/

















































































































































































































































































































Автор: Данилкин А.А.
Теги: artiodactyla парнокопытные систематика животных специальные зоологические науки биология
ISBN: 5-89118-228-09
Год: 2002




Текст
МЛЕКОПИТАЮЩИЕ
РОССИИ
И СОПРЕДЕЛЬНЫХ
РЕГИОНОВ
А.А. ДАНИЛКИН
СВИНЫЕ
ГЕОС
РОССИЙСКАЯ АКАДЕМИЯ НАУК
Институт проблем экологии и эволюции им. А. Н. Северцова
* * *
Московское областное общество охотников и рыболовов
МЛЕКОПИТАЮЩИЕ
РОССИИ
И СОПРЕДЕЛЬНЫХ
РЕГИОНОВ
А.А. ДАНИЛКИН
СВИНЫЕ
(SUIDAE)
Москва
ГЕОС
2002
УДК 599.731
ББК 28.69
Д 18
Свиные (Suidae) / А.А. Данилкин. — М.: ГЕОС, 2002. — 309 с.
(Млекопитающие России и сопредельных регионов).
ISBN 5-89118-228-09
Современный обзор систематики и биологии свиных фауны России и со-
предельных регионов. Рассматриваются филогения, изменчивость, таксоно-
мия, динамика ареала, численность и факторы, ее определяющие, среда оби-
тания, питание, размножение, структура популяций, использование ресурсов,
управление популяциями. Для зоологов, экологов, этологов, охотоведов и
специалистов по охране природы.
Табл. 51. Ил. 80. Библ. 1169.
Рецензенты: доктор биологических наук В.А. Кузякин
кандидат биологических наук Г.И. Иванова
Pigs (Suidae) I А.А. Danilkin. Moscow: GEOS, 2002. 309 p.
The book is the research into systematics and biology of wild boars of the fauna
of the Russia and contiguous districs. The phylogeny, systematics, variability,
range and numbers and its underlying factors, environment, feeding, breeding,
population structure, utilization of their resources are given. For zoologists, ecolo-
gists, ethologists, game biologists.
Tabl. 51. Fig. 80. Ref. 1169.
ISBN 5-89118-228-09
© А. А. Данилкин, 2002
©ГЕОС
Предисловие
Монография “Свиные” - очередная книга в серии “Млекопитающие России и
сопредельных регионов”, продолжение и дополнение издания ’’Млекопитающие
Советского Союза” (1961-1976), основанного В.Г. Гептнером. Ранее были опуб-
ликованы “Зайцеобразные” (1994), “Усатые киты” (1994), “Тушканчикообраз-
ные” (1995), “Оленьи” (1999).
Каждый том серии - всестороннее и сравнительно полное описание опреде-
ленной систематической группы млекопитающих, обитающих на территории
России и сопредельных стран. Разумеется, достичь всеобъемлющей характери-
стики биологических объектов не удается, даже если над монографией трудится
большой коллектив ученых. Особенно проблематичными становятся разделы
«Морфология», «Физиология» (во многих монографиях этой серии они отсутст-
вуют) и «Систематика». Недостаточный уровень знаний генофонда и морфомет-
рической изменчивости животных не позволяет разработать удовлетворительную
внутривидовую таксономию. Из-за отсутствия мечения во многом неясны про-
странственная структура популяций и закономерности перемещения зверей.
В последнее десятилетие в условиях глубокого экономического кризиса по-
левые териологические исследования в России практически, за редким исключе-
нием, прекращены. Значительно уменьшились возможности и в подготовке фун-
даментальных монографий. Однако необходимость в них, особенно по промы-
словым животным, и свиным в частности, не отпала.
После всеобъемлющего тома «Парнокопытные и непарнокопытные» [Гепт-
нер и др., 1961] отечественных сводок по семейству свиных не было. Имеющиеся
многочисленные литературные сведения о единственном представителе этого
семейства на территории России и сопредельных республик - кабане разрознены
и нуждаются в обобщении и анализе. Этот труд автор взял на себя, полагая, что
монография «Свиные», наряду с опубликованной ранее сводкой «Оленьи»
(1999), станет полезным справочным пособием для зоологов, экологов, специа-
листов по охране природы и охотоведов.
Кабан, на мой взгляд, заслуживает самого пристального внимания не только
как интереснейший биологический вид и предок домашней свиньи, но и как важ-
ный охотничий объект, имеющий огромные потенциальные возможности. При-
способленность дикой свиньи к экстремальной и измененной человеком среде
обитания поразительна, что в сочетании с высокой продуктивностью позволяет
этому зверю быстро увеличивать численность и занимать обширный ареал. Как
активный член биоценоза кабан в значительной мере способствует преобразова-
нию природных экосистем, а при высокой плотности населения способен нано-
сить существенный вред сельскому хозяйству. В России ресурсы диких свиней, к
сожалению, скудны и используются крайне нерационально. Поголовье этих ко-
пытных вполне реально увеличить в несколько раз при минимизации вреда ок-
ружающей среде, но для разумного управления популяциями необходимо знать
элементарные биологические основы их функционирования.
Структура книги требует некоторых пояснений. В ней, как и в предыдущих
томах, не рассматриваются вымершие роды и виды. Для краткости изложения
все сравнительные сведения объединены в таблицах, что позволяет использовать
монографию в качестве справочного пособия и значительно облегчает поиск
нужной информации. Статистическая обработка материала, к сожалению, оказа-
лась невозможной из-за отсутствия стандартных математических вычислений в
3
большинстве анализируемых работ. Из-за недостатка места отдельные слова (об-
ласть, край, республика) в таблицах опущены или сокращены: заповедник (з-к),
Центрально-Черноземный (Ц.-Черноземный), северо-восток (с.-в.) и т.п. Назва-
ния республик не всегда совпадают, что вызвано изменениями в административ-
но-территориальном устройстве бывшего СССР и России. Прочерки в таблицах
или знаки вопроса, как правило, - отсутствие сведений или информация, вызы-
вающая сомнение.
Краткая морфометрическая характеристика семейства, рода, вида и синони-
мика, касающаяся главным образом отечественных форм, основаны на анализе
соответствующих разделов известных фундаментальных работ [Ellerman,
Morrison-Scott, 1951; Соколов И.И., 1959; Гептнеридр., 1961; Громов и др., 1963;
Соколов В.Е., 1979; Барышников и др., 1981; Павлинов, Россолимо, 1987] с необ-
ходимыми изменениями и дополнениями. Ссылки на литературные источники,
приведенные в синонимике, указаны в вышеназванных публикациях и не вклю-
чены в список литературы. В связи с заданной структурой серийного издания и
диагностическим подходом в отдельных разделах неизбежно незначительное
«дублирование» материала, что, однако, оказывается полезным при использова-
нии книги в качестве справочного пособия.
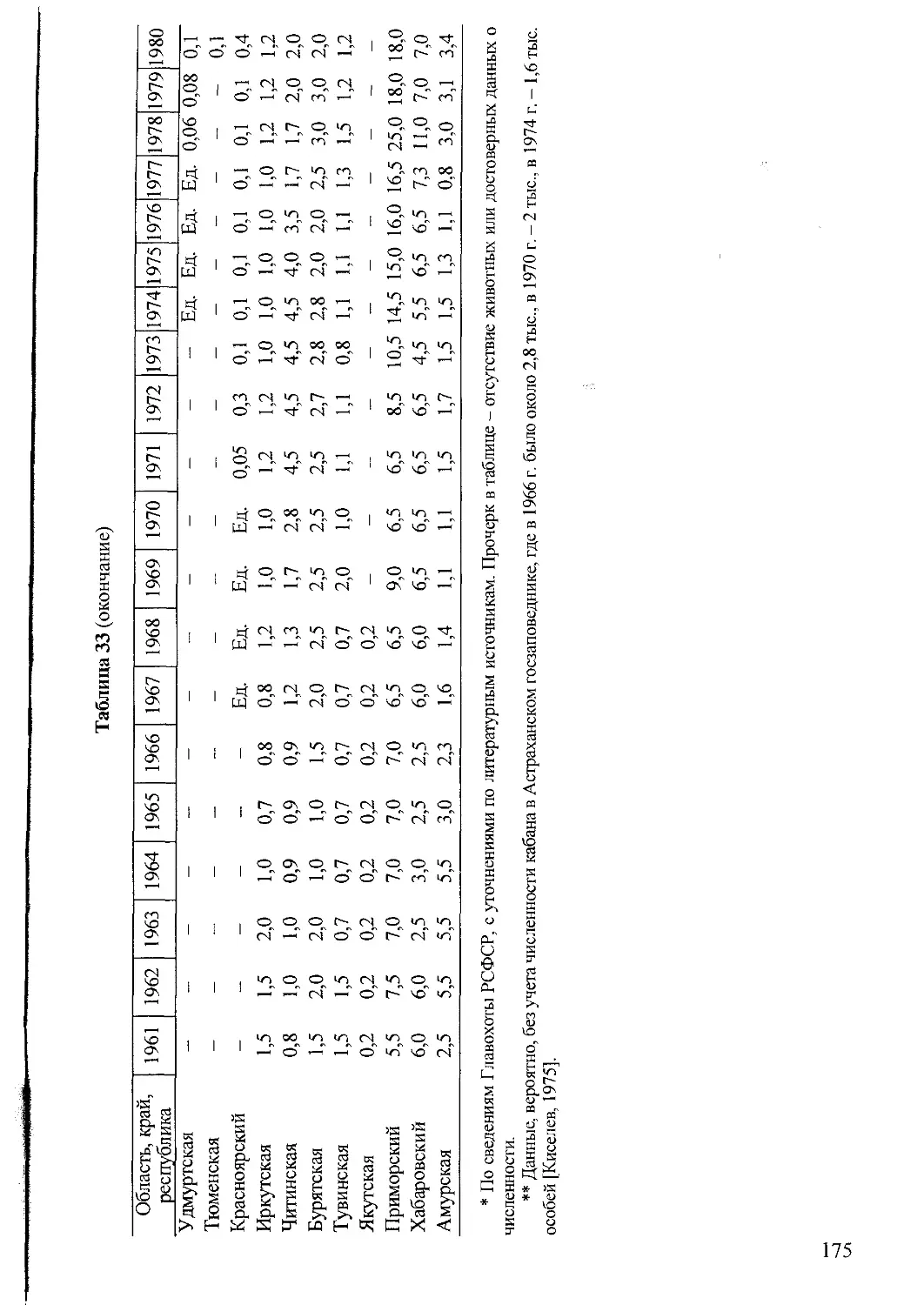
Материалы, касающиеся современного распространения, численности и добы-
чи животных, обобщены по литературным источникам и сведениям Департамента
по охране и рациональному использованию охотничьих ресурсов Минсельхозпро-
да РФ (Главохоты РСФСР, Охотдепартамента РФ), его Центральной научно-
исследовательской лаборатории (ЦНИЛ) и Государственной службы учета охот-
ничьих ресурсов, Ассоциации «Росохотрыболовсоюз» и их территориальных под-
разделений. Из-за сложностей учета все данные о численности зверей сугубо при-
близительные, но они вполне отражают тенденцию динамики популяций.
Автор весьма признателен за помощь в работе над рукописью коллегам: Л.Л.
Данилкиной, В.А. Кузякину, Г.И. Ивановой, М.В. Куликовой, И.Я. Павлинову, а
также многим зоологам и работникам охотничьего хозяйства, участвовавшим в
полевой работе. Оригинальные рисунки выполнены В.М. Гудковым и
А.А.Варшавским. Фотографии любезно предоставлены редакцией журнала
«Охота и охотничье хозяйство»; их авторам, а также главному редактору О.К.
Гусеву я приношу глубокую благодарность. На обложке книги использованы фо-
тографии зверей, сделанные А.Ф. Дигилевичем.
Замечания, уточнения или дополнения прошу направлять по адресу: 117071,
Москва, Ленинский проспект, 33. Институт проблем экологии и эволюции им.
А.Н. Северцова.
4
Семейство СВИНЫЕ
Suidae Gray, 1821
Характеристика семейства. Свиные - парнопалые, коротконогие нежвачные
животные среднего и мелкого размеров с относительно редким волосяным покро-
вом. Передняя часть головы длинная, конусовидная, оканчивающаяся круглым,
плоским, лишенным волос подвижным «пятачком», на поверхности которого от-
крываются ноздри. Остовом пятачка служит непарная хоботковая кость (os rostri),
расположенная между передними краями межчелюстных и носовых костей.
Череп удлиненный, клиновидной формы. Лицевая часть вытянутая. Лобноте-
менная поверхность плоская, постепенно поднимающаяся к затылку. В отличие
от других парнопалых, крыловидная ямка (fossa pterygoidea) хорошо выражена.
Глазница незамкнутая. Слуховая капсула вздута, грушевидной формы. Отверстие
носослезного канала в слезной кости двойное. Максимально (у рода Sus) 44 зуба:
i 3/3, с 1/1, р 4/4, m 3/3, но их число весьма изменчиво. У представителей боль-
шинства родов исчезают боковые резцы или передние предкоренные, иногда те и
другие. Заднекоренные зубы бугорчатые (бунодонтные). Клыки сильно развиты
и с постоянным ростом, их корни открыты. Верхние клыки изгибаются кверху. В
месте их выхода верхнечелюстные кости утолщены незначительно или сильно.
На конечностях по 4 пальца, боковые пальцы сдвинуты назад и значительно
короче средних. Метаподии как на передних, так и на задних конечностях не
слиты; локтевая и малая берцовая кости самостоятельные. Фаланги пальцев (в
отличие от гиппопотамов) в роговых чехлах.
Желудок простой, не разделенный на отделы, с левосторонним выпячивани-
ем в кардиальной части. Слепая кишка и желчный пузырь имеются. Плацента
диффузная. Брюшных или паховых сосков более 2 пар.
Кариотип представителей отряда Suiformes, и семейства Suidae в частности,
резко отличается от такового у Tylopoda и Ruminantia [Трошина, 1975; Графодат-
ский, 1989].
Всеядные формы, предпочитающие области с повышенным увлажнением.
Полигамны. Скороспелы и плодовиты, приносят в год один или два помета. Де-
тенышей в помете от 1 до 15.
Традиционные объекты охоты.
Распространение. Естественный ареал: Европа, Азия, включая Цейлон и
острова Малайского архипелага, Африка, о. Мадагаскар. В последние столетия
отдельные представители семейства акклиматизированы в Северной и Южной
Америке, Австралии, Новой Зеландии, Тасмании, Новой Гвинее.
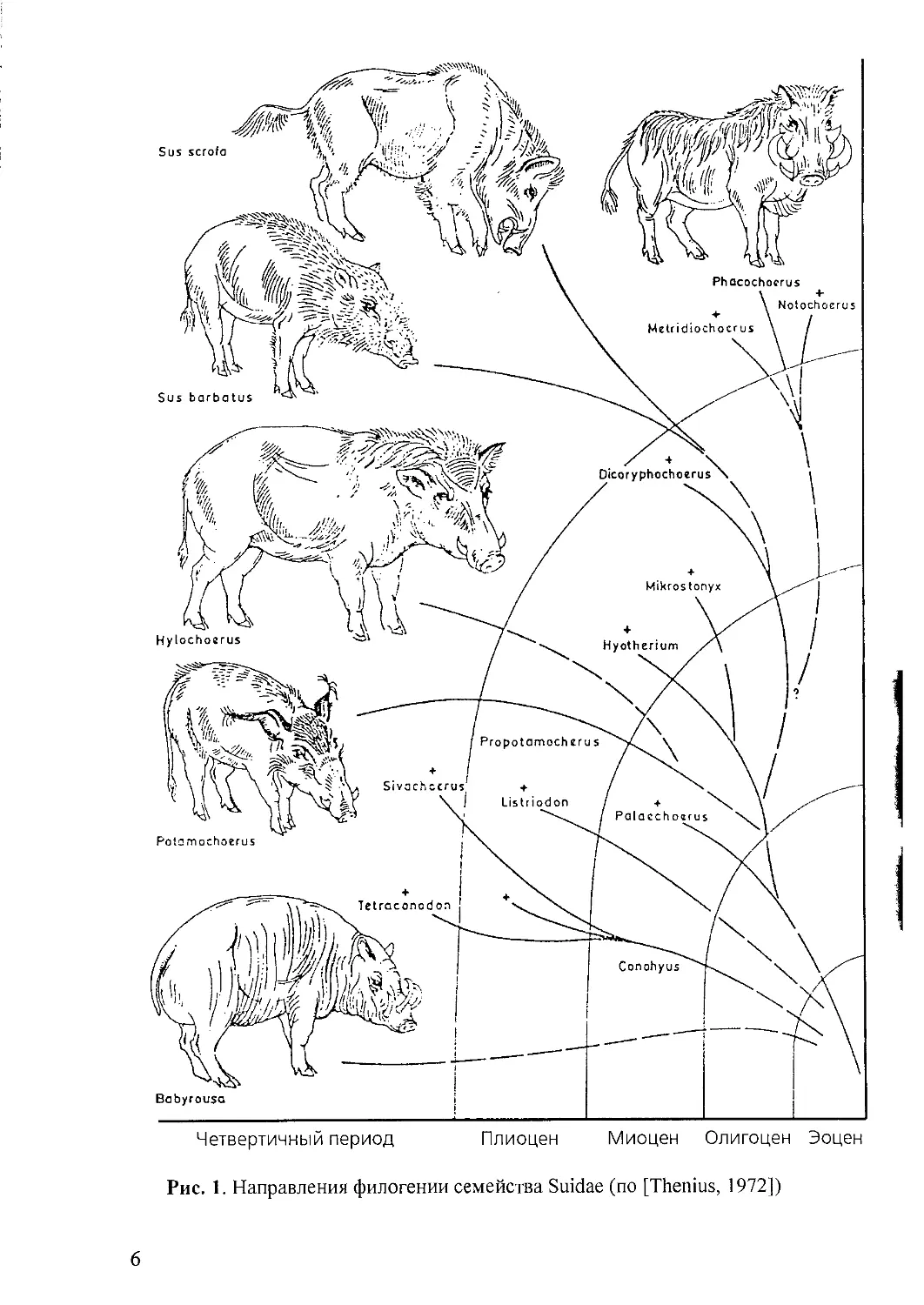
Эволюция. Свиные, вероятно, являются одной из ветвей верхнеэоценовых
палеодонтов (рис. 1), имевших короткую лицевую часть и четырехбугорчатые
коренные зубы. У нижнеолигоценовых представителей рода Propalaeochoerus
удлиняются нижние клыки, у миоценовых Palaeochoerus и Hyotherium они пре-
вращаются в постоянно растущие, усложняются коренные зубы, удлиняются ли-
цевые кости черепа.
Представители подсемейства Suinae Zittel, 1893 появляются в позднем мио-
цене Старого Света. Большая их часть затем вымерла, не оставив потомков [Со-
колов, 1959; Гептнер и др., 1961; Thenius, 1970, 1972]. Роды Propotamochoerus и
Sus известны из отложений эоплейстоцена. Propotamochoerus provincialis Gervais,
5
Четвертичный период
Плиоцен Миоцен Олигоцен Эоцен
Рис. 1. Направления филогении семейства Suidae (по [Thenius, 1972])
6
1859 дожил в лесах Южной Европы и Закавказья до древнего плейстоцена. Со-
временные потомки этой группы, относимые к роду Potamochoerus, сохранились
в Западной Африке до наших дней [Алексеева, 1977; Верещагин, Барышников,
1980а; Барышников и др., 1981].
Основное направление эволюции семейства - приспособление к рытью поч-
вы и добыванию подземных частей растений и почвенных животных, всеядность.
Состав. Свиные (Suidae Gray, 1821), пекариевые (Tayassuidae Palmer, 1897) и
бегемотовые (Hippopotamidae Gray, 1821) составляют подотряд Свинообразных
(Suiformes Jaeckel, 1911).
Семейство Suidae объединяет 34 вымерших и 5 современных родов [Соколов,
1979; Барышников и др., 1981]:
Sus Linnaeus, 1758 - собственно Свиньи (Евразия, север Африки);
Potamochoerus Gray, 1854 - Кистеухие, или речные свиньи (Африка к югу от
Сахары и остров Мадагаскар);
Babyrousa Perry, 1811 - Бабируссы (острова Борнео и Целебес);
Phacochoerus F. Cuvier, 1817 - Бородавочники (Африка к югу от Сахары);
Hylochoerus Thomas, 1904 - Лесные свиньи (зона африканских экваториаль-
ных лесов).
В фауне России и сопредельных стран семейство представлено 7 вымершими
и одним современным родом Sus.
Род СВИНЬИ
Sus Linnaeus, 1758
Sus Linnaeus, 1758. Syst. Nat., ed. 10, 1:49. Германия.
Aper Pallas, 1766. Misc. Zool.:16. Nom. nov. pro.Su.v Linnaeus.
Scrofa Gray, 1868. Proc. Zool. Soc. London:38.
Sinisus Heude, 1892. Mem. Hist. Nat. Emp. Chin., 2:102.
Характеристика рода. Свиньи - животные средней величины, за исключени-
ем карликовой свиньи (S. salvanius), длина тела которой 50-65 см, высота в холке
25-30 см, длина черепа 150-160 мм. Взрослые самцы обычно заметно крупнее са-
мок, в холке они выше, чем в крестце. Голова непропорционально большая, клино-
видная. Вытянутое рыло оканчивается притупленным пятачком, снабженным ося-
зательными ворсинками. Шея короткая. Глаза маленькие. Уши относительно ко-
роткие, широкие. Хвост короткий, подвижный или рудиментарный (у S. salvanius).
Череп узкий и удлиненный в лицевой части, без сагиттального гребня (в от-
личие от Babyrousa). Затылочная область сильно приподнята, задняя ее поверх-
ность вертикальна или скошена вперед. Полный зубной ряд (см. выше) не у всех
форм. Крайняя пара резцов и первый верхний предкоренной выпадают у моло-
дых животных большинства видов. Первый переднекоренной зуб не имеет мо-
лочного предшественника. Утолщений верхнечелюстной кости у основания
верхних клыков нет, или они слабые. Клыки разной формы, постоянно растущие:
нижний - трехгранный, концевые части верхних скошены в результате постоян-
ного стачивания нижними. У карликовой гималайской формы последний корен-
ной зуб верхней челюсти редуцирован.
Волосяной покров (жесткая щетина и пуховые волосы) относительно хорошо
развит, на шее и по хребту удлиненный. У отдельных южных форм отсутствует
подшерсток, у других на боках головы развиты парные бородавчатые выросты
кожи. Окраска детенышей, за редким исключением, продольно-полосатая, у
взрослых однотонная, от темной до светлой и рыжеватой, вдоль хребта тянется
темная полоса.
7
Брюшных и паховых сосков от 6 (у карликовых свиней) до 12. У домашних
свиней в результате направленного отбора их число увеличилось до 17-20 [Гра-
чев, Галанцев, 1973].
Обитатели влажных биотопов и зарослей. Имеют важное промысловое зна-
чение. Родоначальники домашней свиньи.
Распространение. Большая часть Европы, юг Азии, включая многочислен-
ные крупные и мелкие острова Тихого океана, север Африки (рис. 2). Действи-
тельный естественный ареал, скорее всего, меньше, поскольку на многие тихо-
океанские острова свиньи могли быть завезены человеком. На север Африки жи-
вотные проникли, видимо, из Южной Азии по так называемому арабо-
палестинскому коридору. В последние века представители рода акклиматизиро-
ваны в Северной и Южной Америке и Австралии.
Рис. 2. Восстановленный ареал рода Sus (по [Гептнер и др., 1961])
Область распространения Sus scrofa ограничена сплошной линией, S. barbatus - пунктиром. В
центральной части Азии имеется пробел в ареале
Эволюция. Род Sus прослеживается с верхнего миоцена - нижнего плиоцена
Евразии от примитивных миоценовых Dicoryphochoerus Pilg. (см. рис. 1), от кото-
рых отличается усложненными и удлиненными коренными зубами и прогресси-
рующим ростом клыков [Пидопличко, 1951, 1954; Соколов, 1959; Thenius, 1970,
1972; Groves, 1981]. Род наименее специализирован, однотипен и наиболее близок
к древним формам. Основные направления эволюции те же, что и для семейства.
Состав. Классификация рода крайне запутана [Thenius, 1972; Groves, 1981,
1997], что связано как с несовершенством описаний отдельных видов, так и с
влиянием доместикации (смешением диких и домашних форм и одичанием по-
следних на островах между Азией и Австралией). В последние десятилетия из 9-
8
12 выделяемых ранее видов систематики принимают чаще лишь три [Гептнер и
др., 1961; Соколов, 1979]:
S. scrofa L., 1758 (= leucomystax, cristatus, vittatus, falzfeini, orientalis и др.) - кабан,
или дикая свинья (естественный ареал - большая часть Евразии, север Африки);
S. barbatus Muller, 1838 (= verrucosus, celebensis и др.) - бородатая свинья
(Малакка, Суматра, Ява, Калимантан, Сулавеси, Филиппинские острова);
S. salvanius Hodgson, 1847 - карликовая свинья (Восточные Гималаи, Непал,
Бутан).
Однако вряд ли такой видовой состав рода окончательный. Некоторые систе-
матики настаивают на обособлении Sus verrucosus Muller, 1838, а реликтовую
карликовую свинью нередко выделяют в отдельный подрод или род Porcula.
В фауне России и сопредельных регионов один вид.
КАБАН, или ДИКАЯ СВИНЬЯ
Sus scrofa Linnaeus, 1758
Sus scrofa Linnaeus, 1758. Syst. Nat., ed. 10, 1:49. Германия.
Sus setosus Boddaert, 1785. Elench. Anim., 1:157. Nom. nov. pro Sus scrofa Linnaeus.
Sus setosus aper Boddaert, 1785. Ibid., 1:157.
Sus ewopaeus Pallas, 1811. Zoogr. Rosso-Asiat., 1:265. Nom. nov. pro Sus scrofa Linnaeus.
Sus scrofa var. nigripes Blanford, 1875. J. Asiat. Soc. Bengal., 44, 2:112. Китай, Синьцзян, Тянь-
Шань в Кашгарии.
Sus ussuricus Heude, 1888. Mem. Hist. Nat. Emp. China, 2:54. Приморский кр., p. Уссури.
Sus leucomystax var. continentalis Nehring, 1889. Sitzungsb. Ges. Nat. Fr. Berlin, 7:141. Примор-
ский кр., Владивосток.
Susgigas Heude, 1892. Mem. Hist. Nat. Emp. China, 3:191. Приморский кр., Владивосток.
Sus attila Thomas, 1912. Proc. Zool. Soc. London, 1:393 (Abstr. March 12:13). Трансильвания (Ру-
мыния), Колошвар (ныне Клуж).
Sus falzfeini Matschie, 1918. Sitzungsb. Ges. Nat. Fr. Berlin, 8:5. Белоруссия, 75 км к западу от
Минска, Налибоки.
Sus leucomystax sibiricus Staffe, 1922. Arb. Lehrkanz. Tier. Hochschule Bodenkultur Wien, 1:51. Бу-
рятская АССР, хр.Тункинские Гольцы.
Sus scrofa raddeanus Adlerberg, 1930. Докл. АН СССР, 4:95. Монголия, Южный Хэнтэй, Согу-нур.
Характеристика вида. Кабан - наиболее крупная форма рода Sus (рис. 3).
Половой диморфизм четко выражен. Длина тела взрослых самцов в популяциях в
северной части ареала в среднем 150-187 (до 221) см, самок 141-174 (181) см;
высота в холке 80-109 см и 72-97 см; обхват туловища 119-157 см и 101-143 см;
длина ступни 30-37 см и 29-35 см; дайна уха 12-18 см; длина хвоста 24-28 см и
20-29 см. Масса тела в среднем 114-202 (до 350) кг у самцов и 68-144 (до 186) кг
у самок. На юге и западе ареала размеры животных несколько меньше.
Форма тела клиновидная и уплощенная с боков. В холке звери чуть выше, чем
в крестце. Голова непропорционально большая, шея короткая, толстая, с мощной
мускулатурой. Рыло вытянутое, с овальным пятачком, верхняя половина которого
голая, нижняя - с редкими короткими осязательными волосками (рис. 4). Изо рта
торчат клыки, особенно хорошо заметные у взрослых самцов. Глаза маленькие,
глубоко посаженные. Уши сравнительно короткие (11-20 см), широкие. Хвост
(14-35 см) не достигает скакательного сустава, тонкий, подвижный, с удлинен-
ным волосом на конце.
Ноги короткие; боковые пальцы (2-й и 5-й) сравнительно хорошо развиты,
при ходьбе касаются земли. Роговая кромка копыт 3-го и 4-го пальцев острая, что
наряду с вогнутой их поверхностью препятствует скольжению, предоставляя ка-
9
Рис. 3. Кабан, или дикая свинья (фото Н. Власова)
бану, в отличие от других копытных, возможность движения по льду. На мягком
грунте все пальцы широко раздвигаются; это увеличивает площадь опоры и по
зволяет зверю относительно свободно перемещаться по болотистой местности
Длина шага у спокойно идущих животных разная в зависимости от возраста i
размера тела: у полугодовалых поросят - 25-30 см, подсвинков - 28-37 см, двух
годовалых самок - 39-47 см, самцов этого же возраста - 45-54 см, у взрослы:
самок - 50-65 см и у взрослых самцов - 60-75 см [Бромлей, 1964; Чичикин, Во
робьев, 1967].
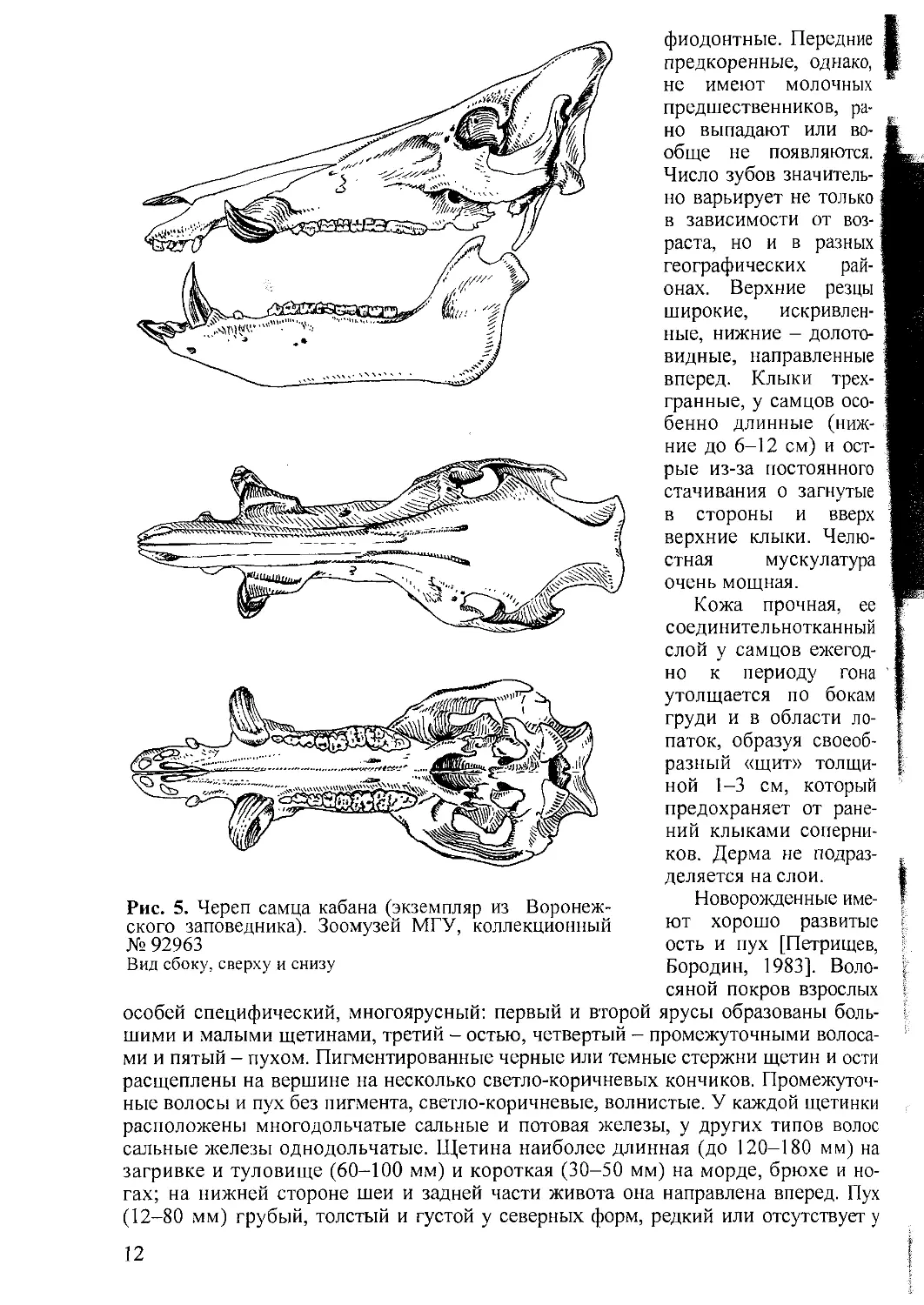
Череп длинный и относительно узкий (рис. 5). У самцов в разных популяция;
в северной части ареала его максимальная длина в среднем 384-466 (до 510) мм
у самок 362^116 (460) мм; скуловая ширина 149-173 мм и 139-161 мм; высот
190-250 мм и 170-227 мм; длина верхнего ряда коренных зубов 110-144 мм
109-137 мм. На юге и западе ареала размеры черепа меньше. Зубы крепкие, ди-
10
Рис. 4. Рыло кабана (фото И. Мухина)
11
Рис. 5. Череп самца кабана (экземпляр из Воронеж-
ского заповедника). Зоомузей МГУ, коллекционный
№ 92963
Вид сбоку, сверху и снизу
фиодонтные. Передние
предкоренные, однако,
не имеют молочных
предшественников, ра-
но выпадают или во-
обще не появляются.
Число зубов значитель-
но варьирует не только
в зависимости от воз-
раста, но и в разных
географических рай-
онах. Верхние резцы
широкие, искривлен-
ные, нижние - долото-
видные, направленные
вперед. Клыки трех-
гранные, у самцов осо-
бенно длинные (ниж-
ние до 6-12 см) и ост-
рые из-за постоянного
стачивания о загнутые
в стороны и вверх
верхние клыки. Челю-
стная мускулатура
очень мощная.
Кожа прочная, ее
соединительнотканный
слой у самцов ежегод-
но к периоду гона
утолщается по бокам
груди и в области ло-
паток, образуя своеоб-
разный «щит» толщи-
ной 1-3 см, который
предохраняет от ране-
ний клыками соперни-
ков. Дерма не подраз-
деляется на слои.
Новорожденные име-
ют хорошо развитые
ость и пух [Петрищев,
Бородин, 1983]. Воло-
сяной покров взрослых
особей специфический, многоярусный: первый и второй ярусы образованы боль-
шими и малыми щетинами, третий - остью, четвертый - промежуточными волоса-
ми и пятый - пухом. Пигментированные черные или темные стержни щетин и ости
расщеплены на вершине на несколько светло-коричневых кончиков. Промежуточ-
ные волосы и пух без пигмента, светло-коричневые, волнистые. У каждой щетинки
расположены многодольчатые сальные и потовая железы, у других типов волос
сальные железы однодольчатые. Щетина наиболее длинная (до 120-180 мм) на
загривке и туловище (60-100 мм) и короткая (30-50 мм) на морде, брюхе и но-
гах; на нижней стороне шеи и задней части живота она направлена вперед. Пух
(12-80 мм) грубый, толстый и густой у северных форм, редкий или отсутствует у
12
южных рас [Сафаров, 1965; Соколов, 1973; Гунчак, 1982; Соколов, Темботов, 1993;
Соколов, Петрищев, 1997]. Толстая прочная щетина, прикрывающая слой пуха, на-
ряду со слоями подкожного (до 3-5 см) и внутриполостного жира, способствует
термоизоляции, скатыванию воды с тела и облегчает передвижение зверей по гус-
тым зарослям.
Окраска взрослых бурая с оттенками от почти черного до серебристо-серого,
соломенного или рыже-бурого, что определяется в основном цветом расщеплен-
ных концов щетины. Передняя часть головы темнее туловища, иногда наоборот.
Пятачок, губы, подбородок, уши, низ конечностей, копыта и кончик хвоста тем-
ные, нередко черные. В целом окраска покровительственная, типа «агути», с ха-
рактерной темной полосой вдоль хребта и светлеющая на боках, с серовато-
белыми участками на животе и в паху, черными передними поверхностями ко-
нечностей. Летом во время единственной линьки с потерей пуховых волос каба-
ны становятся темными или черными. Молодые особи обычно светлее. Для дете-
нышей характерна полосатая окраска - светлые продольные полосы на рыже-
буром фоне.
Хорошо развиты специфические железы: карпальные, межпальцевые (мешки
отсутствуют), гардерова над клыками в верхней губе, подбородочная, на пятачке,
циркуманальные и половые. Мейбомиевы железы, типичные для других предста-
вителей Suidae и домашних свиней, не развиты, а их место занято толстым мы-
шечным тяжем, вытянутым вдоль века. Нет у кабана и четко выраженных пред-
глазничных желез. Верхняя стенка препуция образует впячивание в виде мешка
размером с яйцо, который заполнен секретом сальных и апокриновых трубчатых
желез и сшелушивающимся (или секретирующим по голокриновому типу) эпи-
телием стенок, а также конкрециями с сильным запахом застоявшейся мочи.
Размеры препуциального мешка в период гона увеличиваются в два раза. Его
секрет вместе с мочой образует половой феромон. Запах секрета карпальной же-
лезы также служит половым аттрактантом [Соколов, Чернова, 2001].
У диких свиней тонкое обоняние, хороший слух, но слабое зрение. В экспе-
риментах звери четко различают синий, пурпурно-синий и пурпурный цвета, ху-
же-зеленый и серый и не различают красный и серый [Eguchi et al., 1997].
Эти копытные всеядны и явно приспособлены к собиранию пищи на земле
или в верхнем ее слое. Благодаря сильной шейной мускулатуре легко роют зем-
лю, разламывают смерзшиеся пласты до 9 см толщиной, рвут толстые (до 3 см)
корни, выворачивают пни, лежащие деревья и крупные камни. Благодаря под-
вижной нижней губе могут выбирать из земли мелкие пищевые объекты величи-
ной с зерно. После рытья почвы регулярно прочищают носовые ходы, с силой
выдыхая воздух.
Несмотря на коротконогость, звери весьма подвижны, перемещаются на
большие расстояния, хорошо плавают и форсируют широкие водные преграды.
«Обтекаемая» форма тела позволяет легко проходить через густые заросли. Пе-
редвигаются шагом, рысью и галопом (максимальная скорость до 40 км в час на
коротком расстоянии), способны прыгать в длину до 4 м и в высоту до 1,5-2 м.
Взрослые особи могут преодолевать рыхлый снег высотой до 40-50 см. При
большей его высоте животные передвигаются исключительно по набитым тро-
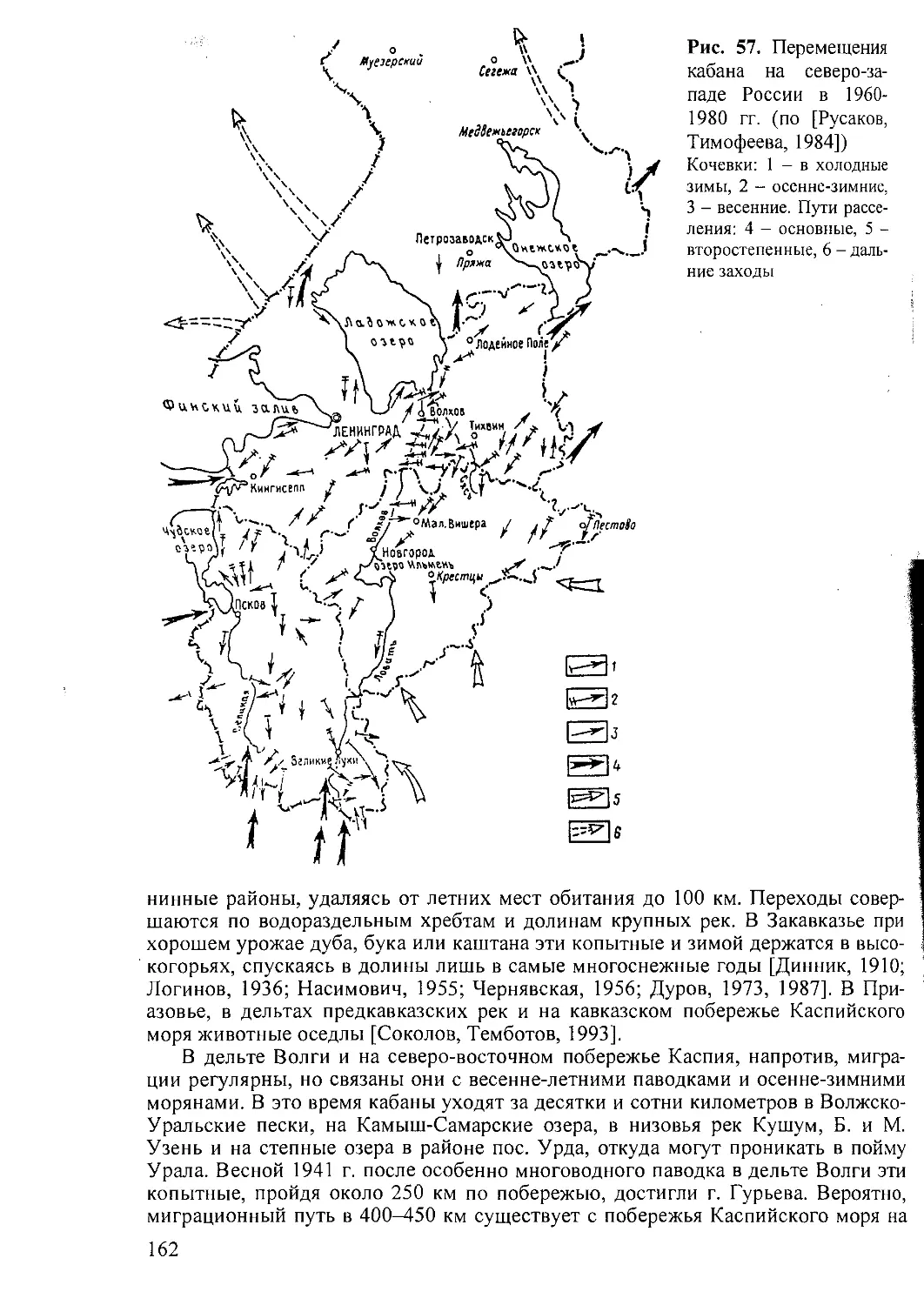
пам. Во многих районах выражены сезонные миграции.
При опасности кабаны спасаются бегством, но могут эффективно обороняться,
используя в качестве «оружия» зубы. В определенные периоды жизни агрессивны.
Образ жизни преимущественно семейно-групповой и скрытный - сумереч-
ный и ночной. При слабом беспокойстве активны и в дневное время. Очень хо-
рошо развито чувство времени, что подтверждается точным (до минут) появле-
нием зверей на подкормочных площадках.
Весьма плодовиты - до 15 эмбрионов, среднее число поросят в выводке - 5-7.
13
Продолжительность жизни в неволе около 20 лет, но в природе редко превы-
шает 10-12 лет.
Родоначальник домашней свиньи, с которой легко скрещивается. Процесс одо-
машнивания шел одновременно в нескольких частях ареала, вероятно, с мезолита.
Важный охотничий вид. При высокой плотности может приносить вред сель-
скому хозяйству и существенно влиять на биоценозы.
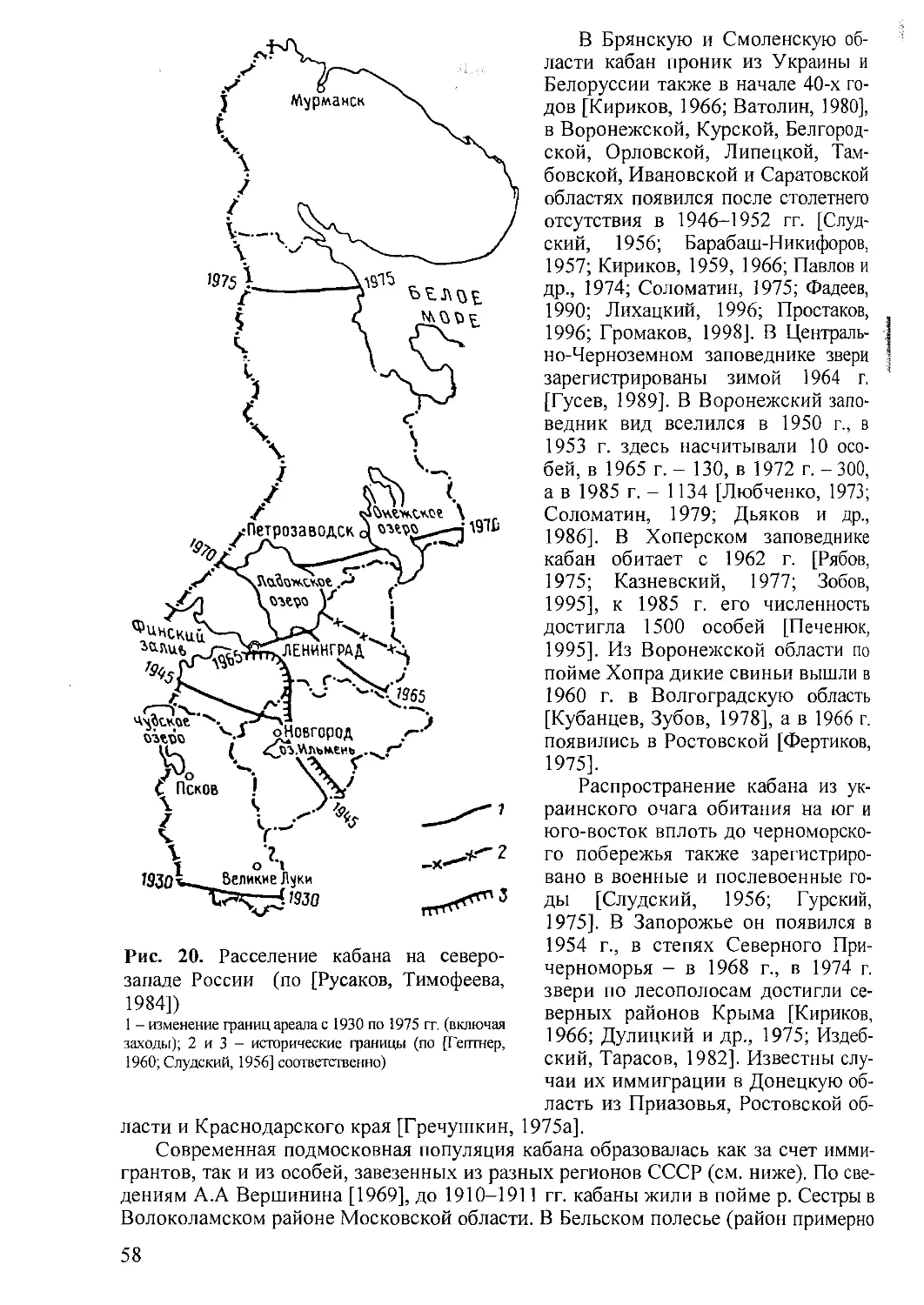
Распространение. Большая часть Европы и Азии, включая многие острова
Тихого океана, север Африки (см. рис. 2). Интродуцирован в Северной и Южной
Америке и Австралии.
Эволюция. В Евразии ископаемые предковые формы дикой свиньи (вероят-
но S. prisons') появляются в позднем плиоцене. На западе Европы, в Сирии, Китае
и Юго-Восточной Азии формы, близкие к современному кабану, обнаруживают-
ся в раннечетвертичных слоях. На территории бывшего СССР они найдены в ок-
рестностях Самарканда, в Таджикистане и в составе хапровской фауны в Приазо-
вье. В других районах Средней Азии, в Закавказье, Предкавказье, Приазовье,
Крыму и Южном Приморье прослеживаются с плейстоцена [Смирнов, 1940;
Громов, 1948; Пидопличко, 1951, 1954; Верещагин, 1959; Соколов, 1959; Биби-
кова, 1960; Батыров, 1969; Байгушева, 1971; Шарапов, 1972а; Алексеева, 1977;
Верещагин, Громов, 1977].
Одна из известных эволюционных ветвей — европейский раннеплейстоценовый
Sus strozzii F. Major, 1881 (возможно, = S. tamanensis N. Verestchagin, 1957) и сред-
неплейстоценовый апшеронский S. apscheronicus Burtschak-Abramovitsch et
Dzjafarov, 1946 из Восточного Закавказья [Бурчак-Абрамович, Джафаров, 1948;
Верещагин, 1957, 1959; Верещагин, Барышников, 1980а; Барышников и др., 1981].
Эволюция вида шла по пути уменьшения размеров тела, усложнения строе-
ния зубов и возникновения морфофизиологических и поведенческих адаптаций к
меняющейся среде обитания. При этом за последние сотни тысячелетий морфо-
логический облик диких свиней изменился незначительно.
Состав. Описано более 30 рас S. scrofcr. scrofa, setosus, aper, europaeus,
celtica, castilianus, boeticus, attila, falz-feini, nigripes, raddeanus, continentalis,
leucomystax, meridionalis, reiseri, majori, barbatus, sennariensis, lybicus, cristatus,
vittatus, floresianus, timorensis, nicobaricus, andamanensis, taivanus, chirodonlus,
coreanus, riukianus, moupinensis, papuensis, niger, aruensis и другие, а количество
имен близко к 50 [Адлерберг, 1933; Филипченко, 1933; Ellerman, Morrison-Scott,
1951; Соколов, 1959; Гептнер и др., 1961; Groves, 1981; Sjarmidi, Gerard, 1988].
Большинство из них выделены исходя, скорее, из географического местоположе-
ния, нежели из морфологического принципа классификации. Действительное
число форм, которые можно было бы признать подвидовыми, несомненно, гораз-
до меньше. Требуется всесторонняя и тщательная ревизия таксономии вида.
На территории России и сопредельных регионов систематики выделяют до 9
форм, из которых 5 имеют статус подвидов. О географической и индивидуальной
(включая развитие и рост) изменчивости и таксономии диких свиней более под-
робно пойдет речь в следующей главе.
Изменчивость и таксономия
Размеры и масса тела
Новорожденные кабаны сравнительно хорошо развиты, зрячие, с плотным воло-
сяным покровом (рис. 6). Их масса колеблется в очень больших пределах в зависи-
мости от числа эмбрионов и физиологического состояния самок - от 500 до 1680 г,
составляя в среднем около 1 кг. Длина тела - 22-42 см, высота в холке - 16-26, об-
14
a
й
Рис. 6. Новорожденный (фото А. Стрелкова)
хват туловища - 23-26, длина стопы - 8-10, длина уха - 4-5, длина хвоста - 5-7 см.
В первые дни после рождения масса поросят несколько уменьшается за счет расхо-
дования эмбрионального жира. К концу первого месяца жизни она достигает 3-6 кг,
второго - 9-15, третьего - 17-22, четвертого - 26, пятого - 30, на шестом - примерно
33-39 кг. Зимой рост сеголеток прекращается, и к весне их масса уменьшается
[Бромлей, 1964; Козло и др., 1974; Козло, 19746, 1975; Курносов, Петрищев, 1977;
Гунчак, 1978; Петрищев, 1980; Русаков, Тимофеева, 1984; Петрищев, Бородин, 1985;
Briedermann, 1986; Недзельский, 1994; Простаков, 1996J.
Сеголетки в возрасте 7-12 месяцев имеют следующие параметры, относительно
близкие у самцов и самок: длина тела в среднем 100—110 см, высота в холке - 55-67
см, обхват туловища-72-91 см, длина стопы -22-28 см, длина уха-9-12 см, длина
хвоста- 14-21 см (табл. 1). Их масса колеблется в пределах 29-40 (до 65) кг (табл. 2).
У растущих диких свиней значительно меняются пропорции тела и относи-
тельные размеры внутренних органов, причем динамика этих изменений у сам-
цов и самок неодинакова и заметно отличается от таковой у домашних животных
[Иваненко, 1956; Сафаров, 1965; Смирнова, 1972; Гунчак, 1982; Trnkova, 1984;
Давлетова, 1986, 1992].
Рост и развитие детенышей существенно зависят от условий питания, а также
от уровня зараженности гельминтами. В вольерах при обильном и регулярном
питании масса особей почти в 1,5 раза больше, чем у их ровесников в природе
[Иваненко, 1956]. В Беловежской пуще при урожаях дуба и бука средняя масса
сеголеток 35-37 кг, длина тела 107-111 см, в неурожайные годы - 27-28 кг и
100-102 см [Козло, 1970а, 1975], в Украинских Карпатах соответственно 40-50
кг и 30-40 кг [Гунчак, 1978, 1982]. На Северном Кавказе в урожайные годы масса
отдельных 8-месячных поросят достигает 60,7 кг, в неурожайные уменьшается на
15
Таблица 1. Размеры тела (см) 7-12-месячных особей
Район п М lim | Источник информации
Германия 1326 Длина тела 100-106* 71-129 Briedermann, 1986
Беловежская пуща 19 101-105* 87-125 Козло, 19706, 1975
Березинский з-к 30 110-109* 96-125 Козло и др., 1974
Литва 52 103-108* 70-122 Янулайтис, 1988; Янулайщс,
Украинские Кар- 18 111-109* 96-131 Козла, 1988 Гунчак, 1978, 1982
паты Юг Украины — 118-142 Щербак, 1986
Ц. Черноземье 124 102 76-105 Простаков, 1994,1996 |
Кавказский з-к 4 113 103-120 Дуров, Александров, 1968 1
Азербайджан 16 103-111* 99-125 Сафаров, 1965 Я
Низовья р. Или 12 100,5 92-115 Слудский, 1956 1
Ю. Киргизия 5 111 105-118 Чичикин, Воробьев, 1967 J
Таджикистан 13 102-126 — Иваненко, 1956 Я
Предбайкалье 21 100-104* 89-112 Недзельский, 1994 Я
Сихотэ-Алинь 26 107-108* __ Бромлей, 1964 1
Беловежская пуща 19 Высота 59-64* в холке 55-75 Козло, 1975 Я
Березинский з-к 30 66* 56-74 Козло и др., 1974 Я
Литва 52 60-62* 40-78 Янулайтис, 1988; Янулайщс, I
Украинские Кар- 18 68-67* 54-79 Козла, 1988 1 Гунчак, 1978, 1982 I
паты Юг Украины — — 71-87 Щербак, 1986 1
Ц. Черноземье 124 62 44-68 Простаков, 1994, 1996 1
Кавказский з-к 4 67 62-73 Дуров, Александров, 1968 1
Азербайджан 16 61-62* 48-76 Сафаров, 1965 1
Низовья р. Или 14 55-57* 48-63 Слудский, 1956 I
Ю. Киргизия 5 64 59-68 Чичикин, Воробьев, 1967 ।
Таджикистан 13 55-65 — Иваненко, 1956 ।
Предбайкалье 21 57-62* 50-67 Недзельский, 1994 |
Сихотэ-Алинь 26 62-65* - Бромлей, 1964 |
Беловежская пуща 19 Обхват туловища 73-79* 65-84 Козло, 1975 :
Березинский з-к 30 91-88* 76-108 Козло и др., 1974
Литва 52 75-78* 48-96 Янулайтис, 1988; Янулайщс,
Украинские Кар- 18 78* — Козла, 1988 Гунчак, 1978
паты Юг Украины — — 83-102 Щербак, 1986
Ц. Черноземье 124 76 57-82 Простаков, 1994, 1996
Кавказский з-к 4 84 73-98 Дуров, Александров, 1968
Азербайджан 16 74-81* 66-90 Сафаров, 1965 s
16
Таблица 1 (окончание)
Район | п | М | lim | Источник информации
Предбайкалье 21 72-77* 64-84 Недзельский, 1994
Сихотэ-Алинь 26 73-80* Длина стопы Бромлей, 1964
Беловежская пуща 19 24-26* — Козло, 1975
Березинский з-к 30 28-27* 19-31 Козло и др., 1974
Литва 52 25-26* — Янулайтис, 1988; Янулайщс, Козла, 1988
Ц. Черноземье 124 25 24-27 Простаков, 1996
Кавказский з-к 4 24 23-25 Дуров, Александров, 1968
Низовья р. Или 8 22 20-24 Слудский, 1956
Сихотэ-Алинь 26 26-27* Длина уха Бромлей, 1964
Беловежская пуща 19 10-11* 8-11 Козло, 1973, 1975
Березинский з-к 30 11* 10-13 Козло и др., 1974
Литва 52 10-11* — Янулайтис, 1988; Янулайщс, Козла, 1988
Украинские Кар- паты 18 12-12,7* — Гунчак, 1978
Ц. Черноземье 124 10,5 9-12 Простаков, 1994, 1996
Кавказский з-к 4 11 11-12 Дуров, Александров, 1968
Низовья р. Или И 9,5 8-11 Слудский, 1956
Предбайкалье 21 9* 8-10 Недзельский, 1994
Сихотэ-Алинь 26 9,5 Длина хвоста Бромлей, 1964
Беловежская пуща 19 16-17* 12-17 Козло, 1973, 1975
Березинский з-к 30 20-21* 17-28 Козло и др., 1974
Литва 52 18-19* — Янулайтис, 1988; Янулайщс, Козла, 1988
Украинские Кар- паты 18 18* — Гунчак, 1978
Ц. Черноземье 124 16 12-17 Простаков, 1994, 1996
Кавказский з-к 4 16 13-19 Дуров, Александров, 1968
Низовья р. Или 10 13,5 10-15 Слудский, 1956
Предбайкалье 21 14-15* 13-16 Недзельский, 1994
Сихотэ-Алинь 26 15 — Бромлей, 1964
* У самок и самцов соответственно.
Прочерк - отсутствие сведений.
47%, а размеры - на 4-5% [Дуров, Александров, 1968; Дуров, 1987]. В нацио-
нальном парке «Завидово» в зависимости от условий обитания средняя масса се-
голеток варьирует от 33,4 кг до 38,6 кг в разные годы [Фертиков и др., 1999]. В
Подмосковье здоровые 10-12-месячные поросята весят 28-35 кг, зараженные
гельминтами - 20-23 кг [Овсюкова, Бельский, 1980], в Таджикистане соответст-
венно 30-34 кг и 18-20 кг [Мельникова, 1968].
2. Заказ №2528.
17
Условия роста и развития сеголеток в значительной мере сказываются на
дальнейшем формировании особей. Как и у оленьих, из поросят-«заморышей»
вряд ли вырастут хорошие производители.
Таблица 2. Масса тела (кг) 7-12-месячных особей
Район 1 п | М | lim | Источник информации
Беловежская пуща 34 30-33* 15-55 Козло, 19706, 1975
Березинский з-к — 35-36* 20-50 44
44 30 34-32* 20-50 Козло и др., 1974
Украинские Карпа- 18 36-39* 27-53 Гунчак, 1978, 1982
ТЫ Юг Украины — — 14-65 Щербак, 1986
“Завидово” 152 33,4-38,6 10-58 Фертиков и др., 1999
Подмосковье 19 — 20-35 Овсюкова, Бельский, 1980
Ц. Черноземье 108 35,6** 26-49** Простаков, 1996
44 — 37 — Сухорослов, 19726
Кавказский з-к 4 45,3 33-61 Дуров, Александров, 1968
Кировская область 5 — 42-51 Павлов, Зарубин, 1982; Павлов,
Азербайджан 16 37-43* 32-56 1999а Сафаров, 1965
Дельта Волги — 35^40 — Лавровский, 1962
Низовья р. Или 9 22,5** 16-30** Слудский и др., 1984
Ю. Киргизия >5 38-39,5 30-45 Чичикин, Воробьев, 1967; Во-
Таджикистан 11 35,3-34,9* — робьев, 1968 Иваненко, 1956
Тува — 20 -25 Смирнов, 2001
Предбайкалье 21 29,5-32,5 20-42 Недзельский, 1994
Сихотэ-Алинь 26 29-32* Бромлей, 1964
* У самок и самцов соответственно.
** Масса без желудка, кишечного тракта и крови.
Летом годовалые особи быстро растут, причем самцы - интенсивнее [Perin et
al., 1987]. К следующей зиме длина тела у самцов увеличивается в среднем до
122-155 см, у самок - до 118-148 см; высота в холке - до 72-95 см и 62-82 см
соответственно (табл. 3), а масса почти удваивается, достигая 52-82 кг и 48-76 кг
(табл. 4). Половой диморфизм у этой группы уже четко выражен, а экстерьерные
различия все более увеличиваются с возрастом. У самцов становятся более мас-
сивными голова и передняя часть туловища, и они заметно выше в холке.
Рост самок продолжается до 4-5 лет, самцов - до 6-7 лет, но темпы прироста
с третьего года жизни у первых и с четвертого - у вторых уже незначительны.
Размеры взрослых особей максимальны в 6-8 лет, после чего происходит сниже-
ние массы тела. Зимой она прогрессивно уменьшается у всех возрастов и незави-
симо от пола, что связано с отрицательным энергетическим балансом. Даже при
благоприятных условиях зимовки это сокращение достигает 20-30%, при небла-
гоприятных - 40-50% [Jezierski, Myrcha, 1975; Козло и др., 1974; Козло, 19746,
1975; Гунчак, 1978, 1982; Бромлей, Кучеренко, 1983; Янулайтис, 1988; Недзель-
ский, 1994]. При потере более 50-55% массы дикие копытные неминуемо гибнут,
что нередко случается и в популяциях кабана.
18
Таблица 3. Размеры тела (см) 1,5-2-годовалых кабанов-подсвинков
Самцы Самки Район (источник информации)*
п М lim п 1 м | lim
Длина тела
292 132 95-150 232 128 92-146 Германия (14)
37 125 104-140 50 120 102-137 Беловежская пуща (3)
8 136 130-142 15 134 98-146 Березинский з-к (2)
15 131 124-138 15 126 119-134 Литва (12)
- 153 145-160 — 148 137-154 С.-з. России (1)
26** 155 142-163 26** 143 126-156 Украинские Карпаты (11)
2 127 124-130 4 128 125-132 Закавказье(6)
4 124 121-127 2 112 104-120 Низовья р. Или (5)
3 136 135-137 2 134 134-135 Ю. Киргизия (8)
17 134 — 8 129 — Таджикистан (9)
7 122 115-128 11 118 109-125 Предбайкалье (13)
4 137 — 3 131 — Сихотэ-Алинь (4)
Высота в холке
- 95 — 88 — С.-з. России (1)
15 78 67-85 15 75 68-79 Литва (12)
37 78 69-84 50 76 57-84 Беловежская пуща (3)
8 85 82-87 15 82 63-91 Березинский з-к (2)
26** 83 78-86 26** 79 72-95 Украинские Карпаты (11)
2 74 71-77 4 76 74-80 Закавказье(6)
6 76 72-80 2 68 61-75 Низовья р. Или (5)
3 82 82-83 2 80 79-81 Ю. Киргизия (8)
17 72 — 8 62 — Таджикистан (9)
7 76 69-82 11 75 67-81 Предбайкалье (13)
4 78 — 3 77 — Сихотэ-Алинь (4)
Обхват туловища
128 — — 115 — С.-з. России (1)
15 100 92-107 15 97 86-108 Литва (12)
37 92 76-109 50 90 73-105 Беловежская пуща (3)
8 114 107-132 15 114 86-124 Березинский з-к (2)
26** 124 — 26** 96 — Украинские Карпаты (11)
2 90 88-92 4 91 86-98 Закавказье (6)
7 91 82-99 И 89 80-97 Предбайкалье (13)
4 98 — 3 97 - Сихотэ-Алинь (4)
В Длина стопы
V 37 30 — 50 29 — Беловежская пуща (3)
8 33 32-35 15 31 25-34 Березинский з-к (2)
15 31 — 15 29 — Литва (12)
3 27 27-28 2 26 24-27 Низовья р. Или (5)
4 32 - 3 29 - Сихотэ-Алинь (4)
19
2»
Таблица 3 (окончание)
Самцы Самки Район (источник информации)*
п 1 м | lim п 1 м | lim
Длина уха
37 13 — 50 12 — Беловежская пуща (3)
8 13 12-14 15 13 10-15 Березинский з-к (2)
15 12 15 12 — Литва (12)
26** 13 — 26** 12 — Украинские Карпаты (11)
2 11,5 11-12 4 13 12-14 Закавказье(6)
4 12 11-12 2 11,5 11-12 Низовья р. Или (5)
7 11,5 10-12 И 13,7 12-14 Предбайкалье (13)
4 11 — 3 И — Сихотэ-Алинь (4)
Длина хвоста
37 25 — 50 24 — Беловежская пуща (3)
8 24 20-28 15 22 16-31 Березинский з-к (2)
15 24 — 15 22 — Литва (12)
26** 32 — 26** 31 — Украинские Карпаты (11)
2 21 20-23 4 21 16-23 Закавказье(6)
4 18 14-20 1 __ 21 Низовья р. Или (5)
7 18 17-19 И 17 15-18 Предбайкалье (13)
4 20 — 3 19 — Сихотэ-Алинь (4)
*1-Русаков, Тимофеева, 1984; 2-Козло и др., 1974; 3-Козло, 1975; 4-Бромлей, 1964; 5-Слуд-’
ский, 1956; 6-Сафаров, 1958а; 7-Дуров, Александров, 1968; 8-Чичикин, Воробьев, 1967; 9-Ива-
ненко, 1956; 10-Лавровский, 1962; 11-Гунчак, 1978, 1982; 12-Янулайтис, 1988; Янулайщс, Коз-
ла, 1988; 13-Недзельский, 1994; 14-Briedermann, 1986.
** Число самцов и самок.
Таблица 4. Масса тела (кг) 1,5-2-го довалых кабанов-подсвинков
Самцы Самки Район (источник информации)*
п М lim п М 1 lim
34 54 -82 50 50 — Беловежская пуща (3)
8 72 65-80 15 76 59-89 Березинский з-к (2)
15 72*** — 15 72*** — Литва (12)
26** 82 69-91 26** 76 62-89 Украинские Карпаты (11)
5 76 57-106 3 70 44-90 Кавказский з-к (7)
2 63 57-69 4 58 54-66 Закавказье (6)
— 70-75* * * — — *** — Дельта Волги (10)
4 — -54 2 — -44 Низовья р. Или (5)
3 70 68-73 2 62 60-65 Ю. Киргизия (8)
5 63 — 2 67 — Таджикистан (9)
— 36*** -50*** — *** *** Тува (15)
7 52 42-60 11 48 40-54 Предбайкалье (13)
4 70 — 3 61 — Сихотэ-Алинь (4)
* Обозначения см. в таблице 3. 15 - Смирнов, 2001.
** Число самцов и самок.
*** Средние значения для самцов и самок.
20
Рост и развитие диких свиней неодинаковы даже в пределах одного региона.
В березинской популяции, например, животные развиваются быстрее и они
крупнее, чем в беловежской (табл. 1—6), что, вероятно, связано с чрезмерной
плотностью населения копытных и с продолжительным выборочным отстрелом
наиболее крупных особей в Беловежской пуще [Козло, 19706, 1975].
Географическая изменчивость размеров и массы тела диких свиней на террито-
рии России и сопредельных регионов менее значительна, чем можно было ожидать.
Наиболее крупные особи - в популяциях Карпат, севера Белоруссии и дельты Волги,
самые мелкие - на западе Европы, в Закавказье и Средней Азии. Дальневосточные
кабаны не самые крупные, как обычно считают охотоведы, а сибирские по результа-
там сравнительного анализа не могут относиться к мелким (табл. 5 и 6).
Таблица 5. Размеры тела (см) взрослых (старше 3 лет) кабанов в различных районах
России и сопредельных регионах
Самцы Самки Район (источник информации)*
п М | Нт п 1 м | lim
Длина тела
53 150 139-178 44 141 127-160 Германия (6)
34 157 — 86 152 — Чехословакия (6)
47 162 145-174 56 142 127-155 Беловежская пуща (9)
14 171 161-182 14 159 150-167 Березинский з-к (29)
19 164 156-175 12 151 146—164 Литва (24)
— — -189 — — -176 С.-з. России (2)
42 162 130-183 62 149 127-174 Ц. Черноземье (1)
101 162 132-202 98 156 132-179 Воронежский з-к (7)
- — -193 — — — Приднепровье (3)
2 — 200-220 — — — Донецкая область (5)
18** 187 174-196 18*4 174 157-181 Украинские Карпаты
(23)
12 169 150-205 23 144 129-169 Кавказский з-к (13)
6 156 109-176 7 152 142-160 -“-(15)
— — -198 — — -180 С. Кавказ (4)
7 164 152-180 — — -168 -“-(20)
5 154 146-163 7 144 136-161 Закавказье (14)
- 200*** 198-215 — 180*** 174-199 Дельта Волги (22)
и 160 148-174 12 144 127-157 Низовья р. Или (10)
5 — 182-217 4 — 149-160 Ю. Киргизия (19)
24 164-172 -184 35 150-154 — Таджикистан (21)
12 173 163-182 21 160 150-169 Предбайкалье (31)
1 — 163 — — — Хамар-Дабан (17)
18 171 137-221 17 149 135-178 Дальний Восток (8)
Высота в холке
11,27 79-81 — 43 72 — Германия (6)
35 93 — 87 88 — Чехословакия(6)
47 101 86-110 56 87 81-93 Беловежская пуща (9)
14 103 99-107 14 93 82-96 Березинский з-к (29)
19 91 83-103 12 86 89-94 Литва (24)
— — -111 — — -97 С.-з. России (2)
42 90 79-105 62 86 68-97 Ц. Черноземье (1)
21
Таблица 5 (продолжение)
Самцы Самки Район (источник информации)*
п М | lim п 1 м | lim
101 88 73-104 98 84 72-95 Воронежский з-к (7)
— — -124 — — - Приднепровье (3)
18** 102 90-104 18** 92 89-105 Украинские Карпаты (23)
12 103 93-120 23 75 61-96 Кавказский з-к (13)
6 101 91-106 7 88 84-92 -“-(15)
5 90 83-98 7 82 72-90 Закавказье (14)
— 86 — 81 — Грузия (28)
— 109 95-120 — 97 89-106 Дельта Волги (22)
13 85 74-102 13 77 71-86 Низовья р. Или (10)
4 87 85-90 4 80 78-83 -“-(32)
5 — 105-116 4 — 92-98 Ю. Киргизия (19)
24 84 — 35 82 — Таджикистан (21)
12 94,5 85-103 21 87,5 79-96 Предбайкалье (31)
1 90 — — — — Хамар-Дабан (17)
18 93 75-118 17 86 63-94 Обхват туловища Дальний Восток (8)
11,26 119 — 53,43 101-105 — Германия (6)
34 130 — 87 122 Чехословакия (6)
47 140 118-151 56 115 95-123 Беловежская пуща (9)
14 156 128-172 14 133 120-150 Березинский з-к (29)
19 127 116-150 12 120 112-129 Литва (24)
— — -170 — — -152 С.-з. России (2)
— — -170 — — — Приднепровье (3)
18** 157 — 18** 143 — Украинские Карпаты (23)
42 140 98-154 62 116 86-146 Ц. Черноземье (1)
100 139 100-198 97 135 102-166 Воронежский з-к (7)
6 137 120-160 7 125 108-144 Кавказский з-к (15)
5 130 115-150 7 106 98-110 Закавказье (14)
— 125 — — 114 — Грузия (28)
— 149 127-163 — 123 111-143 Дельта Волги (22)
4 128 124-135 4 116 112-120 Дельта Или (32)
12 130 113-147 21 112 98-126 Предбайкалье (31)
18 126 114-183 17 118 Длина стопы 90-131 Дальний Восток (8)
17 32 — 33 30 — Германия (6)
33 35 — 33 30 — Чехословакия (6)
47 33-35 — 55 31-32 — Беловежская пуща (9)
14 36 35-38 14 33 26-35 Березинский з-к (29)
19 34 — 12 34 — Литва (24)
42 32 29-36 62 30 20-35 Ц. Черноземье (1)
101 30 25-35 98 29 25-39 Воронежский з-к (7)
6 35 34-37 7 32 29-34 Кавказский з-к (15)
- - -37 - - — Приднепровье (3)
9 31 28-34 9 28,5 25-33 Низовья р. Или (10)
22
Таблица 5 (окончание)
Самцы Самки Район (источник информации)*
n 1 М | lirn п 1 м | lim
17 37 28^18 16 35 28-39 Дальний Восток (8)
Длина уха
17 14 — 33 13 — Германия (6)
34 14 — 89 13 — Чехословакия(6)
47 15 13-20 56 14 13-15 Беловежская пуща (9)
14 17 15-20 14 15 13-17 Березинский з-к (29)
19 15 12 14 — Литва (24)
42 14 13-21 59 14 12-18 Ц. Черноземье (1)
101 15 12-20 97 14 11-17 Воронежский з-к (7)
— — -15 - — — Приднепровье (3)
18** 18 — 18** 17 — Украинские Карпаты (23)
12 16 15-18 23 18 14-20 Кавказский з-к (13)
6 13 11-15 7 13 11-17 -“-(15)
5 16 13-18 7 15 12-17 Закавказье (14)
- 16 15-18 — 15 13-18 Дельта Волги (22)
11 14 13-16 11 13 12-14 Низовья р. Или (10)
12 16 14-19 21 15 14-19 Предбайкалье (31)
18 12 10-14 17 12 9-13 Дальний Восток (8)
Длина хвоста
37,24 24 — 39,41 21-23 — Германия (6)
35 24 — 89 23 — Чехословакия(6)
28 27 21-33 21 24 22-33 Беловежская пуща (9)
14 28 21-34 14 29 25-35 Березинский з-к (29)
19 28 — 12 26 — Литва (24)
42 25 18-33 59 25 17-33 Ц. Черноземье (1)
100 25 17-31 96 23 17-35 Воронежский з-к (7)
- — -29 — — — Приднепровье (3)
18** 27 — 18** 24 — Украинские Карпаты (23)
12 24 19-30 23 23 16-32 Кавказский з-к (13)
6 25 18-30 7 20 14-25 -“-(15)
5 25 22-28 7 24 20-26 Закавказье (14)
— 25 23-28 — 22 19-28 Дельта Волги (22)
11 24,5 20-29 10 22 19-26 Низовья р. Или (10)
12 24 22-28 21 21 18-24 Предбайкалье (31)
18 25 20-33 17 22 21-26 Дальний Восток (8)
*1 - Простаков, 1994, 1996; 2 - Русаков, Тимофеева, 1984; 3 - Евтушевский, 1975; 4 - Дуров,
1983; 5-Павлов и др., 1974; 6 - Briedermann, 1986; 7-Дьякова, 1989; 8 - Бромлей, 1964; 9-Козло,
1975; 10 - Слудский, 1956; 11 - Слудский и др., 1984; 12 - Калниньш, 1950; 13 - Донауров, Теплов,
1938; 14-Сафаров, 1958а, 1965; 15-Дуров, Александров, 1968; 16 - Сержанин, 1961; 17-Фети-
сов, 1956; 18 - Иванова, Молоканов, 1969; 19 - Чичикин, Воробьев, 1967; 20 - Динник, 1910; 21 -
Иваненко, 1956; 22 - Лавровский, 1962; 23 - Гунчак, 1978, 1982; 24 - Янулайтис, 1988; Янулайщс,
Козла, 1988; 25-Ишадов, 1965; 26-Сухорослов, 19726; 27-Мануш, Николаев, 1995; 28 - Хеладзе,
1964; 29-Козлоидр., 1974; 30-Бабкин, 1997; 31-Недзельский, 1994; 32 - Бутарин и др., 1955.
** Число самцов и самок.
*** Не исключено, что в общую длину тела включена длина хвоста.
23
Таблица 6. Масса тела (кг) взрослых (старше 3 лет) кабанов в различных районах России
и в сопредельных регионах
Самцы Самки Район (источник информации)*
п | М lim п 1 м | lim
47 135 90-176 56 106 85-140 Беловежская пуща (9,29)
14 161 104-234 14 109 74-151 Березинский з-к (29)
— — -256 — — — Белоруссия (16)
— — -236 — — — Латвия (12)
— 135 — — 120 — Ярославская область (18)
2 — 151-145 3 ПО 96-123 “Завидово “ (27)
3 92 90-96 3 108 102-110 Кировская область (5)
— 172 69-229 — 144 106-172 Среднее Поволжье (30)
47 134 83-173 59 102 68-147 Ц. Черноземье (1)
— 124 — — 111 — -“-(26)
126 118 62-285 137 110 71-186 Воронежский з-к (7)
— — -250 — — — С.-з. России (2) и Приднепро-
вье (3)
18** 202 160-250 18** 133 105-152 Украинские Карпаты (23)
2 — 250-320 — — — Донецкая область (5)
12 166 64-178 23 68 48-109 Кавказский з-к (13)
6 144 105-181 7 117 82-148 -“-(15)
— — -250 — — -150 С. Кавказ (4)
5 114 90-142 7 86 58-99 Закавказье(14)
— — 80-320 — — — Грузия (28)
— 185 103-243 — 111 86-142 Дельта Волги (22)
(до 296)
— 123 95-240 — 83 65-118 Казахстан (10,11)
5 — 130-243 4 — 90-120 Ю. Киргизия (19)
22 114 79-144 33 96 70-123 Таджикистан (21) 1
— — -224 — — - Туркменистан (25) 1
— 74 -120 — 57 -70 Тува (33) |
12 118 103-139 21 85 64-104 Предбайкалье (31) |
18 120 69-267 17 86 53-142 Дальний Восток (8) ’
* Обозначения см. в таблице 5. 33 - Смирнов, 2001.
Судя по костным остаткам, средне - и позднеголоценовые особи из различ-
ных районов Европы, видимо, были крупнее, чем современные [Пидопличко,
1951; Цалкин, 1958, 1962; Верещагин, 1959; Паавер, 1965; Тимченко, 1972;
Jonsson, 1986]. Сохранились также свидетельства очевидцев, регистрировавших
зверей «чудовищного» роста и массы (до 15-22 пудов или до 240-350 кг) в Мол-
давии, Белоруссии, Литве, на Украине, Северном Кавказе, в Закавказье, на севере
Каспийского побережья, в Западном и Южном Казахстане, Забайкалье и на
Дальнем Востоке [Кириллов, 1902; Карцев, 1903; Динник, 1910; Шиллингер,
1928; Марков, 1932; Донауров, Теплов, 1938; Слудский, 1956; Лавровский, 1962;
Абрамов, 1963; Цалкин, 1963; Янулайтис, 1988].
Измельчание вида могло происходить в результате постоянной охотничьей
антиселекции - выборочной добычи самых крупных особей, что очевидно также
для популяций косуль, благородного оленя и лося [Цалкин, 1951; Данилкин,
1999а, 2000а]. Замечу, что крупные кабаны изредка встречаются и сейчас (см.
табл. 5 и 6), но их было бы гораздо больше, если бы охотники позволяли зверям
доживать до зрелого возраста.
24
Краниометрическая характеристика
У новорожденных кабанов череп округлый с короткой лицевой частью: мозго-
вой отдел составляет около 55%, лицевой — 45%. Его рост в постэмбриональный
период более интенсивен в длину, нежели в ширину и высоту. У годовалых особей
размер черепа примерно в 2-3 раза больше, чем у новорожденных, а у двухлетних
больше еще на 30%. Половые различия в этих возрастных группах четко не выра-
жены, но по линейным характеристикам самцы несколько превосходят самок. К
четырем годам жизни особи лицевой отдел вытягивается до 71-73% размера чере-
па (рис. 7). Рост костей продолжается примерно до 5 лет, но с 3-летнего возраста ли-
нейные изменения уже незначительные, что позволяет при морфометрическом ана-
лизе включать животных старше трех лет в категорию взрослых. Масса черепа также
существенно увеличивается с возрастом: у 6-10-месячных самцов - 450-700 г, у го-
довалых - 560-1190 г, от 2 до 3 лет - 1200-2040 г, у зверей старше 3 лет - 1500-
2700 г. У самок старше 3 лет масса черепа не превышает 1900-2100 г и
значительно меньше в линейных размерах, чем у самцов [Иваненко, 1956;
Сафаров, 19586, 1965; Лавровский, 1962; Бромлей, 1964; Козло, 1973, 1975;
Гунчак, 1978; Русаков, Тимофеева, 1984; Соколов, Темботов, 1993].
В верхнем своде мозговой коробки кабана с возрастом образуется пазуха в ре-
зультате расслаивания лобных, теменных и затылочных костей. Она отсутствует у
сеголеток, у годовалых составляет 6-16 мм, у двухлетних - 12-20 мм, у взрослых
достигает 28-32 мм на лбу и до 64 мм в затылочной области. Это образование явля-
ется, скорее всего, своеобразной адаптацией, предохраняющей головной мозг от
проникающих ранений при драках и от сильных ударов по черепу при быстром дви-
жении зверей в густых зарослях [Иваненко, 1956].
Географическая изменчивость краниометрических показателей весьма значи-
тельна (табл. 7). Наименьшие линейные размеры черепа у особей из популяций
Западной Европы, Закавказья, Средней Азии и Восточной Сибири, наиболее
крупные - в Карпатах, на севере Белоруссии, в дельте Волги и на Дальнем Вос-
токе. Северокавказские и закавказские кабаны явно разнокачественные не только
по размерам и массе тела (табл. 5 и 6), но и по размерам черепа, что тоже не ук-
ладывается в рамки общепринятых представлений о подвидовой таксономии.
Таблица 7. Морфометрическая характеристика черепа (мм) взрослых (старше 3 лет)
кабанов
Самцы Самки Район (источник информации)*
п 1 М 1 lim п м I lim
Наибольшая длина
20 398 366-437 8 379 364-385 Германия (16)
49 427 396-457 57 382 358^413 Беловежская пуща (2)
15 456 438^485 13 389 383^402 Березинский з-к (2)
18** 466 434-510 18** 416 383^446 У крайне кие Кар паты
(13)
12 436 410-467 23 387 344^421 Кавказский з-к (5)
52 427-437 403^470 39 394-414 383^440 -“-(7)
22 410-418 380^448 13 387-399 365460 Центральный Кавказ (7)
7 392^420 370-436 — — — Закавказье(7)
12 414 357-461 12 379 353^423 -“-(9)
9 427 417^442 5 396 387^406 -“-(Ю)
- 444 412^490 — 404 387^431 Дельта Волги (12)
11 402 369^436 2 — 342-358 Казахстан (3)
25
Таблица 7 (продолжение)
Самцы Самки Район (источник информации)*
п 1 м | lim п 1. м lim
3 411 391^131 3 371 342-391 Средняя Азия (10)
24 408 365^435 33 381 342-412 Таджикистан (11)
— — 367-381 1 — 340 Монголия (4,14)
— 384 370-400 — 362 355-375 Восточная Сибирь (8)
3 355 344-366 2 344 340-347 -“-(Ю)
12 406 371^141 21 364 337-390 Предбайкалье (15)
6 441 431-451 4 — 338-361 Дальний Восток (1)
4 - 424-442 2 - 388^125 -“-(14)
Скуловая ширина
49 161 141-172 57 147 132-157 Беловежская пуща (2)
15 173 158-188 13 149 148-150 Березинский з-к (2)
18** 162 154-176 18** 157 150-168 Украинские Карпаты (13)
12 166 157-180 23 148 135-158 Кавказский з-к (5)
47 158-166 138-180 36 146-154 137-163 -“-(7)
20 156-166 142-190 10 146-150 139-174 Центральный Кавказ (7)
7 147-163 134-168 — — — Закавказье (7)
9 156 147-175 5 151 146-157 -“-(6,10)
9 163 141-173 2 — 133-138 Казахстан (3)
24 163 153-178 34 149 135-163 Таджикистан (11)
— 149 145-152 — 139 131-144 Восточная Сибирь (8)
12 152 144-158 21 143 130-154 Предбайкалье (15)
6 168 166-171 4 161 160-163 Дальний Восток (1)
Высота с нижней челюстью
49 232 218-258 57 206 185-220 Беловежская пуща (2)
15 250 238-277 13 218 209-225 Березинский з-к (2)
18** 234 — 18** 227 — Украинские Карпаты
(13)
12 . 238 216-258 23 204 187-232 Кавказский з-к (5)
45 220-229 214-249 26 196-203 128-221 -“-(7)
22 191-203 157-219 12 167-172 153-206 Центральный Кавказ (7)
7 183-201 172-206 — — — Закавказье (7)
12 220 184-249 13 197 158-222 -“-(9)
— — 239-274 — — 205-207 Дельта Волги (12)
10 229 196-258 2 — 151-181 Казахстан (3)
— 225 220-232 — 198 190-206 Восточная Сибирь (8)
12 226 206-245 21 199 169-209 Предбайкалье (15)
6 — 232-268 4 - 225-230 Дальний Восток (1)
Длина лицевой части
49 322-330 300-349 38 297-309 286-328 Кавказский з-к (7)
22 312-330 285-365 14 296-330 280-360 Центральный Кавказ (7)
7 299-319 283-326 — — — Закавказье(7)
9 282 275-290 5 259 252-264 -“-(Ю)
11 317 294-338 2 265 260-270 Казахстан (3)
26
Таблица 7 (окончание)
Самцы Самки Район (источник информации)*
п 1 м | lim п | М lim
24 269 232-290 34 251 223-272 Таджикистан (11)
Длина носовых костей
20 215 — 8 206 — Германия (16)
18** 229 — 18** 210 — Украинские Карпаты (13)
12 218 195-240 23 195 159-209 Кавказский з-к (5)
45 225-237 206-260 33 202-213 184-221 -“-(7)
20 202-226 173-248 13 182-206 110-244 Центральный Кавказ (7)
7 203-219 190-228 — — — Закавказье(7)
10 208 183-237 11 193 172-217 -“-(9)
5 - 200-222 4 - 208-214 Дальний Восток (1)
Длина неба
20 250 — 8 238 — Германия (16)
49 266 243-283 57 235 214-252 Беловежская пуща (2)
15 290 279-302 13 241 230-258 Березинский з-к (2)
12 256 241-279 23 220 188-240 Кавказский з-к (5)
12 257 238-275 21 229 211-240 Предбайкалье (15)
6 - 264-272 4 — 242-247 Дальний Восток (1)
Длина нижнего ряда коренных зубов
- 164 __ — — — Венгрия (10)
9 151 141-159 5 146 142-154 Азербайджан (10)
3 147 142-153 3 145 142-150 Средняя Азия (10)
3 133 130-139 2 128 127-129 Восточная Сибирь (10)
6 167 166-168 4 156 155-158 Дальний Восток (1)
Длина верхнего ряда коренных зубов
- 135 — — - — Венгрия (10)
12 136 125-146 23 131 122-138 Кавказский з-к (5)
12 130 116-142 13 135 113-135 Закавказье (9)
9 133 -138 5 133 -137 Азербайджан (10)
— по 108-112 — 109 107-112 Восточная Сибирь (8)
12 114 107-120 21 112 109-115 Предбайкалье (15)
6 — 141-147 4 — 135-141 Дальний Восток (1)
* 1 - Бромлей, 1964; 2 - Козло, 1975; 3 - Слудский и др., 1984; 4 - Банников, 1954; 5 - Донау-
ров, Теплое, 1938; 6 - Сафаров, 19586; 7 - Соколов, Темботов, 1993; 8 - Фетисов, 1956; 9 - Анучин,
1938; 10-Сафаров, 1965; 11 -Иваненко, 1956; 12-Лавровский, 1962; 13-Гунчак, 1978, 1982; 14-
Филипченко, 1933; 15-Недзельский, 1994; 16-Briedermann, 1986.
** Число самцов и самок.
У кабана, как считают систематики, существенную таксономическую значи-
мость имеют вогнутость профиля черепа у стыка лицевой и носовой костей и
форма слезной кости. Однако к такому выводу исследователи пришли на основа-
нии анализа единичных экземпляров черепов или очень малых выборок,
У восточноазиатских форм диких свиней профиль черепа обычно слегка вы-
пуклый, тогда как у европейских рас, включая кавказских и нижневолжских, он
чаще вогнутый у самцов и почти прямой у самок [Адлерберг, 1930, 1933; Филип-
пенко, 1933; Анучин, 1938; Соколов, 1959; Лавровский, 1962; Козло, 1975; Кня-
27
Рис. 7. Схема возрастных изменений черепа у самцов (А, Б) и самок (В, Г) кабана
(по [Иваненко, 1956])
А и В - по боковому профилю, Б и Г - по верхнему профилю. Заливкой обозначено развитие
мозговой полости
28
4
Рис. 8. Географическая изменчивость профиля черепа и формы слезной кости у самцов ка-
бана (по материалам Зоомузея МГУ)
Районы: А - Сихотэ-Алинь, коллекционный № 69179; Б - Забайкалье, № 54856; В - Монголия,
№43659; Г-Копетдаг, № 5500; Д- Казахстан, № 35148; Е - Астраханская область, № 36872; Ж -
Беловежская пуща, № 54968; 3 - Румыния (Карпаты), № 63986
29
зев, Тихонов, 1985; Соколов, Темботов, 1993]. У особей из Азербайджана он в ос-
новном прямой, а вогнутость обнаруживается лишь на некоторых экземплярах из
низменных районов [Сафаров, 1965]. У среднеазиатской расы и тот, и другой тип
[Иваненко, 1956; Слудский, 1956; Чичикин, Воробьев, 1967]. Индекс ширины и вы- |
соты черепа у азиатских подвидов выше, чем у европейских, а индекс слезной кости, ;
напротив, ниже [Knyazev, Tikhonov, 1998]. :
По моим исследованиям, у всех палеарктических форм кабана встречаются
все рассматриваемые типы профиля черепа, но с разной частотой, и очевидна
клинальность в направлении с востока ареала на запад (рис. 8). На Кавказе
(Предкавказье, Центральный Кавказ и Закавказье), как и в Средней Азии, наблю-
Рис. 9. Изменчивость профиля черепа и формы слезной кости у самцов кавказского
кабана (по материалам Зоомузея МГУ)
Коллекционные номера: 35099, 35143, 35085, 35092, 75116 и 35097
30
Рис. 10. Типичный профиль черепа и формы слезной кости у самок кабана (по материалам
Зоомузея МГУ)
Районы: А- низовья Сырдарьи, коллекционный № 35140; Б - Астраханская область, № 36868;
В - Азербайджан, № 111326
31
дается почти такой же ряд изменчивости (рис. 9). У самок географическая вариа-
бельность выражена менее четко, нежели у самцов; профиль их черепа чаще бли-
зок к прямому (рис. 10). Этот таксономический признак, следовательно, значим
лишь при достаточно больших выборках и, преимущественно, при сравнении по-
пуляций с западной и восточной окраин ареала.
Форма слезной кости у кабана тоже клинально изменяется с востока ареала
на запад (см. рис. 8) - от широкой и почти квадратной у восточных рас до удли-
ненной у западных [Адлерберг, 1930, 1933; Фетисов, 1956]. Длина нижнего края
и высота слезной кости у особей из Беловежской пущи (36-42 и 23-26 мм) близ-
ки к таковым у зверей из Азербайджана (37-40 и 25 мм) и дельты Волги (41,5 и
25 мм). У особей среднеазиатской формы длина ее нижнего края уменьшается
(33 и 24,7 мм), и еще короче она у дальневосточных (32 и 29,8 мм) и сибирских
(23,8 и 27,2 мм) зверей при одновременном увеличении высоты. Отношение дли-
ны нижнего края к высоте изменяется, соответственно, от 1:0,5 до 1:1,2 (табл. 8).
Индивидуальная изменчивость слезной кости, однако, настолько велика, что у
всех выделяемых рас и даже в одной популяции (в Беловежской пуще, например)
встречаются особи как с удлиненным «европейским» типом, так и с квадратным
«азиатским» [Филипченко, 1933; Лавровский, 1962; Сафаров, 1965; Козло, 1975].
У гибридов кабана с домашней свиньей доминирует «азиатский» тип слезной
кости [Князев, Тихонов, 1985]. Следовательно, этот признак тоже не может быть
определяющим в таксономии вида.
Таблица 8. Изменчивость слезной кости у разных форм кабана*
Форма Длина, мм Высота, ММ Соотношение длин
НИЖНИЙ край верхний край по центру нижний край: высота центр: высота
5. s. scrofa 36-42 58-68 54 23-26 1:0,5-0,7 1:0,4—0,5
5. s. attila 37^12 63 48 (до 28) 23-25 1:0,5-0,7 1:0,5
5. S. s. nigripes s. sibiricus (до 44) 33(26-43) 24 58 46 44 29-31 25 (до 32) 27-29 1:0,8 1:1,1-1,2 1:0,6 1:0,9-1,0
S. s. ussuricus 32 (до 36) 50 44 30(27-36) 1:0,9 1:0,7
* По [Адлерберг, 1933; Филипченко, 1933; Фетисов, 1956; Лавровский, 1962; Сафаров, 1965;
Козло, 1975; Недзельский, 1994], с изменениями.
В одном и том же районе, но в разных биотопах, у кабана может быть разной ши- '
рина темени (расстояние между височными гребнями). На Вахшском хребте у осо- 5
бей, добытых в горах, ширина теменной площадки составляет 20-30 мм, тогда как в ]
предгорьях - 40-61 мм. Неодинаковое развитие черепа у особей горных и предгор- •
ных группировок, видимо, связано различными условиями питания и кормодобыва-
ния и разной нагрузкой на отдельные мышцы в онтогенезе. Кроме того, у горных ка-
банов короче и прочнее плечевая и бедренная кости, кости предплечья и голени и
заметна разница в длине тела и высоте в холке [Иваненко, 1956].
Это — наглядный пример существенной морфометрической дифференциации
особей на популяционном уровне в зависимости от условий обитания, причем
различия между равнинными и горными популяциями могут достигать «подви-
довых» величин [Genov et al., 1991].
Дикие свиньи отличаются от домашних большими размерами, рядом морфо-
логических особенностей черепа и посткраниального скелета и большей плотно-
стью трубчатых костей, что позволяет идентифицировать их среди костных ос-
татков на древних стоянках человека [Адлерберг, 1933; Громова, 1948; Хеладзе,
1959; Цалкин, 1970; Нинов, Хрусталева, 1985].
32
Изменчивость зубов
У кабана резцы, клыки, 2-4-е предкоренные зубы дифиодонтные, все задне-
коренные не имеют молочных предшественников, первый предкоренной также
не меняется. Последовательность развития зубной системы отражена в таблице 9.
У новорожденных 2 молочных зуба, у детенышей в возрасте нескольких дней
их уже 8. Формирование молочных зубов завершается к 6 месяцам жизни. 7-12-
месячные особи имеют 36 зубов: отрастает первый коренной, идет смена молоч-
ных на постоянные, число которых колеблется от 12 до 16. К 14-15 месяцам пер-
вый резец заменяется постоянным, отрастает второй коренной. У 18-24-
месячных подсвинков 40 44 зуба: завершается замена молочных резцов, второй
коренной развит полностью, формируется третий, предкоренные сформированы
и имеют стертые вершины, развивается третий коренной. Нижние клыки самцов
вырастают до 4 см, у самок они короче. У 3-летних зверей зубная система сформи-
рована полностью: 44 зуба, резцы и первый и второй коренные слабо стерты, пред-
коренные стерты сильнее, третий коренной полный, клыки трехгранные, направ-
ленные вверх. У 4-летних особей следы стертости на всех зубах, у 5-летних - уко-
рочены резцы и сильно сношены поверхности передних и задних коренных, ост-
рые клыки начинают загибаться назад. Стертость зубов еще более выражена у 6-
7-летних зверей, клыки у них достигают максимальных размеров - более 10 см у
самцов и около 3,5 см — у самок. У особей старше 8 лет заметно разрушение и
выпадение зубов, прогрессирующее с возрастом, клыки также постепенно сти-
раются и обламываются. У животных старше 10 лет задние коренные зубы стер-
ты до десен, а концы верхних клыков, если не сломаны, загибаются к верхней гу-
бе [Иваненко, 1956; Сафаров, 19586, 1965; Лавровский, 1962; Бромлей, 1964;
Козло, Никитенко, 1967; Козло, 1973, 1975; Иванова, Овсюкова, 1976; Русаков,
Тарасов, 1988]. У отдельных секачей клыки верхней челюсти загибаются на-
столько, что достигают поверхности носовой кости и иногда пробивают ее [Дин-
ник, 1910].
Интересно формирование многобугорчатых зубов. Первый верхний коренной у
5-месячных особей закладывается пятью самостоятельными конусами, у 6-
месячных три передних сливаются, образуя переднюю часть зуба, а два других -
заднюю, у 9-месячных все элементы соединяются, у 10-месячных минерализуются
и выходят на поверхность десны. Первый нижний коренной образуется из четырех
конусов, которые соединяются у 7-месячных поросят. Самые большие третьи ко-
ренные закладываются у 9-месячных особей из 11 конусов, из которых три, шесть
и два образуют три сегмента, сливающиеся к концу второго года жизни животных,
а в начале третьего года поверхность зуба выходит наружу [Иваненко, 1956].
Изменчивость зубов (рис. 11) служит критерием определения возраста кабана
[Козло, 1973, 1975]. Однако по внешним признакам, стертости эмали, изменени-
ям корней зубов или размерам клыков определить точный возраст животных
старше 2-3 лет невозможно, даже имея эталонные серии нижних челюстей, по-
скольку многие параметры зависят от наследственности, характера пищи и ее ка-
чества, состава и твердости почвы и др. Более надежен подсчет годовых слоев на
срезах зубов, но у диких свиней линии прироста дентина и цемента значительно
менее контрастны, чем, например, у оленьих [Iff, 1983; Moller, 1984; Meynhardt,
1985с; Клевезаль, 1988].
Точное определение пола особей по форме черепа обычно не вызывает про-
блем. У самцов верхнечелюстная кость в месте выхода клыка разрастается, обра-
зуя своеобразный выступ (см. рис. 5), отсутствующий у самок. Клыки самцов и
самок тоже различаются не только по размерам, но и по форме и структуре. У
3. Заказ №2528.
33
Таблица 9. Развитие зубов у кабана (по [Сафаров, 1965], с изменениями)
Возраст 11 I1 12 I2 1з I3 С, с1 Р1 р1 р2 р2 Рз р3 р4 р4 м, м1 м2 м2 м3 м3
Новорожд. м м
До 5 дней +М +м м м м м +м +м
5-10 дней +М +м м м м м +м +м
3^1 мес. м м м м м м м м м м м м м м +п
7 мес. м м м м м +п +п м м м м м м м м м п п
9 мес. м м м м +п п п +п м м м м м м м м п п
13 мес. м ПМ м м п п п п м м м м м м м м п п +п +п
15 мес. +п +п м м п п п п м м м м м ПМ ПМ ПМ п п п п
16 мес. п п м м п п п п м м +п +п +п +п +п +п п п п п
20-21 мес. п п +п +п п п п п м м п п п п п п п п п п
23-24 мес. п п +п +п п п п п м м п п п п п п п п п п +п
26 мес. п п п +п п п п п м м п п п п п п п п п п +п +п
30 мес. п п п +п п п п п м м п п п п п п п п п п +п +п
32 мес. п п п п п п п п м м п п п п п п п п п п +п +п
33 мес. п п п п п п п п м м п п п п п п п п п п +п +п
Примечание: М - молочный зуб, П - постоянный зуб, ПМ - растет постоянный зуб, но молочный еще не выпал, + - прорезывающийся зуб.
Рис. 11. Возрастные изменения зубов у кабана (по [Козло, 1975])
А - в возрасте 9-11 месяцев, Б - в 2 года, В - в 3 года, Г - в 6-7 лет, Д - в 10-12 лет
первых в верхней челюсти они трапециевидные и с эмалью на корнях, у вторых
треугольные и без эмали [Mayer, Brisbin, 1988].
Следует отметить, что размеры клыков кабана, приводимые в литературных
источниках, существенно разнятся. При трофейной оценке их длина у самцов
составляет около 160-250 мм. В зоологических промерах длина нижних клыков
самцов - 60-125 мм, верхних - 45-120 мм, у самок - 24-38 мм и 18-39 мм [До-
науров, Теплов, 1938; Бромлей, 1964; Чичикин, Воробьев, 1967; Козло, 1975;
Слудский и др., 1984; Простаков, 1996]. Несоответствие возникает из-за разных
методик измерения. Зоологи чаще промеряют лишь видимую часть клыка, вы-
ступающую наружу за альвеолярный край челюсти, тогда как охотоведы - весь
зуб после извлечения из черепа.
3*
35
Существенной географической изменчивости в размерах клыков не усматри-
вается, тогда как частота появления особей с крупными клыками в разных попу-it
ляциях неодинакова. Это во многом зависит не только от условий обитания, но и *
от уровня охотничьего преследования и срока жизни взрослых особей. Клыки
кабана в горной части Карпат относительно длинные (до 234 мм) и узкие, в отли-1
чие от коротких и широких клыков у секачей из Германии, и они существенно ж
больше, чем в Чехии и Моравии. Трофеи, оцененные золотой медалью, напри-1
мер, в Восточно-Словацкой области встречаются с частотой 1 на 190 экземпля-
ров, в Словакии - среди 480, на территории Чехословакии соответственно среди
5,6 тыс. особей. Размеры клыков здесь возрастают с юго-запада ареала на северо-
восток, а также по мере увеличения высоты над уровнем моря. В горах, вероятно,
клыки длиннее и тоньше, чем на равнине [Hell, 1985]. В ГДР один “золотой”
трофей обнаруживался среди 8 тыс. особей [Briedermann, 1986], в Литве - среди
1 тыс. С возрастом доля секачей с “медальными” клыками здесь возрастает и у
особей старше 9 лет достигает 53% [Петялис, 1988]. В Польше в 1923-1985 гг,
медалями отмечено 1147 трофеев, в том числе золотыми - 504 [Uloth, 1990].
Число зубов у кабана заметно варьирует не только в зависимости от возраста.
Полный их набор в Березинском заповеднике имеют лишь 57%, в Центрально-
черноземном - 53% взрослых животных. Одного резца на верхней и нижней челю-
стях недостает у 9-12 % особей, двух - у 11-15%. На обеих челюстях отсутствует
один зуб у 11-19% зверей, два - у 17-24%, три - у 4-7% и четыре - у 4-6%. В за-
падных и центральных областях России полный набор зубов обнаружен только у
63 (54-80)% животных. Гиподонтия у диких свиней, скорее всего, имеет генетиче-
скую природу, а ее высокая частота, вероятно, результат массового близкородст-
венного скрещивания по линии «брат-сестра» [Кораблев, 1986а, 1989].
В географическом аспекте наблюдается постепенное усложнение структуры
жевательной поверхности зубов MJ (появление дополнительных бугорков и раз-
витие пятки) в направлении с востока ареала на запад, что позволяет говорить об
относительной примитивности азиатских рас, из которых среднеазиатская форма
занимает промежуточное положение. Однако для всех форм эти признаки не аб-
солютны, и нередки отступления от общего правила [Адлерберг, 1930, 1933; Со-
колов, 1959; Лавровский, 1962]. *
Окраска и линька
Типичная окраска новорожденных - полосатая: 5-6 светлых и 7 рыжевато-
бурых полос тянутся от головы до хвоста. Морда палевая или рыжевато-бурая,
конец рыла - черный. Область вокруг глаз, боковые части шеи и конечности
светло-бурые, брюхо грязновато-серое. Индивидуальная окраска поросят весьма
изменчива, и даже у особей из одного помета. Иногда рождаются не только по-
лосатые, но и пятнистые детеныши. Со второго месяца жизни продольные поло-
сы постепенно тускнеют (рис. 12), полностью исчезая к 3-4 месяцам. С конца ле-
та - начала осени окраска становится однотонно светло-серой, серовато-рыжей
или бурой из-за отрастающего к зиме густого сероватого подшерстка. Тем не ме-
нее в ноябре и даже в декабре иногда встречаются полосатые поросята из позд-
них выводков. После осенней линьки сеголетки заметно светлее, чем темно-
бурые подсвинки, у которых волосы на холке заметно длиннее. Полная смена
ювенильного волосяного покрова происходит в возрасте 12-14 месяцев [Бром- ]
лей, 1964; Чичикин, Воробьев, 1967; Шостак, 1969; Козло, 1975]. Пятнистые осо- ’
би сохраняют свою окраску до конца жизни. :
Линяет кабан раз в год, постепенно, с марта - апреля по июнь - август. Смена
зимнего волоса начинается с головы и ног, в последнюю очередь он выпадает на
верхней части боков (рис. 13 и 14). В пределах обширного ареала сроки линьки
36
Рис. 12. Со второго месяца жизни продольные
полосы у поросят тускнеют (фото из архива
журнала «Охота и охотничье хозяйство»)
неодинаковы. В Предбайкалье
она начинается в конце апреля
и заканчивается к началу июля.
В Казахстане линька заверша-
ется во второй половине апреля
- начале мая, но у истощенных
особей и кормящих самок длит-
ся до начала июня; в Киргизии,
соответственно, в конце мая - в
июне и в июле - начале августа;
в Белоруссии - к 1-10 июня, у
лактирующих самок - к 25-30
июня. В дельте Волги самцы ме-
няют волос в июне, самки - в
конце июля - начале августа. На
срок линьки существенно влия-
ет общее состояние организма.
Звери с запаздывающей линь-
кой - это больные или имею-
щие ранения или травмы [Ива-
ненко, 1956; Слудский, 1956;
Лавровский, 1962; Бромлей,
1964; Сафаров, 1965; Чичикин,
Воробьев, 1967; Козло, 1975;
Гунчак, 1982; Недзельский,
1994].
Летом животные на один -
два месяца почти лишены во-
лосяного покрова и подкожно-
го жира, что способствует тер-
морегуляции и предохраняет
организм от перегрева, осо-
бенно в южных широтах, где температура достигает +40-50°С. От солнечных
ожогов, в отличие от домашней свиньи, их предохраняет темная кожа. В конце
лета окраска зверей за счет темной подпуши и темной щетины чаще черно-бурая
или почти черная.
Зимний волос начинает отрастать с середины лета, рост пуховых волос продол-
жается до октября. Часть волос находится на стадии роста и зимой [Соколов, Пет-
рищев, 1997]. Зимняя окраска зверей темная или темно-бурая (рис. 15). Зимой длин-
ная ость постепенно сечется, концы ее светлеют, отчего к весне общий тон окраски
становится грязно-охристым, сероватым или белесым при темно-бурых голове и но-
гах, где ость короче (рис. 16). Волосяной покров молодых животных менее густой и
прочный, чем у взрослых, и к концу зимы бывает посечен ветвями кустарников или
стеблями тростников вплоть до образования плешин на боках. Следовательно, моло-
дым особям сложнее переносить сильные морозы и по этой причине.
Изменчивость окраски диких свиней весьма велика - от черной до почти бе-
лой и пятнистой (пятна белые или черные). В редких случаях встречаются аль-
биносы и меланисты. Пятнистую окраску многие охотоведы и охотники связы-
вают с гибридизацией кабана и домашней свиньи, что не совсем верно. Это гене-
тический признак гомозиготных по данному фактору особей [Andrzejewski,
1974] Наиболее часто он проявляется у европейского кабана на западе ареала. В
суровых условиях обитания естественный отбор не благоприятствует выжива-
нию более заметных пятнистых особей, их в первую очередь и отстреливают
37
охотники. Взрослые пятнистые
самки, однако, нормально раз-
множаются и, как правило, прино-
сят потомство с типичной окра-
ской, тогда как от пятнистых сам-
цов получаются как полосатые,
так и пятнистые поросята. Инте-
ресно, что потомство одичавших
белых домашних свиней в Амери-
ке со временем также приобрело
черную (60%) и пятнистую (32%)
окраску, причем у черноокрашен-
ных особей череп стал длиннее и
уже [Brown, 1985].
В Западной и Центральной Ев-
ропе пятнистые дикие свиньи
обычны [Briedermann, 1986]. Я
неоднократно встречал животных
с такой окраской в Белгородской,
Тульской и Тверской областях и
однажды - в Челябинской облас-
ти. В группировке зверей запо-
ведника «Лес на Ворскле» Белго-
родской области зимой 1974 г.
пятнистые особи составляли 37%
[Тимофеева, 1975]. Их добывали
также в Брянской, Владимирской,
Калужской, Московской, Смолен-
ской и Ростовской областях [Ха-
хин, 1980; Казаков, Дапиков,
1983]. В Хоперском заповеднике
наряду с черными и пятнистыми
особями встречаются белые каба-
ны [Антонец, 1987].
Размах изменчивости окраски
высок даже в пределах локальных
районов. Окраска зверей в Бере-
зинском заповеднике, например,
черно-бурая, отклонения редки
(2,9% - меланисты и 0,6% - пестрые), тогда как в Беловежской пуще 72% особей
темно-бурые с переходом к рыжему и серому оттенкам, 11% — черные с серова-
то-бурым оттенком и 17% — черно-белые [Шостак, 1969; Козло, 19706,в, 1973,
1975]. В Германии в середине зимы темная или почти черная окраска наблюдает-
ся у 58% самцов и 30% самок, серая или серебристо-серая, соответственно, у 22 и
44%, промежуточная темно-коричневая - у 12 и 19%, светлая до желто-
коричневой - у 4 и 7% и пятнистая - у 4% [Briedermann, 1986].
На Западном Кавказе у 80% зверей щетина черная, у 15% - светло-серая на
боках и шее [Дуров, 1987]. На Центральном Кавказе темноокрашенных живот-
ных меньше-10%, столько же их с беловатой щетиной (в дистальной части),
20% - с буроватыми кончиками волос, остальные имеют промежуточные типы
окраски. Пятнистые особи не замечены [Динник, 1910; Соколов, Темботов,
1993]. В Закавказье окраска зверей варьирует от грязно-белой и светло-рыжей до
темно-бурой, коричневой и почти черной. Дикие свиньи, живущие в низмеппо-
38
Рис. >4. Разные стадии линьки у диких свиней. У лактирующих самок линька за-
держивается (фото А. Германавичуса)
стях по камышам, более светлые [Марков, 1932; Хеладзе, 1964; Сафаров, 1965]. В
дельте Волги они, напротив, зимой однообразно бурые с черными ушами, подбо-
родком и передней частью морды, иногда встречаются черные или светлые (пес-
чаные) экземпляры [Лавровский, 1962]. На Среднем Поволжье окраска зверей
темно-бурая и черная, но есть светлые и пестрые особи [Бабкин, 1997].
Среднеазиатские кабаны, в отличие от европейских форм, светлоокрашены:
светло-бурые, песчано-бурые, песчано-серые, соломенно-желтые с темными или
бурыми конечностями, иногда встречаются темные экземпляры [Флеров, 1935;
Слудский, 1956; Чичикин, Воробьев, 1967]. В Таджикистане окраска животных
коричнево-бурая с серым налетом на боках и голове, у горных особей темнее,
чем в предгорьях [Иваненко, 1956]
Восточноазиатские формы диких свиней имеют темно-бурую или почти чер-
ную окраску с характерной серой или белесой полосой из грязно-серой ости дли-
ной 10-25 см, тянущейся от угла рта к ушным раковинам [Банников, 1954; Фети
сов, 1956; Бромлей, 1964; Недзельский, 1994]. В Туве примерно 70% взрослых
самцов черные, остальные - темно-бурые или темно-серые. У взрослых самок
преобладает темно-бурая окраска (32%), несколько реже встречаются черные
(25%), темно-серые и серые особи. Подсвинки в основном (75%) темно-бурые и
бурые, сеголетки бурые (80%) [Смирнов, 2001].
Неоднородность окраски зверей значительно затрудняет использование этого
признака в таксономии.
39
Piic. 15. Типичная зимняя окраска кабанов (фото А. Дигилевича)
Рис. 16. К весне окраска взрослых диких свиней становится сероватой или беле-
сой (фото А. Щеголева)
40
Генетическая изменчивость
Отличительная видовая особенность кабана - значительный хромосомный
полиморфизм (2 п = 36, 37 и 38), никак не связанный с гибридизацией диких и
домашних особей. Он четко выражен не только у разных форм (табл. 10), но и в
отдельных популяциях [Bosma, 1976]. В Белоруссии 66,6% исследованных осо-
бей имеют 36 хромосом, 16,7% - 37 и столько же - 38. У животных из Воронеж-
ского заповедника и в закавказской популяции (Азербайджан) кариотип 2п = 36
не обнаружен, 11% и 20% исследованных зверей были с 37 и остальные - с 38
хромосомами. У среднеазиатской формы (Киргизия) кариотип 2п = 38 не найден,
64,9% особей имеют 36 и 35,1% - 37 хромосом. У дальневосточных зверей (При-
морский и Хабаровский края, Амурская область) 37 хромосом обнаружено толь-
ко у одной особи, у остальных - 38. Такой же набор (2 п = 38) у зверей из Тувы.
Все 38-хромосомные кариотипы особей из европейских и азиатских популяций
одинаковы по характеру дифференциальной окраски и полностью соответствуют
кариотипу (2п = 38) домашних свиней. Полиморфизм обусловлен робертсонов-
скими слияниями акроцентрических хромосом 15-й и 17-й у европейских и 16-й
и 17-й у азиатских форм, а также у особей в закавказской популяции [Тихонов и
др., 1972; Тихонов, Трошина, 1974, 1977; Орлов, Булатова, 1983; Трошина и др.,
1983; Трошина, Тихонов, 1985; Горелов, 1994].
Таблица 10. Цитогенетическая и иммуногенетическая изменчивость кабана
(по [Горелов, 1994], с изменениями)
При- знак* Формы кабана
центрально-евро- пейская** (n = 32-370) кавказ- ская*** (п = 8-138) среднеазиат- ская (п = 20-83) сибир- ская**** (п = 7-35) уссурий- ская (п = 11-90)
36 Хромосомный полиморфизм, % 33,3 0 64,9 0 0
37 13,8 25,0 35,1 0 5,0
38 52,8 75,0 0 100 95,0
А 0,72 Встречаемость антигенов 0,59 0,46 0,74
Da . 2,68 8,54 36,00 92,32 64,77
Db 100 100 96,00 15,38 87,50
Fa 0 0 0 0 13,33
Ga 100 62,50 57,83 0 23,33
Gb 0 83,93 97,59 100 100
Da 0,0134 Концентрация аллелей 0,0427 0,20 0,8846 0,3864
Db 0,9866 0,9573 0,8 0,1154 0,6136
pedghkmnp 0,5686 0,5345 0,5317 0,7307 0,8055
gedghjmn 0,2577 0,4655 0,4683 0,11153 0,1556
gbdgkmp 0,1737 0 0 0 0
gaeglns 0 0 0 0,1540 0,0389
Fa 0 0 0 0 0,0667
Fb 1 1 1 1 0,9333
41
Таблица 10 (окончание)
При- знак* Формы кабана
центрально-евро- пейская** (n = 32-370) кавказ- ская*** (n = 8-138) среднеазиат- ская (п = 20-83) сибир- ская**** (п = 7-35) уссурий- ская (п= 11-90)
1?°В' 0,4783 0,4018 0,6024 0,6548 0
Lbdfl 0,5011 0 0,0602 0 0
Ijtgiin 0 0,2946 0 0,0760 0,1111
j^adhi 0,0067 0,3036 0,3374 0,1154 0,6445
j^dhjk 0 0 0 0 0,1611
j^adhjl 0,0139 0 0 0,1538 0,0833
Ga 1 0,3929 0,3012 0 0,1167
Gb 0 0,6071 0,6988 Частота генотипов 0,8833 1
Daa 0 0 4,00 84,62 12,50
Dab 2,68 8,54 32,00 7,70 52,27
Dbb 97,32 91,46 64,00 7,68 35,23
Fab 0 0 0 0 13,33
Fbb 100 100 100 100 86,67
Gaa 100 16,07 2,41 0 0
Gab 0 46,43 55,42 0 23,33
Gbb 0 37,50 Частота 42,17 аллотипов 100 76,67
AM3’4 1 1 0,50 0,50 0,37
AM4’5 0 0 0,50 0,50 0,60
Lpr1+ 0,09 0,11 0,50 1 1
IgG1+ 1 1 0 1 0
Lpb3 Gll+ 0,38 0,16 1 0 1
0,29 0 0 Частоты трансферрина 1 0,67
TfA 0,219 0,133 0,109 0 0
TfB 0,781 0,867 0,891 0 0,909
Tf 0 0 0 1 0,091
* Число хромосом, антиген, аллель, генотип, аллотип, трансферрин.
** Материал в основном из Воронежского и Березинского заповедников и Беловежской пущи.
*** Материал преимущественно из Закавказья.
**** Забайкальский подвид (материал из Тувы).
Кариотип кабана из Японии такой же, как и у восточноазиатских рас: 2п = 38
[Muramoto et al., 1965]. Африканские звери имеют 37 и 38 хромосом, особи в За-
падной и Центральной Европе — от 36 до 38. У потомков диких свиней, завезен-
ных из Германии в Северную Америку, обнаружено 36 хромосом [McFee et al.,
1966; Groop et al., 1969; Rittmannsperger, 1971; Zivkovic et al., 1971; Bosma, 1976;
Нечипоренко, Першин, 1976; Бавин, 1979; Василько, 1984; Горелов, 1994].
Хромосомный набор кабана относительно сходен с таковым у бабирусс (2п = 38),
но существенно отличается от набора африканских бородавочников (2п = 34), у ко-
торых его уменьшение произошло за счет слияния 13-й и 16-й, 15-й и 17-й хро-
мосом. Кариотип американских пекари (2п = 30) преобразован за счет сложных
42
транслокаций и перицентрических инверсий [Орлов, Булатова, 1983; Тихонова,
Трошина, 1985]. Тем не менее по некоторым хромосомным фрагментам пекари
сходны с дикими свиньями, что указывает на возможность происхождения их от
общего олигоценового «предка» [Тихонов и др., 1983а, 1985].
Иммуногенетический полиморфизм диких свиней, как и хромосомный, доста-
точно высок (табл. 10). По данным И.Г. Горелова [1986, 1994], частота эритроци-
тарного антигена А у европейского кабана 0,72, у закавказского и среднеазиатско-
го соответственно - 0,59 и 0,46, у сибирского (из Тувы) — 0,74. Частота антигена Da
у европейской формы всего 2,86, у кавказской - 8,54, у среднеазиатской - 36,0, то-
гда как у сибирской - 92,32 и у уссурийской - 64,77. По антигену Gb четко вы-
ражена клинальная изменчивость - его нет у центральноевропейских популяций
(Беловежская пуща), у восточноевропейских (Воронежский заповедник) его
встречаемость приближается к 5%, в закавказской и северокавказской популяци-
ях- 84-93%, среднеазиатской - 98%, дальневосточных - 100%. У уссурийского
кабана, в отличие от других форм, обнаружен антиген Fa, который обычен у до-
машних свиней азиатского происхождения [Тихонов и др., 1975а, 1978, 1983а,
1985]. У него же, в отличие от центральноевропейской, кавказской (румынской)
и среднеазиатской рас, есть аллотип А-14, но отсутствуют Lpp3, А-11, A-13G, а
также А-23 сывороточного альфа-глобулина, которого нет и у среднеазиатского
подвида. В направлении с запада на восток ареала вида аллотипы А-15 и А-16
встречаются с большей частотой [Князев, 1983; Тихонов, Князев, 1985]. У забай-
кальского (тувинского) кабана, в отличие от остальных рас, обнаружен полимор-
физм по эстеразе D и гемоглобину [Горелов, 1987, 1994]. Полиморфизм по сис-
теме Е у диких свиней разных рас незначителен, тем не менее по некоторым ал-
лелям восточноевропейские особи отличаются от закавказских и среднеазиат-
ских [Тихонов и др., 1985, 1992].
Достаточно высок и межпопуляционный полиморфизм. Несмотря на географи-
ческую близость областей распространения, воронежские и белорусские кабаны
достоверно различаются по частоте антигенов Аа, Eb, Ej, На, Ка, Kb, Lc, Lg, по от-
дельным аллелям системы Е и другим иммуногенетическим параметрам. Анало-
гичные различия были обнаружены у особей северокавказской и закавказской по-
пуляций. Иммуногенетически северокавказские кабаны стоят ближе к восточноев-
ропейским, нежели закавказские [Тихонов и др., 19756, 1978, 1983а, 1985].
Локусы групп крови, сывороточные антигены и изоферменты явно имеют
адаптивное значение в эволюции, которая, возможно, идет за счет изменения ген-
ных частот признаков и новых комбинаций полиморфных генов. Например, анти-
ген Fa, отсутствующий у кабанов Восточной Сибири, с высокой частотой встреча-
ется у южноазиатских свиней, давая им преимущества в условиях жаркого климата
и высокой солнечной радиации [Горелов, Савина, 1994]. В группировке диких сви-
ней национального парка Гауя в Латвии после суровой зимы 1986-1987 гг. про-
изошел резкий сдвиг аллельных частот в локусе Е [Stass, 1992].
В целом цито- и иммуногенетическая изменчивость кабана (табл. 10) носит
клинальный характер с тенденцией обособления европейской и восточноазиат-
ской групп рас [Тихонов и др., 1975а, 1978, 1983а; Князев, 1983; Князев и др.,
1985]. Такого рода дифференциация, являющаяся результатом длительной гео-
графической изоляции, характерна также для косуль, лося и благородного оленя
[Данилкин, 1999а]. Азиатские домашние свиньи по иммуногенетическим показа-
телям имеют азиатское происхождение, а свиньи из Европы и Америки - евро-
пейское [Suzuki et al., 1984].
43
Гибридологические особенности
Гибриды, полученные при скрещивании 36-хромосомных кабанов с 38-
хромосомными домашними свиньями, имеют 37 хромосом, кариотип потомства
при возвратном скрещивании - 2п = 36-38. Жизнеспособность поросят с хромо-
сомным набором 2п = 37 снижена. Плодовитость свиней, осемененных гибрид-
ными хряками, имеющими кариотип 2п = 37, на 0,78 поросенка выше, чем от
гибридов с нормальным азиатским набором хромосом 2п = 38 [Тихонов, Троши-
на, 1977; Тихонов и др., 1983в; Василько, 1984].
У гибридных новорожденных в толщине кожи и развитии волосяного покро-
ва доминируют признаки кабана. Они имеют плотную ость, но у них гораздо
слабее развита подпушь [Петрищев, Бородин, 1983, Петрищев, 1992], что умень-
шает их шансы на выживание в природе.
У гибридов также очевидно проявление гетерозиса. Потомство первого и вто-
рого поколений крупнее диких особей, начинает размножение на год раньше, бо-
лее плодовито, отличается от домашних свиней крепостью конституции, большей
жизнестойкостью и устойчивостью к стрессам, лучше переносит высокие летние и
низкие зимние температуры, у него лучше качество мяса. Третье поколение по
своим параметрам более сходно с кабаном [Бутарин и др., 1955; Иваненко, 1956;
Журовский и др., 1970; Тихонов и др., 19836,в, 1985; Капралова, 1992].
Генофонд кабана широко используется селекционерами для совершенствова-
ния домашних пород и выведения новых [Бутарин и др., 1955; Ладан, 1960; Му-
хамедгалиев и др., 1970; Тихонов и др., 1981, 19836, 1985; Тихонов, 1990; Горе-
лов, 1993]. Одной из последних в результате скрещивания домашней свиньи со
среднеазиатским кабаном создана семиреченская темно-бурая порода, отличаю-
щаяся высокой продуктивностью [Ли и др., 1981; Джангельдина, 1985; Калдыба-
ев, Еськов, 1992]. От ландрас-кабаньих гибридов к 1982 г. получено более 500
тыс. потомков [Тихонов и др., 1983в].
Скрещивания кабана с домашними свиньями на воле нередки, особенно если
домашние стада находятся на вольном выпасе. В дельте Волги, например, в до-
машнем стаде с. Коклюя 21 поросенок из 100 имел полосатую окраску [Лавров-
ский, 1962]. В Грузии такого рода гибридизация происходит во многих районах
[Хеладзе, 1964].
Межвидовое скрещивание кабана с бородатой свиньей (S. barbatus) также
возможно. Получаемые гибридные особи плодовиты [Соколов, 1959].
Таксономия
Как показано выше, морфометрические и генетические признаки Sus scrofa
закономерно и относительно постепенно изменяются с востока на запад. В этом
направлении, вероятно, и происходило расселение и дальнейшее формообразо-
вание вида. Особи европейских форм на ранних возрастных стадиях сохраняют
отдельные примитивные черты восточноазиатских рас [Адлерберг, 1933], что
подтверждает общность их происхождения.
По морфометрическим и, особенно, по цито- и иммуногенетическим пара-
метрам европейские расы кабана весьма сходны, но заметно отличаются от вос-
точноазиатских форм; среднеазиатская занимает промежуточное положение. Та-
кая дифференциация свидетельствует в пользу гипотезы о двух центрах формо-
образования диких свиней [Тихонов и др., 1975а, 1983а; Князев, 1983]. Однако
это разделение может быть и следствием длительной географической изоляции
европейских и азиатских рас, что имеет место у европейской и сибирской косуль,
европейских и азиатских форм благородного оленя и лося [Данилкин, 1999а]. Ев-
ропейские и азиатские расы диких свиней, в свою очередь, стали родоначальни-
44
ками по меньшей мере двух типов домашних свиней (европейской и восточно-
азиатской), а также смешанных форм.
На территории бывшего СССР систематики обычно выделяют 5 подвидов каба-
I на [Адлерберг, 1930, 1933; Филипченко, 1933; Фетисов, 1936, 1956; Анучин, 1938;
Бобринский и др., 1944, 1965; Слудский, 1956; Соколов, 1959; Гептнер и др., 1961;
Громов и др., 1963; Барышников и др., 1981; Орлов, Мазин, 1992]:
западноевропейский (= центральноевропейский) - S. s. scrofa;
румынский (= кавказский) - S’, s. attila;
среднеазиатский - S. .s\ nigripes',
забайкальский (= сибирский) - S. 5. sibiricus (= raddeanus)’,
уссурийский (= дальневосточный) - S. s. ussuricus (= continentalis).
Подвидовые диагнозы, приводимые в первоописаниях и упомянутыми выше
авторами, однако, настолько нечетки, что по ним весьма проблематично отли-
чить одну форму от другой. Эта таксономическая «расплывчатость» имеет свои
причины - малые выборки (1-2 черепа или шкуры) при описании форм, значи-
тельное сходство рас по морфометрическим параметрам, огромная вариабель-
ность животных в окраске даже в пределах одной популяции и отсутствие доста-
точного материала для сравнительного географического анализа. Подвид S.
nigripes, например, выделен Бленфордом [Blanford, 1875] на основании более
темной, чем у S. s. scrofa, окраски конечностей и некоторых заметных различий в
строении черепа (без уточнения каких именно), а у S. s. attila Thomas, 1912 об-
щий тон окраски несколько светлее, чем у номинативного подвида.
В таксономии копытных наибольший вес имеют краниометрические показа-
тели, поскольку они менее вариабельны в зависимости от среды обитания по
сравнению с размерами тела. По Томасу [Thomas, 1912] именно длина (452 мм) и
высота (271 мм с нижней челюстью) черепа отличает S. s. attila от S. s. scrofa
(380-410 мм и до 210 мм соответственно). Если исходить из этих показателей, то
популяции Кавказа (см. табл. 7) более соответствуют S. s. scrofa, нежели S. 5.
attila, к которым их обычно причисляют.
Ю.А. Филипченко [1933] и А.В. Анучин [1938] в результате детального срав-
нительного краниометрического исследования разных географических рас, не-
смотря на некоторые методические ошибки, сделали несколько интересных
предположений, из которых внимания заслуживает близость S. s. scrofa, S. s.
attila и S. s. nigripes. Позднее к такому же выводу пришли И.И. Соколов [1959] и
М.А Сафаров [1965].
Восточноазиатские расы (S. s. sibiricus и S. s. ussuricus) диких свиней, в отли-
чие от европейских, имеют некоторые примитивные черты (иное строение ко-
ренных зубов и иную форму черепа и слезной кости [Адлерберг, 1930, 1933; Со-
колов, 1959]). Однако объединять их в качестве самостоятельного вида Sus
orientalis [Филипченко, 1933] или относить к Sus vittatus [Анучин, 1938], как от-
метил еще Г.П. Адлерберг [1933], нет веских оснований. Замечу также, что по
правилам зоологической номенклатуры подвиды S. s. raddeanus Adlerberg, 1930 и
S. s. continentalis Nehring, 1889 должны выступать в качестве младших синони-
мов S. s. sibiricus Staffe, 1922 и S. s. ussuricus Heude, 1888 соответственно, по-
скольку описаны позднее, примерно из тех же районов и не различаются прин-
ципиально. Путаница с русскими наименованиями подвидов легко устраняется
при учете места первоописания.
Забайкальский кабан (S. 5. sibiricus) заметно отличается от соседних форм
меньшими размерами черепа (см. табл. 7), что позволяет, хотя и не совсем четко,
обозначить границы его распространения. Причина возникновения существен-
ных краниометрических различий при отсутствии географической изоляции от
соседних рас, однако, не совсем ясна. На мой взгляд, измельчание этой формы
могло произойти в результате длительного негативного воздействия мощного
45
экологического фактора - постоянного недостатка зимних кормов на северном
пределе ареала в Восточной Сибири.
Кабаны с южного и северного склонов Главного Кавказского хребта также замет-
но различаются между собой по морфометрическим параметрам (см. табл. 6 и 7), по
телосложению, окраске [Дуров, 1983], краниометрическим признакам [Сафаров,
1965] и иммуногенетически [Тихонов и др., 1985]. Закавказская группировка ди-
ких свиней по многим показателям более сходна со среднеазиатской формой ви-
да, нежели с европейской. Скорее всего, на определенном этапе эволюции Закав-
казье заселили (или проникли туда) особи из Средней Азии, что несложно было
сделать вдоль южного побережья Каспийского моря или южнее. Обмен особями
между среднеазиатской и европейской расами также вполне возможен и в районе
к северу от Каспийского моря, но в очень давнее время этому, видимо, препятст-
вовали его трансгрессии. Отсюда, с учетом склонности кабана к перемещению на
дальние расстояния, становится понятной происхождение своеобразной кли-
нальности и относительной схожести рассматриваемых форм.
Накопленных сведений по географической изменчивости кабана все же не-
достаточно для прояснения его таксономии [Sjarmidi, Gerard, 1988]. Новейшие
охотоведческие преобразования, к сожалению, еще больше запутали системати-
ков. В XX в. в европейской части России, Зауралье, Западной и Восточной Сиби-
ри, а также на Украине, в республиках Закавказья и на Северном Казахстане в ре-
зультате искусственного расселения (более 9 тыс. особей) образованы многочис-
ленные смешанные популяции (см. ниже), а подмосковная группировка включает
особей всех подвидов, известных на территории СССР [Кораблев, 19866]. Соот-
ветственно, с учетом очень значительной подвижности зверей и панмиксии,
морфометрический и генетический материал из этого огромного региона нельзя
считать «чистым» и использовать в таксономии.
Фактически в Восточной Европе и в Сибири в результате масштабных непро-
думанных действий советских охотоведов образована новая «форма» кабана со
смешанным генофондом и неопределенным статусом, которой уместно дать обо-
значение Sus scrofa indet. Было бы чрезвычайно интересно проследить за измене-
нием ее генетических и морфометрических параметров, особенно в популяциях
на северном пределе ареала.
Итак, на территории России в настоящее время можно выделить лишь три
формы, из которых одна смешанная, одна (забайкальская) частично засорена чу-
ждым генофондом и одна (уссурийская) - пока еще «чистая».
Sus scrofa indet. - Смешанная “форма”
Статус не определен. “Форма” образована в результате искусственного сме-
шения разных рас с преобладанием европейских особей.
Распространение. Восточная Европа, Урал, юг Западной Сибири, Северный
Казахстан, юг Восточной Сибири (на восток до Красноярского края включитель-
но).
Sus scrofa sibiricus Staffe, 1922 - Забайкальский кабан :
Сравнительно мелкая форма. Череп короткий, его профиль чаще слегка вы- <
пуклый в лобной области. Наибольшая длина черепа у самцов 344-441 мм, у са-
мок 337-390 мм, высота 206-245 мм и 169-209 мм, скуловая ширина 144-158 мм
и 130-154 мм, длина верхнего ряда коренных зубов 107-120 мм. Слезная кость
укороченная, почти квадратной формы. Третий верхний коренной зуб довольно
широкий, талон слабо развит и упрощен. Окраска тела от темно-бурой до черной.
46
От заднего края рта к ушным раковинам тянется серая или белесая полоса. Горло
и паха серебристо-серые.
Распространение. Саяны, Прибайкалье, Забайкалье, Монголия, Китай к за-
паду от Большого Хингана.
Sus scrofa ussuricus Heude, 1888 - Уссурийский кабан
Более крупная форма. Профиль черепа выпуклый. Наибольшая его длина у
самцов 424-451 мм (до 510 мм [Гептнер и др., 1961]), у самок 338-425 мм; высо-
та 232-268 мм и 225-230 мм, скуловая ширина более 160 мм, длина верхнего ря-
да коренных зубов более 135 мм. Слезная кость относительно широкая, по форме
ближе к квадратной. Последний верхний коренной широкий, талон почти не раз-
вит. Уши чуть короче, чем у других форм. Окраска черно-бурая с более светлыми
участками по низу головы, шеи, тела, хребту и области вокруг глаз. От угла рта к
уху проходит белая полоса.
Распространение. Бассейн Уссури и Амура на запад до Большого Хингана,
Северо-Восточный Китай, Корея.
Распространение
Дикие свиньи - теплолюбивые и влаголюбивые животные с особой пищевой
специализацией, что в целом определяет их распространение в южных и умерен-
ных широтах, где возможно круглогодичное добывание корма в почве и призем-
ном слое. Однако на протяжении многих тысячелетий границы ареала вида су-
щественно расширялись и изменялись, особенно на северном пределе обитания.
Восстановленный ареал
Распространение в плейстоцене и голоцене
Область распространения кабана в плейстоцене была значительно меньше,
чем в голоцене (рис. 17). Костные остатки плейстоценового возраста обнаружены
лишь в Англии, Бельгии, Швейцарии, Южной Германии, Франции, Испании,
Италии, Югославии, Чехии, Венгрии и, возможно, Болгарии, в Крыму, Приазо-
вье, на Северном Кавказе, в Закавказье, Средней Азии, Китае и в Приморье
[Шарффъ, 1918; Пидопличко, 1951, 1954; Бибикова, 1960, 1975; Паавер, 1965;
Kurten, 1968; Батыров, 1969; Верещагин, Громов, 1977; Верещагин, Барышников,
1980а; Давид, 1980; Алексеева, 1982; Давид и др., 1990]. В Северной и на боль-
шей части Восточной Европы, включая Украину и Молдавию, диких свиней, как
считалось, не было [Бибикова, 1960]. Тем не менее позднее их остатки найдены в
сингальском комплексе первой половины плейстоцена на Нижнем Поволжье
[Алексеева, 1990], в средне - позднеплейстоценовых отложениях волжского ост-
рова Хорошевский в Саратовской области [Хромов и др., 2000] и, возможно, на
Самарской Луке [Стрижева, 1991].
В конце плейстоцена - раннем голоцене на большей части Северной Евразии
(в границах бывшего СССР) в фауне доминируют мамонт, шерстистый носорог,
бизон, большерогий и гигантский олени, обычные виды - сайгак, лошади, лось,
косули, благородный и северный олени. Ареал кабана к этому времени явно рас-
ширяется, особенно в верхнем палеолите. Его костные остатки найдены на мно-
гих древних стоянках человека: в Западной Европе, Молдавии, на Украине, в
Крыму, Предкавказье, Закавказье, Иране, Палестине, Сирии, на Нижнем По-
волжье, в Казахстане, Средней Азии и на юге Дальнего Востока [Бируля, 1930;
47
20 10 О 10 20 30 40 SO 60
Рис. 17. Распространение кабана в четвертичном периоде (по [Бибикова, I960]) <
1 и 2 - места находок костных остатков в плейстоценовых и голоценовых отложениях (непол- ;
ные сведения. - Л.Д.), 3 и 4 - северная граница ареала вида в плейстоцене и голоцене соответ- 1
ственно. Цифры - обозначения мест находок плейстоценовых остатков (по первоисточнику).
Бадер, 1940; Беляева, 1940; Громов, 1948; Пидопличко, 1951, 1954, 1956; Слуд- j
ский, 1956; Верещагин, 1959, 1971; Бибикова, 1953, 1960, 1975; Верещагин, Ба-
тыров, 1967; Kurten, 1968; Давид, Чемыртан, 1976; Кожамкулова, 1977, 1981;
Алиев, 1978; Верещагин, Барышников, 19806; Давид, 1982, 1986; Батыров, 1986;
Алексеева, 1990; Худайбердиев, 1990; Турубанова, 1999]. Обнаружены они также
в Белоруссии [Калиновский, 1983], в бассейне Дона [Громов, 1948; Пидопличко,
1954; Верещагин, 1975; Верещагин, Барышников, 1980а] и в долине Оки непода-
леку от Рязани [Антипина, Маслов, 1990].
В фауне плейстоцена Сибири дикие свиньи, в отличие от лося, благородного
оленя и сибирской косули, вероятно, отсутствуют, что связано с неблагоприят-
ными ландшафтно-климатическими условиями, и прежде всего - с остепнением
и глубоким промерзанием почвы. Обширной зоны тайги в Северной Азии прак-
тически не существовало: тундра плавно переходила в зону холодных степей и
полупустынь [Бибикова, 1960; Равский и др., 1964; Вангейгейм; 1977; Алексеева,
1980]. Возможно, именно эти факторы сдерживали распространение вида и в
Восточной Европе.
В голоцене (в последние 12-10 тыс. лет) происходит потепление климата, расши-
ряется зона лесов и очевидна экспансия ареалов многих видов диких копытных на се-
вер. В бореальный период раннего голоцена дикие свиньи заселяют Ирландию, Да-
нию, Южную Швецию и Западную Прибалтику. В атлантический период среднего го-
лоцена (5-2 тыс. лет до н. э.), отличавшийся мягким климатом и обширной зоной ши-
роколиственных лесов, они занимают Южную Норвегию и распространяются по всей
Прибалтике. На стоянках человека на северо-западе бывшего СССР среди костных ос-
татков диких копытных доля кабана составляет от 9 до 100%, но в позднем голоцене
(при похолодании) снижается до 3-54% [Паавер, 1965]. В это время вид становится
обычным и в Белоруссии [Цалкин, 1956,1962; Сержанин, 1961].
48
На Украине и на всем юге Восточной Европы в нижнем голоцене кабан срав-
нительно редок. Многочисленным он становится примерно за 4,5 тыс. лет до н. э.
В неолитических поселениях человека в Причерноморье, Поднепровье и Молда-
вии его костные остатки доминируют (до 80% остатков среди охотничьих зверей
в отдельных захоронениях) наряду с благородным оленем, косулей, лосем и ту-
ром. В конце I тысячелетия до н. э. этот вид, вероятно, был одним их самых мно-
гочисленных среди копытных восточноевропейской лесостепи. В начале нашей
эры поголовье диких свиней несколько сокращается, но вплоть до XVIII в. н. э.
они остаются одним из основных объектов промысла [Кириков, 1952, 1966, 1983;
Корнеев, 1953, 1970; Пидопличко, 1954, 1956; Цалкин, 1962, 1966а, 19696, 1970,
1972а; Бибикова, 1963, 1975; Тимченко, 1972, 1974; Журавлев, 1981; Давид,
1982]. Так же обильно в среднем и позднем голоцене дикие свиньи населяли
Крым, Предкавказье, Кавказ и Каспийское побережье, включая дельту Волги
[Пидопличко, 1954; Верещагин, 1959; Формозов, 1972].
Наибольший интерес вызывает распространение кабана на севере Восточной
Европы. А.Н. Формозов [1946] полагал в свое время, что в области верхнего и
среднего течения Волги кабана в историческое время не было. Сообщения о
встречах зверей севернее изолинии высоты снежного покрова в 50 см, в частно-
сти в Костромской губернии [Дюбюк, 1920], он относил к неправдоподобным.
А.А. Слудский [1956] проводил границу ареала вида в Восточной Европе север-
нее: от Ладожского озера через Рыбинское водохранилище, Кострому, чуть се-
вернее Волги на низовья Ветлуги и к устью р. Белой, что вызывало сомнение у
многих ученых и не получило поддержки.
За основу обычно принимались сведения В. Г. Гептнера, который понятие
“восстановленный” ареал трактовал в сравнительно узком “историческом” смысле,
отбрасывая сомнительные, на его взгляд, сообщения. Северную границу восстанов-
ленного ареала (включая заходы) диких свиней (рис. 18) он проводил от восточной
оконечности Финского залива и юго-западного побережья Ладожского озера (район
Мги) на Новгород, Осташков, Калинин, Москву, Калугу, Тулу, Рязань, Пензу, Сама-
ру, по р. Бузулук и верховьям Белой и далее, в азиатской части, - на Магнитогорск,
Миасс, Куртамыш, Ишим, Усть-Тару, оз. Чаны, Новосибирск, захватывал юг Алтая
и уводил в Китай [Гептнер, 1960; Гептнер и др., 1961]. Новые данные, однако, позво-
ляют отодвинуть эту условную линию значительно севернее.
В среднем - позднем голоцене на северо-западе России северная граница
распространения диких свиней, вне всякого сомнения, достигала Ладожского и
Онежского озер и захватывала юг Карелии. Костные остатки этих зверей обна-
ружены, как минимум, на 23 стоянках человека в лесной зоне: в Ленинградской,
Псковской, Новгородской, Ярославской, Вологодской, Костромской, Тверской,
Московской, Смоленской и Рязанской областях (рис. 18). Вместе с лосем, благо-
родным и северным оленями, косулей, волком и медведем кабан был одним из
основных охотничьих видов в добыче местных племен [Иностранцев, 1882;
Формозов, 1946; Слудский, 1956; Цалкин, 1956, 1962; Юргенсон, 1956; Марвин,
1959; Андреева, Петренко, 1976; Верещагин, Русаков, 1979]. Особо отмечу на-
ходки остатков кабана на мезолит-неолитической стоянке «Ивановское-7» в
Ярославской области [Кириллова, 1999], на р. Шоше (каменный век) в Тверской
области [Степанова, Степанов, 1971, 1974], на нескольких стоянках 1II-II тысяче-
летия до н. э.: Языковской (Кашинский район Тверской области), Ладожской и
Кубенино (Онежский край), Модлонской (Чарозерский район Вологодской об-
ласти), Федоровской (Чухломское озеро, Костромская область), а также в Холмо-
горском (?!) могильнике под Архангельском [Юргенсон, 1956].
В I тысячелетии - первой половине II тысячелетия н. э. ареал кабана на запа-
де и юге Восточной Европы был не менее обширным. Костные остатки этих жи-
вотных присутствуют во многих археологических памятниках Прибалтийских
4. Заказ №2528.
49
республик, Белоруссии, Украины, Молдавии и России: в Пскове, Новгороде,
Старой Ладоге, Брянске и Брянской области, окрестностях г. Кимры Тверской
области, на территории Москвы, в Можайском и Раменском районах Московской
области, в Калужской области, в Старой Рязани, в нескольких селищах в бассей-
не р. Оки, в большинстве городищ Подонья, Поднепровья и Северного Кавказа.
Особенно обильны они в поселениях человека в Воронежской, Орловской и Кур-
ской областях [Третьяков, 1941; Громова, 1948; Ефименко, Третьяков, 1948;
Алексашина, 1950; Цалкин, 1956, 1961, 1962, 1963, 1966а, 1968, 1969; Барабаш-
Никифоров, 1957; Верещагин, 1971; Андреева, 1972; Верещагин, Барышников,
1980а; Александрович, 1999]. В этот период, как и прежде, вместе с дикой свинь- ’
ей жили косули, благородный олень, лось и волк.
В Волжско-Камском крае, как считалось ранее, кабана не было [Павлов, 1978, ;
1999а]. Интереснейшие археологические находки [Прокошев, 1940; Збруева,
1952, 1954; Цалкин, 1958; Андреева, Петренко, 1976; Петренко, 1984] в большин-
стве своем остались незамеченными или проигнорированными зоологами и охо- ;
товедами. Тем не менее остатки диких свиней обнаружены: ,
на мезолитической (8-6 тыс. лет до н. э.) Деуковской-2 стоянке на правом
берегу р. Ик, в городище Гремячий ключ (раннее железо, I тысячелетие до н. э.)
на правом берегу Камы, в ананьинских (VIII—III вв. до н. э.) городищах Сорочьи
горы (Лаишевский район) и Свиногорское (устье р. Вятки) в Татарии;
на мезолитической стоянке Баринка-1 на р. Кильмезь и Усть-Лудяны (эпоха
ранней бронзы, конец III - начало II тысячелетия до н. э.) на левобережье Вятки в
Удмуртии;
в ананьинских городищах Усть-Нечкинское (Сарапульский район) на правом
берегу Камы, Лебяжье (Советский район), Белоглазовское (Нолинский район),
Ройское, Буйское (Уржумский район) и Аргыжское (Малмыжский район) по р.
Вятке в Кировской области;
в Левшинском поселении (ранняя бронза) в устье р. Чусовой, селище Поло-
винное-1 (XI-I вв. до н. э.) в Сальском районе и Горюхалихинском городище (IV
в. до н. э. - VI в. н. э.) в бассейне р. Очер в Пермской области;
в Богородском городище [Альбицкий, Пчелкин, 1931] в б. Ивановской про- ।
мышленной области (р. Ветлуга, Нижегородская область); 1
на стоянках эпохи ранней бронзы Мариер на правом берегу р. Б. Кокшаги и i
Баркужерское-3 в бассейне р. Илеть в республике Марий-Эл;
на стоянках поздней бронзы Инзелга (правый берег р. Белой, Гафурийский
район) и Идрисово [Смирнов и др., 1990] в Салаватовском районе, а также в Бик-
тимировском городище (раннее железо) в Бирском районе Башкирии;
на Гулькинской стоянке (бронза) в Старомайнском районе Ульяновской об-
ласти и, возможно, в поселении Моечное Озеро-1 на берегу р. Усы в Шигонском
районе б. Куйбышевской области.
Более поздние остатки кабана зарегистрированы в нескольких поселениях V-
VII вв. н. э.: Щербетьское, Балымерское (Куйбышевский район), Именьковское
(Лаишевский район), I Троицко-Урайское (Рыбно-Слободской район) в Татарии и
в Выползовском селище в Волжском районе Куйбышевской области. Найдены они
также и в нескольких археологических памятниках II тысячелетия н. э.: Муранском
селище (XII-XIV вв.) в Шигонском районе Куйбышевской области; Чебоксарском
городище (XIV-XVIII вв.); Казанском кремле (VI-XIV вв.); городищах VIII—XIV
вв. Анюшкар (правый берег Иньвы, приток Камы) в Ильинском районе и Редикор-
ское на правом берегу Вишеры в Чердынском районе Пермской области; городище
Кудымкар (IX-XV вв.) в Коми-Пермяцком национальном округе.
Следовательно, палеонтологами подтверждено распространение диких сви-
ней (а также сибирской косули и благородного оленя в отдельных пунктах) на
Среднем Поволжье и в Вятско-Камском крае минимум до Чебоксар и Йошкар-
50
Рис. 18. Распространение кабана в голоцене на севере Восточной Европы
1 - северная граница восстановленного ареала; 2 - то же (по [Гептнер и др., 1961]); 3 - места
находок ископаемых остатков (см. текст)
Олы, по бассейну Вятки - почти до Кирова, по Каме - до ее северной излучины
(до р. Вишеры), по р. Белой - до Уфы (рис. 18). Звери обитали здесь, в историче-
ском понимании, непрерывно с эпохи мезолита и бронзы до второй половины II
тысячелетия н. э. Они были немногочисленны, однако в отдельных памятниках
приказанской культуры кости кабана, наряду с костными остатками зайца, со-
ставляют основу [Попов, 1964]. Последующее исчезновение диких свиней из это-
го обширного региона, как справедливо считал В.И. Цалкин [1958], - «несомнен-
ный результат истребительной и хозяйственной деятельности человека». В Тата-
рии они уничтожены, вероятно, лишь в XVIII в. [Попов, 1978].
Отмечу также, что в Волжско-Камском крае, как и во всей лесной зоне к се-
веру от Верхней Волги, широколиственные леса в прошлом были распростране-
ны гораздо шире, чем в настоящее время, а подзона смешанных лесов простира-
лась примерно до широты Архангельска [Кириков, 1966]. Северная граница вос-
становленного ареала кабана (см. рис. 18) почти совпадает с границей распро-
странения дуба [Долгошов, 1962; Любимова, 1962]. Следовательно, есть все ос-
нования полагать, что склонный к дальним перемещениям вид при отсутствии
существенных географических барьеров проникал гораздо севернее тех районов,
где его остатки обнаружены на месте главнейших древних поселений человека
[Брюсов, 1954; Петренко, 1984], т. е. вполне мог достигать южных районов Ар-
хангельской области и Республики Коми.
4*
51
На Нижней Волге голоценовые остатки кабана найдены в черте г. Саратова и
г. Энгельса, на Варфоломеевской стоянке (V-IV вв. до н. э.) в Новоузенском рай-
оне, в двух пунктах Вольского района и у с. Трубино в Красноармейском районе
Саратовской области [Хромов и др., 2000].
На восточном склоне Южного и Среднего Урала, в Зауралье и на юге Запад-
ной Сибири дикие свиньи появляются, как и в Предуралье, в среднем голоцене,
хотя лось, сибирская косуля и благородный олень известны здесь с плейстоцена
[Цалкин, 19726; Смирнов, 1976а, б; Верещагин и др., 1977; Смирнов и др., 1990;
Варов, Косинцев, 1996]. По сведениям П.А. Косинцева [1986, 1988, 1996], в степ-
ной и лесостепной зонах Среднего Урала и Западной Сибири остатки кабана об-
наружены более чем в 20 древних поселениях человека. Интересен факт его на-
ходок в Шигирском торфянике близ Нижнего Тагила, в поселении Верхняя Са-
нарка в Пластовском районе Челябинской области, а также на неолитической
стоянке в районе Ильменского государственного заповедника [Дворников, 1986;
Матвеев, Бакунин, 1994]. В Омском музее, по сообщению А.А. Слудского [1956],
хранятся кости дикой свиньи из Омской стоянки, датируемой XV в. до н. э. Эта
датировка вызывает некоторое сомнение, поскольку подавляющее большинство
находок зверей в Южной Сибири средне- и позднеголоценового происхождения.
Нельзя не отметить также и случай обнаружения черепа кабана на берегах Те-
лецкого озера на Алтае [Залесский, 1934; Гептнер и др., 1961].
Южнее костные остатки кабана найдены на местах поселений человека эпохи
бронзы на севере и востоке Казахстана, на мезолит-неолитической стоянке Кара-
унгур в Чимкентской области, под Хорезмом, в бассейне Амударьи, в слоях 11
тысячелетия до н. э. в Ферганской долине, в нижнем течении Зеравшана, Южном
Узбекистане, Южном Таджикистане (5-4 тыс. лет до н. э.) и других районах
Средней Азии [Ишадов, 1965; Цалкин, 19666; Верещагин, Батыров, 1967; Баты-
ров, 1969; Кожамкулова, 1969, 1976, 1977, 1981; Шарапов, 19726; Макарова,
1976; Верещагин, Барышников, 1980а; Кожамкулова, Костенко, 1984].
В Предбайкалье на стоянках Ангарской группы кабан появляется лишь в ме-
золите [Вангейгейм, 1977] или, вероятно точнее, - в неолите [Ермолова, 1978].
Его клыки обнаружены в нескольких могильниках эпохи неолита и бронзы не
только на Ангаре, но и в Старом Качуге на Верхней Лене [Окладников, 1950]. В
Приморье вид обитал непрерывно с плейстоцена. Число его костей в Сучанских
пещерах (5-4 тыс. лет до наших дней) уступает лишь кабарге [Верещагин, Ба-
рышников, 1980].
В некоторых источниках упоминается о былом обитании диких свиней на
Сахалине, где они исчезли, видимо, еще в доисторическое время [Абрамов, 1954;
Бромлей, 1964; Беньковский и др., 1989].
Изменения ареала во II тысячелетии н. э.
Распространение кабана в позднем голоцене было, вероятно, почти неизмен-
ным вплоть до середины II тысячелетия н. э. Существенные изменения ареала и
численности очевидны лишь с XV-XVIII вв. в связи усилением антропогенного
воздействия на животных и среду их обитания.
В Англии диких свиней истребили в 1617 г., а последующие попытки интро-
дукции проваливались из-за противодействия крестьян. С 1660 г. кабана не стало
в Нидерландах (отдельные особи, вероятно, жили здесь до начала XIX в. [Pelzers,
1988]) и на островах в Дании. В Ютландии два последних зверя убиты в 1801 и
1803 гг. В этот же период животные уничтожены в Тюрингии, Саксонии, Бава-
рии и в других районах Германии, а также в Швейцарии, и их не было здесь
вплоть до 1940 г. [Briedermann, 1986]. В Западной Европе отдельные очаги оби-
тания кабана сохранились лишь на Пиренеях, Арденнах и Апеннинах [Prossinagg,
52
1985]. Вид полностью перестал существовать не только на Британских островах
и Скандинавском полуострове, но и в Египте и Судане.
В Прибалтике поворотным периодом в послеледниковой истории кабана стал
XVIII в. - звери уцелели в небольшом количестве только в Литве. Их последую-
щее исчезновение из региона отчасти совпадает с уничтожением широколист-
венных лесов и ухудшением кормовой базы, но главная причина - неограничен-
ная охота и исключительное обилие волка. В XIX в. регистрировались лишь ред-
кие заходы из стран, расположенных южнее, но мигрантов сразу же убивали
[Линг, 1955; Паавер, 1965; Кириков, 1966].
В Белоруссии в XII—XVIII вв. дикие свиньи, судя по костным остаткам и го-
сударственным актам, были многочисленными и доминировали в фауне вместе с
благородным оленем, зубром, косулей и лосем [Цалкин, 1951; Кириков, 1960,
1966]. Вплоть до 1915 г. они были распространены на большей части республики,
занимая в Борисовском и Мозырском округах 60-86%, в Минском, Бобруйском и
Слуцком - треть, а в остальных округах - от 1 до 10% территории. К 1925 г. пло-
щадь ареала сократилась в 2-3 раза, в Калининском округе их не стало. Все виды
диких копытных в это время находились «на пути к окончательному истребле-
нию» [Федюшин, 1928; Сержанин, 1961, 1970].
В Киевской Руси в XVI в., по описаниям М. Литвина [1890], «диких зверей:
зубров, диких лошадей и оленей такое множество в лесах и на полях, что за ними
охотятся только ради кожи, а мясо, по причине большого его изобилия, бросают,
за исключением филейных частей; ланями и дикими кабанами совсем гнушают-
ся». Резкое снижение численности копытных происходит с середины XVIII в. В
1750-1764 гг. специальная команда стрельцов, снабжавшая дичью царский двор,
смогла добыть в Запорожье лишь пять кабанов. В южных губерниях охота на
этот вид была прекращена. Однако в тростниковых зарослях по берегам Буга и
Днепра, как и в междуречье, дикие свиньи оставались многочисленными до кон-
ца столетия. К середине XIX в. в связи с усилением охотничьего пресса они поч-
ти исчезают в лесостепных и степных районах, сохраняясь лишь в самых трудно-
доступных местах, но их еще много по р. Пруту. В Киевской губернии последний
кабан убит в 1889 г. [Туркин, 1899]. С начала XX в. и до 30-40-х годов он полно-
стью отсутствует в степном Причерноморье, на юго-востоке республики и редок
на северо-востоке [Браунер, 1923; Мигулин, 1927; Корнеев, 1970; Кириков, 1976;
Турский, 1975]. В Прикарпатье и Карпатах, напротив, звери плотно населяли
почти все лесные и горные районы, а в послевоенные годы в отдельных местах
их объявляли вредителями со всеми вытекающими отсюда последствиями [Со-
кур, 1952]. В горном Крыму вид истреблен в середине XIX в. [Гептнер, 1960].
В Молдавии вплоть до начала XX в. дикие свиньи были распространены по-
всеместно, в некоторых районах их было «великое множество», а вред посевам,
особенно кукурузе, от нашествия зверей был «огромен». В середине XX столе-
тия, в военные и послевоенные годы, они были почти полностью уничтожены
человеком, и лишь единицы их сохранились в Кодрах и в плавнях Прута и Дне-
стра [Аверин, 1960а,б; Лозан, Лозан, 1981в].
В Карелии диких свиней уничтожают после славянской колонизации, начав-
шейся в XI в. [Марвин, 1959]. В X-XIV вв. на них еще охотились вокруг Новго-
рода и Пскова. Позднее кабан исчезает из северных районов северо-запада Рос-
сии и не появляется там вплоть до 30-х годов XX столетия. Соответственно се-
верная граница ареала отодвигается к югу на 300-400 км [Кириков, 1953, 1966;
Цалкин, 1956; Гептнер, 1960; Гептнер и др., 1961; Верещагин, Русаков, 1979; Ру-
саков, Тимофеева, 1984; Русаков, 1989]. Однако в Костромском наместничестве
дикие свиньи обитали вплоть до конца XVIII в. [Дюбюк, 1920], а в Ивановской
промышленной области встречались еще в 1868 г. [Альбицкий, Пчелкин, 1931].
В Подмосковье в XVIII в. кабан обычен, отдельные забеги регистрировались
53
до середины XIX в., но к концу столетия зверей не стало совершенно [Данилов,
1947; Кириков, 1953; Соколов, 1959]. На Брянщине, в Молого-Шекснинском ме-
ждуречье и на территории Тверской области этот вид обитал вплоть до конца
XIX в. [Исаков, 1949; Шапошников и др., 1959; Кириков, 1966].
В XVII в. кабан еще был охотничьим видом в смоленских и рязанских лесах,
водился по рекам Цне, Хопру и Вороне. В XVIII в. он числился в «реестре зве-
рей» Воронежской губернии, Пензенской и Симбирской провинций и Малорос-
сии и отмечался в Орловской губернии. В «Примечаниях к генеральному меже-
ванию» (1776-1780 гг.) кабан, лось, олень и косуля уже не упоминаются в Туль-
ской губернии, а в Смоленской относятся к крайне редким зверям [Кириков,
1953, 1960, 1966; Слудский, 1956].
В низовьях Дона и Приазовье кабаны исчезли в 1812 г. [Зверозомб-
Зубровский, 1923], на Среднем Подонье - в первой половине XIX в. [Барабаш-
Никифоров, 1957]. К концу XIX в. они, а также косули и олени, полностью ис-
треблены на Тамбовщине [Громаков, 1998]. На Среднем Поволжье их не было
уже в начале XIX в. [Богданов, 1871], в северных районах Нижнего Поволжья
[Кубанцев, Зубов, 1978] и Курской лесостепи [Гусев, 1989] - в начале XX в.
В благодатном климате Предкавказья, Северного Кавказа и Закавказья дикие
свиньи были широко распространены и многочисленны вплоть до середины XIX в. -
до заселения Кавказа русскими. Наиболее быстро человек уничтожил их на Армян-
ском нагорье - при отсутствии религиозного запрета на употребление мяса этих жи-
вотных и при интенсивном обезлесении горных склонов. В христианской Грузии
данный негативный процесс также очевиден с XIX в., но здесь звери сохранились
благодаря обилию лесов и труднодоступных заболоченных участков. Сокращению
ареала вида в Предкавказье, несомненно, способствовало строительство со второй
половины XVIII в. кавказской военной линии, для чего тугайные леса на равнине и в
предгорьях намеренно выжигали и вырубали. К началу XX в. казаки истребляют ка-
бана под Ставрополем, на Кубани (немного животных уцелело близ станиц Гривен-
ской и Петровской, где они жили еще в 1932 г.), Тереке и Сулаке, за исключением
отдельных районов. Тем не менее в первом десятилетии XX в. этот вид еще обитал
во всех горных лесах Северного Кавказа, а северная граница его ареала проходила
близко к линии Анапа - Краснодар - Майкоп - Нальчик - Моздок - Владикавказ -
Махачкала - побережье Каспия. Особенно плотно были заселены низовья Кубани,
Терека и Кумы, непроходимые заросли возле станции Буйнак, морское побережье от
Петровска до Дербента, Черноморская и Кутаисская губернии, долины Куры и Ала-
зани. В Закавказье кабаны обитали везде, где были леса, густые заросли кустарника
и камыши. Еще в начале XX в. недалеко от Тбилиси за день охоты можно было до-
быть до 50 голов. В 30-х годах зверей истребляют в лесах северо-восточного Караба-
ха и на северных склонах Малого Кавказа вплоть до Грузии, позднее - почти повсе-
местно, включая равнины и предгорья Дагестана. Отдельные малочисленные груп-
пировки сохраняются лишь в труднодоступных убежищах и в заповедниках [Дин-
ник, 1910; Марков, 1932; Верещагин, 1959; Кириков, 1966; Сафронова, 1973; Сарки-
сов, Хохлов, 1982; Соколов, Темботов, 1993].
Каспийское побережье, лесостепную и степную зоны Предуралья, Зауралья,
Южной Сибири и Казахстан дикие свиньи устойчиво заселяли вплоть до середи-
ны XIX в. Во многих районах они обитали «в необъятном множестве» и «вели-
кими стадами», а побережье Каспия и дельты впадающих в него рек характери-
зовались в охотничьей литературе как «царство» вида. Обилие кабана и других
копытных, включая лося, косуль и благородного оленя, здесь, как и по всему югу
европейской части России, не было чем-то необычным. Несколько веков назад
эта территория была более облесенной, особенно по поймам полноводных рек и
многочисленных озер, и, главное, малолюдной [Кириков, 1952, 1955, 1959, 1966,
1983; Слудский, 1956; Марвин, 1969; Варов, Косинцев, 1996].
54
В конце XVIII - первой половине XIX вв. кабан еще встречался в башкирских
широколиственных лесах (по р. Белой), а в Зауралье заходил до Троицкой и Че-
лябинской крепостей [Формозов, 1946; Кузнецов, 1948; Слудский, 1956]. П.С.
Паллас [1773, 1786] неоднократно упоминал его как типичного представителя
фауны Оренбургского края и юга Западной Сибири. В Тоболо-Ишимской лесо-
степи множество зверей обитало на озерах Копчи и Мангут, изредка попадались
они в Барабинской степи и в предгорьях Алтая. В 1748 г., по сообщению пра-
порщика П. Старцева, этот вид, вместе с маралом, лосем и косулей, водился
«близ р. Алей, где бор от Иртыша пришел концом» [Формозов, 1946]. Топогра-
фический отряд под начальством инженер-прапорщика Зеленого в 1764 г. отме-
чал присутствие диких свиней в степной зоне по берегам Иртыша между устьями
Нарыма и Курчума [Кириков, 1963а, 1966]. В XVIII в., по свидетельствам оче-
видцев, их было много на озерах Чаны и Абышкино, в Исилькульском районе, на
юго-западе Барабы и в Центральной Кулунде, в XIX в. - в Кокчетавском районе
Казахстана [Залесский, 1934].
К концу XIX в. кабан был уничтожен человеком (в основном русскими пересе-
ленцами) почти повсеместно, и о его существовании долгое время напоминали
лишь «кабаньи» названия отдельных рек, озер, урочищ и населенных пунктов. Зве-
ри уцелели только в дельте Волги, на территории Южного Казахстана и Средней
Азии, а также в отдельных местах Тоболо-Ишимского междуречья и Заишимья. С
территории Тюменской области они полностью исчезли после необычно суровой
зимы 1913-1914 гг., когда были вынуждены выходить к населенным пунктам
[Азаров, Климов, 1990; Азаров, 1996]. В конце XIX - начале XX в. северная грани-
ца ареала вида на Урале и в Западной Сибири сдвинулась к югу на сотни километ-
ров, и дикая свинья уже не упоминается в составе местной фауны [Словцов, 1892;
Зверев, 1937; Куклин, 1938; Янушевич, Благовещенский, 1952].
В XIX - начале XX вв. кабан изредка встречался в предгорьях и на юге Алтая
(Западный Алтай, Чуйская высокогорная степь с окрестностями, северный склон Ка-
тунских гор, долина р. Бухтармы). Отдельные заходы зверей еще дальше на север
региона обычно относили к сомнительным случаям, но они имели место неодно-
кратно [Яблонский, 1904; Селевин, 1930; Залесский, 1934; Кузнецов, 1948; Насимо-
вич, 1949; Слудский, 1956; Соколов, 1959; Гептнер и др., 1961; Собанский, 1992].
Сокращение ареала вида в XIX в. заметно и в Восточной Сибири (в нижних
течениях рек Каи, Бирюсы и Чуны), но еще в начале XX в. звери жили в бассейне
Ангары и заходили к вершинам Лены. В Северо-Восточном Забайкалье (в Бодай-
бинском районе, в верховьях р. Чары и близ оз. Нечатка) последние кабаны ис-
треблены, вероятно, в многоснежные зимы 1920-1922 гг. Не стало их и в Тим-
птонском районе Якутии [Скалой и др., 1941; Копылов, 1950; Скалой, 1951;
Слудский, 1956].
В свете имеющихся и новых данных, северная граница восстановленного ареа-
ла диких свиней на западе России захватывает юг Карелии, Вологодскую и Кост-
ромскую области, большую часть Кировской и Пермской областей (по средним те-
чениям рек Ветлуги, Вятки и Камы на север до Вишеры), достигая как минимум
60° с. ш., а заходы зверей вероятны до широты Архангельска (рис. 18). Далее на
восток она идет близко к линии Нижний Тагил - Тюмень - Усть-Ишим - Тара - оз.
Чаны - Новосибирск - Бийск - Телецкое озеро, южнее Западного Саяна, пересе-
кает Восточный Саян в районе р. Кан, через Тайшет выходит на р. Ангару север-
нее Братска (примерно на 57° с. ш.), захватывает верховья Лены в районе Усть-
Кута, огибает оз. Байкал с юга, поднимаясь по восточному побережью на север
до Бодайбо, а по р. Чаре - до юга Якутии, проходит по южным отрогам Станово-
го хребта, вклиниваясь по р. Тимптон в Якутию, и через верховья Бурей выходит
к низовьям Амура, Татарскому проливу и о. Сахалин (рис. 19).
55
Рис. 19. Распространение кабана в России и сопредельных регионах (в границах бывшего СССР)
1 - северная граница восстановленного ареала (там, где она не совпадает с современной границей); 2 — минимальная область обитания в начале
XX в. (по [Гептнер и др., 1961], с изменениями); 3 — максимальный ареал в конце XX в. (включая периодические заходы); 4 — крайние северные
пункты заходов
В последние столетия II тысячелетия н. э. ареал диких свиней, как и других
диких копытных, неуклонно сокращался. Популяции зверей в плотно населенных
человеком лесостепной и степной зонах Европы и Западной Сибири, а также на
северном пределе распространения вида были практически истреблены.
Динамика ареала в XX веке
Наименьшим и разорванным на отдельные части (западную, кавказско-
прикаспийскую и азиатскую) ареал стал в конце XIX - начале XX вв. (см. рис. 19).
Его северная и восточная границы в Европе в период наибольшей депрессии в конце
20-х - начале 30-х годов XX в. отступили до Риги, Великих Лук, Витебска, Моги-
лева, Гомеля, Киева и устья Днестра. В кавказском очаге северная условная линия
границы шла от Азовского моря по р. Кубани на Краснодар, Армавир, Минеральные
Воды, Нальчик, по р. Терек до Каспийского побережья, и далее на север - узкой
полосой вдоль моря до устья р. Белой. На восточном побережье Каспия зверей не
было вплоть до Аральского моря. Южнее и восточнее они занимали лишь Копетдаг,
Большие Балханы, бассейны Теджена и Мургаба и районы к востоку от Амударьи и
Аральского моря, включая низовья Сарысу, Телекольские озера, долину р. Чу, озера
Балхаш и Алаколь, Тарбагатай и Зайсанскую котловину. К северу от этой линии в
Казахстане, возможно, сохранились небольшие островные популяции на озерах
Челкар-Тенгиз, Тенгиз, Кургальджин, в низовьях Тургая, Иргиза и Узеня и на
Камыш-Самарских озерах. На Южном Алтае регистрировали заходы зверей из
Монголии. Несколько сократилась область обитания кабана в Присаянье,
Прибайкалье, Забайкалье и, совсем незначительно, на Дальнем Востоке [Силантьев,
1898; Туркин, 1899; Бобринский и др., 1944; Формозов, 1946; Слудский, 1956;
Гептнер, 1960; Гептнер и др., 1961].
На западе Европы, однако, видовой ареал имел тенденцию к расширению уже
в начале XX в., что связано с уменьшением преследования животных. Из сохра-
нившихся в Польше и Литве локальных популяций дикие свиньи расселяются с
1910 г. по Латвии [Калниньш, 1950], в 20-е годы проникают в Эстонию, в 30-е -
достигают ее северных районов, но в начале 40-х из-за суровых зим исчезают и
вновь появляются здесь в 1945 г. [Линг, 1955, 1978]. В 40-е годы они распростра-
няются по всей Прибалтике [Паавер, 1965]. В Белоруссии расширение ареала за-
метно с 1927 г., и к середине столетия вид занимает все районы республики
[Сержанин, 1961, 1970]. Отсюда, а также из небольших группировок, уцелевших
на Украине, звери начинают расселяться в западные области России.
В Псковскую область дикие свиньи проникли из Эстонии в 1943 г., в Ленин-
градской зарегистрированы в 1945 г., в Новгородской - в 1946 г. До начала 60-х
годов их распространение на северо-западе европейской части России было моза-
ичным: в Ленинградской области животные обитали, видимо, лишь в Кингисепп-
ском районе, встречались в Оредежском, Тосненском и Гатчинском районах, зани-
мали южную часть Новгородской области и более двух третей Псковской. К нача-
лу 70-х кабан расселился в этих областях почти повсеместно, но с разной плотно-
стью (рис. 20). На Карельский перешеек звери вышли в 1967 г., на юге Карелии за-
регистрированы в нескольких пунктах в 1969-1970 гг., а к 1974-1975 гг. достигли
северных районов республики - Беломорского и Лоухского [Данилов, 1974; Вере-
щагин, Русаков, 1979; Фадеев, 1979; Марковский, 1995]. К 80-м годам мигранты из
Ленинградской области и Карелии распространились по территории Финляндии
(первая встреча здесь зарегистрирована в 1956 г.) почти до Полярного круга [Ива-
нов, Русаков, 1970; Русаков, 1975; Сиивонен, 1979; Erkinaro et al., 1982].
57
Рис. 20. Расселение кабана на северо-
западе России (по [Русаков, Тимофеева,
1984])
1 - изменение границ ареала с 1930 по 1975 гг. (включая
заходы); 2 и 3 - исторические границы (по [Гептнер,
1960; Слудский, 1956] соответственно)
В Брянскую и Смоленскую об-
ласти кабан проник из Украины и
Белоруссии также в начале 40-х го-
дов [Кириков, 1966; Ватолин, 1980],
в Воронежской, Курской, Белгород-
ской, Орловской, Липецкой, Там-
бовской, Ивановской и Саратовской
областях появился после столетнего
отсутствия в 1946-1952 гг. [Слуд-
ский, 1956; Барабаш-Никифоров,
1957; Кириков, 1959, 1966; Павлов и
др., 1974; Соломатин, 1975; Фадеев,
1990; Лихацкий, 1996; Простаков,
1996; Громаков, 1998]. В Централь-
но-Черноземном заповеднике звери
зарегистрированы зимой 1964 г.
[Гусев, 1989]. В Воронежский запо-
ведник вид вселился в 1950 г., в
1953 г. здесь насчитывали 10 осо-
бей, в 1965 г. - 130, в 1972 г. - 300,
а в 1985 г. - 1134 [Любченко, 1973;
Соломатин, 1979; Дьяков и др.,
1986]. В Хоперском заповеднике
кабан обитает с 1962 г. [Рябов,
1975; Казневский, 1977; Зобов,
1995], к 1985 г. его численность
достигла 1500 особей [Печенюк,
1995]. Из Воронежской области по
пойме Хопра дикие свиньи вышли в
1960 г. в Волгоградскую область
[Кубанцев, Зубов, 1978], а в 1966 г.
появились в Ростовской [Фертиков,
1975].
Распространение кабана из ук-
раинского очага обитания на юг и
юго-восток вплоть до черноморско-
го побережья также зарегистриро-
вано в военные и послевоенные го-
ды [Слудский, 1956; Гурский,
1975]. В Запорожье он появился в
1954 г., в степях Северного При-
черноморья - в 1968 г., в 1974 г.
звери по лесополосам достигли се-
верных районов Крыма [Кириков,
1966; Дулицкий и др., 1975; Издеб-
ский, Тарасов, 1982]. Известны слу-
чаи их иммиграции в Донецкую об-
ласть из Приазовья, Ростовской об-
ласти и Краснодарского края [Гречушкин, 1975а].
Современная подмосковная популяция кабана образовалась как за счет имми-
грантов, так и из особей, завезенных из разных регионов СССР (см. ниже). По све-
дениям А.А Вершинина [1969], до 1910-1911 гг. кабаны жили в пойме р. Сестры в
Волоколамском районе Московской области. В Бельском полесье (район примерно
58
от г. Невеля до ст. Мостовой) с 1914 по 1950 гг. их встречали не менее 16 раз, но
им редко удавалось закрепиться здесь надолго [Юргенсон, 1959]. В начале 30-х го-
дов зверь был убит в Пеновском районе Западной (ныне Тверской) области [Стро-
ганов, 1934]. В Солотчинском районе Рязанской области животные выпущены в
1948-1950 гг., в 1949 г. они обнаружены в Клепиковском районе, в 1950 г. - в Ок-
ском заповеднике, в 1951 г. - в Тумском и Бельковском районах [Козлов, 1958; Зы-
кова, 1965; Любченко, 1969]. В Тульской области дикие свиньи зарегистрированы
в 1953 г. [Миллер, Скалой, 1990], в Калужской - в 1957 г. [Воронин, 1975а]. В ок-
рестностях г. Владимира крупный зверь убит в 1950 г. [Сысоев, 1970].
В Вологодской области первый кабан появился в 1952 г. [Насимович, 1955], но
массовая иммиграция из соседних районов Ленинградской, Новгородской, Калинин-
ской и Ярославской областей имела место лишь в начале 60-х годов [Семенов, 1976;
Варнаков, 1978а,б]. В Дарвинском заповеднике звери замечены зимой 1967/68 гг.
Устойчивая группировка формировалась здесь в течение пяти лет [Тупицина, 1988].
К 1975 г. дикие свиньи заняли все районы Вологодской области, а их численность
достигла 1,5 тыс. особей [Киселев, Каплин, 1987]. Костромскую область они стали
заполнять с 1966-1967 гг., заходя из Ивановской, Ярославской и, возможно, Горь-
ковской областей [Фадеев, 1973в; Сапоженков, 1975].
Случай добычи кабана 21 января 1953 г. близ с. Одина Ровдинского района
Архангельской области (за 500-700 км от области обитания) в свое время стал
зоологической сенсацией. Однако он, видимо, не был первым в этом регионе. По
сообщению Ф.Я. Павлова, его отец, сплавляя лес по р. Кяхте весной 1916 г., убил
багром взрослого зверя в 40 км к югу от Архангельска [Холостое, 1956; Гаскон-
ский, Паровщиков, 1957; Паровщиков, 1959]. Председатель Каргопольского рай-
онного общества охотников зарегистрировал драку кабанов с домашним хряком
летом 1940 г. [Плешак, Миняев, 1986]. Совокупность этих сведений, считавших-
ся ранее неправдоподобными [Гептнер и др., 1961], позволяет по-иному оценить
потенциальные возможности расселения вида.
Южные районы Архангельской области дикие свиньи стали заселять с 1967 г.,
заходя сюда из Карелии и Вологодской области. Во второй половине 70-х годов
места их зимовок зарегистрированы близ г. Березник и в пойме Северной Двины.
Северная граница распространения вида в европейской части России (рис. 21) в
этот период проходила от Северного Прионежья по условной линии Няндома -
Березник - Котлас - Мураши (Кировская область) - верховья рек Вятки и Камы -
Нытва (Пермская область) - к западу от Уфы и Кумертау [Губарь, 1980; Фадеев,
1980, 1981, 1998; Кузякин, Сурбанос, 1984].
В 1977-1978 гг., выйдя к берегам Онежской губы и в Мезенский район, кабан
достиг Полярного круга. В 1981 г. самец в возрасте двух-трех лет добыт возле
маяка Инцы на берегу Белого моря у 66° с. ш.. Зимой 1982/83 гг. зверей видели
нар. Койде [Плешак, Миняев, 1986]. Это на сегодня самые северные пункты рас-
пространения вида в Восточной Европе (рис. 22), хотя потенциальные возможно-
сти для расселения, вероятно, еще не исчерпаны. К 1992 г. его численность в Ар-
хангельской области достигла максимума — 2,9 тыс. (см. табл. 34), лицензионная
добыча ведется с 1977 г.
Заселение восточных областей европейской части России происходило следую-
щим образом. В Горьковской области дикие свиньи зарегистрированы в начале 50-х
годов на границе с Рязанской областью в Выксунском лесхозе [Павлов и др., 1974].
В 1966-1967 гг. они появились также в юго-западных, а в 1971-1972 гг. - в цен-
тральных районах Кировской области. В середине 70-х годов кабан обитал в 15 рай-
онах, а в 80-е распространился по всей области. С 1977 г. этот вид стал объектом
охоты в вятских лесах, а в 1991 г. его численность достигла 4,5 тыс. голов [Злобин,
19756; Григорьев, 1978; Павлов, 1978, 1999а,б; Фадеев, 1979; Павлов, Зарубин, 1982].
59
Рис. 21. Динамика ареа-
ла кабана в Восточной
Европе (по [Фадеев,
1981])
1 - граница ареала в ЗО-е го-
ды, 2 - в середине 50-х, 3 - в
1970 г„ 4 - в 1974 г.. 5 - к
1980 г.; 6 - места выпуска; 7
- рубеж объединения рассе-
ляющегося европейского ка-
бана с гибридной группи-
ровкой интродуцентов; 8 -
крайние пункты встреч
В западные районы Пермской области животные впервые зашли в 1971-1973 гг.,
вероятно, из Кировской области. В 1976-1977 гг. их отмечали в 450-470 км севернее
Перми, а позднее они заняли большую часть территории вплоть до Гайнского, Чер-
дынского и Красновишерского районов [Фадеев, 1979, 1981; Неганов и др., 1989;
Павлов, 1999а]. В Удмуртии дикие свиньи зарегистрированы в 1974 г. Сюда они,
скорее всего, вселились из Татарии, где появились в 1969-1970 гг. [Попов, Попов,
1980; Попов, 1986; Горшков, 1988], и Кировской области.
В южных районах Коми АССР первые кабаны замечены в 1972-1974 гг. [Се-
менов, 1976; Тюрнин, 1989], к 1980 г. они достигли верховьев Вычегды [Фадеев,
1986, 1987], к 1982 г. - бассейна Верхней Печоры, с 1984 г. ежегодно регистри-
руются в Печоро-Илычском заповеднике [Нейфельд, 1989]. По сообщению пред-
седателя правления республиканского Общества охотников и рыболовов
С.В.Лифиренко, в октябре 1987 г. животные обнаружены близ с. Ижма, в 1988 г. -
в 30 км севернее г. Печоры. К середине 90-х годов звери заселили 13 админист-
ративных районов на юге и западе республики [Тюрнин, 1996], но к 2000 г. менее
100 особей сохранились лишь в южных районах - Прилузском, Сысольском,
Койгородском и Сыктывдинском.
В Мордовии дикие свиньи живут с 1964 г. [Павлов и др., 1974], в Мордовском
заповеднике - с 1970 г. [Филонов, 1989], в Чувашии и Марийской АССР - с 1966 г.
[Любченко, 1969]. В 1975 г. они достигли Жигулевского заповедника [Вехник, Сак-
сонов, 1987]. В конце 70-х годов животные появились в Башкирии, в 1981 г. вошли в
состав фауны Башкирского заповедника [Филонов, 1989], но на хр. Кыркты впервые
после долгого отсутствия зарегистрированы в 1956 г. [Кириков, 1966].
Нижневолжские области заселялись кабаном с юга и запада, а левобережные
60
х / СЭ 2 ----J
Рис. 22. Северные районы распространения ка-
бана в Архангельской области (по [Плешак,
Миняев, 1986])
1 - крайние северные пункты встреч; 2 - очаги оби-
тания; 3 - северная граница области постоянного
обитания
районы Волго-Уральского меж-
дуречья и Камыш-Самарские
озера в частности заполнялась
иммигрантами с Западного Ка-
захстана с середины XX в.
[Слудский, 1956; Кириков, 1966;
Фадеев, 19706, 1981]. Их рассе-
лению вверх по Волге способст-
вовали весенние паводки, кото-
рые вытесняли животных из
дельты. В 1947 г. звери замече-
ны в 150 км севернее Астрахани,
осенью 1950 г. одна свинья уби-
та в Палласовском районе Вол-
гоградской области и группа за-
мечена по р. Горькой [Лавров-
ский, 1962].
Очевидно, что из западного
(Прибалтика, Белоруссия, Ук-
раина) очага обитания кабан
расселялся по территории Евро-
пейской России в трех основных
направлениях (см. рис. 21): по
верховьям Днепра через водо-
раздел в район Верхней Волги и
по бассейну Западой Двины в
Невско-Ильменский бассейн; из
бассейна Десны через водораз-
дел в бассейны Оки и Волги; по
левобережным притокам Днепра
в бассейн Дона [Фадеев, 1973а,в,
1975, 1978, 1981]. Четвертое на-
правление - южное - по долине Днепра и лесополосам в Причерноморье и Крым.
Постоянные островные поселения иммигрантов формировались сначала на
низменностях и по долинам рек, затем звери занимали прилежащие возвышенно-
сти и распространялись далее. За 20-25 лет (начиная с 1930 г.) граница ареала
продвинулась на восток на 250-600 км [Гептнер, 1960; Гептнер и др., 1961; Фа-
деев, 1981], за 50 лет - более чем на 1000 км. Средняя скорость расселения каба-
на колебалось от 20-30 км в год в северном направлении и до 45-90 км - в вос-
точном и южном [Григорьев, 1978; Попов, 1986; Червонный, 1990а].
В начале 60-х годов европейский, кавказский и азиатский очаги обитания ди-
ких свиней практически соединились. В 70-80-е годы вид почти полностью вос-
становил свой былой ареал в Восточной Европе, а к концу XX в. его северная
граница достигла исторического максимума (рис. 19).
Южный (кавказско-прикаспийский) очаг обитания вида, в отличие от запад-
ного и азиатского, не стал заметным источником расселения. Тем не менее в се-
редине XX в. отмечено появление зверей в равнинной части Краснодарского края
и на Ставрополье [Федоров, 1954; Лавровский, 1962]. В высокогорных районах
Тебердинского заповедника дикие свиньи зарегистрированы в 1947 г. [Аренс,
1957; Бобырь, 1975].
Во второй половине XX в. из-за чрезмерного преследования и интенсивного
хозяйственного освоения земель ареал диких свиней на Северном Кавказе суще-
ственно сократился и стал очаговым, приуроченным к охраняемым территориям
61
или к угодьям с хорошими укрытиями, таким, как камышовые и тростниковые
крепи по рекам и горные леса с густым подлеском и подростом. На северо-западе
региона в 60-70-е годы северная граница ареала сместилась к югу в предгорья в
среднем на 50 км. Звери исчезли в пойме Кубани и на Таманском полуострове. В
1969-1972 гг. кабана интродуцировали в островные леса степных районов Став-
ропольского края [Саркисов, Хохлов, 1982]. В 80-е годы вновь отмечено расши-
рение ареала в Краснодарском крае и в Ставрополье, где к концу этого десятиле-
тия животные заняли 28 районов из 34. На севере Восточного Кавказа они обита-
ли в плавнях Терека, Аксая, Акташа, Сулака, по Терско-Кумской низменности, в
предгорных и горных районах Чечни, Ингушетии и Дагестана [Котов, Рябов,
1963; Дуров, 1973, 1987; Гинеев, 1989; Соколов, Темботов, 1993].
В Грузии ареал кабана особенно заметно уменьшился в период с 1915 по
1920 гг. в результате прямого истребления и хозяйственного освоения земель. С
1958 г. охота на этот вид была запрещена. В начале 60-х годов его распростране-
ние ограничивалось Лагодехским, частично Кварельским, Сигнахским, Гарда-
банским и Болнисским районами, локальные группировки сохранились в Абха-
зии, в Очамирском и Гудаутском районах. Значительно реже звери встречались в
других местах на западе и северо-западе республики [Хеладзе, 1964]. В 70-е годы
их регистрировали в Телавском и Ахметском районах, в 80-е - в пойме рек Куры
и Алазани, в Вашлованском и Лагодехском заповедниках, Ахалцихском районе
Аджарии и в ущелье Гумиста в Абхазии [Арабули, 1970, 1989]. В Армении дикие
свиньи уцелели лишь в хорошо охраняемых и в отдельных труднодоступных
угодьях. На Малом Кавказе их распространение во второй половине XX в. было
спорадичным, но более обширным, чем в 30-е годы [Соколов, Темботов, 1993]. В
Куба-Хачмасской зоне звери (около 700 голов) уцелели в трех лесных массивах
на побережье Каспия, в Кусарском и Кубинском районах [Алекперов, Кулиев,
1987]. В Кызыл-Агачском заповеднике, по моим наблюдениям, в 70-е годы они
были многочисленными, а в отдельных стадах насчитывали от 18 до 52 особей.
В Казахстане первые признаки расширения ареала заметны в 1914-1918 гг.,
когда промысел диких свиней резко сократился. В 20-х годах животные обитали
на оз. Зайсан, по долине Черного Иртыша, в горах Тарбагатая, на озерах Балхаш,
Ала-Куль и Салык-Куль, появились в долине Иртыша, окрестностях Семипала-
тинска, Усть-Каменогорска, ст. Локоть, в ленточных борах на широте г. Павло-
дара, но после возвращения охотников с фронтов быстро исчезли из многих мест.
В середине 30-х годов в связи с коллективизацией упраздняется «Охотсоюз», что
привело к снижению числа охотников. Сроки добычи кабана в законодательном
порядке ограничиваются тремя месяцами, запрещается охота с собаками, унич-
тожавшими большое число поросят. Резкое сокращение промыслового пресса,
особенно в военные (1941-1945) годы, и благоприятный водный режим стали ос-
новными причинами бурного роста численности вида и быстрого (до 100 км за
год) расширения ареала. Уже в начале 50-х годов его северная граница достигла
линии Уральск - Кустанай - Курган (по р. Тобол [Федюшин, 1963]) - Петропав-
ловск — оз. Чаны, т. е. продвинулась на север на 300-500 км. Расселение живот-
ных шло в основном по долинам рек и сети озер. За 20 лет область распростране-
ния кабана увеличилась почти в три раза. В 70-е годы он практически полностью
восстановил свой ареал [Селевин, 1930; Слудский, 1956; Никифоров, 1959; Соко-
лов, 1959; Фадеев, 1978; Слудский и др., 1984].
В граничащих с Казахстаном районах Оренбургской, Челябинской, Курган-
ской и Омской областей дикие свиньи после столетнего отсутствия стали появ-
ляться в 1946-1954 гг. [Слудский, 1956; Кириков, 1959, 1966]. В феврале 1959 г.
зверь был убит в 40 км к западу от Омска, в 1961 г. остатки обнаружены на оз.
Тобол-Кушлы б. Саргатского района [Федюшин, 1963]. Челябинская область, од-
нако, заселялась также с севера и запада (из Свердловской области и Башкирии).
62
В 1978 г. животные зарегистрированы в Ильменском заповеднике [Дворников,
Дворникова, 1989], в 1979 г. - в Каслинском районе, в 1980 г. - Нязепетровском,
на следующий год - в Уйском, в 1982 г. - в Красноармейском районе, в 1983 г. -
возле г. Карабаша, в 1984 г. - близ г. Верхнего Уфалея, в 1985 г. - в Карталин-
ском районе, в 1987 г. - в Октябрьском районе и в 1988 г. - возле г. Пласта. К
концу 80-х годов кабан расселился по всей территории области, образовав устой-
чивые группировки, насчитывающие от 130 до 750 голов в Аргаяшском, Ашин-
ском, Каслинском, Кусинском, Нязепетровском и Чебаркульском районах [Мат-
веев, Бакунин, 1994].
На Средний Урал (в Артинский район Свердловской области) кабан проник в
1969 г. из Башкирии. В 1970 г. животные зарегистрированы в Шалинском рай-
оне, позднее в Ачитском и Красноуфимском. В 1978-1984 гг. в Сысертском, Ка-
мышловском, Туринском и Алапаевском районах выпустили 834 зверя. Отсюда
они расселялись в основном на север и восток. В 1978 г. дикие свиньи обнаруже-
ны недалеко от г. Ревды, в 1979-1981 гг. - севернее г. Качканар, в Ново-
Лялинском и Серовском районах, близ г. Ивделя, за 450-500 км от ближайшего
места выпуска, а также в Байкаловском, Ирбитском, Талицком, Таборинском и
Тугулымском районах. Северо-восточные районы, вероятно, пополнялись и за
счет животных, акклиматизированных в Тюменской области. Часть животных из
Свердловской области уходила в северные районы Челябинской и на запад Кур-
ганской областей [Борискин, 1982; Киселев, 1986; Марков, 1995, 1997а, 1999;
Павлов, 1999а].
В южной части Тюменской области кабаны замечены в 1979-1980 гг. [Бакла-
нов, 1980; цит. по: Фадеев, 1987]. В 1980-1984 гг. в Нижнетавдинском, Армизон-
ском, Омутинском, Исетском и Ишимском районах произведены выпуски зверей.
К 1986 г. дикие свиньи заняли еще 9 районов. Несколько лет они зимовали в Ва-
гайском, Тобольском и Уватском районах, а отдельные особи и группы проника-
ли в Ханты-Мансийский автономный округ: до Кондинска, р. Мулымья, посел-
ков Советский, Березово, в Ханты-Мансийский и Октябрьский районы и, воз-
можно, до р. Надым (см. рис. 19). В Октябрьском районе периодические заходы
групп до 6-7 особей регистрировали как на левобережье, так и на правобережье
Оби. При высоком снежном покрове они становились легкой добычей браконье-
ров [Азаров, Деков, 1990; Азаров, Климов, 1990; Азаров, 1996; Павлов, 1999].
Восточная часть Саратовской, Оренбургская, юг Челябинской и Курганская
области в основном заселены эмигрантами из Северного Казахстана, тогда как в
Свердловской, Тюменской, Новосибирской и Омской областях популяции диких
свиней сформированы преимущественно в результате искусственного расселения
в 80-е годы. В Омской области северный очаг обитания ограничен Муромцев-
ским, Тарским, Седельниковским, Тевризским, Большереченским районами, юж-
ный - Называевским, Тюкалинским, Москаленским, Люблинским и Иссык-
Кульским районами [Федотов, 1990]. На востоке Новосибирской области они
расселены в Каргатском, Сузинском и Колывановском районах, западный очаг
сформирован из мигрантов из Омской области [Баранов, 1990].
В 90-х годах XX в. северная граница распространения кабана в Зауралье и
Западной Сибири (включая заходы) проходила от Ивделя по р. Конде до Оби, где
звери зарегистрированы по обеим ее берегам, а также в Ханты-Мансийском и
Октябрьском районах [Павлов, 1999а] и далее по Иртышу выходила к границам
Омской области (см. рис. 19).
В Средней Азии динамика ареала кабана существенно отличалась от таковой
в других частях ареала. Во второй половине XX в. область распространения под
влиянием человека резко сократилась и стала очаговой. В Западной Туркмении в
70-х годах малочисленные популяции сохранились лишь в Больших Балканах,
пойме Западного Узбоя и по рекам на границе с Ираном [Ишадов, 1972; Зархид-
63
Рис. 23. Распростра-
нение кабана на Ал-
тае (по [Собанский,
1992])
1 - область обитания;
2 - районы с более вы-
сокой плотностью на-
селения
зе, 1980]. На Юго-Западном Копетдаге поголовье уменьшилось в начале 80-х го-
дов [Карпинский, Лукаревский, 1987]. В Узбекистане этот зверь уцелел только в
заказниках и заповедниках в лесном поясе Пскемского, Чаткальского, Гиссарско-
го, Туркестанского и Нуратинского хребтов, в горах Кугитанга, Бабатага, Бай-
сунтау, в пойме Амударьи и на юго-востоке Кызылкумов [Ишунин, 1980]. В Се-
верной Киргизии он обычен в горах, окружающих оз. Иссык-Куль: в Кунгей- и
Терскей-Ала-Тоо, Чон-Кемине, на северных склонах Киргизского хребта и на
Таласском хребте; в горных районах его численность в последнее время растет
[Катаевский, 1987]. В Таджикистане четко выделяются две обособленные попу-
ляции диких свиней: тугайная (на юго-западе республики) и горная (в централь-
ной и северной ее частях). Ареал включает Туркестанский, Зеравшанский, Гис-
сарский, Дарвазский и Вахшский хребты, на востоке его граница доходит до гор-
ного узла Академии наук. Звери встречаются по долинам рек Вахш, Ванч,
Пяндж, в тугаях заповедника «Тигровая балка». К середине 70-х годов XX в. из-
за хозяйственного освоения земель область обитания кабана в этом регионе со-
кратилась на 10-15% [Сапожников, 1976]. j
На западе и юге Алтая (Змеиногорский и Кошагачский районы, среднее тече- !
ние р. Бухтармы, р. Чуя, Курайская степь) отдельные заходы диких свиней с тер- [
ритории Восточного Казахстана и Монголии, как упоминалось ранее, регистри- ’
ровали вплоть до 30-40-х годов XX в. [Залесский, 1934; Соколов, 1959]. Осенью
1953 г. кабан был убит у слияния Бии и Катуни [Слудский, 1956]. Заселение этим
видом Чуйской долины произошло в начале 60-х годов. Позднее иммиграция бы-
ла наиболее интенсивной в бассейн р. Чулышман из прилегающих районов Тувы,
а со второй половины 70-х - начала 80-х годов - с территории Восточного Казах-
стана в западные и юго-западные районы республики. К 1983 г. звери заселили
более 30% территории региона, а к концу 80-х - большую его часть (рис. 23),
64
Рис. 24. Распространение кабана в Монголии (по [Соколов и др., 1982])
1 - районы постоянного обитания; 2 - заходы
достигнув окрестностей Горно-Алтайска и северного берега Телецкого озера
[Собанский, 1986, 1990а, б, 1992].
Южнее, в Монголии, в первой половине XX в. кабан обитал почти повсеме-
стно на Монгольском Алтае и в северной части страны [Банников, 1954]. К 80-м
годам, по нашим данным, на западе он сохранился лишь в пограничных с Китаем
районах на побережьях озер Хотон-Нур и Хоргон-Нур, в верховьях р. Сагсай (за-
ходы отмечаются западнее р. Булган-Гол), западнее оз. Ачит-Нур, по берегам
крупных озер Хара-Ус-Нур, Хара-Нур и Убсу-Нур. В Заалтайском Гоби дикие
свиньи, вероятно, исчезли. На Хангае и Хэнтэе они распространены во всех лес-
ных массивах, откуда выходят в лесостепные и степные районы (рис. 24). В бас-
сейне р. Халхин-Гол звери обитают не только по берегам рек, но и занимают кус-
тарниковые заросли по барханам и заболоченные участки в степи за десятки ки-
лометров от ближайших зарослей [Соколов и др., 1982].
В Туве кабан был обычен даже в период максимальной депрессии вида в на-
чале XX в. [Смирнов, 1989]. Отсюда звери эмигрировали на север, заселяя За-
падный Саян. На правобережье Енисея они впервые обнаружены в 1960 г. (в бас-
сейне р. Христафорихи, на склонах хр. Мирского и в долине р. Таловки), позднее
- в Ермаковском и Шушенском районах Красноярского края, в 1967 г. - в Саяно-
Шушенском заповеднике, в начале 70-х годов - в верховьях р. Кан [Лавов, 1975;
Соколов, 1979; Завацкий, 1987, 1989; Филонов, 1989; Смирнов, Бриллиантов,
1990]. К концу 70-х годов дикие свиньи широко расселились по Хакасии, наибо-
лее часто встречаясь в бассейнах рек Кантегир, Малый Абакан, Абакан, Джебаш,
на Джойском и Джебашском хребтах. В середине 80-х они достигли Аскизского
района, стыка Абаканского хребта и Кузнецкого Алатау, в конце десятилетия их
регистрировали в верховьях Белого Июса и в Ширинском районе. Летом 1987 г.
животные отмечены в верховьях Уса в Кемеровской области [Прокофьев, 1992].
В 1982-1987 гг. образован искусственный очаг обитания вокруг Красноярска в
Емельяновском, Балахтинском и Рыбинском районах [Павлов, 1999а].
В Предбайкалье в начале XX в. одиночные заходы кабанов отмечались лишь
до Ангары [Бобринский и др., 1944; Насимович, 1955]. Во второй половине сто-
летия ареал сместился к северу на 250 км. Звери достигли Нижнеудинска, Тай-
5.Заказ № 252S.
65
Рис. 25. Распространение
кабана на юге Дальнего
Востока (по [Бромлей,
Кучеренко, 1983])
1 - граница ареала; 2 - ти-
пичные места обитания; 3 -
лучшие биотопы; 4 - пункты
заходов
шета и р. Туманшет, но их распространение повсеместно было мозаичным [Не-
дзельский, 1994]. Выпуски мелких партий зверей имели место в Качугском и Ир-
кутском районах [Павлов, 1999а].
В Бурятии диких свиней можно причислить к горным животным. Они обитают
на Восточном Саяне, Хамар-Дабане, Улан-Бургасе, Джидинском, Малханском, За-
ганском и, изредка, Икатском хребтах, на Цаган-Хуртэе, Витимском плато, где до-
ходят до подножья Южно-Муйского хребта, в Баргузинской котловине, по берегам
Байкала и в других районах [Москвитин, Смирнов, 1975; Смирнов, 1978].
В Северо-Восточном Забайкалье во второй половине XX в. северная граница
ареала вида проходила по Витиму, нижнему течению р. Калар, верховьям рек
Олекма и Тунгир [Байдавлетов, 1980]; в Читинской и Амурской областях - по
левобережному бассейну Амура примерно в 50-150 км севернее государственной
границы; в Хабаровском крае - севернее Комсомольска-на-Амуре, затем по за-
падным склонам Сихотэ-Алиня уходила к Бикину, откуда поворачивала на север
и по побережью Японского моря достигала Советской Гавани (рис. 25). На Даль-
нем Востоке наиболее дальние заходы зверей к северу зарегистрированы в 100
км южнее Тынды [Фадеев, 1987], в устье Селемджи, южнее Чегдомына и север-
нее Амгуни [Бромлей, 1964; Кучеренко, 1973, 1976; Бромлей, Кучеренко, 1983].
В Якутии (в бассейне р. Алдан) кабан отсутствует с 1968 г. [Иванова, 1976].
Итак, наиболее существенные изменения ареала дикой свиньи произошли во
второй половине II тысячелетия н. э. После максимальной депрессии, имевшей ме-
сто в XVIII - начале XX вв., происходит резкая трансформация его границ. С сере-
дины XX в. кабан быстро расселяется от западных и южных окраин России насе-
66
вер и восток и к концу столетия достигает севера Карелии и Архангельской облас-
ти (на севере доходит почти до Полярного круга, на востоке - до р. Мезень), вос-
точных районов Республики Коми (до Печоры и Уральских гор). За Уралом услов-
ная северная линия ареала по Сосьве доходит до Оби (район пос. Березово), затем
опускается до Иртыша, по нему выходит к восточным границам Омской области,
далее идет севернее Новосибирска, по верховьям р. Ус в Кемеровской области, се-
вернее Красноярска, через Тайшет и севернее Ангары выходит к оз. Байкал (см.
рис. 19). В Забайкалье и на юге Дальнего Востока ареал с середины XX в. сущест-
венных изменений не претерпел. На большей его части животные немногочислен-
ны и обитают отдельными очагами.
Территориальную экспансию кабана в XX в., тем не менее, вряд ли следует
считать «небывалым» в истории «эпохальным» событием и сравнивать с «наше-
ствием» [Павлов, 1978, 1999а]. Этот вид, так же как европейская и сибирская ко-
сули и лось [Данилкин, 1999а], всего лишь восстановил свой видовой ареал и не-
сколько увеличил его при благоприятных обстоятельствах. В Восточной Сибири
и на Дальнем Востоке, где дикие копытные не были полностью истреблены,
расширения границ областей распространения не происходило, хотя периодиче-
ские их пульсации при изменении численности имели место.
Причины изменения ареалов и численности диких копытных весьма дискус-
сионны.
Основные факторы, определяющие область
распространения и численность (исторический аспект)
В антропогене на территории бывшего СССР обитали по меньшей мере 96
видов копытных животных, из них 20 видов непарнокопытных и 76 - парноко-
пытных, включая 6 мозоленогих. К голоцену уцелел 31 вид: 5 непарнокопытных
и 26 парнокопытных (2 мозоленогих) [Верещагин, Барышников, 1980а]. В наши
дни сохранились лишь 1 вид непарнокопытных и 19 парнокопытных (с учетом
разделения косуль на два вида - 20). Мозоленогих животных в составе рассмат-
риваемой дикой фауны теперь нет. В Южной Сибири в конце палеолита жили 36
видов крупных млекопитающих, к настоящему времени остались только 16. Ви-
довое разнообразие особенно уменьшилось среди степных и лесостепных живот-
ных. Средняя скорость исчезновения крупных млекопитающих в каждое тысяче-
летие была примерно следующей: поздний палеолит - мезолит - 0,7, мезолит -
бронзовый век - 0,3, бронзовый - железный века - 0, железный век - средние ве-
ка - 2,4, средние века - новое время - 4,4, новое время - современность - 20
[Смирнов, 1994, 1999]. В Северной Америке, которая была заселена человеком
сравнительно поздно (около 40 тыс. лет назад), в голоцене не стало представите-
лей 17 родов крупных зверей, но уцелели овцебыки и бизоны, «вымершие» в
Старом Свете [Martin, 1967]. Только за исторический период с планеты исчезло
120 видов крупных млекопитающих и 150 видов птиц [Дорст, 1968].
Споры исследователей о значении тех или иных факторов среды в динамике
ареалов и населения диких копытных животных, и кабана в частности, не утиха-
ют до сих пор. При этом четко выделяются два «лагеря». «Солнцепоклонники»
отстаивают ведущую роль климата и сукцессий, а «антропогенисты» считают
доминирующим влияние человека и крупных хищников.
Абиотические факторы
Климат. Кабан, по существу, южный вид, большая часть ареала которого за-
нимает тропическую и теплую зоны Европы и Азии, и лишь незначительная часть
5!
67
находится в умеренно холодной зоне [Гептнер, I960]. В многовековом аспекте ди-
намика области его распространения, как и у большинства других крупных млеко-
питающих, несомненно, определялась климатическими факторами. Ареалы ко-
пытных в плейстоцене и голоцене явно расширялись в периоды потеплений и
смещения лесной зоны и заметно сужались во время похолоданий и сопутствую-
щих им изменений растительности [Пидопличко, 1951; Паавер, 1965; Алексеева,
1990]. Соответственно изменялась и численность животных.
Следует отметить, что в отдельные периоды плейстоцена климатические ус-
ловия были более суровыми, а их изменения более резкими, чем в голоцене. Тем
не менее крупные звери сравнительно благополучно переносили их и, несмотря
на ландшафтно-климатические катастрофы, не исчезли в ледниковое время.
Масштабное сокращение доминирующих видов мамонтовой фауны происходит
лишь в последние 13-10 тыс. лет на грани плейстоцена и голоцена как в резуль-
тате изменения (потепления) климата и ландшафтов, так и по мере роста населе-
ния человека и усиления его воздействия [Громов, 1948; Пидопличко, 1951, 1954,
1963; Верещагин, 1963, 1971, 1975; Алексеева, 1990]. С позднего палеолита кли-
мат становится не первопричиной, а, скорее, лишь фоном, на котором проявля-
лась истребительная деятельность человека [Алексеева, 1980].
Глобальное потепление климата в Арктике и в умеренных широтах, начав-
шееся с середины XIX в. [Паавер, 1965] и усилившееся в XX в., несомненно, спо-
собствовало восстановлению ареала кабана и других копытных. Однако связы-
вать этот процесс только с этим фактором было бы абсолютно неверно. Напом-
ню, что максимальное сокращение ареала и населения диких копытных на терри-
тории России в начале XX в. и резкое уменьшение их численности в самом конце
этого столетия произошли именно в период потепления.
Температура, снежный и водный режим. Эти мощнейшие экологические
факторы, несомненно, играли и продолжают играть немаловажную роль в фор-
мировании ареалов, морфофизиологических адаптаций и сезонного ритма жиз-
недеятельности животных, отсеивая формы, непригодные к существованию.
Ледниковый период, как крайнее их выражение, стал для многих видов экстре-
мальной эпохой [Формозов, 1946].
Весовая нагрузка на площадь следа у кабана, даже при дополнительной опоре
на копыта боковых пальцев и заднюю часть подошвы, в зависимости от возраста
и массы составляет около 250-500 г на 1 см2. У взрослых животных она пример-
но в 2-3 раза выше по сравнению с северным оленем, соответствует таковой у
косуль, лося и благородного оленя и ниже, чем у пятнистого оленя. У волка она
меньше, чем у кабана, минимум в 1,5-3 раза.
Зоологи были убеждены, что грузный коротконогий зверь, у которого рас-
стояние от земли до груди в зависимости от возраста колеблется от 29 до 50 см,
не может жить там, где высота снежного покрова превышает критический уро-
вень в 40-50 см [Формозов, 1946; Насимович, 1955; Бромлей, 1964]. При боль-
шей высоте снега перемещение зверей становится проблематичным, поросята
практически не могут идти, выбиваются из сил и нередко замерзают прямо на
тропе. Особенно сложно перемещаться животным при настах - они сильно ранят
ноги. В многоснежье существенно усиливается преследование и истребление
бедствующих копытных людьми и крупными хищниками.
Этим обстоятельством и объяснялась прежде всего северная граница историче-
ского ареала кабана, которую проводили близко к изолинии высоты снега в 40-50 см
[Слудский, 1956; Гептнер и др., 1961]. Однако во второй половине XX в., опроверг-
нув воззрения зоологов, кабан, как, впрочем, и косули, занял некоторые районы, где
средняя высота снежного покрова, лежащего более 200 дней в году, превышает 50-
60 см, абсолютная - 100—110 см, минусовые зимние температуры достигают 40°С, а
в отдельные дни опускаются до 50°С [Фадеев, 1980, 1981; Данилкин, 1992, 1999а].
68
Рис. 26. Зимой кабаны перемещаются по набитым тропам (фото А. Щеголева)
В таких экстремальных условиях дикие свиньи выживают исключительно
благодаря комплексу физиологических, экологических и поведенческих адапта-
ций. При регулярном передвижении они набивают тропы (рис. 26), а накоплен-
ные с осени жировые запасы, активная «гнездостроительная» деятельность и
коллективная терморегуляция смягчают влияние низких температур (см. ниже).
Снежный покров для кабана можно рассматривать и как благоприятный фак-
тор. Благодаря малой теплопроводности снег предохраняет почву от сильного
выхолаживания, что позволяет зверям в мягкие зимы, даже многоснежные, до-
бывать подземный корм и воду. Гораздо сложнее уцелеть им в малоснежные мо-
розные зимы с настами, когда почва и вода глубоко промерзают. Именно в такие
годы регистрируются массовые падежи животных.
Следовательно, северная граница обитания этого вида определяется не высо-
той снежного покрова и не низкой температурой (хотя существенное влияние
этих факторов бесспорно), а наличием и доступностью влажных зимних кормов,
а также преследованием человеком и хищниками. В полной мере данный вывод
относится практически ко всем оленьим, и прежде всего к косулям.
Большой прогиб северной границы ареала лося в Западной Сибири А.Н. Фор-
мозов [1946] и другие зоологи тоже долгое время объясняли многоснежьем. Од-
нако, несмотря на эти воззрения, при улучшении охраны и сокращении чрезмер-
ного промысла вид во второй половине XX в. заселил всю енисейскую тайгу
вплоть до тундры [Данилкин, 1999а].
Распространение кабана и косуль определенно коррелирует с продолжитель-
ностью залегания снежного покрова. В затяжные бескормные зимы наблюдается
повышенная смертность животных, полностью израсходовавших жировые запа-
сы. После нескольких следующих подряд экстремальных зим северные области
обитания этих копытных смещаются к югу на сотни километров. После серии
мягких, сравнительно малоснежных зим звери из областей переживания снова
69
проникают в северные районы и заселяют их. Соответственно граница ареала по-
стоянно «пульсирует».
В высокогорных районах продолжительный и высокий снежный покров ста-
новится значительной преградой и для кабана, и для косуль [Данилкин, 1992]. На
Дальнем Востоке проникновение животных на север ограничено хребтом Туку-
рингра [Ильяшенко и др., 1982], в Восточной Сибири - системой Станового на-
горья и Становым хребтом. Многоснежные осевые хребты Сихотэ-Алиня (север-
ной части), Саян, Алтая, Тянь-Шаня, Урала и Кавказа [Дуров, 19706] тоже прак-
тически непригодны для их жизни.
Распространение диких свиней на юг в степные, полупустынные и пустын-
ные районы ограничивает недостаток влажного корма и воды, а также укрытий.
Их расселение повсеместно идет по руслам рек и заселяют они прежде всего
поймы рек и берега крупных озер. Здесь же обычно наиболее интенсивна хозяй-
ственная деятельность человека и хорошо развито сельское хозяйство.
На Нижнем Днепре кабан появился вновь после создания лесополос и интен-
сивного орошения полей в Северном Причерноморье [Издебский, Тарасов, 1982].
В песках Кызылкумов он стал жить после образования артезианских водоемов с
сопутствующими им гидрофильным растительным комплексом и густыми тама-
рисковыми зарослями по берегам [Реймов, Карабеков, 1975; Сабилаев, 1978]. В
глубь Каракумов звери заходят на 250 км, но это случается лишь поздней осе-
нью, зимой и ранней весной, когда в вегетирующих растениях содержится наи-
большее количество влаги и доступна вода в редких источниках [Ишадов, 1986].
В зоне пустынь в их распространении большую роль играет Каракумский канал,
протяженность которого составляет около 1000 км [Нургельдыев и др., 1982]. В
равнинной части Казахстана и Западной Сибири при засухах и изменении водно-
го режима происходит усыхание озер, что приводит к сокращению кормовых и
защитных угодий диких свиней и массовой их эмиграции [Слудский, 1956].
Среди рассмотренных факторов, определяющих границы распространения
диких свиней, на мой взгляд, ведущий - кормовой: наличие и доступность влаж-
ных кормов. Он оказывается одним из основных и в динамике популяций. Одна-
ко трансформация ареала, как и динамика численности вида в целом, находится в
теснейшей связи с антропогенным воздействием и влиянием хищников.
Влияние хищников
Кабан, как и другие дикие копытные, многие тысячелетия сосуществовал
вместе с крупными хищниками. Их популяционные взаимоотношения обсужда-
ются ниже в специальном разделе. Здесь же имеет смысл заострить внимание
лишь на историческом аспекте «волчьей» проблемы в связи с усиливающейся
тенденцией охраны и «разведения» этого хищника в некоторых странах Западной
Европы.
На территории России волк почти всегда был в изобилии, а его численность в
периоды пиков, вызванных социальными неурядицами и войнами [Сабанеев,
1877], вполне сопоставима с населением отдельных массовых видов диких ко-
пытных. Вред, причиняемый им диким и домашним животным, а также человеку,
огромен, и об этом не следует забывать. Вот некоторые любопытные и поучи-
тельные факты, отражающие к тому же пищевое предпочтение хищника.
В Европейской России волк был особенно многочисленным в середине XIX в.
- возможно, около 180-200 тыс. особей. В 1873 г. в 45 губерниях им уничтожено
179 тыс. голов крупного и 562 тыс. голов мелкого рогатого скота, и не меньше-
диких копытных. Гибель людей исчислялась сотнями [Лазаревский, 1876]. В
1897 г. в Российской Империи хищниками истреблено 148 тыс. лошадей, 122
тыс. коров, 585 тыс. голов мелкого рогатого скота и свиней, 560 тыс. домаш-
70
них птиц, а всего - более 1,4 млн. голов. Число задавленных диких животных,
вероятно, больше почти на порядок [Туркин, Сатунин, 1902]. В 1896-1897 гг.
съедено около 2,5 тыс. человек [Бутурлин, 1924]. В Калужской области в 1883 г.
потеряно по этой причине 17,6 тыс. голов домашнего скота, в 1902 г. - более 2,4
тыс. [Васильев, Воронин, 1979]. В Беловежской пуще в 1829 г. волки зарезали
более 200 голов крупного рогатого скота, 18 лошадей, 344 овцы, 121 гуся, 128
свиней и немало диких копытных [Карцов, 1903].
Человек, как мог, боролся со своим хищным конкурентом. В Беловежской
пуще в 1869-1882 гг. добыт 471 зверь [Карцов, 1903]. На юге России казаки ре-
гулярно устраивали грандиозные облавы. В степном Подонье и Новороссии в
1843 и 1845 гг. лишь за несколько дней апреля убито более 3000 волков. В 1896 г.
в Актюбинском, Иргизском, Кустанайском и Тургайском уездах Тургайской об-
ласти добыто 2595 хищников [Кириков, 1959, 1983]. Шуйский аптекарь г-н Ва-
левский, предложивший способ их истребления стрихниновыми пилюлями, в пе-
риод с 1861 по 1875 гг. только в 10 верстах вокруг г. Шуи уничтожил 184 зверя
[Лазаревский, 1876]. В Забайкалье казачьи караулы в отдельные годы затравли-
вали таким образом до 1600 особей [Туркин, 1898].
К концу XIX в. с большим трудом удалось уменьшить численность этого
хищника, что содействовало росту поголовья домашних и диких копытных. В
начале XX в. в России насчитывалось не более 50 тыс. волков при среднегодовой
добыче в 15 тыс. [Бибиков и др., 1992]. В годы послевоенной и послереволюци-
онной разрухи их число вновь заметно увеличилось. С.А. Бутурлин и В.Я. Гене-
розов, на основании анализа динамики заготовок шкур в 20-е годы (10-28 тыс. в
год), определяли волчье население в РСФСР в 62-82 тыс. (до 100 тыс.) [Дементь-
ев, 1933]. В СССР в период с 1925 по 1939 гг. их заготовка находилась в преде-
лах 20-39 тыс. [Миленушкин, 1957].
В 1920 г. лишь в Череповецкой губернии хищники погубили более 4,5 тыс.
голов скота, а в 1921 г. в Тарском уезде Омской области - 35 тыс., что составляет
около 15% поголовья [Бутурлин, 1924]. В 1924/25 гг. в Российской Федерации, на
Украине и в Узбекистане ими задавлено 865 тыс. голов скота, среди которых до-
машние свиньи составляли 7,2% [Дементьев, 1933]. В Белоруссии в этот год по-
теряно более 71 тыс. голов, в том числе 9655 свиней [Сержанин, 1961], в Горно-
Алтайской автономной области - 6% всего поголовья лошадей, 2,5% крупного и
4% мелкого рогатого скота, 8,5% маралов и около 1% свиней. В Хакасии за ян-
варь - март 1923 г. уничтожено 10,8 тыс. домашних животных: из них лошади
составляли 38,2%, крупный рогатый скот - 18,7%, мелкий скот - 42,9%, свиньи -
0,2% [Бондарев, 1989]. В Адлерском районе вблизи Кавказского заповедника в
1935 г. хищники зарезали 11 лошадей, 46 голов крупного рогатого скота и 967
свиней [Теплое, 1938].
Эта вспышка численности волка, также оказавшая значительное негативное
воздействие на популяции диких копытных и способствовавшая сокращению их
ареалов, большими усилиями ликвидирована к середине 30-х годов. Однако
вскоре, в период второй мировой войны 1939-1945 гг., борьба с хищником ос-
лабла (в 1942 г. добыли всего 4,1 тыс.), и Россия пережила новую волну подъема
численности волка минимум до 150 тыс. (см. монографию «Волк», 1985). Ее уда-
лось подавить только к концу 50-х годов, чему способствовали высокие премии
за уничтоженных хищников. В 1944-1946 гг. в России ежегодно истребляли 40-
44 тыс., в СССР - до 62 тыс. зверей [Миленушкин, 1957; Павлов, 1990; Бибиков и
др., 1992], и лишь в одной Воронежской области в 1947 г. - 1,1 тыс. [Фадеев,
1990]. В конце 40-х - 50-е годы добыча постепенно уменьшилась с 38 до 14 тыс.
В военные и послевоенные годы потери домашних и диких животных от вол-
ка на территории России также исчисляются сотнями тысяч голов. И лишь после
резкого сокращения его численности они уменьшились, но все-таки были ощу-
71
тимыми и оценивались в десятки тысяч [Приклонский, 1985]. В Калужской об-
ласти в 1951 г. зарезано 68 лошадей, 50 коров, 715 овец и 54 свиньи [Васильев,
Воронин, 1979]. В Брянской области, по сведениям Госстраха, в 1964 г. хищники
уничтожили 2 лошади, 3 коровы, 262 овцы и 24 свиньи [Ватолин, 1979]. В 1975-
1976 гг. среди 19,7 тыс. голов животных, истребленных ими в 30 областях
России, доля домашних свиней составила 1,5% [Осмоловская, Приклонский,
1979], в 1978-1982 гг. при примерно таком же количестве погибшего домашнего
скота в РСФСР - 1,2% [Бондарев, 1989]. В Белоруссии эта цифра выше - 1,5-
11,8% в разных районах [Козло, Банад, 1985], а в Северо-Западном
Причерноморье - около 7% [Гурский, 1985].
Среди домашних животных доля потерь свиней от волка в целом невелика,
что объясняется, однако, не столько пищевым предпочтением хищника, сколько
стойловым содержанием жертвы.
Во второй половине 60-х - первой половине 70-х годов, в период самой низ-,
кой в XX в. численности волка (около 7,5-11 тыс. особей по сведениям Главохо-j
ты РСФСР) в России, наблюдается бурный рост населения и восстановление'
ареалов диких копытных, и кабана в частности (см. рис. 63). В последующие го-
ды на природоохранной волне борьба с хищником снова ослабляется, а в 90-е го-
ды на фоне социальных и экономических неурядиц очевиден очередной пик его
численности (около 50 тыс. особей), также в немалой степени способствовавший
уменьшению ресурсов копытных [Данилкин, 1999а]. В Астраханской области,
например, население волка увеличилось с нескольких особей почти до тысячи,
что сравнимо теперь с местным поголовьем диких свиней. >,jm|
Антропогенное воздействие
В историческое время динамика ареалов и численности диких копытных жи-
вотных определялась в основном антропогенными факторами.
Изменение биотопов. Антропогенное преобразование ландшафта, и особенно
вырубку лесов, обычно считают отрицательным фактором по отношению к диким
копытным, и кабану в частности, хотя такие утверждения не всегда справедливы.
Леса, прежде всего дубовые, ранее всего (в середине II тысячелетия) человек
стал интенсивно осваивать в прибрежных странах Западной Европы в связи с ко-
раблестроительством. На изготовление одного корабля требовалось около 4000
дубов, и лишь Испания на постройку флота израсходовала более полумиллиона
этих деревьев. Ландшафт очень быстро стал почти безлесным [Благосклонов и
др., 1967], что, несомненно, способствовало истреблению копытных.
Лес повсеместно вырубали также для освобождения земель под пашню и для
строительства поселений. Много древесины требовалось на топливо и для полу-
чения древесного угля, необходимого многочисленным рудникам и заводам.
Частые пожары захватывали огромные пространства, причиняя огромный вред
лесным зверям [Кириков, 1959, 1960, 1966, 1983]. В центре европейской части
России дубовые леса были почти полностью сведены к середине - концу XIX в.,
а лесопокрытая площадь уменьшилась во многих районах до 5-20%. В Европей-
ской России за два последних века процент лесистости изменился с 52,7 до 35,9
[Данилов и др., 1966], но в целом в Российской Федерации в конце XX в. он ос-
тался на сравнительно высоком уровне — 45,3% (Гос, доклад о состояниии окру-
жающей природной среды Российской Федерации в 1998 году. М., 1999).
С потерей дубовых, буковых, а позднее и кедровых лесов значительно
уменьшилась кормовая емкость угодий, сократились запасы осенне-зимних кор-
мов, что, безусловно, отразилось на численности диких свиней. Однако лесохо-
зяйственная деятельность, сопровождаемая уменьшением площади сплошных
массивов и образованием мозаичности лесных угодий, как и распашка степей и
72
осушение болот, приводящие к увеличению доли сельскохозяйственных полей, в
целом благоприятствуют жизни этих зверей [Козло, Емельянова, 1987; Толкач,
1987; Хорин, Горшков, 1989]. Плотность их населения во многих районах Запад-
ной и Центральной Европы, где дубовые и буковые леса сведены полностью,
весьма высока, а порой и чрезмерна. Поэтому сетования на снижение численно-
сти вида или его исчезновение в результате трансформации лесных биотопов
обычно спекулятивны. Расширение площади полей в Восточной Европе и в За-
падной Сибири в значительной мере способствовало расселению кабана в XX в.
Интенсификация земледелия и скотоводства привела к обострению отноше-
ний человека с дикими копытными, которых стали преследовать и как вредите-
лей посевов, и как пастбищных конкурентов. Однако наибольший урон они не-
сли все же как охотничьи виды.
Охота. Древние люди, судя по применяемым орудиям, способам добычи и
результатам, были весьма умелыми и удачливыми охотниками, что и стало глав-
ной причиной «вымирания» крупных зверей [Алексеева, 1980]. Они исчезали не
одновременно: сначала - виды с низкой плодовитостью, жившие в холодном
климате, где служили основной пищей человека, для которого охота была едва
ли не единственным источником существования десятки тысяч лет. У археологов
этот ранний период истребления животных человеком получил название плей-
стоценового перепромысла [Флинт, 2000].
Об успешности охот первобытного человека свидетельствуют «кладбища»
костей животных. К примеру, в овраге близ Амвросиевки на Украине убиты
копьями 983 зубра [Пидопличко, 1951]; в местечке Пшедмости в Чехословакии -
около 1 тыс. мамонтов; в Гаронне во Франции - до 30 тыс. северных оленей; у
селения Солютре в Бургундии добычей охотников стали более 100 тыс. лошадей;
в пещерах горных районов Швейцарии и Венгрии зарегистрированы остатки 1-2
тыс. пещерных медведей [Благосклонов и др., 1967; Дежкин, 1983; Смирнов,
1996]. В Молдавии в слоях небольшого трипольского поселения Флорешты об-
наружены остатки 139 кабанов, 84 благородных оленей, 37 косуль, 31 тура и 11
речных бобров; в слоях селетской культуры в гроте у с. Брынзены - 194 лошадей,
117 северных оленей, 21 бизона, 8 благородных оленей, 6 шерстистых носорогов;
в мадленском слое стоянки Старые Дуруиторы - 91 лошади, 235 северных оле-
ней, 5 бизонов и много костей других видов млекопитающих [Давид, 1982].
По расчетам Н.К. Верещагина [1971], население Крыма и юга Русской равни-
ны в позднем палеолите вряд ли превышало 15 тыс. человек, но для пропитания
им требовалось около 10500 т мяса в год; это или 120 тыс. северных оленей, или
80 тыс. лошадей, или 30 тыс. бизонов, или 10 тыс. мамонтов. А стоянок перво-
бытных охотников на территории Евразии было достаточно много!
Процесс истребления животных резко активизировался во второй половине II
тысячелетия н. э., особенно в последние века. С 1600 по 1975 гг., по данным Ме-
ждународного союза охраны природы и природных ресурсов, вымерло 63 вида
зверей и 74 вида птиц, из них по вине человека - 75% и 86% соответственно
[Флинт, 2000]. В XIX в. исчезло столько же видов млекопитающих (33), сколько
за весь предшествующий исторический период, а потери только за первую поло-
вину XX в. были еще большими - около 40 видов. По естественным причинам
исчезли 25%, тогда как под воздействием человека - 75%, в том числе в резуль-
тате охоты - 33%, или 35 видов [Дежкин, 1977]. Лишь на территории Латвии
уничтожены 8 видов охотничьих млекопитающих [Паавер, 1965], в Литве в XV-
XIX вв. - не менее 10 [Блузма, 1999]. На Кавказе за последнюю тысячу лет при
содействии человека вымерло не менее 9 видов крупных зверей [Верещагин,
1959]. Опасность полного исчезновения грозит еще многим видам, существую-
щим пока лишь благодаря принятым мерам охраны.
Скорость исчезновения охотничьих животных вполне соотносится с совершен-
73
ствованием орудий добычи [Верещагин, 1959, 1971, 1975]. В верхнем палеолите
преобладают самоловные и облавные способы охоты. В каменном веке использу-
ются самоловы, топоры, дротики, копья и появляется новое мощное оружие - лук,
который постоянно совершенствуется, видоизменяясь в арбалет. В средние века
техника изготовления луков достигла совершенства. Хороший лучник поражал
цель на расстоянии до 150 м, выпуская в минуту до 12 тяжелых стрел. Арбалет по-
сылал стрелы с хорошей точностью до 200 м. Охотники на крупного зверя исполь-
зовали лошадей, множество собак и были вооружены длинными копьями с широ-
кими и острыми лезвиями. Жертвами облав становились преимущественно олени,
косули и кабаны. Собственноручно прикончить свирепого вепря холодным оружи-
ем считалось высшей доблестью. Известны и печальные рекорды. Герцог Георг I,
правитель Саксонии и Магдебурга, с 1611 по 1655 гг. убил 116906 диких живот-
ных. В одной из охот, устроенных веймарским герцогом Августом, приняло уча-
стие 373 конных охотника с 1100 собаками [Дежкин, 1977, 1983].
В Германии с 1611 по 1680 гг. одними только саксонскими курфюрстами добыто
более 50 тыс. кабанов. В 1728 г. король Фридрих Вильгельм I убил 3586 особей, а в
1782 г. герцог Карл Вюртембергский лишь на одной охоте затравил 2600 [Брем, Рос-
смесслер, 1867; Brehm, 1925; Kiessling, 1925]. Во второй половине XIX в. из-за резко-
го снижения численности добыча этих зверей здесь достигла нижнего предела - в
1865 г. убит 91 кабан, в 1881/82 гг. - 131 [Schwartz, 1988]. В Чехии в 1758 г. лишь в
районе замка «Опочно» за 18 дней охоты добыто 1710 диких свиней, в 1935 г. по
всей республике - 5, в 1950-1953 гг.-200-500 особей [Вольф, 1985].
Особенно впечатляющими были средневековые облавные охоты на копыт-
ных. В Средиземноморье в них участвовало до десяти тысяч человек, а кольцо
окружения достигало 18—20 км. При этом истреблялись тысячи крупных живот-
ных [Верещагин, 1959, 1963]. В азиатских и европейских степях татаро-
монгольские отряды, насчитывавшие десятки тысяч всадников, окружали еще
большее пространство и несколько дней, а иногда и в течение двух-трех месяцев,
гнали дичь, постепенно сжимая кольцо до такого размера, когда можно было
убивать множество животных. В Монголии и Китае для задержки и направления
зверей в определенное место применялись волосяные веревки с птичьими перья-
ми (своеобразные «флажки») или земляные валы, протяженность которых изме-
рялась сотнями (до 800) километров. Остатки этих грандиозных земляных со-
оружений кое-где заметны и по сей день. Массовая охота была одним из методов
обучения войск и способом добывания провианта для огромных армий [Кириков,
1959, 1966; Смирнов, 1994; Жаргалов, 1996]. Позднее истребительные облавные
охоты устраивали степные кочевники, казаки и казахи. На степных озерах зимой
поджигали тростник, и спасающихся от огня кабанов догоняли на лошадях, ко-
лоли пиками или стрелами из луков, или выгоняли на заранее вбитые заострен-
ные колья [Туркин, 1899; Слудский, 1956].
Истребление копытных резко усиливалось во время военных сборов и похо-
дов - войска нужно было кормить. Перед походом на рыцарей Тевтонского орде-
на (1409 г.) князь литовский Ягелло близ Гродно (район Беловежской пущи) за-
готавливал впрок мясо диких копытных для своей 100-тысячной армии, сплавляя
по рекам бочки с солониной на склад в Полоцк. В 1429 г. на большой съезд вель-
можных особ в Луцке на Украине в течение шести недель доставлялось по сотне
зубров, сотне лосей, сотне кабанов, а также другие дикие и домашние животные.
В 1431 г. на прокорм знати и свиты, кроме домашнего скота, пошло 100 зубров,
100 лосей и 500 кабанов. Как следствие неумеренной круглогодичной охоты, уже
с XV в. на западе Европы значительно сокращается численность зубра, в XVI в.
истреблен кулан, в 1627 г. в Янтровском лесу на границе Литвы и Белоруссии
убит последний тур, в 1882 г. в Таврической губернии - последний тарпан. В
этот же период с территории Европы исчезают кулан и сайгак, а многочисленные
74
I
прежде благородные олени и косули во многих местах сохраняются лишь в по-
мещичьих парках [Бутурлин, 1924; Пидопличко, 1951; Корнеев, 1953; Цалкин,
1956; Сержанин, 1961; Кириков, 1966, 1976; Тимченко, 1972].
Промысел зверей и птиц играл очень большую роль в жизни российских кре-
стьян и в экономике княжеств и губерний. Обычная повинность монастырских и
помещичьих крестьян - ходить «на ловы». Охота на копытных имела настолько
большое значение, что с XVI в. сведения о животных и местах их обитания
включались в государственные акты и переписи [Кириков, 1960].
В Сибири и на Дальнем Востоке чрезвычайно масштабным и круглогодич-
ным был самоловный промысел. Тропы копытных оснащали многочисленными
петлями и самострелами, на десятки километров тянулись ловчие изгороди с
ямами [Корнилов, 1856; Пржевальский, 1870; Кириллов, 1902; Туркин, Сатунин,
1902]. Лишь в Забайкалье городьба превышала 2 тыс. верст, и здесь были выко-
паны более 20 тыс. ям [Бионкур, 1901].
Истребление крупных копытных еще более усилилось с появлением и совер-
шенствованием дальнобойного огнестрельного оружия. Наиболее впечатляющий
пример варварского отношения человека к природным ресурсам - уничтожение
бизона в Северной Америке, численность которого в XVIII в. достигала, вероятно,
60-75 млн. голов. К 1830 г. поголовье сократилось до 40 млн., а в 1888 г. (через 58
лет!) в живых осталось всего лишь 26 особей в штате Вайоминг. По расчетам, в
период с 1868 по 1874 гг. ежегодная добыча составляла 1,5-7 млн. голов. Зверей
убивали зачастую только ради шкур и языков [Уатт, 1971; Силантьев, 2000].
Неограниченные возможности для добычи зверей на дальнем расстоянии дало
изобретение четырехлинейной винтовки Бердана, а затем и русского варианта - вин-
товки Мосина, с помощью которых завершилось уничтожение диких копытных
во многих районах. В частности, как упоминалось выше, популяции копытных
значительно поредели во время продолжительных войн на Северном Кавказе, а в
Закавказье (в Ленкорани) в начале XX в. лишь за два года армянами и русскими
добыты около 5 тыс. диких свиней [Верещагин, 1959]. Последний кавказский
зубр уничтожен в 1927 г., а чуть раньше - кавказский лось. И если бы не религи-
озные запреты на употребление мяса свиней мусульманами, такая участь ожида-
ла бы и кабана, которого истребляли и как вредителя посевов. По р. Кубани в
Краснодарском крае во второй половине XIX в. с ним велась «настоящая война»
[Флеров, Баландин, 1931; цит. по: Пидопличко, 1951].
В конце XIX - начале XX в. в России ежегодно промышляли до 50 млн. зве-
рей, в том числе более 1,2 млн. копытных. В Восточной Сибири добывали около
100 тыс. изюбрей и сайги, в Западной Сибири - до 50 тыс. маралов. Добыча ко-
суль достигала 500 тыс., кабарги - 10-15 тыс., лося - 300 тыс. (объем добычи, на
мой взгляд, завышен. -А.Д.), козлов и баранов - до 200 тыс., кабана вместе с ку-
ланом, джигетаем и шакалом - до 100 тыс. [Туркин, 1899; Туркин, Сатунин,
1902]. Перед первой мировой войной в результате охоты изымали из популяций
копытных более 410 тыс. особей: лося - около 80 тыс., изюбря и марала - 25
тыс., дикого северного оленя - 20 тыс., косуль - 200 тыс., кабана - 50 тыс. кабар-
ги - 25 тыс., а также немало баранов и сайгаков. Лишь в зыряновском крае на р.
Ижме завод ежегодно перерабатывал до 120 тыс. оленьих шкур [Бутурлин, 1924].
Известный путешественник В.К. Арсеньев [1914] в одной из китайских фанз в
горах Сихотэ-Алиня обнаружил 40 пудов оленьих жил, для заготовки которых,
по его подсчетам, потребовалось убить около 1600 оленей.
По мнению большинства ученых, именно масштабный нерегулируемый про-
мысел привел к полному или почти полному уничтожению большинства видов
крупных млекопитающих или резкому сокращению численности и ареалов остав-
шихся в живых [Богданов, 1871; Данилов, 1947; Кириков, 1952, 1953, 1959, 1960,
1966; Слудский, 1956; Цалкин, 1956, 1958; Кириков, Исаков, 1959; Формозов, 1959;
75
Гептнер, 1960; Гептнер и др., 1961; Фадеев, 1978, 1981, 1986; Верещагин, Барыш-
ников, 1980а; Давид, 1982; Косинцев, 1986, 1988; Смирнов, 1994, 1996; Лихацкий,
1996; Граков, 1997; Граков, Сафонов, 1997]. В начале XX в. в европейской части
России и на юге Западной Сибири исчезли не только дикие свиньи, но и косули,
благородный и северный олени, до предела сократился ареал лося [Данилкин,
1999а], в Крыму не осталось козерога, в Средней Азии - тугайного оленя и тигра
[Верещагин, Батыров, 1967; Верещагин, Барышников, 1980а].
Наибольший пресс на копытных всегда возникал во время социальных катак-
лизмов: в годы войн и революций и после них, при неурожаях и голоде, в период
неудачных реформ и обнищания населения, когда вооруженные люди устремля-
лись в леса. Массовое истребление зверей в европейской части России имело ме-
сто в 1905-1906 и 1914-1921 гг., когда браконьерство многократно возросло при
полном отсутствии охраны [Гортынский, 1914; Саблинский, 1914; В.П., 1928;
Кулагин, 1932; Юргенсон и др., 1935; Данилов, 1947; Покровский, Щадилов,
1962; Кириков, 1966; Тимофеева, 1974а; Данилкин, 1999а]. В Казахстане, Сред-
ней Азии и Сибири в годы войн численность зверей, напротив, увеличивалась в
связи с уходом большинства охотников на фронт, но сразу же уменьшалась с их
возвращением [Слудский, 1956].
Социальные неурядицы, кроме того, всегда имели важнейшее следствие -
резкое увеличение численности волка, который, как показано выше, также вно-
сил немалую лепту в сокращение поголовья своих жертв, включая кабана, спо-
собствуя их исчезновению из многих районов.
Роль организованного охотничьего хозяйства. Искусственное расселе-
ние. Восстановление ареалов кабана, лося, косуль и благородного оленя в Европе
и Западной Сибири во второй половине XX в. тесно связано с изменением орга-
низации охотничьего хозяйства: с улучшением охраны, подкормки (рис. 27) и с
резким сокращением поголовья волка в послевоенные годы [Любченко, 1969;
Фадеев, 1973в,г, 1975, 1978а,б, 1980, 1981, 1987, 1998; Иванова, Овсюкова, 1976;
Иванова, 1977; Новиков, Тимофеева, 1980]. В Восточной Сибири и на Дальнем
Востоке, где в этот же период охотничье хозяйство отставало в своем развитии и
где не удалось существенно снизить численность хищников, ареал и поголовье
копытных сокращались или, в лучшем случае, были относительно стабильными
[Лавов, 1980а; Данилкин, 1999а]. Кризис охотничьего хозяйства в конце XX сто-
летия [Дежкин, 1997а,б] незамедлительно негативно отразился и на численности
животных.
Процессу естественного восстановления ареала диких свиней, безусловно, спо-
собствовало искусственное расселение, которое, однако, имеет свою историю.
В начале XVI в. этот вид из Испании интродуцирован в Панаму. Отсюда
часть зверей в 1530 г. переселена в Перу, а в 1541 г. - в Чили [Skewes-Ramm,
1994]. В 1890 г. европейский кабан выпушен в Нью Гемпшире, в 1912 г. несколь-
ко голов завезены из Германии в Северную Каролину (США), и позднее - в Ар-
гентину [Пелтон, Конли, 1974; Bratton, 1975]. Сведения об интродукции в 1909 г.
в Северную Каролину 14 особей, отловленных якобы в предгорьях Урала [Ива-
нов, 1962; Павлов и др., 1974], весьма сомнительны, поскольку дикие свиньи к
этому времени на Урале были истреблены. В Америке животные быстро размно-
жились и стали серьезными вредителями сельскохозяйственных посевов, как и в
Австралии, куда их вселили в 1800 г. Попытка реакклиматизации вида имела ме-
сто также в южных районах Швеции и Норвегии, однако впоследствии зверей
уничтожили [Сиивонен, 1979].
В России в последние два столетия кабана, как и других диких копытных, из-
редка выпускали в царские охотничьи угодья и в некоторые поместья. Плано-
мерное расселение вида началось лишь с середины XX в. С 1928 по 1940 гг. в
СССР реакклиматизировали 25 зверей, в 1941-1955 гг. - 258, 1956-1965 гг. - 843,
76
Рис. 27. Сохранению поголовья кабана в немалой степени способствовала зимняя
подкормка, организованная в охотничьих хозяйствах (фото И. Мухина)
1966-1975 гг. - 4821, 1976-1977 гг. - 147 особей [Павлов и др., 1974; Павлов,
1978]. Более всего диких свиней завезли в центральные области европейской час-
ти России. Здесь в 1946-1968 гг. выпустили около 1420 голов [Вершинин, 1969],
до 1972 г. - 3178 [Гаврин, 1973]. К 1988 г. в Подмосковье расселено более 4 тыс.
особей (табл. 11).
Таблица 11. Искусственное расселение кабана в России и республиках бывшего СССР*
Место выпуска (источ- ник информации)* Годы выпуска Выпу- щено, голов Место отлова
Московская область (9, П, 12) 1947-1984 577 Средняя Азия, Дальний Восток, Северный Кавказ, Беловежская пуща, Березинский, Воронежский и Окский з-ки, внутриобластное расселение
Калининская область (11,12) 1935-1985 1616 К.-Балкария, Латвия, Литва, Эсто- ния, Киргизия, Таджикистан, Ка- захстан, Дальний Восток, Кали- нинградская, Горьковская, Астра- ханская, Ярославская, Калужская, Смоленская, Московская и Влади- мирская области. Беловежская пу- ща, Воронежский з-к
Владимирская область (11,12,14) 1954-1988 223 К.-Балкария, Беловежская пуща, Дальний Восток, Воронежская, Горьковская и Смоленская области
Новгородская область (10) Псковская область (12) 1971 22 К.-Балкария
1986 7 Внутриобластное расселение
Калужская область (11,12) 1964-1981 670 Дальний Восток, Казахстан, Бело- вежская пуща, Калининградская, Брянская, Воронежская, Смолен- ская и Горьковская области, Воро- нежский, Окский и Березинский з-ки, К.-Балкария
Смоленская область (Н,12) 1966-1973 265 Воронежский з-к, К.-Балкария, Эс- тония
77
Таблица 11 (продолжение)
Место выпуска (источ- ник информации)* Годы выпуска Выпу- щено, голов Место отлова
Ярославская область (11, 12, 16) 1961-1986 802 Беловежская пуща, К.-Балкария, Казахстан, Приморский край, Мо- сковская и Владимирская области, Воронежский з-к
Вологодская область (12) 1971 50 К.-Балкария
Рязанская область (7,12) 1948-1987 97 Средняя Азия, Смоленская область
Ивановская область (12) 1988 14 Внутриобластное расселение
Горьковская область (11) 1963-1969 51 Беловежская пуща, Белоруссия, Дальний Восток
Кировская область (12) 1975-1976 28 Горьковская область
Татарская АССР (11, 12) 1972-1972 30 it
Ульяновская область (11) 1969 18 а
Пензенская область (11,12) 1970-1973 63 Ct
Тамбовская область (6) 1969 19 Беловежская пуща
Курская область (11,12) 1971-1974 9 Внутриобластное расселение
Белгородская область (12' 1987 40 а
Воронежская область (12) 1984-1986 69 сс
Ростовская область (8, И, 12) 1970-1973 185 К.-Балкария, Воронежский з-к
С. Осетия (И, 12) 1966-1986 400 К.-Балкария, внутриобластное рас- селение
Ставропольский край (И, 12, 17) 1969-1992 88 К.-Балкария, К.-Черкессия, внут- риобластное расселение
Краснодарский край (12) 1979-1985 38 Воронежский з-к, внутриобластное расселение
Самарская область (12) 1974-1982 29 сс
Саратовская область (И,12) 1970-1973 75 Воронежский з-к, Брянская об- ласть
Волгоградская область (Н) 1969 26 Приморский край
Астраханская область (11) 1969-1972 30 Внутриобластное расселение
Оренбургская область (2, И, 12) 1971-1988 463 Воронежская, Калужская, Брян- ская и Московская области, Кирги- зия
Башкирия (12) 1987-1988 10 Внутриобластное расселение
Свердловская область (4, 12) 1978-1984 834 Киргизия, К.-Балкария, Примор- ский край, Брестская, Смоленская, Калужская и Владимирская облас- ти, Беловежская Пуща
Тюменская область (5, 12, 15) 1980-1984 326 Московская (?), Калужская, Брянская и Смоленская области, Киргизия, Бе- лоруссия (?), Воронежский з-к
Новосибирская область (1,12) 1981-1987 >118 Московская, Брянская и Смолен- ская области, Северный Казахстан (?)
78
Таблица 11 (окончание)
Место выпуска (источ- ник информации)* Годы выпуска Выпу- щено, голов Место отлова
Омская область 1982-1987 172 Московская область
(1,12,18,19) Иркутская область (12) 1981-1983 12 Внутриобластное расселение
Красноярский край (12) 1982-1987 223 Московская область
О. Сахалин (13) 1980 6 Хабаровский край
Итого в России Украина (11, 12) 1935-1988 1957-1978 7705 676 К.-Балкария, Дальний Восток,
Белоруссия(11) 1953-1961 19 Средняя Азия, Воронежский з-к, Белоруссия, Казахстан, внутрирес- публиканское расселение Беловежская пуща
Латвия (11) 1956-1958 17 Неизвестно
Литва (11) 1965-1959 4 сс
Эстония (11) 1966 8 а
Армения (11) 1969-1972 404 Дальний Восток, Воронежский з-к,
Грузия (11) 1960-1971 164 Окский з-к, Эстония, Киргизия Узбекистан, внутриреспубликан-
Казахстан (11,12) 1961-1986 56 ское расселение Западная Украина, Дальний Вос-
Киргизия (11,12) 1971-1979 232 ток, Северный Балхаш, Киргизия Внутриреспубликанское расселе-
Всего 1935-1988 9285 ние
* 1 - Баранов, 1990; 2 - Самигуллин, 1990а,б; 3 - Фертиков и др., 1999; 4 - Киселев, 1986; Марков,
1999; 5 - Воронцов, 1982; 6-Синицын, 1982; 7 - Зыкова, 1965; 8-Фертиков, 1975; 9-Иванова, 19756;
10-Русаков, 1978; И - Павлов и др., 1974; 12-Павлов, 1999а; 13-Беньковский и др., 1989; 14-Сысоев,
1970; 15-Азаров, Климов, 1990, Азаров, 1996; 16-Ильинский, 1966; 17-Саркисов, Хохлов, 1982; 18-
Назарова и др., 1982; 19-Околелов, Косачев, 1987.
Наибольшее число зверей выпущено на юге Калининской области в хорошо
организованных охотничьих хозяйствах [Макарова, Хохлов, 1971]: Скнятинском
(1947, 1952 гг. - 26 голов), Калининском (1962, 1964, 1969 гг. - 83), Безбородов-
ском (1960, 1961, 1965 гг. - 50), в Тургиновском (1967 г. - 14), Кушалинском
(1967 г. - 8) и особенно много - в Завидовском, куда в 1935-1971 гг. завезено
1179 особей из многих районов СССР (табл. 12), что привело к смешению разных
форм и массовому расселению «гибридного» потомства.
Первый завоз кабана в Завидовское охотничье хозяйство связан с именами
наркома обороны СССР К.Е. Ворошилова и первого секретаря Кабардино-
Балкарского обкома ВКП(б) Б. Калмыкова, договорившихся о выпуске зверей в
Подмосковье после одной из охот в районе Нальчика. В 1935 г. доставлено 7
особей, в 1936 г. - еще 9. В августе 1937 г. животные (4 взрослых самца, 5 свиней
и 16 поросят и подсвинков) были выпущены на волю. В первый год они уходили
не далее 6 км от загона, к концу 1938 г. встречались в радиусе 17 км от него [Ко-
лодяжный, Зворонос, 1979].
Из этого охотничьего хозяйства звери постепенно расселялись естественным
путем во все стороны, но, в основном, на север и запад. К 1941 г. их регистриро-
79
Таблица 12. Искусственное расселение кабана в Завидовском заповедно-охотничьем
хозяйстве (по [Колодяжный, Зворонос, 1979; Фертиков и др., 1999])
Год выпуска Выпущено, голов Место отлова
1935-1936 7,9 Нальчик
1941 12 Латвия
1946-1949 7,2,4 Р. Сырдарья
1960 62 Беловежская пуща
1961 17 Казахстан '
1963, 1964 18, 22 7
1965 7,77 Дальний Восток, Киргизия, Воронежская область
1966 25, 30, 19,13,22, Беловежская пуща, Дальний Восток, Воронеж-
17 ская область, Киргизия
1967 25,29, 30, 7 Киргизия, Воронежская область, Таджикистан
1968 25, 28, 23, 30, 14, Беловежская пуща, Дальний Восток, Астраханская
16, 8, 1 область, К.-Балкария, Калининградская область
1969 48, 60, 68, 62, 88, Калининградская область, К.-Балкария, Кирги-
6, 11 зия, Беловежская пуща, Литва, Казахстан
1970 17, 23, 25, 43,28, Литва, Дальний Восток, Беловежская пуща,
28, 11 Киргизия
1971 6, 27, 10,26 Киргизия, Казахстан
1935-1971 48 партий, 1179
особей
вали под Волоколамском, в Высоковском, Лотошинском и Ново-Петровском
районах, в 1945 г. - в Шаховском, позднее в Рузском и Можайском районах Мос-
ковской области. В 1946 г. они замечены в Тургиновском охотхозяйстве, в 1948
г. - в Старицком районе, 1950 г. - Ржевском, 1963-1964 гг. - Торжокском, Лихо-
славльском, Спировском и Вышневолоцком, 1966 г. - Весьегонском, 1967-1968
гг. - в Удомельском, Бежецком и Краснохолмском районах Калининской облас-
ти. В 1968-1969 гг. эти копытные заселили всю территорию области и отсюда,
вероятно, расселялись в Вологодскую область [Данилов, 1947; Дормидонтов,
1969; Юргенсон, 1969; Макарова, Хохлов, 1971; Полуйко, 1980].
Крупные партии кабанов были выпущены также в Переславском государст-
венном лесоохотничьем хозяйстве [Иванова, Молоканов, 1969] Ярославской об-
ласти (489 особей), в Вяземском охотхозяйстве Смоленской области (240), в
«Барсуках» Калужской области (272 особи) [Павлов, 1999а].
Наибольший вклад в расселение диких свиней внесли: Зоообъединение Гла-
вохоты РСФСР, передавшее в охотничьи хозяйства в 1950-1980 гг. 2,6 тыс. голов
[Шурупов, 1982]; Воронежский заповедник, где только в 1966—1978 гг. отловлено
1240 особей [Кестер, 1978]; Беловежская пуща и Нальчикское государственное
лесоохотничье хозяйство.
В 70-80-е годы зверей наиболее активно расселяли на Урале и в Сибири: в
Оренбургской области - 463 особи, Свердловской - 834, Тюменской - 326, Ново-
сибирской - более 118, Омской - 172, в Красноярском крае - 223 (см. табл. 11).
Выпуск кабанов на о. Сахалин оказался неудачным [Беньковский, 1989].
Всего в XX в. на территории СССР реинтродуцировано около 9,3 тыс. диких
свиней (см. табл. 11), в т. ч. в России - 7,7 тыс. На Украину до 1972 г. завезено 505
особей, а к 1978 г. - 676. В Крыму в 1957 г. выпустили 35 животных, доставленных
из Приморского края и одну самку - из Черниговской области [Кормилицын, Ду-
80
лицкий, 1972], а позднее, в 1973-1977 гг., еще 45 особей, отловленных в разных об-
ластях Украины. В Армению, где кабан был фактически уничтожен человеком, в
1969-1972 гг. вселили 404 зверей с Дальнего Востока, из Центральной Европы,
Прибалтики и Средней Азии. В других республиках выпуски были менее мас-
штабными (табл. 11): в Грузии - 164 зверя, в остальных - от нескольких особей
до нескольких десятков голов. В Киргизии проводилось лишь внутриреспубли-
канское расселение из Сары-Челекского заповедника [Павлов и др., 1974; Пав-
лов, 1999а].
Искусственное расселение привело к образованию мощных очагов обитания
кабана во многих областях России, что отрадно. К сожалению, есть и существен-
ный негативный результат этой работы - смешение генофонда разных подвидов
и образование в Восточной Европе и Сибири формы (или форм) с новыми при-
знаками, практически еще не изученными. Массовые выпуски нескольких видов
диких копытных способствовали формированию стойких очагов клещевого эн-
цефалита и других заразных болезней в Восточной Европе. Далеко не всегда ин-
тродукция охотничьих животных была осмысленной и необходимой, особенно в
тех областях, где плотность их населения оставалась достаточно высокой.
Роль сельского хозяйства. Как уже отмечалось выше, антропогенное преоб-
разование ландшафта - увеличение площади полей в лесной зоне, лесопосадок в
степной зоне и образование искусственных водоемов [Фадеев, 1986] - оказалось
весьма благоприятным для диких свиней и других копытных. Однако стреми-
тельное расселение и закрепление кабана на северном пределе ареала, где не раз-
вито организованное охотничье хозяйство, вряд ли было бы возможным без ори-
гинального социалистического сельского хозяйства России. Огромные площади с
зерновыми культурами и картофелем, некачественно убранными или полностью
оставленными на корню в зиму, открытое складирование урожая на полях и воз-
ле ферм, многочисленные неутилизированные трупы домашних животных на
скотомогильниках - все это давало пищу для диких свиней. Фактически с сере-
дины XX в. подкормка зверей проводилась в небывалом ранее «государствен-
ном» масштабе.
Итак, очевидно, что в историческом аспекте динамику ареалов и численности
диких копытных определяют как климатические, так и антропогенные факторы.
Они неодинаковы по направленности, продолжительности и силе воздействия, но
теснейшим образом переплетаются и взаимосвязаны. В плейстоцене и раннем-
среднем голоцене изменения границ распространения животных в значительной
мере совпадают со сдвигами климатических и растительных зон - расширяются
при потеплениях и продвижении лесов на север и сокращаются при похолодани-
ях и смене фитоценозов. В позднем голоцене и особенно со второй половины II
тысячелетия н. э. роль антропогенного воздействия становится доминирующей и
в целом истребительной, что приводит к исчезновению многих видов крупных
млекопитающих или резкому уменьшению ареалов оставшихся в живых.
Быстрое восстановление областей распространения диких копытных, и каба-
на в частности, во второй половине XX в., на мой взгляд, не связано напрямую
ни с солнечной активностью и погодно-климатическими флуктуациями, ни с
сукцессиями и изменениями продуктивности фитоценозов, ни с мифическими
«волнами жизни» [Павлов, 1999а]. Этот процесс - закономерное следствие раз-
вития организованного охотничьего хозяйства: уменьшения охотничьего пресса,
улучшения охраны и подкормки, многократного снижения численности волка,
образования мощных искусственных очагов обитания зверей при изобилии кор-
ма на сельскохозяйственных полях. Немаловажно, что дикие свиньи, как и оле-
ньи, смогли адаптироваться к антропогенным преобразованиям среды обитания.
6 Заказ №2528.
81
Среда обитания
Среда обитания кабана чрезвычайно разнообразна: от зон с мягким и влаж-
ным климатом до таежной и лесотундровой зон с длительным периодом низких
температур, мощным и продолжительным снежным покровом и глубоким про-
мерзанием почв. Однако далеко не везде на протяжении обширнейшего ареала
условия благоприятны для его жизни.
Биотопы. Региональные и сезонные особенности
биотопического распределений
В Южной Прибалтике кабан живет повсеместно, однако его распределение
неравномерно. Основные пастбища животных - сельскохозяйственные угодья
(рис. 28), лес играет первостепенную роль лишь в качестве защитной среды
[Янулайтис, Падайга, 1985; Бальчяускас, 1988].
Рис. 28. Основные пастбища диких свиней — сельскохозяйственные угодья (фото А.
Щеголева)
В Белоруссии основным местом обитания диких свиней служат дубово-
грабовые, сосновые и елово-широколиственные леса, ольшаники, зарастающие
вырубки, пойменные луга и сельскохозяйственные поля [Сержанин, 1961; Гатих,
1975]. Плотность населения определяется долями ельников, черноольховников,
82
Рис. 29. На северо-
западе ареала из всех
типов леса для каба-
на наиболее важны
ельники (фото А.
Щеголева)
спелых и приспевающих лесов [Бабинок и др., 1989]. Ключевые зимние стации -
слабо промерзающие заболоченные и пойменные торфяниковые экосистемы
[Козло, 2001]. В Беловежской пуще звери круглогодично встречаются во всех
биотопах, но в осенне-зимний период все же предпочитают хвойные и хвойно-
широколиственные леса, где отыскивают подземный корм, вечнозеленые побеги
черники и желуди, весной и летом перемещаются в елово-дубово-грабовые леса,
ольшаники и поймы, где есть сочная растительная и животная пища. Сельскохо-
зяйственные поля они посещают с весны и до поздней осени [Северцов, Саблина,
1953; Саблина, 1955; Лебедева, 1956а; Козло, 1975].
В зоне Чернобыльской АЭС в первые годы после аварии на атомной станции
и отселения людей кабаны держались в типичных местах обитания: во влажных
лесных массивах, на пойменных лугах и в перелесках. С 1988 г. они стали осваи-
вать заброшенные сельхозугодья и населенные пункты. В начале лета звери кон-
центрируются в лиственных лесах и на зарастающих полях; в июле - августе
значительная часть их переходит в поселки, где основным кормом становятся
фрукты, зерновые культуры и почвенные беспозвоночные; в сентябре - октябре
животные перемещаются в лиственные (в основном с примесью дуба) леса, где
проводят зиму. В заброшенных поселениях человека дикие свиньи проникают в
амбары с запасами зерна и дома, где переворачивают и разламывают стеллажи и
ящики с сыпучими продуктами, салом и консервами С 1992 г. отмечается их
6
83
Рис. 30. Лучшие стации кабана - разнотравные кустарниковые луга в поймах рек и
опушки смешанных лесов (фото А. Щеголева)
обитание непосредственно в г. Чернобыле [Легенда, Панов, 1993; Дунин и др.,
1998; Козло и др., 1998; Воронецкий и др., 1999].
На северо-западе России более половины встреч зверей регистрируется в лесах.
Из всех их типов наиболее важны ельники (рис. 29), особенно окраины, и смешан-
ные леса с примесью ели. В ельниках зимой почва промерзает меньше, что позволяет
кабану рыться в верхнем ее слое, и, кроме того, привлекает отсутствие ветра и воз-
можность устраивать утепленные лежки из подроста и нижних ветвей. Летом эти
стации посещаются в основном в дождливую погоду. Смешанные и мелколиствен-
ные леса и дубравы используются примерно с одинаковой частотой. Менее привле-
кательны чистые сосновые леса, лишенные подроста. Тем не менее весной животные
находят здесь массу личинок майского хруща и совок. В бесснежный период они
предпочитают сырые и заболоченные участки, труднопроходимые для человека,
дубравы, густые заросли кустарников, зарастающие вырубки и гари, лесные поляны,
разные типы лугов. В жаркое время возрастает роль водоемов, заболоченные берега
которых покрыты густым ивняком и разнообразными болотными растениями. Здесь
кабаны нередко устраивают грязевые купалки. Наиболее часто они посещают травя-
ные и тростниковые болота, реже - моховые. В период массового сбора грибов и
ягод основными убежищами крупных копытных становятся тростниковые заросли
по берегам водоемов. Непромерзающие участки болот, заросшие тростником и хво-
щами, очень важны и зимой. В течение года на сырые стации приходится примерно
треть всех встреч зверей и почти столько же - на сельскохозяйственные угодья [Ти-
мофеева, 19746, 1976; Верещагин, Русаков, 1979; Русаков, Тимофеева, 1984; Кисе-
лев, Каплин, 1987; Тупицина, 1988].
Проанализировав размещение диких свиней на северо-западе России, О.С.
Русаков и Е.К. Тимофеева выделяют 5 классов бонитета. Первый класс (лучшие
угодья) - заросшие травой и кустарниками поймы рек, опушки смешанных лесов,
84
Рис. 31. Спелые лиственные и смешанные леса - удовлетворительные угодья для
кабана (фото А. Щеголева)
облесенные окраины болот, разнотравные луга поблизости от леса (рис. 30). В
таких угодьях плотность населения зверей в позднезимний период - не менее 5
особей на 1000 га. Второй класс (хорошие угодья) - разреженные лиственные ле-
са, молодняки смешанных лесов, суходольные злаковые луга; плотность - до 4
особей. Третий класс (удовлетворительные угодья) - спелые лиственные и сме-
шанные леса (рис. 31); плотность - до 3 особей. Четвертый класс (посредствен-
ные угодья) — хвойные леса; плотность — до 2 особей. Пятый класс (плохие уго-
дья) - обширные низинные, кочкарниковые и моховые болота; животные здесь
редки [Русаков, 1983; Русаков, Тимофеева, 1984]. Такая классификация, в прин-
ципе, пригодна и для других регионов севера Европы.
Наиболее типичные стации диких свиней в центральных областях европей-
ской части России - смешанные и лиственные переувлажненные леса пойменно-
го и припойменного типа (рис. 32), примыкающие к полям, дубравы с наличием
оврагов и ручьев, ельники, травяные болота и поляны с влаголюбивой раститель-
ностью [Фадеев, 1970а, Иванова, 1974, 1982; Лаптев, 1983; Миллер, Скалой,
1990]. «Завидовский» кабан довольно равномерно распределен по угодьям, но
предпочитает лесолуговые ландшафты, особенно в сочетании с поймами ручьев
и речек, избегает обширных верховых болот и суходольных сосновых молодня-
ков, в летне-осенний период концентрируется на сенокосах, вблизи полей зерно-
вых и на прудах рыбхозов, где подбирает снулую рыбу [Дормидонтов, 1969; Ма-
нуш, Николаев, 1995].
На северном пределе ареала в Вятско-Камском крае обитание этого парноко-
пытного всецело связано с еловыми и елово-пихтовыми крепями в смешанных
лесах, изобилующих муравейниками, с пойменными лесами и сельскохозяйст-
венными полями, возле которых при неполной уборке урожая звери остаются на
зимовку [Павлов, Зарубин, 1982: Фадеев, 1998; Павлов, 1999а,б]. Основной тип
местообитаний на Среднем Поволжье - хвойно-широколиственные и мелколист-
венные островные леса [Фадеев, 1992]. В Татарии животные предпочитают сме-
85
Рис. 32. Наиболее типичные
стации диких свиней в цен-
тральных областях европей-
ской части России - смешан-
ные и лиственные переув-
лажненные леса (фото А.
Щеголева)
шанные леса (плотность - 7 особей на 1 км2), поймы рек (4,2), лесные овраги
(3,9) и молодые сосняки (3,1) [Горшков, 1988, 1989].
На Нижнем Поволжье и в бассейне Дона группировки локализованы в закры-
тых биотопах вблизи от источников воды - преимущественно в пойменных лесах
[Фадеев, 1993]. В Центральном Черноземье кабан живет в тех лесах, где есть вы-
ходы грунтовых вод, корма и укрытия, предпочитая сомкнутые участки дубрав,
молодые (5-15 лет) посадки сосны и степные лога. Зимой он встречается чаще
всего в пойменных и нагорных дубравах, на заболоченных участках и в смешан-
ных лесах. Основные кормовые стации - сельскохозяйственные поля и фрукто-
вые сады, примыкающие к лесам. Концентрация зверей очевидна на неубранных
плантациях кукурузы, картофеля, сахарной свеклы и конопли. Весной они обыч-
ны в поймах и заболоченных лесах; летом обитают вблизи от сельскохозяйствен-
ных полей и водоемов; осенью тяготеют к заболоченным участкам леса и пой-
менным дубравам. Однако в течение года животные бывают практически во всех
биотопах [Сухорослов, 1970а, 1971, 19726; Любченко, 1973; Тимофеева, 1975;
Гусев, 1989; Простаков, 1996].
На Украине и в Молдавии [Чсгорка, 1984] дикие свиньи обитают почти по-
всеместно, но предпочтение отдают дубравам и буковым лесам, фруктовым са-
дам и агроценозам. Наиболее благоприятные условия они находят в предгорной
зоне Карпат, где широколиственными лесами с густым подлеском занято около
30% территории, а непродолжительный снежный покров колеблется в пределах
от 7 до 33 см. Весной и летом звери концентрируются в горно-лесном районе,
зимой - в предгорье [Гунчак, 1982]. В Причерноморье кабан населяет днепров-
ские плавни, леса нижнеднепровских песков, обычен по побережью Каховского
86
Рис. 33. На юге ареала кабан живет в широколиственных лесах (фото О. Габузова)
водохранилища и в сельскохозяйственных угодьях степных районов, где много-
числен в островных лесах и заросших оврагах. В период вегетации звери держат-
ся в высокостебельных культурах - кукурузе, сорго и на орошаемых участках
полей [Издебский, Тарасов, 1982; Щербак, 1986].
Излюбленные стации диких свиней в Предкавказье - камышовые и тростни-
ковые заросли в руслах и, особенно, в низовьях крупных рек и на побережье
Азовского моря. На Западном Кавказе, и в Кавказском заповеднике в частности,
они занимают также равнинные и горные леса, предпочитая жить в предгорных и
горных долинах, в дубовых, буковых и фруктовых массивах и в субальпийском
криволесье на высоте от 500 до 2500 м над уровнем моря (рис. 33). Летом значи-
тельная часть популяции концентрируется на субальпийских лугах. С конца ав-
густа звери из высокогорий откочевывают на 20-30 км в предгорья, фруктарники
и орехоносы, а зимой держатся там, где есть корм и высота снежного покрова не
превышает 30-40 см [Динник, 1910; Насимович, 1936, 1955; Донауров, Теплое,
1938; Чернявская, 1956; Костоглод, 1969; Дуров, 1973, 1980а, 1986]. В Тебердин-
ском заповеднике весной большая часть животных обитает в широколиственных
и темнохвойных лесах. В начале лета здесь остаются в основном самки с порося-
тами, а остальные откочевывают в субальпийскую зону и на альпийские луга (до
2800 м над у. м.). В июле к ним присоединяются самки с выводками В начале
зимы стада спускаются на малоснежные склоны [Бобырь, 1975]. В Дагестане ка-
бан распространен повсеместно от равнин до горной зоны, но основное поголо-
вье сосредоточено все же на низменностях [Шахмарданов, Спасская, 1975; При-
луцкая, Пишванов, 1988].
В Грузии в окрестностях Лагодехского заповедника летом дикие свиньи под-
нимаются на высоту' до 3000 м над у. м. [Хеладзе, 19б4]. На востоке Кавказского
перешейка его излюбленные биотопы - крепи тугайных и низовых лесов, со-
87
стоящие из лоха, карагача, ивы, зарослей ежевики и тростника [Верещагин,
1959]. К числу благоприятных угодий следует отнести также субтропические
низменности Закавказья и непролазные кустарниковые чащи Абхазии и Аджарии
[Марков, 1932; Соколов, Темботов, 1993].
Наиболее благоприятные условия для обитания кабана имеются на побережье
Каспийского моря и в дельте Волги: сравнительно малоснежные зимы, обильный
корм и прекрасные убежища в массивных тростниковых и ивняковых зарослях,
откуда звери могут выходить в степи [Лавровский, 1962; Киселев, 1975; Литви-
нов, Коновалова, 1979].
В Оренбургской области этот вид занимает леса в поймах рек Урал, Сакмара,
Илек, Иртек, Киндель и в отрогах Южного Урала, предпочитая дубравы [Сами-
гуллин, 1990а, б]. На Южном Урале, по моим наблюдениям, он концентрируется
в обширных кустарниковых и тростниковых болотах, откуда совершает набеги
на окрестные сельскохозяйственные поля. На Среднем Урале его распределение
положительно коррелирует с долей агроландшафта и чернозема и отрицательно -
с высотой снега и долей темнохвойных лесов. В летний и осенний периоды для
диких свиней наиболее значимы разнотравные луга, в осенне-зимний - мелколи-
ственные леса и тростниковые болота. Сезонная динамика биотопического рас-
пределения вида полностью связана с обилием и доступностью кормов. Зависи-
мость численности и плотности его населения от климатических факторов незна-
чительна [Марков, 19976, 1999].
Основные места обитания кабана в Западной Сибири - ельники, сосняки, бе-
резовые и осиновые колки, осоковые и ивняковые болота и тростниковые зай-
мища на озерах. В агрикультурном ландшафте его зимнее существование сопря-
жено с искусственной подкормкой и остатками урожая на полях [Азаров, Кли-
мов, 1990; Баранов, 1990]. В Горном Алтае звери держатся в смешанных разре-
женных лесах с подлеском, полянами, заболоченными участками и водоемами. В
кедровниках в урожайные годы живут круглогодично. Летом обычны в зоне суб-
альпийских лугов и кустарниковых тундр, но выше 2,2-2,4 тыс. м над у. м. не
поднимаются. В горных тундрах и степях концентрируются лишь вблизи водо-
емов. В местах интенсивного выпаса скота животные редки. Зимой они избегают
темнохвойных многоснежных лесов и сухих горных степей [Собанский, 1992].
В равнинной части Казахстана дикие свиньи обитают в зарослях тростника и
кустарников в озерных котловинах и поймах рек и прилежащих к водоемам хол-
мистых степях и пустынях. В мелкосопочнике они держатся в заболоченных кус-
тарниковых долинах и березово-осиновых лесах. В горной части юга и юго-
востока республики распространены повсеместно до субальпийского пояса (до
3000 м над у. м.), предпочитая обильно заросшие ущелья лесного и лугостепного
поясов [Слудский, 1956; Слудский и др., 1984; Ержанов, 2000]. На Северном
Тянь-Шане в ноябре с выпадением снега звери спускаются с гор в предгорья и
долину р. Или, а весной идут в обратном направлении до субальпийских и аль-
пийских лугов. В Таласском Алатау, Терскей-Алатау и Джунгарском Алатау их
жизнь в большей степени связана с долинами рек и арчовниками [Федосенко,
1975; Грачев, Смирнова, 1980; Федосенко, Жиряков, 1984]. На Западном Тянь-
Шане зимой кабан держится в низкогорной степи и в поясе луговой и древесно-
кустарниковой растительности (1000-2000 над у. м.), предпочитая тугаи, каньо-
ны и солнечные склоны. Летом с наступлением засухи и появлением гнуса он
поднимается в высокогорье. Основные кормовые стации - горные луга и остеп-
ненные участки, щебнистые и глинистые склоны с мягкой почвой. Убежищами
служат заросли древовидной и стелющейся арчи и кустарники [Обидина, 1982].
По мнению А.А. Слудского [1956], наиболее оптимальные условия для оби-
тания вида в Казахстане имеются в тростниковых зарослях на юге республики, на
северном побережье Каспия, в долинах рек Сырдарьи и Чу, в низовьях Или, Ка-
88
Рис. 34. Заросли тростника - оптимальные кормовые и защитные стации кабана во
многих районах (фото В. Рашек)
ратала, Ак-су, на озерах Иссык-Куль и Зайсан, в дельте Черного Иртыша и в го-
рах Западного Тянь-Шаня (рис. 34).
Распространение диких свиней в Средней Азии мозаичное. В Приаралье и в
пойме Амударьи звери живут в тростниках и тугайных лесах [Воложенинов, 1974;
Реймов, Карабеков, 1975; Реймов и др., 1989]. На территории Узбекистана они
обитают постоянно практически во всех местах, где есть вода, включая бугристые
пески пустыни Кызылкум. Сюда же в поисках зимнего незамерзающего корма за-
ходят мигранты из поймы Амударьи [Ишунин, 1972]. Отсюда звери проникают и в
Каракумы, где многочисленны по берегам Каракумского канала и на фильтраци-
онных озерах с обильной прибрежной растительностью [Зинченко, Черногаев,
1989]. В Туркмении кабан — единственный представитель фауны копытных, насе-
ляющий все ландшафтные зоны республики. Основные биотопы - кустарниковые
и тростниковые заросли в долинах рек, на побережьях озер и вокруг других вод-
ных источников, включая искусственные, а также горные леса, особенно фисташ-
ковые. Осенью значительная часть животных Зимует в полупустынях и пустынях:
здесь выше температура, меньше замерзает почва и легче добывать луковицы и
корневища растений. Е1 малоснежные зимы и при хороших урожаях фруктов и
орехов они остаются в зоне лесов. Весной звери постепенно поднимаются в горы
на субальпийские луга до высоты 2500 м над у. м., где держатся на сглаженных
участках и в ущельях, заросших густым кустарником [Ишадов, 1965, 1983; Горе-
лов, 1970; Ишадов, Клюшкин, 1975; Кулибаба, Пакулин, 1985]. Лучшие места оби-
тания диких свиней в Таджикистане - тугайные леса из туранги и лоха, заросли
тростника, тамариска, эриатуса, фисташки. Весной с появлением зеленой эфемер-
89
ной растительности звери выходят на жировку в пустыню. В горах они придержи-
ваются древесно-кустарниковых зарослей и арчовников, поднимаясь летом до суб-
альпийской и альпийской зон, причем самцы вне периода гона занимают верхние
стации. Излюбленные места кормежки - лесные плодовые сады [Флеров, 1935;
Иваненко, 1956; Сапожников, 1976]. В Киргизии этот вид занимает большую часть
территории, за исключением высокогорий, куда заходит лишь летом, и Чуйской,
Кочкорской и Джумгальской долин, где выбит. Более 90% поголовья сосредоточе-
но на юге республики в Ошской области. Наибольшая плотность его населения в
Сары-Челекском заповеднике - осенью до 150 голов на 1000 га [Чичикин, Воробь-
ев, 1967; Катаевский, 1987].
В Монголии кабан обитает в темнохвойной тайге, где обычен в кедровниках
и в лиственничных лесах, в долинах рек и ручьев, зарослях тростника и кустар-
ника по берегам озер, на болотах, в ерниках и гольцах у верхней кромки леса.
Нередко звери проникают в степь и оазисы пустынь за десятки и сотни километ-
ров от южной границы леса [Банников, 1954; Соколов и др., 1982].
В Западных Саянах дикие свиньи хорошо приспособились к горному релье-
фу. Они легко карабкаются по склонам и живут даже в многоснежных (50-60
см), труднодоступных местах, где обитают козероги. Весной звери держатся в
поймах рек; летом поднимаются в средний и верхний пояса гор, доходя до верх-
ней границы леса и субальпийских лугов; осенью в урожайные годы сосредота-
чиваются в кедрачах; зимой перемещаются в долины рек южного макросклона и
на остепненные косогоры [Соколов, 1970; Завацкий, 1989]. В Предсаянье живот-
ные обычны в разнотравных смешанных лесах, на лугах и болотах, зимой зани-
мают малоснежные сосновые и елово-пихтовые леса и березовые колки, весной
переходят в черемушники [Беньковская, Беньковский, 1989]. В Туве они обитают
в озерных и речных долинах, лиственничной и кедровой тайге, где плотность их
населения может достигать 10-20 особей на 1000 га. Летом кабаны нередко вы-
ходят на субальпийские луга, а во второй половине зимы - на каменистые солн-
цепеки [Ткаченко, 1992; Горелов, 1994; Смирнов, 2001].
В Предбайкалье дикие свиньи зимой предпочитают кедрово-лиственничные
леса, подножья склонов, увалы и редины; весной - солнцепечные склоны и пой-
менные леса; летом - сырые поймы рек и ручьев, безлесные склоны и распадки
северной экспозиции, где меньше гнуса; осенью - зрелые и переспелые кедрачи
[Недзельский, 1994]. Основные их стации в Прибайкалье и Бурятии - листвен-
ничные, сосново-лиственничные и лиственные леса с подростом и подлеском,
заросли кустарников в речных долинах, остепненные склоны хребтов. В осенне-
зимний период скопления зверей отмечаются в кедрачах и в зоне кедрового стла-
ника [Смирнов, 1978; Субботин, 1980]. Остепненные склоны, кедрачи, поймы
рек, заросли кустарников - типичные биотопы этих животных и в Читинской об-
ласти [Баранов, Иванова, 1989].
Распространение кабана на Дальнем Востоке тесно связано с хвойно-
широколиственными лесами, и прежде всего с формациями маньчжурской фло-
ры. Лучшие стации - дубовые, кедровые и осиново-липовые леса, пади и долины
рек. В елово-пихтовых, лиственничных и мелколиственных лесах, а также на от-
крытых пространствах звери не держатся, за исключением временных заходов
летом и при переходах. В неурожайные годы места зимовок приурочены к сы-
рым ельникам с зарослями хвоща зимующего. Нередко животные выходят на
сельскохозяйственные поля, предпочитая овсяные, кукурузные и картофельные.
На Сихотэ-Алине они круглогодично обитают в долинах, предгорьях и нижней
части среднегорья, а их численность уменьшается по мере обеднения раститель-
ности маньчжурского типа по направлению к главной оси хребта и к северу. Ре-
гулярное перемещение зверей из одних районов и стаций в другие происходит в
зависимости от урожайности основных кормов и высоты снежного покрова [Ра-
90
ков, 1956; Бромлей, 1964; Кучеренко, 1976; Бромлей, Кучеренко, 1983; Дарман,
1990; Зайцев, Зайцева, 1990; Антонов, 1999].
Факторы, влияющие на биотопическое размещение
Кабан находит свою пищу в основном на земле и в ее верхнем слое. Соответ-
ственно, как уже отмечалось, чем выше плодородие почв, тем разнообразнее и
богаче растительная и животная кормовая база, тем выше, как правило, плот-
ность населения животных [Падайга, 1975; Янулайтис, Падайга, 19846]. Немало-
важны также состав и влажность почв: на каменистых, тяжелых и сухих землях
роющая деятельность зверей затруднена, а корма меньше, чем в мягких и влаж-
ных грунтах. Сухая пылящая почва, вероятно, забивает носовые ходы, что за-
трудняет дыхание и нарушает обоняние, поэтому дикие свиньи рыть ее избегают
[Иваненко, 1956] и не задерживаются в таких биотопах.
Кабан - ярко выраженный гигрофил. Несмотря на эвритопность и удивитель-
ную экологическую пластичность, повсеместно очевидно его тяготение к водо-
емам, пойменным, озерным, прибрежным и болотистым комплексам, заросшим
водно-болотной растительностью и труднопроходимыми кустарниками, что и
определяет в основном ленточное и очаговое распространение вида (см. рис. 22).
В засушливый период он повсеместно концентрируется в поймах рек и ручьев, в
ольшаниках, на пойменных лугах и болотах, где наряду с сочным растительным
кормом сосредоточены основные животные корма - дождевые черви и личинки
насекомых.
Кормовой и защитный факторы наиболее удачно сочетаются в тростниковых за-
рослях на юге ареала и на непромерзающих зимой лесных травянистых болотах на
севере, что делает эти стации излюбленными для диких свиней и самыми ценными.
Из всех типов леса кабан предпочитает влажные пойменные леса с обильной
почвенной фауной, а также широколиственные (дубовые и буковые) и кедровые
как наиболее кормные в урожайные годы, и еловые как лучшие защитные угодья.
Об избирательности ельников свидетельствует и тот факт, что звери при выпус-
ках прежде всего заселяют их или смешанные леса с густым еловым подростом
[Иванова, Рыковский, 1968а,б; Иванова, 1970, 1974, 1976; Воронин, 1975а; Фаде-
ев, 1977, 1982; Царев, 1979; Тупицина, 1988]. Спелые темнохвойные леса благо-
приятны для них и по другой причине - густые кроны задерживают большую
часть снега, и внизу остаются малоснежные участки, облегчающие передвижение
и доступ к «резервному» корму - влажным зимнезеленым полукустарникам,
хвощам и мху. Вырубок, мелколесий и хвойных молодняков, где снежный по-
кров существенно выше, они избегают.
Дикую свинью, тем не менее, нельзя причислять к исключительно лесным
видам животных, что обычно делается. Преимущественно лесным жителем ее
вынудил стать человек, веками преследующий копытных. При отсутствии или
слабом факторе беспокойства этот зверь не боится открытых мест и обычен в
степях, полупустынях, заходит далеко в пустыни.
В Литве плотность населения вида находится в прямой зависимости от пло-
дородия почв и доли лиственных пород в составе насаждений и обратно пропор-
циональна проценту лесистости территорий и площади лесных массивов [Падай-
га, Янулайтис, 1980, 19846]. В соседнем северо-западном регионе России такой
закономерности не выявлено, но наивысшие плотности зарегистрированы, тем не
менее, не в сплошных лесных массивах, а в районах с оптимальным сочетанием
лесов и полей [Русаков, Тимофеева, 1984]. На севере Русской равнины этот зверь
более тесно связан с полями, чем на юге [Червонный, 1987]. Очевидно, в лесной
зоне он предпочитает территории, где доли лесов и полей примерно равны, тогда
как в лесостепи частота его поселений напрямую коррелирует с лесистостью
91
Рис. 35. В Подмосковье зимнее размещение диких свиней тесно связано с местами
их подкормки в охотничьих хозяйствах (фото А. Щеголева)
территории [Фадеев, 1982, 1989]. В лиственных лесах Западной Сибири весьма
важный лимитирующий фактор в жизни дикой свиньи - дефицит материала для
устройства зимних лежек [Баранов, 1990].
На большей части ареала кабан стал фоновым видом в агрикультурном
ландшафте. Высокая пластичность позволяет ему не только уживаться рядом с
человеком, но и выживать за счет него. Пространственное размещение животных
практически круглогодично находится в тесной зависимости от хозяйственной
инфраструктуры человека: расположения сельскохозяйственных полей, главным
образом зерновых и картофеля, картофелехранилищ, силосных и зерносенажных
ям, животноводческих ферм, могильников, свалок.
В Подмосковье распределение кабана по угодьям тесно коррелирует не толь-
ко с плодородием почв и долей зерновых полей, но и с интенсивностью его под-
кормки в охотничьих хозяйствах [Иванова, 1982], причем зимнее размещение
существенно зависит от расположения и числа подкормочных площадок (рис.
35). В Кубринском лесничестве Переславского охотничьего хозяйства, например,
где было оборудовано 13 подкормочных площадок, плотность его населения со-
ставляла 18 голов на 1000 га, в Плещеевском на 6 площадках - 12, в Дмитров-
ском при наличии одного пункта подкормки - 1,8 [Иванова, 1974].
На биотопическое размещение диких копытных в немалой степени влияет
охотничий пресс. При постоянном преследовании, особенно с собаками и при ре-
гулярных загонных охотах, звери покидают свои участки обитания и скаплива-
ются в наиболее спокойных и защищенных угодьях [Слудский, 1956; Абрамов,
1963; Чичикин, Воробьев, 1967; Тимофеева, 1975, 1976]. Убежищами для них
обычно становятся заповедники и заказники. В Центрально-Черноземном запо-
веднике с началом осеннего охотничьего сезона численность кабанов и лосей
ежегодно увеличивается в 2-3 раза [Гусев, 1989].
При надлежащей охране и отсутствии постоянного преследования дикие сви-
ньи могут постоянно обитать в густонаселенной человеком (и охотниками!) ме-
стности, вблизи населенных пунктов, шумных автострад и аэропортов. В 70-80-е
годы XX в. они были многочисленны в окрестностях Москвы. По моим наблю-
92
дениям, на окраинах Зеленограда хвойный лес был буквально испещрен следами
и тропами этих животных. Отдельные семьи жили в Лосиноостровском лесопар-
ке, в районе Сходни и Внукова. Зарегистрированы десятки случаев проникнове-
ния зверей в черту Москвы, Киева, Орла и других крупных городов [Фадеев,
1973в, 1987; Иванова, 19756; Фельдман, 1989].
Причиной смены мест обитания кабана нередко становятся стихийные бедст-
вия: паводки, наледи, пожары [Воложенинов, 1974], усыхание водоемов, что
также прямо или косвенно затрагивает проблему кормодобывания. В отдельных
районах смена стаций связана с кровососущими насекомыми [Слудский, 1956].
Итак, биотопы диких свиней существенно различаются в разных регионах и
заметно меняются по сезонам года. Биотопическое размещение этих зверей оп-
ределяется комплексом абиотических и биотических факторов, среди которых,
как и при формировании границ ареала, важнейшая роль принадлежит обилию
доступных кормов, наличию убежищ, водоемов и влиянию человека. Этот вид
прежде всего занимает лучшие в кормовом и защитном отношениях угодья; в
них плотность населения животных, как правило, выше, что может служить
своеобразным критерием оценки их качества. С другой стороны, в заповедниках
и высокоорганизованных охотничьих хозяйствах его население может быть бо-
лее высоким даже в худших по качеству угодьях за счет хорошо налаженной ох-
раны и искусственной подкормки (см. раздел «Плотность населения»).
При высокой плотности населения кабан и сам существенно воздействует на
среду обитания. Эта проблема особенно интересна в практическом отношении, и
она обсуждается после анализа питания вида. !
Питание
Кабан - всеядный зверь, что во многом обеспечивает выживание вида даже в
экстремальных условиях. Его корма можно условно разделить на две группы:
растительные (подземные и надземные части растений, плоды и семена) и жи-
вотные.
Растительные корма
В период вегетации большую часть (от 40 до 90%) рациона зверей составля-
ют надземные зеленые (влажные) части различных видов трав (рис. 36), масса
которых на одном квадратном метре может достигать нескольких килограммов.
Запасы доступного для диких свиней растительного корма оцениваются в 2,5-5
тыс. кг/га, из которых около 20% сконцентрировано в подземной части; зимой
они могут уменьшаться до 10 кг/га [Лебедева, 1956а; Гусев, 1986, 1989].
Подземные части различных растений (корни, корневища, клубни, луковицы
и зимующие побеги) кабан использует круглогодично. Их масса существенна и
варьирует от 20 до 600 г/м2 [Воробьев, 1968; Тимофеева, 1980; Русаков, Тимо-
феева, 1984; Слудский и др., 1984]. Такие запасы, даже с учетом пищевой изби-
рательности зверей, вполне обеспечивают существование популяций, обитающих
на побережьях крупных и мелких озер, морских заливов, в дельтах и поймах рек
[Слудский, 1956; Лавровский, 1962; Соломатин, 1972; Издебский, Тарасов, 1982;
Слудский и др., 1984; Анненков, 1989].
Основной весенне-летний корм диких свиней - зеленые проростки, молодые
корневища и луковицы различных водно-болотных растений, включая тростник,
рогоз, камыш, стрелолист, водяной орех, вахту, ирис, осоки и хвощи. Из лесных,
луговых и степных растений наиболее предпочитаемы следующие виды: иван-
чай, манжетка, подорожник, лютик, земляника, одуванчик, клевер, ветреница,
93
Рис. 36. В период вегетации большую часть рациона кабана составляет зеленая права
(фото А. Германавичуса)
лапчатка, хохлатка, пролеска, звездчатка, пушица, кровохлебка лекарственная
гравилат, горец, гречишник, купена, ятрышник, копеечник, канареечник, вейник
солодка, герань, морковник, тмин, василистник, скереда, пижма, борщевик, дуд
ник, сныть, бутень, крапива жгучая и двудомная, щавель, лебеда, молочай, осо'
болотный, гусиный лук, саранка, кандык, крокус, ферула, тюльпан. Охотно по
едают кабаны и ядовитые растения: белокрыльник, водосбор, калужницу, копы
тень, орляк обыкновенный, аконитум, анемону, купену лекарственную и др. Мо
лодые побеги и корни деревьев и кустарников (осины, ивы, дуба, бука, березы
лещины, ели, вереска, терна, бересклета, яблони и др.) также нередки в их пищ|
[Банников, 1954; Саблина, 1955; Лебедева, 1956а,б; Слудский, 1956; Лавровский
1962; Корнеев, Кричевська, 1967; Дормидонтов, 1969; Козло, 1969, 1975; Дуров
1970а; Соломатин, 1972; Воложенинов, 1974; Иванова, 1974; Тимофеева, 1975
Иванова, Овсюкова, 1976; Верещагин, Русаков, 1979; Гунчак, 1980; Никулин
1980; Genov, 1981а,b; Бромлей, Кучеренко, 1983; Вехник, Саксонов, 1987; Гусев
1989; Смирнов, 2001].
Упомянутые виды растений отличаются, как правило, повышенным содержа
нием протеина, что важно для накопления животными жировых запасов. В otoi*
списке немало и рудеральных видов. Часть из них (крапиву, щавель, лебед\
осот, молочай) издавна с успехом скармливают домашним свиньям в крестьян
ских хозяйствах.
Важнейшие позднелетние и осенние корма - плоды и семена: желуди, орехи
фрукты, ягоды и листья земляники, брусники, черники, голубики и клюквы, гри
бы (не везде), а также корни и корневища, луковицы, зеленая растительност
[Саблина, 1955; Слудский, 1956; Козло, 1969, 1975; Бобырь, 1975; Ишадов
Клюшкин, 1975; Кучеренко, 1976; Собанский, 1992; Горелов, 1994; Смирнов
94
2001]. Эти корма, однако, неравноценны по своей питательности и привлека-
тельности для диких свиней. В широколиственных лесах излюбленной пищей
являются орешки бука, содержащие до 22% белка и до 42% жира, а также грец-
кий орех (8,7% и 51% соответственно). У плодов дуба и каштана доля питатель-
ных веществ значительно меньше (до 4,7 и 3,8% и 4 и 2,5%), гораздо ниже и их
энергетическая ценность. Фрукты звери выбирают с большей сахаристостью -
грушу, сладкие яблоки [Донауров, Теплое, 1938], спелые ягоды винограда, кизил
[Кормилицин, Дулицкий, 1972]. В хвойных лесах наиболее предпочитаемый
корм - орехи кедра. В Казахстане немаловажное место в питании кабана зани-
мают плоды стелющегося кустарника селитрянки [Слудский, 1956; Злобин,
1969], в Средней Азии - лоха [Воложенинов, 1974]. Плоды орехоносов хорошо
сохраняются под снегом, поэтому звери используют их вплоть до весны. Обиль-
ные урожаи, как и полные неурожаи, случаются примерно раз в 3-5 лет, что в
значительной мере сказывается на благополучии популяций.
В агрикультурном ландшафте с весны и до осени, а нередко почти круглого-
дично, наибольшее кормовое значение имеют поля кукурузы, гороха, вики, пше-
ницы, ячменя, ржи, овса, сои, риса, картофеля, свеклы, моркови, топинамбура,
турнепса, клевера, а также бахчи [Флеров, 1935; Логинов, 1936; Бромлей, 1964;
Дормидонтов, 1969; Козло, 1969; Иванова, 1974; Соломатин, 1975; Тимофеева,
1975, 1976; Шуев, 1975; Саркисов, Хохлов, 1982; Гусев, Елисеева, 1983; Казаков,
Даликов, 1983; Варнаков, 1986; Иванова, Овсюкова, 1986; Genov, 1987; Гусев,
1989; Простаков, 1996]. В Подмосковье звери выходят также на плантации капус-
ты [Фадеев, 1968], в Центральном Черноземье посещают посевы конопли [Сухоро-
слов, 1970а], в Беловежской пуще, Карпатах и Казахстане - гречихи и проса [Лебе-
дева, 1956а; Слудский, 1956; Гунчак, 1980], в Березинском заповеднике - сладкого
люпина [Козло, 19726], в Ростовской и Херсонской областях - подсолнечника и
сорго [Казаков, Даликов, 1983; Щербак, 1986], в Киргизии - люцерны, эспарцета,
хлопка [Чичикин, Воробьев, 1967; Воробьев, 1968; Ишадов, Клюшкин, 1975], в
Таджикистане - льна [Иваненко, 1956]. В Грузии в их рационе присутствуют фа-
соль, редис, чеснок, гоми (Setaria italica), тыква [Хеладзе, 1964].
Основной позднеосенний и зимний растительный корм диких свиней не от-
личается большим разнообразием: плоды и семена различных видов деревьев,
упавшие на землю, влажные водно-болотные и зимнезеленые растения (корни,
корневища, луковицы, стебли) и остатки урожая культивируемых растений. Дре-
весно-веточный корм, наиболее доступный для копытных в зимнее время, кабан
употребляет лишь изредка и в небольших количествах, да и то преимущественно
в самый критический период (см. ниже). Поэтому в экстремальных условиях при
многоснежье и глубоком промерзании почвы он имеет гораздо меньше шансов
выжить по сравнению с древесноядными видами животных.
Успешная зимовка вида во многом зависит от накопления жировых резервов.
Для этого требуются высокобелковая пища, и животные корма в частности.
Животные корма
Животные корма имеют весьма существенное значение в питании кабана, со-
ставляя порой до 30% видового разнообразия рациона.
Весной и летом важный компонент их пищи - различные беспозвоночные,
среди которых наибольшее значение имеют дождевые черви, слизни, личинки
майского хруща и жука-плавунца [Дормидонтов, 1969; Иванова, Ильинский,
1969; Сухорослов, 1970а, 1972а,б; Гунчак, 1980]. На 1 м2 влажной почвы в ее
верхнем слое могут жить от нескольких десятков до тысячи червей [Сухорослов,
1972а; Иванова, 1974; Зеньков и др., 1979], биомасса которых составляет 16-34
кг/га [Гусев, 1989]. В сосновых молодняках, суходольных сосняках и смешанных
95
лесах на 1 м2 приходится от 14 до 27 г почвенных беспозвоночных, в поймах, i
елово-дубовых лесах и ольшаниках-от 34 до 61 г [Козло, 1969, 1975].
Беспозвоночные - прекрасный белковый корм. В теле дождевых червей со-
держится около 8% протеина и 1,2% жира, а в насекомых - 21 и 3,8% соответст-
венно [Русаков, Тимофеева, 1984]. В зонах пустынь, полупустынь и степей в пе-
риоды массового размножения саранчи дикие свиньи питаются исключительно
ею и быстро жиреют [Слудский, 1956; Лавровский, 1962].
Основная животная пища кабанов, живущих по берегам водоемов, - лягушки,
змеи, моллюски, ракообразные, крупные личинки различных водных беспозво-
ночных, яйца и птенцы водоплавающих и околоводных птиц, околоводные гры-
зуны (водяная крыса в частности) и рыба, которую они могут ловить на мелково-
дье. Особенно обильный рыбный корм звери находят после летних и зимних за-
моров, нереста и при усыхании водоемов [Лавровский, 1952, 1962; Слудский,
1956; Бромлей, 1964; Козло, 19726; Соломатин, 1972; Саркисов, Хохлов, 1982;
Простаков, 1996]. В Казахстане, где распространен способ добычи диких свиней
на приваде, их специально подкармливают рыбой и ондатровыми тушками [Зло-
бин, 1969, 1975а]. Нередко эти копытные разоряют птичьи гнезда не только на
земле, но и на кустарниках - подминают грудью ветви и стряхивают яйца или
птенцов [Лавровский, 1962]. На оз. Арал они подбирают подранков водоплаваю-
щих птиц, невзирая на выстрелы охотников [Соломатин, 1972].
Кабаны никогда не проходят мимо «гнезд» и «складов» бурундуков, пестру-
шек, полевок, желтогорлых мышей, слепушонок, туркестанских крыс и других
норных зверьков, съедая попутно хозяев, а также мимо захоронок лисиц: остат-
ков зайцев, мышевидных грызунов и птиц [Банников, 1954; Чичикин, Воробьев,
1967; Соломатин, 1972; Тимофеева, 1976; Федосенко, Жиряков, 1984].
В неволе звери нередко съедают домашних птиц, случайно зашедших в их за-
гоны. Из предлагаемой пищи они выбирают в первую очередь животную: мышей
и полевок, мясные отходы, кости, павших птиц, червей, насекомых, лягушек. 4-
месячный поросенок может съесть за день до 30-40 полевок, игнорируя при этом
землероек и кротов [Северцов, Саблина, 1953; Саблина, 1955, 1969]. На скотомо-
гильниках, в окрестностях ферм и чабанских стоянок эти копытные находят, от-
капывают и поедают трупы павших сельскохозяйственных животных или отходы
от убитых.
При троплении кабанов мне не раз приходилось видеть остатки съеденных
ими животных (ондатр, бобра, лисицы, зайца, косуль, уток), пойманных в капка-
ны и петли охотниками и браконьерами или погибших от огнестрельных ран. В
лесу они нередко подчищают все, что остается на месте отстрела и разделки ко-
пытных животных (внутренности, головы, ноги, шкуры, кости). В Арсеньевском
районе Тульской области группа диких свиней из 13 особей с помощью хищных
птиц за 5 январских дней практически полностью утилизировала тушу взрослого
лося. Причем это стадо нашло подранка, пройдя более 1,5 км по его следам. Со-
держимое желудка звери доели в конце марта. На Урале в многоснежные зимы
обильным их кормом становятся погибшие сибирские косули.
По сообщению Т.П. Карцева [1903], в Беловежской пуще кабаны не только
разоряли гнезда тетеревиных птиц, пожирали новорожденных оленьих и трупы
павших сородичей, но и охотились за подранками, уподобляясь хищникам. В
1902 г. за 20 ч. они полностью съели тушу подстреленного накануне благородно-
го оленя. В Воронежском заповеднике эти звери поедали трупы оленей в много-
снежном 1963 г. Позднее их специально подкармливали погибшими копытными,
что привело к развитию хищнических наклонностей. Они стали нападать на ос-
лабевших оленей, сбивали их с ног и разрывали на куски [Соломатин, 1979]. В
Казахстане и Средней Азии зарегистрированы случаи нападения диких свиней на
овец [Слудский и др., 1984]. В Завидовском заповедно-охотничьем хозяйстве они
96
с удовольствием пожирали волчьи туши, выложенные в качестве подкормки
[Дормидонтов, 1977]. В Центрально-Черноземном заповеднике в конце зимы и
весной эти животные охотно кормились оттаявшими трупами лося, косули и сво-
их собратьев [Гусев, Елисеева, 1983; Гусев, 1989]. В Хоперском заповеднике в
суровую зиму 1984/85 гг. 25 трупов кабана из 61 в той или иной степени были
объедены сородичами [Антонец, 1987].
Каннибализм кабанов зарегистрирован также в Кавказском заповеднике
[Чернявская, 1956], в дельте Волги [Лавровский, 1962], на Тургайском плато
(многоснежной зимой 1968/69 гг. здесь обнаружено 15 растерзанных зверей [Со-
ломатин, 1972]), в Заилийском Алатау [Федосенко, Жиряков, 1984], на Дальнем
Востоке [Раков, 1956; Пикунов и др., 1989] и в других районах. Иногда сильные
особи нападают на ослабленных сеголеток и подсвинков, разрывают и съедают
их [Капланов, 1948; Слудский, 1956; Абрамов, 1963; Простаков, 1996].
И все-таки каннибализм у диких свиней, несмотря на их всеядность, проявля-
ется далеко не всегда. Мне не раз приходилось видеть замерзших на лежках по-
росят, которых члены семейной группы не ели, даже будучи голодными.
В районах, где этот вид появился сравнительно недавно или где восстанови-
лась численность ранее истребленного волка, отношения хищника и жертвы от-
личались от традиционных. В Воронежском, Хоперском, Жигулевском, Окском,
Дарвинском, Кавказском и Саяно-Шушенском заповедниках дикие свиньи на ка-
кое-то время становились облигатными комменсалами хищников, доедали их до-
бычу, следуя за стаями в ожидании новых жертв. В Дарвинском заповеднике они
даже отгоняли волков от только что задавленного лося. В Саяно-Шушенском за-
поведнике эти копытные, найдя недоеденную хищниками тушу марала или козе-
рога, сооружали рядом убежище и жили до тех пор, пока не подчищали все. При
этом они переносили части туши в «гнездо» и охраняли добычу от своих со-
братьев, птиц, соболей, росомах, рыси, волка, а иногда не подпускали даже чело-
века. На Дальнем Востоке отдельные семьи благополучно перезимовывают бла-
годаря остаткам «трапез» тигра [Бромлей, 1964; Казневский, 1979; Матюшкин и
др., 1981; Филонов, Калецкая, 1985; Варнаков, 1986; Вишневский, 1986; Завац-
кий, 19866, 1987, 1989; Кудактин, 1986].
Кормовая ниша всеядного кабана существенно шире, чем у других копытных,
что позволяет ему достаточно эффективно конкурировать с ними. Однако видо-
вой набор пищи не отличается большим разнообразием.
Региональные и сезонные особенности питания
Количественный и качественный состав корма диких свиней неодинаков на
протяжении обширного ареала и существенно меняется по сезонам года. В пре-
делах локальных территорий их пища состоит примерно из 40-150 видов (или
групп видов) растений и животных. На севере ареала она скуднее при отсутствии
группы плодов, и зимой к тому же многие виды растений становятся недоступ-
ными из-за промерзания почвы и высокого снежного покрова. Менее разнооб-
разна пища и на тростниковых побережьях морей и озер, в дельтах рек, а также в
высокогорных районах (табл. 13).
В Польше кабан употребляет около 130 видов и групп растений и животных: на
северо-востоке и на западе страны - 41 вид трав, 18 видов кустарников, 14 - куль-
турных растений, 45 - беспозвоночных и 12 видов позвоночных; на Дунайской
равнине соответственно 27, 15, 24 вида растений и 22 вида или таксона животных.
В 89% исследованных желудков обнаружены культурные растения, масса которых
составляет 71% всей съеденной пищи. В Болгарии в число 148 поедаемых им ви-
дов включены 40 культивируемых растений [Genov, 1981а, 1987, 1994].
В Карпатах в состав корма диких свиней входят 54 вида травянистых, 17 дре-
7. Заказ №2528. 97
Таблица 13. Количественный состав растительного и животного корма кабана
Район Число потреб- ляемых видов или групп Источник информации
Польша 130 Genov, 1981а
Дунайская равнина 89 Genov, 1987
Болгария 148 Genov, 1994 >
Беловежская пуща >120 Северцов, Саблина, 1953; Саблина, 1955; Лебедева, 1956а; Козло, 1969,1975
Украинские Карпаты >81 Гунчак, 1980, 1982
Молдавия 120 Лозан, 1983
Северо-запад России 80 Верещагин, Русаков, 1979
Вологодская область 39 Варнаков, 1986
“Завидово” >72 Дормидонтов, 1969
Ц.-Черноземный з-к 131 Гусев, 1986, 1989
З-к “Лес на Ворскле” >49 Тимофеева, 1975
Ц. Черноземье 116 Простаков, 1996
Воронежский з-к >149 Любченко, 1973; Дьяков и др., 1986
Кавказский з-к >74 Донауров, Теплов, 1938; Лебедева, 19566
Грузия ПО Хеладзе, 1964
Куба-Хачмасская зона >41 Алекперов, Кулиев, 1987
Дельта Волги 73 Лавровский, 1952, 1962
Тургайское плато >45 Соломатин, 1972
Низовья р. Или 38 Слудский и др., 1956
Северный Тянь-Шань 48 Жиряков, 1982
Западный Тянь-Шань 75 Обидина, 1982
Заилийское Алатау >80 Слудский и др., 1984; Федосенко, Жиря- ков, 1984
Вахшский хребет >60 Иваненко, 1956
Туркмения 95 Ишадов, 1965, 1969
Западный Копетдаг >43 Кулибаба, Пакулин, 1985
Предбайкалье 202 Недзельский, 1994
Юг Дальнего Востока >100 Бромлей, 1964; Бромлей, Кучеренко, 1983
весно-кустарниковых растений, 10 сельскохозяйственных культур и различные
животные. Весной и летом в пище преобладают надземные части растений, осе-
нью и зимой - плоды и семена бука, дуба, орехоносов и фруктарников (73%), в
неурожайные годы - подземные (16%) и зеленые (8%) части растений и живот-
ные (3%) [Гунчак, 1980].
Беловежский кабан довольствуется минимум 73 видами травянистых расте-
ний, 13 - деревьев и кустарников, 10 - культурных растений, а также нескольки-
ми видами мхов, лишайников, грибов, омелой и примерно 21 видом беспозво-
ночных и позвоночных. Подземные части растений составляют в пище в среднем
56%, зеленые части - 24%, плоды - 7,6% и животные корма - 12,4%. В разные
сезоны и годы это соотношение меняется [Лебедева, 1956а; Саблина, 1955; Коз-
98
ло, 1969, 1975]. У животных, обитающих в центральной части Беларуси, набор
кормов примерно такой же, но в осенне-зимний период большую часть рациона
составляют картофель и зерноотходы, выкладываемые в качестве подкормки, и
выше доля кормов животного происхождения [Емельянова, 1999].
Основная пища диких свиней на северо-западе России растительная: подзем-
ная (корни и клубни) - 24,2% и надземная - 48,5%, в том числе травянистые рас-
тения - 39,6%, древесные - 7,8%, грибы, мхи и лишайники - 1,1%. Животные
корма составляют 27,3%, из них почвенные насекомые - 14% и черви - 12,2%
[Верещагин, Русаков, 1979]. Очень большую роль здесь играют сельскохозяйст-
венные культуры [Тимофеева, 1976]. В Завидовском заповедно-охотничьем хо-
зяйстве этот зверь употреблял в пищу 61 вид растений и 11 наименований жи-
вотных кормов [Дормидонтов, 1969].
В Центральном Черноземье в числе 116 кормовых видов наиболее высока до-
ля травянистых и кустарничковых растений - 61,8%, древесно-кустарниковые
составляют 14,5%, культурные - 10%, низшие - 6,4%, животные - 7,3% [Проста-
ков, 1996]. В Центрально-Черноземном заповеднике в рационе зверей 67 травя-
нистых, 15 древесно-кустарниковых, 13 культивируемых видов и 36 видов по-
звоночных и беспозвоночных [Гусев, 1986, 1989]. В заповеднике «Лес на Ворск-
ле» пища менее разнообразна: 35 видов травянистых растений, 8 древесно-
кустарниковых пород и несколько видов животных. Доля растительных кормов
максимальна зимой (89-96%), летом снижается до 41% [Тимофеева, 1975].
Кормом диких свиней в Кавказском заповеднике служат 57 видов трав, пло-
ды и семена бука, дуба, каштана, лещины, грецкого ореха, груш, яблонь, алычи,
черешни, культурные растения (кукуруза, картофель, свекла), земляные черви,
насекомые, мышевидные грызуны, иногда птицы и зайцы [Донауров, Теплов,
1938; Лебедева, 19566; Дуров, 1970а]. В Грузии в их рационе 110 видов, из Них
26 - культурные растения, 45 - травянистые, 39 - древесные, а также папоротни-
ки, грибы и 16 видов животной пищи. Здесь звери потребляют плоды и семена 49
видов растений, зеленые части - 58, корни и корневища - 11 видов. Из плодов,
помимо перечисленных выше, они едят мушмулу, мандарин, апельсин, лимон,
айву, инжир, персик и др. [Хеладзе, 1964].
В список кормов кабана дельты Волги А.А. Лавровский [1952, 1962] включает 43
вида растений и 30 видов животных, однако в рационе преобладают чилим (водяной
орех), рогоз, тростник, сусак зонтичный, стрелолист, камыш морской и рыба.
На равнинной части Казахстана и Западной Сибири около 6-7 месяцев в году
звери питаются в основном подземными частями растений, главным образом тро-
стника и рогоза, 2 месяца - преимущественно травой, в оставшийся период пища
смешанная - растительная и животная [Злобин, 1969; Слудский и др., 1984]. В го-
рах состав корма иной. На Северном Тянь-Шане в него входят 16 травянистых, 13
древесно-кустарниковых, 5 сельскохозяйственных видов растений, один вид ли-
шайника и 13 видов или групп животной пищи [Жиряков, 1982]. В Заилийском
Алатау в списке поедаемых растений 63 травянистых и древесно-кустарниковых
вида [Федосенко, Жиряков, 1984], на Западном Тянь-Шане - также около 60 видов
травянистых растений, 4 - древесно-кустарниковых и 11 видов животных [Обиди-
на, 1982]. На Вахшском хребте в Таджикистане в желудках среднеазиатского каба-
на обнаружены фрагменты 54 видов растений и несколько групп животных кор-
мов. В весенне-летний период в рационе преобладает трава, осенью - плоды и се-
мена диких фруктов и орехоносов, зимой и ранней весной - подземные органы
(луковицы, корневища, клубни) растений [Иваненко, 1956].
Питание диких свиней на северном пределе ареала в Восточной Сибири и
Монголии представляет особый интерес. Казалось бы, в резко континентальном
климате с низкими зимними температурами, при глубоком промерзании почвы и
высоком снежном покрове, при отсутствии широколиственных лесов и плодовых
7
99
деревьев их рацион должен быть наиболее скудным. Однако, как показали иссле-
дования, в Предбайкалье эти звери потребляют более 200 видов кормов расти-
тельного и животного происхождения. Подземные и зеленые части растений со-
ставляют в среднем по 38%, семена кедра, кедрового стланика, сосны, ели и яго-
ды - 10%, животные корма - 10%, остальное - примеси (ветошь, кора, хвоя,
мхи). Объемное соотношение кормов меняется по сезонам года, но даже зимой
доля подземных и зеленых частей растений остается высокой. Наибольшая эф-
фективность зимнего кормодобывания отмечена на малопромерзающих заболо-
ченных участках в долинах рек, ручьев и ключей [Недзельский, 1994]. Животные
корма (мыши, полевки, моллюски, лягушки, дождевые черви, личинки, куколки и
имаго беспозвоночных) составляют значительную часть их рациона в весенне-
летний период. Основная пища в это время все же травянистая. Особо они выби-
рают корневища пионов, луковицы саранок, бобовые и зонтичные растения, кро-
вохлебку лекарственную, кипрей, подорожник, молодую осоку, черемшу и кан-
дык сибирский. В конце лета и осенью звери наиболее охотно поедают семена
хвойных, ягоды (бруснику, жимолость и др.) и грибы (на Западном Саяне). В аг-
рикультурных биотопах они выходят на посевы зерновых и поля картофеля [Ко-
пылов, 1950; Банников, 1954; Черкасов, 1962; Смирнов, 1978; Завацкий, 1987,
1989; Никулин, 1980].
На юге Дальнего Востока звери потребляют более 90 видов растений и 10
групп животных кормов. На Южном Сихотэ-Алине зеленые корма составляют
около 31% рациона, желуди - столько же, орехи кедра - 16%, корни, беспозво-
ночные животные - 10, хвощ зимующий - 5,5, маньчжурские орехи - 4, орехи
лещины - 0,9, орешки липы - 0,6% [Бромлей, 1964; Бромлей, Кучеренко, 1983].
Количественный набор кормов, тем не менее, не может служить мерилом
пищевого предпочтения и благополучия. При отсутствии высококалорийной пи-
щи звери начинают поедать разнообразные второстепенные и случайные мало-
питательные корма, тогда как при изобилии излюбленных кормов длительное
время могут обходиться 1-4 видами. В многоснежный период их рацион также
ограничивается несколькими доступными видами.
Географическая изменчивость зимнего питания диких свиней достаточно вы-
сока. Основные корма на юге и западе Восточной Европы - плоды и семена бука,
каштана, дуба, грецкого ореха, лещины и различных фруктовых деревьев, куль-
тивируемые растения, корни и корневища травянистых, преимущественно водно-
болотных растений, корни сосны, ели, осины, черники, вереска [Донауров, Теп-
лое, 1938; Лебедева, 1956а; Козло, 1969, 1970в, 19726; Гунчак, 1980; Щербак,
1986].
В Германии особенно значима роль картофеля, что объясняется большими
площадями этой культуры и механизированной уборкой, при которой много
клубней остается в земле. Однако далеко не все сорта по вкусу зверям, что дока-
зано экспериментальным путем [Meynhardt, 1983а; Майнхардт, 1986].
В Прибалтике, европейской части России, в Зауралье и на юге Сибири зимов-
ки кабана особенно тесно связаны с остатками урожая на полях. В Центрально-
черноземном регионе, например, доля культурных растений в общей массе по-
требленной пищи достигает 70-89% у взрослых особей и 42-64% у сеголеток.
Наиболее предпочитаемый корм - корнеплоды сахарной свеклы [Гусев, 1989]. В
Литве сельскохозяйственные культуры по степени встречаемости в рационе рас-
полагаются так: пшеница - 33-78%, овес и смеси с ним - 4-58%, картофель - 2-
36%, рожь - 0-35% [Янулайтис, Падайга, 1985]. В этих регионах немаловажна
также роль незамерзающих болот и ельников, где животные отыскивают корне-
вища водно-болотных растений, древесные корни, подснежные зеленые стебли
осок, злаков, молодые побеги черники, брусники и вереска [Иванова, 1974; Вар-
наков, 1986; Горшков, 1988].
100
По всему Каспийскому побережью, в дельте Волги, на равнинной части Ка-
захстана и Южной Сибири важнейший зимний растительный корм - корни тро-
стника и рогоза. При сильных морозах и промерзании почвы почти единственной
пищей становятся молодые стебли тростника. Нередко дикие свиньи разрывают
тростниковые хатки ондатр и поедают остатки их корма [Слудский, 1956; Лав-
ровский, 1962; Злобин, 1969].
Основная пища кабана в пустынях Средней Азии в осенне-зимний период -
корневища ферул и луковицы тюльпанов. В горных районах он, помимо корней,
корневищ, луковиц и зеленых проростков, подбирает плоды фисташки, алычи,
инжира, миндаля, абрикоса, грецкого ореха, боярышника, шиповника, арчи, яб-
лони, груши, вишни и рябины [Антипин, 1941; Слудский, 1956; Воробьев, 1968;
Соломатин, 1972; Кулибаба, Пакулин, 1985]. В Таджикистане на Вахшском хреб-
те звери научились стряхивать зрелые плоды ударами клыков по деревьям, и по-
этому многие стволы искорежены на высоте 50-110 см [Иваненко, 1956]. На юге
Киргизии обилие плодов в лесах таково, что даже при плотности 150 голов на
1000 га эти копытные не съедают их полностью за зиму и только грецкий орех
выбирают полностью и быстро при содействии других животных [Чичикин, Во-
робьев, 1967]. На Тянь-Шане основу зимнего питания составляют корни и луко-
вицы травянистых растений (44%), яблоки (35%) и вегетативные части травяни-
стых растений (33%) [Жиряков, 1982; Федосенко, Жиряков, 1984]. На Западном
Копетдаге значительная часть животных кормится на посевах зерновых и на бах-
чах [Кулибаба, Пакулин, 1985].
В Восточной Сибири и Монголии зимнее обитание животных особенно тесно
связано с кедрачами, и при неурожаях популяции нередко находятся на грани
вымирания. На Дальнем Востоке зимняя кормовая база диких свиней значитель-
но богаче, но и здесь их благополучие определяется урожаями дуба монгольско-
го, кедра корейского, ореха маньчжурского и лещины. В неурожайные годы ос-
новными кормами становятся хвощ зимующий, богатый влагой и растворимыми
углеводами, корни леспедецы и рябинолистника, побеги желтого клена, актини-
дии и других кустарников [Капланов, 1948; Абрамов, 1954; Раков, 1956; Бромлей,
1964; Кучеренко, 1973, 1976; Пикунов и др., 1989; Дарман, 1990; Антонов, 1999].
На северном пределе ареала зимнее выживание кабана зависит главным обра-
зом от наличия искусственной подкормки в охотничьих хозяйствах. Эта пробле-
ма подробно обсуждается ниже.
Потребление кормов и воды. Минеральное питание
Потребление кормов. Детеныши примерно до 3-месячного возраста кормят-
ся молоком матери. Зеленые травянистые растения в очень небольшом количест-
ве они потребляют с середины первого месяца жизни, корешки растений и жи-
вотную пищу (преимущественно дождевых червей) - на втором месяце, но рыть
почву начинают со второго дня жизни [Иваненко, 1956]. Подражая взрослым
особям, они поедают тот же самый корм, постепенно обучаются ловить мелких
животных и находить грибы [Лозан, 1982; Лозан, Лозан, 1982]. С осени у сеголе-
ток набор пищи практически такой же, как и у взрослых зверей.
Потребность кабана в корме существенно колеблется в зависимости от его
доступности и питательности, сезона года и возраста особей. Масса содержимого
желудка исследованных зверей чаще составляет 2-4 (от 0,1 до 7) кг [Донауров,
Теплов, 1938; Саблина, 1955; Чичикин, Воробьев, 1967; Козло, 1975; Шуев, 1975;
Гунчак, 1980], однако, с учетом быстрого переваривания, взрослое животное, ве-
роятно, может потреблять до 12 кг пищи в сутки [Бромлей, 1964].
В Польше суточный рацион взрослого самца в среднем 4,3 кг, самки - 3 кг,
подсвинка - 2,2 кг [Genov, 19816]. В Предбайкалье у самок масса содержимого
101
желудков также гораздо меньше (1,4-1,6 кг, в сухом виде - 125 г), чем у самцов
(2,3-2,5 кг и 429 г) [Недзельский, 1994]. В Вологодской области в желудках
взрослых особей летом и осенью находят 2,6-2,8 кг, у подсвинков - 1,4-2 кг, у
поросят - 0,2-0,8 кг пищи. Зимой эти показатели уменьшаются соответственно в
три (0,9 кг), 4,5 (0,5 кг) и 1,5 (0,5 кг) раза [Варнаков, 1986]. Следовательно, сего-
летки, кормящиеся рядом с матерями, зимой добывают примерно столько же
пищи, сколько и подсвинки. В неволе 8-месячные особи съедают 2 кг ячневой
крупы, 1,5 кг пшеничной муки, 1 кг хлеба и выпивают 3 л столовых отходов и 2-
2,5 л воды, в 10-месячном возрасте - 3 кг ячневой крупы, 0,6 кг свеклы, 8 л сто-
ловых отходов и 2-3 л воды [Иваненко, 1956].
Очень часто желудки диких свиней заполнены всего лишь одним-тремя
предпочитаемыми в данный сезон видами пищи. В Воронежской области осенью
они содержат от 0,8 до 4,7 кг желудей. Весной в ольховниках животные потреб-
ляют за сутки до 4 кг дождевых червей [Сухорослов, 1972а,б]. В желудках сред-
неазиатских кабанов находили до 1,5 кг саранчи [Слудский, 1956]; у крымских-
1,6 кг личинок пластинчатоусых насекомых [Кормилицин, Дулицкий, 1972]; в
дельте Волги - до 10 кг водяного ореха (чилима), до 3 кг клубеньков стрелоли-
ста, 2 кг воблы, 1,5 кг сазана, около 500 г личинок плавунца [Лавровский, 1962];
на севере Монголии - от 1,8 до 6,7 (в среднем 5,5) кг поедей кедровых шишек
[Банников, 1954]; на Украине - до 20 серых полевок [Корнеев, Кричевская, 1967].
В желудках (п = 11) зверей, добытых в Тверской области в декабре - январе,
я регистрировал от 0,6 до 2,7 кг зеленых побегов хвоща топяного, до 2,2 кг кор-
ней и стеблей осок и других водно-болотных растений, до 370 г корней ивы, а у
крупного секача - 6,7 кг картофеля. Судя по свежим следам после ночной поро-
ши, он кормился в разрытом бурте в 2,3 км от места лежки. Другой пищи в же-
лудке не было, что свидетельствовало о предварительном голодании зверя.
За одну кормежку кабан собирает до 1 тыс. желудей и до 500 плодов водяно-
го ореха, а за сутки - до 600 грецких орехов, 4000 маньчжурских орехов или до
300 кедровых шишек, в которых зимой сохраняется лишь часть семян. При этом
он потребляет лишь около 6-15% урожая кедра и не более 1% - других плодов.
Остальное используют многочисленные конкуренты: птицы, грызуны и копыт-
ные [Донауров, Теплов, 1938; Раков, 1956; Лавровский, 1962; Бромлей, 1964; Чи-
чикин, Воробьев, 1967].
На 1 м2 зверь выкапывает от 8 до 132 луковиц и корневищ, обкусывает до 44
растений и потребляет до 65% их массы, а также съедает от 30 до 79% почвен-
ных беспозвоночных. С этой площади он получает от 28 до 548 г растительных и
животных кормов [Лебедева, 1956а; Соломатин, 1972; Козло, 1975; Тимофеева,
1975; Верещагин, Русаков, 1979]. Летом дикие свиньи потребляют до 7-19 кг/га,
что составляет всего лишь 0,3-0,6% общих кормовых запасов. Зимой потребле-
ние кормов снижается до 1,6—4,9 кг/га, но использование доступных ресурсов
увеличивается до 16-33% (табл. 14). При высокой плотности населения (80—100
особей на 1000 га) использование корма возрастает до 2,3% и 70,5% соответст-
венно, при этом резко усиливается негативное воздействие зверей на среду оби-
тания [Гусев, 1986, 1988, 1989].
На отдельных озерах Казахстана животные утилизируют около 3% доступ-
ных запасов корма, в том числе - до 30% корневищ рогоза и 1% корневищ и по-
бегов тростника [Соломатин, 1972]. На лугах Кавказского заповедника они по-
требляют около 50% общих запасов кормовых растений, при этом наиболее пол-
но (на 69%) используют корневища иван-чая [Дуров, 1970а].
102
Таблица 14. Использование доступных запасов корма и воздействие кабана на среду
обитания при различной плотности населения (Центрально-Черноземный заповедник,
по [Гусев, 1988])
Плотность населения, особей на 100 га леса Период года
теплый холодный
исполь- зование корма, % площадь нарушений, м2/га отчуждение раститель- ности, % использо- вание корма, %
порой тропы на пороях на тропах
1 0,2 80,5 73,5 0,7 0,6 15,5
1-2 0,3 120,0 132,7 1,1 1,2 16,3
2-3 0,3 150,0 138,4 1,7 1,3 18,1
3^ 0,5 290,0 204,4 2,0 2,2 20,3
4-5 0,9 250,0 213,7 3,1 2,4 25,5
5-6 Нет данных 33,3
6-7 Нет данных 44,1
7-8 Нет данных 47,8
8-10 2,3 760,8 415,6 6,7 4,3 70,5
В Беловежской пуще среднее количество корма, добываемое кабаном в тече-
ние года с 1 м2 достигает следующих величин: подземные части растений - 288 г
(зимой до 481 г), зеленые части растений - 87 г, плоды - 56 г, животные корма -
43 г. Зимой он вырывает до 400 г корней черники и сосны, 344 г - белокрыльни-
ка, 159 г - хвоща, 140 г - осоки, 68 г - тростника и скусывает до 200 г зеленых
побегов черники; в вегетационный период — до 444 г стеблей и листьев морков-
ника, 127 г - осота болотного и от 4 до 95 г - других видов трав. При сильном
промерзании почвы и высоком снежном покрове полнота использования корма с
единицы площади резко снижается. На 1 м2 звери также находят и съедают от 30
до 200 экземпляров червей, получая до 70 г высокобелковой пищи. Зимой черви
уходит в глубь земли, поэтому повышается относительное значение насекомых и
их личинок, зимующих в верхнем слое почвы, в гнилых пнях и упавших стволах
деревьев, которые дикие свиньи активно разыскивают и разрушают. Однако мас-
са такого корма ничтожна. Доля дождевых червей в животной пище составляет
62% в количественном отношении и 74% - в весовом, доля личинок насекомых
почти в два раза меньше - 32% и 26% соответственно [Лебедева, 1956а].
Порой кабана, как отмечают многие исследователи, обычны лишь в тех мес-
тах, где сконцентрировано наибольшее количество беспозвоночных или излюб-
ленная растительная пища. Звери явно отыскивают ее с помощью обоняния (рис.
37). Обнаружению корма в разрытой почве способствуют осязательные ворсин-
ки, расположенные на кончике рыла (см. рис. 4). Однако при снежном покрове
выше 40-50 см животные обычно прекращают поиски желудей, орехов и кедро-
вых шишек, что связано, скорее всего, с невозможностью учуять их или с боль-
шими энергозатратами на раскопку.
Шишки, орехи и желуди дикие свиньи обычно разгрызают, шелуху и скорлу-
пу выплевывают. У созревающих колосьев зерновых они нередко высасывают
содержимое, также выплевывая комки шелухи. У кукурузы отламывают и по-
едают початки; иногда, покидая поле, звери уносят их в зубах и доедают на
опушке леса [Муравицкий, 1988].
Потребление воды. Вода играет большую роль в жизни диких копытных,
103
Рис. 37. Кабан отыскивает пищу в земле с помощью обоняния (фото Н Немнонова)
чем принято считать. Она составляет 3/4—4/5 веса тела и до 75% от всех выде-
ляемых организмом веществ, а ее недостаток ведет к тяжелым расстройствам и
смерти. Массовая гибель копытных зимой чаще всего происходит не от недос-
татка корма вообще (у большинства погибших зверей желудок наполнен), а
именно от нехватки волы или влажной пищи. Этот фактор - один из ведущих
при выборе животными корма и мест обитания, он нередко становится причиной
миграций и эмиграции и во многом определяет границы областей распростране-
ния видов [Данилкин, 1989, 1996, 1999а].
У домашней свиньи, как и у дикой, потребность в воде почти в два раза
выше, чем у жвачных животных. На каждый килограмм потребленного сухого
вещества корма ей требуется до 7-8 л воды, тогда как корове - 4-6 л, лошади
и овце - 2-3 л. За сутки свинья выделяет 2-6 л мочи, т. е. примерно столько
же, сколько лошадь, и в 2-4 раза больше, чем овца или коза. Образование во-
ды в результате окисления жира, крахмала или белка ничтожно, поэтому «ме-
таболическая» вода не может удовлетворить потребности организма [Викто-
ров, 1939]. В условиях неволи кабаны выпивают за сутки около 6,5-7 л [Ива-
ненко, 1956].
Практически вся пища диких свиней достаточно влажная. В килограмме зер-
на содержится 120-150 г воды, в картофеле - около 750 г. В содержимом желуд-
ков (п = 11) исследованных мной зверей во все сезоны года находилось не менее
79% воды. Летом животные получают воду в основном с зеленой растительной и
животной пищей и регулярно посещают водопои. Зимой, как и другие копытные,
разыскивают наиболее влажную пищу, сокращают выделение воды с мочой и ка-
лом и, при необходимости, поедают снег. Однако снег почти лишен минеральных
примесей и усугубляет минеральное голодание, а при его плавлении и нагрева-
нии до температуры тела энергии теряется больше, чем может быть получено с
104
пищей. Поэтому именно из-за потребности в свободной воде кабаны зимой тяго-
теют к незамерзшим болотам, ручьям и речкам, где нередко тонут в полыньях.
Грубую волокнистую пищу звери не заглатывают, а жуют, высасывают и затем
выплевывает пережеванные комочки, а у корней деревьев обгладывает лишь
наиболее влажную часть - кору. Это тоже своеобразные адаптации к получению
дополнительной влаги.
В степной, полупустынной и пустынной зонах кабаны концентрируются
лишь поблизости от водных источников. В Таджикистане, например, 214 из 219
зверей были добыты не далее 1-1,5 км от водоема [Иваненко, 1956]. В засушли-
вый период они нередко сами устраивают искусственные водопои, вырывая на
дне высохших рек, ручьев и озер глубокие ямы, в которых скапливается вода.
Иногда они пьют слабосоленую воду [Слудский, 1956; Чичикин, Воробьев, 1967;
Соломатин, 1972].
Минеральное питание. Предпочтение животными тех или иных кормов мо-
жет определяться не только их доступностью, обилием или питательностью, но и
составом минеральных веществ и микроэлементов. В отличие от многих других
копытных, дикие свиньи в достаточной мере получают их с животной пищей и
поэтому не посещают естественные и искусственные солонцы во многих районах
[Гептнер и др., 1961; Бромлей, 1964]. Однако на Урале я неоднократно регистри-
ровал их у искусственных натриевых солонцов (в смеси с глиной), и чаще всего
там, где звери кормились на неубранных зерновых полях или на подкормочных
площадках зерном или зерноотходами, - вероятно, сказывался недостаток жи-
вотной пищи. Довольно часто в течение всего года кабаны посещают солонцы в
Кировоградской области [Ярмоленко и др., 1980]. На Северном Кавказе макси-
мум выходов на них осенью — в это время их следы встречаются на солонцах так
же часто, как и оленьи; зимой потребности в дополнительном минеральном пи-
тании здесь нет [Насимович, 1938].
Летом после продолжительных дождей порой кабанов обычны на глинистых
участках. По моим наблюдениям, они не ищут здесь дождевых червей или личи-
нок насекомых, а поедают глину, выгрызая небольшие ямы, что, возможно, свя-
зано с избыточным потреблением влажной пищи. В данном случае, по-
видимому, глина используется как необходимый элемент для нормализации пи-
щеварения и формирования каловой массы.
Адаптации к критическим условиям. Энергосбережение и обмен
веществ
Один из основных способов добывания дикими свиньями пищи, как было по-
казано выше, - роющая деятельность. Однако взрослые особи могут разрывать
слой почвы, промерзший на глубину не более 7-8 см. После сильных морозов
подземные корма и свободная вода становятся недоступными. В многоснежье
(свыше 50-70 см) поиск и добывание корма значительно затруднены или невоз-
можны из-за резкого ограничения перемещения или непомерных энергозатрат.
Следует учесть также, что при длительных переходах по снегу животные нередко
ранят ноги до крови ледяной коркой, нижняя часть тела, наименее защищенная
волосяным покровом, постоянно переохлаждается, у отдельных свиней вымер-
зают соски, что сказывается на воспроизводстве. Поросята, не имеющие запасов
жира, подвержены простудным заболеваниям.
Как же выживает кабан в экстремальные зимы, особенно при неурожаях ду-
ба, орехов и других плодов? Его адаптационные особенности настолько интерес-
ны, что требуют специального рассмотрения.
Прежде всего, поражает способность этих копытных находить сравнительно
малоснежные и малопромерзающие участки с запасами зимнего корма и неза-
105
мерзшей водой. Для многих группировок характерны регулярные, вероятно мно-
говековые, осенне-зимние миграции к таким кормовым стациям. Весьма эффек-
тивной оказывается и социальная организация популяций, в основе которой ле-
жит семейно-групповой образ жизни с лидерством наиболее опытных и сильных
особей (см. ниже). Молодые животные выживают исключительно благодаря
взрослым, способным торить тропы в снегу, устраивать утепленные «гнезда», ак-
тивно защищать потомство от хищников, находить и выкапывать подземный
корм.
При замерзшей почве дикие свиньи раскапывают снег и поедают лишь влаж-
ные зеленые побеги черники, брусники, осок, хвощей, зеленую прикорневую по-
росль некоторых злаков и других трав, молодые стебли тростника. В отсутствие
зимнезеленых растений они начинают потреблять древесные и наземные лишай-
ники, мхи, травяную ветошь, побеги, ветви, кору, хвою и доступные корни со-
сны, ели, пихты, кедра, лиственницы, осины, ивы, граба, дуба, бука, ясеня, липы,
клена, рябины, березы, вереска, караганы, спиреи, жимолости, богульника и дру-
гих видов деревьев и кустарников, упавшие на землю листья и шишки, труху
гнилых пней и гнилую древесину, древесные грибы-трутовики [Донауров, Теп-
лое, 1938; Северцов, Саблина, 1953; Банников, 1954; Насимович, 1955; Слудский,
1956; Динесман, 1961; Бромлей, 1964; Котов, 1969; Козло, 19726, 1975; Солома-
тин, 1972; Смирнов, 1978; Гунчак, 1980; Никулин, 1981; Завацкий, 1989; Смир-
нов, 2001; и др.]. Эта пища, замечу, хотя и грубая и малокалорийная, но весьма
влагоемкая. На Алтае отмечены случаи поедания сена, запасенного пищухами
[Собанский, 1992]. В Карелии кабан с декабря до конца марта прожил на свежей
вырубке, кормясь корой сваленных осин [Данилов, 1974].
В бескормицу звери, как уже отмечалось выше, пытаются отыскать остатки
урожая на полях, выходят к окраинам населенных пунктов и животноводческим
фермам, посещают овощехранилища, бурты картофеля и свеклы, силосные и
зерносенажные ямы, могильники, свалки, помойки, бродят по дорогам, собирая
сено и конский навоз [Зыкова, 1965; Фадеев, 1968; Сухорослов, 19726; Тимофее-
ва, 1976; Саркисов, Хохлов, 1982; Плешак, Миняев, 1986; Смирнов, 2001], утили-
зируют трупы, остатки волчьих трапез, изредка нападают на живых животных,
подчас - на своих собратьев. В критических ситуациях они заходят в населенные
пункты, где чаще получают картечь, чем помощь. Однако известны случаи [Ру-
саков, Тимофеева, 1984], когда люди подкармливали ослабевших свиней, и те ос-
тавались жить у человека.
Кабан, как ни один другой вид фауны России и сопредельных регионов, не
впадающий в спячку или зимний сон, способен накапливать значительные жиро-
вые резервы. Восстановление потерянных за зиму белков и жиров происходит
очень быстро в связи с обильным потреблением весной и летом животной пищи,
и прежде всего земляных червей. Осенью питание этих зверей носит характер
гиперфагии с явным избытком потребления пищи, которую организм преобразу-
ет в жир, распределяя его по всему телу. У некоторых особей жировые запасы
составляют около 20-25% живой массы, подкожный слой на хребте достигает 6
см, а внутриполостной - до 7-9 кг.
Жир представляет собой ценнейший энергетический резерв, который живот-
ные постоянно носят «с собой», и, кроме того, он прекрасно изолирует организм
от переохлаждения. Взрослые упитанные особи, особенно самки с хорошо разви-
тым волосяным покровом (рис. 38), в суровые зимы имеют гораздо больше шан-
сов на выживание по сравнению с сеголетками, у которых жировые запасы весь-
ма скудны по причине быстрого расходования их растущим организмом, а шер-
стный покров гораздо беднее.
106
Рис. 38. У взрослых
свиней, имеющих высо-
кую упитанность и хо-
рошо развитый волося-
ной покров, в суровые
зимы гораздо больше
шансов на выживание
(фото А. Богданова)
Низкая упитанность зверей в весенне-летний период, возможно, не только ре-
зультат зимнего истощения, но и своеобразная адаптация к высоким летним тем-
пературам, которые особи, не имеющие значительных жировых отложений, пе-
реносят легче [Козло, 19746].
Зимой диким свиньям жизненно важно максимально снизить энергозатраты,
что достигается различными способами. В особенно морозные и ветреные дни
они резко сокращают переходы, строят утепленные «гнезда» и большую часть
времени проводяз в них вместе, согревая друг друга, или забираются в стога,
копны соломы или сена. На жировку звери выходят нередко в середине дня, ко-
гда температура воздуха существенно повышается (рис. 39). Перемещаются они
исключительно по набитым тропам гуськом вслед за вожаком, что также значи-
тельно снижает потери энергии.
Поросята зимой практически не в состоянии самостоятельно добывать корм и
в основном подбирают остатки пищи на пороях взрослых особей. Соответствен-
но каждый взрослый зверь способствует выживанию 1-2 сеголеток, что необхо-
димо учитывать в практике охотничьего хозяйства. Интересно также, что при
ранних (март) опоросах, в отличие от родов в более теплое время, взрослые сви-
ньи не прогоняют от себя прошлогодних детей, используя преимущества коллек-
тивной терморегуляции для сохранения нового потомства [Царев, 1989в, 2000]
Снижение уровня метаболизма, прекращение роста, уменьшение потребленья
корма, максимально экономное расходование жировых запасов, «гнездострои-
тельство» и «коллективная терморегуляция» — основная, преимущественно гипо-
биозная, стратегия выживания кабана зимой. В нее не вписываются лишь круп-
ные энергетические потери взрослых самцов во время гона и ранние опоросы са-
107
Рис. 39. Зимой звери выходят на кормежку в середине дня (фото А. Щеголева)
мок в холодный период. Однако первое обстоятельство характерно практически
для всех видов копытных и приводит к повышенной смертности самцов, а по-
следнее компенсируется особенно высоким жироотложением у самок и способ-
ностью к деторождению в утепленных убежищах. Сеголетки из ранневесенних
пометов достигают к зиме большей массы, что немаловажно для благополучия
популяций и вида в целом.
Во многих отношениях (всеядность, обильное жирозапасание, снижение об-
мена веществ, прекрашение роста, раннее размножение в утепленном убежище)
дикая свинья очень сходна с некоторыми зимоспящими хищниками: медведем,
барсуком и енотовидной собакой, достигшими большого прогресса в адаптациях
к суровым зимним условиям. Не исключено, что на севере ареала у нее могут
возникать отдельные элементы «зимнего сна», близкие к временному оцепене-
нию. Зарегистрированы случаи непрерывного (до 3 суток) пребывания этих зве-
рей на лежках [Соколов, Баскин, 1984]. Я также несколько раз поднимал кабанов
из «гнезд» и стогов соломы, которые они не покидали 1-3 дня, что было видно
по отсутствию следов после выпавшего снега.
Отдельные утверждения о непрерывном нахождении животных на лежке до
месяца и дольше пока что умозрительны По свидетельству известного охотоведа
М.П. Павлова [1999а, б], в Кировской области одна из семей не покидала участок
ельника, где на 200-метровой тропе имелось четыре логова, с середины ноября
по апрель. Звери прожили всю зиму ... не кормясь, и при этом не ослабли даже
сеголетки, которые, как известно, не накапливают запасов жира, достаточных для
выживания. Вятские кабаны, по этому описанию, стали «зимоспящими» живот-
ными! Данный факт весьма любопытен, но, на мой взгляд, очень сомнителен, по-
скольку постоянного слежения за семьей организовано не было.
Тем не менее было бы очень интересно провести биотелеметрические иссле-
дования физиологических параметров дикой свиньи на северном пределе ареала
в экстремальных зимних условиях.
108
Пищевая активность и поведение
Кабана, как и других диких копытных, обычно относят к сумеречным и ноч-
ным животным. Однако этот тип активности не следует считать врожденным и
генетически закрепленным. Он явно приобретен под воздействием человека и
проявляется лишь там, где преследование зверей носит регулярный характер.
Яркий пример - изменение поведения всех видов диких копытных в зоне
Чернобыльской АЭС после отселения людей. Если в 1986-1987 гг. на полях в
светлое время суток их не было, то в 1991 г. днем кормились около 35% учтен-
ных кабанов и косуль, в 1993 г. - 45%, в 1996 г. - более 50%. При этом у боль-
шинства животных исчез страх перед человеком [Дунин и др., 1998].
Мне, как и другим исследователям, также неоднократно приходилось наблю-
дать группы диких свиней днем в «глухих» местах, в заповедниках, заказниках и
на территориях хорошо организованных охотничьих хозяйств [Динник, 1910; Ло-
гинов, 1936; Донауров, Теплов, 1938; Раков, 1956; Слудский, 1956; Лавровский,
1962; Чичикин, Воробьев, 1967; Тимофеева, 1975; Смирнов, 1978; Царев, 19806].
В вольерах, при регулярном кормлении в светлое время суток, кабанята активны
исключительно днем, а ночью лежат неподвижно [Иваненко, 1956]. У европей-
ского кабана, акклиматизированного в США, по данным радиопрослеживания, во
все сезоны года преобладает дневная активность; ночная деятельность несколько
возрастает лишь летом, что связано с терморегуляцией [Kurz, Marchinton, 1972].
Зимой на севере ареала и в горах дикие свиньи кормятся, как отмечалось вы-
ше, в основном в дневные часы при более высокой температуре, что дает им воз-
можность уменьшать потери энергии при передвижении и раскопке более рыхло-
го снега, а также получать в оттепели свободную воду, замерзающую ночью. Се-
мьи, в которых есть маленькие поросята, пасутся исключительно днем. На под-
кормочных площадках звери появляются в сумерках, голодные - нередко днем,
сразу же после подвоза корма. Секачи более осторожны и обычно предпочитают
кормиться в темное время суток. Летом в жару и при массовом появлении крово-
сосущих насекомых ритм питания сдвигается на ночные и утренние часы. Днем
животные отлеживаются в грязи, холодных ключах, высокой траве или в специ-
ально оборудованном из травы «логове».
Наиболее масштабное региональное исследование суточной и сезонной ак-
тивности кабана проведено в Беловежской пуще (рис. 40). Звери здесь наименее
подвижны в декабре - феврале (2,4 следа на 1 км). С весны подвижность посте-
пенно возрастает (2,9-3,5 следа), и ее пик зарегистрирован в сентябре, что может
быть связано с формированием зимних стад. Суточный ритм активности в апреле
трехфазный: вечером - с 19 до 21 ч, ночью - с 23 до 5 ч, днем - с 15 до 17 ч; в
мае пасущиеся животные наблюдаются почти круглосуточно, кроме небольшого
перерыва с 13 до 15 ч. Максимальная частота встреч отмечена в утренние (54,4%)
и ночные (41,9%) часы. В июне дикие свиньи кормятся с восхода солнца до по-
лудня (с 5 до 13 ч), в июльскую жару - с 7 до 9 ч (62,8% встреч) и с наступлени-
ем темноты. Дифазный ночной ритм (75,9% встреч) характерен для августа - с 21
до 9 ч утра с небольшим перерывом на отдых. Максимум следов (38,5%) отмечен
на рассвете - с 5 до 7 ч. В сентябре при монофазном характере активности (с 21
до 13 ч) возрастает число дневных перемещений с максимумом с 7 до 9 ч
(65,8%). Таким образом, в вегетационный период дикие свиньи активны практи-
чески в любое время суток, за исключением отрезка времени с 13 до 15 ч; пики
приходятся на утренние (с 5 до 9) и ночные (с 23 до 3) часы (66,5% встреч).
Дневной ритм питания и перемещений более характерен для весеннего периода
[Кочко и др., 1988].
Радиопрослеживание семейной группы кабанов, проведенное нами в Пере-
славском государственном охотничьем хозяйстве в Ярославской области в авгу-
109
Рис. 40. Гистограмма суточной активности кабана (по
[Кочко и др., 1988])
Темное время суток обозначено штриховкой
сте 1983 г., показало, что
звери активны около 13-
15 ч в сутки, в основном
ночью. Иногда они вста-
вали с лежек в середине
дня, что, возможно, бы-
ло вызвано беспокойст-
вом, недолго (до 40-50
мин) ходили рядом, а за-
тем снова ложились в то
же или новое место не-
подалеку. Примерно за
1,5 ч до заката солнца
вся группа поднималась
с лежек и неторопливо
перемещалась к опушке,
где с продолжительны-
ми остановками бродила
по кромке леса. Ближе к
закату темп хождения
«взад-вперед» ускорял-
ся. Отдельные годова-
лые особи выскакивали
на край пшеничного по-
ля, хватали колоски и
быстро возвращались к
стаду. За 20-30 минут до
захода солнца на край
посевов выходили от-
дельные группы под-
свинков. Свиньи с поро-
сятами, секачи и круп-
ные стада, образовав-
шиеся на опушке, появ-
лялись на зерновых по-
сле захода солнца, но
сразу же устремлялись в
глубь поля. Эти набеги
сопровождались шумом,
чавканьем, хрюканьем и
визгом поросят, на что
собаки из окрестных де-
ревень отзывались ин-
тенсивным лаем. Ночью,
судя по стабильному сигналу радиопередатчика, кабаны могли около часа отды-
хать прямо на поле или на опушке. В лес звери уходили на рассвете, устраиваясь
на лежку неподалеку от ручья в 0,4-1,2 км от опушки.
После нескольких выстрелов по животным на рассвете и закате их поведение
изменилось. Хождение вдоль опушки сопровождалось более длительными оста-
новками и продолжительным прислушиванием; на поля группы выходили в но-
вых местах, с большей осторожностью и не ранее 30-50 минут после захода
солнца, а в лес уходили за 1-2 часа до рассвета. Места лежек переместились на
3-5 км в глубь леса. Тем не менее ежедневные набеги на поля не прекратились.
110
Рис. 41; На подкормочных площадках первыми еще засветло и примерно в одно и то
же время появляются свиньи с поросятами и подсвинки (фото И. Мухина)
Рис. 42. Между взрослыми особями порой возникают стычки за корм (фото А. Ще-
голева)
111
Рис. 43. Слабых молодых особей обычно оттесняют от корма (фото А. Щеголева)
На подкормочных площадках первыми еще засветло и примерно в одно и то
же время (иногда с точностью до минут) появляются свиньи с поросятами и под-
свинки (рис. 41). Перед выходом на них звери около 10—40 минут отстаиваются в
ближайших зарослях, убеждаясь в безопасности. Наиболее нетерпеливые порося-
та, как и на полях, выскакивают к кормушкам, хватают картофелины или зерна и
убегают назад. Отдельные группы настолько привыкают к человеку, регулярно
выкладывающему корм, что выходят на площадку даже в его присутствии Меж-
ду взрослыми особями порой возникают стычки за корм (рис 42), но такие слу-
чаи все же относительно редки. Меньше всего пищи достается более слабым по-
росятам и подсвинкам, которых взрослые особи оттесняют от корма (рис. 43).
Секачи появляются на подкормке лишь в сумерках, но сразу же становятся до-
минантами; семьи при их появлении чаще всего уходят с площадки или держатся
на ее краях. Взрослые особи, несмотря на чавканье и шум, регулярно поднимают
голову вверх, прислушиваются и принюхиваются. Наибольшими раздражителя-
ми, обращающими их в бегство, служат треск ветки, шорох и запах человека.
Итак, сделаем некоторые выводы. Продолжительность непрерывного периода
кормежки диких свиней чаще колеблется в пределах 1-5 часов в зависимости от
обилия и доступности корма. В течение суток обычно регистрируют от 1 до 5 пе-
риодов пищевой активности. Следовательно, кабан - это вид с полифазным ти-
пом активности, смещающимся к сумеречному, ночному и монофазному под
воздействием антропогенного фактора.
Его простой желудок (не разделенный на камеры), в отличие от желудка жвачных
копытных, не приспособлен к перевариванию грубой пищи с большим содержанием
клетчатки. Поэтому даже при ее обилии звери быстро худеют и погибают. Повсеме-
стно предпочтение отдается влажному, концентрированному, богатому протеином и
легкопереваримому корму: корневищам болотных растений, луковицам, зеленым
частям травянистых растений, плодам орехоносов и дикорастущих фруктарников и
животной пище. К сожалению, урожаи основных кормов - дуба, бука, кедра и фрук-
тарников - бывают не на всем ареале и далеко не каждый год. Зимой из-за много-
112
снежья упавшие на землю плоды и семена часто становятся недоступными, как и
подземные корма при глубоком промерзании почвы.
Дикие свиньи (теплолюбивые, в сущности, животные) выживают и весьма
успешно выкармливают своих детенышей даже при низких температурах и в
многоснежных районах с крайне неустойчивой кормовой базой благодаря всеяд-
ности в сочетании с быстрым накоплением и отложением жировых резервов и
экономным их расходованием при специфической «коллективной» терморегуля-
ции и гнездостроительстве.
Кабан сумел приспособиться и к антропогенному преобразованию ландшаф-
тов, и к жизни рядом с человеком, что позволяет надеяться на дальнейшее про-
цветание этого древнего вида при отсутствии чрезмерного преследования. Одна-
ко он оказывает существенное воздействие на биогеоценозы, что нередко приво-
дит к конфликтам. ...
Воздействие на биогеоценозы
Воздействие кабана на среду обитания при высокой плотности населения, по-
жалуй, более заметно по сравнению с другими дикими копытными из-за его
роющей деятельности. В поисках животного корма, семян, корней и корневищ
растений он переворачивает лесную подстилку и разрывает верхний слой земли
на значительной (до нескольких гектаров) площади, выворачивает камни весом
до 100 кг [Иваненко, 1956]. В результате этот зверь приводит в негодность сено-
косные луга и пастбища, изменяет состав растительности, выбирает из почвы по-
лезных в целом дождевых червей. Весьма существенный вред он наносит и агро-
ценозам, и зооценозам.
В конце 70-х годов XX в. на страницах журнала «Охота и охотничье хозяйст-
во» была развернута специальная дискуссия о вреде и пользе кабана. Приводи-
лись веские аргументы за и против его существования, вплоть до исключения
этого «чуждого», плодовитого и трудно регулируемого вида из состава биогео-
ценозов среднерусских лесов [Благовещенский, 1977; Дормидонтов, 1977; Ива-
нова, 1977; Фадеев, 1977, 19786; Ханжин, 1977; и др.]. Эта полемика еще не за-
вершена, хотя не только наука, но и практика показала, что дикая свинья не та-
кой уж «страшно вредный» зверь, каким его представляли, особенно для леса.
Кроме того, численность вида быстро научились «регулировать» вплоть до нуля,
научились также и существенно ослаблять негативное воздействие зверей на аг-
роценозы. И все же эта проблема по-прежнему актуальна.
Влияние на почвенные ценозы и растительность
Повреждения почвы дикими свиньями могут быть поверхностными, сплошными,
диффузными и точечными. Одно животное за час способно «вспахать» 25-30 м2, за
день - от 2,5 до 120 м2, а за год - до 4-5 га. Глубина покопок чаще колеблется в
пределах 8-20 (1-40) см, но при добывании корней или кладовых грызунов они
роют ямы, напоминающие воронки от снарядов (до 100-120 см глубиной и до
100-200 см в диаметре) [Северцов, Саблина, 1953; Лебедева, Качанова, 1955; Ле-
бедева, 1956а; Бромлей, 1964; Чичикин, Воробьев, 1967; Козло, 1969, 1970г,
1975; Дуров, 1970а; Соломатин, 1972; Лавов, 1981; Лаптев, 1983; Лобачев, Пету-
хов, 1984; Гусев, 1989].
В Беларуси кабан перекапывает от 0,2% площади в сосняках и до 65% - в дубра-
вах. В Беловежской пуще под его воздействием находится около 20 (2-36)% терри-
тории, на которой он перемещает 130-430 тыс. м~ почвы. Площадь сплошных
пороев на отдельных участках достигает 4 га, а общие повреждения за
8. Заказ № 2528.
113
год превышают 1300 тыс. м2. На разрытых местах масса травяного покрова
уменьшается на 30—65%, а видовое разнообразие сокращается на 10-60%. При
этом звери поедают только 1-2% растительной массы, а 20-60% погибает в ре-
зультате механических повреждений. Отчуждение фитомассы в ельнике достига-
ет 48%, в луговых сообществах - 67%, в смешанных широколиственных лесах -
86%. Через 2-3 года растительность восстанавливается, при этом ее биомасса
возрастает на 80-150% по сравнению с невзрыхленными участками. Порой
обычно зарастают новыми цветковыми и сорными видами растений [Лебедева,
Качанова, 1955; Лебедева, 1956а; Козло, 1969, 1970г, 1972а, 1975; Козло, Став-
ровская, 1979; Емельянова, 1986, 1989, 1990, 1999].
В степной зоне Украины эти копытные при плотности населения 18-25 особей
на 1000 га «перепахивают» до 40-50% площади пойменных и до 15% - плакорных
лесов, в результате чего существенно изменяются физические свойства и химиче-
ский состав почвы: твердость уменьшается на 8,8-25,4%, влажность увеличивается
на 28—35%, аэрационные свойства — на 8-13%, а водопроницаемость - на 70-75%.
Кабан возвращает на поверхность 70-226 кг марганца, 19-33 кг хрома, 7-13 кг
никеля, 0,3—0,7 кг молибдена, 4—9 кг ванадия и 0,1-1,4 кг меди. Процесс минера-
лизации подстилки ускоряется на 14-16%, что приводит к обогащению почвы
гумусом. Заметно изменяется и растительный состав в местах пороев. Дикие
свиньи уничтожают до 50% клубней и корней растений, что приводит к исчезно-
вению 60-70% трав. Полное восстановление растительности занимает 4-6 лет.
Разрыхление земли, однако, способствует лесовозобновлению - всхожесть семян
древесных пород увеличивается на 14-35%. Роющая деятельность затрагивает и
почвенную мезофауну: ее биомасса уменьшается на 27-39%, но уже через 2-3
недели вспаханные участки интенсивно заселяются дождевыми червями и дру-
гими животными, плотность населения которых увеличивается и превышает
прежние показатели [Булахов, 1975].
В лесостепной дубраве в Белгородской области кабан при высокой плотности
населения (до 50-150 особей на 1000 га в зимний период) перерывает в отдель-
ных кварталах до 50-70% поверхности земли, выкапывая и съедая на 1м2 от 8 до
132 луковиц, клубней и корневищ преимущественно эфемероидов. Видовой со-
став травянистых растений на пороях здесь также существенно изменяется, а
всхожесть семян древесных пород увеличивается в 2,8-7 раз [Тимофеева, 1980,
1982]. Значительное увеличение всхожести дуба достигается благодаря взрыхле-
нию земли и зарыванию на глубину от 3 до 8 см от 1 до 9 желудей на 1 м* пороев.
Без этого лежащие на поверхности почвы семена чаще всего сгнивают [Сухоро-
слов, 19726].
Средняя площадь повреждений почвы в Хоперском и Центрально-
черноземном заповедниках составляет 7-101 (до 435 в отдельные годы) м2 на 1
га леса, а при высокой плотности зверей достигает 761 м2. Наиболее интенсивна
роющая деятельность диких свиней в дубравах (до 2,3% их площади ежегодно),
где они наносят некоторый вред основной лесообразующей породе - дубу, но
способствуют семенному возобновлению терна, бересклета, черемухи, клена и
ясеня. Доля изъятия излюбленного корма - пролески достигает 50,8% по количе-
ству и 47,4% по массе; на некоторых участках это растение деградировало. На
пороях здесь также существенно изменились микроклимат, гидротермический
режим и физические свойства почвы [Ходашева, Елисеева, 1967; Гусев, 1986,
1988, 1989; Ковалева, Антонец, 1995; Антонец, 1997].
В Жигулевском заповеднике, где площадь сплошных нарушенных лесных
участков достигает 1-2 тыс. м2, уничтожению подвергаются редкие виды расте-
ний, а сорные виды, напротив, расселяются [Вехник, Саксонов, 1987]. Реальна
угроза исчезновения ряда редких видов растений и в Черноморском биосферном
заповеднике [Уманец, Селюнина, 1987]. Регулярное перекапывание дикими
114
свиньями почвенного покрова в Воронежском заповеднике (взрыто до 22% тер-
ритории), наиболее интенсивное в апреле и октябре, привело к снижению общей
численности и отдельных групп эпигеобионтной мезофауны (дождевых червей и
многоножек-землянок) [Дьякова, Дьяков, 1989; Емец, Емец, 1995].
При плотности населения в 6-12 особей на 1000 га кабан повреждает от 2 до
12% площади лугов. Заметный ущерб причинен им уникальным лугам Окской
поймы и сенокосам в Московской области [Иванова, Овсюкова, 1976; Иванова,
1977; Лаптев и др., 1987].
На Южном Урале в Ильменском заповеднике каждый зверь для удовлетворе-
ния суточной потребности в корме разрывает около 40,2 м2 почвы. Запасы над-
земной фитомассы на пороях сокращаются в 2,4 раза, потеря корневой массы
достигает 22,4%, клубни и корневища зонтичных используются на 96,7%. Из
травостоя выпадают редко встречающиеся растения: кровохлебка лекарственная,
купена лекарственная, подмаренник озерный, таволга вязолистная, ракитник рус-
ский, но появляются новые виды: княжик сибирский, горец вьющийся, лебеда
садовая. Через два года бобовые восстанавливаются на 13,3%, злаки на - 55,4%,
разнотравье - на 81,6%, зонтичные - на 125,6% [Дворников, 1986; Дворников,
Дворникова, 1989].
На Тянь-Шане у верхней границы леса звери разрывают сплошь до 190 тыс.
м2 почвы [Жиряков, 1982; Федосенко, Жиряков, 1984]. На Западном Копетдаге
повреждается 25-30% площади горно-долинных лесов, 10-15% - сухих склонов
и 1% - горных степей [Кулибаба, Пакулин, 1985].
В целом в экосистемах в результате роющей деятельности кабана и вытапты-
вания отчуждается соответственно не более 1,8% и 0,8% травянистой раститель-
ности, которая восстанавливается на третий-четвертый год. При этом в травяни-
стых ассоциациях доминирующие виды заменяются рудеральными и улучшается
естественное лесовозобновление. Наиболее быстро восстанавливаются луговые
виды растений на открытых полянах в лесу, медленно - многолетние растения с
разветвленной корневой системой [Howe et al., 1981] и субальпийские и альпий-
ские лишайниковые виды (до 15 лет - [Онипченко, Голиков, 1996]). Видовой со-
став, численность и биомасса почвенных беспозвоночных на пороях и тропах ди-
ких свиней уменьшаются в 2-13 раз, что обусловлено главным образом исчезно-
вением группы дождевых червей. Однако их восстановление происходит очень
быстро - на второй год [Дуров, 1970а; Гусев, 1983, 1986, 1989, 1995; Кулибаба,
Пакулин, 1985; Вехник, Саксонов, 1989].
Интересна роль роющей деятельности кабана в 30-километровой зоне Черно-
быльской АЭС, где им «перепахано» по меньшей мере 30% территории бывших
полей. В этих местах гамма-фон в 2 раза ниже, чем на неповрежденных участках,
более интенсивна гумификация растительной ветоши, что снижает пожароопас-
ность территории, продолжительнее сроки вегетации травянистой растительно-
сти. Зарастание пороев древесными растениями способствует ускорению процес-
са облесения полей на 15-20 лет. Естественная рекультивация и интенсивное ле-
совозобновление стали альтернативой идеи формирования в зоне отчуждения ис-
кусственных сосновых насаждений. Увеличение видового разнообразия расти-
тельности и повышение продуктивности приводит к интенсивному поступлению
в почву различных комплексообразующих соединений, способных снижать под-
вижность радионуклидов [Легейда, 1993; Козло и др., 1998].
Воздействие на лесные ценозы
Вред, причиняемый дикими свиньями древесной растительности (сдирание
коры при погрызах, почесах, ударах клыками, повреждения подроста для устрой-
ства лежек, разрывание и поедание корней и однолетних побегов) несравненно
8*
115
меньший, чем от других копытных, хотя в отдельных сосновых посадках [Валь-
кович, 1986] он может быть существенным. В дубравах даже при очень высокой
плотности населения кабан повреждает в среднем 38 экземпляров деревцев (в ос-
новном бересклет, рябину и черемуху) на 1 га, что составляет не более 2% всего
подроста и подлеска [Тимофеева, 1975, 1980, 1982; Гусев, 1989; Гусев, Гусева,
1989]. В Беловежской пуще его воздействие на древесный подрост при недостат-
ке корма, вероятно, более существенно [Саблина, 19536].
Жалобы лесничеств, в основном, сводятся к разрушению этим зверем мура-
вейников, разрыванию сенокосов, поеданию семян дуба, бука, орехоносов и
фруктарников и выкапыванию посевов. В Окском заповеднике из 269 зарегист-
рированных жилищ муравьев оказались разрушенными по разным причинам 108,
на 68 из них обнаружены лежки кабанов, впоследствии восстановлены лишь око-
ло 55% [Червонный, 1978, 19906]. В Верхне-Клязьминском лесхозе Московской
области дикие свиньи повредили 104 муравейника из 200; ежегодно здесь оказы-
ваются поврежденными от 16 до 26% этих сооружений [Захаров, Суворов, 1988].
В заповеднике «Лес на Ворскле» на небольшой площади от кабанов пострадали
133 жилища муравьев [Тимофеева, 1975].
В годы урожая дуба 50-80 особей потребляют около 20-30 т желудей [Тимо-
феева, 1980]. В Хоперском заповеднике, поедая желуди, дикие свиньи снижают
количество подроста более чем в 2 раза [Мусиевский, Ильин, 1986], а в нацио-
нальном парке «Припятский» уничтожают почти все семена [Емельянова, 1999].
В Кавказском заповеднике они собирают примерно 1-3% урожая буковых орехов
[Дуров, 1980а]. Семья за 5 суток может выкопать до 45% посеянных желудей на
площади 3 га [Сухорослов, 19726]. В Бадхызе один зверь выбирает до 3 кг фис-
ташковых орешков на 2-3-километровой борозде, а вместе животные сводят уси-
лия лесоводов на нет [Соломатин, 1972].
Следует отметить, что потребителями семян и подроста дуба, бука и кедра явля-
ются не только дикие свиньи, но и многие другие виды млекопитающих и птиц (см.
раздел «Конкуренты»), что при общей высокой их плотности приводит к деградации
широколиственных лесов [Толкач, Будниченко, 1986]. В отдельных районах Запад-
ной Европы регулярный сбор дикими копытными всего урожая семян способствовал
замене дубово-букового древостоя на березовый [Динесман, 1961].
В орехово-плодовых лесах, и особенно кедрачах, исследуемый вид является
мощным конкурентом человеку (как, впрочем, и человек животным). В урожай-
ные годы такие леса буквально «истолчены» дикими свиньями. Как и медведь,
они при случае утилизируют собранные и складированные людьми шишки кедра,
даже затаренные в мешки [Смирнов, 1978]. Однако, на мой взгляд, это не повод
объявлять их вредителями леса и урожая. С таким же успехом «вредителем»
можно назвать человека, который «грабит» зверей в их исконных угодьях, обре-
кая на голодную смерть или провоцируя выход на сельскохозяйственные поля.
Обитание кабана в лесу, как уже подчеркивалось выше, благоприятно с точки
зрения лесовозобновления и особенно полезно во влажных хвойных массивах,
где толстый слой мха препятствует попаданию семян в почву. На пороях через
несколько лет появляется обильный самосев - до 30 экземпляров подроста на 1
м2 [Корелов, 1947; Лебедева, 1956а; Слудский, 1956; Динесман, 1961; Абрамов,
1963; Корнеев, Кричевская, 1967; Чичикин, Воробьев, 1967; Козло, 1970г; Сухо-
рослов, 19726]. В Карпатах, например, на местах 2-3-летних повреждений почвы
подроста бука в 6 раз больше, чем на нетронутых участках, на 60% взрыхленной
территории появляется береза, на лесокультурных участках вред компенсируется
образованием группировок из ели, пихты и березы [Гунчак, 1982]. В Дарвинском
заповеднике в разных типах леса количество однолетних древесных побегов на
пороях в 4-30 раз больше, чем на неповрежденных участках [Завьялова, 1997].
Необходимо также учитывать, что большая часть семян, проходящих через
116
желудочно-кишечный тракт диких свиней, не теряет всхожести [Слудский и др.,
1984]. Желуди перевариваются почти полностью, косточки вишен повреждаются
на 83%, семена яблок и груш - на 41%, винограда и рябины - на 21%, семена
диаметром менее 4 мм сохраняются полностью [Genard, Lescourret, 1985]. Эти
звери, следовательно, способствуют их рассеиванию с экскрементами, удобряя
одновременно почву. Таким же образом они разносят на значительные расстоя-
ния и споры многих видов грибов [Genard et al., 1986].
Роль в агроценозах
В агроценозах кабан - один из основных вредителей посевов (рис. 44). Более
всего от него страдают плантации кукурузы, гороха, овса, вики, проса, пшеницы,
ячменя, ржи, риса, сои, картофеля и топинамбура (не везде), турнепса, сахарной
свеклы и бахчевые культуры [Динник, 1910; Марков, 1932; Сержанин, 1961; Во-
робьев, 1968; Фадеев, 1973в; Варнаков, Мошева, 1975; Соломатин, 1975; Хан-
жин, 1977; Andrzejewski, Jezierski, 1978; Янулайтис, Падайга, 19846; Варнаков,
1986; Генов, 1986; Горшков, 1989]. Набеги животных на сельскохозяйственные
посевы начинаются ранней весной с момента появления озимых из-под снега,
усиливаются во время посадки гороха, кукурузы, картофеля и колошения зерно-
вых и продолжаются вплоть до уборки урожая. Однако и зимой они тоже могут
причинять немалый вред.
Мне, как и другим исследователям [Иванов, Русаков, 1970; Тимофеева, 1976;
Миллер, Скалой, 1990], в 60-80-е годы XX в. не раз приходилось видеть в разных
областях России раскрытые дикими свиньями промерзшие бурты картофеля и
свеклы. В каждом из них находились десятки тонн испорченного семенного ма-
териала. В охотничьем хозяйстве «Барсуки», например, эти звери в 1966 г. «за-
морозили» 90 т картофеля, а в 1967 г. - 233 т [Вершинин, 1969]. В Можайском
районе Московской области только за период с 5 по 26 января 1976 г. они повре-
дили 37 буртов [Лаптев, 1987]. В целом потери складированных овощей в мас-
штабе страны могли исчисляться многими тысячами тонн.
В вегетационный период лишь за одну ночь 5-7 особей могут выбрать клуб-
ни только что высаженного картофеля на площади в 200-300 м2 или вытоптать
0,1 га посевов хлебных культур и до 0,7 га кукурузы [Раков, 1956; Бромлей, 1964;
Верещагин, Русаков, 1979; Русаков, Тимофеева, 1984]. В Карпатах, по расчетам,
каждый кабан повреждает в среднем около 0,2 га сельскохозяйственных культур
[Гунчак, 1982].
При низкой плотности населения диких свиней ущерб сельскохозяйственным
посевам, однако, невелик. На территории 5 районов Белгородской области, где на
полях кормилось не более 15—20% поголовья, в 1969 г. стравлено всего 2,3 га
урожая [Сухорослов, 19726]. При высокой численности или временной концен-
трации зверей потравы резко увеличиваются.
В Первоуральском районе Свердловской области в 1987 г. стадо за ночь при-
вело в негодность поле овса на площади 6 га [Дунаев, 1991]. В Московской об-
ласти группы из 15-17 особей ранней весной уничтожали посевы кукурузы на
площади 12-22 га. Летом наибольший ущерб от них был на полях, примыкаю-
щих к лесным массивам или находящихся среди леса. Зерновые на площади до
10 га животные стравливали полностью, а на более крупных участках поврежда-
ли преимущественно 30-метровую полосу вдоль опушки. Они не покидали поля
и после уборки урожая, подбирая остатки. Рядом с не полностью убранной кар-
тофельной плантацией площадью около 12 га всю зиму могли держаться около
120 особей [Лаптев и др., 1987]. В Беловежской пуще осенью 1948 г. при неуро-
жае желудей эти копытные большими стадами выходили на картофельные поля,
невзирая на ночную охрану, а после сбора урожая устремлялись на посевы ози-
117
Рис. 44. Поле пше-
ницы после набегов
диких свиней (фото
П. Яровицкого)
мых, которые приходилось пересеивать по несколько раз [Саблина, 1955]. На юге
Дальнего Востока столетие назад в конце лета и осенью дикие свиньи шли на по-
ля крестьян, как «войска на приступ», не пугаясь криков, шума и выстрелов
[Бромлей, Кучеренко, 1983].
По моим наблюдениям, в Переславском охотничьем хозяйстве в Ярославской
области летом и осенью 1983 г. кабаны выходили на посевы каждый вечер. В
июне животных больше всего привлекали клевер и овес, в июле - рожь и пшени-
ца, достигшие молочной спелости, в августе и сентябре - овес. В конце июля -
начале августа на одном из пшеничных полей, примыкающих к лесу, регулярно
кормились около 140-160 особей. В нескольких стадах я насчитывал от 28 до 67
голов, причем их число не изменялось на протяжении 4-9 дней. Утром звери
уходили в ближайший ельник, где ложились весьма компактно, но каждая се-
мейная группа - отдельно. Вечером группы снова объединялись в крупные стада
на опушке леса перед выходом на поле. Десятки гектаров зерновых были факти-
чески уничтожены ими на корню, причем свиньи не столько съедали урожай,
сколько вытаптывали его. На некоторых ближайших к лесу участках осенью
убирать было нечего. Картофельные плантации, напротив, уцелели. Кабаны раз-
рывали, обычно после дождей, десятки кустов, но при этом много клубней оста-
вались несъеденными. Скорее всего, они искали здесь дождевых червей. После
уборки урожая крупные группировки в этом районе уже не встречались.
118
В Белоруссии на полях овса, примыкающих к лесу, уборка также нередко не
требовалась. В 1987 г. в Витебской области из 83 колхозов и совхозов от этих ко-
пытных пострадали 32 хозяйства. В Городокском районе повреждено около 30%
посевов зернобобовых, 3 тыс. га овса, 1,5 тыс. га кукурузы, 500 га картофеля.
Однако вред посевам, возможно, преувеличен - так нерадивые руководители
обычно списывают издержки собственной бесхозяйственности [Муравицкий,
1988].
В Литве при плотности населения диких свиней от 4 до 82 особей на 1000 га
угодий потери урожая пшеницы составляют от 3,4 до 62%, овса - 4—55,3%, кар-
тофеля - 0,6-39,1, ржи - 0,2-6,6, ячменя - 0,05-2,2 и свеклы - до 0,5% [Янулай-
тис, Падайга, 1984а,б, 1985]. В Польше в 70-е годы ущерб урожаю выражался в
следующих цифрах: овес - 0,18%, рожь - 0,06%, пшеница - 0,05%, картофель -
0,11% [Максимов, Романов, 1976].
В некоторых странах Западной и Центральной Европы на кабана списывают
до 5-7% потерь урожая отдельных культур, а ущерб сельскому хозяйству оцени-
вают в миллионы марок, франков или долларов [Briedermann, 1977a,b; Норр,
1979; Gunther, 1980]. В Корее население вынуждено постоянно охранять планта-
ции от непрекращающихся набегов. В Австралии и Пакистане борьба с кабаном,
а также с одичавшими домашними свиньями, ведется всеми способами, включая
отравленные приманки.
Любопытные расчеты ущерба от диких свиней в российском Нечерноземье
сделаны Г. Манушем [1989]. По его мнению, каждому зверю в год требуется
около 1 т корма. В совокупности эти копытные ежегодно «приворовывают» у
сельского хозяйства не менее 100 тыс. тонн кормов. Если их скормить домашним
животным, то можно получить 20 тыс. тонн свинины, тогда как охотники, добыв
16 тыс. кабанов, получают лишь 1 тыс. тонн дикого мяса, что не покрывает вре-
да. Следовательно, эти животные - «бедствие, от которого надо избавляться».
В этих расчетах, к сожалению, допущены существенные натяжки не в пользу
диких свиней. Большую часть корма они находят все же не на сельскохозяйст-
венных полях, а в лесу. Кроме того, отражена только официальная добыча, тогда
как не меньше зверей отстреливают браконьеры. При более разумном ведении
охотничьего хозяйства можно было бы ежегодно добывать около 70-100 тыс. го-
лов, а также существенно снизить вред посевам. Следует учесть и себестоимость
продукции: для диких животных не надо строить свинарники и капитальные хра-
нилища кормов, не нужен ежедневный уход, все трудозатраты многократно ни-
же, а качество дикого мяса выше.
В ГДР в свое время при такой же численности (около 100 тыс. особей) еже-
годно отстреливали 100-120 тыс. голов. Размер причиненного кабаном ущерба
здесь не превышал 10-15% дохода от добычи [Briedermann, 1979, 1986]. Этот
весьма успешный опыт управления многочисленными популяциями позволяет
более оптимистично оценивать шансы вида на существование в агрикультурном
ландшафте.
Замечу также, что совокупный вред сельскохозяйственным посевам от врано-
вых, голубей и мышевидных грызунов обычно гораздо выше, чем от диких ко-
пытных животных.
Воздействие на зооценозы
Дикие свиньи не проходят мимо земноводных и пресмыкающихся, уничтожают
рыбу на мелководьях [Харченко и др., 1980] и перекапывают нерестилища [Варна-
ков, 1986], принося вред рыбному хозяйству. Они разрушают норы выхухоли [Рябов,
Семаго, 1974; Хахин, 1974; Казневский, 1977] и причиняют существенный прямой
(поедание зверьков) или косвенный (разрушение и поедание хаток) ущерб ондатро-
119
Рис. 45. Динамика
численности глуха-
ря (1), тетерева (2) и
кабана (3) в Зави-
довском заповедно-
охотничьем хозяй-
стве в 1954 1978 гг.
(по [Кручинин,
1979])
вому хозяйству [Анненков, 1989]. В Прибалхашье, Южном Приаралье и в дельте р.
Или эти копытные уничтожают около 30-40%, а в отдельных местах - до 90% хаток
ондатры, из-за чего продуктивность угодий снижается на 30-40% [Слудский, 1956;
Злобин, 1969; Ширяев, 1975; Нуратдинов, 1990].
В местах интенсивных жировок кабана редки выводки зайца [Фадеев, 19786;
Кручинин, 1979]. При высокой численности он становится существенным пище-
вым конкурентом барсука и даже уничтожает его физически, что в совокупности
приводит к снижению поголовья последнего [Горшков, 1988].
Большой вред дикие свиньи наносят птицам, особенно гнездящимся на земле.
В плавнях Приазовья они поедают кладки чаек, крачек, чибисов, уток, шилоклю-
вок, тиркушек и других видов [Харченко и др., 1980], а в Кызыл-Агачском запо-
веднике разоряют колонии цапель, караваек и бакланов [Лобачев, Петухов, 1984].
На Арале, по наблюдениям, 5 зверей уничтожили 37 утиных кладок, на трех из
них были съедены взрослые птицы. Урон водоплавающим птицам от этих ко-
пытных достигает здесь 20% всех потерь от хищников [Соломатин, 1972]. По
сведениям А. Ханжина [1977], на отдельных участках Переславского государст-
венного охотничьего хозяйства дикие свиньи истребляют до 50% кладок глухаря,
82% - тетерева и 33% - рябчика. Доля тетерок без выводков достигает 70-76%
при обычном соотношении 24-32%, доля глухарок без птенцов - 57-60% при
нормальном соотношении 25-49%.
В Завидовском заповедно-охотничьем хозяйстве на островах Шошинского
плеса эти звери разоряют утиные кладки полностью. Из 30 кладок тетерева, 6 -
глухаря и 9 - уток, находившихся под контролем, они истребили соответственно
И, 5 и 8 [Кручинин, 1979]. Динамика численности глухаря, тетерева и кабана
здесь находится в относительной противофазе (рис. 45), на что обычно указыва-
ют противники разведения этого вида в охотничьих хозяйствах.
Замечу, однако, что прогрессирующее снижение населения боровой дичи в
Завидовском охотничьем хозяйстве началось задолго до заметного роста поголо-
вья диких свиней (рис. 45), и вряд ли их влияние можно считать основным нега-
тивным фактором в этом процессе. В Лапландском заповеднике, где нет кабана,
за 52 года наблюдений численность глухаря тоже сократилась в 3 раза, рябчика -
в 4 раза [Семенов-Тян-Шанский, 1989].
Следует учитывать также и косвенный вред, наносимый боровой дичи. При
высокой плотности звери неизбежно распугивают и разбивают выводки, а по-
вреждая черничник и брусничник, могут снижать урожай ягодных кормов [Ива-
нова, 1977; Аношин, 1988].
120
Справедливости ради следует заметить, что ущерб от кабана выводковым
птицам может быть сильно преувеличен, поскольку разорять кладки могут и дру-
гие звери и птицы, что не всегда учитывается. В специальных опытах, проведен-
ных американскими учеными, установлено, что в угодьях, плотно и редко засе-
ленных дикими свиньями, сохраняется примерно одинаковое число кладок (21,4
и 27,3%). Эти копытные разоряют всего 2,4% гнезд, остальные - лисицы, еноты,
скунсы, а также ужи и другие змеи (15%). В аналогичных опытах в Беловежской
пуще найдено, что кабаны уничтожают всего 4,7% гнезд, тогда как енотовидная
собака - 28,1%, куница - 10,9%, ласка и горностай - 4,7%. В разорении гнезд
принимают участие также лисица, барсук, ворон, сойка и, возможно, другие жи-
вотные [Боровик, 1973].
В последнее десятилетие поголовье кабана во многих районах России сведено
к минимуму, но заметного увеличения численности боровой и водоплавающей
дичи не происходит. Низкая плотность населения глухаря, тетерева, рябчика и
уток разных видов в охотничьих угодьях может быть вызвана совершенно иными
причинами. Как известно, гораздо больший прямой и косвенный урон птицам
наносят гельминтозы, химизация в сельском и лесном хозяйствах, масштабное
уничтожение лучших биотопов, чрезмерная охота, хищники, зимы с плотными
настами и затяжные холодные весны с обильными дождями, периодические не-
урожаи ягод и беспокойство, причиняемое человеком и домашними животными
[Борщевский, 1989; Захарова, 1989; Лоскутова, 1989; Лыхварь, 1989; Семенов-
Тян-Шанский, 1989]. В Московской области, например, с увеличением плотности
людского населения с 5—20 до 100 человек/км2 численность глухаря сократилась
в 6 раз, тетерева - в 4, рябчика - в 2 раза [Кузьмин, 1987].
Итак, дикую свинью нельзя всецело расценивать как вредителя, особенно в
лесном хозяйстве, где вред и польза от этого вида примерно равны [Янулайтис,
Падайга, 1984а]. Ее роющая деятельность, как показано выше, оказывает благо-
творное влияние на физические и химические свойства почвы, освещенность,
температуру, воздушный режим и водопроницаемость, а взрыхление земли при
последующем затаптывании и зарывании множества семян способствует лесово-
зобновлению. Травянистая растительность и почвенная фауна в местах повреж-
дений животными почвенного покрова довольно быстро (за 2-4 года) восстанав-
ливаются, а биомасса увеличивается. Примерно через 3 года полностью восста-
навливаются и сенокосы.
Кабаны явно приносят пользу, пожирая множество личинок, куколок и имаго
почвенных беспозвоночных — потенциальных вредителей леса: майского хруща,
златки, дубовой плодожорки, сосновой совки, соснового шелкопряда, проволоч-
ника, пилильщика, щелкуна и др. [Лебедева, 1956а; Корнеев, Кричевская, 1967;
Чичикин, Воробьев, 1967; Сухорослов, 19726; Тимофеева, 1975; Горшков, 1989].
В Польше, например, в апреле - мае дикие свиньи уничтожают до 41% коконов
соснового пилильщика [Genov, 1981b], Несомненна польза и от истребления
большого количества саранчи [Слудский, 1956; Лавровский, 1962; Слудский и
др., 1984]. В лесу и на полях этот вид поедает немало мышевидных грызунов, со-
храняя тем самым семена и урожай.
Зимой дикие свиньи облегчают передвижение многим млекопитающим. В не-
которых многоснежных районах европейская и сибирская косули выживают лишь
благодаря развитой сети кабаньих троп. Ими охотно пользуются также благород-
ный и пятнистый олени, иногда - лоси [Бальчяускас, Таугинас, 1983], хищники
(волк, лисица, енотовидная собака) и зайцы. В дельте Волги зимние покопки диких
свиней спасают от бескормицы и гибели фазана и куропаток [Лавровский, 1962].
Кабан, к тому же, представляет важное звено в трофической цепи - он один из ос-
новных источников пищи для крупных хищников: тигра, волка, медведя, леопарда.
Трупы погибших особей утилизируются животными-некрофилами.
121
При разумном подходе негативное воздействие диких свиней на среду обита-
ния и ущерб от них сельскому, лесному и охотничьему хозяйствам можно суще-
ственно снизить (см. «Управление популяциями»). И, конечно же, в пылу поле-
мики не следует забывать, что этот вид - не «чуждый» для фауны России. После
более чем векового отсутствия он занял свою экологическую нишу, снова став
неотъемлемым элементом биоценозов и важнейшим охотничьим видом.
Размножение
Половая зрелость
Кабан - полигамное, раннеспелое и очень плодовитое животное, что во мно-
гом способствует процветанию вида и его выживанию даже в неблагоприятных
условиях.
Часть самок становится половозрелыми уже на первом году жизни [Oloff,
1951; Иваненко, 1956; Бромлей, 1964; Бридерманн, 1970; Козло, 1975; Щербаков,
Стародынова, 1979; Червонный, 19806; Meynhardt, 1982, 1984а,с; Volokh, 1998:
Волох, 1999]. Вероятность успешного покрытия сеголеток зависит от кормовых
условий и их массы к периоду гона [Gaillard et al., 1993] - позднорожденные,
слабые и плохо упитанные особи не принимают участия в размножении. У боль-
шинства самок первая течка проявляется в возрасте полутора лет [Лавровский,
1962; Мануш, 1969; Щербаков, Стародынова, 1979]. В Предбайкалье все иссле-
дованные свиньи становились беременными лишь на 19-20-м месяце жизни [Не-
дзельский, 1994].
Самцы вступают в размножение обычно позднее самок. У сеголеток семенники
еще не полностью развиты, и даже на втором году жизни они составляют примерно
треть массы и объема семенников взрослых особей. Однако в популяциях с высокой
плотностью населения в размножении, вероятно, принимают участие самцы всех
возрастов, включая сеголеток старше 9 месяцев, которые кроют чаще своих сестер
[Майнхард, 1983; Царев, 19866, 1989в, 2000; Spitz, 1987].
Спаривание и половое поведение
Гон у кабана на территории России и сопредельных регионов весьма растя-
нут по срокам и длится преимущественно с октября по февраль (табл. 15). Наи-
более интенсивен он в ноябре - декабре, когда покрываются более 80% самок.
Однако, судя по срокам опоросов (табл. 17), самцы способны оплодотворять сви-
ней практически круглогодично.
Сроки гона определяются степенью упитанности животных, зависят от погод-
ных условий и возраста самок. Первыми покрываются взрослые свиньи и спустя
2-3 недели - подсвинки. В неурожайные годы течка задерживается на 1-2 меся-
ца, в урожайные - начинается раньше, но у сильно разжиревших свиней может
запаздывать, или они остаются яловыми [Oloff, 1951; Лебедева, 1956а; Слудский,
1956; Романов, Козло, 1977; Meynhardt, 1983b; Perin et al., 1987]. В Беловежской
пуще при обильном урожае желудей первые спаривания отмечаются в середине
сентября, в неурожайные годы - в середине декабря [Северцов, Саблина, 1953,
1955]. При раннем похолодании гон начинается на 5-10 дней раньше [Логинов,
1936; Недзельский, 1994].
Ежегодно к периоду спаривания отдельные органы взрослых самцов претер-
певают существенные изменения. У секачей в сентябре - октябре в 2-3 раза уве-
личиваются масса (от 150-300 г до 430-600 г, с придатками - до 875 г) и услов-
ный объем семенников. Сперма в их придатках обнаруживается с сентября - ок-
122
Таблица 15. Сроки гона кабана
Район Начало | Конец Источник информации
Германия Октябрь Февраль (апрель) Meynhardt, 1983b; Майн- хардт, 1986
Беловежская пуща 3 ноября Начало января Козло, 1975
а 19 ноября Январь Лебедева, 1956а
а Середина октября (15 сентября) Начало января Северцов, Саблина, 1953; Саблина, 1955
Карпаты Ноябрь Декабрь Гунчак, 1982
Юг Украины Декабрь Середина января Щербак, 1986
25 октября 1 февраля Volokh, 1998
Латвия Конец ноября Январь Калниньш, 1950
Литва Ноябрь Декабрь Даугела, Падайга, 1980
С.-запад России Середина октября Середина января Русаков, Тимофеева, 1984
Брянская обл. Декабрь С< Петрищев, Бородин, 1985
Подмосковье Середина ноября tc Иванова, 1975а
“Завидово” Декабрь Февраль Мануш, 1969
Ц. Черноземье 3 ноября 2 января Простаков, 1996
Белгородская обл. Ноябрь Апрель Царев, 2000 |
Воронежский з-к а Декабрь Дьяков и др., 1986 1
Кавказский з-к 5 ноября Январь Донауров, Теплое, 1938
Северный Кавказ 15 ноября 25 января Логинов, 1936
Закавказье Декабрь Январь Марков, 1932 ;
Грузия Середина ноября 15 января Хеладзе, 1964 |
Дельта Волги Конец ноября 4-20 января Лавровский, 1962
Тургайское плато 1 ноября Начало января Соломатин, 1972
Низовья р. Или 5 ноября Середина января Слудский, 1956
Заилийское Алатау Конец октября Конец декабря
Тянь-Шань Ноябрь Декабрь Обидина, 1982
Алакуль а (С Анненков, 1989
Туркменистан а а Ишадов, 1965
3. Копетдаг а а Кулибаба, Пакулин, 1985
Юг Киргизии Конец ноября Январь Чичикин, Воробьев, 1967
Таджикистан 24 ноября 2 января Иваненко, 1956 I
Алтай Ноябрь Декабрь Собанский, 1992
Монголия Конец ноября Середина января Банников, 1954
Тува Ноябрь Январь Горелов, 1994; Смирнов, 2001
Предбайкалье Декабрь Начало января Копылов, 1950
Конец ноября а Недзельский, 1994
Забайкалье Ноябрь Декабрь Кириллов, 1902; Смирнов, 1978 ।
Хинганский з-к 30 ноября 11 декабря Дарман, 1990
Дальний Восток 14 ноября 18 декабря Бромлей, 1964
а Конец ноября Начало января Абрамов, 1963
тября до марта, т. е. самцы потенциально способны оплодотворять самок почти
полгода. В конце гона сперматогенез затухает, масса и объем семенников умень-
шаются, и с апреля до августа они находятся в стадии покоя [Слудский, 1956;
Лавровский, 1962; Козло, 1975; Русаков, Тимофеева, 1984].
123
К периоду гона у самцов
появляются и специфические
морфологические адаптации:
кожный покров в области ло-
паток утолщается до 2-3 см, а
иногда - и до 4 см [Динник,
1910], становясь своеобраз-
ным «щитом», в значитель-
ной мере предохраняющим
жизненно важные органы от
повреждения клыками со-
перников (рис. 46). Кожа в
этом месте настолько прочна,
что с большим трудом режет-
ся ножом, а пробить его лез-
вием почти невозможно. На
«калканах», тем не менее,
обнаруживаются глубокие
порезы и даже проколы от
ударов клыками, свидетель-
ствующие о мощи противни-
ков. Бои нередко заканчива-
ются ранениями, увечьями
или гибелью одного из сека-
чей [Донауров, Теплов, 1938;
Иваненко, 1956; Лавровский,
1962; Козло, 1975; Проста-
ков, 1996].
Рис. 46. Схема расположения «кожного щита» у
взрослого самца кабана
Образование кожного «щита» к периоду спаривания - явление не редкое среди
копытных. У косуль он, как правило, выполняет и иную функцию: сальные и пото-
вые железы в этом месте увеличиваются в несколько раз, продуцируя пахучий
секрет, необходимый для маркировки территории [Соколов, Данилкин, 1981].
В гон секачи чаще, чем обычно, трутся о стволы преимущественно хвойных
деревьев и ударами клыков делают на них своеобразные засечки. В этот период
особенно пахучими становятся выделения их половых желез и моча, желто-
оранжевые пятна которой нередко видны на следах. Даже человек с его слабым
обонянием способен почувствовать специфический мускусный запах феромона
за несколько сот метров. Пахучая жидкость выдавливается из особого кожистого
мешочка, расположенного возле выходного отверстия пениса, который увеличи-
вается осенью в объеме почти в два раза. Ольфакторно-оптические метки, остав-
ляемые на следах и на деревьях (рис. 47), служат для маркировки территории и
сигнализируют самкам и противникам о местонахождении самца. Резкий и не-
приятный для человека запах имеет и мясо секачей, содержащее следы полового
феромона [Karg, 1988]. Этот феромон явно стимулирует самок. В животноводст-
ве, как известно, его распыление способствует более полному искусственному
осеменению свиноматок.
Свиньи в период гона тоже проявляют маркировочное поведение. За две не-
дели до начала течки самка-вожак группы начинает грызть стволы деревьев и
тереться о них глазной впадиной, оставляя пахучие выделения слюнных и слез-
ных желез, затем это делают другие свиньи (рис. 48). Самцы, таким образом, по-
лучают информацию о готовности свиней к спариванию [Meynhardt, 1983b,
1984b], Отмечу, однако, что у кабанов предглазничные железы не развиты [Со-
124
Рис. 47, Ольфакторная маркировка (фото А. Щеголева)
колов, Чернова, 2001], поэтому не исключено, что мечение производится секре-
том всего комплекса желез головы.
Перед гоном взрослые самцы широко бродят в поисках самок, проходя за
день до 20 км [Слудский, 1956; Бромлей, 1964]. Присоединившись к семье или
семейной группе, секач ревностно охраняет свой «гарем» от соперников. Он бес-
престанно находится в движении, преследует и кроет самок, пришедших в охоту,
сражается с противниками. При этом самец издает хрюкающие звуки, щелкает
клыками, изо рта на землю и растительность истекает обильная пена. Своими
действиями он постоянно тревожил группу и не дает ей спокойно кормиться. На
месте преследования и покрытия самок остаются утоптанные круговые площад-
ки, испещренные следами поросят, бегающих за матерью. Изредка возбужден-
ный секач наносит клыками порезы и даже глубокие раны самкам. Одна из них,
как мне пришлось наблюдать в Тульской области, после такого «ухаживания»
истекала кровью двое суток. К сожалению, ее дальнейшая судьба осталась невы-
ясненной. Поскольку в период спаривания самцы почти не едят, они быстро ху-
деют, теряя до 20-25% массы.
Взрослые особи относительно оседлы (см. ниже), поэтому секачи в течение не-
скольких лет покрывают одних и тех же самок, в том числе и своих дочерей. Однако
у диких свиней известен промискуитетный тип половых отношений: полигамия, по-
лиандрия, факулыативная моногамия и инбридинг. В группах в период гона чаще
всего находится один взрослый самец, в составе крупных гаремов - несколько. Об-
разование многосамцовых репродуктивных группировок отмечается даже в тех по-
пуляциях, где соотношение полов близко к 1:3 в пользу самок. В Белгородской облас-
ти, например, в 59 гаремах был один секач, в 13 - два, в 2 - три, в 4 - четыре и в 1 -
125
Рис. 48. Свиньи в пе-
риод гона тоже про-
являют маркировоч-
ное поведение (фото
Н Немнонова)
шесть взрослых самцов одновременно. Их число в стадах постоянно изменяется в
зависимости от количества и половой активности самок и от агрессивности секача-
доминанта. Самцы-субдоминанты обычно держатся на периферии гарема, но все же
имеют возможность отбивать и покрывать свиней, иногда поочередно и до 4-5 раз
во время течки. Секачи покидают гарем, когда самки не проявляют признаков течки,
и активно ищут другие группы [Царев, 1989в].
Беременность и эмбриональное развитие
Кабан - моноцикличный вид. Тем не менее в Западной Европе и Северной
Америке зарегистрированы редкие случаи полиэстральности, приводящие к по-
явлению двух выводков в году, что сходно с циклом размножения домашней
свиньи. Обычно это бывает, если первый помет рано и полностью погибает
[Cloth, 1967; Gunther, 1980, Майнхардт, 1986; Gaisford, 1991]. Вероятно, отчасти
именно этим обстоятельством может объясняться появление поросят в неурочное
время осенью или зимой и в разных регионах России.
У диких свиней, как и у домашних, течка длится, видимо, 2-3 дня, следую-
щая проявляется через 21-23 дня. Молодые самки, спаривающиеся позднее
взрослых, соответственно и позднее рожают. Все члены семейной группы обыч-
но покрываются почти одновременно с разницей в 1-8 дней.
126
Рис. 49. Изменение внешних морфологических признаков эмбрионов кабана (по
[Курносов, Петрищев, 1977])
Зародыши: 1 - 19-суточные, 2 - 19-20 и 3 — 20-21-суточные. Эмбрионы: 4 - 34-35-
суточные, 5 - 39-40, 6-41-42, 7-42-43, 8-48-50 и 9-73-75-суточные
Период беременности у самок кабана по сравнению с другими дикими ко-
пытными сравнительно короткий - 114-140 дней, в среднем около 120 дней
[Бромлей, 1964; Сухорослов, 19726; Meynhardt, 1983b; Слудский и др., 1984;
Дарман, 1990]. У рано покрытых (в октябре - ноябре) свиней срок беременности,
вероятно, чуть продолжительнее, чем у особей, спаривающихся в декабре - янва-
ре. При неблагоприятной зимовке и плохом питании развитие эмбрионов у них,
как и у некоторых видов оленьих [Данилкин, 1999а]. скорее всего, может задер-
живаться, что сказывается на сроках родов.
Средняя масса эмбрионов в январе - феврале варьирует в пределах 0,2-330 г,
в марте - апреле - от 300 до 1000 г, но при поздней беременности в апреле может
составлять всего лишь 0,2-0,3 г, а в мае - около 51—53 г [Банников, 1954; Ива-
ненко, 1956, Слудский, 1956; Лавровский, 1962; Бромлей, 1964; Чичикин, Во
127
робьев, 1967; Щербаков, Стародынова, 1979; Давлетова, 1992; Недзельскийч
1994; Простаков, 1996]. |
Чем меньше эмбрионов у самки, тем меньше разница в их массе и более рав-
номерен рост, что отличает диких свиней от домашних [Давлетова, 1985]. У мно-
гоплодных самок масса эмбрионов различается в 1-4 раза, причем неодинаковое
развитие плодов обнаруживается уже на ранней стадии беременности. Из-за это-
го часть новорожденных заметно отстает в росте, и слабейшие детеныши быстро
погибают [Щербаков, Стародынова, 1979].
Возрастная изменчивость массы и линейных параметров тела эмбрионов дос-
таточно детально прослежены при исследовании диких свиней в охотхозяйстве
«Нерусса» Брянской области. Показатели, приведенные в таблице 16, и признаки,
показанные на рисунке 49, могут быть полезны исследователям и охотоведам
при определении стадий развития эмбрионов.
Таблица 16. Возрастная изменчивость массы и длины тела эмбрионов кабана
(по [Петрищев, Бородин, 1985])
Возраст, дни п Масса, г Длина тела, см
М lim М lim
18 6 — — 0,5 —
19 5 0,16 — 0,7 —
19-20 5 0,16 0,15-0,17 0,5 —
20-21 6 0,21 0,17-0,25 0,8 —
25-26 4 0,75 0,6-0,9 0,7 0,6-0,9
34-35 4 3,6 3,4-3,8 3,9 3,9-4,0
40 5 10,0 9,6-10,5 4,8 4,5-5,1
40-41 5 12,8 10,3-15,3 5,6 5,3
41-42 5 13,0 11,7-15,6 5,9 5,8-6,0
42-43 6 14,3 13,8-15,4 7,3 6,0-8,4
43 4 17,4 16,4-18,5 6,7 6,5-7,0
45 И 26,0 25,0-28,5 8,3 7,5-9,0
50 6 38,0 37,7-38,2 8,8 8,5-9,0
60 9 87,3 83,0-91,6 11,5 11,0-12,0
62 3 107,5 104-111 12,0 12,0-12,0
70 8 221,0 218-224 15,2 15,0-15,5
72-74 7 308,0 283-329 17,2 15,5-18,5
80 5 400,0 390-410 18,0 18,0-18,0
90 5 770,6 697-830 23,3 22,0-24,0
100-105 6 852,3 816-900 23,6 23,0-24,0
Новорожденные 5 986,2 835-1090 36,2 33,0-39,5
В зародышевый период в 18-суточном возрасте происходит разделение го-
ловного мозга на передний, средний и задний, закладываются сердце, печень и
мезонефрос, тело имеет 26-28 сомитов, эмбрион находится в амнионе, желточ-
ный мешок большой; в 35 суток образуются закладки больших полушарий сред-
него и заднего мозга, наружное ухо и веки; на 40-е сутки плод полностью рас-
прямляется, на ногах уже хорошо различимы пальцы, можно определить пол
(рис. 49). 60-суточный эмбрион имеет зачатки верхних клыков; на 70-е сутки на
копытцах формируется роговой чехол; к 80-му дню появляются ресницы, волосы
на надбровных дугах, носу, подбородке и в области плечевого пояса; у 90-
суточных клыки достигают 1 см, пробиваются волосы на остальных участках те-
ла; к 100-105 суткам на боках формируются желтые и бурые, а по хребту - тем-
но-коричневая продольные полосы шириной 0,8—1,5 см, характерные и для но-
ворожденных. Удельная скорость линейного роста эмбрионов максимальна в за-
родышевом периоде, резко снижается к началу предплодного периода, составляя
1 см на 1 г массы, а в плодном периоде уменьшается до долей миллиметра.
Удельное изменение массы, напротив, максимально на протяжении плодного пе-
риода, при этом оно замедляется за 1,5-2 недели до родов и затем резко возрас-
тает в последние дни беременности самок, т. е. масса плодов быстро достигает
необходимой физиологической нормы практически без увеличения линейных
размеров тела. Такой процесс развития эмбрионов позволяет самкам быть под-
вижными на протяжении всего срока беременности, что весьма важно для их
выживания при постоянном прессе хищников и беспокойстве людьми [Курносов,
Петрищев, 1977; Петрищев, 1980; Петрищев, Бородин, 1985].
Эмбрионы кабана отличаются от таковых у домашних свиней полосатым во-
лосяным покровом, более ранним (на 10 суток) его формированием, длинноры-
лостью, узкотелостью и взгорбленностью, неодинаковой интенсивностью роста
скелета и плотностью костей, меньшей массой тела и большей длиной кишечни-
ка в позднеплодный период [Курносов, Петрищев, 1977, 1978; Давлетова, 1985,
1986; Петрищев, 1985; Петрищев, Бородин, 1985; Саркисова, Хрусталева, 1985].
Опорос и выкармливание детенышей |
Роды у самок кабана в северном полушарии, как правило, регистрируются в
период с марта по июнь (табл. 17), изредка чуть позже этих сроков и в исключи-
тельных случаях - в декабре - феврале [Oloff, 1951; Саблина, 1969; Сухорослов,
19706; Шостак, 1971; Белов, 1972; Козло, 1972в; Евтушевский, 1975; Неганов и
ДР, 1989].
Поздняя беременность и поздние опоросы более характерны для однолетних
или отдельных больных и раненых самок. Доля поздних родов в разные годы и в
разных районах неодинакова и колеблется от 0,2 до 18%. Увеличение их числа
заметно в группировках с повышенной плотностью населения [Козло, 1972, 1975;
Щербаков, Стародынова, 1979], но они есть и в малочисленных популяциях. В
Архангельской области, например, зарегистрирован случай опороса в ноябре
[Плешак, Миняев, 1986].
В Центральном Черноземье кабан размножается почти круглогодично. Роды
отмечаются во все месяцы года, за исключением ноября. Этот феномен связан,
возможно, с нехваткой самцов-производителей в разреженных популяциях в ост-
ровных лесах среди степи. Поздние детеныши, как правило, погибают, особенно
у молодых самок, которые не всегда устраивают утепленные родильные «гнезда»
[Сухорослов, 1970а, 19726].
Пик опоросов диких свиней в разных частях ареала приурочен обычно к нача-
лу вегетации растительности. В Беловежской пуще он приходится на конец марта
и первую половину апреля, но отдельные особи приносят детенышей в феврале и
далее вплоть до августа. Растянутость сроков деторождения возрастает в годы, не-
благоприятные по кормовым и погодным условиям [Саблина, 1955; Козло, 1975].
На северо-западе России массовые роды (50% встреч выводков) регистрируются в
мае, 29% самок приносит поросят в апреле, 16% - в июне и 5% - в марте [Русаков,
19806; Русаков, Тимофеева, 1984]. В Казахстане пик опоросов отмечен в апреле
[Слудский и др., 1984].
На значительной части видового ареала в северном полушарии в период оп-
росов (в марте - мае) еще лежит снег, а ночные температуры минусовые. В этих
условиях поросята могут выжить лишь в специальных укрытиях - родильных
«гнездах», устраиваемых свиньями перед опоросом (см. раздел «Инфраструктура
участков обитания»).
В первые 2-8 дней жизни, в зависимости от погоды, детеныши практически
9. Заказ№2528. 129
Таблица 17. Сроки опороса кабана
Район | Начало Конец | Источник информации
Испания Февраль Август Saez-Royuela, Telleria, 1987
Германия Март Сентябрь Майнхардт, 1986
Польша 14 марта 19 апреля Журовский и др., 1970
Карпаты Март Апрель Гунчак, 1982
Юг Украины Апрель Сентябрь Щербак, 1986
44 Февраль Июнь Volokh, 1998
З-к “Кодры” Март Апрель Чегорка, 1984
Литва 20 марта 4 4 Даугела, Падайга, 1980
Белоруссия 15 января Август Козло, 1975
Беловежская пуща Март Июнь Лебедева, 1956а; Шостак, 1971
44 Февраль Август Саблина, 1953; Козло, 1975
(январь)
С.-запад России Март Июль Русаков, Тимофеева, 1984
Подмосковье Апрель Июнь Иванова, 1975а
“Завидово” 24 марта и Дормидонтов, 1969; Мануш, 1969;
(август) Щербаков, Стародынова, 1979
Ц. Черноземье Круглогодично Сухорослов, 1970а, 19726
(С 7 января Август Простаков, 1996
Воронежский з-к Март 44 Дьяков и др., 1986
Кавказский з-к Конец марта Середи- Донауров, Теплое, 1938
на мая
Грузия Март Апрель Хеладзе, 1964
Дельта Волги 22 марта 10 мая Лавровский, 1952,1962
Тур гайское плато Март — Соломатин, 1972
Низовья р. Или 18 февраля Май Слудский, 1956
Заилийское Алатау 20 февраля 44 44
Тянь-Шань Март 4 4 Обидина, 1982
Алакуль 44 Слудский, 1956; Анненков, 1989
Туркменистан 44 44 Ишадов, 1965
3. Копетдаг Конец февраля Март Кулибаба, Пакулин, 1985
Юг Киргизии Конец марта Апрель Чичикин, Воробьев, 1967
Таджикистан Март 44 Иваненко, 1956
Алтай Апрель — Собанский, 1992
Монголия 44 Май Банников, 1954
Тува а а Смирнов, 2001
Предбайкалье и 44 Копылов, 1950; Недзельский, 1994
Забайкалье и 44 Кириллов, 1902; Смирнов, 1978
Хинганский з-к 24 марта 25 мая Дарман, 1990
Дальний Восток 9 марта Апрель Бромлей, 1964
не покидают логово. При низкой температуре около суток может не выходить из
него и самка, обогревающая своим теплом поросят. Затем, нуждаясь в воде и
пище, она начинает кормиться вблизи логова, возвращаясь в него каждые 2-3 ча-
са. До месячного возраста поросята питаются в основном молоком. В первые дни
после рождения их кормление происходит по инициативе матери примерно через
каждые 40 минут, позднее - когда проголодавшиеся детеныши начинают повиз-
гивать [Майнхардт, 1986]. После короткого «массажа» вымени каждый сосет мо-
локо из своего соска около 30 секунд. Его жирность у свиней колеблется в пре-
делах 3,8-16,2%, содержание белка- 5-7,6% [Грачев, Галанцев, 1973].
130
Рис. 50. Свинья с выводком (фото В. Пескова)
У самок кабана обычно 5 пар сосков, реже - 4 пары, но передняя пара практиче-
ски не функционирует. При рождении более 8 детенышей не все они могут получать
молоко одновременно. Более сильные оттесняют слабых, и те рано или поздно поги-
бают. Однако отдельным свиньям все же удается выкармливать большие выводки
(рис. 50). Неиспользуемые молочные железы постепенно уменьшаются в размерах и
деградируют. По числу развитых сосков у самок можно определить число поросят в
выводке [Иваненко, 1956; Лавровский, 1962; Бромлей, 1964; Щербаков, Стародыно-
ва, 1979].'
Выживанию детенышей способствует семейно-групповой образ жизни диких
свиней. Опоросы у самок семейной группы происходят почти одновременно.
Они рожают детей в разных местах, но через несколько дней или недель собира-
ются вместе. Активность всех особей стада в значительной мере синхронизиро-
вана. Кормление поросят молоком также происходит одновременно и рядом. Де-
теныши узнают свою мать, скорее всего, по специфическому запаху и при кор-
межке в первую очередь устремляются к ней, но затем в поисках свободного со-
ска могут перебегать от одной свиньи к другой. В двухнедельном возрасте каж-
дый поросенок знает свой сосок и не допускает к нему собратьев [Meynhardt,
1984b, 1985b].
9
131
По наблюдениям С.В. и А.И. Шостак [1988], содержащиеся в вольере бере-
менные самки-сестры устраивали одно родильное логово на двоих, где пороси-
лись рядом брюхом к брюху и головами в разные стороны. Новорожденные ока-
зывались между ними и могли получать молоко от обеих. Когда одна самка ухо-
дила из «гнезда», вторая оставалась с выводками. В крупных семейных группах
на страже чаще всего остается самка-вожак, которая отправляется на кормежку
лишь после возвращения какой-либо из самок [Майнхардт, 1986].
Близкородственные свиньи, следовательно, иногда кормят и чужих детены-
шей, а при гибели одной особи ее выводок теоретически могут выхаживать дру-
гие. Однако число функционирующих молочных сосков у самки соответствует
числу ее поросят, и поэтому она практически не в состоянии выкармливать си-
рот. Кроме того, в разреженных популяциях большая часть свиней рожает и вос-
питывает детенышей отдельно от других, не образуя семейные союзы. Поросята,
потерявшие мать, обычно обречены.
Плодовитость
Дикие свиньи — самые плодовитые животные среди копытных. Среднее число
эмбрионов у беременных самок в исследованных популяциях колеблется от 4,3
до 7,6, но у отдельных особей может достигать 11-15 (табл. 18). В пометах доля
из 4-8 новорожденных также максимальна (табл. 19).
Успешность покрытия и плодовитость свиней увеличиваются с возрастом,
достигая пика в 5-7 лет, и остаются достаточно высокими до старости (табл. 20 и
21). В Германии самки, покрытые в годовалом возрасте, приносят в среднем 4,2
поросенка, тогда как двухгодовалые - 5,6, а более взрослые - 6,5 [Бридерманн,
1970]. При стабильно хороших кормовых условиях у 1—2-летних свиней по 3,2-
6,4 поросенка, у взрослых-7-8 [Meynhardt, 1982, 1984а]. В Окском заповеднике
плодовитость взрослых особей выше, чем у молодых, в среднем на 27% [Червон-
ный, 19806], в Беловежской пуще-в 1,5 раза [Козло, 1970а, 1975].
Самки-сеголетки, несмотря на их способность к зачатию, не вносят большого
вклада в воспроизводство. В Западной Европе даже в самые благоприятные годы
покрываются не более 50% сеголеток, а при слабом урожае кормов они полно-
стью исключаются из размножения [Oloff, 1951; Pedone et al., 1991]. В Германии
в годовалом возрасте приносят потомство от 10 до 45% самок [Бридерманн, 1970;
Meynhardt, 1982, 1984а]. В центральных областях европейской части России при
хороших летне-осенних кормовых условиях в размножении участвует до 18% се-
голеток [Иванова, 1986], в Белгородской области - от 8 до 42% [Царев, 19866,
1989в, 2000]. У молодых самок к тому же высока резорбция эмбрионов, они не-
редко приносят мертворожденных или нежизнестойких поросят [Иваненко,
1956]. Раннеспелые животные, как правило, отстают в росте и развитии и не все-
гда приносят потомство на следующий год.
Годовалые свиньи также нередко остаются прохолоставшими. На северо-
западе России почти 60% двухлетних и 45% трехлетних самок яловые, тогда как
среди особей старше 5 лет их менее трети [Русаков, Тимофеева, 1984]. В Бело-
вежской пуще в размножении участвуют в среднем 32-35% двухлетних, 55-61%
трехлетних, 68-72% 4-5-летних, 75-82% 6-7-летних особей и 67-70% зверей
старшего возраста, в Березинской популяции соответственно 50, 67, 76-78, 84 и
74% [Козло, 1970а, 1975, 2001; Романов, Козло, 1977].
132
Таблица 18. Потенциальная плодовитость диких свиней
Район Исследовано самок Эмбрионов у самок Источник информации
всего, п беремен- ных, % яловых, % в сред- нем lim
Болгария — — — 6,4 — Генов, Коцаков, 1986
Италия — — 4,9 Pedone et al., 1991
Беловежская 85 54,1 45,9 5,7 1-11* Козло, 19706,1975
пуща Березинский з-к — — — 6-6,1 — Лавов, 1981
С.-запад России 306 58,8 41,2 6,7 1-14 Русаков, Тимо- феева, 1984
Вологодская обл. 18 83,0 до 17,0 4,9 — Варнаков, 1985
Брянская обл. 23 100 __ 5,0 2-7 Петрищев, Боро- дин, 1985
“Завидово” 304 100 — 5,1 1-11 Щербаков, Ста- родынова, 1979
Ярославская обл. 116 68,6 31,4 5,7 1-15 Иванова, 1985
Окский з-к 27 100 — 6,0 1-10 Уваров, 1986
Ц. Черноземье 99 62,6 37,4 5,9 — Простаков, 1996
З-к “Лес на 116 46,6 53,4 — Царев, 2000
Ворскле” Воронежский з-к 23 100 — 7,5 (7,2) 5-10 Дьяков и др., 1986
Ростовская обл. 121 100 — 6,1 1-12 Казаков, Дали- ков, 1983
Северный Кавказ 8 100 — 7,6 7-10 Динник, 1910
Кызы л-А гач. з-к 14 100 — 4,4 — Литвинов, 1980
Дельта Волги - 100 __ 6,0 4-10 Лавровский, 1952, 1962
Низовья р. Или 8 100 — 6,0 -7 Слудский, 1956
Тянь-Шань 15 — __ 3,3-4,3 -5 Федосенко, Жи- ряков, 1984
Юг Киргизии 17 47 53 5,8 3—11 *** Чичикин, Во- робьев, 1967
Таджикистан 21 95,2 4,8 5,2(4,3**) 1-9 Иваненко, 1956
Предбайкалье 56 70-75 25-30 >5,4 -8 Недзельский, 1994
Дальний Восток 116 63,8 36,2 5—7*** 2—9*** Бромлей, Куче- ренко, 1983
(С 8 — — 6,4 4-8 Н. Раков [Слуд- ский, 1956]
* Сведения о числе эмбрионов по [Шостак, 1971].
** У двух самок резорбирующиеся зародыши.
*** Включая новорожденных.
Прочерк - отсутствие сведений.
133
Таблица 19. Размер помета у диких свиней
Район (источник информации)* Самок, п Число эмбрионов и детенышей и их доля в помете, % Средний размер помета, п
1 2 3 4 5 6 7 8 9 10 и 12 13-15
С.-запад России (1) 316 0,9 2,5 5,1 9,8 12,0 13,6 17,4 18,0 10,8 6,0 2,8 0,6 0,3 6,6
-“-(1) 296 — 1,7 3,4 13,8 13,8 26,0 19,3 10,8 5,1 3,7 1,4 0,7 0,3 6,2**
“Завидово” (2) 304 0,9 3,6 10,2 18,1 29,9 18,1 13,2 3,9 1,0 0,3 0,6 — — 5,1
Ярославская обл. (7) 99 2,0 з,о 4,0 18,2 17,2 23,2 17,2 8,1 2,0 4,0 — — 2,0 5,7
Брянская обл. (5) 23 — 4,3 4,3 17,4 39,1 30,4 4,3 — — — — — — 5,0
Казахстан (3) 15 — — — 40,0 — 13,3 26,7 6,7 6,7 6,7 — — — 6,1
Низовья р. Или (3) 40 — — — 45,0 17,5 12,5 12,5 7,5 2,5 2,5 — — 5,4**
Хинганский з-к (6) 19 — — — 5,2 21,1 15,8 26,3 21,1 5,2 5,2 — — 6,7**
Сихотэ-Алинь (4) 8 — - — 12,5 12,5 25,0 25,0 25,0 - — - - — 5,4
* 1 - Русаков, Тимофеева, 1984; 2 - Щербаков, Стародынова, 1979; 3 - Слудский, 1956; 4 - Слудский, 1956 (сведения Н. Ракова); 5 - Петрищев, Бородин,
1985; 6-Дарман, 1990; 7-Иванова, 1985.
** Число детенышей.
Таблица 20. Успешность покрытия диких свиней в зависимости от возраста
Самок, п Доля (%) беременных самок в разном возрасте (лет) Источник информации
1-2 3 1 4 I 5 6 7+
306 40,5 55,3 67,9 65,3 72,4 72,2 Русаков, Тимофее- ва, 1984
99 48,1* * 75,6** ** ** 63,2 Простаков, 1996
85 30,7 55,1 68,7* * 78,1 66,6 Козло, 1970а, 1975;
230 32 61 72* * 82 82-67 Романов, Козло, 1977
116 20,5 59,8* * * * * Царев, 2000
* Среднее значение для возрастной группы.
** То же для другой возрастной группы.
Таблица 21. Плодовитость диких свиней в зависимости от возраста
Самок, п Среднее число (п) эмбрионов и детенышей у самок разного возраста (лет) Источник информации
1-2 з 4 5 6 7+
180 5,5 6,0 6,8 7,0 7,7 6,5 Верещагин, Ру- саков, 1984
— 4,2 5,6 6,5* * * * Бридерманн, 1970
212 3,2-6,4 7,1 7,2 7,5 7,4 8,0 Meynhardt, 1982, 1984а
— 3,6-5,5 5,6* * * * * Pedone et al., 1991
470 4,2-4,5 5,5-6,4 6-7 6,5-7,1 6,7-7,4 6,3-7,3 Козло, 1975,2001
13 2,3 4,5 7,3* * * * Иванова, 1975а
— 4,0 4,2 6,0 6,0* * * Мануш, 1969
304 4,0-5,2 5,1* * * * * Щербаков, Ста- родынова, 1979
99 * 4,9* ** 7,1** ** 6,6 Простаков, 1966
27,31 2-5,8 3,4-6,6 3,8-5,5 3-6,2 5,3 6,5-4 Уваров, 1986
10 4,1 — 8,0 5,5 — — Иваненко, 1956
11 5,2 6,5 7,5 7,3 — — Бромлей, 1964
* Среднее значение для возрастной группы.
** То же для другой возрастной группы.
Прочерк - отсутствие сведений
Исследования кабана в Завидове дали иной результат. У немногих беремен-
ных годовалых самок обнаружили в среднем по 4,0 эмбриона, у двухлетних - 5,2,
у животных старше 3 лет - 5,1, т. е. их число у свиней старше 2 лет было при-
мерно одинаковым, как и максимальное количество (11) зародышей. Оказалось
также, что при почти 12-кратном росте поголовья (с 420 до 5000 особей) и увели-
чении плотности населения с 8 до 43 особей на 1000 га угодий плодовитость при
регулярной подкормке не только не уменьшилась, но и возросла с 4,8 до 5,5 эм-
брионов на самку (табл. 22) [Щербаков, Стародынова, 1979].
Уровень плодовитости самок кабана, как и домашних свиней, вероятно, зави-
сит главным образом от условий питания. При недокорме у домашних свинома-
ток могут резорбироваться до 41-46% зародышей. У диких свиней в зависимости
от возраста и упитанности разница между числом желтых тел и нормально раз-
витых эмбрионов может достигать в Средней Азии 1,9-4,4 [Иваненко, 1956], в
135
Восточной Сибири - 30-Л0%. При дополнительной подкормке в зимнее время
число детенышей в помете увеличивается с 5,4 до 7,3 [Недзельский, 1994].
Таблица 22. Плодовитость диких свиней при разных уровнях плотности их населе-
ния в Завидовском заповеднике (по [Щербаков, Стародынова, 1979], с изменениями)
Год Численность, особей Плотность, особей на 1000 га* Исследовано самок, п Эмбрионов на самку в среднем
1965 420 7,9 19 4,8
1966 447 8,3 18 ' 5,2
1967 667 5,8** 18 4,6
1968 902 7,8 16 4,5
1969 1200 10,4 25 4,6
1970 2210 19,2 26 4,5
1971 2750 23,9 32 5,6
1972 3420 29,7 57 5,5
1973 3890 33,8 31 5,4
1974 5000 43,4 62 5,5
* Расчетные данные (см. табл. 39).
** Уменьшение плотности в связи с присоединением новой территории.
При ранних и обильных снегопадах, в неурожайные годы и при плохих усло-
виях питания в летне-осенний период в гоне участвуют только взрослые особи,
он проходит вяло, сильно растянут или же, напротив, быстро заканчивается. В
результате яловость существенно возрастает, а плодовитость резко снижается
[Насимович, 1955; Слудский, 1956; Соломатин, 1972]. При благоприятных кор-
мовых условиях, напротив, яловость уменьшается, в размножении принимают
участие и молодые особи, число поросят в помете увеличивается, а их выживае-
мость повышается. Яловость и средний размер выводков, следовательно, наибо-
лее лабильные параметры в размножении кабана (см. табл. 18).
Приведу некоторые характерные примеры. В дельте Волги при всегда благоприят-
ных кормовых условиях самки без поросят почти не встречаются [Лавровский, 1962],
тогда как на Тургайском плато в голодные годы в отдельных группировках приплода
весной не бывает вовсе [Соломатин, 1972]. В Подмосковье в зависимости от погоды,
условий питания и продолжительности гона яловость изменяется от 7 до 50%, состав-
ляя в среднем 30-35% [Иванова, 1985, 1986]. В заповеднике «Лес на Ворскле» Белго-
родской области этот показатель варьирует у сеголеток в пределах 57-92%, у самок
старше двух лет - от 35 до 50%, составляя в среднем 40,2% [Царев, 1989в]. В Окском
заповеднике при благоприятных кормовых условиях число эмбрионов у молодых са-
мок увеличивается с 4,4 до 5-6,5 [Червонный, 19806]. В Хинганском заповеднике в не-
урожайные и урожайные на желуди годы яловость изменяется в пределах 7-76% (в
среднем 34,2%), средний размер выводков варьирует от 1,3 (после голодной зимовки
1986/87 гг.) до 6,4, а доля сеголеток в популяции - от 12-24 до 70% [Дарман, 1990]. В
Карпатах соответственно яловость колеблется от 47,6% до 68,4%, а средняя величина
выводка — от 3,8 до 5,9 [Гунчак, 1982]. На южной стороне Кавказского хребта из-за
лучшей обеспеченности плодами каштана, бука, грецкого ореха и дуба средний размер
выводка (6,1) на 2,2 поросенка больше, чем на северной [Дуров, 19706, 1980а]. В Бело-
вежской пуще в урожайные годы беременными становятся практически все самки;
число поросят в помете составляет 5,8 в среднем, в неурожайные - 3,1 и лишь у круп-
ных старых самок, а после голодной зимовки в 1952 г. - только 1,5 [Саблина, 1955; Ле-
бедева, 1956а; Сержанин, 1961].
Примерно такая же амплитуда колебаний яловости диких свиней известна для
136
Таблица 23. Количество поросят в выводках диких свиней в весенне-летний
и раннеосенний периоды
Район Семей, п Поросят, п Поросят на самку Источник информации
в среднем lim
Германия 257 1401 5,5 — Meynhardt, 1988
Болгария — — 5,6-6,1 — Генов, Коцаков, 1986
Беловежская пуща 388 2285 5,9 (6,7-4,7) 2-10 Саблина, 1955
а 810 5926 6,2 (7,0-4,8) -10 Лебедева, 1956а
и — >10000 6,3-5,2 — Козло, 1975
Березинский з-к — — 6 — Лавов, 1981
Карпаты — — 5,9-3,8 2-11 Гунчак, 1982 1
Юг Украины 286 1497 7,2-6,3 1-13 Волох, 1999
Крым 99 372 3,7 2-10 Кормилицин, Ду- лицкий, 1972
С.-запад России* 296 1853 6,2 2-14 Русаков, Тимофеева, 1984
* 31 178 5,7 2-12 С ч
Архангельская обл. 13 53 4,1 2-8 Плешак, Миняев, 1986
Дарвинский з-к — — 5,1-5,3 — Тупицина, 1988
Подмосковье — — 5,6-6,0 — Иванова, 1975а
Тверская область (вольера) 7 42 6,0 4-8 Личное сообщение А. Матвеева
Окский з-к 95 432 4,5 Червонный, 19806
Ц. Черноземье — — 7,2 — Сухорослов, 1970а, 19726
Воронежская обл. 8 60-48 7,5-6,0 4-10 Простаков, 1996
Воронежский з-к — 5,4 — Дьяков и др., 1986
Кавказский з-к 75 277 3,7 (4,4—3,0) — Донауров, Теплое, 1938
— — 6,1 — Дуров, 19706
Куба-Хачмасская зона — — 4,1 — Алекперов, Кулиев, 1987
Дельта Волги 63 318 5,0 (6,1-4,5) — Лавровский, 1962
Тургайское плато 11 84 7,6 5-10 Соломатин, 1972
Низовья р. Или 40 215** 5,4 4-10 Слудский, 1956
Тянь-Шань 24 — 4,3-3,2 2-5 Слудский и др., 1984
Алакуль — — 5,9 3-9 Анненков, 1989
3. Копетдаг — — 5,8 — Кулибаба, Пакулин, 1985
Таджикистан 6 31 5,2 3-7 Иваненко, 1956
Тува — — 6,2 2-11 Смирнов, 2001
Предбайкалье 267 — 7,3-5,4 -8 Недзельский, 1994
Забайкалье — — — 4-10 Кириллов, 1902
Хинганский з-к 64 342 5,3 1-10 Дарман, 1989, 1990
Еврейская АО — — 6,2 — Ю. Салмин [Бром- лей, 1964]
Сихотэ-Алинь 16 68 5,2 3-7 Бромлей, 1964
Дальний Восток — — 6 4-12 Абрамов, 1963
* В 1960-1977 гг. и 1977-1982 гг. соответственно.
** Включая эмбрионы.
Прочерк - нет сведений.
137
Рис. 51. Во многих семьях к весне сохраняется лишь один поросенок (фото А. Диги-
левича)
Западной Европы - от 25 до 80%, число поросят в выводках в урожайные годы дос-
тигает 8-9, в бескормные - 3-4 при среднем многолетнем (за 50 лет) уровне 5,5
[Oloff, 1951]. В Кампиносском национальном парке в Польше приплод был мини-
мальным (1,4 детеныша на самку) после многоснежной зимы 1969/70 гг., макси-
мальным (5,7) - после урожайного на желуди 1967 г. [Andrzejewski, Jezierski, 1978].
В Германии в годы обилия желудей и плодов красного бука в помете в среднем 6 де-
тенышей, при недостатке корма - 5 [Бридерманн, 1970]. В отдельных районах США
при неурожаях выводки у кабана отсутствуют [Пелтон, Конли, 1974].
В южных районах ареала уровень плодовитости диких свиней (при лучшей
обеспеченности кормом) иногда может быть ниже, чем на севере. В горах Тянь-
Шаня в выводках насчитывают в среднем 3,3-4,3 детеныша [Федосенко, Жиря-
ков, 1984], в Испании на беременную самку приходится всего 4,3 (1-6) эмбриона
[Saez-Royuela, Telleria, 1987]. В благоприятные годы среднее число эмбрионов
или детенышей на самку, как это ни парадоксально, также может снижаться по
сравнению с обычными за счет размножения большего числа менее продуктив-
ных молодых свиней [Иванова, 1985, 1986].
Число поросят в выводках в весенне-летний период (табл. 23) не намного
меньше регистрируемого числа эмбрионов (см. табл. 18), что свидетельствует о
сравнительно высоком уровне их выживаемости в первые месяцы жизни. Причем
детеныши достаточно неплохо сохраняются даже в больших пометах.
Тем не менее в течение года число поросят в семьях неуклонно снижается
(рис. 51), особенно зимой (табл. 24 и 25). На юге Украины, например, потенци-
альная плодовитость свиней составляет 7,2 детеныша на самку. Чаще всего
138
Таблица 24. Изменение размера выводков у диких свиней в течение года
Район (источник информации) Число вывод- ков Число поросят в них Средний размер выводка в месяцу В среднем на самку
IV V VI VII VIII IX X XI XII I II III
Беловежская 1004 5926 7,0* * 6,7 6,4 5,7 5,7 4,8 5,4 4,2 4,8 4,3 * 5,9
пуща (1) Кавказский з-к (2) 84 295 4,4 3,9 3,5 з,з з,з з,о 2,0 3,5
Дельта Волги (3) 82 380 6,1 5,6 5,0 4,9 4,5 4,5 3,8 — — 3,2 2,8 — 4,6
Низовья р. Или (4) 137 578 5,4 4,5* * 3,9** ** 3 6*** *** 3 3**** **** 2 g***** ***** — 4,2
Дальний Восток (5) 26 101 6,0 5,2 5,0 5,0 5,0 3,0 — 3,0 3,0 1,0 2,5 2,3 3,9
Источник информации: 1-Лебедева, 1956а; 2-Донауров, Теплов, 1938; 3-Лавровский, 1962; 4-Слудский, 1956; 5-Бромлей, Кучеренко, 1983.
Звездочки - средние значения для выборок.
встречаются выводки из 5-7 (56,1%), 8-9 (28,1%), реже - из 10-13 (10,5%) и 1-4
(5,3%) поросят. К октябрю их размер уменьшается до 6,3, к январю (после сезона
охоты) - до 4,4 и к марту - до 3,1 [Волох, 1999]. В низовьях р. Или из 5,4 дете-
нышей в пометах к февралю остается 2,8, а до возраста подсвинка доживают
лишь 2,1 [Слудский, 1956]. В Монголии в апреле - мае преобладают самки с
одним уцелевшим зимой поросенком и очень редко регистрируются с тремя
[Банников, 1954]. На Дальнем Востоке, где потенциальная плодовитость самок
6,5 эмбрионов, до октября сохраняется около 5 сеголеток, а к весне остается в
среднем 2,6 [Бромлей, 1964].
i 'j
Таблица 25. Среднее количество поросят в семьях диких свиней зимой и ранней весной
Район Семей, п Поросят, п Поросят на самку Источник информации
в среднем | lim
С.-запад России* 73 336 4,6 — Русаков, Тимофеева, 1984
44 * 21 60 2,8 1-6 44
Беловежская пуща 52 235 4,5 — Лебедева, 1956а
44 — — 4,6-5,6 — Козло, 1975
Подмосковье — — 5,5-5 — Иванова, 1975а
44 — — 4-5 -9 Иванова, Рыковский, 19686
“Завидово” 88 405** 4,6 — Дормидонтов, 1969
Окский з-к 144 662 4,6 — Червонный, 19806
Воронежская область 8 37 4,6 3-6 Простаков, 1996
З-к “Лес на Ворскле” — — 5 3-7 Тимофеева, 1975
Юг Украины — — 4,3-3,1 1-8 Волох, 1999
Крым — — 2-2,5 — Кормилицин, Дулиц- кий, 1972
Болгария — — 3,1 — Генов, Коцаков, 1986
Кавказский з-к 9 18 2,0 — Донауров, Теплое, 1938
Дельта Волги 14 43 3,1 — Лавровский, 1962
Низовья р. Или 43 141 3,5-2,8 — Слудский, 1956
Бурятия 6 19 3,2 2-5 Смирнов, 1978
Хинганский з-к 37 133 3,6 1-8 Дарман, 1990
Сихотэ-Алинь 13 34 2,6 1-4 Бромлей, 1964
* В 1960-1976 гг. и 1977-1982 гг. соответственно.
** По расчетам.
Итак, дикие свиньи способны размножаться уже на первом году жизни. Половоз-
релые особи почти ежегодно приносят 5-7 поросят, из которых большинство не до-
живает до следующей весны. Уровень воспроизводства в значительной мере сопря-
жен с обеспеченностью кормом в осенне-зимний период, максимален у средневозра-
стных животных и остается высоким вплоть до конца их жизни. Высокая плодови-
тость вида, вероятно, - одна из эволюционных адаптаций, направленных на компен-
сацию повышенной смертности.
140
Популяционная структура
Половой и возрастной состав
Соотношение полов. По исследованиям В.П. Теплова [1954], у большинства
видов млекопитающих рождается больше самцов, а у крота и, возможно, у каба-
на - самок. Это предположение, основанное для диких свиней на малой выборке
(п = 16), в дальнейшем неоднократно цитировалось многими специалистами в
утвердительной форме, хотя оно и не соответствует действительности.
Соотношение полов среди эмбрионов у кабана, как и у других копытных, в
репрезентативных выборках близко 1:1 (табл. 26), но в разные годы в одной и той
же популяции может заметно сдвигаться в ту или другую сторону - от 1:3,5 до
1:0,6 [Иванова, 1985]. Эмбрионы в матках чаще всего разного пола, однако
встречаются и однополые; в Завидове они обнаружены у 6,9% самок [Щербаков,
Стародынова, 1979].
Таблица 26. Соотношение полов у эмбрионов и новорожденных кабана '
Район Всего, п Самцы, % Самки, % Соотношение самцы: самки Источник информации
С.-запад России 48 33,3 66,7 1:2 Русаков, Тимо- феева, 1984
Германия 1110 46,5 53,5 1:1,1 Meynhardt, 1982, 1984а
Ярославская обл. 370 49,5 50,5 1:1 Иванова, 1985
Окский з-к 306 55-41 45-59 1:0,8-1,4 Червонный, 1980а
“Завидово” 304 46,5 53,5 1:1,2 Щербаков, Ста- родынова, 1979
Ц. Черноземье 103 43,7 56,3 1:1,3 Простаков, 1996
а — 42 58 1:1,4 Сухорослов, 19726
Воронежский з-к — — — 1:1 Дьяков и др., 1986
Карпаты — 33,6 66,4 1:2 Гунчак, 1982
Кавказский з-к 16 31 69 1:2,2 Донауров, Теп- лое, 1938
Дельта Волги 23 39,1 60,9 1:1,6 Лавровский, 1962
Низовья р. Или 19 36,8 63,2 1:1,7 П.Еськов [Слуд- ский, 1956]
Таджикистан 52 57,7 42,3 1:0,7 Иваненко, 1956
Предбайкалье 64 51,5 48,5 1:0,9 Недзельский, 1994
Сихотэ-Алинь 71 52,1 47,9 1:0,9 Бромлей, 1964
Соотношение полов среди сеголеток близко к таковому у эмбрионов и ново-
рожденных, но в разных популяциях и в разные годы тоже заметно варьирует в
ту или другую сторону. В Завидове из 1006 отловленных для мечения детенышей
в возрасте 3-6 месяцев самцы составляли 55% [Мануш, Николаев, 1995]. В Кар-
141
Таблица 27. Соотношение полов в популяциях кабана (по визуальным наблюдениям)
Район Всего встреч,п Самцы, % Самки, % Соотношение самцы : самки Источник информации
Литва 2638 39,7 60,3 1:1,5 Янулайтис, Па- дайга, 1985
Беловежская пуща 25000 — — 1:1-1,6 Козло, 1975
598 62,9 37,1 1:0,6 Северцов, Саб- лина, 1953
Карпаты 2110 44 56 1:1,3 Гунчак, 1982
Крым — — — 1:1,2 Кормилицин, Дулицкий, 1972
Дарвинский з-к — — — 1:0,9-1,4 Завьялова, 1997
Ярославская обл. — 49 51 1:1 Иванова, Овсю- кова, 1976
и — 46-32 54—68 1:1,2-2,1 Иванова, Моло- канов, 1982
“Завидово” 205 57 43 1:0,8* Дормидонтов, 1969
Окский з-к 300 30 70 1:2,3 Червонный, 1980а
Юг европейской части России 348 44,1 57,9 1:1,3 Иванова, 1985
Ц. Черноземье — 39 61 1:1,6 Сухорослов, 19726
З-к “Лес на Ворскле” 155 25,2 74,8 1:3 Царев, 1989а
Воронежская обл. 1484 53,2 46,8 1:0,9 Простаков, 1996
Воронежский з-к — — — 1:1,1-1,5 Дьяков и др., 1986
Липецкая обл. 2071 44,4 55,6 1:1,2 Побединский, 1997
Кавказский з-к 1101 48 52 1:1,1 Донауров, Теп- лое, 1938
Куба-Хачмасская — 37,8 62,2 1:1,5 Алекперов, Ку-
зона лиев, 1987
Дельта Волги 536 47,5 52,5 1:1,1 Лавровский, 1952, 1962
Низовья р. Или 269 49,1 50,9 1:1 Слудский, 1956
Тянь-Шань 57 31,6 68,4 1:2,2 Слудский и др., 1984
С.-Челекский з-к 2621 32-25 68-75 1:2,1-3 Титов, Шинга- реев, 1970
Ю. Киргизия 479 44,9 55,1 1:1,2 Чичикин, Во- робьев, 1967
Алтай 67 52,2 47,8 1:0,9 Собанский, 1992
Саяны 118 55,9 44,1 1:0,8 Завацкий, 1989
Предбайкалье 137 — — 1:1 Недзельский, 1994
Сихотэ-Алинь 23 39 61 1:1,6 Антонов, 1999
* Результат выборочной охоты на свиней.
142
патах среди сеголеток и полувзрослых особей преобладали самки - 65% и 56%
[Гунчак, 1982]. На северо-западе России в 1981 и 1982 гг. среди 62 добытых се-
голеток и 70 подсвинков (пол молодых кабанов визуально почти неразличим и,
следовательно, эти промысловые выборки можно считать репрезентативными
для популяции) было больше самцов - 1:0,8 и 1:0,6 [Русаков, Тимофеева, 1984].
В Центрально-Черноземном регионе, напротив, в числе добытых сеголеток
больше самок (1:1,3), но у подсвинков соотношение полов примерно одинаково
[Простаков, 1996]. В Сары-Челекском заповеднике среди 38 отловленных под-
свинков самок было 63% [Титов, Шингареев, 1970].
В большинстве популяций, по визуальным наблюдениям, самок среди взрос-
лых животных несколько больше, чем самцов, что связано с более высокой есте-
ственной смертностью последних и выборочным отстрелом (см. табл. 48). Одна-
ко и в этом случае соотношение полов в целом все-таки остается близким 1:1
(табл. 27).
Возрастной состав. Возрастной состав популяций кабана в России и в сопре-
дельных регионах примерно следующий: сеголетки - около 40-50 (10-80)%,
подсвинки - 10-20 (3-28)%, взрослые особи - 30-40 (17-73)% (табл. 28). Однако
он очень существенно меняется как на протяжении года, так и в разные годы, что
связано прежде всего с цикличностью размножения и неодинаковыми уровнями
воспроизводства и смертности, и различается в разных группировках.
Таблица 28. Возрастной состав популяций кабана (по визуальным наблюдениям)
Район Всего встреч,п Сеголет- ки, % Годова- лые, % Взрослые, % Источник информации
Литва 5779 54,4 ** 45,6** Янулайтис, Падай- га, 1985
Беларусь 46520 44,8 17,6 37,6 Козло, 2001
Беловежская пуща 2859 59,0 8,9 32,1 Северцов, Саблина, 1953
6835* 21-41 6,5-20 44-73 Саблина, 1955
25000 30-70 7 70-30 Козло, 1975
Березинский з-к 1065 56,8 7 43,2 Лавов, 1981
Чернобыль (зона АЭС) — 54-60 18-28 25-18 Козло и др., 1993; Дунин и др., 1998
З-к “Кодры” 100 45 24 31 Чегорка, 1984
Карпаты 2110 4720* 46,5 (35-60) 21,5 32 Гунчак, 1982
Крым — 57,6 7 42,4 Кормилицин, Ду- лицкий, 1972
С.-запад России 254 51,3 15,4 33,3 Русаков, Тимофее- ва, 1984
107 45,8 9,3 44,9
Вологодская обл. - 40-67 3-25 17-37 Варнаков, 1985
- 64 ** 36** Киселев, Каплин, 1987
Дарвинский з-к — 34-42 4-8 55-58 Завьялова, 1997
“Завидово” 660 62,0 7,1 30,9 Дормидонтов, 1969
2138* 42,3 ** 57,7** Мануш, 1969
143
Таблица 28 (окончание)
Район Всего встреч,п Сеголетки, % Годова- лые, % Взрослые, % Источник информации
36850* 50,1 (34-58) ** 49 9** Мануш, Николаев, 1995
Ярославская обл. 516* 49,8 (33-55) 24,6 (12-26) 25,6 (55-19) Иванова, Молока- нов, 1969,1982
— 54,8 22 23,2 Юргенсон, 1969
Окский з-к 300 63-70 11-15 17-26 Червонный, 1980а
Ц.-Черноземный з-к — 43^16 * * 57-54** Гусев, 1989
Воронежская обл. 4290* 53,1 12,2 34,7 Простаков, 1996
Белгородская обл. 432 64,1 10,9 25,0 Царев, 1989а, 2000
Липецкая обл. 2071 46,5 13,5 40,0 Побединский, 1997
Ц. Черноземье — 55,8 ** 44,2** Сухорослов, 19726
Кавказский з-к — 15-57 15 50-28 Дуров, 1980а,б
Дельта Волги 1830 65 ** 35** Лавровский, 1962
Европейская часть России 27527* 45-54 ** 55^16 Любченко, Мама- ев, 1988
а — 66-34 12-27 26-47 Иванова, 1985
Низовья р. Или 734 49,9 13,5 36,6 Слудский, 1956
Тянь-Шань 306* 43,8 20,6 35,6 Слудский и др., 1984
а 140 47-57 ** 53-43** Есипов, 1990,1991
Алтай 220* 50,4 19,1 30,5 Собанский, 1992
Тува - 68-37 ** 32-63 Смирнов, 2001
Саяно- Шушенский з-к 269* 56,2 ** 43,8** Завацкий, 1989
Предбайкалье 137(681*) 28МЗ 16-22 41-50 Недзельский, 1994
Хинганский з-к 240 (1557*) 56(46-61) 12-16 23М2 Дарман, 1989, 1990
Амурская обл. 141 32-53 12-23 55-24 а
Сихотэ-Алинь 42 45,3 ** 54,7 Антонов, 1999
сс — 46-52 (10-80) 10-12 36-44 Бромлей, Кучерен- ко, 1983
* Всего особей.
* * Доля взрослых и годовалых особей.
В группировке в Переславском охотничьем хозяйстве Ярославской области
доля поросят составляет в среднем 55%, подсвинков - 20-22%, свиней - 10-13%,
взрослых самцов - 12-13% [Иванова, Рыковский, 19686], в Липецкой области -
46, 14, 22 и 18% [Побединский, 1997], в заповеднике «Кодры» в Молдавии - 45,
24, 20 и 11% соответственно [Чегорка, 1984]. В литовской популяции диких сви-
144
ней регистрируют 54-59% сеголеток, 15-18% самцов старше года и 25-28% са-
мок того же возраста [Янулайтис, Падайга, 19846, 1987]. В Беларуси на разных
фазах динамики численности в популяциях 40,3-52,3% сеголеток, 14,8-23,2%
двух-трех-летних животных, 8,1—20,8% взрослых самцов и 16,2-24,0% взрослых
самок. По средним многолетным данным возрастной состав популяций следую-
щий: сеголетки - 44,8%, двухлетние особи - 17,6%, трехлетние - 12,5%, 4-5 лет-
10,2%, 6-7 - 8,7%, 8-10 -2,8%, 11-12 лет и страше - 3,0% [Козло, 2001].
В Кавказском заповеднике доля сеголеток после урожайных и неурожайных
лет колеблется от 15 до 57%, в сентябре она составляет в среднем около 30%, к
весне сокращается до 17%, а при высокой численности волка - до 8,6%. Соответ-
ственно изменяется соотношение и в других возрастных группах [Дуров,
1980а,б]. На Дальнем Востоке при благоприятных кормовых условиях в популя-
циях 75-80% сеголеток, при недостатке зимних кормов - 10-15% [Бромлей, Ку-
черенко, 1983].
Социальная организация
Основа социальной организации популяций кабана, как и у многих других ко-
пытных, семейная и семейно-групповая. Единой ее классификации не существует
[Баскин, 1976; Лозан, 1980а, 1983; Царев, 19806, 1982, 19896,в, 2000; Лозан, Лозан,
1981а,б; Майнхард, 1983; Обидина, 1987; Простаков, 1996; Козло, 2001]. Наиболее
типичны, на мой взгляд, следующие внутрипопуляционные группировки:
семья - самка с детьми;
семейная группа (союз) — объединение из двух и более родственных семей;
гурт (гарем) - одна из названных группировок в период гона или, реже, в
другое время, включающая доминирующего взрослого самца (секача) или не-
скольких самцов;
компания - временное объединение нескольких особей одинакового или раз-
ного пола и возраста. Среди таких компаний выделяют годовалых особей - груп-
пы подсвинков;
стадо - крупное кратковременное объединение групп и особей разного пола
и возраста.
Кроме того, в популяции довольно много одиночных животных. Это обычно
самцы и самки старше 1,5 лет, отделившиеся от семьи и не входящие ни в одну
из указанных выше группировок, или отдельные особи, уцелевшие после сезона
охоты и трудной зимовки.
Объединения диких свиней образованы, как правило, кровными родственни-
ками по материнской линии. Они относительно стабильны и «закрыты» для про-
никновения в них неродственных группировок и особей, за исключением поло-
возрелых самцов [Meynhardt, 1988]. В вольерах, тем не менее, группировки могут
формироваться из животных, не связанных родством.
Сезонные изменения. Социальная организация популяций закономерно,
циклически и существенно изменяется в течение года [Olof, 1951; Козло, 1975;
Царев, 1980а,б, 1986а, 1989а-в, 1990, 1991, 2000; Лозан, 1983; Майнхард, 1983,
1986; Hirotani, Nakatani, 1987; Meynhardt, 1988; Зайцев, 1989].
Один из важнейших этапов - весенний (март - май), когда из-за возрастания
агрессивности и антагонизма происходит распад семей и групп. Перед родами
беременные самки отделяются от группы и прогоняют от себя прошлогодних де-
тей, к которым агрессивны и после опороса. Значительная часть подсвинков по-
кидает материнский участок весной, а летом от образовавшейся компании отпоч-
ковываются самцы. Оставшихся самцов свиньи изгоняют из больших семейных
групп в возрасте 15—18 месяцев, и затем они живут поодиночке [Meynhardt,
10. Заказ №2528.
145
Рис. 52. Взрослые сам-
цы ведут преимущест-
венно одиночный об-
раз жизни (фото А.
Дигилевича)
1988]. Яловые самки не агрессивны к своему прошлогоднему потомству, но к
концу лета и возле них остаются лишь отдельные дочери.
Через несколько дней после опороса, когда агрессивность, связанная с охра-
ной нового потомства, несколько затухает, свиньи с новыми выводками присое-
диняются к семье родственной самки-вожака, образуя семейное объединение.
Позднее в него могут вливаться подсвинки, преимущественно самки, и взрослые
яловые особи. Семейные союзы, в состав которых порой входят до 10 свинома-
ток с потомством, иногда достигают очень больших размеров - до 102 особей.
При благоприятных условиях такие формирования сохраняются до весны, но
крупные в конце лета обычно дробятся. Часть свиней покидает союз, уводя своих
детенышей, включая потомство прошлых лет. Отделившиеся семьи живут на но-
вых участках обособленно или образуют новую родственную семейную группу
[Майнхард, 1983, 1986].
Взрослые самцы, ведущие в основном одиночный образ жизни (рис. 52), при-
соединяются к семьям и семейным группам лишь в период гона. Однако отдель-
ные из них остаются в гуртах вплоть до весны, а некоторые сопровождают семьи
и летом [Саблина, 1955, 1969; Царев, 2000].
Во второй половине зимы социальная организация популяций кабана практи-
146
чески не меняется, за исключением уменьшения числа особей в группах под воз-
действием промысла, хищников и других факторов смертности.
Состав группировок. Группы диких свиней численностью в 5-10 особей, как
правило, - семьи. В объединениях, насчитывающих от 10 до 30 особей, тоже чаще
близкородственный состав: 1-4 взрослые свиньи с поросятами, подсвинки, яловые
самки и, иногда, самцы. Более крупные стада, если это не семейные союзы, фор-
мируются спонтанно из соседних групп на ограниченных местах кормежек и при
опасности, когда напуганные особи увлекают за собой остальных. Такие искусст-
венные формирования быстро распадаются на родственные субъединицы.
Одиночные особи в разных популяциях и в разные периоды года составляют
от 18 до 60% встреч. К ним относятся самцы старше 1,5 лет, взрослые свиньи,
потерявшие или не имеющие поросят, подсвинки-самки и поросята, отставшие от
семей или уцелевшие после сезона охоты и зимовки. Молодые животные чаще
все же встречаются вместе по 2-5 особей. Доля таких мелких групп довольно
значительна - от 4 до 26 % (табл. 29). В них регистрируют также самок с 1-4 по-
росятами, взрослых самцов или самок, взрослого самца с подсвинком или с сего-
летком, подсвинка с 1-3 поросятами и др.
Частота встреч групп разного состава и одиночных зверей в разных районах
(см. табл. 29), как и в разные сезоны и годы, неодинакова, что свидетельствует о
лабильности и неоднородности социальной структуры популяций.
На северном пределе ареала в Архангельской области по 60 зарегистриро-
ванным встречам одиночные звери составляют 26,7%, пары - 21,7%, группы из
3-4 особей - 23,3%, из 5-11 - 28,3% [Плешак, Миняев, 1986].
В Беловежской пуще доля одиночных особей в популяции в течение года из-
меняется незначительно - от 25% в июне до 36% в январе. Доля групп из 2-4 го-
лов минимальна в сентябре (16%) и максимальна (34%) в феврале; группы из 5-9
особей чаще встречаются в июне и июле (42-45%), реже (25%) - в январе и мае.
Группировки, насчитывающие более 20 голов, обычны в мае - июне, что связано
с объединением семей, и ранней осенью - в наиболее кормных биотопах [Козло,
1974а, 1975].
Зимой, на удивление, очень мало одиночных животных и пар в Литве. При
1025 встречах 5898 кабанов они составили всего 1,9-4,4% и 1,9—2,4%. Макси-
мальное число зверей (8-12%) находится в группах из 4-8 особей [Янулайтис,
Падайга, 19846]. В низовьях р. Или, напротив, необычайно часто встречаются
одиночные особи - 59,8%. Существенное изменение социальной структуры по-
пуляции здесь связано с интенсивной охотой с применением собак, которые да-
вят в первую очередь молодых особей и разгоняют остальных [Слудский, 1956].
В популяции диких свиней Центрально-Черноземного региона доля одиноч-
ных особей 32%, семей - 43,7%, стад - 11,2%, одновозрастных компаний - 7,6%,
гуртов - 5,2%. Наиболее часто (40,4%) встречаются группы из 5-8 особей [Про-
стаков, 1996]. В Воронежском заповеднике в 1981-1985 гг. одиночные звери со-
ставляли четвертую часть поголовья [Дьяков и др., 1986]. В группировке запо-
ведника «Лес на Ворскле» Белгородской области, насчитывающей 93—127 осо-
бей, отдельно живущие взрослые свиньи зарегистрированы лишь дважды, сека-
чей здесь около 7-9, самцов-подсвинков - 1-3, семей - 2-5 и 6-7 семейных групп
[Царев, 1991].
В популяции Кавказского заповедника выделено 8 структурных единиц: самки с
выводками - 33%; семьи, включающие поросят и подсвинков - 7%; взрослые свиньи
с подсвинками - 7%; взрослые самки без молодняка - 6%; взрослые самцы - 19%
(18% - одиночные особи и 1% - по 2-3 вместе); группы до 10 самок и самцов с мо-
лодняком - 4%; 1-6 подсвинков - 6% и 1-6 поросят- 2% [Дуров, 1986].
В группировке Хинганского заповедника семьи составляют в среднем за год
27%, но от весны к зиме их доля уменьшается с 46 до 15%. Отдельные семьи на-
10!
147
00
Таблица 29. Количественный состав групп кабана
Район (источник информации) Число встреч Число особей Одиночных особей и особей в группе, % от числа встреч
1 2 3 4 5 6 7 8 9 10 11-15 16-20 21-30 31-50 >'
С.-запад России (1) 840 4924 26,4 7,3 7,4 6,5 7,6 8,9 7,7 5,9 4,3 4,9 7,8 2,9 1,6 0,5 о,
Беловежская пуща (2) 4774 22130 35,7 26* * * 25,4** ** ** ** ** у 9*** *** 3,6 । 2**** **** -
-“-(6) 731 2898 35,6 14,5 10,7 6,8 7,9 6,4 5,5 2,1 2,3 2,3 3,8 1,2 0,5 0,3 —
Хоперский з-к (8) 186 796 32,3 12,4 11,8 9,1 6,4 7,0 4,3 6,4 1,6 2,7 5,9* * * * -
Кавказский з-к (3) 273 2195 18,3 14,7 20,5 17,9 17,6 8,4 8,8 7,3 4,0 4,4 7,0 2,9 3.3 1,5 -
Низовья р. Или (4) 314 1127 59,8 5,7 6,4 7,3 8,0 6,4 4,8 4,1 2,2 1,3 2,2 0,6 0,3 0,6 —
Саяны (7) 190 — 27,9 7,4 43,2* * * 15,3** ** ** ** ** 6 3*** *** *** —
Бурятия (9) 62 — 58,1 9,8 6,4 6,4 12,9* * — — — — 6,4 — — — —
Хинганский з-к (5) 1078 5176 44,3 11,3 8,8 4,9 4,1 4,4 2,9 2,6 2,0 3,0 5,7 2,6 2,1 1,2 0,1
Источник информации: 1 - Русаков. Тимофеева, 1974; 2 - Козло, 1975; 3 - Донауров, Теплов, 1938; 4 - Слудский, 1956; 5 - Дарман, 1990; 6 - Северцов, Саблина
1953; 7-Завацкий, 1989; 8-Антонец, 1993; 9-Смирнов, 1978.
Звездочки - средние значения для выборок.
чинают объединяться в апреле (8% встреч), а к июню доля групп возрастает до
16,5%, что приводит к росту показателя стадности в это время. В ноябре - декаб-
ре в состав групп входят 1-2 секача (5,6%). Доля зимних стад колеблется в пре-
делах 11-22%. Одиночных особей здесь около 44% (или 9% от числа встречен-
ных животных), групп из 2-4 молодых зверей - 9%, групп из 3-5 взрослых осо-
бей - 16%. Большая часть кабанов объединена в группы из 6-10 (24%) и 11-20
членов (26%). Крупные стада в 50-75 голов отмечаются в мае - июне и ноябре
[Дарман, 1990].
На Сихотэ-Алине гурты во главе с секачем зарегистрированы в 12% встреч,
семейные группы - в 16%, семьи - 18%, группы подсвинков - 18%, сеголетки -
1,3% и одиночные особи - в 35% встреч. Число свиней в семейных группах не
превышает 4-6 [Зайцев, Зайцева, 1990; Зайцев, 1991; Антонов, 1999].
Размер групп. Размер групп у многих видов копытных определяется биоло-
гическими циклами: увеличивается при появлении молодняка и в период гона и
уменьшается перед сезоном деторождения. Однако немаловажны также общая
численность зверей и степень их концентрации в угодьях в определенный сезон.
В первой половине XX в. в широколиственных лесах Приморья нередко встреча-
лись стада в 60-100 голов [Капланов, 1948; Бромлей, 1964], в 60-х годах - по 50-
60 особей [Кучеренко, 1973], а в конце столетия при сокращении численности -
до 20 [Антонов, 1999].
Среднее число особей в группе (показатель, или коэффициент, стадности) в
разных популяциях кабана и в разные годы колеблется от 2,2 до 14, максималь-
ный размер стад достигает ста голов (табл. 30).
В Кавказском заповеднике максимальный коэффициент стадности у кабана в
декабре в разгар гона (10,2), в сентябре (7,2) и марте (8,0), т. е. в периоды кон-
центрации животных на локальных участках орехоносов и дикорастущих фрук-
тарников [Донауров, Теплов, 1938]. В Хинганском заповеднике этот показатель
изменяется от 4,8 в обычные зимы до 7,2 в голодные и до 3,0 - после них [Дар-
ман, 1990].
В Беловежской пуще в 1946-1953 гг. данный коэффициент варьировал по ме-
сяцам в среднем в пределах 3-5,2, в разные годы - от 2,2 до 5,1, в 1956-1965 гг. -
от 3,7 до 5,3 и 3,3 до 5,0 соответственно. В 70-е годы в период обильной зимней
подкормки и при максимальной плотности, достигавшей 17-28 особей на 1000
га, он увеличился в 2-3 раза (табл. 31). Зависимость показателя стадности от
плотности населения прослеживалась и в отдельных лесничествах: 3,4 при плот-
ности менее 10 особей на 1000 га, 4-4,6 - до 20 и 4,9-5,3 - свыше 20 голов. В
многоснежные и голодные зимы стадность заметно уменьшалась [Козло, 1971,
1974а, 1975]. В 50-е годы при неурожаях желудей она была почти в два раза
меньше, чем в годы урожаев, а максимальное число особей в группе сокращалось
с 22-27 особей до 7-19 [Саблина, 1969].
В Центральном Черноземье показатель стадности сравнительно невысокий (в
среднем 4,7-6,7), что определяется мозаичностью и малокормностью угодий
степной зоны и низкой численностью животных. В отдельных охотничьих угодь-
ях при плотности 20 особей на 1000 га он составляет 4,9, а при 5 - всего 1,8 [Су-
хорослов, 1970а, 19726; Простаков, 1996]. В Переславском лесоохотничьем хо-
зяйстве в Ярославской области при плотности населения 9 особей на ту же пло-
щадь этот коэффициент близок к 4, а при 18 - к 6 [Иванова, Овсюкова, 1976]. В
Калужской области в период с 1961 по 1970 гг. по мере увеличения численности
кабана на порядок среднее число особей в группе изменялось от 1,0 до 10,6 [Во-
ронин, 19756].
149
Таблица 30. Коэффициент стадности и максимальный размер групп у кабана
Район Число особей в группе Источник информации
среднее максимальное
Литва 5,7 28 Янулайтис, Падайга, 1985
О. Сааремаа 6-14 44 Валдманн, 1990
Беловежская пуща 2,2-15 42 Северцов, Саблина, 1953; Лебеде-
ва, 1956а; Козло, 1975; Балюк и
др, 1987
Березинский з-к 4,1 — Козло, 1975
Чернобыль (зона АЭС) 4,7-7,3 — Дунин и др,1998
С.-запад России 5,9 57 Русаков, Тимофеева, 1984
Архангельская обл. 3,4 И Плешак, Миняев, 1986
Кировская обл. — 30 Павлов, 1999а
Ярославская обл. 5,8 67 Личные наблюдения
Ц. Черноземье 6,7 — Сухорослов, 19726
(С 4,7 43 Простаков, 1996
З-к “Лес на Ворскле” 5,7-6,6 18 Тимофеева, 1975
Воронежский з-к — 70 Дьяков и др, 1986
З-к “Кодры” 3,9-4,4 27 Чегорка, 1984
Кавказский з-к 5,9 40 Донауров, Теплое, 1938
Куба-Хачмасская зона 5,1-13,3 — Алекперов, Кулиев, 1987
Низовья р. Или 3,1 (5,8) 35 Слудский, 1956
Тянь-Шань 6,4 24 Есипов, 1990, 1991
Сары-Челекский з-к — 70 Чичикин, Воробьев, 1967
Тюменская область — 46 Азаров, 1996
Алтай 3,1 14 Собанский, 1992
Саяно-Шушенский з-к 3,4 32 Завацкий, 1987,1989
Предбайкалье 4,9 10 Недзельский, 1994
Монголия — 60 Банников, 1954
Бурятия 2,8 48 Смирнов, 1978
Хинганский з-к 4,8 75 Дарман, 1990
Сихотэ-Алинь — 60-100 Капланов, 1948; Раков, 1956
3,2 20 Антонов, 1999
Приморский край 5-7 42-60 Метельский, 1975; Кучеренко,
1979
И все же, несмотря на вариабельность показателя стадности в зависимости от
плотности населения и кормовой ситуации, его многолетние (табл. 31) и сезон- j
ные (табл. 32) изменения у кабана в целом невелики по сравнению с другими ди- '
кими копытными.
150
Таблица 31. Изменения показателя стадности кабана в Беловежской пуще (по [Лебедева, 1956а; Козло, 1975; Балюк и др., 1987], с изменениями)
Год Месяц В среднем за год Плотность, особей/1000 га**
1 2 з 4 5 6 7 8 9 10 11 12
1946 — 1,9 3,5 3,2 2,2 3,5 1,0 4,3 4,8 3,5 4,3 2,5 3,3 5,2
1947 2,8 1,6 3,6 3,6 4,8 5,0 3,3 2,5 3,4 2,4 2,5 3,5 3,4 7,1
1948 4,3 3,1 5,1 4,6 7,6 6,3 6,4 6,3 5,9 4,9 5,0 3,2 5,1 10,1
1949 4,9 4,3 3,8 2,7 4,1 3,9 3,3 2,9 3,5 3,4 3,1 2,1 3,5 11,8
1950 2,4 3,4 4,7 5,1 5,9 4,4 4,3 3,7 5,5 3,2 4,6 3,8 4,2 7,2
1951 3,6 3,5 3,5 4,4 4,2 4,2 4,7 4,4 3,6 3,8 5,6 2,9 4,0 16,1
1952 2,8 2,3 2,3 2,8 3,4 2,3 1,7 2,0 2,6 1,9 2,4 2,2 2,2 12,5
1953 2,0 1,3 4,0 8,6 10,2 6,0 4,6 6,4 5,3 6,4 4,2 1,0 5,0 5,3
1956 3,2 3,5 3,3 1,9 5,0 5,7 3,8 3,7 3,7 3,2 3,4 2,2 3,3 2,5
1957 4,4 3,0 3,9 4,7 3,0 4,7 6,5 5,4 3,8 5,2 3,6 5,9 4,7 2,8
1958 3,2 4,2 4,7 4,6 3,4 3,1 2,5 3,5 3,0 2,8 2,6 2,0 3,4 6,4
1959 2,0 2,1 2,9 3,5 5,5 6,0 4,3 4,9 5,0 2,9 4,6 6,1 4,8 17,7
1960 3,5 5,0 5,2 4,3 3,9 5,8 4,9 5,0 4,4 5,4 5,2 4,5 4,9 18,3
1961 4,0 3,6 3,2 5,0 7,2 5,9 5,6 5,6 6,1 4,5 5,2 4,6 5,1 17,5
1962 5,4 4,1 4,3 3,5 3,6 4,4 3,2 3,7 4,1 5,0 4,4 3,5 4,3 17,2
1963 4,1 5,6 6,3 4,0 3,9 6,2 4,4 4,0 6,2 5,0 3,4 4,9 5,0 18,4
1964 3,8 4,8 4,9 7,0 7,0 4,5 3,1 2,0 4,3 3,1 2,9 2,5 3,3 7,0
1965 3,6 4,0 4,1 4,6 6,6 6,2 5,7 4,6 4,5 4,9 5,0 4,7 4,7 9,2
1974 10,4* 15,4* 8,8* 7,7* 3,5 15,8
1975 8,0* 14,0* 14,7* 9,4* 3,8 17,7
1976 12,4* 17,0* 12,4* 9,4 4,3 25,9
1977 12,5* 16,3* — — — — — 27,8
1978 13,9* 17,9* 16,5* 11,5* 5,0 25,8
1979 16,1* 10,7* 8,8* 7,5* 3,6 18,6
Среднее: 1946-1953 3,5 3,0 3,6 4,2 5,2 4,4 4,3 4,0 4,3 3,6 4,6 3,0 3,9 5-16
1956-1965 3,7 4,0 4,3 4,3 4,9 5,3 4,4 4,2 4,5 4,2 4,0 4,1 4,3 3-18
1974-1979 12,6* 15,1* 11,4* 9,0* 9-15 16-28
Примечание. Число встреч в 1946-1956 гг.-9167, число особей-39470; в 1973-1979 гг. - 8390 и 104193 соответственно.
* В среднем за сезон.
** Сведения о плотности населения по [Шостак, 1978; Балюк и др., 1987].
Таблица 32. Сезонные изменения показателя стадности кабана
Район (источник информации)* Число встреч Число особей Среднее число особей в группе
весна лето осень зима
Ленинградская обл. (1) 214 1166 4,2 5,8 6,4 5,1
О. Сааремаа (2) — 3306 — 6-14 7-9 6-9
Беловежская пуща (6) 17577 143663 3,6-15 4-15 3,6-11,4 3-12,6
“Завидово” (4) 457 — 2,5-3,9 4,7-5,9 4,7-6,6 4,0
Кавказский з-к (8) 273 2195 4-8 3-4,5 4,9-7,2 4-10,2
Низовья р. Или (7) 314 1127 2-5,6 3,2-3,8 2,9—4,1 1,5-3,1
Алтай (5) 83 260 4,0 6,5 10,0 1,6
Предбайкалье (9) 137 681 3,7 4,3 6,3 5,5
Хинганский з-к (3) 1078 5176 4,5 5,3 4,9 4,8
*1 - Русаков, Тимофеева, 1984; 2-Валдманн, 1990; 3-Дарман, 1990; 4 - Дормидонтов, 1969; 5
-Собанский, 1992; 6 - Север цов, Саблина, 1953; Лебедева, 1956а; Козло, 1975; Балюкидр., 1987; 7
-Слудский, 1956; 8-Донауров, Теплов, 1938; 9-Недзельский, 1994.
Поведение. Структурная целостность группировок кабанов и взаимоотноше-
ния особей в них базируются на четко выраженной линейной иерархии [Лозан,
Лозан, 1981а,б; Bartschi, 1983; Лозан, 1983; Meynhardt, 1983b, 1985b; Майнхардт,
1983, 1986; Schnebel, Griswold, 1983; Цыганов, 1985; Rascalou, Teillaud, 1988].
Иерархический статус животных существенно зависит от их возраста и силы.
В семьях и семейных группах доминирует вожак - наиболее крупная старая сам-
ка, субдоминанты - другие взрослые самки, имеющие поросят, затем самцы и
яловые самки. Низшие позиции в иерархической лестнице занимают подсвинки и
сеголетки, которые при приближении взрослых особей вынуждены уходить с
места кормежки. В противном случае те безжалостно отшвыривают их от корма,
и лишь мать позволяет своим детям находиться рядом. При передвижениях за
вожаком чаще следуют ее поросята, затем другие самки с их выводками, другие
взрослые особи и подсвинки. Иногда все поросята идут за лидером. Раненые и
больные особи теряют свой ранг и могут быть изгнаны из группы, а выздоровев-
шие снова становятся полноправными членами. После гибели самки-вожака сре-
ди других самок возникает борьба за лидерство, что приводит к постоянному
беспокойству особей в течение нескольких недель и, в конечном счете, к распаду
и дроблению семейного союза, перераспределению территории, повышенной ги-
бели потомства, снижению упитанности и ухудшению состояния особей.
В гуртах также складывается доминантно-иерархический тип взаимоотноше-
ний, но с наличием двойного управления стадом со стороны самца-вожака и сам-
ки-лидера [Лозан, 1984; Майнхардт, 1986]. Такое объединение отличается наи-
большей жизнестойкостью. Самка-лидер консолидирует жизнедеятельность
группы, определяет время активности, направление движения, место кормежки и
лежки, устанавливает причину тревоги и степень опасности, принимает решение
о бегстве или подает соответствующий сигнал отмены тревоги. Роль секача тоже
многогранна. Он не только выполняет функции «стимулятора» самок во время
гона и основного производителя, но и нередко первым обнаруживает опасность и
активно защищает членов группы. При бегстве самец следует последним, зачас-
тую предохраняя стадо от нападения хищников лишь одним своим присутствием.
Зимой порой лишь только он своей мощью способен пробить траншею в глубо-
ком снегу или расковырять рылом пласты смерзшейся земли, предоставляя место
для кормежки более слабым особям.
Иерархический ранг особей формируется к 10-месячному возрасту в зависи-
152
Рис. 53. Агрессия
(фото В. Алешко)
мости от уровня развития и, видимо, типа нервной системы, что в дальнейшем
способствует разделению их функций в группировках [Meynhardt, 1983b]. Вожа-
ками-лидерами чаще становятся более темные, крупные и сильные животные;
бегство от опасности инициируют более слабые; низкоранговые звери, отличаю-
щиеся повышенной осторожностью, обычно исполняют роль «сторожей» и «раз-
ведчиков». Гетерогенность особей и синхронизация их поведения - одни из ос-
новных элементов, обеспечивающих устойчивость группировок кабана [Зайцев,
1989, 1992, 1999].
В одновозрастных и часто однополых «компаниях» молодых зверей, насчи-
тывающих от 2 до 10 особей, взаимоотношения также строятся на основе ритуа-
лизации и слабой иерархии. Эти группировки не привязаны к определенным уча-
сткам и наиболее подвижны. Они, по сути, представляют биологический резерв
популяции, способствующий диффузному заполнению территории и расшире-
нию границ ареала вида [Лозан, 1980а].
Иерархические отношения в группах регулируются посредством демонстра-
ций угроз (низко опущенное рыло, вздыбленные волосы на загривке, прижатые
уши, приподнятый неподвижный хвост - рис. 53) и имитации полового акта со
стороны доминантов или «умиротворяющих» поз и подчинения (низко опущен-
ная голова и шея и приподнятое вверх рыло) со стороны низкоранговых особей,
сопровождаемых соответствующими звуковыми сигналами. Агрессивное пове-
дение развивается на ювенильной стадии онтогенеза, когда игровое поведение
перерастает в борьбу (рис. 54). Незнакомые однополые животные весьма враж-
дебны друг к другу. Интенсивность агрессии зависит от дистанции между ними:
максимальна на расстоянии 1-4 м и почти не проявляется на расстоянии свыше
12 м. При опасности агрессивность к сородичам блокируется. В критических си-
туациях, например при преодолении препятствий, высокоранговые особи могут
нападать на субдоминантов, вынуждая их идти первыми [Лозан, Лозан, 1978а;
Лозан, 1981, 1983; Майнхардт, 1986; Dardaillon, Teillaud, 1987].
Свиньи особенно активно защищают свои выводки после родов, бросаясь на
любой объект, приблизившийся к логову, и даже на человека. Через одну-две не-
дели агрессивность затухает и заменяется стремлением скрыться от опасности,
153
Рис. 54. Игровое поведение кабанят (по [Зайцев, 1991], с изменением)
а - подход к партнеру в позе «бычья голова», б - удары в ноги и бок противника, в - «марш»
с высоко поднятой головой, г — игра с предметом перед партнером, д — поза «бычья голова»
перед стычкой, е - стычка, толкание, удары, ж - параллельное шествие, з - завершение
борьбы, и - бег за лидером, к - прекращение игры сменой деятельности, л - сексуальные
проявления
154
однако инстинктивные элементы защиты потомства проявляются практически
круглогодично.
При нападении хищников группа кабанов нередко занимает круговую оборо-
ну, а одиночные особи прижимаются задом к дереву или выворотню, забиваются
в густые заросли кустарников или заломы тростника. При удобном случае эти
копытные контратакуют, недолго преследуя врага. Молодые особи, в отличие от
взрослых, обычно не оказывают сопротивления хищникам и спасаются бегством.
От человека животные убегают почти всегда, но вожаки изредка могут демонст-
рировать угрозы и кратковременные выпады, а раненые - затаиваться и нападать;
тогда они представляют реальную опасность. Мне приходилось наблюдать, как
свинья-подранок пыталась достать зубами охотника, взобравшегося на тонкое
накренившееся дерево, и самому отпрыгивать в сторону от раненого секача, рез-
ко изменившего направление движения. Взрослые самцы, содержащиеся в воль-
ерах, нередко становятся весьма агрессивными к людям, что следует учитывать
при работе с животными в неволе.
Поведение кабанов в значительной мере ритуализировано. При встрече разно-
полых взрослых особей самец поворачивается боком, затем партнеры соприкаса-
ются носами и обнюхивают друг друга. Выделено три типа обонятельных контак-
тов: назо-назальные, обнюхивание ног и глаз [Meynhardt, 1980]. При стычках про-
тивники становятся лицом к лицу и рылом ударяют один другого в щеку, шею, ло-
патку или бок. Обычно этого достаточно, чтобы выявить сильнейшего. Однако не-
редко, и особенно у самцов, в ход идут клыки; и лишь специальные адаптации в
виде «кожного щита» на лопатках защищают их от сильнейших увечий.
Агрессивность и нетерпимость, тем не менее, носит ярко выраженный харак-
тер лишь в отдельные периоды жизни и на местах кормежек. В целом же взаимо-
отношения в близкородственных стадах относительно миролюбивые и иногда
игривые. У поросят игровые элементы поведения (борьба, параллельные шест-
вия, гонка за лидером, перекатывание предметов, взбрыкивания, прыжки и сек-
суальные упражнения) проявляются с первых суток после рождения и обычны
вплоть до зимы (см. рис. 54). Порой все члены семьи или стада участвует в свое-
образной игре, длящейся до 15 мин, - стремительном беге кругами и в разные
стороны с резкими разворотами и кружением на месте [Слудский и др., 1984;
Федосенко, Жиряков, 1984; Зайцев, 1991].
Выживанию кабана способствует достаточно обширный набор врожденных и
приобретенных элементов поведения. К врожденным реакциям относятся: при-
слушивание и принюхивание, звуковые сигналы, сбор особей в случае опасности
для круговой обороны, «маятниковые» движения при сборе информации и бес-
покойстве, затаивание и замирание, а также следование за лидером и поиск парт-
неров в случае их потери. Модель поведения (бегство, затаивание или оборона)
выбирается на основе жизненного опыта лидеров, что позволяет избежать чрез-
мерного беспокойства животных. На своем участке, и особенно в центральной
его части, звери более смелы и могут не только активно обороняться, но и напа-
дать на врага [Лозан, Лозан, 1978а,б, 1981а,б; Лозан, 19806,в, 1983, 1984].
Звуковая сигнализация диких свиней весьма разнообразна. Выделяют 3 груп-
пы сигналов (контактные, предостережения и тревоги и связанные с борьбой и
обороной), содержащие 8-10 типов звуков [Meynhardt, 1980; Майнхардт, 1983,
1986; Briedermann, 1986; Лозан, 1988]:
- контактный сигнал, издаваемый поочередно взрослыми членами группы
и свиноматками во время кормления поросят;
- поисковый (призывный) сигнал свиноматок и поросят после затаивания;
- сигнал удовлетворения - протяжный высокий звук при открытой пасти
особей, нашедших лакомый корм или при «приятных» контактах;
155
- сигнал тревоги - резкий гортанный звук взрослых особей при обнаруже-
нии явной опасности;
- сигнал угрозы - резкий гортанный звук при борьбе соперников, ранжи-
ровании в группе или при встрече с врагом;
- исследовательский сигнал - частый носовой звук при обнюхивании воз-
духа или нового объекта;
- визг голодных особей;
- резкий визг при испуге или болевых ощущениях.
Самки особенно бурно реагируют на визг испуганных детенышей, без разду-
мий нападая на хищника или человека. При повизгивании своих голодных поро-
сят, которых они отличают по звуку, ложатся на бок, подставляя соски.
Наиболее характерные реакции диких свиней при опасности - затаивание и
бегство. При внезапной тревоге самка издает специфический звук, после которо-
го поросята стремглав бросаются врассыпную и ложатся, прижимаясь к земле,
или неподвижно застывают на месте в зарослях. Затаивание может длиться часа-
ми до подхода матери. На хрюкающий призывный сигнал выводок сбегается к
ней и сразу же припадает к соскам. Нередко после продолжительного ожидания
голодные детеныши сами перемещаются в том направлении, куда убежала мать,
издавая урчащие звуки. С осени подросшие сеголетки при опасности убегают
вместе с группой.
Затаивание характерно и для взрослых животных. Мне не раз приходилось
наблюдать кабанов, особенно одиночных, затаившихся в нескольких метрах от
кричащих загонщиков или стоящих на линии охотников. На одной из охот близ
Серпухова секач, скрытый густыми еловыми ветвями с налипшим снегом, заме-
рев, отстаивался в 6 м от меня около часа, но так и не вышел на открытое место,
хотя загонщик прошел в 9 м от него.
При опасности эти копытные затаиваются и на лежках. Один из крупных
самцов, за которым мне пришлось наблюдать в Тульской области в январе 1974
г., два метельных дня отлеживался в стожке соломы, не выходя кормиться. Не
поднялся он и при криках людей, подошедших вплотную к стогу. Только после
того, как человек забрался на стог и несколько минут топтался над ним, секач как
бы нехотя появился на свет. В Тверской области в декабре 1983 г. самка с поро-
сятами и подсвинками около 15 мин. отлеживалась в куче соломы, невзирая на
шум подъехавшей машины и голоса людей в 60 м от них, и выскочила лишь по-
сле случайного выстрела.
Уровень настороженности и готовности зверей к тому или иному действию
можно распознать по положению хвоста и ушей. У диких свиней уши относительно
короткие, что не всегда позволяет определить их положение. Хвост же у спокойного
животного опущен вниз (см. рис. 30) и постоянно находится в движении, у встрево-
женного - занимает горизонтальное положение (см. рис. 26), а поднятие его вверх у
замерших в ориентировочно-исследовательской позе особей (рис. 55) означает го-
товность к бегству, которое обычно следует после короткого звукового сигнала.
Дикие свиньи - очень понятливые копытные, способные адекватно оценивать
обстановку. Там, где их постоянно и интенсивно преследуют, они очень осто-
рожны и всячески избегают встреч с людьми, но при умеренном охотничьем
прессе или его отсутствии смелы и назойливы, регулярно и даже днем выходят
на посевы рядом с поселками, невзирая на запахи, шум и лай собак.
Итак, социальная организация в популяциях кабана, даже при достаточно
сложных и закономерно меняющихся на протяжении года взаимоотношениях
между особями, сравнительно проста и эффективна. Семейный и семейно-
групповой образ жизни в значительной мере способствует выживанию и процве-
танию вида: взрослые особи обеспечивают воспроизводство популяции, воспи-
тание и защиту потомства, поиск и добывание корма; молодые выживают ис-
156
Рис. 55. Ориентировочно-
исследовательская реак-
ция (фото А. Щеголева)
ключительно благодаря взрослым, перенимая их жизненный опыт и передавая
его из поколения в поколение. Особенно важна роль самок-лидеров, консолиди-
рующих жизнь стада, наиболее активно защищающих ее членов, прокладываю-
щих путь группе в снегу, что существенно снижает воздействие негативных фак-
торов. Повышенная элиминация взрослых особей при охотничьем промысле (см.
ниже), к сожалению, приводит к разрушению социальной организации популя-
ций и массовой гибели молодняка от истощения и хищников.
Пространственная структура н перемещения
Локальные перемещения и участки обитания
По данным массового мечения кабанов (п = 3344), проведенного в ГДР в
1976-1985 гг., установлено, что все сеголетки находятся в радиусе 2,5 км от мес-
та маркировки. Большая часть самок-подсвинков (65%) также остается в районе
рождения (остальные особи покидают его в июне - июле и в октябре), самцы-
подсвинки перемещаются в среднем на 8 км (пики эмиграции в апреле - мае и в
октябре - декабре). У самок старше двух лет эмиграция не выражена в отличие
157
от самцов [Stubbe, 1987]. По сведениям X. Майнхардта, ни одна семья с вывод-
ком не уходит далее 4-5 км. Самцы удаляются на 20-250 км, но некоторые сека-
чи к началу гона возвращаются назад; один из них переместился на 250 км всего
за 4 недели. Однако, большинство самцов старше двух лет все же относительно
оседлы [Meynhardt, 1982, 1983b, 1988].
Большая часть из маркированных в госкомплексе «Завидово» сеголеток (око-
ло 69%) была отстреляна на тех же площадках мечения спустя 1-3 года, а одна {
свинья — через 6 лет. Годовалых самок добывали в радиусе 6-7 км от мест бирко-
вания, годовалых самцов - на расстоянии 19-24 км (один зверь зарегистрирован
за 200 км), двухлетних особей - за 32-42 км, шестилетняя свинья убита в 24 км
[Мануш, Николаев, 1995].
Максимальная дистанция перемещения радиомеченных диких свиней на юге
Франции - 40 км, но большинство из них не уходят далее 20 км, а самки с вывод-
ками -далее 5 км [Dardaillon, Beugnon, 1987; Spitz, 1989]. Скорость перемещения
животных от убежища к кормовому участку и обратно составляет 1-2,5 км/час
зимой и 0,4-1,2 км/час в бесснежный период, на кормежке - 0,2-0,5 км/час. По-
сле кормежки самки обычно (80%) возвращаются к старому убежищу, самцы
(88%) используют новую лежку [Janeau, Spitz, 1984].
В вегетационный и малоснежный периоды протяженность суточных перехо-
дов отдельных особей и групп кабанов чаще находится в пределах 2-5 (до 20) км
по прямой, в глубокоснежье - 0,2-8,5 км. Поскольку путь их извилист, он обыч-
но в 3-5 раз длиннее [Банников, 1954; Саблина, 1955; Лебедева, 1956а; Раков,
1956; Бромлей, 1964; Дормидонтов, 1969; Тимофеева, 1975; Ватолин, 1980; Руса-
ков, Тимофеева, 1984; Кулибаба, Пакулин, 1985; Жиряков, 1989; Зайцев, Зайцева,
1990; Зайцев, 2000].
В Белоруссии по 325 троплениям зверей установлено, что почти 48% из них
проходят в сутки не более 3 км, 10% - менее 1 км, 17% - по 1-2 и 3-4 км, 25% -
4-8 и 10% - более 8 км [Козло, 1975]. В отдельных горных районах Таджикиста-
на они ежедневно кормятся за 16-18 км от убежища, преодолевая путь длиной в
32-36 км, а при снегопаде за одну ночь могут переместиться на 70 км [Иваненко,
1956]. При усилении охотничьего пресса подвижность животных увеличивается,
и они не задерживаются на одном месте более 3-4 дней [Антонов, 1999], а при
преследовании могут уходить на расстояние до 50-60 км [Слудский, 1956; Сухо-
рослов, 19726].
При отсутствии обильной пищи кабаны перемещаются от одной кормовой
точки до другой, проходя за ночь до 10-15 км и устраиваясь на дневки между
этими пунктами. Однако через день, два или более они все равно замыкают мар-
шрут. Кормовые площади и убежища, на которых животные держатся несколько
дней или недель подряд, соединенные сетью троп, образуют участки обитания, а
их совокупность составляет сезонный или годовой участок (рис. 56).
Сезонные участки обитания одиночных особей, семей, семейных групп и
стад существенно перекрываются и используются совместно или попеременно.
Нередко места кормежек и убежищ у многих из них одни и те же, особенно зи-
мой. Но на кормежке группы держатся обособленно и ложатся они отдельно, хо-
тя иногда расстояние между ними может быть весьма коротким [Тимофеева,
1975; Червонный, 1980в].
Взрослые самки, вырастившие потомство в данном районе, и самцы старше 4
лет более привязаны к определенным территориям, нежели другие особи. В Бел-
городской области, например, на месте мечения в последующие годы регистри-
ровали более 60% взрослых зверей. Центром участка обитания самок становится
место рождения и выращивания молодняка. Его площадь ежегодно наращивается
примерно на 1,1 км2 до 4-летнего возраста животных. Размер зимних участков
158
Рис. 56. Схема
структуры сезон-
ного (А) и годово-
го (Б) участков
обитания групп
кабана (по [Царев,
2000])
-----Границы сезонных участков обитания
EZ22 Временные кормовые участки
/•_ Места отдыха
' ♦ Пути переходов (локальные перемещения)
•• • • Границы годового участка обитания
обитания здесь около 1,2 (0,5-4,6) км2, осенних - 0,6 (0,2-1,4), многолетних - 3,7
(3,1-4,4) км [Царев, 1986а, 1989а,в, 1999, 2000].
На юге Нечерноземья зимой для семей кабана более характерен 1-2-
недельный цикл перемещения по участку. В кормных угодьях эти копытные за-
держиваются 1-2 дня, затем переходят на другие по соединяющим их тропам.
При частом беспокойстве длина суточных переходов увеличивается с 10-12 до
15-25 км, и в этом случае стадо может осваивать территорию до 15-20 тыс. га.
Благодаря системе троп размер участка особей и групп здесь не уменьшается за-
метно даже при высоком снежном покрове. Секачи после гона живут оседло, ис-
пользуя одну-три лежки, соединенные тропами. Их ночные маршруты короче,
чем у семей, и составляют в среднем 4,2 км [Воронин, 1980].
Размеры суточных и сезонных участков обитания диких свиней существенно
варьируют в разных регионах и биотопах, но при этом очевидна тенденция их
сокращения в летний и позднезимний периоды.
Летом в низовьях р. Или при отсутствии беспокойства семьи держатся на
площади в 200-300 га [Слудский, 1952]. Примерно такие же размеры сезонных
участков обитания зверей в Молдавии. В заповеднике «Кодры» участки отдель-
ных гуртов, территориальность которых выше, чем у других группировок, пере-
крываются на 20-50%. Минимальное расстояние между их центрами - 1,5-2 км.
Кормовые угодья, как правило, общие для соседних групп [Лозан, Лозан, 1981а;
Лозан, 1984].
В Подмосковье в малоснежье участки обитания составляют около 500 га [Ива-
нова, Овсюкова, 1976], но при высоком снеге резко уменьшаются. В Окском запо-
веднике их площадь во второй половине зимы сокращается почти в два раза - с 45
до 26 га. Одна самка с выводком более месяца находилась на территории в 6 га.
После снегопадов семья на протяжении недели не отходила от единственной
лежки далее 100 м. Размеры суточных участков колеблются здесь от нескольких
до нескольких десятков гектаров [Червонный, 1980в, 19906]. В Беловежской пу-
ще при подкормке звери всю зиму живут на площади не более 250 га [Лебедева,
1956а]. В Ленинградской области стадо несколько недель не выходило за преде-
лы территории в 15 га, имея обширную сеть троп, соединяющих постоянные
лежки с местами жировок [Царев, 1980а,б]. Такие же небольшие зимние участки
у животных и в Иркутской области [Никулин, 1981]. В Кировской области в мно-
159
госнежье семья с ноября по апрель обитала на 200-метровом участке ельника
[Павлов, 1999а]. В Завидовском заповедно-охотничьем хозяйстве в марте 1960 г.
некоторые особи не отходили от своей лежки далее 30 м, питаясь исключительно
мхом [Дормидонтов, 1969].
По моим наблюдениям, во многих областях России кабаны зимой как бы «при-
вязаны» к одному или нескольким кормовым пунктам (полям с остатками урожая,
подкормочным площадкам, буртам картофеля, силосным ямам, скотомогильникам,
свалкам), от которых набитые тропы уходят до ближайшего укрытия в лесу или на
болоте. В случаях, когда подкормочные площадки или неубранные поля находятся
на краю густого елового леса или болота, расстояние от места кормежки до лежек
исчисляется сотнями метров. При удобном расположении кормовых угодий и убе-
жищ звери неделями не покидают участки, площадь которых не превышает не-
скольких или нескольких десятков гектаров. При отсутствии вблизи от мест кор-
межки подходящих убежищ они проходят до места лежки до 8-13 км, причем рас-
стояние увеличивается в зависимости от степени беспокойства.
Нередко в многоснежье эти копытные скапливаются в наиболее кормных
угодьях. В Апшеронском районе Краснодарского края зимой 1969/70 гг. группи-
ровка в 120 особей в течение месяца держалось на площади в 200 га [Гинеев,
19756]. В низовьях р. Или на участке размером 6 км на 3,5 км регистрировали до
100 голов [Слудский, 1956], а на оз. Б. Аксаут большое стадо в феврале обитало
на площадке 100 м на 300 м [Соломатин, 1972]. На Сихотэ-Алине при урожаях
дуба и кедра отдельные особи и группы могут жить и осенью на территории раз-
мером в 100-400 га, осваивая за сутки всего 2-5 га [Раков, 1956; Зайцев, 2000].
В Италии, по данным радиопрослеживания, размеры суточных участков оби-
тания кабанов 1,1—3,9 км2, перекрывающиеся у самок и более обособленные у
самцов [Boitani et al., 1994]. На юге Франции они составляют 60-74 га, площадь
годовых участков самок достигает 4-6 тыс. га, самцов - 12-15 тыс. га. Наимень-
шие и стабильные на протяжении нескольких лет участки у самок с поросятами -
300-600 га, наибольшие у подсвинков - до 20 тыс. га. Каждая группа или особь
на локальной территории использует 5-10 лежек. Возвращение к логову преды-
дущего дня происходит часто, но непредсказуемо. Локальная группировка раз-
мещается на площади в 700 км2 [Mauget, 1979; Janeau, Spitz, 1984; Spitz et al.,
1984; Spitz, Pepin, 1985; Spitz, 1987; Spitz, Janeau, 1988, 1990]. Эти сведения близ-
ки к данным, полученным при исследовании пространственной структуры диких
свиней в Центральной России и на Дальнем Востоке [Зайцев, 2000].
В США сезонные участки обитания акклиматизированного европейского ка-
бана составляют в среднем 300-350 га, но в неурожайные на желуди годы увели-
чиваются до 1070 га, а суточная активность возрастает почти в 5 раз. И здесь
большинство зверей в течение суток замыкают маршрут и возвращаются к леж-
кам, которые используют от 3 до 7 дней подряд. Около 75% взрослых особей -
оседлые [Singer et al., 1981]. Участки обитания самок минимальны (17-30 га) в
последней фазе беременности и в первые недели после опороса. Среднее рас-
стояние между двумя максимально удаленными местоположениями зверей за су-
тки - 0,6 км у самок и 1,1 км у самцов [Kurz, Marchinton, 1972].
Характерная черта пространственной структуры популяции кабана в горных,
предгорных, пустынных и степных районах - раздробленность на мелкие груп-
пировки (микропопуляции), границами обитания которых становятся хребты,
ущелья, горные реки, безводные и безлесные участки местности, железные и
шоссейные дороги [Дуров, 19706; Сухорослов, 1970а].
В Кавказском заповеднике горные склоны фактически поделены между ло-
кальными группировками, занимающими 1—2 ущелья, и отдельными гуртами ди-
ких свиней. Индивидуальные участки секачей площадью 100-500 га включают
фрагменты 3-4 растительных поясов. В случае гибели владельца его территорию
160
в течение года занимают другие самцы. Группы свиней держатся на участках 1-2
взрослых самцов. Часть самок и молодняк - кочующие [Дуров, 1986].
На Сихотэ-Алине в кедровниках самки с выводками занимают участки оби-
тания в 300-400 га, секачи - 700-1000 га. У групп и отдельных особей их площа-
ди еще больше - до 2-3 тыс. га [Зайцев, Зайцева, 1990]. Во второй половине зи-
мы при высоком снежном покрове их размеры уменьшаются до 17-50 га [Раков,
1956; Антонов, 1999], а иногда и до 1 га [Пикунов и др., 1989]. Однако постоян-
ные участки обитания для уссурийского кабана все-таки не характерны. При не-
достатке корма и беспокойстве звери легко перемещаются из одного района в
другой. За сутки они способны проходить по 30-40 км, за 10-15 дней - до 200-
300 км [Бромлей, 1964; Бромлей, Кучеренко, 1983].
Один из основных факторов, влияющих на подвижность диких свиней, - охо-
та. По данным радиопрослеживания 59 особей, проведенного во Франции в
1987—1996 гг., до периода отстрела, разрешенного с октября по январь, участки
обитания у самцов составляют в среднем 692 га (до 1128 га), у семейных групп -
554 га. В сезон охоты их размеры увеличиваются соответственно до 3327 и 2502
га, а после него снова сокращаются до 500-700 га [Brandt et al., 1988].
Миграции
На северо-западе России осенне-зимние кочевки кабана обычны в южном и
юго-западном направлениях (рис. 57). В районах, где есть непромерзающие бо-
лота с островками хвойного леса, перемещения чаще локальные. Размах сезон-
ных переходов достигает десятков километров, отдельные звери уходят на сотни
километров. Весной они возвращаются назад примерно по тем же экологическим
«руслам» (долинам рек, заболоченным участкам, низменным угодьям), которые
использовались ими и при восстановлении ареала [Верещагин, Русаков, 1979;
Русаков, Тимофеева, 1984].
В Вологодской и Архангельской областях осенью большая часть диких сви-
ней также откочевывает на юг и юго-запад, где останавливается в островных ле-
сах и на незамерзающих болотах. Назад они возвращаются после схода снега
[Варнаков, 1978а; Плешак, Миняев, 1986]. В Дарвинском заповеднике массовые
выселения животных происходят в многоводные и многоснежные годы [Тупици-
на, 1988].
Звери, помеченные в госкомплексе «Завидово», регулярно уходят в примы-
кающие крупные лесные массивы Клинского, Волоколамского и Лотошинского
районов Московской области, Конаковского и Вышневолоцкого районов Твер-
ской области. Основное направление перемещений - с юго-запада на северо-
восток и, реже, - с юга на север. Крупные автострады и железная дорога не ста-
новятся здесь существенной преградой для эмигрантов. Оставшиеся стада зимой
концентрируются в центре охраняемой территории [Мануш, Николаев, 1995].
Сезонные высотные миграции кабана четко выражены в Карпатах: весной
большая часть популяции поднимается в верхние пояса гор, осенью спускается в
предгорье [Гунчак, 1982]. В Беловежской пуще сезонные перемещения зверей
сопряжены с урожаем дуба. Массовая эмиграция очевидна в годы неурожаев при
повышенной плотности населения [Козло, 1975].
На Северном Кавказе осенняя миграция начинается задолго до выпадения
снежного покрова. Она связана в большей степени с поспеванием фруктов и пло-
дов орехоносов и их распределением. Высоко в горах на южных склонах остают-
ся лишь отдельные семьи и крупные одиночные особи, которые покидают эти
места с выпадением снега и, напротив, по мере его таяния идут вверх. В урожай-
ные и малоснежные годы диапазон высотных перемещений зверей не превышает
1000-1500 м, но в неурожайные и многоснежные они уходят в предгорные и рав-
11. Заказ №2528.
161
Рис. 57. Перемещения
кабана на северо-за-
паде России в 1960-
1980 гг. (по [Русаков,
Тимофеева, 1984])
Кочевки: 1 - в холодные
зимы, 2 - осенне-зимние,
3 - весенние. Пути рассе-
ления: 4 - основные, 5 -
второстепенные, 6 - даль-
ние заходы
нинные районы, удаляясь от летних мест обитания до 100 км. Переходы совер-
шаются по водораздельным хребтам и долинам крупных рек. В Закавказье при
хорошем урожае дуба, бука или каштана эти копытные и зимой держатся в высо-
когорьях, спускаясь в долины лишь в самые многоснежные годы [Динник, 1910;
Логинов, 1936; Насимович, 1955; Чернявская, 1956; Дуров, 1973, 1987]. В При-
азовье, в дельтах предкавказских рек и на кавказском побережье Каспийского
моря животные оседлы [Соколов, Темботов, 1993].
В дельте Волги и на северо-восточном побережье Каспия, напротив, мигра-
ции регулярны, но связаны они с весенне-летними паводками и осенне-зимними
морянами. В это время кабаны уходят за десятки и сотни километров в Волжско-
Уральские пески, на Камыш-Самарские озера, в низовья рек Кушум, Б. и М.
Узень и на степные озера в районе пос. Урда, откуда могут проникать в пойму
Урала. Весной 1941 г. после особенно многоводного паводка в дельте Волги эти
копытные, пройдя около 250 км по побережью, достигли г. Гурьева. Вероятно,
миграционный путь в 400 450 км существует с побережья Каспийского моря на
162
Рис. 58. Перемещения кабана в Казахстане (по [Слудский, 1956])
1 - направления сезонных миграций, 2 - пути массовых эмиграций
Аральское через Устюрт. Через пески Большие Барсуки они проникают в низовья
Иргиза, а на п-ове Мангышлак заходят в горы Каратау и Актау. Из поймы Сыр-
дарьи осенью часть популяции откочевывает на 150-200 км на юг в пески Кы-
зылкум, где рассредоточивается по бывшим кустарниковым руслам рек, в бугри-
стых песках и саксаульниках. В конце декабря - феврале с началом сильных мо-
розов и глубокого промерзания почвы звери вновь возвращаются в тростниковые
заросли долины Сырдарьи (рис. 58). В Кызылкумы уходит и часть группировки,
обитающей в долине Амударьи. Не исключено, что между сырдарьинской и аму-
дарьинской популяциями может существовать регулярный обмен особями, осо-
бенно по побережью Аральского моря. Осенние откочевки диких свиней при-
мерно на 100 км отмечены также из низовьев р. Талас через пески Маюнкум в
долину р. Чу, откуда они выходят на 30 40 км в саксаульники и бугристые пески.
Возвращение животных происходит в марте. В отдельные годы летом во время
массового вылета кровососущих насекомых часть группировки из долины Сыр-
дарьи уходит на хр. Каратау, а с низовий р. Чу — через Чу-Илийские горы к ни-
зовьям р. Или, преодолевая расстояние в 50—150 км. Из дельты Или звери могут
переходить на Каратал икр. Аксу. Летние перемещения вдоль южного берега оз.
Балхаш регулярны. Группировки, зимующие в среднем течении Или и в низовьях
горных рек, в мае перемещаются в горы Заилийского и Джунгарского Алатау и
Кетмень, достигая субальпийского и альпийского поясов, а осенью спускаются
вниз. В малоснежные годы значительная часть популяции остается зимовать у
верхней границы леса и в арчовых зарослях в субальпийском поясе. Широко ко-
чуют кабаны и в Таласском Алатау, откуда могут уходить на зимовку, вероятно,
на хр. Каратау, расположенный на расстоянии 100-150 км, а отсюда в много-
снежные зимы - в долину Сырдарьи [Лавровский, 1952, 1962; Слудский, 1956;
Ишунин, 1972; Киселев, 1976; Слудский и др., 1984].
В горах Средней Азии в бесснежный период дикие свиньи предпочитают
жить в верхней трети лесного пояса, преимущественно в арчовниках, заходя на
альпийские луга до высоты 3500 м. На зиму они откочевывают вниз до предго-
11
163
рий, где концентрируются в камышах и тугаях по долинам рек, в ореховых лесах
и фруктарниках. Однако часть популяции остается в лесном поясе гор до января,
а в малоснежные годы проводит здесь всю зиму [Флеров, 1935; Насимович, 1955;
Федосенко, 1975; Обидина, 1982, 1987]. В Таджикистане с Вахшского хребта звери
уходят в долины рек Вахша и Пянджа, назад они возвращаются по мере таяния
снега [Иваненко, 1956]. В Туркменистане, помимо регулярных высотных миграций
в горах, выражены также осенне-зимние перемещения зверей на 30-180 км из гор-
ных районов и речных долин в пустыни Бадхыза, Каракумы и другие пески. Ос-
новной миграционный путь проходит по Унгузу, где имеются водосборные такы-
ры. Благодаря этим перемещениям осуществляется обмен особями в группировках
из низовий Амударьи, Мургаба, Теджена, Узбоя и Тахтинского района. С наступ-
лением жары животные возвращаются в прибрежные и горные тугаи. Часть свиней
приносит поросят в песках и мигрирует после того, как поросята окрепнут. После
строительства Каракумского канала и обводнения обширных территорий отдель-
ные группировки стали жить в пустыне постоянно [Ишадов, 1965, 1969, 1983].
Осенью в период дождей в фисташковые заросли Теджена подкочевывают стада из
Афганистана и Ирана [Соломатин, 1972].
На Алтае, где также отмечены высотные миграции, кабаны нередко задержи-
ваются в горах до снегопадов и переваливают через довольно высокие хребты,
преодолевая снежный покров в 50-90 см [Собанский, 1992]. На севере Монголии
их массовые перемещения связаны с урожаем кедра [Банников, 1954]. На Вос-
точном Саяне многоснежной зимой 1954/55 гг. наблюдалась перекочевка живот-
ных из бассейна р. Хойто-Ока на юг Окинского аймака. Протяженные переходы
зверей регистрировались и в бассейне р. Каа-Хем [Смирнов, 1978, 2001].
На Дальнем Востоке перемещения диких свиней также находятся в тесной
зависимости от урожая дуба и кедра. На Сихотэ-Алине осенние кочевки имеют
общее направление к главной оси хребта и особенно ярко выражены в тех рай-
онах, где мало или нет хвоща зимующего. При неурожаях стада мигрируют в по-
исках корма на десятки и сотни (до 700) километров. Массовое вынужденное пе-
реселение зверей на побережье зарегистрировано в 1932 г., в 1933 г. они эмигри-
ровали с западных склонов на восточные. Известны также перемещения этих ко-
пытных из бассейна р. Бикин в бассейн р. Иман и другие [Раков, 1956; Абрамов,
1963; Руковский, 1963; Бромлей, 1964; Кучеренко, 1973; Бромлей, Кучеренко,
1983; Антонов, 1999]. В Хинганский заповедник в урожайные годы в августе -
ноябре подкочевывают стада с сопредельных территорий, в неурожайные, напро-
тив, характерны две волны эмиграции: в октябре — ноябре и в январе - феврале
[Дарман, 1990].
Инфраструктура участков обитания
Каждый участок обитания кабанов имеет свою инфраструктуру, складываю-
щуюся из кормовых площадей (см. «Питание» и «Среда обитания»), сети троп,
«купалок», «чесалок», временных или добротно устроенных лежек, которые
охотники и натуралисты называют «логовами», «лежбищами», «гнездами» или
«гайнами». В дальнейшем я буду употреблять эти синонимы без кавычек.
Тропы. При высокой плотности населения диких свиней места их обитания
буквально испещрены взрытыми участками земли или снега, следами и многоки-
лометровыми тропами.
В весенне-летний и малоснежный (до 15-20 см) периоды звери в спокойном
состоянии обычно идут врассыпную (см. рис. 39), при опасности и в многосне-
жье — один за другим (рис. 59). Для формирования летней тропы нужно около
120 проходов животных [Бальчяускас, 1988]. Зимой тропа образуется при одно-
кратном перемещении группы, но она периодически заносится снегом, и ее при-
164
L/l
Рис. 59. Выживание кабанов зимой во многом зависит от наличия в группах мощных и сильных лидеров, способных торить тропы
(фото Ф. Тверитина)
Рис. 60. На лежке в бес-
снежный период (фото
В. Зайцева)
ходится каждый раз восстанавливать, особенно после обильных снегопадов.
Передвижение массивного низкорослого кабана в многоснежье, как уже подчер-
кивалось выше, представляет большую проблему. При высоте снега более 40 см
взрослые звери буквально вынуждены бороздить его грудью, оставляя за собой
траншею, а молодые пробиться через его толщу самостоятельно не могут. Зимнее
благополучие группировок, следовательно, во многом зависит от наличия в них
сильных особей, способных торить тропы, - взрослых самцов и самок-лидеров
(см. рис. 59). По набитым тропам-траншеям животные, включая сеголеток, спо-
собны перемещаться даже при 1-1,5-метровом слое снега.
При постоянном обитании диких свиней на участках и высокой плотности их
населения создается своеобразная «дорожная» сеть, связывающая, как правило,
кормовые стации с убежищами, где располагаются лежки. В определенных
местах на основных тропах и на расстоянии нескольких десятков или сотен
метров от долговременных лежек постепенно накапливаются кучи экскрементов
со специфическим «свиным» запахом, хорошо различимым даже человеком с его
слабым обонянием. Тропы, а также ольфакторно-оптические метки на деревьях,
однако, обычны и на границах участков обитания групп, что косвенно свидетель-
ствует о возможности целенаправленной их маркировки и наличии отдельных
элементов «патрулирования», что свойственно «территориальным» видам ко-
пытных.
Лежки. В теплое время года кабаны предпочитают отдыхать в защищенных
166
Рис. 61. Зимние
«гнезда» кабаны ча-
ще всего сооружа-
ют под низко опу-
щенными ветвями
крупных елей (фо-
то С. Лычагина)
сырых местах: в зарослях тростника и других высокостебельных околоводных
растений, в высокотравной растительности в лесу, преимущественно еловом
(рис. 60). Высокотравье и заросли позволяют укрываться от назойливых кровосо-
сущих насекомых. Возможно, по этой причине летом звери нередко днюют и на
хлебных полях.
В бесснежный период лежки чаще представляют углубления или вырытые
ямы в земле овальной формы без подстилки. Свиньи, имеющие маленьких поро-
сят, сгребают в кучи листья или устилают логово высокостебельиыми растения-
ми: тростником, папоротником, малиной, крапивой, борщевиком, ветками де-
ревьев, мхом. Летом земляные лежки-ямы способствуют теплоотдаче и предо-
храняют покрытые редкой щетиной живот и бока от укусов кровососущих насе-
комых. В жаркие дни взрослые звери в качестве убежища используют также ку-
палки, что тоже связано с терморегуляцией и защитой от кровососов.
Дикие свиньи, живущие на юге и западе ареала, зимой практически не уст-
раивают закрытых утепленных гнезд. В Беловежской пуще, например, все логова
без крыши [Саблина, 1955; Лебедева, 1956а; Щербак, 1986]. В Центральном Чер-
ноземье долговременные лежки они делают лишь в исключительных случаях и
обычно спят во временных [Сухорослов, 19726].
На севере ареала в снежный период кабан проявляет особенно активную
«гнездостроительную» деятельность (рис. 61). При устройстве логова разрывает-
ся снег, по возможности разрыхляется верхний слой почвы или вырывается не-
большая яма. Затем животные захватывают пастью ближайшие растения, срыва-
ют их или отламывают ветки и переносят к лежке, где иногда измельчают и ук-
ладывают, поправляя рылом. Дно гнезда чаще всего выстилается нижними вет-
вями рядом стоящих елей или еловым подростом, вырванным с корнем. В каче-
стве подстилки звери используют также ветки сосны, пихты, кедра и, реже, лист-
венных деревьев, тростник, папоротник и другие высокостебельные растения,
травяную ветошь, мох, собранные вокруг гнезда на площади 60-290 м~. Каждая
семья скусывает и обламывает до 130-320 экземпляров подроста деревьев тол-
щиной 0,5-3 см на расстоянии 10-25 м от гнезда, диаметр которого может дости-
167
Рис. 62. В морозные и ветреные дни кабаны устраивают лежки в стожках соломы
или сена (фото А. Дигилевича)
гать 5-6,5 м. В логовах одиночных особей площадь подстилки составляет 0,5-1 м2, в
групповых ее слой достигает толщины 10-42 см на площади 4-10 м~. Число ис-
пользованных веток или стеблей колеблется от 10-15 до 2 тыс. [Лавровский,
1962; Тимофеева, 1975; Русаков, 1980а; Гусев, 1989; Гусев, Гусева, 1989; Завац-
кий, 1989; Червонный, 19906; Смирнов, 2001].
В устройстве логова, как мне приходилось наблюдать, принимает участие вся
семья, включая подросших сеголеток. Попытки уставших детей лечь в недостро-
енном гнезде немедленно пресекаются самкой, отшвыривающей их в сторону.
Она же нередко прогоняет из готового сооружения подсвинков, которые вынуж-
дены устраивать новое лежбище неподалеку или ложиться в разрытом снегу на
тонком слое подстилки по двое головами в разные стороны. В сильные морозы
поросята стараются переместиться в середину группы, что сопровождается виз-
гом и грызней. На край выталкиваются более слабые особи, нередко замерзаю-
щие здесь же.
Большое стадо обычно делает несколько гнезд в нескольких десятках метров
одно от другого Секачи в период гона устраивают свои лежки в нескольких мет-
рах от групповых, в другое время - на значительном расстоянии. Их число у
взрослых самцов больше, чем у остальных животных, а слой подстилки в них не-
редко толще.
«Капитальные» гнезда кабаны чаще всего сооружают в хвойных лесах, преиму-
щественно еловых или в еловых вкраплениях в смешанном лесу, в мелколесье, около
стволов крупных деревьев, под низко опущенными ветвями (см. рис. 61) или упав-
шими деревьями. Они пользуются ими неоднократно и постоянно обустраивают.
Особенно часто приходится «ремонтировать» валик по краям. В морозные и ветре-
ные дни животные предпочитают зарываться в стожки сена (рис. 62) или соломы
[Иванова, 1974] Одиночные звери нередко в качестве лежек используют мура-
вейники и ондатровые хатки, разрытые по центру. Не исключено, что тяга к му-
равейникам связана не только с простым устройством в них гнезд, но и с репел-
168
лентными свойствами муравьиной кислоты. Во время переходов уставшие звери
не делают гнезд, а ложатся на снег или на малоснежное место под нависшими
ветвями елей, вырыв, если это возможно, небольшую ямку в земле, которую ни-
чем не выстилают.
Региональные особенности в распределении и степени обустроенности гнезд
весьма существенные.
В Ленинградской области, по сведениям С.А. Царева [1979], лежки без под-
стилки, в форме ям и купалок, характерные для бесснежного периода, составля-
ют соответственно 21, 18 и 34%, зимние лежбища в муравейниках - 8% и гайна-
19%. Основной гнездостроительный материал - ветки хвойных пород, тогда как
в Белгородской области - ветви желтой акации. На северо-западе России из 115
осмотренных зимних лежек 19% сооружены в муравейниках, 10% - в стогах се-
на, 15% - в завалах леса и кучах хвороста и тростника. Из них 29% имеют под-
стилку из еловых веток, тростника и трав, 16% - своеобразные стены и крыши, и
только 12% открыты и без подстилки [Русаков, 1980а]. В Хоперском заповеднике
более половины (56,6%) из 108 гнезд с подстилкой из тростника, рогоза и вето-
ши, 18,5% выстланы ветками древесных пород и столько же - без подстилки,
6,5% находятся в стогах сена [Антонец, 1987]. В Южном Подмосковье из 248
лежек 25% расположены в хвойных посадках, 28% - в мелколесье, 10% - в жерд-
няках, столько же — в копнах соломы и сена и 13% - в муравейниках [Воронин,
1975а]. В Брянской области почти половина из 106 логов устроена в густых лист-
венных молодняках с примесью ели, 21% - в жердняках, 13% - в сосновых по-
садках, 10% - в спелых смешанных лесах и небольшая часть - в стожках сена и
соломы [Ватолин, 1980]. В Окском заповеднике из 407 зарегистрированных гнезд
63% сооружены под елями с солнечной стороны [Червонный, 1980в, 19906]. В
Беловежской пуще большая их часть (61%) тоже у стволов елей или в еловом
подросте (17%) [Лебедева, 1956а]. В Черноморском биосферном заповеднике
звери предпочитает устраивать лежки в молодых сосняках (82%) и, реже, в дубо-
вых колках и зарослях терна [Москаленко, 1999]. На Северном Кавказе они
обычны у стволов пихт, которые дают защиту от осадков, в густых зарослях ро-
додендрона, азалии и самшита, а летом - в густых папоротниках, под нависшими
кустами смородины и в высокотравье [Донауров, Теплов, 1938]. В Хинганском
заповеднике половина гнезд расположены в зарослях лещины в дубняках, 28% -
в густом подросте дуба и по 11% - в липово-осиновых и черноберезовых лесах
[Дарман, 1990], на Западном Сихотэ-Алине почти все они (94%) устроены у
стволов кедра на склонах южных экспозиций [Антонов, 1999].
В горах Казахстана и Средней Азии кабаны выбирают для лежек пологие мес-
та, а небольшой уклон выравнивают, скапывая почву. Весной они предпочитают
отдыхать под валунами на прогреваемых южных склонах, летом в жаркую пого-
ду - на теневых склонах, под елями, между корнями деревьев, в зарослях кустар-
ников, причем нередко вырывают глубокие (до 1 м) ямы. В период массового
вылета кровососущих насекомых звери зарываются в подстилку, набрасывая на
себя землю и хвою. Зимой животные устраиваются на отдых на южных склонах
гор или под скалами, где температура может быть выше на 10-20°С. Предвари-
тельно они собирают лесную подстилку в большую кучу и разгребают ее в сере-
дине. На равнине, где зимой температура значительно ниже, чем в горах, свиньи
делают теплые гнезда из сухой травы, стеблей тростника, рогоза и осок, исполь-
зуют разрытые ондатровые хатки или забираются в стога сена или соломы. Если
животных не тревожат, то такие утепленные лежки они используют многократно
и длительно (до 20 дней), постоянно подновляя их [Флеров, 1935; Иваненко,
1956; Слудский, 1956; Ишадов, 1965; Чичикин, Воробьев, 1967; Злобин, 1969;
Слудский и др., 1984; Кулибаба, Пакулин, 1985; Жиряков, 1989].
Родильные гнезда. Их самки устраивают незадолго до опороса в укромном и
169
защищенном месте: в густых ельниках под нависшими ветвями, которые нередко
служат крышей, в тростниках, кустарниках, островках на болотах, в малинниках,
муравейниках.
Почти все осмотренные мной места родов свиней (п = 31) находились в пре-
делах 70-600 м от ближайшего водоема, и лишь три из них были на расстоянии
около 1-3 км. Очевидно, в период лактации самки испытывают большую по-
требность в воде, чем обычно. Однако если учесть, что большинство опоросов
приходится на ранневесенний период, когда вода повсеместно в изобилии, то на-
личие водоема, видимо, не обязательное условие при выборе самкой места для
гнезда.
Родильные гнезда почти не отличаются от обычных зимних сооружений, но
все же они, как правило, тщательнее обустроены и слой подстилки в них толще.
Дно выстлано более тонкими ветками, сухой травой, древесной трухой. Крыша и
стены, закрывающие логово, сооружены из веток, подроста хвойных деревьев,
хвороста, сухой травы, тростника. Материал для гнезда берется в радиусе не-
скольких метров, что нарушает его маскировку. Площадь сооружения варьирует
от 3 до 5 м2 при высоте до 1-1,5 м. Масса подстилки порой составляет около 30
кг [Червонный, 1980в], а толщина некоторых обломанных веток достигает 5 см.
Нередко самки рожают в стожках соломы или используют этот материал в каче-
стве подстилки. Иногда свинья сооружает на небольшой площади два-три лого-
ва, из которых затем выбирает одно.
В теплом климате родильные гнезда более просты, чем в холодном. В Герма-
нии они чаще устроены из сухой травы: примерно за 3 часа работы свинья со-
бирает ее в копну, куда потом заползает для опороса. Температура в таком со-
оружении, однако, сохраняется высокой - около +26°С при 8-градусном морозе
снаружи [Майнхардт, 1986]. В Молдавии выводковое логово представлено уг-
лублением в земле размером 120x30 см, чаще всего без подстилки, покрытое
сверху хворостом [Лозан, Лозан, 1981а; Лозан, 1984]. На северо-западе России
утеплены 73% родильных гнезд: половина из них с подстилкой, стенами и кры-
шей, 27% - без крыши, 9% располагаются в муравейниках и 27% - на земле без
подстилки, стен и крыши. Обычные размеры гнезд: длина 90-110 см, ширина 45-
50 см, высота 20-25 см [Русаков, 1980а; Русаков, Тимофеева, 1984].
Все 14 выводковых логов, осмотренных мной в Северной Монголии и на Ура-
ле, были хорошо утеплены, с толстым слоем подстилки, 7 имели крышу и 4 рас-
полагались в стожках соломы. На более тщательное их устройство в зонах с низ-
кими весенними температурами, в частности в Восточной Сибири и на Дальнем
Востоке, обращают внимание и другие исследователи [Сысоев, 1952; Банников,
1954; Черкасов, 1962; Смирнов, 1978].
Купалки и чесалки. В бесснежный период, как отмечают многие исследова-
тели, дикие свиньи охотно купаются, предпочитая стоячую или медленно теку-
щую воду, и часто валяются в жидкой грязи в лужах на дорогах, в ямах с водой,
на окраинах болот, в мелких ручьях, в зарослях ольшаника и тростника, у выво-
ротов корней деревьев. Поросята начинают заходить в грязь, видимо, на 3-м ме-
сяце жизни. Иногда звери устраивают и пылевые ванны, зарываясь в субстрат на
дорогах или на полях. С наступлением холодов «грязевые» процедуры становит-
ся редкими. Тем не менее взрослые особи, как я не раз регистрировал при троп-
лении следов, ложатся в незамерзающие болота, ручьи и лужи в оттепели даже в
ноябре - январе, если их длительно преследуют охотники с собаками. В Грузии
[Хеладзе, 1964] и Бурятии [Смирнов, 1978] кабаны посещают грязевые купалки в
любое время года.
Назначение купалок, вероятно, полифункциональное, прежде всего - термо-
регуляторное, направленное на охлаждение тела, что особенно необходимо упи-
танным зверям с большим слоем подкожного жира при высокой температуре
170
среды, а также при длительных переходах и быстром беге, сопровождаемых зна-
чительным выделением энергии. В опытах (принудительный бег на горизонталь-
ном треке при температуре среды +16...+18°С и скорости перемещения около 8
км/час) сублетальный уровень температуры тела (41,5°С) достигается на 39-й
минуте после 5,4 км пробега, при скорости около 5 км/час - на 96-й минуте после
9,6 км [Bunger и. а., 1976]. Купание в грязи и пыли, видимо, способствует и уда-
лению отмерших клеток эпителия кожи [Иваненко, 1956], эктопаразитов и по-
лезно как лечебная (грязь) процедура. Кроме того, грязевая корка защищает тело
животных от укусов кровососущих насекомых.
После грязевых «ванн» звери обычно чешутся о стволы хвойных деревьев,
кора которых предварительно ударами клыков или зубами превращается в подо-
бие гребенки. Со временем ствол сглаживается, затекает смолой, но на нем по-
стоянно появляются новые насечки от клыков. В Окском заповеднике 74% «че-
салок» обнаружено на елях, 23% - на соснах и лишь 3% - на березах [Червон-
ный, 19906]. В южных районах звери используют для этой цели ребристые или
корявые стволы грецкого ореха, арчи, боярышника, яблонь [Чичикин, Воробьев,
1967]. Предпочтение хвойных пород в качестве чесалок может быть связано с ос-
вобождением от кровососущих насекомых - смола способствует прилипанию эк-
топаразитов и служит природным репеллентом.
На многих деревьях-чесалках, тем не менее, нет следов грязи. Это свидетель-
ствует о том, что они служат не только для счесывания образовавшейся на теле
корки. На Северном Тянь-Шане возле купалок находятся лишь 38% помеченных
деревьев, остальные обнаружены вдали от воды на склонах (44%) и на водораз-
дельных хребтах (18%), из них хвойные породы составляют 67% [Жиряков,
1982]. На Алтае дикие свиньи нередко для чесания используют те деревья, на ко-
торых есть отметины медведя [Собанский, 1992]. Скорее всего, такие деревья
служат своеобразными ольфакторно-оптическими метками (рис. 48), о чем гово-
рилось выше. В репродуктивный период группы кабанов посещают места марки-
ровки чаще, чем обычно, здесь же происходят и спаривания [Царев, 1988].
Основные выводы по данному разделу следующие. Популяции кабана явно
гетерогенны по своему составу: в них есть как оседлые, так и мигрирующие осо-
би и группировки. Число тех и других в разных районах неодинаково, что в зна-
чительной мере определяется экологическими условиями. Недостаток кормов и
постоянное преследование вынуждают животных перемещаться в новые районы,
а при благоприятных кормовых и защитных условиях и в отсутствие чрезмерного
беспокойства они, напротив, ведут оседлый образ жизни на сравнительно не-
больших по размерам участках. Оседлость, следовательно, может служить ус-
ловным показателем благополучия отдельных группировок.
Сезонные миграции имеют место практически на всем ареале кабана в России и
сопредельных регионах. Особенно четко они выражены в горных областях, где име-
ют высотную направленность: зимой звери спускаются в нижние пояса гор, а летом
поднимаются в высокогорья вплоть до субальпийских и альпийских лугов. Нередко
кочевки возникают в связи со стихийными бедствиями: паводками, засухами, пожа-
рами и гололедицами. Массовые перемещения приводят к сезонному перераспреде-
лению численности и, соответственно, плотности населения в отдельных районах,
что необходимо учитывать в практике охотничьего хозяйства.
Размножающиеся самки и взрослые самцы наиболее «привязаны» к своим
участкам обитания. Молодые звери, особенно годовалые самцы, напротив, весь-
ма подвижны и склонны к эмиграции. По данным мечения, около 60-80% пере-
зимовавших поросят, изгнанных матерями весной незадолго до появления нового
потомства, навсегда покидают место своего рождения. На следующий год эмиг-
рантами становятся более половины оставшихся подсвинков, включая тех, кото-
рые ранее сохраняли связь с матерью. В районе рождения оседают всего лишь
171
около 10% молодых самок, но и они не наследуют участки обитания своих мате-
рей. Дистанция расселения молодых особей - от 5 до 250 км [Andrzejewski,
Jezierski, 1978; Майнхардт, 1983; Dardaillon, 1986; Meynhardt, 1988; Царев,
1989а,в, 1990, 2000; Мануш, Николаев, 1995; Уваров, 1995].
Миграционный процесс в целом весьма полезен для вида, поскольку проис-
ходит регулирование численности в плотно населенных популяциях, оставшимся
особям достается больше корма, а разобщенность позволяет сохранить какую-
либо часть группировки даже в экстремальные зимы. Кроме того, благодаря зна-
чительной подвижности зверей в популяциях сравнительно высок уровень пан-
миксии. За счет эмиграции и иммиграции их население ежегодно обновляется
почти наполовину [Andrejewski, Jezierski, 1978], а гетерозис, как известно, повы-
шает жизнестойкость вида. Эта особенность очень важна, поскольку у диких
свиней довольно часто происходит близкородственное скрещивание по линии
“отец-дочь” и “брат-сестра”.
Суточный маршрут особей и групп кабанов чаще представляет собой замкну-
тую кривую от убежища до мест кормежки и обратно. При благоприятных кор-
мовых условиях размеры сезонных участков обитания находятся в пределах от
нескольких десятков до нескольких сотен гектаров и почти совпадают на протя-
жении года. При неблагоприятных условиях они резко увеличиваются или, на-
против, уменьшаются до сотен метров, а зимние и летние местообитания могут
разделять десятки и сотни километров.
Дикие свиньи не охраняют активно свои участки обитания, хотя отдельные
элементы «патрулирования», мечения и «гнездового» обустройства территории
явно выражены. Этим копытным более свойственен так называемый «подвиж-
ный территориализм», при котором основными охраняемыми объектами стано-
вятся члены семьи или группы и место вокруг них.
Численность и динамика населения
Динамика ареала и населения кабана в голоцене, тесно взаимосвязанные,
прослежены выше. Как уже отмечалось, в историческое время этот вид был од-
ним из основных охотничьих объектов, что, в конечном счете, и привело к рез-
кому сокращению численности или полному уничтожению его человеком во
многих районах. Роль климата в данном процессе второстепенна [Кириков, 1952,
1953; Слудский, 1956; Аверин, 1960а,б; Гептнер, 1960; Гептнер и др., 1961; Ди-
несман, 1961; Лавров, Хахин, 1974].
Численность в XX веке
Динамика населения вида в России. Пик сокращения ресурсов кабана при-
ходится на начало XX в. В середине столетия происходит медленный рост пого-
ловья в европейской части России, чему в немалой степени способствовала эмиг-
рация зверей на восток из западного (Прибалтика, Белоруссия, Украина) очага
обитания (см. выше). Резкий подъем численности в локальных группировках от-
мечен при достижении ими плотности 0,2-0,3 особи на 1000 га, а при плотности
свыше 1 особи на ту же площадь они становились устойчивыми к воздействию
неблагоприятных факторов среды [Фадеев, 1973а,г].
Динамика населения вида во второй половине XX в. в областях, краях, рес-
публиках и в целом по Российской Федерации отражена в таблицах 33-35. С се-
редины 60-х годов очевиден неуклонный подъем численности (с 36 тыс. до 300
тыс.), но последнее десятилетие века завершается очередной депрессией, к сча-
стью, не принявшей катастрофического характера (табл. 35).
172
Таблица 33. Динамика численности кабана (тыс. особей) в областях, краях и республиках России в 1961-1980 гг.*
Область, край, республика 1961 1962 1963 1964 1965 1966 1967 1968 1969 1970 1971 1972 1973 1974 1975 1976 1977 1978 1979 1980
Калининградская 1,8 2,6 1,3 1,4 1,4 1,4 1,4 1,6 1,8 1,8 1,7 1,7 2,0 2,5 з,о 2,9 3,5 з,з з,з 2,9
Архангельская — — — — — — Ед. Ед. Ед. Ед. Ед. Ед. Ед. 0,01 0,03 0,05 0,1 0,4 0,8 1,0
Вологодская Ед. Ед. Ед. Ед. Ед. Ед. 0,01 0,01 0,04 0,1 0,2 0,4 0,6 1,4 1,6 1,9 2,9 3,6 3,8 3,8
Карельская __ — — — — — Ед. Ед. Ед. Ед. 0,01 0,03 0,06 0,1 0,1 0,5 0,8 0,8 1,1
Коми — — — — — — — — — — — Ед. Ед. Ед. 0,02 0,02 0,03 0,03 0,03 0,04
Ленинградская 0,2 0,2 0,25 0,3 0,35 0,35 0,4 0,4 0,5 0,65 1,85 1,9 2,3 2,8 5,0 6,9 7,9 9,0 7,0 4,3
Новгородская 0,2 0,2 0,2 0,2 0,2 о,з 0,4 0,6 1,0 1,5 2,4 4,6 4,6 7,0 5,6 6,7 6,8 6,2 4,9 4,8
Псковская 0,65 0,7 0,5 0,8 0,95 1,0 1,1 2,0 4,4 5,5 6,0 6,6 7,2 8,0 9,0 15,2 13,5 10,0 10,1 6,5
Брянская 1,5 1,0 2,1 2,1 2,1 3,4 1,1 2,6 2,6 4,0 4,8 5,5 6,5 3,5 4,9 4,0 3,5 4,2 4,2 з,о
Владимирская 0,01 0,02 0,02 0,02 0,02 0,04 0,05 0,07 0,1 0,3 0,6 0,9 1,1 1,2 3,0 2,5 2,1 2,3 2,0 1,5
Ивановская — — — Ед. 0,03 0,04 0,08 0,1 0,15 0,25 о,з 0,4 0,7 0,75 0,8 0,8 0,8 0,9 1,7
Калининская 0,3 0,5 0,6 0,6 0,6 0,6 0,6 1,1 1,5 з,о 4,5 7,0 6,0 7,5 6,0 6,0 6,0 6,0 5,0 7,7
Калужская 0,2 0,2 0,2 0,2 0,55 0,5 0,7 1,0 з,о 2,0 2,6 3,2 4,0 5,6 5,9 4,2 з,о 2,2 2,5 2,3
Костромская — — — — — Ед. Ед. Ед. Ед. Ед. Ед. 0,05 0,1 0,1 0,2 0,7 0,6 1,0 1,3 1,6
Московская 0,6 0,7 0,8 0,7 0,8 1,1 1,15 1,1 1,9 2,5 3,4 4,0 4,6 5,2 5,8 5,9 з,о 4,8 6,9 7,4
Орловская 0,06 0,05 0,09 0,1 0,25 0,12 0,15 0,5 0,6 0,4 0,45 0,3 0,7 1,1 1,7 1,6 1,1 2,2 0,8 0,7
Рязанская Ед. 0,03 0,03 0,03 0,04 0,06 0,06 0,15 0,2 0,5 0,7 0,8 0,8 1,1 1,1 1,4 2,1 2,0 3,2 2,5
Смоленская о,з 0,35 0,5 0,5 0,65 0,9 1,0 1,4 1,9 3,5 2,4 3,6 4,8 6,2 4,0 5,8 5,0 6,0 4,0 4,3
Тульская 0,06 0,2 о,з 0,3 о,з 0,4 0,4 0,4 0,6 1,0 0,5 0,6 0,9 0,9 1,3 3,0 3,2 2,4 1,3 1,4
Ярославская 0,02 0,1 0,2 0,4 0,4 0,4 0,2 0,4 1,0 2,0 1,5 3,0 з,о 3,5 4,0 2,0 2,0 2,0 2,5 2,5
Г орьковская Ед. Ед. Ед. Ед. 0,02 0,05 0,08 0,2 0,04 0,2 0,4 1,0 1,2 0,9 1,9 2,2 2,0 2,4 2,4 2,4
Кировская — — — — — Ед. Ед. Ед. Ед. 0,02 0,03 0,07 0,1 0,2 0,4 0,4 0,4 1,2 0,8 1,6
Марийская — — — — — Ед. Ед. Ед. Ед. 0,04 0,05 0,04 0,1 0,2 0,3 0,4 0,4 0,4 0,6 0,4
Мордовская — — — Ед. Ед. Ед. Ед. 0,05 0,15 0,09 0,2 0,3 0.4 0,9 0,8 1,1 1,3 1,2 1,2 0,8
Чувашская — — — - — Ед. 0,01 0,01 0,01 0,01 0,01 0,01 0,02 0,1 0,2 0,4 0,5 0,5 0,6 0,6
Таблица 33 (продолжение)
Область, край, республика 1961 1962 1963 1964 1965 1966 1967 1968 1969 1970 1971 1972 1973 1974 1975 1976 1977 1978 1979 1980
Белгородская 0,03 0,04 0,06 0,06 0,06 0,08 0,2 о,з 0,35 0,7 0,7 0,8 0,7 1,6 1,0 1,6 1,5 1,0 1,3 1,3
Воронежская 0,08 0,05 0,06 0,08 0,08 0,2 0,1 0,1 0,5 0,8 1,4 1,4 1,0 2,2 1,5 1,3 1,5 1,5 1,9 2,3
Курская 0,3 0,3 о,з 0,4 0,4 0,5 0,5 1,0 1,5 1,4 1,5 1,6 2,5 2,7 2,8 2,1 2,3 2,3 2,5 1,5
Липецкая 0,02 0,04 0,08 0,06 0,07 0,03 0,03 0,01 0,06 0,2 0,2 0,4 0,7 1,4 1,2 1,9 1,9 1,3 1,9 1,1
Тамбовская — — — — Ед. 0,02 0.02 0,16 0,2 0,3 0,4 0,5 0,5 0,6 0,8 1,3 1,5 1,3 1,4 1,2
Астраханская** 0,9 1,0 1,0 0,5 0,5 0,5 0,5 0,3 0,3 0,3 0,9 1,0 0,8 0,8 0,5 1,0 1,1 1,3 1,0 0,4
Волгоградская Ед. Ед. Ед. Ед. Ед. Ед. Ед. 0,04 0,05 0,15 0,2 0,4 0,6 1,4 1,6 1,8 2,9 3,6 3,8 з,з
Куйбышевская - Ед. 0,05 0,07 0,1 0,1 0,1 0,3 0,3 0,4 0,3
Пензенская — — — — — — — 0,02 0,06 0,25 0,5 0,6 0,6 1,9 2,2 2,6 3,4 2,4 2,2
Саратовская — — — — — — Ед. 0,01 0,01 0,08 0,08 0,15 0,25 0,7 0,8 1,3 2,2 2,3 2,3
Ульяновская — — — — — — — — Ед. Ед. 0,04 0,07 0,4 0,6 1,0 0,8 1,2 1,0 1,0 1,1
Калмыцкая 0,5 0,5 0,3 0,1 0,2 0,2 0,2 0,06 0,07 0,1 0,1 0,1 0,08 0,1 0,1 0,2 о,з 0,4 0,4 0,2
Татарская — — — — — — Ед. Ед. 0,01 0,02 0,1 0,2 о,3 0,4 1,5 0,6 0,8 1,0 0,9
Краснодарский 3,0 6,0 7,0 3,0 3,0 3,0 3,0 7,0 11,0 7,8 7,8 9,0 И,7 13,0 10,5 9,1 8,6 9,1 10,1 6,5
Ставропольски й 0,3 0,4 0,5 0,6 1,0 1,0 1,0 2,5 2,5 2,0 2,5 2,7 3,2 3,4 3,8 4,2 4,2 4,0 4,1 4,1
Ростовская — -- — — — Ед. Ед. Ед. 0,1 0,2 о,з 0,6 0,9 1,7 2,3 2,5 3,6 5,5 5,5 5,2
Дагестанская 7,0 7,4 1,8 1,1 2,0 1,5 3,0 1,6 3,0 1,3 3,о 2,7 2,8 з,о 3,5 3,7 7,1 7,2 4,2 4,2
К.-Балкарская 2,0 2,0 2,0 1,7 2,1 2,3 2,8 з,о 2,5 2,7 2,5 2,7 3,2 3,8 4,0 4,4 5,4 6,0 6,0 6,0
С.-Осетинская 0,5 0,6 0,5 0,5 0,9 1,1 1,3 2,0 2,0 2,8 1,9 1,3 1,3 1,4 1,4 1,6 1,4 1,7 1,7 1,7
Ч.-Ингушская 1,2 0,8 0,8 0,8 0,8 0,8 1,4 1,0 1,0 0,9 1,0 0,8 1,0 1,0 1,0 0,9 1,2 1,3 1,4 1,5
Курганская — — — - Ед. Ед. Ед.
Оренбургская 0,1 0,1 0,1 0,1 0,1 0,1 0,1 0,1 0,2 0,15 0,15 0,1 0,2 0,1 0,2 0,2 0,3 0,1 0,2 0,2
Пермская — — — — — — — — — - Ед. Ед. Ед. 0,05 0,1 0,1 о,1 0,1 0,1 0,5
Свердловская — — — — — — - — Ед. Ед. Ед. Ед. Ед. Ед. Ед. Ед. Ед. 0,1 0,2 0,2
Челябинская Ед. Ед. Ед.
Башкирская — — — — — - - - - - - - - Ед. Ед. Ед. Ед. Ед. 0,1 0,2
Таблица 33 (окончание)
Область, край, республика 1961 1962 1963 1964 1965 1966 1967 1968 1969 1970 1971 1972 1973 1974 1975 1976 1977 1978 1979 1980
Удмуртская - - - - - - - - - - - - - Ед. Ед. Ед. Ед. 0,06 0,08 0,1 Тюменская - - - - - - - - - - - -------- о 1 Красноярский - - ----- Ед. Ед. Ед. Ед. 0,05 0,3 0,1 0,1 0,1 0,1 0,1 0,1 0,1 0,4 Иркутская 1,5 1,5 2,0 1,0 0,7 0,8 0,8 1,2 1,0 1,0 1,2 1,2 1,0 1,0 1,0 1,0 1,0 1,2 1,2 1,2 Читинская 0,8 1,0 1,0 0,9 0,9 0,9 1,2 1,3 1,7 2,8 4,5 4,5 4,5 4,5 4,0 3,5 1,7 1,7 2,0 2,0 Бурятская 1,5 2,0 2,0 1,0 1,0 1,5 2,0 2,5 2,5 2,5 2,5 2,7 2,8 2,8 2,0 2,0 2,5 3,0 3,0 2,0 Тувинская 1,5 1,5 0,7 0,7 0,7 0,7 0,7 0,7 2,0 1,0 1,1 1,1 0,8 1,1 1,1 1,1 1,3 1,5 1,2 1,2 Якутская 0,2 0,2 0,2 0,2 0,2 0,2 0,2 0,2 - - - _________ Приморский 5,5 7,5 7,0 7,0 7,0 7,0 6,5 6,5 9,0 6,5 6,5 8,5 10,5 14,5 15,0 16,0 16,5 25,0 18,0 18,0 Хабаровский 6,0 6,0 2,5 3,0 2,5 2,5 6,0 6,0 6,5 6,5 6,5 6,5 4,5 5,5 6,5 6,5 7,3 11,0 7,0 7,0 Амурская 2,5 5,5 5,5 5,5 3,0 2,3 1,6 1,4 1,1 1,1 1,5 1,7 1,5 1,5 1,3 1,1 0,8 3,0 3,1 3,4
* По сведениям Главохоты РСФСР, с уточнениями по литературным источникам. Прочерк в таблице - отсутствие животных или достоверных данных о
численности.
** Данные, вероятно, без учета численности кабана в Астраханском госзаповеднике, где в 1966 г. было около 2,8 тыс., в 1970 г. - 2 тыс., в 1974 г. - 1,6 тыс.
особей [Киселев, 1975].
Таблица 34. Динамика численности кабана (тыс. особей) в областях, краях и республиках России в 1981-2000 гг. *
Область, край, республика 1981 1982 1983 1984 1985 1986 1987 1988 1989 1990 1991 1992 1993 1994 1995 1996 1997 1998 1999 2000
Калининградская 2,7 1,5 1,4 2,1 1,5 2,3 4,5 2,8 5,9 4,5 2,4 3,6 4,1 3,4 2,7 2,0 1,9 2,4 2,5 2,9
Архангельская 0,8 1,0 0,9 0,9 1,7 1,8 1,0 2,7 1,8 1,7 2,4 2,9 1,8 1,0 0,6 1,3 1,1 0,8 0,4 0,2
Вологодская з,з 3,2 2,7 2,7 3,0 2,9 2,5 4,0 5,0 8,8 10,6 8,4 5,0 4,4 3,4 3,5 2,2 3,7 2,1 2,6
Карельская 0,5 0,5 0,4 о,з 1,2 0,9 2,7 0,9 1,6 2,8 1,8 1,4 1,3 1,2 0,8 0,7 0,8 0,7 0,5 0,5
Коми 0,05 0,1 0,1 о,1 0,3 0,1 0,1 0,2 0,2 0,3 0,03 0,08 0,1 0,08 0,03 0,06 0,1 0,2 0,1 0,02
Ленинградская. 3,9 з,з 3,7 2,9 4,2 3,8 4,8 4,0 5,2 5,5 6,9 8,7 7,1 4,8 3,0 1,6 1,3 1,4 1,1 1,4
Новгородская 4,1 3,1 3,8 4,0 3,2 2,7 3,2 4,8 5,8 7,9 9,6 9,0 8,3 6,2 3,8 2,9 2,8 3,1 2,8 2,1
Псковская 4,2 4,2 5,0 4,2 3,5 5,1 7,6 6,8 9,4 10,9 13,9 13,8 12,7 11,0 6,2 4,6 4,0 4,7 5,0 4,8
Брянская 2,7 2,8 2,5 3,2 2,8 2,8 3,5 3,5 3,4 3,4 4,5 2,9 3,4 2,3 1,5 1,2 1,6 1,8 2,0 1,8
Владимирская 2,4 1,8 1,8 3,2 1,7 2,6 2,3 2,0 2,1 з,з 5,1 2,9 2,3 1,0 1,2 1,0 1,2 1,6 1,8 2,3
Ивановская 1,7 1,8 2,1 2,5 1,5 2,1 2,1 3,8 3,1 3,6 3,2 1,7 1,2 0,7 0,6 0,5 0,7 1,1 1,3 0,9
Калининская 7,6 7,1 8,4 10,1 6,9 8,7 8,7 8,2 8,3 13,7 17,9 13,5 14,2 10,2 7,1 5,7 5,4 5,9 6,4 6,4
Калужская 2,4 2,1 3,4 4,0 з,з 2,9 2,9 3,7 5,1 5,3 5,3 2,8 2,8 2,4 1,6 1,5 3,2 з,о з,з з,з
Костромская 1,4 1,6 2,0 2,4 2,0 1,8 2,8 2,5 2,5 4,1 3,4 2,9 3,4 1,8 1,0 1,1 1,2 1,8 1,0 1,2
Московская 7,8 5,7 5,6 8,4 7,9 8,1 8,1 7,2 6,4 7,0 7,2 4,3 3,6 2,8 2,9 3,3 3,9 4,6 4,9 5,0
Орловская 0,9 0,6 0,6 0,8 1,0 1,9 1,6 1,5 1,8 2,5 2,7 1,7 1,2 1,4 1,1 0,9 0,9 1,2 1,3 1,3
Рязанская 2,9 2,7 2,7 2,8 1,3 1,5 1,0 2,0 2,5 3,8 з,о 1,8 1,0 0,9 0,5 0,9 1,5 1,6 2,1 2,0
Смоленская 6,9 5,1 5,7 5,1 6,3 7,0 7,0 8,7 8,7 8,1 9,8 6,5 4,4 4,1 4,0 5,2 4,0 4,7 6,8 5,0
Тульская 1,7 1,1 1,4 1,5 0,9 1,4 1,8 1,9 1,9 4,5 2,7 2,1 1,9 2,0 1,8 1,9 2,4 2,2 3,4 2,6
Ярославская 2,6 2 2,6 4,0 2,4 2,8 5,0 3,5 4,3 8,0 7,8 5,9 3,8 1,9 1,9 2,1 2,3 3,2 3,0 2,6
Горьковская 2,3 2,5 2,8 2,5 4,7 1,7 2,5 2,1 1,7 2,7 2,4 2,5 1,6 1,2 0,6 0,7 1,2 1,3 1,4 1,7
Кировская 2,0 1,8 2,0 3,7 4,4 4,8 4,9 6,1 3,7 4,0 4,5 3,8 3,2 2,2 2,3 2,6 2,7 3,2 2,1 2,1
Марийская 0,5 0,5 0,6 0,9 0,5 0,5 0,5 0,5 0,8 0,6 0,6 0,6 0,4 0,2 0,2 0,2 о,з 0,6 0,5 0,4
Мордовская 1,1 1,7 1,9 1,3 0,9 0,2 0,3 0,4 1,4 0,6 0,5 0,4 0,4 0,3 0,4 0,4 0,5 0,5 0,7 0,5
Заказ№ 2528.
Таблица 34 (продолжение)
Область, край, республика 1981 1982 1983 1984 1985 1986 1987 1988 1989 1990 1991 1992 1993 1994 1995 1996 1997 1998 1999 2000
Чувашская 0,9 0,8 1,1 1,1 1,4 0,9 0,9 0,6 0,9 0,9 0,7 0,3 о,з 0,5 0,5 0,5 0,4 0,6 0,3 0,4
Белгородская 1,3 1,7 1,7 2,8 2,1 2,6 2,5 з,о 2,8 3,5 3,8 2,9 2,5 2,2 з,о 2,6 2,3 2,6 3,6 3,9
Воронежская 2,3 3,8 4,7 4,7 3,1 4,0 2,9 2,9 2,7 2,6 2,7 1,6 1,3 1,8 1,3 1,9 1,7 2,0 1,6 2,5
Курская 1,1 0,7 0,8 0,6 0,9 0,9 1,2 1,2 0,9 1,2 3,1 2,2 1,7 1,7 1,7 1,5 1,0 1,5 1,9 1,5
Липецкая 1,0 0,9 1,2 1,3 0,9 1,2 1,5 0,9 1,5 2,3 1,2 0,7 0,6 0,9 0,6 0,5 0,8 1,3 1,1 1,0
Тамбовская 1,3 0,9 0,9 0,7 1,0 0,6 0,6 0,8 0,8 1,4 1,9 1,2 1,1 1,0 0,5 0,8 1,0 1,0 1,3 1,0
Астраханская 1,3 1,1 0,9 0,8 1,3 0,3 0,7 0,7 0,7 0,8 0,4 0,5 0,6 0,7 1,1 2,2 1,5 1,2 1,0 1,5
Волгоградская 2,5 1,4 2,1 1,4 1,2 2,7 1,8 2,1 3,8 4,4 3,2 з,з 3,0 2,4 1,9 1,7 2,0 2,9 2,4 1,8
Куйбышевская 0,5 0,5 0,7 0,9 1,5 1,2 0,7 1,2 0,8 2,7 2,8 2,4 2,5 2,1 2,0 2,2 2,4 2,4 2,8 2,2
Пензенская з,о 2,8 2,7 2,3 1,9 1,7 0,7 0,7 2,5 1,9 1,6 1,9 2,1 1,6 1,2 1,4 2,0 2,2 2,0 2,0
Саратовская 2,6 з,о 2,9 2,7 1,9 2,3 1,8 2,0 з,з 3,1 5,0 4,6 4,2 2,9 2,8 3,1 3,1 4,2 6,0 6,0
Ульяновская 2,3 1,7 2,1 3,2 3,1 0,8 1,4 1,5 1,9 1,9 2,8 2,6 2,3 2,4 2,4 2,3 1,8 1,8 1,2 1,5
Калмыцкая 0,2 0,2 0,2 0,2 0,2 0,1 0,2 0,2 0,2 0,2 0,3 о,з 0,2 0,2 0,2 0,1 0,3 0,3 0,3 0,5
Татарская 1,4 1,5 1,4 1,4 1,8 1,3 2,9 1,6 1,7 1,8 2,5 2,7 1,5 1,0 0,6 1,6 1,8 2,3 2,5 2,3
Краснодарский 7,7 7,6 6,0 5,7 5,3 7,2 7,2 7,2 6,6 5,7 8,9 7,4 6,3 4,9 4,7 5,4 5,7 5,9 5,8 5,7
Ставропольский 4,1 4,1 4,0 4,0 1,0 2,1 2,6 3,7 2,7 2,9 з,о 3,1 з,о 2,5 2,6 2,5 2,7 2,9 2,9 2,7
Ростовская 5,2 5,2 5,2 5,2 2,8 2,3 2,3 2,3 2,5 2,3 4,0 3,4 з,о 2,9 2,1 1,6 1,5 1,7 1,9 1,7
Дагестанская 6,3 8,3 2,1 2,3 4,9 1,6 3,3 з,з 3,7 3,7 з,з 3,4 з,з 3,2 3,4 3,1 2,4 2,6 2,5 2,4
К.-Балк арская 4,0 1,6 1,8 1,3 1,5 0,4 0,5 1,3 1,5 5,3 з,о 2,9 2,0 1,0 1,0 1,1 1,4 1,7 1,7 2,3
С.-Осетинская 1,7 1,4 1,4 1,0 0,4 1,2 0,9 1,1 1,6 3,2 2,6 2,5 2,0 1,6 1,5 1,8 1,4 1,4 1,5 1,6
Ч.-Ингушская 2,0 1,9 2,8 3,3 2,4 2,2 3,3 1,9 2,0 2,6 3,1 3,4 3,3 ? ? 0,8 1,6 1,8 2,0 1,7
Курганская Ед. Ед. Ед. Ед. 0,1 0,2 0,4 0,5 0,8 0,9 4,6 з,з 3,6 2,7 2,4 1,8 2,3 2,6 2,3 2,9
Оренбургская 0,6 0,4 0,6 0,9 1,1 0,9 0,6 1,0 1,9 1,5 3,6 3,9 4,3 2,8 2,5 2,1 2,0 2,5 1,8 1,4
Пермская 0,7 0,9 1,2 1,5 1,1 2,9 3,0 3,4 2,7 2,0 з,з 2,7 4,6 3,2 1,4 2,4 3,2 2,4 1,2 1,1
Свердловская 0,2 0,2 о,з 0,5 1,1 3,2 3,0 3,8 3,5 2,8 3,3 2,4 1,5 2,2 2,5 2,5 2,5 2,9 2,5 2,3
Таблица 34 (окончание)
Область, край, республика 1981 1982 1983 1984 1985 1986 1987 1988 1989 1990 1991 1992 1993 1994 1995 1996 1997 1998 1999 2000
Челябинская Ед. 0,05 0,07 0,1 0,2 0,3 0,5 0,3 0,5 1,2 1,9 2,4 2,2 2,4 3,2 3,4 з,з 3,2 3,0 2,6
Башкирская 1,2 1,8 1,5 1,6 2,2 1,6 2,0 1,8 1,6 2,2 2,5 2,7 3,9 з,о 3,0 2,4 2,3 2,6 1,8 1,0
Удмуртская 0,2 0,2 0,6 1,0 0,9 0,7 0,7 1,0 0,7 0,4 0,8 1,0 1,2 1,2 0,8 0,9 0,6 1,1 1,0 1,0
Алтайский — — 0,2 0,1 — — 0,1 — — 0,5 0,7 0,9 1,1 0,5 0,7 1,о 1,0 1,1 2,1
Новосибирская — — — — 0,1 0,1 0,1 0,1 0,3 0,3 0,7 0,6 0,3 0,5 0,4 0,3 0,2 0,3 0,2
Омская — — — — — 0,5 0,5 0,5 0,5 0,5 1,4 1,0 1,1 1,0 0,9 0,4 0,3 о,з 0,4 0,05
Тюменская 0,2 0,2 0,2 0,9 1,2 1,6 1,6 1,7 1,6 1,6 2,0 2,6 2,6 4,2 1,9 2,7 2,1 2,4 1,8 1,8
Красноярский** 0,1 0,1 0,2 о,з 0,6 2,4 2,4 2,4 2,4 10,1 5,1 3,9 2,8 2,4 2,4 з,о 2,5 1,2 1,4 1,0
Иркутская 1,5 5,1 5,8 5,8 4,7 4,5 4,5 4,5 4,5 5,0 4,5 4,4 3,2 2,6 2,5 1,9 3,7 2,9 2,7 3,3
Читинская 2,0 2,6 4,0 4,6 5,2 13,9 6,3 6,3 7,5 19,3 13,3 12,4 8,9 5,8 8,7 8,1 7,0 6,3 9,4 7,8
Бурятская 2,0 з,о 3,5 2,5 4,0 6,3 5,0 5,0 4,2 8,6 12,4 10,3 6,8 5,0 3,7 3,1 4,3 4,1 3,7 4,7
Тувинская 1,2 1,2 1,2 1,2 2,4 2,4 2,4 2,4 4,5 2,4 3,2 2,3 1,7 1,2 1,4 1,9 4,4 4,3 4,3 4,3
Приморский 19,6 19,8 13,6 12,8 3,5 15,5 9,0 19,0 17,3 31,1 21,9 18,8 15,7 12,6 19,1 24,0 20,7 18,3 19,3 22,4
Хабаровский 7,0 8,0 9,0 9,2 3,5 16,1 4,0 4,0 5,3 4,0 22,7 13,8 17,7 14,7 14,4 14,0 12,1 10,0 7,3 10,0
Амурская 2,6 2,7 2,9 з,о 3,1 15,8 2,3 з,о 10,7 П,7 4,3 6,4 3,6 5,2 4,6 3,2 3,1 4,8 5,2 3,6
* По сведениям Главохоты РСФСР (Охотдепартамента РФ), Государственной службы учета охотничьих ресурсов, а также по [Фонд охотничьих угодий и
численность основных видов диких животных в РСФСР, 1992; Овсюкова, 1996; Ломанов, 2000], с уточнениями по литературным источникам.
** Вместе с бывшими автономными областями и округами.
Таблица 35. Приблизительная численность (тыс. особей) кабана в России во второй
половине XX в.*
Год Кол-во Год Кол-во Год Кол-во Год Кол-во
1961 41 1971 83 1981 158 1991 300
1962 51 1972 99 1982 153 1992 250
1963 42 1973 НО 1983 155 1993 223
1964 36 1974 132 1984 165 1994 179
1965 36 1975 138 1985 140 1995 161
1966 38 1976 151 1986 194 1996 164
1967 42 1977 155 1987 172 1997 165
1968 53 1978 177 1988 188 1998 173
1969 71 1979 161 1989 206 1999 174
1970 73 1980 150 1990 275 2000 170
*По сведениям Главохоты РСФСР (Охотдепартамента РФ), Службы учета охотничьих ресур-
сов, а также по [Иванова, Овсюкова, 1976; Фонд охотничьих угодий и численность основных видов
диких животных в РСФСР, 1992; Овсюкова, 1996], с уточнениями.
Рис. 63. Динамика населения кабана (1) и волка (2) в России
Как и у других копытных, начало интенсивного роста поголовья кабана сов-
падает с периодом максимальной депрессии численности волка (рис. 63), а пе-
риоды стабилизации и сокращения ресурсов коррелируют с увеличением населе-
ния этого хищника [Кузьмин, Хахин, 1980; Лавов, 19866; Лавов, Воронова, 1986;
Побединский, 1989, 1997; Фадеев, 1990; Данилкин, 1999а].
Динамика населения диких свиней, однако, не была синхронной в разных ре-
179
гионах. В Ленинградской, Новгородской, Псковской областях и Карельской
АССР в 1960-1976 гг. поголовье увеличилось с 850 голов до 28,9 тыс. В благо-
приятные годы оно удваивалось, в неблагоприятные не росло или же сокраща-
лось (до 12,8 тыс. в 1982 г.) [Русаков, Тимофеева, 1984]. В Северо-Западном и
Центральном районах пик численности зарегистрирован в 1975-1976 гг. В Цен-
трально-Черноземном районе в 70-е годы численность была сравнительно ста-
бильной. В это же время в Волго-Вятском и Поволжском районах группировки
бурно росли, а в Астраханской области и Калмыцкой АССР поголовье уменьша-
лось. Для популяций Северного Кавказа издавна характерен тип динамики насе-
ления с частыми и резкими колебаниями (табл. 33). За десятилетие ресурсы вида
увеличились здесь всего на 12% [Иванова, 1976, 1980а,б; Фадеев, 1989].
На востоке ареала динамика численности диких свиней имела иной характер.
В Восточной Сибири в 60-70-е годы ресурсы вида находились на стабильно низ-
ком уровне, а их рост заметен лишь в 80-е годы. На юге Дальнего Востока мно-
гочисленное прежде поголовье [Пржевальский, 1870] резко сократилось после
особенно многоснежной зимы 1914 г. В дальнейшем, несмотря на многократные
«пики», оно уже никогда не достигало прежнего уровня, а в середине XX в. каж-
дые 10 лет уменьшалось примерно на 10-12% [Бромлей, 1964; Кучеренко, 1976;
Бромлей, Кучеренко, 1983]. Судя по официальным сведениям (табл. 33 и 34), в
60-е годы здесь обитало в среднем около 15 тыс. особей, в 70-е годы численность
увеличилась до 24 тыс., в 80-е - до 29 тыс. и в 90-е - до 37 тыс.
Основные ресурсы кабана в России в 70-80-е годы были сосредоточены в
Центральном (около 40 тыс.), Дальневосточном (25-29 тыс.) и Северо-
Кавказском (24 тыс.) районах [Сицко, 1986]. Максимальная численность (300
тыс. особей) зарегистрирована в 1991 г., после чего поголовье сократилось почти
в 2 раза. К 1995 г. на севере европейской части России и в Центральном регионе
оно уменьшилось в 3 раза, в Северо-Западном и Волго-Вятском - в 2,3, Черно-
земном и Северо-Кавказском - 1,8, Поволжском - 1,5, Уральском - 1,3, Западно-
Сибирском - 1,1, Восточно-Сибирском - 2,1 и Дальневосточном - в 1,3 раза
(табл. 34). Этот негативный процесс имел место в разных природных зонах, при
разных кормовых и климатических условиях и был наиболее интенсивным там,
где выше плотность людского населения и ниже плотность населения зверей.
Резкое снижение численности диких свиней в начале 90-х годов XX в., как и
большинства других видов диких и домашних копытных, на мой взгляд, не свя-
зано напрямую с климатическим и трофическим факторами [Ломанов, 1995,
2000; Овсюкова, 1996; Ломанов, Ломанова, 2000]. Данный процесс, вне всякого
сомнения, - следствие экономических и социальных преобразований в России,
приведших к снижению жизненного уровня населения и кризису в сельском и
охотничьем хозяйствах. В этот период заметно ухудшились охрана диких живот-
ных и их подкормка, существенно выросла численность волка (рис. 63), лицензи-
онная добыча была максимально высокой и крайне нерациональной (см. рис. 69),
резко возрос уровень браконьерства, что не могло не отразиться на состоянии
популяций [Глушков и др., 1997; Данилкин, 1997, 1998а,б, 1999а, 20006]. По тем
же самым причинам сократились ресурсы копытных практически во всех постсо-
ветских республиках [Квакин, 1997; Tonisson, 1997; Козло, 1999, 2001; Муравиц-
кий, 2001].
В самом конце XX в. численность кабана в России стабилизировалась на низ-
ком уровне с тенденциями сокращения поголовья на севере европейской части, в
Волго-Вятском регионе и на Урале и относительно устойчивого роста в цен-
тральных и южных областях, в Восточной Сибири и на Дальнем Востоке [Лома-
нов, 2000].
Динамика населения вида в ареале. В Литве в 1934 г. жили 280 кабанов, в
1943 г. - 1 тыс., 1950 г. - 3,8 тыс., в 1969 г. - 12 тыс., в 1984 г. - 19 тыс. [Baleisis
180
u. a., 1987]. В Латвии в 1940 г. их насчитывалось менее 1 тыс., в 1960 г. - 2,4 тыс.,
в 1970 г. - 8,2 тыс., в 1974 г. - 13,6 тыс. [Сакс, 1975]. В Эстонии в 1950 г. обитало
около 100 голов [Tonisson, 1974], в 1955 г. - 600, в 1974 г. - 6 тыс., а ежегодная
добыча достигла 1-1,6 тыс. голов [Линг, 1978]. К 1987-1989 гг., по сведениям
Министерства сельского хозяйства (МСХ) СССР, в Литве численность возросла
до 22-24 тыс., в Латвии - до 22-26 тыс., в Эстонии - до 11-13 тыс.
В Белоруссии поголовье диких свиней растет с 1927 г. В Речицком Полесье в
1930-1931 гг. регистрировали не более двух десятков кабанов, в 1950 г. - около
100, в 1965 г. - 1365, в 1974 г. - более 2100 [Сержанин и др., 1975]. Во всей рес-
публике в 1936 г. насчитывали 1,5 тыс. особей, в 1948 г. - 5 тыс., в 1967 г. - 15, в
1972-1978 гг. - около 27-30, в 1984 г. - 31 ив 1988 г. - 33 тыс. [Сержанин, 1970;
Козло, 1975; Романов, Козло, 1977; Савицкий, 1980; Вадковский, 1986, 1988]. В
1990 г. численность достигла 39,3 тыс., но к 1997 г. сократилась до 25,3 тыс.
[Козло, 1999; 2001].
На Украине стремительный рост поголовья происходит в 40-е военные годы,
вероятно, в связи с благоприятными кормовыми условиями и меньшим пресле-
дованием человеком. В местах обитания единичных особей появляются крупные
стада до 35 голов, и звери распространяются по 18 областям. В послевоенные го-
ды из-за неограниченной охоты, гололедиц и эпизоотий население этих копыт-
ных снова резко (на 90%) сокращается [Корнеев, 1970]. Новый подъем численно-
сти, связанный с развитием охотничьего хозяйства и улучшением охраны живот-
ных, отмечается с начала 60-х годов. С 1959 по 1974 гг. поголовье возрастает с
7,5 тыс. до 64 тыс. при одновременном увеличении добычи более чем в 8 раз. В
конце 80-х годов оно находится на уровне 55-60 тыс. особей [Болденков, Край-
нев, 1975, 1980; Евтушевский, 1988; сведения Управления охотничьего хозяйства
республики].
В Карпатах в начале 60-х годов насчитывают 2-4 тыс. кабанов, в 70-е годы -
около 10 тыс. при высоком уровне добычи [Керечун, 1980; Гунчак, 1982]. В са-
мой густонаселенной (190 человек на 1 км2) Донецкой области, где доля лесов
составляет не более 6,5% территории, с 1965 по 1977 гг. их численность увеличи-
вается с 13 до 2200 голов [Гречушкин и др., 1975а,б; Харченко и др., 1980].
Среднеприднепровская популяции в 1963-1971 гг. вырастает в 5 раз [Евтушев-
ский, 1975]. В степные районы Северного Причерноморья звери зашли в 1968 г„
в 1970 г. здесь было всего 20 особей, в 1980 г. стало 1500 [Издебский, Тарасов,
1982]. Крымская популяция, образованная в результате выпуска нескольких де-
сятков особей в 1957 г., к 1968 г увеличивает свою численность до 2400 голов
[Кормилицин, Дулицкий, 1972]. В Молдавии в 1987-1989 гг., по данным МСХ
СССР, ресурсы вида не превышали 4,4-5,0 тыс. особей.
В Закавказье, Казахстане и в Средней Азии во второй половине XIX - начале
XX в. население диких свиней резко сократилось, главным образом из-за нерегу-
лируемой охоты и хозяйственной деятельности человека. С середины XX в. ох-
рана животных несколько улучшилась, в большинстве республик были установ-
лены сроки охоты и введена лицензионная добыча. Эти меры позволили сохра-
нить ресурсы, но далеко не везде из-за резко усилившегося браконьерства. В
Южном Приаралье, например, в 50-х годах поголовье составляло около 20 тыс.
особей, к концу 80-х оно уменьшилось до 3 тыс. [Реймов и др., 1989].
В Армении численность кабана в 70-е годы оставалась на стабильно низком
уровне (1-1,3 тыс.), в Азербайджане колебалась от 4 до 17 тыс., в Грузии умень-
шилась с 10 тыс. до 0,9 тыс. В Казахстане в 1973 г. насчитывали около 10 тыс., в
Киргизии - 8 тыс., Таджикистане - 7 тыс., Узбекистане - 23 тыс., в 1978 г. соот-
ветственно - 5,5, 9,6, 7 и 14,5 тыс., в Туркмении - 2,3 тыс. особей [Максимов,
1974; Иванова, Овсюкова, 1976; Фадеев, 1987; Арабули, 1989]. По сведениям
МСХ СССР, в 1987-1989 гг. ресурсы вида в этих республиках были следующи-
181
ми: Армения - 3,7-4,4 тыс., Азербайджан - 7,6-5,9 тыс., Грузия - 4,9-5,6 тыс.,
Казахстан - 7-13 тыс., Киргизия - 4-7,5 тыс., Таджикистан и Узбекистан - около
4 тыс., Туркмения - 1,1—4,0 тыс. В Казахстане в конце XX в. обитали около 10
тыс. особей, в том числе 6 тыс. — в равнинной части республики [Бекенов и др.,
1989; Бекенов, 1999].
В странах Западной и Центральной Европы поголовье диких свиней заметно
увеличивается после первой мировой войны, но особенно быстро - с середины
XX в. В ГДР в период с 1955 по 1984 гг. оно выросло с 29 тыс. до 60-65 тыс. (при
одновременном росте добычи с 12 тыс. до 100-128 тыс.), в Болгарии с 1960 по
1975 гг. - с 13 до 41 тыс. [Briedermann, 1986], в Венгрии с 1980 по 1989 гг. - с 20
до 37 тыс. [Nagasy, Takacs, 1994].
В конце 70-х годов численность вида в Европе оценивалась в 668 тыс., в 80-е
- в 774 тыс. при ежегодной добыче в 409 и 522 тыс. голов соответственно. В Се-
верной Америке ресурсы акклиматизированного европейского кабана составляют
около 160 тыс. голов [Дежкин, 1980, 1983, 1989]. В бывшем СССР в 1989 г. оби-
тало около 460 тыс. особей (табл. 36).
Таблица 36. Приблизительная численность кабана (тыс. особей) в разных странах Евра-
зии и в Северной Америке в конце XX в.*
Страна | Численность | Страна | Численность
Австрия 5 США и Канада 160
Бельгия 9 Япония 30
Болгария 40 Монголия 50
Венгрия 37 СССР (бывший): 460
Г ермания 110 Россия 300-170
Греция 10 Украина 60-50
Дания 1 Белоруссия 39-25
Испания 20 Молдавия 5
Люксембург 1 Литва 24
Нидерланды 2 Латвия 26
Польша 64-74 Эстония 13
Румыния 50 Грузия 5
Словакия 42 Армения 4
Чехия 15 Казахстан 10
Финляндия 1-3 Киргизия 8
Франция 50-70 Таджикистан 4
Швейцария 0,6 Узбекистан 4
Швеция 0,3 Туркмения 4
Югославия 50
* По сведениям Главохоты РСФСР, Службы учета охотничьих ресурсов РФ, Управлений охот-
ничьим хозяйством бывших республик СССР, а также по [Соколов и др., 1982; Кузякин, 1985;
Uloth, 1988, 1990; Климова, 1989] и источникам, приведенным в тексте.
Периодичность в динамике населения. В популяциях диких копытных
численность изменяется циклично лишь на протяжении года. Поздней весной по-
сле рождения молодняка она максимальна, затем с каждым месяцем уменьшает-
ся, и тем быстрее, чем выше смертность. Самый низкий ее уровень — перед появ-
лением нового потомства. Многолетняя динамика населения, определяемая ком-
плексным воздействием многих природных и антропогенных факторов, тоже пе-
182
7000
Рис. 64. Динамика численности кабана в Беловежской пуще (1) и национальном
парке «Завидово» (2)
риодична, однако интервалы между пиками нечетки, непостоянны и неодинако-
вы в разных районах.
При анализе динамики населения кабана многолетнюю цикличность не уда-
ется выявить ни в целом по России (см. табл. 35, рис. 63), ни в отдельных облас-
тях (см. табл. 33 и 34) и регионах [Иванова, Овсюкова, 1976; Русаков, Тимофеева,
1984], ни в локальных группировках (рис. 64, табл. 39 и 40).
На юге Дальнего Востока некая устойчивая периодичность, связанная глав-
ным образом с урожаем дуба и кедра, проявляется через 5-8 лет [Бромлей, 1964],
в приазовских плавнях - через 11, в предгорьях Кавказа - через 12 [Бакеев, Кос-
тоглод, 1975], а на юго-западе Болгарии - через 5 и 18 лет [Генов, Коцаков,
1986]. В разных областях Центрального и Северо-Западного районов устойчивый
рост численности кабана во второй половине XX в. имел место на протяжении
12—30 лет. Заметное уменьшение населения зверей, однако, произошло почти
одновременно в 1976-1977 гг. из-за неблагоприятных зимовок и затяжных весен,
усугубленных эпизоотиями чумы, увеличением численности волка и нерациональ-
ным промыслом [Иванова, 1982]. В Центрально-Черноземном регионе динамика
популяций различалась даже в соседних областях: в Воронежской и Тамбовской их
рост длился 9 лет, в Курской - 10, Липецкой - 13, Белгородской - 14 лет. В целом
по региону с 1966 по 1995 гг. рост и уменьшение численности чередовались сле-
дующим образом: +12, -1, +1, -3, +3, -2, +1,-1, +2, -4 [Простаков, 1996]. В России
в период с 1961 по 2000 гг. это соотношение было таким (табл. 35): +1, -2, 0, +13,
-2, +1, -1, +2, -1, +1, -1, +4, -4, +4, -1.
183
Плотность населения
Плотность населения кабана в европейской части России в 70-80-е годы воз-
росла в несколько раз, достигнув в отдельных южных и западных областях 6-12
особей на 1000 га охотничьих угодий. Однако на юге Сибири она по-прежнему
оставалась на очень низком уровне - не более 0,4 экземпляра на ту же площадь, а
на юге Дальнего Востока колебалась в пределах 0,4-2,8 особей (табл. 37). Рас-
пределение плотности населения этого вида в России в 1981—1995 гг. отражено
на рисунке 65.
Рис. 65. Среднемноголетняя плотность населения (особей на 1000 га лесной площа-
ди) кабана в России в 1981- 1995 гг. (по [Овсюкова, 1996])
1-0,06-0,3; 2-0,31-0,5; 3-0,51-1,0; 4-1,01-2,3; 5-2,31-4,0; 6-4,01-12,0; 7-более12,0
На периферии ареала эта величина обычно ниже 0,3 особей на 1000 га. Ее ли-
митирует, как правило, единственный фактор среды, находящийся в минимуме.
На севере это недостаток влажных кормов или ограничение доступа к ним при
многоснежье и глубоком промерзании почвы. Именно по этой причине животные
могут полностью исчезать из северных районов и появляться здесь вновь спустя
несколько лет за счет иммигрантов. На юге основным лимитирующим фактором
становится дефицит воды и, следовательно, недостаток влажных растений и
скудность почвенной фауны.
В северной части ареала распределение плотности населения кабана носит
как бы ступенчатый широтный характер, в определенной мере совпадающий с
высотой и продолжительностью залегания снежного покрова. Даже в пределах
отдельных областей на севере плотность обычно ниже, чем на юге. В Архангель-
ской области и в Предбайкалье она ниже почти на порядок [Плешак, Миняев,
1986; Недзельский, 1994]. В 70-е годы в юго-западных районах Псковской облас-
ти на 1000 га угодий обитали 7-8 диких свиней, в северо-восточных - 3-4. С та-
кой же густотой были заселены и юго-западные районы Новгородской и Ленин-
градской областей, в то время как в северных районах было менее 1 особи [Руса-
ков, Тимофеева, 1984]. В этот же период в соседней Литве средняя плотность на-
селения вида достигала 10 голов при высокой численности других диких копыт-
184
ных: лося - 6, благородного оленя - 13, косули - 25 особей на 1000 га [Падайга,
1982; Baleisis и. а, 1987]. В юго-западных областях европейской части России
плотность составляла 6-10, в западных и центральных - 2-6, в северных - 1-2
особи [Иванова, 1976].
Таблица 37. Средняя плотность населения кабана во второй половине XX в.
Район Особей на 1000 га Источник информации
Литва 10-11 Падайга, 1982; Baleisis и. а., 1987
Польша 5-11 (до 21,5) Uloth, 1990
Белоруссия 3,2-3,6 Вадковский, Костин, 1978;
1,9-7,2 Козло, Дунин, 1989; Козло, 2001
Речицкое Полесье 8,4 Сержанин и др., 1975 <
Украина 2,5-19 Иванова, Овсюкова, 1976
Карпаты 6,9 Гунчак, 1982 *
Россия: С.-Западный р-н 1-8 Русаков, Тимофеева, 1984
Центральный р-н 8 Иванова, 1980
Ц.-Черноземный р-н 6-11 44
Поволжский р-н 6-10 и
Северо-Кавказский р-н 3-10 Иванова, 1976; Дуров, 1973 :
С.-Восточный Кавказ 3,6 Равкин, 1976 .
Подмосковье 1,8-4,2 Приклонский, 1999
Европейские области 0,1-4,3 Фадеев, 1973г, 1978, 1982
Оренбургская область 0,4-2 Самигуллин, 1990а
Алтай 0,3 Смирнов, 1994
Юг Красноярского края 0,06 а
Тува 0,3 и
Иркутская область, 0,3-0,4 и
Бурятия Читинская область 0,2 и
Забайкалье 0,1-0,4 Смирнов, 1978; Байдавлетов, 1980
Юг Дальнего Востока 0,4-2,8 Метельский, 1975; Кучеренко, 1973,
Тянь-Шань 5-25 1976; Бромлей, Кучеренко, 1983 Жиряков, 1982
Каракумский канал 8-12 Зинченко, Черногаев, 1989
Дельта Амударьи 6-16 Реймов, Карабеков, 1984; Реймов и
Монголия 1-2 др., 1989 Соколов и др., 1982
Однако в ряде регионов плотность населения копытных выше не на юге, где
выше температура, меньше снежный покров, короче продолжительность его за-
легания и богаче растительность, а на севере. В Белоруссии, например, плотность
населения кабана и лося увеличивается в направлении с юга на север в 2,5 и 5 раз
соответственно [Козло, Дерябина, 1986; Козло, Дунин, 1989; Козло, 2001]. То же
характерно и для Подмосковья, где для диких свиней более значимы благоприят-
ное сочетание пойменных угодий, богатых естественным кормом, ельников или
их вкраплений в лесах, служащих убежищами, а также специализация сельского
хозяйства. Концентрация животных наблюдается в районах с наиболее плодо-
родными почвами, где развито зерноводство, и в местах интенсивного ведения
185
охотничьего хозяйства, где образованы кормовые поля и подкормочные площад-
ки [Иванова, 19756, 1982].
В зоне кедрово-широколиственных лесов юга Дальнего Востока плотность на-
селения кабана тесно коррелирует с плодоношением дуба и кедра и при урожаях
почти всегда существенно возрастает за счет подтока мигрантов. При средней мно-
голетней плотности населения в 3-4 особи на 1000 га в урожайные годы и после
них насчитывают от 6 до 12 голов на ту же площадь [Кучеренко, 1976; Бромлей,
Кучеренко, 1983]. Наивысшие плотности (до 50-100 голов) регистрируются в оре-
хово-плодовых лесах Кавказа и Средней Азии [Иванова, Овсюкова, 1976].
Плотность населения диких свиней на особо охраняемых территориях - в за-
поведниках, заказниках и образцовых охотничьих хозяйствах (табл. 38) несрав-
ненно выше, чем в других угодьях (табл. 37). При этом замечу, что во многих за-
поведниках обитают крупные хищники, нет подкормки, и молодые особи в массе
эмигрируют отсюда в соседние, менее заселенные территории. Очевидно, что в
настоящее время ведущую роль в восстановлении и сохранении вида играет его
охрана.
Таблица 38. Плотность населения кабана на особо охраняемых территориях
Заповедник, заказник, охотхозяйство Особей на 1000 га Источник информации
Беловежская пуща 7-27 Толкач, 1975; Шостак, 1975,1978
Березинский 11-15 Лавов, Воронова, 1986; Лавов, 1987;
Филонов, 1989
Припятский 8 Гатих, 1975
Полесский 8,5-36 Дунин и др., 1998
Дарвинский 4,4-5,3 Завьялова, 1997
“Завидово” 10-58 Фертиков и др., 1999
“Переславское” 9-18 Иванова, 1976
Приокско-Террасный 12 Иванова, 19756
Воронежский 36-37 Дьяков и др., 1986
Центр.-Черноземный 16-144 Гусев, 1986,1989
“Лес на Ворскле” 80-150 Тимофеева, 1982; Царев, 1989в
Хоперский 96 Ковалева, Антонец, 1995
Жигулевский 5,5-6,5 Вехник, Саксонов, 1987
Астраханский 27-75 Лавровский, 1952,1962
и 10-36 Киселев, 1976
Кавказский 19-60 Дуров, 1980а
Тебердинский 10 Бобырь, 1975
Хинганский 4-25 Дарман, 1986, 1989,1990
Сихотэ-Алинский 14 Мысленков, Волошина, 1989
Кызыл-Агачский 35-37 Литвинов, 1980, 1981
Закатал ьский 80 Иванова, Овсюкова, 1976
Сары-Челекский 60-70 U
“Цуманское” 21 U
Алма-Атинский 5 Жиряков, 1986
С.-Хасардагский 8-30 Кулибаба, Пакулин, 1985; Лукарев-
ский, 1986
Чаткал ьский 11 Есипов, 1991
Келифский 24 Зинченко, Черногаев, 1989
Исмаиллинский 144 Кулиев, 1989
Карадагский 14 Курочкина, 1999
186
В связи с таким заключением весьма любопытно проследить многолетние
изменения численности и плотности населения кабана и других копытных в не-
которых заповедниках и заповедно-охотничьих хозяйствах, где хорошо налажена
охрана животных.
В Березинском заповеднике после его закрытия в 1951 г. численность косули
сократилась почти в 7 раз, кабана - в 2,6 раза. К моменту восстановления запо-
ведного статуса в 1969 г. плотность населения копытных, включая лося, здесь
была такой же, как и в соседних угодьях. Затем при режиме охраны и истребле-
нии волка за 8-10 лет плотность населения лося увеличилась в 7,5, а диких сви-
ней - в 8,5 раз и достигла соответственно 19 и 12 особей на 1000 га лесной пло-
щади, что в несколько раз выше, чем в соседних охотничьих угодьях, где кормо-
вые условия лучше [Лавов, 1987]. Снижение численности копытных в 90-е годы
тесно связано с усилением легального и браконьерского отстрела и ростом пого-
ловья волка [Каштальян, 1997].
В Сары-Челекском заповеднике с момента его организации в 1960 г. всего за
6 лет население кабана выросло почти на порядок - с 80 до 720 особей, а плот-
ность достигла 60-70 голов на 1000 га [Воробьев, 1968]. В 30-километровой зоне
Чернобыльской АЭС после отселения людей и снижения антропогенного пресса
его поголовье за 4—5 лет увеличилось в 8-15 раз, а плотность возросла с 0,6 до 36
особей [Панов, Легейда, 1993; Козло и др., 1998].
В Завидовском заповедно-охотничьем хозяйстве после выпуска двух десят-
ков диких свиней в 1937 г. (см. выше) их поголовье к 1940 г. выросло до 100 осо-
бей. Отсутствие подкормки в военные (1941-1945) годы стало причиной эмигра-
ции зверей и снижения стада до 47 особей. В послевоенное десятилетие числен-
ность увеличивалась весьма медленно из-за ликвидации хозяйства в 1951-1953
гг., ряда многоснежных зим и недостаточного объема биотехнических работ. С
середины 60-х годов, несмотря на чередование малоснежных и многоснежных
зим, наблюдается бурный рост поголовья, что напрямую связано с образованием
кормовых полей, увеличением объема подкормки и полным истреблением (в
1964 г.) волка (табл. 39, рис. 64). В это же время выпускали новые партии каба-
нов, но этот искусственный прирост нивелировался отстрелом. К 1975 г. плот-
ность группировки достигла 58 особей на 1000 га угодий, что потребовало уве-
личения добычи. В начале 90-х гг. в период изменения общественных и эконо-
мических отношений в СССР в «Завидово» имел место массовый отстрел копыт-
ных, в основном взрослых особей. За три года добыто около 5 тыс. кабанов и 1
тыс. пятнистых и благородных оленей. Плотность населения диких свиней была
снижена до 9,5 особей на 1000 га [Юргенсон, 1969; Дормидонтов, 1969; Щерба-
ков, 1971; Фертиков и др., 1999]
Таблица 39. Динамика численности и плотности населения диких копытных в «Завидо-
во» (по [Юргенсон, 1969; Колодяжный, 1971; Щербаков, Стародынова, 1979; Фертиков и
др., 1999])*
Год Кабан Лось Марал Пятнистый олень
1 2 1 1 2 1 2 1 2
1933 69 2,3 7 —
1934 183 з,з
1936 8 0,1 — —
1937 25 0,5 — —
1939 50 0,9 — —
1940 100 1,8 — —
1947 47 0,8 271 5,0
1948 80 1,5 211 3,9
1949 144 2,7 384 7,1 14
187
Таблица 39 (окончание)
Год Кабан Лось Марал Пятнистый олень
1 2 1 1 2 1 2 1 1 2
1950 133 2,5 482 8,9 14 —
1951 — — 593 11,0 17 —
1954 188 3,5 400 7,4 24 —
1955 237 4,4 372 6,9 31 —
1956 160 з,о 406 7,5 20 —
1957 274 5,1 523 9,6 34 —
1958 155 2,9 619 11,2 29 —
1959 246 4,6 552 10,2 50 —
1960 175 з,з 586 10,8 43 —
1961 250 4,7 580 10,7 60 —
1962 292 5,5 592 10,9 59 —
1963 297 5,6 621 11,5 77 —
1964** 265 5,0 769 14,2 84 —
1965 420 7,9 989 18,3 116 —
1966 440 8,3 713 13,2 139 —
1967*** 667 5,8 736 7,0 178 1,6 50 0,4
1968 902 7,8 436 4,1 482 4,4 71 0,6
1969 1200 10,4 460 4,4 496 4,6 121 1,0
1970 2210 19,2 500 4,8 550 5,0 220 1,9
1971 2750 23,9 1000 9,5 750 6,9 260 2,2
1972 3420 29,7 980 9,3 830 7,6 300 2,5
1973 3890 33,8 990 9,4 935 8,6 344 2,9
1974 5000 43,4 1000 9,5 1110 10,1 400 3,4
1975 6680 58,0 800 7,6 1258 11,5 486 4.1
1976**** 4000 34,7 880 8,3 1400 12,8 520 4,4
1977 4500 39,1 900 8,5 1300 11,9 510 4,3
1978 5260 45,7 990 9,4 1429 13,1 532 4,5
1979 6710 58,2 900 8,5 1552 14,2 592 5,0
1980 5800 50,3 789 7,5 1715 15,7 652 5,5
1981 5850 50,8 800 7,6 1568 14,3 700 5,9
1982 5890 51,1 890 8,4 1780 16,3 750 6,3
1983 4500 39,1 620 5,9 1250 Н,4 800 6,8
1984 5990 52,0 750 7,1 1570 14,4 1100 9,3
1985 6500 56,4 650 6,2 1800 16,5 1800 15,2
1986 6500 56,4 600 5,7 1700 15,6 2100 17,8
1987***** 5050 43,8 500 4,7 1100 10,1 2100 17,8
1988 2700 23,4 570 5,4 1270 Н,6 1440 12,2
1989 2060 17,9 500 4,7 850 7,8 1100 9,3
1990 1100 9,5 500 4,7 900 8,2 1000 8,5
1991 1100 9,5 500 4,7 900 8,2 1100 9,3
1992 1300 и,з 500 4,7 800 7,3 930 7,9
1993 1600 13,9 480 4,6 600 5,5 950 8,0
1994 1500 13,0 410 3,9 856 7,8 850 7,2
1995 2300 20,0 400 3,8 719 6,6 1000 8,5
1996 2500 21,7 300 2,8 1100 10,1 1000 8,5
1997 2400 20,8 294 2,8 1051 9,6 950 8,0
1998 2110 18,3 268 2,5 1070 9,8 924 7,8
* 1 - численность (особей), 2 - плотность населения (особей на 1000 га); прочерк - отсутствие
сведений, пустая графа - отсутствие животных.
** Добыт последний волк.
*** Присоединена новая территория.
**** Эпидемия чумы свиней.
***** Массовый отстрел кабанов в годы «перестройки».
188
Трудно переоценить роль этого заповедника и как природного резервата, в
значительной мере способствовавшего насыщению дикими копытными Русской
равнины, и как научного стационара. «Завидово», на мой взгляд, великолепный
образец для отечественного охотничьего хозяйства, дающий представление о по-
тенциальных возможностях в разведении крупных зверей. На площади около 120
тыс. га поголовье кабана составляло 6,5 тыс. (плотность - 18-58 особей на 1000
га), лося - 1 тыс. (5-10), благородного оленя - 1,8 тыс. (7-16) и пятнистого оленя
- 2,1 тыс. (2-18 особей). Общая численность копытных здесь не меньше, а пого-
ловье диких свиней выше, чем во многих областях России!
Многолетние сведения о динамике численности копытных Беловежской пу-
щи также весьма интересны как в плане потенциальных возможностей охотничь-
его хозяйства, так и в связи с конкурентными отношениями между видами и воз-
действием различных факторов среды (рис. 64). В 60-е годы XX в. на площади 79
тыс. га численность и плотность населения достигала следующих величин: кабан
- 2,2 тыс. (26 особей на 1000 га); благородный олень - 2,9 тыс. (36); косуля - 2,0
тыс. (24) при наличии нескольких десятков лосей и зубров. Еще больше копыт-
ных было в 1914 г.: на 130 тыс. га обитали 6,8 тыс. оленей (52 особи), 5 тыс. ко-
суль (38), 1,5 тыс. ланей (12), 2,3 тыс. диких свиней (18) и 785 зубров (6), или
всего более 16 тыс. голов при плотности около 126 особей на 1000 га (табл. 40).
Население кабана, которого не лимитируют древесно-веточные корма, изме-
нялось здесь до определенного уровня независимо от оленьих и зубра, а ампли-
туда колебаний была самой высокой среди копытных - в отдельные годы оно
уменьшалось или увеличивалось вдвое. Рост поголовья обуславливался хороши-
ми урожаями желудей, интенсивной зимней подкормкой, уничтожением крупных
хищников. Снижение численности и исчезновение отдельных видов происходило
в военные, послевоенные и неурожайные годы, в периоды усиления браконьер-
ства, увеличения населения волка, отсутствия подкормки и при эпизоотиях [Саб-
лина, 1953а,б; Северцов, Саблина, 1953; Козло, 1970а, 1975; Романовский, Коча-
новский, 1971; Шостак, 1978; Балюк и др., 1987].
Таблица 40. Динамика численности и плотности населения диких копытных в Беловеж-
ской пуще (по [Карцов, 1903; Шостак, 1978])*
Год Кабан Олень Косуля Лось Зубр
1 1 2 1 1 2 1 2 1 2 1 1 2
1890 329 2,5 4 — 541 4,1 375 2,9 391 3,0
1891 900 6,9 28 0,2 729 5,6 535 4,1 464 3,6
1892 758 5,8 200 1,6 434 з,з 430 3,3 476 3,7
1893 890 6,9 250 1,9 685 5,3 460 3,5 465 3,6
1894 1000 7,7 373 2,9 835 6,4 404 3,1 474 3,7
1895 — — 665 5,1 959 7,4 490 3,8 535 4,1
1896 1026 7,9 717 5,5 1345 10,4 500 3,9 533 4,1
1897 1100 8,5 849 6,5 1690 13,0 545 4,2 605 4,7
1898 653 5,0 1046 8,1 2178 16,8 660 5,1 630 4,9
1899 750 5,8 1742 13,4 2960 22,8 730 5,6 640 4,9
1900 1500 11,5 2100 16,2 4500 34,6 700 5,4 710 5,5
1901 2000 15,4 2600 20,0 5100 39,2 700 5,4 730 5,6
1902 1800 13,9 2500 19,2 5000 38,5 450 3,5 655 5,1
1903 1140 8,8 2530 19,5 4560 35,1 370 2,9 703 5,4
1904 1033 8,0 2935 22,6 5495 42,3 392 3,0 708 5,5
1905 1500 11,5 3500 26,9 6000 46,2 340 2,6 651 5,0
1906 1200 9,2 3750 28,8 4300 33,1 250 1,9 653 5,1
1907 1639 12,6 5054 38,9 5329 41,0 222 1,7 741 5,7
1908 1412 10,9 4769 36,7 3582 27,6 144 1,1 696 5,4
1910 1842 14,2 5000 38,5 — — — — 600 4,6
189
Таблица 40 (окончание)
Год Кабан Олень Коса /ля Лось Зубр
1 2 1 1 2 1 ' 2 1 2 1 2
1914 2320 17,9 6800 52,3 4966 38,2 58 0,5 785 6,0
1915 — — 3000 23,1 3000 23,1 — — 739 5,7
1916 — — 1500 И,5 1063 8,2 — — 216 1,7
1917 — — 1769 13,6 — — — — 167 1,3
1928 250 1,9 210 1,6 2000 15,4 — —
1935 1240 9,5 687 5,3 2400 18,5 — —
1939 1900 14,6 1700 13,1 2938 22,6 9 0,1
1945 600 8,5 380 5,3 547 7,7 — —
1946 367 5,2 274 3,9 497 7,0 2 —
1947 506 7,1 310 4,4 371 5,2 4 0,1
1948 720 10,1 360 5,1 480 6,8 5 0,1
1949 840 11,8 510 7,2 670 9,4 9 0,1
1950 510 7,2 680 9,6 850 12,0 7 0,1
1951 1140 16,1 770 10,8 850 12,0 9 0,1
1952 890 12,5 560 7,8 710 10,0 9 0,1
1953 400 5,3 570 7,6 710 10,0 И 0,2
1954 325 4,3 680 9,1 750 10,0 31 0,4
1955 310 4,1 760 10,1 550 7,3 34 0,4
1956 180 2,5 540 8,5 480 6,4 32 0,4
1957 210** 2,8 700 9,3 330 4,4 19 0,2
1958 480 6,4 820 10,9 270 3,6 33 0,4
1959 1330 17,7 900 12,0 550 7,3 41 0,5
1960 1370 18,3 1100 14,7 660 8,8 56 0,7
1961 1380 17,5 1250 15,8 750 9,5 60 0,8
1962 1360 17,2 1330 16,8 850 10,8 — —
1963 1450 18,4 1600 20,3 1000 12,7 — —
1964 550*** 7,0 1700 21,5 1110 13,9 — — 22 о,з
1965 724 9,2 1630 20,6 1030 13,0 — — 28 0,4
1966 990 12,5 1670 21,1 1100 13,9 60 0,8 49 0,6
1967 2100 26,6 2880 36,5 1800 22,8 43 0,5 51 0,7
1968 2200 26,6 2550 30,7 1650 19,9 62 0,8 58 0,7
1969 1900 22,9 2650 31,9 1950 23,5 40 0,5 60 0,8
1970 1450 17,1 2090 24,6 620 7,3 79 0,9 63 0,8
1971 1440 17,0 2290 27,0 570 6,7 83 0,9 63 0,8
1972 1370 16,1 2430 28,6 610 7,2 60 0,8 66 0,9
1973 1690 19,9 2290 27,0 800 9,4 70 0,8 77 0,9
1974 1370 15,8 2100 24,1 690 7,9 73 0,8 82 1,0
1975 1535 17,7 2120 24,4 880 10,1 144 0,8 90 1,0
1976**** 2230 25,9 — — — — — — — —
1977 2396 27,8 — — — — — — — —
1978 2219 25,8 — — — — — — — —
1979 1600 18,6 — — — — — — — —
1980 2190 25,4 - - - - - - - -
* 1 - численность (особей), 2 - плотность населения (особей на 1000 га угодий); прочерк - отсутствие
сведений, пустая графа - отсутствие животных.
** Начало регулярной подкормки зверей.
*** После эпизоотии чумы.
**** Сведения с 1976 г. по [Балюк и др., 1987].
В Кавказском заповеднике в середине 60-х годов насчитывали около 40 тыс.
голов копытных: до 3 тыс. диких свиней, более 10 тыс. благородных оленей, 16
тыс. туров, 10 тыс. серн, 700 косуль и 700 зубров [Голгофская и др., 1979]. Для
190
популяции кабана здесь предельна плотность в 60 особей на 1000 га [Дуров,
1980а]. В отдельных угодьях Краснодарского края в 70-е годы плотность населе-
ния вида достигала на короткое время 400-800 голов на 1000 га [Гинеев, 19756].
В Хоперском заповеднике на площади в 16,2 тыс. га в >1985 г. обитали 2800 пят-
нистых оленей и 1500 кабанов [Печенюк, 1995]. В конце 80-х годов плотность
населения диких свиней составляла в нем 100 особей на 1000 га [Ковалева, Ан-
тонец, 1995].
Разумеется, в отдельных заповедниках и высокоорганизованных охотничьих
хозяйствах плотность населения диких копытных явно чрезмерна, но при этом
весьма показательна в отношении реальной емкости угодий и потенциальных
возможностей в разведении животных.
В заключение подчерну следующее. Ресурсы диких свиней в России в целом
скудны, а в конце XX в. они снова существенно сократились. Плотность населе-
ния животных почти повсеместно многократно ниже возможной. Вместе с тем
очевидно, что при хорошей охране, отсутствии крупных хищников и обильной
подкормке дикие копытные могут образовывать предельно высокие плотности
даже в многоснежных районах. Многолетней цикличности в динамике населения
вида не усматривается.
Основные факторы, определяющие динамику популяций
Факторы, определяющие популяционную динамику кабана, разнообразны и
весьма интересны как с теоретической, так и с практической точек зрения.
Воспроизводство
Уровень воспроизводства — один из ключевых факторов динамики популяций
диких копытных. Он определяется числом участвующих в размножении живот-
ных, процентом беременных самок, числом родившихся и выживших детенышей
и длительностью их полового созревания, долей самок в приплоде и популяции.
При благоприятных условиях обитания и оптимальном половом и возрастном
составе группировок уровень воспроизводства в них сравнительно высок и отно-
сительно стабилен. Скорость роста большинства популяций, как и динамика в
целом, в большей степени зависит от факторов, вызывающих смертность.
Кабан, как показано выше, весьма плодовит. Потенциальный биологический
прирост у дикой свиньи, приносящей в среднем по 5—7 поросят, близок к 500-700%
от числа беременных самок и к 100-200% от весенней численности популяции. Ре-
альный годичный прирост, однако, обычно гораздо меньше и нередко бывает отри-
цательным. Средний размер ежегодного прироста у этого вида, как это ни странно,
не намного выше, чем у других видов копытных животных, главным образом из-за
низкого процента беременных самок и высокой смертности сеголеток.
Доля самок старше 1 года в его группировках - около 25-30%, но большая
часть годовалых свиней еще не приносит потомства, а часть взрослых остается
яловой. Потенциально могут размножаться лишь около 10-25% особей, реально -
еще меньше. В популяции Беловежской пущи доля беременных свиней составля-
ет 4,8-15,6% [Саблина, 1955; Козло, 1970а,1975], в Березинском заповеднике -
17,9% [Лавов, 1981], на юге Киргизии - около 7% [Чичикин, Воробьев, 1967]. По
расчетам Г.И. Ивановой [1983], в группировке из 100 голов потомство приносят
лишь 8-12 свиней.
При интенсивном отстреле крупных взрослых особей доля размножающихся
самок обычно уменьшается до предела. После массовой гибели сеголеток (что
нередко случается и приводит к изменению возрастного состава популяции) и
191
выборочном отстреле взрослых самцов (что приводит к сдвигу соотношения по-
лов) она, напротив, может быть максимальной - около 25-30%. Плодовитость
свиней неодинакова в урожайные и голодные годы и различается у молодых и
взрослых животных (см. выше). Все это существенно отражается на размере го-
дачиого прироста..
В Беловежской пуще в период с 1890 по 1975 гг. максимальный прирост, превы-
шающий 100% (до 177%), отмечался 5 раз. Гораздо чаще он был в пределах 1—45%, а
в отдельные годы (более 20 раз) регистрировалась убыль популяции, достигающая
62%. Среднегодичный прирост, с учетом убыточных лет, в общем оказался невы-
соким - около 15% (табл. 40). При средних кормовых и метеорологических усло-
виях максимальное пополнение популяции наблюдается лишь при плотности 10-
15 особей на 1000 га [Саблина, 19536; Козло, 1975]. В годы после урожаев дуба
прирост увеличивается в 1,5-2 раза [Романовский, Кочановский, 1971].
В Литве коэффициент годичного прироста кабана - 1,3-1,5. При нерегуляр-
ной подкормке зверей он минимальный — 0,9, но при обильном кормлении воз-
растает до 1,7, а доля молодняка увеличивается с 47% до 64%. При равном соот-
ношении полов в популяции коэффициент близок к 1,1, доля сеголеток составля-
ет 51,5%, но при соотношении 1:2 в пользу самок эти показатели увеличиваются
до 1,6 и 62,6% соответственно. Связь между уровнем прироста и плотностью на-
селения животных здесь не прослеживается, видимо, из-за хорошей кормовой ба-
зы [Янулайтис, Падайга, 1984а,б, 1985].
На юго-западе Болгарии среднемноголетний прирост популяции диких сви-
ней невелик - 22,7% [Генов, Коцаков, 1986]. В Чехословакии он гораздо выше-
70-130% [Skultety et al., 1974], а в Германии даже в неблагоприятные годы дости-
гает 110-130%, в нормальные - 140%, а в благоприятные годы - 150-200% [Бри-
дерманн, 1970; Briedermann, 1971, 1979; Meynhardt, 1984с; Stubbe, 1987].
В европейской части России в 60-70 годы прирост варьировал в пределах 10-
91% [Фадеев, 1982], составляя в охотничьих хозяйствах Подмосковья от 5 до
100% [Фадеев, 1978], в областях Центрального Черноземья - от 1 до 150% (в
среднем 22-56%) [Простаков, 1996], в Хоперском заповеднике - 82% [Казнев-
ский, 1977]. В Завидовском заповедно-охотничьем хозяйстве (табл. 39) в 40-50-е
годы при отсутствии или недостаточной подкормке годичный прирост стада был
на уровне 13%, в 60-е - 88%, позднее при обильном кормлении увеличился до
130-170% [Юргенсон, 1969; Колодяжный, 1971]. В областях северо-запада Рос-
сии этот показатель изменялся от 10 до 33% [Верещагин, Русаков, 1979]. В Хин-
ганском заповеднике в Амурской области его размер в зависимости от урожай-
ных и неурожайных на желуди лет варьировал от 49 до 232% соответственно,
расчетный потенциальный прирост определяется здесь в 158% [Дарман, 1989,
1990]. На юге Киргизии он составляет около 40% [Чичикин, Воробьев, 1967]. В
рекомендациях по охотустройству средний размер годичного прироста у кабана
оценивается в 50% [Данилов и др., 1966], Е.В. Фадеев [1987] определяет его в
размере 30-40%.
Очевидно, что годичный прирост в популяциях кабана очень изменчив. Од-
нако такие существенные различия - отчасти следствие разных методов его рас-
чета.
По моим расчетам, среднегодичный прирост населения кабана в России в пе-
риод относительного роста поголовья с 1961 по 1991 гг. составляет около 8%, в
период с 1961 по 2000 гг. - менее 5% с вариациями от +38% до -20% при средне-
годовом уровне лицензионной добычи около 11% от численности. Следователь-
но, при потенциальном биологическом приросте вида в 100% непромысловые
потери достигают как минимум 84%.
Рассмотрим основные факторы смертности, влияющие на динамику популяций.
192
Детская смертность
Причины смертности детенышей кабана различны и в основном сопряжены с
погодой, кормами, хищниками, болезнями и влиянием человека.
Часть поросят из ранних (март - апрель) выводков при минусовых ночных
температурах неизбежно погибает от переохлаждения в плохо утепленных гнез-
дах, при передвижении по пропитанному водой снегу и форсировании многочис-
ленных канав и ручьев с ледяной водой, в которых они к тому же тонут [Бакеев,
Костоглод, 1975; Верещагин, Русаков, 1979; Майнхардт, 1986]. Много их гибнет
в поймах рек и тростниках озер во время мощных весенних паводков и пожаров,
а также при нападении хищников [Слудский, 1956; Лавровский, 1962; Анненков,
1989]. При высокой плотности населения кабана довольно высока смертность де-
тенышей от гельминтозов [Фертиков и др., 1999].
Судя по разнице между средним числом эмбрионов и числом детенышей в
выводках (табл. 18 и 23), в весенне-летний период погибает около 5-22% сеголе-
ток, что подтверждается некоторыми региональными исследованиями [Лебедева,
1956а; Слудский, 1956; Лавровский, 1962; Дормидонтов, 1969; Andrzejewski,
Jezierski, 1978; Червонный, 19806; Лавов, 1981; Кудряшова, 1982; Иванова, 1986;
Простаков, 1996]. В Карпатах, например, летние потери составляют 10-14%,
осенние - 15-17%, зимние - 19-21% [Гунчак, 1982].
Наиболее существенна убыль сеголеток зимой (табл. 24 и 25). Естественный
отбор не благоприятствует поздно родившимся (июнь - июль) детенышам: они
не набирают к зиме необходимой массы и первыми замерзают на лежках или в
местах кормежки. Поросята, родившиеся в нормальные сроки, также почти не
накапливают жира с осени, поскольку все эйергетические резервы расходуют на
рост. При плохой зимней кормежке они быстро теряют упитанность и массу тела.
По моим наблюдениям (п = 16), в средней (полосе России гибель поросят в мо-
розные зимы происходит при исхудании до 21—27 кг. Величина критического по-
рога, вероятно, во многом зависит от температуры воздуха.
Сеголетки - легкая добыча для крупных хищников по сравнению с взрослы-
ми животными: они не в состоянии активно обороняться, в семьях идут послед-
ними и чаще подвергаются нападению. В Березинском заповеднике при высокой
численности волка в выводках свиней к весне остается 1-3 поросенка, а в Кав-
казском заповеднике их число уже в октябре сокращается до 2. В охотничьих хо-
зяйствах Подмосковья и в Беловежской пуще, напротив, при отсутствии крупных
хищников и регулярной зимней подкормке размер приплода у кабана в течение
года уменьшается незначительно - до 5,5 и 5,6—4,3 соответственно [Донауров,
Теплов, 1938; Саблина, 1955; Лебедева, 1956а; Иванова, Рыковский, 19686; Коз-
ло, 1975; Иванова, Овсюкова, 1976].
Изъятие хищниками низкоранговых особей, однако, не приводит к разруше-
нию структуры группировок. Наибольший урон детенышам и, в конечном счете,
популяциям наносят охотники и браконьеры. Заинтересованные в добыче круп-
ного животного, они, как правило, стреляют лидера группы - свинью. Поросята,
оставшиеся без матери, в суровых зимних условиях России обречены на гибель,
поскольку не в состоянии самостоятельно добывать корм и не имеют «гнездо-
строительных» навыков. В масштабах страны ежегодная добыча взрослых сви-
ней исчисляется тысячами голов. Соответственно масштаб смертности сеголеток,
оставшихся без матерей, может исчисляться десятками тысяч.
Уровень смертности детенышей в пределах обширного ареала различен
(табл. 41), но в большинстве районов достигает 40-70 (100)%. На северо-западе
России он находится в пределах 34-70%, а в экстремальные зимы может быть и
выше [Русаков, Тимофеева, 1984]. В Вологодской области доля сеголеток в по-
пуляции к весне снижается почти в 3 раза [Киселев, Каплин, 1987]. Уровень по-
13. Заказ № 2528.
193
терь в этой возрастной группе составляет 60-80% на севере европейской части
страны и 40-60% в центральных областях. В Подмосковье данный показатель
колеблется в пределах 13—36%, но в лучших охотничьих хозяйствах при налили
обильной подкормки он не превышает 8-15%. В отдельные суровые зимы падеж
существенно увеличивается [Иванова, 1975а, 1983, 1985]. В Центрально-
черноземном регионе даже при относительно благоприятных условиях обитания
гибнет 27-38% молодняка [Гусев, 1989; Простаков, 1996], в Крыму - 32% [Кор-
милицин, Дулицкий, 1972], на юге Украины до годовалого возраста доживают
только 30% [Волох, 1999]. В Беловежской пуще до 1953 г. отход сеголеток дос-
тигал 36% [Лебедева, 1956а], но при регулярной зимней подкормке и отстреле
волка с 1957 г. сократился до 6,2-19,4%. До 19-20-месячного возраста здесь до-
живает около 30-40% особей от числа родившихся [Козло, 1975].
Таблица 41. Приблизительный уровень смертности сеголеток в течение года в разных
районах
Район Уровень смертности, % Источник информации
Польша 48 Jezierski, 1977
Беловежская пуща 36 Лебедева, 1956а
6-19* Козло, 1975
Березинский з-к 60-70**
Карпаты 44-52 Гунчак, 1982
Юг Украины 70 Волох, 1999
Крым 32 Кормилицин, Дулицкий, 1972
Северо-запад России 34-70 Русаков, Тимофеева, 1984
60-80 Иванова, 1983
Центр европейской части РФ 40-60 (С
Костромская область 54,5 Зайцев, 2000
Подмосковье 8-36 Иванова, 1975а, 1985
Окский з-к 21-52 Червонный, 19806; Кудряшова, 1982
Ц.-Черноземный з-к 27-35 Гусев, 1989
З-к «Лес на Ворскле» 55 Царев, 2000
Кавказский з-к 60-91 Дуров, 1980а
Дельта Волги 50-60 Лавровский, 1952,1962
Тюменская область 20-80 Азаров, 1996
Монголия 60 Банников, 1954
Предбайкалье 40-46 (24-100) Недзельский, 1994
Хинганский з-к 74 (до 90) Дарман, 1989, 1990
Юг Дальнего Востока 48-62 Абрамов, 1963; Бромлей, 1964; Куче- ренко, 1976; Бромлей, Кучеренко, 1983
Алакуль 65 Анненков, 1989
* При регулярной подкормке с 1957 г. и истреблении крупных хищников.
** При наличии волка.
В сравнительно благоприятных условиях Кампиновского национального пар-
ка в Польше 15—20% поросят гибнет через 3 месяца после рождения, к концу го-
да - 48%, на второй год - еще 36%. До трех лет доживают всего 8% особей при
максимальной продолжительности жизни отдельных животных до 8-9 лет
[Jezierski, 1977]. В Германии в группировке, находившейся под постоянным на-
блюдением, отход сеголеток в первые 4 недели жизни составлял 21,8%, за 8 ме-
сяцев - 38,6%. Сырая погода была более губительной для них, нежели сухая и
холодная и снегопады [Meynhardt, 1982, 1984а, с].
Очень высок уровень отхода детенышей в Кавказском заповеднике: к маю -
194
около 11%, к августу - 25%, к ноябрю - 55% [Донауров, Теплов, 1938; Теплов,
1938]. В низовьях р. Или к октябрю гибнет 32%, к февралю - 48% сеголеток, а
возраста подсвинка достигают лишь 40% из них [Слудский, 1956]. Примерно
столько же (40-50%) молодняка остается в дельте Волги. В годы высоких павод-
ков он здесь погибает почти полностью [Лавровский, 1952, 1962]. В Алакульской
котловине отход сеголеток к осени колеблется в разные годы в пределах 25-48%,
до годовалого возраста доживают не более 35%, а количество взрослых самок, не
участвующих в размножении или потерявших потомство, изменяется от 29 до
65% [Анненков, 1989]. В Монголии в течение года гибнет около 60% приплода
[Банников, 1954]. В Хинганском заповеднике до полутора лет доживают лишь
11% от числа рожденных [Дарман, 1990].
Масштаб потерь сеголеток особенно отчетливо виден при анализе возрастно-
го состава популяций (см. табл. 28), в которых доля подсвинков находится в пре-
делах 3-28%.
Корма и погода
Корма и погода - взаимосвязанные важнейшие факторы, влияющие не только
на воспроизводство и динамику, но и на половозрастную структуру популяций.
Массовая гибель истощенных животных, преимущественно сеголеток, происхо-
дит главным образом из-за недоступности или недостатка пищи как в годы с вы-
соким и плотным снежным покровом и настами, так и в морозные малоснежные
зимы, отличающиеся глубоким промерзанием почвы и обледенениями. После го-
лодных зимовок всегда значительно снижается плодовитость (см. выше).
По расчетам М.Д. Перовского [1998], доля гибели кабана от истощения на
территории России составляет 4,3%, что, на мой взгляд, не соответствует дейст-
вительности. Эта величина в разных районах достигает в среднем примерно тре-
ти от всех потерь и колеблется в пределах 3-96% (табл. 42). Однако и эти данные
неполны. Гибель истощенных зверей особенно высока поздней зимой и ранней
весной, но в многоснежье и распутицу обнаружить погибших животных в глухих
лесных массивах весьма сложно [Лавов, 19806; Червонный, 19806; Иванова,
1985]. Поэтому большинство случаев смертности от истощения, как правило, не
реги стрируется.
Во многих районах Прибалтики в суровые зимы кабаны исчезают почти полно-
стью или полностью, как это случилось в Эстонии в 1939/40 и 1941/42 гг. [Кал-
ниньш, 1950; Линг, 1955, 1978]. В Беловежской пуще в суровые зимы 1946, 1950,
1952, 1956, 1970 и 1976 гг. популяция диких свиней снижалась на 45-55%. При этом
погибал почти весь молодняк и до 20—30% годовалых особей [Козло, 2001].
На северо-западе России доля истощения в общей внепромысловой смертно-
сти составляет 52,6% [Русаков, Тимофеева, 1984]. Массовая гибель зверей заре-
гистрирована в отдельных районах Псковской, Новгородской и Ленинградской
областей в 1954, 1957, 1962, 1965—1967, 1969, 1972, 1975 и 1982 гг. В Новгород-
ской области суровой зимой 1974/75 гг. поголовье сократилось на 25%. Погибали
преимущественно поросята, иногда до 8-25 трупов находили в одном месте. В
Карелии массовый падеж отмечен в многоснежные зимы 1980-1982 и 1993—1995
гг. [Марковский, 1995].
В Вологодской области массовая гибель от истощения (до 17,5% поголовья в
Дарвинском заповеднике) зарегистрирована в 1973, 1976, 1980 и 1985 гг. Здесь
зверей также обнаруживали замерзшими на лежках семьями по 3-12 особей, а
доля умерших сеголеток достигала 55-83% в разные годы. Периоды замедления
роста численности или снижения поголовья следовали после многоснежных зим
или глубокого промерзания почвы [Варнаков, 19786; Киселев, Каплин, 1987; Ту-
пицина, 1988].
13*
195
40
04
Таблица 42. Основные причины непромысловой гибели (%) кабана
Район (источник информации) Всего п погибших, п Браконьеры Волк Медведь Исто- щение Болезни Травмы** Утонули Ране- ния Дру- гие
С.-запад России (1) 720 ; ? 10,7 0,9 52,6 4,6 1,8 14,1 4,8 10,5
145 30,3 8,3* * 31,8 — 15,2 П,7 — 2,7
Архангельская 21 ? — 23,8 19,1 — — 9,5 — 47,6
обл. (12) Березинский з-к (2) 156 ? 39,7 7,7 15,4 3,8 21,8 9,6 2,0
Дарвинский з-к (6) 51 ? 6 4 86* * — 4 — —
--(8) 116 1,4-2,2* 9,9 7,0 72-96 — * 2,2-2,8 * —
“Завидово” (5) 49 26,5 — — -- 26,5 10,2 6,1 30,6
Центральный р-н 658 20,4 38,3 — 19,9 4,8 7,4 5,6 ? 3,6
(Ю) Ц.-Черноземный р-н 141 34,0 9,9 15,6 1,4 20,7 — 18,4
(Ю) “Лес на Ворскле” (15) 25 ? — 68,0 — — — 28,0 4,0
Ц. Черноземье (7) 233 15,9 9,9 — 9,9 5,6 11,2 6,0 37,3 4,2
Самарская обл. (9) 32 ? — — 3,1 — 62,5 25,0 — 12,5
Волго-Вятский р-н 164 20,5 8,3* * 52,1 — 8,3 9,1 — 1,7
(Ю) С.-Кавказский р-н 95 76,9 1,0 4,2 8,5 4,2 5,2
(Ю) Европейская часть 1389 27,9 21,4* * 24,6 3,5 9,8 7,0 ? 5,8
РФ (10) Тува (16) ? 11,8* * 75,7 4,2 0,7 1,4 ? 6,2
Предбайкалье (14) 47 41,7 9,3* * 28,4 — — — — 20,7
Бурятия (13) 20 ? 15 5 10 40 — 20 — 10
Приамурье (11) 834 ? 6,9 21,8 49,3 13,4 0,5 0,8 — 7,3
Хинганский з-к (3) 129 9,3 31,0 4,7 17,8 27,1 — 0,8 — 9,3
РСФСР (4) 714 38,2 28,4 2,2 — — — — 14,8 16,4
Источник информации: 1 - Русаков, Тимофеева, 1984; 2 - Лавов, 1986а-в; 3 - Дарман, 1990; 4 - Глушков и др., 1989; 5 - Щербаков, 1971; 6 - Филонов, 1989; 7 - Простаков,
1996; 8 - Тупицина, 1988; 9-Ковалев, Кармаев, 1997; 10-Иванова, 1985; 11 - Раков, 1970; 12-Плешак, Миняев, 1986; 13-Смирнов, 1978; 14 - Недзельский, 1994; 15 - Царев,
2000; 16-Смирнов, 2001.
* Среднее значение для выборки.
** В основном гибель от транспорта.
В Завидове в 1958 г. погибло 119 особей из 274. Зимой 1966/67 гг. в 11 подмос-
ковных охотничьих хозяйствах найдено около 70 трупов, в основном молодых жи-
вотных [Фадеев, 1968]. В Брянской области массовая гибель наблюдалась в много-
снежные (более 80 см) зимы 1967 и 1968 гг. [Ватолин, 1980]. В 1968 г. большая часть
зверей погибла в Воронежском заповеднике [Любченко, 1973]. В Удмуртии в первые
годы естественного расселения кабана при отсутствии подкормки зимняя смертность
доходила до 40-80% [Попов, Попов, 1980]. В суровую зиму 1978/79 гг. в северных
областях европейской части России не стало значительной части локальных группи-
ровок [Фадеев, 1981]. В Татарии многоснежной зимой 1986 г. из-за бескормицы по-
головье сократилось в 6—10 раз [Горшков, 1989]. Этой же и следующей зимой в Тю-
менской области погибло от 30 до 80% молодняка, но в местах подкормки отход не
превышал 20-30% [Азаров, Климов, 1990].
Массовая гибель зверей от истощения в глубокоснежье регистрируется даже
в регионах с благоприятным климатом и в зоне широколиственных лесов: в Кар-
патах [Пидопличко, 1951; Гунчак, 1980, 1999], Ставропольском [Двойченко,
1955] и Краснодарском [Гинеев, 19756] краях и Воронежской области [Антонец,
1987]. На Северном Кавказе значительный падеж зверей наблюдался в 1902—
1904, 1906, 1908, 1927-1929, 1932 и 1937 гг. [Динник, 1910; Донауров, Теплов,
1938], в Закавказье - в 1910 и 1925 гг. [Марков, 1932]. В Кавказском и
Закатальском заповедниках зимой 1931/32 гг. погиб почти весь приплод
[Насимович, 1955]. Многоснежные зимы 1948-1950, 1951/52 и 1966-1968 гг.
также унесли немало жизней диких копытных животных в этом регионе [Дуров,
Александров, 1968; Котов, Рябов, 1963; Котов, 1969].
В низовьях Сырдарьи и Амударьи в морозные малоснежные и с настами зи-
мы 1949/50, 1950/51 и 1953/54 гг., когда почва промерзала особенно глубоко, на-
ходили десятки трупов, вмерзших в лежки. Массовый падеж сеголеток и под-
свинков в низовьях рек Или и Сырдарьи зарегистрирован и в многоснежные зи-
мы 1939/40 и 1949/50 гг., что было связано с оттепелями и обледенением. В горах
Казахстана особенно губительными были зимы 1949, 1951, 1960, 1966 и 1969 гг.,
после которых во многих урочищах звери исчезали полностью [Слудский, 1956;
Слудский и др., 1984]. В Сары-Челекском заповеднике зимой 1968/69 гг. от бес-
кормицы погибло 2/3 популяции [Титов, Шингареев, 1970].
Практически на всем ареале и всеми исследователями в той или иной мере
отмечается зависимость уровней смертности и плодовитости кабанов от урожая
дуба, бука, кедра, лещины, фисташки, яблонь. Пики численности диких свиней
наблюдаются после урожайных лет, резкий спад, соответственно, после неуро-
жайных и голодных. Особенно масштабной смертность бывает в неурожайные
годы, когда животные не успевают накопить запасы жира.
Особенно сложно выживать диким свиньям на юге Восточной Сибири, где
основным источником корма служат кедрачи, плодоносящие далеко не регуляр-
но. Ежегодная смертность зверей в Восточном Предсаянье и Предбайкалье, в ос-
новном из-за нехватки зимних кормов, колеблется от 28 до 42% [Беньковская,
Беньковский, 1989; Недзельский, 1994]. В Туве доля кабанов, погибших от исто-
щения достигает в общей смертности 75,7%. Сеголетки среди погибших состав-
ляют 53-90% [Смирнов, 2001]. В Бурятии массовый отход животных зарегистри-
рован в 1960/61 и 1972/73 гг. [Смирнов, 1978].
В Приамурье в умеренно снежную зиму 1959/60 гг. в бассейне р. Малая Далма
на площади 60 кмгобнаружено 37 трупов сеголеток, 3 - подсвинков и 1 - секача.
Большинство животных замерзли ночью в логовах [Раков, 1970]. На Сихотэ-
Алине многоснежной (100—150 см) зимой 1914 г. кабаны погибли почти полно-
стью. Сохранились лишь наиболее сильные взрослые особи, пожиравшие моло-
дых и более слабых собратьев. Минимальной численность была здесь и в 1935 г.
после высоких (94 см) снегов и интенсивного хищнического уничтожения, ко-
197
гда много убитых зверей остались не вывезенными из тайги [Капланов, 1948]. С
1928 по 1978 гг. одновременные неурожаи желудей и орехов кедра случались 15
раз, эпизоотии - четырежды, «завальные» (более 1 метра) снега - 5 раз. В эти 50
лет падежи отмечались 18 раз, т. е. в среднем 3-4 в 10 лет. После очередной мно-
госнежной зимы 1987/88 гг. уцелела лишь третья часть популяции. В первую
очередь погибали сеголетки и подсвинки [Пикунов и др., 1989а,б].
При очень высоком снежном покрове животных не спасает от гибели даже
обильный урожай орехов и желудей. В Приамурье и Приморье после урожайных
малоснежных лет поголовье увеличивается до 20 и 40 тыс. соответственно, а по-
сле неурожайных и особенно многоснежных сокращается до 1-3 тыс. [Бромлей,
1964; Раков, 1970; Кучеренко, 1973, 1976; Бромлей, Кучеренко, 1983; Дарман,
1990]. В Украинских Карпатах в мягкие зимы после хорошего урожая бука и ду-
ба численность кабана возрастает в среднем на 35 (18-52)%, при среднем - на
13%, при плохом урожае и в трудные зимы снижается на 20%, а при хорошем
урожае в многоснежные годы (1995-1997) здесь погибает более 50% популяции
(Гунчак, 1999).
Суровые многоснежные зимы и гололедицы были существенными регулято-
рами численности зверей и раньше. По описанию путешественника Броневского,
в середине XVI в. в южноукраинских степях погибло множество зубров, сайга-
ков, диких лошадей и кабанов. Бедственными здесь были также зимы 1788, 1812,
1824, 1833 и 1849 гг. [Кириков, 1955, 1959]. В Беловежской пуще массовый па-
деж диких свиней имел место неоднократно. В 1840 г. находили замерзшие груп-
пы по 20-30 особей. В 1882 г. после нескольких гололедиц в ноябре и декабре
почти все животные ушли из пущи, а оставшиеся к весне казались живыми ске-
летами [Карцов, 1903]. В суровую зиму 1897 г. отход достиг 2-3 тыс. голов [Тур-
кин, 1899].
Гибель кабанов в сравнении с оленьими [Данилкин, 1999а], однако, не столь
значительна, как можно было бы ожидать. В Латвии, например, в 1930-1940 гг.
ежегодно находили от 1 до 12 павших особей, в то время как число погибших ко-
суль в зависимости от суровости зим колебалось от 121 до 8969 [Калниньш,
1950]. По нашим наблюдениям, в Курганской области многоснежной зимой и
весной 1998 г. от истощения погибли десятки тысяч сибирских косуль [Данил-
кин, Останин, 1998; Данилкин и др., 2000]. При этом дикие свиньи уцелели и вы-
глядели довольно упитанными исключительно благодаря способности поедать
трупы. В Центрально-Черноземном заповеднике в многоснежную зиму 1974/75
гг. они также выжили в основном благодаря значительному падежу косуль [Гу-
сев, Елисеева, 1983; Гусев, 1989]. На Алтае эти звери сохранялись в периоды
массовой смертности маралов и косуль [Собанский, 1992].
Следует отметить, что в многоснежные зимы истощенные копытные стано-
вятся более легкой добычей охотников, браконьеров и хищников, более воспри-
имчивы к заболеваниям и заражению гельминтами, что усугубляет снижение
численности. Не благоприятствуют их жизни и засухи, приводящие к усыханию
водоемов и растительности и, в конечном счете, к голодной смерти.
Хищники
Крупные хищники оказывают на популяции кабана весьма существенное
воздействие (см. табл. 42). Наибольшие потери они несут от волка, тигра и буро-
го медведя.
Волк. Этот хищник весьма избирателен в отношении своих жертв. В ряде
районов он давит чаще благородного оленя и косуль, затем лося, а дикие свиньи
занимают последнее место (табл. 43). В некоторых областях европейской
части России, где кабан по численности не уступает лосю, хищник режет его
198
вдвое реже [Приклонский, 1999]. В Дарвинском заповеднике в период с 1973 по
1986 гг. соотношение погибших кабанов и лосей еще выше - 1:11 [Нестеренко,
1988].
Таблица 43. Избирательность добычи диких копытных животных крупными
хищниками, %*
Район, хищник Число регистра- ций, п** Ка- бан Лось Оле- ни Косули Другие ВИДЫ Источник информации
Карпаты: 336 3,0 — 21,1 75,9 — Керечун, 1975
В т. ч. волк 144 12,5 — 46,5 41,0 — Керечун, 1979
рысь 149 4,5 — 30,9 64,4 — U
медведь 19 10,5 — 89,5 —- — а
Карпаты 1636 14,0 — 5,0 56,0 25,0 Алмэшан и др., 1970
Прикарпатье 350 27,1 — 42,9 30,0 — Слободян и др., 1975
Причерноморье 49 32,7 — — 67,3 — Курский, 1985
Беловежская пуща 560 48,9 0,2 14,3 36,6 — Гаврин, До- науров, 1954
it >186 10-14 1,8 47-55 22-24 13 Буневич, 1986,1988
Белоруссия 729 28-46 13- 72 8 1-39 — Козло, Банад, 1985
Березинский з-к 299 41 54 4 1 — Филонов, 1989
а 456 33,6 55,7 8,3 2,4 — Лавов, 19866****
В т. ч. волк 262 22,5 65,3 Н,1 1,1 — и
рысь 7 — 28,6 — 71,4 — а
медведь 44 27,3 72,7 — — — сс
Припятский з-к 115 76,5 3,5 — 20,0 Литвинов и др., 1979
а — 36,5 15,4 — 19,2 28,9 Кучмель, 1999
Дарвинский з-к 472 25 74,9 — 0,1 Филонов, 1989
Окский з-к 269 15 73 12 — а
Мордовский з-к 217 8 45 47 — — а
Алтайский з-к 348 9 4 69 5 13 а
Припятский з-к >47 57 31 — 12 — а
С.-Шушенский з-к >367 9 3 45 1 42 а
С.-Алинский з-к >62 12 7 26 16 39 а
Лазовский з-к >81 19 — 51 16 14 а
Ц.-Лесной з-к 184 22,8 77,2 — — — Кочетков, Со- колов, 1979
Хоперский з-к 280 2,8 6,8 90,4 — — Печенюк, 1995
Кавказский з-к 131 44,3 — 19,1 8,4 28,2 Теплов, 1938
сс 77 15,6 — 74,0 — 10,4 Кудактин, 1982
с 678 16,7 — 47,8 — 35,5 Кудактин, 1986
812 13,9 — 37,4 — 48,7 Филонов, Ка- лецкая, 1985
199
Таблица 43 (окончание)
Район, хищник Число регистра- ций, п** Ка- бан Лось Оле- ни Косули Другие ВИДЫ Источник информации
Кызыл-Агач- ский з-к 454 66,7 — — — 33,3 Литвинов, 1980
44 75 82,9 — — — 17,1 Лобачев, Пе- тухов, 1984
Подмосковье 60 26,7 21,7 51,6 — — Кузьмин, Ха- хин, 1980
Брянская обл. 179 14,5 29,6 3,4 52,5 — Ватолин, 1979
Южный Урал ? 23,2 47,1 29,7 — Гордиюк, 1996
Р. Или ? 23,7 — — 35,9 40,4 Слудский, 1970
Тянь-Шань 134 13,3 — 2,1 27,0 42,4 Жиряков, 1990
Джунгарский Алатау 571 4-37 — 17-54 3-20 4-20 Федосенко, Жиряков, 1979
Таджикистан 50 48,0 — 12 — 40,0 Соков, 1985
3. Копетдаг*** 130 61,5 — — — 38,5 Лукаревский, 1995
Западный Саян 396 1,8 — 66,2 3,0 29,0 Завацкий, 1986а,б
Приамурье 72 6,9 6,9 5,6 73,6 7,0 Дымин, 1975
Хинганский з-к 332 28,0 — 6,5 61,9 3,6 Дарман, 1986
Дальний Восток: 1621 18,1 6,2 24,2 40,2 п,з Раков, 1979
В т. ч. волк 828 6,9 2,3 39,1 50,9 0,8 а
рысь 210 6,7 — 2,8 89,0 1,5 41
медведь 296 62,1 26,3 11,6 — — 44
* В основном сведения по хищничеству волка.
** Найдено жертв или исследовано желудков и экскрементов.
*** Жертвы леопарда.
**** О потерях копытных в Березинском и Припятском заповедниках см. также [Лавов, 19806;
Литвинов и др., 1975, 1979, 1981].
В Беловежской пуще, однако, волк предпочитал охотиться на кабана (48,9%)
и косулю (36,6%), а оленя при такой же численности давил в 3 раза меньше
(14,3%). Хищничество усиливалось в годы, когда в популяции было больше мо-
лодых особей и при неурожаях желудей, когда истощенные животные ослаблены
[Гаврин, Донауров, 1954]. Позднее, в 80-е годы, основным объектом хищника
стала косуля (коэффициент избирательности 4,4), затем олень (3,2), лось (2,7), и
последнее место (1,2) занимал кабан [Буневич, 1988].
В Полесском заповеднике (в зоне Чернобыльской АЭС) в первые годы после
его образования в рационе волка преобладал лось (50-60%), затем кабан (20-
30%) и реже встречалась косуля (5-7%). При увеличении плотности населения ко-
пытных основной добычей хищника стали дикие свиньи, но после сокращения их
поголовья от чумы в 1992 г. — лось. С 1998 г. кабан, восстановивший числен-
ность, снова составляет основу рациона волка (48,7%), затем идет косуля
(27,4%), а доля лося сократилась значительно [Воронецкий и др., 1998, 1999; Во-
ронецкий, Тышкевич, 1998; Кучмель, 1999]. В Припятском [Литвинов и др., 1979,
1981] и Кавказском [Теплов, 1938] заповедниках дикие свиньи также преоблада-
ли в добыче хищника лишь в отдельные периоды.
Очевидно, что в питании волка чаще доминирует все же наиболее многочис-
200
ленный вид копытных. Меньшая уязвимость диких свиней в некоторых районах,
возможно, отчасти связана с их активной и, порой, эффективной защитой. Убить
взрослого зверя не всегда удается даже стае, а нападения на секачей иногда за-
канчиваются гибелью хищника [Гаврин, Донауров, 1954; Слудский, 1956; Кова-
лев и др., 1975; Дарман, 1990]. Однако успешность охот волка все же достаточно
высока, особенно на молодых животных, и составляет примерно 30-35%, что
близко к эффективности добычи других видов копытных [Кудактин, 1982; Гор-
диюк, 1996; Приклонский, 1999].
При вселении диких свиней в новые районы хищник сравнительно продол-
жительное время (иногда до 10-12 лет) игнорирует потенциальную добычу. Эта
особенность отмечена многими исследователями [Дормидонтов, 1969; Лавов,
1980а; Варнаков, 1986; Завацкий, 19866, 1987, 1989; Киселев, 1986; Нестеренко,
1989; Филонов, 1989; Стахеев и др., 1999]. На северо-западе России, например,
до конца 70-х годов XX в. случаи нападения волка на проникшего сюда кабана
были сравнительно редки. Он предпочитал привычную добычу - лося (до 53%
состава пищи) и зайца-беляка (до 26%). С 1982 г. встречаемость остатков лося в
экскрементах хищника снизилась до 14,6%, зайца-беляка - до 6,3%, а доля каба-
на возросла до 47,5% [Русаков, Тимофеева, 1984].
Говоря о постепенном привыкании волка к новой жертве, не следует забы-
вать, что кабан сосуществует вместе с ним многие тысячелетия. Поэтому в эво-
люционном аспекте взаимные адаптации у них весьма давние, и вряд ли они бы-
ли утеряны за несколько поколений.
Большую роль в формировании пищевой специализации волка играет тот
вид, которым щенки выкармливаются летом. В Дарвинском заповеднике, где
места воспитания молодняка приурочены к концентрациям лося, в их пище в
дальнейшем доминирует этот вид (98,1%), тогда как на сопредельных территори-
ях - кабан, доля которого среди жертв достигает 73,4% [Нестеренко, 1989].
Основная причина сравнительно низких регистрируемых потерь диких сви-
ней от волка и других хищников, скорее всего, кроется в другом. Остатки этих
копытных, предпочитающих жить в хорошо защищенных местах, сложнее обна-
ружить. Кроме того, крупные хищники съедают сеголеток практически полно-
стью. Я наблюдал в Тульской области, как три волка в декабре утилизировали
крупного поросенка за одну трапезу, оставив лишь раздробленные лопаточные
кости. Реальная смертность вида от этого фактора, на мой взгляд, гораздо выше
фиксируемой.
Любопытные сведения собраны в заповедниках. В Кавказском доля кабана в
пище волка составляет 14-16%, но в годы эпизоотий, вероятно, достигает 50%
[Донауров, Теплов, 1938], в Припятском, Кызыл-Агачском и Хинганском - 28-
83% (см. табл. 43), в Уссурийском в отдельные годы (1945-1948) он служит осно-
вой рациона [Абрамов, 1963]. В Алма-Атинском хищники изымают за год около
30-60% местной популяции диких свиней [Жиряков, 1986, 1990]. В Березинском
после резкого ограничения численности волка с 1959 г. за 8 лет поголовье кабана
выросло со 114 до 820 особей и, напротив, в 1979-1983 гг. размножившийся
хищник сократил популяцию жертвы на 38% [Лавов, 1981, 1986а,б; Лавов, Воро-
нова, 1986]. Уменьшение группировки в Полесском заповеднике в 1996-1997 гг.
также обусловлено мощным прессом волка (Козло и др., 1998). В Дарвинском в
1984/85 гг. волк и медведь изъяли 12-14% популяции [Филонов, 1989]. В Кызыл-
Агачском в 1976-1979 гг. 60-70 волков полностью изымали годичный прирост,
стабилизируя поголовье жертвы примерно на уровне 700 голов. Из-за быстрой
утилизации остатков добычи шакалами, лисицами, барсуками и воронами им
приходится добывать копытных больше, чем нужно для прокорма [Литвинов,
Коновалова, 1979; Литвинов, 1980, 1981; Лобачев, Петухов, 1984]. В Припятском
201
и Центрально-Лесном заповедниках остатки диких свиней в экскрементах волка
встречаются круглогодично [Гатих, 1979; Кочетков, Соколов, 1979].
Особенно уязвимы молодые животные (табл. 44). В Кавказском заповеднике
при высокой численности волка до годовалого возраста доживают всего лишь
8,6% поросят [Дуров, 1980а]. В долине р. Или хищник давит до 50% сеголеток, а
в многоснежные годы уничтожает их и подсвинков почти полностью [Слудский,
1956, 1970]. Основная добыча кызыл-агачских волков - также сеголетки (69,3%),
которыми они выкармливают свои выводки [Лобачев, Петухов, 1984]. В горных
экосистемах Узбекистана по этой причине в живых остается лишь 50% молодня-
ка копытных [Абдуназаров, Тарянников, 1991; Есипов, 1991]. Доля сеголеток в
добыче этого хищника в Приамурье еще выше - 76,2%о [Раков, 1970].
Таблица 44. Половой и возрастной состав погибших кабанов, %
Район Исследо- вано, п Сего- летки Годо- валые Взрос- лые Причина гибели Источник информации
Дарвинский з-к 116 55-83 2-10 14-35 Истощение Тупицина, 1988
Белоруссия 65 56,9 23,0 20,1 Волк Козло, Банад, 1985
Березинский з-к 59 59,3 * 40,7* а Козло, 19866
Полесский з-к — 64,1 26,8 9,1 и Воронецкий и др., 1999
Дельта Вол- ги 1187 75,8 8,8 15,4 Утонули Киселев, 1976
Кызыл-Агач- ский з-к 68 50,0 13,2 36,8 Волк Литвинов, 1980, 1981
Приамурье и 21 76,2 9,5 14,3 (С Раков, 1970
151 53,5 8,6 37,7 Медведь
и 33 36,4 15,1 48,5 Тигр и
а 5 40 40 20 Леопард а
а 14 100 — Рысь а
Хинганский з-к 32 71,9 9,3 18,8 Волк Дарман, 1990
а 6 16,7 — 83,3 Медведь а
а 23 78,3 17,4 4,3 Истощение а
а 35 34,3 20,0 45,7 Болезни а
а 7 42,9 14,2 42,9 Браконьеры а
Липецкая обл. — 40 * 60* а Побединский, 1997
* Доля годовалых и взрослых особей.
В Белорусии волк изымает из популяций диких копытных до 7-8% особей
[Вадковский, Костин, 1978; Кононов, 1988]. По расчетам, 2,5 тыс. хищников в
течение года убивают здесь около 8 тыс. кабанов, 6 тыс. лосей и 4 тыс. европей-
ских косуль, что в 2-6 раз больше, чем добывают охотники [Козло, 2001]. В Ук-
раинских Карпатах от волка и медведя ежегодно гибнет около 10-20% диких
свиней [Гунчак, 1982, 1999].
В Московской области отдельные группы волков буквально «пасут» кабанов,
предпочитая питаться парным мясом. На дневки хищники устраиваются непода-
леку от лежек своих жертв [Михайлов, Клюкин, 1996]. У 5 из 9 волков, отстре-
лянных в марте в центральных районах европейской части России, в желудках
обнаружены остатки новорожденных поросят. В 1977 г. в Смоленской области от
202
этих хищников погибло (зафиксировано) более 100 диких свиней [Кузьмин, Ха-
хин, 1980]. В Брянской области в 1975-1976 гг. только на территории трех охот-
ничьих хозяйств они задавили 53 лося, 94 косули, 6 оленей и 26 кабанов [Вато-
лин, 1979].
В Свердловской области в отдельных районах волк истребляет до 10% попу-
ляции диких свиней. Стая из 9 хищников за 5 дней способна полностью уничто-
жить группу из 6 особей [Киселев, 1986]. Урон от него довольно ощутим и в Бу-
рятии [Смирнов, 1978]. В Предбайкалье на каждого волка приходятся всего лишь
два кабана [Недзельский, 1994], что сдерживает рост поголовья копытных.
В равнинной части Казахстана остатки диких свиней обнаружены в 22% экс-
крементов и поедей (п = 1052) этого хищника, в горных районах этот показатель
достигает порой 37%. В течение года каждый зверь уничтожает до 50-80 особей,
в основном поросят. Пара волков за день съедает и растаскивает подсвинка. В
многоснежные зимы 1945/46 и 1947/48 гг. в низовьях р. Или, в Балхаш-
Алакульской котловине и на оз. Зайсан хищники истребили почти всю косулю и
более половины местной популяции кабана - сначала сеголеток и подсвинков,
затем свиней и секачей [Слудский, 1956, 1970; Слудский и др., 1984; Федосенко,
1986]. На Тургае каждую зиму волки изымают 10-15% поголовья кабана [Соло-
матин, 1972]. В Таджикистане в период щенения они устраивают логова в местах
сосредоточения беременных и лактирующих свиней [Иваненко, 1956].
В условиях глубокоснежья этот хищник не только «режет» копытных боль-
ше, чем может съесть, но и постоянным преследованием разгоняет их, что нега-
тивно сказывается на зимовке [Бромлей, 1964; Керечун, 1979; Лавов, 1986в].
Следует отметить, что в результате безобразных, на мой взгляд, методов и спо-
собов добычи копытных в России и применения неэффективного оружия и снаря-
дов рацион волка в охотничий сезон существенно дополняется ранеными или по-
гибшими от ран животными, а также сеголетками, оставшимися без матерей.
Тигр. Кабан - одна из основных жертв амурского тигра [Капланов, 1948; Аб-
рамов, 1963; Бромлей, 1964; Кучеренко, 1973, 1976; Раков, 1979]. Доля разных
видов копытных в питании этого хищника (табл. 45) примерно соответствует со-
отношению их численности [Животченко, 1976, 1979, 1981; Салькина, 1993,
1994; Мысленков, Микуэлл, 1997], но в Сихотэ-Алинском заповеднике в послед-
ние десятилетия тигры чаще добывали изюбрей [Матюшкин, 1992], а в Уссурий-
ском — пятнистых оленей [Поддубная, Ковалев, 1993].
Успешность охоты на кабана - около 50-55%, на изюбря - 29-37%. При вы-
соком снежном покрове доля успешных нападений на диких свиней существенно
возрастает (до 73%), тогда как на оленя остается практически неизменной [Юда-
ков, Николаев, 1977, 1987; Матюшкин, 1991]. Замечено, что взрослые тигры-
самцы предпочитают охотиться на изюбря, самки - на кабана и косуль [Пикунов,
1981, 1983]. Обнаружив стадо диких свиней, хищник постепенно уничтожает
всех, причем нередко давит их ночью на лежках [Раков, 1970].
Суммарная годовая потребность в пище каждого тигра - 70-100 голов диких
копытных, из них 40-47 кабанов, 25-34 изюбря, некоторое количество косуль и
кабарог, а также до 3 медведей и несколько собак. В Хабаровском крае за год
этот хищник изымает не менее 30% поголовья кабана и 7—10% - изюбря, в При-
морье - 24-35% и 10-20% соответственно, что близко к величине годового при-
роста копытных [Юдаков, 1973, 1974; Кучеренко, 1977а,б, 1993, 1994; Пикунов,
1983; Дунишенко, 1985, 1987; Пикунов и др., 1985; Юдаков, Николаев, 1987;
Pikunov, 1988].
По расчетам, в Амуро-Уссурийском крае тигры ежегодно добывают 3,8-4,5
тыс. диких свиней, бурые медведи -3-4 тыс., волки и рыси - около 1,5 тыс., а
203
вместе - около 10 тыс. особей в год, что значительно больше добычи охотников
[Бромлей, Кучеренко, 1983].
Таблица 45. Избирательность добычи диких копытных животных тигром, %*
Заповедник, район Число регист- раций, п* Кабан Лось Оле- ни Косули Другие виды Источник информации
Уссурий- ский 15 40,0 — 60,0 — — Данченко, 1981
С i 91 33,0 — 57,1 7,7 2,2 Поддубная, Ковалев, 1993
Болыпехех- цирский 27 37,0 — 29,6 3,7 29,7 Ткаченко, 1996
Лазовский 362 31,8 — 50,1 5,3 2,9 Животченко, 1981
а 48 8,3 — 85,4 6,3 — Салькина, 1994
Сихотэ- Алинский 61 23,0 1,6 72, 1 3,3 — Матюшкин и др., 1981
а а 144** 292 28,5 31,8 4,1 57,6 54,5 3,5 6,3 13,7 Матюшкин, 1992 Смирнов, 1993
а 53 32,1 — 54,7 9,4 3,8 Смирнов и др., 1996
а 552 29,5 — 54,3 — 16,2 Miquelle et al., 1996
а — 36,0 — 57,0 — 7 q*** Мысленков, Микуэлл, 1997
Сихотэ- Алинь 720 55 __ 37 7 Pikunov, 1988
64 54,4 — 25,0 И,9 8,7 Юдаков, 1974; Юдаков, Ни- колаев, 1987
Дальний Восток 98 56 — 27 6 И Кучеренко, 1972
(С 83 50,0 3,5 23,0 23,5 — Раков, 1979
* Найдено жертв или исследовано желудков и экскрементов.
** Совокупные сведения за 1937-1977 гг.
*** вИд жертвы не определен.
Бурый медведь. Этот зверь нападает на кабана в основном весной и в начале
зимы [Бобырь, 1980; Варнаков, 1986; Смирнов, 2001] и в целом наносит популя-
циям этого вида все же меньший урон, чем волк и тигр.
В горной части Казахстана остатки диких свиней в экскрементах этого хищ-
ника составляют 0,1-5,9% [Слудский и др., 1984]. На Дальнем Востоке в зоне
хвойно-широколиственных лесов медведь, напротив, специализируется на добы-
че кабана. Его доля достигает здесь 21,8% в общей смертности вида и 61-62% в
гибели от хищников. Применяемые медведем способы добычи копытных разно-
образны - скрадывание, подкарауливание и длительное (до 20 км) преследова-
ние, причем он легко справляется даже с секачами [Раков, 1970, 1979].
Один медведь способен за месяц уничтожить 3-8 диких свиней, шатуны в
204
районе своего зимнего обитания истребляют стада полностью, начиная с поро-
сят. Суммарные потери от этого хищника только на Дальнем Востоке оценива-
ются в 6 тыс. голов в «голодные» годы [Бромлей, 1964; Кучеренко, 1973, 1976].
Другие хищники. Рысь из копытных животных явно предпочитает косуль
(табл. 43) и на кабанов нападает сравнительно редко, выбирая из засады некруп-
ных особей, чаще - поросят [Бромлей, 1964; Раков, 1970; Русаков, Тимофеева,
1984; Дарман, 1990; Аромов, Вашетко, 1999]. В ее экскрементах остатки диких
свиней составляют в разных районах от 4 до 16 % [Донауров, Теплов, 1938;
Слудский, 1956; Козло, 1975; Желтухин, 1979; Федосенко, 1980].
В Приамурье [Раков, 1970] и в заповеднике «Кедровая падь» [Коркишко,
1986] отмечены случаи смертности кабана, в том числе секачей, от леопарда. В
Западном Копетдаге в Туркмении дикие свиньи составляют заметную долю (до
27%) в его питании [Кулибаба, Пакулин, 1985; Лукаревский, 1986, 1995; Кузне-
цов, 1999]. Зарегистрирована также гибель этих копытных от снежного барса
[Донауров, Теплов, 1938; Слудский, 1956; Федосенко, 1986] и харзы [Бромлей,
1964; Кучеренко, 1976].
Новорожденных и мелких поросят могут пожирать енотовидная собака [Ру-
саков, 1979] и лисица [Иваненко, 1956; Слудский, 1956; Козло, 1975; Есипов,
1991]. Я наблюдал в Челябинской области попытку лисовина напасть на отстав-
шего от стада месячного поросенка. На пронзительный визг детеныша стремглав
выскочила мать, обратив хищника в бегство.
В годы высокой численности кабана и в морозные многоснежные зимы, ко-
гда ослабевшие звери выходят к населенным пунктам, на них нападают бродячие
и деревенские собаки, которые легко давят поросят и подсвинков. Иногда собаки
объединяются в стаю и совершают набеги на копытных в их типичных местооби-
таниях [Соломатин, 1972; Щербак, 1986]. В Прибалхашье в одном из урочищ 5
брошенных рыбаками собак лишь за сентябрь уничтожили 2 выводка, насчиты-
вающие 9 поросят [Злобин, 1969]. В Ставрополье в 1979 г. зафиксировано И
случаев гибели диких свиней от бродячих собак [Саркисов, Хохлов, 1982].
В Ярославской области мне довелось видеть работу собаки-лайки одного из
браконьеров, специализировавшегося на добыче кабана. Она по снежной тропе
легко настигала группу и останавливала лидера. Свинья принимала оборони-
тельную позу и временами бросалась на собаку. Во время поочередного пресле-
дования и кружения группа рассыпалась и сходила с тропы. Заметив отбившего-
ся сеголетка, собака оставляла свинью и, как только та выходила на тропу, при-
давливала поросенка, держа зубами за ухо. Если хозяина долго не было, лайка
давила жертву, затем находила человека и короткими пробежками вперед-назад
призывала его следовать к месту добычи.
Однако гораздо чаще охотничьи собаки настигают стадо, душат поросят, а затем
преследуют взрослых особей, не возвращаясь к растерзанным сеголеткам [Злобин,
1969], или съедают их еще до подхода людей. Эти особенности следует учитывать в
охотничьем хозяйстве при выборе оптимальных способов промысла копытных.
Конкуренты
Конкурентов у кабана сравнительно много, но межвидовые отношения не та-
кие напряженные, как, например, у оленьих. Тем не менее другие виды живот-
ных могут опосредованно оказывать существенное влияние на популяции диких
свиней, сокращая запасы их пищи.
Антагонизм между копытными заметен лишь на подкормочных площадках,
где благородные олени вытесняют диких свиней. В урожайные годы оленьи ста-
новятся массовыми потребителями желудей и плодов фруктовых деревьев. Кро-
ме того, они ежегодно сокращают запасы влажной надземной растительности,
205
снижая кормовую емкость угодий для кабана. В Беловежской пуще при общей
очень высокой численности оленьих и зубра плотность населения диких свиней
не поднималась выше 27 особей на 1000 га угодий даже при наличии подкормки
[Шостак, 1978]. В Средней Азии их поголовье сокращалось из-за трофической
конкуренции с домашним скотом [Андреенков, 1980]. С другой стороны, кабан,
при высокой плотности его населения, оказывается более весомым конкурентом
и источником беспокойства для оленьих, чем они для него. В Саянах маралы
обычно покидают свои участки обитания через несколько дней после появления
большой группы диких свиней [Завацкий, 1989].
В совместном обитании разных видов копытных на одной и той же территории
есть и преимущества. Зимой оленьи часто перемещаются по тропам диких свиней,
экономя энергию и получая доступ к новым кормовым участкам, кабан же нередко
использует их тропы, выложенную для них подкормку и трупы погибших особей.
Наиболее значимые пищевые конкуренты диких свиней - барсук, в меньшей
степени - бурый и гималайский медведи, имеющие сходный тип питания, и ено-
товидная собака. Однако летом и осенью запасы пищи практически не ограниче-
ны, а зимой эти виды животных находятся в спячке или малоактивны. Тем не ме-
нее замечено, что при высокой плотности населения кабана резко снижается чис-
ленность барсука [Горшков, 1988; Хорин, Горшков, 1989].
Осенью дикие свиньи нередко посещают те же поля овса, на которых кор-
мится бурый медведь [Плешак, Миняев, 1986], и даже вытесняют его с них [Вар-
наков, 1986], но чаще все же сами первыми уходят при появлении хищного кон-
курента или не выходят на кормежку до тех пор, пока тот находится на поле. Зи-
мой, как отмечалось выше, крупные хищники нередко «предоставляют» кабанам
остатки своей добычи, способствуя их выживанию, но при этом свиньи сами час-
то становятся их пищей.
Мелкие млекопитающие (мышевидные грызуны, белки, бурундуки, сони,
зайцеобразные, куньи) и птицы (кедровки, сойки, дятлы, поползни, дубоносы,
дрозды, рябчики, фазаны и многие др.) - потребители желудей, орехов, плодов,
ягод, семян и корневищ повсеместно и весьма быстро уменьшают запасы корма,
растаскивая его даже на подкормочных площадках. Крот в отдельных стациях
может значительно снижать численность дождевых червей. Зимой главными пи-
щевыми конкурентами кабана становятся ворона, сорока, лисица, куницы, со-
боль, быстро утилизирующие трупы животных.
На водоемах пищевыми конкурентами могут быть ондатра, бобр, водяная по-
левка, гуси, лысуха и другие водоплавающие, поедающие водно-болотную рас-
тительность. Однако кормовые ниши этих животных все же разные. Ондатра,
как, впрочем, пищуха и бурундук в лесах, чаще выступает в роли своеобразного
заготовителя корма для диких свиней, уничтожающих их запасы, остатки пищи,
хатки и самих хозяев.
Человека, как потребителя плодов и преобразователя биотопов, также можно
причислить к основным трофическим конкурентам кабана. Но он же оказывает
ему одновременно и неоценимую помощь, «предоставляя» корм на сельскохо-
зяйственных полях, в овощехранилищах, на скотомогильниках, подкормочных
площадках и после охот (подранки, остатки после разделки туш).
Как видим, трофические связи диких свиней в биоценозе очень сложны.
Важно, что многие пищевые конкуренты, включая хищников, одновременно
весьма способствуют их выживанию.
Гибель в водоемах
При недостатке кормов дикие свиньи перемещаются на десятки и сотни ки-
лометров, что особенно отчетливо выражено на Дальнем Востоке в неурожайные
206
годы (см. выше). Соответственно в одних районах их поголовье резко уменьша-
ется, в других - увеличивается. Однако общая численность крупных группировок
при этом существенно не изменяется, хотя часть животных неминуемо гибнет
при переходах, и особенно часто - в водоемах.
Дикие свиньи хорошо плавают, однако они нередко тонут в полыньях при
форсировании рек по тонкому льду. Иногда на лед и в воду их выгоняют волки
[Насимович, 1955], но чаще так они гибнут в осенне-зимний сезон охоты при
преследовании человеком или собаками. При этом больше тонет крупных осо-
бей, имеющих большую массу тела и, следовательно, легче проламывающих лед.
В Ленинградской области в январе 1974 г. только на 3-километровом участке р.
Долгой зарегистрирована гибель 5 взрослых особей, а на р. Тигоде утонули 7
весьма упитанных зверей [Русаков, Тимофеева, 1984]. Подобные случаи зареги-
стрированы и в других районах [Слудский, 1956; Раков, 1970; Простаков, 1996].
Очень много животных гибнет в половодья и в паводки, особенно мощные в
низовьях Днепра, в дельте Волги и на всех крупных реках и озерах Казахстана, а
также на северном побережье Каспия, где из-за сильного ветра в осенне-зимний
период образуются нагонные волны, замерзающие в тростниках. В отдельные го-
ды дикие свиньи, живущие на равнинных водоемах, в большом количестве гиб-
нут в наледях и в ледяных пустотах [Слудский, 1956; Слудский и др., 1984]. Вы-
сокая вода к тому же выгоняет зверей в прибрежные степи и пустыни, где они
становятся легкой добычей человека и хищников.
В дельте Волги массовая гибель кабана зарегистрирована в 1910, 1923, 1928,
1946 и 1947 гг. [Лавровский, 1952, 1962]. В 1966 г. здесь во время весеннего по-
ловодья нашли около 800 трупов, из них 82 взрослых особей и более 700 поросят,
в 1974 г. - около 900 особей, в том числе в Астраханском заповеднике - более
400 голов (25% популяции). В их числе 101 самка, 104 подсвинка и 200 сеголет-
ков [Киселев, 1975, 1976].
Доля этого фактора в общей смертности вида сравнительно высока и колеб-
лется в разных районах в пределах 0,8-25% (табл. 42). М.Д. Перовский [1998]
оценивает долю потерь кабана из-за гибели в воде в 6,4%.
Болезни и травмы
Болезни. Роль болезней в динамике популяций диких свиней исследована не-
достаточно. В общей зарегистрированной смертности зверей доля этого фактора
составляет в разных районах от 1 до 40%, но все же он гораздо меньше уровня
гибели от истощения (см. табл. 42).
Кабаны восприимчивы ко многим болезням, характерным для других диких и
домашних копытных. Зарегистрированы классическая чума свиней, ящур, рожа,
туляремия, бруцеллез, пастереллез, пироплазмоз, паратиф, туберкулез, колибак-
териоз, сибирская язва, болезнь Ауески, чесотка, ложное бешенство, сальмонел-
лез, лептоспироз и др. [Донауров, Теплов, 1938; Слудский, 1956; Горегляд, 1971;
Кормилицин, Дулицкий, 1972; Стародынова, 1974а,б, 1979; Козло, 1975; Ивано-
ва, Овсюкова, 1976; Литвинов, 1980; Седов и др., 1992].
Сравнительно высока гибель поросят из ранних выводков от пневмонии.
Другая причина гибели или отставания в росте молодняка - острые желудочно-
кишечные заболевания: катаральные, катарально-геморрагические, геморрагиче-
ские энтериты и гастроэнтериты, вызываемые употреблением недоброкачествен-
ного корма [Стародынова, 1974а, 1979].
Особенно высоки потери диких свиней от чумы. Это острое вирусное заболева-
ние поражает животных всех возрастов с летальностью до 80%. Типичный признак
чумы - красные пятна на коже живота и в паховых складках, на ушах и морде, а
также гнойные истечения из глаз и носа [Иванова, Овсюкова, 1976]. Заболевшие жи-
207
вотные теряют аппетит, неохотно поднимаются с лежек, у них характерная сидячая
поза и шаткая походка [Фертиков, Егоров, 1997], при беге они отстают от группы.
Лихорадка сопровождается жаждой, что побуждает зверей пить воду. Поэтому тру-
пы обычно находят у водных источников [Шиков, Гришок, 1995].
Источником инфекции чаще всего становятся неблагополучные по этому за-
болеванию свиноводческие хозяйства [Равилов и др., 1992]. В природу возбуди-
тель заносят при выбрасывании павших животных. Иногда в поисках пищи или в
период гона кабаны забредают на свинофермы, где также могут заразиться от
домашнего скота или заразить его.
В последние годы стало обозначаться участие кабана в эпизоотическом про-
цессе классической чумы свиней и как резервуара возбудителя, и как источника
инфекции. Периодичность вспышек болезни составляет 4-7 лет (частота 55%) и
12-13 лет (22%), но отдельные ее проявления отмечаются через 2-36 лет. Эти пе-
риоды, как правило, совпадают со временем вспышек чумы среди домашних i
свиней. Наибольшая продолжительность эпизоотии на большом пространстве ;
(60% территории Молдавии, треть Украины и 7 областей России) - 7 лет [Коло- !
мыцев и др., 1995, 1999].
В Беловежской пуще вспышки чумы у кабанов имели место в 1906, 1911-1912,
1927 и 1963-1964 гг. В 1964 г. поголовье сократилось в 2,6 раза [Врублевский,
1908; Горегляд, 1971; Козло, 1975; Шостак, 1978]. В Белоруссии эпидемия зареги-
стрирована и в 1992-1993 гг. [Каштальян, Медведев, 1999]. В Карпатах эта болезнь
проявлялась в 1946 г., а рожа-в 1951-1952 гг. [Гунчак, 1982]. В Крыму падеж ди-
ких свиней от чумы и пастереллеза отмечался в 1965, 1968 и 1971 гг. [Мельникова,
1968; Павлов и др., 1974]. На Украине и юге России мощные вспышки чумы были
в 1974-1976 и 1985-1994 гг., причем эпизоотия 1985 г. возникла в Молдове [Ши-
ков и др., 1992; Коломыцев и др., 1995, 1999; Шиков, Гришок, 1995]. В Черкасской
области в 1972-1974 гг. погибла треть поголовья [Евтушевский, 1975]. На Север-
ном Кавказе эта болезнь наблюдалась в 1935-1936 гг. и, по-видимому, в 1947 г. В
1974-1975 гг. в черноморских лесах погибло около 7% популяции [Донауров, Теп-
лов, 1938; Котов, Рябов, 1963; Дуров, 1980].
На западе России (Брянская, Смоленская, Тверская, Калужская, Московская,
Воронежская и Курская области) классическую чуму свиней регистрировали в
1966-1967, 1974-1976, 1991-1992 и 1996 гг. В Калининской и Московской об-
ластях зимой 1975/76 гг. пало около 2 тыс. особей [Фадеев, 1977; Фомушкин,
Максимов, 1992]. В «Завидово» по этой причине в 1976 г. группировка диких
свиней уменьшилась значительно (табл. 39). В 1991 г. благодаря вакцинации па-
ло не более 70 голов [Егоров, 1992]. С октября 1990 г. по апрель 1991 г. в Мос-
ковской области обнаружено 318 павших и отстреляно 245 кабанов с признаками
болезни [Коломыцев и др., 1995, 1999]. Общие их потери в Московской и Твер-
ской областях в 1976 и 1992 гг. оцениваются в несколько тысяч голов [Фертиков,
Егоров, 1997].
Гибельные эпизоотии чумы свиней в разных районах Бурятии и на западе Чи-
тинской области наблюдались в 1950-1960 гг. [Смирнов, 1978]. В Приамурье падеж
имел место в 1945-1946, 1953-1955, 1957-1959 гг. [Раков, 1970] и 1984-1985 гг.
[Юсупов и др., 1992], на юге Дальнего Востока- в 1936, 1941, 1955 и 1957 гг. [Куче-
ренко, 1973, 1976]. В Хинганском заповеднике вспышка чумы отмечена в 1968 г.,
массовая гибель - в 1980 г. Из 500 особей выжило лишь 130-150 голов [Дарман,
1990]. В Приморье эпизоотии регистрируются через 5-6 лет, унося иногда до 70%
локальных популяций или более 10 тыс. особей в отдельные годы. На восстановле-
ние поголовья требуется минимум 3-4 года при условии подтока мигрантов и хоро-
ших урожаев естественных кормов [Бромлей, 1964].
В Казахстане массовый падеж зверей от чумы отмечался в разных районах: 1936 г.
- в Семиречье, 1937 г. - в долине Сырдарьи, 1950 г. - в пустыне Кызылкум, 1951—
208
1952 гг. - на северном побережье Каспия. В июле 1939 г. мощная эпизоотия зареги-
стрирована в дельте Амударьи [Слудский и др., 1984]. На юге Киргизии кабан стал
исключительно редким животным после эпизоотии 1931/32 гг. Поголовье постепен-
но восстановилось только к 1940 г. [Чичикин, Воробьев, 1967].
За пределами бывшего СССР чуму регистрировали в Германии (в 1949, 1990,
1993, 1995 и 1997 гг.), Италии и Австрии [Шиков, Гришок, 1995; Depner и. а., 1997].
В Беловежской пуще массовая гибель кабанов и других копытных (7,6% по-
головья) при геморрагической септицемии зарегистрирована в 1910 г. Заболева-
ния паратифом и свиной рожей не оказали заметного влияния на эту популяцию
[Козло, 1970а].
Вспышка ящура, занесенного крупным рогатым скотом в 1952 г., была осо-
бенно губительной для диких свиней в Южной Киргизии. Весной 1953 г. по мере
стаивания снега в ущельях находили большое количество трупов [Чичикин, Во-
робьев, 1967].
Из болезней алиментарного происхождения наиболее известна остеодистрофия
диких свиней, вызываемая недостатком минеральных веществ, чаще всего - каль-
ция и фосфора. В коллекциях черепов биологических институтов Санкт-
Петербурга доля особей с признаками остеодистрофии достигает 35%, что гораздо
выше, чем у других копытных. Наиболее часто это заболевание встречается у
взрослых животных на северном и южном пределах ареала вида - вероятно, вслед-
ствие пессимальных условий обитания, регулярного голодания и истощения. Для
профилактики этой болезни в охотничьих хозяйствах эффективными могут быть
добавки в корм мела, костной муки и пережженных костей, монокальцийфосфата и
диаммонийфосфата [Русаков и др., 1981; Русаков, Тимофеева, 1984].
Паразиты. Кабан - объект разнообразных и многочисленных паразитов, на-
считывающих около 110 видов. В их числе 12 видов простейших (амебы, инфу-
зории, кокцидии, гемоспоридии, трихомонады, саркоспоридии, токсоплазмы), 48
видов гельминтов и паразитических аннелид (нематоды, трематоды, цестоды,
скребни, пиявки), 30 видов паразитических паукообразных (иксодовые, аргасо-
вые и чесоточные клещи), вши и 19 видов двукрылых кровососущих насекомых.
Каждому возрасту зверей свойственен свой комплекс паразитов, достигающих
наибольшего разнообразия у взрослых особей, особенно у самок, ведущих, в от-
личие от секачей, групповой образ жизни. Дикие свиньи инвазированы круглого-
дично, но по сезонам происходит заметное колебание численности паразитов и
смена одних видов и групп другими. Интенсивность инвазии существенно зави-
сит от плотности населения зверей и их биотопического распределения [Мельни-
кова, 1968].
Фауна гельминтов обычно общая для диких и домашних копытных, населяю-
щих конкретную территорию, и чаще всего именно домашний скот становится ис-
точником инвазии [Говорка и др., 1988]. Однако в ряде районов доминирующая
роль диких копытных в этом процессе неоспорима. В Таджикистане, например, где
свиноводство стало развиваться лишь в 30-х годах XX в., кабан был и остается ос-
новным носителем и резервентом паразитофауны [Мельникова, 1968].
В разных районах бывшего СССР у диких свиней регистрируют от 1 до 22
видов паразитических червей. В кишечнике локализуются 15, в желудке - 7, в
бронхах - 5, в мозгу и мускулатуре - 2, в легких, мочевом пузыре и языке - по 1
виду [Овсюкова, 1975]. Среди гельминтозов обычны метастронгилез, трихинел-
лез, дикроцелиоз, фасциолез, эхинококкоз, аскаридоз, цистицеркоз (финноз), ре-
же встречаются тонкошейный саркоцистоз, макрокантаринхоз, спарганоз, глобо-
цефалез, трихоцефалез и др. [Белопольская, 1952; Слудский, 1956; Мельникова,
1968; Горегляд, 1971; Карасев, 1974; Назарова, Стародынова, 1974а; Стародыно-
ва, 1974а; Козло, 1975; Литвинов, 1975, 1980а; Шималов и др., 1975; Иванова,
Овсюкова, 1976; Литвинов, Карасев, 1977; Ромашов, 1979; Закариев, 1980; Слуд-
14. Заказ №2528.
209
Рис. 66. Схема разви-
тия метастронгилу-
сов у кабана (по [Го-
регляд, 1971])
ский и др., 1984; Говорка и др., 1988; Егоров, 1992; Горелов, 1994; Пенькевич,
1998а; Фертиков и др., 1999].
Наибольшее воздействие на популяции кабана оказывает метастронгилез.
Это легочно-глистное заболевание, вызываемое Metastrongylus elongatus, М.
pudendotectus, М. confusus, М. salmi и М. pulmonalis, известно на всем ареале вида
[Овсюкова, 1974]. Возбудители - белые, тонкие и круглые черви длиной до 5 см,
паразитирующие в бронхах зверей. Самки метастронгилид выделяют яйца со
сформировавшейся личинкой, скрученной восьмеркообразно. При кашле яйца
попадают в носоглотку, оттуда в пищеварительный тракт и на землю вместе с
фекалиями. Промежуточные хозяева паразита - более десятка видов дождевых
червей, заглатывающих рассеянные в почве яйца метастронгилусов (рис. 66).
Яйца паразита устойчивы к воздействию внешних факторов и в сухой почве
жизнеспособны до 20 месяцев, а во влажной сохраняются более двух лет. Из яиц
появляются личинки, которые паразитируют в дождевом черве до конца его жиз-
ни. Один экземпляр может быть носителем сотен личинок. В метастронгилезных
очагах зараженность червей достигает 80%. От их обилия напрямую зависит эпи-
зоотическое значение разных типов охотничьих угодий. В годы с повышенной
влажностью заболеваемость наиболее высока, в сухие годы, напротив, сокраща-
ется. Заражение диких свиней возможно в течение всего бесснежного периода.
Проглоченный дождевой червь переваривается, личинки освобождаются и через
стенки кишечника проникают в лимфатические узлы, а затем через лимфатиче-
ские и кровеносные сосуды внедряются в легкие. Здесь они проникают в альвео-
лы, потом в бронхиолы и бронхи, где быстро растут и через 25-35 дней достига-
ют половозрелости. Продолжительность жизни метастронгилусов в легких жи-
вотного - около одного года [Иванова, Овсюкова, 1976; Зеньков и др., 1979; Ста-
родынова, Сыроежкин, 1979; Говорка и др., 1988].
Кабан, следовательно, - основной хранитель инвазии в природе. Ее уровень
зависит от возраста и плотности населения этого вида, численности дождевых
червей, сезона года, погоды, биотопа. Новорожденные обычно свободны от пара-
210
зитов, но с каждым месяцем жизни их инвазированность увеличивается, а у
взрослых, напротив, становится меньше. В Подмосковье заражены 40-100% се-
голеток и подсвинков, 14-72% 4-летних и 33% 7-летних особей [Овсюкова, 1974,
1975], в Березинском заповеднике - 78,6% сеголеток, 62,4% подсвинков и 46-
47% взрослых животных [Карасев, 1974], в разных зонах Беларуси - 21-76%
зверей [Пенькевич, 1997; Одинцова, 1998].
В легких отдельных молодых особей находят до 9,8 тыс. экземпляров паразита,
чаще - около 200-500. Зараженные животные становятся более восприимчивыми к
другим заболеваниям, острее реагируют на недостаток кормов, хуже переносят не-
благоприятную погоду. Рост поросят при высоком уровне инвазии задерживается
или совершенно прекращается, а их масса обычно в 1,5-2 раза меньше, чем у здо-
ровых. Отход инвазированных сеголеток, особенно в сырые годы, может достигать
8-30% [Морозов, Назарова, 1962; Мельникова, 1968; Стародынова, 1974а; Старо-
дынова, Сыроежкин, 1979; Литвинов, 1980а; Овсюкова, Бельский, 1980; Варнаков,
Михайлова, 1982; Пенькевич, 19986; Фертиков и др., 1999].
Эпизоотическое значение имеет и трихинеллез, в формировании и поддержа-
нии природных очагов которого ведущую роль играют хищники и грызуны [Па-
шук, 1970]. Кабан чаще всего заражается, поедая мышевидных грызунов и трупы
хищных животных, в том числе выбрасываемые охотниками после снятия шкур.
Доля зараженных особей в популяциях диких свиней сравнительно невелика. В
Московской и Тверской областях, например, она составляет 0,01-0,09%, в то время
как обследованные волки поражены на 20%, лисицы и енотовидные собаки - на
11-12% [Исаев, Иванова, 1979; Егоров, 1992]. В Березинском заповеднике экс-
тенсивность инвазии этих видов выше: 0,66, 35,7 и 34,4% соответственно, у ено-
товидной собаки - 38,4%, медведя - 18,1, рыси - 29, собак - 12,5, кошек - 8,4,
черной крысы - 16, обыкновенной бурозубки - 2,7%. Уровень зараженности ли-
чинками эхинококка достигает 16% [Карасев, Литвинов, 1974; Литвинов, Кара-
сев, 1977]. В Таджикистане трихинеллез у кабана встречается еще чаще - 1,3%.
Кроме того, этот вид играет большую роль в поддержании природных очагов эхи-
нококкоза, описторхоза, гнатостоматоза, гонгилонематоза и других болезней
[Мельникова, 1968]. В Туве уровень зараженности диких свиней трихинеллезом
составляет 2%, лисицы - 5%, волка - 7%, медведя - 12%, колонка - 15% [Gorelov,
1998]. Наиболее поражены трихинеллами дикие животные на Северном Кавказе и
в Предкавказье: кабан - 2,5—15,8%, волк - 60%, шакал - 83%, медведь - 15- 89%,
лисица - 20-37%, енотовидная собака - 14-75%, куница - 27%, серая крыса -
1,8-3,4% [Пиголкин, Бокун, 1992].
Непосредственная гибель копытных от гельминтозов, за исключением метас-
тронгилеза, наблюдается сравнительно редко. Тем не менее паразиты играют
важную роль в поддержании стабильности биоценозов как регулирующий фак-
тор [Говорка и др., 1988]. Ущерб от гельминтов выражается в снижении уровня
воспроизводства и выживаемости ослабленного молодняка, увеличении смертно-
сти от других факторов среды, уменьшении массы тела и упитанности и, в целом,
в снижении продуктивности популяции, ее трофейной ценности и качества мяс-
ной продукции. Зараженность и потери тем значительнее, чем выше плотность
населения зверей [Козло, 1975; Рыковский, Маклакова, 1999].
В национальном парке «Завидово», например, от гельминтозов гибнет до 20%
молодняка диких свиней. При ветеринарно-санитарной экспертизе выбраковывается
и уничтожается до 2% туш, пораженных трихинеллами, и до 3,4% - в связи с зара-
жением спарганумами (личиночной формой Spirometra erinaceieuropaei\ а также до
60% печеней, пораженных фасциолой [Фертиков и др., 1999].
Немаловажно, что инвазированные животные, в свою очередь, становятся ис-
точником заражения человека, употребившего их мясо.
14
211
На кабане паразитируют клещи родов Haemaphysalis, Dermacentor, Ixodes,
Rhipicephalus, Boophilus и свиные вши рода Haematopinus. Общее число эктопа-
разитов на теле отдельных зверей, особенно весной и летом, может исчисляться
сотнями. Кровососы явно доставляют беспокойство, и дикие свиньи пытаются
избавиться от них во время грязевых купаний и чесания о деревья. Как и другие
виды копытных, они, и особенно сеголетки, страдают и от гнуса (мошки, мокре-
цов, комаров, слепней и оводов), стараясь проводить жаркие дни в укрытиях, во-
де, грязи, поднимаются высоко в горы, уходят в пески, степи или на продуваемые
ветром места.
Как прокормитель клещей и других кровососов кабан играет существенную
роль в формировании и поддержании природных очагов возбудителей и пере-
носчиков паразитарных и инфекционных болезней. Из многих видов паразити-
рующих на нем клещей выделены возбудители лихорадки Ку, туляремии, клеще-
вого энцефалита, бруцеллеза и других болезней. Гибель самих зверей от эктопа-
разитов сравнительно редка. Случаи зудневой чесотки с летальным исходом за-
регистрированы у молодых особей в Сары-Челекском заповеднике в Киргизии
[Мельникова, 1968]. Зудневая эпизоотия имела место также в Приамурье в 1927—
1928, в 1933-1934 и в 1961 гг. [Раков, 1970].
Травмы. Травмы, вызванные причинами внутривидового характера, чаще
всего отмечаются в двух возрастных группах - среди поросят и взрослых самцов.
Сеголетки обычно получают их на местах кормежки, особенно на небольших по
площади подкормочных площадках. Подсвинки, в отличие от сравнительно ми-
ролюбивых к детям взрослых особей, нередко увечат их, ломая ребра, кости че-
репа, конечности, позвоночник. Взрослые кабаны, несмотря на чрезвычайную
живучесть, порой погибают от ранений клыками в период гона. Смерть наступа-
ет спустя длительное время от осложнений, плеврита и перикардита [Раков, 1970;
Стародынова, 1974]. Нередко звери при бегстве от опасности калечатся о сучья и
стволы поваленных деревьев, иногда распарывают животы. Раны им могут нано-
сить и хищники.
Доля этого фактора в общей смертности в целом сравнительно невелика и
вряд ли превышает 2%. Гораздо выше смертность от огнестрельных ран (см. раз-
дел «Использование ресурсов»).
Антропогенные факторы
Травматизм на дорогах. На автодорогах ФРГ от столкновения с транспортом
ежегодно погибают 0,5-1,5 тыс. кабанов [Икерманн, 1970; Фокина, Размахнин,
1989]. В Центральном Черноземье доля погибших при наездах автотранспорта и
поездов достигает 8-11% [Простаков, 1996; Побединский, 1997].
Отравления. На северо-западе России зарегистрировано 35 случаев (4,5% в
общей смертности) отравления диких свиней минеральными удобрениями, ос-
тавляемыми в кучах на полях [Русаков, Тимофеева, 1984]. В 1967 г. в Лискин-
ском районе Воронежской области погибли 8 особей, в 1968 г. в Долгоруковском
районе Липецкой области - 2 секача [Сухоруков, 19726], в Каракулинском рай-
оне Удмуртии в августе 1977 г. — 6 зверей [Попов, Попов, 1980]. В Завидовском
заповедно-охотничьем хозяйстве на картофельных полях в 1969 г. пали 2 зверя от
отравления суперфосфатом [Щербаков, 1971], а в 1971 г. - 6 голов, предположи-
тельно после опыления полей гербицидами. Здесь же имел место случай гибели
животного после поедания кусочков соли-лизунца [Стародынова, 1974]. В Рос-
товской области из 87 зарегистрированных случаев смертности кабана на отрав-
ления приходится 30% [Казаков, Даликов, 1983], в Липецкой области - 19,1°Z
[Побединский, 1997].
Казахстанские популяции диких свиней в середине XX в. сильно пострадали
212
от ядов (мышьяковокислого или мышьяковистокислого натра и белого мышья-
ка), применяемых при истреблении саранчи [Слудский, 1956; Сыроечковский,
Рогачева, 1975]. Отмечены случаи гибели этих копытных от стрихнина и фтор-
ацетата бария при поедании отравленных приманок и трупов затравленных вол-
ков, лисиц и ворон [Бромлей, 1964; Собанский, 1992; Смирнов, 2001]. В марте
1974 г. в Каякентском районе Дагестана обнаружен 21 труп кабанов, собравших
разбросанное по снегу зерно, обработанное фосфидом цинка в целях борьбы с
мышевидными грызунами [Метелкин и др., 1974].
Уничтожение биотопов. Уменьшение ареала и численности диких копыт-
ных в России нередко связано с сокращением площади лучших местообитаний и,
соответственно, кормов и убежищ. Мелиорация, осушение плавней, хозяйствен-
ное освоение пойменных земель, вырубка тугаев, широколиственных, плодовых
и орехоносных лесов повсеместно не благоприятствуют и жизни кабана [Гинеев,
1975а; Иванова, 1976, 1982; Карпинский, Лукаревский, 1987; Реймов и др., 1989].
Приведу некоторые примеры.
В неосвоенных человеком плавнях Приазовья в 50-е годы свежие тропы ка-
бана попадались «чуть ли не на каждом шагу», а обсыхающие мелководья были
так изрыты, что походили на вспаханные поля. В 60-е годы большие участки бы-
ли отведены под рисовые поля и нерестово-вырастные рыбные хозяйства. Экска-
ваторами прорыто множество каналов, по которым днем и ночью стали курсиро-
вать моторные лодки. Для охотников (и браконьеров) стали доступны самые глу-
хие места. Все это привело к резкому снижению численности и площади обита-
ния местной группировки диких свиней [Костоглод, 1969].
В тростниковых крепях и тугаях дельты Волги, Каспийского побережья, Ка-
захстана и Средней Азии нередки пожары, длящиеся несколько дней и охваты-
вающие огромные пространства. Масса кабанов, особенно молодняка, гибнет в
огне или калечится, в результате чего их поголовье значительно сокращается.
Причина большинства пожаров - антропогенная. Палы пускают пастухи ради
улучшения кормовых угодий для домашнего скота, рыбаки - для лучшего проез-
да и прохода, браконьеры - сборщики яиц, а также охотники - с целью выгона
зверей на открытое место [Слудский, 1956; Лавровский, 1962; Соломатин, 1972].
Весеннее выжигание тростника и массовая заготовка молодых побегов на
больших площадях до двух раз за лето лишают диких зверей естественных укры-
тий, существенно увеличивая негативное влияние на популяции браконьеров и
хищников. Выпас домашнего скота в типичных биотопах диких свиней также
повсеместно не благоприятствует их обитанию [Киселев, 1975; Шахмарданов,
Спасская, 1975; Слудский и др., 1984; Анненков, 1989]. В дельте Волги тростник
на значительном пространстве выкашивали ранее в качестве сырья для Астра-
ханского целлюлозно-картонного комбината, что приводило к уничтожению
лучших стаций [Мошонкин, 1987].
На Дальнем Востоке уменьшение ресурсов кабана, помимо браконьерства и
увеличения численности волка, вызвано сокращением площади хвойно-
широколиственных лесов и изменением их облика после обширных пожаров, а
также масштабных вырубок кедрачей [Бромлей, 1964].
Охота. Один из важнейших факторов динамики популяций диких свиней -
прямое воздействие человека: охота легальная и браконьерская. Использованию
ресурсов вида посвящена следующая глава.
В заключение замечу, что быстрый рост численности в популяциях кабана
обычно определяется сочетанием нескольких позитивных факторов, в то время
как резкое уменьшение, как правило, зависит от единственного неблагоприятно-
го, чаще - от недостатка зимних кормов и болезней. В России в динамике попу-
ляций этого вида наиболее значимый фактор - высокая смертность сеголеток от
истощения и хищников. Характер колебаний численности может быть изменен
213
активным воздействием человека - от максимального сохранения животных до
их полного уничтожения.
Использование ресурсов
Кабан - важнейший объект охотничьего промысла со времен палеолита (см.
выше). Добывают его в основном ради мяса и сала. Из всех съедобных видов ди-
ких животных мясо диких свиней, как и медведя, наиболее питательно - до 5000
калорий в килограмме, содержит 14-20% белка и 0,5-37% жира, доля мышечной
ткани - 57-65% [Белоносов, 1965; Житенко и др., 1987]. Выход мясной продук-
ции в зависимости от возраста и упитанности составляет около 45-70% (в том
числе сала - 10-30%) от массы тела животного. Средняя масса туш (без головы,
шкуры, внутренностей и части ног) в осенне-зимний охотничий сезон примерно
следующая: сеголетки - 20-25 кг, подсвинки - 40-45 кг, взрослые особи - 55-85
кг, крупные самцы - 130-150 кг. Во время гона мясо секачей приобретает резкий
неприятный запах, но его можно частично устранить, если после добычи сразу
же вырезать семенники вместе с дивертикулом препуциального мешка [Петской
и др., 1974]. В народной медицине применяется желчь кабана. Шкуры использу-
ют в кустарном и промышленном производстве для выделки кож, из которых
шьют обувь, коврики, седла, сумки, ремни, стельки. Щетина и пух - ценное сы-
рье для изготовления щеток, кистей, набивки сидений и матрасов. Из пуха вяжут
теплые носки и варежки. В последние десятилетия из-за бесхозяйственности
шкуры диких копытных чаще всего выбрасывают на месте отстрела.
Рис. 67. Клыки кабана -
трофей охотника (фото Ю.
Елькина)
С XX в. в России все большее распространение получает спортивная и, осо-
бенно, трофейная (с целью получения клыков - рис. 67) охота. Добыча этого чут-
214
Рнс. 68. Кабан - зверь стремительный, трудно добываемый (фото И. Бесарабы)
кого, осторожного, проворного, стремительного (рис. 68) и порой свирепого зве-
ря, вооруженного мощными клыками, на мой взгляд, более увлекательна и инте-
ресна по сравнению с охотой на другие виды диких копытных и требует от охот-
ника большей выдержки, сноровки, быстроты реакции.
Охота на диких свиней сопряжена с риском и в прошлые века считалась
весьма опасной. Случаи ранений охотников кабанами, как самцами, так и самка-
ми, отмечаются поныне [Динник, 1910; Шиллингер, 1928; Черкасов, 1962; Мура-
вицкий, 1983]. В Окском заповеднике на 270 охотах зарегистрировано 6 таких
случаев, причем однажды на человека напал не раненный зверь, а самка, имев-
шая поросят [Червонный, 1980]. Тем не менее при нынешнем вооружении стрел-
ков у зверя остается совсем немного шансов остаться в живых.
Добывание (промысел) животных - это, собственно, работа по «сбору урожая»
со своей спецификой, так или иначе влияющая на популяцию. От того, как, когда и
при каких условиях она выполняется, во многом зависит благополучие локальных
группировок, их биологическая продуктивность и эффективность охотничьего хо-
зяйства в целом.
Объем добычи
Объем добычи в XIX - первой половине XX вв. В дореволюционной России
ежегодно добывали, вероятно, не менее 50 тыс. диких свиней, в основном на Кавка-
зе, Каспийском побережье и на юге Дальнего Востока [Гептнер и др., 1961].
На юге Дальнего Востока в XIX в. отдельные охотники убивали за зиму до
40-100, а за свою жизнь - до 1000 голов. Даже во второй половине XX в. добыча
одного охотника за зимний сезон в годы высокой численности достигала 35 осо-
бей, в обычные годы - 8, в годы депрессии - до 4 особей [Бромлей, 1964; Куче-
ренко, 1973; Бромлей, Кучеренко, 1983]. В глубокоснежье кабаны истреблялись
поголовно, и городские базары в такие зимы были завалены их мясом [Туркин,
1899]. По неполным данным, в 1909 г. лишь в б. Приморской области добыто
около 4 тыс. диких свиней и свыше 12 тыс. косуль [Сухомиров, 1968]. В середине
прошлого столетия колхоз «Ударный охотник» района им. Лазо сдавал заготови-
тельным организациям более 10 т кабанятины [Сысоев, 1950].
215
В Забайкальской области в конце XIX - начале XX вв. ежегодная добыча ди-
ких свиней составляла около 500 голов, изюбря - 1,7 тыс. и косули - 31 тыс.
Промышляли их у Байкала, по Чикою, в верховьях Ингоды и по Онону. В Читин-
ском округе добывали около 100 голов кабана, 11,6 тыс. косуль и 2,3 тыс. изюб-
рей, в Верхнеудинском соответственно 219, 7,5 тыс. и 0,7 тыс., в Баргузинском-
57, 2,9 тыс. и 0,1 тыс., в Троицкосавском округе - 25 голов, 1,2 тыс. и 0,15 тыс., в
Иркутской губернии-до 100 особей [Туркин, 1899; Кириллов, 1902, 1904], в Ту-
ве - около 300 голов [Смирнов, 2001]. В дельте Волги в начале XX в. за сезон от-
стреливали 300-350 особей [Лавровский, 1952].
Масштабы промысла диких свиней в Казахстане прослежены А.А. Слудским
[1956]. В начале XX в. добыча здесь исчислялась десятками тысяч голов. У оз.
Зайсан отставной казак Илья Антонов, выезжавший на охоту со сворой собак,
семилинейной винтовкой и двумя рогатинами в некоторые дни добывал по 10-15
зверей, в зиму 1894/95 гг. за три недели - 199, в 1895/96 гг. - 179, в 1896/97 гг. -
167, в 1898/99 гг. - 175. В зиму 1923 г. 4 человека с 27 собаками в районе г.
Кзыл-Орда добыли 61 кабана, из них 46 крупных, а на Кыз-Балинских островах
зимой 1924 г. - 80 свиней. Близ оз. Балхаш зимой 1922 г. один охотник застрелил
120 зверей. В сезон 1923/24 гг. 28 промысловых бригад, имевших около 600 со-
бак, только в низовьях р. Или заготовили 1800 взрослых кабанов (поросят в этот
период заготовительные организации не принимали). При этом своры собак за-
давили не менее 7200 сеголеток. Следовательно, этой зимой только в одном рай-
оне было истреблено около 9000 диких свиней. В 1936 г. здесь же добыто около
2500 голов (в конце 60-х годов в этом районе отстреливали не более 500 особей в
год [Злобин, 1969]). В 1928/29 гг. лишь «Кожсиндикатом» принято 19763 шкуры
кабана и «козлов». В 1930 и 1931 гг. в Казахстане заготавливали около 7 тыс.
шкур диких свиней ежегодно, что составляло лишь небольшую часть от десятков
тысяч добываемых животных. В сезон 1941 г. в низовьях р. Чу отстреляно 500
особей. На Камыш-Самарских озерах в 1951-1953 гг. один из охотников убил 70
зверей.
Не менее интенсивным промысел был в Киргизии. По сообщению Н.А. Се-
верцова, «в окрестностях Чимкента кабанов в зиму 1865-1866 годов убили не-
сколько сот - попадались до 70 на три ружья в двухдневную охоту». Позднее, в
1928-1929 гг., только Джалал-Абадский союз охотников изготавливал из мяса
этих копытных до 300 ц колбасы. Отстрел зверей резко сократился лишь с лик-
видацией в 1931 г. Охотсоюза, после чего дикие свиньи заметно размножились. В
30-х годах их причислили к категории вредных животных и начали борьбу с ни-
ми всевозможными способами, а на юге республики даже выплачивали премии
за уничтожение. Основанием для выплаты служили хвосты убитых зверей, а мя-
со зачастую оставалось на месте отстрела. Такие меры быстро привели к резкому
сокращению ресурсов [Чичикин, Воробьев, 1967].
В Таджикистане по установившейся традиции кабана уничтожали как вредите-
ля круглогодично всеми доступными средствами и при любых обстоятельствах. В
1950 г. в одном из колхозов Сарыхасорского района с помощью собак 7-8 семей
выгнали из кустарника на высокий берег реки, и все поросята (около 50 голов) бы-
ли сброшены с обрыва в реку. В результате повсеместного истребления дикие сви-
ньи стали редким видом [Иваненко, 1956]. В середине XX в. их добыча достигала 5
тыс. особей [Гептнер и др., 1961]. В Туркмении в агроценозах разрешена круглого-
дичная безлицензионная охота, поэтому установить число добываемых животных
здесь практически невозможно [Кулибаба, Пакулин, 1985].
В Закатальском округе в 1896 г. промышлялось около 600 зверей, в Кубан-
ской области - до 300 в год, столько же волков и 100 оленей [Туркин, 1899]. На
Кавказе в XIX в. кабан был приравнен к хищным животным. В 40-х годах XX в.
здесь отстреливали до 8 тыс. особей, в том числе 2 тыс. на севере региона, в Да-
216
1
1
Рис. 69. Динамика численности (1)
и добычи (2) кабана в России
>
гестане - 1,5 тыс., в Азербайджане - 4 тыс., в Грузии и Армении - около 200-300
голов [Верещагин, 1959]. Е.Л. Марков [1932] круглогодичную добычу кабана в
Закавказье в 20-е годы оценивал в 4 тыс. голов. С 1930 г. Азербайджанский Нар-
комзем ввел запрет на добычу свиней с 1 марта по 1 октября и нормировал от-
стрел - не более 15 голов за одну охоту.
Лицензионная добыча. В России отстрел кабана по лицензиям ведется с
1958 г. Ежегодный объем добычи во второй половине XX в. составлял 0,8-48
тыс. голов, максимальным он был в 1988-1991 гг. (табл. 46). В последующие го-
ды в связи с сокращением численности, в том числе и из-за чрезмерного и нера-
ционального промысла, уровень добычи уменьшился в 5 раз - до 9,3 тыс. в
1996/97 гг. (рис. 69). С 1958 по 2000 гг. официально отстреляно около 680 тыс.
особей. Уровень неофициальной добычи, скорее всего, больше в несколько раз.
Таблица 46. Лицензионная добыча кабана (тыс. особей) в России*
Годы | Добыча | Годы | Добыча | Годы | Добыча | Годы | Добыча
1958/59 0,82 1969/70 3,83 1980/81 23,3 1991/92 44,6
1959/60 2,12 1970/71 4,37 1981/82 22,1 1992/93 36,4
1960/61 1,96 1971/72 5,55 1982/83 22,3 1993/94 20,6
1961/62 2,59 1972/73 8,17 1983/84 25,7 1994/95 14,2
1962/63 1,37 1973/74 12,5 1984/85 27,2 1995/96 10,6
1963/64 1,43 1974/75 14,8 1985/86 28,3 1996/97 9,34
1964/65 2,37 1975/76 18,0 1986/87 27,0 1997/98 10,3
1965/66 1,73 1976/77 17,5 1987/88 30,2 1998/99 11,9
1966/67 2,07 1977/78 19,1 1988/89 40,3 1999/00 Н,4
1967/68 2,43 1978/79 22,3 1989/90 44,5
1968/69 3,38 1979/80 23,4 1990/91 47,8
* По сведениям Главохоты РСФСР (Охотдепартамента РФ), Государственной службы учета
охотничьих ресурсов, а также по [Дежкин и др., 1975].
217
В 70-80-е годы в центральных и центрально-черноземных областях европей-
ской части России, в Поволжье и Волго-Вятском районе официальная добыча
достигала 20-50% поголовья. В Северо-Кавказском регионе отстреливали не бо-
лее 13% от общей численности, но здесь особенно велики потери от браконьер-
ства [Иванова, 19756, 1976, 1980а,б; Иванова, Овсюкова, 1976; Иванова, Молока-
нов, 1978; Сицко, 1986; Горшков, 1988].
В целом ежегодная лицензионная добыча кабана в России в 1958-2000 гг., по
моим расчетам, находилась в пределах 2,7-21,6% (в среднем 10,8%) от после-
промысловой численности, тогда как в Польше, для сравнения, отстреливали
106-142% от учтенного весной поголовья [Uloth, 1990]. В России на 1000 га
площади ареала в среднем добывали менее 0,1, на такой же лесной площади - не
более 0,2—0,3 головы, тогда как в ГДР - 14 и 40-50 особей соответственно
[Briedermann, 1986].
Максимальная годовая добыча диких свиней в большинстве областей Россий-
ской Федерации (до 1-2 тыс. особей) сопоставима с уровнем отстрела зверей лишь в
одном госкомплексе «Завидово» [Фертиков и др., 1999]. Это сравнение наводит на
грустные мысли об огромных потенциальных возможностях отечественного охот-
ничьего хозяйства, остающихся нереализованными до сих пор.
Официальный объем добычи диких копытных животных в России составляет
лишь небольшую часть от всех потерь, которую можно сравнить с надводной частью
айсберга. Масштаб смертности кабана из-за нерационального использования ресур-
сов, браконьерства, от хищников и недостатка зимних кормов огромен - по прибли-
зительным расчетам, около 100-200 тыс. голов ежегодно!
В других республиках бывшего СССР объем лицензионной добычи был сле-
дующим. На Украине в 1965 г. официально отстреляно 0,7 тыс. голов, в 70-е годы
ежегодно - 6-8 тыс. [Болденков, Крайнев, 1975, 1980]. Этот уровень изъятия сохра-
нился и в 80-е годы. Примерно таким же он был и в Белоруссии: 5,8-6,4 тыс. В Литве
в 1985 г. отстреляно 14,6 тыс. [Baleisis и. а., 1987]. В 1987-1989 гг., по данным МСХ
СССР, ежегодная добыча находилась здесь в пределах 18,2-23,0 тыс., в Латвии -
14,7-18,1 тыс., Эстонии - 5,9-6,8 тыс., Молдавии - 161-243 особей, в Армении она
не велась, в Грузии исчислялась единицами и десятками голов, в Азербайджане,
Таджикистане и Туркмении - 2-4 сотнями. В Казахстане в период с 1975 по 1980 гг.
официально отстреляно всего 1,4 тыс., в 1981 г. - 196 голов [Слудский и др., 1984], в
1987-1989 гг. - 574, 274 и 215 соответственно. В Киргизии в 1963-1966 гг. по лицен-
зиям добывали до 1,5 тыс. [Чичикин, Воробьев, 1967], в 1976 и 1977 гг. - 810 и 762
[Айзин, 1980], в 1987-1989 гг. - 745, 914 и 576 особей.
В СССР объем лицензионной добычи диких свиней в 1973 г. составил 39,2
тыс. [Максимов, 1974], в 1977 г. - 52,4 тыс., в 1978 г. - 53,5 тыс. [Висящев, 1979].
По сведениям Министерства сельского хозяйства, в 1987 г. отстреляно 82,9 тыс.,
в 1988 г. - 102,1 тыс. и в 1989 г. - 106,1 тыс.
Уровень ежегодной лицензионной добычи диких свиней в Прибалтийских рес-
публиках находился в пределах 20-75% от учтенной численности [Максимов, 1974;
Иванова, Овсюкова, 1976; Падайга, 1982; Мазин, 1988; Климова, 1989]. В Белорус-
сии и на Украине он был примерно таким же, как и в России, - в среднем около 10-
12% (в отдельные годы до 19-24%) [Романов, Козло, 1977; Вадковский, 1988; Евту-
шевский, 1988; Кононов, 1988; Романовский, Корочвич, 1989; Козло, 2001].
Добыча в других странах. В Австрии в XIX в. ежегодная добыча кабана
достигала 3 тыс. голов [Туркин, 1899]. В 1927 г. здесь отстреляли всего 99 осо-
бей, в 1935 г. - 156, 1957 г. -711, 1961 г,- 1352, 1971 г. - 2670, в 1981 г. - 4428
[Prossinagg, 1985], в 1989 г. - 10058 [Sternath, 1998]. В Пруссии в XIX в. добыва-
ли около 10 тыс. особей [Туркин, 1899]. Столько же их отстреливали в Германии
в 1936—1939 гг. В ФРГ добыча в 1960 г. составила 25 тыс., в 1984 - 66 тыс.; в ГДР
в 1954 г. - 12 тыс., в 1960 - 27 тыс., в 1981 - 128 тыс., в 1988 г. - 150 тыс., а так-
218
же 155 тыс. косуль, 28 тыс. благородных оленей, 18 тыс. ланей и 3,7 тыс. муфло-
нов. В Чехословакии в период с 1950 по 1983 гг. отстрел диких свиней возрос с
1,3 до 23 тыс., в Польше с 1949 по 1980 гг. - с 16 до 80 тыс., в Венгрии с 1970 по
1990 гг. — с 9 до 40 тыс., в Югославии с 1970 по 1980 гг. - с 5 до 15 тыс., во
Франции с 1974 г. за 10 лет - с 37 до 70 тыс. [Wotchikowsky, 1981; Meynhardt,
1984с; Briedermann, 1986; Liess, Horstmann, 1989; Kohalmy, 1991]. В Италии, где к
началу XIX в. кабан был почти полностью уничтожен, в конце XX в. его добыча
приблизилась к 30 тыс. голов [Pedone et al., 1991].
В целом в Западной и Центральной Европе с конца 60-х до начала 80-х годов
отстрел этих копытных увеличился со 115 до 400 тыс. особей [Дежкин, Петров-
ская, 1970; Дежкин, 1985]. Приблизительный объем добычи кабана в разных
странах мира отражен в таблице 47.
Таблица 47. Приблизительный уровень официальной добычи кабана (тыс. особей) в раз-
ных странах Евразии и Северной Америки в конце XX в.*
Страна Добыча Страна Добыча
Австрия 10 СССР 106
Бельгия 4-7 Россия 48-10
Болгария 7-20 Украина 6-8
Венгрия 40 Белоруссия 5-6
Германия 160-200 Молдавия 0,2-1
Греция 8 Литва 23
Испания 30-35 Латвия 18
Италия 30 Эстония 7
Люксембург 0,5 Армения —
Нидерланды 1 Грузия 0,1 1
Польша 80 Азербайджан 0,2
Румыния 10 Казахстан 0,2-0,6
Словакия 9 Киргизия 0,6-0,8
Финляндия 0,1 Таджикистан 0,3-0,4
Франция 50-70 Туркмения 0,1-0,6
Чехия 14 Узбекистан 0,2
Швейцария 0,6 Япония 42 ,
Югославия 15 США, Канада 120
Монголия 1
* По [Дежкин, 1983, 1985, 1989], с уточнением и добавлением по литературным источникам и
сведениям охотничьих организаций стран.
Браконьерство. В России и сопредельных регионах, к сожалению, это мас-
совое, широко распространенное и почти неискоренимое явление, имеющее глу-
бокие социальные корни. Как справедливо заметили в свое время С.В. Кириков и
Ю.А. Исаков [1959], «браконьерство в нашей стране приняло размеры государст-
венного бедствия». Даже лицензии зачастую служат лишь прикрытием для от-
стрела большого числа животных.
На северо-западе России браконьеры ежегодно изымают 5-10% населения
кабана [Русаков, Тимофеева, 1984]. Они наносят ощутимый урон популяциям и в
Центральном Черноземье [Сухорослов, 19726]. В Липецкой области потери дос-
тигают 10% поголовья диких свиней и 25% - европейской косули [Побединский,
1997]. В заповеднике «Лес на Ворскле» Белгородской области лишь в весенне-
летний период от них гибнет около 10% местной группировки [Царев, 1989а]. В
Предкавказье этих копытных добывают гораздо больше, чем планируют офици-
ально [Саркисов, Хохлов, 1982]. На Северном Кавказе круглогодичной охотой
219
изымается не менее 30% поголовья, т.е. практически весь годовой прирост [Кс
тов, Рябов, 1963; Дуров, 1973].
Случаи браконьерства, когда семьи кабанов уничтожаются полностью, не
редки на Урале [Киселев, 1986]. На Алтае размер этого ущерба сравним с поте
рями от волка [Собанский, 1992]. В Прибайкалье нелегальный отстрел превыше
ет лицензионный в 10 раз [Недзельский, 1994]. На Дальнем Востоке в 60-70-е гс
ды ежегодно официально добывали 1-2 тыс. голов, но с учетом браконьерской
отстрела - в несколько раз больше. Интенсивная охота нередко велась со свора
ми собак, с нарушением норм добычи и сроков охоты. Общий объем добычи ди
ких свиней оценивается здесь в 4-12 тыс. особей [Бромлей, 1964; Сухомиров
1968; Войлочников, 1972; Кучеренко, 1973, 1976; Бромлей Кучеренко, 1983].
На Украине, так же как и в других республиках бывшего СССР, основно!
“урожай” диких копытных собирают браконьеры [Евтушевский, 1988]. В Карпата:
они изымают более 10% популяции, отстреливая преимущественно взрослых особе!
[Гунчак, 1982, 1999]. В Белоруссии истинный размер добычи, вероятно, в 3-6 ра
выше официального [Муравицкий, 1983, 2001]. В республиках Закавказья, как мн
довелось наблюдать, браконьерство распространено повсеместно, а урон животные
велик даже в заповедниках, несмотря на хорошо вооруженную и многочисленнун
охрану. По сведениям В.С. Лобачева и А.Г. Петухова [1984], в Кызыл-Агачском за
поведнике с осени до весны выстрелы звучат каждые 30 секунд.
В Казахстане нелегальный отстрел тоже значительно превышает лицензион
ный. С середины XX в. широчайшее распространение получило автобраконьерст
во. Тростник на озерах повсюду примят колесами автомашин и тракторов. За оды
выезд добывают до 10-15 животных. Зверей массово истребляют здесь и с самоле
тов и вертолетов, оснащенных пулеметами [Слудский, 1956; Соломатин, 1972
Слудский и др., 1984]. В низовьях Амударьи браконьеры отстреливают их в 3-
раз больше, чем по лицензиям [Реймов, Карабеков, 1984]. В Киргизии фактическа
добыча минимум втрое больше официальной [Чичикин, Воробьев, 1967]. В Турк
мении в 50-60-е годы различные изыскательские экспедиции почти полностью ис
требили популяцию кабанов в Восточных Каракумах и тугаях Амударьи [Ишадо!
1969, 1983]. В Монголии официально добывают около 1 тыс. особей, а с учето,
браконьерского отстрела - несколько тысяч [Соколов и др., 1982].
Иногда браконьерство носит варварский характер. В Ярославской области
обнаружил 5 трупов кабанов с животами, распоротыми о ножи-подрезы, вмонтг
рованные в поваленное дерево, лежащее поперек зимней тропы. Кишки волоч?
лись за ранеными животными несколько сот метров.
В конце XX в. браконьерство заметно усилилось в связи с экономически!
кризисом, обнищанием населения, развалом организованного охотничьего хозя!
ства, недальновидной политикой охотничьих чиновников, непомерно увеличр
вающих стоимость лицензий и путевок на отстрел копытных, отсутствием эл(
ментарной социальной справедливости при распределении лицензий. Массовс
вооружение населения дальнобойными карабинами и применение быстроходно
снегоходной техники оставляет зверям все меньше шансов уцелеть.
Немаловажный побочный результат браконьерской охоты - особенно бол1
шое число недобранных подранков.
Способы промысла, их эффективность и
влияние на группировки
Существует немало способов добычи кабана, как разрешенных, так и запр(
щенных, но все же до сих пор применяемых браконьерами: облавы, загоны (ри
70), с подхода, с собакой или со сворой собак (рис. 71), из засидок или специал,
но оборудованных вышек на подкормке, на приваде, посевах и тропах, с подъе:
220
Рис. 70. На кабана (фото А. Дигилевича)
да на лошади, автомашине, тракторе, снегоходе, с вертолета и самолета, ночью
из-под фар, стационарными и переносными ловушками, сетями, а также загани-
вание по насту, ямами на тропах, капканами, петлями, ножами-подрезами, пика-
ми, самострелами и др. [Динник, 1910; Каверзнев, 1932; Марков, 1932; Сысоев,
1950; Лавровский, 1962; Черкасов, 1962; Абрамов, 1963; Чичикин, Воробьев,
1967; Костоглод, 1969; Злобин, 1975а; Чегодаев, 1975; Муравицкий, 1983; Малов,
1989; Русанов, 1991; Блюм, 1995; Варнаков, 1997; Слепых, 1998]. В Южной Си-
бири большая часть диких свиней добывается с подхода [Смирнов, 1994, 2001], в
Казахстане - с собаками [Слудский, 1956], в европейской части России - на за-
гонных охотах, из засидок и с вышек на подкормке.
Далеко не каждая легальная охота завершается добычей зверя. В 1983-1987 гг.
в охотничьих хозяйствах Росохотрыбловсоюза за 7618 дней встречено 27527 диких
свиней, из которых отстреляно только 6064 [Любченко, Мамаев, 1988], т. е. в
среднем менее одной особи за день охоты.
Трудоемкость промысла кабана сравнительно высока и неодинакова в зави-
симости от способа охоты и плотности его населения. Трудозатраты на добычу
взрослых особей обычно в 1,5-2 раза выше, чем на отстрел молодых. Наиболее
успешна охота с собакой (4,2 человеко-дня на отстрел 1 зверя), из засидки (4,5),
наименее - с подхода (8,0) и загоном (23,9). В среднем на отстрел одного зверя
тратится около 12-19 человеко-дней [Русаков, Тимофеева, 1984; Варнаков,
19886; Воронин, Заикин, 1990]. В раннеосенний период добыча наиболее эффек-
тивна на засидках (48,5%) и с подъезда (42%), менее успешна - с подхода (9%)
[Варнаков, 1985].
Загонная охота, сопровождаемая шумом, имеет крайне негативное следствие,
если проводится регулярно - большая часть зверей покидает свои участки обита-
ния. Эмиграция особенно очевидна при низкой численности и очаговом распре-
221
Рис. 71. Охота на кабана с собаками (фото П. Яровицкого и А. Дигилевича)
делении животных. Кабаны «не держатся» в тех охотничьих хозяйствах, где име-
ет место этот мощный фактор беспокойства. И далеко не случайно численность
копытных в заповедниках резко увеличивается (иногда вдвое [Гусев, 1986]) по-
сле начала охотничьего сезона.
222
Еще большее воздействие на группировки диких свиней оказывает охота со
сворой собак. Псы давят и нередко съедают поросят еще до подхода людей. Ста-
да и семьи разбиваются, социальная структура популяции разрушается. Остав-
шиеся в живых свиньи находятся в стрессовом состоянии и уводят выводки в
другие районы. При продолжительном преследовании отмечались случаи заболе-
вания зверей пневмонией зимой и гибели, вероятно от перегрева, летом [Иванен-
ко, 1956; Слудский, 1956; Абрамов, 1963; Бромлей, 1964; Чичикин, Воробьев,
1967; Кучеренко, 1973, 1976; Бромлей, Кучеренко, 1983]. Этот способ охоты,
безусловно, должен быть в числе запрещенных.
В приазовских плавнях в 60-е годы охота на кабанов осуществлялась с верто-
летов. За 3 сезона добыто 247 голов, по неофициальным источникам - много
больше. Отстреливали лишь взрослых животных, а без них молодняк в массе по-
гибал от истощения. Оставшиеся в живых звери в панике покидали обжитые мес-
та, нередко проваливаясь сквозь тонкий лед лиманов. Некогда богатейшие угодья
оказались опустошенными [Костоглод, 1969].
Отстрел с вышек и из засидок оказывает наименьшее воздействие на группи-
ровки и позволяет производить селективный выбор особей, что весьма важно в
охотничьем хозяйстве. Тем не менее при интенсивной охоте на подкормочных
площадках звери временно прекращают посещать их [Иванова, Молоканов, 1969].
Во второй половине XX в. в европейской части России на диких свиней охоти-
лись в основном загонным способом, а отстрел велся из гладкоствольных ружей кар-
течью и, реже, пулей. При этом для добычи одного зверя делалось более 10 выстре-
лов в среднем [Русаков, Тимофеева, 1984]. Успешность таких охот невелика, а коли-
чество раненых и недобранных животных превышает число отстрелянных.
Судя по моим дневниковым записям, на 76 загонных охотах, в которых я при-
нимал участие, были добыты 72 и ранены 127 кабанов, из которых затем добраны
22. На одной из коллективных охот в Калининской области в декабре 1981 г. по
животным стреляли 23 раза. В результате убит один подсвинок и еще 4 свиньи
ранены, но не добраны ни в этот, ни на следующий день.
А.П. Варнаков [1985,1988а] считает, что в отдельных областях России доля
подранков достигает 70% от числа отстрелянных. На Украине на одного добыто-
го зверя приходится 2-3 раненных [Евтушевский, 1975]. На Алтае их много из-за
применения на охоте малокалиберного оружия [Собанский, 1992]. Доля гибели
диких свиней от ран в общей смертности составляет в некоторых районах 4,8—
37,3 % (см. табл. 42).
Избирательность добычи по полу и возрасту
В зависимости от выбранного способа охоты избирательность добычи каба-
нов по полу и возрасту может быть разной.
На северо-западе России на загонных охотах и с подхода чаще добывают
взрослых животных (73,5% и 81,8% соответственно), преимущественно самцов
(55,8% и 66,7%); с собакой - взрослых особей (73%) и самок (53,5%). Из засидок
можно выбрать зверя любого пола и возраста, но и в этом случае охотники пред-
почитают отстреливать крупных особей (81,4%), чаще самцов (57,1%). На про-
мысловой охоте добывают больше взрослых самцов (69,3%), на спортивной -
самок (60,6%), имеющих лучшие вкусовые качества [Русаков, Тимофеева, 1984].
В Воронежской области с собакой берут больше (до 65%) самцов, но и здесь под
выстрел попадают в основном взрослые звери [Соломатин, 1975].
По статистике, охотники чаще добывают самцов (табл. 48). Это можно объяс-
нить большей осторожностью самок, а также трофейной направленностью спор-
тивной охоты. Однако нередко отстрел взрослых самок скрывается, и в лицензии
вместо них указываются самцы, что существенно искажает реальную картину.
223
Таблица 48. J Юловой состав кабанов, добытых по лицензиям
Район Годы Иссле- довано, п Самцы, % Самки, % Источник информации
Белоруссия 1972 2400 57 43 Вадковский, К<
1974 3360 63 37 тин, 1978
Литва 80-е 15464 60 40 Янулайтис, Г
С.-запад России 1975-1982 3951 61,9 38,1 дайга, 1985 Русаков, Тик
Вологодская обл. 1975-1976 54-63 46-37 феева, 1984 Варнаков. 1978(
1977/78 154 61,0 39,0 Варнаков, 1985
и 1983/84 475 54,7 45,3 44
Окский з-к 1970-1978 532 37,0 63,0 Червонный, 198
Московская обл. 1967-1972 2060 56,3 43,7 Иванова, 19756
и 1974-1978 — 59,0 41,0 Иванова, 19806
Ярославская обл. сс — 61,8 38,2 _44_
Ивановская обл. It — 66,0 34,0 44
Псковская обл. 44 — 62,0 38,0 _4<_
Новгородская обл. 44 — 63,9 36,1 __44
Волгоградская обл. 44 — 62,9 37,1 _44_
Ставропольский край 44 — 64,9 35,1
Брянская обл. 1976-1977 321 65,7 34.3 Ватолин, 1980
Ярославская обл. 1981/82 500 62,0 38,0 Дан-Чин-Ю, 19
Хоперский з-к 1980-1984 177 41.8 58,2 Антонец, 1993
Калужская обл. 1985-1987 2227 69-65 31-35 Воронин, Заик1
Европейская часть 1983-1987 6064 63,5 36,5 1989 Любченко, IV
России Свердловская обл. 1991-1997 2099 68,5 31,5 маев, 1988 Марков, 1999
Рис. 72. Планомерное уничтожение крупных взрослых особей - репродуктив
ного ядра популяций — одна из основных причин снижения поголовья дики:
копытных в России (фото С. Чевнорева)
224
При любом способе охоты в России отстреливают преимущественно взрос-
лых особей (табл. 49), и лишь в самом конце XX в. под давлением ученых наме-
тилась тенденция к увеличению доли отстреливаемых сеголеток (до 45%).
Таблица 49. Возрастной состав кабанов, добытых по лицензиям
Район Годы Исследо- вано, п Сеголет- ки, % Г одова- лые, % Взрос- лые, % Источник информации
С.-запад России 1971-1975 1511 5,0 14,7 80,3 Вереща- гин, Руса- ков, 1979
(С 1981/82 404 15,3 17,3 67,4 Русаков, Тимофеева, 1984
Московская обл. 1967-1972 2060 14,3 25,5 61,2 Иванова, 19756
it 1974-1976 — 24,0 20,0 56,0 Иванова, 19806
Ярославская обл. (6 — 6,1 15,7 78,2 Сс
Ивановская обл. it — 5,0 19,0 76,0 it
Псковская обл. (С — 25,5 23,5 51,0 ii
Новгородская обл. ii — 14,8 19,3 65,9 к
Волгоградская обл. U it — 12,5 19,7 67,8
1977/78 154 14,0 17,0 69,0 Варнаков, 1985
it 1983/84 475 20,0 19,7 60,3 it
Брянская обл. 1976-1977 321 4,6 12,7 82,7 Ватолин, 1980
Ростовская обл. 1978-1981 2831 14,0 * 86,0 Казаков, Даликов, 1983
Хоперский з-к 1980-1984 492 46 18 36 Антонец, 1993
Калужская обл. 1985-1987 2227 44-58 * 56- Воронин,
42* Заикин, 1989
Европ. часть 1983-1987 6064 35,7 * 64,3* Любченко,
России Мамаев, 1988
Свердловская обл. 1991-1997 2099 26,4 * 73,6* Марков, 1 ООО
Тува 1961-1999 - 36 * 64* Смирнов, 9001
Россия 1995/96 6762 45,4 * 54,6* Служба учета охот-
ничьих
ii 1996/97 7170 36,0 * 64,0* ресурсов
а 1997/98 9143 42,4 * 57,6* сс
а 1998/99 6177 44,6 * 55,4* U
Литва 80-е 15464 58,2 * 41,8* Янулайтис, Падайга, 1985
Польша 80-е — 60-70 15-20 10-15 Uloth, 1990
Франция 90-е — 70 30* * Маньен,
% 1999
< * Доля годовалых и взрослых особей.
225
15. Заказ №2528.
Изъятие из группировки более 50% половозрелых самок на следующий год
приводит к снижению ее численности вдвое [Варнаков, 1985]. В Баларуси вслед-
ствие излишней добычи охотниками взрослых самок приплод у диких свиней
ежегодно снижается на 5-6 тыс. особей [Козло, 2001]. Сеголетки, оставшиеся без
матерей, как показано выше, в массе гибнут зимой. Планомерное уничтожение
репродуктивного ядра популяций (рис. 72) - одна из основных причин медленно-
го роста или снижения поголовья диких копытных [Данилкин, 1999а], и кабана в
частности, в России и сопредельных регионах.
Сроки охоты
Сроки охоты на диких свиней по действующим типовым правилам непомер-
но растянуты - с 1 июня (на самок, имеющих приплод текущего года, - с 1 ок-
тября) по 15 января. Нередко они продляются до 1 февраля, что не оправдано ни
с биологической, ни с экономической точек зрения.
Многомесячное преследование копытных не благоприятствует сохранению и
увеличению их ресурсов. При регулярном зимнем беспокойстве, как уже отмеча-
лось выше, звери уходят из привычных мест обитания за десятки километров,
растрачивая при переходах по снегу последние энергетические запасы. Нередко в
сезон охоты они появляются в новых районах, далеко не лучших для их жизни,
где также подвергаются преследованию или гибнут от бескормицы.
Большая часть кабанов добывается в конце сезона охоты, обычно в период
многоснежья, когда они находятся в бедственном положении [Павлов, Зарубин,
1982]. В Московской области, например, в 1967-1972 гг. в октябре добыто 6,3%, в
ноябре — 25,3%, в декабре - 28,4, а за 15 дней января - 40% животных. В сезон
1973/74 гг. в ноябре отстреляно 15,7%, в декабре - 38,3%, а за две недели января -
46% [Иванова, 19756; Иванова, Овсюкова, 1976]. На северо-западе России в
1971-1975 гг. добыча 1511 особей происходила следующим образом: июль-
1,3%, август - 3, сентябрь - 2,7, октябрь - 7,9, ноябрь - 10, декабрь - 19, январь -
35,7 и февраль - 20,4% [Верещагин, Русаков, 1979]. В Кировской области путем
анализа 409 лицензий, сданных после сезона охоты в 1980/81 гг., определено, что
3,4% зверей добыто в сентябре, 3,7 - в октябре, 16,1 - в ноябре, 25,2 - в декабре,
51,1 - в январе и 0,5% - в феврале [Павлов, 1999а]. В Туве 70-80% лицензий так-
же реализуется в декабре-январе [Смирнов, 2001].
К январю масса копытных значительно уменьшается, соответственно снижа-
ется продуктивность. По исследованиям О.С. Русакова [Верещагин, Русаков,
1979], с октября до января - февраля масса туш самцов уменьшается в среднем
на 25 кг, самок - на 17 кг. В масштабах страны потери мясной продукции при
позднем отстреле кабана исчисляются сотнями тонн, что эквивалентно потере
нескольких тысяч животных.
Ранний летний отстрел, производимый из засидок и чаще в сумерках, также
имеет свои негативные стороны. Он приводит к элиминации из популяции ис-
ключительно взрослых особей, в числе которых немало самок, имеющих поро-
сят, несмотря на запрет их добычи до 1 октября, и лучших самцов-
производителей, изымаемых непосредственно перед сезоном спаривания.
Итак, очевидно: охота была и остается важнейшим фактором, не только оп-
ределяющим динамику населения, но и существенно влияющим на половозраст-
ную и пространственную структуру популяций кабана. Ресурсы вида в России
используются крайне нерационально, и это - одна из основных причин низкой
численности и продуктивности популяций.
226
Управление популяциями
В конце II тысячелетия проблема охраны диких копытных и среды их обита-
ния снова стала наиболее актуальной в охотничьем хозяйстве России. Во многих
областях из-за резкого снижения численности охота на лося, европейскую и си-
бирскую косуль, благородного оленя, кабана и другие виды резко ограничена или
закрыта на ряд лет. На мой взгляд, за последнее десятилетие это единственное
верное, хотя и сильно запоздавшее решение чиновников контролирующего охот-
ничьего ведомства в аспекте управления популяциями.
Абсолютная охрана копытных, тем не менее, не может быть гарантией их со-
хранения. В конечном счете она не приносит пользы ни животным, увеличиваю-
щим свое население и рано или поздно погибающим от недостатка корма, ни био-
ценозу, который под их чрезмерным воздействием деградирует и видоизменяется,
ни человеку, безвозвратно теряющему ежегодный «биологический урожай». Бес-
предельное наращивание численности, как давно стало ясно, так же вредно и недо-
пустимо, как и неограниченная добыча зверей. Следовательно, задача их охраны,
как это ни парадоксально, сводится к рациональной эксплуатации.
Охота, по сути, должна стать одной из форм охраны животных, направленной
на долговременное использование биологического ресурса, а охотничье хозяйство,
по определению известного российского охотоведа Н.Ф. Томкевича, собственно и
«есть деятельность человека, направленная на разведение и сохранение различных
животных, служащих объектом охоты» [Томкевич, 1914 (по изд. 1992)].
Увеличение народонаселения планеты и быстрое сокращение пахотных зе-
мель неизбежно приведут к глобальной проблеме лимитированности белковой
продукции, к необходимости повышения биологической продуктивности естест-
венных угодий, не вовлеченных в сельскохозяйственный оборот, к необходимо-
сти инвестиций в охотничье хозяйство, где приоритетными становятся мясодич-
ное и трофейное направления. Немаловажно, что затраты труда на единицу про-
дукции здесь во много раз меньше, чем в смежных отраслях.
Воздействие человека на диких животных должно быть экологически осоз-
нанным и направленным на оптимизацию численности, сокращение непроизво-
дительных потерь, увеличение продуктивности и сохранение среды обитания.
Элементарным объектом охотничьего хозяйства и промысла должна быть попу-
ляция [Шварц, 1969, 1974а,б]. Этот тезис в настоящее время ни у кого не вызыва-
ет сомнений. Споры не утихают лишь по поводу стратегии и тактики, методов и
способов управления популяциями охотничьих животных.
Теоретические основы управления популяциями диких
копытных. Парадоксы теории и практики
Управление популяциями - это, по моему мнению, система взаимосвязанных
научно обоснованных мер, направленных на охрану, воспроизводство и рацио-
нальное использование ресурсов и получение максимума продукции при мини-
мизации ущерба окружающей среде и самим популяциям. Единой трактовки это-
го термина, однако, нет, и его используют или в очень узком (только использова-
ние ресурсов), или примерно в таком же широком смысле.
Генеральная цель управления - оптимизация, или выбор лучшего варианта из
возможных [Павлов, 1989]. Чем совершеннее управление популяциями, тем эф-
фективнее охрана животных и среды их обитания и использование ресурсов.
Позитивным примером может служить управление популяциями кабана в
ГДР в 70-80-е годы. Ежегодная добыча (около 100 тыс. голов) здесь была в 1,6
раза выше имеющегося весеннего поголовья (около 60 тыс.) при относительно
227
15
низком ущербе сельскому хозяйству [Briedermann, 1986]. В России, напомню, в
последние 30 лет при среднегодовой численности в 170 тыс. особей добыча со-
ставляет около 22 тыс., что почти в 8 раз ниже имеющегося поголовья, и при
этом довольно высок уровень вреда биотопам. Разница в продуктивности попу-
ляций при пересчете на единицу площади достигает минимум двух порядков!
Ярчайший пример крайне неразумного использования ресурсов и неумелого
управления популяциями копытных в России - массовая (десятки тысяч голов, пре-
имущественно - сеголеток) гибель сибирской косули в Курганской области от голода в
конце XX в. [Данилкин, 1995; Останин, 1996; Данилкин, Останин, 1998; Данилкин и
др., 2000]. Этот ущерб можно было бы свести к минимуму предлагаемыми мерами:
увеличением объема и изменением структуры добычи, а также внедрением новой эф-
фективной системы подкормки зверей во всех охотничьих хозяйствах области. Одна-
ко, прекрасно зная о прогнозируемом учеными массовом падеже косуль [Данилкин,
1995], «ответственные» охотничьи чиновники не предприняли никаких мер для изме-
нения ситуации. Более того, впоследствии они всячески пытались скрыть масштабы
произошедшей трагедии. И никто из них не получил даже малейшего административ-
ного взыскания, хотя «управленческий» ущерб животному миру многократно превы-
сил многолетние потери от всех браконьеров и хищников вместе взятых!
Тем не менее даже после такого урока кардинальных решений по оптимиза-
ции управления популяциями диких копытных животных не последовало ни на
федеральном, ни на территориальном уровнях, что неминуемо приведет к новым
катастрофам!
Анализ состояния и использования ресурсов диких копытных животных в
России не оставляет сомнений в том, что их скудность, низкая продуктивность
популяций и регулярная массовая гибель зверей - следствие ошибочной страте-
гии управления.
Рассмотрим некоторые наиболее очевидные парадоксы теории и практики в
управлении популяциями.
Использование ресурсов диких копытных в современном охотничьем хозяй-
стве должно базироваться не только на сведениях о численности, но и на глубо-
ком знании структуры популяций и роли в них отдельных половых и возрастных
групп животных.
Важнейшее свойство популяции - биологическая продуктивность, опреде-
ляемая совокупностью абиотических и биотических факторов среды. Конкретное
проявление популяционного гомеостаза - интенсификация размножения и сни-
жение смертности молодняка при уменьшении численности [Наумов, 1963, 1965;
Шварц, 1965, 1970, 1974а,б; Шилов, 1977].
Принцип «компенсации» (промысел как один из факторов смертности со-
кращает естественную гибель и замещает потери от других факторов, стимули-
руя при этом повышенное воспроизводство популяции) стал одним из основных
в теории управления популяциями охотничьих животных, а определение опти-
мального режима изъятия некоего «экологического резерва» стало важнейшей
практической задачей охотоведения. Слабое опромышление снижает рентабель-
ность охотничьего хозяйства и может привести к нежелательному увеличению
населения дичи. Чрезмерная эксплуатация ведет к уменьшению численности,
продуктивности, негативным изменениям половой, возрастной и генетической
структуры группировок и, в конечном счете, к их биологической деградации. Це-
ленаправленным воздействием на половой и возрастной состав можно сущест-
венно увеличивать или уменьшать биологическую продуктивность популяций
[Шварц, 1969; Юргенсон, 1973; Банников, 1974; Дежкин, 1976, 1985; Шварц,
Михеева, 1976; Язан, 1979; Глушков и др., 1989; Данилкин, 1989, 1999а; Глуш-
ков, Байбиков, 1998; Козло, 2001].
228
Однако даже самая правильная научная идея на практике может быть извра-
щена и доведена до абсурда, что особенно заметно в охотничьем и сельском хо-
зяйствах. Замечательный в целом принцип «компенсации» был подхвачен прак-
тиками как установка на максимизацию отстрела копытных. Этот тезис на «язы-
ке» охотоведов, проповедующих максимизацию добычи [Павлов, 1999а], звучит
уже примерно так: «чем больше добудешь, тем больше будет!»
Естественная смертность копытных при повышении объема добычи действи-
тельно может несколько сокращаться, однако далеко не адекватно и не всегда.
При существующих условиях использования ресурсов усиливается браконьерст-
во (под прикрытием большего числа лицензий), увеличивается доля подранков,
возрастает промысловая нагрузка на репродуктивное ядро, что ведет к омоложе-
нию популяции, снижению ее воспроизводственного потенциала и большим по-
терям сеголеток, оставшихся без матерей, и, в конечном счете, к уменьшению
поголовья. На оставшуюся часть популяции усиливается пресс хищников и бра-
коньеров. При плотности (численности), упавшей ниже критического уровня,
даже максимальный для данного вида прирост не компенсирует естественной
убыли, и популяция гибнет.
«Компенсационная» идея как теоретическая основа промысла животных, сле-
довательно, замечательна лишь при некоторых непременных условиях: достаточно
высокой плотности населения и при умеренном (меньше годичного прироста) изъ-
ятии из популяции половых и возрастных групп животных, наиболее подвержен-
ных естественной смертности. На практике же, к сожалению, все наоборот.
Планирование и нормирование добычи диких копытных животных в России
ведется от «достигнутой» численности, а не от оптимального ее уровня. Группи-
ровка с любой численностью (100000, 10000, 1000 или 100 особей) считается
промысловой. Применение одинаковых для крупных регионов норм добычи при-
водит к перепромыслу животных в популяциях с низкой плотностью населения
или их недоопромышлению при высокой плотности.
Теоретически не обоснованы, противоречат здравому смыслу, но воплощены
в ведомственных инструкциях Главохоты РСФСР «О порядке добычи диких ко-
пытных животных по разрешениям (лицензиям) ...» и реализованы на практике
запрет или ограничения на добычу сеголеток и установка на преимущественный
отстрел наиболее крупных взрослых особей, «дающих больше мяса». Основные
мотивы ограничения добычи сеголеток - они должны подрасти, а также якобы
эффективнее, чем взрослые животные, используют корма.
Зимой, когда запасы кормов минимальны, детеныши, отличающиеся повы-
шенным обменом веществ, потребляют пищи почти столько же, сколько и взрос-
лые, но при этом практически не растут, а масса их тела значительно уменьшает-
ся. В отличие от взрослых особей, молодые практически не накапливают жиро-
вых запасов, у них выше теплоотдача, и они обычно погибают в суровые зимы.
Большая эффективность использования первичной (растительной) продукции се-
голетками, следовательно, иллюзорна, а потери животной биомассы, как показа-
но выше, реальны и огромны.
Ежегодный массовый отстрел взрослых особей приводит к уничтожению ре-
продуктивного ядра группировок, сдерживая их рост. Длительный выборочный
отстрел наиболее крупных особей стал главной причиной деградации многих по-
пуляций копытных. К сожалению, масштабная антиселекция не только не пре-
кращается, но и имеет тенденцию к усилению в связи с развитием трофейной
охоты. Целенаправленная элиминация элитных самцов осуществляется повсеме-
стно, и даже появились браконьеры, специализирующиеся на добыче лучших
трофеев [Данилкин, 1999а, 2000а].
На мой взгляд, изъятие из популяции преимущественно взрослых особей (ре-
продуктивного ядра) допустимо лишь при чрезмерной численности и необходи-
229
мости ее резкого снижения. «Пропорциональная» добыча разных половых и воз-
растных групп, эквивалентная их доле в группировке, экономически оправдана
только в тех случаях, когда плотность населения достигла оптимальной величи-
ны и требуется поддерживать ее стабильное состояние. Сейчас же популяции ко-
пытных в России, за редким исключением, настолько малочисленны, что многие
из них находятся на грани выживания. Именно поэтому стратегия управления
ими должна быть совершенно иной, направленной на максимальное сохранение
репродуктивного ядра, увеличение поголовья и улучшение качества животных.
В период увеличения численности диких копытных в России многие специа-
листы призывали к увеличению норм их добычи, и кабана в частности [Фадеев,
19706, 1978а,б; Воронин, 19756; Иванова, 19756, 1976, 1977, 1978; Мелехин,
1983; Русаков, Тимофеева, 1984; Варнаков, 1988а; Павлов, 1999а]. Нормы были
увеличены, но выделяемые квоты стали хронически недоосваиваться, особенно
при очередном резком уменьшении ресурсов.
На этой основе в недрах управления отечественным охотничьим хозяйством
был «рожден» «коэффициент успешности охоты», позволяющий территориаль-
ным органам добавлять к выделенному лимиту добычи копытных недоиспользо-
ванную часть прошлогодней квоты. Чем ниже этот коэффициент, тем относи-
тельно больше выдается дополнительных разовых лицензий на отстрел зверей.
Соответственно большее число охотников имеют потенциальную («бумажную»)
возможность добыть зверя, а контролирующее охотничье ведомство получает
больший доход от продажи большего числа лицензий. Охотничий пресс на ко-
пытных в разреженных популяциях при этом значительно возрастает. Более того,
при малоуспешной охоте охотничьи организации обычно настаивают на продле-
нии сроков добычи и заявляют о невозможности отстрела сеголеток из-за малой
их доли в популяциях. При большом количестве неосвоенных лицензий сроки
охоты, как правило, продляются, а в добыче преобладают взрослые особи. При
многомесячной охоте под прикрытием лицензий усиливается браконьерство, ко-
пытные подвергаются длительному стрессу и вытесняются непрерывными «заго-
нами» в худшие местообитания, где в массе гибнут от голода и хищников.
При хроническом недоосвоении квот численность копытных в России в по-
следние годы не растет или же снижается, чего не должно быть. Эта ситуация,
однако, легко объяснима - практически повсеместно ресурсы не недоиспользу-
ются, а, напротив, очевиден ... перепромысел, усугубленный целенаправленным
истреблением репродуктивного ядра популяций, непомерным уровнем браконь-
ерства и хищничеством волка!
В связи с таким выводом, кардинально отличающимся от ведомственной точ-
ки зрения, напомню некоторые признаки, позволяющие определить избыточную
эксплуатацию популяций [Уатт, 1971]: снижение процента беременных самок
(увеличение яловости); снижение продуктивности и выживаемости животных
(сокращение прироста); уменьшение относительной эффективности промысла
(добычи за единицу времени на одной и той же территории); неспособность по-
пуляции быстро возвращаться в норму после прекращения промысла или сниже-
ния интенсивности эксплуатации. И все эти признаки характерны для отечест-
венных «недоопромышляемых» популяций диких копытных!
Напомню также замечательное высказывание одного из основоположников
теории научного управления популяциями охотничьих животных академика С.С.
Шварца [1969], забытое охотоведами: «Популяция не может справиться с одно-
временным снижением численности (результат промысла) и нарушением струк-
туры (результат ошибочного планирования промысла). При этом падение чис-
ленности внешне может производить впечатление гибели животных от естест-
венных причин. В подобных случаях не всегда удается установить, что перво-
причиной краха популяции является непродуманная система промысла. Поэтому
230
нарушение структуры популяции в результате промысла чрезмерной интенсив-
ности есть основание назвать абсолютным перепромыслом».
Отечественные охотоведы оказались невосприимчивыми к урокам прошлого.
А ведь на протяжении последних веков поголовье копытных при увеличении их
добычи без уменьшения воздействия других факторов среды уже неоднократно
стремилось к нулю. Многие виды (тур, зубр, лось, благородный, пятнистый и се-
верный олени, европейская и сибирская косули, сайга, кабан и др.) исчезли со
значительной части своих ареалов на столетия или совсем не из-за естественной
цикличности, а из-за чрезмерного промысла.
Весьма интересное заключение в свое время сделал Ю.П. Язан [1979]: «Нор-
мирование промысловых нагрузок, лишенное, как правило, серьезных научных
обоснований, и преимущественный отстрел взрослых крупных особей обоего по-
ла - вот наиболее распространенные погрешности современного охотничьего хо-
зяйства. Выравнивание этих погрешностей и есть главная задача управления по-
пуляциями охотничьих зверей».
Думаю, что одна из основных задач управления популяциями охотничьих жи-
вотных заключается все же не в вечном «выравнивании» этих погрешностей, а пре-
жде всего - в их недопущении.
В последние годы в охотничьем хозяйстве, в довершение многих бед, усили-
вается антибиотехническая тенденция. Известный российский охотовед М.П.
Павлов [1999а] считает, например, что кабан зимой вполне может обходиться без
подкормки и даже ... без еды. Такого рода утверждения вряд ли полезны для жи-
вотных - дикие свиньи, как показано выше, не зимоспящий вид. Они, несомнен-
но, выживают даже в экстремальные зимы, но сколько их уцелеет, сколько навсе-
гда эмигрирует из района обитания, какова будет плодовитость выживших осо-
бей и продуктивность популяции?
В.В.Петрашов [1999] пришел к выводу о неэффективности биотехнических
мероприятий для копытных и на этом основании призывает к отказу от управле-
ния природными комплексами. Основные аргументы: с помощью биотехнии не
удается остановить снижение численности кабанов, лосей и других видов, хотя
емкость угодий далеко не заполнена, и, к тому же, в тех областях, где она не про-
водится, показатели динамики численности лучше.
На мой взгляд, в этой диссертационной работе явно перепутаны причинно-
следственные связи. Главная причина очередного снижения населения диких ко-
пытных животных связана отнюдь не с биотехнией, а с прямым их уничтожени-
ем человеком и не в меру размножившимся волком. Кроме того, в суровых мно-
госнежных условиях России значительная часть зверей гибнет даже при низкой
плотности населения вовсе не от избытка подкормки, а от недостатка или полно-
го отсутствия кормов. Для специалистов давно не секрет, что во многих охот-
ничьих хозяйствах зимняя подкормка проводится лишь «на бумаге» или же зве-
рям выкладывают пищу, от которой больше вреда, чем пользы, например сухие
лиственные веники и злаковое сено для косуль при отсутствии свободной воды
[Данилкин, 1989, 1996а, 1999а; Данилкин и др., 1999]. И все это - результат не-
эффективного в целом устройства отечественного охотничьего хозяйства на фоне
огромного позитивного мирового опыта, в том числе и в биотехнии.
К сожалению, до предела извращена и замечательная идея охраны охотничьих
животных в России путем создания многочисленных заказников. Они далеко не все-
гда выполняют функции резерватов. Плотность населения диких зверей и птиц в них
нередко ниже, чем в соседних охотничьих угодьях [Русаков, 1987; Побединский,
1999], что связано не только со слабой охраной, но и с отсутствием или меньшим
объемом биотехнических работ, более высокой численностью волка и легальным от-
стрелом охраняемых животных. В конце XX в. большинство заказников охотничьи
чиновники превратили в ведомственные охотничьи хозяйства.
231
Магистральный путь управления популяциями диких копытных очевиден
[Глушков, 1988; Папонов, 1988; Размахнин, 1988; Граков, Глушков, 1989; Данил-
кин, 1989, 1999а; Русаков, 1989; Управление популяциями охотничьих живот-
ных, 1999; Козло, 2001; Муравицкий, 2001]:
- формирование высокопродуктивных средневозрастных группировок оп-
тимального размера;
- массовая добыча малопродуктивных, наиболее уязвимых и склонных к
эмиграции особей младших возрастных групп;
- выборочный селекционный и трофейный отстрел среди половозрелых
особей, улучшение качества популяций;
- проведение эффективных биотехнических мероприятий.
Собственно, это и есть основные задачи охотоведения и охотоведов на бли-
жайшие десятилетия в отношении диких копытных животных. Однако при суще-
ствующей системе отечественного охотничьего хозяйства, действующих прави-
лах и сроках охоты и «Инструкции о порядке добычи диких копытных ...» (кото-
рая, по моему мнению, в большей степени стала инструкцией по их уничтоже-
нию!) эффективное управление популяциями и рациональное использование ре-
сурсов невозможны в принципе.
Методы и способы управления популяциями. Повышение
продуктивности
Определение оптимальной плотности (численности). Для каждой популя-
ции существует сугубо своя, специфическая оптимальная плотность населения,
зависящая от многих факторов, среди которых ведущее место занимает пища
[Тимофеев-Ресовский и др., 1973]. Ее оценка весьма сложна, и к тому же она
осуществляется не с позиций самой популяции, стремящейся к гомеостазу, а с
«целесообразной» (хозяйственной) точки зрения человека, что не одно и то же.
Ясно также, что уровни оптимальной плотности населения зверей не могут быть
стандартными, поскольку неодинаковы среда их обитания и экономические воз-
можности охотничьих хозяйств.
При низкой численности зверей кормовые ресурсы недоиспользуются и про-
дуктивность популяций гораздо ниже возможной. При чрезмерной плотности
быстро истощаются запасы корма, что ведет к голоданию и истощению особей,
падежу, снижению плодовитости, массовой эмиграции и усилению ущерба.
Определение оптимальной численности у копытных, и кабана в частности,
оказалось неразрешимой задачей для отечественного охотоведения, что во мно-
гом вызвано как скудностью знаний экологии животных, среды их обитания и
реакций популяций на изменения условий существования, так и «шаблонностью»
подходов к этой проблеме.
Оптимальную плотность населения копытных охотоведы чаще всего пыта-
ются вычислить исходя из известного экологического «правила минимума» [Да-
нилов и др., 1966]. К примеру, если запас летних кормов для кабана допускает
обитание 100 голов на 1000 га, а запас зимних кормов - только 5, то естественная
емкость угодий, по этому правилу, будет определяться фактором среды, имею-
щимся в минимуме, т.е. равняться 5 особям.
Традиционная бонитировка и расчеты плотности населения животных в зави-
симости от естественной кормовой емкости угодий оказываются, однако, недос-
таточно точными. К тому же искусственная подкормка позволяет значительно
увеличить емкость угодий. Именно поэтому реальная плотность населения ко-
пытных в лучших охотничьих хозяйствах, как правило, в несколько раз выше ре-
комендованной при охотустройстве.
232
Более перспективно, видимо, определение неких «экологических норм» чис-
ленности (плотности) для каждого района и охотничьего хозяйства на основе объ-
ективных многолетних учетных данных, изучения тенденций динамики популяций
[Кузякин, 1989] и изменения среды обитания, с учетом возможного ущерба лесно-
му и сельскому хозяйствам, т. е. на основании эмпирического опыта.
Оптимальные нормы плотности, рекомендуемые специалистами для разных
районов, приведены в таблице 50. Несмотря на существенные различия, все же
улавливается некая закономерность. В большинстве районов эти показатели на-
ходятся в пределах 10-20 голов на 1000 га.
Таблица 50. Оптимальная (допустимая) плотность населения кабана
Район | Особей на 1000 га | Источник информации
Германия 3-15 (до 35) Briedermann, 1977a,b, 1979, 1986
Польша 2-10 Haber et al., 1975
а 30-40* Andrzejewski, Jezierski, 1978
Болгария 20 Атанасов, 1981
Беловежская пуща 10-15, 18-19 Саблина, 1955; Лебедева, 1956а;
Козло, 1970а, 1975; Шостак, 1978
Белорусское Полесье 10-12 Козло, Емельянова, 1987
Украина 5-8 Корнеев, 1970
Карпаты 5-7*, 10-12 Гунчак, 1982, 1999
Молдова 20 Лозан, Лозан, 1981 в
Литва 4-30(7-15) Янулайтис, Падайга, 1984а,б 1985,1987
Россия: 1-20 (по бонитету) Данилов и др., 1966
С.-западный р-н 2-3,5 Иванова, 1978
Центральный р-н 4-5
Поволжский р-н 4-5 —Горшков, 1988
Северный Кавказ 6-8 it
Зона южной тайги 30-35** Говорка и др., 1988
Зона хвойно-широко- 20**
лиственных лесов
Лесостепная зона 20-30 Гусев, 1986, 1989, 1995
Ц. Черноземье 6-10 Простаков, 1996
а До 18-20 Сухорослов, 19726
“Завидово” 10 Фертиков и др., 1999
Кавказский з-к 19 Дуров, 1980
Краснодарский край 15-20 Котов, Рябов, 1963
Киргизия 15-50 Воробьев, 1968
* На 1000 га лесопокрытой площади.
* * С ветеринарной точки зрения.
Г.И. Иванова [1983] полагала, что для угодий зоны южной тайги нормальной
можно считать плотность 1,5-3, в смешанных лесах - 3-6, в лиственных, байрач-
ных, горных лесах, лесостепи и степи - 4-8 особей на 1000 га. Для лесостепной
зоны России, однако, вполне допустимо обитание 20-30 особей на 1000 га уго-
дий. При такой плотности дикие свиньи съедают лишь около 0,3% доступных за-
пасов корма летом и до 18% зимой, а степень повреждения фитомассы находится
в пределах 1,3-1,7% [Гусев, 1986, 1988, 1989, 1995].
В Беловежской пуще при очень высокой плотности населения других копытных
воспроизводительная способность популяции кабана наиболее высокая при 10-15
особях на ту же площадь [Козло, 1975]. Польские специалисты после тщательного
анализа вреда, наносимого этим видом сельскому хозяйству, пришли к заключению,
233
что при интенсивной подкормке плотность его населения можно поддерживать в
пределах 30-40 особей на 1 000 га лесопокрытой площади, а не 10 на 10ОО га, как
считалось ранее [Andrzejewski, Jezierski, 1978]. Примерно такую же величину (30
особей) указывают и французские ученые [Spitz et al., 1984].
Вспышка метастронгилеза у диких свиней возможна при плотности более 35
особей на 1000 га [Пенькевич, 1997]. Предельный уровень плотности населения с
ветеринарной точки зрения: в широколиственных и хвойно-широколиственных
лесах - 20, в южной тайге - 30-35, а при комплексных биотехнических, селекци-
онных и профилактических мероприятиях - до 50 особей на 1000 га [Романов,
Козло, 1977; Говорка и др., 1988; Фертиков и др., 1999]. Эти значения, на мой
взгляд, могли бы быть ориентировочными при определении допустимой плотно-
сти населения в высокоорганизованных охотничьих хозяйствах.
Помимо определения неких пределов оптимальной или предельно допусти-
мой плотности (численности), существует другая немаловажная проблема - оп-
ределение минимального уровня, при котором следует прекращать промысел. Ра-
зумеется, и этот показатель не может быть одинаковым для всех районов и охот-
ничьих хозяйств. Е.М. Недзельский [1994] считает, например, что в Восточной
Сибири при плотности населения кабана менее 1 особи на 1000 га нарушается
соотношение полов, в результате чего резко снижается биологическая продук-
тивность популяции. В Киргизии Г. Воробьев [1968] рекомендует запрещать охо-
ту, если плотность меньше 5-10 особей на 1000 га, но для ряда областей России
эта величина может быть близкой к оптимальной.
Имитационное математическое моделирование, выполненное с учетом спе-
цифических социально-экономических проблем в отечественном охотничьем хо-
зяйстве, привело нас к общему выводу: добычу копытных не следует вести, если
плотность их населения стала менее 40-50% от оптимального уровня, и до его
скорейшего достижения не следует добывать более 1-5% от учтенной (после-
промысловой) численности.
Как быть, если плотность населения (или численность) копытных в области,
крае, республике или районе меньше необходимого для открытия охоты мини-
мального уровня, тогда как в отдельных хорошо работающих охотничьих хозяй-
ствах она гораздо выше? В этом случае охотничьи хозяйства, несомненно, долж-
ны иметь возможность пользоваться плодами своего труда.
Формирование зимнего «стада». Прокормить сотни или тысячи кабанов и дру-
гих копытных в конкретном охотничьем хозяйстве в течение длительного зимнего
периода весьма накладно экономически и трудно физически. Следовательно, зимняя
численность зверей должна быть минимально необходимой, но это поголовье долж-
но обладать максимальным воспроизводственным потенциалом.
В зиму желательно оставлять в основном репродуктивное ядро популяции -
взрослых беременных самок и крупных самцов-производителей. Такой подход к
формированию зимнего «стада» диктуется и другими причинами.
Отстрел лидеров - катастрофа для семей и семейных групп. Убив взрослую
беременную самку, мы теряем сразу же более 10-15 зверей: саму свинью, 5-7
будущих поросят и примерно столько же детенышей этого года, которые неми-
нуемо погибнут зимой без матери. При расчете потенциальных потерь популя-
ции с учетом воспроизводства потомков самки за период ее жизни это значение
увеличивается на порядки.
При отстреле крупных секачей мы лишаемся лучших производителей и
«стимуляторов» самок во время гона, лучших «охранников» стад от крупных
хищников, лучших «пахарей» и прокладчиков троп в глубокоснежье. В их отсут-
ствие молодые самцы не всегда способны покрывать крупных самок.
Кроме того, бессмысленно кормить всю зиму всех сеголеток, значительная
часть которых все равно погибнет к весне, поскольку их растущий организм тре-
234
бует гораздо больше энергии, чем получает с пищей. На севере ареала даже при
постоянной опеке со стороны свиньи и секача к весне сеголетки истощены до
предела [Царев, 19806]. Они к тому же - самая инвазированная возрастная груп-
па, что способствует массовой гибели. Весьма уязвимы зимой и подсвинки, из
которых далеко не все участвуют в размножении, плодовитость их заметно ниже,
чем у взрослых, свое потомство они сохраняют хуже.
Нормирование добычи. Продуктивность группировок кабана в разные годы
неодинакова, как и не безграничны возможности охотничьих хозяйств в их со-
хранении зимой или в возмещении ущерба, причиняемого животными. Следова-
тельно, нормы добычи не должны быть строго фиксированными даже в пределах
одного региона.
При чрезмерной численности они могут превышать уровень прироста попу-
ляции, а ее репродуктивное ядро необходимо уменьшать. При низкой плотности
населения диких свиней, напротив, охота на них должна быть закрыта или суще-
ственно ограничена при запрете отстрела взрослых особей. При оптимальной
численности ежегодная добыча должна быть несколько ниже величины годично-
го прироста популяции, в зависимости от масштаба естественной смертности и
браконьерства.
Отечественные охотничьи чиновники, ратующие за увеличение норм добычи
кабана, обычно ссылаются на опыт отдельных европейских стран, где отстрел
планируется в объеме имеющейся весенней численности или больше. Макси-
мальные нормы добычи (до 100-200%) в действительности установлены лишь в
тех странах, где имеющаяся плотность населения кабана намного превышает ем-
кость угодий, а естественная смертность в благоприятных климатических усло-
виях, при отсутствии крупных хищников и изобильной подкормке, низка. Кроме
того, здесь особое внимание уделяют структуре добычи и сохранению репродук-
тивного ядра популяций. У нас же, за исключением нескольких хорошо органи-
зованных охотничьих хозяйств, проблема прямо противоположная - как увели-
чить или хотя бы сохранить то, что имеем. Поэтому повышать нормы отстрела
при сложившейся практике преимущественной добычи наиболее продуктивных
взрослых особей и при значительных зимних потерях зверей, по меньшей мере,
непрофессионально.
Большинство специалистов считают, что добыча кабана при низкой его плот-
ности должна находиться в пределах 5-25%, а при оптимальной и чрезмерной -
от 20 до 100% от весенней численности (табл. 51).
В странах Западной Европы с благоприятным для кабана климатом и при вы-
сокой численности вида специалисты рекомендуют отстреливать 60-80% сеголе-
ток, 10-15% подсвинков и не более 5-15% взрослых особей от общего объема
добычи при обязательной селекции. На практике доля изымаемых детенышей не-
сколько ниже рекомендуемой, но все же повсеместно выше 50% при высокой
(иногда до 40%) доле отстрела подсвинков [Briedermann, 1977b, 1979; Groger,
1979; Hopp, 1979, 1980; Gunther, 1980; Menzel, 1981; Blume, Hopp, 1982, 1986; Ге-
нов, Коцаков, 1986; Remmele, 1986; Spitz, 1987; Говорка и др., 1988].
В России, которая отличается длительными, многоснежными и суровыми зи-
мами, промысловая нагрузка на сеголеток, практически не участвующих в раз-
множении и наиболее подверженных естественной смертности, должна быть еще
выше. Преимущественный их отстрел важен и с ветеринарной точки зрения -
именно молодые особи наиболее инвазированы и часто гибнут от метастронгилеза.
На необходимости более интенсивной эксплуатации молодых животных и
сохранении репродуктивного ядра популяций давно настаивают многие отечест-
венные ученые [Телишевский, 1972, 1990; Козло, 1975; Фадеев, 1975, 1978а;
Иванова, Овсюкова, 1976; Хахин, 1980; Иванова, 1983, 1985; Русаков, Тимофее-
ва, 1984; Кононов, 1988; Русаков, 1989; Приклонский, 1999]. На практике, к со-
235
Таблица 51. Рекомендации по нормированию добычи кабана
Объем добычи (% от весенней численности) Сеголетки, %* Подсвин- ки, %* Взрослые, %* Источник информации
15-25 Преимуще- ственно — — Козло, 1970а, 1975
20-50 30-50 ** 50-70** Иванова, Молоканов, 1978
10-60 50-60 25-35 10-20 Иванова, 1983, 1985
75 15 10 Телишевский, 1972, 1990; Briedermann, 1979; Hopp, 1980; Blume, Hopp, 1982; Meynhardt, 1984c
20-25 60-70 15-25 10-20 Русаков, 1989
— 80-90 ** 10-20** Кононов, 1988
— 90** ** 10 Фадеев, 1987
20*** 50-60 ** 50-40** Смирнов, 2001
60-100 70-80 15-20 5-10 Янулайтис, Падайга, 1984а,б, 1985, 1987
__ 70 15 15 Маньен, 1999
10-40 70-75** ** 25-30 Варнаков, 1985
60-80 10-30 10 Говорка и др., 1988
5-10 — — Приклонский, 1999
100 60-80 10-30 До ю Фертиков и др., 1999
До 20-60 в разных районах До 60 ** >40** Инструкция Главохоты РСФСР, 1984
* От общего объема добычи, %.
** Среднее значение для выборки.
*** От осенней численности !
Прочерк — нет сведений. .
жалению, все наоборот: при очень низкой плотности населения диких свиней и в
многоснежные голодные годы основная промысловая нагрузка падает на взрос-
лую репродуктивную часть популяции (см. табл. 49), что ведет к ее омоложению,
уменьшению плодовитости и снижению темпов воспроизводства, увеличению
смертности молодых особей и, в конечном счете, к снижению численности.
По моему мнению, доля сеголеток кабана в объеме добычи, до достижения
оптимальной плотности населения, должна быть не менее 95-100%. В неурожай-
ные годы с ранней многоснежной зимой и гололедицами и при недостатке искус-
ственных кормов число молодых животных следует сокращать особенно резко.
При таком подходе существенно упрощается проблема выбора охотником того
или иного зверя - разрешается отстрел только детенышей, хорошо отличающих-
ся от взрослых особей меньшими размерами и коричневой (в европейской части
ареала) окраской (рис. 73).
Организовать массовую добычу сеголеток и подсвинков невозможно без j>
дифференциации стоимости лицензий на отстрел взрослых и молодых особей, а
также без применения нарезного оружия. Впрочем, в настоящее время ни то, ни
другое уже не представляет большой проблемы.
Оптимальные сроки добычи. Длительность охоты на кабана и других ди-
ких копытных, по моему разумению, должна находиться в тесной зависимости от
плотности их населения. При низкой численности и, соответственно, мизерном
объеме добычи вполне достаточно двух недель или одного месяца. При плотно-
сти, близкой к оптимальной, срок охоты может достигать двух-трех месяцев, при
чрезмерном поголовье и ощутимом вреде сельскому хозяйству он может быть
еще увеличен, но только не за счет продления зимнего сезона.
236
Рис. 73. Сеголетки легко различимы среди взрослых особей по размерам и окраске,
что упрошает их выборочный отстрел (фото П. Яровицкого)
Наиболее подходящие сроки массовой добычи кабана в разных регионах Рос-
сии, как и в Белоруссии [Козло, 1975], - сентябрь - ноябрь (в отдельных районах -
до середины декабря). В этот период животные наиболее упитанны. Позднее
масса тела взрослых особей резко снижается, а рост сеголеток прекращается (см.
раздел «Изменчивость и таксономия»). При раннеосеннем массовом отстреле со-
кращается число потребителей корма, и его больше останется особям, выжившим
после сезона охоты. При сравнительно коротких сроках охоты животных меньше
беспокоят, они затрачивают гораздо меньше энергии на вынужденные переме-
щения и, при наличии подкормки, не уходят из охотничьих хозяйств. С 15 декаб-
ря (1 января при высокой плотности) охота на копытных должна быть закрыта
практически на всей территории страны. В Сибири ученые рекомендуют пре-
кращать промысел диких свиней с 15 ноября или 1 декабря [Смирнов, 1978, 2001;
Недзельский, 1994].
Замечу, что в Киргизии эти рекомендации уже частично воплощены. С 1966 г.
охота здесь разрешена в течение двух месяцев - с 1 октября до 1 декабря [Чичи-
кин, Воробьев, 1967], в 80-е годы - с 15 сентября по 15 декабря, но предлагается
открывать ее с 1 сентября по 15 ноября [Катаевский, 1987].
При оптимальной плотности населения выборочная охота на кабана возмож-
на с конца лета (предпочтительнее - не ранее 15 августа). Установка на интен-
сивную его добычу в летний период [Варнаков, 1988а] явно неприемлема, по-
скольку, как показывает практика, приводит к массовому легальному и нелегаль-
ному отстрелу взрослых животных, включая лактирующих самок. Поросята, ос-
тавшиеся без матерей, затем гибнут или отстают в росте и развитии [Варнаков,
1985]. Кроме того, упитанность зверей в это время низка, что приведет к значи-
тельным потерям мясной продукции. При низкой численности копытных охота
на них в летние месяцы (на потравах) должна быть запрещена.
В странах Западной Европы, где плотность населения диких свиней намного
превышает емкость угодий, охота ведется большую часть года или круглогодич-
но. Возможно, при значительном увеличении ресурсов круглогодичную охоту
можно будет внедрять и у пас, но пока что для этого нет абсолютно никаких
предпосылок
237
Рис. 74. Схема стационарной живоловушки для кабанов (по [Новиков, 1987])
Размеры - в сантиметрах
Для минимизации нелегальных потерь срок действия разовых лицензий на
отстрел зверя желательно максимально ограничивать и не допускать неконтро-
лируемой охоты. Оптимальный вариант - все лицензии должны находиться у ад-
министрации охотничьих хозяйств и егерей.
Оптимальные способы добычи. Видимо, нет смысла подробно рассматри-
вать все существующие и давно известные специалистам и охотникам способы
добычи кабана. Отмечу лишь еще раз наиболее эффективные из них: из засидок
(с вышек), с собакой, а также живоловушками и сетями. При всех этих способах
возможен выборочный селекционный отстрел животных.
От загонных охот, при всех их спортивно-массовых достоинствах, в интен-
сивном охотничьем хозяйстве придется отказаться или существенно уменьшить
масштаб, применяя только в тех местах, где нет подкормки и постоянных убе-
жищ зверей. Однако при чрезмерной численности загонные охоты могут быть
полезными, так как будут способствовать изгнанию из угодий «лишних» особей.
В любом случае необходимо культивировать собак, работающих по кровяному
следу, что существенно уменьшит долю недобранных подранков.
При высокой и чрезмерной численности перспективна добыча диких свиней
живоловушками и сетями. Эти способы необходимы также для отлова их с целью
расселения, мечения, в ветеринарных и селекционных целях.
Отлов живоловушками. Ловушки (стационарные или переносные) просты по
конструкции (рис. 74) и могут быть без значительных затрат сооружены в любом
охотничьем хозяйстве - небольшой, предпочтительнее ромбовидный, загон (из мел-
коячеистой сетки, частокола, жердей или бревен) диаметром 4-15 м, высотой 2-
2,5 м, с одной или двумя опадными дверями и длинным (до 5 м), узким (90-60 см) и
затемненным коридором (стрункой). Этот коридор, оборудованный двумя или тремя
выдвижными или откидными (только в одну сторону) дверцами, служит для фикса-
ции и перегона пойманных зверей в транспортные клетки (длина - 165-180 см, ши-
рина- 85-90 и высота - 80-105 см в зависимости от возраста животных), которые
ставят одну за другой к внешнему его концу. Иногда для предотвращения травмиро-
вания кабанов сооружают два коридора или внутрь загона помещают автоматически
захлопывающиеся транспортные клетки. Ловушку оснащают автоматической или
приводимой в действие с помощью троса из наблюдательного пункта насторожкой.
Для этого на верхнем конце опадной двери делают петлю из толстой проволоки, ку-
да вставляют сторожок с тросом, перекинутым через перекладину. Второй конец
238
Рис. 75. Живоловушка для сеголеток (фото П. Мануша и В. Николаева)
троса привязывают к колышку-насторожке, скрывая его под корытом с кормом. По-
едая корм, звери сталкивают корыто с насторожки, трос ослабляется, сторожок вы-
скакивает, и дверь падает. Животные начинают метаться по загону в поисках выхода
и тыкаются рылом в нижние жерди. Нажав на откидную дверь, они поднимают ее и
сами, без принуждения, заходят в струнку, где не могут развернуться, и затем прохо-
дят в транспортные клетки. Для мечения свиней пол в струнке делают приподнятым
над землей и подвижным (опускающимся на тросах с помощью ворота) перед выпу-
скной дверцей. Над полом по центру прокладывают жердь диаметром около 12 см
для надежной фиксации зверей, которые повисают на ней без опоры и теряют под-
вижность [Комов, 1972; Козло, 1973; Чегорка, 1984; Царев, 19866,в, 2000; Новиков,
1987]. Во Франции этих копытных отлавливают с помощью переносных ловушек
(2 х 0,7 х 1,7 м), поставленных по 2-3 через каждый километр [Mauget, 1979].
Корм (зерно, силос, зерносенаж), выкладываемый в корыта в ловушке и для
приманки (небольшими кучками или россыпью в виде дорожек) с внешней ее
стороны, можно пропитывать растворами успокаивающих препаратов (например,
15 мг аминазина или 100 мг хлоралгидрата на 1 кг массы зверя). При обездвижи-
вании зверей ромпуном и эторфином возможна гибель самок на последних ста-
диях беременности [Царев, 19866,в, 2000].
Для отлова сеголеток на подкормочной площадке достаточно соорудить ло-
вушку размером 3x4 м высотой 1,5-1,7 м, с настороженной опадающей жердью и
дощатым полом, на который высыпается корм (рис. 75). Для ограничения про-
никновения взрослых особей высота входного проема не более 20-30 см от зем-
ли. Размеры настила должны быть на 0,5—0,7 м короче размеров забора. Поросята
при кормлении сбивают насторожку и остаются внутри вольеры. При появлении
людей они спрыгивают с настила и бегают по периметру забора, что облегчает их
239
Рис. 76. Отлов кабанят сачком-конусом (фото П Мануша и В. Николаева)
поимку изготовленным из металлических прутьев сачком-конусом, скрепленным
по бокам кольцами (рис. 76). В приподнятом вертикально сачке удобно прово-
дить мечение и ветеринарные манипуляции [Мануш, Николаев, 1995]
Отлов сетями. Этот способ можно применять круглогодично. Кабан - сильный
зверь, поэтому сети необходимо применять прочные, капроновые, с толщиной ни-
ти не менее 3 мм (рис. 77). Оптимальные ее размеры: высота - 2,5 м, длина - до 1
км, длина отдельных звеньев - 40-50 м, размер ячеи 15x15 см. Дель крепится
без усадки на шнур диаметром 8 мм только с одной стороны. Шнур должен быть
на метр длиннее с каждой стороны отрезка дели. Эти концы служат для связыва-
ния собранной сети. Для маскировки снасть окрашивают синтетическими краси-
телями в темный цвет.
Как показала наша практика, зверь надежно запутывается, если его голова про-
ходит в ячейку, при этом грудь и передние ноги становятся своего рода ограничи-
телями, удерживающими его в упавшей снасти (рис. 78). Сеть с ячеей 15 х 15 см
весьма уловиста для сеголеток (с августа) и подсвинков. Крупные самцы, у кото-
рых голова не проходит в ячею, нередко выпутываются до подхода ловцов. Для
отлова крупных животных необходима ячея 20 х 20 см, но такая сеть не удержи-
вает мелких особей.
Техника установки снасти следующая. Ее развешивают на двухметровые ко-
лышки диаметром 3-4 см. Для крепления шнура в их верхние концы заранее
вбивают гвозди, острие которых на 2-3 см выступает наружу, или же подбирают
колышек с сучком в верхней части. Острия сучков или гвоздей на установленных
колышках располагают с противоположной стороны от предполагаемого хода
зверей. Шнур ни в коем случае нельзя привязывать к колышкам или к деревьям -
он должен свободно соскакивать с креплений при прикосновении к сети. Нижний
'40
Рис. 77. Линия сетей для отлова диких свиней
Рис. 78. Зверь надежно запутывается в сети, если его голова проходит в ячейку
16. Заказ №2528.
241
ее край и отдельные секции тоже не закрепляют, низ немного подворачивают в
сторону предполагаемого захода зверя, как это делается при отлове зайцев. Кабан
наступает на сеть, просовывает голову в ячею, дергает снасть и сбрасывает ее
верхнюю часть на себя. При этом падает лишь одно звено, а цельность остальной
линии не нарушается.
Диких свиней отлавливают сетями активным (загоном) или пассивным спосо-
бами, или их комбинацией [Данилкин, 1984; Варнаков, 19886]. Наиболее эффек-
тивный и вместе с тем наименее трудоемкий - пассивный способ отлова, осно-
ванный на доминировании пищевого рефлекса над другими реакциями.
Сетью зверям преграждают доступ к корму (кормовым полям или к подкор-
мочным площадкам). У голодных зверей, видящих или чующих корм, по-
видимому, несколько притупляется инстинкт самосохранения — они пытаются
прорваться сквозь преграду. Самыми нетерпеливыми и менее осторожными ока-
зываются молодые животные, чаще других попадающие в сеть.
Этот способ особенно полезен при отлове животных, повреждающих посевы.
В этом случае линию сетей ставят на опушках в местах выхода зверей на поле
(см. рис. 77) и не снимают несколько дней, проверяя 2-4 раза в сутки. Иногда за
одну августовскую ночь в 300-500-метровую снасть попадают до 8 особей. Наши
исследования показали, что группы зверей, отдельные члены которых были пой-
маны сетями и оснащены радиопередатчиками, не пугаются таких ловушек, что
позволяет без особых усилий отлавливать, если необходимо, всех.
Следует учесть, что попавшие в сеть кабаны при появлении человека иногда
бросаются на него и пытаются схватить зубами или ударить клыком. Поэтому
подходить к ним следует весьма осторожно. Если звери нужны живыми, то их
фиксируют, прижав к земле длинным шестом с двух сторон, шприцем вводят
препарат для обездвиживания и только после этого выпутывают из сетей.
Селекция. Одна из основных задач охотничьего хозяйства - улучшение ге-
нофонда диких животных. К сожалению, как уже упоминалось выше, сейчас в
России прогрессирует охотничья антиселекция, приводящая к деградации попу-
ляций копытных [Данилкин, 2000а].
Тем не менее селекцию копытных, и кабана в частности, необходимо вести круг-
логодично. Принципы отбора весьма просты. На племя оставляют самых жизнестой-
ких животных с максимальным репродуктивным потенциалом - крупных секачей и
самок, сохранивших ранние выводки. Безусловному отстрелу подлежат самки с
поздними выводками или позднорожденные, мелкие (с массой до 25 кг), полосатые
зимой, кашляющие, сгорбленные, с приподнятой на спине шерстью поросята, а
также сильно истощенные, ослабленные, больные, травмированные или раненые
особи и звери с аномальной окраской [Козло, 1972в, 1973, 19746; Телишевский,
1972]. В природе такие животные обычно вымирают сами, однако в охотничьих
хозяйствах при наличии обильной подкормки выживают и оставляют потомство
далеко не лучшего качества, к тому же часто зараженное паразитами или ставшее
носителем инфекций.
Потери сеголеток за год почти повсеместно составляют более половины от
числа родившихся, поэтому максимальное изъятие обреченных на естественную
гибель особей выгодно для любого охотничьего хозяйства и, при выборочном от-
стреле, полезно для популяции. Элиминация старых особей, о чем обычно упо-
минают при обсуждении принципов селекционного отстрела, вряд ли возможна.
В природе их практически не удается различить, и, кроме того, опытные особи,
уцелевшие при очень непростых условиях жизни, на мой взгляд, весьма полезны
для группировок, причем плодовитость старых самок не меньше, чем у молодых.
Известную рекомендацию [Данилов и др., 1966] преимущественного отстрела
наиболее крупных и массивных секачей, которые отгоняют других особей от корма
242
и якобы имеют проблемы при оплодотворении мелких самок, не выдерживающих их
во время садок, я считаю биологически не обоснованной и крайне вредной.
Агрессивность взрослых самцов на подкормочных площадках выражена го-
раздо меньше, чем, например, у подсвинков по отношению к сеголеткам. Только
крупные секачи способны покрывать массивных самок, они же кроют большую
часть молодых, а для остальных всегда имеется достаточное количество разно-
возрастных самцов. Роль мощных самцов заключается не только в оставлении
лучшего по качеству потомства. Они выполняют и другие важные социальные
функции в группировке, о которых говорилось выше. Секачи, наряду с взрослы-
ми самками, вне всякого сомнения, - самые жизнеспособные и наиболее ценные
члены популяции, и их следует максимально сохранять, а не убивать.
Уменьшение негативного воздействия на биоценозы. С изменением эко-
номических отношений, произошедшим в России в конце XX в., важной пробле-
мой для охотничьего хозяйства становится не только повышение продуктивности
популяций диких копытных и угодий в целом, но и снижение негативного воз-
действия животных на среду обитания и, в частности, уменьшение ущерба сель-
скому хозяйству, теперь уже в значительной мере частному и фермерскому.
Нужно отчетливо сознавать, что кабан был, есть и будет одним из многих
«умных» и «умелых» вредителей, с которыми трудно бороться. Его тяга к сель-
скохозяйственным посевам, где сконцентрирован лучший белковый корм, неис-
требима. Почти невозможно предотвратить и роющую деятельность в лесу, на
сенокосных полянах и лугах.
Огораживание полей - дорогостоящая и малоэффективная мера. Отпугивание
с помощью сторожей, костров, собак, привязанных на длинной проволоке, шу-
мовых эффектов и т.п. имеет кратковременное действие [Слудский, 1956; Абра-
мов, 1963; Бромлей, 1964; Саблина, 1969]. Электрошоковые приспособления не
становятся реальным препятствием для крупных копытных [Гусев, 1988, 1989;
Хахин, Гончаров, 1989]. Тем не менее электрические изгороди в какой-то мере
могут сдерживать набеги диких свиней на посевы [Gasparik et al., 1993], а сигна-
лы тревоги, записанные на магнитофон и передаваемые через репродуктор, спо-
собствуют их изгнанию с полей [Майнхардт, 1986]. При высокой плотности по-
пуляции отстрел нескольких особей на потравах тоже не дает положительного
эффекта [Варнаков, 19786]. Кабаны выходят на поле в разных местах или пере-
ходят на другие участки.
Проблема сокращения ущерба от этого вида существует во многих странах,
но успеха добились только там, где от борьбы перешли к хозяйственному ис-
пользованию ресурсов на научной основе и в сотрудничестве с сельским хозяй-
ством [Briedermann, 1986].
Замечу, что человек разумный обычно довольно быстро, в течение одного-
двух десятилетий, решает вопрос спасения от диких свиней запасов картофеля,
овощей и зерна путем сооружения капитальных хранилищ или расположения
буртов в населенных пунктах. Поля, примыкающие к лесу, стали засевать плохо
поедаемыми культурами: травами, льном, остистым ячменем и специальными
сортами картофеля. Кукурузные, гороховые, пшеничные, овсяные, картофельные
и свекольные плантации в местах обитания этих копытных обычно располагают
не ближе 1,5-2 км от опушки [Иванова, Рыковский, 1968а; Варнаков, Мошева,
1975; Фадеев, 19786; Майнхардт, 1986]. Новые репелленты также оказались
весьма эффективными, и в настоящее время довольно широко применяются в за-
падноевропейских странах для отпугивания кабанов от посевов. Кроме того, до-
вольно просто, огораживанием, решается проблема сохранения муравейников в
лесу [Ханжин, 1977; Янулайтис, 1984] и посадок в лесопитомниках.
Ущерб можно снизить и путем сохранения лучших естественных кормовых
стаций кабана, но все же основное направление - комплексные биотехнические
16
243
мероприятия, в числе которых размещение в лесах отвлекающих кормовых полей
и регулярная искусственная подкормка. Даже применение в качестве своеобразных
кормушек больших стеклянных бутылок, наполненных зерном, которые звери ча-
сами катают по земле, позволяет долгое время удерживать их в лесу [Воробьев,
1968; Иванова, Рыковский, 19686; Сухорослов, 19726; Фадеев, 19736; Козло, 1975;
Briedermann, 1977a,b, 1986; Романов, Козло, 1977; Groger, 1979; Гунчак, 1980;
Genov, 1981b; Генов, 1986; Козло, Дунин, 1989; Телишевский, 1990].
По «Рекомендациям по предотвращению ущерба, наносимого кабанами сель-
скому хозяйству» (М., 1986), площадь отвлекающих кормовых полей должна со-
ставлять 0,3-0,5 га на каждые 1000 га угодий при плотности населения диких
свиней на ту же площадь в южной тайге 1-4, в смешанных лесах - 2-10, в широ-
колиственных и горных лесах и лесостепи - 6-15 голов. На 15-20 особей необхо-
дима одна подкормочная площадка. Норма выкладки - 500 г картофеля и 300 г
зерноотходов на каждого зверя в сутки. В этой методической разработке подчер-
кивается также, что охотничьи животные — государственная собственность (об-
щенародное достояние), поэтому «мероприятия по предотвращению ущерба, на-
носимого кабанами сельскому хозяйству, проводятся не только охотничьими хо-
зяйствами, но и сельскохозяйственными организациями, на которых возлагаются
функции охраны посевов».
Все эти меры позволят также уменьшить повреждаемость лугов и пастбищ, :
сократят поисковую активность зверей, что будет благоприятствовать сохранно-
сти тетеревиных птиц.
И все же главное условие уменьшения ущерба - поддержание численности
диких свиней на оптимальном уровне, приемлемом как для охотничьего, так и 1
для сельского хозяйства. При этом немаловажно правильно регулировать поло- I
вой и возрастной состав. Уже давно замечено, что наибольший ущерб посевам 1
причиняют не взрослые особи, а детеныши, оставшиеся без матерей, и группы "
подсвинков. Меньше всего вредят посевам старые звери, знакомые с опасностью
выхода на поля. Следовательно, молодые животные должны быть основным объ-
ектом добычи и с этой точки зрения.
Подкормка. Как показано выше, численность диких копытных неоднократно
резко сокращалась во многих районах из-за недостатка зимних кормов, и затем
требовались годы для восстановления ресурсов. «Предоставленный самому себе, |
кабан влачит довольно жалкое существование, почти совсем исчезая после тяже- I
лых снежных зим и несколько оправляясь после двух-трех благоприятных зим I
сряду. По иному обстоит дело там, где его зимой не подкармливают, а кормят» - 1
вывод, сделанный П.Б. Юргенсоном [1969] по результатам исследований в Зави-
довском заповедно-охотничьем хозяйстве, вряд ли когда-либо потеряет свою ак-
туальность в суровых климатических условиях России.
Опыт Беловежской пущи и Березинского заповедника отчетливо продемон-
стрировал, что без участия человека сохранить высокое поголовье этого вида
нельзя. Лишь благодаря интенсивному кормлению здесь прекратилась массовая
гибель животных, что нередко случалось ранее. Подкормку посещают до 70% !
особей [Лебедева, 1956а; Козло, 1969, 1970, 19726, 1975; Романовский, Кочанов-
ский, 1971; Романов, Козло, 1977]. Необходимость этого мероприятия убеди-
тельно подтверждена и многолетней практикой охотничьего хозяйства [Саблина,
1955; Иванова, Рыковский, 19686; Фадеев, 1968, 1969, 1970а,б, 1973б,д; Верши-
нин, 1969; Киселев, 1986; Варнаков, Млодиков, 1988; Козло, Дунин, 1989; Аза-
ров, Климов, 1990; Недзельский, 1994; Козло, 2001], хотя затраты на подкормку и
продуктивность популяций не всегда адекватны в разных угодьях и при разных
приемах управления популяциями [Мамаев, 1983]. Не менее важная задача, ко-
торая решается подкормкой, — концентрация зверей в нужном месте и в нужное
244
Рис. 79. Подкормка необходима также для концентрации зверей в нужном месте и в
нужное время (фото А. Богданова)
время, удержание на ограниченной территории и предотвращение миграций за
пределы охотничьего хозяйства (рис. 79).
Посредством искусственного повышения кормовой емкости угодий происходит
«смягчение» основного экологического фактора, находящегося в минимуме. Однако
кабану нужен не всякий корм. В природе для него особенно важна высококалорий-
ная белковая пища (земляные черви, насекомые, зерновые и бобовые культуры, пло-
ды), обеспечивающая максимальное накопление жировых резервов (см. выше).
В охотничьих хозяйствах для подкормки зверей обычно выкладывают овес,
ячмень, пшеницу, рожь, зерноотходы, кукурузу, подсолнечник, люпин, карто-
фель, свеклу, морковь, топинамбур, яблоки, груши, желуди, буковые орешки,
комбикорма, жмых, отходы различных пищевых предприятий, мясокостную му-
ку, мясные и рыбные отходы и павших животных. Нередко такую пищу остав-
ляеют в кучах под открытым небом с осени, что приводит к ее порче. На подкор-
мочных площадках мне нередко приходилось видеть пораженные гнилью желуди
и кукурузу, испорченный картофель, заплесневелые кучи зерна, зерноотходов и
комбикормов, протухшие животные корма.
Дикие свиньи, несмотря на относительную всеядность, поедают далеко не все
предлагаемые им корма, а, как показывает практика, лишь доброкачественные,
высокопитательные и, преимущественно, влажные. В большинстве случаев зве-
ри, у которых хорошо развит инстинкт самосохранения, к испорченной пище не
подходят до тех пор, пока имеют возможность находить другую. В голод поеда-
ется и такой корм, но последствия могут быть печальными как для животных, так
и для охотничьих хозяйств. Случаи отравления кабанов, особенно сеголеток, не-
качественной пищей регистрируются повсеместно.
Картофель обычно считают лучшим кормом для свиней, хотя это не совсем
так Он богат углеводами, но содержа! мало протеина, поэтому эту пищу можно
расценивать лишь как «поддерживающую». Во всех отношениях гораздо ценнее
топинамбур, мало вымерзающий в земле и входящий в состав рациона зверей до
8 месяцев в году, а в мягкие зимы и в южных районах — почти круглогодично.
245
Следует отметить, что кабаны очень осторожно относятся к новому корму и,
даже будучи голодными, не сразу поедают его. В неволе свеклу, морковь и ка-
пусту они начинают употреблять после двухнедельного опробывания, яблоки -
на четвертый день [Иваненко, 1956], а к топинамбуру не подходят до месяца. В
Иркутской области животные, прежде незнакомые с подкормкой, не притрагива-
лись к ней три недели [Никулин, 1981].
Овощи и картофель быстро замерзают на морозе и становятся несъедобными
для молодых особей (взрослые кабаны с их мощными челюстями разгрызают и
мерзлые клубни), поэтому их лучше скармливать осенью и весной, а зимой -
лишь в оттепели или вывозить небольшими порциями для скармливания в тот же
день. Морковь, капуста и турнепс, по моим наблюдениям, поедаются плохо, в
измельченном виде - более охотно.
В каждом охотничьем хозяйстве необходимо иметь собственные поля, на ко-
торых высеваются растения, потребляемые всеми видами копытных: люцерна,
козлятник, эспарцет, горохо-вико-овсяная смесь и др. Из однолетних культур ди-
кие свиньи предпочитают кукурузу, горох и овес. Следует учитывать, что план-
тации этих растений, находящиеся в непосредственной близости от мест концен-
трации зверей, посещаются в первую очередь и уничтожаются полностью на
корню еще до зимы. Кукурузу и горох животные выбирают и на подкормочных
площадках. Зерновые культуры они охотно поедают со времени колошения.
Сладкий люпин посещают лишь после первых заморозков, и, следовательно, этот
вид может быть одним из основных на зимних подкормочных полях.
При необходимости быстрого увеличения и сохранения поголовья кабана нет
смысла делать ставку на топинамбур как основную подкормочную культуру, по-
скольку звери очень долго привыкают к нему. По моим наблюдениям и сведени-
ям других исследователей [Чегорка, 1984], плантации с этой культурой они мо-
гут игнорировать несколько лет, пока не найдется голодная особь, которой рас-
тение придется по вкусу. Постепенно все большее число животных начинают по-
сещать поля, выбирая клубни.
Во многих охотничьих хозяйствах урожай на собственных кормовых полях
обычно оставляют в зиму на корню. Исследования показывают, что с таких не
скошенных полей копытные чаще всего используют от 1 до 20% биомассы, тогда
как убранные и складированные (в копнах, стогах, под навесами) корма утилизи-
руют зимой на 70-100%. Соответственно мои рекомендации сводятся к необхо-
димости полной или почти полной уборке урожая с полей, за исключением клуб-
ней топинамбура. В противном случае зимняя подкормка становится эффектив-
ной лишь для мышевидных грызунов.
Необмолоченные стожки гороховой, пшеничной, ячменной, овсяной или го-
рохо-вико-овсяной соломы нередко даже более привлекательны для диких сви-
ней по сравнению с зерном или с зерноотходами этих культур. Вероятно, в сто-
гах зерно более влажное, и, кроме того, солома необходима зверям в качестве на-
полнителя. В них же свиньи устраивают зимние и родильные «гнезда». Поэтому
подвоз необмолоченных стожков, сложенных при стоговании на волокуши
(срубленные ветвистые деревья), к местам убежищ зверей может стать одним из
основных приемов подкормки, поскольку при этом значительно снижаются тру-
дозатраты на выкладку кормов.
При возможности выбора копытные всегда отдают предпочтение более
влажному белковому корму. На мой взгляд, наиболее перспективный влагоемкий
корм для всех видов диких копытных в отечественном охотничьем хозяйстве -
силос или зерносенаж (утрамбованная смесь трав и зерновых нередко с добавле-
нием подсолнечника). Технология силосования сравнительно проста и хорошо
отработана в сельском хозяйстве. Косули, однако, весьма неохотно едят кукуруз-
ный силос. На заросших озерах силос можно готовить из молодых стеблей и кор-
246
невищ тростника и рогоза [Слудский, 1956]. В некоторых районах дикие свиньи с
удовольствием потребляют зеленый рапс, скошенный после мороза и складиро-
ванный в кучах на подкормочных площадках.
Заслуживают внимания исследования Е.М. Недзельского [1994], предлагаю-
щего для снижения затрат использовать в качестве подкормки измельченную ве-
точно-травяную смесь (хвоя, побеги осины, березы, ивы и разнотравье в соотно-
шении 1:3) и измельченные пищевые отходы (очистки картофеля, хлеб, свекла,
морковь, капуста, брюква, мясо-рыбные продукты, иногда комбикорма). Для
приготовления россыпи и брикетов корма пригодны типовые измельчители и
гидравлические прессы. Подача корма в кормушки осуществляется по лоткам из
бункеров объемом 1-1,5м3. Пища для сеголеток готовится из комбикорма (90%)
и концентрата растительного белка из веточно-травяной массы. Доступ взрослых
особей на «детские» площадки ограничивается забором. Эксперименты по под-
кормке кабана в Восточной Сибири белковой пищей дали следующий результат:
— повышение плодовитости в среднем на 25%, или на 2,1 детеныша;
— увеличение массы новорожденных на 2-4%;
— увеличение массы и размеров тела сеголеток в среднем на 5-13 кг и 3-10 см;
— увеличение упитанности и массы взрослых особей на 5% (до 7 кг);
— большую сохранность животных в зимний период.
Расчетный срок подкормки диких свиней - 70-165 дней в зависимости от клима-
тических условий, суточная норма выкладки - 1—3 кг на голову в зависимости от ви-
да корма и суровости зим [Данилов и др., 1966; Ильинский, 1966; Иванова, Рыков-
ский, 1968а,б; Фадеев, 19736; Козло, Дунин, 1989; Никифоров, Бринза, 1991]. Годо-
вая норма подкормки в «Завидово» - 100-110 кг картофеля и около 7 кг гороха на
особь, что недостаточно в многоснежные зимы. В январе - марте норма выкладки
увеличивается до 2-3,5 кг на животное [Дормидонтов, 1969; Мануш, 1969]. В Бело-
вежской пуще и в Березинском заповеднике на одного зверя в сутки выкладывают от
0,5 (ноябрь) до 2-4 кг (до марта). В морозные дни суточный рацион увеличивают до
3-^4 кг на особь [Козло, 19726, 1975]. Фактически в снежный период для каждого ка-
бана требуется около 300-500 кг корма.
Как показано выше, стратегия выживания диких свиней зимой основана на
максимальной экономии энергозатрат. Это важное обстоятельство, к сожалению,
не учитывается в отечественном охотничьем хозяйстве. Повсеместно зверей под-
кармливают у охотничьих вышек, расположенных, как правило, вблизи населен-
ных пунктов, где живет егерь и есть удобный подъезд. Животные вынуждены
почти ежедневно совершать многокилометровые переходы от убежищ до мест
подкормки, затрачивая при этом энергии гораздо больше, чем получают со скуд-
ной пищей. При такой подкормке они быстро худеют и погибают от истощения.
Кроме того, на переходах их подстерегают браконьеры, волки, собаки. Людям
невдомек, что, располагая кормовые пункты вдали от основных убежищ, они не
сохраняют, а губят копытных.
Биотехнические сооружения следует приурочивать к стациям переживания:
болотам, зарослям в поймах рек, ельникам или их вкраплениям в лиственные ле-
са, дубравам. Подкормочные площадки, по возможности, желательно оборудо-
вать поблизости от воды и болот, но на сухом месте, предпочтительнее в сосня-
ках. Необходимо учитывать, что кабану, как и другим копытным, круглогодично
нужна свободная вода. При поедании сухих кормов потребность в ней сущест-
венно возрастает. По расчетам известного охотоведа С.Г. Мануша [1969], диким
свиньям на 1 кг сухого корма требуется 7 л воды, что невозможно получить, по-
едая снег. Добавлю, что на превращение снега в воду и нагревание ее до темпе-
ратуры тела звери иногда затрачивают энергии больше, чем получают с кормом.
Игнорирование животными сухой подкормки нередко объясняется лишь отсут-
ствием свободной воды. Именно из-за ее дефицита зимой нельзя скармливать ко-
247
пытным соленую рыбу и другие подсоленные корма. Звери с жадность поедают
их, получая отравляющие дозы натрия. Однако «глиняные» солонцы или соль-
лизунец на подкормочных площадках необходимы.
При регулярной подкормке кабаны обычно не уходят далее 0,5-2 км от под-
кормочной площадки. При привычных звуках, связанных с раздачей пищи, они
могут выходить кормиться в дневное время и даже в присутствии человека.
Взрослые особи всегда более осторожны. Секачи посещают места кормежки ча-
ще с наступлением сумерек.
Взрослые самцы довольно агрессивны, особенно по отношению к молодняку.
Поэтому на каждой подкормочной площадке желательно раскладывать корм как
можно шире или оборудовать из жердей отдельные загоны, ограничивающие
доступ к корму взрослым животным. Эта мера поможет обеспечить сеголеток
пищей, снизит возможность их заражения болезнями, существенно уменьшит
травматизм, обеспечит возможность проведения дегельминтизации.
Корма не должны гнить в кучах под открытым небом. Их следует хранить в спе-
циальных сараях или под навесами. Наиболее удобны закрытые стационарные со-
оружения бункерного типа в виде сарая или вышки, с несколькими корытами (для
молодых и взрослых животных) у наружной стены. Корм, по мере его употребления,
под действием силы тяжести автоматически поступает в корыта. Очень неплохо ра-
ботают стационарные (или передвижные, установленные на полозья [Дунаев, 1991])
бункерные кормушки, сделанные из металлических бочек емкостью до 250 л. Таких
кормушек на площадке лучше делать несколько, иначе возникают постоянные драки
за корм. Сараи с открытым доступом к корму также эффективны, но корм здесь за-
грязняется свиньями, и на ограниченном пространстве между животными иногда
возникают драки, приводящие к серьезным травмам.
Из закрытых хранилищ корм можно высыпать на снег. При соприкосновении
со снегом он становится более влажным, и звери прекрасно находят его даже по-
сле порош. При раннеосенней подкормке сыпучие корма предпочтительнее вы-
кладывать в длинные корыта (рис. 80) или на площадки из досок, что предотвра-
щает затаптывание их в грязь и снижает опасность заражения гельминтами [Шу-
рин-Юхкум, 1994].
Подкормка должна быть регулярной и ежегодной, с выкладкой кормов задол-
го до установления снежного покрова. Иначе в глубокоснежье далеко не все зве-
ри могут найти пищу. При отсутствии корма хотя бы в течение одной суровой
зимы кабан погибает или откочевывает в другие районы [Иванова, 19756]. В за-
сушливых районах не обойтись без устройства искусственных водоемов.
Следует учитывать и негативные последствия искусственной подкормки:
чрезмерную концентрацию животных, повышенное заражение гельминтами и
быстрое распространение инфекционных заболеваний, выживание ослабленных
зверей, насыщение популяции менее жизнестойкими особями. Этому необходи-
мо активно противодействовать путем селекционного отстрела и проведения
профилактических ветеринарных мероприятий.
Профилактика заболеваний. Чем выше уровень ведения охотничьего хо-
зяйства, тем обычно больше плотность населения диких копытных и их концен-
трация у водопоев, кормовых полей, подкормочных площадок и солонцов, и тем
меньше воздействие на популяцию таких факторов естественного отбора, как
крупные хищники, корма и погода. При высокой плотности резко возрастает
возможность циркуляции инвазии и зараженность животных и увеличивается ве-
роятность их гибели. Для профилактики болезней и гельминтозов необходимо
регулировать численность и круглогодично вести выборочный отстрел особей с
признаками болезни [Говорка и др., 1988]. ।
248
Рис. 80. При раннеосенней подкормке сыпучие корма предпочтительнее выклады-
вать в длинные корыта или на площадки из досок (фото И. Мухина)
Наибольшие потери поголовья в охотхозяйствах - от чумы свиней и метаст-
рогилеза. Профилактика чумы заключается в регулярной вакцинации диких и
домашних животных [Фертиков, Егоров, 1997], ограничении контактов между
ними (недопущении домашних свиней в лес и отстреле зверей, посещающих
свиноводческие фермы) и тщательной утилизации трупов павших животных.
При вспышках болезни необходимо сократить численность (минимум до 20 осо-
бей на 1000 га леса [Depner и. а., 1997]) и выборочно отстреливать животных, от-
стающих от стада, не убегающих от опасности, истощенных, с шаткой походкой,
помятой щетиной, а также наладить сбор и утилизацию (сжигание) трупов. В не-
благополучных районах объявляется карантин [Мельникова, 1968; Горегляд,
1971; Комаров, Богацкий, 1980]. При отстреле не следует применять загонные
методы охоты и собак, что распугивает зверей и приводит к еще большему рас-
пространению болезни.
Профилактика гельминтозов заключается в ограничении контактов кабанов с
домашними свиньями и в организации выборочного отстрела поросят (кашляющих,
исхудавших, отстающих в росте, с задержанной линькой), а также в организации
обильной подкормки, включающей белковые корма, в сухих угодьях с целью отвле-
чения этих копытных от мест с обилием дождевых червей. В местах концентрации
зверей желательны посевы кормового люпина и других растений с антигельминт-
ными свойствами [Мельникова, 1968; Иванова, 1970; Рухлядев, 1970].
При высокой плотности населения диких свиней и угрозе вспышек гельмин-
тозов рекомендованы:
- массовая дегельминтизация;
- ассенизация (дезинфекция) подкормочных площадок после схода снега
(очистка и сжигание мусора, подсыпка свежего грунта, а также обработка не-
гашеной известью из расчета 2 кг на 1 м“);
249
- сокращение численности промежуточных хозяев гельминтов путем приме-
нения механических либо химических средств (что весьма сложно сделать и
обычно не делается. -А.Д.у,
- плановая дегельминтизация охотничьих собак;
- ограничение численности основных паразитоносителей - хищных зверей.
Дегельминтизация сеголеток проводится дважды в году (в мае - июле и сен-
тябре — ноябре) на подкормочных площадках высокоэффективным препаратом
нилвермом или его аналогами: тетрамизолом, тибензолом, мебендазолом (а так-
же панакуром или фенбендазолом) в дозе 10—15 мг на 1 кг массы животных, ко-
торая летом составляет около 8-10 кг, осенью - около 30 кг. Расчетное потребле-
ние корма поросятами: 0,3 кг весной и 1 кг - осенью. При дозе нилверма более 50
мг/кг возможно отравление препаратом. Химиотерапию желательно проводить в
специальных загонах с дощатым настилом, на который высыпают зерна кукуру-
зы или ячменя, пропитанные нилвермом (из расчета 10-12 кг зерна на 1 л водно-
го раствора препарата). Для предотвращения проникновения взрослых особей
площадку огораживают жердями: нижнюю планку устанавливают на земле, меж-
ду ней и второй жердью промежуток должен быть не более 16-18 см летом и до
25 см - осенью. Через 20-30 мин после употребления лечебного корма детеныши
начинают усиленно кашлять, освобождаясь от паразитов [Назарова и др., 1974,
1982; Литвинов, 1975, 19806; Шостак, Василюк, 1975; Иванова, Овсюкова, 1976;
Стародынова, 1979; Стародынова, Сыроежкин, 1979; Говорка и др., 1988; Пень-
кевич, 1998а, б; Фертиков и др., 1999]. При высокой зараженности необходима
дегельминтизация всего или почти всего поголовья на подкормочных площадках
из расчета 0,01 г препарата на 1 кг массы животного [Овсюкова, 1975].
Профилактика борьбы с трихинеллезом заключается главным образом в со-
кращении численности распространителей гельминтозов: волка, лисицы, еното-
видной собаки, бродячих собак и кошек и утилизации трупов. В целях предот-
вращения заболевания человека обязателен ветеринарный осмотр и трихинелло-
скопия туш добытых кабанов и сжигание их в случае обнаружения трихинелл.
Отношение к крупным хищникам. Крупные хищники, как и другие виды
животных, безусловно, имеют право на существование и защиту. Однако в охот-
ничьем хозяйстве их поголовье, особенно волка, должно жестко регулироваться.
Замечу, что плодовитость этого хищника близка к таковой у кабана - он в со-
стоянии ежегодно увеличивать свою численность почти в два раза [Смирнов,
1992], что требует особенно пристального контроля его популяций. Селективная
и «санитарная» роль волка в отношении диких и домашних копытных весьма
проблематична [Данилкин, 1999а], а вред очевиден и огромен.
Тигр, несомненно, нуждается в жесткой охране, однако подчинять все охот-
ничье хозяйство Дальнего Востока служению одному виду, на мой взгляд, не со-
всем разумно. Беспредельное наращивание его численности при ограниченных
кормовых ресурсах рано или поздно может закончиться катастрофой для охра-
няемого вида. В особенно многоснежные и неурожайные годы массовая гибель
диких копытных (основного корма) при современном иррациональном ведении
охотничьего хозяйства неминуема. Для сохранения тигра было бы полезнее ог-
раничить его численность в рамках, определенных учеными с учетом естествен-
ной кормовой емкости среды обитания, и направить дальнейшие усилия на соз-
дание устойчивой кормовой базы - увеличение поголовья копытных, и кабана в
частности.
Бродячих собак в охотничьих угодьях, безусловно, быть не должно.
Учет численности. Учет поголовья — одна из обязательных составляющих
управления популяциями.
Основной метод учета кабана и других диких копытных в России - зимний
маршрутный. Однако он проводится в самом конце зимы - начале весны в мно-
250
госнежье, когда активность зверей минимальна, а переходы нередко измеряются
сотнями метров. По этой причине следы животных часто не регистрируются.
В большинстве охотничьих хозяйств зверей с гораздо большей точностью
можно подсчитать на подкормке, добавляя определенное число особей, которые
не посещают подкормочные площадки.
Охотничьи «фермы» («ранчо»). Для охотничьих целей во многих странах
издавна практикуется разведение копытных в загонах. Их добыча внутри ограды
далека от спортивной и порой смахивает на узаконенное убийство - есть люби-
тели, которые с вышки отстреливают за один-два дня до сотни голов. Однако
«ранчо» приносят неплохой доход их владельцам, а спрос на охоту в них растет
не только за рубежом, но и в России. Они необходимы и для разведения диких
животных в целях последующего их выпуска в окрестные угодья.
В вольеры завозят только здоровых зверей после тщательного ветеринарного
контроля. Перевозку и передержку лучше всего переносят одно-двухлетние осо-
би. Однако наличие взрослых животных весьма желательно, поскольку они легче
адаптируются к новым условиям и быстрее формируют группы. При вселении
новой партии между взрослыми особями возникает борьба, иногда заканчиваю-
щаяся смертью притесняемых низкоранговых чужаков.
Величина поголовья зависит от площади загона и возможности круглогодич-
ного обеспечения животных кормами. Желательно, чтобы на одного зверя при-
ходилось не менее 1 га [Hell, 1986]. Чем больше территория, тем больше копыт-
ных она может вместить, и тем более естественными будут казаться условия
«охоты», что тоже немаловажно.
Типичные размеры «ранчо» - от 30 до 3000 га. Они обнесены чаще 1,5-2-
метровым сетчатым забором (толстостенным снизу при содержании кабанов), обыч-
но без заглубления сетки в землю. Внутри загона обязательно должен находиться
источник воды, желательно незамерзающий, густой кустарник или густой ельник,
кормовые поля пшеницы, овса, люцерны, топинамбура и других культур. Часть по-
лей огораживают и стравливают по мере необходимости. Здесь же оборудуют под-
кормочные площадки, закладывают силосные ямы и сооружают убежища.
В качестве убежищ для свиней желательно построить из жердей навесы раз-
мером 3 м на 8-10 м, одна из сторон которых отстоит от земли на 0,5 м, другая -
на 1,5 м. Жерди плотно накрывают еловым лапником, под навес укладывают со-
лому или сено [Ильинский, 1966]. Искусственные лежки делают также под гус-
тыми еловыми ветвями или в плотном кустарнике в виде площадки из елового
лапника размером около 150x150 см и высотой около 50 см. Рядом желательно
выкладывать нарубленный еловый лапник, который животные укладывают в
гнезда по своему усмотрению. Через три года эти лежки лучше сжигать, устраи-
вая новые [Варнаков, 1995]. По моему опыту, лучшими зимними убежищами
обычно становятся стожки необмолоченной (или обмолоченной) пшеницы, овса,
гороха и люцерны, завезенные в типичные места отдыха зверей. Они одновре-
менно служат и кормовыми пунктами.
Одно из основных условий успешной работы с животными в вольерах - ре-
гулярные ветеринарно-профилактические мероприятия.
В больших по площади вольерах возможно отстреливать кабанов не только с
вышек, но и с подхода, и даже с собакой. В этом случае, при наличии нескольких
секачей в вольере и мелькающих повсюду зверей, острота охотничьих ощущений
вполне сравнима с таковыми при охоте в естественных условиях.
Кабаньи, косулиные или многовидовые (сибирская косуля, благородный или
пятнистый олени, лань и небольшое число диких свиней) «ранчо», на мой взгляд,
- один из наиболее перспективных путей развития отечественного охотничьего и
фермерского хозяйства. В загоне площадью около 1000 га при обильном кормле-
нии можно содержать репродуктивное стадо примерно в 100-300 голов, ежегод-
251
но отстреливая несколько сотен сеголеток и несколько взрослых самцов на тро-
фей. Добыча только в одном «ранчо» может быть не меньше, чем в настоящее
время в опустошенных охотничьих угодьях многих областей России.
Заключение
Анализ представленного выше материала не оставляет даже малейших со-
мнений в том, что в последние столетия динамика ареала и населения кабана, как
и других диких копытных животных, в большей степени определялась антропо-
генными факторами, нежели природными. Именно человек разумный уничтожил
многие виды крупных млекопитающих и резко сократил численность оставшихся
в живых, и он же, благодаря охране и развитию охотничьего хозяйства, восста-
новил и даже частично увеличил их ресурсы в XX в., что особенно характерно
для стран Западной и Центральной Европы и Северной Америки.
Охотничья история России также изобилует примерами массового истребле-
ния зверей на огромных территориях, вплоть до их полного исчезновения и пре-
вращения некогда богатейших угодий в пустынные. В начале XX в. известней-
ший зоолог С.А. Бутурлин [1924] вынужден был обратиться к своим современ-
никам со следующим призывом в «Настольной книжке охотника», изданной мас-
совым тиражом: «И в других странах звери и птицы сильно истреблялись, но во
многих из них давно взялись за ум и стали относиться к этим богатствам береж-
но, и они снова стали размножаться. И нам нужно твердо помнить, что не только
на наших внуков и детей, но и на наш век птицы и зверя уже не хватает. Надо
браться за ум и беречь их, если не хотим остаться не только без охоты, но и без
пушнины, без мяса, без кож. ... Главный враг всякого зверя и птицы, в том числе
и промысловых животных, - сам человек, и это надо твердо помнить».
Уроки истории, однако, остались невыученными. Отечественные охотоведы,
в большинстве своем, так и не «взялись за ум» и не научились грамотно работать
с популяциями диких копытных животных. Управление ими в третьем тысячеле-
тии нашей эры все так же находится в вопиющем противоречии с научным под-
ходом и здравым смыслом. При этом свои многочисленные ошибки и «провалы»
охотничьи чиновники весьма умело прикрывают рассуждениями о цикличности
динамики численности копытных, вызванной изменением солнечной активности,
температуры, влажности, трофическим и другими сравнительно малозначимыми
в настоящее время факторами.
Ресурсы диких копытных в России крайне скудны, сокращаются или же не
растут (за исключением северного оленя, на которого заметно уменьшился про-
мысловый и браконьерский пресс), но по-прежнему используются крайне нера-
ционально. Это во многом связано с неэффективным устройством отечественно-
го охотничьего хозяйства, многолетний «застой» которого превратился в затяж-
ной «кризис». При существующих методах и способах его ведения, при приме-
няемых методах планирования и «регулирования» численности копытных нельзя
не только достичь максимальной биологической продуктивности, но и сохранить
сами популяции!
О скудности ресурсов мы будем с горечью говорить до тех пор, пока не будет
изменено охотничье законодательство и реформировано управление охотничьим
хозяйством, пока не будут решены элементарные проблемы охраны, подкормки,
оптимизации поголовья и половозрастной структуры популяций, нормирования
добычи, селекции копытных и регулирования численности волка [Данилкин,
19966, 19996, 2000а].
252
Наукой разработаны и мировой практикой успешно апробированы иные
принципы и методы управления популяциями диких копытных. Эффект от вне-
дрения научных разработок порой превосходит ожидаемый, что видно на приме-
ре западноевропейских стран, где продуктивность угодий многократно выше,
чем у нас.
Потенциальная численность диких копытных животных в России может быть
почти на порядок выше существующей [Шварц, Михеева, 1976; Данилкин,
1999а]. При разумном подходе их ресурсы и добычу в ближайшие десятилетия
реально увеличить в несколько раз, что позволило бы превратить охотничье хо-
зяйство в высокоэффективную отрасль производства белковой продукции. Это
особенно важно в аспекте продовольственной безопасности страны, большая
часть сельскохозяйственных угодий которой находится в зоне «рискованного»
земледелия.
253
Литература
Ч
Абдуназаров Б.Б., Тарянников В.И. Волк в Западном Тянь-Шане // Экология, ох-
рана и акклиматизация млекопитающих в Узбекистане. Ташкент: Изд-во
ФАН, 1991. С. 42-48.
Абрамов К.Г. Копытные звери Дальнего Востока. Хабаровское кн. изд-во, 1954.128 с.
Абрамов К.Г. Копытные звери Дальнего Востока и охота на них. Владивосток:
Приморское кн. изд-во, 1963. 131 с.
Аверин Ю.В. Главнейшие изменения в видовом составе фауны млекопитающих и
птиц Молдавии в XVII-XX столетиях // Бюл. МОИП. Отд. биол. 1960а. Т. 65,
вып. 2. С. 5-12.
Аверин Ю.В. Некоторые изменения в составе охотничье-промысловой фауны Молда-
вии за последние столетия // Охрана природы Молдавии. 19606. Вып. 1. С. 125-132.
Адлерберг Г.П. Предварительный обзор русских и монгольских кабанов // Докл.
АН СССР. 1930. № 4. С. 91-96.
Адлерберг Г.П. К вопросу о происхождении домашних свиней // Проблема про-
исхождения домашних животных. Л.: Изд-во АН СССР, 1933. С. 185-209. (Тр.
лаборатории генетики АН СССР. Вып. 1).
Азаров В.И. Редкие животные Тюменской области и их охрана. Амфибии, реп-
тилии, птицы, млекопитающие. Тюмень: Вектор Бук, 1996. 238 с.
Азаров В.И., Деков В.М. Дикие копытные Тюменской области и задачи их ра-
циональной эксплуатации // Ресурсы животного мира Сибири. Новосибирск:
Наука, 1990. С. 181-184.
Азаров В.И., Климов Ю.П. Кабан в Тюменской области // Там же. 1990. С. 187—
189.
Айзин Б.М. Ареалы копытных Киргизии // Копытные фауны СССР. М.: Наука,
1975. С. 32-33.
Айзин Б.М. Охрана и использование диких копытных Киргизии // Копытные
фауны СССР. М.: Наука, 1980. С. 58-59.
Алекперов Х.М., Кулиев С.М. Некоторые данные по экологии кабана и косули в
Куба-Хачмасской зоне Азербайджана // Экология и охрана горных видов мле-
копитающих. М., 1987. С. 8-9.
Алексашина КС. Остатки фауны из некоторых городищ Верхнего Поволжья //
Материалы и исследования по археологии СССР. 1950. № 13. С. 148-151.
Алексеева Л.И. Териофауна раннего антропогена Восточной Европы. М.: Наука,
1977. 214 с. (Тр. геол, ин-та. Вып. 300).
Алексеева Л.И. Фаунистический комплекс млекопитающих времени последнего
межледниковья Русской равнины // III съезд Всесоюз. териол. о-ва. М., 1982.
Т. 1. С. 7-8.
Алексеева Л.И. Териофауна верхнего плейстоцена Восточной Европы (крупные
млекопитающие). М.: Наука, 1990. 109 с. (Тр. Геол, ин-та. Вып. 455).
Алексеева Э.В. Млекопитающие плейстоцена юго-востока Западной Сибири. М.:
Наука, 1980. 187 с.
Александрович Н.П. Охотничья териофауна из раскопок средневекового Турова
// Биологическое разнообразие национального парка «Припятский». Туров-
Мозырь, 1999. С. 229-231.
Алиев С.Д. Фауна Азыхской пещеры - объект охоты первобытного человека // II
съезд Всесоюз. териол. о-ва. М.: Наука, 1978. С. 3-4.
254
Алмэшан X., Скэрлатеску Дж., Нестеров В., Манолаке Л. О режиме питания
волка (Canis lupus) в Румынских Карпатах // Тр. IX Междунар. контр, биоло-
гов-охотоведов. М., 1970. С. 529.
Алъбицкий Ф.А., Пчелкин В.М. Растительный и животный мир области // Ивановская
промышленная область. М.-Иваново-Вознесенск: Госиздат, 1931. Вып. 2. 48 с.
Андреева Е.Г. Изучение костных остатков животных из раскопок пронской экс-
педиции // Основные проблемы териологии. М.: Наука, 1972. С. 82-96. (Тр.
МОИП. Отд. биол. Т. 48).
Андреева Е.Г., Петренко А.Г. Древние млекопитающие по археозоологическим
материалам Среднего Поволжья и Верхнего Прикамья // Из археологии Вол-
го-Камья. Казань, 1976. С. 137-189.
Андреенков В.И. Условия обитания копытных в районах интенсивного пастбищ-
ного животноводства Киргизии // Копытные фауны СССР. М.: Наука, 1980. С.
59-61.
Анненков Б.П. Кабан в Алакульской котловине // Экология, морфология, исполь-
зование и охрана диких копытных. М., 1989. Ч. 2. С. 191.
Аношин Р.М. Кабан и боровая дичь: к проблеме совместного обитания // Вопро-
сы интенсификации охотничьего хозяйства. М., 1988. С. 170-177.
Антипин В.М. Млекопитающие Казахстана. Копытные. Алма-Ата: Казгосиздат,
1941. Т.З. 107 с.
Антипина Е.Е. Антропогенное воздействие как фактор формирования современ-
ной териофауны Крыма // III съезд Всесоюз. териол. о-ва. М., 1982. Т. 1. С.
93-94.
Антипина Е.Е., Маслов С.П. Фауна палеолитической стоянки Шатрище-П И V
съезд Всесоюз. териол. о-ва АН СССР. М., 1990. Т. 1. С. 4-5.
Антонец Н.В. Некоторые особенности экологии кабана хоперской популяции //
Влияние антропогенной трансформации ландшафта на население наземных
позвоночных животных. М., 1987. Ч. 1. С. 174-176.
Антонец Н.В. Об особенностях параметров дикого кабана Хоперского заповед-
ника // Материалы VI совещания «Вид и его продуктивность в ареале». СПб.:
Гидрометеоиздат, 1993. С. 35-37.
Антонец Н.В. Особенности влияния дикого кабана на эфемероиды в дубравах степ-
ной и лесостепной зон //Заповщ. справа в Украшь 1997. Т. 3, № 1. С. 55-57.
Антонов А.Л. Состояние популяций копытных кедрово-широколиственных лесов
Приамурья при антропогенном воздействии: Автореф. дис. ... канд. биол. наук.
Владивосток, 1999. 23 с.
Анучин А.В. Видообразование и систематика (к происхождению диких свиней) //
Тр. зоол. сектора Грузинского фил. АН СССР. 1938. Т. 2. С. 5-93.
Арабули А.Б. Распространение и численность некоторых охотничье-
промысловых млекопитающих в Восточной Грузии // Зоол. журн. 1970. Т. 49,
вып. 3. С. 418^121.
Арабули А.Б. Современное состояние редких видов копытных Грузии // Эколо-
гия, морфология, использование и охрана диких копытных. М., 1989. Ч. ГС.
28-29.
Аренс Л.Е. О влиянии заповедности на охотничье-промысловую фауну Тебер-
динского заповедника и прилегающих районов // Тр. Тебердинского гос. запо-
ведника. 1957. Т. 1. С. 113-140.
Арамов В., Вашетко Э.В. Туркестанская рысь в Гиссарском заповеднике И VI
съезд териол. о-ва. М., 1999. С. 13.
Арсеньев В.К. Китайцы в Уссурийском крае // Зап. Приамурского отд. рус. геогр.
о-ва. 1914. Т. 10, вып. 1. 203 с.
Атанасов П. Състояние на едрия дивеч в Смядовското горско стопанство // Гор-
ско стопанство. 1981. Т. 37, № 11. С. 46-47.
255
Бабинок В.В., Романов В.С., Мухуров Л.И. Экологические связи копытных в лес-
ной экосистеме // Экология, морфология, использование и охрана диких копыт-
ных. М., 1989. Ч. 1.С. 5-6.
Бабкин Г. Банк биологических данных дикого кабана для селекции и гибридиза-
ции в свиноводстве Среднего Поволжья // Свиноводство. 1997. № 4. С. 18-20.
Бавин В.Г. Идентификация кариотипа диких и домашних свиней методом фазо-
во-контрастной микроскопии // Генетические и биохимические основы повы-
шения воспроизводительной способности сельскохозяйственных животных.
Л., 1979. С. 59-66. (Тр. ВНИИ разведения и генетики с/х. животных. № 27).
Бадер О.Н. Новые данные по палеолиту горного Крыма // Бюл. Комиссии по
изучению четвертичного периода. 1940. №6-7. С. 85-88.
Байгушева В.С. Ископаемая териофауна Ливенцовского карьера (Северо-
Восточное Приазовье) // Материалы по фаунам антропогена СССР. Л.: Наука,
1971. С. 5-29. (Тр. Зоол. ин-та. Т. 49).
Байдавлетов Р.Ж. Распределение и численность копытных в Северо-Восточном
Забайкалье //Копытные фауны СССР. М.: Наука, 1980. С. 63-64.
Бакеев Ю.Н., Костоглод В.Е. Причины и особенности вспышки численности ка-
бана в Краснодарском крае в 1973-1974 гг. // Копытные фауны СССР. М.:
Наука, 1975. С. 37-38.
Балъчяускас Л.П. Пространственное распределение копытных в антропогенном
ландшафте Литвы: Автореф. дис. ... канд. биол. наук. М., 1988. 23 с.
Балъчяускас Л.П., Таугинас ИС. Совместное использование зимних троп различны-
ми видами копытных//Прикладная этология. М.: Наука, 1983. С. 42-43.
Балюк С.С., Вакула В.А., Буневич А.Н. Динамика численности и стадность каба-
нов Беловежской пущи // Заповедники Белоруссии. 1987. Вып. 11. С. 99-108.
Банников А.Г. Млекопитающие Монгольской Народной Республики. М.: Изд-во
АН СССР, 1954. 669 с.
Банников А.Г. Проблема охраны охотничьих и редких животных // Охотоведе-
ние. М.: Лесн. пром-сть, 1974. С. 295-304.
Барабаш-Никифоров И.И. Звери юго-восточной части черноземного центра. Во-
ронежское кн. изд-во, 1957. 370 с.
Баранов П.В. Акклиматизация кабана в Западной Сибири // V съезд Всесоюз. те-
риол. о-ва АН СССР. М., 1990. Т. 3. С. 75-76.
Баранов П.В., Иванова В.А. Крупные хищники и копытные звери Сохондинского
заповедника // Экологические исследования в заповедниках Южной Сибири.
М., 1989. С. 39-50.
Барышников Г.Ф., Гарутт В.Е., Громов И.М. и др. Каталог млекопитающих
СССР (плиоцен - современность). Л.: Наука, 1981. 456 с.
Баскин Л.М. Поведение копытных животных. М.: Наука, 1976. 295 с.
Батыров Б.Х. Плейстоценовые копытные Узбекистана // IV съезд Всесоюз. те-
риол. о-ва. М., 1986. Т. 1. С. 8-9.
Бекенов А.Б. Ресурсы охотничье-промысловых зверей в Казахстане и их исполь-
зование // VI съезд териол. о-ва. М., 1999. С. 23.
Бекенов А.Б., Байдавлетов Р.Ж., Кыдырбаев Х.К. Копытные Казахстана: ресур-
сы и перспективы использования // Экология, морфология, использование и
охрана диких копытных. М., 1989. Ч. 1. С. 31-32.
Белов С. Редкий случай // Охота и охотничье хоз-во. 1972. № 12. С. 44-45.
Белоносов В. Мясо кабана // Охота и охотничье хоз-во. 1965. № 12. С. 38.
Белопольская М.М. Паразитофауна кабана (Sus scrofa continentalis) Приморья И
Тр. Ленинградского о-ва естествоиспыт. 1952. Т. 21, вып. 4. С. 3-9.
Беляева Е.И. О фауне млекопитающих из палеолитической пещеры Вирхова
(Сакажия) // Бюл. Комиссии по изучению четвертичного периода. 1940. № 6-
7. С.105-107.
256
Бенъковская И.Л., Беньковский Л.М. Некоторые особенности популяций копыт-
ных Восточного Предсаянья // Экология, морфология, использование и охрана
диких копытных. М., 1989. Ч. 1. С. 35—36.
Беньковский Л.М., Мартынов В.А., Бенъковская ИЛ. Есаулов Н.П. Некоторые во-
просы интродукции копытных на Сахалине // Там же. М., 1989. Ч. 1. С. 33-34.
Бибиков Д.И., Губарь Ю.П., Филимонов А.Н. Волк в СССР: состояние и управле-
ние популяциями // Крупные хищники. М., 1992. С. 14-23.
Бибикова В.И. Фауна раннетрипольского поселения Лука-Врублевецкая // Мате-
риалы и исследования по археологии. 1953. № 38. С. 411-458.
Бибикова В.И. О распространении дикого кабана в четвертичном периоде // Бюл.
Комиссии по изучению четвертичного периода. I960. № 25. С. 107-112.
Бибикова В.И. Из истории голоценовой фауны позвоночных в Восточной Европе
// Природная обстановка и фауны прошлого. Киев: Изд-во АН УкрССР, 1963.
Вып. 1. С. 119-146.
Бибикова В.И. О смене некоторых компонентов фауны копытных на Украине в
голоцене // Бюл. МОИП. Отд. биол. 1975. Т. 80, вып. 6. С. 67-72.
Бионкур А.А. О состоянии ружейного дела в России // Природа и охота. М., 1901.
Кн. 8. С. 10-16.
Бируля А.А. Предварительное сообщение о хищниках (Carnivora) из четвертич-
ных отложений Крыма//Докл. АН СССР. 1930. № 6. С. 139-144.
Благовещенский В. Какому лесу нужен кабан? // Охота и охотничье хоз-во. 1977. №
5. С. 10-11.
Благосклонов К.Н., Иноземцев А.А., Тихомиров В.Н. Охрана природы. М.: Высш,
шк., 1967. 442 с.
Блузма П. Состояние охотничьей териофауны Литвы: динамика, использование
и охрана популяций // VI съезд териол. о-ва. М., 1999. С. 27.
Блюм А. На кабанов по чернотропу // Охота и охотничье хоз-во. 1995. № 9. С.
14-17.
Бобринский Н.А., Кузнецов Б.А., Кузякин А.И. Определитель млекопитающих
СССР. М.: Сов. наука, 1944. 440 с.; М.: Просвещение, 1965. 382 с.
Бобырь Г.Я. Распределение и численность кабана в Тебердинском заповеднике //
Копытные фауны СССР. М.: Наука, 1975. С. 65-66.
Бобырь Г.Я. Взаимоотношения бурого медведя с копытными в горах Северного
Кавказа // Копытные фауны СССР. М.: Наука, 1980. С. 126-128.
Богданов М. Птицы и звери черноземной полосы Поволжья и долины Средней и
Нижней Волги // Тр. о-ва естествоиспыт. при Казанском ун-те. 1871. Т. 1, отд.
1. С. 3-226.
Болденков С.В., Крайнев Е.Д. Динамика численности и использование поголовья
кабана, косули и лося в Украинской ССР // Копытные фауны СССР. М.: Нау-
ка, 1975.С. 213-214.
Болденков С.В., Крайнев Е.Д. Парнокопытные фауны Украины // Копытные фау-
ны СССР. М.: Наука, 1980. С. 75-76.
Бондарев А.Я. Избирательная добыча волком домашних копытных животных //
Экология, поведение и управление популяциями волка. М., 1989. С. 24-33.
Борискин ГФ. Акклиматизация копытных на Среднем Урале // Обогащение
фауны и разведение охотничьих животных. Киров, 1982. С. 67.
Боровик А.А. Влияние хищников и копытных на популяцию глухаря в период
размножения // Беловежская пуща. 1973. Вып. 7. С. 196-201.
Борщевский В.Г. К проблеме использования ресурсов глухарей в РСФСР // Тете-
ревиные птицы в заповедниках РСФСР. М., 1989. С. 5-20.
Браунер А.А. Сельскохозяйственная зоология. Киев: Укриздат, 1923. 436 с.
Брем А., Россмесслер Е. Лесные животные. Звери, птицы и пресмыкающиеся. С.-П.,
1867.790 с.
17. Заказ №2528.
257
Бридерманн Л. Значение кабана (Sus scrofa) для повышения продуктивности
охотничьего хозяйства // Тр. IX Междунар. конгр. биологов-охотоведов. М.,
1970. С. 810-811.
Бромлей ГФ. Уссурийский кабан. М.: Наука, 1964. 107 с.
Бромлей Г.Ф., Кучеренко С.П. Копытные юга Дальнего Востока СССР. М.: Нау-
ка, 1983. 305 с.
Брюсов А.Я. Предки народов Зауралья // По следам древних культур. От Волги
до Тихого океана. М.: Гос. изд-во культ.-просвет. лит-ры, 1954. С. 131-162.
Булахов В.Л. Влияние роющей деятельности кабана на физико-химические и био-
геоценотические свойства почв лесных биогеоценозов И Копытные фауны СССР.
М.: Наука, 1975. С. 159-161.
Буневич А.Н. Влияние волка на поголовье копытных в Беловежской пуще // IV
съезд Всесоюз. териол. о-ва. М., 1986. Т. 2. С. 40—41.
Буневич А.Н. Динамика численности и питание волка в Беловежской пуще // За-
поведники Белоруссии. 1988. Вып. 12. С. 108-113.
Бурчак-Абрамович Н.И., Джафаров Р.Д. Остатки дикого кабана из бинагадин-
ских Кировых отложений (Апшерон) // Тр. Ест.-ист. музея АН АзССР. 1948.
Вып. 1-2. С. 76-136.
Бутарин Н.С., Еськов П.А., Милецкий Д.М., Багровская Н.Н. Отдаленная гибриди-
зация диких свиней с домашними. Алма-Ата: Изд-во АН КазССР, 1955. 113 с.
Бутурлин С.А. Настольная книжка охотника. Свердловск: Издание Уральского
обл. к.-п. союза охоты, 1924. 256 с.
Вадковский В. Б. К управлению популяциями копытных в Белоруссии путем
промысла // IV съезд Всесоюз. териол. о-ва. М., 1986. Т. 2. С. 243-244.
Вадковский В.Б. Охотничье хозяйство Белоруссии: стратегия, тактика // Научно-
технический прогресс - в практику перестройки охотничьего хозяйства. М.,
1988. С. 3-6.
Вадковский В.Б., Костин В.А. Пути улучшения эксплуатации популяций копыт-
ных в Белоруссии // Пути и методы рациональной эксплуатации и повышения
продуктивности охотничьих угодий. М., 1978. С. 175-177.
Валдманн XX. Стадность кабана и использование ее для определения численно-
сти популяций // V съезд Всесоюз. териол. о-ва АН СССР. М., 1990. Т. 3. С.
77-78.
Валъкович В.М. Повреждение сосны копытными животными в Саратовской об-
ласти // Роль крупных хищников и копытных в биоценозах заповедников. М.,
1986.С. 138-141.
Вангейгем Э.А. Палеонтологическое обоснование стратиграфии антропогена Се-
верной Азии (по млекопитающим). М.: Наука, 1977. 172 с.
Варнаков А.П. К вопросу о продвижении кабана на север // Пути и методы ра-
циональной эксплуатации и повышения продуктивности охотничьих угодий.
М., 1978а. С. 7-9.
Варнаков А. Кабан в Вологодской области // Охота и охотничье хоз-во. 19786. №
ЕС. 4-5.
Варнаков А.П. Опыт селекционного отстрела кабана // Управление популяциями
диких копытных животных. М., 1985. С. 122-133.
Варнаков А.П. Питание кабана и его роль в биоценозах Молого-Шекснинского
междуречья // Роль крупных хищников и копытных в биоценозах заповедни-
ков. М., 1986. С. 86-94.
Варнаков А.П. Об изменении сроков охоты на кабана // Научно-технический про-
гресс - в практику перестройки охотничьего хозяйства. М., 1988а. С. 100-102.
Варнаков А.П. Добыча кабана переносными сетевыми ловушками // Там же. М.,
19886. С. 171-174.
Варнаков А. Лежки для кабанов // Охота и охотничье хоз-во. 1995. № 8. С. 3.
258
Варнаков А. Летняя охота на кабана// Там же. 1997. № 8. С. 12-13.
Варнаков А.П., Михайлова Е.П. Метастронгилез кабана в Саратовской области //
III съезд Всесоюз. териол. о-ва. М., 1982. Т. 1. С. 166-167.
Варнаков А.П., Млодиков Н.Ф. О зимней подкормке кабана в центральных об-
ластях европейской части РСФСР // Научно-технический прогресс - в практи-
ку перестройки охотничьего хозяйства. М., 1988. С. 199-202.
Варнаков А.П., Мошева Т.С. Роль кабана в сельском хозяйстве Московской об-
ласти // Копытные фауны СССР. М.: Наука, 1975. С. 161-162.
Варов А.И., Косинцев П.А. Крупные млекопитающие Приуралья в позднем голо-
цене // Материалы и исследования по истории современной фауны Урала.
Екатеринбург: Изд-во «Екатеринбург», 1996. С. 110-117.
Васильев А.Л., Воронин А.А. Волк в Калужской области // Экологические основы
охраны и рационального использования хищных млекопитающих. М.: Наука,
1979. С.93-94.
Василька М. Кариотипическое исследование при разведении домашних свиней и
их гибридов с диким кабаном в норме и при некоторых наследственных забо-
леваниях: Автореф. дис. ... канд. биол. наук. М., 1984. 16 с.
Ватолин Б.А. О популяции волка в Брянской области и регулировании его чис-
ленности // Экологические основы охраны и рационального использования
хищных млекопитающих. М.: Наука, 1979. С. 91-93.
Ватолин БА. Размещение, численность и влияние экологических факторов на каба-
на Брянской области //Копытные фауны СССР. М.: Наука, 1980. С. 131-133.
Верещагин Н.К. Остатки млекопитающих из нижнечетвертичных отложений
Таманского полуострова // Тр. Зоол. ин-та АН СССР. 1957. Т. 22. С. 9-74.
Верещагин Н.К. Млекопитающие Кавказа. М.-Л.: Изд-во АН СССР, 1959. 704 с.
Верещагин Н.К. Основные черты формирования териофаун Голарктики в антро-
погене//Зоол. журн. 1963. Т. 42, вып. И. С. 1686-1698.
Верещагин Н.К Охоты первобытного человека и вымирание плейстоценовых
млекопитающих в СССР // Материалы по фаунам антропогена СССР. Л.: Нау-
ка, 1971. С. 200-232. (Тр. Зоол. ин-та АН СССР. Т. 49).
Верещагин Н. Охота наших предков // Охота и охотничье хоз-во. 1975. № 2. С.
39-41.
Верещагин Н.К, БарышниковГ.Ф. Ареалы копытных фауны СССР в антропогене //
Млекопитающие Восточной Европы в антропогене. Л., 1980а. С. 3-20. (Тр. Зоол.
ин-та АН СССР. Т. 93).
Верещагин Н.К, Барышников Г.Ф. Млекопитающие предгорного Северного
Крыма в эпоху палеолита // Там же. 19806. С. 26—49.
Верещагин Н.К, Батыров Б. Фрагменты истории териофауны Средней Азии //
Бюл. МОИП. Отд. биол. 1967. Т. 72, вып. 4. С. 104-115.
Верещагин Н.К, Громов И.М. Формирование териофауны СССР в четвертичном
периоде // Успехи современной териологии. М.: Наука, 1977. С. 32-М5.
Верещагин Н.К, Громов И.М., Ермолова Н.М., Паавер К.А. Основные черты
формирования териокомплексов Северной Евразии в голоцене // История био-
геоценозов СССР в голоцене. М.: Наука, 1976. С. 101-116.
Верещагин Н.К, Русаков О.С. Копытные северо-запада СССР. Л.: Наука, 1979.
309 с.
Вершинин А.А. Расселение охотничьих животных в Подмосковье // Вопр. повы-
шения продуктивности охотничьих угодий. М.: Колос, 1969. С. 50-98.
Вехник В.П., Саксонов С.В. Роющая деятельность кабана в условиях Жигулев-
ского заповедника // Влияние антропогенной трансформации ландшафта на
население наземных позвоночных животных. М., 1987. Ч. ЕС. 176-178.
17
259
Вехник В.П., Саксонов С.В. Опыт оценки воздействия кабана на фитоценозы ка-
менистой степи Жигулевского заповедника // Экология, морфология, исполь-
зование и охрана диких копытных. М., 1989. Ч. 2. С. 192-193.
Викторов К.Р. Физиология домашних животных. М.: Сельхозгиз, 1939. 518 с.
Висящее Г. Итоги 1978 года // Охота и охотничье хоз-во. 1979. № 12. С. 4-5.
Вишневский Ю.Н. Влияние волка на диких копытных Дарвинского заповедника
// Проблемы охраны генофонда и управления экосистемами в заповедниках
лесной зоны. М., 1986. Ч. 2. С. 44-46.
Войлочников А.Т. Ресурсы промысловых животных, их размещение и использо-
вание в лесных угодьях Амуро-Уссурийского края // Зоологические проблемы
Сибири. Новосибирск: Наука, 1972. С. 366-368.
Воложенинов Н.Н. К некоторым вопросам экологии кабана на Арал-Пайгамбаре
// Узб. биол. журн. 1974. № 4. С. 45-46.
Волох А.М. Репродуктивный потенциал популяций дикого кабана // VI съезд те-
риол. о-ва. М., 1999. С. 53.
Вольф Р. Развитие на запасите на дивата свиня в Чехословакия // Дивечьт и
околната среда. София: Изд-во наОтеч. фронт, 1985. С. 51-57.
Воробьев Г. Кабаны Киргизии // Охота и охотничье хоз-во. 1968. № 10. С. 22.
Воронецкий НН, Дунин В.Ф., Одинцова Т.М., Парейко О.А. Волк в Полесском
радиационно-экологическом заповеднике //10 лет Полесскому гос. радиаци-
онно-экологическому заповеднику. Минск, 1998. С. 142-150.
Воронецкий Н.Н., Дунин В.Ф., Пискунов В.С., Киреенко К.М. Численность и био-
топическое распределение диких животных в Полесском радиационно-
экологическом заповеднике // Биологическое разнообразие национального
парка «Припятский». Туров-Мозырь, 1999. С. 312-315.
Воронецкий Н.Н., Одинцова Т.М., Парейко О.А. Радиоэкологический анализ
хищников в ПГРЭЗ // Структурно-функциональное состояние биологического
разнообразия животного мира Беларуси. Минск, 1999. С. 40-42.
Воронецкий Н.Н., Тышкевич В.Е. Копытные (зубр, кабан, косуля, благородный
олень, лось) и хищники (волк, рысь, лисица) Полесского ГРЭЗ И 10 лет По-
лесскому гос. радиационно-экологическому заповеднику. Минск, 1998. С.
151-159.
Воронин А.А. Адаптация кабана к условиям Южного Подмосковья // Копытные
фауны СССР. М.: Наука, 1975а. С. 72-73.
Воронин А.А. Структура популяции кабана по результатам наблюдений и про-
мысла // Охрана природы и совершенствование биогеоценозов. Тула, 19756.
Вып. 3. С. 49-52.
Воронин А.А. Суточная активность кабана на юге Нечерноземья // Копытные
фауны СССР. М.: Наука, 1980. С. 285-287.
Воронин А.А., Заикин С.М. Анализ промысловых выборок кабана в Калужской
области // Экология, морфология, использование и охрана диких копытных.
М„ 1989. Ч. 2. С. 194-195.
Воронин А.А., Заикин С.М. Стратегия промысла копытных // V съезд Всесоюз.
териол. о-ва АН СССР. М„ 1990. Т. 3. С. 81-82.
Воронцов Я.Н. Акклиматизация промысловых зверей в Тюменской области //
Обогащение фауны и разведение охотничьих животных. Киров, 1982. С.74.
Врублевский К.И. Чума диких кабанов//. Архив ветеринарных наук. 1908. Кн. 10.
С. 943-959.
В.П. Охрана природы. Лоси // Охота и природа. 1928. № 5. С. 12.
Гаврин В.Ф. Акклиматизация и реакклиматизация охотничьих животных в СССР
в 1963-1973 гг. // Акклиматизация и реакклиматизация животных. М.: Колос,
1973. С. 3-5.
260
Гаврин В.Ф., Донауров С.С. Волк в Беловежской пуще // Зоол. журн. 1954. Т. 33, №
4. С. 904-924.
Гасконский А., Паровщиков В. Кабан в Архангельских лесах // Охота и охотни-
чье хоз-во. 1957. № 4. С. 25.
Гатих В.С. Распространение и численность диких копытных на территории При-
пятского заповедника//Копытные фауны СССР. М.: Наука, 1975. С. 73-74.
Гатих В.С. Роль волка в биогеоценозах Белорусского Полесья // Экологические
основы охраны и рационального использования хищных млекопитающих. М.:
Наука, 1979. С. 94-95.
Генов П. Ролята на дивата свиня в горските биоценози и агроценозите на Смо-
лянски окръг // Науч. тр. Пловд. ун-та. Биол. 1986. Т. 24, № 1. С. 287-295.
Генов П.В., Коцаков Л. Динамика на числеността на дивата свиня (Sus scrofa at-
tila Thomas) в Югозападна България // Фауна Югозап. Българии. София, 1986.
Ч. 1. С. 44-60.
Гептнер В.Г. Динамика ареала некоторых копытных и антропокультурный фактор
(материалы к проблеме ареала) //Вопросы географии. 1960. Сб. 48. С. 24-54.
Гептнер В.Г, Насимович А.А., Банников А.Г. Млекопитающие Советского Сою-
за. Парнокопытные и непарнокопытные. М.: Высш, шк., 1961. Т. 1. 776 с.
Гинеев А.М. Численность и распространение копытных в плавнях Дагестана И
Копытные фауны СССР. М.: Наука, 1975а. С. 164-165.
Гинеев А.М. Промысловое освоение и охрана запасов кабана в Краснодарском
крае //Там же. 19756. С. 219-220.
Гинеев А.М. Динамика численности копытных в плавнях Терека // Экология, мор-
фология, использование и охрана диких копытных. М., 1989. Ч. 1. С. 40-42.
Глушков В.М. К обоснованию стратегии управления популяциями диких копыт-
ных животных // Научно технический прогресс - в практику перестройки
охотничьего хозяйства. М., 1988. С. 108-110.
Глушков В.М., Байбиков Е.В. К вопросу об управлении популяциями лося мето-
дом регулируемого отстрела (сообщение первое) // Современные проблемы
охотничьего хозяйства. М., 1998. С. 49-59.
Глушков В.М., Граков Н.Н. Теория динамики численности, концепция циклично-
сти и реальность идеи управления популяциями лося // Управление популя-
циями диких копытных животных. Киров, 1989. С. 14-35.
Глушков В.М., Пиминов В.Н., Пономарев В.П. Зимняя смертность и резервы про-
мысла диких копытных // Управление популяциями диких копытных живот-
ных. Киров, 1989. С. 81-92.
Глушков В.М., Пиминов В.Н., Синицын А.А. К вопросу о программе восстановле-
ния ресурсов диких копытных // Вопросы прикладной экологии (природо-
пользования), охотоведения и звероводства. Киров, 1997. С. 86-88.
Говорка Я., Маклакова Л.П., Митух Я. и др. Гельминты диких копытных Вос-
точной Европы. М.: Наука, 1988. 208 с.
Голгофская К.Ю., Кудактин А.Н., Бибиков ДИ. К проблеме изучения трофиче-
ских связей хищники-копытные-пастбища на Северо-Западном Кавказе И
Экологические основы охраны и рационального использования хищных мле-
копитающих. М.: Наука, 1979. С. 25-27.
Гордиюк НМ. Особенности взаимоотношений копытных и крупных хищников
Южного Урала: Автореф. дис. ... д-ра биол. наук. М., 1996. 48 с.
Горегляд Х.С. Болезни диких животных. Минск: Наука и техника, 1971. 302 с.
Горелов И.Г. Изучение полиморфизма групп крови диких кабанов сибирского
подвида// Сиб. вести, с.-х. науки. 1986. № 3. С. 107-110.
Горелов И.Г Иммуногенетические и биохимические особенности популяции ди-
ких кабанов Тувинской АССР // Исчезающие, редкие и слабо изученные рас-
261
тения и животные Алтайского края и проблема их охраны. Барнаул, 1987. С.
55-59.
Горелов И.Г. Незаменимый объект исследований // Наука в Сибири. 1993. № 4.
С. 6-7.
Горелов ИГ Биология сибирского кабана. Новосибирск, 1994. 82 с.
Горелов И.Г., Савина М.А. Генетические маркеры как показатель адаптационных
способностей диких кабанов Сибири (Sus scrofa sibiricus) // Междунар. симп.
«Молекулярная генетика и биотехнология в оценке и изменении геномов
сельскохозяйственных животных». СПб.; Пушкин, 1994. С. 107-108.
Горелов Ю.К. Факторы, определяющие территориальные группировки копытных
Бадхыза // Популяционная структура вида у млекопитающих. М., 1970. С. 31-33.
Гортынский В.П. Экономическое значение охоты в Лужском уезде С.-
Петербургской губернии // Материалы к познанию русского охотничьего де-
ла. СПб., 1914. Вып. 3. С. 3-72.
Горшков П.К. Экология кабана в Татарии // Экология, охрана и воспроизводство
животных Среднего Поволжья. Казань, 1988. С. 126-133.
Горшков П.К. Экология и биоценотическая роль кабана в Татарии // Экология, мор-
фология, использование и охрана диких копытных. М., 1989. Ч. 2. С. 195-196.
Граков Н.Н. Значение охоты, сокращения и трансформации лесов в сокращении
численности охотничьих животных // Вопросы прикладной экологии (приро-
допользования), охотоведения и звероводства. Киров, 1997. С. 90-92.
Граков Н.Н., Сафонов В.Г. К истории промысла бобра на Руси // Там же. 1997. С.
92-93.
Графодатский А.С. Цитогенетические аспекты филогении Artiodactyla// Экология,
морфология, использование и охрана диких копытных. М., 1989. Ч. 1. С. 13.
Грачев ИМ., Галанцев В.П. Физиология лактации. Л.: Наука, 1973. 590 с.
Грачев Ю.А., Смирнова Э.Д. Вертикальное распространение и кочевки копытных
в Таласском Алатау И Копытные фауны СССР. М.: Наука, 1980. С. 82-83.
Гречушкин В.С., Харченко В.И., Молодан Г.Н., Мглинец С.М. Современное со-
стояние популяций копытных Донецкой области // Копытные фауны СССР.
М.: Наука, 1975а. С. 78-79.
Гречушкин В.С., Харченко В.И., Молодан Г.Н., Никаноров А.Г. Перспективы уве-
личения норм изъятия из популяций диких копытных Донбасса И Там же.
19756. С.237-238.
Григорьев Г.П. Заселение кабаном Кировской области И Акклиматизация охот-
ничьих животных в СССР. Минск: Ураджай, 1978. С. 132-134.
Громаков НА. Страницы истории охоты в Тамбовском крае. Тамбов, 1998. 180 с.
Громов В.И. Палеонтологическое и археологическое обоснование стратиграфии
континентальных отложений четвертичного периода на территории СССР
(млекопитающие, палеолит) // Тр. Ин-та геол. наук. 1948. Вып. 64. Геол, се-
рия, № 17. 521 с.
Громов И.М., Гуреев А.А., Новиков ГА. и др. Млекопитающие фауны СССР. М,-
Л.: Изд-во АН СССР, 1963. Ч. 2. С. 641-1101.
Громова В.И. Остатки млекопитающих из раннеславянских городищ вблизи г.
Воронежа И Материалы и исследования по археологии СССР. 1948. № 8. С.
113-123.
Губарь Ю.П. Копытные Архангельской области И Копытные фауны СССР. М.:
Наука, 1980. С. 84-85.
Гунчак Н.С. Морфометрическая характеристика кабана (Sus scrofa) Украинских
Карпат // Зоол. журн. 1978. Т. 57, вып. 12. С. 1870-1877.
Гунчак Н.С. Питание дикой свиньи (Sus scrofa L.) в Украинских Карпатах //
Вести, зоологии. 1980.№ 5. С. 73—78.
262
Гунчак Н.С. Морфологические и экологические особенности кабана Украинских
Карпат: Автореф. дис.....канд. биол. наук. Минск, 1982. 22 с.
Гунчак Н.С. Факторы, определяющие численность и воспроизводство дикого ка-
бана в Украинских Карпатах И Структурно-функциональное состояние биоло-
гического разнообразия животного мира Беларуси. Минск, 1999. С. 45-46.
Гурский И.Г. Кабан, косуля, лось и благородный олень в северо-западном При-
черноморье И Копытные фауны СССР. М.: Наука, 1975. С. 79-80.
Гурский ИТ. Украина и Молдавия // Волк. М.: Наука, 1985. С. 487-492.
Гусев А.А. Питание и роющая деятельность кабана в Центрально-Черноземном
заповеднике // Периодические явления в жизни животных. М., 1983. С. 37-46.
Гусев А.А. Функциональная роль диких копытных животных в заповедных био-
геоценозах // Роль крупных хищников и копытных в биоценозах заповедни-
ков. М„ 1986. С. 94-105.
Гусев А.А. Допустимая плотность населения диких копытных животных и опыт
ее поддержания в Центрально-Черноземном заповеднике // Популяционные
исследования животных в заповедниках. М.: Наука, 1988. С. 114-128.
Гусев А.А. Животные на заповедных территориях. Воронеж: Центр.-Черноземное
кн.. изд-во, 1989. 208 с.
Гусев А.А. Антропогенное влияние на наземные экосистемы лесостепной зоны
европейской части России: Автореф. дис. ... д-ра с.-х. наук. Курск, 1995. 39 с.
Гусев А.А., Гусева НА. Повреждение подроста и подлеска кабанами в дубравах
европейской лесостепи И Экология, морфология, использование и охрана ди-
ких копытных. М., 1989. Ч. 2. С. 197-198.
Гусев А.А., Елисеева В.И. Особенности зимнего питания кабана в лесостепи И
Периодические явления в жизни животных. М., 1983. С. 46-51.
Давид А.И. Териофауна плейстоцена Молдавии. Кишинев: Штиинца, 1980. 186 с.
Давид А.И. Формирование териофауны Молдавии в антропогене. Кишинев:
Штиинца, 1982. 151 с.
Давид А.И. Териофауна позднего плейстоцена - раннего голоцена Молдавии И
IV съезд Всесоюз. териол. о-ва. М., 1986. Т. 1. С. 12.
Давид А.И., Татаринов К.А., Свистун В.И. Хищные, хоботные и копытные ран-
него плейстоцена юго-запада СССР. Кишинев: Штиинца, 1990. 131 с.
Давид А.И., Чемыртан ГД. История развития териофауны Молдавии в голоцене
// История биогеоценозов СССР в голоцене. М.: Наука, 1976. С. 207-213.
Давлетова Л.В. Рост желудочно-кишечного тракта у диких и домашних свиней //
Морфология и генетика кабана. М.: Наука, 1985. С. 167-183.
Давлетова Л.В. Экологические и доместикационные особенности в морфофунк-
циональной характеристике развития желудочно-кишечного тракта у диких и
домашних свиней // Влияние экологических факторов на морфофункциональ-
ное состояние внутренних органов животных. М., 1986. С. 89-94.
Давлетова Л.В. Рост желудочно-кишечного тракта гибридных семиреченских
свиней в течение эмбрионального развития // Морфология и генетика гибрид-
ных свиней. М.: Наука, 1992. С. 144-160.
Данилкин А. Отлов кабанов сетями И Охота и охотничье хоз-во. 1984. № 3. С. 14-15.
Данилкин А.А. Экология и систематика косуль Евразии (биологические основы
рационального использования и охраны): Дис. ... д-ра биол. наук. М., 1989.
670 с.
Данилкин А.А. Ареал //Европейская и сибирская косули. М.: Наука, 1992. С. 64-85.
Данилкин А. Российская программа "Косуля": итоги, проблемы и перспективы //
Охота и охотничье хоз-во. 1995. № 2. С. 14-17.
Данилкин А.А. Зимняя подкормка копытных: биологический аспект проблемы //
Там же. 1996а. № 4. С. 12-14.
263
Данилкин А. Охотничье законодательство и дикие копытные животные И Там же.
19966. № 10. С. 4-6.
Данилкин А. О цикличности в динамике численности лося // Там же. 1997. № 6.
С. 12-16.
Данилкин А. Лось в России: использование ресурсов. Исторический аспект про-
блемы // Там же. 1998а. № 4. С. 4-9.
Данилкин А. Лось в России: использование ресурсов. Биологический аспект про-
блемы И Там же. 19986. № 9. С. 8-11.
Данилкин А.А. Оленьи (Cervidae). М.: ГЕОС, 1999а. 552 с.
Данилкин А. Каким путем идти? // Охота и охотничье хоз-во. 19996. № 6. С. 4-6.
Данилкин А. Трофейная охота как фактор антиселекции // Там же. 2000а. № 6. С.
16-17.
Данилкин А.А. Дикие копытные животные востока европейской части России:
анализ динамики и состояния популяций И Охотник и рыболов Поволжья.
20006. № 7-8. С. 2-3.
Данилкин А., Останин В. Массовая гибель косули // Охота и охотничье хоз-во.
1998. №7. С. 6-8.
Данилкин А., Останин В., Стрекаловских В. Подкормка косули: нужны коренные
изменения И Там же. 1999. № 2. С. 1-3.
Данилкин А.А., Останин В.А., Стрекаловских В.А. Демографические параметры
популяции сибирской косули в Зауралье и основные факторы, их определяю-
щие И Экология. 2000. № 6. С. 432-437.
Данилов Д.Н. Размещение и численность лося, косули и кабана в Московской
области И Очерки природы Подмосковья и Московской области. М.: Изд-во
МОИП, 1947. № 4. С. 85-95.
Данилов Д.Н., Русанов Я.С., Рыковский А.С. Методическое руководство по внут-
рихозяйственному устройству охотничьих хозяйств «Росохотрыболовсоюза».
М.: Лесн. пром-сть, 1965. 137 с.
Данилов Д.Н., Русанов Я.С., Рыковский А.С. и др. Основы охотустройства. М.:
Лесн. пром-сть, 1966. 331 с.
Данилов П.И. Появление кабана {Sus scrofa L.) и косули {Capreolus capreolus L.)
в Карелии // Вопросы экологии животных. Петрозаводск, 1974. С. 158-150.
Данилов П.И. Распространение и численность копытных в Карелии // Копытные
фауны СССР. М.: Наука, 1975. С. 80-81.
Данченко В.В. Амурский тигр в Уссурийском заповеднике И Редкие и исчезаю-
щие животные суши Дальнего Востока СССР. Владивосток, 1981. С. 168-170.
Дан-Чин-Ю В. Эффект экономический // Охота и охотничье хоз-во. 1983. № 1. С.
3-4.
Дарман Ю.А. Волк и копытные в Хинганском заповеднике И IV съезд Всесоюз.
териол. о-ва. М., 1986. Т. 2. С. 45-46.
Дарман Ю.А. Копытные Хинганского заповедника И Экология, морфология, ис-
пользование и охрана диких копытных. М., 1989. Ч. 1. С. 52-53.
Дарман Ю.А. Млекопитающие Хинганского заповедника. Благовещенск, 1990.
164 с.
Даугела А., Падайга А. К вопросу о размножении дикого кабана // Диагностика,
лечение и профилактика болезней с.-х. животных. Вильнюс: Мокслас, 1980.
С. 81-85. (Науч. тр. Литовской вет. акад. Т. 13).
Двойченко Г.Г. Пушные и промысловые звери Ставрополья. Ставропольское кн.
изд-во, 1955. 88 с.
Дворников М.Г. Кабан в Ильменском заповеднике И Проблемы охраны генофон-
да и управления экосистемами в заповедниках лесной зоны. М., 1986. Ч. 2. С.
66-68.
264
Дворников М.Г., Дворникова Н.П. О влиянии роющей деятельности кабана на
травянистую растительность лесных биогеоценозов Ильменского заповедника
// Экология, морфология, использование и охрана диких копытных. М., 1989.
Ч. 2. С. 200-201.
Дементьев Г.П. Волк. М.-Л.: Внешторгиздат, 1933. 52 с.
Дежкин В.В. Пути и методы поддержания оптимальной продуктивности экс-
плуатируемых популяций охотничьих животных // Итоги науки и техники.
Зоология позвоночных. М., 1976. Т. 8. С. 8-67.
Дежкин В.В. Охота и охрана природы. М.: Физ-ра и спорт, 1977. 104 с.
Дежкин В. Дикие копытные в Европе // Охота и охотничье хоз-во. 1980. № 12. С.
26-27.
Дежкин В.В. Охота и охотничье хозяйство мира. М.: Лесн. пром-сть, 1983. 358 с.
Дежкин В.В. Управление популяциями диких копытных // Итоги науки и техни-
ки. Зоология позвоночных. М., 1985. Т. 13. С. 66-137.
Дежкин В.В. Дикие копытные в Европе (численность и добыча) И Управление
популяциями диких копытных животных. Киров, 1989. С. 35-47.
Дежкин В.В. Кризис охотничьего хозяйства России // Вопросы прикладной эко-
логии (природопользования), охотоведения и звероводства. Киров, 1997а. С.
350-352.
Дежкин В. Кризис охотничьего хозяйства // Охота и охотничье хоз-во. 19976. №
8. С. 1.
Дежкин В.В., Кондратьева Л.С., Менькова Н.В. Промысел и заготовки диких
копытных в РСФСР // Охотоведение. М.: Лесн. пром-сть, 1975. С. 126-172.
Дежкин В., Петровская Н. Охотничья продукция стран Европы И Охота и охот-
ничье хоз-во. 1970. № 12. С. 38-41.
Джангельдина З.Н. К возрастной и сравнительной морфологии гонад семиречен-
ских свиней И Биоморфология сельскохозяйственных и промысловых животных.
Алма-Ата, 1985. С. 42-43.
Динесман Л.Г. Влияние диких млекопитающих на формирование древостоев. М.:
Изд-во АН СССР, 1961. 166 с.
Динник Н.Я. Звери Кавказа. Ч. 1. Китообразные и копытные. Тифлис, 1910. (Зап.
Кавказского отд. Имп. Рус. геогр. о-ва. Кн. 27, вып. 1). 248 с.
Долгошов В.П. Географические особенности плодоношения дуба и его спутни-
ков на Русской равнине // Исследования географии природных ресурсов жи-
вотного и растительного мира. М.: Изд-во АН СССР, 1962. С. 101-108.
Донауров С.С., Теплое В.П. Кабан в Кавказском заповеднике // Тр. Кавказского
гос. заповедника. 1938. Вып. 1. С. 191-226.
Дормидонтов Р.В. Опыт и результаты интродукции кабанов в Завидовском за-
поведно-охотничьем хозяйстве в период 1935-1961 гг. И Тр. Завидовского за-
поведно-охотничьего хозяйства. М.: Воениздат, 1969. Вып. 1. С. 209-250.
Дормидонтов Р. Кабан нужен культурному лесу // Охота и охотничье хоз-во.
1977. №5. С. 10-11.
Дорст Ж. До того как умрет природа. М.: Прогресс, 1968. 145 с.
Дулицкий А.И., Кормилицин А.А., Кормилицина В.В. Изучение копытных Крыма
И Копытные фауны СССР. М.: Наука, 1975. С. 88.
Дунаев Ю. Кабаны на Урале (опыт подкормки в зимний период) // Вестник Росо-
хотрыболовсоюза. М.: Росагропромиздат, 1991. Вып. 1. С. 17-22.
Дунин В.Ф., Парейко О.А., Одинцова Т.М. Оценка изменения состояния диких
копытных в Полесском радиационно-экологическом заповеднике И 10 лет По-
лесскому гос. радиационно-экологическому заповеднику. Минск, 1998. С.
123-135.
265
Дуншиенко Ю.М. К вопросу охраны тигра в Хабаровском крае // Изучение и ох-
рана редких и исчезающих видов животных фауны СССР. М.: Наука, 1985. С.
62-65.
Дунишенко Ю.М. Амурский тигр // Они нуждаются в защите. Редкие животные
Хабаровского края. Хабаровск: Хабаровское кн. изд-во, 1987. С. 135-148.
Дуров В.В. Влияние кабана на состояние его кормовых растений в районе Кав-
казского заповедника // Средообразующая деятельность животных. М.: Изд-во
Московского ун-та, 1970а. С. 35-36.
Дуров В. В. Пространственные границы популяции кабана Западного Кавказа //
Популяционная структура вида у млекопитающих. М., 19706. С. 36-37.
Дуров В.В. Размещение, миграции и численность кабана в Краснодарском крае //
Вопр. экологии позвоночных животных. Краснодар, 1973. С. 99-136.
Дуров В.В. Репродуктивность и плотность популяции кабана Западного Кавказа
// Копытные фауны СССР. М.: Наука, 1980а. С. 148-150.
Дуров В.В. Объем изъятия и возрастная структура популяции кабана на Запад-
ном Кавказе // Там же. 19806. С. 239-241.
Дуров В.В. Морфологические особенности кавказского кабана И Популяционная
изменчивость вида и проблемы охраны генофонда млекопитающих. М., 1983.
С. 48-49.
Дуров В.В. Социальная структура популяции кавказского кабана // IV съезд Все-
союз. териол. о-ва. М., 1986. Т. 1. С. 199-200.
Дуров В.В. Кабан Западного Кавказа: Автореф. дис. ... канд. биол. наук. М.,
1987. 20 с.
Дуров В.В., Александров В.Н. Размеры и вес кабанов в Кавказском заповеднике И
Тр. Кавказского гос. заповедника. 1968. Вып. 10. С. 294-301.
Дымин В.А. Воздействие хищников на диких копытных Верхнего Приамурья //
Копытные фауны СССР. М.: Наука, 1975. С. 194-195.
Дьяков Ю.В., Дьякова ГА., Комов НМ. Современное состояние популяции кабана в
Воронежском биосферном заповеднике И Проблемы охраны генофонда и управ-
ления экосистемами в заповедниках лесной зоны. М., 1986. Ч. 2. С. 71-73.
Дьякова ГА. Некоторые экстерьерные особенности кабанов усмано-воронежской
популяции // Экология, морфология, использование и охрана диких копытных.
М., 1989. Ч. 2. С. 201-203.
Дьякова Г.А., Дьяков Ю.В. Методы изучения интенсивной роющей деятельности
кабанов в Воронежском заповеднике в различные сезоны года И Там же. 1989.
Ч. 2. С. 204-205.
Дюбюк Е. Из материалов к истории зверовой охоты в Костромском крае // Тр.
Костромского науч. о-ва. 1920. Вып. 18. 27 с.
Евтушевский Н.Н. Состояние среднеприднепровской популяции кабана И Ко-
пытные фауны СССР. М.: Наука, 1975. С. 93-94.
Евтушевский Н.Н. О резервах охотничьих хозяйств Украины И Научно-
технический прогресс - в практику перестройки охотничьего хозяйства. М.,
1988. С. 21-24.
Егоров А.Н. Опыт борьбы с болезнями диких животных и профилактика эпизо-
отий на охраняемой территории // Болезни и паразиты диких животных. М.,
1992. С. 50-56.
Емельянова Л.Г. Влияние кормодобывающей деятельности кабана (Sus scrofa L.)
// IV съезд Всесоюз. териол. о-ва. М., 1986. Т. 1. С. 203-204.
Емельянова Л.Г. Изменение продуктивности надпочвенного покрова на пороях ка-
бана (Sus scrofa L.) в лесных и луговых экосистемах // Динамика зооценозов, про-
блемы охраны и рационального использования животного мира Белоруссии. VI
зоол. конф. Минск, 1989. С. 204-205.
266
Емельянова Л.Г. Оценка процесса восстановления фитоценозов на пороях кабана в
смешанных лесах Белоруссии И V съезд Всесоюз. териол. о-ва АН СССР. М.,
1990. Т. 2. С. 276-277.
Емельянова Л.Г. Осенне-зимнее питание кабана в центральной части Беларуси И
Структурно-функциональное состояние биологического разнообразия живот-
ного мира Беларуси. Минск, 1999а. С. 50-51.
Емельянова Л.Г. Оценка роющей деятельности кабана в естественных экосисте-
мах национального парка «Припятский» // Биологическое разнообразие на-
ционального парка «Припятский». Туров-Мозырь, 1999. С. 241-243.
Емец В.М., Емец Н.С. Многолетняя динамика численности эпигеобионтной ме-
зофауны в дубняках Воронежского биосферного заповедника, в различной
степени нарушенных деятельностью кабана // Проблемы изучения и охраны
заповедных природных комплексов. Воронеж: Изд-во Воронежского ун-та,
1995. С.136-137.
Ержанов Н.Т. Млекопитающие Казахского мелкосопочника (фауна, биология,
экология, цитогенетика, охрана и рациональное использование): Автореф.
дис. ... д-ра биол. наук. Алматы, 2000. 47 с.
Ермолова Н.М. Териофауна долины Ангары в позднем антропогене. Новоси-
бирск: Наука, 1978. 222 с.
Есипов А.В. Кабан на Тянь-Шане // V съезд Всесоюз. териол. о-ва АН СССР. М.,
1990. Т. 2. С. 74-75.
Есипов А.В. К экологии кабана Западного Тянь-Шаня // Экология, охрана и акк-
лиматизация млекопитающих в Узбекистане. Ташкент: Изд-во ФАН, 1991. С.
73-76.
Ефименко П.П., Третьяков П.И. Древнерусские поселения на Дону И Материалы
и исследования по археологии СССР. 1948. № 8. С. 5-112.
Жаргалов Д. Облавные охоты до и после Чингизхана. Иркутск: Имя, 1996. 176 с.
Желтухин А.С. Зимнее питание и охотничье поведение рыси в Центрально-
Лесном заповеднике // Экологические основы охраны и рационального ис-
пользования хищных млекопитающих. М.: Наука, 1979. С. 266-267.
Животченко В.И Амурский тигр И Охота и охотничье хоз-во. 1976. № 7. С. 16-19.
Животченко В.И. О величине годовой добычи копытных тиграми одной семей-
ной группы // Экологические основы охраны и рационального использования
хищных млекопитающих. М.: Наука, 1979. С. 246-247.
Животченко В.И. О питании амурского тигра // Хищные млекопитающие. М.,
1981. С. 64-75.
Жиряков В.А. Питание и воздействие кабана на растительность в Северном
Тянь-Шане // III съезд Всесоюз. териол. о-ва. М., 1982. Т. 1. С. 199-200.
Жиряков В.А. О взаимоотношениях хищных и копытных млекопитающих в Ал-
ма-Атинском заповеднике (Северный Тянь-Шань) // IV съезд Всесоюз. тери-
ол. о-ва. М., 1986. Т. 1. С. 211-212.
Жиряков В.А. Распределение лежек кабана по сезонам в Заилийском Алатау И
Экология, морфология, использование и охрана диких копытных. М., 1989. Ч.
2. С. 205-206.
Жиряков В.А. Роль волка в биоценозах Алма-Атинского заповедника (Северный
Тянь-Шань) И V съезд Всесоюз. териол. о-ва АН СССР. М., 1990. Т. 2. С. 278-279.
Житенко И.В., Белякова Н.П., Чебакова Г.В., Хритинина Г.А. Морфологический
и химический состав мяса дикого кабана // Теория и практика товароведения
и рациональное использование сырья и продуктов животноводства. М., 1987.
С. 98-100.
Журавлев О.П. Дикие млекопитающие эпохи бронзы Нижнего Дона // Эколого-,
морфологические особенности животных и среда их обитания. Киев: Наукова
Думка, 1981. С. 20-23.
267
Журовский В., Сиудова Г., Галка Б. Влияние добавления крови домашней свиньи
на популяцию местных диких кабанов (Sus scrofa} И Тр. IX Междунар. конгр.
биологов-охотоведов. М., 1970. С. 238.
Завацкий Б.П. Избирательность зимних охот волков на Западном Саяне // IV
съезд Всесоюз. териол. о-ва. М., 1986а. Т. 1. С. 212-213.
Завацкий Б.П. Роль волка в биоценозах Саяно-Шушенского заповедника // Роль
крупных хищников и копытных в биоценозах заповедников. М., 19866. С. 35-54.
Завацкий Б.П. Кабан как новый вид Западного Саяна И Экология и охрана гор-
ных видов млекопитающих. М., 1987. С. 66-68.
Завацкий Б.П. Кабан в Саяно-Шушенском заповеднике И Экологические иссле-
дования в заповедниках Южной Сибири. М., 1989. С. 34-39.
Завьялова Л. Ф. Тема: Биоценотическая роль кабана в Дарвинском заповеднике и
его значение в соседних сельхозугодьях. 1992, 1993 гг. // Научные исследова-
ния в заповедниках и национальных парках России (Федеральный отчет за
1992-1993 г.). М„ 1997. С. 99-101.
Зайцев В.А. Формирование групп кабанят. Синхронизация деятельности и лиди-
рование И Экология, морфология, использование и охрана диких копытных.
М„ 1989. Ч. 2. С. 206-208.
Зайцев В.А. Игры кабанов И Бюл. МОИП. Отд. биол. 1991. Т. 96, вып. 1. С. 38-48.
Зайцев В.А. Синхронизация поведения и индивидуальные дистанции в группах
кабанов (Sus scrofa L.) И Журн. общей биол. 1992. Т. 53, № 2. С. 243-257.
Зайцев В.А. Вожаки, лидеры и разведчики у кабанов И Бюл. МОИП. Отд. биол.
1999. Т. 104, вып. 1. С. 10-17.
Зайцев В.А. Использование участков обитания и пространственная структура по-
пуляций у лесных копытных Artiodactyla // Зоол. журн. 2000. Т. 79, № 4. С.
397-411.
Зайцев В.А., Зайцева В.К. Структура групп кабана И V съезд Всесоюз. териол. о-
ва АН СССР. М., 1990. Т. 3. С. 90.
Закариев А.Я. Дикие парнокопытные Северного Кавказа как носители паразити-
ческих червей И Копытные фауны СССР. М.: Наука, 1980. С. 159-160.
Залесский П.М. Прошлое и современное распространение копытных в Западно-
Сибирском крае И Охотник Сибири. 1934. № 7-8. С. 26-28.
Зархидзе В.А. Копытные Западной Туркмении И Копытные фауны СССР. М.:
Наука, 1980. С. 88-90.
Захаров А.А., Суворов А.А. Влияние кабанов на рост и развитие муравейников //
Биологические основы использования полезных насекомых. М., 1988. С. 40-42.
Захарова Л.С. Закономерности и факторы динамики численности тетеревиных
птиц в заповеднике «Кивач» // Тетеревиные птицы в заповедниках РСФСР.
М„ 1989. С. 30-46.
Збруева А. В. История населения Прикамья в ананьинскую эпоху // Материалы и
исследования по археологии СССР. № 30. Материалы и исследования по ар-
хеологии Урала и Приуралья. Т. 5. М.: Изд-во АН ССР, 1952. С. 5-319.
Збруева А.В. Население берегов Камы в далеком прошлом //По следам древних
культур. От Волги до Тихого океана. М.: Гос. изд-во культ.-просвет. лит-ры,
1954. С. 95-130.
Зверев М.Д. Млекопитающие Новосибирского района И Тр. Новосибирского зоо-
сада. Звери и птицы Западной Сибири. Новосибирск: Зап.-Сиб. краевое, изд-
во, 1937. Т. 1. С. 7-30.
Зверезомб-Зубовский Е.В. К познанию фауны млекопитающих Донской области.
Ростов-на-Дону, 1923. 29 с.
Зеньков А.В., Пенькевич В.А., Пенькевич А.А. Некоторые вопросы эпизоотологии
метастронгилеза диких кабанов И Заповедники Белоруссии. 1979. Вып. 3. С.
76-80.
268
Зинченко Е.И., Черногаев Е.А. Численность копытных в равнинной части Чард-
жоуской области ТССР // Экология, морфология, использование и охрана ди-
ких копытных. М., 1989. Ч. 1. С. 53-54.
Злобин Б. Кабан в Прибалхашье // Охота и охотничье хоз-во. 1969. № 2. С. 22-23.
Злобин Б.Д. Использование кабаньего стада в Прибалхашье И Копытные фауны
СССР. М.: Наука, 1975а. С. 220-221.
Злобин Б.Д. Редкие виды копытных Кировской области // Там же. 19756. С. 264-265.
Зобов А.И. Основные этапы деятельности Хоперского государственного запо-
ведника // Проблемы изучения и охраны заповедных природных комплексов.
Воронеж: Изд-во Воронежского ун-та, 1995. С. 5-11.
Зыкова Л.Ю. Кабан и пятнистый олень в Окском заповеднике // Охотничье-
промысловые звери. Биология и хозяйственное использование. М.: Россельхозиз-
дат, 1965. Вып. 1.С. 120-134.
Иваненко И.Д. Постнатальное развитие кабана в связи с условиями внешней
среды. Сталинабад, 1956. 233 с.
Иванов И. Уральские кабаны в США // Охота и охотничье хоз-во. 1962. № 12. С. 59.
Иванов П.Д., Русаков О.С. Кабан И Охотничьи звери и их промысел. М.: Лесн.
пром-сть, 1970. С. 156-159.
Иванова Г.И. Экологические основы профилактики матастронгилеза кабана в
Подмосковье // Тр. IX Междунар. конгр. биологов-охотоведов. М., 1970. С.
612-615.
Иванова Г.И. Размещение кабанов в северных районах их выпусков И Охотове-
дение. М.: Лесн. пром-сть, 1974. С.58-69.
Иванова Г.И. Особенности размножения кабанов Подмосковья // Копытные
фауны СССР. М.: Наука, 1975а. С. 95-96.
Иванова Г.И. Размещение и численность кабана в Московской области И Охото-
ведение. М.: Лесн. пром-сть, 19756. С. 307-314.
Иванова Г.И. Распространение и численность кабана в РСФСР // Охотоведение.
М.: Лесн. пром-сть, 1976. С. 71-82.
Иванова Г. Строго регулировать численность // Охота и охотничье хоз-во. 1977.
№9. С. 10-12.
Иванова Г.И. К вопросу о рациональном использовании кабана в РСФСР И Пути
и методы рациональной эксплуатации и повышения продуктивности охот-
ничьих угодий. М., 1978. С. 180-184.
Иванова Г.И. Динамика численности кабана в европейской части РСФСР И Ко-
пытные фауны СССР. М.: Наука, 1980а. С. 161-162.
Иванова Г.И. Состояние промысла кабана в европейской части РСФСР // Там же.
19806. С.242-243.
Иванова Г.И. Размещение кабана (Sus scrofa L.) в европейской части РСФСР И
Промысловые звери РСФСР (пространственные и временные изменения насе-
ления). М„ 1982. С. 9-32.
Иванова Г. Нормы добычи кабана И Охота и охотничье хоз-во. 1983. № 1. С. 10-11.
Иванова Г.И. Эколого-биологические предпосылки рационального использова-
ния популяции кабана в европейской части РСФСР // Управление популяция-
ми диких копытных животных. М., 1985. С. 133-147.
Иванова Г.И. Влияние особенностей размножения кабана на прирост И IV съезд
Всесоюз. териол. о-ва. М., 1986. Т. 2. С. 230.
Иванова Г.И., Ильинский В.О. Размещение кабана по лесным угодьям в районе
выпуска И Науч. тр. ВНИИ лесоводства и механизации лесного хоз-ва. 1969.
Вып. 2. С. 64-83.
Иванова Г.И, Молоканов Б.В. Результаты выпуска копытных в Переславском го-
сударственном лесоохотничьем хозяйстве // Вопр. повышения продуктивно-
сти охотничьих угодий. М.: Колос, 1969. С. 99—110.
269
Иванова Г.И., Молоканов Б.В. Использование кабана в европейской части
РСФСР И Акклиматизация охотничьих животных в СССР. Минск: Ураджай,
1978. С.129-130.
Иванова Г.И., Молоканов Б.В. Половозрастная структура и динамика численно-
сти кабана// III съезд Всесоюз. териол. о-ва. М., 1982. Т. 2. С. 220-221.
Иванова Г.И., Овсюкова И.И. Кабан И Охота на копытных (биологические осно-
вы промысла). М.: Лесн. пром-сть., 1976. С. 103-161.
Иванова Г.И., Рыковский А.С. Кабан в Подмосковье // Проблемы охраны и ра-
ционального использования промысловых животных. М., 1968а. С. 90-93.
Иванова Г.И., Рыковский А.С. Хозяйство на кабана в Подмосковье // Передовой
опыт в охотничьем хозяйстве. 19686. Вып. 1. С. 61-65.
Издебский В.М., Тарасов И.А. Кабаны в таврических степях Северного Причер-
номорья И III съезд Всесоюз. териол. о-ва. М., 1982. Т. 2. С. 221-222.
Икерманн Е. Гибель охотничьих животных на путях транспорта в ФРГ // Тр. IX
Междунар. конгр. биологов-охотоведов. М., 1970. С. 165-166.
Ильинский В. Кабаны в Ярославской области // Охота и охотничье хоз-во. 1966.
№ 10. С. 4-5.
Илъяшенко В.Ю., Костенко В.А., Родионов СИ., Юдин В.Г. Хребет Тукурингра
как зоогеографический рубеж // III съезд Всесоюз. териол. о-ва. М., 1982. Т. 1.
С.115-116.
Иностранцев А.А. Доисторический человек каменного века побережья Ладож-
ского озера. СПб., 1882. 241 с.
Исаев В.А., Иванова Т.Е. К вопросу изучения распространения трихинеллеза
среди диких животных в лесах Калининской и Московской областей // Тр. За-
видовского заповедника. М.: Воениздат, 1979. Вып. 4. С.158-165.
Исаков Ю.А. Краткий очерк фауны млекопитающих и птиц Молого-
Шекснинского междуречья до образования водохранилища// Тр. Дарвинского
гос. заповедника. 1949. Вып. 1. С. 137-171.
Ишадов И. Эколого-географические особенности некоторых копытных Туркме-
нии: Автореф. дис. ... канд. биол. наук. Ашхабад, 1956. 22 с.
Ишадов Н. Некоторые вопросы экологии кабана в Туркмении // Учен. зап. Турк-
менского гос. ун-та им. А.М. Горького. 1969. Вып. 53. С. 59-61.
Ишадов И. Распространение и современное состояние численности копытных
животных в Северо-Западной Туркмении // Териология. Новосибирск: Наука,
1972. Т. 1. С. 417-419.
Ишадов И. Особенности миграции кабана в Каракумах // Прикладная этология.
М.: Наука, 1983. С. 210-211.
Ишадов И. Особенности водопоя промысловых зверей в аридной зоне Туркме-
нистана//IV съезд Всесоюз. териол. о-ва. М., 1986. Т. 1.С. 231-232.
Ишадов И., Клюшкин Е.А. Копытные животные Кутитанга и прилегающей рав-
нины // Копытные фауны СССР. М.: Наука, 1975. С. 98-99.
Ишунин Г.И. Популяция кабана (Sus scrofa nigripes Blanford) в Кызылкумах //
Териология. Новосибирск: Наука, 1972. Т. 1. С. 412-413.
Ишунин Г.И. Дикие копытные Узбекистана // Копытные фауны СССР. М.: Наука,
1980. С.91-93.
Каверзнев В.И. Охота на кабанов. М.: КОИЗ, 1932. 56 с.
Казаков Б., Даликов Р. Кабан в Ростовской области И Охота и охотничье хоз-во.
1983. № И. С. 22-23.
Казневский П.Ф. Дикие копытные животные Хоперского заповедника и пробле-
мы управления их популяциями И Охотничье хозяйство и заповедное дело.
М.: Россельхозиздат, 1977. С. 36-42.
270
Казневский П.Ф. Волк в Хоперском заповеднике И Экологические основы охра-
ны и рационального использования хищных млекопитающих. М.: Наука,
1979. С. 110-112.
Калдыбаев С. У., Еськов П.А. Популяция свиней гибридной семиреченской поро-
ды с окраской агути // Морфология и генетика гибридных свиней. М.: Наука,
1992. С.23-42.
Калиновский П.Ф. Териофауна позднего антропогена и голоцена Белоруссии.
Минск: Наука и техника, 1983. 154 с.
Калниньш А.И. Охота и охотничье хозяйство в Латвийской ССР. Рига: Латгосиздат,
1950.540 с.
Катанов Л.Г. Тигр, изюбрь, лось. М.: Изд-во МОИП, 1948. 128 с.
Капралова Л.Т. Рост и гистогенез мышечной ткани гибридных семиреченских сви-
ней // Морфология и генетика гибридных свиней. М.: Наука, 1992. С. 71-80.
Карасев П.Ф. Метастронгилез кабанов Березинского заповедника // Березинский
заповедник. 1974. Вып. 3. С. 55-59.
Карасев Н.Ф., Литвинов В.Ф. Эпизоотология трихинеллеза животных Березин-
ского заповедника // Там же. 1974. Вып. 3. С. 62-65.
Карпинский Ю.С., Лукаревский В.С. Распространение кабана на Юго-Западном
Копетдаге // Влияние антропогенной трансформации ландшафта на население
наземных позвоночных животных. М., 1987. Ч. 1. С. 178-179.
Карцов Г.П. Беловежская пуща. Ее исторический очерк, современное охотничье
хозяйство и высочайшие охоты в Пуще. СПб., 1903. 414 с.
Катаевский В.Н. Численность, рациональное использование и охрана копытных
в горных районах Северной Киргизии И Экология и охрана горных видов мле-
копитающих. М., 1987. С. 84-86.
Каштальян А.П. Состояние фаунистических комплексов копытных Березинско-
го биосферного заповедника // Охраняемые природные территории и объекты
Белорусского Поозерья: современное состояние, перспективы развития. Ви-
тебск, 1997. С. 87-88.
Каштальян А.П., Медведев И.Г. Динамика численности копытных в Березин-
ском биосферном заповеднике в 1966-1998 годах и факторы, ее определяю-
щие // Структурно-функциональное состояние биологического разнообразия
животного мира Беларуси. Минск, 1999. С. 52-54.
Квакин В.И. Современное состояние, проблемы охраны и дальнейшего исполь-
зования копытных животных в охотхозяйствах Белорусского общества охот-
ников и рыболовов И Вопросы прикладной экологии (природопользования),
охотоведения и звероводства. Киров, 1997. С. 106-108.
Керечун С. Ф. Влияние хищников на карпатскую популяцию копытных // Копыт-
ные фауны СССР. М.: Наука, 1975. С. 197-198.
Керечун С.Ф. Влияние хищников на поголовье копытных в зоне Карпат И Эколо-
гические основы охраны и рационального использования хищных млекопи-
тающих. М.: Наука, 1979. С. 43-44.
Керечун С.Ф. Прирост и использование копытных в Карпатах И Копытные фау-
ны СССР. М.: Наука, 1980. С. 93-94.
Кестер Б.В. Некоторые итоги и перспективы работ Воронежского заповедника
по отлову и расселению оленя, кабана и речного бобра И Акклиматизация
охотничьих животных в СССР. Минск: Ураджай, 1978. С. 53-54.
Кириков С.В. Исторические изменения животного мира нашей страны в XIII—
XIX веках // Изв. АН СССР. Сер. геогр. 1952. № 6. С. 31-48.
Кириков С.В. Исторические изменения животного мира нашей страны в XIII—
XIX веках. Сообщение 2-е И Изв. АН СССР. Сер. геогр. 1953. № 4. С. 15-27.
Кириков С.В. Исторические изменения животного мира нашей страны в XIII—
XIX веках. Сообщение 3. // Изв. АН СССР. Сер. геогр. 1955. № 1. С. 32-40.
271
Кириков С.В. Изменения животного мира в природных зонах СССР (XIII—XIX
вв.); Степная зона и лесостепь. М.: Изд-во АН СССР, 1959. 175 с.
Кириков С. В. Изменения животного мира в природных зонах СССР (XIII—XIX
вв.). Лесная зона и лесотундра. М.: Изд-во АН СССР, 1960. 157 с.
Кириков С.В. О «смешанной» фауне в степной и лесной зонах // Изв. АН СССР.
Сер. геогр. 1963а. № 1. С. 29-35.
Кириков С.В. Промысловые животные, природная среда и человек. М.: Наука,
1966.348 с.
Кириков С.В. Звери и птицы северного степного Причерноморья и их среда обита-
ния в XVIII и первой трети XIX в. // История биогеоценозов СССР в голоцене.
М.: Наука, 1976. С. 193-207.
Кириков С.В. Человек и природа степной зоны. Конец X - середина XIX в. (ев-
ропейская часть СССР). М.: Наука, 1983. 128 с.
Кириков С.В., Исаков Ю.А. Запасы наземных промысловых животных; динамика
промысла и перспективы использования. Л., 1959. 50 с.
Кириллов Н. Охотничье хозяйство в Забайкальской области И Природа и охота.
1902. Янв. С. 31-40.
Кириллов Н.В. Охотничье хозяйство в Забайкальской области И Там же. 1904.
Кн. 9. С. 21-25.
Кириллова И.В. Охотничья добыча с мезолит-неолитического памятника Ива-
новское-7 (Переславский р-н Ярославской области) как отражение изменений
среды обитания И VI съезд териол. о-ва. М., 1999. С. 117.
Киселев А. Кабан на Среднем Урале // Охота и охотничье хоз-во. 1986. № 11. С.
12-13.
Киселев В.Е., Каплин В.В. Кабан - новый вид в таежных биоценозах // Влияние
хозяйственного освоения лесных территорий Европейского Севера на населе-
ние животных. М., Наука, 1987. С. 3-11.
Киселев Н.П. Факторы, влияющие на численность кабанов в дельте Волги И Ко-
пытные фауны СССР. М.: Наука, 1975. С. 103-104.
Киселев Н. Судьба кабана в дельте Волги // Степные просторы. 1976. № 4. С. 47—48.
Клевезалъ Г.А. Регистрирующие структуры млекопитающих в зоологических ис-
следованиях. М.: Наука, 1988. 285 с.
Климова В.Н. Ресурсы основных видов копытных и их использование в СССР //
Экология, морфология, использование и охрана диких копытных. М., 1989. Ч.
1. С. 59-60.
Князев СП. Использование полиморфизма аллотипов белков сыворотки крови
для изучения генофонда кабанов разных регионов СССР И Популяционная
изменчивость вида и проблемы охраны генофонда млекопитающих. М., 1983.
С. 85-87.
Князев С.П., Тихонов В.Н. Краниологические особенности европейских и азиат-
ских диких кабанов и их гибридов с домашними свиньями // Морфология и
генетика кабана. М.: Наука, 1985. С. 33 48.
Князев С.П., Тихонов В.Н., Сузуки С. и др. Генетические особенности диких и до-
машних свиней (Suidae) Евразии по сывороточным полиморфным системам. 2.
Полиморфизм по системам трансферрина, церулоплазмина и амилазы И Зоол.
журн. 1985. Т. 64, №11. С. 1712-1717.
Ковалев А.А., Кармаев В.Н. Гибель копытных животных в Ставропольском рай-
оне Самарской области в 1968-1996 гг. // Вопросы прикладной экологии
(природопользования), охотоведения и звероводства. Киров, 1997. С. 116-117.
Ковалев А.К, Ковалева И.Н., Голдырев Ф.Н. К взаимозависимости численности ка-
бана и волка в Таджикистане // Копытные фауны СССР. М.: Наука, 1975. С. 199.
Ковалева Ж.В., Антонец Н.В. Роющая деятельность кабана и ее влияние на лесо-
возобновление в Хоперском заповеднике // Проблемы изучения и охраны за-
272
поведных природных комплексов. Воронеж: Изд-во Воронежского ун-та,
1995. С. 73-74.
Кожамкулова Б. С. Антропогеновая ископаемая териофауна Казахстана. Алма-
Ата: Наука, 1969. 149 с.
Кожамкулова Б.С. Голоценовая фауна копытных Казахстана И История биогео-
ценозов СССР в голоцене. М.: Наука, 1976. С. 260-267.
Кожамкулова Б.С. Зоогеографический анализ состава и распределения антропо-
геновой фауны Казахстана в пространстве и времени // Мезокайнозойская
фауна и флора Северо-Западного Казахстана. Алма-Ата: Наука, 1977. С. 89-
104. (Материалы по истории фауны и флоры Казахстана. Т. 7).
Кожамкулова Б.С. Позднекайнозойские копытные Казахстана. Алма-Ата: Нау-
ка, 1981. 144 с.
Кожамкулова Б.С., Костенко Н.Н Вымершие животные Казахстана (палеогео-
графия позднего кайнозоя). Алма-Ата: Наука, 1984. 103 с.
Козло П.Г. Питание кабана и сезонная смена его корма // Беловежская пуща.
1969. Вып. З.С. 132-144.
Козло П.Г. Факторы, определяющие динамику численности кабана (Sus scrofa) в
Беловежской пуще // Зоол. журн. 1970а. Т. 49, вып. 3. С. 422-430.
Козло П.Г. Морфологические и экологические особенности популяций кабана
Беловежской пущи и Березинского заповедника И Популяционная структура
вида у млекопитающих. М., 19706. С. 117-119.
Козло П.Г. Внутрипопулянионная изменчивость морфологических признаков
кабана Беловежской пущи //Там же. 1970в. С. 119-121.
Козло П.Г. Роющая деятельность дикого кабана И Средообразующая деятель-
ность животных. М.: Изд-во Московского ун-та, 1970г. С. 79-80.
Козло П.Г. Сравнительная характеристика роющей деятельности дикого кабана
(Sus scrofa L.) в Березинском заповеднике и Беловежской пуще // Березинский
заповедник. 1972а. Вып. 2. С. 130-133.
Козло П.Г. О составе осенне-зимних кормов дикого кабана (Sus scrofa L.) в Бере-
зинском заповеднике // Там же. 19726. Вып. 2. С. 133-140.
Козло П. Селекция кабана И Охота и охотничье хоз-во. 1972в. № 2. С. 12-14.
Козло П.Г. Определение возраста, селекция и отлов дикого кабана. Минск: Урад-
жай, 1973. 55 с.
Козло П.Г. Динамика структуры стад и внутристадных отношений между особя-
ми дикого кабана Беловежской пущи И Березинский заповедник. 1974а. Вып.
3. С. 92-98.
Козло П.Г. Изменчивость веса тела дикого кабана, обитающего в Беловежской
пуще И Там же. 19746. Вып. 3. С. 98-103.
Козло П.Г. Дикий кабан. Минск: Ураджай, 1975. 223 с.
Козло П.Г. Направленность динамики численности охотничьих макромаммалий
в Беларуси И Биологические ритмы. Брест, 1999. С. 101-103.
Козло П.Г. Морфофизиологические адаптации и структурно-функциональный ана-
лиз динамики популяций парнокопытных (Artiodactyla), проблемы их охраны и
рационального использования в Беларуси: Дис. ... докт. биол. наук в форме науч-
ного доклада. Минск, 2001. 68 с.
Козло П.Г, Банад Э.Б. Белоруссия // Волк. М.: Наука, 1985. С. 483-487.
Козло П.Г., Дерябина Т.Г. Ресурсно-пространственная оценка размещения ко-
пытных как основа разработки стратегии оптимизации ведения охотничьего
хозяйства в Белоруссии И IV съезд Всесоюз. териол. о-ва. М., 1986. Т. 2. С.
231-232.
Козло П.Г., Дунин В.Ф. Биотехнические мероприятия для копытных зверей как
основа повышения эффективности охотничьего хозяйства Белоруссии. Минск,
1989. 49 с.
18. ЗаказN>2528.
273
Козло П.Г, Емельянова Л.Г. Особенности изменения ресурсов копытных в Бело-
русском Полесье в связи с трансформацией естественных экосистем // Влия-
ние антропогенной трансформации ландшафта на население наземных позво-
ночных животных. М., 1987. Ч. 1. С. 205-207.
Козло П.Г., Емельянова Л.Г., Кучмель С.В., Парейко О.А. Экологические особен-
ности популяции кабана, обитающей в Полесском радиологическом заповед-
нике // 10 лет Полесскому гос. радиационно-экологическому заповеднику.
Минск, 1998. С. 130-135.
Козло П.Г, Никитенко М.Ф. Методика определения возраста дикого кабана //
Экология млекопитающих и птиц. М.: Наука, 1967. С. 209-221.
Козло П.Г., Сидоренко О.Н., Дунин В.Ф. Изучение закономерностей изменения
численности и структуры популяций и сообществ охотничье-промысловых
млекопитающих в 30-километровой зоне ЧАЭС // Радиобиологический съезд.
Пущино, 1993. Ч. 2. С. 473.
Козло П.Г., Ставровская Л.А. Влияние роющей деятельности кабана (Sus scrofa
L.) на травянистую растительность // Заповедники Белоруссии. 1979. Вып. 3.
С. 91-99.
Козло П.Г, Хацкевич А.И., Пустошило Р.П, Автушко ГВ. Возрастные изменения
веса и размеров тела кабана Березинского заповедника // Березинский заповед-
ник. 1974. Вып. 3. С. 3-15.
Козлов В.В. Кабаны в Рязанской области // Зоол. журн. 1958. Т. 37, вып. 1. С.
142-143.
Колодяжный ПК. Некоторые вопросы динамики численности животных охот-
ничьей фауны Завидовского заповедно-охотничьего хозяйства. 1966-1970 гг.
// Тр. Завидовского заповедно-охотничьего хоз-ва. М.: Воениздат, 1971. Вып.
2. С. 8-35.
Колодяжный И.К, Зворонос Г.И. Страницы истории Завидовского хозяйства //
Тр. Завидовского заповедника. М.: Воениздат, 1979. Вып. 4. С.6-52.
Коломыцев А.А., Вишняков И.Ф., Жестерев В.И., Калабеков М.И. Классическая
чума свиней среди диких кабанов в России И Вести, ветеринарии. 1999. № 13
(2). С. 52-56.
Коломыцев А.А., Вишняков И.Ф., Семенихин А.Л. и др. Классическая чума свиней
среди кабанов на территории бывшего СССР И Актуальные вопр. ветеринар-
ной вирусологии. Покров, 1995. С. 35-36.
Комаров Б.А., Богацкий Е.К Профилактика классической чумы среди диких ка-
банов // Ветеринария. 1980. № 8. С. 37-39.
Комов Н. Отлов кабанов в Воронежском заповеднике // Охота и охотничье хоз-
во. 1972. № 1.С. 22-23.
Кононов С.И. Хозяйственное использование копытных в БССР и пути увеличе-
ния их добычи // Научно-технический прогресс - в практику перестройки
охотничьего хозяйства. М., 1988. С. 124—127.
Копылов ИЛ. Дикие копытные животные Иркутской области. Иркутское обл.
гос. изд-во, 1950. 76 с.
Кораблев П.Н. Изменчивость зубной формулы млекопитающих // IV съезд Все-
союз. териол. о-ва. М., 1986а. Т. 2. С. 63-64.
Кораблев П.Н. К вопросу об охране и изучении генофонда кабана // Проблемы
охраны генофонда и управления экосистемами в заповедниках лесной зоны.
М„ 19866. 4.2. С. 112-114.
Кораблев П.Н. Изменчивость зубной формулы кабана // Экология, морфология,
использование и охрана диких копытных. М., 1989. Ч. 1. С. 174-176.
Корелов М.Н. О значении кабана в жизни моховых ельников Тянь-Шаня // Веста.
АН КазССР. 1947. № 3 (24).
274
Коркшико В.Г. Экологические особенности и поведение дальневосточного лео-
парда: Автореф. дис. ... канд. биол. наук. М., 1981. 26 с.
Кормилицин А.А., Дулицкий А.И. К реакклиматизации свиньи дикой (Sus scrofa
L.) в Крыму И Вести, зоологии. 1972. Вып. 1. С. 38-44.
Корнеев А.П. История промысла диких зверей на Украине. Изд-во Киевского
гос. ун-та им. Т.Г. Шевченко, 1953. 37 с.
f Корнеев А.П. Колебания численности дикого кабана на Украине и рациональные
нормы плотности его поголовья в охотничьих хозяйствах // Тр. IX Междунар.
конгр. биологов-охотоведов. М., 1970. С. 812-814.
Корнеев А., Кричевская Ц. Кабан в лесах Украины // Охота и охотничье хоз-во. 1967.
№ 4. С. 22.
Корнеев О.П., Кричевська Ц.Ю. Особливост! живлення дикого кабана на Украш!
И В1стн. Кшвськ. ун-ту. Сер. бюл. 1967. № 9. С. 150-154.
Корнилов И.П. Олени Восточной Сибири. Козуля И Вести, естеств. наук. 1856. Т.
3, № 16. С. 494-504.
Косинцев П.А. Териофауна Урала и Западной Сибири в голоцене И IV съезд Все-
союз. териол. о-ва. М., 1986. Т. 1. С. 16-17.
Косинцев П.А. Голоценовые остатки крупных млекопитающих Западной Сибири
// Современное состояние и история животного мира Западно-Сибирской
низменности. Свердловск, 1988. С. 32-51.
Косинцев П.А. Фауна крупных млекопитающих Северного Урала в позднем
плейстоцене и голоцене // Материалы и исследования по истории современ-
ной фауны Урала. Екатеринбург, 1996. С.84-109.
Косинцев П.А., Ражее Д.И. Новые данные по териофауне Южного Урала и За-
уралья в голоцене // Млекопитающие в системе природопользования на Ура-
ле. Свердловск, 1985. С. 17-18.
Костоглод В.Е. Кабан в Приазовских плавнях // Сб. науч.-техн, информ.
ВНИИЖП. Охота-пушнина-дичь. Киров, 1969. Вып. 24. С. 23-27.
Котов В.А. Трудная для диких копытных зима в Кавказском заповеднике // Бюл.
11 МОИП. Отд. биол. 1969. Т. 74, вып. 1. С. 57-61.
Котов В.А., Рябов Л.С. Промысловые и ценные млекопитающие предгорных и
горных районов Краснодарского края // Тр. Кавказского гос. заповедника.
1963. Вып. 7. 239 с.
Кочетков В.В., Соколов А.А. Питание волка в Центрально-Лесном государствен-
ном заповеднике И Экологические основы охраны и рационального использо-
вания хищных млекопитающих. М.: Наука, 1979. С. 112-114.
Кочко Ф.П., Буневич А.Н., Вакула В.А. Суточная и сезонная активность кабана
Беловежской пущи//Заповедники Белоруссии. 1988. Вып. 12. С. 113-118.
Кручинин В.Д. Динамика численности тетеревиных в Завидовском государственном
научно-опытном заповеднике И Тр. Завидовского заповедника. М.: Воениздат,
1979. Вып. 4. С. 85-106.
Кубанцев Б.С., Зубов В.Г. Акклиматизация и реакклиматизация некоторых копыт-
ных в северных районах Нижнего Поволжья И II съезд Всесоюз. териол. о-ва. М.:
Наука, 1978. С. 248-250.
Кудактин А.П. Соотношение численности копытных и волка в Кавказском запо-
веднике И Копытные фауны СССР. М.: Наука, 1975. С. 199-200.
Кудактин А.П. Волк Западного Кавказа (экология, поведение, биоценотическое
положение): Автореф. дис. ... канд. биол. наук. М., 1982. 22 с.
Кудактин А.П. Влияние волка на копытных в Кавказском заповеднике // Роль круп-
ных хищников и копытных в биоценозах заповедников. М., 1986. С. 21-35.
Кудряшова Л.М. Воздействие некоторых факторов среды на выживаемость сего-
леток кабана И III съезд Всесоюз. териол. о-ва. М., 1982. Т. 2. С. 227-228.
18
275
Кузьмин И.Ф. Основные черты формирования фауны охотничьих животных Мо-
сковской области за столетний период // Влияние антропогенной трансформа-
ции ландшафта на население наземных позвоночных животных. М., 1987. Ч.
1. С. 104-106.
Кузьмин И.Ф., Хахин ГВ. Влияние волка на копытных в центре европейской час-
ти СССР И Копытные фауны СССР. М.: Наука, 1980. С. 171-172.
Кузнецов Б.А. Млекопитающие Казахстана. М.: Изд-во МОИП, 1948. 226 с. (Ма-
териалы к познанию фауны и флоры СССР. Новая сер. Отд. зоол. Вып. 13).
Кузнецов В.И. Леопард на Западном Копетдаге и пути его сохранения // VI съезд
териол. о-ва. М., 1999. С. 135.
Кузякин В.А. Состояние ресурсов диких копытных И Итоги науки и техники.
Зоология позвоночных. М.: ВИНИТИ, 1985. Т. 13. С.5-65.
Кузякин В.А. К оценке качества и емкости охотничьих угодий // Современные
проблемы охотничьего хозяйства. М., 1989. С. 15-27.
Кузякин В.А., Сурбонос Н.Э. Северо-восточная граница ареала кабана в европей-
ской части РСФСР // VIII Всесоюз. зоогеографическая конф. М., 1984. С. 81.
Куклин С.А. Звери и птицы Урала и охота на них. Свердловск, 1938. 244 с.
Кулагин Н.М. Лоси СССР И Тр. Лаб. прикладной зоологии. Л.: Изд-во АН СССР,
1932. 120 с.
Кулибаба В.В., Пакулин В.А. Материалы по экологии кабана бассейна р. Сумбар
// Растительность и животный мир Западного Копетдага. Ашхабад: Ылым,
1985. С.100-107.
Кулиев С.М. Численность, половой и возрастной состав копытных в Исмаиллин-
ском заповеднике и заказнике // Экология, морфология, использование и ох-
рана диких копытных. М., 1989. Ч. 1. С. 60-62.
Курносов К.М., Петрищев Б.И. Эмбриогенез кабана Sus scrofa L. И II съезд Все-
союз. териол. о-ва. М.: Наука, 1978. С. 62-63.
Курносов К.М., Петрищев Б.И. Эмбриогенез кабана {Sus scrofa). Состояние
внешних морфологических признаков у эмбрионов кабана разных стадий раз-
вития // Зоол. журн. 1977. Т. 56, вып. 3. С. 412 419.
Курочкина О.Г. Влияние роющей деятельности кабана на биоценозы Карадаг-
ского заповедника // VI съезд териол. о-ва. М., 1999. С. 136.
Кучеренко С.П. К экологии амурского тигра И Охота и охотничье хоз-во. 1972.
№ 1. С. 18-20.
Кучеренко С.П. Уссурийский кабан И Там же. 1973. № 1. С. 20-22.
Кучеренко С.П. Копытные млекопитающие Амуро-Уссурийского региона // Жи-
вотный мир и охотничье хозяйство Дальнего Востока. Владивосток, 1976. С.
97-125.
Кучеренко С.П. Воздействие амурского тигра на популяции его жертв И Редкие
виды млекопитающих и их охрана. М.: Наука, 1977а. С. 133-134.
Кучеренко С.П. Регулировать численность хищников И Охота и охотничье хоз-
во. 19776. № 10. С. 16-19.
Кучеренко С.П. Цена охраны тигра И Там же. 1993. № 2. С. 16-19.
Кучеренко С.П. Амурский тигр // Там же. 1994. № 2. С. 14-18.
Кучмель С.В. К оценке изменчивости питания волка И Структурно-
функциональное состояние биологического разнообразия животного мира Бе-
ларуси. Минск, 1999. С. 75-78.
Лавов М.А. Распространение и примерная численность копытных зверей в Крас-
ноярском крае // Вопросы экологии. Тр. гос. заповедника «Столбы». 1975.
Вып. 10. С. 339-347.
Лавов М.А. Вероятные причины расширения кабана в СССР // Копытные фауны
СССР. М.: Наука, 1980а. С. 27-28.
276
Лавов М.А. Причины гибели копытных // Охота и охотничье хоз-во. 19806. № 7.
С. 16-17.
Лавов М.А. Динамика и регулирование численности кабана в Березинском запо-
веднике // Заповедники Белоруссии. 1981. Вып. 5. С. 93-98.
Лавов М.А. Волк, медведь, рысь и их роль как хищников в Березинском заповед-
нике И IV съезд Всесоюз. териол. о-ва. М., 1986а. Т. 2. С. 14-16.
Лавов М.А. Управление популяциями диких копытных в Березинском заповед-
нике // Проблемы охраны генофонда и управления экосистемами в заповедни-
ках лесной зоны. М., 19866. Ч. 2. С. 130-132.
Лавов М. Волк в Березинском заповеднике // Охота и охотничье хоз-во. 1986в. №
11. С. 10-11.
Лавов М.А. Изменение качества угодий и численности охотничьих животных
под влиянием режима заповедности И Влияние антропогенной трансформации
ландшафта на население наземных позвоночных животных. М., 1987. Ч. 1. С.
121-123.
Лавов М.А., Воронова Т.Н. Влияние заповедного режима на состояние популя-
ций копытных и хищных зверей И Проблемы охраны генофонда и управления
экосистемами в заповедниках лесной зоны. М., 1986. Ч. 2. С. 133-135.
Лавров Н.П., Хахин ГВ. Результаты расселения охотничьих животных в Москов-
ской области И Тр. каф. биол. педвузов РСФСР. М., 1974. Вып. 30. С. 144-152.
Лавровский А.А. Кабан в дельте Волги. Астрахань: Изд-во «Волга», 1962. 67 с.
(Тр. Астраханского заповедника. Вып. 7).
Ладан П.Е. Межвидовая гибридизация в свиноводстве // Тр. Новочеркасского
зоовет. ин-та. 1960. Вып. 2. С. 47-53.
Лазаревский В.М. Об истреблении волком домашнего скота и дичи и об истреб-
лении волка. Владимир на Клязме, 1876. 106 с.
Лаптев С.П. Стадиальное распределение и роющая деятельность кабана в лесных
охотничьих угодьях Смоленской области И Биологические основы охотничьего
дела. М„ 1983. С. 71-77.
Лаптев С.П., Кузьмин И.Ф., Гасевич С.Д. Кабан в условиях интенсивного сель-
скохозяйственного производства И Влияние антропогенной трансформации
ландшафта на население наземных позвоночных животных. М., 1987. Ч. 1. С.
179-181.
Лебедева Л.С. Экологические особенности кабана Беловежской пущи // Учен. зап.
Моск. гор. пед. ин-та им. В.П. Потемкина. 1956а. Т. 61, вып. 4-5. С. 105-271.
ЛебедеваЛ.С. О составе летних кормов кабана в высокогорной зоне Кавказского
заповедника// Там же. 19566. Т. 61, вып. 4-5. С. 273-289.
Лебедева Л.С., Качанова А.А. О кормах и роющей деятельности кабана в запо-
веднике Беловежская пуща // Там же. 1955. Т. 38, вып 3. С. 141-146.
Легейда ИС. О роли роющей деятельности кабана в зоне ЧАЭС И Радиобиоло-
гический съезд. Пущино, 1993. Ч. 2. С. 585-586.
Легейда И.С., Панов ГМ. Некоторые особенности биологии кабана в зоне ЧАЭС
И Там же. 1993. Ч. 2. С. 587-588.
Ли В.А., Калдыбаев С. У., Тян П.Е. Семиреченская порода свиней. Алма-Ата: Кай-
нар, 1981. 139 с.
Линг ХИ. Изменения в распространении кабана в Советской Прибалтике за по-
следние столетия // Ежегодник о-ва естествоиспыт. при АН ЭстССР. Таллин,
1955. Т. 48. С. 176-200.
ЛингХ.И. Структура и динамика популяции промысловых млекопитающих: Ав-
тореф. дис. ... докт. биол наук. Л., 1978. 37 с.
Литвин М. О нравах татар, литовцев и московитян. Извлечение из сочинения Ми-
хаила Литвина (1550) // Мемуары, относящиеся к истории Южной Руси. К1ев,
1890. Вып. 1. (XVI ст.). С. 1-58.
277
Литвинов В.П. Влияние волка на численность кабана в Кызыл-Агачском запо-
веднике // Копытные фауны СССР. М.: Наука, 1980. С. 173-174.
Литвинов В.П. Волк (Canis lupus) и кабан (Sus scrofa) в Кызыл-Агачском запо-
веднике//Зоол. журн. 1981. Т. 60, вып. 10. С. 1588-1591.
Литвинов В.П., Коновалова НА. Численность и размещение кабана (Sus scrofa
L.) в угодьях Кызыл-Агачского государственного заповедника И Изв. АН
АзССР. Сер. биол. наук. 1979. № 1. С 70-73.
Литвинов В.И, Литвинов В.Ф., Тиханский А.Д. Питание волка в Березинском и
Припятском заповедниках// Заповедники Белоруссии. 1981. Вып. 5. С. 98-102.
Литвинов В.Ф. Болезни лося и кабана И Копытные фауны СССР. М.: Наука,
1975.С. 150-151.
Литвинов В. Ф. Паразитарные болезни лося и кабана // Копытные фауны СССР.
М.: Наука, 1980а. С. 174-175.
Литвинов В. Ф. Лечебно-профилактические мероприятия при паразитарных бо-
лезнях диких копытных // Там же. 19806. С. 176-177.
Литвинов В.Ф., Ильясевич В.А., Дунин В.Ф. Роль хищников в гибели диких ко-
пытных И Экологические основы охраны и рационального использования
хищных млекопитающих. М.: Наука, 1979. С. 44-46.
Литвинов В. Ф., Карасев И. Ф. Эхинококкоз животных Березинского заповедника
//Заповедники Белоруссии. 1977. Вып. 1. С. 134-136.
Литвинов В.Ф., Пачкаев В.И., Ильясевич В.А. Влияние хищников на поголовье
диких копытных // Копытные фауны СССР. М.: Наука, 1975. С. 201.
Лихацкий Ю.П. Экологические основы функционирования, принципы охраны и
эксплуатации сообществ копытных животных русской лесостепи: Автореф.
дис. ... д-ра биол. наук. М., 1966. 40 с.
Лобачев В.С., Петухов А.Г. Численность, размещение и питание крупных хищ-
ных млекопитающих в Кызыл-Агачском заповеднике // Бюл. МОИП. Отд.
биол. 1984. Т. 89, вып. 4. С. 45-54.
Логинов В.В. Новые данные к познанию биологии кавказского кабана (Sus scrofa
attila Thomas) // Бюл. МОИП. Отд. биол. 1936. Т. 45, вып. 1. С. 10-21.
Лозан А.М. Социальная организация популяции кабана // Копытные фауны
СССР. М.: Наука, 1980а. С. 293-294
Лозан А.М. Роль врожденных и приобретенных компонентов ориентационного
поведения дикого кабана И Там же. 19806. С. 294-296.
Лозан А.М. Сложные формы групповой защиты дикого кабана // Фауна, эколо-
гия и физиология животных. Кишинев, 1980в. С. 39-43.
Лозан А.М. Агрессивно-оборонительное поведение дикого кабана и его адаптив-
ное значение // Экология птиц и млекопитающих Молдавии. 1981. С. 106-124.
Лозан А.М. Об адаптациях дикого кабана И Эколого-этологические и физиологи-
ческие исследования животных. Кишинев: Штиинца, 1982. С. 23-32.
Лозан А.М. Дикий кабан (Sus scrofa L.) в Молдавии. Его этология и практическое
использование: Автореф. дис. ... канд. биол. наук. Киев, 1983. 23 с.
Лозан А.М. Структура популяции и территориальность дикого кабана (Sus scrofa
L.) //Природа заповедника «Кодры». Кишинев: Штиинца, 1984. С. 88-93.
Лозан А.М. Передача и прием информации у диких свиней И Адаптация птиц и
млекопитающих к антропогенному ландшафту. Кишинев: Штиинца, 1988. С.
81-86.
Лозан М.Н., Лозан А.М. Агрессия и защита у кабана И II съезд Всесоюз. териол.
о-ва. М.: Наука, 1978а. С. 223-224.
Лозан М.Н., Лозан А.М. Пространственная ориентация дикого кабана И Там же.
19786. С. 224-225.
Лозан М.Н., Лозан А.М. Пространственно-этологическая структура популяции
дикого кабана в весенне-летний период И Экология, структура популяций и
278
внутривидовые коммуникативные процессы у млекопитающих. М.: Наука,
1981а. С. 151-181.
Лозан М.Н., Лозан А.М. Пространственная структура популяции дикого кабана и
ее адаптивное значение И Новые данные об экологии и охране флоры и фауны
СССР. Докл. МОИП. Зоология и ботаника. 1979 г. М., 19816. С. 26-28.
Лозан М., Лозан А. Дикий кабан: вчера, сегодня, завтра И Сельское хоз-во Мол-
давии. 1981 в. № 8. С. 47.
Лозан М.Н., Лозан А.М. О возникновении поведенческих признаков у высших
позвоночных и их роли в эволюции // Эколого-этологические и физиологиче-
ские исследования животных. Кишинев: Штиинца, 1982. С. 3-12.
Ломанов И.К. Закономерности динамики численности и размещения населения
лося в европейской части России. М., 1995. 60 с.
Ломанов И.К. Кабан // Охотничьи животные России. 2000. Вып. 2. С. 24-31.
Ломанов И.К., Ломанова Н.В. Лось И Там же. 2000. Вып. 2. С. 13—23.
Лоскутова Н.М. Изменение численности тетеревиных в Башкирском заповедни-
ке // Тетеревиные птицы в заповедниках РСФСР. М., 1989. С. 50-55.
Лукаревский В.С. Леопард в антропогенном ландшафте Юго-Западного Копетда-
га // IV съезд Всесоюз. териол. о-ва. М., 1986. Т. 2. С. 72-73.
Лукаревский В.С. Современное состояние популяций крупных хищных млекопи-
тающих Западного Копетдага и прилежащих территорий: Автореф. дис. ...
канд. биол. наук. М., 1995. 23 с.
Лыхваръ В.П. Об изменениях численности и видового соотношения у тетереви-
ных птиц в бассейне р. Малая Сосьва, произошедших со времен ликвидации
Кондо-Сосьвинского заповедника // Тетеревиные птицы в заповедниках
РСФСР. М„ 1989. С. 56-60.
Любимова ЕЛ. О распространении широколиственных пород на Русской равни-
не в историческое время (по данным топонимики) // Исследования географии
природных ресурсов животного и растительного мира. М.: Изд-во АН СССР,
1962. С.95-100.
Любченко О.В. Современное распространение кабана в средних и северных об-
ластях европейской части СССР И Бюл. МОИП. Отд. биол. 1969. Т. 74, вып. 2.
С. 17-25.
Любченко О.В. Кабан в Воронежском заповеднике // Бюл. МОИП. Отд. биол.
1973. Т. 78, вып. З.С. 17-29.
Любченко О.В., Мамаев Ю.Ф. Анкетный метод контроля за состоянием популя-
ции копытных и составом промысловой пробы (на примере кабана) // Научно-
технический прогресс - в практику перестройки охотничьего хозяйства. М.,
1988. С. 142-144.
Мазин Л.Н. В охотничьих хозяйствах Латвии И Научно-технический прогресс - в
практику перестройки охотничьего хозяйства. М., 1988. С. 31-33.
МайнхардтX. Моя жизнь среди кабанов. М.: Лесн. пром-сть, 1983. 129 с.
МайнхардтX. Рассказы о диких кабанах. Минск: Ураджай, 1986. ПО с.
Макарова Л.А. Дикие млекопитающие из археологических памятников эпохи
неолита и бронзы Казахстана // История биогеоценозов СССР в голоцене. М.:
Наука, 1976. С. 268-270.
Макарова О.А., Хохлов А.М. О распространении кабана в Калининской области
(Sus scrofa L.) // Тр. Завидовского заповедно-охотничьего хоз-ва. М.: Воениз-
дат, 1971. Вып. 2. С. 69-73.
Максимов И. Итоги 1973 г. // Охота и охотничье хоз-во. 1974. № 11. С. 6-7.
Максимов И., Романов С. Разведение кабанов в социалистических странах И
Охота и охотничье хоз-во. 1976. № 6. С. 44-45.
Малов О. С собакой на кабана // Охота и охотничье хоз-во. 1989. № 11. С. 18-21.
279
Мамаев Ю. Подкормка кабанов: всегда ли она нужна? И Охота и охотничье хоз-
во. 1986. № 11. С. 14-16.
Мануш Г. Кабанья напасть И Нечерноземье. 1989. № 4. С. 42-44.
Мануш П., Николаев В. Мечение - перспективный метод изучения кабана И Охо-
та и охотничье хоз-во. 1995. № 6. С. 12-15.
Мануш С.Г. Новые данные о кабане и ведении хозяйства на этого зверя в Зави-
довском заповедно-охотничьем хозяйстве // Тр. Завидовского заповедно-
охотничьего хозяйства. М.: Воениздат, 1969. Вып. 1. С. 251-261.
Маньен Ф. Франция: на каких кабанов охотиться? И Охота. 1999. № 9-10. С. 10-12.
Марвин М.Я. Млекопитающие Карелии. Петрозаводск: Гос. изд-во Карельской
АССР, 1959. 239 с.
Марвин М.Я. Фауна наземных позвоночных Урала. Свердловск, 1969. Вып. 1.
Млекопитающие. 155 с.
Марков Е.Л. Дикие свиньи Закавказья. Тифлис: Изд. Заккнига, 1932. 40 с.
Марков Н.И. К вопросу о путях формирования популяции кабана на Среднем
Урале И Механизмы поддержания биологического разнообразия. Екатерин-
бург, 1995. С. 98-100.
Марков Н.И. Современное состояние популяции кабана в Свердловской области
И Вопросы прикладной экологии (природопользования), охотоведения и зве-
роводства. Киров, 1997а. С. 158-159.
Марков Н.И. Динамика численности кабана в Свердловской области и ее связь с
климатическими факторами // Экология. 19976. № 4. С. 305-310.
Марков Н.И. Биология кабана (Sus scrofa L.) на Среднем Урале: Автореф. дис. ...
канд. биол. наук. Екатеринбург, 1999. 16 с.
Марковский В.А. Копытные Карелии (современное состояние популяций): Авто-
реф. дис. ... канд. биол. наук. Петрозаводск, 1995. 32 с.
Матвеев А.С., Бакунин В.А. Промысловые звери и птицы Челябинской области.
Челябинск, 1994. 383.
Матюшкин Е.Н. Приемы охоты и поведение у добычи амурского тигра И Бюл.
МОИП. Отд. биол. 1991. Т. 96, вып. 1. С. 10-27.
Матюшкин Е.Н. Тигр и изюбрь на приморских склонах Среднего Сихотэ-Алиня
//Там же. 1992. Т. 97, вып. 1. С. 3-20.
Матюшкин Е.Н., Астафьев А.А., Зайцев В.А., Костоглод Е.Е. и др. История, со-
временное состояние и перспектива охраны тигра в Сихотэ-Алинском запо-
веднике//Хищные млекопитающие. М., 1981. С. 76-118.
Мелехин А. Добыча лосей и кабанов И Охота и охотничье хоз-во. 1983. № 9. С.
17.
Мельникова Т.Г. Паразиты кабана (Sus scrofa L.) Средней Азии: Автореф. дис. ...
докт. биол. наук. М.: МГУ, 1968. 31 с.
Метелкин А.И., Захарченко Н.М., Михалько Н.С. и др. Отравлены фосфидом
цинка // Охота и охотничье хоз-во. 1974. № 9. С. 29.
Метельский А.П. Численность диких копытных в Приморском крае // Копытные
фауны СССР. М.: Наука, 1975. С. 55-56.
Мигулин О. Щкидни та корисни звири УкрашЁ Харьюв: Рад. Селянин, 1927. 166 с.
Миленушкин Ю. Предисловие И Волки и их истребление. М.: Воениздат, 1957. С.
3-8.
Миллер И.Д., Скалой О.В. О состоянии популяции кабана в Тульской области //
V съезд Всесоюз. териол. о-ва АН СССР. М., 1990. Т. 3. С. 109-110.
Михайлов А.М., Клюкин НК. Волки Подмосковья. М., 1996. 193 с.
Морозов Ю.Ф., Назарова Н.С. К вопросу о гельминтозах диких копытных Бело-
вежской пущи ИII зоол. конф. БССР. Минск, 1962. С. 168-170.
280
Москаленко Ю.А. Использование территории дикими кабанами (Sus scrofa L.) в
регионе лесостепных участков Черноморского биосферного заповедника // VI
съезд териол. о-ва. М., 1999. С. 168.
Москвитин В.П., Смирнов М.Н. Ареал, ресурсы, использование и охрана диких
копытных Бурятии // Копытные фауны СССР. М.: Наука, 1975. С. 115-116.
Мошонкин Н.Н. Значение для кабана низовьев дельты Волги некоторых элемен-
тов антропогенного ландшафта // Влияние антропогенной трансформации
ландшафта на население наземных позвоночных животных. М., 1987. Ч. 1. С.
182-183.
Муравицкий А. Лось и кабан в Белоруссии // Охота и охотничье хоз-во. 2001. №
3. С. 1-3.
Муравицкий В. Облавные охоты на кабана И Там же. 1983. № 1. С. 6-9.
Муравицкий В.А. Во что обходятся дикие кабаны? // Сельское хоз-во Белоруссии.
1988. № 12. С. 34-35.
Мусиевский А.Л., Ильин А.М. Плотность копытных и состояние пойменных дуб-
равных фитоценозов Хоперского государственного заповедника // Проблемы
охраны генофонда и управления экосистемами в заповедниках лесной зоны.
М., 1986. Ч. 2. С. 163-164.
Мухамедгалиев Ф.М., Исенжулов А.И., Еськов П.А., Ли В.А. Отдаленная гибри-
дизация в свиноводстве //Вести, с.-х. науки. 1970. № 3. С. 171.
Мысленков А.И., Волошина ИВ. Авиаучеты копытных в Сихотэ-Алинском запо-
веднике // Экология, морфология, использование и охрана диких копытных.
М„ 1989. Ч. 1. С. 66-67.
Назарова НС., Околелое В.И., Косачев Н.Н. Кабаны - новоселы Сибири // Земля
сибирская, дальневосточная. 1982. № 8. С. 47.
Назарова Н.С., Стародынова А.К. Гельминты диких парнокопытных в лесах Ка-
лининской и Московской областей И Тр. Завидовского гос. научно-опытного
заповедника. М.: Воениздат, 1974. Вып. 3. С. 173-180.
Назарова Н.С., Стародынова А.К., Сыроежкин Ю.М. Лечение кабанов от метас-
тронгилеза в угодьях Завидовского государственного научно-опытного запо-
ведника // Там же. 1974. Вып. 3. С.181-184.
Насимович А.А. О некоторых закономерностях зимнего распространения копыт-
ных в горах западного Кавказа // Бюл. МОИП. Отд. биол. 1936. Т. 45, вып. 1.
С. 5-9.
Насимович А.А. К познанию минерального питания диких животных Кавказско-
го заповедника // Тр. Кавказского гос. заповедника. 1938. Вып. 1. С. 103-152.
Насимович А.А. Промысловые звери Центрального Алтая // Пушные богатства
СССР. М.: Заготиздат, 1949. Вып. 1. С. 127-148.
Насимович А.А. Роль режима снежного покрова в жизни копытных животных на
территории СССР. М.: Изд-во АН СССР, 1955. 403 с.
Наумов Н.П. Экология животных. М.: Высш, шк., 1963. 618 с.
Наумов Н.П. Некоторые проблемы популяционной биологии и охотничье хозяй-
ство//Вопр. охотничьего хозяйства СССР. М.: Колос, 1965. С. 13-18.
Неганов В.Г., Никулин В.Ф., Ладыгина ГД. и др. Отряд Парнопалые (Artiodactyla) //
Животный мир Прикамья. Пермское кн. изд-во, 1989. С. 170-174.
Недзельский Е.М. Кабан Прибайкалья Sus scrofa sibiricus Straffe, 1922 (черты
биологии, охрана и хозяйственное использование): Автореф. дис. ... канд.
биол. наук. М., 1944. 28 с.
Нейфелъд Н.Д. Состояние населения копытных в Северном Предуралье И Эколо-
гия, морфология, использование и охрана диких копытных. М., 1989. Ч. 1. С.
67-69.
Нестеренко В.В. Роль волка в природных комплексах заповедников // Популяцион-
ные исследования животных в заповедниках. М.: Наука, 1988. С. 139-144.
281
Нестеренко В.В. Формирование специализации питания волка на примере его
взаимоотношений с кабаном в зоне южной тайги И Экология, поведение и
управление популяциями волка. М., 1989. С. 96-102.
Нечипоренко В.Х., Першин М.М. Карютип европейських диких свиней // Свинар-
ство. 1976. Вып. 24. С. 76-79.
Никифоров В., Бринза В. Эффективность подкормки кабанов в Осташовском
охотхозяйстве Московского общества охотников и рыболовов // Вестник Ро-
сохотрыболовсоюза. М.: Росагропромиздат, 1991. С. 22-24.
Никифоров Л.П. Опыт биосъемки населения млекопитающих Тоболо-Ишимской
лесостепи // География населения наземных животных и методы его изучения.
М.: Изд-во АН СССР, 1959. С. 7-22.
Никулин В.С. Состав растительных кормов в питании кабана Иркутской области
// Воспроизводство и организация использования промысловых зверей и птиц
в Сибири и на Дальнем Востоке. Иркутск, 1980. С. 74-78.
Никулин В.С. Опыт зимней подкормки кабанов в Иркутской области // Биология,
охрана и хозяйственное использование зверей и птиц в Восточной Сибири.
Иркутск, 1981.
Нинов Л.К., Хрусталева И.В. Морфология скелета домашних и диких свиней в
постнатальном онтогенезе // Морфология и генетика кабана. М.: Наука, 1985.
С.122-130.
Новиков ГА., Тимофеева Е.К. Урбанизация природных ландшафтов и синантропи-
зация лесных копытных // Копытные фауны СССР. М.: Наука, 1980. С. 38-39.
Новиков И. Живоловушки для кабанов // Разведение дичи и рыбы. М.: Россель-
хозиздат, 1987. С. 22-25.
Нуратдинов Т. Влияние кабана на поселения ондатры в водоемах Южного При-
аралья//V съезд Всесоюз. териол. о-ва АН СССР. М., 1990. Т. 3. С. 110-111.
Нургелъдыев О.Н., Щербина Е.И., Маринина Л.С. Влияние Каракумского канала
на состав и распределение млекопитающих И III съезд Всесоюз. териол. о-ва.
М„ 1982. Т. 1. С. 258.
Обидина В.А. Материалы по экологии дикого кабана заповедника Аксу-
Джабаглы (Западный Тянь-Шань) // III съезд Всесоюз. териол. о-ва. М., 1982.
Т. 1. С. 259.
Обидина В.А. К вопросу о социальной структуре дикого кабана Таласского Ала-
тау // Экология и охрана горных видов млекопитающих. М., 1987. С. 129-131.
Овсюкова Н.И. О зараженности кабанов Подмосковья метастронгилидами //
Охотоведение. М.: Лесн. пром-сть, 1974. С. 99-102.
Овсюкова Н.И. Эколого-фаунистический обзор гельминтофауны кабанов Под-
московья и методы дегельминтизации И Охотоведение. М.: Лесн. пром-сть,
1975. С.267-295.
Овсюкова Н.И., Бельский А.В. Селекционный отстрел поросят // Копытные фау-
ны СССР. М.: Наука, 1980. С. 248-249.
Овсюкова Н.Э. Кабан // Ресурсы основных видов охотничьих животных и охот-
ничьи угодья России (1991-1995 гг.). М., 1996. С. 77-98.
Одинцова Т.М. Особенности динамики гельминтозов диких копытных в услови-
ях ПГРЭЗ //10 лет Полесскому гос. радиационно-экологическому заповедни-
ку. Минск, 1998. С. 221-225.
Окладников А.П. Неолит и бронзовый век Прибайкалья. Историко-
археологическое исследование // Материалы и исследования по археологии
СССР. 1950. №18. С. 5-412.
Околелое В.И, Косачев Н.Н. Сибирская популяция кабанов // Земля сибирская,
дальневосточная. 1987. № 7. С. 8-9.
282
Онипченко В.Г., Голиков К.А. Демутационные смены после пороев кабанов на
альпийских лишайниковых пустошах в Тебердинском заповеднике // Бюл.
МОИП. Отд. биол. 1996. Т. 101, вып. 4. С. 49-54.
Орлов В.Н., Булатова Н.Ш. Сравнительная цитогенетика и кариосистематика
млекопитающих. М.: Наука, 1983. 405 с.
Орлов В.Н., Мазин С.М. О некоторых проблемах систематики Sus scrofa L. //
Морфология и генетика гибридных свиней. М.: Наука, 1992. С. 48-52.
Осмоловская В.И., Приклонский С.Г. К оценке ущерба от волка животноводству
в европейской части РСФСР // Экологические основы охраны и рационально-
го использования хищных млекопитающих. М.: Наука, 1979. С. 129-131.
Останин В. Курганская популяция сибирской косули: проблемы управления //
Охота и охотничье хоз-во. 1996. № 6. С. 24-27.
Паавер К.Л. Формирование териофауны и изменчивость млекопитающих При-
балтики в голоцене. Тарту, 1965. 510 с.
Павлинов И.Я., Россолимо О.Л. Систематика млекопитающих СССР. М.: Изд-во
МГУ, 1987. 285 с.
Павлов Б.К. Управление популяциями охотничьих животных. М.: Агропромиз-
дат, 1989. 144 с.
Павлов М. Нашествие кабанов И Охота и охотничье хоз-во. 1978. № 12. С. 18-19.
Павлов М.П. Состояние и перспективы акклиматизации охотничьих животных в
СССР // Акклиматизация охотничьих животных в СССР. Минск: Ураджай,
1978. С. 3-15.
Павлов М.П. Волк. М.: Агропромиздат, 1990. 352 с.
Павлов М.П. Акклиматизация охотничье-промысловых зверей и птиц в СССР.
Киров, 1999а. Ч. 3. Копытные. 666 с.
Павлов М. Кабан: почему он прижился в вятской тайге И Охота и охотничье хоз-
во. 19996. №7. С. 8-11.
Павлов М.П., Зарубин Б.Е. Кабан в таежных лесах Кировской области И Обога-
щение фауны и разведение охотничьих животных. Киров, 1982. С. 95.
Павлов М.П., Корсакова И.Б., Лавров Н.П. Акклиматизация охотничье-
промысловых зверей и птиц в СССР. Киров: Волго-Вятское кн. изд-во, 1974.
4.2. 459 с.
Падайга В.И. Биохимический состав кормов // Охотничье хозяйство в интенсив-
ном комплексном лесном хозяйстве. Каунас: Гирионис, 1975. С. 18-19.
Падайга В.И. Особенности разведения копытных в интенсивном охотничьем и
лесном хозяйстве // Обогащение фауны и разведение охотничьих животных.
Киров, 1982. С. 187.
Падайга В.И., Янулайтис З.П. Кабан в современном культурном ландшафте И
Хозяйственная деятельность и охотничья фауна. Киров, 1980. Т. 1. С. 172-173.
Паллас П.С. Путешествие по разным провинциям Российской Империи. СПб.,
1773. Кн. 1. 551 с. (1809. Ч. 1. 657 с.)
Паллас П.С. Путешествие по разным местам Российского государства по пове-
лению Санкт-Петербургской Императорской академии наук. СПб., 1786.
(1770. Ч. 2, Кн. 2). 571 с.
Панов Г.М., Легейда И.С. Динамика ресурсов кабана в 30-км зоне ЧАЭС в послеава-
рийный период // Радиобиологический съезд. Пущино, 1993. Ч. 2. С. 761.
Папонов В.А. Совершенствовать хозяйство на лося // Научно-технический про-
гресс - в практику перестройки охотничьего хозяйства. М., 1988. С. 152-153.
Паровщиков В.Я. Изменения ареалов и новые данные о границах распростране-
ния некоторых млекопитающих севера европейской части СССР // География
населения наземных животных и методы его изучения. М.: Изд-во АН СССР,
1959. С.217-226.
283
Пашук В.П. Материалы изучения инвазированности трихинеллами охотничьих
животных в Белоруссии И Тр. IX Междунар. конгр. биологов-охотоведов. М.,
1970. С.700-708.
Пелтон М.Р., Конли Р.Х. Статус Sus scrofa L. (Artiodactyla) в Северной Америке
ИI Междунар. конгр. по млекопитающим. М.: ВИНИТИ, 1974. Т. 2. С. 110.
Пенькевич В.А. Эколого-гельминтологическая характеристика биотопов дикого
кабана Беларуси // Охраняемые природные территории и объекты Белорус-
ского Поозерья: современное состояние, перспективы развития. Витебск,
1997.С. 110-111.
Пенькевич В.А. Гельминты и гельминтозы дикого кабана // Ветеринарная наука-
производству. Минск: Белор. изд. тов-ство «Хата», 1998а. С. 151-158.
Пенькевич В.А. Эффективность некоторых антигельминтиков при нематодозах
дикого кабана//Там же. 19986. С. 159-163.
Перовский М.Д. Методы управления популяциями ценных охотничьих живот-
ных России: Автореф. дис. ... д-ра биол. наук. М., 1998. 59 с.
Петрашов В.В. Воспроизводство ресурсов охотничьих животных и концепция ко-
эволюции природы и общества: Автореф. дис. ... д-ра биол. наук. М., 1998. 44 с.
Петренко А.Г. Древнее и средневековое животноводство Среднего Поволжья и
Предуралья. М.: Наука, 1984. 174 с.
Петрищев Б.И. Линейно-весовой рост эмбрионов дикой свиньи (Sus scrofa) //
Вести, зоологии. 1980. Вып. 6. С. 63-67.
Петрищев Б.И. Эмбриональное развитие кожно-волосяного покрова кабана (Sus
scrofa) И Морфология и генетика кабана. М.: Наука, 1985. С. 105-114.
Петрищев Б.И. Сравнительная морфофункциональная оценка кожи новорож-
денных поросят различного происхождения И Морфология и генетика гиб-
ридных свиней. М.: Наука, 1992. С. 63-71.
Петрищев Б.И., Бородин А.Л. Особенности формирования волосяного покрова
гибридных свиней И Популяционная изменчивость вида и проблемы охраны
генофонда млекопитающих. М., 1983. С. 152-154.
Петрищев Б.И., Бородин А.Л. Развитие эмбрионов кабана популяции широколи-
ственных лесов Брянской области // Морфология и генетика кабана. М.: Нау-
ка, 1985. С. 87-104.
Петской П., Бекеш П, Фролов М. Как устранить неприятный запах мяса кабана
//Охота и охотничье хоз-во. 1974. № И. С. 14.
Петялис КА. Развитие и качество клыков у кабанов Южной Литвы И Териоло-
гические исследования в Литве. Вильнюс, 1988. С. 54-58.
Печенюк А.Д. Волк в Хоперском заповеднике // Проблемы изучения и охраны
заповедных природных комплексов. Воронеж: Изд-во Воронежского ун-та,
1995. С. 70-73.
Пиголкин А.У., Бокун В.А. Трихинеллез диких животных на Северном Кавказе //
Болезни и паразиты диких животных. М., 1992. С. 164-170.
Пидопличко П.Г. О ледниковом периоде. Киев: Изд-во АН УкрССР, 1951. Вып.
2. 264 с.
Пидопличко П.Г. О ледниковом периоде. Киев: Изд-во АН УкрССР, 1954. Вып.
3. 220 с.
Шдогипчко 1.Г. Матер1али до вивчення минулих фаун УРСР. 1956. Вып. 2. 235 с.
Пидопличко П.Г. Современные проблемы и задачи изучения истории фаун и
среды их обитания И Природная обстановка и фауны прошлого. Киев: Изд-во
АН УкрССР, 1963. Вып. 1. С. 9-30.
Пикунов Д.Г. Размеры хищничества амурских тигров И Редкие и исчезающие
животные суши Дальнего Востока СССР. Владивосток, 1981. С. 71-75.
Пикунов Д.Г. Амурский тигр и его влияние на диких копытных Приморья // Ред-
кие виды млекопитающих СССР и их охрана. М., 1983. С. 128-130.
284
Пикунов Д.Г., Базылъников В.И., Рыбачук В.В. Методы изучения и экологические
основы охраны тигра в Приморье И Изучение и охрана редких и исчезающих
видов животных фауны СССР. М.: Наука, 1985. С. 70-74.
Пикунов Д.Г., Коваленок С.И., Фоменко П.В. О гибели диких копытных в Сихо-
тэ-Алине после многоснежной зимы 1987-1988 гг. И Экология, морфология,
использование и охрана диких копытных. М., 1989а. Ч. 1. С. 69-70.
Пикунов Д.Г, Фоменко П.В., Коваленок С.И. Адаптация уссурийских кабанов к
условиям многоснежной зимы // Там же. М., 19896. Ч. 2. С. 211-213.
Плешак Т., Миняев А. Кабан в Архангельской области И Охота и охотничье хоз-
во. 1986. №7. С. 14-15.
Побединский Г.А. Начальная реакция популяций копытных на снижение числен-
ности волка // Экология, морфология, использование и охрана диких копыт-
ных. М„ 1989. Ч. 1.С. 71.
Побединский Г.Д. Антропогенное воздействие на динамику численности копыт-
ных Центрального Черноземья (на примере Липецкой области): Автореф. дис.
... канд. биол. наук. Воронеж, 1997. 19 с.
Побединский Г.Д. Роль охотничьих заказников в сохранении фауны копытных И
VI съезд териол. о-ва. 1999. С. 200.
Поддубная Н.Я., Ковалев В.А. Тигр в Уссурийском заповеднике: состояние и пер-
спективы сохранения // Бюл. МОИП. Отд. биол. 1993. Т. 98, вып. 3. С. 54-62.
Покровский В.С., Щадилов Ю.М. Влияние деятельности человека на изменение
ареала и численности лося И Охрана природы и заповедное дело в СССР. Бюл.
АН СССР. 1962. № 7. С. 57-65.
Полуйко Ю.В. Кабан в Калининской области // Фауна Нечерноземья, ее охрана,
воспроизведение и использование. Калинин, 1980. С. 87-90.
Попов В.А. Наземные позвоночные животные Волжско-Камского края (история
формирования, современное состояние, перспективы развития и использова-
ния): Докл. дис. ... докт. биол. наук. М., 1964. 43 с.
Попов В.А. Редкие и исчезающие виды животных Татарии (позвоночные). Ка-
зань: Тат. кн. изд-во, 1978. 96 с.
Попов Ю.К. О заселении кабаном Удмуртской АССР И IV съезд Всесоюз. тери-
ол. о-ва. М„ 1986. Т. 2. С. 210.
Попов Ю.К, Попов Н.Ю. Кабан в Удмуртской АССР И Копытные фауны СССР.
М.: Наука, 1980. С. 196-197.
Пржевальский Н.М. Путешествие в Уссурийском крае в 1868-1869 годах. СПб.,
1870.297 с.
Приклонский С.Г. Воздействие волка на домашних животных // Волк. М.: Наука,
1985. С.374-377.
Приклонский С.Г. Научные основы мониторинга охотничьих животных на больших
территориях: Дис. в виде научного доклада ... д-ра биол. наук. М., 1999. 52 с.
Прилуцкая Л.И., Пишванов Ю.В. Современная численность и размещение охот-
ничье-промысловых животных Дагестана // Ресурсы животного мира Север-
ного Кавказа. Ставрополь, 1988. С. 136-140.
Прокофьев С.М. Фауна и состояние численности охотничьих млекопитающих в
Хакасии // Экология промысловых животных Сибири. Изд-во Красноярского
ун-та, 1992. С. 20-37.
Прокошев Н.А. К вопросу о неолитических памятниках Камского Приуралья II
Материалы и исследования по археологии СССР. 1940. № 1. С. 11-40.
Простаков Н.И. Особенности становления фауны диких парнокопытных Сред-
него Подонья // Состояние и проблемы экосистем Усманского бора. Воронеж,
1992. Вып. 2. С. 45-58. (Тр. биол. уч.-научн. базы Воронежского гос. ун-та
«Веневитово»),
285
Простаков Н.И. Эколого-морфологическая характеристика популяции кабана
Центрального Черноземья И Там же. 1994. Вып. 4. С. 79-83.
Простаков Н.И. Копытные животные Центрального Черноземья. Воронеж, 1996.
375.
Равилов А.З., Юсупов Р.Х., Верхолетов В.А., Ахметов Р.М. К эпизоотологии за-
разных болезней, распространяемых дикими животными // Болезни и парази-
ты диких животных. М., 1992. С. 27-31.
Равкин Е.С. Ресурсы диких копытных животных Северо-Восточного Кавказа //
Охотоведение. М.: Лесн. пром-сть, 1976. С. 82-95.
Равский Э.И., Александрова Л.П., Вангенгейм Э.А и др. Антропогеновые отложе-
ния юга Восточной Сибири // Тр. Геол, ин-та. 1964. Вып. 105. 280 с.
Размахнин В.Е. Совершенствовать систему использования ресурсов охотничье-
промысловых видов диких копытных // Научно-технический прогресс - в
практику перестройки охотничьего хозяйства. М., 1988. С. 158-160.
Раков Н.В. Некоторые особенности условий существования кабана на Сихотэ-
Алине // Бюл. МОИП. Отд. биол. 1956. Т. 61, вып. 1. С. 13-22.
Раков Н.В. О факторах смертности кабана и его взаимоотношениях с хищниками
в Приамурье // Зоол. журн. 1970. Т. 49, вып. 8. С. 1220-1228.
Раков Н.В. О роли хищников в ограничении численности копытных в Приамурье
и Приморье // Экологические основы охраны и рационального использования
хищных млекопитающих. М.: Наука, 1979. С. 58-59.
Реймов Р., Карабеков М. Состояние популяции копытных Каракалпакии И Ко-
пытные фауны СССР. М.: Наука, 1975. С. 276-277.
Реймов Р., Карабеков М. Распространение и численность кабана в низовьях
Амударьи // Вести. Каракалп. фил. АН УзбССР. 1984. № 2. С. 31-36.
Реймов Р., Карабеков М., Нуратдинов Т. Современное состояние популяции ка-
бана и сайгака в Южном Приаралье // Экология, морфология, использование и
охрана диких копытных. М., 1989. Ч. 2. С. 213-214.
Рекомендации по предотвращению ущерба, наносимого кабанами сельскому хо-
зяйству. М., 1986. 14 с.
Романов В.С., Козло П.Г. Зимняя подкормка кабана // Охота и охотничье хоз-во.
1955. №2. С. 20-21.
Романов В.С., Козло П.Г. Копытные звери Белоруссии, их изучение, охрана и хо-
зяйственное использование // Лесоведение и лесное хоз-во. 1977. Вып. 12. С.
144-155.
Романовский В.П., Корочвич В.И. Пути рационального использования Госохот-
фонда республики // Динамика зооценозов, проблемы охраны и рационально-
го использования животного мира Белоруссии. VI зоол. конф. Минск, 1989. С.
217-218.
Романовский В.П., Качановский С.Б. Влияние биотехнических мероприятий на чис-
ленность копытных Беловежской пущи // Беловежская пуща. Минск: Урожай,
1971. Вып. 4. С. 152-167.
Ромашов В.А. Фауна гельминтов копытных животных в Усманском бору // Ве-
дение заповедного хозяйства в лесостепной и степной зонах СССР. Воронеж,
1979. С. 40-46.
Руковский Н.Н. Влияние периодичности урожая кедра на млекопитающих Вос-
точного Сихотэ-Алиня // Зоол. журн. 1963. Т. 42, вып. 11. С. 1741-1742.
Русаков О.С. История, современное состояние и перспективы искусственного
расселения копытных на северо-западе СССР // Акклиматизация охотничьих
животных в СССР. Минск: Ураджай, 1978. С. 38-40.
Русаков О.С. Хищники как фактор динамики численности копытных на северо-
западе СССР // Экологические основы охраны и рационального использова-
ния хищных млекопитающих. М.: Наука, 1979. С. 61-62.
286
Русаков О.С. Аспекты строительной деятельности кабана на северо-западе
СССР И Копытные фауны СССР. М.: Наука, 1980а. С. 198-199.
Русаков О.С. Особенности размножения кабана в северо-западных областях
СССР // Там же. 19806. С. 199-200.
Русаков О.С. Особенности стадиального распределения и бонитировка угодий
кабана на северо-западе СССР // Фауна и экология птиц и млекопитающих се-
веро-запада СССР. Петрозаводск, 1983. С. 139-149.
Русаков О.С. Значение охотничьих заказников в динамике численности живот-
ных в условиях густонаселенных и хозяйственно освоенных территорий, пути
повышения их эффективности // Влияние антропогенной трансформации
ландшафта на население наземных позвоночных животных. М., 1987. Ч. 1. С.
126-128.
Русаков О.С. Об экологических и охотхозяйственных аспектах рационализации
промысла копытных на северо-западе СССР И Управление популяциями ди-
ких копытных животных. Киров, 1989. С. 48-59.
Русаков О.С., Савельев В.Д., Никаноров А.С. Остеодистрофия у кабана // Экология
наземных позвоночных северо-запада СССР. Петрозаводск, 1981. С. 167-175.
Русаков О С, Тарасов С.А. Рентгенологический метод определения возраста и
некоторые особенности строения зубной системы и скелета кабана (5. scrofa}
и лося (A. alces} // Рационализация методов изучения охотничьих животных.
Киров, 1988. С. 72-85.
Русаков О.С., Тимофеева Е.К. Кабан (экология, ресурсы, хозяйственное значение
на северо-западе СССР). Л.: Изд-во Ленингр. ун-та, 1984. 207 с.
РусановЯ.С. На кабанов // Охота и охотничье хоз-во. 1991. № 12. С. 18-20.
РухлядевД.П. Гельминтозы диких парнокопытных в районах Крыма и Кавказа//
Тр. IX Междунар. конгр. биологов-охотоведов. М., 1970. С. 605-608.
Рыковский А.С. Маклакова Л.П. Воздействие зараженности гельминтами на ус-
тойчивость и продуктивность популяций копытных И Взаимоотношения пара-
зита и хозяина. М., 1999. С. 88-92.
Рябов Л.С. Копытные, лес и выхухоль в бассейне среднего Хопра И Бюл. МОИП.
Отд. биол. 1975. Т. 80, вып. 5. С. 11-22.
Рябов Л., Семаго Л. Кабан против выхухоля // Советская Россия. 1974. 4 апреля.
Сабанеев Л.П. Волк // Природа. 1877. Кн. 2. С. 227-331.
Сабилаев А.С. О причинах изменения численности и ареалов некоторых млеко-
питающих Каракалпакии под воздействием человека И II съезд Всесоюз. те-
риол. о-ва. М.: Наука, 1978. С. 188-189.
Саблина Т.Б. Прошлое и настоящее лося в заповеднике «Беловежская пуща» // Тр.
Ин-та морфологии животных им. А.Н. Северцова. 1953а. Вып. 9. С. 206-216.
Саблина Т.Б. Перспективы разведения копытных в широколиственных и сме-
шанных лесах европейской части СССР // Там же. 19536. Вып. 9. С. 217-230.
Саблина Т.Б. Копытные Беловежской пущи // Там же. 1955. Вып.. 15. 191 с.
Саблина Т.Б. Об этологии кабана в широколиственных лесах // Поведение жи-
вотных и проблема одомашнивания. М.: Наука, 1969. С. 57-62. (Тр. МОИП.
Отд. биол. Т. 35).
Саблинский А.К. Опыт учета состояния и состава лосиных стад в Царскосель-
ском уезде С.-Петербургской губернии // Материалы к познанию русского
охотничьего дела. СПб., 1914. Вып. 3. С. 73-107.
Савицкий Б.П. Дикие копытные в антропургических очагах клещевого энцефа-
лита//Копытные фауны СССР. М.: Наука, 1980. С. 201-202.
Сакс К.А. О распространении кабана в Северной Прибалтике // Охотничье хозяй-
ство в интенсивном комплексном лесном хозяйстве. Каунас, 1975. С. 37-73.
Салъкина Т.П. Современное состояние популяции тигра на юго-востоке Сихотэ-
Алиня // Бюл. МОИП. Отд. биол. 1993. Т. 98, вып. 3. С. 45-53.
287
Салькина ГП. Тигр в Лазовском заповеднике И Природоохранные территории и
акватории Дальнего Востока и проблемы сохранения биологического разно-
образия. Владивосток, 1994. С. 98-102.
Самигуллин ГМ. Распределение и численность кабана в Оренбургской области //
V съезд Всесоюз. териол. о-ва АН СССР. М., 1990а. Т. 3. С. 113—114.
Самигуллин Г.М. Распространение и численность копытных в Оренбургской области //
Ресурсы животного мира Сибири. Новосибирск: Наука, 19906. С. 184-186.
Сапоженков Ю.Ф. Копытные Костромской области и их промысел // Копытные
фауны СССР. М.: Наука, 1975. С. 249-250.
Сапожников ГН Дикие копытные Таджикистана // Редкие млекопитающие
фауны СССР. М.: Наука, 1976. С. 113-124.
Саркисов В., Хохлов А. Кабан Центрального Предкавказья И Охота и охотничье
хоз-во. 1982. №3. С. 14-15.
Саркисова Т.Б., Хрусталева И.В. Скелет диких и домашних свиней в раннем он-
тогенезе // Морфология и генетика кабана. М.: Наука, 1985. С. 114-122.
Сафаров М.А. Морфологические особенности дикого кабана Азербайджана (экс-
терьер и интерьер) // Изв. АН АзССР. Сер. биол. и с.-х. наук. 1958а. № 3. С.
45-56.
Сафаров М.А. Морфологические особенности дикого кабана И Там же. 19586.
Вып. 6. С. 51-61.
Сафаров М.А. Морфологические особенности и систематическое положение ди-
кого кабана Азербайджана // Фауна и экология наземных позвоночных Азер-
байджана. Баку: Изд-во АН АзССР, 1965. С. 3-68.
Сафронова Ж.И. О формировании современных ареалов копытных на Северном
Кавказе // Вопр. экологии позвоночных животных. Краснодар, 1973. С. 81-93.
Северцов С.А., Саблина Т.Б. Олень, косуля и кабан в заповеднике Беловежская
пуща // Тр. Ин-та морфологии животных им. А.Н. Северцова АН СССР. 1953.
Вып. 9. С. 140-205.
Седов В.А., Ведерников В.А, Черниченко С.А. Важнейшие инфекции диких пар-
нокопытных животных (состояние проблемы в СНГ) И Болезни и паразиты
диких животных. М., 1992. С. 4-11.
Селевин В.А. Кабан Западной Сибири // Охотник. 1930. № 4. С. 14-15.
Семенов Б.Т. Естественное расселение кабана и современная граница его ареала
в Вологодской, Архангельской областях и Коми АССР // Биологические про-
блемы севера. Зоология. 1976. С. 297-299.
Семенов-Тян-Шанский О.И. Колебания численности тетеревиных птиц за полве-
ка и их причины // Тетеревиные птицы в заповедниках РСФСР. М., 1989. С.
20-30.
Сергеев Е.Б. Ресурсы копытных в Кировской области и их использование: Дис. ...
канд. биол. наук. М., 2000. 148 с.
Сержанин И.Н. Млекопитающие Белоруссии (второе издание). Минск: Изд-во
АН БССР, 1961.318 с.
Сержанин Ю.И. Географическое распространение и состояние естественных запа-
сов охотничье-промысловых млекопитающих в Белорусской ССР. Минск: Нау-
ка и техника, 1970. 124 с.
Сержанин Ю.И., Курское А.Н., Рождественский В.В. Распространение и дина-
мика численности копытных на территории Речицкого Полесья // Копытные
фауны СССР. М.: Наука, 1975. С. 119.
Сиивонен Л. Млекопитающие Северной Европы. М.: Лесн. пром-сть, 1979. 231 с.
Силантьев А.А. Обзор промысловых охот в России. СПб., 1898. 619 с.
Силантьев В. Бизоны в Америке И Охота и охотничье хоз-во. 2000. № 9. С. 42-43.
Синицын А.Н. Акклиматизация охотничьих зверей в Тамбовской области // Обо-
гащение фауны и разведение охотничьих животных. Киров, 1982. С. 107.
288
Сицко А.В. Состояние и перспективы использования ресурсов диких копытных
животных в РСФСР ИI Всесоюз. совещ. по проблемам зоокультуры. М., 1986.
Ч. 1. С. 187-190.
Скалон В.Н. О некоторых промысловых млекопитающих Бодайбинского района
Иркутской области//Бюл. МОИП. Отд. биол. 1951. Т. 56, вып. 1. С. 27-32.
Скалон В., Щербаков И., Базыкин М. Новые материалы по распространению
промысловых млекопитающих в Якутии И Природа и соц. хоз-во. 1941. Сб. 8,
ч. 2. С. 266-285.
Слепых В. Охота на кабана в Ставрополье // Охота и охотничье хоз-во. 1998. №
1. С. 14-16.
Слободян А.А., Олейник Я.В. Влияние хищников на популяцию копытных в ле-
сах Прикарпатья // Копытные фауны СССР. М.: Наука, 1975. С. 203-204.
Словцов И.Я. Позвоночные Тюменского округа и их распространение в Тоболь-
ской губернии. М., 1882. 78 с.
Слудский А.А. Кабан (морфология, экология, хозяйственное и эпизоотологиче-
ское значение, промысел). Алма-Ата: Изд-во АН КазССР, 1956. 220 с.
Слудский А.А. Проблема «хищник-жертва» в охотничьем хозяйстве Казахстана//
Тр. IX Междунар. конгр. биологов-охотоведов. М., 1970. С. 467-470.
Слудский А.А., Байдавлетов Р.Ж., Бекенов А., Жиряков В.А. и др. Млекопитаю-
щие Казахстана. Парнокопытные (оленьи, кабарговые, свиные) и непарноко-
пытные (лошадиные). Алма-Ата: Наука КазССР, 1984. Т. 3, ч. 4. 232 с.
Смирнов В.П. Череп ископаемого Sus scrofa L. из окрестностей Самарканда // Тр.
Узб. гос. ун-та. Новая сер. № И. Геология. 1940. Вып. 1. С. 1-6.
Смирнов В.С. Численность и истребление волка на Урале за 50 лет И Крупные
хищники. М., 1992. С. 23-34.
Смирнов Е.Н. Группировка тигров Сихотэ-Алинского заповедника и сопредель-
ной территории // Бюл. МОИП. Отд. биол. 1993. Т. 98, вып. 3. С. 37—44.
Смирнов Е., Микуэлл (Микэл) Д., Николаев И. и др. Проект “Амурский тигр” //
Охота и охотничье хоз-во. 1996. № 6. С. 16-18.
Смирнов М. Кабан в Бурятии И Охота и охотничье хоз-во. 1978. № 6. С. 16-17.
Смирнов М.Н. Дикие копытные на юге Сибири (исторические изменения населения,
проблемы использования) // Управление популяциями диких копытных живот-
ных. Киров, 1989. С. 59-75.
Смирнов М.Н. Крупные промысловые млекопитающие Южной Сибири: Авто-
реф. дис. ... докт. биол. наук. М., 1994. 68 с.
Смирнов М.Н. Охотники и охотничье хозяйство в доземледельческий период.
Красноярск, 1996. 65 с.
Смирнов М.Н. К вопросу снижения видового разнообразия крупных промысло-
вых млекопитающих Южной Сибири И VI съезд териол. о-ва. М., 1999. С. 238.
СмирновМ.Н. Кабан в центре Азии //Охота и охотничье хоз-во. 2001. № 8. С. 12-15.
Смирнов М.Н., Бриллиантов А.В. Современное состояние численности и пер-
спективы использования диких копытных в Красноярском крае // Ресурсы
животного мира Сибири. Новосибирск: Наука, 1990. С. 240-243.
Смирнов НГ. Некоторые особенности голоценового этапа развития териофауны
Урала // История биогеоценозов СССР в голоцене. М.: Наука, 1976а. С. 213-221.
Смирнов Н.Г. Динамика териофауны и изменчивость некоторых млекопитаю-
щих Среднего, Южного Урала и Зауралья в голоцене: Автореф. ... дис. канд.
биол. наук. Свердловск, 19766. 20 с.
Смирнов Н.Г., Большаков В.Н., Косинцев НА. и др. Историческая экология жи-
вотных гор Южного Урала. Свердловск, 1990. 244 с.
Смирнова Ю.Г. К вопросу о морфофизиологических показателях развития диких
свиней в сравнении с домашними // Тр. Моск. вет. академии. 1972. Т. 61 (Ве-
теринария). С. 170-171.
19. Заказ №2528.
289
Собанский ГГ Расширение ареалов и рост численности копытных на Алтае И
Бюл. МОИП. Отд. биол. 1986. Т. 91, вып. 1. С. 25-30.
Собанский ГГ Ресурсы копытных Горного Алтая И Ресурсы животного мира
Сибири. Новосибирск: Наука, 1990а. С. 254-257.
Собанский ГГ Заселение кабаном Горного Алтая И Экологические и экономиче-
ские аспекты охраны и рационального использования охотничьих животных и
растительных пищевых ресурсов Сибири. Шушенское, 19906. С. 146-148.
Собанский Г.Г. Копытные Горного Алтая. Новосибирск: Наука, 1992. 257.
Соков А.И. Таджикистан //Волк. М.: Наука, 1985. С. 510-513.
Соколов В.Е. Кожный покров млекопитающих. М.: Наука, 1973. 487 с.
Соколов В.Е., Баскин Л.М. Задачи прикладных экологических и этологических
исследований промысловых наземных позвоночных в лесной зоне // Живот-
ный мир южной тайги. М.: Наука, 1984. С. 6-38.
Соколов В.Е., Данилкин, А.А., Дуламцэрэн С. Современное распространение и
численность лесных копытных в Монголии // Зоологические исследования в
МНР. М.: Наука, 1982. С. 37-56.
Соколов В.Е., Петрищев Б.И. Кожный покров домашних млекопитающих (ко-
пытные). М., 1997. 288 с.
Соколов В.Е., Темботов А.К. Позвоночные Кавказа. Млекопитающие. Копыт-
ные. М.: Наука, 1993. 527 с.
Соколов В.Е., Чернова О.Ф. Кожные железы млекопитающих. М.: ГЕОС, 2001.
648 с.
Соколов ГА. Млекопитающие кедровых лесов Сибири. Новосибирск: Наука,
1979. 256 с.
Соколов И.И. Копытные звери (отряды Perissodactyla и Artiodactyla). Фауна
СССР. Млекопитающие. М.-Л.: Изд-во АН СССР, 1959. Т. 1, вып. 3. 640 с.
Сокур I.T. 3Bipi Радянских Карпат i ix господарське значения. Кшв: Видавн. АН
УРСР, 1952. 68с.
Соломатин А О. Кабан Тургайского плато и природная среда // Основные про-
блемы териологии. М.: Наука, 1972. (Тр. МОИП. Отд. биол. Т. 48). С. 238-251.
Соломатин А.О. Состояние кабана в Воронежской области // Копытные фауны
СССР. М.: Наука, 1975. С. 128-129.
Соломатин А.О. Экологическая ниша волка в Усманском бору // Экологические
основы охраны и рационального использования хищных млекопитающих. М.:
Наука, 1979. С. 140-142.
Стародынова А.К. Болезни лосей, маралов и кабанов в лесных угодьях Кали-
нинской и Московской областей // Тр. Завидовского гос. научно-опытного за-
поведника. М.: Воениздат, 1974а. Вып. 3. С. 147-172.
Стародынова А.К. Локализация и степень проявления туберкулезных пораже-
ний у диких свиней И Там же. 19746. Вып. 3. С. 185-194.
Стародынова А.К. Некоторые вопросы ветеринарной службы в Завидовском госу-
дарственном научно-опытном заповеднике // Там же. 1979. Вып. 4. С. 122-134.
Стародынова А.К, Сыроежкин Ю.М. Опыт борьбы с метастронгилезом диких
свиней // Там же. 1979. Вып. 4. С. 148-157.
Стахеев В.А., Сонникова А.Е., Завацкий Б.П., Желтухина Т.П. и др. Саяно-
Шушенский заповедник И Заповедники Сибири. М.: ЛОГАТА, 1999. Т. ЕС.
116-127.
Степанова Л.И., Степанов Н.М. Первобытная охота на Шоше // Тр. Завидовско-
го заповедно-охотничьего хоз-ва. М.: Воениздат, 1971. Вып. 2. С. 271-279.
Степанова Л.Н., Степанов Н.М. Стоянки первобытных охотников на реке Шо-
ше // Тр. Завидовского гос. научно-опытного заповедника. М.: Воениздат,
1974. Вып. З.С. 195-208.
290
Стрижева И.М. Костные остатки крупных млекопитающих в районе Самарской
Луки // Самарская Лука. 1991. № 2. С. 197-205.
Строганов С. Заметки по фауне промысловых млекопитающих северо-
восточной части Западной обл. //Учен. зап. Моск. гос. ун-та. 1934. Вып. 2. С.
293-310.
Субботин А.М. Распространение и биотопическое размещение парнокопытных
(Artiodactyla) и хищных (Carnivora) в горах Хамар-Дабана (Прибайкалье) // Фауна
и экология позвоночных Сибири. Новосибирск: Наука, 1980. С. 115-122.
Сухомиров Г.И. Хозяйственное использование копытных животных в Хабаров-
ском крае // Сб. науч.-техн, информ. ВНИИ животного сырья и пушнины.
1968. Вып. 22. С. 95-101.
Сухорослов М.С. Об особенностях популяции кабана Центрального Черноземья
И Популяционная структура вида у млекопитающих. М., 1970а. С. 93-95.
Сухорослов М.С. Случаи раннего размножения диких кабанов в Белгородской
области // Вопр. зоологии, физиологии и биофизики. Воронеж: Изд-во Воро-
нежского ун-та, 19706. С. 3-4. (Тр. Воронежского гос. ун-та. Т. 79).
Сухорослов М.С. Особенности размещения кабанов в Центрально-Черноземной
полосе // Вопр. зоологии и физиологии. Воронеж: Изд-во Воронежского ун-та,
1971. С. 64-67. (Тр. Воронежского гос. ун-та. Т. 93).
Сухорослов М.С. О роли дождевых червей в питании кабана (Sus scrofa L.) в пе-
риод весеннего половодья // Териология. Новосибирск: Наука, 1972а. Т. 1. С.
413-414.
Сухорослов М.С. Экологические особенности и хозяйственное значение кабана
ЦЧО: Автореф. дис. ... канд. биол. наук. Воронеж, 19726. 18 с.
Сыроечковский Е.Е., Рогачева Э.В. Животный мир СССР. М.: Мысль, 1975. 439 с.
Сысоев В. Охота в Хабаровском крае. Дальневосточ. гос. изд-во, 1950. 144 с.
Сысоев Н.Д. Животный мир Владимирской области (охотничье-промысловые
звери). Ярославль, 1970. 291 с.
Телишевский Д. Критерий селекционного отстрела диких копытных // Охота и
охотничье хоз-во. 1972. № 5. С. 18-19.
Телишевский Д.А. Дикие кабаны: формирование стада, способы предотвращения
ущерба //Лесное хоз-во. 1990. № 1. С. 51-52.
Теплое В.П. Волк в Кавказском заповеднике // Тр. Кавказского гос. заповедника.
1938. Вып. 1. С. 343-366.
Теплое В.П. К вопросу о соотношении полов у диких млекопитающих // Зоол.
журн. 1954. Т. 33, вып. 1. С. 174-179.
Тимофеева Е.К. Лось. Л.: Изд-во ЛГУ, 1974а. 167 с.
Тимофеева Е.К. Экология Sus scrofa L. (Artiodactyla) на северном пределе его
обитания ИI Междунар. конгр. по млекопитающим. М.: ВИНИТИ, 19746. Т. 2.
С. 259-260.
Тимофеева Е.К. К экологии кабана в лесостепных дубравах Белгородской облас-
ти И Бюл. МОИП. Отд. биол. 1975. Т. 80, вып. 5. С. 23-34.
Тимофеева Е.К. Стациальное распределение и питание кабана на северном пре-
деле его распространения И Биоценотические отношения организмов. Л.: Изд-
во Ленинградского ун-та, 1976. С. 107-120. (Тр. Петерг. биол. ин-та. № 24).
Тимофеева Е.К. Влияние кабана на растительность лесостепных дубрав И Ко-
пытные фауны СССР. М.: Наука, 1980. С. 207-208.
Тимофеева Е.К. Роль кабана в биоценозе лесостепных дубрав И III съезд Всесо-
юз. териол. о-ва. М., 1982. Т. 1. С. 307.
Тимофеев-Ресовский Н.В., Яблоков А.В., Глотов Н.В. Очерк учения о популяци-
ях. М.: Наука, 1973. 277 с.
Тимченко Н.Г. К истории охоты и животноводства в Киевской Руси (Среднее
Поднепровье). Киев: Наукова Думка, 1972. 204 с.
19
291
Тимченко Н.Г. Промысловые и домашние млекопитающие Правобережной Ук-
раины в I тысячелетии нашей эры И I Междунар. конгр. по млекопитающим.
М.: ВИНИТИ, 1974. Т. 2. С. 262-263.
Титов Э.К., Шингареев Р.С. Особенности формирования полового состава по-
пуляции кабана Сары-Челекского заповедника // Популяционная структура
вида у млекопитающих. М., 1970. С. 95-96.
Тихонов В.Н. Генетика мини-свиней. Новосибирск: ИЦиГ СО АН СССР, 1990.
207 с.
Тихонов В.Н., Бобович В.Е., Солодуха К.В. Использование генофонда европей-
ских и среднеазиатских кабанов для изучения связи системы Е групп крови с
продуктивностью и ее хромосомной локализации И Морфология и генетика
гибридных свиней. М.: Наука, 1992. С. 17-23.
Тихонов В.Н., Горелов ИГ, Бобович В.Е. Различия по группам крови кабанов
разных подвидов, обитающих на территории СССР // Популяционная измен-
чивость вида и проблемы охраны генофонда млекопитающих. М., 1983а. С.
192-193.
Тихонов В.Н., Горелов И.Г., Бобович В.Е. Иммуногенетические особенности не-
которых форм диких свиней Европы, Азии, Африки и Америки // Морфология
и генетика кабана. М.: Наука, 1985. С. 3-17.
Тихонов В.Н., Кестер Б.В., Трошина А.И., Горелов И.Г. Иммуногенетические
особенности популяции кабанов Воронежского заповедника И Восстановле-
ние и изучение природных экосистем в Воронежском государственном запо-
веднике. Воронеж: Изд-во Воронежского ун-та, 1978. С. 63-76.
Тихонов В.Н., Князев С.П. Аллотипические особенности крови центральноевро-
пейского, среднеазиатского, уссурийского и кавказского кабанов // Морфоло-
гия и генетика кабана. М.: Наука, 1985. С. 28-32.
Тихонов В.И, Никитин С.В., Травушкин В.А. Генетические особенности европейско-
го, среднеазиатского и дальневосточного кабанов при гибридизации с домашней
свиньей // Популяционная изменчивость вида и проблемы охраны генофонда
млекопитающих. М., 19836. С. 193-195.
Тихонов В.Н., Попов В.И, Никитин С.В. и др. Использование генофонда европей-
ского, среднеазиатского и уссурийского кабанов для укрепления конституции и
улучшения мясных и откормочных качеств домашних свиней // Морфология и
генетика кабана. М.: Наука, 1985. С. 56-72.
Тихонов В.Н., Травушкин В.А., Бобович В.Е. Продуктивность ландрас-кабаньих
гибридов на промышленных свинокомплексах И Особенности разведения
сельскохозяйственных животных в условиях Сибири. Новосибирск, 1983с. С.
40-46.
Тихонов В.Н., Трошина А.И. Идентификация хромосом и их перестроек в карио-
типах подвидов дикого кабана Sus scrofa L. методом дифференциальной окра-
ски И Докл. АН СССР. 1974. Т. 214, № 4. С. 932-935.
Тихонов В.И, Трошина А.И. Плодовитость и жизнеспособность трех поколений
гибридов, полученных при скрещивании домашних и диких свиней с разным
числом хромосом //Генетика. 1977. Т. 13, № 4. С. 627-640.
Тихонов В.Н., Трошина А.И., Горелов И.Г. Цитогенетическое исследование даль-
невосточных и среднеазиатских диких кабанов и домашних свиней И Изв. СО
АН СССР. Сер. биол. наук. 1972. № 10, вып. 2. С. 87-93.
Тихонов В.Н., Трошина А.И, Горелов И.Г. Иммуногенетические особенности ди-
ких кабанов разных подвидов И Копытные фауны СССР. М.: Наука, 1975а. С.
24-25.
Тихонов В.Н., Трошина А.И., Горелов И.Г. Дифференциация по антигенам групп
крови популяций кабана {Sus scrofa) из разных регионов СССР И Зоол. журн.
1978. Т. 57, вып. 7. С. 1074-1082.
292
Тихонов В.Н., Трошина А.И., Горелов И.Г. и др. Создание ландрас-кабаньих гиб-
ридов для промышленных свинокомплексов. Новосибирск: ИциГ СО АН
СССР, 1981. 25 с.
Тихонов В.Н., Трошина А.И., Кестер Б.В. Дифференциация популяций кабанов в
пределах некоторых подвидов по антигенам групп крови И Копытные фауны
СССР. М.: Наука, 19756. С. 25-26.
Тихонов В.Н., Трошина А.И., Сруога А.А. Сравнительное изучение кариотипа ом-
ских серых свиней и дикого кабана // Зоологические проблемы Сибири. Ново-
сибирск: Наука, 1972. С. 481-482.
Ткаченко В.А. Распространение и динамика численности копытных в Юго-
Западной Туве // Экология промысловых животных Сибири. Изд-во Красно-
ярского ун-та, 1992. С. 50-57.
Ткаченко КН Тигр, Panthera tigris (Carnivora, Felidae) в Большехехцирском за-
поведнике (Хабаровский край) // Зоол. журн. 1996. Т. 75, вып. И. С. 1729—
1736.
Толкач В.Н. Роль древесных и кустарниковых пород в питании копытных Бело-
вежской пущи // Копытные фауны СССР. М.: Наука, 1975. С. 185-186.
Толкач В.Н. Нарушенность природных территориальных комплексов и плот-
ность диких копытных в условиях режима заповедно-охотничьего хозяйства И
Влияние антропогенной трансформации ландшафта на население наземных
позвоночных животных. М., 1987. Ч. 1. С. 125-126.
Толкач В.Н, Будниченко Н.Н. Влияние диких копытных на возобновление дубрав
И Проблемы охраны генофонда и управления экосистемами в заповедниках
лесной зоны. М., 1986. Ч. 2. С. 215-217.
Томкевич НФ. Об организации правильного охотничьего хозяйства (практиче-
ское элементарное руководство для охотников и охотничьих обществ) И Охо-
та и охрана природы. М., 1992. Ч. 1. С. 13-50.
Третьяков П.Н. К истории племен Верхнего Поволжья в первом тысячелетии н.
э. // Материалы и исследования по археологии СССР. 1941. № 5. С. 5-150.
Трошина А.Н. Изучение иммуногенетического и хромосомного полиморфизма
диких и домашних свиней в связи с полиморфизмом: Автореф. дис. ... канд.
биол. наук. Новосибирск, 1975. 150 с.
Трошина А.И., Тихонов В.Н. Цитогенетические особенности некоторых диких
свиней Европы, Азии, Африки и Америки // Морфология и генетика кабана.
М.: Наука, 1985. С. 17-28.
Трошина А.И., Тихонов В.Н., Астахова Н.М. Цитогенетические особенности ка-
банов, обитающих на территории СССР, в связи с их филогенезом // Популя-
ционная изменчивость вида и проблемы охраны генофонда млекопитающих.
М„ 1983. С. 195-197.
Тутщина Л.Ф. Динамика численности и размещения кабана (Sus scrofa) в Дар-
винском заповеднике И Популяционные исследования животных в заповедни-
ках. М.: Наука, 1988. С. 128-139.
Туркин Н. Исторический очерк деятельности Императорского Общества раз-
множения охотничьих и промысловых животных и правильной охоты за 25-
летний период существования, 1872-1897 // Природа и охота. 1898. Кн. 1.
Приложение. С. 1-115.
Туркин Н.В. Промысел на кабанов в области их распространения в России И
Природа и охота. 1899. Июль. С. 1-8.
Туркин Н.В., Сатунин КА. Звери России. М.: Изд. Н.В. Туркина, 1902. 506 с.
Турубанова С.А. Изменение ареалов ключевых видов животных в голоцене на
территории бывшего СССР//VI съезд териол. о-ва. М., 1999. С. 261.
Тюрнин Б.Н. Современное состояние копытных в Коми АССР И Экология, мор-
фология, использование и охрана диких копытных. М., 1989. Ч. 1. С. 81-82.
293
Тюрнин В.Н Животный мир Республики Коми. Сыктывкар, 1996. 46 с.
Уатт К. Экология и управление природными ресурсами. М.: Мир, 1971.463 с.
Уваров Н.В. Материалы по размножению кабана в питомнике Окского заповедни-
ка// I Всесоюз. совещ. по проблемам зоокультуры . М., 1986. Ч. 1. С. 199-201.
Уваров Н.В. Первые итоги мечения кабанов в Окском биосферном заповеднике//
Проблемы изучения и охраны заповедных природных комплексов. Воронеж:
Изд-во Воронежского ун-та, 1995. С. 127-129.
Уманец О.Ю., Селюнина З.В. Влияние антропогенного изменения растительно-
сти на увеличение численности кабана в Черноморском государственном био-
сферном заповеднике // Влияние антропогенной трансформации ландшафта
на население наземных позвоночных животных. М., 1987. Ч. 1. С. 183-185.
Управление популяциями охотничьих животных. Киров, 1999. 210 с.
Фадеев Е.В. Кабаны Центральной России // Охота и охотничье хоз-во. 1968. № 7.
С. 12-13.
Фадеев Е.В. Результаты искусственного расселения некоторых видов диких ко-
пытных в охотничьих хозяйствах России // Бюл. МОИП. Отд. биол. 1969. Т.
74, вып. 1. С. 37-46.
Фадеев Е.В. Кабан в Подмосковье // Охота и охотничье хоз-во. 1970а. № 8. С.
20-21.
Фадеев Е.В. Естественное и искусственное расселение кабана в европейской
части РСФСР //Науч. докл. высш. шк. Биол. науки. 19706. № 1. С. 28-34.
Фадееве Е.В. Динамика численности кабана (Sus scrofa L.) в Европейской России
//Зоол. журн. 1973а. Т. 52, вып. 8. С. 1214-1219.
Фадеев Е. Разведение кабана в охотничьих хозяйствах Центральной России //
Передовой опыт в охотничье-рыболовном хоз-ве. 19736. Вып. 6. С. 67-76.
Фадеев Е.В. К экологии кабана Центральной России И Вести. Моск, ун-та. Био-
логия, почвовед. 1973в. № 5. С. 5-28.
Фадеев Е.В. Динамика численности кабана (Sus scrofa) в Европейской России //
Зоол. журн. 1973г. Т. 52, вып. 8. С. 1214-1219.
Фадеев Е.В. Разведение кабана в охотничьих хозяйствах Центральной России //
Передовой опыт в охотничье-рыболовном хоз-ве. 1973д. Вып. 6. С. 67-76.
Фадеев Е. Кабан в европейской части СССР И Охота и охотничье хоз-во. 1975.
№2. С. 16-17.
Фадеев Е. Кабан в Подмосковье // Там же. 1977. № 5. С. 12-13.
Фадеев Е.В. Кабан // Крупные хищники и копытные звери. М.: Лесная пром-сть,
1978а. 256-293.
Фадеев Е. Итоги дискуссии И Охота и охотничье хоз-во. 19786. № 1. С. 6-8.
Фадеев Е. Динамика ареала кабана И Там же. 1979. № 2. С. 14-15.
Фадеев Е.В. Современное состояние североевропейской части ареала кабана //
Копытные фауны СССР. М.: Наука, 1980. С. 49-50.
Фадеев Е.В. О динамике северной границы ареала кабана в Восточной Европе //
Биол. науки. 1981.№ 9. С. 56-64.
Фадеев Е.В. Размещение и динамика численности кабана на восточноевропей-
ской окраине ареала И Там же. 1982. № 3. С. 53-57.
Фадеев Е.В. Экологические аспекты охраны и рационального использования
охотничьих животных И Там же. 1985. № 4. С. 15-23.
Фадеев Е.В. Динамика фауны копытных лесов Русской равнины И Там же. 1986.
№9. С. 15-23.
Фадеев Е.В. Кабан И Охота и охотничье хоз-во. 1987. № 2. С. 10-13.
Фадеев Е.В. Специфика заселения кабаном северной части нечерноземного цен-
тра России // Вести. Моск, ун-та. Сер. 16. Биология. 1989. № 3. С. 45-50.
Фадеев Е.В. К восстановлению фауны копытных в лесах Среднего Подонья И
Там же. 1990. № 1. С. 47-53.
294
Фадеев Е.В. Кабан лесостепи Среднего Поволжья И Там же. 1992. № 2. С. 28-32.
Фадеев Е.В. Пространственная структура населения кабана в Саратовском По-
волжье // Там же. 1993. № 3. С. 30-36.
Фадеев Е.В. Размещение населения кабана в Волго-Камском крае во второй по-
ловине XX столетия // Там же. 1998. № 2. С. 22-26.
Федоров С.М. Млекопитающие (Mammalia) Ставропольского края: видовой со-
став, значение и использование // Матер, по изуч. Ставропольского края. 1954.
Вып. 6. С. 177-192.
Федосенко А.К. Сезонные кочевки копытных в Северном Тянь-Шане и Джунгар-
ском Алатау И Копытные фауны СССР. М.: Наука, 1975. С. 27-29.
Федосенко А.К. Марал. Алма-Ата: Наука, 1980. 198 с.
Федосенко А.К. Взаимоотношения крупных хищников и копытных в горах Юго-
Восточного Казахстана и Южной Сибири И Роль крупных хищников и копыт-
ных в биоценозах заповедников. М.: 1986. С. 4-21.
Федосенко А.К, Жиряков В.А. Взаимоотношения хищников и диких копытных в Се-
верном Тянь-Шане и Джунгарском Алатау И Экологические основы охраны и ра-
ционального использования хищных млекопитающих. М.: Наука, 1979. С. 72-74.
Федосенко А.К, Жиряков В.А. Экология и поведение кабана в горах юга и юго-
востока Казахстана И Бюл. МОИП. Отд. биол. 1984. Т. 89, вып. 4. С. 36^15.
Федотов О.А. Первые итоги акклиматизации кабана в Омской области И Интен-
сификация воспроизводства ресурсов охотничьих животных. Киров, 1990. С.
234-235.
Федюшин А.В. Динамика и географическое распространение охотничьей фауны
БССР. Изд. “Паляунщы БеларуД”, 1928-29. 81 с.
Федюшин А.В. Новые данные о расселении дикого кабана в Западной Сибири И
Изв. Омского отд. географ, о-ва Союза ССР. 1963. Вып. 5 (12). С. 127-129.
Фельдман В. Лоси и кабаны на улицах Москвы // Охота и охотничье хоз-во. 1989. №
3. С. 24-25.
Фертиков В.И. Реакклиматизация и акклиматизация охотничьих животных в
Ростовской области //Охотоведение. М.: Лесн. пром-сть, 1975. С. 244-254.
Фертиков В., Егоров А. Как спасти кабанов от чумы И Охота и охотничье хоз-во.
1997. №10. С. 15.
Фертиков В.И., Сонин М.Д., Рыковский А.С., Егоров А.Н. Гельминты диких ко-
пытных национального парка «Завидово». Тверь, 1999. 80 с.
Фетисов А.С. Материалы по систематике и географическому распространению
млекопитающих Западного Забайкалья // Изв. Гос. противочум. ин-та Сибири и
ДВК. 1936. Т.З.С. 86-119.
Фетисов А.С. Систематическое положение дикого кабана из Восточной Сибири
и Северо-Западной Монголии И Изв. биол.-геогр. НИИ при Иркутском гос.
ун-те им. А.А. Жданова. 1956. Т. 16, вып. 1-4. С. 203-205.
Филипченко Ю.А. Краниологическое исследование диких видов свиньи // Про-
блема происхождения домашних животных. Л.: Изд-во АН СССР, 1933. С.
157-184. (Тр. лаборатории генетики АН СССР. Вып. 1).
Филонов К.П.. Калецкая М.Л. Воздействие волка на диких копытных животных
//Волк. М.: Наука, 1985. С. 336-354.
Филонов КП. Копытные животные и крупные хищники на заповедных террито-
риях. М.: Наука, 1989. 254 с.
Флеров КК Копытные звери (Ungulata) Таджикистана И Звери Таджикистана.
М.-Л.: Изд-во АН СССР, 1935. С. 93-130. (Тр. Таджикской базы. Т. 1. Зооло-
гия и паразитология).
Флеров КК, Баландин В.Н. Степи Северо-Кавказского края. Изд. «Сев. Кавказ»,
1931.
295
Флинт В.Е. Стратегия сохранения редких видов в России: теория и практика. М.:
ГЕОС, 2000. 328 с.
Фокина В.Д., Размахнин В.Е. Использование ресурсов диких животных в СССР и
за рубежом. М., 1989. 59 с.
Фомушкин В.М., Максимов А.А. Распространение болезней диких животных и меры
профилактики//Болезни и паразиты диких животных. М., 1992. С. 12-19.
Фонд охотничьих угодий и численность основных видов диких животных в
РСФСР. М., 1992. 97 с.
Формозов А.А. К истории древнейшего скотоводства на юге СССР // Основные про-
блемы териологии. М.: Наука, 1972. С. 19-25. (Тр. МОИП. Отд. биол. Т. 48).
Формозов А.Н. Снежный покров в жизни млекопитающих и птиц СССР. М.:
Изд-во МОИП, 1946. 141 с.
Формозов А.Н. О движении и колебании границ распространения млекопитаю-
щих и птиц // География населения наземных животных и методы его изуче-
ния. М.: Изд-во АН СССР, 1959. С. 172-196.
Ханжин А. О вреде кабана // Охота и охотничье хоз-во. 1977. № 9. С. 12-13.
Харченко В.И., Молодан Г.Н., Кормачев НА. Современное состояние популяций
диких копытных Донецкой области И Копытные фауны СССР. М.: Наука,
1980. С. 111-112.
Хахин ГВ. Влияние роющей деятельности Sus scrofa L. на местообитание Des-
тапа moschata L. И I Междунар. конгр. по млекопитающим. М.: ВИНИТИ,
1974. Т. 2. С. 303.
Хахин ГВ. Ресурсы копытных Нечерноземного Центра И Биологические основы
охотничьего дела. М., 1980. С. 57-66.
Хахин Г.В., Гончаров В.Л. Охрана сельскохозяйственных объектов от диких ко-
пытных И Экология, морфология, использование и охрана диких копытных.
М, 1989. Ч. ЕС. 25-26.
Хеладзе П. К краниологическому изучению дикого кабана, водящегося в Грузии,
и примитивной сванской домашней свиньи И Тр. Тбилисского гос. ун-та им.
Сталина. Сер. биол. наук. 1959. Т. 70, вып. 1. С. 147-162. (На груз. яз.).
Хеладзе П.С. Биологические особенности кабана, перспективы охраны и раз-
множения его в условиях Грузии: Автореф. дис. ... канд. биол. наук. Тбилиси,
1964. 19 с.
Ходашева К.С., Елисеева В.И. Участие позвоночных животных - потребителей
веточных кормов в круговороте веществ в лесостепных дубравах И Структура
и функционально-биогеоценотическая роль животного населения суши. М.:
Изд-во АН СССР, 1967. С. 81-84.
Холостое В.Г. Кабан в Архангельской области // Бюл. МОИП. Отд. биол. 1956.
Т. 61, вып. 4. С. 82.
Хорин С.И., Горшков П.К. Сравнительная экология кабана и роль его в биоцено-
зах Северного Кавказа. Экологичесие проблемы Ставропольского края и со-
предельных территорий. Ставрополь, 1989. С. 331-333.
Хромов А.А., Архангельский М.С., Иванов А.В. Местонахождения крупных чет-
вертичных млекопитающих Саратовского Поволжья. Саратов: Научная книга,
2000. 156 с.
Худайбердиев Т.Н. Промысловые виды животных многослойной палеолитиче-
ской стоянки Кульбулак // V съезд Всесоюз. териол. о-ва АН СССР. М., 1990.
Т. ЕС. 33-34.
Цалкин В.И. Об аборигенном олене Западной Белоруссии И Докл. Акад. наук.
1951. Т. 77, №3. С. 521-523.
Цалкин В.И. Материалы для истории скотоводства и охоты в древней Руси И Ма-
териалы и исследования по археологии СССР. М.: Изд-во АН СССР, 1956. №
51. 186 с.
296
Цалкин В.И. Фауна из раскопок археологических памятников Среднего Повол-
жья (материалы для истории скотоводства и охоты в СССР) И Там же. 1958. №
61. С. 221-281. (Тр. Куйбышевской археолог, экспедиции. Т. 2).
Цалкин В.И. Млекопитающие бассейна Оки и Верхней Волги в начале нашей
эры //Бюл. МОИП. Отд. биол. 1961. Т. 66, вып. 1. С. 23-39.
Цалкин В.И. К истории животноводства и охоты в Восточной Европе // Мате-
риалы и исследования по археологии СССР. М.: Изд-во АН СССР, 1962. №
107. 129 с.
Цалкин В.И. К истории млекопитающих восточноевропейской лесостепи И Бюл.
МОИП. Отд. биол. 1963. Т. 68, вып. 2. С. 43-62.
Цалкин В.И. Животноводство и охота племен восточноевропейской лесостепи в
раннем железном веке // Материалы и исследования по археологии СССР.
1966а. № 135. С. 3-107.
Цалкин В.И. Фауна древнего Хорезма в свете данных археологии // Там же.
19666. № 135. С. 108-157.
Цалкин В.И. Фауна из раскопок городищ скифского времени на Среднем Дону И
Население Среднего Дона в скифское время. М.: Наука, 1968. С. 128-136.
Цалкин В.И. Фауна Танаиса // Античные древности Подонья-Приазовья. М.:
Наука, 1969а. С. 273-285.
Цалкин В.И. Фауна из раскопок Борщевских и Роменских городищ И Советская
археология. 19696. № 4. С. 91-101.
Цалкин В.И. Древнейшие домашние животные Восточной Европы. М.: Наука,
1970.280 с.
Цалкин В.И. Фауна из раскопок славянских памятников Молдавии // Основные про-
блемы териологии. М.: Наука, 1972а. С. 38-65. (Тр. МОИП. Отд. биол. Т. 48).
Цалкин В.И. Фауна из раскопок андроновских памятников в Приуралье // Там
же. 19726. С. 66-81.
Царев С.А. Сезонная изменчивость лежек кабана// Вести. ЛГУ. № 21. Биология.
1979. Вып. 4. С. 22-28.
Царев С.А. Внутривидовые отношения кабанов на северном пределе распро-
странения // Копытные фауны СССР. М.: Наука, 1980а. С. 321-322.
Царев С.А. Поведение кабана на северной границе ареала // Вести. ЛГУ. Биоло-
гия. Вып. 2. 19806. №9. С. 15-21.
Царев С.А. Социальная структура населения кабана И III съезд Всесоюз. териол.
о-ва. М, 1982. Т. 2. С. 189-190.
Царев С.А. Изучение пространственно-этологической структуры популяций ка-
бана И IV съезд Всесоюз. териол. о-ва. М., 1986а. Т. ЕС. 369-370.
Царев С.А. Опыт отлова и мечения кабанов в заповеднике «Лес на Ворскле» //
Роль крупных хищников и копытных в биоценозах заповедников. М., 19866.
С. 141-147.
Царев С.А. Опыт отлова и группового мечения кабанов в заповеднике «Лес на
Ворскле» // Комплексные исследования биогеоценозов лесостепных дубрав.
Л., 1986в. С. 131-140.
Царев С.А. Коммуникативное и территориальное поведение кабана И Коммуника-
тивные механизмы регулирования популяционной структуры у млекопитающих.
М., 1988. С. 182-185.
Царев С.А. Механизмы расселения кабанов // Современные проблемы охотничь-
его хозяйства. М., 1989а. С. 83-94.
Царев С.А. Циклические модификации социальной структуры группировок ка-
банов И Экология, морфология, использование и охрана диких копытных. М.,
19896. Ч. 2. С. 116-218.
Царев С.А. Сезонные изменения социальной структуры и территориального по-
ведения кабана (Sus scrofa L.): Автор, дис. ... канд. биол. наук. М., 1989в. 22 с.
20. Заказ №2528.
297
Царев С.А. Механизмы расселения кабанов И V съезд Всесоюз. териол. о-ва АН
СССР. М., 1990. Т. 3. С. 68.
Царев С.А. Циклические изменения социальной структуры группировок кабанов
(Sus scrofa) И Зоол. журн. 1991. Т. 70, вып. 11. С. 97-109.
Царев С.А. Развитие пространственных отношений кабанов (Sus scrofa L.) // VI
съезд териол. о-ва. М., 1999. С. 270.
Царев С.А. Кабан. Социальное и территориальное поведение И Охотничьи жи-
вотные России. М., 2000. Вып. 3. 113 с.
Цыганов А. Вожаки в сообществе кабанов // Охота и охотничье хоз-во. 1985. №
10. С. 28.
Чегодаев Е. Охота на кабанов в Азербайджане // Охота и охотничье хоз-во. 1975. №
6. С. 22-23.
Чегорка П.Т. К экологии копытных // Природа заповедника «Кодры». Кишинев:
Штиинца, 1984. С. 75-87.
Червонный В.В. К вопросу о повреждаемости муравейников кабанами // II съезд
Всесоюз. териол. о-ва. М.: Наука, 1978. С. 210.
Червонный В.В. Структура популяции кабана в Окском заповеднике И Копытные
фауны СССР. М.: Наука, 1980а. С. 114-115.
Червонный В.В. Особенности размножения кабанов в Окском заповеднике И Там
же. 19806. С. 215-216.
Червонный В.В. Индивидуальный участок обитания кабанов зимой И Там же. 1980в.
С. 216-217.
Червонный В.В. Особенности поведения кабана в Окском заповеднике И Там же.
1980г. С. 323-324.
Червонный В.В. О длине суточного хода лося и кабана в Белоруссии // III съезд
Всесоюз. териол. о-ва. М., 1982. Т. 1. С. 323-324.
Червонный В.В. О роли некоторых антропогенных элементов ландшафта в жиз-
ни копытных Русской равнины И Влияние антропогенной трансформации
ландшафта на население наземных позвоночных животных. М., 1987. Ч. 1. С.
204-205.
Червонный В.В. Особенности динамики ареала кабана и косули в европейской
части РСФСР за последние 20 лет // V съезд Всесоюз. териол. о-ва АН СССР.
М„ 1990а. Т. 1.С. 152-153.
Червонный В.В. Особенности структуры и размещения участков обитания кабана
в зимний период (Окский заповедник) // Биологические основы учета числен-
ности охотничьих животных. М., 19906. С. 103-120.
Черкасов А.А. Записки охотника-натуралиста. М.: Изд-во АН СССР, 1962. 504 с.
Чернявская С.И. Сезонное размещение и кочевки диких копытных и медведя в районе
Кавказского заповедника в связи с распределением урожая плодов фруктарников и
орехоносов // Бюл. МОИП. Отд. биол. 1956. Т. 61. Вып. 4. С. 7-21.
Чичикин Ю.Н., Воробьев ГГ. Дикий кабан юга Киргизии. Фрунзе: Изд-во Кыргыз-
стан, 1967. 80 с.
Шапошников Л., Головин Л., Сорокин М., Тараканов А. Животный мир Калинин-
ской области. Калинин: Кн. изд-во, 1959. 459 с.
Шарапов Ш. Обзор местонахождений плиоцен-нижнеплейстоценовых наземных
млекопитающих Таджикистана И Вопр. зоологии Таджикистана. Душанбе: До-
ниш, 1972а. С. 216-231.
Шарапов Ш. Остатки млекопитающих из неолитической стоянки Сай-Сайед
(Южный Таджикистан) //Там же. 19726. С. 232-236.
Шарффъ Р.Ф. Европейсюя животныя, ихъ геологическая история и географиче-
ское распространеше. М.: Природа, 1918. 233 с.
Шахмарданов З.А., Спасская Т.Х. Популяция и промысел кабана в Дагестане //
Копытные фауны СССР. М.: Наука, 1975. С. 141-142.
298
Шварц С.С. Возрастная структура популяций животных и проблемы микроэво-
люции (теоретический анализ проблемы) И Зоол. журн. 1965. Т. 44, вып 10. С.
1443-1453.
Шварц С. Популяционная экология - теоретическая основа охотничьего хозяй-
ства // Охота и охотничье хоз-во. 1969. № 4. С. 16-17.
Шварц С.С. Эколого-популяционные основы ведения охотничьего хозяйства //
Тр. IX Междунар. конгр. биологов-охотоведов. М., 1970. С. 74-77.
Шварц С. Популяция - элементарный объект охотничьего хозяйства // Охота и
охотничье хоз-во. 1974а. № 10. С. 16-17.
Шварц С.С. Биологические основы охотничьего хозяйства И Современное состоя-
ние и пути развития охотоведческой науки в СССР. Киров, 19746. С. 9-11.
Шварц С.С., Михеева К.В. Теоретические основы рационального использования
охотничье-промысловых животных // Итоги науки и техники. Зоология по-
звоночных. М., 1976. Вып. 8. С. 8-67.
Шиков А.Т, Балинский В.П., Федулов О.В. Классическая чума свиней у диких каба-
нов наУкраине //Болезни и паразиты диких животных. М., 1992. С. 101-102.
Шиков А.Т., Гришок Л.П. Экологические и эпизоотологические особенности клас-
сической чумы диких кабанов // Актуальные вопр. ветеринарной вирусологии.
Покров, 1995. С. 50-51.
Шиллингер Ф.Ф. Кабан. М.: Изд-во Московского комунальн. хоз-ва, 1928. 39 с.
Шилов И.А. Эколого-физиологические основы популяционных отношений у жи-
вотных. М.: Изд-во МГУ, 1977. 261 с.
Шималов В.Т., Дорощук Л.П., Касач Л.А., Бурак М.Н. и др. Хищничество и эпи-
зоотии как факторы, обуславливающие численность диких копытных в Бело-
руссии // Копытные фауны СССР. М.: Наука, 1975. С. 209-211.
Ширяев В.В. Влияние кабана на поселения ондатры в дельте р. Или И Копытные
фауны СССР. М.: Наука, 1975. С. 187-188.
Шостак С.В. Возрастные изменения в окраске и расцветке волосяного покрова
дикой свиньи (Sus scrofa L.) // Беловежская пуща. М.: Урожай, 1969. Вып. 3.
С.144-146.
Шостак С.В. Особенности размножения дикой свиньи в Беловежской пуще //
Там же. 1971. Вып. 4. С. 185-186.
Шостак С.В. Комплекс биотехнических мероприятий в Беловежской пуще И
Копытные фауны СССР. М.: Наука, 1975. С. 256-257.
Шостак С.В. Численные соотношения европейского благородного оленя с дру-
гими копытными // Заповедники Белоруссии. Минск: Ураджай, 1978. Вып. 2.
С.130-139.
Шостак С.В., Василюк И.Ф. Мероприятия по сохранению диких копытных в Бе-
ловежской пуще //Копытные фауны СССР. М.: Наука, 1975. С. 257-259.
Шостак С.В., Шостак А.И. Особенности поведения дикой свиньи при размно-
жении и их роль в расселении вида И Коммуникативные механизмы регули-
рования популяционной структуры у млекопитающих. М., 1988. С. 194-195.
Шуев В.А. Поведение кабана в Псковской области И Копытные фауны СССР. М.:
Наука, 1975. С. 340.
Шурин-Юхкум А. Подкормка копытных // Охота и охотничье хоз-во. 1994. № 2.
С. 1-2.
Шурупов И.И. Роль «Зоообъединения» в расселении охотничьих животных И
Обогащение фауны и разведение охотничьих животных. Киров, 1982. С. 116.
Щербак В С. Некоторые особенности экологии кабана Херсонской области //
Воспроизводство, использование и охрана диких зверей и птиц. Пермь, 1986.
. С.40-45.
299
Щербаков В. Причины, сдерживающие процент продуктивного разведения ос-
новных видов охотничьей фауны // Тр. Завидовского заповедно-охотничьего
хоз-ва. М.: Воениздат, 1972. Вып. 2. С. 74-80.
Щербаков В.П., Стародынова А.К. Плодовитость и некоторые вопросы размно-
жения кабанов в угодьях Завидовского заповедника // Тр. Завидовского запо-
ведника. М.: Воениздат, 1979. Вып. 4. С. 76-84.
Юдаков А.Г. О влиянии тигра на численность копытных // Редкие виды млеко-
питающих фауны СССР и их охрана. М.: Наука, 1973. С. 93-94.
Юдаков А.Г. Экология Panthera tigris altaica //1 Междунар. конгр. по млекопи-
тающим. М.: ВИНИТИ, 1974. Т. 2. С. 354-355.
Юдаков А.Г, Николаев И.Г. Снег как экологический фактор в жизни амурского ти-
гра// Редкие виды млекопитающих и их охрана. М.: Наука, 1977. С. 133-134.
Юдаков А.Г, Николаев И.Г. Экология амурского тигра. По зимним стационар-
ным наблюдениям 1970-1973 гг. в заповедной части Среднего Сихотэ-Алиня.
М.: Наука, 1987. 152 с.
Юргенсон П.Б. Очерки по сравнительному изучению соболя и куниц // Сб. мате-
риалов по результатам изучения млекопитающих в гос. заповедниках. М.:
Изд-во МСХ СССР, 1956. С.33-71.
Юргенсон П.Б. Роль алиментарного и других факторов в тенденции к расширению
границ ареала и в распределении популяций // География населения наземных
животных и методы его изучения. М.: Изд-во АН СССР, 1959. С. 7-22.
Юргенсон П.Б. Состояние и перспективы охотничьих ресурсов // Тр. Завидовского
заповедно-охотничьего хозяйства. М.: Воениздат, 1969. Вып. 1. С. 79-208.
Юргенсон П.Б. Биологические основы охотничьего хозяйства в лесах. М.: Лесн.
пром-сть, 1973. 173 с.
Юргенсон П.Б., Капланов Л.Г., Книзе А.А. Лось и его промысел. М.: Изд-во
Главпушнины НКВТ, 1935. 155 с.
Юсупов Р.Х., Ильясов Г.Х., Хамдеев Р.Х. Случай выделения вируса классической
чумы свиней у диких кабанов // Болезни и паразиты диких животных. М.,
1992. С.103-104.
Яблонский Н.И. Алтай // Природа и охота. 1904. Кн. 9. С. 14-20.
Язан Ю.П. Искусственное регулирование численности и структур популяций
охотничьих млекопитающих // II съезд Всесоюз. териол. о-ва. Пленарные док-
лады. М., 1979. С. 146-151.
Янулайтис З.П. Влияние дикого кабана на состояние муравейников // Весщ АН
БССР. Сер. б!ял. н. 1984. №2. С. 112-113.
Янулайтис З.П. Определение массы тела кабана по экстерьерным признакам //
Териологические исследования в Литве. Вильнюс, 1988. С. 44-53.
Янулайтис 3., Падайга В. Рациональное использование популяции кабана в
Южной Прибалтике. Каунас, 1984а. 8 с.
Янулайтис З.П., Падайга В.И. Продуктивность популяции и управление числен-
ностью кабана в Литовской ССР // Вопр. лесного охотоведения и недревесной
продукции леса. М., 19846. С. 36-48.
Янулайтис З.П., Падайга В.И. Управление численностью популяции кабана в
Южной Прибалтике // Управление популяциями диких копытных животных.
М„ 1985. С. 111-122.
Янулайтис З.П., Падайга В.И. Управление популяцией кабана в Литовской ССР
// Вопросы лесозащиты в Литовской ССР. Вильнюс: Мокслас, 1987. С. 163—
176. (Сб. тр. ЛитНИИЛХ. Т. 27).
Янулайщс З.П., Козла П.Г. Марфалапчная характарыстыка папуляцый кабана
(Sus scrofa} Л1твы i Беларуа // Весщ АН БССР. Сер. б!ял. навук. 1988. № 3. С.
87-92.
300
Янушевич А., Благовещенский И. Промысловые звери и птицы Западной Сибири.
Новосибирск: Обл. гос. изд-во, 1952. 191 с.
Ярмоленко Б.Н., Козлова А.З., Самарский С.Л. Минеральная подкормка копыт-
ных в Среднем Приднепровье // Копытные фауны СССР. М.: Наука, 1980. С.
223-224.
Andrzejewski R. Spotty mutation of the wild boar Sus scrofa Linnaeus, 1758 // Acta
Theriol. 1974. Vol. 19,N l.P. 159-163.
Andrzejewski R., Jezierski W. Management of a wild boar population and its effects on
commercial land//Acta Theriol. 1978. Vol. 23, N 19-30. P. 309-339.
Baleisis R., Bluzma P., Balciauskas L. Lietuvos kanopiniai zverys. Vilnius: Mokslas,
1987. 199 s.
Bartschi R. Das Familienleben der Wildschweine // Osterr. Tierschutz. 1983. N 5. S.
16-21.
Blume K, Hopp P.-J. Zur Hege und Bejagung des Schwarzwildes. Dritter Bericht aus
dem Spessart // Wild und Hund. 1982. Bd. 85, N 10. S. 36—40.
Blume K, Hopp P-J. Die Saujagd im Spiegel der Streiken // Ibid. 1986. Bd. 89, N 12.
S. 52-56.
Boitani L., Mattei L., Nonis D., Corsi F. Spatial and activity patterns of wild boars in
Tuscany, Italy // J. Mammal. 1994. Vol. 75, N 3. P. 600-612.
Bosma A.A. Chromosomal polymorphism and G-banding patterns in the wild boar
(Sus scrofa L.) from Netherlands // Genetica. 1976. Vol. 46, N 4. P. 391-399.
Brandt S., Vissant J, Jullien J.-M. Damaine vital diurne des sangliers en foret de
Chateauvillain - Arc-en-Barrois // Bui. Mens. 1988. N 234. P. 4-11.
Bratton S.P. The effect of the European wild boar, Sus scrofa, on Grey Beech Forest
in the Great Smoky Mountains // Ecology. 1975. Vol. 56. P. 1356-1359.
Brehm A. Die Saugetiere. Berlin: Bibliogr. Inst, in Leipzig, 1925. Bd. 4. 714 s.
Briedermann L. Zur Reproduktion des Schwarzwildes in der Deutschen
Demokratischen Republik // Tagungsber. Dt. Akad. Landwirtschaftswiss. 1971. N
113. S.169-186.
Briedermann L. Zu den Grundsatzen der Schwarzwildbewirtschaftung // Unsere Jagd.
1977a. Bd. 27, N l.S. 8-9.
Briedermann L. Moglichkeiten zur Feldschadenverminderung durch Verbesserung der
Ernahrungsbedingungen fur das Schwarzwild // Ibid. 1977b. Bd. 27, N U.S. 332—
333.
Briedermann L. Zasady a efektivnost obhospodarovania diviacej zveri v NDR // Folia
Venator. (CSSR). 1979. N. 9. P. 241-248.
Briedermann L. Schwarzwild. B.: VEB Dt. Landwirtschaftsverlag, 1986. 539 s.
Brown L.N. Elimination of a small feral swine population in an urbanising section of
central Florida // Fl. Sci. 1985. Vol. 48, N 2. P. 120-123.
Bunger U, Steihardt M„ Riehm G., Lyhs L. Verhalten der Rektaltemperatur bei
Laufbandbelastungen von Europaischen Wildschweinen (Sus scrofa scrofa L.) //
Biol. Rdsch. 1976. Bd. 14, N 6. S. 373-376.
Dardaillon M. Seasonal variations in habitat selection and spatial distribution of wild
boar (Sus scrofa) in the Camargue, southern France // Behav. Process. 1986. Vol.
13,N3.P. 251-268.
Dardaillon M., Beugnon G. The influence of some environmental characteristics on
the movements of wild boar Sus scrofa II Biol. Behav. 1987. Vol. 12, N 2. P. 82-92.
Dardaillon M., Teillaud H. Ethogramme du sanglier adulte et du Marcassin (Sus
scrofa L.) // Monit. Zool. Ital. 1987. Vol. 21, N l.P. 41-68.
Depner K.R., Granzow H, Kern B., u. a. Schweinepest - Uneingeschrankte Jagd ist
kontrapdoduktiv // Wild und Hund. 1997. Bd. 100, N 15. S. 34-39.
Eguchy Y., Tanida H, Tanaka T, Yoshimoto T. Colour discrimination in wild boars //
J. Ethol. 1997. Vol. 15, N 1. P. 1-7.
301
Ellerman J.R., Morrison-Scott T.C.S. Checklist of Palaearctic and Indian mammals
1758 to 1946. L.: British Mus. (Nat. Hist.), 1951. 810 p.
Erkinaro E., Heikura K., Lindgren E„ Pulliainen E. Occurrence and spread of the wild
boar (Sus scrofa) in eastern Fennoscandia // Mem. Soc. Fauna et Flora Fenn. 1982.
Vol. 58, N 2. P. 39-47. j
Gaillard J.M., Brandt S., Jullien J.-M. Body weight effect on reproduction of young
wild boar (Sus scrofa) females: a comparative analysis // Folia Zool. 1993. Vol. 42,
N 3. P.204-212.
Gaisford M. Breeding wild boar: Danish experience pays off // Farmers Week. 1991.
N 26. P. 32. |
Gasparik J., Hell P., Sabados K. Ochrana kultury kukurice elektrickym oplotenim
proti skodam sposobenym raticovou zverou // Folia Venator. 1993. N 23. S. 89-98.
Genard M., Lescourret F. Le sanglier (Sus scrofa scrofa L.) et les diaspores dans le
sud de la France // Rev. Ecol. 1985. Vol. 40, N 3. P. 343-353.
Genard M., Lescourret F., Durrieu G. Mycophagie chez le sanglier et dissemination
des spores de champignons hypoges // Gaussenia. 1986. N 2. P. 17-23.
Genov P. Food composition of wild boar in north-eastern and western Poland // Acta
Theriol. 1981a. Vol. 26, N 8-15. P. 185-205.
Genov P. Significance of natural biocenoses and agrocenoses as the source of food for
wild boar (Sus scrofa L.) // Ecol. Pol. 1981b. Vol. 29, N 1. P. 117-136.
Genov P. V. Food composition of the wild boar (Sus scrofa attila Thomas) in the
Danubian plan // Екология. 1987. Vol. 20. P. 47-57.
Genov P.V. Nourriture du sanglier (Sus scrofa attila Thomas, 1912) dans les
montagnes de Bulgarie // Ibid. 1994. Vol. 26. P. 51-67.
Genov P., Nikolov H, Massei G., Gerasimov S. Craniometrical analysis of Bulgarian
wild boar (Sus scrofa) populations // J. Zool. 1991. Vol. 225, Pt. 2. P. 309-322.
Gorelov LG. Trichinosis in wild boars in East Siberia // Euro-Amer. Mammal. Congr.
Santiago de Compostella, 1998. P. 398-399.
Groger H. Erfahrungen und Ergebnisse bei der Bewirtschaftung von Schwarzwild im
Kreis Niesky // Unsere Jagd. 1979. Bd. 29, N 6. S. 176-177.
Groop A., Giers D., Tettenborn U. Das Chromosome komplement des wildschwein
(Sus scrofa) // Experientia. 1969. Bd. 25. S. 778.
Groves C. Ancestors for the pigs: taxonomy and philogeny of the genus Sus // Aust.
Nat. Univ. Dept. Prehist. Techn. Bull. 1981. N 3. P. 1-90.
Groves C.P. Taxonomy of wild pigs (Sus) of the Philippines // Zool. J. Linn. Soc.
1997. Vol. 120, N 2. P. 163-191.
Gunther W. Erfahrungen mit der Schwarzwildbewirtschaftung im Odenwald // Allg.
Forstzeitschrift. 1980. Bd. 35, N 18. S. 478-480.
Haber A., Paslawski T, Zaborowski S. Gospodarstwo lowieckie. Warszawa: Panstw.
Wydawn. Nauk., 1975. 368 s.
Hell P. Zhodnotenie trofejovej kvality divaiacej v Ceskoslovensku // Folia Venator.
1985.N 15. S.125-139.
HellP. Koncepcia diviacich zvernic v SSR//Folia Venator. 1986. N 16. S. 137-155.
Hell P., Paule L. Systematische Stellung des westkarpatischen Wildschweines Sus
scrofa II Prirodoved. Pr. Ustavu CSAV Brne. 1983. Bd. 17, N 3. 54 s.
Hirotani A., Nakatani J. Grouping-patterns and inter-group relationships of Japanese
wild boars (Sus scrofa leucomystax) in the Rokko Mountain areas // Ecol. Res.
1987. Vol. 2, N 1. P. 77-84.
Hopp P.-J. Schweinereien Feststellungen, Beispiele und Vorschlage zur Bejahgung
des Schwarzwilds // Wild und Hund. 1979. Bd. 82, N 10. S. 236-239.
Hopp P.-J. Zur Situation der Schweine//Ibid. 1980. Bd. 83, N 6. S. 129-133.
302
Howe T.D., Singer FI, Ackerman B.B. Forage relationships of European wild boar
invading northern hardwood forest // J. Wildlife Manag. 1981. Vol. 45, N 3. P.
748-754.
IffU. Altersbestimmung und -schatzung beim Schwarzwild // Wild und Hund. 1983.
Bd. 86, N U.S. 26-30.
Janeau G., Spitz F. L’espace chez le sanglier (Sus scrofa scrofa L.). Occupation et
mode d’utilisation // Gibier Faune Sauvage. 1984. N 1. P. 73-89.
Jezierski W. Longevity and mortality rate in a population of wild boar // Acta Theriol.
1977. Vol. 22, N 24. P. 337-348.
Jezierski W., Myrcha A. Food requirements of a wild boar population // Pol. Ecol.
Stud. 1975. Vol. 1,N2. P. 61-83.
Jonsson L. From wild boar to domestic pig - a reassessment of neolithic swine of
northwestern Europe // Striae. 1986. Vol. 24. P. 125-129.
Karg H. Pheromones in mammals with special emphasis on the boar scent steroids,
also occuring in humans and truffles // Verh. Kon. Acad. Geneesk. Belg. 1988. Vol.
50, N 2. P. 121-138.
Kiessling W. Das Schwarzwild und seine Jagd. Berlin: Verlag von J. Neumann in
Neudamm, 1925. 368 s.
Knyazev S„ Tikhonov V. Sus scrofa phylogenesis: genogeographic and hybridologic
analysis of cranial parameters // Abstr. Euro-Amer. Mammal. Congr. Santiago de
Compostela, 1998. P. 284-285.
Kohalmy T. Assessment of wild boar stocks in Hungary // Trans. XX Intern. Congr.
Union Game Biol. Godollo, 1991. Pt. 1. P. 99.
Kurten B. Pleistocene mammals of Europe. Chicago: Aldine Publ. Comp., 1968. 318 p.
Kurz J.C., Marchinton R.L. Radiotelemetry studies of feral hogs in South Carolina // J.
Wildlife Manag. 1972. Vol. 36, N 4. P. 1240-1248.
Liess C., Horstmann H.-D. Stand und Entwicklung unserer Schalenwildbestande.
Analyse der Jagdstrecke 1988 und der Wildbestandsermittlung 1989 // Unsere Jagd.
1989. Bd. 39, N 9. S. 258.
Martin P.S. Prehistoric overkill // Proc. VII Congr. INQUA. New Haven; L.: Yale
Univ. Press, 1967. Vol. 6. P. 75-120.
Mauget R. Mise en evidence, par captures-recaptures et radiotracking, du domaine
vitat chez le sanglier (Sus scrofa L.) en foret de Chize // Biol. Behav. 1979. Vol. 4,
N 1. P. 25-^41.
Mayer J. J., Brisbin L.I. Sex identification of Sus scrofa based on canine morphology //
J. Mammal. 1988. Vol. 69, N 2. P. 408-M12.
McFee A.F., Banner M.W., Рагу J.M. Variation in chromosome number among
European wildpigs //Cytogenetics. 1966. Vol. 5. P. 75-81.
Menzel K. Frischlinge, Uberlaufer, grobe Sauen // Wild und Hund. 1981. Bd. 84, N 16.
S. 386-389.
Meynhardt H. Untersuchungen zur akustischen, olfaktorischen und visuellen
Kommunikation des Europaischen Wildschweines (Sus scrofa L.) // Beitr.
Forstwirt. 1980. Bd. 14, N 2. S. 72-82.
Meynhardt H. Zehn Jahre verhaltensbiologische Untersuchungen an freilebenden
Wildschweinen // Unsere Jagd. 1982. Bd. 32, N 11. S. 336-337.
Meynhardt H. 10 Jahre unter Wildschweinen. II. Nahrungs-auswahl und
Sinnesleistungen // Wild und Hund. 1983a. Bd. 86, N 14. S. 44-48.
Meynhardt H. 10 Jahre unter Wildschweinen. III. Rangordnung und Rauschzeit // Ibid.
1983b. Bd. 86, N 18. S. 53-56.
Meynhardt H. 10 Jahre unter Wildschweinen. IV. Geschlechterverhaltnis beim
Schwarzwild // Ibid. 1984a. Bd. 86, N 21. S. 28-30.
Meynhardt H. 10 Jahre unter Wildschweinen.VI. Die Aufzucht der Frischlinge // Ibid.
1984b. Bd. 87, N 6. S. 42-45.
303
Meynhardt Н. 10 Jahre unter Wildschweinen.VIH // Ibid. 1984c. Bd. 87, N 18. S. 58-62.
Meynhardt H. 10 Jahre unter Wildschweinen. IX. Das Ansprechen des Schwarzwildes
//Ibid. 1985a. Bd. 87, N 22. S. 31-35.
Meynhardt H. Leitoder Fubrungsbachen-ihre Bedeutung in Schwarzwildrotten // Ibid.
1985b. Bd. 87, N25. S. 30-32.
Meynhardt H. Vierzehn Jahre verhaltensbiologische Untersuchungen an freilebenden
Wildschweinen // Unsere Jagd. 1988. Bd. 38, N 8. S. 236-237.
Miquelle D.G., Smirnov E.N., Quigley H.G., et. al. Food habits of Amur tiger in
Sikhote-Alin Zapovednik and the Rusian Far East, and implications for
conservation //Wildl. Research. 1996. Vol. 1,N 2. P. 138-147.
Moller D. Beitrage zur Altersbestimmung des Schwarzwildes // Unsere Jagd. 1984.
Bd. 34, N 8. S. 232-233; N ll.S. 330-331; N12. S. 358-359.
Muramoto J., Makino S., Ishikawa T., Kanagawa H. On the chromosomes of the wild
boar and boar-pig hybrids // Proc. Japan Acad. 1965. Vol. 41. P. 236-239.
Nagasy M., Takacs A. Possibilities of decreasing game damages by individual defence
// XXXXVI Intern. Symp. Crop. Prot. Meded. Fac. Landbouwetensch. Univ. Gent.
1994. Vol. 59, N 28. P. 741-744.
Oloff H.B. Zur Biologie und Okologie des Wildschweines // Beitr. z. Tierkunde u.
Tiererz.. 1951. Bd. 2. S. 1-95.
Pedone P., Mattioli L., Mattioli S. et al. Body growth and fertility in wild boars of
Fuscany, Central Italy // Trans. XX Intern. Congr. Union Game Biol. Godollo,
1991. Pt. 2. P. 604-609.
Pelzers E. Het wilde zwijn Sus scrofa in Nederland voor de tweede wereldoorlog //
Lutra. 1988. Vol. 31, N 2. P. 145-152.
Perin D., Spitz F., Janeau G., Valet G. Dynamics of reproduction and development of
weight in the wild boar (Sus scrofa) in South-west France // Z. Saugetierk. 1987.
Vol. 52, N 1. P. 21-30.
Pikunov D.G. Eating habits of the Amur tiger (Pantera tigris altaica) in the wild //
Proc. V World Conf, on breeding endangered species in captivity. Cincinnati,
1988. P. 185-190.
Prossinagg H. Das Schwarzwild // Osterr. Weidwerk. 1985. N 2. S. 21-23, 26.
Rascalou S., Teillaud P. Plasticite du comportement parental chez le sanglier (Sus
scrofa) en captive//Bull. SFECA. 1988. Vol. 3,N l.P. 157-159.
Remmele W. Gerechtigkeit fur unsere “Armen Schweine”! Kritik der gultigen
Jagdzeiten fur Schwarzwild // Wild und Hund. 1986. Bd. 89, N 3. S. 70-73.
Rittmannsperger C. Chromosome investigation in wild and domestic pigs // Ann.
Genet. Sol. Anim. 1971. N 3. P. 105.
Saez-Royuela C., Telleria J.L. Reproductive trends of the wild boar (Sus scrofa) in
Spain//Folia Zool. 1987. Vol. 36, N l.P. 21-25.
Schnebel E.M., Griswold J.G. Agonistic interactions during competition for different
resources in captive European wild pigs (Sus scrofa) // Appl. Anim. Ethol. 1983.
Vol. 10, N 4. P. 291-300.
Schwartz E. Uber den Verlauf der Bestandsentwicklung von Rot,- Dam,- Reh- und
Schwartzwild in den letzten 120 Jahren // Beitr. Jagd- und Wildforschung. 1988.
Bd. 15. S. 59-68.
Singer F.J., Otto D.K., Tipton A.R., Habie C.P. Homes ranges, movements, and
habitat use of european wild boar in Tennessee // J. Wildlife Manag. 1981. Vol. 45,
N2. P. 343-353.
Sjarmidi A., Gerard J.F. Autor de la systematique et la distribution des suides //
Monit. Zool. Ital. 1988. Vol. 22, N 4. P. 415-448.
Skewes-Ramm O. Schwarzwild im Chile: Ph. D-r. Dissert. Institut fur Wildbiol. und
Jagdkunde. Goettingen, 1994.
304
Skultety J., Ladziansky A., Katreniak J., Hell P. Studie polovnickej rajonizacie a
bonitacie polovnynych revirov Slovenska. Polovnicke Study, 2. 1974. 104 s.
Spitz F. Aspects demographiques de la strategic adaptative du sanglier (Sus scrofa L.)
// Actes Colloq. Biol. Populat. Lion, 1987. P. 585-589.
Spitz F. Mortalite et dispersion chez le sanglier (Sus scrofa) de Camarque // Gibier
Faune Sauvage. 1989. N. 6. P. 27-42.
Spitz F., Janeau G. Strategies spatiales du sanglier dans diverses situation
demographiques // Comportement biol. popul. Bull. SFECA. 1990. Vol. 5, N 2. P.
27-35.
Spitz F., Janeau G., Valet G. Elements de demographic du sanglier (Sus scrofa) dans
la region de Gresigne // Acta Oecol. Oecol. Appl. 1984. Vol. 5, N 1. P. 43-59.
Spitz F., Pepin D. Occupation de 1’espace par le sanglier en zone de grande culture //
Trans. XVII Congr. Int. Union Game Biol. Brussels, 1985. Pt. 2. P. 953-959.
Stass V. Genetical studies of wild sows in the Baltic // Eksp. Biol. 1992 (1993). N 3-4.
P. 130-131.
Sternath M. Osterreichs jagd statistik 1989 // Osterr. Weidwerk. 1990. N U.S. 21-24.
Stubbe C. Lebensraumnutzung und Populationsumsatz des Schwarzwildes in DDR -
Ergebnisse der Wildmarkierung // Unsere Jagd. 1987. Bd. 37, N 8. S. 228-230.
Suzuki S„ Tanaka K., Kurosawa Y., Tikhonov V.N., Khyazev S.P. Geographic
distribution of serum amylase type electrophoretic variations among Sus scrofa wild
pig populations in eastern European and Asian regions // Proc. Jap. Acad. 1984.
Vol. B60, N 10. P. 418-422.
Thenius E. Zur Evolution und Verbreitungsgeschichte der Suidae // Z. Saugetierk.
1970. Bd. 35, N 6. S. 321-342.
Thenius E. Grundzuge der Verbreitungsgeschichte der Saugetiere. Jena: VEB Gustav
Fischer-Verlag, 1972. 345 s.
Thomas O. The races of the European wild swine // Proc. Zool. Soc. London. 1912. Pt.
1. P.390-393.
Tonisson I. Metsiga meie looduses //Jant ia Ulukid. Tallin, 1974. S. 125-140.
Tonisson Ju. The moose research and management in Estonia // IV Intern. Moose
Symp. University of Alaska. Fairbanks, 1997. P. 70.
Trnkova J. Prispevek к poznani rustu prasete dikokeho, Sus scrofa L., 1758 //
Vertebratol. Zpr. 1984. S. 102-106.
Uloth W. Zur Einburgerungsgeschichte europaischen Schwarzwildes in Nordamerika
//Unsere Jagd. 1967. Bd. 17, N 11. S. 344.
Uloth W. Bestand, Dichte und Strecke des Schwarzwildes sowie die Qualitat seiner
Trophaen in der VR Polen // Ibid. 1990. Bd. 40, N 6. S. 199.
Volokh A. The structure and reproduction of the wild boar population of the Ukrainian
steppe // Euro-Amer. Mammal. Congr. Santiago de Compostella, 1998. P. 246.
Wotchikowsky U. Sind die Sauen degeneriert? // Jager. 1981. Bd. 99, N 2. S. 39—42.
Zivkovic S., Jovanovic V, Isakovic I., Milosevic M. Chromosome complement of the
European wild pig (Sus scrofa L.) // Experientia. 1971. N 27. P. 224.
305
Оглавление
Предисловие......................................................з
Семейство Свиные.................................................5
Род Свиньи ...................................................7
Кабан, или Дикая свинья....................................9
Изменчивость и таксономия ...............................14
Размеры и масса тела .................................14
Краниометрическая характеристика......................25
Изменчивость зубов ...................................33
Окраска и линька......................................36
Генетическая изменчивость ............................41
Гибридологические особенности.........................44
Таксономия............................................44
Распространение .........................................47
Восстановленный ареал.................................47
Распространение в плейстоцене и голоцене............41
Изменения ареала во II тысячелетии н. э.............52
Динамика ареала в XX веке.............................57
Основные факторы, определяющие область распространения и
численность (исторический аспект) .......................67
Абиотические факторы..................................67
Влияние хищников......................................70
Антропогенное воздействие.............................72
Среда обитания ..........................................82
Биотопы. Региональные и сезонные особенности биотопического
распределения.........................................82
Факторы, влияющие на биотопическое размещение ........91
Питание .................................................93
Растительные корма....................................93
Животные корма........................................95
Региональные и сезонные особенности питания...........97
Потребление кормов и воды. Минеральное питание .......101
Адаптации к критическим условиям. Энергосбережение и обмен
веществ..............................................105
Пищевая активность и поведение........................109
Воздействие на биогеоценозы .............................113
Влияние на почвенные ценозы и растительность .........113
Воздействие на лесные ценозы .........................115
Роль в агроценозах ..................................117
Воздействие на зооценозы..............................119
Размножение .............................................122
Половая зрелость......................................122
Спаривание и половое поведение .......................122
Беременность и эмбриональное развитие ................126
306
Опорос и выкармливание детенышей ....................129
Плодовитость.........................................132
Популяционная структура ................................141
Половой и возрастной состав..........................141
Социальная организация ..............................145
Пространственная структура и перемещения.............157
Локальные перемещения и участки обитания ..........157
Миграции ..........................................161
Инфраструктура участков обитания ..................164
Численность и динамика населения .......................172
Численность в XX веке................................172
Плотность населения .................................184
Основные факторы, определяющие динамику популяций........191
Воспроизводство .....................................191
Детская смертность...................................193
Корма и погода.......................................195
Хищники .............................................198
Конкуренты...........................................205
Гибель в водоемах....................................206
Болезни и травмы ....................................207
Антропогенные факторы ...............................212
Использование ресурсов .................................214
Объем добычи.........................................215
Способы промысла, их эффективность и влияние на группировки 220
Избирательность добычи по полу и возрасту.............223
Сроки охоты...........................................226
Управление популяциями...................................227
Теоретические основы управления популяциями диких копытных.
Парадоксы теории и практики ..........................227
Методы и способы управления популяциями. Повышение продук-
тивности .................................232
Заключение.........................................252
Литература ........................................254
Contents
Preface
з
Family Suidae ..........................................................5
Genus Sus ..........................................................7
Wild pig ........................................................9
Variability and taxonomy .....................................14
Body dimensions and weight ................................14
Craniometric characteristics...............................25
Variability of tooth...............................................33
Coloration and shedding............................................36
Genetics variability ..............................................41
Hybridological peculiarity.........................................44
Taxonomy ..........................................................44
Distribution...........................................................47
Historical range ..................................................47
Distribution in pleistocene and holocene ......................47
Range variations in II millennium ours era.....................52
Range variations in XXth century ..................................57
Factors which determine distribution and numbers (historical
aspect)............................................................67
Abiotic factors................................................67
Influence of predators ........................................70
Antropogenic influence ........................................72
Environment .......................................................82
Habitats ......................................................82
Factors affecting habitat distribution.........................91
Feeding ...........................................................93
Vegetative forages.............................................93
Animal forages ................................................95
Regional and seasonal peculiarity nutrition....................97
Nutrition and water consumption. Mineral nutrition............101
Metabolism.....................................................105
Forage activity and behaviour ................................109
Influence on biogeocenosis ........................................113
Influence on soil cenosis and vegetation ........................ 113
Influence on forest cenosis ..................................... 115
Role in agriculture.............................................. 117
Influence on zoocenosis .......................................119
Reproduction ......................................................122
Puberty .......................................................122
Mating and sexual behaviour ...................................122
Pregnancy and embryogeny ......................................126
Birth .........................................................129
Fertility......................................................132
Population structure ..............................................141
308
Sex-age structure.............................................141
Social organization...........................................145
Spatial structure and movement ...............................157
Local movements and home range ............................157
Migrations ................................................161
Home range structure.......................................164
Numbers and population dynamics .................................172
Numbers in XXth century ......................................172
Density ......................................................184
Numbers factors..................................................191
Reproduction .................................................191
Children’s mortality .........................................193
Forages and weather...........................................195
Predators.....................................................198
Competitors...................................................205
Mortality in water............................................206
Diseases and wounds...........................................207
Antropogenic factors .........................................212
Use of resources ................................................214
Volume of use resources ......................................215
Methods of hunting ...........................................220
Hunting selection ............................................223
Period of hunting.............................................226
Population management ...........................................227
Theoretical principles of the ungulate population management. Parado-
xes of theory and practice....................................227
Methods of population management. Rise of productivity .......232
Conclusion.................................................................252
References ................................................................254
Научное издание
Алексей Алексеевич Данилкин
СВИНЫЕ
(SUIDAE)
Серия “Млекопитающие России
и сопредельных регионов”
Утверждено к печати
Ученым советом Института
проблем экологии и эволюции
им. А.Н. Северцова
Российской академии наук
Редактор М.В. Куликова
Художник Е.Н. Руденко
Макет Е.Н. Руденко
Подписано к печати 15.11.2001.
Формат 70x100 1/8. Бумага офсет № 1, 80 г/см2.
Гарнитура Таймс. Печать офсетная.
Уч.-изд. л. 33,0. Тираж 2000 экз.
Заказ № 2528.
Издательство ПК ГЕОС
Изд. лицензия ИД № 01613 от 19.04.2000.
125315, Москва, 1-й Амбулаторный пр., 7/3-114.
Тел.: (095) 230-80-92. Факс: (095) 951-04-43
Отпечатано с готовых пленок издательства
в РГУП «Чебоксарская типография № 1».
428019, г. Чебоксары, пр. И. Яковлева. 15.