/









































































































































































































































































































Автор: Данилкин А.А.
Теги: ruminantia (ruminants) систематика животных специальные зоологические науки география млекопитающие фауна
ISBN: 5-87317-231-5
Год: 2005




Текст
МЛЕКОПИТАЮЩИЕ РОССИИ
И СОПРЕДЕЛЬНЫХ РЕГИОНОВ
А.А. ДАНИЛКИН
ПОЛОРОГИЕ
Москва ❖ 2005
РОССИЙСКАЯ АКАДЕМИЯ НАУК
Институт проблем экологии и эволюции им. А.Н. Северцова Программа фундаментальных исследований Президиума РАН «Научные основы сохранения биоразнообразия России»
МЛЕКОПИТАЮЩИЕ РОССИИ
И СОПРЕДЕЛЬНЫХ
РЕГИОНОВ
А.А. ДАНИЛКИН
ПОЛОРОГИЕ
(BOVIDAE)
Товарищество научных изданий КМК Москва ❖ 2005
УДК 599.735.5
ББК 28.69
Полорогие (Bovidae) / А.А. Данилкин. М.: Т-во научных изданий КМК, 2005. 550 с. (Млекопитающие России и сопредельных регионов)
Сведения о полорогих животных фауны России и сопредельных регионов: филогения, систематика, динамика ареала, численность и факторы, ее определяющие, среда обитания, питание, размножение, структура популяций, использование ресурсов и охрана. Для зоологов, экологов, этологов, охотоведов и специалистов по охране природы.
Табл. 96. Ил. 85. Фото на вклейках 61. Библиогр. 2378.
Рецензенты: доктор биологических наук Л.В. Жирнов доктор биологических наук В.Н. Орлов
Hollow-horned ruminants (Bovidae) I A.A. Danilkin. Moscow: KMK Scientific Press, 2005. 550 p.
The book is the research of hollow-homed ruminants of the fauna of the Russia and contiguous districs. The phylogeny, systematics, variability, range and numbers and its underlying factors, environment, feeding, breeding, population structure, utilization of their resources and preservation are given. F or zoologists, ecologists, ethologists, game biologists.
Tabl. 96. Fig. 85. Foto 61. Ref. 2378.
ISBN 5-87317-231-5
© A.A. Данилкин, текст, иллюстрации, 2005
© Товарищество научных изданий КМК, 2005
Посвящается выдающимся ученым -исследователям диких копытных животных: Н.А. Северцову, Н.В. Насонову, И.И. Соколову, В.Е. Соколову и Н.К. Верещагину
Предисловие
Монография “Полорогие” — очередная книга в серии “Млекопитающие России и сопредельных регионов”, продолжающей и дополняющей издание ’’Млекопитающие Советского Союза” [1961-1976]. Ранее были опубликованы “Зайцеобразные” [1994], “Усатые киты” [1994], “Тушканчикообразные” [1995], “Оленьи” [1999] и “Свиные” [2002].
Каждый том этой серии - всестороннее рассмотрение в пределах определенного семейства существующих ныне родов и видов млекопитающих, обитающих на территории России и сопредельных стран. Основная цель сводок - обобщение ценнейшего научного материала, накопленного отечественной и мировой наукой за последние десятилетия.
Структура книг стандартна. После краткой характеристики семейства, подсемейств и родов по единой схеме и без излишней детализации приведены видовые или подвидовые (для неэндемичных видов) очерки. Научные сведения, которые можно рассматривать в сравнительном аспекте, для краткости изложения объединены в таблицах. В связи с диагностическим подходом в отдельных разделах неизбежно незначительное “дублирование” материала, что, однако, оказывается полезным при использовании монографий в качестве справочного пособия.
В монографии “Полорогие”, как и в других книгах серии, характеристика таксонов основана, главным образом, на критическом анализе соответствующих разделов известных фундаментальных работ [Simpson, 1945; Ellerman, Morrison-Scott, 1951; Цалкин, 1951; Громова, 1953; Соколов И.И., 1953, 1959, 1962; Гептнер и др., 1961; Громов и др., 1963; Haltenorth, 1963; Соколов В.Е., 1979; Барышников и др., 1981; Павлинов, Россолимо, 1987] с необходимыми изменениями и дополнениями. Литературные источники, приведенные в синонимике, указаны в вышеназванных публикациях и не включены в список литературы.
Наиболее проблематичная часть монографии - систематика. Она основательно запутана, а существующая видовая и внутривидовая таксономия полорогих, во многих случаях, совершенно неприемлема. Решение противоречивых вопросов упирается в извечную проблему критерия таксонов, и, прежде всего, вида и подвида. Развитие систематики невозможно без новейших иммунологических, биохимических и генетических методов, но их результаты нередко оказываются далеко не тождественными морфологическим построениям и спорными. Поэтому на данном этапе исследований пришлось взять за основу все же традиционную (морфологическую) классификацию Bovidae с существенной корректировкой по результатам последних молекулярно-генетических исследований и с учетом возможного параллелизма в филогении животных. Предлагаемая ниже таксономия, тем не менее,
отражает всего лишь сегодняшний уровень накопленных знаний и по-прежнему остается несовершенной. По этой причине в монографии не приводятся традиционные характеристики (диагнозы) подвидовых форм, но необходимый для этого сравнительный материал приведен достаточно полно.
Материалы, касающиеся распространения, численности и добычи зверей, обобщены по литературным источникам и сведениям Министерства сельского хозяйства СССР, Департамента по охране и рациональному использованию охотничьих ресурсов Минсельхозпрода РФ (Главохоты РСФСР, Охотдепартамента РФ) и Службы учета охотничьих ресурсов. Эти собранные по крупицам данные - ценный вклад в исторический мониторинг фауны России и сопредельных регионов.
Большую помощь в поиске необходимой литературы оказала Л.Л. Данилкина. Ценные замечания сделаны рецензентами Л.В. Жирновым и В.Н. Орловым. Специальные исследования филогении, систематики и генетической изменчивости наиболее сложных групп полорогих животных новейшими методами провели мои сотрудники М.В. Кузнецова, Е.Ю. Казанская и Т.П. Сипко. В сборе полевого материала принимали участие многие зоологи, охотоведы, егеря и охотники. Оригинальные рисунки выполнены В.М. Гудковым и А.А. Варшавским. Фотографии любезно предоставлены Т.В. Баженовым, А.П. Вербером, А. Волковым, А.Ф. Дигилевичем, Р.В. Дормидонтовым, Д.Г. Медведевым, С.С. Романовым, Т.П. Сипко, П.А. Сорокиным, С.А. Тишкевичем и А.Н. Хохловым. Мной использованы также некоторые, имеющие научное значение, снимки животных, опубликованные в журналах “Охота и охотничье хозяйство”, “Сафари”, “Охотник за трофеями”, в Интернете и других изданиях. На обложке книги помещены фотографии копытных, сделанные С.А. Тишкевичем и П.А. Сорокиным. Всем коллегам, так или иначе внесшим вклад в подготовку монографии “Полорогие”, приношу глубокую благодарность.
Замечания, уточнения или дополнения прошу направлять по адресу: 119071, Москва, Ленинский проспект, д. 33. Институт проблем экологии и эволюции им. А.Н. Северцова.
Семейство ПОЛОРОГИЕ Bovidae Gray, 1821
Характеристика. Разнообразная группа жвачных парнокопытных животных мелких, средних и крупных размеров с полыми рогами. Сложение от легкого и стройного до тяжелого и массивного. Половой диморфизм у большинства видов незначителен.
Волосяной покров густой и относительно длинный у северных и горных видов, короткий и редкий - у южных. Окраска чаще однотонная - от почти черной до белой, на голове и туловище могут быть контрастные узоры. Хвостовое “зеркало” имеется или отсутствует.
Залобная часть черепа сильно изогнута. Теменные кости заходят на затылочную плоскость черепа. Теменные гребни широко расставлены. Лицевая часть слезной кости с углублением для преорбитальной железы или без него; ее орбитальный край с одним слезным каналом по внутреннему краю. Верхнечелюстная кость простирается на лицевую часть. Носовые отростки межчелюстных костей сравнительно короткие. У многих видов кости черепа сильно пневматизированы. Зубы тетраселенодонтные, щечные - гипсодонтные и столбчатые, их призмы переходят в корни почти без шейки. Верхние клыки закладываются, но за редким исключением (у карликовой антилопы Madoqua) не прорезываются. Зубная формула типичная для копытных животных: i 0/3, с 0/1, р 3/3, m 3/3 = 32, но иногда в нижней и верхней челюсти только два премоляра.
Рога эпидермического происхождения. В отличие от рогов оленьих (Cervidae), они полые внутри, не сбрасываются ежегодно как у вилорогов (Antilocapridae) и не ветвятся. Их костные пневматизированные стержни отрастают в вершине, тогда как роговые чехлы циклически растут в основании. В течение жизни, однако, происходит частичная смена рогового чехла: под ним нарастает новый слой, прободающий концы чехлов первых рогов у взрослых самцов. У самок нарастающий новый чехол сталкивает первый. Форма рогов разнообразная: есть массивные, закрученные спирально, длинные копьевидные и саблеобразные, короткие и толстые, направленные в стороны, вниз и назад. На поперечном разрезе они округлой, овальной или треугольной формы, на поверхности - гладкие или чаще с буграми, поперечными складками, кольцами и продольными ребрами. У большинства видов рога имеют как самцы, так и самки, но у последних они значительно меньше в размерах.
Боковые пальцы обычно с рудиментарным скелетом и небольшими копытами. Метакарпальные и метатарзальные кости II и V пальцев редуцированы: от первых остаются лишь проксимальные части, вторые в значительной мере срастаются с метатарзальными костями III и IV пальцев. Из фаланг II и V пальцев сохраняются лишь две или одна, иногда они отсутствуют.
Волосяной покров у большинства представителей семейства состоит из ости и пуха, у Bovinae имеются направляющие волосы. Специфические кожные железы чаще хорошо развиты и расположены перед глазом, на затылке, в пахах, у корня хвоста, на
запястье и между копытами. Их расположение - важный диагностический признак в систематике и филогении. Молочная железа двухдолиатая или четырехдольчатая.
Желудок сложный. Желчный пузырь есть почти у всех. Плацента поликотиле-донная. У многих видов penis с нитевидным отростком на конце.
Кариотип разнообразный - 2п = 30-60, у примитивных представителей - 2п = 60, NF = 60. Не исключено, что 60-хромосомомный набор с акроцентрическими аутосомами был предковым для всех жвачных [Графодатский, 1989].
Экологически очень разнообразная группа. Преимущественно стадные травоядные животные, обитатели степей, полупустынь, пустынь и гор (встречаются до высоты в 5500 м над уровнем моря), но некоторые виды живут в лесах, в тундре или в прибрежных зарослях.
Большинство видов - полигамы. Продолжительность беременности неодинакова - от 4 до 11 месяцев. В помете от 1 до 5 детенышей. Продолжительность жизни - до 15-30 лет.
Важные охотничьи или редкие и исчезающие виды. Некоторые из них стали родоначальниками домашних животных.
Распространение. Африка, Европа, Азия и большая часть Северной Америки. Отсутствуют в Австралии, Центральной и Южной Америке. Отдельные виды акклиматизированы в Новой Зеландии. В одомашненном состоянии обитают повсеместно.
Эволюция и филогения. Bovidae, возможно, произошли от мелких олигоценовых оленьков (Tragulidae) типа Eotragus около 15-20 млн лет назад (рис. 1). Древнейшие представители семейства прослеживаются с начала миоцена в Африке (Namibiomeryx senuti - [Morales et al., 1995]), с конца раннего миоцена - в Европе [Gentry, 1996], с верхнего миоцена - в Азии [Pilgrim, 1939,1947] и с плейстоцена - в Северной Америке. Судя по последним находкам в Монголии (роды Palaeohypsodontus и Hanhaicerus), время су-
Рис. 1, Предполагаемая радиация представителей семейства Bovidae по морфологическим данным (по [Thenius, Hofer, 1960])
Рис. 2. Схема базальной радиации Ruminantia по молекулярным данным (по [Кузнецова и др., в печати])
ществования древних полорогих в Азии охватывает интервал раннего - позднего олигоцена и раннего миоцена. Высококоронковый па-леогипсодошус, возможно, стоял в основе ветви, приведшей к появлению козлиных, газелевых и коровьих антилоп, а низкокоронковый хангайцерус - к группам лошадиных, винто-рогих антилоп и к бычьим [Дмитриева, 2002].
Bovidae четко отграничены от остальных семейств Ruminantia. Наиболее близки к ним, как считается, вилороговые (Antilo-capridae) [Gentry, Hooker, 1988] и жирафовые (Giraffidae) [Janis, Scott, 1988; Gentry, 1994]. Наши филогенетические построения, основанные на молекулярных исследованиях (рис. 2), отличаются от традиционных морфологических схем. От базальной группы Tragulidae ответвляется эволюционная линия жирафовых или, иногда, вилороговых, но место Antilocapridae в схеме остает
ся неясным. Семейство Giraffidae является внешней группой к общей кладе Cervidae, Moschidae и Bovidae. Оленьи оказываются сестринской кладой по отношению к Bovidae, а кабарговые явно сближаются с полорогими. При таком построении очевидна несостоятельность общепринятого состава надсемейства Cervoidea, включающего только Antilocapridae (или Giraffidae), Cervidae и Moschidae. В его состав должно быть включено и семейство Bovidae [Кузнецова и др., в печати].
Полорогие являются самой молодой из современных групп копытных. Для них характерна быстрая радиация и ускоренное эволюционное развитие. С верхнетретичного времени и поныне - это наиболее многочисленное и процветающее семейство. Основное направление эволюции: переход лесных форм животных к жизни в степи и горах; потребление исключительно растительной, преимущественно жесткой травянистой пиши с примесью пыли и песка; приспособление к быстрому бегу на равнине или к специфическому передвижению в горах, что повлекло за собой увеличение и укрепление заднекоренных зубов, частичную редукцию предкоренных, усложнение и дифференцирование желудка и изменение строения конечностей. В социальном плане очевиден переход к стадному образу жизни и активным способам защиты, что во многом привело к преобразованию и увеличению рогов, сдвигу их оснований вместе с участками лобных костей к вершине черепа, укорочению заро-говой части и излому его оси, и к соответствующему изменению, укреплению и пневматизации костей черепа [Соколов, 1953]. Большие рога, тем не менее, не могут служить бесспорным показателем прогрессивности тех или иных групп животных.
Неоднородность среды обитания на обширном ареале обусловила разнообразный внешний вид и размеры этой группы животных - от зверей величиной с кролика, таких как африканские карликовые антилопы (Neotragini), до гигантов вроде зубра, бизона, буйвола и уничтоженного тура, и от стройных стремительных газелей до коротконогих мускусных быков.
Состав. Систематика семейства основательно запутана не только в связи с большим разнообразием видов, но также из-за слабой изученности многих из них. Исследователи, базирующиеся на традиционных морфологических методах, включали в состав семейства от 2 до 16 подсемейств, в последнее время чаще 5-8: хохлатых (Cephalophinae) и карликовых (Neotraginae) антилоп, газелевых (Antilopinae, = Gasellinae), козлиных (Caprinae), бычьих (Bovinae), а также лошадиных (Hippotraginae), винторогих (Tragelaphinae) и коровьих (Alcelaphinae) антилоп. В них насчитывали 140-174 рода, в том числе около 120 вымерших и 43-54 современных, а число принимаемых видов превышало 200. По последним оценкам, в семействе около 104—120 родов (современных 43-48), что составляет примерно 70% видов Ruminantia и 60% Artiodactyla. Более половины из них африканские, треть - евразийские, 3 вида - североамериканские и 1 - евразийскоамериканский [Simpson, 1945; Ellerman, Morrison-Scott, 1951; Соколов И.И., 1953, 1959, 1962; Haltenorth, 1963; Соколов В.Е., 1979; Барышников и др., 1981; Durst et al., 1982; Gentry, 1992; Тихонов, 1999; Павлинов, 2003].
Новые биохимические и генетические исследования позволили уточнить статус ряда групп полорогих, но не дали однозначных результатов даже на уровне подсемейств [Wurster, Benirschke, 1968; Lowenstein, 1986; Georgiadis et al., 1990; Allard et al., 1992; Chikuni et al., 1995; Groves, Shields, 1996, 1997; Modi et al., 1996; Глазко и др., 1997, 2002; Glazko, 1998; Hassanin, Douzery, 1998, 1999a,b; Hassanin et al., 1998; Klungland et al., 1999; Matthee, Robinson, 1999; Rebholz, Harley, 1999; и др.]. Результаты многих биохимических исследований в систематике позднее были признаны артефактами.
Молекулярно-генетические методы, включая использование феномена геномного полиморфизма и анализ последовательностей ядерной и митохондриальной ДНК, оказались более достоверными [Рысков и др., 1993,1994; Потапов и др., 1994,1997; Engel et al., 1996; Потапов, 1998; Семенова и др., 1999; и др.]. Однако результаты, полученные с их помощью, тоже далеко не всегда согласуются с традиционной классификацией. В частности, на основании анализа последовательностей митохондриального гена цитохрома b у 51 вида, семейство Bovidae разделено на две клады: к первой отнесены быки и буйволы (Bovini), нильгау (Boselaphini) и лесные антилопы (Tragelaphini), ко второй - газели (Antilopini), козлы и бараны (Caprini) и представители остальных триб [Hassanin, Douzery, 1999а]. Подсемейство Hippotraginae при исследованиях с использованием в качестве маркеров митохондриальных генов рибосомных РНК распадается на отдельные, не связанные между собой, трибы. Триба Reduncini входит в кластер Cephalophinae, в который вписываются также Oreotragus, Aepyceros и Neotragus, относимые прежде к Antilopinae. Такое же несоответствие очевидно и по другим таксонам полорогих (см. ниже). В целом геносистематики определяют в семействе 5 монофилетических групп на уровне подсемейств: Bovinae, Antilopinae, Caprinae, Alcelaphinae и Cephalophinae [Gatesy et al., 1992, 1997; Кузнецова, 2002; Кузнецова и др., 2002].
Классификация обширного семейства Bovidae в общем виде приведена И.Я. Павли-новым [2003] в сводке “Систематика современных млекопитающих”, что избавляет меня от такой необходимости, за исключением отдельных уточнений и дополнений. В целом систематику полорогих нельзя признать завершенной даже на уровне подсемейств и триб, не говоря уже о таксонах более низкого ранга, о чем не раз упоминается ниже.
В фауне России и сопредельных регионов (в пределах бывшего СССР) 3 подсемейства: Bovinae, Antilopinae и Caprinae, включающие около 30 вымерших и 8 современных родов.
Подсемейство БЫЧЬИ Bovinae Gray, 1821
Характеристика. Бычьи - животные средних и крупных размеров, тяжелого массивного сложения с короткой и толстой шеей и низко посаженой головой. У многих видов развит подгрудок. Морда большая с широко расставленными ноздрями, ее носовая часть голая. Хвост средней длины или длинный, достигающий скакательного сустава, обычно с кистью на конце. Конечности короткие, в их скелете пястная кость короче лучевой, а иногда и плечевой кости. Окраска однотонная, темная с разными оттенками.
Лобные кости черепа большие, затылочная кость расширена в задней части. Зароговая часть сильно укорочена. Преорбитальное вдавление на слезной кости развито слабо или отсутствует, как и этмоидальное отверстие. Слуховые пузыри вздуты и сжаты с боков. Пневматизация костей черепа хорошо выражена. Роговые стержни с внутренними полостями, доходящими почти до вершины. Резцы широкие. Верхние коренные зубы обычно с широкими коронками, гипсодонтные, заднекоренные (нижние, по крайней мере) - с добавочными бугорками или столбиками.
Рога изогнутые, гладкие, имеются у самцов и самок, но у последних меньше в размерах. Обычно их основания расположены позади глазниц близко к затылочной части черепа и направлены в стороны. В сечении рога круглые или иногда треугольной формы.
Волосяной покров короткий, представлен 3 категориями волос: направляющими, остевыми и пуховыми [Соколов, 1973]. Специфические железы чаще отсутствуют. Молочная железа четырехдольчатая.
Кариотип - 2п = 46-60, NF = 58-62. Аутосомы акроцентрические, Х-хромосома субметацентрическая, Y-хромосома варьирует по форме [Wurster, Benirschke, 1968; Koulischer et al., 1972].
Обитатели лесов, лесостепи и степи, реже - гор и болот. Иногда образуют большие стада и скопления.
Охотничьи, охраняемые и домашние животные.
Распространение. Европа, Азия, Африка и Северная Америка. Домашние виды есть повсеместно.
Эволюция и филогения. Древнейшие ископаемые формы Bovinae появляются в нижнем - позднем миоцене Евразии (Leptobos) и, возможно, Африки. В Америке эта группа животных возникает в плейстоцене. Типичные Bovini с гладкими, относительно прямыми рогами обнаруживаются в среднем плиоцене Южной Азии [Barry et al., 1985; Раутиан и др., 2000]. В основе их родословного древа, возможно, находитсяParabos [Groves, 1981].
По морфологическим и генетическим параметрам бычьи четко отличаются от остальных Bovidae [Wurster, Benirschke, 1968; Gentry, 1992; Gatesy et al., 1997]. Основные направления эволюции - травоядность и увеличение размеров и массы тела.
Состав. Состав подсемейства окончательно не определен, но все же он более однозначен по сравнению с остальными таксонами полорогих [Соколов, 1953,1959; Groves, 1981], и даже по молекулярно-генетическим исследованиям близок к классификации Симпсона [Simpson, 1945]. В нем три трибы: Bovini с родами Bos, Poephagus, Bison, Bibos, Bubalus, Anoa, Syncerus и новыми вьетнамскими родами и видами - Pseudoryx nghetinhensis [Dung et al., 1993] и Pseudonovibos spiralis [Peter, Feiler, 1994]; Boselaphini с родами Boselaphus и Tetracerus'. Tragelaphini c Tragelaphus и Taurotragus (нередко эти группы объединяют) [Кузнецова, 2002; Павлинов, 2003].
Принадлежность рода Pseudoryx к бычьим, тем не менее, оспаривается - его предлагали причислить к козлиным [Thomas, 1994]. Еще более неясна ситуация с Pseudonovibos spiralis - по последним молекулярно-генетическим исследованиям образцы этого животного идентифицируются как филогенетически близкие к буйволам [Kuznetsov et al., 2001; Кузнецов и др., 2002], или как принадлежащие домашнему скоту [Brandt et al., 2001; Hassanin et al., 2001; Thomas et al., 2001; Galbreath, Melville, 2003].
Классификация трибы Bovini по-прежнему дискуссионна. Предложение оставить в ней только два (или три) рода: Bos, включая Bison, Poephagus и Bibos, и Bubalus с Syncerus и Anoa [Bohlken, 1958; Stormont et al., 1961; Орлов, Чудиновская, 1979; Соколов, 1979] не лишено оснований - линии быков и буйволов прослеживаются не только по морфологическим признакам, но и по результатам геномной дактилоскопии, таксономического типирования и рестрикционного картирования рибосомной РНК [Wall et al., 1992; Рысков и др., 1993, 1994; Потапов и др., 1994; Семенова и др., 1999]. Буйволы, скорее всего, отпочковались от общей ветви подсемейства раньше других плиоценовых родов, и их обособленность, в отличие от Syncerus и Апоа, не вызывает сомнений. Уровень репродуктивной дивергенции Bos и Bison весьма существенный, что не позволяет объединить эти роды. Як (Poephagus) занимает промежуточное положение между Bos и Bison (рис. 3), но по генетическим показателям он более близок к последнему [Бадагуева, Сулимова, 1994; Бадагуева, 1995; Modi et al., 1996; Агаджанян, Раутиан, 1997; Zimmermann u. а., 1998; Burzynska et al.,
Рис. 3. Филогенетические отношения форм трибы Bovini (по [Раутиан и др., 2000])
1999; Раутиан, Агаджанян, 1999; Раутиан и др., 2000; Раутиан, Калабушкин, 2003].
В составе дикой фауны России и сопредельных регионов один современный род — Bison. Тур (Bos primigenius Bojanus, 1827) и дикий як (Bos mutus Przewalski, 1883) истреблены здесь сравнительно недавно -в XVII-XVIII вв. В домашнем состоянии сохранились потомки тура (крупный рогатый скот - Bos taurus), яка (Bos grun-niens) и завезены потомки индийского буйвола (Bubalus bubalis).
Род БИЗОНЫ
Bison Н. Smith, 1827
Bison Н. Smith, 1827. Griffith’s Cuvier Animal Kingd., 5:373. Тип рода Bos bison Linnaeus, 1758 (Сев. Америка).
Bonasus Wagner, 1844. Schreber Saugeth., Suppl., 4:515. Bos bison H. Smith.
Характеристика. Бизоны - крупные животные с массивной головой, мощной передней частью туловища и высокой холкой в виде горба, который появляется вследствие удлинения остистых отростков 3-5 грудных позвонков. Длина тела достигает 330 см, высота в холке - 210 см, масса порой превышает 1000 кг. Хвост длинный, доходит до пяточного сустава, с кистью на конце. Уши короткие. Имеется подгрудок.
Окраска варьирует от серо- и темно-бурой до рыжеватой. Волосяной покров густой. На нижней стороне головы и шеи длинный волос образует своеобразный подвес.
Череп широкий, относительно короткий, с трубкообразными глазницами. Его лобные кости и зароговая часть сравнительно хорошо развиты. Межчелюстные кости не достигают носовых. Межроговой валик отсутствует. Слезные кости без ямок, этмоидальных отверстий нет. Кости черепа сильно пневматизированы. Поверхность роговых стержней не имеет продольных ребер, что отличает бизонов от буйволов. Верхние коренные зубы резко гипсодонтные с дополнительной призмой с внутренней стороны.
Рога небольшие (меньше, чем у видов рода Bos), толстые в основании, с загнутыми внутрь концами, гладкие. Они имеются как у самцов, так и у самок (меньших размеров). Их поперечное сечение круглое. Роговое вещество серо- или черно-бурое.
Специфических желез, вероятно, нет.
Диплоидное число хромосом - 60, что сближает их с родом настоящих быков Bos (2 п = 58-60), но отделяет от азиатских (Bubalus) и африканских (Sinceros) буйволов, у которых 2 п = 46-50 и 2 п = 52-54 соответственно [Wurster, Benirschke, 1968; Koulischer et al., 1972; Графодатский и др., 1990].
Степные, лесостепные, лесные и горнолесные жители. Образ жизни преимущественно групповой и стадный. У степных бизонов четко выражены сезонные миграции. Живут до 24—30 лет.
Охотничьи или строго охраняемые в последнее столетие виды.
Распространение. Верхнеплиоценовые и раннеплейстоценовые представители Bison прослеживаются в фауне Индии (В. sivalensis), Китая (В. palaeosinensis), юга Сибири, Казахстана и юга Европы (В. suchovi, В. tamanenis, В. schoetensacki) [Кулагин, 1919; Пидопличко, 1951, 1954; Верещагин, 1957, 1959а; Соколов, 1959; Флеров, 1976; Алексеева, 1977; Верещагин, Барышников, 1980а; Кожамкулова, 1981; Бурчак-Абрамович, Векуа, 1982; Хозацкий, 1986; Тлеубердина и др., 1989; Давид и др., 1990; Форонова, 2001]. В нижнем плейстоцене примитивные формы бизонов достигают северо-западных окраин Европы и широко распространяются по Сиби-
Рис. 4. Распространение Bison в Евразии (по [Флеров, 1979])
1-3 - реальная, предполагаемая и неизвестная границы восстановленного ареала рода Bison, соответственно; 4 - местонахождения остатков древнейших бизонов подрода Eobison', 5 - вероятные пути расселения Bison в плиоцене и плейстоцене; 6 - ареал Bison bonasus в голоцене и в раннеисторическое время
ри (рис. 4). В минделе они сменяются более прогрессивными формами, которые занимают почти всю Палеарктику, но исчезают в Южной и Юго-Восточной Азии. Восточные популяции проникают через Беренгийский перешеек в Северную Америку в среднем плейстоцене, вероятно, неоднократно [Флеров, Заблоцкий, 1961], и расселяются к югу (рис. 5).
География районов обитания первобытных бизонов в России и сопредельных республиках обширна. Их остатки известны из плейстоценовых отложений Приазовья, Крыма, Северного Кавказа, Краснодарского края, Ростовской, Астраханской, Оренбургской, Волгоградской, Саратовской, Самарской, Пензенской, Ульяновской, Воронежской, Орловской, Курской, Брянской, Московской, Рязанской, Тверской, Владимирской, Ленинградской, Новгородской, Псковской, Вологодской, Архангельской, Кировской и Пермской областей, Республики Коми, Татарии, Башкирии, Мордовии, из многих районов Среднего и Южного Урала, Зауралья, Западной Сибири (Прииртышье, Новосибирская и Томская области, Кузнецкая котловина, Алтай), Восточной Сибири (окрестности Красноярска, бассейны Енисея, Бирюсы, Ангары, Нижней Тунгуски, Селенги, Никоя, Алдана и Вилюя, окрестности Севе-робайкальска и Читы, среднее и нижнее течение Лены, низовья Яны и Анадыря, бассейн Колымы, о. Б. Ляховский) и Южного Приморья, а также из Молдавии, Украины, Беларуси, Казахстана, Киргизии, Узбекистана и всех республик Закавказья [Кулагин, 1918, 1919; Громова, 1935а; Верещагин, 1947, 1954, 1959а; Громов, 1948; Бибикова, 1950, 1975, 1985; Пидопличко, 1951, 1954; Трофимов, 1955; Ван-генгейм, 1961,1977; Александрова и др., 1963; Беляева, Курдюков, 1963; Соколов, 1963; Кузьмина, 1966, 1971, 1975; Верещагин, Оводов, 1968; Межлумян, 1968; Ко-жамкулова, 1969,1972,1981; Савинов, 1972; Ларичев, 1976; Смирнов, 19766; Алек-
Рис. 5. Распространение Bison в Северной Америке (по [Флеров, 1979])
1 - границы восстановленного ареала; 2 - границы ареала Bison bison в XV в.; 3 - южная граница ареала Bison priscus athabascae-, путь миграции Bison в плейстоцене
сеева, 1977, 1980, 1982, 1990а,б; Оводов, Орлова, 1977; Ермолова, 1978; Верещагин, Русаков, 1979; Галкина, Форонова, 1980; Давид, 1980, 1986, 1990, 2000; Лазарев, 1982, 1990; Батыров, 1986; Бурчак-Абрамович, 1986; Каспаров, 1986;Кожам-кулова, Пак, 1986; Татаринов, 1986, 1995; Худайбердиев, 1986; Белан, 1989; Тлеуберди-на и др., 1989; Антипина, Маслов, 1990; Белан, Старкин, 1990; Давид и др., 1990; Оводов, Ивлева, 1990; Смирнов и др., 1990; Абрамова и др., 1991; Бобковская, Косинцев, 1991; Байгушева, Тишков, 1995; Барышников, 1995; Васильев, 1995, 2003а-в; Калмыков, 1995; Кузьмина, Синицына, 1995; Малеева и др., 1995;Манасерян, 1995,1997а; Логвиненко, 2001; Пономарев, 2001, 2003; Форонова, 2001; Хромов и др., 2001; Бай-
гушева, 2003; Бобковская, 2003; Косинцев, 2003; Косинцев, Бобковская, 2003; Подопригора, 2003; Раутиан, Пузаченко, 2003; Улитко, 2003].
В конце вюрма ареал Bison сокращается и начинает разрываться. В начале голоцена эти копытные исчезают в Северной Европе, затем на большей части Восточной Сибири и на крайнем севере Америки [Флеров, Заблоцкий, 1961; Флеров, 1972, 1976, 1979; Барышников, 1978]. На Северном Урале они вымерли 9-10 тыс. лет назад [Пономарев, 2001, 2003], в Зауралье и Западной Сибири - во П-1 тысячелетиях до н. э. [Смирнов, 19766; Косинцев, 1982, 1986, 1988]. Не исключено, что в голоцене бизон еще населял Кузнецкую котловину [Алексеева, 1980]. В Забайкалье он дожил вместе с куланом до эпохи камня и ранней бронзы. “Сибирский зубр” всего лишь тысячу лет назад (VIII-X вв. н. э.) входил в состав фауны лесостепного Предбайкалья [Ермолова, 1978; Окладников, Кириллов, 1980; Смирнов, 1983]. В последние столетия отдельные очаги обитания представителей рода Bison сохранялись только в Европе и Северной Америке.
Эволюция. Корни рода Bison не совсем ясны, но очевидно, что эти копытные близки к Bos и Bibos, с которыми могут гибридизировать. Филогенетическая ветвь бизонов отделилась от ствола подсемейства, по-видимому, еще в миоцене.'Наиболее известная ранняя форма - Protobison kuschkunensis из Азербайджана [Бурчак-Абрамович, Векуа, 1980], но более реальный центр происхождения большинства родов трибы Bovini — Индостан [Соколов, 1953].
В процессе эволюции морфологический облик животных заметно менялся. Бизоны позднего плиоцена (Bison sivalensis, В. palaeosinensis') из Индии и Китая, эоп-лейстоценовый В. tamanensis и другие древние формы имели сравнительно мелкие размеры, резко отогнутые назад основания небольших роговых стержней, узкий затылок и мозговой отдел, слабо трубкообразные глазницы. Позднее у них изгибается ось черепа, его лобная и мозговая части расширяются и утолщаются, орбиты становятся резко трубкообразными, зубная система видоизменяется в связи с переходом животных на питание более плотными кормами. В миндельское время обширный ареал (от Западной Европы до Средней Сибири) занимал Bison schoetensacki, который имел морфологические адаптации, характерные для лесных обитателей. В плейстоцене в открытых холодных ландшафтах Северной Евразии сформировались формы гигантских размеров (Bison priscus и др.) с длинными широко расставленными (более 2 м между концами) рогами, которые затем постепенно мельчали и становились короткорогими. Приблизительно в вюрме дифференцируются экологически и морфологически линии зубра (bonasus) и бизона (bison). Достоверные остатки собственно зубра известны лишь из голоценовых слоев Европы, включая Кавказ, и, возможно, Зауралья и юга Западной Сибири. В Западной Сибири на протяжении всего четвертичного периода, видимо, существовали две формы. Одна из них была более приспособлена к жизни в лесных биотопах, другая - к открытым пространствам. Лесной бизон (форма athabascae), скорее всего, населял редколесья и светлохвойную тайгу северо-востока Сибири и Канады, а в прериях Северной Америки жил В. bison [Громова, 1935а, 19486; Громов, 1948; Верещагин, 1959а; Соколов, 1959, 1962, 1963; Guthrie, 1970; Шер, 1971, 1986; Флеров, 1972, 1976, 1979; Ермолова, 1978; Алексеева, 1980; Верещагин, Барышников, 1980а, 1985; Косинцев, 1982, 1986, 1988; Gienc, 1994; Зудин, Фороно-ва, 1995; Раутиан, Сипко, 1997; Форонова, 2001; Раутиан, Пузаченко, 2003].
В целом для рода характерны два основных типа экологической специализации: обитание в лесах и питание листвой, веточным кормом и травой; и обитание в степи, где основной корм травянистый, что привело к возникновению определенных морфологических адаптаций. У первых (ветвь зубров) передний отдел межчелюстных костей не расширен; узкий симфиз нижней челюсти; резцы прямые, долотообразные; коренные и предкоренные зубы не имеют дополнительных образований; конечности более длинные; грудная клетка уплощена; остистые отростки позвонков менее развиты. У вторых (ветвь бизонов) передний отдел межчелюстных костей расширенный; симфиз нижней челюсти широкий; резцы с расширенными коронками; коренные и предкоренные зубы имеют дополнительные образования, противодействующие стиранию коронок при пережевывании жесткой травы; конечности относительно короткие; ребра длинные и изогнутые; грудная клетка бочкообразная; остистые отростки позвонков крупные и по размерам близкие к таковым у ископаемых бизонов; хвостовой и крестцовый отделы позвоночника укороченные [Соколов, 1971; Флеров, 1972, 1979; Решетов, 1974; Верещагин, Громов, 1977].По фенам черепа В. bison близокк В. priscus [Пузаченко и др., 1999].
Процесс формообразования зубра и бизона и становления их ареалов, к сожалению, не ясен до сих пор, несмотря на многочисленные исследования по этому поводу. Скорее всего, адаптация к лесному и степному образу жизни у них не абсо
лютна, морфологические отличия менее значительны, а родственные отношения более тесные, чем предполагалось ранее.
Состав. Единого мнения о составе рода среди систематиков нет. В нем признают от одного (Bison bison L., 1758) [Bohlken, 1967] до трех близкородственных видов: степного и лесного американских бизонов (В. bison L., 1758 и В. athabascae Rhoads, 1897) и зубра (В. bonasus L., 1758). Лесного и степного бизонов все же чаще считают подвидами В. bison [Флеров, 1932, 1935а, 1965, 1979; Соколов И.И., 1959, 1971; Гептнер и др., 1961; Громов и др., 1963; Haltenorth, 1963; Соколов В.Е., 1979; Krumbiegel, 1980; Groves, 1981]. Однако по последним молекулярно-генетическим и фенетическим исследованиям дифференциация степного и лесного бизонов кажется еще менее существенной - на уровне, характерном для географических популяций [Bork et al., 1991; Zyll de Jong et al., 1995; Wilson, Strobeck, 1999; Раутиан, Калабушкин, 2003].
Бизон (фото 1), как считается, отличается от зубра (фото 2 и 3) красновато-коричневым оттенком окраски, черной окраской молодых особей, более удлиненными темными волосами на лбу и предплечьях, более короткими рогами и хвостом, некоторыми особенностями строения скелета, скелетной мускулатуры, черепа и зубов [Флеров, 1965; Соколов, 1971; Kobryn, 1973; “Зубр”, 1979; Раутиан и др., 2000]. Однако морфологическую дифференциацию этих копытных вряд ли можно считать безусловно достигшей “видового” уровня (табл. 1), что наглядно показали Зилл де Енг [Zyll de Jong, 1990] и, несмотря на иной вывод, Г.С. Раутиан с соавторами [2003].
По результатам биохимических, цитогенетических (см. ниже) и молекулярногенетических исследований зубр и бизон тоже или весьма близки, или почти идентичны [Schreiber, Goltenboth, 1990; Wall et al., 1992; Рысков и др., 1993, 1994; Потапов и др., 1994; Сипко и др., 1994, 1996, 1997а,б; Удина и др., 1994; Потапов, 1998; Ломов и др., 1999а; Семенова и др., 1999, 2000; и др.]. Основные черты экологии, популяционной динамики, социальной структуры и поведения этих копытных сходны [Дерягина, 1972; Фабри, Дерягина, 1972; Krasinski, 1978]. Репродуктивные барьеры полностью отсутствуют, что установлено экспериментально и подтверждено в природе на примере процветавшей до недавнего времени (до усиления браконьерского пресса) кавказской зубробизоньей группировки. Все это убедительно свидетельствует в пользу точки зрения об одновидовой структуре рода.
Тем не менее на некоторых генетических дендрограммах зубр и бизон заметно обособлены не только от других видов Bovini, но и друг от друга, особенно по частотам антигенов крови [Агаджанян, Раутиан, 1997; Дымань и др., 1999, 2000; Раутиан, Агаджанян, 1999; Глазко и др., 2002; Раутиан, Калабушкин, 2003; Раутиан, Немцев, 20036]. Это, отчасти, может быть связано с гибридизацией бизона с домашним скотом: в некоторых популяциях обнаружен гаплотип мтДНК, характерный для крупного рогатого скота [Polziehn et al., 1995; Janecek et al., 1996; Ward et aL, 2001].
По мнению Раутиан и Калабушкина [2003], “современные зубры и бизоны представляют собой разные виды, занимающие специфические экологические ниши и существенно различающиеся морфологически. Однако они дивергиро-вали недавно (в геологическом масштабе времени), чем объясняется сохранение высокого генетического сходства по различным характеристикам: свободной межвидовой гибридизации, идентичности таксопринтов и кариотипа, гомологии групп
крови и отдельных эритроцитарных антигенов и т. п.”. Скорость дивергенции гипервариабельных последовательностей между зубром и бизоном, по расчетам, составляет 78,5% за миллион лет [Burzynska et al., 1999].
Таксономия субъективна, и, чаще всего, мнения ученых радикально расходятся даже при оценке одних и тех же результатов. Проведенные исследования сами по себе не могут дать однозначного ответа на вопрос о статусе зубра и бизона, но при всестороннем анализе чаша весов пока склоняется в пользу монотипичной структуры рода Bison с двумя (или тремя) подвидами: В. bison bison, (В. b athabascae) и В. b. bonasus. Такая система рода позволила бы решить многие практические проблемы, связанные с угрозой увеличения инбридинга и вырождения зубра и бизона, с гибридным происхождением западнокавказской группировки, расселением животных и восстановлением ареала (см. ниже). Замечу также, что эволюционная история, уровень морфологической и генетической дифференциации и таксономическая структура рода Bison весьма сходна с таковой у монотипичного рода A Ices, благородного оленя [Данилкин, 1999] и снежного барана (см. ниже).
Разногласия по статусу зубра, бизона и зубробизона (см. ниже) - один из ярких примеров кризиса в систематике, который возник, во многом, из-за несоответствия терминов “биологический вид” и “подвид” реалиям, особенно в части репродуктивной изоляции. Без решения возникших проблем невозможна дальнейшая разработка стратегии сохранения и восстановления представителей рода Bison в природе. Однако принять ту или иную систему можно лишь коллективно на специальном форуме, тщательно взвесив все аргументы.
В современной фауне России - только зубр, которого до специального обсуждения я рассматриваю пока в качестве самостоятельного вида В. bonasus. В последнее время созданы серьезные предпосылки и для реакклиматизации лесного бизона.
ЗУБР
Bison bonasus Linnaeus, 1758
Bos bonasus Linnaeus, 1758. Syst. Nat., 10 ed., 1:71.
Bos urus Boddaert, 1785. Elench. Anim., 1: 151.
Bos bison H. Smith, 1827. Griffith’s Cuvier Animal Kingd., 4: 398. Nom. praeocc., non Linnaeus, 1758. Urus nostras Bojanus, 1827. Nova Acta Physiol.-med. Acad. Carol.-Leopold., 13: 413. Беловежская Пуща.
Bison europaeus von Meyer, 1832. Ibid. (1835), 17: 131.
Bison europaeus Owen, 1849. Proc. Zool. Soc. London (1848): 126. Nom. nov. pro Bos bonasus Linnnaeus.
Bos bonasus caucasicus Satunin, 1903. Зап. Кавк. отд. Имп. Русск. геогр. о-ва. 24: 63. “Кавказ”. Nom. nudum.
Bos bonasus caucasicus Turkin et Satunin, 1904. Звери России, 4, 9: 754. Северо-Зап. Кавказ, Кавказский заповедник.
Bison bonasus var. caucasica Greve, 1906. Zool. Beobacht., 47: 270. Кавказ. Nom. praeocc., non Turkin and Satunin, 1904.
Bison bonasus hungarorum Kretzoi, 1946. Annal. Hist. Nat. Mus. Nation. Hung. 39, 5: 105. Карпаты, Трансильвания.
Характеристика. Зубр - крупный зверь тяжелого сложения с большой головой (фото 2 и 3). Быки заметно крупнее самок и отличаются от них массивной передней частью туловища с высокой холкой. Длина тела самцов - 332, самок - 290 см, высота в холке - 210 и 197 см, масса -более 1000 и 630 кг соответственно (табл. 1). Хвост длинный - до 92 см, с концевыми волосами достигает скакательного сустава. Уши короткие (15-18 см) широкие и частью скрыты в шерсти. Глаза относительно небольшие. Конец морды голый.
Голова, шея и передняя часть туловища покрыта густым, отчасти курчавым волосом, особенно длинным (до 20-26 см) на темени, между рогами, под нижней челюстью и низу шеи. По верху шеи и на холке имеется пышная грива. Большая оволоснен-ность передней части тела - адаптация к защите от холода и, особенно, от ветра, к которому животные обычно поворачиваются головой. На остальной части туловища мех короткий. Волосы разделяются на три категории: изогнутые направляющие, слабоизвитые остевые с сердцевиной и сильноизвитые пуховые. Зимой окраска меха темно-бурая и буро-коричневая, более светлая на шее и плечах. Ноги и живот черно-бурые. Летняя шерсть короче и светлее. Линька одна, весенняя.
Рис. 6. Череп самца зубра (экземпляр из Хоперского заповедника; Зоомузей МГУ, коллекционный № 102005) Вид сбоку, сверху и снизу
Череп относительно короткий в лицевой части и узкий в лобной (рис. 6). Передние концы intermaxillaria малы или не имеют выступающих лопастей. Носовые кости широкие. Орбиты направлены вперед и чуть в стороны. Слезные кости узкие с выемкой впереди, в их передних частях имеется широкая предглазничная впадина. Теменная и затылочная области опущены ниже лба. Верхний край затылочного гребня ниже основания роговых стержней. Слуховые барабаны большие. Резцы долотообразные со слабым расширением и изгибом коронок наружу.
Роговые чехлы черно-бурые, округлые, гладкие, суживающиеся к концу, д линой около 30-49 см. Их основания у взрослых особей отходят от вершины черепа в стороны,
затем загибаются вверх и внутрь, у молодых зверей рога почти прямые. У самок они менее массивные, сравнительно тонкий стержень занимает лишь треть чехла, тогда как у самцов более толстые роговые стержни входят в чехлы примерно на 2/3.
Специфические железы отсутствуют. Мускусный запах, свойственный как самцам, так и самкам, скорее всего, обусловлен бактериями [Соколов и др., 1990].
Современный зубр обитает в лесостепи, равнинных и горных лесах. Образ жизни оседлый, у самок групповой, у самцов - чаще одиночный. В спокойном состоянии животные медлительны, передвигается шагом и не способны к длительному бегу. При опасности и в сильном возбуждении стремительны и ловки, почти без разбега преодолевают двухметровые препятствия, легко бегут галопом по метровому снегу. Хорошо плавают и не боятся водных преград. Из органов чувств хорошо развиты обоняние и слух, зрение слабое. Известная продолжительность жизни в неволе: самцов - 23 года, самок - 28 лет (сообщения о жизни зубров до 50 лет, видимо, ошибочны).
Почти истребленный, а в настоящее время охраняемый и искусственно расселяемый вид, который постепенно увеличивает свою численность и восстанавливает ареал.
Распространение. В историческое время - Центральная и Южная Европа, в последние столетия - отдельными очагами в Европе.
Состав. Две слабо различимые расы: беловежская, или европейская (В. bonasus bonasus L., 1758) и вымершая кавказская (В. bonasus caucasicus Turkin et Satunin, 1904). Восстановление почти уничтоженного вида идет по трем линиям — беловежской, беловежско-кавказской и гибридной с бизоном.
Изменчивость и таксономия
Размеры и масса тела. Масса новорожденных зубрят - 19-29 кг. Примерно такой же вес (25-30 кг) и у новорожденных бизона [Agabriel et al., 1996]. Через полгода самочки весят 84-110 кг, самцы - 110-130 кг. Интенсивный рост длится примерно до 4 лет, затем замедляется, но продолжается до 8-10 лет [Калугин, 1968; Киселева, 1974, 1986а; Перерва, 1992; Сипко и др., 2002].
Длина тела взрослых самцов современных линий разведения составляет в среднем 281-289 (225-332) см, высота в холке - 180-183 (162-210) см, масса -615-631 (310-920) кг, самок - 240-262 (188-290) и 169-174 (123-197) см и 424-439 (до 633) кг соответственно (табл. 1). Столетие назад, возможно, существовали гораздо более крупные экземпляры. И.С. Башкиров [1940], ссылаясь на литературные источники, указывает рекордную массу беловежского зубра в 1900 кг, а кавказского - более 1 т (75-80 пудов). Измельчание животных, как считают некоторые ученые, связано с ухудшением условий обитания [Кулагин, 1919; Цалкин, 19516] или с изменением генофонда.
Большинством исследователей беловежский зубр признается более крупным, по сравнению с кавказским. Иное мнение имел И.И. Соколов [1971], который, вслед за Болкеном [Bohlken, 1967], справедливо подчеркивал, что “...представление о том, что исконный кавказский зубр был более мелким, чем беловежский, лишено оснований”. Достоверные размеры вымершего кавказского зубра следующие: длина вместе с хвостом - 307, высота в холке - 160, окружность груди -
Таблица 1. Размеры тела (см) и масса (кг) взрослых особей зубра и бизона
Самцы Самки Форма, линия разведения, район (источник информации)*
п М lim п М lim
1 2 3 4 5 6 7
Длина тела
1 - 295 1 — 262 Беловежский зубр (10)
54 289 260-332 47 262 245-290 Беловежская линия (9)
1 — 306 1 306 (с хвостом) Кавказский зубр (10)
2 287 285-289 2 242 234-249 Б.-кавказская линия (3)
45 281 225-330 36 240 188-280 -“-(9)
43-72 279 260-300 32 225 192-263 Зубробизон (2, 7, 9)
Высота в холке
20 164 132-195 18 147 123-166 Беловежский зубр до 1921 г. (4, 7)
16 183 166-204 5 174 161-197 Беловежская линия (4, 7)
81 183 171-192 64 169 155-187 -“-(9)
3 167 160-180 — - — Кавказский зубр до 1921 г. (4)
39 177 151-210 33 174 123-190 Б.-кавказская линия (4, 7)
2 180 163-197 2 170 166-173 -“-(3)
45 180 162-210 36 174 136-197 -“-(9)
23 — 160-188 3 164 160-167 Зубробизон (1)
43 175 167-180 32 160 154-165 -“- (2, 7)
72 171 148-187 32 158 148-167 -“- (4, 7, 9)
6 161 134-172 3 141 133-148 Бизон, Аскания-Нова (6)
Обхват туловища
16 227 159-262 15 186 146-237 Беловежский зубр до 1921 г. (4, 7)
81 245 225-270 64 212 190-240 Беловежская линия (9)
1 253 3 202 198-206 Кавказский зубр до 1921 г. (4)
38 234 146-312 32 209 130-240 Б.-кавказская линия (4, 7)
45 236 178-312 36 207 152-240 -“-(9)
2 235 230-240 2 215 200-230 -“-(3)
23 — 212-253 3 200 191-210 Зубробизон (1)
43 248 219-269 32 204 189-213 -“- (2, 7)
28 235 169-269 14 205 180-244 -“-(4)
72 235 180-269 32 204 169-244 -“-(9)
6 250 198-270 3 203 196-210 Бизон, Аскания-Нова (6)
Длина уха
44 16 14-18 46 16 14-19 Беловежская линия (9)
45 17 14-19 36 18 14-21 Б.-кавказская линия (9)
2 16 15-16,5 2 15 15 -“-(3)
43-72 16 16-17 32 15 14-16 Зубробизон (2, 7, 9)
6 18 15-21 — - — Бизон, Аскания-Нова (6)
Длина хвоста
1 — 63 1 — 58 Беловежский зубр до 1921 г. (10)
44 62 54-74 46 56 49-61 Беловежская линия (9)
45 60 42-92 36 62 45-86 Б.-кавказская линия (9)
2 56 53-58 2 51 49-53 -“-(3)
43-72 51 38-64 32 60 47-72 Зубробизон (2, 7, 9)
6 57 48-65 3 46 44-46,5 Бизон, Аскания-Нова (6)
Масса
16 540 368-760 17 322 123-560 Беловежский зубр до 1921 г. (4,7)
8 683 500-920 1 — 530 Беловежская линия (4)
50 615 485-850 46 430 305-540 -“-(9)
Таблица 1 (окончание)
1 1 2 1 з 1 4 1 5 | 6 1 7
2 449 448-450 — — Кавказкий зубр до 1921 г. (4)
16 625 310-860 23 424 101-633 Б.-кавказская линия (4, 7)
19 631 408-860 24 439 291-633 -“-(9)
2 742 625-860 1 __ 550 -“-(3)
30 421-1150 1 - 602 Зубробизон (1)
— — 550-1150 — — — -“-(5)
33 621 398-1150 - — — -“-(9)
6 935 880-990 3 705 690-720 Бизон, Аскания-Нова (6)
- - 700-800 - - 450-500 -“-, на фермах Франции (8)
* J - Калугин, 1966; 2-Немцев, 1988в; 3 - Киселева, 1974; 4-Сипко и др., 2002; 5 -Камьянов, 1969; 6 - Лобанов, 1968; 7 - Немцев и др., 2003; 8 - Agabriel et al., 1996; 9 - личное сообщение Т.П. Сипко; 10 -Соколов, 1959.
207, длина рогов - 33-42 см [Динник, 1910]. Размеры некоторых чучел больше, но уверенности в точном соответствии их размера оригиналу нет. Судя по немногочисленным данным (табл. 1), кавказский зубр очень близок к параметрам беловежского, жившего с ним в одно время.
Размеры особей современных (инбредных) беловежской и беловежско-кавказской линий зубра практически одинаковы (табл. 1), хотя отдельные ученые считают последних более крупными. Линейные размеры и масса зубробизонов (фото 4) из Кавказского заповедника на 10-30% меньше, за исключением обхвата туловища, длины конечностей, длины и ширины головы и т. д. Наряду с легкостью склада и стройным широким туловищем, у горных особей сравнительно крупная и короткомордая голова, длинный с большой кистью хвост и характерный для вымершего кавказского подвида изгиб рогов [Немцев, Киселева, 1987; Сипко и др., 2002; Немцев и др., 2003]. Авторы подчеркивают эволюционный тренд и адаптивный характер изменений экстерьера “горной” формы и близость ее к уничтоженному кавказскому подвиду.
Это заключение, при внимательном рассмотрении, сомнительно, поскольку отдельные выборки крайне малы (по Кавказу до 1921 г. всего 1-3 особи), анализируемые параметры весьма изменчивы, при их измерениях обычна большая погрешность, в сравнительном анализе задействованы разновозрастные животные, среди которых много выращенных в неволе и выбракованных по состоянию здоровья и измененному экстерьеру и, кроме того, современная “горная” форма гибридная.
По линейным размерам, включая длину хвоста, зубр не отличается существенно от бизона (табл. 1).
Краниометрическая характеристика. Размеры черепа и рогов у самцов зубра и бизона существенно крупнее, чем у самок (табл. 2). Половой диморфизм краниометрических признаков у этих форм и вымершего В. priscus перекрывает размах географической и временной изменчивости. Возрастные изменения некоторых структур черепа продолжаются до конца жизни особей [Пузаченко, Раутиан, 2001; Раутиан и др., 2003].
Наибольшая длина черепа у самцов колеблется в пределах 465-612, у самок -390-503 мм, ширина - 290-350 и до 284 мм соответственно. Большинство краниологических признаков у беловежского и кавказского зубров, бизона и зуброби-
Таблица 2. Размеры черепа (мм) взрослых особей зубра и бизона
Самцы Самки Форма, линия разведения, район (источник информации)*
п М lim п М lim
1 2 3 4 5 6 7
Наибольшая длина
- - 500-580 — — Беловежский зубр (1)
52 521 - 49 489 Беловежская линия (5)
- - 490-560 - - - Кавказский зубр (1)
19 501 — 11 479 “ (5)
23 587 465-612 24 498 390-503 Б.-кавказская линия (5)
31 510 460-542 71 472 390-485 Зубробизон (5)
4 586 — 2 517 — Бизон (5)
— 540 490-570 - — — Степной бизон (1)
- 575 560-600 - - - Лесной бизон (1)
Основная длина
9 462 457-480 9 441 419-452 Беловежский зубр (2)
24 470 - 24 450 — Беловежская линия (3)
12 468 456-490 10 442 428-460 Кавказский зубр (2)
9 463 — 4 435 — -“-(3)
23 514 446-563 24 450 369^478 Б.-кавказская линия (5)
13 474 - 15 451 — Зубробизон (3)
31 469 442-500 71 432 360—458 -“-(5)
3 501 490-517 4 461 438—4-81 Бизон (2)
4 513 - 2 464 - Степной бизон (3)
Максимальная ширина
— 320 290-350 — — — Беловежский зубр (1)
54 310 — 47 264 — Беловежская линия (3)
34 297 - 21 243 - Кавказский зубр (3)
13 309 — 12 268 — Б.-кавказская линия (3)
23 319 309-334 24 275 268-284 -“-(5)
8 334 — 9 284 Окский питомник (3)
15 307 - 19 258 Зубробизон (3)
8 348 — 4 281 — Степной бизон (3)
>29 317 270-345 — — — -“-(1,4)
>4 355 325-385 — — — Лесной бизон (1,4)
Межглазничная ширина
53 241 — 47 208 — Беловежская линия (3)
22 236 — 9 197 - Кавказский зубр (3)
23 270 199-325 24 223 199-312 Б.-кавказская линия (5)
15 241 19 201 - Зубробизон (3)
15 241 195-315 71 236 191-291 -“-(5)
8 274 — 4 225 - Степной бизон (3)
Длина носовых костей
26 186 — 23 181 — Беловежская линия (3)
5 195 - 4 181 - Кавказский зубр (3)
23 201 179-238 24 186 131-235 Б.-кавказская линия (5)
9 203 - 10 193 - Зубробизон (3)
31 189 176-215 71 180 128-220 -“-(5)
4 204,5 2 202 — Степной бизон (3)
Максимальная ширина носовых костей
53 95 — 46 79 — Беловежская линия (3)
9 93 — 8 75 - Кавказский зубр (3)
1 1 2 | 3 1 4 1 5 1 6 1 7
23 102 76-231 24 82 61-91 Б.-кавказская линия (5)
9 94 - 12 76 - Зубробизон (3)
31 89 70-112 71 72 60-86 -“-(5)
7 103 - 4 83 - Степной бизон (3)
Длина межчелюстных костей
25 156 — 24 149 — Беловежская линия (3)
9 179 - 8 156 - Кавказский зубр (3)
23 167 112-172 24 166 103-172 Б.-кавказская линия (5)
13 159 - 15 152 - Зубробизон (3)
31 153 100-176 71 145 113-158 -“- (5)
4 178 - 2 177 - Степной бизон (3)
Длина верхнего зубного ряда
54 143 — 48 137 — Беловежская линия (3)
24 142 — 20 140 — Кавказский зубр (3)
23 153 138-157 24 148 122-161 Б.-кавказская линия (5)
15 142 - 19 137 — Зубробизон (3)
31 137 130-146 71 134 121-142 -“-(5)
8 152 - 4 150 — Степной бизон (3)
Длина рогового стержня
19 277 — 22 156 — Беловежская линия (3)
8 246 - 9 191 - Кавказский зубр (3)
13 250 — 12 156 — Б.-кавказская линия (3)
23 277 250-318 24 173 156-196 -“-(5)
8 318 — 9 196 , Окский питомник (3)
14 254 — 12 166 — Зубробизон (3)
31 219 123-327 71 174 112-280 -“-(5)
4 208 4 131 — Степной бизон (3)
>29 233 - — — — -“- (4)
>4 255 — — — — Лесной бизон (4)
Расстояние между концами роговых стержней
- 610 525-690 — — — Беловежский зубр (1)
21 579 — 23 440 — Беловежская линия (3)
8 559 5 367 — Кавказский зубр (3)
13 611 — 12 458 — Б.-кавказская линия (3)
23 648 611-705 24 490 458-536 -“-(5)
8 705 9 536 — , Окский питомник (3)
14 608 - 11 438 — Зубробизон (3)
31 510 360-800 71 402 195-490 -“-(5)
4 560 — 4 451 — Степной бизон (3)
— 580 485-665 — — — (13)
— 665 585-740 - — — Лесной бизон (1,4)
Обхват основания рогового стержня
55 226 — 51 164 — Беловежская линия (3)
32 229 — 23 161 — Кавказский зубр (3)
23 231 133-322 24 152 138-216 Б.-кавказская линия (5)
15 209+18 — 15 160 — Зубробизон (3)
31 203 123-301 71 161 130-205 -“-(5)
16 240 — 6 156 — Степной бизон (3)
Диаметр основания рогового стержня
— 80 65-90 - - - Беловежский зубр (1)
55 74 51 50,5 Беловежская линия (3
Таблица 2 (окончание)
1 1 2 1 з 1 4 1 5 1 6 1 7
32 77 — 23 50 - Кавказский зубр (3)
13 73 — 12 49 — Б.-кавказская линия (3)
23 81 62-92 24 51 44-83 (5)
8 82 9 60 — , Окский питомник (3)
15 69 — 16 48 - Зубробизон (3)
31 63 52-70 71 50 42-81 -“-(5)
4 82 — 4 50 - Степной бизон (3)
— 75 65-95 — - (1)
>29 78 — — - - (4)
... 95 80-110 - - - Лесной бизон (1)
>4 92 — — — — (4)
* 1 - Флеров, 1965; 2 - Соколов, 1971; 3 - Раутиаи и др., 2003; 4 - Skinner, Kaisen, 1947 (по: [Раутиан и др., 2003]); 5 - личное сообщение Т.П. Сипко.
зона широко трансгрессируют (табл. 2) [Флеров, 1965; Bohlken, 1967; Соколов, 1971; Немцев, 19826, 1985а; Немцев, Киселева, 1987; Раутиан и др., 2000, 2003; Сипко, 2003]. По исследованиям Г.С. Раутиан с соавторами [2000, 2003], “горный” зубр близок к беловежской и кавказской расам (что и следовало ожидать исходя из его происхождения), но отличается от них по некоторым промерам (длине носовых костей, затылочному гребню, боковой длине лба, диаметру роговых стержней). В вольных стадах изменчивость признаков сравнима с различиями между линиями. Зубры и бизоны, живущие в неволе, по краниометрическим характеристикам существенно отличаются от животных из природных популяций.
Для дальнейших таксономических выводов важно отметить и другое обстоятельство. У инбредных видов, прошедших через “бутылочное горлышко”, как известно, возможны значительные морфологические отклонения от нормы, включая аномалии зубной системы, которые ярко выражены и у зубра, и у бизона [Crooks, Van Vuren, 1994; Раутиан, 20036]. Изменчивость краниометрических признаков уничтоженной кавказской расы, судя по сохранившимся черепам, была незначительной [Соколов, Темботов, 1993]. Современные линии зубров отличаются от предковых повышенным уровнем асимметрии фенов черепа. Этот показатель максимальный у особей беловежской (особенно -плесской) линии, что, видимо, связано с наибольшим уровнем инбридинга, и у животных, выращенных в питомниках в связи с более высокими темпами роста [Makowiecka, 1994; Baranov et al., 1997; Раутиан и др., 1997, 1998; Пузаченко и др., 1999; Раутиан, 2003а].
Окраска и линька. Современные беловежские самцы зубра однотонно бурые (фото 2) с характерной челкой, беловежско-кавказские - рыжие (фото 3) и без челки [Перерва, 1992]. Бока головы, нижняя часть шеи, брюхо и нижние части конечностей темно-коричневые. Уши, челка, переносье, бока шеи, холка, плечи и бока охристо-бурые или рыже-коричневые. Окраска вымершего кавказского зубра, возможно, была более темной и однотонной - коричнево-бурой с кофейным оттенком. Самки той и другой формы светлее самцов. Наиболее темной окраска бывает сразу после линьки, когда мех короткий. Зимний мех хорошо развит, с густым подшерстком. Ость мягкая и неломкая, и к весне, в отличие от оленьих, почти не снашивает-
ся. Молодые особи в зимнем меху чаще серо-бурые с шоколадными потемнениями на морде, вдоль хребта, на брюхе и конечностях. Новорожденные пепельно-серые с охристым оттенком [Флеров, 1979].
Зубр и зубробизоны линяют один раз в году, но этот процесс сильно растянут, особенно в неволе (от 100 до 250 дней) - с января - марта до сентября - ноября, а у части животных - до декабря. Волос выпадает сначала на шее, передней части туловища и боках, затем на спине, задней трети туловища, брюхе, и дольше всего он сохраняется на хвосте. Взрослые самцы начинают и завершают линьку раньше, менее упитанные лактирующие самки и больные звери - позже [Калугин, 1958; Киселева, 1974, 1978а; Перерва, 1981, 1982, 19836].
Генетическое разнообразие и происхождение современных линий. Кариотип зубра содержит 60 хромосом, NF = 62. Все аутосомы акроцентрические, X — крупный субметацентрик, Y-хромосома мелкая и, по данным разных авторов, суб-мета-, субтело- или акроцентрическая с почти редуцированными р-плечами. У бизона при таком же наборе Y-хромосома обычно акроцентрическая, варьирующая до субмета- и субтелоцентрической. Кариотип зубробизона (кавказского “горного” зубра) такой же, как и у исходных форм. Хромосомный набор зубра и бизона в целом сходен с таковым у крупного рогатого скота, за исключением незначительных отличий в морфологии Y-хромосомы, которая у скота субметацентрическая, и иного расположения ядрышкообразующего района на 27 паре аутосом. У гибридов зубра и бизона с домашним скотом половые хромосомы резко гетероморфны, Y-хромосома акроцентрическая [Wurster, Benirschke, 1968; Орлов, Чудиновская, 1979; Шумов, 1980, 1994; Ясинецкая, 1982а,б; Стекленев, Ясинецкая, 1989, 1990; Графодатский и др., 1990; Соколов, Темботов, 1993; Треус, 1993, 1994; Треус, Ясинецкая, 1993; Треус и др., 1994, 1997; Шаршов, 1994; Дзуев, 1995, 1998; Раутиан, Калабушкин, 2003; Sipko et al., 2004а,с].
Современный зубр происходит от 12 уцелевших особей-основателей (рис. 7), причем в становлении чистокровной беловежской линии принимали участие только 7 особей, из которых максимальный генетический вклад внесли самцы-производители № 42 и № 45 (по классификации European Bison Pedigree Book - ЕВРВ). Беловежско-кавказская линия сформирована из сходного набора “базовых” особей-основателей I и II, а кавказское происхождение, видимо, имело только животное № 100. Степень участия “кавказского” генома в формировании генофонда современных группировок составляет 1,8-4,8%. Так называемая “горная” форма происходит от 5 зубробизонов П-Ш генераций поглотительного скрещивания с использованием тех же самцов беловежско-кавказской линии и особей №№ 50, 51, 65. Гены зубров-основателей №№ 87, 147 и 15 утрачены или почти утрачены в процессе разведения (табл. 3). В целом степень сохранности исходного генофонда в беловежской и беловежско-кавказской линиях составляют 46,7 и 42,0% (табл. 4) [Slatis, 1960; Флинт, Перерва, 1986; Olech, 1989, 1998, 1999; Pucek, 1991; Белоусова, 1993а, 1999; Сипко, 1995, 2003; Сипко и др., 1995, 1999; Белоусова и др., 1999; Кудрявцев, Белоусова, 1999; Раутиан, Немцев, 2003а; Раутиан и др., 2003].
Из трех кавказских особей, содержавшихся в зоосадах в начале XX в., потомство, скорее всего, оставил лишь самец № 100 по кличке “Кавказ”, пойманный на Северном Кавказе теленком в мае 1907 г. и доживший в.Бойценбурге до февраля 1925 г. Его прямой потомок - бык “Бодо” (№ 193) из “Аскании-Нова”. Не исключе-
о (X
л & S ч ю el
*
s & S
Ь4 S & S о Ё С
о л S к к
X л
о л S к к
X 2 & 5 сх<
S л к
CQ
* *
*
* бк - беловежско-кавказская линия, б - беловежская линия, г - горная линия. ** В среднем по всем питомникам мира [Olech, 1989].
*** Мировая совокупность вольноживущих популяций [Белоусова и др., 2003].
1-3 - группы западной ветви разведения с низким (<30%), средним (30-60%) и высоким (>60%)
вкладом “основателсй-П”, ие представленных у животных вольных стад, соответственно
Таблица 4. Генетическая характеристика современного зубра (по [Кудрявцев, Белоусова, 1999])
Характеристика Линия
беловежская беловежско-кавказская
Число основателей 7 11
Степень сохранности исходного генофонда, % 46,7 42,0
Сохранилось геномов основателей 3,27 4,61
Сохранилось исходного генетического разнообразия, % 67,2 75,0
Показатель среднего сходства геномов особей 0,32 0,25
Численность на 1994 г., особей 760 1110
но, что среди родоначальников мог быть и кавказский зубр “Казан”, завезенный в Беловежскую пущу в 1899 г. Этот малорослый зверь не считался полноценным производителем, но он все же мог быть отцом или дедом зубров-основателей №№ 85, 86, 89, 95, 96 и 123, происходящих от неизвестных родителей [Раутиан и др., 2003; Сипко, 2003].
В западноевропейских странах в начале XX в. содержали преимущественно чистокровных беловежских зубров, но в 20-50-е годы зубриц нередко спаривали с беловежско-кавказскими самцами. В результате число животных с примесью “кавказской” крови увеличилось с 8 в 1921 г. до 82 в 1950 г. Число чистокровных “беловеж-цев” в этот период осталось почти неизменным - 45-46 особей [Заблоцкий, 1957а]. В Восточной Европе доминирует беловежско-кавказская линия зубров. В СССР в
1957 г. среди 69 чистокровных зубров беловежских было только 24. Большинство стад происходит от зверей, выращенных в советской части Беловежской пущи, и, частично, от особей, разводимых в “Аскании-Нова”. Всего в мире в 1994 г. числилось 760 вольных беловежских зубров и 1100 - беловежско-кавказских [Фортунатов, 1922,1924, 1929; Башкиров, 1939а; Заблоцкий, 1939,1947, 1956, 1957а, 1963; Север-цов, 1940а; Соколов, 1959; Гептнер и др., 1961; Корочкина, 1963; и др.].
В “Асканию-Нова” в 1897 и 1900 гг. завезены две пары бизонов, и с 1902 по 1938 гг. - 8 чистокровных зубров (5 самцов и 3 самки), от которых родилось 15 зубрят, из них 12 погибли. Последующий завоз бизонов (5 самцов и 3 самки) имел место в 1911-1914 гг., и по одному самцу ввезли в 1921, 1933 и 1950 гг. Зубров скрещивали с бизоном, серым украинским скотом и другими породами. Всего получено 295 гибридов. В 1940 г. 5 асканийских гибридов зубра и американского степного бизона (из них, возможно, один самец и две самки с примесью крови кавказских зубров) вселены в Кавказский заповедник. В 1948-1957 гг. сюда же завезли 15 чистокровных беловежско-кавказских самцов из Беловежской пущи, Приокско-Террасного и Хоперского заповедников, из которых заметное участие в воспроизводстве приняли только два [Салганский и др., 1963; Лобанов, 1968; Тре-ус, 1968; Заблоцкий, Заблоцкая, 1986а].
Эта интродукция и дальнейшая племенная работа (только за 19 лет выбраковано более 60 низкокровных самцов) позволили частично изменить генофонд гибридной кавказской группировки. Кровность стада по зубру в 1964 г. составляла 81% для самцов и 67% для самок [Крайнова, 1947; Калугин, 1968, 1970]. Г.С. Раутиан с соавторами [2003] оценивают генетическую составляющую бизона в 6,4-5,9%. По мнению М.А. и М.М. Заблоцких [1986, 1987], группировка Кавказского заповедника, разводимая “в себе”, преимущественно зубробизонья, что представляет угрозу для генофонда чистокровной беловежско-кавказской популяции, созданной в Северной Осетии, Чечено-Ингушетии и Карачаево-Черкессии [Заблоцкий, 1983]. Генофонд кавказского “горного” зубра, следовательно, существенно отличается от генофонда истребленной кавказской расы.
Геномы зубра, зубробизона и американского степного бизона, исследованные методом таксономического фингерпринта ДНК, весьма близки, но обособлены от таковых домашнего скота (см. выше). У зубра, в сравнении с бизоном и домашним скотом, низкие показатели генетического разнообразия и высокие индексы внутригруппового сходства [Рысков и др., 1994; Удина, 1994; Сипко и др., 1996, 1997а,б; Ломов и др., 19996]. Средняя гетерозиготность по всем изученным локусам - 4,1%, по ферментным локусам - 2,75%. Генетическое разнообразие минимально у беловежской линии (1,3 по mu-критерию) и несколько выше у беловежско-кавказской и горной (1,45 и 1,50 соответственно). У первых найдено 33 антигена (66% вариантов, выявленных у вида), у вторых - 41 (82%), у третьих - 36 (70%). Средний уровень гетерозиготности в расчете на локус составляет, соответственно, 0,038, 0,044 и 0,038 (табл. 5). В целом генетические различия животных этих линий примерно такие же, как между популяциями одного вида или сравнимы с уровнем различий между породами крупного рогатого скота, или даже стадами одной породы [Сипко, Черкашенко, 1989; Сипко, 1990, 1995, 2002, 2003; Перерва, 1992; Сипко и др., 1993, 1994, 1995, 1996, 1997а,б, 1999; Раутиан и др., 1994; Удина и др., 1994; Udina et al., 1994; Бадагуева, 1995; Удина, 1996; Раутиан, Сипко, 1997; Сипко, Раутиан,
Таблица 5. Характеристика генетического разнообразия зубра (по [Сипко, 2002, 2003])
Параметр Линии разведения
беловежская беловежско-1 кавказская | горная
Кровь. Количество антигенов: в линии 33 (66%) 41 (82%) 35 (70%)
на особь 11,2 (5-17) 13,4 (5-18) 12,9 (6-26)
Разнообразие (mu-критерий) 1,31 1,45 1,50
Биохимический полиморфизм. Средняя
гетерозиготность (21 локус) 0,038 0,044 0,038
ДНК полиморфизм. Гетерозиготность 0,07 0,09 -
Доля полиморфных локусов 0,42 0,57 -
19976, 1999; Белоусова, 1999; Ломов и др., 1999а; Семенова и др., 2000; Раутиан, Калабушкин, 2003; Nowak et al., 2004].
Зубр входит в число видов, относительно устойчивых к инбредной депрессии, однако при длительном близкородственном скрещивании животных этот процесс прогрессирует. Коэффициент инбридинга в беловежской линии в мире составляет 0,324, в России- 0,375, в беловежско-кавказской линии - 0,193 и 0,269 соответственно [Перерва, 1986; Olech, 1987, 1989; Белоусова, 19936, 1999; Раутиан, Сипко, 1997; Белоусова и др., 1999; Сипко и др., 1999; Belousova, Kudryavtsev, 2000; Раутиан, Калабушкин, 2003; Раутиан, Немцев, 20036].
Гибридизация. Помеси зубра и бизона плодовиты неограниченно. Гибридные самки, полученные при скрещивании и зубра, и бизона с разными породами домашнего скота, также плодовиты, имеют фенотип диких предков, скороспелы, сильны и выносливы. Однако есть большие проблемы в осеменении коров из-за отсутствия или слабо выраженной половой реакции на них у самцов зубра и бизона, у них высокая яловость, значительная эмбриональная смертность (около 30%) из-за избыточного количества околоплодной жидкости, затруднены отелы из-за крупного гибридного плода, и они не могут повторно приносить гибридное потомство. Гибридные самцы I—II поколений бесплодны, как считается, из-за разных Y-хромосом и нарушения процесса мейоза, что ведет к стерильности животных [Фортунатов, 1922, 1924, 1929; Соколов, 1959; Лобанов, 1968; Треус, 1968; Рубайлова, 1980; Шумов, Рубцова, 1980; Стекленев, 1982,1983а,б, 1986; Krasinska, 1988а; Стекленев, Ясинецкая, 1990; Шумов, 1994; Немцев, Мироненко, 2003]. А.С. Графодатский с соавторами [1990], однако, считают попытку связи проблем гибридизации зубра и бизона с домашним скотом с межвидовыми различиями в морфологии Y-хромосом неоправданной, поскольку гибридные самцы Bos taurus х В. indicus, имеющие разные Y-хромосомы плодовиты, а самцы В. taurus х В. gaurus стерильны при идентичной Y-хромосоме.
Таксономия. Номенклатура зубра и статус кавказской зубробизоньей группировки окончательно не определены, что стало существенной проблемой при выработке стратегии сохранения биоразнообразия, реинтродукции и использования ресурсов.
Ведущие отечественные зоологи [Туркин, Сатунин, 1904; Динник, 1910; Флеров, 1932, 1965; Соколов, 1959; Гептнер и др., 1961; и др.] принимали две формы
зубра: беловежскую и полностью уничтоженную человеком кавказскую. К.К. Флеров [1979] считал подвидом и мелкую трансильванско-карпатскую горную расу (В. bonasus hungarorum Kretzoi, 1946), но эта точка зрения, основанная на ничтожном материале (одного скелета и зарисовок с натуры), не нашла поддержки.
Основные отличительные признаки беловежской и кавказской форм, приводимые в вышеназванных литературных источниках, следующие. Высота самцов беловежского зубра в холке достигает 185 см, масса - 850-1000 кг, наибольшая длина черепа - 580 мм. Лоб выпуклый. Роговые стержни низко опущены. Верхняя сторона их оснований у самцов лежит ниже лобной поверхности на 30 55 мм. Предглазничная ямка выражена слабо. Мыщелки округлены и без рудиментов переднего загиба. Небные отростки дорзальной поверхности почти плоские. Рога у коров слабо изогнуты внутрь, концы их широко расставлены. У самцов рога изогнуты спирально: наружу, вверх, внутрь и назад. Конец кисти хвоста достигает пяточного сустава. Копыта удлиненные. Окраска однотонная буро-коричневая. Волосы слабо завитые.
Высота в холке самцов вымершего кавказского зубра - 160-180 см, масса достигает 700-800 кг, наибольшая длина черепа - 490-560 мм. Лоб почти плоский. Роговые стержни слабо опущены. Верхняя сторона их оснований у самцов лежит на уровне лобной поверхности или чуть ниже. Предглазничная ямка выражена четко. Мыщелки часто с рудиментами переднего загиба. Небные отростки по дорзальной поверхности вогнуты продольно. Рога у коров сильно изогнуты внутрь, концы их близко сходятся. У самцов рога изогнуты крутой спиралью: наружу, вперед, вверх, внутрь и назад. Конец кисти хвоста немного не достигает пяточного сустава. Копыта короткие, округлые. Окраска коричнево-бурая с кофейным оттенком, более темная, чем у беловежской расы. Вдоль спины тянется темная полоса. Волосяной покров короче, чем у беловежского зубра, и более извитой.
Недавно Г.С. Раутиан с соавторами [2000] выделили новый “горный” подвид (В. bonasus montanus) на базе кавказской гибридной зубробизоньей группировки, что вызвало недоумение у зоологов. Напомню, что ранее ученые тщательно избегали придания ранга таксонов смешанным группировкам животных, и поэтому не признавали кавказских зубробизонов зубрами и не вносили их в международные списки поголовья вида, а М.А. и М.М. Заблоцкие [1983,1986а-в, 1987] даже настаивали на их полной элиминации. Это нововведение может привести к серьезным последствиям в систематике, и его нельзя оставить без внимания, особенно после выхода в свет монографии А.С. Немцева с соавторами [2003] “Зубр на Кавказе”, где делается очередная попытка теоретического и практического обоснования такого шага.
Основанием для выделения горного подвида зубра послужили малозначимые различия в отдельных краниометрических признаках потомков зубробизонов в сравнении с особями инбредной беловежской линии и некоторое морфометрическое сходство с вымершей кавказской формой. Замечу, что исследователи не включили в анализ более крупные черепа животных, выращенных в питомниках, хотя их изменчивость вряд ли могла выходить за рамки видовой “нормы реакции”. Позднее в “Зубре на Кавказе” они фактически опровергли свои таксономические доводы, показав, что по краниометрическим параметрам беловежский, беловежско-кавказский и горный зубры образуют единую группу, противостоящую кластеру абори
генной кавказской формы, и, следовательно, горная и беловежско-кавказская группировки, обитающие на Кавказе, по словам авторов, “безусловно по основным характеристикам ближе к беловежскому, чем к аборигенному подвиду” (с. 70) [Раутиан и др., 2003].
Отличительные морфологические особенности горных зубров, по мнению Раутиан с соавторами [2000,2003], “могут быть интерпретированы как результат адаптации равнинных зубров к условиям гор”. На мой взгляд, природа морфометрических отличий особей горной (зубробизоньей) линии от других в большей мере не адаптивно-экологическая и не эволюционная, а генетическая. Гибриды млекопитающих, включая гибридов зубра и бизона с домашним скотом, и их потомки, как известно, отличаются от своих предков по форме и размерам черепа, размерам и массе тела и окраске, но отдельные признаки они наследуют [Соколов, 1959; Шумов, Рубцова, 1980; Krasinska, Suminski, 1981; Krasinska, 1988a,b; Стрекозов и др., 2001; Шилова и др., 2001]. Установлено, что если беловежский зубр имеет даже менее 10% примеси крови кавказского, то его скелет и череп имеют особенности, свойственные последнему [Kobrynczuk, 1985]. Отчетливое сходство горного зубра с бизоном усматривается, например, в чрезмерно широко расставленных роговых стержнях [Раутиан, Немцев, 20036].
Собственно генетическую природу морфометрической изменчивости “горного” зубра признают и Раутиан с соавторами [2003] (цит.): “современные зубры Кавказа занимают центральное положение между всеми формами, что объясняется гибридным происхождением кавказско-беловежских и горных зубров, среди предков которых были представители как беловежского, так и кавказского подвидов” (с. 70). “Горные, кавказско-беловежские и беловежские зубры, населяющие ныне леса Кавказа и Беловежскую пущу, оказались похожи друг на друга больше, чем современные беловежцы на своих предшественников - уничтоженных в начале века обитателей Пущи, не говоря уже о зубрах кавказского подвида. Это сходство неудивительно, имея в виду близкое кровное родство (в буквальном смысле слова) между всеми современными зубрами и сравнительно малый срок независимого развития каждой из популяций” (с. 75).
Генетические различия беловежской, беловежско-кавказской и “горной” линий зубра, как показано многими учеными, примерно такие же, как между популяциями одного вида или сравнимы с уровнем различий между породами крупного рогатого скота или даже стадами одной породы (см. выше). Этот вывод подтверждается и исследованиями Раутиан и Калабушкина [2003]: генетические расстояния между анализируемыми линиями зубров колеблются в пределах 0,013-0,119, а по частотам полиморфных аллелей белковых локусов они отличаются статистически незначимо.
Следует обратить внимание и еще на один принципиально важный аргумент цитируемых исследователей. Раутиан и Немцев [2003] считают, что называть современных горных зубров гибридами некорректно с биологической точки зрения, поскольку в БЭС [1991] гибридом называется “организм, полученный в результате скрещивания генетически различающихся родительских форм (видов, пород, линий и др.)”. Следовательно, современные зубробизоны Кавказского заповедника, как и потомки беловежско-кавказского зубра, не подходят под это определение, поскольку родились в результате скрещивания животных, относящихся не к раз
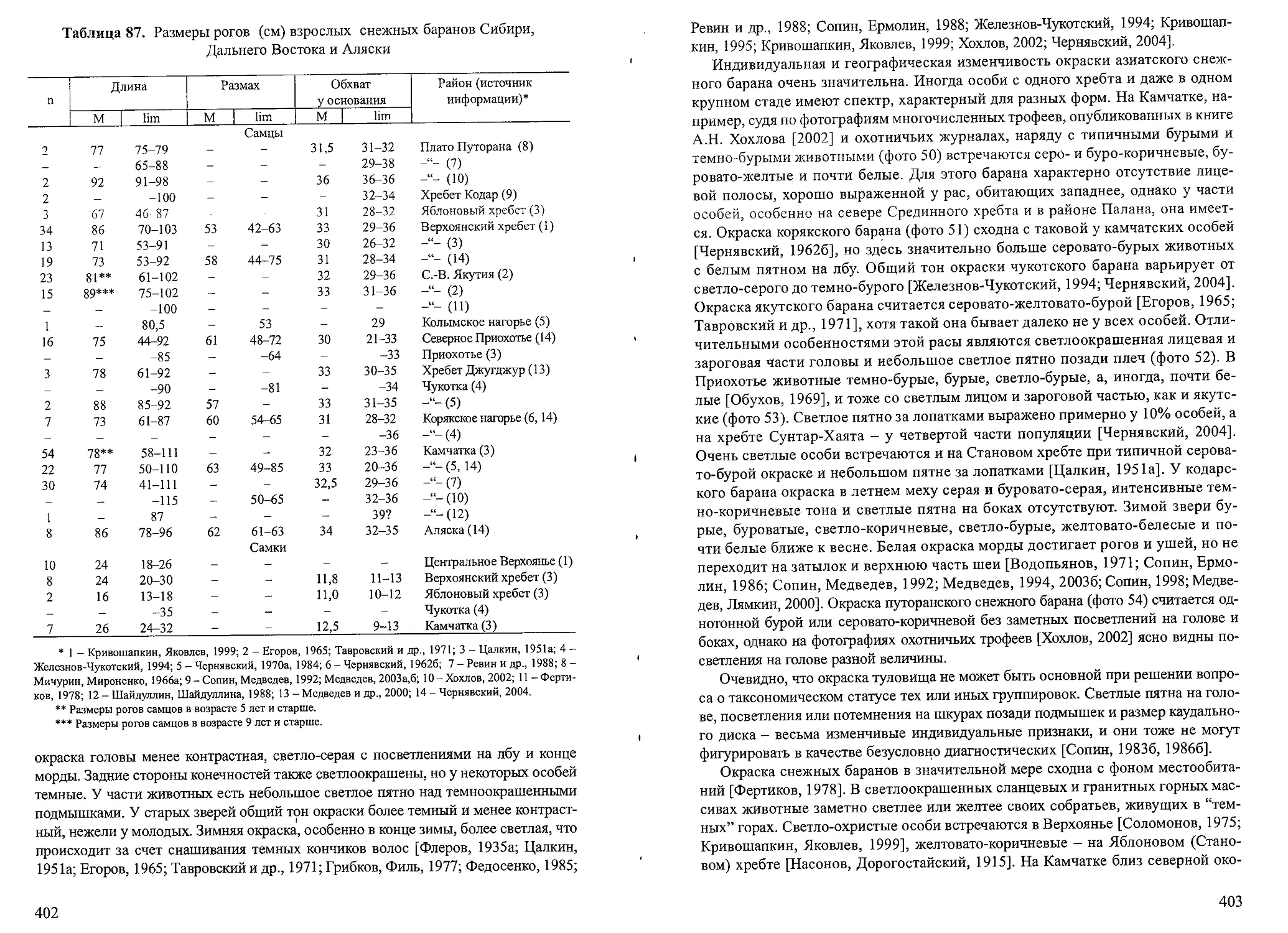
ным, а к одной и той же форме, длительно существующей в природе. При таком понимании термин “гибрид” утрачивает свое существование и становится синонимом термина “особь” (с. 11-12).
Вынужден заметить в этой связи, что в систематике существуют свои правила, которые прописаны нс в БЭС, а в Международном кодексе зоологической номенклатуры. В нем дается иное толкование термина “гибрид”, смысл которого, к сожалению, меняется от издания к изданию. В третьем издании Кодекса на русском языке [1988] гибрид - это “потомок двух особей, относящихся к разным видам. Потомок двух особей, относящихся к разным подвидам одного и того же вида, не является гибридом”. В четвертом издании Кодекса [2000] гибрид - “потомок двух особей, относящихся к разным таксонам”. Более того, согласно ст. 17.2 последнего издания “пригодность названия не затрагивается, даже если оно было применено к таксону, гибридное происхождение которого было известно или выяснилось впоследствии”. Следовательно, пригодность названия, предложенного Раутиан с соавторами [2000] для кавказской “горной” группировки, не может оспариваться в отличие от всего остального.
Данное положение Кодекса и таксономический прецедент, когда группировке гибридного происхождения (кавказскому зубробизону) придан подвидовой статус через несколько поколений после ее образования, открывает перед палеонтологами и зоологами широчайшие возможности к дальнейшему “видодробительству”. Подвидами, видимо, можно будет признавать группы особей, получаемых в результате скрещивания разных видов, пород, линий и др. сразу же при разведении “в себе”. Подвидом, например, по логике следовало бы считать не только горного зубра, но и беловежско-кавказскую линию, поскольку она существует в природе под контролем естественного отбора еще дольше, чем горная, а отдельные стада адаптированы к жизни не только на равнине, но и в горах Кавказа (цейская группировка в частности). Подвидами, в таком случае, придется признать многочисленные прекрасно приспособленные к жизни в природе помесные популяции кабанов, образовавшиеся во многих регионах России после выпусков особей разных форм в одних и тех же районах. К бесспорным ssp. nova следует отнести и аска-нийского гибридного оленя с острова Бирючьего, имеющего роскошные рога, помесные группировки благородного и пятнистого оленей, а также помеси многих других видов и подвидов, особенно на стыках ареалов.
Стремление отдельных ученых выделить свой “уникальный” подвид и затем придать ему статус особо охраняемого, к сожалению, может привести не только к дополнительным проблемам в таксономии, но, главное, к бережному сохранению ублюдков в природе и даже специальному выведению новых гибридных форм, подобно сотням пород собак, кошек, овец и других домашних животных. Помимо классификации придется пересмотреть и темпы эволюции крупных млекопитающих, поскольку, следуя логике Г.С. Раутиан с соавторами, подвиды могут образовываться в течение нескольких или нескольких десятков лет, что никак не увязывается с известной скоростью формообразования, на что требуются многие тысячелетия.
Считать кавказского зубробизона горным подвидом зубра (В. bonasus montanus) при достоверно известной родословной вряд ли уместно. Более оправдано, видимо, выделение у зубра трех генетических линий: беловежской, беловежско-кавказской и горной. Замечу также, что проблемы, связанные с гибридным происхожде
нием кавказского зубробизона и его таксономическим статусом, автоматически снимаются, если род Bison будет признан одновидовым.
Распространение
Динамика ареала. В голоцене зубр занимал обширный ареал - от Атлантического побережья на западе и, видимо, до Западной Сибири на востоке и Закавказья на юге. Северную границу его распространения в историческое время в бывшем Советском Союзе В.Г. Гептнер [Гептнер и др., 1961] проводил от Рижского залива по восточной окраине Литвы и западной части Белоруссии, к югу от Москвы и Рязани до Дона, и далее на юго-запад до побережья Азовского и Черного морей. От этой части ареала он отделял кавказский участок (рис. 8). Все сведения о распространении зверей далее к северу и востоку автор относил за пределы исторического времени или считал ошибочными и основанными на недоразумении. Тем не менее, он допускал, что ареал зубра мог быть несколько большим: на север доходить до Оки, а на восток - до Волги, т. е. совпадать с границами ареалов зоогеографических аналогов - благородного оленя, косуль и кабана. В числе мест возможного обитания животных за пределами очерченного ареала В.Г. Гептнер ука-
Рис. 8. Распространение зубра на территории бывшего СССР в историческое время (по [Гептнер и др., 1961])
1 - VIII-XIII вв.; 2 - XV-XVI вв.; 3 - XVII в.; 4 - XVIII в.; 5 - в разное время до XVIII в; 6 - XIX и начало XX вв.; 7 — область вероятного соединения южнорусской части ареала с кавказской; 8 - предполагаемая граница распространения за историческое время (неполные сведения - см. текст); 9 - места содержания в неволе или выпуска животных в заповедники в середине XX в.
пивал Енотаевск и Дубровку на Нижней Волге, р. Иргиз, Сызрань, Пензу, Спасск Рязанской губернии, Орел, Тверь, Новгород, Вологду, Нижегородскую область, Вятку, Пермь, а также донские степи, что подтверждается новыми находками.
В Западной Европе зубр был широко распространен вплоть до исторического времени, хотя, не исключено, что к этому периоду его не стало уже в Англии и Испании. В I - начале II тысячелетия н. э. человек полностью истребил этот вид в Болгарии, Австрии, Югославии, Голландии, Бельгии, Швеции, Франции, в XIV в. - в Чехии, в 1762 г. - в Румынии, в 1793 г. - в Германии (в Пруссии - в 1755 г.) [Брем, Россмесслер, 1867;Bochenek, 1955; Соколов, 1959; Гептнер и др., 1961;Паавер, 1965; Винников, Рустамов, 1977; Флеров, 1979]. В Восточной Европе голоценовые остатки лого копытного, близкого к современной форме, по более крупного, обнаружены но многих районах Прибалтики, включая Эстонию, на южном берегу Ладожского озера, под Нарвой, возле Пскова и Новгорода, в Белоруссии, Молдавии, на Украине, и Крыму, Приазовье, Предкавказье и Закавказье, в Волгоградской области. Поздне-юлоценовые находки известны из древней Ольвии с территории Николаевской обчисти (начало н. э.), Молдавии (XIII-XIV вв.), прибалтийских городищ X-XVI вв., древних Гродно, Витебска (X-XVIII вв.), Турова, Киева, Вышгорода. Очень часто имеете с зубром встречаются кости тура [Громова, 1935а; Пидопличко, 1951, 1954, 1956; Цалкин, 19516, 1956, 1962, 1963, 1966, 1972а; Верещагин, 1959а; Соколов, 1959; Соколов, Рафалович, 1961; Паавер, 1965; Тимченко, 1972, 1974; Верещагин и др., 1976; Давид, Чемыртан, 197б;Чемыртан, 1986; Александрович, 1989,1999; Алексеева, 19906; Манасерян, 1995, 1997а]. На Северном Кавказе и в Закавказье зубр жил непрерывно до последних веков, о чем свидетельствуют находки его костей и черепов в осетинских святилищах-дзуарах (XTV-XIX вв.), в Дигории, на севере Армении (III тысячелетие до н. э., бронза), в Грузии и Азербайджане [Динник, 1910; Верещагин, Наниев, 1949; Наниев, 1956; Межлумян, 1968]. В средневековье кавказская группировка, скорее всего, была тесно связана с равнинными европейскими популяциями через лесистую долину Дона. Во II—I тысячелетиях до н.э. этот вид, возможно, обитал в лесостепи и на севере степной зоны Зауралья и Западной Сибири вместе с туром, сайгой, тарпаном, северным и благородным оленем, сибирской косулей, лосем и кабаном [Косинцев, 1982, 1986, 1988].
В Великом Княжестве Литовском зубр был охотничьим видом до XV в. Позднее он стал цениться дороже остальных копытных, и за его отстрел полагался большой денежный штраф. В XVI в. он еще обитал в Киевском воеводстве. На Десне этих зверей кололи на “плави” во время сезонных перекочевок. В украинской лесостепи их стада и стада диких коней и оленей мешали сторожевой службе, затаптывая следы проникавших сюда татарских отрядов. Южнее зубры населяли причерноморские степи. В степях между Днестром и Южным Бугом они водились вплоть до второй половины XVI в. В XVII в. в исторических источниках уже не упоминается об изобилии этих животных, но они еще встречались в лесостепной Украине по границе с Московским государством и на Дону. В 1696 г. из левобережной Украины в подарок царскому двору вместе с лосями, кабанами и оленями был послан зубр. В одной из книг Денежного стола Разрядного приказа есть сведения о добыче в 1627 г. пяти молодых зверей в Курском уезде. До конца XVII в. зубры еще сохранялись в лесостепном и степном Подонье, откуда казаки вывозили для продажи их шкуры, а также шкуры лосей, бобров и
выдр. В воронежской лесостепи этих копытных видели в 1709 г. (здесь же в это время жили и тарпаны [Гмелин, 1806]). В отдельных местах зубры, возможно, дожили до 30-х годов этого столетия, а последний зверь был убит в конце века в лесах по Аксаю и Дону неподалеку от Верхне-Курмоярской станицы. В первой половине XVIII в. их еще встречали в степном Причерноморье, во второй - в Украинских Карпатах, Буковине, Беловежской пуще и смежных лесах [Литвин, 1890; Карцов, 1903; Пидопличко, 1951; Кириков, 1952, 1959, 1960, 1966, 1976, 1979, 1983; Соколов, 1959; Гептнер и др., 1961; Аверин, 1976].
По сообщению лесничего Далматова, в 1840 г. зубр был убит в Лыковской казенной даче Семеновского уезда Нижегородской губернии. В 1855 г. он видел свежую шкуру и рога этого животного у кожевника села Лыково. В 1895 г., по сведениям А. А. Ширинского-Шахматова, 5 зверей были встречены в Устюжинском уезде Новгородской губернии, причем один из них был убит [Кулагин, 1919].
В начале XX в. этот вид сохранился только в Беловежской пуще и на Кавказе (рис. 9). Первые письменные упоминания о кавказских зубрах относятся к концу XIII - началу XIV вв. Рашид-ад-дин описал облавную охоту воинов Га-зан-хана в горах Талыша в 1302 г., во время которой внутрь огромного загона сгонялись “горные буйволы”, дикие козы, ослы, волки, медведи и другие звери. Позднее, в XVI в., “дикие буйволы” отмечены в других источниках как обита-
Рис. 9. Распространение зубра на Кавказе (по [Соколов Темботов, 1993])
1-27 - районы обитания в конце XIX и XX вв. (см. в оригинале). На врезке - распространение в прошлом (по [Верещагин, 1959а]): 1 и 2 - верхний и средний плейстоцен, 3 - нижний плейстоцен и верхний плиоцен; 4 - начало XX в.; 5 - место гибели последних зубров; 6 - XIV-XVIII вв. (по коллекционным экземплярам);
7 - XIV-XIX вв. (по литературным сведениям); 8 - ранний голоцен
тели Черкесии и Абхазии [Верещагин, 1959а]. Из сообщения академика Ловила в Петербургскую Академию наук, обнаруженного в архивах П. Паппасом и опубликованного в его Zoographia [Pallas, 1831. Т. 1], известно, что в 1776 г. в верховьях Кумы убит огромный зубр. Сведения о распространении этого копытного в зоне между р. Кубанью и р. Псиб и в Прикубанье содержатся в публикации А. Нордманна [Nordmann, 1838, цит. по: Кириков, 1979]. Академик Гюльденштедт, исследовавший фауну Кавказа в 1770 г., нашел в пещере близ р. Урух 14 зубровых черепов. В 1830 и в 1836 гг. в Академию наук из этого региона были доставлены шкуры зверей, которые затем были утрачены, но сохранилось описание одной из них, сделанное академиком Бэром. Позднее о существовании зубра на Кавказе не раз сообщали русские поселенцы, офицеры русской армии, воевавшей с черкесами, и исследователи. На одном из черепов, найденном в осетинском святилище, сохранилась ясная надпись “1833”, возможно, свидетельствующая о дате добычи одного из последних зверей в Центральной части Северного Кавказа [Верещагин, Наниев, 1949]. В 1835 г. главнокомандующий кавказскими войсками барон Розен прислал в Академию наук Санкт-Петербурга еще одну шкуру этого копытного [Сатунин, 1914]. В середине XIX в. зубры были обычными лишь в горных лесах Северо-Западного Кавказа. В 1848 г. один зверь был убит в прикумской степи [Ребров, 1849], в 1866 и 1868 гг. -самец и самка в верховьях Урупа [Виноградов, 1870], в 1883 г. - на р. Хосте близ Сочи. В это время стада еще жили в бассейнах рек Б. и М. Лабы, по Уруш-тену, Кише, Абаго и в других районах [Васильев, 1896]. Прежнее распространение вида усматривали в границах от Белой до Терека и Военно-Грузинской дороги с выходом на южные склоны хребта в область истоков Мзымты и Бзыби [Сатунин, 1914].
В конце XIX - начале XX в. появилось особенно много сообщений о кавказском зубре, компактная группировка которого в этот период еще сохранилась в горах Кубанской области в междуречье Белой и Малой Лабы, откуда отдельные особи заходили в соседние районы. В августе 1891 г. Литтлдаль добыл в лесах по р. Кише быка и корову, шкуры и скелеты которых отправлены в Британский музей. В 1895 г. Великим князем Сергеем Михайловичем и его спутниками убиты бык и корова, а в 1897 г. - два зубра. Доктор Рейнер убил быка, корову и ранил телка [Радде, 1899; Шильдер, 1901; Динник, 1910]. В 1866, 1899 и 1907 гг. три зубренка были отправлены в Москву и Беловежскую пущу. Зимой 1909/10 гг. Д. Филатов [1910, 1912] добыл старых самца и самку для зоомузея Академии наук в бассейне рек Киши и Шиши. В 1915 г. один зверь был убит вблизи Сухуми, в 1921 г. - у станицы Линейной в 50 км к западу от г. Майкопа, в 1925 или 1926 гг. - в верховьях Кодора. К 1927 г. зубр на Кавказе был истреблен полностью [Северцов, 1926; Башкиров, 1939а; Верещагин, 1959а].
Факторы, определяющие область распространения и численность. Причины вымирания или резкого сокращения численности и ареалов крупных плейстоценовых млекопитающих, среди которых много видов полорогих, включая зубра, бизона, тура, овцебыка и сайгу, недостаточно ясны и по-прежнему дискуссионны. Есть сторонники климатической гипотезы, и есть противники, отстаивающие ведущую роль антропогенных факторов [Флеров,
1976; Верещагин, 1977; Верещагин, Барышников, 1985; Алексеева, 19906, 1995; Буровский, 1995; Маслов, Антипина, 1995; Праслов, 1995; Пучков, 1995; Шуле, 1995; Маслов, 1997]. Эти же две позиции доминируют и при рассмотрении факторов современной динамики населения копытных животных.
Процесс изменения численности и ареалов крупных млекопитающих на границе плейстоцена и голоцена (15-10 тыс. лет назад) нельзя объяснить одной причиной - он, скорее всего, обусловлен как изменением климата и растительности, так и воздействием антропогенных факторов. Многие виды так называемого “мамонтового комплекса”, обитавшие в обширной зоне холодной гундростепи, видимо, не смогли выжить в условиях прогрессирующего повышения температуры на больших пространствах заболоченных тундр и сфагновых болот, а также в районах с высоким снежным покровом при периодических гололедицах. Ареалы животных в этот период существенно сокращались, группировки становились изолированными. Быстрому уменьшению оставшегося поголовья способствовал человек, стремительно увеличивающий свою численность. После значительного сокращения населения крупных фитофагов степная и лесостепная растительность постепенно заменялась лесными ценозами, что, в свою очередь, ограничивало распространение степных животных и давало преимущество типичным лесным обитателям.
Об успешности охот древних людей, начиная с конца среднего плейстоцена (ангельской культуры камня), свидетельствуют “кладбища” костей животных в удобных местах добычи или стоянок с многочисленными следами или обломками применяемых орудий. В процессе загонных охот нередко истреблялись целые стада [Верещагин, 1971, 19756; Бибикова, 1975; Ермолова, 1978; Верещагин, Русаков, 1979; Алексеева, 1980; Верещагин, Барышников, 19806; Аникович, Анисюткин, 1995; Пучков, 1995]. К примеру, в овраге близ Амвросиевки на Украине убиты копьями 983 зубра [Пидопличко, 1951]; в местечке Пшедмости в Чехословакии - около 1 тыс. мамонтов; в Гаронне во Франции - до 30 тыс. северных оленей; у селения Солютре в Бургундии добычей охотников стали более 100 тыс. лошадей; в пещерах горных районов Швейцарии и Венгрии зарегистрированы остатки 1-2 тыс. пещерных медведей [Благосклонов и др., 1967; Дежкин, 1983; Смирнов, 1996]. В Молдавии в слоях небольшого трипольского поселения Флорешты обнаружены остатки 139 кабанов, 84 благородных оленей, 37 косуль, 31 гура и 11 речных бобров; в слоях селетской культуры в гроте у с. Брынзены - 194 лошадей, 117 северных оленей, 21 бизона, 8 благородных оленей, 6 шерстистых носорогов; в мадленском слое стоянки Старые Дуруиторы - 91 лошади, 235 северных оленей, 5 бизонов и много костей других видов млекопитающих [Давид, 1982]. В кухонных отбросах раннепалеолитических поселений человека в Волыно-Подолии доминируют кости мамонта, ископаемой лошади, северного, благородного и гигантского оленей и первобытного зубра [Татаринов, 1986]. На Средней Оби тоже больше костей мамонта (39%), бизона (35%) и лошади (23%) [Алексеева, 1980]. На Енисее и Ангаре в верхнем палеолите люди специализировались на добыче северного оленя, остатки которого достигают 50% среди других животных, в Крыму - сайги, на Десне - овцебыка и мамонта, в Александровке - зайцев, которых убито более 3 тыс. [Громов, 1948].
Исчезновение крупных травоядных животных ускоряется по мере совершенствования орудий добычи [Верещагин, 1959а, 1971, 1975а; Смирнов, 1965; Дежкин, 1977, 1983] и достигает пика с появлением дальнобойного огнестрельного оружия. Истребление копытных особенно резко усиливается во время военных сбо
ров и походов (войска нужно кормить), а также во время массовых охот знати, что подтверждается многочисленными историческими свидетельствами.
Перед походом на рыцарей тевтонского ордена литовский князь Ягелло в 1409 г. заготавливал в Беловежской пуще впрок мясо диких копытных для своей стотысячной армии, сплавляя на плотах в Полоцк ежедневно по 200 т солонины в бочках. В 1429 и 1431 гг. на съезд вельможных особ в Луцке на Украине в течение шести-семи недель доставлялось по сотне зубров, лосей и кабанов. В середине XVI в. зубровую солонину, которая ценилась очень высоко, из Киевского воеводства отсылали в Литву. На одной из охот, устроенной в Беловежской пуще польской королевской знатью 27 сентября 1752 г., убито 42 зубра. Численность этого вида из-за неумеренной круглогодичной охоты заметно сокращается с XV в., в 1627 г. в Янтровском лесу на границе Литвы и Белоруссии убит последний тур, в 1882 г. в Таврической губернии - последний европейский тарпан (в Казахстане тарпан дожил до начала XX в.). В этот же период с территории Европы исчезают кулан, а многочисленные прежде благородные олени и косули сохраняются во многих районах лишь в помещичьих парках. Охота на зубра продолжалась и в XIX в., когда счет оставшимся животным шел на сотни. Беловежской популяции особенно большой урон нанесли браконьеры, осевшие в лесу после подавления польских восстаний 1831 и 1863 гг. [Карцев, 1903; Бутурлин, 1924; Пидопличко, 1951; Кириков, 1952, 1959, 1966, 1976, 1979; Корнеев, 1953; Сержанин, 1955; Цалкин, 1956; Кестер, Шостак, 1968; Тимченко, 1972; Дежкин, 1983].
И все же наиболее впечатляющий пример варварского отношения человека к природным ресурсам - истребление бизона в Северной Америке, численность которого в XVIII в. достигала, вероятно, 60-75 млн. К 1830 г. охотники сократили поголовье до 40 млн, а в 1888 г. (через 58 лет) в живых осталось всего лишь 26 особей в штате Вайоминг. По расчетам, в период с 1868 по 1874 гг. ежегодная добыча составляла 1,5-7 млн голов. Зверей убивали зачастую только ради шкур и языков, а туши оставляли гнить в прериях [Башкиров, 1940; Уатт, 1971; Силантьев, 2000]. Среди людей были своеобразные “чемпионы”, уничтожавшие в среднем по 4 тыс. бизонов ежегодно [Дежкин, 1977].
Замечу, что начиная с раннего - среднего палеолита, наряду с истреблением, шло вытеснение зверей вследствие вырубки лесов, распашки пастбищ и увеличения поголовья домашнего скота. В историческое время численность домашних копытных исчислялась миллионами. В 20-х годах XIX в. только в одной Воронежской губернии было 570 тыс. лошадей и 600 тыс. голов рогатого скота [Кириков, 1966]. Домашние копытные, как справедливо подчеркивает И.И. Пидопличко [1951], во многих районах заменили диких.
В последние тысячелетия, вне всякого сомнения, антропогенный фактор стал доминирующим. Массовое истребление и вытеснение диких копытных имеет место и в наше время, в результате чего немало видов полорогих стали редкими или исчезающими.
Среда обитания
Биотопы. Среди ведущих зоологов не сложилось единого мнения об экотипе зубра. В.Г. Гептнер с соавторами [1961] причислял его к типичным обитателям
разреженных лиственных лесов с полянами и открытыми пространствами, лесостепи и даже степи с уремными и водораздельными лесами. Л.М. Баскин [1979], на основании анализа литературы, сделал заключение, что основная экологическая ниша вида - опушки широколиственных лесов. К.К. Флеров [1979] был убежден, что он возник и всегда существовал как настоящее лесное животное. Представление об обитании этих зверей в степях он считал ошибочным, и особо выделял горные формы. М.А. Заблоцкий [1965а] и И.И. Соколов [1971], напротив, относили истребленного кавказского зубра к типичным равнинным жителям, плохо приспособленным к горным условиям.
В историческом плане прослеживается постепенный переход степных форм Bison к жизни в лесах. В плейстоценовых отложениях Восточной Европы и Сибири В. prisons обычно находится вместе с мамонтом, сайгой, лошадью, шерстистым носорогом, северным оленем и овцебыком, сурками, сусликами, леммингами и песцом, т.е. с типично степными или тундровыми видами. Среди костных остатков конца этого периода он все чаще встречается вместе с лесостепными и лесными видами: туром, благородным оленем, косулей, лосем и кабаном. Судя по пыльцевым остаткам, ландшафты Европы не раз меняются от степных и тундростепных до лесных, и снова становятся степными при аридизации климата. В микулин-ское межледниковье территория Русской равнины была гораздо менее облесенной, чем запад Европы. В валдайское время доминировала перигляциальная фауна [Кулагин, 1918; Пидопличко, 1954; Алексеева, 19906].
На грани плейстоцена и голоцена происходят существенные изменения климата, увеличивается снежный покров, меняется растительность [Флеров, Заблоцкий, 1961; Флеров, 1976]. В голоцене очевидна широчайшая экспансия лесов, что в значительной мере связано с уменьшением численности копытных, подавлявших появление лесной растительности. Древний бизон был вынужден адаптироваться к лесной жизни, измельчал и стал близким к современной форме В. bonasus, но тяготение к открытым биотопам осталось. В историческое время зубр обитал не только в лесах, но и в лесостепи и степной зоне, которая в прошлом отличалась от нынешней. В открытом ландшафте убежищем для крупных животных служили многочисленные пойменные леса, боры, густые заросли кустарников и камышовые крепи по берегам полноводных рек и озер [Кириков, 1959,1963], что, тем не менее, не спасало их от человека. Оптимальными биотопами для зубра, судя по всему, были лесостепь и разреженные широколиственные сообщества. Ареал беловежского и кавказского зубров несколько столетий назад, скорее всего, был единым и приуроченным преимущественно к равнинам [Туркин, Са-тунин, 1904; Верещагин, 1959].
Распределение в современных стациях и влияние на среду обитания. Наиболее полные сведения по стадиальному размещению зубра накоплены в Беловежской пуще, представляющей собой заболоченный массив леса с холмистыми возвышенностями, покрытыми сосновым лесом с примесью дуба, березы и ели, и пологими участками, занятыми лиственным лесом. Эти стации никак нельзя назвать оптимальными для животных, и поэтому их приходилось сезонно менять.
В XIX - начале XX вв. с середины лета до поздней осени зубры держались в сырых низинах (ольсах), где растительность долго не увядала; осенью охотно выходили на сенокосные луга, где отрастала зеленая отава; зимой жили в густом подлеске и подро
сте, на вырубках, в буреломе, где много лишайников, а в многоснежье концентрировались у сенных сараев, переходя от одного к другому по набитым тропам; весной переходили на пойменные луга, где раньше пробивалась зеленая трава, а потом уходили в сосновый лес. Смена стаций определялась наличием доступного качественного корма и, прежде всего, выбором животными влажного травянистого корма и молодой листвы. Особую роль играли дубравные формации, но с увеличением плотности населения копытных дубравы практически утратили кормовое значение, особенно зимой [Карпов, 1903; Врублевский, 1912; Кулагин, 1919, 1940].
Во второй половине XX в. беловежские зубры предпочитали елово-дубово-со-сновый орляково-кисличный природный территориальный комплекс [Толкач, 1989]. Степень использования ими различных стаций определяется, главным образом, их кормностью. Кормовые возможности лесов Беловежской пущи из-за высокой плотности населения копытных и уничтожения ими подроста, подлеска и даже травянистой растительности весьма ограничены, что вынуждает часть зверей выходить за пределы заповедника. Лесные угодья чаще являются убежищем, куда зубры уходят на отдых. Основные стации этого вида весной и осенью - кормовые поляны сеяных трав, сельскохозяйственные поля, низкополнотные участки леса, вырубки с хорошо развитой травянистой растительностью и сенокосы. Летом животные все же чаще держатся в дубравах и ельнике, а зимой концентрируются в местах подкормки [Корочкина, 1971в, 1973а,б, 1977; Корочкина, Кочко, 1980, 1981а,б; Буне-вич, 19896, 1991]. Основные факторы, определяющие плотность их размещения-наличие дубрав, грабняков и других твердолиственных пород; доля разреженных насаждений, полян и редин; доля спелых и перестойных лесов; количество опушек и лесных сенокосов [Бабинок, Зубко, 1990]. В местах концентрации звери оказывают существенное воздействие на формирование древесно-кустарниковой растительности - из состава древостоя постепенно выпадают предпочитаемые виды: ива, ясень, рябина, дуб и клен. Приемлемая плотность населения зубра при высокой численности других копытных - 4-5 особей на 1 000 га [Корочкина, 1970,1971 а].
В других районах Беларуси, на Украине и в Польше зубры также предпочитают лиственные и смешанные леса, особенно дубово-грабовые. В период вегетации они держатся на полянах, вырубках, лесных и пойменных сенокосах, регулярно выходят на посевы зерновых, кукурузы, рапса, люпина и сеяных трав, посещают сады; в осенне-зимний период предпочитают отаву многолетних трав, концентрируются у силосных ям и неогороженных буртов свеклы и картофеля, у стогов сена, становятся “нахлебниками” животноводческих ферм. Лес, преимущественно разреженный, и здесь служит им больше в качестве убежища [Лавов, Чичикина, 1980; Лавов, Воронова, 1982; Лавов, 1983; Лихотоп, Сологор, 1983; Колиснык, 1989; Дунин, Ставровский, 1990; Krasinski, Krasinska, 1994; Тышкевич, 20026].
Группа зверей, выпущенная на волю в пойме Припяти, где богатейшая кормовая база, тем не менее, откочевала в мае из-за обилия кровососущих насекомых в менее кормные, но хорошо продуваемые сосновые и сосново-лиственные леса. В районе ее обитания, несмотря на регулярную зимнюю подкормку, заметны негативные изменения лесных фитоценозов: угнетение подлеска и подроста, выпадение из состава древостоев вяза и ясеня, сильные повреждения дуба, сосны, клена, липы и граба. При истощении кормовой базы участились случаи выхода животных в сельскохозяйственные угодья [Углянец, Домашкевич, 1999а,б; Угиянец и др., 2001; Ушянец, 2003].
В Приокско-Террасном заповеднике летом вольное стадо держалось в лиственном лесу, сосняках широкотравных, на вырубках, опушках, в поймах, на лугах и сенокосных полянах, где был зеленый влажный корм (фото 5). Зимой основными кормовыми стациями становились зарастающие вырубки, редины, опушки полян и места с ветровальными деревьями. Существенный вред вольные звери наносили лишь малоценным породам: иве, бересклету и подросту ольхи, но в загонах и возле них были окольцованы и погибли многие деревья [Заблоцкая, 1957; Заблоцкий, 1957а]. Со временем из состава местной флоры полностью выпали предпочитаемые кормовые виды растений [Шовкун, 2003].
В Крыму переселенцы регулярно выходили из леса и паслись на полях, в садах, где губили немало фруктовых деревьев, и в степи. Быки заходили на фермы и в поселки, где иногда нападали на автомашины и людей, что послужило поводом для переселения 16 голов в Черниговскую область. Примерно такая же ситуация имела место в Хоперском заповеднике, где звери паслись на колхозных полях. В Мордовском заповеднике их зимнее благополучие полностью зависйло от возможности кормиться у стогов сена. В Сары-Челекском заповеднике в Киргизии эти звери приносили большой вред орехово-яблоневым садам [Кузнецов, 1969; Прусаков, 1989; Павлов, 1999].
На Кавказе в конце XIX - начале XX вв. зубр зимой обитал преимущественно в широколиственных предгорных лесах и в пихтарниковом поясе. Летом животные перемещались в верхний пояс леса и на субальпийские пастбища, откуда выходили на альпийские луга [Виноградов, 1870; Динник, 1910; Филатов, 1912; Сатунин, 1914; Касимович, 1939]. Зубробизоны, вселенные на Кавказ во второй половине XX в., были выращены в неволе в равнинных условиях, и оказались не приспособленными к горной местности. Они не могли перепрыгивать через ветровальные деревья, неосторожно шли по осыпям, нередко оступались и падали с обрывов. Однако навыки, характерные для горных животных, появились у них сравнительно быстро. В Кавказском заповеднике в период вегетации растительности звери держатся преимущественно на полянах в зоне пихтово-буковых лесов, у верхней границы леса в субальпийском и вейниковом пихтарниках, кленовниках, злаковых и разнотравных березняках и выходят на субальпийские и альпийские луга (фото 6). Зимой эти копытные концентрируются в злаковых осинниках с обширными полянами, а также в овсяницевых и разнотравно-ожиновых букняках и овсяницевых пихтарниках. Повсеместно наиболее предпочитаемы смешанно-лиственные (24% встреч) и смешанные (18,5%) леса и ольшаники (9%). Выход в открытые биотопы чаще носит временный характер, особенно в последние годы, что связано с усилением браконьерства с вертолетов, звук которых вызывает паническое бегство зверей в лес. При умеренной плотности населения они не приносят существенного вреда фитоценозам, хотя нередко “окольцовывают” молодые деревья, которые затем засыхают. Наибольшие повреждения животные причиняют, устраивая “каталки”, где вытаптывают почву и полностью выбивают растительный покров на площади до 200-300 м2 (фото 7). При высокой плотности их влияние на лесные ассоциации более ощутимо. Из древостоя выпадают поврежденные и затем зараженные энтомовредителями, бактериями и грибами ильм, пихта, бересклет, сильно подавляется возобновление груши, черешни и других видов. Леса в местах обитания зубробизонов стали более осветленными, сменились и домини
рующие виды травяного яруса. Оптимальная плотность зубра на Северном Кавказе - около 10 особей на 1000 га [Калугин, 1958, 1968, 1973; Дыренков и др., 1990; Немцев и др., 2003].
В Тебердинском заповеднике зубры встречаются повсеместно от лесного и горно-лугового поясов до альпийского. Зимой они чаще держатся на нижней половине склонов гор, в долинах рек и лесных полянах, летом - у верхней границы леса и на высокогорных лугах. В лесной зоне животные предпочитают пойменный лиственный лес, смешанные лиственно-хвойные участки на относительно пологих склонах и конуса выноса снежных лавин, заросшие древесно-кустарниковой растительностью. В непогоду они укрываются в хвойном лесу под густыми пихтами и елями [Тарасов, 1975, 1980; Бобырь, 1992]. Места обитания этих копытных в Кабардино-Балкарии сходны: пояс горных лесов и субальпийская и альпийская зоны вблиз опушек. Наиболее характерный биотоп - смешанный лес с хорошо развитым подлеском и наличием полян [Дзуев, 2000а].
В Северной Осетии зубр обитает в буково-грабовых лесах и на субальпийских высокотравных лугах Пастбищного и Лесистого хребтов на высоте от 650 до 1500 м. В местах концентрации его плотность достигает 50-100 особей на 1000 га. Негативное воздействие на лесные фитоценозы проявляется только зимой, когда звери включают в свой рацион кору и древесно-кустарниковые побеги. Особенно сильно они повреждают липу, клен, бузину (до 80%), бересклет (73), граб (57), ильм и тис ягодный (100%), а из фруктовых деревьев - дикие яблони, грецкий орех и алычу. Величина изъятия фитомассы на летних пастбищах колеблется в пределах 13-30%, на зимних, за исключением полян верхней части южных склонов и лесных редин, не превышает 3-8% запаса. В целом трофическая деятельность этого копытного не представляет угрозы для лесов Центрального Кавказа, хотя и может привести к деградации отдельных древесных видов [Литвинова, 1980; Вейнберг, 19866,в; Казьмин, 1988; Казьмин и др., 1992; Казьмин, Смирнов, 1992]. Тем не менее при наличии в местах обитания зубра других видов копытных совокупный вред коренным фитоценозам может быть значительным.
По результатам исследований зубра в разных частях ареала, Л.В. Заблоцкая [1957] заключила, что для него жизненно важно наличие обильного и разнообразного лиственного подлеска и подроста, развитого травяного покрова, лесных речек и ручьев с луговой и древесно-кустарниковой растительностью, выкашиваемых лесных полян и зарастающих вырубок. Хозяйственная деятельность человека (рубка леса и сенокошение) в целом способствует улучшению условий обитания вида.
Питание
Корма. Беловежского зубра в XIX - начале XX в. ученые считали более травоядным, нежели древесноядным зверем, но единого мнения по этому поводу все же не было [Далматов, 1849; Бобровский, 1863, Усов, 1865; Карцов, 1903; Врублевский, 1912]. Дальнейшие исследования позволили выяснить многие интересные особенности его питания.
В польской части Беловежской пущи доля злаков, осок и разнотравья в рационе зверей, как оказалось, достигает 90%, веточных кормов - около 10%, мхов, хвощей, папоротников и грибов - 0,1-0,7% [Gebczynska et al., 1991]. В белорусской
части Пущи и окрестностях основу питания зубра с мая по октябрь тоже составляет травянистый корм: 331 вид растений, относящихся к 42 семействам. Из этого разнообразия животные чаще включает в рацион 85 видов, преимущественно бобовых, злаковых, сложноцветных и осоковых, растущих на хорошо освещенных открытых местах и имеющих высокую питательную ценность. Из них наиболее предпочитаемы: овсяницы овечья и гигантская, мятлик луговой, ежа сборная, лисохвост луговой, бор развесистый, золотая розга, мелколепестник, бодяки, серпуха красильная, тысячелистник, пижма, ястребинки, осот полевой, кульбаба, скереда, одуванчик лекарственный, клевера, чина луговая и лесная, горошки, сочевичник весенний, астрагал солодколистный, лядвенец рогатый, ветреницы, калужница, перелеска, купальницы, водосбор обыкновенный, василистник, лютики и др. Древеснокустарниковые корма в вегетационный период составляют до 30% суточного рациона, поздней осенью и ранней весной они доминируют в питании, но зимой при обильной подкормке их доля незначительна. Звери потребляют почти все виды древесной, кустарниковой и кустарничковой растительности Пущи: ясень, клен, осину, иву, дуб, граб, вяз, лещину, крушину, рябину, липу, бересклет, сосну, реже ель и березу, можжевельник, ракитник, сирень, вереск, омелу, дрок, чернику, малину, ежевику, смородину, костянку, толокнянку. Очень любят желуди и плоды яблонь и груш. Они охотно кормятся на сельскохозяйственных полях, где поедают клевер, озимые рожь и пшеницу, кукурузу, овес, картофель, топинамбур, свеклу, зимой потребляют сено и даже солому, силос, иногда разрывают бурты картофеля [Кулагин, 1919,1940; Корочкина, 1966, 1970, 1971в,г, 1972а, 19736, 1974].
В национальном парке “Припятский” эти копытные выбирают 33 вида травянистых растений, преимущественно из семейств злаковых (чаще молинию, вейник, ежу сборную, бор развесистый), бобовых (ракитник русский, дрок красильный), норичниковых (марьянник, веронику дубравную), вересковых (чернику) и сложноцветных (золотарник обыкновенный, мицелис стенной). Из древесно-кустарниковых видов они предпочитают бересклет бородавчатый, осину, рябину, липу, граб, дуб и крушину. Наиболее богаты излюбленным кормом дубравы и сосняки, наименее - смешанные лиственные леса [Козло и др., 1999]. С ноября основу зимнего рациона составляет подкормка [Углянец, 1999а].
В пище зубра в Цуманской пуще на Волыни зарегистрировано более НО видов растений, из них 84 - травянистых и 30 - древесно-кустарниковых. Летом травянистая растительность составляет 80% рациона, древесно-кустарниковая - 20%, зимой преобладает древесная растительность, ежевика, а также сено и сочные корма (на подкормочных площадках). Здесь животные сильно повреждают кору деревьев [Татаринов, Дякун, 1969; Колиснык, 1989].
В Приокско-Террасном заповеднике эти копытные употребляют в пищу свыше 200 видов растений, из них около 180 травянистых, 36 деревесно-кустарнико-вых и полукустарниковых и 6 видов грибов, что составляет около 20% местной флоры. В число основных кормов входит около 30 видов трав и 9 древесных и кустарниковых пород. Особенно охотно звери поедают сочные высокостебельные травы (растущие в местах повышенного и среднего увлажнения, на ранних фазах вегетации или в виде отавы): осоку волосистую, иван-чай, золотую розгу, купену лекарственную, ландыш, гравилат, бодяк, таволгу вязолистную, клевер, верхушки молодого тростника, а также вейники, щучку дернистую, молинию,
лапчатку и сныть, весной - первоцветы. Ряд предпочитаемых древесно-кустарниковых кормов следующий: ивы, осина, бересклет, рябина, черная смородина, клен, черемуха, дуб, крушина ломкая, липа, ольха, береза, ель, сосна, что соответствует уровню содержания в этих растениях протеина и, отчасти, кальция. Летом листва, кора и древесно-кустарниковые побеги диаметром до 1,5-2 см составляют не менее 50% массы поедаемого корма, в другие сезоны их доля еще выше. Кора употребляется круглогодично, но в большем объеме - с ранней осени и до весны. В конце лета в рационе обычны грибы (белые, подберезовики, лисички и сыроежки), осенью - желуди в урожайные годы. Сено, особенно клеверное, заготовленное на поздних стадиях вегетации, поедается плохо [Заблоц-кая, 1957].
Список растений, поедаемых зубробизоном в Кавказском заповеднике, еще обширнее: более 325 видов, из которых 59 - древесно-кустарниковые, 196 - разнотравье, 41 - злаки, 16 - бобовые, 6 - осоки и ситники. Основной зимний корм - зеленые подснежные растения (ежевика, ожина, овсяница горная, осока лесная), ветошь трав, древесные побеги, кора деревьев и кустарников, сохранившиеся с осени желуди, древесный лишайник. Звери употребляют в пищу практически все растущие здесь древесно-кустарниковые виды, предпочитая ильм, осину, козью иву, лещину, бересклет, пихту, в меньшей мере - дуб, граб, ольху, грушу, алычу, черешню и тис. При содержании в загонах едят сено, концентрированные корма и корнеплоды. В ранневесеннем питании доминируют зеленые травы: пролеска, черемша, вейник, ежа, мятлик, пырей, типчак, подмаренник, воронец, ясколка, позднее - злаки, осоки, щавель, чемерица, клевер, козлятник восточный, сочевичник, а также молодые листья и побеги деревьев и кустарников. Летом основным кормом становится вы-сокотравье верхней части лесного, субальпийского и, отчасти, альпийского поясов, где они выбирают борщевик, лигустикум крылатый, мятлики, вейники, ежу сборную, овсяницы, щучки, козлятник восточный, головчатку, купену, мать-мачеху, сныть, чину луговую, тимофеевку альпийскую и др. Древесно-кустарниковый корм в этот период становится второстепенным. Осенью в рационе возрастает доля мелкотравья и отавы, но особенно охотно звери поедают плоды груш, яблонь, алычи и шиповника, выходят на картофельные, свекольные и кукурузные плантации [Крайнова, 1951; Александров, 1958; Калугин, 1968; Дыренков и др., 1990; Немцев и др., 2003]. Примерно таким же был корм и аборигенного кавказского зубра в начале XX в. [Филатов, 1912].
В Тебердинском заповеднике зимой и ранней весной основной корм зубра -ветки и кора ильма шершавого, бересклета, черемухи, осины, клена, лещины, рябины, кора и хвоя молодых пихт и сосны, побеги рододендрона, ежевики, черники, смородины, древесные лишайники, ветошь вейника и других злаков, клевер, манжетка, хвощ. Для подкормки зверей здесь ежегодно заготавливают сено. В вегетационный период в пище преобладают травянистые растения, из которых чаще поедается овсяница пестрая, ежа сборная, вейник, пырей, щавель, головчатка кавказская, борщевик, козлятник, отава на сенокосах, а также листья деревьев, листья и побеги смородины и шиповника. Осенью рацион дополняется плодами фруктовых деревьев [Бобырь, 1992].
В Северной Осетии на Лесистом хребте в список кормов этого копытного входят 47 видов травянистых растений и 22 - древесно-кустарниковых. Сходный набор кор
мов и на Пастбищном хребте, где летом звери потребляют около 50 видов растений из 18 семейств. Основу травянистого питания составляют сложноцветные, губоцветные и бобовые. Злаки, папоротники и хвощи поедаются гораздо реже. Из древесно-кустарникового и кустарничкового корма и летом, и зимой зубры предпочитают иву козью, ежевику, бузину черную и травянистую, липу, клен, ильм, граб, вербу, осину, тис ягодный и лещину. Особенно охотно животные едят молодые побеги и кору, но при температуре ниже -10... 15 °C кора выпадает из рациона. При небольшом снежном покрове в пище доминируют побеги ежевики (22% потребленного корма), зимнезеленые травянистые растения, ветошь и папоротники (38%). С увеличением слоя снега потребление ежевики и травянистых растений уменьшается с 59 до 6% при одновременном росте доли коры (с 39 до 79%) и древесно-кустарниковых побегов (с 2 до 15%). Величина изъятия фитомассы на летних пастбищах колеблется в пределах 13-30%, на зимних не превышает 3-8% запаса и не подрывает кормовой базы, за исключением полян верхней части южных склонов и лесных редин, где использование кормов достигает 24% [Литвинова, 1980; Вейнберг, 19866; Казьмин и др., 1992; Казьмин, Смирнов, 1992]. Замечено, что питание твердыми буковыми видами ускоряет стирание зубов и приводит к раннему (в 13-15 лет) старению животных [Варзиев, Курятников, 1980].
Итак, по типу питания зубр явно относится к траво-древесноядным животным: летом основу его рациона составляет травянистая растительность, преимущественно злаки и бобовые, зимой - древесно-кустарниковый корм, дополняемый искусственной подкормкой. В питании характерны следующие черты: эврифагия; высокая пластичность в использовании местных кормов; предпочтение высокостебельной травянистой мезофильной и гидрофильной растительности и трав на ранних стадиях вегетации; большой удельный вес лиственных древесных кормов (выше, чем у бизонов), используемых круглогодично. Зимой древесно-кустарниковый корм незаменим даже при подкормке сеном и концентратами [Заблоцкая, 1957; Заблоцкий, 1957в].
Бизона, однако, тоже нельзя считать чисто травоядным. Он, как и зубр, предпочитает злаки и осоки, а на Аляске у животных, выпасающихся в долинах рек, доля веточных кормов в питании достигает 94% [Campbell, Hinkes, 1983; Van Vuren, 1984; Waggoner, Hinkes, 1986].
Потребление кормов и воды. Минеральное питание. В Беловежской пуще экспериментально установлено, что 2-3-летние зубры летом поедали 19-28,5 кг, зимой - 4—6 кг корма, взрослые быки, соответственно, 32 и 7 кг [Gebczynska, Krasinska, 1972]. М.В. Холодова с соавторами показала, что 1,5-3-летние животные летом потребляют 5-8 кг сухого вещества, 4-8-летние - 10-15 кг, или, соответственно, 20-25 кг и 40-50 кг сырого корма на особь. Переваримость кормов варьирует от 46 до 79% в зависимости от содержания в них клетчатки. Пороговая величина переваримости, не приводящая к дефициту энергии в организме, у взрослых не размножающихся особей составляет около 55%, у беременных и лактирую-щих самок и растущего молодняка - 75%. При уменьшении запаса травянистой растительности наблюдается снижение интенсивности ее потребления. Зимой уровень потребления кормов уменьшается в 1,4—2 раза и быстро возрастает с апреля, что связано с изменением метаболизма [Холодова, 1989; Холодова, Белоусова, 1989а,б; Холодова и др., 1990]. Примерно такую же величину потребления корма (30—45 кг, в среднем 36 кг травы и до 4 кг веточного корма) приводит для зуброби
зонов С.Г. Калугин [1958, 1968]. Он опытным путем нашел также, что в вегетационный период на каждого зверя должно приходиться не менее 3,5 га пастбищ при соотношении полян и леса 2:1,5. В Приокско-Террасном заповеднике в опытах звери за сутки съедали: в ноябре - 7 кг сена и 15 кг древесно-веточного корма, в феврале и апреле - по 6 кг этих кормов, в марте - 4 и 11 кг соответственно [Заблоцкая, 1957]. В Тебердинском заповеднике пришли к заключению, что для зимней подкорки одной особи требуется около 330 кг сена ежегодно [Бобырь, 1992].
Экспериментально установлено большое сходство зубра и бизона в энергетических потребностях и уровне потребления и усвоения кормов. При этом зубр, видимо, все же лучше адаптирован к усвоению древесной клетчатки [Холодова, Белоусова, 1993].
Зубр нуждается в большом объеме свободной воды. Летом в неволе взрослая особь выпивает за сутки 30-60 л воды, а иногда - до 13 ведер. В жару звери приходят к водным источникам дважды в день, в пасмурные и дождливые дни - раз в 2-3 суток. Они предпочитают пить свежую проточную воду, к застойной не подходят, зимой используют снег [Сержанин, 1955; Калугин, 1958, 1968].
На Кавказе копытные круглогодично посещают естественные и искусственные солонцы, потребляя в среднем от 10 до 53 г соли в сутки и от 8 до 13 кг на особь за год. Потребность в минеральных веществах особенно высока в вегетационный период. Пики посещений солонцов приходятся на весенние месяцы и август [Динник, 1910; Филатов, 1912; Касимович, 1938; Башкиров, 1939а; Калугин, 1958,1968; Бобырь, 1992; Немцев и др., 2003].
В зубропитомнике Окского заповедника в каждом загоне звери выгрызали песчаный грунт в 2-6 местах. Размер одной из ям - 7 х 6 м и 0,6 в глубину. Молодые животные поедали грунт в бесснежный период, взрослые - в конце зимы - начале весны, а в другое время года использовали его только при желудочно-кишечных заболеваниях. Ранней весной зубры ели также трухлявые пни и строительный материал муравейников. В качестве минеральной подкормки им давали мел и соль-лизунец, которую они использовали особенно интенсивно в июле-октябре. Среднегодовой расход соли на особь здесь был гораздо меньше, чем на Кавказе - 2,5-4,5 кг [Киселева, 1974, 19786].
Свои особенности минерального питания были и в Приокско-Террасном заповеднике. Здесь звери охотно посещали площадки с солью-лизунцом, комбинированные солонцы из смеси поваренной соли и красной глины, а также известняковые выбросы у ям и колодцев. Во второй половине зимы они разрывали снег и поедали гумусный слой почвы по склонам ручьев и оврагов, между корнями деревьев и на кочках [Заблоцкая, 1957].
Пищевая активность. Суточный ритм активности зубра полифазный, но без строгих циклов. В Беловежской пуще в вегетационный период звери больше времени затрачивают на пастьбу, нежели на отдых. Продолжительность пастьбы здесь в значительной мере определяется состоянием естественной кормовой базы. Суточная периодика у разных половых и возрастных групп животных сходна и во многом зависит от воздействия абиотических факторов среды. Зимой питание занимает около четверти времени суток, отдых - 60%, переходы - 10%, остальные формы поведения - 5% [Корочкина, 19726; Cabon-Raczynska et al., 1983]. На Кавказе летом животные тратят на пастьбу и отдых примерно равное время, вес-
ной дольше пасутся, а в снежный период дольше остаются на лежках. В жаркие дни они находятся на пастбище с вечера до утра, а в пасмурную и дождливую погоду - и в дневное время. Взрослые самцы обычно кормятся меньше времени, чем самки, и дольше отдыхают. Непосредственно на пастьбу звери затрачивают в среднем около 5 ч в сутки, лактирующие самки зимой - до 9 ч. Суточный ритм членов группы или стада относительно синхронизирован [Калугин, 1958, 1968, 1977; Казьмин и др., 1992; Немцев, 2003].
Размножение
Половая зрелость. Самки и самцы зубра и бизона, как показала практика, становятся половозрелыми в возрасте 2 лет, но большая часть вступает в размножение позднее, а отдельные особи - в 5-6 лет. Сперматогенез начинает затухать после 12 лет жизни, но способность к воспроизводству сохраняется до 16-24-летнего возраста [Заблоцкий, 1957а; Jaczewski, 1958; Krasinski, Raczynski, 1967; Стекленев, 1983а, 1996; Agabriel et al., 1996; Kirkpatrick et al., 1996; Czykier et al., 1999].
В Беловежской пуще большинство самок зубра (65%) приносили первое потомство в 4 года, 16% - в 3, 13% - в 5 и 5% - в 6 лет. Особи беловежской линии позднеспелые (первый отел в среднем в возрасте 4,5 лет) по сравнению с беловежско-кавказскими (3,9 года) [Корочкина, 19716]. В Окском заповеднике 50-60% самок телились в 3-летнем возрасте, 25-28 - на четвертом году жизни, 17-20 - в 5 и 5% - в 6 лет [Киселева, 1973, 1974, 1977, 1978а]. В вольерах Кавказского заповедника первый отел зубробизонов происходил в 3-^1-летнем возрасте, на воле - в 4—5 лет. Здесь же зарегистрирован случай успешного оплодотворения самки полуторагодовалым самцом [Калугин, 1968]. Позднее в кавказской группировке самки приносили потомство в возрасте 3-6 лет [Немцев, 1987а; Немцев и др., 2003].
Гон. Гон у животных, содержащихся в вольерах, растянут со второй половины июля по март, с пиком в сентябре. В природе он обычен в июле - сентябре, и лишь немногие особи спариваются раньше или позже (табл. 6). У большинства зубриц первая течка отмечается в июле - сентябре, а у впервые пришедших в охоту - в августе - сентябре [Перерва, 1992], что служит дополнительным указанием на ви-доспецифичность этого срока. Примерно в это же время был гон у зубра и в XIX -начале XX вв. - с июня по октябрь включительно [Виноградов, 1870; Карцов, 1903; Кулагин, 1919; Башкиров, 1939а]. У бизона, как и у зубра, гон протекает с последней декады июля до октября, но часть особей спаривается вплоть до конца апреля, а в загонах размножение может идти практически круглогодично с типичным сезонным пиком [Стекленев, 1983а, 1996; Agabriel et al., 1996].
Сильная растянутость сроков гона у зубра вызвана, скорее всего, длительным разведением животных в неволе, где быки постоянно находятся вместе с самками. В природе в местах зимней концентрации зверей (чаще на подкормочных площадках), несмотря на агрегацию групп самок и взрослых самцов, гон тоже имеет место даже в разгар зимы, что отражается на сроках рождения телят. Самок нередко покрывают молодые самцы, которых в обычных условиях к ним не допустили бы взрослые особи.
Длительным срокам спаривания способствует и другие обстоятельства. У зубриц в течение года бывает 2-3 течки, у некоторых - до 5. Каждая течка длится 1-2, 46
Таблица 6. Сроки гона у современного зубра
Район, заповедник Начало Конец Источник информации
Беловежская пуща Июнь 27 июня Польша Август Припятский з-к Середина августа Полесский з-к 27 июля Волынь (Украина) Начало октября Окский з-к (питомник) Вторая половина Октябрь 20 августа Сентябрь Середина сентября 4 октября Начало марта Март (апрель) Кулагин, 1940 Сержанин, 1955 Jaczewski, 1958 Углянец, 1999а Тышкевич, 2000 Татаринов, Дякун,1969 Киселева, 1973, 1974,
Приокско-Террасный з-к (питомник) Кавказский з-к июля Июль Август Конец июля Январь Февраль Октябрь (единично -до февраля) 1978а Перерва, 1981 Белоусова, 1986 Калугин, 1958, 1968
реже - от 3 до 8 дней. При неоплодотворении они наступают в интервале от 9 до 103, чаще - через 20-30 дней. При позднем эструсе гон сдвигается на позднезимние месяцы, что, в свою очередь, отодвигает срок родов. В обычных условиях интервал между отелом и первой в текущем году течкой - около 100-120 дней, но в неволе он сокращается до 80-22 дней, и изредка приводит к успешному оплодотворению. Старые самки, которых много в вольерах, чаще оплодотворяются на месяц позднее молодых, хотя в природе приходят в охоту раньше [Калугин, 1919, 1968; Jaczewski, 1958; Киселева, 1974, 1978а]. Половая активность животных резко снижается при похолоданиях в октябре - ноябре, и возрастет при потеплениях, что тоже удлиняет гон [Перерва, 1992].
Сроки гона и отела у разных генетических линий зубра неодинаковы. В Центральном зубровом питомнике гон у животных в первые годы их разведения проходил в два этапа - с июля по сентябрь и с ноября по январь. Беловежские зубры в эти два периода спаривались примерно с равной интенсивностью, кавказско-беловежские -в соотношении 4:1, а гибриды зубров с бизонами и серым украинским скотом - 7:1. Со временем под влиянием среды обитания у всех произошло выравнивание сроков гона до соотношения 1,8-2,4:1 [Перерва, 1983а].В 1981-1985 гг. на август-сентябрь приходилось 56% покрытий, на октябрь - 16, на остальные месяцы до февраля включительно -по 6-8% [Белоусова, 1986].
В период гона взрослые самцы, обычно держащиеся по одному или по 2-3, присоединяются к группам самок. В этот период они мало едят, издают хоркающие звуки и мускусный запах, становятся агрессивными и порой нападают не только на других самцов, но и на зубрят, взрослых самок, и весьма опасны для человека. Иногда они заходят на животноводческие фермы, где могут покрывать коров [Киселева, 1974; Бобырь, 1992]. Сильные быки, вошедшие в состав маточного стада, вытесняют молодых 2-3-летних самцов. Эта особенность весьма важна для популяций, поскольку уменьшает вероятность близкородственного скрещивания [Заблоцкий, 19756; Немцев, 1980а]. В любом случае в природе потомство оставляют самые мощные самцы зрелого возраста, спаривающиеся, прежде всего, с высокоранговыми самками [Перерва, 1989а].
В брачных скоплениях кавказских зубробизонов насчитывается несколько десятков особей (фото 8). В августе 1991 г. в одном из стад зарегистрировано 97 голов, включая 30 быков. Наиболее сильные самцы занимали центральное положение, более слабые - периферийные участки [Немцев, 2003]. Ранее присутствие нескольких взрослых самцов в брачной группе считалось артефактом, связанным с гибридным происхождением группировок и длительным разведением животных в неволе [Калугин, 1968].
В ритуале ухаживания самцов за самками обычны следующие элементы: обнюхивание и флемование; неотступное преследование; глухое урчание; ассистирование в положении сбоку и чуть сзади от самки (у бизонов - и спереди), сопровождаемое угрожающими кивками, грумингом и флемованием; вскидывание головы на спину самке, дополняемое звучным сопением; вспрыгивание на самку и копуляция в положении стоя (у бизонов - в движении). Бык может копулировать до четырех раз в сутки, или же после акта теряет интерес к самке и отходит от нее. Нередко у самок, особенно у старых, отмечается нимфомания, усиливающаяся при импотенции самцов [Перерва, 1992].
Беременность. Беременность самок зубра в Беловежской пуще длится 265 (257-272) дней [Jaczewski, 1958; Корочкина, 19716], в Окском и Приокско-Тер-расном заповедниках-268 (217-297 и 255-286 соответственно) [Киселева, 1973, 1974, 1978а; Белоусова, 1986]. Зубробизоны в Кавказском заповеднике вынашивают плод 261-283 дня [Калугин, 1958, 1968], бизоны, разводимые в Аска-нии-Нова на Украине и в вольерах во Франции, - в среднем 265 и 270 дней [Лобанов, 1968; Agabriel et al., 1996], что соответствует природным срокам.
Деторождение и выкармливание молодняка. Отел у зубра происходит с апреля до декабря (табл. 7). Его пик в вольерах обычен в июле - августе, а на воле -в мае-июле [Виноградов, 1870; Калугин, 1958, 1968; Киселева, 1973, 1974, 1978а; Буневич, Кочко, 1988; Буневич, 1999в]. Растянутость времени отела вызвана измененными сроками гона, но в природе этот процесс постепенно нормализуются до эволюционно обусловленного периода, приходящегося, видимо, на конец апреля -июнь. Именно в это время чаще телятся прохолоставшие и впервые рожающие самки [Перерва, 1992]. Самки бизона тоже приносят телят в апреле - июне, но значительная часть их, особенно в неволе, рожает в летне-осенние и даже зимние месяцы [Стекленев, 1983а, 1996; Rutberg, 1984; Agabriel et al., 1996].
Перед отелом вольные беременные самки зубра обычно покидают стадо, уединяясь в густых лесных зарослях. В загонах звери могут телиться на виду у членов стада и человека, и нередко стельную самку к месту отела сопровождают ее подруги. Самки бизона в стадах, обитающих в облесенной местности, тоже уединяются для отела в зарослях, а живущие в открытой местности рожают в стаде [Lott, Galland, 1985а].
Самка телится стоя или лежа. Процесс отела длится от 30 до 130 мин. Мать съедает околоплодную оболочку и тщательно вылизывает детеныша. Через час после рождения теленок стоит на ногах, а спустя полчаса может, спотыкаясь, следовать за матерью, которая уводит его с места родов. Если теленок не идет за ней, то она остается на месте отела еще несколько дней. В отсутствие матери, уходящей на кормежку, детеныш затаивается, и при опасности прижимается к земле. Самка каждый раз ищет его для кормления, издавая при этом свое-
Таблица 7. Сроки отела у зубра
Район,заповедник | Начало Конец Источник информации
Беловежская пуща Конец мая Ноябрь Кулагин, 1919, 1940
U 31 марта 9 ноября Сержанин, 1955
к Апрель Декабрь Буневич, Кочко, 1988
Припятский заповедник Середина мая Июнь Углянец, 1999
Волынь Июнь Декабрь Татаринов, Дякун, 1969
Окский з-к (питомник) Апрель Ноябрь Киселева, 1973, 1974, 1978а
Приокско-Террасный Май Октябрь Перерва, 1981
заповедник (питомник)
Кавказ Апрель Май Динник, 1910
— Май Начало июня Сатунин, 1914
Кавказский заповедник Апрель Ноябрь Калугин, 1958, 1968
Тебердинский заповедник Май Август Тарасов, 1975
образные звуки, напоминающие хрюканье. Ее голос теленок различает на 3-5-й день жизни, но уверенно идет на него примерно с месячного возраста, радостно мыча в ответ. В первые месяцы зубрята сосут мать до 12-15 раз в сутки, в 5-6-месячном возрасте - 3-5, в последний месяц лактации - 1-2 раза. Продолжительность разового кормления молоком, жирность которого достигает 8-9%, составляет 1-7 мин. Лактация длится от 5 до 11 мес. в зависимости от времени отела, а если мать остается яловой, до года и даже, в редких случаях, до двух лет [Карцов, 1903; Кулагин, 1919; Киселева, 1974; Daleszczyk, Krasinski, 2001].
Голодный теленок очень требователен, и может бодать мать, заставляя ее встать с лежки, но чаще зубрица сама определяет время его кормления по наполнению вымени молоком. Самки обычно кормят только своих телят (иногда одновременно и прошлогодних), но некоторые не отказывают в молоке и чужим детенышам, и известны случаи выкармливания телят-сирот. Лактирующие самки осторожны и агрессивны. В первые месяцы они активно защищают свое потомство, демонстрируя нападения на приближающихся людей, крупных хищников и даже своих сородичей, что существенно снижает детскую смертность. В холодные зимние дни самки не позволяет своим телятам долго лежать на снегу, или стремятся перевести их на подстилку из сена [Сержанин, 1955; Калугин, 1958, 1968; Киселева, 1974, 1986а; Тарасов, 1975; Заблоцкий, 1976; Перерва, 1992].
Плодовитость. В природе при хорошем питании большинство самок зубра приносит одного теленка ежегодно, при плохом - через 1-4 года, а в среднем - через 1,5-2 года. Двойни рождаются крайне редко, всего около десятка случаев за всю историю разведения вида. В Кавказском заповеднике у зубробизона двоен не зарегистрировано ни разу при 631 отеле [Калугин, 1968]. Периодичность отелов и уровень плодовитости зверей в неволе существенно зависит не только от питания, но и от состояния здоровья, возраста и иерархического ранга особей [Кулагин, 1919; Заблоцкий, 1957а; Киселева, 1973, 19866; Перерва, 1981, 1989а; Krasinski et al., 1994; Немцев и др., 2003].
В Беловежской пуще до 1971 г. ежегодно телилось 78% способных к воспроизводству самок с интервалом в 1 год и 3 месяца в среднем [Корочкина, 19716,в, 1973; Krasinski, 1978; Буневич, Кочко, 1988]. В 1971-1995 гг. впольской части Пущи потомство приносили только 51% половозрелых самок, в белорус
ской - 39 (21-55)%. За 34 года наблюдений (1964-1997) плодовитость варьировала в пределах 86-16%. Самки наиболее полно (более 50%) реализовали свои репродуктивные возможности в 1965, 1968, 1971, 1976, 1978 и 1990 гг., следующие за обильными урожаями желудей, депрессия (16-27%) имела место в 1966, 1981 и 1992 гг. после неурожая желудей и при некачественной зимней подкормке. В целом воспроизводственные показатели польских зубров были на 13% выше белорусских при аритмии годовых уровней плодовитости, что обусловлено, прежде всего, кормовым фактором [Krasinski etal., 1994; Буневич, 1999а,б; Козло, 2001].
В Кавказском заповеднике показатель плодовитости самок зубробизона в первые десятилетия после выпуска варьировал от 50 до 100%, яловость чаще находилась в пределах 25-30%. Позднее доля яловых самок в отдельные годы возрастала до 53%, а рождаемость после тяжелых зимовок падала до 12-18%. При низкой плотности популяции самки размножались почти ежегодно, при высокой интервал между отелами достигал 2-3 лет [Калугин, 1968; Немцев и др., 2003].
В вольере Окского заповедника в 60-е годы размножались 85% (75-100% в разные годы) половозрелых самок. Большинство из них приносили потомство каждый год или с редкими годичными перерывами, вызванными заболеваниями. В 70-е годы приплод получен от 69, а в 80-е - от 44% самок [Киселева, 1973, 1974, 1977, 1978а, 19866]. В питомнике Приокско-Террасного заповедника доля яловых особей в отдельные периоды достигала 30-50%. Воспроизводственный потенциал стада здесь сокращался при многолетнем содержании самок с одним и тем же самцом. В первый год потомство приносили 67% самок, в следующем - 62, затем 47, 58 и 38%. Следовательно, для увеличения плодовитости необходимо чаще менять самцов, что и происходит в природе [Перерва, 1981, 1983а; Петров, 2003].
Самцы наиболее активны в возрасте 7-11 лет, в гоне участвуют до 14—15 лет. Самки наиболее плодовиты в возрасте от 3-4 до 12-14 лет. Позднее репродуктивные возможности уменьшаются в 2-^1 раза [Заблоцкий, 1957а; Krasinski, Raczynski, 1967; Корочкина, 19716; Белоусова, 1986; Киселева, 19866].
Плодовитость бизона примерно такая же, как и у зубра, но в природе у самок все же чаще проявляется 3-летний цикл при больших индивидуальных вариациях, а в неволе при хорошем кормлении они могут рожать ежегодно. В целом репродуктивная стратегия, сроки размножения и половое поведение этих животных сходны [Стекленев, 1983а, 1996; Lott, Galand, 1985b; Maher, Byers, 1987; Schaw, Carter, 1989; Перерва, 1992; Agabriel et al., 1996; Kirkpatrick et al., 1996; Towne, 1999].
Структура группировок
Половой и возрастной состав. Судя по накопленным сведениям, соотношение полов у детенышей зубра во всех наблюдаемых природных группировках близко 1:1с незначительным варьированием в ту или иную стороны в разные годы. При содержании зверей в неволе оно изменяется в широких пределах - от 1:0 до 1:5. Замечено, что доля новорожденных самочек несколько увеличивается при хорошем кормлении зверей, и их чаще рожают зрелые самки [Заблоцкий, 1957а,б; Jaczewski, 1958; Корочкина, 1970, 19716; Киселева, 1974, 1977, 1978а; Krasinski, 1978; Корочкина, Кочко, 1983; Белоусова, 1986; Pucek et al., 1996; Буневич, 1999в].
Таблица 8. Половозрастной состав (%) и соотношение полов в современных группировках зубра
Район, заповедник (источник информации)* Сеголетки 1-3-летние Взрослые Самцы : самки
самки | самцы
Беловежская пуща (1) 6-17 23 38 (33-42) 18-24 1:1,1-2,4
-“-(7) 14 25 41 20 1:2
(5) 13,6 24,6 37,7 24,4 1:1,5
(2) 15,5 23,4 35,7 25,5 1:1,4
Припятский з-к (б) 12-25 25-48 29-44 4-12 1:1,5-2.6
Кавказский з-к (3) 11 49 25 15 1:1,2
-“-(8) 12,8 10,1** 42,6 34,5 1:1,2
Тебердинский з-к (4) 7-16 15-27,5 31-39 28-44 1:1
* 1 - Буневич, Кочко, 1988; 2 - Krasinski et al., 1994; 3 - Калугин, 1968,1970; 4 - Бобырь, 1992; 5 - Козло, 2001; 6-Углянсц, 19996; 7 - Буневич, 1999а; 8-Немцев, 1987а.
** Годовалые особи.
В XIX - начале XX вв. в стадах беловежского зубра самцов, возможно, было больше, чем самок [Кулагин, 1919], но по учету, проведенному 1 марта 1909 г., последние преобладали при соотношении полов 1:1,3 [Wroblewski, 1927]. В последние десятилетия в Беловежской пуще на одного взрослого самца приходится в среднем 1,5 (1,1-2,4) самки. Самок несколько больше и в составе других современных вольных стад (табл. 8), а также среди животных, содержащихся в неволе [Заблоцкий, 1957а,б], что связано с повышенной гибелью самцов. В группировке Кавказского заповедника доля самок составляла в среднем 52,5% (43-59% в разные годы) [Немцев и др., 2003].
Демографическая структура большинства стад сформирована (или формируется) под непрерывным воздействием человека, и далека от оптимального уровня. Доля сеголеток в разных группировках варьирует в пределах 6-25%, 1-3-летних особей - 10-49, взрослых самок - 25-44, взрослых самцов - 4-44% (табл. 8).
Социальная организация и поведение. В группировках зубра исследователи выделяют семьи (мать с теленком), маточные стада (несколько взрослых самок с детьми), группы самцов, смешанные (разнополые и разновозрастные) стада и одиночных особей. Основа социальной организации - семьи и маточные стада, далеко не всегда объединенные кровным родством. Последние по уровню интеграции приближаются к объединениям закрытого типа. Взрослые и молодые самцы большую часть года живут в одиночестве, реже - парами или втроем, или изредка образуют группы численностью до 7-15 особей. Первую неделю жизни зубрята находятся рядом с матерями, а со второй недели более 50% времени проводят со сверстниками, держась в стаде отдельной группой, которую нередко возглавляет младшая из отелившихся самок. Эта особенность в значительной мере способствует снижению травматизма сеголеток, особенно в присутствии взрослого самца. Летом в горах и зимой в равнинной части ареала звери концентрируются на ограниченных пастбищах или у подкормочных площадок, образуя скопления [Кулагин, 1919; Заблоцкий, 19756, 1976; Krasinski, 1978; Немцев, 1980а, 2003; Перерва, 1992; Krasinska, Krasinski, 1994, 1995; Daleszczyk, 2002].
В XIX - начале XX вв. наиболее крупные стада зубров в Беловежской пуще состояли из 15-20, а в редких случаях кратковременно (2-5 дней) - из 25-60 голов,
но зимой иногда отмечали скопления до 200 особей. Обычно же животные держались мелкими группами от 5 до 15 экземпляров [Далматов, 1849; Бобровский, 1863; Усов, 1865; Карцов, 1903; Кулагин, 1919, 1940; Wroblewski, 1927]. Во второй половине XX в. в восточной части Беловежской пущи чаще всего регистрировали группы из 2-5 особей. Крупные объединения, насчитывающие 60-140 голов, редки. Они формируются лишь в экстремальных ситуациях при нехватке естественных кормов. Средний размер групп в течение года - 14,6, в вегетационный период -21,3 (табл. 9). В современных группировках преобладают смешанные стада (72% встреч), чаще состоящие из 6-30 особей. Отдельные самцовые и самочьи группы, численностью от 2 до 9 голов, обычны с мая по июль (70-80%). Доля одиночных особей - 12,5% [Буневич, Кочко, 1988; Буневич, 1989а]. В польской части Пущи показатели стадности меньше - в среднем 12, максимально - 38 особей [Krasinski, 1978]. Установлено также, что размер стад в обеих частях Пущи зависит от уровня лесистости и кормности территории: в лесных биотопах в группах обычно 7-15 голов, вблизи полей и сенокосных полян - 40-70 [Krasinski et al., 1994; Красинска и др., 1999]. Примерно такой же показатель стадности (12) зарегистрирован и в Борецкой пуще. Группы менее 20 особей составляли в этой группировке 87%, одиночных самцов регистрировали в 59% встреч [Krasinski, Krasinska, 1994].
На Кавказе, судя по историческим сведениям, в прошлые столетия в группах насчитывали не больше десятка особей (чаще 4—7), стада из 14-60 голов были редки, взрослые самцы жили обособленно [Радде, 1899; Шильдер, 1901; Динник, 1910; Филатов, 1910, 1912; Сатунин, 1914; Башкиров, 1939а]. Зубробизоны, вселенные в XX в. в Кавказский заповедник, сначала держались одним общим стадом, а когда оно увеличилось до 12 голов, сильный бык в период гона вытеснил других самцов, которые стали жить отдельно. Когда стадо увеличилось до 29 голов, из него стали уходить зубрицы старших возрастов - по две-три вместе с молодняком. Периодически все звери объединялись. Постепенно выделились несколько семейных групп с примкнувшими к ним другими животными, и группа, состоящая из взрослой самки и 6 молодых самцов. Позднее стали появляться объединения молодых разнополых особей, а также одиночки или пары взрослых. По мере расширения области обитания контакты между отдельными группами становились все реже. В первой половине 60-х годов большинство животных держались по 3-10 (29%), 11-20 (27%) и
Таблица 9. Среднее и максимальное число особей в группах зубра в XX в.
Район,заповедник Число особей в группе Источник информации
среднее | максимальное
Беловежская пуща 5-13 60 Кулагин, 1919, 1940
сс 14,6 140 Буневич, Кочко, 1988
U 12 38 Krasinski, 1978
Борецкая пуща 12 - Krasinski, Krasinska, 1994
Цуманская пуща 18-20 65 Лихотоп, Сологор, 1983; Колиснык, 1989
Кавказ 6-8 27 Динник, 1910
Кавказский з-к 15,8-9,5 124 Калугин, 1968, 1973
Тебердинский з-к Чаще 2-16 40 Бобырь, 1992
Заказник “Цейский” 3,6-7 зимой, 15.8 летом 78 Казьмин и др., 1992
21 40 (22%) голов. Одиночных зубров и пары регистрировали в 16% встреч. Стада более 40 голов были редки (5%). Они образовывались только на ограниченных пастбищах и быстро распадались. В период гона на отдельных субальпийских и альпийских лугах иногда скапливалось более сотни голов. После гона взрослые самцы покидали стада. Средний показатель стадности в момент формирования природной группировки достигал 15,8, варьируя в разные сезоны года от 14 до 17, а затем, по мере одичания животных, снизился до 9,5 [Калугин, 1968,1973]. В дальнейшем одиночные самцы и самки составляли здесь 13% населения, семьи (самка с 1-3 молодыми) - 6, маточные стада — 28, непрочные группы самцов — 10, смешанные стада - 36 и скопления - 7%. Однополые группы взрослых животных включали от 4 до 12 особей, стада самок с молодняком - 13-35 [Немцев, 19806, 1990, 2003].
В Тебердинском заповеднике по мере увеличения численности состав и число особей в группах постоянно изменялись. Животные большую часть года держатся группами от 2 до 16 особей, самцы-чаще отдельно от самок, а зимой они объединяются в большое стадо, насчитывающее, порой, до 40 голов [Бобырь, 1992]. Примерно такая же величина групп зверей (5-15, максимально 30) в заказнике “Цейский” в Северной Осетии [Липкович, 1985].
Стада зубров в Беловежской пуще, Кавказском и Приокско-Террасном заповедниках и других районах начинали делиться, если число особей в них приближалось к двум десяткам. Показатель стадности в 12-15 особей, видимо, является видоспецифичным и оптимальным. Однако размеры групп, как и вся социальная организация, существенно изменяются в гон, во время отела, в экстремальные периоды жизни, а также при содержании животных в неволе и на местах зимней подкормки в природе.
Социальные отношения в группировках зубра основаны на линейной иерархии [Калугин, 1968; Дерягина, 1972; Заблоцкий, 19756; Немцев, 19806, 2003; Жесткова, 1988; Перерва, 1992]. В смешанных стадах и в скоплениях чаще доминируют сильные самцы, в маточных группах - крупные (опытные) самки старшего возраста. Следующие ступени в иерархической лестнице распределяются в соответствии с возрастом и размерами - чем мельче зверь, тем ниже его ранг. Эта особенность характерна даже для детенышей. Самка-доминант во многом определяет ритм активности всех членов группы, занимает лучшие места на пастбище, у кормушек и на отдыхе, лидирует при переходах с одного места на другое, у нее чаще первой наступает течка. Для поддержания иерархического положения доминанту обычно достаточно демонстраций - раскачивания головой и бросков, а также вспрыгиваний на подчиненную особь, которая проявляет покорность и убегает или начинает сражение, если хочет повысить свой социальный статус. Лидером, однако, далеко не всегда бывают самые сильные, крупные или агрессивные особи, занимающие высшие ступени социальной иерархии.
При установлении социального, ранга между зубрицами происходят ожесточенные схватки, во время которых члены стада возбуждаются и чаще конфликтуют между собой, что приводит к повышению травматизма и смертности. Наиболее конфликтные особи - стареющие особи, теряющие свой ранг. При объединении двух групп между вожаками всегда возникают стычки. При вхождении в стадо новой самки происходит своеобразный ритуал знакомства, который, как правило, завершается взаимными угрозами, дракой с субдоминантам и оттеснением пришельца на последнее
место в иерархии или изгнанием. Индивидуальные дистанции в самочьих группах относительно постоянны - 4,5-5 м. К взрослым самцам самки не агрессивны. В стаде самец нередко выступает в роли “миротворца” - хрюкнув и подняв хвост, он вклинивается между дерущимися самками, и разъединяет их. Поднятый хвост у зубра служит своеобразным индикатором возбуждения.
Стычки взрослых самцов между собой учащаются в период гона. Они проходят с демонстрациями угроз, противостоянием, проявлениями элементов маркировочного поведения, но не всегда заканчиваются непосредственными столкновениями. Иногда соперники ограничиваются демонстрациями и яростным боданием предметов, после чего слабейший уходит. Бои между равными по силам быками яростные, с фронтальным столкновением лбами, остервенелым толканием друг друга и боковыми ударами острыми и незагнутыми, как у зубриц, рогами. От серьезных ранений их, в какой-то мере, предохраняют разросшиеся кости черепа и толстая плотная кожа на голове и передней части туловища, но удары в бок и брюхо противника бывают смертельными. Иногда несколько молодых самцов, изгнанных доминантом, объединяются и нападают на него, забивая до смерти.
Зубр на воле и в вольерах обычно не боится человека. Зимой звери нередко выходят к жилым поселкам, дорогам, фермам, стогам сена и сеновалам. При охотничьем преследовании они становятся осторожными, и после испуга проходят без остановок несколько километров.
Структура группировок и социальные отношения у зубров, бизонов и их гибридов, за исключением отдельных специфических особенностей, в целом сходны. Самцы бизонов, в отличие от зубра, в неволе чаще держатся вместе с самками, хотя в природе - отдельно. При совместном содержании зубров, бизонов и их гибридов, в группах доминируют бизоны и зубробизоны, а самый низкий ранг у тригибридов (зубробизонокоров) [Дерягина, 1972; Robitaille, Prescott, 1985; Rutberg, 1983, 1986; Перерва, 1989в, 1992].
Пространственная структура и перемещения. Зубр - оседлое животное, весьма привязанное к району рождения или выпуска. Даже в переуплотненных группировках не происходит широкого освоения соседних угодий и образования новых очагов обитания за счет эмиграции. За полвека работ по реакклиматизации исключением стало возникновение кнышынского очага в Польше [Пуцек и др., 1996] и соединение областей обитания буковинского и зубровицкого стад на Украине [Перерва и др., 1991].
Часть взрослых самцов и молодые особи, тем не менее, весьма подвижны и склонны к эмиграции, что в целом полезно для вида в связи с повышением вероятности “дрейфа” генов в неродственные стада. В Кавказском заповеднике отдельные эмигранты уходили на 90 км от его границ, в Хоперском заповеднике - на 160 км, в Крыму - на 100 км от места выпуска в степные районы, на Волыни в Цуманской пуще -на 40 км [Колиснык, 1989; Павлов, 1999; Немцев и др., 2003]. Зарегистрированы случаи перемещения отдельных особей на 500-600 км [Заблоцкий, 19756].
Беловежский зубр столетие назад был оседлым, но часть самцов мигрировала за пределы Пущи [Карцов, 1903; Кулагин, 1919]. Позднее звери тоже, в основном, держались в ее пределах. Взрослые самцы и группы самок с молодняком в бесснежный период занимали относительно небольшие для крупного зверя участки обитания, которые существенно перекрывались и были близки по размерам - 70 (29-152) и 69 (45-97) км2 соответственно, у молодых быков - 44 км2. Зимой
площадь участков самцов и самок сокращалась до 11 и 8 км2 [Krasinska, Krasinski, 1995; Буневич, 1999а; Красинска и др., 1999; Krasinska et al., 2000, 2004]. С 1965 г. в восточной части Пущи участились выходы одиночных взрослых самцов за пределы заповедника, но они носили кратковременный характер - 1-2 мес. в период гона. К 1975 г. число таких животных, среди которых были и молодые самцы, возросло до 20, причем они стали уходить группами по 2-5 особей, и жили на расстоянии 2-10 км от заповедника сравнительно долго - от 2-3 мес. до 1-2 лет. Отдельные звери эмигрировали на десятки, а иногда - на сотни километров, преимущественно по одним и тем же путям, к границе с Литвой. В 1968 г. взрослый самец, прошедший за осенние месяцы 350 км, был отловлен на территории Литвы. Наиболее вероятные причины ухода самцов из стад - вытеснение более сильными особями, ухудшение кормовой базы из-за перенаселенности угодий копытными и проявление древних миграционных инстинктов. В отличие от чистокровных беловежских зубров особи беловежско-кавказской линии не выходили за пределы границ Пущи [Корочкина, 1971 в, 1973а; Корочкина, Вакула, 1976; Корочкина, Кочко, 1981]. В польской части Пущи выходы отдельных взрослых самцов за пределы района зимовки группировки зарегистрировали в 1963 г., а в 80-е годы доля таких животных возросла до 67%. Расселение зверей из заповедника было вызвано истощением кормовых ресурсов и пересыханием водоемов [Krasinska, Krasinski, 1994].
На Кавказе в середине XIX в., судя по сохранившимся историческим сведениям, у зубра были четко выражены сезонные миграции. Летом звери держались в горах, а на зимовку спускались в предгорья вплоть до Псебея, станиц Новосвободной, Преградной и Сторожевой на севере и почти до черноморского побережья на западе. Основная причина осенних откочевок - высокие снега в горах. Тем не менее значительная часть популяции оставалась зимой в обширных долинах на Главном хребте и боковых отрогах. По мере колонизации и хозяйственного освоения региона сезонные миграции стали затухать [Башкиров, 1939а].
В Кавказском заповеднике пространственная структура группировки и поведение зубробизонов менялось и усложнялось по мере роста численности и одичания животных. На начальном этапе вольного разведения звери после принудительного выгона из загонов на высокогорные пастбища стремились вернуться в парк. Потребовались десятилетия, чтобы они освоили площадь в 520 км2. Их расселение шло по долинам рек, пологим склонам и по субальпийским и альпийским лугам вдоль верхней границы леса. Постепенно стали формироваться высотные миграции в диапазоне 400-3000 м над уровнем моря. Зимой зубры держались в широколиственном лесу, весной и в начале лета поднимались на субальпийские и альпийские луга, следуя за отступающей границей снега и ковром зеленой травы и спасаясь от кровососущих насекомых. В сентябре - октябре, по мере увядания травянистой растительности, они откочевывали вниз в лесную зону. Ежедневные переходы обычно не превышали 6-7 км. Протяженность сезонных путей миграции, проходящих по долинам рек и ручьев и пологим склонам, - до 50 км. В период гона отдельные самцы уходили за 25-30 км от своих сезонных участков обитания, но возвращались к местам зимовок. В районах обитания животных со временем появились хорошо набитые тропы к солонцам, переходам через горные реки, хребты и ущелья, а также “каталки” (площадки на склонах, с которых лежащие на
боку звери сползают вниз - фото 7) и “чесала” (стволы деревьев и выступы каменных глыб, о которые они регулярно чешутся) [Калугин, 1968, 1970].
Миграционные традиции зубробизона в Кавказском заповеднике сохранились по сей день. Основная причина сезонных переходов за пределы заповедника в предгорья - снежный покров. Сигналом к началу осенних перемещений служит увеличение его высоты до 30-^10 см. Северную границу заповедника в октябре - декабре первыми пересекают самки с телятами, самцы уходят с гор в январе. Скорость перемещения невелика - от 0,7 до 10 км в сутки. При небольшой снежности характерны кратковременные откочевки назад с образованием встречных потоков. Весенние кочевки вверх более кратковременные. Размах сезонных миграций не превышает 30-40 км. В ключевые периоды жизни (во время гона, лактации и зимовок) звери оседлы. Размеры летнего участка обитания у семьи близки к 300 га. У самцов они намного больше, у некоторых особей - до 1600 га. Зимой их площадь уменьшается в 2-3 раза, а в глубокоснежье стада по нескольку дней концентрируются на участках, не превышающих сотни гектаров. Суточный ход в этот период сокращается порой до 150-300 м, т. е. животные кормятся практически на одном месте, что приводит к сильному повреждению древостоя [Немцев, 1990, 1994, 2003; Немцев и др., 2003].
В Тебердинском заповеднике стадо более 20 лет держалось на одном участке горной долины р. Кизгич и прилегающих урочищах, занимая площадь около 20 тыс. га. За пределы этой зоны изредка выходили (на 15-30 км) быки-одиночки, группы взрослых самцов и небольшие группы самок, но через несколько дней или месяцев они возвращались назад. Максимальные дистанции эмиграции взрослых самцов -70 и 150 км. Диапазон высотных перемещений животных - от 1200 до 2800 м. При переходах они легко преодолевали бурные реки и участки с высоким (до 1-1,5 м) снежным покровом, следуя друг за другом [Тарасов, 1975, 1980; Бобырь, 1992]. В Цейском заказнике в Северной Осетии зимой средняя протяженность суточного хода зубров находилась в пределах 1,9-1,2 км, сокращаясь почти на 30% при повышении высоты снежного покрова [Казьмин, Смирнов, 1992].
Существующие тропы, лежки, каталки и чесала - это своеобразные ориентиры и ольфакторно-оптические метки, сигнализирующие другим особям о занятости участка. Пришельцы обычно обнюхивают предметы, на которых остается запах животных, и чешутся о них головой (чаще лбом и окологлазничной областью), шеей, грудью и даже животом, оставляя свою метку. Хозяева территорий постоянно подновляют метки, особенно на границах участков, предпочитая маркировать хвойные деревья, запах которых, возможно, возбуждает их. Возбужденные звери нередко бодают деревья и кусты, взрыхляют почву рогами и как бы разметают ее бородой. Возле чесал они уринируют, а затем разбрасывают землю передними копытами, пятясь назад. Со временем у таких меток-чесал появляются ямы до 20 см глубиной и до 4 м в диаметре, в которые звери уринируют и затем валяются в образовавшейся грязи. Выйдя из “купалок”, зубры счесывают грязь о деревья, оставляя на них специфический запах. Сходный способ маркировки участков обитания характерен отчасти для кабана и благородного оленя [Данилкин, 1999, 2002]. Молодые самцы, вытесняемые взрослыми особями из стад, стремятся занять и маркировать свободные участки, на которые со временем приходят смешанные группы молодых животных или отпочковавшиеся от стад взрослые самки с потомством
[Перерва, 1988, 1992]. Такое поведение самцов в сочетании с оседлостью самок позволяет говорить о зубре как о территориальном животном, несмотря на то, что защиты семейного или стадного участка у них нет. Обычно “неприкосновенной” становится лишь индивидуальная дистанция - около 4-5 м, при нарушении которой следует угроза [Немцев, 19806]. Другие функции катания в грязи и снегу и почесывания о деревья - избавление от кожных кровососущих насекомых и, как считал Н.М. Кулагин [1919], своеобразный массаж кожи.
В Окском заповеднике проведено исследование размещения лежек зубра. Установлено, что они устраивают их обычно (43-73%) на возвышенных продуваемых ветром местах, преимущественно в сосняках и дубняках вблизи от мест кормежки. В светлое время суток животные ложатся от 1 до 27 раз, чаще летом (12-27 раз) и реже - зимой (1-6 раз). Летом они с большой охотой “купаются” в песке (10-23 раза в день), что, скорее всего, связано с линькой и необходимостью избавления от кровососущих насекомых. Расстояние между отдельными лежащими животными - 0,5-3 м. Иногда они лежат “цепочкой”, но чаще стадо разбивается на 2-3 группы, располагающиеся на отдых в 12-50 м одна от другой. Телята ложатся рядом с матерью или образуют “детский сад”. Поскольку одни и те же места отдыха используются многократно, со временем образуются большие участки без растительности площадью до 655 м2 [Киселева, 1978в].
Существующую пространственную структуру и некоторые элементы поведения зверей, видимо, пока еще нельзя считать видоспецифичными “зубровыми” не только в гибридных группировках, но и в чистокровных стадах, подвергшихся одомашниванию.
Американский степной бизон, в отличие от зубра, типичный сезонный мигрант, однако в районах, где сезонные условия существования относительно однообразны, он оседлый, хотя участки обитания у стад сравнительно большие - около 56 км2 в среднем [Lott, Minta, 1983]. У лесного бизона размеры участков еще больше - у самцов в среднем 433, у самок - 335 км2 [barter, Gates, 1990]. В национальном парке Вуд-Буффало установлено, что перемещение животных на значительное расстояние может быть связано с хищничеством волка. В одном случае после нападения хищников стадо за сутки переместилось на 86 км [Carbyn, 1997].
Численность
Численность в XIX - начале XX вв. Беловежский зубр сохранился в природе до XX в., вне всякого сомнения, лишь благодаря строгой охране, введенной в Пуще в 1538 г. королем польским Сигизмундом I и поддерживаемой позднее знатью в целях успешной охоты. Известно, что 27 сентября 1752 г. король польский Август III со свитой убили 42 зверя. В 1798 г. Беловежская пуща отходит к России. В 1802 г. Александр I издает указ “О сохранении зубров”, и с этого времени ведется их учет (табл. 10), показывающий неуклонный рост поголовья. В 1810 г. здесь жили 310 особей, в 1840 г. — 817, в 1857 г. - 1898. Затем заметно снижение численности, что отчасти связано с охотой. В 1860 г. во время царской “потехи” Александра II в Беловежской пуще за два дня (6 и 7 октября) убито 28 зубров и десятки других видов копытных, а перед этим много зверей погибло при загоне в зверинец. В 1860-1888 гг. 34 зверя застрелены для пополнения зоологических
Таблица 10. Динамика численности зубра в Беловежской пуще*
Год Особей Год Особей Год Особей Год Особей Год Особей Год Особей
1809 350 1838 906 1868 559 1893 494 1953 7 1977 102
1810 310 1839 932 1869 541 1894 496 1954 13 1978 114
1813 363 1840 817 1870 542 1895 561 1955 16 1979 132
1816 483 1841 946 1871 528 1896 561 1956 19 1980 143
1817 497 1843 984 1872 528 1897 637 1957 27 1981 159
1818 546 1844 993 1873 527 1898 662 1958 30 1982 156
1819 504 1845 1025 1874 536 1899 661 1959 28 1983 159
1820 528 1846 1095 1875 558 1900 727 1960 30 1984 196
1821 508 1848 1264 1876 561 1901 747 1961 28 1985 206
1822 477 1849 1354 1877 559 1902 665 1962 33 1986 204
1823 515 1850 1560 1878 565 1903 703 1963 37 1987 226
1824 572 1851 1642 1879 571 1904 708 1964 22 1988 242
1826 653 1852 1748 1880 579 1905 651 1965 28 1989 255
1827 690 1853 1802 1881 574 1906 653 1966 49 1990 315
1828 711 1854 1824 1882 600 1907 741 1967 51 1991 315
1829 711 1855 1824 1883 592 1908 696 1968 58 1992 295
1830 772 1856 1771 1884 384 1910 600 1969 60 1993 308
1831 657 1857 1898 1885 433 1912 700 1970 63 1994 290
1832 770 1858 1434 1886 427 1914 727 1971 63 1995 280
1833 768 1860 1575 1887 438 1916 216 1972 66 1996 270
1834 810 1861 1447 1889 380 1917 167 1973 77 1997 260
1835 845 1862 1251 1890 403 1918 68 1974 82 1998 232
1836 858 1863 874 1891 479 1921 - 1975 87 1999 238
1837 860 1865 724 1892 491 1952 — 1976 90
* По: [Карцов, 1903; Кутепов, 1911; Кулагин, 1919, 1940; Wroblewski, 1927; Северцов, 19406; Корочкина, 1958, 1961, 1971в, 1973а, 1975; Шостак, 1978, 19896; Корочкина, Кочко, 1983; Буневич, Кочко, 1988; Буневич, 1997а, 1999а,в; Шостак, Шохалсвич, 1997; Козло, 2001]. Данные разных авторов могут не совпадать. Со второй половины XX в. численность только в восточной части Беловежской пущи.
коллекций. После очередного польского восстания, вспыхнувшего в 1863 г., и последующего массового браконьерства в Пуще недосчитались 377 голов. В последние десятилетия XIX - начале XX вв. численность зубра долгое время находилась в пределах 400-700 особей (табл. 10), что было следствием возросшей пищевой конкуренции с другими видами диких копытных. В конце 60-х - начале 70-х годов XIX в. в Пущу завезли благородных оленей (полностью уничтоженных в XVIII в.), которые быстро размножились (до 5-7 тыс. в 1905-1914 гг.) и вместе с косулями и ланями почти уничтожили лиственный подрост. В отсутствии полноценных древесных кормов звери нередко гибли от воспаления кишечного тракта и чаше заражались паразитами. Часть их умерла от заразных болезней, занесенных крупным рогатым скотом, прогоняемым через Пущу. Отстрел копытных не прекращался и в начале XX в. В 1900 г. на одной из царских охот убито 690 животных, в том числе 45 зубров, но большая часть их поголовья истреблена в 1914-1915 гг., в военное время, 15 зверей вывезли в Восточную Пруссию. В 1916 г. в Пуще оставалось 216 особей, в 1917 г. - 167, в 1918 г. - 68, к концу 1920 г. - одна самка, которая была убита 9 февраля 1921 г. бывшим лесником Шпаковичем. По другим данным, зубр был уничтожен полностью уже в 1919 г. К этому времени полностью истребили ланей, а счет оленям, косулям и каба
нам шел на единицы [Карцов, 1903; Кутепов, 1911; Врублевский, 1912; Север-цов, 1933, 1940а,б; Башкиров, 1939а, 1940; Кулагин, 1940; Заблоцкий, 1948; Корочкина, 1958; Кестер, Шостак, 1968; Перерва, 1992; Буневич, 1999а].
На Кавказе в 60-70-х годах XIX в. учитывали до 2 тыс. зубров. Из-за неумеренного истребления к концу первого десятилетия XX в. в междуречье Белой и Лабы сохранилось около 300-600 особей, да и то лишь благодаря организации в 1888 г. Кубанской великокняжеской охоты. Несмотря на запрет охоты, браконьерство процветало. Отдельные охотники убивали за свою жизнь до 20 зверей, а “знаменитые” - до 80. В годы революции и гражданской войны 1917-1920 гг. началось безудержное истребление копытных местным населением, почти поголовно вооруженным трехлинейками. Часть животных, вероятно, погибла от эпизоотии, занесенной в горы домашним скотом в 1919 г. К 1920 г. осталось около 50-100 особей, к 1924 г. - не более 10-20. Уцелевшие зубры были полностью уничтожены браконьерами в 1925-1927 гг. уже на территории Кавказского государственного заповедника, учрежденного 12 мая 1924 г. По сообщению С.А. Северцова [1926], “в архиве отдела охраны природы Главнауки есть данные, что один охотник, за короткое время, просто из спортивного интереса убил 18 зубров. При этом даже мясо и шкура не были использованы”. Последний свежий след видели в бассейне Б. Лабы на Псыше осенью 1927 г. [Виноградов, 1870; Радде, 1899; Динник, 1910; Филатов, 1910,1912; Воронцов-Вельяминов, 1929; Башкиров, 1939а; Верещагин, 1947,1959а; Крайнова, 1947; Заблоцкий, 1948; Калугин, 1968, 1977].
В 1927 г. нигде в мире не осталось зубров, живущих в природе. Этот зверь, как и тур, мог навсегда исчезнуть, если бы его не содержали в зверинцах, зоопарках и охотничьих парках мира.
В 1914 г. в неволе и полувольных условиях жило до 300 особей. Наиболее крупное стадо сохранялось в одном из охотничьих парков князя Плесе в Верхней Силезии. После очередного завоза в 1915 г. 22 особей из Беловежской пущи, здесь весной 1918 г. обитали 74 зверя. В 1919 г. после передела государственных границ этот парк перешел к Польше и стал называться Пшинским. К 1921 г. браконьеры истребили почти всех зубров, за исключением двух быков и одной самки, от которой постепенно развели новое стадо в 16 голов. До революции стадо беловежских зубров содержали в Гатчине под Петроградом (36 особей в 1917 г.), в охотничьем парке “Пилявин” на Западной Украине (22 зубра и 9 бизонов). В 1913 г. 1 беловежского быка и 3 зубрицы вселили в Крым. В 1917 г. стадо выросло до 9 голов. Практически все эти животные позднее погибли во время военных действий или от браконьеров [Фортунатов, 1922; Житков, 1934; Башкиров, 19396; Северцов, 1940а; Соколов, 1959; Заблоцкий, 1960; Гептнер и др., 1961; Перерва, 1992].
Восстановление поголовья. В 1923 г. в Париже на Международном конгрессе деятелей охраны природы основано Международное общество сохранения зубра, впоследствии внесшее важный вклад в создание питомников в разных странах, инвентаризацию поголовья и организацию племенной работы [Заблоцкий, 1957а, 1960].
В 1927 г. в зоопарках и зверинцах мира числилось 48 (52) чистокровных зубров. В 1938 г. в двух десятках питомников насчитывали 97 голов, в том числе в Германии - 35, Польше - 30, Швейцарии и Англии - по 9, Голландии - 8, Швеции - 2, Венгрии, Австрии и СССР - по 1. К 1944 г. поголовье увеличилось до 146 особей, но в 1945 г. во время военных действий сократилось до 84, а число питомников уменьшилось до
семи. В СССР до 1941 г. жили 126 зубров, бизонов и их гибридов, в 1946 г. после войны сохранилось всего 29 особей: 1 зубр, 8 бизонов, 13 зубробизонов и 7 сложных гибридов [Фортунатов, 1929; Заблоцкий, 1938, 1939, 1957а,б, 19656, 1971, 1975а; Башкиров, 1940; Размахнин, 1977а].
Основная работа по восстановлению поголовья зубра выполнена Польшей и СССР. Подробные обзоры реакклиматизации вида в республиках СССР в разное время сделаны П.Б. Юргенсоном [1956], М.А. Заблоцким [1960] и М.П. Павловым [1999], поэтому в настоящей монографии я освещаю лишь основные результаты этой деятельности с необходимыми уточнениями и дополнениями.
Особый вклад сделан Беловежской пущей. В 1929 г. Польша закупила 4 (2) зверей в Германии у Гагенбека, позже пару в Швеции, 1 - в княжестве Плесе в Силезии и 3 - в питомнике графа Арнима в Бойтценбурге. В 1929-1930 гг. в Пущу завезли 7 особей из Познаньского зоосада. В 1936 г. здесь насчитывали 15, а в 1938 г. - 13 зубров и 14 зубробизоньих гибридов. Осенью 1939 г. часть территории Пущи вместе с 16 зубрами отошла к Белоруссии. В 1941 г. здесь жил 21 зверь. В войну поголовье уменьшилось на 8 особей. В 1945 г. при установлении новых государственных границ зубропарк вместе с 17 уцелевшими животными оказался на территории Польши. В советскую часть Пущи 10 зубров завезли в 1946 и 1949 гг. (по 3 самца и 2 самки). В 1951 г. в питомнике содержали 17 особей. С 1946 по 1962 гг. родились 95 зубрят, из которых сохранено 92%. В 1962 в Пуще находилось 82 зверя [Башкиров, 1939а, 1940; Заблоцкий, 1939, 1947, 1948, 1957а, 1974; Северцов, 1940а; Сержанин, 1955; Корочкина, 1958, 1963, 1970; Соколов, 1959; Гептнер и др., 1961; Кестер, Шостак, 1968; Корочкина, Кочко, 19836; Krasinski, 1994; и др.].
Вольное разведение зубра в польской части Беловежской пущи началось с 1952 г. К началу 1961 г. стадо выросло до 34, а в 1973 г. - до 253 особей. В белорусской части Пущи выпуск 7 (5) животных состоялся в 1953 г. Летом 1962 г. стадо состояло из 56 экземпляров. В 1976 г. в природе жили около 90 зверей, и 118, преимущественно беловежско-кавказской линии (114 особей), вывезли в другие районы. Оставшееся стадо пополнили 49 животными беловежской линии из Польши и Приокско-Террасного заповедника. В середине 80-х годов в местной группировке было более 200 чистокровных особей (табл. 10). Плотность населения достигла 3, а в некоторых районах - 12 голов на 1000 га [Корочкина, 1961, 1963, 1970, 1973в, 1977; Шостак, 1977, 1989а; Krasinski, 1978, 1983; Буневич, Кочко, 1988; Перерва, 1992]. В начале 90-х годов в беловежском стаде в Беларуси насчитывали 315 голов, в 1994 г. - 290, 1998 г. - 232, 1999 г. - 238, 2002 г. - 275, а на территории Польши - около 349 [Буневич, 19976, 1999а; Красинска и др., 1999; Коз-ло, 2001; ЕВРВ, 2002].
В Беларуси, помимо беловежской популяции, к концу XX в. образовано несколько мелких вольных группировок зубра в Борисовском, Воложинском, Гродненском и Осиповичском лесхозах, Полесском заповеднике, национальном парке “Припятский”, в лесоохотничьем хозяйстве “Лясковичи” с числом особей от 11 до 41 [Козло и др., 1996; Тышкевич, 20026]. В национальный парк “Припятский” 13 особей из Беловежской пущи и Приокско-Террасного заповедника завезены в 1987-1988 гг. и выпущены на волю в 1990 г. К концу 2002 г. в двух стадах было более 40 особей. В 2000 г. в левобережной части поймы создано еще одно стадо из 14 особей [Козло, Углянец,
1999; Углянец, 2003]. Всего в Беларуси в 2002 г., по данным ЕВРВ [2002], находилось 534 зверя, в Польше - 804.
Далеко не все выпуски зубра в СССР были успешными. Например, из 11 зверей (5 самцов и 6 самок), завезенных в 1974 и 1976 гг. в Березинский заповедник, 5 погибли в первые два года. К 1988 г. стадо выросло лишь до 18 особей [Лавов, Чичикина, 1980; Лавов, Воронова, 1982; Лавов, 1983; Дунин, Ставровс-кий, 1990]. Основные проблемы в этом регионе - гибель копытных от браконьеров и истощения в малокормных лесах, выход на сельскохозяйственные поля, к фермам и жилью человека. Не вполне удачными были выпуски животных в Литве, осуществлявшиеся с 1969 г. В 1990 г. здесь находились 35 особей (столько же их было в 1977 г. [Мицкус, Турса, 1978]), из них 12 оставались в вольерах. В Казахстане (в многоснежном Заилийском Алатау) все выпущенные звери погибли. В Киргизии в Сары-Челекском заповеднике, куда завезли 13 особей, численность стада за 25 лет разведения не превысила 18 голов [Янушевич, 1978; Прусаков, 1989; Павлов, 1999].
Наиболее успешным было разведение зубра на Украине. В “Аскании-Нова”, охотничьем парке “Пилявин” и в Крыму [Башкиров, 19396; Дулицкий, 1973], как отмечено выше, реинтродукция началась еще в дореволюционное время, продолжалась в предвоенные годы, но во время войны животные были практически уничтожены. В послевоенные годы в республике расселено более 160 зверей, преимущественно беловежско-кавказской линии. В 1968 г. в 4 областях обитали 53 зубра, в 1977 г. - 154 в 7 областях, в 1988 г. -476 в 10 пунктах [Болденков, Крайнев, 1980; Перерва и др., 1991]. К 1990 г. в 9 областях жили 578 зверей (284 самки, 229 самцов и 65 зубрят), в том числе 424 в Волынской, Черновицкой и Львовской. В 1992— 1994 г. на Украине насчитывали 640-660 зубров [Павлов, 1999; Kryzhanovskii, 2004]. Самая крупная группировка образована в Цуманской пуще на Волыни, куда в 1965 г. выпущены 15 беловежско-кавказских особей [Татаринов, Дякун, 1969; Лихотоп, Сологор, 1983]. В 1989 г. здесь обитали 128 зверей (с плотностью 10 голов на 1000 га в отдельных лесничествах) [Колиснык, 1989], в 1989 г. - 185 [Перерва и др., 1991], в 1992 г. - 202. К 1995 г. их число уменьшилось до 170 в результате выбраковки и гибели от болезней [Демчук и др., 1997]. В 90-е годы много животных погибло в результате браконьерства и охот иностранных туристов. К началу 2004 г. на Украине осталось около 320 особей, т. е. за десятилетие утрачено более половины поголовья [Мерзликин и др., 1999; Kryzhanovskii, 2004].
В России основными центрами расселения вида стали питомники, образованные в Приокско-Террасном и Окском заповедниках. В Центральный питомник Приокско-Террасного заповедника пару беловежских и пару беловежско-кавказских зубров завезли из польской части Беловежской пущи в конце 1948 - начале 1949 гг. Позднее к ним добавили животных из Голландии, Германии и Швеции, самца и двух самок бизона из Беловежской пущи и Голландии и гибридную “зубробизонокорову” из Алма-Атинского зоопарка. Животных разных линий здесь содержали раздельно. К 1960 г. стадо в питомнике увеличилось до 50 голов, позднее - до 60-70. В 1981-1985 гг. здесь было 6 племенных групп зубра общей численностью около 30 голов. В 2000 г. осталось 23 зверя. К 2003 г. в питомнике родилось около 480 зубрят, из них выращены и перевезены в более чем 25 пунктов 343 особи [Заблоцкий, 1960, 1983; Белоусова, 1986; Заблоцкий, Заблоцкая, 1990; Перерва, 1992; Петров, 2003; Требогано-
ва, 2003а]. В питомник Окского заповедника в 1959-1974 гг. завезли 14 беловежско-кавказских зубров из Приокско-Террасного заповедника и Беловежской пущи. На 1 января 1970 г. здесь находилось 44 зверя (16 самцов и 28 самок), в 1976 г. - 45, в 1986 г. - 31. За сорокалетний период родилось 327 зубрят, из них 196 выпущены на волю [Киселева, 1974, 1977, 19866; Цибизова, 2003]. В 1982-1984 гг. 11 беловежских зубров завезены на Алтай в экспериментальное хозяйство СО РАН (с. Черга). К 1990 г. вольерное поголовье увеличилось до 38 особей [Павлов, 1999]. Позднее здесь содержали около 20 животных.
На равнинной территории России зубров и зубробизонов выпускали в Завидовском заповедно-охотничьем хозяйстве (2 зубробизона в 1954 г.), в Скнятинс-ком охотхозяйстве (27 беловежско-кавказских из Окского заповедника), Хоперском (13 беловежских, беловежско-кавказских и гибридных в 1955-1978 гг.) и Мордовском (19 чистокровных и гибридных особей в 1956—1967 гг.) заповедниках. Эти выпуски не были результативными. В Скнятинском охотхозяйстве численность искусственно поддерживали на уровне 30-40 голов. В Хоперском заповеднике в 70-е годы, когда стадо увеличилось до 30 голов, звери стали разбредаться по степным дубравам и стравливать озимые посевы. В 1977 г. их заключили в капитальную ограду. В начале 80-х годов здесь осталось только 2 зубра, в 1984 г. не стало и их. В Мордовском заповеднике к 1978 г. полувольное стадо выросло до 28 голов, во второй половине 80-х годов гибридная группировка насчитывала около 35 особей, затем 6 зверей погибли в ледяной ловушке, другие пали от пневмонии или от рук браконьеров.
На Среднерусской возвышенности на территориях Орловской, Брянской и Калужской областей планируется создание многочисленной (до 1000 голов) популяции зубра. В национальном парке “Орловское полесье” в 1966-2001 гг. выпущены 65 особей. К 2002 г. 11 зверей погибли, но родилось 24 детеныша. К 2003 г. в 4 группировках насчитывали 78 голов [Белоусова и др., 2003; Казьмин, 2003а,б; Перерва и др., 2003]. Из 11 зверей, завезенных в заповедник “Брянский лес”, к сожалению, в живых остались только три [Цибизова, 2003]. Реализуется также программа создания группировок во Владимирской (Великоозерское охотхозяйство) и Вологодской областях [Гусаров и др., 1998а,б; Перерва, Перерва, 2003; Сипко, 2003].
Удачным, хотя и дискуссионным, оказался выпуск зубра и зубробизона на Кавказе. В Кавказский заповедник 5 асканийских зубробизонов вселили в 1940 г., а позднее в 1948-1957 гг. завезли 15 чистокровных беловежско-кавказских самцов из Беловежской пущи и Приокско-Террасного заповедника. До 1945 г. животных содержали в загонах, затем перешли к подкормочно-выпасной системе, а с 1953 г. -к вольному выпасу. К 1957 г. численность выросла до 110 голов, к 1966 г. - до 449. В зоне летнего обитания плотность достигла 8-10, а зимняя - 40-50 особей на 1000 га. В 1969 г. здесь насчитывали 630 зверей [Крайнова, 1947; Калугин, 1958, 1968, 1970; Заблоцкий, 1960]. В 70-е годы в Кавказском заповеднике и окрестностях жили 700-800 зубров, в 80-е - начале 90-х годов - около 1000-1300 особей [Равкин, 1976; Немцев, 1983, 1985, 19876, 1988а; Липкович, 1989; Дыренков и др., 1990; Перерва, 1992]. Затем из-за резко возросшего браконьерства и охоты иностранцев по научным лицензиям к 1996 г. поголовье сократилось до 450 особей, а в самом заповеднике осталось около 300-350 особей [Кудактин, 1996; Павлов, 1999].
Из Кавказского заповедника в 1959 г. 4 зубробизона переселены в Нальчикское государственное лесоохотничье хозяйство. В 1960-1967 гг. к ним добавлены 30
беловежско-кавказских особей (из Приокско-Террасного и Окского заповедников и Беловежской пущи). К концу 80-х годов поголовье выросло до 288 особей [Лип-кович, 1989; Павлов, 1999]. В конце столетия в Кабардино-Балкарии осталось около 100-150 зверей [Дзуев, 2000а].
В Северной Осетии 46 зубров беловежско-кавказской линии и 1 - беловежской (из Беловежской пущи и Приокско-Террасного заповедника) выпущены в 1964— 1968 гг. в Цейском государственном заказнике и 9 - в военно-охотничьем хозяйстве “Сунженское” в 1987 г. В Цейском заказнике к 1978 г. группировка увеличилась до 115 особей, к 1990-1991 гг. - до 270-287. Плотность населения этих копытных в местах обитания достигла предельных величин: в среднем 14—17 особей на 1000 га, в местах зимовок - до 70 особей. К 1995 г. из-за ухудшения условий существования, эмиграции животных и браконьерства в заказнике осталось 117 особей. В 1998-2002 гг. в Северной Осетии уцелело всего около 50-67 зубров [ЕВРВ, 2000, 2002], а плотность их населения уменьшилась до 3 особей на 1000 га [Варзи-ев, Курятников, 1980; Курятников, Варзиев, 1983; Липкович, 1985,1988,1989,1993; Казьмин, 1988, 1989; Казьмин и др., 1992; Казьмин, Смирнов, 1992; Вейнберг, Комаров, 1999; Павлов, 1999; Комаров, Попов, 2003; Немцев и др., 2003].
В Тебердинский заповедник 16 беловежско-кавказских зубров (по 8 особей из Приокско-Террасного и Окского заповедников) завезли в 1968 г. Две самки вскоре погибли в горах в многоснежную зиму. Первый приплод получен в 1971 г. В 1978 г. скда выпустили еще 7 особей из Окского питомника. Стадо росло медленно из-за значительной смертности и низкой плодовитости, и за 20 лет увеличилось до 57 особей. В конце XX — начале XXI вв. эта группировка, в которой осталось 45 зверей [ЕВРВ, 2002], занимала территорию Главного Кавказского хребта от водораздела рек Кизгич и София на западе до истоков р. Бугойчат и Зеленчукской обсерватории на востоке, с выходом на север до окрестностей пос. Нижний Архыз и к верховьям рек Карачайки и Пхия [Тарасов, 1973, 1980; Бобырь, 1989, 1992, 1999; Павлов, 1999].
В 1969 г. 7 беловежско-кавказских зубров из Окского заповедника переселили на территорию Чечено-Ингушетии. Вскоре два зверя сорвались с обрывов и разбились. До 1975 г. сюда ввезли еще 39 особей, из которых два погибли в результате несчастных случаев, двух задавили волки, пять перешли на территорию Северной Осетии. Часть животных, видимо, погибла в экстремальную зиму 1972/73 гг. и от браконьеров. К концу 80-х годов здесь насчитывали около 30 экземпляров [Рашкевич, 1973; Брагин, 1987; Липкович, 1988, 1989; Павлов, 1999].
Всего в середине 70-х годов на Северном Кавказе обитали около 1000-1100 особей, из них до 200.чистокровных беловежско-кавказских [Калугин, 1975, 1977; Равкин, 19756, 1976]. В 80-е годы общее поголовье увеличилось до 1,5-1,7 тыс. особей, а площадь обитания превысила 250 тыс. га [Язан, Немцев, 1985; Немцев, 1987, 1988а; Гинеев и др., 1988; Павлов, 1999; Сипко и др., 1999].
Выпуск 12 зубров в Азербайджане не дал результата. По официальной версии звери погибли от воспаления легких и отравления листьями тиса, по неофициальной- выбиты браконьерами [Павлов, 1999]. В 1961 и 1962 гг. 7 сложных гибридов и 3 чистокровных самца выпущены в Алма-Атинской области Казахстана. Эти зоо-парковские, большей частью - туберкулезные, животные не были приспособлены к жизни в горах, и вскоре пали или были отстреляны [Слудский, Афанасьев, 1964]. В 1963-1964 гг. 13 особей из Центрального зубрового питомника выпущены в Сары-
Челекском заповеднике в Киргизии. Часть из них травмировалась в горах, но впоследствии звери успешно акклиматизировались [Чичикин, Чичикина, 1965; Кузнецов, 1969; Прусаков, 1989].
Численность во второй половине XX в. В 1951 г. в Польше содержали 65 зубров, в Швеции - 23, СССР - 22, Западной Германии - 18, Голландии - 8, Дании 3, Англии - 2, Чехословакии и Швейцарии - по одному. К 1962 г. мировое поголовье вида достигло полутысячи особей: в Польше - 176, СССР — 140, ФРГ - 50, Швеции - 28, ГДР и Голландии - по 18, Дании - И, Чехословакии - 9, Венгрии и Бельгии - по 6, Румынии, Болгарии, Франции, Швейцарии и Финляндии - по 4, Австрии - 3, Англии - 2, в Китае и Италии - по одному зверю. В 1968 г. насчитывали 918 особей, в 1971 г. в 28 странах обитали 1166 чистокровных животных, а к началу 80-х годов поголовье превысило 2 тыс. Около 70% мирового поголовья было сконцентрировано в трех странах: Польше (более 500 особей), ФРГ (около 300) и СССР. На территории СССР в 1951 г. помимо 22 чистокровных зубров имелось 12 бизонов и 46 гибридов, в 1956 г. - 57, 19 и 97 особей соответственно. В 1965 г. здесь содержали 231 чистокровного зубра, в 1968г.-314,в 1973 г.-436, а гибридных особей было более 800. В 1983 г. насчитывали 783 зубра, из них 264 - беловежских и 519 - беловежско-кавказских, в том числе на воле 201 и 405 соответственно [Заблоцкий, 1956, 1957а, 1960, 1963, 1971, 1974а,б, 1981, 1983; Юргенсон, 1956; Соколов, 1959; Раз-махнин, 1977а; Puzek, 1984; Пуцек, 1987].
В конце 80-х годов мировое поголовье этих копытных, разводимых в 236 пунктах, увеличилось до 3,2 тыс. В СССР насчитывали 1345 чистокровных особей, из них 1145 свободе в 20 группировках. В Польше около 600 зверей находились в 5 районах [Krzeminski, Dobrowolski, 1981; Заблоцкая, Заблоцкий, 1986; Заблоцкая, 1990; Перерва, 1992]. К 1994 гг. мировые запасы зубра достигли 3,3-3,4 тыс.: 1470— 1698 особей содержали в неволе и около 1720-1800 находились в природе. Численность зверей беловежской линии составляла 1,1 тыс., беловежско-кавказской -2,2 тыс. [Сипко, 1995; Пуцек и др., 1996; Сипко и др., 1999]. В Международной племенной книге [ЕВРВ, 2000] учтено 2864 чистокровных особи, из них 1154 в неволе в 192 пунктах и 1710 в 33 вольных стадах. По ЕВРВ [2002] на воле числилось 1893 зубра, в вольерах - 1186, а всего - 3079 особей, не считая кавказских зубробизонов.
Динамика численности зубра и его гибридов с бизоном в России отражена в таблице 11. На 1 января 1994 г. здесь учтено 53 беловежских и 346 беловежско-кавказских зверя, из них 121 - в питомниках [Красная книга РФ. 2001]. Максимальным общее поголовье было в конце 80-х - начале 90-х годов (2,2 тыс.), затем оно сократилось в 3 раза - до 0,7 тыс. Число чистокровных особей уменьшилось в 2 раза: с 569 особей в 1992 г. до 277 в 2003 г. При этом следует учесть, что в 1996-2001 гг. из зоопарков и питомников западноевропейских стран в Россию ввезено 65 чистокровных зубров. Вместе с этими животными к началу 2002 г. здесь осталось 324 особи, из них 218 вольных и 106 в неволе [Перерва, 1997; Перерва, Перерва, 2003]. В питомнике Приокско-Террасного заповедника содержали 34 зверя, в Окском заповеднике - 24 и на Алтае в “Чегре” - 25 [ЕВРВ, 2002]. Остальные вольерные животные рассредоточены по разным зоопаркам, зоосадам и частным зверинцам по одиночке или, нередко, мелкими однополыми группами, и практически не участвуют в размножении.
Таблица 11. Динамика численности зубра и зубробизона (особей) в России*
Год В России В том числе на Северном Кавказе
всего** чистокровные всего чистокровные зубробизоны
1 2 3 4 5 6
1940 7 0 7 0 7
1941 6 0 6 0 6
1942 9 0 9 0 9
1943 9 0 9 0 9
1944 10 0 10 0 10
1945 12 0 12 0 12
1946 16 0 16 0 16
1947 14 0 14 0 14
1948 18 0 18 0 18
1949 21 7 21 0 21
1950 36 8 28 0 28
1951 43 И 32 0 32
1952 62 14 48 0 48
1953 77 16 61 0 61
1954 90 18 72 0 72
1955 111 21 90 0 90
1956 139 29 ПО 0 110
1957 169 33 136 0 136
1958 210 37 173 0 173
1959 250 43 207 0 207
1960 286 50 236 0 236
1961 340 59 281 0 281
1962 396 70 326 0 326
1963 458 85 373 0 373
1964 529 101 441 13 428
1965 587 НО 496 19 477
1966 676 129 570 23 547
1967 790 142 696 48 648
1968 731 164 640 73 567
1969 785 176 686 77 609
1970 828 177 743 92 651
1971 886 197 796 107 689
1972 893 218 797 122 675
1973 924 197 834 107 727
1974 1032 239 935 142 793
1975 1099 255 1005 161 844
1976 1086 262 985 161 824
1977 1183 273 1072 162 910
1978 1232 301 1106 175 931
1979 1302 291 1194 183 1011
1980 1455 341 1339 225 1114
1981 1531 365 ' 1404 238 1166
1982 1590 360 1461 231 1230
1983 1687 366 1546 225 1321
1984 1812 403 1660 251 1409
1985 1943 446 1760 263 1497
1986 2201 494 2018 311 1707
1987 1989 521 1808 340 1468
Таблица 11 (окончание)
1 2 3 4 1 5 6
1988 2180 521 1999 340 1659
1989 1952 524 1768 340 1428
1990 1696 525 1511 340 1171
1991 1612 507 1445 340 1105
1992 1588 569 1403 384 1019
1993 1359 409 1182 232 950
1994 1218 410 1033 225 808
1995 972 417 784 229 555
1996 816 418 626 228 398
1997 700 303 520 123 397
1998 667 303 487 123 364
1999 696 306 513 123 390
2000 718 308 533 123 410
2001 758 319 551 112 439
2002 786 298 567 79 488
2003 757 277 525 45 480
* По сведениям из Международных родословных книг зубров, литературным данным (см. выше), сведениям заповедников, Службы учета охотничьих ресурсов и материалам Т.П. Сипко (личное сообщение). Отдельные данные из разных источников могут не совпадать.
** Включая кавказских зубробизонов.
Особенно сильно пострадали группировки зубра и зубробизона на Кавказе (табл. 11), где численность уменьшилась почти в 4 раза (с 2 до 0,5 тыс.), причем в “горной” линии - в 3 раза (с 1,5 до 0,4-0,5 тыс.), в чистокровной - в 8 раз (с 384 до 45 особей). Группировка Кавказского биосферного заповедника и окрестностей сократилась с 1400 до 480 (по сведениям А.С. Немцева и др. [2003] - до 380) голов, Цейского государственного заказника и Северо-Осетинского заповедника - с 287 (1991 г.) до 50-70 особей, Нальчикского государственного охотничьего хозяйства-с 380 до 8 голов [Сипко и др., 1999; Т.П. Сипко (личное сообщение)]. В Адыгее в 1996 г. в охотничьих хозяйствах обитали 58 зубробизонов, зимой 2002 г. их по результатам учета не зарегистрировано [Шебзухова, 2003]. Стада Ассинского заказника, Сунженского охотничьего хозяйства и Исмаиллинского заповедника уничтожены полностью [Павлов, 1999].
Основная причина столь резкого изменения численности зубра, как и других видов копытных, в России - непомерный уровень браконьерства при слабой государственной охране в сочетании с другими негативными факторами.
Основные факторы, определяющие динамику группировок
Воспроизводство, прирост. Ежегодный прирост в группировке зубров Беловежской пущи в период с 1852 по 1909 гг. составлял 6-24% (в среднем 11-15%), а в последние десятилетия этого отрезка времени - 2,7-4,4%. Примерно таким же он был в питомниках и зоосадах в 1921-1951 гг. [Кулагин, 1919; Северцов, 19406; Заблоцкий, 1957а; Корочкина, Кочко, 1982]. В западной части Пущи в 1965-1973 гг. прирост стада был на уровне 13%, затем уменьшился до 4%. В восточном стаде показатель плодовитости в 40-60-е годы достигал 21-24%, а в 1971-1996 гг. вос
производство заметно снизилось: приплод - до 14,7% (6-27% в разные годы), а среднегодовой прирост - до 7%, что в значительной мере связано с ухудшением состояния естественной кормовой базы [Корочкина, 1963, 1970, 1971 в, 1975; Krasinski, 1978; Корочкина, Кочко, 1982, 1983а; Буневич, Кочко, 1988; Буневич, 1997а, 1999а,в].
В группировке Кавказского заповедника в 40-70-е годы XX в. ежегодный приплод составлял около 18%, отход сеголеток - 10-15, отход взрослых особей - 4, ежегодный прирост - 10-15%. До 1965 г. сюда ввезено 20 особей, в приплоде 631 зубренок, отстреляно 43, вывезено 20, погибло по разным причинам 139 зверей, из них 66 пали в детском возрасте. При загонном содержании здесь ежегодно погибала третья часть всех детенышей, на воле - 7,7%. Основные причины смертности: инфекции и инвазии (18%), убиты взрослыми животными (12), умерли от ушибов и ран и утонули (12), родились мертвыми (7,5), задавлены волком (3%) [Калугин, 1968, 1973, 1977; Немцев, 1982а]. В 1950-1982 гг. группировка увеличивалась ежегодно в среднем на 6,9%. К 1983 г. прирост в многочисленной популяции снизился до 4,6%, плодовитость самок уменьшились почти в 2 раза. В отдельные годы этого десятилетия группировка не только не росла, но даже сокращалась. В суровые зимы смертность неполовозрелых особей достигала 80%, а яловость самок после таких зим составляла 60-70%. В целом годовой прирост в этот период снизился до 0,8%. В 90-е годы популяция сокращалась ежегодно в среднем на 6% преимущественно из-за резко возросшего браконьерства [Немцев, 1985,1987а,б, 1988а; Немцев и др., 2003]. В Тебердинском заповеднике в 80-е годы прирост группировки колебался в пределах 7,5-15,2%, в 90-е- был на уровне 7% [Бобырь, 1989, 1992, 1999], а в Цейском заказнике в период с 1969 по 1987 гг. составлял 9-11%, что втрое выше, чем в чечено-ингушском стаде [Брагин, 1987; Казьмин, 1989].
Максимальным уровень плодовитости стада зубров, видимо, был в Окском заповеднике в 1960-1976 гг. - 29,5%, но здесь достаточно велик и ежегодный отход -18,7%. В 80-е годы прирост уменьшился до 9% [Киселева, 1974, 1977, 1978а, 19866]. Высоким уровнем воспроизводства отличалось и стадо национального парка “Припятский”: приплод - 24%, смертность - 7, убыль - 10, прирост - 14% [Углянец, 19996, 2003].
Уровень смертности. Уровень смертности неодинаков в разных возрастных группах. Отход среди сеголеток во многих стадах, судя по литературным источникам, колеблется от 7 до 20%, и существенно зависит от времени рождения телят. В Окском заповеднике из числа детенышей, родившихся весной, погибали 2%, а из осеннего приплода - 10% [Киселева, 1986а]. В питомнике Приокско-Террасного заповедника доля мертворожденных и умерших в первые дни детенышей достигала 42% в общей гибели [Перерва, 1981]. По исследованиям М.А. Заблоцкого [1957а], средний процент отхода среди годовалых особей составляет 15,8, среди 2-8-летних животных - 9,5, в группе от 9 до 15 лет - 11,9, 16-21 года - 22,8, 22-25 лет - 19,5, 26-27 лет - 83,3. Близкие цифры приводит и Т.П. Сипко [2003], считающий уровень смертности зубра весьма высоким во всех возрастных классах из-за инбредной депрессии. У самцов он все же выше, нежели у самок, из-за большей подвижности и склонности к поединкам [Корочкина, 1975; Корочкина, Кочко, 1982, 1983а].
В целом в питомниках и в вольных стадах зубра в СССР среднегодовая смертность детенышей в первое десятилетие разведения не превышала 17%, среднего-
Таблица 12. Изменение уровня смертности молодых особей и интервала между отелами самок у зубров беловежской (б) и беловежско-кавказской (бк) линий в Приокско-Террасном (ПТ) и Окском (О) питомниках (по [Сипко, 2003])
Годы рождения Особей, доживших до репродуктивного возраста Интервал между отелами
ПТ (б) ПТ (бк) О (бк) ПТ (б) ПТ (бк) О(бк)
п | % п | % П 1 % п | мес. п | мес. п | мес.
1950-1965 56 60.7 75 80,0 28 89,3 85 16,4 114 14,5 141 14,5
1966-1975 51 72,5 58 82,8 120 80,8 40 18,0 51 16,0 56 17.1
1976-1985 55 63,6 68 66,2 93 71,0 47 20,0 31 18,8 58 20,7
1986-1996 45 53,3 47 48,9 60 55,0 - —
довой прирост составлял около 15, а в лучшие годы достигал 22-25% [Заблоцкий, 1957а,б]. В последние десятилетия и в питомниках (табл. 12), и в вольных стадах и в России, и по всему миру очевидна тенденция к увеличению детской смертности, интервала между отелами у самок и к снижению темпов воспроизводства. Среднегодовой прирост мирового стада уменьшился с 11% (в период с 1950 по 1970 гг.) до 9%, что связано с ростом инбридинга, обеднением кормовой базы, болезнями и старением племенных стад [Raczynsky, 1980; Puzek, 1984, 1985, 1987; Киселева, 19866; Перерва, 1986; Сипко, Раутиан, 1997а; Сипко, 2002].
Погода и корма. Смертность зубра (преимущественно сеголеток) от истощения проявляется в основном в Беловежской пуще, где составляет около 6% от числа павших. Влияние погоды и недостатка кормов в большей мере отражается на уровне плодовитости группировки, которая в экстремальные годы снижается в несколько раз [Буневич, Кочко, 1988]. В борисовском стаде близ границ Березинского заповедника по этой причине пали 5 молодых зверей [Дунин, Ставров-ский, 1990].
Массовую гибель зубра от истощения в многоснежные зимы не раз регистрировали и на Кавказе [Динник, 1910; Филатов, 1910; Башкиров, 1939а]. В Кавказском заповеднике зимой 1967/68 гг. погибло свыше 120 особей, в основном молодых, или 24% популяции. В 1971/72 и 1975/76 гг. отход был меньше - около 12% [Немцев, 1985; Немцев и др., 2003]. В Тебердинском заповеднике зарегистрирована гибель 7 зверей [Бобырь, 1992], в Цейском заказнике в Северной Осетии многоснежной зимой 1972/73 гг. пали 10 особей [Павлов, 1999]. Снижение численности отмечено и после зим 1967/68, 1971/72, 1975/76, 1986/87 и 1991-1993 гг. [Немцев, 1988а; Липкович, 1993; Перерва, 2001].
Хищники. Зубр и бизон - крупные животные, вооруженные рогами и имеющие отработанную веками тактику отражения атак хищников и защиты своих телят, при опасности занимающих центральное положение в стаде. Из хищников лишь волк и медведь нападают на молодых животных, и иногда их жертвами становятся взрослые больные и ослабленные особи, отбившиеся от стада. В Беловежской пуще в 1846 г. один медведь “заел” за лето 5 зубров [Кутепов, 1911]. Рысь и леопард тоже охотятся на зубрят, но такие случаи крайне редки [Башкиров, 1939а; Калугин, 1968; Рашкевич, 1973; Бобырь, 1980, 1992]. И все же групповая защита детенышей у этих копытных недостаточно эффективна, и потери от хищников порой могут быть очень существенными, что видно на примере бизона [Haynes, 1984; Carbyn, Trottier, 1987].
Конкуренты. Очевидные пищевые и территориальные конкуренты зубра - европейская косуля, благородный и пятнистый (акклиматизированный в некоторых районах) олени и лось, занимающие порой одни и те же стации и потребляющие сходный набор кормов, и, в меньшей мере, кабан, который потребляет желуди и вредит лесовозобновлению [Саблина, 1955; Заблоцкая, 1957; Корочкина, 1971г; Данилкин, 1999, 2002]. На Кавказе к ним добавляются кавказские козлы и серна, а вне заповедных территорий - домашний скот.
Наиболее остро конкуренция разных видов копытных за пищу проявилась в Беловежской путце, где при общей высокой или чрезмерной плотности их населения очевидна деградация естественной кормовой базы. Это привело к ухудшению состояния и качества группировки зубра и других видов, измельчанию животных, снижению плодовитости, увеличению смертности молодняка и эмиграциям на сопредельные неохраняемые территории [Карцов, 1903; Врублевский, 1912; Кулагин, 1919; Цалкин, 19516; Корочкина, 1971в,г, 1973, 1974, 1977]. На Кавказе при изобилии кормов конкурентные отношения между копытными выражены слабо, да и то лишь в зимний период [Башкиров, 1939а; Калугин, 1968; Немцев и др., 2003].
Болезни, паразиты, травмы. Практически все зарегистрированные болезни зубра (ящур, пастереллез, сибирская язва, бруцеллез, туберкулез, эмфизематозный карбункулез, паратиф, пневмония, столбняк, бешенство, актиномикоз и др.) типичны для многих других видов диких и домашних копытных, от которых он чаще всего и заражается. Судя по литературным сведениям, наиболее губительными для вида были эпизоотии ящура, пастереллеза и сибирской язвы. Именно эти эпизоотии унесли в конце XIX - начале XX вв. значительную часть поголовья в Беловежской пуще и на Кавказе [Северцов, 1933; Башкиров, 1939а]. Стада североамериканских бизонов чаще поражены бруцеллезом и туберкулезом, и являются одним из источников этих инфекций наряду с другими дикими и домашними животными [DeYoung, 1973; Wisent, 1973; Tessano, 1986, 1989; Tessano et al., 1990].
Основные причины гибели ныне живущих зубров, тем не менее, - травмы и неинфекционные болезни. Травмы чаще связаны с нападением взрослых животных на молодых и столкновениями одноранговых особей, особенно во время гона и на подкормочных площадках, а также ранениями за пределами охраняемых зон. Среди погибших преобладают взрослые самцы. В горных районах звери нередко срываются и падают со скал, ломают ноги при переходах через осыпи, попадают в снежные лавины. Гибель от болезней большей частью вызвана поражением желудочно-кишечного тракта, мочеполовой системы, слизистых оболочек глаз, воспаления легких и мозга. Часть зубриц гибнет при родах или рожает мертвых детенышей [Корочкина, 1970; Василюк, 1973; Калугин, 1973; Киселева, 1974, 1977; Перерва, 1981; Корочкина, Кочко, 1982,1983а; Киселева, Ибрагимов, 1986а,б; Буневич, Кочко, 1988; Бобырь, 1992]. В восточной части Беловежской пущи в 1981-1998 гг. вынужденно отстреляно 250 безнадежно больных зверей и более 118 погибли от болезней сами [Козло, Пикулик, 1999; Козло, 2001]. В Сары-Челекском заповеднике 7 особей изъято из-за травмирования конечностей [Прусаков, 1989].
Иногда болезнь или смерть наступают от отравления. На Кавказе зарегистрированы случаи гибели зубров от тимпанита из-за неумеренного потребления овса и свеклы и отравления ядовитыми травами (чемерицей белой и лютиком едким) [Калугин, 1968]. В Белоруссии взрослый самец отравился минеральными удобре
ниями [Дунин, Ставровский, 1990]. В Окском заповеднике болезни желудочно-кишечного тракта были вызваны употреблением недоброкачественного комбикорма и сена, пораженного грибком Aspergillus nlger [Киселева, Ибрагимов, 1986а].
У зубра и его гибридов зарегистрировано, по меньшей мере, 43 вида гельминтов, а в разных стадах и группировках - от 7 до 24 видов. Состав гельминтофауны в целом сходен с таковым у бизона и других представителей Bovidae и, в меньшей мере, Cervidae, обитающих на той же территории. Специфичных для зубра гельминтов не обнаружено. Зубры, переселенные в новые районы, теряют часть видов, но начинают участвовать в циркуляции местной паразитофауны, преимущественно свойственной крупному рогатому скоту [Беляева, 1959; Матевосян, 1964; Рухлядев, 1964; Гагарин, Назарова, 1966; Назарова, 1973; Киселева, 1974; Козлов, Назарова, 1979; Перерва, 1980; Москвин, Багрова, 1993; Багрова и др., 1995; Горохов и др., 1995; Кочко, 1999; Требоганова, 20036; Немцев и др., 2003].
Заболевания, вызываемые гельминтами, могут приводить к снижению продуктивности и гибели зубров, однако такие случаи единичны. Зарегистрирована смертность животных от фасциолеза, дикрацелиоза, диктиокаулеза и ценуроза мозга. Сильно зараженные гельминтами зубрята отстают в росте и развитии от сверстников. При дегельминтизации эффективны 20% тетрамизол гранулят и панакур гранулят [Корочкина, 1958; Заблоцкий, 1960; Татаринов, Дякун, 1969; Назарова, Киселева, 1980; Киселева, 1990; Киселева, Цибизова, 2003].
Из эктопаразитов на зубре обычны иксодовые клещи, слепни, мошки, мокрецы, мухи-жигалки и другие кровососы. На Кавказе у него не найдено личинок кожного и носоглоточного оводов [Калугин, 1968].
Антропогенные факторы. Антропогенная трансформация среды обитания, несомненно, сыграла отрицательную роль в жизни и зубра, и бизона, однако главной причиной их вымирания, как показано выше, было прямое истребление.
В XIX-XX вв. у охраняемого зубра, в отличие от многих других копытных, динамика численности определялась, в основном, естественными факторами (за исключением военных и революционных десятилетий начала XX в., когда вид был почти уничтожен человеком). В Беловежской пуще доля естественной смертности в 1832-1902 гг. достигала 91%, антропогенной - 8, в 1971-1980 гг. - 74 (из них болезни и истощение - 55, травмы - 45) и 7% соответственно. В Кавказском заповеднике в 1940-1965 гг. из 117 погибших животных от естественных причин пали 70% (в том числе от травм - 54, от болезней и истощения - 7, утонули - 6, от хищников - 2), от антропогеных - 8% [Филонов, 1989]. Благодаря минимизации антропогенных потерь численность вида неуклонно росла, постепенно восстанавливался и ареал.
В последние десятилетия XX в. ситуация кардинально изменилась: из-за социально-экономических преобразований в республиках бывшего СССР ослабла государственная охрана и резко усилилось браконьерство даже на особо охраняемых территориях. В Беларуси браконьерами застрелены 5 зубров в борисовском стаде [Дунин, Ставровский, 1990]. В заповеднике “Брянский лес” уничтожены почти все выпущенные звери [Цибизова, 2003]. На Кавказе браконьерство достигло беспрецедентного размаха. Несколько стад уничтожены полностью (см. выше). В особо охраняемом Кавказском биосферном заповеднике в 1989-1997 гг. найдено 40, а за его пределами - 144 убитых или погибших от ран животных. Судя по резкому па
дению численности, еще большее число убитых зверей не обнаружено. До 78% жертв браконьеров составляют самки, откочевавшие из заповедника в окрестные угодья на зимовку. С 1996 г. горный зубр отнесен к охотничьим объектам, что еще более усугубило ситуацию [Кудактин, 1996; Павлов, 1999; Немцев, Мироненко, 2003; Немцев и др., 2003].
Проблемы сохранения и использования ресурсов
В прошлые столетия, как показано выше, зубр был одним из основных охотничьих видов. Кроме мяса, прочной шкуры и теплой зимней шерсти высоко ценились рога, которые использовали в качестве кубков на пирах, ими украшали шлемы средневековые рыцари, из них изготавливали фонарики, применявшиеся в интерьерах домов. Охота, главным образом, и привела к почти полному уничтожению этого зверя, как и многих других крупных копытных. С другой стороны, зубр все же уцелел именно благодаря своему статусу важного охотничьего вида - “королевского”, “княжеского” или “царского”. Если бы не запрет на его добычу в разных районах Европы, введенный еще в XIII-XVI вв., и строгую охрану, вряд ли мы могли бы сейчас созерцать это могучее животное.
В XX в. международному сообществу с большими усилиями удалось не только спасти зубра от полного исчезновения, но и вернуть его в природу. Этот мощный зверь снова стал неотъемлемой частью лесных и лесостепных биоценозов Центральной и Восточной Европы. К концу столетия поголовье увеличилось настолько, что проблема сохранения этого вида стала уже менее актуальной, хотя еще не решен ряд принципиальных вопросов, связанных с реинтродукцией животных в современные преобразованные человеком экосистемы и сохранением генетического разнообразия.
Сохранение генетического разнообразия. Все современные вольноживущие группировки зубра, за исключением гибридной кавказской, невелики и изолированы: в Беловежской пуще не более 320 особей, в большинстве других - от 2 до 50 голов. В связи с крайне малым числом особей-основателей генофонд стад обеднен, а возможность генетического обмена между изолированными группировками практически отсутствует. Во многих стадах очевидны признаки вырождения, снижается уровень воспроизводства, высокая смертность и низкие темпы прироста. Существующая система разведения животных в зубропитомниках, к сожалению, тоже не совершенна и приводит к безвозвратной утрате части генофонда. Без специальных мер по подержанию генотипической целостности и предотвращению автогенетических процессов, вызванных, прежде всего, дрейфом генов, редкий вид теряет жизнеспособность. Очевидно, что стратегия работы с зубром должна быть переориентирована на сохранение генетического разнообразия и образование полноценных природных популяций [Балюк и др., 1986; Флинт, Перерва, 1986; Перерва, 1987; Пуцек, 1987; Сипко, 1990, 1995; Белоусова, 1993а-в, 1999; Кудрявцев, Сафронова, 1993; Сипко, Белоусова, 1993; Кудрявцев, Белоусова, 1997, 1999, 2000; Белоусова и др., 1999; Сипко, Раутиан, 1999; Сипко и др., 1999; Орлов, 2000; Колотова, 2002; Раутиан, Калабушкин, 2003; Раутиан, Немцев, 20036; Sipko, Kazmin, 2004b].
Единой международной программы сохранения генетического разнообразия зубра, тем не менее, нет. Для гарантированного долговременного сохранения вида
специалисты рекомендуют создавать природные группировки с минимальной эффективной численностью не менее 100-500 голов [Soule, 1980; Заблоцкий, Заблоцкая, 19866; Пуцек и др., 1996; Pucek et al., 1996; Белоусова, 1993в; Сипко, 2002; Pucek, 2004]. В Беларуси принята “метапопуляционная” стратегия, предусматривающая создание 10-12 микропопуляций с численностью до 50 особей и более каждая. В таких микрогруппировках предлагается раз в поколение проводить искусственную “деизоляцию” посредством “отлова - выпуска” взрослых самцов. Генетическая изменчивость в совокупности микропопуляций может быть выше, нежели в крупной единой группировке. Кроме того, разным субгюпуляциям придается разный статус: “страховой” (в заповедниках и национальных парках) и “охотничьей” (на территориях общего пользования при достижении определенного потолка численности) [Козло, 1999а,б, 2001; Козло, Пикулик, 1999; Романов, 2001]. Собственно, в Беларуси реализовано предложение М.А. и М.М. Заблоцких [1986в] о создании регулируемых популяций, численность которых искусственно поддерживается на определенном уровне. Р. Грачик [Graczyk, 1988] в целях пространственной изоляции, снижения опасности эпизоотий и уменьшения вреда лесу тоже считал возможным реинродуцировать в природу небольшие семейные группировки (6-12 особей), численность которых следовало регулировать путем отстрела.
В “Стратегии сохранения зубра в России” [2002] главной целью определено сохранение этого копытного как вида с максимально возможным генетическим разнообразием и функционирующего как естественный компонент природных экосистем. Основные стратегические задачи: сохранение основных вольно живущих стад, создание не менее двух популяций, численностью около 1000 особей каждая, и обеспечение разведения животных в неволе как части мировой мегапопуляции. К сожалению, государство, провозгласившее этот курс, как показано выше, пока не способно наладить должную охрану вольных стад, а состояние зубровых питомников даже в принятой “Стратегии ...” оценивается как неблагополучное. Питомники фактически не могут выполнять функцию сохранения страхового генофонда не только из-за ненадлежащих условий содержания животных, но и из-за малой численности племенного поголовья, что неизбежно приводит к эффекту основателя, генетическому дрейфу и инбредной депрессии [Сипко, 2003].
Особенно дискуссионной оказалась проблема сохранения в чистоте генофонда беловежской и беловежско-кавказской генетических линий, тесно связанная с расселением животных в природу. М.А. и М.М. Заблоцкие настаивали на жестком географическом районировании и разведении беловежско-кавказского зубра только на Кавказе, в Крыму, Карпатах, Молдавских Кодрах и на Тянь-Шане, а беловежского - в равнинных лесах Европы, предварительно переселив на Кавказ или в другие горные районы местных беловежско-кавказских особей [Заблоцкий, Криницкий, 1974; Заблоцкий, 1975а, 1978, 1981, 1983; Заблоцкая, 1990; Заблоцкий и др., 1999]. Эти рекомендации частично выполнены. Из Беловежской пущи были вывезены все беловежско-кавказские звери [Буневич, 19976]. Другие ученые полагали, что морфы должны иметь обособленные области обитания между которыми возможны зоны контакта и обмен генами, но контакт с кавказской гибридной формой должен быть исключен [Перерва, 1982, 1987; Флинт, Перерва, 1986; Флинти др., 1986; Брагин, 1988]. Есть и более радикальное предложение - прекратить раздельное разведение генетически
близких линий, поскольку для вида, прошедшего через “бутылочное горлышко” отбора и вновь воссозданного из нескольких особей-основателей, при высоком уровне инбридинга крайне необходим обмен генофондом [Сипко и др., 1993, 1995; Сипко, Раутиан, 1999; Сипко, 2003]. “Перемешивание” генофонда полезно и при формировании зубра как охотничьего животного [Бабипок, Зубко, 1990].
После всестороннего анализа результатов исследований, я тоже не сторонник линейного разведения зубра и считаю это ошибкой, которую рано или поздно придется исправлять. Следует осознать, что попытка сохранить вид на базе ничтожного остатка единственной популяции с крайне обедненным генофондом весьма рискованна она может привести к его потере из-за негативных последствий инбридинга. Конечная цель рабогы заключается не в восстановлении отдельных генетически чистых липин зубра, а зоологического вида в целом.
Нет единой позиции и в отношении гибридных группировок зубробизона, или так называемого “горного” зубра, обитающего в Кавказском государственном заповеднике и Нальчикском лесоохотничьем хозяйстве. Этих животных многие специалисты не признают зубрами и не вносят в международные списки поголовья [Липко-вич, 1988]. М.А и М.М. Заблоцкие [1983, 1986, 1987] настаивали на их полной элиминации. С.Г. Калугин [1977], напротив, считал, что на Северо-Западном Кавказе нужно наращивать поголовье гибридных зубров до 1,5 тыс. при дальнейшем совершенствовании их фенотипа и генотипа путем селекционного отстрела и без завоза равнинных чистокровных особей, а также продолжать расселение животных в новые районы. Ю. Язан и А. Немцев [1985] предлагали придать гибридной группировке особый правовой статус и название “чистопородный горный зубр, или кавказский горный зубр (чистопородный)”. Позднее этой форме был оформлен, как отмечено выше, подвидовой статус В. bonasus montanus [Раутиан и др., 2000].
В связи с возникшей проблемой напомню о мечте специалистов в 30-е годы XX в.: ... “Мы надеемся, что зубр на Кавказе будет вновь. Пусть вначале это будет равнинный и полуодомашненный беловежец, содержащий даже небольшую примесь крови бизона: умелым подбором производителей, селекцией, через ряд лет можно получить приспособленное к горным условиям животное” [Башкиров, 1939а]. Эта мечта фактически осуществилась: в природе появилась мощная, адаптированная горная группировка гибридогенного происхождения, которая вряд ли заслуживает истребления [Флинт, 2000].
В “Стратегии сохранения зубра в России” [2002] говорится о необходимости сохранения этой уникальной формы и поддержан ее подвидовой статус. Тем самым горная (гибридная) форма как бы признана “зубром”, однако вместе с тем подчеркивается необходимость обеспечения независимого развития горной, беловежско-кавказской и беловежской линий до тех пор, “когда зубры займут устойчивое положение в природных экосистемах и их развитие будет контролироваться естественными эволюционными факторами”. Г.С. Раутиан и А.С. Немцев [2003а,б], напротив, считают горного зубра “источником генетического материала для “улучшения породы” других вольно живущих популяций. Поэтому следует бояться не гибридизации чистых зубров кавказско-беловежской линии разведения с “нечистым” горным подвидом, а того, что этого может не произойти”.
Судя по результатам молекулярно-генетических исследований, зубр и бизон находятся в гораздо более близком родстве, чем считали ранее - они, скорее, не виды,
а разные формы одного вида (см. выше). Если это так, то, по моему мнению, кавказского зубробизона следует признать одной из видовых генетических линией со всеми вытекающими отсюда последствиями в плане охраны и реинтродукции.
Реинтродукция и реконструкция ареала. Проблематичной оказывается и реинтродукция зубра, и реконструкция его ареала в связи сильным антропогенным прессом. Этот вид вряд ли следует считать типично лесным и пытаться вселять его в густые леса, особенно таежного типа, и в многоснежные районы. Как подчеркивалось выше, оптимальные ландшафтные зоны для него, скорее всего, лесостепь и разреженные широколиственные сообщества. Однако именно в таком ландшафте мало крупных массивов, способных вместить значительную (более 1000 особей) группировку животных, и велика плотность людского населения, что чревато постоянными конфликтами с реинтродуцентами, которые губят деревья, выходят пастись на посевы, съедают и разметывают рогами копны сена и стога, выходят к фермам и поселкам, теряют страх перед человеком и изредка нападают на него. По этой причине далеко не многие заповедники, заказники или охотничьи хозяйства хотели бы иметь этих животных на своей территории. Кроме того, зубр при вселении в новые районы становится пищевым конкурентом аборигенным видам копытных: лосю, благородному оленю и косулям, разведение которых в охотничьем хозяйстве менее затратное.
Тем не менее зубр должен стать полноправным видом дикой фауны России. В европейской части подходящие для него лесные массивы сохранились в Брянской, Калужской, Смоленской, Орловской, Тульской, Рязанской, Владимирской, Новгородской, Вологодской областях и Мордовии [Перерва, 1982, 1997; Сипко и др., 1999; Сипко, 2002; Стратегия ..., 2002]. К этому перечню я добавил бы еще крупные лиственные и смешанные леса Ярославской и Тверской областей. М.М. Заблоцкий [1963] предлагал разводить беловежско-кавказского зубра в порядке эксперимента в Башкирии, малоснежных районах Южного Алтая и приморских лесах Дальнего Востока. Далеко не исчерпаны возможности Кавказского региона, где могли бы жить вольные стада общей численностью не менее 5000 особей [Немцев, Мироненко, 2003; Раутиан, Немцев, 2003а].
Представителей рода Bison вполне реально реакклиматизировать и в Сибири. По мнению К.К. Флерова, ныне живущие в Канаде лесные бизоны почти не отличаются от исчезнувшего сибирского “зубра” [Флеров, Заблоцкий, 1961; Флеров, 1979]. Их вселение возможно в Центральную Якутию [Егоров, 1965; Тихонов и др., 1996], а также в лесостепные районы Приангарья и Хакассии, где эти копытные обитали сравнительно недавно [Ермолова, 1978; Смирнов, 1983], и малоснежные районы Читинской области [Заблоцкий, 1963].
Я полностью поддерживаю эту идею. Отмечу, что такого рода эксперимент спонтанно проведен в конце XX в. на Северном Урале, где бизоны, выращенные в зоопарке, вырвались из загона на свободу. Более 3 лет они прожили в очень суровых условиях в нетипичных биотопах. Затем все звери были уничтожены человеком (устное сообщение Т.П. Сипко). Бизоны, как известно, без проблем живут и размножаются в “Аскании-Нова” на Украине, в питомниках в европейской части России, а гибриды зубра и бизона - на Западном Кавказе.
Канадского лесного бизона в 2005 г. планировали реакклиматизировать в Якутии (Программа “Реакклиматизация бизона в лесной зоне Якутии”), где животные
первоначально будут размещены в обширной по площади вольере. В России появилась реальная возможность восстановить еще один полностью истребленный вид (или подвид). По предварительным расчетам, угодья, пригодные для его обитания в азиатской части страны, могут вместить десятки, если не сотни тысяч особей. Единственная проблема, которую я вижу в этом проекте случайный завоз животных, зараженных туберкулезом и бруцеллезом. В данном случае нужен очень тщательный ветеринарный осмотр племенного стада и дальнейший его контроль после выпуска в загон.
Зубр как охотничий вид. В конце XX в. этот редкий “краснокнижный” вид фактически стал охотничьим объектом. В Польше в последние годы отстреляны более 40 зубров, с соблюдением традиционного охотничьего церемониала. В Беловежской пуще на территории Беларуси ежегодно изымают до 10 особей. Отстрел в трофейных целях одного старого или травмированного самца, “не представляющего ценности для дальнейшего воспроизводства”, эквивалентен в денежном отношении добыче 5-6 самцов благородного оленя. Максимальных трофейных качеств самцы достигают в возрасте 12-15 лет, к старости роговые чехлы у животных сильно стираются [Буневич, 2002; Тышкевич, 2002а].
Предварительные расчеты показывают, что селекционный отстрел 1-2 особей возможен, если стадо достигло численности в 60 голов, а в группировке из 100 особей можно ежегодно изымать до 6-7 зверей, а всего при численности в 2000 особей - около 70 [Флинт, Перерва, 1986; Перерва, 19896; Перерва и др., 1991; Перерва, Жирнов, 1995]. На необходимости охотничьего регулирования поголовья зубробизона в районе Кавказского биосферного заповедника посредством селекции и под контролем ученых в свое время настаивал А.С. Немцев [19886, 1994]. Вместе с И.В. Мироненко [2003] он считал, что в кавказской группировке при достижении промысловой численности (не менее 1500 особей) без ущерба для воспроизводства можно ежегодно изымать до 10% поголовья в незаповедных частях ареала и в сжатые сроки в ноябре - декабре.
С 1997 г. гибридные формы (зубробизон и гибриды зубра с домашним скотом) исключены из Красной книги Российской Федерации (приложение 2 к приказу Госкомэкологии РФ от 19.12.1997 № 569) и отнесены к объектам охоты (постановление Правительства Российской Федерации от 30 июля 1998 г. № 859). Этим постановлением Правительства, как признается в “Стратегии сохранения зубра в России” [2002], утвержденной Министерством природных ресурсов РФ, фактически узаконена добыча “горного” зубра, который и есть гибрид. С другой стороны, в этой же “Стратегии ...” зубробизон считается “уникальной” формой зубра - подвидом Bison bonasus montanus, который следует охранять. В последнее издание Красной книги Российской Федерации [2001] включен “беловежский подвид зубра и внутривидовые гибридные формы”.
Отнесение “горного” зубра к охотничьим видам еще больше стимулировало добычу не только зубробизонов, но и чистокровных особей по всему ареалу. Отстрел этих копытных на Кавказе превысил все разумные пределы [Павлов, 1999]. Охотничья “выбраковка” мало похожа на селекцию - скорее этот процесс стал откровенным истреблением зверей, а отстрел зубра на трофей в вольерах вызвал острейшую дискуссию об охотничьей этике в охотничьей и природоохранной прессе.
Результат совместной деятельности основных государственных природоохранных организаций (Министерства природных ресурсов и Охотдепартамента МСХ
РФ), охраняющих зубра, в последнее десятилетие XX в. весьма плачевен. Как показано выше, численность этого зверя резко уменьшилась, а на территориях охотничьих хозяйств и государственных заказников Кавказа и “горный”, и чистокровный беловежско-кавказский зубр почти уничтожены, значительно поредели и группировки заповедников.
И тем не менее, в условиях рыночных отношений процесс превращения зубра из редкого и охраняемого в охотничий вид, в том числе и в вольерах, неизбежен, и ему нужно не противодействовать, а направлять в правовое русло, не забывая при этом об основной цели - восстановлении биоразнообразия фауны России.
Перспективы гибридизации и фермерского разведения. Эксперименты по гибридизации зубра и бизона с домашним скотом, проводившиеся в разных странах, включая Россию, пока не привели к созданию новых высокопродуктивных пород домашнего скота ни мясного, ни жирномолочного направления. Основное препятствие - частичная биологическая несовместимость представителей разных родов животных. Несмотря на реальную возможность получения и разведения гибридов на фермах [Долгоброд, 1985; Шумов, 1994; Стрекозов и др., 2001], эта работа оказалась весьма сложной и трудоемкой. Уверенность в получении новых гибридных пород животных, которые могли бы обитать в полувольных условиях или на воле в лесных районах, используя преимущественно веточный корм, тоже в значительной мере утрачена. Более перспективно, судя по мировой тенденции, полу-вольное разведение бизона в качестве фермерского охотничьего и рекреационного вида.
Подсемейство ГАЗЕЛЕВЫЕ Antilopinae Gray, 1821
Характеристика. Животные мелких (карликовые антилопы Madoqua Ogilby) и средних размеров, легкого сложения, с тонкими высокими ногами, нередко удлиненной и высоко поставленной шеей. Подгрудка нет. Ноздри сближены. На конце морды безволосый участок небольшой или отсутствует. Хвост короткий, обычно не достигает скакательных суставов и без кисти на конце.
Окраска варьирует от однотонной светлых, желтых и коричневых тонов до полосатой и пятнистой на голове, туловище и конечностях. Околохвостовое зеркало чаще четко выражено. Волосяной покров короткий и относительно равномерной длины.
Зароговая часть черепа хорошо развита. Лобные кости короткие. Передние края глазниц расположены на уровне коренных зубов верхней челюсти. Предглазнич-ные ямки й этмоидальные щели имеются, но не у всех видов. Межчелюстные кости на конце не расширены. Основная затылочная кость узкая в передней части. Слуховые пузыри вздутые. Пневматизация черепа слабая или ее нет. Роговые стержни без внутренних полостей. Средняя пара резцов расширена в форме асимметричных лопаток; остальные резко сужаются от первой пары к клыкам. Последний предкоренной нижней челюсти, за редким исключением, с открытой передней или задней долинкой. Заднекоренные зубы, как правило, без добавочных столбиков и бугорков.
У большей части видов рога имеют только самцы. Их основания сближены и расположены над глазницами на передней поверхности лобных костей. Форма рогов разнообразная: прямые, лировидные, загнутые назад и вперед, спиральные, они могут быть короткими и длинными, гладкими и гофрированными. Продольных ребер на поверхности роговых чехлов нет. Окраска рогов чаще светлая.
Волосяной покров представлен одними остевыми волосами с сильно развитым сердцевинным слоем. Специфические кожные железы (предглазничные, паховые, межпальцевые, метакарпальные, иногда спинные, зароговые, препуциальные и цир-куманальные) хорошо развиты, но имеются не у всех видов [Соколов, 1973; Соколов, Чернова, 1988]. Сосков две или, реже, одна пара.
Диплоидное число хромосом варьирует от 30 до 60, NF = 60.
Обитатели саванн, степей, полупустынь и пустынь, прибрежных зарослей и, изредка, высокогорий. Образ жизни преимущественно групповой и стадный, иногда образуют скопления. Очень подвижные животные, склонные к миграциям.
Охотничьи или охраняемые виды.
Распространение. Африка, Передняя, Средняя и Южная Азия, некоторые районы Южной Сибири, юг Европы.
Эволюция и филогения. Предковая группа животных, давшая начало Antilopinae, неизвестна. Древнейшие формы газелей прослеживаются с верхнего миоцена - нижнего плиоцена в Азии, с плиоцена - в Европе и с плейстоцена - в
Африке. В Северной Евразии, как считают, жило не менее 14 видов [Верещагин, Барышников, 1980а], однако часть из них, скорее всего, выделена ошибочно [Дмитриева, 1974].
Ближайшие признанные родственники - Caprinae и Hippotraginae. Несколько неожиданной представляется обнаруженная по некоторым молекулярно-генетическим маркерам близость ряда видов антилоп к непарнокопытным, нежели к полорогим [Глазко и др., 2002].
По новейшим исследованиям, дивергенция Antilopinae очевидна, по меньшей мере, на 3-5 триб: Antilopini и Neotragini, близость которых подтверждается на молекулярном уровне [Rebholz, Harley, 1999], Gazellini, Saigini и, возможно, Procaprini [Кузнецова, 2002; Кузнецова, Холодова, 2003].
Наиболее примитивная группа семейства с длинной зароговой частью черепа и просто устроенными рогами. Основное направление эволюции - постепенное удлинение и увеличение массивности роговых стержней, разнообразие их формы, возрастание гипсодонтности зубов и дифференциация Р/4 (от премоляров с замкнутой задней долинкой к таковым с замкнутой передней или с открытыми долинками) [Дмитриева, 1974], приспособление к жизни в открытом пространстве, быстрому бегу и высоким прыжкам, что обусловило легкость сложения. Кариотипическая эволюция газелей шла, в основном, по пути центрического слияния одноплечих хромосом [Vassart et al., 1995].
Состав. Дискуссионный. В подсемейство включают от 12 до 24 родов, в которых насчитывают 34-48 видов [Simpson, 1945; Ellerman, Morrison-Scott, 1951; Соколов И.И., 1953, 1959, 1962; Гептнер и др., 1961; Haltenorth, 1963; Соколов В.Е., 1979; Барышников и др., 1981; Durst et al., 1982; Павлинов, Россолимо, 1987; Gentry, 1992; Барышников, Крахмальная, 1994; Тихонов, 1999; Павлинов, 2003].
Наибольшую сложность для классификации представляет род Saiga. Исследователи относили его к разным подсемействам полорогих, и даже выделяли в отдельное подсемейство Saiginae, но в последнее время трибу Saigini Gray, 1872 все же чаще причисляли к Caprinae. К козлиным род отнесен и в недавней фундаментальной сводке “Сайгак” [1998].
По экологическим (равнинный вид) и ряду морфологических показателей (удлиненная зароговая часть черепа, не пневматизированные лобные кости и роговые стержни, наличие открытой передней долинки на последнем нижнем предкорен-ном зубе, отсутствие рогов у самок, строение метаподий, блоковидный тазобедренный сустав, форма спермиев, отсутствие пуховых волос [Берггрин, 1948; Егоров, 1955; Гаспарян, 1969; Стекленев, 1978а]) сайга, однако, стоит гораздо ближе к Antilopinae, нежели к Caprinae. По результатам молекулярно-генетических исследований она четко кластеризуется с антилопами, причем находится с ними в большем родстве, нежели дзерен (Ргосарга) и карликовые антилопы (триба Neotragini) [Gatesy et al., 1992,1997; Groves, Shields, 1996; Hassanin, Douzery, 1998; Hassanin et al., 1998; Глазко и др., 2002; Кузнецова, 2002; Кузнецова и др., 2002; Kuznetsova, Kholodova, 2002; Кузнецова, Холодова, 2003; Lei et al., 2003].
Морфологически сайга сходна с представителями рода оронго (Pantholops), которые обитали в Южной Сибири с плейстоцена и, возможно, до VII-VIII в. н. э. [Смирнов, 1989, 1994]. Их иногда объединяли в одну трибу, полагая, что эта группа через предковые формы могла быть связующим звеном Antilopinae с Caprinae.
Тем не менее по результатам молекулярного анализа Pantholops относится к козлиным [Kuznetsova, Kholodova, 2002; Кузнецова и др., 2002] и, следовательно, его родственные связи с сайгой сомнительны.
В составе фауны России два рода газелевых - Ргосарга и Saiga. Из списка представителей фауны я вынужден, к глубокому сожалению, исключить джейрана (Gazella subgutturosa), жившего в историческое время в степных районах Северного Кавказа [Верещагин, 1959а] и до середины XX в. обитавшего или заходившего в степи Юго-Восточного Алтая. С 70-х годов достоверных сообщений о его нахождении в этих или других районах нет.
Род ДЗЕРЕНЫ
Procapra Hodgson, 1846
Procapra Hodgson, 1846. J. Asiat. Soc. Bengal., 15:334. Тип рода Procapra picticaudata Hodgson. Зап. Тибет, обл. Гундес.
Prodorcas Рососк, 1918. Ann. Mag. Nat. Hist., 2:130. Antilope gutturosa Pallas.
Характеристика. Животные мелких (до 25 кг у тибетского Procapra picti-caudala} и средних размеров, плотного сложения, с относительно крупной головой, широкой грудью и не длинной шеей. Конец морды с большими ноздрями, оволосненный у тибетского дзерена или с узким голым вертикальным участком у Р gutturosa (монгольского дзерена). Глаза выпуклые. Хвост короткий, не более 16 см, снизу голый.
Половой диморфизм выражен. Самки мельче в размерах и безрогие. У самцов сильно развита гортань, которая выступает снизу на шее в виде зоба. Именно из-за этой морфологической особенности дзерен в некоторых сводках назван “зобатой газелью”.
Окраска верха тела летом рыжеватая, зимой - песчано-серых тонов. Околохво-стовое зеркало белое, заходит на круп выше корня хвоста. Волос густой, мягкий, зимой удлиненный на лбу, затылке, ушах, горле и задней части спины. По бокам верхней губы жесткие длинные волосы образуют подобие усов.
Лицевая часть черепа относительно массивная, лобно-носовая область выпуклая (рис. 10). Носовые кости удлиненные, без вырезки на переднем крае, их концы клинообразные. Носовые отростки межчелюстных костей обычно касаются носовых костей. Преорбитальное вдавление на слезной кости очень мало или его нет. Этмоидальное отверстие имеется. Слуховые барабаны небольшие. Коренные зубы брахиодонтные. Первый верхний предкоренной появляется поздно, и у Р. picticaudata он имеет тенденцию к рудиментации. Последний премоляр нижней челюсти с замкнутой передней долинкой [Соколов, 1959], что существенно отличает дзерена от других антилоп, и рода Gazella в частности.
Основания рогов тонкие, расположены над глазами. Рога небольшие, сближенные в основании, лировидные, загнутые назад, с кольцевыми утолщениями в нижней части и гладкие - в верхней. Их концы загнуты немного внутрь и вверх. Цвет рогов темно-серый или черный.
Предглазничные железы редуцированы, запястные - развиты очень слабо или отсутствуют, межпальцевые — небольшие. Зароговая железа имеется. У Р. gutturosa, в отличие от Р picticaudata, есть слаборазвитая паховая железа. Молочная железа двудольная.
Обитатели степей, полупустынь, пустынь и высокогорных плоскогорий. Стадные животные.
Промысловые или строго охраняемые в некоторых районах звери.
Распространение. Центральная Азия (Южный Алтай, юг Тувы и Забайкалья, западная часть Китая, Монголия, Тибет, Гималаи).
Эволюция. Древнейшие газели линии Procapra известны из миоплиоценовых отложений Бэгэрской котловины юго-запада Монголии и провинций Шаньси и Гань
Рис. 10. Череп самца дзерена (экземпляр из Монголии; Зоомузей МГУ, коллекционный № 39841) Вид сбоку, сверху и снизу
су Китая. Возможный филогенетический ряд: верхнемиоценовые Gazellapaotehensis, среднеплиоценовые G. blacki, позднеплиоценовые и нижнеплейстоценовые G. sinensis, G. paragutturosa, G. parasinensis -Procapra [Дмитриева, 1982].
По морфологическим признакам Procapra сходны с представителями рода Gazella, однако цито- и молекулярно-генетические исследования отвергают их близкое родство, в том числе и с джейраном, у которого иной кариотип (2п = 30-32, NF = 60) [Соколов и др., 1975; Орлов и др., 1978;VassartetaL, 1993; Schreiber, Hegel, 1999]. Дзерены представляются самостоятельной монофилетической ветвью (триба Рго-caprini), отщепившейся от общего ствола антилоп, видимо, в верхнем плиоцене прежде рода газелей. Во всяком случае, Procapra стоит ближе к карликовым антилопам рода Mado-qua, нежели к Gazella [Кузнецова, 2002; Кузнецова, Холодова, 2003; Lei et al., 2003].
Эволюция Procapra шла по пути редукции преорбитальных ям, образования на последнем премоляре нижней челюсти замкнутой передней долинки, сохранения комолости у самок. Наличие слаборазвитых паховых желез, характерных для Gazella, лишь у одного вида дзеренов (у Р gutturosa) свидетельствует скорее не о прогрессивной эволюции этого органа, как принято считать, а о рудиментарном его характере. В экологическом плане очевидно приспособление животных к стадной жизни и быстрым перемещениям на дальние расстояния в равнинной местности.
Состав. Состав рода не совсем ясен. В нем признают 2 или 3 вида: Р picticaudata Hodgson, 1846 (Гималаи и Тибет), Р przewalskii Buchner, 1891 (Китай от Цинхая до Внутренней Монголии) и Р gutturosa [Строганов, 1949; Соколов И.И., 1959; Гептнер и др., 1961; Wang et al., 1963; Соколов В.Е., 1979; Durst et al., 1982]. По молекулярным данным форма przewalskii более близка к монгольскому, нежели к тибетскому дзерену [Lei et al., 2003]. К сожалению, она находится на грани уничтожения. К концу XX в. сохранилось лишь несколько стад общей численностью, по разным оценкам, от 0,2 до 3 тыс. особей в провинции Цинхай [Jiang et al., 1995; Джиган, Дикян, 1998; Lei et al., 2001; Ли, Цзян, 2002]. В фауне России и сопредельных территорий - Р gutturosa.
МОНГОЛЬСКИЙ ДЗЕРЕН
Procapra gutturosa Pallas, 1777
Antilope gutturosa Pallas, 1777. Spicil. Zoo!., fasc. 12: 46, tab. 2. Вост. Забайкалье, верхи, течение р.
Онон.
Antilope orientalis Erxieben, 1777. Syst. Regn. Anim.: 288. Nom. nov. pro Antilopegutturosa Pallas. Antilope altaica Hollister, 1913. Smiths. Misc. Collect., 60, 19:1. Монгольский Алтай, плато Суок у перевала Баян-Цаган.
Характеристика. Длина тела взрослых самцов - 105-148 см, самок - 110-121 см, высота в холке, соответственно, 62—84 и 54—74 см, длина уха - 10-15 см, хвоста - 9-16 см, масса 24—52 и 20-37 кг.
Половой диморфизм и географическая изменчивость в окраске не выражены. Окраска головы в зимнем меху преимущественно желтовато-серая, от носа ко лбу тянется коричневатая полоса. Верх тела песчано-серый, нижняя и задняя части туловища светлые. Шея самцов в области гортани, выдающейся в виде зоба, во время гона темно-серая с синим оттенком. К концу зимы, вследствие обламывания верхушек волос и выгорания, окраска становится белесой. Летом верх головы и туловища рыжеватый и желтовато-песчаный, нижние части тела и крупа - белые с сероватым или желтоватым оттенком (фото 9 и 10). Хвост белый, его кончик бурокоричневый, а по центру нередко проходит узкая рыжая полоса, заходящая на круп и разделяющая зеркало на две половины. Волос между копытами светло-желтый. Зимние волосы сравнительно длинные, 30-50 мм, слегка волнистые, удлиненные на лбу, темени и щеках. Летний волос короткий, 20-30 мм, редкий. Выпадает он в мае - первой половине июня, зимний волос растет с конца августа до ноября.
Череп с удлиненной предглазничной и вздутой ростральной частью (рис. 10). Наибольшая его длина у самцов 236-263 мм, у самок 222-251 мм. Самки безрогие. У взрослых самцов рога лировидной формы, темно-серые или черные длиной 19-29 см с 4-19 кольцевыми утолщениями в нижней части [Лукашкин, 1927; Банников, 1954; Гептнер и др., 1961; Жирнов, Винокуров, 1975; Соколов, Орлов, 1980; Кирилюк, 2000а, 2002].
Предглазничные и запястные железы рудиментарные, межпальцевые небольшие на всех ногах. Паховая железа развита, но слабо. Зароговая железа имеется. Увеличены сальные и апокриновые железы половых губ самок и циркуманальные и препуциальные у взрослых самцов. Препуциальный мешок очень крупный, и летом содержит секрет, похожий на ушную серу с “козлиным” запахом [Соколов, Чернова, 1988,2001].
Диплоидное число хромосом равно 58 (60?), NF = 60. В наборе крупные акроцентрические хромосомы, включая половые, и одна (две?) пары мелких точечных матацентриков [Соколов и др., 1975; Орлов и др., 1978; Орлов, Булатова, 1983; Soma, Kada, 1984]. Уровень генетического разнообразия (Н) составляет 0,36-0,43 (в отдельных группировках - 0,09-0,32). Нуклеотидная изменчивость по фрагменту D-петли весьма высокая - 6,37% [Лущекина и др., 1983; Холодова и др., 2002; Сорокин и др., 2004].
Стадный мигрирующий житель степей, полупустынь и пустынь. Прекрасный бегун, передвигающийся со скоростью около 25—30, а при преследовании — 60—80 км/ч
[Лукашкин, 1927; Банников, 1954; Гамбарян, 1972]. Хорошо развито зрение и обоняние. В природе живет недолго - самки примерно до 10 лет, самцы - до 6 лет.
В России редок и включен в Красную книгу как вид, находящийся под угрозой исчезновения, в Монголии — объект промысла.
Распространение. Монголия, соседние районы Китая, юг Тувы и Забайкалья, Южный Алтай.
Состав. Монотипичный вид. Форма altaica Hollister, 1913 отвергнута А.Г. Банниковым [1954].
Распространение
Древнейшие формы газелей линии Procapra населяли юго-запад Монголии и провинции Шаньси и Ганьсу Китая с верхнего миоцена [Дмитриева, 1982]. В раннем антропогене они занимали степные районы Киргизии, Казахстана (на запад до Балхаша), Алтая, Тувы, Забайкалья, Монголии, Китая и, видимо, Приморья. Позднепалеолитические и неолитические остатки дзерена известны из степного Забайкалья, в том числе из поселения Сухотино-4 в черте Читы [Громов, 1948; Верещагин, 1954,19756; Александрова и др., 1963; Кожамкулова, 1969,1976,1981,1982; Ермолова, 1978; Верещагин, Барышников, 1980а; Окладников, Кириллов, 1980; Каспаров, 1986; Кольцов, Медведев, 1989; Тлеубердина и др., 1989].
В голоцене ареал дзерена существенно сократился, особенно в западной и восточной частях, но минимальным он стал в наше время. В XIX - начале XX вв. его северная граница проходила южнее лесной и горной зон Алтая, Тувы и юга Забайкалья; западная и южная - по склонам Монгольского и Гобийского Алтая, севернее пустыни Гоби на восток до р. Хуанхе; восточная - по склонам Большого Хингана. В Северо-Восточном Китае звери обитали на плоскогорье Барги, в степях Маньчжурской равнины и в Содовой степи. До постройки китайской восточной железной дороги мигрирующие стада доходили до р. Сунгари и Харбина, позднее - не далее линии железной дороги Цицикар - Аньда [Пржевальский, 1875 (по изд. 1946); Лукашкин, 1927, 1975; Банников, 1954; Жирнов, 1977а].
До середины XX в. дзерен обитал почти по всей территории Монголии, за исключением пустынной юго-западной и лесной северной части, куда животные забегали лишь изредка (рис. 11). Летом южная граница ареала совпадала с южным пределом
Рис. 11. Распространение монгольского дзерена в Центральной Азии (по [Луще-кина, 1998])
1 - в середине XX в. (по [Банников, 1954]), 2 - в конце XX в., 3 - основные направления сезонных миграций
Рис. 12. Распространение монгольского даерена на Алтае (по [Собанский, 1992]) 1 - в начале XX в., 2 - в 30-50-е годы, 3 - пункты встреч в 1974, 1982 и 1989 гг.
сухих степей, а западная проходила по склонам Хангая и Монгольского Алтая, сдвигаясь зимой к югу в зону пустынь. На востоке этот вид был распространен до рек Хал-хин-Гол и Дэге-Гол, на севере - почти до окраины лесостепной зоны, а в степных районах - до государственной границы с Россией [Банников, 1951, 1954]. С середины XX в. за 30 лет площадь ареала сократилась с 780 до 100-170 тыс. км2, что стало одной из основных причин уменьшения притока копытных в смежные районы России. В 70-80-е годы крупные группировки сохранились только на востоке Монголии и сопредельной территории Китая. Отдельные мелкие стада уцелели в предгорьях Арц-Богдо, в районе озер Хара-Нур, Хара-Ус-Нур, Дургэн-Нур, близ сомонов Манхай (Кобдоский аймак), Цаган-Хайрхан, Хувсгэл, Сулан-Хэр, Улан-Бадрах, Ундэр-Шил, Гоби-Угтал. В 90-е годы ареал в значительной мере восстановлен естественным путем по мере увеличения численности [Даш, 1970; Буяндэлгэр и др., 1977; Жирнов, 1978, 1982; Соколов, Орлов, 1980; Соколов и др., 1982, 1992; Лущекина и др., 1983, 1986; Даш, Соколов, 1986; Лущекина, 1990,1998; Milner-Gulland, Lhadvasuren, 1998].
На Алтае многочисленные наскальные изображения дзерена в отрогах хребтов, окружающих Чуйскую степь, датируются VII-IV вв. до н.э. [Ешелкин, 1972]. Вплоть до 30-х годов XX в. этот вид был обычным в фауне региона. В Чуйской степи во время миграций наблюдали тысячи зверей. Многие стада жили постоянно не только здесь, но и населяли урочища хребтов Сайлюгема, Курайского и Чихачева. Их распростране
ние на север ограничивал Курайский хребет, на запад - Северо- и Южно-Чуйский хребты (рис. 12). Позднее поголовье копытных в этом районе сильно уменьшилось из-за резко возросшего браконьерства с применением автотранспорта, особенно при сооружении Чуйского тракта [Булавин, 1934; Колосов, 1938, 1939]. В военные (1941-1945) годы дзерена, как и архара, в массе добывали для нужд армии и местного населения, что окончательно подорвало ресурсы. Тем не менее, и в 50-е годы стада, насчитывающие порой до 200 голов, регулярно заходили из Монголии на юг Алтая, проникая к северу до истоков р. Чулышман, в отроги Сайлюгема и выходили в Чуйскую степь. В 60-е - начале 70-х годов отмечали
Рис. 13. Районы обитания монгольского дзерена в Читинской области в конце XX в. (по [Кирилюк, 2000а])
лишь спорадические заходы мелких групп численностью не более 10-12 особей, возможно, из Синьцзян-Уйгурского автономного района Китая. Последние встречи зверей (по одной в год) зарегистрированы в 1974, 1982 и 1989 гг. [Савинов, 1964; Ешел-кин, 1973; Собанский, 1988,1992]. Западнее дзерен обитал, видимо, в долине Черного Иртыша, в район гор Катутау, Актау, Илийской долине, на юго-западе Джунгарского Алатау, куда заходил из соседней Джунгарии [Антипин, 1941; Кожамкулова, 1981].
В Туве этот вид населял полупустынные и степные районы южнее Восточного Танну-Ола вплоть до середины XX в. В периоды высокой численности (в 30-е -начале 40-х годов XX в.) большие стада переходили через этот хребет в окрестностях Самагалтая. Северная граница ареала доходила до озер Чагытай, Хадын и Че-дер в центральной части республики. В послевоенное время из-за перепромысла с применением автотранспорта численность этих копытных резко сократилась. В 50-е годы поголовье оценивали в 5 тыс., а в начале 60-х годов в Убсунурской котловине еще встречали стада по 100 и более особей [Сулимов, 1977]. Позднее ежегодные миграции зверей из Монголии прекратились. В последние десятилетия одиночных животных и мелкие группы (до 10 голов) наблюдали лишь спорадически в районе озер Торе-холь, Дус-холь, Шара-Нур и по правобережью р. Тес-хем [Янушевич, 1952; Никифоров, 1973; Очиров, Башанов, 1975; Никифоров и др., 1977; Смирнов, 1978, 1983, 1988, 20026; Соколов, 1988].
В степях Забайкалья в XVIII - первой половине XIX вв. путешественники наблю
дали многотысячные стада антилоп, которые во время осенне-зимних кочевок уходили из Монголии и Китая на север до лесостепных районов и Нерчинска, а в экстремальные годы заходили и в таежные районы. Летом они “водились и плодились здесь во множестве”. К середине XIX в. .численность местной группировки и область ее обитания стали сокращаться, чему способствовали высокие снега и охотничий промысел. Буряты добывали этих животных на облавных охотах сотнями. Заходы огромных стад зимой в Даурские степи, тем не менее, были регулярными вплоть до середины XX в., но летом здесь оставались лишь мелкие группы и одиночные особи. Позднее миграции стали затухать и почти прекратились. Зверей, остававшихся после кочевок, быстро уничтожали моторизованные браконьеры. Последние случаи размножения антилопы
в Читинской области имели место в 70-х годах. Позднее мелкие группы и одиночных особей изредка отмечали лишь в приграничных районах [Черкасов, 1867 (по изд. 1990); Pallas, 1776; Дементьев, 1935;Г1одаревский, 1936; Фетисов, Хрусцелевский, 1948; Павлов, 1949; Лавов, 1966; Варнаков, 1973; Баранов, Бойко, 1988; Смирнов, 1989; Соколов, Парнес, 1993]. В 1985 г. Ю.Г. Швецов зарегистрировал группу из 11 зверей на юге Бурятии юго-западнее оз. Барун-Торей [Швецов и др., 1984; Швецов, 1987]. Причиной прекращения миграций стало не только резкое уменьшение численности копытных, но и сооружение протяженной линии инженерных заграждений вдоль государственной границы.
Регулярные заходы тысячных стад животных из Монголии в Забайкалье возобновились лишь в 90-е годы, что способствовало образованию небольшой оседлой группировки в Кыринском, Ононском, Борзинском и Забайкальском районах Читинской области и Агинском Бурятском округе (рис. 13). С 1992 г. в заповеднике “Даурский” вновь зафиксировано размножение дзерена [Кирилюк, 1997а,б, 2001а, 2002; Кирилюк, Черепицын, 1998; Кирилюк, Лущекина, 2002].
Среда обитания
Область обитания дзерена находится в зоне дерновинно-злаковых и, частично, пустынных степей Центральной Азии. Этот регион отличается суровым, резко континентальным климатом, большим перепадом температур, частыми засухами, коротким периодом вегетации растений, низкой продуктивностью фитоценозов, и мало пригоден для обитания других видов копытных, что уменьшает конкуренцию.
Оптимум современного ареала приходится на зону даурско-монгольских равнинных сухих мелкодерновинно-злаково-тырсовых степей, находящихся на высотах от 800 до 1000 м над уровнем моря, со среднегодовым количеством осадков 200-300 мм, мало затронутых антропогенным воздействием. Лучшие биотопы - ковыльные, ковыльковые, ковыльково-луковые, ковыльково-солянковые степи и полупустыни волнистых равнин, холмов и низкогорий, широких межгорных долин и котловин. Разнотравные и горные степи, злаково-солянковые полупустыни и солянковые пустыни служат временными пастбищами преимущественно зимой [Банников, 1951, 1954; Буяндэлгэр и др., 1977; Жирнов, 1977а, 1978, 1982; Лущекина и др., 1986; Лущекина, 1990].
Узких долин, каменистых склонов, песчаных холмов, зарослей кустарников и лесов стада избегают. На юго-востоке Монголии, тем не менее, летом мы неоднократно встречали мелкие группы этих копытных в песчаных холмах и кустарниках вместе с сибирской косулей. В Китае под Хайларом в многоснежные зимы стада держались в сосновых рощах [Лукашкин, 1927]. В Забайкалье дзерен, обитая в степной зоне, нередко заходит в лесополосы, лесные колки, приозерные кустарники и пойменные заросли. В многоснежном 2001 г. несколько мигрирующих стад прошли через широкий (15-20 км) Цасучейский сосновый бор, и затем долго оставались на опушке, ще не было наста [Кирилюк, 2001а, 2002].
Северно-восточная популяция монгольского дзерена, проникающая в Забайкалье, населяет холмистые равнины, озерные и речные долины бассейнов рек Ульдзы и Керулена. В летний период животные концентрируются в степях с разнотравно-
мелкодерновинно-злаковыми, разнотравно-леймусовыми и злаково-разнотравными ассоциациями. Спектр зимних стаций гораздо шире, а зимовочная территория в десятки раз превышает площадь летовок. Места обитания этого вида не приурочены к водным источникам. Тем не менее, с наступлением холодов, животные в поисках влаги идут на север к кромке снежной зоны [Слудский, 1963], а летом крупные стада все же чаще держатся вблизи пресных и слабосоленых водоемов. Здесь, помимо воды, более сочные корма и большая безопасность, поскольку такие места трудно проходимы для автомашин [Лущекина и др., 1986; Кирилюк, 1997а,б; Кирилюк, Лущекина, 2002, 2004].
Снежный покров - один из основных факторов, определяющих не только биото-пическое размещение, но и северную границу распространения вида. Животные имеют узкие копыта, поэтому весовая нагрузка на след весьма высока - около 1 кг/см2, что сильно затрудняет передвижение по снегу. Проблематичной становится и тебеневка, в связи с чем звери практически не могут потреблять растения, укрытые снегом. В степи с низкорослой растительностью критическим для них становится снежный покров высотой более 10 см, и особенно, с настом. Наиболее оптимальные условия для дзерена в районах, где снежный покров неплотный, пятнистый и не более 7-8 см высотой [Кирилюк, 1997а, 2001а].
Кочевки зверей в Монголии напрямую связаны и с состоянием пастбищ. Однообразные низкоурожайные центральноазиатские степи не в состоянии прокормить массы зверей. По этой причине группировки, обитающие на востоке ареала, кочуют к северу в более продуктивные разнотравные степи, а центральные и западные перемещаются южнее в ковыльковые пустынные степи [Банников, 1951].
Питание
Дзерен употребляет в пищу несколько десятков видов растений, но основных всего около десятка: ковыль тырса (до 60% содержимого желудков), ковылек гобийский, вострец, тонконог, прутняк, лапчатка, луки, полыни, карагана, осоки и солянки [Банников, 1951, 1954; Жирнов, 1977а, 1982; Лущекина, 1990; Кирилюк, 2002].
Лучшие пастбища этого вида - мелкодерновинно-злаково-тырсовые степи с участием востреца, луков, караганы и дэриса. Урожайность в степных сообществах колеблется в пределах 1,5-6,9 ц/га сухой массы. Массовый отел копытных обычно совпадает с наивысшей питательностью травостоя. Содержание протеина в основных весенне-летних нажировочных кормах колеблется от 8 до 26%. Зимой на пастбищах сохраняется от 20 до 50% урожая травостоя, в котором сохраняется не более 40% протеина от летних значений [Соколов и др., 1982; Лущекина и др., 1986].
На пастбищах дзерены никогда не стравливают растения под корень. Пастьба происходит на ходу: звери скусывают лишь верхушки растений, что стимулирует их к кущению, повторной вегетации и приводит к увеличению биомассы. Осенью семена растений втаптываются в разрыхляемую копытами дерновину, что способствует их прорастанию в удобренной животными почве. При наличии снежного покрова они также скусывают лишь верхушки растений, и тебенюют лишь в исключительных случаях.
На поля с зерновыми культурами копытные обычно не заходят, но при недостатке естественных кормов могут повреждать посевы, что отмечалось в некоторых районах Китая. Гораздо больше их привлекают старые залежи, заросшие сор-
ними растениями и полынью. Зимой при недостатке корма звери едят сено из стогов и копен [Кирилюк, 2001а].
Воду весной и летом дзерен в достаточной мере получает с растительным кормом, особенно утром в виде росы, и очень экономно расходует ее. Поэтому животные встречаются за десятки и сотни километров от открытых водоемов. При наличии водоемов животные посещают их раз в несколько дней, а при засухах - ежедневно. Зимой звери поедают снег или грызут и лижут лед [Банников, 1951, 1954; Кирилюк, Цэвээнмядаг, 1999; Кирилюк, 2002].
Величина потребления корма (570 г/особь) и его переваримость (82-87%) у детенышей дзерена выше, чем у сайги (350 г и 52-55%, соответственно) [Магомедов и др., 1989].
Суточная активность полифазная. Наиболее активны звери вечером и в начале ночи, а также утром до 10-11 ч дня. В июне - июле они кормятся и днем с небольшими перерывами (30-40 мин) на отдых, причем в стаде постоянно происходит смена кормящихся и отдыхающих животных. Большую часть ночи дзерены отдыхают [Банников, 1951, 1954; Кирилюк, 1997а].
Размножение
Половая зрелость. Самки монгольского дзерена принимают участие в спаривании на втором году жизни, самцы - на третьем [Жирнов, 1977а; Кирилюк, 2002].
Гон. Гон проходит с конца ноября до начала - середины января (табл. 13). Наиболее интенсивен он с середины до конца декабря. Спаривание обычно длится не более двух недель.
Перед гоном наиболее сильные самцы отбивают от стада часть самок и удерживают их на определенной территории 2-3 недели. В результате образуются несколько крупных гаремных групп (до 60 особей), по периферии которых распределяются мелкие гаремы (от 2 до 30 особей), одиночные самцы и стада животных (от нескольких десятков до нескольких сот особей), не принимающих участия в размножении. Расстояние между территориальными самцами составляет около 80-500 м. Свою территорию самцы маркируют, оставляя кучи помета, обрызганного мочой, и охраняют от соперников, постоянно перемещаясь по участку с громкими рявкающими звуками. Драки между взрослыми самцами редки - для выявления силы обычно достаточно демонстраций. Самцы пытаются не только удержать самок на своем участке, но и завлечь новых. Часть самок, включая оплодотворенных, уходит из гарема или их отбивают другие самцы, но вместо них появляются другие. В результате состав гарема постоянно меняется. При отсутствии самок самцы перемещаются на другие участки.
Таблица 13. Сроки гона у монгольского дзерена
Район 1 Начало I Конец I Источник информации
Забайкалье Конец ноября Начало января Кирилюк, 2002
Монголия С( Январь Банников, 1954
а (( Середина января Жирнов, 1977а
4 декабря — Львов, 1982
(С 10 декабря Начало января Кирилюк, 1997а
Наиболее стабильны территории при высокой плотности населения. В гон самцы мало едят, теряют много энергии и сильно истощаются [Жирнов, 1977а, 1982; Львов, 1982; Кирилюк, 1997а, 2002].
Беременность и деторождение. Продолжительность беременности самок монгольского дзерена - 187-191 день [Кирилюк, 2002].
Отел происходит в июне-июле (табл. 14), его пик обычен в конце июня — начале июля, причем 90% самок рожают в течение 4—7 дней [Лущекина, 1990]. Сроки родов, видимо, смещаются в разные годы, но районы отела остаются почти неизменными. Беременные самки непременно возвращаются туда, где родились сами.
Таблица 14. Сроки отела у монгольского дзерена
Район Начало Конец Источник информации
Забайкалье 15 июня 15 июля Кирилюк, 2000а, 2002
Монголия 8 июля — Банников, 1954
сс 16 июня Июль Жирнов, 1977а
сс Конец июня Начало июля Лущекина, 1990
(4 15 июня 10-15 июля Кирилюк, 1997а
Рожают самки чаще в высокой траве. Новорожденные дзерена крупны и хорошо развиты. Их масса составляет в среднем 3,7-4 (2,9-4,6) кг. Они встают на ноги и кормятся молоком через 20-60 мин после родов. Через 2-3 дня телята способны быстро бегать, но все же не следуют за матерью, а затаиваются в характерной позе среди густой степной растительности. Встают они только при подходе матери, которая кормится на расстоянии 50-300 м от них. Мать находит своих детенышей среди многочисленного потомства по голосу и запаху. В случае опасности они некоторое время бегут за матерью, затем резко отворачивают в сторону, с разбега падают на землю и затаиваются. Траву детеныши начинают срывать в недельном возрасте. Лактация продолжается до ноября - начала декабря. В “родильных домах” группировки остаются 6-12 дней, после чего стада с подросшими детенышами (фото 9) покидают район отела. Однако здесь еще остается часть беременных и только что отелившихся самок. Поэтому период родов в каждой группировке длится около месяца [Жирнов, 1977а; Кирилюк, 1997а, 2002; Кирилюк, Черепицын, 1998].
Плодовитость. Самки дзерена сравнительно поздно для газелей вступают в размножение и рожают преимущественно одного теленка, двойни редки - 2-8 (до 20)%. В благоприятные годы потомство приносит 60-85% самок, в неблагоприятные - почти половина из них не имеет детей [Банников, 1954; Шагдарсурен, 1974; Жирнов, 1977а, 1982; Milner-Gulland, Lhadvasuren, 1998; Кирилюк, 1997а, 2002]. Соответственно потенциальный уровень плодовитости близок к 1 детенышу на половозрелую самку, но фактически он существенно ниже и гораздо меньше, чем у сайги (табл. 15).
Популяционная структура
Половой и возрастной состав. Соотношение полов у новорожденных монгольского дзерена примерно равное, но, в связи с высокой смертностью самцов, оно с возрастом изменяется в пользу самок вплоть до соотношения 1:2—5 [Жирнов, 1977а;
Таблица 15. Число детенышей у самок монгольского дзерена
Район (источник информации)* Встречено самок, п Самок с детенышами, % Половозрелых самок без телят, % Детенышей на самку
с одним I с двумя
Монголия (1) - 97 3 - -
-“-(3) — 96,2 3,8 32 -
-“-(5) Монголия и 342 96,5 3,5 (2,5-8,2) 47 0,6
Забайкалье (2) - 96-98 2-4 2-4 1,0
Забайкалье (4) 333 - - 43-47 -
* 1 - Жирнов, 1977а; 2 - Кирилюк, 2002; 3 - Банников, 1951; 4 - Кирилюк, 1997а: 5 - Лущекина. 1990.
Таблица 16. Половозрастной состав (%) и соотношение полов в популяциях монгольского дзерена
Район (источник информации)* Встречено животных, п Сеголетки Годовалые Взрослые Самцы: самки
самки самцы
Монголия (1) 279 групп — - - - 1:2,3
-“-(2) 572 особи — — — — 1:1,1
-“-(3) - 11-31 4-7 - 1:1,4-1,7
-“-(5) 7210** 19,5 — 41,6 38,9 1:1,07
-“-(6) 774 24,9 - 44,2 30,9 1:1,4
Забайкалье(4) 2282 26-29 — 52-54 19-20 1:2,7
* 1 - Жирнов, 1977а; 2 - Банников, 1951, 1954; 3 - Кирилюк, 1997а, 1999; 4 - Кирилюк, 2001а; 5 -Соколов и др., 1982; 6 - Лущекина, 1990.
** По результатам элиминации стад в декабре 1977 г. (данные Я. Даша).
Кирилюк, 2002]. Доля сеголеток в популяциях колеблется в пределах 11-31%, годовалых особей меньше - 4-7, взрослых самок — 41—54, взрослых самцов — 19-39% (табл. 16). В группировках в Северо-Восточной Монголии осенью 1994-1998 гг. доля сеголеток достигала 19-31%, но после эпизоотии 1998 г., унесшей около 20-32 тыс. особей, сократилась до 10,7% [Кирилюк, 1999]. В Китае в 1988 г. доля молодняка в популяции составляла 40% [Jiang et al., 1993].
Социальная организация. Монгольский дзерен - стадное животное (фото 9), но крупные стада, достигающие 2-10 тыс. голов, характерны лишь для многочисленных центральных популяций. На периферии ареала и при депрессии преобладают группы до 10-20 особей, стада из 30-200 голов и много одиночных животных. В периоды миграций образуются кратковременные многотысячные (до 10-40 тыс.) скопления. Еще большими (до 100 тыс. особей) скопления бывают на местах отелов. Во время отела крупные стада распадаются, и животные держатся поодиночке или группами, состоящими только из самок, самцов или подростков. После родов самки живут отдельно от других животных на выбранных участках, контактируя с телятами только во время кормления молоком. По мере взросления детенышей отдельные семьи постепенно группируются, а с конца августа “маточные” группы начинают пополняться самцами и годовалыми животными. Во время гона взрослые самцы агрессивны к сеголеткам, и те начинают жить самостоятельно или же после гона снова присоединяются к матерям, оставаясь с ними до следу-
тощего отела [Пржевальский, 1875 (по изд. 1946); Дементьев, 1935; Банников, 1951, 1954; Шагдарсурен, 1974; Жирнов, 1977а, 1978, 1982; Лущекина и др., 1982, 1986; Соколов и др., 1982; Кирилюк, 2001а, 2002].
Средний размер групп в нижнеульдзинской популяции и в группировке заповедника “Даурский” существенно варьирует в течение года, но изменение этого показателя стадности в целом сходно: в июне - июле - 1,3-2,5, в августе - 5-7, в сентябре - 8, в октябре во время миграции - 33-58, в декабре во время гона - 19, на зимовке в январе - марте - от 21 до 266 в разные годы, в апреле во время весенней миграции - 42, в мае перед отелом - 18-24 особи [Кирилюк, 1997а; Кирилюк, Че-репицын, 1998].
Ориентировочно-исследовательское и оборонительное поведение, а также звуковая сигнализация дзерена существенно отличается от такового у ближайших родственников - сайги и джейрана. Особь, заметившая опасность, принимает ориентировочную позу, резко выдувает воздух через нос, издавая “чмыхающие” звуки, и нередко топает передними ногами. Члены стада настораживаются, и некоторые повторяют сигналы тревоги [Кирилюк, 2002]. В высокой траве или в кустарниковых зарослях отдельные звери, как мне приходилось видеть, поднимаются на задние ноги и вытягивают шею для лучшего обзора местности. При бегстве животные, чаще самцы, высоко подпрыгивают, что типично для африканских газелей. Лидируют в стадах взрослые самки. При преследовании стада могут разбиваться на отдельные группы со своими лидерами.
Пространственная структура и перемещения. Дзерен - типичный сезонно мигрирующий вид, но менее номадный и территориально более консервативный по сравнению с сайгой [Жирнов, Винокуров, 1975; Жирнов, 1977а, 1982; Кирилюк, 2002; Кирилюк, Лущекина, 2002].
До середины XX в. группировки дзерена в Монголии летом концентрировались в основном в центральной и восточной части ареала. В сентябре стада начинали постепенно перемещаться по одним и тем же из года в год миграционным коридорам к периферии ареала: восточные группировки двигались на северо-восток в направлении нижнего течения р. Ульдзы и через государственную границу к оз. Далай-Нур и р. Аргунь, а центральные и западные уходили на юг. К началу зимы животные рассредоточивались на огромной территории, в десятки раз превышающей летнюю площадь обитания. Весенняя миграция начиналась в марте. Звери сравнительно медленно возвращались назад к местам отела, постоянным на протяжении многих лет, и к летним пастбищам. В многоснежные и гололедные зимы стада проникали далеко на юг в пустынные районы и на север в степные и лесостепные районы и долины горных лесов. В такие зимы они заполняли даурские и маньчжурские степи, предгорные районы Хангая и Монгольского Алтая, долину р. Тэс, где значительная часть их гибла [Банников, 1951, 1954]. Иногда звери появлялись за сотни километров от границ ареала в нетипичных стациях. Зимой 1911 г., например, взрослый самец дзерена был убит в лесу под Иркутском [Лукаш-кин, 1927; Дементьев, 1935]. С середины столетия, по мере сокращения численности, хозяйственного освоения территории и в связи с постройкой железной дороги и проволочный заграждений на участке Улан-Батор — Дзамын-Уд, сезонные миграции стали затухать, и для животных стали более характерны местные кочевки. Тем не менее в Восточной Монголии сезонные перемещения стад были четко выраже
ны в районе Тухмийн-Гоби и юго-восточнее Эрдэнэ-Цаган-сомона [Лущекина, 1990].
Более детальные сведения о пространственной структуре восточных популяций дзерена получены в самом конце XX - начале XXI вв. [Кирилюк, 1997а,б, 2000а, 2001а, 2002; Кирилюк, Черепицын, 1998; Кирилюк, Цэвээнмядаг, 1999; Кирилюк, Лущекина, 2002]. Крупная северокеруленская популяция дзерена ежегодно с середины августа - начала сентября перемещается на 200—380 км в западном, северном и северо-восточном направлениях, и лишь небольшая ее часть, включая нижнеульдзинскую группировку, зимует в местах летнего обитания. Животные идут шагом или легкой рысью, периодически отдыхая. Средняя скорость перемещения - 6-10 км в сутки, иногда до 24-29 км. В июле 1979 г. скопление дзерена (около 40 тыс. голов) в период массового выплода кровососущих насекомых из района развалин Улан-Цирик переместилось за сутки более чем на 100 км к югу [Лущекина, 1990]. Озера звери обходят, а реки переплывают или переходят по льду. Железная дорога Чойбалсан - Борзя разделяет потоки мигрантов, но не препятствует перемещению стад, которые при необходимости пересекают полотно и тянущиеся по сторонам невысокие заграждения из проволоки. Общая продолжительность осенней миграции составляет около 2-2,5 мес. На зимовках плотность населения зверей на отдельных участках достигает 600-900 особей на 1 км2. Такие концентрации все же кратковременны (одна — две недели), после чего животные перемещаются в новый район. При благоприятных условиях стада подолгу остаются на одном месте.
На севере ареала мигрирующие животные регулярно достигают границ России, а на отдельных участках переходят государственную границу, минуя инженерные сооружения. В Забайкалье в последние десятилетия такие заходы зарегистрированы в 1989/90, 1991/92, 1993-1995 гг. В декабре 1999 г. - январе 2000 г. многотысячные стада проникли вглубь Читинской области на 18-20 км к западу от Торейских озер, а в январе 2001 г. - почти на 100 км (рис. 14). В следующем году мигранты удалялись на север от границы всего на 10-15 км. Весенние миграции стад в обратном направлении обычны в феврале - мае. В места отела животные возвращаются к середине - концу мая, где рассредоточиваются по территории с высокой плотностью - от 20 до 40 особей на 1000 га, а нередко и выше.
Размножающиеся самки с мая и до конца июля живут поодиночке на небольших родовых участках площадью около 2-3 км2, а затем объединяются в группы и постепенно расширяют участок обитания. Высокая плотность копытных в местах размножения вдали от водоемов имеет свои преимущества, поскольку уменьшается вероятность их гибели от волка, который в это время выкармливает потомство поблизости от водоемов и привязан к логовам. Кроме того, в крупном скоплении родившие самки смело атакуют и преследуют лисицу, корсака и низколетящих пернатых хищников, защищая не только своих, но и чужих телят. Во время гона плотность населения животных также значительна, что гарантирует почти поголовное покрытие самок.
При обилии корма стада дзерена держатся на ограниченной территории, при его недостатке их перемещения становятся беспорядочными и стремительными, и они могут появляться далеко от типичных районов обитания в несвойственных
Рис; 14. Пространственная структура северокеруленской популяции монгольского дзерена и ее размещение в Забайкалье на рубеже XX-XXI вв. (по [Кирилюк, 2001 а]) 1 - основной район отела, 2 - направления осенней миграции, 3 - размещение на российской территории в январе - марте 2000 и 2001 гг., 4 - места встреч отдельных групп в начале 2001 г., 5 - районы вероятных заходов в 2001 г.
угодьях. Эмиграции приводят к значительной гибели животных, но именно эта стратегия позволяет не только сохранить часть популяций, но и восстанавливать и расширять ареал за счет оседания отдельных групп в новых районах, в результате чего образуются периферийные очаги обитания. В них животные тоже сравнительно оседлы. Расселение в таких очагах идет, в основном, за счет эмиграции годовалых особей.
Численность и основные факторы, ее определяющие
Численность. Динамика ареала и численности дзерена на территории России до 90-х годов XX в. отражена выше (см. раздел “Распространение”)- К этому времени вид практически исчез из состава фауны страны, а заходы мелких групп животных из Монголии в приграничные районы были крайне редкими и нерегулярными.
В Монголии до 40-х годов XX в. обитали, видимо, несколько миллионов дзеренов. В 1941-1945 гг., в результате массовых заготовок, численность сократилась до 300-400 тыс. [Shagdarsuren, 2002]. В послевоенные годы ресурсы этого вида оценивали почти в 1 млн [Банников, 1954]. В конце 60-х годов поголовье снова уменьшилась до 300-400 тыс. [Даш, 1970], в 70-е годы, возможно, -до 150-200 тыс. (по более оптимистическим оценкам в этот период оставалось около 400-500 тыс.). В 1980 г. после массового падежа численность снизилась, возможно, до 180 тыс., но уже к 1982 г. выросла до 300-400 тыс., и в дальнейшем колебалось в этих пределах [Буяндэлгэр и др., 1977; Жирнов, 1982; Лущекина и
др., 1982, 1983, 1986; Соколов и др., 1982, 1992; Даш, Соколов, 1986; Лущекина, 1990]. В 90-е годы поголовье стало быстро расти, чему способствовало уменьшение браконьерства в связи с выводом российских войск и отъездом многочисленных гражданских специалистов. В Китае в середине XX в. ресурсы дзерена, возможно, достигали 2 млн голов [Tan, 1979]. К концу столетия осталось около 200-400 тыс. особей [Gao et al., 1996; Zhigang, Diqiang, 1996]. Общая численность вида в конце XX - начале XXI в., видимо, находилась в пределах 1,9-3,8 млн особей [Milner-Gulland, Lhadvasuren, 1998; Reading et al., 2002]. He исключено, что эти цифры существенно завышены [Лущекина, 1998].
Поголовье северокеруленской популяции, находящейся в Монголии близ границы с Россией, выросло до 300 тыс. Следствием этого процесса стало появление мигрирующих зверей в пограничных районах Читинской области после многолетнего отсутствия (см. выше) и образование здесь в 90-е годы оседлой группировки численность около 20-80 особей. В конце декабря 1999 г. - начале января 2000 г. на территорию России зашли 10-12 тыс. особей, а в январе 2001 г. - около 45-70 тыс. особей. В январе - феврале 2002 г. наблюдали кратковременные заходы отдельных стад общей численностью от 3-5 до 12-15 тыс. Часть мигрантов осталась жить в Забайкалье. В июне 2001 г. в Читинской области насчитывали 550-800, в 2002 г. -400-500 оседлых особей [Кирилюк, 1997а, 2000а, 2001а, 2002; Кирилюк, Лущекина, 2001, 2002, 2003].
Основные факторы, так или иначе влияющие на динамику популяций дзерена, следующие.
Воспроизводство, прирост. Несмотря на позднее вступление самок в размножение и сравнительно низкую плодовитость по сравнению с сайгой, уровень воспроизводства в популяциях дзерена в благоприятные годы все же высок - до 76-96 детенышей на 100 самок после сезона деторождения и до 55-64 сеголеток в весенней популяции на то же количество самок. Однако чаще годовой прирост существенно ниже, чем у сайги - 12-29% от исходного поголовья, в неблагоприятные годы - 6-7%, а после экстремальных зим он может быть отрицательным [Жирнов, Винокуров, 1975; Жирнов, 1977а, 1982; Лущекина, 1990; Кирилюк, 1997а, 2001а, 2002; Кирилюк, Черепицын, 1998; Milner-Gulland, Lhadvasuren, 1998].
Детская смертность. По сведениям В.Е. Кирилюка [2002], в первую неделю жизни погибает от 2 до 10% детенышей, до полуторамесячного возраста - еще 16-22%, а в осенне-зимний период их отход составляет 1-1,5% в месяц. Основные причины гибели - врожденные аномалии, родовые и послеродовые травмы, хищники и истощение в случае потери матерей.
Погода и корма. Массовая гибель дзерена от истощения при недостатке корма в многоснежные зимы и при настах и гололедице - типичное явление по всей области обитания. В Монголии такая ситуация возникала зимой 1922, 1925/26, 1935/36, 1944/45, 1947/48, 1951/52, 1957/58 и 1967/68 гг., и усугублялась отсутствием запрета охоты на бедствующих животных. Именно по этой причине копытные исчезли в большинстве районов Западной Монголии. Смертность особенно высока среди самцов, теряющих жировые запасы во время гона. Летние засухи обычно не приводят к гибели животных, но они действуют на них опосредованно, вынуждая больше кочевать и затрачивать больше энергии на добывание скудного корма, что способствует истощению организма. В засушливые годы часты пожары, которые полнос
тью уничтожают степную растительность на огромных площадях [Банников, 1954; Даш, 1970; Жирнов, Винокуров, 1975; Жирнов, 1977а; Соколов и др., 1982; Кирилюк, 1997а, 2002].
В Забайкалье смертность истощенных животных была необычайно высокой в январе 2001 г., особенно в полосе между государственной границей и инженерно-техническими заграждениями. Только в Забайкальском, Борзинском и Ононском районах было собрано для утилизации около 5 тыс. трупов, а общее число погибших, вероятно, превышало 6 тыс. Основные причины гибели - искусственная преграда на пути мигрирующих стад, что вызвало скопление зверей на ограниченной территории в условиях многоснежья при низкой температуре и недостатке корма, массовое браконьерство, приведшее к большому числу подранков, и хищничество волка и собак. Среди павших несколько выше была доля сеголеток и взрослых самцов. Численность куруленской группировки после этой зимы сократилась на 40-45%, или на 90-100 тыс. особей [Кирилюк, 2001а,б].
Хищники. Стадный образ жизни и быстрый бег спасают дзерена от хищников, однако они все же наносит значительный урон популяциям. Волки в период отелов копытных легко давят беременных самок и новорожденных, убивая их до 3-5 в день на каждую волчью семью с потомством. С осени их стаи могут кочевать за стадами дзерена, подбирая подранков, больных или истощенных животных, особенно самцов после гона, и легко настигают их в глубокоснежье. Нередко хищники устраивают на копытных засады или облавы. На востоке Монголии в районах зимовок дзерена плотность размещения волков в 6 раз выше, чем других районах. Доля этого хищника в общей непромысловой смертности копытных достигает 54%, от истощения и болезней погибают до 14% [Жирнов, 1977а; Кирилюк, 1997а, 2001, 2002].
По наблюдениям известного путешественника М.В. Певцова [1951], во второй половине XIX в. потери дзерена от волка, которого в Монголии было “множество”, гораздо превышали урон от промысла. По расчетам А.А. Слудского [1962], основанных на ежегодных заготовках шкур волков (до 10 тыс.) и уровне ежегодного потребления жертв, в Монголии и Китае копытные должны были гибнуть от этого хищника сотнями тысяч. Во Внутренней Монголии зимой 1958/59 гг. охотники за один месяц добыли свыше 10 тыс. голов дзерена и более тысячи волков. Наименьший урон волк наносил дзерену в 80-е годы XX в., когда его численность была резко снижена, благодаря массовой борьбе с ним [Лущекина, 1990].
Большой вред копытным причиняют и пастушьи собаки, которые обычно добывают пропитание в природе. На детенышей могут нападать также манул и беркут. Корсак, лисица, канюк и коршун чаще довольствуются падалью [Лукашкин, 1927; Лущекина, 1990].
Конкуренты. Острой конкуренции дзерена с другими дикими и домашними копытными не возникает из-за ярко выраженной номадности. Антилопы нередко пасутся вместе с коровами и овцами или рядом с группами сибирских косуль. И все же в местах интенсивного животноводства этих копытных, как правило, нет, что связано не только со скудностью стравленного растительного покрова, но, в большей мере, с преследованием человеком и собаками.
Болезни, паразиты, травмы. Массовый падеж дзерена от эпизоотий неясной этиологии имел место в Монголии в 1871, 1923 и 1926 гг. [Пржевальский, 1875; Лукашкин, 1927; Дементьев, 1935]. В 1964-1965, 1974, 1978, 1980, 1982,
1994 и 1998 гг. от инфекционных болезней (чаще от пастереллеза и некробакте-риоза) смертность достигала 25-30% поголовья, или 20-140 тыс. Причиной некоторых эпизоотий, возможно, был ящур, что достоверно зарегистрировано лишь в 1963-1964 гг. В 2001 г. почти у всех исследованных зверей, мигрировавших из Монголии в Россию, были выявлены антитела к вирусу ящура, что свидетельствует о перенесенной ранее болезни [Жирнов, 1977а; Дуламцэрэн и др., 1981; Соколов и др., 1982; Даш, Соколов, 1986; Лущекина и др., 1986; Лущекина, 1990; Кирилюк, 1999, 2001а, 2002]. Не исключено также участие дзерена в поддержании циркуляции вируса гриппа типа А I [Неронов и др., 1982].
У дзерена найдено 23 вида гельминтов, в том числе на территории Тувы - 8 [Сулимов, 1973]. Сильно изнуряют животных кровососки, подкожный и носоглоточный овода. Личинками последнего они поражены почти поголовно [Лущекина и др., 1986]
Массовое травмирование зверей, приводящее к гибели, постоянно имеет место в пограничных инженерно-технических сооружениях [Кирилюк, 2001а].
Антропогенные факторы. Исчезновение дзерена с территории России в XX в. вызвано исключительно антропогенными факторами: круглогодичной неумеренной охотой, резко усилившейся с появлением дальнобойного нарезного оружия и автотранспорта, а также распашкой степей и зимним выпасом домашнего скота в типичных районах обитания вида [Янушевич, 1952; Ешелкин, 1973; Никифоров, 1973; Баранов, Бойко, 1988; Собанский, 1992; Кирилюк, 2000а, 2002; Собанский, Часовских, 2002].
Воздействие хозяйственной деятельности человека (выпаса скота, распашки степей и фактора беспокойства) на группировки дзерена в Монголии все же ничтожно по сравнению с охотой и, отчасти, степными пожарами. Одной из важных причин сокращения численности в середине XX в. могла быть постройка железной дороги Улан-Батор - Дзамын-Уд, которая, как отмечалось выше, перерезала сложившиеся веками пути миграции. Непреодолимым препятствием для животных стали проволочные заборы вдоль этой ветки, а также мощные заграждения вдоль многих участков государственной границы [Эрегдэндагва, Сосорбарам, 1969; Жирнов, 1978; Соколов и др., 1982, 1992; Кирилюк, 1997а, 2001а, 2002; Лущекина, 1998].
Использование ресурсов и охрана
Дзерен издавна был одним из основных промысловых видов у народов, населяющих Центральную Азию. Его добывали ради вкусного и жирного в осенний период мяса и шкур, из которых изготавливали дохи, замшу, прочные сыромятные ремни и коврики. Рога шли на всевозможные поделки и применялись в традиционной китайской медицине. На их основе синтезирован лечебный препарат “прокаприн” [Хайдав и др., 1986]. В последнее время возросла и трофейная ценность рогов.
В степях Забайкалья, Тувы и Алтая охота на этот вид была массовой лишь во время зимних миграций, но она носила варварской характер. Вот как описывает ее А.А. Черкасов [1867 (по изд. 1990)]: “... Валовый выход зеренов из Китая в наши пределы быстрой и радостной вестью проносится по всей границе, из одного селения в другое, из улуса в улус, из стойбища в стойбище, из уст
в уста, и все охотники от мала до велика, караульцы и туземцы, выждав удобный случай, подметив где-либо большой табун зеренов, собираются по нескольку человек вместе, садятся на лихих скакунов и едут на промысел бить, резать, душить появившихся животных. Ход всей этой бойни состоит в том, чтобы загнать целое стадо зеренов, состоящее иногда из 500 и более особей, на лед, на котором животные, попадав, волей-неволей попадаются под ножи охотников. ... Караульцы так ловко производили эту охоту, что загнав все стадо на озеро, вырезали несчастных животных до одной головы и сотнями пудов привозили домой их жирное мясо. Вся поверхность небольшого озерка покрывалась кровью, которая застывала и впоследствии долго напоминала побоище и служила пищею хищным зверям и воронам”. В многоснежные зимы животных загоняли на лошадях и убивали палками и нагайками, иногда сотнями за загон. Промысловый пресс на вид постепенно усиливался по мере увеличения людского населения в степной зоне и с появления дальнобойного огнестрельного оружия. С 30-х годов XX в. стали практиковать преследование зверей на автомобилях, истребляя за раз по нескольку десятков голов [Булавин, 1934; Дементьев, 1935]. Введение государственных заготовок диких копытных на мясо и резкое увеличение воинского контингента в районах обитания дзерена привели к полному его уничтожению во многих районах.
Запрет охоты на вид введен в Западносибирском крае в 1933 г. [Булавин, 1934 (ред. уточнение); Жирнов, 1977а], но в Забайкалье промысел был разрешен и в послевоенные годы с 15 ноября по 1 февраля [Павлов, 1949]. Нелегальная круглогодичная и круглосуточная добыча животных с применением быстроходного авто-и мототранспорта и нарезного оружия велась и ведется до сих пор. В Читинской области, например, в январе 2001 г. браконьерство на мигрирующих зверей было массовым - убиты около 2-6 тыс. особей, а следы огнестрельных ранений имели 26% осмотренных трупов [Кирилюк, 2001а,б, 2002; Кирилюк, Лущекина, 2001].
В Монголии дзерен издавна является традиционным объектом промысла местного населения. Способы охоты, применяемые монголами в XIX в. (засидки, загоны, с помощью верблюда, ножными ловушками и др.), подробно описаны Н.М. Пржевальским [1875]. Позднее зверей добывали преимущественно из нарезного оружия и с помощью автотранспорта. Государственные заготовки, в том числе для нужд армии и на экспорт, ведутся с 20-х годов XX в. В 1932 г. заготовлено 13,6 тыс. туш, 1933 г. - 30 тыс., 1935 г,- 44 тыс., 1940 г. - 60 тыс. В годы Второй мировой войны ежегодно добывали до 100-150 тыс. особей. В 50-е и первой половине 60-х годов ежегодный отстрел не превышал 70 тыс., позднее сократился до 10-30 тыс., но в середине 70-х вырос снова почти до 80 тыс. С 1932 по 1970 гг. только официально было заготовлено около 1,3 млн голов, а с учетом потребления населения - много больше. В отдельные годы из популяций изымали почти весь прирост, что, наряду с неблагоприятными погодными условиями и гибелью от эпизоотий, стало одной из основных причин сокращения ресурсов и потребовало принятия мер по их охране и восстановлению. К концу 70-х годов объем государственных заготовок уменьшился до 10-12 тыс. В 80-е годы официальная добыча не превышала 30 тыс. голов [Банников, 1954; Даш, 1970; Жирнов, 1977а, 1982; Лущекина, 1990, 1998].
Замечу, что к “дзерену” монгольские охотничьи чиновники нередко относили и заготавливаемую на мясо сибирскую косулю. Следовательно, приводимые 5 - 1475 97
официальные данные по добыче антилопы, возможно, нуждаются в корректировке, однако реальные цифры, с учетом браконьерской добычи, гораздо выше. Нелегальная охота на этот вид, особенно моторизованная, как нам приходилось наблюдать в 70—80-е годы, велась повсеместно и круглогодично. Редкий водитель не брал с собой в дорогу оружия. Особенный урон стадам наносили армейские браконьеры, применявшие для отстрела копытных автоматы и пулеметы. После их ночных охот в степи оставались десятки искалеченных животных.
В Китае, как и в Монголии, издавна применялись изощренные способы охоты на дзерена, которого особенно “нещадно били” под Хайларом [Лукашкин, 1927]. В 1956-1961 гг. в стране официально добыто 2,5 млн голов, в том числе 670 тыс. -во Внутренней Монголии в 1960-1961 гг. [Tan, 1979]. В 60-70-е годы ежегодная добыча сократилась до 70 тыс., в 80-е - возросла до 100 тыс. без учета браконьерской добычи, уровень которой тоже был высоким [Zhang, 1991; Tong, 1993]. Группировки, мигрирующие в Китай с территории Монголии, обратно практически не возвращались [Даш, 1970].
В настоящее время дзерен включен в Красные книги Российской Федерации, Республики Тыва, Читинской области и Агинского Бурятского автономного округа. В Китае он тоже отнесен к охраняемым животным, в Монголии по-прежнему имеет промысловое значение.
В XXI в. восстановить утраченную часть ареала вида реально не только в Монголии, где нужно создать сеть заповедников и заказников [Лущекина, 1990], но и в Забайкалье, где современное состояние среды обитания практически не лимитирует расселение и воспроизводство популяции. Единственный сдерживающий фактор здесь - нелегальная охота, но, при функционировании заповедника “Даурский” и принятых дополнительных мерах охраны, она имеет тенденцию к сокращению [Кирилюк, Черепицын, 1989; Кирилюк, Лущекина, 2002]. Реакклиматизация вида возможна в Убсу-Нурской и Тувинской котловинах [Смирнов, 20026], а степной части Южого Алтая, а также в полупустынной и пустынной зонах Казахстана [Слудский, Афанасьев, 1964].
Для сохранения генофонда в случае экстремальной ситуации, видимо, целесообразно полувольное разведение дзерена на специальных фермах. Его детеныши, как показывает опыт, легко приручаются и живут рядом с человеком, но осенью стремятся уйти к диким сородичам. В Китае этих копытных в прошлом содержали в больших огороженных парках [Кирилюк, 2002]. Вольерных зверей можно использовать и для восстановления некогда истребленных популяций, а также в коммерческих целях: организации туризма и платной охоты, получения мясной и кожевенной продукции и изготовления лекарственного препарата “Прокаприна” из рогов [Лущекина, 1990].
Род САЙГИ
Saiga Gray, 1843
Saiga Gray, 1843. List Mamin. Brit. Mus. London: XXVI, 160. Тип рода - Capra tatarica Linnaeus.
Colus Wagner, 1844. Schreber’s Saugeth., Suppl., 4: 419. Antilope saiga Pallas. Nom. praeocc., non Humphrey, 1797.
Характеристика. Животные среднего размера, сравнительно легкого сложения, с бочкообразным туловищем и горизонтально расположенной шеей. Конечности тон
кие, короткие, но с удлиненными плюсневой и пястной костями [Гаспарян, 1969,1976;
Мельник, Клыкова, 1989]. Хвост короткий, не более 18 см, снизу голый. Голова боль
шая, необычной формы. Морда горбоносая, нос в виде короткого подвижного хобота с тесно сближенными широкими трубкообразными ноздрями на конце (фото 11 и 12). Наружная поверхность верхней губы покрыта волосами, за исключением узкой срединной полоски голой кожи. Хоботообразный нос, вероятно, - адаптация к длительному перемещению животных в экстремальных условиях: зимой— в облаке снежной пыли, летом - в сплошной пылевой пелене, образуемой многочисленным стадом. В хо
боте развита сеть кровеносных сосудов и желез, выделяющих слизь на внутренних стенках полости пред дверия носа. Сама полость покрыта волосками, которые задерживают пыль. Здесь, как в респираторе, воздух очищается, увлажняется
и прогревается зимой. Кроме того, у сайги высоко адаптирована и дыхательная система [Масенов, 1966].
Череп уникального строения. Носовые кости сильно укорочены, приподняты вверх и полностью слиты с лобными костями (рис. 15). Носовой вход очень длинный и сильно расширенный в вентральной части. Его край образо
Рис. 15. Череп самца сайги (экземпляр из Калмыкии; Зоомузей МГУ, коллекционный № 60850) Вид сбоку, сверху и снизу
10 см
—।
ван носовыми, межчелюстными, верхнечелюстными и увеличенными слезными костями, что необычно для полорогих. Передняя часть верхнечелюстных костей сильно удлинена. Сошник не срастается с небными костями. Глазницы большие, трубкообразные, заметно выступают в стороны и вверх над лбом. Предглазничные ямки неглубокие. Этмоидальных щелей нет. Задние части сагиттального лобного и венечного швов с выступающими гребнями. Основания рогов расположены почти над глазницами. Синусы в роговых отростках, в отличие от козлиных, не развиты. Резцы долотовидные. Предкоренные сильно редуцированы, в нижней челюсти их обычно два, и изредка сохраняется второй молочный зуб. Последний нижний предкоренной с открытой передней долинкой (в результате смыкания метастилида с энтостили-дом). Заднекоренные зубы гипсодонтные, с узкими стилями на наружной поверхности в верхней челюсти.
Рога есть только у самцов. У самок на их месте обозначены лишь костные бугорки, не более 5 мм, и иногда образуются прямые короткие роговые стержни или уродливые наросты. Рога самцов сравнительно длинные, до 42 см, поставлены почти вертикально, слегка лировидные, с загнутыми внутрь кончиками, нередко темными, часто асимметричные. Число кольцевых утолщений на поверхности роговых чехлов у взрослых самцов доходит до 22, но чаще их 12-14. Вершины рогов гладкие. Поперечное сечение округлое. Роговое вещество необычное среди полорогих - полупрозрачное, воскового цвета.
Окраска зверей однотонная, летом желтовато-рыжая, зимой - белесая с потемнениями на голове, шее и спине у самцов. Борода не образуется. Кожа вокруг ануса и в пахах голая круглогодично. Волос не подразделяется на категории и не образует групп и пучков [Соколов, 1973] в отличие от козлиных, у которых меховой покров состоит из ости и пуха, растущего пучками и группами пучков. Соотношение и количество волос на единицу площади почти не меняется по сезонам, но зимний волос почти в 2 раза толще и в 2-3 раза длиннее (до 6-7 см на туловище, а на щеках и шее достигает 19 см) летнего, извитой и имеет воздушную полость. У самок на хребте, боках и брюхе он на 25-30% длиннее, чем на тех же участках у самцов. Зимой волосяной покров очень плотный и устойчив против раздувания ветром. Хорошо развиты предглазничные, межпальцевые, карпальные и паховые железы [Рашек, 1963, 1965, 1974; Соколов, Чернова, 1988, 2001]. Молочная железа с 4 сосками, но передняя пара недоразвита.
Кариотип примитивный: 2п = 60, NF = 60.
Степные и полупустынные животные, ведущие стадный кочевой образ жизни. Прекрасные бегуны. Скороспелые и очень плодовитые звери. Срок жизни непродолжительный - 5-7 лет у самцов и 12-13 лет у самок.
Важный промысловый вид, периодически почти истребляемый, затем охраняемый и восстанавливаемый.
Распространение. В плейстоцене - Европа, Центральная и Северная Азия вплоть до Новосибирских островов, северо-запад Северной Америки. В голоцене - юг Восточной Европы и Центральная Азия. В последние столетия ареал очаговый.
Эволюция. Происхождение сайги неясно. Ее древние формы четко прослеживаются лишь со среднего плейстоцена Северо-Востока Сибири и Аляски [Шер, 1967, 1971], хотя отпочкование предковой ветви, судя по архаичным морфологическим особенностям, должно было произойти еще в плиоцене. Безуспешность поиска родственников
сайги, па мой взгляд, во многом объясняется неверным предположением о ее родстве с козлиными, тогда как предка логичнее искать среди ископаемых газелей Азии.
Эволюция сайги почти на протяжении миллиона лет происходила в суровых климатических условиях арктических тундростепей, вероятно, при необходимости быстрых перемещений крупных стад. Скорее всего, эти обстоятельства способствовали формированию своеобразных адаптаций, не имеющих аналогов среди копытных.
По краниометрическим признакам прослеживаются две древние линии рода. Череп плейстоценовой европейской сайги небольшой, с прямыми, короткими и слабо разваленными стержнями рогов. Восточноазиатская форма отличается большим развитием зароговой части черепа, более длинными носовыми костями, слабым развалом рогов и прямизной стержней, сильно сплющенных с боков. Верхнеплейстоценовые S. tatarica prisca, S. t. borealis и S. t. binagadensis имеют характерный для современных животных развал рогов и изогнутость стержней во фронтальной плоскости. К голоцену роговые стержни заметно удлиняются, череп становится более широким, а его зароговая часть укорачивается [Верещагин, 1959а; Шер, 1967; Алексеева, 1980; Барышников, Крахмальная, 1994; Лущекина, Дулам-цэрэн, 1997; Барышников и др., 1998].
Состав. Род включает единственный вид Saiga tatarica L., 1766. В 1946 г. А.Г. Банников описал новый вид Saiga niongolica, основываясь на более мелких размерах тела и черепа, более тонких и коротких рогах, слабой горбоносости, небольшом переднем носовом отверстии и серо-бурой окраске животных в зимнем меху. Позднее монгольская форма признана подвидом S', tatarica mongolica [Банников, 1946,1954; Ellerman, Morrison-Scott, 1951; Соколов, 1959; Гептнер и др., 1961; Барышников, Крахмальная, 1994].
САЙГА
Saiga tatarica Linnaeus, 1766
Ibeximberbis Gmelin, 1760. Nov. Comment. Acad. Sci. Petropol., 5: 345. Омская обл.,Тарана Иртыше. Nom. nudum.
Capra tatarica Linnaeus, 1766. Syst. Nat., 12 ed., 1: 97. Казахстан, Уральские степи.
Antilope saiga Pallas, 1766. Misc. Zool.: 6. Pro Ibex imberbis Gmelin. Nom. nudum.
Antilope scythica Pallas, 1767. Spicil. Zool., 1: 9. Pro Ibex imberbis Gmelin. Nom. nudum. Capra sayga Forster, 1768. Philos. Trans., 57: 344. “Поволжье”.
Cernas colus Oken, 1816. Lehrb. Naturg., 3, 2: 736. Pro Ibex imberbis Gmelin. Nom. nudum.
Saiga mongolica Bannikov, 1946. Докл. АН СССР, 51,5: 397. 3. Монголия, Шаргин Гоби, Духмэн-тала, 150 км к зап. от Баян-сомона.
Характеристика. См. характеристику рода.
Распространение. То же, что и для рода.
Состав. Два очевидных подвида: S. t. tatarica L., 1766 и S. t. mongolica Bannikov, 1946. В составе фауны России - S. t. tatarica.
Изменчивость
Размеры и масса. Длина тела новорожденных колеблется в пределах 42-69 см, масса - 1,5-4,5 кг (до 5,2-5,5 кг в исключительных случаях), чаще 2,9-4,2 кг. Сай-
Таблица 17. Размеры (см) взрослых (старше 2 лет) особей сайги
Самцы Самки Район (источник информации)*
п 1 м lim п 1 М 1 lim
31 132 Длина 123-146 25 116 108-125 Калмыкия (4)
53 126 114-138 63 но 104-118 -“-(2)
182 133-140 123-150 195 116-121 104-134 Казахстан (1)
49 140 124-156 51 126 114-135 -“-(2)
144 - 134 (5)
50 138 126-150 50 117 109-127 Бетпакдала (8)
- 138 131-140 - 124 120-127 О. Барсакельмес (9)
3 115 110-116 1 — 110 Монголия (3)
- - -120 - - - -“-(7)
53 81 Высота в холке 75-87 63 73 66-77 Калмыкия (2)
31 73 69-79 25 68 57-73 -“-(4)
182 73 66-78 195 68 60-78 Казахстан (1)
49 85 75-92 51 81 65-88 -“-(2)
- 71 - - 69 - -“-(5)
50 73 66-78 50 68 52-72 Бетпакдала (8)
- 74 69-80 — 67 63-70 О. Барсакельмес (9)
3 64 60-67 1 — 63 Монголия (3)
53 81 Обхват туловища 76-89 63 71 66-76 Калмыкия (2)
49 88 79-96 51 79 71-85 Казахстан (2)
50 93 80-106 50 81 75-92 Бетпакдала (1,8)
— 33,5 Длина ступни 32,1 — О. Барсакельмес (6)
53 7,1 6-8 Длина уха 63 6,8 6-8 Калмыкия (2)
31 11 8-12 25 8,5 7-11 -“-(4)
182 8-9 7-12 195 8-9 6-11 Казахстан (1)
49 6,9 6-8 51 6,6 6-7,5 -“-(2)
50 9 7-12 50 9,2 6-12 Бетпакдала (8)
- 10,2 10-11 - 8,2 8-8,5 О. Барсакельмес (9)
3 8,3 8-8,5 1 - 8 Монголия (3)
53 12,7 Длина хвоста 11-15 63 11,9 9-18 Калмыкия (2)
31 10 8-12,5 25 8,5 6-10 -“-(4)
182 12 9-17 195 11 7-17 Казахстан (1)
49 И,2 9-14 51 10,6 8-14 -“-(2)
50 11,7 9-14 50 12 9-15 Бетпакдала (8)
- 12 12-12 - п,з 10-12 О. Барсакельмес (9)
3 11 8-13 1 - 13 Монголия (3)
* 1 - Фадеев, Слудский, 1982; 2 - Соколов и др., 1998; 3 - Банников, 1954; 4 - Банников и др., 1961; 5 -Слудский, 1955; 6 - Рашск, 1965, 1974; 7 - Соколов н др., 1996; 8 - Слудский и др., 1983; 9 - Басенко, 1950.
гачата с массой 1,5-2,8 кг обычно гибнут в первые дни жизни. Детеныши быстро растут: среднесуточный прирост массы в первый месяц составляет около 65 г, в 1,5-месячном возрасте - 118 г в неволе и 292 г в природе. В июне их масса достигает 9-11 кг, в июле — 17-18 кг, в октябре — 18—25 кг. Масса внутренних органов
близка к максимальным значениям уже к 4 месяцам жизни. Половой диморфизм у детенышей почти не выражен [Васенко, 1950; Бакеев, Формозов, 1955;Раков, 1956; Даль и др., 1958; Фандеев, 1960, 1965; Проняев, 1978; Петрищев, Холодова, 19806; Присяжнюк, Проняев, 1982; Матвиенко, Треус, 1986; Треус, 1993; Петрищев, 1996, 1997а,б; Близнюк, Букреева, 2000]. Зимой рост сеголеток замедляется и возобновляется лишь с началом вегетации растительности. С мая по сентябрь масса годовалых животных увеличивается примерно на 7-10 кг. Молодые звери достигают минимальных размеров взрослых к 18-19 мес., после чего самки практически не растут, а у самцов рост продолжается до 3-4 лет [Слудский, 1955; Банников и др., 1961; Фадеев, Слудский, 1982; Слудский и др., 1983; Проняев, Фандеев, 1998; Субботин, 2001; Букреева, 2002]. Рост и развитие сайги наиболее полно отражены в монографии А.Г. Банникова с соавторами [1961] и книге “Европейский сайгак” [1997].
Длина тела взрослых самцов колеблется в пределах 110-156, самок - 104-135 см, высота в холке, соответственно, 60-92 и 52-88 см, масса - 26-60 и 15-41 кг (табл. 17 и 18). После благоприятных летних сезонов масса самцов на 18, а самок - на 13 кг выше, чем в неблагоприятные годы [Лещенко, 1997]. Во время гона самцы быстро теряют накопленные жировые отложения.
Таблица 18. Масса (кг) взрослых (старше 2 лет) особей сайги
Самцы Самки Район (источник информации)*
п I М I lim п 1 м 1 lim
53 40 32-50 63 25 20-30 Калмыкия (2)
29 43 32-51 25 31 21-41 -“-(4)
21 43 31-58 23 27 22-31 -“-(12)
96 40 26-44 75 24,5 15-28 -“-(Ю)
182 36-44 28-49 195 27-31 21-37 Казахстан (1)
26 47 — 16 30,5 - -“-(6)
— 46 37-60 - 36 34-37 -“-(5)
156** 47 - ** 33 - -“-(7)
50 44 37-49 50 31 22-37 Бетпакдала (9)
- 37 35-39 - 29,5 27-32 О. Барсакельмес (11)
3 31 29-32 1 — 26 Монголия (3)
- - -35 -“-(8)
* 1 - Фадеев, Слудский, 1982; 2 - Соколов и др., 1998; 3 - Банников, 1954; 4 - Банников и др., 1961; 5 -Слудский, 1955; 6-Раков, 1956; 7 - Присяжнюк, Проняев, 1980; 8 - Соколов и др., 1996; 9- Слудский и др., 1983; 10-Лещенко, 1997; 11 -Васснко, 1950; 12 - Присяжнюк, Проняев, 1982.
** Самцы и самки вместе.
Географическая изменчивость отчетливо выражена. Особи европейской популяции в среднем несколько мельче, чем в казахстанских группировках (уральской, устюртской и бетпакдалинской), но существенно крупнее особей монгольской формы (табл. 17 и 18). Фенотипическая дивергенция европейской и казахстанских популяций, видимо, возникла сравнительно недавно в связи с ухудшением условий обитания и перепромыслом первых [Проняев, 1985а,б, 1998].
Краниометрическая характеристика. Новорожденные сайгачата имеют молочные резцы, клыки и предкоренные зубы, покрытые тонкой слизистой оболочкой. Верхние клыки выпадают вскоре после рождения и больше не отрастают, мо-
Таблица 19. Размеры черепа (мм) взрослых особей (старше 2 лет) сайги
Самцы Самки Район (источник информации)*
п 1 м | lim п | М | lim
Наибольшая длина
23 248 239-258 50 222 205-234 Калмыкия (2)
50 245 232-266 50 222 211-230 Казахстан (2)
16 260 256-263 11 238 229-242 Междуречье Волги и Урала (1)
45 249 233-271 45 225 213-235 Бетпакдала (1)
25 240-245 226-261 25 229 221-235 Барсакельмес (1)
3 255 243-267 2 240 - Джунгария (3)
5 217 203-237 2 203 201-204 Монголия (3)
Максимальная ширина
23 130 123-136 50 111 104-116 Калмыкия (2)
50 129 122-137 50 113 105-118 Казахстан (2)
Межглазничная ширина
23 90 82-98 50 74 67-80 Калмыкия (2)
50 90 81-102 50 76 68-83 Казахстан (2)
16 95 92-102 11 81 78-82 Междуречье Волги и Урала (1)
45 90-93 72-100 45 78 72-84 Бетпакдала (1)
25 89 82-98 25 76 74-81 Барсакельмес (1)
11 95 83-100 9 75 70-80 Бетпакдала (4)
Длина лицевой части
23 206 197-217 50 179 165-192 Калмыкия (2)
50 201 188-221 50 180 166-196 Казахстан (2)
16 209 205-221 11 189 183-192 Междуречье Волги и Урала (1)
45 197 188-204 45 178 169-196 Бетпакдала (1)
25 196 177-208 25 181 176-185 Барсакельмес (1)
5 125** 110-141 2 ПО** 107-114 Монголия (3)
Длина мозговой части
23 94 89-98 50 81 76-84 Калмыкия (2)
50 95 91-100 50 82 78 Казахстан (2)
5 90 87-100 2 79 76-81 Монголия (3)
Длина носовых костей
23 105 97-112 50 89 84-97 Калмыкия (2)
50 104 93-113 50 91 82-103 Казахстан (2)
Длина верхнего ряда зубов
23 68 63-74 50 67 60-72 Калмыкия (2)
50 69 66-74 50 68 59-75 Казахстан (2)
3 82** 80-83** 2 70** - Джунгария (3)
5 68 63-75 2 68 67-69 Монголия (3)
* 1 - Фадеев, Слудский, 1982; 2 - Соколов и др., 1998; 3 - Банников, 1954; 4 - Басенко, 1950.
** Измерения, скорее всего, сделаны по иной методике.
лочные резцы и премоляры меняются на постоянные в возрасте 13-18 мес. Полного развития зубная система достигает к 20 мес., а к 6-7 годам зубы сильно изнашиваются, что и определяет срок жизни животных [Соколов, Рашек, 1961 ]. Рост черепа у самок завершается к двум годам жизни, у самцов - к трем, но пропорции,
характерные для взрослых особей, формируются к 19 мес. [Банников и др., 1961; Фандеев, 1965; Проняев, Фандеев, 1998].
Максимальная длина черепа взрослых самцов - 203-271, самок - 201-242 мм; максимальная ширина достигает 137 и 118 мм соответственно. Отличия особей европейской и казахстанских популяций по размерам черепа в целом незначительные, хотя и выражены по отдельным показателям [Проняев, 1985а; Присяжнюк и др., 1986]. Линейные параметры особей из Монголии заметно меньше, но длина верхнего ряда зубов у исследованных форм одинакова, что для копытных является надежным видоспецифическим признаком. В целом вид представляется фенотипически однородным с прерывом постепенной изменчивости лишь в монгольской части ареала (табл. 19).
Изменчивость рогов. Костные шипики на лобных костях становятся заметными уже у двухнедельных сайгачат. У месячных детенышей бугорки вырастают до 1-2 см. У самцов рога прорезываются к исходу восьмой недели, иногда—раньше. К 6—7-месячному возрасту они отрастают до 8—13 см и начинают очищаться от эпидермальных чехлов, постепенно светлея и приобретая восковую окраску и 1-2 кольцевых утолщения. К 20-21 мес. эти образования почти полностью развиты, число колец приближается к 12-14. У 1,5-летних особей в нижней трети роговых чехлов заметно отслоение наружного слоя, похоже на шелушение. У взрослых животных длина рогов обычно варьирует в пределах 19-38 см, а число колец доходит до 22 [Адольф, 1954; Соколов, Рашек, 1961; Рашек, 1963; Черченко, 1975; Проняев, Фандеев, 1998]. Средняя масса одного рога близка к 192 (140-240), масса рогового чехла - около 100 (60-130) г [Размахнин и др., 1976; Размахнин, Рывкин, 1976]. Средний вес пары рогов составляет около 400-440, роговых чехлов - 217-235 г [Слудский, 1955; Раков, 1956; Присяжнюк, Проняев, 1982].
Несмотря на грандиозный промысел сайги в последние десятилетия, когда животных добывали сотнями тысяч, а рога вывозили из страны буквально вагонами, и при многочисленности ученых-охотоведов, работающих с этим видом, я, к удивлению, не встретил ни одной заслуживающей внимания публикации, посвященной географической изменчивости рогов. Весь имеющийся скудный материал укладывается в небольшую таблицу (табл. 20). Рога монгольской сайги, видимо, несколько отличаются от рогов особей номинативной формы: они меньше по размерам, тоньше, кольца на них выражены слабее [Банников, 1954].
Таблица 20. Размеры рогов (см) взрослых особей сайги
Длина- Район (источник информации)*
П | М | lim
53 30 26-38 Калмыкия (2)
31 32,5 28-38 -“-(4)
182 29-31 25-35 Казахстан (1)
16 35 33-37 Междуречье Волги и Урала (1)
45 30 19-37 Бетпакдала(1)
25 28 24-34 Барсакельмес (1)
50 31 27-36 ?, (5)
- - -22 Монголия (3)
* 1 - Фадеев, Слудский, 1982; 2 - Соколов и др., 1998; 3 - Соколов и др., 1996; 4 - Банников и др., 1961;
5 - Размахнин, Рывкин, 1976.
Окраска и линька. Летом окраска сайги рыжая или желтовато-рыжая с более светлой окраской нижней части тела. Зимний мех на туловище буровато-песчаного цвета с буро-коричневым пятном на крестце (фото 13). Шея темная, контрастная с окраской туловища. Низ туловища, внутренние части ног и круп белые, без резкого перехода от темных к светлым участкам. На щеках отрастают пучки длинных темных волос, образующие “бакенбарды”, а у самцов они есть и на задней кромке ушной раковины (фото 12). У взрослых самцов в зимнем наряде больше черного цвета, чем у самок, а бока шеи, щеки и пространство вокруг глаз - темно-серые. К весне мех выцветает и становится почти белым. Молодые особи имеют более темную окраску. С возрастом она светлеет из-за многочисленных белых остевых волос, выступающих над шерстным покровом. Иногда в стадах встречаются альбиносы (ягнята) и меланисты. Альбиносы, видимо, менее жизнеспособны, и не доживают, за редким исключением, до взрослого состояния. Географическая изменчивость в окраске не выявлена.
Ювенильный покров у детенышей начинает меняться на постоянный через 2-3 недели после рождения, и линька у них заканчивается в июле-августе. Взрослые особи линяют дважды в году: в конце марта-июне и сентябре-ноябре. На востоке ареала этот процесс начинается позднее на 10-15 дней, нежели на западе. Максимальная продолжительность смены волоса - 60-75 дней, но весенняя линька (фото 13) обычно протекает гораздо интенсивнее и почти в два раза короче осенней. Смена волоса начинается с головы и ног и заканчивается на боках, спине и брюхе. В разные годы сроки линьки могут сдвигаться на месяц в зависимости от погоды и кормовых условий. Молодые особи меняют волос примерно на неделю позже взрослых [Васенко, 1950; Даль и др., 1958; Банников и др., 1961; Рашек, 1963, 1965, 1974; Фадеев, Слудский, 1982; Слудс-кий и др., 1983].
Генетическая изменчивость. Кариотип сайги - один из самых примитивных среди Bovidae: 2п = 60, NF = 60. Все хромосомы, включая Y-хромосому, акроцентрические [Wurster, Benirschke, 1968; Проняев, 1985а; Проняев и др., 1986]. В публикации В.Н. Орлова и Н.Ш. Булатовой [1983] Y-хромосома описана как маленький метацентрик. Географической изменчивости кариотипа не обнаружено.
В прикаспийской и казахстанских популяциях не выявлено также каких-либо аллельных вариантов миоглобина. Гетерозиготность особей в них практически одинакова - 0,231 и 0,223 соответственно. Генетическое расстояние по Нею незначительное (0,02), что свидетельствует о слабой дивергенции популяций и возможном обмене генами между ними [Проняев, 1985а, 1986; Соколов и др., 1998]. В прикаспийской популяции, однако, в 80-е гг. XX в. на порядок (до 0,02) сократилось разнообразие редких аллелей и резко уменьшилась гетерозиготность, что связано с чрезмерной промысловой элиминацией [Проняев, Рожков, 1988; Проняев, 1998].
Генетическая изменчивость изолированной и малочисленной группировки монгольской сайги (S. t. mongolica) еще ниже. Различия между нуклеотидными последовательностями в 10 гаплотипах не превышают 3%, тогда как доля вариабельных сайтов в том же фрагменте мтДНК у номинативного подвида (S. t. tatarica) достигает 24%. Уровень изменчивости гаплотипов монгольской группировки составляет 0,75, в прикаспийской популяции - 0,87-0,93, индекс нуклеотидного разнообра
зия - около 3%. Ни один из выделенных у монгольской сайги специфических гаплотипов не встречается у номинативной формы, что однозначно свидетельствует об их генетической обособленности [Kholodova et al., 2000; Luschekina et al., 2000; Кузнецова, Холодова, 2001; Холодова и др., 2001, 2002, 2004].
Распространение
Восстановленный ареал. Древнейшие среднеплейстоценовые и нижнеплейстоценовые костные остатки сайги обнаружены на Аляске близ Фербенкса, в Восточной Сибири (в низовьях Колымы и Индигирки, на берегах рек Адыча и Большая Чукочья), в Западной Сибири на р. Тобол у с. Худяково, в Павлодарской области Казахстана, в Поволжье недалеко от Саратова и у с. Райгород Волгоградской области. В конце среднего плейстоцена эти копытные расселяются на запад вплоть до Атлантики, заселяют степной Крым, Предкавказье и проникают в Восточное Закавказье при сильном усыхании Каспия. Находки позднеплейстоценовых остатков особенно многочисленны и обширны по географии (рис. 16): Аляска, Северо-Западная Канада, Восточная Сибирь (включая Новосибирские и Ляховские острова, низовья Лены, Оленека, Яны и Колымы, бассейны Вилюя, Селенги, Ангары, Бирюсы и Енисея, окрестности Нижнеудинска, Абакана, Минусинска и Красноярска), Западная Сибирь (Кузнецкая котловина, окрестности Новосибирска и Омска, Алтай, Тюмень, низовья Иртыша и Оби), Северный, Средний и Южный Урал, Пре-
Рис. 16. Динамика ареала сайги (по [Барышников и др., 1998])
1 - восстановленный ареал за исторический период, 2 - основные очаги обитания в конце XIX - начале XX вв., 3 - область обитания в 50-60-с годы XX в., 4 - то же в 1975-1993 гг., 5 - места ископаемых находок (неполные сведения, см. текст)
дуралье, Казахстан, Северный Узбекистан, Восточная и Западная Европа до Британских островов и Греции включительно, но без Скандинавии, Пиренейского и Апеннинского полуостровов. В Восточной Европе остатки сайги найдены в Молдавии, на Южной Украине к северу до Киева и Харькова, в Крыму, на Кубани, в Предкавказье и Восточном Закавказье, на Среднем и Нижнем Поволжье, в бассейне Дона, близ Воронежа и Владимира, на берегах Оки, Камы и Верхней Печоры, в Пермском и Южном Приуралье. На палеолитических стоянках ее остатки часто встречаются вместе с костями мамонта, лошади, шерстистого носорога, овцебыка, бизона, лося, северного и благородного оленей, сибирской косули, кабана, кулана, волка, росомахи, песца, бобра, байбака и суслика, что свидетельствует о смешении тундровой, лесной, лесостепной и степной фаун [Громова, 1941, 1948а,б; Адольф, 1948, 1950; Громов, 1948; Верещагин, 1949, 1954, 1959а, 1963, 19756; Пидоплич-ко, 1951, 1954; Алекперова, 1953, 1955; Трофимов, 1955; Верещагин, Колбутов, 1957; Джафаров, 1960; Банников и др., 1961; Вангенгейм, 1961, 1976; Бибикова, 1963,1985; Кузьмина, 1966,1971,1975; Шер, 1967, 1971; Кожамкулова, 1969,1976, 1981; Цалкин, 1970; Демеуов, 1971; Савинов, 1972; Давид, Чемыртан, 1976; Ларичев, 1976; Смирнов, 1976а,б; Алексеева, 1977,1980,1990а,б; Оводов, Орлова, 1977; Галкина, Форонова, 1980; Верещагин, Барышников, 19806; Давид, 1980; Лазарев, 1982, 1990; Delpech, 1983; Слудский и др., 1983; Маслов, Антипина, 1984; Батыров, 1986; Бурчак-Абрамович, 1986; Худайбердиев, 1986; Барышников и др., 1989, 1990, 1998; Тлеубердина и др., 1989; Смирнов и др., 1990; Абрамова и др., 1991; Бобковская, Косинцев, 1991; Kahlke, 1991; Барышников, Крахмальная, 1994; Барышников, 1995; Васильев, 1995, 2003а,в; Малеева и др., 1995; Косинцев, 1996, 2003; Пономарев, 2001,2003; Форонова, 2001; Хромов и др., 2001; Косинцев, Бобковская, 2003; Подопригора, 2003; Улитко, 2003; Боескоров, 2004].
В плейстоцене, как считалось, сайга отсутствовала в восточной части “аридного пояса” Азии: в Забайкалье, Монголии и Китае [Барышников, Крахмальная, 1994; Барышников и др., 1998]. Этот факт долго не находил разумного объяснения. Скорее всего, зоологи не обратили внимания на сообщения об обнаружении позднепалеолитических и неолитических остатков вида в Забайкалье (в бассейне Селенги вместе с винторогой антилопой Spirocerus kiakhtensis и дзереном, на стоянке Сухо-тино-4 в черте Читы, на Ононе в поселении каменного века Чиндант недалеко от устья р. Борзи и в поселении Арын-Жалга), а также в Китае в пещерах Чжоукоу-дянь неподалеку от Пекина [Громов, 1948; Верещагин, 1954; Макеев, 1963; Окладников, Кириллов, 1980; Каспаров, 1986]. Со временем плейстоценовые остатки сайги, скорее всего, будут найдены и в Монголии. Голоценовые остатки вида здесь уже извлечены из погребений древних монголов (XII - начало XIII вв. н. э.) на юго-востоке страны в Дариганге [Динесман и др., 1986], что далеко за пределами современного очагового ареала. В монгольских горных степях сохранилось и много наскальных рисунков, относящихся к бронзовому веку, что подтверждает прежнее широкое распространение этих копытных (рис. 17) [Лущекина, 1997; Лущекина, Дуламцэрэн, 1997].
Резкое сокращение огромного ареала вида, занимавшего почти всю Северную и Центральную Азию, Европу и северо-западную часть Америки, происходит на рубеже плейстоцена и голоцена, около 10—15 тыс. лет назад. В раннем голоцене сайга уцелела лишь на юге Восточной Европы и в Центральной Азии [Барышни-
Рис. 17. Распространение сайги в Монголии (по [Лущекина, Дуламцэрэн, 1997])
1 и 2 - районы обитания и пункты забегов в середине XX в., соответственно (по [Банников, 1954]); 3 - место обнаружения голоценовых костных остатков (по [Динссман и др., 1986]); 4 - места обнаружения петроглифов с изображениями сайги (по сообщениям Л.Г. Динссмана)
ков, Крахмальная, 1994; Барышников и др., 1998], но численность продолжала уменьшаться. В археологических памятниках Крыма и Причерноморья с позднего плейстоцена и до конца раннего голоцена доля ее костей среди других копытных животных снижается в несколько раз [Бибикова, 1975, 1984, 1985]. Тем не менее в голоцене сайга населяла еще почти все лесостепные, степные и полупустынные районы юго-востока Европы от Воронежской области до Крыма и Северного Кавказа, Приуралья и Зауралья, юга Западной и Восточной Сибири (на севере - до окрестностей Красноярска), Казахстана, Средней Азии, Монголии и северо-запада Китая. В IV тысячелетии до н. э. этот вид жил на юге Туркмении, во II - в Ферганской долине. В Х-Ш вв. до н. э. он был одним из основных охотничьих зверей в Забайкалье и предгорьях Алтая. На Юго-Восточном Алтае его наскальные изображения имеются в урочище Туюк-Гобо [Ешелкин, 1972]. В Минусинской котловине кости сайги найдены в погребениях человека XVIII-XIV вв. до н. э. В Западной Сибири в III—I тысячелетиях до н. э. северная граница ареала доходила до Омска и включала Приишимскую лесостепь и Барабинские степи. В Зауралье в VI-IV вв. до н.э. вид был распространен на север до оз. Аргази в Аргаяшском районе Челябинской области. Эпизодически звери заходили и в южные районы лесной зоны Среднего Урала [Пидопличко, 1951, 1954; Кузьмина, 1966; Верещагин, Батыров, 1967; Деме-уов, 1968, 1971; Цалкин, 19726; Смирнов, 1976а,б; Косинцев, 1982, 1986, 1988; Слудский и др., 1983; Косинцев, Ражев, 1985; Каспаров, 1990; Смирнов и др., 1990; Жирнов и др., 19986].
В Восточной Европе в последние тысячелетия до н. э. -1 тысячелетие н. э. область обитания сайги простиралась к западу до Карпат, к северу, как минимум, - до Киева, Курска, Тамбова, Самары и Уфы, к востоку - до р. Урал, к югу - до Черного и Азовского морей, Крыма, предгорий Кавказа и Восточного Закавказья [Пщошпчко, 1956; Цалкин, 1958,1960,1966,1969; Верещагин, 1959а; Банников и др., 1961; Биби
кова, 1963; Давид, Чемыртан, 1974,1976; Шилов, 1975; Антипина, 1982]. Ее костные остатки, датируемые V-XI вв. н. э., найдены при раскопках под Киевом, Харьковом, Воронежем, в Старой Рязани, а также в городище Балымеры на левом берегу Волги севернее Ульяновска и в Именьковском городище близ Казани (в Куйбышевском и Лаишевском районах Татарии) [Цалкин, 1956, 1958, 1963; Шрамко, 1962; Андреева, 1972; Тимченко, 1972; Андреева, Петренко, 1976; Петренко, 1984]. Позднее сайга исчезает из приокских степей, а в начале XVIII в. ее не стало в Молдавии (последние сообщения о встречах зверей относятся к 1716 г.). Тем не менее в первой половине XVIII в. область распространения вида еще обширна: северная граница шла от низовий Дуная и предгорий Карпат на Киев, Курск, Тамбов, Самару, Уфу, Оренбург, Орск, южная - по берегам Черного и Азовского морей, по правобережью Кубани, низовьям Терека и побережью Каспийского моря до Дербента. Особенно много зверей было на землях запорожского казачества. В 60-80-х годах сайгу во множестве добывали в степном Крыму и степях Таврии. В 1769 г., по описанию П.С. Палласа [1809], многочисленные стада доходили до р. Бузулук и переходили за Самару. Позднее численность резко сократилась после многоснежных буранных зим 1788/89 и 1805 гг. К началу XIX в. этих копытных практически полностью истребили в междуречье Днестра и Буга, затем в причерноморских степях, а к середине столетия - на всей Украине, где вскоре исчез и тарпан. К востоку от Дона звери еще жили “огромными стадами”, особенно в степях и полупустынях Нижнего Поволжья. Тысячные стада переправлялись через Волгу против г. Камышина и г. Саратова. К северу животные проникали до рек Большой Иргиз, Самара и вглубь Башкирии, южная граница ареала шла по кромке степей в Прикаспии и Предкавказье. В январе 1850 г. в прикумских степях зверей “можно бы сотни тысяч насчитать”, но после этой суровой зимы большая часть европейской популяции погибла. Возможно, отчасти поэтому до 80-х годов сайга была сравнительно редкой и в Волго-Уральском междуречье, хотя Н.А. Северцов [1861] считал, что она вытеснена отсюда человеком [Богданов, 1871; Анучин, 1875; Дементьев, 1935; Адольф, 1948, 1957; Бакеев, Формозов, 1955; Раков, 1956; Кириков, 1959, 1960, 1966, 1976, 1983; Банников и др., 1961; Жирнов, 1982; Динесман, 1998; Жирнов и др., 1998а].
В Казахстане в историческое время сайга обитала почти повсеместно, а изменения границ ареала вплоть до XIX в. были незначительными [Черников, 1960; Демеуов, 1968, 1971; Кожамкулова, 1969, 1976, 1977, 1981; Макарова, 1970, 1973, 1976; Фадеев, Слудский, 1982; Слудский и др., 1983; Динесман, 1998]. На юге ее распространение ограничивалось восточным берегом Кара-Бугаз-Гола, нижним течением Амударьи, средним течением Сырдарьи, предгорьями Каратау, Киргизского и Заилийского Алатау и р. Или. На севере линия распространения проходила близко от границы с Россией (рис. 18). На востоке весной стада переплывали через Ишим и Иртыш и перекочевывали на лето в барабинские степи, множество животных жило в притобольской степи и вблизи Омской крепости, а южнее они достигали предгорий Алтая. Во второй половине XIX в. северная граница обитания сместилась к югу на линию Уральск - Тургай - Баян-Аул - хр. Чингизтау - оз. Зайсан, южная осталась почти без изменений [Паллас, 1786; Эверсман, 1850; Карелин, 1883; Зарудный, 1897; Антипин, 1941; Адольф, 1948; Васенко, 1950; Слудский, 1955; Банников и др., 1961; Кириков, 1959, 1961, 1963, 1966; Цалкин, 1966; Жирнов, 1982; Жирнов и др., 1998а].
Рис. 18. Распространение сайги в Заволжье, Оренбургских степях и Казахстане в XVIII - середине XX вв. по историческим сведениям (по [Кириков, 1966])
1 - места встреч в 60-70 годы XVIII в.; 2 и 3 - то же летом и зимой, соответственно, в первой половине XIX в.; 4 и 5 - то же в 1959 г.
В степном Алтае, а также в аргуно-ононских, селенгинских и даурских степях Забайкалья сайга, возможно, дожила до конца XVIII в., причем в селенгинских степях путешественники видели “превеликие стада”, а в даурских ее численность была небольшой, и здесь водилось много дзеренов и куланов [Кириков, 1959,1966; Смирнов, 1978, 1983, 1994]. Не исключено, что сайгу путешественники путали с дзереном, но факт ее прежнего обитания на юге Сибири очевиден. В XIX - начале XX вв. этот вид еще считался обычным в Зайсанском крае, был многочисленным в степях и предгорных районах Джунгарии и Монгольского Алтая и встречался близ Гобийского Алтая и в Западной Гоби [Грум-Гржимайло, 1922; Певцов, 1951; Банников, 1954].
Динамика ареала в XX в. В начале XX в. становится заметным сокращение численности и области распространения сайги, на что обращают внимание многие исследователи, но она еще была фоновым видом на севере Средней Азии и в Казахстане, а в Европе обитала в калмыцких, астраханских, кумо-манычских и прикумских степях [Динник, 1910; Сатунин, 1914].
Минимальным ее ареал (рис. 16) был во втором и третьем десятилетиях [Банников и др., 1961; Фадеев, 1974; Жирнов, 1982; Жирнов и др., 1998], но, как показывает анализ литературы, он все, же шире, чем принято считать. В европейской части эти животные встречались не только в калмыцкой степи. В 1922-1928 гг. их отмечают в небольшом количестве в разных районах Самарской губернии, по Малому Иргизу, в караногайской степи, в Астраханском заповеднике, в волго-уральских песках, а в волго-кумских степях они были обычными [Берков, 1927; Сытин, 1927; Воробьев, 1929; Дементьев, 1935; Гептнер, Формозов, 1941; Адольф, 1948; Слудский, 1955; Кириков, 1966]. В Азии эти копытные уцелели на Устюрте, по р.
Рис. 19. Область обитания европейской популяции сайги в 1950-1994 гг. (по [Жирнов и др., 19986])
1 - зона обитания в 1950-1965 гг.; 2 - границы распространения в периоды засух и многоснежных зим; 3 - зона обитания в 1988-1994 гг.; 4-6 - районы массового отела в 1957-1961, 1966-1975 и 1990-1994 гг., соответственно
Сарысу, в Бетпакдале, южной части Семипалатинской губернии, в Акмолинской и Актюбинской губерниях, на острове Барсакельмес, низовьях р. Или, в Прибалхашье, по Иртышу, в причуйских степях (по сведениям Н.А. Зарудного, здесь звери были многочисленны зимой 1915 г.), к северо-западу от хребта Каратау, у Алакуля, оз. Зайсан, к востоку от Ашхабада и в других районах, а также в отдельных очагах в Западной Монголии и в Джунгарии [Барышев, 1925; Горчаковский, 1925; Шпота, 1928; Дементьев, 1935; Тимофеев, 1934; Антипин, 1941; Адольф, 1948; Кузнецов, 1948; Слудский, 1955; Раков, 1956].
В 30-е годы ареал постепенно восстанавливается, а в 40-е отмечено появление мелких стад на северо-востоке Ставрополья, к югу от р. Кумы в Грозненской
области, в Ростовской области, возле Сталинграда и Вольска. В Астраханской области стали образовываться сотенные и тысячные стада. Особенно многочисленны они были в приманычских и прикумских степях. В 1950 г. границы ареала еще более расширяются в результате массового выселения сайги из традиционных мест обитания после сильного бурана, случившегося 9-11 января. В северные районы Грозненской области зашло до 3000 особей. Сюда же десятки тысяч животных откочевали многоснежной зимой 1953/54 гг. Отдельные мигрирующие стада по побережью Каспия достигли территории Дагестана (г. Махачкалы), пересекли Аграханский полуостров и по льду вышли на острова Чечень и Тюлений. В результате этого вселения образовались новые очаги обитания в Кизлярском и Крайновском районах, на Аграханском полуострове и на востоке Сулакс-ко-Терской низменности, впоследствии исчезнувшие из-за массовой браконьерской охоты [Адольф, 1948, 1950, 1957; Бакеев, Формозов, 1955; Слудский, 1955; Рак, 1956; Жирнов, 1982; Жирнов и др., 19986].
К середине 50-х годов XX в. на правобережье Волги сайга занимает почти все пригодные стации, включая район Ергеней, сальские и кумо-манычские степи, а к концу десятилетия область распространения европейской популяции достигает максимума в 100-120 тыс. км2 (рис. 19). В 60-е - начале 70-х годов площадь ареала сокращается в 2-3 раза (до 46-30 тыс. км2) вслед за уменьшением популяции, и затем по мере ее роста снова расширяется до 87 тыс. км2 к 1979 г., охватывая территорию Калмыкии, запад Астраханской области, северо-восток Ставропольского края, восток Ростовской области и север Дагестанской АССР [Близнюк, 1980, 1982]. В Дагестане в 70-80-е годы насчитывали от 5 до 20 тыс. особей [Равкин, 1976; Газалиев, 1980а; Спасская, Абдулманапов, 1980; Прилуцкая, Пиш-ванов, 1988]. В 80-е годы западная и северная границы снова сдвинулись на 100— 200 км, что опять-таки связано с очередным сокращением численности, и к середине 90-х площадь зоны обитания вида уменьшилась более чем в 7 раз - до 17 тыс. км2. В конце XX - начале XXI вв. животные концентрировались в юго-восточной части Калмыкии в районе Черных земель. Большая часть популяции размещалась в единственном относительно безопасном месте - в заповеднике “Черные земли” (рис. 20) [Близнюк, 1995; Лещенко, 1997; Букреева, 1998, 2002; Жирнов и др., 19986].
В Волго-Уральском междуречье и на всей азиатской части, за исключением Монголии, изменение ареала было примерно сходным. Процесс его восстановления очевиден с 30-х годов. В Урало-Эмбенской степи зимой 1933-1934 гг. отмечено массовое появление зверей. В группах мигрантов насчитывали от 20 до 150 особей [Дементьев, 1935]. В середине 40-х повсеместно стали встречаться тысячные стада, и к началу 50-х годов северная граница распространения вида проходила уже по линии Саратов - Уральск - Орск - Атбасар - Киевка - горы Кент - хр. Чингиз-Тау - оз. Зайсан, южная - через Мангышлак, по северу Туркмении и Узбекистана, предгорьям Тянь-Шаня, Джунгарского Алатау и долине Черного Иртыша [Басенко, 1950; Соколов, 1951; Слудский, 1955; Залетаев, 1956; Раков, 1956; Ишадов, 1966, 1972, 1975а, 1999а; Фадеев, 1974]. В первой половине 60-х годов сайга появляется в Актюбинской, Кустанайской, Тургайской и Целиноградской областях и других районах Казахстана. В этот период ее ареал приближается к максимальному, хотя из-за возрастающей хозяйственной деятельности человека животные не
смогли “закрепиться” в Зай-санской котловине, на левобережье Иртыша и в Алакульс-кой котловине [Фадеев, Слуд-ский, 1982; Жирнов и др., 19986].
В дальнейшем область обитания вида уменьшается на севере из-за распашки целинных степей. В 80-е годы обособляются 3 очага: волгоуральский, устюртский и бет-пакдалинский. В конце XX -начале XXI вв. ареал сайги в Казахстане стремительно сокращается. Животных не стало в Северном и Южном Прибалхашье, Чуйских Муюнку-мах, на большей части При-аральских Каракумов и восточном побережье Каспийского моря, а уцелевшие группировки обособились еще более территориально (рис. 21) и раздробились на мелкие стада [Bekenov et al., 1998;
Рис. 20. Область обитания европейской популяции сайги в Lushchekina Struchkov 2001 ’ конце XX в. (по [Кузнецов, Лущекина, 2002]) Лущекина, 2002; Грачев, Беке-
1 - граница распространения во время засух и суровых зим, 2 - пааоз
основная область обитания в 1990-2000 гт., 3 - административ- НОВ, ZUUJJ.
ная граница Калмыкии, 4 - основные места отелов МОНГОЛЬСКИЙ участок аре-
ала оказался изолированным, видимо, еще в XIX в. В первой половине XX в. сайга обитала в котловине Больших Западных озер: Хиргиз-Нур, Хара-Нур, Хараусу-Нур, Дургэ-Нур и южнее - в Шар-гин-Гоби и Хуйсин-Гоби. Отдельные нерегулярные заходы отмечались к юго-востоку от этих районов и на север в южную часть котловины Убсу-Нур [Банников, 1946, 1951, 1954]. В 60-80-е годы происходит резкое сокращение области обитания. Животные уцелели лишь на двух небольших участках (рис. 17): в северных предгорьях Монгольского Алтая в Шаргин-Гоби и в районе Манхан-сомона [Цэвэгмид, 1970; Соколов, Орлов, 1977, 1980; Сапожников, Дуламцэрэн. 1982; Соколов, 1986; Соколов и др., 1986, 1996; Дуламцэрэн, Амгалан, 1995; Лущекина, Дуламцэрэн, 1997; Жирнов и др., 1998]. В июле 1981 г. группа из 10 особей обнаружена в котловине Барун-Хурай [Жирнов, Ильинский, 1985]. К концу 80-х годов площадь ареала сайги в Монголии сократилась примерно до 2-5 тыс. км2. В начале 90-х в связи с ростом поголовья очевидна тенденция расширения области обитания, площадь которой к 1998 г. достигла 10 тыс., а в 2000 г. - 13,3 тыс. км2, в том числе в Шаргин-Гоби - 4,1, Хуйсин-Гоби - 5,9, в районе Дургун-Чандмани и Манхана - 2,9 и 0,4 тыс. км2. Зимой
Рис. 21. Ареал сайги в конце XX в. (по [Кузнецов, Лущекина, 2002])
1 - области обитания популяций: I - европейской, Па-с - волго-уральской, устюртской и бетпакдалинской, соответственно, Ша,Ь - монгольских; 2 - граница распространения вида в историческое время (по [Соколов, Жирнов, 1998])
1991 г. двух зверей видели намного восточнее - в северных предгорьях хр. Гурван-Сайхан [Соколов и др., 1996]. В самом начале XXI в. численность монгольской группировки снова резко уменьшилась [Dulamtseren, Amgalan, 2002], что может привести к очередному сокращению ареала. В Китае сайга полностью истреблена еще в 50-е годы XX в. [Джиган, Дикян, 1998].
Факторы, определяющие область распространения. Резкое сокращение ареала сайги в постплейстоценовое время исследователи связывают с потеплением и увлажнением климата Евразии, заменой тундростепных сообществ лесными, заболачиванием и повсеместным увеличением высоты снежного покрова [Пидоп-личко, 1953; Верещагин, 1971; Бибикова, 1975; Верещагин, Барышников, 1985; Барышников и др., 1998]. В этот период, как известно, вымирают многие представители “мамонтовой” фауны, а уцелевшие степные животные вынуждены отступать или сезонно мигрировать к югу. Обилие сайги в “кухонных отбросах” палеолитических стоянок человека в Крыму может косвенно свидетельствовать о концентрации вида здесь в зимнее время [Громов, 1948; Раков, 1963].
В раннем и среднем голоцене она населяет ксерофитные ландшафты. Проникновение на запад и на север Русской равнины, видимо, ограничивают высокий снежный покров, леса и высокотравная степная растительность. Некоторому расширению ареала в I тысячелетии н. э. могли способствовать ксерофитизация степей и их зональная пастбищная дигрессия в связи с развитием скотоводства, а также периодические эмиграции стад на север и запад в засушливые годы [Динесман, 1982а,б; Жирнов, 1982; Жирнов и др., 1998].
В XVII-XIX вв. на юге Восточной Европы быстро растет людское оседлое население, и бурно развивается земледелие и скотоводство. Лесостепи и степи интенсивно распахиваются и выбиваются скотом, естественные биотопы сайги уничтожаются, резко возрастает охотничий пресс [Кириков, 1963, 1983; Близнюк, 1995]. Такие же экологические тенденции, усугубляемые джутами, характерны и для азиатской
части ареала вида. Все это негативно отражается и на численности, и на области распространения копытных. Основной причиной катастрофического сокращения поголовья в начале XX в., помимо неумеренного охотничьего преследования (см. ниже), стали джутовые зимы 1917/18 и 1927/28 гг. Восстановление ареала и численности в 20-50-е годы XX в. явно связано с комплексом благоприятных факторов [Бажанова, 1955; Слудский, 1955, 1974; Раков, 1956; Банников, 1958; Верещагин, 1959а; Смирнов, 1965; Жирнов, 1982; Цаплюк, 1982; Слудский и др., 1983]:
- запретом охоты (декрет от 27 мая 1919 г. “О сроках охоты и о праве на охотничье оружие” и постановление Президиума ВЦИК РСФСР от 24 августа 1921 г. “Правила производства охоты, ее сроки и способы”);
- изъятием у населения нарезного оружия и улучшением охраны животных;
- ликвидацией контрабанды и уменьшением спроса на рога (их некуда было сбывать);
- сокращением хозяйственной деятельности и домашнего скота в годы коллективизации;
- переходом кочевников на оседлый образ жизни;
- уменьшением уровня браконьерства в годы войны, когда большинство охотников были на фронтах;
- сокращением численности волка в послевоенные годы;
- мягкими зимами без джутов;
— исчезновением кожного овода (Pallasiomyia antilopum Pall.) после сокращения поголовья сайги в 20-е годы.
В середине XX в. сайга вновь заняла почти все пригодные для жизни пустынные, полупустынные и степные стации. Дальнейшее ее расселение и в европейской, и в азиатской части ареала ограничивали районы интенсивного земледелия с густой сетью населенных пунктов. Сокращение области обитания вида в 70-е годы вызвано, главным образом, усилением антропогенного пресса. В 80-е годы границы распространения существенно не менялись, но площадь пригодных биотопов значительно уменьшилась из-за распашки степей, огораживания пастбищ, строительства поселков, оросительных каналов, дорог, военных и хозяйственных объектов. В 90-е годы в результате резкого снижения численности ареал снова заметно сократился и стал очаговым. Причины этого явления обсуждаются ниже.
Основные факторы, определяющие динамику ареала вида в Монголии в последние столетия, практически такие же, что и в бывшем СССР - нелегальная охота и хозяйственное освоение территории в сочетании с гибелью зверей в многоснежные зимы.
Среда обитания
Сайга - типичный обитатель аридных ландшафтов, но мнения исследователей в отношении ее зонального экотипа сильно расходятся. Этих копытных считают степными животными [Кучерук, 1959], жителями северных пустынь [Васенко, 1950; Афанасьев, 1960] или полупустынь [Берггрин, 1948; Банников, 1954; Слудский, 1955; Банников и др., 1961; Жирнов, 1982, 1998а,б].
На мой взгляд, экологический тип вида не укладывается в определенные “зональные” рамки. В эволюционном плане в его ареале прослеживается смена ландшафтов - от тундростепей и северных пустынь в плейстоцене до разнотравно-тип-
Рис. 22. Районы сезонного обитания и основные пути миграций казахстанских группировок сайги (по [Жирнов, 1982])
1 - районы летовок; 2 - районы зимовок; 3 - основные пути миграций; 4-8 - северные границы степной зоны, зоны полупустынь, пустынь умеренного пояса, субтропических пустынь и горных областей, соответ-
ственно
чаково-ковыльных степей в голоцене и сухих степей, полупустынь и пустынь в историческое время. Кроме того, сезонно мигрирующие животные регулярно переходят из одной зоны в другую (рис. 22).
Зону пустынь все же вряд ли можно отнести к основным биотопам сайги в связи с коротким периодом вегетации растительности, малой продуктивностью и дефицитом водопоев. Тем не менее азиатские популяции 4-8 мес. в году обитают в пустынях, а на о. Барсакельмес - круглогодично. Здесь животные в течение года используют практически все стации, что свидетельствует о значительной широте нормы реакции вида [Рашек, 1974]. Степные пастбища с господством рано заканчивающих вегетацию дерновинных злаков и разнотравья тоже характеризуются низким кормовым качеством и используются копытными лишь в короткий весенний и раннелетний периоды. Наиболее оптимальны для сайги пустынно-степные фитоценозы, распространенные в зоне полупустынь. Они отличаются большим разнообразием (более 100 видов) и обилием хорошо поедаемых и длительно вегетирующих двудольных растений (трав и полукустарничков), что обеспечивает почти круглогодичное присутствие зеленой фитомассы с высоким содержанием протеина. В таких фитоценозах даже сильное пастбищное воздействие домашнего скота не вызывает ощутимого ухудшения их качества для сайги. Привлекательными для нее оказываются и ценозы, формирующиеся на распаханных землях, где обильно разрастаются длительно вегетирующие виды сор
ных растений, охотно потребляемые животными [Субботин, Абатуров, 1999; Субботин, 2001; Абатуров, 2004].
В европейской части ареала основные биотопы сайги находятся в равнинной Прикаспийской низменности. С осени до весны эти копытные живут преимущественно в зоне полупустынь, летом заселяют сухие и дерновинно-злаковые степи. В Казахстане сезонные скопления животных и пути их миграций в большей степени приурочены к равнинам и сильно сглаженному рельефу полупустынь Устюрта, Бетпакдалы, Иргиз-Тургайского междуречья, района оз. Тенгиз. Появление зверей в высокотравной лесостепи, бугристых песках, зарослях кустарников и тростника, предгорьях, горах, горных ущельях и плато, вблизи городов или на льду и островах Каспия обычно вызвано неблагоприятными условиями жизни: недостатком корма и воды, обилием насекомых, пожарами, засухами, глубокоснежьем, буранами, гололедицами [Раков, 1956; Банников и др., 1961; Капитонов, 1980; Жирнов, 1982]. В предгорных районах Монгольского Алтая весьма велик верхний предел вертикального диапазона распространения сайги - 1900-2000 м над уровнем моря. Область ее обитания здесь лежит в границах зон петрофитных ковыльково-солян-ковых, солянково-ковыльковых и баглурово-луково-ковыльковых степей, а западнее Дарви животные заходят в караганово-ковыльковые степи. Ранее они держалась и в злаково-солянковых полупустынях и солянковых пустынях [Банников, 1954; Сапожников, Дуламцэрэн, 1982; Соколов и др., 1986].
Одна из характерных черт экологии вида - сезонная смена участков обитания. Основные факторы, влияющие на распределение стад по территории в весенне-летнее время, - наличие влажного корма и водопоев. В засушливый период животным приходится широко перемещаться в их поисках. Поскольку более влажные корма находятся на севере ареала, то вполне понятной становится причина весенних миграций в северном направлении. На традиционных путях миграций звери нередко попадают в сельскохозяйственные зоны, образованные на распаханных целинных землях, где наносят некоторый ущерб (часто преувеличенный) лесозащитным полосам и вегетирующим культурам. Это неизбежно приводит к усилению преследования копытных [Бакеев, Формозов, 1955; Банников и др., 1961; Бакеев, 1964; Голованова, Анохина, 1976; Жирнов, 19776, 1982; Жирнов, Калецкий, 1976; Жирнов и др., 1977; Фадеев, Слудский, 1982; Слудский и ДР-, 1983].
Зимой размещение стад во многом определяется высотой и плотностью снежного покрова, а также силой ветра. Перед снежными буранами и в сильный ветер они стремятся скрыться в зарослях тростника и кустарников или уходят в бугристые пески. Сайга не способна быстро передвигаться и добывать пищу при плотном снеге высотой более 20 см. Критический уровень при рыхлом снеге - 35-40 см, когда раздвигание его мордой и тебеневка уже невозможны. В типичных зимних местах обитания снежный покров обычно не превышает 10-15 см, но даже такой слой при его уплотнении и, особенно, при гололеде, делает недоступными низкорослые после засух растения. Удельная нагрузка на след у этих копытных сравнительно велика - 530-860 г/см2 в зависимости от возраста и пола. При перемещении по глубокому снегу звери вынуждены идти цепочкой друг за другом по тропам, что не характерно для них, или использовать дороги, проложенные людьми [Слудский, 1963; Рашек, 1965, 1974; Фадеев, Слудский, 1982]. Малоснежные
кормные участки находятся на юге ареала, поэтому причина осенних миграций стад в южном направлении также легко объяснима.
Многовековые сезонные миграции животных с юга на север весной и обратно осенью, скорее всего, уже закреплены на генетическом уровне, тогда как локальные кочевки и стремительные перемещения в разных направлениях спонтанны и вызваны неблагоприятными условиями обитания. Постоянные перемещения в значительной мере предохраняют пастбища от сильного стравливания, что обеспечивает устойчивое существование экосистемы “копытные - растительность” [Кучерук, 1963; Фадеев, Слудский, 1982; Абатуров, 1984].
В последние столетия, в результате интенсивной хозяйственной деятельности человека, значительная часть лучших пастбищ утрачена: распахана, перегорожена, изрыта многочисленными каналами, покрыта дорогами и всевозможными сооружениями. Мощным пастбищным конкурентом и источником беспокойства стал домашний скот. В результате постоянного охотничьего преследования сайгу не только в массе истребляют, но и вытесняют в худшие биотопы (см. ниже). В целом среда обитания этого вида, к сожалению, стремительно ухудшается, что негативно отражается на состоянии популяций.
Питание
Корма. Основу питания сайги составляет пустынно-степное разнотравье, эфемеры, эфемероиды и пустынные полукустарнички. Список кормовых растений невелик - по моим подсчетам, не более 160 видов на всем ареале. Для отдельных популяций он меньше почти в два раза, а по сезонам количество потребляемых видов сокращается до 1-3 десятков. Более 13% видов растений из этого списка -ядовитые, горькие, с избытком солей или резким запахом, которые не поедает или плохо поедает домашний скот [Банников и др., 1961; Жирнов, 1982; Слудский и др., 1983; Абатуров и др., 1987; Жирнов и др., 1998].
В Северо-Западном Прикаспии сайга из 125 поедаемых видов растений выбирает преимущественно прутняк, грудницу, люцерну, кермек, горец, черную и белую полыни, эфедру, анабазис и, реже, пырей, костер, полевичку малую, мятлики, солодку, типчак, житняк, комфорсому, солянки, ковыли, марь белую, лапчатку двувильчатую, лук гусиный и молочай [Адольф, 1950, 1954; Бакеев, Формозов, 1955; Лебедева, 1959, 1960; Банников и др., 1961; Абатуров, Петрищев, 1998; Близнюк, Бакташева, 2001]. Набор ее кормов в Казахстане также небольшой -немногим более сотни растений, из которых основные примерно те же, что и на европейской части ареала с преобладанием полыней, биюргуна, эфедры, прутняка, солянок, кохии, ромашника и саксаула в осенне-зимний период и некоторых злаков - в летний. На острове Барсакельмес эти копытные из 130 видов местной флоры потребляют всего 26, из которых 20 относятся к 3 семействам: маревым, злаковым и сложноцветным. Наиболее предпочитаемы следующие растения: кохия, эбелек, лебеда, солянки, мятлик, пыреи, ковыли, полыни, горчак, изень, би-юргун, сарсазан и саксаулы. Состав пищи сайги здесь близок к таковому у верблюда [Басенко, 1950; Демченко, 1950; Соколов, 1951; Раков, 1956; Рашек, 1965, 1974; Мамбетжумаев, 1969; Демеуов, 1971; Фадеев, Слудский, 1982; Слудский и др., 1983; Фадеев, Иванов, 1988]. В Монголии эти животные поедают около 20 видов, пре
имущественно гобийский ковылек, луки, баглур и терескеп [Банников, 1954], но эти сведения, видимо, не полные.
В рационе сайги во все сезоны преобладают несколько видов растений, доминирующих в растительном покрове: летом - прутняк, грудница, люцерна, горец, солодка, солянки, эфедра и верблюжья колючка, сохраняющие наибольшее количество влаги; в осенне-зимний период - прутняк, солянки, эфедра, полыни (иногда до 60-70% содержимого желудков), зеленые листья мятлика, а в пустынях - веточки саксаула и кусты анабазиса. Наиболее предпочитаемы длительно вегетирующие двудольные растения пустынно-степных фитоценозов. Злаки, господствующие в полупустынях, составляют все же небольшую по объему долю пищи, а большинство видов не входит в состав качественных кормов. В весеннем питании на большей части ареала наиболее важны зеленые эфемеры, среди которых доминируют тюльпаны (до 89% в рационе) [Соколов, 1951; Субботин, Абатуров, 1999; Субботин, 2001; Абатуров, 2004].
При недостатке корма животные вынуждены поедать тростник, рогоз и даже побеги и листья деревьев и кустарников в лесополосах: терескена, тамариска, лоха, дуба, вяза. В джуты они подходят к стогам или собирают сено, рассыпанное на дорогах, заходят на деревенские огороды, где обгрызают капустные кочерыжки, едят картофельные и помидорные плети. В засухи эти копытные заходит на зеленые поля кукурузы, зерновых культур, люцерны, эспарцета и горчицы, частично поедая и вытаптывая урожай. Однако они избегают посевов выше 15-20 см и на зерновых полях охотнее поедают сорняки [Бакеев, Формозов, 1955; Слудский, 1955; Рак, 1956; Банников и др., 1961; Динесман, 1961; Бакеев, 1964; Голованова, Анохина, 1976; Жирнов, Калецкий, 1976; Фадеев, Слудский, 1982; Слудский и др., 1983].
Потребление кормов и воды. Минеральное питание. Сайгачата начинают срывать траву в 3-4-дневном возрасте, а через 15 дней растительный корм составляет около 95% объема желудка. В 3-месячном возрасте состав потребляемых ими растений примерно такой же, как и у взрослых животных, но меньше по объему - 170-430 г сухого корма в сутки; 4—5-месячные особи потребляют около 700-800 г; взрослые -1,2-1,7 (0,5-2,0) кг в среднем в сухом виде и от 2 до 7 кг - в сыром. Зимой потребление корма уменьшается в 1,4—2,4 раза (в пересчете на сухой вес), что связано со снижением интенсивности метаболизма. У беременных самок потребление корма на 15-30%, а у лактирующих - на 50-60% выше, чем у других особей. Самцы во время гона почти не кормятся. Пороговая величина переваримости, не приводящая к дефициту энергии в организме взрослых не размножающихся особей, составляет 55-59%, у беременных и лактирующих самок и растущего молодняка - 68-75%. Для поддержания жизнедеятельности животных переваримость корма должна быть не ниже 59%, содержание протеина - не менее 7,7, кремния - не более 1, а для роста и лактации - 61—68, 14 и 0,8-1% соответственно [Раков, 1956; Рашек, 1963; Абатуров и др., 1980, 1982а,б, 1998; Фадеев, Слудский, 1982; Холодова, 1989; Абатуров, 1999, 2000; Субботин, 2001].
Необходимое качественное питание достигается посредством высокой избирательности высококалорийных растений и их наиболее энергоемких частей: верхушек, соцветий, листьев и молодых побегов. Эфедра, солянки, прутняк, биюргун, саксаул, боя-лыч, эбелек и Польши даже зимой имеют от 7 до 17% белка и от 1 до 6% жира [Басенко, 1950; Соколов, 1951; Фадеев, Иванов, 1988]. Переваримость основных кормов ва
рьирует от 50 до 77% в зависимости от содержания в них клетчатки. Поступление протеина с кормом максимально ранней весной (до 18%), когда животные поедают проростки тюльпанов, затем оно снижается до 13%. Содержание остальных питательных веществ относительно постоянно в разные сезоны: жира - 2,6-2,7%, клетчатки -27-33, БЭВ - 44—48%. Поступление влаги с весны снижается с 81 до 32%. Зимой потребность организма в энергии в 1,5 раза превышает ее поступление, в результате чего масса животных уменьшается [Абатуров и др., 1998; Субботин, 2001].
При разной плотности населения и в разных биотопах сайга потребляет от 11 до 261 кг фитомассы с 1 га пастбищ, изымая от 1 до 20% (чаще не более 5%) валового урожая растительности при допустимой нагрузке в 60-70%. Однако при высокой численности она вместе с домашним скотом и грызунами быстро стравливает наиболее питательные растения, биомасса которых в фитоценозе невелика. Ухудшение качества пастбищ отрицательно сказывается на состоянии популяций, а при снижении переваримости корма до критической величины воспроизводство популяции прекращается [Абатуров, 1979, 1980, 1984; Субботин, Джапова, 1984; Абатуров и др., 1987, 1998; Субботин, 2001].
Немаловажен и водный баланс. Большую часть воды животные получают с кормом. Весной в злаках и эфемерах содержится до 60-90% влаги, что полностью удовлетворяет потребность зверей, включая лактирующих самок. Летом растения усыхают, содержание влаги в них не превышает 30%. Сайга в этом случае переключается на питание влагосодержащими растениями, такими как солянки, прутняк, ягоды эфедры, избирательно выискивая их на пастбищах. При недостатке влажных кормов звери кочуют в их поисках или к водным источникам. На острове Бар-сакельмес животные, включая полумесячных детенышей, пьют воду из Аральского моря (концентрация солей в ней в 3 раза меньше обычной морской и сульфаты преобладают над хлоридами), но и здесь они предпочитают посещать пресноводные озерцы и лужи после дождя. Зимой основным источником воды становится снег, изморозь и лед на растениях. Поэтому далеко не случайно в осенне-зимний период стада держатся на кромке снежной линии [Тимофеев, 1934; Адольф, 1950, 1954; Васенко, 1950; Раков, 1956; Банников и др., 1961; Рашек, 1963, 1965, 1974; Слудский, 1963; Исмагилов, 1970; Фадеев, Слудский, 1982; Слудский и др., 1983; Жирнов, Холодова и др., 1998].
В жару стада не уходят далее 10-15 км от водоемов и посещают их чаще с 10 до 14 ч, в пасмурную погоду утоляют жажду в любое время дня, а в дождливый период на водопой не приходят. К водоему звери подходят осторожно, и при малейшей опасности лидирующая самка уводит стадо прочь. Пьют они очень быстро, утоляя жажду за 1,5—4 минуты, а весь процесс водопоя у стада обычно занимает не более 20 минут. Иногда во время водопоя стадо или ее часть переплывает на противоположный берег, теряя на переправах сеголеток, отставших от матерей [Фадеев, 1972а; Жирнов и др., 1976].
В аридной зоне вода и растения содержат много солей, поэтому животные не испытывают дефицита в минеральных веществах. После отела самки нередко поедают глинистый грунт. Следует отметить, что светлокаштановые почвы и солонцы Прикаспия отличаются малым содержанием меди и избытком ее антагониста -микроэлемента бора, который накапливается в полынях и других растениях в больших дозах, способных вызвать тяжелые расстройства желудочно-кишечного трак
та у овец. Сайга, в отличие от овец, аккумулирует медь в организме, чему способствуют перемещения по разным пастбищам [Лихачев, 1974].
Пищевая активность. Для сайги во время отела характерны два периода повышенной активности: утренний - с 4 до 9-10 ч и вечерний - с 16-17 до 1 ч ночи. Ночной отдых длится 2-3 ч, дневной - 5-6 ч. Часть животных активна в любое время суток. Летом пики активности, как и у других копытных, сдвинуты на раннеутренние, поздневечерние и ночные часы, что связано с изменением влажности воздуха и увеличением содержания влаги в растениях и на них в виде росы. В пасмурную и дождливую погоду, а также зимой, звери активны и днем. Ранней весной при самых низких запасах пищи они пасутся в 1,5 раза дольше. Отдых лежа во все сезоны составляет 50-60% суточного бюджета времени. Кочуют стада в любое время суток, но чаще ночью [Васенко, 1950; Рашек, 1965, 1974; Банников и др., 1961; Жирнов, 1982; Фадеев, Слудский, 1982; Слудский и др., 1983; Жирнов, Холодова и др., 1998; Субботин, 2001].
Размножение
Половая зрелость и половые циклы. Сайга - скороспелое животное. Большая часть самок вступает в размножение в 7-8-месячном возрасте, остальные - в 1,5-летнем. Самки находятся в охоте от 12 ч до 3 суток. Повторная течка у небпло-дотворенных особей возможна через 16-19 дней. Самцы физиологически способны к размножению с 1,5-летнего возраста, и успешно покрывают самок, но полного развития их половая система достигает к 2,5 годам. Пик сперматогенеза у взрослых особей отмечается в ноябре - январе, в феврале этот процесс затухает, затем в конце этого месяца активизируется, достигает максимума в мае - июне, уменьшается в июле и возобновляется в августе. Самцы, следовательно, могут оплодотворять самок большую часть года. Наличие двух половых циклов (с чем не согласен А.А. Фандеев [1965]) позволяет предположить существование у предковых форм сайги двух сезонов размножения, что типично для тропических и субтропических животных. В измененном климате весенний половой цикл утратил свое значение [Цаплюк, 1962, 1966,1968, 1970, 1972, 1982; Рашек, 1963, 1974]. Всплеск весеннелетней половой активности характерен также и для некоторых других видов евроазиатских копытных, в частности для джейрана, сибирского козла, европейской и сибирской косуль, у предков которых тоже могли быть два периода размножения [Цаплюк, 1974].
Замечу, что у косуль характер размножения принципиально иной: гон проходит в летние или раннеосенние сроки, но затем следует уникальная задержка в развитии эмбрионов вплоть до зимы, что позволяет самкам рожать в обычные для копытных Евразии сроки. Более вероятно, что у них сдвиг сроков спаривания с осенне-зимних месяцев на летние произошел в результате развития территориализма у самцов, что обусловило преждевременное (летом, а не осенью) достижение ими пика половой потенции [Данилкин, 1990,1999]. Не исключено, что и у сайги, джейрана и сибирского козла раньше тоже было не два половых цикла в году, как считает О.Э. Цаплюк, а в процессе эволюции и изменения климата мог произойти сдвиг сроков единственного в году периода спаривания с весенне-летних на осенне-зимние месяцы.
Гон и половое поведение. Гон у сайги проходит в конце ноября - декабре (табл. 21). Отдельных неоплодотворенных самок самцы повторно кроют в январе. Всплеск половой активности отмечается и в конце весны — начале лета, когда молодые и, изредка, взрослые самцы преследуют самок. Отдельные случаи весенне-летнего спаривания, судя по регистрации беременных самок и новорожденных в период с августа по ноябрь, могут быть результативными [Банников и др., 1961; Цаплюк, 1962, 1966, 1968, 1982; Букреева, Кравчук, 2004].
Таблица 21. Сроки гона у сайги
Район | Начало | Конец 1 Источник информации
С.-З. Прикаспий Конец ноября 20 декабря Адольф, 1954
и а Конец декабря Банников и др., 1961
и Начало декабря 4 4 Близнюк, 1982
с< 3 декабря 30 декабря Жирнов и др., 1998в
- Середина января Лещенко, 1997
<л Начало декабря — Лущекина, 2002
44 5 декабря Середина января Букреева, 2002
Казахстан 2 декабря Конец декабря Раков, 1956
и Середина ноября 44 Слудский, 1955
и 25 декабря 19 января Слудский, Фадеев, 1974
Бетпакдала 10 декабря 1 января Соколов, 1951
44 Декабрь 27 декабря Цаплюк, 1966
и 20 декабря Начало января Слудский и др., 1983
О. Барсакельмес Середина ноября Конец декабря Васенко, 1950
Монголия Вторая половина ноября Середина января Банников, 1954
Аскания-Нова Вторая половина декабря — Салганский и др., 1963
(в вольерах)
В течку первыми обычно приходят средневозрастные самки, последними - старые и молодые [Рашек, 1963, 1974; Фандеев, 1965]. Продолжительность гона в разных районах достигает 40 дней, но большинство самок оплодотворяется в течение 5-10 дней. Сроки спаривания на протяжении обширного ареала имеют тенденцию к сдвигу в направлении с запада на восток (табл. 21), и могут изменяться год от года в зависимости от условий обитания. В европейской популяции, например, судя по анализу развития эмбрионов, в 1985 г. гон длился с 3 декабря до 1 января, в 1986 г. -с 19 декабря до 20 января [Петрищев, 19976]. В последнее десятилетие XX в. массовое спаривание имело место как с 5 по 10 декабря, так и с 28 декабря по 8 января, причем в отдельные годы гон растягивался до середины января, а общая продолжительность спаривания достигала 90 дней [Лещенко, 1992; Букреева, 2002].
У взрослых самцов к периоду гона происходит существенная морфофизиологическая перестройка организма: семенники достигают максимального размера и массы, увеличиваясь почти в 3 раза; окраска становится контрастной; ниже глаз и по бокам шеи отрастает длинный волос, образуя бакенбарды и гриву; увеличивается мягкая часть хоботообразного носа (фото 12); резко усиливается секреция паховых, предглазничных и карпальных желез. Выделения паховых желез вместе с пахучей мочой, которой самец часто орошает шею, низ головы и передние ноги, принимая характерную позу, образуют резкий специфический запах. Сильный запах, наряду с изменившейся от орошения мочой и ставшей темно-бурой окраской меха, скорее
всего, привлекает и стимулирует самок. У последних, принимавших участие в гоне, белый мех вокруг полового отверстия тоже окрашивается в темно-бурый цвет [Банников и др., 1961; Рашек, 1963, 1965, 1974; Фандеев, 1965; Соколов, Чернова, 2001].
В вольерах признаки предгонного состояния у самцов заметны уже с ноября: они становятся возбужденными и агрессивными, издают громкие “храпящие” звуки, бодают предметы и сражаются друг с другом. Бои иногда заканчиваются смертельным исходом. Активность зверей резко возрастает при низкой температуре и в вечерние сумерки. Днем доминант собирает самок в плотную группу и удерживает их до наступления ночи. Половое поведение проявляется в частом замирании в одной позе, обнюхивании гениталий самок, мочевых пятен и экскрементов на земле. Самок самец кроет многократно и нескольких в один день [Цаплюк, 1966, 1968, 1982; Кокшу-нова, 2003аб; Петрищев, Арылов, 2003а].
В природе в брачный период взрослые самцы находятся в крупных стадах. Они отбивают группы самок, насчитывающие порой до 50-60 особей, и пытаются удерживать их возле себя. Расстояние между гаремами измеряется десятками, а, порой, несколькими метрами. При передвижении группа идет гуськом, а доминант шествует сзади и постоянно контролирует движение, не позволяя своим самкам уйти в сторону. Половозрелые самцы, не имеющие самок, стремятся отбить их, что наталкивается на противодействие владельцев гаремов. Драки соперников весьма ожесточенные. Удары рогами настолько мощны, что у противников зачастую оказываются сломанными ребра, кости предплечья, таза и конечностей. В неволе звери легко пробивают рогами трехслойную фанеру. Перераспределение самок длится более месяца, что к концу гона приводит к уменьшению размеров брачных групп до 5-15 особей. Гаремы сохраняются и после гона, но с января - февраля резко увеличивается доля одиночных взрослых самцов и их скоплений [Васенко, 1950; Адольф, 1954; Слудский, 1955; Раков, 1956; Фандеев, 1961, 1965; Рашек, 1963, 1974; Цаплюк, 1972; Близнюк, 1982]. В 1999-2001 гг. при очень малом числе самцов в европейской популяции гон проходил в больших скоплениях, гаремы не образовывались, турнирных боев не наблюдалось. Взрослые самки не допускали молодых самок к самцам, изгоняя их из стад, и те чаще оставались неоплодотворенными [Букреева, 2002].
Пришедшую в течку самку самец преследует, высоко задрав морду, прогнув спину и часто перебирая ногами. Если она отбегает, самец в такой же позе передвигается в сторону и опускается на землю. Пролежав 5-10 с, он вскакивает, подбегает к самке, и, если та не принимает его, снова падает на землю, и так - многократно [Рашек, 1963]. Спаривание происходит круглосуточно. Садок бывает несколько, каждая из них длится 2-3 с [Цаплюк, 1968]. Один самец за день покрывает две-три самки, за период гона - 10-15. Самцы в гон очень подвижны, постоянно сражаются, плохо питаются (желудочно-кишечный тракт у них пуст или наполнен не более чем на 20%) и быстро истощаются. Ослабленные и травмированные в постоянных драках особи после гона становятся легкой добычей хищников или в массе погибают в сильные морозы и снегопады [Адольф, 1954; Слудский, 1955, 1963].
Беременность и развитие плода. Продолжительность беременности сайги составляет 131-145 (в среднем 138) дней [Адольф, 1954; Салганский и др., 1963; Цаплюк, 1966, 1968; Жирнов и др., 1998в] за редким исключением (в неволе одна из самок вынашивала плод около 6 месяцев [Васенко, 1950]). Зарегистрировано
несколько случаев внесезонной беременности [Банников и др., 1961; Фадеев, Слудский, 1982; Соколов и др., 1998].
Через месяц после оплодотворения зародыш вырастает до 5 см и весит около 5 г, через два месяца - до 12 см и 60-70 г, через три - до 23 см и 420-525 г, через четыре -до 40-43 см и 1,1-1,8 кг. К последнему месяцу беременности длина плодов достигает 44—54 см, масса - 2,2-2,9 кг. В последние недели перед рождением скорость прироста массы снижается. Размеры и масса плодов во многом зависят от срока покрытия и кормовых условий. В конце марта в одной и той же группировке о. Барсакельмес масса эмбрионов может колебаться от 0,01 г до 406 г [Рашек, 1974]. При недостаточном питании самок в последний месяц беременности часть новорожденных имеет малую массу, отличается пониженной жизнеспособностью и гибнет вскоре после рождения [Даль и др., 1958; Чагиров, Литвинова, 1960; Петрищев, 1987а, 1997а,б, 1998].
Деторождение и выкармливание молодняка. Отел у сайги на обширном ареале происходит в разгар вегетации растительности во второй половине апреля -первой половине мая, однако его сроки, как и сроки гона, имеют тенденцию к временному сдвигу с запада ареала на восток (табл. 22).
Таблица 22. Сроки отела у сайги
Район | Начало Конец Источник информации
С.-З. Прикаспии 25 апреля 20 мая Адольф, 1954
и 27 апреля 11 мая Даль и др., 1958
25 апреля 27 мая Фандеев, 1960, 1965; Банников и др., 1961
и 15 апреля 20 мая Перовский, 1974; Жирнов и др., 1982, 1998в
и 25 апреля 5 мая Петрищев, 1987а
3 мая 20 мая Близнюк, Букреева, 2000
и 26 апреля - 14 мая 16 мая - 8 июня Букреева, 2002
Казахстан Конец апреля 1 июня Слудский, 1955
сс 1 мая Конец мая Раков, 1956
30 апреля 29 мая Слудский, Фадеев, 1974
Бетпакдала 18 апреля 10 июня Цаплюк, 1982
с с 9 мая - Корнеев и др., 1986а
10 мая 1 июня Соколов, 1951
Устюрт 5 апреля Конец мая Фадеев, 19756
Узбекистан Начало мая — — Мамбетжумаев, 1966;Реймов, Карабеков, 1980; Мельниченко и др., 1989
О. Барсакельмес Последняя декада мая — Васенко, 1950
и 16 мая Июнь Рашек, 1963, 1974
Аскания-Нова (в вольерах) Май Май Салганский и др., 1963
Монголия Конец мая Первая половина июня Банников, 1954
В Северо-Западном Прикаспии пик отела обычен в первой половине мая, но иногда он сдвигается на последнюю декаду апреля или мая. Рождение молодняка длится около месяца, но около 80% самок рожают в сжатые сроки - за 3-7 дней.
Годовалые самки приносят телят позднее, чем взрослые [Близнюк, 1982; Жирнов и др., 1982, 1998в]. В 90-е годы XX в. отел проходил в более поздние сроки по сравнению с 70-80-ми годами, и заканчивался лишь в первой декаде июня [Букреева, 2002]. В 2001-2003 гг. из-за недостаточного количества взрослых самцов в популяции отел был растянут вплоть до сентября - октября, что указывает на возможность спаривания отдельных особей в мае - июне [Букреева, Кравчук, 2004]. В Западном Казахстане массовый отел имеет место в первой половине мая, на острове Барсакельмес и на востоке Казахстана - во второй половине этого месяца [Слудский, 1955; Раков, 1956; Цаплюк, 1968; Фадеев, 19756; Рашек, 1963; Фадеев, Слудский, 1982; Корнеев и др., 1986а]. В Монголии сайга приносит потомство в конце мая - начале июня [Банников, 1954].
Сдвиг сроков отела связан как с погодными условиями, так и с интенсивностью промысла и беспокойством зверей в период гона, что, как правило, приводит к задержке спаривания [Жирнов, 1982]. В заповедной популяции о. Барсакельмес, где антропогенное влияние минимальное, годовые колебания сроков массового ягнения не превышали трех дней [Рашек, 1963].
Самки выбирают для родов участки с ровным рельефом и с редкой низкорослой растительностью. “Родильные дома” обычно одни и те же из года в год. В Западном Казахстане весной 1955 г. сайга приносила потомство на посевах пшеницы в урочище, которое до распашки служило им родильным домом. Места отела чаще не приурочены к водоемам, поскольку в период весенней вегетации достаточное количество влаги животные получают с кормом. Кроме того, вдали от водоемов всегда меньше волка, собак и людей, т. е. меньше беспокойства и потерь новорожденных. Однако в засушливые весны стада размещаются вблизи водоемов [Слудский, 1956, 1962; Фандеев, 1960, 1965; Рашек, 1963].
В период отелов самки осторожны, но концентрация тысяч особей сопровождается значительным шумом. Роды, вероятно, чаще происходят утром, что благоприятствует выживанию новорожденных при оптимальной дневной температуре [Корнеев и др., 1986а]. Процесс родов занимает от нескольких минут до нескольких часов, в среднем - около получаса. Непосредственно перед ним некоторые самки разрыхляют землю, делая небольшое углубление в земле - “родовую” лежку. Рожают они чаще лежа, чем стоя [Адольф, 1954; Раков, 1956; Фадеев, 19726]. Новорожденных матери энергично вылизывают, освобождая от околоплодных оболочек. Сайгачата рождаются хорошо сформированными, и сразу же пытаются вставать, а с 10—12 минуты жизни способны сосать молоко, ходить и даже пытаются бегать, но первые три дня после кормлений предпочитают лежать, вытянувшись на земле в характерной позе (фото 14). Через несколько часов после родов мать старается увести их в другое место, но сайгачата, родившиеся в вечернее и ночное время, остаются на лежке до утра. Через трое суток детеныши уже быстро бегают и кратковременно следуют за матерями, 4-5-дневные неотступно держатся с ними, а 10-дневные в беге не отстают от взрослых особей. Самки кормят молоком только своих малышей, которых отличают от других по запаху, но за ними иногда пытаются следовать несколько детей (в неволе кормление самками чужих детенышей обычно [Rubin, Michelson, 1994]). Реакция затаивания проявляется у сайгачат примерно до 7-10-дневного возраста. Через две недели после рождения во время игр они сбиваются в группы, образуя свое
образные “детские сады” [Васенко, 1950; Фандеев, 1960, 1965; Рашек, 1963; Жирнов, 1982; Хахин, 1983].
Детеныши в первые дни жизни питаются исключительно молоком матери, которое сосут несколько раз в день по 5-20 сек. Лактация продолжается до середины июля - сентября. У некоторых сайгачат в желудках нет молока уже в середине июня, но отдельных кормящих самок регистрируют вплоть до второй половины октября [Васенко, 1950; Соколов, 1951; Слудский, 1955; Даль и др., 1958; Банников и др., 1961; Рашек, 1963, 1974; Фадеев, Слудский, 1982; Цаплюк, 1982].
Плодовитость. В благоприятные годы в размножении участвуют почти все самки, включая сеголеток. Яловость среди взрослых особей низка, и в многочисленных популяциях с ненарушенным половозрастным составом варьирует в пределах 1-4%. В матках взрослых беременных самок обычно находят два, реже - одного и изредка (до 3%) - три эмбриона, у сеголеток почти всегда бывает лишь один плод. В среднем на одну самку приходится 1,3-1,8 эмбриона, в неблагоприятные годы -меньше 1 (табл. 23).
Таблица 23. Потенциальная плодовитость сайги
Район (источник информации)* Годы Самок, п Самок, % Эмбрио-нов на самку, п
с двумя эмбрионами С одним эмбрионом яловых
взрослые сего- летки взрослые сего- летки взрослые сеголетки
1 2 3 4 5 6 7 8 9 10
С.-З. При- 1955 25 88,8 25,0 11,2 75,0 4,5 1,2
каспий (1)
-“-(1, 14) 1958 433 73,8 5,0 26,2 95,0 3,8 15 1,4
-“-(14) 1959 - - — - - 13,7 44,8 -
(2, 3) 1973-
1978 167 71,6 25,4 3,0 1,7
-“-(3) 1979 - 19,4 55,6 25,0 0,9
-“-(3) 1980 — 36,4 56,0 7,6 1,3
-“-(13) 1983,
1986 450 — — — 10-13 1,3, 0,9
-“-(Ю) 1986 164 17,7 82,3 10,4 1,1
1987 99 7,1 92,9 20,2 0,9
-“-(6, 7) 1993 24 62,5 37,5 — 1,5
(С 1994 32 32,3 - 68,7 3,1 - 1,3
с< 1995 85 39,5 2,4 60,5 97,6 0 12,8 1,2
11 1996 65 80,0 0 20 100 2,0 12,5 1,4
1997 86 78,7 0 21,3 100 0 16,7 1,4
а 1998 29 62,1 0 37,9 100 0 42,9 1,4
и 1999 70 40,9 • 0 59,1 100 — - 1,1
2000 88 66,1 0 33,9 100 8,8 50 1,3
и 2001 103 62,5 0 37,5 100 75,1 97,2 0,3
Казахстан 1964 40 81,0 5,3 19,0 89,4 0 5,3 1,4
(8)-“- 1965 87 80,7 0 19,3 93,1 0 6,9 1,5
<< 1966 90 64,0 ** 0 32,0 95,0 4,0 5,0 1,2
а 1967 86 83,0* 4,5 14,6 93,2 2,4 2,3 1,4
а 1969 63 90,0 0 10,0 100 0 0 1,6
Таблица 23 (окончание)
1 1 2 1 3 1 4 | 5 1 6 1 7 1 8 1 9 1 10
я 1970 73 82,6 0 17,4 100 0 0 1.6
к 1972 90 87,6 ** 4,0 9,2 88,0 3,2 8,0 1,6
к 1974 142 92,7 ** 11,9 6,4 73,8 0,9 14,3 1,6
я 1975 18 72,7 0 18,1 100 9,2 0 1,4
я 1976 42 70,0 4,6 30,0 63,6 0 31,8 | £)***
я 1977 63 83,0** 0 15,1 80,0 1,9 20,0 1,5
я 1978 89 60,1 ** П,2 36,5 88,8 2,9 0 1,5
-“-(4) 1964- 1,5
1978 892 80,6 ** 4 17,7 88,3 1,7 7,7 (1-1,6)
Бетбак- 1960-
дала (5) 1967 303 76 2-8 24 92-98 2 4,5 -
-“-(2) 1986-
1994 1030 72-76 ** 9-18 15-26 51-63 3,7-7,1 20-31 1,5-1,8
О. Барса- 1953-
кельмес (12) 1966 164 48,4 14,3 51,6 85,7 5,2 87,9 1,0
-“-(5) 1960-
1967 151 59 — 41 - 8,5 87,5 -
Монголия
(15) 1981 - 5 95 - -
* 1 - Банников и др., 1961; 2 - Жирнов и др., 1998в; 3 - Близнюк, 1982; 4 - Фадеев, Слудский, 1982; 5 -Цаплюк, 1966, 1970; 6 - Букреева, 2002, 2003; 7 - Близнюк, Букреева, 2000; 8 - Слудский и др., 1983; 9 -Даль и др., 1958; 10 - Петрищев, 19976; 11 - Мамбстжумасв, 1966; 12 - Рашек, 1974; 13 - Максимук, Проняев, 1986; 14 - Фандеев, 1965; 15 - Сапожников, Дуламцэрэн, 1982.
** Включая самок с тремя эмбрионами.
*** После засухи 1975 г.
Наиболее плодовиты самки, как считалось, в возрасте 2-5 лет [Соколов, 1951; Рашек, 1963; Фадеев, 1980а]. Исследования А.И. Близнюка [1999] показали, однако, что число эмбрионов на самку остается стабильно высоким почти до конца жизни. Уровень плодовитости в значительной мере зависит и от размера самок -чем крупнее особь, тем более вероятно рождение двоен (табл. 24). Этот вывод весьма важен в плане планирования добычи сайги. Резкое уменьшение доли самок с двумя эмбрионами в европейской популяции в последние годы во многом связано с неумеренной и длительной промысловой элиминацией более крупных животных [Проняев, 1998].
Таблица 24. Изменение плодовитости самок сайги в зависимости от размера особей (по [Рожков, Проняев, 1994])
Косая длина туловища, см Доля самок с двумя эмбрионами, %
73-78 0
79-84 0,05
85-87 0,13
88-90 0,37
91-93 0,67
При неблагоприятной погоде и нехватке кормов и воды яловость самок существенно увеличивается (до 75% у взрослых и почти до 100% у молодых), и у
большинства особей регистрируют только по одному эмбриону. Особенно заметно уменьшается плодовитость после интенсивных охот в период гона, когда преследуемые днем и ночью звери вынуждены постоянно перемещаться и не могут нормально спариваться [Цаплюк, 1966, 1968], кормиться и отдыхать. В европейской группировке при острейшем дефиците самцов (3,5-0,9% во время гона), уничтоженных охотниками, воспроизводство особенно резко снижалось в 1959, 1979 и 2001 гг. (до 0,3 эмбрионов на самку) [Фандеев, 1965; Близнюк, 1982; Букреева, 2002, 2003]. Такая же ситуация наблюдалась и в казахстанских популяциях (см. ниже).
Примером изменения плодовитости этого вида из-за нехватки кормов может служить группировка о. Барсакельмес. До 1945 г. 70% самок приносили здесь по два детеныша, позднее - чаще по одному. В 60-е годы общее поголовье сайги и джейрана на острове достигало 4—5 тыс., что привело к истощению пастбищ. В результате время наступления половой зрелости у большинства самок увеличилось на год. И у самок, и у молодых самцов были недоразвиты половые гонады. Резко возросло число холостых самок, хотя сроки гона и отела не изменились. Здесь же замечено, что уровень воспроизводства группировки коррелирует с количеством осадков весной предыдущего года и, следовательно, с летними кормовыми условиями. В засушливые годы большинство или все молодые самки не вступают в размножение [Рашек, 1963, 1974; Цаплюк, 1966, 1970; Исмагилов, 1970]. В материковых казахстанских популяциях после засух 1964—1967 гг. и джута 1971— 1972 гг. яловость возросла до 4-8% и еще до 4—10,5% эмбрионов рассасывалось, что привело к снижению среднего числа эмбрионов на самку [Слудский, Фадеев, 1974], но не такому резкому, как после перепромысла. В Монголии А.Г. Банников [1954] регистрировал без детенышей до 87% самок.
Популяционная структура
Половой и возрастной состав. Половозрастной состав, как известно, один из важнейших инструментов механизма регуляции численности и показателей состояния популяций, во многом зависящий от воздействия как природных, так и антропогенных факторов.
Соотношение полов среди эмбрионов и новорожденных сайги близко 1:1 в разных группировках и в разные годы (табл. 25). При депрессии несколько возрастает доля самок, при пиках численности - доля самцов. У сеголеток соотношение сдвигается в пользу самок, что связано с повышенной смертностью самцов [Банников и др., 1961; Присяжнюк, Проняев, 1982; Слудский и др., 1983; Жирнов и др., 1998]. Среди особей старше года явно преобладают самки, особенно в старших возрастных группах (табл. 26). В европейской популяции в 1978-1990 гг. соотношение полов у полуторалетних было 1:1,2-1,8, у зверей старше 2,5 лет - 1:2,4-6,0 (в среднем - 1:3,3), в казахстанских популяциях в 1981-1993 гг. среди особей старше года - 1:3,6 [Жирнов и др., 1998].
Доля самок старше года в популяциях варьирует от 30 до 78%, чаще - 33-55%. Половозрелых самцов в норме около 18-30%, но их доля изменяется в разные годы от 35 до 1% (табл. 27). Резкое снижение их числа обычно вызвано высокой смертностью после гона в очень суровые зимы или чрезмерным отстрелом. Относительная
Таблица 25. Соотношение полов у эмбрионов и новорожденных сайги
Район Годы Исследовано, п Самцы, % Самки, % Самцы : самки Источник информации
С.-З. Прикаспий 1941 23 52,2 47,8 1:0,92 Адольф, 1954
(С 1955 31 51,6 48,4 1:0,93 Даль и др., 1958
с< 1957 3905* 50,1 49,9 1:0,99 Фандеев, 1960, 1965;
<< 1958 10135* 52,5 47,5 1:0,90 Банников и др., 1961 <<
сс 1959 4067*' 50,1 49,9 1:1,0 сс
к 19571977 26442* 50,6 49,4 1:0,98 Жирнов, 1982
4 С 1986-1987 2618 44,4 55,6 1:1,25 Петрищев, 1997а
1995 220* 52,8 47,2 1:0,89 Лещенко, 1997
1994-1998 2776 48,9 51,1 1:1,04 Близнюк, Букреева,
Казахстан 40-е 25* 56,0 44,0 1:0,79 2000 Соколов, 1951
50-е 47* 51,1 48,9 1:0,96 Слудский, 1955
44 1963-1965 1017 53 47 1:0,90 Демеуов, 1971
44 1964-1978 1360 47,3 52,7 1:1,12 Фадеев, Слудский,
44 44 18781* 51,8 48,2 1:0,93 1982; Слудский и др., 1983 а
44 1983-1994 1447 49,2 50,8 1:1,03 Жирнов и др., 1998
44 44 17114* 52,2 48,8 1:0,93 сс
О. Барсакельмес 1953-1966 157 51,9 48,1 1:0,93 Рашек, 1974
44 2581* 52,7 47,3 1:0,90 С<
Узбекистан, Устюрт 1965 74 55,4 44,6 1:0,80 Мамбетжумаев, 1966
* Новорожденные.
Таблица 26. Соотношение полов в популяциях сайги*
Район Годы Исследовано, п Самцы, % Самки, % Самцы : самки Источник информации
С.-З. Прикаспий 50-е 28 72 1:2,6** Банников и др., 1961
СС 1971 66354 35 65 1:1,9 Перовский, 1974
Cl 1978-1990 57172 23 77 1:3,3 Жирнов и др., 1998
1988-1998 64344 27 73 1:2,7 Букреева, 2002
Cl 1999-2001 4427 9 91 1:9,9 сс
Казахстан 1966-1967 1760 32 68 1:2,2** Фадеев, Слудский, 1982
с с 1973 19291 25 75 1 ’3** СС
сс 1981-1993 224403 22 78 1:3,6** Жирнов и др., 1998
Монголия 40-е 176 22 78 1:3,7 Банников, 1951, 1954
1981-1982 — — — 1:2,8 Соколов и др., 1986
1981 101 — - 1:7,1 Сапожников, Дуламцэрэн, 1982
* По осенним визуальным наблюдениям и данным промыслового отлова сетевыми коралями.
** Соотношение полов среди особей старше 1 года.
Таблица 27. Возрастной и половой состав популяций сайги, %
Район (источник информации) Годы Учтено особей, п Сеголетки Особи старше года
самцы | самки
С.-З. Прикаспий (11) 1949 937 42-45 22-24 33-34
сс 1950 — — 10-14** —
-“-(10) 1955 882* 57 5 40
-“-(1) 1958 1828 34,9 18,3 46,8
-“-(1, 15) 1959 - - -
-“-(8) 1966-1979 58000 42,4 18,9 (16-24) 38,7
-“-(2) 1978 17622**** 33,7 20,0 46,3
-“-(8) 1979 - 26,2 30,4 43,4
(3) 1981 1604 39,1 18,9 40,7
-“-(4) 1982-1990 37710 25-47 18-20 33-55
-“-(6) 1988 7897 38,3 16,6 45,1
1989 9435 35,0 20,6 44,4
сс 1990 15130 35,8 16,7 47,5
1991 33405 30,6 24,3 45,1
tt 1992 7098 38,7 6,4 54,9
с с 1993 3352 45,5 8,2 46,3
сс 1994 4508 33,9 10,3 55,8
и 1995 9163 40,9 12,3 46,8
СС 1996 4336 45,6 14,4 40,0
<4 1997 3802 47,6 12,3 40,1
СС 1998 2592 40,9 11,0 48,1
СС 1999 1925 38,2 7,5 54,3
СС 2000 2663 42,7 3,5 53,9
2001 2068 17,3 8,2 74,6
-“-(16) 2002 — — <1 —
Казахстан (7) 50-е — - 20 (35-4,2) —
-“-(18) 1963-1965 — 35-41 34-4,2 30-65
-“-(5) 1966-1967 3050 47-33 16-22 36,6-45
-“-(13) 1972-1980 — 35-44 17-27 37-40
-“-(4) 1981-1993 >242000 21-41 26-7 35-66
СС 1992-1993 81965 18-47 2,3-9,1 48-78
-“-(17) 1994-2001 - - 10-1,7 -
О. Барсакельмес (14) 1956-1959 — 47 (37-57) — —
Узбекистан (12) 1966 1000 36,5 31,9 31,6
Монголия (9) 1981-1982 — 43—46 7-14 50-40
-“-(19) 1981 101 42,6 6,9 50,5
1 - Банников и др., 1961; 2 - Присяжнюк, Проняев, 1982; 3 - Жирнов, 1982; 4 - Жирнов и др., 1998; 5 -Фадеев, Слудский, 1982; 6 - Букреева, 2002; 7-Слудский, 1955; 8 - Близнюк, 1982; 9-Соколов и др., 1986; 10 - Даль и др., 1958; 11 - Адольф, 1950, 1952, 1954; 12 - Мамбетжумаев, 1966; 13 - Слудский и др., 1983; 14 - Рашек, 1974; 15 - Фандеев, 1965; 16 - Сидоров, 2003; 17 - личное сообщение Ю.А. Грачева на VII съезде Териологического о-ва (2003); 18-Демеуов, 1971; 19 - Сапожников, Дуламцэрэн, 1982.
* На местах отела.
** После суровой зимы 1949/50 гг.
*** Результат перепромысла.
**** Промысловая выборка.
стабильность их доли в европейской популяции в 70-е годы связана с нормированием добычи [Адольф, 1954; Раков, 1955; Слудский, 1955, 1963; Банников и др., 1961; Фандеев, 1965; Рашек, 1974; Близнюк, 1982, 1983; Жирнов, 1982]. Осенью сеголет
ки составляют примерно треть группировок, но из года в год их соотношение колеблется в широких пределах (от 17 до 57%) в зависимости от уровня плодовитости самок в текущем году и детской смертности. Соответственно ежегодно значительно изменяется и доля особей старше года (табл. 27).
С 90-х годов очевидна тенденция снижения числа взрослых самцов практически во всех популяциях (табл. 27), что, безусловно, связано с усилением выборочного браконьерского пресса (см. ниже). В Северо-Западном Прикаспии в 1992 г. их доля сократилась до 4,6%, а в 1998-2002 гг. в период гона составляла всего 3,5-0,9% [Язан, Лещенко, 1995; Букреева, 1998,2002,2003; Сидоров, 2000,2003; Milner-Gulland et al., 2003]. Резкое изменение полового состава, в свою очередь, - одна из основных причин снижения воспроизводственного потенциала вида.
В связи с промыслом и высокой естественной смертностью состав популяций сайги обновляется почти полностью за 3-5 лет. По данным мечения, животные старше 5,5 лет составляют в них всего несколько процентов [Фадеев, 1980а; Фадеев, Слудский, 1982]. В интенсивно эксплуатируемых популяциях средний возраст половозрелых самцов обычно не превышает 2 лет, самок - 2,7 года (с учетом сеголеток - 1,3 и 2 года соответственно), что примерно в 2 раза меньше, чем в неэксп-луатируемых или слабо опромышляемых популяциях в конце 50-х годов. Лишь отдельным особям удается дожить до 8-10 лет [Жирнов, 1982; Рожков, Проняев, 1994; Жирнов и др., 1998].
В европейской популяции сайги В.Е. Соколов с соавторами [1998] выделили, на мой взгляд, весьма спекулятивно, две биологические формы (“типичную и нетипичную экоморфы”), формирующие разноразмерные стада, имеющие разную половозрастную и пространственную структуру, иной тип динамики численности и другие особенности. Большая или меньшая частота тех или иных признаков в разных группировках, находящихся под воздействием промысла, характерна практически для каждого охотничьего вида. При прекращении или ослаблении промыслового пресса такого рода “экоморфы” быстро приходят в “типичное” состояние.
Социальная организация и поведение. Сайга, пожалуй, - самый общественный вид из всех отечественных копытных. Почти вся жизнь особи с рождения и до смерти проходит в стаде в тесном взаимодействии с другими ее членами (фото 15). Животные вместе пасутся, отдыхают, перемещаются, спасаются от опасности, соблюдая при этом определенную индивидуальную дистанцию (в летний период на отдыхе - в среднем 2,7, при бегстве - 0,3, пастьбе - 5,0 м) и индивидуальный ритм активности [Баскин, 1976]. Наиболее прочная связь у членов семьи (самки с детенышами), хотя, в отличие от многих других видов копытных, она кратковременна -обычно только в период лактации.
Величина и состав группировок сайги весьма изменчивы на протяжении года. Чаще всего исследователи регистрируют стада численностью от 20 до 200 особей. В определенные периоды формируются более крупные объединения, а в районах концентраций (на местах отелов, на лучших пастбищах, у водопоев) и при миграциях образуются многотысячные скопления. При снижении численности стадность заметно уменьшается. На периферии ареала преобладают одиночные особи, пары и мелкие группы [Адольф, 1950, 1954; Васенко, 1950; Соколов, 1951; Бакеев, Формозов, 1955; Раков, 1955; Слудский, 1955; Рак, 1956; Даль и др., 1958; Фандеев, 1960, 1965; Банников и др., 1961; Рашек, 1963; Исмагилов, 1970; Перовский, 1974;
Близнюк, 1982; Жирнов, 1982,1998в; Сапожников, Дуламцэрэн, 1982; Фадеев, Слудский, 1982; Слудский и др., 1983].
Мелкие группы (преимущественно до 20 особей) чаще встречаются в декабре во время гона, когда взрослые самцы формируют гаремы. В этот период обособленно держатся лишь отдельные самцы и группы неполовозрелых особей. В январе — феврале гаремы сливаются в смешанные стада, насчитывающие сотни и тысячи особей. После гона часть взрослых самцов отделяется, и вскоре они образуют группы из 3-40 особей. При неблагоприятных погодных условиях формируются самцовые стада, которые весьма подвижны и обычно первыми приходят на весенние пастбища. В ранневесенний период большая часть (до 75-95%) мигрирующих животных находится в крупных стадах, насчитывающих при высокой численности тысячи или десятки тысяч особей. Эти временные объединения в предродовой период концентрируются в определенных районах, образуя скопления в сотни тысяч особей. В так называемых “родильных домах” на территории в несколько тысяч гектаров плотность животных достигает 300-12000 голов/км2. Плотность распределения детенышей варьирует от 0,1—2 до 153 особей на 1 га. Взрослые самцы в таких скоплениях составляют не более 2-5%. Рано родившие самки чаще держатся обособленно от остальных.
Концентрацию самок на местах отелов А.А. Слудский [1955, 1962] рассматривает как приспособление к снижению гибели молодняка от хищников. Действительно, при известной весенней территориальности волка, на участках “родильных домов”сайги не может быть более одной семьи этого хищника. Тем не менее ущерб от него все же велик из-за наличия периферийных мест отелов [Филимонов, 1979].
В мае самки с подросшими детенышами начинают кочевать. В этот период несколько увеличивается число мелких групп, однако основная масса животных находится в крупных группировках. К началу июня скорость перемещения маточного поголовья и самцовых групп становится примерно одинаковой, что способствует формированию смешанных стад. Летом стадность сайги резко уменьшается. Крупные группировки распадаются на мелкие и средние, в которых насчитывают одну-две сотни голов. При засухах преобладают мелкие стада и группы из нескольких особей, но на отдельных пастбищах с сочным кормом держатся большие стада. В конце лета небольшие группы образуют сеголетки. С сентября стада объединяются, достигая многотысячных размеров в период осенних миграций в октябре - ноябре. Во время гона эти скопления вновь разбиваются на мелкие группы.
В “джутовые” годы многотысячные скопления характерны для сайги и вне обычных сезонов миграций. В январе 1951 г. в Чимкентской области пилоты наблюдали скопление кочующих стад животных в полосе шириной около 15-18 км. Фланги этой группировки, насчитывающей 120-140 тыс. особей, не были видны с высоты 500 м. В третьей декаде декабря 1953 г. в междуречье Урал - Кушум стада держались в четырех группировках, которые в январе 1954 г. слились в одну, насчитывающую около 20-25 тыс. голов [Раков, 1956].
Стадная жизнь в открытом ландшафте более безопасна для этого весьма пугливого и осторожного животного, у которого хорошо развито зрение и обоняние, но относительно слабый слух. При опасности обеспокоенные особи, после типичной ориентировочной позы и короткого перемещения с твердой постановкой копыт,
прыгают вперед и вверх, предупреждая остальных членов стада. Волна сигнальных прыжков прокатывается по всему стаду, которое с топотом и ревом сбивается в более плотное объединение перед бегством. При преследовании звери развивают скорость до 60-80, отдельные особи - до 85 км/час (фото 16). Преобладающий тип бега - иноходь [Соколов, 1951; Слудский, 1955, 1962].
В обычных условиях сайга избегает кустарников и гор. При постоянном беспокойстве, однако, животные могут укрываться в горах, быстро и ловко перемещаясь по склонам, усеянных глыбами камней [Капитонов, 1980]. При миграциях стада нередко преодолевают крупные водные преграды. На переправах животные весьма осторожны, не решаются переправляться сходу и неоднократно в течение нескольких часов подходят к хорошо просматриваемым береговым участкам. Затем несколько особей отделяются, заходят в воду и увлекают за собой остальных. Звери, находящиеся в хвосте стада, теснят передних, давят молодняк, теряют осторожность и издают громкие звуки, слышные за 2 км. Особенно большой отход детенышей имеет место в случае паники животных при опасности [Жирнов и др., 1976].
В стадах сайги нет постоянных вожаков, но всегда находятся лидеры, обычно старые опытные самки. В самцовых стадах в их роли чаще оказываются молодые особи. В гон “вожаком” временно становится владелец гарема.
Во время гона, как отмечено выше, самцы весьма агрессивны. Столкновения с соперниками, однако, чаще ограничиваются демонстрациями угроз: при сближении с противником самец вытягивает шею, приподнимает хвост и периодически закидывает голову вверх или опускает ее. Бои непродолжительны. Слабейший из них убегает сразу же после демонстраций или после столкновения лбами и рогами; сильнейший кратковременно преследует его рысью или галопом. Бои равных по силам противников могут быть ожесточенными, и, нередко, заканчиваются гибелью одного из них [Банников и др., 1961; Рашек, 1963]. Агрессивное поведение вне периода гона выражено сравнительно слабо. В стадах доминируют взрослые самцы, которые летом, как и в период гона, проявляют отдельные элементы территориального поведения. В вольерах в группах самцов четко выражена иерархия, особенно во время кормления. Иерархический статус определяется в боях [Кок-шунова, 2003а].
Обычно сайга молчалива, за исключением периода родов, когда повсюду в стадах раздается блеяние ягнят и призывное “мекание” самок, и периода гона, когда самцы издают “хрюкающие” и “рычащие” звуки. При опасности и самцы и самки фыркают.
Пространственная структура и перемещения. Сайга - типичный кочевник, привязанный к определенному участку только на короткий период, связанный с появлением потомства, однако основные направления перемещений и сезонное пространственное распределение ее группировок относительно традиционны.
Северо-Западный Прикаспий. До середины XX в. большая часть европейской популяции в конце марта - начале апреля находилась в местах зимовок на юге Прикаспийской низменности, но отдельные стада держались в ее центральных и северных районах, а также на Восточном Маныче и на восточных склонах Южных Ергеней. В первой половине апреля мелкие группы и многотысячные стада южной группировки смещались на 300-850 км на север Прикаспия, где концентрировались в восточной части Сарпинской низменности на наиболее влажных и
кормных злаково-эфемерных пастбищах. Здесь, вдали от поселений человека и выпасающихся отар овец, обычно происходил отел. Иногда после многоснежных зим “родильные” дома размещались между Яшкулем и Сарпой и на правобережье Кумы западнее Бакресса. Мелкие группы могли останавливаться на отел на северном берегу Восточного Маныча, в прикумских степях, на Черных землях и на побережье Каспия.
Весенние миграции сайги отличались строгой направленностью на север. В отдельные годы скопления (до 100-200 тыс. особей) животных двигались ночью и в первой половине дня почти непрерывным многокилометровым потоком в полосе до 1,5 км шириной с средней скоростью от 3-6 до 10-20 км/час в зависимости от уровня беспокойства, проходя до 40-60 км в сутки. Во второй половине дня звери обычно задерживались на кормежку и отдых.
До середины 70-х годов стада самцов появлялись в Ергенях и приманычских степях в конце апреля - начале мая, самки с молодняком - в третьей декаде этого месяца. Отсюда животные кочевали еще дальше на запад в сальские степи, достигая густонаселенных человеком районов с развитым земледелием, а затем уходили на север вдоль западных склонов Ергеней и к Сарпинским озерам, где больше доступных водоемов, богаче растительный покров и растянуты сроки вегетации растений. После июльских дождей и возобновления вегетации растительности стада перемещались по равнинам Прикаспийской низменности и заходили в полупустынные районы, а в засушливые годы (летом 1949 г.) особенно широко разбредались, проникая в Ростовскую область и выходя к Дону и Волге. В целом летние стации обитания вида, по сравнению с весенними, сдвигались к западу и юго-западу на 120-200 км.
С конца лета сайга начинала перемещаться на восток в Сарпинскую низменность и центральные районы Калмыкии, где концентрировалась в октябре и ноябре. В отдельные годы животные скапливались также в приманычских степях и на Южных Ергенях. Иногда многотысячные стада выходили на водопой к Волге, Восточному Манычу, на оз. Сарпа. В засушливые годы в ноябре животные уходили на юг, в Кумо-Манычское междуречье и на северное побережье Кумы, при снегопадах могли кочевать на запад в Северные Ергени и юго-западнее. Зимой группировка располагалась, в основном, к северу от р. Кумы в окрестностях оз. Сарпы и на Ергенях, в западных и юго-западных районах Прикаспийской низменности, где стада кочевали чаще в меридиональном направлении. При неблагоприятных погодных условиях, глубокоснежье (более 10 см) и малокормных пастбищах звери покидали эти районы и стремительно перемещалась на юг, юго-запад и юго-восток на сотни километров, достигая в отдельные годы западной части Ставрополья, северных районов Ингушетии и Чечни (р. Терек и г. Хасавюрта), севера Дагестана, побережья Каспия и песков Приволжья (рис. 19). При потеплениях часть группировок возвращалась назад. Отдельные стада по льду пересекали Волгу и выходили на ее левый берег, но чаще мигрировали в противоположном направлении. В особенно многоснежные и суровые зимы мигранты появлялись в Волгоградской (Сталинградской) области, в бассейне р. Медведицы южнее Саратова, на льду Каспийского моря далеко от берегов [Ребров, 1849; Воробьев, 1929; Адольф, 1950, 1955; Лавровский, 1950; Бакеев, Формозов, 1955; Насимович, 1955; Даль и др., 1956, 1958; Рак, 1956; Фандеев, 1960; Банников и др., 1961; Жирнов, 1962, 1982; Слудс-
кий, 1963; Бакеев, 1964; Фандеев, 1965; Близнюк, 1982; Жирнов и др., 1998; Жирнов, Максимук, 1998а].
В конце XX в. пространственная структура европейской популяции сайги претерпела существенные изменения в связи с хозяйственной деятельностью человека и неумеренным преследованием. К 90-м годам пастбища деградировали из-за перевыпаса домашнего скота и оказались изрезанными разветвленной сетью оросительных каналов, дорог и проволочных заграждений. Северные районы Прикаспийской низменности практически утратили значение для вида, вытесненного в зону полупустынь и пустынь. В 1990 г. в Калмыкии на Черных землях организован заповедник. Улучшение охраны и прекращение выпаса скота усилило привлекательность этого района для сайги, которая стала обитать именно здесь (рис. 20). В отдельные многоснежные зимы часть стад откочевывала на юг вплоть до Северного Дагестана, а в летнюю засуху доходила на север до южной окраины Сарпинских озер. Дальнейшее продвижение зверей на север стало проблематичным из-за сети оросительных каналов на рисовых полях. Основные “родильные дома” и весенние пастбища в Сарпинской низменности оказались безвозвратно утраченными, что не замедлило сказаться на выживаемости молодняка и воспроизводстве европейской популяции. В 80-90-е годы отел сайги проходил в центральной части Черных земель и на юге современной зоны обитания. Регулярные сезонные миграции животных практически прекратились. Последняя массовая откочевка, вызванная тяжелыми условиями зимовки, имела место в 1998/99 гг. Часть стад ушла на территорию Дагестана, откуда из-за массового браконьерства возвратились, видимо, лишь немногие особи. Популяция перешла от сезонно-кочевого к полуоседлому образу жизни на ограниченной территории, в основном в Черноземельском и Яшкульс-ком районах Калмыкии (рис. 20), где в сильнопересеченной местности с барханами и колеями автомобильных дорог затруднено преследование копытных человеком [Близнюк, 1995; Лещенко, 1997; Жирнов, Максимук, 1998а; Жирнов и др., 1998; Букреева, 2002].
Казахстан, Узбекистан, Туркменистан. В Казахстане сайга живет более или менее постоянно в одних и тех же районах с ноября до середины февраля - марта, в мае и с июня до середины сентября, а в остальное время кочует, преодолевая за год до 3-4 тыс. км [Соколов, 1951; Раков, 1956, 1957; Кириков, 1959, 1966; Фадеев, 1974, 19756; Фадеев, Слудский, 1982, 1983; Слудский и др., 1983; Фадеев, Иванов, 1988; Грачев, Бекенов, 1993; Бекенов, Грачев, 1998; Bekenov et al., 1998].
Летом звери размещаются близко к северной границе степей, в дождливые годы граница ареала смещается к югу. Небольшое количество животных остается на лето в Южном Прибалхашье, в междуречье Или - Топар, в Северных Кызылкумах и на Устюрте. Стада самцов приходят на летние пастбища в мае - первой половине июня примерно на 10-30 дней раньше самок с детенышами. Общая площадь лето-вок сайги достигает 300-350 тыс. км2, а плотность распределения по территории составляет 5-20 (до 80) особей/км2. В сентябре - ноябре группировки перемещаются на юг и зимуют в зоне пустынь, где высота снежного покрова находится в пределах 5-17 см (рис. 22).
На территории Казахстана, как отмечено выше, обособлены 3 популяции: уральская, устюртская и бетпакдалинская (рис. 21), контакт между которыми затруднен многочисленными барьерами (каналами, проволочными ограждениями, заборами, различ
ными сооружениями), воздвигнутыми человеком на путях миграций [Фадеев, 1980а; Слудский и др., 1983; Фадеев, Иванов, 1988; Грачев, Бекенов, 1993]. Тем не менее обмен генофондом между ними возможен.
Уральская популяция круглогодично находится в междуречье Волги и Урала, размещаясь зимой на северном побережье Каспийского моря. Часть ее (несколько тысяч особей) в сентябре — начале октября с восточной кромки Волго-Уральских песков перемещается по Приуральской равнине к р. Урал. Реку стада форсируют на протяжении 50 км в лесистой пойме между поселками Ак-тогай и Орлик, пополняя устюртскую популяцию [Трощенко, 1986]. Весной звери возвращаются назад. Этот миграционный путь отмечал также Э. Эверсман [1850], говоривший о “неимоверном количестве” кочующих животных. Еще ранее С.Г. Гмелин [1777] сообщал, что “около Астрахани, но только зимою, во время снеговых вьюг, а особливо когда долго дует восточный ветер, бывают те дикие козы, коих татары на своем языке сайгаками называют. Они не принадлежат собственно к зверям Волжским, но приходят в сии страны с реки Уралу, однако часто большими стадами”. Часть уральской популяции периодически подходит к Волге и, видимо, иногда форсирует ее и в наше время. В этом районе возможны контакты и обмен генофондом с прикаспийской группировкой.
Устюртская группировка зимует на плато Устюрт, включая территории Узбекистана и Туркмении. В Каракалпакию ежегодно приходило до 35-80, а иногда и до 200 тыс. особей. Отдельные скопления достигали 26 тыс. Здесь же находились и места отела, на которых порой концентрировалось до 60 тыс. самок [Мамбетжу-маев, 1966; Павленко, Тарянников, 1975; Реймов и др., 1975, 1987;Ишунин, 1980; Реймов, Карабеков, 1980, 1986; Реймов, 1994; Мельниченко и др., 1998]. В Северо-Западную Туркмению ежегодно в ноябре - феврале заходили от нескольких до нескольких десятков тысяч особей, а в отдельных скоплениях насчитывали до 11 тыс. Весенне-летние миграции зарегистрированы в 1975, 1979 и 1984 гг. Летом 1984 г. в Северном Прикарабогазье держалось около 10 тыс., а на Южном Устюрте - 17 тыс. голов. Максимальное число животных, мигрирующих на Крас-новодский полуостров - 700, в Заузбойский район - 12 тыс., на Южный Устюрт -25 тыс., в Северное Прикарабогазье - 15 тыс. [Зархидзе, 1980; Горбунов, 1986а,б, 1987, 1995а]. В многоснежные годы часть стад мигрировала в Сарыкамышскую впадину и южнее к Ашхабаду, проходя по территории Туркмении до 360 км. В декабре 1974 г. здесь держалось 123 небольших стада от 8 до 300 особей в каждом. Плотность копытных в Сарыкамышской впадине достигала 10-20 голов на 1000 га. Отдельные группы задерживались здесь до лета и размножались [Иша-дов, 1965, 1966, 1972, 1975а, 1999а].
Бетпакдалинская группировка зимует севернее р. Чу, в Алакульской котловине и Северном Прибалхашье, в центральной части Бетпакдалы, а в многоснежные годы - в Муюнкумах и Саксаулдале [Корнеев и др., 1975]. Отдельные стада иногда достигают гор Заилийского Алатау, где поднимаются до высоты 2400 м над уровнем моря.
Весной стада перемещаются на север к местам отела, которые не постоянны. В Волго-Уральском междуречье они размещаются на территории примерно 200x300 км, в междуречье Урала и Эмбы - 300 х 300, а в Центральном Казахстане - 400x700 км. На этих участках скапливаются сотни тысяч животных с плотностью 15-400 особей/км2, а на отдельных участках - до 100-120 голов/га.
Миграционные пути животных в Казахстане, как и на юго-востоке европейской части России, проходят по равнинным, наиболее кормным, участкам с наличием водоемов. В Волго-Уральском междуречье есть два основных направления с зимовок на север: западное - по линии Новый Уштоган - Азгир - Суюндук - Урда -Хакисор - Аралсора - Нижинские разливы и восточное - по линии Новобогатинс-кое - Тас - Ахмедшкола - Камышсамарские озера - оз. Едильсор. Устюртская популяция перемещается на север параллельно восточному побережью Каспийского моря по нескольким путям в зависимости от расположения зимовок. Часть стад достигает р. Урал и иногда форсирует ее, соединяясь с уральской группировкой [Фадеев, Иванов, 1988]. Бетпакдалинская популяция придерживается двух “коридоров”: один тянется от оз. Балхаш до р. Сарысу, пересекает Бетпакдалу, выходит к железной дороге Джесказган - Жарык и оз. Тенгиз, другой пролегает между пос. Байконур и Приаральскими Каракумами на северо-запад к рекам Тургай, Иргиз, Улькояк, а в отдельные годы простирается на север до пос. Баянаул Павлодарской области, Целинограда, Актюбинска и Северо-Казахстанской области (рис. 22).
Осенью сайга перемещается в обратном направлении примерно по тем же путям, что и весной, но часть животных, как показало мечение, возвращается на зимовку иным маршрутом или уходит в другие районы за 300—900 км от места мечения. Звери, зимовавшие вдоль р. Чу, например, на следующий год останавливаются в Арыскуме или Приаралье. Скорость мигрирующих копытных в нормальных условиях составляет 5-20 км в сутки, при неблагоприятных увеличивается до 40-45 км. Расстояние сезонных миграций по прямой линии - 200-1200 км.
При неблагоприятных погодных факторах (высокий снежный покров, гололедицы, сильные засухи, длительные бураны) животные кочуют на десятки и сотни километров в разных направлениях и иногда появляются в нетипичных местах обитания, подходят вплотную к населенным пунктам. Причиной откочевок могут быть также пожары, обилие кровососущих насекомых и антропогенные факторы: преследование стад охотниками, концентрация людей, техники и домашнего скота на пастбищах, шум низко летящих вертолетов и самолетов.
В аномальные годы учащаются переходы стад из Казахстана на правобережье Волги и обратно и заходы далеко на север или юг ареала. Многоснежной зимой 1942 г. сайга в массе скопилась в нижнем течении р. Урал, откуда крупные стада уходили по льду в Каспийское море на сотни километров от берега [Формозов, 1946; Слудский, 1963]. В январе 1952 г. сотенные стада снова появились на льду моря в 80 км от Гурьева, а в июне животные были обнаружены рыбаками еще южнее, на о. Кулалы [Раков, 1956]. В суровую зиму 1928 г. из Актюбинской и Акмолинской губерний эти копытные откочевали в Туркестанский уезд Сырдарьинс-кой области, где стали легкой добычей “двуногих и четвероногих хищников” [Шпо-та, 1928]. В многоснежные зимы 1950/51 и 1953/54 гг. стада, зимующие в пустыне Далялык и в Бетпакдале, ушли на юг к линии железной дороги Кзыл-Орда- Аральское море, где звери гибли в большом количестве. Часть животных откочевала в Кызылкумы, часть дошла до предгорий Каратау и Чу-Илийских гор и проникла в Отарскую и Илийскую долины, где тоже гибли тысячами. Зимой 1976/77 гг. мигранты появились на землях Семиречья, возле Куртинского водохранилища, Капча-гая и близ г. Алма-Аты [Цаплюк, 1982]. В 1954 г. два сайгака зашли на окраину г. Кзыл-Орды [Антипин, 1955]. В Туркмении зимой 1965/66 гг. стада дошли к югу
до Кизил-Арвата, а в 1968/69 и 1972/73 гг. из Сарыкамышской впадины часть животных мигрировала севернее Дарвазы и юго-восточнее Северного Завода, откуда несколько особей проникли в Центральные Каракумы. В феврале 1973 г. истощенный самец был пойман в 90 км севернее Ашхабада [Ишадов, 1972, 1975а]. В особенно суровые зимы известны заходы зверей из Бетпакдалы на юг вплоть до окрестностей г. Ташкента [Быкова, Есипов, 2004].
В Казахском нагорье в обычные годы мелкие группы сайги встречаются лишь на южной окраине. В засушливое лето 1974 и 1975 гг. животные продвинулись необычно далеко к северу, где держались не только в открытом ландшафте, но и в зарослях кустарников, на вершинах и гребнях холмов и таких гор как Актау, Кызылтау и Узунтау, на высоте около 1000 м, вблизи от пасущихся архаров. Стада в 200-300 особей дошли до южных окраин гор Темирши, Кызылрай, Жаксы-тагылы, а мелкие группы - до Сарытау и Кызыл-тас и оз. Сарыкуль [Капитонов, 1980]. В 1975 г. зверей регистрировали во многих районах кустанайской и петропавловской лесостепи, самый северный пункт встреч - оз. Щучье в 5 км от южной границы Курганской области [Дробовцев, 1977].
В самом конце XX - начале XXI вв., по сообщению Ю.А. Грачева и А.Б. Беке-нова [2003] на VII съезде териологического общества, традиционные места отела, летовок, зимовок и путей миграций сайги в Казахстане еще сохранялись, но крупных скоплений животных уже не было даже во время отела, мелкие группы широко рассеяны по территории и нигде не задерживаются подолгу.
Численность
Во второй половине II тысячелетия поголовье сайги, судя по сохранившимся историческим сведениям и описаниям путешественников-первопроходцев, исчислялось миллионами. На землях Запорожского казачества в первой половине XVIII в. эти копытные “покрывали всю степь”, а в Нижнем Поволжье (от Астрахани до Царицына), Предкавказье, Волго-Уральском междуречье и Казахстане в том же столетии водились в “громадном, неимоверном количестве”, “великом множестве”, “неисчислимыми табунами”, “великими и несметными стадами” [Рычков, 1772; Гмелин, 1777; Адольф, 1948; Раков, 1956; Кириков, 1959, 1966, 1976, 1983; Банников и др., 1961; Демеуов, 1968; Фадеев, Слудский, 1982; Жирнов и др., 1998а].
Прогрессирующая тенденция сокращения ареала и ресурсов вида в Европе заметна с конца XVIII в., но вплоть до середины XIX в. он был многочисленным. В степном Прикумье многоснежной зимой 1848/49 гг. А. Ребров оценил поголовье в несколько сот тысяч [Кириков, 1983]. Позднее численность быстро сокращается, что вызвано периодическими гибельными джутами, хозяйственной деятельностью и неумеренным истреблением этих копытных резко возросшим людским населением [Адольф, 1948; Бакеев, Формозов, 1955; Слудский, 1955, 1956; Раков, 1956; Близнюк, 1995].
Чрезмерное преследование быстро поставило вид на грань исчезновения. В 20-е годы XX в. в самых глухих местах Калмыкии, Казахстана и Монголии сохранилось, как считается, всего лишь несколько сот особей или максимум 1 тыс. [Слудский, Страутман, 1955; Банников, 1958; Банников и др., 1961; Слудский, 1974; Бе-кенов, Грачев, 1998]. На мой взгляд, судя по анализу литературы, уцелело, скорее
всего, несколько десятков тысяч зверей, что подтверждается сравнительно широким распространением сайги в это время и весьма существенным объемом нелегальной заготовки рогов (см. ниже). В калмыцких и волго-кумских степях в середине 20-х годов ее было “много”, и с 1926 г. на нее опять начали усиленно охотиться. В 20-е годы в Прикаспии встречали не только одиночных животных и мелкие группы, но и стада до 100 экземпляров и более [Берков, 1927; Сытин, 1927; Орлов, 1928; Гептнер, Формозов, 1941]. В Балхашской котловине до 1925 г. этот вид обитал в “большом количестве” [Горчаковский, 1925], а из южной части Семипалатинской губернии был большой сбыт рогов в Китай [Барышев, 1925]. В 30-е годы в Китае на рынке традиционной медицины ежегодно продавали около 50 тыс. пар рогов [Джиган, Дикян, 1998].
Рост поголовья стал особенно заметным в 30-е и бурным в 40-е, военные, годы, когда большинство охотников находилось на фронтах, а на большей части пастбищ не было домашнего скота. В середине 40-х годов тысячные стада регистрировали по всему ареалу. В конце десятилетия весенняя численность европейской популяции оценивалась примерно в 100 тыс. голов, а в Казахстане превысила 600 тыс. [Дементьев, 1935; Антипин, 1941; Слудский, 1955; Исаев, 1959].
После многоснежной зимы 1950 г. часть прикаспийской группировки погибла [Лавровский, 1950], ноужев 1952 г. она увеличилась до 180 тыс.,в 1953 г.-до 250-300 тыс., а затем после массовой гибели в исключительно суровую зиму 1953/54 it. снова сократилась почти в два раза [Даль и др., 1956; Рак, 1956]. В Казахстане в это время насчитывали около 900 тыс. В 1958 г. европейская группировка достигла пика численности в 540 тыс., а население казахстанских популяций, заходящих на территорию Узбекистана и Туркмении, к концу этого десятилетия приблизилось к 1,5 млн. Общее поголовье вида оценивалось в 2 млн [Слудский, 1955; Банников и др., 1961; Бекенов, Грачев, 1998; Жирнов, Максимук, 1998а].
Дальнейшая динамика населения сайги в России и Казахстане отражена в таблице 28. Очередной пик численности европейской популяции был в 1978 г. (720 тыс., плотность - около 9 особей/км2), а ее постепенное уменьшение очевидно с 1979 г. Сокращение поголовья казахстанских популяций произошло после трудной зимовки 1975/76 гг., а затем, после очередного подъема и пика примерно в 1 млн в начале 90-х годов, оно снова резко уменьшается. В 2003 г. в Северо-Западном Прикаспии уцелело около 17 тыс., в Казахстане - 21 тыс. особей. В Узбекистане, судя по недавнему сообщению Е.А. Быковой и А.В. Есипова [2004], сохранилось 30-35 тыс. особей, во что верится с трудом. Миллионное поголовье быстро, всего за 7 лет (1994— 2002 гг.), сократилось почти в 20 раз. Вид снова оказался на грани выживания.
В Монголии до середины XX в. сайга была обычным видом в котловине Больших Западных озер. К концу 70-х - началу 80-х годов в результате нелегальной охоты, вытеснения скотом и после многоснежных зим поголовье сокращается до 300-750 особей в Шаргин-Гоби и до нескольких десятков особей в районе сомона Манхан. В последующие годы охрана копытных улучшилась, и их численность стала возрастать, даже несмотря на отдельные многоснежные зимы. В 1982 г. запасы вида оценивали в 900-950 голов, в 1988 г. - в 1700, а к 2001 г. группировка увеличилась до 5200 особей. В 2002 г. из-за неблагоприятных погодных условий и усиления нелегальной охоты (особенно на самцов) ее население снова сокращается до 1 тыс. [Соколов, Орлов, 1977, 1980; Сапожников, Дуламцэрэн, 1982; Соко-
Таблица 28. Динамика популяций сайги (тыс. особей) во второй половине XX - начале XXI вв. в России и Казахстане*
Год Россия, Прикаспий Казахстан, популяции
весной осенью уральская устюртская бетпакдалинская всего
1 2 3 4 5 6 7
1950 100 - - — -
1951 180 - - - 900
1952 180 - - — —
1953 300 80 - - 900
1954 100 85 - 560 830
1955 - 300 - - - -
1956 400 - - - - -
1957 450 500 - - — —
1958 540 811 - - - 1500?
1959 348 522 - - - -
1960 186? 297? — - 690 1200-1300
1961 342 513 - — - 1300
1962 267 400 — - — >650
1963 125 185 - - - >620
1964 115 270 - — >700
1965 75 118 — 64 412 >480
1966 105 215 — 581 590
1967 130 230 — — — —
1968 80 116 — 34 519 550-600
1969 — 213 - — - -
1970 190 - - — -
1971 150 246 50 56 995 1100
1972 - 213 - — 850 900
1973 260 324 - — - -
1974 240 425 70 199 956 1200
1975 340 500 - — - -
1976 390 650 45 117 480 640
1977 420 660 116 — 484 600-700
1978 — 720 120 - 280 500
1979 — 430 100 120 290 510
1980 - 380 120 170 400 690
1981 — 430 160 190 470 820
1982 - 395 180 190 480 850
1983 220 280 150 180 440 770
1984 185 265 40 190 340 570
1985 165 222 50 190 400 640
1986 176 240 70 150 250 470
1987 119 145 100 140 300 540
1988 131 157 90 207 368 665
1989 80 149 135 265 323 723
1990 — 160 ' 138 202 361 700
1991 133 168 236 232 357 825
1992 — 152 298 254 375 927
1993 — 148 250 216 510 976
1994 113 142 274 254 282 810
1995 125 220 - — 212
1996 196 300 (336)? - 214 248 -
Таблица 28 (окончание)
1 1 2 1 3 1 4 1 5 1 6 1 7
1997 270? 380? — —
1998 180 - 104 246 120 470
1999 55 - 84 200 64 348
2000 25 - 17 116 15 148
2001 18 - 9 58 12 79
2002 19 - 7 19 4 30
2003 17 — — — 1,8 21
* По: [Слудский, 1955; Раков, 1956, 1957; Сорока, 1957; Исаев, 1959; Банников и др., 1961; Жирнов, 1961, 1963, 1982; Слудский, Шубин, 1961, 1963; Жирнов и др., 1965; Смирнов, 1970; Дсмсуов, 1971: Близнюк, 1975, 1978; Фадеев, 1975а; Фадеев, Шаад, 1978; Близнюк и др., 1977, 1980; Фадеев, Слудский, 1982; Слудский и др., 1983; Дежкин, 1985; Жирнов, Максимук, 1994, 1998а; Бекенов, Грачев, 1998; Букреева, 1998; Грачев и др., 1999; Сидоров, 2000, 2003, 2004; Бекенов н др., 2002; Кузнецов, Лущекина, 2002; Лущекина, 2002; Грачев, Бекенов, 2003; Шаймуханбстов, 2004], сведениям Охотдспартамента РФ (Главохоты РФ) н Службы учета охотничьих ресурсов с изменениями и дополнением. Прочерк - учет нс проводился или нет достоверных сведений. Отдельные учетные данные в разных источниках не совпадают.
лов, 1986; Соколов и др., 1986, 1992, 1996; Лущекина, 1997; Лущекина, Дуламцэ-рэн, 1997; Лущекина, Неронов, 1997; Жирнов и др., 1998; Lushchekina et al., 1999, 2000; Dulamtseren, Amgalan, 2002].
Самые значительные количественные изменения ресурсов в XX в., как показывает анализ, вызваны чрезмерной охотничьей добычей и массовой гибелью зверей в многоснежные или гололедные зимы, при сильных засухах и эпизоотиях, что подробнее рассматривается в следующих разделах монографии.
Основные факторы, определяющие динамику популяций
Динамика популяций копытных определяется взаимодействием различных факторов среды, влияющих как на плодовитость, так и вызывающих смертность животных.
В этом плане очень показательно изменение населения сайги на о. Барсакельмес, куда животные зашли после одной из суровых зим в XIX в. В 1881 г. на острове обитали около 500-700 голов. К 1915 г. большую часть копытных здесь и всю группировку на о. Николая истребили охотники, исключительно ради получения рогов. В 1918 г. на Барсакельмес завезли несколько семей казахов, которые в том же году убили около 60 самцов. Оставшихся животных прикончил заведующий рыбными промыслами о. Возрождения (бывшего о. Николая) Гольцев, который в 1921 г. отловил и вывез на баркасе 50 особей. Из-за задержки в пути при отсутствии корма все они, кроме двух, погибли. В 1922 г. убит последний самец, и на острове уцелело не более 5-10 самок. Двух самцов, завезенных в 1927 г., вскоре застрелили местные жители. В 1929 г. местное охотничье-про-мысловое хозяйство снова завезло одного самца и двух самок, а год спустя еще одного самца и 4 самок. В 1931 г. на острове жили 5 взрослых самок с двумя детенышами и три самца, в 1932 г. - 10-11 зверей. В 1936 г., после уничтожения волка и лисицы, стадо выросло до 65 особей, а к 1938 г. - до 2 сотен. В 1938 г. большая часть копытных была уничтожена или выгнана на лед борзыми собаками, и к 1939 г. группировка уменьшилась до 50-60 голов. В этом же году террито-
рия острова стала заповедной, что не замедлило сказаться на численности популяции. В 1940 г. насчитывали 85-110 особей, в 1948 г. — 1059. Не исключено, что в холодные зимы мог быть небольшой подток животных с материка по льду. После многоснежных зим 1948-1950 гг. осталось 297 особей. В 1951 г. поголовье увеличилось до 349, в 1955 г. - до 647, в 1957 г. - до 1167 и в 1959 г. - до 3400 особей. В этот год, по отчетам, отстреляно 217 особей, на следующий - 218 (по другим сведениям - 457), в 1961 г. добыты и отловлены 309 (388) зверей, в 1962 г. - 264 (300). Скорее всего, добыча здесь велась в больших масштабах, поскольку к 1961 г. популяция уменьшилась до 1,5 тыс. особей. В 1962 г. на острове насчитали 2,5 тыс. После засух 1962 и 1963 гг. прохолостовали все самки, рожденные в 1962 г., число двоен в пометах сократилось до 50% и увеличилась смертность. Зимой 1964 г. отмечался падеж сайги и кулана. В результате популяция уменьшилась вдвое -до 1700 голов. Во второй половине 60-х годов общая численность сайги (3,5 тыс.) вместе с джейраном, куланом и сотней голов крупного рогатого скота и лошадей достигала 4—5 тыс., что при частых засухах привело к истощению пастбищ и, как показано выше, к резкому уменьшению плодовитости и сокращению популяции до 600 голов [Тимофеев, 1934; Антипин, 1941; Слудский, 1948, 1955; Васенко, 1950; Соколов, 1958; Слудский, Афанасьев, 1964; Исмагилов, 1970; Цаплюк, 1970; Соломатин, 1973]. В 70-е годы условия жизни копытных на острове ухудшились из-за повышения концентрации солей в Аральском море и уменьшения водоснабжения из пресных источников. В 1971 г. отмечен большой падеж сайги, но на острове оставалось около 700 голов [Цаплюк, 1982]. В 80-е годы здесь снова появились хищники - волк и лисица, мясом диких копытных стало питаться и местное население. В 1990 г., по одним источникам, часть популяции по льду ушла на материк, по другим - сюда пришли около 200 особей. В 1991 г. на Барса-кельмесе уцелело всего около сотни животных [Павлов, 1999].
Примерно такие же факторы среды воздействуют и на материковые популяции, периодически вызывая резкие изменения численности.
Воспроизводство, прирост. Сайга - один из самых плодовитых видов среди копытных. Скороспелость и рождение преимущественно двоен, а также преобладание в популяциях самок, позволяют виду в благоприятные годы увеличивать весеннюю численность почти в 2 раза. Из-за высокой детской смертности осенью на самку в лучшем случае приходится 1,0-1,4 детеныша, в худшем - 0,3-0,6. Соответственно осенний прирост популяций в разные годы колеблется от 13 до 85%, оставаясь чаще на уровне 40-50% от весенней численности. В период промысла и зимой происходит дальнейшее сокращение доли молодняка, и к весне в популяциях обычно остается менее трети сеголеток [Слудский, 1955; Раков, 1956; Банников и др., 1961; Рашек, 1963, 1974; Перовский, 1974; Жирнов, 1978, 1982; Близнюк, 1982, 1983; Фадеев, Слудский, 1982; Слудский и др., 1983; Лещенко, 1997; Жирнов и др., 1998в].
Особенно негативное воздействие на воспроизводство оказывает неумеренный выборочный отстрел взрослых самцов и взрослых самок, что приводит к резкому изменению полового и возрастного состава и омоложению популяций, снижению числа двоен в пометах и увеличению яловости самок. В интенсивно эксплуатируемых группировках самцы участвуют в размножении всего лишь 1-2 раза, самки - 3-4, и лишь немногие из них приносят детенышей более 4 раз [Фадеев, Слудский, 1982; Жирнов и др., 1998].
Л.В. Жирнов с соавторами [1998] выделяют у сайги два морфотипа - “рано- и поздноразмножающихся” животных, и на этом основании предлагают оригинальную интерпретацию популяционной динамики вида, что, на мой взгляд, совершенно не приемлемо. Островные и мелкоочаговые группировки, особенно на периферии ареала, чаще находятся в худших условиях обитания, и поэтому особи в них имеют меньшие размеры, массу и низкие репродуктивные показатели. На о. Барсакельмес и в небольших относительно изолированных стадах, например, при недостатке качественного корма, как отмечалось выше, сеголетки медленнее растут и до 80-100% самок не приходят в течку на первом году жизни. Крупных рано созревающих самок-сеголеток больше (до 96%) в многочисленных и широкомигрирующих казахстанских популяциях, имеющих хорошую кормовую базу практически круглогодично. Однако в неблагоприятные годы их доля резко уменьшается [Васенко, 1950, Даль и др., 1958; Банников и др., 1961; Рашек, 1963; Цаплюк, 1966, 1970; Исмагилов, 1970; Фадеев, Слудский, 1982]. Зависимость времени полового созревания от размеров и массы животных не является характерной особенностью только сайги, а известна для многих видов копытных.
При взаимодействии нескольких негативных абиотических или биотических факторов и, особенно, при чрезмерной добыче годичный прирост популяций сайги нередко оказывается отрицательным, а численность вида, несмотря на высокий уровень воспроизводства, стремится к нулевым значениям.
Детская смертность. Часть сайгачат рождается мертвыми, часть гибнет от травм, и особенно много их уничтожают хищные звери и птицы. Они гибнут также при заморозках, холодных дождях, от града и от голода, поскольку в крупных “родильных домах” далеко не все самки могут отыскать и накормить своих детей молоком. Смертность детенышей в первые дни жизни обычно составляет 5-16%, что типично для стадных животных, и лишь в редких случаях при ухудшении условий обитания и ранних засухах этот показатель достигает 18-24% [Раков, 1956; Даль и др., 1958; Соколов, 1951; Банников и др., 1961; Жирнов и др., 1965, 1998а; Фандеев, 1965; Филимонов, 1979; Жирнов, 1982; Фадеев, Слудский, 1982; Хахин, 1983; Максимук, Проняев, 1986; Лещенко, 1997; Близнюк, Букреева, 2000; Сидоров, 2000].
В благоприятные годы в весенне-летний период отход сайгачат близок к 20%, но при засухах возрастает в 2-3 раза. Из-за недостатка сочных кормов и воды нарушается режим активности животных и уменьшается продолжительность лактации самок. Стада в это время концентрируются у водоемов, близ которых обычно расположены поселения человека. Специальные исследования в районе отелов у Сар-пинских озер в Прикаспии в 1959 г. показали, что 48% из числа погибших телят были задавлены собаками, 13% - погибли под колесами автомашин, 4% - умерли от ран, полученных при переходах через скошенный тростник, т. е. большинство детенышей лишились жизни в результате антропогенных причин. До 10% сеголеток гибнет во время кочевок: их затаптывают взрослые особи на водных переправах, они отстают от матерей, тонут в воде, вязнут в иле, погибают от хищников [Фандеев, 1965; Жирнов, 1982; Жирнов и др., 1976, 1977; Петрищев, 1987а].
В европейской популяции детская смертность заметно увеличилась во второй половине XX в. по мере сокращения площади пастбищ и ухудшения питания. В конце 50-х годов их отход к августу составлял около 22%, а в 1972-1978 гг. он возрос до 29%. Максимальные потери (87% от числа погибших) приходились на
периоды отелов. По естественным причинам погибли 76% сайгачат, еще 20% - от наезда транспорта и около 5% - в оградах пастбищ, каналах, задавлены собаками или застрелены охотниками [Близнюк, 1982, 1983]. В 1993 г. отход сеголеток в отдельных районах достигал 35% [Петрищев, 1996]. Однако, по сведениям О.М. Букреевой [1998, 2002], в 90-е годы смертность среди новорожденных была в пределах нормы (4,6-16,8%), как и число детенышей на самку осенью (0,6-1,2). При этом нельзя не отметить высокий уровень плодовитости (1,1-1,5 эмбриона на самку), что свидетельствует о достаточно хороших условиях обитания копытных в последнее десятилетие XX в. (табл. 29).
Таблица 29. Уровень воспроизводства сайги в Северо-Западном Прикаспии в конце XX - начале XXI вв. (по [Букреева, 1998, 2002]; с изменениями и дополнением)
Год Доля самцов в популяции, % Число эмбрионов на самку Смертность новорожденных, % Число сеголеток на самку осенью
1983 20,0 — - 1,39
1985 16,8 — 0,78
1986 14,9 — - 0,89
1987 15,1 — - 0,45
1988 16,6 — — 0,85
1989 20,6 — - 0,79
1990 16,7 — — 0,75
1991 24,3 — - 0,68
1992 6,4 - - 0,70
1993 8,2 1,50 8,4 0,98
1994 10,3 1,28 16,8 0,61
1995 12,3 1,23 (1,39)** 5,4 0,87
1996 14,4 1,44 4,8 1,14
1997 12,3 (5)* 1,43 4,6 1,19
1998 10,9 (3,5)* 1,37 12,8 0,85
1999 7,5 (2,5)* 1,06 15,8 0,70
2000 3,5 (0,9)* 1,28 23,4 0,79
2001 8,2 0,26 6,5 0,23
2002 (<!)* । 13*** 6 5*** 0,91***
2003 9,1 (6,5)* - з*** -
* В скобках - доля самцов в период гона (по [Lushchekina, Struchkov, 2001; Букреева, 2003, 2004; Сидоров, 2003]).
** По сведениям А.П. Лещенко [1997].
*** По [Сидоров, 2004].
Осенью и зимой естественный отход сеголеток редко превышает 10%. Значительно большие потери они несут от охотников. В целом смертность молодняка ежегодно достигает 50-80% от числа родившихся.
Погода и корма. Судя по данным мечения, обычно в осенне-зимний период по естественным причинам гибнет около 5% поголовья сайги. В аномальные годы (засушливые, многоснежные, морозные, буранные и гололедные) животные особенно широко кочуют, в массе погибая от голода, браконьеров, волков и собак. Число погибших исчисляется десятками и сотнями тысяч. На севере аридной зоны сильные засухи бывают 5-6 раз, а многоснежье (более 20 см) и гололедицы -
1-3 раза в 10-12 лет. В засушливые годы от обезвоживания чаще гибнут кормящие самки и молодняк. При остром недостатке зимнего корма происходит массовое (20-30%) абортирование самок и резорбция эмбрионов, что сказывается на уровне воспроизводства популяций. Зимняя смертность особенно высока среди взрослых самцов, истощенных после гона. В многоснежные 1948-1951 гг. находили десятки тысяч их трупов [Слудский, 1955, 1963; Раков, 1956]. На о. Барсакельмес в отдельные зимы гибнет до 60% местной группировки. В 1953— 1964 гг. среди погибших особей взрослые самцы составляли 70,4%, самки - 17,2, молодняк - 12,4% [Рашек, 1963, 1965, 1974]. Массовая зимняя элиминация самцов приводит к резкому сдвигу полового и возрастного состава популяций [Лавровский, 1950; Банников и др., 1961; Близнюк, 1982, 1983; Фадеев, Слудский, 1982; Горбунов, 1986а; Жирнов и др., 1998а].
В европейской части ареала исследуемого вида многоснежные и буранные зимы, во время которых гибло множество голов домашнего скота и, скорее всего, диких копытных, были в 1560, 1578, 1756, 1787, 1790, 1798, 1802, 1805, 1812, 1823, 1833, 1841, 1844, 1848-1849, 1875-1876, 1910, 1923, 1926, 1928, 1939, 1948-1949,1953 и 1955 гг. [Кириков, 1959, 1976, 1983; Слудский, 1963]. Массовую гибель (до 40-50%) сайги регистрировали в суровые зимы 1848-1850, 1949/50, 1953/54 (более 150 тыс. особей), 1959/60 и 1972 гг. и в сильные засухи 1949, 1957 и 1959 гг. [Ребров, 1849; Лавровский, 1950; Бакеев, Формозов, 1955; Кириков, 1955, 1966; Даль и др., 1956; Рак, 1956; Исаев, 1959; Близнюк, 1982, 1995; Жирнов, Максимук, 1998а]. Неблагоприятные условия зимовки были здесь также в 1967/68, 1978/79, 1986/87, 1993/94и 1998/99 гг., а засухи-в 1984,1987, 1994и 1998-2000 гг. [Букреева, 2000], но значительного отхода копытных в эти годы не отмечено. 6 мая 1971 г. много животных погибло от переохлаждения и травм после продолжительного ливня с градом [Близнюк, 1983].
В разных районах Казахстана высокую смертность сайги после джутов отмечали в 1826/27, 1840/41, 1848/49, 1851/52, 1855-1857, 1866/67, 1879/80, 1888/89, 1891/92, 1903/04, 1917/18, 1919/20, 1921/22, 1927/28, 1940-1946, 1948-1951, 1953/54, 1958/ 59,1963/64, 1968/69,1971-1973, 1975-1977,1983-1985, 1987/88, 1993/94, 1996/97 гг. После суровой зимы 1826/27 гг. эти звери почти на 30 лет исчезли в междуречье Волги и Урала. Зимой 1949/50 и 1975/76 гг. поголовье этого вида в республике уменьшилось вдвое, в 1971/72 гг. погибло около 400 тыс., а в - 100 тыс. особей. В многоснежную зиму 1993/94 гг. бетпакдалинская группировка сократилась с 510 тыс. до 280 тыс., а уральская понесла значительные потери (десятки тысяч особей) в 1996/97 г. В устюртской популяции падеж зарегистрирован в 1972/73 и 1975/76 гг. [Соколов, 1951; Слудский, 1955, 1956, 1963, 1974; Раков, 1956; Кириков, 1959; Рашек, 1963,1965; Корнеев и др., 1975; Фадеев, 1975а, 19806; Слудский, Фадеев, 1977; Фадеев, Слудский, 1982; Слудский и др., 1983; Цаплюк, 1982; Рей-мов, Карабеков, 1986; Грачев, Бекенов, 1993; Бекенов, Грачев, 1998; Bekenov et al., 1998; Бекенов и др., 2002].
Хищники. Один из основных потребителей сайги - волк. Этот хищник не в состоянии догнать здоровое животное, быстро развивающее высокую скорость, но его охота на беременных самок, новорожденных, раненых, травмированных и истощенных особей, а также у водопоев, в местах переправ при миграциях, по глубокому снегу, при настах и группой с применением загонов и засад весьма ус
пешна и истребительна. Группы волков обычно сопровождают мигрирующие стада, подбирая отставших особей, и держатся в местах сезонной концентрации копытных. Зимой остатки сайги встречаются в 90-100% исследованных желудков и экскрементов хищников. В отдельных районах волк истребляет до 60-70% истощенных и травмированных во время гона самцов, влияя, следовательно, не только на количественный, но и на половозрастной состав популяций. Нередко этот хищник давит копытных гораздо больше, чем может съесть. В местах отела 1 зверь за 1 час может загрызть 5-6 сайгачат, а по насту стая из 12-15 особей уничтожает в стаде за раз до 2(М10 голов. При обилии пищи волк часто объедает у жертвы лишь жир, как бы оставляя мясо “про запас”. По некоторым оценкам, видимо завышенным, каждый зверь за год давит до 50-100 сайгаков. В совокупности этот хищник истребляет до 20-25% популяции, на всем ареале - десятки и сотни тысяч (при высокой численности) особей ежегодно [Формозов, 1946; Соколов, 1951; Адольф, 1954; Раков, 1955, 1956; Слудский, 1955, 1962, 1970; Банников и др., 1961; Филимонов, Лаптев, 1975; Слудский и др., 1981, 1983; Фадеев, Слудский, 1982; Лещенко, 1997].
Во все времена волк был охотничьим конкурентом человеку, который, понимая это, как мог, боролся с ним. В России особенно резко его численность была сокращена в середине XX в. в послевоенные годы. В Калмыкии с появлением мотоциклов и автомашин хищник был почти полностью истреблен к 1954 г. Особенно эффективно его добывали в засушливый период года возле колодцев и в зарослях кустарника. В Центральной Калмыкии последний волк убит в 1969 г. В первой половине 70-х годов хищника почти полностью уничтожили на юге европейской части страны: в областях, краях и республиках счет ему шел на единицы, что, несомненно, способствовало быстрому росту населения сайги. На каждого уцелевшего волка в этот период приходилось по нескольку тысяч копытных. В 80-е годы борьба с ним ослабла, его поголовье в областях стало исчисляться десятками, а в 90-е годы - сотнями голов. В Калмыкии и Астраханской области в 60-70-е годы весной учитывали от 5 до 40 волков, в середине 80-х годов - 100-170, в 2000 г. - от 700 до 1200-1400 [Зуев, Хахин, 1982а,б; Букреева, 2000; Букреева, Убушаев, 2003]. Осенняя численность хищников, как минимум, на 50-70% больше. Следовательно, в конце XX - начале XXI вв. в этом регионе находилось около 2 тыс. волков, и на каждого из них приходилось в среднем не более 10 голов сайги.
В Казахстане в начале 50-х годов XX в. в местах концентрации сайги нередко наблюдали стаи волков, насчитывающие от 18 до 42 голов. В зоне пустынь и полупустынь обитали примерно 25 тыс. зверей [Слудский, 1955, 1962]. В 1955 г. в республике добыли 21,2 тыс. хищников, а в целом во второй половине этого десятилетия - около 80 тыс., что значительно ослабило пресс на копытных и ускорило рост их популяций [Банников и др., 1961; Слудский, 1974]. В 70-е годы здесь насчитывали около 30 тыс. волков [Фадеев, 1981]. В этот период на каждого зверя приходилось в среднем 20-40 голов сайги. В конце XX в. борьба с волком почти прекратилась, и его численность, вероятно, приблизилась к 40 тыс. [Бекенов, Лобачев, 2003] (по другим данным - к 90-100 тыс. [Бекенов, 1999]), что значительно превышает население сайги.
Потери и европейской, и казахстанских популяциях от волка в последнее десятилетие могут оцениваться в десятки тысяч голов. Следовательно этот хищник,
наряду с человеком, стал одним из главных негативных факторов динамики населения копытных, численность которых сократилась до критического уровня.
Этот вывод противоречит мнению отдельных ученых [Бибиков, Жирнов, 1975; Жирнов, 1982; Проняев, 1985а; Бибиков и др., 1998], полагающих, что волк не способен ограничить рост численности сайги, поскольку основная часть его добычи приходится на дефективную часть группировок (травмированных, больных и ослабленных животных, находящихся в “хвосте” стад), и он, скорее, является санитаром, оздоровляющим популяции копытных.
Действительно, волк не может остановить рост многочисленных популяций сайги, насчитывающих сотни тысяч особей, что было заметно в середине XX в., когда промысловая эксплуатация группировок была минимальной. Более того, в этот период он, видимо, мог оказывать определенную селективно-оздоровительную роль. Однако его негативное воздействие на популяции жертв многократно усиливается при депрессиях, вызванных, как правило, чрезмерным промыслом или массовой гибелью в суровые зимы и при эпизоотиях. И именно в это время чрезмерно размножившийся хищник, наряду с браконьерами, способен полностью истребить локальные группировки. Поэтому роль волка в динамике популяций копытных нельзя недооценивать.
Существенный урон сайге наносят и собаки. В местах ее обитания только в Калмыкии численность пастушьих собак, не считая дворовых, достигает 15 тыс. Они давят копытных почти круглогодично, но чаще - в период отела, миграций, в многоснежье, при джутах и засухах, когда животные выходят к населенным пунктам и водоемам. В 1957-1970 гг. до 50% случаев гибели молодняка в некоторых районах приходилось на их долю [Бибиков и др., 1998]. В 90-е годы их жертвами чаще становились взрослые самцы после гона (65%), реже взрослые самки (23%) и молодые особи (12%). Собаки “контролировали” периферийные районы обитания копытных, где не было волка [Лещенко, 1997]. В Бетпакдале 1 собака за неделю отлавливала до 11 сайгачат, а всего в Казахстане, как и в Калмыкии, от этого хищника ежегодно гибнут тысячи особей [Слудский, 1962; Слудский и др., 1983].
На сайгачат нападают также лисицы, корсаки и крупные птицы (беркут, орел-могильник, европейский степной орел, орлан-белохвост, коршун, ворон, серые и черные вороны, серебристые чайки), но в целом их роль в динамике населения исследуемого вида, за исключением, пожалуй, лисицы, несущественна [Васенко, 1950; Бакеев, Формозов, 1955; Раков, 1955; Слудский, 1962; Рашек, 1963, 1974; Исмагилов, 1970; Жирнов, 19776, 1982; Фадеев, Слудский, 1982; Слудский и др., 1983; Хахин, 1983; Лещенко, 1997].
Конкуренты. Пастбища сайги и домашнего скота, зачастую, одни и те же, но дикие копытные поедают многие растения, которые домашние животные не потребляют, и при номадном образе жизни не составляют им значительной конкуренции [Демеуов, 1971]. Напротив, оседлый домашний скот обычно сильно стравливает и выбивает пастбища, которые впоследствии становятся непригодными для диких животных. При высокой плотности населения участие в этом деструктивном процессе принимают и дикие копытные.
Домашние животные стали серьезными конкурентами сайги и за источники воды, хотя следует отметить и положительный фактор. В последние десятилетия для скота сооружено много искусственных водопоев, которыми в ночные и раннеутренние часы пользуются дикие копытные.
Из других растительноядных конкурентов можно отметить лишь джейрана (локально) и суслика-песчаника на юге ареала [Васенко, 1950].
Болезни, паразиты, травмы. Среди болезней наибольшее влияние на динамику населения сайги оказывают ящур и пастереллез, эпизоотии которых в отдельные годы приводят к гибели 10-20% группировок. Источником инфекции, как правило, служат домашние копытные, но затем мигрирующие стада сами становятся разносчиками заболеваний [Курченко, 1995; Бекенов и др., 1998]. Важно, что они могут быть потенциальным переносчиком возбудителей чумы, которой звери заражается от блох и клещей во время миграций через колонии больших песчанок. На Мангышлаке зарегистрирован случай заболевания человека чумой от сайги [Пей-сахис и др., 1979].
В Северо-Западном Прикаспии эпизоотии ящура регистрировали в 1957, 1958, 1972 и 1977 гг. Особенно высокой (более 40 тыс. особей) смертность была засушливым летом 1957 г. Среди погибших преобладали сеголетки и самки [Исаев, 1959; Банников и др., 1961; Жирнов, 1962; Фандеев, 1965; Жирнов, Максимук, 1998а; Жирнов и др., 1998а]. В Казахстане эпизоотии имели место в 1955-1956, 1958, 1967, 1969, 1971 и 1974 гг. В Центральном Казахстане летом 1967 г. погибло около 50 тыс. сеголеток, падеж среди взрослых особей был незначительным. Взрослые самцы более стойкие к этой болезни, но у переболевших зверей шелушатся роговые чехлы и отпадают их верхние части, поэтому рога становятся короткими. В некоторые годы в стадах находятся до 20% пораженных ящуром особей [Фадеев, Слудский, 1982; Слудский и др., 1983; Бекенов, Грачев, 1998].
Вспышки пастереллеза отмечены в Казахстане в 1981, 1984 и 1988 гг. В Тургай-ской области в 1981 г. погибло около 70 тыс., в 1988 г. - 270 тыс., в междуречье Волги и Урала в 1984 г. - более 100 тыс. особей [Фадеев, Слудский, 1982; Слудский и др., 1983; Фадеев, 1986; Грачев, Бекенов, 1993; Бекенов, Грачев, 1998].
У сайги найдены также сибирская язва, бруцеллез, некробактериоз, токсоплазмоз, энтероксемия, колибактериоз, диплококкоз и другие болезни [Ременцова, 1962; Галузо и др., 1963; Горегляд, 1971; Петров и др., 1979; и др.], однако они не играют существенной роли в динамике ее населения.
У этого копытного обнаружено также 55 видов эндопаразитов (из которых около 85% общие с домашними животными) и 10 видов эктопаразитов [Банников и др., 1961; Горегляд, 1971; Фадеев, Слудский, 1982; Слудский и др., 1983; Петров, 1985; Звегинцова, Треус, 1986; Прядко и др., 1993]. В целом роль паразитарных инвазий в популяционной динамике незначительна, что отчасти объясняется большой подвижностью животных и частой сменой мест обитания.
Кожный овод сильно изнуряет сейчас только монгольские группировки. В европейской и казахстанских популяциях он исчез в конце 20-х годов XX в., когда поголовье сократилась до минимума [Слудский, 1955; Бекенов и др., 1998]. В том, что овод паразитировал на них ранее, нет сомнений. Вот что сообщает С.Г. Гмелин [1777] о животных, добытых в районе Астрахани в 70-е годы XVIII в.: “Мясо молодых сайгаков вкусом не неприятно, и козы гораздо вкуснее козлов, да и об старых сайгаках не думали столь худо, если бы некоторый особливый род червей, коих у них между кожей и мясом превеликое множество бывает не причинял чрезвычайного омерзения”. Примерно то же пишет и П.С. Паллас [1809]: “... мясо старых сайгаков весьма гнусно; ибо вся спина под кожею так усыпана чирьями и толсты
ми червяками оленьих шершней, что едва можно найти такое место, в которых бы оных не было”. О сильной пораженности сайги личинками овода свидетельствуют также Э. Эверсман [1850], Н.А. Северцов [1861] и Д. Анучин [1875].
Значительная часть самцов погибает от травм, полученных при драках во время гона. На о. Барсакельмес их отход по этой причине достигал 10-20% [Васенко, 1950; Рашек, 1963, 1965, 1974]. В междуречье Урал-Кушум в период с 21 декабря по 3 января 1953 г. Н.В. Раков [1955, 1956] обнаружил 318 трупов сайги, из них 300 взрослых самцов, из которых часть была со следами ранений, и наблюдал 62 живых зверя с признаками увечий. Большинство из них хромали на одну из передних ног, что, видимо, было вызвано не только травмами, но и ослаблением роговых чехлов копыт при непрерывном перемещении самцов вокруг самок по промерзшей земле [Слудский, 1955].
Гибель на водных переправах. Тысячи кочующих животных гибнут ежегодно на водных переправах, особенно при переходах по тонкому льду рек и озер. Под лед иногда уходят стада, насчитывающие сотни особей [Цаплюк, 1982]. Весной высока смертность сайгачат, особенно на переправах через “рукотворные” каналы (см. ниже).
Антропогенные факторы. Негативное антропогенное воздействие на популяции сайги достигло апогея во второй половине XX в., когда существенно увеличилось людское население, резко выросло поголовье домашнего скота (до 4 млн только в Калмыкии) и усилилась трансформация среды обитания. Интенсификация животноводства потребовала дополнительной распашки земель под кормовые культуры, их мелиорации и сооружения сети оросительных каналов, протяженность которых в местах обитания европейской группировки достигла 1300 км. Параллельно шло строительство населенных пунктов, автодорог с твердым покрытием, искусственных водопоев, трубопроводов, военных сооружений, кошар для содержания скота и огораживание пастбищ (только в 1973-1980 гг. огорожено около 300 тыс. га). На Черных землях кошары были расположены через 3-4 км одна от другой, и с любой точки их можно было видеть от 5 до 12. Между ними по степи днем и ночью курсировали автомашины, представлявшие не только источник постоянного беспокойства для диких копытных, но и прямую угрозу для их жизни. Сотни автомашин имели многочисленные геологоразведочные партии, предприятия по добыче и перекачке нефти и газа и вновь возникшие крупные населенные пункты [Жирнов и др., 1965; Близнюк, 1977, 1982, 1995; Дежкин, Менькова, 1981; Жирнов, 1982; Зуев, Хахин, 1982а,б; Хахин, 1986; Максимук и др., 1989].
В результате интенсивной хозяйственной деятельности человека в Прикаспии в 60-80-е годы сайга лишилась большей части жизненно важных мест обитания. Более того, антропогенное преобразование естественного ландшафта и стационарный выпас скота привели к опустыниванию (до 75-83% площади ареала) и засолению пастбищ, что стало масштабным экологическим бедствием как для копытных, которым уже не хватало качественных кормов, так и для человека в связи с сокращением посевных площадей и поголовья домашнего скота. В изменившихся условиях обитания сайга была вынуждена заходить на орошаемые земли, посевы зерновых культур и кормовых трав и выходить к искусственным водоемам в густонаселенных людьми и собаками районах, где их неимоверно истребляли. Несоизмеримо возросло и беспокойство животных. Их практически круглогодично пре
следовали на автомашинах и мотоциклах, и особенно интенсивно - в период промысла. В результате этого у копытных изменилось поведение - едва заслышав шум моторов, они стали панически срываться с места. Сверхподвижный образ жизни вел к нарушению процесса питания и размножения, изменению суточной активности, сроков и путей миграций, мест отела, сроков гона и, в конечном счете, к увеличению яловости самок и снижению уровня воспроизводства. Беременные самки нередко рожали “на ходу”, что не способствовало сохранению молодняка, отход которого увеличился в несколько раз. При расширении контакта с домашними животными участились случаи массовой гибели зверей в результате заболеваний ящуром и пастереллезом [Жирнов и др., 1977; Близнюк, 1982; Зуев, Хахин, 1982а,б; Максимук, Проняев, 1986; Петрищев, 1987а,б; Максимуки др., 1989; Жирнов, Максимум 1994; Лещенко, 1995, 1997; Букреева, 1998; Жирнов, 1998г; Близнюк, Бак-ташева, 2001].
В Казахстане преобразование среды обитания сайги достигло максимума в середине XX в., когда были распаханы огромные площади целинных степей, где копытные выпасались летом. В 1880-1928 гг. пашня занимала 4,5-5, в 1949 г. - 6,8, а в 1956 г. - 27,8 млн га [Бажанова, 1955; Слудский и др., 1962]. Мигрирующие животные были вынуждены пересекать сельскохозяйственные зоны, где их всячески преследовали браконьеры и легальные охотники даже летом, после чего на полях оставались многочисленные трупы и осиротевшие детеныши. Многократно усилился фактор беспокойства, что отражалось на упитанности и плодовитости зверей [Голованова, Анохина, 1976; Фадеев, Исмагилов, 1987]. Деградация пастбищ была очевидной и на севере Узбекистана. Устюрт в 70-80-е годы пересекали более 780 дорог, протяженность каждой достигала 200-450 км, что привело к заметному разрушению почвенного слоя, уничтожению растительности на больших площадях и препятствовало нормальному ходу миграций животных [Реймов и др., 1986].
Существенно возросла смертность этих копытных в сооружениях человека и от наезда транспорта. В Казахстане, Узбекистане и Туркмении тысячи зверей ежегодно гибли в траншеях и коллекторах нефте-, газо- и водопроводов с крутыми берегами [Ишадов, 1975а; Реймов и др., 1986; Фадеев, Исмагилов, 1987; Жирнов, 1998г]. В Калмыкии в конце 70-х годов при переправах через Яшкульский распределительный канал гибли десятки тысяч особей, преимущественно детеныши в возрасте 3-10 дней. На одном километре канала насчитывали до 25 тыс. трупов. Летом 1975 г. в Черноземельском и Яшкульском районах в проволочных ограждениях пастбищ на 1 км ограды обнаруживали 1-3 трупа, большей частью - самок. Гибель животных от умышленного наезда транспорта в 1972-1978 гг. достигала 9% от зарегистрированных случаев непромысловой смертности [Близнюк, 1977, 1982, 1983; Жирнов и др., 1977; Зуев, Хахин, 1978,1982а,б; Зуев, 1980; Петрищев, 1987а]. В начале 80-х годов доля антропогенных факторов (без лицензионной добычи) в общей смертности сайги в РСФСР составляла 83% (Перовский, 1988).
Самым мощным негативным антропогенным фактором, однако, была охота. Ее роль в динамике населения вида рассматривается ниже в специальном разделе.
Основные причины сокращения ресурсов в конце XX в. Причину резкого уменьшения поголовья сайги в последнее десятилетие XX в., также как лося и других копытных [Ломанов, 1995], российские ученые-охотоведы и охотничьи чиновники связывают с “циклической закономерностью развития” популяций и “кли
матическими и негативными флористическими изменениями условий среды обитания, оказавшими влияние на снижение репродуктивного потенциала и выживаемость животных” [Сидоров, Букреева, 1999; Сидоров, 2000, 2003, 2004; Букреева, 2002]. Однако эта “климатическо-циклическая” гипотеза не согласуется с имеющимися фактами.
“Циклической закономерности” в динамике населения сайги, при внимательном анализе, не усматривается. Многолетняя динамика казахстанских группировок в отдельные периоды находится в противофазе по отношению к европейской популяции (табл. 28). Монгольская группировка в 90-е годы и вовсе бурно увеличивала свою численность в то время, когда другие быстро сокращались. Популяционные “циклы”, выделяемые учеными-охотоведами, разномасштабные у разных группировок и не связаны с климатом. Они, в большей мере, “рукотворные”: популяции быстро депрессируют после неумеренного промысла (рис. 23) и постепенно увеличивают свою численность после запрета охоты, что типично для всех эксплуатируемых видов копытных. Более того, говорить о существовании “закономерности” на основании всего двух периодов изменения известной численности животных, по меньшей мере, непрофессионально.
Истинные причины, давшие начало выделяемым охотоведами “естественным циклам” во второй половине XX в., иные. Первое масштабное сокращение населения европейской группировки, достигшей максимума (540 тыс.) в 1958 г., произошло после рекордной добычи в самом конце 50-х годов (см. ниже). Цитирую И. Дремова [1963]: “...работники Главохоты, в частности заместитель начальни-
Годы
Рис. 23. Динамика населения и добычи сайги в России
1 - численность, 2 - добыча сайги, 3 - численность волка (х 100) (по материалам Главохоты РСФСР, Охотде-партамента РФ и Государственной службы учета охотничьих ресурсов)
ка Главохоты тов. Круторогов, оправдывая создавшееся положение (резкое уменьшение численности -А.Д.), заявляют, что они сознательно, в интересах развития местного животноводства и нужд сельского хозяйства, стремятся снизить, довести до минимума численность основного поголовья сайгаков”.
Очередной пик численности был в 70-е годы (табл. 28). Животные снова стали наносить определенный вред сельскому хозяйству. Цитирую очевидца и участника событий охотоведа А. Максимука [1982]: “Некоторые республиканские организации предложили объявить сайгака “сорным видом” и начать компанию по его уничтожению на пастбищах сельскохозяйственных животных. Учитывая тревожную ситуацию, было решено провести редукционный промысел сайгаков и привести их численность в соответствие с емкостью угодий (450-500 тыс. животных). В промысловый сезон 1978 г. Астраханский и вновь созданный Калмыцкий госпромхозы добыли в общей сложности 200 тыс. сайгаков, на 40 тыс. больше, чем в предыдущем сезоне. Пришлось применить одновременно два способа добычи сайгаков - ночной, из-под фар и метод кольцевого нагона (с использованием мотоциклов). Но даже при таких мерах интенсификации промысел затянулся до декабря, когда у сайгаков обычно уже начинается гон. Популяция пришла к гону в неудовлетворительном состоянии, что отразилось на ее половозрастной структуре и воспроизводстве. Весной 1979 г. было учтено 360 тыс. сайгаков”.
Нет прямой связи и с “негативными флористическими изменениями”. Напротив, в 90-е годы кормовые условия для европейской популяции сайги были в целом лучше, чем в предыдущие десятилетия, поскольку поголовье домашнего скота в Калмыкии сократилось почти в 5 раз (с 3,65 млн голов в 1989 г. до 755 тыс. в 1999 г.). Продуктивность пастбищ возросла в 3-4 раза - с 1,5-2 до 6-8 ц сухой массы на гектар. Масса новорожденных сайгачат в это десятилетие, за исключением 1994 г., превышает показатели 80-х годов, что свидетельствует о хорошем физиологическом состоянии самок. Гибель детенышей в первые дни жизни близка к норме [Петрищев, 1997а,б; Букреева, 1998,2002; Близнюк, Букреева, 2000; Близнюк, Бакташева, 2001; Кузнецов, Лущекина, 2002].
Заметного снижения репродукционного потенциала европейской группировки в 90-е годы, по сравнению с предыдущими десятилетиями, тоже не было. Доля беременных самок и число эмбрионов на самку оставались в пределах нормы вплоть до 2000 г. (табл. 23 и 29). Эти показатели резко уменьшились позднее, когда в популяции в период гона осталось менее 1% самцов. Не усматривается до 2000 г. и заметного уменьшения числа сеголеток осенью (табл. 29). Плодовитость самок во всех трех казахстанских популяциях в 1991-1996 гг., по сообщению Ю.А. Грачева и А.Б. Бекенова на VII съезде териологического общества (2003 г.), как и в европейской группировке, было высоким (в среднем 1,7-1,8 эмбрионов на самку старше года и 0,9-1,0 - на молодую) даже при низкой (3,7-10%) доле самцов (табл. 30). В уральской популяции нормальное воспроизводство продолжалось до 1999 г. (в среднем 1,4 детеныша на самку), в бетпакдалинской плодовитость резко уменьшилась с 1999 г., когда осталось менее 2% самцов. В устюртской популяции в мае 2001 г. на одну родившую самку приходилось 1,4 детеныша, но из-за малой доли самцов резко возросла яловость, и показатель плодовитости составил всего 0,6.
Таблица 30. Уровень воспроизводства популяций сайги в Казахстане в 90-е годы XX в.*
Популяции Год Число эмбрионов на самку Число детенышей на самку осенью Смертность молодняка, % Прирост популяций, %
Бетпакдалинская 1990 1,57 0,69 56,1 48
1991 1,60 0,75 53,1 50
1992 1,40 0,39 72,2 23
— — 1993 1,37 0,97 29,2 73
Устюртская 1990 1,52 0,59 61,2 36
а 1991 1,20 0,44 63,3 26
— — 1992 1,72 0,68 60,5 72
<1 1993 1,34 0,23 82,9 13
а 1999 0,55** - — -
Уральская 1992 1,16 0,59 49,2 36
и 1993 1,44 0,65 54,9 42
1999 1,40** - - -
* По ([Жирнов и др., 1998в]; с дополнениями).
** Число детенышей на самку в период отела (по личному сообщению Ю.А. Грачева).
Резкое сокращение уровня плодовитости сайги, следовательно, очевидно лишь с 1999-2000 гг., но вызвано оно не “климатическими и флористическими явлениями”, а почти полным уничтожением браконьерами взрослых самцов, доля которых в популяциях сократилась с 15-20% до 3,3%, а к периоду гона - до 1% (табл. 27 и 29). В европейской популяции такая же ситуация уже имела место в 1958-1959 гг., когда в результате чрезмерного отстрела доля взрослых самцов уменьшилась к периоду гона до 1—2%, а яловость самок, соответственно, возросла почти в 3 раза [Банников и др., 1961; Жирнов, 1963; Фандеев, 1965].
Погода, несомненно, внесла немалую лепту в динамику группировок. Однако в Северо-Западном Прикаспии в 80-90-е годы катастрофических зим, губительных засух и эпизоотий не было. Многоснежной зимой 1998/99 г. значительная часть популяции мигрировала на территорию Дагестана и не вернулась обратно не из-за изменения климата и растительности, а из-за того, что была уничтожена браконьерами. В Казахстане бетпакдалинская популяция уменьшилась в два раза в многоснежную зиму 1993/94 гг., уральская несколько сократилась зимой 1996/97 гг., но устюртская потерь не несла (табл. 28). Многоснежные зимы, уносившие десятки и сотни тысяч особей, в Казахстане неоднократно случались и раньше, но после них популяции быстро восстанавливались. Массовой смертности копытных от болезней в 90-е годы в республике тоже не отмечено.
На мой взгляд, “климатическо-циклическая” гипотеза динамики численности копытных, “рожденная” Охотдепартаментом РФ и его Контрольным информационноаналитическим центром, отвечающими за состояние ресурсов животного мира - это или недоразумение, или попытка переложить масштабные потери животных, возникшие в результате собственного неумелого управления популяциями и слабой государственной охраны, на климат.
Главная причина очередной масштабной депрессии популяций сайги, как и у других видов [Данилкин, 1999, 2002, 2003; Глушков, 2003], несомненно, иная -
антропогенно-хищническая. Если к уровню официальной и браконьерской добычи животных (см. ниже) и другого антропогенного ущерба добавить не менее значимый урон от волка, численность которого на постсоветском пространстве в конце XX в. выросла в несколько раз, то места для потерь от “климатическо-флористических” явлений практически не остается.
Очевидно же, что в последнем десятилетии XX в. в России, в период изменения социально-экономических отношений, многократно усилилось нелегальное истребление копытных местным обнищавшим населением, а также моторизованными браконьерами, специализирующимися на их добыче ради дорогостоящих рогов и мяса [Кузнецов, Лущекина, 2002; Лущекина, 2002]. Охрана угодий госохотнадзором значительно ухудшилась. Борьба с волком почти прекратилась, что привело к увеличению его численности в Калмыкии и Астраханской области, как отмечено выше, с 37 особей в 1980 г. до 2 тыс. к концу XX в. (рис. 23). В зоне обитания сайги ежедневно находилось около 1 тыс. оседлых и кочующих за стадами хищников [Букреева, Убу-шаев, 2003]. По наблюдениям, в 1988 г. каждый волк резал в сутки в среднем 0,17 особей, в 1995 г. - 0,23 [Лещенко, 1997], а за год, следовательно, - 60-80 экземпляров. Потери популяции от волка, собак и браконьеров, помимо официальной добычи, исчисляются десятками тысяч [Данилкин, 2003]. Именно в результате этого, а не “климатических и флористических явлений”, европейская группировка утратила промысловое значение.
В Северо-Западном Прикаспии примерно такая же ситуация уже имела место в конце 50-х - первой половине 60-х годов, когда из-за ослабления охраны, роста браконьерства и чрезмерной лицензионной добычи (преимущественно - взрослых самцов) численность сайги уменьшилась в несколько раз [Жирнов и др., 1965; Демьянова, Любаев, 1986].
Основная причина сокращения населения этого вида в Казахстане та же, что и в России: “круглогодичное браконьерство во всех районах обитания сайги, практически при полном отсутствии охраны”. Те же и социальные корни браконьерства: “бедность и отсутствие работы у местного населения (особенно сельского), нашедшего в охоте на сайгу источник заработка и пропитания”. Кроме того, вплоть до 1999 г. этих животных добывали легально (табл. 31). Снижение воспроизводства популяций так же вызвано “целенаправленным истреблением самцов” ради получения дорогостоящих рогов [Бекенов и др., 2002; Грачев, Бекенов, 2003; Шай-муханбетов, 2004]. Главные причины уменьшения численности вида в Туркменистане и Узбекистане в конце XX в. точно такие же - хищнический промысел и браконьерство [Ишадов, 1999а; Быкова, Есипов, 2004].
Факты крайне нерационального использования ресурсов приведены в следующем разделе.
В заключение замечу, что проблема резкого сокращения поголовья европейской популяции сайги была рассмотрена учеными Российской академии наук 3 июня 1993 г., когда ее ресурсы составляли около 120 тыс. особей. Участниками совещания было принято специальное обращение в адрес руководства Российской Федерации о немедленном принятии необходимых мер по сохранению этого вида, а также зубра и лошади Пржевальского [Кудрявцев, Сафронова, 1993]. Глас ученых, как в пустыне, остался без внимания. Спустя десятилетие численность сайги уменьшилась еще в 7 раз (до 17 тыс. особей), а поголовье зубра - в 3 раза!
Использование ресурсов
Сайга - важнейший объект охоты человека в аридной зоне Евразии, дающий диетическое мясо (15-30 кг), жир (2-8 кг), шкуры и ценные рога, препараты из которых издавна применяются в восточной медицине. Их целебные свойства были экспериментально подтверждены специалистами Калмыцкого государственного университета. Изготовленный из роговых чехлов препарат сайтарин обладает обезболивающим, противосудорожным и успокаивающим эффектом. В народной медицине использовали также мясо, семенники, предглазничные железы и копыта этих животных [Соколов, 1951; Слудский, 1955; Нестеренко, 1971; Демеуов, 1972; Размахнин и др., 1976; Размахнин, Рывкин, 1976; Размахнин, 1986; Жирнов, Максиму», 19986].
В прошлые века сайгу добывали круглогодично, всевозможными способами, что не способствовало ее сохранению. Она была одним из основных видов степной дичи, “кормившей” огромные армии. Монголы и татары кололи ее во множестве во время многодневных, а, порой, и многонедельных облавных охот, в которых одновременно участвовали десятки тысяч всадников. Степные кочевники и казаки, помимо облав, практиковали загоны зверей на гладкий лед, в ямы, топи и на срезанный тростник, в массе истребляли в многоснежье и по насту, а на “плави” (во время переправ животных через реки) били дубинами и резали “сколько кому угодно”. В низовьях р. Чу при загонах на срезанный тростник за один день добывали до 12 тыс. голов, и еще тысячи уходили искалеченными [Эверсман, 1850; Грум-Гржимайло, 1922; Барышев, 1925; Тимофеев, 1934; Адольф, 1948, 1957; Бажанова, 1955; Кириков, 1955, 1959, 1966, 1983; Слудский, 1955; Смирнов, 1965; Демеуов, 1971].
Масштаб заготовок и стоимость рогов. Промысловое воздействие на этот вид усиливалось по мере увеличения людского населения на юге России и в Казахстане, а также с появлением огнестрельного оружия и увеличением спроса на рога, которые стали предметом торговли еще в конце XVIII в. В Крыму их продавали от 3 до 20 “пара” за пару [Кириков, 1966, 1983]. Спрос на них особенно возрос с середины XIX в. в связи с экспортом в Китай. В 1840-1850 гг. бухарские и хивинские купцы скупили в киргизской степи и ввезли 345 тыс. пар. В 1852-1882 гг. экспортировано более 4 млн пар (вместе с изюбриными рогами, которые составляли небольшую долю), или ежегодно в среднем 112 тыс. Только через Петропавловскую таможню в 1857-1861 гг. каждый год переправляли от 7,7 тыс. до 41,7 тыс. пар, и примерно столько же - через другие таможни. В киргизской степи в это время ежегодно заготавливали от 9 до 61 тыс., а в отдельные годы с убитых, павших от джута и загрызенных волками копытных снимали до 100 тыс. пар [Эверсман, 1850; Небольсин, 1855; Мейер, 1865; Красовский, 1868; Силантьев, 1898; Туркин, Сатунин, 1902; Антипин, 1941]. На небольшом о. Николая в Аральском море, где до середины XIX в. сайга не видела человека, один из промышленников в 1887 г. заготовил, по сведениям Л.С. Берга, 1500 пар рогов [Дементьев, 1935].
Следует особо отметить, что рекордное количество рогов (437 тыс. пар) вывезено в Китай в 1854 г., и после такой массовой заготовки поголовье сайги во многих азиатских областях заметно сократилось, а ряде районов она исчезла. Промысловый пресс на европейскую популяцию в этот период был, вероятно, слабее.
В конце XIX в. цена за пару сайгачьих рогов на рынках России колебалась в пределах 1-5 рублей, а в начале XX в. доходила до 20-25 рублей, что равнялось стоимости хорошей лошади или верблюда [Силантьев, 1898; Слудский, 1955,1956; Слудский, Страутман, 1955]. Высокая стоимость рогов, несомненно, стимулировала не только круглогодичную, но и избирательную охоту на самцов, что не способствовало воспроизводству популяций.
По сведениям Д.К. Соловьева [1922], несмотря на чрезвычайно резкое сужение ареала, в первые десятилетия XX в. промысел сайги велся в кумо-манычских и киргизских степях, к востоку от Каспийского моря, на Устюрте, возле Аральского моря, близ Сырдарьи и на восток до Семиречья и Алтая. Объем добычи стал заметно меньше: если в XIX в. ее добывали сотнями тысяч в год, то “теперь - только тысячами”. Избирательная охота на самцов процветала также в Восточной Джунгарии и на Гобийском Алтае [Грум-Гржимайло, 1922].
Как уже отмечалось, к 20-м годам XX в. в результате неумеренного истребления вид оказался на грани уничтожения. Несмотря на запреты добычи, введенные в 1919 и 1921 гг., нелегальный отстрел животных и контрабандная торговля их рогами продолжались до начала 30-х годов. В 1922 г. во времена НЭПа цены на эту продукцию выросли в 10-15 раз, поскольку в Монголии и Китае, где не принимали советские дензнаки, рога стали служить своеобразной валютой при продаже скота. В 20-е годы цена за пару рогов сайги колебалась в пределах 35-80, а в 1926 г. в Астрахани и Махачкале за них давали 100-150 рублей. Высокие цены привели к еще более беспощадному истреблению зверей [Барышев, 1925; Жемчужников, 1926; Берков, 1927; Сытин, 1927; Тимофеев, 1934; Гептнер, Формозов, 1941; Слудский, 1955, 1956]. В 1922 г. в Алма-Ате задержали партию рогов весом в 1,5 пуда, и в этом же году через Кульджинское отделение Внешторга переправлено в Китай около 10 пудов, для чего потребовалось убить около 1500-1840 самцов [Дементьев, 1935; Антипин, 1941].
Необходимо заметить, что правилами охоты в Туркестанской республике (приказ Народного Комиссариата Земледелия от 10 мая 1921 г., № 102) охота была запрещена круглогодично только на самок и телят копытных, а на самцов, включая сайгу, - лишь с 1 января по 15 августа (на самцов кабана - с 15 февраля по 15 июня) [Смирнов, 1965], т.е. в 20-е годы в Казахстане добыча самцов сайги была разрешена официально.
Перерыв в поступлении рогов этого копытного на международный рынок из-за “железного занавеса” на границах СССР вызвал увеличение спроса на них и, соответственно, цен. В 1933 г. на рынках Китая 1 кг стоил 480-640 рублей золотом [Тимофеев, 1934]. Тем не менее в 30-е годы объем продаж здесь был значительным - около 50 тыс. пар в год. Сокращение поставок сырья из-за рубежа стимулировало добычу самцов сайги и в самом Китае, где этот вид был полностью уничтожен в 50-е годы [Джиган, Дикян, 1998]. В середине XX в. стоимость трех пар первосортных рогов была эквивалентной стоимости шкурки баргузинского соболя или трех шкурок серебристо-черной лисицы, или 10-12 норок [Размахнин, 1969; Размахнин, Рывкин, 1976].
Лицензионная добыча. В СССР для восстановления поголовья сайги до промыслового уровня потребовалось почти три десятилетия (20-40-е годы) жесткой охраны. Лицензионная добыча разрешена в Калмыкии с 1951 г. (10 тыс. голов), в Казахстане - с 1954 г. (50 тыс.).
В начальный период лицензионного освоения ресурсов основными способами добычи были дневные загоны животных на стрелков и стрельба с автомашин, в том числе по методу “замкнутого кольца”, предложенного Т. Солдатовым. В дальнейшем практиковались ночная охота с автомашин с применением мощных фар или прожекторов, ас 1976 г. внедрен кольцевой метод добычи с использованием спортивных мотоциклов [Жирнов и др., 1965; Демеуов, 1971; Близнюк, 1978; Жирнов. 1982; Перовский, 1985; Размахнин, 1985]. В 1954 г. А.А. Слудским [1955] предложена достаточно продуманная система организации промысла этих копытных специализированными охотничьими хозяйствами и эффективная технология добычи, в том числе с применением загородок-ловушек. Позднее примерно такой же способ отлова в ловушки с направляющими “крыльями” и “воронкой”, по типу практики добывания северных оленей, предлагал В.Н. Скалой [1975]. Этот метод, успешно апробированный при отлове сайгиВ.Л. Рашеком на о. Барсакельмес [1959], был усовершенствован и в дальнейшем получил название “корально-сетевой” [Максимук, 1981, 1982, 1986].
Легальный промысел копытных бурно развивался, и уже к концу 50-х годов его масштабы превысили разумные пределы (табл. 31). В Северо-Западном Прикаспии с 1951 по 1955 гг. отстреляно 218 тыс. особей, причем в 1951-1952 гг. была разрешена летняя охота, которая велась варварскими способами. В 1956 г. создано специализированное охотничье хозяйство - Астраханский госпромхоз, который в 1956-1963 гг. добывал за сезон от 35 до 180 тыс. животных. К этой цифре следует добавить тысячи погибших после охот подранков, немалую долю спортивных охотничьих хозяйств и браконьерскую добычу, исчислявшуюся десятками тысяч голов. В отдельные годы из популяции единовременно изымали до 250 тыс. преимущественно взрослых особей. Чрезмерный промысловый пресс очевиден даже по результатам массового мечения сайгачат в 1957-1959 гг.: 77% из погибших стали жертвами промысловых охотников, 11% уничтожили браконьеры, и только 12% пали от других причин, включая ранения [Бакеев, Формозов, 1955; Банников и др., 1961; Жирнов, 1962, 1982; Жирнов и др., 1965; Максимук, 1981; Перовский, 1974; Жирнов, Максимук, 1994, 1998а].
В 1956-1960 гг. в Северо-Западном Прикаспии официально отстреляно 612 тыс. (122 тыс. в год), в 1961-1965 г. - еще 347 тыс. особей. Объем лицензионной добычи в 1959/60 и 1962/63 гг. достигал 31-34% предпромысловой численности и 47-52% весенней [Жирнов и др., 1965], т. е. был эквивалентен весеннему приросту популяции или превышал его. Эти масштабные акции, наряду с другими факторами смертности, привели к значительному сокращению популяции (до 75 тыс. к 1965 г.) и к ее депрессии (рис. 23).
В 1965, 1968-1970 и 1972 гг. охота на европейскую сайгу была запрещена, что, несомненно, способствовало росту ее численности. В 1971-1975 гг. ежегодные размеры заготовок не превышали 70 тыс. голов, но с образованием в 1977 г. Калмыцкого госпромхоза добыча снова резко выросла и в 1976-1980 гг. достигла рекордного уровня - 653 тыс., или в среднем 130 тыс. в год. В 1977 г. отстреляно 170 тыс., в 1978 г. - 201 тыс. (табл. 31). Среди отстрелянных животных преобладали взрослые особи (до 80-95%), причем доля самцов достигала порой 60% [Жирнов и др., 1965, 1977; Близнюк, 1977; Максимук, 1981; Жирнов, 1982; Фадеев, Слудский, 1982; Жирнов, Максимук, 19986].
Таблица 31. Динамика официальной добычи сайги (тыс. голов)*
Годы | Россия I Казахстан | Годы | Россия I Казахстан
1951-1955 218 - 1979 81 36
1955 ? 2,3 1980 71,5 150
1956 106 16,5 1981 91,4 193
1957 128 2,6 1982 81,6 223
1958 158** 16,9 1983 41,5 205
1959 180** 71 1984 Запрет 123
1960 38** 138 (175) 1985 20,7 157
1961 119** 203 1986 15,3 87
1962 125** 176 1987 Запрет 100
1963 40** 165 1988 — — 63
1964 40 ПО 1989 ПО
1965 Запрет 73 1990 11,3 95
1966 22 75 1991 Запрет, 0,2 112***
1967 30 61 1992 0,2 108
1968 Запрет 85 1993 , 0,3 62
1969 и 33 1994 , 0,3 37,5
1970 11 161 1995 , 0,5 28,6
1971 15,5 195 1996 14,5 30
1972 Запрет 265 1997 Запрет, 0,3 12,8
1973 33 297 1998 , 0,93 7,3
1974 66 344 1999 , 0,26 Запрет
1975 87 501 2000 , 0,23 сс
1976 138 321 2001 -“-,0,12 сс
1977 163 351 2002 сс **** сс
1978 201 100 2003 сс сс
* По [Банников, 1958; Банников и др., 1961; Слудский, 1961; Жирнов и др., 1965; Смирнов, 1965; Фан-десв, 1965; Круторогое, 1975; Менькова, 1976; Максимук, 1981; Жирнов, 1982; Фадеев, Слудский, 1982; Фадеев, 1986; Дежкин, 1989; Жирнов, Максимук и др., 1998] и ведомственным материалам Охотдепарта-мента РФ (Главохоты РСФСР).
** Заготовка сайги Астраханским госпромхозом.
*** Объем добычи сайги в Казахстане с 1991 г. - по сообщению Ю.А. Грачева и А.Б. Бекенова на VII съезде Териологического о-ва (Москва, 2003).
**** В 2002 г. охотуправление Республики Калмыкия планировало изъятие из популяции 170 голов, в том числе 70 половозрелых самок в марте.
Такого промыслового пресса, дополняемого спортивной и браконьерской охотой и огромным числом оставленных и погибших позднее подранков, европейская группировка снова не смогла выдержать. Ее численность, начиная с 1979 г., сокращается (табл. 28), но промысел, хотя и в меньшем объеме, продолжается (табл. 31). В 1987 г. вновь вводится запрет охоты сроком на 5 лет, но в 1990 г. все же отстреляно 11,3 тыс. Всего с 1951 по 1990 гг. из популяции официально изъято более 2,2 млн особей [Зуев, Хахин, 1982а,б; Максимук, 1982, 1986; Жирнов, Максимук и др., 1998].
Прекращение массового лицензионного промысла позволило в дальнейшем поддерживать летнее поголовье европейской группировки на относительно стабильном уровне в 140-160 тыс. и даже несколько увеличить его. Однако при масштабном браконьерстве и неуклонном росте волчьего населения популяция не смогла выйти из депрессии. Ее численность, как и в начале XX в., сократилась до критического уровня (табл. 28). В 1987-1989, 1991-1995 гг. и с 1997 г. промысел сайги запрещен, но по так называемым “научным”, “котловым” и другим лицензиям про
должали официально изымать из популяции сотни особей (табл. 31), и устраивались трофейные охоты на самцов [Зверев, 2001].
В Казахстане в 1955-1960 гг. ежегодно отстреливали в среднем 43 (3-138) тыс., в 1960-1964 гг. - 160 (110-202) тыс., позднее до 1969 г. - 64 тыс., в 1972-1977 гг.-345 (264—501) тыс. Уровень официальной добычи сайги здесь в целом был ниже, чем в России (10-20% от числа учтенных), но в отдельные годы, как и в европейской популяции, достигал 36-39% поголовья, причем в 1966, 1970-1976 гг. практиковался и летний (с 1 июля), и позднезимний (в январе) отстрел. Всего в 1958— 1968 гг. добыто 1198 тыс. [Демеуов, 1971], в 1971-1976 гг. - 1906 тыс. голов, причем госпромхозами в этот период заготовлено 1655 тыс. шкур и 961 тыс. пар рогов, из них экспортировано 128,9 т [Фадеев, Шаад, 1978]. Доля взрослых самцов в объеме добычи в этот период, следовательно, достигала 58%. Браконьерская добыча тоже исчислялась сотнями тысяч особей [Жирнов, Калецкий, 1976]. Чрезмерный охотничий пресс, сопровождаемый преимущественным отстрелом взрослых самцов (до 60-69% в отдельные годы), неминуемо должен был привести к подрыву ресурсов, что и произошло. После рекордной добычи (501 тыс. в 1975 г.) поголовье в 1976 г. сократилась вдвое, чему способствовала также суровая зима 1975/76 гг. В 80-е годы доля взрослых самцов в добыче составляла 15-20%, самок - 70-75, сеголеток - 10, и только в конце этого десятилетия объем добычи сеголеток увеличен до 30-40% [Грачев, Бекенов, 1993]. В 1979-1993 гг. среднегодовая добыча находилась на уровне 122 (36-223) тыс. голов. Из числа детенышей, помеченных в 1986-1993 гг. и добытых позже, 52% - сеголетки, 25 - годовалые, 15 - в возрасте 2,5 лет, 4 - в 3,5 года и 2-0,5% - в возрасте 4,5-9 лет. Доля животных, отстрелянных в возрасте 0,5-2,5 лет, достигала 92% [Грачев, Исмагулов, 1999], что свидетельствует о чрезмерной интенсивности промысла. Всего в 1955-1993 гг. добыто более 5,5 млн особей. С 1965 по 1981 гг. экспортировано более 330 трогов. Устюртскую популяцию, кроме того, с 1966 г. промышляли на территории Узбекистана и Туркмении, куда стада заходили во время осенней миграции [Банников и др., 1961; Мамбетжумаев, 1966; Слудский, Фадеев, 1975, 1977; Голованова, Анохина, 1976; Жирнов и др., 1977, 1998; Фадеев, Шаад, 1978; Ишунин, 1980; Фадеев, Слудский, 1982; Слудский и др., 1983; Фадеев, 1986; Фадеев, Иванов, 1988; Бекенов, Грачев, 1998; Жирнов, Максимук и др., 1998].
С 1998 г. в Бетпакдале, и с 1999 г. на всей территории Казахстана введен временный запрет добычи сайги в связи с катастрофическим сокращением численности [Шаймуханбетов, 2004]. В Узбекистане запрет охоты объявлен более своевременно - в 1990 г. и вплоть до 2010 г. [Быкова, Есипов, 2004], что, во многом, позволило предотвратить уничтожение вида.
В целом в России и Казахстане за 40-летний период эксплуатации восстановленного вида официально добыто около 8 млн голов и получено более 122 тыс. тонн мяса, миллионы шкур и экспортировано более 500 т рогов (более 2 млн пар -А.Д.) по цене около 50 американских долларов за 1 кг [Соколов и др., 1991; Жирнов, Максимук, 1994; Жирнов, Максимук и др., 1998], что стало несомненной экономической заслугой охотоведов и охотничьих организаций. Однако эксплуатация ресурсов, как показано выше, проходила с многочисленными грубыми ошибками, не раз негативно отражавшимися на состоянии популяций. Считаю необходимым более детально рассмотреть их, дабы на повторять в будущем.
Основные ошибки при эксплуатации ресурсов. Главной причиной резкого снижения численности и депрессии популяций не раз становился перепромысел. Моделирование динамики ясно показывает, что при 40% объеме добычи численность быстро сокращается даже при прочих благоприятных условиях. В благоприятные годы допустимо добывать лишь до 30% осеннего поголовья, а при расширенном воспроизводстве - не более 20% [Жирнов, 1982; Заикин, Жирнов, 1989]. При численности группировки ниже 400 тыс. особей норма добычи не должна быть более 10% [Близнюк, 1982]. Очень высокая доля самцов в добыче в модели тоже неминуемо приводит к резкому сокращению популяции из-за уменьшения плодовитости самок при недостатке самцов-производителей [Milner-Gulland, 1994].
Интенсивно эксплуатируемым популяциям наносился существенный качественный урон, который можно расценивать как масштабную антиселекцию. Из них ежегодно изымали преимущественно репродуктивное ядро - наиболее крупных и плодовитых самок и самцов-производителей, что приводило к уменьшению уровня воспроизводства, к сокращению генотипического и фенотипического разнообразия, к уменьшению размеров животных, изменению их поведения и снижению жизнестойкости группировок [Проняев, 1985а,б, 1990, 1998; Проняев, Аксенов, 1988; Рожков, Проняев, 1994]. В отдельные периоды диспропорция полов, вызванная перепромыслом взрослых самцов, принимала размеры, угрожающие существованию сайги как биологического вида. Как отмечалось выше, в 1959 г. их доля в европейской популяции уменьшилась до 1-2%, а яловость самок в последующие годы возросла почти в 3 раза. То же повторилось в самом конце XX - начале XXI вв., когда из-за резко возросшего браконьерского отстрела и в европейской, и в казахстанских популяциях самцов почти не осталось.
Сроки промысла были непомерно растянуты (в отдельные периоды - с 15 июня до 20 февраля). Массовым отстрел был перед гоном и в период гона. Постоянное охотничье преследование нарушало ход размножения копытных со всеми вытекающими отсюда негативными последствиями. Зверей отстреливали картечью с автомашин и мотоциклов днем и ночью (из-под фар) после длительной погони на предельной скорости. Обессиленные животные падали сотнями. Часть их гибла от отека легких, патологических изменений желез внутренней секреции, появления инфильтрата, самки — от абортов, а ушедшие от погони долго не могли восстановить нормальное физиологическое состояние. В степях после таких охот оставалось множество подранков. Их доля достигала 10-18% от числа добытых, а иногда и много больше (до 70%). В Калмыкии в 1978 г. специальная бригада из двух человек ежедневно за 3-4 ч подбирала от 42 до 50 еще живых зверей. По расчетам, только в Калмыкии за время промысла до 1981 г. потеряно более 300 тыс. подранков. В отдельные годы во время массового отстрела, из-за отсутствия холодильных установок, до половины продукции портилось [Бакеев, Формозов, 1955; Сорока, 1957; Дремов, 1963; Демеуов, 1971; Фадеев, Шаад, 1978; Близнюк, 1978,1982,1983; Максимук, 1981,1982,1986; Зуев, Хахин, 19826; Перовский, 1985; Размахнин, 1985; Зуев, 1986].
С 1979 г. в практику внедрен усовершенствованный корально-сетевой способ добычи. В сетевую ловушку, имеющую форму “улитки”, при помощи мотоциклов направляли все или большую часть стада, которое затем полностью утилизировали. При этом практически не оставалось подранков, в два раза увеличивалась про
изводительность труда охотников, в два-три раза сокращался период промысла, исключалась избирательность добычи взрослых животных и значительно улучшалось качество мясной продукции и кожевенного сырья [Житенко, 1974, 1986; Максимум 1981, 1982, 1986; Размахнин, 1985]. Достоинства этого метода были настолько очевидны, что не были просчитаны все негативные последствия.
Стада сайги разнокачественные по составу: есть преимущественно состоящие из взрослых самок с детенышами, из самцов, и есть смешанные. Первыми мигрируют наиболее подвижные животные, среди которых преобладают самцы. Массовый отстрел или отлов в корали авангардных стад мог нанести непоправимый ущерб популяциям, поскольку значительно нарушал половой состав. Кроме того, при отлове всего стада происходило уничтожение мигрирующих в определенное время и в определенном направлении животных, т. е. безвозвратно исчезал их генофонд.
Замечено, что при раннем массовом промысле стадных мигрирующих копытных, таких как сайга и северный олень, нередко происходит резкая смена путей их миграций, летних и зимних мест обитания. Это смещение некоторые исследователи пытались объяснить многолетними циклическими “маятниковыми” колебаниями в популяциях [Сыроечковский, 1986]. На самом же деле все гораздо проще -после нескольких лет интенсивного промысла по традиционным путям и к типичным местам летовок и зимовок практически некому идти. Уцелевшие животные обычно примыкают к другим стадам, кочующим в ином направлении, или становятся оседлыми. Сезонные миграции возобновляются лишь спустя много лет, когда группировка вновь станет многочисленной, и если в ней найдутся особи, сохранившие генетическую память о путях перемещений.
Нерациональный в целом лицензионный промысел копытных в значительной мере дополнялся нелегальной охотой, масштабы которой в XX в. менялись в зависимости от уровня охраны и социально-экономических явлений.
Нелегальная добыча. В Северо-Западном Прикаспии браконьерство стало массовым в 50-е - начале 60-х годов с началом официального промысла сайги. На нее охотились преимущественно ночью из-под фар ради мяса и шкур, которые выделывали кустарным способом. Нелегальную добычу этого зверя специалисты оценивали в 25-50 тыс. голов, что составляло 10-14% от объема лицензионного промысла [Рак, 1956; Банников и др., 1961; Жирнов, 1962, 1963; Фандеев, 1965]. В Казахстане в середине XX в. браконьеры уничтожали этих копытных десятками тысяч. Некоторые группы в годы войны добывали до 1500-2000 особей за сезон, заготавливая до 8 т мяса [Слудский, 1955; Тихонов, 1956; Адольф, 1957]. В 60-е годы Ю.А. Смирнову [1965] “не удалось встретить ни одного человеческого жилья, где не было бы улик охоты на сайгу или же самих виновников этой охоты -браконьеров”. Нелегальная добыча в 70-е годы достигала здесь 200 тыс. особей [Жирнов, Калецкий, 1976].
С середины 60-х годов в Российской Федерации значительно улучшилась охрана животных. В 1965 г. на базе Астраханского госпромхоза организован мобильный отряд по охране сайги (с 1969 г. он придан Калмыцкой госохотинспекции [Зуев, Хахин, 1982а]), внесший значительный вклад в сохранение этого вида и сокращение уровня нелегальной охоты, порой ценой жизни егерей и охотоведов. Тем не менее, браконьерство оставалось массовым и, порой, принимало варварский характер. Например, в Астраханской области на территории совхоза “Прикаспийс
кий” браконьеры загнали на еще неокрепший лед озера Джурак большое стадо зверей. Под их тяжестью лед провалился. Прибежавшие люди вытаскивали находившихся в беспомощном состоянии животных, и ... тут же перерезали им горло. “Мясо в поселок возили на тракторах с прицепами, автомашинах и солили в бочках. Работниками охотнадзора только на берегу и в поселке было найдено несколько сотен туш сайгаков”. В Калмыкии в ноябре 1971 г. браконьеры из Волгограда для добычи копытных использовали две грузовые и одну легковую автомашины и мотоцикл. Лишь в одной из автомашин обнаружено 77 туш [Жаворонков, Зуев, 1976]. В феврале 1962 г. ночью милицией города Астрахани задержана автомашина, на которой группа штатных охотников госпромхоза везла 96 “нелегальных” туш [Дремов, 1963]. И таких примеров массового уничтожения копытных браконьерами можно привести сотни.
В Северо-Западной Туркмении в 70-80-е годы хищнический характер носила промысловая охота: сайгу стреляли не только “из-под фар”, но и сбивали машинами. Браконьерские “экспедиции” нередко состояли из нескольких машин. В суровые зимы охотники истребляли половину местного поголовья [Горбунов, 1986а, 1995а].
Очередная волна нелегальной охоты стала нарастать в 80-е годы с увеличением в сельской местности числа автомашин и мотоциклов, которые применяли для преследования копытных. Однако невиданный ранее размах браконьерство приобрело в конце 80-х и в 90-е годы - с началом “перестройки” и изменения социально-экономических отношений, после ликвидации государственной монополии на внешнюю торговлю и в связи с повсеместным ослаблением охраны животных и всеобщим законодательным нигилизмом. Местное население получило к тому же мощный экономический стимул к браконьерству. К 1989 г. цена 1 кг рогов сайги на внешнем рынке достигла 500-600 американских долларов, а на местном - 30 долларов, что привело к “валютной лихорадке”. Многочисленные “заготовители” из многих областей России, Прибалтики и Средней Азии, помимо нелегальной добычи, скупали рога копытных у местного населения, и даже не гнушались раскапывать могильники, извлекая трупы зверей, павших от эпизоотий в 1985-1987 гг. В 1988-1990 гг. из Казахстана в Россию вывезено около 77 т рогов, а всего собрано более 200 т. Такое количество рогов можно получить, лишь добыв около 500-700 тыс. самцов. После такой акции их доля в стадах снизилась до 3-7% [Соколов и др., 1991; Жирнов, Максимук, 1994; Павлов, 1999].
Об увеличении нелегальной и избирательной добычи сайги в конце XX в. говорят и другие факты. Степи Калмыкии и Казахстана в этот период были усеяны черепами с отпиленными рогами. Массовым явлением стала индивидуальная и групповая добыча зверей днем на мотоциклах, развивающих скорость до 130 км/ час: после 5-6 минут погони животные умирали от разрыва сердца [Кузнецов, Лущекина, 2002; Лущекина, 2002]. В 1991 г. охотинспекция во время рейдов в охотничьих угодьях Калмыкии выявляла в среднем за сутки около 16 голов копытных, добытых незаконным путем, что в 2 раза больше, чем в 1987 г. Среди отстрелянных животных доля самцов достигала 89%. Среди задержанных браконьеров процент мотоциклистов увеличился с 29 в 1988 г. до 72 в 1992 г., а их доля в общем объеме нелегального промысла зверей возросла в эти годы с 13 до 72% [Лещенко, 1997]. В
Джезказганском районе Казахстана почти в каждом из 12 обследованных осенью 2002 г. населенных пунктов имелись браконьерские бригады мотоциклистов, включающие 15-25 человек. Каждый из этих браконьеров был способен за день охоты добыть 5-7 рогачей [Шаймуханбетов, 2004].
В 90-е годы объем нелегальной добычи этих копытных, преимущественно взрослых самцов, в Калмыкии составлял не менее 15-25, а в Казахстане - 100-150 тыс. голов [Соколов и др., 1991; Жирнов, 1998г; Кузнецов, Лущекина, 2002], что значительно превышало объем лицензионного промысла. Браконьерство фактически стало определяющим фактором динамики и структуры популяций.
В целом очевидно, что сайга является ценнейшим возобновляемым биологическим ресурсом, который при разумном управлении мог бы быть неистощимым вечно! К глубокому сожалению, в XX в. этот вид варварски эксплуатировался охотничьими организациями и армией браконьеров. В результате - снова приходится говорить о неотложных мерах по его сохранению.
Сохранение вида
Как показано выше, в последние десятилетия XX в. динамику популяций сайги определяли, в основном, антропогенные факторы, и в первую очередь - официальный промысел, массовое браконьерство, сокращение жизненно важных мест обитания, а также многократно увеличивший свою численность волк. Соответственно в деле сохранения и восстановления ресурсов этого реликтового копытного, чудом уцелевшего до наших дней, на первый план выходит охрана животных и среды их обитания.
Охрана животных и среды их обитания. Одна из первоочередных и вынужденных мер при чрезмерном снижении поголовья вида - включение его в Красные книги и списки, что уже сделано Монголией (1997) [Lushchekina et al., 2000], Туркменистаном (1999) и МСОП (2002). Жизненно необходимо значительно расширить площадь последнего прибежища европейской популяции - заповедника “Черные земли” и организовать несколько крупных хорошо охраняемых территорий в основных районах обитания сайги в Казахстане. Эксплуатация популяций, находящихся в депрессии, безусловно, должна быть прекращена полностью, включая отстрел животных в научных, культурных и других целях, что часто служит удобной “лазейкой” для браконьеров, особенно наделенных властными полномочиями. В России в 90-е годы XX в. под таким прикрытием в период запрета охоты легально добывали сотни (табл. 31), а нелегально - тысячи особей ежегодно. Охрана популяций должна быть мобильной и эффективной. Для этого нужно материально-технически оснастить службу охраны, ужесточить наказание за нелегальную добычу сайги, разработать меры поощрения лиц, задержавших браконьеров с поличным, а также перекрыть нелегальные каналы вывоза рогов на экспорт в соответствие с решениями IX конференции СИТЕС (1994 г.), запретить торговлю не только рогами, но и мясом сайги. В ближайшие годы крайне необходимо свести к минимуму численность волка, сократить число беспривязных дворовых и пастушьих собак и исключать их проникновение в места отелов копытных. Нестандартные подходы к этой проблеме - повышение трудовой занятости, улучшение благосостояния и просвещение местного населения [Кузнецов, Лущекина, 2002].
После восстановления поголовья копытных до промыслового уровня следует пересмотреть нормы, сроки и способы добычи, обеспечив щадящий режим эксплуатации популяций при максимально возможном сохранении репродуктивного ядра - взрослых животных. При корально-сетевом способе добычи недопустимо уничтожение стад полностью - здесь нужна выборочная элиминация сеголеток и старых зверей с определенной корректировкой в формировании необходимого соотношения полов в популяциях. Наиболее безопасной стратегией, скорее всего, является промысел животных только в благоприятные годы, при умеренной доле добываемых самцов [Milner-Gulland, 1994].
Многие научно-практические разработки, касающиеся охраны среды обитания сайги, сводятся к рекомендациям нежелательности вмешательства человека в сложившиеся экосистемы, хотя они уже до предела трансформированы. Эта позиция, как справедливо считают Л.В. Жирнов и его соавторы [1976, 1977,1982], ошибочна. Напротив, человек обязан принять все необходимые меры по восстановлению и увеличению продуктивности пастбищ. В современных условиях гарантированное сохранение популяций сайги реально, если будут разработаны специальные региональные комплексные программы восстановления ее ресурсов, интегрированные в перспективные планы хозяйственно-экономического развития областей. В них, помимо конкретных мер по охране животных, должно быть предусмотрено районирование угодий (выделение животноводческой и “сайгаководческой” зон, зоны совместного использования угодий и резерватов [Дежкин, Менькова, 1981; Дежкин, 1986]), исключение ключевых мест обитания сайги из сферы хозяйственного использования [Кузнецов, Лущекина, 2002] и недопущение промышленного или сельскохозяйственного освоения земель без предварительной экологической экспертизы проектов. Южнее границы зоны земледелия желательно образовать сеть подкормочных полей и искусственных водопоев с целью улучшения кормовой емкости угодий, предотвращения потрав сельскохозяйственных кулыур и проникновения копытных в густонаселенные человеком районы. Подкормку животных следует организовать и в районах их зимнего обитания. На местах отелов, в миграционных коридорах, в местах переправ через водные преграды и на постоянных водопоях нужно исключить беспокойство зверей, создавая здесь временные “зоны покоя” (заказники, воспроизводственные участки и т. п.), где запрещается проезд транспорта, выпас скота и другая хозяйственная деятельность на период нахождения стад.
Необходимо усовершенствовать мониторинг популяций. Перепромысел сайги в локальных районах нередко происходил из-за отсутствия сведений о численности вида или неточного учета поголовья, а также из-за отсутствия контроля за уровнем воспроизводства, который заметно снижался при резком нарушении половозрастного состава группировок. Мониторинг нужен постоянный, причем он может выполняться с помощью аэрофотосъемки и слежения со спутников, что уменьшит беспокойство копытных [Демьянова, Любаев, 1986; Соколов и др., 1998].
Одна из предлагаемых мер сохранения генофонда вида - разведение животных в неволе [Кудрявцев, Сафронова, 1993; Жирнов, Максимук, 1994; Лущекина, Неронов, 1997]. Безусловно, она заслуживает внимания, но вряд ли может стать основной в восстановлении ресурсов. Рассмотрим эту проблему подробнее.
Разведение в неволе и перспективы реинтродукции. Разведение сайги в неволе практикуется издавна. Известно, что запорожские казаки не убивали бере
менных самок, и они рожали близ хуторов и жили там вместе с ягнятами, легко приручаясь. В Прикаспии в казачьих и казахских крепостях и поселениях крестьяне и охотники тоже нередко выкармливали сайгачат, и те свободно ходили с домашним скотом и к вечеру возвращались домой. Иногда удавалось приручать и обессиленных в голодные годы взрослых животных, попадавших в руки человека [Паллас, 1809; Северцов, 1861; Тимофеев, 1934; Васенко, 1950; Соколов, 1951; Бакеев, Формозов, 1955; Кириков, 1959].
В XIX в. попытку разведения сайги в неволе по поручению Русского общества акклиматизации предпринял саратовский биолог К. Глич. Однако все отловленные капканами и петлями взрослые особи погибли от травм. Из 49 новорожденных сайгачат, содержавшихся в тесном дощатом загоне, погибли 36. В Московский зоопарк доставлены лишь 9 молодых особей [Цаплюк, 1982]. Д. Анучин [1875] видел в Московском зоологическом саду целое стадо молодых особей, пойманных около Сарапеты. Все они погибли, но 6-7 штук успели отправить за границу.
Эксперименты по разведению сайги в Аскании-Нова на Украине ведутся с 1887 г. За 70 лет до 1958 гг. сюда поступило 174 зверя, и вместе с родившимися было 340 особей, из которых 300 пали, 18 проданы и 19 выпущены на о. Бирючьем, откуда разбежались и погибли. Продолжительность жизни зверей в неволе в среднем не превышала трех лет. Большинство из них умирали на первом и втором годах жизни из-за болезней внутренних органов и нарушения обмена веществ, травм, инфекций, инвазий и отравления, и только единицы доживали до 5-7 лет. В 1969 и 1978 гг. сюда завезли 73 зверя из Казахстана и 50 из Калмыкии, но почти половина первой партии вскоре погибла. И все же, несмотря на неудачи, здесь доказана реальная возможность разведения сайги в полувольных условиях. От 39 оставшихся в живых животных к 1997 г. получено 545 детенышей. Ежегодный приплод достиг значительной величины - 49 особей, при этом ежегодно 2М сайгаченка рождались мертвыми или слабыми. В 1998 г. в двух загонах площадью 807 и 1550 га содержали 212, в 2004 г. - 134 особей. 20 зверей отсюда отправлены в Чехию и 6 завезены в заповедник “Еланецкая степь” [Салганский и др., 1963; Треус, 1968, 1986, 1990, 1993, 1997, 1998; Павлов, 1999; Треус, Смаголь, 2004].
В зоопарках Европы сайгу содержат с 1864 г., в США - с 1934 г., в Китае в полувольных условиях - с 1989 г. В бывшем СССР сайгу пытались разводить в Москве, Санкт-Петербурге, Новосибирске, Ростове-на-Дону, Таллине, Алма-Ате и Чемкенте. Большинство особей здесь тоже не жили больше 3 лет, и лишь отдельные из них достигали 5-10-летнего возраста. В последние десятилетия размножение этих копытных в зоопарках, в результате накопленного опыта работы с видом, идет более успешно, чем раньше, что весьма радует в плане сохранения генофонда [Dolan, 1977; Цаплюк, 1982; Kamsay et al., 1992; Rubin, Michelson, 1994; Соколов, Холодова, 1996; Холодова, Неронов, 1996; Букреева, Арылов, 1998]. В 60-70-е годы опыты по выращиванию сайги в неволе проводили научно-исследовательские противочумные институты [Черченко, 1975; Корнеев и др., 19866] и Институт зоологии АН Казахстана, где самки приносили потомство в течение нескольких лет [Цаплюк, 1982]. Разведение этого копытного в парке экоцентра “Джейран” в Бухарской области Узбекистана, насколько мне известно, не было успешным.
В последние годы вольерное разведение вида практикуется в Калмыкии, где к 2004 г. численность зверей на ферме достигла 82 особей, и начато в Астраханском
госохотхозяйстве. В неволе лучше выживают крупные детеныши (с массой не менее 2,6 кг), пойманные в природе, с “лояльной” оборонительной реакцией на человека (не сопротивляются, когда их берут на руки). В вольерах их содержат небольшими группами (до 10 особей), выпаивая до 3^1-месячного возраста заменителем цельного молока, применяемым для искусственного выращивания овец и телят в животноводстве. Кормление заменителем молока происходит от 7 до 1 раза в сутки в зависимости от возраста сайгачат. С 4-го дня молочное питание сочетают с выпасом. По мере стравливания растительности в вольере, сюда привозят свежескошенную траву, из которой детеныши выбирают полынь, типчак, солодку, марь белую, люцерну. С третьего месяца в их рацион включают дробленый ячмень и комбикорм. В вольере должна находиться вода и соль-лизунец. В сутки на одно животное требуется 2,1-3,9 кг сырого или 1,3-2,0 сухого корма [Абатуров, Петрищев, 1978, 1986, 1987а; Петрищев, Холодова, 1980а; Петрищев и др., 1998, 2004; Букреева, Арылов, 1998; Арылов, Букреева, 1999; Петрищев, 2000; Петрищев, Арылов, 20036]. Приемы выращивания сайги в неволе подробно освещены в работах Б.И. Петрищева и соавторов [1997] и Ю.Н. Арылова [2002].
Выращенных в искусственных условиях животных, как считают некоторые специалисты, можно и нужно выпускать в природу для пополнения популяций. На мой взгляд, эти выпуски, скорее всего, будут обречены на неудачу - вольерные особи быстро станут жертвами волка, собак и браконьеров. Напомню, что выпуски сайги в Сюгатинской долине Заилийского Алатау (38 особей), на островах Булла (20 и 58) и Глиняном (53) Бакинского архипелага, в ширванской степи (35 голов) и Кызыл-Агачском заповеднике Азербайджана не дали результата, несмотря на начавшееся размножение и расселение животных, большей частью именно по этой причине [Алиев, 1961, 1962, 1970; Банников и др., 1961; Слудский, Афанасьев, 1964; Павлов, 1999]. Возрождение популяции о. Барсакельмес (см. выше) стало возможным лишь благодаря организации заповедника и хорошей охране.
Без устранения главных причин, вызвавших очередную депрессию природных популяций, сайгу такими мерами не спасти, а при действенной охране и резком сокращении численности крупных хищников вряд ли понадобится масштабная и дорогостоящая работа по разведению зверей в неволе и их реинтродукции в природу.
Подсемейство КОЗЛИНЫЕ Caprinae Gray, 1821
Характеристика. Животные мелких, средних и довольно крупных размеров, крепкого и тяжелого сложения, с короткими и относительно толстыми конечностями, где пястная кость короче лучевой. Носовое зеркало, если имеется, небольшое. Ноздри сближены. Хвост короткий и не достигает скакательного сустава.
Окраска чаще слабоконтрастная, преимущественно бурых и коричневых тонов, с потемнениями или посветлениями на отдельных участках. У многих видов на передней части тела развиты волосяной подвес и борода. Хвостовое зеркало не выражено. Волосяной покров густой и состоит из остевых и пуховых волос, растущих пучками.
Зароговая часть черепа несколько укорочена. Основная затылочная кость короткая, уплощенная, слабо сужена спереди (кроме рода Budorcas). Носовые кости без вырезки на передних концах. Глазницы, как правило, трубкообразные, выступающие в стороны. Предглазничные ямки и этмоидальные щели часто отсутствуют. Слуховые пузыри небольшие, сжатые с боков. Роговые стержни в нижней трети, а иногда и выше, с внутренними полостями. Основания рогов расположены над глазницами или сдвинуты назад за их края. Резцы узкие, долотообразные, постепенно убывающие в ширине от средней пары к клыкам. Коренные зубы гипсодонтные, добавочные бугорки и столбики на них отсутствуют. Последний предко-ренной в нижней челюсти с замкнутой передней лункой.
Рога чаше имеют животные обоих полов, но у самок они меньше. Форма рогов различная: серповидная, дугообразная, улиткообразная, с двойным изгибом, в виде загнутых на вершине крючков. Поверхность роговых чехлов снабжена одним-тремя продольными ребрами (килями).
Копыта некоторых горных видов имеют твердый край и мягкую подушку внутри, что противодействует скольжению на скальном грунте.
Кожные железы (предглазничные, зароговые, паховые, хвостовые, запястные, межпальцевые) хорошо развиты и с заглубленными выводными протоками [Соколов, 1973; Соколов, Чернова, 1988]. У разных видов они присутствуют в разной комбинации. Их секрет у самцов нередко с сильным специфическим запахом. Молочная железа с 2 или 4 сосками.
Кариотип весьма изменчив: 2п = 42-60, но число плеч почти у всех видов, кроме Nemorhaedus, равняется 60 [Wurster, Benirschke, 1968; Bunch, Nadler, 1980; Орлов, Булатова, 1983; Soma, Kada, 1984].
Преимущественно высокоспециализированные горные и высокогорные животные, приспособленные к жизни в холодном климате. Большей частью ведут групповой или стадный образ жизни.
Охотничьи или охраняемые виды. Одомашненные коза и овца стали важнейшими сельскохозяйственными животными.
Распространение. Южная Европа, Азия, Северная Африка, северная и западная части Северной Америки. В одомашненном состоянии живут почти повсеместно.
Эволюция и филогения. Наиболее древними капринами ранее считали ойоце-роса (Qioceros) - миоцен-плиоценового козлообразного животного Евразии с относительно крупными гомонимными рогами овального сечения. По новейшим исследованиям, к древнейшим Caprinae относят раннемиоценовых Gobiocerus (Монголия), у которых зубы не достигают уровня гипсодонтности и не имеют “козьей” складки, а также Turcocerus (Китай), Hypsodontus и Kubanotragus (Кавказ) и Beniceros (Морокко) [Дмитриева, 2003]. Среднемиоценовые представители козлиных Samotraguspilgrimi и Aragoral mudejar известны из Средиземноморья [Alcala, Morales, 1997; Azanza et al., 1998]. Предковой формой для Caprinae и, видимо, для сестринской группы Hippotraginae мог быть среднемиоценовый род Caprotragoides [Thenius, 1979; Gentry, 1994; Kohler et al., 1995]. В любом случае непосредственный предок каприн еще не определен.
Расцвет группы козлиных имел место в плиоцене и раннем плейстоцене не только в Центральной Азии [Соколов, 1953], но и в Африке [Gentry, 1996; McKenna, Bell, 1997]. В Новом Свете остатки древних видов прослеживаются лишь с плейстоцена. Из современных триб наиболее древние корни имеют Ovibovini и Caprini [Тихонов, 1999]. Родственные отношения представителей каприн с вымершими раннеплейстоценовыми Soergelia, распространенными от Западной Европы до Северо-Восточной Сибири [Тлеубердина и др., 1989], остаются неясными.
Предполагаемая схема эволюции родов подсемейства, получившая наибольшее признание ученых во второй половине XX в., отражена на рис. 24, но сейчас очевидно, что она нуждается в уточнении. По исследованиям митохондриальной ДНК, козлиные близки к Antilopinae, но еще в большем родстве они находятся с представителями трибы Alcelaphini из подсемействаHippotraginae [Allard et al., 1992; Gatesy et al., 1997; Hassanin, Douzery, 1998].
Рис. 24. Предполагаемая радиация представителей подсемейства Caprinae по морфологическим данным (по [Thenius, Hofer, 1960])
Caprinae считают монофилетической группой полорогих, хотя и не бесспорно [Соколов, 1953, 1959; Halthenort, 1963; Gentry, 1992; Gatesy et al., 1997; Hassanin et al., 1998]. При молекулярно-генетическом анализе в подсемействе прослеживаются две эволюционные линии - овцебыков (Ovibos), горалов (Nemorhaedus) и сероу (Capricornis) с одной стороны, и остальных родов - с другой [Кузнецова, 2002].
Род Capra, как предполагалось ранее, находится в тесном родстве с Ovis. У них, несмотря на разное число хромосом у большинства представителей, одинаковое число плеч (NF = 60) и идентичная картина исчерченности дифференциально окрашенных хромосом [Мамедов, 1978, 1985; Шарипов, 1989]. Однако по биохимическим [Hartl et al., 1990] и молекулярным исследованиям [Кузнецова, 2002] их родство оказывается гораздо более далеким, чем представлялось и по кариологи-ческим, и по морфологическим исследованиям, хотя морфологические отличия в строении черепа, рогов, посткраниального скелета [Андреева, 1933], желез, окраске и структуре волос, при внимательном сравнении, тоже весьма значительны.
Гибридизация козлов с овцами и их дикими предками, несмотря на отдельные неподтвержденные сообщения и успешные опыты с редкими породами домашних животных, практически невозможна из-за генетической несовместимости и гибели гибридных плодов. В экспериментах не получено потомства и при скрещивании представителей козлов и баранов с Ammo tragus, хотя у коз, в отличие от овец, гибридные плоды развивались до конца 3-го - начала 5-го месяца эмбриогенеза, и даже получена гибридная самка. Гривистый баран, по результатам гибридизации, следовательно, тоже филогенетически обособлен от козлов, но все же стоит к ним гораздо ближе, чем к баранам, несмотря на большее сходство кариотипа с последними [Стекленев, 1972, 1975, 1978а, 1980, 1986; Мамедов, 1978; Шарипов, 1989].
Предковые формы Caprinae, судя по костным остаткам, скорее всего, были некрупными лесными животными. Основное экологическое направление эволюции представителей подсемейства - адаптация к обитанию в суровом холодном климате в горах, холмистой местности, бореальных степях, полупустынях и тундре, где они, как и другие представители семейства, перешли на питание относительно жесткой травянистой растительностью, заняли свободную нишу и достигли расцвета при групповом или стадном образе жизни. Внутривидовая борьба способствовала развитию полового диморфизма у части родов и отбору более крупных особей с мощными рогами, что не могло не отразиться на строении черепа.
Морфологическая эволюция шла по пути изменения структуры волосяного покрова (развитию густого подшерстка); строения метаподий; укорочения зароговой части черепа с одновременным сдвигом оснований рогов за уровень глазницы, вследствие чего основная затылочная кость становится уплощенной и почти квадратной; большого угла излома оси черепа; пневматизации роговых стержней. Зубной ряд укорачивается; зубные коронки удлиняются (гипсодонтия), что противодействует быстрому стиранию зубов; ребра на верхних коренных и дополнительные столбики на заднекоренных редуцируются; на Р4 метастилид смыкается с па-растилидом и формируется типичная “козья” складка [Geist, 1966, 1987; Тихонов, 1999; Дмитриева, 2003].
Эволюция кариотипа у козлиных происходила путем слияния равно- или разноразмерных акроцентрических хромосом с образованием метацентрических или суб
метацентрических хромосом [Evans et al., 1973; Nadler et al., 1973; Орлов, 1974; Орлов, Булатова, 1983; Шарипов, 1989].
Состав. Систематика подсемейства особенно сложна и запутана. Число триб, включаемых в него, варьирует от 2 до 9, а количество современных родов - от 9 до 14 [Simpson, 1945; Ellerman, Morrison-Scott, 1951; Соколов И.И., 1953,1959; Thenius, Hofer, 1960; Haltenorth, 1963; Соколов B.E., 1979; Thenius, 1980; Durst et al., 1982; Geist, 1987; Hartl et aL, 1990; Gentry, 1992; Gatesy et al., 1997; Hassanin et al., 1998; Hassanin, Douzery, 1999а].
A.H. Тихонов [1999], проведя ревизию Caprinae преимущественно традиционными методами, выделил 5 триб, 12 родов и 28 видов. Однако и его достаточно аргументированная классификация оказалась далеко не идеальной, особенно в отношении рода сайги, который, как показано выше, заметно отличается от козлиных и стоит гораздо ближе к Antilopinae, чем к Caprinae. Структура подсемейства, предлагаемая И.Я. Павлиновым [2003] с учетом новых данных, на мой взгляд, выглядит более привлекательной. Она включает 3 трибы с 12 родами: Rupicaprini (Rupicapra, Capricornis, Nemorhaedus и Oreamnos), Ovibovini (Ovibos и Budorcas) и Caprini (Hemitragus, Capra, Pseudois, Ammotragus и Ovis). Род Pantholops вынесен им за пределы выделенных триб в связи с неясными родственными связями.
Место овцебыков в системе полорогих, видимо, прояснилось окончательно. Ovibos в переводе с латинского - “баранобык”. Это словосочетание само по себе отражало неясность в происхождении и классификации животных. До XIX в. этих копытных относили к роду Bos подсемейства бычьих, затем де Блэнвилл выделил его в самостоятельный род. В дальнейшем систематики причисляли его или к бычьим, или к козлиным, или считали отдельным подсемейством. По морфологическим признакам родство Ovibos усматривалось с южноазиатскими такинами, хотя были очевидны и существенные отличия между ними, позволявшие некоторым систематикам выделять Budorcas даже в отдельное подсемейство. С середины XX в. овцебыков все же чаще рассматривали в качестве монотипичного рода трибы Ovibovini подсемейства Caprinae [Simpson, 1945; Соколов, 1959; Гептнер и др., 1961]. Позднее их родство с козлиными подтверждено на молекулярно-генетическом уровне [Кузнецова, 2002].
Результаты сравнительных молекулярных исследований козлиных в целом оказались парадоксальными. Представители Capra, Pseudois и Ammotragus иногда группируются вместе, а все виды Ovis образует другой кластер. Ammotragus, возможно, является формой, произошедшей от древних козлов и настоящих баранов, a Capra и Pseudois образуют свою эволюционную линию [Ludwig, Fischer, 1998]. По мнению М.В. Кузнецовой [2002], роды Budorcas и Oreamnos должны быть исключены из трибы Ovibovini и помещены в трибу Caprini вместе с Capra, Pseudois, Hemitragus, Ovis и Pantholops, a Ammotragus представлен в трибе Rupicaprini. He подтверждается также видовой статус многих животных, в частности в родах Capra и Ovis (см. ниже) и даже в “ясном” роде Pseudois, в который включали всего два вида: Р. nayaur и Р. schaeferi. Судя по анализу митохондриального гена цитохрома Ь, голубые бараны монотипичны и разделяются, по меньшей мере, на 5 групп, рассматриваемых в качестве географических популяций [Zhou et al., 2003]. Замечу, что сравнительно недавно голубой баран обнаружен на Памире в пределах территории бывшего СССР [Сапожников, 1984, 1985].
По накопленным молекулярно-генетическим сведениям, классификация Caprinae может выглядеть следующим, но далеко не окончательным образом:
Триба Ovibovini Gill, 1872
Род Ovibos Blainville, 1816 - овцебыки
Ovibos moschatus Zimmermann, 1780
Род Nemorhaedus H. Smith, 1827 - горалы
Nemorhaedus gora/Hardwicke, 1825
Nemorhaedus caudatus Milne-Edwards, 1867
Nemorhaedus baileyi Pocock, 1914
Род Capricornis Ogilby, 1837 - cepoy
Capricornis sumatraensis Bechstein, 1799
Capricornis crispus Temminck, 1845
Capricornis swinhoei Gray, 1862
Триба Rupicaprini Brookes, 1828
Род Rupicapra Blainville, 1816 - серны
Rupicapra rupicapra L., 1758
Rupicapra pyrenaica Bonaparte, 1845
Род Ammotragus Blyth, 1840 - гривистые бараны
Ammotragus lervia Pallas, 1777
Триба Caprini Gray, 1821
Род Capra L., 1758 - козлы
Capra ibex L., 1758
Capra sibirica Pallas, 1776
Capra aegagrus Erxleben, 1777
Capra caucasica Guldenstaedt et Pallas, 1783
Capra nubiana F. Cuvier, 1825
Capra pyrenaica Schinz, 1838
Capra falconeri Wagner, 1839
Capra cylindricornis Blyth, 1841
Род Hemitragus Hodgson, 1841 - тары
Hemitragus jemlahicus Smith, 1826
Hemitragus hylocrius Ogilby, 1838
Hemitragus jayakari Thomas, 1894
Род Budorcas Hodgson, 1850 - такины
Budorcas taxicolor Hodgson, 1850
Род Oreamnos Rafmesque, 1817 - снежные козы
Oreamnos americanus Blainville, 1816
Род Ovis L., 1758 - бараны
Ovis ammon L., 1758
Ovis canadensis Shaw, 1804
Род Pseudois Hodgson, 1846 - голубые бараны
Pseudois nayaur Hodgson, 1833
Род Pantholops Hodgson, 1834 - оронго
Pantholops hodgsoni Abel, 1826
В составе фауны России 5 родов козлиных, в числе которых Ovibos.
Род ОВЦЕБЫКИ
Ovibos Blainville, 1816
Ovibos Blainville, 1816. Bull. Sci. Soc. Philom. Paris: 76. Тип рода - Bos moschatus Zimmermann, 1780.
Bosovis Kowarzik, 1911. Zool. Anz., 37: 107.
Характеристика. Животные крупного размера, плотного тяжелого сложения, с горбом, массивной головой и широкой мордой, на конце которой голый участок кожи (фото 17). Половой диморфизм выражен. Длина тела самцов достигает 262, высота в холке - 152, обхват туловища - 195 см, у самок - 204, 123 и 169 см соответственно. Хвост короткий - 6 см, скрыт в волосах. Длина уха - 12-15 см.
Масса новорожденных около 7-13 кг. Они быстро растут и в 2-месячном возрасте весят около 40—45 кг, в 4 мес. - 70-75 и в 6-12 мес. - 80-95 кг, в два года - около 140-180 кг. Полного физического развития особи достигают к 5-7 годам. Масса взрослых самцов в природе колеблется в пределах 200-408 кг в зависимости от времени года и упитанности, а на фермах достигает 658 кг. У самок этот показатель изменяется в пределах 150-188, на фермах - до 300 кг. Летом и осенью животные накапливают значительные жировые запасы, которые полностью расходуют зимой, теряя при этом и мышечную массу, а их вес снижается на треть [Чернявский, 1984; Lent, Devis, 1993; Якушкин, 1998].
Новорожденные бурые или серо-бурые с серебристо-желтыми “чулками” на ногах. Окраска взрослых особей темно-бурая, темно-коричневая или почти черная с желтыми подпалинами. На горбу у самцов рыжеватая грива. На подбородке и горле имеется черная борода. Поясница и нижние части ног белесые или светло-кремовые. У молодых особей (фото 18) и взрослых самок белый волос есть также на лбу. Светлая окраска головы более характерна для арктических островных животных Канадского архипелага и Гренландии, которых называют “белоголовыми”. Волос длинный (10-62 см) по всему телу, особенно по низу шеи, растущий, в отличие от других представителей козлиных, не образуя пучков и групп. Шерстный покров густой, четырехьярусный с многочисленными пуховыми волосами. Соотношение остевых и пуховых волос составляет 1:37, что в два раза больше, чем у лучших пород овец. Теплоизоляционные свойства меха, пожалуй, самые высокие среди копытных. Линька одна, протекающая с мая до июля. У лактирующих самок смена волоса запаздывает на 3-4 недели. С августа до октября происходит увеличение длины и густоты меха [Тепег, 1965; Якушкин, Олькова, 1988; Соколов и др., 1992; Якушкин, 1992, 1998].
Череп массивный, с хорошо развитой лицевой частью (рис. 25). Зароговая и лобная поверхности расположены в одной плоскости. Основная затылочная кость почти квадратная. Межчелюстные кости не соприкасаются с носовыми. Сошник делит носовую полость пополам, срастаясь с небными костями. Глазницы трубкообразные, сильно выступающие в стороны. Предглазничные ямки развиты. Этмоидальные щели отсутствуют. Слуховые барабаны небольшие. Резцы брахиодонт-
Рис. 25. Череп самца овцебыка (экземпляр из Московского зоопарка; Зоомузей МГУ, коллекционный № 136379)
Вид сбоку, сверху и снизу
ные. Коренные зубы относительно узкие, на верхних иногда имеется базальный столбик. Наибольшая длина черепа самцов достигает 502 мм, ширина - 259, длина носовых костей - 156, верхнего ряда зубов -150 мм, у самок -421, 215, 144 и 132 мм соответственно [Якушкин, 1998]. Размеры черепов у современных овцебыков Таймыра несколько мельче, чем у ископаемых, но их пропорции почти идентичны [Груздев и др., 1999].
Рога имеют животные обоих полов, но у самцов они заметно больше, чем у самок. Рога появляются на первом году жизни, полного развития достигают к шести годам. У самцов их основания сильно утолщены, сближены и у старых особей почти смыкаются на лбу. От основания чехлы загибаются резко в стороны между глазом и ухом вниз по боковой части головы, а затем плавно поднимаются вверх. Синусы находятся только в основаниях роговых
отростков. Форма сечения рогов округлая. У молодых особей на конце они черные, у старых - темно-серые. Длина рогов у самцов на Таймыре достигают 79 см, размах - 61, диаметр в основании - 14 см, у самок - 34, 48 и 7 см соответственно [Якушкин, 1998], но, скорее всего, встречаются и более крупные экземпляры.
Копыта широкие, округлые с острыми нижними краями и с широкой прочной “подушкой” снизу, что препятствует скольжению на склонах и позволяет копытить плотный снег при добывании корма. Боковые пальцы развиты слабо.
Предглазничные железы хорошо развиты, межпальцевые увеличены, но “мешки” у них отсутствуют, паховых и специфических “мускусных” железистых образований нет. В коже препуция самцов имеются многодольчатые сальные, а у самок развиты циркуманальные железы [Соколов, Чернова, 1988, 2001]. Сосков 2 пары.
Диплоидный набор хромосом овцебыка - 2п = 48, NF = 60, что заметно отличает его от большинства представителей бычьих, кариотип которых 2п = 60, но сближает с индийским буйволом Bubalus buffelus, имеющим такой же хромосомный набор. При G-окрашивании акроцентрические хромосомы бычьих и овцебыка практически идентичны, но, в то же время, сходны и с отдельными представителями козлиных [Teitz, Teal, 1967; Heck et al., 1968; Wurster, Benirschke, 1968; Desaulniers et al., 1989; Biltueva et al., 1995].
Рис. 26. Распространение Ovibovini (по [Шер, 1971])
Места находок ископаемых остатков представителей различных родов: 1 - Ovobos, 2 - Simbos, 3 - Praeovibos, 4 — Bootherium, 5 - Preptoceras, Euceratherium, 6 — Boopsis, 7 — Ovibos в Евразии (неполные сведения, см. текст). Ареал Ovibos moschatus'. 8 - в середине XX в., 9 - исторический
Овцебыки живут группами или небольшими стадами на равнинных и скальных участках арктической тундры. Звери медлительны, передвигаются шагом или галопом со скоростью до 35 км/ч. При опасности группируются и активно обороняются. Все органы чувств достаточно хорошо развиты. Предельная продолжительность жизни - 23-25 лет, но в природе самки живут не более 18 лет.
Преимущественно охраняемый вид, имеющий значительные перспективы для расширения ареала, увеличения численности и фермерского разведения.
Распространение. Ovibovini в плейстоцене населяли большую часть Евразии и Северной Америки (рис. 26). В историческое время представители рода Ovibos истреблены повсеместно, за исключением северо-запада Канады и Гренландии. В наше время реакклиматизированы на Аляске, Шпицбергене, а также в Норвегии, Швеции и России.
Европа Азия Америка
Голоцен
Верхний плейстоцен
Средний плейстоцен
Нижний плейстоцен
Верхний плиоцен
Рис. 27. Стратиграфическое положение родов Ovibovini (по [Шер, 1971])
Эволюция. Ovibovini, возможно наряду с вымершими в начале плиоцена Oiocerini [Masini, Thomas. 1989], - наиболее древняя евроазиатская группа козлиных. Предковая форма линии Ovibos не ясна, предполагаемая - род Hypsodontus Sokolov, 1949 из среднемиоценовых отложений Восточной Европы. Род Praeovibos прослеживается в Северной Азии и Европе с верхнего плиоцена - раннего плейстоцена, род Ovibos - с нижнего плейстоцена. В Северной Америке в плейстоцене существовали и другие архаичные роды Ovibovini, в частности Symbos и Bootherium (рис. 27). Типичный позднеплейстоценовый овцебык Евразии - Ovibos pallantis Н. Smith, 1827, несколько отличающийся по морфологии черепа от Ovibos moschatus [Шер, 1971; Кожамкулова, 1981; McDonald, 1985; McDonald, Davis, 1989; Тихонов, 1994, 1999; Kirkland, Hilliard, 1996; Форонова, 2001; Косинцев, Бобковская, 2003].
Группа возникла и развивалась в условиях равнинного и сравнительно низкогорного ландшафта и сухого холодного климата, что привело к возникновению специфических экологических и морфологических адаптаций, связанных с проблемой терморегуляции и обороны от хищников.
Состав. Род монотипичен.
ОВЦЕБЫК
Ovibos moschatus Zimmermann, 1780
Bos moschatus Zimmermann, 1780. Geogr. Gesch. Thiere, 2: 86. Канада, междуречье Сил и Черчилл.
Характеристика. См. характеристику рода.
Распространение. То же, что и для рода.
Состав. Исследователи, на основании различий в окраске головы и небольшой разницы в размерах животных, чаще признают у О. moschatus два подвида: номинативный О. т. moschatus Zimmermann, 1780 (= niphoecus Elliot, 1905) (Северная Канада) и О. т. wardi Lydekker, 1900 (острова Канадского архипелага и Гренландия). Тем не менее вид, скорее всего, монотипичный, в пользу чего свидетельствует морфологическое и генетическое сходство, отсутствие непреодолимых географических барьеров и тесный контакт популяций [Флеров, 1935а; Тепег, 1965; Fleischman, 1986; Groves, 1995].
Распространение
Восстановленный ареал. В Евразии ископаемые остатки представителей рода Ovibos чаще датируются второй половиной плейстоцена, но часть их - из Западной и Средней Европы, находки на Волге у Сормова, в черте Москвы, на верхней Оби, в Павлодарской области Казахстана, в Кузнецкой котловине, на Вилюе, долине р. Хилок в Забайкалье и в бассейне Индигирки - может быть отнесена к более раннему времени, включая нижний плейстоцен [Вангенгейм, 1961; Шер, 1971; Алексеева, 1977; Тлеубердина и др., 1989; Оводов, 2000; Форонова, 2001].
Ареал плейстоценовых овцебыков был обширным. Эти животные населяли север Америки и Гренландию, острова Новосибирские, Фаддеевский и Врангеля, мыс Челюскин, Северо-, Восточно- и Западно-Сибирскую низменности, бассейны
Колымы, Алдана и Вилюя, Кузнецкую котловину, Таймыр, Ямал, Северный и Средний Урал и значительную часть Европы (рис. 26). В Европе их остатки найдены на многих древних стоянках человека: в Болыпеземельской тундре, Республике Коми, в Пермской, Архангельской и Вологодской областях, в Карелии, на окраинах Москвы и Казани, в Московской, Смоленской, Воронежской, Нижегородской, Саратовской и Астраханской областях, бассейне Дона, Десны и Среднего Днепра, близ Киева и Чернигова, на Скандинавском полуострове, Британских островах, во Франции, Германии, Румынии, Чехии, Словакии, Венгрии и Польше [Громова, 1935в; Громов, 1948; Пидопличко, 1951, 1954; Трофимов, 1955; Верещагин, 1959а, 19756, 2002; Марвин, 1959; Вангенгейм, 1961; Кузьмина, 1966, 1971, 1975; Шер, 1971; Кожамкулова, 1972; Алексеева, 1977, 1980, 19906; Верещагин, Громов, 1977; Верещагин, Русаков, 1979; Галкина, Форонова, 1980; Верещагин, Кузьмина, 1982; Девятова, 1982; Лазарев, 1982, 1990; Белан, 1989; Ревин, 1989; Бобковская, Косинцев, 1991; Смирнов, 1994; Тихонов, 1994; Малеева и др., 1996; Косинцев, 1996, 2003; Пономарев, 2001, 2003; Хромов и др., 2001; Косинцев, Бобковская, 2003; Подопригора, 2003]. Недавно плейстоценовый овцебык обнаружен на территории Монголии в районе Ноинулина [Алексеева, 1997]. Эта находка, вместе с забайкальской, позволяет значительно сдвинуть границы его древнего ареала в Азии к юго-востоку.
В позднем плейстоцене область распространения овцебыка быстро сокращается. В начале голоцена эти копытные сохранилось лишь на северном пределе ареала, а затем практически исчезли с территории Евразии, за исключением Крайнего Севера Сибири, где жили еще около трех тысячелетий назад [Верещагин, Барышников, 1985]. Сообщение И.Г. Пидопличко [1951] (со ссылкой на Маркуса и Кузнецова) об обитании этого вида в начале нашей эры в Монголии пока не нашло подтверждения. К началу XX в. небольшие стада сохранились только в Гренландии, в материковой тундре на севере Канады и на островах Канадского архипелага [Верещагин, 2002].
Причины сокращения ареала и численности. Главной причиной сокращения ареала и численности овцебыка и других крупных травоядных млекопитающих Евразии на рубеже плейстоцена и голоцена считается, как упоминалось выше, глобальное потепление климата, вызвавшее размораживание Полярного бассейна, заболачивание тундростепей, увеличение влажности и повышение уровня и плотности снежного покрова. На севере Евразии в это время исчезло около 30, а в Северной Америке - 32 вида [Трофимов, 1955; Флеров, 1955; Верещагин, Заблоцкий, 1961; Верещагин, Русаков; 1961; Томирдиаро, 1977; Верещагин, Барышников, 1985]. Однако, некоторые из вымерших животных, включая мамонта и носорога, жили в условиях уже измененного климата тысячи лет и окончательно исчезли по эволюционным меркам совсем недавно - 9-8 тыс. лет назад, а отдельные, например - тур и тарпан, всего сотни лет назад. Типичные представители “мамонтовой” фауны -сайга, северный олень, косули, зубр, бизон и овцебык и поныне живут в сравнительно теплом климате.
К полному исчезновению многих видов, резко сокративших свои ареалы при смене климата, несомненно, причастен человек, которому крупные звери служили источником пищи и одежды. Антропогенный пресс неуклонно усиливался по мере прогрессирующего увеличения людского населения, совершенствования орудий и
способов добычи зверей и появления транспортных средств - одомашненных лошадей и северных оленей. Соответственно ускорялись и темпы исчезновения животных.
Овцебык тоже был в числе основных охотничьих объектов человека. Его добывали ради мяса и жира (до 200-250 кг с одного животного) и длинноволосой шкуры, из которой шили теплую одежду, обувь и спальные мешки, что и стало одной из основных причин уничтожения многих группировок в последние столетия. Многочисленные арктические экспедиции, китобои и охотники за пушниной с помощью луков или нарезного оружия зачастую полностью выбивали стада, занимавшие круговую оборону. Драматическую роль в судьбе этого вида сыграло появление лаек, которые удерживали животных от бегства [Верещагин, 19596,1979,2002].
В Сибири овцебыка полностью уничтожили около 2800 лет до н. э., но на Таймыре, скорее всего, гораздо позднее. На Аляске последнее стадо истреблено в окрестностях мыса Хоп в середине XIX в. В материковой тундре Канады охота на этот вид велась вплоть до 1917 г. В 1860-1915 гг. компания Гудзонова залива и другие торговцы заготовили около 23 тыс. шкур. В Гренландии зверей промышляли до 50-х годов XX в. За сотню лет добыто около 12-16 тыс., и лишь в сезон 1929/ 30 гг. приезжими охотниками отстреляно 1,2 тыс. [Лент, 1970; Успенский, 1976; Burch, 1977; Barr, 1995; Якушкин, 1998].
В начале XX в. молодых животных охотно закупали зоопарки мира, что только усилило истребление оставшихся популяций. Телят нельзя было отловить, не уничтожив взрослых, которые инстинктивно защищали потомство. В период с 1900 по 1925 гг. из Гренландии вывезли около 250 молодых овцебыков, для чего пришлось истребить до 1200-1500 взрослых особей. Звери, помещенные в зоопарки, не имели иммунитета ко многим болезням и большей частью гибли. Неоправданно большие потери стали причиной отказа зоопарков от дальнейшего приобретения этих копытных, что способствовало сохранению оставшихся стад. В 1917 г. введен запрет охоты на вид в Канаде, а с середины столетия эффективная охрана налажена и в Гренландии [Прасолов, 1973; Якушкин, 1979; Alendal, 1980а].
Восстановление ареала в XX в. Планомерное восстановление ареала и численности овцебыка осуществляется с начала XX в. В 1900-1901 гг. 6 молодых животных из гренландской популяции переселили в горную часть Швеции, в 1929-1930 гг. 14 особей - в Исландию. Эти выпуски не принесли успеха. Более удачной была интродукция в Швецию 20 особей в 1980-1981 гг. В Норвегию 43 зверя завезли в 1924 г. В 1932 г. к ним добавили 10 и в 1938 г. - еще двух особей. В годы Второй мировой войны все животные были истреблены браконьерами. Повторные выпуски 37 особей осуществлены в 1947-1953 гг. Несколько десятков их потомков живут здесь до сих пор [Успенский, 1976; Alendal, 1980а,b; Железнов, 1990;Якушкин, 1998].
В 1929 г. 17 зверей завезли на Шпицберген. К 1942 г. стадо насчитывало около 70 голов, затем большая его часть была уничтожена во время войны. К середине 60-х годов здесь обитали 150 особей, спустя десятилетие - около 200. В 1974 г. на архипелаге зарегистрировано лишь 35 экземпляров, в 1978 - одна пара, в 1981 г. - 1 особь. Исчезновение овцебыка на Шпицбергене связывают с возникновением пищевой конкуренции с северным оленем, климатическими и антропогенными факторами [Успенский, Чернявский, 1966; Прасолов, 1973; Успенский, 1976; Alendal, 1980b; Якушкин, 1998].
На Аляске первая партия животных (17 годовалых особей и 17 сеголеток из Гренландии) выпущена в 1930 г. в загоны в районе г. Фербенкса, где вскоре 10 особей погибли. В 1935-1936 гг. 31 зверя перевезли, с целью защиты от нападения медведя, на о. Нунивак в Беринговом море. К 1938 г. стадо выросло до 50 голов, к концу 60-х годов - до 800. В 1987 г. нунивакская популяция насчитывала 587 экземпляров. С 60-х годов животных отсюда переселяли на другие арктические острова и на материковую часть Аляски, где они успешно закрепились и стали расселяться естественным путем. Группировка полуострова Сьювард, куда в 1970 г. ввезено 36 особей, в 1988 г. насчитывала 527 голов. Всего в XX в. на зарубежном Севере образовано 9 достаточно крупных популяций, включая лабрадорскую в Канаде [Лент, 1970; Успенский, 1976; Smith, 1989; Якушкин, 1998].
На необходимости реинтродукции овцебыка в Северную Азию (на Таймыр, Северный Урал, в Якутию, на северо-восток Сибири, Новосибирские и Медвежьи острова, острова Врангеля, Белый, Северную и Новую Земли, Землю Литке) зоологи настаивали с начала XX в. [Бутурлин, 1928а,б; Верховский, 1928; Насимович, 1933; Иванов, 1934; Подаревский, 1936; Зубков, 1937; Лебедев, 1958; Егоров, 1963; Успенский, Чернявский, 1965, 1966; Успенский, 1966; Прасолов, 1973]. В Арктическом институте в 30-е годы был разработан конкретный план реакклиматизации этого вида, однако реальные меры предприняты только в 70-е годы при улучшении международных отношений. В 1974 г. правительство Канады и в 1975 г. - США приняли решение подарить СССР стада овцебыков. В дальнейшем огромную работу по завозу, сохранению и восстановлению поголовья этих копытных в России проделали сотрудники Института сельского хозяйства Крайнего Севера [Успенский, 1975а,б, 19826; Верещагин, 2002; Забродин, 2002], самоотверженный труд которых должен быть рано или поздно оценен по достоинству.
В 1974 г. 10 полуторагодовалых особей с канадского о. Банкс и в 1975 г. 20 зверей с о. Нунивак (Аляска) завезли на Таймыр, где до 1979-1981 гг. их содержали в загонах в долине р. Бикады, а затем выпустили на волю. До 1979 г. от бронхопневмонии и травм погибли 13 особей, но родилось 24 детеныша, из которых выжили 20. Эти копытные стали ядром новой, быстро растущей популяции, постоянно расширяющей область своего обитания, площадь которой на Восточном Таймыре к концу столетия достигла 11 млн га (рис. 28). В начале XXI в. животные начали заселять тундры Центрального Таймыра и появились в предгорьях северо-восточной части плато Путорана [Якушкин, 1975, 1976, 1978, 1992, 1998, 2001; Успенский, 1976; Кацарский и др., 1982; Алабугин и др., 1984; Кацарский, Якушкин, 1986; Кацарский, 1989; Кравец, 2003; Сипко, 2004].
20 овцебыков (6 годовалых самцов, 12 самок и двухлетние самец и самка с о. Нунивак) 7 мая 1975 г. выпущены на о. Врангеля в среднем течении р. Мамонтовая после 20-дневной передержки в вольере. Звери сразу же разделилось на группы из 3, 6 и 11 голов и начали широко перемещаться по острову, уйдя от места выпуска на 150-200 км. В первые 3 года после выпуска погибли 11 особей, но оставшиеся в живых адаптировались к местным условиям и закрепились в северной части - в Тундре Академии [Тархов, 1976; Чернявский и др., 1980, 1981; Сташкевич, Казьмин, 1984; Чернявский, 1984; Казьмин, 1986, Ковалев, 1989, 1990а,б, 1991; Чернявский, Ковалев, 1995].
В октябре 1996 г. 24 молодых таймырских овцебыка перевезены в Булунский улус Республики Саха (Якутия), в охранную зону заповедника “Усть-Ленский”. Вскоре
Рис. 28. Размещение овцебыков на полуострове Таймыр (по [Якушкин, 2001])
1 - границы летнего ареала в 2000 г.; 2 - районы обитания отдельных субпопуляций; 3 - места встреч холостых самцов
два зверя погибли (от травмы и стресса), а выпущенные на волю образовали два стада. К 2002 г. в обеих группах насчитывали 39 голов, и еще 2 самца жили поодиночке. В 1997 г. 28 особей переселили в Анабарский улус. И здесь сразу же в первый день вселения погибли самец (от стресса) и самка (утонула в реке). В 2000 г. сюда же доставили еще 15 и в 2001 г. -10 животных. В Аллайховский улус вселили 11 особей в 2000 г., на о. Б. Бегичев в море Лаптевых - 25 в 2001-2002 гг. Всего в 1996-2002 гг. в Якутию завезли 124 овцебыка из Восточного Таймыра и 23 - с о. Врангеля. Первый приплод получен в 1999 г. - 6 телят от 11 самок. В 2002 г. здесь насчитывали 180 голов. Через 8 лет, как показывают расчеты, группировка вырастет до 1000 экземпляров [Тихонов и др., 1997, 2002; Царев, 1997, 2002; Царев, Лякин, 2001; Горохов, Якшина, 2002; Тихонов, 2002, 2003а; Тихонов, Егоров, 2002; Груздев и др., 20036].
В 1997 г. 15 таймырских зверей выпущены в Приуральском районе Ямало-Ненецкого АО. На следующий год они вышли к побережью Карского моря в сотне километров от места выпуска. В 1998 г. в тот же район завезли еще 15 особей с Таймыра и двух самок из Московского зоопарка, а в 2001 г. группировку пополнили 16 животными [Ширшов и др., 1999; Тихонов и др., 2002, 2003а].
В перспективе планируется расселение овцебыка и в других областях России (см. ниже).
Среда обитания
Овцебык - обитатель арктических и субарктических тундровых зон, отличающихся длительными низкими температурами и малым количеством осадков. Многоснежных стаций звери избегают, хотя и способны добывать корм при рыхлом слое высотой до 40-50 см. Зимой они обычно держатся на горных плато, пологих склонах и верхних участках речных террас, на малопродуктивных, но малоснежных (до 20-30 см) пастбищах, а в вегетационный период выбирают участки с богатой растительностью в долинах рек и озер и понижениях в тундре [Тепег, 1965; Успенский, Чернявский, 1965; Успенский, 1976; Якушкин и др., 1980, 1998; Верещагин, 2002].
Канадские овцебыки переселены на Восточный Таймыр на ту же широту (74°) и в аналогичную подзону южных арктических тундр в сходные экологические условия. Американские животные выпущены на Таймыр и о. Врангеля на 11-15" севернее их типичных местообитаний в более суровую климатическую зону и менее продуктивные угодья, что привело к гибели части интродуцентов. Однако уцелевшие звери адаптировались сравнительно быстро [Якушкин, 1979].
По исследованиям В.В. Рапота [1979], на Таймыре эти копытные, помещенные в обширные загоны, круглогодично использовали бугорковатые, мелкобу-горковатые, пятнисто-бугорковые, бугорковые и щебнистые растительные группировки арктических тундр, с весны до поздней осени — болотно-тундровые комплексы и арктические луговины. Излюбленные летние и осенние пастбища -разнотравно-ивово-дриадовые тундры (лесотундры), гигрофитные луговины, обводненные мочажины и заболоченные озерки с сообществами арктофилы рыжеватой. Зимой звери выпасались в низкоивняковых осоково-моховых и бугор-коватых тундрах, где дернина имеет рыхлую структуру и не смерзается благодаря переплетению ветвей ив. В мелкобугорковых тундрах они выбирали участки с преобладанием лишайников. Весной животные выходили на проталины на возвышенностях и затем, по мере таяния снега, переходили на болотно-тундровые комплексы.
В местах обитания овцебыков на воле в бассейне р. Бикады выделено 7 типов пастбищ. Малопродуктивные арктические кустарничковые и разнотравно-кустарничковые тундры, также как и субарктические кустарничково-моховые и кустарничково-осоково-моховые ассоциации, овцебыки используют в основном весной и летом. На высокопродуктивных субарктических кустар-никово-осоково-пущициево-моховых тундрах звери выпасаются круглогодично и сильно стравливают их. Наиболее продуктивны субарктические разнотравно-кустарничково-моховые тундры в сочетании с другими ассоциациями: они имеют сплошной растительный ковер, но занимают небольшие участки по берегам рек и крупных ручьев, и служат основными пастбищами лишь в весенне-раннелетний период. Кустарниковая и луговая растительность речных и озерных долин и днищ оврагов обильна, но доступна только в бесснежный и малоснежный сезоны. Зимой животные чаще держатся на малокормных полигональных и плоскобугристых болотах и в болотно-тундровых комплексах [Поспелова, 1989].
На о. Врангеля в снежный период группы зверей выпасаются в основном на малоснежных возвышенностях с крутыми террасированными склонами, с которых снег сдувается ветром; в нижних частях южных склонов гор в кочкарных ивняково-кустарничковых и осоково-пушициевых тундрах, где снежный покров не превышает 25 см; в долинах рек и ручьев, где выше снежный покров, но травянистая растительность (осоковые) уходит под снег в зеленом состоянии. Летом животные держатся на северных склонах гор и в долинах рек, предпочитая кочкарные осоково-пушициевые трещиноватые и пятнистые мелкокустарничковые тундры. При необходимости они с легкостью поднимаются по крутым склонам и каменистым осыпям на щебнистые платообразные вершины гор [Ковалев, 1991; Железнов-Чукотский и др., 1996; Царев, 1999].
Питание
В набор основных кормов овцебыка на всем ареале входят различные виды кустарниковых ив и берез, ольха, осоки, злаки, пушица, камнеломка, голубика, смородина, разнотравье, хвощи и, в незначительном количестве, лишайники и мхи [Успенский, Чернявский, 1965; Тепег, 1965; Klein, Вау, 1990].
На Таймыре эти копытные потребляют более сотни видов сосудистых растений (почти половину все видов местной флоры). Весенний рацион состоит из 23 основных кормовых видов, летний - из 43, осенний и зимний - из 32 и 20 соответственно. Весной они предпочитают зеленые проростки пушицы, осоки, новоси-версии ледяной, остролодочника Миддендорфа, астрагала альпийского и различные виды ив. Основу рациона в летний, осенний и раннезимний периоды составляет гигрофильная растительность - пушица узколистная, осока прямостоячая и арктофила рыжеватая, а также дюпонция голоцветковая, мытник судетский, кисличник, щавель и лисохвост. Зимой и ранней весной в пище преобладают ветошь однодольных трав, побеги ив, дриада и лишайники, из которых предпочитается кладония и пельтигера пупырчатая. В загонах зимой звери охотно поедали прессованное сено, комбикорм и травяную муку в гранулах [Рапота, 1979, 1981; Якушкин, 1998]. На о. Врангеля животные используют в пищу 72 вида растений, преимущественно ивовые, злаковые и бобовые [Чернявский и др., 1981; Железнов-Чукотский и др., 1996].
В бесснежный период овцебыки выбирают и скусывают отдельные излюбленные растения, а при их сплошном покрытии на луговинах и болотных мочажинах, медленно передвигаясь и почти не поднимая головы, “подстригают” пастбища узкими лентами размером 10 х 300 см на высоте 1-4 см от земли, изымая 95% биомассы. Зимой, в поисках растений, тонкий и рыхлый слой снега они разгребают мордой, плотный раскапывают широкими и прочными копытами, оставляя лунки площадью 0,1-2,6 м2 или глубокие борозды [Рапота, 1979]. Суточная площадь выпаса стада из 20-25 особей в зимний период колеблется в пределах 1,3—4,6 га. За 8 снежных месяцев оно осваивает 5-6 тыс. га пастбищ, но непосредственно под выпас использует 1,2-1,5 тыс. На одну особь, следовательно, приходится 240-250 га общей площади и около 60 га кормовой [Якушкин, 1998].
Степень утилизации растительного корма овцебыком гораздо выше, чем северным оленем, живущим в сходных условиях. Как показали опыты в загонах на Аляске, первые используют до 84% растений в предлагаемом корме, тогда как вторые -до 57%, а из предлагаемой порции разных видов лишайников - 52 и 37% соответственно [Успенский, Чернявский, 1965].
На Таймыре содержавшиеся в загонах звери охотно грызли и лизали поваренную соль. На воле они быстро нашли естественные солонцы с явным преобладанием солей Na, и в бесснежный период стали регулярно, не реже одного раза в неделю, посещать их [Якушкин, 1986, 1998].
Суточный ритм активности полифазный. Летом овцебыки чередуют кормежку и отдых 6-9 раз. В прохладные и ветреные дни они дольше пасутся и отдыхают, в теплую и тихую погоду при повышенной активности кровососущих насекомых периоды отдыха короче. Осенью резко увеличивается продолжительность и пастьбы, и отдыха, в связи с чем суточный цикл становится четырехфазным. Зимой дви
гательная активность снижается, а периоды отдыха увеличиваются, особенно при сильном морозе и в пургу. Весной количество фаз возрастает до 5-6. Синхронизация пастбищной активности более характерна для отдельных возрастных групп, нежели для всего стада, которое на подъем или укладывание нередко затрачивает более 1 ч. На отдых животные ложатся обычно на пастбище, кучно, спиной к ветру, а самцы и самки-лидеры - головой к ветру для контроля обстановки. В пургу выбирают защищенные от ветра участки склонов и ложбины. Телята прижимаются к матерям и сворачиваются калачиком. Летом звери отдыхают на сырых участках или поднимаются на высокие сопки, обдуваемые ветром [Рапота, 1979; Якушкин, 1981, 1998].
Размножение
Половая зрелость. В условиях Арктики, как считалось, самки впервые приходят в течку в трехлетием возрасте, в Субарктике - в двухлетнем [Успенский, Чернявский, 1965; Успенский, 1976]. Позднее выяснилось, что и в материковых, и арктических островных популяциях большинство самок становятся половозрелыми в двухлетнем возрасте, а при хороших кормовых условиях до 30% из них оплодотворяются на 15-17 месяце жизни. Размножение завершается в 11-14 лет, в редких случаях -в 15-17 лет. Самцы способны успешно спариваться с самками с 2-3-летнего возраста, однако их участию в гоне препятствуют взрослые самцы-доминанты [Alendal, 1971b; Кацарский, 1987; Якушкин, 1989, 1996, 1998; Ковалев, 1990а].
Гон и половое поведение. Гон у овцебыка в разных частях ареала, на фермах и в зоопарках весьма растянут - с конца июля - начала августа до середины октября, а иногда сдвигается на сентябрь - декабрь. Сдвиг в сроках наступления течки связан с погодными и кормовыми условиями. Продолжительность астрального цикла у самок 20-21 день, разовой течки - 12 ч [Тепег, 1965; Успенский, Чернявский, 1965; Гавердовский, 1980; Чернявский, 1984; Reinhardt, 1985; Орджоникидзе, Хить, 1989; Железнов-Чукотский и др., 1996; Якушкин, 1998].
По наблюдениям Г.Д. Якушкина [1982, 1983, 1989, 1998], на Таймыре у овцебыков в середине апреля - первой половине мая проявляется ложный гон. Самцы преследуют самок и пытаются делать садки. Нередко соперники сталкиваются между собой, но бои носят чаще демонстрационный характер с целью выявления иерархического статуса. Перед гоном в конце июля - начале августа в группу самок с молодняком вливается мощный самец, который вытесняет из нее молодых самцов и образует гарем. В 1982-1995 гг. средний размер гаремных групп достигал в июле 18 (3—47), в августе - 20 (3-69) особей. Доминант в период гона почти не кормится, преследует самок в эструсе, многократно кроет их и, во избежание конкуренции с соперниками, пытается удерживать группу в укромном месте. Гаремный самец может покрыть до двух десятков самок. В крупных стадах, однако, вместе с ним находятся до 4 самцов-субдоминантов и в стороне держатся до 8 молодых (2-3-летних) особей.
В половом поведении самцов выделено 14 элементов: 1) ориентация на самку в эструсе; 2) подход в позе угрозы; 3) удар передней ногой или толчок мордой в корпус или зад самки; 4) обнюхивание гениталий; 5) флемен; 6) обоюдное потирание телом; 7) обоюдное кружение; 8) позы контакта; 9) “подбрасывание” передней
части тела; 10) давление подбородком о круп самки; 11) подготовка к садке; 12) заскок; 13) садка с интромиссией и эякуляцией; 14) соскок и подготовка к очередной садке. Брачная пара держится вместе один-два дня, но в пик гона самец за 9 ч может успешно покрыть 3^4 самки.
Беременность. Беременность длится около 8-8,5 месяцев [Тепег, 1965; Орджоникидзе, Хить, 1989; Якушкин, 1996, 1998]. В Норвегии, по наблюдениям за парой овцебыков, установлены более точные сроки вынашивания плода - 249-252 дня [Alendal, 1971а]. В Московском зоопарке детеныш родился через 244 дня после спаривания родителей [Гавердовский, 1980].
Деторождение и выкармливание молодняка. Деторождение у овцебыков в разных частях ареала и в неволе происходит в апреле - июне. На Таймыре оно обычно с 15 апреля по 25 июня, более 85% самок рожает с конца апреля до середины мая. На о. Врангеля этот процесс растянут с 20 апреля до середины июля, но новорожденные чаще появляются с середины мая до середины июня [Тепег, 1965; Успенский, Чернявский, 1965; Якушкин, 1982, 1983, 1987; 1996, 1998; Чернявский, 1984; Кацарский, 1987; Орджоникидзе, Хить, 1989; Железнов-Чукотский и др., 1996].
Самки овцебыка приносят детенышей в стаде или рядом на снегу при устойчивой ночной минусовой температуре, достигающей 30-35 °C. Роды длятся 10-110 мин. Самка при этом неоднократно ложится, встает и кружится на месте. Рожает она в положении лежа, иногда - стоя, и в первые минуты интенсивно вылизывает новорожденного, который конечно же страдает от гипотермии. Однако у овцебы-чат, как и у детенышей других видов копытных, большая потеря тепла компенсируется за счет теплообразования при реакции окисления бурого жира. Новорожденный встает на ноги через 5-40 мин после появления на свет, и через 30-50 мин начинает сосать вымя. В первые два дня мать кормит его до 20 раз за сутки и от 1 до 9 мин за период кормления, давая за день до 3-5 л молока. После насыщения детеныш ложится. На третьи сутки число кормлений снижается до 6-9, к 15-20 дням жизни -до 2-6, в месячном возрасте - до 1—4, к двум месяцам - до 0-2 и к четырем - до 0-1. Месячный овцебычок опустошает вымя за 20-30 с. Молозивное молоко содержит до 5-12% жиров и 5-13% белков, что обеспечивает интенсивный рост сеголеток. Ветошь трав они пробуют с недельного возраста, а через месяц переходят на подножный зеленый корм, дополняемый порциями молока. Лактация заканчивается на пятом месяце их жизни, но некоторые позднородившие и прохолоставшие самки кормят детей до года и больше. Продолжительная лактация не препятствует наступлению беременности [Якушкин, 1982, 1983, 1998; Кацарский, 1986; Lent, 1991; Adamczewski et al., 1997].
В Московском зоопарке детеныш со второго дня жизни пил воду, копал ногами землю и поедал ее. После случайного захода в загон самца, мать не приняла его и перестала кормить. В дальнейшем этого овцебычонка выпаивали искусственно овечьим молоком и молочными смесями. В трехнедельном возрасте он начал потреблять фрукты и овощи, в 2,5 месяца - траву и веточный корм [Гавердовский, 1980].
Плодовитость. Самки овцебыка приносят обычно одного детеныша, двойни редки. Процент беременных 18-35-месячных самок не превышает 25, а у 3,5-летних и старше достигает 63. При хорошем питании взрослые звери рожают ежегодно, но особи старше 10 лет приносят детенышей через год [Тепег, 1965; Якуш-
кин, 1983, 1996, 1998; Кацарский, 1987; Adamczewski etal., 1997]. На Таймыре яловость самок составляет 4-15%, а на каждую из рожавших приходится в среднем 0,8-0,9 теленка. В аномальном по климатическим условиям сезоне 1989/90 гг. яловость была необычайно высокой - 37%, а в 1999 г. достигала 43% [Якушкин, 1989, 1998, 2001]. На о. Врангеля в 80-е годы плодовитость особей старше 3 лет была довольно высокой и стабильной - 91 (75-100 в разные годы) детеныш на 100 самок, а у двухлетних этот показатель варьировал в пределах 25-100 (62,5 в среднем) [Ковалев, 1990а].
Популяционная структура
Половой и возрастной состав. Соотношение полов среди эмбрионов и детенышей овцебыка в разных частях ареала близко 1:1с незначительным преобладанием в ту или другую сторону в разные годы. На Таймыре несколько чаще рождались самцы, но в популяции при регистрации преобладали самки [Якушкин, 1989, 1998], что, скорее всего, вызвано недоучетом широко кочующих самцов. На о. Врангеля в 80-е годы среди телят (п = 87) было больше самцов (55%) [Ковалев, 1990а], соотношение полов в группировке составляло 1:0,9 [Груздев и др., 2003а]. Доля сеголеток в группировках колеблется в пределах 16-27%, годовалых особей - около 6-20, одиночных самцов - 6-30, самцов в стадах - 10-18, самок - 30—40% (табл. 32).
Таблица 32. Половозрастной состав (%) группировок овцебыка
Район (источник информации)* Сеголетки Годовалые особи Одиночные самцы Самцы в стадах Самки
О. Врангеля (1) 17,6 5,9 29,9 10,9 35,7
Таймыр (1) 22,6 15,4 6 17,9 38,1
-“-(3) 20-26 22 7 11 40
-“-(2) 16-27 17-20 14-17 10-13 30-36
* 1 - Груздев и др., 2003а; Сипко и др., 2003; 2 - Якушкин, 1998; 3 - Кацарский, Ликонцсв, 1990.
Социальная организация и поведение. Овцебыки - высокоорганизованные стадные животные (фото 17) с разнообразным социальным поведением и эффективным набором оборонительных реакций.
В осенне-зимний период звери образуют смешанные стада, включающие половозрелых самок с приплодом, годовалых особей, 2-3-летних самцов и одного-двух половозрелых самцов. При переизбытке самцов могут образовываться их однополые группы, насчитывающие до 11-13 особей. Весной стада распадаются. В период отела и после него самки с молодняком держатся отдельно от взрослых самцов, живущих поодиночке, по два-три йли вместе с другими особями разного возраста. В отдельных небольших группах взрослые самцы остаются с самками весной и летом, но выпасаются и отдыхают обычно в стороне и присоединяются к ним только на переходах [Якушкин, 1987, 1998; Ковалев, 1989, 19906, 1991].
На Таймыре средние размеры групп в зимний, весенний и раннелетний периоды достигают 22-29 особей, в июле - сентябре - 17-20. В крупных стадах насчи
тывают до 50-80 голов, во временных скоплениях на нагульных пастбищах - до 100 [Кацарский, 1989; Якушкин, 1989, 1998, 2001; Царев и др., 1999; Сипко и др., 2003]. На о. Врангеля максимальный размер смешанного стада - 53 экземпляра, но чаще звери держатся группами от 2-6 до 10-15 особей, а взрослые самцы - поодиночке или небольшими компаниями [Ковалев, 1989; Железнов-Чукотский и др., 1996].
Отдельные элементы полового, агрессивного и оборонительного поведения овцебыка более сходны с таковыми у представителей подсемейства Bovinae, нежели козлиных [Лент, 1974]. Отношения особей в стадах строятся по линейному иерархическому принципу с главенствующей ролью самца-доминанта, который нередко остается с самками и молодыми самцами и после гона. При переходах и при бегстве от опасности чаще лидирует высокоранговая самка, а доминант и другие самцы обычно замыкают группу. За каждой самкой следует ее теленок. Иерархия поддерживается демонстрациями, столкновения сравнительно редки даже в период гона. Демонстрационное поведение самцов включает сближение противников с твердой поступью, рев, параллельные марши, позы угрозы, потирание предглаз-ничной железой о выставленную вперед ногу, рытье земли копытами и рогами. Столкновения лбами происходит на большой скорости с расстояния в 10-15 м, а иногда - 30-50 м. Таких сильнейших ударов в одном бою может быть до 40, Порой у противников после столкновения из ноздрей и ушей течет кровь, и, оглушенные, они несколько минут стоят неподвижно. Иногда ожесточенные лобовые удары или удары рогами в бок поверженного на землю соперника бывают смертельными [Железнов-Чукотский и др., 1996; Якушкин, 1998; Забродин, 2002].
Оборонительное поведение овцебыков по отношению к человеку и хищникам отличается. При приближении волков или собак стадо сразу же образует плотный круг, внутрь которого оттесняются молодые животные, и активно обороняется. Хищников чаще атакует самец-доминант. После выпада на 5-10 м он, пятясь, возвращается в круг или же к нему приближаются члены стада. При появлении людей или техники ближе 400-500 м животные группируются и пытаются сблизиться, чтобы рассмотреть объект, а затем организуют оборону или, чаще, быстро уходят плотной группой. В загонах при постоянной зимней подкормке животные утрачивают страх перед человеком, но весной и летом оборонительные реакции проявляются и здесь. В период гона самцы агрессивны и опасны для человека [Якушкин, 1979, 1983а, 1998; Ковалев, 1991; Царев, 1997, 1999; Царев, Лякин, 2001].
Из средств общения звери больше используют зрение. Звуковая сигнализация развита слабо. Взрослые самцы ревут при подходе к стаду и при турнирных схватках. При тревоге животные фыркают и громко сопят. Детеныши, отставшие от матерей, призывно блеют, и матери идут на их зов.
Пространственная структура и перемещения. Овцебык - сравнительно оседлое животное, однако в местах интродукции растущие группировки интенсивно осваивают новые районы, а многие стада становятся сезонными мигрантами.
На о. Врангеля осенью звери кочуют в Тундре Академии с запада на восток к побережью, весной - перемещаются обратно вдоль горной цепи на расстояние до 120 км. Отел самок происходит в районах, достигнутых во время миграции. После отела стада около месяца оседлы. Зимой животные размещаются на возвышенностях между реками Красный Флаг и Насхок, на южных и юго-западных склонах Безмянных гор, в верховьях рек Ледниковой и Равнинной, летом - в западной и
северной части Тундры Академии. Площадь зимнего участка каждого стада не превышает 50 км2, размер годового участка достигает 200 км2. Одиночные самцы и их группы перемещаются гораздо шире, нежели самки, освоив почти всю территорию острова [Ковалев, 1991; Железнов-Чукотский и др., 1996].
Таймырская популяция состоит из трех группировок. Одна из них размещается в подзоне типичных тундр в долине р. Бикады (на месте интродукции), вторая - на северных склонах гор Бырранга в подзоне арктических тундр, третья - в долинах рек Большая Балахня и Гусиха в подзоне кустарниковых тундр. В последние годы центральная и северная группировки объединились в низкогорной части Бырранга, третья разбита на мелкие группы в результате отлова молодняка для расселения. Из-за чрезмерного антропогенного пресса многие стада покинули бассейн Бикады. Расширение области обитания популяции идет интенсивно во всех направлениях, кроме южного (рис. 28). Холостые самцы особенно подвижны и встречаются почти по всей тундровой зоне Таймыра. Максимальная дистанция эмиграций - 600-800 км. Отдельные особи на севере вышли на побережье Ледовитого океана и мыс Челюскин, на западе преодолели крупные реки Пясину и Пуру, на юге достигли зоны редколесья [Якушкин, 1989, 1996, 1998, 2001, 2002а,б, 2003].
Отел обычно происходит на местах зимовок, после чего стада с уже окрепшими телятами в конце мая - начале июня (до начала снеготаяния) уходят за десятки километров на более богатые кормом летние пастбища. Самки, задержавшиеся с отелом, приносят телят во время кочевки. В половодье стада выбирают путь по водоразделам и ищут менее широкие русла рек и ручьев и спокойное течение воды. Телята стремятся преодолевать ручьи прыжками. За 1 ч быстро передвигающиеся звери могут пройти по снегу до 5 км, однако такая скорость быстро утомляет, и они останавливаются на отдых. За сутки в период кочевок и летом в период интенсивного нагула стада обычно проходят не более 8-10 км, в обычной ситуации - 2-4 км, а в период гона, стопоримые гаремными самцами, долго остаются на одном небольшом участке. На зимовках животные появляются к концу октября [Якушкин, 1998].
Овцебыки не терпят постоянного беспокойства со стороны людей, включая чрезмерное преследование с земли и с воздуха с целью отлова, - стада покидают район обитания, репродуктивный потенциал животных при этом снижается, а стадность возрастает [Якушкин, 1990; Кравец, 2003].
Численность и основные факторы, ее определяющие
Численность. В 30-е годы XX в. мировые ресурсы овцебыка оценивали в 10-12, в середине столетия - в 18-20 тыс. В 1970 г. на планете обитали 25 тыс. особей, из них 18 - в Гренландии, 1,5 - в Северо-Западной Канаде, 3,5 тыс. - на арктических островах, 800 особей - на Аляске и прилегающих островах, 150 - на Шпицбергене. К середине 90-х годов миррвая популяция увеличилась до 170-180 тыс., в том числе в Канаде - до 140, включая 45-тысячную группировку о. Виктория и 45-65-тысячную о. Банкс [Тепег, 1965; Успенский, Чернявский, 1965; Якушкин, 1974, 1998, 2001; Adamczewski et al., 1997; barter, Nagy, 2001].
В российских группировках также очевиден бурный рост поголовья. В 1979 г. на о. Врангеля жили 10 особей, 1989 г. - 88,1996 г. - около 400. К 2001 г. популяция выросла до 650 голов, в 2002 г. учтено 684 взрослых особей и 156 детенышей. На
острове ощущается недостаток пастбищ и уже необходима регуляция численности [Железнов, 1990; Железнов-Чукотский и др., 1996; Груздев и др., 2001, 2003а,б]. На Таймыре в 1981 г. насчитывали 51 особь, в 1982 г. - 60, 1985 г. - 132, 1986 г. - 175, 1987 г. - 230, 1988 г. - 290, 1989 г. - 370-390. К 1996 г. группировка увеличилась, по оценкам разных исследователей, до 1000-1600, 1997 г. - 1260-1800, 1998 г. - 1450— 2150 особей, а к 2002 г. - до 2,5-3,6 тыс. (последняя цифра, скорее всего, завышена) [Якушкин, 1983а, 1989, 1996, 2001; Кацарский, Якушкин, 1986; Кацарский, 1989; Кацарский, Ликонцев, 1990; Царев и др., 1999; Царев, 2002; Kolpashikov, Layshev, 2003]. По сведениям Службы учета охотничьих ресурсов в России в 2001— 2003 гг. обитали около 2,5-3 тыс. овцебыков.
Динамика популяций этих копытных определяется совокупностью многих экологических факторов.
Воспроизводство, прирост. Уровень воспроизводства в популяциях овцебыка, на удивление, высок, особенно в благоприятные по кормовым условиям годы. На Таймыре 100 половозрелых самок приносят 86-87 телят. Естественный годовой отход не превышает 4—7%. В 1982-1985 гг. прирост популяции колебался в пределах 23-29% в год, сохранность телят составляла 89-100%. В 1989/90 гг. из-за аномальной погоды показатель прироста оказался минимальным - около 16%, а в последующие годы он был близким к 23 (17-29)% [Кацарский, 1987; Якушкин, 1989, 1998, 2001, 20026; Царев, 1999; Царев и др., 1999; Сипко и др., 2003]. За 20 лет поголовье увеличилось почти в 30 раз. Ежегодный прирост группировки о. Врангеля в 80-е годы был почти таким же, как и на Таймыре - около 25 (17-33)%, а смертность во всех возрастных группах, за исключением старшей, не превышала 5% [Казьмин, 1986; Ковалев, 1990а; Железнов, 1990; Железнов-Чукотский и др., 1996; Сипко, 2003]. В многочисленной популяции о. Банкс на Канадском архипелаге в 1986-1999 гг. на 100 самок приходилось от 31 до 56 телят, выживаемость которых в разные годы составляла 23-83% [barter, Nagy, 2001].
Погода и корма. Благодаря прекрасно развитому шерстному покрову, овцебык легко переносит холодные, малоснежные и ветреные зимы, но мягкие, с оттепелями, дождями и гололедом, нередко заканчиваются массовой гибелью животных от бескормицы и переохлаждения - в заморозки намокший мех смерзается и превращается в ледяной панцирь, утрачивая теплозащитные свойства. После аномальной зимы 1953/54 гг. в Восточной Гренландии погибли несколько тысяч особей, популяция сократилась почти наполовину, а на ее воссстановление потребовалось около 20 лет. В 60-80-е годы на некоторых арктических островах Канадского архипелага отход достигал 70-80%. При сильных морозах в сочетании с пургой, несмотря на превосходные теплоизоляционные свойства меха, ослабленные от недостатка пищи животные простужаются и тоже умирают [Успенский, Чернявский, 1965; Якушкин, 1998]. Демографические показатели популяции о. Врангеля, однако, более всего зависят от погодных условий в осенний период [Груздев, Сипко, 2002].
Хищники. Главный враг овцебыка - волк, в меньшей мере, бурый и белый медведи, а также росомаха. Эти копытные, как уже отмечалось выше, могут сравнительно эффективно обороняться, используя “защитный круг”, однако хищники нападают на одиночных животных и телят в случае испуга и бегства стада. На Таймыре экологическая связь “хищник - жертва” установилась очень быстро. В июле 1981 г. зарегистрирован первый случай нападения волка на стадо, в мае
1982 г,- первый случай гибели новорожденного. Пресс хищников неуклонно возрастает и, для предотвращения потерь овцебыков, волков пришлось отстреливать с вертолета и снегоходов [Успенский, Чернявский, 1965; Якушкин, 1979, 1998; Алабугин и др., 1984; Кацарский, Ликонцев, 1987; Забродин, 2002].
Конкуренты. К потенциальным трофическим конкурентам исследуемого вида относят северного оленя, леммингов, зайца-беляка и гусей. Однако в районы обитания овцебыка на Таймыре северный олень приходит только летом, когда корма изобильны, да и то в небольшом количестве, а гуси и заяц-беляк не оказывают существенного влияния на пастбища. Более влиятельные конкуренты - сибирский и копытный лемминги, подстригающие растительность в годы высокой численности “под корень” [Якушкин, 1979, 1998]. На о. Врангеля трофическая конкуренция овцебыка с северным оленем, резко увеличившим численность в 90-е годы, уже ощущается, и дальнейшее увеличение поголовья копытных может нанести непоправимый ущерб местной флоре [Царев, 1999].
Болезни, паразиты. У овцебыков зарегистрирован вирус контагиозной экти-мы (“овечьей оспы”), псевдотуберкулез, бруцеллез, инфекции центральной нервной системы и бронхопневмония, от которой на о. Врангеля в 1976 г. погибли 6 из 20 выпущенных зверей [Чернявский, 1984]. Бронхопневмония как фактор смертности имела место и в таймырской популяции, но простудные заболевания, несмотря на суровость климата, сравнительно редки. Здесь же у животных обнаружено 7 видов гельминтов, из них 5 характерны и для северного оленя. В других частях ареала у овцебыка найдено более 20 видов нематод [Якушкин, 1998].
Антропогенные факторы. Антропогенный пресс в районах реинтродукции овцебыка в Северной Азии минимален. При факторе беспокойства, частых авиаполетах в частности, стада, как отмечалось выше, покидают районы своего обитания [Якушкин, 1989]. В последние годы на Таймыре участились случаи нелегального отстрела животных [Царев, 1999; Кравец, 2003].
Перспективы восстановления и использования ресурсов
В XX в. усилиями природоохранных и охотничьих организаций удалось не только сохранить это крупное млекопитающее, но, как показано выше, и значительно увеличить поголовье, а также частично восстановить былой ареал. В Канаде аборигенные популяции вновь стали эксплуатировать с 1969 г. В конце столетия ежегодная квота на добычу достигла 2-3 тыс. голов. Возрожденная нунивакская популяция на Аляске промышляется с 1975 г., в других стадах ограниченная трофейная охота открыта с начала 80-х годов [Якушкин, 1998; Царев, Лякин, 2001]. Трофейная охота практикуется также в Гренландии.
В перспективе овцебык может стать одним из важнейших охотничьих видов и в России. В 2001 г. на Таймыре впервые выделена квота на отстрел 10 взрослых самцов, но она не была использована полностью. В дальнейшем возможен трофейный отстрел десятков и сотен особей [Якушкин, 2002а]. Популяция о. Врангеля уже сейчас нуждается в регуляции. Сократить ее можно селективным отстрелом, но гораздо целесообразнее ежегодно отлавливать часть зверей и переселять в другие регионы [Царев, 1999]. С другой стороны, объявление овцебыка охотничьим видом уже привело к активизации браконьеров [Кравец, 2003].
Овцебык, как показала практика, вполне может жить даже в районах со скудной растительностью, которые избегает северный олень. В Северной Азии экологическая ниша вида остается пустующей во многих малоснежных районах Восточной Сибири и Приполярного Урала. Его дальнейшее расселение возможно на Западном Таймыре, севере полуострова Ямал, Гыданском полуострове, в тундрах Якутии, Шмидтовском, Певекском, Анадырском и Чукотском районах Чукотского АО, в Магаданской области, горных районах Сибири и Дальнего Востока, на Новой и Северной Земле, о. Сибирякова, Вайгаче, Новосибирских островах и в других местах [Бутурлин, 1928а; Насимович, 1933; Егоров, 1963; Сыроечковский, 1995в; Железнов-Чукотский и др., 1996; Якушкин, 1998; Царев, 1999, 2002; Тихонов, 2003а; Тихонов и др., 2003].
По расчетам С.М. Успенского [1982а], в Российской Арктике могли бы обитать несколько десятков тысяч овцебыков. Однако эта цифра уже нуждается в корректировке. В тундровой зоне России, площадь которой оценивается в 300-400 млн га [Андреев, 1975], могут жить около 0,5 млн особей [Царев, 1999], а с учетом остальной пригодной территории эту цифру следовало бы увеличить еще как минимум наполовину. Лишь в Якутии поголовье вида можно увеличить до 100-150 тыс. без ущерба домашнему оленеводству, что позволит ежегодно добывать 20-30 тыс. особей и получать до 4000 т мяса (столько же продукции дает добыча 80-100 тыс. диких северных оленей) [Тихонов и др., 2002; Тихонов, 2003а]. Существуют серьезные предпосылки и для разведения овцебыка на фермах.
Фермерское разведение. В 30-х годах XX в. специальная комиссия, созданная правительством Канады, рассмотрев эту идею, признала ее экономически нерентабельной по сравнению с промыслом вольно живущих животных. Тем не менее в 1954-1955 гг. в США в штате Вермонт, а десятилетие спустя на Аляске близ Фербенкса были образованы опытные фермы (фото 19). В 70-е годы они появились в Канаде и Норвегии, где успешно функционируют до сих пор, хотя их рентабельность невелика. Всего их в мире пока чуть больше десятка. Овцебыки легко поддаются приручению, могут круглогодично жить на подножном корму, дают жирное молоко, вкусное мясо, кожевенное сырье и прекрасный тонкий и длинный пух (около 2,5-3 кг от одной особи в период линьки), из которого вяжут свитера, платья и шарфы. Из 1 кг пуха можно получить нить длиной до 30 км, а на платье или теплый свитер требуется всего 100-150 г [Успенский, Чернявский, 1965; Успенский, 1976; Тихонов и др., 2002; Царев, 2002; Тихонов, 20036].
В последние годы в мире явно наметилась тенденция к дальнейшему развитию фермерской индустрии [Lewis, 2000]. Одомашнивание этого вида вполне реально и в России, и не только на Крайнем Севере, но и в горных районах Средней и Южной Сибири. Небольшие фермы уже начали создавать в Якутии [Тихонов, Егоров, 2002; Тихонов и др., 2002; Царев, 2002; Груздев и др., 20036; Тихонов, 20036]. И все же, на мой взгляд, более перспективно не фермерское, а полувольное разведение животных по типу домашнего оленеводства.
Род ГОРАЛЫ Nemorhaedus Smith, 1827
Nemorhaedus H. Smith, 1827. Griffith’s Cuver Animal Kingd., 5: 352. Тип рода - Antilope goral Hardwicke, 1825. Непал, Гималаи.
Kemas Ogilby, 1837. Proc. Zool. Soc. London (1836): 138.
Caprina Wagner, 1844. Schreber’s Saugeth., Suppl., 4, 9: 457.
Urotragus Gray, 1871. Ann. Mag. Nat. Hist., 8: 372.
Nemorhaedus Flower et Garson, 1884. Cat. Osteol. Mus. College Surg., 2: 254.
Cernas Blanford, 1891. Fauna Brit. India. Mamm.: 516. Nom. praeocc., non Gioger, 1841. Non Oken,
1816.
Характеристика. Размеры животных небольшие, сложение плотное, шея утолщенная, конечности короткие (фото 20). Длина тела достигает 135 см, высота в холке - 90 см, масса - 48 кг.
Хвост отрастает до 19 см, с длинным волосом на конце. Носовое зеркало хорошо развито. Половой диморфизм выражен слабо.
Окраска варьирует от серой до рыжей и темно-бурой. Шерстный покров относительно одинаковой длины по всему телу и с небольшой гривой по верхней части шеи, четырехъярусный, зональный. Вершины направляющих, остевых, промежуточных и пуховых волос не перекрывают другой ярус. Мех очень густой за счет большого числа пуховых волос, и по теплоизоляционным свойствам сравним с шерстью овцебыка [Бромлей, 1963; Соколов и др., 1992]. Бороды нет.
Череп с выпуклым профилем лобно-лицевой части (рис. 29). За-роговая часть хорошо развита. Теменные гребни сближены. Носовые кости несколько укорочены и не соприкасаются с верхнечелюстными и предчелюстными. Глазницы не трубкообразные и не выступают заметно в стороны. Слезная кость достигает носового от-
10 см
Рис. 29. Череп самца горала (экземпляр из Судзухин-ского заповедника; Зоомузей МГУ, коллекционный № 39796)
Вид сбоку, сверху и снизу
Рис. 30. Ареал рода Nemorhaedus (по [Тихонов, 1999])
верстия. Предглазничные ямки и этмоидальные щели отсутствуют. Роговые стержни расположены позади глазниц и наклонены назад. Основания роговых отростков сближены, в поперечном сечении они округлые. Синусы имеются лишь в нижней их трети. Резцы брахиодонтные. Предкоренные зубы не редуцированы, в верхней челюсти они со слабо развитыми стилями. Коренные слабо гипсодонтные.
Рога имеют животные обоих полов. Роговые чехлы черные, короткие (до 23 см), кольца в их нижней трети хорошо выражены.
Предглазничные железы редуцированные, паховые отсутствуют, межпальцевые и зароговая хорошо развиты, препуциальные имеются [Соколов и др., 1983; Соколов, Чернова, 1988, 2001]. Молочная железа четырехдольчатая.
Спермин горала четко отличаются от таковых у других представителей козлиных мелкими размерами и лопатообразной формой головки, что косвенно указывает на невозможность его скрещивания с другими видами подсемейства [Стекленев, 1978а].
Диплоидный набор - 2п = 55-56. Все хромосомы акроцентрические. Кариотип несколько отличается от остальных козлиных по числу плеч хромосом - NF = 56 [Soma et al., 1980, 1987; Орлов, Булатова, 1983; Soma, Kada, 1984].
Типичные обитатели скалистых участков, ведущие оседлый семейно-групповой образ жизни. Имеют отличное обоняние и слух, хорошее зрение. Максимальная продолжительность жизни в вольерах 18 лет.
Охраняемые животные, внесенные в Красные книги Международного союза охраны природы и России.
Распространение. Южная Азия. Современный ареал очаговый. Западный участок включает горы Северного Пакистана, Кашмира, южные склоны Гималаев на севере Индии и Непала, юго-восток Тибета, северную часть Бирмы, южные и центральные провинции Китая, восточный участок - северо-восток Китая, Корею и юг Дальнего Востока (рис. 30).
Эволюция. Древнейшие формы горала известны из плиоценовых отложений Западной Европы и нижнеплейстоценовых - Китая. В раннем плейстоцене эти животные вымерли в Европе, но на Сардинии, вероятно, дожили до голоцена. На Дальнем Востоке они прослеживаются с плейстоцена [Верещагин, Барышников, 1979, 1980а; Алексеева, 1997]. Происхождение рода неясно. Родство усматривается с Ovibos и Capricornis.
Самобытная высокоспециализированная группа животных, адаптированная к жизни в малоснежных скально-луговых биотопах.
Состав. Окончательно не определен. В роде насчитывают до 6 видов: Nemorhaedus caudatus, N. goral, N. baileyi, N. griseus, N. hodgsoni и N. arnouxianus, или всех их сводят к одному-трем [Lydekker, 1913; Ellerman, Morrison-Scott, 1951; Соколов, 1959; Громов и др., 1963; Гептнер и др., 1961; Павлинов, 2003]. Существование репродуктивной изоляции доказано лишь между Nemorhaedus caudatus и N. goral [Volf, 1976]. В фауне России - Nemorhaedus caudatus.
АМУРСКИЙ ГОРАЛ
Nemorhaedus caudatus Milne-Edwards, 1867
Antilope (Caprina) crispa Radde, 1862. Reisen Sud. Ost-Sibir., 1: 262. Буреинский xp. Nom. praeocc., non Temminck, 1845.
Antilope caudatus Milne-Edwards, 1867. Ann. Sci. Nat. Zool., 5, 7: 377. Буреинский xp.
Nemorhaedus griseus Milne-Edwards, 1871. Nouv. Arch. Hist. Nat. Paris, 7, 93. Мупин, Сычуань, Китай.
Kemas raddeanus Heude, 1894. Mem. Hist. Nat. Emp. Chin., 2: 240. Буреинский xp.
Характеристика. См. характеристику рода.
Распространение. Китай, Корея, юг Дальнего Востока России, Бирма. Ареал очаговый.
Состав. Не ясен. И.И. Соколов [1959] выделял три подвида амурского, или китайского горала: N. с. caudatus Milne-Edwards, 1867 (Западный, Центральный и Северный Китай), N. с. griseus Milne-Edwards, 1871 (юго-запад Китая и Бирма) и N. с. raddeanus Heude, 1894 (Дальний Восток, Корея и, вероятно, Северо-Восточный Китай). В.Г. Гептнер [1961] считал raddeanus синонимом caudatus.
Изменчивость
Размеры и масса. Средняя масса новорожденных амурского горала 3,2 кг (2,2-4,0, а у одного из мертворожденных - 5,1 кг), в месячном возрасте - 5-7,7, в полугодовалом - 17, в годовалом - 22 и в двухлетнем - 35 кг. К двум годам рост завершается, а весовые изменения носят только сезонный характер. Половой диморфизм по экстерьерным показателям не выражен (табл. 33). Длина тела взрослых особей колеблется в пределах 106-135, высота в холке -69-85 см. Масса осенью достигает 40-48 кг, а зимой при истощении снижается до 24 кг. Особи с буреинских гор и гималайские горалы, судя по единич-
Таблица 33. Размеры (см) и масса (кг) взрослых особей амурского горала на юге Дальнего Востока
Самцы Самки Источник информации
п 1 М 1 lim n 1 М lim
7 113,5 109-118 Длина 3 112 106-117 Бромлей, 1977а; Бромлей,
11 128 123-132 5 130 126-135 Кучеренко, 1983 Волошина, Мысленков, 1992а
5 73,2 Высота в холке 69-78 3 73,3 71-75 Бромлей, 1963
10 81 77-84 3 81 74-85 Волошина, Мысленков, 1992а
11 78,7* 75-82 6 76,7* 70-83 Соломкина, 1992
4 86 Обхват туловища 78-98 2 74 70-77 Бромлей, 1963
И 79 70-87 4 76 72-82 Волошина, Мысленков, 1992а
11 89,6* 85-98 6 86,3* 76-94 Соломкина, 1992
7 31 Длина ступни 30-32 3 29 28-30 Бромлей, 1977а; Бромлей,
10 33 32-34 4 32 31-33 Кучеренко, 1983 Волошина, Мысленков, 1992а
7 14 Длина уха 13-15 3 14 13-15 Бромлей, 1977а; Бромлей,
10 14 13-14 4 14 13-15 Кучеренко, 1983 Волошина, Мысленков, 1992а
4 17* 14-19 4 16* 15-18 Соломкина, 1992
7 16 Длина хвоста 15-18 3 15,5 15-16 Бромлей, 1977а; Бромлей,
9 15 14-18 3 16 16-17 Кучеренко, 1983 Волошина, Мысленков, 1992а
7 34 30-39 Масса 3 30 24-32 Бромлей, 1977а; Бромлей,
11 33 26-41 5 32 27-45 Кучеренко, 1983 Волошина, Мысленков, 1992а
И 40,3* 37-48 7 40,4* 38-43 Соломкина, 1986, 1992
* Вольерные животные.
ным промерам тела, сделанным известным зоологом Г.И. Радде, близки по размерам. Особи южной лазовской популяции в длине тела и высоте в холке, вероятно, несколько мельче, чем в урочище Абрек [Бромлей, 1963; Соломки-на, 1975а, 1980а, 1986, 1990, 1992; Волошина, Мысленков, 1992а].
Краниометрическая характеристика. Новорожденные имеют 20 молочных зубов: прорезавшиеся резцы и намечающиеся клыки, которые к 10-14 дням жизни достигают размеров резцов. В это время прорезаются первые коренные. Смена молочных зубов на постоянные начинается в 1,5-летнем возрасте и заканчивается к 2 годам. Стирание коренных зубов заметно на пятом году, разрушение-в 14-15 лет [Бромлей, 1963; Срломкина, Хохряков, 1993].
Размеры черепа самцов и самок практически одинаковы. Наибольшая его длина колеблется в пределах 204—223 мм, ширина - 98-118, длина костных стержней рогов - 55-90 мм (табл. 34). Наибольшее диагностическое значение при определе-
Таблица 34. Размеры черепа (мм) взрослых особей амурского горала на юге Дальнего Востока
Самцы Самки Источник информации
п М I lim п М I lim
Наибольшая длина
14 214,4 206-223 14 212,1 Максимальная ширина 204-220 Соломкина, Хохряков, 1993
14 107,1 99-117 14 103,9 98-118
1 - 111 Межглазничная ширина - Юргенс, 1963
14 70,1 64-78 14 67,4 Длина лицевой части 64-70 Соломкина, Хохряков, 1993
14 147,9 139-159 14 151,6 142-157 а
1 - 164 Длина носовых костей - Юргенс, 1963
14 70,7 65-78 14 69,6 65-73 Соломкина, Хохряков, 1993
Длина костных стержней рогов
14 65-90 14 55-90 сс
14 Окружность оснований стержней рогов 60-85 14 - 55-70 «с
Таблица 35. Размеры рогов (см) взрослых особей амурского горала на юге Дальнего Востока
п Длина Размах Обхват у основания Источник информации
М lim М 1 lim М 1 lim
12 16,1 14-18,3 7,8 Самцы 7,0-9 9,8 8-12 Соломкина, 1992;
25 8,2 5,6-10,7 9,9 9,4-10,8 Соломкина, Хохряков, 1993 Тихонов, 1995
13 15,8 13,2-22 5,1 Самки 3,5-6,5 8,8 8-10 Соломкина, 1992
18 — — 4,9 4,4-5,6 8 7,5-8,6 Соломкина, Хохряков, 1993 Тихонов, 1995
нии пола имеет расстояние между основаниями роговых стержней: у самцов оно в среднем 67,3 (62,4-71,5), у самок - 59,5 (55,4-63,4) мм [Тихонов, 1995].
Изменчивость рогов. Рожки у сеголеток, самцов и самок, появляются в 1,5-2-месячном возрасте и интенсивно растут до 2 лет, после чего их концы начинают расщепляться и стираться. В 6 месяцев их длина достигает 6-10, через год - 13-14 см. В 16-18-месячном возрасте ювенильные темно-оливковые роговые чехлы заменяются постоянными черными. Рога у взрослых особей сравнительно тонкие, короткие, примерно одинаковой длины у самцов и самок, но у первых они массивнее в основании, шире поставлены, сильнее загнуты назад и с большим числом колец. Их длина составляет в среднем 16 (13-22) см, размах - 5-8 (3,5-10,7), окружность основания - 9-10 (8-12) см (табл. 35). Число неполных кольцевых утолщений на рогах у самцов доходит до 29, у самок - до 23 [Бромлей, 1963,1977а; Мысленков,
1976; Соломкина, 1977, 19806, 1990, 1992; Бромлей, Кучеренко, 1983; Соломкина, Хохряков, 1993; Волошина, Тихонов, 1995].
Окраска и линька. Окраска горала варьирует от серой и белой с охристым, палевым или землистым оттенками до темно-бурой, но чаще встречаются серые и буро-палевые животные с бурыми или черными полосами, тянущимися от носа через верх головы по спине до конца хвоста. Белые пятна присутствуют на груди, обеих губах, вокруг основания хвоста, белые полосы - на передних и задних ногах. Половой диморфизм в окраске выражен слабо. У самцов крупнее горловое пятно, всегда имеется беловатая шерсть на губах, а шерсть на лбу более однородная и темная, чем у самок, у которых контрастная темная полоса проходит посредине. Новорожденные светло-серые или молочно-кофейные с черными гривой и ремнем на спине [Соколов, 1959; Абрамов, 1963; Бромлей, 1963, 1977а; Бромлей, Кучеренко, 1983; Соколов и др., 1992].
Линька одна, продолжительная. Зимний волос выпадает с апреля до августа, новый отрастает к концу октября [Бромлей, 1977а]. Во время линьки, как и у овцебыка, идет интенсивная смена пуховых волос, а кроющие меняются постепенно [Соколов и др., 1992].
Распространение
На рубеже плиоцена и плейстоцена ареал горалов, вероятно, был трансконтинентальным и простирался от Восточного Китая до Западной Европы включительно. В плейстоцене область распространения резко уменьшается, а в голоцене ограничивается Южной Азией. Амурский горал дожил до наших дней лишь на горных хребтах российского Дальнего Востока, Корейского полуострова и Северо-Восточного Китая [Верещагин, Оводов, 1968; Верещагин, Барышников, 1977, 1979].
И.И. Соколов [1959] и В.Г. Гептнер [Гептнер и др., 1961], ссылаясь на заметки путешественников-первопроходцев Шренка, Маака и Радде, северную границу восстановленного ареала амурского горала вели от южной оконечности Буреинского хребта до устья Амура и правобережья Амурского лимана с включением Северного Сихотэ-Алиня. Далее к югу, по мнению В.Г. Гентнера, эта линия шла по западным склонам Сихотэ-Алиня вплоть до южной оконечности, и затем по возвышенностям к югу и западу от оз. Ханка. Пределом распространения вида на востоке служит океан. Г.Ф. Бромлей [1963] с соавторами [1978, 1983], однако, считали, что горал в силу своих биологических особенностей вряд ли мог обитать в многоснежных нижнеприамурских районах севернее 50° с.ш.
Современный ареал вида на территории России разбит на три очага, видимо, связанных между собой через Восточно-Маньчжурскую горную страну. Западный участок, который считали исчезнувшим [Соколов, 1959; Гептнер и др., 1961], включает северо-восточные отроги Малого Хингана и южную оконечность Буреинского хребта. Местное население периодически встречает, а порой и добывает этих копытных на скалистом побережье Амура севернее железной дороги Биробиджан - Облучье, южнее курорта Кульдур и В|бассейне р. Хор в Хабаровском крае [Абрамов, 1963; Раков, 1981; Дунишенко, 1983, 1985]. Существование этой микропопуляции, видимо, поддерживается за счет подтока иммигрантов из сопредельных районов Китая, как и на другом небольшом по площади участке в районе Черных
гор и Пограничного хребта на юго-западе Приморского края [Бромлей, 19776; Бромлей, Кучеренко, 1983; Нестеров, 1992].
Самая крупная группировка находится на Сихотэ-Алине. До середины XX в. северным пунктом распространения вида считалась гора Айгач на правом берегу р. Тумнин несколько южнее 50° с.ш., в середине 60-х годов - приток Бикина ключ Килой, куда они могли заходить из района р. Макси-мовки, т. е. с восточного склона хребта. Южнее они держались в горах Яковлевского района Приморского края и по берегам р. Ар-сеньевки. На побережье Японского моря горал был обычен на кручах притоков р. Максимовки и по берегам р. Таежной в Тернейском районе, на склонах г. Абрек на территории Сихотэ-Алинского госу
Рис. 31. Распространение амурского горала на юге Дальнего Востока (по [Бромлей, Кучеренко, 1983]) 1 - места обитания отдельных групп
дарственного заповедника, встречался на мысе Бринера, а также во многих скалистых прибрежных районах. Значительная группиров
ка сохранялась в Лазовском заповеднике. Небольшие очаги обитания имелись на Партизанском хребте [Абрамов, 1963; Бромлей, 1963, 1977а,б; Костоглод, 1973].
Распространение горала (рис. 31) на север в 70-80-е годы прослеживалось по морскому побережью до мыса Белкина, по восточному макросклону Сихотэ-Алиня - до р. Амги, по западному - до бассейна р. Хор, куда звери проникали, скорее всего, из верховьев Бикина и Максимовки. Несколько животных жили на горе Змеиной в Уссурийском заповеднике. На юго-западе Приморья животные по-прежнему обитали в Черных горах и в истоках р. Комиссаровка на Пограничном хребте. Южный пункт их встреч на морском побережье - мыс Разградского. Основное поголовье сосредоточено в Лазовском, Ольгинском, Дальнегорском, Тернейском, Партизанском, Анучинском, Чугуевском, Кавалеровском и Хасановском районах Приморского края, более чем в 20 локальных участках плошадью от 10-15 га до 40 тыс. га [Волошина и др., 1977; Глебов, 1981; Нестеров, 1981, 1992; Бромлей, Кучеренко, 1983; Дунишенко, 1983].
В формировании границ и структуры ареала вида первостепенное место занимают характер рельефа И растительных формаций и режим снежности, а также пресс хищников и антропогенное воздействие. В последние столетия северная граница распространения этого копытного вряд ли выходила за пределы зоны хвой
но-широколиственных лесов, которая имела тенденцию отступления на юг.
Среда обитания
Амурский горал - обитатель горных широколиственных и хвойно-широколиственных лесов, предпочитающий скально-луговые биотопы и склоны большой крутизны (60-80°) в диапазоне высот от 0 до 800 м над уровнем моря. Важнейшие факторы, оказывающие влияние на биотопическое распределение животных, - рельеф и снежный покров. Вид населяет районы гранитных интрузий, которые расположены отдельными островами на морском побережье и в верховьях рек. Там, где отсутствуют крупные скальные массивы и где устойчивый снежный покров свыше 35-40 см, нет и горала. Соответственно очаги его обитания удалены один от другого на десятки километров, что и определяет мозаичность ареала. Если участки гранитных разломов невелики, то небольшими оказываются и группировки населяющих их животных. Значение имеют также высота и конфигурация скал и наличие ущелий. Лучшие стации - сильно расчлененные прибрежные скальные массивы длиной 5—10 км, не прерываемые бухтами. Наибольшее количество гранитных интрузий имеется в Лазовском и Партизанском районах Приморского края. Здесь сосредоточена и большая часть поголовья [Юргенс, 1961, 1963; Бромлей, 1963; Вейнгер, 1963; Бромлей, Кучеренко, 1983; Волошина, 1988; Волошина, Нестеров, 1992].
Исследователи выделяют два основных типа местообитаний горала - прибрежный и материковый. На скалистых прибрежных участках лучше защитные условия, более благоприятный режим снежного покрова, который под воздействием солнца и ветра сходит через несколько дней после снегопада, меньше хищников, из-за сильных ветров меньше кровососущих насекомых, больше запас травянистой растительности, сроки вегетации которой сильно растянуты. В материковых стациях выше снежный покров, больше гнуса, короче сроки вегетации растительности, но по разнообразию и доступности корма материковые пастбища не уступают таковым на морском побережье [Нестеров, 1985]. Однако в поисках пищи животным приходится спускаться со скал, что увеличивает вероятность встреч с хищниками на пологих склонах или на равнинной местности, где эти мелкие копытные с короткими конечностями быстро становятся их жертвами, особенно зимой [Бромлей, 1963, 1977а; Юргенс, 1963; Волошина и др., 1976; Бромлей, Кучеренко, 1983].
Растительный покров пастбищ горала на протяжении тысячелетий формировался под его непосредственным воздействием и претерпел большие изменения. На травяных склонах при пастьбе животные разрыхляют дернину и войлок и устраняют подрост некоторых древесно-кустарниковых пород. По опушкам дубняков, где чаще всего пасутся эти копытные, обычны рыхлозадернованные разнотравно-злаковые фитоценозы, отличающиеся большой продуктивностью (до 25 ц/га) и быстрорастущим высокорослым травостоем. В местах их обитания постоянно подстригаемые древесно-кустарниковые растения приобретают приземистую форму, и многие из них не имеют генеративных побегов. При высокой плотности населения на тропах и крутых склонах заметно оголение почвы и эрозия, поскольку звери выдирают плотную дерновину злаков и осок и разгребают землю, добывая корневища и луковицы. При мечении территории они рогами обдирают кору, повреждая до 1% деревьев на участках обитания [Шаульская, 1992].
Питание
Набор кормовых растений амурского горала весьма богат. Только в прибрежной полосе среднего Сихотэ-Алиня он состоит из 268 видов (58% от общего числа растений местной фауны), из которых в разные сезоны травы составляют 76-89%, деревья - 3-5, кустарники - 7-15, мхи, грибы и лишайники - менее 3%. Летний рацион (более 223 видов) разнообразнее зимнего почти втрое. Летом и ранней осенью эти копытные предпочитают травянистые растения, бобовые в особенности: вику, горец широколистный, ветреницу, кровохлебку, шпороцветник, веронику, клевер, щавель, зубровку, мятлики, купену, термопсис бобовый, мытник, бубенчики, герань, ясменник, звездчатку, очиток, черемшу, луки, различные виды осок и полыней, а также листья и побеги дуба монгольского, леспедецы двухцветной, лещины разнолистной, чубушника, барбариса амурского, липы, клена, ив, амурского винограда, корневища красноднева съедобного, кровохлебки аптечной и луковицы лука густого. Зимой, наряду с древесно-веточным кормом, в пище доминируют полыни, осоки и разнотравная ветошь. На осыпях и обвалах они раскапывают корневища колосняка мягкого, в лесу подбирают желуди, объедают кору деревьев и древесные лишайники. Весной охотно потребляют зеленые проростки лука густого, осок, злаков и молодые древесные листья. Вольерные звери выбирают леспедецу, дуб и его желуди, ягоды лимонника китайского, боярышника, актинидий и плоды дикой яблони, едят морковь, свеклу, капусту, картофель, брюкву, тыкву, сухофрукты, овсянку, комбикорм, хлеб и сено [Юргенс, 1961,1963; Бромлей, 1963,1977а; Вейнгер, 1963; Соломкина, 1975, 19836,в; Валова, 1978; Шаульская, 1980, 1992; Соломкина, Глебов, 1981; Бромлей, Кучеренко, 1983; Мысленков, Волошина, 1989].
В отсутствии конкурентов и при невысокой численности горалы сравнительно неплохо обеспечены кормом в течение всего года. Летом запасы пищи практически не лимитированы - 29,5 ц/га фитомассы в воздушно-сухом состоянии, зимой они уменьшаются до 5-8 ц/га. В типичных прибрежных местообитаниях даже при высокой плотности населения (около 40 особей на 100 га) общий запас зимних кормов превышает потребности животных минимум в 1,5 раза [Вейнгер, 1963; Шаульская, 1992]. Тем не менее в многоснежные зимы на небольших участках обитания доступная растительность бывает почти полностью стравлена, и звери вынуждены голодать [Бромлей, 1963].
Горалы, как и другие копытные, нуждаются в минеральных веществах. На морских побережьях они иногда жуют выброшенные на берег ламинарию и взморник морской (возможно, случайно), лижут кристаллы соли, образовавшиеся на различных выбросах и предметах, а в глубине материка регулярно посещают сухие солонцы и минерализованные водные источники. Водопоями служат многочисленные родники и горные ключи, зимой звери едят снег [Бромлей, 1963, 1977а; Вейнгер, 1963; Волошина, Мысленков, 1989; Мысленков, 1989].
Суточный ритм полифазный. Летом животные пасутся 4-5 раз и столько же отдыхают. В жаркие дни периоды пастьбы сдвигаются на сумеречные и ночные часы. В дождливую погоду горалы более подвижны. Время пастбищной активности летом не превышает 50% светлого времени суток. Пик активности отмечен в июне, что связано с вегетацией растительности и физиологическим состоянием животных. Зимой в течение дня два-три длительных периода кормежки, пастьба
занимает 62-78% дневного времени. На пастбище звери передвигаются медленно (60 м/ч зимой и до 240 м/ч весной), часто меняя направление движения. Нередко они встают на задние ноги, чтобы дотянуться до верхних побегов, или срывают растения, стоя на коленях, если находятся на крутом склоне. При выборе корма используют зрение и обоняние. Пережевывание пищи происходит на лежке или, реже, стоя. Продолжительность жвачки увеличивается к концу лета, что связано с огрублением кормов. Пастбищная активность членов одной группы не отличается синхронностью: часть их может отдыхать, когда другие кормятся [Бромлей, 1963; Вейнгер, 1963; Юргенс, 1963; Волошина и др., 1976, 1992; Волошина, 1988].
Размножение
Половая зрелость. Самки и самцы горала вступают в размножение в возрасте 2,5 лет. В вольерах в гоне участвуют и 1,5-летние самцы, но в природе молодых зверей не допускают к самкам взрослые особи [Соломкина, 1986; Мысленков, Волошина, 1989].
Гон и половое поведение. Гон у амурского горала, судя по наблюдениям и срокам деторождения, весьма растянут - с сентября - второй половины октября до конца декабря, что связано с полиэстральностью самок. Его разгар приходится на середину ноября [Юргенс, 1961; Бромлей, 1963; Вейнгер, 1963]. У особей тибетско-гималайской формы (N. g. griseus), содержавшихся в Пражском зоопарке, спаривание имело место в конце сентября - октябре. В норме у самок один астральный цикл, но в случае неоплодотворения в вольерах наблюдали до 4-5 течек, следующих через 18-25 дней. Самец держится с самкой от 4 до 6 дней. В период гона он может покрыть до 6 самок [Мысленков, 1975; 1976, 1982а; Мысленков, Волошина, 1978, 1989; Соломкина, 1989].
Половое поведение стереотипно. Приподняв хвост, самец быстро побегает к самке, вытягивается и передней ногой или попеременно обеими ногами 3-5 раз “поглаживает” разные части ее тела, затем пятится назад, опускает хвост и замирает на 20-40 с. Самка в ответ принимает позу угрозы или убегает. Самец образует пару с наименее агрессивной партнершей, продолжая ритуал ухаживания и фле-муя после ее уринации и дефекации. Эти элементы полового поведения нередко проявляются и вне периода гона, особенно весной: самец обнюхивает свежую мочу самки и иногда устремляется в погоню за ней. Если в моче обнаруживаются вещества, указывающие на близость эструса, он, путем ухаживания и многочисленных ударов передней ногой, вынуждает самку мочиться, после чего долго флемует. Сигналом о готовности самки к спариванию служит частое принудительное мочеиспускание и поднимание хвоста, и бурный флемен партнера. Резкое уменьшение числа флемований - признак начала копуляции. Перед копуляцией самец вылизывает аногенитальную область самки и ее ноги до копыт в местах, куда стекает астральная жидкость. При этом для партнера типичны следующие элементы поведения: замирание позади партнерши, быстрое подбегание к ней, поглаживания ногой, вылизивание аногенитальной области, флемен, вспрыгивание, движения тазом, вылизывание спины, интромиссия, эякуляция. Максимальное число копуляций за день - 17. Садки следуют в среднем через 100 с, и каждая из них длится 2-5 с. Самка в промежутке между садками нередко пасется, и пара при этом переме
щается по кругу. Течка длится 10-16 ч, изредка до 30 ч. Если эструс у нескольких самок наступает одновременно, то самец пытается крыть их поочередно [Волошина, 1976а, 1988; Мысленков, Волошина, 1978, 1989; Мысленков, 1982а].
Беременность и развитие эмбрионов. Беременность у горала длится 212-218 дней [Соломкина, 1986, 1989], по другим данным - 225-231 (в среднем 228) день [Волошина, 1986, 1988; Мысленков, Волошина, 1989], но не 5-8 месяцев, как считалось ранее. В конце февраля зародыши вырастают до 20 см, но еще лишены волоса. К концу апреля их масса достигает 800 г при длине тела до 33 см, к началу июня - 1,9 кг и 43 см [Бромлей, 1963, 1977а].
Деторождение и выкармливание молодняка. В природе деторождение происходит с середины мая до конца июня, но иногда новорожденных регистрируют в июле - сентябре [Юргенс, 1961; Бромлей, 1963, 1977а; Вейнгер, 1963; Волошина, 1986, 1988]. Самый ранний срок появления детенышей на свет в вольерах - 22 апреля, поздний - 28 августа, а массовые роды обычны в мае - июне [Соломкина, 1989; Мысленков, Волошина, 1989].
Перед родами самка уединяется и приносит потомство в скальных убежищах. В вольерах большинство самок остаются дружелюбными даже в период родов. Некоторые из них вытягивали у рожениц послед, вместе съедали его, вылизывали новорожденных и вместе кормили их [Бромлей, 1977а; Мысленков, Волошина, 1989; Соломкина, 1989].
Мать кормит детеныша молоком от 6 до 1 раз в сутки в зависимости от возраста последнего: по своей инициативе — в период его затаивания, который длится около 24-35 дней, а затем по инициативе гораленка, неотступно следующего за ней. Частота актов сосания в день колеблется от 28 до 1. Одноразовым кормление становится через 1,5—2-месяца после родов. Лактация длится около 100 дней. Траву молодняк начинает потреблять с 10-12-дневного возраста [Соломкина, 1977; Волошина, 1986, 1988; Мысленков, Волошина, 1989].
Плодовитость. Самки горала обычно приносят одного детеныша. Сообщалось также о рождении двух и, в исключительных случаях, трех горалят [Юргенс, 1961; Бромлей, 1963, 1977а; Вейнгер, 1963], однако прямых подтверждений многодетности этих копытных в природе нет. В вольерах самки также рожают по одному детенышу, и лишь в Пражском зоопарке из 10 приплодов однажды зарегистрирована двойня, и два теленка родились в вольере Лазовского заповедника. Яловость не превышает 5%. Потенциальная плодовитость вида близка к 1 детенышу на половозрелую самку [Мысленков, 1982а; Мысленков, Волошина, 1989; Соломкина, 1989].
Популяционная структура
Половой и возрастной состав’. В вольерах Лазовского заповедника горалы принесли 11 самочек и 13 самцов [Соломкина, 1989]. Соотношение полов у детенышей, следовательно, близко 1:1. По наблюдениям в Судзухинском заповеднике в 1937 г. и 1944—1947 гг., молодые звери составляли в местной группировке 38-39, взрослые самки - 39-42, взрослые самцы — 18-22%. Соотношение полов среди взрослых особей -1:1,8-2,3 [Бромлей, 1963]. Половозрастной состав популяции Сихотэ-Алинского запо
ведника следующий: взрослые самки — 31, взрослые самцы - 13, двухлетние особи -12, годовалые - 18 и сеголетки - 26% [Мысленков, 1982а, 1986].
Социальная организация и поведение. Горал - один их немногих видов полорогих с семейно-групповым образом жизни. Семья состоит из самки с детьми этого и, нередко, прошлого года, и самца, который значительную часть года живет обособленно. Зимой соседние семьи, хорошо знающие друг друга и, как правило, имеющие близкое родство, могут кратковременно образовывать группы, насчитывающие до 5-10 и, изредка, до 11-16 голов, в которых могут находиться несколько самцов. Иногда группы до 5 голов образуют 1-3-летние звери. Взаимоотношения особей строятся по возрастному иерархическому типу с доминированием взрослого территориального самца и старых самок [Волошина и др., 1976; Мысленков, 1976, 1982; Мысленков, Волошина, 1978,1989,1997; Бромлей, Кучеренко, 1983; Волошина, 1988]. В социальном отношении образ жизни горала сходен с таковым у косуль [Данилкин, 1999], за исключением большей роли самца в семейной жизни. Стадность существенно возрастает в период гона: в октябре - 2,9, ноябре - 4,6 и декабре - 6,1 особей в группе в среднем [Мысленков, Волошина, 1975]. Зимой одиночных зверей регистрируют в 48-71% встреч, пары составляют 12-20%, группы из трех экземпляров - 15% [Глебов и др., 1980].
Горалы в природе сравнительно миролюбивы. Выяснение иерархических отношений обычно ограничивается демонстрациями. Конфликты, происходящие между взрослыми самцами на границах территорий, не простираются дальше взаимных угроз на расстоянии. Драки редки, столкновения фронтальные как у антилоп [Мысленков, Волошина, 1978, 1989; Мысленков, 1982а]. В неволе, напротив, между самцами нередки ожесточенные драки, иногда заканчивающиеся летально. Они также весьма агрессивны к домашним животным и людям [Соломкина, 19756,1989].
Эти копытные весьма доверчивы и не пугливы, подпускают человека в среднем на 88 (25-200) м, а к молодым особям можно подойти почти вплотную. При беспокойстве животные принимают ориентировочную позу, поднимают “гриву” и хвост, ударяют передними ногами о землю, издают своеобразный шипящий звук и прыжками удаляются в скальные убежища, обычно не далее 100-150 м. Лидирует при бегстве чаще взрослая самка. Основное направление бегства - вверх по склону. Отличительная особенность ориентировочно-исследовательского поведения - поиск источника беспокойства медленным шагом с твердой постановкой передних ног (“топание”), короткими высокими прыжками и с длительным стоянием (до получаса) в замершей позе на трех ногах. Нередко такое замирание происходит и после бегства. При неожиданном обнаружении явной опасности звери затаиваются: если находятся на лежке, то прижимаются к земле, если стоят, то пытаются спрятаться за любое укрытие или быстро ложатся, вытянув голову в сторону опасности [Бромлей, 1963, 1977а; Мысленков, 1982а,б; Бромлей, Кучеренко, 1983; Мысленков, Волошина, 1989; Волошина и др., 1992].
Звуковая сигнализация у горала развита относительно слабо, звуки негромкие, но разнообразные. А.И. Мысленков и И.В. Волошина [1980, 1989] выделяют 9 типов сигналов: “хав-хав-хав” - у взрослых особей и резкий дребезжащий звук “чшав-чшав-чшав” - у молодых, издаваемые круглогодично сериями из 2-40 криков при тревоге стоя или на бегу; “э-э-э-э-э” - у взрослых самцов с мая по ноябрь при мечении территории и при половом возбуждении; “ме-е-е-е” - у детенышей и самок с мая по октябрь при потере визуального контакта; “пшш” - у самцов как элемент агони
стического поведения; визг “ааааа” - у субдоминантой при нападении доминанта; “вопли”, “стоны” и “скрежет зубами” у пойманных животных и “топание”.
В спокойном состоянии эти копытные медлительны, но при возбуждении или опасности ловки и быстры. При снежном покрове до 35 см прыгают в длину на 2-3 м, в высоту без разбега - на 2 м. В скалах передвигается стремительно, как бы рикошетируя от выступов [Бромлей, 1963; Вейнгер, 1963; Бромлей, Кучеренко, 1983].
Пространственная структура и перемещения. Горал считается оседлым и малоподвижным видом. Каждая группировка имеет свой район обитания, поделенный самцами на охраняемые территории площадью 22-55 га, на которых находятся 2^4 взрослых самки с детенышами и несколько полувзрослых особей. Участки обитания самок существенно меньше (в среднем около 6 га) и широко перекрываются, а некоторые включают части территорий соседних самцов. Самки, в отличие от самцов, не охраняют свои участки, но их границы поддерживаются взаимным избеганием контакта. Территориальные самцы в период гона явно имеют больший репродуктивный успех, нежели нетерриториальные. Зимой территориальная система отчасти утрачивает свое значение. Меченые в природе, выпущенные на свободу или сбежавшие из вольер животные обычно не перемещаются далее 2 км. Тем не менее вид расселяется, и отдельных зверей регистрируют за десятки километров от мест обитания, особенно осенью в период гона. Эмигрантами чаще всего становятся молодые самцы, которых вытесняют взрослые особи [Юргенс, 1963; Мысленков, 1982а; Бромлей, Кучеренко, 1983; Волошина, Мысленков, 19926].
После пожаров и в особенно многоснежные зимы животные могут покидать свои участки обитания. В апреле 1946 г. Г.Ф. Бромлей [1963] наблюдал переход горалов из-за пожара со склонов бухты Пашагоу на гору Горал, где образовалось укрупненное стадо. Он же отметил проникновение одного зверя с материка по льду на о. Петрова и возврат его назад через 15 дней. При высоте снега в 50 см зарегистрирована перекочевка группы из 17 особей с западного склона Партизанского хребта на восточный. По опросным сведениям, с морского побережья по первому снегу эти копытные уходят вверх по долинам рек Зеркальная и Аввакумовка на 50-70 км [Нестеров, 1992].
Свои территории горалы (самцы и самки) маркируют секретом кожных желез, в том числе специфической зароговой, при трении лбом, теменем и участком у основания рогов о деревья, кустарники и скальные выступы. Пахучими метками служат также места уринации и дефекации на земле. Порция пахучих экскрементов, оставляемая в постоянных “уборных”, обычно значительно меньше, чем при обычном опорожнении кишечника. Зароговая железа используется для мечения территории круглогодично. Вне периода гона самки метят ей в 1,5-2,6 раза чаще, чем самцы. На стволах тонких деревьев рогами сдирается кора на высоте 25^47 см от земли (что ниже, чем у деревьев, маркируемых в тех же биотопах самцами косуль). Такие деревья служат не только ольфакторной, но и оптической меткой. На следах зверей, скорее всего, остается и секрет межпальцевых желез, который обильно выступает на удлиненных волосах межпальцевого углубления. Секрет этой железы самцы, возможно, целенаправленно используют и для мечения самок, проводя передней ногой по их телу. Интенсивность мечения территории резко возрастает во время гона и после дождей. Плотность меток порой достигает 6 на 1 м2. На 5 м тропы может размещаться до 15 меток на деревьях. В некоторых местах кучи экскрементов располагаются через 5-15 м, мочевые пятна — через 5-30 м, а между пятнами остаются
капельки мочи. На 1 га территории приходится от 33 до 66 уборных, размеры которых порой достигают 2,5 м2 при толщине слоя экскрементов до 10 см. Иногда один и тот же ствол дерева могут поочередно маркировать несколько членов группы. Если на метке обнаружен запах самки, то самец флемует. При мечении самцы издают негромкий звук, напоминающий меканье [Бромлей, 1963; Волошина, 19766,1982,1988; Мысленков, Волошина, 1978, 1989; Волошина и др., 1982; Мысленков, 1982а; Соколов и др., 1992; Соколов, Чернова, 2001].
Лежки горалов располагаются чаще на открытых склонах сопок, на скальных выступах с хорошим обзором, а в непогоду - в нишах, изредка - на лугах под деревьями и в густых зарослях. Многие лежки долговременные, и ими пользуются разные особи, но приоритет имеют доминанты. В удобных укрытиях экскременты лежат слоем до 15-18 см на площади в 1-2 м2. На лежке звери многократно меняют позы, нередко встают и разворачиваются на 180° [Вейнгер, 1963; Юргенс, 1963; Волошина и др., 1992].
Численность и основные факторы, ее определяющие
Численность. До середины XIX в. численность горала на Дальнем Востоке определялась “многими тысячами”. В конце XIX - начале XX вв. здесь обитали, вероятно, не менее 2 тыс. животных. Группы из 20-30 особей не были редкостью вплоть до 20-х годов. К середине XX в. по мере роста людского населения и браконьерства, участившихся пожаров, периодического многоснежья и появления волка поголовье вида сократилось до 400-500 экземпляров. Наиболее крупная группировка сохранялась в Судзухинском заповеднике, где в 1937-1959 гг. регистрировали от 116 до 200 особей [Юргенс, 1961, 1963; Бромлей, 1963; Кучеренко, 1976; Бромлей и др., 1978]. В 60-80-е годы на юге Дальнего Востока насчитывали около 500-750 голов. В Сихотэ-Алинском заповеднике жили 60-150, в Лазовском и его окрестностях - 120-210, в скальниках у истоков рек Бикин, Уссури, Арсеньевка, Максимовка, Малиновка, Милоградовка и др. - 120, в Хасановском районе Приморского края - 30, на Малом Хингане - 10-15 зверей. В труднодоступных районах отдельные группы, возможно, остались не учтенными [Вейнгер, 1963; Костог-лод, 1973; Пикунов и др., 1973; Смирнов, 1973; Бромлей, 1977а,б; Глебов и др., 1980; Глебов, 1981; Нестеров, 1981; Волошина и др., 1986; Мысленков, 1986; Волошина, Мысленков, 1990]. В 90-е годы население горала на Дальнем Востоке, скорее всего, увеличилось до 700-900 голов. Локальные группировки в Сихотэ-Алинском и Лазовском заповедниках насчитывали 180 и 250 экземпляров соответственно [Волошина, Мысленков, 2001], но в большинстве других районов они не превышали 10-50 голов.
Численность горала в Северном Китае и Северной Корее неизвестна. В Южной Корее к началу XXI в. осталось около 250 особей (Lee, Rhim, 2002).
Воспроизводство. Уровень воспроизводства у этого вида относительно стабилен - чуть меньше 1 детеныша на взрослую самку. Зимой на каждую из них приходится 0,8 сеголетка, что свидетельствует о сравнительно низкой детской смертности; До года доживает 75% сеголеток [Мысленков, 1982а; Мысленков, Волошина, 1989].
Погода и корма. Один из основных факторов смертности горала - высокий снежный покров. При его высоте в 35-40 см эти копытные становится беспомощными, не
могут передвигаться и добывать корм, и сильно истощаются. В многоснежные зимы, которые в ареале вида, к счастью, не часты, в отдельных группировках регистрировали гибель до 8% особей, из них до 50% - взрослые самцы. Массовые падежи копытных отмечены в 1914/15, 1941/42 и 1947/48 гг. [Бромлей, 1963, 1977а; Мысленков, Волошина, 1989].
Хищники. Горал, несмотря на обитание в труднодоступных скалах, зимой довольно часто становится жертвой волка при выходе на кормежку на пологие склоны. В некоторых группировках от этого хищника гибнет от 3 до 18% особей. При появлении волков копытные обычно стараются не покидать скальных убежищ. Иногда они становятся жертвами леопарда, тигра, рыси, харзы, белохвостого и белоплечего орланов и беркута. В Сихотэ-Алинском заповеднике доля хищников в общей смертности вида составляет около трети, причем наибольший урон причиняет рысь, примерно столько же приходится на болезни и 14% - на браконьерство [Абрамов, 1963; Бромлей, 1963, 1977а; Вейнгер, 1963; Юргенс, 1963; Кучеренко, 1976; Бромлей, Кучеренко, 1983; Мысленков, Волошина, 1989].
Конкуренты. Своеобразные скальные местообитания горала непригодны для других копытных Приморья, поэтому кормовых конкурентов у него зимой практически нет, а летом при обилии корма конкуренция не ощутима. В дубняках и на пологих травянистых склонах за нижней границей леса он контактирует с изюбрем и пятнистым оленем, а в некоторых местах занимает те же участки, что и сибирская косуля и кабарга [Бромлей, 1963; Волошина и др., 1976, 1982; Бромлей, Кучеренко. 1983].
Болезни, паразиты, травмы. Массовая гибель горала от инфекционных болезней в естественных условиях не отмечалась, что вполне объяснимо как обособленностью его скальных местообитаний, малочисленностью и отсутствием тесного контакта с другими животными, так и малоизученностью. У вида зарегистрированы следующие болезни: бронхопневмония, инфекционный пиелонефрит, цистит, мочекаменная болезнь, рак почки, энтероколит, спайки кишечника, цирроз печени, воспаление головного мозга, эмфизема плода, эндометрит, сепсис, остеодистрофия и др. Среди них наиболее губительна бронхопневмония, от которой в Московском зоопарке умерли девять из десяти особей, в Пражском - три из восьми. Иногда звери погибают от травм, полученных при падении со скал. Большие потери от травм, миопатии и паралича конечностей случаются и при отлове животных [Бромлей, 1963; Волошина, Соломкина, 1992].
В природе у этого копытного отмечен 21 вид гельминтов, но интенсивность инвазии невелика (1-150 экземпляров) в отличие от особей, содержащихся в зоопарках, у которых эндопаразиты найдены в большом количестве (до 817 экземпляров) и разнообразии (26 видов), включая экзотические. Всего список гельминтов содержит 30-33 вида, из которых 9 присущи только горалу. Наиболее патогенны легочные стронгиллиды, способствующие проявлению бронхопневмонии [Бромлей, 1963; Бромлей, Кучеренко, 1983; Волошина, 1988; Волошина, Мысленков, 1989; Волошина, Хрусталев, 1992].
Эктопаразитами (клещи родов Ixodes, Dermacentor и Haemaphysalts, власоед, оленья кровососка и другие кровососущие насекомые) горал в силу специфичности местообитаний поражен в меньшей степени, нежели другие виды копытных [Бромлей, 1963, 1977а], исключая, пожалуй, клеща Haemaphysalis japonica, самцы
которого зимуют на нем. Максимальное количество клещей на одном звере в марте (282), минимальное - в сентябре (45) [Волошина и др., 1980].
Антропогенные факторы. Наибольший вред популяциям причиняет человек. На юге Дальнего Востока горал издавна был в числе излюбленных объектов охоты. Зверей добывали, главным образом, ради красивых пышных шкур, высококачественного мяса и в целях изготовления ценных лекарственных препаратов из рогов и отдельных частей тела. В тибетской медицине находила применение вся туша, из которой варили лекарственный “холодец”.
В СССР из-за крайне низкой численности отстрел горала запрещен с 1924 г. Однако нелегальная добыча имела место всегда и везде. Основной урон виду браконьеры наносят вблизи населенных пунктов и на морских побережьях. В 1944 г. вскрыт случай незаконного отстрела сразу 6 особей. Во многих местах эти доверчивые звери уничтожены полностью [Бромлей, 1963, 1977а,б; Вейнгер, 1963; Волошина, Мысленков, 1977].
Хозяйственная деятельность человека пока что почти не сказывается на труднодоступных скальных биотопах копытных.
Перспективы сохранения
Амурский горал занесен в Красный список МСОП, Приложение 1 СИТЕС, Красную книгу РФ (2001). Перспективы его сохранения как вида фауны России достаточно высоки. Он неплохо охраняется в 3 заповедниках и 8 заказниках Дальнего Востока [Нестеров, Смирнов, 1981; Волошина, Мысленков, 2001], но в ближайшие десятилетия его численность вряд ли заметно увеличится из-за ограниченности охраняемых мест обитания, наличия волка и браконьерства.
При малой численности горал не будет объектом охоты, хотя многие коллекционеры хотели бы иметь такой трофей. Более важен он как неотъемлемый элемент фауны Дальнего Востока, как объект научных исследований и демонстраций в зоопарках и зверинцах.
Этот вид, к сожалению, плохо переносит содержание в неволе. В 1937 г. в Московском зоопарке через 8 месяцев погибли все 10 особей, доставленных из Судзу-хинского заповедника. В 1984 г. из 4 зверей, отловленных в Лазовском заповеднике, через год погибли 2. В Сихотэ-Алинском заповеднике из 16 особей вскоре после отлова погибли 5, но уцелевшие принесли потомство. В Лазовском заповеднике в 1973-1990 гг. содержали 47 горалов. Здесь из 39 детенышей погибли 15, из взрослых - 8 [Бромлей, 1963; Волошина, Соломкина, 1992].
К настоящему времени методики отлова [Глебов, Соломкина, 1983] и разведения этих копытных в вольерах, сооруженных в природе в ареале вида, разработаны весьма неплохо. Специализированные фермы, в какой-то мере, могли бы быть гарантами сохранения генофонда, а парковые животные - материалом для реинтродукции [Соломкина, 1977,1980, 1981, 1983а-в; Волошина, 1981, 1988; Соломкина, Глебов, 1981; Храмцов, 1986; Мысленков, Волошина, 1989; Волошина, Мысленков, 19926,2001]. ,
Род СЕРНЫ
Rupicapra Blainville, 1816
Rupicapra Frisch, 1775. Natur-Syst. vierf.
Thiere: 2. Nom nudum.
Rupicapra de Blainville, 1816. Bull. Sci.
Soc. Philom. Paris: 75. Тип рода -Capra rupicapra Linnaeus.
Capella Keyseriing et Blasius, 1840.
Wirbelth. Europas, 4: 28.
Cernas Gloger, 1841. Naturgesch., 1: 153.
Nom. nov. pro Rupicapra de Blainville. Non Oken, 1816.
Caprina Wagner, 1844. Schreber Saugeth., Suppl., 4: xi, 457. Nom. praeocc., non d’Orbigny, 1822.
Характеристика. Серны -мелкие стройные животные с тонкой длинной шеей (рис. 32). Длина тела достигает 135 см, высота в холке - 90 см, масса -65 кг. Конечности относительно высокие за счет удлинения костей голени, плюсны и берцовой кости, что не типично для козлиных [Гаспарян, 1969, 1976]. Хвост короткий, около 12 см, с безволосой нижней поверхнос
Рис. 32. Серны (фото А. Щеголева)
тью. Носовое зеркало неболь-
шое. Половой диморфизм выражен слабо.
Окраска контрастная, темно- или черно-бурая с посветлениями, летом с желтизной или ржаво-рыжая. Голова заметно светлее туловища. На морде от угла рта до основания уха тянется темная полоса. Самцы несколько темнее самок. Бороды и околохвостового зеркала нет. Волосяной покров одинаковой длины по всему телу, за исключением полоски удлиненного зимнего меха по хребту.
Череп с большим углом излома оси и сильно вогнутым профилем лицевой части (рис. 33). Мозговая часть хорошо развита. Верхнечелюстные кости соприкасаются с носовыми, межчелюстные не достигают носовых. Слезные кости не доходят до входа в носовое отверстие. Сошник иногда срастается с небными костями в передней трети. Орбиты трубкообразные. Предглазничные ямки отсутствуют, этмоидальные щели с возрастом зарастают. Кости черепа слабо пневматизированы. Роговые стержни вертикальные, расположены над глазницами, их основания сближены. Синусы имеются лишь в нижней трети. Резцы слабо гипсодонтные. Третий предкоренной
Рис. 33. Череп самца серны (экземпляр из Кавказского заповедника; Зоомузей МГУ, коллекционный № 38008)
Вид сбоку, сверху и снизу
зуб присутствует в верхней и нижней челюстях. Стили на верхних коренных выражены слабо.
Рога у представителей обоих полов, но у самцов они чаще несколько длиннее - до 28-34 см. Основания рогов прямые, вершины загнуты назад и вниз. Кольца на чехлах слабо выражены, вершины гладкие, кили и ребра отсутствуют. В поперечном сечении рога округлые. Цвет их черный.
Копыта своеобразные: с жестким и острым краем, опущенной вниз передней частью и слегка вогнутой мягкой подошвой, что препятствует скольжению. Половинки передних копыт могут раздвигаться более чем на 90°, что, вероятно, способствует торможению на спусках. Боковые пальцы сильно развиты.
Имеется только зароговая и межпальцевые железы [Соколов, Чернова, 2001]. Сосков 2 пары.
Диплоидное число хромосом -58, NF = 60.
Горные животные, предпочитающие скальные участки альпийской зоны и верхнюю границу леса. Ведут групповой образ жизни. Обладают прекрасным обонянием, слухом и зрением. Необычайно ловки, быстры и смелы при передвижении в скалах и осыпях. Прекрасные прыгуны в длину (до 11,3 м [Гамбарян, 1972]) и высоту. На ровной местно-
сти скорость бега невелика - около 30-40 км [Гаспарян, 1969], что делает этих
копытных сравнительно легкой добычей хищников. Максимальная продолжительность жизни - 17-20 лет.
Преимущественно охотничьи звери.
Распространение. Горные области Центральной и Южной Европы и Малой Азии. Современный ареал очаговый: Пиренеи, Альпы, центральная часть Апеннин, Балканы, Карпаты, горы в Турции, Кавказ (рис. 34). Акклиматизированы в Новой Зеландии.
Эволюция. Предковой формой серн считается верхнеплиоценовый род Procamptoceras Schaub, 1923, представители которого имели вертикально поставленные рога и сходное строение посткраниального скелета. Происхождение рода, возможно, азиатское, откуда через горы Эльбурс звери распространились до Кав-
Рис. 34. Ареал рода Rupicapra (по [Lovari, Scala, 1980]) Цифрами обозначена локализация разных форм
каза, и через Малую Азию и существующий перешеек Дарданеллы - Босфор вышли в Западную Европу, где в ледниковое время происходило интенсивное формообразование [Деннлер, 1975; Lovari, Scala, 1980]. В Европе ископаемые формы Rupicapra известны из среднеплейстоценовых отложений [Kurten, 1968]. Здесь же обычны позднеплейстоценовые и голоценовые находки в горных и предгорных районах, включая Бельгию, Францию, Германию и Италию. Позднеплейстоценовые остатки этих животных найдены также в Карпатах, в бассейне Днестра (?) и в Закавказье. На Северном Кавказе и в Малой Азии пока известны лишь голоценовые остатки [Громов, 1948; Громова, 19486; Верещагин, Наниев, 1949; Пидоплич-ко, 1951, 1954; Верещагин, 1959а; Соколов, 1959; Baryhsnikov, Tikhonov, 1993].
Род Rupicapra четко обособлен в подсемействе козлиных. Ближайшая родственная группа - Caprini.
Своеобразные животные, адаптированные к обитанию в скальниках, однако могут жить почти на равнинных участках.
Состав. Род считается монотипичным. Тем не менее в нем иногда выделяют 2 вида: Rupicapra pyrenaica Bonaparte, 1845 (Пиренеи и Апеннины) и Rupicapra rupicapra L., 1758 [Masini, Lovari, 1988]. Обособленность этих копытных подтверждается молекулярно-генетическими исследованиями: средняя нуклеотидная дивергенция гаплотипов составляет 0,56%, время дивергенции мтДНК - около 280 тыс. лет [Hammer et al., 1995]. Не исключено, однако, что этот уровень расхождения форм всего лишь подвидовой.
СЕРНА
Rupicapra rupicapra Linnaeus, 1758
Capra rupicapra Linnaeus, 1758. Syst. Nat., 10 ed., 1: 68. Швейцария.
Rupicapra tragus Gray, 1843. ListMamm. Brit. Mus.: 167. Nom. nov. pro Capra rupicapra Linnaeus.
Rupicapra capella Bonaparte, 1845. Atti Sci. Ital. Milano, 6: 377. Norn. nov. pro Capra rupicapra Linnaeus.
Rupicapra europaea Comalia, 1871. Fauna Ital., 1: 53. Nom. nov. pro Capra rupicapra Linnaeus.
Rupicapra dorcas Schulze, 1897. Helios, 14: 81. Nom. nov. pro Capra rupicapra Linnaeus.
Rupicapra tragus caucasica Lydekker, 1910. Ward’s Rec. Big Game, 6 ed.: 338. Краснодарский кр., Кавказский заповедник.
Rupicapra rupicapra carpatica Couturier, 1938. Le Chamois: 369. Румыния, Южные Карпаты (“Трансильванские Альпы”), горы Ретезат.
Характеристика. См. характеристику рода.
Распространение. То же, что и у рода, возможно, за исключением Пиренеев и Апеннин.
Состав. В мировой фауне насчитывают до 11 подвидов Rupicapra rupicapra (рис. 34): rupicapra, caucasica, parva, asiatica, balcanica, olympica, cartusiana, carpatica, tatrica, а также pyrenaica и ornata, которых иногда относят к R. pyrenaica [Соколов,
Таблица 36. Размеры (см) и масса (кг) взрослых особей кавказской серны
Самцы Самки Район, заповедник (источник информации)*
п М lim n 1 м lim
3 118 Длина тела 115-121 4 121 118-130 Краснодарский край (2)
8 120,7 115-132 — — — Кавказский заповедник (4)
5** 116 111-127 ** — Закатальский заповедник (5)
2 81 Высота в холке 80-82 3 82 78-86 Кавказ (3)
3 84 81-90 4 80 77-84 Краснодарский край (2)
8 88 — 3 77 — Кавказский заповедник (4)
1 - 90 — - — Тебердинский заповедник (1)
5** 83 80-87 ** — Закатальский заповедник (5)
1 — Обхват туловища 83 I 80 Краснодарский край (2)
1 — 79 — — — Тебердинский заповедник (1)
8 90 — 3 80 - Кавказский заповедник (4)
3 37 Длина ступни 34—40 4 38 36-40 Краснодарский край (2)
2 13 Длинауха 12-14 3 13,5 12,5-14 Кавказ (3)
3 13 13-13,5 4 13 13-14 Краснодарский край (2)
4** 13,5 12,5-14,6 ** — Закатальский заповедник (5)
3 10 Длина хвоста 9-11 4 11 9-12 Краснодарский край (2)
— — 35-40 Масса — — Кавказ (3)
2 29,0 27-31 2 36 31-41 Краснодарский край (2)
8 36,2 — — — — Кавказский заповедник (4)
2 — 31-43 3 30 26-34 -“-(6)
1 — 41 — — Тебердинский заповедник (1)
*1 - Лопырин и др., 1960; 2 - Котов, Рябов, 1963 (включая данные И.И. Соколова); 3 - Динник, 1910; Верещагин, 1947; 4 - Котов, 1964в; 5 - Попкова, 1967; 6 - Ромашин, 2001.
** Самцы и самки вместе.
И.И. 1959; Гептнер и др., 1961; Громов и др., 1963; Haltenorth, 1963; Деннлер, 1975; Соколов В.Е., 1986]. В действительности число подвидов гораздо меньше.
В фауне России - кавказская серна (Rupicapra rupicapra caucasica).
Изменчивость
Размеры и масса. Половой диморфизм у кавказской серны выражен слабо. Самки лишь немного уступают самцам в размерах и массе тела. Длина тела взрослых особей достигает 132 см, высота в холке - 90 см, масса - 43 кг (табл. 36). Кавказская серна, возможно, несколько мельче европейских рас. Масса тела у R. rupicapra carpatica, например, достигает 60-65 кг [Koubek cl al., 1985]. Однако имеющийся морфометрический материал слишком мал для однозначных выводов.
Краниометрическая характеристика. Молодые самцы и самки практически не отличаются по краниометрическим показателям. Черепа взрослых самцов в среднем чуть длиннее и шире в лобной части. Наиболее весомый половой отличительный признак-длина лицевого отдела [Попкова, 1967; Koubek, Hrabe, 1983; Koubek et al., 1985, 1989; Тихонов, 1995].
Максимальная длина черепа кавказской серны не превышает 226, максимальная ширина - 118, длина верхнего ряда зубов - 64 мм (табл. 37). Эта форма по некоторым показателям достоверно отличается от номинативной R. г. rupicapra, однако краниологические различия между ними в целом весьма незначительны [Koubek, Hrabe, 1983]. Последняя, в свою очередь, отличается по ряду средних краниометрических показателей от расы carpatica, максимальная длина черепа которой достигает 230, кондилобазальная - 205, межгпазничная - 120 мм, и от tatrica [Hrabe, Koubek, 1984; Koubek et al., 1985]. Наименьшие размеры черепа и сближенные основания рогов
Таблица 37. Размеры черепа (мм) взрослых особей кавказской серны
Самцы Самки Источник информации
п М | lim п | М | lim
14 216,7 Наибольшая длина 210-222 5 212,6 211-214 Соколов, Темботов, 1993
26 216,5 206-226 11 210,3 201-220 Koubek, Hrabe, 1983
14 111,5 Максимальная ширина 104-118 5 106,6 105-107 Соколов, Темботов, 1993
28 110,0 102-118 И 106,7 98-117 Koubek, Hrabe, 1983
15 77,2 Межглазничная ширина 73-82 5 73,5 70-77 Соколов, Темботов, 1993
28 79,3 75-85 12 74,7 71-80 Koubek, Hrabe, 1983
14 122,7 Длина лицевой части 122-133 5 125,1 125-126 Соколов, Темботов, 1993
27 126,3 117-136 11 121,3 115-127 Koubek, Hrabe, 1983
15 72,9 Длина носовых костей 63-78 5 72,3 70-76 Соколов, Темботов, 1993
28 73,4 62-82 11 70,5 65-78 Koubek, Hrabe, 1983
15 58,6 Длина верхнего ряда зубов 53-64 5 56,9 54-60 Соколов, Темботов, 1993
30 59,0 52-64 13 59,7 55-62 Koubek, Hrabe, 1983
Таблица 38. Размеры рогов (см) у взрослых особей кавказской серны
п Длина Размах Обхват у основания Источник информации
М | lim М lim М I lim
Самцы
- 20-22 -24 - 4-10 7,5 9,3 Динник, 1910
12-15 20,8 19-24 7,7 5-10 8,2 6-9 Соколов, Темботов, 1993
22 20,3 17-24 8,2 7-9 Koubek, Hrabe, 1983
12* 20 — — — - — Попкова, 1967
Самки
- - -23 - - 6- Динник, 1910
4-5 19,3 18-20 7,2 6-8 7,0 6-9 Соколов, Темботов, 1993
10 18,9 15-22 7,3 6-9 Koubek, Hrabe, 1983
* Самцы и самки.
характерны для форм pyrenaica и omata с Апеннин, a cartusiana занимает промежуточное положение между ними и другими подвидами [Lovari, Scala, 1980; Scala, Lovari, 1984]. Популяции из Болгарии по краниометрическим признакам относят к R. г. balcanica [Markov et al., 1998]. При внимательном анализе приведенных в литерату
Рис. 35. Черепа серн с уродливым и одиночным рогами (фото С. Тишкевича)
ре данных, прослеживается кли-нально-зональная изменчивость вида, и очевидна несостоятельность выделения многих подвидов.
Изменчивость рогов. В первый и второй годы жизни рога у серны отрастают на 6-7, в третий - на 3-4, в четвертый - на 1,5-2, в последующие годы-на 0,5—0,1 см. У самцов они в среднем чуть длиннее и массивнее, чем у самок, однако их длина, развал, а также диаметры роговых чехлов и стержней могут служить д ля определения пола только у одновозрастных животных. Размеры рогов особей из разных регионов Европы варьируют, но географическую изменчивость нельзя назвать существенной. Длина рогов у кавказской серны не превышает 24 см (табл. 38), тогда как у альпийской достигает 28-34 см [Динник, 1910; Попкова, 1967;Koubek,Hrabe, 1983, 1984; Lovari, Scala, 1984; Hrabeetal., 1986;Masseietal., 1994].
Рога у серн, несмотря на маленькие размеры, очень прочные. Они часто остаются целыми даже
после падения животных с большой высоты [Динник, 1910]. Тем не менее в природе иногда встречаются звери с уродливыми после травм рогами, и, в исключительных случаях, однорогие экземпляры (рис. 35).
Окраска и линька. Половой диморфизм в окраске выражен слабо, но возрастная, сезонная, индивидуальная и географическая [Деннлер, 1975] изменчивость серн весьма высока. Окраска зверей контрастная, неодинаковая зимой и летом, что связано с неравномерным ростом разных категорий волос во время продолжительной линьки, протекающей с марта - апреля до июля - августа. Новый волос достигает максимальной длины к ноябрю.
Зимний наряд кавказской серны черно-бурый, грива черная. Голова и шея светлее туловища. Горловое светло-рыжее пятно отделено черно-бурой полосой. Глаза обрамляет бело-желтое кольцо. Грудь, задняя сторона бедер и основание хвоста светло-бурые, живот светло-рыжий. Летняя окраска ржаво-рыжая. По спине от затылка до конца хвоста тянется черно-бурая полоса. Грудь коричневая или темнобурая, брюхо светло-желтое или ржаво-желтое, круп желто-рыжий, светлее боков, передние ноги спереди почти черные, задние - бурые. Окраска головы контрастная. Горловое пятно большое, заходящее на нижнюю челюсть, светлое. От угла рта до основания ушей идет узкая темно-бурая полоса. Половой диморфизм практически не выражен, хотя у самцов в целом окраска несколько темнее и более очерчены светлые полосы на морде. Молодые животные серовато-бурые или рыжеватые с темным ремнем по спине. Цвет меха у них менее яркий и контрастный по сравнению с взрослыми. Лицевой узор, тем не менее, отчетливо выражен [Динник, 1896, 1910; Гептнер и др., 1961; Попкова, 1967; Дормидонтов, Блохин, 1977; Соколов, Темботов, 1993].
Генетическая изменчивость. В кариотипе 58 хромосом, число плеч аутосом -58. Одна пара аутосом метацентрическая, остальные и Х-хромосома - акроцентрические. Y-хромосома точечная двуплечая [Орлов, Булатова, 1983]. Уровень генетической изменчивости у серны один из наиболее высоких среди крупных млекопитающих. В 5 альпийских популяциях полиморфными оказались 10 локусов из 42. Их доля варьировала от 0,095 до 0,167, составляя в среднем около 0,140, среднее число аллелей на локус - 1,192. Средняя гетерозиготность на особь равна 0,041 [Miller, Hartl, 1986, 1987].
Гибридизация. Достоверных случаев гибридизации серны с другими видами козлиных не известно. Опыты скрещивания самца этого вида с козами и овцами в Те-бердинском заповеднике дали отрицательный результат. Лишь у одной из овец была обнаружена яйцеклетка с вклинившимися спермиями [Лопырин и др., 1959, 1960].
Распростр анение
В историческое время кавказская серна была распространена по всему Главному Кавказскому хребту, его отрогам и горам Малого Кавказа. Еще столетие назад западная граница ее ареала на Главном хребте проходила по перевалу Гойтх и горе Семешхо в верховьях Пшиша, полосе Черных гор и прибрежных скал. В многоснежные годы животные появлялись вблизи Новороссийска и Сочи (на горе Ахун) [Васильев, 1896]. Крайний пункт распространения на востоке - гора Бабадаг в Азербайджане. Многочисленные места обитания этих животных в прошлом тщательно
перечислены в публикациях замечательных ученых: Н.Я. Динника [1896, 1910], Г.И. Радде [1899], А.А. Насимовича [1936а,б, 1939] и Н.К. Верещагина [1959а].
В XX в. ареал кавказской серны постепенно уменьшался, становился очаговым (рис. 36), а некогда фоновый вид превратился в редкий [Гинеев и др., 1988; Соколов, Темботов, 1993]. На Северо-Западном Кавказе численность и область обитания этого копытного заметно сократились в период гражданской войны, но в 30-е годы животных еще регистрировали на Черноморском побережье, в районе Бакишеевско-го и Грачевского перевалов на Главном хребте, на горах Аутль и Бзыч, в верховьях Дагомыса и Агуа, на правобережье р. Ажу, по р. Бзыч, на горе Амуко, хребтах вблизи Мацесты и Ахну, на горах Кепши, Сапун, по берегам р. Чвежипсе, на горе Индюк, у перевала Гойтх, в окрестностях Псебая и Бесленеевской и в других районах. В середине столетия северная граница распространения вида проходила (с востока на запад) близ сел и станиц Соленое, Бугунжа, Даховская, Самурская, Безымянное, Крепостная, Эриванская и Шапсугская, южная - у населенных пунктов Адербиевка, Михайловский перевал, Пшада, Дефановка, Георгиевское, Чвежипсе. Иногда зверей встречали и даже убивали в районе Майкопа и Псебая [Насимо-вич, 1936а,б, 1939; Верещагин, 1959а; Котов, Рябов, 1963]. В 60-70-е годы северная граница ареала в Краснодарском крае сместилась к югу на 25-30 км. Животные исчезли в бассейнах рек М. Лаба, Уруштен, Цице и Белая. Фактически сохранились лишь три немногочисленные группировки, находящиеся на расстоянии 100— 130 км одна от другой: в Кавказском и Тебердинском заповедниках и окрестнос-
Рис. 36. Распространение серны на Кавказе в XX в. (по [Соколов, Темботов, 1993]) Точки с цифрами - зафиксированные районы обитания. На врезке - высотное распределение
тях, а также в Пшада-Геленджикском районе [Дубень, 1977, 1983; Дуров, 1977]. В многоснежные зимы отдельные животные заходят в низкогорье на побережье. В 80-90-е годы их наблюдали в ущелье Ах-цу на р. Мзымта, у и. Марьино Лазаревского района, в Адлерском и Геленджикском районах [Ромашин, 2001].
На Центральном Кавказе в XIX - начале XX в. этот вид был обычен в пределах среднегорья и даже низкогорья. В Приэльбрусье граница ареала до последнего десятилетия XX в. проходила по северным лесистым отрогам Скалистого хребта. К концу столетия в верховьях р. Малка она отступила вверх до Бокового хребта. На Меловом и Скалистых хребтах серна уже не встречается, но по Баксанскому ущелью еще обычна от Тырныауза до Эльбруса, живет в верховьях Баксана, Чегема и Черека, а также очагами в Дигории и Осетии [Темботов, 1960, 1972; Амирханов и др., 1981; Дзуев, 20006].
Восточнее небольшая популяция оставалась в горной части Ингушетии в ущельях Шони, Ами и Гулой-лом [Точиев, 1975]. В Дагестане В.Г. Гептнер и А.Н. Формозов [1941] отмечали серну в высокогорьях Андийского и Гунибского округов, на Богосском хребте и в верховьях Самура, а Н.К. Верещагин [1959а] - на лесистом хребте Черных гор до среднего течения Аргуни, в верховьях Уруха и Ардона и на восточном склоне Шахдага. В 80-е годы этот вид населял лишь высокогорные районы республики по Главному Кавказскому хребту: Тляратинский, Чародинский, Советский, Агульский, Цумадинский и Цунтинский. Изредка животные появлялись в горах хребтов Нукатль, Дюльтыдаг и Диклосмта [Гинеев и др., 1988; При-луцкая, Пишванов, 1988].
В Грузии в начале 70-х годов серна обитала на Гагрском, Бэйбском, Кодорском, Сванетском, Двалетском и смежных с ними хребтах, в районе Хевсуретии, Борба-лийском массиве и южных склонах Кахетинского Кавкасиони на Большом Кавказе, а также в пределах Аджаро-Имеретинского хребта на Малом Кавказе в высокогорных Адигенском, Чохатаурском, Маяковском и Хулойском районах [Капанадзе, 1973, 1974]. К концу 80-х годов она занимала верхний пояс лесов и безлесные субальпы, а в Лагодехском и Боржомском заповедниках, Саирме и Окуни спускалась до низа лесного пояса [Арабули, 1989].
В Азербайджане в середине XX в. этот вид был распространен по южным склонам Большого Кавказа от притоков Гирдыманчая до границ Грузии. В 70-е годы копытные населяли преимущественно Шеки-Закатальскую зону. Небольшие очаги сохранялись на северо-востоке республики на хребтах Шахдаг и Бабадаг и на Муровдагском хребте Малого Кавказа [Алиев, 1961; Алекперов и др., 1973, 1976].
В XIX в. серна жила на севере Армянского нагорья, где исчезла, видимо, в 30-х годах XX в. [Верещагин, 1959а].
Среда обитания
Кавказскую серну считают типичным обитателем высокогорий, что не совсем верно. Ее местообитания простираются почти от уровня моря (нижних зон леса) и до 4000 м. В недалеком прошлом она в изобилии водилась в среднегорных и встречалась в низкогорных лесах, откуда была вытеснена в высокогорные стации.
На Северном Кавказе летом эти копытные встречаются во всех поясах гор, но основное поголовье сосредоточено в альпийской и субальпийской зонах и в верх
ней кромке лесной зоны, на высотах от 1500 до 2500 м. Осенью из многоснежных районов Главного Кавказского хребта звери уходят на зимовку в боковые отроги и под защиту хребтов-“ширм”, где живут в темнохвойных лесах на крутых склонах, в скальных выходах и в скалистых участках речных долин на высоте 1000-1600 м. Часть их спускается ниже, в широколиственные леса, вплоть до долин или остается на малоснежных участках в субальпийской зоне. Весной, по мере таяния снега, животные снова поднимаются в высокогорье. Типичные места обитания - крутые, но не сильно расчлененные, склоны гор, заросшие редким лесом и горные плато со скальными выходами, мелкощебнистыми осыпями, мезофитными лугами и лужайками. Лучшие биотопы - задернованные склоны с крутизной в 20-40° и скальные обнажения с карнизами и нишами, дополняемые наличием естественных солонцов [Радде, 1899; Динник, 1910; Насимович, 1936а, 1939, 1941, 19496, 1955; Аренс, 1957; Жарков, 1959; Котов, Рябов, 1963; Котов, 1969; Тарасов, 1976; Дуров, 1977; Кудактин, 1977; Амирханов и др., 1981; Вейнберг, 19836; Дубень, 1985; Бобырь, 1999; Дзуев, 20006; Ромашин, 2001].
В Грузии и Азербайджане осенью, зимой и ранней весной серна концентрируется на малоснежных склонах в среднем и верхнем поясах леса и в субальпийской зоне. Летом верхняя граница высотного размещения достигает верхнего пояса альпийской зоны, но нижняя остается почти неизменной. В заповедниках звери зимой спускаются до нижнего уровня лесного пояса и предгорий, и обычны на мезофит-ных лугах, расположенных вблизи крутых осыпей и скал [Эквтимишвили, 1952; Енукидзе, 1953; Капанадзе, 1967, 1973, 1975, 1976; Попкова, 1967; Арабули, 1989; Кулиев, 1989; Гасанов, 1990].
У серны весовая нагрузка на след (200 г/см2) меньше, чем у других козлиных, что позволяет ей относительно свободно передвигаться по плотному снегу. Животные без особого труда преодолевают рыхлый снег высотой около 30 см, замедляя движение при его слое в 40-45 см [Насимович, 1955]. В отличие от горных козлов, они нередко не откочевывают из многоснежных районов, не избегают лесов и более тесно связаны с мезофитной луговой растительностью.
Питание
Кавказская серна употребляет в пищу более 250 видов растений. Основной корм в весенне-летний период - разнотравье, из которого звери выбирают подорожник, анемоны, манжетки, овсяницы, мятлики, лютики, копеечник, белокопытник, прострел лиловый, тмин, бутень, девясил, бодяк, мытники, валериану, колокольчики. Осенью они предпочитают хлопушки, анемоны, купальницы, бутень, борщевик, мытники, цефалярии, зеленые листья буквицы и горечавки, охотно едят семена трав, ягоды черники, малины, ежевики, смородины, желуди, орешки бука и каштана, плоды фруктовых деревьев, грибы. Зимой животные копытят зимнезеленую траву и ветошь, если слой рыхлого снега не превышает 20 см. В рационе в этот период доминируют овсяницы, мятлики, щучки, иглица подлистная, ожина, ежевика, черника, омела, плющ, лавровишня, а в многоснежье - веточные корма (ива, бук, липа, ильм, граб, клен, медвежий орех, дуб, ясень, рябина, хвоя сосны, пихты и ели, шиповник, рододендрон, можжевельник и др.), древесные лишайники и мхи. В целом серны в достаточной мере обеспечены кормом практически круглогодич
но. Суточная потребность - 5-6 кг на особь. В рубцах обычно находится от 250 до 4400 г пищи [Динник, 1910; Насимович, 1939, 19496, 1955; Жарков, 1959; Попкова, 1967; Капанадзе, 1974; Дубень, 1985; Ромашин, 2001].
В Тебердинском заповеднике сернята, выращиваемые в вольерах, начинали питаться зеленым кормом с месячного возраста. Молодые звери поедали не менее 42 видов растений, наиболее охотно - листья дикой яблони, клевер, щавель кислый, клубнику, ежу сборную, костер береговой, манжетку, лабазник и язвенник [Иняко-ва, 19576].
Серны регулярно, и даже днем, выходят на естественные и искусственные солонцы, посещая их чаще в весенне-летний период, реже - поздней осенью и изредка - зимой. Динамика солонцевания согласуется с сезонным изменением питания. Тяга к минеральным источникам вызвана острым дефицитом хлористого натрия, кальция, фосфора и, возможно, железа и йода в растениях горнолуговой зоны [Насимович, 1938, 19496; Розмахов, 1940; Попкова, 1967].
Водные источники звери чаще всего посещают лишь в жару. Летом они получают достаточно воды вместе с сочным альпийским кормом, покрываемым к тому же утром и вечером обильной росой. Кроме того, в горах нередки дожди, а в высокогорье лежат снега, которые можно использовать круглогодично [Динник, 1910].
Летом эти копытные пасутся наиболее интенсивно с рассвета и до 7-11 ч, после чего уходят на дневной отдых в скальные укрытия, ниши и пещеры, в тень деревьев или на снежники. Животные, находящиеся в лесу, периодически кормятся и днем. На вечернюю пастьбу серны выходят несколько позже козлов (в 17-18 ч) и проводят на пастбище часть ночи. В пасмурную и дождливую погоду они остаются на открытых склонах весь день, чередуя пастьбу с отдыхом. Зимой звери активны преимущественно в дневное время. Суточный ритм может значительно меняться под воздействием выпаса скота и при регулярном охотничьем преследовании. По сравнению с кавказским и безоаровым козлами, серна более подвижна: на лежке животные чаще меняют позы, встают и кратковременно кормятся поблизости, чем и объясняется довольно высокий показатель дневной активности (до 40% от встреченных в это время особей) [Динник, 1910; Насимович, 1941, 19496; Жарков, 1959; Попкова, 1967].
Размножение
Сведения о размножении кавказской серны весьма скудны и однообразны [Динник, 1896, 1910; Насимович, 1939, 19496; Эквтимишвили, 1952, 19546; Инякова, 19576; Попкова, 1967]. Самки вступают в размножение чаще на втором году жизни и первого теленка приносят в возрасте 3 лет. Для альпийской серны на большом материале было показано, что в двухлетнем возрасте становятся половозрелыми 63% особей, позднее - 92,5% [Houssin et al., 1993]. В конце сентября - октябре у взрослых самцов увеличиваются зароговые железы, секрет которых имеет резкий, специфический запах. Звери становятся беспокойными, агрессивными и подвижными: они часто перебегают от одной группы самок с сеголетками и подростками к другой, сражаются с соперниками, издают звуковые сигналы, напоминающие глухое “блеяние”. Спаривание происходит с середины октября до конца ноября - конца декабря. Продолжительность беременности - около 165-170 дней. Рожают серны с середины апреля до начала июня. В Кавказском заповеднике в 1991 г. зареги
стрирован случай появление детеныша в сентябре [Ромашин, 2001]. Перед отелом самки отделяются от стада и некоторое время держатся отдельно в скалах лесной и субальпийской зон, вблизи от пастбищ и минеральных источников. Детеныши рождаются хорошо развитыми, и уже через несколько дней способны передвигаться за матерями по скальным кручам. Лактация продолжается до начала гона.
Самка приносит чаще одного детеныша, двойни редки. В Кавказском заповеднике в 1991 г. встречено 8 и в 1996 г. - 6 самок с двумя сеголетками. Изменение плодовитости, отчасти, циклическое (3,5 года), но в большей мере оно связано с интенсивностью трофической конкуренции с другими видами диких копытных [Ромашин, 2001].
Популяционная структура
Половой и возрастной состав. В популяциях серн доля сеголеток в сезон деторождения близка к 20%, годовалых особей - 7-15, взрослых самок - около 30-50, взрослых самцов - 15-38%. Соотношение полов колеблется в значительных пределах - от 1:0,8 до 1:3,4 (табл. 39).
Таблица 39. Половозрастной состав (%) и соотношение полов в популяциях кавказской серны
Район (источник информации)* Встречено животных, п Сеголетки Годо-валые Взрослые Самцы: самки
самки | самцы
Кавказский з-к (3) 3074 17-20 — — —
-“-(1) >28000 20-21 10 70-79 —
-“-(6) >7000 13-29 7-14 5-11**, 44-56 1:3,3
-“-(7) - 19(14-28) 15 51 15 1:3,4
Тебердинский з-к (5) - 23,8 — -
Исмаилинский з-к (2) — 21 10,5 31,1 37,4 1:0,8
Азербайджан (4) - 22 15,4 62,6 1:0,9
* 1 - Котов, 19606, 1969; 2 - Кулиев, 1989; 3 - Насимович, 1941, 19496; 4 - Кулиев, 1997; 5 - Тарасов, 1973; 6 - Дубснь, 1985; 7 - Ромашин, 2001.
** Полувзрослые особи.
Социальная организация. Самки вскоре после деторождения формируют группы, куда постепенно вливаются другие особи, включая молодых самцов. В таких
Таблица 40. Размер групп у кавказской серны
Район Число особей в группе Источник информации
среднее i максимальное
Кавказ Чаще 15-20 50-60 и более Динник, 1896, 1910
Кавказский заповедник 7-16 60-93 Насимович, 1939, 1941,
19496
а — 106-201 Котов, Рябов, 1963
Грузия 10(5-15) 40-50 Капанадзе, 1967, 1973
Лагодехский заповедник - ' 70-100 Верещагин, 1947
и Чаще 3-7 50 Енукидзе, 1953
Закатальский заповедник 5,7 46 Попкова, 1967
Рис. 37. Стадо серн (фото В. Котова)
смешанных стадах (рис. 37) порой бывает до 200 особей (табл. 40). Большая часть взрослых самцов с апреля по октябрь ведет одиночный образ жизни. В гон крупные стада дробятся на группы, чаще состоящие из 3-7 (до 22) самок с молодняком, в которые входят взрослые самцы. Эти объединения сохраняются и зимой. Отдельные группы взрослых самцов встречаются очень редко [Динник, 1910; Насимович, 1939, 1941, 19496; Попкова, 1967].
В отличие от кавказских козлов, объединения серн не отличаются большим постоянством. Стада в течение нескольких дней распадаются на мелкие группы и формируются вновь в другом составе, чему в немалой степени способствует своеобразное оборонительное поведение - при опасности эти копытные не бегут в одном направлении, как козлы, а бросаются врассыпную, соединяясь позднее в случайные группы.
Размер групп в значительной мере зависит от плотности населения. В XIX в., когда серна была многочисленной, в высокогорных стадах насчитывали сотни особей [Радде, 1899]. В Кавказском заповеднике в первые годы его существования (1924-1926) в них было в среднем 7 особей, при увеличении поголовья в 1937 г. -16 (до 93), а в начале и средине XX в. при максимальной численности в отдельных стадах регистрировали до 100-200 особей [Насимович, 1941, 1496; Котов, Рябов, 1963]. В 30-х годах по 70-100 голов в стаде наблюдали и в Лагодехском заповеднике [Верещагин, 1947].
Вожаков или даже ярко выраженных лидеров у серн нет. Иерархия линейна, ранг коррелирует с возрастом, массой тела и размерами рогов особи. Агрессивные взаимодействия обычны, но столкновения взрослых зверей между собой сравнительно редки [Locati, Lovari, 1990, 1991].
Эти копытные очень осторожны, особенно при постоянном охотничьем преследовании. На солонцах они, в отличие от кавказских козлов, часто прислушива
ются и оглядывают окрестности. В глубинных районах заповедников звери, однако, мало пугливы, любопытны и подпускают человека на 20-50 шагов. При опасности они издают протяжный шипящий свист, непохожий на свист других горных животных. В иных ситуациях иногда слышно “блеяние” детенышей и самок [Дин-ник, 1910; Насимович, 1938; Жарков, 1959; Котов, 1960в]. Сернята в неволе быстро привыкают к человеку, но с возрастом становятся все более осторожными и агрессивными [Инякова, 19576].
Пространственная структура и перемещения. Серна - весьма консервативный вид в отношении мест обитания. Территории половозрелых самцов остаются неизменными практически в течение всей жизни, а их конфигурация не меняется и при смене владельца. Летом животные не уходят со своих участков и при интенсивном выпасе скота, присутствии людей и преследовании охотниками. В малоснежных районах лесной зоны они живут практически оседло круглогодично, но в многоснежных вынуждены ежегодно покидать летние участки и переходить в другие места. И с северного, и с южного макросклонов Главного Кавказского хребта, где высота снежного покрова достигает 2 м, животные в октябре - ноябре спускаются вниз и откочевывают в боковые отроги. Диапазон сезонных высотных кочевок достигает 2000 м, их пространственный размах - 10-25 км, а иногда и больше. Суточные перемещения стада в пределах лесной зоны обычно не превышают 2-3 км. Индивидуальные размеры участков обитания достигают 5-18 км2, но чаще они меньше [Насимович, 1938,1941, 19496, 1955; Енукидзе, 1953; Жарков, 1959; Капанадзе, 1967; Попкова, 1967; Дубень, 1985].
По данным мечения и радиопрослеживания серн в Альпах и Пиренеях установлено, что взрослые самцы в конце весны занимают из года в год одни и те же территории, размеры которых, как правило, не превышают 100 гектаров. В период гона значительная их часть концентрируется в зонах, особенно привлекательных для самок, куда они приходят, порой, за 20 км. Молодые особи до двухлетнего возраста держатся на участках их матерей. Летние и зимние участки у зверей обычно разные, и при высокой плотности населения они меньше 20 га [Harm, 1984; Levet, Pepin, 1994; Hardenberg, 2000]. При реинтродукции в новые горные районы почти все особи (85%) остаются в местах выпуска, осваивая большие участки. У самок их размеры достигают 2,2 тыс. га, у самцов — 8,2 (до 26) тыс. га [Michallet, Toigo, 2000].
Численность и основные факторы, ее определяющие
Численность. В XIX - начале XX вв. кавказская серна была многочисленной на всем ареале [Радде, 1899], несмотря на обилие волка, рыси и медведя. На Западном Кавказе в верховьях Белой, Уруштена и Малой Лабы за день встречали до 200-400 особей [Динник, 1896,1910; Насимович, 1939,19496], а ее поголовье здесь, видимо, приближалось к 40-50 тыс. [Дубень, 1985]. Общие ресурсы вида на Кавказе, скорее всего, были много больше 100 тыс. Позднее, из-за неумеренного отстрела, вытеснения с лучших пастбищ и эпизоотий, занесенных домашним скотом, поголовье неуклонно сокращалось.
В середине XX в. на Северном Кавказе сохранилось, видимо, не более 15 тыс. особей. В 70-80-х годах в Дагестане жили около 250-350 серн, Чечено-Ингушетии -300, Северной Осетии - около 400-600, Кабардино-Балкарии - 400, Ставропольском
крае - 1-1,2 тыс., Краснодарском крае - около 6-7 тыс., а всего около 9-10 тыс. [Рав-кин, 19756, 1976; Точнее, 1975; Дубень, 1977, 1985; Амирханов и др., 1981; Курятников, Варзиев, 1983; Гинеев и др., 1988; Прилуцкая, Пишванов, 1988]. В конце столетия в Дагестане учли около 350 особей [Магомедов, Ахмедов, 1999], в Кабардино-Балкарии численность “очень низка” [Дзуев, 20006]. В Адыгее с 1996 г. по 2003 гг. ресурсы уменьшились в 2 раза - до 400 голов [Шебзухова, 2003]. По данным Службы учета охотничьих ресурсов, в 2000-2003 гг. в Дагестане обитали около 420-460 особей, Кабардино-Балкарии - 600-700, Краснодарском крае (без Кавказского заповедника) -700-550, Северной Осетии - 570, в Карачаево-Черкесии - 1600-1900. Общая численность вида в России, с учетом зверей в заповедниках, следовательно, близка к 5-6 тыс.
Основные ресурсы серны, как и кавказских козлов, сконцентрированы на особо охраняемых территориях, где регистрируют наиболее высокие плотности их населения (табл. 41). В первые десятилетия XX в. к западу от р. Лабы эти копытные были почти уничтожены - сохранилось не более 1 тыс. После образования Кавказского заповедника в 1936/37 гг. численность вида определена в 4,4 тыс., в 1939-1940 гг. - в 12 тыс. [Жарков, 1940; Насимович, 1941; Крайнова, 1947]. В годы войны и в 1951— 1957 гг., когда площадь заповедника уменьшилась почти в 3 раза, из-за браконьерства и интенсивного хозяйственного использования территории поголовье снова сократилось до 3^4 тыс. В 60-е - начале 80-х годов здесь насчитывали от 8 до 4 тыс. На отдельных участках в центральной части заповедника и в поясе открытых лугов плотность населения достигала 150, а на периферии - 3-6 экземпляров на 1000 га [Котов, 19606,д; Калугин, 1968; Дуров, 1974, 1977, 1978; Равкин, 19756; Дубень, 1977,1983; Кудактин, 1977; Дубень, 1985; Гинеев и др., 1988]. В последнее десятилетие XX в. в результате “невиданного” истребления зверей численность уменьшилось до 3-1 тыс. [Ромашин, 1994, 2001; Кудактин, 1996; Немцев и др., 1999]. В 2002-2003 гг., по данным отчетов, здесь жили около 1650-1700 особей.
Таблица 41. Плотность населения кавказской серны
Район, заповедник Особей на 1000 га Источник информации
Краснодарский край 0,1-10,5 Котов, Рябов, 1963
Кавказский заповедник 9-131 на разных участках Котов, 19606, 1969
3-148 Дубень, 1977
и 16 Ромашин, 2001
Тебердинский заповедник 17-6 Тарасов, 1973, 1976, 1977, 1983
и 2,3-4,4 Бобырь, 1999
Дагестан 7,2 Магомедов, Ахмедов, 1999
Грузия 2-50 на разных участках Капанадзе, 1973, 1974, 1975
к 10-15 Арабули, 1986
Закатальский заповедник 26 Попкова, 1967
Исмаилинский заповедник 18 Кулиев, 1989
Исмаилинский заказник . 1
В Тебердинском заповеднике в 1936-1952 гг. насчитывали около 2-3,6 тыс. серн [Аренс, 1957], в 1955 г. - 1 тыс. [Инякова, 1957а], в первой половине 70-х - 600-770 особей, позднее - около 300, а в конце 90-х их осталось не более 100. Плотность населения, соответственно, снизилась с 17 до 2-4 голов на 1000 га [Тарасов, 1973, 1976, 1977, 1983; Дуров, 1977; Поливанова, 1990; Бобырь, 1999]. В Северо-Осе
тинском заповеднике в 70-80-е годы находилось около 100-130 голов [Вейнберг, 1977; Амирханов и др., 1990]. На 1 января 1990 г. в трех из четырех кавказских заповедников учли всего 2750 особей [Филонов, Шадрина, 1996].
В Грузии ресурсы вида были значительно подорваны в первой половине XX в., но все же оставались значительными. В 1950 г. только в Лагодехском заповеднике насчитывали около 1 тыс. [Енукидзе, 1953]. В 1973 г. в республике, по официальным данным, обитало до 20 тыс. серн, но эта цифра, скорее всего, завышена. На незаповедной территории жили около 2,3 тыс., а всего, видимо, несколько тысяч [Капанадзе, 1973, 1975, 1976, 1978]. В конце 80-х годов осталось не более 5 тыс. [Арабули, 1989]. В Боржомском заповеднике уцелело 20-30 особей [Бараташвил-ли, 1980].
В Азербайджане в 30-х годах только на территории Закатальского заповедника держалось до 1 тыс. серн [Марков, Млокосевич, 1935]. В 50-е - начале 60-х годов в республике насчитывали 2-2,5 тыс. [Алиев, 1961, 1962; Попкова, 1967]. В 70-80-е годы 700-800 зверей жили на Большом Кавказе (в том числе 450-500 на территории Закатальского заповедника) и 150 в Исмаилинском заповеднике. Они встречались также в северо-восточной части республики и на Муровдагском хребте [Алекперов и др., 1973, 1976; Кулиев, 1989]. К концу XX в. осталось всего около 300-600 голов [Кулиев, 1997].
В СССР общую численность серны в 60-80-е годы оценивали в 20-35 тыс. особей, причем более 50% ресурсов находилось в Грузии [Банников, 1962, 1965, 1978; Климова, 1989]. К началу XXI в., по экспертной оценке, на той же территории сохранилось менее 10 тыс. Угроза исчезновения этого вида из фауны Кавказа, и фауны России в частности, реальна как никогда раньше.
В Европе в 70-80-е годы XX в. обитали более 400 тыс. серн, в том числе в Австрии - 110-160, Швейцарии - 55-70, Италии - 60-63, Франции - 36-54, Испании - 32, Югославии - 24, Германии - 16, Румынии - 8,5, Чехословакии - 1,6 и Болгарии - 1,5 тыс. [Дежкин, 1983, 1989].
Воспроизводство. Уровень воспроизводства у серны невысок. На половозрелую самку приходится менее 1 теленка. В течение года погибает от 33 до 70% сеголеток [Попкова, 1967;Кулиев, 1989; Allaineetal., 1990]. Определенная цикличность в уровне плодовитости практически не сказывается на динамике популяций, поскольку та определяется, в большей мере, уровнем смертности [Ромашин, 2001].
Погода и корма. Кавказская серна, как считалось, меньше других копытных страдает от бескормицы в многоснежные зимы, чему способствует рассредоточение зверей по угодьям мелкими группами [Насимович, 1939]. Однако, по исследованиям А.В. Ромашина [2001], этот вид более чувствителен к суровости зим, по сравнению с другими копытными, в связи с меньшими размерами тела. У пиренейской серны тоже обнаружена четкая корреляция уровня смертности с суровостью зим [Gonzalez, Crampe, 2001]. В Кавказском заповеднике в июне 1986 г. зарегистрирована гибель почти половины новорожденных после сильного похолодания и выпадения снега (до 40 см) [Ромашин, 2001].
Хищники. Основной враг серны - рысь, легко передвигающаяся по крутым склонам [Насимович, 1939; Попкова, 1967]. Колебания численности этого хищника и жертвы в Тебердинском заповеднике “зеркальны” по отношению друг к другу [Тарасов, 1976]. Не менее опасен для нее и волк, которому более доступен молодняк. В
Кавказском заповеднике остатки серны встречаются в 3-16% экскрементов и желудков волка и в 18% - рыси. В 1966-1977 гг. доля волка в общей смертности жертвы достигала 39% [Тсплов, 1938; Котов, 1958; Жарков, 1959; Кудактин, 1977, 1986; Ромашин, 2001]. На Боковом хребте в Северной Осетии, где численность серны мала, ее доля в пище рыси не превышает 4% [Вейнберг, 1988]. К потенциальным врагам следует отнести также медведя, лисицу, одичавших собак и крупных пернатых хищников, однако их роль в динамике численности жертвы невелика.
Конкуренты. Серна - мелкое копытное с небольшими рогами, и поэтому вынуждена уступать место на пастбищах и солонцах более крупным животным - козлам, косулям и оленям и занимать лишь “свободные” биотопы [Насимович, 1949а; Котов, 1968; Соколов, Кузнецов, 1978; Вейнберг, 19836]. В Восточных Альпах реинтродукция и последующее увеличение численности альпийского козла привело к вытеснению серны в результате перекрывания пространственной ниши на 80-90% и трофической на 40% [Schroder, Kofler, 1985]. На Кавказе совместное сосуществование нескольких многочисленных видов копытных до последнего столетия не вызывало сильного угнетения кого-либо из них, хотя конкурентное давление, безусловно, имело место. Тем не менее, не исключено, что именно трофическая конкуренция была причиной выселения кавказской серны с летних пастбищ альпийской зоны в лесной пояс и стала одним из основных факторов сокращения ее численности [Ромашин, 1994, 1995, 2001].
Конкуренция кавказской серны с домашним скотом, особенно с овцами и козами, особенно остра, поскольку последние занимают лучшие пастбища и стравливают около 30-60% излюбленных кормовых растений. Однако она, как и кавказские козлы, приспособилась к выпасу домашних животных, выходя на пастбища из высокогорных убежищ ночью или осваивая бедные участки горных лугов и небольшие лужайки среди осыпей и скал [Жарков, 1959; Попкова, 1967]. В последнее время серьезным конкурентом для нее стали яки [Дзуев, 20006], которые поднимаются в высокогорье, вытесняя из последних прибежищ.
Болезни, паразиты, травмы. Массовый падеж кавказской серны от эпизоотий, возможно, имел место в 60-х - начале 70-х годов XIX в. и в 1919 г. [Насимович, 19496]. Позднее гибель животных от болезней не регистрировали.
Число видов гельминтов, паразитирующих на серне в пределах всего ареала, около 50, в том числе в Западной Европе - более 30 [Котрлы, Котрла, 1970]. На Кавказе у этих копытных также зарегистрировано более трех десятков видов паразитов, преимущественно сосальщиков и ленточных червей, но их патогенное значение в целом невелико. Большая часть из гельминтов характерны также для кавказского козла, европейской косули и домашнего скота. Чаще всего у серны обнаруживают цистокаулез. Источником паразитарных заболеваний становится домашний скот, выпасаемый на горных пастбищах. По другой версии, серна сама является резервентом гельминтов, типичных для домашних животных [Асадов, 19596; Жарков, 1959; Рухлядев, 1959, 1964, 1970; Попкова, 1967; Закариев, 1980].
Гибель от травм, получаемых при падении со скал, в осыпях или при драках с соперниками во время гона, видимо, обычна, несмотря на необычайную ловкость и живучесть животных. Охотники нередко добывают зверей со старыми следами переломов ног, ребер и рогов.
Гибель в лавинах. Гибель серны в лавинах случается несколько реже по сравнению с кавказскими козлами [Попкова, 1967; Тарасов, 1976], но чаще, чем у других копытных, поскольку она нередко зимует вблизи лавинных русел. В районе Кавказского заповедника весной находили до 10-17 трупов сразу, а гибель мелких групп регистрировали неоднократно [Динник, 1910; Насимович, 1939, 1955]. Доля этого фактора в общей смертности вида достигает здесь 39-48% [Ромашин, 2001].
Антропогенные факторы. Расширение рубок леса, прокладка дорог и неумеренное использование высокогорных лугов под выпас скота способствовали исчезновению копытных во многих районах [Бараташвилли, 1977; Дубень, 1977; Дуров, 1977]. В последние десятилетия с развитием туризма фактор беспокойства, на который серна реагирует более остро, нежели кавказские козлы, резко усилился даже в заповедниках [Тарасов, 1977; Ромашин, 2001]. Однако главной причиной сокращения ареала и численности серн во многих районах, несомненно, стал человек-охотник.
Использование ресурсов и охрана
Серна издавна была одним из излюбленных охотничьих животных, особенно в европейских странах. Стреляют ее обычно не для получения мяса, которое не отличается высокими вкусовыми качествами, и не из-за шкур, хотя они весьма прочны и красивы, а ради трофея и забавы. Принц Август Саксон Кобургский, например, в своих владениях в Верхней Штирии убил более 3400 особей. Во второй половине XX в. объем добычи этого копытного в европейских странах был близок к 60 тыс. голов [Дежкин, 1977, 1980, 1989].
На Кавказе охотничий пресс на серну и других копытных заметно усилился в XIX в., когда местные охотники и пастухи сменили кремневые ружья на более современные. Человек стал главным истребителем этих животных, и уже к началу XX в. выбил или заметно сократил их население во многих районах [Динник, 1910; Гептнер, Формозов, 1941]. О масштабах промысла и былой численности копытных свидетельствуют следующие факты. 1 сентября 1897 г. в Кубанской охоте на горе между Аспидной и Уруштеном граф П.М. Стенбок добыл 10 серн, побив рекорд Е.П. Демидова, который 1 сентября 1895 г. убил 7 зверей [Шильдер, 1901]. Охотник Баба Гасанов из селения Геок-Таша в бассейне Геок-чая в Исмаилинском районе Азербайджана добыл в 1900-1940 гг. около 500 кабанов, 200 оленей, 400 косуль и 15 серн, а лесничий Боржомского и Лагодехского районов Грузии Л. Млокосевич за 40 лет с конца XIX в. - 828 кабанов, 157 оленей, 475 косуль, 249 кавказских козлов, 6 безоаровых козлов и 215 серн. Конечно же не все охотники были такими удачливыми, но их число (только зарегистрированных) на Кавказе в 30-е годы XX в. достигало 130000. Общая добыча серны была не менее 1500 голов в год [Верещагин, 1947], и лишь в Азербайджане отстреливали не менее 500-600 особей [В-н, 1936].
Несмотря на учреждение ряда заповедников и усиление охраны животных в годы советской власти, ресурсы вида, восстановить не удалось. Более того, как показано выше, они постепенно таяли из-за усиления хозяйственной деятельности человека и непрекращающегося браконьерства по всему Кавказу [Насимович, 19366; Попкова, 1967; Арабули, 1989; Кудактин, 1996; Немцев и др., 1999; Дзуев,
20006]. В последние десятилетия XX в. браконьерство резко возросло даже на особо охраняемых территориях. В Кавказском заповеднике в 1990 г. браконьерский выстрел в лесу регистрировали в среднем через 25 дней, в 1991-1994 гг. - через 12-16, в 1996 г. - через 2,5 дня. Доля этого фактора в общей смертности серны достигла 36% [Ромашин, 2001].
Кавказскую серну необходимо незамедлительно включать в Красную книз у Российской Федерации.
Со временем, при улучшении охраны и сокращении числа хищников, потребуется реакклиматизация вида в утраченных частях ареала на Кавказе. Возможна также его акклиматизация в горах Крыма, Средней Азии и в других горных областях [Гептнер и др., 1961]. Эффект от интродукции может быть значительным. В Новой Зеландии, куда в 1907 г. завезли всего 7 особей, численность в отсутствии хищников выросла до 10 тыс. [Деннлер, 1975].
Считается, что серна плохо живет и размножается в неволе [Соколов, 1959], хотя при хорошем уходе ее разведение вполне реально [Инякова, 19576].
Род КОЗЛЫ
Capra Linnaeus, 1758
Capra Linnaeus, 1758. Syst.Nat., 10ed., 1: 68. Тип рода-Capra hircus Linnaeus (домашняя коза).
Hircus Brisson, 1762. Regn, Anim.: 12. Nom. nudum.
Ibex Pallas, 1776. Spicil. Zool., 2: 52. Ibex sibiricus Pallas.
Aries Link, 1795. Zool. Beytr., 1, 2: 96. Nom. nov. pro Capra Linnaeus. Non Brisson, 1762.
Tragus Schrank, 1798. Fauna Boica, 1: 78. “Tragus aegagrus".
Ibex Hodgson, 1847. Asiat. Soc. Bengal., 16: 700.
Orthaegoceros Trouessart, 1905. Cat. Mamm., Suppl.: 738. Capra falconeri Wagner.
Turus Hilzheimer, 1916. Brehm’s Tierleben, 4 ed., Sauget., 4: 273. Capra caucasica Guldenstaedt et
Pallas.
Euibex Camerano, 1916. Atti Acad. Torino, 51: 338. Capra ibex Linnaeus.
Eucapra Camerano, 1916. Ibid., 51: 338. Capra sibirica Meyer.
Turocapra de Beaux, 1949. Atti Soc. Ital. Sci. Nat. Milano, 88: 17. Capra pyrenaica Schinz.
Характеристика. Животные средних размеров, плотного тяжелого сложения с относительно короткими и толстыми конечностями. Самки заметно меньше самцов. Хвост небольшой, с плоской голой нижней поверхностью. На верхней губе узкая вертикальная полоска голой кожи. Ноздри сближены.
Окраска самок и молодняка преимущественно одноцветная, серых и коричневых тонов. Темные полосы вдоль морды характерны для С. aegagrus и С. falconeri, по спине - для первых и С. sibirica. У взрослых самцов этих видов и С. nubiana резко выражены темные полосы вдоль хребта, на лопатках, груди, боках и ногах,
С. aegagrus
С. cylindricomis
Взрослые самки
Сеголетки
Взрослые самцы
С. aegagrus С. falconeri С. nubiana С. sibirica
C-aegagrus С. falconeri (fmibiana С. sibirica C^ytinfincomis
C. aegagrus C. falconeri C. pyrenaica c nubiana
. caucasica cylindricomis
Рис. 38. Отличительные признаки в окраске видов рода Capra (по [Вейнберг, 1993]) Окраска: а - летом, б - зимой, в-в период гона
особенно заметные в период гона и в зимнем меху (рис. 38). Остальные (ibex, caucasica, cylindricomis') окрашены относительно монотонно, но не одинаково [Вейнберг, 1993]. Шерстный покров относительно короткий, равномерной длины, густой, с пучками пуховых волос. У некоторых видов бороду имеют не только самцы, но и самки. Хвостового зеркала нет. Линька одна - весенне-летняя.
Зароговая часть черепа хорошо развита, ее профиль выпуклый. Межчелюстные кости обычно соприкасаются с носовыми. Носовые кости без выемок спереди и отделены щелью от близлежащих по сторонам костей. Сошник не срастается с небными костями. Глазницы трубкообразные. Слезные кости слабо изогнуты. Теменные гребни сближены. Предглазничные ямки отсутствуют, этмоидальные щели имеются. Кости черепа пневматизированы. Основания роговых отростков расположены над глазницами и слабо наклонены. Роговые стержни внутри с системой крупных полостей, достигающих верхней трети. Их форма округлая, близкая к треугольной или скрученная в спираль (рис. 39). Зубы гипсодонтные. Предкоренные сильно редуцированы, но третий из них, за редким исключением, присутствует. Верхние коренные со стилями. На лабиальной поверхности параконида нижних коренных имеется типичная складка.
Рога - у животных обоих полов, но у самцов они более массивные. Форма роговых чехлов разнообразная: чаще они длинные саблевидные, реже со спиральным изгибом, а у винторогого козла (С. falconeri) - круто спиральные. На их передней стороне обычны валики, кили или бугры. Окраска чаще черная.
Специфических кожных желез немного: межпальцевые (без “мешков”), у некоторых видов - зароговая, а также подхвостовая и препуциальная, секрет которых смешивается с мочой, обуславливая резкий “козлиный” запах. Молочная железа, как правило, двудольная.
Рис. 39. Форма роговых стержней у представителей рода Capra (по [Цалкин, 1950])
Слева направо: черепа самцов сибирского (С. sibirica), безоарового (С. aegagrus) и винторогого (С. falconeri) козлов
Рис. 40. Восстановленный ареал рода Capra (по [Гептнер и др., 1961]; с дополнением)
Кариотип Capra, видимо, наиболее примитивный среди представителей Сарппае: 2п = 60, NF = 60 (у Ammotragus - 2п = 58, Ovis - 52-58, Pseudois - 54, Hemitragus - 48). Аутосомы акроцентрические, Х-хромосома крупная акроцентрическая, Y-хромосома мелкая субметацентрическая или точечная [Heck et al., 1968; Schmitt, Ulbrich, 1968; Wurster, Benirschke, 1968; Мамедов, 1978; Bunch, Nadler, 1980; Орлов, Булатова, 1983; Шарипов, 1989; Мамедов, Агаев, 1990].
Горные и высокогорные скальные животные, ведущие групповой и стадный образ жизни. Скорость бега не велика - 20-45 км/час, но в горах они стремительны и делают большие прыжки в длину и высоту [Гаспарян, 1969; Гамбарян, 1972]. Продолжительность жизни около 15-20 лет.
Важные охотничьи или охраняемые звери, находящиеся под угрозой уничтожения. Домашняя коза стала одним из важнейших сельскохозяйственных животных.
Распространение. Горные системы Европы, Передней и Центральной Азии и северо-востока Африки (рис. 40). Современный ареал очаговый.
Эволюция. Происхождение рода неясно. Древнейшие ископаемые остатки Capra (С. prisca) известны с верхнего плиоцена Европы и Азии (из Индии). Ствол Ibex прослеживается с миоцена. Наиболее примитивными представителями рода считаются формы aegagrus, затем sibirica, ibex и nubiana, сохранившие ряд архаичных черт, наиболее специализированными - восточнокавказский (cylindricornis) и винторогий (falconer!) козлы [Громова, 19486, 1953]. Судя по молекулярным исследованиям, расхождение всех видов рода было практически одновременным (в эволюционном масштабе времени) [Klanceau et al., 1999].
Эволюция рода шла по пути адаптации животных к жизни в скалистых горах (обеспечивающих защиту от хищников), что особенно отразилось на строении опорно-двигательного аппарата. У козлов укороченные широкие метаподии, шаровид
ная головка бедренной кости, широко раздвигающиеся копыта с твердыми быстро растущими краями и мягкой подушкой внутри. Угол подъема копытной фаланги у них в два раза выше по сравнению с равнинными видами, что позволяет им ставить конечности почти вертикально [Громова, 1953; Тихонов, 1999]. Эти особенности повышают устойчивость и препятствуют скольжению зверей на крутых скалистых склонах. Суровые климатические условия обитания в высокогорье привели к максимальному развитию пуховых волос. Стадный образ жизни способствовал усилению полового диморфизма и развитию у самцов мощных рогов в качестве турнирного оружия и как средства защиты от хищников [Северцов, 1951; Егоров, 1955]. Форма чехлов рогов у самцов усложнялась от плоской в сечении с передним или задним ребром (aegagrus и falconer!) до овальной с бугорчатой передней гранью (ibex) и треугольно-округлой (caucasica, cylindricornis), а изгиб из саблевидного превращался в спиральный (рис. 41). У самок сечение рога осталось уплощенным с передним ребром у винторогого козла, каплевидным с передним ребром у безоа-рового и округлым без ребер у остальных форм [Вейнберг, 1993].
Состав. Особенно дискуссионный. Систематики чаще выделяли 2 подрода: Capra, включающий от 1 до 9 видов, и Ammotragus с единственным видом Л. lervia [Lydekker, 1913; Schwarz, 1935; Ellerman, Morrison-Scott, 1951; Соколов И.И., 1953, 1959; Гепт-нер и др., 1961; Громов и др., 1963; Haltenorth, 1963; Ansell, 1972; Соколов В.Е., 1979; Durst et al., 1982; Honacki et al., 1982; Uerpmann, 1987; Grubb, 1993; Wild Sheep and Goats ..., 1997; Тихонов, 1999]. Гривистый баран, как показано выше, имеет весьма отдаленное родство с козлами, и его приходится исключить не только из состава рода Capra, но и трибы Caprini.
Сеголетки
Взрослые самки
Молодые самцы
Взрослые самцы
С.aegagrus C.falconeri C.pyrenaica C.nubiana C.ibex C.sibirica Ccaucasica C.cylindricornis
Рис. 41. Форма сечения и изгиба роговых чехлов у представителей рода Capra (по [Вейнберг, 1993]) Стрелки показывают, что форма изгиба или сечения сохраняется у следующих таксонов; в рамке - схема развития рога самца в онтогенезе
Таксономия рода Capra не стала яснее даже после многомерного анализа довольно большой выборки черепов (п = 112) разных форм по 55 признакам [Fandos et al., 1993]. При исследовании краниометрических признаков самок однозначно различаются формыpyrenaica, nubiana и ibex, тогда как falconeri, caucasica и sibirica образуют перекрывающиеся зоны. Среди самцов выделяются ibex, pyrenaica и falconeri. Наиболее дифференцирован С. pyrenaica, наименее - формы, включаемые в группу ibex.
В аналогичном исследовании рода Capra, основанном на многомерном анализе 380 черепов по 80 параметрам (рис. 42), установлено, что винторогий козел четко
Самки
С. aegagrus
С. sibirica
С. ibex
С. caucasica
С. cylindricornis
С. falconeri
Root 1
Самцы
° С. aegagrus
о С. sibirica
« С. ibex а С. caucasica
• С. cylindricornis
я С. falconeri
Рис. 42. Распределение видов рода Capra в системе координат с каноническими переменными по краниальным признакам взрослых особей (по [Казанская, 2004])
Root 1 и 2 - первые канонические переменные дискриминантного анализа
rtf:
caucasica (Карачаево-Черкесия) 55
caucasica (Карачаево-Черкесия) 26
caucasica (Карачаево-Черкесия) 25
caucasica (Карачаево-Черкесия) 6
caucasica (Карачаево-Черкесия) 8
caucasica (Карачаево-Черкесия) 5
caucasica (Карачаево-Черкесия) 54
caucasica (Карачаево-Черкесия) 53
ibex A Fl 72367*
ibex AF034735*
pyrenaica AJ010054*
pyrenaica AJ010049*
pyrenaica AJ010056*
pyrenaica A JO10053*
falconeri AJ293418*
falconeri AB044309*
falconeri (Таджикистан) 30
falconeri AF034736*
falconeri D84202*
cylindricornis (Азербайджан) 19
caucasica (?) AF034738*
cylindricornis AF034737*
cylindricornis (Кабардино-Балкария) 9
cylindricornis (Кабардино-Балкария) 7 cylindricornis (Кабардино-Балкария) 10 cylindricornis (Кабардино-Балкария) 11 cylindricornis (Кабардино-Балкария) 42 hircus AB004075*
hircus AB00470*
aegagrus cretica AF217255*
aegagrus D84204*
aegagrus AB004069*
nubiana AF217256*
nubiana AF034740*
sibirica (Киргизия) 29
sibirica (Киргизия) 27
sibirica (Киргизия) 28
sibirica AF034734*
sibirica (Алтай) 43
sibirica (Гобийский Алтай) 41
sibirica (Алтай) 15
sibirica (Алтай) 16
Ammotragus lervia AF034731*
Рис. 43. Филогенетическое древо рода Capra, построенное на основании анализа последовательностей митохондриального гена цитохрома b методом минимальной эволюции (по [Казанская и др., в печати])
Цифры слева - уровень поддержки каждого кластера (%), справа - номера образцов. Звездочкой обозначены образцы из генетического банка данных
обособлен от других видов, которые объединяются в две неясно выражение группы: кавказских козлов (формы caucasica и cylindricomis) и группу, состоящую из sibirica, ibex и aegagrus. При этом наблюдается частичное перекрывание параметров (дистанций), особенно при анализе черепов самцов, между формами caucasica и cylindricomis, а также между sibirica и ibex, sibirica и aegagrus (см. табл. 63). Наибольшие дистанции выявлены между aegagrus и cylindricomis, а также sibirica иfalconeri, т. е. между формами, ареалы которых перекрываются [Казанская, 2004].
При молекулярном исследовании рода кластеризация образцов в целом соответствует принятой видовой принадлежности, однако взаимное расположение клад отличается от морфологических построений (рис. 43). Сибирский козел оказывается в отдельном кластере, образцы остальных форм группируются в другом. Ба-зоаровый козел с хорошей поддержкой объединяется в одной кладе с домашней козой, что подтверждает их тесное родство. Западнокавказский козел (caucasica) формирует кладу, четко обособленную от восточнокавказского козла (cylindricomis) и примыкающую к европейским формам (ibex и pyrenaica). Генетическое различие западнокавказских и восточнокавказских козлов, о котором упоминали также В. Манко с соавторами [Manceau et al., 1999], крайне интересно, поскольку эта группа образует клинальный ряд морфологических форм, что служит основной причиной таксономических разногласий (см. ниже).
Судя по имеющимся данным, представители рода более близкородственны, чем представлялось ранее (и что подтверждается отсутствием репродуктивной изоляции), и, видимо, более заслуживают ранга подвидов, нежели видов. Тем не менее винторогий, сибирский, безоаровый, пиренейский, альпийский и нубийский козлы обособлены географически и имеют несомненные отличия по форме рогов. Самостоятельность безоарового козла подтверждается многовековым совместным существованием с кавказскими козлами. Существование двух разных форм кавказских козлов (caucasica и cylindricomis) несомненно. Вопрос о статусе эфиопского (wallie) и синайского (sinaitica) козлов тоже остается открытым из-за отсутствия материала. Необходимость дальнейшей таксономической ревизии рода Capra очевидна.
БЕЗОАРОВЫЙ КОЗЕЛ
Capra aegagrus Erxleben, 1777
Capra aegagrus Erxleben, 1777. Syst. Regn. Anim.: 260. Дагестан, вост, часть Большого Кавказа.
Capra caucasica Gray, 1843. ListMamm. Brit. Mus.: 167. “Кавказ”. Nom. praeocc., non Guldenstaedt et Pallas, 1783.
Capra blythi Hume, 1875. Proc. Asiat. Soc. Bengal, 1874: 240. Синд, Юго-Вост. Пакистан.
Capra aegagrus turcmenicus Zalkin, 1950. Докл. АН СССР, 20,2: 325. Туркмения, Центр. Копетдаг, сев. склоны хр. Тава, 25 км южнее ст. Гяуре.
Характеристика. Размеры сравнительно некрупные, сложение легкое (фото 21). Хвост короткий, менее 20 см, с удлиненным волосом, но без кисти на конце. Половой диморфизм выражен.
Окраска летом рыжеватобурая, зимой серовато-бурая. У самцов по хребту, вокруг основания шеи, по ее низу, на груди и ногах имеются черно-бурые полосы. Голова темно-бурая. Нижняя часть тела бело-вато-серая, хвост черный. Борода длинная (до 20 см), густая, черная, за что вид нередко называют “бородатым”. У самок бороды чаще нет. У сеголеток и самок темные полосы тянутся вдоль морды, маскируя глаза; у последних темная полоса и вдоль хребта. На передней стороне запястных суставов расположены округлые роговые мозоли, над которыми есть характерный белый перехват.
Череп вытянутый, неширокий, с удлиненной зароговой частью (рис. 44). На лобных кос
Рис. 44. Череп самца безоарового козла (экземпляр с Кавказа; Зоомузей ЗИН РАН, коллекционный № 958) Вид сбоку, сверху и снизу
тях у основания роговых стержней возвышения. Носовые кос-
ти относительно широкие и короткие. Роговые стержни слабо
сдвинуты назад, уплощены с боков, с одним острым передним ребром.
Роговые чехлы у самцов длинные (до 130-140 см), сравнительно узкие и сближенные в основании, саблевидно изогнутые в одной плоскости или со сходящимися
вершинами, и с крупными, сжатыми поперечными валиками по ребру. Поперечное сечение рога каплевидное, линзовидное или грушевидное. У самок рога короткие (до 39 см), тонкие, округлой формы, без бугров, мелкоморщинистые, слабо изогнутые или почти прямые. Роговое вещество серо-бурого или буро-черного цвета.
Хорошо развиты межпальцевые и препуциальные железы [Соколов, Чернова, 2001].
Специализированный обитатель скалистых гор. Предпочитает держаться небольшими группами на облесенных склонах. В зоопарках живет до 17 лет.
Родоначальник домашней козы.
Распространение. Острова Греческого архипелага, Кавказ, Малая Азия, Ирак, Иран, Туркменистан, Афганистан, Восточный Оман, Индия, Пакистан (рис. 45). Успешно акклиматизирован в штате Нью-Мексико США.
Состав. Исследователи чаще выделяют 4 подвида безоарового козла: С. a. aegagrus Erxleben, 1777 (Малая Азия, Иран и Кавказский перешеек), С. a. cretica Schinz (острова Греческого архипелага), С. a. turcmenicus Zalkin, 1950 (Копетдаг и Балха-
ны в Туркменистане, Туркмено-Хорасанские горы в Иране) и С. a. blythi Hume, 1875 (Индия и Пакистан) [Соколов, 1959]. Туркменского козла нередко относят к последней расе. Помимо них описаны формы neglecta (возможно, синоним blythi), а также dorcas и pictus, которые, скорее всего, помесные с домашней козой [Гептнер и др., 1961]. По результатам молекулярно-генетических исследований критский козел (cretica) четко входит в кластер домашних коз [Bar-Gal et al., 2002].
Все подвиды описаны на основании ничтожного материала. Требуется таксономическая ревизия.
В фауне России - номинативная форма С. a. aegagrus.
Изменчивость
Размеры и масса тела. Масса безоаровых козлят, рожденных в неволе, составляет 2,7-3 кг в среднем. В первые месяцы ежемесячный прирост массы достигает 1-2,5 кг, и через полгода самцы весят около 14, а самки - 10 кг [Антонян, Эгнато-сян, 1984].
Длина тела взрослых самцов с Кавказа колеблется в пределах 136-160 см, высота в холке - 81-90, длина уха - 11, хвоста - 12-13 см [Даль, 1951; Кулиев, 1981]. Экстерьерные показатели особей из Туркменистана практически идентичны [Мориц, 1930]. Масса самцов варьирует от 35 до 70 кг [Точиев, 1973; Ишадов, 1965; Коршунов, 1995а]. Самки заметно мельче самцов.
Размеры черепа. Максимальная длина черепа самцов - 280, самок - 268 мм. Размеры черепа особей кавказской и туркменской форм безоарового козла сходны (табл. 42), что ранее заметил В.И. Цалкин [1950а]. При многомерном краниометрическом анализе самки этих форм не отличаются друг от друга, различия самцов незначительные [Казанская, 2004].
Изменчивость рогов. У самцов рога в первый год жизни вырастают до 10 см, позднее их среднегодовой прирост колеблется в пределах 6-15 см. Максимальный он на втором году, минимальный - к концу жизни. Число бугров на рогах у взрослых особей достигает 7-9 [Гаспарян, 1975; Коршунов, 1995а]. Размеры рогов кор-
Таблица 42. Размеры черепа (мм) взрослых особей безоарового козла
Самцы Самки Район (источник информации)*
п м 1 lim 1 п 1 м 1 lim
Наибольшая длина
4 257,8 241-279 4 224,1 218-233 Кавказ (1)
11 263,4 248-278 6 224,2 215-233 -“-(2)
2 — 245-251 - - -“-(4)
— — 240-277 — - 220-245 -“-(5)
1 — 258 - - - -“-(7)
1 - 260 6 226,5 218-233 Туркменистан (1)
и 261,6 250-279 6 227,5 224-234 -“-(2)
10 254,8 232-280 18 228,3 213-268 Кавказ и Туркменистан (6)
Максимальная ширина
4 131,7 123-139,5 4 117,4 112-125 Кавказ (1)
2 - 129-134 - - - -“-(4)
— — 115-154 - — 109-127 -“-(5)
1 - 131 - - -“-(7)
7 124,1 115-133 7 117,4 112-122 Туркменистан (1)
19 130,1 120-157 25 122,7 109-141 Кавказ и Туркменистан (6)
Межглазничное расстояние
4 92,0 86-97,5 4 79,2 76-83 Кавказ (1)
— — 76-110 — — 70-93 -“-(5)
7 87,3 79-99 8 81,7 78-85 Туркменистан (1)
21 89,1 79-99 27 81,1 72-100 Кавказ и Туркменистан (6)
Длина лицевой части
10 183,9 169-203 18 172,6 155-206 Кавказ и Туркменистан (6)
Ширина мозговой коробки
21 73,4 68-84 27 68,1 63-75 Кавказ и Туркменистан (6)
Длина носовых костей
4 74,9 69-86 5 72,1 62-84 Кавказ (1)
— - — — - 60-78 -“-(5)
1 - 71 — — - -“-(7)
6 74,1 61-85 7 71,6 63-83 Туркменистан (1)
18 77,4 62-88 25 73,7 61-92 Кавказ и Туркменистан (6)
Длина верхнего ряда зубов
4 75,0 72-78 5 70,0 66-72 Кавказ (1)
1 — 71 — — - -“-(7)
7 72,9 71-75 8 69,6 60-74 Туркменистан (1)
21 73,7 68-80 28 70,0 63-75 Кавказ и Туркменистан (6)
Длина костных стержней
— 378 320-565 — 102 70-140 Кавказ (5)
5 386 250-560 15 130 90-180 Кавказ и Туркменистан (6)
Обхват основания костных стрежней
— 172 165-205 — 84,0 — Кавказ (5)
6 165 146-200 16 81,1 67-113 Кавказ и Туркменистан (6)
* 1 - Соколов, Темботов, 1993; 2 - Цалкин, 1950а; 3 - Коршунов, 1995а; 4 - Даль, 1944; 5 - Манасерова, 1989; 6 - Казанская, 2004; 7 - Пфиценмайер, 1915.
релируют с возрастом. Их длина у старых самцов достигает 128 (у отдельных особей, видимо, 140), размах - 67, обхват у основания - 26 см. У самок длина рогов обычно не превышает 39 см (табл. 43).
Таблица 43. Размеры рогов (см) у взрослых особей безоарового козла
п Длина Размах Обхват у основания Район (источник информации)*
М | lim М I lim м lim
Самцы
— — — 115 — 35-55 - Кавказ (7, 3)
45,37 - — - - 22,4 20-25 -“-(2)
1 83 31 20,5 -“-(5)
36 __ 42-128 - — - 18-26 -“-(8)
67 — -127 — - 21,5 18-25 Туркменистан (4)
- 60-90 - - - — (6)
37 - - - - 19,8 17-23 (2)
21 73 42-122 28 8,6-67 Самки 21,0 18-25,5 Кавказ и Туркменистан (9)
- - 12-30 - - — — Кавказ (8)
— - -25 - — - - -“-(1)
— — -25 - — - — Туркменистан (4)
— — 15-16 — — — — -“-(6)
26 25 12-39 14 6-29 9,8 7,5-13 Кавказ и Туркменистан (9)
*1 - Разсвиг, 1904; 2 - Цалкин, 1950а; 3 - Радде, 1899; 4 - Коршунов, 1995а; 5 - Пфицснмайср, 1915; 6 - Мориц, 1930; 7 - Даль, 1944, 1951; 8 - Манасерова, 1989; 9 - Казанская, 2004.
По исследованиям В.И. Цалкина [1950а], кавказские безоаровые козлы отличаются от туркменских и индийских более толстыми основаниями рогов. У первых их окружность составляет в среднем 22,4 (20-25) см, у вторых - 19,8 (18-23) см. Судя по новым сведениям (табл. 43), различение форм по этому признаку практически невозможно. Следовательно, оснований для выделения кавказской и туркменской форм нет ни по краниометрическим признакам, ни по признакам рогов.
Окраска и линька. Сеголетки безоарового козла летом серые, зимой - темнопесочные с темными продольными полосами на лбу, от глаз к носу и на передней поверхности кисти и стопы. Молодые и взрослые самки летом песочно-рыжие, зимой - темно-песочные с теми же полосами. Окраска самцов варьирует от рыжей до песочно-серой и седой летом, а зимой - от темной до буровато-серой, светло-бурой, серебристо-белой и даже белой у старых особей с черными контрастными полосами на ногах, хребте, боках, лопатках и нижней стороне шеи (фото 21). Хвост черноватобурый. Изменчивость окраски у особей в значительной мере является результатом различных комбинаций растущего, снашивающегося или выпадающего волоса. Линька наблюдается с конца апреля до июня. К зиме развивается подпушь и удлиняется ость [Даль, 1951; Гаспарян, 1969, 1975; Вейнберг, 1999]. Окраска безоаровых козлов Кавказа и Туркмении сходна, а особи индийской расы, видимо, более светлые с интенсивно окрашенной головой [Цалкин, 1950а].
Генетическая изменчивость. Диплоидное число хромосом у безоарового козла - 2п = 60, NF = 60. Все хромосомы акроцентрические, за исключением мелкой точкообразной Y-хромосомы. Кариотцп идентичен диплоидному набору восточнокавказского козла и домашней козы как по числу и морфологии хромосом, так и по картине их исчерченности при дифференциальной окраске [Орлов, 1974; Кулиев, Мамедов, 1974, 1975; Мамедов, 1978, 1982; Мамедов, Агаев, 1990]. Генетичес-
Рис. 46. Форма рогов самцов безоарового и кавказских козлов и гибридных особей (по [Пфицснмай-ер, 1915]; с изменением) Верхний ряд - типичные рога С. aegagrus, С. Caucasian и С. cylindricornis, ниже - рога гибридов
ки (PCR-RFLP анализ) безоаровый козел очень сходен с домашней козой [Takada et al., 1998], что подтверждает его непосредственное участие в становлении последней. Уровень генетической изменчивости не известен.
Гибридизация. Гибриды безоаровых козлов с домашними козами плодовиты, по экстерьеру близки к дикой форме и имеют большую жизнестойкость. Плодовито и потомство, полученное при скрещивании безоарового козла с кавказскими и винторогим козлами. Гибридные самцы безоарового и кавказских козлов имеют оригинальную форму рогов (рис. 46) и отличаются от родителей в окраске [Радде, 1899; Пфиценмайер, 1915; Бутарин, 1947; Верещагин, 1947,1959а; Саркисов, 1953; Лопырин и др., 1959; Соколов, 1959; Айрумян, Гаспарян, 1986].
Распространение
Судя по ископаемым остаткам, в плейстоцене безоаровый козел населял обширную территорию, включающую Ирландию, Францию, Италию, Болгарию, Греческий архипелаг, Кавказ, Малую Азию, Иран, Ирак, Афганистан, Туркменистан, Восточный Оман, Индию и Пакистан. На Кавказ этот вид проник, скорее всего, в позднем плейстоцене из Передней Азии и распространился на север до Дагестана (до окрестностей г. Буйнакса), Кабарды, и на запад по северному склону Главного хребта, возможно, до бассейна Кубани. Костные остатки и наскальные изображения зверей периода неолита - средневековья особенно обильны в Кабристане к западу от Баку, в Армении и Туркменистане [Верещагин, 1947,1949,1959а; Пидопличко, 1951;Гептнеридр., 1961; Верещагин, Батыров, 1967; Сапоженков, 1969; Алиев, 1978; Барышников, 1978; Верещагин, Барышников, 1980а; Межлумян, Оганесян, 1986; Манасерова, 1986; Гинеев и др., 1988; Манасерян, 19976; Manaseryan, 2001]. В историческое время область распространения сильно уменьшилась и стала очаговой.
Рис. 47. Распространение безоарового козла на Кавказе в XX в. (по [Соколов, Темботов, 1993]) Точки с цифрами - зафиксированные районы обитания (см. в оригинале). На врезке - высотное распределение
На территории России находится самая северная часть современного ареала вида (рис. 47). В конце XIX - первой половине XX вв. безоаровый козел занимал восточную часть Главного Кавказского хребта от Военно-Грузинской дороги и до восточных склонов, примыкающих к Каспийскому морю, а также центральную и восточную части Малого Кавказа от Имеритинского хребта на западе (где его добывали близ Боржоми) и до Талышских гор на востоке [Радде, 1899; Динник, 1910; Гептнер, Формозов, 1941; Верещагин, 1959а; Гептнер и др., 1961]. Я. Васильев [1896] сообщал, видимо ошибочно, что бородатый козел водится и в пределах зоны обитания серны в Кубанско-Черноморском районе, но он здесь редок. Во второй половине XX в. его ареал резко сократился и стал мелкоочаговым. Самый западный пункт обитания животных - верховья р. Аргун [Темботов, 1972]. В Чечено-Ингушетии до 70-х годов вид сохранялся в районе от р. Ассы вдоль Скалистого хребта до верховий Чанты-Аргуна и Шаро-Аргу-на. В конце 80-х годов звери достоверно жили только по Чанты-Аргуну, в ущельях Кериго и Маисты-хи, на горе Байсты-лам и по Шаро-Аргуну в ущельях Цеси и Хулан-дой [Точиев, 1973; Батхиев, 1980,1989а,б; Точиев, Батхиев, 1988]. В Дагестане до 80-х годов безоаровый козел обитал в верховьях рек Аварского, Андийского и Казикумух-ского Койсу, Каракойсу и Самура, позднее - только в Аварском и Андийском Койсу (рис. 48) в высокогорных Ботлихском, Ахвахском, Цумадинском, Цунтинском, Тляро-тинском и Советском районах [Гептнер и др., 1961; Шахмарданов, Рамазанов, 1980;
Рис. 48. Распространение безоарового козла в Дагестане в конце XX в. (по [Вейнберг, 1999])
Закариев, 1982; Гинеев и др., 1988; При-луцкая, Пишванов, 1988,1989,1990; Вейнберг, 1999; Магомедов, Ахмедов, 1999].
В Грузии небольшая его группировка сохранилась на Главном Кавказском хребте в районе Хевсуретии и Мтагушетского высокогорья [Капанадзе, 1975], а также на Малом Кавказе на Аренанском хребте. Общая площадь области обитания не превышает 11 тыс. га [Арабули, 1989]. Не исключено, однако, что в последнем районе копытных уже нет [Соколов, Темботов, 1993].
На территории Азербайд жана этот вид в начале XX в. занимал почти все районы Малого Кавказа и был обычным на Большом Кавказе. Во второй половине столетия он уцелел, главным образом, в Нахичевани на хребтах Неграмдаг, Дарыдаг, Иландаг, на Шахдагском, Муровдагском
и Карабахском хребтах, а на Большом Кавказе - лишь в верховьях Гирдыманчая [Алекперов и др., 1973, 1976; Кулиев, 1977, 1981, 2004; Алекперов, Кулиев, 1981].
В недавнем прошлом безоаровый козел был широко распространен и многочислен в Армении. Сотенные стада здесь были обычными. Еще в середине XX в. его ареал простирался с юга и юго-востока от гор Мегринского, Кафанского, Го-риского и Сисианского районов на север до р. Азат и водораздельных хребтов Севанского бассейна. Звери встречались на Памбакском и Сарайбулагском хребтах, а чуть раньше, и в бассейне оз. Севан. К 70-м годам в результате интенсивного хозяйственного освоения территории область обитания существенно сократилась и раздробилась на отдельные очаги, а в северной части республики животные исчезли полностью. На Армянском нагорье ареал искусственно разделен на две части пограничными сооружениями [Даль, 1944, 1948,1951, 1954; Гаспарян, 1969, 1975; Айрумян, Гаспарян, 1973, 1976]. Тем не менее в конце XX - начале XXI вв. эти копытные еще населяли Гегамский, Урцский, Варденисский и Вайоцдзорский хребты и районы, примыкающие к Нахичевани [Казарян, 2003].
Кавказский участок ареала, несомненно, связан с малоазиатским и иранским, и через него - со среднеазиатским. В Туркменистане безоаровый козел до XX в. “во множестве” населял почти все горы. В 40-е - начале 50-х годов животные были уничтожены в Бадхызе, в 70-е годы - в Западном Копетдаге. Здесь лишь изредка регистрируют отдельных особей, вероятно мигрантов. Полностью истреблены звери также на Гязгедыке, в Кара-Калинском этрапе и Кубадаге, на грани исчезновения они на Бегарслане, Малом и Большом Балханах. В Центральном Копетдаге популяция еще многочисленна, но площадь обитания и здесь уменьшилась почти в 2,5 раза и оказалась раздробленной на отдельные участки [Карелин, 1883; Мориц, 1930; Шукуров, 1962; Кибакин и др., 1964; Ишадов, 1965, 1972,1977, 19996, 20036; Сапоженков, 1969; Горелов, 1970, 1973; Зархидзе, 1980; Коршунов, 1986, 1987, 1995а].
Повсеместно основная причина резкого сокращения ареала и численности безоарового козла - неумеренное преследование, расширение выпаса домашних животных и усиление хозяйственной деятельности человека в горах [Точиев, 1973; Саблина, 1983; Коршунов, 1995а].
Среда обитания
Безоаровый козел - горный житель, но его, как и серну, нельзя причислить к высокогорным видам. Диапазон высотного распределения весьма широк - от уровня моря на Кубадаге у Красноводска и хребте Большой Балхан и до 4500 м на Главном Кавказском хребте. На Северном Кавказе эти копытные живут обычно в лесной зоне ниже типичных мест обитания кавказских козлов на высоте 1200-2500 м, но при постоянном охотничьем преследовании держатся в высокогорье вблизи ледников и вечных снегов. Повсеместно они предпочитают невысокие скалистые горы с ксерофитной растительностью и среднегорье, где скальные выходы соседствуют с пологими безлесными участками и древесно-кустарниковыми зарослями [Радде, 1899; Динник, 1910; Гептнер, Формозов, 1941; Насимович, 1955; Верещагин, 1959а; Капанадзе, 1975; Закариев, 1982; Ахмедов, 1997а].
Типичные места обитания безоарового козла в Чечено-Ингушетии - труднодоступные изрезанные скальные массивы и гребни с наличием пещер и глубоких ниш и небольшими участками кустарника и можжевелового редколесья по травянистым склонам [Точиев, 1973; Батхиев, 1980, 1989а,б]. В Дагестане животные тяготеют к скалистым участкам лесной зоны, где самки чаще держатся на склонах гор в диапазоне 1550-1900 м, самцы - на высоте 1800-2400 м. Их биотопическое распределение в значительной мере коррелирует с расстоянием до ближайшей реки или ее притока, крутизной скалистых местообитаний (предпочитают склоны с крутизной 35-50°), служащих убежищами, а зимой приурочено к ксерофитным участкам среднего пояса гор и склонам южной экспозиции [Гептнер, Формозов, 1941; Ахмедов, 1997а, 1999; Вейнберг, 1999; Яровенко, Насрулаев, 1999; Магомедов и др., 2002].
В Азербайджане эти копытные заселяют верхнюю часть леса и субальпийские и альпийские луга [Алиев, 1962; Кулиев, 1974]. В Армении козлы уцелели лишь в больших труднодоступных горных массивах, где живут среди скал, осыпей и нагромождения камней. Летом они встречаются на высотах от 500 до 3900 м, зимой находятся ниже зоны сплошного снежного покрова, а в малоснежных районах 1фуглогодично держатся в одних и тех же местах [Даль, 1944; Гаспарян, 1969,1975; Айрумян, Гаспарян, 1976]. В Туркменистане звери преимущественно обитают в труднодоступных скалистых участках гор, покрытых зарослями арчи, в каменистых ущельях, а на горные плато выходят на короткое время только для кормежки. В Малых Балханах, однако, они живут в безлесных горах в поясе полупустынь [Мориц, 1930; Шукуров, 1962; Ишадов, 1965, 19756, 19996; Коршунов, 1995а].
Безоарового козла считают экологически более пластичным по сравнению с кавказскими козлами. Он живет в более разнообразных биотопах и сравнительно легко пересекает открытые пространства, что способствует расширению ареала. Тем не менее этот вид не смог занять западную часть Кавказа, что не находит пока разумного объяснения. Можно лишь предположить, что в более доступных горах он был полностью уничтожен человеком.
Питание
В Чечено-Ингушском регионе безоаровый козел потребляет более 160 видов растительности, преимущественно травянистой. Древесно-кустарниковые корма играют значимую роль лишь в зимнем питании [Точиев, 1973; Точиев, Батхиев, 1980]. В Дагестане в состав пищи входят не менее 104 видов травянистых растений (главным образом из семейств злаковых, бобовых, крестоцветных, зонтичных, сложноцветных, камнеломковых и осоковых) и 8 видов деревьев и кустарников [Закариев, 1982; Яровенко, Насрулаев, 1999], однако этот список далеко не полный. В Армении рацион зверей состоит из 159 видов [Гаспарян, 1964; Айрумян, Гаспарян, 1976], в Азербайджане - из 72, из них 84% - травы, 10 - кустарнички и 6% - кустарники [Кулиев, 1978, 1981]. В Туркменистане звери поедают более 38 видов растений, из которых излюбленной пищей служит арча, злаки (61%), горный клен, барбарис, эфедра и шиповник [Мориц, 1930; Ишадов, 1965; Коршунов, 1995а].
На Кавказе весной и летом звери наиболее охотно едят маки, василистник, очиток, колокольчики, беллевалию, луки, пролеску, спаржу, купырь лесной, вейники, мятлики, овсяницы, клевера, кобрезию, герани, манжетки, камнеломки, молодило кавказское; вику, ромашку, смолевку, желтушник, терескен, козлобородник, чистяк, козелец, дубровник, подмаренник, лабазник, васильки, марь, эспарцет, микро-лофус, синеголовник, птицемлечник, резеду, астрагалы и др., а также шиповник, вишню, спирею, крушину, кизильник и миндаль. Зимой в рационе преобладает ветошь трав и древесно-кустарниковая растительность: ива, ольха, береза, дуб, шиповник, жимолость, можжевельник, рододендрон, боярышник, крушина и др. [Даль, 1944, 1951; Гаспарян, 1964,1975; Кулиев, 1978, 1981; Батхиев, 1989а,б; Яровенко, Насрулаев, 1999]. Доля веточных кормов в содержимом рубцов достигает 40-65% [Магомедов и др., 2002]. Уровень переваримости кормов в экспериментах составляет 49-68% [Яровенко и др., 2003].
Обычно животные довольно долго пасутся на кормном участке среди каменистых россыпей, тщательно объедая растения и порой выгрызая их до земли. При высоком снежном покрове, в отличие от кавказских козлов, они не раскапывают снег в поисках пищи, а довольствуются древесно-кустарниковым кормом [Мориц, 1930; Ахмедов, 1997а]. Солонцы они посещают круглогодично, особенно интенсивно - весной [Даль, 1951; Кулиев, 1978]. В жару остро нуждаются в водопоях, которые могут посещать несколько раз в день [Коршунов, 1995а].
Летом звери кормятся с 4—6 ч утра до 8-11 ч, затем уходят на отдых, а вечером активны с 16-19 ч до темноты. При постоянном беспокойстве пасутся преимущественно в сумерках и ночью. Такой тип активности более характерен для взрослых самцов. Зимой козлы активны чаще в дневное время [Даль, 1951; Гаспарян, 1964, 1975; Кулиев, 1974, 1981].
Размножение
Половая зрелость. Самцы безоарового козла становятся половозрелыми на третьем году жизни, но в размножении принимают участие не ранее 4—5-летнего возраста, поскольку во время гона вытесняются из стад более сильными соперниками.
Часть самок первое потомство приносит в двухлетнем возрасте, но большинство - в трехлетием [Мориц, 1930; Коршунов, 1995а].
Гон. К периоду гона, который начинается в ноябре и завершается обычно к середине декабря (табл. 44), группы самцов и самок объединяются. При высокой плотности популяции в смешанных стадах, состоящих из нескольких десятков особей, находятся несколько крупных козлов, которые постепенно изгоняют неполовозрелых и слабых самцов, и покрывают самок. При низкой плотности в группах самок с молодняком, редко превышающих 10 особей, находится один крупный самец и, иногда, молодой, не принимающий участие в размножении [Коршунов, 1995а].
Таблица 44. Сроки гона у безоарового козла
Район Начало Конец Источник информации
Кавказ Ноябрь Начало декабря Динник, 1910
Ч.-Ингушетия 10 декабря Начало января Батхиев, 1989а
Дагестан Середина ноября Середина декабря Закариев, 1982
17 декабря 21 января Вейнберг, 1999
Грузия Середина ноября Середина января Эквтимишвили, 1954а
Армения Вторая декада — Гаспарян, 1975
декабря
Азербайджан Начало ноября Вторая половина Кулиев, 1974,1981
декабря
Туркменистан Конец ноября Первая половина Мориц, 1930; Ишадов, 1965
декабря
Ц. Копетдаг 1 ноября 15 декабря Коршунов, 1988а
Беременность и деторождение. Судя по срокам гона и отела, беременность у самок безоарового козла длится около 5 месяцев. Деторождение на Северном Кавказе происходит в мае - июне, в Закавказье начинается в апреле, а в Туркменистане наиболее ранние окоты зарегистрированы в конце марта - начале апреля, самые поздние - в июле (табл. 45). Перед родами самки отделяются от стада и уходят в наиболее безопасные места в скальниках. Детеныши рождаются хорошо развитыми. В первые дни они затаиваются, а затем неотступно следуют за матерями по горным тропам [Динник, 1910; Мориц, 1930; Вейнберг, 1999].
Таблица 45. Сроки деторождения у безоарового козла
Район | Начало | Конец | Источник информации
С. Кавказ Май Июнь Динник, 1910
Закавказье Апрель - (С
Дагестан Вторая половина июня Первая половина июля Вейнберг, 1999
Грузия Середина апреля Конец июня Эквтимишвили, 1954а
Азербайджан 18 апреля Конец мая Кулиев, 1974, 1981
Армения Середина мая (15-20 апреля) ' Середина июня Саркисов, 1944; Даль, 1951, 1954; Гаспарян, 1975
Туркменистан Апрель (конец марта) Май (июль) Мориц, 1930; Шукуров, 1962; Коршунов, 1995а
Плодовитость. Самки безоарового козла весьма плодовиты. В благоприятные годы они часто приносят двух козлят а, изредка, и тройни [Динник, 1910; Мориц, 1930; Эквтимишвили, 1954а; Шукуров, 1962; Ишадов, 1965; Кулиев, 1974, 1981; Вейнберг, 1999]. На Северном Кавказе к концу периода деторождения на каждую взрослую самку приходится 1,2-1,6 козленка. Самки с одним детенышем составляют 44-47%, с двойнями - 32—40, с тройнями - до 6, яловые - 13-18% [Вейнберг, 2001; Магомедов и др., 2004]. В Копетдаге потенциальный приплод на самку - 1,2, яловость не превышает 20% [Коршунов, 1983, 1995а]. В Армении значительная часть самок остается непокрытыми из-за недостатка самцов в период гона в районах с интенсивным выпасом скота и в местах браконьерских охот [Гаспарян, 1975]. В течение года повсеместно регистрируют не более 0,5-0,9 детеныша на самку, что свидетельствует о высокой детской смертности (табл. 46).
Таблица 46. Число детенышей на одну самку в популяциях безоарового козла (по визуальным наблюдениям)
Район Взрослых самок, и Сеголеток, п Число детенышей на самку Источник информации
Восточный Кавказ 2429 1360 0,6 (0,5-0,7) Магомедов и др., 2001
Дагестан 48 45 0,9 Ахмедов, 19976, 1999
а 159 124 0,8 Магомедов и др., 2004
Азербайджан 273 167 0,6 Кулиев, 1981
Ц. Копетдаг 572 287 0,5* Коршунов, 1988а, 1995а
ч 594 295 0,5** сс
* В период от отела до осени.
** В период с осени до весны.
Популяционная структура
Половой и возрастной состав. Соотношение полов у молодых особей безоарового козла, как и у других козлиных, близко 1:1, но с возрастом оно меняется чаще в сторону преобладания самок, что, как правило, вызвано преимущественным отстрелом браконьерами взрослых самцов. В составе группировок чаще от 15 до 30% сеголеток, 15-18% годовалых особей, 22-50% взрослых самок и 22-32% взрослых самцов (табл. 47).
Социальная организация. Образ жизни безоарового козла преимущественно групповой. Самки с детьми и молодые животные обоих полов с весны и до осени обычно держатся отдельно от групп взрослых самцов, которые встречаются и по одному. В гон образуются смешанные стада. После гона самцы отделяются, но часть их, особенно молодые, остаются с самками (рис. 49). Состав стад не постоянен.
В отличие от кавказских козлов, безоаровый козел редко образует многочисленные стада. Средний размер групп чаще не превышает десяти особей, при высокой плотности - двух десятков, а максимальный (в Туркменистане) - не более сотни особей (табл. 48).
В горах Чечни и Дагестана большинство животных держится в группах из 2-5 особей. Средний показатель стадности не превышает 4,2, максимальный размер
Таблица 47. Половозрастной состав (%) и соотношение полов в популяциях безоарового козла
Район (источник информации)* Встречено животных, п Сеголетки Годовалые Самки Самцы Самцы : самки
Чечня (1) 99 10 — 65 25 1:2,6
Дагестан (8) 148 30,4 14,9 32,4 22,3 1:1,45
(3) 469 25-27 16-18 34 22-23 1:1,5
(7) 502 25 16 31 28 1:1,1
Азербайджан (4) 1194 15 26 (9,2) 22 28 1:0,7
Армения (5) — — — 61-72 28-39 1:1,8-2,2
(6) 354 36,4** ** 31,4 32,2 1:1,0
Ц. Копетдаг(2) 2835 26,2 — 49,9 23,8 1:2,1
* 1 - Точиев, Батхиев, 1988; 2 - Коршунов, 1988а, 1989,1990,1995а; 3 - Ахмедов, Насруллаев, 2003; Магомедов и др., 2004; 4 - Кулиев, 1977; 5 - Гаспарян, 1975; 6 - Даль, 1951; 7 - Вейнберг, 1999; 8 - Ахмедов, 1999.
** Сеголетки и годовалые вместе.
группы зимой - 15 голов [Точиев, Батхиев, 1988; Вейнберг, 1999]. В Нахичевани число животных в стаде колеблется от 7 до 30, достигая максимума к периоду гона и минимума во время деторождения [Кулиев, 1974]. В Центральном Копетдаге на заповедной территории средняя величина стад относительно одинакова в течение года - около 15 особей, и лишь в зимне-весенний период увеличивается до 19. Среднегодовой показатель стадности (16 особей) резко снижается при уменьшении плотности популяции и факторе беспокойства, и в эксплуатируемой группировке он в два раза меньше (7,5 особей), чем в заповедной. В популяции почти круглогодично преобладают смешанные стада, доля которых максимальна (63%) в зимне-весенний период и минимальна (31%) в период окота. Самцовые группы
Рис. 49. Стадо безоаровых козлов (фото А. Белова)
Таблица 48. Размер групп у безоарового козла
Район Число особей в группе Источник информации
среднее максимальное
Ч.-Ингушетия 4,2 15 Точиев, Батхиев, 1988; Батхиев, 1989а,б
Дагестан 2,6-4,1 14 Вейнберг, 1999
Азербайджан 7-11 42 Кулиев, 1974, 1977, 1981
Армения 10 35 Айрумян, Гаспарян, 1976
<< Чаще 2-6 31 Даль, 1951
Туркменистан 8,5 25 Ишадов, 1965, 19756
Копетдаг 16 18*, 99 Коршунов, 1988а, 1990, 1995а
Б. Балханы - 10 Шукуров, 1962
— - 70 Зархидзе, 1980
* В группах самцов.
составляют около 2% зимой и ранней весной и 12-15% в летне-осенний период. Беременные самки покидают стада перед родами, но вскоре после окота объединяются сначала в мелкие, а затем и в крупные группы, в состав которых постепенно входят нерожавшие самки и самцы. Доля отдельных групп самок с молодняком летом в три раза выше (34%), чем зимой. В период гона большинство животных находятся в смешанных стадах. Одиночные звери редки во все сезоны года. Особи в группах обычно родственные или хорошо знакомые, что способствует стабилизации их состава. Лидирует в смешанных стадах взрослая самка. За ней следуют самки с молодняком и молодые самцы, затем самый крупный самец-доминант, за ним самцы-субдоминанты, второстепенные самцы и молодые самки. При опасности лидером чаще становится опытная самка или молодой самец, а старые самцы замыкают стадо, нередко останавливаясь и оценивая ситуацию. На водопое первым утоляет жажду доминантный самец и его окружение, затем взрослые самки с потомством и последними - молодые особи [Коршунов, 1988а, 1995а].
Безоаровый козел, несмотря на хорошо развитые обоняние, слух и зрение, в осторожности все же уступает кавказским козлам [Динник, 1910], и, обитая в лесистой местности на меньшей высоте, чаще становится жертвой охотников. По сравнению с кавказскими козлами, у него более сложный комплекс импонирования, и он чаще использует звуковые сигналы. Соотношение составляющих элементов поведения распределяется следующим образом: голосовые сигналы - 28%, резкое опускание головы - 17, высовывание языка - 23, удар ногой - 28, спурт - 3,8, поза “вытянувшись” -0,2% [Эрнитс, 1982].
Пространственная структура и перемещения. Безоаровый козел - оседлое животное, живущее чаще в пределах одного-двух ущелий. Перемещения, за исключением периода гона, носят локальный характер. Суточные переходы короткие - от нескольких сот метров до 1-2 км по горизонтали и до 100-200 м в высотном диапазоне из скальных убежищ на пастбище или к водопоям и обратно. Сезонные миграции, которые, скорее всего, раньше имели место, теперь не выражены. Стадиальная половозрастная сегрегация, в отличие от кавказских козлов, незначительна, что, возможно, связано с ограниченностью пригодных для обитания угодий. Тем не менее в Дагестане самки, в отличие от самцов, предпочитают северные склоны с большей крутизной в летний период, и с меньшей - в зимний. В горах Азербайджана самки с
потомством и годовалые особи держатся на высоте 900-1200 м, прохолоставшие самки и молодые самцы - в диапазоне 1800-2500 м, а взрослые самцы - выше 3500 м. Нижняя граница высотного распределения, особенно у взрослых самцов, в значительной мере зависит от уровня антропогенного пресса [Гаспарян, 1975, 1981; Коршунов, 1988а; Вейнберг, 1999; Магомедов и др., 2002].
В спокойной ситуации звери флегматичны и малоподвижны. При бегстве по относительно ровной местности развивают скорость до 46 км/час, но очень быстро устают. При опасности стремятся скрыться в скалах, где могут перепрыгивать пропасти шириной в 13,5 м, а в высоту способны запрыгнуть на 2-3 м [Гаспарян, 1969; Гамбарян, 1972].
Численность и основные факторы, ее определяющие
Численность. Общая численность безоарового козла в Чечено-Ингушетии в 70-е годы XX в. оценивалась в 250-600 голов, в 80-е - в 350-360. В местах обитания средняя плотность составляла 6-9 особей на 1000 га [Точиев, 1973, 1975; Рав-кин, 19756; Батхиев, 1980,1989а; Саблина, 1983]. В Дагестане в 70-80-е годы держалось около 1 тыс. особей: в Ахвакском районе - 40-60, Цумадинском - 200-300, Советском - 200, Тляротинском - 60, Ботлихском - 40-60, на Богосском хребте в Цунтинском районе - 600 [Равкин, 1976; Прилуцкая, Пишванов, 1988,1989,1990]. К концу 90-х годов область обитания сократилась, но в Аварском и Андийском Койсу сохранилось примерно столько же зверей с плотностью 9-21 экземпляров на 1000 га [Магомедов, Ахмедов, 1999; Магомедов и др., 2004], а, возможно, и больше-до 1,5 тыс. с плотностью 130-170 особей на 1000 га лесных угодий [Вейнберг, 1999, 2001]. В начале 2004 г. здесь насчитали 1156 голов [Кричевский, 2004]. Всего на Северном Кавказе в конце XX в. жили около 1,3-1,6 тыс. безоаровых козлов [Гинеев и др., 1988; Вейнберг, 1997]. В последние годы очевидна тенденция к дальнейшему сокращению площади обитания и численности этих копытных [Магомедов и др., 2002].
В Грузии в 70-80-е годы сохранилось около 250-300 особей [Капанадзе, 1975, 1978; Соколов, 1986; Арабули, 1989]. В Армении тенденция быстрого сокращения ресурсов вида стала очевидной с середины XX в., но в некоторых районах плотность его населения в этот период достигала 5-53 экземпляра на 1000 га [Даль, 1944, 1951; Гамбарян и др., 1963]. В 70-е годы здесь насчитывали около 400-700 особей (с учетом заходящих из Нахичевани) [Айрумян, Гаспарян, 1973, 1976; Гаспарян, 1975]. По последним данным, в Армении и Карабахском нагорье обитают около 2-2,5 тыс. козлов [Казарян, 2003]. В Азербайджане в первой половине столетия нередко встречали стада в 100—150 особей [Верещагин, 1947]. Позднее поголовье держалось на уровне 1-2,4 тыс. при средней плотности около 8, а в Нахичевани - до 15-17 экземпляров на 1000 га [Алиев, 1961; Алекперов и др., 1973, 1976; Кулиев, 1977, 1981, 1997; Алекперов, Кулиев, 1981]. В начале XXI в. здесь осталось всего около 600-650 зверей [Кулиев, 2004].
В Туркменистане к середине XX в. безоаровый козел был истреблен во многих районах (см. выше). В 60-е годы насчитывали 2-2,5 тыс. особей. Позднее в результате улучшения охраны численность выросла. В Центральном Копетдаге популяция увеличилась до 6,6 тыс., в том числе до 2 тыс. - в Копетдагском заповеднике. В начале 90-х годов на Малом Балхане оставалось 20 голов, на главном хребте Большо
го Балхана-не более 100. Всего в республике обитало примерно 8 тыс. особей [Иша-дов, 1965, 19996; Коршунов, 1986, 19886, 1995а; Вейнберг, 1997]. Позднее ситуация резко ухудшилась, и на Центральном Копетдаге осталось не более 2-2,5 тыс. [Лука-ревский, 1996].
Ресурсы этого вида в СССР в середине XX в. оценивали в 20-25 тыс. [Банников, 1962], в конце столетия на той же территории осталось не более 10 тыс., а в России - 1-1,5 тыс. Примерно такая же численность (около 10 тыс.) реинтродуци-рованного в 1914—1921 гг. безоарового козла в Швейцарии, где он является охотничьим видом [Lups, 1985]. Мировые запасы, видимо, достигают 50 тыс. особей [Вейнберг, 2001].
Воспроизводство. Воспроизводство в стадах безоарового козла относительно стабильно. Динамика популяций в большей мере определяется уровнем смертности, зависящим от многих факторов.
Погода и корма. Зимой часть сеголеток погибает от недостатка кормов [Магомедов и др., 2004]. Гибель зверей и откочевка части группировок обычна и в особенно засушливые годы [Коршунов, 1995а].
Хищники. Основные враги этого копытного на Кавказе - волк, рысь, для детенышей - беркут и орел-ягнятник [Закариев, 1982; Батхиев, 1989а], в Средней Азии — леопард, на долю которого приходится до 86% от числа погибших особей, и потенциально - гиена, лисица и рысь. Леопард чаще давит самцов (68%), существенно влияя на половой состав популяции жертвы [Мориц, 1930; Коршунов, 1989, 1995а].
Конкуренты. У безоарового козла, населяющего труднодоступные скальные участки, конкуренция с дикими баранами и кавказскими козлами, выпасающимися на более пологих участках, сведена к минимуму. Большее значение имеет, видимо, беспокойство, причиняемое домашним скотом, собаками и людьми, которые фактически уже вытеснили его с лучших пастбищ.
Болезни, паразиты, травмы. Болезни безоарового козла практически не изучены, и их роль в динамике популяций не ясна. На Кавказе у него обнаружено более 20 видов гельминтов - нематод и цестод, и зараженность ими сравнительно высокая [Закариев, 1982; Батхиев, 1989а]. В Туркменистане на этих копытных паразитирует 24 вида гельминтов из 7 семейств, личинки подкожного и носоглоточного овода, кровососки и 6 видов клещей [Кибакин и др., 1964; Коршунов, 1995а]. В целом па-разитофауна козла достаточно обширная, но вряд ли она оказывает существенное влияние на популяции.
В многоснежные годы эти звери, как и другие горные копытные, гибнут под обвалами и в лавинах.
Антропогенные факторы. Безоаровый козел, наряду с кавказскими козлами, издавна был одним из основных охотничьих объектов, добываемых ради вкусного мяса и прочной кожи. В Европе и Передней Азии на него охотились и ради минерализованных образований в желудках (“безоаров”), которым приписывались магические свойства [Гмелин, 1785]. Отсюда, видимо, и происходит название козла-безоаровый. Рога животных использовали для украшения жилищ, а отдельные племена туркмен устанавливали их над могилами предков [Коршунов, 1995а].
В 30-е годы XX в. его добыча по всему Кавказу достигала 4 тыс. особей [Верещагин, 1947]. В Туркменистане в начале 40-х годов ежегодно отстреливали тысячи
зверей. В Копетдаге одна бригада охотников с ноября по февраль добывала до 362-374 козлов [Цалкин, 1948; Шукуров, 1962].
Неумеренная добыча стала главной причиной сокращения численности вида. Уровень современной нелегальной охоты наглядно характеризуют следующие сведения. В Дагестане в 2001-2002 гг. из 72 погибших безоаровых козлов жертвами браконьеров стали 68, и только 4 зверя пали от естественных причин. Копытных отстреливают круглогодично, независимо от пола и возраста, но выбирают все же крупных самцов [Кричевский, 2004; Магомедов и др., 2004]. С середины XX в. охота на этих копытных запрещена во всех республиках бывшего СССР. Безоаро-вый козел внесен в Красные книги МСОП (1996), Российской Федерации (2001), Туркменистана (1985, 1999) и других республик, однако его поголовье не восстанавливается, главным образом, из-за браконьерства, вытеснения людьми и домашним скотом с лучших пастбищ и вырубки древесно-кустарниковой растительности в типичных местах их обитания [Даль, 1951; Гамбарян и др., 1963; Гаспарян, 1964, 1975; Алекперов и др., 1973; Батхиев, 1980, 1989а; Точиев, Батхиев, 1988; Коршунов, 1995а; Вейнберг, 1999, 2001; Ишадов, 19996; Казарян, 2003].
Перспективы сохранения. При неизменной ситуации этот древнейший вид будет неизбежно уничтожен! Сохранение животных реально только на особо охраняемых территориях, которых явно недостаточно, и в специализированных питомниках, где они, как показала практика, вполне нормально живут и размножаются [Антонян, Эгнатосян, 1984]. При улучшении охраны (во что верится с трудом) возможна масштабная реинтродукция копытных во все утраченные участки ареала.
ЗАПАДНОКАВКАЗСКИЙ И ВОСТОЧНОКАВКАЗСКИЙ КОЗЛЫ*
Capra caucasica Guldenstaedt et Pallas, 1783 Capra cylindricornis Blyth, 1841
Capra caucasica Guldenstaedt et Pallas, 1783. Acta Acad. Sci. Petropol. (1779), 3, 2: 273. Кабардино-Балакария, водораздел рек Малка и Баксан.
Ovis cylindricornis Blyth, 1841. Proc. Zool. Soc. London (1840): 68. Грузия, Главный Кавказский xp., г. Казбек.
Aegocerospallasii Rouillier, 1841. Bull. Soc. Nat. Moscou, 14: 910. “Кавказ, Дживки”. Nom. praeocc., non Schinz, 1838.
Capra severtzowi Menzbier, 1888. Proc. Zool. Soc. London (1887), 6: 618. Карачаево-Черкесия, горы к зап. от Эльбруса и к югу от Теберды.
Capra raddei Matschie, 1901. Sitzungsb. Ges. Naturf. Freunde. Berlin, I: 32. Грузия, верхи, течение p. Ингури.
Capra dinniki Satunin, 1905. Zool. Anz., 29, 11: 344. Адыгея, верхи, течение p. Белая.
* Кавказские козлы рассматриваются вместе в целях сравнения в связи с дискуссионным таксономическим статусом. Их также нередко называют “турами”, что приводит к путанице с собственно туром (Bos primigenius Boianus, 1827), истребленным в XVII в. В научной литературе это наименование желательно нс употреблять.
Рис. 50. Череп самца западнокавказского козла (экземпляр из б. Кубанской области;
Зоомузей МГУ)
Вид сбоку, сверху и снизу
Характеристика. Козлы крупных размеров, массивного сложения с толстой недлинной шеей и коротким (до 17 см) хвостом. Максимальная длина тела самцов -196, самок - 164 см, высота в холке, соответственно, НО и 93 см, масса - 155 (192) и 72 кг.
Окраска летом рыжеватая или бурая, зимой - серо- или темно-бурая. Темного ремня на спине и у основания шеи нет. Борода у самцов зимой сравнительно широкая и длинная (до 12-18 см). Волос на лбу удлиненный. Роговые мозоли на тыльной стороне запястных суставов отсутствуют.
Зароговая часть черепа укороченная, без вогнутости на верхней поверхности (рис. 50). Максимальная длина черепа самцов 326 мм, самок - 265 мм. Предкоренные зубы заметно редуцированы. У самцов костные роговые стержни относительно короткие (до 51 см), толстые в основании (до 31 см в окружности), широко расставленные.
Рога разной формы. Роговые чехлы массивные, с широким основанием (до 36 см в обхвате), широкой лобовой поверхностью и закругленными ребрами. У западнокавказского козла они круто дугообразно изогнуты в одной плоскости, а их концы направлены внутрь или с небольшим отклонением наружу (фото 22), у восточнокавказского изогнуты пологой спиралью со сближающимися концами (фото 23). Между этими крайними типами рогов есть ряд переходов (рис. 54). В основании
рога расходятся в стороны под углом более 45°. Поперечное сечение оснований округлое у восточнокавказской расы животных или со слабо выраженным передним ребром у западнокавказской, концы округло-треугольной формы. На передней поверхности чехлов крупные складки или волнообразные морщины с небольшими буграми в основании. У самок рога тонкие и короткие (до 33 см), со слабым загибом, эллиптической формы в сечении.
Хорошо развиты подхвостовые железы, усиленно функционирующие в период гона [Соколов, Чернова, 2001].
Горные стадные животные, предпочитающие жить в скальных выходах в лесном, субальпийском и альпийском поясах. Весьма медлительные и флегматичные звери, обычно передвигающиеся шагом или неуклюжей рысью, при испуге переходят в тяжелый галоп. Скорость бега по ровному месту невелика - до 30-40 км/ч [Гаспарян, 1969], но в скалах козлы исключительно ловки, точны и быстры. Обладают прекрасным зрением, обонянием и слухом. Продолжительность жизни - 20-23 года, но в природе редко доживают до 15 лет.
Охотничьи или охраняемые звери.
Распространение. Главный Кавказский хребет и его боковые хребты (рис. 51).
Состав. Особенно дискуссионный (см. ниже). Достаточно отчетливо выделяются лишь крайние формы: западнокавказская, или кубанская (caucasicd), и восточнокавказская, или дагестанская (cylindricornis).
Рис. 51. Ареалы восточнокавказского (1)'и западнокавказского (2) козлов (по [Гептнер и др., 1961]).
3 - крайний западный пункт обитания животных в середине XX в., 4 - места прежнего обитания, 5 - место выпуска (в Боржоми)
Изменчивость и таксономия
Размеры и масса. Масса новорожденных кавказских козлов колеблется в пределах 3-4,7 кг. Ее удвоение происходит через 40 дней, последующее - через 65-70. Зимой прирост массы замедляется до 0,1 кг в месяц. Длина тела увеличивается за месяц примерно на 3 см. Половые различия в линейных размерах животных заметны на втором году жизни, в массе - на третьем. В Кавказском заповеднике и окрестностях длина тела двухлетних самцов достигает 115-156 (138 в среднем), высота в холке - 69-93 (83), у самок - 115-130 (124) и 71-79 (74,5) см [Котов, 1968]. На Восточном Кавказе у их сверстников-самцов длина около 110, высота в холке - 68, а у самок - 100 и 65 см соответственно [Магомедов и др., 2001]. Рост сацок прекращается к 4-5 году жизни, тогда как у самцов линейные размеры относительно стабилизируются к 6-7 годам, а масса - в 10-11 -летнем возрасте (рис. 52) [Лопырин и др., 1960; Залиханов, 1967, 1968; Ахмедов, Яровенко, 1991; Магомедов и др., 2001].
Масса самцов восточнокавказского козла в возрасте 4—6 лет составляет в среднем 71 кг, в 6-11 лет - 99, в 12-16 лет - 139 кг; у самок от 2 до 4 лет - 36, от 6 до 9 лет- 56 кг [Абдурахманов, 1973, 1977]. Масса самцов старше 6 лет западнокавказской формы колеблется в пределах 123-155 (в среднем 143) кг, взрослых самок - 58-71 (66) кг [Котов, 19646, 1968]. Вес отдельных самцов, в исключительных случаях, возможно достигает 192 кг [Воронцов-Вельяминов, 1929].
В.И. Цалкин [1955] полагал, что заметных различий в размерах тела между западне- и восточнокавказским козлами нет. Действительно, судя по имеющимся сведениям, эти звери почти одинаковы и по линейным размерам (табл. 49), и по массе (табл. 50). Наиболее крупные особи встречаются в заповедниках, а в районах интенсивной охоты они крайне редки. Это обстоятельство следует учитывать при сравнении средних показателей животных из разных районов.
Рис. 52. Изменение массы (А) и длины тела (Б) восточнокавказского козла с возрастом (по [Магомедов и др., 2001])
Самцы Самки Район, заповедник (источник информации)**
п 1 М 1 lim п 1 м 1 lim
Длина
23 178,7 159-196 63 145-150 136-164 Западный Кавказ (3)***
16 171,7 147-187 9 139,0 128-146
2 158,5 152-165 2 141,0 140-142 Кавказский з-к (4)
- 172 — - 143 - Кабардино-Балкария (7)
2 160,5 143-178 1 - 146 Северная Осетия (2)
- - -170 6 — -119? Дагестан (1)
- 189,8 - - 138,0 - (8)
- - - - 146,6 - Азербайджан (10)
Высота в холке
28 101,3 93-108 63 84-87 78-92 Западный Кавказ (3)
16 100,3 101-110 9 87,0 82-93 -“-(3)
2 95,5 92-99 2 88,0 87-89 Кавказский з-к (4)
1 — 77,5 - - - Тебердинский з-к (5)
- 104 — - 82 - Кабардино-Балкария (7)
2 90,0 81-99 1 - -83 Северная Осетия (2)
2 100,0 99-101 — — — -“-(6)
10 91,7 86-90 5 76,4 71-80 Дагестан (1)
- 104,8 — — 85,2 — -“-(8)
Обхват туловища
28 118,5 106-128 63 96-101 92-114 Западный Кавказ (3)
16 136,3 128-150 9 104 92-112 -“-(3)
2 124,0 123-125 - - — Кавказский з-к (4)
1 — 130 — - — Тебердинский з-к (5)
2 105,5 97-114 - - — Северная Осетия (2)
2 116,5 116-117 — — __ -“-(6)
12 114,2 99-130 6 92 82-101 Дагестан (1)
- 148,2 - - 98,7 — -“-(8)
1 - 116 - — - Азербайджан (9)
- — - — 95,7 — -“-(Ю)
Длина ступни
28 37,8 36—41 63 33 31-35 Западный Кавказ (3)
16 38,1 36-42 9 32 30-33 -“-(3)
12 35,5 32-37 6 31 27-32 Дагестан (1)
1 — 35 — - - Азербайджан (9)
Длина уха
28 12,3 10-14 63 11,5 10-13 Западный Кавказ (3)
16 и,з 10-13 9 11,0 9-12 -“-(3)
2 12,2 11-13,5 2 11,5 11-12 Кавказский з-к (4)
2 11,5 11-12 - — - Северная Осетия (6)
7 12,4 12-13 6 12,2 12-13 Дагестан (1)
Длина хвоста
28 14,3 12-17 63 12,0 8-14 Западный Кавказ (3)
16 14,6 13-16 9 12,0 11-14 -“-(3)
2 13,5 13-14 2 11,5 10-13 Кавказский з-к (4)
2 14,5 14-15 — — — Северная Осетия (6)
8 13,5 11-17 6 11,8 10-13 Дагестан (1)
3 14,5 13-16 - - Азербайджан (9)
* Взрослые особи: самки - 4 года и старше, самцы - 5-6 лет и старше.
**1 - Магомедов и др., 2001; 2 - Вейнберг, 1984; 3 - Котов, 19646, 1966а,б, 1968; 4 - Цалкин, 1955; 5 - Лопырин и др., 1960; 6 Наниев, 1958; 7 - Залиханов, 1967; 8 - Абдурахманов, 1973; 9 - Верещагин. 1938; 10-Кулиев, 1981.
*** Размеры отловленных и отстрелянных особей.
Таблица 50. Масса (кг) взрослых особей кавказских козлов
Самцы Самки Район, заповедник (источник информации)*
п м 1 lim п м 1 lim
14 143 123-155 9 65,5 58-71 Западный Кавказ (3)
1 — 79 2 - 42-51 Кавказский з-к (8)
2 - 113-114 - - - Тебердинский з-к (4, 7)
- 125 -140 - 75 - Кабардино-Балкария (5)
14 99 62-148 8 51,5 39-68 Дагестан (1)
— 99-139** -149 - 56 -72 -“-(2)
1 91 - - 54 Азербайджан (6)
* 1 - Магомедов и др., 2001; 2 - Абдурахманов, 1973, 1977; 3 - Котов, 1966а, 1968; 4 - Лопырин и др., 1960; 5 - Залиханов, 1967; 6 - Верещагин, 1938; 7 - Мишарев, 1969; 8 - Ромашин, 2001.
** В разных возрастных группах.
Верхнеплейстоценовые кавказские козлы были, скорее всего, крупнее современных, что видно при сравнении размеров метаподий, однако основные морфофизиологические черты древних и современных животных сходны [Верещагин, 1959а].
Краниометрическая характеристика. Зубная система кавказских козлов достигает полного развития к 4—5 годам [Верещагин, 1938], к этому времени в основном завершается и рост костей черепа.
По основным линейным признакам черепа западная и восточная формы кавказских козлов весьма близки (табл. 51). Дискретных отличий не удается выявить даже при многомерном анализе выборок (рис. 42). Тем не менее дистанция Маха-лонобиса, разделяющая эти две формы, не меньше, чем аналогичные дистанции между С. sibirica, С. ibex и С. aegagrus [Казанская, 2004].
Н.К. Верещагин [1959а] нашел, что шов теменной и лобной костей у западнокавказского козла образует прямую линию, тогда как у восточнокавказского он имеет форму тупого угла. З.И. Члаидзе [1966] обнаружил в черепе восточнокавказского козла добавочную ромбовидную кость в промежутке между носовыми и лобными костями. Чуть реже эта косточка встречается у западнокавказского козла, а у безоарового обнаруживается даже чаще, чем у восточнокавказского. У С. sibirica, С. ibex и С. falconeri она, однако, отсутствует [Казанская, 2004].
Изменчивость рогов. У кавказских козлов рога начинают расти вскоре после рождения. К концу первого месяца жизни детенышей их длина достигает 1 см, к 4 мес. - 7 см. У самцов они растут быстрее, чем у самок. В летний период ежемесячный прирост у первых составляет 2 см, у вторых - 0,85 см, зимой - 0,46 и 0,07 см соответственно [Ахмедов, Яровенко, 1991; Магомедов и др., 2001]. У самок рост рогов замедляется с 3-летнего возраста, что, возможно, связано с их участием в размножении [Вейнберг, 2002]. У самцов они интенсивно растут до 7-летнего возраста, затем их прирост составляет 2-3 см в год [Котов, 1968].
Определение пола и возраста животных в полевых условиях по форме и размерам рогов не представляет сложности, за исключением идентификации са-
Самцы Самки Район,заповедник (источник информации)*
п | М lim п 1 м 1 lim
Наибольшая длина
9 292,5 284-310 10 252,0 240-265 Западный Кавказ (1)
18 283,5 266-326 10 250,2 227-265 (5)
- 284 -295 - 249 - Кабардино-Балкария (4)
17 298,7 280-310 13 249,8 235-265 Восточный Кавказ (1)
- 263-267 - - 243,4 - -“-(2)
15 294,5 273-320 8 254,3 244-267 (5)
Максимальная ширина
34 155,3 137-170 13 136,4 122-143 Западный Кавказ (5)
27 158,3 140-177 13 135,1 128-144 Восточный Кавказ (5)
Межглазничное расстояние
34 111,6 94-125 12 95,7 89-103 Западный Кавказ (5)
28 114,8 93-131 11 93,2 88-100 Восточный Кавказ (5)
Расстояние от основания рога до края глазницы
34 21,1 12-28 14 31,6 22-39 Западный Кавказ (5)
28 18,1 12-22 11 31,7 28-35 Восточный Кавказ (5)
Длина лицевой части
18 215,0 196-234 11 193,6 178-208 Западный Кавказ (5)
14 223,1 207-241 8 199,3 182-213 Восточный Кавказ (5)
Ширина мозговой коробки
33 84,7 75-96 14 73,0 67-78 Западный Кавказ (5)
27 87,0 78-95 11 74,6 72-77 Восточный Кавказ (5)
Длина носовых костей
29 111,7 96-124 12 100,6 86-116 Западный Кавказ (5)
34 107-123 - - - — Центральный Кавказ (3)
26 116,4 91-149 9 101,2 87-106 Восточный Кавказ (5)
Длина верхнего ряда зубов
32 79,2 71-87 14 75,2 72-82 Западный Кавказ (5)
- 75-78 - - - - Центральный Кавказ (3)
27 81,0 75-86 11 74,0 64-80 Восточный Кавказ (5)
- 74-84 - - 74,2 - -“-(2)
Длина костного стержня рогов
— — 300-400 Западный Кавказ (1)
24 314,2 185—430 10 91,0 80-110 -“-(5)
— — -510 Восточный Кавказ (1)
- — -421 -“-(2)
24 368,8 210—480 8 86,3 60-105 -“-(5)
Обхват костного стержня рогов
32 219,7 185-265 12 97,8 60-110 Западный Кавказ (5)
23 248,6 184-310 8 94,4 85-110 Восточный Кавказ (5)
* 1 - Цалкин, 1955; 2 - Абдурахманов, 1973; 3 - Темботов, 1974; 4 - Залиханов, 1968; 5 - Казанская, 2004.
мок и 2-3-летних самцов. Однако у последних рога все же больше отклонены в стороны, а их основания толще. У 4-5-летних самцов концы рогов широко расставлены и загнуты внутрь, с 7 года ^сизни они начинают загибаться вверх, а к 11-12 годам утолщаются, затупляются и стираются (рис. 53). Нередко встречаются особи со сломанными или неправильно сросшимися рогами, у многих обломаны концы, а у отдельных самцов есть дыры на стенках оснований рого-
Рис. 53. Возрастная изменчивость формы рогов у восточнокавказского козла (по [Магомедов и др., 2001])
А - вид спереди, Б - вид сбоку; а - взрослая самка; б-ж - самцы (б - 2-3 года, в - 4—5 лет, г -6-7, д - 8-9, е - 10-12 лет, ж - более 13 лет)
вых чехлов. Происхождение дыр, возможно, связано с заражением животных личинками носоглоточного овода, что вынуждает животных постоянно чесаться об острые кромки скал и стволы деревьев [Бобырь, Бобырь, 19996].
Форма рогов у взрослых самцов на западе и востоке ареала неодинакова. У особей западнокавказской формы (от Зеленчука до бассейна р. Белой) они преимущественно саблеобразно изогнуты в одной плоскости (в отдельных стадах - бокаловидные), на передней поверхности бугристые, складчатые или морщинистые (у некоторых животных - почти гладкие [Котов, 19646]), их сечение округло-треугольной формы. У восточнокавказской расы они изогнуты пологой спиралью вокруг своей оси примерно на 180°, гладкие, округлые в сечении у взрослых особей, но у молодых - округло-треугольные, как и у западнокавказских особей. На Центральном Кавказе (от верховьев Малки до Черека) преобладает промежуточный тип рогов (рис. 54) [Радде, 1899; Динник, 1909, 1910; Верещагин, 1938; Цалкин, 1955; Темботов, 1960, 1974; Темботов и др., 1980а,б; Кудакгин, 1985; Вейнберг, 1993; Соколов, Темботов, 1993].
Рис. 54. Географическая и популяционная изменчивость рогов у кавказских козлов (по [Соколов, Темботов, 1993])
Районы добычи животных: 1 - верховья Андийского Койсу; 2-4 - Черек-ское, Чегемское и Баксанское ущелья, соответственно; 5 - верховья Те-берды; 6 - бассейны Зеленчука и Лабы
Западный и восточный тип рогов у кавказских козлов прослеживается со средневековья, причем в некоторых капищах Северной Осетии в бассейне Ардона и в алтарях горной части Западного Закавказья в истоках Ингура их черепа присутствуют вместе. В святилище Реком в Цейском ущелье найдено 292 пары рогов восточного типа и 6 - западнокавказского. Современные черепа с рогами восточнокавказского типа известны из Теберды, переходного типа - с Баксана и Ингура, обеих форм - из ущелий Псыгансу, Безингийского и Балкарского Черека [Верещагин, 1959а].
В.И. Цалкин [1955] полагал, что рога самцов восточной формы заметно длиннее, чем западной. Средняя длина 4 сегментов (от второго до пятого) у первых (46-77 см) почти на 10 см больше, чем у вторых. Однако при анализе большого материала очевидно сходство этих форм и по этому, и по другим параметрам (табл. 52). Средняя длина рогов взрослых самцов — 59-88, максимальная - 114 см, расстояние между концами колеблется в пределах 16-91 см, обхват у основания - 22-36 см. У самок эти признаки находятся в пределах 10-33, 13-32 и 11-13 см соответственно.
К.Р. Айунц [1992] на основании биометрических исследований определил, что форма рогов взрослых самцов кавказских козлов клинально изменяется по длинной оси ареала, где существенные географические барьеры для этого копытного отсутствуют. Индекс “скрученности” роговых чехлов коррелирует с климатическими характеристиками высокогорий: сопряженность наиболее высока с радиационными
п Длина Расстояние между концами Обхват у основания Район (источник информации)**
М | lim м | lim м | lim
Самцы
26 76 61-111 50 42-70 30,0 26-33 Западный Кавказ (4)
49 66 46-81 - - 28,3 22-35 -“-(6)
14 68 40-95 - - 29,0 22-33 -“-(7)
- - 53-85 26-31 -“-(8)
22 83-88 65-104 — — 31,6 27-33 (9)
41 77 - __ - - -“-(В)
29-32 59*** 30-95 51 36-88 27,0 23-32 -“- (14)
— - 69-87 — 51-91 - -30 Район от р. Белой до
Эльбруса (8)
5 86 — 81 — 33,4 - Р. Теберда (12)
4 69 — 58 — 30,0 — Р. Баксан (12)
5 65 - 56 — 28,2 - Р. Чегем (12)
19 72 - 54 — 28,8 - Р. Черек (12)
— — -91 — 39-70 — -32 Кабардино-Балкария (10)
— - 75-90 — — — - Северная Осетия (3)
32 68 42-91 — — 28,3 25-32 Центральный Кавказ (7)
— — -85 — 50-62 — -31 -“-(8)
2 77 77-78 51 46-56 29,0 29-29 Сванетия (11)
19 73 58-88 — — 28,4 25-32 Восточный Кавказ (7)
39 77 57-102 29,9 26-35 -“-(6)
— — -75 — 26-35 - -25 -“-(8)
8 73 55-92 - — - — -“-(1)
— — -88 — -64 — -30 -“-(2)
— — 70-103 — — — 28-32 -“-(5)
5 67 63-75 33 26-39 27,2 26-29 -“-(И)
8 80 - 36 29,7 -“-(12)
25-26 67 52-88 41 26-55 29,7 22-36 -“-(14)
25 78 53-100 42 16-70 30,2 27-34 Восточное Закавказье (11)
Самки
63 20-24 16-33 19-22 15-31 12,5 11-16 Западный Кавказ (4)
— — -25 11-12,5 -“-(6)
4-6 16,6 10-22 23,0 19-25 12,2 12-12,5 -“-(14)
- - -21 — - — -12 Кабардино-Балкария (10)
— — -21 — — — 11 Восточный Кавказ (8)
6 19,5 15-21 - — - — -“-(1)
— - -24 — — — -12 -“-(2)
8 17,6 14-20 20,8 13-32 12,0 11-13 -“- (14)
* Самцы преимущественно старше 6 лет.
**1 - Магомедов и др., 2001; 2 - Абдурахманов, 1973, 1977; 3 - Вейнберг, 1984; 4 - Котов, 1966а, 1968; 5 - Хохлов, 2002; 6 - Цалкин, 1955 (расчет по материалам); 7 - Соколов, Темботов, 1993 (расчет по материалам); 8 - Динник, 1910; 9 - Кудактин, 1985; 10 - Залиханов, 1968; 11 - Верещагин, 1938; 12 - Темботов, 1974; 13 - Ромашин, 2001; 14 - Казанская, 2004.
*** В анализе особи 4-х лет и старше.
параметрами и первичной биопродуктивностью ландшафтов. Начало формирования внутривидовой генотипической изменчивости отнесено к эпохе среднечетвертичного оледенения [Айунц, Коломыц, 1986]. Ранее подверженность толщины, массивнос
ти и степени закрученное™ рогов у самцов сильной популяционной изменчивости отмечал М.Г. Абдурахманов [1973].
Окраска и линька. Детеныши кавказских козлов светло-коричневые (фото 24). Окраска взрослых особей в летнем наряде варьирует от сероватой, рыжевато-серой и рыжевато-бурой до бурой и коричневой. Нижняя часть туловища более светлая и грязно-белая или желтовато-серая на брюхе и в пахах. Хвост темно-бурый или буровато-коричневый. Голова и ноги несколько темнее туловища, щеки светлые. Зимой западнокавказские козлы грязновато-белые, серовато-бурые, реже -желтовато-каштаново-бурые, центрально- и восточнокавказские - каштаново-бурые или темно-бурые. Борода у самцов коричневая, темно-коричневая, темно-бурая или желтовато-бурая. Зеркало грязно-белое, серое или буроватое, верхняя сторона хвоста черная. К концу зимы окраска животных светлеет и даже на востоке ареала становится охристой, грязно-желтой и светло-желтой. Иногда встречаются альбиносы [Динник, 1909, 1910; Цалкин, 1955; Наниев, 1958; Залиханов, 1967, 1968; Котов, 1968; Абдурахманов, 1973, 1977; Вейнберг, 1984; Соколов, Темботов, 1993; Магомедов и др., 2001]. Популяционная изменчивость в окраске столь велика, что этот признак нельзя считать диагностическим [Темботов, 1974].
Линька сильно растянута. На Западном Кавказе смена волоса у большинства особей начинается с конца марта - апреля и заканчивается в начале - середине июля, у отдельных старых самцов - в первых числах августа [Котов, 19666, 1968]. На Центральном и Восточном Кавказе самки и молодые особи завершают линьку в мае -июне, самцы - в июне-июле. Зимний мех отрастает с сентября до конца ноября [Верещагин, 1938; Залиханов, 1967, 1968; Абдурахманов, 1973; Вейнберг, 19816, 1984].
Различие в сроках линьки животных разных половых и возрастных групп и в разных частях ареала пока не находит достаточно убедительного объяснения. На мой взгляд, это связано все же с климатическими условиями и обитанием животных на разных высотах. На западе ареала летом снеговая линия значительно ниже, чем на востоке [Котов, 1968]. Старые самцы обычно держатся на большей высоте, чем самки и молодые особи. В высокогорье климат гораздо холоднее, что и обуславливает задержку их линьки. В тех районах, где летние места обитания самцов и самок относительно сходны, отличия в сроках линьки менее заметаы. В тоже время нельзя сбрасывать со счетов и возможную изменчивость, связанную с разным генотипом.
Генетическая изменчивость. Кариотап кавказских козлов в западной, центральной и восточной частях ареала одинаков, и он практически идентичен таковому домашней козы, безоарового, винторогого и сибирского козлов и европейского ибекса [Мамедов, Кулиев, 1974, 1975; Мамедов, 1978, 1982; Мамедов, Агаев, 1990; Дзуев, 1995, 1998]. По последовательностям фрагмента контрольного региона мтДНК и митохондриального гена цитхрома Ъ (рис. 43) западнокавказские и восточнокавказские формы четко различаются, причем у С. cylindricomis из Кабардино-Балкарии обнаружена уникальная делеция в 18 п.н., которая не встречается ни у С. caucasica, ни у других видов [Казанская, 2004; Казанская и др., в печата].
Гибридизация. Восточнокавказские и западнокавказские козлы легко скрещиваются между собой, с безоаровым и сибирским козлами, а также с домашними козами, но не могут гибридизировать с домашними овцами [Разевиг, 1904; Пфицен-майер, 1915; Верещагин, 1938,1947,1959а; Бутарин, 1947; Саркисов, 1953; Инякова, 1957в; Лопырин и др., 1959, 1960; Соколов, 1959; Алиев, 1970]. Гибриды с домашни
ми козами отличаются скороспелостью (самки вступают в размножение в 7-месяч-ном возрасте), большой массой (до 108 кг у самцов), высоким качеством пуха, значительным удоем жирного молока (1-2,1 л в сутки), длительной лактацией (до 8 мес.) и устойчивостью к низким температурам [Джанашвили, 1943; Верещагин, 1947; Банников, 1959; Залиханов, 1967; Мишарев, 1969; Котов, Рябов, 1963; Котов, 1968]. Это явление гетерозиса хорошо знают горцы, выгоняющие осенью стада домашних коз в горы в надежде на то, что их покроют дикие козлы, или проводящие скрещивание в домашних условиях [Абдурахманов, 1973; Вейнберг, 1984].
Таксономия. На Кавказе выделены 5 видов козлов: Capra caucasica, С. cylindri-cornis, С. severtzowi, С. raddei и С. dinniki (см. синонимиму), не считая безоарового козла. Это, как справедливо заметил В.И. Цалкин [1955] в прекрасном очерке истории классификации группы, единственный в своем роде случай даже в богатой курьезами систематике копытных.
Дискуссия о таксономии кавказских козлов, чаще называемых турами, ведется более 200 лет. Основные причины разногласий - разная форма рогов животных и неясность происхождения и родственных связей. По строению черепа и рогов западнокавказская раса близка к Capra ibex, и поэтому далеко не случайно Е. Шварц [Schwarz, 1935] предлагал объединить ее и альпийского и сибирского козлов в один вид, а Еллерман и Моррисон-Скотт [Ellerman, Morrison-Scott, 1951] отнесли западнокавказского козла к ибексу. Г.И. Радде [1899], изучив обширную коллекцию рогов в Тифлисском музее, не нашел отличий Capra severtzowi от С. caucasica. В.А. Разе-виг [1904] выделял 3 вида козлов: западно-, центрально- и восточнокавказкого. Н.Я. Динник [1909, 1910] считал, что на Кавказе обитают четыре вида. Вместе с К.А. Сатуниным он сделал также заключение об ошибочном выделении профессором Матчи С. raddei. В.И. Цалкин [1955] формы козлов из западной части Кавказа свел к С. caucasica, но решительно возражал против объединения его с С. cylindricornis, с чем были согласны другие ученые [Верещагин, 1959а; Гептнер и др., 1961; Schaller, 1977; Соколов В.Е., 1979; Павлинов, Россолимо, 1987; Grubb, 1993; Павлинов и др., 1995].
Границу между этими видами ученые проводили в районе гор Эльбрус и Казбек, а в Кабарде и Сванетии предполагалось их совместное существование и гибридизация (рис. 51). М.Ч. Залиханов [1967, 1968] на основании краниометрических исследований нашел, однако, что зона гибридизации находится между Баксаном и Череком-Безенгийским, четких видовых границ распространения не существует, а высокая популяционная изменчивость зависит от условий обитания. На территории Грузии на южных склонах Кавказского хребта западным пунктом распространения восточнокавказской формы считается Мамисонский перевал, центральнокавказской - Марухский перевал, западнокавказской - гора Ажра [Чла-идзе, 1974].
По мнению ряда других специалистов, на Кавказе реально существует лишь один полиморфный вид, объединяющий два, три или четыре подвида: severtzowi и dinniki (западная часть Главного Кавказского хребта к востоку до р. Теберды или Эльбруса), caucasica (район от Теберды до р. Черек) и cylindricornis (восточная часть Кавказского хребта от р. Черек) [Соколов И.И., 1959; Громов и др., 1963; Темботов, 1970, 1972, 1973, 1974; Темботов и др., 19806, 1982; Барышников и др., 1981; Соколов, Темботов, 1993; Абдурахманов, 2003].
К.Р. Айунц [1992] пришел к еще более радикальному выводу - на Кавказе цепь незначительно отличающихся между собой популяций козлов образует единый вид С. caucasica, деление которого на подвиды по форме рогов не обосновано. А.В. Ромашин [2001], изучивший фенотипическую изменчивость роговых чехлов кавказских козлов, также не нашел прерыва в кливальной изменчивости. Чуть меньшую длину рогов у особей на Центральном Кавказе он логично связывает с большим антропогенным воздействием на популяции и меньшей продолжительностью жизни животных, живущих в пессимальных условиях.
Е.Ю. Казанская [2004] убедительно показала, что деление кавказских козлов на отдельные таксоны по краниометрическим признакам, как и по форме рогов, затруднено даже при многомерном анализе, что говорит в пользу монотипичности вида. Однако по последовательностям фрагмента контрольного региона мтДНК и по митохондриальному гену цитохрома b они четко делятся на две группы, соответствующие формам caucasica и cylindricomis, причем западнокавказский козел генетически близок к западноевропейским расам (рис. 43).
Распространение
Ископаемые остатки кавказских козлов, датируемые средним - поздним плейстоценом, голоценом и историческим временем, обычны во всей горной части Северного Кавказа и Закавказья, и особенно обильны в Западном и Центральном Закавказье [Громов, 1948; Громова, 19486; Верещагин, Наниев, 1949; Пи-допличко, 1951, 1954; Верещагин, 1959а; Цалкин, 1970; Формозов, 1972; Барышников, 1978; Барышников, Хварцкая, 1990]. Очевидно, что эти звери жили здесь непрерывно с плейстоцена, соседствуя с безоаровым козлом, серной, зубром, оленем, косулей, лосем и кабаном.
В последние столетия границы ареала кавказских козлов уменьшились, однако не так сильно, как у серны и безоарового козла. В начале XIX в. эти копытные, как считалось, были уничтожен в Пятигорье, к концу столетия - на Фиште и Оштене, в 1908 г. - на горе Ачишхо у Красной Поляны и примерно в этот же период - на Кинжале и Бармамыте, в 40-х годах - в Дигории [Насимович, 1949а; Верещагин, 1959а]. Однако позднее (в 60-80-е годы XX в.) на массиве Фишт - Оштен, Ачишхо, в Лазаревском районе, на Кинжале и в малодоступных местах Сванетии встречали одиночных особей и отдельные мелкие группы, видимо, изредка проникающие на окраину былого ареала [Залиханов, 1967; Ромашин, 2001].
В XIX-XX вв. область распространения кавказских козлов (рис. 55) простиралась узкой полосой вдоль Главного Кавказского, Бокового и Скалистого хребтов. Крайние пункты их обитания на западе - горы Абаго, Атамажи и Чугуш, на востоке - горы Базардюзи в Дагестане, Бабадаг и Гюмишлю в Азербайджане [Васильев, 1896; Разевиг, 1904;Динник, 1909,1910; Верещагин, 1938,1959а; Насимович, 1939,1949а, 1955; Гептнер, Формозов, 1941; Наниев, 1956, 1958; Котов, Рябов, 1963; Залиханов, 1967, 1968; Котов, 1968; Темботов, 1972; Абдурахманов, 1973, 1982; Члаидзе, 1974; Точиев, 1975; Алекперов и др., 1976; Кулиев, 1977, 1981; Арабули, 1989; Соколов, Темботов, 1993; Ромашин, 2001].
Ширина современной области распространения зверей на западе Кавказа - около 15 км, в центре в районе Эльбруса - 55 км, на востоке 30-35 км, общая протяжен-
Рис. 55. Районы обитания кавказских козлов в XX в. (по [Соколов, Темботов, 1993])
Точки с цифрами - зафиксированные районы обитания (см. оригинал). На врезке - высотное распределение
ность ареала - 800 км. На этом пространстве кавказские козлы заселяют не более 25% площади [Темботов и др., 1980а]. Размещение группировок преимущественно очаговое [Котов, 19666, 1970]. В Дагестане животные распределены с разной плотностью в полосе дайной около 250 км в верховьях Андийского, Аварского и Казику-мухского Койсу, Каракойсу, Самура, Ахтьгчай, в горах и урочищах Базардюзи, Шал-буздага, Деавгия, Алахундага, Дюльтыдага, Тактика, Хултайдага, Тутона, Шавикла-де, Диклосмта, на талибском, Нукатлинском и Богосском хребтах. Площадь обитания составляет здесь 4—4,7 тыс. [Абдурахманов, 1973; Ахмедов, 1995; Магомедов и др., 2001], в Чечне и Ингушетии - 0,8 тыс., в Северной Осетии - 1 тыс. [Равкин, 1975а], в Кабардино-Балкарии - 1,2 тыс. км2 [Темботов и др., 19806].
Среда обитания
Западнокавказский козел обитает в высотном диапазоне от 1000 до 3800 м в пихтово-еловых лесах и в субальпийской, альпийской и субнивальной зонах. В бесснежный период большинство животных (92-99%) держится в альпийском и субальпийском поясах в местах, где доминируют осочково-разнотравные и разно-травно-осочковые пастбищные ассоциации (фото 25). В верхних ярусах гор они обитают и зимой, выбирая малоснежные участки на склонах южной и юго-восточной экспозиций, но часть, преимущественно группы самок с молодняком, переходит в скалистые участки лесной зоны. Запас зимних кормов в районах, где нет
выпаса домашнего скота, значителен - до 1315 кг сухой массы на 1 га. В апреле -мае копытные концентрируются в верхней границе леса, представленного пихтовыми, еловыми и сосновыми участками и березовым криволесьем, на хорошо ин-солируемых склонах, где раньше появляется зеленая трава. По мере таяния снега они поднимаются выше и широко расходятся по склонам гор. Летом животные особенно тесно связаны с субальпийскими высокотравными лугами, а в жару тяготеют к участкам гор, где сохраняются снежники и ледники, есть зеленая трава и меньше кровососущих насекомых. Во всех биотопах и во все сезоны они предпочитают скалистые склоны, а на пологих участках появляются лишь при переходах [Динник, 1909, 1910; Насимович, 1936а, 1939, 1949а, 1955; Аренс, 1957; Котов, 1960а, 19666, 1968, 1971; Котов, Рябов, 1963; Ромашин, 1995, 2001; Бобырь, 1999, 2002].
В центральной части Кавказского хребта большую часть года звери живут в субальпийской и лесной зонах, а альпийскую используют в основном летом. В Кабардино-Балкарии козлы обычны в высотном диапазоне от 1200 до 3400 м. Зимой они предпочитают крутые облесенные склоны, “выгрева” и “выдува”, летом - высокогорные альпийские луга выше березового криволесья, а в леса заходят в основном для солонцевания. В районах выпаса скота они концентрируются в глубоких ущельях с разреженной лесной растительностью и высоким травостоем, а платообразных хребтов избегают [Темботов, 1961; Залиханов, 1967, 1968]. В Северной Осетии козлы обитают в пределах высотной зоны 1200-4000 м. И здесь они тоже интенсивно используют лесной пояс. Основными местами зимовок служат глубокие горные долины между Главным, Боковым и Скалистым хребтами и их отрогами, отличающиеся своеобразными климатическими условиями. Снег на крутых склонах сдувается ветром, обнажая сухую растительность, которая служит животным кормом. В поисках пищи и водопоев они нередко переходят из одного ущелья в другое [Наниев, 1958; Вейнберг, 1977, 1984].
Основные места обитания восточнокавказского козла приурочены к высокогорным районам до высоты 4500 м, где звери предпочитают альпийские и субальпийские луга со скалистыми участками (фото 26). В многоснежные зимы нижняя граница их размещения совпадает с верхней границей леса. Здесь они находят древесный и лишайниковый корм и убежища в виде навесов и пещер. Весной, пр мере появления зеленой растительности, животные переходят в альпийскую зону, где наиболее характерны следующие типы лугов: разнотравные рыхлодернинные (альпийские ковры), осоково-овечье-овсянициевые плотнодернинные, высокогорные пестроовсяни-циевые с альпийским мелкотравьем и субальпийским разнотравьем и кобрезиевые альпийские с урожайностью фитомассы от 27,5 до 13 ц/га. На этих пастбищах они изымают от 8 до 51% растительной продукции, а вместе с домашними овцами - до 77%, что нередко приводит к существенному изменению не только биомассы, но и структуры фитоценозов. В целом только в пределах дагестанского участка ареала козлы ежегодно утилизируют до 11000 т фитомассы. Зимой площадь доступных для них пастбищ резко уменьшается и составляет в разных районах от 12 до 45% размера летних стаций. В зимне-весенний период (до прихода отар овец) звери используют, в основном, высокогорные пестроовсянитщевые луга на высотах 2500-2800 м по склонам южной экспозиции и кобрезиевые альпийские луга в пределах 2400-2700 м. Естественные зимние пастбища при отсутствии интенсивного выпаса домашнего скота в состоянии прокормить до 31-35 особей/км2 [Абдурахманов, 1973, 1977, 1982; Рав-
кин, 1975а; Яровенко, 1990; Магомедов, Ахмедов, 1994; Ахмедов, 1995, 1997; Магомедов, Яровенко, 1998; Магомедов и др., 2001].
На южном склоне Главного Кавказского хребта козлы тоже типичные обитатели субальпийского и альпийского поясов. В заповедниках отдельные особи и группы держатся и в лесной зоне на высоте 900-1500 м. Субнивальную зону животные используют лишь для отдыха. На зимовку они спускаются в менее многоснежный лесной пояс до высоты 800-1000 м, но предпочитают все же его верхнюю часть и субальпийское редколесье, где живут в скальниках на малоснежных южных и юго-восточных склонах [Марков, 1934; Верещагин, 1938; Эквтимишвили, 1952, 1953; Енукидзе, 1953; Насимович, 1955; Капанадзе, 1975, 1976; Кулиев, 1981].
Во многих районах Кавказа пространственное размещение кавказских козлов определяется основным конкурентом - домашним скотом, пригоняемым весной на горные пастбища. Отары вытесняют диких животных в ущелья, на вершины хребтов и в труднодоступные скалы, откуда они спускаются лишь в сентябре -октябре после ухода скота на зимовку, или ежедневно выходят на пастбища ночью [Верещагин, 1938; Темботов, 1973; Абдурахманов, 1977; Соколов, Темботов, 1993].
Очевидно, что кавказских козлов нельзя считать специфическими обитателями высокогорий, что нередко встречается в научной и научно-популярной литературе. Тяготение этих копытных к высокогорным биотопам в отдельных районах, скорее всего, вторично и вызвано не только вытеснением их с пастбищ домашним скотом, но и с неумеренным охотничьим преследованием, что Н.Я. Динник [1909, 1910] отмечал еще столетие назад. Пребывание зверей в лесной зоне тоже скорее правило, чем исключение. Более того, лес для них служит одной из основных стаций переживания зимой и убежищем при резких похолоданиях и затяжных дождях в летне-осенний период, а субальпийский и альпийский пояса используются преимущественно в благоприятное время или, вынужденно, в многоснежье.
Древние формы этих копытных, видимо, возникли в лесном поясе, а затем освоили высокогорье, что подтверждается некоторыми особенностями экологии и поведения современных форм. Кавказские козлы чаще спариваются и рожают в лесном поясе; звери интенсивно маркируют деревья ольфакторно-оптическими метками; они предупреждают сородичей об опасности звуковым сигналом; у детенышей длительная реакция затаивания-как у типичных лесных обитателей [Верещагин, 1938,1959а; Залиханов, 1967; Вейнберг, 19816, 1984]. А.В. Ромашин [2001], однако, считает мнение о первичной связи этих животных с лесом ошибочным.
Жизненно необходимое условие для кавказских козлов, как, впрочем, и для других горных копытных, - наличие скальных образований, которые служат убежищем. В скальниках звери передвигаются с большой скоростью и вверх, и вниз, эффективно используя едва заметные выступы, по которым не способен передвигаться ни один крупный хищник. Здесь же животные спасаются и от преследования человека, благодаря чему уцелели до сих пор. Во многом именно поэтому плотность их населения достигает максимальных значений в скалистой местности и на склонах большой крутизны.
Немаловажен также снежный фактор. Слой снега высотой в 40-50 см становится серьезной преградой для относительно коротконогих и грузных животных [Насимович, 1955], но наибольшие проблемы с кормодобыванием возникают все же в сильные метели, сопровождаемые гололедицей. Из-за мощного снежного покрова
эти копытные, за редким исключением, не зимуют на южном макросклоне Главного Кавказского хребта, и сосредоточиваются на северном, где большая часть их держится на южных склонах на участках, где есть выдувы и проталины, или снежный покров не превышает 30-35 см [Котов, 1968]. В вегетационный период распределение козлов в большей мере определяется наличием зеленой травы.
Питание
Корма. Список растений, потребляемых кавказскими козлами, весьма обширный. В Кавказском заповеднике звери использует в пищу не менее 195 видов [Котов, 19666, 1968], в Тебердинском заповеднике - 145 [Бобырь, Бобырь, 1999а; Бобырь, 2002], в Кабардино-Балкарии - более 108 [Залиханов, 1967], Чечне и Ингушетии - 205, Дагестане - 256 [Абдурахманов, 1973, 1977], Закатальском заповеднике - 99 [Кулиев, 1978], Грузии - 80 [Эквтимишвили, 1953], а всего в ареале -более 442, в том числе 388 - травянистые, 37 - древесно-кустарниковые и не менее 17 - низшие растения. Из них злаки составляют 80% массы, бобовые - 2,5 и древесный корм - 4,5% [Абдурахманов, 2003].
Замечено, что набор кормовых растений козлов минимален при изобилии корма и при обитании зверей в верхних поясах гор в пессимальных условиях, и максимальный при жесткой трофической конкуренции с другими видами диких и домашних копытных, когда вынужденно потребляются не самые качественные растения. Разнообразие кормов закономерно повышается весной, несколько снижается летом и существенно обедняется в осенне-зимний период [Ромашин, 2001]. В Дагестане основу питания составляют около 60 видов растений, из которых в первую очередь поедаются лишь 20 [Магомедов, Яровенко, 1998].
В вегетационный период рацион животных состоит преимущественно из трав и включает многие растения, считающиеся ядовитыми. Наиболее часто копытные скусывают астрагалы, эспарцеты, тмины, пупавки, овсяницы, овсец, клевер, пырей, душистый колосок, ячмень фиолетовый, тонконог кавказский, кобрезию, вейник, полевицу, костер, кострец, мятлики, лисохвост, гречишник, герани, анемоны, лютики, камнеломки, козлобородник, подорожники, манжетки, одуванчик Стивена, лапчатку, горошек альпийский, бодяк, подмаренник, колокольчики, белокопытник, гравилат, веронику, примулы, осоки, а также иву козью. Осенью рацион дополняется грибами. Зимой звери поедают все доступные корма, но наиболее значимы травяная ветошь и подснежные зеленые травы (преимущественно злаки и осоки), а также кустарнички, кустарники и древесные виды (ивы, березы, осина, рябина, клен, липа, черемуха, ольха, рододендрон, бересклет, шиповник, сосна, пихта, черника, ежевика, брусника, плющ), доля которых в рационе возрастает до 40%. Значительно меньшую роль в зимнем питании играют папоротники, лишайники и мхи [Верещагин, 1938; Насимович, 1939, 1949а, 1955; Эквтимишвили, 1953; Залиханов, 1967, 1968; Точиев, 1970; Абдурахманов, 1973, 1977; Вейнберг, 1984; Ахмедов, 1995, 1997; Магомедов, Яровенко, 1998; Бобырь, Бобырь, 1999а; Магомедов и др., 2001; Ромашин, 2001].
Ботанический анализ экскрементов козлов в пещерах Северной Осетии показал, что на протяжении 4000 лет их основным кормом служили злаки, разнотравье и древесно-кустарниковые растения, из которых наибольшую роль играла сосна [Князев, Савинецкий, 1992]. Злаки остаются основной пищей и по сей день. В
Кавказском заповеднике они составляют до 30-90% содержимого желудков [Ромашин, 2001], на востоке ареала - около 30-80%. В летний период в рационе доминирует разнотравье (до 70%), зимой и ранней весной возрастает роль бобовых (до 80%) и веточных кормов (до 40%) [Абдурахманов, 1973; Яровенко, 1997; Магомедов и др., 2001].
Детеныши начинают потреблять растения в месячном возрасте, а к концу второго месяца жизни включают в свой рацион почти все основные виды, который поедают взрослые особи [Ахмедов, Яровенко, 1991; Магомедов и др., 2001].
Потребление кормов и воды. Минеральное питание. Масса содержимого желудков у козлов составляет 5-8 кг зимой и 8-14 кг летом [Залиханов, 1967, 1968]. Уровень среднесуточного потребления корма находится в пределах 1000-2400 г сухого вещества на особь в зависимости от сезона, а зимой порой снижается до 375-410 г. Суточная потребность в энергии - 16000-17500 кДж на особь, получаемая энергия - 12000-25000 кДж. Уровень потребления возрастает с увеличением подвижности животных. Молодые растущие особи отличаются более высокой интенсивностью зимнего питания. На пастбищах разной продуктивности среднесуточное потребление кормов почти одинаково, что достигается увеличением или уменьшением времени пастьбы и изменением видового состава используемых в пищу растений. Содержание протеина в потребленных кормах варьирует от 20% летом до 10% зимой. Переваримость пищи колеблется в пределах 41-81% в зависимости от доли сырой клетчатки. Высокая питательность рациона во многом связана с постоянным стравливанием пастбищ - в отаве содержание белков и жира в 1,5-2,5 раза выше, чем в сформированных растениях. Самки, выпасающиеся на более влажных склонах северных экспозиций, потребляют более питательную пищу по сравнению с самцами. Избыточное потребление энергии в летне-осенний период приводит к быстрому увеличению массы животных и накоплению жирового резерва. Дисбаланс в поступлении и расходовании энергии проявляется зимой и, особенно, в конце весны, что нередко приводит к гибели истощенных зверей, чаще молодых [Магомедов, Яровенко, 1997а,б, 1998; Магомедов и др., 2001]. В Кавказском заповеднике после холодной зимы и затяжной весны 1987 г. у козлов регистрировали капрофагию [Ромашин, 2001].
В связи с ограниченностью зимних пастбищ, нагрузка на них весьма велика. В Тебердинском заповеднике за зимний период козлы изымают более 70% надземной растительности. Это, однако, не только не приводит к деградации экосистем альпийских лугов и пустошей, а, напротив, стимулирует их продуктивность, поскольку удаление отмерших частей растений улучшает световой режим для новых побегов, а экскременты животных служат источником дополнительного минерального питания [Бобырь и др., 1999; Бобырь, 2002].
При добывании корма козлы нередко встают на задние ноги, опираясь передними о скалу или дерево, или балансируют ими в воздухе. Иногда они грудью прижимают тонкие деревца к земле, или залезают на наклонные деревья. Снег раздвигают мордой или копытят его, если высота снежного слоя не превышает 30 см. В поисках сочных корней они могут разгребать ногами подстилку и тонкий слой земли [Насимович, 1955; Вейнберг, 1983а, 1984].
Кавказские козлы регулярно посещают солонцы как естественные (сухие и водные, включая серные), так и искусственные, которые устраивают для них в охотни-
чьих хозяйствах и браконьеры (фото 27). Здесь этих копытных обычно отстреливают или отлавливают для расселения в другие районы. Сюда они выходят чаще в весенне-летний период (наиболее активно - в мае - июне), причем, в отличие от оленей, и в дневное время. На соль и глину набрасываются с жадностью, отталкивая друг друга. На отдельных солонцах скапливается до 200 животных. Поскольку места для всех не хватает, часть их лежит или пасется рядом. Детеныши начинают поедать субстрат с двухмесячного возраста. На искусственных солонцах при высокой плотности населения козлов и серн в альпийском поясе среднесуточный расход соли в июне - июле достигает 2,9-3,б кг (более 30 г на голову) [Динник, 1910; Верещагин, 1938; Насимович, 1938, 1949а; Котов, 1960в,г, 1964а, 19666, 1968; Залиханов, 1967, 1968; Абдурахманов, 1973; Кулиев, 1981; Магомедов и др., 2001; Ромашин, 2001; Бобырь, 2002].
В Кавказском государственном заповеднике насчитывается более 175 водных и несколько сотен естественных сухих “туровых” солонцов, занимающих большую площадь. Как установил А.А. Насимович [1938], последние образуются следующим образом. На склоне пастбищ отдельные звери выбивают копытами небольшую ямку и съедают верхний слой почвы. При длительном обитании в одном районе козлы постепенно выедают землю в ямках на площади в несколько квадратных метров на глубину до полуметра, и за многие годы расширяют солонцы до нескольких гектаров. Площадь “Большого турового солонца” в субальпийской зоне на Тыбге достигает 8 га. Солонцы звери интенсивно используют с февраля до конца сентября. Их зимние и ранневесенние экскременты нередко состоят сплошь из серо-пепельной глинистой массы.
Проблем в потреблении воды у кавказских козлов нет. Летом достаточно влаги содержится в зеленом корме, и у снежников всегда есть вода, зимой животные обычно довольствуются снегом, содержащимся на поедаемых растениях, или, как описывает В.И. Наниев [1958], посещают незамерзающие минеральные источники.
Пищевая активность. У кавказских козлов, как и у многих других видов копытных, полифазный тип активности с пиками в раннеутренние и вечерние часы, но разный у самцов и самок. Летом, в жару, самцы почти весь день проводят на лежках, и пасутся в сумеречные и ночные часы, тогда как значительная часть лактирующих самок, у которых повышенные энергетические потребности, кормится в любое время суток, чередуя пастьбу с отдыхом. Самки кормятся чаще, чем самцы, но последние в темное время суток пасутся дольше и потребляют больше корма. В утренние часы масса содержимого рубца у взрослых самцов составляет в среднем около 20 кг, тогда как у самок - в два раза меньше. В дождливую и пасмурную погоду все звери активны в равной мере. Зимой они пасутся преимущественно днем [Динник, 1909, 1910; Насимович, 1949а; Котов, 1960а, 19666, 1968; Котов, Рябов, 1963; Залиханов, 1967, 1968; Абдурахманов, 1973, 1977, 1982; Вейнберг, 19816, 1982а, 1984; Кулиев, 1981; Ахмедов, Магомедов, 1996; Магомедов и др., 2001].
Разный ритм суточной активности у самцов и самок обычно связывают с неодинаковыми физиологическими потребностями полов. На мой взгляд, более выраженная сумеречная и ночная активность у взрослых самцов - это и своеобразная адаптация животных, имеющих роскошные рога, к длительному выборочному охотничьему воздействию. Как показывают исследования, в районах с интенсивным браконьерством суточный ритм пастьбы практически всех видов диких копытных
резко нарушается и становится преимущественно ночным у всех половых и возрастных групп.
Размножение
Половая зрелость. Самки кавказских козлов вступают в размножение на втором (до 40%) или третьем году жизни [Верещагин, 1938; Инякова, 1957в; Залиханов, 1967, 1968; Котов, 19666, 1968]. Самцы начинают ухаживать за ними в 4-6-летнем возрасте и способны покрывать их, но более зрелые и сильные козлы обычно вытесняют молодых из стада. Поэтому потомство оставляют, как правило, самцы старше 6 лет [Вейнберг, 19816, 19826].
Гон и половое поведение. Гон у кавказских козлов имеет место в ноябре - январе (табл. 53). Его сроки и интенсивность, вероятно, зависят от температурного режима. Он начинается раньше в более холодной высокогорной области, после резкого похолодания и ранних снегопадов. Однако в особенно холодные дни этот процесс прекращается, звери часами стоят неподвижно, и самцы не обращают внимания на самок. В морозные зимы спаривание затягивается до конца января [Залиханов, 1967; Абдурахманов, 1973, 1982; Вейнберг, 1984; Магомедов и др., 2001]. В неволе у особей восточнокавказской формы гон более растянут, чем у западнокавказской, а отдельные элементы полового поведения наблюдаются круглогодично [Эрнитс, 1980].
Гон проходит на местах зимовок. К его началу группы взрослых самцов распадаются. С начала - середины ноября одиночные особи и пары козлов держатся вблизи от групп самок, а затем входят в их состав или переходят от одного стада к другому. В этот период самцы весьма активны, агрессивны и менее осторожны, почти не кормятся, у них постоянно поднят хвост, а специфические железы издают сильный мускусный запах, что типично для всех видов козлов. Драки за самок сравнительно редки, но порой они носят ожесточенный характер - борьба продолжается несколько часов, а победитель долго преследует противника [Залиханов, 1967; Абдурахманов, 1973].
По наблюдениям П.И. Вейнберга [1980, 19816, 19826, 1984], при наличии признаков эструса у какой-либо из самок, самец-доминант не отходит от нее ни на шаг
Таблица 53. Сроки гона у кавказских козлов
Район, заповедник | Начало | Конец | Источник информации
Западный Кавказ Вторая половина ноября Конец декабря Котов, 19666, 1968
Кавказский з-к Декабрь Начало января Насимович, 1949а
Тебердинский з-к (вольера) Начало ноября 3 февраля Инякова, 1957в
Кабардино-Б алкария Середина ноября Начало января Залиханов, 1967, 1968
Северная Осетия Конец ноября Конец января (начало февраля) Вейнберг, 1980, 1984
Дагестан а Конец декабря -января Абдурахманов, 1979,1982; Магомедов и др., 2001
«С Вторая половина декабря Начало января Газалиев, 19806
Грузия Начало ноября Конец декабря Эквтимишвили, 1952
Азербайджан Декабрь Декабрь Верещагин, 1938
Конец декабря Конец января Кулиев, 1981
и не подпускает близко других козлов. Самки явно предпочитают спариваться с сильными взрослыми козлами, которых в крупных смешанных стадах несколько. Пяти-шестилетние самцы получают доступ к еще непокрытым самкам только в конце гона, когда истощенные доминанты покидают группы.
Ухаживание самца за самкой включает 7-10 элементов: горизонтальное вытягивание головы и шеи, резкие повороты головы, периодическое высовывание языка, демонстрацию профиля головы, гребущие движения передней ногой, назо-генитальные и назо-назальные контакты, касания лежащей самки копытом, флемование и урина-цию себе на морду для усиления запаха. В последнем случае самец наклоняет голову и мочится на нее стоя, а иногда и сидя в присутствии самки или других взрослых самцов. Демонстрации и сильнейший “козлиный” запах, содержащий половые феромоны, явно стимулируют самок. Молодые (3-6-летние) самцы демонстрируют те же позы, что и взрослые, за исключением показа профиля головы, но с меньшей интенсивностью. Их ухаживание, к тому же, содержит элементы агрессии, что не способствует успешному спариванию, вызывая ответные атаки самок или их бегство.
Агонистическое поведение взрослых самцов во время гона весьма разнообразно и содержит элементы, часть из которых сходна с брачным поведением: демонстрацию соперникам профиля тела; скребущие движения ногой; резкое опускание и вытягивание головы, сопровождаемое высовыванием языка; угрозы наклоном головы и броски вперед; уринацию на морду; вставание на задние ноги перед столкновением; удары рогами и теснение противника; удары в корпус в позиции “голова к хвосту”; прижимание головы противника к земле; блокировку подхода соперника к самке. Молодые самцы подвергаются атакам взрослых особей почти в 4 раза чаще, чем одновозрастные соперники. При подходе взрослого козла молодые обычно опускают голову или отворачиваются и, иногда, начинают торопливо пастись, или обнюхивают его, облизывают и трутся мордой и рогами, а затем один из них или оба уринируют себе на морду.
В неволе половая активность особей восточнокавказского козла выше, чем у западнокавказского. Самцы последнего при импонировании чаще высовывают язык, делают выпады головой и обнюхивают голову самки по сравнению с самцами первого, которые чаще обнюхивают бока самки и выбрасывают вперед ноги. Готовая к спариванию самка встает сбоку и касается передними ногами спины самца, тогда как самка западнокавказской формы дотрагивается до паховой области самца и кружится вместе с ним [Эрнитс, 1980].
Беременность, деторождение и выкармливание молодняка. Беременность длится 150-165 дней [Верещагин, 1938; Инякова, 1957в; Газалиев, 19806; Абдурахманов, 1982].
На северных склонах Главного Кавказского хребта самки рожают в основном в мае - первой половине июня, на южных - на месяц раньше. Нередки случаи появления детенышей в июле и даже в августе (табл. 54).
Деторождение происходит преимущественно в скалах северных склонов в субальпийском поясе. Перед родами и в первые дни после них самки держатся обособленно и отдельно от затаивающихся телят. Вскоре часть семей группируется, образуя своеобразные “детские сады”,, в которых наблюдались случаи кормления самками чужих детей. Козлята быстро растут, и в недельном возрасте уже могут ловко прыгать по скалам [Котов, 1968; Абдурахманов, 1973, 1977, 1982; Магомедов и др., 2001]. Лактация продолжается до ноября - декабря, известны случаи
Таблица 54. Сроки деторождения у кавказских козлов
Район, заповедник 1 Начало I Конец ; | Источник информации
Кавказский з-к 3 мая Начало июля Котов, 1966, 1968
17 мая Вторая половина июня Насимович, 1949а
Тебердинский з-к 25 апреля 10 августа Эквтимишвили, 1953,19546
-“- (вольера) 18 мая 26 июля Инякова, 1957в
К.-Балкария 26 апреля Начало июля Залиханов, 1967
С.-Осстинский з-к Конец апреля Август Вейнберг, 1984, 1989
Восточный Кавказ 25 мая Конец августа Магомедов и др., 2001
Дагестан 20 мая Конец июля Абдурахманов, 1973,
1977, 1982
-- Конец мая - Газалиев, 19806
Грузия Апрель Май Капанадзе, 1976
Азербайджан 20 мая 20 июня Верещагин, 1938
а Конец апреля Середина июня Кулиев, 1981
нахождения молока в вымени в январе. Самка дает за сутки 635-900 г молока 5% жирности [Верещагин, 1938; Залиханов, 1967; Вейнберг, 19816, 1984].
Плодовитость. Самки кавказских козлов способны размножаться до 15-18 лет, принося ежегодно одного или, изредка (1-5%), двух детенышей. Уровень яловости колеблется в пределах 3-14%, но иногда без козлят (возможно погибших) бывает до 40% самок [Верещагин, 1938; Насимович, 1949а; Эквтимишвили, 19546; Залиханов, 1967, 1968; Котов, 19666, 1968; Абдурахманов, 1973, 1977, 1979, 1982; Кулиев, 1981; Магомедов и др., 2001].
Потенциальный уровень воспроизводства, следовательно, близок к 1 детенышу на одну половозрелую самку. Наблюдатели обычно регистрируют 0,5-0,9 сеголеток на самку (табл. 55), что свидетельствует все же не о низкой плодовитости этих копытных, а скорее указывает на значительный уровень детской смертности.
Таблица 55. Число детенышей на самку в популяциях кавказских козлов
Район, заповедник Самок, п Сеголеток, п Число детенышей на 1 самку Источник информации
Кавказский з-к 21935 10393 0,5 Котов, 1968
С.-Осетинский з-к >1279 >738 0,6 (0,3-0,9) Вейнберг, 1984, 2002
Восточный Кавказ 2429 1360 0,6 (0,5-0,7) Магомедов и др., 2001
Азербайджан - - 0,8-0,9 Кулиев, 1981, 1989
Уровень воспроизводства в популяциях кавказских козлов существенно зависит от условия питания самок в период, предшествующий размножению, и снижается по мере увеличения плотности населения животных и уменьшения запасов качественного корма [Ахмедов, 1995; Магомедов и др., 2001].
Популяционная структура
Половой и возрастной состав. В популяциях кавказских козлов чаще 11-22% сеголеток, 8-18% подростков, 32-37% взрослых самок и 23-36% взрослых самцов (табл. 56). Соотношение полов среди сеголеток и годовалых зверей близко 1:1.
Среди взрослых животных на заповедных территориях чаще регистрируют больше самцов, что, видимо, связано с большей скрытностью самок. В эксплуатируемых популяциях это соотношение смещается до 1:1,6-2 в пользу самок, а среди особей старше 13 лет оно достигает 1:2,6 из-за избирательного отстрела козлов. Взрослые самцы преобладают в многочисленных популяциях (при плотности населения, превышающем 70 особей на 1000 га), в которых доля сеголеток обычно ниже из-за высокой смертности и уменьшения плодовитости самок [Абдурахманов, 1973, 1982, 1987; Ахмедов, Магомедов, 1995, 2000; Магомедов и др., 2001]. Изменение доли сеголеток слабо связано с динамикой солнечной активности и с температурным режимом, но достоверно - с урожайностью злаков на высокогорных пастбищах [Ромашин, 2001].
Таблица 56. Половозрастной состав (%) и соотношение полов в популяциях кавказских козлов
Район, заповедник (источник информации)* Встречено животных, п Сеголетки Под-ростки Взрослые Самцы: самки
самки | самцы
Кавказский з-к (4) 64584 16(11-18) 9 34 41 1:0,8
-“-(Н) - 13 (7-18) 9 31 31 1:1,1
Тебердинский з-к (3) - 14-15 - — — -
Кабардино-Балкария (8) 19792 15 28 32 25 1:1,3
Северная Осетия (2) — 17 - 45** 38** 1:1,2
С.-Осетинский з-к (5) 7511 19(16-20) 8-11 34 36 1:0,8
-“- 1988-2000 гг. (5) 2030 14 — — 23-5 1:0,5
Дагестан (1) 10287 22 18 37 23 1:1,6
-“-(7) 7856 28** ** 45 28 1:1,6
Исмаилинский з-к (6) - 22 13 32 33 1:0,9
Закатальский з-к (9) 2928 — - — — 1:0,5
* 1 - Ахмедов, 1995; Магомедов и др., 2001; 2 - Равкин, 1975а; 3 - Тарасов, 1973; 4 - Котов, 1968; 5 -Вейнберг, 1984, 2002; 6 - Кулиев, 1986; 7 - Абдурахманов, 1973, 1982; 8 - Залиханов, 1967, 1968; 9 - Верещагин, 1938; 10-Кулисв, 1981; 11 -Ромашин, 2001.
** Вместе с подростками.
Социальная организация и поведение. Кавказские козлы - стадные звери, но взрослые разнополые животные вне периода гона предпочитают жить отдельно друг от друга. Относительно обособлены объединения самок с детенышами, включающие также подростков обоего пола (фото 28), группы взрослых козлов (фото 22 и 23) и группы молодых самцов (фото 29), но большая часть последних обычно примыкает к тем и другим. Зимой существуют как однополые, так и смешанные стада. Последние обычно формируются на основе самочьих групп, в которые перед гоном и после него входят взрослые и молодые самцы. Иногда в отдельные группы объединяются травмированные особи, которые не в состоянии следовать за быстро убегающим среди скальных нагромождений стадом [Вейнберг, 1981а, 1984]. Объединившись, они получают несомненную выгоду, меньше привлекая хищников и чувствуя себя в большей безопасности среди других членов группы.
Из всех объединений наиболее стабилен состав групп самок, которые, скорее всего, состоят из близкородственных или хорошо знакомых особей. Объединения самцов нередко одновозрастные. В Северной Осетии в составе группы из 10 голов, погиб
шей в лавине, семь зверей были 15-летними, три - 18-летними; в группе из 12 особей - пять 15-летних, четыре - 17-летних и три - в возрасте 3 лет [Наниев, 1962].
Средний размер групп у кавказских козлов обычно варьирует от 6 до 35 экземпляров (табл. 57). Это один из самых высоких показателей стадности среди отечественных видов горных копытных. Стада, содержащие до сотни особей, не редкость, а ранее по всему ареалу регистрировали объединения, насчитывающие до 200-500 (1000) голов [Радде, 1899; Динник, 1909, 1910; В-н, 1936; Верещагин, 1938, 1947, 1959а; Насимович, 1939, 1949а]. Крупные смешанные объединения чаще образуются при скоплениях животных на ограниченных по площади пастбищах. Они кратковременны и легко распадаются на мелкие субъединицы.
Таблица 57. Показатель стадности и максимальный размер групп у кавказских козлов
Район, заповедник Число особей в группе Источник информации
в среднем | максимально
Кавказский з-к 11-16 152 Насимович, 1949а
и 11-20 300 Котов, 1960а,д, 1961, 19666,
1968, 1971
К.-Балкария 15 (4-24) 534 Залиханов, 1967, 1968
К.-Балкарский з-к 20-30, 10-15* > 100 Аккиев, 2001
Северная Осетия 6-9 85 Вейнберг, 1984
Ч.-Ингушетия 22 81 Равкин, 1975а
Дагестан 16-34 180 сс
с с 14-35 194 Магомедов и др., 2001
Грузия - 200 Капанадзе, 1976
Лагодехский з-к - 125, 500, 1000* Верещагин, 1938
Азербайджан 76 205 Кулиев, 1981
* В группах самок, самцов и смешанных стадах соответственно.
С увеличением численности и плотности популяции показатель стадности, как правило, возрастает до определенного предела, видимо близкого к 30-35 (табл. 57). В некоторых районах группы самцов крупнее, чем самок. Ранней весной и осенью стада больше, чем зимой и летом. В местах интенсивной охоты стадность заметно меньше [Залиханов, 1967; Котов, 1968; Темботов, 1973; Вейнберг, 1983г, 2002; Ромашин, 2001].
В Кавказском заповеднике в период 1938-1959 гг. средний показатель стадности достигал 17 (14-21 в разные годы): летом в среднем 17, весной и осенью - 15, зимой - 10. Стада в 150-300 голов здесь не редкость. В июне 1954—1955 гг. на солонцах на горе Тыбга регистрировали скопления самок с сеголетками и подростками в 400 и 700 голов [Савельев, 1960; Котов, Рябов, 1963].
Средний размер разных половозрастных групп кавказских козлов в центральной части ареала в течение года (3-10 особей) заметно меньше, чем в западной и восточной. Стада, превышающие 30-40 особей, тоже относительно редки и более часты встречи одиночных взрослых самцов. Эти различия, возможно, связаны с обитанием значительной части животных в лесной зоне, в которой группи-
Таблица 58. Среднее число особей в группах козлов в Северо-Осетинском заповеднике (по [Вейнберг, 1984])
Группы Месяцы В течение года
XI-I | II-IV | V-VII | VIII-X
Взрослых самцов 3,3 8,8 16,0 10,4 9,7
Молодых самцов 1,7 1,6 2,6 3,8 2,6
Самок с детьми 5,4 6,4 7,7 8,7 6,8
Смешанные 10,0 2,7 10,0 21,0 9,8
ровки копытных всегда меньше, чем в открытых биотопах. Показатель стадности здесь существенно изменяется по сезонам, неодинаков в разных половых и возрастных группах (табл. 58), отличается в лесном и субальпийском поясах и максимален в смешанных стадах осенью и весной. Вне периода гона в смешанных стадах доля взрослых самцов не превышает 0,9-14%, 16% молодых самцов образуют самостоятельные группы, 51% - входят в состав групп самок и треть держится вместе с взрослыми козлами [Залиханов, 1967; Вейнберг, 19816, 1982а, 1984].
В Дагестане летом наиболее часто наблюдаются группы самок и группы самцов (26% и 27% встреч), насчитывающие 11—20 особей, но наибольшее число зверей сосредоточено все же в стадах, состоящих из 41-80 (28% самок и 43% самцов) и 81-120 голов (24 и 14% соответственно). В зимний период чаще регистрируют группы самок и самцов из 6-20 особей (60 и 50% встреч), но наибольшее количество самок (46%) держатся в стадах из 21-40 голов, тогда как 49% самцов предпочитают жить в более крупных объединениях. Смешанные группы обычно состоят так же из 21-40 (28%) и 41-80 (38%) зверей. В конце зимнего периода максимальный размер стад самок может достигать 194, самцов - 125 особей [Магомедов и др., 2001].
Социальное поведение кавказских козлов не отличается существенно от других видов горных козлов. Ярко выраженных вожаков у них нет, а лидеры меняются даже на пути следования на кормежку и обратно. В смешанных группах лидирует какая-либо взрослая самка, за ней идут самки с козлятами, а замыкают группу молодые самцы и яловые самки. В период гона последним шествует взрослый самец. Активность особей в мелких группах более синхронизирована, нежели в крупных стадах. Как и у большинства горных копытных, самцы молчаливы, и в стадах слышно лишь блеяние самок и сеголеток. При опасности звери настораживаются, свистят и издают своеобразный фыркающий звук [Динник, 1910; Залиханов, 1967; Котов, 1968; Аккиев, 2001]. У козлят свист впервые регистрируется на 18-й день после рождения [Ахмедов, Яровенко, 1991].
Детеныши игривы, в играх доминируют элементы брачного и агонистического поведения. Иногда животные играют на снежниках, разбегаясь и скользя по ним, как на лыжах. Агонистическое поведение у взрослых самцов после гона проявляется редко. Стычки чаще наблюдаются среди самок. Иерархия линейна по размерности рогов и тела. Низшие ступени в иерархической лестнице занимают сеголетки, подростки и самки, высшие - молодые и взрослые самцы. В поведении всех возрастных и половых групп животных присутствуют элементы маркировки территории: звери обдирают рогами кору на деревьях и кустах и трутся об оголенные участки затылком и лбом, оставляя на растениях не только визуальные, но и оль
факторные метки, которые затем обнюхивают. Другие особи, обнюхав метку, весьма возбуждаются и, подняв хвост, сами метят то же или другое растение [Вейнберг, 19816, 1984]. Соотношение составляющих элементов при импонировании у особей дагестанской и кубанской форм распределяется так: голосовые сигналы - 9 и 17%, резкое опускание головы - по 17%, высовывание языка - 25 и 35%, удар ногой - 35 и 4%, спурт - 4 и 1%, поза вытянувшись - 10 и 26% [Эрнитс, 1982].
В отсутствие интенсивного преследования козлы не боятся человека. В Лаго-дехском заповеднике стадо взрослых самцов подпускало открыто идущего человека на 50 м, изредка издавая короткий свист [Верещагин, 1938]. На 30-40 м можно было подходить к животным в Кабардино-Балкарском заповеднике [Аккиев, 2001]. В Тебердинском заповеднике несколько групп козлов сами подходили к людям на 10-15 м, ожидая подкормки хлебом [Баскин, 1980]. В Цейском ущелье животные почти утратили страх перед многочисленными туристами и альпинистами - из всех ориентировочно-исследовательских реакций остались только разглядывание и свист [Вейнберг, 19816]. В районах постоянных охот звери, однако, весьма осторожны, особенно зимой, и убегают при малейшем подозрении на опасность, находящуюся за сотни метров [Динник, 1909, 1910; Насимович, 1939]. Тем не менее при современном дальнобойном нарезном оружии это не спасает их от истребления.
Пространственная структура и перемещения. Кавказские козлы ведут преимущественно оседлый групповой образ жизни на ограниченных участках обитания, но их стации, как показано выше, неодинаковые в разные сезоны года. Взрослые самцы и самки с молодняком большую часть года живут обособленно, что снижает конкуренцию за пищу и беспокойство.
Сезонные кочевки зверей обычны в пределах одного хребта или горного отрога, в редких случаях - с одного горного массива на другой, при этом козлы могут заходить далеко в лес, пересекать водораздельные долины и глубокие ущелья иногда вблизи от населенных пунктов. С южного многоснежного макросклона Главного Кавказского хребта часть зверей в октябре - декабре, видимо, уходит на зимовку в область снежной тени на северный макросклон, где сосредоточивается в 10-25 км от водораздельного гребня. Размах сезонных кочевок здесь, следовательно, может превышать 25 км, но во многих районах он значительно меньше [Верещагин, 1938; Насимович, 1939, 1949а, 1955]. По мнению З.С. Эквтимишвили [1952], такие миграции не имеют места. Протяженные переходы козлов, достигающие 10-25 км, обычны в сентябре - первой половине января в Кабардино-Балкарии [Залиханов, 1967, 1968]. В Дагестане известны миграции самцов в Богосском массиве [Абдурахманов, 1973, 1982].
Размах ежедневных горизонтальных перемещений кавказских козлов, как правило, не превышает нескольких километров. Однако на солонцы звери ходят за 10-12 км, стекаясь на отдельные из них с площади до 10 тыс. га. Перемещения животных в высотном диапазоне обычны не только в разные сезоны, но и разные дни и в течение суток. Размах ежедневных высотных перемещений групп взрослых самцов колеблется в пределах 400-2000 м. У самок он гораздо меньше, особенно летом, что связано с приуроченностью их участков обитания к скальным выходам - убежищам, где появляется потомство, для которого ежедневные переходы по горной местности непосильны и небезопасны. Взрослые самцы в бесснежный период менее уязвимы для хищников и поэтому могут позволить себе регулярные переходы с мест дневок
на пастбища и обратно [Котов, 1964а, 1968; Залиханов, 1967, 1968; Абдурахманов, 1982; Вейнберг, 1984; Магомедов и др., 2001].
В Северной Осетии сроки высотных миграций групп взрослых самцов и самок разные: первые спускаются в лесной пояс осенью на месяц позже, а во второй половине зимы на 1-2 месяца раньше вторых покидают его. Фактически самцы живут вместе с самками в одном поясе только во время гона. Большинство козлов входит в этот период в состав самочьих групп, остальные широко разбредаются в поисках свободных самок, нередко выходя далеко за пределы типичных зимних стаций. Территориальность отсутствует даже во время гона, но элементы маркировки сохраняются, хотя и носят агонистический характер [Вейнберг, 1980, 1982а; 1984].
На северном макросклоне Большого Кавказа зимой большинство зверей концентрируется на склонах южной экспозиции. В весенне-летний период самки и молодые особи предпочитают северные склоны субальпийского и альпийского поясов, где прохладнее и имеются зеленые пастбища и водопои. Взрослые самцы, у которых преобладает сумеречная и ночная активность, не проявляют избирательности в отношении определенной экспозиции склонов. Они ежедневно перемещаются в высотном направлении до 1-1,5 км: кормятся на богатых субальпийских и альпийских пастбищах, а дневки устраивают в субнивальном поясе [Насимович, 1949а; Наниев, 1958; Абдурахманов, 1977; Залиханов, 1967,1968; Котов, 1968; Вейнберг, 1981а, 1984; Магомедов и др., 2001]. На южном макросклоне летом козлы, напротив, нередко не спускаются, а поднимаются на альпийские пастбища от верхней границы леса и из субальпийской зоны, что связано с иной структурой рельефа: изрезанные скальные массивы (каньонообразные ущелья) и русла рек, служащие убежищами, находятся здесь ниже основных пастбищ на сглаженном высокогорье. Немалую роль в их пространственном размещении играет и домашний скот, выпасаемый в альпийской зоне [Верещагин, 1938; Эквтимишвили, 1952].
Самцы, как подчеркивают многие исследователи, в горах обычно обитают не только выше самок, но и в более открытых биотопах. На востоке ареала в летний период свыше 90% самок держится в диапазоне высот от 2500 м до 3250 м на склонах средней крутизны (около 36°), богатых убежищами; самцы - в пределах 3250-3500 м в зоне с крутизной склонов от 28 до 46°. Зимой животные выпасаются, в основном, в высотном поясе от 2500 до 3000 м. Ниже этого уровня встречается больше самок, выше - больше самцов. Зимой звери концентрируются на менее крутых склонах (до 35°), где больше площадь пастбищ [Магомедов, Ахмедов, 1994; Ахмедов, 1995; Магомедов и др., 2001]. В Северной Осетии летом в скальниках субальпийского и альпийского поясов держатся 63% взрослых самок и лишь 18% самцов, которые пасутся чаще на открытых склонах [Вейнберг, 1982а].
Места обитания кавказских козлов пронизаны густой сетью хорошо заметных, отполированных копытами троп, поскольку животные ежедневно по нескольку раз перемещаются из одних и тех же мест отдыха к пастбищам и обратно цепочкой друг за другом. При опасности животные стремятся кратчайшим путем уйти в скалы, что спасает их от хищников, но не от охотников, знающих такую особенность поведения. Лежки звери устраивают на открытых местах с хорошим обзором или под скалами, на каменистых склонах и осыпях, под карнизами, в пещерах и нишах. Перед тем как лечь, отгребают передней ногой лишние камни, и, если есть возможность, делают небольшое углубление в земле. На крутых
склонах отдыхают сидя. На постоянных лежках постепенно скапливается большое количество помета, слой которого порой достигает 37 см [Князев, Савинец-кий, 1992]. Зимой они нередко отдыхают прямо на пастбище, ложась на снег или на пучки сухой травы и землю, или в лесу у стволов [Насимович, 1939; Котов, 1968].
Численность
По оценке А.Г. Банникова [1962, 1965 1974, 1978], видимо сильно заниженной, в первой половине XX в. численность кавказских козлов в СССР составляла 20-25 тыс., в 60-70-е годы - 25 40 тыс. голов. По более достоверным сведениям, в 60-80-е годы только на Северном Кавказе жили 30-50 тыс., в том числе в Дагестане -8-10 тыс., Чечено-Ингушетии - 2 тыс., Северной Осетии - около 4 тыс., Кабардино-Балкарии - 9-5,5 тыс., Ставропольском крае - 3,5 тыс., Краснодарском крае -26 тыс. [Залиханов, 1967, 1968; Абдурахманов, 1973, 1977, 1979, 1982; Равкин, Овчаренко, 1973; Круторогов, 1975; Равкин, 1975а,б, 1976; Точиев, 1975; Айунц, 1983; Прилуцкая, Пишванов, 1988]. В Азербайджане насчитывали 7-8 тыс. особей [Алиев, 1961, 1962; Алекперов и др., 1973, 1976; Кулиев, 1981], в Грузии 12-13,6 тыс. [Капанадзе, 1975, 1978], а в конце 80-х годов - около 2 тыс. [Арабули, 1989]. Ресурсы вида в СССР в эти десятилетия, скорее всего, колебались в пределах 60-70 тыс. [Климова, 1989]. Поголовье восточнокавказской формы на северном и южном макросклонах было примерно равным-по 15-16 тыс., всего около 30 тыс., а в 90-е годы - 25 тыс. [Вейнберг, 1984]. Численность западнокавказской формы достигала такого же уровня. В. Котов [1964г], однако, оценивал ее в 100 тыс., а П. Вейнберг [1997] позднее - в 6-10 тыс.
В конце XX - начале XXI вв. ресурсы кавказских козлов в России составляли около 50 тыс. особей: в Дагестане - 18-20 тыс. [Ахмедов, 1995; Магомедов, Ахмедов, 1994, 1999; Магомедов и др., 2001], в Кабардино-Балкарии - 18-21,5 тыс., Северной Осетии - 1,8-3,5, Карачаево-Черкесии - 4-5 тыс. и в Краснодарском крае - 0,3-0,1 тыс. (без поголовья Кавказского заповедника) [Ломанов и др., 2000; Сидоров, 2000, 2004].
Основные запасы сосредоточены в заповедниках и заказниках. В Кавказском заповеднике в 1927 г. насчитано 0,8 тыс., в 1935-1940 гг. - 7-8, в 40-50-е годы - 8-13, в 60-80-е - от 13-16 до 9-11, в 90-е. - около 3-4 тыс. [Насимович, 1940, 1949а; Верещагин, 1947; Крайнова, 1947; Котов, 1960а,д, 1961, 19666, 1968, 1969, 1971; Котов, Рябов, 1963; Равкин, Овчаренко, 1973; Дуров, 1974, 1978; Равкин, 19756; Кудактин, 1977, 1985,1996; Ромашин, 1989, 1994,2001]. В последние годы, судя по отчетам, здесь учитывают около 1-1,2 тыс. В Тебердинском заповеднике в 1936— 1941 гг. обитали 2-2,5 тыс., 1949-1952 гг. - 2,6-3,6 [Аренс, 1957], в 1954—1955 гг. -2,3-2,5 [Инякова, 1957а], в 70-80-е годы-3,6-1,8 [Тарасов, 1973, 1976, 1977, 1982, 1983; Равкин, 19756; Бобырь, Онипченко, 1990], в 90-е - 0,9-1,1 [Бобырь, 1999, 2002], в 2003 г. - 0,8 тыс. (по отчету). В охотничьих угодьях и заказниках Краснодарского и Ставропольского краев, на площади в 6 раз большей, чем в Кавказском и Тебердинском заповедниках, жило около 100 особей [Дуров, 1978]. В Кабардино-Балкарском заповеднике, организованном в 1976 г. на площади в 74 тыс. га, в 1982-1987 гг. размещались 5,2 тыс. [Айунц, Шалыбков, 1990], в 2003 г., судя по
данным отчета, - 6,4 тыс. В Северо-Осетинском заповеднике в 70-90-е численность колебалась в пределах 1-2 тыс. С начала 90-х заметно сокращение поголовья, а плотность населения уменьшилась с 85-90 до 40 особей/1000 га [Вейнберг, 2002]. На 1 января 1990 г. в 4 российских заповедниках находилось около 14 тыс. кавказских козлов [Филонов, Шадрина, 1996], позднее - менее 10 тыс.
В Закавказье козлы также сохраняются в основном в заповедниках: в Закаталь-ском во второй половине XX в. обитали 4,5-2,5 тыс. [Верещагин, 1947,1959а; Алиев, 1961,1962; Попкова, 1967; Абдурахманов, 1977; Кулиев, 1977,1981; Гасанов, 1990], Тушетском - 70-120 голов [Чиковани и др., 1990а], Исмаилинском - 1,1 тыс. [Кулиев, 1989], Лагодехском - 3-3,9 тыс. [Енукидзе, 1953; Алекперов и др., 1976], в 80-е годы - не более 1,2 тыс. [Чиковани и др., 1990в].
Плотность населения кавказских козлов (табл. 59) весьма высока даже в сравнении с другими массовыми видами диких копытных. В разных частях ареала этот показатель превышает 5 особей на 1000 га, на особо охраняемых территориях заповедников достигает 100-400, а зимой на отдельных участках - 600 голов. В Белоканской лесной даче, площадью 10 тыс. га, в Азербайджане, когда она находилась в аренде у Демидовых-Сан-Донато и хорошо охранялась, перед 1914 г. жило не менее 3000 козлов [Витович, 1928]. Допустимая (расчетная) плотность еще выше - 700-800 экземпляров на 1000 га пригодных угодий [Котов, 1964г, 1968; Абдурахманов, 1973, 1979; Вейнберг, 1977, 1984].
Таблица 59. Плотность населения кавказских козлов
Район, заповедник | Особей на 1000 га | Источник информации
Кавказский з-к 30-200 Жарков, 1940, 1959
и 75 (12-342) Котов, Рябов, 1963
сс 170-190 (до 441 зимой) Котов, 1960а,д, 1961, 1964г, 1968, 1971
(( 38 в луговой зоне Ромашин, 2001
Западный Кавказ До 33 в лучших угодьях Котов, 1968
Тебердинский з-к 40-60 (до 181 зимой) Тарасов, 1973, 1976, 1977, 1982, 1983; Бобырь, Онипченко, 1990; Бобырь, 1999
сс 48-114 Айунц, 1980
Кабардино-Балкария 27—400 Залиханов, 1967, 1968
а 52-62 Темботов и др., 19806
Северная Осетия 37 Равкин, 1973, 1975а
C.-Осетинский з-к 80-180 Вейнберг, 19816, 1984
с с 40 Вейнберг, 2002
Ч.-Ингушетия 25 Равкин, 1973, 1975а
Дагестан 18-30 «С
(( 51 (18-73) Ахмедов, 1995; Ахмедов, Магомедов, 2000; Магомедов и др., 2001
а 86 (до 175 зимой) Абдурахманов, 1973
Грузия 5-20 Члаидзе, 1974; Капанадзе, 1975
в заповедниках 17-190 сс
30-50 Арабули, 1986
вне заповедников 5-6 it
Азербайджан 15-59 Кулиев, 1981, 1989
Закатальский з-к 120-175 (250) ' Верещагин, 1938, 1947; Попкова, Попков, 1965*; Кулиев, 1977, 1981; Гасанов, 1990
* По [Равкин, 1975а].
Плотность населения козлов на Западном Кавказе в лучших угодьях в среднем не более 33 особей на 1000 га угодий [Котов, 1968], на Центральном в ландшафтах первого класса бонитета - 40-70 голов [Айунц, 1980], на Восточном этот показатель достигает 50 даже вне заповедных территорий. Наивысшие плотности населения животных отмечены в горно-луговой зоне в отрогах крупных высокогорных образований с наличием протяженных скальных выходов (в сочетании хороших кормовых и защитных условий) средней части Главного Кавказского и Бокового хребтов, наименьшие - ближе к границам ареала в районах со сглаженным среднегорным рельефом. В местах выпаса овец плотность населения диких копытных обычно в 2-3 раза ниже [Равкин, 1975а; Ахмедов, Магомедов, 2000; Магомедов и др., 2001].
Основные факторы, определяющие динамику популяций
Воспроизводство, прирост. Годовой прирост в популяциях кавказских козлов при потенциальном уровне воспроизводства в 25-30% невелик: в Дагестане - 8-12% [Абдурахманов, 1973, 1977, 1979, 1982; Магомедов и др., 2001], Кабардино-Балкарии - 5-14 [Залиханов, 1967, 1968], Северо-Осетинском заповеднике - 3-11 [Вейнберг, 1984, 2002], Тебердинском - 11 [Бобырь, 2002], Кавказском - 8% [Котов, 19666, 1968, 1971].
Детская смертность. Динамика популяций этих копытных в значительной мере определяется уровнем детской смертности. На Восточном Кавказе ежегодно погибает около 40-50% сеголеток. Из оставшихся в живых зверей до двухлетнего возраста доживает около 93%. Хорошая сохранность годовалых особей объясняется тем, что они долго держатся вместе с матерями, неплохо питаются и не участвуют в размножении. В дальнейшем выживаемость самок остается высокой (до 90%) до старости, у самцов этот показатель с возрастом снижается до 70% [Абдурахманов, 1977, 1979, 1982; Ахмедов, 1995; Ахмедов, Магомедов, 2000; Магомедов и др., 2001]. На Западном и Центральном Кавказе уровень детской смертности примерно такой же, как и на востоке - 35-50%, а иногда и выше. В отдельные годы половина детенышей гибнет уже в первый месяц после рождения, а до годовалого возраста доживают не более 25% [Теплов, 1938; Котов, Рябов, 1963; Залиханов, 1967; Котов, 1968; Ромашин, 2001]. Смертность животных в большей мере связана со следующими факторами.
Погода и корма. Часть новорожденных погибает при резких похолоданиях и поздних снегопадах. У сеголеток в осенне-зимний период сохраняется высокий уровень метаболизма, что при недостатке пищи приводит к быстрому истощению организма и повышенной смертности. При дефиците кормов сравнительно высока смертность и истощенных после гона самцов. Зимой животные концентрируются на ограниченных по площади пологих пастбищах, более доступных для хищников и охотников [Ахмедов, Магомедов, 2000; Магомедов и др., 2001]. В особенно многоснежные зимы имеет место массовая гибель зверей, что наблюдалось на Центральном Кавказе в 20-е годы, а так же в 1962/63 гг. [Залиханов, 1967]. В Северо-Осетинском заповеднике в 1986/87 гг. погибло до 5% популяции, а серия снежных зим 1987-1993 гг. стала, видимо, одной из причин резкого сокращения численности копытных на Центральном и Западном Кавказе [Вейнберг, 2002]. Кормовой фак
тор, в значительной мере связанный с климатическим, является не только одним из ведущих в популяционной динамике козлов [Ромашин, 1997, 2001], но и опосредованно, через повышенную смертность тех или иных половых и возрастных групп, определяет структуру группировок.
Хищники. Основной враг кавказских козлов, уничтожающий ежегодно сотни особей, - волк. В летний период от него особенно страдают самки с детенышами. Для взрослых самцов одиночные хищники менее опасны, поскольку группа успешно защищается, сбиваясь в круг и постепенно отходя в таком построении к скалам. Однако в период гона, когда группы распадаются, и зимой, когда волки объединяются в крупные стаи, истощенные и менее проворные самцы погибают чаще, чем самки [Залиханов, 1967, 1968; Бобырь, 1977; Кудактин, 1982; Ахмедов, Магомедов, 2000; Магомедов и др., 2001].
В Кавказском заповеднике, где соотношение волка и козла близко 1:140-300, i остатки жертв отмечается в 12-56% экскрементов и содержимого желудков хищников. От волка здесь порой погибает до 60% приплода копытных, а в 70-е годы доля сеголеток в популяции козлов сократилась до 4%. В 1986-1997 гг. смертность от этого фактора достигала 20% в общей гибели [Теплов, 1938; Насимович, 1939, ,
1949а; Котов, 19666; Дуров, 1974; Кудактин, 1975, 1977, 1986; Ромашин, 2001]. В Тебердинском заповеднике волк ежегодно изымает до 4% поголовья [Бобырь, Онип-ченко, 1990]. В Дагестане на его долю приходится до 20% от числа погибших [Абдурахманов, 1973].
Медведь представляет меньшую угрозу для козлов, поскольку большая часть хищников залегает на зиму в спячку. Однако в горах Кавказа этот зверь распространен повсюду до высоты 3000 м. Остатки задавленных копытных и их шерсть в экскрементах медведя отмечают все исследователи. Нередко хищник устраивает засады и давит козлов у солонцов, но чаще все же питается трупами животных, погибших в лавинах [Бобырь, 1979, 1980].
Рысь относительно немногочисленна и в целом не играет регулирующей роли в популяционной динамике козлов. Однако этот хищник может круглогодично жить в условиях сильно расчлененного рельефа в типичных местах обитания самок с ягнятами, нанося им значительный локальный урон. На Боковом хребте доля этих копытных в пище рыси достигает 66% [Залиханов, 1967; Вейнберг, 1984, 1988], в Кавказском заповеднике - 19% [Котов, 19666]. В прошлом более серьезным врагом был барс [Динник, 1910; Верещагин, 1938; Насимович, 1939]. Козлята иногда становятся жертвами лисицы, но гораздо чаще погибают от беркута и бородача-ягнятника. Естественная их реакция на появление хищников -затаивание, что, однако, не спасает от нападения. С трехмесячного возраста ягнята начинают активно обороняться от хищных птиц ногами и рогами, но до 5-месячного возраста при их появлении все же стремглав бросаются к матерям и ' прячутся под них. Несмотря на такое оборонительное поведение, потери сеголеток от хищных птиц, вероятно, составляют около 5-7% [Залиханов, 1967; Абдурахманов, 1973, 1977; Ахмедов, Магомедов, 2000; Магомедов и др., 2001].
Конкуренты. К пастбищным конкурентам кавказских козлов обычно относят всех высокогорных копытных и грызунов. Однако эти стадные животные доминируют среди диких копытных, включая и более крупного благородного оленя, и нередко оттесняют их с пастбищ и солонцов [Насимович, 1949а; Котов, 1968; Ромашин, 1995].
Основной их конкурент на протяжении последних 4 тыс. лет, и, особенно, в конце II тысячелетия н. э. [Савинецкий, 1992] - домашний скот, выпасаемый летом на горных лугах, который потребляет растительной продукции не меньше, а в ряде районов гораздо больше, чем дикие копытные, и, кроме того, вытесняет последних с лучших пастбищ. Звери осваивают их в темное время суток, но чаще держатся в местах, недоступных для домашних животных. Более того, диким копытным приходится зимовать на уже стравленных пастбищах, что не благоприятствует их жизни [Насимович, 1949а; Залиханов, 1967; Абдурахманов, 1973, 1982; Бобырь, 1977; Вейнберг, 1984]. И чаще всего именно поэтому в местах интенсивного выпаса скота плотность населения кавказских козлов в несколько раз ниже возможной.
Болезни, паразиты, травмы. Смертность кавказских козлов от болезней в целом невелика. Зарегистрированы случаи падежа зверей от чесотки, у некоторых животных обнаружена копытная гниль [Верещагин, 1938; Ахмедов, Магомедов, 2000; Магомедов и др., 2001]. В конце XIX в. в бассейне р. Белой видели “громадную шайку туров штук в 200, все окончательно хромые” [Шильдер, 1901]. Возможна также их гибель от чумы рогатого скота и ящура [Динник, 1909, 1910; Насимович, 1949а]. При вселении козла в низкогорные леса Кубани часть животных погибла из-за нарушений пищеварительной системы [Гинеев и др., 2002].
На Западном Кавказе у козлов найдено 18 видов гельминтов, на Центральном - 24, на Восточном - 53. Общими для разных форм этих копытных оказались лишь 9 видов паразитических червей из 60, тогда как с домашним скотом - 38, с серной - 32, с муфлоном и косулей - 27, благородным оленем -21. Уровень зараженности козлов паразитами в целом невелик, хотя он выше, чем у серны, а гибель от них, в том числе от личинок носоглоточного овода, носит единичный характер. Тем не менее наличие паразитов способствуют исхуданию животных, и они, видимо, чаще погибают в тяжелый для жизни зимний период [Грунин, 1948; Асадов, 1959а; Рухлядев, 1959, 1964; Котов, 19666, 1968; Залиханов, 1967; Абдурахманов, 1973, 2003; Закариев, 1980; Газаев, Аккиев, 1997; Бобырь, 2002].
Иногда в желудках кавказских козлов находят “безоары” - минерализованные образования, состоящие из шерсти животных и тонких корешков растений, которые у домашних животных становятся причиной их гибели [Котов, 19666; Залиханов, 1967, 1968].
Более частое явление - травматизм, который присущ всем горным копытным. Нередко животные срываются со скал и ломают ребра, конечности, рога или погибают. Во время драк сильные самцы могут сбрасывать соперников вниз [Бобырь, Бобырь, 19996]. Покалеченные, хромые и с обломанными и уродливыми рогами звери встречаются повсеместно.
Гибель в лавинах. При переходах вдоль склона или при пересечении лавинного русла стадо животных, идущее цепочкой, как бы “подрезает” снежный пласт, что может вызвать его сход. Звери, Возможно, знают лавиноопасные места на своих участках обитания и стараются их избегать. Тем не менее этот фактор смертности весьма высок, особенно на Западном и Центральном Кавказе среди взрослых самцов [Динник, 1910; Насимович, 1955; Наниев, 1962; Залиханов, 1967, 1968; Котов, 1968; Тарасов, 1976,1982; Бобырь, 1977,1979; Вейнберг, 19816,1984,2002]. В Кавказском заповеднике каждый год в лавинах находят 20-50 особей. Общие ежегодные потери достигают здесь 100-300 экземпляров, или до 9 голов на 1000
га в местах зимовок. В общей смертности доля этого фактора составляет 31% [Котов, 19666, 1968; Кудактин, 1977; Ромашин, 1989, 2001]. В Кабардино-Балкарии погибает не менее 370 особей [Залиханов, 1967], а в Тебердинском заповеднике -2,2-3,3% популяции [Бобырь, 1999]. На востоке такого рода потери регистрируются реже [Ахмедов, Магомедов, 2000; Магомедов и др., 2001], но и здесь иногда находят большие группы животных, погребенные под снегом [Перовский, 1988]. В Дагестане доля таких потерь в общей смертности козлов достигает 12% [Абдурахманов, 1973]. В целом на Кавказе под лавинами, видимо, ежегодно гибнет более тысячи особей.
Антропогенные факторы. П.И. Вейнберг [1984] выделяет четыре основных фактора антропогенного воздействия на кавказских козлов: выпас скота, промышленное освоение горных районов, туризм и альпинизм, браконьерство.
Скот, как отмечено выше, стравливает пастбища и вытесняет диких копытных в высокогорье. Кроме того, он является переносчиком опасных заболеваний и гельминтов. Не меньшее зло - пастухи, обычно вооруженные, и их собаки, постоянно находящиеся в угодьях. В результате постоянного беспокойства у горных копытных изменяется суточный ритм активности, и они выходят на пастбища лишь ночью, а днем держатся на перевалах и осыпях или в лесу [Гептнер, Формозов, 1941; Верещагин, 1959а; Котов, 1968; Абдурахманов, 1973].
В последнее время все большее беспокойство причиняют туристы, горнолыжники и альпинисты, которые проникают даже в самые труднодоступные районы. Тем не менее козлы мирятся с присутствием человека, если оно не сопровождается их уничтожением и изменением среды обитания, и не покидают своих участков, даже если рядом проходят оживленные автомобильные трассы или туристические тропы. При частых встречах с людьми у них ослабевает реакция на опасность, что повышает успех охоты на них хищников и браконьеров [Тарасов, 1977; Вейнберг, 1984].
В конце XX в. в связи с военными действиями на Кавказе, антропогенный пресс, вызванный передвижением войск, прокладкой коммуникаций и локальными боями, стал масштабным, что негативно отражается на популяциях диких копытных [Магомедов и др., 2001]. И все же гораздо более значимый фактор - охота.
Использование, охрана и перспективы восстановления ресурсов
Кавказские козлы - эндемики Кавказа, символ горных народностей и излюбленный объект охоты, дающий прекрасное по вкусовым, питательным и диетическим свойствам мясо [Абдурахманов, 1973]; ценнейшие рога, из которых делают кубки и настенные украшения; мягкие и прочные шкуры, которые идут на пошив теплой одежды, обуви, бурдюков, мешков и покрытия седел. Жир широко используют в народной медицине.
Неумеренное охотничье преследование стало главной причиной резкого сокращения ресурсов копытных на Кавказе, которое началось с появлением русских поселений в предгорьях в период кавказских войн, с завозом четырехлинейных винтовок Бердана, а затем и русских трехлинеек Мосина, что позволяло отстреливать зверей на дальних дистанциях. Вооруженность населения достигла пика после войны 1914 г., революции 1917 г. и гражданской войны. Кавказских и безоаровых козлов добывали в любое время года, отстреливая с подхода, из засад, подкарауливая на солонцах, у водо
поев и на тропах или отлавливая капканами [Разевиг, 1904; Динник, 1909; Каверзнев, 1933; Верещагин, 1959а]. Наибольший урон причиняли пастухи, “которые пасут своих баранов на высоких альпийских лугах и отправляются туда с винтовкой в руках, чтобы иметь возможность ежедневно охотиться за турами, сернами или оленями, добывать себе даровое мясо и сохранять за летние месяцы лишний десяток своих баранов или коз. От этих истребителей дичи на Кавказе погибают ежегодно сотни туров” [Динник, 1910]. Группа из 4-5 пастухов задень отстреливала до 10-12 козлов. Помимо пастухов их неумеренно истребляли зверопромышленники и ветеринарные стражники [Витович, 1928]. Охотники за свою жизнь добывали несколько сотен, а отдельные -более тысячи животных. В первой половине XX в. ежегодная добыча составляла по всему Кавказу около 4—8 тыс. особей, в том числе 0,8-1 тыс. в Азербайджане [В-н, 1936; Гептнер, Формозов, 1941; Верещагин, 1947, 1959а].
Известный зоолог Н.Я. Динник [910] полагал в свое время, что кавказские козлы могут совершенно исчезнуть с лица земли через несколько десятков лет. К счастью, его предположения не сбылись. Эти звери по-прежнему являются промысловыми, но их легальная добыча невелика. В начале 60-х годов в СССР отстреливали до 1000 особей [Банников, 1965]. В 1981 г. на весь Северный Кавказ выдано 175 лицензий [Вейнберг, 1985]. С 1980 по 1984 гг. официально добыто всего 270 особей [Перовский, 1985], в 1986 г. - 91 [Дежкин, 1989], в конце XX - начале XXI вв. ежегодно 115-166 голов, или от 3 до 87 экземпляров в разных областях [Ломанов и др., 2000; Сидоров, 2000, 2004].
Нелегальный отстрел многократно выше и, по экспертной оценке, достигает нескольких тысяч особей в год. Браконьерство распространено повсеместно и носит массовый характер. Особенно большой урон этим копытным, как и прежде, наносят вооруженные пастухи, жители горных селений, а также егеря, лесники, работники турбаз и альпийских лагерей. Нередко браконьеры действуют артелями, а в местах охоты оборудуют коптильни и салотопки [Котов, 1960а, 19666; Залиханов, 1967]. В Дагестане на их долю приходится не менее 43% от числа всех погибших животных [Абдурахманов, 1973].
Судя по личным сообщениям охотоведов, незаконный промысел диких копытных на Кавказе особенно расцвел в последнее десятилетие XX в., когда возрос спрос на трофейную охоту, правовой нигилизм достиг максимума, а местное население в результате военных действий получило на руки огромное количество автоматического нарезного оружия. Немалую лепту в нелегальный отстрел животных вносят многочисленные армейские, пограничные и милицейские подразделения, дислоцирующиеся в горах. Браконьерский пресс заметно усилился даже в заповедниках, причем в Кавказском - в 10 раз. Здесь для охот на копытных широко применяются вертолеты, что делает доступными даже самые удаленные угодья. Много зверей потеряно из-за прекращения выкладки соли и эмиграции их за пределы охраняемой территории [Кудактин, 1996; Бобырь, 1999,2002; Ромашин, 2001; Вейнберг, 2002].
В целом наметилась тенденция к резкому сокращению численности кавказских козлов, причем даже в высокогорьях, где очевидна постепенная изоляция отдельных групп [Аккиев, 2001]. Наибольший урон от браконьеров повсеместно несут взрослые самцы, на которых выборочная охота ведется практически круглогодично. Уничтожение преимущественно крупных и сильных производителей, по мне
нию М.Г. Абдурахманова [1973], приводит к включению в размножение молодых особей, ослаблению полового отбора, обеднению генофонда популяций и снижению жизнеспособности животных.
Учитывая низкий годовой прирост популяций, норма допустимого охотничьего изъятия кавказских козлов, даже при условии полной ликвидации браконьерства, не должна превышать 5% от численности [Вейнберг, 1984]. Поскольку браконьерство на Кавказе масштабно и, на мой взгляд, практически неискоренимо, то эксплуатация козлов должна быть очень ограниченной, но при этом максимально доходной, что возможно только при хорошо организованной трофейной охоте. Без ущерба для популяций на Северном Кавказе, видимо, можно ежегодно добывать до сотни самцов старше 10 лет, что при высоких ценах на трофеи приносило бы немалый доход. В районах трофейной охоты в целях создания и поддержания высокой плотности населения диких копытных необходимо ограничивать или полностью прекращать выпас домашнего скота, и полностью запрещать охоту на другие виды дичи.
При условии реализации хотя бы небольшой части средств, полученных от трофейной охоты, на охрану и биотехнические мероприятия, общее поголовье козлов реально увеличить до 150-200 тыс. Только в Дагестане на значительно сокращенном ареале этих копытных может быть не менее 56 тыс. [Абдурахманов, 1973], в Кабардино-Балкарии - около 40^17 тыс. [Залиханов, 1968], а на Западном Кавказе численность могла бы быть выше минимум в 5 раз [Котов, 19666]. При таких ресурсах возможно значительно увеличить добычу, что, в свою очередь, способствовало бы развитию охотничьего хозяйства, охотничьего туризма, сферы обслуживания и занятости местного населения. Рациональное использование диких горных копытных для трофейной охоты в принципе экономически гораздо выгоднее пастбищного животноводства.
По расчетам М.Г. Абдурахманова [1973, 1979, 1980, 1982], кавказских козлов можно реинтродуцировать на территории, где они были уничтожены, достигающей 36% площади современного ареала. К сожалению, в настоящее время при массовом браконьерстве эта работа не будет успешной. Все усилия сейчас целесообразнее направить на сохранение оставшихся животных, которые служат пищей не только охотникам, но и таким редким хищникам как переднеазиатский леопард, кавказский бурый медведь, белоголовый сип, черный гриф, беркут и бородач-ягнятник.
Эти копытные заслуживают гораздо большего внимания животноводов и селекционеров как материал для выведения новых пород домашних коз и создания высокопродуктивных гибридных стад. Их, как показывает опыт, можно разводить и на охотничьих фермах. В 1888-1890 гг. 29 дагестанских козлов выпустили в большой загон в Боржомском великокняжеском имении. К 1917 г. здесь насчитывали около 500 особей, а в 1919 гг. все звери были истреблены. В 1934 г. в другой загон поместили трех и в 1941 г. еще 8 особей, но война помешала реализации этого мероприятия [Динник, 1910; Чиковани и др., 19906]. В 1913 г. козлов завезли в горную часть Крыма, где их сначала содержали в вольере, а потом выпустили на волю. Позднее все животные были уничтожены [Гептнер и др., 1961]. В 1982-1986 гг. 14 особей из Кавказского заповедника вселили в низкогорные леса Геленджикского района (лесоохотничье хозяйство “Кубанское”). В 1993 г. в полувольных условиях содержали около 20 зверей, а к 1995 г. и здесь все переселенцы были истреблены [Павлов, 1999; Гинеев и др., 2002].
СИБИРСКИЙ КОЗЕЛ Capra sibirica Pallas, 1776
Ibex sibiricus Pallas, 1776. Spicil. Zool., 11: 52. Вост. Саян, xp. Мунку-Сардык.
Capra sibirica Meyer, 1794. Zool. Annal., 1: 397. Саяны, xp. Мунку-Сардык.
Caprapallasii Schinz, 1838. N. Denkschr. Schweiz. Ges. Naturwiss., 2: 9. Nom nov. pro Ibex sibiricus
Pallas. Nom. praeocc., non Roulier, 1841.
Capra sakeen Blyth., 1842. J. Asiat. Soc. Bengal., 5, 11: 283. Тибетский склон Гималаев.
Capra sibirica lydekkeri Rotschild, 1900. Novit. Zool., 7: 277. Горный Алтай, Катунский xp.
Capra altaica Noak, 1902. Zool. Anz., 25: 623. Казахстан, “Прииртышский Алтай”.
Capra fasciata Noak, 1902. Ibid., 25: 623. Горный Алтай, Телецкое оз.
Capra alaiana Noak, 1902. Ibid., 25: 624. Киргизия, ср. часть Алайского xp.
Capra sibirica hagenbeckii Noak, 1903. Ibid., 26: 384. Монголия, Кобдо.
Capra sibirica lorenzi Satunin, 1905. Природа и охота, 5: 27. Иркутская обл., 300 км к югу от Нижнеудинска.
Capra sibirica merzbacheri Leisewitz, 1906. Zool. Anz., 29: 655. Киргизия, верхи, течение p. Ha-рын.
Capra sibirica almasyi Lorenz, 1906. Denkschr. K. Akad. Wiss. Wien, 80: 89, 98. Киргизия, xp. Tep-скей Алатау, Аксу.
Capra sibirica transalaiana Lorenz, 1906. Ibid., 80: 90,103. Таджикистан, Горный Бадахшан, южн. склон Заалайского хр., “Сарык-могул”.
Capra sibirica typica Lorenz, 1906. Ibid., 80: 95. Бурятия, Вост. Саян, Тункинские Гольцы.
Capra sibirica formosovi Zalkin, 1949. Бюл. МОИП. Отд. биол., 54, 2: 20. Киргизия, Киргизский (Александровский) хр., “ущ. Кыяматгу, в 26 км ю.-з. ст. Ново-Дмитриевской”.
Capra sibirica dementjevi Zalkin, 1949. Ibid., 54,2: 21. Зап. Куэн-лунь, горы Тохта-хон ю.-з. Яркенда.
Характеристика. Козел крупных размеров (фото 30), плотного сложения, с короткой шеей и относительно длинным среди горных козлов хвостом (до 23 см). Половой диморфизм значителен. Длина тела самцов достигает 171 см, самок - 145 см, масса - 123 (в исключительных случаях, возможно, 150-180) и 64 кг.
Окраска серовато- или коричневато-бурая, с потемнениями или посветлениями на разных участках головы и туловища в зимнем меху. У самцов на хребте темная полоса. Борода длинная, до 20 см, черная или бурая, у самок ее нет или она представлена тонким пучком грубых волос. Подвеса из удлиненных волос на шее и передней части туловища нет. Хвост с удлинными волосами. Зимняя шерсть длинная (до 10 см), густая и грубая, подпушь хорошо развита. Летний мех короткий и без подпуши.
Зароговая часть черепа относительно длинная. Мозговая коробка широкая. Носовые кости заметно выдаются вперед. Возвышения на лобных костях у оснований роговых стержней отсутствуют. Костные стержни рогов относительно короткие, загнуты в одной плоскости, сильно сжаты с боков и в поперечном сечении имеют форму равнобедренного треугольника, основанием которого служит широкая и слегка выпуклая передняя поверхность (рир. 56). Предкоренные зубы иногда заметно редуцированы.
Рога у самцов типично “козлиные”: длинные (до 148 см), массивные, сжатые с боков, саблевидно изогнутые в одной плоскости, с двумя четко обозначенными ребрами на широкой передней поверхности, снабженной поперечными валиками. От основания они расходятся под небольшим (менее 45°) углом. Роговой чехол в сечении близок к треугольной форме с выпуклым основанием. У самок рога корот-
Рис. 56. Череп и рог самца сибирского козла (экземпляры из Восточной Сибири; Зоомузей ЗИН РАН, коллекционные №№ 482 и 7653) Вид черепа сбоку, сверху и снизу
кие (до 45 см), тонкие, с пологим загибом назад, без ребер на передней округлой поверхности. Сечение их эллиптической формы. Роговое вещество серо-вато-бурое.
Хорошо развиты только препуциальная и подхвостовая специфические железы. Железистое поле последней окружает и устье заднего прохода [Соколов, Чернова, 1988, 2001].
Стадные малоподвижные животные. Обитают в крупных малоснежных скалистых массивах с наличием остепненной растительности. Обладают острым зрением. В неволе живут до 18-22 лет, в природе редко доживают до 13-15 лет.
Имеют важное охотничье значение.
Распространение. Горные системы Центральной Азии (рис. 57). Современный ареал очаговый.
Состав. Выделено более 13 подвидов, большинство из которых, безусловно, неприемлемы, поскольку при внимательном рассмотрении их характеристики совпадают или неудовлетворительны (см. ниже). Необходима тщательная таксономическая ревизия.
Изменчивость и таксономия
Размеры и масса. Новорожденные сибирского козла достаточно крупные (от 2,6 до 4,8 кг, в среднем около 3 кг) и хорошо развитые. Рост наиболее интенсивен в первый и второй год жизни, но у самок он заметно мень-
Рис. 57. Восстановленный ареал сибирского козла (по [Гептнер и др., 1961 ]; с дополнением)
ше уже в годовалом возрасте. С третьего года рост замедляется, и у самок прекращается к 5 годам, а самцы растут до 8-10 лет [Шнаревич, 1948; Егоров, 1953; Савинов, 1962; Федосенко, Бланк, 1982; Слудский и др., 1983].
Взрослые самцы значительно крупнее самок. Длина тела у первых достигает 171, у вторых - 145 см, высота в холке, соответственно, 106 и 92 см (табл. 60), масса - 123 и 64 кг (табл. 61). Вес крупных самцов на Алтае и в Джунгарском Алатау, возможно, близок к 150-180 кг [Слудский, Афанасьев, 1964; Размахнин, 1977; Собанский, 1992], однако эти сведения основаны на сообщениях охотников и требуют подтверждений.
Судя по приведенным в таблицах данным, существенных отличий в размерах и массе тела у особей их разных географических районов нет. Центральноазиатские козлы по размерам не превосходят алтайских, как нередко ошибочно считают [Федосенко, 2003].
Краниометрическая характеристика. Половой диморфизм по признакам черепа у взрослых особей сибирского козла четко выражен. Наибольшая его длина у самцов составляет в среднем 279 (232-310), у самок - 250 (222-269) мм; ширина— 150 и 134 мм, длина костных стержней рогов - 319 и 108 мм (табл. 62).
Индивидуальная и географическая изменчивость также значительна. В.И. Цалкин [1949, 19506] нашел, что у сибирского козла в северной части ареала (Алтай, Саяны, Монголия) размеры черепа заметно меньше, чем в южной (Тянь-Шань и Памир), но наиболее мелкие размеры имеют животные с Западного Куэнь-луня. Особи с Гобийского Алтая и Заалтайской Гоби по строению черепа сходны с ал-
Таблица 60. Размеры (см) взрослых особей сибирского козла*
Самцы Самки Район (источник информации)**
п М | lim п 1 М lim
Длина тела
8 151,6 138-160 5 141,0 132-145 Горный Алтай (1)
- 160,2 - 132,7 — Предбайкалье (8)
14 155,4 141-171 30 126,5 122-134 Джунгарский Алатау (3)
17? 155,1 140-171 55? 127,1 111-140 -“-(6)
10 143,7 130-165 3 117,7 115-120 Тянь-Шань (4)
1 - 169 - - - Заилийский Алатау (7)
1 - 144 1 124 Восточный Памир (7)
Высота в холке
8 94,7 86-102 5 80,2 78-85 Горный Алтай (1)
- 99,3 - 87,3 — Предбайкалье (8)
14 98,7 94-102 30 80,9 77-85 Джунгарский Алатау (3)
17 98,8 88-104 55 82,0 65-92 -“-(6)
11 91,6 78-102 6 70,5 67-74 Тянь-Шань (4)
4 89,0 — 2 68,7 — Сарытау, Заилийский Алатау (5)
1 - 106 — — — Заилийский Алатау (7)
14 93,2 86-99 - - — Терскей Алатау и В. Памир (2)
1 - 101 1 — 69 Восточный Памир (7)
— — 95-102 — — — Балтистан (7, по сведениям
Stockley, 1926)
Обхват туловища
8 123,0 116-130 5 93,5 90-97 Горный Алтай (1)
14 115,8 103-127 30 90,6 87-100 Джунгарский Алатау (3)
11 105,8 94-124 6 84,2 74-89 Тянь-Шань (4)
4 110,0 - 2 87,0 - Сарытау, Заилийский Алатау (5)
14 104,0 99-115 - — - Терскей Алатау и В. Памир (2)
Длина ступни
14 35,6 35-37 30 31,2 30-32 Джунгарский Алатау (3)
17 35,8 34-38 55 31,5 29-35 -“-(6)
2 34,0 34-34 3 30,5 29-32,5 Тянь-Шань (4)
14 31,1 29-32 - — — Терскей Алатау и В. Памир (2)
Длина уха
8 15,3 14-17 5 14,0 13-15 Горный Алтай (1)
14 14,5 14-15 30 14,7 14-16 Джунгарский Алатау (3)
17 14,7 14-16 55 14,7 13-18 -“-(6)
И 15,3 14-16 6 14,8 14-16 Тянь-Шань (4)
1 - 16 1 - 13 Восточный Памир (7)
Длина хвоста
8 14,7 13-17 5 15,1 14-16 Горный Алтай (1)
14 19,1 18-20 30 16,1 15-17 Джунгарский Алатау (3)
17 19,1 16-23 55 16,0 13-22 -“-(6)
1 - 18 3 11,0 10-12 Тянь-Шань (4)
1 — 17,5 1 — 13 Восточный Памир (7)
* Самки преимущественно старше 4 лет, самцы - старше 5 лет.
** 1 - Собанский, 1992; 2 - Егоров, 1953***; 3 - Савннов, 1962***; 4 - Цалкин, 19506 (по Денисову и Демнну)***; 5 - Бутарин, 1947; 6- Слудский н д[>., 1983***; 7 - Федосенко, 2003; 8 - Недзельский, Недзель-ский, 2003.
*** Расчет по материалам.
Таблица 61. Масса (кг) взрослых особей сибирского козла
Самцы Самки Район (источник информации)*
п м 1 lim п м 1 lim
8 86 70-103 5 56 50-64 Горный Алтай (1)
— 81 - - 56 - Предбайкалье (8)
13 78 71-100 37 41 38-45 Джунгарский Алатау (3)
17 76 58-114 55 43 27-56 -“-(6)
13 71 64-77 4 50 40-56 Терскей Алатау (2)
2 78 65-90 3 35** 29-41 Тянь-Шань (9, 4)
4 — -123 2 - -59 Сарытау, Заилийский Алатау (5)
1 113 - - - Заилийский Алатау (7)
3 74 70-77 3 40 35-44,5 Памир (2)
1 - - 1 - 51 -“-(7)
1 — -120 1 — 60 -“-(Ю)
3 87 85-89 — — — Балтистан (7, по данным
Stockley, 1926)
* Источники информации (1-8) см. в таблице 60; 9 - Денисов, 1935; 10 - Одннашоев, 1986.
** Весеннее взвешивание.
тайско-монГольскими. Эти выводы послужили основой для предложенной им таксономии вида.
Действительно, на основании имеющегося в наличии скудного материала (табл. 62), другого вывода сделать невозможно. При многомерном краниологическом анализе материала из зоомузеев МГУ и Зоологического института, из общей выборки обособляются лишь самки с Западного Тянь-Шаня (форма formosovi) [Казанская, 2004]. Следовательно, выделение нескольких подвидов сибирского козла по краниометрическим признакам нельзя признать обоснованным.
Таблица 62. Размеры черепа (мм) взрослых особей сибирского козла
Самцы Самки Район (источник информации)*
п м lim п М lim
1 2 3 4 5 6 7
Наибольшая длина
5 269,0 258-275 5 246,0 238-249 Горный Алтай (1)
17 270,5 256-293 11 247,0 239-260 Алтай, Саяны, Монголия (2)
— 281,4 - — 256,7 - Предбайкалье (4)
2 265,0 261-269 2 245,5 245-246 Гобийский Алтай, Гоби (2)
20 292,2 276-306 11 254,0 248-262 Тянь-Шань, Памир (2)
7 293,0 286-302 3 254,0 249-258 3. Тянь-Шань (2)
- - - 2 231,0 230-232 3. Куэнь-лунь (2)
48 281,1 236-310 39 250,2 230-269 В ареале (3)
Максимальная ширина
5 146,0 140-153 5 137,8 132-140 Горный Алтай (1)
14 146,6 138-164 6 135,7 126-146 Алтай, Саяны, Монголия (2)
— 159,2 - - 141,9 - Предбайкалье (4)
2 148,5 148-149 2 134,0 131-137 Гобийский Алтай, Гоби (2)
12 156,4 149-170 6 136,7 130-142 Тянь-Шань, Памир (2)
7 155,7 150-159 3 135,7 132-140 3. Тянь-Шань (2)
- - - 2 125,0 125-125 3. Куэнь-лунь (2)
Таблица 62 (окончание)
1 | 2 1 3 1 4 1 5 | 6 1 7
65 151,2 131-182 39 135,9 123-150 В ареале (3)
Межглазничное расстояние
5 107,0 100-113 5 97,7 94-104 Горный Алтай (1)
14 103,2 94-119 6 93,7 86-101 Алтай, Саяны, Монголия (2)
2 104,5 101-108 2 92,5 90-95 Гобийский Алтай, Гоби (2)
12 113,2 102-127 6 91,8 82-99 Тянь-Шань, Памир (2)
7 110,6 101-118 3 95,3 94-97 3. Тянь-Шань (2)'
— - - 2 84,0 82-86 3. Куэнь-лунь (2)
64 107,4 88-127 40 92,5 82-107 В ареале (3)
Длина лицевой части
5 206,0 202-211 5 188,0 186-192 Горный Алтай (1)
14 201,1 188-211 6 193,2 183-207 Алтай, Саяны, Монголия (2)
2 199,5 195-204 2 190,5 188-193 Гобийский Алтай, Гоби (2)
12 217,4 208-234 6 201,3 195-208 Тянь-Шань, Памир (2)
7 215,4 211-220 3 195,3 193-197 3. Тянь-Шань (2)
— — — 2 179,5 178-181 3. Куэнь-лунь (2)
50 207,6 180-234 38 194,6 181-209 В ареале(3)
Ширина мозговой коробки
13 82,2 76-88 6 71,8 70-75 Алтай, Саяны, Монголия (2)
2 80,5 78-83 2 72,0 72-72 Гобийский Алтай, Гоби (2)
11 85,6 79-97 6 72,8 70-75 Тянь-Шань, Памир (2)
7 86,1 83-89 3 74,7 73-76 3. Тянь-Шань (2)
— - — 2 68,0 67-69 3. Куэнь-лунь (2)
61 81,9 70-90 40 72,6 67-78 В ареале (3)
Длина носовых костей
5 85,4 82-89 5 81,0 70-85 Горный Алтай (1)
14 85,9 78-100 6 93,0 85-100 Алтай, Саяны, Монголия (2)
2 94,5 94-95 2 80,0 80-80 Гобийский Алтай, Гоби (2)
18 103,9 90-120 6 101,7 96-113 Тянь-Шань, Памир (2)
7 105,7 94-115 3 95,0 88-106 3. Тянь-Шань (2)
— — - 2 85,5 81-90 3. Куэнь-лунь (2)
59 96,5 78-120 37 92,9 74-113 В ареале (3)
Длина альвеолярного ряда зубов
5 70,6 68-74 5 66,5 63-71 Горный Алтай (1)
14 71,2 62-75 6 65,2 62-69 Алтай, Саяны, Монголия (2)
2 71,0 69-73 2 72,5 71-74 Гобийский Алтай, Гоби (2)
12 75,9 71-83 6 75,0 71-78 Тянь-Шань, Памир (2)
7 77,4 73-85 3 71,7 67-74 3. Тянь-Шань (2)
- - - 2 69,0 68-70 3. Куэнь-лунь (2)
64 74,1 61-85 40 70,1 55-79 В ареале(3)
Длина костных стержней рогов
7 292,8 260-330 2 80,0 75-85 Алтай, Саяны, Монголия (2)
4 430,0 350-510 — — - Тянь-Шань, Памир (2)
— — — 1 — 80 3. Тянь-Шань (2)
21 336,9 205-510 19 109,7 70-220 В ареале (3)
Обхват оснований стержней рогов
7 176,4 155-200 2 85,0 80-90 Алтай, Саяны, Монголия (2)
5 208,0 180-240 — — 1 - Тянь-Шань, Памир (2)
23 203,9 152-270 19 91,3 70-110 В ареале (3)
* 1 - Собанский, 1992; 2 - Цалкнн, 1949, 19506 (расчет по материалам); 3 - Казанская, 2004; 4 - Не-дзельскнй, Недзельский, 2003.
В целом по краниологическим параметрам сибирский козел очень близок к бе-зоаровому и альпийскому козлам, что ранее отмечали В.И. Цалкин, [19506] и В.И. Громова [1953]. Различить эти и другие виды рода Capra реально только по средним значениям некоторых признаков в репрезентативных выборках, а по отдельным экземплярам черепов (без роговых стержней и чехлов) вряд ли можно достоверно определить даже видовую принадлежность (табл. 63).
Таблица 63. Размеры черепа (мм) представителей рода Capra
Самцы Самки Вид (источник информации)*
п м lim п м lim
1 2 3 4 5 6 7
Наибольшая длина
— 269-293 236-310 — 246-254 222-269 С. sibirica (1)
7 278 270-287 7 235 224-262 С. falconeri (2)
- 255-263 232-280 — 224-229 213-268 С. aegagrus (1)
- 263-299 273-320 - 250-254 235-267 С. cylindricomis (1)
— 284-292 266-326 — 250-252 227-265 С. caucasica (1)
9 253 221-276 2 225 217-233 С. ibex (2)
- 253 — - 230 - С. pyrenaica (3)
Максимальная ширина
— 146-156 138-182 — 136-138 125-150 С. sibirica (1)
9 148 141-159 9 127 118-139 С. falconeri (2)
- 124-132 115-157 - 117-123 109-141 С. aegagrus (1)
27 159 140-177 11 136 128-144 С. cylindricomis (2)
37 155 137-170 14 136 122-143 С. caucasica (2)
16 133 115-149 4 119 115-124 С. ibex (2)
- 143 - - 129 - С. pyrenaica (3)
Межглазничное расстояние
— 103-113 88-127 — 92-98 82-107 С. sibirica (1)
9 101 96-109 8 83 73-92 С. falconeri (2)
- 87-92 76-110 - 79-82 70-100 С. aegagrus (1)
27 115 101-131 11 93 88-100 С. cylindricomis (2)
34 112 94-125 12 97 89-103 С. caucasica (2)
16 95 79-107 4 80 77-85 С. ibex (2)
- 97 - - 88 - С. pyrenaica (3)
Длина лицевой части
— 201-217 180-234 — 188-201 178-209 С. sibirica (1)
7 205 195-218 7 177 171-183 С. falconeri (2)
10 184 169-203 ,18 173 155-206 С. aegagrus (2)
14 223 207-241 8 199 182-213 С. cylindricomis (2)
18 215 198-234 11 194 178-208 С. caucasica (2)
9 190 167-209 2 - 167-186 С. ibex (2)
Ширина мозговой коробки
— 82-86 70-97 — 72-73 67-78 С. sibirica (1)
8 83 77-99 8 71 68-74 С. falconeri (2)
21 73 68-84 28 68 63-75 С. aegagrus (2)
27 87 78-95 11 75 72-77 С. cylindricomis (2)
33 85 75-96 14 73 67-78 С. caucasica (2)
15 74 62-82 4 66 63-68 С. ibex (2)
Длина носовых костей
- 85-106 78-120 - 81-102 70-113 С. sibirica (1)
Таблица 63 (окончание)
1 1 2 3 4 5 6 7
8 103 85-121 8 88 80-103 С. falconeri (2)
— 74-77 61-88 - 72-74 60-92 С. aegagrus (1)
26 116 91-149 9 101 87-106 С. cylindricornis (2)
29 111 90-124 12 101 86-116 С. caucasica (2)
12 94 79-101 3 83 77-87 С. ibex (2)
- 88 83 Длина верхнего ряда зубов С. pyrenaica (3)
— 71-77 61-85 — 65-75 55-79 С. sibirica (1)
9 80 76-85 8 73 69-77 С. falconeri (2)
- 73-75 68-80 - 70 60-75 С. aegagrus (1)
27 81 75-86 11 74 64-80 С. cylindricornis (2)
32 79 71-87 14 75 72-82 С. caucasica (2)
16 67 56-73 4 65 62-68 С. ibex (2)
- 71 67 Длина костных стержней рогов С. pyrenaica (3)
— 292-43 205-510 — НО 70-220 С. sibirica (1)
3 314 310-322 4 96 45-140 С. falconeri (2)
- 378-386 250-565 — 102-130 70-180 С. aegagrus (1)
21 369 210-480 8 86 60-105 С. cylindricornis (2)
24 314 185-430 10 91 80-110 С. caucasica (2)
6 306 155-380 4 102 80-120 С. ibex (2)
- 275 78 Обхват оснований стержней рогов С. pyrenaica (3)
— 176-208 152-270 — 91 70-110 С. sibirica (1)
6 189 160-215 4 86,5 65-112 С. falconeri (2)
— 165-171 146-205 — 81-84 67-113 С. aegagrus (1)
23 249 184-310 8 94 85-110 С. cylindricornis (2)
32 220 185-265 12 98 60-110 С. caucasica (2)
6 183 135-205 4 95 85-108 С. ibex (2)
- 145 - 75 - С. pyrenaica (3)
* 1 - обобщенные табличные материалы (см. выше); 2 - Казанская, 2004; 3 - Fandos, Vigal, 1993.
Изменчивость рогов. Рога у сибирского, альпийского и нубийского козлов почти одинаковые: саблевидные, овальные в сечении и с бугорчатой передней гранью. У молодых особей их форма близка к таковым у безоарового козла [Вейнберг, 1986а].
Форма рогов сибирского козла, судя по коллекционным материалам, сильно варьирует. Есть рога слабо изогнутые и почти параллельные, есть сильно изогнутые и с широко разведенными концами. Каких-либо географических различий в форме рогов не усматривается [Цалкин, 19506]. У старых самцов рога потрескавшиеся, местами раскрошенные или обломанные в скалах или в турнирных боях. В Саянах дефектные роговые чехлы имеют 7,5% взрослых особей [Федосенко и др., 1992].
Темпы роста рогов и их размеры зависят от пола и возраста животных и, во многом, определяются условиями питания. У самцов максимальный прирост сегментов бывает в первые 5 лет жизни - до 11-1 / см (в дальнейшем до старости их ежегодный прирост не превышает 4-9 см) и в годы с ранней весной и теплой продолжительной осенью при хорошей кормовой базе. В этом случае на передней стенке рога вырастают три поперечных бугра вместо обычных двух или даже одного при поздней весне
Таблица 64. Размеры рогов (см) у взрослых особей сибирского козла
п Длина Размах** Обхват у основания Район (источник информации)*
М I lim М I lim м I lim
Самцы
11 87,3 54-110 50 28-80 23,0 20-28 Алтай (1)
23 105 74-126 61 30-103 25,0 22-29,5 (4)****
1 — -108 - - - 27 Западный Саян (2)
11 92,0 55-117 — - - - -“-(Н)
11 82,0 58-103 42 29-63 21,6 18,5-26 Алтай, Саяны, Монголия (5)
72,3 - - - - Предбайкалье (13)
2 80,0 76-84 48,5 47-50 20,5 20-21,5 Гобийский Алтай, Гоби (2)
14 92,4 56-116 - - - - Джунгарский Алатау (7)
30, 12 101,1 56-134 — — 22,9 20-25 -“-(8)
— - 87-140 - — — — Заилийский Алатау (10)
>53 — 100-147 — — — — Национальный парк “Алтын Эмель”, Казахстан (12)****
23 131 114-148 95 45-148 26,0 20-29 Средняя Азия (4)****
9-6 100,2 80-136 47,2 30-68 25,7 23-29 Киргизия (10)
43-12 101,4 74-132 47,8 22-68 23,0 19-29 Тянь-Шань, Памир (5)
7 101,2 78-124 62,9 36-89 22,9 21,5-25 Западный Тянь-Шань (5)
31, 19 109,4 74-137 - - 22,4 21-28 Северный Тянь-Шань (8)
25 82,0 45-116 — - - - Внутренний Тянь-Шань (11)
19 -132 22-25 - Памир (6)
20 88,1 61-119 — — — — Восточный Памир (11)
1 — 76 — 38 — 23 Бадхыз (3)
— — — — — 25,2 21-31 Индия, Гималаи (5)
3, 1 122,8 102-140 - 64 - -26,5 Гималаи (11, по данным Roberts, 1997)
Самки
5 37,0 30-41 20 13-32 13,5 12-15 Алтай (1)
3 30,3 28-34 21,2 19-24 11,2 10-13 Алтай, Монголия (5)
- 32,6 - - - — — Предбайкалье (13)
2 27,8 27,5-28 — 19 10,5 10-11 Гобийский Алтай, Гоби (5)
30 21,3 19-29 — - — - Джунгарский Алатау (7)
55,45 23,0 13-39 — — 10,0 8,5-12 -“-(8)
— - -45,5 - — — Заилийский Алатау (10)
6-5 25,0 24-26 10,5 8-13 10,8 9,5-12 Тянь-Шань, Памир (5)
4,3 23,2 16-36 8,2 6-11 10,5 9-13 Киргизия (9)
3,2 23,7 23-25 9,0 6-12 10,2 9,5-10,5 Западный Тянь-Шань (5)
5,4 28,2 24-37 — — 11,7 11-12,5 Северный Тянь-Шань (8)
— — -28 — — — — Гималаи (11, по данным Roberts, 1997)
* 1 - Собанский, 1992; 2- Размахнин, 1977; 3 - Горелов, 1961; 4 - В.А. Часовских (личное сообщ.); 5 -Цалкин, 1949, 19506***; 6 - Егоров, 1955; 7 - Савинов, 1962***; 8 - Слуцскнй и др., 1983***; 9 - Денисов, 1935; 10 - Шнаревнч, 1948; 11 - Федосенко, 2003***; 12 - Мельников, 2003; 13 - Недзельскнй, Недзельс-кий, 2003.
** Измерения по разным методикам (расстояние между концами или максимальное расстояние между рогами).
*** Расчет по материалам.
**** Трофейная избирательная добыча.
и ранней зиме. У зараженных паразитами и травмированных зверей рост сегментов и формирование валиков задерживается. В условиях зоопарков при хорошем кормлении рост сегментов рогов не прекращается и зимой, в результате чего они могут отрастать в первый год до 25-30 см, почти не имеют годовых перемычек, и в течение жизни рога у самцов вырастают до 2 м. Их среднегодовой прирост достигает 10-12 см, у самок 2-4 см. У яловых самок рога растут более быстро, чем у лактирующих [Шнаревич, 1948; Цалкин, 19506; Егоров, 1955].
Длина рогов у самцов сибирского козла колеблется в пределах 45-148 см, размах - 22-148, обхват у основания - 18-31 см. У самок рога гораздо короче и с меньшим расстоянием между концами (табл. 64).
В.И. Цалкин [1949, 19506] считал, что рога сибирского козла в северной части ареала (Алтай, Саяны, Северная Монголия) отличаются небольшими размерами и не достигают 100 см даже у очень крупных старых животных при обхвате оснований, не превышающем 24 см. На юге ареала (Тянь-Шань, Памир) они обычно длиннее 100 см, у старых особей отрастают до 135-140 см и до 29 см в основании, а в Индии и Гималаях у отдельных особей - до 31 см. Последующие исследования опровергли эту точку зрения. Г.Г. Собанский [1992] определил максимальную длину рогов взрослых алтайских козлов в ПО см, полагая, что у старых особей она может доходить до 125-130 см. В.А. Часовских (личное сообщение) на трофейном материале из этого же региона установила, что даже средняя длина рогов составляет 105 см, а максимальная достигает 126 см при обхвате основания в 29,5 см. Эти показатели все же чуть меньше трофейных характеристик среднеазиатских козлов, длина рогов которых колеблется в пределах 114-148 см.
Параметры рогов козлов из разных географических районах близки (табл. 64), несмотря на существенное варьирование средних значений, которые, как показывает анализ, в большей мере определяются не генотипом животных, а величиной разновозрастных выборок - чем больше в них доля старых самцов, тем выше средние значения. Размеры рогов зверей в значительной мере зависят и от кормовой базы, тесно связанной с количеством осадков. О.В. Егоров [1955] нашел, что на Восточном Памире в группировках, обитающих в хороших условиях, рога самцов обычно бывают более 100 см, в плохих - менее 100 см.
Окраска и линька. Окраска сибирского козла подвержена существенной половой, возрастной, сезонной, индивидуальной и географической изменчивости (рис. 58), пожалуй, большей, чем у других представителей рода. Зимняя окраска самцов на Алтае сероватая или бледно-охристая с темно-бурыми участками на нижней стороне шеи и груди у старых особей и относительно однотонная у молодых. По средней линии спины до конца хвоста тянется бурая или темная полоса, такого же цвета и борода. Живот и паха бледно-серые или желтоватые. Летний мех с преобладанием желтоватых тонов. Самки, в отличие от самцов, не имеют потемнений на плечах, груди и бедрах, и темных полос на боках и по средней линии спины, характерных для зимнего наряда самцов. Окраска молодых сходна с таковой у самок. Изредка встречаются альбиносы [Собанский, 1992]. Обший тон окраски самцов в Саянах варьирует от коричневато- и темно-бурого до пегого, светлого и песчано-желтого. В Гобийском Алтае и Заалтайской Гоби темные полосы у зверей выражены слабо, и в зимнем наряде они имеют однотонную серовато-коричневую окраску. На Восточном Тянь-Шане и Памире спина и бока взрослых самцов в зимнем наряде коричне-
Рис. 58. Типичная зимняя окраска (схематично) у самцов выделяемых форм сибирского козла (по [Цалкин, 19506])
Слева направо: Capra sibirica sibirica, С. s. alaiana, C. s. sakeen
во-бурые, на верху спины и шеи расположено светлое пятно разного размера и тональности, темные полосы на спине и на нижней части боков хорошо развиты и имеются потемнения на передних и задних конечностях. В Джунгарском Алатау и на Тарбагатае светлое “седло” у животных сливается с шейным пятном и занимает верхнюю часть тела. На Западном Тянь-Шане оно у самцов отсутствует. В Северной и Южной Киргизии крайние цветовые формы, в Центральной - промежуточная. В целом на севере ареала и в Гималаях животные более светлые и с более контрастным узором [Денисов, 1935; Цалкин, 1949, 19506; Филь, 1964; Вейнберг, 1993; Федосенко, 2003].
Линька очень продолжительная: начинается в конце марта - мае и длится до конца июня - середины июля. Лактирующие самки, больные, зараженные паразитами и травмированные особи сохраняют остатки старого волоса на хребте и загривке иногда до конца августа. Зимний мех полностью отрастает в ноябре [Шнаре-вич, 1948; Егоров, 1955; Савинов, 19756; Слудский и др., 1983; Федосенко, 2003].
Генетическая изменчивость. Кариотип, как и у других видов козлов, содержит 60 хромосом, NF = 60. Все аутосомы и половые хромосомы акроцентрические [Wurster, Benirschke, 1968; Орлов и др., 1978]. Генетическая изменчивость у сибирского козла не исследована, но теоретически все показатели у относительно многочисленного вида должны быть существенно выше, чем у ибекса. В альпийских популяциях Capra ibex, неоднократно проходивших через стадии низкой численности и восстановленных от нескольких особей-основателей, генетическая изменчивость крайне мала: доля полиморфных локусов колеблется от 5,3 до 15,1%, среднее число аллелей на локус - 1,18, средняя гетерозиготность - от 0,007 до 0,061 [Stuwe, Scribner, 1989; Tosi et al., 1991]. Генофонд Caprapyrenaica значительно разнообразнее [Marquez et al., 1998]. По последовательностям митохондриального гена цитохрома b сибирский козел чето обособлен от других видов (рис. 43).
Гибридизация. Сибирский козел легко скрещивается с домашними козами. Гибриды плодовиты и по мужской, и по женской линиям. По фенотипу они более сходны с диким предком. Гибридизация сибирского козла возможна и с кавказскими, и с винторогим козлами, но в последнем сочетании иногда происходят выкидыши и ма
церация второго гибридного плода [Бутарин, 1947; Соколов, 1959; Салганский и др., 1963; Треус, 1968; Стекленев, 1972, 1978а; Одинашоев, 1986; Собанский, 1992].
Таксономия. Видовая самостоятельность сибирского козла подвергается сомнению со времен П.С. Палласа, описавшего его в Spicilegia в 1766 г. по экземплярам, добытым на Восточном Саяне в междуречье Уды и Бирюсы, как форму европейского горного козла {Capra alpium sibiricorum), а в 1776 г. как Ibex sibiricus. К альпийскому козлу его относили и некоторые известные систематики [Schwarz, 1935; Ellerman, Morrison-Scott, 1951; Haltenorth, 1963]. Большинство ученых считали сибирского козла все же отдельным видом, выделяя при этом длинный ряд подвидов (см. синонимику) большей частью по географическому принципу - на каждом хребте “своя” форма. Многие расы (или виды) были описаны по единичным экземплярам шкур, рогов или черепов, порой, неизвестного происхождения. Как справедливо заметил Б.А. Кузнецов [1948а], диагнозы большинства подвидов составлены небрежно, не обоснованы научно, материалы скудны и не этикетированы точно, и все эти описания вряд ли имеют большую научную ценность. Именно по этой причине я не привожу в синонимике и на рассматриваю в этом и других разделах совершенно непригодные с точки зрения таксономии формы.
К.К. Флеров [19356], одним из первых усомнившийся в реальности многообразия форм сибирского козла, 11 из 13 известных ему рас свел в синонимику, оставив лишь С. sibirica sibirica (Южная Сибирь и Монголия) и С. s. sakeen (остальная часть ареала). Особи типичной сибирской формы, по его мнению, имеют крупный рост, массивный череп и рога, короткие носовые кости (около 90 мм) и контрастную окраску. Особи другой формы отличаются меньшими размерами тела, удлиненными (около 110 мм) носовыми костями, однотонной окраской со светлым пятном на спине у многих особей. Этот диагноз, однако, явно не точен, особенно в отношении размеров черепа и рогов (табл. 62 и 64).
В.И. Цалкин [1949, 19506], проведя ревизию вида, определил иной подвидовой состав С. sibirica, который был поддержан И.И. Соколовым [1959]: sibirica (Русский и Монгольский Алтай, Саяны, Северная Монголия), hagenbeckii (Гобийский Алтай и Заалтайская Гоби), alaiana (Восточный и Центральный Тянь-Шань, Памир, Алайский и Заалайский хребты), formosovi (Киргизский хребет, Таласский Алатау и другие хребты Западного Тянь-Шаня, исключая Каратау), dementjevi (Западный Куэн-лунь), а также, видимо, sakeen (Кашмир и Гималаи). Замечу, что, по мнению Цалкинa, formosovi отличается от остальных рас по отсутствию у самцов зимой светлого седловидного пятна (такое пятно, хотя и слабое, у части особей имеется [Гептнер и др., 1961] и отсутствует у некоторых типичных alaiana [Федосенко, 2003]), а для dementjevi характерны мелкие размерами черепа взрослых самок, что определено по измерению всего лишь двух экземпляров. Форма hagenbeckii выделена Ноаком на основании однородной серовато-коричневой окраски взрослых самцов в зимнем наряде.
И.М. Громов с соавторами [ 1963] в пределах СССР различали sibirica, alaiana и formosovi. В.Г. Гептнер [1961] принял, с некоторыми оговорками, точку зрения К.К. Флерова, выделив лишь алтайского (С. г. sibirica) и среднеазиатского (С. 5. alaiana) козлов. Последнюю форму он отделял от С. s. sakeen чисто условно, подчеркивая недостаточный уровень знаний.
Судя по приведенным выше сведениям, особи сибирской формы вида (Южная Сибирь, Северная Монголия) по размерным показателям не отличаются достоверно от среднеазиатских и близки к ним генетически (рис. 43).
Распространение
Восстановленный ареал. Область распространения сибирского козла, скорее всего, - лишь небольшая часть некогда обширнейшего ареала группы Ibex, простиравшегося в плейстоцене непрерывно от горных районов запада Европы до гор Средней и Центральной Азии и Африки, включая Карпаты, Крым, Кавказ, Аравию и Южный Урал [Громова, 19356, 19486; Громов, 1948; Цалкин, 1970; Барышников, 1978; Верещагин, Барышников, 1980а,б].
Костные остатки сибирского козла обнаружены на многих палеолитических стоянках человека в горах Южной Сибири, где они, нередко, доминируют среди остатков других диких копытных: бизона, архара, благородного и северного оленей, лося, сибирской косули, лошади, кулана, сайги, шерстистого носорога и мамонта. Наиболее древним из них в восточных отрогах Кузнецкого Алатау более 30 тыс. лет [Ларичев, 1976; Оводов, Орлова, 1977]. Этот козел в прошлом плотно заселял Алтай, включая среднегорье и северные и северо-западные предгорья [Оводов, Ивлева, 1990], горные массивы Саян и их отроги в приенисейских котловинах, окрестности Абакана, Красноярска и Иркутска, берега Бирюсы и, возможно, скалистые откосы Приморского и Байкальского хребтов (рис. 59). В Забайкалье его остатки найдены в устье Селенги, на правобережье Уды, в поселении “Студеное” на р. Чикой далеко от границ современного ареала [Громов, 1948; Цалкин, 1950; Ермолова, 1978; Васильев, 1986; Абрамова и др., 1991; Смирнов, Ткаченко, 1992].
Наскальные изображения сибирского козла, датируемые главным образом серединой I тысячелетия до н. э. - серединой I тысячелетия н. э., встречаются по всему югу Сибири, включая Западный Саян и Забайкалье (в бассейнах Селенги и Онона, в устье Джиды), но особенно многочисленны они на Алтае, в Туве и Хакасии [Окладников, Запорожская, 1970; Шер, 1980; Гришин, 1981; Хамзина, 1982;
Рис. 59. Распространение сибирского козла в Южной Сибири (по [Смирнов, Ткаченко, 1992])
1 - предполагаемая граница распространения вида в верхнем палеолите, 2 - местонахождения петроглифов н ископаемых остатков, 3 - районы современного обитания
нию, две тысячи лет назад козел заселял всю южную часть региона, а северная граница его ареала проходила южнее Горно-Алтайска [Ешелкин, 1972; Собанский, Ешелкин, 1975; Собанский, 1992].
В горах Средней Азии в мустьерскую эпоху и позднее он тоже был одним из основных охотничьих видов, наряду с диким бараном [Верещагин, Батыров, 1967; Батиров, 1986; Батыров, 1986; Коробков, 1989; Худайбердиев, 1990]. В палеолите, раннем и среднем голоцене сибирский козел населял почти все горные системы
Рис. 60. Распространение сибирского козла в Казахстане в голоцене (по [Соколов, 1964])
Точки с номерами - пункты находок наскальных изображений (см. в оригинале); предполагаемая область обитания заштрихована
Казахстана (рис. 60), включая Каратау, Гиссарский хребет и Чу-Илийские горы, что подтверждается не только наличием костных остатков, но и многочисленными наскальными изображениями зверей, относимыми к I тысячелетию до н. э. [Громов, 1948; Соколов, 1964; Кожамкулова, 1969, 1976, 1981; Макарова, 1976; Тлеу-бердинаидр., 1989].
Эти животные, судя по костным остаткам, найденным в пещерах по берегам р. Сим в Челябинской области, с позднего плейстоцена и до конца голоцена жили и на Южном Урале [Громов, 1948; Смирнов и др., 1990].
Фактически сибирский козел обитает непрерывно с плейстоцена до наших дней в обширном ареале, простирающемся от гор Южной Сибири на севере до Гималаев на юге, включая горные системы Алтая, Саян, Хангая, Гоби, Тянь-Шаня, Памира, Гиндикуша, Парапомиза и Куэн-луня [Цалкин, 19506; Егоров, 1955; Размахнин, 1977]. ,
Ареал в XX в. Детальное описание распространения вида до середины XX в. содержится в замечательных монографиях В.И. Цалкина [19506], И.И. Соколова [1959] и В.Г. Гептнера с соавторами [1961]. Эти книги изданы массовыми тиражами, и нет смысла еще раз перечислять все многочисленные пункты обитания зверей. 296
Рис. 61. Ареал сибирского козла во второй половине XX в. (по [Федосенко, 2003])
[1959] и В.Г. Гептнера с соавторами [1961]. Эти книги изданы массовыми тиражами, и нет смысла еще раз перечислять все многочисленные пункты обитания зверей.
Во второй половине XX в. ареал сибирского козла несколько сжался, во многих районах стал мелкоочаговым, но все же остался достаточно обширным (рис. 61).
На Алтае в XX в. его площадь уменьшилась вдвое, а численность животных - в десятки раз. Этот вид полностью исчез из северных и юго-восточных районов, и к концу столетия сохранился лишь в разрозненных очагах на Северо- и Южно-Чуйском хребтах, Сайлюгеме, хребтах Чихачева, Шапшальском, Катунском и Чулыш-манском нагорье (рис. 62). Основные причины сокращения ареала - развитие скотоводства, в результате чего значительно уменьшилась площадь пастбищ и возникли эпизоотии, изменение климата, сопровождающееся увеличением снежности и оттепелей среди зимы, хищничество, но, главное, - неумеренная охота [Дмитриев, 1938; Колосов, 1938, 1939; Малков, Малков, 1972; Собанский, Ешелкин, 1975; Собанский, 1988, 1992].
До 70-х годов сибирский козел оставался фоновым видом в приенисейской части Западного Саяна, хотя и здесь область его обитания сократилась значительно из-за антропогенного воздействия. В конце столетия звери населяли хребет Сайлыг-хем-Тайга, верховья рек М. и Б. Абакан, Она, бассейны Джебеш, Кантегир, Ус, Б. и М. Уры, Казыр-Суг, Хемчикский и западную часть Куртушибинского хребта, берега Енисея в районе скальных выходов саянских хребтов (рис. 63). В Хакасии основной очаг обитания находится в горной степи в бассейне р. Каратош. Северная граница распространения проходит по р. Тепсель (приток Енисея в 40 км ниже устья р. Ус) [Соколов, Балагура, 1973; Соколов, 1979, 1988; Гущин, Завацкий, 1980; Сыроечковский, Рогачева, 1980; Завацкий, 1989,1990; Смирнов, Бриллиантов, 1990; Прокофьев, 1992; Смирнов, Ткаченко, 1992; Федосенко и др., 1992; Сыроечковский, 1995].
Рис. 62. Распространение сибирского козла на Алтае (по [Собанский, 1992])
1 - восстановленный ареал по наскальным изображениям, 2 - современное распространение (номерами обозначены локальные районы обитания), 3 -основные места скопления наскальных изображений
Рис. 63. Распространение сибирского козла в приенисейской части Западного Саяна (по [Соколов, 1979]) 1 - во второй половине XX в., 2 - в прошлом
В Туве этот вид до 70-х годов населял все крупные хребты: Западный и Восточный Танну-Ола, Сангилен, Цаган-Шибэту, Шапшальского, Кантегирский, Хемчик-ский, Уюкский, Куртушибинский, Эргак-Таргак-Тайга, Академика Обручева, южные отроги Западного Саяна [Янушевич, 1952; Никифоров, Шурыгын, 1973; Никифоров и др., 1977]. К концу века мелкие очаги отмечены в Монгун-Тайгинском районе, в окрестностях озер Шара-Нур, Дус-холь, Тере-холь и в юго-восточной части хребта Сангилен. На юго-западе региона, скорее всего, существуют три обособленные группировки: на хр. Чихачева, Монгун-Тайгинская и занимающая хребты Цаган-Шибэту, Шапшальского и Чингекатское плато [Смирнов, Ткаченко, 1992; Ткаченко, 1992; Смирнов, 1994]. На севере мелкие группировки сохранились на Уюкском и Хемчикском хребтах и Алашском плато. Они, возможно, связаны с популяциями хребтов Сайлыг-хем-Тайга и Главного Саянского [Соколов, 1988].
Севернее, на Восточном Саяне козел населяет его центральные части: Удинский хребет и Ергак-Таргак-Тайга, где встречается в верховьях рек Казыр и Уды, по долине которой звери иногда спускаются до Алыгджера, живет в верховьях Оки, Иркута, Китоя, Урика, по Сархою, Хоре, Диби, Тиссе, Сенце, Хойто-Оке, в районе оз. Хара-Нур и Тункинских белках [Филь, 1964, 19776; Лавов, 1966; Опрышко, 1975; Богородский, Литвинов, 1988; Смирнов, 19886; Медведев, Аюпов, 1993; Борисова и др., 2001]. В Байкальском бассейне эти копытные встречается исключительно в приграничных районах [Швецов и др., 1984]. Местообитания в Восточном Саяне, Тункинских Альпах, на хр. Академика Обручева и в смежных районах Монголии составляют, скорее всего, общий очаг [Соколов, 1988], но связь группировок весьма слабая. Ранее В.Б. Подаревский [1936] сообщал, видимо ошибочно, что горный козел известен для Бодайбинского района, где живет возле оз. Орон.
В Монголии козел занимает горные системы Прихубсугулья, Хангая, Монгольского и Гобийского Алтая и возвышенностей Заалтайской и Западной Гоби [Колосов, 1939; Цалкин, 19506; Банников, 1954; Соколов, Орлов, 1980]. Здесь его ареал, как, впрочем, и в других регионах, во многом совпадает с областью распространения архара.
На территории Казахстана козел живет на Курчумском и Нарымском хребтах, Сарымсакты и Южном Алтае, обычен в Джунгарском Алатау и на большинстве хребтов Тянь-Шаня. На северо-востоке республики в Дельбегетейских горах и к юго-западу от Семипалатинска (в горах Коконь и, вероятно, Ортау) этот вид уничтожен, скорее всего, после 1925 г. В горах Саура, Каратау и на Тарбагатае он крайне редок, а в Чу-Илийских горах истреблен [Антипин, 1941; Цалкин, 19506; Савинов, 1964, 1975а,6; Слудский и др., 1983]. В Киргизии козел занимает все осевые горные хребты Тянь-Шаня и Памир-Гиссара: Терскей, Кунгей, Киргизский, Таласский, Чаткальский, Нарынский, Актабашинский, Кок-Шаалу, Ферганский, Алайский и Заалайский, а также горы близ озер Сары-Чилек, Сон-Куль, верховья Сары-Джаза, хребты Барколдой, Ак-Шийрак, Куйлютоо и др. [Кузнецов, 1948а; Айзин, 1969,1975]. В горах Таджикистана он распространен очагами практически по всем крупным хребтам Памира, но более многочислен на западе [Мекленбурцев, 1949; Сапожников, 1976а]. В Узбекистане козел населяет Угамский, Пскемский, Чаткальский, Туркменский и Гиссарский хребты [Ишунин, 1980]. Из Узбекистана этот вид, видимо, проникает на хребет Кугитанг на территорию Туркменистана, где встречается также на юго-востоке республики в горах междуречий Кушки и Кашана.
Ранее он входил в состав фауны Бадхыза. В южные районы звери заходят, скорее всего, с севера Афганистана [Горелов, 1961,1970; Ишадов, 1977; Коршунов, 19956]. В Афганистане сибирский козел занимает северную и центральные части, в Пакистане и Индии водится только на севере, в Китае - в районах, примыкающих к Памиру, Тянь-Шаню, Монгольскому Алтаю и Тибету (рис. 61).
Целенаправленной работы по восстановлению ареала рассматриваемого вида в XX в. не было. Немногочисленные выпуски были случайными. В 1947 г. 7 особей, оставшихся после съемок кинофильма, выпустили в Крыму. Часть этих животных вскоре погибла от браконьеров. Оставшиеся размножались, но, видимо, не выжили в многоснежную зиму 1954/55 гг. Не увенчался успехом и завоз 27 голов из Киргизии в Щучинский район Кокчетавской области в 1963-1965 гг. Через 10 лет здесь сохранились лишь 2 особи [Ткаченко, 1963; Слудский, Афанасьев, 1964; Павлов, 1999].
Среда обитания
Сибирский козел - преимущественно высокогорный житель, населяющий труднодоступные верхние части лесной и кустарниковых зон и горные тундры вплоть до снежников [Шнитников, 1936; Вырыпаев, 1973; Соколов, Балагура, 1973; Жиряков, 1976; Федосенко, 2003]. Однако его потенциальная экологическая ниша гораздо обширнее. Столетия назад он населял среднегорья и даже низкогорья, о чем свидетельствуют многочисленные костные остатки и наскальные рисунки. В Заалтайской Гоби этот вид встречался в мелкосо-почнике и даже в котловинах, если поблизости находились спасительные скалы [Банников, 1954], в Казахстане занимал невысокие хребты Б. Богуты, Ту-райгыр, Матай, Коконь, Дельбегетей, Джамбыл, Чулак и др. [Антипин, 1941, 1946]. Обитание в высокогорье, следовательно, - явление во многом вторичное, связанное с истреблением вида человеком и вытеснением домашним скотом [Егоров, 1955]. Скальные массивы в связи этим были и остаются жизненно важными стациями.
На Алтае исследователи выделяют два типа биотопов сибирского козла: высокогорные, где обитает 3/4 поголовья, и среднегорные. На северо-востоке и юге региона звери живут преимущественно в гольцовой зоне, глубоко расчлененной, с цирками, каньонами, ущельями, обрывами и россыпями на высотах от 1700 до 3200 м. Здесь они предпочитают горные тундры: кустарниковые, лишайниковые и каменистые, где находят пищу на альпийских лужайках и полянах среди кустарника, скал и россыпей. Зимой животные держатся на лишенных снега или малоснежных остепненных склонах и в верхнем поясе леса, где снег рыхлый и его легче копытить. Летом они не заходят ниже верхней границы леса, а участки тайги пересекают лишь во время переходов. В бассейнах рек Башкаус, Аргут и других местах козлы живут на труднодоступных для человека и хищников склонах каньонообразных долин, но нередко спускаются в долины до 500-700 м. В ущельях кое-где есть отлогие участки с хорошим травостоем, зарослями кустарников (спиреи, крыжовника, караганы, ив, берез) и куртинами леса, где звери укрываются в непогоду. Нередко они выходят кормиться к подножиям гор: на равнину и в широкие долины верхних течений рек
[Дмитриев, 1938; Насимович, 1955; Малков, Малков, 1972; Собанский, 1988, 1992].
В Саянах у них более обширный спектр стаций - от сухостепных и горностепных растительных ассоциаций до участков лиственничников, альпийских лугов и горных тундр с наличием скальных выходов. Летом животные преимущественно обитают в верхней части долин рек и ключей и на субальпийских и альпийских разнотравных лугах. Зимой они откочевывают ниже, в пояса сухих и горных полынно-злаковых и злаково-разнотравных степей, занимают островные лиственничники и полоски кустарников по склонам гор, живут в верхней и средней части лесного пояса и даже в разреженной кедрово-лиственничной тайге [Насимович, 1955; Соколов, 1979; Ткаченко, 1992]. В Саяно-Шушенском заповеднике сибирский козел населяет в основном пояс горной степи, где держится ниже типичных мест обитания лося и северного оленя, занимающих подгольцовый и гольцовый пояса гребневой части осевого хребта. Зимует он вместе с маралом на малоснежных открытых солнцепечных склонах, заросших травами и кустарником, перемежающихся с лиственничным редколесьем и скалами [Гущин, Завацкий, 1980; Завацкий, Гущин, 1980; Федосенко и др., 1992; Федосенко, 2003].
В Джунгарском Алатау и на хребтах и отрогах Тянь-Шаня высотная зона обитания вида простирается от 400-500 до 3500-4000 м. Летом звери живут в альпийском поясе, после первых снегопадов спускаются в лесную зону, где концентрируются на остепненных склонах южной экспозиции. К наиболее характерным биотопам относят альпийские и субальпийские луга, типчаково-разнотравные и высокогорно-типчаковые степи и крутые склоны с обязательным наличием скал, на которых нередко растет арча. В скальниках с редкой растительностью и возле ледников и снежников чаще держатся старые самцы, скалы с альпийскими лужайками предпочитают молодые самцы и часть холостых самок и самок с козлятами, субальпийские луга с обильными скальными выходами занимают самки с приплодом. От скальных массивов звери не уходят дальше 1-2 км [Кузнецов, 1948а; Шульпин, 1948; Насимович, 1955; Савинов, 1961,1964,1975а,б; Филь, 1970; Плешак, 1973; Грачев, Савинов, 1975; Жиряков, 1976; Бургело, 1980; Слудский и др., 1983].
На Памире эти копытные встречаются в диапазоне от 500 до 5500 м в скальных массивах лиственных горных лесов, арчевников и субальпийской и альпийской зон. На востоке региона звери многочисленны на сглаженных хребтах с небольшими участками легко доступных скал. На юго-западе Таджикистана они обитают и в низкогорье в поясе фисташкового редколесья [Мекленбурцев, 1949; Егоров, 1955; Сапожников, 1976].
Очевидно, что при весьма высокой экологической пластичности сибирский козел - узкоспециализированный вид. Его биотопическое размещение определяется, главным образом, наличием убежищ в виде скальных массивов и бесснежных или малоснежных (не более 30 см) пастбищ вокруг них. В многоснежных районах, в отсутствие скал и в скалах, лишенных растительности, он практически не живет. Распределение животных по территории в немалой степени зависит от влияния хищников, кровососущих насекомых и фактора беспокойства, вызванного выпасом домашнего скота, туризмом и охотой.
Питание
Основная пища сибирского козла в горах Южной Сибири - злаки, осоки, луки, полыни, гречишник, анемона, осот разнолистный, горькуша, левзея и другие травы на альпийских и субальпийских лугах и наземные лишайники в горных тундрах. В кустарниковом поясе звери поедают молодые побеги ив, берез, акаций, спиреи, караганы, таволги, жимолости, кизильника, рододендрона, красной и черной смородины, рябины и черемухи. Летом в рационе преобладают травянистые растения, зимой их ветошь и древесно-кустарниковые растения: ива, ольха, осина, береза, можжевельник и кедр, а также лишайники и мхи. Весной животные предпочитают злаки и осоки, осенью - бобовые [Дмитриев, 1938; Филь, 1964; Гущин, Завацкий, 1980; Собанский, 1992; Недзельский, Недзельский, 2003; Федосенко, 2003]. На севере Монголии набор их кормовых растений схож, но на юге в рационе доминируют луки, ковылек, ковыль, баглур, миндаль, житняк, остролодочники, вострец и осоки [Банников, 1954].
В Джунгарском Алатау и на Северном Тянь-Шане список кормовых растений козла наиболее обширен - около 140 видов растений, в том числе 26 древеснокустарниковых. Летом и осенью в рационе преобладают злаки и разнотравье; зимой к травяной ветоши добавляется полынь, морозоустойчивые и вегетирующие зимой очиток гибридный, молочай алатавский и хвойник хвощевой, а также веточный корм - молодые побеги спиреи и листья эфедры. При изменении условий обитания звери легко переключаются с одной группы растений на другие, и даже в самые суровые зимы не страдают от бескормицы, чему в значительной мере способствует тебеневка [Савинов, 1964,19756; Слудский и др., 1983; Федосенко, 2003]. В Терскей Алатау эти копытные живут в основном в альпийском поясе и предпочитают кормиться на лугах двух растительных формаций: манжетковой и кобрези-евой, где выбирают мятлики, манжетку, чемыр, гречиху, кобрезию, осоки, типец, астрагал и луки, а у границ снежников и в истоках ручьев обкусывают соцветия примулы. Осенью они охотно едят ягоды рябины, шиповника, смородины, побеги и листья ивы. На Памире к этим видам растений добавляются лисохвост, вейник, кольподиум, копеечник, бескильницы, пижма, ячмень, полыни, лапчатки, клине-лимус, остролодочник, регнерия, ковыли и терескен. В многоснежные зимы возрастет роль кустарниковой растительности, особенно ив, в малоснежные доминируют осоки и злаки. В неволе звери зимой едят разнотравное сено, яблоки, картофель, морковь, корки дынь и арбуза [Егоров, 1955; Одинашоев, 1986]. В Таласском Алатау в их рационе более 77 видов растений. Ранней весной они поедают хвою можжевельника и веточки арчи, затем побеги тюльпанов, эремусов и молодые древесные листья. Летом основу питания составляют эфемероиды, мезофитное разнотравье и полукустарники, зимой - хвоя, ветки и кора кустарников и ветошь трав [Шульпин, 1948; Ковшарь, Янушко, 1965; Бургело, 1980]. В Заилийском Алатау в содержимом желудков почти круглогодично присутствуют можжевельник, эфедра, карагана, вишня, таволга, шиповник (осенью - плоды), типец, ковыль, дикий овес, волосянец, люцерна, гречиха. В неволе животные выбирают люцерну, клевер, злаки, разнотравье и практически те же виды древесно-кустарниковой растительности, что и в природе [Шнаревич, 1948]. В таджикском заповеднике “Рамит” для зимней подкормки козлов заготавливают сено в копнах [Андреенков, 1977].
При анализе питания сибирского козла складывается впечатление, что в его рационе весьма скудно представлены древесные, и, особенно, хвойные растения, которые доминируют в питании его альпийского и пиренейского собратьев [Martinez et al., 1985; Schroder, Kofler, 1985; Martinez, 1988].
Масса содержимого желудков у самок и молодых животных достигает 3-5 кг, у крупных самцов - 8 кг. Количество потребляемой ими пищи за сутки, следовательно, летом должно быть примерно в 2 раза больше [Егоров, 1955].
В горах козлы, как и другие горные копытные, обычно не испытывают недостатка в воде летом, а зимой едят снег. Солонцы они посещают охотно во многих районах, деля их с маралом, сибирской косулей и горными баранами или используя в высокогорье единолично. Наиболее интенсивно солонцевание с апреля по октябрь с пиком в июне - августе. В Алма-Атинском заповеднике на солонцы приходят в основном самки с молодняком и только в дневное время, затрачивая на минеральное питание около часа (от 5 мин до 3 ч). На Алтае и в Тофаларии звери предпочитают желто-серо-белую глину, выедая в ней траншеи, зимой лижут минеральные выходы в известняках [Дмитриев, 1938; Филь, 1964; Савинов, 19756; Жиряков, 1976, 1989; Слудский и др., 1983; Собанский, 1992; Федосенко, 2003].
Сибирскому козлу присущ полифазный тип активности с более частыми периодами летом, когда звери кормятся с рассвета до 8-9 ч, днем с 11-12 ч до 1-2 ч и с 17-18 ч до темноты, а также ночью. В жару они весь день проводят на лежках, которые устраивают в тени скал, кустов, на ледниках, под обрывами и в пещерах, где спасаются также и от кровососущих насекомых. В пасмурную погоду пищевая активность возрастает, но заметно снижается при обильных осадках и сильном ветре. Осенью и зимой кормежка занимает почти весь световой день с небольшими перерывами на отдых. Тем не менее в любой час можно видеть как кормящихся, так и отдыхающих животных. Весной они тратят на кормежку до 11 ч в день, на отдых - 4 ч, на посещение солонцов - около 1 ч. При охотничьем преследовании, выпасе скота и беспокойстве, причиняемом жалящими насекомыми, время активности смещается на сумеречные и ночные часы [Шнаревич, 1948; Егоров, 1955; Ковшарь, Янушко, 1965; Слудский и др., 1983; Федосенко, 19836, 2003; Грачев, 1988; Собанский, 1992].
Размножение
Половая зрелость и половые циклы. Часть самок сибирского козла (около 25%) впервые приходит в течку в 1,5-летнем возрасте. С 2,5 лет в размножении участвуют практически все особи. Самцы могут преследовать самок уже в годовалом возрасте, но полноценно участвуют в гоне лишь с 5 лет [Шнаревич, 1948; Егоров, 1955; Савинов, 1962, 19756; Слудский и др., 1983; Федосенко и др., 1992]. Сперматогенез во всех стадиях, но низкой интенсивности, впервые проявляется у них с 1,5 лет, а в 2,5 года он такой же, как и у взрослых особей. Отчетливо фиксируются два периода сперматогенной активности: в апреле -мае и в июле - первой половине ноября. Придатки семенников наполнены сперматозоидами с сентября по январь, и не содержат их совсем в марте и июне. Весенний период повышенной половой активности характерен и для самок, но он ановуляторный и, видимо, является затухшим звеном в половой ритмике этого
вида, который в далеком прошлом вполне мог размножаться дважды в году [Цаплюк, Савинов, 1973].
Два сезонных всплеска половой активности характерны и для некоторых других видов козлов, что отличает их от баранов. Домашние козы нередко приносят козлят почти круглогодично, одичавшие - обычно один раз в год, но, в некоторых стадах в Калифорнии гон проходит до 4 и более раз в год с интервалом в 2-4 мес. [Coblentz, 1980].
Гон и половое поведение. Гон у сибирского козла длится с октября до конца января (табл. 65), но сроки его начала и окончания сильно варьируют в разные годы.
К периоду гона группы самцов разбиваются, и звери входят в состав стад самок. В крупных смешанных стадах, насчитывающих до сотни особей, в гон могут находиться до 8-20 взрослых самцов, в группах из 10-20 особей - 1-3, в гареме у единственного самца чаще содержатся 2-3 самки с козлятами и, в исключительных случаях, на две самки приходится 3 козла. В спаривании, как правило, участвуют самцы старше 5-6 лет. Они преследуют молодых козлов, которые вынуждены держаться поблизости или уходить в поисках свободных самок. За самкой в течке могут следовать несколько самцов, но сильнейший не подпускает к ней остальных. Иногда после нескольких часов ухаживаний доминант покидает самку, и она остается с молодыми самцами [Савинов, 1962; Айзин, 1969; Филь, 1970; Слудский и др., 1983].
В гон взрослые самцы мало кормятся, много передвигаются и сильно истощаются, становясь позднее легкой жертвой хищников. Столкновения между козлами в этот период обычны. Демонстрации (параллельная пастьба, показ профиля головы и туловища, уринация на морду) нередко заканчиваются дракой: соперники,
Таблица 65. Сроки гона у сибирского козла
Район,заповедник | Начало | [ Конец | Источник информации
Горный Алтай Вторая половина ноября Декабрь Дмитриев, 1938
25 октября- 15 ноября 20 ноября -10 декабря Собанский, 1992
Западный Саян Середина октября - Федосенко и др., 1992
и Октябрь Ноябрь Завацкий, 1990
Восточный Саян Ноябрь Декабрь Смирнов, 19886
Предбайкалье Конец сентября Начало декабря Недзельский, Недзельский, 2003
Тянь-Шань Середина ноября Декабрь Федосенко, Орлов, 1976
Северный Тянь-Шань Конец октября Конец января Федосенко, Бланк, 1982
Терскей Алатау Вторая половина ноября Конец декабря Айзин, 1969
Заилийский Алатау Ноябрь Декабрь Филь, 1970
Таласский Алатау 25 ноября 10 декабря Шульпин, 1948
З-к “Аксу-Джабаглы” Октябрь Декабрь Ковшарь, Янушко, 1965
Джунгарский Алатау и Ноябрь Савинов, 1962
Заилийский Алатау Начало ноября Начало декабря Шнаревич, 1948
Алма-Атинский з-к Ноябрь ' Декабрь Жиряков, 1976
Памир Декабрь Январь Егоров, 1953, 1955
Гималаи <4 Schaller, 1977; Fox et al., 1992
привстав на задние ноги, сталкиваются рогами, теснят друг друга, наносят удары в бок или другие части тела, что иногда приводит к серьезным травмам и падениям со скал [Шнаревич, 1948; Егоров, 1955; Савинов, 1962; Федосенко и др., 1992].
Половое поведение складывается из следующих элементов (рис. 64). Самец с вертикально поднятым или трясущимся хвостом приближается к самке, затем, горизонтально вытянув шею и голову, замирает в неподвижной позе, нередко с поднятой ногой, иногда встряхивает головой или поворачивает ее, пытаясь коснуться туловища или морды самки, или делает скребущие движения передней ногой. Если самка отходит, он снова сближается с ней и замирает в той же позе, совершая червеобразные движения языком и обнюхивая морду и половые органы. При этом самец нередко приподнимает хвост самки, облизывает ее гениталии и флемует. Такое следование за партнершей длится до 15-20 мин, и, если она в течке, завершается короткой садкой, длящейся около 5-10 с. Чаще же она угрожает ему рогами, убегает и отстаивается на карнизе скалы. При следовании за самкой самцы нередко уринируют и даже эякулируют спонтанно: высовывают пенис, изгибаются и берут его в рот, затем выпрямляются, долго жуют губами и поднимают морду вверх, видимо, флемуя. Молодые самцы, непрерывно следующие за парой, более суетливы, неосторожны и, в отсутствии взрослого самца, пытаются крыть партнершу после кратковременного ухаживания, что обычно заканчивается ее бегством в скальные укрытия. Однако они настойчивы и пытаются делать садки даже на бегу. Брачный репертуар сибирского козла в целом сходен с таковым у других видов козлов, однако у него, возможно, отсутствуют такие элементы как загребание или удар передней ногой [Федосенко, Орлов, 1976; Федосенко, Бланк, 1982; Слудский и др., 1983: Федосенко и др., 1992; Федосенко, 2003].
Рис. 64. Элементы полового поведения сибирского козла (по [Слудский и др., 1983])
а - подход самца к самке, б - урннирование самки и обнюхивание мочи самцом, в - прикосновение к туловищу самки, г - эякулирование
Беременность и развитие эмбрионов. Беременность длится 152-180 дней [Шнаревич, 1948; Недзельский, Недзельский, 2003]. Эмбрионы развиваются сравнительно быстро. Масса плодов в марте колеблется в пределах 1-3,2 кг, длина тела - 38-55 см, высота в холке - 22-39 см, в апреле, соответственно - 1,9-3,6 кг, 46-63 и 27-38 см. Темпы роста плодов и размеры и масса новорожденных в большей мере определяются характером питания самок в последние месяцы беременности, нежели возрастом животных [Савинов, 1962].
Деторождение и выкармливание молодняка. Деторождение у сибирского козла происходит в апреле - июне (табл. 66). На Алтае встречена самка с козленком в конце марта [Собанский, 1992]. На Восточном Памире массовое появление молодняка обычно в середине июня [Егоров, 1955].
Таблица 66. Сроки деторождения у сибирского козла
Район, заповедник 1 Начало 1 Конец 1 Источник информации
Горный Алтай Вторая половина Май Собанский, 1992
апреля
Западный Саян Середина апреля а Завацкий, 1990; Федосенко и др., 1992
Монголия 5 апреля - Enkhbujin, Amgalan, 2002
Тянь-Шань 11 апреля Начало июня Федосенко, Бланк, 1982
Таласский Алатау Конец мая - Шульпин, 1948
З-к “Аксу-Джабаглы” Конец апреля Май Ковшарь, Янушко, 1965
Алма-Атинский з-к Май Июнь Жиряков, 1976
Джунгарский Алатау Апрель Май Савинов, 1962; Анненков, Литун,
1989; Слудский и др., 1983
Заилийский Алатау 30 апреля Начало июня Шнаревич, 1948
Памир 13 июня 25 июня Егоров, 1953, 1955
«< Июнь Июнь Мекленбурцев, 1949
Гималаи Середина июня — Fox et al., 1992
Незадолго до родов самка отделяется от группы и уединяется в труднодоступном месте в скалах, где приносит потомство. В первые дни козлята подолгу лежат в отсутствие пасущихся матерей, которые кормят их молоком через 35-50 мин, а с августа - через 1,5-2 ч. Продолжительность сосания в первые дни составляет в среднем 27-34 с, с августа - в 2-3 раза меньше. Растут детеныши очень быстро, и уже через 5-7 дней стремительно бегают и ловко прыгают. С этого периода лак-тирующие самки могут объединяться в группы, однако у каждой из них сохраняется свое время кормления. Лактация длительная - до октября - конца декабря. Сирот могут выкармливать другие самки, которые водят порой до 3-5 козлят. Траву детеныши начинают употреблять рано - в 8-10-дневном возрасте [Шнаревич, 1948; Егоров, 1953, Савинов, 1962; Федосенко, Бланк, 1982; Слудский и др., 1983; Завацкий, 1990; Собанский, 1992; Федосенко, 2003].
Плодовитость. На Алтае в июле - августе 65-70% лактирующих самок имеют по одному козленку, остальные - по два, и в исключительных случаях - три [Собанский, 1992]. В Западном Саяне двойни регистрируют не ежегодно [Завацкий, 1989; Федосенко и др., 1992]. В Предбайкалье почти половина самок с двумя эмбрионами, а яловых - не более 9% [Недзельский, Недзельский, 2003]. У четырех беременных самок на Тянь-Шане и двух - на Памире О.В. Егоров [1953, 1955]
обнаружил только по одному эмбриону, а по сообщению охотников, заготовлявших зверей на мясо во время войны, из 50 самок с двумя эмбрионами были только две. В Джунгарском Алатау из каждых 100 самок в размножении ежегодно участвуют 72-85, из них 9-25% (старших возрастов) приносят по два детеныша [Савинов, 1962; Слудский и др., 1983; Анненков, Литун, 1989]. Двойни зарегистрированы и в Аскании-Нова, где сибирского козла разводили в неволе [Салганский и др., 1963]. Следовательно, потенциальная плодовитость этого вида - более 1 детеныша на взрослую самку, но их число из-за высокого уровня детской смертности сокращается с каждым месяцем (табл. 67).
Таблица 67. Плодовитость самок сибирского козла
Район, заповедник (источник информации)* Самок, п Самок с детенышами (эмбрионами), % Детенышей (эмбрионов) на самку
с одним | с двумя
Алтай (1) - 65-70 25-30 >1
Западный Саян (4) 384 - - 0,6
-“-(6) - - - 0,8-1,1**
Предбайкалье (10) — 53 (52) 47 (48) —
Алма-Атинский з-к (2) 25 92 8 1,1
-“-(3) - 70 30 >1
З-к “Аксу-Джабаглы” 138 93 7 1,1
Джунгарский Алатау (5) - 91-75 9-25 1-0,5
-“-(7) 70 43 (82) (86) (18) (14) (1,2) (1,4)
Тянь-Шань, В. Памир (8) 6 (ЮО) - (1)
* 1 - Собанский, 1992; 2 - Жнряков, 1976; 3 - Жнряков, 1977; 4 - Федосенко и др., 1992; 5 - Анненков, Литун, 1989; 6 - Завацкий, 1989; 7 - Савинов, 1961, 1962; 8 - Егоров, 1953, 1955; 9 - Ковшарь, Янушко, 1965; 10 - Недзельский, Недзельский, 2003.
** На рожавшую самку.
Популяционная структура
Половой и возрастной состав. Соотношение полов у эмбрионов и новорожденных сибирского козла, как и у других видов копытных, близко 1:1 [Савинов, 1962; Жиряков, 1973; Слудский и др., 1983; Недзельский, Недзельский, 2003]. Примерно таким же оно остается у взрослых особей в заповедниках и в труднодоступных районах, тогда как в охотничьих угодьях резко меняется в пользу самок из-за выборочной элиминации самцов. В большинстве группировок доля сеголеток достигает 20-25%, годовалых особей обычно менее 18, взрослых самок - 33^15, взрослых самцов - от 9 до 42% (табл. 68). В заповедных и слабооп-ромышляемых группировках доля самцов старше 5 лет чаще составляет 10-18%, а среди всех самцов - от 37 до 62% [Слудский и др., 1983]. В местах интенсивной охоты зрелых самцов в популяциях остается не более 2%, и во время гона на одного производителя приходится до 30 самок, тогда как обычно - около 6 [Егоров, 1953, 1955].
Социальная организация и поведение. Типичный образ жизни сибирского козла - стадный, хотя крупные объединения, насчитывающие несколько сотен особей, редки и кратковременны. Осенью, в период гона, животные находятся в смешанных ста-
Таблица 68. Половозрастной состав (%) и соотношение полов в популяциях сибирского козла
Район,заповедник (источник информации) Особей, n Сеголетки Годова-лые Взрослые Самцы : самки
самки самцы
Горный Алтай (3) >2000 19,9 (13-30) 8,6 (5-Ю) 39,5 32,1 1:1,2
Тува (4) 104 9,6 14,4 25,0 51,0 1:0,5
Западный Саян (6) 9549 24,5* (10-42) * 48,1 (29-65) 27,4 (21-37) 1:1,8
Саяно-Шушенский з-к (13) 473 18-19,8 * 40,4* 39,8^41,6 1:1
Джунгарский Алатау (9) 308 21,4 * 55,8* 15,7 (22,8)* 1:2,5
-“-(11) 92 27,1 25,0** 32,7 15,2 1:2,2
Терскей Алатау (8) 460 - * 59,5* 9,3 (40,5)* 1:1,5
Т аласский Алатау (11) 387 16,5 14,9 40,2 28,4 1:1,4
Заилийский Алатау (12) 989 38,5* * 34,3 27,2 1:1,3
-“-(И) 1067 25,6 12,2 40,3 21,9 1:1,8
Чаткальский з-к (1) 1791 18-25 - — 28-46* -
З-к “Аксу-Джабаглы” (2) 1320 23,6 * 48,3* 28,1* 1:1,7
-“-(Ю) 1352 20,1 * 43,3* 36,6* 1:1,2 .
Алма-Атинский з-к (5) Природный парк — 14-20 * 43,8* 42,2* 1:1
“Алтын-Эмель” (7) — — 6,3-17,7 — — 1:2,9-3,6
Восточный Памир (8) (13, по данным 603 - * 57,5* 9,8 (42,5)* 1:1,4
П.И. Вейнберга) Китай (13, по [Schaller 179 24,6 16,2 44,7 14,6 1:3,1
et al., 1987]) Индия (13, по [Fox 201 24,9 * 36,3* 38,8* 1:0,9
et al., 1991]) 302 25,0 10,9 33,1 30,1 1:1,1
1-Вырыпасв, 1973; 2 - Плешак, 1973; 3 - Собанский, 1992; 4-Смирнов, Ткаченко, 1992; 5 - Жиряков, 1976, 1977; 6 - Завацкий, 1990; 7 - Бекенов н др., 1999; 8 - Егоров, 1953, 1955; 9 - Савинов, 1962; 10 -Ковшарь, Янушко, 1965; 11 - Слудский н др., 1983; 12 - Филь, 1970; 13 - Федосенко, 2003.
* Вместе с годовалыми особями.
** Полувзрослые особн.
дах, после чего взрослые самцы отделяются и формируют самцовые группы. Этот процесс растянут до июня, а отдельные их группы входят в самочьи стада уже с конца июня, поэтому смешанные стада встречаются практически круглогодично, особенно при ограниченных кормовых или защитных стациях. В районах интенсивной охоты взрослые козлы весной и летом чаще живут отдельно от самок и присоединяются к их стадам только осенью. С началом ягнения самок молодые особи тоже могут образовывать самостоятельные группы, однако они быстро сливаются со стадами самок с детенышами, среди которых находятся и яловые самки. Некоторые молодые самцы держатся вместе со старыми самцами или с яловыми самками. Одиночные звери - это, как правило, молодые особи или взрослые самцы, переходящие от одной группы к другой [Дмитриев, 1938; Цалкин, 19506; Егоров, 1953, 1955; Савинов, 1962, 19756; Филь, 1970; Слудский и др., 1983; Федосенко, 2003]. Примерно такая же социальная структура группировок и у альпийского ибекса [Toigo et al., 1995].
Средний размер групп сибирского козла в разных районах и в разные месяцы варьирует в пределах 2-24 особей, максимальный - достигает 200-300 голов (табл. 69}.
Таблица 69. Размер групп у сибирского козла
Район, заповедник Число особей в группе Источник информации
среднее, чаще | максимальное
Алтай 8-12 200, 25* Собанский, 1992
и 11 16 Федосенко, 2003
Западный Саян 2-9 140, 19* Соколов, 1979
Саяно-Шушенский заповедник 9-15* 25, 27* Федосенко и др., 1992
7,5 150 Завацкий, Гущин, 1980; Завацкий, 1989
и — 160, 2000** Сашко, Смазнов, 1999
С1 6-12, 6-14* 40, 44* Федосенко, 2003
Восточный Саян 6-15 22 Филь, 19776
Тофалария - 56 (89?) Филь, 1964
Иркутская область — 40 Медведев, Аюпов, 1993
Тува 3-10 50 Никифоров, Шурыгын, 1973
и 4,6 35 Смирнов, Ткаченко, 1992
Монгольский Алтай 3-7 100-150 Банников, 1954
Гобийский Алтай - 20 4 С
Джунгарский Алатау 2-10 150-200 Грачев, Савинов, 1975; Савинов, 1961, 1962, 1975а
и — 50 Махмутов, 1999
4-12 60 Анненков, Литун, 1989
Горы юга Казахстана 4-26, 2-13* 60, 19* Слудский и др., 1983
Тянь-Шань 14-24 - Кошкарев, Нестеров, 1989
<< - 70 Егоров, 1953, 1955
Заилийский Алатау 10-15 200-300 Филь, 1970
Алма-Атинский заповедник 2-10 150 Жиряков, 1973, 1976
Чаткальский заповедник — 50 Вырыпаев, 1973
Заповедник “ Аксу-Джабаглы” 2-10 75 Ковшарь, Янушко, 1965; Плешак, 1973
Природный парк “Алтын-Эмель” 6-10 __ Бекенов и др., 1999
Северная Киргизия - 100 Катаевский, 1987
Памир 5-30, 5-15* 100 Мекленбурцев, 1949
Восточный Памир Заповедник “Рамит” 13-22 88 Федосенко, 2003
(Таджикистан) - 26 Андреенков, 1977
Пакистан 9 26, 33* Schaller, 1977
Синьцзян 18 74 Schaller et al., 1987 (по [Федосенко, 2003])
Гималаи 11 40 Fox et al., 1992
* В группах самцов.
** При миграции.
Размеры стад в значительной мере зависят от плотности населения, сезона года и типа биотопа - в сильно расчлененных горах и при низкой численности стадность обычно меньше.
В Горном Алтае летом взрослые самцы образуют группы от 3 до 25 особей, самки - от 4 до нескольких десятков голов. В состав маточных групп иногда входят молодые самцы и изредка 2-4 взрослых. Молодые самцы (до 2^1 лет) чаще все же держатся отдельно в объединениях, насчитывающих от 3 до 15 голов. Эти группи
ровки наиболее подвижны и нередко уходят из мест рождения. В сентябре, перед гоном, самцы присоединяются к самкам. В зимний период максимальный размер стад обычно не превышает 50-60, и лишь в исключительных случаях достигает 200 особей [Дмитриев, 1938; Собанский, 1992]. В разных районах Тувы чаще встречаются группы из 3-10, и гораздо реже - из 15-50 голов [Никифоров, Шурыгын, 1973].
В Западном Саяне в приенисейской популяции летом типичны группы из 2-12 самок с ягнятами, зимой преобладают смешанные группы до 30 голов. Группы самцов (до 19 особей, чаще 2-3) и одиночные звери встречаются круглогодично. В июле 1972 г. на левобережье Б. Уры зарегистрировано 8 стад общей численностью около 500 особей. В самом большом из них было до 100 голов. В сентябре 1967 г. встречено смешанное стадо из 140-150 особей [Соколов, 1979]. В Саяно-Шушенском заповеднике взрослые самцы (старше 4 лет) держатся чаще в группах по 3-11, но иногда объединяются в стада из 18-27 особей, в которых находятся не более 3 дней. Состав их групп меняется ежедневно, тогда как объединения самок с детенышами и примкнувших к ним 1-3-летних животных обоего пола более постоянны. Изменения в них происходят лишь за счет ухода или присоединения молодых зверей [Завацкий, 1990; Федосенко и др., 1992]. В июле - августе, в жару, в нижней части долин у воды образуются формирования самок с детенышами и молодыми особями, в которых насчитывается до 160 экземпляров. Крупные скопления зверей здесь зарегистрированы и в период миграции - в одном стаде от нескольких сотен до 2 тыс. [Сашко, Смазнов, 1999].
В Алма-Атинском заповеднике чаще встречаются группы из 2-10 особей (около 60-70%), осенью - из 6-10 (40%). Одиночные звери (1,3-4,6% в разные сезоны) -это преимущественно взрослые самцы. Максимальный размер стада - 150 голов [Жиряков, 1973, 1976]. Примерно такой же размер групп и в заповеднике Аксу-Джабаглы. Здесь круглогодично преобладают смешанные стада (более 50%), реже регистрируют группы самцов (18-24%), самок с молодняком и холостых самок (24%) [Плешак, 1973]. В Чаткальском заповеднике максимальное количество особей в объединениях в разные годы колебалось от 20 до 50 [Вырыпаев, 1973].
Отношения животных в стадах основаны на иерархии. Смешанные и самочьи группы возглавляет опытная самка, самцовые - наиболее мощный самец с крупными рогами, за которым следуют другие крупные особи. При кормежке доминанты держатся компактно в центре стада, субдоминанты - на периферии. Самец, занимающий в крупном стаде подчиненное положение, может стать вожаком в небольшой группе. В гон брачную группу возглавляет самец. Вожаки обычно занимают лучшие лежки, а взрослые звери нередко сгоняют с удобных лежек молодых. Иерархия поддерживается демонстрациями угроз, для чего обычно достаточно наклона головы с выставленными вперед рогами. Драки вне периода гона редки [Егоров, 1953, 1955; Schaller, 1977; Слудский и др., 1983; Федосенко и др., 1992; Федосенко, 2003].
У сибирского козла, по сравнению с кавказским, более сложный комплекс импонирования, и он реже других видов козлов использует звуковые сигналы и чаще находится в позе вытянувшись. Соотношение составляющих элементов поведения следующее: голосовые сигналы - 7%, резкое опускание головы - 22, высовывание языка - 18, удар ногой - 18, спурт - 4, поза вытянувшись - 31% [Эрнитс, 1982]. По сравнению с другими видами горных копытных, он игрив (особенно
козлята и молодые особи), любопытен и не очень осторожен. Звери способны обнаружить человека или хищника за 2-3 км, когда их с трудом различаешь в бинокль, но на охраняемых территориях, где их не беспокоят, подпускают человека на 60-80 м [Махмутов, 1999]. Слух у них тоже острый, но обоняние, видимо, слабое. При опасности козлы издают громкий и резкий свист, после которого группа настораживается, пытаясь установить источник тревоги, и затем обращается в бегство, обычно вверх в направлении ближайшего скального отстоя. Животные панически боятся шума вертолета, хотя спокойно реагируют на пролетающий самолет.
Пространственная структура и перемещения. Сибирский козел - сравнительно оседлое животное. Места пастьбы и отдыха у зверей обычно одни и те же или расположены в сотнях метров. Ежедневные “горизонтальные” переходы в несколько километров представляются исключительными, тогда как высотные в несколько сот метров обычны [Цалкин, 1950; Банников, 1954; Гептнер и др., 1961; Филь, 1964,1970; Johnsingh et al., 1999; Федосенко, 2003]. Тем не менее и для этого вида характерны сезонные миграции в ряде районов.
Как упомянуто выше, козлы, живущие в альпийской и гольцовой зонах из-за многоснежья вынуждены ежегодно в сентябре - ноябре спускаться к верхней кромке леса или переходить в лесную зону. Переход в обратном направлении происходит в мае - июне с появлением гнуса или домашних животных. Размах сезонных высотных перемещений порой превышает 2000 м, а протяженность миграций из одного района в другой достигает нескольких десятков километров [Насимович, 1955].
В Горном Алтае в конце ноября при снежном покрове высотой 30-50 см звери группами от 2-4 до 7-8 особей переходят из верховий р. Еринат на правобережье Чульчи, преодолевая расстояние в 30 км и пересекая при этом два водораздела и каньонообразную долину р. Кыги, заросшую лесами таежного типа. Миграции зарегистрированы также на Айгулакском и Курайском хребтах и на Сайлюгеме, где на зиму козлы перемещаются на склоны южной экспозиции. В феврале - марте 1975 г. около 400 особей перешли через хребет Табын-Богдо-Ола из Китая, что, возможно, было связано со взрывными работами на сопредельной территории. Наиболее подвижные в популяциях молодые самцы. Их нередко регистрируют на расстоянии 30-100 км от ближайших типичных мест обитания вида [Собанский, 1992]. На хребте Цаган-Шибэту в Туве после установления снежного покрова часть группировки уходит с северного макросклона на южный, где концентрируется в нижней трети южных склонов или на водораздельных участках [Смирнов, Ткаченко, 1992].
На Западном Саяне самки с молодняком большую часть года занимают скалистые участки в нижней части склонов гор, изредка удаляясь от них на 1-1,5 км. Взрослые самцы выпасаются отдельно от них в верхней части остепненных склонов на относительно пологих участках. Обычно они не уходят дальше 2 км от постоянных убежищ, но осенью перед гоном встречаются на всех высотах и иногда спускаются в пойму. В нижние части гор самцы уходят и после обильных снегопадов, где остаются до тех пор, пока не растает снег на прогреваемых солнцем участках [Федосенко и др., 1992]. Летом на берегах Енисея самки с козлятами и молодые самцы ночью пасутся в нижних частях склонов, а с рассветом поднимаются вверх к каменистым и скальным убежищам. Сезонные миграции не протяженные - до 5-10 км. Начало осенней перекочевки на зимние стойбища вблизи Енисея связано с выпадением снега. Зимой площадь обитания животных сокращается в 2-3 раза [Соколов, 1979]. В отдельные годы
отмечаются массовые перемещения зверей. По сведениям Ю.К. Ошарова, в начале 70-х годов на правобережье Енисея напротив устья Хемчика осенью мигрировали сотни особей. Исключительный переход козлов зарегистрирован 14 декабря 1998 г. у кордона Базага - утром стадо из 1800-2000 голов, растянувшись на 1,5 км, прошло из долины р. Узунсук вниз к долине Енисея [Сашко, Смазнов, 1999].
В Северном Тянь-Шане и Джунгарском Алатау высотные перемещения козла весной и осенью совпадают с началом прихода или откочевки скота. В малоснежные годы звери остаются в верхних поясах гор до декабря - января, после чего уходят к верхней границе леса или еще ниже. В Джунгарском Алатау они спускаются с водоразделов с середины октября. Весенняя откочевка в обратном направлении начинается в середине апреля и завершается через месяц. Часть копытных регулярно сезонно переходит с главного хребта и хребта Тастау в горы Аксай и Теректылы и обратно, покрывая расстояние в 30-100 км. Осенью сюда первыми приходят небольшие группы самцов, затем самки с молодняком и, после снегопада, крупные смешанные стада в 50-100 голов. Весной самки мигрируют позже самцов - в конце мая - начале июля, когда окрепнут козлята. К летним пастбищам козлы идут медленно, преодолевая за 11 дней около 20 км. Около трети самок с молодняком живут в горах Аксай и Теректылы круглый год. С хребта Барлык звери на зимовку уходят, пересекая широкую (10 км) долину, в малоснежные полупустынные горы Кату, возвышающиеся над равниной всего на 200 м. Отсутствие водных источников и раннее выгорание растительности вынуждает их весной возвращаться назад. В Терскей Алатау размах сезонных кочевок достигает 15-50 км. Примерно на такое же расстояние животные уходят при массовых охотах на сыртах [Егоров, 1955; Савинов, 1961, 1962, 1964; Филь, 1970; Федосенко, 1975; Слудский и др., 1983].
В Заилийском Алатау особенно четко выражено высотное распределение козлов по полу и возрасту. На высотах до 2500 м концентрируются самки до 5-летнего возраста и самцы до 3^4 лет. На высотах 2500-3000 м живут средневозрастные животные: самки 6-9 лет и самцы 4-12 лет. Особи старше 10-12 лет, преимущественно старые самцы и яловые самки, держатся выше 3000 м. Дифференцированное размещение зверей в горах во многом связано с уровнем их защиты от хищников - старые менее быстрые особи предпочитают жить в худших кормовых условиях, но в отсутствии волка [Филь, 1970].
На Памире с наступлением зимы копытные перемещаются на малоснежные участки пониженных склонов южной экспозиции, но на востоке региона, где господствуют сильные ветры, сдувающие снег, они не спускаются ниже 3800^4000 м. Летом и осенью самки держатся в горах ниже самцов в более теплых местах, что связано с деторождением и воспитанием молодняка [Егоров, 1955].
Для отдыха сибирские козлы выбирают осыпи и небольшие, относительно горизонтальные, площадки в скальных выходах. Их число ограничено, поэтому одни и те же лежки используются постоянно, и вокруг них скапливается помет. В отсутствии беспокойства звери отдыхают в местах кормежки, причем самцы летом чаще ложатся на открытых местах, обдуваемых ветром, что спасает от назойливых насекомых. В непогоду и жару они укрываются в нишах, пещерах, трещинах, за скалами и большими камнями, в зарослях кустарника или в лесу. Зарегистрирован случай долговременного обитания животных в домах заброшенного людьми горного поселка [Савинов, 1962; Собанский, 1992; Федосенко и др., 1992].
Численность
Численность сибирского козла стала быстро уменьшаться с конца XIX - начала XX вв., особенно заметно в районах интенсивного промысла и скотоводства [Цалкин, 19506]. В Южной Сибири, по экспертной оценке, поголовье вида в историческое время сократилось в десятки раз [Смирнов, Ткаченко, 1992], в XX в. - в 15 раз [Соколов, 1988]. В 80-е годы XX в. в России обитали около И тыс. особей (табл. 70) при средней плотности около 4 зверей на 1000 га ареала [Смирнов, 1994]. Примерно столько же животных (10-15 тыс.) учитывали и в конце этого -начале следующего столетия [Сидоров, 2000, 2004; Федосенко, 2003].
В Горном Алтае в 70-80-е годы плотность населения сибирского козла в разных очагах составляла от 5 до 80 особей на 1000 га, а численность в большинстве из них колебалась в пределах 20-250 голов. Основные ресурсы вида (2-2,5 тыс.) сосредоточены на Северо- и Южно-Чуйском хребтах и в бассейне р. Аргус. На территории Алтайского заповедника учтено немногим более 700 голов [Собанский, Ешелкин, 1975; Собанский, 1988, 1992]. По сведениям С.В. Сидорова [2004], в 2000-2003 гг. в Республике Алтай находилось 8,2-9 тыс. козлов, во что верится с трудом. В Туве около 300 голов жили на хр. Чихачева, в горном массиве Монгун-Тайга, на Хемчикском хребте и его окружении, по 500 - на хребте Цаган-Шибэту и Сангилене, свыше 200 - на Танну-Ола, а всего примерно в 20 группировках было около 3-тыс. особей при средней плотности 2,4 экземпляра на 1000 га угодий. За последние 40 лет ресурсы уменьшились в 2 раза [Опрышко, 1975; Смирнов, 1988а; Смирнов, Ткаченко, 1992; Ткаченко, 1992]. В Красноярском крае общая численность этого вида - около 2,5 тыс., из них большая часть поголовья (1,5-2 тыс.) сосредоточена в Саяно-Шушенском заповеднике [Завацкий, 1989, 1990; Смирнов, Бриллиантов, 1990; Сашко, Смазнов, 1999; Сидоров, 2004]. В Хакасии поголовье неуклонно сокращается. В 1990 г. здесь осталось около 150 зверей [Прокофьев, 1992]. В Иркутской области [Филь, 1964, 19776; Медведев, Аюпов, 1993] и Бурятии [Смирнов, 1988а] численность вряд ли превышает 1 тыс.
Средняя плотность населения козла в очагах обитания на юге Сибири во второй половине XX в. колебалась в пределах 2-8, в Монгольском и Гобийском Алтае - от 8 до 17,5, в отдельных районах Памира достигала 26 голов на 1000 га (табл. 70). В Алма-Атинском заповеднике в 60-70-е годы XX в. она составляла 10-13 [Жиряков, 1973, 1976], в заповеднике “Аксу-Джабаглы” - 56 [Плешак, 1973], на отдельных участках Саяно-Шушенского заповедника в 80-е годы - 300 особей [Федосенко и др., 1992].
Ресурсы сибирского козла в бывшем СССР в 60-е годы XX в. оценивали в 200-550 тыс., в 70-е годы - в 100 тыс. [Банников, 1962, 1965, 1978], в 80-е - в 110-150 тыс. [Климова, 1989]. Основные его запасы были сосредоточены в Таджикистане (несколько десятков тысяч) [Сапожников, 1973, 1976], Киргизии (от 10 [Айзин, 1969] до 60-70 тыс. [Федосенко, 2003]) и горной части Казахстана (26-27 тыс.) [Савинов, 1975а,б]. В конце 90-х годов в Казахстане насчитывали 20-25 тыс. [Бекенов, 1999]. Общие запасы вида в республиках СНГ в этот период были близки к 100 тыс. при очевидной тенденции к дальнейшему сокращению из-за браконьерства [Вейнберг, 1997].
В Монгольском и Гобийском Алтае X. Сухбат [1975] в 70-е годы насчитал около 9 тыс. особей. Позднее В. Груздев и X. Сухбат [1982] численность козла в
Таблица 70. Плотность населения и численность сибирского козла на территории России и в сопредельных регионах во второй половине XX - начале XXI вв.
Район, заповедник Плотность, особей/10 км2 Численность, тыс. особей Источник информации
Алтай 5-8 (до 80 на отдельных участках) 2-4,5 Собанский, Ешелкин, 1975; Собанский, 1977, 1988, 1992; Смирнов, 1994
(С - 3-9 Сидоров, 2000, 2004
Саяны 6 2 Смирнов, 1994
Саяно-Шушенский з-к До 300 на отдельных участках 1,7-2 Федосенко и др., 1992; Сашко, Смазнов, 1999
Красноярский край - 4-2,5 Сидоров, 2000
Тува (Тыва) 3-4 2-3 Сулимов, 1977; Смирнов, 1980, 1994; Смирнов, Ткаченко, 1992
(< — 2,5-3,1 Сидоров, 2000
Тофалария — 1,0 Филь, 1964
Прибайкалье и Забайкалье 2 1,8 Смирнов, 1994
Казахстан 4-25 26-17 Слудский и др., 1983
а — 15-20, 20-25 Бекенов и др., 1998; Бекенов, 1999
сс — 26-27 Савинов, 1975 а,б
Джунгарский Алатау Киргизский и Таласский 13 13 а
Алатау и окрестности 9 4,0 (<
Тянь-Шань 9 5,1 <<
СС 4-58 - Смоляр, 1987
Терскей Алатау 6-110 - Егоров, 1955
З-к “Аксу-Джабаглы” Природный парк 56 1,3 Плешак, 1973
“Алтын-Эмель” 40-50 1,4-2,8 Бекенов и др., 1999
Алма-Атинский з-к 10-14 05-0,7 Жиряков, 1975, 1976, 1977, 1990
Киргизия 5-7 - и
СС 0,2-19 >10 Айзин, 1969
Северная Киргизия 5-23 - Катаевский, 1987
Таджикистан До 26 Несколько десятков тысяч Сапожников, 1976
сс — 40 Фролов, 1983 (по [Федосенко, 2003])
СССР Монгольский — 110-150 Дежкин, 1983; Климова, 1989
и Гобийский Алтай 8-17,5 8,6 Сухбат, 1975
Монголия - 45 Груздев, Сухбат, 1982
Монголии определили в 45 тыс. В конце века монгольские зоологи называли цифру в 80 тыс., что, скорее всего, далеко от истинного значения. В Афганистане, Пакистане и Индии ресурсы, видимо, близки к 10 тыс., в Китае оцениваются в несколько десятков тысяч [Schaller, 1977; Федосенко, 2003].
Запасы ближайших родственников - пиренейского и альпийского козлов в Европе невелики. В 70-90-е годы XX в. в Испании насчитывали 30 тыс., Австрии -4-4,5, Франции - около 2-3, Швейцарии - 8,5-12,5, Италии - 3,5-6,7, Югосла
вии и Германии - 0,1-0,4 тыс., Чехословакии и Румынии - по нескольку десятков особей. В большинстве стран козлы реинтродуцированы в природу после продолжительного разведения в неволе [Дежкин, 1980, 1983, 1989; Estive, Villaret, 1989; Stuwe, Scribner, 1989; Stuwe, Nievergelt, 1991; Bauer, 1992].
Основные факторы, определяющие динамику популяций
Воспроизводство, прирост. При потенциальной плодовитости, близкой к 1 детенышу на самку, годичный прирост в популяциях сибирского козла относительно невысок. В Западном Саяне в летне-осенние месяцы в разных урочищах на 100 взрослых самок приходится от 60 до 84 козлят [Завацкий, 1989; Федосенко и др., 1992]. В Терскей Алатау в июне - июле детенышей имеют только 40-54% самок, в октябре - около 30%, а на Восточном Памире уже в конце июля 71% самок ходят без них [Егоров, 1953, 1955]. В Джунгарском и Заилийском Алатау летом на каждую самку приходится в среднем около 0,8 сеголеток, осенью - 0,4-0,5, а до годовалого возраста доживают лишь 25-30% от числа родившихся [Савинов, 1962; Слудский и др., 1983; Анненков, Литун, 1989]. На Алтае к концу года не имеет детенышей каждая вторая самка [Собанский, 1992]. Примерно такая же ситуация и в популяциях в Гималаях, где с начала зимы и до начала лета число козлят на 100 самок снижается с 85 до 52 [Fox et al., 1992]. Сравнительно низкий прирост связан, прежде всего, с высокой детской смертностью, вызванной ливневыми дождями, снегопадами, низкой температурой и хищниками, а также с высокой смертностью животных в многоснежные зимы.
Погода и корма. Сибирский козел, в сравнении с архаром, живет в относительно благоприятных условиях [Савинов, 19756], но и его популяции иногда существенно “регулируются” многоснежными зимами. В Горном Алтае, по сообщению Г.Г. Собан-ского [1988, 1992], в одной из группировок, насчитывавшей около 300 особей, после суровой зимы 1965/66 гг. осталось всего 25-30. Много животных погибло также в бассейне р. Чульча, на Курайском хребте и в других районах. В многоснежье звери вынуждены переселяться в низкогорные угодья, где резко усиливается их преследование охотниками и хищниками, увеличивается гибель в лавинах. При переходах они нередко тонут в бурных реках [Филь, 19776]. Суровые многоснежные зимы 1965/66, 1968/69 и 1970/71 гг. значительно уменьшили группировки в горах юго-востока Казахстана [Федосенко, Грачев, 1975]. Падеж этих копытных случается и в Саяно-Шушенском заповеднике [Завацкий, 1989,1990]. Гололедной зимой 1943/44 гг. большое количество козлов погибло на Гобийском Алтае [Дагва, 1958]. На Восточном Памире от бескормицы чаще гибнут старые самцы [Егоров, 1955].
Хищники. Основные враги сибирского козла - волк, снежный барс и рысь. Доля этих хищников в общей смертности вида, однако, невелика и не превышает нескольких процентов, хотя локально каждый из них может наносить значительный ущерб популяциям жертвы [Шнаревич, 1948; Савинов, 19756; Жиряков, 1976, 1977; Завацкий, Гущин, 1980; Федосенко, 1986, 2003; Собанский, 1992; Захарченко, 2003].
Скальные убежища козла недоступны для волков, и они подстерегают копытных на переходах или скрадывают на кормежке. На Памире в их экскрементах обычно присутствует зимняя шерсть взрослых животных и, изредка, копытца козлят. Боль
шинство жертв - самцы старше 6 лет [Егоров, 1955]. В Алма-Атинском заповеднике за год один хищник убивает в среднем 2-3 козлов, а все изымают из популяции около 30—45 экземпляров, или 4-5% [Жиряков, 1990]. Случаи нападения волка на копытных нередки и в заповеднике “Аксу-Джабаглы” [Ковшарь, Янушко, 1965]. В горах Джунгарского Алатау среди жертв хищника доля сибирского козла, наряду с маралом, наиболее высокая - до 19-34% в отдельные годы [Федосенко, Жиряков, 1979; Анненков, 1988]. В Саяно-Шушенском заповеднике за 10 лет найдены 152 задавленных козла, в том числе 40 (26,4%) самок в возрасте от 2 до 12 лет и 103 (67,8%) самца в возрасте от 5 лет до 21 года [Завацкий, 1986а,б, 1989, 1990]. В Туве в 1979 г. обнаружено 135 животных, убитых волками [Смирнов, 2002а]. В первой половине XX в. эти хищники вместе с человеком почти уничтожили популяцию козлов на Восточном Саяне в бассейне р. Уды. Возродилась она лишь после 1944 г. в результате истребления волка [Филь, 1964].
Снежный барс и рысь, в отличие от волка, могут проникать в самые труднодоступные места обитания сибирского козла, где давят копытных круглогодично [Кузнецов, 1948а; Савинов, 1962; Слудский и др., 1983; Федосенко, 1986, 20003; Смирнов, Ткаченко, 1992; Ишадов, 2003а]. По наблюдениям О.В. Егорова [1955], на территории семьи барсов 82% самок не имеют козлят уже в августе. Из взрослых животных этот хищник чаще убивает самцов (до 73%). И далеко не случайно ареал барса в Южной Сибири и Средней Азии почти полностью совпадает с областью распространения жертвы.
Ущерб от медведя, красного волка [Егоров, 1955], росомахи и лисицы имеет больше теоретический, нежели практический интерес в силу малочисленности этих зверей в типичных биотопах сибирского козла. Самки этого вида довольно эффективно охраняют своих детенышей от нападения крупных пернатых хищников, хотя случаи их гибели все же имеют место [Дмитриев, 1938; Слудский и др., 1983; Федосенко, 2003].
Конкуренты. Местообитания сибирского козла настолько специфические, что он почти избавлен от присутствия других видов копытных животных, и в первую очередь - оленьих. Косули и маралы заходят в скальники лишь кратковременно, и только в сглаженные участки, а лось практически не поднимается в высокогорье. Кабарга обычно не покидает таежных стаций. Некоторую конкуренцию составляют архары, но они в типичных местах обитания козла малочисленны, и не могут оказывать существенного влияния на пастбища в отличие от домашнего скота, выпасаемого летом в горах.
Домашний скот не только сильно стравливает пастбища, но и вытесняет диких копытных в высокогорья в худшие места обитания. Яки, овцы и, особенно, домашние козы легко проникают в типичные биотопы сибирских козлов, уничтожая скудную высокогорную растительность и заражая пастбища яйцами гельминтов [Анд-реенков, 1980; Слудский и др., 1983; Собанский, 1992; Ткаченко, 1992].
Болезни и паразиты. Из видовых болезней известны пастереллез, некроба-циллез, инфекционная плевропневмония, злокачественная катаральная горячка и другие. Массовая гибель сибирского крзла может быть вызвана поражением легких протостронгилидами [Урсова, 1985], но наибольшее влияние на численность оказывают, вероятно, эпизоотии саркоптоза, возбудителем которой является клещ Acarus siro саргае. В 1917-1920 гг. эта болезнь опустошила многие популяции на
Алтае [Дмитриев, 1938]. В 1959-1963 гг. массовый падеж зарегистрирован на северо-востоке Джунгарского Алатау [Савинов, 19756; Слудский и др., 1983]. В 1968-1972 гг. зудневой чесоткой была поражена популяция копытных заповедника “Аксу-Джабаглы” в Заилийском Алатау, которой она, видимо, заразились от домашнего скота [Плешак, 1973; Федосенко, 1983]. Значительный падеж козлов от саркоптоза в эти годы имел место и в западной части Чаткальского хребта в Узбекистане. На Угамском хребте эпизоотия началась раньше - в 1966 г. [Вырыпаев, 1973; Вырыпаев, Литвинов, 1974].
У сибирского козла обнаружено более 30 видов гельминтов [Геп тнер и др., 1961; Слудский и др., 1983]. В Туве очаги гельминтозной инвазии, включающие 15 видов, формируются в результате контактов с домашними овцами и козами на пастбищах [Сулимов, 1973]. Из эктопаразитов более всего досаждают зверям носоглоточный овод, слепни, комары, мошки и мухи-кровососки. На них встречаются также блохи, вши и, в небольшом количестве, клещи [Дмитриев, 1938; Шнаревич, 1948; Грунин и др., 1955; Егоров, 1955; Дагва, 1958; Савинов, 19756], но эти паразиты вряд ли становятся причиной смертности копытных.
Травматизм, гибель в лавинах. Гибель козлов от травм, полученных в драках, редка. Гораздо чаще они травмируются (ломают конечности, ребра и рога) при быстром беге по крупнокаменистым осыпям и при неудачных прыжках со скал. Гибель групп животных происходит в лавинах, при камнепадах и снежных обвалах. Их остатки обычно находят у основания лавинных лотков и у подножий скал. В Заилийском Алатау на лавины приходится 34% от числа всех зарегистрированных случаев гибели. На Западном Памире в многоснежные зимы под обвалами остается, возможно, до 10% поголовья. Много зверей гибнет здесь после землетрясений под обломками скал [Егоров, 1955; Жиряков, 1976, 1977, 1979; Соколов, 1979; Собанский, 1992].
Антропогенные факторы. На популяции козла, как отмечено выше, существенное воздействие оказывает фактор беспокойства, вызванный интенсивным выпасом домашнего скота, который вытесняет зверей из типичных мест обитания. Большой ущерб молодняку причиняют многочисленные пастушьи собаки, постоянно рыскающие в поисках пропитания по угодьям. Часть детенышей гибнет, отставая от убегающих матерей, по вине туристов, беспокоящих зверей в их местообитаниях [Дмитриев, 1938; Вырыпаев, 1973; Федосенко, 1975,2003; Жиряков, 1976; Слудский и др., 1983; Смоляр, 1987; Собанский, 1992; Ткаченко, 1992; Медведев, Аюпов, 1993]. И все же, как и у других видов копытных, несравненно больший урон популяциям причиняют браконьеры, нередко полностью уничтожающие локальные группировки.
Использование ресурсов и охрана
Сибирский козел издавна служит объектом охоты населения Южной Сибири и Центральной Азии, о чем свидетельствуют многочисленные наскальные рисунки с изображением сцен поимки зверей в ямы и ловушки, загонов их всадниками и собаками на засады пеших лучников. Костные остатки этого вида сохранились в древних жилищах человека во многих районах (см. выше). Мясо козлов очень вкусное, но несколько жестковатое по сравнению с мясом горных баранов. Зимние
шкуры обычно используют для пошива теплой и прочной верхней одежды и обуви, в качестве ковриков и одеял, летние чаще выбрасывают, а из шкур молодых особей выделывают шевро и изготавливают бурдюки для хранения и перевозки кумыса. Рога применяют в качестве украшений жилищ, как груз для установки капканов, используют на ручки, рукоятки ножей, а в последнее время они стали желанным трофеем.
Основные способы охоты на козлов - облавы, преследование по глубокому снегу, добыча с помощью луков-самострелов, капканов, сплавом с плотов и лодок на прибрежных скалах, из засадок и с подхода с помощью современного оружия и собак [Каверзнев, 1933; Цалкин, 19506; Егоров, 1955; Дагва, 1958; Смирнов, 1983; Смирнов, Ткаченко, 1992].
Никаких ограничений в сроках и способах добычи зверей не существовало вплоть до 30-50-х годов XX в. Неумеренный хищнический промысел и стал главной причиной резкого сокращения ареала и численности вида. На Алтае сибирский козел взят под охрану в 1935 г. [Дмитриев, 1938], в Бурятии - в 1949 г. Примерно в это же время была запрещена охота в Красноярском крае, чуть позже - в Иркутской области, в 1979 г. - в Туве (в период с 1971 по 1978 г. здесь официально добывали около 50 особей ежегодно) [Никифоров и др., 1977].
В первой половине XX в. добыча сибирского козла в Южной Сибири, вероятно, исчислялась тысячами голов. Только в Кош-Агачском аймаке на Алтае ежегодно промышляли свыше 200 особей [Колосов, 1939]. С 1975 г. за семь охотничьих сезонов официально отстреляно всего 92 козла, или 13 экземпляров ежегодно [Перовский, 1985]. В 80-е годы среднегодовой легальный объем добычи на юге Сибири составлял около 30 особей [Смирнов, 1994], а в конце XX - начале XXI вв. не превышал 50-76 экземпляров [Сидоров, 2004].
Фактическая добыча несопоставима с лицензионной. В Горном Алтае она близка к 200-300 экземплярам. В последние десятилетия козлы полностью исчезли с берегов Телецкого озера, в верховьях Чарыша и Чуи, по левобережью Катуни и в других местах. Особенно большой вред горным копытным причиняют геологические отряды, базирующиеся в труднодоступных районах и имеющие в своем распоряжении вертолеты, без проблем доставляющие охотников в нужные пункты. В середине 70-х годов в верховьях р. Большой Абакан небольшой отряд геологов с помощью одного армейского карабина и двух лаек всего за 1,5 месяца уничтожил изолированную еринатскую группировку (60-70 голов) сибирских козлов [Собанский, Ешелкин, 1975; Собанский, 1988, 1992]. Такая же ситуация и в Туве, где из-за неумеренного браконьерства перестали существовать многие мелкие и некоторые крупные группировки, насчитывающие до 1 тыс. особей [Смирнов, 1988а, 1989; Смирнов, Ткаченко, 1992]. По Енисею процветает “мото-лодочное” браконьерство - зверей отстреливают с подъезда [Федосенко, 2003]. В Бурятии нелегальный отстрел не прекратился и после полного запрета охоты на вид [Смирнов, 19886].
О масштабах промысла козла в Средней Азии и его последствиях свидетельствуют следующие факты. В Терскей Адатау только одной бригадой из 3 человек в 1947 г. отстреляно более 200 особей [Егоров, 1955]. В Джунгарском Алатау в долине р. Теректы и прилежащих районах чабаны, снабженные малокалиберными винтовками, сократили численность козла с 1 тыс. в 1959 г. до 90 особей в 1963 г.
[Грачев, Савинов, 1975а]. В 1963 г. бригада охотников Алма-Атинского общества с трудом отстреляла здесь с десяток зверей, и почти все они были со следами прежних ранений малокалиберными пулями [Смирнов, 1965]. Из-за браконьерства, доля которого в общей смертности животных достигает трети, и вытеснения скотом поголовье козла заметно уменьшилось и на Тянь-Шане, но здесь он все же остался фоновым видом [Жиряков, 1977; Смоляр, 1987]. На Памире сильное сокращение численности произошло во время и после войны в связи с интенсивным промыслом [Кузнецов, 1948а; Егоров, 1955]. В Киргизии даже в 70-е годы козлов добывали больше (1,4—1,6 тыс.), чем кабана и косуль [Айзин, 1980]. Всего в СССР в середине XX в. ежегодная добыча сибирского козла достигала 20-25 тыс., в Монголии - 5 тыс. [Банников, 1954, 1965].
В последнее время сибирский козел стал пристальным объектом трофейной охоты, но и она, в большей мере, ведется нелегально. Интенсивное изъятие лучших самцов в национальном парке “Алтын-Эмель” (Казахстан), где ежегодно добывают по 50-70 особей, уже привело к заметному снижения качества популяции - максимальна длина рогов уменьшилась со 147 до 107-123 см, а средняя - со 114 до 100-103 см [Мельников, 2003].
Сохранить этих копытных становится все сложнее из-за доступности высокогорий с помощью вертолета и отсутствия действенной охраны. Спасти вид от полного истребления во многих регионах можно лишь расширив сеть особо охраняемых природных территорий [Смирнов, 19886]. Как показывает практика, при охране сибирский козел способен быстро увеличивать численность. В пограничной зоне Тувы его поголовье заметно выросло после запрещения охоты в 1981—1983 гг. [Ткаченко, 1992]. За 10 лет существования Саяно-Шушенского заповедника популяция увеличилась в 2,3 раза, а плотность достигла 19 особей на 1000 га, что в 7 раз выше, чем в соседних охотничьих угодьях [Смирнов, Ткаченко, 1992]. В дальнейшем в восстановлении и увеличении ресурсов вида, скорее всего, будет возрастать роль организованного охотничьего хозяйства трофейного типа.
При улучшении охраны угодий сибирского козла желательно реакклиматизи-ровать в прителецкой части Алтайского заповедника, где он обитал вплоть до 40-х годов XX в. [Собанский, 1977, 1988, 1992]; в Тувинской котловине; на левобережье Енисея между Красноярском и устьем Бирюсы; в скалистых массивах в месте пересечения Енисеем Батеневского кряжа и гор Оглахты; по берегам р. Белый Июс [Смирнов, 1994]; в горных степях Минусинской котловины. Расселять зверей реально из мощной популяции Саяно-Шушенского заповедника. Для получения племенного материала желательно также создать питомник в степной или лесостепной части Кузнецкого Алатау [Прокофьев, 1992]. Большие перспективы в восстановлении ареала козла имеются и в Казахстане [Слудский, Афанасьев, 1964].
Разведение этого вида в неволе и полувольных условиях тоже вполне возможно. Опыт успешного воспроизводства зверей в степи накоплен в Аскании-Нова [Треус, 1968, 1970]. На Памире местные жители отлавливали детенышей и содержали их долгое время в домашних условиях. Известен случай, когда выращенная в неволе и прирученная самка ежегодно весной перед родами уходила в горы за 3-4 км от поселка, а в середине августа вместе с козленком возвращалась к людям. Ее козлята 3-5 лет жили в качестве домашних животных, а затем уходили в горы и больше не возвращались [Одинашоев, 1986].
Род БАРАНЫ Ovis Linnaeus, 1758
Ovis Linnaeus, 1758. Syst. Nat., 10ed., 1: 70. Тип рода- Ovis aries Linnaeus, 1758 (домашняя овца). Швеция.
Aries Brisson. 1762. Regn. Anim.: 12. Nom. nudum.
Musimon Pallas, 1776. Spicil. Zool., 2, fasc. 2: 8.
Musmon Schrank, 1798. Fauna Boica, 1: 78. Nom. nudum.
Aegoceros Pallas, 1811. Zoogr. Rosso-Asiat., 1: 224.
Ammon de Blainville, 1816. Bull. Soc. Philom. Paris: 76. Nom. nudum.
Oegoceros Lesson, 1827. Man. Mamm.: 399. Pro Capra ammon Linnaeus.
Caprovis Hodgson, 1847. J. Asiat. Soc. Bengal., 16: 702. Ovis musimon Pallas.
Argali Gray, 1852. Cat. Ungulate Mamm. Brit. Mus., 3: 174. Aegoceros argali Pallas.
Pachyceros V. Gromova, 1936. Neue Forsch. Tierzucht. und Abstammung. (Festschr. Dr. Durst): 84.
Ovis nivicola Eschscholtz.
Характеристика. Копытные мелких, средних и крупных размеров, крепкого сложения, с относительно высокими и тонкими ногами и коротким оволосненным хвостом. Половой диморфизм и географическая изменчивость значительны. Длина тела самцов разных форм колеблется от 110 до 210 см, высота в холке - от 60 до 135 см, масса - от 30 до 216 кг (табл. 71).
Окраска баранов разнообразная: от черной до белой, однотонная или контрастная (см. ниже). У самцов муфлона (фото 31) и уриала (фото 33 и 34) зимой на груди и шее образуется подвес из длинных волос: у первых он черного и коричневочерного цвета, у вторых - буровато-черный на груди и белый у подбородка или светлый. У архара (фото 37-39) подвес чаще не развит, и в виде светлой манишки обычен только у южных рас. У снежного барана (фото 50-54) подвеса нет, окраска преимущественно бурая, но есть формы с почти черным, белым и светло-рыжим мехом. Борода у баранов, в отличие от козлов, отсутствует, хвостовое зеркало имеется.
Волосяной покров у баранов, как и у других представителей Caprinae, представлен остью и пухом, но пуховых волос меньше, чем у Capra. Летний волос относительно короткий и равномерной длины, зимний - пневматизированный и удлиненный. Линька одна - весенняя. Развиты межпальцевые (с “карманами”), предглазничные (образованы только потовыми железами) и паховые железы [Соколов, 1973; Соколов, Чернова, 2001]. Сосков одна пара, вторая пара недоразвита.
Череп относительно широкий, с укороченной зароговой частью, удлиненным и суженным лицевым отделом. Его размеры значительно варьируют у разных форм (табл. 71). Кости сильно пневматизированы. Межчелюстные кости не всегда достигают носовых. Теменные гребни широко расставлены. Предглазничные ямки имеются или почти не выражены, этмоидальных щелей чаще нет. Слуховые барабаны небольшие. В нижней челюсти с каждой стороны три или два предкоренных зуба. Роговые стержни длинные. Их основания расположены за глазницами, и у снежного барана сильно утолщены. Синусы роговых отростков обычно не достигают верхней трети. Размеры черепа коррелируют с размерами рогов.
Таблица 71. Морфологическая характеристика разнохромосомных баранов Евразии
Признаки (у взрослых самцов) Муфлон* Уриал* Архар** Снежный баран**
Число хромосом 54 58 56 52
Длина тела, см 110-144 120-160 144-210 140-188
Высота в холке, см 60-89 77-98 91-135 76-112
Масса, кг 30-65 (95) 39-96 75-216 56-150
Наибольшая длина черепа, мм 225-264 233-297 260-395 256-296
Максимальная ширина черепа, мм 114-154 118-155 138-218 139-189
Высота черепа, мм 115-154 124-158 138-244 116-170
Длина носовых костей, мм 71-101 82-119 70-170 67-106
Длина верхнего ряда зубов, мм Число предкоренных зубов в 60-78 68-84 69-104 65-81
нижней челюсти 3 3 3 3-2
Длина костных стержней рогов, см Обхват костных стержней рогов 20-45 25-48 26-52 13-30
у основания,см 12,5-20 17-23 (27) 18,5-39,5 18-33,5
Длина роговых чехлов, см Обхват роговых чехлов 37-104 51-104 51-190 41-115
у основания, см 18-27 21-30 23-55 20-38
Тип рогов Гетеронимные, изредка первер-тированные Перветиро-ванные, реже гомонимные Гомонимные Гомонимные
Окраска животных От черной до рыжей, со светлым пятном От рыже-бурой От буро-до песочной коричневой до светлосерой От бурой до светлой
Подвес из удлиненных зимних волос на груди Черный, темный От буроваточерного до светлого Светлый или отсутствует Отсутствует
*По [Насонов, 1923; Цалкин, 19486, 1951а; Даль, 1954; Дулицкий, Кормилицын, 1970; Дулицкий, 1976; Сапожников, 1971, 19766; Бекенов, Савинов, 1981; Hoefs, 1982; Ишунин, 1983а,в; Слудский и др., 1983; Сопин, 1984а,6; Авруян, 1970; Минасян и др., 1986а; Манассрова, 1986, 1988; Sabados, Holy, 1992; Хохлов, Харебов, 2001; Федосенко, 2002а; Хохлов, 2002; Woloch, 2003].
** Обобщенные сведения (см. ниже).
У самок рога короткие (до 56 см), тонкие, сжатые с боков, немного загнутые назад, или у значительной части отсутствуют (на западе ареала). У самцов они длинные (до 190 см), массивные, широкие в основании и постепенно сужающиеся к вершине. Роговые чехлы без валиков и бугров на передней поверхности, но с глубокими бороздами, возникающими ежегодно в зимние месяцы в период пре-рыва их роста из-за неблагоприятных условий существования, и с поперечными складками-морщинами, образование которых связывают с тем, что рост стержня не поспевает за ростом чехла [Сопин, 19756]. Существует и другая гипотеза. Поперечные кольцевые утолщения на рогах у домашних и диких баранов возникают с интервалом в 6-7 и 13-14 дней в периоды антициклональной погоды с повышенным атмосферным давлением и пониженной геомагнитной напряженностью и, возможно, отражают биоритмы изменений уровня обмена веществ в орга
низме [Маликов, 1978]. Цвет рогов варьирует от светло-охристого и желтовато-серого до темно-бурого.
Рога баранов разнообразны, но с запада на восток и, отчасти, с юга на север ареала четко прослеживается увеличение их размеров, усложнение формы и изменение изгиба от отрицательного (гетеронимного) у муфлона к нейтральному (первертированному) у уриала и положительному (гомонимному) у аргали-образных форм и снежного барана. Исключение представляют некоторые пустынные расы архара, имеющие небольшие рога [Цалкин, 1951а], что, возможно, связано с худшими условиями обитания. В наименьшем развитии рога слабо изогнуты назад, а их концы направлены внутрь или слабо отогнуты вниз, в максимальном - закручены назад, вниз и в стороны.
Муфлоны, уриалы, архары и снежные бараны различаются не только по размерам и форме рогов и окраске, но и по темпам роста, пропорциям тела и черепа, некоторым особенностям посткраниального скелета [Северцов, 1873а,б; Насонов, 1923; Sushkin, 1925; Gromova, 1936; Цалкин, 19486,1951 а; Чернявский, 1962в, 1963, 19646; Явруян, 1970; Сапожников, 19766, 1977; Сопин, 1980а, 19836, 1984а,б; Сопин, Улоть, 1980; Минасян и др., 1986а,б; Ревин и др., 1988], а также имеют разный хромосомный набор, что обычно служит основанием для разделения их на виды. Тем не менее географическая изменчивость морфологических признаков у разнохромосомных форм, за редким исключением, клинальна. Основные параметры снежного барана не выходят за рамки варьирования признаков горных баранов, что свидетельствует о их близком родстве (табл. 71).
Бараны - преимущественно горные животные. Образ их жизни групповой и, в определенные периоды, стадный. Хорошие бегуны. Максимальная продолжительность жизни и в природе, и в неволе обычно не превышает 12—14 лет, но отдельные особи, судя по годовым кольцам рогов и результатам мечения, доживают до 17-20 лет.
Охотничьи или охраняемые виды. Домашняя овца (Ovis aries) стала важнейшим для человека животным.
Распространение. Юг Европы (в плейстоцене на запад до Англии), включая острова Средиземного моря, Передняя, Средняя и Центральная Азия, Южная и Восточная Сибирь, запад Северной Америки (рис. 65). Современный ареал очаговый, приуроченный к горным системам.
Эволюция. Группа по происхождению азиатская. Наиболее древние верхнеплиоценовые ископаемые представители рода (Ovis zdanskyi Bohlin, 1938 и Ovis schantungensis de Chardin, Piveteau, 1930) известны из Китая. В Европе архароподобные бараны появились, видимо, в конце этого периода, в Южной Сибири - в эоплейстоцене.
Для Ovis характерна высокая адаптивная радиация. Дивергенция архароподобных, муфлонообразных и снежных баранов прослеживается со среднего плейстоцена. Темпы эволюционных преобразований у отдельных представителей рода необычайно велики. Корсиканский муфлон, расселенный в Центральной Европе, изменился в размерах и окраске всего за столетие, чему, скорее всего, способствовала его гибридизация с домашней овцой. Снежные бараны в морфологическом отношении более консервативны - современные формы близки к четвертичным.
Бараны - наиболее специализированная группа среди Caprini и, пожалуй, среди Caprinae. Замечательный “продукт” их эволюции - колоссальные размеры рогов,
являющихся “турнирным оружием” у самцов [Северцов, 1951] и средством защиты от хищников. Завершающее звено эволюционного ряда, идущего от примитивных Capra через промежуточные формы к Ovis, - собственно архар (О. аттоп аттоп). С другой стороны у архара сохраняются отдельные архаичные черты, в частности рудимент пятой пястной кости, что не позволяет однозначно считать его “венцом” эволюционных преобразований [Сапожников, 19766, 1977].
Снежные бараны заметно уклонились от основного направления эволюции рода, представляющего адаптацию к жизни в сравнительно открытой холмистой местности или пологих частях гор и к быстрому продолжительному бегу (чему более соответствуют типы муфлона и уриала), и приспособились к обитанию в расчлененном скалистом рельефе и к передвижению по крутым склонам, подобно козлам. Это привело к перестройке опорно-двигательного аппарата и морфологической и экологической дифференциации от предковых форм. По строению черепа и рогов они наиболее примитивные среди Ovis, а по ряду морфологических признаков близки к Capra [Громова, 1936, 1953].
По цитогенетическим показателям 52-хромосомный азиатский снежный баран, напротив, является конечным звеном эволюции группы, а наиболее примитивным в этом плане представляется 58-хромосомный уриал. В процессе эволюции, скорее всего, происходило постепенное уменьшение числа хромосом (2п = 58-56-54—52) в результате транслокационных соединений отдельных пар акроцентрических хромосом и преобразования их в метацентрические (при неизменном числе плеч), что весьма распространено у млекопитающих при генных мутациях. По полифилетической гипотезе робертсоновские транслокации могли возникать независимо в разных группах баранов. Не исключена и возможность увеличения числа хромосом путем центрического разделения [Воронцов и др., 1972; Bunch, 1978;Мамедов, 1978, 1985,1987; Орлов, 1978; Сопин, 1980, 1983в; Орлов, Булатова, 1983; Шарипов, 1989; Ляпунова и др., 1997; Bunch et al., 2000; Шайдуллин, 2001].
Свободное скрещивание разнохромосомных баранов в экспериментах и возникновение гибридных группировок в зоне наложения их ареалов (см. ниже) свидетельствует об отсутствии репродуктивной изоляции и монофилетическом происхождении этой группы копытных.
Бараны, обитающие в многоснежных районах на севере ареала и в высокогорье в суровом холодном климате, как правило, крупнее, чем на юге и в относительно равнинных и малоснежных районах, и такая морфологическая изменчивость укладывается в правило Бергмана. Известно также, что у животных, постоянно живущих высоко в горах, увеличиваются размеры грудной клетки, ее форма, гипертрофируется сердце и легкие, меняется картина красной крови [Булатова, 1962], что показывает возможный путь дифференциации горных и снежных баранов на ряд форм.
Состав. Размах географической изменчивости признаков у баранов максимальный среди копытных животных, что дает повод для выделения более 50 форм (рис. 65). Состав рода Ovis остается особенно дискуссионным до сих пор. В нем принимают до 4 подродов, но чаще 2: собственно Ovis s. str. с 1-5 видами и Pachyceros Gromova, 1936 с 1-3 видами [Северцов, 1873а,б; Насонов, 1923; Sushkin, 1925; Флеров, 1935а,б; Ellerman, Morrison-Scott, 1951; Цалкин, 1951а; Соколов И.И., 1959; Гептнер и др., 1961; Громов и др., 1963; Haltenorth, 1963; Соколов В.Е., 1979; Барышников и др., 1981; Durst et al., 1982; Valdez, 1982; Co-
пин, 1983а; Павлинов, Россолимо, 1987; Ludwig, Вепеске, 2003; Павлинов, 2003; Чернявский, 2004; и др.]. История классификации баранов весьма интересна и достойна отдельного рассмотрения.
История и проблемы классификации. Морфологический этап. Основоположником систематики рода Ovis следует считать К. Линнея, описавшего типичного представителя — домашнюю овцу (Ovis aries Linnaeus, 1758) и горного барана (Capra аттоп L., 1758). Н.А. Северцов [1873а,б] первым систематизировал этих копытных, разделив их, преимущественно по признакам рогов, на два рода: арка-ров (Ovis) с 11 видами в двух группах (северной и южной) и муфлонов (Musimon) с 6 видами. Он же выделил путоранского барана (О. borealis) и несколько новых туркестанских видов; О. coIlium, О. karelini, О. heinsii и О. nigrimontana. Уровень таксономических исследований того времени характеризует хотя бы описание О. coIlium-. “Не виденный мной и никем не описанный; по словам Г. Карелина отличен от Чуйских аркаров. An varietas О. Karelini? Сомнительный вид”. Однако позднее эти и другие формы, описанные им практически без сравнения, перекочевывали из одной таксономической сводки в другую.
Р. Лидеккер [Lydekker, 1898, 1913] в единственном роде Ovis определил 3 подрода: собственно Ovis с 8 видами и 12 подвидами, североафриканских гривистых баранов (Ammotragus) и центральноазиатских голубых баранов (Pseudois). Затем он пересмотрел свою точку зрения в отношении родства горных баранов с Ammotragus и Pseudois и уменьшил число видов Ovis до 6 (О. musimon, О. orientalis, О. laristanica, О. vignei, О. аттоп и О. canadensis), но число подвидов довел до 42.
Академик Н.В. Насонов [1910, 1911, 19136, 1914, 1915, 1916, 1919, 1920] после многолетних капитальных исследований в фундаментальной сводке “Географическое распространение диких баранов Старого Света” [1923] выделил, главным образом по форме и изгибу рогов, 36 географических рас 9 видов: О. musimon, О. orientalis, О. ophion, О. gmelini, О. laristanica, О. vignei (группа Mufloniformes) и О. polii, О. аттоп и О. nivicola (группа Argaliformes). Он же дал детальный обзор имеющейся литературы по систематике и географическому распространению рода и заключил, что группа Mufloniformes имеет более примитивные черты по сравнению с Argaliformes, а наиболее примитивный вид - О. ophion и его кипрская раса О. о. ophion. Замечу также, что академик первым из исследователей отнес кызылкумского барана, или барана Север-цова (О. polii severtzovi) к группе аргали.
Известнейшие систематики Дж. Эллерман и Т. Моррисон-Скотт [Ellerman, Morrison-Scott, 1951] включали в два подрода (Pachyceros и Ovis) 5 видов: О. canadensis, О. аттоп (с 15 подвидами), О. orientalis, О. laristanica и О. musimon. Однако в целом их классификация близка к предыдущей.
К.К. Флеров [19356] справедливо считал различия баранов по форме рогов “чрезвычайно туманными”, как, впрочем, и по окраске тела. Позднее В.И. Цалкин [19486, 1951а] пришел к однозначному выводу о значительном сходстве муфлона и архара (аргали) и их отличии от толсторога (снежного барана). Он нашел, что в направлении с запада на восток и, отчасти, с юга на север ареала у баранов четко выражена клинальная изменчивость признаков в соответствии с правилом Бергмана и с небольшими отклонениями от него у пустынных форм. В постэмбриональном развитии аргали как бы проходит стадии, характерные для муфлона, но с дальнейшим гиперморфизмом. Усложнения в строении черепа связаны с разрастани
ем и утяжелением рогов. Прерыв постепенности изменения признаков очевиден лишь у граничащих популяций архара и толсторога. Замечу, что эти утверждения справедливы и по сей день (табл. 71), несмотря на новые тенденции в систематике. Соответственно, все формы баранов Евразии В.И. Цалкин свел к двум аллопатрическим видам: архару (О. аттоп) с 24 подвидами и снежному барану (О. canadensis) с 4 расами (см. ниже). Эта классификация в целом поддержана В.Г. Гентнером [Гептнер и др., 1961] и В.Е. Соколовым [1979].
И.И. Соколов [1953, 1959], напротив, рассматривал род Ovis в широком объеме и включал в его состав 4 подрода: Ovis, Pachyceros, Ammotragus и Pseudois. В пределах СССР к подроду настоящих баранов он отнес 3 вида: европейского муфлона (О. musimon), азиатского муфлона (О. orientalis) и архара (О. аттоп) с 15 подвидами (см. ниже), к подроду Pachyceros - снежного барана (О. nivicola). Эта же система отражена и в сводке И.М. Громова с соавторами [1963].
Наиболее радикальная точка зрения - объединение всех баранов Голарктики в один вид [Haltenorth, 1963] не нашла поддержки у большинства ученых, хотя аргументы автора, на мой взгляд, не лишены оснований.
Цитогенетический этап. Наиболее масштабная ревизия систематики рода произошла в период развития кариологии. В подроде Ovis цитогенетики определили 4 вида [Schmitt, Ulbrich, 1968; Воронцов и др., 1972а,б; Nadler et al., 1973; Korobitsina et al., 1974; Воронцов, 1975; Коробицина, Воронцов, 1975; Ляпунова и др., 1997]:
домашнюю овцу (О. aries L., 1758) - диплоидный набор хромосом 2п = 54;
муфлона (О. orientalis Gmelin, 1774) - 2n = 54 (юг Европы, Малая Азия, Закавказье, северо-запад Ирана);
уриала {О. vigneiBlyth, 1841)-2п = 58 (юго-западКазахстана, Узбекистан,Туркменистан, юго-запад Таджикистана, Афганистан, Пакистан, Кашмир, восток Ирана, север Омана);
архара (О. аттоп L., 1758) - 2п = 56 (горы и возвышенности Средней и Центральной Азии).
В подроде Pachyceros по кариотипу обособлены 2 вида [Wurster, Benirschke, 1968; Nadler, 1971; Korobitsina et al., 1974; Коробицина, Воронцов, 1975]:
азиатский снежный баран (О. nivicola Eschscholtz, 1829) - 2n = 52 (Северо-Восточная Азия);
американский (канадский) толсторог (О. canadensis Shaw, 1803) - 2n = 54 (запад Северной Америки).
Несмотря на этот вывод, в подроде Pachyceros, помимо О. canadensis и О. nivicola, некоторые исследователи признают барана Далла (О. dalli Nelson, 1884), который имеет такой же кариотип, как и у канадского толсторога, но отличается размерами тела, черепа и белой или почти черной окраской меха, ограничивая его ареал Аляской и горами Британской Колумбии в Канаде [Cowan, 1940; Чернявский, 1963, 2004; Сопин, 1980, 1983а, 19866; Durst et al., 1982; Ревин и др., 1988; Павлинов, 2003].
Цитогенетики, несомненно, внесли весомый вклад в таксономию диких животных, однако число и морфология хромосом не стали бесспорным аргументом в систематике. Многие виды копытных, как оказалось, имеют не только одинаковое их число (чаще 2п = 60 и 2п = 70), но и разное: лось (2п ~ 68-70), сибирская косуля (2п = 70+1-14), пятнистый олень (2п = 64-68), благородный олень (2п =
66-68), кабан (2n = 36-38). Виды с варьирующим кариотипом обычны и в других систематических группах млекопитающих [Орлов, Булатова, 1983].
Не стали исключением и бараны. У домашней овцы при типичном 2п = 54 нередки гиподиплоидные клетки с числом хромосом от 50 до 53, а в некоторых породах (у ромнейской в Новой Зеландии) преобладают 52-хромосомные особи. Однако, судя по морфологической структуре кариотипа, 52-хромосомные снежные бараны вряд ли могли быть предками последних [Мамедов, 1978, 1985; Шарипов, 1989]. В зоне наложения ареалов муфлона и уриала на хребте Эльбурс в Иране существует группировка животных (фото 35) с хромосомным набором 2п = 54—58. Морфологические признаки эльбурского барана заметно варьируют: в западной части преобладают черты муфлона, в восточной - уриала [Nadler et al., 1971; Воронцов и др., 1972; Valdez et al., 1978]. Разнохромосомные особи, скорее всего, существуют и на стыке областей распространения других форм горных баранов.
Кариотип европейского и азиатского муфлонов, американского толсторога и домашней овцы оказался одинаковым как по числу (2п = 54), так и по морфологии хромосом. Такой же диплоидный набор и у голубого барана (Pseudois), но при иной морфологии [Nadler, 1971; Кулиев, Мамедов, 1974, 1975; Коробицина, Воронцов, 1975; Мамедов, 1985, 1987; Шарипов, 1989; Мамедов, Агаев, 1990]. Можно предположить, что американские снежные бараны и муфлон некогда имели общего 54-хромосомного предка [Nadler et al., 1973; Сопин, 1983в] или в процессе эволюции утратили некоторые хромосомы независимо один от другого, что ближе
- горный баран
- муфлон (2п = 54)
- уриал (2п = 58)
- архар (2п = 56)
- снежный баран
- снежный баран (2п = 52)
к истине.
Сомнения в правомерности “хромосомной” классификации Ovis привели известного зоолога и цитогенетика В.Н. Орлова [1978] к выводу о двухвидовом составе рода (О. аттоп и О. canadensis) и полувидовом (semispecies) ранге других форм баранов. Предлагаемая им система рода весьма логична:
Ovis аттоп
musimon (semisp.) vignei (semisp.) аттоп (semisp.) Ovis canadensis
nivicola (semisp.)
canadensis (semisp.) - канадский баран (2n = 54).
Гибридизация как таксономический критерий. Цитогенетическое сходство и теоретически, и практически позволяет свободную гибридизацию домашней овцы, американского толсторога и муфлона между собой [Пфиценмайер, 1915; Фортунатов, 1924; Gray, 1954; Соколов, 1959; Треус, 1968; Стекленев, 1972; Треус, Лобанов, 1973,1976; Blunt et al., 1977; Piegert, 1980; Айрумян, Гаспарян, 1986; Минасян и др., 1986а]. Муфлон считается непосредственным предком домашней овцы. Их скрещивание особенно легко приводит к получению гибридов. Наиболее известный продукт гибридизации - порода “горный меринос” [Иванов, 1949; Иванов, Гребень, 1949].
Гибридизация разнохромосомных уриала (2п = 58), архара (2п = 56) и азиатского снежного барана (2п = 52) с домашней овцой и муфлоном (2п = 54), как оказалось, тоже возможна, но ее результаты неодинаковы [Пфиценмайер, 1915; Саркисов, 1953; Салганский и др., 1963; Треус, 1968; Лобанов, Треус, 1978; Шарипов,
1989]. Гибриды архара с овцой и муфлоном имеют 55 хромосом, гибриды уриала -56 [Савицкий и др., 1974; Шарипов и др., 1980; Шарипов, 1989], а в Бадхызе получен ряд плодовитых гибридов местного уриала с каракульской овцой с хромосомным спектром от 54 до 58 [Орлов и др., 1980]. При скрещивании бухарского уриала с гор Бабатага с серой каракульской овцой выведен новый тип домашней овцы (бабатагский), отличающийся повышенной жизнестойкостью молодняка [Гигиней-швили, 1959,1983; Гигинейшвили, Чариев, 1980]. В результате гибридизации тянь-шаньского архара с тонкорунной овцой выведен казахстанский архаромеринос с кариотипом 2п = 54. Не представляет также проблем скрещивание архара с грубошерстными и курдючными овцами [Румянцев и др., 1935; Бутарин, 1949, 1959, 1964; Мухамедгалиев, Савицкий, 1974; Стамбеков и др., 1987]. Плодовитые и высокопродуктивные гибриды получены и при скрещивании с каракульскими овцами кызылкумского и каратауского баранов [Рахимов, 1959, 1966; Шарипов и др., 1980; Шарипов, 1989].
Гибриды азиатского снежного барана с разными породами овец тоже плодовиты и по мужской, и по женской линиям, но результативность гибридизации значительно хуже. Новорожденные в связи с более коротким сроком беременности овец (144—158 дней), по сравнению с диким видом (170 дней), имеют небольшую массу, и большинство их погибает в постнатальный период, а особи, достигшие половой зрелости, отличаются пониженной плодовитостью. Все гибриды F1 имеют промежуточное число (53) хромосом [Стекленев, 1972; Давлетова, 1986; Смирнов и др., 1987; Шайдуллин, 1992, 1994а,б, 2001; Шайдуллин и др., 1994; Живалев и др., 1996]. Эти данные однозначно свидетельствуют в пользу гипотезы о более существенной таксономической дифференциации снежного барана по сравнению с остальными формами.
Отсутствие репродуктивных барьеров и постепенность морфологической (табл. 71) и кариологической изменчивости ставит видовую самостоятельность разнохромосомных форм баранов, за исключением, пожалуй, снежного, под сомнение. Впрочем, легко скрещиваются между собой и многие другие “ясные” и признанные систематиками виды копытных, давая плодовитые гибриды с разным числом хромосом, что, в принципе, противоречит биологической концепции вида.
Новые версии. В последних отечественных таксономических справочниках (“Каталог млекопитающих СССР” [Барышников и др., 1981 ] и “Систематика млекопитающих СССР” [Павлинов, Россолимо, 1987]) в качестве видов указаны европейский (О. musimon) и азиатский муфлоны (О. orientalis), уриал (О. vignei), архар (О. аттоп) и снежный баран (О. nivicola). Позднее И.Я. Павлинов [2003] все же свел О. musimon к О. orientalis.
Специалистами по капринам (IUCN/SSC Caprinae Specialist Group) недавно предложена новая версия классификации рода Ovis, в которой выделены 5 видов: О. orientalis, О. аттоп, О. canadensis, О. dalli и О. nivicola (табл. 72). В этой системе муфлон и уриал объединены под названием О. orientalis Gmelin, 1774, а общее число принимаемых подвидов сокращено до 33.
Очевидно, что таксономия баранов Старого и Нового Света, несмотря на двухсотлетнюю историю, по-прежнему далека от ясности. Морфологический и цитогенетический подходы к проблеме классификации этой группы животных полностью исчерпаны, так и не дав однозначного результата. В систематике до сих пор
Таблица 72. Классификация рода Ovis, предложенная специалистами по группе каприн (по [Wild sheep and goats and their relatives. 1997]; с изменениями)
Вид Подвид Распространение
Ovis orientalis Gmelin, 1774 areal Eversmann, 1850 bochariensis Nasonov, 1914 cycloceros Hutton, 1842 (= blanfordi) gmelini Blyth, 1841 isphahanica Nasonov, 1910 laristanica Nasonov, 1909 musimon Schreber, 1782 ophion Blyth, 1841 orientalis Gmelin, 1774 punjabiensis Lydekker, 1913 severtzovi Nasonov, 1914 vignei Blyth, 1841 Иран, Киргизстан, Туркменистан, Узбекистан Таджикистан, Туркменистан, Узбекистан Афганистан, Иран?, Пакистан, Туркменистан Армения, Азербайджан, Иран, Ирак?, Турция Иран (С Корсика, Сардиния Кипр Иран Пакистан (Пенджаб) Узбекистан Индия (Джамму и Кашмир), Пакистан
Ovis аттоп Linnaeus, 1758 аттоп Linnaeus, 1758 coIlium Severtzov, 1873 darwini Przewalski, 1883 hodgsonii Blyth, 1840 (= dalai-lamae, adametzi) jubata Peters, 1876 karelini Severtzov, 1873 (= littledalei, sairensis) nigrimontana Severtzov, 1873 polii Blyth, 1841 Китай, Монголия, Россия Казахстан Китай, Монголия Китай, Индия, Непал Китай Китай, Киргизия, Казахстан Казахстан Афганистан, Китай, Пакистан, Таджикистан
Ovis canadensis Shaw, 1804 auduboni Merriam, 1901 californiana Douglas, 1829 canadensis Haw, 1804 cremnobates Elliot, 1903 mexicana Merriam, 1901 nelsoni Merriam, 1897 weemsi Goldman, 1937 США Канада, США сс Мексика, США Северо-Западная Мексика, США США Мексика
Ovis dalli Nelson, 1884 dalli Nelson, 1884 stonei Allen, 1897 Канада, США (Аляска) Канада
Ovis nivicola alleni Matschi, Г907 Eschscholtz, 1829 (= tschuktschorum, koriakorum) borealis Severtzov, 1873 lydekkeri Kowarzik, 1913 nivicola Eschscholtz, 1829 Россия (Амурская область, Хабаровский край, Магаданская область) Россия (Красноярский край) Россия (Якутия) Россия (Камчатка)
господствует все тот же принцип - что ни автор, то “система”. А зоологическая система на сегодняшний день такова, что европейского муфлона, например, авторы обозначают как О. аттоп musimon, О. gmelini musimon и О. orientalis musimon [Beitrage zur Jagd- und Wildforschung. 2003. Bd. 28]. Помимо весьма противоречивой зоологической номенклатуры существует также и “трофейная” классификация баранов, базирующаяся на первой, но со своими специфическими элементами, что вконец запутывает всех, кто пытается разобраться в таксономии рода.
Новые подходы к классификации этой группы копытных связаны с молекулярными методами.
Молекулярно-генетический этап. Паттерны фрагментов геномов разнохромосомных диких баранов и домашней овцы, исследованные методом таксономического фингерпринта ДНК и PCR-RAPD, оказались очень близкими. Более того, найдено значительно большее сходство азиатских пород овец с архаром и уриалом, нежели с муфлоном [Ертаев, Турарбеков, 1987; Мельникова, 1995; Мельникова и др., 1995; Мельникова, Медников, 1996; Ломов и др., 19996], что подтверждает мнение С.Н. Боголюбского [1956] о возможном участии в становлении домашней овцы трех или четырех диких подвидов. Замечу, что задолго до Боголюбского об этом же говорил Н.А. Северцов [1873а], причем его замечания интересны и с таксономической точки зрения:... “Весьма вероятно, что домашние овцы не составляют один вид - О. aries, а происходят от порабощения нескольких видов, принадлежащих к обоим родам, Ovis и Musimon. ...Если считать всех домашних овец одним видом, О. aries, то и все виды обоих родов, Ovis и Musimon, должны считаться породами одного вида, так как у домашних овец встречаются признаки обоих родов” (стр. 156-157). Различия паттернов рестрикционных фрагментов ДНК между архаром и муфлоном, от-
Аргалиобразные
2п=56
Горные бараны
100
100
Муфлонообразные
2п=54, 58
Снежные -------- Азиатские
бараны 100 2п=52
---- Американские
2п=54
---------------------- Внешняя группа (Capricornis)
Рис. 66. Схема филогенетических связей в роде Ovis по результатам анализа последовательностей фрагмента контрольного региона мтДНК 100 особей (по [Кузнецова и др, в печати]) Возле каждого узла даны значения индексов бутстрепа
носимым к разным видам, оказалась такого же порядка, как между породами домашней овцы [Турарбеков и др., 1988]. При анализе ND5 гена митохондриальной ДНК выявлено сравнительно близкое родство архара и уриала [Tuya et al., 2002].
По результатам нашего анализа последовательностей контрольного региона мтДНК, исследованные представители рода Ovis разделяются на две крупные группировки. В одну из них входят отдельными кластерами 52- и 54-хромосом-ные азиатский и американский снежные бараны, имеющие специфические делении, в другую, тоже двумя кластерами, - архар и уриал с муфлоном и домашней овцой (рис. 66).
Молекулярные исследования, как и другие методы, не могут дать однозначного ответа о ранге таксона, и полученные результаты можно трактовать по-разному. На наш взгляд, они, как и морфологические, свидетельствуют в пользу точки зрения о существовании лишь двух близкородственных политипических видов баранов - Ovis аттоп и Ovis canadensis, включающих в качестве подвидов разнохромосомные формы.
АРХАР
Ovis аттоп Linnaeus, 1758
Capra аттоп Linnaeus, 1758. Syst. Nat., 10 ed., 1: 70. Монголия, Монгольский Алтай.
Ovis orientalis Gmelin, 1774. Reise d. Russl., 3: 432. Иран, Мазендеран, зап. часть Эльбурса, Решт, Musimon asiaticus Pallas, 1776. Spicil. Zool., fasc., 2: 8. Казахстан, верхи, течение p. Иртыш.
Ovis argali Boddaert, 1785. Elench. Animal.: 147. Сев. Азия.
Aegoceros musimon Pallas, 1811. Zoogr. Rosso-Asiat., 1: 230. Вост, побережье Каспийского моря.
Ovis polii Blyth, 1841. Proc. Zool. Soc. London (1840): 62. Памир, окрести, оз. Серы-Куль (Зор-куль).
Ovis hodgsonii Blyth, 1841. Ibid.: 65. Непал.
Ovis gmelini Blyth, 1841. Ibid.: 69. Турция, Эрзурум.
Ovis vignei Blyth, 1841. Ibid.: 70. Индия, Джамму и Кашмир, Астор.
Ovis ophibn Blyth, 1841. Ibid.: 73. Кипр.
Ovis ammonoides Hodgson, 1841. J. Asiat. Soc. Beng., 10: 230. Тибет.
Ovis cycloceros Hutton, 1842. Calcutta J. Nat. Hist., 2: 514. Афганистан, Кандагар, горы Хазара.
Ovis arkal Eversmann, 1850. Естеств. история Оренбург, края, 2: 276. Узбекистан, Сев. Устюрт. Ovis arcar Brandt, 1852. Beitr. z. Kenntn. d. Russ. Reiches, 17: 310. Замена arkal Eversm.
Ovis nigrimontana Severtzov, 1873. Изв. Имп. о-ва любит, естеств., антропол., этногр., 8, 2: 84, 87,
154. Хр. Каратау.
Ovis karelini Severtzov, 1873. Ibid.: 84, 86, 150, 154. Тянь-Шаньская горная система.
Ovis heinsii Severtzov, 1873. Ibid.: 84, 87, 150, 154. Зап. Тянь-Шань и верховья Аму.
Ovis argali daurica Severtzov, 1873. Ibid.: 151, табл. IV. Забайкалье.
Ovis argali mongolica Severtzov, 1873. Ibid.: 154. Вост. Монголия и Забайкалье.
Ovis argali altaica Severtzov, 1873. Ibid.: 154. Алтай, преимущественно степные хребты.
Ovis coIlium Severtzov, 1873. Ibid.: 154. Степные низкие хребты к сев. от оз. Балхаш.
Ovis brookei Ward, 1874. Proc. Zool. Soc. London: 14. Ладак, окрести. Леха.
Ovis jubata Peters, 1876. Monatsberichte Akad. Berlin: 177. Горы к сев. от Пекина.
Ovis darwini Przevalski, 1883. Третье путешествие в Центр. Азию: 463. Юго-Вост. Гоби, горы Хара-Нарын-Ула.
Ovis dalai-lamae Przevalski, 1888. Четвертое путешествие в Центр. Азию: 274. Московский хребет. Ovis аттоп typica Lydekker, 1898. Wild oxen, sheep and goats of all lands: 177. Алтай.
Ovis sairensis Lydekker, 1898. Ibid.: 185. Cayp.
Ovis polii typica Lydekker, 1898. Ibid.: 192. Памир.
Ovis sairensis littledalei Lydekker, 1902. Proc. Zool. Soc. London: 2, 83. Вост. Тянь-Шань, p. Или к ю.-в. от Кульджи.
Ovis vignei varenzovi Satunin, 1905. Зап. Кавк. отд. Ими. Рус. геогр. о-ва, 25, 3: 11. Копетдаг.
Ovis аттоп humei Lydekker. Cat. Hume Bequest.: 6. Тянь-Шань к сев. от Кашгара.
Ovis аттоп kozlovi Nasonov, 1913. Изв. Акад, наук: 621. Южн. Гоби, горы Ябарай.
Ovis polii karelini melanopiga Nasonov, 1914. Ibid.: 699. Заилийский Алатау, Суук-Тюбе. Непригодно (инфраподвидовое название).
Ovis severtzovi Nasonov, 1914. Ibid.: 761. Нуратау.
Ovis vignei bochariensis Nasonov, 1914. Ibid.: ИЗО. Таджикистан, верховья p. Кызылсу, Бальджу-ан.
Ovis ophion armeniana Nasonov, 1921. Изв. Росс. Акад, наук: 1215. Турция, окрести. Баязета к югу от Арарата.
Ovispolii coIlium albula Nasonov, 1923. Геогр. распростр. горн, баранов Старого Света: 82. Казахстан, “Акмолинск”. Непригодно (инфраподвидовое название).
Ovispolii coIlium obscura Nasonov, 1923. Ibid.: 82. Казахстан, “горы Аркат” (к с.-в. от хр. Чингиз-тау). Непригодно (инфраподвидовое название).
Ovis аттоп przewalskii Nasonov, 1923. Ibid.: 118. Алтай, Сайлюгем.
Ovis polii nasonovi Laptev, 1929. Определитель млекопит. Ср. Азии: 76. Таласский Алатау.
Характеристика. См. характеристику рода.
Дополнительные отличительные черты следующие. Череп сравнительно вытянутый, с короткой мозговой коробкой (рис. 67). Межчелюстные кости крупные, всегда достигают носовых и соприкасаются с ними или внедряются между ними и верхними челюстными костями. Носовые кости длинные, носовое отверстие широкое. Очень часто встречаются вставочные косточки между межчелюстными, носовыми и верхнечелюстными костями. Углубления на слезной кости выражены отчетливо. Длина роговых стержней у самцов, в отличие от снежного барана, больше длины окружности их основания. В нижней челюсти с каждой стороны по три предкоренных зуба.
Рога массивные, изогнутые, в крайнем выражении дважды спирально закрученные. Передненаружное ребро на роговых чехлах хорошо выражено, передняя грань плоская. Первый годовой сегмент сохраняется всю жизнь. Самки с серповидно изогнутыми морщинистыми рогами длиной около 23-56 см или без них. Цвет рогов у самцов варьирует от светло-желтого до грязно-бурого и темно-серого, у самок темно-серый.
Околохвостовое зеркало не простирается выше хвоста. Предглазничные и паховые специфические железы крупные, межпальцевые имеются у обоих полов на всех конечностях. У взрослых самок увеличены апокриновые железы в коже половых губ, у самцов - сальные железы в циркуманальной области [Соколов, Чернова, 1988, 2001]. У некоторых форм на затылке имеются зароговые “мозоли”, функциональное значение которых неясно [Спивакова и др., 1999].
Горно-степные стадные животные с широкой экологической нишей. Способны к длительному бегу с высокой скоростью (до 65 км/ч). В природе живут до 15 лет.
Преимущественно охотничий вид. Некоторые его формы, находящиеся на грани уничтожения, охраняются.
Распространение. Острова Средиземного моря, горы Передней, Средней и Центральной Азии на север до Закавказья, Казахского нагорья и Южной Сибири, на юг до Гималаев, на восток до Большого Хингана включительно (рис. 65). Одна из
Рис. 67. Череп и рог самца архара (экземпляр с Горного Алтая; Зоомузей ЗИН РАН, коллекционный № 26638) Вид сбоку, сверху и снизу
форм (муфлон) расселена во многих странах Западной, Центральной и Южной Европы, США, Аргентине, Новой Зеландии, на Гавайских островах в Тихом океане и о. Кергелен в Индийском океане.
Состав. Насчитывается более 40 подвидовых имен. Действительное число подвидов гораздо меньше и по-прежнему дискуссионно (см. раздел “Таксономия”). В фауне России - номинативный подвид архара (О. аттоп аттоп L., 1758), представленный алтайским бараном. Соответственно ниже рассматривается преимущественно группа собственно архаров, или аргалиобразных баранов.
Изменчивость и таксономия
Размеры и масса. Масса новорожденных у аргалиобразных баранов варьирует от 2,6 до 6,3 кг. Архарята в среднем несколько крупнее и на первом году жизни растут почти в два раза быстрее, чем детеныши уриала и муфлона. К 1,5-2 месяцам жизни они весят 10-24 кг, к 6 месяцам - 25-40, в 1,5 года - 35-72 кг. В пустынных низкогорных популяциях детеныши, однако, растут и развиваются медленнее, чем в высокогорных, что, скорее всего, связано с недостатком молока у самок в
Таблица 73. Размеры (см) взрослых аргалиобразных баранов
Самцы Самки Район (источник информации)*
n M | lim n L М lim
Длина тела
4 176 172-180 3 170,5 167-174 Южная Сибирь (3)
30 179 165-201 5 155 147-165 Казахское нагорье (6)
58 167 144-202 — — - (10)
3 - 172-190 2 166,5 166-167 Таласский Алатау (9)
1 — -190 — — -164 Тянь-Шань (4)
1 - -180 - — - Памир (4)
3 164 160-168 2 148 — -“- (7)
1 - 210 - - - Западный Тибет (12)
2 - 152-167 1 — 148 Гоби, Тибет (12)
- - -170 — — - Внутренняя Монголия (5)
Высота в холке
4 117 115-120 3 108 100-114 Южная Сибирь (3)
- - -105 — — — Южная Монголия (2)
— - 110-115 - — Внутренняя Монголия (5)
30 120 109-135 5 92 83-104 Казахское нагорье (6)
58 112 103-126 — — -“- (10)
3 - 103-115 2 102,5 100-105 Таласский Алатау (9)
— - -130 - - - Киргизия (8)
4 - 115-118 — — 95-105 Памир (1)
1 — -120 (4)
3 104 102-106 2 95 - -“- (7)
3 118 115-120 — — 99-112 Гоби, Тибет (4, 12)
1 - 91 — — — Каратау (11)
Обхват туловища
4 135 126-140 3 114 110-118 Южная Сибирь (3)
30 137 127-152 5 107 102-114 Казахское нагорье (6)
3 112 108-116 2 107 — Памир (7)
3 140 135-144 1 — 109 Гоби, Тибет (12)
I 113 - — Каратау (11)
Длина ступни
4 46 45-48 3 — 45 Южная Сибирь (3)
3 44 44-45 2 39 — Памир (7)
2 - 43-50 — — — Гоби, Тибет (12)
Длинауха
4 13 12-14 3 11 10-12 Южная Сибирь (3)
30 12,3 10-14 6 11,5 10-12 Казахское нагорье (6)
3 10,4 10-11 2 10,4 — Памир (7)
2 — 10-15 — — — Гоби, Тибет (12)
Длина хвоста
4 11 10-12 3 10,7 10-12 Южная Сибирь (3)
25 14,7 10-18 6 10,7 10-12 Казахское нагорье (6)
3 10,3 9-11 2 10,5 — Памир (7)
2 - 10-18 1 - 8 Гоби, Тибет (12)
*1 - Мсклснбурцев, 1948; 2 - Соколов и др., 1996; 3 - Сопин, 19756, 1976в; 4 - Федосенко, 2000; 5 -Сопин, 1982а; 6 - Бербер, 1999; 7 - Сапожников, 19^66; 8 - Хохлов, 2002; 9 - Ковшарь, Янушко, 1965; 10 -Байдавлетов, 1997; 11 - Шарипов, 1989; 12 - Bunch et al., 2000.
засушливых районах и с более коротким сроком лактации. Темпы роста самок замедляются со второго года жизни, что вызвано их участием в размножении, и сравнительно слабо увеличивается позднее. Самцы муфлона и уриала достигают типичных размеров взрослых особей к 4 годам, аргали интенсивно растут до 7-9 лет (рис. 68 и 69) [Сапожников, 1971, 19766, 1977; Сопин, 19776; Федосенко, 1983а, 2000; Антонян, Эгнатосян, 1984; Минасян и др., 1986а,б; Шакула, 1989в; Бербер, 1999].
Географическая, индивидуальная и половая изменчивость линейных размеров и массы горных баранов очень значительны. Наиболее крупные из них - аргалиоб-разные животные. Максимальная длина тела взрослых самцов варьирует в пределах 152-210 см, самок - 147-174 см, высота в холке, соответственно, 91-135 и 83-114 см. Масса самцов почти в два раза больше, чем у самок: у первых 75-180, у вторых - 47-100 кг (табл. 73 и 74). Отдельные особи, вероятно, крупнее 200 кг. В Хорогском краеведческом музее выставлено чучело 7-8-летнего самца, добытого в ноябре 1960 г. на Памире в окрестностях оз. Кара-Куль, с отметкой массы в 216 кг (Сапожников, 1971, 19766). После зимовки вес животных обычно на 20-40% меньше, чем осенью. Длина тела взрослых самцов муфлона колеблется в пределах 110— 144 см, высота в холке - 60-89 см, масса - 30-65 (до 95) кг. Соответствующие показатели уриала: 120-160 см, 77-98 см и 39-96 кг (табл. 71).
По анализируемым экстерьерным признакам при имеющихся скудных сведениях (табл. 73 и 74) обозначить какой-либо подвид среди аргалиобразных баранов нереально.
Таблица 74. Масса (кг) взрослых аргалиобразных баранов
Самцы Самки Район (источник информации)*
п 1 м 1 lim п 1 м 1 lim
4 134 101-160 3 90 80-100 Южная Сибирь (3)
— - 100-160 - — — Казахстан (4)
25 138 118-165 5 65 58-80 Казахское нагорье (5)
58 148 113-180 - - - -“-(8)
3 - 60-104 2 52 50-54 Таласский Алатау (7)
4 113 97-152 13 54 47-66 Тянь-Шань (2)
- - -180 - — - Киргизия (6)
3 108 83-130 1 — 59 -“-(9)
- - 101-160 — - 56-70 Таджикистан, Памир (1)
1 — 124 — — — -“-(2)
— - 98-105 - - -68 Тибет (2)
3 113 98-146 — - - Гоби, Тибет (12)
- - 110-125 - - - Нуратау (10)
1 — 75 — — — Каратау (11)
* 1 - Сапожников, 1971,19766, 1977; 2 - Федосенко, 1983а, 2000; 3 - Сопин, 19756, 1976в; 4 - Антипин, 1947; 5 - Бербер, 1999; 6 - Хохлов, 2002; 7 т- Ковшарь, Янушко, 1965; 8 - Байдавлстов, 1997; 9 - Румянцев и др., 1935; Денисов, 1935; 10-Рахимов, 1966; 11 - Шарипов, 1989; 12-Bunch et al., 2000.
Краниометрическая характеристика. Основные пропорции черепа архара устанавливаются к 2-летнему возрасту, но рост костей и формирование зубов продолжается до 5 лет, после чего их прирост незначительный. Стержни рогов растут до конца
Н—I—I—I—I—I—I—I—I—I—I—I
1 2 3 4 5 6 7 8 9 10 11 12
12 34567 89 10 11 12
Возраст, лет
Рис. 68. Возрастная изменчивость размеров тела самцов (А) и самок (Б) североказахстанского барана (по [Бербер, 1999]) '
I - длина тела, II - высота в холке, III - обхват груди, IV - длина передней ноги, V -косая длина туловища, VI - длина хвоста, VII - длина уха. Штриховкой обозначена область варьирования значений
жизни, максимально увеличиваясь на втором году. Чем сильнее развиты костные стержни и роговые чехлы, тем больше разрастаются лобные кости и поперечный гребень на затылочной кости. У старых особей обычна облитерация швов между костями черепа [Цалкин, 1951а; Сопин, 19756; Сапожников, 19766, 1977].
Половой диморфизм выражен резко. Географическая изменчивость краниометрических признаков горных баранов явно клинальна (табл. 71 и 75). Ее размах тоже значителен
180
0 т—।—।—।—।—।—।—।—।—।—।—।—।—।—।—।
0 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15
Возраст, лет
Рис. 69. Возрастная изменчивость массы тела самцов (А) и самок (Б) североказахстанского барана (по [Бербер, 1999]) Точкой отмечены средние значения, прямоугольником - предельные значения для особей
при еще большем уровне индивидуальной изменчивости, на что В.И. Цалкин [1951а] обратил особое внимание. Отдельно взятые черепа взрослых особей практически
невозможно достоверно отнести к той или иной форме. Среди выделяемых рас собственно архара максимальные размеры черепа у алтайского аргали (наибольшая длина - 325-395 мм), минимальные - у особей из Южной Гоби (305-337), с хребта Каратау (285-315) и, особенно, из Наратау и Актау (260-292 мм). Последние четко выделяются и по многим другим признакам, включая длину зубного ряда. По средним значениям репрезентативных выборок реально различимы лишь крупные черепа аргали и мелкие уриала и муфлона (табл. 75).
Таблица 75. Размеры черепа (мм) взрослых аргалиобразных баранов и граничащих форм уриала и муфлона
Самцы Самки Район, форма (источник информации)*
п М lim п м lim
1 2 3 4 5 6 7
Наибольшая длина
25 363,0 345-385 10 318,0 305-337 Южная Сибирь (10)
14 361,7 345-385 5 322,6 305-337 Алтай, С.-З. Монголия (1)
26 368,0 350-395 — — — Монгольский Алтай (4)
27 339,0 325-357 - — — Гобийский Алтай (4)
3 315,7 305-337 1 __ 285 Южная и Ю.-В. Гоби (1)
2 305,0 305-305 2 281,5 278-285 Южная Гоби (4)
24 345,7 335-361 12 318,4 297-330 Казахстан (2)
6 319,2 308-335 5 290,6 286-297 -“-(1)
58 328,0 302-344 - - — -“-(7)
3 - 309-318 2 - 245-290 Таласский Алатау (6)
19 324,7 303-338 1 — 275 Терскей и Заилийский Алатау (1)
17 327,0 315-338 - — - Тянь-Шань (4)
Таблица 75 (продолжение)
Таблица 75 (продолжение)
1 2 1 3 4 1 5 6 7 1 1 2 1 з 4 5 6 1 7
13 342,6 330-355 3 295,3 283-305 Памир (1) 28 180,0 166-196 - — — -“-(4)
29 338,0 320-355 - — - 24 182,1 172-193 и 124,9 117-135 -“-(11)
10 338,3 328-351 9 298,7 292-307 -“-(И) 1 - 182 1 - 137 Тибет (1)
— — 305-355 — — 288-305 -“-(14) 6 162,3 157-168 2 115,5 114-117 Каратау (1)
1 — 354 1 - 302 Тибет (1) 2 141,5 140-143 - - - Нуратау, Актау (1)
6 300,2 285-315 2 266,0 258-274 Каратау (1,4) 9 146,0 138-157 -“-(3)
2 265,0 260-270 - - - Нуратау, Актау (1) 31 147,0 136-156 - - - Форма cycloceros (3)
9 269,0 260-279 - — — -“-(3) 14 136,0 124-144 2 102,5 93-112 vignei (11)
— 283,0 270-292 - — — -“-(12) 11 134,0 124-143 — - — -“- bochariensis (3)
15 267,4 255-283 10 240,8 230-249 Форма area! (8) 46 128,7 117-139 •“ gmelini (5)
31 273,0 257-297 — - — -“- cycloceros (3) Межглазничная ширина
4 248,0 242-254 2 228,5 217-240 -“- vignei (11) 25 140,0 132-162 10 124,0 121-130 Южная Сибирь (10)
11 247,0 233-255 - - - -“- bochariensis (3) 14 146,8 138-155 5 123,2 112-128 Алтай, С.-З. Монголия (1)
— 253,0 233-272 — 226,0 217-240 -“-(Н) 26 152,0 135-184 — — Монгольский Алтай (4)
46 244,4 228-264 - 229,0 — -“- gmelini (5) 27 143,0 132-155 — — — Гобийский Алтай (4)
40 242,0 225-264 - 226,5 213-238 -“-(13) 3 125,3 116-136 1 - 115 Южная и Ю.-В. Гоби (1)
Максимальная ширина 2 120,0 116-124 2 109,0 103-115 Южная Гоби (4)
25 185,0 170-208 10 172,0 164-180 Южная Сибирь (10) 40 159,7 132-183 16 126,6 120-130 Казахстан (2)
14 195,7 190-203 5 171,2 164-180 Алтай, С.-З. Монголия (1) 6 133,5 130-136 5 112,8 109-116 -“-(1)
26 196,0 175-218 — - — Монгольский Алтай (4) 18 130,0 120-150 — — Тянь-Шань (4)
27 177,0 171-190 — - — Гобийский Алтай (4) 19 129,1 114-150 1 — 111 Терскей и Заилийский Алатау (1)
3 174,7 166-178 1 — 161 Южная и Ю.-В. Гоби (1) 13 135,9 127-147 3 112,3 105-112 Памир (1)
2 168,5 166-171 2 155,5 150-161 Южная Гоби (4) 29 131,0 113-147 - — — -“-(4)
22 181,7 165-190 15 160,9 150-170 Казахстан (2) 23 130,8 124-142 11 111,2 105-118 -“-(И)
6 183,5 178-188 4 160,8 152-164 -“-(1) 1 143 1 — 120 Тибет (1)
18 176,0 160-191 — — — Тянь-Шань (4) 6 117,8 114-122 2 98,5 98-99 Каратау (1)
19 175,5 160-191 1 - 146 Терскей и Заилийский Алатау (1) 5 101,6 98-110 - - - Нуратау, Актау (1)
13 184,5 178-198 3 157,7 152-163 Памир (1) 9 103,0 94-114 — — — -“-(3)
29 181,0 166-198 — — — -“-(4) 15 101,4 97-105 10 84,6 80-88 Форма areal (8)
23 181,3 171-193 и 156,5 150-162 -“-(И) 18 111,8 95-130 10 95,4 90-100 -“-(9)
1 — 194 1 - 165 Тибет (1) 31 103,0 93-115 - - - -“- cycloceros (3)
6 161,7 158-164 2 144,5 142-147 Каратау (1) 13 94,5 85-101 2 83,0 78-88 -“- vignei (11)
5 141,4 138-147 - — - Нуратау, Актау (1) И 93,0 81-100 - - — bochariensis (3)
9 142,0 138-148 -“-(3) 46 90,1 81-102 — - — -“- gmelini (5)
15 141,9 136-146 10 124,8 122-132 Форма areal (8) Длина лицевой части
31 145,0 137-155 — — - -“- cycloceros (3) 14 282,1 273-304 5 259,2 241-267 Алтай, С.-З. Монголия (1)
14 130,6 120-140 2 123,0 115-131 -“- vignei (11) 26 283,0 260-305 — — Монгольский Алтай (4)
11 130,0 118-140 — — — -“- bochariensis (3) 27 257,0 247-265 — - — Гобийский Алтай (4)
46 131,2 117-154 - — -“- gmelini (5) 3 251,0 239-269 1 — 225 Южная и Ю.-В. Гоби (1)
Высота** 2 242,0 239-245 2 223,0 221-255 Южная Гоби (4)
25 194,0 181-220 10 136,0 126-146 Южная Сибирь (10) 16 256,2 241-265 8 237,5 232-242 Казахстан (2)
14 185,5 171-200 5 129,2 125-135 Алтай, С.-З. Монголия (1) 6 246,5 235-261 5 233,2 230-237 -“-(1)
26 215,0 193-244 — - - Монгольский Алтай (4) ' 17 260,0 245-287 — — — Тянь-Шань (4)
27 191,0 182-200 - — - Гобийский Алтай (4) 18 258,7 230-287 1 — 226 Терскей и Заилийский Алатау (1)
3 157,0 144-173 1 — 118 Южная и Ю.-В. Гоби (1) 11 266,2 257-284 3 237,0 228-244 Памир (1)
2 149,0 144-154 2 117,0 116-118 Южная Гоби (4) 25 264,0 244-284 — — — -“-(4)
22 180,7 170-205 12 134,5 132-137 Казахстан (2) 11 260,4 248-272 9 241,0 237-246 -“-(П)
5 171,0 168-175 4 123,2 121-129 -“-(1) 1 - 283 1 — 240 Тибет (1)
17 176,0 165-186 — — — Тянь-Шань (4) 6 228,2 216-239 2 211,5 206-217 Каратау (1)
18 175,2 153-186 — — - Терскей и Заилийский Алатау (1) 4 198,5 192-208 - - — Нуратау, Актау (1)
12 159,9 120-187 3 125,7 119-129 Памир (1) 9 203,0 192-212 - - - -“-(3)
1 2 3 1 4 5 1 6 7
31 209,0 197-224 — - - Форма cycloceros (3)
3 185,3 180-190 2 182,0 171-193 -“- vignei (11)
11 215,0 202-225 — — — -“- bochariensis (3)
46 185,2 174-197 - - - -“- gmelini (5)
Длина носовых костей
25 138,0 124-158 10 118,0 108-125 Южная Сибирь (10)
14 141,3 127-158 5 118,6 108-125 Алтай, С.-З. Монголия (1)
26 147,0 125-170 — — — Монгольский Алтай (4)
27 127,0 112-146 - — — Гобийский Алтай (4)
3 109,3 98-122 1 - 77 Южная и Ю.-В. Гоби (1)
2 103,0 98-108 2 82,0 77-78 Южная Гоби (4)
26 122,7 111-140 12 108,3 100-117 Казахстан (2)
6 120,3 109-138 5 107,6 97-117 -“-(1)
19 124,1 108-140 1 - 103 Терскей и Заилийский Алатау (1)
11 133,0 115-152 3 111,0 103-112 Памир (1)
16 135,0 113-154 — — — -“-(4)
9 137,7 114-154 9 123,6 119-128 -“-(П)
1 — 136 — — — Тибет (1)
6 116,5 102-129 2 105,5 103-108 Каратау (1)
5 83,4 70-93 — — — Нуратау, Актау (1)
9 84,0 70-93 — — — -“-(3)
15 102,2 98-108 10 90,0 84-96 Форма areal (8)
15 103,5 90-115 4 90,0 90-90 -“-(9)
31 104,0 93-119 — - — -“- cycloceros (3)
7 94,4 90-100 2 75,0 74-76 -“- vignei (11)
11 93,0 86-103 • — — — -“- bochariensis (3)
46 88,5 71-100 - - - -“- gmelini (5)
Ширина носовых костей
14 53,3 47-58 5 41,0 34-49 Алтай, С.-З. Монголия (1)
3 38,0 35-40 1 — 35 Южная и Ю.-В. Гоби (1)
22 58,4 50-70 16 43,3 38-48 Казахстан (2)
6 51,2 47-54 5 38,2 36-42 -“-(1)
19 46,9 42-57 1 - 36 Терскей и Заилийский Алатау (1)
И 53,1 42-63 3 39,7 36-43 Памир (1)
20 54,6 49-60 10 42,4 40-46 -“-(И)
1 - 49 — — Тибет (1)
6 42,8 39-47 2 29,5 28-31 Каратау (1)
5 33,0 30-37 — - — Нуратау, Актау (1)
12 34,5 28-40 2 29,0 26-32 Форма vignei (11)
Длина верхнего ряда зубов
25 97,0 91-104 10 92,0 6-97 Южная Сибирь (10)
12 96,8 91-101 5 91,6 86-96 Алтай, С.-З. Монголия (1)
26 98,0 82-112? — - - Монгольский Алтай (4)
27 91,0 80-99 — - — Гобийский Алтай (4)
3 94,3 93-96 1 — 79 Южная и Ю.-В. Гоби (1)
2 95,0 94-96 2 84,5 79-90 Южная Гоби (4)
62 87,9 80-95 22 84,2 77-92 Казахстан (2)
6 88,0 78-91 5 82,2 75-92 -“-(1)
19 91,8 83-102 1 - 83 Терскей и Заилийский Алатау (1)
11 91,7 87-95 3 89,3 87-92 Памир (1)
Таблица 75 (окончание)
1 1 2 3 1 4 1 5 6 7
27 91,0 87-97 - - -“-(4)
21 89,8 79-98 11 86,1 84-91 -“-(11)
1 — 97 1 — 88 Тибет (1)
5 85,0 83-89 2 83,5 82-85 Каратау (1)
5 73,0 69-76 - - - Нуратау, Актау (1)
9 76,0 69-88 — — — -“-(3)
31 77,0 71-81 - - - Форма cycloceros (3)
12 72,3 68-77 2 68,0 68-68 vignei (11)
11 73,0 69-80 — — — bochariensis (3)
46 72,7 - - gmelini (5)
Длина костного стержня рогов
25 385,0 320-460 10 137,0 120-170 Южная Сибирь (10)
9 393,3 360-440 5 141,0 120-170 Алтай, С.-З. Монголия (1)
3 318,3 260-355 1 - 120 Южная и Ю.-В. Гоби (1)
4 367,5 350-420 4 111,2 100-125 Казахстан (1)
8 374,4 300-395 - - - Терскей и Заилийский Алатау (1)
7 428,6 325-515 - — — Памир (1)
1 — 360 — — — Тибет (1)
4 337;5 315-370 2 85,0 80-90 Каратау (1)
3 300,0 260-350 — — — Нуратау, Актау (1)
46 342,0 200-450 - — — Форма gmelini (5)
18 339,0 200-435 - 50,0 - -“-(13)
Обхват костного стержня рога у основания
25 334,0 280-395 10 142 130-160 Южная Сибирь (10)
9 340,6 300-370 5 143 130-160 Алтай, С.-З. Монголия (1)
9 340,4 310-374 — — - -“-(15)
3 253,3 210-310 1 — 130 Южная и Ю.-В. Гоби (1)
4 288,8 270-320 4 120 110-130 Казахстан (1)
10 278,5 240-320 - — — Терскей и Заилийский Алатау (1)
9 312,8 275-345 - — - Памир (1)
1 — 325 — — — Тибет (1)
4 237,5 225-250 2 100 90-110 Каратау (1)
3 190,0 185-195 - — - Нуратау, Актау (1)
19 187,0 125-205 — 68,8 - Форма gmelini (13)
* 1 - Цалкин, 1951а (расчет по “Приложению”); 2- Бербер, 1999; 3 - Сопин, 1984а,б; 4 - Сопин, 19826; 5-Явруян, 1969 (по: Соколов, Темботов [1993]), 1970; 6-Ковшарь, Янушко, 1965; 7 - Байдавлетов, 1997; 8 -Горбунов, 19956; 9 - Слудский и др., 1983; 10-Сопин, 19756; 11 - Сапожников, 19766 (расчет по “Приложению”); 12-Рахимов, 1966; 13-Манассрова, 1988; 14-Сопин, 2003; 15-Боескоров, 2001. Разные авторы нередко используют одни и те же экземпляры черепов из зоомузеев Зоологического института РАН и МГУ.
** Методика измерений данного признака не одинаковая у разных авторов, что приводит к значительной вариабельности значений.
Л.В. Сопин [19826, 1984а], проанализировав репрезентативные выборки, пришел к выводу, что краниометрические различия между памирскимpolii, тянь-шаньским karelini и казахстанским coIlium значительно меньше принятого подвидового уровня CD по Майру [1971], а отличия этой группы от алтайского аттоп достигают его. Популяции с Русского и Монгольского Алтая, несмотря на некоторые различия, близки, но существенно отличаются от группировок из Гобийского Алтая (форма darwini) и, особенно, Южной Гоби (форма kozlovi) [Сопин, Сухбат, 1980,
1983]. Материал из Западного и Восточного Тибета и Гималаев (формы dalai-lamae, jubata, hodgsonii) крайне скуден, и не позволяет сделать однозначные заключения. Каратауский nigrimontana имеет переходные черты между казахстанским архаром и уриалом (табл. 75) и вполне может иметь гибридное происхождение. Кызылкумский баран, или баран Северцова (форма severtzovi) с гор Нуратау и Актау, считавшийся ранее уриалом или промежуточной формой между уриалом и архаром [Цалкин, 1951а; Гептнер и др., 1961; Сапожников, 19766], по краниометрическим показателям стоит явно ближе к граничащим формам туркменского (cycloceros, arkal, arcar) и бухарского (bochariensis) уриала, нежели к архару (табл. 75), хотя и отличается от первых по числу хромосом. К уриалу очень близок и закавказский (арме-нийский) муфлон [Явруян, 1970].
При детальной ревизии аргалиобразных баранов на основе признаков 368 черепов с использованием современных методов статистики выявлены 3 краниологических типа: severtzovi, который отличается от других форм сильно сближенными теменными гребнями и степенью наклона зароговой части черепа относительно плоскости затылка, что сближает его с уриалом и муфлоном; собственно аттоп, включающий монгольские расы аттоп и darwini, имеющие широко расставленные теменные гребни; и промежуточный тип, состоящий из форм nigrimontana, polii, karelini и coIlium. Формаpolii все же выделяется из них по признакам рогов [Капитанова, 2004]. Д.В. Капитанова фактически подтвердила вывод Л.В. Сопина [19826, 1984а] о краниологической близости киргизско-казахстанских баранов (еще ранее к такому заключению пришли Н.В. Насонов [1923] и В.Г. Гептнер [1961]) и отметила уникальность и морфологическую примитивность барана Северцова, несущего отдельные “капроидные” черты. Она заключила, что формы аттоп и darwini, также как казахстанские и тянь-шаньские coIlium и karelini, не достигают уровня подвидов, и поэтому первых можно свести к аттоп, а вторых к karelini. Близкое родство аттоп и darwini, как показано ниже, подтверждено и на молекулярном уровне, чего нельзя сказать о coIlium и karelini, поскольку последний объединяется с формой polii.
Изменчивость рогов. Рога у самок архара значительно меньше, чем у самцов (табл. 76), у которых они растут всю жизнь, исключая зимние периоды. Величина прироста роговых чехлов существенно колеблется в зависимости от условий питания [Ишунин, 1968] и возраста животных. В первые 5-6 лет ежегодный прирост рогов у самцов достигает 15-20 см, и максимальным , бывает чаще на втором и третьем году жизни (до 60 и 45 см у отдельных особей). Начиная с четвертого года каждый последующий сегмент прогрессивно укорачивается, и у старых зверей не превышает 1-3 см. Толщина рогов также постоянно увеличивается с возрастом. У старых особей их окружность в основании иногда превышает 40 см. У 1,5-2-летних самцов рога по форме и размерам почти такие же, как у взрослых самок. У 2,5-3-летних особей они достигают четверти круга, в 3—5 лет — половины, в 5-7 лет - три четверти, а у самцов старше 8 лет составляют полный круг. У старых баранов концы рогов обычно стерты, поскольку лежащие животные упираются ими в землю [Горелов, 1986], или обломаны, а поверхность бывает повреждена во время турниров и при случайных ударах [Цалкин, 1951а; Сопин, 19746; Жиряков, Федосенко, 1983; Федосенко, 1983а; Бербер, 1999].
Географическая изменчивость рогов горных баранов весьма велика (табл. 76), но более характерны три основных типа (рис. 70) [Цалкин, 1951а; Гептнер и др., 1961]:
Таблица 76. Размеры рогов (см) взрослых аргалиобразных баранов
п Длина Размах Обхват у основания Район (источник информации)*
М lim М lim м lim
1 2 3 4 5 6 7 8
25 110 82-158 Самцы** 84 66-95 43 34-50 Южная Сибирь (7,14)
6 133 123-146 80 75-87 51 45-55 Алтай (3, 6)
10-21 116 95-168 74 63-87 46 42-55 -“-(2)
13 108 82-125 — — 45 40-50 С.-З. Монголия (12)
3 115 100-131 - -87 44 41-46 Монгольский Алтай (2)
- 136 -176 - 48 -53 Монголия (18)
— -87 - — - -38 Южная Монголия (17)
1 — 112 - - - 42 Нань-Шань (12)
2 74 61-88 — — 28 26-30 Южная Гоби (1)
174-91 116 82-150 76 50-105 38 32-44 Казахстан (10)
6 106 97-115 - - 37 35-38 -“-(12)
58 117 85-140 71 49-103 39 35-44 -“-(13)
17 103 94-138 - - - - Таласский Алатау (2)
18 103 74-127 — - 35 30-40 Терскей и Заилийский
-180 — -39 Алатау (12) Тянь-Шань (11)
58-40 120 100-154 95 69-119 36 32-47 Аксай (2)
12 125 100-148 - -109 — Джунгарский Алатау (2)
10 104 69-134 — — — -38 Калканы (2, 19)
2 95 - - - 28 - Алайский хребет (4)
39-35 129 107-156 104 73-124 38 35-43 Памир (2)
29-27 126 89-190 105 70-130 37 35-41 -“-(1)
1 4 - 190 138-142 39 -43 -“-(11) -“- (5)
11 130 105-150 — - 38 35-40 -“-(12)
- 139-165 - - — — -“-(23)***
22 118 106-142 — - 38 32-40 , Киргизия (14)
— — 89-145 — — — 43 Тибет (2)
— — 120-143 — — 42-49 Гималаи (12)
- — . 117-130 — — — 47-50 С.-В. Китай (9)
11 107 95-117 — - - Ю.-В. Каратау (2)
5 91 77-102 — — 31 28-33 Каратау (12)
— — -120 — — 31 28-34 -“- (22)
2 82 82-83 49 43-55 24 24-26 Нуратау (8)
5 64 55-73 — — 24 23-26 -“-(12)
14 75 51-98 — — - -29 -“-(15)
— 98,5 (Ю5) 80-107 — — 29 28-30,5 -“-(16)
- - 86-95 — 48-52 — 23-28 -“-(1)
10 47 40-56 Самки** 41 37-47 18 16-19 Южная Сибирь (7)
1 — 39,5 — 43,5 — 18,5 Алтай (6)
8 45 39-51 42 39-46 17 17-18 -“-(2)
5 48 42-53 — 17 16-19 С.-З. Монголия (12)
2 38 32-44 — — 14,5 14-15 Южная Гоби (12)
1 - 44 - - - 15 -“- (1)
Таблица 76 (окончание)
1 2 3 4 5 6 7 8
23 33 28-40 40 37-44 14 12-14 Казахстан (10)
4 31 29-33 - - 14 12-15 -“-(12)
17 30 26-34 — - - — Тянь-Шань (2)
1 - 28 - — — 14 -“-(12)
7 33 30-37 - - - - Джунгарский Алатау (20)
7 30 26-34 -34 — 12-13 Калканы (19, 20)
2 31 30-32 __ 14,8 14-15 Памир (12)
9 35 32-39 36 30-48 14,4 14-16 -“- (1)
5-1 33 29-38 - - - 13,5 -“-, Киргизия (14)
— — 36—46 — — — -19 Тибет (2)
2 24 23-25 - - 12 11-13 Каратау (12)
* 1 - Сапожников, 19766, 1977; 2 - Федосенко, 1983а, 2000; 3 - Собанский, 1992; 4 - Мскленбурцсв, 1962; 5 - Мекленбурцсв, 1948; 6 - Филус, 1992; 7 - Сопин, 19756; 8 - Bunch et al., 1998; 9 - Сопин, 19826; 10 - Бербер, 1999; 11 - Хохлов, 2002; 12 - Цалкин, 1951а (расчет по “Приложению”); 13 - Байдавлстов, 1997; 14 - Федосенко и др., 1995; 15-Ишунин, 1968; 16-Рахимов, 1966; 17 - Соколов и др., 1996; 18-Груздев, Сухбат, 1982; 19-Жиряков, Федосенко, 1983; 20 - Слудский и др., 1983; 21 -Манассрова, 1988; 22-Сопин, 2003; 23 - Мельников, 2003.
** Самцы, в основном, старше 5-6 лет, самки - старше 4 лет.
*** Охотничьи трофеи.
тип муфлона - примитивные, небольшие (чаще не более 90 см в длине по изгибу), сравнительно тонкие в основании, гетеронимные, отходящие от черепа в стороны и вверх с загнутыми назад к шее и внутрь навстречу друг другу концами. Однако у отдельных особей они длинные (до 104 см) с дополнительным загибом в стороны (рис. 71);
тип уриала - средние по размерам (до 110 см), чаще первертированные, отходящие от черепа вверх и в стороны, серповидные без загиба концов вверх и в стороны. Тем не менее у некоторых особей они с загибом как у архара;
тип архара - высокоспециализированные, массивные (до 22 кг вместе с черепом у алтайского аргали), длинные (до 190 см), отходящие вверх, вбок и назад, гомонимные, спирально закрученные (правый направо, левый налево) с направленными в стороны концами. Их размах может превышать 140 см. В индивидуальном развитии рога архаров как бы проходят стадии, аналогичные развитым рогам других форм баранов.
Наиболее длинные и максимально закрученные рога с относительно узким основанием характерны для баранов западной части ареала - Памиро-Алайской и Тянь-Шаньской горных систем [Насонов, 1923; Цалкин, 1951а]. Аргали с Алтая и Заал-тайской Гоби имеют массивные рога (у самок они и самые длинные - до 56 см) с широким основанием округлой формы без четких внутренних и наружных граней (рис. 70). У других рас архара они чаще ближе к треугольной форме. Форма рогов барана Северцова явно промежуточная между типичными рогами архара и уриала [Ляпунова и др., 1997], а по размерам они ближе к последним (табл. 76), причем могут быть крупно- и мелкоморщинистыми (фото 39). Длина роговых чехлов у муфлона варьирует в пределах 37-104 см, у .уриала - 51-110 см (табл. 71). Длина рекордных рогов самцов алтайского аргали - 178 см, окружность основания - 50 см, а у памирского архара - 190,5 см при окружности 43 см. Рога тянь-шаньского барана, найденные в 1921 г. в долине р. Юлдаш, впадающей в р. Или (Кыргызстан), достига-
Рис. 70. Географическая изменчивость и типы рогов горных баранов (по [Цалкин, 1951а; Гептнер и др., 1961]; с изменениями)
Типы рогов: 1 - гетеронимный (форма ophion)-, 2-3 -первертированный (urmiana и orientalis)-, 4 - резко выраженный первертированый (cycloceros)-, 5 - гомоним-ный (cycloceros)-, 6 - гомонимный (karelini)-, 7-8 - резко выраженный гомонимный (polii и аттоп)
Рис. 71. Рекордный трофей (длина большего рога 104 см) европейского муфлона (фото из архива А. Хохлова)
ли 179,7 см в длину, окружность у основания - 39,4 см. В марте 2001 г. в Нарынс-кой области Кыргызстана добыт тринадцатилетний самец с рогами длиной в 171 см [Хохлов, 2002]. По трофейным каталогам рога памирского барана обычно не менее 55 дюймов (140 см), у тянь-шаньского - 49 (125 см) [Кошкарев, 2002].
Очевидно, что изменчивость рогов горных баранов тоже клинальна. Их размеры в значительной мере коррелируют с размерами тела. Крупные животные, населяющие наиболее холодные и многоснежные районы на севере ареала и высокогорье, имеют и самые мощные рога. Группировки, живущие в пессимальных условиях при недостатке корма и воды, отличаются сравнительно слаборазвитыми рогами. Статус большинства подвидов баранов, выделенных на основании отличий в размерах и форме рогов, при анализе больших выборок не подтверждается.
3
2
1
Окраска и линька. Новорожденные архарята серо-голубые или светло-коричневые с серо-голубыми или грязно-белыми нижними частями тела. У подросших сеголеток окраска темно-бурая в верхней части головы и туловища и более светлая по низу тела и на конечностях (фото 36). У взрослых животных более темный тон окраски верхней части тела сохраняется. Зимняя окраска самцов в разных частях ареала варьирует от буро-коричневой до бледно-бурой и светло-серой. Нижняя, а иногда и верхняя, части шеи, грудь, брюхо, задняя часть туловища и ноги до колен желтовато-белые или почти белые, как и большая часть морды. У молодых самцов и самок окраска чаще светло-серая или серо-коричневая. Самки в летнем меху желтовато-бурые или коричневатые с белым зеркалом, самцы - бурые, рыжеватые, коричневатые и красноватые, морда белесая, шея светлая, передние и задние конечности светло-кремовые, зеркало белое или кремовое. Линька сильно растянута и протекает с апреля до конца июня, у лактирующих самок (на Памире) затягивается до июля - начала августа. Зимний волос появляется в августе, а в октябре мех достигает полного развития [Антипин, 1947; Егоров, 1955; Савинов, 19756; Сопин, 19756; Сапожников, 19766, 1977; Слудский и др., 1983; Федосенко, 1983а, 2000; Собанский, 1992; Филус, 1992; Федосенко и др., 1995; Бербер, 1999].
Географическая, сезонная и индивидуальная изменчивость окраски горных баранов значительна. У европейского муфлона она рыжевато-красноватая, темно-коричневая или почти черная со светлым пятном на боках и спине (фото 31). У закавказского муфлона (фото 32) и эльбурского красного барана (фото 35), считающегося гибридом муфлона и уриала, светлое пятно на туловище отсутствует или слабозаметно. У уриала окраска желтовато-буроватая и песочная (фото 33 и 34).
У аргалиобразных баранов окраска чаще бурая в верхней части тела и светлая в нижней. У алтайского аргали она варьирует от темной до светлой, преобладающий цвет коричневато-бурый зимой и желтовато-бурый летом. У южногобийского барана при сходной окраске на груди развит подвес [Цалкин, 1951а; Федосенко, 2000]. Особенно большой он и в виде манишки у особей в Северном Тибете [Пржевальский, 1875]. Некоторые самцы из Заалтайской Гоби и Юго-Восточной Монголии пятнистые (в бурых яблоках) или со светлым пятном на крупе [Соколов и др., 1996]. Самцы памирского архара в летнем меху бурые и крапчато-коричневые, с кремовой или белой мордой, грудью, нижней частью тела и светлым околохвостовым зеркалом. Зимняя окраска туловища разнообразная - от коричневато-бурой до бледно-бурой и даже белесой. Морда белая или кремовая. У некоторых зверей на пояснице и задней части боков кремово-белые пятна 2-4 см в диаметре. На нижней части шеи и груди имеется небольшой желтовато-серый или желтовато-белый подвес. В целом зимний мех самцов и самок светлее летнего в нижней части туловища и на конечностях, и темнее в верхней части тела. Граница между светлыми и более темными частями окраски на боках обычно хорошо выражена (фото 37). Изредка встречаются альбиносы [Мекленбурцев, 1948; Федосенко и др., 1995; Хохлов, 2002]. Североказахстанский баран (фото 38) несколько темнее, чем памирский. Буро-коричневая или серо-вато-бурая окраска верхней части головы и туловища у него чаще резко отделена от белого низа, а по низу фронтальной частр задних ног проходит темная полоса [Слудский и др., 1983; Бербер, 1999]. У тянь-шаньского барана буроватая или бурая верхняя часть туловища тоже обычно отграничена темной полосой от белого брюха [Се-верцов, 18736]. На севере хребта Каратау бараны буровато-коричневые или серова
то-бурые с небольшими рогами. Сверху по середине хвоста и на задних ногах (у некоторых особей - и на передних) спереди, от копыт до колен, проходит темная полоска, губы и нос белые, морда темная. Волосы на груди и по низу шеи грязноватого тона. У особей, живущих на юге этого же хребта, окраска и форма рогов, как мне довелось видеть на снимках охотничьих трофеев, иные, такие же как у тянь-шаньской расы. У кызылкумского барана Северцова основной тон окраски серовато-коричневый, серовато-бурый или буровато-коричневый, в нижней части - грязно-вато-белый или с коричневым оттенком (фото 39). Слабозаметный подвес сероватобелый, но у отдельных особей он хорошо развит и снежно-белый [Рахимов, 1966; Сапожников, 19766; Ишунин, 19836; Сопин, 19846; Ляпунова и др., 1997].
Генетическая изменчивость. Кариотип собственно архара, как отмечалось выше, - 2п = 56, NF = 60. В наборе 2 пары крупных метацентрических хромосом, 25 пар имеют акро- и субтелоцентрическое строение, Х-хромосома - крупный акро-центрик, Y-хромосома мелкая субметацентрическая, метацентрическая или акроцентрическая. Хромосомные наборы тянь-шаньского karelini, казахстанского coIlium, а также форм аттоп, darwini, przewalskii, jubata, dalai-lamae, hodgsonii из Монгольского и Гобийского Алтая, Заалтайской Гоби, Ганьсу и Тибета, severtzovi из Нуратин-ских гор Узбекистана и nigrimontana из Каратау практически идентичны [Schmitt, Ulbrich, 1968; Воронцов и др., 1972а,б; Орлов и др., 1975, 1978; Орлов, 1978; Шарипов и др., 1980; Bunch et al., 1990, 2000; Шакула и др., 1994; Ляпунова и др., 1997; Сливинский, 1999]. Диплоидный набор устюртского и бухарского баранов, граничащих с архаром, оказался типичным для уриала - 2п = 58, NF = 60. Аутосомы представлены одной парой крупных метацентриков и 27 парами ароцентрических хромосом, Х-хромосома - крупный акроцентрик [Воронцов и др., 1972а; Сливинский и др., 2003].
Наименее изменчивым в генетическом отношении среди баранов (и даже среди копытных) является европейский муфлон (доля полиморфных локусов равна 0,022, средняя гетерозиготность — 0,011), что связано с прохождением его через “бутылочное горлышко” эволюционного отбора еще на островах Средиземного моря и с акклиматизацией в Центральной и Восточной Европе малого числа особей-основателей [Hartl, 1990]. Бараны из Китая и Средней Азии, напротив, генетически высоко вариабельны (2,4—11,5%, в среднем - 7,7% по исследованиям последовательностей D-петли мтДНК). Формы nigrimontana и severtzovi, скорее всего, отделились от общего ствола Ovis раньше других. Они значительно отличаются от китайских баранов, среди которых генетически близки dalai-lamae и hodgsonii, а также sairensis и adametzi. Форма darwini в этом кластере образует третью кладу [Wu et al., 2003]. Алтайская и гоби-хангайская группировки генетически однородны. Уровень их различия по ND5 гену митохондриальной ДНК скорее популяционный, нежели подвидовой. Форма darwini близка к алтайскому аргали [Reading et al., 2002; Tuya et al., 2002]. По PCR-RAPD полиморфизму геномной ДНК выявлены достаточно отчетливые различия между формами coIlium и karelini. Бараны, населяющие Джунгарский Алатау (littledalei), и Чу-Илийская группировка животных имеют, возможно, гибридное происхождение [Сливинский и др., 2002].
Результаты анализа последовательностей фрагмента контрольного региона мтДНК 60 особей разных форм горных баранов, проведенного нами, не вполне укладываются в рамки известных представлений об их родственных связях (рис. 72).
Capricornis crispus Ovis canadensis AY116621* O. c. nivicola 72 O. c. nivicola 73 O. ariesbN582№* Туркмения AY091490* О. musimon AY091488* musimon O. musimon AY091487* + aries”
O. musimon AF039579* 0. aries AF039580* Туркмения,”42 \
Туркмения,53 Туркмения, 47 Туркмения, 48 Туркмения, 52
Плато Усть-Урт AY091489* Плато Усть-Урт, 43 О. vigney АХ565593* Туркмения, 46 Туркмения, 49 Туркмения, 50 Туркмения, 51______
Таджикистан, 44 Казахстан, 27 Казахстан, 33 Казахстан, 22 Казахстан, 36 Казахстан,63 Казахстан, 23 Казахстан, 37 Казахстан, 28 Казахстан, 24 Казахстан, 26 Казахстан, 25 Казахстан, 34 Казахстан, 65 Казахстан, 64 Казахстан, 32 Казахстан, 35^ Таджикистан, 45~\ Таджикистан, 16 Таджикистан, 20 Таджикистан, 86 Таджикистан, 66 Таджикистан, 67 Таджикистан, 15 Таджикистан, 17 Таджикистан, 19 Таджикистан, 87 Таджикистан, 18 Таджикистан, 88 Таджикистан, 21 Таджикистан, 14 Таджикистан, 54у Киргизия, 11 * Киргизия, 13 Киргизия, 38 Киргизия, 41 Киргизия, 40 Киргизия, 39 Монголия AF242348* Китай AJ238301* Китаи AJ238303* Китай AJ238305*
У “collium
>-“karelini
> аттоп
Рис. 72. Филогенетическое дерево горных баранов, построенное по результатам анализа последовательностей фрагмента контрольного региона мтДНК 60 особей (по [Кузнецова и др., в печати]) Цифры слева - уровень поддержки каждого кластера (%), полученный методом бутстрсп-анализа, справа -номера образцов. Звездочками обозначены образцы из генетического банка данных
Исследованные образцы обособляются в двух кластерах. В одном из них, включающем всех аргалиобразных 56-хромосомных баранов, компактные клады образуют североказахстанские (соответствующие обычно выделяемой форме coIlium'), кир-гизско-таджикистанские (karelini, polii) и монгольско-китайские (аттоп, darwini) расы. В другой кластер входят 54 и 58-хромосомные муфлонообразные бараны: европейский муфлон, домашняя овца и уриалы.
Гибридизация. Все формы горных баранов гибридизируют между собой и с домашними овцами беспрепятственно (см. выше).
Таксономия. История систематики баранов, и архара в частности, кратко отражена выше, поэтому в данном разделе вряд ли следует еще раз приводить весьма противоречивые воззрения ученых.
В середине XX в. зоологи чаще всего принимали классификацию настоящих баранов Ovis s. str. И.И. Соколова [1959], который, следуя Н.В. Насонову [1923], выделил 14 подвидов архара, или аргали (аттоп, hodgsonii, dalai-lamae, jubata, kozlovi, mongolica, przevalskii, nigrimontana, karelini, littledalei, coIlium, polii, heinsii и sairensis), добавив к ним форму intermedia, а также считал отдельными видами европейского и азиатского муфлонов. К последнему он причислил 14 форм (anatolica, armeniana, urmiana, isphaganica, orientalis, erskinei, cycloceros, dolgopolovi, areal, punjabensis, blanfordi, vignei, bochariensis и severtzovi). He менее популярной была классификация В.И. Цалкина [19486, 1951а], относившего к Ovis аттоп следующие подвиды: musimon, ophion, anatolica, gmelini, urmiana, isphaganica, laristanica, punjabiensis, blanfordi, vignei, bochariensis, orientalis, cycloceros, areal, severtzovi, nigrimontana, karelini, poli (polii), coIlium, аттоп, kozlovi, jubata, dalai-lamae и hodgsonii. В.Г. Гептнер [Гептнер и др., 1961] считал все же самостоятельность некоторых рас сомнительной и в пределах СССР определил всего 10 автохтонных подвидов. К группе собственно архаров он причислял кара-тауского (nigrimontana), тянь-шаньского (karelini, syn. heinsii, nasonovi, melanopyga), казахстанского (coIlium, syn. sairensis, albula, obscura), памирского (polii, syn. poli, typica, littledalei, humei) баранов; алтайского барана (аттоп, syn. argali, altaica, daurica, mongolica, przevalskii, asiaticus) относил к группе аргали; закавказского (gmelini, syn. armeniana) - к ophion; ладакского (vignei) и бухарского (bochariensis) - к vignei; туркменского (cycloceros) и кызылкумского (severtzovi) — к группе cycloceros. В сводки “Млекопитающие фауны СССР” [Громов и др., 1963] и “Каталог млекопитающих СССР” [Барышников и др., 1981] включены 5 подвидов архара: аттоп (= altaica, daurica, mongolica, typica, przevalskii), nigrimontana, karelini (= melanopyga, nasonovi), coIlium (= albula, obscura) и polii (= littledalei, humei). Подвид gmelini в “Каталоге ...” отнесен к азиатскому муфлону (Ovis orientalis), a varentzovi, areal, severtzovi и bochariensis причислены к уриалу (Ovis vignei).
Л.В. Сопин [1982а,б, 19836, 1984а,б, 2003] после морфометрической ревизии собственно архара указывал, не раз меняя точку зрения, от 8 до 11 подвидов: nigrimontana, polii (= heinsii, sairensis, littledalei, humei), karelini, coIlium, аттоп (= argali, altaica, daurica, mongolica, typica, przevalskii), darwini, jubata (= commosa), koslovi (= intermedia), dalai-lamae, hodgsonii и severtzovi. Фактически он восстановил в ранге подвида барана Дарвина, описанного Н.М. Пржевальским [1875], и вслед за Н.В. Насоновым [1914] причислил к архарам барана Северцова. Формы мелких по размерам баранов (кызылкумскую, каратаускую и южногобийскую) он,
не без оснований, считал реликтовыми, наиболее примитивными среди архаров (праархарами) и промежуточными между архаром и уриалом. В группе уриала им выделен новый оманский подвид (Ovis vignei arabica Sopin et Harrison, 1986) из Юго-Восточной Аравии, имеющий относительно крупные размеры, красноватые пятна на щеках, белый узкий подвес, окаймленный темно-коричневыми полосами, малые величины базиллярной длины черепа и верхнего ряда коренных зубов [Сопин, Харрисон, 1986].
В. Гайст [Geist, 1991], положив в основу классификации группы архаров особенности брачного наряда самцов, определил 7 подвидов: nigrimontana, hodgsonii, аттоп, darwini, jubata, polii и karelini, которого объединил с coIlium. Баран Северцова, однако, им не был причислен к архару. Специалисты по группе каприн (IUCN/SSC Caprinae Specialist Group) дробят аргали на те же подвиды, но выделяют еще и coIlium, а к Ovis orientalis относят 12 других рас баранов, включая кызылкумского (табл. 72). А.К. Федосенко [2000, 2002а] отнес к архару все перечисленные В. Гайстом подвиды, барана Северцова и казахстанского coIlium, а к уриалу причислил 7 подвидов: устюртского (areal), туркменского, или афганского (cycloceros), таджикского (bochariensis), ладакского (vignei), пенджабского (punjabensis), белуджистанского (blanfordi) и оманского (arabica) баранов.
Судя по приведенным выше морфологическим и молекулярно-генетическим данным, среди горных баранов обособлены лишь две “ясные” систематические группы (аргалиобразная и муфлонообразная), которые более соответствуют статусу подвидов (О. a. аттоп и О. a. orientalis), нежели видов, а все выделенные ранее многочисленные расы заслуживают ранга популяций, если считать популяцией совокупность особей одного вида, обладающих общим генофондом и занимающих определенную территорию. Каратауский баран и баран Северцова явно занимают промежуточное положение между аргалиобразными и муфлонообразными баранами и примыкают к одной или другой группе в зависимости от анализируемых признаков. В целом таксономия горных баранов требует дальнейшей разработки, особенно на молекулярном уровне.
Распространение
Восстановленный ареал. В прошлом ареал крупных архароподобных баранов был гораздо шире современного и простирался от Атлантического до Тихого океана. В верхнем плиоцене и раннем антропогене эти животные населяли Китай, Индию, Забайкалье, Среднюю и Переднюю Азию, Казахстан, Закавказье, Таманский полуостров, Крым и Южную Европу, включая Италию, Францию и Моравию, где в некоторых районах жили вплоть до верхов плейстоцена. Остатки мелких муфлонообразных баранов появляются в более поздних отложениях: в Западной Европе - с плейстоцена, в Крыму и вблизи Адлера в Ахштырской пещере - в мустьерских и верхнепалеолитических слоях, в Дагестане - с мезолита, в Южном Закавказье -с палеолита, в Узбекистане - со среднего плейстоцена, но в голоцене в этих районах уцелели только они. Удивительными представляются находки остатков мелких баранов на равнине вдали от гор - в палеолитической стоянке Мезинской на р. Десне и в низовьях р. Урал [Громова, 19356, 19486, 1953; Громов, 1948; Пидопличко, 1951, 1954; Верещагин, 1959а; Джафаров, 1960; Кожамкулова, 1969, 1981; Цал-
кин, 1970; Верещагин, Барышников, 1980а; Манасерова, 1986; Межлумян, Оганесян, 1986;Тлеубердинаидр., 1989; Барышников, 1995;Манасерян, 19976;Manaseryan, 2001].
В плейстоцене архар был обычен в Казахстане, Средней и Центральной Азии и Южной Сибири, где населял северные и северо-западные предгорья Алтая, окрестности Усть-Канска и Барнаула, восточные склоны Кузнецкого Алатау, окрестности Омска, Томска, Абакана, Минусинска, Красноярска, Братска, Иркутска и Балаганс-ка, Минусинскую и Тувинскую котловины, устье Бирюсы, верховья Уды, долину Ангары, о. Ольхон на Байкале и Западное Забайкалье, включая бассейны Селенги и Джиды, Баргузинскую долину, район оз. Баунт, окрестности Сретенска, Кяхты и Улан-Удэ, Южную Якутию (рис. 73). Его многочисленные остатки найдены также в позднепалеолитическом поселении Сухотино-4 в черте Читы вместе с сайгой, дзере-ном, бизоном, северным оленем, лошадью, сибирской косулей, благородным оленем, лосем и носорогом. В меж- и послеледниковье архар, видимо, вместе с сибирским козлом жил на Южном Урале, проникая в Башкирии до р. Белой, а на востоке — до камышловской террасы [Черский, 1861; Павлова, 1911; Насонов, 1923; Громо-
Рис. 73. Распространение аргали в Сибири (по [Боескоров, 2001])
я - современный ареал; б - границы ареала в XVIII - середине XIX вв.; в-д - ископаемые находки, относящиеся к эоплейстоцену, плейстоцену и голоцену, соответственно. Номерами обозначены конкретные пункты находок (см. в оригинале)
Рис. 74. Распространение архара в Казахстане в голоцене (по [Соколов, 1964])
Точки с номерами - пункты находок наскальных изображений (см. в оригинале); предполагаемая область обитания заштрихована
ва, 1941; Громов, 1948; Цалкин, 1951а; Верещагин, 1954; Вангенгейм, 1961, 1977;
Александрова и др., 1963; Ко-жамкулова, 1969,1981; Савинов, 1972; Сопин, 1974а,
1998; Антощенко-Оленев,
1975; Ларичев, 1976; Оводов, Орлова, 1977; Барышников, 1978; Алексеева, 1980,19906;
Верещагин, Барышников, 1980а; Батиров, 1986; Батыров, 1986; Каспаров, 1986а,6;
Кожамкулова, Пак, 1986; Тле-убердина и др., 1989; Оводов, Ивлева, 1990; Смирнов и др., 1990; Худайбердиев, 1990; Абрамова и др., 1991; Калмы-
ков и др., 1992; Калмыков, 1995; Малеева и др., 1995; Боескоров, 2001; Косинцев, 2003].
На юге Сибири, в Казахстане (рис. 74), Средней и Центральной Азии этот баран был широко распространен и в голоцене, что видно по обилию ископаемых остатков [Верещагин, Батыров, 1967; Кожамкулова, 1969, 1976, 1981; Алексеева, 1980; Тлеубердина и др., 1989] и наскальных рисунков, датируемым Ш-П тысячелетиями до н.э. - Х-ХШ вв. н.э. [Соколов, 1964; Окладников, Запорожская, 1970; Ешелкин, 1972; Смирнов, 1990]. В Забайкалье его черепа найдены в хуннском погребении Ш-П в. до н. э. в долине Джиды, а костные остатки периода бронзового и раннего железного веков - в Шилкинской пещере в 18 км от Усть-Карска. Всего три столетия назад путешественники-первопроходцы видели сотенные стада животных в предгорьях Алтая, Чуйской степи и на хребтах Прибайкалья. На Джидинском хребте, Малом Хамар-Дабане и по р. Хилок архар дожил до начала XIX в. В этот период он населял также бассейны Селенги, Ингоды и Онона, верховья Чикоя и Шилки, степные горы Даурии и горно-степные районы Восточной Монголии. Сравнительно недавно этот вид занимал почти всю территорию Тувы, сотню лет назад жил в Минусинской котловине, а в восточной части Восточного Саяна (бассейн рек Тисса и Казас) встречался до середины XX в. [Северцов, 1873а,б; Насонов, 1923; Колосов, 1938, 1939; Цалкин, 1951а; Соколов, 1959; Гепт-неридр., 1961;Гришин, 1975,1981;Сопин, 19756; Макарова, 1976; Смирнов, 1978, 1983,1989,1990; Федосенко, 1983а,2000; Швецовидр., 1984; Соколов, 1988; Смирнов, Бриллиантов, 1990; Прокофьев, 1992; Борисова и др., 2001].
Главной причиной исчезновения аргали в Восточной Сибири, и Забайкалье в частности, А. Хлебников [1877] справедливо считал появление человека с винтовкой. Выпавшие в конце 30-х годов XIX в. глубокие снега “доконали” этот вид окончательно, а последних особей истребили казаки. В Северной Маньчжурии (на плоскогорье Барги) архар исчез, скорее всего, одновременно с баранами Забайкалья и Восточной Монголии после особенно многоснежной и суровой зимы 1831/32 гг. На
Хэнтэе единичные животные дожили до начала XX в. [Лу-кашкин, 1975; Груздев, Сухбат, 1982].
Ареал в XX в. Отечественные ученые различали от 8 до 15 форм аргалиобразных баранов, “привязывая” их к определенным географическим районам [Север-цов, 1873а,б; Насонов, 1923; Цалкин, 1951а; Соколов, 1959; Гептнер и др., 1961; Воронцов и др., 1972а,б; Федосенко и др., 2000; Сопин, 2003, и др.]. Однако границы распространения выделяемых рас нечетки и весьма дискуссионны, как и сама
Рис. 75. Локализация форм аргалиобразных баранов (по [Федосенко, 2000])
Формы: 1 - алтайская, 2 - казахстанская, 3 - тянь-шаньская, 4 -каратауская, 5 - кызылкумская, 6 - памирская, 7 - тибетская, 8 -северокитайская, 9 - гобийская
таксономия баранов. Тем не менее попытаемся обозначить их и проследить, где
это возможно, тенденцию динамики современного ареала.
Алтайский аргали. Алтайский аргали населяет горы юга Сибири, севера и запада Монголии и северо-запада Китая (рис. 75). Современный ареал очаговый. Районы обитания удалены один от другого на 30-160 км и более [Анчифоров, 1990; Анчифоров, Фалеев, 1993; Федосенко, 20026].
На юге Сибири северная граница его распространения в связи с усиливающейся хозяйственной деятельностью человека и охотой заметно сдвинулась к югу в XIX в., но вплоть до 70-х годов XX в. он населял восточные отроги Катунского хребта, хребты, окружающие Чуйскую степь, северные склоны хребта Южный Алтай, южные части Шапшальского и Чулышманского хребтов, хребет Чихачева и Талдуаир. В конце столетия эти копытные уцелели только на юго-востоке Горного Алтая (рис. 76) в приграничных районах с Китаем (на плато Укок и прилегающих районах), Монголией и Тувой (на хр. Чихачева, в массиве Талдуаир и на хр. Сай-люгем). На Южном Алтае на территории Казахстана несколько десятков аргали обитали в верховьях р. Бухтармы и в южных отрогах Курчумского хребта. Северная граница ареала вида проходит в Алтайском заповеднике по бассейну р. Богояш и водоразделу с р. Башкаус [Колосов, 1938, 1939; Цалкин, 1951а; Ирисов и др., 1973; Сопин, 1973, 19756, 1976, 1977в; Федосенко и др., 1987; Филус, Золотухин, 1987; Собанский, 1988, 1989, 1992; Собанский, Федосенко, 1990; Филус, 1992; Федосенко, 2000, 2001; Пальцын, Спицын, 2002; Абатуров и др., 2004]. Все оставшееся на Алтае поголовье диких баранов равно лишь одному крупному стаду, каких здесь ранее было много.
Область распространения аргали в Туве особенно резко сократилась к середине XX в. К 40-м годам его не стало на Хемчикском хребте и в междуречье Казыра и Кизира. Во второй половине столетия он населял высокогорья хребтов Танну-Ола, Сангилена, Шапшальского, Цаган-Шибэту и восточные склоны хр. Чихачева.
Рис. 76. Распространение аргали на Русском Алтае и пограничных территориях (по [Абатуров и др., 2004]) А - районы постоянного обитания; В - места периодических или единичных встреч баранов или следов их пребывания за последние 100 лет; С - северная граница Дауро-Монгольской степной области; D, Е-номсра и границы округов Монголо-Алтайской высокогорно-степной подпровинции и ботанико-географических районов (см. в оригинале); F- границы государств; С - административные границы субъектов Российской Федерации
А
. в ч_/ с xivj D "й : Е
ч._., - G
Изолированные очаги были на хребтах Западном Танну-Ола, Хан-Тайга, Ергак-Таргак-Тайга и Хазарама. На западе республики к концу столетия мелкие группировки уцелели лишь на восточном склоне хр. Чихачева (20-30 особей), северном макросклоне массива Монгун-Тайга (около 50-100 особей) и на западной окраине Западного Танну-Ола. Небольшой очаг есть в истоках р. Оны. Ежегодно в Туву мигрируют несколько десятков особей из прилежащих районов Монголии и Горного Алтая [Янушевич, 1952; Сопин, 19756, 1977в; Никифоров и др., 1977; Смирнов, 1983, 1988а, 1990, 2002в; Швецов и др., 1984; Соколов, 1988; Ткаченко, 1992; Федосенко, 2000, 2001].
В Красноярском крае эти животные могли сохраниться до настоящего времени в междуречье Кизира и Казыра. В начале 1980 г. несколько самцов барана видели на хр. Крыжина [Сыроечковский и др., 2000]. В Хакасии аргали регистрировали в высокогорном поясе в верховьях рек Он, Малый и Большой Абакан и в бассейне р. Еринат в типичных местах обитания сибирского козла. Возможно, сюда звери заходят с сопредельной территории Алтая и Тувы. В первой половине XX в. небольшие группы баранов встречали в верховьях рек Большие и Малые Уры. В 60-е годы их изредка отмечали в предгорьях Кузнецкого Алатау [Смирнов, Бриллиантов, 1990; Прокофьев, 1992].
В Бурятии в начале XX в. аргали жили в истоках Иркута, в середине столетия -в окрестностях озера Хара-Нур, в бассейне р. Тиссы, во второй половине века они в небольшом числе заходят на крайний запад республики из приграничных районов Монголии [Смирнов, Сопин, 1988]. В Читинской области последний архар,
видимо зашедший из Восточной Монголии, убит близ государственной границы к югу от ст. Мангут в 1918 г. [Гептнер и др., 1961]. После этого достоверных встреч не было вплоть до середины 90-х годов. В 1994—1997 гг. три барана жили на левом скалистом берегу Онона ниже устья р. Или [Кирилюк, 20006].
В Монголии алтайского аргали исследователи отмечают только на севере и северо-западе в Прихубсугулье, Хангае, Монгольском и Гобийском Алтае. Юг Монголии (хребты Байтаг-Богди, Тахийн-Шар, Заалтайскую Гоби на восток до Дорноговь аймака и Замын-Уудэ) и Внутреннюю Монголию, как считалось, населяет форма darwini [Банников, 1954; Соколов, Орлов, 1980; Жирнов, Ильинский, 1985; Соколов и др., 1996]. К существенным географическим барьерам относили Алакнурскую депрессию [Сопин, Сухбат, 1983], хотя она вряд ли служит значительным препятствием для этих подвижных животных. В октябре 1976 г. три самки и самец, например, замечены на хребте Дархан Хэнтэйского аймака далеко от границ обитания ближайших популяций [Груздев, Сухбат, 1982]. Генетически алтайская и гоби-хангайская группировки аргали, как показано выше, однородны. Современное распространение архара в Монголии преимущественно мелкоочаговое [Amgalandaatar et al., 2002].
Казахстанский баран. В XIX в. границы распространения этой формы простирались на север до Кокчетава, Экибастуза, Семипалатинска, на восток - до Иртыша и Черного Иртыша, на запад - до гор Улытау и Ортау, на юг - до Балхаша, Сасыколя и Алаколя [Селевин, 1928; Антипин, 1941;Цалкин, 1951а; Кириков, 1963, 1966; Бербер, 1999]. В первой половине XX в. ареал сократился почти в 2 раза. В середине столетия благодаря запретам охоты и улучшению охраны, приведшим к снижению браконьерства, а также в связи с переходом населения от кочевого к оседлому образу жизни область обитания баранов стала быстро расширяться, особенно на севере республики. Они появились во многих горах, а в Карагандинской области их насчитывали более тысячи. В 1948 г. 3 зверя зашли в Шучинский район Кокчетавской области. В 1952 г. значительное их количество регистрировали в Ма-кинском районе Акмолинской области и в Майском районе Павлодарской области в 120 км севернее ближайшего хребта Чингизтау, а одного самца видели гораздо севернее - в Максим-Горьковском районе. В 1953 г. звери отмечены северо-восточнее Баян-Аула [Слудский, 1956; Пакиж, 1958].
Во второй половине XX в. ареал казахстанского барана, несмотря на естественное расселение животных, так и не восстановился полностью. Северный его участок в Казахском нагорье (горы Койтас, Ерментау и Нияз) находится в отрыве от основной области обитания. Северная граница ареала проходит по горным массивам Бая-наул, Кыргытас, Тотия, Кызылтау, Аркалык, Едрей, Муржик, Дегелен; западная - по горам Шантимес, Топай, Улькенаюл, Ортау и др. и по сглаженным остепненным массивам западнее оз. Балхаш; южная тянется с востока на запад через Котан-Эмэль, Колмак-Эмель, Бектауата, Шунак, Караунгир, Тюлькили, Жанет и Аркарлы севернее Балхаша; восточная включает приграничные горные районы северо-запада Китая, Тарбагатай, Саур и Калбинский Алтай (рис. 75). Баран редок повсеместно, за исключением заповедных и приграничных территорий [Савинов, 19756; Капитонов, Махмутов, 1977; Капитонов, 1980; Слудский и др., 1983; Федосенко, 1983а, 2000; Федосенко, Жиряков, 1987; Бербер, 1999; Байдавлетов, 20036; Бербер, Ержанов, 2003].
Тянь-шаньский баран. Обитает на большинстве хребтов Тянь-Шаня, расположенных от пика Хан-Тенгри на востоке до Ферганского хребта и Алайской долины
на западе, преимущественно в приграничной с Китаем полосе, где звери занимают лишь труднодоступные высокогорные участки (рис. 75). В зону обитания помимо Киргизского, Угамского и Пскемского хребтов входит Джунгарский Алатау, Бо-ролдай, горы южнее юго-восточной и юго-западной оконечностей оз. Балхаш, Чу-Илийские горы, Заилийский, Таласский, Терскей и Кунгей Алатау, Кетмень, юго-восточная часть Каратау, горы Бетпакдалы, Калканы и Каржантау [Северцов, 1873а,б; Чернавин, 1916; Кузнецов, 1948а; Цалкин, 1951а; Шапошников, 1956; Айзин, 1975, 1977, 1980; Савинов, 19756; Танюшин, 1977; Капитонов, Лобачев, 1977; Федосенко, 1977, 2000; Ишунин, 1983г; Слудский и др., 1983; Сопин, 1986а]. В северных отрогах Алайского и Туркестанского хребтов, возможно, тоже обитает “karelini" [Мекленбурцев, 1962] или другая форма, отличающаяся небольшими размерами рогов.
Памирский баран. Населяет Памиро-Алайскую горную систему в пределах Таджикистана и соседних районов Киргизии, Афганистана и Китая (рис. 75). Имеются также сообщения о встречах этих животных в Пакистане в районе пересечения Ки-лика и Кхунджераба с р. Хунза [Хохлов, 2002]. В Таджикистане архар распространен по всем высокогорным хребтам к востоку от Бадахшана. Граница области обитания на западе проходит от Ленгара через озера Яшилкуль и Сарезское до восточной части ледника Федченко, на севере - по Заалайскому хребту, на востоке и юге - за государственной границей. В Киргизии он занимает высокогорную территорию близ государственной границы от Алайской долины и Ферганского хребта на западе до пика Хан-Тенгри на востоке, включая долины рек Кек-Су, Ой-Тал (Карадарья), южную оконечность Ферганского хребта, окрестности оз. Чатыркуль, западную часть хребта Кокшаалтау и бассейны Аксая и Узенгикуша. Акшийрак, видимо, населяет смешанная форма, а на Сарыджасе уже обитает тянь-шаньский баран [Флеров, 19356; Айзин, 1969; Сапожников, 19766; Федосейкой др., 1995; Федосенко, Вейнберг, 1999, 2001; Федосенко, 2000]. Е. Кошкарев [2002] считает, что границей распространения памирского и тянь-шаньского баранов служит скорее многоснежный Ферганский хребет, нежели р. Нарын. В последние годы отмечен подток крупных светлоокрашенных животных с большими рогами (которых называют бараном “Марко Поло”) с территории Киргизии в Терскей Алатау [Шубин и др., 2002]. В Таджикистане на Ваханском, Ишкашимском, Шахдаринском, Шугнанском хребтах и западных отрогах Южно-Аличурского хребта ареал архара перекрывается или соприкасается с областью обитания уриала [Сапожников, 1976а,б, 1977; Сопин, 1986а].
Каратауский баран. Долгое время исследователи, вслед за Н.А. Северцовым [1873а], считали, что эта форма распространена по всему хребту Каратау. Позднее выяснилось, что она занимает лишь северо-западную часть (Сырдарьинское Каратау), где живет оседло (рис. 75), а на юго-востоке хребта и на Боролдае обитает тянь-шаньский баран, который сезонно мигрирует в западную часть Таласского Алатау [Шапошников, 1956; Грачев, 1982].
Кызылкумский баран, или баран Северцева. Область современного обитания (рис. 77) - хребет Нуратау и отдельные горы-останцы Центрального Кызылкума. Ранее этот баран населял, возможно, [Туркестанский, Зеравшанский, Гиссарский и Алайский хребты и БаЙсунские горы [Насонов, 1923; Цалкин, 1951а; Ишунин, 1960, 1967, 1977, 19836; Мекленбурцев, 1962; Сапожников, 1976а; Присяжнюк, 1984; Реймов, 1994; Федосенко, 2000].
Рис. 77. Распространение кызылкумского барана (по [Присяжнюк, 1984]) Современная область обитания и распространение в прошлом выделены частой и редкой штриховкой соответственно; отдельное современное место обнаружения зверей обозначено черным кружком
Тибетский баран. Эта группировка занимает Тибет, Нань-Шань, Кунь-лунь, Алтын-таг (Китай), Ладак (Индия), Непал вдоль границы с Тибетом и, вероятно, северо-восток Бутана [Schaller, 1998; Сопин, 2003]. Границы распространения не ясны (рис. 75).
Северокитайский баран (фото 49). Область обитания - Внутренняя Монголия и север Центрального Китая (рис. 75), однако границы распространения тоже приблизительны [Федосенко, 2000; Сопин, 2003].
В целом очевидно, что область обитания аргалиобразных баранов на территории бывшего СССР заметно сократилась за счет исчезновения северной и северо-восточной частей из-за чрезмерного преследования человеком, дополняемого гибелью зверей в многоснежные зимы. Значительно уменьшились и границы распространения муфлонообразных баранов (см. ниже).
Среда обитания
Архара можно причислить к эвризональным и эврибионтным экологически пластичным видам [Северцов, 1873а,б; Насонов, 1923; Антипин, 1947; Цалкин, 1945, 1951а]. Он живет практически на любой высоте: в высокогорье, среднегорье, низ-когорье и даже на мелких возвышенностях, занимая при этом, в отличие от горных козлов и снежных баранов, более открытые, ровные и пологие участки с расположенными неподалеку убежищами - изрезанными скалистыми участками, где звери отдыхают и где рожают самки. На большей части ареала условия обитания животных весьма суровы: резко континентальный сухой и холодный климат, высокая инсоляция, малое количество осадков даже в зимнее время, сильные ветра и короткий вегетационный период. Среднегодовые температуры воздуха во многих районах отрицательные, а абсолютный минимум порой превышает -50...60 °C.
Эти звери при большой массе тела имеют относительно узкие копыта, и больше других копытных зависят от высоты снежного покрова. Обязательное условие для их жизни - малоснежье, обусловленное либо малым количеством осадков, либо выдуванием снега сильными ветрами. Бараны раскапывают рыхлый слой снега мордой и копытами лишь при острой необходимости, и обычно прекращают тебе
невку если его высота превышает 20-25 см. При снеге выше 30 см движение рысью становится затруднительным, они переходят на прыжки или передвигаются один за другим по тропам. Весовая нагрузка на след очень высокая (около 700 г/ см2 у самок и 1000 у самцов), и при беге она еще увеличивается. Критическая высота для относительно высоконогого аргали близка к 70 см, для других форм она меньше. Высота снежного покрова, во многом, определяет северную границу ареала [Насимович, 1955; Сопин, 19756, 1983г; Слудский и др., 1983].
Распределение животных зимой также в большей мере зависит от высоты снежного покрова, а летом - от наличия зеленого корма и беспокойства кровососущими насекомыми. Зимуют архары в малоснежных районах чаще в низкогорье, а весной перемещаются вверх за отступающей линией снежного покрова. В высокогорье эти копытные пасутся на субальпийских и альпийских лугах, в нивальном поясе встречаются редко. Излюбленные биотопы - сочетание пастбищ и щебнистых участков с хорошим обзором, где они устраиваются на отдых. Убежищами для них служат также отвесные скалы, останцы, каменные россыпи, ниши и пещеры. Летом звери нередко заходят в осиново-березовые колки, сосновые боры и заросли кустарников [Насимович, 1955; Сапожников, 1977; Грачев, Смирнова, 1980; Федосенко, 1979, 1983а, 2000].
В вегетационный период самки горных баранов почти повсеместно держатся на меньших высотах по сравнению с самцами, хотя условия обитания здесь нередко хуже: напряженнее конкурентные отношения с домашним скотом, выше охотничий пресс, меньше воды, суше растительность, больше хищников и кровососущих насекомых. Гипотез, объясняющих это явление, выдвигалось немало, но внимания, на мой взгляд, заслуживает лишь “физиологическая”. Новорожденные горных баранов, как и домашних овец, не приспособлены к обитанию на больших высотах, и их адаптация к условиям высокогорья происходит постепенно с месячного возраста. Эту особенность можно рассматривать и как одно из доказательств низкогорного происхождения горных баранов [Цалкин, 1945, 1951а]. Их современное обитание в труднодоступных местах на больших высотах, скорее всего, вторично, и связано, как справедливо заметил еще Н.А. Северцов [18736], с вытеснением домашним скотом и человеком из холмогорий и низкогорий.
В Горном Алтае сотни лет назад аргали заселял не только горные, но и предгорные районы и плоскохолмистые межгорные котловины (Чуйскую, Курайскую и Уймонскую), откуда был вынужден уйти на вершины хребтов на высоты в 2000-3000 м над уровнем моря. Основные современные местообитания животных - ос-тепненные луговые осоково-злаковые и кобрезиевые тундры, горные степи, альпийские луга и лужайки, склоны хребтов крутизной не более 20-30°, крутые щебнистые осыпи и скалы, служащие убежищем. Ерники и луговые тундры с переувлажненными осоково-кобрезиевыми и лишайниково-моховыми сообществами, занимающие большие площади, для них почти не пригодны [Колосов, 1939; Сопин, 19756, 1976а, 1982в; Филус, Золотухин, 1987; Собанский, 1988, 1992; Федосенко, 1989, 2000; Федоткина, 2002; Абатуров и др., 2004]. В Алтайском заповеднике летом животные предпочитают травяно-дриадо-лишайниковые, травяно-лишайниковые, каменистые и щебнисто-полигональные тундры и альпийские луга, осенью и зимой - тундры, луго-тундры, остепненные луга, кобрезиевники и ерники [Филус, 1992]. На хребте Сайлюгем бараны чаще (82% встреч) держатся на крутых скло
нах горных вершин, что, возможно, связано не только с удобным сочетанием убежищ и пастбищ [Петрищев, 2003; Петрищев, Петросян, 2003], но и с неумеренным охотничьим преследованием. В Туве аргали круглогодично обитают в высокогорье на субальпийских и альпийских лугах, в кустарниковых и щебнистых тундрах с участками субальпийского разнотравья и альпийской растительности в типичных биотопах горных козлов [Ткаченко, 1992]. В целом для архара Южной Сибири наиболее характерны следующие стации: мелкотравные злаковые степи; мелкотравные, кобрезиевые и злаково-осоковые альпийские луга; травянистые, кустарниковые, мохово-лишайниковые и каменисто-щебнистые тундры [Сопин, 19756].
В Монголии горные бараны населяют обширнейшую территорию от высокогорной тайги на севере до полупустынь и пустынь на юге, предпочитая холмистые плоскогорья, предгорные склоны и высокогорья со сглаженным рельефом. На Хан-гае эти копытные держатся преимущественно в кобрезиево-осоковых горных степях и на остепненных склонах, на Монгольском и Гобийском Алтае - в горных степях с альпийским разнотравьем. В Заалтайской и Джунгарской Гоби они с высокой плотностью заселяют горы-останцы (100-300 м) в пустынях, мелкосопоч-ник, изредка их регистрируют на пустынных равнинах, поросших саксаулом, солянками и эфедрой, в типичных местах обитания кулана и джейрана. Зимой они концентрируются у подножий гор и возвышенностей и в полупустынных степях, летом поднимаются на высокогорные альпийские луга. В Южной Гоби звери обычны в мелкосопочнике, на склонах небольших гор, щебнисто-каменистых плато, увалах и межгорных котловинах. Важное условие их обитания - наличие водных источников [Банников, 1954; Дагва, 1958; Львов, 1981; Жирнов, Ильинский, 1985; Соколов и др., 1996; Федосенко, 2000; Amgalandaatar et al., 2002].
Биотопы баранов в Казахстане представляют особый интерес. В Казахском нагорье даже самые высокие горы, где они обитают, не более 1500 м над уровнем моря, а в мелкосопочнике животные населяют возвышенности, редко превышающие 500 м (фото 40). Подножья гор и межгорные понижения представлены полынно- и ковыльно-типчаковыми степями и полупустынями, склоны остепненные или заросшие кустарником (казацким можжевельником, ивами) и березо-осиновыми и сосновыми лесами, или в виде каменистых осыпей и выходов скал. Звери не избегают леса и кустарниковых зарослей. Самки и молодые особи предпочитают кварцитовые горы со сложным рельефом и богатой растительностью, самцы - гранитные, менее доступные для человека и хищников. Зимой животные держатся лишь на малоснежных участках. Летом в местах зимовок остается не более 10-40% особей, остальные расходятся по межгорным понижениям, сглаженным сопкам, холмистым равнинам и даже встречаются на сельскохозяйственных полях. На Тарбагатае и Сауре архары обитают, в основном, на высокогорных субальпийских и альпийских лугах [Антипин, 1947; Капитонов, Махмутов, 1977; Слудский и др., 1983; Бе-кенов и др., 1999; Бербер, 1999; Федосенко, 2000].
На Памире и Внутреннем Тянь-Шане эти копытные живут в сложном ландшафтном горном комплексе на высотах от 3000 до 6000 м, где предпочитают сглаженные хребты, пересеченные широкими приозерными и речными долинами и крутыми ущельями, с наличием мелкощебнистых склонов, скальных или крупнощебнистых убежищ, ксерофитных растительных группировок, альпийских лужаек, ручьев и родников. Зимние места их обитания обычно на 500-600 м ниже летних, при
чем самки держатся на склонах ниже взрослых самцов (фото 41). Здесь особенно низки среднегодовые температуры, ничтожное количество осадков, сильные ветры, интенсивная солнечная радиация и очень короткий вегетационный период. Субальпийский пояс представлен преимущественно пустынями и степями с разреженной растительностью и бедным видовым составом: терескеном, злаками и полынями, на лугах типичны кобрезия и осоки. В альпийском поясе преобладают растения-подушечники, на альпийских лугах - осоки, луки, гречишниковые и овсяницы, в степях - беломятликовые, типчаковые и овсецовые сообщества [Северцов, 18736; Мекленбурцев, 1948; Айзин, 1969; Сапожников, 1976а,б; Абдусаля-мов, 1997; Федосенко, 2000].
Тянь-шаньские бараны занимают как низкие пустынные и малоснежные горы на севере области обитания, так и высокогорья (до 3000 м) [Северцов, 1873а,б], отличающиеся многоснежьем и низкими температурами. Выживают они здесь во многом благодаря миграциям на зимовку в малоснежные периферийные части хребтов и низкогорья (см. ниже) и сильным ветрам, сдувающим снег с отдельных участков, а также благодаря сильной изрезанности ландшафта и многочисленности убежищ.
На северном макросклоне Сырдарьинского Каратау баран живет в кустарниковых зарослях среди скал, ущелий и осыпей (фото 42). Более пригодные для жизни участки хребта и плато заняты домашним скотом [Федосенко, 2000].
Основное поголовье барана Северцова сосредоточено в охраняемом Нуратинс-ком заповеднике, где мало домашнего скота и есть родники. Звери свободно выпасаются в горных степях и на травянистых и кустарниковых склонах на высоте 1000-1800 м (фото 43). Несколько хуже условия их обитания на Актау, где суше климат и мало водных источников. При отсутствии осадков растительность усыхает уже в конце мая, и баранам приходится большую часть года использовать “сено на корню”. Зимой они выдерживают сильные морозы и штормовые ветры при крайне скудном корме [Ишунин, 1960, 1968, 1977, 19836; Коршунов, Коршунова, 1983; Шакула, 1989а; Федосенко, 2000].
Питание
Корма. По моим подсчетам по литературным источникам, аргалиобразные бараны на обширном ареале поедают более 300 видов растений, однако их количество в отдельных регионах обычно не превышает 70-110 и, кроме того, рацион существенно меняется по сезонам года.
Весной звери предпочитают листья и бутоны тюльпанов и маков, побеги ревеня, луков, пырея ползучего, мятликов, лапчатки, ячменя луковичного, костра, эги-лопса, осок, тонконога, зопника, гречишника и других зеленых растений. Летом в рационе доминируют бобовые, разнотравье и злаки: астрагалы, люцерна, эспарцет, копеечники, мытники, горноколосник, кипрей, герани, пижма, горец змеиный и живородящий, лапчатка, водосбор, примулы, аконит, молочай, кобрезия, спирея, очиток гибридный, клевера, подорожники, лисохвост, овсяницы, мятлики, ежа сборная, одуванчики, горчак, солянки, луки, ковыли, побеги и листья смородины, кизильника, жимолости, шиповника и спиреи. Осенью в пище обычны также мытники, горечавки, житняк, гречишники, щавель, сон-трава, осоки, полыни и эфедра. Типичная зимняя пища - ветошь злаков (на Памире и Алтае до 59-99% содержи
мого рубцов), кобрезии, осоки, полыни, ковыли, типчак, очиток гибридный, зоп-ник горный, альфредия снежная, порезник, лишайники, мхи, побеги ивы, осины, березы, таволги, терескена, спиреи, эфедры, малины, жимолости, шиповника, кизильника и арчи. В пустынях к ним добавляются ковыльки, баглур, прутняк, миндаль, караганы, вяз, саксаул, тамариск, солянки и другие растения. Нередко животные раскапывают корни крестоцветных и зонтичных растений. В целом высокогорные популяции архара более травоядные по сравнению с низкогорными, в питании которых немаловажную роль играют листья и побеги кустарников и кустарничков (до 51% содержимого рубцов животных в Северном Казахстане) [Антипин, 1947; Мекленбурцев, 1948; Банников, 1954; Дагва, 1958; Ковшарь, Янушко, 1965; Мамбетжумаев, 1974; Капитонов, Кубыкин, 1975; Сопин, 19756; Сапожников, 19766, 1977; Бургело, 1980; Плешак, 1980; Ишунин, 19836; Слудский и др., 1983; Федосенко, 1983а, 2000; Жирнов, Ильинский, 1985; Филус, Золотухин, 1987; Филус, 1992; Анчифоров, Фалеев, 1993; Бербер, 1999; Федоткина, 2002].
Питание уриала и арменийского муфлона в целом сходно с таковым у архара [Мамбетжумаев, 1969; Явруян, 1970; Сапожников, 19766; Горелова, 1980; Слудский и др., 1983; Жесткова, 1999], а европейский муфлон, возможно, более древес-ноядный [Heroldova, 1988; Rodriquez et al., 1988; Homolka, 1991].
Потребление кормов и воды. Минеральное питание. Детеныши начинают пробовать растительную пищу на 2-16-й день жизни, пастись - на 18-19, продолжительная жвачка отмечена на 24—25, поедание рыхлой земли - с 6 по 40-й день [Шакула, 1989в; Анчифоров, Фалеев, 1993].
За сутки в период вегетации взрослые самцы потребляют до 19 кг, самки - до 16 кг растительного корма [Сапожников, 19766, 1977; Бербер, 1999]. В осенне-зимний период масса содержимого рубцов гораздо меньше - около 3-4,5 кг в отдельных районах [Мамбетжумаев, 1974]. Весной в начале вегетации и летом животные быстро передвигаются по пастбищу в поисках излюбленных растений или их частей. Зимой их движения замедлены, они подолгу остаются на одном месте, тщательно выедая растения на выдувах или на местах тебеневки.
Архары, как и другие копытные, нуждаются в минеральном питании. В весеннелетний и раннеосенний периоды во многих районах они регулярно посещают сухие и водные солонцы, содержащие соли кальция, магния и натрия, и иногда выгрызают ямы до 3 м в диаметре. Помет животных в эти периоды может состоять почти полностью из глинистой массы. Бараны активно используют и искусственные солонцы из поваренной соли, сделанные чабанами для домашнего скота или специально устроенные в охотничьих хозяйствах и заповедниках. Однако в некоторых пустынных горах они на солонцы не выходят, что, возможно, связано с эрозией почвы при сильных ветрах и потреблением солей, оседающих на растениях [Слудский и др., 1983; Федосенко, 1983а, 2000; Бербер, 1999; Бербер, Ержанов, 2002].
Воду горные бараны получают с влажным кормом, но в жару они нуждаются в водопоях (рис. 78). Поскольку число водоемов в засушливых районах ограничено, зверям приходится ходить к ним за несколько (иногда за 10-15) километров или выходить в высокогорье, где близ снежников много ручьев и родников [Жиряков, 1976; Слудский и др., 1983; Федосенко, Жиряков, 1987]. В отличие от многих диких и домашних копытных, бараны могут употреблять сильно минерализованную воду. В частности, уриал в Бадхызе и на Устюрте пьет воду, содержа-
Рис. 78. Горные бараны на водопое (фото Г. Левенштейна)
щую до 23 г солей на литр, а в засушливые годы - даже до 57-195 г/л. Молодые животные иногда гибнут здесь же от солевого отравления, однако такие случаи редки [Горелов, 1964; Горбунов, 1986в, 1987, 19956]. После дождей звери несколько дней утоляют жажду из луж и котлованов, образуемых в скальных углублениях. Зимой они поедают снег. В местах водопоев копытные особенно осторожны: на подход к воде иногда затрачивают до часа времени, часто останавливаются, осматриваются и прислушиваются, а начав пить, нередко стремглав выскакивают на открытое место.
Пищевая активность. Суточная активность архара - полифазная. В весеннелетний и раннеосенний периоды, как и большинство других копытных, они наиболее интенсивно и продолжительно пасутся в раннеутренние и вечерние часы, чередуя кормежки с кратковременным отдыхом, а жаркий день проводят в укрытиях в тени или на снежниках. В пасмурные дни животные выходят на пастбища и днем. Зимой они пасутся в основном в дневное время, что связано с терморегуляцией. При постоянном беспокойстве, вызванном выпасом домашнего скота или охотничьим преследованием, их активность становится преимущественно ночной. Во время кормежки звери через каждые 3-4 мин поднимают голову и осматриваются. Каждая фаза активности длится около часа, после чего бараны примерно столько же времени отдыхают на пастбище. Суточный ритм обычно заметно изменяется в периоды гона и лактации. В апреле, с появлением ягнят, самки пасутся в среднем около 46 мин, отдыхают всего 13 мин, и почти каждый час кормят молоком детенышей. К концу апреля продолжительность отдыха увеличивается до 40 мин, в
мае - до 1 ч. В июне в периоды активности они тратят собственно на кормежку 82% бюджета времени, на передвижение - 7, на осматривание - 11%. В конце лета и осенью пастьба занимает около 60% времени суток. У самцов меньше периодов активности, чем у самок, но они дольше пасутся и отдыхают. При недостатке корма время, затрачиваемое на перемещение и кормодобывание, существенно увеличивается. До 15% времени звери тратят на посещение солонцов и водопоев. Активность баранов на обширном ареале в одни и те же периоды неодинакова в связи с разным климатом и средой обитания [Антипин, 1947; Мекленбурцев, 1948; Банников, 1954; Ковшарь, Янушко, 1965; Сопин, 19756, 19766; Сапожников, 19766, 1977; Плешак, 1980; Dawaa и. а., 1983; Слудский и др., 1983; Федосенко, 1983а,б, 2000; Федосенко, Жиряков, 1987; Грачев, 1988; Шакула, 19896; Федосенко, Скуратов, 1990; Собанский, 1992; Филус, 1992; Федосенко и др., 1995; Бербер, 1999].
Размножение
Половая зрелость. Самки аргали приносят первое потомство в трехлетием возрасте, и лишь изредка - в двухлетнем. Самцы становятся половозрелыми в 2,5 года. В вольере на Алтае и в Карагандинском зоопарке бараны в этом возрасте результативно покрывали самок, в том числе 1,5-летних [Анчифоров, Фалеев, 1993; Бербер, 1999]. Однако в природе самцы обычно не принимают участие в размножении до 4,5-5,5 лет, поскольку их вытесняют из гаремов взрослые соперники [Мекленбурцев, 1948; Сопин, 19776; Федосенко, 1983а]. У самок и самцов муфлона половая зрелость наступает раньше - в 1,5 года [Стекленев, 19786, 1995]. У уриала, как и у аргали, большая часть самок приносит потомство в трехлетием возрасте, а бараны могут участвовать в гоне с третьего года жизни [Горбунов, 19956; Коршунов, 1995в; Жесткова, 1999].
Физиологически бараны способны к оплодотворению самок только в осенне-зимний период. Половые гонады достигают максимального развития в октябре — ноябре. С конца ноября сперматогенез затухает, в феврале - марте в семенниках и их придатках признаки глубокой инволюции, в июне - июле половые железы находятся в стадии относительного покоя, с августа активизируется сперматогенез, а с сентября - спермиогенез [Спивакова, Байдавлетов, 1997; Спивакова и др., 2002]. Половая активность и у самцов, и у самок сохраняется до глубокой старости.
Гон и половое поведение. Гон у аргалиобразных баранов обычен в октябре -декабре. В пустынных и низкогорных районах он начинается раньше (в октябре), в высокогорных - на 1-1,5 месяца позже, но проходит в более сжатые сроки (табл. 77).
Примерно в эти же сроки (с конца октября - ноября до декабря - начала января, в неволе - с августа до конца декабря) спариваются уриал [Мамбетжумаев, 1969; Сапожников, 19766; Ишунин, 1983а,в; Слудский и др., 1983; Горбунов, 1987,19956; Коршунов, 1988а; Горелов, Ишадов, 1999; Жесткова, 1999] и муфлон (октябрь -декабрь, в неволе - с сентября до февраля) [Даль, 1954; Салганский и др., 1963; Треус, 1968; Rougeot, 1969; Дулицкий, Кормилицын, 1970; Явруян, 1970; Треус, Лобанов, 1976; Стекленев, 1977, 19786, 1995]. У снежного барана гон чаще начинается позднее (см. ниже).
Спаривание в осенне-зимние месяцы, следовательно, типично для всех представителей рода Ovis. Начало сроков гона, видимо, каким-то образом связано с
Таблица 77. Сроки гона у аргалиобразных баранов
Район, заповедник Начало Конец Источник информации
Алтай Конец октября Начало декабря Сопин, 19776,в
Монголия Октябрь Ноябрь Соколов и др., 1996
15 ноября - Груздев, Сухбат, 1982
Казахское нагорье 16 октября Начало декабря Бербер, 1999
Тянь-Шань Ноябрь — Вырыпаев, 1983а
ч Октябрь Ноябрь Кузнецов, 1948а
ч Середина ноября - Федосенко, 2000
Таласский Алатау 20 октября 5 декабря Ковшарь, Янушко, 1965
Конец ноября Федосенко, 1983а
Калканы Середина октября ч Федосенко, 1979, 1983а
Алма-Атинский з-к Ноябрь Ноябрь Жиряков, 1979
Нуратау Конец октября Начало декабря Ишунин, 19836
ч 20 ноября Шакула, 1989в
Памир Декабрь Конец декабря Сапожников, 1977
ч ч Январь Мекленбурцев, 1948
сс Ноябрь Декабрь Айзин, 1969
ч Конец ноября — Федосенко, 2000
Тибет Вторая половина Январь ч
декабря
укорочением светового дня. В экспериментах с искусственно укороченным дневным временем (8 ч в сутки) самки корсиканского муфлона приносили потомство дважды в год [Rougeot, 1969]. Сроки гона могут заметно сдвигаться в ту или другую сторону и в зависимости от погоды. В Казахском нагорье спаривание у архара чаще начинается с 16-20 октября, но теплой осенью 1995 г. брачные пары стали образовываться только 11-12 ноября [Бербер, 1999].
Гон архара, как правило, проходит на зимовках. В конце сентября - октябре взрослые самцы начинают подтягиваться к районам обитания самок с детенышами. Мелкими группами и поодиночке они медленно бродят по холмам и гребням и подолгу стоят в местах с хорошим обзором, срываясь с них при приближении самок, которых стремятся обнюхать. В этот период между соперниками обычны демонстрации и турнирные бои [Слудский и др., 1983; Федосенко, 1983а, 2000].
Непосредственно в гоне участвуют самцы старше 5 лет, которые входят в состав самочьих групп и формируют “гаремы”. При низкой плотности населения при одном баране находится 1-2 две самки с ягнятами, при высокой - до 8-15. Реже в смешанном стаде в гон бывает два самца, а иногда, кратковременно, - до 10. После выяснения иерархического статуса слабейшие из них покидают группу. Молодые самцы остаются в гареме, или, преследуемые взрослыми баранами, поодиночке или по 2-5 держатся на некотором расстоянии [Сопин, 19756; Сапожников, 19766; Бербер, 1999].
Взрослые самцы охраняют свой гарем от посягательств соперников. При высокой плотности популяции поединки меду ними нередки и в разгар гона. При борьбе противники пытаются оттеснить друг друга или, реже, встают на дыбы перед ударом и с силой сталкиваются рогами. Победитель остается с самками, постоянно контролируя их состояние и проявляя типичное половое поведение: подходит к ним “вытянувшись” с хриплыми звуками, сгоняет с лежки постукиванием ноги,
Рис. 79. Половое поведение аргали (по [Филус, 1992])
теснит грудью, трогает бока и живот передней ногой, обнюхивает половые органы и мочу, флемуя при этом, и пытается запрыгивать на круп (рис. 79). Перед обнюхиванием самец обычно широко расставляет передние ноги и делает характерный поворот головой, направляя основание одного рога вверх, другого - вниз. Самка, не пришедшая в охоту, принимает позу угрозы или убегает, а находящаяся в эстру-се позволяет обнюхивать морду и другие части тела и облизывать половые органы, затем приседает и мочится. Баран подставляет морду под струю, флемует и, если самка остается на месте, шеей и грудью касается ее крупа, передней ногой - живота и ног, издает хриплый урчащий звук, иногда теснит ее, кладет голову на холку и круп, и, став сзади, делает садку, длящуюся 5-15 с. Садки следуют одна за другой с интервалом в несколько минут. Если самка все же убегает, то самец неотступно преследует ее, если она ложится, то он может поднять ее ударом передней ноги [Сапожников, 19766, 1977; Сопин, 19766, 19776; Федосенко, Орлов, 1976; Федосенко, 1979, 2000; Вырыпаев, 1983а; Слудский и др., 1983; Филус, 1992; Бербер, 1999].
Беременность. Срок беременности самок архара, по данным разных исследователей, колеблется в пределах 155-180 дней. По уточненным сведениям, включающим наблюдения в зоопарках, он находится в пределах 160-168 дней [Са-
Таблица 78. Сроки деторождения у аргалиобразных баранов
Район,заповедник | Начало | Конец | Источник информации
Алтай Конец апреля Май Сопин, 19776,в
Монголия Апрель Начало мая Соколов и др., 1996
Монгольский Алтай 15 апреля - Груздев, Сухбат, 1982
Южная Монголия Конец апреля Май Жирнов, Ильинский, 1985
Казахское нагорье 25 марта Начало мая Бербер, 1999
З-к “Аксу-Джабаглы” Конец марта 5 мая Ковшарь, Янушко, 1965
Джунгарский Алатау Середина апреля - Федосенко, 1983а, 2000
Калканы Таласский Алатау 11 апреля — (С
и Чу-Илийские горы Заилийский Алатау Конец марта — и
и Терскей Алатау Третья декада апреля - и
Нуратау Начало апреля Май Ишунин, 19836; Коршунова, Коршунов, 1983
и 10 апреля Конец мая Шакула, 1989в
Каратау Вторая половина февраля Конец марта Антипин, 1947
Боролдай 8 апреля Плешак, 1980
Алма-Атинский з-к Конец марта Апрель Жиряков, 1976
Тянь-Шань Март С( Кузнецов, 1948а
Памир Конец марта — Айзин, 1969
Киргизия Май — Федосенко и др., 1995
Тибет Конец мая Июнь Федосенко, 2000
пожников, 1977; Сопин, 19776; Федосенко, 1979, 1983а, 2000; Шакула, 1989в; Бербер, 1999].
Деторождение и выкармливание молодняка. Сроки ягнения самок аргали в разных частях ареала и горных массивах неодинаковы (табл. 78). В низкогорье и среднегорье они рожают с конца марта - начала апреля, в высокогорье - с середины апреля, а на Памире - с конца мая, что связано с разным временем прекращения морозного периода и начала вегетации растительности. В высокогорных популяциях деторождение, как и гон, проходит в более сжатые сроки. Судя по цитированным выше литературным источникам, примерно такие же сроки родов характерны для уриала и муфлона (в неволе они более растянуты), а у снежного барана (табл. 90) они сдвинуты на 1-2 месяца.
Рожают самки поодиночке, в укромных местах, в сильно пересеченном рельефе: в скалах, на склонах каньонов и ущелий, в пещерах, где особенно благоприятный температурный режим, вблизи останцов, в крупнокаменистых осыпях, по краю кустарников. Роды длятся около 40 мин. Новорожденные хорошо развиты. Через 3-5 ч после рождения они способны карабкаться по пологим склонам и уходить на 10-20 м от самок. При опасности они затаиваются, но через несколько дней неотступно следуют за матерями. Лактация продолжительная - до октября -декабря, в высокогорных районах - др января - февраля и, в исключительных случаях, до мая, причем детенышей могут кормить молоком и беременные самки. В пустынных районах период лактации короче, и детеныши получают молока меньше, что негативно отражается на их развитии и росте [Мекленбурцев, 1948;
Сапожников, 1977; Сопин, 19755, 19776; Федосенко, 1979, 1983а, 2000; Слудский и др., 1983; Федосенко, Жиряков, 1987; Анчифоров, Фалеев, 1993; Бербер, 1999].
Плодовитость. Самки агали рожают чаще одного теленка, но двойни все же обычны, а в низкогорных и среднегорных популяциях два ягненка регистрируют почти у трети самок (табл. 79). В Карагандинском зоопарке три самки рожали 17 раз, и в 13 случаях (76,5%) приносили двойни [Бербер, 1999].
Таблица 79. Число детенышей у самок архара
Район (источник информации)* 1 Самок с детенышами, % Самок без ягнят, % Детенышей на самку
| с одним | с двумя
Алтай (3) 28 62 -
-“-(7) 88 - 12 0,5-0,7
Казахстан (10) 65 35 34 1,3**
Казахское нагорье (8) 71 29 - 1,3**
-“-(9) 83 17 50 1,2**, 0,6
Джунгарский Алатау (2, 6) 89 11 35 1 3**
Калканы (7) 86,7 13,3 34,1 0,6-0,75
Таласский Алатау (7, 13) 100 - - 0,6
88,2 11,8 — 1,1
Боролдай (4) 82,4 17,6 51,4 1 2**
С.-З. Каратау (7) 86,7 13,3 9,1 1,0**
Нуратау (5) 83,7 16,3 — 0,8
-“-(12) 90,5 9,5 (14) 23,5
Памир (1) 92 8 - -
-“-(7) 100 — — 0,3-0,5
Тянь-Шань (7) 100 — — 0.4-0.7
*1 - Сапожников, 19766, 1977; 2 - Федосенко, 1979; 3 - Сопин, 1977; 4 - Плешак, 1980; 5 - Вейнберг, 2003а; 6 - Анненков, Литун, 1989; 7 - Федосенко, 2000; 8 - Капитонов, Махмутов, 1977; 9 -Бербер, 1999; 10 - Антипин, 1947; 11 - Ковшарь, Янушко, 1965; 12-Ишунин, 1967,19836; 13 - Федосенко, Жиряков, 1987.
** Детенышей на родившую самку.
Потомство ежегодно приносят около 70-85% самок архара [Ишунин, 1967; Федосенко, 1983а, 2000; Анненков, Литун, 1989], но при недостатке самцов - менее трети [Сопин, 19756, 19776]. Число ягнят на одну взрослую самку, с учетом появления двоен, в обычные годы близко к 1. Наблюдатели часто показывают гораздо меньшее их число (табл. 79), поскольку часть детенышей ко времени регистраций уже погибает.
Плодовитость уриала и арменийского муфлона, видимо, чуть выше, чем у аргали. В приплоде обычно один, реже - два (11-32%) и, изредка, три ягненка. Каждая половозрелая самка приносит в среднем 1,2-1,7 детеныша. Яловость в разные годы колеблется от 7 до 37% [Йшадов, 1965; Мамбетжумаев, 1969; Явру-ян, 1970; Сапожников, 19766; Ишунин, 1983в; Слудский и др., 1983; Минасян и др., 1986а; Горбунов, 1987, 19956; Горелов, Ишадов, 1999; Жесткова, 1999]. У самок европейского муфлона в неволе преобладают двойни [Даль, 1954; Треус, Лобанов, 1976].
Популяционная структура
Половой и возрастной состав. Соотношение полов у эмбрионов и новорожденных аргали, также как у муфлона и уриала, близко 1:1, а у взрослых особей в разных группировках изменяется в ту или другую сторону в значительных пределах в зависимости от условий обитания и степени охотничьего воздействия.
Замечу, что при исследованиях популяционной структуры баранов особенно важна репрезентативность выборок. Самки с детенышами и взрослые самцы большую часть года живут раздельно и, нередко, в разных районах и на разных высотах, и поэтому при малом числе регистрируемых животных в узколокальных районах возможны значительные искажения половозрастного состава популяций.
В большинстве группировок аргали доля сеголеток колеблется в пределах 13-33%, годовалых особей чаще 10-16, половозрелых самок - 30-60, самцов - 20-40%, в том числе взрослых от 0,6 до 29% (табл. 80). В заповедных и слабо опро-мышляемых популяциях соотношение полов близко 1:1-2, в эксплуатируемых самцов обычно в несколько раз меньше, чем самок. Доля сеголеток к весне следующего года в большинстве районов снижается минимум в 2 раза, что свидетельствует о значительных потерях в этой возрастной группе.
На Памире в середине XX в. среди самцов было около 14% 2-3-годовалых особей, 40% - 4-5-летних и 46% - старше 6 лет [Егоров, 1955]. В 60-е годы доля взрослых баранов в популяциях достигала 33% [Сапожников, 1977], а в 90-е годы во многих районах их число уменьшилась до 0,6-19,5% [Федосенко и др., 1995; Федосенко, 2000]. В Киргизии в конце 80-х годов в разных группировках насчитывали 19-28% самцов, из них особей старше 10 лет - около 8%. К 1999 г. в результате неумеренной валютной охоты их число уменьшилось, соответственно, до 6 и 1% [Кошкарев, 2002]. Среди самцов архара в Южной Гоби 1-3-годова-лые особи составляли 24%, 3-4-летние - 13, 5-7 летние - 50 и старше 7 лет -13% [Valdez, Frisina, 1994; Reading et al., 1997 (по: Федосенко, 2000)].
У туркменского уриала в Центральном Копетдаге половозрастной состав популяции примерно такой же, как и у аргалиобразных баранов: сеголетки - 22, самки - 58 и самцы - 20%, соотношение полов на заповедной территории - 1:2,9, а в эксплуатируемых стадах - 1:5,6 [Коршунов, 1988а, 1995в]. В Бадхызском заповеднике доля сеголеток достигает 28%, годовалых - 13, самцов - 24 и самок -34% [Жесткова, 1997, 1999], у устюртского уриала, соответственно без годовалых особей, - 27, 19 и 54% [Горбунов, 19956]. По сведениям А.К. Федосенко с соавторами [2000], у туркменского уриала в разных группировках доля сеголеток колеблется в пределах 22-40%, самок - около 45-49 и самцов - 14-37%, в том числе старше 5 лет - 0,5-10%. У закавказского барана в Азербайджане доля сеголеток и годовалых особей оказалась очень низкой - 8 и 5% соответственно. Соотношение самцов и самок в популяции близко 1:1 [Кулиев, 1997]. У европейского муфлона в Крыму половозрастной состав не отличается существенно от такового у архара [Дулицкий, 1992].
Социальная организация и поведение. Горные бараны - стадные животные. Их популяции представляют собой совокупность разных половых и возрастных объединений, включающих: отдельные семьи (самку с одним или двумя ягнятами сразу после родов, а позднее - с молодняком прошлого года); семейные группы
Таблица 80. Половозрастной состав (%) в популяциях аргалиобразных баранов
Район, заповедник п Сего- Годо- Полово- Самцы Самцы :
(источник информации)* летки валые зрелые молодые взрослые самки
самки
Алтай (1) 227 20 8? 48 24 1:2
- 8-12 - 52 40 1:1,3
хр. Чихачева (13) — 25 — 54 21 1:2,6
-“-(4) 155 15-18 — 42-50 35-40 1:1,1-1,4
Сайлюгем (4) 2251 19-33 - 40-53 8-27 2-16 1:1-3,7
-“-(17) 248 15-20 - 44-46 ? 16-18 1:2,7
Монгольский Алтай (17) _. 20 26 - - - - 1:1,5
-“-(4) 764 20-22 - 52-72 11-15 10 1:0,5-9,1
Заалтайская Гоби (10) — 8 - 52 10 1:1,3
Южная Гоби (4) 673 13-22 - 30-61 16-57 1:0,5-3,7
Казахстан (4) 446 17-27 - 40—44 11-13 18-29 1:1-1,5
-“-(14) 3231 18-29 - 34-51 24-37 1:1,1-2
-“-(18) 4236 23 - 39 22 16 1:1
Тарбагатай (18) - 24 - 38 20 16 1:1
(20) 95 20 12 30 18 1:0,8
Терскей Алатау (4) 118 18 - 23 5 9 1:0,4
-“-(21) 120 21 28 51 1:0,5
Калканы (4) 209 11 49 32 9 1:1,2
С.-З. Каратау (8) 35 17 6 37** 9 14 1:1,6
-“-(4) 139 32 — 35 3 2 1:1,1
Боролдай (7) - 19 - 33** 48 1:0,7
Таласский Алатау (15) 4161 19(6-26) - 41,5 39,6 1:1,05
-“-(4) 478 19-27 — 44-61 6-22 7-14 1:1,5-3
Алма-Атинский з-к (2) 269 16 - 40** 44 1:0,9
-“-(3) 295 7 27 29 12 25 1:0,8
Киргизия (6) - - - — 19-28, 6*** -
-“- (22) 565 15 10 36 38 1:1
Нуратау (11) 2180 33 - 40 27 1:1,5
(23) - 34 - 45** 21 1:2,1
Памир (5) — - - - 33 -
ю.-в. часть (4) 4015 21-23 — 46-58 25-10 8-7 1.1,4-3
с.-в. часть (4) 492 24 - 62 11,6 0,6 1:4,6
Горный
Бадахшан (9, 24) 5033 16-24 14-16 40 20-30 1:1,3-2
Киргизия (16) 460 15 10 36 24 14 1:1,1
Тянь-Шань (8) — 9 — 32 ? 13 —
-“-(4) 986 27 - 40 17 16 1:1,2
-“-(4) 460 15 - 41 29 14 1:1
Центр. Тянь-Шань (24) 544 22 13 42 2 1:1,9
Тибет (12) 202 20,5 - 50 29,5 1:1,7
*1 - Собанский, 1992; 2 - Жиряков, 1976; 3 - Федосенко, Жиряков, 1977; 4 - Федосенко, 2000; 5 -Сапожников, 1977; 6 - Кошкарев, 2002; 7 - Плсшак, 1980; 8 - Байдавлстов, 2003а,б; 9 - Магомедов и др., 2003; 10 - Жирнов, Ильинский, 1985; 11 - Ишунин, 1967; 12 - Schaller, 1998; 13 - Пальцын, Спицын, 2002; 14 - Бербер, 1999; 15 - Ковшарь, Янушко, 1965; 16 - Федосенко и др., 1995; 17 - Анчифоров, Фалеев, 1993; 18 - Байдавлстов, 1996, 20036; 19 - Сопин, 1977в; 20 - Капитонов, Махмутов, 1977; 21 -Ганюшин, 1977; 22 - Fedosenko et al., 1995; 23-Коршунова, Коршунов, 1983; 24 - Magomedov et al., 2003.
** Вместе с молодыми особями.
*** После трофейной охоты.
(объединения нескольких семей); группы взрослых самцов (фото 44); группы яловых самок нередко вместе с молодыми самцами; самочьи и смешанные стада и скопления. Одиночные особи в группировках редки - обычно не более 4-15% от числа встреч [Ишунин, 1967; Сопин, 19756, 1976а; Баскин, 1976; Жиряков, 1976; Сапожников, 1976а, 1977; Зархидзе, 1980; Слудский и др., 1983; Федосенко, 1983а, 2000; Филус, 1992; Бербер, 1999; Федосенко, Вейнберг, 2001].
Смешанные стада формируются во время гона и сохраняются некоторое время спустя. В многоснежные зимы самцы и самки могут жить на ограниченных участках вместе вплоть до весны. Чаще же в конце декабря - январе взрослые бараны отделяются и образуют одновозрастные или разновозрастные группы, в которых живут до осени, т. е. до очередного периода гона. В стадах самок обычно находится приплод и молодые особи, включая самцов до 2-3 лет. Мелкие самочьи группы, как правило, состоят из животных, связанных родством, и наиболее устойчивы. Распадаются стада примерно за две недели до отела, когда взрослые самки уединяются для родов. Молодые особи держатся поблизости от мест отела, нередко вместе с яловыми самками. Через 2-3 недели после родов самки с детенышами начинают объединяться в группы, в которые со второй половины июня вливаются молодые и яловые звери. Однако отдельные одиночные самки с ягнятами встречаются до сентября - октября.
Аргали чаще образуют группы от 4 до 15 особей. Максимальные размеры стад самок с детенышами достигают 1-3 сот голов, в группах самцов регистрируют по нескольку десятков экземпляров (табл. 81). Скопления, насчитывающие сотни, а в многоснежье на ограниченных пастбищах - тысячи особей (на Памире), редки и кратковременны. Состав крупных группировок и групп молодых животных особенно непостоянен и может изменяться даже в течение суток.
Среднее и максимальное число особей в стадах выше в открытых биотопах, при высокой плотности популяции и при минимальном уровне охотничьего пресса и пастбищного животноводства. Низкая стадность может служить показателем неблагополучного состояния группировок. В Северном Казахстане, например, даже в открытых биотопах среднегодовой показатель стадности не превышал 5, изменяясь по месяцам в пределах 3-7, и только в гон в смешанных стадах насчитывали до 62 особей. Доля одиночных особей здесь максимальная для вида - 26% [Бербер, 1999].
У уриала и муфлона примерно такая же социальная организация, как и у аргали, и такой же размер групп: обычно от 4-5 (в семьях и эксплуатируемых стадах) до 30 голов, максимально - до 300. Зимой все самцы находятся в смешанных стадах. Отдельные самцовые группы образуются лишь весной и сохраняются до гона, но и в этот период значительная часть взрослых и молодых самцов находится вместе с самками и ягнятами [Цалкин, 1948а; Ишадов, 1965; Явруян, 1970; Сапожников, 19766; Зархидзе, 19836; Слудский и др., 1983; Коршунов, 1988а, 1995в; Ду-лицкий, 1992; Ciucci et al., 1998; Жесткова, 1999; Федосенко и др., 2000].
Лидирует в смешанных или самочьих стадах горных баранов старая самка, в группах самцов - самый крупный баран. В смешанных стадах за самкой-лидером цепочкой идут другие самки с молодняком, молодые самцы до 3 лет, затем доминирующий самец в сопровождении других взрослых самцов. Замыкают шествие второстепенные самцы и самки без ягнят. Самки обычно быстрее самцов реагируют на опасность и более осторожны, у самцов сравнительно дольше ориентиро-
Район, заповедник Число особей в группе Источник информации
среднее | максимальное
Алтай 9(5-12) 39 Сопин, 19756, 1976а, 1977в
— — 10 40 Собанский, 1992
Чаган-Бургазы 7-34, 6-12* 92, 27* Федосенко, 2000
Сайлюгем 9-14 28, 12* Петрищев, Петросян, 2003
Алтайский з-к 7,5 37, 11* Филус, Золотухин, 1987; Филус,
1992
Тува 9-10 30(40), 6* Смирнов, 1990
Монголия 240, 40* Груздев, Сухбат, 1982
— 240, 24* Dawaa и. а., 1983
з-к “Хохцир” 30-39, 12* 200, 28* Dzieciolowski et al, 1980
(по: Федосенко, 2000)
48, 13* 78, 29* Федосенко, 1983а, 2000
Восточная Гоби 7 30 Львов, 1981
Южная Гоби 4-6 29 Reading et al., 1997
(по: Федосенко, 2000)
Казахское нагорье 5, 4* 62, 13* Бербер, 1999
сс 7(4-9) 50 (100) Капитонов, Махмутов, 1977
сс 2-9 33, 5* Федосенко, 1983а, 2000
Сырдарьинский Каратау 3-4, 2* 11, 3* с с
Таласский Алатау 10-16, 7* 52, 26* сс
Заилийский Алатау 4-8, 2* 19, 3* сс
Терскей Алатау 4-9, 3-6* 28, 18* Ганюшин, 1977
Джунгарский Алатау 2-5, 2-3* 15, 9* Анненков, Литун, 1989
Алма-Атинский з-к 5 28 Жиряков, 1976
Природный парк
“Алтын-Эмель” 4 — Бекенов и др., 1999
Боролдай Чаще 1-5 20 Плешак, 1980
Нуратау 9 - Вейнберг, 2003а
5-10 54, 33* Ишунин, 1967
Тянь-Шань 9-26, 6-9* 69, 51* Федосенко, 2000
Центральный Тянь-Шань 12 - Кошкарев, Нестеров, 1989
Западный Тянь-Шань — 17* Чернавин, 1916
Памир 7-12 300 Мекленбурцев, 1948
СС 5-10 Несколько Сапожников, 1976, 1977
тысяч, 40*
сс — 35 Соков, 19776
с с 15-17 50 Одинашоев и др., 1990
Киргизия 10, 4* >20, 15* Федосенко и др., 1995
сс — 40 Абдусалямов, 1997
сс 13-42, 4-11* 135 (500), 58* Федосенко, 2000; Федосенко,
Вейнберг, 2001
сс — 300-400 Хохлов, 2002
Северный Тибет 5-15 30 Пржевальский, 1875
Тибет 10, 4* 31, 9* Schaller, 1998
* В группах самцов.
вочно-исследовательская реакция. При появлении мнимой или реальной угрозы самец-доминант один или со своим сопровождением нередко выдвигается в сторону опасности и принимает решение об отражении атаки или бегстве в определен-
ном направлении, увлекая за собой остальных животных, а иногда поднимает лежащее стадо, “набегая” на ее членов или “подгоняет” отставших или находящихся в стороне особей. Доминирующий самец во многом определяет поведение всего стада не только при опасности, но и на водопое, где имеет явное преимущество при подходе к воде [Баскин, 1976; Сапожников, 19766; Сопин, 19766; Коршунов, 1988а; Филус, 1992].
На пастбищах и на отдыхе доминируют сильные взрослые особи, прогоняющие более слабых с мест кормежки или с лежек. В самочьих стадах доминантами иногда становятся 2-3-летние самцы. Одноранговые животные нередко выясняют отношения с помощью рогов. Особенно часты стычки в стадах одновозрастных самцов, но они, как правило, не ожесточенные. Один из соперников отбежав, машет головой, встает на задние ноги и идет к ближайшему самцу. Тот тоже встает или подставляет рога под удар, или же отворачивается, прерывая таким образом атаку противника. Иногда в стычках участвуют сразу несколько баранов [Федосенко, 1979, 1983а, 2000].
Звуковые сигналы у архара редки: самки призывают телят блеянием, самцы издают своеобразные звуки в период гона; животные, заметившие опасность, с силой ударяют копытом передней ноги о землю и иногда издают шипящий звук, что настораживает других особей. Телята в первые дни жизни затаиваются, в неволе такое поведение характерно и для взрослых особей [Абрамов, Фалеев, 1996]. Самки активно защищают детенышей при нападении пернатых хищников, выставляя рога и подпрыгивая вверх. Ягнята при этом прячутся под животом матери [Слудский и др., 1983]. Раненые самцы агрессивны и порой пытаются напасть на человека.
В спокойной обстановке звери передвигаются шагом, при опасности — галопом и, реже, рысью, и всегда стремятся скрыться за ближайшим гребнем. Они весьма быстры и выносливы. У горных баранов хорошо развиты все органы чувств. Человека они замечают за 2,5-3 км и убегают, но при отсутствии постоянного беспокойства могут подпускать его на 300-800 м, постепенно сбиваясь в группу. По запаху человека распознают с расстояния около 1 км, звук слышат с 500 м [Мекленбурцев, 1948; Сапожников, 19766]. Если баранов не преследуют, они пасутся вместе с домашним скотом рядом с жильем человека [Пржевальский, 1875].
Пространственная структура и перемещения. Архар - преимущественно оседлый вид, хотя и более подвижный по сравнению с другими горными копытными. В малоснежных районах звери совершают лишь незначительные ежедневные и сезонные высотные перемещения или переходят с одного склона хребта на другой. Как отмечалось выше, весной по мере таяния снега животные, зимующие в нижних поясах гор, перемещаются в субальпийский и альпийский пояса, а осенью спускаются вниз. При временных потеплениях или похолоданиях обычны перемещения “вверх-вниз” [Грачев, Смирнова, 1980]. Со второй половины зимы и до осени взрослые самцы и самки держатся в разных стациях и, порой, в разных горах, при этом бараны предпочитают верхние пояса и более ровные участки рельефа, самки - нижние и изрезанные скалистые, что связано с необходимостью защиты молодняка от хищников и непогоды.
Популяции, находящиеся в многоснежных или засушливых районах, вынуждены сезонно мигрировать на десятки километров. Основные причины перемещений - не
доступность корма или его отсутствие, появление кровососущих насекомых, пересыхание водоемов, а также беспокойство со стороны человека и домашнего скота.
Группировка, зимующая на севере Монгольского Алтая, с середины апреля перемещается в Горный Алтай на хребет Сайлюгем, в основном, в верховья р. Баян-Чаган, где лучше летние пастбища. Отсюда часть животных спускается вниз по долине реки и, не доходя до ее устья, переваливает через отрог хребта в долину р. Саржематы. Протяженность миграционного пути - 10-25 км, продолжительность перехода - 1,5-2 недели. Летом звери, объединившись в крупные стада, перемещаются через каждые 3-4 дня на 3-8 км вверх-вниз по долинам этих рек. Откочевка зверей на юг в малоснежные районы Монголии происходит в октябре после пригона домашнего скота на зимние стоянки. С южной части хребта Лихачева аргали уходят зимовать на юг на сопредельную территорию Монголии и на запад в холмистую равнину у отрогов Курайского хребта. Из бассейна р. Богояш и верховьев р. Калбакан они кочуют на 25-30 км на восток в приграничные районы Тувы. На Талдуаире животные перемещаются с северных и северо-восточных склонов на южные и дальше на хребет Лихачева [Сопин, 19756, 1976а; Федосенко, 1989; Собанский, 1992]. В районе Алтайского заповедника звери кочуют, видимо, в пределах бассейнов рек Богояш, Баш-каус, Кобакая и Буйлюкем [Филус, Золотухин, 1987].
В Монголии сезонные перекочевки баранов отмечены на Монгольском и Гобийском Алтае, Хангае, в Заалтайской и Джунгарской Гоби. На севере республики они вызваны многоснежьем, на юге - недостатком воды [Груздев, Сухбат, 1982]. В Туве мигранты из Монголии регулярно появляются на южном макросклоне хребта Монгун-Тайга, но их численность с 70-х годов XX в. сократилась со 100 или более особей до 20-40. Еще меньше зверей приходит в районы, граничащие с Горно-Алтайской республикой [Ткаченко, 1992].
Очень подвижны архары в Казахском нагорье. Из Ерментау они откочевывают на зиму к востоку в горные кряжи Желтау, Бекежан-Акшокы и Акшокы, а в многоснежье - в Торежал и Керегетас. Летом, с появлением кровососущих насекомых, животные с гор уходят в мелкосопочник и межгорные понижения. С горных массивов Кошубай, Коныр-Темирши, Мыржик, Аркалык, Едрей и др. перемещается до 60-80% популяции. Эти кочевки, однако, более характерны для самок и менее присущи взрослым самцам, которые приходят в места их обитания к периоду гона, преодолевая в некоторых районах до 50-100 км. Пути миграций постоянны из года в год, причем переходы нередки и по ровной местности. Передвигаются звери неторопливо, но при опасности могут бежать со скоростью 55-60 км/ч [Слудский и др., 1983; Бекенов и др., 1999; Бербер, 1999]. В сильные засухи и пожары часть популяции в середине лета может мигрировать на север к горам Балхаш-Иртышс-кого водораздела или в другие горные массивы. В особенно засушливое лето 1974 г. из-за высыхания водоемов почти все животные покинули горы Калмак-Эмель, Бектаута и Таятхан и вернулись назад лишь в октябре. Оставшиеся бараны были вынуждены пить из источников вместе с овцами, на виду у людей [Капитонов, 1980, 1983]. Из Тарбагатая и Саура часть зверей перемещается в приграничные районы Китая [Федосенко, 2000].
Очень четко выражен миграционный путь животных из Таласского Алатау и Каратау. В конце сентября - декабре большая часть группировки спускается вниз
и идет по северным склонам Джабаглинского хребта, пересекает долины Аксая и Коксая, и у перевала Куюк поворачивает на север в Каратау. Мигранты из заповедника “Аксу-Джабаглы” за 2-3 дня преодолевают 40-70 км, форсируя при этом р. Терс и линию железной дороги восточнее станции Бурное. Весенняя миграция в обратном направлении начинается в марте - апреле и растягивается на месяц, причем отдельные группы, преимущественно самцовые, возвращаются назад через восточную оконечность Боролдайских гор, долину р. Арысь и Джабаглинс-кий хребет. Небольшая часть животных остается зимовать в Джабаглинских горах [Шапошников, 1956; Ковшарь, Янушко, 1965]. С сокращением поголовья кочевки стали малозаметными и бывают не ежегодно, также как в Заилийском Алатау и Терскей Алатау [Слудский и др., 1983].
В Заилийском Алатау из района Тау-Чилик звери в ноябре - декабре уходили на зимовку в восточном направлении к горам Большие и Малые Богуты, пересекая в долине р. Сюгаты 18-километровый степной участок. Размах сезонных кочевок в этом районе достигал нескольких десятков километров [Насимович, 1955]. Животные, обитающие в Сарытау и в верховьях Чилика, зимой появляются в малоснежных районах Кши-Тургеня и Ассинской долине, где остаются до мая. В Джунгарском Алатау бараны, зимовавшие в Калканах и Катутау, в конце мая -начале июня с появлением кровососущих насекомых и увяданием растительности переходят в соседние горы Алтын-Эмель, Матай и Чулак, пересекая населенную человеком равнину протяженностью в 15-20 км. Кочевка в обратном направлении происходит в сентябре - первой половине октября. Из восточной части Джунгарского Алатау архары в середине октября перемещаются за 10-30 км в Теректы, где протекает гон. С появлением домашнего скота на зимовочных пастбищах они откочевывают в верхние части окружающих гор [Антипин, 1947; Жиряков, 1973, 1976; Федосенко, 1975, 1977, 1979, 1983а, 2000; Слудский и др., 1983; Федосенко, Жиряков, 1987].
На Памире в особенно многоснежные зимы зарегистрированы откочевки архаров из окрестностей Кызылрабата в малоснежный район Тохтамыша на расстояние в 50 км [Сапожников, 19766, 1977]. В других районах перемещения баранов носят локальный характер и связаны в большей мере с вытеснением их из типичных местообитаний домашним скотом [Федосенко, 2000].
В высокогорье постоянные места отдыха животных обычно находятся выше пастбищ и к ним идут хорошо набитые тропы (фото 45). Лежки звери чаще всего устраивают на щебнистых участках склонов гор, под отдельными валунами (фото 46) или скалами, в нишах и пещерах. Многие из них архары используют в течение десятилетий, что приводит к скоплению по краям и вокруг толстого слоя (до 20-30 см) экскрементов, который осыпается вниз по склону, оставляя длинный и хорошо заметный шлейф. В пещерах и нишах самки нередко рожают, а затем долго держатся поблизости, заходя сюда вместе с ягнятами на отдых или в непогоду. Место лежки звери каждый раз очищают царапающими движениями передних ног. Эти движения у самцов носят скорее ритуальный характер, но самки чистят место для отдыха очень энергично и тщательно. Лучшие лежки занимают высокоранговые животные. Ложатся звери обычно так, чтобы в их поле зрения находилось если не все стадо, то несколько особей (фото 47). При таком расположении они имеют круговой обзор и быстро реагируют на поведение какой-либо особи, резко вскочившей
при опасности [Сопин, 19766; Капитонов, Махмутов, 1977; Слудский и др., 1983; Федосенко, Жиряков, 1987; Бербер, 1999; Федосенко, 2000].
Во многих местах на участках обитания животных находятся отполированные крупные камни и выступы скал, о которые они часто чешутся разными частями тела, пытаясь освободиться от эктопаразитов. Иногда им в этом помогают сороки и альпийские сойки, которые садятся на них и выклевывают клещей.
Численность
Динамика населения архара в XX в. На большей части ареала, за исключением малонаселенных и высокогорных районов, население горных баранов существенно уменьшилось еще в XIX в., а в начале XX в. эти животные почти полностью исчезли из легко доступных для человека холмо-, низко- и среднегорий [Слудский, 1956]. Благодаря запрету охоты, введенному в отдельных республиках в 20-30-е годы, удалось не только остановить падение численности, но и увеличить поголовье. Очередное резкое снижение (в несколько раз) ресурсов вида произошло в 60-80-е годы, прежде всего в связи с расширением пастбищного животноводства, а также из-за возросшего антропогенного пресса и браконьерства в частности. Общую численность архара в СССР в 60-80-е годы оценивали от 33 тыс. [Федосенко, 1983а, 1988] до 40-60 тыс. [Климова, 1989], но, по мнению Л.В. Сопина [1986а], здесь оставалось не более 15-20 тыс.
В 90-е годы за распадом СССР последовал глубокий экономический кризис. Поголовье домашнего скота, мощного конкурента архара, значительно сократилось во всех республиках (в Казахстане более чем в 10 раз [Байдавлетов, 2002]). На летних пастбищах стало меньше не только скота, но и людей и собак. Все это благоприятно отразилось на группировках горных баранов, однако далеко не повсеместно. В некоторых регионах очевидна противоположная тенденция, что связано с усилением браконьерства, возросшей численностью волка и валютной трофейной охотой.
Алтайский аргали. Численность аргали в России в начале 70-х годов была близка к 1 тыс. особей, из них 600 голов жили на Алтае (табл. 82). Во второй половине 70-х годов в этом регионе насчитывали 200 особей, в 1980 г. - 100, позднее -несколько десятков [Сопин, 1973, 19756, 1977а,в; Бондарев, 1982; Юдин, Собанский, 1984]. Авиаучеты и наземные наблюдения, проведенные в 1986 г., дали более точную цифру - 215 экземпляров, а с учетом пропуска зверей - около 280-300. Причиной недоучета животных в прошлые годы стала их откочевка на зимовку в соседние районы Монголии и Тувы [Федосенко и др., 1987;. Федосенко, 1989; Собанский, Федосенко, 1990; Собанский, 1992]. В Алтайском заповеднике и на смежной территории в верховьях рек Колбакая, Башкаус и Буйлюкем держатся 80-100 особей [Филус, 1992]. В конце 90-х годов на Алтае зимой насчитывали около 200 особей, летом за счет иммигрантов из Монголии и Тувы численность увеличивалась до 450-500 [Федосенко, 2000]. В Туве в 70-80-е годы обитали 200-270 голов при плотности около 2 особей на 1000 га угодий [Сулимов, 1977; Смирнов, 1980, 1988а, 1990, 1994], в 90-е годы численность сократилась до 100-150 экземпляров с учетом мигрантов [Ткаченко, 1992; Федосенко, 2000]. По сведениям М.Ю. Пальцы-на и С.В. Спицына [2002а,б], в 1999 г. в южной части хребта Чихачева и на Талдуаи-
ре в пределах Республики Алтай жили 140-150 баранов, примерно столько же - в северной части хребта в Туве и около 100 животных держалось в монгольской части этого горного массива. В Алтайском заповеднике осталось не более 40 особей. Общее поголовье аргали на данном участке ареала определено в 400 особей. По последней оценке А.К. Федосенко [20026], на территории Алтая и Тувы живет около 650-700 аргали. В Хакасии [Прокофьев, 1992], Бурятии [Смирнов, Сопин, 1988] и Читинской области [Кирилюк, 20006] регистрируют лишь редкие встречи этих животных.
Таблица 82. Численность аргалиобразных баранов (тыс. особей) в России и сопредельных республиках в XX в.*
Форма архара Страна, район Годы
50-60-е | 70-80-е | 90-е - начало XXI в.
Алтайская Россия 1,0 1-0,5 0,6-0,7
Алтай 0,6 0,2-0,3 0,4-0,5
Тува 0,4 0,25 0,1-0,15
Казахстан, Ю. Алтай 0,1 0,1 0,07-0,05
Монгольский Алтай - — Несколько
(Китай) сотен особей
С.-з. Монголии — 6 4-5
Гобийская Монголия, Китай - - 11**
Казахстанская Казахстан 16 7-10 12-6
Тянь-шаньская Киргизия — 20-4 10-15
Казахстан — 2-2,5 2
Китай — - 4,5-5
Каратауская Казахстан - 0,15 0,1
Кызылкумская Узбекистан 0,1-1,0 0,8-1,2 1,4-2,5
Памирская Таджикистан 70-80 20-10 5-14
Киргизия Несколько Несколько 6-7
десятков ТЫСЯЧ
тысяч
Тибетская Китай и соседние районы Тибета — — 7
Северокитайская Внутренняя Монголия — 0,6-0,7 —
* По приведенным в тексте сведениям. Прочерк - отсутствие достоверных сведений.
** По экспертной оценке.
По сведениям, собранным А.К. Федосенко [2000], в последнем десятилетии XX в. на Южном Алтае (на территории Казахстана) жили около 70 особей алтайского аргали (по данным Р. Байдавлетова [20036] - 50-55 голов); на западных склонах Монгольского Алтая в Китае - несколько сот; на северо-западе Монголии - около 4-6 тыс., из них 2,5-5 тыс. - на Монгольском Алтае, около 500-800 голов - в районах, примыкающих к Туве и 60-70 особей - в Прихубсугулье. Численность аргали на северо-западе Монголии в последние годы быстро сокращается из-за резкого увеличения поголовья скота и бесконтрольного использования пастбищ даже в заповедниках.
В Гобийском Алтае и Южной Гоби в 70-90-е годы насчитывали примерно по 4 тыс. особей, на территории Большого Гобийского заповедника - 900 голов, в Китае в
Заалтайской и Джунгарской Гоби - 1,5-2,0 тыс., во Внутренней Монголии - 300-400 особей [Федосенко, 2000].
Общую численность архара в Монголии в 1970 г. оценивали в 40 тыс., в последующие десятилетия - в пределах 12-20 тыс. [Сухбат, 1975; Груздев, Сухбат, 1982; Сухбат, Груздев, 1986; Lushchekina, Dulamtseren, 2000]. По другой оценке, скорее всего, сильно завышенной, в конце века здесь обитали около 50-60 тыс. особей [Reading et al., 2002].
Североказахстанский баран. В Казахском нагорье в начале 60-х годов XX в. насчитывали около 16 тыс. голов [Смирнов, 1965]. В 60-70-е годы популяция уменьшилась, вероятно, до 7-5 тыс. [Савинов, 1974,19756; Федосенко, Грачев, 1975; Слудский и др., 1983; Федосенко, 1983а], но позднее отмечен рост. В конце 80-х - начале 90-х годов здесь учли около 10-12 тыс., в том числе в Карагандинской области - 7 тыс., Джезказганской - 2 тыс. В Тарбагатае жили около 600 особей, в Сауре - не более 100. С середины последнего десятилетия заметно снижение ресурсов (в отдельных районах в 1,5-2 раза) из-за возросшей численности волка, массового браконьерства, пожаров и трофейной охоты. В конце XX - начале XXI вв. в Карагандинской области учтено 3,1 тыс., на Тарбагатае - 1,1 тыс., в горных массивах Чингизстау, Канчингиз и Акштау - 300, в Монраке - 130, Сауре - 100, Колбинском Алтае - 50 и не менее 1420 голов - в других горных массивах, а всего около 6,2 тыс. Вместе с особями других форм (см. ниже) в Казахстане обитает до 11 тыс. баранов [Байдавлетов, 1996,20036; Бекенов и др., 1999; Бербер, 1999; Федосенко, 2000].
Каратауский баран. В горах Каратау в 1976 г. было около 750 голов, в конце 70-х - 400, из них 150 - каратауского подвида и 250 - тянь-шаньского [Грачев, 1982]. В 1980-1982 гг. численность каратауского барана уменьшилась до нескольких десятков, и даже предполагали, что он исчез [Соколов, 1986]. А.К. Федосенко [1997], однако, оценивал его запасы в 300 голов. По исследованиям Р.Ж. Байдавле-това [2003а,б], в конце XX в. в западной части Каратау жили около 100 зверей на площади 220 тыс. га со средней плотностью 0,45 экземпляра на 1000 га.
Кызылкумский баран, или баран Северцова. К 1959 г. в Актау и Нуратау сохранилось, как считали, по нескольку десятков особей (скорее всего, оставалось несколько сотен -АД). После образования Нуратинского заказника поголовье быстро увеличилось. В 60-90-е годы в Узбекистане насчитали от 800 до 2500 особей, в том числе в Нуратинском заповеднике - 550-1200 [Ишунин, 1967, 19836; Коршунов, Коршунова, 1983; Коршунова, Коршунов, 1983; Шакула, Снитько, 1995; Федосенко, 1997; Абдуназаров, 2003]. Из хребтов Нуратау и Актау звери стали заходить на территорию Казахстана в массивы Карактау и Койтас [Байдавлетов, 20036].
Тянь-шаньский баран. Численность тянь-шаньского барана была высокой до середины 50-х годов XX в. В предгорных районах Казахстана сотенные стада выходили на кормежку в степь, а охотники привозили их туши телегами [Федосенко, 1983а]. В 60-80-е годы ресурсы стала уменьшаться по мере роста поголовья домашнего скота, браконьерства и эпизоотий зудневой чесотки в локальных районах, а часть периферийных группировок полностью исчезла. Сокращение поголовья произошло даже в заповедниках. В “Аксу-Джабаглы” в Казахстане в 1933-1935 гг. насчитывали 60-80 особей, 1943 г. - 281, 1950 г. - 670, 1972 г. - 331, 1978 г. - 91, 1981 г. - 47 экземпляров [Шульпин, 1948; Ковшарь, Янушко, 1965; Ковшарь, 1977; Мельников, 1980; Бургело, 1986]. В казахстанской части ареала в конце 80-х годов обитали около 2-2,5 тыс. [Фе
досенко, 2000]. В начале XXI в. на Тянь-Шане осталось около 900 голов, из них 150 - на Западном, 200 - Северном, 400 - Центральном и 150 - в Чу-Илийских горах. В Джунгарском Алатау современная численность близка к 800 особей [Байдавлетов, 20036].
В Киргизии сокращение численности горных баранов стало заметным в конце 50-х годов, что связано с начавшимся хозяйственным освоением высокогорных сыртов и круглогодичной охотой [Айзин, 1969]. Однако до начала 80-е годов их плотность оставалось высокой. К концу десятилетия из-за антропогенного пресса и огромной пастбищной нагрузки (выпасали до 10 млн домашних овец) ресурсы вида уменьшились с 20 до 4,2-6 тыс. В 90-е годы поголовье домашнего скота резко сокращается, а численность архара увеличивается до 10-15 тыс. [Кошкарев, 2002]. В Терскей Алатау группировка из нескольких десятков особей выросла до 1 тыс. [Шубин и др., 2002]. На Восточном Тянь-Шане и в горах, граничащих с Джунгарскими воротами (Китай), в начале 90-х годов находилось около 4,5-5 тыс. [Федосенко, 2000]. Общая численность тянь-шанского барана в начале XXI в., видимо, близка к 15-20 тыс.
Памирский баран. На Памире в 30-50-е годы XX в. поголовье архара было настолько велико, что многочисленные стада полностью стравливали высокогорные пастбища. В некоторых долинах одновозрастные группы животных в утренние и вечерние часы покрывали все нижние участки склонов [Егоров, 1955]. Т.Н. Сапожников [1973, 1976а,б, 1977] в 60-е годы видел здесь тысячные стада и оценивал популяцию в 70-80 тыс. А.И. Соков [1975, 1977а,б] считал эти цифры сомнительными, определяя численность в 18-20 тыс. В 1977 г. насчитывали не более 18 тыс., в конце 80-х- 12-10 тыс. [Одинашоев и др., 1977; Соков, 1989]. К 1991 г. ресурсы вида в Таджикистане сократилось до 9,4 тыс., в 1993 г. - до 5, а в 2000-2001 гг. - до 3 тыс. [Абдусалямов, 1997, 2003]. Однако, по исследованиям А.К. Федосенко [2000], в середине 90-х годов здесь жили около 9-9,5 тыс. баранов, а к концу века, благодаря резкому сокращению скота, поголовье архара увеличилось до 13-14 тыс. Позднее М.-Р.Д. Магомедов с соавторами [2003] определили, что только в Горно-Бадахшанской области обитает около 14,5 тыс. с плотностью 14-22 экземпляра на 10 км2.
На киргизской части ареала ранее население памирского барана исчислялось десятками тысяч, в 70-х годах - несколькими тысячами, в том числе около 2 тыс. обитали в бассейне р. Аксай [Айзин, 1977] и до 3,5 тыс. - во Внутреннем Тянь-Шане [Андреенков, 1983]. В начале 90-х годов в нескольких локальных группировках, расположенных вдоль границы с Китаем, насчитывали около 6-7 тыс. особей, в том числе в ферганской - 1,8 тыс., аксайской - 3,1, мюдюрюмской - 0,5-0,9, узенгикуш-ской - 1,3 тыс. [Андреенков, 1991; Федосенко и др., 1995; Федосенко, 2000].
До 60-65% поголовья вида в Таджикистане и Киргизии находится за линией инженерных проволочных сооружений, протяженностью около 300 км и шириной от нескольких километров до 30 км, в приграничной зоне с Китаем. В этом своеобразном резервате площадью около 500 тыс. га, куда ограничен доступ местных жителей и скота, плотность популяции баранав 6 раз выше, чем вне его [Федосенко, 1997,2000; Федосенко, Вейнберг, 1999].
Общая численность памирского барана близка к 20 тыс. [Федосенко, Вейнберг, 2001; Fedosenko, 2003].
Таблица 83. Плотность населения аргалиобразных баранов
Район,заповедник | | Особей на 1000 га | | Источник информации
Алтай 3-8 Сопин, 1977в
и 0,02-0,16 Анчифоров, Фалеев, 1993
и 1,3-3,5 Абатуров и др., 2004
Тува 2,3 Смирнов, 1990
Монгольский Алтай 0,09-0,6 tl
Заалтайская Гоби 0,5-0,6 Соколов и др., 1996
Джунгарская Гоби 0,8 а
Монголия 0,5 (0,1-15) Сухбат, Груздев, 1986
Памир 2,6 Соков, 1975, 1977а,б
разные районы 36-80 Сапожников, 19766, 1977
приграничные районы 25-40 Федосенко, Вейнберг, 1999
Горный Бадахшан 14-22 Магомедов и др., 2003
Ю. Киргизия 25 Fedosenko etal., 1995
Казахское нагорье 1,8-2,2 (0,3-12,4 на разных участках) Капитонов, Махмутов, 1977; Савинов, 1974; Федосенко, 1988, 2000; Бербер, 1999
Центральный Казахстан 6,1 (0,4-11,2) Бекенов и др., 1999; Байдавле-тов, 20036
Тарбагатай 1,2-1,6 (0,3-6,8) Федосенко, 1988; Байдавлетов, 20036
Каратау 1,5-2,4 Грачев, 1982
Западная часть Каратау 0,45 Байдавлетов, 2003 а
Терскей Алатау 1,1-2,1 Ганюшин, 1977
Заповедник “Аксу-Джабаглы” 17-25 Вырыпаев, 1983а
Джунгарский Алатау 0,8-11,9 Байдавлетов, 20036
Горы Ю.-З. Прибалхашья 0,6-7 Федосенко, 2000
Алма-Атинский заповедник 8-10 Жиряков, 1976
Нуратинский заказник 50 Ишунин, 1967
Тибетский баран. Численность этих копытных, по одним источникам, не превышает 7 тыс., по другим - достигает 29-36 тыс., в том числе в Синьцзяне и Ганьсу -19-23, в Тибетском автономном районе - 7-8 тыс. [Piao, 1996; Schaller, 1998; Федосенко, 2000].
Северокитайский баран. По сведениям китайских исследователей, общая его численность не более 600-700 особей [Федосенко, 2000].
Плотность населения архара в большинстве районов обитания не превышает 3 особей на 1000 га угодий, но в хорошо охраняемых заповедниках и на труднодоступных участках достигает 10-80 голов на ту же площадь (табл. 83).
Численность муфлона. Резкие изменения численности, вызванные, прежде всего, истреблением, наблюдались не только в популяциях архара, но и в популяциях муфлона и уриала.
Европейский муфлон, ранее обитавший во всех горных областях Европы, к XX в. почти исчез, за исключением островов Корсики, Сардинии и Кипра (куда, возможно, был завезен в античное время из Малой Азии) и резервата Конья-Боздач в Турции в 170 км от Анкары. От некогда многотысячных популяций сохранились от 15 особей до 1-3 сотен, а на Сардинии - около 650-750 голов [Reinhardt, Schenk, 1969]. И лишь масштабное искусственное расселение позволило не только вос
становить, но и значительно расширить ареал этих копытных. Звери, реинтро-дуцированные в Западной и Центральной Европе и акклиматизированные в Северной Америке, размножились и заняли не только горные, но и относительно равнинные районы. Европейский муфлон снова стал массовым охотничьим объектом. В 1968 г. его поголовье (вместе с гибридами) оценивали в 32 тыс., в 70-80-е годы-в 60-70 тыс. [Дежкин, 1980, 1983; Кузякин, 1985; Соколов, 1986; Стенин, 1986; Лобанов, 1988; Briedermann, 1989]. К концу века в 23 европейских странах насчитывали 94 тыс. особей (в том числе в Чехии - 20, Германии - 19, Венгрии - 11, Австрии, Франции и Словакии - 6-7,5, Болгарии, Италии и Испании - 3-4 тыс.), в США - около 4 тыс., мировая популяция превышает 100 тыс. [Piegert, 2003].
В пределах бывшего СССР муфлона неоднократно, начиная с 1894 г. и до середины 80-х годов прошлого века, вселяли в разные области Украины и Крым, в 60-80-е годы - в Белоруссию, Литву и Латвию. В 1982 г. 16 особей из Чехословакии завезли в Россию в Адлерское гослесоохотничье хозяйство. Во многих районах звери прижились и дали потомство, однако в целом вольное разведение признано не перспективным, за исключением горных районов Крыма. Но и здесь за весь многолетний период разведения (с 1913 г.) группировка, главным образом, из-за браконьерства не превышала 0,5 тыс. Общая численность этого копытного во всех республиках СССР в конце XX в. составляла 800-900 особей [Ткаченко, 1963; Ду-лицкий, Кормилицын, 1970; Керечун, 1974; Треус, Лобанов, 1976; Мицкус, Турса, 1978; Болденков, Крайнев, 1980; Перовский, 1984; Лобанов, 1988; Паршинцев, 1992; Павлов, 1999; Piegert, 2003; Woloch, 2003].
Численность арменийского муфлона, обитающего в пределах бывшего СССР, скорее всего, не превышает 1,5 тыс. В Грузии в 1985 г. оставалось всего около 40 особей в Кедском районе Аджарии в нейтральной зоне на границе с Турцией [Арабули, 1989]. В Армении, где в первой половине XX в. встречались стада в 300 и более особей, насчитывают всего 350-400 голов с учетом мигрантов из нахичеванской части ареала. Оседлых животных из-за истребительной и хозяйственной деятельности человека сохранилось всего несколько десятков, несмотря на запрет охоты с 1936 г. [Явруян, 1970, 1975; Айрумян, Гаспарян, 1976; Гаспарян, Маркарян, 1977]. В Нахичевани в 70-90-е годы обитали 1-1,2 тыс. особей на площади в 68 тыс. га, что на 60-70% меньше, чем в середине столетия. В 1998 г. уцелело около 450-500 особей в Ордубадском национальном парке, на Неграмдаге, Дарыдаге и Иландаге. Основная причина сокращения ресурсов и ареала этой формы - интенсивное освоение территории под пастбища для домашнего скота и массовое браконьерство [Алекперов и др., 1976; Алекперов, Кулиев, 1981; Кулиев, 1997, 2004].
Численность уриала. На территории Арало-Каспийского водораздела выделяют 3 группировки устюртского уриала (фото 34): казахстанскую (от Северного чинка Устюрта до уступов Каясанирек на юге), северотуркменскую (от Южного чинка до хребта Большой Балхан) и почти исчезнувшую каракалпакскую (от Восточного чинка до Сарыкамышской котловины и впадины Акчакая). В прошлом этот участок ареала был сплошным, а “аркаров” было много [Рычков, 1762 (по изд. 1999); Карелин, 1883; Ишадов, 1965; Мамбетжумаев, 1969; Ишунин и др., 1981; Ишунин, 1983а; Горбунов, 19956; Горелов, Ишадов, 1999; Плахов, 1999, 2003а].
В Казахстане в середине XX в. обитали 7-10 тыс. голов устюртского уриала, в 60-70-е годы его численность из-за неумеренного антропогенного воздействия сократились до 2-3 тыс., в 1982 г. - до 1,5 тыс., но позднее, по мере снижения негативного пресса, выросла и в 90-е годы достигла 5,5-6,5 тыс. В 1992-2000 гг. проведен эксперимент по трофейной эксплуатации баранов - добыт 91 взрослый самец. К 2002 г. из-за резко возросшего браконьерства и большого количества волка группировка снова уменьшилась до 2-2,5 тыс. [Савинов, Бекенов, 1977; Бекенов, Савинов, 1980, 1983; Слудский и др., 1983; Бекенов, 1997; Плахов, 1997, 2003аб; Федосенко, 2002а]. На территории Узбекистана (в Каракалпакии) уриала уже нет на Восточном чинке Устюрта, а на Южный чинок до 100-350 особей регулярно заходят из Туркмении. Основная причина сокращения популяции та же - хозяйственное освоение пустынных земель и браконьерство [Ишунин, 1983а; Реймов и др., 1989; Реймов, Реймов, 1997; Плахов, 1999]. Поголовье северотуркменской группировки из-за браконьерства к середине 80-х годов сократилось с 5-7 до 1,4 тыс., позднее - до 1 тыс., а область обитания уменьшилась втрое [Зархидзе, 1983а; Горбунов, 1983, 1987, 19956; Зархидзе, Горбунов, 1983; Плахов, 2003а].
Более многочислен туркменский баран (“cycloceros ”), но и его ресурсы и ареал заметно уменьшились к середине XX в. [Сапоженков, 1969]. Охота на него велась круглогодично без ограничений, причем добывались и беременные и лактирую-щие самки и ягнята, что стало основной причиной резкого снижения поголовья во многих районах [Цалкин, 1948в; Шукуров, 1962]. В 60-е годы на Балканах осталось около 200 особей, а к 1996 г. эта группировка практически исчезла. На Сюнт-Хасардаге в 90-е годы популяция уменьшилось вдвое (до 100 особей). В Бадхызе в довоенные годы насчитывали до 3-4 тыс. баранов, в 60-80-е - 1,5-2,5 тыс., в 1995 г. - 1210 особей, в 1998 г. - около 700 голов. Здесь он является объектом трофейной охоты. На приграничных хребтах на правом берегу р. Теджен и в куш-ко-мургабском междуречье тысячная группировка практически истреблена [Горелова, 1980; Соколов, 1986; Горелов, 1999; Горелов, Ишадов, 1999; Федосенко и др., 2000]. На Западном Копетдаге в отдельных очагах осталось от 15 до 200 особей, а на центральном участке заповедника - не более 60-80 [Лукаревский, 1989,1999]. В Центральном Копетдаге в первой половине XX в. популяция была особенно многочисленной. За день не составляло труда встретить до полутысячи зверей. В начале 40-х годов бригада охотников за сезон (с ноября по февраль) добывала до 430-480 баранов и безоаровых козлов, или до 106-120 голов на человека. Общая добыча уриала достигала примерно 1000 голов в год, что привело к подрыву запасов и к омоложению популяции. Из 906 исследованных особей только 2 экземпляра были старше 7 лет [Цалкин, 1948а,в]. В середине XX в. здесь жили около 3,5-4 тыс. баранов. К 1984 г., благодаря учреждению Копетдагского заповедника, поголовье в этом регионе увеличилось до 8 тыс. На Восточном Копетдаге насчитывали 1,4 тыс. особей. Общие ресурсы этого копытного в Туркмении в 70-80-е годы оценивали примерно 10-12 тыс. [Ишадов, 1965, 1977; Коршунов, 1986, 19886,1995в; Федосенко, 1997]. К 1996 г. популяция в Центральном Копетдаге уменьшилась до 2 тыс., заметно сократилась она и в восточной части хребта. Основная причина уменьшения ареала и ресурсов - браконьерство и валютная охота в заповедниках и заказниках [Горелов, Ишадов, 1999; Лукаревский, 1999]. В связи с сокращающейся чис
ленностью уриал занесен в Красные книги СССР (1984), Туркменистана (1985), МСОП (1995) и списки CITES (1997).
Поголовье туркменского уриала в Иране в середине 70-х годов достигало 20 тыс., но после исламской революции и войны уменьшилось в несколько раз. В Афганистане обитает, видимо, несколько тысяч особей [Федосенко, 2002а].
Численность бухарского уриала (“bochariensis ”) в Узбекистане, распространенного от хребта Кугитанг на восток до гор правобережья Амударьи и нижнего Пянджа, а на севере - до Заравшана, заметно сократилась в 70-е годы, но он еще встречался в Бабатаге и на Кугитанге [Султанов, 1980; Ишунин, 1982а,б, 1983в]. В 80-е годы его считали исчезнувшим [Реймов и др., 1989], но затем насчитали около 200 особей [Абдуназаров, 2003]. В Кугитанском заповеднике сохранилось не более сотни животных [Федосенко и др., 2000]. В исчезающей (около 20 голов в 1968— 1989 гг.) туркменской группировке к 1990 г. выжила лишь одна самка с ягненком [Коршунов, 1995в; Горелов, Ишадов, 1999]. В Таджикистане бухарский баран (фото 33) до середины XX в. повсеместно населял низкогорье (фото 48), был многочисленным, а в его стадах регистрировали сотни голов. По воздействием антропогенного фактора таджикская группировка в 70-90-е годы сократилась до 1,4-1,2 тыс., а к концу века еще не менее чем вдвое [Флеров, 19356; Соков, 1977а, 1989; Фролов, Голуб, 1983; Соколов, 1986; Федосенко, 1997, 2002а]. В долине Пянджа в 2002 г. уриал был очень редок [Вейнберг, 2003].
Современная численность ладакского и пенджабского уриалов близка к 1-1,5 тыс., белуджистанского - к 3 тыс., поголовье оманской популяции не определено [Федосенко, 2002а]. В целом население уриала продолжает неуклонно сокращаться.
Основные факторы, определяющие динамику популяций
Воспроизводство, смертность, прирост. В 80-90-е годы XX в. на Алтае на 100 взрослых самок приходилось в среднем 50-68 ягнят, в Южной Гоби - 37-44, на Памире - 32-58, на Тянь-Шане - 37-71 [Федосенко, 1989, 2000; Fedosenko et al., 1995; Федосенко, Вейнберг, 2001], в Казахском нагорье - 58 (23-106) [Бербер, 1999]. В Казахском нагорье этот показатель в северной группировке составлял 42-67, в южной - 22-46. Разница в уровне воспроизводства была связана с неодинаковыми кормовыми условиями и прессом трофейной охоты [Бекенов и ДР-, 1999].
Ягнята архара весьма жизнестойки, и их отход в первые недели после рождения незначителен - около 2% в общей смертности [Байдавлетов, 2002]. Летом гибель сеголеток существенно возрастает, и у алтайского аргали к осени может достигать 12-36%, к марту остается 46% от числа родившихся, к июлю - 37, к двум годам - 25%. У тянь-шаньского барана летом погибает до 36% ягнят, у памирского за год - 51-69 [Федосенко, 1983а, 1989, 2000; Федосенко, Вейнберг, 2001]. По данным М.Ю. Пальцына и С.В. Спицына [2002], на хребте Лихачева до полугодовалого возраста доживают 72% ягнят, до 1 года - 50-55. В Джунгарском Алатау до 6-месячного возраста сохраняются 75% сеголеток [Анненков, Литун, 1989]. В Центральном Казахстане в июле - августе на одну самку приходится 0,6 детеныша, в сентябре - октябре - 0,3, в феврале - марте - 0,2 и практически не остается двоен. Отход молодняка за год достигает 72% [Бербер, 1999]. Годовой прирост популя
ций горных баранов в разных районах колеблется от 5 до 35% [Сопин, 19756; Dawaa и. а., 1983], в неблагоприятные годы его значения отрицательные.
Уровень воспроизводства и детской смертности у уриала примерно такой же, как и у архара. В Центральном Копетдаге осенью на одну самку приходится 0,4 ягненка, а к весне остается 0,35 [Коршунов, 1988, 1995в], на Западном Устюрте в течение года погибает до 70% приплода [Федосенко, 19866]. У муфлона, судя по литературным сведениям, ежегодный прирост существенно выше.
Погода и корма. Наибольшие потери популяции горных баранов несут в многоснежные зимы. На Алтае они повторяются через 9-10 лет [Сопин, 1973]. О высокой смертности зверей от истощения в районах, граничащих с Монголией, во второй половине XIX в. упоминал путешественник П.М. Певцов [1880, по изд. 1951]. Основной причиной полного вымирания архара в Забайкалье и Хэнтэе, помимо истребления человеком, считается аномальная зима 1831/32 гг. В Северо-Западной Монголии значительный падеж животных был в 70-е годы XIX в., в 1983/84 и 1992/ 93 гг. Высокая смертность аргали и других копытных отмечена зимой 1912, 1965/ 66 и 1968/69 гг. Трупы истощенных баранов и “кладбища” (десятки пар рогов) регистрировали наМонгольском Алтае, в Заалтайской и Южной Гоби [Колосов, 1939; Формозов, 1946; Собанский, 1992; Соколов и др., 1996; Федосенко, 2000].
В Казахстане большие джуты бывают каждые 10-12 лет, и от них страдают не только степные, но и горные животные. Массовая гибель архара от истощения имела место зимой 1950/51 гг. в Чу-Илийских горах и в Казахском нагорье [Слудский, 1956], в 1961/62 - в восточной части Джунгарского Алатау, в 1965/66 - по всему ареалу, 1966/67 - на Чингизтау и Тарбагатае, 1968/69 - в Калканах и Ерментау, 1971/72 - в Сырдарьинском Каратау, в 1975/76 гг. - в Таласском Алатау [Савинов, 19756; Капитонов, Махмутов, 1977; Федосенко, 1983а, 2000]. Долю погибших от истощения баранов в их общей смертности Р. Байдавлетов [2002] оценивает в 8%, что в три раза меньше гибели от волка и браконьеров. На Памире многоснежные зимы редки, и поэтому потери здесь меньше, нежели в других районах. Тем не менее в суровую зиму 1985/86 гг. отмечена гибель части поголовья [Федосенко, 2000; Федосенко, Вейнберг, 2001].
Хищники. Основной потребитель горных баранов - волк. Его влияние на динамику популяций жертв особенно заметно в Казахстане, где гибель архара от этого хищника с 1993 г. стала большей (40-73%), нежели от браконьеров (20-37%). В заповеднике “Аксу-Джабаглы” остатки архара содержатся в 7% экскрементов волка, в Калканах - в 35%. В Сырдарьинском Каратау и Казахском нагорье, где соотношение хищник-жертва составляет 1:8-21, доля погибших от волка баранов достигает 24-35% в общей смертности. В январе-марте 1991-1997 гг. только на хребтах Мыржик, Коныр-Темирши и Кошубай на площади 500 км2 обнаружены останки 109 архаров, из которых 84 (77%) были задавлены волком. Среди этих жертв взрослые самцы составляли 84%. Весной этот хищник чаще давит ягнят. Их свежие останки обнаружены у 19 из 39 осмотренных логов в Казахском нагорье. Свою лепту в смертность копытных вносят также рысь (1,4-1,8%), снежный барс (0,8), лисица (1,1), крупные хищные птицы (3,2) и бродячие собаки (7,9%) [Ганюшин, 1977; Слудский и др., 1983; Федосенко, 1983а,в, 1986а, 2000; Федосенко, Жиряков, 1987; Байдавлетов, 1996, 2002; Бекенов, Байдавлетов, 1997; Бербер, 1999]. На Устюрте волк с не меньшим успехом давит уриала [Федосенко, 19866].
Волк - главный враг архара и на Центральном Тянь-Шане, где встречаемость жертвы в рационе хищника в осенне-зимний период достигает 36% [Вырыпаев, 1980, 19836]. На Памире урон от него и от малочисленного снежного барса составляет 7% в общей смертности вида. В некоторых урочищах после многоснежных зим находили останки 60-80 баранов (до 20% группировки), убитых волками. Среди погибших доля взрослых самцов наиболее высокая (72-78%). Самки гибнут гораздо реже (около 3%), что, возможно, связано с большей скоростью их бега (до 60 км/ч) по сравнению с массивными взрослыми самцами (до 50 км/ч), оснащенными тяжелыми рогами. Грузные бараны гораздо медленнее самок передвигаются и по снегу с настом, что позволяет хищникам давить сразу нескольких особей. Черепа самок и молодых животных чаще разгрызаются и растаскиваются хищниками, поэтому в местах традиционных охот волков со временем образуются скопления черепов преимущественно взрослых самцов. К числу второстепенных хищников в этом регионе относят красного волка, медведя, лисицу, рысь, собак, беркута, ягнятника и грифа-бородача [Мекленбурцев, 1948; Егоров, 1955; Айзин, 1969; Сапожников, 1977; Федосенко и др., 1995; Федосенко, 2000; Федосенко, Вейнберг, 2001; Кошкарев, 2002].
Ущерб от волка ощутим и на Алтае, и в Монголии, особенно в многоснежные годы и при сокращении поголовья домашнего скота. И здесь он давит чаще взрослых самцов и молодняк. Снежный барс, рысь, росомаха, бурый медведь, лисица и крупные хищные птицы или малочисленны, или наносят гораздо меньший урон популяциям. Более губительным хищником являются вездесущие пастушьи собаки [Сопин, 19756,1976г, 1977в; Груздев, Сухбат, 1982; Собанский, 1992; Соколов и ДР-, 1996].
Конкуренты. В высокогорных районах бараны конкурирует за пищу с горными козлами, но стации этих копытных все же отличаются. Часть пастбищ заметно стравливают сурки и суслики, однако эти потребители растительных кормов впадают в спячку и в зимний период выпадают из числа конкурентов.
Главный конкурент архара - домашний скот, особенно в зимнее время. Яки, овцы, козы, лошади зачастую выпасаются на тех же малоснежных или бесснежных участках, где концентрируются дикие копытные. На Алтае в некоторых долинах находится одновременно до 6 отар овец и коз по 800-1000 голов в каждой [Собанский, 1988]. На зимних пастбищах в Сарджементы и Баян-Чагане плотность домашнего скота такова, что весной их помет остается на каждом квадратном метре [Федосенко, 2000]. При перевыпасе горные растительные сообщества деградируют, происходит разрушение дерновины и эрозия почвенного слоя. В верховьях Чаган-Бургазы проективное покрытие летом составляет всего 19-28%, зимой 6-12%, а запас наземной фитомассы (воздушно-сухой) -30-50 г/м2, что в 2-3 раза ниже, чем в сходных биотопах Монголии [Анчифоров, Фалеев, 1993].
Домашний скот не только сильно стравливает пастбища, но и вытесняет диких копытных в худшие биотопы. В засушливые годы он скапливается у родников и других водных источников, затрудняя получение воды. В результате звери вынуждены эмигрировать в другие районы или уходят в зиму истощенными, что существенно увеличивает смертность. Кроме того, в результате постоянного контакта с домашним скотом дикие копытные заражаются как гельминтами, так и инфекционными болезнями от больных и павших животных, трупы которых в горах практически не
утилизируют [Сопин, 1973, 19756; Федосенко, Жиряков, 1977; Андреенков, 1980, 1983; Вырыпаев, 1980, 19836; Груздев, Сухбат, 1982; Коршунов, Коршунова, 1983; Федосенко, 1983в, 1988, 2000; Собанский, 1992]. В этой связи В.И. Цалкин [1945, 1951а] совершенно справедливо подчеркивал, что наряду с человеком “злейший враг дикого барана есть баран домашний”.
Болезни и паразиты. Влияние болезней на динамику населения архара изучено слабо, а сведения об эпизоотиях фрагментарны. На Памире массовый падеж животных от сибирской язвы наблюдался в 1895-1898 гг. В 1886-1887 гг. какая-то эпизоотия, поражавшая легкие, унесла половину поголовья [Мекленбурцев, 1948]. Не исключено, что некоторые эпизоотии могли быть вызваны протостронгилидами [Ур-сова, 1985]. Известны случаи гибели зверей от некробациллеза, пастереллеза, пироплазмоза и зудневой чесотки [Федосенко, 1983а]. Эпизоотия зудневой чесотки, сопровождавшаяся гибелью и сибирского козерога, и архара, отмечена в заповеднике “Аксу-Джабаглы” в Таласском Алатау в 1968-1971 гг. [Ивашенко, 1973].
В Казахстане у архара найдено более 35 видов гельминтов, из которых большинство типичны для домашнего скота и человека. Видоспецифичных паразитов у него не обнаружено [Слудский и др., 1983; Федосенко, 1983а, 2000; Белякова, Байдавлетов, 1997; Бербер, 1999; Дзержинский и др., 2003]. В Казахском мелкосопочнике зарегистрирован мощный очаг дикроцелиоза и эхинококкоза, в котором архар служит, в первом случае, окончательным, во втором - промежуточным хозяином. Рост инвазии связан с увеличением численности волка и бродячих собак [Белякова, Байдавлетов, 1999]. У казахстанских баранов зарегистрировано также 7 видов эктопаразитов: иксодовые и чесоточные клещи, овод, власоед и во множестве - муха-кровососка, которая особенно досаждает животным [Белякова, Ахметова, 2002].
На Памире, а также в Джунгарском Алатау и Каратау бараны сильно поражены кожным оводом. В других районах Казахстана этих копытных особенно изнуряет носоглоточный овод. В жару животные нередко стоят в угнетенной позе, опустив голову в собственную тень или уткнув ноздри в песок. Время от времени они бьют передней ногой, стараясь поднять пыль, чтобы отогнать оводов или лишить их активности хотя бы на короткое время [Мекленбурцев, 1948; Сапожников, 19766, 1977; Федосенко, 1983а, 2000].
Травматизм, гибель в лавинах. У архара, как и у других горных животных, нередки травмы конечностей. Изредка животные срываются со скал [Филус, 1992]. Значительное их количество погибает в лавинах. В 1942 г. в заповеднике “Аксу-Джабаглы” в одной из них обнаружено сразу 18 баранов [Насимович, 1955]. По наблюдениям Р. Байдавлетова [2002], в многоснежные годы на Тянь-Шане и в Джунгарском Алатау доля зверей, погибших от этого фактора, достигает 20-25%.
Антропогенные факторы. Динамика населения разных форм и группировок баранов во второй половине XX в. не была цикличной и в большей мере определялась антропогенными факторами, нежели климатическими.
Архар, пожалуй, один из самых притесняемых человеком и домашними животными видов, о чем упоминалось выше. Хозяйственное освоение предгорных и горных районов, особенно распашка межгорных участков, геологоразведочные и взрывные работы, добыча полезных ископаемых с применением тяжелой техники и вертолетов, демаркация государственных границ и строительство пограничных со
оружений, дорог и населенных пунктов также внесли немалую лепту в усиление беспокойства копытных и сокращение их численности. В степных районах в последние десятилетия участились пожары, которые приводят к вынужденному выселению зверей или гибели [Жирнов и др., 1975; Жиряков, 1976; Айзин, 1977; Вырыпаев, 1980, 19836; Федосенко, 1983а, 2000; Собанский, 1992; Абдусалямов, 1997; Бекенов и др., 1999; Бербер, 1999].
В большинстве районов среди всех факторов смертности архара все же доминирует охота. Охотники преследуют баранов повсеместно. Особенно интенсивно добывают их чабаны, геологи, пограничники, охотники за сурками, а в последние годы, по моим сведениям, охотники за трофеями. Автотранспорт и вертолет сделали доступными практически все угодья, а дальнобойное нарезное оружие оставляет все меньше шансов для зверей. Далеко не случайно на Алтае, в Туве, Киргизии и в Таджикистане группировки баранов уцелели в основном в приграничных районах, где антропогенный пресс ослаблен в связи с пограничным режимом. Масштабы охотничьего воздействия на популяции рассматриваются ниже.
Использование ресурсов и охрана
Охота на архара ведется со времен палеолита. Его добывают, в основном, ради вкусного мяса. Шкуры недолговечны, и используются населением для подстилки в юртах, хотя они могут служить ценным сырьем для выделки кож, которые идут на изготовление верхней части обуви [Денисов, 1935]. Черепа и рога самцов в безлесных районах применяют в качестве строительного материала для заборов и стен, а также в качестве настенных украшений и сувениров. В последнее время приоритетной становится трофейная охота.
Использование ресурсов. Объем добычи архара в целом весьма значителен. Во многих горных районах местное население почти половину всего мяса получало и получает до сих пор за его счет.
В XIX - начале XX в. в горах Центральной и Средней Азии путешественников поражали груды черепов взрослых самцов с рогами, оставляемых охотниками в традиционных местах [Чернавин, 1916]. Позднее воинские подразделения во многом обеспечивали себя продовольствием за счет диких баранов, применяя для их отстрела дальнобойное нарезное и автоматическое оружие, вплоть до станковых пулеметов [Цалкин, 1945]. В Алайской долине, например, бойцы пограничной заставы Бор-даба за зиму 1933/34 гг. заготовили 7 т архарьего мяса, а рога весной с трудом вывезли на 1,5-тонном грузовике [Кузнецов, 1948а]. В Киргизии в 1934 г. заготовлено 28480 шкур архаров и сибирских козлов. Учитывая, что около 10% шкур осело у местного населения, общее число добытых животных превышает 31 тыс. [Денисов, 1935].
На Памире в 30-50-е годы XX в. ежегодно заготавливали около 3,5^4- тыс. шкур, при этом еще больше их оставалась у местного населения или выбрасывалось. Охотники убивали за год в среднем 20-25 баранов, а лучшие из них - до 50. Большую роль на охоте играли собаки, загонявшие зверей на стрелков или на отстой. Несмотря на запрет охоты, вводимый в 50-х годах, добыча архара не прекращалась никогда, и доля охотников в его общей гибели достигала 93% [Мекленбурцев, 1948; Айзин, 1969; Сапожников, 19766, 1977]. В 70-80-е годы местному населению ежегодно разрешалось добыть 100-120 баранов, нелегально отстреливали около 1000
особей. Погранзаставы проводили заготовку мяса для личного состава с применением автоматов, добывая баранов десятками [Хохлов, 2002]. С 1987 г. памирский баран включен в Красные книги. Тем не менее с 1992 г. браконьерство резко усилилось в связи с ослаблением пограничного режима и появлением вооруженных беженцев. Нелегальная охота имела место и в приграничной полосе. Здесь же был открыт промысел сурка, что привело к вытеснению баранов к государственной границе. Доля человека в гибели самцов памирского архара в последнем десятилетии XX в. достигала 62,5%, причем 78% добытых особей были старше б лет, что свидетельствует об избирательности охоты. В середине 90-х годов часть оружия у местного населения изъяли, что способствовало уменьшению браконьерства. Однако охрана диких копытных па Памире, как и в других районах, практически отсутствует, за исключением участков, где проводится трофейная охота [Федосенко, 2000; Федосенко, Вейнберг, 2001].
В Казахстане охота на самок и сеголеток архара и других копытных запрещена с 1921 г., но этот запрет повсеместно не соблюдался [Слудский, 1956; Смирнов, 1965; Ганюшин, 1977] и не соблюдается до сих пор. В Казахском нагорье нелегальная охота особенно развита среди сельского населения. Нередко в ней участвуют от 3 до 25 всадников и 7^40 собак. О масштабах браконьерства говорят следующие цифры: 6 из 10 самцов барана, добытых иностранными охотниками в 1992 г., были со свежими (1-2-месячной давности) огнестрельными ранениями [Бербер, 1999]. В Северном Прибалхашье эти копытные встречаются не ближе 30-50 км от поселков, хотя еще в середине столетия держались рядом с населенными пунктами [Шубин, 1975]. Во второй половине XX в. браконьерство в общей смертности архара в Центральном Казахстане составляло 24-42% [Байдавлетов, 1996, 2002], а в горах юго-востока республики - две трети. Следы нелегальных охот и останки животных встречаются повсеместно, в том числе в заповедниках. В феврале 1978 г. в пустынной части Алма-Атинского заповедника в Калканах в выходные дни можно было услышать до 10 выстрелов за утро. Местные жители регулярно охотятся и на каратауского барана, занесенного в I приложение СИТЭС как форма, которой угрожает реальная опасность исчезновения. Более того, в 90-е годы здесь было организовано несколько трофейных охот для иностранцев [Федосенко, 1977, 1983а, 2000; Слудский и др., 1983; Федосенко, Жиряков, 1987]. Основная причина уменьшения численности барана Северцова в Узбекистане та же - браконьеры (58% в общей смертности), волк и пастушьи собаки [Рахимов, 1966; Ишунин, 19836; Коршунов, Коршунова, 1983; Федосенко, 1983а, 2000].
В Монголии резкое сокращение численности архара и других копытных произошло в 30-50-е годы по всему ареалу с появлением современного нарезного оружия. Охотником в этот период был почти каждый мужчина. В Кобдосском аймаке некоторые из них с сентября по декабрь 1944 г. добывали до 60 аргали и сотни дзеренов и джейранов. Ситуация несколько улучшилась лишь с 1953 г. после запрета охоты на копытных [Груздев, Сухбат, 1982].
На Алтае запрет на отстрел аргали введен с 1934 г., но и здесь он тоже не соблюдался никогда. Браконьерская охота ведется повсеместно, в том числе и в Алтайском заповеднике и заказниках, и даже после внесения барана в Красные книги МСОП и Республики Алтай. Основной пресс нелегальной охоты приходится на летние месяцы, т. е. на период выпаса домашнего скота и экспедиционный сезон.
Доля браконьеров в общей смертности животных составляет не менее 40%. В последнее десятилетие XX в. усилилось “армейское, начальственное и трофейное” браконьерство. Аналогичная ситуация в Туве и Монголии в отсутствии реальной охраны [Сопин, 1973,19756,1976г, 1977в; Собанский, 1992; Федосенко, 1989,2000, 20026; Соколов и др., 1996; Пальцын, Спицын, 2002; Калмыков, 2003; Собанский, Часовских, 2002]. Браконьеры отстреливают преимущественно взрослых самцов. Среди остатков черепов (п = 37) на скотоводческих стоянках в верховьях Чаган-Бургазы на Алтае доля самок 2-10-летнего возраста составляет около 16%, самцов 3-4 лет - 6%, 5-8-лет - 78% [Анчифоров, Фалеев, 1993].
Трофейная охота. В последние десятилетия в России и сопредельных странах одним из ведущих факторов динамики численности баранов стала трофейная охота. В Монголии она практикуется для иностранных охотников с 1967 г. Лицензия на отстрел архара стоит около 23-25 тыс. долларов США. До 1989 г. официально добыто 1630 взрослых самцов, за отстрел которых в казну страны поступило около 20 млн долларов. Доход от продажи лицензий распределяется следующим образом: 70% - в госбюджет, 10 - в бюджет местного административного органа, на территории которого производится отстрел животных, и 20% получает фирма, организующая охоту. В центральной части Монгольского Алтая в 1978-1992 гг. добыт 201 взрослый самец аргали, что привело к заметному снижению в популяции баранов с крупными рогами [Груздев, Сухбат, 1982; Федосенко, 2000].
В Центральном Казахстане ограниченная трофейная охота на “краснокнижного” барана начата с 1990 г. С 1992 г. здесь ежегодно добывали от 6 до 19 взрослых самцов, а всего в 1990-1997 гг. - 86 экземпляров, за которых иностранные охотники заплатили более миллиона долларов (официально государству). Это позволило осуществить ряд мероприятий, направленных на сохранение животных, а 20% средств направлено на научно-исследовательские работы. К сожалению, охоту здесь чаще проводили в период гона, изымая лучших производителей. При неизбежном факторе беспокойства самцы откочевывали, и часть самок в районах трофейной охоты оставались непокрытыми. В результате в группировках значительно уменьшилась доля баранов с мощными рогами. На двух экспериментальных трофейных участках с 1991 г. по 1995 г. прирост популяции снизился с 16,8 и 27,4% до 9,6 и 17,9% соответственно [Бекенов, Байдавлетов, 1997; Бекенов и др., 1999; Бербер, 1999; Федосенко, 2000]. В связи с негативным влиянием трофейной охоты на популяции архара Правительство Республики Казахстан приняло постановление, согласно которому с 1998 г. запрещено проводить охоту на баранов в период гона и изымать более 12 взрослых самцов [Бербер, 2001].
В Таджикистане трофейная охота на памирского барана началась с 1987 г. (в год внесения его в Красную книгу -А.Д.), в Киргизии - с 1990 г. Ежегодная лицензионная добыча составляла 15-60 особей. К 1999 г. легально отстреляно около 460 взрослых самцов (285 - на Памире и 175 - на Тянь-Шане), но, судя по изменению возрастной структуры популяций и сообщениям отдельных охотове-' дов, много больше. На Памире в районах трофейной по форме и антиселекцион-ной по сути охоты доля молодых самцрв в группировках, по мере убывания взрослых баранов, увеличилась с 29-39% до 55% [Абдусалямов, 1997; Федосенко, Вейнберг, 1999; Федосенко, 2000]. В Киргизии в начале 90-х годов цена двухнедельной охоты на тянь-шаньского барана, который эксплуатировался под маркой
барана Марко Поло, доходила до 45 тыс. долларов. Позднее из-за массового предложения она снизилась до 12 тыс. Официальная ежегодная квота отстрела самцов увеличилась с 20 до 40 голов, неофициально их добывали в 2-3 раза больше. Нередко практиковалась охота с вертолета. В результате неумеренной добычи доля половозрелых самцов в популяциях уменьшилась в 3-5 раз, а трофейных (старше 10 лет) - в 12-18 раз. Элитных особей в районах интенсивных трофейных охот практически не осталось [Кошкарев, 2002]. В окрестностях Нуратинс-кого заповедника в Узбекистане трофейная охота (5-6 экземпляров ежегодно) на барана Северцова ведется с 1992 г. Поскольку на него охотятся и местные жители, то из-за неумеренной добычи рост численности в последнее время прекратился. Средства, получаемые от трофейных охот, практически не используются для улучшения охраны и восстановления ресурсов [Федосенко, 2000].
Очевидно, что трофейная охота, которая обычно существенно дополняется браконьерским отстрелом, чаще приводит к негативным для популяции результатам. Положительные примеры редки, и они приводятся, как правило, бизнесменами от охоты. По сообщению В.В. Мельникова [2003], в Мургабе (Таджикистан) благодаря хорошо организованной охране численность и трофейные качества памирского барана возросли. Средняя длина рогов добытых животных увеличилась с 1997 по 2002 гг. со 139 до 146 см. На мой взгляд, в этом районе быстро прогрессирует не улучшение трофейных качеств популяции, а усиление выборочного охотничьего пресса на лучших самцов. И при такой “селекции” через ряд лет ситуация, скорее всего, будет иной.
Охрана. Охрана горных баранов неудовлетворительна по всему ареалу, несмотря на занесение многих форм в Красные книги. Очевидно, что для их сохранения необходимо не только улучшение охраны и расширение сети реально охраняемых территорий, но и снижение численности волка, закрепление охотничьих угодий за конкретными пользователями и экологическое просвещение населения. Не менее важная задача - ограничить масштабы животноводства в ключевых районах обитания группировок горных копытных или хотя бы применять систему “чересполосного выпаса” [Андреенков, 1980; Федосенко, 1983а, 2000, 20026; Бербер, 1999; Байдавлетов, 2002; Пальцын, Спицын, 2002].
Особую тревогу вызывает ситуация с алтайским аргали на территории России, который находится на грани исчезновения. Для его охраны необходимо создать заповедник на хребте Сайлюгем в верховьях Чаган-Бургазы, несколько заказников и запретить выпас скота в основных районах обитания. На южном макросклоне Сайлюгема в Монголии особо охраняемая территория, скорее всего, будет образована [Сопин, 1975а,б; Филус, Золотухин, 1987; Собанский, 1992; Федосенко, 2000, 2001, 20026; Пальцын, Спицын, 2002]. Охраняемая территория нужна также в Туве на хребте Монгун-Тайга [Федосенко, 20026] и в Бурятии на северных склонах Джи-динского хребта [Смирнов, Сопин, 1988]. Очень нужен заповедник и в Западном Каратау для охраны чудом уцелевшего сотенного поголовья каратауского барана. Не будет лишним создание резерватов в горах-останцах Кызылкума [Ишунин, 19836] и в Южной Гоби [Соколов и др., 1996].
Перспективы реакклиматизации. Плановая работа по расселению архара в СССР не велась. В 1961-1969 гг. 57 особей, отловленных в Чу-Илийских горах и в Киргизии, выпустили в специализированное охотничье хозяйство Боровское в Кок-четавской области Казахстана. Впоследствии все звери погибли большей частью в
вольерах из-за гельминтозов, и позднее, в природе, от волка и рыси [Слудский, Афанасьев, 1964; Павлов, 1999].
Восстановление ареала горных баранов искусственным путем, тем не менее, необходимо. Аргали реально реакклиматизировать на Приморском хребте, в Иркутском и Ольхонском районах Иркутской области, горах Адун-Чолон, на Джи-динском хребте, в Сохондинском заповеднике, Минусинской котловине и на Хэн-тэе в Монголии [Мельников, Сопин, 1978; Смирнов, 1983, 1994; Смирнов, Сопин, 1988; Прокофьев, 1992; Соколов и др., 1996; Сопин, 1998]. Для него вполне пригодны малоснежные горы в Хакасии. Казахстанского барана можно расселить во многих районах прежнего обитания [Бербер, 1999]. Аргалиобразных баранов, как считал Н.К. Верещагин [1959а], следует выпустить в Центральном Дагестане в районе Гуниба, Ботлиха и Хунзага. Однако при современном уровне браконьерства и высокой численности волка реинтродукция этих копытных в ближайшие годы вряд ли целесообразна во всех районах.
Отлавливать архаров в природе для расселения можно с помощью капроновых сетей и живоловушек различной конструкции. В сравнительно открытой местности эффективен отлов с мотоцикла арканом с петлей диаметром 80-90 см на 4-5-метровом шесте [Львов, 1978].
Разведение в неволе. В целях сохранения генофонда вида в зоопарках и на специальных фермах желательно образовать резервный фонд из баранов разных форм.
Аргали в настоящее время успешно разводят в зоопарках Германии, Голландии, США и Канады [Соколов и др., 1996]. В конце 80-х годов их воспроизводство пытался наладить Биологический институт СО РАН. В 1991 г. здесь получено первое потомство, но затем два взрослых самца погибли при иммобилизации. Замечено, что на состояние взрослых зверей угнетающе действует повышение температуры среды выше 23-24 °C, поэтому рекомендовано содержать этих копытных в условиях, максимально приближенным к естественным [Анчиферов, Фалеев, 1993]. Казахстанского барана с 1986 г. пытались разводить в Карагандинском зоопарке. Из 5 отловленных ягнят три самки пали, не достигнув половой зрелости. От оставшихся в живых самца и самки в 1989-1997 гг. получено И детенышей, из них 7 погибли. В 1990 г. стадо пополнили двумя новыми самками. В 1993-1997 гг. от них получено 11 ягнят, из которых не выжили 5. Позже часть животных была отправлена в другие зоопарки, часть убита собаками и человеком. В 1998 г. в вольерах оставалось 9 особей [Бербер, 1999].
Для охотничьих ферм более перспективен муфлон. Размножение этого копытного в неволе идет без проблем, но часть молодняка все же погибает в первый год жизни от болезней. Наименее жизнеспособно потомство инбредных животных [Треус, 1968; Антонян, Эгнатосян, 1984; Минасян и др., 1986а,б].
СНЕЖНЫЙ БАРАН
Ovis canadensis Shaw, 1804
Ovis canadensis Shaw, 1804. Naturalist’s Misc.', 15: 610. Канада, Альберта.
Ovis nivicola Eschscholtz, 1829. Zool. Atlas, 1:11. Камчатка.
Aegoceros montanus Middendorff, 1853. Sibir. Reise, 2, 2: 116. Становой xp., p. Половинная (приток Уды). Nec Schreber, 1804.
Ovis borealis Severtzov, 1873. Изв. Имп. о-ва любит, естеств., антропол., этногр., 8, 2: 153. Горы в верховьях Пясины и Хатанги.
Ovis аттоп storcki J. Allen, 1904. Bull. Amer. Mus. Nat. Hist., 20: 293. Зап. Камчатка, окрести, форта Тигил.
Ovis alleni Matschie, 1907. Niedieck’s Kreuzfahrten im Beringsmeere: 236. П-ов Тайгонос.
Ovis аттоп middendorfi Kowarzik, 1913. Zool. Anz., 41, 10: 439. Окрести, p. Удд.
Ovis borealis lydekkeri Kowarzik, 1913. Ibid., 41: 439. С.-з. часть Верхоянского хр., в 40 милях от устья Яны.
Ovis lenaensis Kowarzik, 1914. Peterm. Geogr. Mitt. Jrg., 1: 4. Бассейн, p. Лены. Nom. nudum. Ovis nivicola potanini Nasonov, 1915. Изв. Имп. Акад, наук, 9: 1599. Ю.-з. часть Станового хр. Ovis nivicola lydekkeri albula Nasonov, 1923. Геогр. распростр. горн, баранов Старого Света: 133.
Верхоянский хр. Непригодно (инфраподвидовое название).
Ovis nivicola lydekkeri obscura Nasonov, 1923. Ibid.: 133. Якутия, хр. Тас-хаяхтах. Непригодно (инфраподвидовое название).
Ovis nivicola koriakorum Tchemyavsky, 1962. Докл. АН СССР, 145, 5: 1174. Корякское нагорье, верховья р. Ачай-ваям.
Ovis nivicola tschuktschorum Zheleznov, 1990. Дикие копытные Северо-Востока СССР: 285. Анадырское плоскогорье, хр. Пекульней.
Ovis nivicola kodarensis Medvedev, 1994. Байкал - природная лаборатория для иссл. изменений окр. среды и климата, 5: 37. Центр, часть хр. Кодар, верховья р. Мускунах.
Характеристика. Размеры средние. Сложение плотное, коренастое, голова небольшая, шея и ноги относительно короткие и толстые (фото 50). Хвост и уши короткие, до 14 и 11 см, соответственно. Длина тела самцов в азиатской части ареала достигает 188 см, самок - 179 см, высота в холке - 112 и 100 см, масса - 150 и 68 кг.
Окраска азиатского снежного барана варьирует от темно-бурой и почти черной до светло-серой и белой (см. ниже). У американских форм вида тоже преобладает бурый цвет меха, более темный на севере ареала и светлый с песочным и красноватым оттенками на юге. У барана Далла, как отмечено выше, окрас зимой чисто белый и с желтоватым оттенком летом, а у барана Стоуна нередко почти черный [Cowan, 1940; Сопин, 19866]. Околохвостовое зеркало у азиатского снежного барана не простирается выше хвоста, у американского толсторога заходит далеко на круп. Подвес на груди и шее не образуется. Зимний мех длинный (3-15 см), густой, с хорошо развитым подшерстком, обладает высокими теплоизоляционными свойствами, к весне заметно светлеет.
Череп широкий и низкий, с укороченным лицевым отделом и относительно длинной мозговой коробкой (рис. 80). Максимальная его длина у самцов азиатского снежного барана - 296 мм, у самок - 270 мм, максимальная ширина - 189 и 162 мм. Межчелюстные кости не всегда достигают носовых и соприкасаются с ними. Носовые кости короткие и широкие. Углубления на слезной кости выражены слабо. Костные стержни рогов относительно короткие и толстые; их длина, в отличие от рогов самцов архара, меньше обхвата основания стержня. Передний премоляр в нижней, а у самцов, изредка, и в верхней челюсти, отсутствует или рудиментарный, однако едва ли не у половины зверей сохраняются 3 предкоренных зуба, как и у горных баранов. У некоторых особей в нижней челюсти с одной стороны 3, с другой 2 премоляра.
Посткраниальный скелет, в сравнении с архаром, имеет укороченный шейный и удлиненный поясничный отделы позвоночника; относительно укороченные конечности; удлиненные передние и задние пальцы и плечевую и бедренную кости; более массивные лучевую кость, метаподии и фаланги пальцев; более округлую
Рис. 80. Череп и рога самца азиатского снежного барана (экземпляр с Камчатки;
Зоомузей МГУ, коллекционный № 1406)
Вид сбоку, сверху и снизу
форму суставной головки плеча и бедра; расширенную лопатку; узкий таз в сочетании с удлиненной подвздошной костью, что сближает (конвергентно) снежного барана с высоко специализированными горными животными - козлами [Громова, 1936, 1953; Егоров, 1955; Чернявский, 1963, 19646; Ревин и др., 1988].
Роговые чехлы у самцов снежного барана однотипные, круто спирально изогнутые, относительно длинные (до 115 см у азиатского и 147 см у североамериканского барана Стоуна), с толстыми основаниями (до 38 см в окружности), широкой и плоской лобной поверхностью. Внутреннее и наружное ребра хорошо выражены, нижнее, как правило, отсутствует, лобная поверхность нависает над височной. Поверхность рога мелкоморщинистая без поперечных возвышений. Первый годовой сегмент слабо развит и у взрослых особей исчезает. У самок рога короткие, с расходящимися в стороны концами. Цвет рогов варьирует от желтовато-серого до темнобурого.
Узкоспециализированный холодолюбивый горный вид, предпочитающий ос-тепненные скалистые участки. Занимает сходную с козлами, но более широкую экологическую нишу. Животные хорошо бегают по открытой местности подобно архару, ловко лазают и прыгают по скалам, подобно козлам, и умело маскируются, сливаясь с окружающим фоном местности. Имеют острое зрение, прекрасное обоняние и слух. Живут сравнительно долго - до 15-20 лет.
Охотничий или охраняемый в отдельных районах вид.
Распространение. Горные массивы Восточной Сибири и Дальнего Востока и Северной Америки. Ареал очаговый (см. ниже).
Состав. Число подвидовых названий близко к 20, что, как и у архара, излишне. По результатам последних исследований вид представляется более однородным. В нем четко выделяются лишь разнохромосомные азиатские и американские бараны.
Ниже рассматривается преимущественно азиатский снежный баран - эндемик фауны России.
Изменчивость и таксономия
Размеры и масса. Новорожденные азиатского снежного барана хорошо развиты и крупны (3-4,5 кг). Такая же масса (3-4,3 кг) и у новорожденных американских баранов, сходны и темпы роста и развития ягнят [Hansen, 1965; Bunnell, Olsen, 1976; Hass, 1995]. В 4-месячном возрасте сеголетки весят 20-25 кг. Зимой их рост практически прекращается, и вновь бурно продолжается с весны до следующей зимы. За этот период их масса удваивается. Половой диморфизм в линейных размерах проявляется на втором году жизни. Полного развития самки достигают к З-Л-летнему возрасту, самцы - к 5-летнему, но медленный рост продолжается до 9-10 лет [Чернявский, 1962а, 1984; Егоров, 1965; Бромлей, Кучеренко, 1983; Кри-вошапкин, Яковлев, 1999]. Отличительные половозрастные признаки животных отражены на рис. 81.
Самец, 5 лет старше
Сеголеток 1 год Самка, Самка,
2 года ad
Самец, Самец, 2 года 3-4 года
Рис. 81. Половозрастные различия азиатского снежного барана (по [Кривошапкин, Яковлев, 1999])
Таблица 84. Размеры (см) взрослых снежных баранов Сибири, Дальнего Востока и Аляски
Самцы Самки Район (источник информации)*
п М lim п М lim
1 2 3 4 5 6 7
Длина
2 152 151 153 - — — Плато Путорана (8)
— -155? - - -“-(4)
2 - 162-171 2 - 139-144 Хребет Кодар (12)
9 152 140-161 7 135,5 128-156 Верхоянский хребет(1)
— - -170 — — - Горные системы Якутии (4)
- - - 2 168 - Хребет Сунтар-Хаята (4)
— - 150-175 — - -168 Приохотье (4)
6 158 152-163 - - - Северное Приохотье (15)
5 182 178-188 5 176 169-179 Чукотка (4)
1 - 152 — — — -“-(5)
8 150 148-154 9 134 126-142 Корякское нагорье (2)
4 168 162-178 10 134 127-145 Камчатка (6)
9 165 -168 2 134 127-141 -“-(9)
— 160 - — 140 — -“-(10)
6 163 155-174 - - - Аляска (15)
Высота в холке
- -111? — - — Плато Путорана (4)
2 - 94-103 2 84 — Хребет Кодар (12)
9 94 85-102 7 84 80-89 Верхоянский хребет (1)
3 103 93-108 5 89 87-94 С.-В. Якутия (7)
— - -112 — - -100 -“-(Н)
— — - 2 90 — Хребет Сунтар-Хаята (4)
- 100 - — 90 - Приохотье (4)
6 100 89-104 — — — Северное Приохотье (15)
5 104 102-106 5 91 87-100 Чукотка (4)
1 — 96 — — — -“-(5)
8 98 95-100 9 89 86-91 Корякское нагорье (2)
5 97 90-105 3-7 86 76-94 Камчатка (3, 6)
9 86 76-94 - 85 - -“-(9, 10)
1 — 106 — - - -“-(13)
6 103 98-109 — — — Аляска (15)
Обхват туловища
2 по 108-111 — — - Плато Путорана (8)
9 115 106-122 7 95 90-103 Верхоянский хребет (1)
— — — 2 102 — Хребет Сунтар-Хаята (4)
6 118 110-126 — - - Северное Приохотье (15)
5 146 138-150 5 119 110-130 Чукотка (4)
1 — 140 — — — -“-(5)
8 106 103-111 9 96 92-99 Корякское нагорье (2)
1 — 137 — — — Камчатка (13)
6 116 110-124 — - - Аляска (15)
Длина ступни
2 37 37-37 — - Плато Путорана (8)
9 39,5 35-41 7 38 36-39 Верхоянский хребет (1)
- — — 2 38 36^40 Хребет Сунтар-Хаята (4)
5 42 40-43 5 38 37^41 Чукотка (4)
Таблица 84 (окончание)
1 1 2 1 з 1 4 | 5 1 6 1 7
1 39 - - -“-(5)
8 39 38-41 9 37 37-39 Корякское нагорье (2)
6 40 37-42 Длина уха — — Аляска (15)
2 — 7,5-8,5 — — - Плато Путорана (8)
8 8,2 2 - 7,8-8,1 Хребет Кодар (14)
9 8,9 7,5-9 7 9,6 8-11 Верхоянский хребет(1)
- - - 2 9,8 9,5-10 Хребет Сунтар-Хаята (4)
- 10 - - - - Приохотье (4)
6 9,5 8,5-10,5 — - Северное Приохотье (15)
5 9,3 9-10,3 5 9,6 9-10 Чукотка (4)
1 - 9 - — - -“-(5)
8 8,6 7,5-9 9 8,6 7,5-9,5 Корякское нагорье (2)
5 9,5 9-10 3-10 8,7 8-10 Камчатка (3, 6)
9 9 -9 2 10 9-11 -“-(9)
1 — 9 — - — -“- (13)
6 8,9 8,5-9,2 Длина хвоста — — Аляска (15)
2 - 6,5-7 - — - Плато Путорана (8)
2 — 11-12 2 - 8,8-9,5 Хребет Кодар (14)
9 11 10-12 7 12 10-13 Верхоянский хребет (1)
- - — 2 11,5 - Хребет Сунтар-Хаята (4)
- — -12 — - — Приохотье (4)
6 9,2 8,5-10 - - - Северное Приохотье (15)
4 12,5 12-13 5 11 9-13 Чукотка (4)
1 — 11 — — — -“-(5)
8 10,3 9,5-11,5 9 10 7,5-10 Корякское нагорье (2)
5 12 10-14 3-10 10 8-14,5 Камчатка (3, 6)
9 9 -11 2 10 -10 -“-(9)
1 - 9 - — - -“-(13)
6 8,7 7-10 — - - Аляска (15)
*1 - Кривошапкин, Яковлев, 1999; 2 - Чернявский, 19626, 1964а; 3 - Ревин и др., 1988; 4 - Железнов-Чукотский), 1990, 1994; 5 - Чернявский, 1970а; 6 - Аверин, 1948, 1951; 7 - Егоров, 1965; 8 - Мичурин, Мироненко, 1966а; 9 - Гаврилов, 1947; 10 - Грибков, Филь, 1977; 11 - Тавровский и др., 1971; 12-Мсдвс-дсв, 1994, 20036; Медведев, Лямкин, 2000; 13 - Шайдуллин, Шайдуллина, 1988; 14 - Сопин, Медведев, 1992; 15 - Чернявский, 2004.
Длина тела взрослых самцов варьирует в пределах 140-188 см, высота в холке- 76-112 см, масса - 56-150 кг, у самок, соответственно, 126-179, 76-100 см и 33-68 кг. Наиболее крупные особи населяют Камчатку и Чукотку. Бараны Корякского нагорья, Верхоянской горной системы, Приохотья, Южной Якутии, Забайкалья и плато Путорана весьма сходны по своим параметрам (табл. 84 и 85). По экстерьеру азиатский снежный баран очень близок к американским сородичам. Длина тела самцов барана Далла составляет 140-160 см, масса - 75-90 кг, максимальные показатель канадского толсторога - 190 см и 140 кг соответственно [Bunnell, Olsen, 1976; Сопин, 19866].
Возрастная и индивидуальная изменчивость снежного барана в размерах тела и массе значительна, и поэтому экстерьерные показатели не могут быть определяющими в систематике, как, впрочем, и размеры рогов, которые растут в течение жиз-
Таблица 85. Масса (кг) взрослых снежных баранов Сибири, Дальнего Востока и Аляски
Самцы Самки Район (источник информации)*
п 1 М 1 lim п | м I lim
2 108 106-109 - 55 - Плато Путорана (8, 4)
- - 80-120 - - - Хребет Кодар (12, 14)
9 68 56-81 7 48 42-50 Верхоянский хребет (1)
— 85-97 — — — -“-(16)
- - - 1 - -64 С.-В. Якутия (11)
— - - 2 58 57-60 Хребет Сунтар-Хаята (4)
- -98 - -68 Приохотье (3)
2 103 102-105 - - - Северное Приохотье (15)
4 99 86-105 5 62 45-68 Чукотка (4)
1 - 78 - - -“-(5)
8 73 59-81 9 42 33-49 Корякское нагорье (2)
5 93 79-109 3-8 49 46-54 Камчатка (3, 6)
9 114 -128 2 46 44-18 -“-(9)
— 135 -150 - 56 - -“-(Ю)
1 - 105 - - -“-(13)
6 75 74-82 - - Аляска (15)
* Обозначения 1-15 см. в таблице 84; 16 - Фертиков, 1978.
ни полорогих копытных. К концу зимы звери обычно теряют около 35-40% массы, а в аномальные годы - еще больше. По мнению Л.В. Сопина [19866], размеры тела снежных баранов во многом зависят от высоты местности над уровнем моря и, что особенно важно, от состояния кормовой базы. Животные, обитающие в некоторых районах при постоянном дефиците зимних кормов, могут быть мельче, чем в группировках, находящихся в более благоприятных условиях. Для канадского толсторога установлено заметное снижение массы молодых особей при увеличении плотности популяции [King et al., 1993].
Краниометрическая характеристика. Формирование зубной системы и костей черепа у снежного барана завершается к 4 годам. Полное снашивание зубов происходит, в сравнении с архаром, поздно (в 14-18 лет), что связано, видимо, с поеданием менее грубой пищи и с меньшим количеством минеральных частиц (пыли), попадающих на растения в Северной Азии [Цалкин, 1951а; Чернявский, 1962а, 1963; Егоров, 1965].
Географическая изменчивость размеров черепа у азиатского снежного барана в репрезентативных выборках, в отличие от архара, незначительна. Эту особенность заметил еще В.И. Цалкин [1951а], полагая все же, что бараны Камчатки отличаются от верхоянско-анадырских более крупными костными стержнями рогов, более короткой мордой и менее широким черепом в области орбит. Однако на большем материале эти особенности практически не выявляются ни у самцов, ни и у самок, за исключением, пожалуй, максимальной длины костных стержней у самцов. Отдельные средние линейные параметры баранов Корякского нагорья чуть мельче, чем у соседних группировок. В целом по краниометрическим показателям азиатские популяции вида почти идентичны (табл. 86).
По недавним исследованиям Д.Г. Медведева [1994, 2003а,б], череп кодарского барана существенно отличается от такового у других форм. У молодых особей от-
Таблица 86. Размеры черепа (мм) взрослых снежных баранов Сибири, Дальнего Востока и Аляски
Самцы Самки Район (источник информации)*
п М lim п м lim
1 2 3 4 5 6 7
Наибольшая длина
5 281,0 274-294 - - - Плато Путорана (8)
2 274,5 274 -275 — — -“-(9)
1 - 288 - - - Хребет Кодар (8)
— — -288 — — -254
4 275,0 262-286 2 242,5 233-252 Становой хребет (2, 8)
6 279,8 275-286 10 251,1 245-262 Центральное Верхоянье (1)
13 277,2 258-291 10 258,2 242-270 Верхоянский хребет(2)
16 277,2? 256-290 - — — -“-(13)
6 280,0 269-295 6 258,0 252-262 С.-В. Якутия (5)
13 277,0 258-291 9 260,0 250-270 Якутия (8)
- - 275-280 - — 265-270 Чукотка (3, 4)
1 — 276 — - — Колымское нагорье (4)
2 278,5 276-281 — — - Охотское побережье (12)
- - 256-280 - - 245-256 Приохотье (3)
18 272,4 256-283 — - - Северное Приохотье (13)
8 267,3 256-277 5 246,0 240-252 Корякское нагорье (6, 13)
43 278,7 261-296 11 256,9 247-263 Камчатка (2)
23 280,1 266-291 - — - -“- (13)
12 283,3 238-297 - — - Аляска (13)
Максимальная ширина
6 171,0 160-182 — — Плато Путорана (8)
2 164,5 162-167 — — — -“-(9)
1 — 186 — - - Хребет Кодар (8)
— — -189 — — -162 -“-(Ю)
4 164,0 155-173 2 140,0 138-142 Становой хребет (2)
6 174,5 165-184 10 151,1 144-161 Центральное Верхоянье (1)
13 169,0 158-178 10 153,4 145-161 Верхоянский хребет(2)
16 168,3 139-184 — — — -“-(13)
6 173,8 152-189 6 153,2 150-157 С.-В. Якутия (5)
13 169,0 156-177 9 154,0 146-161 Якутия (8)
2 166,0 159-173 - — - Охотское побережье (12)
18 166,2 150-176 - - — Северное Приохотье (13)
— — 166 - - - Чукотка (3, 4)
8 167,2 163-177 - - - Корякское нагорье (13)
43 169,7 153-187 11 154,6 146-159 Камчатка (2)
23 170,8 155-186 - - - -“-(13)
12 162,1 151-174 - - - Аляска (13)
Высота
3 163,0 155-170 - - - Плато Путорана (8)
1 - 167 - - - Хребет Кодар (8)
4 142,5 136-153 2 105,5 101-110 Становой хребет (2)
6 148,2 143-154 10 104,8 103-108 Центральное Верхоянье (1)
13 144,5 138-153 10 106,0 99-110 Верхоянский хребет (2)
16 141,7 116-153 — — — -“-(13)
13 145,0 138-153 9 107,0 105-110 Якутия (8)
2 155,5 151-160 - - - Охотское побережье (12)
Таблица 86 (продолжение)
1 1 2 1 3 1 4 1 5 1 6 1 7
18 146,3 128-157 Северное Приохотье (13)
1 — 147 - - - Чукотка (4)
8 143,3 130-153 - — — Корякское нагорье (13)
43 147,8 130-161 11 107,6 103-113 Камчатка (2)
23 148,2 129-161 - - - -“-(13)
12 142,9 115-161 - - — Аляска (13)
Межглазничная ширина
6 119,0 110-130 - - - Плато Путорана (8)
2 117,0 113-121 — — — (9)
1 — 138 - - - Хребет Кодар (8)
4 120,5 111-132 2 96 92-100 Становой хребет (2)
6 121,3 113-127 10 105,6 99-114 Центральное Верхоянье (1)
13 119,3 114-135 10 110,9 102-117 Верхоянский хребет (2)
16 127,1 99-136 — — — -“-(13)
13 127,0 114-135 9 111,0 102-117 Якутия (8)
2 119,0 119-119 - - - Охотское побережье (12)
18 118,7 102-129 - - - Северное Приохотье (13)
8 120,7 118-127 - - - Корякское нагорье (13)
43 126,0 113-138 11 111,8 104-117 Камчатка (2)
23 126,7 113-138 9 108 104-111 -“-(4, 13)
12 118,2 110-129 - - - Аляска (13)
Длина лицевой части
2 207,5 207-208 - - - Плато Путорана (8)
1 - 214 - - - Хребет Кодар (8)
4 208,5 201-216 2 188,5 181-196 Становой хребет (2)
6 211,2 207-214 10 197,2 191-206 Центральное Верхоянье (1)
13 208,8 197-217 10 200,1 189-209 Верхоянский хребет (2)
16 204,3 190-215 — — — (13)
13 209,0 197-217 9 201,0 195-209 Якутия (8)
1 - 203 — — - Колымское нагорье (4)
2 - 202-205 - - - Охотское побережье (12)
- — 190-200 - - - Приохотье (3)
18 202,8 191-214 - - Северное Приохотье (4, 13)
1 - 206 — - - Чукотка (4)
8 196,9 188-203 - - - Корякское нагорье (13)
43 206,7 196-219 11 197,5 190-203 Камчатка (2)
23 207,0 195-216 - - - -“-(13)
12 215,3 206-224 - - - Аляска (13)
Длина мозговой коробки
4 131,0 123-136 - - - Плато Путорана (8)
2 134,0 132-136 — — — -“-(9)
1 - 134 - - - Хребет Кодар (8)
4 123,2 118-129 2 106,0 104-108 Становой хребет (2)
6 126,5 118-135 10 109,3 104-115 Центральное Верхоянье (1)
13 126,5 117-133 10 113,2 106-118 Верхоянский хребет (2)
16 125,6 114-133 — - — -“-(В)
13 126,0 117-132 9 114,0 112-118 Якутия (8)
2 122,0 118-126 - - - Охотское побережье (12)
18 128,7 114-140 - - - Северное Приохотье (13)
1 - 126 - - - Чукотка (4)
8 124,4 120-129 - - - Корякское нагорье (13)
Таблица 86 (продолжение)
1 | 2 1 3 1 4 1 5 | 6 1 7
43 129,1 115-144 11 115,8 111-121 Камчатка (2)
23 130,2 115-140 — - - -“-(В)
12 125,4 111-142 - - - Аляска (13)
Длина носовых костей
1 — 85 - - - Плато Путорана (8)
1 — 90 - - - Хребет Кодар (8)
4 89,7 82 94 о 74,0 73-75 Становой хребет(2)
6 92,8 85-102 10 80,2 72-85 Центральное Верхоянье (1)
12 90,2 81-98 10 81,3 71-89 Верхоянский хребет(2)
16 88,1 80-104 - — — -“-(13)
5 93,4 91-95 5 81,4 74-88 С.-В. Якутия (5)
13 90,0 81-98 9 82,0 72-89 Якутия (8)
1 — 89 — — — Колымское нагорье (4)
2 85,5 83-88 — - — Охотское побережье (12)
18 85,6 76-106 - - - Северное Приохотье (4, 13)
8 80,7 77-83 — - — Корякское нагорье (13)
41 88,8 76-100 и 78,1 67-87 Камчатка (2)
23 87,2 67-96 9 80 76-87 -“-(7, 13)
12 87,9 75-100 - - __ Аляска (13)
Длина верхнего зубного ряда
5 76,0 73-80 — — - Плато Путорана (8)
2 74,5 73-76 — — — • -“-(9)
1 — 80 - - - Хребет Кодар (8)
4 77,2 73-81 1 — 79 Становой хребет (2)
6 73,5 70-77 10 71,3 66-83 Центральное Верхоянье (1)
12 75,4 70-80 10 73,1 70-78 Верхоянский хребет (2)
16 72,5 66-78 — — — -“- (13)
6 74,0 72-76 6 72,8 71-74 С.-В. Якутия (5)
13 75,0 70-80 9 73,0 70-78 Якутия (8)
1 — 73 - — — Колымское нагорье (4)
2 71,5 70-71 — - — Охотское побережье (12)
18 73,6 68-80 - - - Северное Приохотье (4, 13)
1 — 79 - - — Чукотка (4)
8 68,2 65-73 — - — Корякское нагорье (13)
36 74,9 68-80 9 73,2 71-76 Камчатка (2)
15 71,6 65-79 9 71,0 65-74 -“-(7, 13)
12 76,6 54-89 - - — Аляска (13)
Длина костного стержня рогов
— — 140-190 — - — Плато Путорана (8)
— — -250 - — - -“-(3)
4 178,8 150-205 2 60 50-70 Становой хребет (2)
6 181,3 135-230 - - - Центральное Верхоянье(1)
9 175,6 150-210 3 66,7 60-70 Верхоянский хребет (2)
19 169,3 130-200 — - - -“-(13)
9 211,3 195-231 — - — Верхоянье (11)
2 197 180-215 — - - С.-В. Якутия (5)
13 175,0 150-190 - - - Якутия (8)
1 190 — - - Колымское нагорье (4)
16 200,0 160-230 — - — Северное Приохотье (4, 13)
— - -200 - - - Чукотка (3)
2 - -220 — - - -“-(4)
Таблица 86 (окончание)
1 1 2 1 3 1 4 1 5 1 6 1 7
7 186,0 165-205 Корякское нагорье (13)
— — -270 — — — -“-(3)
38 208,0 155-270 11 62,2 50-75 Камчатка (2)
8 237,1 201-296 - - -“-(И)
22 210,6 150-270 — — — -“-(13)
8 222,5 200-230 - - - Аляска (13)
Обхват костного стержня рогов у основания
- - 250-300 - - - Плато Путорана (8)
— — -330 — — — -“- (3)
3 268,3 230-290 - - Становой хребет (2)
6 282,3 250-315 - - - Центральное Верхоянье (1)
9 284,4 230-290 3 90 85-95 Верхоянский хребет (2)
19 251,4 180-280 — — — -“-(13)
9 301,7 280-330 - - - Верхоянье (11)
2 310,0 290-330 — — — С.-В. Якутия (5)
13 260,0 230-290 - - - Якутия (8)
1 271 - - - Колымское нагорье (4)
16 266,0 230-290 - - — Северное Приохотье (4, 13)
- - -330 — — — Приохотье (3)
— — -300 — — Чукотка (3)
2 285,0 280-290 — - - -“-(4)
7 281,0 265-305 — - — Корякское нагорье (13)
38 291,6 210-335 11 95,5 90-120 Камчатка (2)
8 300,6 274-316 — — — -“-(Н)
22 277,0 220-330 — — — -“-(13)
8 280,0 270-290 - - - Аляска (13)
* 1 - Кривошапкин, Яковлев, 1999; 2 - Цалкин, 1951а (расчет по “Приложению”); 3 - Железнов-Чукотский, 1994; 4 - Чернявский, 1970а, 1984; 5 - Егоров, 1965; 6 - Чернявский, 19626; 7 - Чернявский, 1962в; 8 - Ревин и др., 1988; 9 - Мичурин, Мироненко, 1966а; 10 - Медведев, 2003а,б; 11 - Босскоров, 2001; 12 - Медведев и др., 2000; 13 - Чернявский, 2004.
сутствует шов между предклиновидной и клиновидной костями, слезная кость очень узкая и у взрослых особей отделена от носовых костей соединением отростков верхнечелюстной и лобных костей. У них узкие слуховые проходы, длинный и мощный ряд зубов. У самцов почти овальное сечение роговых стержней в отличие от якутских и охотских баранов, у которых оно округло-треугольное. Ширина черепа максимальна для снежных баранов Азии при сравнительно короткой его длине.
Резкая изменчивость консервативного в целом вида в узколокальном неизолированном районе настораживает. Известно, что пропорции черепа копытных могут заметно меняться под воздействием мощного внешнего воздействия, например, радиоактивного излучения, что обнаружено мной у сибирской косули на месте радиоактивного “следа” в Челябинской области. Замечу также, что еще В.И. Цалкин [1951а] и В. Громова [1953] обратили внимание на значительную географическую и индивидуальную изменчивость отдельных костей черепа и исключительно сильное срастание черепных швов у баранов, имеющих тяжелые рога. У взрослых особей зачастую практически невозможно различить границы между отдельными костями. С возрастом щели между носовыми и межчелюстными, слезными и частью верхнечелюстных костей у большинства животных смыкаются.
Между носовыми и верхнечелюстными костями нередко внедряются межчелюстные кости или образуются особые вставочные косточки, которые прирастают к носовой или верхнечелюстной кости соответственной стороны. Уникальность отдельных признаков кодарского барана не подтверждается и при сравнительном морфометрическом анализе. Ширина черепа и длина зубного ряда особей этой популяции не выходят за рамки изменчивости других форм (табл. 86).
Краниометрические показатели североамериканских баранов несколько отличаются от таковых у азиатского сородича [Сопин, 19866; Чернявский, 2004], но вряд ли эти различия следует считать достигшими видового уровня. Размеры костных остатков азиатских плейстоценовых снежных баранов сходны с параметрами современных особей [Ревин и др., 1988; Боескоров, 2001], а ископаемые бараны Северной Америки близки к современному канадскому толсторогу [Stock, Stokes, 1969], что свидетельствует о низких темпах изменчивости этих животных.
Изменчивость рогов. У ягнят снежного барана рост рогов начинается в июле и длится около 2,5-3 месяцев. Продолжительность роста первого годового сегмента примерно на месяц меньше следующих, и поэтому он всегда существенно короче. Со второго года жизни рога у самцов растут гораздо быстрее, чем у самок. Максимальный прирост сегментов, в отличие от архара, регистрируется на четвертый -шестой годы жизни, и он остается высоким до глубокой старости. У самцов старше 8 лет лобная поверхность рогов нередко сильно выщерблена в результате поединков с соперниками, а концы рогов изношены, расщеплены или обломаны, встречаются и однорогие звери. Обломанные рога регистрируют и у части самок [Цалкин, 19486, 1951а; Егоров, 1965; Тавровский и др., 1971; Филь, 19736; Фертиков, 1978; Кривошапкин, Яковлев, 1999].
Рога азиатских и американских снежных баранов практически идентичны по строению и форме и близки по размерам. В.И. Цалкин [1951а] считал, что у камчатских баранов они длиннее и толще в основании, чем у верхоянских и анадырских, и неотличимы от особей с Джугджура и Яблонового хребта. Позднее это утверждение не раз цитировали другие специалисты, хотя оно не вполне соответствует реалиям. По линейным параметрам рога зверей из разных районов Сибири и Дальнего Востока практически одинаковы. Их средняя длина у взрослых особей варьирует в пределах 71-89 (41-115), размах - 53-63 (51-85), обхват у основания - 30-36 (21-38) см. У самок рога значительно меньше, чем у самцов, и обычно их длина не превышает 35, а обхват у основания - 13 см (табл. 87).
Окраска и линька. Окраска новорожденных азиатского снежного барана контрастная: голова, шея, грудь, плечи и передняя часть конечностей темно-серые; спина и бока пепельно-серые; на лбу выражено белое пятно; брюхо, задняя сторона конечностей и ягодицы белые или серовато-белые; по спине тянется темно-бурая полоса, которая на хвосте становится черной [Чернявский, 1962а; Тавровский и др., 1971].
Окраска взрослых особей варьирует от буро-коричневой и черновато-бурой до кремовой, светло-охристой, светло-серой и почти белой. Нижняя часть шеи, грудь и плечи в летнем меху несколько темнее туловища, брюхо - светлее. Хвост темный, хвостовое зеркало белое. У самок вдоль хребта заметна темная полоса, у самцов она нередко отсутствует. Конец морды, лоб, межроговая и зароговая части у самцов чаще светло-серого или чисто белого цвета, на носу выражена темная полоса. У самок
Таблица 87. Размеры рогов (см) взрослых снежных баранов Сибири, Дальнего Востока и Аляски
п Длина Размах Обхват у основания Район (источник информации)*
М lim М | lim М 1 lim
2 77 75-79 — Самцы 31,5 31-32 Плато Путорана (8)
— — 65-88 — — — 29-38 (7)
2 92 91-98 - - 36 36-36 -“- (10)
2 — -100 - - - 32-34 Хребет Кодар (9)
3 67 46- 87 - 31 28-32 Яблоновый хребет (3)
34 86 70-103 53 42-63 33 29-36 Верхоянский хребет (1)
13 71 53-91 — — 30 26-32 -“- (3)
19 73 53-92 58 44-75 31 28-34 (14)
23 81** 61-102 - - 32 29-36 С.-В. Якутия (2)
15 75-102 — — 33 31-36 (2)
— — -100 - - — - -“- (И)
1 — 80,5 — 53 - 29 Колымское нагорье (5)
16 75 44-92 61 48-72 30 21-33 Северное Приохотье (14)
- — -85 - -64 - -33 Приохотье (3)
3 78 61-92 — — 33 30-35 Хребет Джугджур (13)
- — -90 - -81 — -34 Чукотка (4)
2 88 85-92 57 — 33 31-35 -“-(5)
7 73 61-87 60 54-65 31 28-32 Корякское нагорье (6,14)
— — — — — — -36 -“-(4)
54 78** 58-111 — — 32 23-36 Камчатка (3)
22 77 50-110 63 49-85 33 20-36 -“-(5, 14)
30 74 41-111 — - 32,5 29-36 -“-(7)
- — -115 - 50-65 - 32-36 -“-(Ю)
1 — 87 - — — 39? -“-(12)
8 86 78-96 62 61-63 34 32-35 Аляска (14)
10 24 18-26 — Самки — — Центральное Верхоянье (1)
8 24 20-30 — — 11,8 11-13 Верхоянский хребет (3)
2 16 13-18 — — 11,0 10-12 Яблоновый хребет (3)
- — -35 - — - — Чукотка (4)
7 26 24-32 - - 12,5 9-13 Камчатка (3)
* 1 - Кривошапкин, Яковлев, 1999; 2 - Егоров, 1965; Тавровский и др., 1971; 3 - Цалкин, 1951а; 4 -Железнов-Чукотский, 1994; 5 - Чернявский, 1970а, 1984; 6 - Чернявский, 19626; 7 - Ревин и др., 1988; 8 -Мичурин, Мироненко, 1966а; 9- Сопин, Медведев, 1992; Медведев, 2003а,б; 10-Хохлов, 2002; 11 - Фертиков, 1978; 12 - Шайдуллин, Шайдуллина, 1988; 13 - Медведев и др., 2000; 14 - Чернявский, 2004.
** Размеры рогов самцов в возрасте 5 лет и старше.
*** Размеры рогов самцов в возрасте 9 лет и старше.
окраска головы менее контрастная, светло-серая с посветлениями на лбу и конце морды. Задние стороны конечностей также светлоокрашены, но у некоторых особей темные. У части животных есть небольшое светлое пятно над темноокрашенными подмышками. У старых зверей общий тон окраски более темный и менее контрастный, нежели у молодых. Зимняя окраска, особенно в конце зимы, более светлая, что происходит за счет снашивания темных кончиков волос [Флеров, 1935а; Цалкин, 1951а; Егоров, 1965; Тавровский и др., 1971; Грибков, Филь, 1977; Федосенко, 1985;
Ревин и др., 1988; Сопин, Ермолин, 1988; Железнов-Чукотский, 1994; Кривошапкин, 1995; Кривошапкин, Яковлев, 1999; Хохлов, 2002; Чернявский, 2004].
Индивидуальная и географическая изменчивость окраски азиатского снежного барана очень значительна. Иногда особи с одного хребта и даже в одном крупном стаде имеют спектр, характерный для разных форм. На Камчатке, например, судя по фотографиям многочисленных трофеев, опубликованных в книге А.Н. Хохлова [2002] и охотничьих журналах, наряду с типичными бурыми и темно-бурыми животными (фото 50) встречаются серо- и буро-коричневые, буровато-желтые и почти белые. Для этого барана характерно отсутствие лицевой полосы, хорошо выраженной у рас, обитающих западнее, однако у части особей, особенно на севере Срединного хребта и в районе Палана, она имеется. Окраска корякского барана (фото 51) сходна с таковой у камчатских особей [Чернявский, 19626], но здесь значительно больше серовато-бурых животных с белым пятном на лбу. Общий тон окраски чукотского барана варьирует от светло-серого до темно-бурого [Железнов-Чукотский, 1994; Чернявский, 2004]. Окраска якутского барана считается серовато-желтовато-бурой [Егоров, 1965; Тавровский и др., 1971], хотя такой она бывает далеко не у всех особей. Отличительными особенностями этой расы являются светлоокрашенная лицевая и зароговая Части головы и небольшое светлое пятно позади плеч (фото 52). В Приохотье животные темно-бурые, бурые, светло-бурые, а, иногда, почти белые [Обухов, 1969], и тоже со светлым лицом и зароговой частью, как и якутские (фото 53). Светлое пятно за лопатками выражено примерно у 10% особей, а на хребте Сунтар-Хаята - у четвертой части популяции [Чернявский, 2004]. Очень светлые особи встречаются и на Становом хребте при типичной серовато-бурой окраске и небольшом пятне за лопатками [Цалкин, 1951а]. У кодарс-кого барана окраска в летнем меху серая и буровато-серая, интенсивные темно-коричневые тона и светлые пятна на боках отсутствуют. Зимой звери бурые, буроватые, светло-коричневые, светло-бурые, желтовато-белесые и почти белые ближе к весне. Белая окраска морды достигает рогов и ушей, но не переходит на затылок и верхнюю часть шеи [Водопьянов, 1971; Сопин, Ермолин, 1986; Сопин, Медведев, 1992; Медведев, 1994, 20036; Сопин, 1998; Медведев, Лямкин, 2000]. Окраска путоранского снежного барана (фото 54) считается однотонной бурой или серовато-коричневой без заметных посветлений на голове и боках, однако на фотографиях охотничьих трофеев [Хохлов, 2002] ясно видны по-светления на голове разной величины.
Очевидно, что окраска туловища не может быть основной при решении вопроса о таксономическом статусе тех или иных группировок. Светлые пятна на голове, посветления или потемнения на шкурах позади подмышек и размер каудального диска - весьма изменчивые индивидуальные признаки, и они тоже не могут фигурировать в качестве безусловно диагностических [Сопин, 19836, 19866].
Окраска снежных баранов в значительной мере сходна с фоном местообитаний [Фертиков, 1978]. В светлоокрашенных сланцевых и гранитных горных массивах животные заметно светлее или желтее своих собратьев, живущих в “темных” горах. Светло-охристые особи встречаются в Верхоянье [Соломонов, 1975; Кривошапкин, Яковлев, 1999], желтовато-коричневые - на Яблоновом (Становом) хребте [Насонов, Дорогостайский, 1915]. На Камчатке близ северной око-
нечности Срединного хребта в желто-красных скальниках обитают животные, имеющие охристо-ржавый оттенок меха [Грибков, Филь, 1977].
Замечено, что на востоке Камчатки во второй половине зимы экскременты снежного барана сходны по цвету с окрестными хребтами и скалами, возможно, из-за того, что звери тщательно съедают растения вместе с корнями и приставшими к ним частицами почвы [Аверин, 1951]. На Яблоновом хребте они регулярно посещают солонцы, представленные выходами туфа красновато-бурого цвета [Насонов, Доро-гостайский, 1915]. Не исключено, что окраска зверей на протяжении тысячелетий каким-то образом могла изменяться, и стала покровительственной, отчасти, через питание. Тем не менее природа изменчивости окраски, в большей мере, генетическая - она определяется и контролируется несколькими локусами, слагающимися из ряда аллелей, вызывающих ослабление или усиление признака. У белого барана Далла преобладает локус С; у барана Стоуна он выпадает, а черную окраску определяют аллели группы D; у азиатского снежного барана в контроле окраски помимо С участвует локус S, вызывающий пятнистость у северных популяций. У горных баранов доминирует локус В - ген бурой (рыжей) окраски [Сопин, 1985а].
Сезонная смена волосяного покрова изменчива по срокам в зависимости от возраста и пола животных и географического района. Выпадение зимней шерсти на юге ареала азиатского снежного барана начинается в мае, на севере - чаще в июне и продолжается до конца июля. Первыми вылинивают взрослые самцы, последними - лактирующие самки. Линька начинается с лопаток и шеи, затем зимний волос выпадает на животе, боках, задних конечностях, голове и спине. В летний период пуховой волос на шкуре отсутствует, и начинает подрастать только со второй половины августа. Завершается линька в ноябре [Аверин, 1951; Чернявский, 1961,1963; Портенко и др., 1963; Егоров, 1965; Тавровский и др., 1971; Кищинский, 1975а].
Генетическая изменчивость. В хромосомном наборе азиатского снежного барана (2п = 52) 4 пары крупных субметацентрических и 21 пара акроцентрических аутосом, типичных для Ovis. Х-хромосома - крупный акроцентрик, Y - мелкий метацентрик [Korobitsina et al., 1974; Коробицина, Воронцов, 1975; Смирнов и др., 1987; Шайдуллин, 19946; Шайдуллин и др., 1994].
Уровень генетической изменчивости снежного барана, даже у относительно давно изолированной путоранской формы, весьма высок. У животных выявлен полиморфизм по 5 локусам: TF, GPI, MPI, АК и СК. Доля полиморфных локусов составляет суммарно 16,7%, в расчете на локус — 7,3%, средняя гетерозиготность — 0,073 (0,104). В целом генетическая изменчивость путоранского барана близка к уровню, характерному для домашней овцы. Генетическое расстояние по Нею между снежным бараном и разными породами домашней овцы составляет 0,875-0,891 [Glazko et al., 1998; Сипко и др., 1999; Дымань и др., 2000; Глазко и др., 2001]. Такое расстояние у копытных характерно для “надежных” видов.
Проведенный нами [Кузенцова и др, в печати] филогенетический анализ последовательностей фрагмента контрольного региона мтДНК (длина участка 713 нуклеотидов) 40 особей снежного барана показал четкое их разделение на азиатскую и американскую группы. Азиатские бараны, в свою очередь, делятся на две крупные группировки. В первую входят особи с Камчатки и Корякского нагорья, во вторую -особи с плато Путорана, хребта Кодар, из Якутии, со Станового хребта и Чукотки (рис. 82). Генетическая вариабельность американских снежных баранов (образцы из
Ovis аттоп
Камчатка, № 2
Камчатка, № 3
Корякское нагорье, № 68
Камчатка, № 7
Камчатка, № 5
Камчатка, № 10
Камчатка, № 69
Камчатка, № 6
Корякское нагорье, № 70
Камчатка, № 4
Корякское нагорье, № 56
Корякское нагорье, № 57
Кодар, №55
Кодар, № 8
Кодар, № 58
Плато Путорана, № 60
Плато Путорана, № 61
Плато Путорана, № 62
Кодар, № 59
Верхоянский хребет, № 75
Верхоянский хребет, № 76
Верхоянский хребет, № 81
Верхоянский хребет, № 82
Момский хребет, № 79
Становой хребет, № 1
Становой хребет, № 95
Становой хребет, № 96
Чукотка, № 71
Чукотка, № 72
Чукотка, № 73
США, AF076914*
США, AF076913*
США, AF076917*
США, AF076911*
США, AF076915*
Канада, AY116621*
США, AF076916*
США, AF076913* Канада, AY116623* Мексика, AY091486*
Рис. 82. Филогенетическое дерево снежных баранов, построенное по результатам анализа последовательностей фрагмента контрольного региона мтДНК 40 особей (по [Кузнецова и др., в печати]) Цифры слева - уровень поддержки кластеров, %, справа - номера образцов. Звездочками обозначены образцы из генетического банка данных
Канады, США и Мексики) оказалась ниже, чем в любой азиатской группировке, а генетическая дистанция между американскими и азиатскими группами (0,068-0,082), почти такая же, как между азиатскими группировками (0,036-0,076).
Гибридизация. См. выше.
Таксономия. Ведущие российские специалисты выделяли 4-5 азиатских подвидов снежного барана [Насонов, 1923; Флеров, 1935а; Цалкин, 1951а; Соколов, 1959; Гептнер и др., 1961]:
камчатский {nivicola, syn. storcki) - Камчатка;
охотский {alleni, syn. montanus, middendorfi, potanini) - Становой и Яблоновый хребты, Джугджур, юго-западные части Колымского хребта, горы полуострова Тайгонос. И.И. Соколов [1959] отделял барана Станового хребта {potanini), описанного Н.В. Насоновым [Насонов, Дорогостайский, 1915], от alleni по наличию белого пятна позади плеч;
якутский {lydekkeri, syn. lenaensis, albula, obscura) - Верхоянский хребет, хребет Черского, Момского, северные части Колымского хребта, Анадырский край;
таймырский, или путоранский {borealis) - горы Путорана.
Замечу, что диагнозы подвидов, приведенные этими признанными систематиками, не отличаются строгостью и касаются, в основном, особенностей окраски. Форма alleni, например, отличается от nivicola заметным посветлением позади плеч. У баранов формы lydekkeri, помимо такого же светлого пятна и более светлой окраски, костные стержни рогов и роговые чехлы несколько короче и тоньше. Точный диагноз путоранского подввда {borealis) и вовсе отсутствует. Н.А. Северцов [1873а] выделил его на том основании, что .. .“экземпляры этого вида, доставленные Г. Шмидтом в музей Академии наук, с гор у вершин Пясины и Хатанги, показались мне средней формой между О. nivicola и О. argali, и ближе к первой, с которой этот сомнительный вид может быть и тождествен; роговые углы не измерены; от О. argali отличается меньшими рогами, меньшим ростом и беловатым брюхом”. Н.В. Насонов [1923] отмечал у этого барана сильное удлинение шерсти над пахами, но К.К. Флеров и В.И. Цалкин считали данный признак сомнительным, а Л.Н. Мичурин и О.Н. Мироненко [1966а] такой особенности не обнаружили вовсе.
Позднее Ф.Б. Чернявский [19626, 1963, 1964, 1967, 1970а, 1984] выделил корякский подвид - koriakorum (в Красную книгу РСФСР [1983] он включен под названием “чукотский снежный баран” -А.Д.) и по единственному экземпляру сделал заключение о самобытности барана Чукотки, у которого чуть длиннее костные стержни рогов и отсутствуют светлые пятна позади подмышек. Чукотской расе Н.К. Железнов [1990] дал название tschuktschorum на основании тех же диагностических признаков и имеющегося в его распоряжении, но не приведенного в публикации, краниометрического материала. В сводке “Млекопитающие фауны СССР” [Громов и др., 1963] и “Каталоге млекопитающих СССР” [Барышников и др., 1981] приняты nivicola, alleni, lydekkeri, borealis и koriakorum. Совсем недавно Д.Г. Медведев [1994] описал кодарскую форму {codarensis).
В сводках Н.К. Железнова-Чукотского [ 1994,2003] даны диагнозы 6 подвидов снежного барана (путоранского, якутского, охотского, чукотского, корякского и камчатского), но по ним, на мой взгляд, отличить одну форму от другой практически невозможно. Эта таксономия, тем не менее, поддержана А.А. Кривошапкиным и Ф.Г. Яковлевым [1999] с замечаниями о границах распространения отдельных форм.
Таблица 88. Основные диагностические признаки выделяемых азиатских рас снежного барана
Признаки самцов Камчатская Корякская* Чукотская Якутская Охот-кая Кодарс-кая Путоранс-кая*
Длина тела, ем 162-178 148-154 152-188 140-170 150-175 162-171 151-153
Высота в холке, см 76-106 95-100 96-106 85-112 89-104 94-103 -111
Масса, кг Наибольшая 79-150 59-92 78-105 56-97 -98 80-120 106-109
длина черепа, мм Длина стержней 261-296 256-277 275-280 258-295 256-283 288 274-294
рогов, мм Обхват стержней 150-296 165-270 -220 130-231 160-230 140-250
рогов, мм 210-335 265-305 280-330 180-330 230-330 — 250-330
Длина рогов, см 41-115 61-87 85-92 53-103 44-92 84-100 65-98
Обхват рогов, см 20-36 28-36 31-35 26-36 21-35 32-34 29-38
Окраска От темно- От светло- От светло- Серо- От светло- Буро- Бурая,
бурой серой серой желто- серой вато- серо-
до светлой до бурой до темно- вато- до темно- серая корич-
бурой бурая бурой невая
Посветления Есть у Есть у Есть Есть Есть Есть Есть
на голове части части
особей особей
Посветления Нет Нет Нет Есть Есть Нет Нет
позади плеч у части особей
Окраска низа Бурая, Светло- Серая От свет- От свет- Светло- Светло-
живота светло- серая, ЛОЙ ЛОЙ серая серая
серая грязно- до до
белая бурой бурой
Хвостовое зеркало Белое Белое Белое Белое Белое Белое Белое
* Нерепрезентативные выборки
О.В. Егоров справедливо указывал на недостаточную обоснованность выделения якутского барана в качестве подвида и на идентичность его и охотской формы [Тавровский и др., 1971]. П.Ф. Грибков и В.И. Филь [1977] тоже не нашли достоверных морфологических различий между охотским, якутским и камчатским подвидами, хотя и отметили иную форму рогов у последнего. Л.В. Сопин [19866] пришел к выводу о монотипичности азиатского снежного барана, показав, что краниометрические различия между группировками, включая популяции Корякского нагорья и Чукотки, не достигают подвидового уровня. Позднее, говоря об отсутствии оснований для разделения азиатского снежного барана на подвиды, он вместе с соавторами все же выделил три формы (путоранскую, якутскую и камчатскую) исключительно по принципу давней географической изоляции, а статус кодарского барана был снижен до популяционного уровня [Сопин, Ермолин, 1986; Ревин и др., 1988]. Ф.Б. Чернявский [2003, 2004] все известные формы свел к четырем подвидам: lydekkeri, koriakorum, nivicola и borealis. В Перечне (списке) объектов животного мира, занесенных в Красную книгу РФ по состоянию на 1 ноября 1997 г. [Красная книга России: правовые акты. М., 2000] и Красной книге Российской Федерации [2001] значатся путоранский подвид и якутский подвид Ovis nivicola lydekkeri
(чукотская популяция), т. е. чукотский подвид низведен до ранга популяции. Специалисты по группе каприн (IUCN/SSC Caprinae Specialist Group) чукотского и корякского баранов относят к форме alleni (табл. 72).
Как видим, таксономия азиатского снежного барана по-прежнему не ясна, но прослеживаются две разнонаправленные тенденции - дробление и стремление к монотипизации.
Основные диагностические морфологические признаки выделяемых азиатских подвидов снежного барана, приводимые разными авторами, сведены мной в таблице 88 с дополнением новых сведений (см. табл. 84—87). При анализе параметров этих форм очевидно, что существенных различий между ними, за редким исключением, нет, а выявленные отличия нельзя рассматривать только в аспекте географической или популяционной изменчивости - это, скорее, следствие очень малых и разновозрастных выборок. Именно по этой причине чаще всего возникают ошибочные выводы и рождаются “новые” виды и подвиды.
В целом азиатский и американский снежные бараны очень близки. И один, и другой слабо дифференцированы и по морфологическим, и по генетическим показателям. Деление снежных баранов на множество подвидов представляется несостоятельным. В связи с этим логично, на мой взгляд, принять более простую таксономическую систему, выделив азиатскую и американскую группы в качестве подвидов Ovis canadensis nivicola и Ovis с. canadensis единого политипического вида.
Распространение
Восстановленный ареал. Плейстоценовые и голоценовые ископаемые остатки снежного барана в Азии скудны, однако по районам их находок можно очертить примерные границы былого ареала. Они обнаружены в Кузнецкой котловине (р. Барзас), на Салаирском кряже, на р. Кара-Чумыш в Кемеровской области, в Хакасии, по берегам Красноярского водохранилища, в Предбайкалье (стоянка Мальта на р. Белой, окрестности Иркутска, верховья Уды, Приморский хребет), на о. Ольхон на Байкале, в Баргузинской котловине, на р. Витим, в окрестностях Бодайбо, Верхневилюйска, Северо-Байкальска и оз. Баунт, в Усть-Оборе и долине р. Хилок в Западном Забайкалье, а также на берегах р. Лены в окрестностях пос. Качикатцы и г. Якутска, р. Буотаме, в низовьях р. Юнюгян, в верхнем течении Алдана и Индигирки, на Колыме и Яне, в Дежневском древнеэскимосском поселении на Чукотке и на Сахалине. Во многих районах Южной Сибири наряду с костями снежного барана присутствуют ископаемые остатки архара [Черский, 1891; Павлова, 1911; Громова, 1941, 1947; Громов, 1948; Вангенгейм, 1961, 1977; Алексеева, 1970, 1980, 1995, 1997; Кищинский, 1972; Ермолова, 1978; Ревин и др., 1988; Сопин, Ермолин, 1988; Ревин, 1989; Динесман и др., 1996; Сопин, 1998; Оводов, 1999, 2000; Боескоров, 2001].
Можно с уверенностью говорить, что в плейстоцене ареал снежного барана в Азии простирался от Енисея и Кузнецкого Алатау до Камчатки, Сахалина и северных островов Курильской и Алеутской гряд, включая Средне-Сибирское плоскогорье, все Становое нагорье, Становой хребет и прилегающие районы (рис. 83).
Возраст известных ископаемых остатков азиатского снежного барана -10-40 тыс. лет, а из долины р. Хилок, возможно, 100 тыс. лет, американского толсторога - 25-
Рис. 83. Распространение азиатского снежного барана (по [Боескоров, 2001]) а - современный ареал, б - границы распространения в XVIII - начале XX вв.; в - ископаемые находки, относящиеся к плейстоцену; г - ископаемые находки, относящиеся к голоцену (по литературным источникам). Цифрами обозначены конкретные районы ископаемых находок (см. в оригинале)
100 тыс. лет [Hansen, 1982; Ревин и др., 1988; Оводов, 2000; Чернявский, 2004]. Дифференциация снежных баранов, проникших по Беренгийскому мосту из Азии в Америку в конце плиоцена - начале плейстоцена [Cowan, 1940], произошла, возможно, еще до исчезновения перешейка между материками, и их древние формы эволюционировали независимо. Этот вывод, подтверждаемый результатами исследований фенетики [Сопин, 19856], противоречит гипотезе об обратной миграции толсторога из Северной Америки в Азию [Северцов, 1873а,б; Насонов, 1923] и не согласуется с предположением о существовании двух волн обмена териофауной через Беренгийс-кий мост в плейстоцене [Флеров, Заблоцкий, 1961].
Современный ареал. Сведения о распространении снежного барана в Азии в историческое время тщательно собраны В.И. Цалкиным [1951а] и затем дополнены другими учеными [Соколов, 1959; Гептнер и др., 1961; Егоров, 1965; Чернявский, 1974, 1984; Ревин и др., 1988; Железнов-(Чукотский), 1990, 1994, 2003; Кри-вошапкин, Яковлев, 1999; и др.]. В генерализованном виде его ареал, общая площадь которого около 1,4 млн км2 [Ревин и др., 1988], представлен на рис. 83. В последние столетия он оказался разорванным на две части - западную и восточную, а затем стал очаговым.
Западный участок ареала снежного барана - горы Путорана (800-1700 м над уровнем моря) на Средне-Сибирском плоскогорье. В XIX - начале XX вв. его северная граница, видимо, вплотную подходила к северному пределу плато Путора
на; южная шла почти по широте Северного полярного круга и включала верхние части бассейнов правых притоков Нижней Тунгуски; западная - по водоразделу рек Пясина - Хантайка (горы Сыверма); восточная граница определялась началом сглаженных форм рельефа. Площадь области обитания, скорее всего, достигала 200 тыс. км2. Рог барана был найден в низовьях Енисея, и оттуда академиком Шмидтом доставлены в Академию наук череп и шкура. Эти копытные, возможно, населяли междуречье Хатанга - Анабар и горы Бырранга на полуострове Таймыр [Сидоров, 1881; Наумов, 1934; Флеров, 1935а; Подаревский, 1936; Цалкин, 1951а; Ревин и др., 1988].
Сообщения об обитании снежного барана в горах Таймыра И.И. Соколов [1959] и В.Г. Гептнер [Гептнер и др., 1961] считали ошибочными, хотя я не нахожу веских причин для такого заключения - звери могут перемещаться на десятки и сотни километров и по равнинной, и по лесистой местности (см. ниже). По сведениям Н.К. Железнова-Чукотского [1994], в ноябре 1975 г. баран был встречен на северном побережье Гыданского полуострова напротив острова Оленьего, а свежесло-манный рог найден оленеводами еще западнее - в северной части полуострова Мамонта. Эти пункты находятся в тундровой зоне за 650-700 км от современных западных границ ареала вида, что свидетельствует о реальной возможности проникновения этих животных на север Таймыра и в другие районы, которые прежде считались недосягаемыми.
К середине XX в. снежные бараны сохранились в основном в центральной и северо-восточной возвышенной части плато Путорана [Мичурин, Мироненко, 1966а,б]. Современная северная граница ареала проходит по водоразделам рек Микчанды, Калтамы и устью Хибарбы, южная - вблизи озер Агата и Дюпкун, западная - по горам в окрестностях озер Лама, Кета и Хантайское, на востоке - по горам высотой более 1000 м над уровнем моря (рис. 84). Площадь этого участка, состоящего из мелких разрозненных очагов, но имеющего тенденцию к расширению в последние десятилетия в результате прекращения выпаса стад домашних оленей и улучшения охраны животных, составляет примерно 70-120 тыс. км2, что уже близко к его размерам в XIX - начале XX вв. [Павлов, 1977; Боржонов и др., 1979, 1988; Ларин, 19876, 1989, 1990; Ларин и др., 1988; Сыроечковский, 19956; Ларин, Сипко, 2004].
Восточный участок ареала снежного барана (рис. 83), расположенный примерно в 1300 км от западного, включает Верхоянскую горную систему, где северная точка распространения - устье р. Кенгдей на правобережье Лены (изредка звери заходят на левобережный отрог хребта Туора-Сис), горные массивы хребтов Сунтар-Хаята, Черского и Момского, Охотско-Колымское нагорье, почти всю Чукотку вплоть до восточной оконечности полуострова, Корякское нагорье, горные хребты и скалистые прибрежные районы Камчатки. На юге современный ареал ограничен Становым нагорьем, северной частью Яблонового хребта, Становым хребтом и его обрамлением: Ал-дано-Учурским и Сутамо-Гонамским хребтами, Токкинским Становиком, Джугджу-ром и далее Майским и Прибрежным хребтами. Южной границей распространения вида считаются р. Удд и верховья р. Селемджи [Насонов, 1923; Флеров, 1935а; Аверин, 1948,1951; Цалкин, 1951а; Соколов, 1959; Гептнер и др., 1961; Чернявский, 1961, 1968, 1970а, 1974, 1984; Портенко и др., 1963; Егоров, 1965; Кищинский, 19676; Обухов, 1969; Тавровский и др., 1971; Вершинин, 1972; Метельский, 1973; Грибков, Филь,
Рис. 84. Распространение снежного барана на плато Путорана (по [Ларин, 1990])
1 - основные очаги обитания; 2 - граница высокогорий, нс пригодных для обитания; 3 - места встреч локальных групп и одиночных особей; 4 - граница восстановленного ареала
1977; Чернявский, Железнов, 1977; Кривошеев, Чернявский, 1981; Ревин, 1982а, 1989; Бромлей, Кучеренко, 1983; Мосолов, 19866,1992; Ревин и др., 1988; Железнов-(Чукот-ский), 1990, 1994, 2003; Чернявский и др., 1990; Кривошапкин, Яковлев, 1999; Мым-рин, 2003].
Область обитания вида на Корякском нагорье с севера и запада отделена низменностью Анадырско-Пенжинской депрессии, а от Камчатки - протяженным (около 300 км) Парапольским долом [Чернявский, 19626; Портенко и др., 1963]. Тем не менее, учитывая возможность перемещения животных на десятки и сотни километров даже по открытой местности, этот и другие очаги, на которые раздробился восточный участок ареала в последнее столетие, нельзя считать полностью изолированными.
Всего две-три сотни лет назад снежный баран жил на Курильских и Алеутских островах, но затем человек полностью истребил его [Флеров, 1935а; Воронов, 1974, 1982, 1983].
В связи с выделением кодарского снежного барана в качестве подвида особый интерес вызывает юго-западный участок ареала. В Предбайкалье (по Ангаре) снежный баран обитал, вероятно, до XVIIв. включительно [Гептнер и др., 1961]. В Х1Х-начале XX вв. он плотно населял Витимо-Олекминский район (окрестности оз. Орон, бассейны Витима, Чары и Олекмы) и Становой хребет [Поляков, 1873; Насонов, Дорогос-тайский, 1915; Цалкин, 1951а]. В первой половине XX в. самым юго-западным местом распространения был Бодайбинский район Иркутской области, где звери жили близ оз. Ничатка в верховьях р. Эльгер, в районе верхних притоков р. Чары, а также в бассейне оз. Леприндо. Нередко встречались они и в верховьях рек М. и Б. Оконон, у озера Токко и на хребте Кодар. В верховьях р. Токко барана истребили к 40-м годам [Флеров, 1935а; Подаревский, 1936; Туров, 1937; Павлов, 1949; Скалой, 1949,1951]. Во второй половине века эти копытные сохранились лишь на хребте Кодар. Ядро
Рис. 85. Распространение снежного барана на хребте Кодар (по [Ревин и др., 1988])
немногочисленной популяции сосредоточено в междуречьях Сюльбана, Верхнего, Среднего и Нижнего Сакукана. На востоке область распространения ограничена р. Чара, на юге - горами Сюльбана, на севере - р. Сень (рис. 85). В Иркутской области баран уцелел в верховьях рек Левой Сыгыкты, Куда Малая в Муйской котловине, по р. Талая (правый приток Витима) и в верховьях р. Ледниковая на территории Витимского заповедника. Животные изредка встречаются также близ оз. Ничатка и на хребтах Удокан и Калар [Лавов, 1966; Водопьянов, 1971; Сопин, Ермолин, 1986, 1988; Медведев и др., 1991; Сопин, Медведев, 1992; Сопин, 1993; Медведев, 1997, 20036; Медведев, Лямкин, 2000]. Кодарская популяция, несомненно, ранее имела связь с соседними группировками, истребленными или исчезнувшими в относительно недавнее время, и поэтому оснований считать ее “древним изолятом” нет.
Основные факторы, определяющие область распространения. В плейстоцене, судя по районам находок ископаемых остатков, азиатский снежный баран заселял не только высокогорье и среднегорье, но и малоснежное низкогорье и хол-могорье. На рубеже плейстоцена и голоцена увеличение высоты снежного покрова, вызванное потеплением и увлажнением климата, скорее всего, привело к его исчезновению на плоскогорьях и к разрыву прежней области обитания на ряд очагов. В дальнейшем человек-охотник и крупные хищники завершили формирование ареала, сделав его исключительно горным.
Современные границы распространения снежного барана определяются комплексом факторов, и, прежде всего, наличием убежищ (скалистых участков), доступностью зимнего корма и охотничьим прессом. Он не живет там, где высота снега превышает 40 см и в районах с плотным снежным покровом. Равнинные малоснежные участки, не имеющие убежищ, ему не позволяют осваивать крупные хищники и человек.
Факторы, ограничивающие южную границу распространения этого животного не ясны. Барьером, возможно, служит высокий снежный покров (более 40 см), что типично для лесной и лесостепной зон и гряды высоких хребтов Южной Сибири и юга Дальнего Востока. Однако на некоторых участках южная граница ареала выходит за многоснежный рубеж, и вид, казалось бы, должен был плотно заселить малоснежные районы юга Забайкалья, где он, судя по ископаемым остаткам, обитал ранее.
Определенную роль в ограничении южных границ ареала, возможно, играет температурный фактор, но его роль совершенно не исследована. Американский пустынный толсторог на юге США и в Мексике живет в гораздо более теплом климате, чем южные популяции снежного барана в Азии. Отдельные исследователи, однако, отмечают худшую приспособленность этих копытных к аридным условиям обитания
по сравнению с другими крупными млекопитающими: они более уязвимы из-за продолжительной беременности, растянутого срока рождения детенышей, редкости двоен в помете, дневной активности в летнюю жару, оседлости и неманевренности [Bailey, 1980]. Другие, напротив, считают пустынную форму хорошо адаптированной к ксе-рофитным условиям: животные выносливы к обезвоживанию; способны противостоять чрезмерным тепловым нагрузкам, ведя ночной образ жизни, используя тень в дневное время и устраивая лежки в светлом и сыром грунте; сроки рождения детенышей у них сдвинуты и приурочены к самому благоприятному времени вегетации растительности; они живут небольшими группами, рационально используя немногочисленные разрозненные пастбища и водоемы [Hansen, 1982].
Не исключено, что в давние времена имела место конкуренция азиатского снежного барана с архаром, северная граница восстановленного ареала которого перекрывается с южной границей распространения первого [Оводов, 2000]. И все же основная причина ограничения его расселения на юг, скорее всего, иная - крупные хищники, пресс которых на юге ареала гораздо выше, чем на севере, а также более интенсивная охотничья деятельность человека.
Среда обитания
Основные биотопы. Снежный баран - типичный обитатель горных массивов Северной Азии и Северной Америки. Он живет на разных высотах (от 0 до 3000 м над уровнем моря) в низкогорье, среднегорье и высокогорье и в разных растительных зонах (от лесной до альпийской), что свидетельствует о его высокой экологической пластичности.
В северных горах имеются хорошие защитные условия, обилие корма и воды в вегетационный период, а зимой звери могут добывать пищу на отдельных склонах, с которых ветром сдувается снег. Прекрасные теплоизоляционные свойства меха и запасы жира, в сочетании с оптимальным выбором укрытий, позволяют этим копытным выживать даже при очень низких (до -50-60 °C) температурах и сильных ветрах.
В ареале азиатского снежного барана Н.К. Железнов [1990, 1994] выделяет 11 типов местообитаний: пологоовальные склоны (гряды холмов, отроги низкогорных цепей) высотой до 500 м над уровнем моря, водораздельные склоны, горные плато, речные и морские скалистые берега, склоны и отроги горных цепей, кряжей и хребтов, кары (понижения на склонах гор), горные цирки и гряды, глубокие вулканические врезы. А.В. Ревин с соавторами [1988] все местообитания сводит к трем основным типам: склоны и вершины в альпийском и субальпийском поясах в крупных горных массивах; плато вулканического происхождения с выраженной террасированностью склонов (характерные для плато Путорана, Корякского нагорья и Камчатки); скалистые каньоны и береговые обрывы в руслах рек и уступы морских побережий, локализованные на относительно низких высотах и чаще в пределах пояса высокоствольной растительности.
На плато Путорана летом большая часть популяции живет в гольцовом и подгольцовом поясах преимущественно на склонах гор у верхней кромки леса, на безлесных и облесенных террасах и уступах бортов долин (фото 55). На периферии ареала звери держатся в крутосклонных долинах с выходами базальтов, которые
служат укрытиями. Растительные комплексы террассированных речных и озерных долин на высотах от 200 до 1000 м являются своеобразными экологическими зонами, обеспечивающими существование баранов. С установлением снежного покрова животные концентрируются в подгольцовой зоне, а по мере увеличения высоты снега спускаются в лесной пояс. Гон проходит в нижней части подгольцовой зоны, после чего звери перемещаются в малоснежную гольцовую зону, где выпасаются на выдувах. Весной большая часть группировки спускается к границе леса на участки дриадово-травяных и кустарничково-моховых тундр. В гольцах остаются только беременные и лактирующие самки, которые занимают верхние части цирков ручьев в нагромождениях базальтовых выходов, соседствующих с ровными площадками щебнистых тундр. В июле сюда подтягиваются и другие половые и возрастные группы животных [Боржонов и др., 1979; Ларин, 1983, 19876, 1990; Федосенко, 1985; Ларин, Шелковникова, 1988; Ларин, Сипко, 2004]. Иногда бараны встречаются в биотопах с сильно сглаженным рельефом, где скалы отсутствуют [Мичурин, Мироненко, 1966а], что убедительно подтверждает возможность их былого обитания почти на всем Средне-Сибирском плоскогорье.
На многочисленных хребтах в Якутии летом эти копытные чаще держатся у снежников в зоне горных тундр на высоте 1900-2300 м, где меньше гнуса. Излюбленные пастбища - альпинотипные лужайки в скалистых верховьях горных рек, ручьев и у подножий горных цирков и каров, а также участки сухой тундростепи и увлажненные луговины по берегам ледниковых озер (фото 56). Осенью они мигрируют в малоснежные низкогорья (700-1500 м) восточного и северного макросклонов или остаются на зимовку в горно-тундровом и гольцовом поясах (1200-2000 м), лишенных кустарниковой растительности, предпочитая гладкие или частично террасированные склоны с обширными участками мелкощебнистых осыпей. Весной звери концентрируются на крутых южных склонах, которые раньше освобождаются от снега. Характерно, что наряду с безлесными горно-тундровыми биотопами снежные бараны Якутии населяют пояс кедрового стланика, лиственничники и пойменные угодья или круглогодично заходят в них [Егоров, 1965; Тавровский и др., 1971; Фертиков, 1978, 1979; Ревин, 19826; Кривошапкин, 1995; Кривошапкин, Яковлев, 1999].
В Северном Забайкалье они летом и осенью встречаются по всему Центральному Кодару, а зимой переходят на восточный макросклон хребта, пригреваемый солнцем. Основные типы местообитаний - плато, гребни и вершины гольцов, кары и цирки, долины рек и ключей и склоны гор с луговыми ассоциациями в альпийском и субальпийском поясах (фото 57). Изредка звери встречаются в лесной и нивальной зонах, а в малоснежные годы заходят и в редколесье [Водопьянов, 1971; Сопин, Ермолин, 1988; Сопин, 1993; Медведев, 1997].
На Охотско-Колымском нагорье и Чукотке летними пастбищами снежных баранов служат горные плато, пологие склоны холмов и нагорные террасы, где господствует дриадовая тундровая растительность, а также площадки с влаголюбивыми растениями в местах тектонических разрывов, разреженные ольховники с низкорослыми ивами, увлажненные луговины в речных долинах и на приводораздельных склонах (фото 58). Иногда звери пасутся на границе лесной и кустарниковой зон и непосредственно в лесу (фото 59). С началом зимы они обычно поднимаются на пастбища подгольцового и гольцового поясов, где придерживаются труднодос
тупных скальных выходов и склонов с неравномерным залеганием снежного покрова. На побережье Охотского моря зимой животные обитают на значительном удалении от населенных пунктов на скалистых склонах с задернованными участками до высоты 400-500 м, летом поднимаются до 1500 м [Кищинский, 1967а,б; Чернявский, 1968, 1970а; Железнов, 1990, 1994; Чернявский и др., 1990].
Па Корякском нагорье снежный баран населяет горно-тундровой пояс большинства хребтов. Наиболее типичные местообитания - труднодоступные отроги горных хребтов со скалистыми вершинами, на склонах которых скальные выходы и мелкощебнистые осыпи чередуются с лужайками горно-тундровой растительности на задернованных склонах (фото 60); горные цирки и кары, днища и нижние части склонов которых служат пастбищем, а ближайшие скалы - убежищем; скалистые речные каньоны и базальтовые плато вулканического происхождения, с поверхности которых зимой сдувается снег. На Олюторских горах животные выходят на низкие приморские скалы. Зимой они чаще держатся на малоснежных высокогорных пастбищах, весной перемещаются на склоны южной экспозиции, где ранее всего появляется зеленая трава, а с августа, по мере ее увядания, уходят на северные склоны. Летом звери предпочитают хребты средней высоты, где самцы живут и в лесном поясе [Чернявский, 1961, 1963; 19676, 1984; Портенко и др., 1963; Железнов-Чукотский, 1994].
На Камчатке местообитания снежного барана (фото 61) преимущественно двух типов: высокогорные (горные хребты и изрезанное вулканическое высокогорье до 2600 м) и прибрежные (береговые уступы с обрывистыми скалами, тянущиеся узкой полосой по побережью). Некоторые группировки населяют скалистые участки невысоких хребтов или обрывы глубоких речных долин между высокогорьем и побережьем, но они более доступны для людей, и охотники уничтожают их здесь в первую очередь. Лучшие летне-осенние пастбища расположены в карах ледникового происхождения и цирках, в которых долго сохраняется влага, на лужайках скалистых выступов на морском побережье и в кустарниковом поясе, а также на разнотравных альпийских лугах. Зимой они охотно пасутся на моренных холмах с кустарничково-лишайниковыми ассоциациями и разнотравьем, на выдувах в голь-цовом поясе и на приморских террасах [Гаврилов, 1947;Аверин, 1948, 1951;Гриб-ков, 1970; Вершинин, 1972; Вяткин, Филь, 1975; Филь, 1977в; Стенченко, 1980; Мосолов, 1986а, 1992].
Факторы, влияющие на биотопическое размещение. Ведущую роль в био-топическом размещении снежного барана играют защитный и трофический факторы, во многом взаимосвязанные. Эти копытные предпочитают открытые верхние пояса гор - подгольцовый и гольцовый, где выбирают расчлененные скальные образования и скалистые обрывы, пригодные в качестве убежищ от хищников и укрытий в непогоду, с расположенными поблизости пологими участками с лишайнико-травянисто-кустарничковыми сообществами. От ближайших убежищ они обычно не уходят на кормежку далее 400-500 м, а при опасности бегут к скалам даже из леса и кустарников через открытые места.
Зимой распределение зверей по территории в большей мере определяется наличием и доступностью корма. При высоком или плотном снежном покрове и гололеде они вынуждены перемещаться в высотном направлении или переходить на малоснежные склоны или в другие районы. При многоснежье доступными чаще
всего остаются лишь скудные арктические щебнистые тундры на выдуваемых местах в гольцовом поясе. Выбор летних стаций зависит не только от наличия зеленого корма и убежищ, но и от воздействия кровососущих насекомых. В последние десятилетия в биотопическом распределении баранов значительную роль играет антропогенный фактор.
В выборе биотопов и приспособленности к среде обитания снежные бараны как бы сочетают свойства горных баранов и козлов, что и позволило им освоить обширнейшие и в целом малопригодные для других видов копытных горные пространства Северной Азии и Северной Америки. В целом пространственное размещение вида отличается ярко выраженной мелкоочаговостью и относительной изолированностью отдельных групп и группировок.
Питание
Корма. Список кормовых растений азиатского снежного барана весьма обширный - более 320 видов по моим подсчетам по литературным источникам, причем он достаточно велик в каждом районе (от 40 до 215 видов) и близок к фоновому спектру растительности в местах обитания. Более 90% пищи представлено высшими растениями, остальное - лишайники и зеленые мхи. В рубцах животных чаще находят от 1 до 6 видов, составляющих до 80% объема пищи, и изредка - до 24-34 [Капитонов и др., 1975].
В вегетационный период эти копытные отдают предпочтение травянистой растительности (злаковым, бобовым, осоковым, гречишным и сложноцветным),, составляющим до 70-95% рациона. Ведущую роль в весенне-летнем и раннеосеннем питании играют зеленые травы с повышенным содержанием протеина. Излюбленными растениями в этот период становятся пушица влагалищная, доминирующая по биомассе и отличающаяся высокой питательностью молодых побегов, а также астрагалы, остролодки, копеечники, горцы, кислица, мытники, ветреница сибирская, иван-чай, клевера, зеленые проростки осок, ситника, вейника, мятлика, овсяницы, молодые побеги и листья ив и берез. В видовом списке нередко встречаются лекарственные и ядовитые растения, такие как родиола розовая, пижма, одуванчик, толокнянка, прострел раскрытый, живокость. Раннеосенний рацион животных дополняется лишайником, ягодами и грибами. Они охотно едят подберезовики, маслята и сыроежки, особенно старые, напичканные личинками грибных паразитов, что служит существенной белково-витаминной добавкой в период окончания вегетации растительности. В снежный период массовым кормом служат наземные лишайники (до 30-65% рациона и до 65-90% объема содержимого рубцов), ветошь трав (иногда до 50% содержимого рубцов), осоки и злаки, уходящие зелеными под снег или растущие под его покровом, полыни, зеленые хвощи, мхи (около 4-16%), плауны, кустарниковые и кустарничковые растения: карликовые ивы, березы, вереск, багульник, дриада, голубика, брусника, Кассиопея и арктери-ка. В содержимом рубцов встречаются корневища мытника и астрагала, корни остролодочника и копеечника, кора и хвоя лиственницы и кедрового стланика. Нередко эти копытные разоряют стожки северной пищухи [Гаврилов, 1947; Аверин, 1948, 1951; Федосенко и др., 1961; Чернявский, 1961, 1963, 1967а,б, 1968, 1974, 1984; Портенко и др., 1963; Егоров, 1965; Мичурин, Мироненко, 1966а,б; Тавровс-
Фото 1. Бизон в Йеллоустонском национальном парке (фото И. Середкина)
Фото 3. Беловежско-кавказский зубр (фото Т. Сипко)
Фото 4. Кавказский зубробизон (фото С. Трепета)
Фото 5. Типичный биотоп зубра разреженный лиственный лес (фото А. Дигиле-вича)
Фото 6. Места обитания зубробизона в Кавказском заповеднике (фото С. Трепета)
Фоiо 7. Зубробизон у "каталки" (фото С. Трепета)
Фото 9. Стадо монгольского дзерена (фото В. Кирилюка)
Фото 10. Детеныш монгольского дзерена (фото П. Сорокина)
Фото 11. Отличительная особенность сайги - хоботообразный ное (фото П. Сорокина)
Фото 12. Окраска головы и форма носа самца сайги во время гона (фото П. Сорокина)
Фото 13. Самцы сайги в летнем (вверху) и зимнем (в центре) наряде и во время весенней линьки (фото А. Лущекиной. П. Сорокина и Р. Дор-мидонтова)
Фото 14. Новорожденный сайгачснок (фото Т. Баженова)
Фото 15. Стадо сайги (фото Т. Баженова)
Фото 16. При преследовании сайга развивает скорость до 60-80 км/ч (фото Р. Дормидонтова)
Фото 17. Овцебыки (фото С. Царева)
Фото 18. Детеныши овцебыка (фото В. Тихонова)
Фото 19. Овцебыки на ферме (фото В. Тихонова)
Фото 21. Безоаровый козел (фото Й. Хласека)
Фото 20. Амурский горал (фото П. Романова)
Фото 22. Западнокавказские козлы (фото С. Тпшксвича)
Фото 23. Восточнокавказские козлы (фото С. Тишкевича)
Фото 25. Типичные места обитания козлов на Западном Кавказе (фото С. Тишкевича)
Фото 26. Восточнокавказские козлы предпочитают держаться на скалистых участках (фото С. Тишкевича)
Фото 24. Детеныш восточнокавказского козла (фото С. Тишкевича)
Фото 27. Западнокавказские козлы на солонце (фото С. Тишкевича)
Фото 28. Группа самок восточнокавказского козла с детенышами и подростками (фото С. Тишкевича)
Фото 29. Группа молодых восточнокавказских козлов (фото С. Тишкевича)
Фото 31. Европейский муфлон (фото С. Ерошенко)
Фото 32. Закавказский муфлон (Г. Казаряна)
Фото 34. Устюртский уриал (фото из архива А. Хохлова)
Фото 35. Эльбурский баран, считающийся гибридом муфлона и уриала (фото из архива Л. Хохлова)
Фото 36. Изменчивость окраски архарят (фото А. Бербера и С. Тишкевича)
Фо।о 37. Памирский архар (фото С. Тишкевича)
Фото 38. Североказахстанский баран (фото С. Тишкевича)
Фото 39. Баран Северцова (горы Нуратау, фото С. Тишкевича)
Фото 40. Типичные биотопы архара в Казахском нагорье (фото А. Бербера)
Фото 41. На Памире самки архара держатся в нижних частях горных массивов (фото С. Тишкевича)
Фото 42. Места обитания баранов в Каратау (фото С. Тишкевича)
Фото 43. Стации барана Северцева в горах Нуратау (фото С. Тишкевича)
Фото 45. Тропы (вверху) и лежка баранов в Казахском мелкосопочнике (фото А. Бербера)
Фото 46. Отдыхающий памирский архар (фото С Тишкевича)
Фото 47. Типичное расположение архаров на отдыхе (фото С. Тишкевича)
Фото 48. Стации уриала в Таджикистане (фото С Тишкевича)
Фото 49. Северокитайский баран (Внутренняя Монголия. фото С. Тишкевича)
Фото 50. Камчатский снежный баран (фото А. Дигилевича)
Фото 52. Типичная окраска молодого и взрослого самцов якутского снежного барана (фото А. Кривошапкина)
Фото 53. Охотский снежный баран (фото С. Тишкевича)
Фото 54. Путоранский снежный баран (фото А. Волкова)
Фото 55. Места обитания снежного барана на плато Путорана (фото А. Волкова)
Фото 56. Места обитания снежного барана в Центральном Верхоянье (фото Ф. Яковлева)
Фото 57. Места обитания снежного барана на Центральном Кодаре (фото Д. Медведева)
Фото 58. Места обитания охотского снежного барана (фото С. Тишкевича)
Фото 59. Охотский снежный баран, выпасающийся в лесу (фото С. Тишкевича)
Фото 60. Места обитания снежного барана в Корякском нагорье (фото С. Тишкевича)
Фото 61. Места обитания снежного барана на Камчатке (фото С. Тишкевича)
кий и др., 1971; Капитонов и др., 1975; Грибков, Филь, 1977; Железнов, 1981а, 1990, 1994; Ларин, 1983, 1987а, 1990; Соскин, Глушнев, 1983; Тишков, 1985; Федосенко, 1985; Мосолов, 1986а; Ларин, Шелковникова, 1988; Ревин и др., 1988; Чернявский и др., 1990; Колесник, Головушкин, 1994; Медведев, 1997; Кривошапкин, Яковлев, 1999; Ларин, Сипко, 2004].
Большую долю лишайников в зимнем питании снежных баранов исследователи связывают с большим содержанием в них углеводов [Ревин и др., 1988], а зеленых мхов - с повышенным содержанием железа [Кривошапкин, Яковлев, 1999]. На мой взгляд, основная причина выборочного поедания этих групп растений, как и растений, уходящих под снег в зеленом состоянии, кроется в большом содержании в них воды.
Масса содержимого желудков взрослых зверей, добытых летом и ранней осенью, когда запасы корма не ограничены, достигает 6-12 кг, зимой - в два раза меньше [Аверин, 1948, 1951; Чернявский, 1961; Капитонов и др., 1975]. За лето полувзрослые и взрослые животные, в отличие от растущих сеголеток, быстро накапливают жировые резервы. Зимой почти повсеместно очевиден дефицит пищи. При снежной толще в 10 см бараны начинают раскапывать растения передними ногами, а критический уровень, при котором тебеневка прекращается, - 25-30 см. При плотном снеге или гололеде добывание пищи становится невозможным. К концу зимы ближайшие к убежищам пастбища обычно сильно стравливаются, и бараны вынуждены уходить все дальше от них и в более многоснежные места. Из-за недостатка корма звери, особенно сеголетки, быстро истощаются и нередко погибают.
В экспериментах показано, что уровень потребления кормов канадским толсторогом в зимний период с октября до февраля - марта уменьшается почти вдвое. Скорость метаболизма резко снижается в феврале и восстанавливается в марте - апреле. Метаболизм у самцов выше, чем у самок, и значительно ускоряется при температуре ниже -20 °C и сильном ветре [Chappel, Hudson, 1978].
Почти все исследователи отмечают регулярное посещение снежными баранами твердых глинистых или грязевидных солонцов и минеральных водных источников в бесснежный период с пиком посещений в июне - июле и всплеском активности в сентябре [Насонов, Дорогостайский, 1915; Черемкин, 1959; Чернявский, 1967а,б; Водопьянов, 1971; Федосенко, 1985; Ревин, 1989; и др.]. На хребте Кодар имеются каменноугольные солонцы [Медведев, 20036]. На Камчатке звери поедают выброшенные на берег морские водоросли [Вяткин, Филь, 1975]. Беременные и лактиру-ющие самки и годовалые особи ходят на солонцы чаще, чем взрослые самцы и яловые самки. В период “белых ночей” звери появляются на них круглосуточно, с конца июля - только в светлое время. Средняя продолжительность солонцевания животных в июле достигает 3 ч. Зафиксированы случаи поедания самками свежих остатков зайца-беляка [Кривошапкин, Яковлев, 1999] и костей животных [Keating, 1990], что также может быть связано с недостатком минеральных веществ в организме.
Пищевая активность. Суточный ритм активности полифазный. Летом у снежного барана 3-4 периода кормежки, из которых наиболее продолжительный (4—5 ч) -раннеутренний. В дневную жару звери отдыхают у гребня хребтов, где постоянно дуют ветры, прохладно и нет гнуса. В пасмурную погоду дневная пастбищная активность возрастает. В короткие зимние дни при наличии скудного корма они па
сутся с рассвета и до сумерек. Весной с началом вегетации животные также кормятся всю светлую часть суток с небольшими перерывами на отдых и пережевывание пищи. При кормежке почти треть времени тратится на осматривание местности [Аверин, 1948, 1951; Чернявский, 1961, 1962г; Федосенко, 1985].
Размножение
Половая зрелость. Самки снежных баранов могут вступать в размножение с полутора лет, но большинство созревает на год, а часть - на два года позднее. Зрелая сперма в придатках семенников самцов обнаруживается в возрасте полутора лет, и они успешно кроют самок, однако в природе в гоне участвуют обычно сильные самцы старше 5 лет. При недостатке взрослых баранов шансы молодых самцов на оставление потомства существенно возрастают [Чернявский, 1961, 1962а, 1974, 1984; Егоров, 1965; Тавровский и др., 1971; Филь, 1975, 1978; Грибков, Филь, 1977; Nichols, 1978; Железнов, 1980, 1990, 1994; Ларин, 1989, 1990; Festa-Bianchet et al., 1995]. В вольерах в США зарегистрированы случаи оплодотворения 10-11-месячных самок, которые рожали нормальных детенышей в годовалом возрасте [McCutchen, 1977; Morgart, Krausman, 1983].
Гон и половое поведение. Гон у снежных баранов проходит в ноябре - декабре (табл. 89) в специфических условиях, когда световой день не превышает 4—5 ч или же полярной ночью при минусовых температурах, достигающих 40-50 °C.
В октябре перед гоном разновозрастные стада самцов подтягиваются к традиционным местам обитания самок, где и проходит гон. Здесь стада дробятся: обособляются бараны старше 6 лет, которые вносят основной вклад в продолжение рода; группы 4—6-летних особей, пытающихся оставить потомство, особенно, в отсутствии старых самцов, и группы 3-4-летних зверей, практически не участвующих в спаривании [Оконешников, 2003]. Одиночные взрослые бараны особенно активны. В поисках
Таблица 89. Сроки гона у азиатского снежного барана
Район | Начало I Конец I Источник информации
Плато Путорана Середина ноября Середина декабря Наумов, 1934
н Вторая декада ноября 10 декабря Ларин, 1990
Якутия 13 ноября Начало января Егоров, 1965
it 25 октября 25 ноября Черемкин, 1959
Верхоянье Середина ноября Середина декабря Кривошапкин, Яковлев, 1999
Хребет Кодар 15 ноября 15 декабря Медведев, 1997
Хребет Джугджур Октябрь Ноябрь Рудков, 1979
Хребет Сунтар-Хаята 6 декабря 26 декабря Железнов, 1990, 1994
Чукотка Середина ноября Конец декабря сс
Корякское нагорье Декабрь Начало января Чернявский, 1962а, 1963;
« 3 декабря 29 декабря Портенко и др., 1963 Железнов, 1994
Камчатка Ноябрь , Декабрь Аверин, 1948, 1951
<( 5 ноября 3 января Филь, 1978
Юг Дальнего Востока Середина сентября Начало января Бромлей, Кучеренко, 1983
самок они могут преодолевать десятки километров. Замечено, что в этот период самки подолгу стоят неподвижно на вершинах холмов, откуда хорошо видны их силуэты, на которые самцы реагируют с расстояния до 3 км. Одиночных самок, в свою очередь, “притягивают” силуэты самцов, тогда как самки, находящиеся в стадах, не проявляют активности в поиске половых партнеров. На Крайнем Севере, в условиях полярной ночи, животные разыскивают друг друга, главным образом, с помощью обоняния [Железнов, 1980; Hogg, 1987; Ревин и др., 1988].
Сблизившись, партнеры обнюхивают друг друга, самец при этом флемует, и если самка готова к спариванию, следует несколько садок, длящихся в среднем около 36 с. После копуляции половые партнеры могут кормиться. При встрече с группой самок самец сначала поочередно обнюхивает их, и только потом спаривается с пришедшей в охоту. Если в группе нет особей в течке, то самки стремятся скрыться от приближающегося самца, а он начинает преследовать их [Железнов-Чукотский, 1994], возможно стимулируя таким образом. Среди диких копытных такая стимуляция самок наиболее ярко проявляется у косуль [Данилкин, 1999].
Иную пространственную структуру и половое поведение наблюдал у снежных баранов В.И. Филь [1978] на Камчатке. Самцы перед гоном занимали определенные участки, которые охраняли от соперников и, возможно, маркировали, многократно обходя границы. К периоду гона на них подходили самки, вытесняемые глубокоснежьем из субальпийского пояса. Контакт с самцом стимулировал у самок начало течки. Следовательно, камчатские бараны в период гона становятся, по этим наблюдениям, типичными территориальными животными, что не укладывается в рамки известного об их поведении.
Образовав брачную группу, самец активно защищает ее от соперников. При их приближении хозяин принимает позу угрозы, временами привставая на задние ноги. Демонстраций обычно достаточно, чтобы держать противников на расстоянии. Манера боя во многом зависит от характера местообитаний. На больших площадках бараны сталкиваются лбами, разгоняясь с расстояния в 10-15 м, а на узких заснеженных скаль-никах - вставая на задние ноги и без разбега, что более типично для козлов [Северцов, 1951; Грибков, Филь, 1977; Филь, 1978; Кривошапкин, Яковлев, 1999].
На одного самца старше 5 лет, участвующего в гоне, приходится в среднем 1—2 самки. Общее их количество в брачных группах обычно не превышает 9, чаще их 2-4, но это половое соотношение может меняться едва ли не ежедневно [Чернявский, 1962а; Филь, 1975, 1978; Железнов-(Чукотский), 1980, 1994]. Иногда самку в течке преследуют сразу несколько самцов, а их число при 3-4 половозрелых самках в группе может достигать 12. Доминант за 1 час делает от 3 до 9 садок, длящихся по 2-3 с [Егоров, 1965; Ларин, 1990].
Гон у снежного барана в целом протекает спокойно, без значительных энергетических затрат и резкого исхудания производителей. После гона соседние брачные группы образуют смешанное стадо, из которого затем уходят взрослые самцы, объединившиеся в однополую группу.
Беременность и развитие эмбрионов. Продолжительность беременности самок снежного барана колеблется в пределах 171-200 дней: в Северной Азии -180-190, в Америке - 171-178 [Чернявский, 1962а; Nichols, 1978; Бромлей, Кучеренко, 1983; Shackleton et al., 1984; Ревин и др., 1988; Ларин, 1989,1990; Железнов, 1990, 1994; Hass, 1995; Кривошапкин, 1995; Goodrowe, 1996; Кривошапкин, Яков
лев, 1999]. Запаздывание начала вегетации растительности, возможно, вызывает задержку родов, что сказывается на варьировании сроков беременности. Это явление известно для оленьих [Данилкин, 1999]. Масса эмбриона снежного барана составляет в феврале около 150-300 г, марте - до 650 г, мае - от 500 до 2180 г, перед рождением - свыше 3 кг [Аверин, 1948, 1951; Егоров, 1965; Тавровский и др., 1971; Кривошапкин, Яковлев, 1999].
Деторождение и выкармливание молодняка. На юге ареала ягнение у азиатского снежного барана чаще начинается в мае, на севере - в июне (табл. 90), и оно приурочено к наступлению теплого периода, прекращению ночных заморозков и началу вегетации растительности. Примерно такие же сроки родов и у американских толсторогов: на севере ареала - в мае - июне, южнее - с конца апреля до начала июля, а у пустынной формы детеныши могут появляться в любое время года, но чаще в апреле, что тоже связано с вегетацией растений [Geist, 1971; Nichols, 1978; Bailey, 1980; Bunnell, 1980, 1982; Hansen, 1982; Stewart, 1982; Festa-Bianchet, 1988; Rachlow, Bowyer, 1991].
Таблица 90. Сроки деторождения у азиатского снежного барана
Район | Начало | Конец | Источник информации
Плато Путорана Начало июня — Боржонов и др., 1979; Федосенко, 1985
<< 27 мая 9 июня Ларин, 1989, 1990
Якутия 9 июня Конец июля Егоров, 1965; Тавровский и др., 1971
сс 5 мая 10 июня Черемкин, 1959
Верхоянье Конец мая Июль Кривошапкин, Яковлев, 1999
Хребет Кодар и — Водопьянов, 1971
Хребет Джугджур Апрель Май Рудков, 1979
Колымское нагорье Конец мая Июнь Кищинский, 1967а
Чукотка Вторая половина июня Чернявский, 1970а
Корякское нагорье Конец мая Вторая половина июля Чернявский, 1961, 1963, 1962а, 1984
к 6 июня 28 июня Железнов, 1994
Камчатка Конец мая Июнь Аверин, 1951
За несколько дней до родов беременные самки отделяются от стада и уединяются в труднодоступных скалистых участках, где рожают из года в год в одних и тех же местах, чаще на склонах южной экспозиции, в нишах, расщелинах, пещерах и на карнизах, защищенных от ветра и хищников. Иногда самок к месту родов сопровождают их годовалые дочери. Впервые рожающие самки стремятся приносить потомство неподалеку от матери, что приводит к быстрому объединению семьи и совместному воспитанию детенышей, которых чаще кормит и водит за собой более зрелая самка. Молодая мать через 18-20 дней нередко теряет интерес к своему ягненку и перестает его кормить. Новорожденные хорошо развиты и покрыты густой шерстью. Через час они способны недолго следовать за матерью, а через день легко передвигаются по скалам. В первые дни самки кормит ягнят молоком очень часто — от 2 до 6 раз в час, в июле - раз в 3 ч, в сентябре - раз в сутки. Лактация длится до октября, в
редких случаях - до декабря. Зеленый корм детеныши пробуют спустя неделю, а в месячном возрасте в их рационе доминирует растительная пища [Чернявский, 1962а; Егоров, 1965; Филь, 19736; Капитонов и др., 1975; Федосенко, 1985; Ларин, 1989, 1990; Железнов-Чукотский, 1994; Кривошапкин, Яковлев, 1999].
Плодовитость. Большинство исследованных самок азиатского снежного барана имеют по одному эмбриону, двойни редки (табл. 91). По визуальным наблюдениям, два детеныша следуют за самками тоже не часто (табл. 92). По одному ягненку обычно приносят и самки канадского толсторога [Bunnell, Olsen, 1981]. На плато Путорана два ягненка - обычное явление, что объясняется воспитанием отдельными взрослыми самками детенышей своих дочерей [Боржонов и др., 1979; Ларин, 1989, 1990]. В Якутии в бассейне р. Адычи в благоприятных условиях обитания по 2 ягненка имеют до 17% самок [Егоров, 1965; Тавровский и др., 1971]. В Оймяконском районе осенью 1977 г. В.И. Фертиков [1978, 1979] обнаружил с вертолета 5 самок с двумя и 1 - с тремя сеголетками.
Таблица 91. Потенциальная плодовитость самок азиатского снежного барана
Район (источник нформации)* Исследовано самок, п Самок с эмбрионами, % Эмбрионов на самку,
С одним I с двумя
Плато Путорана (4) 16 100 - 1,0**
Якутия (1) 5 60 40 1,4
Корякское нагорье (2) 3 67 33 1,2
Камчатка (3) 19 100 — 1.0
* 1 - Егоров, 1965, Тавровский и др., 1971; 2 - Чернявский, 1962а; 3 - Аверин, 1951; 4 - Ларин, 1989.
** Включая новорожденных.
Таблица 92. Число детенышей у самок азиатского снежного барана
Район (источник информации)* Встречено самок, п Самок с детенышами, % Половозрелых самок без ягнят, %** Детенышей на самку
С одним I с двумя
Плато Путорана (10) 135 100 - - 0,7
Центр. Верхоянье (1) 974 96,8 3,2(1-13) 22,3 (7,3-35) 1***
Якутия (2) — 83 17 — —
Хребет Кодар (4) Чукотка и Колымское — — — - 0,6
нагорье (5) — - - 0,3-0,4
Хребет Сунтар-Хаята (5) - - — - 0,5-1
Корякское нагорье (3) 21 90,5 - 9,5 0,9
Камчатка (8) 75 — — 0,4
-“-(9) 157 - - — 0,5
-“-(6) — 94 5-6 15 —
(7) 19 • 100 - - 1
-“-(7) 116 - - 7,6 0,9
* 1 - Кривошапкин, Яковлев, 1999; 2 - Егоров, 1965; 3 - Чернявский, 1962а; 4 - Медведев, 1997; 5 -Железнов-Чукотский, 1994; 6- Грибков, Филь, 1977; 7 - Аверин, 1951; 8 - Грибков, 1969; 9 - Филь, 1973а; 10 - Ларин, 1990.
** Самки яловые и потерявшие телят.
*** На лактирующую самку.
По мнению Н.К. Железнова [1994], для снежного барана характерна высокая яловость, поскольку самки, имеющие ягнят, не участвуют в гоне из-за длительной лактации. С таким утверждением никак нельзя согласиться. Самки всех видов диких копытных, лактировавшие до гона и даже лактирующие в период гона (косули в особенности), за редким исключением, приходят в течку. Самки снежного барана обычно прекращают кормить детенышей молоком в сентябре - октябре, т. е. еще до гона. При анализе литературных источников становится ясно, что в категорию яловых многие исследователи включают самок, потерявших детей. Соответственно доля прохолоставших особей в популяциях баранов значительно завышается и не соответствует действительности.
Для самок снежного барана, в неэксплуатируемых популяциях которого высокая доля взрослых самцов (табл. 93), более характерен относительно низкий уровень яловости. На Камчатке, по сообщению А.В. Аверина [1951], из 21 осмотренной после гона половозрелой самки яловыми оказались только 2 (9,5%), а из визуально наблюдаемых до 10 августа 116 самок без детенышей были всего 9 (7,6%), и при этом часть их уже могла потерять ягнят. Высокий уровень оплодотворения самок (85%) отмечает и В.И. Филь [1978]. В Корякском нагорье в июле ягнят имеют 79% взрослых самок, в августе - 61%, в сентябре - только 55% [Чернявский, 1962а]. На плато Путорана детенышей приносят ежегодно 65% самок [Ларин, 1989, 1990]. В эксплуатируемых популяциях воспроизводство может резко снижаться из-за высокой доли самок, прохолоставших из-за отсутствия взрослых самцов. На Камчатке в местах интенсивной охоты на баранов доля самок с потомством уменьшилась в три раза (с 54 до 17%), а число детенышей на одну самку снизилось с 0,85 до 0,4-0,6 [Филь, 1977а, 1981].
Популяционная структура
Половой и возрастной состав. Соотношение полов у новорожденных снежного барана близко 1:1 [Ларин и др., 1988; Ларин, 1990]. Преобладание детенышей-самок в отдельных группировках может быть связано с качественным питанием матерей, что установлено экспериментально для канадского толсторога [Hoefs, Nowlan, 1994].
Половозрастной состав популяций снежного барана существенно различается в разных районах, причем соотношение самцов и самок колеблется от 1:0,2 до 1:7 [Железнов-(Чукотский), 19836, 1990, 1994]. Однако такой большой “разброс” получается, в основном, при анализе малых выборок, когда исследователи учитывают менее сотни особей. В репрезентативных выборках доля сеголеток в группировках составляет 17-29%, годовалых особей - 9-23, взрослых самок - 25—46, молодых самцов — 6—13 и зрелых самцов - от 12 до 33%. Соотношение полов среди особей старше двух лет чаще находится в пределах 1:1-1,6 в пользу самок, и резко меняется лишь в местах интенсивной охоты (табл. 93). На Камчатке, например, в эксплуатируемых группировках оно смещается до 1:3,5-9,4 [Филь, 1977а, 1978, 1981; Мосолов, 19866, 1992].
В средне- и высокогорных группирбвках взрослых самцов нередко больше, чем самок, которые преобладают в низкогорье. Высотная дифференциация полов, в принципе, типична для горных видов копытных. Кроме того, низкогорья
Таблица 93. Половозрастной состав (%) и соотношение полов в популяциях азиатского снежного барана
Район, заповедник (источник информации)* п Сеголетки Годовалые Взрослые самки Самцы Самцы: самки
молодые взрослые**
Плато Путорана (11) 162 26,5 14,8 30,9 27,8 1:1,1
(12) 193 4,1 22,8 42,0 31,1 1:1,4
-“-(1) 387 23,8 15,8 34,9 12,1 13,4 1:1,4
С.-В. Якутия (2) 397 25,4 17,2 34,5 11,3 11,6 1:1,5
Центральное Верхоянье (3' 3003 25,2 12,8 32,4 12,0 17,1 1:1,1
(22-29) (11-15) (29-35) (4-17) (13-27)
Хребет Сунтар-Хаята (4) 107 9,3 11,2 23,4 10,3 45,8 1:0,4
-“-(4) 236 25,7 15,0 25,7 13,3 20,3 1:0,8
-“-(5) 124 12,9 8,1 15,3 2,4 61,3 1:0,2
Хребет Кодар (8) 343 23,3 (6,2)? 41,1 10,5 25,0 1:1,1
Анюйское нагорье (9) 89 20,2 14,2 46,8 10,3 8,2 1:2,5
Колымское нагорье (5) 124 18,5 11,3 29,0 7,3 33,9 1:0,7
Анадырское плоскогорье (4) 315 21,6 15,2 46,0 6,4 10,8 1:2,7
Корякское нагорье (6) 771 19,1 10,6 35,3 12,8 22,2 1:1
-“-(4) 201 17,4 8,9 35,3 13,9 24,5 1:0,9
Камчатка (7) 1024 22,6 10,2 41,1 9,4 16,7 1:1,6
-“-(14) 178 15,2 - 42,1 42,7 1:1
Ю.-В. Камчатка (13) — 7-16 4-11 34-55 4-12 17-31 -
Кроноцкий з-к (10) 635 5,5 4,9 45,3 10,9 33,4 1:1
* 1 - Ларин и др., 1987, 1988, 1990; 2 - Егоров, 1965; 3 - Кривошапкин, Яковлев, 1999; 4 - Железнов, 1990, 1994; 5 - Чернявский и др., 1990; 6 - Чернявский, 1962а, 1963, 1984; 7 - Филь, 1977в; 8 - Медведев, 1997; 9 - Чернявский, 1970а; 10 - Мосолов, 19866; 11 - Боржонов и др., 1979; 12 - Боржонов, 1988; 13 -Мосолов, 1992; 14-Грибков, 1969.
** Старше 5 лет.
более доступны для человека, и взрослые самцы здесь чаще становятся трофеем, что существенно меняет локальную демографическую структуру группировок. Поэтому вряд ли уместно выделять разные “экологические типы” популяций снежного барана по высотному признаку, как предлагает Н.К. Железнов [1987, 1990, 1994].
Социальная организация. Образ жизни снежного барана преимущественно семейно-групповой. При визуальных наблюдениях чаще всего регистрируют семейные группы (объединения двух или нескольких семей, возможно родственных), включающие самок, сеголеток и прошлогодних детей. Число отдельных семей максимально в период отелов, семейных формирований — после отелов. Взрослые самцы большую часть года держатся обособленными группами, а старые бараны нередко живут поодиночке. Молодые самцы в возрасте от 2 до 5 лет летом также образуют компании, насчитывающие до полутора десятков особей. В конце лета они могут присоединяться к семейным группам. Перед гонам группы самцов, как уже отмечалось выше, дробятся, а самки с ягнятами, напротив, объединяются в стада, насчитывающие несколько десятков особей. После гона самцы вновь группируются и отделяются от самок [Чернявский, 1961, 1962а, 19676, 19706; Филь, 1977в; Ларин, 1989, 1990; Медведев, 1997; Кривошапкин, Яковлев, 1999; Оконешников, 2003; Ларин, Сипко, 2004].
Таблица 94. Размер групп у азиатского снежного барана
Район, заповедник Число особей в группе Источник информации
среднее, чаще максимальное
Плато Путорана 2-7 13 Боржонов и др., 1979; Боржонов, 1988
а 8 10*, 40 Ларин, 1983; Ларин, Сипко, 2004
Якутия 2-3*, 5-8 5*, 18 Егоров, 1965; Тавровский и др., 1971
Южное Верхоянье 3,9 - Кривошапкин, Яковлев, 1999
Центр. Верхоянье 3,3*, 4,7 72 Кривошапкин и др., 1997
— 15*, 35 Оконешников, 2003
- - 2,9*, 8,6 7*, 25 Ревин и др., 1988
Северное Верхоянье 4,4*, 6,9 9*, 20
Хребет Черского 3,5*, 8 5*, 11 —
4-8*, 7-12 40*, 62 Фертиков, 1978, 1979
Хребет Кодар 3,7*-6,7 14 Медведев, 1997
Аяно-Майский район 3-5 20 Метельский, 1973
Колыма 3-7 40 Обухов, 1969
Колымское нагорье 6,8 15 Чернявский и др., 1990
Хребет Сунтар-Хаята 3,9 12 и
Анюйское нагорье - 12 Чернявский, 1970а
П-ов Тайгонос — 24 Дронсейко и др., 1983
Корякское нагорье 6,7 (4,5-8,3) 15*, 30 Чернявский, 1961, 1963, 1967а,б
а 8 — Филь, 1977в
Камчатка - 60 Аверин, 1951
4,4 12 Грибков, 1969
сс - 40 Вершинин, 1972
_44 10 34, 42* Филь, 1973 а,б, 1977в
Ю.-В. Камчатка 3*, 6 29 Мосолов, 1992
Кроноцкий з-к 6,3 25 Мосолов, 1986а
* В группах самцов
Азиатский снежный баран, как правило, не образует крупных объединений (табл. 94). Максимальный размер смешанного стада - 72 особи (осенью), самцо-вого - 42, а среднее число зверей в группах в разных районах колеблется чаще от 3 до 8. Группы самок обычно крупнее, чем самцовые. Коэффициент стадности существенно изменяется по сезонам года и коррелирует с уровнем плотности населения [Филь, 1977в], хотя и не всегда. Наиболее высок он осенью, когда образуются смешанные стада, и минимален поздней весной и в начале лета, когда стада дробятся и резко возрастает доля одиночных особей и семей. При постоянном охотничьем преследовании размеры групп животных уменьшаются почти вдвое [Филь, 1973а].
В смешанных стадах лидирует опытная самка, и лишь изредка, при бегстве, впереди оказываются молодые особи. Самцовые группы возглавляет крупный доминант с большими рогами. Конфликты среди баранов редки.
Снежный баран может активно обороняться от хищников с помощью рогов и передних копыт, что снижает потери, особенно от крупных птиц. Скорость бега у этого копытного невелика, но животные весьма эффективно используют недоступные или труднодоступные для хищников отстой. В спокойной обстановке звери медлительны, но при опасности преображаются: они стремительно уносятся в скалы, где, подобно горным козлам, ловко прыгают с уступа на уступ. У них прекрасно развиты все органы чувств. Подойти к баранам с наветренной стороны почти
невозможно. Человека в движении они замечают за 1,5-2 км, и в отсутствие постоянного беспокойства подпускают его на 300-500 м, а при регулярном преследовании-до 800-1000 м [Чернявский, 1961; Филь, 1973а]. В Кроноцком заповеднике, где животных не тревожат, человек может подходить к ним иногда почти вплотную. Особенно доверчивы самки и молодые звери [Аверин, 1951]. Такое же поведение характерно и для якутского и путоранского баранов в местах редкого появления людей [Тавровский и др., 1971; Боржонов и др., 1979; Федосенко, 1985]. При появлении вертолета самцы разбегаются в разные стороны, а самки с молодыми продолжают держаться вместе [Фертиков, 1978]. Снежные бараны молчаливы, однако во время отела нередко слышно протяжное блеяние самок и ягнят. Самцы издают низкие блеющие звуки лишь во время гона [Ларин, 1989; Кривошапкин, Яковлев, 1999].
Пространственная структура и перемещения. Снежные бараны - оседлые животные. Их летние и зимние участки обитания совпадают или смещаются сезонно в высотном направлении и широко перекрываются у разных групп. Летом звери чаще держатся в верхних ярусах гор, в зимний период - ниже, но в ряде районов они, напротив, на зиму перемещаются в верхние зоны, где снег сдувается ветром. Суточные высотные перемещения обычно не превышают 500 м, горизонтальные - нескольких километров. Участки обитания отдельных групп малы - около 20—40 га. Расселение происходит, в основном, за счет молодых особей [Аверин, 1951; Чернявский, 1961, 1963, 1967а,б, 19706; Водопьянов, 1971; Simmons, 1982; Федосенко, 1985; Festa-Bianchet, 1986; Ревин и др., 1988; Ларин, 1989, 1990; Железнов-Чукотский, 1994; Ларин, Сипко, 2004].
Тем не менее в некоторых горных районах Якутии (бассейн р. Адычи, юго-западные отроги Верхоянского хребта, хребты Черского и Туора-Сис, верховья рек Собопол и Тара-Сала и др.) зимние стации баранов расположены за 25-120 км от летних мест обитания. Соответственно стада и мелкие группы ежегодно в конце сентября - ноябре мигрируют, пересекая широкие долины рек и надпойменные террасы в лесном поясе. При этом часть зверей остается зимовать в местах летних концентраций. Начало весенней миграции в обратном направлении совпадает с переходом среднесуточной температуры выше нулевого значения и с началом вегетации на солнцепечных склонах южной экспозиции, что типично для мая. Самки до конца июня задерживаются на границе лесного и подгольцово-кустарнико-вого пояса вблизи скальных выходов, где происходит отел. По мере подрастания ягнят они перемещаются выше в горные тундры. Места обитания взрослых самцов и самок, за исключением периода гона и сезонных миграций, разобщены. Первые занимают верхний пояс гор, в то время как самки с молодняком и часть полувзрослых самцов держатся ниже по склонам, на надпойменных террасах, а также в долинах горных рек, где есть убежища и естественные солонцы. Минеральные солонцы обычно служат центрами летних концентраций животных [Егоров, 1965; Тавровский и др., 1971; Фертиков, 1978; Кривошапкин, Яковлев, 1999].
Сезонные миграции отмечены и в прибрежных районах Чукотки и Камчатки. Весной животные спускаются с гор и переходят на пастбища приморских террас на расстояние до 50-90 км. При этом они иногда пересекают кустарниковые долины рек шириной в 5-19 км. На Камчатке на Срединном хребте зимой самки живут на западных склонах, летом - на восточных. Самцы занимают верхний ярус
зоны обитания, причем старые особи держатся на его верхнем пределе. В горновулканическом районе Кроноцкого заповедника с выпадением снега животные спускаются к границе леса и в зону стлаников. Группировка взрослых самцов, обитающая на вулкане Центральный Семячик, ежегодно к периоду гона переходит на 25 км к подножью Тауншица, минуя зоны горных тундр и участки кедрового стланика. С вулкана Кихпиныч звери уходят в долину р. Гейзерной. На мысе Лопатка двух взрослых самцов в конце августа наблюдали в 30 км от ближайшего горного участка [Гаврилов, 1947; Аверин, 1948,1951; Вершинин, 1972; Грибков, Филь, 1977; Мосолов, 1986а, 1992; Железнов-Чукотский, 1994].
Одиночные бараны и мелкие группы могут перемещаться на десятки и сотни километров вдоль долин рек и по лесным массивам. Нередко они переходят по льду через широкие реки, такие как Лена, проникают в горные отдельности, выходят в открытую тундру. На Чукотском полуострове животных регистрировали на побережье Северного Ледовитого океана в 40 км от ближайших гор [Чернявский, 1972], а путоранский баран замечен в тундре за сотни километров от мест обитания [Железнов-Чукотский, 1994].
Причиной эмиграций зверей могут быть не только ухудшившие кормовые условия, но и высокая активность в период гона, а также беспокойство хищниками и охотниками. Особенно остро снежные бараны реагирует на охотничье преследование зимой: взрослые самцы после стрельбы по ним поднимаются на максимальные высоты, и возвращаются назад спустя полтора—два месяца, самки — быстрее. Зная эту особенность поведения, коренное население в прошлые времена стремилось вести добычу животных лишь в конце лета и осенью [Егоров, 1965; Филь, 1973а; Грибков, Филь, 1977].
Протяженные сезонные и спонтанные перемещения снежных баранов, следовательно, нередки, что позволяет критически относится к рассуждениям отдельных исследователей об “изолированности” группировок, и во многом объясняет относительно слабую географическую изменчивость этих зверей.
Как и у других копытных [Данилкин, 1999,2002], инфраструктура участков обитания групп снежных баранов складывается из пастбищ, солонцов, водопоев, “чесалок” (которыми служат крупные валуны, плиты, скальные выступы, деревья и кусты), мест отдыха и троп к ним. Тропы обычно тянутся по гребням водоразделов и вдоль склонов, как бы “срезая” вершины гор, от седловины к седловине, с зигзагами на крутых участках. Иногда они пересекают долины рек и ручьев, а в лесной зоне идут по их берегам. Регулярно используемые тропы на склонах имеют вид канавок шириной до 1 м. По ним легко определять очаги обитания животных в том или ином районе даже с воздуха [Тавровский и др., 1971; Ревин и др., 1988].
Лежки бараны устраивают чаще всего в мелкоземном грунте на пологих уступах гребней горных распадков и водоразделов и пережимах горных цепей, откуда хорошо просматриваются окрестности, а также на скальных обнажениях и карнизах, где более безопасно. Такие места используются постоянно, особенно в ночное время. Каждый зверь ложится в свою лежку. Самцы, вставая с них, мочатся, маркируя таким образом свое место и участок группы. Постепенно на лежках и вокруг них накапливается слой экскрементов. В ненастную погоду животные нередко используют пещеры с широким входом, ниши и карнизы с навесами, где за длительное время также скапливается 15-20-сантиметровый слой помета. Временные лежки обычны
вблизи пастбищ и нередки в лесном поясе, где летом звери отдыхают в тени деревьев и кустарников, а зимой спасаются от непогоды. Зарегистрированы случаи отдыха баранов в заброшенных домах человека, а также на галечно-песчаных косах крупных рек в нескольких метрах от уреза воды [Чернявский, 1961; Егоров, 1965; Тавровский и др., 1971; Ларин, 1983; Железнов, 1990, 1994].
Численность
В 60-90-е годы XX в. общее поголовье снежного барана в России, по мнению ученых [Банников, 1965; Рудков, 1979; Сопин, 1986; Ревин и др., 1988; Железнов-Чукотский, 1994; Zheleznov-Chukotsky, 2003], находилось в пределах 30-97 тыс. (табл. 95), а по официальным сведениям Главохоты РСФСР, не превышала 25-40 тыс. [Круторогов, 1975; Фертиков, 1978]. В 1997-2003 гг. Государственная служба учета охотничьих ресурсов оценивала запасы в 50-58 тыс. [Ломанов и др., 2000; Сидоров, 2000, 2004].
На плато Путорана в 70-е годы насчитывали около 1-1,5 тыс. снежных баранов, спустя десятилетие — 3,5 тыс., в начале XXI в. — 6-6,5 тыс. Основное поголовье сосредоточено к западу от оз. Аян, в долинах рек Делочи, Дулук, Холокит, Хибар-ба, Оран, Дюгадяк и Хугдякит. Численность вида в этом очаге возрастает и, скорее всего, ее можно увеличить в несколько раз [Павлов, 1977; Фертиков, 1978; Боржо-нов и др., 1979; Ларин и др., 1987, 1988; Ларин, 1990; Сыроечковский, 1995; Ларин, Сипко, 1997, 1999; Kolpashikov, Layshev, 2003].
Запасы барана в Якутии в 60-е годы О.В. Егоров [1965] оценивал, видимо завышая в два раза [Ревин, 1982а; Ревин и др., 1988], в 80-100 тыс. голов, а В.И. Фертиков [1978], занижая, - в 22,5 тыс., а в 70-е годы - в 12 тыс. По данным авиаучета, проведенного в 1990 г., в Верхоянье обитали 21-24 тыс. (в северной части - 5-6 тыс. с плотностью 0,9 голов на 10 км2, в центральной и южной - от 7,5 до 9,5 тыс. при плотности 1,7), в горной системе Черского - 15,5-17 тыс., а всего в Северо-Восточной Якутии - 36,5—41 тыс. особей [Кривошапкин, Яковлев, 1999]. Примерно столько же (40 тыс.) учтено в Саха и в конце XX в. [Сидоров, 2000].
На Чукотке численность этого вида, по оценке Н.К. Железнова-Чукотского [1994], близка к 2,5-3,5 тыс. На восточном и южном морском побережье полуострова в самом конце XX в. насчитывали около 200-250 голов [Мымрин, 2003]. По расчетам Ю.В. Ревина с соавторами [ 1988], на Чукотке поголовье достигает 6-7 тыс., на Камчатке - 12-13 тыс. На юге Дальнего Востока жили около 4-6 тыс. [Бромлей, Кучеренко, 1983]. В области северного побережья Охотского моря обитают 1,5-2 тыс. особей, в том числе в прибрежной полосе - 400-500 экземпляров [Чернявский и др., 1990]. Общее поголовье зверей в Колымском нагорье в 60-е годы А.А. Кищинский [1967а] оценивал в 30-70 тыс., завышая его, скорее всего, в несколько раз. По сведениям Главохоты РСФСР, в Магаданской области в 60-70-е годы было около 10 тыс., на Камчатке - 7-3,5 тыс. [Фертиков, 1978]. Специалисты Государственной службы учета охотничьих ресурсов в 1997-1999 гг. на Камчатке насчитали 3,1-3,2 тыс., в Корякском автономном округе - 2,5 тыс., в Магаданской области -2,1 тыс., в Хабаровском крае - 2 тыс. баранов [Сидоров, 2000].
На хребте Кодар в 80-е годы учтено всего 50-150 особей [Баранов, Бойко, 1984; Сопин, Ермолин, 1986, 1988; Сопин, 1993], в конце десятилетия и по-
Таблица 95. Плотность и численность снежного барана в Северо-Восточной Азии во второй половине XX - начале XXI вв.
Район, заповедник Площадь обитания, тыс. км2 Плотность, особей/10 км2 Численность’ тыс. особей Источник информации
Плато Путорана 120 0,3 (0,1-0,6) 1,4 Боржонов и др., 1979
- — 60 0,2-3,1 3,6 Ларин и др., 1987, 1988; Ларин, 1990
(р. Делочи) - 3-5 0,2 Ларин, Шелковникова, 1988
с с 120 0,05-4,8 5,5-6,5 Ларин, Сипко, 1999, 2004
Якутия 350 2-3 (0,5-15) 80-100 Егоров, 1965; Лавровский и др., 1971
(4 - - 22-12 Фертиков, 1978
44 341 о, 1-2,9 48-50 Ревин, 1982а,б
(4 466 0,01-1,7 39-44 Кривошапкин, Яковлев, 1999
Ч - - 40 Сидоров, 2000, 2004
Южная Якутия Верхоянская горная 27 0,1-0,3 0,3-0,8 Ревин, 1988
система Хребет Черского и 277 0,8-1,5 29-32 Ревин и др., 1988
прилежащие районы Становой хр. и 286 0,5-1,1 25-26,5 44
прилежащие районы 96 0,01-0,3 2,4-2,8 44
Колымское нагорье 156 0,5 7-8 44
Хребет Кодар 12 0,1-0,3 0,3
44 — — 0,5-0,3 Медведев, 1997, 2003а,б
Чукотка — 1,2 5-8 Чернявский, 1970а
44 262 0,25 6-7 Ревин и др., 1988
Ч — 1-1,6 2,8 Железнов-Чукотский, 1994
Чукотский п-ов Северное побережье — — 0,5 Чернявский, Железнов, 1977
Охотского моря — 0,4-8,5 1,5-2 Чернявский и др., 1990
Магаданская область — — .10 Фертиков, 1978
44 — - 2,1 Сидоров, 2000
44 - — 2,1-4 Дубинин, 2002
Корякское нагорье - 1,8-13 — Чернявский, 1961, 19676
44 — 6,7 — Филь, 1977в
44 138 0,5 6-7 Ревин и др., 1988
44 1,8 2,5-3,5 Железнов-Чукотский, 1994
Корякский а. о. — — 2,5-4,8 Сидоров, 2000, 2004
Камчатка — 9-11 (3-14) — Грибков, 1969, 1970
44 — 7-12 5-10 Вершинин, 1972
44 — 6-9 12-15 Филь, 1973а,б, 1977в
44 144 0,9 12-13 Ревин и др., 1988
Камчатская область Восточное побережье — — 3-6 Сидоров, 2000, 2004
Камчатки — 6-7 0,2-0,3 Вяткин, Филь, 1975
Кроноцкий з-к 9,6 0,6 0,6 Мосолов, 1986а
Аяно-Майский район — - 1,0 Метельский, 1973
Юг Дальнего Востока — 4-6 Бромлей, Кучеренко, 1983
Хабаровский край - - 2,0 Сидоров, 2000, 2004
зднее - 300-500 [Ревин и др., 1988; Медведев, 1997; Медведев, Лямкин, 2000], в 2002 г. - около 270-300 [Медведев, 2003а,б].
Средняя плотность населения снежного барана в крупных очагах на северо-востоке Азии, где влияние человека на популяции относительно невелико, лежит в пределах 0,5-1 экземпляра на 10 км2 (табл. 95). Этот показатель определяется, главным образом, погодными факторами и хищниками. В отдельных группировках на хребтах Камчатки и Корякского нагорья он составляет 6-18, а на Колымском нагорье и Чукотке не превышает 1,2-1,5 голов [Чернявский, 19676, 19706; Грибков, Филь, 1977; Филь, 1977в]. В Центральном Верхоянье плотность этого вида в лучших летних стациях достигала 20 особей на 1000 га, но в последние годы в результате интенсивной трофейной охоты наблюдается ее снижение [Кривошапкин, Яковлев, 1999; Охлопков, 2002].
Имеющиеся ресурсы снежного барана в России в несколько раз меньше потенциального уровня. В начале XIX в. его поголовье, скорее всего, исчислялась сотнями тысяч. В Америке в это время лишь пустынного толсторога было около 1,5-2 млн, но к середине XX в., в основном в результате чрезмерного антропогенного пресса, осталось менее 10 тыс. [McCutchen, 1981; Cooperrider, 1985]. Численность барана в путоранском очаге можно увеличить в десятки раз [Боржонов и др., 1979]. В Якутии, по мнению О.В. Егорова [1965], могли бы жить до 500 тыс. особей. Эта цифра, на мой взгляд, несколько завышена, поскольку малая плотность населения копытных на Севере связана не только с отстрелом животных, но и с дефицитом зимних кормов и убежищ. Тем не менее, при значительном улучшении охраны и сокращении числа крупных хищников, существенно увеличить запасы вида вполне реально. Однако сейчас в большинстве районов более очевидна тенденция к дальнейшему уменьшению численности.
Основные факторы, определяющие динамику популяций
Воспроизводство, прирост. По визуальным наблюдениям в Центральном Верхоянье на 100 взрослых самок приходится около 80 (67-96) сеголеток [Кривошап-кин и др., 1997; Кривошапкин, Яковлев, 1999], в Якутии [Егоров, 1965] и Корякском нагорье [Чернявский, 1962а] - 76, на Камчатке - 36—46 [Грибков, 1969; Филь, Вяткин, 1972; Филь, 1973, 1977в, 1978], а в приморских группировках - не более 20 [Мосолов, 1992], на хребте Кодар - 58 [Медведев, 1997], Чукотке - 37, хребте Сунтар-Хаята - 50-100 [Железнов-Чукотский, 1994].
Замечу, что эти регистрации сделаны на протяжении года, когда значительная часть новорожденных уже погибла. В Корякском нагорье, например, в июле без ягнят ходят 24% взрослых самок, в августе - 39, в сентябре - 45%. До годовалого возраста здесь не доживает половина сеголеток, а годовой прирост не превышает 8-11% [Чернявский, 1961, 1962а, 1963, 1974]. В некоторых районах Чукотки после тяжелой зимовки на 100 взрослых самок остается всего 10 ягнят, что, тем не менее, не может служить характеристикой “низкого репродуктивного потенциала” вида [Железнов-Чукотский, 1994]. У снежного барана при типичном для копытных уровне плодовитости (примерно 1 детеныш на самку) низок уровень выживаемости сеголеток, который отдельные исследователи путают с “показателем плодовитости или рождаемости”.
В целом отход молодняка в первый год жизни в разных регионах колеблется в пределах 20-80% (табл. 96). Эти цифры сопоставимы с таковыми по американскому толсторогу: показатель рождаемости составляет около 75%, выживаемость новорожденных - 80%, до одного года доживают 49% [Simmons et al., 1984], причем смертность гораздо выше у позднородившихся детенышей [Festa-Bianchet, 1988].
Таблица 96. Уровень смертности сеголеток азиатского снежного барана
Район I Уровень смертности, % 1 Источник информации
Плато Путорана 25 Ларин и др., 1988
Якутия 30 Егоров, 1965
Центральное Верхоянье 48 (38-59) Кривошапкин, Яковлев, 1999
Чукотка 18-54 Железнов-Чукотский, 1994
Хребет Сунтар-Хаята 24 44
Корякское нагорье 18-37 44
44 50 Чернявский, 1963, 19676, 1974
Камчатка 17-79 Железнов-Чукотский, 1994
с 4 50 Чернявский, 1961, 1962а
и 50 Аверин, 1951
Динамика популяций снежного барана, следовательно, в значительной мере определяется не уровнем плодовитости, а уровнем детской смертности.
Погода и корма. Кормовая база снежного барана в горах Северо-Востока не является основным лимитирующим фактором. Гибель зверей от бескормицы регистрируется лишь во время сильных гололедов и особенно плотного снега. Более обычна массовая гибель детенышей при затяжных холодных дождях и снегопадах в летнее время [Чернявский, 1962а, 1974; Железнов-Чукотский, 1994; Кривошапкин, Яковлев, 1999].
Самцы снежного барана во время гона имеют небольшие гаремы и не особенно агрессивны к соперникам. В гон они обычно кормятся и, в отличие от самцов других видов копытных, не истощаются сильно, поэтому и смертность среди них невысока.
Хищники. Значительный урон популяциям причиняет волк, особенно в низко-горье и холмогорье. Доля гибели от этого хищника в общей смертности баранов иногда достигает 60% [Перовский, 1988], а на Анадырском плоскогорье - 70%. Здесь волк преследует копытных или нападает из засады, и давит независимо от их пола и возраста [Железнов, 1983а, 1990, 1994]. На плато Путорана в его добыче преобладают самцы (42%) [Ларин, 19876, 1990; Ларин, Крашевский, 1989], в Якутии - сеголетки (33) и молодые особи (37%) [Егоров, 1965]. В Верхоянье остатки баранов встречаются в 4,5-8% экскрементов хищника [Кривошапкин, Яковлев, 1999], а на Камчатке в местах зимовок он питается исключительно этими копытными [Аверин, 1948, 1951]. И все же снежный баран в Северной Азии имеет второстепенное значение для волка, которому более доступен северный олень.
Медведь тоже успешно охотится на баранов. М.А. Кречмар в горах Восточной Якутии зарегистрировал гибель от этого хищника 6 полувзрослых особей. Медведь подстерегал копытных на крутом подъеме тропы и бросался на них сверху с расстояния 4-6 м [Чернявский, Кречмар, 2001]. На молодых животных нападают также росомаха и рысь, особенно успешно - на лежках, отстоях, тропах, у солон
цов и в глубоком снегу. Ягнята иногда становятся жертвами песца и лисицы, беркута, орлана-белохвоста, белоплечего орлана и ворона [Аверин, 1951; Чернявский, 1961, 1963; Егоров, 1965; Тавровский и др., 1971; Ларин, 19876, 1990; Ревин и др., 1988; Ларин, Крашевский, 1989; Железнов-(Чукотский), 1990, 1994; Кривошапкин, Яковлев, 1999].
Конкуренты. Пищевыми конкурентами снежного барана считаются все основные растительноядные животные горно-тундровой зоны: северная пищуха, высокогорные полевки, лемминг, беренгийский суслик, черношапочный сурок и заяц-беляк. Однако их население в горах невелико, часть из этих млекопитающих зимой впадает в спячку, и они не могут подорвать кормовые ресурсы. Заяц-беляк, тем не менее, в годы пиков почти полностью уничтожает подрост ивы в отдельных районах, что существенно уменьшает кормовую базу копытных.
Трофическая ниша толсторога частично совпадает с таковой у северного оленя. Стада домашних и диких копытных в локальных районах на плато Путорана, на Камчатке и Чукотке не только сильно вытаптывают и стравливают лучшие зимние лишайниковые пастбища, но и вытесняют баранов в худшие стации. Иногда в многоснежье эти виды копытных вынуждены пастись вместе, при этом бараны охотно пользуются местами тебеневок оленей. В Якутии конкурентом за ягельные пастбища выступает кабарга. Лось занимает иную экологическую нишу и поедает другой корм, поэтому считать его конкурентом толсторога можно лишь с большой натяжкой [Егоров, 1965; Тавровский и др., 1971; Боржонов и др., 1979; Ларин, 1983, 1990; Ревин и др., 1988; Железнов, 1990, 1994; Кривошапкин, Яковлев, 1999].
Болезни, паразиты, травмы. Массовой гибели снежных баранов от инфекционных болезней не отмечено. Простудные заболевания животных в суровом климате Северо-Восточной Сибири, видимо, нередки, и поэтому в аномальные годы возможен значительный отход молодняка от воспаления легких [Кривошапкин, Яковлев, 1999].
Эти копытные весьма незначительно заражены эндопаразитами, чему способствует холодный климат, сравнительная разреженность популяций, жизнь мелкими группами и поедание лекарственных и ядовитых растений. В Якутии у них обнаружено 7 видов гельминтов [Губанов, 1964; Егоров, 1965], на Чукотке - 8, но общим для этих регионов оказался только 1 вид [Железнов, 1990, 1994]. На хребте Кодар у баранов выявлено 11 видов паразитов, но среди них нет общих с гельминтами якутских копытных, и лишь один входит в гельминтофауну чукотского барана [Медведев, 1997]. Разнородность паразитофауны может свидетельствовать об определенной географической изоляции животных, что немудрено в очаговом ареале. Гельминто фауна толсторога во многом сходна с таковой у северного оленя, поскольку они нередко пасутся на одних и тех же пастбищах. Слабо поражены бараны и эктопаразитами, и даже личинками носоглоточного и кожного оводов, хотя во многих районах контактируют с домашними и дикими оленями [Чернявский, 1961; Тавровский и др., 1971].
Часть зверей ежегодно гибнет в лавинах и камнепадах на горных скалах и срывается со скалистых участков [Портенко и др., 1963; Егоров, 1965; Водопьянов, 1971; Рудков, 1979; Мосолов, 1986а; Железнов-Чукотский, 1994]. Однако такие случаи все же редки, поскольку многовековой опыт жизни в горах привил им навыки осторожности.
Антропогенные факторы. Влияние хозяйственной деятельности человека на снежного барана в отдаленных районах Сибири в целом невелико, но оно неуклонно возрастает. Негативное воздействие проявляется, в основном, в браконьерстве местного населения, сотрудников различных экспедиций и туристов. На местах стоянок оленеводов, геологов и старателей повсеместно остаются груды костей, черепов и рогов этих копытных. Особенно быстро зверей истребляют в низкогорных и прибрежных биотопах, вблизи поселков и рудников. В ряде районов вид вытеснен стадами домашнего северного оленя, стравливающего зимние пастбища. Площадь пастбищ барана заметно сокращается и при добыче полезных ископаемых и строительстве всевозможных промышленных комплексов, которые (в частности - Норильский), в свою очередь, начинают оказывать негативное воздействие на среду обитания животных [Егоров, 1965; Чернявский, 1968; Ларин, 1987в, 1990; Ревин и др., 1988; Железнов, 1990, 1994; Кривошапкин, Яковлев, 1999; Ларин, Сипко, 1999; Калякин, 2001].
Использование ресурсов и охрана
Снежный баран, наряду с северным оленем, - ценнейший объект промысла местного населения. Его мясо превосходно на вкус и очень питательно. Шкуры идут на изготовление ковриков, спальных мешков, зимней верхней одежды, рукавиц и детских унтов, которые обладают высокими теплоизоляционными свойствами и легче изделий из оленьего меха. Рога служат украшением жилищ и материалом для всевозможных бытовых поделок.
Охота на этого копытного в Сибири и на Дальнем Востоке не носила массового характера из-за немногочисленности аборигенного населения, высокой трудоемкости добычи и обилия других легко добываемых видов [Ревин, Сопин, 1988; Ревин и др., 1988]. Кроме того, она издавна была относительно регламентированной: охотничьи угодья поделены между семьями; охотники добывали преимущественно взрослых самцов и сеголеток (для детской одежды); охота велась в сжатые сроки в сентябре - октябре в период наивысшей упитанности зверей. У камчадалов за отстрел взрослых самок предусматривалось жесткое наказание - тяжелые и неприятные общинные работы и жизнь вне дома в специальной яме [Грибков, Филь, 1977].
Во второй половине XIX - начале XX вв. добыча снежного барана стала более интенсивной. По сообщению И.С. Полякова [1873], на хребте Кодар “тунгусы” убивали его в большом количестве, а “наиболее ловкие и проворные клали в один день более, чем по полудесятку”. В конце XIX в. здесь ежегодно добывали 100-120 особей [Водопьянов, 1971]. На Яблоновом (Становом) хребте в начале XX в. бараны подвергались особенному преследованию со стороны “ороченов”, пригонявших стада северных оленей в горы на лето [Насонов, Дорогостайский, 1915].
Промысловая ситуация особенно резко изменилась с появлением некоренного населения, для которого не существовало племенных запретов. В 20—30-х годах XX в. в Верхоянском и Колымском округах ежегодно добывали около 2 тыс., а на Дальнем Востоке - 1,6 тыс. баранов. В первой половине этого столетия на территории Якутии отстреливали до 4 тыс., в 50-е годы - около 5-6 тыс. В 1957 г. два охотника в верховьях р. Собопол только за осень добыли для зверофермы около 150 особей. Охотники и оленеводы бывшего Саккырырского района в послевоен
ные годы отстреливали от 2-3 десятков до сотни зверей каждый, на территории колхоза им. Ленина в отдельные годы - свыше 1 тыс., и примерно столько же - на территории двух других колхозов этого района. По нескольку десятков баранов некоторые охотники добывали ежегодно и в Булунском и Томпонском районах, а также на хребтах Черского и Момском [Егоров, 1965; Тавровский и др., 1971; Ревин и др., 1988; Ревин, Сопин, 1988].
На Камчатке в конце XIX - первой половине XX вв. ежегодно отстреливали, видимо, около 1-1,3 тыс. особей. В 1926 г. здесь добыто 937 голов. Зимой 1933/34 гг. охотники только из трех сел убили 46 баранов. В военные 40-е годы этих копытных заготавливали местные жители и специальные команды, что привело к снижению их численности во многих районах. Отдельные охотники добывали до 70 баранов в год [Аверин, 1951; Грибков, 1969; Вершинин, 1972; Филь, 1973а,б].
В послевоенное время охота на снежного барана запрещается везде, кроме Хабаровского края и Камчатки [Вершинин, 1972], с 1956 г. - и на Камчатке [Филь, Вяткин, 1972; Филь, 19736], а типовыми правилами от 10 июля 1961 г. - на всей территории РСФСР [Зуев, 1977]. Запрет охоты и, впоследствии, лицензирование добычи в ряде районов способствовало стабилизации поголовья, однако в целом эта мера оказалась малоэффективной из-за полного отсутствия контроля за охотниками на огромном пространстве. С 1979 по 1983 гг. в России официально изъято около сотни баранов [Перовский, 1985]. В конце XX - начале XXI вв. ежегодно отстреливали от 81 до 185 экземпляров [Ломанов и др., 2000; Сидоров, 2000, 2004]. Нелегальная добыча этого копытного, по экспертной оценке, достигает 3-5 тыс. особей.
На плато Путорана снежный баран был поставлен на грань исчезновения, главным образом, бесконтрольной охотой оленеводов (каждая бригада добывала за год до 15-20 особей) и работников всевозможных экспедиций [Павлов, 1977; Ларин и др., 1988]. В южной части заказника “Путоранский” с пригодных мест обитания зверей вытесняли охотники, использующие снегоходы и моторные лодки. Беспокойство здесь стало основной причиной резкого снижения уровня воспроизводства популяции. Летом 1986 г. среди 148 встреченных особей было только 2 сеголетка [Боржонов, 1988]. Более строгие меры по охране были приняты после включения путоранского подвида в Красные книги СССР [ 1978,1984] и РСФСР [1983] и организации крупного республиканского заказника (в 1985 г.) и заповедника (в 1988 г.). Улучшение охраны и уменьшение браконьерства, особенно со стороны оленеводов, благотворно сказалось на популяции, которая стала быстро расти [Ларин, 1990; Сыроечковский, 19956; Присяжнюк, 2001]. При дальнейшем увеличении численности возможен перевод путоранского барана в разряд охотничьих животных.
В Верхоянье браконьеры ежегодно изымают 1,5-2 тыс. особей или около 7-8% популяции [Кривошапкин, Яковлев, 1999]. Браконьерство - основной лимитирующий фактор популяции снежного барана и на хребте Кодар, где ежегодно отстреливают не менее 30 зверей. В последнее время участились попытки организовать незаконную трофейную охоту на этого копытного [Сопин, Медведев, 1992; Сопин, 1993; Медведев, Лямкин, 2000; Медведев, 20036].
На Чукотке каждая бригада оленеводов отстреливает ежемесячно около 3-5 баранов. Браконьеры ежегодно изымают более 500-600 голов (до 15-20% группировок), что превышает уровень годового прироста. На северо-востоке Сибири осо
бенно большой ущерб стадам наносит преследование с вертолета. Водный туризм по горным рекам и прибрежное рыболовство также нередко завершаются отстрелом животных [Воронов, Воронов, 1973; Железнов-(Чукотский), 19816, 1994; Кривошеев, Чернявский, 1981; Ревин и др., 1988; Чернявский и др., 1990].
Пастухи оленьих стад и промысловики бесконтрольно и круглогодично добывают этих копытных и на Корякском нагорье. На Колымском нагорье браконьеры также круглогодично отстреливают до 200-300 голов. На побережье Охотского моря вблизи поселков звери истреблены полностью, или их численность резко снижена. Ряд субпопуляций прекратили свое существование, а быстрое заселение пустующих районов вряд ли возможно из-за территориального консерватизма вида [Чернявский, 1961, 19676, 1974; Чернявский и др., 1990].
Браконьерство с применением вездеходов и вертолетов процветает и на Камчатке, в чем особенно усердствуют работники различных экспедиций. На западном побережье снежный баран полностью выбит еще в середине XX в. На осевых хребтах при частом преследовании охотниками бараны остаются на зимовку на труднодоступных альпийских пастбищах со скудным кормом, не успевают накопить достаточного запаса жира, что снижает их выживаемость и негативно сказывается на воспроизводстве потомства. Смертность особей при факторе беспокойства возрастает до 50% по сравнению с 12-15% на контрольных участках. При выборочной охоте на взрослых самцов происходит нарушение полового и возрастного состава популяций. В нижнем поясе гор и на периферии горных систем наибольшему преследованию подвергаются самки. В местах интенсивной охоты их доля в популяции снижается в 3 раза [Вяткин, Филь, 1975; Филь, 1973а,б, 1975, 1977а; Грибков, Филь, 1977].
На юге Дальнего Востока этих копытных тоже отстреливают сотрудники различных экспедиций, работники оленеводческих совхозов и колхозов, нередко используя специально натасканных собак. И здесь при постоянном беспокойстве и вынужденном переселении по глубокому снегу в новые районы обитания гибнет до 40-50% поголовья, в основном молодняк [Бромлей, Кучеренко, 1983].
В последнее десятилетие снежный баран стал желанным объектом трофейной охоты. Многие зарубежные охотники-коллекционеры хотели бы иметь его рога, особенно редких форм, что создает определенную угрозу для существования вида. Насколько мне известно, нелегальная добыча животных в трофейных целях ведется повсеместно, круглогодично и с применением вертолетов, в том числе в Путораках и на Кодаре. Есть отдельные фирмы, охотоведы, егеря, и даже летчики, для которых этот вид деятельности стал одним из основных источников дохода.
Тем не менее использование широко распространенного снежного барана в качестве одного из основных объектов охотничьего и экологического туризма в Северной Азии весьма перспективно. Для этого необходимо значительно увеличить численность и плотность его населения в районах трофейной охоты, что невозможно без эффективной охраны зверей, создания сети заказников и заповедников и специализированных охотничьих хозяйств трофейного типа [Сопин, 1986; Ревин, Сопин, 1988; Ревин и др., 1988]. Под особой охраной должны находиться наиболее уязвимые прибрежные и низкогорные популяции, как более доступные для человека и крупных хищников, а также места зимовок и ягнения животных в изолированных и исчезающих очагах. Охоту на снежного барана следует вести в ко
роткие (1-1,5 мес.) раннеосенние сроки [Филь, 1981; Ревин, Сопин, 1988] в верхних ярусах высокогорных угодий, где обычно обитают старые самцы, умеренный отстрел которых не окажет отрицательного влияния на популяции.
При значительном улучшении охраны животных, что в России, видимо, произойдет еще не скоро, можно ускорить восстановление ареала снежного барана путем реинтродукции и даже расширить его. По мнению специалистов, для этого пригодны следующие районы: Командорские и Курильские острова, хребты Бад-жальский, Дуссе-Алинь, Джагды, Икатский, Канарский, Баргузинский и Байкальский, Становое нагорье, Бытантайское плато, долина р. Лены, горы Бырранга на полуострове Таймыр, Средний, Северный, Полярный и Приполярный Урал [Чернявский, 1963; Егоров, 1965; Вершинин, 1972; Воронов, Воронов, 1973; Воронов, 1982; Петрашов, 1982; Терновский и др., 1986; Ревин и др., 1988; Железнов, 1994; Сопин, 1998; Медведев, Лямкин, 2000]. Желательно детально обследовать и малоснежные участки Западного и Восточного Саян, Абаканский хребет и Кузнецкий Алатау, где этот вид обитал в давние времена.
Снежный баран перспективен и в животноводстве для создания новых пород домашних овец, адаптированных к суровым условиям Сибири [Флеров, 1935а; Соколов, 1959]. Такая работа уже ведется на Камчатке, где получено плодовитое гибридное стадо пока с невысоким уровнем воспроизводства из-за значительной гибели ягнят (до 80%) в подсосный период. Во втором поколении потери среди гибридов уменьшаются в два раза [Шайдуллин, Ролдугин, 1986; Шайдуллин, Шайдуллина, 1988; Шайдуллин, 1992, 1994а,б, 2001; Шайдуллин и др., 2000]. Весьма желательно также экспериментально проверить возможность одомашнивания снежного барана. Известны случаи приручения молодых особей и содержания их в стадах вместе с домашними животными, и имеется непроверенное сообщение К. Дит-мара о небольшом стаде баранов, выпасаемом кочевниками близ Ичинской сопки на Камчатке [Вершинин, 1972; Соскин, Глушнев, 1983].
В заключение замечу, что работа по улучшению и выведению новых пород домашнего скота, безусловно, нужна, но я призываю селекционеров, зоологов и охотоведов задуматься о возможных отдаленных негативных последствиях искусственной гибридизации. Домашние породы со свойствами, близкими к дикому виду, становятся для него наиболее опасными конкурентами, вытесняющими с лучших пастбищ и, в конечном счете, способствующими его уничтожению. И вряд ли при увеличении количества и качества домашних пород и общего поголовья скота удастся сохранить диких копытных в природе, сохранить желанное биологическое разнообразие.
Литература
Абатуров Б.Д. Биопродукционный процесс в наземных экосистемах. М.: Наука, 1979. 128 с.
Абатуров Б.Д. Освоение сайгаками растительных кормовых ресурсов // Копытные фауны СССР. М.: Наука, 1980. С. 116-117.
Абатуров Б.Д. Млекопитающие как компонент экосистемы. М.: Наука, 1984. 285 с.
Абатуров БД. Критические параметры качества растительных кормов для сайгаков (Saiga tatarica) на естественном пастбище в полупустыне // Зоол. журн. 1999. Т. 78, вып. 8. С. 999-1010.
Абатуров Б.Д. Качество корма и его влияние на обеспеченность пищей и состояние популяций растительноядных млекопитающих на естественных пастбищах // Арид. экосистемы. 2000. Т. 6, № 11-12. С. 66-76.
Абатуров Б.Д. Сравнительная пригодность различных типов пастбищной растительности для вольноживущих сайгаков // Млекопитающие как компонент аридных экосистем (ресурсы, фауна, экология, медицинское значение и охрана). М., 2004. С. 6-7.
Абатуров БД., Анчифоров П. С., Огуреева ГН. и др. Распространение алтайского горного барана (Ovis аттоп аттоп) на Алтае в связи с особенностями растительного покрова // Зоол. журн. 2004. Т. 83, № 2. С. 241-251.
Абатуров Б.Д, Петрищев Б.И. Опыт вольерного содержания сайгаков И Акклиматизация охотничьих животных в СССР. Минск: Ураджай, 1978. С. 166-167.
Абатуров БД., Петрищев Б.И. Сравнительная оценка рациона свободнопасущегося сайгака (Saiga tatarica) микроскопическим анализом растительных остатков в фекалиях и визуальным подсчетом поедаемых растений И Зоол. журн. 1998. Т. 77, № 8. С. 964-970.
Абатуров БД., Петрищев Б.И., Колесников М.П., Субботин А.Е. Сезонная динамика кормовых ресурсов и питание сайгака на естественном пастбище в полупустыне И Усп. совр. биол. 1998. Т. 118, вып. 5. С. 564-584.
Абатуров Б.Д., Субботин А.Е., Джапова Р.Р. Использование сайгаками запасов пастбищной растительности в Северо-Западном Прикаспии И Экология. 1987. № 5. С. 93-95.
Абатуров БД., Холодова М.В., Субботин А.Е. Переваримость растительных кормов у сайгаков И Копытные фауны СССР. М.: Наука, 1980. С. 117-118.
Абатуров БД., Холодова М.В., Субботин А.Е. Интенсивность потребления и переваримость кормов у сайгаков (Saiga tatarica) И Зоол. журн. 1982а. Т. 61, вып. 12. С. 1870— 1881.
Абатуров БД., Холодова М.В., Субботин А.Е. Потребление и переваримость кормов у сайгака//Млекопитающие СССР. III съезд Всесоюз. териол. о-ва. М., 19826. Т. 1. С. 150-151.
Абдуназаров Б.Б. Состояние численности редких видов млекопитающих Узбекистана И Териофауна России и сопредельных территорий. VII съезд териол. о-ва. М., 2003. С. 11.
Абдурахманов М.Г. Экология, охрана и использование туров Дагестана: Автореф. дис.... канд. биол. наук. Махачкала, 1973. 24 с.
Абдурахманов М.Г. Кавказский тур // Копытные звери. М.: Лесн. пром-сть, 1977. С. 186-200.
Абдурахманов М. Восточнокавказский тур // Охота и охотничье хоз-во. 1979. № 8. С. 10-12.
Абдурахманов М.Г. Современное состояние популяций кавказских туров и задачи их охраны // Копытные фауны СССР. М.: Наука, 1980. С. 225.
Абдурахманов М.Г. Экология восточнокавказского тура в Дагестане И Изв. Сев.-Кавк, науч, центра высш. шк. Естеств. науки. 1982. № 4. С. 48-51.
Абдурахманов М.Г. Некоторые особенности возрастной и половой структуры популяции туров Дагестана// Экология и охрана горных видов млекопитающих. М., 1987. С. 3-4.
Абдурахманов М.Г. Паразитофауна кавказских туров и меры профилактики паразитарных болезней: Автореф. дис. ... докт. биол. наук. Махачкала, 2003. 47 с.
Абдусалямов И.А. Состояние популяции и проблемы охраны памирского архара Ovis аттоп polii Blyth, 1940 в Таджикистане // Редкие виды млекопитающих России и сопредельных территорий. М., 1997. С. 4.
Абдусалямов И.А. Современное состояние некоторых краснокнижных видов парнокопытных Таджикистана // Териофауна России и сопредельных территорий. VII съезд териол. о-ва. М., 2003. С. 11.
Абрамов К.Г Копытные звери Дальнего Востока и охота на них. Владивосток: Приморское кн. изд-во, 1963. 132 с.
Абрамов С.А., Фалеев В.И. Поведение алтайского горного барана - аргали {Ovis аттоп аттоп) при разведении в неволе // Состояние териофауны в России и ближнем зарубежье. М., 1996. С. 5-9.
Абрамова З.А., Астахов С.Н., Васильев С.А. и др. Палеолит Енисея. Л.: Наука, 1991.158 с.
Аверин Ю.В. Наземные позвоночные Восточной Камчатки // Тр. Кроноцкого гос. заповедника. 1948. Вып. 1. С. 3-223.
Аверин Ю.В. О снежном баране на Камчатке // Бюл. МОИП. Отд. биол. 1951. Т. 56, вып. 1. С. 15-26.
Аверин Ю.В. Истребленные и нуждающиеся в охране исчезающие и редкие млекопитающие Молдавии // Редкие млекопитающие фауны СССР. М.: Наука, 1976. С. 43—49.
Агаджанян А.К., Раутиан Г. С. Дифференциация ряда видов Bovinae по частотам групп крови //Редкие виды млекопитающих России и сопредельных территорий. М., 1997. С. 5.
Адольф ТА. Изменения распространения сайги // Охрана природы. М.: Всерос. о-во охраны природы, 1948. Сб. 2. С. 61-93.
Адольф ТА. Сайга в астраханских степях правобережья Волги И Там же. 1950. Сб. 10. С. 73-82.
Адольф Т.А. Количественный учет сайгаков в Астраханских степях И Методы учета численности и географического распределения наземных позвоночных. М.: Изд-во АН СССР, 1952. С. 245-249.
Адольф Т.А. Некоторые данные по биологии сайгака в Астраханских степях И Уч. зап. Моск. гор. пед. ин-та им. В.П. Потемкина. 1954. Т. 28, вып. 2. С. 247-256.
Адольф Т.А. О миграциях сайгаков в Астраханских степях И Там же. 1955. Т. 38, вып. 3. С. 23-27.
Адольф Т.А. Динамика ареала и численности сайги И Охрана природы и заповедное дело в СССР. М.: Изд-во АН СССР, 1957. № 2. С. 125-139.
АйзинБ.М. Копытные//Охотничье-промысловые звери Киргизии. Фрунзе: Илим, 1969. С. 78-110.
Айзин Б.М. Ареалы копытных Киргизии // Копытные фауны СССР. М.: Наука, 1975. С. 32-33.
Айзин Б.М. Распространение, численность, перспективы охраны архара в Киргизии // Редкие виды млекопитающих и их охрана. М.: Наука, 1977. С. 180-181.
Айзин Б.М. Охрана и использование диких копытных Киргизии // Копытные фауны СССР. М.: Наука, 1980. С. 58-59.
Айрумян К.А., Гаспарян К.М. Редкие копытные и хищные млекопитающие Армении // Редкие виды млекопитающих фауны СССР и их охрана. М.: Наука, 1973. С. 6-7.
Айрумян К.А., Гаспарян К.М. Редкие копытные и хищные Армении // Редкие млекопитающие фауны СССР. М.: Наука, 1976. С. 35—42.
Айрумян К.А., Гаспарян К.М. Разведение и гибридизация муфлонов и безоаровых коз в питомнике Института зоологии АН АрмССР //1 Всесоюз. совещ. по пробл. зоокультуры. М„ 1986. 4.2. С. 5-6.
Айунц К.Р. Особенности популяций кавказского тура в Кабардино-Балкарии // Популяционная изменчивость вида и проблем охраны генофонда млекопитающих. М., 1983. С. 3^1.
Айунц К.Р. К систематике кавказских туров // Бюл. МОИП. Отд. биол. 1992. Т. 97, вып. 2. С. 19-25.
Айунц К.Р, Коломыц Э.Г Морфологический показатель изменчивости кавказского тура и его связь с климатом высокогорий // Тр. Высокогорн. геофиз. ин-та. 1986. № 64. С. 3-21.
Айунц К.Р., Шалыбков А.М. Кабардино-Балкарский заповедник И Заповедники СССР. Заповедники Кавказа. М.: Мысль, 1990. С. 34-45.
Аккиев М.И. Некоторые поведенческие аспекты кавказского тура в Кабардино-Балкарском государственном высокогорном заповеднике И III Междунар. конф. “Биологическое разнообразие Кавказа”. Нальчик, 2001. С. 79-81.
Алабугин С., Кацарский О., Никитин В. и др. Овцебык в СССР: уникальный эксперимент продолжается И Охота и охотничье хоз-во. 1984. № 8. С. 4—6.
Алекперов Х.М., Ерофеева С.Н., Рахматулина Н.К Современное состояние некоторых видов млекопитающих Азербайджана в связи с необходимостью их охраны и рационального использования // Редкие виды млекопитающих фауны СССР и их охрана. М.: Наука, 1973. С. 8-10.
Алекперов Х.М., Ерофеева С.Н., Рахматулина И.К. Современное состояние некоторых видов млекопитающих Азербайджана И Редкие млекопитающие фауны СССР. М.: Наука, 1976. С. 28-34.
Алекперов Х.М., Кулиев С.М. Редкие виды копытных Азербайджана и пути их сохранения И Биологические аспекты охраны редких животных. М., 1981. С. 3-5.
Алекперова Н.А. Описание черепа ископаемой бинагадинской сайги (Saiga binagadensis sp. nova) // Изв. АН АзербССР. 1953. № 4. С. 65-70.
Алекперова Н.А. Ископаемая бинагадинская сайга//Тр. Естеств.-ист. музея АН АзербССР. 1955. Вып. 10. С. 10-64.
Александров В.Н. К изучению естественного питания зубров в Кавказском заповеднике И Тр. Кавказского гос. заповедника. 1958. Вып. 4. С. 37-55.
Александрова Л.П., Вангенгейм Э.А., Гербова В.Г. и др. Новые данные о разрезе антро-погеновых отложений горы Тологой (Западное Забайкалье) // Бюл. Комис, по изуч. четвертич. периода. 1963. № 28. С. 85-101.
Александрович Н.П. Фауна из раскопок древнего Витебска // Динамика зооценозов, проблемы охраны и рационального использования животного мира Белоруссии. Минск: Наука и техника, 1989. С. 192.
Александрович Н.П. Охотничья териофауна из раскопок средневекового Турова И Биологическое разнообразие национального парка “Припятский” и других особо охраняемых природных территорий. Туров - Мозырь, 1999. С. 229-231.
Алексеева Л.И. Териофауна раннего антропогена Восточной Европы. М.: Наука, 1977. 216 с. (Тр. Геол, ин-та. Вып. 300).
Алексеева Л.И. Фаунистический комплекс млекопитающих времени последнего межледниковья Русской равнины // Млекопитающие СССР. III съезд Всесоюз. териол. о-ва. М., 1982. Т. 1. С. 7-8.
Алексеева Л.И. Позднепалеолитическая фауна млекопитающих стоянки Сунгирь (г. Владимир) и ее значение для стратиграфии верхнего плейстоцена Восточной Европы // V съезд Всесоюз. териол. о-ва АН СССР. М., 1990а. Т. ЕС. 3—4.
Алексеева Л.И. Териофауна верхнего плейстоцена Восточной Европы (крупные млекопитающие). М.: Наука, 19906. 109 с. (Тр. Геол, ин-та. Вып. 455).
Алексеева Л. К вопросу о вымирании мамонтов // Цитология. 1995. Т. 37, № 7. С. 596.
Алексеева Э.В. Находки ископаемого снежного барана в Кузбассе // Краеведение Кузбасса. 1970. Вып. 2. С. 270-279.
Алексеева Э.В. Млекопитающие плейстоцена юго-востока Западной Сибири. М.: Наука, 1980. 187 с.
Алексеева Э.В. Ископаемые остатки снежного барана на Сахалине И Вести. Дальневост, отд. РАН. 1995. № 6 (64). С. 92-93.
Алексеева Э.В. Редкие млекопитающие плейстоцена Сибири и Дальнего Востока // Редкие виды млекопитающих России и сопредельных территорий. М., 1997. С. 6.
Алиев С.Д. Фауна Азыхской пещеры - объект охоты первобытного человека // II съезд Всесоюз. териол. о-ва. М.: Наука, 1978. С. 3^1.
Алиев Ф.Ф. Современное распространение и динамика численности диких копытных Азербайджанской ССР И I Всесоюз. совещ. по млекопитающим. М.: Изд-во Моск, ун-та, 1961. Т. 1. С. 77-79.
Алиев Ф. Дикие копытные Азербайджана И Охота и охотничье хоз-во. 1962. № 12. С. 28-30.
Алиев Ф.Ф. Результаты и перспективы акклиматизации млекопитающих и птиц на Кавказе И Тр. IX Междунар. конгр. биологов-охотоведов. М., 1970. С. 167-169.
Амирханов А.М., Будун А. С., Вейнберг П.И. и др. Красная книга Северной Осетии. Орджоникидзе: Изд-во “Ир“, 1981. 88 с.
Амирханов А.М., Липкович А.Д., Попов К.П. и др. Северо-Осетинский заповедник И Заповедники СССР. Заповедники Кавказа. М.: Мысль, 1990. С. 50-69.
Андреев В.Н. Состояние кормовой базы северного оленеводства и вопросы использования пастбищ диким северным оленем И Дикий северный олень в СССР. М.: Сов. Россия, 1975. С. 68-79.
Андреева Е.Г. О строении костей некоторых диких и домашних животных // Проблема происхождения домашних животных. Л.: Изд-во АН СССР, 1933. Вып. 1. С. 263-311. (Тр. Лабор. генетики АН СССР. 1933).
Андреева Е.Г. Изучение костных остатков животных из раскопок Пронской экспедиции И Основные проблемы териологии. М.: Наука, 1972а. С. 82-96. (Тр. МОИП. Отд. биол. Т. 48).
Андреева Е.Г., Петренко А.Г. Древние'млекопитающие по археозоологическим материалам Среднего Поволжья и Верхнего Прикамья // Из археологии Волго-Камья. Казань, 1976. С. 137-189.
Андреенков В.И. Редкие виды млекопитающих и проблема их охраны в заповеднике Рамит // Редкие виды млекопитающих и их охрана. М.: Наука, 1977. С. 181-182.
Андреенков В.И. Условия обитания копытных в районах интенсивного пастбищного животноводства// Копытные фауны СССР. М.: Наука, 1980. С. 59-61.
Андреенков В.И. Проблема охраны горных баранов Тянь-Шаня // Редкие виды млекопитающих СССР и их охрана. М., 1983. С. 155-157.
Аникович М., Анисюткин Н. Человек и мамонт в палеолите Восточной Европы // Цитология. 1995. Т. 37, № 7. С. 597.
Анненков Б.ПЛ Экология и поведение млекопитающих Казахстана. Алма-Ата, 1988. С. 69-73. (Тр. Ин-та зоол. Т. 44).
Анненков Б.П., Литун В.И. Некоторые данные по размножению и выживаемости молодняка у диких копытных в Джунгарском Алатау // Экология, морфология, использование и охрана диких копытных. М., 1989. Ч. 1. С. 27-28.
Антипин В.М. Млекопитающие Казахстана. Копытные. Алма-Ата: Казогиз, 1941. Т. 3. 107 с.
Антипин В.М. К экологии дикого горного козла - Capra sibirica // Вести. АН КазССР. 1946. № 11 (20). С. 61-62.
Антипин В.М. Экология, происхождение и расселение диких баранов (Ovis аттоп) Казахстана// Изв. АН КазССР. Сер. зоол. 1947. Вып. 6. С. 3-22.
Антипин В.М. К экологии джейранов и сайгаков И Природа. 1955. № 7. С. 117.
Антипин Е.Е. Антропогенное воздействие, как фактор формирования современной териофауны Крыма // Млекопитающие СССР. III съезд Всесоюз. териол. о-ва. М., 1982. Т. 1. С. 93-94.
Антипина Е.Е., Маслов С.П. Фауна палеолитической стоянки Шатрище-П // V съезд Всесоюз. териол. о-ва АН СССР. М., 1990. Т. 1. С. 4-5.
Антонян А.Ш., Эгнатосян В.А. Рост молодняка муфлонов и безоаровых коз в условиях неволи//Биол. жури. Армении. 1984. Т. 37, № 11. С. 955-958.
Антощенко-Оленев И.В. Кайнозой Джидинского района Забайкалья. Новосибирск: Наука, 1975. 125 с.
Анучин Д. Сайга//Природа. 1875. Кн. 3. С. 201-222.
Анчифоров П. С. К вопросу о структуре ареала алтайского архара // Экологические и экономические аспекты охраны и рационального использования охотничьих животных и растительных пищевых ресурсов Сибири. Шушенское, 1990. С. 4—5.
Анчифоров П. С., Фалеев В.И. Экология алтайского горного барана в связи с вопросами его охраны // Бюл. МОИП. Отд. биол. 1993. Т. 98, вып. 5. С. 15-20.
Арабули А.Б. Роль заповедников Грузии в сохранении генофонда копытных И Проблемы охраны генофонда и управления экосистемами в заповедниках лесной зоны. М., 1986. Ч. 2. С. 11-12.
Арабули А.Б. Современное состояние редких видов копытных Грузии И Экология, морфология, использование и охрана диких копытных. М., 1989. Ч. 1. С. 28-29.
Аренс Л.Е. О влиянии заповедности на охотничье-промысловую фауну Тебердинского заповедника и прилегающих районов // Тр. Тебердинского гос. заповедника. 1957. Т. 1. С. 113-140.
Арылов Ю.Н. Научные аспекты полноценного питания и развития сайгаков в неволе: Автореф. дис.... докт. биол. наук. Дубровицы, 2002. 37 с.
Арылов Ю.Н., Букреева О.М. Выкармливание сайгачат в неволе // Совершенствование технологий производства и переработки сельскохозяйственной продукции в современных условиях. Волгоград, 1999. С. 294-296.
Асадов С.М. Гельминтофауна дагестанского тура (Capra cylindricomis Blyth.) в Азербайджане И Изв. АН АзССР. Сер. биол. и с.-х. наук. 1959а. № 3. С. 35-44.
Асадов С.М. Гельминтофауна серны в Азербайджане И Там же. 19596. № 5. С. 92-96.
Афанасьев А.В. Зоогеография Казахстана. Алма-Ата: Изд-во АН КазССР, 1960. 258 с.
Ахмедов Э.Г. Структурно-функциональная организация популяций дагестанского тура (Capra cylindricomis Blyth.) в условиях Восточного Кавказа: Автореф. дис. ... канд. биол. наук. М., 1995. 23 с.
Ахмедов Э.Г. Распределение дагестанского тура и безоарового козла в районах их совместного обитания И Экология млекопитающих горных территорий. Популяционные аспекты. Нальчик: Изд. центр “Эль-Фа”, 1997а. С. 137-139.
Ахмедов Э.Г. Структура популяции безоарового козла в оптимальных условиях// Редкие виды млекопитающих России и сопредельных территорий. М., 19976. С. 7.
Ахмедов Э.Г. Структура популяции безоарового козла в оптимальных местообитаниях // Редкие виды млекопитающих России и сопредельных территорий. М., 1999. С. 5-10.
Ахмедов Э.Г, Магомедов М.-Р.Д. Зависимость динамики поло-возрастной структуры популяции дагестанского тура (Capra cylindricomis) от условий обитания и плотности популяций // Зоол. журн. 1995. Т. 74, вып. 7. С. 109-118.
Ахмедов Э.Г, Магомедов М.-Р.Д. Пространственно-временная структура популяций дагестанского тура (Capra cylindricomis) в условиях Восточного Кавказа и факторы, ее определяющие И Там же. 1996. Т. 75, вып. 12. С. 1863-1872.
Ахмедов Э.Г., Магомедов М.-РД. Закономерности формирования демографической структуры популяции дагестанского тура (Capra cylindricomis) И Там же. 2000. Т. 79, вып. 4. С. 461-470.
Ахмедов Э.Г., Насруллаев Н.И. Антропогенное влияние на структуру популяций безоарового козла на территории Дагестана // Териофауна России и сопредельных территорий. VII съезд териол. о-ва. М., 2003. С. 25.
Ахмедов Э.Г, Яровенко Ю.А. Рост и развитие молодняка дагестанского тура // Экологические проблемы Прикаспийской низменности. Махачкала, 1991. С. 137-140.
Бабинок В.В., Зубко В.Г. Проблемы реакклиматизации и использования зубров И Интенсификация воспроизводства ресурсов охотничьих животных. Киров, 1990. С. 231-234.
Багрова Н.В., Гордеева Л.Н., Москвин А. С. и др. Ассоциации паразитов у зубров и бизонов при паразитологическом мониторинге в Приокско-Террасном биосферном государственном заповеднике И Ассоциативные паразитарные болезни, проблемы экологии и терапии. М., 1995. С. 16-18.
Бадагуева Ю.Н. Сравнительный анализ полиморфизма гена каппа-казеина у представителей подсемейства Bovidae: Автореф. дис. ... канд. биол. наук. М., 1995. 22 с.
Бадагуева Ю.Н., Сулимова ГЕ. ДНК-полиморфизм гена к-казеина у представителей семейства Bovidae И Генетика. 1994. Т. 30. Прилож. С. 11.
Бажанова М.В. Влияние хозяйственной деятельности человека на распространение и численность охотничье-промысловых зверей Казахстана: Автореф. дис.... канд. биол. наук. Алма-Ата, 1955. 15 с.
Байгушева В. С., Литвиненко В.П., Титов В.В. Позднеплейстоценовая фауна из речных отложений у ст. Вешенской И Териофауна России и сопредельных территорий. VII съезд териол. о-ва. М., 2003. С. 27.
Байгушева В., Тишков М. К истории териофауны Северо-Востока Приазовья И Цитология. 1995. Т. 37, № 7. С. 597-598.'
Байдавлетов РЖ. Современное состояние популяции архара (Ovis аттоп coIlium) в Центральном Казахстане // Состояние териофауны в России и ближнем зарубежье. М„ 1996. С. 21-26.
Байдавлетов РЖ. Экстерьерная характеристика казахстанского архара И Редкие виды млекопитающих России и сопредельных территорий. М., 1997. С. 9.
Байдавлетов РЖ. О факторах смертности архара в Казахстане И Современные проблемы природопользования, охотоведения и звероводства. Киров, 2002. С. 133-135.
Байдавлетов РЖ. Современное состояние популяции каратауского архара // Териофауна России и сопредельных территорий. VII съезд териол. о-ва. М., 2003а. С. 27-28.
Байдавлетов РЖ. Современное состояние популяции архара в Казахстане И Охрана и рациональное использование животных и растительных ресурсов России. Иркутск, 20036. С. 368-374.
Бакеев Н. Вредят ли сайгаки сельскому хозяйству? И Охота и охотничье хоз-во. 1964. № 1. С. 19.
Бакеев Н.Н., Формозов А.Н. Распространение и некоторые черты экологии сайги в западной части Прикаспийской низменности // Материалы по биогеографии СССР. М.: Изд-во АН СССР, 1955. Вып. 2. С. 208-240. (Тр. Ин-та геогр. АН СССР. Вып. 66).
Балюк С. С., Буневич А.Н., Кочко Ф.Ц. Пути сохранения генофонда беловежского зубра // Проблемы охраны генофонда и управления экосистемами в заповедниках лесной зоны. М., 1986. Ч. 2. С. 17-19.
Банников А.Г. Новый вид сайги из Монголии //Докл. АН СССР. 1946. Т. 51, № 5. С. 397-399.
Банников АТ. Материалы к познанию млекопитающих Монголии. V. Антилопы И Бюл. МОИП. Отд. биол. 1951. Т. 56, вып. 3. С. 21^11.
Банников А.Г. Млекопитающие Монгольской Народной Республики. М.: Изд-во АН СССР, 1954. 669 с.
Банников А. Сайгаки и биологические основы их промысла И Охота и охотничье хоз-во. 1958. № 12. С. 23-25.
Банников А. Кавказский тур и его гибриды И Там же. 1959. № 1. С. 25-26.
Банников А. Копытные. Перспективы их хозяйственного использования // Там же. 1962. № 9. С. 26-28.
Банников А.Г. Проблема охраны охотничьих и редких животных // Охотоведение. М.: Лесн. пром-сть, 1974. С. 295-304.
Банников А.Г. Дикие копытные СССР, их использование и охрана//Вопросы рационального использования и охраны диких копытных и других животных. М., 1978. С. 3-6. (Сб. науч. тр. Моск. вет. акад. им. К.И. Скрябина. Т. 97).
Банников А.Г., Жирнов Л.В., Лебедева Л. С., Фандеев А.А. Биология сайгака. М.: Изд-во сельхоз. лит-ры, журн. и плакатов, 1961. 336 с.
Банников А.Г., Рустамов А.К. Охрана природы. М.: Колос, 1977. 207 с.
Баранов П.В., Бойко А.Н. Редкие виды млекопитающих Читинской области И Редкие наземные позвоночные Сибири. Новосибирск: Наука, 1988. С. 13-20.
Бараташвилли Т.К К охране серны в Боржомском заповеднике //Редкие виды млекопитающих и их охрана. М.: Наука, 1977. С. 183-184.
Бараташвилли Т.К. Дикие копытные Боржомского заповедника // Копытные фауны СССР. М.: Наука. 1980. С. 62-63.
Барышев М. Уничтожение сайгаков // Охотник и пушник Сибири. 1925. № 5. С. 36-37.
Барышников Г.Ф. К истории горной териофауны Кавказа// II съезд Всесоюз. териол. о-ва. М.: Наука, 1978. С. 4-5.
Барышников Г. Плейстоценовые млекопитающие из мустъерской стоянки Пролом-2 в Крыму И Цитология. 1995. Т. 37, № 7. С. 598-599.
Барышников Г. Ф., Гарутт В.Е., Громов И.М. и др. Каталог млекопитающих СССР (плиоцен-современность). Л.: Наука, 1981. 456 с.
Барышников Г.Ф., Дмитриева Е.Л., Крахмальная ТВ., Шер А.В. Происхождение, эволюция и систематика// Сайгак. М., 1998. С. 9-24.
Барышников ГФ., Каспаров А.К., Тихонов А.Н. Плейстоценовая сайга Крыма // Экология, морфология, использование и охрана диких копытных. М., 1989. Ч. 2. С. 303-304.
Барышников Г.Ф., Каспаров А.К., Тихонов А.Н. Сайга палеолита Крыма//Тр. Зоол. ин-та АН СССР. 1990. Т. 212. С. 3^18.
Барышников Г.Ф., Крахмальная Т.В. История и систематика сайгаков (Artiodactyia, Bovidae) // Вести, зоол. 1994. № 3. С. 32-38. № 4-5. С. 48-54.
Барышников ГФ., Хварцкая М.Х. Млекопитающие мустьерской стоянки Манагуа в Абхазии // Бюл. МОИП. Отд. биол. 1990. Т. 95, вып. 2. С. 20-31.
Баскин Л.М. Поведение копытных животных. М.: Наука, 1976. 296 с.
Баскин Л.М. Экология и поведение зубра // Зубр. М.: Наука, 1979. С. 442-470.
Баскин Л.М. Использование оборонительного поведения при управлении копытными И Экологические основы управления поведением животных. М.: Наука, 1980. С. 135-143.
Батиров А.Р. Состав и история фауны млекопитающих Узбекистана в послеледниковую эпоху ИIV съезд Всесоюз. териол. о-ва. М., 1986. Т. 1. С. 9-10.
Батхиев А.М. Безоаровый, или бородатый козел в горной Чечено-Ингушетии (экологический анализ географического распространения) И Природа и хозяйство Чечено-Ингушской АССР. 1989а. Вып. 5. С. 103-109.
Батхиев А.М. Современное состояние безоарового козла в Чечено-Ингушской АССР И Редкие и нуждающиеся в охране животные: Материалы к Красной книге. М., 19896. С. 30-34.
Батыров Б.Х. Плейстоценовые копытные Узбекистана ИIV съезд Всесоюз. териол. о-ва. М., 1986. Т. 1. С. 8-9.
Башкиров И. Кавказский зубр (монографический очерк) // Кавказский зубр. М., 1939а (1940). С. 3-72.
Башкиров И. Первый год работы по восстановлению зубра и его акклиматизации в Крымском государственном заповеднике // Там же. 19396 (1940). С. 138-144.
Башкиров И.С. За спасение зубра! // Природа. 1940. № 10. С. 43-50.
Бекенов А. Редкие виды млекопитающих Казахстана и проблемы их охраны И Редкие виды млекопитающих России и сопредельных территорий. М., 1997. С. 10.
Бекенов А.Б. Ресурсы охотничье-промысловых зверей в Казахстане и их использование // VI съезд териол. о-ва. М., 1999. С. 23.
Бекенов А., Байдавлетов РЖ. Научный эксперимент “Архар” И Редкие виды млекопитающих России и сопредельных территорий. М., 1997. С. 10.
Бекенов А.Б., Байдавлетов Р.Ж., Кыдырбаев Х.К. Копытные Казахстана: ресурсы и перспективы использования И Экология, морфология, использование и охрана диких копытных. М., 1989. Ч. 1. С. 31-32.
Бекенов А.Б., Байдавлетов Р.Ж., Федосенко А.К, Вейнберг П.И. О состоянии популяции архара в Карагандинской области И Проблемы охраны и устойчивого использования биоразнообразия животного мира Казахстана. Алматы, 1999. С. 13-14.
БекеновА.Б., ГрачевЮ.А. Казахстан //Сайгак. М., 1998. С. 105-115, 225-227.
Бекенов А.Б., Грачев Ю.А, Бежанов Б., Грачев А.В. Сайга в Казахстане: состояние популяций и проблемы охраны // Зоологические исследования в Казахстане: современное состояние и перспективы. Алматы, 2002. С. 81-83.
Бекенов А.Б., Лобачев Ю.С. О численности волка в Казахстане И Териофауна России и сопредельных территорий. VII съезд териол. о-ва. М., 2003. С. 37-38.
Бекенов А.Б., Плахов К.Н., Бежанов Б., Шаймарданов РТ. Копытные национального парка “Алтын-Эмель” И Проблемы охраны и устойчивого использования биоразнообразия животного мира Казахстана. Алматы, 1999. С. 14-15.
Бекенов А.Б., Поле С.Б., Хахин ГВ., Грачев Ю.А. Гибель от болезней и паразитарных инвазий//Сайгак. М., 1998. С. 247-252.
Бекенов А., Савинов Е. Устюртский муфлон//Охота и охотничье хоз-во. 1980. № 12. С. 8-9.
Бекенов А.Б., Савинов Е.Ф. Азиатский муфлон. Алма-Ата: Кайнар, 1981. 119 с.
Бекенов А., Савинов Е.Ф. Перспективность гибридов азиатского муфлона с домашними овцами // Популяционная изменчивость вида и проблемы охраны генофонда млекопитающих. М., 1983. С. 216-217.
Белан Н.Г. Позднеплейстоценовые копытные Среднего Поднепровья // Экология, морфология, использование и охрана диких копытных. М., 1989. Ч. 2. С. 306-307.
Белан Н.Г, Старкин А.В. Первобытный бизон Северного Причерноморья в позднем плейстоцене // V съезд Всесоюз. териол. о-ва АН СССР. М., 1990. Т. 1. С. 6-7.
Белоусова И.П. Разведение зубров в Центральном зубровом питомнике в 1981-1985 годах //1 Всесоюз. совещ. по пробл. зоокультуры. М., 1986. Ч. 2. С. 11-13.
Белоусова И.П. К вопросу о происхождении современных линий разведения вида зубр (Bison bonasus L.) И К вопросу о возможности сохранения зубра в России. Пущино, 1993а. С. 6-28.
Белоусова И.П. Влияние инбридинга на жизнеспособность зубров в питомниках России И Там же. 19936. С. 29—43.
Белоусова И.П. Вероятность сохранения генетического разнообразия в современных вольных популяциях зубра (Bison bonasus L.); стратегия деятельности по сохранению генетического разнообразия вида И Там же. 1993в. С. 43-56.
Белоусова И.П. Значение и оценка показателей генетического разнообразия для решения проблемы сохранения европейского зубра. Пущино, 1999. 108 с.
Белоусова И.П., Орлов В.Н., Кудрявцев ИВ. Генетическое разнообразие и возможность микроэволюционных изменений европейского зубра (Bison bonasus) И Успехи совр. биол. 1999. Т. 119, № 2. С. 144-150.
Белоусова И.П., Смирнов К.А., Кудрявцев И.В., Казьмин В.Д. Реинтродукция европейского зубра в лесную экосистему национального парка “Орловское полесье” // Проблемы сохранения и восстановления диких копытных животных в Центральном регионе России. Орел: Изд-во ОРАГС, 2003. С. 66-74.
Беляева Е.И. О новых находках ископаемых млекопитающих в Северной Киргизии И Бюл. Комис, по изуч. четвертич. периода. 1963. № 28. С. 76-83.
Беляева М.Я. К познанию гельминтофауны зубра И Работы по гельминтологии к 80-летию акад. К.И. Скрябина. М.: Изд-во АН СССР, 1959. С. 14-16.
Белякова Ю.В., Ахметова Б.А. К фауне эктопаразитов архара (Ovis аттоп Linn.) в Казахстане И Зоологические исследования в Казахстане: современное состояние и перспективы. Алматы, 2002. С. 276-278.
Белякова Ю.В., Байдавлетов РЖ. Паразиты архара (Ovis аттоп L.) Казахского мелкосопоч-ника И Редкие виды млекопитающих России и сопредельных территорий. М.,1997.С.П.
Белякова Ю.В., Байдавлетов Р.Ж. Роль архаров (Ovis аттоп L.) и эпизоотология дик-роцелиоза и эхинококкоза в Казахстане // VI съезд териол. о-ва. М., 1999. С. 24.
Бербер А.П. Горный баран (О. аттоп coIlium) в Центральном Казахстане (биологические основы сохранения): Дис. ... канд. биол. наук. М., 1999. 138 с.
Бербер А. Кто спасет аргали? И Сафари. 2001. № 1. С. 20-23.
Бербер А.П., Ержанов Н. Т. Минеральное питание архаров в Казахстанском нагорье И Зоологические исследования в Казахстане: современное состояние и перспективы. Алматы, 2002. С. 87-88.
Бербер А.П., Ержанов Н. Т. Состояние северной популяции горного барана в Казахском мелкосопочнике // Териофауна России и сопредельных территорий. VII съезд териол. о-ва. М., 2003. С. 40.
Берггрин А.П. К биоморфологии сайги // Тр. Алма-Атинского ветзооинститута. Т. 5. С. 333-345.
Берков. О сайгаке в Калмыцкой автономной области // Охотник. 1927. № 9. С. 18.
Бибиков Д.И., Жирнов Л.В. О роли волков и собак в регулировании численности сайгаков // Копытные фауны СССР. М.: Наука, 1975. С. 190-191.
Бибиков Д.И., Жирнов Л.В., Филимонов А Н., Бекенов А.Б. Гибель сайгаков от хищников // Сайгак. М., 1998. С. 241-247.
Бибикова В.И. О некоторых биологических особенностях первобытного зубра // Бюл. МОИП. Отд. биол. 1950. Т. 55, вып. 5. С. 35—43.
Бибикова В.И. Из истории голоценовой фауны позвоночных в Восточной Европе И Природная обстановка и фауны прошлого. Киев: Изд-во АН УССР, 1963. Вып. 1. С. 119-146.
Бибикова В.И. О смене некоторых компонентов фауны копытных на Украине в голоцене //Бюл. МОИП. Отд. биол. 1975. Т. 80, вып. 6. С. 67-72.
Бибикова В. И. Копытные Северного Причерноморья в раннем голоцене И Проблемы изучения истории современных биогеоценозов. М., 1984. С. 171-203.
Бибикова В.И. Остатки сайгака позднеплейстоценового возраста из раскопок стоянки Анетовка II (Украина) // Вести, зоол. 1985. № 5. С. 47-51.
Благосклонов К.Н., Иноземцев А.А., Тихомиров В.Н. Охрана природы. М.: Высш, шк., 1967. 442 с.
Близнюк А.И. Методика учета и численность сайгаков в Калмыцкой АССР И Копытные фауны СССР. М.: Наука, 1975. С. 39-40.
Близнюк А.И. Сайгак Северо-Западного Прикаспия и перспективы его промысла И Охотничье хозяйство и заповедное дело. М.: Россельхозиздат, 1977. С. 60-68.
Близнюк А.И. Методы промысла и их воздействие на продуктивность популяции сайгаков // Пути и методы рациональной эксплуатации и повышения продуктивности охотничьих угодий. М., 1978. С. 36-37.
Близнюк А.И. Использование пастбищ популяцией сайгаков Северо-Западного Прикаспия // Хозяйственная деятельность и охотничья фауна. Киров, 1980. Т. 2. С. 68-69.
Близнюк А.И. Экология и рациональное использование популяции сайгаков Северо-Западного Прикаспия: Автореф. дис. ... канд. биол. наук. М., 1982. 21 с.
Близнюк А.И. Смертность сайгаков в Северо-Западном Прикаспии И Периодические явления в жизни животных. М., 1983. С. 24-37.
Близнюк А.И. Роль хозяйственного освоения территории в изменении численности калмыцкой популяции сайгака // Биота и природная среда Калмыкии. М. - Элиста, 1995. С. 222-244.
Близнюк А.И. Влияние возраста самок сайгака (Saiga tatarica L.) на плодовитость и пол молодняка// Экология. 1999. № 5. С. 385-387.
Близнюк А.И., Бакташева Н.М. Весеннее питание сайгаков, Saiga tatarica, калмыцкой популяции И Зоол. журн. 2001. Т. 80, № 6. С. 743-748.
Близнюк А.И., Букреева О.М. Плодовитость и отел сайгака, Saiga tatarica, калмыцкой популяции на современном этапе И Там же. 2000. Т. 79, № 9. С. 1124-1132.
Близнюк А.И., Любаева Л.И., Любаев В.Л. Животный мир Калмыкии. Млекопитающие. Элиста: Калмыцкое кн. изд-во, 1980. 127 с.
Бобковская НЕ. Крупные млекопитающие раннего неоплейстоцена севера Западной Сибири // Териофауна России и сопредельных территорий. УП съезд териол. о-ва, М., 2003. С. 50-51.
Бобковская Н.Е., Косинцев П.А. Крупные млекопитающие из аллювиальных местонахождений реки Иртыш //VI Координац. совещ. по изуч. мамонтов и мамонт, фауны. Л., 1991. С. 14-16.
Бобровский П. Зубр И Материалы для географии и статистики России. Гродненская губерния. СПб., 1863. Ч. 1 (22). С. 436-459.
Бобырь Г.Я. Факторы, сдерживающие рост тура в Тебердинском заповеднике // Редкие виды млекопитающих и их охрана. М.: Наука, 1977. С. 185-186.
Бобырь Г.Я. Значение снежных лавин для бурого медведя на Северо-Западном Кавказе // Экологические основы охраны и рационального использования хищных млекопитающих. М.: Наука, 1979. С. 211-212.
Бобырь Г.Я. Взаимоотношения бурого медведя с копытными в горах Северного Кавказа // Копытные фауны СССР. М.: Наука, 1980. С. 126-128.
Бобырь Г.Я. Состояние популяции беловежско-кавказского зубра Тебердинского заповедника И Экологические проблемы Ставропольского края и сопредельных территорий. Ставрополь, 1989. С. 311-312.
Бобырь Г.Я. Кавказско-беловежский зубр в Тебердинском заповеднике И Охрана и изучение редких и исчезающих видов животных в заповедниках. М., 1992. С. 5-20.
Бобырь Г.Я. Редкие виды копытных и хищных Карачаево-Черкессии // Редкие виды млекопитающих России и сопредельных территорий. М., 1999. С. 30-39.
Бобырь Г.Я., Онипченко В.Г. Воздействие крупных хищников на популяцию тура в Тебердинском заповеднике И V съезд Всесоюз. териол. о-ва АН СССР. М., 1990. Т. 2. С. 264—265.
Бобырь КГ О динамике численности западнокавказского тура в Тебердинском заповеднике И Роль заповедников Кавказа в сохранении биоразнообразия природных экосистем. Сочи, 1999. С. 91-94.
Бобырь КГ. Экологические особенности и охрана западнокавказского тура {Capra caucasica Guldenstaedt et Pallas, 1783) Тебердинского заповедника: Автореф. дис.... канд. биол. наук. Ставрополь, 2002. 21 с.
Бобырь КГ, Бобырь Г.Я. Питание западнокавказского тура (Capra caucasica Guldenstaedt et Pallas, 1783) в Тебердинском заповеднике И VI съезд териол. о-ва. М., 1999а. С. 28.
Бобырь КГ, Бобырь Г.Я. Болезни тура (Capra caucasica Guldenstaedt et Pallas, 1783), обусловленные травмами И Там же. 19996. С. 28.
Бобырь К.Г., Бобырь Г.Я. Сравнительный анализ линейных размеров западнокавказского тура Тебердинского и Кавказского заповедников И Териофауна России и сопредельных территорий. VII съезд териол. о-ва. М., 2003. С. 52.
Бобырь К.Г., Онипченко В.Г, Бобырь Г.Я. Опыт оценки питания западнокавказского тура (Capra caucasica Guldenstaedt et Pallas, 1783) на зимних пастбищах в высокогорьях Тебердинского заповедника// Тр. Тебердинского гос. биосф. заповедника. 1999. Вып. 15.
Богданов М. Птицы и звери черноземной полосы Поволжья и долины средней и нижней Волги (био-географические материалы) И Тр. О-ва естествоисп. Казанского ун-та. 1871. Т. 1. С. 3-226.
Боголюбский С.Н. Происхождение домашних животных. М.: Знание, 1956. 32 с.
Богородский Ю.В., Литвинов Н.И. Редкие и исчезающие наземные позвоночные Иркутской области И Редкие наземные позвоночные Сибири. Новосибирск: Наука, 1988. С. 35^11.
Боескоров Г.Г. К систематике и распространению баранов рода Ovis (Artiodactyla, Bovidae) в плейстоцене и голоцене Сибири и Дальнего Востока// Зоол. журн. 2001. Т. 80, № 2. С. 243-256.
Боескоров Г.Г. Сайгак в плейстоцене Якутии И Млекопитающие как компонент аридных экосистем (ресурсы, фауна, экология, медицинское значение и охрана). М., 2004. С. 21-22.
Болденков С.В., Крайнев Е.Д. Парнокопытные фауны Украины // Копытные фауны СССР. М.: Наука. 1980. С. 75-76.
Бондарев А.Я. Состояние численности горных баранов и некоторых других животных на Алтае // Исчезающие и редкие растения и животные Алтайского края и проблемы их охраны. Барнаул, 1982. С. 33-34.
Боржонов Б.Б. Распределение и численность снежного барана в заказнике “Путоранс-кий” // Животный мир плато Путорана, его рациональное использование и охрана. Новосибирск, 1988. С. 14-17.
Боржонов Б.Б., Дорогое В. Ф., Зырянов В.А. и др. Снежный баран гор Путорана // Проблемы охраны и хозяйственного использования ресурсов диких животных Енисейского Севера. Новосибирск, 1979. С. 44-56. (Тр. НИИ сельск. хоз-ва Крайнего Севера. Т. 26).
Борисова Н.Г., Абрамов А.В., Старков А.И. и др. Фауна млекопитающих Республики Бурятия И Тр. Зоол. ин-та РАН. 2001. Т. 288. С. 3-95.
Брагин А.Б. Влияние частного скотоводства на состояние чечено-ингушской популяции зубров И Влияние антропогенной трансформации ландшафта на население наземных позвоночных животных. М., 1987. Ч. 1. С. 185-186.
Брагин А. Куда расселять зубров? // Охота и охотничье хоз-во. 1988. №3. С. 15-17.
Брем А., Россмесслер Е. Лесныя животныя. СПб., 1867. 790 с.
Бромлей ГФ. Биология амурского горала (Nemorhaedus goral caudatus Milne-Edwards, 1867) И Тр. Сихотэ-Алинского гос. заповедника. 1963. Вып. 3. С. 191-267.
БромлейГ.Ф. Горал И Копытные звери. М.: Лесн. пром-сть, 1977а. С. 118-140.
Бромлей Г. Ф. Распространение горала на Дальнем Востоке СССР И Редкие виды млекопитающих и их охрана. М.: Наука, 19776. С. 187-188.
Бромлей Г.Ф., Кучеренко С.П. Копытные юга Дальнего Востока СССР. М.: Наука, 1983. 305 с.
Бромлей Г.Ф., Панкратьев А.Г, Раков Н.В. Распространение амурского горала (Nemorhaedus caudatus) на Дальнем Востоке СССР // Экология и зоогеография некоторых позвоночных суши Дальнего Востока. Владивосток, 1978. С. 86-101.
Букреева О.М. Сохранение сайгака Северо-Западного Прикаспия И Проблемы сохранения биоразнообразия аридных регионов России. Волгоград, 1998. С. 119-121.
Букреева О.М. Европейская популяция сайгака (Saiga tatarica L.) и факторы, определяющие ее состояние: Автореф. дис. ... канд. биол. наук. М., 2002. 19 с.
Букреева О.М. Плодовитость самок европейской популяции сайгака И Териофауна России и сопредельных территорий. VII съезд териол. о-ва. М., 2003. С. 59.
Букреева О.М. Состояние европейской популяции сайгака // Млекопитающие как компонент аридных экосистем (ресурсы, фауна, экология, медицинское значение и охрана). М., 2004. С. 22-23.
Букреева О.М., Арылов Ю.Н. Опыт содержания сайгака И Проблемы сохранения биоразнообразия аридных регионов России. Волгоград, 1998. С. 121-123.
Букреева О.М., Кравчук О. А. Некоторые нарушения моноциклического ритма размножения европейской популяции сайгака И Млекопитающие как компонент аридных экосистем (ресурсы, фауна, экология, медицинское значение и охрана). М., 2004. С. 23-24.
Букреева О.М., Убушаев Б.И. Численность и особенности распространения волка в Калмыкии // Териофауна России и сопредельных территорий. VII съезд териол. о-ва. М„ 2003. С. 59-60.
Булавин А. О некоторых копытных в Кошагаченском аймаке // Охотник Сибири. 1934. №3. С. 24-25.
Буневич А.Н. Характеристика смешанных групп зубров Беловежской пущи // Экология, морфология, использование и охрана диких копытных. М., 1989а. Ч. 2. С. 222-223.
Буневич А.Н. Места обитания зубров в вегетационный период // Динамика зооценозов, проблемы охраны и рационального использования животного мира Белоруссии. Минск: Наука и техника, 19896. С. 196-197.
Буневич А.Н. Итоги расселения зубров по территории Беловежской пущи // Заповеди. Белоруссии. 1991. № 15. С. 98-109.
Буневич А.Н. Воспроизводительные показатели популяции зубров Беловежской пущи // Редкие виды млекопитающих России и сопредельных территорий. М., 1997а. С. 17.
Буневич А.Н. Роль Беловежской пущи в восстановлении зубров // Охраняемые природные территории и объекты Белорус. Поозерья: современное состояние, перспективы развития. Витебск, 19976. С. 45—46.
Буневич А.Н. Результаты реинтродукции зубра (Bison bonasus bonasus) И Биологическое разнообразие национального парка “Припятский” и других особо охраняемых природных территорий. Туров - Мозырь, 1999а. С. 309-310.
Буневич А.Н. Биоритмы репродукции зубра (Bison bonasus bonasus) Беловежской Пущи И Биологические ритмы. Брест, 19996. С. 53-54.
Буневич А.Н. Воспроизводительные показатели популяции зубров Беловежской пущи // Редкие виды млекопитающих России и сопредельных территорий. М., 1999в. С. 72-82.
Буневич АН. Трофейные качества рогов беловежских зубров (Bison bonasus bonasus) // Современные проблемы природопользования, охотоведения и звероводства. Киров, 2002. С. 165-166.
Буневич А.Н., Кочко Ф.П. Динамика численности и структура популяции зубров Беловежской пущи И Популяционные исследования животных в заповедниках. М.: Наука, 1988. С. 96-114.
Бургело Т.Б. Копытные арчевых биоценозов //Копытные фауны СССР. М.: Наука, 1980. С. 128.
Бургело Т.Б. Краткие сообщения об архаре заповедника Аксу-Джабаглы И Редкие животные Казахстана. Алма-Ата, 1986. С. 38.
Буровский А. Палеолитический человек и мамонт в перигляциальных ландшафтах // Цитология. 1995. Т. 37, № 7. С. 602.
Бурчак-Абрамович Н.И., Бурчак-Абрамович Д.Н. Отложения плейстоценовых асфальтов (битумов, киров) - ценные хранилища палеонтологических объектов (млекопитающих, птиц, беспозвоночных, растений и др.) И IV съезд Всесоюз. териол. о-ва. М., 1986. Т. 1.С. 11.
Бурчак-Абрамович Н.И., Векуа А.К. Палеобиологическая история позднекайнозойских быков Кавказа. Тбилиси: Мецниереба, 1980. 92 с.
Бурчак-Абрамович Н.И., Векуа А.К. Важнейшие этапы палеобиологической истории позднекайнозойских быков Кавказа // Млекопитающие СССР. III съезд Всесоюз. териол. о-ва. М., 1982. Т. 1. С. 13-14.
Бутарин Н.С. Хромозомный комплекс архара (Ovis polii karelini Sev.) и курдючного барана И Видовая гибридизация домашних животных с дикими видами. М.-Л., 1935. С. 77-80. (Тр. Киргизск. компл. экспед. 1933-1934 гг. Т. 4, вып. 3).
Бутарин Н. С. Межвидовая гибридизация диких и домашних коз как метод создания новых пород // Веста. АН КазССР. 1947. № 1-2 (22-23). С. 86-91.
Бутарин Н. С. Краткие итоги работы по созданию новой породы овец архаромеринос // Изв. АН КазССР. Сер. биол. 1949. № 71, вып. 5. С. 10-57.
Бутарин Н.С. Некоторые итоги опытов по отдаленной гибридизации животных в Казахстане // Наследственность и изменчивость растений, животных и микроорганизмов. М.: Изд-во АН СССР, 1959. Т. 1. С. 703-708.
Бутарин Н. С. Отдаленная гибридизация в животноводстве (архаромеринос и гибридная свинья). Алма-Ата: Наука, 1964. 210 с.
Бутурлин С.А. Настольная книжка охотника. Свердловск: Издание Уральского обл. к.-п. союза охоты, 1924. 256 с.
Бутурлин С.А. Освоение (акклиматизация) чужеземных зверей // Охотник. 1928а. № 2. С. 9-10.
Бутурлин С.А. К вопросу об акклиматизации овцебыка // Там же. 19286. № 7. С. 21.
Буяндэлгэр У, Даш Я., Шанявски А., Львов И. Учет дзерена в Монголии И Охота и охотничье хоз-во. 1977. № 12. С. 43—45.
Быкова Е.А., Есипов А.В. Современное состояние сайгака в Узбекистане И Млекопитающие как компонент аридных экосистем (ресурсы, фауна, экология, медицинское значение и охрана). М., 2004. С. 28-29.
Валова З.Г Кормовые растения амурского горала в Приморском крае И Раст, ресурсы.
1978. Т. 14, вып. 3. С. 446-454.
Вангенгейм Э.А. Палеонтологическое обоснование стратиграфии антропогеновых отложений Севера Восточной Сибири (по фауне млекопитающих) И Тр. Геол, ин-та. 1961. Вып. 48. 184 с.
Вангенгейм Э.А. Перигляциальная зона и формирование фауны млекопитающих СССР в голоцене //История биогеоценозов СССР в голоцене. М.: Наука. 1976. С. 92-101.
Вангенгейм Э.А. Палеонтологическое обоснование стратиграфии антропогена Северной Азии. М.: Наука, 1977. 169 с.
Варзиев А.Б., Курятников Н.Н. Итоги реакклиматизации зубра в Северо-Осетинской АССР // Копытные фауны СССР. М.: Наука, 1980. С. 229-230.
Варнаков А.П. Редкие виды зверей юга Читинской области // Редкие виды млекопитающих фауны СССР и их охрана. М.: Наука, 1973. С. 109-110.
Васенко Е.П. Экология и распространение сайги (Saiga tatarica L.) И Тр. гос. заповедника Барса-Кельмес. 1950. Вып. 1. С. 38-115.
Васильев С.А. Охота палеолитических обитателей Западного Саяна на сибирского козерога//Палеоэкономика Сибири. Новосибирск: Наука, 1986. С. 133-135.
Васильев С. Фауна млекопитающих последнего межледниковья Новосибирского При-обья // Цитология. 1995. Т. 37, № 7. С. 604-605.
Васильев С.К. Млекопитающие каргинского интерстадиала Новосибирского Приобья И Териофауна России и сопредельных территорий. VII съезд териол. о-ва. М., 2003а. С. 67.
Васильев С.К. Новая находка остатков зоргелии Soergelia sp. на юге Западной Сибири И Там же. 20036. С. 68.
Васильев С.К. Териофауна казанцевского (R-W) времени Новосибирского Приобья И Там же. 2003в. С. 68.
Васильев Я. Предельные линии распространения в Кубанско-Черноморском районе красного зверя и птицы И Природа и охота. 1896. Февраль. С. 1-35.
Василюк И. Ф. Половозрастная характеристика убыли зубров в Беловежской пуще И Редкие виды млекопитающих фауны СССР и их охрана. М.: Наука, 1973. С. 110-111.
Вейнберг П.И. Особенности размещения копытных в Северо-Осетинском заповеднике // Редкие виды млекопитающих и их охрана. М.: Наука, 1977. С. 190-192.
Вейнберг П.И. К поведению дагестанского тура во время гона // Копытные фауны СССР. М.: Наука, 1980. С. 284-285.
Вейнберг П.И. Внутривидовые различия в экологии и разделении взрослых самцов и самок с молодняком у дагестанского тура (Capra cylindricornis Blyth.) // Бюл. МОИП. Отд. биол. 1981а. Т. 86, № 3. С. 23-28.
Вейнберг П.И. Дагестанский тур (Capra cylindricornis Blyth) на Центральном Кавказе (экология, поведение и охрана): Автореф. дис.... канд. биол. наук. М., 19816. 26 с.
Вейнберг П.И. Внутривидовые различия в экологии и сегрегация взрослых самцов от самок с молодняком у дагестанского тура // Млекопитающие СССР. III съезд Всесоюз. териол. о-ва. М., 1982а. Т. 1. С. 167-168.
Вейнберг П.И. К познанию брачного и агонистического поведения дагестанского тура // Там же. 19826. Т. 2. С. 118-119.
Вейнберг П.И. Пищевое поведение дагестанского тура// Редкие виды млекопитающих СССР и их охрана М., 1983а. С. 159-160.
Вейнберг П.И. Местообитания серны в зависимости от наличия конкурирующих видов // там же. 19836. С. 160-161.
Вейнберг П.И. Динамика стадности дагестанского тура в зависимости от репродуктивного цикла // Там же. 1983в. С. 161.
Вейнберг П.И. Дагестанский тур. М.: Наука, 1984. 88 с.
Вейнберг П.И. О формах рогов рода Capra ИIV съезд Всесоюз. териол. о-ва. М., 1986а. Т. 1. С. 47—48.
Вейнберг П.И. Влияние зубров на древесную растительность широколиственных лесов Северо-Осетинского заповедника и заказника Цейский // Роль крупных хищников и копытных в биоценозах заповедников. М., 19866. С. 131-138.
Вейнберг П.И. Влияние зубров на древесную растительность Северо-Осетинского заповедника и заказника “Цейский” // Проблемы охраны генофонда и управления экосистемами в заповедниках лесной зоны. М., 1986в. Ч. 2. С. 41—44.
Вейнберг П.И. Материалы по питанию рыси в Северо-Осетинском заповеднике И Ресурсы животного мира Северного Кавказа. Ставрополь, 1988. С. 35-37.
Вейнберг П.И. Годовая динамика возрастной структуры популяции тура в Северо-Осетинском заповеднике И Экология, морфология, использование и охрана диких копытных. М., 1989. Ч. 2. С. 237-238.
Вейнберг П.И. Анализ формы рогов и окраски волосяного покрова горных козлов (Capra L., 1758; Artiodactyla) // Бюл. МОИП. Отд. биол. 1993. Т. 98, вып. 5. С. 3-14.
Вейнберг П.И. Сведения о современном состоянии популяций горных козлов на территории СНГ И Редкие виды млекопитающих России и сопредельных территорий. М., 1997. С. 18.
Вейнберг П.И. О состоянии популяции и особенностях биологии безоарового козла (Capra aegagrus Erxleben) в Дагестане И Бюл. МОИП. Отд. биол. 1999. Т. 104, вып. 4. С. 12-21.
Вейнберг П.И. Безоаровый козел Capra aegagrus Erxleben, 1777 И Красная книга Российской Федерации (животные). Балашиха: Астрель, 2001. С. 717-718.
Вейнберг П.И. Годовые сегменты рогов самок дагестанского тура (Capra cylindricornis Blyth) как возможный показатель участия в размножении И Бюл. МОИП. Отд. биол. 2002. Т. 107, вып. 5. С. 3-8.
Вейнберг П.И. К состоянию популяции барана Северцова в Нуратауском заповеднике И Териофауна России и сопредельных территорий. VII съезд териол. о-ва. М., 2003а. С. 70-71.
Вейнберг П.И. Зависимость индекса сеголеток от размера и состава групп самок у дагестанского тура // Там же. 20036. С. 71.
Вейнберг П.И. Об уриале на Юго-Западном Памире // Там же. 2003в. С. 71.
Вейнберг П.И. Многолетняя динамика численности, половой и возрастной структуры популяции дагестанского тура (Capra cylindricornis Blyth, 1841) в Северо-Осетинском заповеднике//Бюл. МОИП. Отд. биол. 2002. Т. 107, вып. 2. С. 14—22.
Вейнберг П.И., Комаров Ю.Е. Состояние зубра в Северной Осетии - Алании // VI съезд териол. о-ва. М., 1999. С. 45.
Вейнгер Г.М. Популяция горалов Сихотэ-Алинского заповедника // Тр. Сихотэ-Алинс-кого гос. заповедника. 1963. Вып. 3. С. 269-281.
Верещагин Н.К. Дагестанский тур (Capra cylindricornis Blyth.) в Азербайджане // Тр. Зоол. ин-та Азерб. фил. АН СССР. 1938. Т. 9, вып. 45. С. 5-72.
Верещагин Н.К. Охотничьи и промысловые животные Кавказа. Баку: Изд-во АН АзССР, 1947. 144 с.
Верещагин Н.К. Плейстоценовые реликты Кабристана и Апшеронского полуострова // Бюл. МОИП. Отд. биол. 1949. Т. 54, вып. 4. С. 3-14.
Верещагин Н.К. Байкальский як (Poephagus baikalensis N. Ver., sp. nova, Mammalia) из плейстоценовой фауны Восточной Сибири // Докл. АН СССР. 1954. Т. 99, № 3. С. 455-458.
Верещагин Н.К. Остатки млекопитающих из нижнечетвертичных отложений Таманского полуострова И Тр. Зоол. ин-та АН СССР. 1957. Т. 22. С. 9-74.
Верещагин Н.К. Млекопитающие Кавказа. М.-Л.: Изд-во АН СССР, 1959а. 704 с.
Верещагин Н.К. Овцебык на севере Сибири И Природа. 19596. № 8. С. 105-106.
Верещагин Н.К. Основные черты формирования териофаун Голарктики в антропогене И Зоол. журн. 1963. Т. 42, вып. 11. С. 1686-1698.
Верещагин Н.К. Охоты первобытного человека и вымирание плейстоценовых млекопитающих в СССР//Материалы по фаунам антропогена СССР. Л.: Наука, 1971. С. 200-232. (Тр. Зоол. ин-та АН СССР. Т. 49).
Верещагин Н. Охота наших предков // Охота и охотничье хоз-во. 1975а. № 2. С. 39—41.
Верещагин Н.К. Видовой состав и ареалы копытных четвертичного периода И Копытные фауны СССР. М.: Наука, 19756. С. 7-9.
Верещагин Н.К. Гибель мамонтовой фауны в плейстоцене И Природа. 1977. № 9. С. 90-95.
Верещагин Н.К. Почему вымерли мамонты? Л.: Наука, 1979. 196 с.
Верещагин Н.К. К истории реакклиматизации овцебыка на Таймыре в связи с филогенезом И Овцебык в тундре России: Эксперимент XX века по восстановлению исчезнувшего вида. СПб.: Астерион, 2002. С. 19-29.
Верещагин Н.К, Барышников Г.Ф. Некоторые проблемы изучения истории горных териофаун // Экология, методы изучения и организация охраны млекопитающих горных областей. Свердловск, 1977. С. 6-8.
Верещагин Н.К, БарышниковГ.Ф. Палеозоологическое картирование и ареал млекопитающих СССР ИII съезд Всесоюз. териол. о-ва. Пленарные доклады. М., 1979. С. 14—20.
Верещагин Н.К, Барышников Г.Ф. Ареалы копытных фауны СССР в антропогене И Млекопитающие Восточной Европы в антропогене. Л., 1980а. С. 3-20. (Тр. Зоол. ин-та АН СССР. Т. 93).
Верещагин Н.К, Барышников Г.Ф. Млекопитающие предгорного Северного Крыма в эпоху палеолита И Там же. 19806. С. 26—49.
Верещагин Н.К., Барышников Г. Ф. Вымирание млекопитающих в четвертичном периоде Северной Евразии И Млекопитающие Северной Евразии в четвертичном периоде. Л., 1985. С. 3-38. (Тр. Зоол. ин-та АН СССР. Т. 131).
Верещагин И.К., Батыров Б. Фрагменты истории териофауны Средней Азии // Бюл. МОИП. Отд. биол. 1967. Т. 72, вып. 4. С. 104-115.
Верещагин Н.К., Громов И.М. Формирование териофауны СССР в четвертичном периоде // Успехи современной териологии. М.: Наука, 1977. С. 32-45.
Верещагин Н.К., Громов И.М., Ермолова Н.М., Паавер К.А. Основные черты формирования териокомплексов Северной Евразии в голоцене // История биогеоценозов СССР в голоцене. М.: Наука, 1976. С. 101-116.
Верещагин Н.К., Колбутов А.Д. Остатки животных на мустьерской стоянке под Сталинградом и стратиграфическое положение палеолитического слоя // Тр. Зоол. ин-та АН СССР. 1957. Т. 22. С. 7.5-89.
Верещагин Н.К., Наниев В.И. Прежнее и современное распространение копытных в Северной Осетии. 1. Вымершие виды // Зоол. журн. 1949. Т. 28, вып. 3. С. 277-280.
Верещагин Н.К., Оводов Н.Д. История фауны Приморья // Природа. 1968. № 9. С. 42-49.
Верещагин Н.К., Русаков О. С. Копытные Северо-Запада СССР (история, образ жизни и хозяйственное использование). Л.: Наука, 1979. 309 с.
Верховский. Мускусный бык//Охотник. 1928. № 5. С. 14—15.
Вершинин А.А. Распространение и численность диких копытных в Камчатской области // Охотоведение. М.: Лесн. пром-сть, 1972. С. 109-127.
Виноградов А.Ф. Зубр {Bison europaeus) Северо-Западного Кавказа И Тр. II съезда рус. естествоиспыт. по отд. зоологии, анатомии и физиологии. М., 1870. С. 179-189.
Витович В. Тур и охота на него в Азербайджане // Охотник. 1928. № 11. С. 23.
В-н Н. Охота на крупную дичь в Восточном Закавказье // Боец-охотник. 1936. №11. С.35-37.
Водопьянов Б.Г Снежный баран в Северном Забайкалье и возможности его хозяйственного использования И Пути повышения эффективности охотничьего хозяйства. Иркутск, 1971. Ч. 1. С. 55-57.
Волошина И.В. “Ухаживания” и поведение горалов во время копуляций И Групповое поведение животных. М.: Наука, 1976а. С. 47—49.
Волошина И.В. Маркировочное поведение и его социальные функции у амурского горала - Nemorhaedus goral caudatus И Там же. 19766. С. 49-51.
Волошина И.В. Парковое содержание амурских горалов И Биологические аспекты охраны редких животных. М., 1981. С. 6-9.
Волошина И.В. Маркировочное поведение самок амурского горала И Млекопитающие СССР. III съезд Всесоюз. териол. о-ва. М., 1982. Т. 2. С. 123-124.
Волошина И.В. Взаимоотношения матери и детеныша у амурского горала И IV съезд Всесоюз. териол. о-ва. М., 1986. Т. 2. С. 127-128.
Волошина И.В. Биологические особенности амурского горала в связи с перспективами его реинтродукции. Автореф. дис. ... канд. биол. наук. М., 1988. 25 с.
Волошина И.В., Мысленков А.И. Методы изучения поведения горала {Nemorhaedus goral caudatus) // Групповое поведение животных. М.: Наука, 1976. С. 51-53.
Волошина И.В., Мысленков А.И. Охрана горала на Среднем Сихотэ-Алине И Редкие виды млекопитающих и их охрана. М.: Наука, 1977. С. 194-195.
Волошина И.В., Мысленков А.И. Гельминты амурских горалов // Экология, морфология, использование и охрана диких копытных. М., 1989. Ч. 2. С. 296-298.
Волошина И.В., Мысленков А.И. Современное состояние популяций амурского горала в СССР // V съезд Всесоюз. териол. о-ва АН СССР. М., 1990. Т. 2. С. 64-65.
Волошина И.В., Мысленков А.И. Морфо-физиологические показатели амурского горала И Амурский горал. М.: Изд-во ЦНИЛ охотничьего хоз-ва и заповедников, 1992а. С. 75-83.
Волошина И.В., Мысленков А.И. Теоретические и практические аспекты реинтродукции амурского горала // Там же. 19926. С. 123-131.
Волошина И.В., Мысленков А.И. Амурский горал Nemorhaedus caudatus raddeanus (Heude, 1894) //Красная книга Российской Федерации (животные). Балашиха: Астрель, 2001. С. 715-717.
Волошина И.В., Мысленков А.И., Нестеров Д.А. Распространение горала в Ольгинском районе Приморского края // Редкие виды млекопитающих и их охрана. М.: Наука, 1977. С. 192-194.
Волошина И.В., Мысленков А.И., Обверткина Т.В. Суточная активность горала // Амурский горал. М.: Изд-во ЦНИЛ охотничьего хоз-ва и заповедников, 1992. С.58-70.
Волошина И.В., Мысленков А.И, Смирнов Е.Н. К вопросу об охране горала // Редкие млекопитающие фауны СССР. М.: Наука, 1976. С. 125-133.
Волошина И.В., Мысленков А.И., Тихонова Т.В. Ольфакторно-оптические маркеры маньчжурской косули и амурского горала И Млекопитающие СССР. III съезд Всесоюз. териол. о-ва. М., 1982. Т. 2. С. 121.
Волошина И.В., Мысленков А.И., Шаулъская Н.А. Особенности пищевого поведения го-k ралов (Nemorhaedus goral caudatus} в зимний период И Групповое поведение живот-
» ных. М.: Наука, 1976. С. 53-54.
Волошина И.В., Нестеров Д.А. Характеристика мест обитания горала в Приморском крае И Амурский горал. М.: Изд-во ЦНИЛ охотничьего хоз-ва и заповедников, 1992. С. 21-35.
Волошина И.В., Сагдиева П.Д., Мысленков А.И. Взаимоотношения амурского горала с иксодовыми клещами на Среднем Сихотэ-Алине И Копытные фауны СССР. М.: Наука, 1980. С. 133-134.
Волошина ИВ., Соломкина Н.В. Болезни амурского горала в дикой природе и при содержании в неволе // Амурский горал. М.: Изд-во ЦНИЛ охотничьего хоз-ва и заповедников, 1992. С. 105-119.
► Волошина И.В., Хрусталев А.В. Эндопаразиты: видовой состав, обилие и патогенность // Там же. 1992. С. 119-123.
Воробьев К.А. Сайга в Астраханском заповеднике И Охотник. 1929. № 2. С. 30.
Воронов В.Г. Млекопитающие Курильских островов. Л.: Наука, 1974. 164 с.
Воронов В.Г. Стратегия охраны редких животных на Сахалине и Курильских островах И Редкие виды млекопитающих СССР и их охрана М., 1983. С. 6-9.
Воронов В.Г, Воронов ГА. Реакклиматизация толсторога на северных Курильских островах как мера охраны его в восточной части ареала И Редкие виды млекопитающих фауны СССР и их охрана. М.: Наука, 1973. С. 113.
Воронов Г. А. Акклиматизация млекопитающих на Сахалине и Курильских островах (итоги и перспективы). М.: Наука, 1982. 133 с.
Воронцов Н.Н. О работах по советско-американской программе “Систематика и эволюция голарктических млекопитающих” И Биологические исследования на Дальнем Востоке. Владивосток, 1975. С. 23-28.
Воронцов Н.Н., Коробицына К.В., Надлер Ч.Ф. и др. Хромосомы диких баранов и происхождение домашних овец // Природа. 1972а. № 3. С. 74-82.
Воронцов Н.Н., Коробицина К.В., Надлер Ч.Ф. и др. Цитогенетическая дифференциация и границы видов у настоящих баранов (Ovis s. str.) Палеарктики И Зоол. журн. 19726. Т. 51, вып. 8. С. 1109-1122.
Воронцов-Вельяминов Б. О животных Малого Карачая И Охотник. 1929. № 11. С. 19.
Врублевский К. Теоретическая дифференцировка некоторых жвачных на древеснояд-ных (Fruticivora) и травоядных (Herbivora) и практическое ее значение // Архив ветеринар. наук. 1912. Кн. 8. С. 746-778.
Вырыпаев В.А. Состояние популяции центрально-азиатского горного козла в западной части Чаткальского хребта И Редкие виды млекопитающих фауны СССР и их охрана. М.: Наука, 1973. С. 114-116.
Вырыпаев В.А. Факторы, определяющие численность архара в Центральном Тянь-Шане, и меры по его охране // Копытные фауны СССР. М.: Наука, 1980. С. 233-234.
Вырыпаев В.А. О брачном поведении архара в Тянь-Шане // Прикладная этология. М.: Наука, 1983а. С. 202-203.
Вырыпаев В.А. О факторах, определяющих численность архара в Центральном Тянь-Шане, и мерах по его охране // Взаимодействие биотических компонентов и среды в некоторых экосистемах Тянь-Шаня. Фрунзе, 19836. С. 130-136.
Вырыпаев В.А., Литвинов В. Ф. Влияние саркоптоза на популяцию Capra sibirica Pall. В Западном Тянь-Шане //1 Междунар. контр, по млекопитающим. М.: ВИНИТИ, 1974. Т. 1. С. 131.
Вяткин П.С., Филь В.И. Численность и экология снежного барана восточного побережья Камчатки // Копытные фауны СССР. М.: Наука, 1975. С. 263-264.
Гавердовский М.Д. Опыт искусственного выращивания овцебыка в Московском зоопарке И Копытные фауны СССР. М.: Наука, 1980. С. 234-235.
Гаврилов В. Т. Заметки по биологии снежного барана в Кроноцком заповеднике // Зоол. журн. 1947. Т. 26, вып. 6. С. 561-562.
Гагарин В.Г, Назарова Н. С. Формирование гельминтофауны зубра в связи с его расселением по Советскому Союзу И Гельминты животных Киргизии и сопредельных территорий. Фрунзе: Илим, 1966. С. 62-65.
Газаев М.А., Аккиев М.И. К изучению гельминтозов копытных животных в условиях Центрального Кавказа И Экология млекопитающих горных территорий. Популяционные аспекты. Нальчик: Изд. центр “Эль-Фа”, 1997. С. 85-86.
Газалиев Н.А. Дагестанская популяция сайгаков и перспективы ее рационального использования // Биологическая продуктивность ландшафтов Дагестана. Махачкала, 1980а. Вып. 3. С. 33—40.
Газалиев Н.А. Об экологии и продуктивности дагестанского тура И Там же. 19806. С. 54—57.
Галкина Л.И., Форонова И.В. Антропогеновая териофауна Бачатского карьера Кузнецкой котловины (Кузбасс) // Фауна и экология позвоночных Сибири. Новосибирск: Наука, 1980. С. 176-188.
Галузо И.Г, Голосов В.И., Горбунова З.И. Штамм Toxoplasma gondii “STG", выделенный от сайги И Паразиты сельскохозяйственных животных Казахстана. Алма-Ата, 1963. С. 199.
Гамбарян П.П. Бег млекопитающих. Приспособительные особенности органов движения. Л.: Наука, 1972. 334 с.
Гамбарян П.П., Гаспарян К.М., Григорян В.К., Айрумян В.А. Учет копытных с вертолета в условиях сильно пересеченной местности И Ресурсы фауны промысловых зверей в СССР и их учет. М.: Изд-во Акад, наук СССР, 1963. С. 104-109.
Ганюшин А.Е. О распространении и численности архаров в Центральном Тянь-Шане И Редкие и исчезающие звери и птицы Казахстана. Алма-Ата, 1977. С. 71-74.
Гасанов Х.Н. Закатальский заповедник // Заповедники СССР. Заповедники Кавказа. М.: Мысль, 1990. С. 251-258.
Гаспарян КМ. К кормовому режиму безоаровых коз Capra aegagrus Erxl. на Урцском (Сарайбулагском) хребте // Изв. АН АрмССР. Биол. науки. 1964. Т. 17, № 2. С. 85-100.
Гаспарян К.М. Экология безоарового козла и адаптации к горным условиям некоторых Bovidae: Автореф. дис. ... канд. биол. наук. Ереван, 1969. 19с.
Гаспарян К.М. Экология безоарового козла // Фауна позвоночных животных АрмССР. Ереван, 1975. С. 78-106. (Зоол. сб. Зоол. ин-та АН АрмССР. 1974. Вып. 16).
Гаспарян К.М. К вопросу о строении конечностей некоторых копытных в связи с их образом жизни // Проблемы экологии и морфологии животных. Изд-во Моск, ун-та, 1976. С. 8-11.
Гаспарян К.М., Маркарян Э.Г Перспективы восстановления численности арменийского муфлона И Редкие виды млекопитающих и их охрана. М.: Наука, 1977. С. 196-197.
Гептнер В.Г, Насимович А.А., Банников А.Г. Млекопитающие Советского Союза. Парнокопытные и непарнокопытные. М.: Высш, шк., 1961. Т. 1. 776 с.
Гептнер В.Г, Формозов А.Н. Млекопитающие Дагестана // Тр. гос. зоол. музея МГУ. 1941. Т. 6. С. 3-74.
Гигинейшвили Н.С. Опыт гибридизации каракульской овцы с диким бараном // Наследственность и изменчивость растений, животных и микроорганизмов. М.: Изд-во АН СССР, 1959. Т. 1. С. 709-715.
Гигинейшвили Н.С. Создание нового типа серых каракульских овец с повышенной жизнеус-тойчивостью путем гибридизации с муфлоном И Селекция, гибридизация и акклиматизация сельскохозяйственных животных. М.: Колос, 1983. С. 224-235.
Гигинейшвили НС., Чариев Э.К Результаты гибридизации серых каракульских овец с горным бараном И Вопросы гибридизации копытных. М.: Наука, 1980. С. 40-44.
Гинеев А.М., Абдурахманов М.Г, Спасская ТХ. и др. К современной численности и распространению некоторых редких и исчезающих млекопитающих Северного Кавказа//Ресурсы животного мира Северного Кавказа. Ставрополь, 1988. С. 42-49.
Гинеев А.М., Могильная Г.М., Дудецкий В.И. К решению проблем интродукции - одного из основных и наиболее эффективных путей воспроизводства охотничьих животных И Современные проблемы природопользования, охотоведения и звероводства. Киров, 2002. С. 181-184.
Глазко В.И., Дыманъ Т.Н., Сипко Т.П., Кушнир А.В. Участие маркеров структурных генов и анонимных последовательностей ДНК в генетической дифференциации видов рода Ovis // Докл. Рос. акад. с.-х. наук. 2001. № 2. С. 12-15.
Глазко В.И., Дыманъ Т.Н., Кутузов Д.А., Городня А.В. Участие маркеров ISSR-PCR в межвидовой дифференциации некоторых представителей видов Ungulata И Там же. 2002. № 5. С. 36-40.
Глазко В.И., Зеленая Л.Б., Ясинецкая Н.А. Сравнительный анализ некоторых видов Ungulata по различным молекулярно-генетическим маркерам (белки, RAPD-PCR) И Цитол. и генетика. 1997. Т. 31, № 4. С. 81-88.
Глебов В.В. Места обитания и численность амурского горала в Лазовском и Ольгинском районах Приморского края И Редкие и исчезающие животные суши Дальнего Востока СССР. Владивосток, 1981. С. 173-174.
Глебов В.В., Соломкина НВ. Отлов горалов И Редкие виды млекопитающих СССР и их охранам., 1983. С. 167-168.
Глебов В.В., Шалдыбин С.Л., ЖивотченкоВ.И. Опыт авиаучета горалов и пятнистых оленей в Лазовском заповеднике И Копытные фауны СССР. М.: Наука. 1980. С. 8-10.
Глушков В.М. Экологические основы управления популяциями лося в России: Автореф. дис. ... докт. биол. наук. М., 2003. 45 с.
Гмелин С.Г. Путешествие по России для исследования трех царств естества. СПб.: Им-пер. Акад, наук, 1806 (1771). Ч. 1. 272 с.
Гмелин С.Г. Путешествие по России для исследования трех царств природы. СПб.: Им-пер. Акад, наук, 1777. Ч. 2. 361 с.
Гмелин С.Г. Путешествие по России для исследования всех трех царств в природе. СПб.: Импер. Акад, наук, 1785. Ч. 3, полов. 2. 737 с.
Голованова Э., Анохина Ю. Сайгаки на посевах // Охота и охотничье хоз-во. 1976. № 1. С. 16-17.
Горбунов А.В. Проблемы охраны горного барана в Северо-Западной Туркмении // Популяционная изменчивость вида и проблемы охраны генофонда млекопитающих. М., 1983. С. 221-222.
Горбунов А.В. Численность и промысел сайгака в Северо-Западной Туркмении // IV съезд Всесоюз. териол. о-ва. М., 1986а. Т. 3. С. 141-142.
Горбунов А.В. Миграция сайгаков в Северо-Западную Туркмению // Там же. 19866. С. 142-144.
Горбунов А.В. Водопои копытных и хищных млекопитающих в Северо-Западной Туркмении И Бюл. МОИП. Отд. биол. 1986в. Т. 91, вып. 1. С. 17-24.
Горбунов А.В. Млекопитающие Северо-Западной Туркмении и Южного Устюрта: Авто-реф. дис. ... канд. биол. наук. Киев, 1987. 24 с.
Горбунов А.В. Сайга, или сайгак И Млекопитающие Туркменистана. Хищные, ластоногие, копытные. Ашхабад: Ылым, 1995а. Т. 1. С. 237-243.
Горбунов А.В. Устюртский горный баран // Там же. 19956. С. 268-278.
Горегляд Х.С. Болезни диких животных. Минск: Наука и техника, 1971. 304 с.
Горелов Ю.К. Нахождение сибирского горного козла в Бадхызе (Ю.-В. Туркмения) // Бюл. МОИП. Отд. биол. 1961. Т. 66, вып. 6. С. 146-147.
Горелов Ю.К. Водопои горных баранов Бадхызского заповедника И Там же. 1964. Т. 69, вып. 2. С. 61-70.
Горелов Ю.К. Факторы, определяющие территориальные группировки копытных Бадхы-за // Популяционная структура вида у млекопитающих. М., 1970. С. 31-33.
Горелов ЮАС. Охрана и восстановление численности копытных Бадхыза // Редкие виды млекопитающих фауны СССР и их охрана. М.: Наука, 1973. С. 117-118.
Горелов Ю. Роль рогов в жизни баранов И Охота и охотничье хоз-во. 1986. № 2. С. 8-9.
Горелов ЮАС. Состояние копытных Бадхыза в 1998 г. //VI съезд териол. о-ва. М., 1999. С. 60.
Горелов Ю.К., Ишадов Н. Горный баран (уриал) // Красная книга Туркменистана. Беспозвоночные и позвоночные животные. Ашхабад, 1999. Т. 1. С. 342-343.
Горохов В.В., Багрова Н.В., Гордеева JI.M. и др. Паразитологический мониторинг популяции зубров //Ветеринария. 1995. № 12. С. 32-36.
Горохов Д.Н., Якшина И.А. Реакклиматизация овцебыков на севере Якутии И Современные проблемы природопользования, охотоведения и звероводства. Киров, 2002. С. 192-193.
Горчаковский А.П. Природа Балхашской котловины // Охотник и пушник Сибири. 1925. № 4. С. 12.
Графодатский А. С. Цитогенетические аспекты филогении Artiodactyla// Экология, морфология, использование и охрана диких копытных. М., 1989. Ч. 1. С. 13.
Графодатский А. С., Шаршов А.А., Билтуева Л. С., Alonoe В.А. Кариотип зубра (Bison bonasus L.) И Цитология и генетика. 1990. Т. 24, № 3. С. 34-37.
Грачев Ю.А. Редкие виды млекопитающих заповедника Аксу-Джабаглы и хребта Каратау (Сырдарьинского) И Млекопитающие СССР. III съезд Всесоюз. териол. о-ва. М., 1982. Т. 1. С. 101-102.
Грачев ЮА. О суточной активности копытных в Таласском Алатау (Западный Тянь-Шань) И Экология и поведение млекопитающих Казахстана. Алма-Ата, 1988. С. 48-55. (Гр. Ин-та зоол. Т. 44).
Грачев Ю.А., Бекенов А.Б. Современное состояние популяций сайгака в Казахстане // Зоологические исследования в Казахстане. Алматы, 1993. Ч. 2. С. 165—189.
Грачев Ю.А., Бекенов А.Б. Териофауна России и сопредельных территорий // VII съезд териол. о-ва. М., 2003. С. 97.
Грачев Ю.А., Есжанов Б., Шаймарданов Р. Т. Результаты авиаучетов сайгака в Казахстане//VI съезд териол. о-ва. М., 1999: С. 62.
Грачев Ю.А., Исмагулов Е.Ж. О результатах мечения сайгаков в Казахстане // Проблемы охраны и устойчивого использования биоразнообразия животного мира Казахстана. Алматы, 1999. С. 20-21.
Грачев Ю.А, Савинов Е. Ф. Изменение численности сибирского горного козла и архара в северо-восточной части Джунгарского Алатау // Копытные фауны СССР. М.: Наука, 1975. С. 76-77.
Грачев Ю.А, Смирнова Э.Д. Вертикальное распространение и кочевки копытных в Таласском Алатау // Копытные фауны СССР. М.: Наука, 1980. С. 82-83.
Грибков П.Ф. Состояние поголовья снежного барана {Ovis nivicola) в центральной части Камчатки И Зоол. журн. 1969. Т. 68, № 11. С. 1716-1721.
Грибков П.Ф. Снежный баран на Камчатском полуострове // Краеведч. зап. Петропавловск-Камчатский: Кн. изд-во, 1970. Вып. 2. С. 117-121.
Грибков П.Ф., Филь В.И. Снежный баран // Копытные звери (Редкие животные СССР). М.: Лесн. пром-сть, 1977. С. 222-240.
Гришин Ю.С. Бронзовый и ранний железный века Восточного Забайкалья. М.: Наука, 1975. 135 с.
Гришин Ю.С. Памятники неолита, бронзового и раннего железного веков лесостепного Забайкалья. М.: Наука, 1981. 190 с.
Громов В.И. Палеонтологическое и археологическое обоснование стратиграфии континентальных отложений четвертичного периода на территории СССР (млекопитающие, палеолит) И Тр. Ин-та геол. наук. 1948. Вып. 64. Геол. сер. (№ 17). С. 1-521.
Громов И.М., Гуреев А.А., Новиков ГА. и др. Млекопитающие фауны СССР. М.-Л.: Изд-во АН СССР, 1963. Ч. 2. С. 641-1101.
Громова В.И. Первобытный зубр {Bison priscus Bojanus) в СССР И Тр. Зоол. ин-та АН СССР. 1935а. Т. 2, вып. 2-3. С. 6-204.
Громова В. Об остатках диких баранов и козлов в четвертичных отложениях Крыма // Докл. АН СССР. 19356. Т. 4, № 1-2. С. 97-100.
Громова В. О распространении остатков овцебыка Ovibos moschatus Zimm, в Восточной Европе и Северной Азии // Изв. АН СССР. Отд. матем. и естеств. наук. 7 сер. 1935в. № 1.С. 101-114.
Громова В. Фауна верхнепалеолитической стоянки Мальта близ Иркутска И Докл. АН СССР. 1941. Т. 33, № 1. С. 94-96.
Громова В. К вопросу о прежнем распространении толсторогов {Ovis nivicola Eschsch.) И Там же. 1947. Т. 57, № 5. С. 493-495.
Громова В.И. Остатки млекопитающих из раннеславянских городищ вблизи г. Воронежа И Материалы и исследования по археологии СССР. 1948а. № 8. С. 113-123.
Громова В.И. К истории фауны млекопитающих Кавказа И Изв. АН СССР. Сер. биол. 19486. №5. С. 517-537.
Громова В. Остеологические отличия родов Capra (козлы) и Ovis (бараны) // Тр. Комис, по изуч. четвертич. периода. 1953. Т. 10, вып. 1. 124 с.
Груздев А., Давыдов А., Овсяников Н. и др. Копытные острова Врангеля: сохранять или регулировать? // Охота и охотничье хоз-во. 2001. № 4. С. 6-9.
Груздев А.Р, Сипко Т.П. Зависимость демографических параметров популяции овцебыка (Ovibos moschatus L.) острова Врангеля от погодных условий // Вопросы современного охотоведения. М., 2002. С. 406-Л13.
Груздев А.Р, Сипко Т.П, Гребенков А.Б. Сравнительно-морфологический анализ черепов современных и ископаемых овцебыков (Ovibos moschatus) // VI съезд териол. о-ва. М., 1999. С. 66.
Груздев А.Р, Сипко Т.П., Журба Е.А. Состояние популяции овцебыков на острове Врангеля // Териофауна России и сопредельных территорий. VII съезд териол. о-ва. М., 2003а. С. 101.
Груздев А.Р, Сипко Т.П., Тихонов В.Г., Егоров С. С. Успехи расселения и доместифика-ции овцебыков на Севере России // Охрана и рациональное использование животных и растительных ресурсов России. Иркутск, 20036. С. 393-398.
Груздев В., Сухбат X. Горные копытные Монголии И Охота и охотничье хоз-во. 1982. № 9. С. 42-44.
Грум-Гржимайло ГЕ. Сайга и охота на нее // Ежегодник Всерос. союза охотников. 1922. С. 81-87.
Грунин К.Н. Овод (Oestrus caucasicus sp. nov.), паразитирующий на дагестанском туре (Capra cylindricomis Blyth.) И Докл. АН СССР. Новая сер. 1948. Т. 61, № 6. С. 1125-1127.
ГрунинК.Н., ГребенюкР.В., Сартбаев С.К. Носоглоточный овод (Oestrus caucasicus Grunin) сибирского горного козла// Тр. Зоол. ин-та АН СССР. 1955. Т. 21. С. 365-367.
ГубановН.М. Гельминтофауна промысловых млекопитающих Якутии. М.: Наука, 1964.164 с.
Гусаров И.В., Шумов А.В., Прозоров А.А. Миграции регулируемой популяции зубра в условиях Вологодской области И Актуальные проблемы производства и переработки сельскохозяйственной продукции. Вологда-Молочное, 1998а. С. 40-41.
Гусаров И.В., Шумов А.В., Прозоров А.А. Акклиматизация зубров в Вологодской области Там же. 19986. С. 42-44.
Гущин Н.Н., Завацкий Б.П. Зимние местообитания копытных в Саяно-Шушенском заповеднике // Копытные фауны СССР. М.: Наука, 1980. С. 141-143.
Давид А.И. Териофауна плейстоцена Молдавии. Кишинев: Штиинца, 1980. 186 с.
Давид А.И. Формирование териофауны Молдавии в антропогене. Кишинев: Штиинца, 1982. 151 с.
Давид А.И. Териофауна позднего плейстоцена- раннего голоцена Молдавии ИIV съезд Всесоюз. териол. о-ва. М., 1986. Т. 1. С. 12.
Давид А.И. Остатки млекопитающих из раскопок позднепалеолитической стоянки Ко-соуцы в Молдавии//V съезд Всесоюз. териол. о-ва АН СССР. М., 1990. Т. 1. С. 11.
Давид А.И., Паскарь В.Н. Териофауна многослойной позднепалеолитической стоянки Ко-суэць 1 (Республика Молдова) // Чтения памяти А.А. Браунера. Одесса, 2000. С. 94-98.
Давид А.И., Татаринов К. А., Свистун В.И. Хищные, хоботные и копытные раннего плейстоцена юго-запада СССР. Кишинев: Штиинца, 1990. 131 с.
Давид А.И., Чемыртан ГД. История развития териофауны Молдавии в голоцене И История биогеоценозов СССР в голоцене. М.: Наука, 1976. С. 207-212.
Давлетова Л.В. Использование диких родичей домашних животных для улучшения имеющихся и создания новых пород в целях зоокультуры ИI Всесоюз. совещ. по пробл. зоокультуры. М., 1986. Ч. 1. С. 105-108.
Дагва Д.Э. Аргали на Бага-Богдо Ула (Юго-Западная Монголия, Гобийский Алтай) // Изв. Иркутского с.-х. ин-та. 1958. Вып. 8. С. 90-93.
Далматов Д.Я. История зубра или тура, водящегося в Беловежской пуще Гродненской губернии И Лесн. журн. 1849. № 24. С. 188-191; № 28. С. 220-222.
Даль С.К. Позвоночные животные Сарайбулагского хребта // Зоол. сб. Зоол. ин-та АН АрмССР. 1944. Вып. 3. С. 5^16.
Даль С.К. Позвоночные животные Памбакского хребта // Там же. 1948. Вып. 5. С. 5-68.
Даль С.К. Данные по биологии, распространению, численности и количественному соотношению в стадах безоаровых коз на Урцском хребте И Изв. АН АрмССР. Биол. и с.-х. науки. 1951. Т. 4, № 1. С. 33-40.
Даль С.К. Животный мир Армянской ССР. Ереван: Изд-во АН АрмССР, 1954. Т. 1. Позвоночные животные. 415 с.
Даль С.К., Гусев В.М., Бедный СВ. Влияние зимы 1953/1954 г. на состояние, численность и распределение сайги в Западном Прикаспии И Тр. науч.-иссл. противочумного ин-та Кавказа и Закавказья. 1956. Вып. 1. С. 430-440.
Даль С.К., Гусев В.М., Бедный С.Н. Об экологии и размножении сайги (Saiga tatarica L.) // Зоол. журн. 1958. Т. 37, вып. 3. С. 447—456.
Данилкин А.А. Эмбриональная диапауза у косуль (Capreolus, Cervidae) и этологическая гипотеза ее возникновения И Зоол. журн. 1990. Т. 69, вып. 3. С. 120-124.
Данилкин АЛ. Млекопитающие России и сопредельных регионов. Оленьи. М.: ГЕОС, 1999. 552 с.
Данилкин АЛ. Млекопитающие России и сопредельных регионов. Свиные. М.: ГЕОС, 2002. 309 с.
Данилкин АА. Сайга: стратегия сохранения популяций (альтернативная точка зрения) И Охота и охотничье хоз-во. 2003. № 8. С. 4—7. № 9. С. 14-16.
Данченко В.В. Нахождение амурского горала в Уссурийском заповеднике И Редкие и исчезающие животные суши Дальнего Востока СССР. Владивосток, 1981. С. 175-176.
Даш Я. Проблема восстановления промысловых запасов тарбагана и дзерена в Монгольской Народной Республике // Тр. IX Междунар. контр, биологов-охотоведов. М., 1970. С. 255-257.
Даш Я., Соколов М.Н. Сокращение запасов дзерена в МНР, его возможные причины и задачи по сохранению И Природные условия и биологические ресурсы Монгольской Народной Республики. М.: Наука, 1986. С. 141-143.
Девятова Э.И. Природная среда позднего плейстоцена и ее влияние на расселение человека в северодвинском бассейне. Петрозаводск, 1982. 156 с.
Дежкин В.В. Охота и охрана природы. М.: Физ-ра и спорт, 1977. 104 с.
Дежкин В. Дикие копытные в Европе (численность и добыча) И Охота и охотничье хоз-во. 1980. № 12. С. 26-27.
Дежкин В.В. Охота и охотничье хозяйство мира. М.: Лесн. пром-сть, 1983. 358 с.
Дежкин В.В. Управление популяциями диких копытных //Итоги науки и техники. Т. 13. Зоология позвоночных. М.: ВИНИТИ, 1985. С. 66-137.
Дежкин В.В. Эколого-экономические преимущества разведения диких копытных ИI Всесоюз. совещ. по пробл. зоокультуры. М., 1986. Ч. 1. С. 32-34.
Дежкин В.В. Дикие копытные в Европе (численность и добыча) И Управление популяциями диких копытных животных. Киров, 1989. С. 35—47.
Дежкин В.В., Менькова Н.В. Сравнительные аспекты использования ресурсов диких и сельскохозяйственных копытных животных И Экономика, организация и использование ресурсов охотничьего хозяйства РСФСР. М., 1981. С. 7—40.
ДементьевГ.П. Дзерен, джейран, сайга. М.-Л.: КОИЗ, 1935. 44 с.
Демеуов Д. История ареала и численность сайгаков в Казахстане И Биология и география. Алма-Ата, 1968. Вып. 4. С. 97-104.
Демеуов Д. Сайгаки Казахстана и их хозяйственное использование: Автореф. дис.... канд. с.-х. наук. Алма-Ата, 1971. 20 с.
ДемеуовД. Отношение к сайгакам до Октябрьской революции // Биология и география. Алма-Ата, 1972. Вып. 7. С. 104-110.
Демченко Л.А. Растительность острова Барса-Кельмес как кормовая база копытных // Тр. гос. заповедника Барса-Кельмес. 1950. Вып. 1. С. 6-37.
Демчук В.В., Омелькович Я.А., Сологор Е.А. О динамике численности зубра в Волынской области (Украина) // Редкие виды млекопитающих России и сопредельных территорий. М., 1997. С. 27.
Демьянова О.М., Любаев В.Л. Проблемы и перспективы охраны сайгака в Калмыкии // IV съезд Всесоюз. териол. о-ва. М., 1986. Т. 3. С. 145-146.
Денисов В. Ф. Некоторые данные по изменчивости тау-тэкэ {Capra sibirica М.) и мясной продуктивности архаров (Ovispolii karelini Sev.) //Видовая гибридизация домашних животных с дикими видами. М.-Л., 1935. С. 81-113. (Тр. Киргизск. компл. экспед. 1933-1934 гг. Т. 4, вып. 3).
Деннлер де ла Тур Г. Палеоисторический путь расселения серн (Rupicapra) и образование рас. Палеогенетический этюд И Журн. общ. биол. 1975. Т. 36, № 3. С. 403—413.
Дерягина М.А. Внутристадные взаимоотношения у зубров, бизонов и их гибридов И Зоол. журн. 1972. Т. 51, вып. 3. С. 429^-34.
Джанашвили А.Г. Гибридизация дагестанского тура (Capra cylindricornis Blyth.) и домашней козы (С. hyrcus L.) // Сообщ. АН ГрузССР. 1943. Т. 4, № 8. С. 819-828.
Джафаров Р.Д. Бинагадинские раскопки // Природа. 1960. № 2. С. 99-100.
Джиган Ж., Дикян Л. Антилопы Китая И Изв. РАН. Сер. биол. 1998. № 4. С. 458—461.
Дзержинский В А., Федосенко В.М., Байдавлетов Р.Ж. К изучению паразитофауны архара Ovis аттоп на территории Республики Казахстан И Териофауна России и сопредельных территорий. VII съезд териол. о-ва. М., 2003. С. 111-112.
Дзуев Р.И. Закономерности хромосомной изменчивости млекопитающих в горах Кавказа: Автореф. дис. ... докт. биол. наук. Екатеринбург, 1995. 52 с.
Дзуев Р.И. Хромосомные наборы млекопитающих Кавказа. Нальчик: “Эльбрус”, 1998.249 с.
Дзуев Р.И. Зубр Bison bonasus Linnaeus, 1758 // Красная книга Кабардино-Балкарской республики. Нальчик: Изд. центр “Эль-Фа”, 2000а. С. 44-46.
Дзуев Р.И. Серна Rupicapra rupicapra caucasica Lydekker, 1910/7Тамже. 20006. С. 49-50.
Динесман Л.Г. Влияние диких млекопитающих на формирование древостоев. М.: Изд-во АН СССР, 1961. 166 с.
Динесман Л.Г. История сайгака в послеледниковое время И Сайгак. М., 1998. С. 54-57.
Динесман Л.Г. Голоценовая история биогеоценотического покрова степей Русской равнины // Журн. общ. биол. 1982а. Т. 43, № 3. С. 346-359.
Динесман Л.Г. Изменение численности копытных в степях европейской части СССР в голоцене // Бюл. МОИП. Отд. биол. 19826. Т. 87, вып. 2. С. 3-14.
Динесман Л.Г, Киселева Н.К., Савинецкий А.Б., Хасанов Б.Ф. Вековая динамика прибрежных экосистем Северо-Востока Чукотки. М.: Изд-во “Аргус”, 1996. 186 с.
Динесман Л.Г, Князев А.В., Болд Г. К истории охоты и скотоводства в Монголии И Бюл. МОИП. Отд. биол. 1986. Т. 91, вып. 1. С. 31-37.
Динник Н. Кавказская серна и ее образ жизни И Природа и охота. 1896. Февр. С. 36-59.
Динник Н.Я. Кавказские каменные козлы или туры И Матер, к позн. фауны и флоры Российской империи. Отд. зоол. 1909. Вып. 9. С. 1^17.
Динник Н.Я. Звери Кавказа. Ч. 1. Китообразные и копытные. Тифлис, 1910. 248 с. (Зап. Кавказского отд. Импер. Рус. геогр. о-ва. Кн. 27, вып. 1).
Дмитриев В.В. Копытные звери Алтайского заповедника и прилежащих мест (Восточного Алтая и Западных Саян) // Тр. Алтайского гос. заповедника. 1938. Вып. 1. С. 171-262.
Дмитриева Е.Л. Эволюция прокапр // Млекопитающие СССР. III съезд Всесоюз. териол. о-ва. М., 1982. Т. 1. С. 30-31.
Дмитриева Е.Л. Обзор ископаемых газелей {Gazella) СССР //1 Междунар. конгр. по млекопитающим. М.: ВИНИТИ, 1974. С. 189.
Дмитриева Е.Л. К вопросу о ранней эволюции полорогих // Палеонтол. журн. 2002. № 2. С. 86-88.
Дмитриева Е.Л. К истории древних каприн // Териофауна России и сопредельных территорий. VII съезд териол. о-ва. М., 2003. С. 116.
Долгоброд Н.А. Создание нового типа мясного скота с использованием бизонов И Науч,-техн. бюл. НИИ животноводства лесостепи и полесья УССР. 1985. Т. 41. С. 24-27.
Дормидонтов Р.В., Блохин Ю.Ю. Серна // Копытные звери. М.: Лесн. пром-сть, 1977а. С. 140-153.
Дормидонтов Р.В., Блохин Ю.Ю. Безоаровый козел // Там же. 19776. С. 153-164. Дремов И. В защиту сайгаков // Охота и охотничье хоз-во. 1963. № 9. С. 15-17. Дробовцев В. Сайгаки в лесостепи И Охота и охотничье хоз-во. 1997. № 6. С. 17. Дронсейко Э.Г., Чижов М.М., Зверев П.А. Использование закономерностей поведения снежных баранов при их отлове методом иммобилизации И Прикладная этология. М.: Наука, 1983. С. 43-45.
Дубень А.В. Серна в Краснодарском крае // Редкие виды млекопитающих и их охрана. М.: Наука, 1977. С. 204-205.
Дубень А.В. Актуальные вопросы охраны аборигенной фауны Западного Кавказа И Популяционная изменчивость вида и проблемы охраны генофонда млекопитающих. М., 1983. С. 224-225.
Дубень В.В. Численность и структура популяции серн во взаимосвязи с некоторыми экологическими факторами И Экологические исследования в Кавказском биосферном заповеднике. Изд-во Ростовского ун-та, 1985. С. 31—49.
Дубинин Е.А. Современное состояние и перспективы мониторинга популяций охотничьих животных в Магаданской области И Вопросы современного охотоведения. М., 2002. С. 65-69.
Дуламцэрэн С., Амгалан Л. Современное состояние распространения, размещения и численности монгольского сайгака (Saiga tatarica mongolica Bann.) И Природные условия и биологические ресурсы Большого Гобийского заповедника. Улан-Батор, 1995. С. 46-51.
Дуламцэрэн Ц., Бадарч П., Хухоров В.М. Роль диких животных в эпизоотологии ящура // Матер, симпоз. “Профилактика и эффективная борьба с ящуром, а также создание высокоэффективных противоящурных вакцин”. Владимир, 1981. С. 77-80.
Дулицкий А.И. Редкие звери фауны Крыма // Редкие виды млекопитающих фауны СССР и их охрана. М.: Наука, 1973. С. 13-14.
Дулицкий А.И. О трофейных качествах рогов европейского муфлона (Ovis musimon Pall.) в Крыму // Вести, зоол. 1976. № 6. С. 28-32.
Дулицкий А.И. Некоторые аспекты структуры популяции европейского муфлона в Крыму // Там же. 1992. № 3. С. 50-56.
Дулицкий А.И., Кормилицын А.А. Муфлон европейский (Ovis musimon Pallas, 1811) в Крыму // Там же. 1970. № 4. С. 25-29.
Дунин В. Ф„ Ставровский Д.Д. Экология вольноживущего борисовского стада зубров И V съезд Всесоюз. териол. о-ва АН СССР. М., 1990. Т. 3. С. 147-148.
Дунишенко Ю.М. Находки горала в Хабаровском крае И Редкие виды млекопитающих СССР и их охрана. М., 1983. С. 170-171.
Дунишенко Ю.М. Находки горала в Хабаровском крае // Изучение и охрана редких видов и исчезающих видов животных фауны СССР. М.: Наука, 1985. С. 65-66.
Дуров В. Волки и копытные в Кавказском заповеднике // Охота и охотничье хоз-во. 19746. № 7. С. 12-13.
Дуров В.В. Размещение и численность серны на Северо-Западном Кавказе И Редкие виды млекопитающих и их охрана. М.: Наука, 1977. С. 205-206.
Дуров В.В. Численность и экологические основы сохранения популяций копытных Западного Кавказа// II съезд Всесоюз. териол. о-ва. М.: Наука, 1978. С. 272-273.
Дыманъ Т.Н., Городная А.В., Тарасюк С.И. и др. Участие маркеров структурных генов и анонимных последовательностей ДНК в генетической дифференциации у видов Ovis aries L. и Ovis nivicola borealis // Цитол. и генетика. 2000. Т. 34, № 6. С. 49-58.
Дыманъ Т.Н., Ясинецкая Н.И., Сипко Т.П., Глазго В.М. Полиморфизм биохимических маркеров видов Bovinae И Молекулярно-генетические маркеры животных. Киев: Норд-принт, 1999. С. 13-14.
Дыманъ Т.Н., Ясинецкая Н.И., Глазго В.М. Полиморфизм белков, RAPD-PCR- и ISSR-PCR-маркеров у некоторых видов Ungulata И С.-х. биология. Сер. биол. животных. ' 2000. № 6. С. 29-38.
Дыренков С.А., Голгофская К.Ю, Немцев А. С. Влияние зубров Кавказского государственного биосферного заповедника на лесную растительность: биогенные сукцессии И Общие проблемы биогеоценологии. М.: Наука, 1990. С. 231-252.
Европейский сайгак: развитие, морфология, экология. М.: Наука, 1997. 191 с.
Егоров О.В. Материалы по размножению и росту сибирского горного козла И Бюл. МОИП. Отд. биол. 1953. Т. 58, вып. 3. С. 3-13.
Егоров О.В. Экология сибирского горного козла (Capra sibirica Meyer) И Тр. Зоол. ин-та АН CCCR 1955. Т. 17. С. 7-134.
Егоров О.В. Перспективы акклиматизации копытных в Якутии И Проблемы охраны природы Якутии. Якутск, 1963. С. 99-106.
Егоров О.В. Дикие копытные Якутии. М.: Наука, 1965. 259 с.
Енукидзе Г.П. К фауне млекопитающих Лагодехского государственного заповедника // Тр. Ин-та зоол. АН ГрузССР. 1953. Т. 12. С. 169-183.
Ермолова Н.М. Териофауна долины Ангары в позднем антропогене. Новосибирск: Наука, 1978. 222 с.
Ертаев Е.Е., Турарбеков М.З. Применение метода молекулярной гибридизации ДНК для определения степени генетического сходства некоторых пород овец // Генетические аспекты породообразования. Алма-Ата: Наука КазССР, 1987. С. 122-128. (Тр. Ин-та экс-пер. Биол. АН КазССР. Т. 20).
Ешелкин И.И. Изменение ареалов копытных животных в Юго-Восточном Алтае И Природа и природные ресурсы Горного Алтая. Горно-Алтайск, 1972. С. 76-77.
Ешелкин И.И. Распространение дзерена в Горном Алтае И Редкие виды млекопитающих фауны СССР и их охрана. М.: Наука, 1973. С. 118-119.
Жаворонков Н.М., Зуев ЕЯ. Охрана охотничьих животных в СССР. М.: Лесн. пром-сть, 1976. 152 с.
Жарков ИВ. Результаты учета животных в Кавказском заповеднике в 1939году //Науч, -метод, записки Главн. упр. по заповедникам, зоопаркам и зоосадам. 1940. Вып. 7. С. 138-150.
Жарков ИВ. О взаимоотношениях серн с домашними животными на высокогорных пастбищах Северо-Западного Кавказа И Тр. Кавказского гос. заповедника. 1959. Вып. 5. С. 3-38.
Железнов Н.К. Гон снежных баранов в Корякском нагорье // Копытные фауны СССР. М.: Наука, 1980. С. 289-191.
Железнов Н.К. Корма снежного барана (Ovis nivicola Eschsch.) и их сезонная изменчивость на Чукотке // Экология млекопитающих Северо-Восточной Сибири. М.: Наука, 1981а. С. 115-137.
Железнов Н.К. Проблемы охраны диких животных Чукотки // Проблемы охраны природы. Изд-во Моск, ун-та, 19816. Ч. 1. С. 69—72.
Железнов Н.К. Материалы по экологии волка (Canis lupus L.) на Северо-Востоке СССР // Териологические исследования в Якутии. Якутск, 1983а. С. 28-50.
Железнов Н.К. Структура популяций снежных баранов в некоторых очагах их обитания на Крайнем Северо-Востоке Сибири // Редкие виды млекопитающих СССР и их охрана. М., 19836. С. 174—175.
Железнов Н.К. Дикие копытные Северо-Востока СССР. Владивосток: ДВО АН СССР, 1990. 480 с.
Железнов- Чукотский Н.К. Экология снежных баранов Северной Азии. М.: Наука, 1994. 256 с.
Железнов-Чукотский Н.К., Назаров А.В., Овсяников Н.Г, Успенский С.М. Овцебык на Чукотке и программа его реинтродукции. Анадырь - М., 1996. 62 с. (Препринт).
Жемчужников Н. Сайга и ее значение в охотхозяйстве // Охотник. 1926. № 12. С. 10-11.
Жесткова И. А. Об экологии и поведении зубра в Хоперском заповеднике И Бюл. МОИП. Отд. биол. 1988. Т. 93, вып. 4. С. 30-38.
Жесткова Н.А. Особенности биологии горных баранов Ovis cycloceros Бадхыза И Редкие виды млекопитающих России и сопредельных территорий. М., 1997. С. 34.
Жесткова И.А. Особенности биологии горного барана (Ovis vignei) Бадхыза И Редкие виды млекопитающих России и сопредельных территорий. М., 1999. С. 128-138.
ЖивалевИ.К, КленовицкийП.М., ШайдулинИ.Н. Цитогенетическая характеристика Ovis aries, Ovis nivicola nivicola и их гибридизация Fl // Проблемы сохранения редких пород домашних животных и близкородственных диких видов. Пущино, 1996. С. 19-20.
Жирнов Л.В. Количественный учет сайгаков в Западном Прикаспии И Вопросы организации и методы учета ресурсов фауны наземных позвоночных. М., 1961. С. 29-31.
Жирнов Л.В. Опыт применения массового мечения при изучении размещения и кочевок сайгаков в Западном Прикаспии (предварительное сообщение) // Миграции животных. М.: Изд-во АН СССР, 1962. Вып. 3. С. 40-61.
Жирнов Л.В. Учеты сайгаков с самолета и ресурсы этой дичи в Западном Прикаспии И Ресурсы фауны промысловых зверей в СССР и их учет. М.: Изд-во Акад, наук СССР, 1963. С. 92-98.
ЖирновЛ.В. Дзерен //Копытные звери. М.: Лесн. пром-сть, 1977а. С. 62-78.
Жирнов Л.В. Сайгак // Там же. 19776. С. 79-118.
Жирнов Л.В. Экологические основы охраны копытных аридной зоны Евразии//Вопросы рационального использования и охраны диких копытных и других животных. М., 1978. С. 36-55. (Сб. науч. тр. Моск. вег. акад. им. К.И. Скрябина. Т. 97).
Жирнов Л.В. Возвращенные к жизни (экология, охрана и использование сайгаков). М.: Лесн. пром-сть, 1982. 224 с.
Жирнов Л.В. Среда обитания И Сайгак. М., 1998а. С. 71-79.
Жирнов Л.В. Особенности размещения сайгаков по природным зонам и типы кочевок И Там же. 19986. С. 115-121.
Жирнов Л.В. Этологическая структура и социальная организация И Там же. 1998в. С. 144-155.
Жирнов Л.В. Антропические факторы и их роль в регуляции численности популяций И Там же. 1998г. С. 252-263.
Жирнов Л.В., Бекенов А.Б., Грачев Ю.А. Ареал и его изменение в XVI-XIX вв. Численность сайгаков в прошлые века (XVI-XIX). Абиотические, или погодно-климатические факторы // Там же. 1998а. С. 57-60, 216-219, 233-241.
Жирнов Л.В., Бекенов А.Б., Грачев Ю.А. и др. Ареал и его изменение в XX в. // Там же. 19986. С. 60-67.
Жирнов Л.В., Бекенов А.Б., Грачев Ю.А., Проняев А.В. Размножение // Там же. 1998в. С. 156-179.
Жирнов Л.В., Винокуров А.А. Дзерен // Природа. 1975. № 8. С. 45—49.
Жирнов Л.В., Винокуров А.А., Бычков В.А. Редкие млекопитающие, птицы и их охрана в СССР. М., 1975. 82 с.
Жирнов Л.В., Дуламцэрэн С., Лущекина А.А. Состояние численности популяций сайгака в Китае и Монголии // Сайгак. М., 1998. С. 227-229.
Жирнов Л.В., Ильинский В. О. Большой Гобийский заповедник - убежище редких животных пустынь Центральной Азии. М.: ЦМП ГКНТ СССР, 1985. 130 с.
ЖирновЛ.,КалецкийА. В защиту сайгаков// Охота и охотничье хоз-во. 1976. № 2. С. 1-2.
Жирнов Л. В., Лебедев-Нечаев В.П., Фандеев А.А. Состояние популяции сайгаков и перспективы промысла в Западном Прикаспии И Охотничье-промысловые звери. М.: Россельхозиздат, 1965. Вып. 1. С. 40-56.
Жирнов Л.В., Лопырев Б.Н., Хахин ГВ. Проблемы освоения и охраны ресурсов сайгаков на современном этапе И Охотничье хозяйство и заповедное дело. М.: Россельхозиздат, 1977. С. 46-59.
Жирнов Л.В., Максимук А.В. Стратегия охраны сайгаков! // Охота и охотничье хоз-во. 1994. №5. С. 15-17.
ЖирновЛ.В., МаксимукА.В. Северо-Западный Прикаспий//Сайгак. М., 1998а. С. 80-105, 219-225.
Жирнов Л.В., Максимук А.В. Использование рогов сайгака в китайской медицине. Система управления и рационального использования ресурсов сайгаков И Там же. 19986. С. 267-271, 274-280.
Жирнов Л.В., Максимук А.В., Бекенов А.Б., Грачев Ю.А. Размеры заготовок и продукция сайгачьего промысла// Там же. 1998. С. 264—267.
ЖирновЛ.В., Проняев А.В., Бекенов А.Б. и др. Структура популяций (половой и возрастной состав) И Там же. 1998. С. 180-192.
Жирнов Л.В., Хахин Г.В., Проняев А.В. О поведении сайгаков на переправах через водные преграды и водопоях И Групповое поведение животных. М.: Наука. 1976. С. 116-118.
Жирнов Л.В., Холодова М.В., Бекенов А.Б., Грачев Ю.С. Корма, трофические связи и пищевое поведение И Сайгак. М., 1998. С. 122-143.
Жиряков В.А. К охране редких видов млекопитающих Алма-Атинского заповедника И Редкие виды млекопитающих фауны СССР и их охрана. М.: Наука, 1973. С. 15-17.
Жиряков В.А. Допустимые плотности населения диких копытных в Алма-Атинском заповеднике // Копытные фауны СССР. М.: Наука, 1975. С. 241-242.
Жиряков В.А. Редкие копытные Алма-Атинского заповедника и их охрана И Редкие млекопитающие фауны СССР. М.: Наука, 1976. С. 141-155.
Жиряков В.А. Центральноазиатский козел в Алма-Атинском заповеднике // Редкие виды млекопитающих и их охрана. М.: Наука, 1977. С. 209-210.
Жиряков В.А. Влияние крупных хищников на популяции диких млекопитающих в Алма-Атинском заповеднике И Экологические основы охраны и рационального использования хищных млекопитающих. М.: Наука, 1979. С. 37-39.
Жиряков В.А. Минеральное питание сибирского горного козла в Алма-Атинском заповеднике И Зоопарки и их роль в сохранении диких животных. Алма-Ата, 1989. С. 167-194.
Жиряков В.А. Роль волка в биоценозах Алма-Атинского заповедника (Северный Тянь-Шань) И V съезд Всесоюз. териол. о-ва АН СССР. М., 1990. Т. 2. С. 278-279.
Жиряков В.А., Федосенко А.К. Строение, рост и изменчивость рогов архара из западных отрогов Джунгарского Алатау // Редкие виды млекопитающих СССР и их охрана. М., 1983. С. 175-176.
Житенко П. Мясо сайгака // Охота и охотничье хоз-во. 1974. № 5. С. 20-21.
Житенко П.В. Рациональное использование продуктов сайгачьего промысла ИIV съезд Всесоюз. териол. о-ва. М., 1986. Т. 3. С. 147-148.
Житков Б.М. Акклиматизация животных. М.-Л.: Биомедгиз, 1934. 112 с.
Заблоцкая Л.В. Питание и естественные корма зубров // Тр. Приокско-Террасного гос. заповедника. 1957. Вып. 1. С. 66-143.
Заблоцкая М.М. О положении с зубром в Советском Союзе И N съезд Всесоюз. териол. о-ва АН СССР. М., 1990. Т. 3. С. 148-149.
Заблоцкая М.М., Заблоцкий М.А. Современное размещение и численность чистокровных зубров в СССР И Проблемы охраны генофонда и управления экосистемами в заповедниках лесной зоны. М., 1986. Ч. 2. С. 81-84.
Заблоцкий М.А. Тридцать пять лет разведения зубров и их гибридов в зоопарке Аскания-Нова (1902-1937 гг.) // Кавказский зубр. М., 1939 (1940). С. 73-137.
Заблоцкий М.А. Борьба за сохранение зубра в России // Охрана природы. М., 1948. Сб. 3. С. 22-48.
Заблоцкий М.А. Государственная племенная книга зубров и бизонов (чистокровных, чистопородных и гибридных). М.: Изд-во МСХ СССР, 1956. Т. 1. 116 с.
Заблоцкий М.А. Некоторые биологические особенности зубра и их изменение в условиях загонного содержания // Тр. Приокско-Террасного гос. заповедника. 1957а. Вып. 1. С. 5-65.
Заблоцкий М.А. Система мероприятий, направленных к быстрейшему восстановлению зубра в СССР И Вопр. экологии. Киев, 19576. Т. 2. С. 35—45.
Заблоцкий М.А. Загонное содержание, кормление и транспортировка зубров (инструкция). М., 1957в. 114 с.
Заблоцкий М.А. Восстановление зубров в СССР и за границей И Бюл. Комиссии по охране природы АН СССР. 1960. № 4. С. 52-70.
Заблоцкий М.А. Работы по восстановлению зубра в СССР И Акклиматизация животных в СССР. Алма-Ата: Изд-во АН КазССР, 1963. С. 96-100.
Заблоцкий М.А. Отличительные особенности беловежского и кавказского типов зубра и их помесей между собой И Матер. I и II польско-советской конф, по разведению зубров в Беловежской пуще. Варшава, 1965. С. 37-44.
Заблоцкий М.А. К методике увеличения численности редких видов на примере европейского зубра// Пути повышения эффективности охотничьего хозяйства. Иркутск, 1971. Ч. 1.С. 85-90.
Заблоцкий М.А. Опыт сохранения в СССР редких видов на примере Bison bonasus L. ИI Междунар. контр, по млекопитающим. М.: ВИНИТИ, 1974а. Т. 1. С. 218-219.
Заблоцкий М.А. Размещение и численность в СССР Bison bonasus L. (Artiodactyla) И Там же. 19746. С. 219-220.
Заблоцкий М.А. Принципы, методы и формы охраны зубра // Копытные фауны СССР. М.: Наука, 1975а. С. 242-243.
Заблоцкий МА. Экология поведения зубра и ее значение для сохранения структуры популяций // Там же. 19756. С. 318-320.
Заблоцкий М.А. Материнское поведение у зубров // Групповое поведение животных. М.: Наука, 1976. С. 118-122.
Заблоцкий МА. Реакклиматизация и акклиматизация чистокровных зубров в СССР // Акклиматизация охотничьих животных в СССР. Минск: Ураджай, 1978. С. 48-50.
Заблоцкий М.А. Чистокровные зубры СССР // Биологические аспекты охраны редких животных. М., 1981. С. 9-14.
Заблоцкий МЛ. Сохранение генофонда зубра // Популяционная изменчивость вида и проблем охраны генофонда млекопитающих. М., 1983. С. 62-63.
Заблоцкий МЛ., Заблоцкая М.М. О неправомерности применения наименования Bison bonasus caucasicus Sat. и “чистопородный зубр” к зубробизонам Кавказского государственного заповедника // IV съезд Всесоюз. териол. о-ва АН СССР. М., 1986а. Т. 2. С. 177-178.
Заблоцкий М.А., Заблоцкая М.М. Перспективы дальнейшего развития работ по проблеме восстановления зубра в СССР И I Всесоюз. совещ. по пробл. зоокультуры. М., 19866. 4.2. С. 29-31.
Заблоцкий М.А., Заблоцкая М.М. Создание регулируемых популяций как метод сохранения генофонда и хозяйственного использования восстанавливаемого вида И Там же. 1986в. С. 31-33.
Заблоцкий М.А., Заблоцкая М.М. Реакклиматизация зубра на Кавказе и ее насущные проблемы И Экология и охрана горных видов млекопитающих. М., 1987. С. 64-66.
Заблоцкий МЛ., Заблоцкая М.М. Центральный зубровый питомник Приокско-Террасного заповедника как центр работы по восстановлению зубра в СССР: итоги работы за 40 лет И V съезд Всесоюз. териол. о-ва АН СССР. М., 1990. Т. 3. С. 149-150.
Заблоцкий МЛ., Заблоцкая М.М., Заблоцкая Л.В. Перспективы и стратегия восстановления зубра в России в современных условиях (1999 год) И VI съезд териол. о-ва. М., 1999. С. 95.
Заблоцкий М.А., Криницкий В.В. Зубры в СССР И Сб. науч. тр. Моск. вет. акад. 1974. Вып. 72. С. 112-114.
Забродин ВЛ. О создании природной популяции овцебыков на Таймыре // Овцебык в тундре России: Эксперимент XX века по восстановлению исчезнувшего вида. СПб.: Астерион, 2002. С. 30—43.
Завацкий Б.П. Избирательность зимних охот волков в Западном Саяне // IV съезд Всесоюз. териол. о-ва. М., 1986а. Т. 1. С. 212-213.
Завацкий Б.П. Роль волка в биоценозах Саяно-Шушенского заповедника И Роль крупных хищников и копытных в биоценозах заповедников. М., 19866. С. 35-54.
Завацкий Б.П. Сибирский горный козел Западного Саяна // Экология, морфология, использование и охрана диких копытных. М., 1989. Ч. 2. С. 239-240.
Завацкий Б.П. Экология сибирского горного козла Западного Саяна // Экологические и экономические аспекты охраны и рационального использования охотничьих животных и растительных пищевых ресурсов Сибири. Шушенское, 1990. С. 32-34.
Завацкий Б.П., Гущин Н.Н. Взаимоотношения крупных хищников и копытных Западного Саяна зимой И Копытные фауны СССР. М.: Наука, 1980. С. 158-159.
Заикин С.А., Жирнов JI.B. Прогнозирование динамики калмыцкой популяции сайгаков на основе математической модели // Экология. 1989. № 3. С. 45-52.
Зарудный Н. Заметки по фауне млекопитающих Оренбургского края // Материалы к познанию флоры и фауны Российской империи. Отд. зоол. 1897. Вып. 3. С. 329-372.
Закариев А.Я. Дикие парнокопытные Северного Кавказа как носители паразитических червей И Копытные фауны СССР. М.: Наука, 1980. С. 159-160.
Закариев А.Я. К экологии безоарового козла в Дагестане И Млекопитающие СССР. III съезд Всесоюз. териол. о-ва. М., 1982а. Т. 1. С. 202-203.
Залетаев В.С. Джейран, сайгак и заяц толай на восточном побережье Каспия // Природа. 1956. № 2. С. 98-100.
Залиханов М.Ч. Туры в Кабардино-Балкарии. Нальчик: Каб.-Балк. кн. изд-во, 1967. 103 с.
Залиханов М. Ч. Туры Кабардино-Балкарии, особенности биологии и их хозяйственное значение: Автореф. дис. ... канд. биол. наук. Ростов-на-Дону, 1968. 27 с.
Зархидзе В.А. Копытные Западной Туркмении//Копытные фауны СССР. М.: Наука. 1980. С. 88-90.
Зархидзе В.А. Распространение и численность популяций устюртского горного барана (Ovis vignei агсаГ) на юго-западе ареала И Зоол. журн. 1983а. Т. 62, № Н.С. 1756-1759.
Зархидзе В.А. К экологии южных популяций устюртского горного барана и их охране И Редкие виды млекопитающих СССР и их охрана. М., 19836. С. 176-177.
Зархидзе В.А., Горбунов А.В. Состояние численности и проблема сохранения популяций горных баранов в Западной Туркмении // Популяционная изменчивость вида и проблемы охраны генофонда млекопитающих. М., 1983. С. 227-229.
Захарченко В.Н. Хищничество волка на территории Алтайского заповедника И Териофауна России и сопредельных территорий. VII съезд териол. о-ва. М., 2003. С. 136-137.
Звегинцова Н.С., Треус М.Ю. Зараженность сайгаков гельминтами в условиях полу-вольного содержания в Аскании-Нова// IV съезд Всесоюз. териол. о-ва. М., 1986. Т. 3. С. 148-149.
Зверев С. Сайга - антилопа российских степей // Сафари. 2001. № 3. С. 36-37.
Зубков А.Н. К вопросу об акклиматизации овцебыков И Проблемы Арктики. 1937. № 5. С. 55-65.
Зудин А., Форонова И. О биомеханических особенностях, стратиграфическом и палеогеографическом значении четвертичных бизонов Сибири И Цитология. 1995. Т. 37, № 7. С. 613- 614.
Зуев ЕЯ. Охрана диких животных в СССР // Редкие виды млекопитающих и их охрана. М.: Наука, 1977. С. 11-13.
Зуев ЕЯ. Антропогенное воздействие на сайгаков Калмыкии И Копытные фауны СССР. М.: Наука, 1980. С. 241-242.
Зуев ЕЯ. Проблемы охраны сайгаков Калмыкии ИIV съезд Всесоюз. териол. о-ва. М., 1986. Т. 3. С. 150.
Зуев ЕЯ., ХахинГВ. В защиту сайгаков Калмыкии //Охота и охотничье хоз-во. 1978.№ 1. С. 16-17.
Зуев Е„ Хахин Г. Сайгаки: проблемы остаются И Там же. 1982. № 3. С. 12-14.
Зуев ЕЯ., Хахин ГВ. Пути повышения продуктивности поголовья сайгаков в Калмыцкой АССР // Повышение продуктивности охотничьих угодий. М., 19826. С. 49-62.
Иванов И.М. Мускусный бык и проблема его разведения на советском Севере И Землеведение. 1934. Т. 36. Вып. 2. С. 201-205.
Иванов М.Ф. Опыт создания горного мериноса И Избранные сочинения М.Ф. Иванова. М.: Гос. изд-во с.-х. литературы, 1949. Т. 1. С. 310-329.
Иванов М.Ф., Гребень Л.К. Гибриды от муфлона и домашних овец разных пород // Там же. 1949. Т. 1. С. 266-293.
Ивашенко А.А. О численности архара в заповеднике Аксу-Джабаглы и мерах по улучшению его охраны // Редкие виды млекопитающих фауны СССР и их охрана. М.: Наука, 1973. С. 119-120.
Инякова А.П. К вопросу численности и распределения охотничье-промысловых животных в Тебердинском заповеднике // Тр. Тебердинского гос. заповедника. 1957а. Т. 1. С. 249-255.
Инякова А.П. Опыт содержания серн Rupicapra rupicapra caucasica Lyd в неволе И Там же. 19576. С. 257-268.
Инякова А.П. К вопросу размножения западно-кавказского тура Capra caucasica Guld в неволе // Там же. 1957в. С. 269-270.
Ирисов Э.А., Марин Ю. Ф., Шилов В.А. Некоторые данные о редких видах млекопитающих Алтая // Редкие виды млекопитающих фауны СССР и их охрана. М.: Наука, 1973. С. 18-19.
Исаев Е.М. О состоянии поголовья диких копытных животных не территории РСФСР // Сообщ. Ин-та леса. 1959. Вып. 13. С. 54-57.
Исмагилов М.И. Распространение и динамика численности копытных на острове Бар-са-Кельмес И Популяционная структура вида у млекопитающих. М., 1970. С. 75-77.
Ишадов Н. Эколого-географические особенности некоторых копытных Туркмении: Ав-тореф. дис. ... канд. биол. наук. Ашхабад, 1965. 22 с.
ИшадовН. Сайгак (Saiga tatarica L.) в Туркмении // Зоол. журн. 1966. Т. 45, вып. 1.С. 151.
Ишадов Н. Распространение и современное состояние численности копытных животных в Северо-Западной Туркмении И Териология. Новосибирск: Наука, 1972. Вып. 1. С. 417—419.
Ишадов Н. Миграция и зимовка сайгака в Туркмении // Копытные фауны СССР. М.: Наука, 1975а. С. 96-98.
Ишадов Н. Поведение трех видов копытных в различных условиях их обитания И Там же. 19756. С. 321-322.
Ишадов И. Перспектива разведения и охраны редких и исчезающих зверей Туркмении // Редкие виды млекопитающих и их охрана. М.: Наука, 1977. С. 28-30.
ИшадовН. Сайгак, или сайга Saiga tatarica Linnaeus, 1758//Красная книга Туркменистана. Беспозвоночные и позвоночные животные. Ашхабад, 1999а. Т. 1. С. 336.
Ишадов Н. Бородатый, или безоаровый козел // Там же. 19996. С. 338-339.
Ишадов Н. Охотничье поведение барса // Териофауна России и сопредельных территорий. VII съезд териол. о-ва. М., 2003а. С. 145.
Ишадов Н. Крупные млекопитающие как биоиндикаторы изменения окружающей среды в Юго-Западном Туркменистане И Там же. 20036. С. 145-146.
Ишунин Г.И. О жизни горного барана в горах-останцах Центральных Кызылкумов // Зоол. журн. 1960. Т. 39, вып. 10. С. 1581—1583.
Ишунин Г.И. О состоянии популяции кызылкумского барана на Нуратинском хребте // Там же. 1967. Т. 46, вып. 2. С. 306-308.
Ишунин Г.И. Экология горных баранов в останцах Кызылкума и чинках Устюрта // Экология, методы изучения и организация охраны млекопитающих горных областей. Свердловск, 1977. С. 37-38.
Ишунин Г.И. Новые данные по систематике и морфологии среднеазиатских диких баранов // Бюл. МОИП. Отд. биол. 1968. Т. 73, вып. 6. С. 141-145.
Ишунин Г.И. Дикие копытные Узбекистана// Копытные фауны СССР. М.: Наука, 1980. С. 91-93.
Ишунин ГН. Охрана и управление популяциями редких млекопитающих Узбекистана// Млекопитающие СССР. Ш съезд Всесоюз. териол. о-ва. М., 1982а. Т. 1. С. 117-118.
Ишунин Г.И. Задача сохранения в пустынях Узбекистана уникальных форм азиатского муфлона//Там же. 19826. Т. 1. С. 118-119.
Ишунин Г.И. Устюртский баран, или аркал // Красная книга Узбекской ССР. Позвоночные животные. Ташкент: Изд-во “Фан”, 1983а. Т. 1. С. 41-42.
Ишунин Г.И Кызылкумский баран, или баран Северцова // Там же. 19836. С. 42^14.
Ишунин Г.И. Таджикский, или бухарский баран // Там же. 1983в. С. 44—46.
Ишунин Г.И. Тяньшанский баран, или архар И Там же. 1983г. С. 45—47.
Ишунин Г.И., Бекенов А., Савинов Е. Ф. Современное распространение и охрана устюртского барана//Бюл. МОИП. Отд. биол. 1981. Т. 86, вып. 3. С. 17-22.
Каверзнев В.И. Полорогие фауны СССР и их добывание. М.-Л.: КОИЗ, 1933. 86 с.
Казанская Е.Ю. Таксономия рода Capra-. Дипломная работа. М.: МГУ, 2004. 85 с.
Казарян Г.В. Современное состояние популяции безоарового козла Capra aegagrus и проблемы его охраны в Армении // Териофауна России и сопредельных территорий. VII съезд териол. о-ва. М., 2003. С. 146-147.
Казьмин ВД. Половозрастной состав и динамика численности овцебыков на острове Врангеля И Териология, орнитология и охрана природы. XI Всесоюз. симпоз. “Биол. пробл. Севера”. Якутск, 1986. Вып. 3. С. 35.
Казьмин ВД. Зубр // Ресурсы редких животных РСФСР, их охрана и воспроизводство (Материалы к Красной книге). М., 1988. С. 150.
Казьмин ВД. Динамика численности и современное состояние кавказско-беловежских зубров в Северо-Осетинском заповеднике и заказнике “Цейский” И Экологические проблемы Ставропольского края и сопредельных территорий. Ставрополь, 1989. С. 323-325.
Казьмин ВД. Проблемы создания популяции вольно живущих зубров на Среднерусской возвышенности // Териофауна России и сопредельных территорий. VII съезд териол. о-ва. М., 2003а. С. 147.
Казьмин ВД. Проблемы сохранения и восстановления копытных животных в национальном парке “Орловское полесье” // Проблемы сохранения и восстановления диких копытных животных в Центральном регионе России. Орел: Изд-во ОРАГС, 20036. С. 54-59.
Казьмин ВД., Арбузова М.В., ЗембатоваА.А. Травянистая растительность в летнем питании кавказско-беловежских зубров на Пастбищном хребте Центрального Кавказа // Охрана и изучение редких и исчезающих видов животных в заповедниках. М., 1992. С. 21-28.
Казьмин В.Д., Смирнов КА. Зимнее питание, кормовые ресурсы и трофическое воздействие зубра на лесные фитоценозы Центрального Кавказа И Бюл. МОИП. Отд. биол. 1992. Т. 97, вып. 2. С. 26-35.
Калмыков И. В. Сохранение снежного барса и Алтайского горного барана в Алтай-Саян-ском экорегионе И Охрана и рациональное использование животных и растительных ресурсов России. Иркутск, 2003. С. 426-431.
Калмыков Н. Структура мамонтовой фауны и ландшафты позднего плейстоцена Прибайкалья и Забайкалья // Цитология. 1995. Т. 37, № 7. С. 616.
Калмыков Н.П., Пискунов А. С., Хандуева В.Б. Ископаемые млекопитающие Музея природы Бурятии. Улан-Удэ: Изд-во БНЦ СО РАН, 1992. 70 с.
Калугин С.Г. Зубры в естественных условиях Кавказского заповедника И Тр. Кавказского гос. заповедника. 1958. Вып. 4. С. 4-36.
Калугин С.Г. Восстановление зубра на Северо-Западном Кавказе//Там же. 1968. Вып. 10. С. 3-94.
Калугин С.Г. Территориальные группировки зубров на Северо-Западном Кавказе И Популяционная структура вида у млекопитающих. М., 1970. С. 41-42.
Калугин С.Г. Зубры в горах Северо-Западного Кавказа // Редкие виды млекопитающих фауны СССР и их охрана. М.: Наука, 1973. С. 121.
Калугин С.Г. Зубры на Северном Кавказе И Копытные фауны СССР. М.: Наука, 1975. С. 99-100.
Калугин С.Г. Итоги работы по восстановлению зубров на Кавказе И Тр. Кавказского гос. заповедника. 1977. Вып. 11. С. 30-33.
Калякин В.Н. Современный и прошлый ареалы снежного барана// Природа. 2001. № 6.
С. 47-50.
Камъянов Б. Т. Сравнительное изучение морфологических показателей кавказского зубра в условиях Северного Кавказа: Автореф. дис.... канд. биол. наук. Нальчик, 1969. 17 с.
Капанадзе А.Д. Некоторые вопросы биологии серны в условиях Кахетинского хребта // Материалы к фауне Грузии. 1967. Вып. 2. С. 237-251.
Капанадзе А.Д. Новые данные о распространении серны в Грузии // Редкие виды млекопитающих фауны СССР и их охрана. М.: Наука, 1973. С. 122-123.
Капанадзе А.Д. Серна на Малом Кавказе (в пределах Грузии) // Материалы к фауне Грузии. 1974. Вып. 4. С. 458-480.
Капанадзе А.Д. К изучению поведения серны, тура и безоарового козла в высокогорьях Грузии И Копытные фауны СССР. М., 1975. С. 322-323.
Капанадзе А.Д. Территориальное поведение серн (R. rupicapra caucasica) и туров (Capra caucasica cylindricomis) на южных склонах Кавказа И Групповое поведение животных. М.: Наука, 1976. С. 161-162.
Капанадзе А.Д. Структура и численность популяций высокогорных копытных Кавказа И II съезд Всесоюз. териол. о-ва. М.: Наука, 1978. С. 137-138.
Капитанова Д.В. Подвидовая систематика архара (Ovis аттоп L.) на основе краниологических признаков: Дипломная работа. М.: МГУ, 2004. 93 с.
Капитонов В.И. Влияние летней засухи 1974 и 1975 гг. на копытных Казахского нагорья // Копытные фауны СССР. М.: Наука, 1980. С. 163-165.
Капитонов В.И. Архар в горах Бектауата (Прибалхашье) // Редкие виды млекопитающих СССР и их охрана. М., 1983. С. 177-178.
Капитонов В.И., Кубыкин РА. Питание архара в горах Кошубай И Копытные фауны СССР. М.: Наука, 1975. С. 101.
Капитонов В.И., Лазарев А.А., Борисенко В.А., Грибков П.Ф. Осеннее питание снежного барана на хребте Ганальские Востряки (Камчатка) И Там же. 1975. С. 265-266.
Капитонов В.И., Лобачев Ю. С. Распространение архара на Киргизском и Таласском хребтах (Тянь-Шань) И Редкие и исчезающие звери и ппщы Казахстана. Алма-Ата, 1977. С. 68-71.
Капитонов В.И., Махмутов С.М. О распространении и экологии архара у северного предела ареала в Казахском нагорье // Там же. 1977. С. 7-21.
Карелин Г. С. Путешествия ГС. Карелина по Каспийскому морю // Зап. Импер. Рус. геогр. о-ва по общ. географ. 1883. Т. 10. С. 1—497.
Карцев ГП. Беловежская пуща. Ее исторический очерк, современное охотничье хозяйство и высочайшие охоты в Пуще. СПб., 1903. 414 с.
Каспаров А.К. Остатки млекопитающих из позднепалеолитического поселения Сухоти-но-4 в Забайкалье // IV съезд Всесоюз. териол. о-ва. М., 1986а. Т. 1. С. 15.
Каспаров А.К. Остатки млекопитающих из позднепалеолитического поселения Сухоти-но-4 в Забайкалье // Тр. Зоол. ин-та АН СССР. 19866. Т. 148. С. 98-106.
Каспаров А.К. Фауна млекопитающих из некоторых энеолитических памятников Южной Туркмении // V съезд Всесоюз. териол. о-ва АН СССР. М., 1990. Т. 1. С. 15-16.
Катаевский В.Н. Численность, рациональное использование и охрана копытных в горных районах Северной Киргизии // Экология и охрана горных видов млекопитающих. М., 1987. С. 84-86.
Кацарский О.П. Взаимоотношения матери и новорожденного теленка овцебыков //Науч,-техн. бюл. СО ВАСХНИЛ. 1986. № 33. С. 52-55.
Кацарский О.П. Динамика отела и численность овцебыков на Таймыре И Там же. 1987. Вып. 24-25. С. 53-56.
Кацарский О.П. Состояние популяции овцебыка на Таймыре // Млекопитающие и птицы Севера Средней Сибири. Новосибирск, 1989. С. 74-78.
Кацарский О.П., Ликонцев В.В. Волк и его хищничество в районе обитания овцебыков (Таймыр)//Там же. 1987. Вып. 5. С. 8-11.
Кацарский О.П., Ликонцев В.В. Демографические параметры популяции овцебыка на Таймыре И Экологические и экономические аспекты охраны и рационального использования охотничьих животных и растительных пищевых ресурсов Сибири. Шушенское, 1990. С. 55-57.
Кацарский О.П., Никитин В.А., Матюшенков Н.В., Якушкин ГД. К зимней экологии овцебыков Таймыра // Обогащение фауны и разведение охотничьих животных. Киров, 1982. С. 84.
Кацарский О.П., Якушкин Г.Д. Рост поголовья и расширение ареала овцебыков на полуострове Таймыр // Териология, орнитология и охрана природы. XI Всесоюз. симпоз. “Биол. пробл. Севера»” Якутск, 1986. Вып. 3. С. 37-38.
Керечун С. Ф. Акклиматизация Ovis musimon Pall, в Закарпатье ИI Междунар. конгр. по млекопитающим. М.: ВИНИТИ, 1974. Т. 1. С. 262-263.
Кестер Б.В., Шостак С.В. Прошлое и настоящее Беловежской пущи. М.: Лесн. пром-сть, 1968. 68 с.
Кибакин В.В., Ишадов Н.И., Кибакина Л.Б. К изучению паразитофауны безоарового козла и горного барана (архара) // Изв. АН ТуркмССР. Сер. биол. наук. 1964. Вып. 3. С. 77-79.
Кириков С.В. Исторические изменения животного мира нашей страны в XIII-XIX веках И Изв. АН СССР. Сер. геогр. 1952. № 6. С. 31—48.
Кириков С.В. Исторические изменения животного мира нашей страны в XIII-XIX веках. Сообщение 3 // Там же. 1955. № 1. С. 32-40.
Кириков С.В. Изменения животного мира в природных зонах СССР (XIII-XIX вв.); Степная зона и лесостепь. М.: Изд-во АН СССР, 1959. 175 с.
Кириков С.В. Изменения животного мира в природных зонах СССР (XIII-XIX вв.). Лесная зона и лесотундра. М.: Изд-во АН СССР, 1960. 157 с.
Кириков С.В. Новые данные о размещении млекопитающих в XVII-XIX вв. //1 Всесоюз. совещ. по млекопитающим. М.: Изд-во Моск, ун-та, 1961. Т. 1. С. 114-116.
Кириков С.В. О “смешанной” фауне в степной и лесной зонах // Изв. АН СССР. Сер. геогр. 1963. № 1. С. 29-35.
Кириков С.В. Промысловые животные, природная средаи человек. М.: Наука, 1966.348 с.
Кириков С.В. Звери и птицы северного степного Причерноморья и их среда обитания в XVIII и первой трети XIX в. // История биогеоценозов СССР в голоцене. М.: Наука, 1976. С. 193-207.
Кириков С.В. Распространение зубра на территории Советского Союза в XI-XX вв. И Зубр. М.: Наука, 1979. С. 476-487.
Кириков С.В. Человек и природа степной зоны. Конец X - середина XIX в. (европейская часть СССР). М.: Наука, 1983. 128 с.
Кирилюк В.Е. Редкие виды млекопитающих Юго-Восточного Забайкалья (биологические основы сохранения): Автореф. дис. ... канд. биол. наук. М., 1997а. 27 с.
Кирилюк В.Е. Дзерен низовий р. Ульдзы (Северо-Восточная Монголия) // Редкие виды млекопитающих России и сопредельных территорий. М., 19976. С. 47.
Кирилюк В.Е. Об эпизоотии дзерена {Procapra gutturosa) в Северо-Восточной Монголии И VI съезд териол. о-ва. М., 1999. С. 118.
Кирилюк В.Е. Дзерен Procapra gutturosa (Pallas, 1777) // Красная книга Читинской области и Агинского Бурятского автономного округа. Животные. Чита, 2000а. С. 29-30.
КирилюкВ.Е. Горный баран, или архар Ovis аттоп (Linnaeus, 1758)//Там же. 20006. С. 31-32.
Кирилюк В.Е. Современные миграции дзерена {Procapra gutturosa) в Забайкалье // Тр. Зоол. ин-та РАН. 2001а. Т. 288. С. 136-153.
Кирилюк В.Е. Дзерен возвращается в Забайкалье//Степной бюл. 20016. № 10. С. 24—27.
Кирилюк В.Е. 100 вопросов о дзерене. Чита, 2002. 34 с.
Кирилюк В., Лущекина А. Новогодние гости в России // Охрана дикой природы. 2001. № 4. С. 4-8.
Кирилюк В.Е., Лущекина А.А. Дзерен на севере ареала: исторический обзор, современные миграции и расселение, перспективы восстановления вида в России // Proc. Inst. Biol. (Ulaanbaatar). 2002. N 24. P. 77-84.
Кирилюк B.E., Лущекина А.А. О расселении дзерена Procapra gutturosa в Забайкалье И Тери-офаунаРоссии и сопредельных территорий. VII съезд териол. о-ва. М., 2003. С. 159.
Кирилюк В.Е., Лущекина А.А. Влияние различных факторов на пространственно-временную структуру популяций монгольского дзерена {Procapra gutturosa) // Млекопитающие как компонент аридных экосистем (ресурсы, фауна, экология, медицинское значение и охрана). М., 2004. С. 58-59.
Кирилюк В.Е., Черепицын А.А. Дзерен в Юго-Восточном Забайкалье // Бюл. МОИП. Отд. биол. 1998. Т. 103, вып. 1. С. 21-25.
Кирилюк В.Е., Цэвээнмядаг Н. Пространственно-временная структура и численность популяций дзерена низовий р. Ульдза И Редкие виды млекопитающих России и сопредельных территорий. М., 1999. С. 154-167.
Киселева Е.Г. Материалы о размножении и плодовитости зубров в Окском заповеднике И Редкие виды млекопитающих фауны СССР и их охрана. М.: Наука, 1973. С. 123-125.
Киселева Е.Г. Разведение зубров в питомнике Окского заповедника // Березинский заповедник. 1974. Вып. 3. С. 103-138.
Киселева Е.Г. Результаты и перспективы разведения зубров в Окском заповеднике // Редкие виды млекопитающих и их охрана. М.: Наука, 1977. С. 213-214.
Киселева Е.Г. К биологии зубра в Окском заповеднике ИII съезд Всесоюз. териол. о-ва. М.: Наука, 1978а. С. 140-141.
Киселева Е.Г. Некоторые материалы по минеральному питанию зубров И Там же. 19786. С. 141.
Киселева Е.Г. Размещение и роль лежек зубров // Там же. 1978в. С. 220-221.
Киселева Е.Г. О влиянии различных сроков рождения зубров на их рост и развитие // IV съезд Всесоюз. териол. о-ва. М., 1986а. Т. 2. С. 58-59.
Киселева Е.Г. Зубровый питомник в Окском заповеднике: современное состояние и проблемы ИI Всесоюз. совещ. по пробл. зоокультуры. М., 19866. Ч. 2. С. 41-43.
Киселева Е.Г. Динамика инвазированности зубров Окского заповедника гельминтами Dictyocaulus // V съезд Всесоюз. териол. о-ва АН СССР. М., 1990. Т. 3. С. 199-200.
Киселева Е.Г, Ибрагимов Ю. С. Внутренние незаразные заболевания зубров в питомнике Окского заповедника //1 Всесоюз. совещ. по пробл. зоокультуры. М., 1986а. Ч. 2. С. 43-46.
Киселева Е.Г, Ибрагимов Ю.С. Травматизм зубров в питомнике Окского заповедника / / Там же. 19866. С. 46-47.
Киселева Е.Г, Цибизова Е.Л. Влияние зараженности гельминтами на развитие молодняка зубров // Териофауна России и сопредельных территорий. VII съезд териол. о-ва. М., 2003. С. 161.
Кищинский А.А. Снежный баран в Колымском нагорье // Бюл. МОИП. Отд. биол. 1967а. Т. 72, вып. 4. С. 36-42.
Кищинский А.А. Материалы о распространении диких копытных в Северо-Восточной Сибири//Проблемы Севера. 19676. Вып. 11. С. 142-148.
Кищинский А.А. Формирование горной териофауны Северо-Восточной Сибири И Основные проблемы териологии. М.: Наука, 1972. С. 177-198. (Тр. МОИП. Отд. биол. Т. 48).
Климова В.Н. Ресурсы основных видов копытных и их использование в СССР // Экология, морфология, использование и охрана диких копытных. М., 1989. Ч. 1. С. 59-60.
Князев А.В., Савинецкий А.Б. Изучение отложений ископаемого помета копытных в Северной Осетии // Историческая экология диких и домашних копытных. История пастбищных экосистем. М.: Наука, 1992. С. 147-164.
Ковалев Д.Н. Формирование социальной структуры популяции овцебыков острова Врангеля // Экология, морфология, использование и охрана диких копытных. М., 1989. Ч. 2. С. 278-280.
Ковалев Д.Н. Формирование демографической структуры популяции овцебыков осгроваВран-геля //V съезд Всесоюз. териол. о-ва АН СССР. М., 1990а. Т. 2. С. 161-162.
Ковалев Д.Н. Социально-демографическая структура искусственной популяции овцебыков (Ovibos moshatus) острова Врангеля // Зоол. журн. 19906. Т. 69, вып. 11. С. 120-131.
Ковалев Д.Н. Пространственная организация популяции овцебыков острова Врангеля в 1985-1990 годах // Популяции и сообщества животных острова Врангеля. М., 1991. С. 51-74.
Ковшарь А.Ф. О численности и мерах по сохранению архара в заповеднике Аксу-Джа-баглы // Редкие и исчезающие звери и птицы Казахстана. Алма-Ата, 1977. С. 96-97.
Ковшарь А. Ф., Янушко П.А. Новые данные о млекопитающих заповедника Аксу-Джа-баглы // Тр. гос. заповедника Аксу-Джабаглы. 1965. Вып. 2. С. 203-236.
Кожамкулова Б. С. Антропогеновая ископаемая териофауна Казахстана. Алма-Ата: Наука, 1969. 149 с.
Кожамкулова Б. С. Териофауна антропогеновых отложений Новосибирской области, ее палеогеографическое и стратиграфическое значение И Териология. Новосибирск, Наука, 1972. Т. 1. С. 172-176.
Кожамкулова Б. С. Голоценовая фауна копытных Казахстана И История биогеоценозов СССР в голоцене. М.: Наука, 1976. С. 260-267.
Кожамкулова Б. С. Позднекайнозойские копытные Казахстана. Алма-Ата: Наука, 1981. 144 с.
Кожамкулова Б. С. Позднеплиоценовая териофауна Казахстана // Млекопитающие СССР. III съезд Всесоюз. териол. о-ва. М., 1982. Т. 1. С. 39-40.
Кожамкулова Б.С., Пак Т.К. Комплексная фауна млекопитающих из позднепалеолитической стоянки Жамантуз Кокчетавской области // IV съезд Всесоюз. териол. о-ва. М., 1986. Т. 1. С. 16.
Козло П.Г. Направленность динамики численности охотничьих макромаммалий в Беларуси//Биологические ритмы. Брест, 1999а. С. 101-103.
Козло П.Г. Стратегия и концепции государственной Программы по сохранению, расселению и использованию зубров в Беларуси // VI съезд териол. о-ва. М., 19996. С. 121.
Козло П.Г. Морфофизиологические адаптации и структурно-функциональный анализ динамики популяций парнокопытных (Artiodactyla), проблемы их охраны и рационального использования в Беларуси: Автореф. ... дис. докт. биол. наук. Минск, 2001. 68 с.
Козло П.Г. Прогресс в реинтродукции и разведении зубра в Беларуси // Териофауна России и сопредельных территорий. VII съезд териол. о-ва. М., 2003. С. 166.
Козло П.Г, Емельянова Л.Г, Дерябина Т.Г, Кучмель С.В. Интродукция зубра - важное условие его сохранения в Беларуси // Сохранение биологического разнообразия Белорус. Поозерья. Витебск, 1996. С. 34-35.
Козло П.Г, Емельянова Л.Г, Дерябина Т.Г, Кучмель С.В. Питание зубров и оценка кормовой продуктивности лесных экосистем в национальном парке “Припятский” // Биологическое разнообразие национального парка “Припятский” и других особо охраняемых природных территорий. Туров - Мозырь, 1999. С. 246-254.
Козло П.Г, Пикулик М.М. Состояние и программа сохранения беловежского зубра в Беларуси // Структурно-функциональное состояние биологического разнообразия животного мира Беларуси. Минск, 1999. С. 69-71.
Козло П.Г, Углянец А.В. Оценка состояния припятской микропопуляции зубров, условия обитания и пути их улучшения // Биологическое разнообразие национального парка “Припятский” и других особо охраняемых природных территорий. Туров -Мозырь, 1999. С. 254-260.
Козлов Д.П., Назарова Н.С. Гельминтофауна зубра и пути ее формирования И Зубр. М.: Наука, 1979. С. 471-475.
Кокшунова Л.Е. Иерархия в самцовой группе сайгака в условиях неволи И Териофауна России и сопредельных территорий. VII съезд териол. о-ва. М., 2003а. С. 169-170.
Кокшунова Л.Е. О поведении самцов сайгака в период гона в неволе И Там же. 20036. С. 170.
Колесник А.Д., Головушкин М.И. Результаты первого протистологического обследования снежного барана (Ovis nivicola Eschs) И Вести, зоол. 1994. № 4-5. С. 54.
Колиснык Б.И. Реакклиматизация зубров в лесах Волынской области. Цуманская пуща И Экология, морфология, использование и охрана диких копытных. М., 1989. Ч. 2. С. 225-226.
Колосов А.М. История фаунистических исследований Алтая // Тр. Алтайского гос. заповедника. 1938. Вып. 1. С. 327-393.
Колосов А.М. Звери Юго-Восточного Алтая и смежной области Монголии И Учен. зап. Моск. гос. ун-та. 1939. Вып. 20 (зоол.). С. 123-190.
Колотова М.А. Зубры из Мещеры И Наука в России. 2002. № 3. С. 94-99.
Кольцов Л.В., Медведев Г.И. Мезолит Юга Сибири и Дальнего Востока И Археология СССР. Мезолит СССР. М.: Наука, 1989. С. 174-186.
Комаров Ю., Попов К. Ландшафтный заказник “Цейский” // Охота и охотничье хоз-во. 2003. № 11. С. 4-5.
Корнеев А.П. История промысла диких зверей на Украине. Изд-во Киевского гос. ун-та им. Т.Г. Шевченко, 1953. 37 с.
Корнеев ГА., Карпов А.А., Поле С.Б. Особенности использования сайгой территории в Му-юнкумах и Саксаулдале// Копытные фауны СССР. М.: Наука, 1975. С. 104-105.
Корнеев ГА., Карпов А.А., Путятин В.В. К фенологии ягнения самок сайгаков // IV съезд Всесоюз. териол. о-ва. М., 1986а. Т. 3. С. 151.
Корнеев ГА., Кукин В.М., Карпов А.А., Путятин В.В. Опыт содержания и кормления сайгачат в неволе // Там же. 19866. Т. 3. С. 152-153.
Коробицина КВ., Воронцов Н.Н. Хромосомные наборы диких баранов и пути использования генофонда рода Ovis в отдаленной гибридизации с домашними овцами // Биологические исследования на Дальнем Востоке. Владивосток, 1975. С. 153-158.
Коробков Г. Ф. Мезолит Средней Азии и Казахстана // Археология СССР. Мезолит СССР. М.: Наука, 1989. С. 149-173.
Корочкина Л.Н. Беловежский зубр И Тр. заповед.-охотничьего хоз-ва “Беловежская пуща”. 1958. Вып. 1.С. 7-34.
Корочкина Л.Н. К вопросу о вольном разведении зубров в Беловежской пуще //1 Всесоюз. совещ. по млекопитающим. М.: Изд-во Моск, ун-та, 1961. Т. 3. С. 54—55.
Корочкина Л.Н. Реакклиматизация зубров в Беловежской пуще и ее перспективы // Акклиматизация животных в СССР. Алма-Ата: Изд-во АН КазССР, 1963. С. 114-115.
Корочкина Л.Н. К вопросу о значении древесной растительности в питании зубров Беловежской пущи // Изв. АН БССР. Сер. биол. наук. 1966. № 1. С. 106-111.
Корочкина Л.Н. Древесная растительность в питании зубров Беловежской пущи // Беловежская пуща. Минск: Урожай, 1969. Вып. 3. С. 120-126.
КорочкинаЛ.Н. Влияние зубров на подрост древесной растительности // Там же. 1971а. Вып. 4. С. 167-176.
Корочкина Л.Н. Показатели размножения зубров в Беловежской пуще // Там же. 19716. С. 176-184.
Корочкина Л.Н. Кормовая база и некоторые итоги реакклиматизации зубров в Беловежской пуще И Там же. 1971в. Вып. 5. С. 164-176.
КорочкинаЛ.Н. Взаимоотношения зубров с другими копытными// Там же. 1971г. С. 176-184.
Корочкина Л.Н. Травянистая растительность в питании зубров Беловежской пущи И Там же. 1972а. Вып. 6. С. 110-124.
Корочкина Л.Н. Суточный ритм активности зубров Беловежской пущи // Там же. 19726. С. 125-131.
Корочкина Л.Н. Район обитания и стадиальное размещение зубров в Беловежской пуще И Там же. 1973а. Вып. 7. С. 148-165.
Корочкина Л.Н. Биотехнические мероприятия при разведении зубров в загонах и естественных условиях И Там же. 19736. С. 165-174.
Корочкина Л.Н. Некоторые вопросы вольного разведения зубров в Беловежской пуще // Редкие виды млекопитающих фауны СССР и их охрана. М.: Наука, 1973в. С. 129.
Корочкина Л.Н. Место Bison bonasus bonasus (Bovidae) в экосистеме Беловежской пущи // I Междунар. конгр. по млекопитающим. М.: ВИНИТИ, 1974. Т. 1. С. 293.
Корочкина Л.Н. Методика учета и динамика численности вольноживущих зубров Беловежской пущи // Беловежская пуща. 1975. Вып. 9. С. 91-106.
Корочкина Л.Н. Изменение территориального размещения зубров в связи со снижением кормовой емкости Беловежской пущи // Редкие виды млекопитающих и их охрана. М.: Наука, 1977. С. 214-215..
КорочкинаЛ.Н. Зубр Беловежской пущи (история, восстановление, режим содержания, экология): Автореф. дис. ... канд. биол. наук. Минск, 1970. 27 с.
КорочкинаЛ.Н., Вакула В.А. Миграции зубров Беловежской пущи //Групповое поведение животных. М.: Наука, 1976. С. 182-185.
Корочкина Л.Н., Кочко Ф.П. К вопросу о смертности зубров в естественных условиях Беловежской пущи И Заповедники Белоруссии. 1982. Вып. 6. С. 96-103.
Корочкина Л.Н., Кочко Ф.П. Динамика численности вольноживущих зубров Беловежской пущи // Там же. 1983. Вып. 7. С. 52-59.
Корочкина Л.Н., Кочко Ф.П. Влияние кормовых условий на размещение зубров Беловежской пущи в вегетационный сезон // Копытные фауны СССР. М.: Наука, 1980. С. 170-171.
Корочкина Л.Н., Кочко Ф.П. Зубры в антропогенном ландшафте // Животный мир Белорусского Полесья, охрана и рациональное использование. Гомель, 1981а. С. 29-30.
Корочкина Л.Н., Кочко Ф.П. Значение биотехнических мероприятий при реакклиматизации зубров // Заповедники Белоруссии. 19816. Вып. 5. С. 80-81.
Корочкина Л.Н., Кочко Ф.П. Смертность зубров в естественных условиях Беловежской пущи // Редкие виды млекопитающих СССР и их охрана. М., 1983а. С.180-182.
Корочкина Л.Н., Кочко Ф.П. Роль Беловежской пущи в восстановлении численности зубров//Там же. 19836. С. 182-183.
Коршунов В.М. Состояние редких видов копытных Центрального Копетдага И Биологические основы использования и охраны диких животных. М., 1983. С. 26-33.
Коршунов В.М. Численность и биотопическое распределение туркменского горного барана и туркменского бородатого козла в Центральном Копетдаге // IV съезд Всесоюз. териол. о-ва. 1986. Т. 1. С. 249-250.
Коршунов В.М. Особенности изменения численности и ареалов горных копытных в Центральном Копетдаге в современных условиях// Влияние антропогенной трансформации ландшафта на население наземных позвоночных животных. М., 1987. Ч. 1. С. 208-209.
Коршунов В.М. Структура стада горных копытных Центрального Копетдага // Популяционные исследования животных в заповедниках. М.: Наука, 1988а. С. 79-96.
Коршунов В.М. Опыт проведения учетов численности копытных в Центральном Копетдаге И Изв. АН ТуркССР. Сер. биол. наук. 19886. С. 19-27.
Коршунов В.М. Регулирующее влияние хищников на структуру популяций горных копытных Центрального Копетдага // Экология, морфология, использование и охрана диких копытных. М., 1989. Ч. 1. С. 15-16.
Коршунов В.М. Сравнительная экология горных баранов и бородатых козлов на примере популяций Центрального Копетдага // V съезд Всесоюз. териол. о-ва АН СССР. М., 1990. Т. 2. С. 83-84.
Коршунов В.М. Бородатый, или безоаровый козел И Млекопитающие Туркменистана. Хищные, ластоногие, копытные. Ашхабад: Ылым, 1995а. Т. 1. С. 243-258.
Коршунов В.М. Сибирский горный козел // Там же. 19956. С. 258.
Коршунов В.М. Горный баран, или архар // Там же. 1995в. С. 262-268.
Коршунов Е.Н., Коршунова Е.И. Проблемы охраны кызылкумского барана // Редкие виды млекопитающих СССР и их охрана. М., 1983. С. 184-185.
Коршунова Е.Н., Коршунов Е.Н. Кызылкумский баран в Нуратинском заповеднике // Экология растений и животных заповедников Узбекистана. Ташкент: Изд-во “Фан”, 1983. С. 83-85.
Косинцев ПА. Костные остатки диких млекопитающих из археологических памятников лесной полосы Урала и Западной Сибири (эпоха бронзы - средневековье) // Млекопитающие СССР. III съезд Всесоюз. териол. о-ва. М., 1982. Т. 1. С. 42-43.
Косинцев П.А. Териофауна Урала и Западной Сибири в голоцене // IV съезд Всесоюз. териол. о-ва. М., 1986. Т. 1. С. 16-17.
Косинцев П.А. Голоценовые остатки крупных млекопитающих Западной Сибири // Современное состояние и история животного мира Западно-Сибирской низменности. Свердловск, 1988. С. 32-51.
Косинцев П.А. Фауна крупных млекопитающих Северного Урала в позднем плейстоцене и голоцене // Материалы и исследования по истории современной фауны Урала. Екатеринбург, 1996. С. 84—109.
Косинцев П.А. Крупные млекопитающие Урала в плейстоцене и голоцене И Четвертичная палеозоология на Урале. Екатеринбург: Изд-во Уральского ун-та, 2003. С. 55-72.
Косинцев П.А., Бобковская Н.Е. Крупные млекопитающие неоплейстоцена широтного отрезка Нижнего Иртыша И Там же. 2003. С. 226-231.
Косинцев П.А., Ражее Д.И. Новые данные по териофауне Южного Урала и Зауралья в голоцене // Млекопитающие в системе природопользования на Урале. Свердловск, 1985. С. 17-18.
Костоглод В.Е. Распространение и численность горала на Среднем Сихотэ-Алине // Редкие виды млекопитающих фауны СССР и их охрана. М.: Наука, 1973. С. 130-131.
Котов В.А. Питание рыси в Кавказском заповеднике // Тр. Кавказского гос. заповедника. 1958. Вып. 4. С. 214-217.
Котов В.А. Количественный учет туров в Кавказском заповеднике и некоторые вопросы их экологии // Там же. 1960а. Вып. 6. С. 165-183.
Котов В.А. Количественный учет серн в Кавказском заповеднике // Там же. 19606.
С. 185-189.
Котов В.А. Искусственные солонцы Кавказского заповедника // Там же. 1960в. С. 191-208.
Котов В.А: Самоловы для отлова туров и серн // Там же. 1960г. С. 213-218.
Котов В.А. Туры и серны в Кавказском заповеднике И Природа. 1960д. № 10. С. 113.
Котов В.А. Учет численности туров и динамика их запасов в Кавказском заповеднике // Вопросы организации и методы учета ресурсов фауны наземных позвоночных. М., 1961. С. 36-37.
Котов В.А. Отлов и мечение туров (Capra caucasica) на Западном Кавказе // Бюл. МОИП. Отд. биол. 1964а. Т. 69, вып. 2. С. 54-60.
Котов В.А. Некоторые данные по морфологии западнокавказского тура ИII науч. конф, зоологов пед. ин-тов РСФСР. Краснодар, 19646. С. 180-181.
Котов В.А. Вес и размеры серн Кавказского заповедника // Там же. 1964в. С. 182. Котов В. Кавказский заповедник // Охота и охотничье хоз-во. 1964г. № 5. С. 26-28.
Котов В.А. Вес и размеры кубанских туров//Зоол. журн. 1966а. Т. 45., вып. 8. С. 1229-1235.
Котов В.А. Экология западнокавказского тура (Capra caucasica Guld.): Автореф. дис. ... канд. биол. наук. Воронеж, 19666. 20 с.
Котов В.А. Кубанский тур, его экология и хозяйственное значение // Тр. Кавказского гос. заповедника. 1968. Вып. 10. С. 201-293.
Котов В.А. Плотность населения высокогорных копытных Кавказского заповедника как показатель биологической продуктивности угодий И Естественная производительность и продуктивность охотничьих угодий СССР. Киров, 1969. Ч. 2. С. 86-88.
Котов В.А. Территориальные группировки кубанских туров в Кавказском заповеднике // Популяционная структура вида у млекопитающих. М., 1970. С. 42—43.
Котов В.А. Западно-кавказский тур// Охота и охотничье хоз-во. 1971. №4. С. 24-25.
Котов В. А., Рябов Л. С. Промысловые и ценные млекопитающие пред горных и горных районов Краснодарского края//Тр. Кавказского гос. заповедника. 1963. Вып. 7. С. 1-240.
Котрлы А., Котрла Б. Приспособление гельминтофауны серн к новым биотопам И Тр. IX Междунар. конгр. биологов-охотоведов. М., 1970. С. 601-604.
Кочко Ф.П. Динамика парамфистомозной инвазии зубров в Беловежской пуще // Биол. ритмы. Брест, 1999. С. 162-163.
Кошкарев Е. Валютные бараны и министерские волки Киргизии // Охрана дикой природы. 2002. № 1 (24). С. 26-32.
Кошкарев Е.П., Нестеров Н.А. Индикация состояния популяций горного козла и архара в Тянь-Шане на основе индекса стадности // Экология, морфология, использование и охрана диких копытных. М., 1989. Ч. 2. С. 240-241.
Правей В. Овцебыки возвращаются на Таймыр. М.: “Полярная звезда”, 2003. 156 с.
Крайнова Л. В. Восстановление зубра на Кавказе И 20 лет Кавказского государственного заповедника. М., 1947. С. 61-73.
Крайнова Л.В. Питание зубров Кавказского государственного заповедника // Бюл. МОИП. Отд. биол. 1951. Т. 56, вып. 6. С. 3-17.
КрасинскаМ., КрасинскиЗ., Буневич А.Н. Влияние условий обитания на пространственную структуру зубров различного пола и возраста И Структурно-функциональное состояние биологического разнообразия животного мира Беларуси. Минск, 1999. С. 71-72.
Красная книга России: правовые акты. М., 2000. 134 с.
Красная книга Российской Федерации (животные). Балашиха: Астрель, 2001. 860 с.
Красная книга Туркменистана. Т. 1. Беспозвоночные и позвоночные животные. Ашхабад, 1999. 371 с.
Кривошапкин А.А. Снежный баран Верхоянья: морфология, экология, ресурсы: Автореф. дис. ... канд. биол. наук. М., 1995. 19 с.
Кривошапкин А.А., Лукавцев Ю.С., Яковлев В.Г. К экологии снежного барана в Центральном Верхоянье // Экология млекопитающих горных территорий. Популяционные аспекты. Нальчик: Изд. центр “Эль-Фа”, 1997. С. 151-152.
Кривошапкин А.А., Яковлев В.Г. Снежный баран Верхоянья. Якутск, 1999. 134 с.
Кривошеев В.Г., Чернявский Ф.Б. Редкие и нуждающиеся в охране наземные млекопитающие севера Дальнего Востока СССР И Редкие и исчезающие животные суши Дальнего Востока СССР. Владивосток, 1981. С. 27-35.
Кричевский В. Безоаровые козлы Дагестана страдают от международного терроризма // Российская охотничья газета. 2004. № 17. С. 26.
Круторогое Н. Ф. Хозяйственное использование диких копытных в РСФСР И Копытные фауны СССР. М.: Наука, 1975. С. 225-227.
Кудактин А.Н. Соотношение численности копытных и волка в Кавказском заповеднике // Копытные фауны СССР. М.: Наука, 1975. С. 199-200.
Кудактин А.Н. Влияние волка на численность туров и серн в Кавказском заповеднике // Редкие виды млекопитающих и их охрана. М.: Наука, 1977. С. 216-218.
Кудактин А.Н. Взаимоотношения волка и копытных в Кавказском заповеднике И Экология, охрана и использование хищных млекопитающих в РСФСР. М., 1982. С. 65-70.
Кудактин А.Н. Полиморфизм западнокавказских туров// Фенетика популяций. М., 1985. С. 265-266.
Кудактин А.Н. Влияние волка на копытных в Кавказском заповеднике // Роль крупных хищников и копытных в биоценозах заповедников. М., 1986. С. 21-35.
Кудактин А. Невиданное истребление зверей И Охота и охотничье хоз-во. 1996. № 9. С. 1-2.
Кудрявцев И.В., Белоусова И.П. Принципиальные подходы к оценке современного состояния и перспектив выживания популяций европейского зубра И Редкие виды млекопитающих России и сопредельных территорий. М., 1997. С. 52. М., 1999. С. 188-198.
Кудрявцев И.В., Белоусова И.П. Проблема восстановления и сохранения дикого типа исчезающих & Idquo; краснокнижных & rdquo; видов млекопитающих, существующих в условиях неволи ИII съезд Вавиловского о-ва генет. и селекционеров. СПб, 2000. С. 121-122.
Кудрявцев И.В., Сафронова Л.Д. Совещание по проблеме “Сохранение редких и исчезающих видов - зубр, лошадь Пржевальского, сайгак” // Зоол. журн. 1999. Т. 72, вып. 11. С. 158-159.
Кузнецов Б.А. Звери Киргизии. М.: Изд-во МОИП, 1948а. 210 с. (Материалы по фауне и флоре СССР. Новая сер. Отд. зоол. Вып. 12).
Кузнецов Б.А. Млекопитающие Казахстана. М.: Изд-во МОИП, 19486. 226 с. (Материалы к познанию фауны и флоры СССР. Новая сер. Отд. зоол. Вып. 13).
Кузнецов Б. Сары-Челекский заповедник//Охота и охотничье хоз-во. 1969. № 5. С. 14—15.
Кузнецов В., Лущекина А. Калмыцкий сайгак на грани катастрофы // Степи, бюл. 2002. № 11. С. 28-33.
Кузнецов Г.В., Петров Н.Б., Куликов Е.Е. и др. Таксономический статус и филогенетические отношения нового вида и рода парнокопытного Pseudonovibos spiralis W.P. Peter, A. Feiler, 1994 (Artiodactyla, Bovidae) // Зоол. журн. 2001. T. 80, № 11. С. 1395-1403.
Кузнецова М.В. Реконструкция филогенетических отношений в семействе полорогих (Bovidae; Artiodactyla) на основании ядерных и митохондриальных последовательностей ДНК: Дис. ... канд. биол. наук. М., 2002. 134 с.
Кузнецова М.В., Холодова М.В. Использование RAPD-PCR анализа для идентификации монгольского и калмыцкого подвидов сайгака Saiga tatarica // Памяти Грегора Менделя. М., 2001. С. 66-67.
Кузнецова М.В., Холодова М.В. Ревизия филогенетических отношений в подсемействе Antilopinae на основании анализа последовательностей митохондриальных рРНК и ядерного гена белка 5-спектрин //Докл. Акад. наук. 2003. Т. 391, № 2. С. 281-285.
Кузнецова М.В., Холодова М.В., Лущекина А.А. Филогенетический анализ последовательностей митохондриальных генов 12S и 16S рРНК представителей семейства Bovidae: новые данные И Генетика. 2002. Т. 38, № 8. С. 1115-1124.
Кузьмина И.Е. К истории териофауны Северного Урала и Приуралья в верхнем антропогене И Бюл. МОИП. Отд. биол. 1966. Т. 71, вып. 3. С. 91-102.
Кузьмина И.Е. Формирование териофауны Северного Урала в позднем антропогене // Тр. Зоол. ин-та АН СССР. 1971. Т. 49. С. 44-122.
Кузьмина И.Е. Некоторые данные о млекопитающих Среднего Урала в позднем плейстоцене И Бюл. Комис, по изуч. четвертич. периода АН СССР. 1975. № 43. С. 63-77.
Кузьмина И., Синицына Г. Значение фаунистических остатков для хронологии многослойной стоянки Бирюса на Среднем Енисее // Цитология. 1995. Т. 37, № 7. С. 620-621.
Кузякин В.А. Состояние ресурсов диких копытных И Итоги науки и техники. Т. 13. Зоология позвоночных. М.: ВИНИТИ, 1985. С. 5-65.
Кулагин Н.М. К истории распространения зубров в России И Изв. РАН. 1918. С. 1649-1658.
Кулагин Н.М. Зубры Беловежской пущи. М.: Изд-е Моск. науч, ин-та, 1919. 166 с.
Кулагин Н.М. Фауна БССР. Млекопитающие, копытные. Минск: Изд-во АНБССР, 1940. Т. 1, вып. 4. 63 с.
Кулиев ГК, Мамедов Т.О. Изучение хромосомного набора Ovis orientalis Gmel. с помощью дифференциальной окраски //1 Междунар. конгр. по млекопитающим. М.: ВИНИТИ, 1974. Т. 1. С. 327.
Кулиев Г.К., Мамедов Т.О. Изучение кариотипов копытных животных Азербайджана // Систематика и цитогенетика млекопитающих. М.: Наука, 1975. С. 37-39.
Кулиев К.М., Мамедов Т.О. Изучение хромосомного набора безоарового козла (Capra aegagrus Erxlleben) и дагестанского тура (С. с. cylindricornis Blyth) // Изв. АН АзССР. Сер. биол. наук. 1974. № 6. С. 88-93.
Кулиев С.М. К распространению и эгологии Capra aegagrus Erxl. (Artiodactyla) в Нахичеванской АССР //1 Междунар. контр, по млекопитающим. М.: ВИНИТИ, 1974. Т. 1. С. 327.
Кулиев С.М. О распространении, численности и структурных особенностях популяций безоарового козла и дагестанского тура в Азербайджане // Экология, методы изучения и организация охраны млекопитающих горных областей. Свердловск, 1977. С. 42-43.
Кулиев С.М. К питанию безоарового козла и дагестанского тура в Азербайджане // II съезд Всесоюз. териол. о-ва. М.: Наука, 1978. С. 148-149.
Кулиев С.М. Безоаровый козел и дагестанский тур Азербайджана (Биология, охрана и хозяйственное значение): Автореф. дис. ... канд. биол. наук. М., 1981. 21 с.
Кулиев С.М. Численность, половой и возрастной состав копытных в Исмаиллинском заповеднике и заказнике И Экология, морфология, использование и охрана диких копытных. М., 1989. Ч. 1. С. 60-62.
Кулиев С.М. Редкие виды парнокопытных Азербайджана // Редкие виды млекопитающих России и сопредельных территорий. М., 1997. С. 52.
Кулиев С.М. Современное состояние некоторых парнокопытных аридных экосистем Азербайджана и их охрана И Млекопитающие как компонент аридных экосистем (ресурсы, фауна, экология, медицинское значение и охрана). М., 2004. С. 68-69.
Курченко Ф.П. Об инфекционной патологии сайгаков и их опасности в распространении болезней рогатого скота И Вирусные болезни сельскохозяйственных животных. Владимир, 1995. С. 216.
Курятников Н.Н., Варзиев А.Б. Численность и вопросы охраны млекопитающих Северной Осетии // Редкие виды млекопитающих СССР и их охрана М., 1983. С. 13.
Кутепов Н. Императорская охота на Руси. Конец XVIII и XIX век. СПб., 1911. Т. 4.289 с.
Кучеренко С.П. Копытные млекопитающие Амуро-Уссурийского региона И Животный мир и охотничье хозяйство Дальнего Востока. Владивосток, 1976. С. 97-125.
Кучерук В.В. Степной комплекс млекопитающих в фауне Палеарктики // География населения наземных животных и методы его изучения. М.: Изд-во АН СССР, 1959. С. 45-83.
Кучерук В. В. Воздействие травоядных млекопитающих на продуктивность травостоев степи и их значение в образовании органической части степных почв // Биология, биогеография и систематика млекопитающих СССР. М., 1963. С. 157—194.
Лавов М.А. Распространение и численность диких копытных в Восточной Сибири И Бюл. МОИП. Отд. биол. 1966. Т. 71, вып. 5. С. 30-39.
Лавов М.А. Динамика численности и образ жизни березинского стада зубров // Редкие виды млекопитающих СССР и их охрана. М., 1983. С. 186-188.
Лавов М.А., Воронова Т.Н. Зубры в Березинском заповеднике и прилегающих территориях//Заповедники Белоруссии. 1982. Вып. 6. С. 109-112.
Лавов М.А., Чичикина С.Н. Вольноживущее стадо зубров в Северной Белоруссии // Копытные фауны СССР. М.: Наука, 1980. С. 246-247.
Лавровский А.А. О влиянии летней засухи и суровой зимы 1949/50 г. на численность и размещение сайгаков // Охрана природы. 1950. № 10. С. 83-87.
Лазарев П.А. Мамонтовая фауна Якутии // Млекопитающие СССР. III съезд Всесоюз. териол. о-ва. М., 1982. Т. 1. С. 83-87.
Лазарев П.А. К истории формирования антропогеновой териофауны Якутии // V съезд Всесоюз. териол. о-ва АН СССР. М., 1990. Т. 1. С. 19-20.
Ларин В.В. К экологии снежного барана гор Путорана И Экология и рациональное использование наземных позвоночных Севера Средней Сибири. Новосибирск, 1983. С. 78-86.
Ларин В.В. Снежный баран гор Путорана (некоторые особенности питания) // Экология и охрана горных видов млекопитающих. М., 1987а. С. 103-105.
Ларин В.В. К особенностям обитания снежного барана в горах Путорана // Науч.-техн. бюл. СО ВАСХНИЛ. 19876. № 24-25. С. 16-21.
Ларин В. В. Влияние антропогенного фактора на численность путоранского снежного барана (редкий вид) // Влияние антропогенной трансформации ландшафта на население наземных позвоночных животных. М., 1987в. Ч. 1. С. 198-200.
Ларин В.В. О размножении и материнском поведении путоранского снежного барана // Млекопитающие и птицы Севера Средней Сибири. Новосибирск, 1989. С. 61-68.
Ларин В.В. Снежный баран (Ovis nivicola borealis) (плато Путорана): Автореф. дис. ... канд. биол. наук. М., 1990. 23 с.
Ларин В.В., Боржонов Б.Б., Дорогое В.Ф. и др. Современное состояние подвида путоранского снежного барана и проблемы его охраны И Науч.-техн. бюл. СО ВАСХНИЛ. 1987. № 24-25. С. 56-60.
Ларин В.В., Крашевский О.Р. Хищничество волка и росомахи в отношении путоранского снежного барана // Млекопитающие и птицы Севера Средней Сибири. Новосибирск, 1989. С. 68-74.
Ларин В.В., Павлов Б.М., Дорогое В.Ф., Боржонов Б.Б. Численность и половозрастной состав путоранского снежного барана // Животный мир плато Путорана, его рациональное использование и охрана. Новосибирск, 1988. С. 6-14.
Ларин В.В., Сипко Т.П. Состояние популяции путоранского снежного барана (Ovis nivicola borealis) II Редкие виды млекопитающих России и сопредельных территорий. М., 1997. С. 53. М., 1999. С. 199-203.
Ларин В.В., Сипко Т.П. Баран снежный (Ovis nivicola) И Фауна позвоночных животных плато Путорана. М., 2004. С. 378-398.
Ларин В.В., Шелковникова ТА. Сезонные особенности экологии питания снежных баранов в бассейне р. Делочи // Животный мир плато Путорана, его рациональное использование и охрана. Новосибирск, 1988. С. 17-26.
Ларичев В.Е. У истоков верхнепалеолитических культур и искусства Сибири (к открытию в Кузнецком Алатау поселения Малая Сыя и скульптурного изображения черепахи) И Рериховские чтения. Новосибирск: Наука, 1976. С. 14-26.
Лебедев В. Мускусный овцебык И Охота и охотничье хоз-во. 1958. № 11. С. 27.
Лебедева Л. С. Питание сайгака на правобережье Волги И Бюл. МОИП. Отд. биол. 1959. Т. 64, вып. 5. С. 27-35.
Лебедева Л. С. Материалы к изучению весенних кормов и пастбищ сайгаков правобережья Волги// Зоол. журн. 1960. Т. 39, вып. 9. С. 1438-1442.
Лент П. Овцебык на Аляске // Охота и охотничье хоз-во. 1970. № 4. С. 43.
Лент П. С. Характеристика особенностей поведения представителей рода Ovibos (Bovidae) И I Междунар. конгр. по млекопитающим. М.: ВИНИТИ, 1974. Т. 1. С. 360.
Лещенко А.П. Влияние некоторых антропогенных факторов на состояние и размещение сайгаков в Северо-Западном Прикаспии И ВСХИЗО - агропромышленному комплексу. М., 1995. С. 124-125.
Лещенко А.П. Особенности территориального размещения сайгаков Северо-Западного Прикаспия. Стратегия охраны и использования: Автореф. дис. ... канд. биол. наук. М., 1997. 25 с.
Ли Дицян, Цзян Чжиган. Анализ жизнеспособности популяции дзерена Пржевальского И Экология. 2002. № 2. С. 126-131.
Липкович А.Д. Состояние популяции зубров заказника “Цейский” Северо-Осетинской АССР // Изучение и охрана редких и исчезающих видов животных фауны СССР. М.: Наука, 1985. С. 75-76.
Липкович А.Д. Современное состояние и перспективы восстановления чистокровных зубров на Северном Кавказе // Ресурсы животного мира Северного Кавказа. Ставрополь, 1988. С. 101-104.
Липкович А.Д. Современное состояние и перспективы восстановления чистокровных зубров на Северном Кавказе // Экология, морфология, использование и охрана диких копытных. М, 1989. Ч. 2. С. 226-228.
Липкович А.Д. Зубры на юге России // К вопросу о возможности сохранения зубра в России. Пущино, 1993. С. 82-84.
Литвин М. О нравах татар, литовцев и московитян. Извлечение из сочинения Михаила Литвина (1550) // Мемуары, относящиеся к истории Южной Руси. Kiee, 1890. Вып. 1. (XVI ст.). С. 1-58.
Литвинова Н.А. К питанию зубров в Цейском заказнике И Копытные фауны СССР. М.: Наука, 1980. С. 177-179.
Лихачев А.И. Некоторые черты адаптации Saiga tatarica L. к геохимическим факторам пустыни //1 Междунар. конгр. по млекопитающим. М.: ВИНИТИ, 1974. Т. 1. С. 369-370.
Лихотоп Р.И., Сологор Е.А. Состояние популяции зубра в Волынской области И Редкие виды млекопитающих СССР и их охрана. М., 1983. С. 188-189.
Лобанов Н.В. Бизон, его воспроизводство и гибридизация в зоопарке “Аскания-Нова” // Наследуемость хозяйственно-полезных признаков у сельскохозяйственных животных. Киев, 1968. С. 122-134.
Лобанов Н. Европейский муфлон // Охота и охотничье хоз-во. 1988. № 12. С. 10-11.
Лобанов Н.В., Треус В.Д. Эколого-морфологические особенности диких баранов (роды Ammotragus и Ovis) в Аскании-Нова // Эколого-морфологические особенности диких родичей домашних овец. М.: Наука, 1978. С. 30-36.
Логвиненко В.Н. Зубры (Bison, Artiodactyla) эоплейстоцена Северного Причерноморья Украины // Доп. нац. АН Укр. 2001. № 11. С. 150-155.
Ломанов И.К., Володина О.А., Сиголаева Е.А. Комплексный анализ состояния ресурсов охотничьих животных в Российской Федерации в 1999 г. и прогноз до 2005 г. // Охотничьи животные России. 2000. Вып. 2. С. 122-129.
Ломов А.А., Банникова А.А., Сипко Т.П., Долгов В.А. Использование рестриктаз-ного анализа геномной ДНК для решения проблем систематики и филогении млекопитающих (на примере русской выхухоли и зубра) // Редкие виды млекопитающих России и сопредельных территорий. М., 1999а. С. 204-215.
Ломов А.А., Банникова А.А., ФедороваЛ.В., и др. Использование таксономического фин-герпринта ДНК для решения проблем систематики и эволюции млекопитающих // VI съезд териол. о-ва. М., 19996. С. 143.
Лопырин А.И., Логинова Н.В., Инякова А.П. Опыт скрещивания тура с домашней козой И Наследственность и изменчивость растений, животных и микроорганизмов. М.: Изд-во АН СССР, 1959. Т. 1. С. 709-722.
Лопырин А.И., Логинова Н.В., Инякова А.П. Опыт скрещивания овец и коз с туром и серной // Тр. Тебердинского гос. заповедника. 1960. Вып. 2. С. 217-231.
Лукаревский В. С. Современное распределение, проблемы охраны и восстановления горного барана (уриала) на Западном Копетдаге И Экология, морфология, использование и охрана диких копытных. М., 1989. Ч. 2. С. 242-243.
Лукаревский В.С. Крупные млекопитающие юга Туркмении и проблемы их охраны // Редкие виды млекопитающих России и сопредельных территорий. И., 1999. С. 216-230.
Лукашкин А. С. Монгольская степная антилопа “дзерен” // Тр. о-ва изуч. Маньчжурского края. Зоология. Харбин, 1927. Вып. 1. С. 8-24.
Лукашкин А. С. Монгольские элементы в фауне млекопитающих Северной Маньчжурии // Зоол. журн. 1975. Т. 54, вып. 10. С. 1578-1581.
Лущекина А.А. Эколого-географические основы охраны и рационального использования дзерена (Procapragutturosa) в МНР: Автореф. дис.... канд. биол. наук. М., 1990. 20 с.
Лущекина А. Выживет ли монгольская сайга? И Охота и охотничье хоз-во. 1997. № 7. С. 40-41.
Лущекина А.А. Современное состояние и перспективы международного сотрудничества по охране и устойчивому использованию монгольского дзерена // Изв. РАН. Сер. биол. 1998. № 4. С. 462-466.
Лущекина А. Живая легенда калмыцкой степи // Охрана дикой природы. 2002. № 1 (24). С. 4-11.
Лущекина А.А., Дуламцэрэн С. Монгольская сайга: современный статус и перспективы сохранения // Изв. РАН. Сер. биол. 1997. № 2. С. 177-185.
Лущекина АА., Неронов В.М Современный статус и перспективы сохранения монгольской сайги // Редкие виды млекопитающих России и сопредельных территорий. М., 1997. С. 54.
Лущекина А.А., Неронов В.М., Огуреева Г.Н., Соколова А.Л. Распространение, экология, охрана и рациональное использование дзерена в МНР И Бюл. МОИП. Отд. биол. 1986. Т. 91, вып. 5. С. 73-82.
Лущекина А.А., Неронов В.М., Шурхал А.В. Материалы по структуре ареала и населения дзерена в МНР И Млекопитающие СССР. III съезд Всесоюз. териол. о-ва. М., 1982. Т. 1.С. 123-124.
Лущекина А.А., Неронов В.М., Шурхал А.В. Структура ареала, внутривидовая изменчивость и вопросы рационального использования запасов монгольского дзерена // Прикладные аспекты программы “Человек и биосфера”. М.: ВИНИТИ, 1983. С. 134—146.
Ляпунова Е.А., Банч Т.Д., Воронцов Н.Н., Хоффман Р.С. Хромосомные наборы и систематическое положение барана Северцова (О. аттоп severtzovi) И Зоол. журн. 1997. Т. 76, вып. 9. С. 1083-1093.
Львов И.А. Опыт отлова архаров И Акклиматизация охотничьих животных в СССР. Минск: Ураджай, 1978. С. 146-147.
Львов ИА. К экологии архара из Восточной Гоби И Бюл. МОИП. Отд. биол. 1981. Т. 86, вып. 6. С. 16-19.
Львов И.А. О полигамии дзеренов И Млекопитающие СССР. III съезд Всесоюз. териол. о-ва. М., 1982. Т. 2. С. 146-147.
Магомедов М.-Р.Д., Абатуров БД., Ахмедов Э.Г. и др. Современное состояние популяции памирского архара Ovis аттопpolii И Териофауна России и сопредельных территорий. VII съезд териол. о-ва. М., 2003. С. 205-206. ’
Магомедов М.-Р.Д., Ахмедов Э.Г. Закономерности пространственного размещения и численность дагестанского тура (Capra cylindricomis) на Восточном Кавказе // Зоол. журн. 1994. Т. 73, вып. 10. С. 120-129.
Магомедов М.-Р.Д., Ахмедов Э.Г. Современное состояние горных копытных Дагестана И VI съезд териол. о-ва. М., 1999. С. 154.
Магомедов М.-Р.Д., Ахмедов Э.Г., Насруллаев Н.И. Закономерности пространственной организации популяции безоарового козла, Capra aegagrus (Artiodactyla), в условиях Восточного Кавказа // Зоол. журн. 2002. Т. 81, № 8. С. 1008-1016.
Магомедов М.-Р.Д., Ахмедов Э.Г., Насруллаев Н.И. Демографическая структура популяции безоарового козла, Capra aegagrus (Artiodactyla), и закономерности ее формирования в условиях Восточного Кавказа // Там же. 2004. Т. 83, № 2. С. 234-240.
Магомедов М.-Р.Д., Ахмедов Э.Г., Яровенко Ю.А. Дагестанский тур: популяционные и трофические аспекты экологии. М.: Наука, 2001. 138 с.
Магомедов М.-Р.Д., Петрищев Б.И., Лхагвасурен Б. Интенсивность питания и переваримость кормов у ягнят дзерена // Экология, морфология, использование и охрана диких копытных. М., 1989. Ч. 2. С. 259-260.
Магомедов М.-Р.Д., Яровенко Ю.А. Интенсивность питания и переваримость кормов дагестанского тура {Capra cylindricornis) в неволе // Зоол. журн. 1997а. Т. 76, вып. 2. С. 243-250.
Магомедов М.-Р.Д., Яровенко Ю.А. Питание и энергетическая обеспеченность дагестанского тура в естественных условиях Восточного Кавказа // Экология млекопитающих горных территорий. Популяционные аспекты. Нальчик: Изд. центр “Эль-Фа”, 19976. С. 35-44.
Магомедов М.-Р.Д., Яровенко Ю.А. Трофо-энергетические связи дагестанского тура Capra cylindricornis (Artiodactyla, Bovidae) с пастбищными экосистемами высокогорий Восточного Кавказа И Зоол. журн. 1998. Т. 77, вып. 4. С. 465—473.
Мазин Б.Н. Сайгак на восточном побережье Аральского моря // IV съезд Всесоюз. териол. о-ва. М., 1986. Т. 3. С. 154-155.
Майр Э. Принципы зоологической систематики. М.: Мир, 1971. 454 с.
Макарова Л.А. Предварительное сообщение о животных эпохи бронзы в поселении Чаг-линка И По следам древних культур Казахстана. Алма-Ата: Наука, 1970. С. 269-276.
Макарова Л.А. Териофауна пещеры Караунгур // Археологические исследования в Казахстане. Алма-Ата: Наука, 1973. С. 146-155.
Макарова Л.А. Дикие млекопитающие из археологических памятников эпохи неолита и бронзы Казахстана И История биогеоценозов СССР в голоцене. М.: Наука, 1976. С. 268-272.
Макеев П. С. К вопросу о природных условиях палеолита по археологическим данным // Природная обстановка и фауны прошлого. Киев: Изд-во АН УССР, 1963. Вып. 1. С. 31-65.
Максимук А.В. Совершенствование организационно-экономических основ и техники добычи сайгаков в Калмыцкой АССР И Экономика, организация и использование ресурсов охотничьего хозяйства РСФСР. М., 1981. С. 40-49.
Максимук А. Добыча сайгаков и биотехния И Охота и охотничье хоз-во. 1982. № 7. С. 9.
Максимук А. Промысел сайгака в Северо-Западном Прикаспии // IV съезд Всесоюз. териол. о-ва. М., 1986. Т. 3. С. 156-157.
Максимук А.В., Петрищев Б.И., Хахин ГВ. Динамика численности сайгака Северо-Западного Прикаспия // Экология, морфология, использование и охрана диких копытных. М, 1989. Ч. 2. С. 272-273.
Максимук А.В., Проняев А.В. Влияние преобразованной среды обитания на плодовитость самок сайгака Северо-Западного Прикаспия И I Всесоюз. совещ. по пробл. зоокультуры. М., 1986. С. 164-165.
Малеева А., Стефановский В., Струкова Т, Елькин Ю. Мамонтовая фауна на Урале // Цитология. 1995. Т. 37, № 7. С. 625-626.
Маликов Д.И. Биоритмы отрастания рога у домашних баранов // Бюл. МОИП. Отд. биол. 1978. Т. 83, вып. 2. С. 58-62.
Малков Н.П., Малков Ю.П. К распространению сибирского горного козла в бассейне Ар-гута И Природа и природные ресурсы Горного Алтая. Горно-Алтайск, 1972. С. 75-76.
Мамбетжумаев А. Сайгаки на Устюрте // Охота и охотничье хоз-во. 1966. №6. С. 22-23.
Мамбетжумаев А.М. О питании сайгаков Устюрта в весенне-летний период// Узб. биол. журн. 1969. №3. С. 59-61.
Мамбетжумаев А.М. Новое об устюртском архаре // Природа. 1969. № 8. С. 76-77.
Мамбетжумаев А.М. Об осенне-зимнем питании кызылкумского архара {Ovis аттоп severtzovi Nasonov) на хребтах Тамдытау и Нуратау // Териология. Новосибирск: Наука, 1974. Т. 2. С. 321-327.
Мамедов Т. О. Классификация хромосом млекопитающих рода Capra // Млекопитающие СССР. III съезд Всесоюз. териол. о-ва. М., 1982. Т. 1. С. 53—54.
Мамедов Т. О. Сравнительная кариология диких и домашних полорогих Азербайджана (в цитогенетическом и эволюционном аспектах): Автореф. дис. ... канд. биол. наук. Баку, 1978. 19 с.
Мамедов Т.О. Приложение цитогенетических данных для изучения филогении горных баранов и происхождения домашних овец И Изв. АН АзССР. Сер. биол. наук. 1985. № 6. С. 61-72.
Мамедов Т.О. Цитогенетический подход к изучению филогении баранов // Экология и охрана горных видов млекопитающих. М., 1987. С. 116-117.
Мамедов ТО., Агаев ИГ Сравнительно-кариологические исследования некоторых видов полорогих Азербайджана И V съезд Всесоюз. териол. о-ва АН СССР. М., 1990. Т. 1. С. 84.
Манасерова Н. С. Распространение и хозяйственное использование диких и домашних представителей родов Ovis и Capra // Зоол. сб. Ин-та зоол. АН АрмССР. 19866. Вып. 20. С. 80-98.
Манасерова Н.С. Морфологическая характеристика черепа арменийского муфлона И Биол. журн. Армении. 1988. Т. 41, № 8. С. 668-675.
Манасерова Н.С. Морфологическая характеристика черепа кавказского безоарового козла Capra aegagrus aegagrus Erxleben // Там же. 1989. Т. 42, № 8. С. 727-733.
Манасерян Н. Изменение антропогеновой фауны Армении // Цитология. 1995. Т. 37, № 7. С. 626.
Манасерян Н. У. Териофауна Армении в голоцене // Редкие виды млекопитающих России и сопредельных территорий. М., 1997а. С. 56.
Манасерян Н. У. Бараны и козлы голоцена Армении И Там же. 19976. С. 56.
Манасян Л.Г, Гаспарян К.М., Антонян А.Ш. и др. Особенности роста и развития арме-нийских муфлонов (азиатских горных баранов) в условиях неволи // Зоол. сб. (Ереван). 1986. № 20. С. 48-79.
Марвин М.Я. Млекопитающие Карелии. Петрозаводск: Гос. изд-во Карельской АССР, 1959. 238 с.
Марков Е.Л. Охотничье хозяйство Закавказья. Тбилиси, 1934. 230 с.
Марков Е.Л. Охотничье-промысловые животные Лагодехского заповедника. Тбилиси, 1938. 67 с.
Марков Е.Л., Млокосевич Л.Л. Закатальский заповедник. Баку: Изд. Закавказ. фил. АН СССР, 1935. С. 65 с.
Масенов Т.М. К сравнительной морфологии легких у копытных млекопитающих в связи с их экологией и видовыми особенностями // Изв. АН КазССР. Сер. биол. 1966. № 5. С. 73-81.
Маслов С.П. Антропогенные факторы формирования основ современного биоразнообразия животного мира в позднем плейстоцене // Динамика биоразнообразия животного мира. М., 1997. С. 6-13.
Маслов С.П., Антипина Е.Е. Природная обстановка и биоценотические комплексы среднего течения Оки в конце плейстоцена // VIII Всесоюз. зоогеограф, конф. М., 1984. С. 208-210.
Маслов С., Антипина Е. Гибель мамонтов и судьба мамонтовой фауны // Цитология. 1995. Т. 37, №7. С. 627-628.
Матвиенко А. О., Треус М.Ю. Развитие внутренних органов сайгаков в условиях зоопарка “Аскания-Нова” // IV съезд Всесоюз. териол. о-ва. М., 1986. Т. 3. С. 157-159.
Матевосян Е.М. Гельминты зубро-бизонов Мордовского заповедника//Тр. Мордовского гос. заповедника им. П.Г. Смидовича. 1964. Вып. 2. С. 181-189.
Махмутов С.М. К поведению горного козла в Джунгарском Алатау // Проблемы охраны и устойчивого использования биоразнообразия животного мира Казахстана. Алматы, 1999. С. 32-33.
Медведев Д.Г. Новый подвид снежного барана из хребта Кодар (Витимо-Олекминское нагорье) // Байкал - природная лаборатория для исследований изменения окружающей среды и климата. Иркутск: Изд-во ЛИСНА, 1994. Т. 5. С. 37-38.
Медведев Д.Г. Экология снежного барана (Ovis nivicola Eschscholtz, 1829) в высокогорьях хребта Кодар (Северное Забайкалье): Автореф. дис.... канд. биол. наук. Иркутск, 1997. 26 с.
Медведев Д.Г. Снежный баран хребта Кодар (распространение и морфологические отличия) И Териофауна России и сопредельных территорий. VII съезд териол. о-ва. М., 2003а. С. 219.
Медведев Д.Г. Снежный баран хребта Кодар и перспективы его охраны // Охрана и рациональное использование животных и растительных ресурсов России. Иркутск, 20036. С. 437-441.
Медведев Д.Г., Аюпов В.Г. Сибирский горный козел И Редкие животные Иркутской области (наземные позвоночные). Иркутск, 1993. С. 40—43.
МедведевД.Г, Будлянский А.М., Жарков Л.В. Снежный баран (чубук) Северного Забайкалья И Экология озера Байкал и Иркутской области. Иркутск, 1991. С. 13.
Медведев Д.Г, Лямкин В.Ф. Снежный баран Ovis nivicola Eschscholtz, 1829 // Красная книга Читинской области и Агинского Бурятского автономного округа. Животные. Чита, 2000. С. 32-34.
Медведев Д.Г., Никулина Н.А., Романюк А.А. К изучению охотского подвида снежного барана И Вести. Иркутской гос. с.-х. акад. 2000. Вып. 19. С. 15-16.
Международный кодекс зоологической номенклатуры. Изд. третье. Л.: Наука, 1988.205 с. Изд. четвертое. СПб: Наука, 2000. 221 с.
Межлумян С.К. О распространении зубра в бассейне озера Севан И Уч. зап. Ереванского гос. ун-та. Естеств. науки. Биология. 1968. № 2. С. 84-99.
Межлумян С.К., Оганесян Л.Е. Охота и скотоводство урартов И Биол. журн. Армении. 1986. № 8, С. 664-665.
Мейер Л. Киргизская степь Оренбургского ведомства // Матер, для геогр. и статистики России, собранные офицерами Генерального штаба. СПб, 1865. Т. 14. С. 1-288.
Мекленбурцев PH. Памирский архар (Ovis polii polii Blyth.) // Бюл. МОИП. Отд. биол. 1948. Т. 53, вып. 5. С. 65-84.
Мекленбурцев PH. К экологии сибирского горного козла на Памире И Зоол. журн. 1949. Т. 28, вып. 5. С. 482-483.
Мекленбурцев Р.Н. Об архаре в северных отрогах Алайского хребта // Там же. 1962. Т. 41, вып. 10. С. 1588-1589.
Мельник К.П., Клыкова В. А. Морфология и биомеханические свойства скелета конечностей сайгака//Экология, морфология, использование и охрана диких копытных. М., 1989. Ч. 2. С. 324-325.
Мельников А.В. Динамика численности тяньшанского горного барана в заповеднике Аксу-Джабаглы//Копытные фауны СССР. М.: Наука. 1980. С. 181-182.
Мельников В.В. Влияние охотничьего туризма на состояние популяций охотничьих животных и проблемы его развития: Автореф. дис. ... канд. биол. наук. Киров, 2003. 21 с.
Мельников В. К, Сопин Л.В. Реакклиматизация архаров в Восточной Сибири // Акклиматизация охотничьих животных в СССР. Минск: Ураджай, 1978. С. 65-66.
Мельникова М.Н. Полиморфизм геномной ДНК на видовом и популяционном уровне, исследованной с помощью методов таксопринта и PCR-RAPD (на примере диких, домашних овец и агамных видов рептилий): Автореф. дис. ... канд. биол. наук. М., 1995. 33 с.
Мельникова М.Н., Гречко В.В., Медников Б.М. Исследование полиморфизма и дивергенции геномной ДНК на видовом и популяционном уровнях (на примере ДНК пород домашних овец и диких баранов) // Генетика. 1995. Т. 31, № 8. С. 1120-1131.
Мельникова М.Н., Медников Б.М. Исследование полиморфизма и дивергенции геномной ДНК домашних овец и диких баранов И Проблемы сохранения редких пород домашних животных и близкородственных диких видов. Пущино, 1996. С. 24-25.
Мельниченко А.В., Реймов Р.Р., Карабеков М.К. Места окота сайгака в каракалпакской части Устюрта // Экология, морфология, использование и охрана диких копытных. М., 1989. Ч. 2. С. 274-275.
МеньковаН.В. Заготовки мясной продукции диких копытных // Охотоведение. М.: Лесн. пром-сть, 1976. С. 189-197.
Мерзликин И.Р., Галета В.А., Гетъман Е.Т. Зубр Bison bonasus в Сумской области И VI съезд териол. о-ва. М., 1999. С. 158.
Метельский А.П. Снежные бараны хребта Джугджур и прилегающих отрогов // Редкие виды млекопитающих фауны СССР и их охрана. М.: Наука, 1973. С. 131-133.
Минасян Л.Г., Гаспарян К.М., Антонян А.Ш., Хримян А.Н. К вопросу о сохранении и использовании генофонда азиатского муфлона (арменийского муфлона) Ovis (Ovis) orientalis gmelini Blyth // Биол. журн. Армении. 1986а. № 8. С. 658-664.
Минасян Л.Г, Гаспарян К.М., Антонян А.Ш. и др. Особенности роста и развития арме-нийских муфлонов (азиатских горных баранов) в условиях неволи // Зоол. сб. Ин-та зоол. АН АрмССР. 19866. Вып. 20. С. 48-79.
Мицкус А.В., Турса ГК. Итоги и перспективы акклиматизации охотничье-промысловых зверей в Литовской ССР // Акклиматизация охотничьих животных в СССР. Минск: Ураджай, 1978. С. 22-25.
Мичурин Л.Н., Мироненко О.Н. О толстороге (Ovis nivicola borealis Severtzov, 1873) в горах Путорана// Зоол. журн. 1966а. Т. 45, вып. 11. С. 1736-1738.
Мичурин Л.Н., Мироненко О.Н. Распространение копытных в горах Путорана И Вопросы сельского и промыслового хозяйства Крайнего Севера. Красноярск, 19666. С. 69-75. (Тр. НИИСХ Крайнего Севера. Т. 14).
Мишарев С. С. Тур кавказский и его гибриды с домашними козами // Тр. Всесоюз. НИИ овцеводства и козоводства. 1969. Вып. 29, т. 1. С. 156-165.
Мориц Л. Безоаровый козел в Туркмении // Охотник. 1930. № И. С. 24—25.
Москвин А. С., Багрова Н.В. Гельминтофауна зубров Приокско-Террасного заповедника // К вопросу о возможности сохранения зубра в России. Пущино, 1993. С. 90-95.
Мосолов В. И. Экология копытных горновулканических районов Восточной Камчатки // Экосистемы экстремальных условий среды в заповедниках РСФСР. М., 1986а. С. 38-49.
Мосолов В. И. Численность и половозрастная структура популяции снежных баранов в Кроноцком заповеднике // Териология, орнитология и охрана природы. XI Всесоюз. симпоз. “Биол. пробл. Севера”. 19866. Вып. 3. С. 54-55.
Мосолов В.И. Снежный баран юго-восточного побережья Камчатки И Бюл. МОИП. Отд. биол. 1992. Т. 97, вып. 2. С. 10-18.
Мухамедгалиев Ф.М., Савицкий В. Ф. Кариотип овец породы казахский архаромеринос / / Генетика. 1974. Т. 10, № 4. С. 43-47.
Мымрин Н. Звери Чукотки: миграции и расселение И Охота и охотничье хоз-во. 2003. № 8. С. 18-20.
Мысленков А.И. Групповое поведение у горала (Nemorhaedus goral caudatus) // Групповое поведение животных. М.: Наука, 1976. С. 272-274.
Мысленков А.И. Поведение и внутрипопуляционная структура амурского горала (Nemorhaedus caudatus raddeanus Heude): Автореф. дис.... канд. биол. наук. М., 1982а. 24 с.
Мысленков А.И. Оборонительное поведение горала //Млекопитающие СССР. III съезд Всесоюз. териол. о-ва. М., 19826. Т. 2. С. 151-152.
Мысленков А.И. Структура внутрипопуляционных группировок амурского горала // IV съезд Всесоюз. териол. о-ва. 1986. Т. 1. С. 295.
Мысленков А.И. Поведение копытных во время солонцевания на морском побережье// Экология, морфология, использование и охрана диких копытных. М., 1989. Ч. 1. С. 64—65.
Мысленков А.И., Волошина И.В. Гон у горалов // Копытные фауны СССР. М.: Наука, 1975. С. 272-273.
Мысленков А.И., Волошина И.В. Поведение горала во время гона // Бюл. МОИП. Отд. биол. 1978. Т. 83, вып. 1. С. 30-39.
Мысленков А.И., Волошина И.В. Звуковая сигнализация амурского горала // Копытные фауны СССР. М.: Наука, 1980. С. 297-300.
Мысленков А.И., Волошина И.В. Экология и поведение амурского горала. М.: Наука, 1989. 128 с.
Мысленков А.И., Волошина И.В. Социальное доминирование в группах амурского горала // Редкие виды млекопитающих России и сопредельных территорий. М., 1997. С. 63.
Назарова Н. С. Влияние акклиматизации и доместикации на распространение гельминто-зов // Проблемы общей и прикладной гельминтологии. М.: Наука, 1973. С. 112-116.
Назарова Н. С., Киселева Е.Г. Зараженность гельминтами зубров Окского заповедника и опыт их дегельминтизации И Копытные фауны СССР. М.: Наука. 1980. С. 184-185.
Наниев В.И. Кавказский зубр - домбай (Bison banasus caucasicus Sat.) // Учен. зап. С.-Осет. гос. пед. ин-та им. К.Л. Хетагурова. 1956. Вып. 20. С. 249-276.
Наниев В.И. К биологии кавказского горно-каменного козла (Capra cylindricomis Blyth.) на Центральном Кавказе // Там же. 1958. Т. 23, вып. 1. С. 195-205.
Наниев В.И. О гибели восточнокавказских туров от снежных обвалов во время миграций И Миграции животных. М.: Изд-во АН СССР, 1962. Вып. 3. С. 37-39.
Насимович А.А. О мускусном быке И Сов. Север. 1933. № 2. С. 80-82.
Насимович А.А. О некоторых закономерностях зимнего распространения копытных в горах Западного Кавказа // Бюл. МОИП. Отд. биол. 1936а. Т. 45, вып. 1. С. 3-9.
Насимович А.А. Охота на крупную дичь в горах Черноморского Кавказа // Боец-охотник. 19366. № 11. С. 30-35.
Насимович А.А. К познанию минерального питания диких животных Кавказского заповедника//Тр. Кавказского гос. заповедника. 1938. Вып. 1. С. 103-152.
Насимович А.А. Зима в жизни копытных Западного Кавказа // Вопросы экологии и биоценологии. Л., 1939. Вып. 7. С. 3-91.
Насимович А.А. К методике количественного учета поголовья туров // Науч.-метод, зап. Главн. упр. по заповедникам, зоопаркам и зоосадам. 1940. Вып. 7. С. 23-28.
Насимович А.А. Количественный учет серн и динамика их поголовья в Кавказском заповеднике// Там же. 1941. Вып. 8. С. 178-181.
Насимович А.А. Очерк экологии западнокавказского тура // Тр. Кавказского гос. заповедника. 1949а. Вып. 3. С. 5-38.
Насимович А.А. Новые данные по биологии серны на Западном Кавказе И Там же. 19496. С. 51-64.
Насимович А.А. Роль режима снежного покрова в жизни копытных животных на территории СССР. М.: Изд-во АН СССР, 1955. 403 с.
Насонов Н.В. О диком восточном баране С. Гмелина (Ovis orientalis Pall.) // Изв. Импер. Акад. наук. 1910. С. 681-710. (Отд. оттиск).
Насонов Н.В. Муфлоны и близкие к ним формы диких баранов//Там же. 1911. С. 1267-1296. (Отд. оттиск).
Насонов Н.В. Ovis arcar и близкие к нему форм диких баранов // Там же. 1913а. С. 3-32. (Отд. оттиск).
Насонов НВ. О новом виде дикого барана из Южной Гоби Ovis kozlovi И Там же. 19136. С. 621-626. (Отд. оттиск).
Насонов Н.В. Виды диких баранов Туркестана, описанные Н.А. Северцовым // Там же.
1914. С. 695-726. (Отд. оттиск).
Насонов Н.В. Дополнительные сведения о диком восточном баране С. Гмелина (Ovis orientalis Gmel.) И Там же. 1916. С. 1767-1778. (Отд. оттиск).
Насонов Н.В. О “перверзии” в рогах диких баранов Ovis vignei Blyth, gmelini Blyth и urmiana (Guenther) // Там же. 1919. С. 1215-1246. (Отд. оттиск).
Насонов Н.В. Европейский муфлон (О. musimon Schreb.) по материалам Зоологического музея Академии наук. Там же. 1920. С. 421-452. (Отд. оттиск).
Насонов Н.В. Географическое распространение диких баранов Старого Света. Петроград, 1923. 255 с.
Насонов Н.В., Дорогостайский В. Ч. Диюе бараны (Ovis nivicola potanini) Яблоноваго хребта// Изв. Импер. Акад. наук. 1915. С. 1599-1616. (Отд. оттиск).
Наумов Н.П. Млекопитающие Тунгусского округа // Тр. Полярной комис. АН СССР. 1934. Вып. 17. С. 5-83.
Небольсин П.И. Очерки торговли России с Средней Азией И Зап. Импер. Рус. геогр. о-ва. СПб, 1855. С. Кн. 10. С. 1-445.
Недзельский Е.М., НедзельскийА.Е. Экология, морфология, численность и плотность сибирского горного козла в Предбайкалье. Иркутск, 2003.9 с. (Деп. в ВИНИТИ, № 1325-В2003).
Немцев А.С. Учет численности горных зубров во время яра // Копытные фауны СССР. М.: Наука. 1980. С. 36-38.
Немцев А.С. Формы социальных группировок зубров Кавказского заповедника // Там же. 19806. С. 300-301.
Немцев А.С. Динамика населения зубров Кавказского заповедника // Экология горных млекопитающих. Свердловск, 1982а. С. 83-85.
Немцев А.С. Морфо-экологические особенности черепа восстановленных горных зубров // Там же. 19826. С. 85-87.
Немцев А. С. Сорок лет восстановления зубров в Кавказском заповеднике // Редкие виды млекопитающих СССР и их охрана. М., 1983. С. 191-193.
Немцев А.С. Краниометрия и фенетика чистокровных и гибридных зубров // Фенетика популяций. М., 1985а. С. 266-268.
Немцев А.С. Динамика населения зубров Кавказского заповедника // Экологические исследования в Кавказском биосферном заповеднике. Изд-во Ростовского ун-та, 19856. С. 49-63.
Немцев А.С. Демографическая структура популяции гибридных горных зубров // Экология и охрана горных видов млекопитающих. М., 1987а. С. 122-124.
Немцев А.С. Проблемы восстановления зубров на Северном Кавказе // Там же. 19876.
С. 125-127.
Немцев А.С. Демографический анализ популяции горных зубров // Ресурсы животного мира Северного Кавказа. Ставрополь, 1988а. С. 129-131.
Немцев А.С. Перспективы и особенности охоты на зубров в Краснодарском крае // Там же. 19886. С. 131-133.
Немцев А.С. Особенности биологии, охрана и пути использования популяции горных зубров Северо-Западного Кавказа: Автореф. дис.... канд. биол. наук. М., 1988в. 20 с.
Немцев А.С. Эволюция пространственно-этологической структуры популяции зубров Северо-Западного Кавказа И V съезд Всесоюз. териол. о-ва АН СССР. М., 1990. Т. 2. С. 98-99.
Немцев А.С. Проблемы охраны и рационального использования популяции кавказских горных зубров//Тр. Кавказского гос. биосферного заповедника. 1994. Вып. 15. С. 185-192.
Немцев А.С. Социальная организация и важнейшие особенности образа жизни И Зубр на Кавказе. М. - Майкоп: “Качество”, 2003. С. 213-233.
Немцев А.С., Киселева Е.Г. Сравнительное морфоэкологическое изучение горных и равнинных зубров И Экология и охрана горных видов млекопитающих. М., 1987. С. 127-128.
Немцев А.С., Мироненко И.В. Охрана и использование современных зубров Кавказа // Зубр на Кавказе. М. — Майкоп: “Качество”, 2003. С. 234-248.
Немцев А.С., Пузаченко А.Ю., Раутиан Г. С. Экологическая характеристика зубров Северо-Западного Кавказа И Там же. 2003. С. 150-213.
Немцев А.С., Пузаченко А.Ю., РаутианГ.С., Сипко Т.П. Сравнительная характеристика экстерьера зубров различных подвидов и линий разведения И Там же. 2003. С. 76-97.
Немцев А.С., Раутиан Г. С., Пузаченко А.Ю. Горные зубры - эталон природной популяции зубров И Териофауна России и сопредельных территорий. VII съезд териол. о-ва. М„ 2003. С. 233.
Немцев А.С., Раутиан Г. С., Пузаченко А.Ю. и др. Зубр на Кавказе. М. - Майкоп: “Качество”, 2003. 292 с.
Немцев А.С., Сипко Т.П., Гребенков А.Б. Современное состояние кавказской серны (Rupicapra rupicapra caucasica) // VI съезд териол. о-ва. М., 1999. С. 174.
Неронов В.М., Лущекина А .А., Ямникова С. С. Материалы по эпизоотологической роли дзерена //Млекопитающие СССР. III съезд Всесоюз. териол. о-ва. М., 1982.Т. 1.С. 127-128.
Нестеренко И. Ф. Изучение транквилизирующего действия рогов сайги: Автореф. дис.... канд. биол. наук. Владивосток, 1971. 23 с.
Нестеров Д.А. Горал на Среднем и Южном Сихотэ-Алине // Редкие и исчезающие животные суши Дальнего Востока СССР. Владивосток, 1981. С. 179-180.
Нестеров Д.А. Сравнительная характеристика пастбищ горала - Nemorhaedus goral caudatus Milne-Edwards на Южном Сихотэ-Алине И Сихотэ-Алинский биосферный район: экологические исследования. Владивосток, 1985. С. 141-150.
НестеровД.А. Распространение горала в Приморском крае // Амурский горал. М.: Изд-во ЦНИИ охотничьего хоз-ва и заповедников, 1992. С. 6-20.
Нестеров Д.А., Смирнов Е.Н. Охрана Сихотэ-Алинской популяции амурского горала//Редкие и исчезающие животные суши Дальнего Востока СССР. Владивосток, 1981. С. 177-178.
Никифоров Н.М. Дзерен в Тувинской АССР И Редкие виды млекопитающих фауны СССР и их охрана. М.: Наука, 1973. С. 133-134.
Никифоров Н.М., Шурыгын В.В. Современное распространение и численность сибирского горного козла в Тувинской АССР // Там же. 1973. С. 134—135.
Никифоров Н.М., Шурыгын В.В., Данковцев А.Г. Распространение редких копытных в Тувинской АССР // Редкие виды млекопитающих и их охрана. М.: Наука, 1977. С. 220-222.
Обухов П. Снежные бараны на Колыме // Охота и охотничье хоз-во. 1969. № 2. С. 20-21.
Оводов Н.Д. К истории снежного барана {Ovis nivicola) // VI съезд териол. о-ва. М., 1999. С. 180.
Оводов Н. Судьба снежного барана И Охота и охотничье хоз-во. 2000. № 10. С. 10-11.
Оводов Н.Д., Ивлева Н.Г. Позднеантропогеновая териофауна Алтая // V съезд Всесоюз. териол. о-ва АН СССР. М., 1990. Т. 1. С. 24-25.
Оводов Н.Д., Орлова Н.Г. Фауна млекопитающих восточных склонов Кузнецкого Алатау в позднем плейстоцене - голоцене // Экология, методы изучения и организация охраны млекопитающих горных областей. Свердловск, 1977. С. 97-98.
Одинашоев А.О. Опыт содержания сибирского козерога в условиях неволи //1 Всесоюз. совещ. по пробл. зоокультуры. М., 1986. Ч. 1. С. 174-175.
Одинашоев А. О., Кедамшоев М.К., Темиров Т. Т. Распространение и численность архара на Памире // V съезд Всесоюз. териол. о-ва АН СССР. М., 1990. Т. 3. С. 164-165.
Окладников А.П., Запорожская В.Д. Петроглифы Забайкалья. Л.: Наука, 1970. Ч. 2. 263 с.
Окладников А.П., Кириллов И.И. Юго-Восточное Забайкалье в эпоху камня и ранней бронзы. Новосибирск: Наука, 1980. 176 с.
Оконешников В.В. К зимнему поведению снежных баранов Ovis nivicola И Териофауна России и сопредельных территорий. VII съезд териол. о-ва. М., 2003. С. 240.
Опрышко ИЛ. Ареал и численность сибирского горного козла в восточной части Восточного Саяна // Проблемы охотоведения и охраны природы. Иркутск, 1975. С. 130-131.
Орджоникидзе С.В., Хитъ А.В. Опыт содержания и разведения овцебыков {Ovibos moschatus) в Московском зоопарке И Достижения зоопарков в изучении биологии разведения редких и исчезающих животных. М., 1989. С. 9.
Орлов В.Н. Кариосистематика млекопитающих. Цитогенетические методы в систематике млекопитающих. М.: Наука, 1974. 207 с.
Орлов В.Н. Систематика горных баранов и происхождение домашних овец по кариоло-гическим данным // Эколого-морфологические особенности диких родичей домашних овец. М.: Наука, 1978. С. 5-17.
Орлов В.Н. Зубры вырождаются // Химия и жизнь. 2000. № 2. С. 6-7.
Орлов В.Н., БулатоваН.Ш. Сравнительная цитогенетика и кариосистематика млекопитающих. М.: Наука, 1983.406 с. .
Орлов В.Н., Раджабли С.И., Малыгин В.М. и др. Кариотипы млекопитающих Монголии И География и динамика растительного и животного мира МНР. М.: Наука, 1978. С. 149-164.
Орлов В.Н., ТумуржавМ., Малыгин В.М. Кариотипы горных баранов {Ovis a. przewalskii) и домашних овец Центральной Азии в связи с происхождением последних И Систематика и цитогенетика млекопитающих. М.: Наука, 1975. С. 47-48.
Орлов В.Н., Чудиновская ГА. Хромосомный набор зубра и других быков трибы Bovini // Зубр. М.: Наука, 1979. С. 435-441.
Орлов В.Н., Чудиновская Г.А., Горелов Ю.К. Кариотипы гибридов уриала (Ovis аттоп cycloceros) с каракульской овцой // Вопросы гибридизации копытных. М.: Наука, 1980. С. 89-91.
Орлов Е.И. Материалы к познанию фауны наземных позвоночных Калмыцкой области // Матер, к позн. фауны Нижнего Поволжья. Саратов, 1928. Вып. 2. С. 1-47.
Охлопков И.М. Охотничье-промысловые млекопитающие гор Центрального Верхоянья: ресурсы и перспективы использования // Современные проблемы природопользования, охотоведения и звероводства. Киров, 2002. С. 324—326.
ОчировЮ.Д., БашановКА. Млекопитающие Тувы. Кызыл: Тувинское кн. изд-во, 1975.139 с.
Паавер К.Л. Формирование териофауны и изменчивость млекопитающих Прибалтики в голоцене. Тарту, 1965. 497 с.
Павленко Т.А., Тарянников В.И. Промысел сайгака в Узбекистане // Копытные фауны СССР. М.: Наука, 1975. С. 223-224.
Павлинов И.Я. Систематика современных млекопитающих. М.: Изд-во Моск, ун-та, 2003. 297 с.
Павлинов И.Я., Борисенко А.В., Крускоп С.В., Яхонтов Е.Л. Млекопитающие Евразии. И. Non-Rodentia: систематико-географический справочник. М.: Изд-во Моск, ун-та, 1995. 336 с.
Павлинов И.Я., Россолимо О.Л. Систематика млекопитающих СССР. М.: Изд-во Моск, ун-та, 1987. 285 с.
Павлов Б.М. Снежный баран (толсторог) // Охотничье хозяйство Енисейского Севера. Красноярское кн. изд-во, 1977. С. 87-90.
Павлов Е.И. Промысловые звери Читинской области. Чита: Обл. издат., 1949. 188 с.
Павлов М.П. Акклиматизация охотничье-промысловых зверей и птиц в СССР. Киров, 1999. Ч. 3.666 с.
Павлова М.В. Описание ископаемых остатков млекопитающих Троицкосавско-Кяхтин-ского музея // Тр. Троицкосавско-Кяхтинского отд. Приамур. отд. Рус. геогр. о-ва. 1911. Т. 13, вып. 1. С. 21-59.
Пакиж В. Архары в Казахстане // Охота и охотничье хоз-во. 1958. № 9. С. 24.
Паллас П. С. Путешествие по разным местам Российского государства по повелению Санкт-Петербургской Императорской академии наук. СПб., 1786. (1770). Ч. 2, кн. 2. 571 с.
Паллас П.С. Путешествие по разным провинциям Российской Империи. СПб., 1809 (второе издание). Ч. 1. 657 с.
Палъцын М.Ю., Спицын С.В. Использование геоинформационной системы (ГИС) для определения численности алтайского горного барана аргали в южной части хребта Чихачева // Изучение и охрана природы Алтае-Саянской горной страны. Горно-Алтайск, 2002а. С. 104-106.
Палъцын М.Ю., Спицын С.В. Рекомендации по сохранению группировки алтайского горного барана аргали на хребте Чихачева // Там же. 20026. С. 107-111.
Паршинцев А.В. Современное состояние популяции европейского муфлона в Крымском заповедно-охотничьем хозяйстве И Охрана и изучение редких и исчезающих видов животных в заповедниках. М., 1992. С. 56-64.
Певцов М.В. Путешествия по Китаю и Монголии. М.: Гос. изд. географ, лит-ры, 1951.283 с.
Пейсахис Л.А., Корнеев ГА., Самсонович Л.Г. и др. Экспериментальная чума у сайгаков // X науч. конф, противочумных учрежд. Средней Азии и Казахстана. Алма-Ата, 1979. Вып. 1.С. 148-151.
Перерва В.И. Гельминтофауна зубров Приокско-Террасного заповедника // Копытные фауны СССР. М.: Наука, 1980. С. 191-192.
Перерва В.И. Особенности экологии зубра при содержании в питомниках // Биологические аспекты охраны редких животных. М., 1981. С. 33-36.
Перерва В. И. Разработка программы интродукции зубров в природу с учетом их адаптации к климатическим условиям // Научные основы охраны и рационального использования животного мира. М., 1982. С. 49-57.
Перерва В.И. Изменение экологии зубров под влиянием условий разведения в неволе // Популяционная изменчивость вида и проблемы охраны генофонда млекопитающих. М., 1983а. С. 151-152.
Перерва В.И. Особенности линьки зубров в условиях загонного содержания в Приокс-ко-Террасном заповеднике // Редкие виды млекопитающих СССР и их охрана. М., 19836. С. 195.
Перерва В.И. Инбридинг в разведении зубров в неволе //1 Всесоюз. совещ. по пробл. зоокультуры. М., 1986. Ч. 2. С. 68-70.
Перерва В.И. Проблемы возвращения редких видов животных в трансформированную антропогенным воздействием среду (на примере зубра) // Влияние антропогенной трансформации ландшафта на население наземных позвоночных животных. М., 1987. Ч. 1. С. 140-143.
Перерва В.И. Маркировочное поведение зубров в условиях неволи // Коммуникативные механизмы регулирования популяционной структуры у млекопитающих. М., 1988. С. 132-135.
Перерва В.И. Успешность размножения и иерархический статус зубров И Экология, морфология, использование и охрана диких копытных. М., 1989а. Ч. 2. С. 228-229.
Перерва В.И. Некоторые особенности ведения охотничьего хозяйства по зубру // Там же. 19896. Ч. 2. С. 230-231.
Перерва В.И. Угрожающее поведение зубра (Bison bonasus) в сравнении с бизоном (В. bison) И Зоол. журн. 1989в. Т. 68, вып. 1. С. 124—132.
Перерва В.И. Возвращение зубра. М.: Колос, 1992. 207 с.
Перерва В.И. Современные проблемы сохранения зубра в России // Редкие виды млекопитающих России и сопредельных территорий. М., 1997. С. 71.
Перерва В.И. Зубр Bison bonasus bonasus (Linnaeus, 1758) // Красная книга Российской Федерации (животные). Балашиха: Астрель, 2001. С. 711-713.
Перерва В.И., Жирнов Л.В. Зубр - потенциальный объект охоты (современное состояние и перспективы решения проблемы) // Дичь, охрана окружающей среды, охрана редких видов животных. М., 1995. С. 328-338.
Перерва В.И, Литус И.Е., Крыжановский В.И. Состояние поголовья зубров на Украине и перспективы его рационального использования И Вести, зоологии. 1991. № 5. С. 11-15.
Перерва В.И., Перерва А.В. Зубр - прошлое и настоящее // Териофауна России и сопредельных территорий. VII съезд териол. о-ва. М., 2003. С. 257.
Перерва В.И., Перерва А.В., Верижников Ю.А., Александров А.А. Современное состояние и перспективы формирования вольной популяции зубров на западе европейской части России // Проблемы сохранения и восстановления диких копытных животных в Центральном регионе России. Орел: Изд-во ОРАГС, 2003. С. 75-79.
Перовский М.Д. Учет сайгаков//Охотоведение. М., Лесн. пром-сть, 1974. С. 186-201.
Перовский М.Д. Итоги акклиматизации некоторых копытных в РСФСР // Повышение продуктивности охотничьих угодий. М., 1984. С. 120-133.
Перовский М.Д. Пути улучшения освоения ресурсов мяса диких животных. М., 1985. 49 с.
Перовский М.Д. Роль различных факторов в потерях популяций диких копытных животных в РСФСР И Хронологические изменения численности охотничьих животных в РСФСР. М., 1988. С. 106-126.
Петрашов В.В. Вопросы расселения пятнистого оленя и снежного барана И Повышение продуктивности охотничьих угодий. М., 1982. С. 138-150.
Петренко А.Г. Древнее и средневековое животноводство Среднего Поволжья и Преду-ралья. М.: Наука, 1984. 174 с.
Петрищев Б.И. Принципы содержания сайгаков в неволе и на ограниченной территории// IV съезд Всесоюз. териол. о-ва. М., 1986. Т. 3. С. 159-162.
Петрищев Б.И. Пустыне нужны сайгаки. М.: Мысль, 1987а. 127 с.
Петрищев Б.И. Сайгак в окультуренном ландшафте // Охота и охотничье хоз-во. 19876. №5. С. 16-18.
Петрищев Б.И. Влияние пренатального развития Saiga tatarica на размеры и смертность новорожденных // Состояние териофауны в России и ближнем зарубежье. М., 1996. С. 264-271.
Петрищев Б.И. Эмбриогенез сайгака (Saiga tatarica)-. формирование фенотипических признаков роста // Европейский сайгак: развитие, морфология, экология. М.: Наука, 1997а. С. 5-15.
Петрищев Б.И. Характеристика эмбриогенеза сайгака (Saiga tatarica) и оценка состояния приплода // Зоол. журн. 19976. Т. 76, вып. 2. С. 234—242.
Петрищев Б.И. Эмбриональное развитие// Сайгак. М., 1998. С. 193-202.
Петрищев Б.И. Изменчивость оборонительной реакции сайгаков в онтогенезе // II съезд Вавиловского о-ва генет. и селекционеров. 2000. Т. 1. С. 266.
Петрищев Б.И. Интенсивность использования биотопов аргали Ovis а. аттоп в верховьях Чаган-Бургазы (юго-восток Республики Алтай) в зависимости от времени года // Териофауна России и сопредельных территорий. VII съезд териол. о-ва. М., 2003. С. 259.
Петрищев Б.И, Абатуров БД., Максимук А.В. Методические приемы ранчирования сайгаков // Европейский сайгак: развитие, морфология, экология. М.: Наука, 1997. С. 162-183.
Петрищев Б.И, Арылов Ю.Н. Репродуктивное поведение сайгаков Saiga tatarica при разведении в неволе И Териофауна России и сопредельных территорий. VII съезд териол. о-ва. М., 2003а. С. 258-259.
Петрищев Б.И, Арылов Ю.Н. Значение оборонительного поведения сайгаков Saiga tatarica при содержании в вольерах // Там же. 20036. С. 258-259.
Петрищев Б.И., Арылов Ю.Н., Лущекина А.А., Холодова М.В. Центры разведения сайгаков и их роль в сохранении генофонда и повышении продуктивности различных популяций // Млекопитающие как компонент аридных экосистем (ресурсы, фауна, экология, медицинское значение и охрана). М., 2004. С. 99-100.
Петрищев Б.И., Максимук А.В., Абатуров БД. Методика отлова и содержания молодняка сайгаков // Сайгак. М., 1998. С. 281-289.
Петрищев Б.И, Петросян В.Г. Влияние горного рельефа на размер групп аргали Ovis а. аттоп и их распределение в верховьях Чаган-Бургазы (юго-восток Республики Алтай) // Териофауна России и сопредельных территорий. VII съезд териол. о-ва. М., 2003. С. 259-260.
Петрищев Б.И, Холодова М.В. Выкармливание сайгаков в полевых условиях // Копытные фауны СССР. М.: Наука, 1980а. С. 250-252.
Петрищев Б.И., Холодова М.В. Линейно-весовой рост сайгаков, выращенных в неволе // Там же. 19806. С. 357-358.
Петров В.И. Особенности разведения зубров в Приокско-Террасном заповеднике И Раз-ведение, селекция, генетика и воспроизводство сельскохозяйственных животных. СПб, 2003. С. 75-77.
Петров В. С. Гельминты сайгаков и их значение в эпизоотологии гельминтозов овец: Автореф. дис. ... канд. вет. наук. М., 1985. 24 с.
Петров В.С., Исламов Р.З., Дутов М.М. Некробактериоз у сайгаков //Ветеринария. 1979. № 3. С. 48.
Пидопличко ИГ О ледниковом периоде. Киев: Изд-во АН УкрССР, 1951. Вып. 2. 264 с.; 1954. Вып. 3. 220 с.
Шдогшчко 1.Г. Матер1али до вивчення минулих фаун УРСР. Ки1в: Видавн. АН УРСР, 1956. Вып. 2. 235 с.
Пикунов Д.Г, Абрамов В.К., Скрипчинский К.К. Некоторые особенности распространения и охраны редких видов млекопитающих юга Дальнего Востока И Редкие виды млекопитающих фауны СССР и их охрана. М.: Наука, 1973. С. 25-26.
Плахов К.Н. Устюртский горный баран Ovis аттоп areal в Казахстане // Редкие виды млекопитающих России и сопредельных территорий. М., 1997. С. 74.
Плахов К.Н. Устюртский горный баран Ovis vignei areal // VI съезд териол. о-ва. М., 1999. С. 197.
Плахов К.Н. Устюртский горный баран Ovis vignei areal. Сообщение 1: Современное состояние популяций И Териофауна России и сопредельных территорий. VII съезд териол. о-ва. М., 2003а. С. 262-263.
Плахов К.Н. Устюртский горный баран Ovis vignei areal. Сообщение 2: Охрана и использование ресурсов популяций в Казахстане // Там же. 20036. С. 263.
Плешак Т.В. Центрально-азиатский козел в заповеднике Аксу-Джабаглы и Заилийском Алатау // Редкие виды млекопитающих фауны СССР и их охрана. М.: Наука, 1973. С. 136-137.
Плегиак Т.В. К экологии архара весной И Копытные фауны СССР. М.: Наука, 1980. С. 194-195.
Подаревский В.Б. Проблемы охотхозяйственной акклиматизации в Восточной Сибири. Иркутск: ОГИЗ, 1936. 119 с.
Подопригора И.Н. Териофауна крупных млекопитающих Пермского Приуралья в по-зднеледниковье И Териофауна России и сопредельных территорий. VII съезд териол. о-ва. М., 2003. С. 268.
Поливанова Н.Н. Тебердинский заповедник // Заповедники СССР. Заповедники Кавказа. М.: Мысль, 1990. С. 18-29.
Поляков И.В. Отчет об Олекминско-Витимской экспедиции 1866 года. Зоологические наблюдения И Зап. Импер. Рус. геогр. о-ва по общ. географ. 1873. Т. 3. С. 1-175.
Пономарев Д.В. Крупные млекопитающие Европейского Северо-Востока в позднем плейстоцене и голоцене. Сыктывкар, 2001. 48 с.
Пономарев Д.В. Позднеплейстоцен-голоценовая мегатериофауна Северо-Востока европейской части России И Четвертичная палеозоология на Урале. Екатеринбург: Изд-во Уральского ун-та, 2003. С. 123-132.
Попкова И. Ф. Серна на южных склонах Главного Кавказского хребта (экология, морфология и хозяйственное значение)//Тр. Тебердинского гос. заповедника. 1967. Вып. 7. С. 160-211.
Портенко А.А., Кищинский А.А., Чернявский Ф.Б. Млекопитающие Корякского нагорья. Л.: Изд-во АН СССР, 1963. 131 с.
Поспелова Е.Б. Пастбища овцебыков в бассейне р. Бикада и их продуктивность И Млекопитающие и птицы севера Средней Сибири. Новосибирск, 1989. С. 79-87.
Потапов С.Г. Диагностические возможности молекулярно-генетических маркеров в систематике млекопитающих (на примере грызунов и копытных): Автореф. дис. ... канд. биол. наук. М., 1998. 24 с.
Потапов С.Г., Кудрявцев И.В., Рысков А.П. Использование таксономического типи-рования ДНК для анализа геномной вариабельности представителей Bovidae Gray, 1821 // Генетика. 1994. Т. 30, № 6. С. 858-860.
Потапов С.Г., Токарская О.Н., Семенова С.К. и др. Диагностические возможности мультилокусных маркеров ДНК в систематике диких копытных животных (Artiodactyla) // Генетика. 1997. Т. 33, № 7. С. 961-966.
Праслов Н. Мамонт в жизни палеолитического человека // Цитология. 1995. Т. 37, № 7. С. 634.
Прасолов В. С. Вернуть овцебыка в Евразию // Природа. 1973. № 9. С. 98-101.
Пржевальский Н.М. Монголия и страна тангутов. М.: ОГИЗ, 1946. 333 с. (СПб.: изд. Импер. Рус. геогр. о-ва, 1875. Т. 1).
Прилуцкая Л.И., Пишванов Ю.В. Современная численность и размещение охотничье-промысловых животных Дагестана И Ресурсы животного мира Северного Кавказа. Ставрополь, 1988. С. 136-140.
Прилуцкая Л.И., Пишванов Ю.В. Ареал и численность безоарового козла в Дагестане // Экология, морфология, использование и охрана диких копытных. М., 1989. Ч. 2. С. 243-245.
Прилуцкая Л.И., Пишванов Ю.В. Безоаровый козел и кавказская выдра в Дагестане // V съезд Всесоюз. териол. о-ва АН СССР. М., 1990. Т. 3. С. 112-113.
Присяжнюк В.Е. Кызылкумский горный баран, или баран Северцова Ovis аттоп severtzovi Nasonov, 1914 // Красная книга СССР. М.: Лесная пром-сть, 1984. Т. 1. С. 82.
Присяжнюк В.Е. Снежный баран Ovis nivicola Eschscholtz, 1829 И Красная книга Российской Федерации (животные). Балашиха: Астрель, 2001. С. 721—724.
Присяжнюк В.Е., Проняев А.В. Сравнительный анализ размеров сайгаков Калмыкии и Северного Казахстана И Копытные фауны СССР. М.: Наука, 1980. С. 363-364.
Присяжнюк В.Е., Проняев А.В. Продукция сайгачьего промысла // Научные основы охраны и рационального использования животного мира. М., 1982. С. 57-68.
Присяжнюк В.Е., Проняев А.В. Краниометрическая характеристика сайгаков Калмыкии и Северного Казахстана ИIV съезд Всесоюз. териол. о-ва. М., 1986. Т. 3. С. 162-163.
Прокофьев С.М. Фауна и состояние численности охотничьих млекопитающих в Хакасии// Экология промысловых животных Сибири. Изд-во Красноярского ун-та, 1992. С. 20-37.
Проняев А.В. Изменчивость размеров тела сайгаков Калмыкии // Вопросы рационального использования и охраны диких копытных и других животных. М., 1978. С. 82-84. (Сб. науч. тр. Моск. вет. акад. им. К.И. Скрябина. Т. 97).
Проняев А.В. Фенотипическая, генотипическая характеристики и современное состояние популяций сайгака: Автореф. дис.... канд. биол. наук. М., 1985а. 23 с.
Проняев А.В. Фенотипическая характеристика популяций сайгака на основе количественных признаков экстерьера И Управление популяциями диких копытных животных. М., 19856. С. 43-54.
Проняев А.В. Фенотипическая, генотипическая характеристики популяций сайгака// IV съезд Всесоюз. териол. о-ва. М., 1986. Т. 3. С. 163-165.
Проняев А.В. Микроэволюционная роль антропогенных воздействий в изолированной популяции сайгака Северо-Западного Прикаспия // Молекулярные механизмы генетических процессов. М., 1990. С. 100.
Проняев А.В. Микроэволюционные процессы и рациональное использование популяций животных: Автореф. дис. ... докт. биол. наук. М., 1998. 48 с.
Проняев А.В., Аксенов С.В. Фенотипическая изменчивость изолированной популяции сайгака в ходе изменения численности // Хронологические изменения численности охотничьих животных в РСФСР. М., 1988. С. 143-151.
Проняев А.В., Бакай А.В., Васильев В.Г. Цитогенетическое исследование калмыцкой популяции сайгака И I Всесоюз. совещ. по пробл. зоокультуры. М., 1986. Ч. 1. С. 185-186.
Проняев А.В., Рожков Ю.И. Генетический эффект действия стабилизирующего отбора в изолированной популяции сайгака (Saiga tatarica) // Генетика. 1988. Т. 24, № 7. С. 1256-1261.
Проняев А.В., Фандеев А.А. Постнатальный онтогенез. Возрастная изменчивость черепа// Сайгак. М., 1998. С. 202-207.
Прусаков А.А. К акклиматизации зубров в Сары-Челекском биосферном заповеднике // Экология, морфология, использование и охрана диких копытных. М., 1989. Ч. 2. С. 231-233.
Прядко Э.И, Баитурсинов К.К., Осипов П.П., Беркинбаев О. Трансформация гельмин-тофауны сайги на путях миграции хозяина И Зоологические исследования в Казахстане. Алматы, 1993. Ч. 2. С. 237-258.
Пузаченко А.Ю., Раутиан Г.С. Половой диморфизм представителей рода Bison (Bovidae, Artiodactyla) по признакам черепа и идентификация пола у Bison priscus И Палеонтол. журн. 2001. № 1. С. 85-98.
Пузаченко А.Ю., Раутиан Г.С., Сипко Т.П., Киселева Е.Г. Дифференциация современных и вымерших форм рода Bison по признакам черепа И Редкие виды млекопитающих России и сопредельных территорий. М., 1999. С. 330-338.
Пуцек 3. Европейский зубр: современное состояние и проблемы И Охрана природы, наука и общество. М., 1987. Т. 2. С. 32-35.
Пуцек 3., Белоусова И.П., Лапшов В.А. и др. Зубр (Bison bonasus) — итоги и перспективы исследований: проект международной программы сохранения редкого вида // Зоол. журн. 1996. Т. 75, вып. 12. С. 1873-1883.
ПучковН. Плейстоценовый кризис в Палеарктике И Цитология. 1995. Т. 37, № 7. С. 635-636.
Пфиценмайер Е.В. Несколько интересных ублюдков сем. полорогих из Закавказья И Изв. Кавказского музея. Тифлис, 1915. Т. 8, вып. 3-4 (1914). С. 251-266.
Равкин Е.С. Численность и размещение туров Северо-Восточного Кавказа И Охотоведение. М.: Лесн. пром-сть, 1975а. С. 286-301.
Равкин Е.С. Ресурсы диких копытных на Северном Кавказе и антропогенное воздействие на них И Копытные фауны СССР. М.: Наука, 19756. С. 17-18.
Равкин Е. С. Ресурсы диких копытных животных Северо-Восточного Кавказа // Там же. 1976. С. 82-95.
Равкин Е. С., Овчаренко Г.И. О турах на Северном Кавказе И Редкие виды млекопитающих фауны СССР и их охрана. М.: Наука, 1973. С. 137-139.
Радде Г.И. Коллекции Кавказского музея. Зоология. Тифлис, 1899. Т. 1. С. 521 с.
Разевиг В.А. Кавказские горные козлы. (Туры.) // Псовая и ружейная охота. 1904. № 4. С. 25-38. № 5. С. 82-95. № 6. С. 38-54.
Размахнин В. Рога сайгака и тибетская медицина // Охота и охотничье хоз-во. 1969- № 2. С. 20-21.
Размахнин В.Е. Зубр И Копытные звери. М.: Лесн. пром-сть, 1977а. С. 240-249.
Размахнин В.Е. Сибирский горный козел И Там же. 19776. С. 164—175.
Размахнин B E. Способы промысловой добычи копытных И Итоги науки и техники. Т. 13. Зоология позвоночных. М.: ВИНИТИ, 1985. С. 138-153.
Размахнин В.Е. Рога сайгака - ценное лекарственное сырье // IV съезд Всесоюз. териол. о-ва. М., 1986. Т.З. С. 165-167.
Размахнин В.Е., Рывкин Л.М. Рога диких копытных - ценное лекарственное сырье // Охотоведение. М.: Лесн. пром-сть, 1976. С. 197-266.
Размахнин В., Силаев А., Сухарева Н., Челъцова Г. Рога сайгаков // Охота и охотничье хоз-во. 1976. № 10. С. 16-17.
РакА.С. Сайгаки в Грозненской области//Изв. Грозненского обл. краевед, музея. 1956. Вып. 7-8. С. 230-237.
Раков Н.В. Возможные причины изменений численности сайгаков в палеолите Крыма // Природная обстановка и фауны прошлого. Киев: Изд-во АН УССР, 1963. Вып. 1. С. 147-151.
Раков Н.В. О роли волка и других хищников в ограничении численности сайгака // Тр. Инта зоол. АН КазССР. 1955. Т. 4. С. 56-66.
Раков Н.В. Сайгак в Западном Казахстане И Там же. 1956. Т. 6. С. 28-60.
Раков Н.В. Опыт аэровизуального учета крупных млекопитающих в Центральном Казахстане// Зоол. журн. 1957. Т. 36, вып. 9. С. 1403-1411.
Раков Н.В. Об обитании горалов в левобережье Приамурья И Редкие и исчезающие животные суши Дальнего Востока СССР. Владивосток, 1981. С. 181-184.
Рапота В.В. Пастбищные сезоны и обеспечение овцебыков естественными кормами в условиях изгородного выпаса на Таймыре // Проблемы охраны и хозяйственного использования ресурсов диких животных Енисейского Севера. Новосибирск, 1979. С. 82-96 (Тр. НИИ сельск. хоз-ва Крайнего Севера. Т. 26).
Рапота В.В. Сосудистые растения района р. Бикады (Восточный Таймыр) и их кормовое значение для овцебыков И Экология и хозяйственное использование наземной фауны Енисейского Севера. Новосибирск, 1981. С. 73-93.
Раутиан Г. С. Флуктуирующая асимметрия по признакам черепа // Зубр на Кавказе. М. -Майкоп: “Качество”, 2003а. С. 97-107.
Раутиан Г. С. Нарушение положения щечных зубов в нижней челюсти зубров // Там же. 20036. С. 107-112.
Раутиан Г. С., Агаджанян А.К. Дифференциация ряда видов Bovinae по частотам групп крови // Редкие виды млекопитающих России и сопредельных территорий. М., 1999. С. 339-348.
Раутиан Г. С., Агаджанян А.К., Мироненко И.В. Дифференциация трибы быков и буйволов (Bovini) по морфологическим и генетическим признакам // Палеонтол. журн. 2000. № 5. С. 95-104.
Раутиан Г. С., Калабушкин Б.А. Генетические исследования зубров И Зубр на Кавказе. М. - Майкоп: “Качество”, 2003. С. 111-136.
РаутианГ.С., Калабушкин Б.А., Немцев А.С. Новый подвид зубра Bison bonasus montanus ssp. nov. (Bovidae, Artiodactyla) // Докл. PAH. 2000. T. 375, № 4. C. 563-567.
РаутианГ.С., Немцев А.С. Введение//Зубр на Кавказе. М.-Майкоп: “Качество”, 2003а.
С. 8-14.
Раутиан Г.С., Немцев А.С. Заключение//Там же. 20036. С. 249-253.
Раутиан Г.С., Немцев А.С, Пузаченко А.Ю. Краниологическая характеристика зубров // Там же. 2003. С. 51-76.
Раутиан Г. С., Пузаченко А.Ю. Ископаемые зубры и родственные им формы на Кавказе//Там же. 2003. С. 15-31.
Раутиан Г.С., Пузаченко А.Ю., Буневич А.Н. Участие кавказского подвида в формировании современных беловежских зубров // Териофауна России и сопредельных территорий. VII съезд териол. о-ва. М., 2003. С. 287.
Раутиан Г.С., Пузаченко А.Ю., Сипко Т.П. Асимметрия черепа современных и субре-центных зубров Bison bonasus (Bovidae, Artiodactyla) И Зоол. журн. 1998. Т. 77, вып. 12. С. 1403-1413.
РаутианГ.С., Пузаченко А.Ю., Сипко Т.П., КиселеваЕ.Г. Сравнительный анализ асимметрии фенов и морфометрических признаков у рецентных и субрецентных зубров // Динамика биоразнообразия животного мира. М., 1997. С. 13-18.
Раутиан Г. С., Сипко Т.П. Дифференциация современных и ископаемых форм рода Bison по фенам черепа // Редкие виды млекопитающих России и сопредельных территорий. М., 1997. С. 80.
Раутиан Г.С., Сипко Т.П., Немцев А.С. Происхождение современных популяций зубров Кавказа И Зубр на Кавказе. М. - Майкоп: “Качество”, 2003. С. 32-50.
Раутиан Г.С., Сипко Т.П., Уханов С.В. Генетический полиморфизм зубра // Генетика. 1994. Т. 30. Прилож. С. 131.
Рахимов А.А. О гибридизации серых каракульских овец с архаром // Наследственность и изменчивость растений, животных и микроорганизмов. М.: Изд-во АН СССР, 1959. Т. 1. С. 797-811.
Рахимов А.А. Кызылкумские бараны-архары и их гибриды с каракульскими овцами: Автореф. дис. ... докт. с.-х. наук. Ереван, 1966. 39 с.
Рашек В.Л. Лов сайгаков сетями И Охота и охотничье хоз-во. 1959. № 10. С. 8-10.
Рашек В.Л. Материалы по размножению сайгаков на острове Барса-Кельмес (Аральское море) // Тр. Ин-та зоол. АН КазССР. 1963. Т. 20. С. 164—193.
Рашек В.Л. Зима в жизни сайгаков острова Барса-Кельмес // Охотничье-промысловые звери. М., Россельхозиздат, 1965. Вып. 1. С. 57-81.
Рашек В.Л. Биология сайгаков острова Барса-Кельмес: Автореф. дис. ... канд. биол. наук. М., 1974. 21с.
Рашкевич Н.А. Результаты реакклиматизации зубра в Чечено-Ингушской АССР // Редкие виды млекопитающих фауны СССР и их охрана. М.: Наука, 1973. С. 139-140.
Ребров А. Ф. Перекочевье антилоп, диких коз и лошадей в Кумские камыши // Журн. сельск. хоз-ва и овцеводства. 1849. № 1. С. 17-18.
Ревин Ю.В. О распространении и численности снежного барана (Ovis nivicola Eschsch.) в Якутии // Распространение и экология млекопитающих Якутии. Якутск, 1982а. С. 82-97.
Ревин Ю.В. Опыт авиаучета снежных баранов (Ovis nivicola) в Якутии // Зоол. журн. 19826. Т. 61, вып. 4. С. 593-599.
Ревин Ю.В. Млекопитающие Южной Якутии. Новосибирск: Наука, 1989. 321 с.
РевинЮ.В., СопинЛ.В. Проблемы охраны и рационального использования популяций снежного барана И Редкие наземные позвоночные Сибири. Новосибирск: Наука, 1988. С. 191-197.
РевинЮ.В., СопинЛ.В., ЖелезновН.К. Снежный баран. Новосибирск: Наука, 1988.193 с.
Реймов А., Реймов Р. Антропогенное влияние на состояние популяции редких видов млекопитающих на Каракалпакском Устюрте и Кызылкуме И Редкие виды млекопитающих России и сопредельных территорий. М., 1997. С. 81.
Реймов Р. Млекопитающие Южного Приаралья, их охрана и рациональное использование в условиях деградирующей природной среды: Дис. ... докт. биол. наук в форме науч, доклада. Екатеринбург, 1994. 48 с.
Реймов Р., Карабеков М. Состояние популяции сайгака в Южном Устюрте // Копытные фауны СССР. М.: Наука, 1980. С. 197-198.
Реймов Р.Р., Карабеков М.К. Динамика численности сайгака в Каракалпакской части Устюрта // IV съезд Всесоюз. териол. о-ва. М., 1986. Т. 3. С. 169-170.
Реймов Р.Р, Карабеков М.К., Асенов Г.А. Современное состояние среды обитания сайгака в Южном Приаралье и его охрана // Там же. 1986. С. 171-172.
Реймов Р, Карабеков М., Нуратдинов Т. Динамика численности копытных пустынь Южного Приаралья // Влияние антропогенной трансформации ландшафта на население наземных позвоночных животных. М., 1987. Ч. 1. С. 207-208.
Реймов Р., Карабеков М., Сапарбаев Д. Численность сайгаков Устюрта и их охрана // Копытные фауны СССР. М.: Наука, 1975. С. 248-249.
Реймов Р„ Нуратдинов Т, Карабеков М. Состояние популяций копытных Южного Приаралья, включенных в “Красную книгу СССР” // Экология, морфология, использование и охрана диких копытных. М., 1989. Ч. 1. С. 72-73.
Ременцова М.М. Бруцеллез диких животных. Алма-Ата, 1962. 272 с.
Решетов В.Ю. Сравнительно-остеологический анализ посткраниального скелета бизонов в связи с историей рода Bison // Териология. Новосибирск, Наука, 1972. Вып. 2. С. 57-64.
Рожков Ю.И., Проняев А.В. Микроэволюционный процесс. М., 1994. 364 с.
Розмахов И.Г. К проблеме минерального питания диких животных Кавказского заповедника // Науч.-метод, записки Главн. упр. по заповедникам, зоопаркам и зоосадам. 1940. Вып. 7. С. 201-210.
Романов В. Почти исчезнувший // Лес и охотничье хоз-во. 2001. № 2. С. 46-47.
Ромашин А.В. Современное состояние поголовья западнокавказского тура в Краснодарском крае // Экологические проблемы Ставропольского края и сопредельных территорий. Ставрополь, 1989. С. 330.
Ромашин А.В. Динамика сообщества жвачных животных Кавказского заповедника // Тр. Кавказского гос. биосферного заповедника. 1994. Вып. 15. С. 193-201.
Ромашин А.В. Статистический анализ факторов, детерминирующих летнее размещение копытных в луговом поясе Кавказского заповедника // Экология. 1995. № 1. С. 20-28.
Ромашин А.В. Уточнение численности по данным выборочных учетов с помощью имитационного моделирования и интерпретация ее динамики на примере тура (Capra caucasica Guld.) Кавказского заповедника // Там же. 1997. № 5. С. 382-387.
Ромашин А.В. Эколого-популяционный анализ высокогорных копытных животных Западного Кавказа и их рациональное использование. Сочи, 2001. 184 с.
Рубайлова Н.Г. Вопросы отдаленной гибридизации копытных в СССР и хозяйственного использования гибридов // Вопросы гибридизации копытных. М.: Наука, 1980. С. 3-15.
Рудков А. С. Условия обитания и биология снежного барана // Природа. 1979. № 12. С. 82-85.
Румянцев Б. Ф., Бутарин Н. С., Денисов В. Ф. Опыт гибридизации курдючных овец с диким бараном Тянь-Шаня (Ovispolii karelini Sev.) // Видовая гибридизация домашних животных с дикими видами. М.-Л., 1935. С. 15-58. (Тр. Киргизск. компл. экспед. 1933-1934 гг. Т. 4, вып. 3).
Рухлядев Д.П. Гельминтофауна серн, туров, оленей и косуль в Кавказском заповеднике // Тр. Кавказского гос. заповедника. 1959. Вып. 5. С. 95-124.
Рухлядев Д.П. Гельминтофауна диких парнокопытных животных Крыма и Кавказа в эколого-зоогеографическом освещении. Саратов: Изд-во Саратовского ун-та, 1964. 450 с.
Рухлядев Д.П. Гельминтозы диких парнокопытных в районах Крыма и Кавказа // Тр. IX Междунар. конгр. биологов-охотоведов. М., 19706. С. 605-608.
Рысков А.П., Кудрявцев И.В., Васильев В.А и др. Молекулярно-генетические подходы к анализу генетического разнообразия у представителей Bison и Bos: геномная дактилоскопия и таксономическое типирование ДНК // К вопросу о возможности сохранения зубра в России. Пущино, 1993. С. 74-82.
Рысков А.П., Кудрявцев И.В., Васильев В.А и др. Диагностические возможности молекулярно-генетических подходов к таксономии трибы Bovini // Зоол. журн. 1994. Т. 73, вып. 11. С. 115-123.
Рычков П.И. Топография Оренбургской губернии. Уфа: “КИТАП”, 1999. 309 с. (По изд. 1762).
Саблина Т.Б. Копытные Беловежской пущи // Тр. Ин-та морфологии животных им. А.Н. Северцова АН СССР. 1955. Вып. 15. 191 с.
Саблина Т.Б. Безоаровый козел // Красная книга РСФСР. Животные. М.: Россельхозиз-дат, 1983. С. 116-117.
Савельев П.А. Туры и серны в Кавказском заповеднике // Природа. 1960. № 3. С. 113-114.
Савинецкий А.Б. Вековая динамика пастбищных экосистем центральной части Северного Кавказа за последние тысячелетия // Историческая экология диких и домашних копытных. История пастбищных экосистем. М.: Наука, 1992. С. 165-171.
Савинов В. Звери и птицы Алтая. Алтайское кн. изд-во, 1964. 135 с.
Савинов Е.Ф. Экология сибирского козерога в Джунгарском Ала-Тау И I Всесоюз. со-вещ. по млекопитающим. М.: Изд-во Моск, ун-та, 1961. Т. 2. С. 79.
Савинов Е. Ф. Размножение и рост сибирского козерога в Джунгарском Алатау (Казахстан) // Материалы по фауне и экологии наземных позвоночных Казахстана (млекопитающие). Алма-Ата: Изд-во АН КазССР, 1962. С. 167-182. (Тр. Ин-та зоол. АН КазССР. Т. 17).
Савинов Е.Ф. Кочевки и суточная активность сибирского горного козла в Казахстане // Промысловые и вредные млекопитающие Казахстана. Алма-Ата: Изд-во АН КазССР, 1964. С. 197-207. (Тр. Ин-та зоол. АН КазССР. Т. 23).
Савинов Е. Ф. Результаты авиаучета архара в Казахском нагорье // Современное состояние и пути развития охотоведческой науки в СССР. Киров, 1974. С. 168-169.
Савинов Е.Ф. Распространение и численность сибирского горного козла в Казахстане //Копытные фауны СССР. М.: Наука, 1975а. С. 118-119.
СавиновЕ.Ф. Сибирский горный козел и архар в Казахстане (экология и хозяйственное значение): Автореф. дис. ... канд. биол. наук. Алма-Ата, 19756. 24 с.
Савинов Е.Ф., Бекенов А. Азиатский муфлон и необходимые меры его охраны // Редкие виды млекопитающих и их охрана. М.: Наука, 1977. С. 226-227.
Савинов П. Ф. Общие результаты палеобиологических исследований Павлодарского Прииртышья//Териология. Новосибирск, Наука, 1972. Т. 1. С. 131-142.
Савицкий В. Ф., Шарипов А., Кабир-Биюк-Оглы. Кариотип межвидового гибрида уриала с тонкорунной овцой // Генетика. 1974. Т. 10, № 4. С. 180-181.
СалганскийА.А., Слесъ И.С., Треус В.Д„ Успенский ГА. Зоопарк “Аскания-Нова”. Киев: Гос. изд. с.-х. лит-ры УССР, 1963. 306 с.
Сапоженков Ю. Ф. Очерки по истории охоты и охотничье-промысловых животных Туркмении И Бюл. МОИП. Отд. биол. 1969. Т. 74, вып. 2. С. 33-41.
Сапожников ГН. Форма суставной впадины (cavitas glenoidalis) как критерий отличия родов Capra и Ovis по остаткам os scapula И Природа. 1968. Т. 73, вып. 5. С. 18-20.
Сапожников ГН. Экстерьерные особенности диких баранов Таджикистана И Бюл. МОИП. Отд. биол. 1971. Т. 76, вып. 5. С. 45-53.
Сапожников ГН. Дикие копытные звери Таджикистана И Редкие виды млекопитающих фауны СССР и их охрана. М.: Наука, 1973. С. 140-141.
Сапожников Г.Н. Дикие копытные Таджикистана И Редкие млекопитающие фауны СССР. М.: Наука, 1976а. С. 113-124.
Сапожников Г.Н. Дикие бараны (род Ovis) Таджикистана. Душанбе: Изд-во “Дониш”, 19766. 200 с.
Сапожников Г.Н. Архар // Копытные звери. М.: Лесн. пром-сть, 1977. С. 201-221.
Сапожников Г. Нахур, или голубой баран: новый вид в фауне СССР // Охота и охотничье хоз-во. 1984. № 7. С. 6-7.
Сапожников Г.Н. Голубой баран на Памире! // Природа. 1985. № 2. С. 102-103.
Сапожников Г.Н., Дуламцэрэн С. Экологические особенности и современная численность сайгака - Saiga tatarica mongolica Bannikov в районе Шаргын-Гоби (Монголия) // Изв. АН ТаджССР. Отд. биол. наук. 1982. № 1. С. 49-57.
Саркисов А.А. Острый катар кишок сосунов безоаровых коз {Capra aegagrus Erxl.) И Тр. Ереванского зоол. парка. 1944. Вып. 1-2. С. 133-134.
Саркисов А.А.О помесях полорогих // Природа. 1953. Т. 42. № 2. С. 113.
Сатунин К.А. Кавказский зубр И Очерки родиноведения Кавказа. 1914. № 4. С. 4-16
Сашко В.К., Смазное В.А. О необычной миграции сибирского горного козла // VI съезд териол. о-ва. М., 1999. С. 223.
Северцов Н.А. Звери при-Уральского края // Акклиматизация. М., 1861. Т. 2, вып. 2. С. 49-70.
Северцов Н.А. Вертикальное и горизонтальное распределение туркестанских животных//Изв. Импер. о-ва любит, естествозн., антроп. иэтногр. 1873а. Т. 8, вып. 2. С. 1-157.
Северцов Н.А. Архары (горные бараны) // Природа. 18736. Кн. 1. С. 144-245.
Северцов С.А. Судьба зубров И Охотник. 1926. № 4. С. 7.
Северцов С.А. О падежах диких животных // Сов. Север. 1933. № 5. С. 50-58.
Северцов С.А. Беловежская Пуща // Природа. 1940а. № 10. С. 32-43.
Северцов С.А. Видовые константы размножения беловежского зубра и динамика населения этого вида // Тр. Ин-та эвол. морфол. им. А.Н. Северцова. 19406. Т. 3, вып. 1. С. 3-32.
Северцов С А. Эволюция рогов некоторых парнокопытных как турнирного оружия в боях за самку И Проблемы экологии животных. М.: Изд-во АН СССР, 1951. С. 30-57.
Селевин В.А. Об охотничьей фауне Семипалатинского уезда // Охотник. 1928. № 5. С. 15-16.
Семенова С.К., Васильев В.А., Морозова Е.В. и др. ДНК-фингерпринтинг и генетическое разнообразие европейского зубра {Bison bonasus), американского бизона {Bison bison) и серого украинского скота {Bos taurus) // Генетика. 2000. Т. 36, № Н.С. 1535-1545.
Семенова С.К., ВасильевВ.А., Стекленев Е.П. и др. ДНК-фингерпринтинг представителей подсемейства Быковых (Bovinae) с использованием теломерного маркера (TTAGGG)4//TaM же. 1999. Т. 35, № 1. С. 101-104.
Сержанин И.Н. Млекопитающие Белорусской ССР. Минск: Изд-во АН БССР, 1951.311 с.
Сидоров М.К. Об Ovis nivicola // Изв. Импер. о-ва любит, естествозн., антроп. и этногр. 1881. Т. 37, вып. ЕС. 45.
Сидоров С.В. Снежный баран, туры, сибирский козерог. Сайгак // Охотничьи животные России. 2000. Вып. 2. С. 55-65; 2004. Вып. 6. С. 62-73.
Сидоров С.В. Сайгак Северо-Западного Прикаспия: состояние, перспективы развития // Териофауна России и сопредельных территорий. VII съезд териол. о-ва. М., 2003. С. 315-316.
Сидоров С.В., Букреева О.М. Популяционные циклы сайгака Северо-Западного Прикаспия //VI съезд териол. о-ва. М., 1999. С. 232.
Силантьев А.А. Обзор промысловых охот в России. СПб, 1898. 619 с.
Силантьев В. Бизоны в Америке // Охота и охотничье хоз-во. 2000. № 9. С. 42—43.
Сипко Т.П. Метод оценки генофонда отдельных популяций зубра // Экологические проблемы охраны живой природы. М., 1990 Ч. 1. С. 115-116.
Сипко Т.П. Биологические аспекты сохранения и использования зубра: Автореф. дис. ... канд. биол. наук. М., 1995. 22 с.
Сипко Т.П. Зубр. Популяционно-генетический анализ // Вопросы современного охотоведения. М., 2002. С. 386—405.
Сипко Т.П. Состояние генофонда зубра и подходы к его спасению // Проблемы сохранения и восстановления диких копытных животных в Центральном регионе России. Орел: Изд-во ОРАГС, 2003. С. 30-53.
Сипко Т.П. Овцебык (Ovibos moschatus) И Фауна позвоночных животных плато Путорана. М„ 2004. С. 377-378.
Сипко Т.П., Белоусова И.П. Анализ состояния генофонда зубра в питомниках России // К вопросу о возможности сохранения зубра в России. Пущино, 1993. С. 56-62.
Сипко Т.П., Груздев А.Р., Егоров С.С., Тихонов В.Г. Динамика состояния популяции овцебыка в России // Охрана и рациональное использование животных и растительных ресурсов России. Иркутск, 2003. С. 486-499.
Сипко Т.П., Ларин В.В., Глазго В.И. Фенетическое и генетическое состояние популяции путоранского снежного барана (Ovis nivicola borealis) // Молекулярно-генетические маркеры животных. Киев: Норд-принт, 1999. С. 22-24.
Сипко Т.П., Ломов А.А., Банникова А.А. Сравнение ДНК европейского зубра, американского бизона и их гибридных форм методом рестриктазного анализа // Редкие виды млекопитающих России и сопредельных территорий. М., 1997а. С. 87.
Сипко Т.П., Ломов А.А., Банникова А.А., Медников Б.М. Сравнение ДНК домашних и диких представителей семейства Bovidae методом рестриктазного анализа И Проблемы сохранения редких пород домашних животных и близкородственных диких видов. Пущино, 1996. С. 28-29.
Сипко Т.П., Ломов А.А., Банникова А.А., Медников Б.М. Оценка степени генетической дивергенции некоторых полорогих методом рестриктазного анализа ДНК И Цитол. и генетика. 19976. Т. 31, № 4. С. 76-81.
Сипко Т.П., Пузаченко А.Ю., Буневич А.Н. и др. Изменчивость экстерьерных признаков зубра // Вопросы современного охотоведения. М., 2002. С. 367-385.
Сипко Т.П., Немцев А.С., РаутианГС. Современное состояние горных зубров//VI съезд териол. о-ва. М., 1999. С. 234.
Сипко Т.П., РаутианГС. Динамика факторов роста численности зубров // Редкие виды млекопитающих России и сопредельных территорий. М., 1997а. С. 88.
Сипко Т.П., Раутиан Г.С. Дифференциация и разнообразие популяций зубра по генетическим маркерам И Там же. 19976. С. 88.'
Сипко Т.П., Раутиан Г.С. Стратегические приоритеты сохранения зубра в России И VI съезд териол. о-ва. М., 1999. С. ’234.
Сипко Т.П., Раутиан Г. С., Киселева Е.Г. Состояние зубра и его популяций в европейской части России И Редкие виды млекопитающих России и сопредельных территорий. М., 1999. С. 403-418.
Сипко Т.П., Раутиан Г. С., Удина ИГ, и др. Изучение полиморфизма групп крови у зубров (Bison bonasus) // Генетика. 1995. Т. 31, № 1. С. 93-100.
Сипко Т.П., РаутианГ.С., Удина И.Г, Ракицкая Т.А. Полиморфизм биохимических маркеров зубра {Bison bonasus) // Там же. 1996. Т. 32, № 3. С. 400-405.
Сипко Т.П., Удина И.Г, Бадагуева Ю.Н., Сулимова ГЕ. Сравнительная характеристика полиморфизма ДНК гена каппа-казеина у представителей семейства Bovidae // Там же. 1994. Т. 30, № 2. С. 225-229.
Сипко Т.П., Ужанов С.В., Удина ИГ Иммуногенетический анализ популяций зубра СНГ // К вопросу о возможности сохранения зубра в России. Пущино, 1993. С. 62-73.
Сипко Т.П., Черкашенко В.И. Генетический анализ зубров в СССР // Современные проблемы Красной книги СССР. М., 1989. С. 31-37.
СкалонВ.Н. Об организации Витимского заповедника// Охрана природы. М., 1949. Вып. 8. С. 118-121.
Скалон В.Н. О некоторых промысловых млекопитающих Бодайбинского района Иркутской области //Бюл. МОИП. Отд. биол. 1951. Т. 56, вып. 1. С. 27-32.
Скалон В.Н. Хозяйственное использование сайгаков // Копытные фауны СССР. М.: Наука, 1975. С. 231.
Сливинский ГГ. Кариотип казахстанского архара {Ovis аттоп coIlium) И Проблемы охраны и устойчивого использования биоразнообразия животного мира Казахстана. Алматы, 1999. С. 40—41.
СливинскийГГ.,ЛюдвиковаЕ.К, Мазурова О.С. идр. ИсследованияPCR-RAPD полиморфизма геномной ДНК архара {Ovis аттоп L.) И Зоологические исследования в Казахстане: современное состояние и перспективы. Алматы, 2002. С. 118-119.
Сливинский Г.Г., Тлеулиева Р, Плахов К.Н., Бекенов А.Б. Хромосомный набор устюртского горного барана Ovis vignei areal // Териофауна России и сопредельных территорий. VII съезд териол. о-ва. М., 2003. С. 325.
Слудский А.А. Итоги работ по акклиматизации охотничье-промысловых животных в Казахской ССР за 30 лет советской власти И Изв. АН КазССР. Сер. зоол. 1948. № 51, Вып. 7. С. 3-17.
Слудский А.А. Сайгак в Казахстане И Тр. Ин-та зоол. АН КазССР. 1955. Т. 4. С. 18-55.
Слудский А.А. Фауна Казахстана и ее охрана // Там же. 1956. Т. 6. С. 3-27.
Слудский А.А. Современное состояние запасов охотничье-промысловых зверей Казахстана и задачи их хозяйственного использования ИI Всесоюз. совещ. по млекопитающим. М.: Изд-во Моск, ун-та, 1961. Т. 3. С. 93-96.
Слудский А.А. Взаимоотношения хищников и добычи (на примере антилоп и других животных и их врагов) // Тр. Ин-та зоол. АН КазССР. 1962. Т. 17. С. 24-143.
Слудский А.А. Джуты в евразийских степях и пустынях // Там же. 1963. Т. 20. С. 5-88.
Слудский А.А. Проблема “хищник - жертва" в охотничьем хозяйстве Казахстана // Тр. IX Междунар. конгр. биологов-охотоведов. М., 19706. С. 467-471.
Слудский А.А. Saiga tatarica L. Его настоящее и будущее ИI Междунар. конгр. по млекопитающим. М.: ВИНИТИ, 1974. Т. 2. С. 204.
Слудский А.А., Афанасьев Ю.Г Итоги и перспективы акклиматизации охотничье-промысловых животных в Казахстане И Промысловые и вредные млекопитающие Казахстана. Алма-Ата: Изд-во АН КазССР, 1964. С. 5-74. (Тр. Ин-та зоол. АН КазССР. Т. 23).
Слудский А.А., Бадамшин Б.И., Бекенов А. и др. И Млекопитающие Казахстана. Алма-Ата: Наука, 1981. Т. 3, ч. 1. 244 с.
Слудский А,А., Бекенов А., Жевнеров В.В. и др. // Млекопитающие Казахстана. Алма-Ата: Наука, 1983. Т. 3, ч. 3. 248 с.
Слудский А .А., Страутман Е.И. Охотничье-промысловые звери Казахстана и задачи их хозяйственного использования // Тр. Ин-та зоол. АН КазССР. 1955. Т. 4. С. 3-47.
Слудский А.А., Страутман Е.И., Афанасьев Ю. Т. Пушные ресурсы Казахстана и возможности развития охотничьего хозяйства в республике // Там же. 1962. Т. 17. С. 5-23.
Слудский А.А., Фадеев В.А. Ресурсы и перспективы использования диких копытных Казахстана // Копытные фауны ССССР. М.: Наука, 1975. С. 232-233.
Слудский А.А., Фадеев В.А. Размножение Saiga tatarica Р. в Казахстане И I Междунар. конгр. по млекопитающим. М.: ВИНИТИ, 1974. Т. 2. С. 205.
Слудский А.А., Фадеев В.А. Сайгак в Казахстане и перспективы его промысла И Вести. АН КазССР. 1977. № 3. С. 28-32.
Слудский А.А., Шубин И.Г. Авиавизуальный учет охотничье-промысловых зверей в зоне пустынь Казахстана // Вопросы организации и методы учета ресурсов фауны наземных позвоночных. М., 1961. С. 28-29.
Слудский А.А., Шубин И.Г. Авиавизуальный учет сайгаков, джейранов и ресурсов этой дичи в пустынях Казахстана // Ресурсы фауны промысловых зверей в СССР и их учет. М.: Изд-во Акад, наук СССР, 1963. С. 84-91.
Смирнов Е.Н. К вопросу об охране горала // Редкие виды млекопитающих фауны СССР и их охрана. М.: Наука, 1973. С. 141-143.
Смирнов М.Н. К проблеме восстановления диких копытных животных в степях юга Сибири // Акклиматизация охотничьих животных в СССР. Минск: Ураджай, 1978. С. 63-64.
Смирнов М.Н. Дикие копытные Тувинской АССР // Копытные фауны СССР. М.: Наука, 1980. С. 104-105.
Смирнов М.Н. Дикие животные Южной Сибири // Природа. 1983. № 11. С. 76-83.
Смирнов М.Н. Редкие копытные Бурятии и Тувы И Редкие наземные позвоночные Сибири. Новосибирск: Наука, 1988а. С. 205-211.
Смирнов М.Н. Сибирский горный козел Capra sibirica Pallas, 1776 И Красная книга Бурятской АССР. Улан-Удэ: Бурятское кн. изд-во, 19886. С. 49-51.
Смирнов М.Н. Дикие животные на юге Сибири (исторические изменения населения, проблема использования) // Управление популяциями диких копытных животных. Киров, 1989. С. 59-75.
Смирнов М.Н. Аргали в Туве И Экологические и экономические аспекты охраны и рационального использования охотничьих животных и растительных пищевых ресурсов Сибири. Шушенское, 1990. С. 137-141.
Смирнов М.Н. Крупные промысловые млекопитающие Южной Сибири: Автореф. дис. ... докт. биол. наук в форме науч, доклада. М., 1994. 68 с.
Смирнов М.Н. Охотники и охотничье хозяйство в доземледельческий период. Красноярск, 1996. 65 с.
Смирнов М.Н. Крупные хищные млекопитающие в центре Азии. Красноярск, 2002а. 256 с.
Смирнов М.Н. Дзерен // Красная книга Республики Тыва. Животные. Новосибирск: Изд-во СО РАН (филиал “Гео”), 20026. С. 144-145.
Смирнов М.Н. Алтайский горный баран (аргали) // Там же. 2002в. С. 146-147.
Смирнов М.Н, Бриллиантов А.В. Ресурсы, промысел, охрана и восстановление копытных в Красноярском крае // Экология диких животных и растений и их использование. Красноярск: Изд-во Красноярского гос. ун-та, 1990. С. 74—92.
Смирнов М.Н, Сопин Л.В. Архар, или аргали Ovis аттоп Linnaeus, 1758 // Красная книга Бурятской АССР. Улан-Удэ: Бурятское кн. изд-во, 1988. С. 51-54.
Смирнов М.Н., Ткаченко В.А. Сибирский горный козел в Южной Сибири // Экология промысловых животных Сибири. Изд-во Красноярского ун-та, 1992. С. 71-95.
Смирнов Н.Г. Некоторые особенности голоценового этапа развития териофауны Урала // История биогеоценозов СССР в голоцене. М.: Наука, 1976а. С. 213-221.
Смирнов Н.Г. Динамика териофауны и изменчивость некоторых млекопитающих Среднего, Южного Урала и Зауралья в голоцене: Автореф.... дис. канд. биол. наук. Свердловск, 19766. 20 с.
Смирнов Н.Г., Большаков В.Н., Косинцев П.А. и др. Историческая экология животных гор Южного Урала. Свердловск, 1990. 245 с.
Смирнов О.К., Живалев И.К, Кленовицкий П.М., Кракосевич А.Н. Кариотипические исследования азиатского снежного барана, домашней овцы и их гибридов // биотехнология в животноводстве. Дубровицы, 1987. С. 16-19.
Смирнов ЮА. Влияние охоты на охотничье-промысловую фауну Казахстана. Алма-Ата: Кайнар, 1965. 240 с.
Смирнов Ю.А. Учет сайгака на территории Казахской ССР // Тр. IX Междунар. конгр. биологов-охотоведов. М., 1970. С. 340-344.
Смоляр М.Н. Состояние численности сибирского горного козла в некоторых районах Тянь-Шаня и пути сохранения и увеличения поголовья вида // Экология и охрана горных видов млекопитающих. М., 1987. С. 156-158.
Собанский Г.Г. Горный козел в горах Алтая // Редкие виды млекопитающих и их охрана. М.: Наука, 1977. С. 227-229.
Собанский ГГ Промысловые звери Горного Алтая. Новосибирск: Наука, 1988. 160 с.
Собанский ГГ Алтайский горный баран (аргали) - размещение, состояние численности, миграции // Экология, морфология, использование и охрана диких копытных. М., 1989. Ч. 2. С. 245-246.
Собанский ГГ Копытные Горного Алтая. М.: Наука, 1992.257 с.
Собанский Г, Ешелкин И. Сибирский горный козел на Алтае // Охота и охотничье хоз-во. 1975. №4. С. 12-15.
Собанский Г.Г., Федосенко А.К. Размещение, численность и миграции алтайского горного барана // Ресурсы животного мира Сибири. Охотничье-промысловые звери и птицы. Новосибирск, 1990. С. 243-245.
Собанский Г.Г, Часовских В.А. Антропогенная трансформация фауны крупных млекопитающих на юго-востоке Горного Алтая: история и современность // Бюл. МОИП. Отд. биол. 2002. Т. 107, вып. 3. С. 3-7.
Соков А.И. Влияние антропогенного фактора на популяцию памирского архара в Таджикистане//Копытные фауны СССР. М.: Наука, 1975. С. 279.
Соков А.И. Современное состояние некоторых популяций диких парнокопытных Таджикистана // Редкие виды млекопитающих и их охрана. М.: Наука, 1977а. С. 229-230.
СоковА.И. В защиту памирского архара//Природа. 1977б.№9.С. 129-131.
Соков А.И. Состояние редких видов парнокопытных и методы их охраны в Таджикистане// Экология, морфология, использование и охрана диких копытных. М., 1989. Ч. 1. С. 79-81.
Соколов В.Е. О прошлых ареалах копытных Казахстана по наскальным изображениям // Бюл. МОИП. Отд. биол. 1964. Т. 69, вып. 1. С. 113-117.
Соколов В.Е. Кожный покров млекопитающих. М.: Наука, 1973. 487 с.
Соколов В.Е. Систематика млекопитающих. М.: Высш, школа, 1979. 528 с.
Соколов В.Е. Редкие и исчезающие животные. Млекопитающие. М.: Высш, шк., 1986. 519 с.
Соколов В.Е., Даш Я., Лущекина А.А., Неронов В.М. Современное распространение и количественное размещение дзерена в МНР // Зоологические исследования в МНР. М.: Наука, 1982. С. 7-20.
Соколов В., Жирнов Л., Еремеев А. и др. Валютная лихорадка и сайгаки И Охота и охотничье хоз-во. 1991. №4. С. 1-3.
Соколов В.Е., ЖирновЛ.В., Лущекина А.А. Копытные аридной зоны Евразии (проблемы охраны и рационального использования) И Экология и природопользование в Монголии. Пущино, 1992. С. 180-192.
Соколов В.Е., ЖирновЛ.В., МаксимукА.В. и др. Проблемы охраны сайгаков в современный период // Сайгак. М., 1998. С. 302-314.
Соколов В.Е., Кузнецов ГВ. Суточные ритмы активности млекопитающих. М.: Наука, 1978. 264 с.
Соколов В.Е., Орлов В.Н. Редкие и охраняемые виды млекопитающих Джунгарской и Заалтайской Гоби (МНР) И Редкие виды млекопитающих и их охрана. М.: Наука, 1977. С. 32-34.
Соколов В.Е., Орлов В.Н. Определитель млекопитающих Монгольской Народной Республики. М.: Наука, 1980. 351 с.
Соколов В.Е., Орлов В.Н., Лобачев В.С. Класс млекопитающие (Mammalia) И Редкие животные Монголии (позвоночные). М., 1996. С. 121-182.
Соколов В.Е., Орлов В.Н., Малыгин В.М. Хромосомные наборы двух видов антилоп из Монголии (Antilopinae, Bovidae) // Систематика и цитогенетика млекопитающих. М.: Наука, 1975. С. 46-47.
Соколов В.Е., ПарнесЯ.А. У истоков отечественной териологии. М.: Наука, 1993.412 с.
Соколов В.Е., Проняев А.В., Жирнов Л.В. Таксономия и изменчивость // Сайгак. М., 1998. С. 25-53.
Соколов В.Е., Сапожников Г.Н., Дуламцэрэн С. Состояние популяций сайгаков в Монголии // IV съезд Всесоюз. териол. о-ва. М., 1986. Т. 3. С. 175-176.
Соколов В.Е., Темботов А.К. Позвоночные Кавказа. Млекопитающие. Копытные. М.: Наука, 1993. 527 с.
Соколов В.Е., Ушакова НА., Приходько В.И. и др. Особенности микробных ассоциаций в секретах некоторых кожных желез млекопитающих И Микробиология. 1990. Т. 59, вып. 3. С. 472-481.
Соколов В.Е., Холодова М.В. Сайгаки в зоопарках мира. Сообщ. 2. Опыт содержания сайгаков в зоопарках России и бывших республик СССР И Науч. иссл. в зоол. парках. 1996. № 6. С. 254-272.
Соколов В.Е., Чернова О.Ф. Специфические кожные железы некоторых копытных // Эколого-функциональная морфология кожного покрова млекопитающих. М.: Наука, 1988. С. 91-144.
Соколов В.Е., Чернова О.Ф. Кожные железы млекопитающих. М.: ГЕОС, 2001. 647.
Соколов В.Е., Чернова О.Ф., Волошина И.В. и др. Кожно-волосяной покров амурского горала // Амурский горал. М.: Изд-во ЦНИЛ охотничьего хоз-ва и заповедников, 1992. С. 83-105.
СоколовВ.Е., ЧерноваО.Ф., ВолошинаИ.В., МысленковА.И. Специфическаязароговаяжелеза горала И Редкие виды млекопитающих СССР и их охрана. М., 1983. С. 205-206.
Соколов ГА. Млекопитающие кедровых лесов Сибири. Новосибирск: Наука, 1979. 256 с.
Соколов ГА. Современное состояние популяций некоторых редких и исчезающих видов млекопитающих юга Сибири //Редкие наземные позвоночные Сибири. Новосибирск, Наука, 1988. С. 211-218.
Соколов ГА., Балагура НН. Состояние популяций некоторых редких видов млекопитающих в приенисейской части Западного Саяна // Редкие виды млекопитающих фауны СССР и их охрана. М.: Наука, 1973. С. 31-32.
Соколов И.И. Опыт естественной классификации полорогих (Bovidae) И Тр. Зоол. ин-та АН СССР. 1953. Т. 14. С. 1-295.
Соколов И. В заповеднике Барса-Кельмес//Охота и охотничье хоз-во. 1958. № 9. С. 21-24.
Соколов И.И. Копытные звери (отрады Perissodactyla и Artiodactyla). Фауна СССР. Млекопитающие. М.-Л.: Изд-во АН СССР, 1959. Т. 1, вып. 3. 640 с.
Соколов И.И. Семейство Bovidae Gray, 1821 (=Cavicomia liliger, 1811). Полорогие// Основы палеонтологии. Млекопитающие. М.: Гос. Науч.-техн, изд-во литер, по геологии и охране недр. 1962. Т. 13. С. 383—408.
Соколов И.И. О некоторых интересных находках первобытного зубра (Bison priscus Boj.) на территории СССР //Бюл. Комис, по изуч. четвертич. периода. 1963. № 28. С. 24—32.
Соколов И.И. Посткраниальный скелет представителей рода Bison // Тр. Зоол. ин-та АН СССР. 1971. Т. 48. С. 198-219.
Соколов И.И., Рафалович И.А. О зубре в Молдавии // Бюл. МОИП. Отд. биол. 1961. Т. 66, вып. 3. С. 144-146.
Соколов И.И., Рашек В.Л. Развитие зубов и черепа как показатель возраста у сайги (Saiga tatarica L.) // Бюл. МОИП. Отд. биол. 1961. Т. 66, вып. 6. С. 77-98.
Соколов С. С. Материалы по экологии сайги Saiga tatarica L. // Вести. АН КазССР. 1951. №З.С. 143-151.
Соловьев Д.К. Промыслы копытных зверей и лесной дичи в России // Ежегодник Все-рос. союза охотников. 1922. С. 53-73.
Соломатин А.О. Кулан. М.: Наука, 1973. 145 с.
Соломкина Н.В. Рост и развитие горала // Копытные фауны СССР. М.: Наука, 1975а. С. 279-280.
Соломкина Н.В. Поведение амурского горала в неволе И Там же. 19756. С. 336.
Соломкина Н.В. Опыт вольерного содержания горалов в Лазовском заповеднике // Редкие виды млекопитающих и их охрана. М.: Наука, 1977. С. 230-232.
Соломкина Н.В. Изменения веса тела амурского горала И Копытные фауны СССР. М.: Наука, 1980а. С. 364.
Соломкина Н.В. Развитие рогов вольерных горалов // Там же. 19806. С. 365-366.
Соломкина Н. Питомник горалов // Охота и охотничье хоз-во. 1980в. № 9. С. 16-18.
Соломкина Н.В. Амурские горалы в неволе И Редкие и исчезающие животные суши Дальнего Востока СССР. Владивосток, 1981. С. 185.
Соломкина Н.В. Охрана амурского горала И Популяционная изменчивость вида и проблемы охраны генофонда млекопитающих. М., 1983а. С. 232-233.
Соломкина Н.В. Искусственное выкармливание горалят // Редкие виды млекопитающих СССР и их охрана. М., 19836. С. 207-209.
Соломкина Н.В. Питание вольерных горалов // Там же. 1983в. С. 209-210.
Соломкина Н.В. Разведение горала в питомнике Лазовского заповедника //1 Всесоюз. совещ. по пробл. зоокультуры. М., 1986. Ч. 2. С. 75-76.
Соломкина Н.В. Опыт содержания и разведения горала И Экология, морфология, использование и охрана диких копытных. М., 1989. Ч. 2. С. 283-284.
Соломкина Н.В. Рост и развитие горалов И V съезд Всесоюз. териол. о-ва АН СССР. М., 1990. Т. 1. С. 223-224.
Соломкина Н.В. Рост и развитие горала // Амурский горал. М.: Изд-во ЦНИЛ охотничьего хоз-ва и заповедников, 1992. С. 70-75.
Соломкина Н.В., Глебов В.В. Питомник горалов // Биологические аспекты охраны редких животных. М., 1981. С. 62-64.
Соломкина Н.В., Хохряков С.А. Половой диморфизм и возрастные изменения черепа амурского горала, Nemorhaedus caudatus (Artiodactyla, Bovidae) // Зоол. журн. 1993. Т. 72, вып. 3. С. 153-158.
Соломонов Н.Г. Животный мир Якутии. Якутск: Кн. изд-во, 1975. 184 с.
Сопин Л.В. Дикий баран архар в Алтайском крае // Редкие виды млекопитающих фауны СССР и их охрана. М.: Наука, 1973. С. 145-146.
Сопин Л.В. История ареала дикого барана {Ovis аттоп L.) // I Междунар. конгр. по млекопитающим. М.: ВИНИТИ, 1974а. Т. 2. С. 220.
Сопин Л. Рога архаров // Охота и охотничье хоз-во. 19746. № 7. С. 20-21.
Сопин Л.В. Охрана аргали Южной Сибири // Копытные фауны СССР. М.: Наука, 1975а. С. 280-281.
Сопин Л.В. Дикий баран Южной Сибири: Автореф. дис. ... канд. с.-х. наук. Иркутск, 19756. 23 с.
Сопин Л.В. Некоторые данные по экологии архара хребта Чихачева и горного массива Тал-дуаир // Редкие млекопитающие фауны СССР. М.: Наука, 1976а. С. 134-140.
Сопин Л.В. Некоторые аспекты поведения аргали // Групповое поведение животных. М.: Наука, 19766. С. 370-372.
Сопин Л.В. Экстерьерная характеристика аргали И Экология охотничьих зверей и птиц, технология производства в охотничьем хозяйстве. Иркутск, 1976в. С. 30-35.
СопинЛ.В. Хищники и смертность аргали в Юго-Восточном Алтае//Там же. 1976г С. 36-41.
Сопин Л.В. Состояние сайлюгемской популяции аргали и проблема его сохранения в СССР И Редкие виды млекопитающих и их охрана. М.: Наука, 1977а. С. 232-233.
Сопин Л.В. Размножение аргали // Там же. 19776. С. 233-234.
Сопин Л. Рога архаров И Охота и охотничье хоз-во. 1977в. № 6. С. 14-17.
Сопин Л.В. Изменчивость размеров костей конечностей диких баранов И Копытные фауны СССР. М.: Наука, 1980а. С. 372-373.
Сопин Л.В. Кариотипы и филогенез диких баранов И Воспроизводство и организация использования промысловых зверей и птиц в Сибири и на Дальнем Востоке. Иркутск, 19806. С. 66-74.
Сопин Л.В. К систематике архаров И Млекопитающие СССР. III съезд Всесоюз. териол. о-ва. М., 1982а. Т. 1. С. 77-76.
Сопин Л.В. О внутривидовой структуре архаров Ovis аттоп (Artiodactyla, Bovidae) // Зоол. журн. 19826. Т. 61, вып. 12. С. 1882-1892.
Сопин Л.В. Характеристика местообитаний аргали и их охрана // Исчезающие и редкие растения и животные Алтайского края и проблемы их охраны. Барнаул, 1982в. С. 19-26.
Сопин Л.В. Структура вида и таксономия горных баранов // Популяционная изменчивость вида и проблемы охраны генофонда млекопитающих. М., 1983а. С. 170-172.
Сопин Л.В. Фенетика горных баранов как основа их внутривидовой таксономии // Там же. 19836. С. 173-175.
Сопин Л.В. Половой диморфизм и филогенетические отношения диких баранов И Бюл. МОИП. Отд. биол. 1983в. Т. 88, вып. 6. С. 30-36.
Сопин Л.В. Роль снежного покрова в жизни аргали И Редкие виды млекопитающих СССР и их охрана М., 1983г. С. 211-212.
Сопин Л.В. О таксономическом положении кызылкумского барана {Ovis аттоп severtzovi) // Зоол. журн. 1984а. Т. 63, вып. 5. С. 789-792.
Сопин Л.В. Мелкие формы архаров и их таксономическое положение И Изв. СО АН СССР. Сер. биол. наук. 19846. № 6, вып. 1. С. 85-90.
Сопин Л.В. Генетический контроль окраски горных баранов // Фенетика популяций. М., 1985а. С. 268-269.
Сопин Л.В. Фенентика и филогения снежных баранов // Там же. 19856. С. 270-271.
Сопин Л. Горные бараны: проблемы охраны и рационального использования // Охота и охотничье хоз-во. 1986а. № 5. С. 20-22.
Сопин Л.В. Внутривидовая изменчивость и таксономия снежных баранов (Ovis, Pachyceros) // Охотничье-промысловые ресурсы Сибири. Новосибирск: Наука, 19866. С. 137-145.
Сопин Л.В. Снежный баран И Редкие животные Иркутской области (наземные позвоночные). Иркутск, 1993. С. 37-39.
Сопин Л. Дикие бараны Прибайкалья//Охота и охотничье хоз-во. 1998. № 6. С. 14—15.
Сопин Л.В. Структура вида и систематика аргали И Охрана и рациональное использование животных и растительных ресурсов России. Иркутск, 2003. С. 506-515.
Сопин Л.В., Ермолин А.Б. Снежный баран в Забайкалье может исчезнуть // Природа. 1986. № 7. С. 78-79.
Сопин Л.В., Ермолин А.Б. Состояние и перспективы сохранения кодарского снежного барана И Редкие наземные позвоночные Сибири. Новосибирск: Наука, 1988. С. 222-225.
Сопин Л.В., Медведев Д.Г. Снежный баран Севера Читинской области // Зоологические исследования в Восточной Сибири. Иркутск: ИСХИ, 1992. С. 80-84.
Сопин Л.В., Сухбат X. Географическая изменчивость черепа аргали // Копытные фауны СССР. М.: Наука, 1980. С. 374-375.
Сопин Л.В., Сухбат X. Таксономический ранг аргали Монгольского и Гобийского Алтая И Редкие виды млекопитающих СССР и их охрана М., 1983. С. 213-215.
Сопин Л.В., Улотъ В. Морфофункциональная характеристика посткраниального скелета европейского муфлона // Копытные фауны СССР. М.: Наука, 1980. С. 375-376.
Сопин Л.В., Харрисон Д.Л. О таксономическом положении горного барана из Омана (Ovis, Artiodactyla, Bovidae) //Зоол. журн. 1986. Т. 65, вып. 6. С. 952-955.
Спасская ТХ, Абдулманапов Г.Н. Использование и охрана сайгаков в Дагестане // Копытные фауны СССР. М.: Наука, 1980. С. 264.
Сорока Б. О сайгаках И Охота и охотничье хоз-во. 1957. № 8. С. 17-18.
Сорокин П.А., Кирилюк В.Е., Лущекина А.А., Холодова М.В. Генетическое разнообразие монгольского дзерена (Procapra gutturosa) в Забайкалье и Монголии И Млекопитающие как компонент аридных экосистем (ресурсы, фауна, экология, медицинское значение и охрана). М., 2004. С. 141-142.
Соскин А.А., Глушнев С.В. Доместикация снежного барана с целью гибридизации с домашними овцами И Селекция, гибридизация и акклиматизация сельскохозяйственных животных. М.: Колос, 1983. С. 235-239.
Спивакова Л.В., Байдавлетов РЖ. Микроструктура гонад половозрелых самцов Ovis аттоп coIlium в Центральном Казахстане // Редкие виды млекопитающих России и сопредельных территорий. М., 1997. С. 92.
СпиваковаЛ.В., Байдавлетов РЖ., Плахов К.Н. Динамика репродуктивной активности самцов горных баранов Казахстана // Зоологические исследования в Казахстане: современное состояние и перспективы. Алматы, 2002. С. 119-122.
Спивакова Л.В., Плахов К.Н., Байдавлетов РЖ. Зароговая мозоль у горных баранов // VI съезд териол. о-ва. М., 1999. С. 247.
Стамбеков С.Ж., Середина З.П., Туманбаев В.М. Некоторые результаты скрещивания архара с грубошерстными матками // Селекционно-племенная работа в промышленном животноводстве Сибири. Новосибирск, 1987. С. 88-93.
Сташкевич Л.Ф., Казьмин В.Д. Овцебыки на Чукотке И Магаданский оленевод. 1984. Вып. 36. С. 29-31.
Стекленев Е.П. Отдаленная гибридизация представителей подсемейства козлообразных (Caprinae) И Науч. тр. Укр. НИИ животноводства “Аскания-Нова”. 1972. Т. 15. С. 220-231.
Стекленев Е.П. Современное состояние исследований в области отдаленной гибридизации животных (обзор) // Сельскохоз. биол. 1975. Т. 10, № 3. С. 283—390.
Стекленев Е.П. Особенности размножения европейского муфлона при полуволь-ном содержании // Редкие виды млекопитающих и их охрана. М.: Наука, 1977. С. 234-235.
Стекленев Е.П. Морфология спермиев представителей подсемейства козлообразных (Caprinae) как один из методов определения их генеалогической близости и подбора пар гибридизантов // Эколого-морфологические особенности диких родичей домашних овец. М.: Наука, 1978а. С. 17-23.
Стекленев Е.П. Физиологические особенности размножения европейского муфлона (Ovis musimon Pall.) в связи с его акклиматизацией в условиях юга Украины // Там же. 19786. С. 36-43.
Стекленев Е.П. Основные проблемы в области отдаленной гибридизации животных и перспективы их дальнейшего решения И Вопросы гибридизации копытных. М.: Наука, 1980. С. 15-21.
Стекленев Е.П. Основные показатели акклиматизации и отдаленной гибридизации некоторых видов животных И Науч.-техн. бюл. Укр. НИИ животноводства степных районов. 1982. Вып. 2. С. 10-14.
Стекленев Е.П. Физиологические особенности размножения бизона, акклиматизируемого в степных условиях юга Украины // Селекция, гибридизация и акклиматизация сельскохозяйственных животных. М.: Колос, 1983а. С. 76—81.
Стекленев Е.П. Особенности гибридизации бизона (Bison bison bison L.) с домашней коровой // С.-х. биология. 19836. № 5. С. 81-84.
Стекленев Е.П. Отдаленная гибридизация животных, ее проблемы и перспективы практического использования для целей зоокультуры //1 Всесоюз. совещ. по пробл. зоокультуры. М, 1986. Ч. 1. С. 111-113.
Стекленев Е.П. Характеристика воспроизводительной способности европейского муфлона (Ovis аттоп musimon Pal.), акклиматизируемого в степных условиях юга Украины // Чтения памяти проф. В.В. Станчинского. 1995. № 2. С. 67-70.
Стекленев Е.П. Особенности размножения равнинной формы североамериканского бизона (Bison bison bison), акклиматизируемого на юге Украины И Вести, зоол. 1996. № 4-5. С. 60-69.
Стекленев Е.П., Ясинецкая Н.И. Кариологическая характеристика гибридов бизона с домашней коровой и ее подродовых гибридов с бантенгом И Цитоген. и биотехнология. Л., 1989. С 49-51.
Стекленев Е.П., Ясинецкая Н.И. Кариотипическая характеристика гибридов бизона и домашней коровы//V съезд Всесоюз. териол. о-ва АН СССР. М., 1990. Т. 1. С. 102-103.
Стенин Г. Някои изводи и препоръки за стопанисване на муфлона у нас И Гор. стоп., гор. пром. 1986. Т. 42, № 10. С. 34-35.
Стенченко А.М. Снежные бараны Узон-Гейзерского района Камчатки // Копытные фауны СССР. М.: Наука, 1980. С. 103-104.
Стратегия сохранения зубра в России. М.: WWF, 2002. 45 с.
Стрекозов Н.И., Иолчиев Б.С., Бугрим Л.Н. и др. Биологические особенности гибридов крупного рогатого скота и зубра И Вести. Рос. акад. с.-х. наук. 2001. № 3. С. 63-65.
Строганов СУК.систематике и географическому распространению некоторых антилоп Центральной Азии // Бюл. МОИП. Отд. биол. 1949. Т. 54, вып. 4. С. 15-29.
Субботин А.Е. Экспериментальное изучение питания сайгаков (Saiga tatarica) в связи с особенностями кормовых ресурсов: Автореф. дис.... канд. биол. наук. М., 2001. 24 с.
Субботин А.Е., Абатуров Б.Д. Влияние состава поедаемых растений на энергетический баланс и состояние популяции сайгаков в условиях полупустыни // VI съезд териол. о-ва. М., 1999. С. 250.
Субботин А.Е., Джапова Р.Р. Влияние сайгаков на урожай пастбищной растительности в Калмыкии // IV съезд Всесоюз. териол. о-ва. М., 1986. Т. 3. С. 177-178.
Сулимов А.Д. Гельминты редких зверей Тувы // Редкие виды млекопитающих фауны СССР и их охрана. М.: Наука, 1973. С. 32-33.
Сулимов А.Д. Редкие виды млекопитающих и их охрана в Туве // Редкие виды млекопитающих и их охрана. М.: Наука, 1977. С. 36-37.
Султанов Г. С. Распространение парнокопытных Узбекистана и меры их охраны // Копытные фауны СССР. М.: Наука, 1980. С. 106-107.
Сухбат X. Учет горных копытных в Монгольском и Гобийском Алтае // Проблемы охотоведения и охраны природы. Иркутск, 1975. С. 165-167.
Сухбат X, Груздев В.В. Горный баран-аргали в Монгольской Народной Республике // Природные условия и биологические ресурсы Монгольской Народной Республики. М.: Наука, 1986. С. 196.
Сыроечковский Е.Е. Северный олень. М.: Агропромиздат, 1986. 256 с.
Сыроечковский Е.Е. Сибирский горный козел Capra sibirica Pall. И Красная книга Красноярского края. Красноярск: Красноярское кн. изд-во, 1995а. С. 72-75.
Сыроечковский Е.Е. Путоранский, или норильский снежный баран Ovis nivicola borealis Sev. // Там же. 19956. С. 75-80.
Сыроечковский Е.Е. Овцебык Ovibos moschatus L. // Там же. 1995в. С. 88-93.
Сыроечковский Е.Е., Рогачева Э.В. Животный мир Красноярского края. Красноярское кн. изд-во, 1980. 360 с.
Сыроечковский Е.Е., Рогачева Э.В., Савченко А.П. и др. Красная книга Красноярского края. Красноярск, 2000. 248 с.
Сытин В. О сайгаке в Волго-Кумских степях // Охотник. 1927. № 2. С. 24.
Тавровский В.А., Егоров О.В., Кривошеев В.Г. и др. Млекопитающие Якутии. М.: Наука, 1971. 660 с.
Тарасов М.А. Некоторые данные о динамике численности копытных в Тебердинском государственном заповеднике // Редкие виды млекопитающих фауны СССР и их охрана. М.: Наука, 1973. С. 147-148.
Тарасов М.А. Состояние популяции зубра в Тебердинском заповеднике // Копытные фауны СССР. М.: Наука, 1975. С. 282-284.
Тарасов М.А. Факторы, определяющие динамику численности копытных в Тебердинском заповеднике и прилегающих районах // Редкие млекопитающие фауны СССР. М.: Наука, 1976. С. 156-166.
Тарасов М.А. Влияние рекреации на состояние популяций редких видов в Тебердинском заповеднике // Редкие виды млекопитающих и их охрана. М.: Наука, 1977. С. 235-236.
Тарасов М.А. Структура популяции зубров и использование ими территории в Тебердинском заповеднике//Копытные фауны СССР. М.: Наука, 1980. С. 108-109.
Тарасов М.А. Влияние снежных лавин на популяцию кавказского тура в горах Северо-Западного Кавказа // Экология горных млекопитающих. Свердловск, 1982. С. 118-119.
Тарасов М.А. Редкие виды млекопитающих Северо-Западного Кавказа и их охрана // Редкие виды млекопитающих СССР и их охрана М., 1983. С. 28-30.
Тархов В. С. Новоселы острова Врангеля // Магаданский оленевод. 1976. Вып. 28. С. 41-43.
Татаринов К.А. Зверовая охота - основа трудовой деятельности древнего населения Волыно-Подолии // IV съезд Всесоюз. териол. о-ва. М., 1986. Т. 1. С. 32.
Татаринов К. Антропогеновые позвоночные Приднестровья - индикаторы среды обитания // Цитология. 1995. Т. 37, № 7. С. 644—645.
Татаринов К.А., Дякун Ф.А. Реакклиматизация зубров в лесах Волынской области И Зоол. журн. 1969. Т. 48, вып. 4. С. 612-614.
Темботов А.К. Млекопитающие Кабардино-Балкарской АССР. Нальчик: К.-Балк. кн. изд-во, 1960. 196 с.
Темботов А.К. Распространение и численность туров в Кабардино-Балкарской АССР И I Всесоюз. совещ. по млекопитающим. М.: Изд-во Моск, ун-та, 1961. Т. 1. С. 150.
Темботов А.К. География млекопитающих Северного Кавказа. Нальчик: Эльбрус, 1972. 245 с.
Темботов А.К. О современном распространении и состоянии охраны туров в Кабардино-Балкарской АССР // Редкие виды млекопитающих фауны СССР и их охрана. М.: Наука, 1973. С. 148-149.
Темботов А.К. О географической изменчивости туров (Capra caucasica Guld.) на Северном Кавказе И Фауна, экология и охрана животных Северного Кавказа. Нальчик, 1974. Вып. 2. С. 118-132.
Темботов А .К, КетенчиевХ.А., Дзуев Р.И. Популяционная структура и состояние охраны генофонда кавказских туров // Копытные фауны СССР. М.: Наука, 1980а. С. 266—267.
Темботов А.К., Кетенчиев Х.А., Дзуев Р.И. Центральнокавказский тур - палеоэндемик Большого Кавказа // Памятники природы Кабардино-Балкарии, их выявление, учет и организация охраны. Нальчик: Изд-во “Эльбрус”, 19806. С. 15-23.
Темботов А.К, Шхашамишев XX., Хатуг А. О закономерностях структуры ареалов и географической изменчивости млекопитающих в горах Кавказа // Млекопитающие СССР. III съезд Всесоюз. териол. о-ва. М., 1982. Т. 1. С. 140-141.
Теплое В.П. Волк в Кавказском заповеднике // Тр. Кавказского гос. заповедника. 1938. Вып. 1. С. 343-366.
Терновский Д.В., Терновская Ю.Г., Воронов В.Г., Воронов ГА. Курильские острова— природные резерваты исчезающих и редких видов млекопитающих // Охотничье-про-мысловые ресурсы Сибири. Новосибирск: Наука, 1986. С. 29-34.
Тимофеев В.К. Материалы по биологии и экологии млекопитающих острова Барса-кель-мес в связи с акклиматизацией на нем желтого суслика-песчаника (Citellus fulvus Licht) // Зоол. журн. 1934. Т. 13, вып. 4. С. 731-758.
Тимченко Н.Г. К истории охоты и животноводства в Киевской Руси (Среднее Поднепро-вье). Киев: Наук. Думка, 1972. 204 с.
Тимченко Н.Г. Промысловые и домашние млекопитающие Правобережной Украины в I тысячелетии нашей эры //1 Междунар. контр, по млекопитающим. М.: ВИНИТИ, 1974. Т. 2. С. 262-263.
Тихонов А.Н. Плейстоценовый овцебык (Ovibos pallantis) Урала и Западной Сибири // Четвертичная фауна Северной Азии. СПб., 1994. С. 92-110. (Тр. Зоол. ин-та РАН. Т. 256).
Тихонов А.Н. Половой диморфизм у полорогих трибы Rupicaprini (Artiodactyla, Bovidae) // Исследования по плейстоценовым и современным млекопитающим. СПб., 1995. С. 214-226. (Тр. Зоол. ин-та РАН. Т. 263).
Тихонов А.Н. Систематика полорогих подсемейства Caprinae (Artiodactyla, Bovidae): Дис. ... канд. биол. наук. СПб., 1999. 171 с.
Тихонов В.Г. Опыт переселения овцебыков в Республику Саха (Якутия) // Овцебык в тундре России: Эксперимент XX века по восстановлению исчезнувшего вида. СПб.: Астерион, 2002. С. 49-53.
Тихонов В. Овцебык - будущее жителей Севера // Охота и охотничье хоз-во. 2003а. № 8. С. 1-3.
Тихонов В. Фермы по разведению овцебыков // Там же. 20036. № 11. С. 8-10.
Тихонов В.Г, Егоров С. С. Вопросы реакклиматизации и создания питомников овцебыка в республике Саха (Якутия) // Вопросы современного охотоведения. М., 2002. С. 363-366.
Тихонов В.Г., Луковцев Ю.С., Слепцов М.К. и др. Рекклиматизация лесного бизона в Республике Саха (Якутия) // Проблемы сохранения редких пород домашних животных и близкородственных диких видов. Пущино, 1996. С. 65-66.
Тихонов В.Г, Сипко Т.П., Груздев А.Р. Перспективы расселения овцебыка в Российской Арктике // Овцебык в тундре России: Эксперимент XX века по восстановлению исчезнувшего вида. СПб.: Астерион, 2002. С. 60-62.
Тихонов В.Г, Сипко Т.П., Груздев А.Р., Егоров С.С. Отлов и расселение овцебыков на севере России // Териофауна России и сопредельных территорий. VII съезд териол. о-ва. М., 2003. С. 351.
Тихонов В.Г, Царев С.А., Слепцов М.К. и др. Расселение овцебыка в Якутии // Редкие виды млекопитающих России и сопредельных территорий. М., 1997. С. 94.
Тихонов Ф. В защиту сайгака // Охота и охотничье хоз-во. 1956. № 8. С. 15.
Тишков А.А. Растительноядные млекопитающие в экосистемах тундры // Млекопитающие в наземных экосистемах. М.: Наука, 1985. 296 с.
Ткаченко А. Акклиматизация животных в Крыму//Охота и охотничье хоз-во. 1963. №7. С. 25-26.
Ткаченко В.А. Распространение и численность копытных в Юго-Западной Туве // Экология промысловых животных Сибири. Изд-во Красноярского ун-та, 1992. С. 50-57.
Тлеубердина П.А., Кожамкулова Б. С., Кондратенко Г.С. Каталог кайнозойских млекопитающих Казахстана. Алма-Ата: Наука, 1989. 159 с.
Толкач В.Н. Природные территориальные комплексы - основа выбора региона для реакклиматизации зубра (Bison bonasus) // Динамика зооценозов, проблемы охраны и рационального использования животного мира Белоруссии. Минск: Наука и техника, 1989. С. 225-226.
Томирдиаро С.В. Изменения физико-географической обстановки наравнинах Северо-Восточной Азии на границе плейстоцен - голоцен как основная причина вымирания териофауны мамонтового комплекса // Тр. Зоол. ин-та АН СССР. 1977. Т. 73. С. 64—72.
Точиев Т.Ю. Вопросы экологии и охраны кавказского оленя и безоарового козла в условиях Чечено-Ингушской АССР // Редкие виды млекопитающих фауны СССР и их охрана. М.: Наука, 1973. С. 150-151.
Точиев Т.Ю. Редкие виды диких парнокопытных Чечено-Ингушетии и их охрана // Копытные фауны СССР. М., 1975. С. 284-285.
Точиев Т.Ю., Батхиев А.М. К вопросу о питании безоаровых коз в Чечено-Ингушской АССР // Фауна, экология и охрана животного мира Северного Кавказа. Нальчик, 1980. С. 57-64.
Точиев Т.Ю., Батхиев А.М. Безоаровый козел//Ресурсы редких животных РСФСР, их охрана и воспроизводство (Материалы к Красной книге). М., 1988. С. 151-153.
Требоганова Н.В. О роли Центрального зубрового питомника Приокско-Террасного заповедника в восстановлении поголовья зубров в России // Проблемы сохранения и
восстановления диких копытных животных в Центральном регионе России. Орел: Изд-во ОРАГС, 2003а. С. 92-95.
Требоганова Н.В. Формирование гельминтофауны зубров Центральной России // Териофауна России и сопредельных территорий. VII съезд териол. о-ва. М., 20036. С. 355—356.
Треус В. В. Кариотип бизона (Bison bison) и его сравнение с кариотипом крупного рогатого скота (Bos taurus L.) // Бюл. Всерос. НИИ генетики и разведения с.-х. животных. 1993. Вып. 137. С. 48-50.
Треус В.В. Специфика хромосомной организации представителей семейства Bovidae: Автореф. дис. ... канд. биол. наук. СПб. - Пушкин, 1994. 16 с.
Треус В.В., Павлова В.А., Стекленев Е.П. и др. Изменчивость сателлитной ДНК и гетерохроматиновых районов хромосом у представителей Bovinae и их гибридов // Генетика. 1994. Т. 30. Прилож. С. 159.
Треус В.В., Ясинецкая Н.И. Гетерохроматиновые участки митотических хромосом некоторых представителей семейства Bovidae (зубр, бизон, гаял) и их сравнение с таковыми крупного рогатого скота // Бюл. Всерос. НИИ генетики и разведения с.-х. животных. 1993. Вып. 137. 1993. С. 51-54.
Треус В.В., Ясинецкая Н.И., Треус М.Ю. и др. Гетерохроматиновые и эухроматиновые районы хромосом некоторых представителей подсемейства Bovinae // Генетика. 1997. Т. 33, № 9. С. 1273-1280.
Треус В.Д. Акклиматизация и гибридизация животных в Аскании-Нова. Киев: Урожай, 1968. 316 с.
Треус В.Д. Охрана и разведение охотничье-промысловых и редких животных в Аскании-Нова // Тр. IX Междунар. контр, биологов-охотоведов. М., 1970. С. 155-157.
Треус В.Д., Лобанов Н.В. Разведение редких и исчезающих копытных фауны СССР в Аскании-Нова // Редкие виды млекопитающих фауны СССР и их охрана. М.: Наука, 1973. С. 151-152.
Треус В.Д., Лобанов Н.В. Разведение редких и исчезающих копытных фауны СССР в Аскании-Нова //Редкие млекопитающие фауны СССР. М.: Наука, 1976. С. 103-112.
Треус М.Ю. Реакклиматизация антилопы сайга на юге Украины // IV съезд Всесоюз. териол. о-ва. М., 1986. Т. 3. С. 178-179.
Треус М.Ю. Опыт реакклиматизации антилопы сайга на юге Украины // Науч.-техн. бюл. Укр. НИИ животноводства степных районов. 1990. Вып. 2. С. 48-50.
Треус М.Ю. Реакклиматизация антилопы сайга в зоопарке “Аскания-Нова” // Научные исследования в зоологических парках. М., 1993. С. 76-80.
Треус М.Ю. Состояние популяции сайги (Saiga tatarica) в биосферном заповеднике “Аскания-Нова” // Редкие виды млекопитающих России и сопредельных территорий. М., 1997. С. 94.
Треус М.Ю. Итоги реинтродукции антилопы сайга в Аскании-Нова // Акгуальш питания збереження 1 вщновлення степових екосистем. Аскания-Нова, 1988. С. 100-102.
Треус М.Ю., Смаголь В.А. Многолетняя динамика численности и структура популяции сайгака (Saiga tatarica L.) при полувольном содержании на юге Украине // Млекопитающие как компонент аридных экосистем (ресурсы, фауна, экология, медицинское значение и охрана). М., 2004. С. 151-152.
Трофимов Б.А. К вопросу о происхождении и развитии фаун млекопитающих четвертичного периода умеренной и северной зон // Тр. Комис, по изуч. четвертич. периода. 1955. Т. 12. С. 106-120.
Трощенко Б.В. Об осенних миграциях сайгаков Волго-Уральской популяции через р. Урал//IV съезд Всесоюз. териол. о-ва. М., 1986. Т. 3. С. 179-181.
Турарбеков М.З., Саитбекова Н.Д., Шубина Е.А. и др. Полиморфные повторяющиеся последовательности ДНК в геномах диких и домашних овец // Докл. АН СССР. 1988. Т. 302, № 5. С. 1265-1269.
Туркин Н.В., Сатунин К.А. Звери России. Cervidae. М., 1902. 506 с.
Туров СС. Снежный баран. М.-Л.: КОИЗ, 1937. 112 с.
Тышкевич В.Е. Продолжительность яра (гона), загонного и выпасного периодов у зубра полесской микропопуляции // Проблемы экологии и экологического образования Полесья в постчернобыльский период. Мозырь: РИФ “Белый ветер”, 2000. С. 225-228.
Тышкевич В.Е. Наиболее перспективные направления развития охотничьего хозяйства Беларуси в XXI веке // Современные проблемы природопользования, охотоведения и звероводства. Киров, 2002а. С. 94-97.
Тышкевич В.Е. Зубр в Беларуси - состояние натурализированных популяций // Там же. 20026. С. 393-396.
Уатт К. Экология и управление природными ресурсами. М.: Мир, 1971. 463 с.
У глянец А.В. Особенности годового жизненного цикла припятской микропопуляции зубра//Биологические ритмы. Брест, 1999а. С. 132-134.
Углянец А.В. Структурно-функциональные особенности озеранской микропопуляции европейского зубра // Структурно-функциональное состояние биологического разнообразия животного мира Беларуси. Минск, 19996. С. 100-103.
Углянец А.В. Реакклиматизация европейского зубра в бассейне р. Припять в Беларуси // Териофауна России и сопредельных территорий. VII съезд териол. о-ва. М., 2003. С. 361.
Углянец А.В., Дамашкевич А.М. Повреждения древесных и кустарниковых растений зубрами в условиях Припятского Полесья // Биологическое разнообразие национального парка “Припятский” и других особо охраняемых природных территорий. Туров -Мозырь, 1999а. С. 268-271.
Углянец А .В., Дамашкевич А.М. Цикличность годовой пространственной динамики припятской микропопуляции зубра и факторы ее определяющие // Биологические ритмы. Брест, 19996. С. 130-132.
Углянец А., СтрукД., Дамашкевич А. Прогнозная оценка влияния зубров на древесно-кустарниковую растительность в пойме Припяти // Лес и охотничье хоз-во. 2001. № 2. С. 20-21.
Удина И. Г. Изучение ДНК-полиморфизма генов каппа-казеина и главного комплекса гистосовместимости у зубров // Генетика. 1994. Т. 30. Прилож. С. 161.
Удина И.Г. Генетические аспекты проблемы сохранения зубров // Молекулярно-генетические маркеры животных. Киев: Аграрна наука, 1996. С. 22-23.
Удина И.Г., Соколова С.С., Сипко Т.П., Сулимова ГЕ. Сравнительная характеристика полиморфизма ДНК локусов DQB и DRB главного комплекса гистосовместимости у представителей семейства Bovidae // Генетика. 1994. Т. 30, № 3. С. 356-360.
Улитко А.И. Локальные фауны крупных млекопитающих из отложений пещеры Дыро-ватый камень на реке Серга (Средний Урал) // Четвертичная палеозоология на Урале. Екатеринбург: Изд-во Уральского ун-та, 2003. С. 185-192.
Урсова Л.Е. Роль диких жвачных в эпизоотологии протостронгилидозов овец на Памире // Профилактика протозойных, гельминтозных и незаразных болезней сельскохозяйственных животных. Душанбе, 1985. С. 60-69.
Усов С.А. Зубр. Монография. М., 1865. 114 с.
Успенский С. Овцебыков-в Советскую Арктику// Охота и охотничье хоз-во. 1966. № 8.
С. 17-19.
Успенский С. Овцебык - новосел Таймыра // Там же. 1975а. № 2. С 4—5.
Успенский С. Овцебык в Советской Арктике // Там же. 19756. № 10. С. 22-23.
Успенский С.М. Овцебык в СССР // Природа. 1976. №1. С. 58-63.
Успенский С.М. Овцебыки в СССР, перспективы их дальнейшего расселения и использования // Обогащение фауны и разведение охотничьих животных. Киров, 1982а. С. 111.
Успенский С.М. История завоза и выпуска овцебыков в СССР // Охота и охотничье хоз-во. 19826. №7. С. 29.
Успенский С.М., Чернявский Ф.Б. Овцебык и перспективы его акклиматизации в Советской Арктике//Охотничье-промысловые звери. М.: Россельхозиздат, 1965. Вып. 1. С. 161-177.
Успенский С.М., Чернявский Ф.Б. Обитатель высоких широт // Природа. 1966. № 6. С. 97-99.
Фабри К.Э., Дерягина М.А. О сравнительноэтологической диагностике видотипичного поведения Bivinae// Зоол. журн. 1972. Т. 51, вып. 2. С. 259-266.
Фадеев В А. Поведение сайгаков на водопоях // Поведение животных. Экологические и эволюционные аспекты. М.: Наука, 1972а. С. 195-196
Фадеев В.А. Поведение сайгаков в период массовой миграции и окота // Там же. 19726. С. 197-198.
Фадеев В.А. Миграции Saiga tatarica L. в Казахстане//1 Междунар. конгр. по млекопитающим. М.: ВИНИТИ, 1974. Т. 2. С. 284-285.
Фадеев В А. Динамика численности сайгаков в Казахстане // Копытные фауны СССР.
М.: Наука, 1975а. С. 136-137.
Фадеев В А. Сайгак на Устюрте // Там же. 19756. С. 138-139.
Фадеев В.А. Результаты мечения сайгаков на территории Казахстана // Копытные фауны СССР. М.: Наука, 1980а. С. 210-212.
Фадеев В.А. Влияние джутов на численность сайгаков в Казахстане // Там же. 19806. С. 212-213.
Фадеев В А. Волк (распространение и численность) // Млекопитающие Казахстана. Алма-Ата: Наука, 1981. Т. 3, ч. 1. С. 12-15.
Фадеев В А. Сайгак в Казахстане (численность, промысел) // IV съезд Всесоюз. териол. о-ва. М., 1986. Т. 3. С. 182-183.
Фадеев В А., Иванов А.А. Численность сайгаков в Казахстане в зависимости от кормовых ресурсов // Экология и поведение млекопитающих Казахстана. Алма-Ата, 1988. С. 37-47. (Тр. Ин-та зоол. АН КазССР. Т. 44).
Фадеев В А., Исмагилов Е.Ж. Влияние антропогенных факторов на поголовье сайгаков // Влияние антропогенной трансформации ландшафта на население наземных позвоночных животных. М., 1987. Ч. 1. С. 195-197.
Фадеев В А., Слудский А.А. Сайгак в Казахстане (экология, хозяйственное значение). Алма-Ата: Наука, 1982. 160 с.
Фадеев В.А., Слудский А.А. Сайгак // Млекопитающие Казахстана. Алма-Ата, Наука, 1983. Т. 3, ч. 3. С. 56-92.
Фадеев В., Шаад А. Промысел сайгака в Казахстане // Охота и охотничье хоз-во. 1978. № 10. С. 10-12.
Фандеев А.А. Окот сайги на правобережье Волги // Зоол. журн. 1960. Т. 39, вып. 6. С. 906-911.
Фандеев А.А. Размножение и развитие сайгака в связи с вопросами рационализации промысла: Автореф. дис. ... канд. биол. наук. М., 1965. 21 с.
Федосенко А.К. Сезонные кочевки копытных в Северном Тянь-Шане и Джунгарском Алатау // Копытные фауны СССР. М.: Наука, 1975. С. 27-29.
Федосенко А.К. Архар в Илийской долине И Редкие и исчезающие звери и птицы Казахстана. Алма-Ата, 1977. С. 111-114.
Федосенко А.К. Поведение архаров (Ovis аттоп) в репродуктивный период в Северном Тянь-Шане и Джунгарском Алатау // Зоол. журн. 1979. Т. 58, вып. 6. С. 903-911.
Федосенко А.К. Архар. Алма-Ата: Кайнар, 1983а. 96 с.
Федосенко А.К. О суточной активности архаров и сибирских горных козлов на Тянь-Шане и в Джунгарском Алатау//Периодические явления в жизни животных. М., 19836. С. 5-14.
Федосенко А.К. Состояние поголовья архаров на юго-востоке Казахстана // Редкие виды млекопитающих СССР и их охрана М., 1983в. С. 217-219.
Федосенко А.К. Экология путоранского снежного барана (Ovis nivicola borealis) // Зоол. журн. 1985. Т. 64, вып. 1. С. 107-116.
Федосенко А.К. Взаимоотношения крупных хищников и копытных в горах юго-востока Казахстана и Южной Сибири // Роль крупных хищников и копытных в биоценозах заповедников. М., 1986а. С. 4—21.
Федосенко А.К. О смертности устюртского муфлона на Западном чинке Устюрта // Редкие животные Казахстана. Материалы ко 2-му изданию Красной книги КазССР. Алма-Ата, 19866. С. 33-36.
Федосенко А.К. Современное состояние популяций архара в СССР // Редкие наземные позвоночные Сибири. Новосибирск: Наука, 1988. С. 251-256.
Федосенко А.К. Материалы по экологии алтайского горного барана в Юго-Восточном Алтае // Экологические исследования в заповедниках Южной Сибири. М., 1989. С. 17-31.
Федосенко А.К. Состояние популяций диких баранов на юге СНГ // Редкие виды млекопитающих России и сопредельных территорий. М., 1997. С. 96.
Федосенко А.К. Архар в России и сопредельных странах. М., 2000. 291 с.
Федосенко А.К. Алтайский горный баран Ovis аттоп аттоп Linnaeus, 1758 // Красная книга Российской Федерации (животные). Балашиха: Астрель, 2001. С. 719-720.
Федосенко А.К. Уриал. М., 2002а. 154 с.
Федосенко А.К. Состояние популяций аргали в Алтае-Саянском регионе // Вопросы современного охотоведения. М., 20026. С. 172-176.
Федосенко А.К. Сибирский горный козел в России и прилежащих странах (состояние популяций, экология, поведение, охрана и хозяйственное использование). М.: Изд-во ГУ “Центрохотконтроль”, 2003. 193 с.
Федосенко А.К, Бланк Д.А. О поведении горных козлов (Capra sibirica) в репродуктивный период в Северном Тянь-Шане // Зоол. журн. 1982. Т. 61, вып. 3. С. 428-435.
Федосенко А.К, Вейнберг П.И. Состояние популяций памирского барана // VI съезд териол. о-ва. М., 1999. С. 265.
Федосенко А.К, Вейнберг П.И. О состоянии популяций памирского архара (Ovis аттоп polii) в Таджикистане и Киргизии // Бюл. МОИП. Отд. биол. 2001. Т. 106, вып. 4. С. 5-12.
Федосенко А.К, Вейнберг П.И., Вальдез Р. К экологии памирского барана в Киргизии // Там же. 1995. Т. 100, вып. 1. С. 34—41.
Федосенко А.К, Вейнберг П.И., Вальдез Р.А. О состоянии популяций туркменского и таджикского уриалов и винторогого козла в Туркменистане // Там же. 2000. Т. 105, вып. 3. С. 10-16.
Федосенко А.К, Вейнберг П.И., Скуратов Н.И. Сибирский горный козел в Западном Саяне // Охрана и изучение редких и исчезающих видов животных в заповедниках. М„ 1992. С. 29-44.
Федосенко А.К, Грачев Ю.А. Ресурсы диких копытных в горах юго-востока Казахстана и перспективы их хозяйственного использования // Копытные фауны СССР. М.: Наука, 1975. С. 29-31.
Федосенко А.К., Жиряков В.А. Состояние и охрана поголовья архаров в пустынной части Алма-Атинского заповедника // Редкие виды млекопитающих и их охрана. М.: Наука, 1977. С. 236-237.
Федосенко А.К., Жиряков В.А. Взаимоотношения хищников и диких копытных в Северном Тянь-Шане и Джунгарском Алатау // Экологические основы охраны и рационального использования хищных млекопитающих. М.: Наука, 1979. С. 72-74.
Федосенко А.К., Жиряков В.А. Особенности экологии архара в пустынных горах Илий-ской долины (Казахстан) // Бюл. МОИП. Отд. биол. 1987. Т. 92, вып. 1. С. 31-41.
Федосенко А.К., КолпащиковЛ.А., ПавловБ.М. Материалы по питанию путоранского снежного барана (Ovis nivicola borealis) И Зоол. журн. 1985. Т. 64, вып. 2. С. 315-316.
Федосенко А.К, Орлов Г.И. О брачном поведении архаров и сибирских горных козлов // Групповое поведение животных. М.: Наука, 1976. С. 402-404.
Федосенко А.К, ПолеВ.Б., Байдавлетов Р.Ж. и др. Копытные//Млекопитающие Казахстана. Алма-Ата, 1984. 230 с.
Федосенко А.К, Скуратов Н.И. О суточной активности и питании аргали на Алтае // Бюл. МОИП. Отд. биол. 1990. Т. 95, вып. 2. С. 11-20.
Федосенко А.К, Собанский Г.Г, Скуратов Н.И, Сергеев М.В. Алтайский горный баран // Проблемы охраны редких животных (Материалы к Красной книге). М., 1987. С. 81-84.
Федоткина Н.В. Высокогорные пастбища копытных животных в Юго-Восточном Алтае (хр. Сайлюгем) и проблемы их охраны // Проблемы ботаники Южной Сибири. Барнаул, 2002. С. 71—72.
Фертиков В. Снежный баран // Охота и охотничье хоз-во. 1978. № 7. С. 18-19.
Фертиков В.И. Состояние популяции якутского снежного барана//Природа. 1979. № 12.
С. 86.
Фетисов А.С., Хрусцелевский В.Н. Млекопитающие Юго-Восточного Забайкалья // Тр. Иркутского гос. ун-та им. А.А. Жданова. Сер. биол. 1948. Т. 3, вып. 3. С. 1-15.
Филатов Д. Летняя и зимняя поездки в с.-з. Кавказ в 1909 году для ознакомления с кавказским зубром // Ежегодн. Зоол. муз. Импер. Акад. наук. 1910. Т. 15, № 4. С. 171-217.
ФилатовД. О кавказском зубре//Зап. Импер. Акад. наук. Сер. 8.1912. Т. 30, №8. С. 1-40.
Филимонов А.Н. О возможной селективной роли волка в период отела сайги // Экологические основы охраны и рационального использования хищных млекопитающих. М.: Наука, 1979. С. 142-143.
Филимонов А.Н, Лаптев С.П. Наблюдения за волком и сайгой на юге Актюбинской области // Копытные фауны СССР. М.: Наука, 1975. С. 207-208.
Филонов К.П. Копытные животные и крупные хищники на заповедных территориях. М.: Наука, 1989. 256 с.
Филонов К.П., Шадрина ГД. Крупные наземные млекопитающие в заповедниках России: состояние и перспективы сохранения // Состояние териофауны в России и ближнем зарубежье. М., 1996. С. 343-348.
Филус И.А. Материалы по экологии и поведению аргали в Алтайском заповеднике // Охрана и изучение редких и исчезающих видов животных в заповедниках. М., 1992. С. 44-56.
Филус И.А., Золотухин Н.И. Алтайский горный баран // Проблемы охраны редких животных (Материалы к Красной книге). М., 1987. С. 84-87.
Филь В.Н О популяции сибирского горного козла в Тофаларии (Восточный Саян) // Зоол. журн. 1964. Т. 43, вып. 4. С. 621-622.
Филь В.И. Особенности размещения горных козлов на зимовках в Заилийском Ала-Тау // Сб. науч.-техн. информ. ВНИИОЗ. 1970. Вып. 28. С. 21-25.
Филь В.И. Влияние охоты на популяцию снежного барана в Центральной Камчатке // Редкие виды млекопитающих фауны СССР и их охрана. М.: Наука, 1973а. С. 154-155.
Филь В.И. Снежный баран Южной Камчатки, его охрана и хозяйственное использование // Природа и человек. Владивосток, 19736. С. 244-251.
Филь В.И. Экологические аспекты хозяйственного использования и охраны популяций снежного барана // Копытные фауны СССР. М.: Наука, 1975. С. 281-282.
Филь В.И. Влияние охоты на снежного барана Камчатки // Редкие виды млекопитающих и их охрана. М.: Наука, 1977а. С. 237-239.
Филь В.И. Сибирский горный козел центральной части Восточного Саяна // Там же. 19776. С. 239-240.
Филь В.И. О динамике структуры популяций снежного барана Ovis nivicola nivicola на Камчатке и прилежащих территориях // Зоол. журн. 1977в. Т. 56, вып. 11. С. 1712— 1718.
Филь В.И. Гон снежного барана на Камчатке // Экология. 1978. № 3. С. 98-101.
Филь В.И. О сроках охоты на снежного барана // Проблемы охраны фауны. Изд-во Моск, ун-та, 1982. Ч. 1. С. 133-135.
Филь В.И., Вяткин И. С. Некоторые проблемы изучения копытных на Камчатке // Зоологические проблемы Сибири. Новосибирск: Наука, 1972. С. 488-489.
Флеров К.К. Обзор диагностических признаков беловежского и кавказского зубров И Изв. АН СССР. Отд. матем. и естеств. наук. 1932. № 10. С. 1579-1890.
Флеров К.К. О некоторых географических и исторических изменениях евразийских копытных //Докл. АН СССР. 1934. Т. 4, № 3. С. 167-169.
Флеров К.К. Копытные (Ungulata) арктических стран И Звери Арктики. Л.: Изд-во Глав-севморпути, 1935а. С. 105-263.
Флеров К.К. Копытные звери Таджикистана // Звери Таджикистана. М.-Л.: Изд-во АН СССР, 19356. С. 93-130.
Флеров К.К. Основные черты формирования фауны млекопитающих четвертичного периода в северном полушарии // Тр. Комис, по изуч. четвертич. периода. 1955. Т. 12. С. 121-126.
Флеров К.К. Сравнительная краниология современных представителей рода Bison // Бюл. МОИП. Отд. биол. 1965. Т. 70, вып. 1. С. 40-54.
Флеров К.К. Древнейшие представители и история рода Bison // Териология. Новосибирск: Наука, 1972. Т. 1. С. 81-88.
Флеров К.К. История бизонов и причины изменения их ареалов И История биогеоценозов СССР в голоцене. М.: Наука, 1976. С. 166-175.
Флеров К.К. Систематика и эволюция // Зубр. М.: Наука, 1979. С. 9-127.
Флеров К.К., Заблоцкий М.А. О причинах изменения ареала бизонов // Бюл. МОИП. Отд. биол. 1961. Т. 66, вып. 6. С. 99-109.
Флинт В.Е. Стратегия сохранения редких видов в России: теория и практика. М.: ГЕОС, 2000. 200 с.
Флинт В., Перерва В. Перспективы восстановления численности зубров в СССР // Охота и охотничье хоз-во. 1986. № 3. С. 4—7.
Флинт В.Е., Перерва В.И., Брагин А.Б. Основные принципы расселения зубра //1 Всесоюз. совещ. по пробл. зоокультуры. М., 1986. Ч. 2. С. 93-95.
Формозов А.А. К истории древнейшего скотоводства на юге СССР // Основные проблемы териологии. М.: Наука, 1972. С. 19-25. (Тр. МОИП. Отд. биол. Т. 48).
Формозов А.Н. Снежный покров в жизни млекопитающих и птиц СССР. М.: Изд-во МОИП, 1946. 152 с.
Форонова И.В. Четвертичные млекопитающие юго-востока Западной Сибири (Кузнецкая котловина): филогения, биостратиграфия, палеоэкология. Новосибирск: Изд-во СО РАН (филиал “ГЕО”), 2001. 243 с.
Фортунатов Б.К. Бизоны и зубры в Аскании-Нова//Изв. гос. степи, заповедника Аскания-Нова. 1922. Вып. 1. С. 1-66.
Фортунатов Б.К. Гибридные стада за последние годы // Аскания-Нова. Степной заповедник Украины. М.: Гос. издат., 1924. С. 319-341.
Фортунатов Б.К. Восстановление зубра в Государственном заповеднике “Чапли” (б. Аскания-Нова) //Изв. гос. степи, заповедника “Чапли” (б. Аскания-Нова). 1929 (1928). Т. 7. С. 69-87.
Фролов М.В., Голуб О.Н. Винторогий козел и бухарский горный баран Таджикистана и пути их охраны И Редкие виды млекопитающих СССР и их охрана М., 1983. С. 219-220.
Хайдав Ц„ Варламова С.Т., Цэндэм Ц. и др. Препарат из рогов монгольского дзерена // Природные условия и биологические ресурсы Монгольской Народной Республики. М.: Наука, 1986. С. 206-207.
Хамзина Е.Л. Археологические памятники Бурятии. Новосибирск: Наука, 1982.152 с.
Хахин ГВ. Некоторые особенности поведения сайгаков И Прикладная этология. М.: Наука, 1983. С. 258-259.
Хахин ГВ. Сохранение сайгаков в местах интенсивного природопользования // IV съезд Всесоюз. териол. о-ва. М., 1986. Т. 3. С. 183-184.
Хлебников А. Исчезнувшие каменные бараны Восточной Сибири и обилие лосей в Канском округе//Природа. 1877. Кн. 3. С. 124—126.
Хозацкий Л.И. Новые данные о териофауне плиоцена Молдавии // IV съезд Всесоюз. териол. о-ва. М., 1986. Т. 1. С. 35-36.
Холодова М.В. Потребление и усвоение кормов дикими жвачными умеренных зон: Автореф. дис. ... канд. биол. наук. М., 1989. 17 с.
Холодова М.В., Арылов Ю.Н., Лущекина А.А., Сорокин П.А. Европейский сайгак: динамика численности и генетического разнообразия популяции // Млекопитающие как компонент аридных экосистем (ресурсы, фауна, экология, медицинское значение и охрана). М., 2004. С. 158-159.
Холодова М.В., Белоусова И.П. Период весеннего роста интенсивности потребления кормов у зубров // Экология, морфология, использование и охрана диких копытных. М., 1989а. Ч. 2. С. 233-234.
Холодова М.В., Белоусова И.П. Потребление и усвоение питательных веществ и энергии зубрами (Bison bonasus) И Зоол. журн. 19896. Т. 68, вып. 12. С. 107-117.
Холодова М.В., Белоусова И.П. Изучение эффективности усвоения кормов и энергетики зубров и бизонов в Центральном зубровом питомнике Приокско-Террасного биосферного заповедника // К вопросу о возможности сохранения зубра в России. Пущино, 1993. С. 85-90.
Холодова М.В., Белоусова И.П., Дылис В.Н. Влияние обилия кормов в среде на интенсивность их потребления зубрами И V съезд Всесоюз. териол. о-ва АН СССР. М., 1990. Т. 2. С. 304.
Холодова М.В., Лущекина А.А., НероновВ.М. и др. Сравнительный анализ генетического разнообразия калмыцких и монгольских сайгаков // Докл. Акад. наук. 2001. Т. 376, № 6. С. 1-3.
Холодова М.В., Лущекина А.А., Неронов В.М. и др. Генетическое разнообразие монгольских сайгаков (Saiga tatarica mongolica) // Proc. Inst. Biol. (Ulaanbaatar). 2002. N 24. P. 159-162.
Холодова М.В., Неронов В.М. Сайгаки в зоопарках мира. Сообщ. 1. Опыт содержания сайгаков в зарубежных зоопарках // Науч. иссл. в зоол. парках. 1996. № 6. С. 223-253.
Холодова М.В., Рябинина О.М., Лущекина А.А. и др. Предварительные результаты генетического анализа северной популяции монгольского дзерена (Procapra gutturosa Pallas, 1777) // Proc. Inst. Biol. (Ulaanbaatar). 2002. N 24. P. 163-169.
Хохлов А. Вершины охоты. M., 2002. 424 с.
Хохлов А., Харебов С. Охотничьи (трофейные) животные Европы и Азтт. М.: ФОП, 2001. 688 с.
Храмцов В. С. Перспективы реакклиматизации горалов в Приморье //1 Всесоюз. совещ. по пробл. зоокультуры. М., 1986. Ч. 2. С. 98-99.
Хромов А.А., Архангельский М.С., Иванов А.В. Крупные четвертичные млекопитающие Среднего и Нижнего Поволжья. Дубна, 2001. 254 с.
Худайбердиев Т.Н. Антропогеновая териофауна Восточного Узбекистана ИIV съезд Всесоюз. териол. о-ва. М., 1986. Т. 1. С. 36-37.
Худайбердиев Т.Н. Промысловые виды животных многослойной палеолитической стоянки Кульбулак //V съезд Всесоюз. териол. о-ва АН СССР. М., 1990. Т. 1. С. 33-34.
Цалкин В.И. О вертикальном распространении диких баранов // Бюл. МОИП. Отд. биол. 1945. Т. 50, вып. 1-2. С. 39-51.
Цалкин В.И. Дикий баран в Туркмении // Там же. 1948а. Т. 53, вып. 1. С. 31-48.
Цалкин В.И. О росте рогов и строении черепа у диких баранов // Зоол. журн. 19486. Т. 27, вып. 5. С. 445-464.
Цалкин В.И. О сокращении ареала и численности диких баранов // Охрана природы. М.: Всерос. о-во охраны природы. 1948в. Сб. 2. С. 94—102.
Цалкин В.И. Систематика сибирских горных козлов //Бюл. МОИП. Отд. биол. 1949. Т. 54, вып. 2. С. 3-21.
Цалкин В.И. Новый подвид безоарового козла из Туркмении (Capra aegagrus turcmenicus subsp. nova) // Докл. АН СССР. Новая сер. 1950а. Т. 70, № 2. С. 323-326.
Цалкин В.И. Сибирский горный козел. М.: Изд-во МОИП, 19506. 119 с. (Матер, к позн. фауны и флоры СССР. Новая сер. Отд. зоол. Вып. 21).
Цалкин В.И. Горные бараны Европы и Азии. М.; Изд-во МОИП, 1951а. 344 с. (Там же. Вып. 27).
Цалкин В.И. К истории литовского зубра И Докл. АН СССР. 19516. Т. 77, № 2. С. 349-352.
Цалкин В.И. Изменчивость и систематика туров Западного Кавказа (Caprinae, Artiodactyla) // Бюл. МОИП. Отд. биол. 1955. Т. 60, вып. 4. С. 17-33.
Цалкин В.И. Материалы для истории скотоводства и охоты в древней Руси // Матер, и исслед. по археолог. СССР. М.: Изд-во АН СССР, 1956. № 51. 186 с.
Цалкин В.И. Фауна из раскопок археологических памятников Среднего Поволжья (Материалы для истории скотоводства и охоты в СССР) // Там же. 1958. № 61. С. 221-281. (Тр. Куйбышевской археолог, экспедиции. Т. 2).
Цалкин В.И. Домашние и дикие животные Северного Причерноморья в эпоху раннего железа// Там же. 1960. № 53. С. 7-109.
Цалкин В.И. К истории животноводства и охоты в Восточной Европе // Там же. 1962. № 107. С. 1-129.
Цалкин В.И. К истории млекопитающих восточноевропейской лесостепи И Бюл. МОИП. Отд. биол. 1963. Т. 68, вып. 2. С. 43-62.
Цалкин В.И. Древнее животноводство племен Восточной Европы и Средней Азии // Матер, и исслед. по археолог. СССР. 1966. № 135. С. 1-159.
Цалкин В.И. Фауна из раскопок Боршевских и Роменских городищ // Советская археология. 1969. № 4. С. 91-101.
Цалкин В.И. Древнейшие домашние животные Восточной Европы. М.: Наука, 1970. 280 с.
Цалкин В.И. Фауна из раскопок славянских памятников Молдавии // Основные проблемы териологии. М.: Наука, 1972а. С. 38-65. (Тр. МОИП. Отд. биол. Т. 48).
Цалкин В.И. Фауна из раскопок андроповских памятников в Приуралье // Там же. 19726. С. 66-81.
Цаплюк О.Э. Наступление половозрелое™ и процесс сперматогенеза у самцов сайгака // Материалы по фауне и экологии наземных позвоночных Казахстана (млекопитающие). Алма-Ата: Изд-во АН КазССР, 1962. С. 222-240. (Тр. Ин-та зоол. АН КазССР. Т. 17).
Цаплюк О.Э. Половая цикличность у сайгака // Охотничье-промысловые звери Казахстана. Алма-Ата: Наука, 1966. С. 193-211. (Тр. Ин-та зоол. АН КазССР. Т. 26).
Цаплюк О.Э. Динамика половой активности сайгака: Автореф. дис.... канд. биол. наук. Алма-Ата, 1968. 16 с.
Цаплюк О.Э. Некоторые особенности в размножении сайгаков на острове Барса-Кельмес в Центральном Казахстане // Популяционная структура вида у млекопитающих. М., 1970. С.96-97.
Цаплюк О.Э. Поведение сайгаков в брачный период в условиях неволи // Поведение животных. Экологические и эволюционные аспекты. М.: Наука, 1972. С. 198-199.
Цаплюк О.Э. Атавистические явления в половой ритмике некоторых копытных //1 Междунар. конгр. по млекопитающим. М.: ВИНИТИ, 1974. Т. 2. С. 312-313.
Цаплюк О.Э. Возрождение сайгака. Алма-Ата: Кайнар, 1982. 108 с.
Цаплюк О.Э., Савинов Е.Ф. Половая ритмика сибирского горного козла в Джунгарском Алатау // Промысловые млекопитающие Казахстана. Алма-Ата, 1973. С. 200-212.
Царев С. Переселение овцебыков // Охота и охотничье хоз-во. 1997. № 8. С. 7-9.
Царев С.А. Стратегия восстановления исторического ареала овцебыка на Крайнем Севере России // Охотничьи животные России. 1999. Вып. 1. 83 с.
Царев С.А. Актуальные вопросы расселения и одомашнивания овцебыка на Севере России // Вопросы современного охотоведения. М., 2002. С. 357-362.
Царев С., Лякин Б. Овцебыки: ровесники мамонта // Охота и охотничье хоз-во. 2001. № 6. С. 8-11.
Царев С.А., Сипко Т.П., Гребенков А.Б. Состояние популяции овцебыков на Таймыре // VI съезд териол. о-ва. М., 1999. С. 270.
Цибизова Е.Л. Участие Окского заповедника в создании вольноживущей популяции зубра в Орловско-Брянско-Калужском регионе // Проблемы сохранения и восстановления диких копытных животных в Центральном регионе России. Орел: Изд-во ОРАГС, 2003. С. 87-88.
ЦэвэгмидД. Редчайшие животные Монголии и их современное состояние // Тр. IX Междунар. конгр. биологов-охотоведов. М., 19706. С. 872-874.
Чагиров И.А., Литвинова Л.Д. Некоторые данные о развитии плодов сайгака // Тр. Инта зоол. АН КазССР. 1960. Т. 13. С. 111-124.
Чемыртан ГД. Млекопитающие энеолита Молдавии // IV съезд Всесоюз. териол. о-ва. М., 1986. Т. 1. С. 37-38.
Черемкин Р.Г. Снежный баран в Якутии // Природа. 1959. № 5. С. 107-108.
Черкасов А.А. Записки охотника Восточной Сибири. М.: Физк. и спорт, 1990. 554 с.
Чернавин В.В. Заметка о диких баранах Западного Тянь-Шаня // Ежегодн. Зоол. музея Акад. наук. 1916. Т. 21, №. 2-3. С. 200-213.
Черников С.С. Восточный Казахстан в эпоху бронзы И Матер, и исслед. по археолог. СССР. М.: Изд-во АН СССР, 1960. № 88. 272 с.
Черский И.Д. Описание коллекции послетретичных млекопитающих животных, собранных Новосибирской экспедицией 1885-1886 гг. // Зап. Импер. Акад. наук. 1891. Т. 65. Прилож. № 1. С. 1-706.
Чернявский Ф.Б. Материалы по биологии снежного барана (Ovis nivicola Esch.) в Корякском нагорье // Бюл. МОИП. Отд. биол. 1961. Т. 66, вып. 6. С. 61-76.
Чернявский Ф.Б. О размножении и росте снежного барана // Зоол. журн. 1962а. Т. 41, вып. 10. С. 1556-1566.
Чернявский Ф.Б. Новая форма снежного барана из Корякского нагорья // Докл. АН СССР. 19626. Т. 145, № 5. С. 1174-1176.
Чернявский Ф.Б. О систематических взаимоотношениях и истории снежных баранов Старого и Нового Света // Бюл. МОИП. Отд. биол. 1962в. Т. 67, вып. 6. С. 17-26.
Чернявский Ф. Снежный баран // Охота и охотничье хоз-во. 1962г. № 8. С. 24—27.
Чернявский Ф.Б. Снежный баран Корякского нагорья: Автореф. дис.... канд. биол. наук. Л., 1963.22 с.
Чернявский Ф.Б. Некоторые данные по морфологии снежного барана // Бюл. МОИП. Отд. биол. 1964а. Т. 69, вып. 1. С. 122-126.
Чернявский Ф.Б. О некоторых приспособительных особенностях строения посткраниального скелета снежного барана (Ovis nivicola Esch.) // Зоол. журн. 19646. Т. 43, вып. 2. С. 242-252.
Чернявский Ф.Б. К экологии и промысловому значению снежного барана в Корякском нагорье//Проблемы Севера. 1967. Вып. 11. С. 128-141.
Чернявский Ф. Чукотский толсторог//Охота и охотничье хоз-во. 1968. № 11. С. 12-13.
Чернявский Ф.Б. Снежный баран (Ovis nivicola Esch.) на Чукотке // Бюл. МОИП. Отд. биол. 1970а. Т. 75, вып. 1. С. 92-99.
Чернявский Ф.Б. Опыт учета снежных баранов на Корякском нагорье и на Чукотке И Тр. IX Междунар. контр, биологов-охотоведов. М., 19706. С. 345-349.
Чернявский Ф.Б. Основные экологические и этологические факторы, определяющие структуру и динамику ареалов диких копытных (Artiodactyla, Mammalia) на Крайнем Северо-Востоке Сибири // Териология. Новосибирск: Наука, 1974. Т. 2. С. 106-115.
Чернявский Ф.Б. Млекопитающие Крайнего Северо-Востока Сибири. М.: Наука, 1984.388 с.
Чернявский Ф.Б. Таксономия и история снежных баранов (подрод Pachyceros) // Териофауна России и сопредельных территорий. VII съезд териол. о-ва. М., 2003. С. 380.
Чернявский Ф.Б. О таксономии и истории снежных баранов (подрод Pachyceros, Artiodactyla) // Зоол. журн. 2004. Т. 83, № 8. С. 1059-1070.
Чернявский Ф.Б., Аксенов В.В., Вовченко В.Е. Снежный баран (Ovis nivicola) в области северного побережья Охотского моря // Зоол. журн. 1990. Т. 69, вып. 3. С. 114—119.
Чернявский Ф.Б., Вовченко В.Е., Домнич В.И. Акклиматизация овцебыков на острове Врангеля // Копытные фауны СССР. М.: Наука, 1980. С. 273-274.
Чернявский Ф.Б., Вовченко В.Е., Домнич В.И. Об акклиматизации овцебыков Ovibos moschatus Zimmerman, 1780 // Экология млекопитающих и птиц острова Врангеля. Владивосток: Наука, 1981. С. 97-114.
Чернявский Ф.Б., Железнов Н.К. Распространение, численность и задачи охраны снежного барана на Северо-востоке Сибири И Редкие виды млекопитающих и их охрана. М.: Наука, 1977. С. 241-242.
Чернявский Ф., Ковалев Д. Результаты и перспективы акклиматизации овцебыка (Ovibos moschatus) на Северо-Востоке Азии И Цитология. 1995. Т. 37, № 7. С. 651.
Чернявский Ф.Б., Кречмар М.А. Бурый медведь (Ursus arctos L.) на Северо-Востоке Сибири. Магадан, 2001. 93 с.
Чернявский Ф.Б., Матюшенков Н.В., Никитин В.А. Снежный покров и использование пастбищ овцебыками // Копытные фауны СССР. М.: Наука, 1980. С.275-277.
Черченко Н.И. Сайга в неволе // Копытные фауны СССР. М.: Наука, 1975. С. 339.
Чиковани Т.Г., Вронский Н.Г, Гигаури ГН, Ичуаидзе ГК. Ахметский заповедник И Заповедники СССР. Заповедники Кавказа. М.: Мысль, 1990а. С. 200-210.
Чиковани Т.Г., Гигаури ГН, Дидманидзе Э.А., Курашвили Б.Е. Боржомский заповедник//Там же. 19906. С. 162-172.
Чиковани Т.Г, Гигаури Г.Н., Дидманидзе Э.А. и др. Лагодехский заповедник // Там же. 1990в. С. 210-226.
Члаидзе З.И. О нахождении добавочной кости в черепе восточнокавказского тура // Со-общ. АН ГрузССР. 1966. Т. 41, № 1. С. 181-183.
Члаидзе З.И. Capra caucasica на южном склоне Главного Кавказского хребта //1 Меж-дунар. конгр. по млекопитающим. М.: ВИНИТИ, 1974. Т. 2. С. 321-322.
Чичикин Ю.Н, Чичикина С.Н. Новые виды копытных в Сары-Челекском заповеднике // Тр. Сары-Челекского заповедника. 1965. Вып. 1. С. 89-94.
Шагдарсурен О. К экологии Gazella gutturosa Pall. //1 Междунар. конгр. по млекопитающим. М.: ВИНИТИ, 1974. Т. 2. С. 324-325.
Шайдуллин ИН. Гибриды плодовиты // Овцеводство. 1992. № 3. С. 18-21.
Шайдуллин И.Н. Биологические особенности акклиматизации овец и гибридизации их со снежным бараном Ovis nivicola nivicola в условиях Камчатки: Автореф. дис. ... докг. биол. наук. Дубровицы, 1994а. 41с.
Шайдуллин И.Н. Кариологическая оценка гибридов снежного барана камчатской популяции и домашних овец // Опыт и проблемы зоотехнической науки. Ульяновский с,-х. ин-т, 19946. С. 37-42.
Шайдуллин И.Н. Горные бараны - генный источник повышения жизнестойкости овец // Современные проблемы в зоотехнии. М., 2001. С. 14—17.
Шайдуллин И.Н., Живалев И.К., Шихов ИЯ. Цитогенетическая характеристика гибридов Ovis nivicola nivicola // С.-х. биология. Сер. биол. животных. 1994. № 4. С. 40-42.
Шайдуллин И.Н., Жиряков А.М., Бин Н.Н. Акклиматизация и гибридизация романовских овец в условиях Камчатского региона с целью сохранения и восстановления породы // Там же. 2000. № 4. С. 37-44.
Шайдуллин И.Н., Ролдугин В.Н. Возможна отдаленная гибридизация // Овцеводство. 1986. №4. С. 40-41.
Шайдуллин И.Н., Шайдуллина РГ. Отдаленная гибридизация овец на Камчатке // Там же. 1988. №4. С. 11-13.
Шаймуханбетов О. Сохранение сайги в Центральном Казахстане: общественные организации, государство и местное население // Степи, бюл. 2004. № 15. С. 28-31.
Шакула В.Ф. Использование территории бараном Северцова // Экология, морфология, использование и охрана диких копытных. М., 1989а. Ч. 2. С. 248-249.
Шакула В.Ф. Суточный цикл и активность барана Северцова // Там же. 19896. Ч. 2. С. 249-250.
Шакула В.Ф. Размножение, рост и развитие барана Северцова // Там же. 1989в. Ч. 2.
С. 251-252.
Шакула В.Ф., Каюмов Б.К., Снитько В.П. Кариотип кызылкумского горного барана // Selevinia. 1994. Т. 2, № 2. С. 83-84.
Шакула В. Ф., Снитько В.П. Перспективы охраны и воспроизводства кызылкумских горных баранов // Дичь, охрана окружающей среды, охрана редких видов животных. М., 1995. С. 366-368.
Шапошников Ф.Д. Горные бараны Западного Тянь-Шаня // Природа. 1956. № 1. С. 109— НО.
Шарипов И.К. Кариотипы диких и домашних овец. Алма-Ата: Наука, 1989. 143 с.
Шарипов И.К., Вишневская С.С., Всеволодов Э.Б., Кабир-Биюк-Оглы А. Цитогенетическое исследование межвидовых гибридов овец и свиней // Вопросы гибридизации копытных. М.: Наука, 1980. С. 45-56.
Шаршов А.А. Хромосомные наборы ряда видов домашних и диких парнокопытных Горного Алтая: Автореф. дис. ... канд. биол. наук. Новосибирск, 1994. 16 с.
Шаулъская Н.А. Сезонные изменения поедаемости растений горалом в Сихотэ-Алинском заповеднике//Раст, ресурсы. 1980. Т. 16, вып. 2. С. 177-186. Вып. 3. С. 374—388.
Шаулъская Н.А. Пастбища горала в урочище Абрек и их продуктивность // Амурский горал. М.: Наука, 1992. С. 35-58.
Шахмарданов Э.А., Рамазанов Х.М. Ареалы копытных Дагестана // Копытные фауны СССР. М.: Наука, 1980. С. 113.
Швецов Ю.Г. Дзерен // Проблемы охраны редких животных (Материалы к Красной книге). М., 1987. С. 81.
ШвецовЮ.Г., СмирновМ.Н., Монахов Г.И. Млекопитающие бассейна оз. Байкал. Новосибирск: Наука, 1984. 358 с.
Шебзухова Э.А. Парнокопытные Республики Адыгея // Териофауна России и сопредельных территорий. VII съезд териол. о-ва. М., 2003. С. 385-386.
Шер А.В. Ископаемая сайга на севере Восточной Сибири и Аляске // Бюл. Комис, по изуч. четвертич. периода. 1967. № 33. С. 97-112.
Шер А.В. Млекопитающие и стратиграфия плейстоцена крайнего Северо-Востока СССР и Северной Америки. М.: Наука, 1971. 310 с.
Шер А.В. О систематическом положении бизона “тираспольского гравия” // VI Всесоюз. совещ. по изучению четвертичного периода. Кишинев: Штиинца, 1986. С. 272.
Шилова А.В., Стекленев Е.П., Пилипенко В.П., Елистратова Т.М. Особенности строения черепа межродовых гибридов бизона (Bison bison') с домашней коровой (Bos (Bos) taurusprimigenius) И Изв. ТСХА. 2001. № 3. С. 166—182.
Шильдер В.А. Кубанская охота Его Императорского Высочества Великого Князя Сергея Михайловича в 1898 г. // Природа и охота. 1901. № 5. С. 1-28.
Ширшов С.М., Сипко Т.П., Царев С.А. и др. Расселение овцебыка на Полярном Урале// VI съезд териол. о-ва. М., 1999. С. 283.
Шнаревич И.Д. Материалы по биологии горного козла // Тр. Алма-Атинского зоопарка. 1948. Вып. 1. С. 73-94.
Шнитников В.Н. Млекопитающие Семиречья. M.-JL: Изд-во АН СССР, 1936. 323 с.
Шовкун М.М. Состояние растительности зубрового питомника Приокско-Террасного государственного биосферного заповедника // Проблемы сохранения и восстановления диких копытных животных в Центральном регионе России. Орел: Изд-во ОРАГС, 2003а. С. 107-114.
Шостак С.В. Роль Беловежской пущи в сохранении и восстановлении численности редких копытных фауны СССР // Редкие виды млекопитающих и их охрана. М.: Наука, 1977. С. 242-245.
Шостак С.В. Численные соотношения европейского благородного оленя с другими копытными // Заповедники Белоруссии. 1978. Вып. 2. С.130-139.
Шостак С.В. Стратегия охраны зубра // Экология, морфология, использование и охрана диких копытных. М., 1989а. Ч. 2. С. 235-236.
Шостак С.В. Копытные Беловежской пущи // Динамика зооценозов, проблемы охраны и рационального использования животного мира Белоруссии. Минск: Наука и техника, 19896. С. 227.
Шостак С.В., Шохалевич Г.С. Современное состояние среднеевропейского природного комплекса копытных Беловежской пущи // Междунар. науч.-практ. конф. “Охраняемые природные территории и объекты Белорусского Поозерья: современное состояние, перспективы развития”. Витебск, 1997. С. 145.
Шпота И.К. Появление сайги {Saiga saiga, Pall) // Охотник. 1928. № Н.С. 26.
Шрамко Б.А. Древности Северного Донца. Харьков: Изд-во Харьковского ун-та, 1962. 399 с.
Шилов В.П. Очерки по истории племен Нижнего Поволжья. Л.: Наука, 1975. 215 с.
Шубин И.Г. Численность архара в Северном Прибалхашье // Копытные фауны СССР. М.: Наука, 1975. С. 145.
Шубин В.И., Шаймарданов Р. Т, Спивакова Л.В. Архар в горах Терскей Алатау И Зоологические исследования в Казахстане: современное состояние и перспективы. Алматы, 2002.С. 124-126.
Шукуров Г.Ш. Фауна позвоночных животных гор Большие Балханы (Юго-Западный Туркменистан). Ашхабад: Изд-во АН ТуркмССР, 1962. 158 с.
Шуле В. Причины, механизмы и последствия некомпенсированных вымираний крупных млекопитающих в верхнем плиоцене и четвертичном периоде// Цитология. 1995. Т. 37, № 7. С. 656-657.
Шулъпин Л.М. Материалы по млекопитающим и гадам Таласского Алатау // Изв. АН КазССР. Сер. зоол. 1948. №. 51, вып. 7. С. 65-83.
Шумов А.В. Хромосомы подрода Bos {Bison) и их гибридов с крупным рогатым скотом // Вопросы гибридизации копытных. М.: Наука, 1980. С. 91-94.
Шумов А.В. Исследование отдаленной гибридизации Bovinae в связи с проблемой их хозяйственного использования: Автореф. дис.... докт. биол. наук. СПб., 1994. 31с.
Шумов А.В., Рубцова Р.А. Гибридизация зубра с домашним скотом // Вопросы гибридизации копытных. М.: Наука, 1980. С. 33-40.
Эверсман Э. Естественная история Оренбургского края. Казань, 1850. Ч. 2. 294 с.
Эквтимишвили 3. С. Вертикальное распространение некоторых копытных на южном склоне Кавказского хребта // Сообщ. АН ГрузССР. 1952. Т. 13, № 8. С. 477-484.
Эквтимишвили З.С. Материалы к изучению питания дагестанского тура {Сарпа cylindricornis Blyth.) // Тр. Ин-та зоол. АН ГрузССР. 1953. Т. 11. С. 15-23.
Эквтимишвили 3. С. Материалы к изучению размножения безоарового козла И Там же. 1954а. Т. 13. С. 93-97.
Эквтимишвили З.С. О сроках окота у тура Северцова и серны // Там же. 19546. С. 99-105.
Эрегдэндагва Д„ Сосорбарам Ж. Размещение дзерена в степях Восточной Монголии // Тр. Ин-та биологии АН МНР. 1969. Вып. 4. С. 131-135.
Эрнитс П.Х. Поведение горных козлов в Таллиннском зоопарке // Копытные фауны СССР. М.: Наука, 1980. С. 324-325.
Эрнитс П.Х. О сравнительной этологии горных козлов // Млекопитающие СССР. III съезд Всесоюз. териол. о-ва. М., 1982. Т. 2. С. 192-193.
Юдин Б., Собанский Г. В защиту алтайского архара // Охота и охотничье хоз-во. 1984. № 6. С. 6-7.
Юргенс И.Л. Новые данные по экологии горала //1 Всесоюз. совещ. по млекопитающим. М.: Изд-во Моск, ун-та, 1961. Т. 2. С. 110-111.
Юргенс И.Л. Популяция горала хребта Та-Чинджан // Тр. Сихотэ-Алинского гос. заповедника. 1963. Вып. 3. С. 283-297.
Юргенсон П.Б. Государственная племенная книга зубров и бизонов (чистокровных, чистопородных и гибридных). М.: Изд-во МСХ СССР, 1956. Т. 1. С. 5-12.
Явруян Э.Г. Распространение дикого барана Ovis аттоп Gmelini в Закавказье // Бюл. МОИП. Отд. биол. 1969. Т. 74, вып. 2. С. 26-32.
Явруян Э.Г. Закавказский дикий баран: Автореф. дис.... канд. биол. наук. Ереван, 1970.22 с.
Явруян Э.Г. Численность и методы охраны дикого барана в Армении и Нахичеванской АССР // Копытные фауны СССР. М.: Наука, 1975. С. 285.
Язан Ю.П., Лещенко А.П. Сайгаки в новых экологических условиях Северо-Западного Прикаспия // ВСХИЗО - агропромышленному комплексу. М., 1995. С. 122-124.
Язан Ю„ Немцев А. Кавказские чистопородные горные зубры И Охота и охотничье хоз-во, 1985. № 1. С. 16-17.
Якушкин ГД. Овцебык - новый акклиматизируемый вид в СССР // Копытные фауны СССР. М.: Наука, 1975. С. 147-149.
Якушкин ГД. О начальном этапе интродукции овцебыков на полуострове Таймыр // Биологические проблемы Севера. Петрозаводск, 1976. С. 344-347.
Якушкин Г. Овцебык на Таймыре // Охота и охотничье хоз-во. 1978. № 9. С. 16—19.
Якушкин ГД. Сравнительная экологическая характеристика районов отлова (Канада, Аляска) и выпуска овцебыков (СССР) // Проблемы охраны и хозяйственного использования ресурсов диких животных Енисейского Севера. Новосибирск, 1979. С. 57-81. (Тр. НИИ сельск. хоз-ва Крайнего Севера. Т. 26).
Якушкин ГД Суточная и сезонная активность овцебыков И Экология и хозяйственное использование наземной фауны Енисейского Севера. Новосибирск, 1981. С. 57-72.
Якушкин Г. Рожденные в таймырских тундрах И Охота и охотничье хоз-во. 1982. № 9. С. 20-21.
Якушкин ГД. Оборонительное поведение овцебыков Таймыра // Прикладная этология. М.: Наука, 1983а. С. 56-58.
Якушкин ГД. Репродуктивный цикл овцебыков на полуострове Таймыр // Экология и рациональное использование наземных позвоночных севера Средней Сибири. Новосибирск, 19836. С. 56-72.
Якушкин ГД. Естественные солонцы овцебыков в долине реки Бикады Восточного Таймыра // Науч.-техн. бюл. СО ВАСХНИЛ. 1986. № 33. С. 48-52.
Якушкин Г. Весна в жизни овцебыков // Охота и охотничье хоз-во. 1987. № 7. С. 7-9.
Якушкин ГД. Демографические параметры популяции овцебыка на полуострове Таймыр // Экология, морфология, использование и охрана диких копытных. М., 1989. Ч. 2. С. 292-294.
Якушкин Г. Овцебык на Таймыре И Охота и охотничье хоз-во. 1992. № 5-6. С. 20-23.
Якушкин Г. 20 лет - и 1000 овцебыков: историческое событие свершилось // Охота и охотничье хоз-во. 1996. № 3. С. 16-20.
Якушкин ГД. Овцебыки на Таймыре. Новосибирск, 1998. 236 с.
Якушкин ГД. Состояние популяции овцебыка на Таймыре // Научное обеспечение рационального природопользования Енисейского Севера. Сб. науч. тр. РАСХН. Сиб. отд. Новосибирск, 2001. С. 100-107.
Якушкин ГД. Оценка состояния овцебыка на Таймыре и возможности ее эксплуатации // Современные проблемы природопользования, охотоведения и звероводства. Киров, 2002а. С. 414-415.
Якушкин ГД. Копытные виды в горах Бырранга // Вопросы экологии традиционного природопользования на Крайнем Севере. Новосибирск, 20026. С. 73-81.
Якушкин ГД. Освоение овцебыками гор Бырранга // Териофауна России и сопредельных территорий. VII съезд териол. о-ва. М., 2003. С. 397-398.
Якушкин Г.Д., Олъкова В.В. Морфология и линька волосяного покрова мускусных овцебыков (Ovibos moschatus) И Сиб. вести, с.-х. науки. 1988. № 1. С. 60-67.
ЯнушевичА.И. Фауна позвоночных Тувинской автономной области. Новосибирск, 1952. 143 с.
Янушевич А.И. Итоги и перспективы акклиматизации охотничьих животных в Киргизии // Акклиматизация охотничьих животных в СССР. Минск: Ураджай, 1978. С. 29-31.
Яровенко Ю.А. Характеристика сезонного использования пастбищ дагестанским туром // V съезд Всесоюз. териол. о-ва АН СССР. М., 1990. Т. 2. С. 129-130.
Яровенко Ю.А. Трофическое воздействие дагестанского тура на пастбищные экосистемы Восточного Кавказа // Экология млекопитающих горных территорий. Популяционные аспекты. Нальчик: Изд. центр “Эль-Фа”, 1997. С. 179-180.
Яровенко Ю.А., Насрулаев Н.И. Питание и использование территории безоаровыми козлами // VI съезд териол. о-ва. М., 1999. С. 290.
Яровенко Ю.А., Насрулаев Н.И., Гасанова С.М. Качественные аспекты питания безоарового козла Capra aegagrus И Териофауна России и сопредельных территорий. VII съезд териол. о-ва. М., 2003. С. 399.
Ясинецкая Н.И. Результаты исследования митотических хромосом бизона // Науч.-техн. бюл. Укр. НИИ животноводства степных районов. 1982а. Вып. 2. С. 58-59.
Ясинецкая Н.И. Особенности кариотипа гибридов серого украинского скота с бизоном в сравнении с исходными формами // Там же. 19826. С. 59-60.
AdamczewskiJZ., FloodP.F., Gunn A. Seasonal patterns in body composition and reproduction of female muskoxen (Ovibos moschatus) // J. Zool. 1997. Vol. 241, N 2. P. 245-269.
Agabriel J., Bony J., Micol D. Elevage du bison d’Amerique (Bison bison) // Prod. Anim. 1996. Vol. 9, N 5. P. 379-388.
Alcala L., Morales J. A primitive caprine from the Upper Vallesian of La Roma 2 (Alfambra, Teruel, Aragon, Spain) // C.R. Acad. Sci. Paris. 1997. Vol. 324, Ser. 2a. P. 947-953.
Alendal E. Drektighetspererioden hos moskusfe // Fauna (Norge). 1971a. Bd. 24, N 2. S. 101-105.
Alendal E. Tidlig og god formering hos moskusfe pa Dovrefjell // Ibid. 1971b. S. 96-100.
Alendal E. Overforinger av moskusfe fra Ost-Gronland i perioden 1899-1969 // Medd. Norsk polarinst. 1980b. N 107. S. 3-22.
Alendal E. Overforinger og arsaker til utsettinger av moskusfe i Norge og pa Svalbard // Ibid. 1980b. S. 23-38.
Allaine D., Houssin H., Gaiillard J.M. Etude de la variabilite spatio-temporelle d’un indice de reproduction dens une population de chamois (Rupicapra rupicapra) // Gebier Faune Sauvage. 1990. Vol. 7, mars. P. 85-94.
Allard M. W., Miyamoto M.M., Jarecki L., et al. DNA systematics and evolution of the artiodactyl family Bovidae // Proc. Nat. Acad. Sci. USA. 1992. Vol. 89, N 9. P. 3972-3976.
Amgalanbaatar S., Reading R.P., Dulamtseren S. et al. Assessment of argali sheep (Ovis аттоп) distribution in Mongolia using GIS // Mammalogical studies in Mongolia and its adjacent territories. Ulaanbaatar, 2002. P. 26—35. (Proc. Inst. Biol., N 24).
Ansell W. Order Artiodactyla//The mammals of Africa. Pt. 15. Smithsonian Inst. Press, 1972. P. 1-84.
Azanza B., Nieto M., Morales J. Samotraguspilgrimi n. sp., a new species of Oiocerini (Bovidae, Mammalia) from the Middle Miocene of Spain // C. R. Acad. Sci. Paris. 1998. Vol. 97. P. 377-382.
Bailey J.A. Desert bighorn, forage competition, and zoogeography// Wildlife Soc. Bull. 1980. Vol. 8, N3.P. 208-216.
Baranov A.S., Zakharov V.M., Kiseleva E.G., Pucek Z. Developmental stability of skull morphology European bison, Bison bonasus // Acta Theriol. 1997. Suppl. 4. P. 79-85.
Bar-Gal G.K., Smith P, Tchernov E., et al. Genetic evidence for origin of the agrimi goat (Capra aegagrus cretica) И J. Zool. 2002. Vol. 256, N 3. P. 369-377.
Barr W. The role of independent traders in the rear-extermination of musk-oxen on the mainland tundra of Canada, 1892-1915 // Polar. Rec. 1995. Vol. 31, N 179. P. 425-426.
Barry J.C., Johnson N.M., Raza S.M., Jacobs L.L. Neogene mammalian faunal change in Southern Asia// Geology. 1985. Vol. 13. P. 637-640.
Baryhsnikov G., Tikhonov A. The history and systematic of Rupicapra in the fauna of USSR // Abstr. Symp. “Ungulates, Ongules/91”. Toulouse, France. 1991. P. 8.
Bauer K. Steinwild in Osterreich// Osterr. Weidwerk. 1992. N 6. S. 17-20.
Bekenov A.B., Grachevand I.A., Milner-Gulland E.J. The ecology and management of the Saiga antelope in Kazakhstan // Mammal. Rev. 1998. Vol. 28, N 1. P. 1-52.
Belousova I.P., Kudryavtsev I. V. Genetic characteristics of the modem world population of European bison // Adv. Ethol. 2000. N 35. P. 131.
Biltueva L.S., Sharshov A.A., Graphodatsky A.S. G-banding homologies in musk-ox, Ovibos moschatus, and other bovids // Hereditas. 1995. Vol. 122, N 2. P. 185-187.
Blunt M., Dawson H.A., Thorne E. T. Birth weights and gestation in a captive Rocky Mountain bighorn sheep // J. Mammal. 1977. Vol. 58, N 1. P.106.
Bochenek S. Remains of the European bison, Bison bonasus (L.), from Podhale in Poland // Acta Theriol. 1955. Vol. 1, N 2. P. 15-25.
Bohlken H. Vergleichende Untersuchungen an Wildrindem (Tribus Bivini Simpson, 1945) // Zool. J. Phys. 1958. Bd. 68. S. 113-202.
Bohlken H. Beitrag zur Systematik der rezenten Formen der Gattung Bison H. Smith, 182711 Ztschr. Zool. Syst. Evolutionsforsch. 1967. Bd. 5, N 1. S. 54-110.
BorkA.M., Strobeck C.M., Veh F.C., et al. Genetic relationship of wood and plains bison based on restriction fragment length polymorphism’s // Can. J. Zool. 1991. Vol. 69, N 1. P. 43-48.
Brandt J.H., Dioli M., Hassanin A., et al. Debate on the authenticity of Pseudonovibos spiralis as a new species of wild bovid from Vietnam and Cambodia// J. Zool. Vol. 255, N 4. P. 437-444.
Briedermann L. Der Europaische Mufflon (Ovis аттоп misimon Schreber, 1782) als Objekt wildbiologischer Forschung // Biol. Rdsch. 1989. Bd. 27, N 6. S. 323-330.
Bunch T.D. Fundamental kariotype in domestic and wild species of sheep: identity and ranking of autosomal acrocentrics involved in biarmed formations // Heredity. 1978. Vol. 69. P. 77-80.
Bunch T.D., Mitchell R.M., Maciulis A. G-banded chromosomes of the Gansu argali (Ovis аттоп jubata) and their implications in the evolution of the Ovis kariotype // Ibid. 1990. Vol. 81, N 3. P. 227-230.
Bunch T.D., Nadler C.F. Giemsa-band patterns of the tahr and chromosomal evolution of the tribe Caprini//Ibid. 1980. Vol. 71. P. 110-116.
Bunch T.D., Wang S., Valdez R., et al. Cytogenetics, morphology and evolution of four subspecies of the Giant Sheep argali (Ovis аттоп) of Asia // Mammalia. 2000. Vol. 64, N 2. P. 199-207.
Bunnell F.L. Factors controlling lambing period of Dall’s sheep H Canad. J. Zool. 1980. Vol. 58, N6.P. 1027-1031.
Bunnell F.L. The lambing period of mountain sheep: synthesis, hypotheses, and tests // Ibid. 1982. Vol. 60, N l.P. 1-14.
Bunnell F.L., Olsen N.A. Weights and growth of Dall sheep in Klaune Park Reserve, Yukon Territory // Can. Field. Natur. 1976. Vol. 90, N 2. P. 157-162.
Bunnell F.L., Olsen N.A. Age-specific natality in Dall’s sheep // J. Mammal. 1981. Vol. 62, N 2. P. 379-380.
Burch E.S.J. Muskox and man in the central Canadian subarctic 1689-1974 // Arctic. 1977. N30(3). P. 135-154.
Burzynska B., Olech W, Topczewski J. Phylogeny and genetic variation of the European bison Bison bonasus based on mitochondrial DNA D-loop sequences // Acta Theriol. 1999. Vol. 44, N 3. P. 253-262.
Cabon-Raczynska K, Krasinska M„ Krasinski Z. Behaviour and daily activity rhythm of European bison in winter // Acta Theriol. 1983. Vol. 28, N 9-20. P. 273-299.
Campbell B.H., Hinkes M. Winter diets and habitat use of Alaska bison after wildfire // Wildlife Soc. Bull. 1983. Vol. 11, N 1. P. 16-21.
Carbyn L.N. Unusual movement by bison, Bison bison, in response to wolf, Canis lupus, predation //Can. Field.-Natur. 1997. Vol. Ill, N 3. P. 461-462.
Carbyn L.N., Trottier T. Responses of bison on their calving grounds to predation by wolves in Wood Buffalo National Park // Can. J. Zool. 1987. Vol. 65, N 8. P. 2072-2078.
Chappel R. W, Hudson R.J. Winter bioenergetics of Rocky Mountain bighorn sheep // Can. J. Zool. 1978. Vol. 58, N 11. P. 2388-2393.
ChikuniK, Mori Z, Tabata T, etal. Molecular phylogeny based on the kappa-casein and cytochrome b sequences in the mammalian suborder Ruminantia// J. Mol. Evol. 1995. Vol. 41, N 6. P. 859-866.
Ciucci P, Boitani L., Ricci S. Social patterns of moufflon (Ovis gmelini) in the northern Apennines (Italy) // J. Mammal. 1998. Vol. 62, N 3. P. 442-446.
Coblentz B.E. A unique ungulate breeding pattern // J. Wildlife Manag. 1980. Vol. 44, N 4. P. 929-933.
Cooperrider A. The desert bighorn // Audubon Wildlife Repot. 1985. P. 473-492.
Crooks K, Van Vuren D. Supernumerary teeth in a captive population of American bison Bison bison // Acta Theriol. 1994. Vol. 39, N 3. P. 325-328.
Cowan J. T. Distribution and variation on the native sheep of North America // Amer. Midi. Natur. 1940. Vol. 24, N 3. P. 505-580.
Czykier E., Sawicki B., Krasinska M. Postnatal development of the European bison spermatogenesis // Acta Theriol. 1999. Vol. 44, N 1. P. 77-90.
Daleszczyk K. Reproductive strategies of the European bison // Adv. Ethol. 2002. N 37. P. 30.
Daleszczyk K, Krasinski Z. Parturition behaviour of European bison Bison bonasus living in reserves // Folia Zool. 2001. Vol. 50, N 1. P. 75-78.
Dawaa N., Suchbat Ch., Stubbe M. Beitrag zur Okologie und Morphologie von Ovis аттоп L., 1758 in der MVR // Wiss. Beitr. 1983. N 19. S. 61-65.
Desaulniers D.M., King W.A., Rowell J.E., Flood PF. The banded chromosomes of the muskox (Ovibos moschatus) // Proc. II Intern. Muskox Symp. Saskatoon: National. Research Council of Canada, 1989. P. 1155-1158.
Delpech F. Le temps de 1’Antilope saiga // Le temps de la Prehistoire. 1989. T. 2. P. 48-49.
DeYoungH.G. The bison is beleaguered again //Natur. Hist. 1973. Vol. 82, N 5. P. 48-55.
Dolan J.M. The saiga (Saiga tatarica): a review as a model for the management of endangered species // Intern. Zoo Yearbook. 1977. Vol. 17. P. 25-32.
Dulamtseren S., Amgalan L. Mongolian saiga (Saiga tatarica mongolica) // Mammalogical studies in Mongolia and its adjacent territories. Ulaanbaatar, 2002. P. 64—70. (Proc. Inst. Biol., N 24). Dung V.V., Giao P, Chinh N.N., et al. A new species of living bovid from Vietnam // Nature. 1993. Vol. 363, N 3. P. 398,443^145.
Durst C.R., Gordon D. C„ Grubb P, et al. Family Bovidae // Mammal species of the World: a taxonomic and geographic reference. Lawrence, Kansas, 1982. P. 326-343.
Ellerman J.R., Morrison-Scott T.C.S. Checklist of Palaearctic and Indian mammals 1758 to 1946. L.: British Mus. (Nat. Hist.), 1951. 810 p.
Engel S.R., Linn R.A., Taylor J.F, Davis S.K. Conservation of microsatellite loci across species of artiodactyls: Implications for population studies // J. Mammal. 1996. Vol. 77, N 2. P. 504-518.
Enkhbujin D., Amgalan L. Current situation of ibex population (Capra sibirica Pall., 1776) in Bogdkhan mountain national protected area // Mammalogical studies in Mongolia and its adjacent territories. Ulaanbaatar, 2002. P. 203-209. (Proc. Inst. Biol., N 24).
Esteve R., Villaret J.-Ch. Le bouquetin. Aujord’hui sauve ... Demain gere? // Courr. Nature. 1989. N 119. P. 18-23.
Evans H.J., Buckland R.A., Sumner A.T. Chromosome homology and heterochromatin in goat, sheep and ox studies by banding techniques // Chromosoma. 1973. Vol. 42, N 4. P. 383—402.
Fandos P, Aranda Y, Orueta J.F, Cregut-Bonnoure E. Introduction to skull variability of wild goats (Capra L.) //Folia Zool. 1993. Vol. 42, N 2. P. 111-125.
Fandos P., Vigal C.R. Sexual dimorphism in size of the skull of Spanish ibex Capra pyrenaica // Acta Theriol. 1993. Vol. 38, N 1. P. 103-111.
Fedosenko А.К. Status of the Argali (Archar) Populations in Russia and Republics of Middle Asia // Beitr. Jagd- und Wildforsch. 2003. Bd. 28. S. 143-149.
Fedosenko A. K., Weinberg PL, ValdezR. Argali sheep survey in the headwaters of the Korunduk River, Kirghizstan // J. Mammal. 1995. Vol. 59, N 3. P. 452-455.
Festa-Bianchet M. Seasonal dispersion of overlapping Mountain sheep ewe groups // J. Wildlife Manag. 1986. Vol. 50, N 2. P. 325-330.
Festa-Bianchet M. Birthdate and survival in bighorn lambs (Ovis canadensis) // J. Zool. 1988. Vol. 214, N 4. P. 653-661.
Festa-Bianchet M„ Jorgenson J.T., Lucherini M„ Wischart W.D. Life history consequences of variation in age of primiparity in bighorn ewes // Ecology. 1995. Vol. 76, N 3. P. 871-881.
Fleischman C.L. Genetic variation in musk-oxen (Ovibos moschatus): MSc thesis. Fairbanks, Univers, of Alaska, 1986. 77 p.
Fox J.L., Sinha S.P, Chundawat R. S. Activity patterns and habitat use of ibex in the Himalayan Mountains of India // J. Mammal. 1992. Vol. 73, N 3. P. 527-534.
Galbreath G.J., MelvilleR.A. Pseudonovibos spiralis: Epitaph // J. Zool. 2003. Vol. 259, N 2. P. 169-170.
Gao Z., Jin K„ Guan D., et al. Изучение экологии монгольского дзерена // J. North-East Forest. Univ. 1996. Vol. 24, N 2. P. 37-42.
GatesyJ.,AmatoG„ VrbaE., etal. A cladistic analysis of mitochondrial ribosomal DN A from the Bovidae 11 Mol. Phylogenet. Evol. 1997. Vol. 7, N 3. P. 303-319.
Gatesy J., Yelon D., DeSalle R., Vrba E.S. Phylogeny of the Bovidae (Artiodactyla, Mammalia), based on mitochondrial ribosomal DNA sequences H Mol. Biol. Evol. 1992. Vol. 9, N 3. P. 433-446.
Gebczynska Z„ Gebczynski M., Martynowicz E. Food eaten by the free-living European bison in Bialowieza Forest // Acta Theriol. 1991. Vol. 36, N 3-4. P. 307-313.
Gebczynska Z., Krasinska M. Food preferences and requirements of the European bison // Acta Theriol. 1972. Vol. 17, N 1-11. P. 105-117.
Geist V. The evolution of horn-like organs // Behavior. 1966. Vol. 27. P. 175-214.
Geist V. Mountain sheep. A study in behavior and evolution. Chicago - L.: Univ. Chicago Press, 1971. 383 p.
Geist V. On the evolution of the Caprinae // The biology and management of Capricornis and related mountain antelopes. L., 1987. P. 3-40.
Geist V. On the taxonomy of giant sheep (Ovis аттоп Linnaeus, 1766) // Can. J. Zool. 1991. Vol. 69. P. 706-723.
Gentry A. The subfamilies and tribes ofthe family Bovidae//Mammal. Rev. 1992. Vol. 22,N 1.
P. 1-32.
Gentry A. The Miocene differentiation of Old World Pecora (Mammalia) // Hist. Biol. 1994. Vol. 7. P. 115-158.
Gentry A. A fossil Budorcas from Africa // Paleoecology and paleoenvironments of Late Cainozoic mammals. Toronto: Toronto Univ. Press, 1996. P. 571-587.
Gentry A., Hooker J. The phylogeny of the Artiodactyla // The phylogeny and classification of the tetrapods. 1988. Vol. 2. P. 235-272.
Georgiadis N., Kat P, Oketch H„ Patton J. Allozyme divergence within the Bovidae // Evolution. 1990. Vol. 44, N 8. P. 2135-2149.
Gienc J. Bisonpriscus (Bojanus, 1827) - prazubr czy prabizo? // Prz. Zool. 1994. Vol. 38, N 1-2. P. 63-78.
Glazko V. The comparative analysis of enzyme and RAPD-PCR polymorphisms in some domestic and wild ungulata species // Abstr. Euro-Amer. Mammal. Congr. Santiago de Compostela, 1998. P. 352.
Glazko V.I., Sipko TP, Sholudtchenko S.P Genetic structure of Ovis canadensis in Putoran’s reserve //Adv. Ehtol. 1998. N 33. P. 83.
Gonzalez G„ Crampe J.-P. Mortality patterns in a protected population of isards (Rupicapra pyrenaica) I I Can. J. Zool. 2001. Vol. 79, 11. P. 2072-2079.
Goodrowe K.L., Smak B., Presley N., Monfort S.L. Reproductive, behavioural, and endocrine characteristics of the Dall’s sheep (Ovis dalli dalli) // Zoo Biol. 1996. Vol. 15, NLP. 45-54.
GraczykR. European bison, Bison bonasus bonasus (Linnaeus, 1758), in Wielkopolska // Rocz. AR Pozn. Rozpr. nauk. 1988. N 173. P. 1-35.
GrayA.P. Mammalian hybrids // Commonv. Agr. Bur. 1954. N 10. P. 144.
Gromova V. Uber Kraniologie und Geshichte der Gattung Oves, neue Forsh. in Tierzucht und Abstammungeslehre // Festschz. Z. 60 Geburstag v. prof. Y.U. Deurst. Bern, 1936. S. 48-67.
Groves C.P. Systematic relationship in Bovini (Artiodactyla, Bovidae) // Ztschr. Zool. Syst. und Evolutionforsch. 1981. Bd. 19, N 2. S. 264—278.
Groves P. Muskoxen have little intraspecific variation as indicated by mitochondrial DNA sequences // II Intern. Arctic Ungulate Conf. Fairbanks: Univers, of Alaska, 1995. P. 46-47.
Groves P, Shields G. Phylogenetics of the Caprinae based on cytochrome b sequence // Molec. Phylog. Evol. 1996. Vol. 5, N 3. P. 467-476.
Groves P, Shields G. Cytochrome b sequences suggest convergent evolution of the Asian takin and Arctic muskox // Ibid. 1997. Vol. 8, N 3. P. 363-374.
Grubb P. Bovidae // Mammal species of the World. II ed. Smithsonian Inst. Press, 1993. P. 393-414.
Guthrie R.D. Bison evolution and zoogeography in North America during the Pleistocene // Quart. Rev. Biol. 1970. Vol. 45, N 1. P. 1-15.
Haltenorth T. Klassifikation der Saugetiere: Artiodactyla // Handbuch. der Zoologie. 1963. Bd. 8, H. 32(18). S. 1-167.
Hammer S., Nadlinger K„ Hartl G.B. Mitochondrial DNA differentiation in chamois (genus Rupicapra)-. Implications for taxonomy, conservation, and management // Acta Theriol. 1995. Suppl. 3. P. 145-155.
Hansen Ch.G. Growth and development of desert bighorn sheep // J. Wildl. Manag. 1965. Vol. 29, N2. P. 387-391.
Hansen M.C. Desert bighorn sheep: another view // Wildlife Soc. Bull. 1982. Vol. 10, N 2. P. 133-140.
Hardenberg A., Bassano B., Peracino A., Lovan S. Male alpine chamois occupy territories at hotspots before the mating season // Ethology. 2000. Vol. 106, N 7. P. 617-630.
Harm J. Home range sizes of male chamois, Rupicapra rupicapra, in the Tyrolean Alps, Austria // Acta Zool. Fenn. 1984. N 171. P. 293-296.
Hartl G.B. Genetische Variabilitat beim Muffion (Ovis аттоп musimon) // Ztschr. Jagdwiss. 1990. Bd. 36, N 2. S. 95-103.
Hartl G.B., Burger H, Willing R„ Suchentrunk F. On the biochemical systematics of the Caprini and the Rupicaprini // Biochem. Syst. Ecol. 1990. Vol. 18, N. 2-3. P. 175-182.
Hass C.C. Gestation periods and birth weights of desert bighorn sheep in relation to other Caprinae // Southwest. Natur. 1995. Vol. 40, N 2. P. 139-147.
Hassanin A., Douzery E. Molecular phylogeny of the family Bovidae (Artiodactyla) as inferred from cytochrome В gene sequences // Euro-American Mammal. Congr. Santiago de Compostela, Spain. 1998. P. 347.
Hassanin A., Douzery E.J. The tribal radiation of the family Bovidae (Artiodactyla) and the evolution of the mitochondrial cytochrome b gene//Mol. Phylogenet. Evol. 1999a. Vol. 13, N 2. P. 227-243.
Hassanin A., Douzery E.J. Evolutionary affinities of the enigmatic saola (Pseudoryx nghetinhensis) in the context of the molecular phylogeny of Bovidae // Proc. R. Soc. bond. Biol. Sci. 1999b. Vol. 7, N 266 (1422). P. 893-900.
Hassanin A., Pasquet E., Vigne J-D. Molecular systematics of the subfamily Caprinae (Artiodactyla, Bovidae) as determined from cytochrome b sequences // Mammal. Evol. 1998. Vol. 5, N 3. P. 217-236.
Hassanin A., Seveau A., Thomas H, et al. Evidence from DNA that the mysterious “linh duong” (Pseudonovibos spiralis) is not a new bovid // C. R. Acad. Sci. Ser. 3. 2001. Vol. 324, N l.P. 71-80.
Haynes G. Toots wear rate in northern bison // J. Mammal. 1984. Vol. 65, N 3. P. 487-491.
Heck H., Wurster D„ Benirschke K. Chromosome studies of members of the subfamilies Caprinae and Bovinae, family Bovidae; the muskox, ibex aoudad, Congo buffalo and gaur// Saugetierk. Mitt. 1968. Bd. 33, H. 3. S. 172-179.
Heroldova M. The diet of moufflon (Ovis musimon) outside the growing period 1983-1984 // Folia Zool. 1988. Vol. 37, N 4. P. 309-319.
Hoefs M. Beitrag zur Morphometrie und Wachstumdynamik der Schneckedes Muffelwidders (Ovis аттоп musimon Schreber, 1782) // J. Jagdwiss. 1982. Bd. 28, N 3. 145-162.
Hoefs M„ Nowlan U. Distorted sex ratios in young ungulates: the role of nutrition // J. Mammal. 1994. Vol. 75, N3. P. 631-636.
Hogg J. T. Intrasexual competition and mate choice in Rocky Mountain bighorn sheep // Ethology. 1987. Vol. 75, N 2. P. 119-144.
Homolka M. The diet of moufflon (Ovis musimon) in the mixed forest habitat of the Drahanska vrchovina Highland // Folia Zool. 1991. Vol. 40, N 3. P. 193-201.
Honacki J.H., Kinman K.E., Koeppl J.W. Mammal species of the World. Lawrence, 1982. 694 p.
Houssin H, Loison A., Gaillard J.M. Participation a la reproduction des femelles de Chamois (Rupicapra rupicapra) en fonction de leurage dans une population alpine Chassee // Giber Faune Sauvage. 1993. Vol. 10, mars. P. 39-45.
Hrabe V., KoubekP. Craniometrical characteristics of Rupicapra rupicapra tatrica (Mamm., Bovidae) // Folia Zool. 1984. Vol. 33, N 1. P. 73-84.
Hrabe V., Weber P, Koubek P. The morphometrical characteristics and the dynamics of horn growth in Rupicapra rupicapra carpatica (Mammalia, Bovidae) // Ibid. 1986. Vol. 35, N 1. P. 43-54.
Jaczewski Z. Reproduction of the European bison, Bison bonasus (L.), in reserves // Acta Theriol. 1958. Vol. 1, N 9. P. 333-376.
JanecekL.L., Honeycutt R.L., Adkins R.M., Davis S.K. Mitochondrial gene sequences and the molecular systematics of the artiodactyl subfamily Bovinae // Mol. Phylogenet. Evol. 1996. Vol. 6,N l.P. 107-119.
Janis C, Scott K. The phylogeny of the Ruminantia // The phylogeny and classification of the tetrapods. 1988. Vol. 2. P. 273-282.
Jiang Z., Feng Z., Wang Z., Chen L. Историческое и современное распространение дзерена Пржевальского И Acta Theriol. Sin. 1995. Vol. 15, N 4. P. 241-245.
Jiang Z., Ma I., Gao Z. Изучение структуры популяции и тенденций ее динамики у монгольского дзерена в Китае // Ibid. 1993. Vol. 13, N l.P. 16-20.
Johnsingh A. J, Stuve M., Rawat G.S., et al. Ecology and conservation of Asiatic ibex (Capra ibex sibirica) in Pin Valley National Park, Himachal Pradesh, India. Wildlife Institute of India, 1999. 133 p.
Kahlke R.-D. Pleistocene distributional and evolutionary history of genus Saiga Gray, 1843 (Mammalia, Artiodactyla, Bovidae) in the Palaearctic // Vertebrata PalAsiatica. 1991. Vol. 10. P. 314-322.
KamsayE., Compton B.L., Savage W. Saiga husbandry and management at the Oklahoma City zoo // Zool. Gait 1992. Vol. 62, N 2. P. 93-102.
Keating KA. Bone chewing by rocky mountain bighorn sheep // Great Basin Natur. 1990. Vol. 50,N l.P. 89.
KholodovaM. V., KuznetsovaM. V., LomovA.A., LuschekinaA.A. Searchingformolecularmarkers: RAPD analyses of different populations of saiga antelope (Saiga tatarica L.) // Adv. Ethol. 2000. N 35. P. 138.
King W.J., Festa-Bianchet M„ Jorgenson J. T. Density effects on individual body and horn growth of Bighorn sheep И VI Intern. Theriolog. Congr., Sydney. Rensington, 1993. P. 210-211.
Kirkland H, Hilliard J. Extinct musk ox from Western Oklahoma // Southwest. Natur. 1996. Vol. 42, N 2. P. 190-191.
Kirkpatrick J.F., VcCarthy J.C., Gudermuth D.F., et al. An assessment of the reproductive biology of Yellowstone bison (Bison bison) subpopulations using noncapture methods // Can. J. Zool. 1996. Vol., 74, N 1. P. 8-14.
Klein D.R., Bay C. Foraging dynamics of muskoxen in Peary Land, Northen Greenland H Holarctic Ecol. 1990. Vol. 13. P. 260-280.
Klungland H, Roed K.H., Nesboo C.L. et al. The melanocyte-stimulating hormone receptor (MC1-R) gene as a tool in evolutionary studies of Artiodactyles // Hereditas. 1999. Vol. 131, N l.P. 39-46.
Kobryn H. The thorax, in European bison and other ruminants // Acta Theriol. 1973.Vol. 18, N 7-18. P. 313-341.
Kobrynczuk F. The influence of inbreeding of the shape and size of the skeleton of the European bison // Acta Theriol. 1985. Vol. 30, N 21-31. P. 379-422.
Kohler M., Moya-Sola S., Morales 1. Bovidae and Giraffidae // The vertebrate locality Moramena (Mocedonia, Greece) at the Turolien-Ruscinian boundary (Neogene). Munch. Geowiss. Abh. 1995. Bd. 28. S. 167-180.
Kolpashikov L.A., Layshev K.A. Wild ungulates in Taimyr // Rangifer. 2003. N 7. P. 39.
Korobitsina К. V., Nadler C.F., Vorontsov N.N., Hoffmann R.S. Chromosomes of the Siberian snow sheep, Ovis nivicola, and implications concerning the origin of Amphiberingian Wild sheep (subgenus Pachyceros) II Quatern. Res. 1974. Vol. 4. P. 235-245.
Koubek R, Hrabe V. Craniometrical characteristics of Rupicapra rupicapra caucasica I I Folia Zool. 1983. Vol. 32, N 3. P. 213-223.
KoubekP, Hrabe И Morphometrical characteristics and horn growth dynamics'm Rupicapra rupicapra tatrica (Mammalia, Bovidae) // Ibid. 1984. Vol. 33, N 4. P. 289-302.
KoubekR, Hrabe V, HorakovaM. Postnatal development ofthe skull ofRupicapra rupicapra (Mammalia, Bovidae) // Ibid. 1989. Vol. 38, N 1. P. 31-44.
KoubekR, Weber R, Hrabe V. Craniometrical characteristics of Rupicapra rupicapra carpatica (Mammalia, Bovidae) // Ibid. 1985. Vol. 34, N 3. P. 227-234.
Koulischer L., Tyskens J., Mortelmans J. Mammalian cytogenetics: VI. The chromosomes of a male specimen of Anoa depressicornis quarlesi H Acta Zool. Pathol. Antverp. 1972. Vol. 56. P. 21-24.
Krasinska M. Hybrydy zubra i bydia domowego. Warshawa: Wydown. Polskiej academ. Nauk, 1988a. 192 s.
Krasinska M. Variability of the skull shape in hybrids between European and domestic cattle // Acta Theriol. 1988b. Vol. 33, N 12-25. P. 147-186.
Krasinska M., Krasinski Z.A. Struktura przestrzenna populacji zubrow w polskiej czesci Puszczy Bialowieskiej w Jatach 1976-1993 // Parki Nar. i Rezerw. Przyr. 1994. Vol. 13, N 4. P. 69-87.
Krasinska M., Krasinski Z.A. Composition, group size, and spatial distribution of European bison bulls in Bialowieza Forest И Acta Theriol. 1995. Vol. 40, N 1. P. 1-21.
Krasinska M., Krasinski Z.A., Bunevich A.N. Factors affecting the variability in home range size and distribution in European bison in the Polish and Belorussian parts of the Bialowieza Forest // Acta Theriol. 2000. Vol. 45, N 3. P. 321-334.
Krasinska M., Krasinski Z.A., Bunevich A.N. Spatial distribution of the European bison population in Polish and Belarussian parts of Bialowieza Forest // European bison conservation. Bialowieza, 2004. P. 73.
Krasinska M., Suminski E. Colour phenotypes in hybrids of the European bison and domestic cattle H Acta Theriol. 1981. Vol. 26, N 1. P. 3-22.
Krasinski Z.A. Dynamics and structure of the European bison population in the Bialowieza Primeval Forest // Acta Theriol. 1978. Vol. 23, N 1. P. 3-48.
Krasinski Z.A. Pierwsze 10 lat wolnej hodowli zubrow w Puszczy Bialowieskiej 1952-1961 // Parki Nar. i Rezerw. Przyr. 1983. Vol. 4, N 2. P. 39-50.
KrasinskiZ.A. RestytucjazubrowwBialowiezy wlatach 1929-1952//Ibid. 1994. Vol. 13,N4. P. 3-23.
Krasinski Z.A., Bunevich A.N., Krasinska M. Charakterystyka populacij zubra nizinnego w polskiej i bialoruskiej czesci Puszczy Bialowieskiej // Ibid. 1994. Vol. 13, N 4. P. 25-67.
Krasinski Z.A., Krasinska M. Funkcjonowanie populacji zubra nizinnego w Puszczy Boreckiej // Ibid. 1994. Vol. 13, N 4. P. 89-106.
Krasinski Z., Raczynski J. The reproduction biology of European bison living in reserves and in freedom // Acta Theriol. 1967. Vol. 12, N 29. P. 407-444.
Krumbiegel I. Die unterartliche Trennung des Bisons, Bison bison (Linnne, 1788), und seine Ruckzuchtung // Saugetierk. Mitt. 1980. Bd. 28, N 2. S. 148—160.
Kryzhanovskii V.I. State of contemporary existing European bison populations and the strategy of forming new free-ranging herds in Ukraine // European bison conservation. Bialowieza, 2004. P. 78-82.
Krzeminski Z., Dobrowolski K. Zubr 2000 - sukces i problemy restytucji // Kosrnos (PRL). 1981. Vol. A30, N 4. P. 379-387.
Kurten B. Pleistocene Mammals of Europe. Chicago: Aldine Publ. Co., 1968. 317 p.
Kuznetsov G.V., Kulikov E.E., Petrov N.V., et al. The “Linh Duong” Pseudonovibos spiralis (Mammalia, Artiodactyla) is a new buffalo // Naturwissensch. 2001. Vol. 88. P. 123-125.
Kuznetsova M. И, Kholodova M. V. Phylogenetic reconstruction of bovid evolutionary history based on mitochondrial gene sequences // Adv. Ethol. 2002. Vol. 37. P. 101.
barter N. C., Gates С. C. Home ranges of wood bison in an expanding population // J. Mammal. 1990. Vol. 71, N 4. P. 604-607.
barter N. C., NagyJA. Calf production, calf survival, and recruitment of muskoxen on Banks Island during a period of changing population density from 1986—1999 // Arctic. 2001. Vol. 54, N 4. P. 394-406.
bei R., Jiang Z., Hu Z„ Yang W. Phylogenetic relationships of Chinese antelopes (subfamily Antilopiinae) based on mitochondrial ribosomal RNA gene sequences // J. Zool. 2003. Vol. 261, N3.P. 227-237.
bei R„ Jiang Z., biu B. Group size and group composition in Przewalskis gazelle // Fol. Zool. 2001. Vol. 50, N 2. P. 117-126.
bentP.C. Maternal-infant behaviour in mushoxen// J. Mammal. 1991. Vol. 55, N 1. P. 3-21.
bent PC., Devis W.J. Variables influencing survival in four generations of captivebom muskoxen // Ragifer. 1993. Vol. 13 (2). P. 137-142.
bevit M., Pepin D. Sociabilite et domaine vital d’lsards {Rupicapra pyrenaica) mabs auprintemps a Orlu (Ariede) // Giber Faune Sauvage. 1994. Vol. 11, N. 1. P. 51-64.
bewis C.E. Musk ox: An historical industry looks toward the future // Agroborealis. 2000. Vol. 32, N l.P. 30-32.
bee Woo-Shin, Rhim Sin-Jae. Changes in distribution of arnur goral (Nemorhaedus caudatus) in South Korea // Acta Theriol. Sin. 2002. Vol. 22, N 3. P. 225-227.
bocati M., bovari S. Sexual differences in aggressive behaviour of the Apennine chamois // Ethology. 1990. Vol. 84, N 4. P. 295-306.
bocati M., bovari S. Clues for dominance in female chamois: age, weight, or horn size // Aggress. Behav. 1991. Vol. 17, N 1. P. 11-15.
bott D.F., Minta S. Home ranges of American bison cows on Santa Catalina Island, California // J. Mammal. 1983. Vol. 64, N 1. P. 161-162.
bott D.F., Galland J. C. Parturition in American bison: precocity and systematic variation in cow isolation // Ztschr. Tierpsychol. 1985a. Bd. 69, N 1. S. 66-71.
bott D.F., Galland J.C. Individual variation in fecundity in an American bison population // J. Mammal. 1985b. Vol. 49, N 2. P. 300-302.
bovari S„ Scala C. Revision of Rupicapra Genus. I. A statistical re-evaluation of Couturier’s data on the morphometry of six chamois subspecies // Boll. Zool. 1980. Vol. 47, N 1-2. P. 113-124.
Lovari S., Scala C. Revision of Rupicapra Genus. IV. Hom biometrics ofR. rupicapra asiatica and its relevance to the taxonomic position of R. rupicapra caucasica 11 Ztschr. Saugetierk. 1984. Bd. 49, Hf. 4. S. 246-253.
Lowenstein J Bovid relations based on serum immunology//S. Afr. J. Sci. 1986. Vol. 82. P. 77-78.
Ludwig A., Benecke N. Zur Systematik und Palaontologie der Gattung Ovis mit besonderer Berucksichtigung der Stellung des Europaischen Muffions // Beitr. Jagd- und Wildforsch. 2003. Bd. 28. S. 35-45.
Ludwig A., Fischer S. New aspects of an old discussion - Phylogenetic relationships of A mmotragus and Pseudois within the subfamily Caprinae based on comparison of the 12S rDNA sequences // J. Zool. Syst. and Evol. Res. 1998. Vol. 36, N 4. P. 173-178.
Lups P. Der Alpensteinbock. Symbol fur eine gegluckte Wiedereinburgerung // Natur und Mensch. 1985. Bd. 25, N 3. S. 140-143.
LuschekinaA.A., Kholodova M.V., Milner-Gulland E.J, etal. A preliminary molecular-genetic study of the Mongolian population of the saiga antelope // Central Asian ecosystems. Ulaanbaatar, 2000. P. 90-93.
Lushchekina A., Amgalan L., Neronov V. Conservation problems of the Mongolian saiga {Saiga tatarica mongolica) 11 Species. 2000. N 33. (Без стр.).
Lushchekina A., Dulamtseren S. The status and conservation of argali in Mongolia // Ibid. 2000. (Без стр.).
Lushchekina A., Dulamtseren S„ Amgalan L„ Neronov V.M. The status and prospects for conservation of the Mongolian saiga Saiga tatarica mongolica // Oryx. 1999. Vol. 33, N 1. P. 21-30.
Lushchekina A., Struchkov A. The saiga antelope in Europe: Once again on the brink? // The Open Country. 2001. N 3. P. 11-24.
Lydekker R. Wild oxen, sheep and goats of all lands. L., 1898. 318 p.
Lydekker R. Catalogue of the ungulate mammals in the British Museum. 1. Artiodactyla, Fam. Bovidae. Brit. Mus. Nat. Hist., 1913. Vol. 17. P. 1-249.
Magomedov M-R.D., Akhmedov E. G., Wall W.A., Subbotin A.E. Current Status and Population Structure of Argalis {Ovis аттоп L., 1758) in Central Asia //Beitr. Jagd- und Wildforsch. 2003. Bd. 28. S. 151-163.
Maher C.R., Byers J.A. Age-related changes in reproductive effort of male bison // Behav. Ecol. and Sociobiol. 1987. Vol. 21, N 2. P. 91-96.
Makowiecka M. Usefulness of the chosen qualitative and quantitative features of skull differentiation in three breeding lines of European bison, Bison bonasus (Linnaeus, 1758) // Ann. Warsaw Agr. Univ. - SGGW. Vet. Med. 1994. N 19. P. 3-17.
Manaseryan N. Armenia: Rare hoofed animals in Holocene // J. Morphol. 2001. Vol. 248, N 3. P. 258-259.
Manceau V., Despres L., Bouvert J., Taberlet P. Systematics of the Genus Capra inferred from mitochondrial DNA sequence data // Mol. Phylogen. and EvoL 1999. Vol. 13, N 3. P. 504—510.
Markov G.,Massei G., GenovP. Craniometric characteristics and morphometric analyses of the bulgarian population of the chamois // Acta Zool. Bulg. 1998. Vol. 50, N 2-3. P. 79-86.
Marquez F.J., Soriguer R.C., Granados J.E, et al. Molecular evolutionary dynamics of cytochrome В in Capra pyrenaica И Abstr. Euro-Amer. Mammal. Congr. Santiago de Compostela, 1998. P. 24.
Martinez T. Donnees sur 1’alimentation du bouquetin d’Espagne {Capra pyrenaica) danz la Sierra de Tejeda (Granada) // J. Mammal. 1988. Vol. 52, N 2. P. 284-285.
Martinez T, Martinez E., Fandos P. Composition of the food of the Spanish wild goat in Sierras de Cazorla and Segura, Spain // Acta Theriol. 1985. Vol. 30, N 21-31. P. 461-494.
Masini F, Lovari S. Systematics, phylogenetic relationships and dispersal of the chamois (Rupicapra spp.) // Quaternary Research. 1988. N 30. P. 339-349.
Massei G., Randi E„ Genov P. The dynamics of the horn growth in Bulgarian chamois Rupicapra rupicapra balcanica //Acta Theriol. 1994. Vol. 39, N 2. P. 195-199.
Masini E, Thomas H. Samotragus occidentalis n. sp. a new bovid from the late Messinian of Italy // Boll. Soc. Paleont. Ital. 1989. Vol. 28, N 2-3. P. 307-316.
Matthee C.A., Robinson T.J. Cytochrome b phylogeny of the family Bovidae: resolution within the Alcelaphini, Antilopini, Neotragini, and Tragelaphini // Mol. Phylogenet. Evol. 1999. Vol. 12. N l.P. 31^16.
McCutchen H.E. A minimum breeding age for a desert bighorn ewe // Southwest. Natur. 1977.
Vol. 22, N l.P. 153.
McCutchen H.E. Desert bighorn zoogeography and adaptation in relation to historic land use// Wildlife Soc. Bull. 1981. Vol. 9, N 3. P. 171-179.
McDonald J.N. Symbos cavifronz (Artiodactyla: Bovidae) from delte County, Colorado // Great Basin Natur. 1985. Vol. 45, N 3. P. 455-461.
McDonald N.G., Davis R.A. Fossil muskoxen of Ohio // Can. J. Zool. 1989. Vol. 67, N 5. P. 1159-1166.
McKenna M„ Bell S. Classification of mammals above the species level. N.-Y: Columbia Univ. Press, 1997. 631 p.
MichalletJ, Toigo C. Home ranges of chamois (Rupicapra rupicapra cartusiana) translocated to reinforce a population in the Grande Chartreuse mountain massif Isere // Game and Wildlife Sci. 2000. Vol. 17, N 4. P. 259-272.
Miller G, Hartl G.B. Genetic variation in two alpine populations of chamois, Rupicapra rupicapra L. // Ztschr. Saugetierk. 1986. Bd. 51, N 2. S, 114—121.
Miller C, Hartl G.B. Genetische Variation bei Gemsen der Alpen (Rupicapra rupicapra rupicapra L.) // Ztschr. Jagdwiss. 1987. Bd. 33, N 4. S. 220-227.
Milner-Gulland E.J. A population model for the management of the saiga antelope // J. Appl. Ecol. 1994. Vol. 31, N 1. P. 25-29.
Milner-GullandE.J., Bukreeva O.M., Coulson T, etal. Reproductive collapse in saiga antelope harems // Nature. 2003. Vol. 422. March. P. 135.
Milner-Gulland E.J, Lhadvasuren B. Population dynamics of the Mongolia gazelle Procapra gutturosa-. an historical analysis // J. Appl. Ecol. 1998. Vol. 35, N 2. P. 240-251.
Modi W.S., Gallagher D.S., WomackJ.E. Evolutionary histories ofhighly repeated DN A families among the Artiodactyla (Mammalia) // J. Mol. Evol. 1996. Vol. 42, N 3. P. 337-349.
Morales J., Soria D., Pickford M. Sur les origines de la famille des Bovidae (Artiodactyla, Mammalia)//C. R. Acad. Sci. Paris. 1995. T. 321. P. 1211-1217.
Morgart J.R., Krausman P.R. Early breeding in bighorn sheep // Southwest. Natur. 1983. Vol. 28, N 4. P. 460-461.
Nadler C.F. Chromosomes of the Dall sheep, Ovis dalli dalli (Nelson) // J. Mammal. 1971. Vol. 52, N2. P. 461-463.
Nadler C.F., Hoffman R.S., Woolf A.S. G-band patterns as a chromosomal markers and the interpretation of chromosome evolution in wild sheep (Ovis) 11 Experientia. 1973. Vol. 29, N l.P. 117-119.
Nadler C.F., Korobitsina К. V., Hoffman R.S., Vorontsov N.N. Cytogenetic differentiation, geographic distribution, and domestication in Palearctic sheep (Ovis) // Ztschr. Saugetierk. 1973. Bd. 38, N 2. S. 109-125.
NadlerC.F., LayD.M., HassingerJ.D. Cytogenetic analyses of wild seep population in Northern Iran// Cytogenetics. 1971. Vol. 10, N 2. P. 137-152.
Nasonov N. Uber Ovis severtzovi Nas. und uber die Methode der Untersuchungen der Homer der Wildschafe in systematischer Hinsicht// Изв. Импер. Акад. наук. 1914. С. 761-778. (Отд. оттиск).
Nichols L. Dall sheep reproduction // J. Wildlife Manag. 1978. Vol. 42, N 3. P. 570-580.
Nowak Z., Bukowczyk I., Charon K.M., Olech W. Genetic polymorphism in the European bison population // European bison conservation. Bialowieza, 2004. P. 97.
Olech IV. Analysis of inbreeding in European bison // Acta Theriol. 1987. Vol. 32, N 22. P. 373-387.
Olech W. The participation of ancestral genes in the existing population of European bison // Ibid. 1989. Vol. 34, N 29—43. P. 397-407.
Olech IV. The genetic structure of European bison (Bison bonasus L.) world population // Abstr. Euro-Amer. Mammal Congr. Santiago de Compostela, 1998. P. 275.
Olech W. The number of ancestors and their contribution to European bison (Bison bonasus L.) population // Ann. Warsaw Agr. Univ. - SGGW. Anim. Sci. 1999. N 35. P. 111-117.
Pallas P.S. Reise durch verschiedene Provinzen des Russischen Reichs. T. 3. SPb: Akad. Der Wissenshaften, 1776. В. 1. S. 1-154. B. 2. S. 455-780.
Piao R. Оценка размера популяции горного барана (Ovis аттоп) в Тибете // Acta Ecol. Sin. 1966. Vol. 16, N 3. P. 295-301.
Peter W.P, Feiler A. Eine neue Bovidenart aus Vietnam und Cambodia (Mammalia: Ruminantia) // Zool. Abh. 1994. Bd. 48, N 2. S. 169-176.
PiegertH. Muffelwild - Hausschaf-Blendlinge // Unsere Jagd. 1980. Bd. 30, N 10. S. 299.
Piegert H. Zur Geschichte und Verbreitung des Muffelwildes (Ovis gmelini musimon) auf dem Festland Europas // Beitr. Jagd- und Wildforsch. 2003. Bd. 28. S. 27-33.
Pilgrim G.E. The fossil Bovidae of India // Paleont. Ind. 1939. Vol. 26. P. 1-356.
Pilgrim G. The evolution of the buffaloes, oxen, sheep and goats // J. Linn. Soc. London. 1947. Vol. 41, N 279. P. 272-286.
Polziehn R.O., Beech R., Sheraton J., StrobeckC. Genetic relationships among North American bison populations // Can. J. Zool. 1996. Vol. 74, N 4. P. 738-749.
Polziehn R. 0., Strobeck C., Sheraton J., Beech R. Bovine mtDNA discovered in North American bison populations // Conserv. Biol. 1995. Vol. 9, N 6. P. 1638-1643.
Pucek Z. The European bison: current state and problems of management ИI Intern, biosphere reserve congr. (Minsk, 1983). Paris, 1984. Vol. 2. P. 276-282.
Pucek Z. What to do with the European bison, now saved from extinction? // Acta Zool. Fenn. 1985. N 172. P. 187-190.
Pucek Z. History of the European bison and problems of its protection and management // Trans. XVIIIIUGB Congr. (1987). Krakow - Warszawa, 1991. P. 19-39.
Pucek Z. Current state, threats and conservation strategy for European bison // European bison conservation. Bialowieza, 2004. P. 113-114.
Pucek Z., Udina I., Seal U.S., Miller PS. Population and habitat viability assessment for the European bison (Bison bonasus) // IUSN/SSC Conservation Breeding Specialist Group. Minnesota, 1996. 228 p.
Rachlow J.L., Bowyer R.T. Interannual variation in timing and synchrony of parturition in Dall’s sheep // J. Mammal. 1991. Vol. 72, N 3. P. 487—492.
Raczynsky I. Biologische Grundlagen der Zuchtung und der Restitution des Wisents, Bison bonasus II Zool. Gait. 1980. Bd. 50, N 5. S. 311-316.
Reading R.P., Mix H, Blumer E.S., et al. Wild ungulate conservation in Mongolia H Mammalogical studies in Mongolia and its adjacent territories. Ulaanbaatar, 2002. P. 120-132. (Proc. Inst. Biol., N 24).
Rebholz W., Harley E. Phylogenetic relationships in the bovid subfamily Antilopinae based on mitochondrial DNA sequences // Mol. Phylogenet. Evol. 1999. Vol. 12, N 2. P. 87-94.
Reinhardt V. Courtship behavior among Musk-ox males kept in confinement // Zool. Biol. 1985. Vol. 4, N3. P. 295-300.
Reinhardt S„ Schenk H. Zur Verbreitung der Grossauger auf Sardinien // Bonn. Zool. Beitr. 1969. Bd. 20, N 4. S. 429-436.
Robitaille J-F„ Prescott J. Organisation hierarchique d’un troupeau de bisons (Bison bison) en captivite // Zool. Gart. 1985. Vol. 55, N 2-3. P. 124-128.
Rodriquez J.L., Rodriquez J. C., Ramos M.T. Autumn diet selectivity of the Corsica mouflon (Ovis аттоп musimon Schreber, 1782) on Tenerife (Canary Island) H J. Mammal. 1988. Vol. 52, N 4. P. 475-481.
Rougeot J. Acceleration du rythme de la reproduction chez le mouflon de Corse (Ovis аттоп musimon Schreber, 1782) au moyende cycles photoperiodiques semestriels // Ann. Biol. Anim. 1969. Vol. 9, N 3. P. 441-444.
Rubin E.S., Michelson KJ. Nursing behavior in dam-reared Russian saiga (Saiga tatarica tatarica) at the San Diego Wild Animal Park // Zoo Biol. 1994. Vol. 13, N 4. P. 309-314.
Rutberg A.T. Factors infuencing dominance status in American bison cows (Bison bison) // Z. Tierpsychol. 1983. Vol. 63, N 2-3. P. 206-212.
Rutberg A.T. Birth synchrony in American bison (Bison bison)', response to predation of season? // J. Mammal. 1984. Vol. 65, N 3. P. 418-423.
Rutberg A.T. Dominance and its fitness consequences in American bison cows // Behaviour. 1986. Vol. 96, N 1-2. P. 62-91.
Sabados K., Holy R. Sucasne rozsirenie a zhodnotenie populaciimuflonej zveri na Slovensku // Folia Venator. 1992. Vol. 22. P. 53-74.
Scala C, Lovari S. Revision of Rupicapra Genus. II. A skull and horn statistical comparison of Rupicapra rupicapra ornata and R. rupicapra pyrenaica chamois // Boll. Zool. 1984. Vol. 51, N 3-4. P. 285-294.
Schaller G.B. Mountain monarchs. Wild sheep and goats of the Himalaya. Chicago and London, 1977. 425 p.
Schaller G.B. Wildlife of the Tibetan Steppe. Chicago: Univers. Chicago Press, 1998. 373 p.
SchawJ.H., Carter T.S. Calving patterns among American bison // J. Wildlife Manag. 1989. Vol. 53, N 4. P. 896-898.
Schmitt J., Ulbrich F. Die Chromosomen verschiedener Caprini Simpson, 1945 // Ztschr. Saugetierk. 1968. Bd. 33, N 3. S. 180-186.
Schreiber A., Goltenboth R. The haemoglobins of wild cattle (Bovini Simpson, 1945) // Ztschr. Saugetierk. 1990. Bd. 55, N 4. S. 276—283.
Schreiber A., Hegel G. Genetic variability in goitred gazelle Gazella subgutturosa subgutturosa: Allozymes and kariotypes // Acta Theriol. 1999. Vol. 44, N 1. P. 55-66.
Schroder W, Kofler H. Coexistence and competitive exclusion between ibex Capra ibex L. and chamois Rupicapra rupicapra L. //Acta Zool. Fenn. 1985. N 172. P. 87-88.
Schwarz E. On Ibex and Wild Goat H Ann. and Mag. Nat. Hist. 1935. Vol. 16, ser. 10. P. 433-437.
Shackleton D.M., Peterson R. G., BottrellA. Gestation period in Ovis canadensis // J. Mammal. 1984. Vol. 65, N 2. P. 337-338.
Shagdarsuren O. Mongolian gazelle (Procapra gutturosa) lifestyle and its traditional conservation activity // Proc. Inst. Biol. (Ulaanbaatar). 2002. N 24. P. 178-184.
Simmons N.M. Seasonal ranges of Dall’s sheep, Mackenzie Mountains, Northwest Territories 11 Arctic. 1982. Vol. 35, N 4. P. 512-518.
Simmons N.M., Bayer М.В., Sinkey L. O. Demography of Dall’s sheep in the Mackenzie Mountains, Northwest Territories // J. Wildlife Manag. 1984. Vol. 48, N 1. P. 156-162.
Simpson G.G. The principles of classification and a classification of Mammals. N.Y., 1945. 350 p. (Bull. Amer. Mus. Natur. Hist. Vol. 85).
Sipko T.P, lolchievB.S., Klenovitskii P.M., etal. The comparative characteristics ofmorphology of sex chromosomes of European bison Bison bonasus and cattle Bos taurus, using method of constructing the average optical structures // European bison conservation. Bialowieza, 2004a. P. 118-122.
Sipko T.P, Kazmin V.D. Modem problems of European bison protection and their solution in Russia// Ibid. 2004b. P. 123-128.
Sipko T.P., Strelchenko N.S., Andreeva L. V. Comparative karyotypes of European bison and domestic cattle // Ibid. 2004c. P. 129-134.
Skinner M.F., Kaisen О. C. The fossil bison of Alaska and preliminary revision of the genus // Bull. Amer. Mus. Natur. Hist. 1947. Vol. 89. Art. 3. P. 127-256.
Slatis H.M. An analysis of inbreeding in the European Bison // Genetics. 1960. N 45. P. 275-287.
Smith Т.Е. The role of bulls in pioneering new habitats in an expanding muskox population on the Seward Peninsula, Alaska // Can. J. Zool. 1989. Vol. 67, N 5. P. 1096-1101.
Soma H., Kada H. Evolutionary pathway of chromosomes of the Capricomis // One Med. (Berlin). 1984. P. 109-118.
Soma H., Kada H„ Matayoshi K. Some chromosomal aspects of Naemorhedus goral and Procapra gutturosa // Proc. Japan. Acad. 1980. Vol. 56. P. 273-277.
Soma H„ Kada H„ Matayoshi K. Evolutionary pathways of kariotypes of the tribe Rupicaprini // The biology and management of Capricomis and related mountain antelopes. 1987. P. 62-71.
Soule M.E. Thresholds for survival: criteria for maintenance of fitness and evolutionary potential // Conservation biology: An evolutionary ecological perspective. Sinauer, Sunderland: MA, 1980. P. 151-170.
Stewart S. T. Late parturition in bighorn sheep // J. Mammal. 1982. Vol. 63, N 1. P. 154-155.
Stock A. D., Stokes W.L. A reevaluation of Pleistocene bighorn sheep from the Great Basin and their relationship to living members of the genus Ovis // J. Mammal. 1969. Vol. 50, N 4. P. 805-807.
Stormont C., Miller W.J., Suzuki Y. Blood groups and the taxonomic status of American buffalo and domestic cattle // Evolution. 1961. Vol. 15. P. 196-208.
Stuwe M., Nievergelt B. Recovery of alpine ibex-from near exitinction: the result of effective protection, captive breeding, and reintroductions// Appl. Anim. Behav. Sci. 1991. Vol. 79, N 3-4. P. 379-387.
Stuwe M., Scribner К. T. Low genetic variability in reintroduced alpine ibex {Capra ibex ibex) populations // J. Mammal. 1989. Vol. 70, N 2. P. 370-373.
Sushkin P.P. The wild sheep of the Old World and their distribution // J. Mammal. 1925. Vol. 6, N3. P.145-156.
Takada T, Kikkawa Y, Nomura K, et al. PCR-RFLP analysis of Goat MNC class II DRB gene // J. Agr. Sc. Tokyo Nogyo Daigaku. 1998. Vol. 43, N 1. P. 7-13.
TanB.G. Extinction and protection of wildlife// J. Chinese Nat. 1979. Vol. 2, N 7. P. 425-428.
Teitz W.J., Teal J. J. Chromosome number of the muskox {Ovibos moschatus) // Can. J. Zool. 1967. Vol. 45. P. 235-237.
Tessano S. V. The existing and potential importance of brucellosis and tuberculosis in Canadian wildlife: a review// Canad. Veter. J. 1986. Vol. 27, N 3. P. 119-124.
Tessano S. V. Reveiw of the diseases, parasites and miscellaneous pathological conditions of North American bison // Ibid. 1989. Vol. 30, N 5. P. 416—422.
Tessano S. V., Forbes L.B., Turcotte C. A survey of brucellosis and tuberculosis in bison in and around Wood Buffalo National Park, Canada // Ibid. 1990. Vol. 31, N 3. P. 174—180.
Tener J. S. Muskoxen in Canada: a biological and taxonomic review. Ottawa, 1965. 166 p.
Thenius E. Zur systematischen Stellung und Verbreitung von “Gazella” stehlini (Bovidae, Mammalia) aus dem Miozan Europas // Anz. Oesterr. Acad. Wissen. 1979. Bd. 116. S. 9-13.
Thenius E. Grundzuge der Faunen- und Verbreitungsgeschichte der Saugetiere. Jena: VEB Gustav Fischer, 1980. 375 s.
Thenius E„ Hofer H Stammesgeschichte der Saugetiere. Berlin: Springer-Verlag, 1960. 322 s.
Thomas H. Anatomie cranienne et relations phylogenetiques du nouveau bovide {Pseudoryx nghetinhensis) decouvert dans la cordillere annamitique au Vietnam // J. Mammal. 1994. Vol. 58, N3.P. 453-482.
Thomas H, SeveauA., Hassanin A. The enigmatic new indochinese bovid, Pseu donovibos spiralis: An extraordinary forgery // C. R. Acad. Sci. Ser, 3.2001. Vol. 324, N 1. P. 81-86.
Toigo C, Michallet J, Blanc D. Structure des groupes chez le bouquetin des Alpex {Capra ibex ibex) dans le massif de Belledonne-Sept-Laux (Isere) // Gibier Faune Sauvage. 1995. Vol. 12, N 2. P. 133-146.
Tong Y.C. The changes in abundance of mammal populations in Hulunbeirmeng region // The successional changes of mammals in China under the influences of human activities. Beijing: China Sc. and Techn. Press, 1993. P. 142-145.
Tosi G., Toso S„ Randi E. Demografia e varjabilita genetica in alcune colonie di Stambecco {Capra ibex ibex) // Suppl. Ric. Biol. Selvagg. 1991. Vol. 18. P. 109-122.
Towne E. G. Bison performance and productivity on tailgrass prairie // Southwest. Natur. 1999. Vol. 33, N3.P. 361-366.
Tuya T.S., Reading R., Ramey R.R., et al. New perspectives on the subspecies, population structure, and conservation of mongolian argali sheep // Mammalogical studies in Mongolia and its adjacent territories. Ulaanbaatar, 2002. P. 149-158. (Proc. Inst. Biol., N 24).
Uerpmann H.-P. The ancient distribution of ungulate mammals in the Middle East // Beih. Tubinger Atlas Orients. 1987. P. 1-173.
Udinal.G., SipkoT.P., Rautian G.S., Sulimova G.E. The study ofblood groups andDNA-polimorphism of European bison // Anim. Genet. 1994. Vol. 25, Suppl. 2. P. 40-41.
Valdez R. The wild sheep of the World. New Mexico: Mesilla, 1982. 186 p.
Valdez R., Nadler C.F., Bunch T.D. Evolution of wild sheep in Iran//Evolution. 1978. Vol. 32, N 1. P. 56-72.
Van Vuren D. Summer diets of bison and cattle in Southern Utah // J. Range Manag. 1984. Vol. 37, N 3. P. 260-261.
Vass art M., Greth A., Durand V, Cribin E.P. Chromosomal polymorphism in sand gazelles {Gazella subgutturosa marica) // Heredity. 1993. Vol. 84, N 6. P. 478—481.
Vassart M.,SeguelaA., HayesH. Chromosomal evolution in gazelles//Ibid. 1995. Vol. 86, N 3. P. 216-227.
Volf J. Some remarks on the taxonomy of the genus Nemorhaedus H. Smith, 1827 (Bovidae: Rupicaprinae) // Vestn. Cesk. Spol. Zool. 1976. Vol. 40, N 1. P. 75-80.
Waggoner V, Hinkes M. Summer and fall browse utilization by an alaskan bison herd // J. Wildl. Manag. 1986. Vol. 50, N 2. P. 322-324.
Wall D.A., Davis S.K., Read B.M. Phylogenetic relationships in subfamily Bovidae (Mammalia: Artiodactyla) based on ribosomal DNA // J. Mammal. 1992. Vol. 73, N 2. P. 262-275.
Wang Z., Wang S., Lu C., Zhang R. Taxonomy, distribution and utilization of bovids in China // Biol. Bui. 1963. N 5. P. 24-30.
Wand T.J., SkowL.C., GallagherD.S., etal. Differential introgresion of uniparentally inherited markers in bison populations with hybrid ancestries H Anim. Genet. 2001. Vol. 32, N 2. P. 89-91.
Wilson G.A., Strobeck C. Genetic variation within and relatedness among wood and plains bison populations // Genome. 1999. Vol. 42, N 3. P. 483-496.
Wisent W. Last stand for Yellowstone’s wild bison? // Def. Wildl. News. 1973. N 2. P. 182-185.
Woloch A. Aktueller Stand der Muffelwildpopulationen in der Ukraine // Beitr. Jagd- und Wildforsch. 2003. Bd. 28. S. 135-141.
WroblewskiK. Zubr Puszczy Bialowieskiej //Nakf. OgroduZool. Pozn. 1927. Vol. 25. S. 1-232.
Wu C.H., Zhang Y.P., Bunch T.D., etal. Mitochondrial control region sequence variation within the argali wild sheep (Ovis аттоп): Evolution and conservation relevance // Mammalia. 2003. Vol. 67, N l.P. 109-118.
Wurster D.H., Benirschke K. Chromosome studies in the superfamily Bovoidae // Chromosoma. 1968. Vol. 25, N 2. P. 152-171.
Zheleznov-Chukotsky N.K. Modem Status of Snow Sheep (Ovis nivicola Eschscholtz, 1829) in North Asia // Beitr. Jagd- und Wildforsch. 2003. Bd. 28. S. 165-189.
Zhang Y. Mammalian zoogeography and conservation of the endangered species in arid areas of China // Mammals in the Palaearctic desert: status and trends in the Sahara-Gobian region. M., 1991. P. 269-284.
Zhigang J., Diqiang L. Antelopes in China 11 Russian-Chinese Workshop on study of animal biodiversity and ecosystem function. M., 1996. P. 17-19.
Zhou Cai-Quan, Zhou Kai-Ya, Hu Jin-Chu. Валидность видового статуса китайского барана Pseudois schaeferi - исследование по данным митохондриального гена цитохрома b // Acta Zool. Sin. 2003. Vol. 49, N 5. P. 578-584.
Zimmermann S., Zehner R„ Herzog M.A. Cytochrom b-Sequenz-Vergleiche zwischen Wisent (Bison bison bonasus) und Hausrind (Bosprimigenius f. taurus) IIZ. Jagdwiss. 1998. Bd. 44, N l.S. 26-32.
Zyll de Jong C. G. A systematic study of recent bison with particular consideration of the wood bison (Bison bison athabascae Rhoads, 1898) // Natur. Sci. Ottawa (Canada). 1990. N 6. 130 p.
ОГЛАВЛЕНИЕ
Предисловие ........................................................3
Семейство Полорогие...............................................5
Подсемейство Бычьи.............................................9
Род Бизоны..................................................И
Зубр....................................................16
Изменчивость и таксономия.............................18
Распространение.......................................32
Среда обитания........................................37
Питание...............................................41
Размножение...........................................46
Структура группировок............................... 50
Численность..........................................57
Основные факторы, определяющие динамику группировок..66
Проблемы сохранения и использования ресурсов.........71
Подсемейство Газелевые........................................77
Род Дзерены................................................80
Монгольский дзерен......................................82
Распространение......................................83
Среда обитания.......................................86
Питание..............................................87
Размножение..........................................88
Популяционная структура..............................89
Численность и основные факторы, ее определяющие......93
Использование ресурсов и охрана......................96
Род Сайги..................................................99
Сайга................................................. 101
Изменчивость........................................101
Распространение.....................................107
Среда обитания......................................116
Питание.............................................119
Размножение.........................................122
Популяционная структура.............................129
Численность.........................................139
Основные факторы, определяющие динамику популяций....142
Использование ресурсов..............................156
Сохранение вида.....................................164
Подсемейство Козлиные........................................168
Род Овцебыки..............................................173
Овцебык................................................176
Распространение.....................................176
Среда обитания......................................180
Питание............................................182
Размножение.........................................183
Популяционная структура.............................185
Численность и основные факторы, ее определяющие.....187
Перспективы восстановления и использования ресурсов..189
Род Горалы................................................191
Амурский горал.........................................193
Изменчивость.........................................193
Распространение......................................196
Среда обитания.......................................198
Питание..............................................199
Размножение..........................................200
Популяционная структура..............................201
Численность и основные факторы, ее определяющие .....204
Перспективы сохранения...............................206
Род Серны ...................................207
Серна...................................................209
Изменчивость.........................................211
Распространение......................................213
Среда обитания.......................................215
Питание..............................................216
Размножение..........................................217
Популяционная структура..............................218
Численность и основные факторы, ее определяющие......220
Использование ресурсов и охрана......................224
Род Козлы...................................226
Безоаровый козел........................................232
Изменчивость.........................................234
Распространение......................................237
Среда обитания.......................................240
Питание..............................................241
Размножение..........................................241
, Популяционная структура..........................243
Численность и основные факторы, ее определяющие......246
Западнокавказский и восточнокавказский козлы............248
Изменчивость и таксономия............................251
Распространение......................................260
Среда обитания.......................................261
Питание..............................................264
Размножение..........................................267
Популяционная структура..............................269
Численность..........................................275
Основные факторы, определяющие динамику популяций....277
Использование, охрана и перспективы восстановления ресурсов 280
Сибирский козел.........................................283
Изменчивость и таксономия............................284
Распространение......................................295
Среда обитания.......................................300
Питание..............................................302
Размножение..........................................303
Популяционная структура..............................307
Численность..........................................313
Основные факторы, определяющие динамику популяций....315
Использование ресурсов и охрана......................317
Род Бараны...................................320
Архар...................................................331
Изменчивость и таксономия............................333
Распространение......................................350
Среда обитания.......................................357
Питание..............................................360
Размножение..........................................363
Популяционная структура..............................368
Численность..........................................375
Основные факторы, определяющие динамику популяций....382
Использование ресурсов и охрана......................386
Снежный баран...........................................390
Изменчивость и таксономия............................393
Распространение......................................408
Среда обитания.......................................413
Питание..............................................416
Размножение..........................................418
Популяционная структура..............................422
Численность..........................................427
Основные факторы, определяющие динамику популяций....429
Использование ресурсов и охрана......................432
Литература .........................................................436
CONTENTS
Preface ......................................................................3
Family Bovidae........................................................... 5
Subfamily Bovinae......................................................9
Genus Bison........................................................11
European bison................................................. 16
Variability and taxonomy................................... 18
Distribution................................................32
Environment.................................................37
Feeding.....................................................41
Reproduction................................................46
Population structure........................................50
Numbers.....................................................57
Numbers factors.............................................66
Problem of preservation and use of resources................71
Subfamily Antilopinae................................................77
Genus Procapra.....................................................80
Mongolian gazelle.............................................. 82
Distribution................................................83
Environment.................................................86
Feeding.................................................... 87
Reproduction................................................88
Population structure........................................89
Numbers and numbers factors................................ 93
Use of resources and preservation...........................96
Genus Saiga....................................................... 99
Saiga......................................................... 101
Variability............................................... 101
Distribution.............................................. 107
Environment................................................116
Feeding....................................................119
Reproduction.............................................. 122
Population structure...................................... 129
Numbers................................................... 139
Numbers factors........................................... 142
Use of resources.......................................... 156
Preservation of species................................... 164
Subfamily Caprinae................................................. 168
Genus Ovibos......................................................173
Musk-ox....................................................... 176
Distribution.............................................. 176
Environment............................................... 180
Feeding................................................... 182
Reproduction.............................................. 183
Population structure...................................... 185
Numbers and numbers factors............................... 187
Perspective of recovery and use of resources.............. 189
Genus Nemorhaedus................................................ 191
Goral......................................................... 193
Variability.................................................. 193
Distribution................................................. 196
Environment.................................................. 198
Feeding...................................................... 199
Reproduction..................................................200
Population structure..........................................201
Numbers and numbers factors...................................204
Perspective of preservation...................................206
Genus Rupicapra......................................................207
Chamois..........................................................209
Variability...................................................211
Distribution................................................. 213
Environment...................................................215
Feeding.......................................................216
Reproduction..................................................217
Population structure..........................................218
Numbers and numbers factors...................................220
Use of resources and preservation.............................224
Genus Capra..........................................................226
Bezoar...........................................................232
Variability...................................................234
Distribution..................................................237
Environment...................................................240
Feeding.......................................................241
Reproduction..................................................241
Population structure..........................................243
Numbers and numbers factors...................................246
Westcaueasian and Eastcaucasian tur..............................248
Variability and taxonomy......................................251
Distribution..................................................260
Environment...................................................261
Feeding.......................................................264
Reproduction..................................................267
Population structure..........................................269
Numbers.......................................................275
Numbers factors...............................................277
Use, preservation and perspective of recovery of resources....280
Siberian ibex....................................................283
Variability and taxonomy......................................284
Distribution..................................................295
Environment...................................................300
Feeding.......................................................302
Reproduction..................................................303
Population structure..........................................307
Numbers.......................................................313
Numbers factors...............................................315
Use of resources and preservation.............................317
Genus Ovis...........................................................320
Argali...........................................................331
Variability and taxonomy......................................333
Distribution.................................................350
Environment................................................. 357
Feeding......................................................360
Reproduction.................................................363
Population structure.........................................368
Numbers......................................................375
Numbers factors..............................................382
Use of resources and preservation............................386
Bighorn sheep.................................................. 390
Variability and taxonomy.....................................393
Distribution.................................................408
Environment..................................................413
Feeding......................................................416
Reproduction.................................................418
Population structure.........................................422
Numbers......................................................427
Numbers factors..............................................429
Use of resources and preservation............................432
References .................................................................. 436
Научное издание
Алексей Алексеевич Данилкин
ПОЛОРОГИЕ (BOVIDAE)
Серия “Млекопитающие России и сопредельных регионов”
Утверждено к печати Ученым советом Института проблем экологии и эволюции им. А.Н. Северцова Российской академии наук
Редактирование и макет: Св.В. Найденко
Москва: Товарищество научных изданий КМК. 2005. 550 с.
Для заявок:
123100 Москва а/я 16
ши: kmk2000@online. ги
Отпечатано в ППП “Типография “Наука” 121099 Москва, Шубинский пер., 6.
Подписано к печати 25.08.2005. Формат 70x100 1/16. Бумага офсетная. Гарнитура
Таймс. Печать офсетная. 34,5 +1,5 печ.л. Тираж 1000 экз. Заказ № 1475
МЛЕКОПИТАЮЩИЕ РОССИИ
И СОПРЕДЕЛЬНЫХ РЕГИОНОВ
А.А. ДАНИЛКИН
ПОЛОРОГИЕ
ж к