/




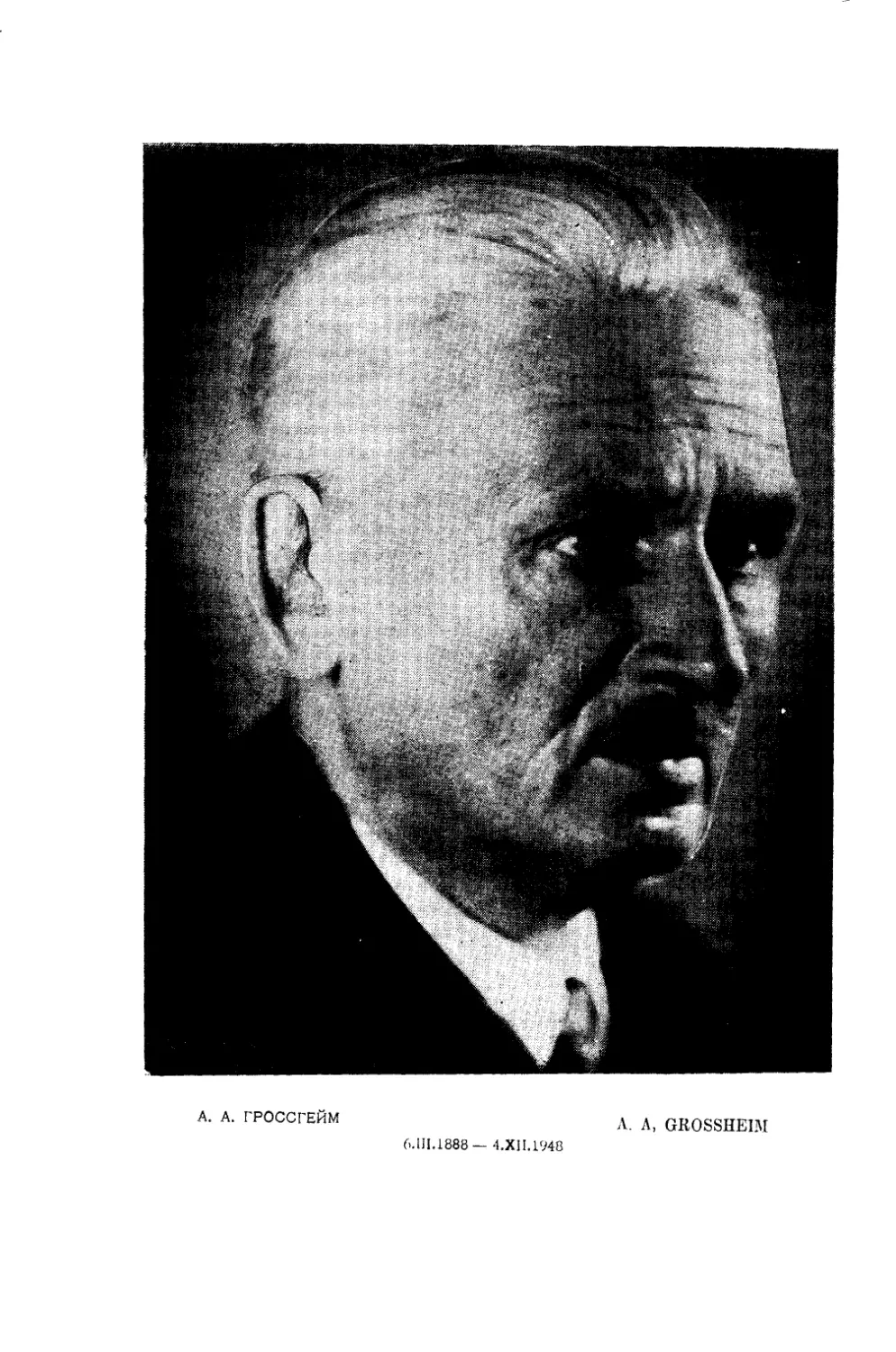

















































































































































































































































Текст
к. А. ГРОССГЕЙМ
дополнений
М. «К САХО КИА
ОБЗОР НОВЕЙШИХ СИСТЕМ
ЦВЕТКОВЫХ РАСТЕНИЙ
A-A. CROSSHEIM
scppi ements
Ol M. F SAKHOKJA
REVIEW OF MBDERR SY5TESS OF FtOWEBIHS PLAHTS
ТБИЛИСИ 1006 THBliUI
bddd60)3oc?(nb ььл аоббообз&дслд д<з№аог>
ACADEMY OF SCIENCES OF THE GEORGIAN SSS
OT&oc?obOb г>(п*дбозоь обь*о*,а*о
TBILISI INSTITUTE OF BOTANY
d абпгььазоао
£0>8a(5)a&i6o
а. ьдьпиодьо
аздзоеоздб аозбдбошд
ажаьо ьоьоааэбоь
аоаоьосзд
9. UdbrojOdb
бдра^оосо (ja ^oSaboftagaradncn
A. A. GROSSHEIM
r
Supplements
of M. F. SAKHOKIA
REVIEW OF MODERN SYSTEMS OF FLOWERING PLANTS
Edited and prefaced
by M. F. Sakhokia
&da(naeaa2?(n&d „асвбообо&д"
(лбосуоЬо 1966 Tbilisi
АКАДЕМИЯНАУК ГРУЗИНСКОЙ ССР
ТБИЛИССКИЙ БОТАНИЧЕСКИЙ ИНСТИТУТ
А. А. ГРОССГЕЙМ
Дополнения
М. Ф. САХОКИА
Обзор
новейших систем
цветковых растений
Редакция и предисловие
М. Ф. Сахокиа
Издательство „Мецниереба
Тбилйси 1966
581.5
582 5/9
Г 896
Книга посвящена научно-крнтнческому анализу филоге-
нетических систем советских (отчасти и русских) и зарубеж-
ных ученых. Оценка анализируемых систем сопровождается, в
частности, сопоставлением их с довлеющими в науке концеп-
циями Энглера и Веттштейна. Показана выдающаяся роль
советских ученых в разработке проблемы филогении цветко-
вых растений. В заключительной части книги изложены вы-
воды, касающиеся современного состояния разработки данной
проблемы; сформулированы основные теоретические положе-
ния и высказаны соображения организациоииого характера,
относительно осуществления дальнейших исследований в’ об-
ласти филогении цветковых растений.
Книга является не только научной сводкой, но и весьма
полезным учебным пособием для студентов-биологов и моло-
дых научных работников. Она представляет интерес и для
широкого круга' биологов.
СОДЕРЖАНИЕ
Стр.:
Предисловие редактора . ............................... IX
Система Г. Га л лира (Д. А. Гроссгейм)............ я . i
Система Ч. Бесси (Д. А. Гроссгейм)................... 3
Система Н. Арбера и Дж. Паркина (А- А. Гроссгейм) ... 5
Система К. Мена (А. А. Гроссгейм)................... 6
Система К. Меца и Г. Цигенспека (А. А. Гроссгейм) . . . 7
Система Дж. Гатчинсона (А. А. Гроссгейм)................. 8
Система А. Гейнце (А. А. Гроссгейм)...................... н
Система Г. Шелленберга (Д. Д. Гроссгейм).......... и
Система Дж. Шаффнера (А. А. Гроссгейм) ...... 13
Система Р. Пула (А. А. Гроссгейм)................ 15
Система Г. Лама (М. Ф. Сахокиа).................. т"
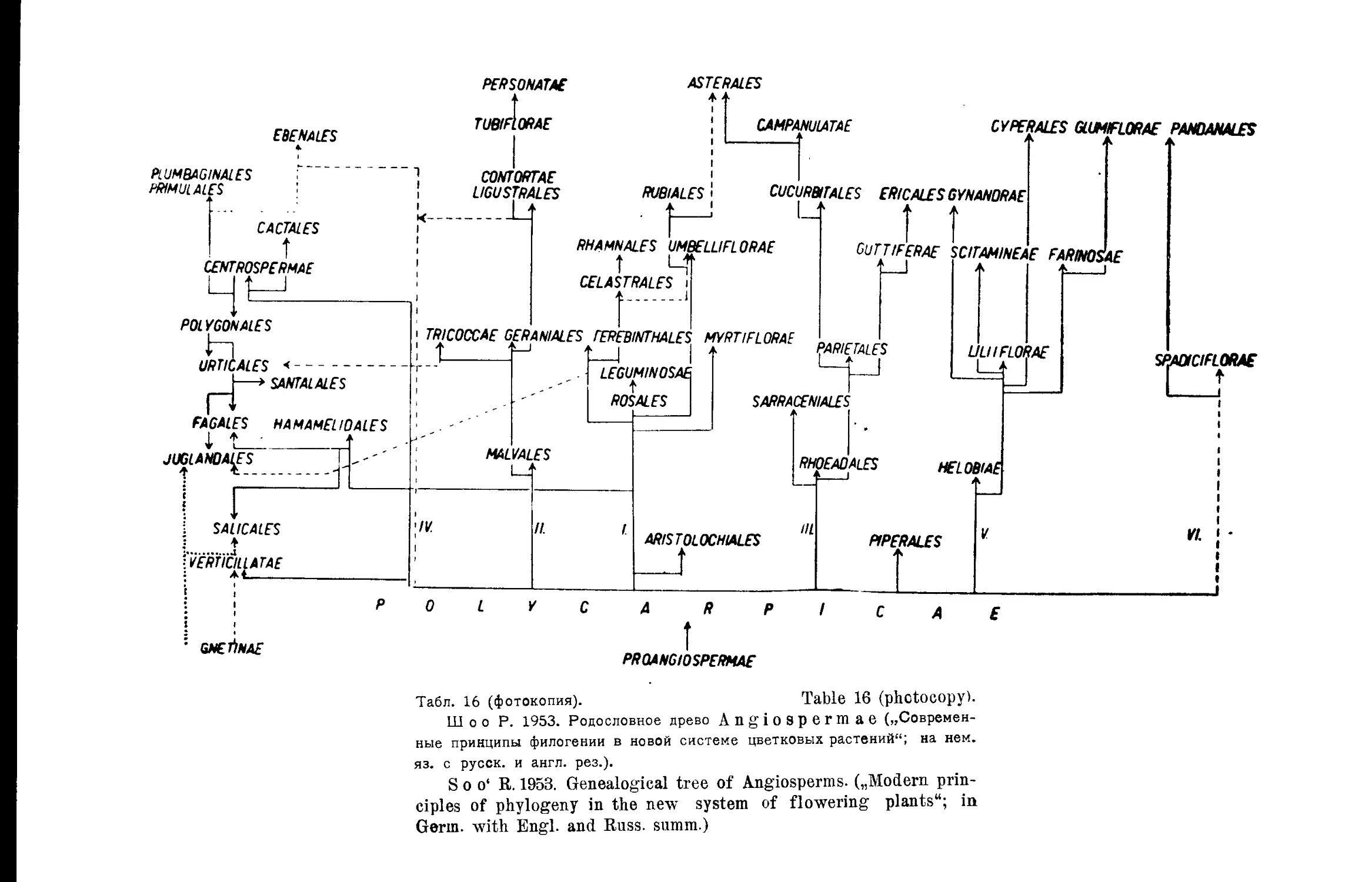
Система Р. Шоо (Ж. Ф. Сахокиа)...........................22
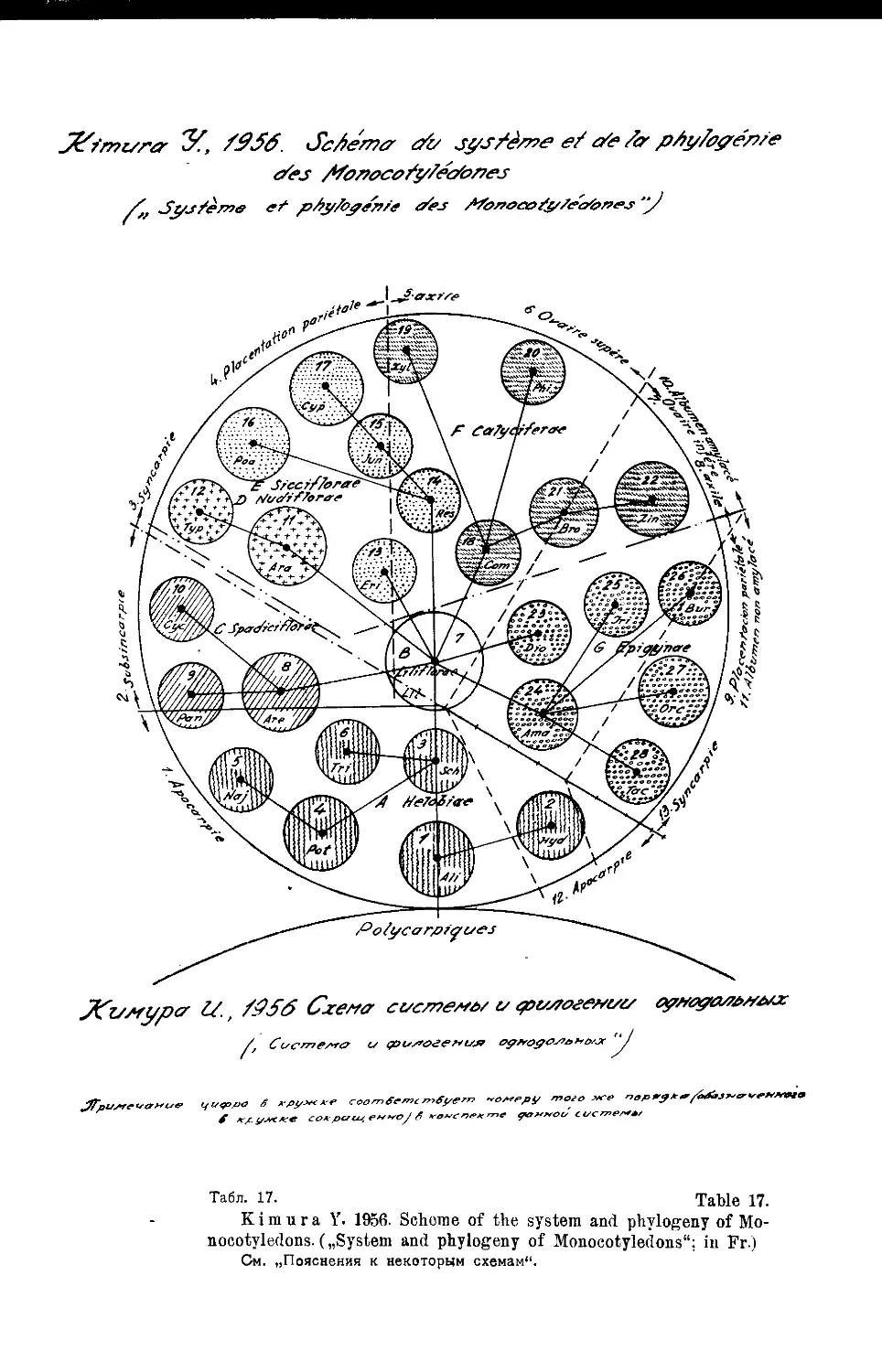
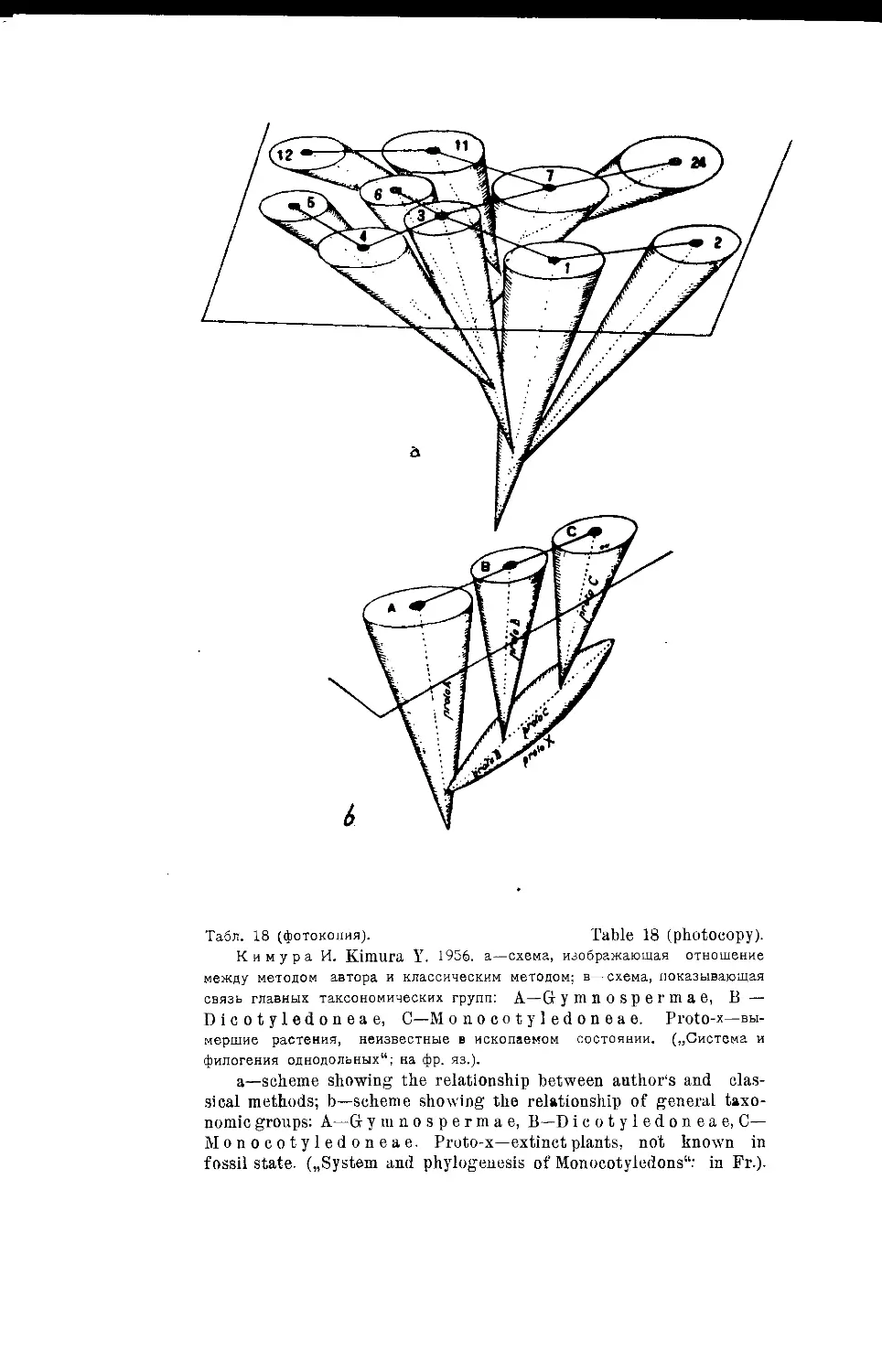
Система однодольных И. Кимуры (М. ф. Сахокиа) .... 23
Система двудольных А. Кронквиста (Л1. Ф, Сахокиа) ... 25
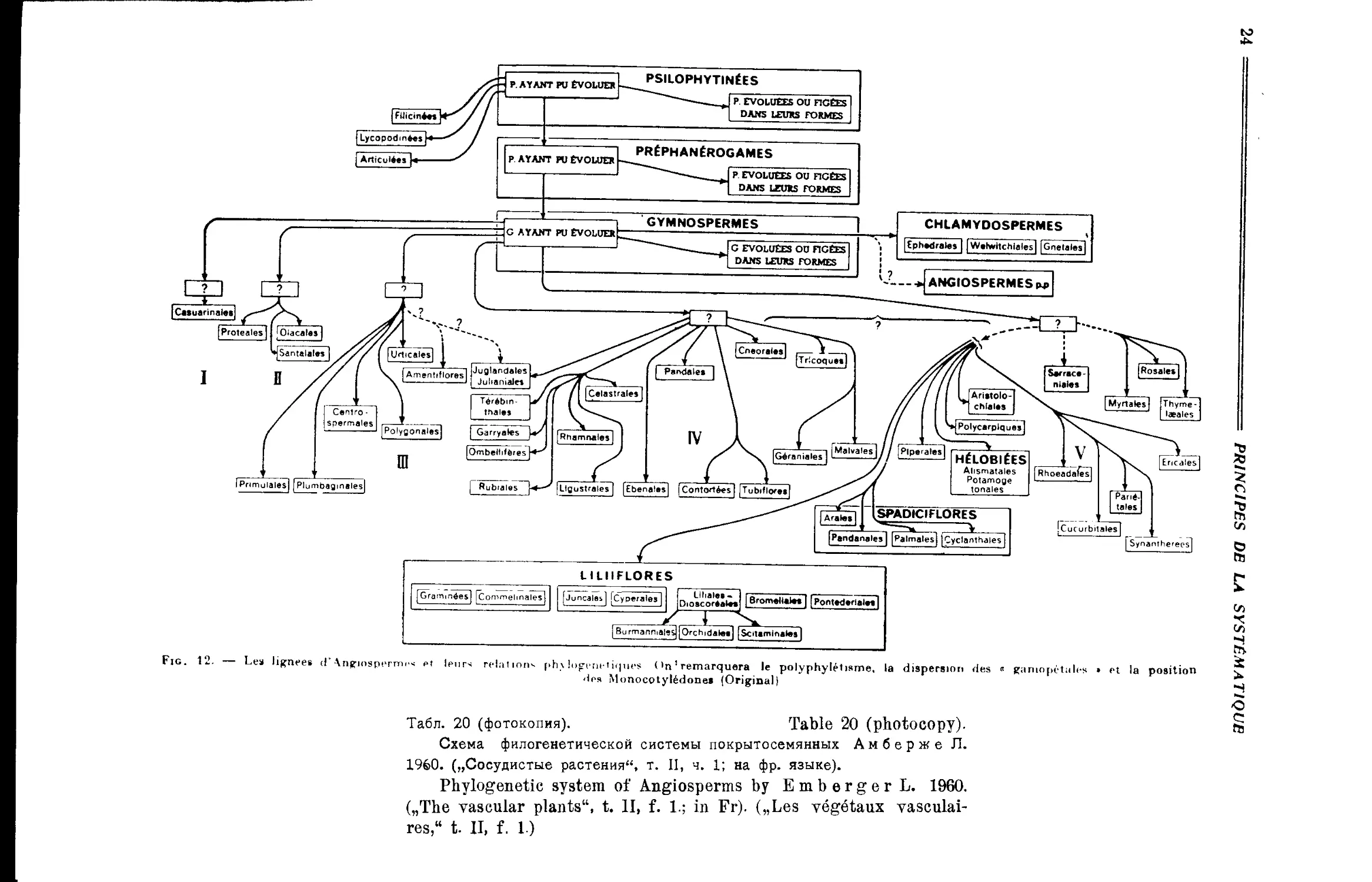
Система Л. Амберже (М. Ф. Сахокиа) ........ 27
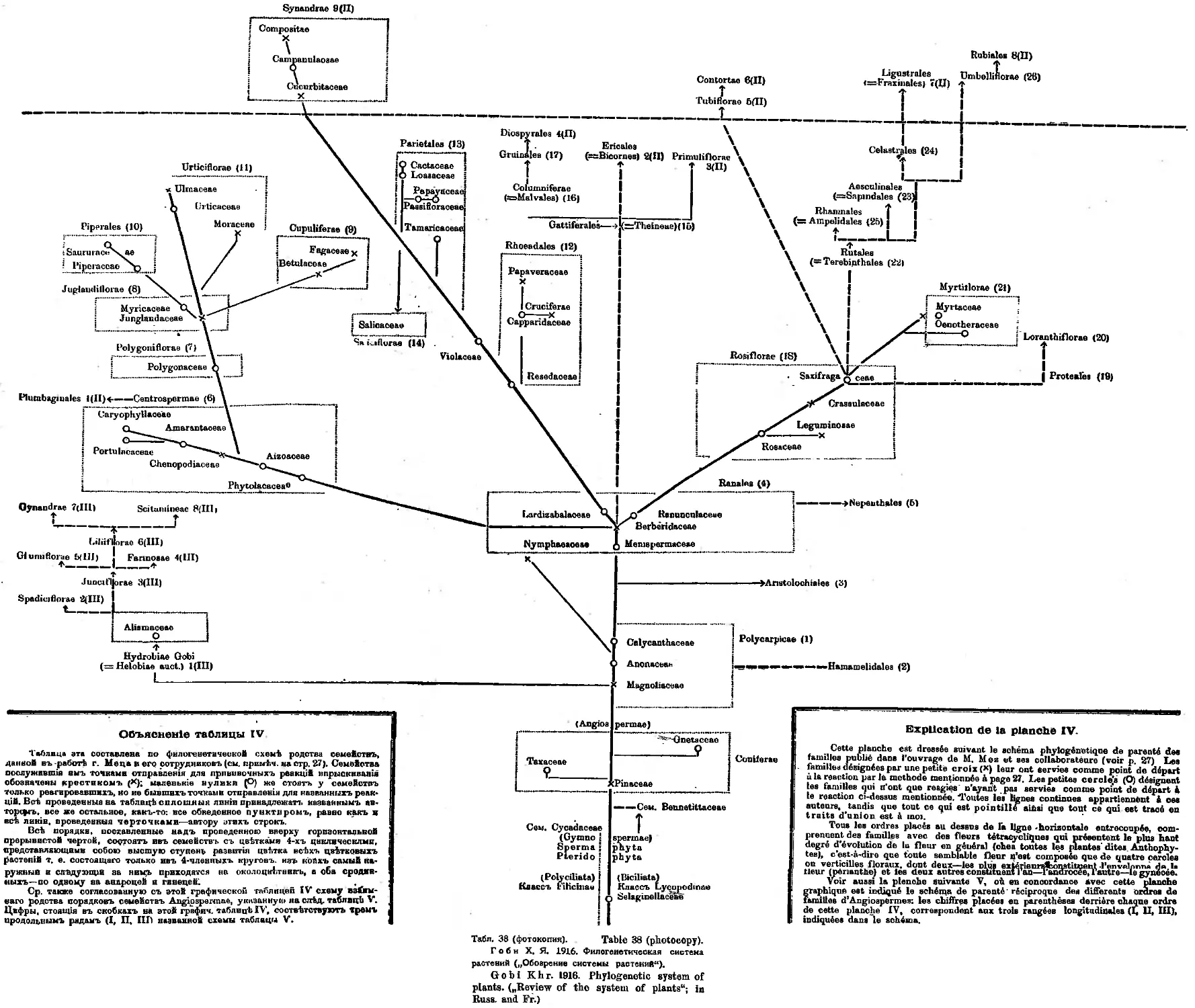
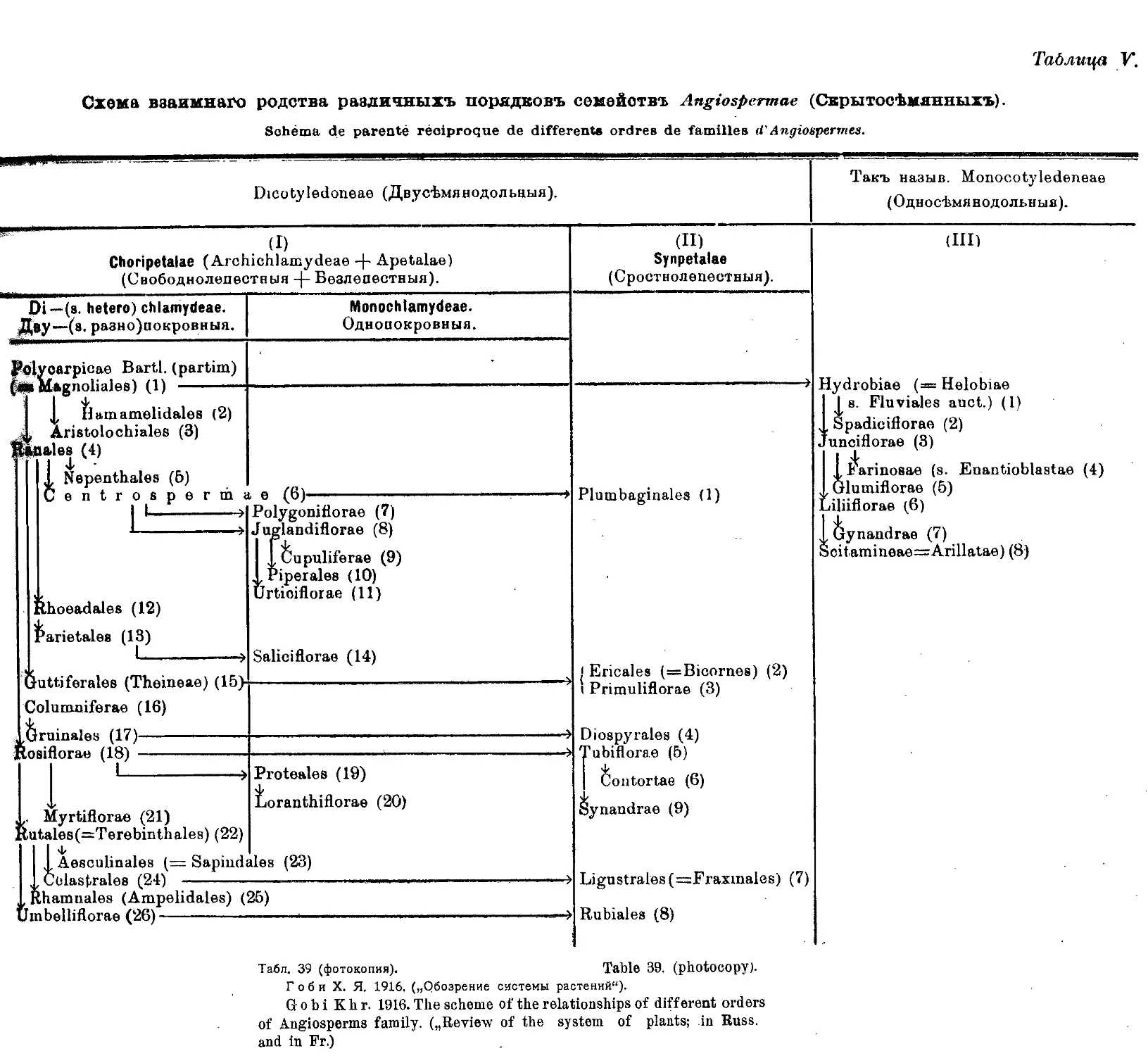
Система X. Я. Гоби (А. А. Гроссгейм) ........ 35
Система Н. И. Кузнецова (А. А. Гроссгейм)............... 37
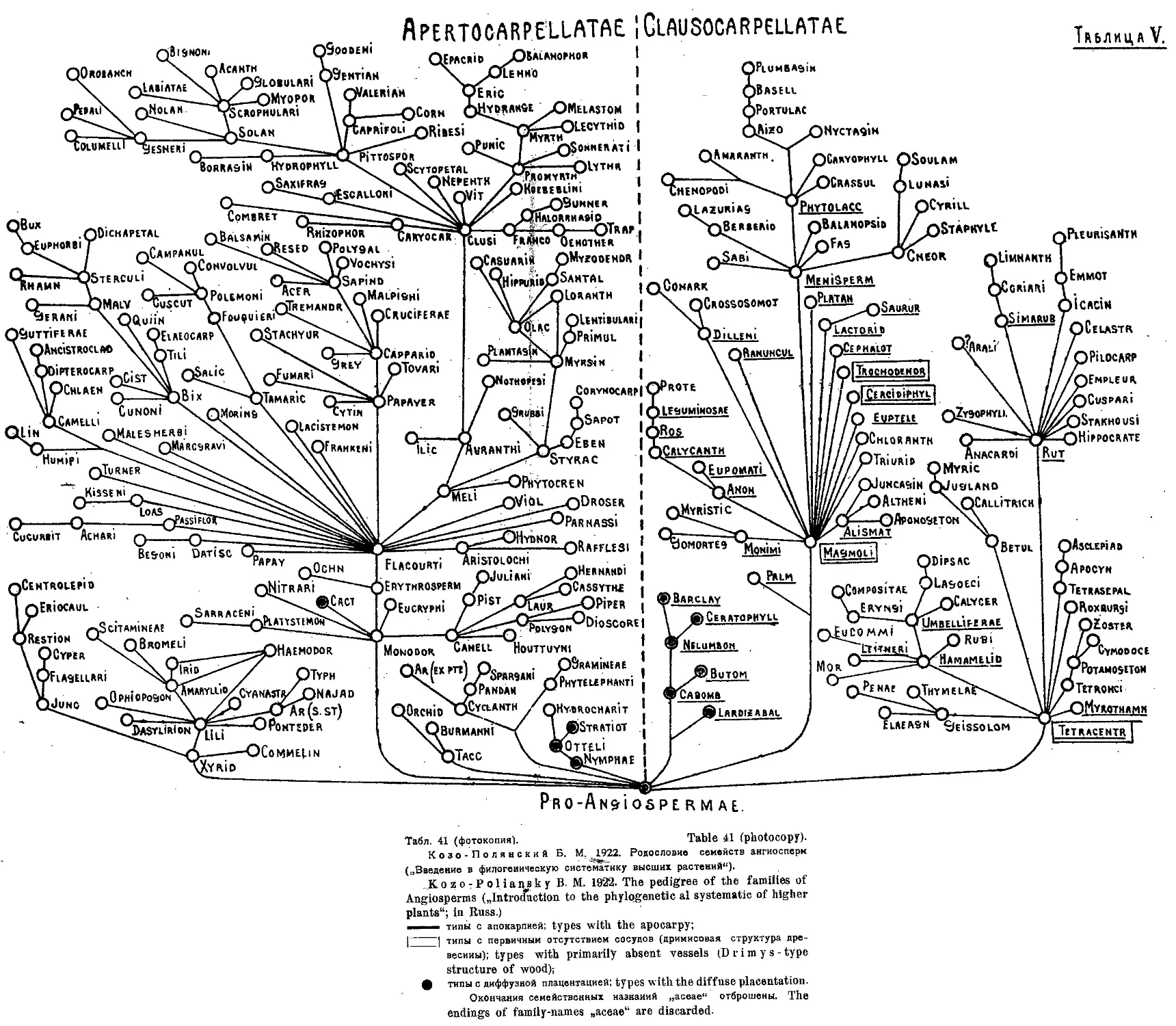
Система Б. М. Козо-Полянского (А. А. Гроссгейм) .... 40
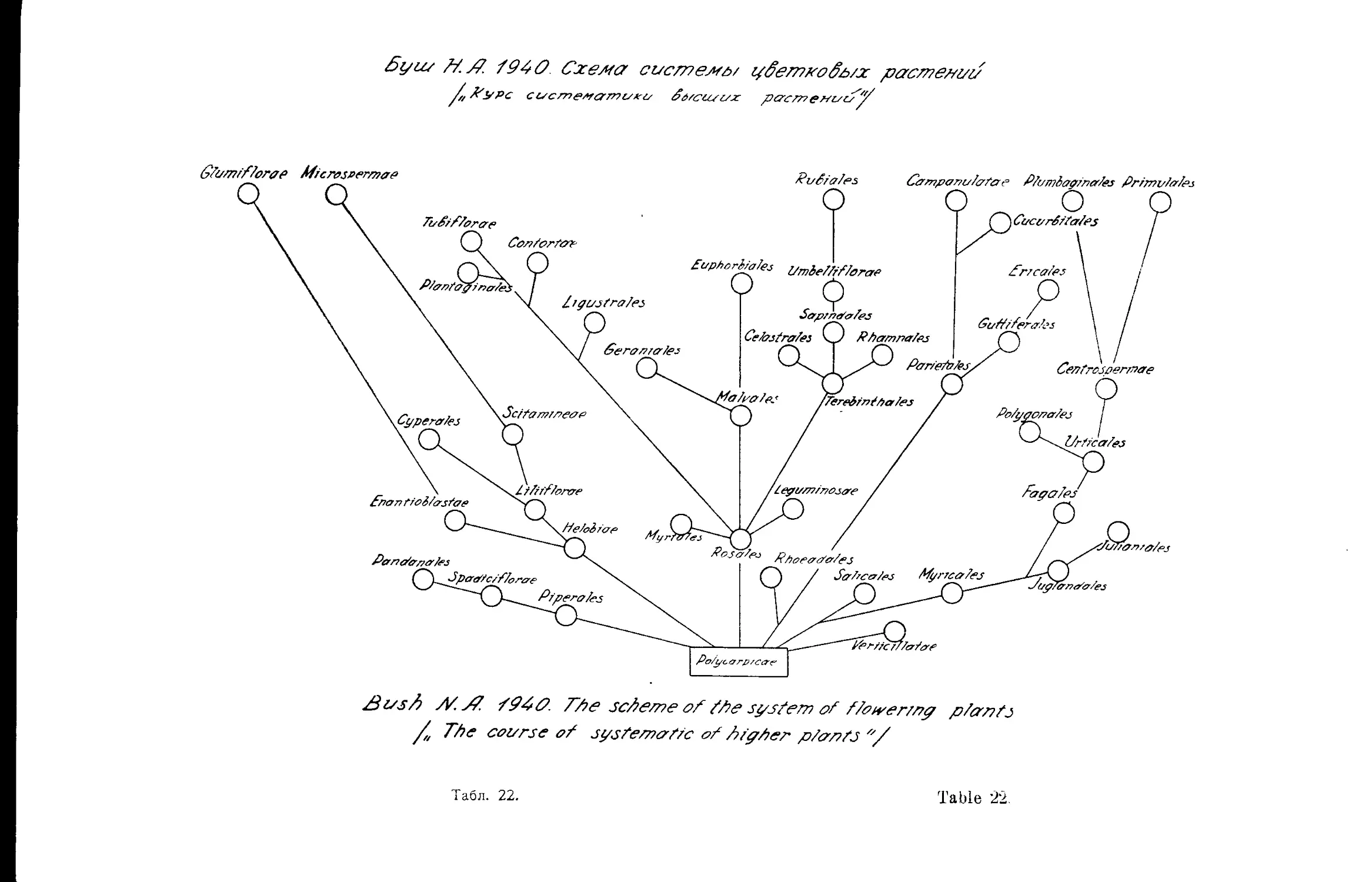
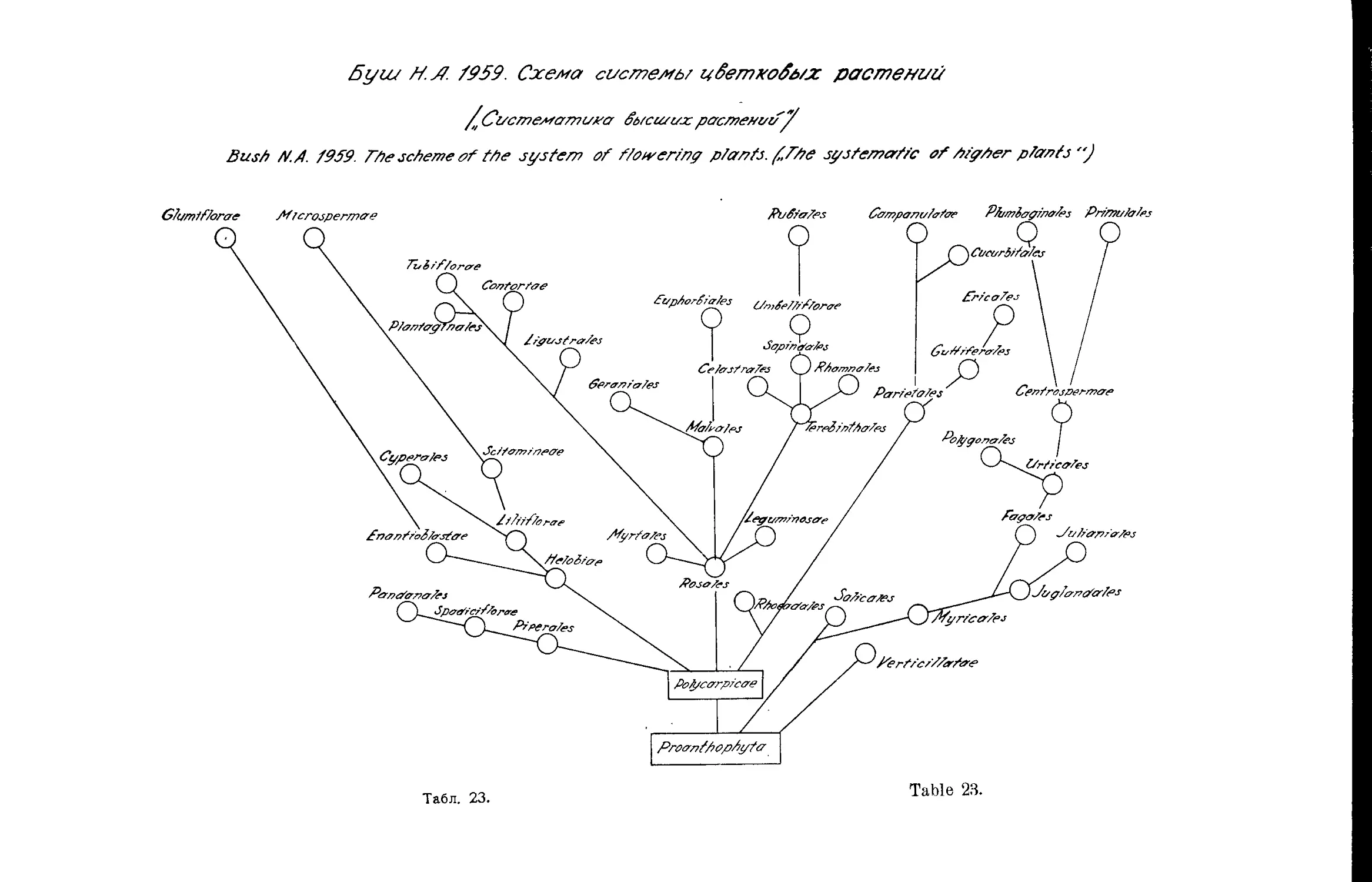
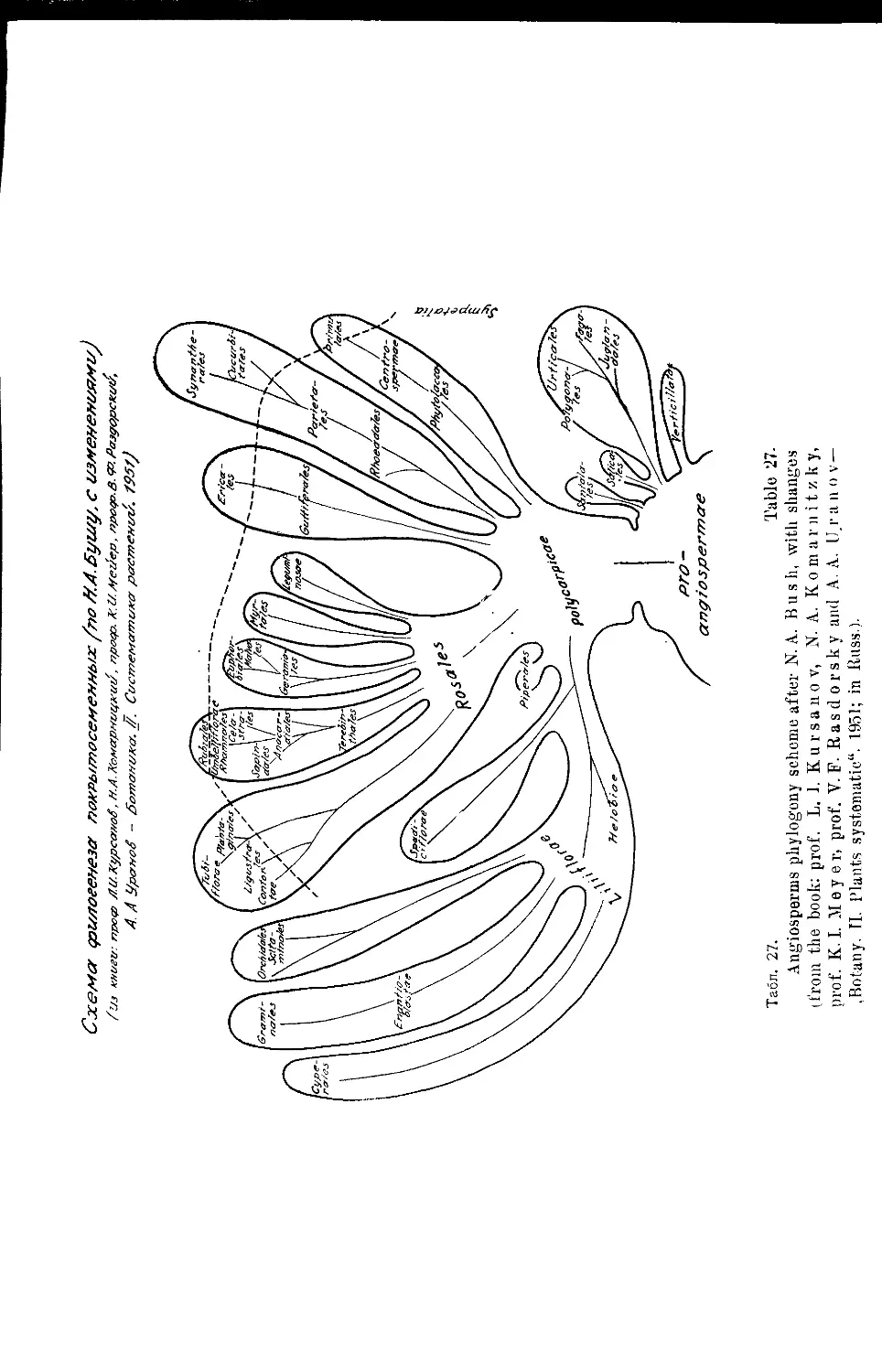
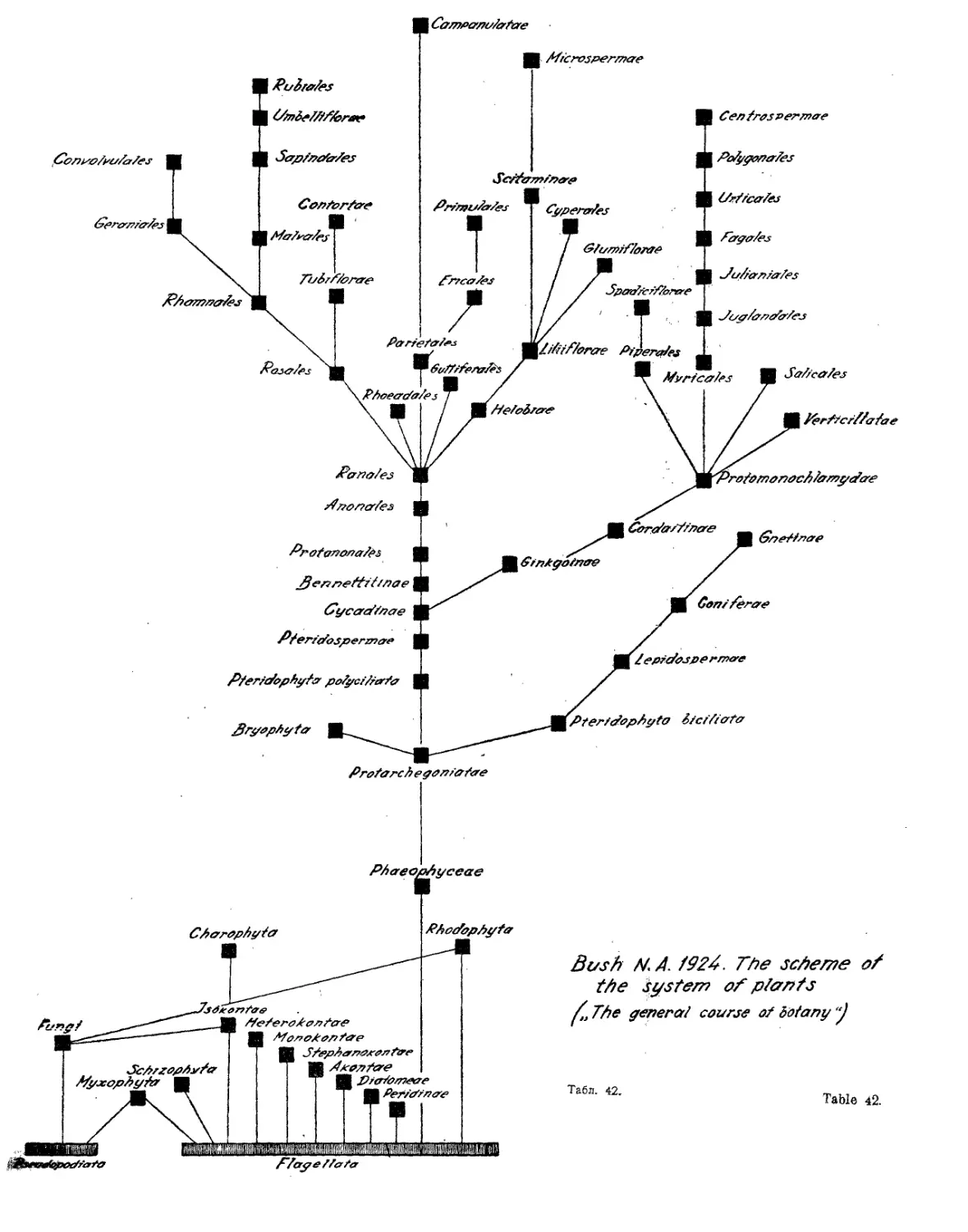
Система Н. А. Буша (Д. А. Гроссгейм).....................43
Система П. М. Жуковского (А. Д. Гроссгейм)...............45
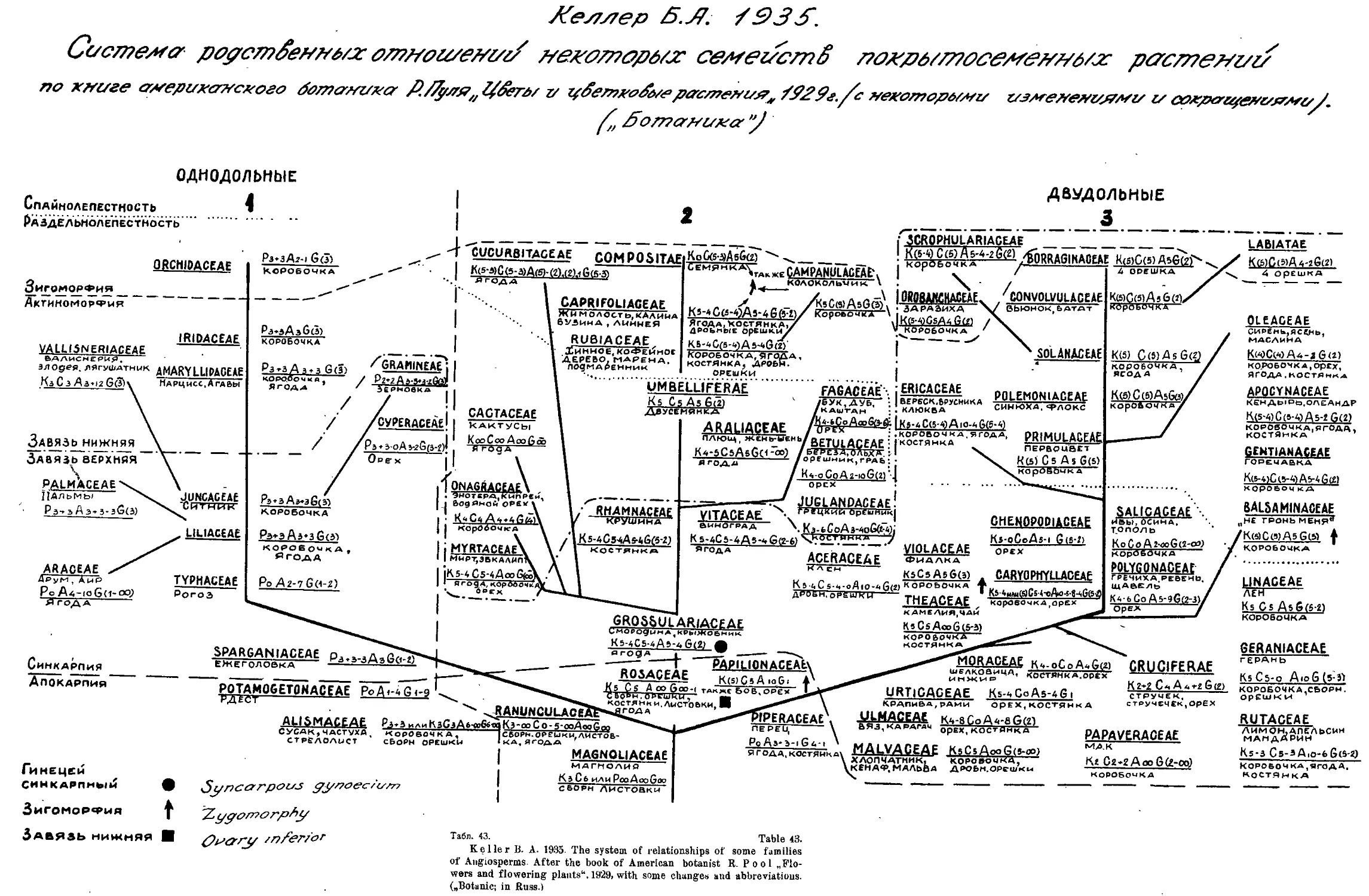
Система Б. А. Келлера (А. А. Гроссгейм) ....... 46
Система М. И. Голенкина (А. А. Гроссгейм)............... 46
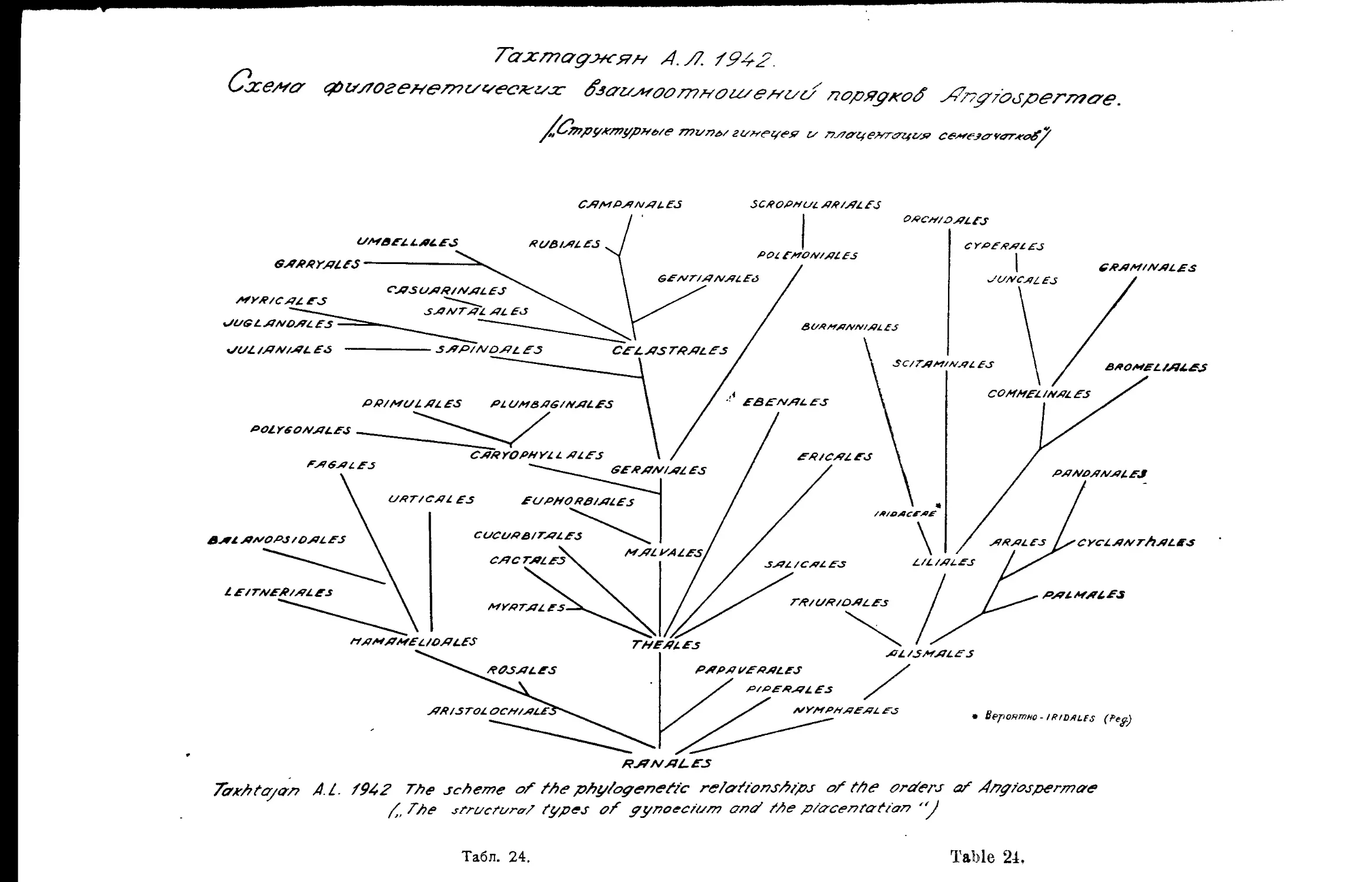
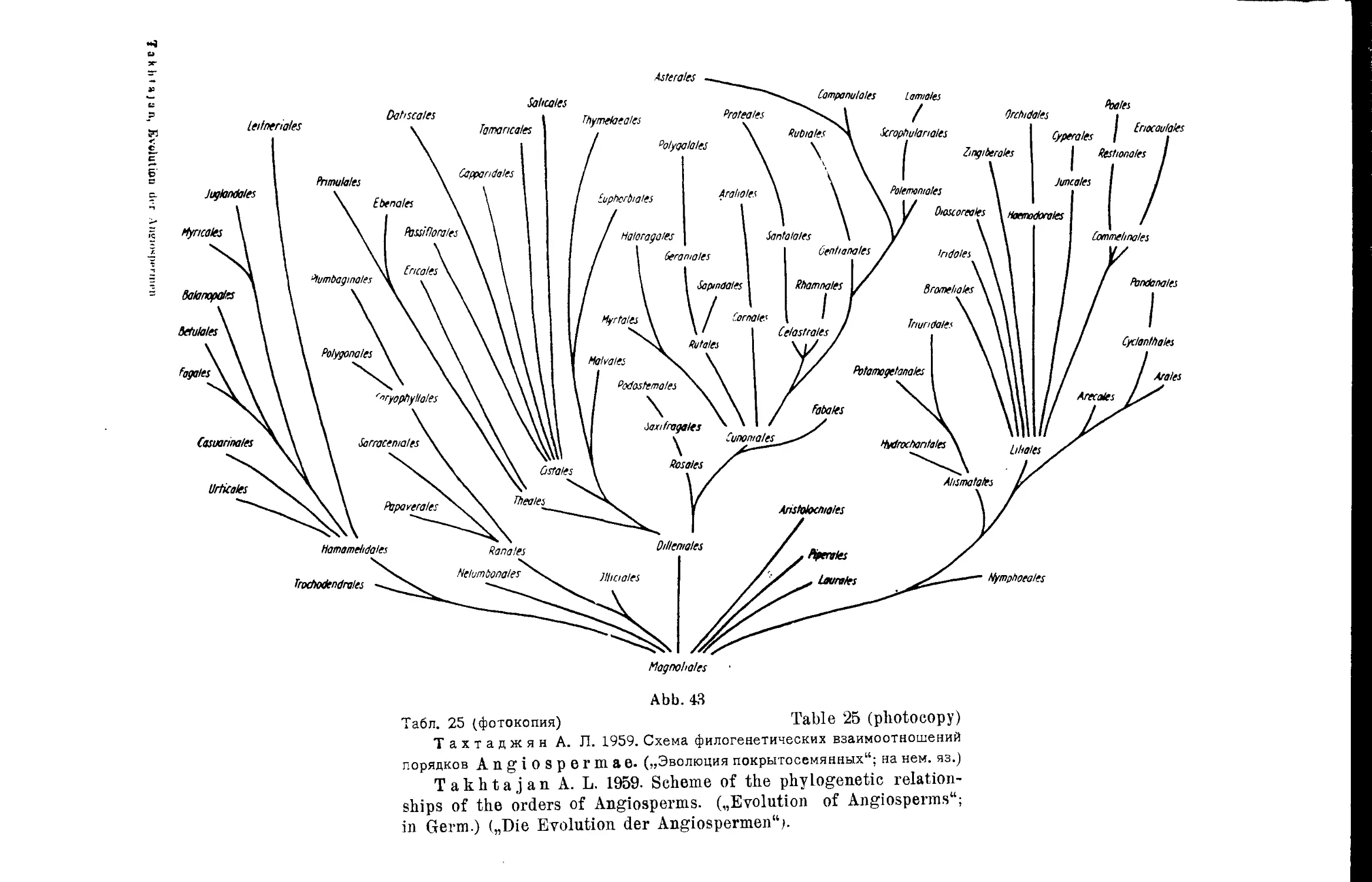
Система А. Л. Тахтаджяна (А. А. Гроссгейм).............. 46
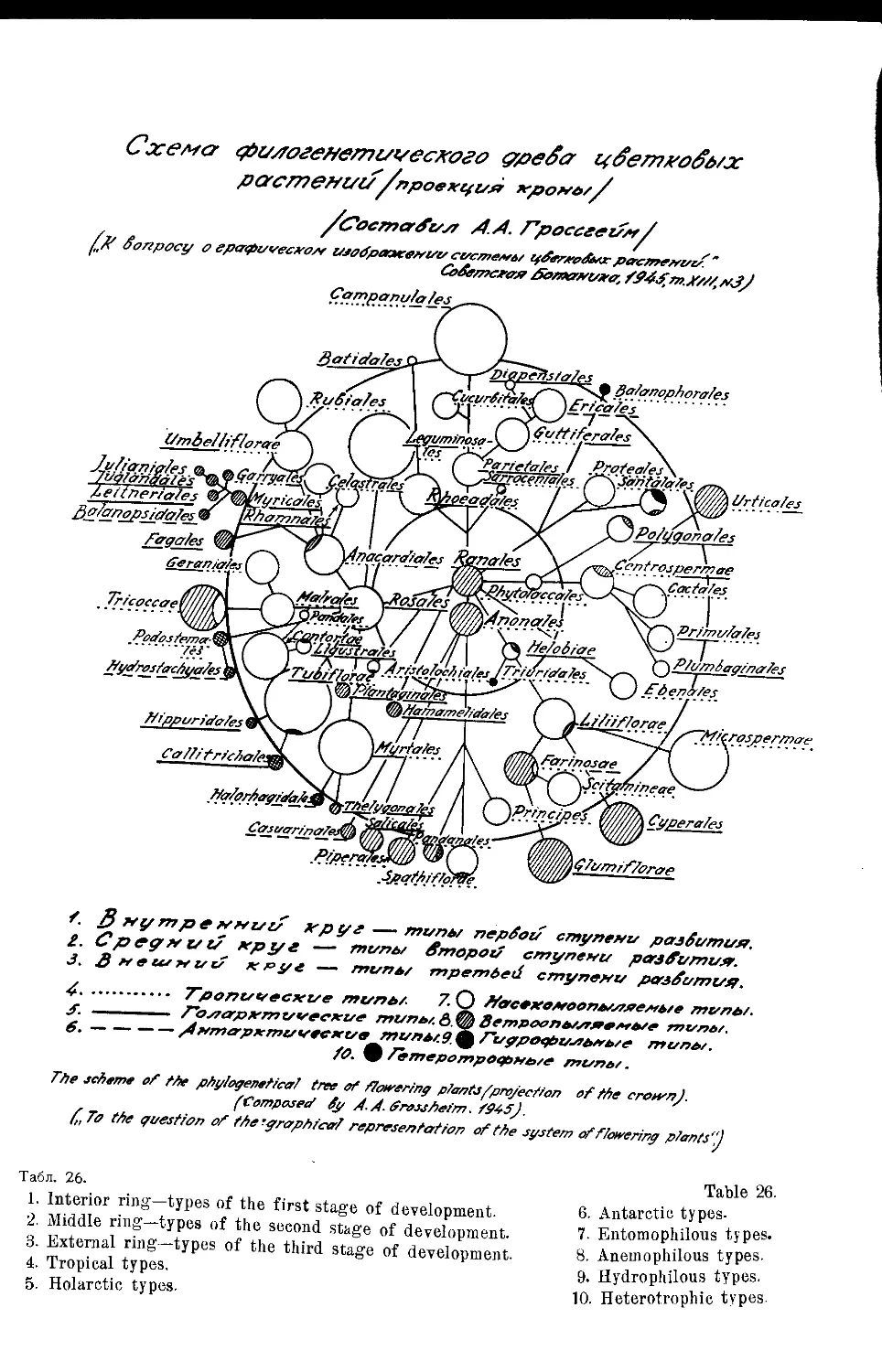
Система А. А. Гроссгейма (А. А. Гроссгейм) . ... . 50
Система Н. А. Комарницкого и А. А. Уранова (Л1. Ф. Сахокиа) 52
Система однодольных Л, А. Куприяновой (Л1. Ф. Сахокиа) $4
Заключение (А. А. Гроссгейм) ............................56
ПРИЛОЖЕНИЯ
(Составление М. Ф. Сахокиа)
А. А. Гроссгейм. К вопросу о графическом изображении
системы цветковых растений............................. 75
VI
Содержание Contents
Стр.:
II
Конспекты (обзоры) ф ил оге н ети ч е с к их систем
Система Р. Веттштейна................................103
„ Ч. Бесси......................................113
„ Дж. Гатчинсона ...............................121
„ А. Гейнце................................... 128
„ Дж- Шаффнера..................................135
, Р. Шоо.......................................138
„ И. Кимуры....................................147
„ А. Кронквиста................................149
„ Л. Амберже................................. 153
„ Б. М. Козо-Полянского........................164
Ш
Литература......................................... 166
IV
Графические схемы филогенетических систем
Т абл.:
Система Г. Галлира............................. г—7
„ Ч. Бесси ............................"... 8—ю
„ Н. Арбера и Дж- Паркина ........ II
„ К- Меца и Г. Цигенспека ...................... 34
„ Дж- Гатчинсона . . . . ................12,31—33
„ Дж. Шаффнера............................... 35,36
“ Р. Пула ..................................... 37
„ Г. Лама.................................. 13—15
,, Р. Шоо . ..............................• . 16
,, И. Кимуры ...... •......................... 17,18
„ А. Кронквиста........................... . 19
„ Л. Амберже.................................... 20
„ X. Я. Гоби .... •....•........... 38,39
,, Н. И. Кузнецова......................... t 40
я Б. М. Козо-Полянского . . . • 21,41
, Н. А. Буша..............•....•............. 22,23,42
„ Б. А. Келлера....................... 45
т А. Л. Тахтаджяна................. 24,25
„ А. А. Гроссгейма ...................... . 26
я Н. А. Комарницкого и А. А. Уранова .... 27,28
„ Л. А. Куприяновой............•...........• . 29
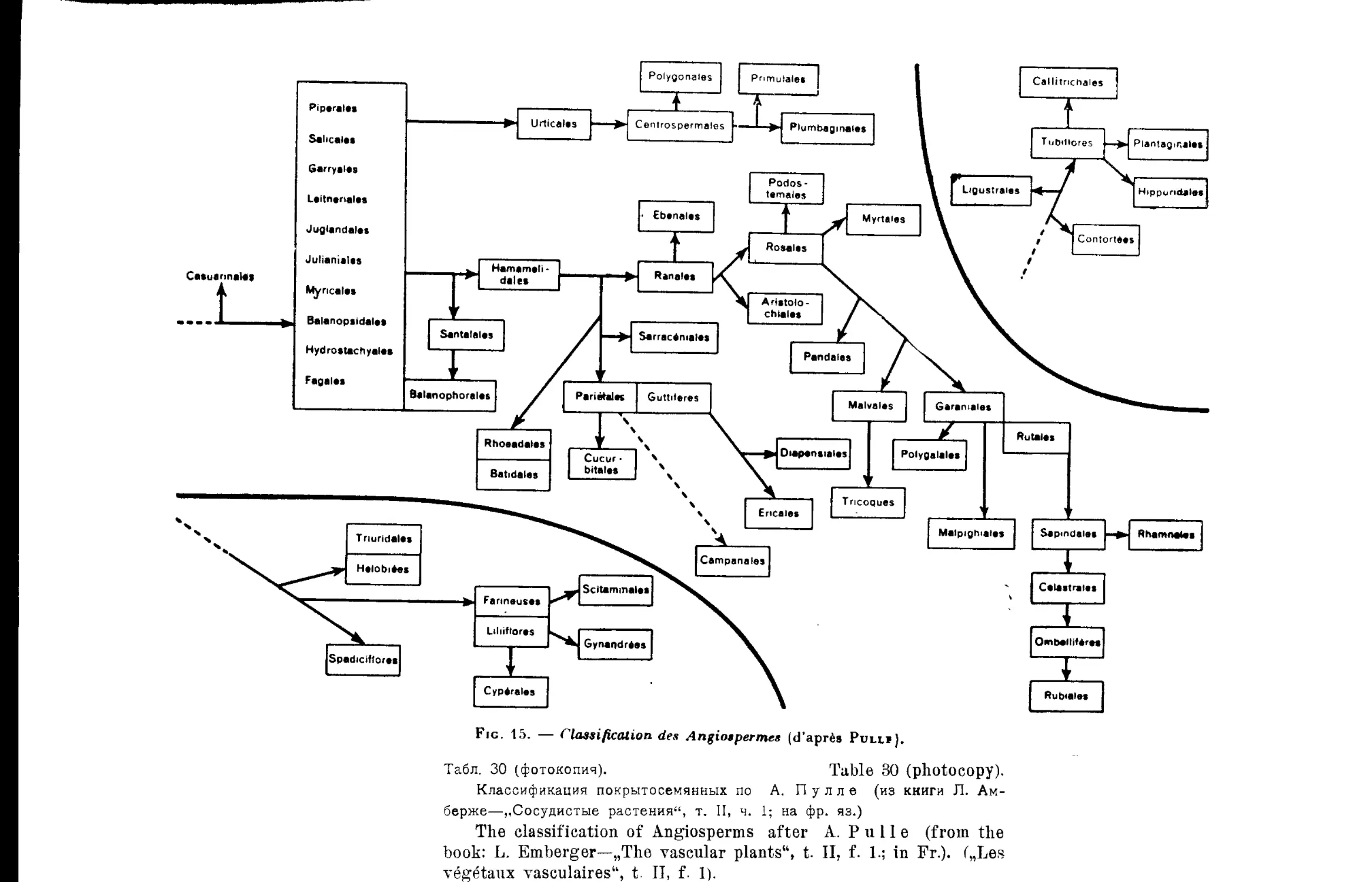
г А. Пулле .......................•............... 30
„ Р. Веттштейна .......................• ' . 44
Пояснения к некоторым схемам............см. после табл. 30
14 графических схем (из 45) приложены в виде отдель-
ных складных таблиц (таблицы № 31—44).
В книге помещены также 8 портретов ботаников-филогенетиков.
CONTENTS
Page!
Editor’s preface.............................................. IX
H. Hallier’s System (A- A- Grossheim).......................... 1
Ch. Bessey’s System (A- A- Grossheim).......................... 3
N. Arber's and J. Parkin’s System (A. A. Grossheim) .... 5
C- Metz’s System (A. A- Grossheim.)........................... 6
C. Metz’s and H. Ziegenspeck’s System (J. A- Grossheim) . . 1
J. Hutchinson’s System (A. A. Grossheim) ...................... 8
A- Heintze’s System (A. A- Grossheim) . . ....... 11
G. Schellenberg’s System (A- A- Grossheim).................... 12
J. Schaffner’s System (A- A’ Grossheim)....................... 13
R. Pool’s System (A- A. Grossheim)................... . . . 15
H. Lam’s System (M- F- Sakhokia).............................. 17
R. Sob’s System (2lf- F- Sakhokia)............................ 22
Y. Kimura’s System of Monocotyledons (Jf. F- Sakhokia) . . 23
A. Cronquist’s System of Dicotyledons (M- F. Sakhokia) . . 25
L. Emberger’s System (№ F- Sakhokia)......................... 27
Khr. Gobi’s System (A. A. Grossheim) ........... 35
N- Kuznetzov’s System (AL. A- Grossheim) ......... 37
B- Kozo-Poliausky’s System (A. A. Grossheim) ...... 40
N. Bush’s System (A. A- Grossheim)............................ 43
P. Zhukovsky’s System (A. A- Grossheim) ......... 45
B. Keller’s System (A- A- Grossheim) ....................... 46
M. Golenkin’s System (A- A- Grossheim)........................ 46
A- Takhtajan’s System (A. A- Grossheim)....................... 46
A. Grossheim’s System (A. A- Grossheim)................• . 50
N- Komarnitzky’s and A. Uranov’s System (2И. F- Sakhokia) . 52
L. Kuprianova’s System (2И- F- Sakhokia)...................... 54
Conclusion (A- A. Grossheim).................................. 56
Supplements
(completed by M, F. Sakhokia)
1
A. A- Grossheim. On the question of graphical representation
of the system of flowering plants.......................... 75
vni
Содержание Contents
Page:
II
Conspectus (reviews) of the phylogenetic Systems
R- Wettstein’s System .................................• • ЮЗ
Ch. Bessey’s System................................ ... 113
J. Hutchinson’s System..........•........................... 121
A. Heintze’s System .................... • . 128
J- Schaffner’s System .... ........................ 135
R. Soo’s System . . . . •..............• . • •................138
Y. Kimura’s System........................................... 147
A. Cronquist’s System..........•............•............... 149
L. Emberger’s System.................................• . . . 153
B. Kozo-Poliansky's System.................................. 164
III
Literature ..................................•............... 166
IV
Graphical Schemes of the phylogenetic Systems
Tables:
H. Hallier’s System ...................... . . . . 1—7
Ch. Bessey’s System................................\ . 8—10
N- Arter’s and J- Parkin’s System 11
C. Metz’s and H. Ziegenspeck’s System . . . • . . . 34
J. Hutchinson’s System............................• . 12,31—33
J. Schaffner’s System............................... 35,36
R. Pool’s System............................................. 37
H. Lam’s System..................................... 13—15
R. Soo’s System..............................• . . . . 16
Y. Kimura’s System....................................... 17,18
A. Cronquist’s System............................... 19
L. Emberger’s System......................................... 20
Khr. Gobi’s System............................ . . . . 38,39
N. Kuznetzov’s System ........................................ 40
B. Kozo-Poliansky's System . . • . •...................... 21,41
N. Bush’s System . ........................•............ 22,23,42
B. Keller’s System........................•.................. 43
A- Takhtajan’s System...................................... 24,25
A- Grossheim’s System . . . ................................. 26
N. Komarnitzky’s and A. Uranov’s System . . • • . . . 27,28
L. Kuprianova’s System . ’.................................... 29
A. Pulle’s System ........................................... 30
R. Wettstein’s System....................................... 44
Explanations to some Schemes.................see after table 30
14 graphical Schemes (out of 45)—in the form of separate
folding tables (tabl. 31—44).
In addition, there are in the book 8 portraits of botanists.
Предисловие редактора
Editor’s preface
В основу издаваемого „Обзор-а" легла, оставшаяся после смерти
академика А. А. ,Г р о с с г е й м а, рукопись объемом в 76 убористо написан-
ных страниц. Текст ее представляет основную часть намеченного им тру-
да; к нему не были приложены схемы и конспекты филогенетических си-
стем и список цитированной литературы; ему не было препровождено
предисловие. Написание текста автором завершено 20 февраля 1948 г.
в г. Баку (начато в июле 1947 г.) Этот труд, вышедший из под плодо-
творного пера А. А. Гроссгейма, и, издаваемый посмертно, безусло-
вно, явится ценным вкладом в литературу по филогении цветковых ра-
стений.
Он написан автором, по-видимому, в качестве конспекта лекций,
читанных им в Ленинградском университете им. Жданова. Вместе с тем, он
является, в сущности, своего рода прологом к его филогенетической
системе, хотя и опубликованной намного раньше (в 1945 г.).
А. А. Гроссгейм категорически придерживается монофилетичес-
кой теории происхождения цветковых растений. Я лично не разделяю
позиции столь абсолютного монофилетизма, но это обстоятельство нис-
колько не отражается на^ обшей композиции публикуемого „Обзорна.
Из систем зарубежных авторов А. А. Гроссгеймом в „Обзор"
были включены системы Г. Га л лира, Ч. Бесси, Н. Арбера и Дж-
Паркина, К. Меда и Г. Цигенспека, Дж- Г ат ч и н с о н а, А.
Гейнце, Т. Шелленберга, Дж- Шаффнера и Р. Пула; из сис-
тем отечественных филогенетиков им рассмотрены системы: Хр. Я. Гоби,
Н. И. Кузнецова, Б. М. К о з о-П о л я н с к о г о, Н. А. Буша, П. М,
Ж у к о в с к о г о, Б. А. Келлера, М. И. Г о л е н к и н а, А. Л. Т а х т а д-
Ж я н а, и, наконец, его же, только что отмеченная система. Ему же при-
надлежит заключительная часть. Кроме того, в рукописи упомянута бы-
ла система О. В. Троицкой (1939), которая, собственно говоря, не
является не только особой системой,' но даже частичным видоизменени-
ем какой-либо из известных к тому времени систем. По поводу нее
А. А. Гроссгейм пишет следующее (цитирую текст целиком): „В своей
Книге „Систематика растений" О. В. Троицкая в отношении покрытосе-
мянных полностью принимает систему Энглера. Из новейших авторов в
тексте упоминаются Н. И. Кузнецов и Галлир". Поэтому, этот раздел ру-
кописи мною полностью опущен. В то же время, А. А. Гроссгейм
новее не приводит опубликованную еще в 1925 году систему Н. Г'ай-
д‘у к о в а, а также малоизвестную систему эстонского ученого Т. Л и п п м а а
(1937)’ В нем не упоминаются и системы некоторых зарубежных исследо-
яателей. Никаких существенных изменений в текст А. А. Гроссгейма
X
Предисловие Preface
мною не внесено. Но, то обстоятельство, что опубликование „Обзор“-а
осуществляется через 17 с лишним лет после его написания, понятно,
вызвало необходимость хотя бы частичного дополнения системами после-
дующих авторов. Помимо этого, я счел целесообразным дополнить „Об-
зор" различными приложениями, с целью повысить его значение, как
учебное, так и научное.
В конечном счете, все внесенные мною дополнения сводятся к сле-
дующему:
i) основная часть „Обзор“-а дополнена характеристикой систем от-
дельных зарубежных авторов, а именно: Г. Лама, Р. Ш о о, И. Киму-
ры, А. Кронк в и ста и Л. Амбер же. Из систем отечественных авто-
ров мною дополнительно включены лишь система Н. А. Комарницко-
го и А. А. У ра н о в а, как дальнейшая разработка системы Н. А. Буша, и
система однодольных Л. А. Куприяновой, напечатанная в малораспрост-
раненном издании. Однако, по независящим от меня обстоятельствам, в част-
ности, из-за ограниченности объема книги, в „Обзор“-е не нашли отражения
труды ряда других ученых, как зарубежных, так и отечественных. Нап-
ример, в него не вошли работы Н. Гайдукова, Л. М. Кречетов и-
ча и И. С. В и н о г р а д о в а, библиографически, впрочем, советскому чи-
тателю более доступные. В частности, исследования Л. М. Кречето-
в и ч а, на мой взгляд, заслуживают самого пристального’ внимания, но
они до сего времени недостаточно оценены. Должен отметить и то,
что я условно ограничил выбор теми системами, которые опубликованы
по i960 год;
2) в „Обзор" включены также графические схемы большинства упо-
минаемых в нем систем, в общей сложности 45 схем. Из них 44 схемы поме-
щены в виде особого приложения в конце книги, а I (система А. Г е-
й н ц е) — включена в текстовую часть (в раздел конспектов). Почти все
они выполнены по первоисточникам; некоторые из них являются фотоко-
пиями оригиналов; это отмечено в подписи каждой соответствующей
схемы (между прочим, среди них имеется и схема системы А. Пу л ле,
хотя А. А. Гроссгейм касается ее лишь вскользь при сопоставлении
взглядов отдельных авторов); другие—представляют собою чертежные ко-
пии подлинников. Прямых указаний на то, что А. А. Гроссгейм пред-
полагал приложить к своему труду, в том или ином виде, схемы обсуж-
даемых им систем—в рукописи нет, если не говорить об одной его поме-
тке на поле рукописи: „привести схему системы Жеффри", желая, ви-
димо, иллюстрировать результаты сравнительно-анатомических исследований
последнего. Включить именно ее, к сожалению, на представилось воз-
можным;
З) . книга дополнена также конспектами (обзорами) филогенетичес-
ких систем отдельных авторов, являющимися перечнями высших таксонов
цветковых растений вплоть до семейств. Количество их пришлось сильно
ограничить- В частности, был изъят включенный вначале мною конспект
Обзор новейших систем цветковых растений
XI
системы Энглера; хотя последняя широко известна и ознакомление с
нею наиболее доступно, все же наличие ее цельного конспекта и графи-
ческой схемы в данном „Обзор“-е представлялось мне желательным, по-
скольку в нем, как правило, сопоставлены те или иные системы именно
с этой наиболее распространенной системой.
Приобщение же графических схем и конспектов вообще мне
представляется необходимым, тем более, что анализ систем в „Обзор“-е
изложен весьма кратко, а порою слишком лаконично; поэтому, эти
приложения, мне думается, послужат, в известной мере, компенсацией;
4) к „Обзор“-у приложена библиография, составленная полностью
мною (около боо названий из иностранной и около 200 из отечественной
литературы). Судя по немногим ссылкам в тексте А. А. Гроссгейм
намеревался снабдить свой труд списком только цитированных им источ-
ников (кстати, малочисленных). Мною же она составлена в более широ-
ком плане. Однако, она не является полной—-эта' цель и не преследова-
лась. Ее назначение—облегчить читателю общую ориентацию в богатой и,
при этом, тематически разнообразной литературе по филогении цветко-
вых растений; иными словами, она имеет лишь общесправочный характер.
В нее вошли публикации до 1963 г. Лишь немногие из позже вы-
шедших в свет трудов удалось включить в процессе напечатания „Об-
зор“-а;
5) наконеп, я счел целесообразным приложить труд А. А. Гросс-
гейм “-а „К вопросу о графическом изображении системы цветковых ра-
стений", в которой изложена его филогенетическая система, также ана-
лизируемая им, как отмечено выше, в данном „Обзор“-е. Она напечатана
была в 1945 г. в № з журнала „Советская ботаника", вышедшем весьма
малым тиражом и тогда же являвшемся библиографической редкостью.
Наряду с указанными выше дополнениями, местами, в написанном
А. А. Гроссгеймом тексте, мною сделаны вставки; в одних слу-
чаях они касаются содержания более поздних изданий упомяну-
тых им же систем, в других—отражают взгляды отдельных ученых по
тем или иным дискуссионным вопросам филогении цветковых растений,
опубликованные за последнее время в разных специальных изданиях. Все
эти вставки заключены в* и в конце отмечены инициалами (М. С-). В дан-
ном случае я стремился к тому, чтобы четко пазграничить тексты авто-
ров и правильно ориентировать читателя в этом отношении.
Полагаю, что даже в столь неполном и сжатом виде „Обзор новей-
ших систем цветковых растений" окажется полезным пособием не только
для студентов-биологов и молодых научных работников, в частности бо-
таников, но представит интерес и для специалистов, поскольку в нем за-
тронуты злободневные вопросы филогении пветковых растений. Хоте-
лось бы надеяться, что издаваемая книга возбудит и усилит интерес
молодых ботаников к филогении покрытосемянных растений, многие явле-
ния которой до сего, времени остаются загадочными. Перед филогенетика-
ми, вооруженными новейшими методами исследования, простирается
благодатное поле деятельности.
Быть может представится возможность переиздать „Обзор" в более
полном виде, в частности, дополнив его системами, не включенными по
различным причинам в данное издание.
XII
Предисловие Preface
Настоящее же издание будет оправдано, если оно в какой-то мере
окажется полезным для ботаников и этим мой труд/ по его подготовке
к печати, явится скромной лептой на алтарь светлой памяти моего учи-
теля, выдающегося ученого и исключительного человека А. А. Гросс-
гейма, творческая мысль которого проникла во многие отрасли бота-
ники.
В заключение считаю своим приятным долгом отметить с чувством
глубокой благодарности содействие отдельных лиц в подготовке к печати
настоящего издания. Я весьма признателен А. А. Уранову за сообще-
ние ценных сведений относительно разработанной им (вначале в соавтор-
стве с Н. А. Комар ницк им) филогенетической системы (in lift.). Ряд
сотрудников Тбилисского ботанического института Академии наук Гру-
зинской ССР оказал мне большую помощь. В частности, выражаю свою
признательность старшему библиотекарю А. В. Павкович, оказавшей
мне содействие в использовании иностранной литературы; ей же прина-
длежат переводы на английский язык. Неоценимую услугу оказал стар-,
ший научный сотрудник, кандидат биологических наук Б. Д. Гаври-
ленко, который со свойственными ему любовью к делу, самоотверженно-
стью и высоким сознанием ответственности, выполнил трудоемкую и слож-
ную работу по полной подготовке чертежей графических схем, требо-
вавшую от него и знания сущности их содержания’(табл. 7—12, 17, 22—
24, 26—27, 31—36, 42—44, а также условные обозначения на русском
языке к схеме 37 и нумерация таксонов на табл. 28).
Ценную помощь оказали кандидаты б/н Л. С. Хинтибидзе и 3. И.
Гвинианидзе и младшие научные сотрудницы и лаборантки: Е. Е.
Мироненко, Л; Я. Абашидзе, М. В. Гаганидзе, М. Ф. Да-
влианидзе, М. А. Иванишвили, Д. И. Мцхветадзе, Л. М. Но-
задзе, Л. Б. С е р д ю к о в а, Н. И. Т у л а ш в и л и и М. И. Эри ст а-
ви, научные сотрудницы Тбилисского государственного университета
Л. М. Алоева и Н. В. Челидзе.
С глубокой признательностью должен отметить любезность доцента
Тбилисского государственного университета Л. К. Гамсахурдия, про-
смотревшей текстовые части на английском языке.
В процессе подготовки и, в особеннности, напечатания книги я по-
стоянно пользовался технической помощью неизменно отзывчивого сотруд-
ника А. Н. Рухадзе.
С благодарностью хочу отметить весьма заботливое отношение ди-
рекции Тбилисского ботанического института к изданию настоящей
книги.
Из-за ограниченности объема книги, в нее не нключены предметный
и другие указатели.
М. Ф. Сахокиа
Тбилисский ботанический институт
XII. 1964 г.
Обзор новейших систем цветковых растений
XIII
Примечание I. По техническим обстоятельствам (главным
образом из-за неимения надстрочных и подстрочных знач-
ков) в приводимых ниже словах не могли быть напеча-
таны соответственные типографские знаки (буквы).
Стр. Строка В слове He мог быть напечатан знак
сверху снизу
177 18 Kobenhavn 0
197 15 dekkroete п
178 14 Celakovsky t.
180 15 bunecuych п
180 15 bunecuych ve
188 8 Nemejc Я
188 10 Nemejc п
189 4 rise
189 4 rocenky п
180 15 vyvoje У
180 15 pribuzenskychvztachu п
184 14 Hruby я
180 ' 15 pribuzenskychvztachu г
182 31 Caile а
182 33 coufatuirilor я
182 13 roslin S
185 27 swiata я
185 27 rosliunego я
19а 27 swietle я
182 13 Gumin ski п
185 27 pokrewienstwa п
182 24 Gusuleac S
182 26 Gusuleac я
182 29 Gusuleac я
182 31 Gusuleac я
182 32 Si
182 33 Bukuresti я
182 29 Consideratiuni t
182 29 drirectiei
182 29 evolutiei
182 32 evolutiei
184 26 utrudnijacych а
189 4 rise 1
XIV
Примечания. Сокращения. Notes. Abbreviations
Примечание 2. Как правило, я везде сохранил принятое тем
или другим цитируемым автором написание латинского названия так-
сонов, хотя разнобой в этом отношений резко бросается в глаза. Но, в
отдельных случаях, более общепринятое написание названия мною от-
мечено следующим образом: в круглые скобки заключены буквы, кото-
рые должны фигурировать в более принятом написании названия; на-
пр., Шаффнер пишет Thymeleales, вместо более распространенного
начертания Thymelaeales, поэтому оно мною дано в виде Tliymel(a)
eales (стр. 137); в квадратные скобки заключены буквы, которые
не должны быть употреблены в более правильно написанном наимено-
вании, напр., у Веттштейна написано Euptelaeaceae вместо Eupte-
leaceae, поэтому последнее название много изложено здесь так:
Euptel[a]eaceae. В менее спорных случаях я придерживался совре-
менного написания названий. Замеченные явные опечатки, допущен-
ные в тексте отдельных авторов, мною исправлены. (Ред.).
ПОЯСНЕНИЕ СОКРАЩЕНИЙ. ABBREVIATIONS.
ЕЖ — „Ботанический журнал". Всесоюзное ботаническое об- щество АН СССР.
БСЭ — Большая Советская Энциклопедия.
Бюлл. МОИП — Бюллетень Московского Общества Испытателей При- роды.
Докл. ТСХА — Доклады Тимирязевской Сел.-Хоз. Академии.
Л. — Ленинград.
м. С. — Инициалы имени и фамилии редактора (ред.).
м. — Москва.
'Сов. Бот. — Журнал „Советская Ботаника". Изд. АН СССР.
Полвека тому назад почти одновременно появились работы двух спе-
циалистов, резко восставших против системы цветковых растений Э н г л е-
ра и его взглядов на филогению покрытосемянных. Этими систематиками
были Ганс Гал лир в Голландии и Чарлз Эдвин Бесси в Север-
ной Америке. Шаг за шагом Гал лир разрабатывал свою систему в
многочисленных статьях, начавших появляться с 1893 года; в наиболее
законченном виде система Галлира была опубликована в 1912:
году. Первые наметки новой системы Б е с с и мы находим в его ранней
работе 1894 года, в более разработанном виде она опубликована в 1897
году и в наиболее полном виде в 1915 году—в год его смерти.
Что же существенно нового внесли Галлир и Бесси в вопрос
филогении покрытосемянных ?
Общим между ними является то, что оба они, в противополож-
ность Энглеру и Веттштейну, считают наиболее древней группой
покрытосемянных растений не Monochlamydeae, а lian ales и Ап о па-
les и, таким образом, переворачивают вверх дном укоренившиеся под
влиянием тяжелого авторитета Энглера и Веттштейна систематиче-
ские представления (см. табл. 44 и конспект системы Веттштейна).
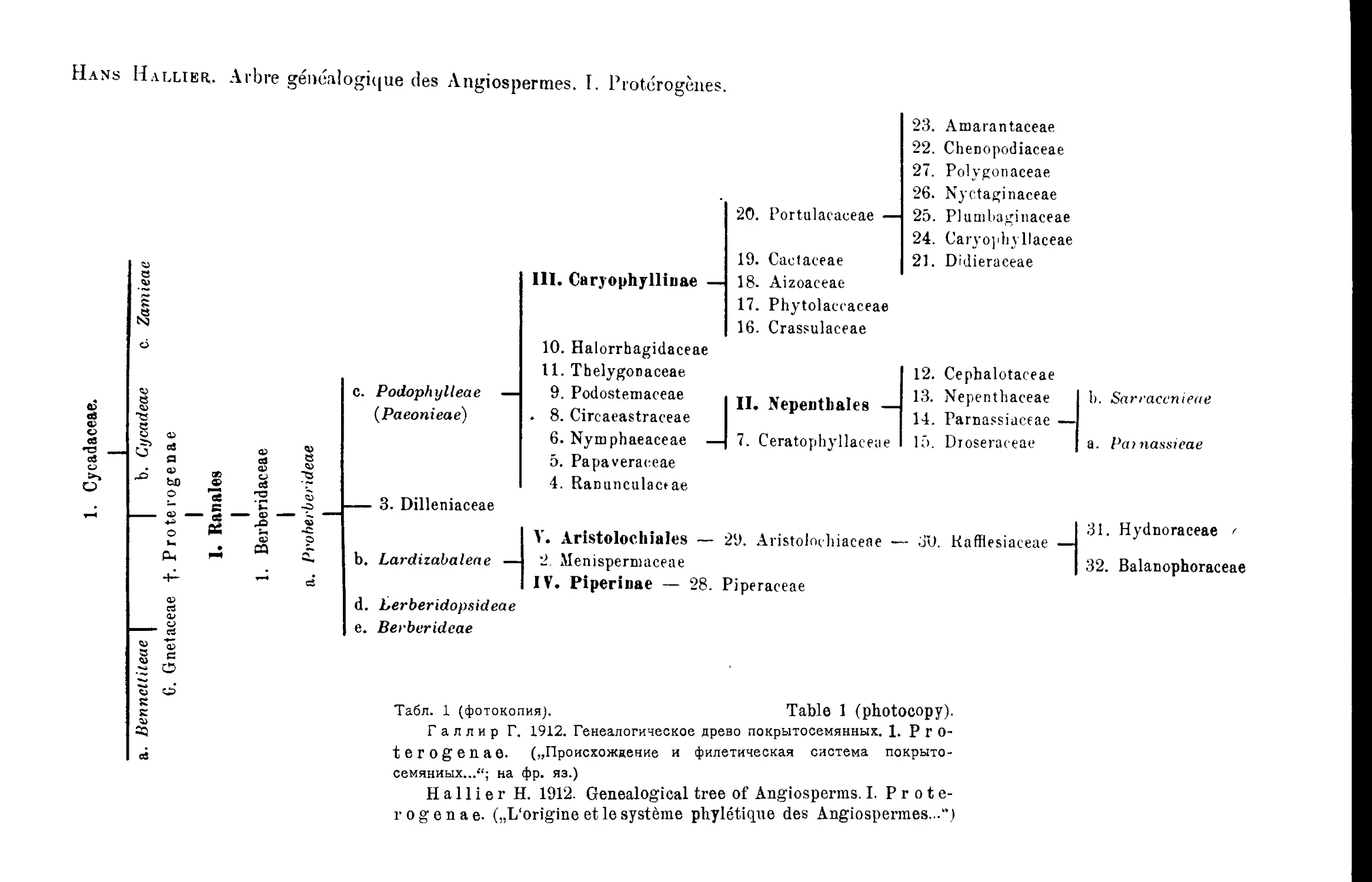
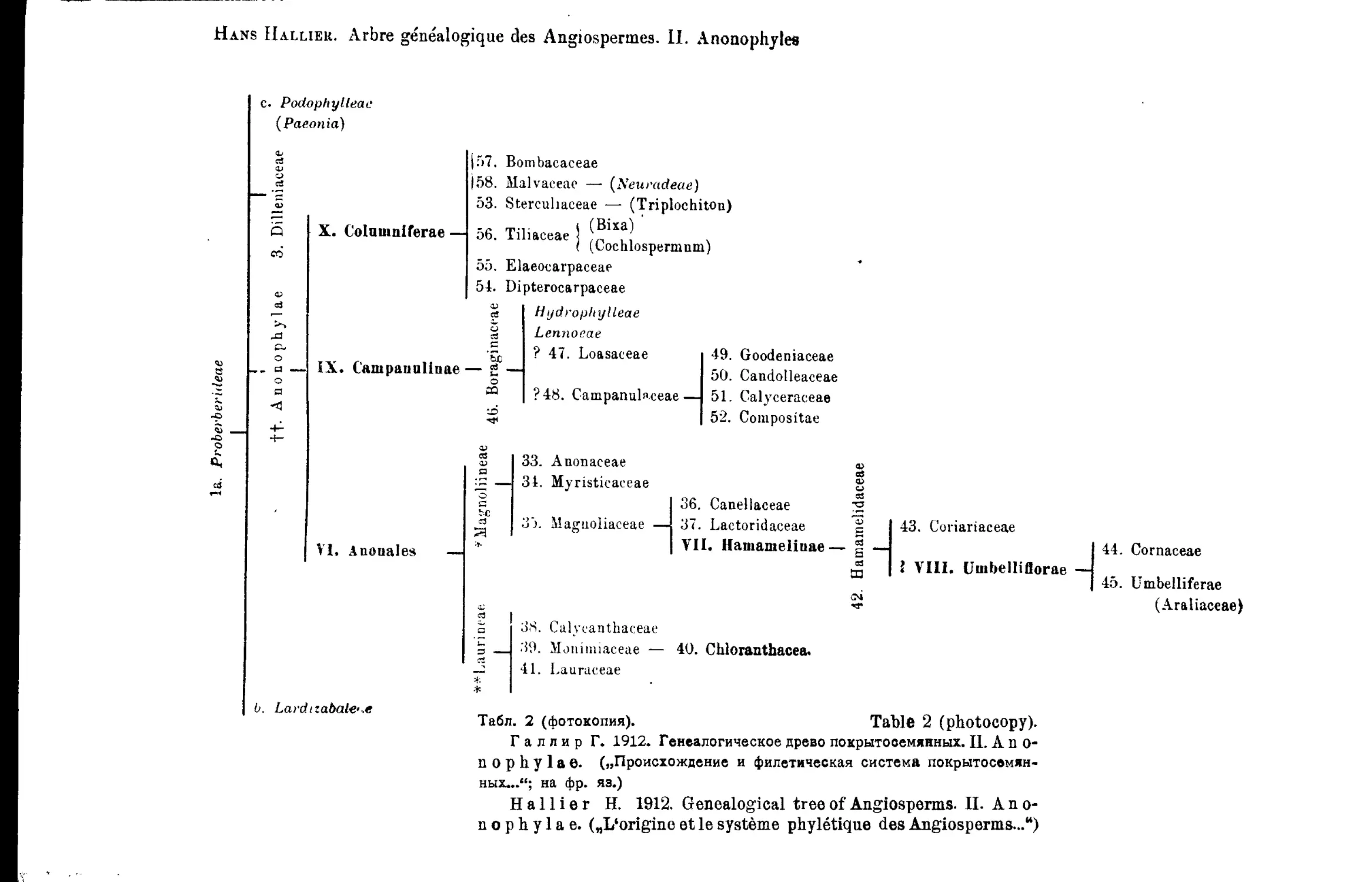
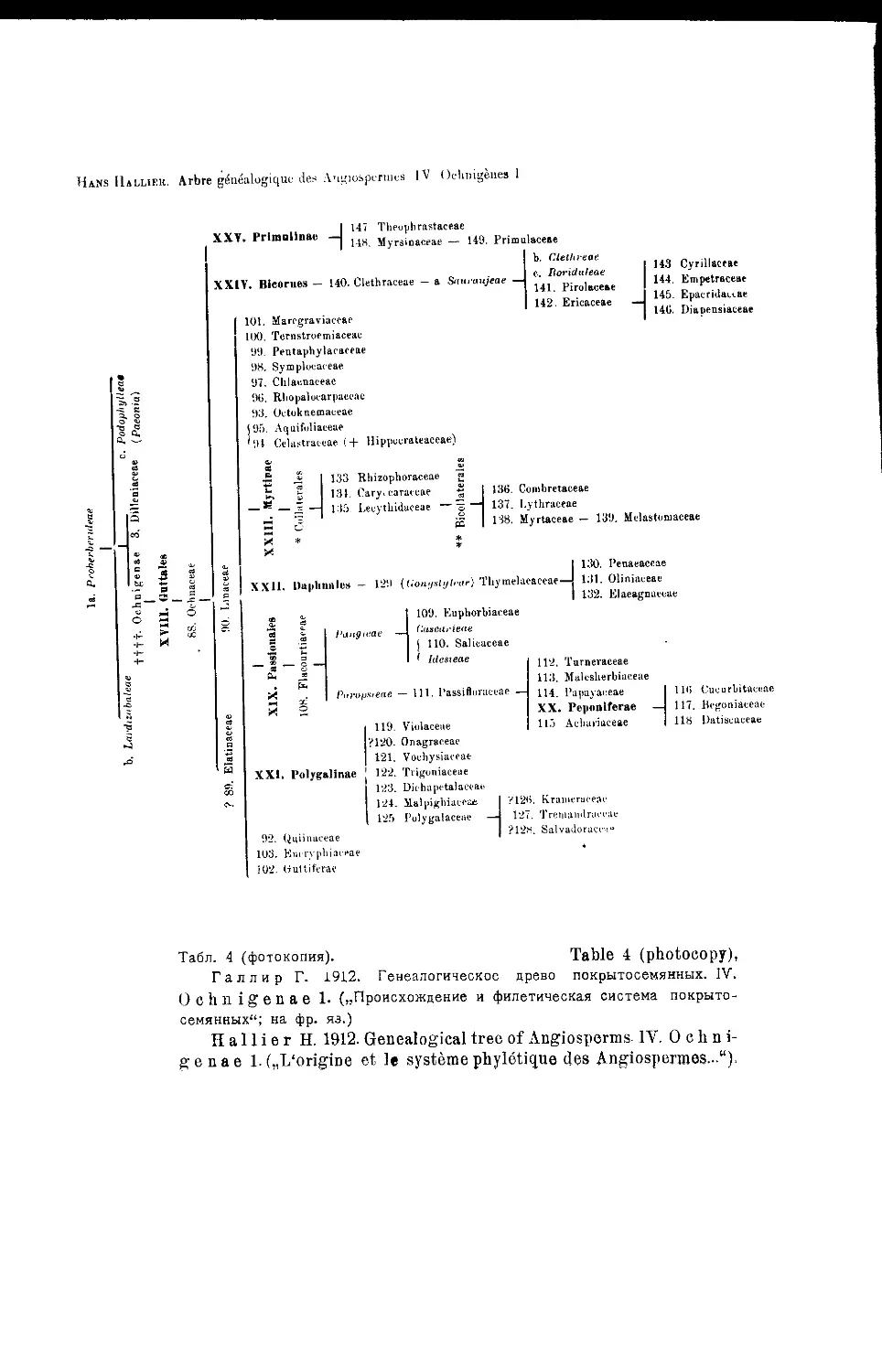
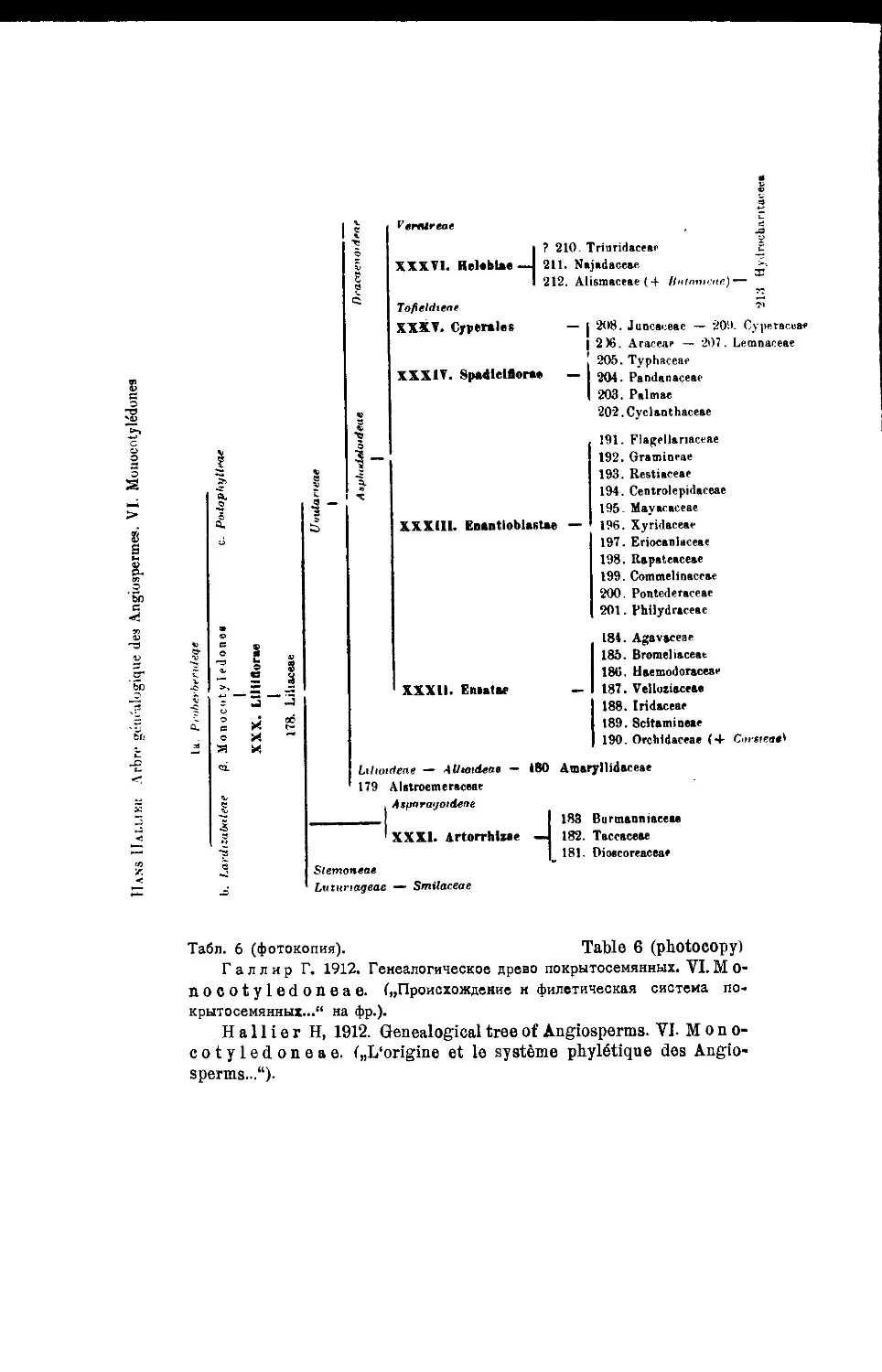
Система Г. Галлира
Н. Нallier’s System
1905; 1912-
Галл ир в 1905 году пишет по этому поводу следующее: „Главней-
шие особенности моей системы следующие: Ангиоспермы—естественная мо-
нофилетическая группа, а не полифилетическая, как это утверждает Эн-
глер... Amentaceae не рассматривается как древний тип, находящийся на
низкой стадии развития со связями или непосредственно происходящий
от Gymnospermae, но, наоборот, как высокий и сильно редуцирован-
ный тип от одной из линий двудольных.
Они и все другие линии развития двудольных развились вследствие
редукции в цветке и плоде от Р olycar picae, последняя же группа прои-
зошла непосредственно от Pennettitaceae или других вымерших Cycadales.
Таким же самым образом Liliiflorае и все другие синкарпные одно-
дольные произошли вследствие срастания плодолистиков, редукции в ко-
личестве частей, развития нижней завязи и других изменений структуры
цветка и плода от поликарпных однодольных (Нelobiae)', последняя же
группа непосредственно произошла от поликарпных двудольных (Р о I у с а-
1. А. А, Гроссгейм и М. Ф. Сахокиа
2 Система Галлира Hallier’s System
pieae и Ran ales)- Среди двудольных раздельнолепестные и сростно-
лепестные являются неестественными группами полифилетического происхо-
ждения".
В 1912 году в наиболее полной и детальной статье о своей системе
Га л лир несколько изменил в деталях эту свою точку зрения, но
сущность ее осталась той же. Как и прежде, он считает примитивной
группой Poly с ar pi с а е и детально мотивирует их примитивность в 32
положениях (каждый раз не забывая в качестве противопоставления под-
черкнуть вторичность однопокровных). Но, он считает теперь, что предка,
ми покрытосемянных являлись подобные низшим, сильно ветвящимся фор-
мам Cycadales (как, например, Wielandiella angustifolia Nath.), не остави-
вшие ископаемых остатков представители типа Berberidaceae, в форме низ-
ких кустов или небольших деревьев с толстыми ветвями с просто или
несколько раз перистыми листьями типа Anomozamites, имевшие вид
Decaisnea insignia, Paeonia Montan, Nandina domestica из Японии и Ma-
honia nepalensis, которые все по Галлиру принадлежат к сем. Berberi-
daceae-
В этой статье Г а л л и р уже не связывает непосредственное проис-
хождение покрытосемянных с беннеттитовыми.
В построенном подробном филогенетическом дереве здесь Г а л л и р
отличает 6 основных линий развития покрытосемянных: I—Proterogenae;
II—Anonophylae; III—Rhodophylae; IV-V—Ochnigenae (две линии) и VI—
Monoc6tyledones (табл, i—6 и 7)1.
Наиболее примитивную из этих линии он ведет от Cycadeae к порядку
R a nates, центральным семейством которого является Berberidaceae и его
примитивная группа a- Proberberideae, от которой и ответвляются
все остальные порядки и семейства этой первой линии. Беннеттитовые по
этой схеме каким-то образом связываются с Gnetaceae, a Zamieae вообще
не имеют потомков. Все остальные линии развития покрытосемянных, в
том числе и однодольные, берут начало в трибе Proberberideae-
Гал лир отказывается здесь от признания Н elobiae примитивной
группой и высказывается в пользу ее вторичного происхождения от при-
митивных Liliaceae’, апокарпию Alismataceae он склонен считать вторично
возникшей. Также отказывается Гал лир здесь и от своих первона-
чальных взглядов на узловое первичное положение сем. Saxifгадасеае',
сейчас он считает, что Saxifгадасеае произошли от Rosaceae-'
Однопокровные сосредоточены Галлиром в семействе Amentaceae
(разбитом, на 5 подсемейств: Quercineae, Myriceae, Coryleae, Casuarineae
л Betuleae), которое помещено в пределах порядка Terebinthinae, не-
посредственно происходящего от Proberberideae-
: В пределах порядка намечается также линия развития: Rutaceae—
Terebinthaceae—Amentaceae-
Всего, Га л лир отличает среди покрытосемянных 36 порядков и 213
семейств.
1 См. также схему его системы в книге: G- Н. М. L a w г е и с е—„Taxonomy
of vascular plants*, 1951 (стр. 132, f. 14). (Ред.).
Обзор новейших систем цветковых растений j
К сожалению, у Г а л л и р а нет графического изображения оконча-
тельно принятого им варианта филогенетического дерева и его построе-
ния, сделанные при помощи типографических средств (1912) очень сло-
жны и мало наглядны.
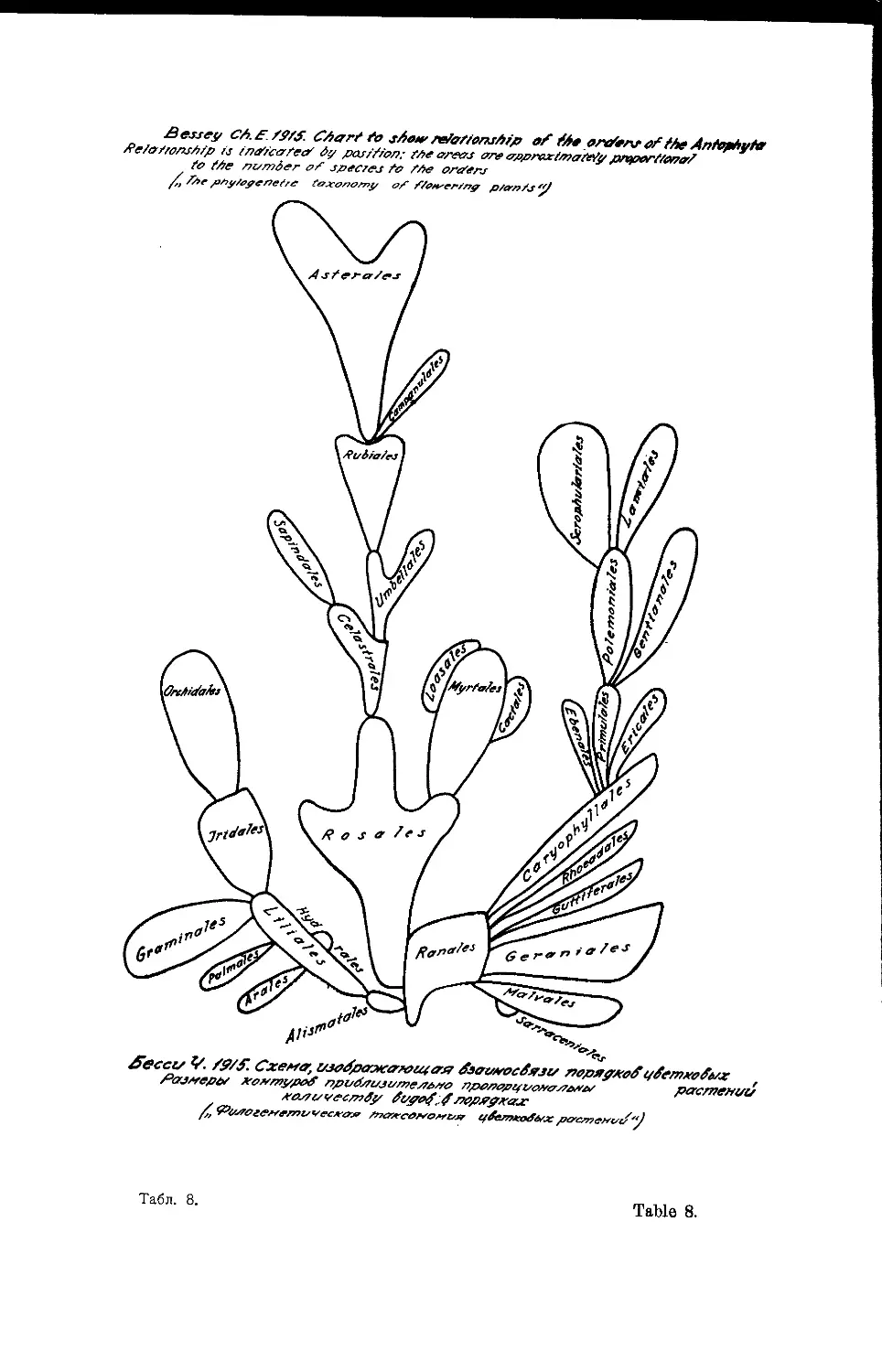
Система Ч- Э- Бесси
Ch. Е. Bessey’s System
1915
Одновременно с Галлиром в Северной Америке выступает с пред-
ложением новых принципов системы цветковых растений Чарлз Бесси.
Если Галлир в своих изысканиях и построениях шел совершенно ори-
гинальным путем, отталкиваясь от филогенетической системы Энглера-
Веттштейна, и резко противопоставлял им свои взгляды, то Бесси
шел иным путем. Его филогенетическая система есть по существу пере-
работка естественной (не филогенетической) системы Бентама и Гу-
кера. За основу своих построений им была взята система Бентама и
Гукера; в 1894 году он несколько видоизменил и улучшил эту систему,
в 1897 году влил в нее филогенетическое содержание, следствием чего
явилась уже новая оригинальная система Бесси, в 1915 году Бесси
придал своей системе наиболее законченный вид (табл. 8; см. также табл.
9 и ю и конспект)2, и дал исчерпывающее обоснование своих основных
принципов филогении цветковых растений (так наз. dicta).
Замечательно то, что и Галлир и Бесси, работая параллельно
над одним и тем же вопросом и не имея общения, пришли к одним и
тем же основным выводам. 28 dicta Бесси (1915) по существу общие с
соответствующими принципами Галлира.
Б е с с и также считает, что наиболее примитивными покрытосемян-
ными нужно признать порядки Ran ales и Al ism at ales- В своих
знаменитых dicta Бесси устанавливает 7 общих принципов эволюции
цветковых растений, 7 принципов эволюций общей структуры пветковых
растений и 14 принципов эволюции структуры цветка. Очень большое
значение Бесси придает положению завязи и основные линии его систе-
мы отражают это явление. Прежде всего цветковые делятся Бесси на
два крупных отдела, соответствующих однодольным и двудольным других
авторов. Первую группу, берущую начало от Alismatales, Бесси
предпочитает называть классом Alternifoliae, вторую группу, происход-
ящую от Ranales, Бесси называет классом Oppositifoliae, избегая ста-
рых названий Monocotyledoneae и Dicotyledoneae, как неясных и ведущих
к ошибкам. У Alternifoliae положение первых листьев (cotyledons) заро-
дыша очередное, у Oppositifoliae—супротивное.
В каждом из этих классов прослеживаются две основные параллель-
ные линии развития: гипогиничная и эпигиничная. У Oppositifoliae ги-
* См. также схему системы Бесси в обработке J. F. Cornman в книге:
G- Н. М- L a w'r е n с е—„Taxonomy of vascular plants"- 1951 (стр. 126, f. 13). (Род.)
4
Система Бесси Bessey’s System
погиничная линия берет начало от Пап ales и приводит к таким совер-
шенным типам как Lamiales, эпигиничная линия начинается среди
Rosales и приводит к Aster ales- У самого Бесси эпигиничная и
гипогиничная линии у Alternifoliae не так ясно обозначены, но на диаг-
рамме его системы, построенной его учеником Раймондом Пулем
(1941), эти две линии выдвигаются с полной ясностью; обе они берут на-
чало от Liliales, гипогиничная заканчивается G г а т in ale s, эпигини-
чная—Orchid ales (табл. ю).
В своих филогенетических построениях Бесси исходит из допуще-
ния стробилярного строения цветка покрытосемянных.
Предложенная Бесси в 1915 году схема филогенетического дерева,
покрытосемянных чрезвычайно наглядна, легко запоминается и стала в-
учебниках ботаники классической.
Таким образом, на рубеже XX века и в первые его десятилетия,,
двумя крупными систематиками, работавшими самостоятельно, без всякой
связи друг с другом, на разных концах мира, было положено начало но-
вым воззрениям на происхождение и развитие цветковых растений, рез-
ко отличающимся от воззрений школы Э н г л ер а-В е тт шт е й н а. Воз-
зрения Г а л л и р а-Б е с с и встретили резкую оппозицию со стороны ста-
рой школы, выражавшуюся, главным образом, в замалчивании и игнори-
ровании, на что неоднократно жалуется Гал лир, и первые десятилетия
нашего' века заполнены борьбой этих двух противоположных направле-
ний. Борьба эта закончилась полной победой и признанием большинст-
вом систематиков воззрения Галлира и Бесси.
Ганс Галлир, ученый-романтик, мятущийся, страстный исследова-
тель, постоянно ищущий, часто меняющий точки зрения, непрерывно пе-
чатающий все новые и новые работы с поправками, оговорками и зачастую
полным изменением того, что было им сказано раньше, с острой поле-
мической направленностью против энглеровской школы, оказал сильней-
шее влияние на европейских ученых.
Чарлз Бесси, спокойный глубокий'исследователь, ярко выражен-
ный тип ученого-классика, публикующий мало работ, но работ закон-
ченных, завершенных, в которых ему не приходится вступать в противо-
речие с самим собою, работ, ровного и спокойного тона, без каких бы то
ни было полемических выпадов, хотя каждая строка работ Бесси спло-
шная полемика против школы Энглера и Веттштейна. Влияние Бес"*
си особенно сказалось в англо-саксонских странах—в США и в Англии*
Продолжателями дела Галлира в Европе были Н. И. Кузне-
цов, Б. М. К о з о-П о л я н с к и й, Н. А. Буш, А. Л. Т акт а джяв,
А. А. Гроссгейм в России и Советском Союзе. Продолжателями дела
Бесси были в Англии Гатчинсон ив США ученик Бесси—Рай-
монд Пул. Но, прежде чем перейти к обзору этих систем, необходимо
остановиться на работах палеоботаников Арбера и Паркина, внесших
существенно новое в вопрос о происхождении цветковых растений. Основ-
ные положения Галлира и Бесси были уже высказаны и сформули-
рованы- когда в 1907 году на помощь им пришли палеоботаники.
Обзор новейших систем цветковых растений
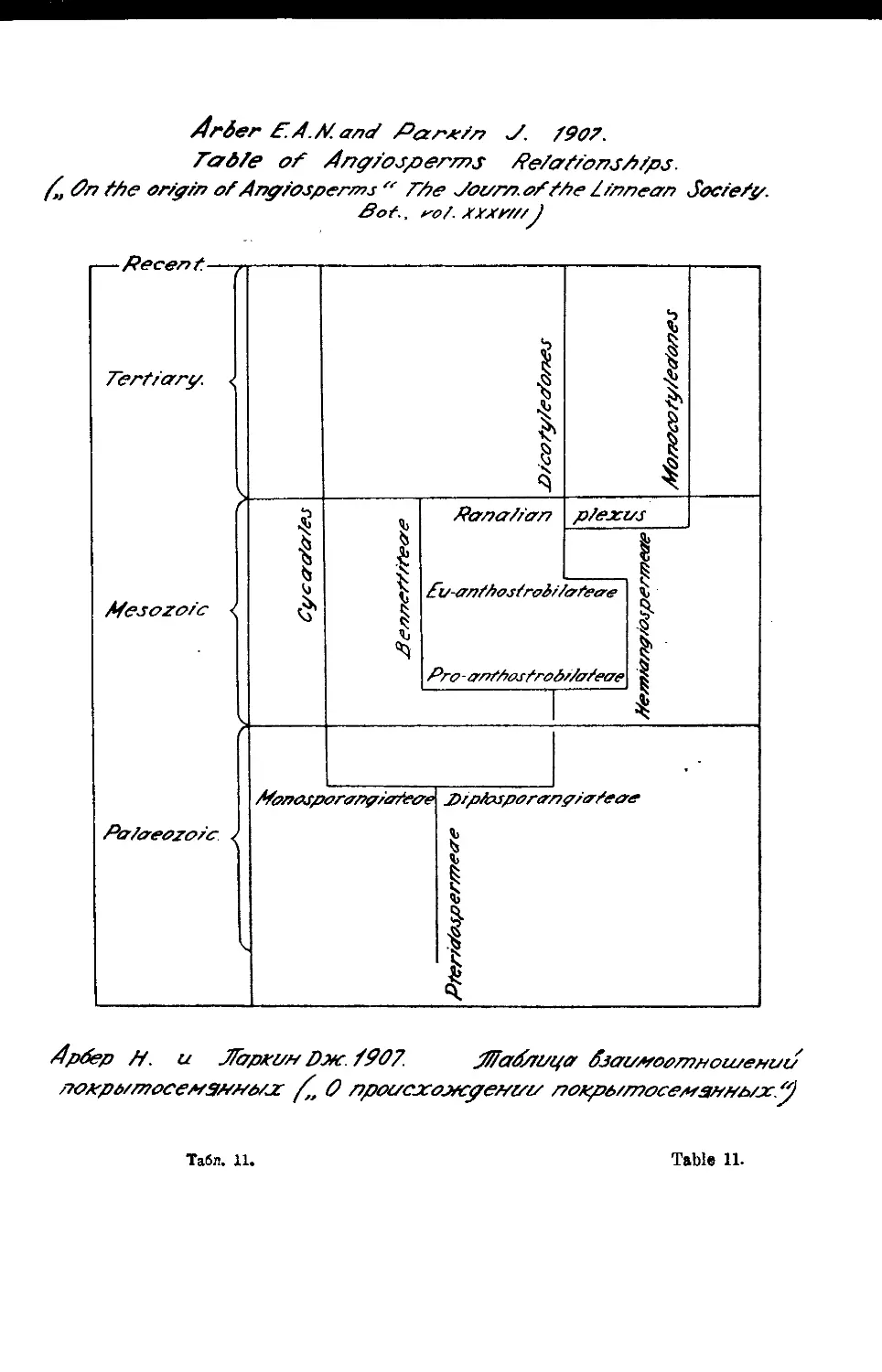
Система Н. Арбера и Дж. Паркина
И. Arber’s and J. Parkin’s System
1907
Мы подчеркиваем то обстоятельство, что и Галлир и Бесси к
своим воззрениям на происхождение цветковых растений пришли совер-
шенно самостоятельным путем до появления в свет стробилярной теории
Ньювелла Арбера и Джона Паркина. Стробилярная теория Ар-
бера и хПар к и на и сближение покрытосемянных с беннеттитовыми,
основанное на данных палеоботаники, прекрасно согласовалось и явилось
подтверждением идей Г ал лира и Бесси, в основном построенных на
данных не палеоботаники, а сравнительной морфологии и систематики.
Галлир принял сближение беннеттитовых с цветковыми, Бесси особен-
но подчеркивал стробилярный путь развития цветковых растений, но об-
щие воззрения обоих реформаторов систематики цветковых растений сло-
жились и развились независимо от воззрений Арбера и Паркина.
Тем не менее, работа и воззрения Арбера и Паркина явились
также определенным этапом в развитии учения о филогении цветковых рас-
тений, этапом, резко противопоставленным воззрениям Энглера и Вет-
тш т ей н а. .
Узловыми моментами учения Арбера и Паркина, как известно,
являются развитая ими стробилярная теория и сопоставление цветковых
растений с вымершими В е n n е t tit а 1 е s (табл. ц). Стробилярная теория,
зачатки которой можно найти и у предшествующих ботаников, развита
Арбером и Паркиным в стройное учение. Евантостробилу цветко-
вых растений эволюционно предшествовал проантостробил полупокрыто-
семянных растений, отличающийся от евантостробила незакрытыми, еще не
приспособленными для улавливания пыльцы макроспоролистиками. К типу
проантостробила относятся и Bennettitales. Стробилярным растениям
предшествовали растения нестробильного строения органов плодоношения.
Стробил явился с самого начала амфиспорангиатным органом. С уса da-
les рассматривается как боковая ветвь развития палеозойского периода
с моноспорангиатным плодоношением. Таким образом, среди Cycadales
нельзя найти предков покрытосемянных растений. Беннеттитовые рассма-
триваются также как боковая мезозойская ветвь стробилярных растений,
тесно связанная с гипотетической также проантостробилярной группой
Hernia в giospermae. Эта последняя группа и дала начало еванто-
стробилярным растениям—покрытосемянным. Евантостробилярия появилась
еще в Мезозое в форме раналиеподобных растений; уже в Мезозое из ра-
налиеподобных развились две основные ветви евантостробилярных расте-
ний—двудольные и однодольные, параллельным развитием которых и за-
полнено все третичное и послетретичное время.
Говоря о причинах подобного пути развития Арбер и Паркин
решающее значение придают развитию насекомоопыляемости. Ветроопыле-
ние по Арберуи Паркину—первобытное явление; переход к вне-
6 Система Арбера и Паркина Arbefs and Parkin’s System
шему типу—насекомоопылению, вызвал смыкание макроспорофилла и об-
разование на его базе улавливающего пыльцу аппарата в форме рыльца,,
столбика и т. д.
Теория Арбера и Паркина заострена против воззрений Энгле-
ра и нанесла им сильнейший удар. Стройно и логически построенная,
подкрепленная фактом существования вымершей гемиангиоспермной груп-
пы беннеттитовых, теория Арбера и Паркина произвела громадное
впечатление на ботаников-систематиков и отголоски ее влияния чувству-
ются во всех последующих филогенетических и классификационных по-
строениях.
Три обстоятельства поколебали авторитет стробилярной теории А р-
бера и Паркина в последующем. Во-первых, были обнаружены и изу-
чены новые палеоботанические материалы, которые также, как и бение-
ттитовые, могли послужить исходными точками для филогенетического
построения; особенное значение в этом смысле имело изучение группы
Psilophytales, меньшее значение—группы Cay toni. ale s-
Затем нужно отметить, что все же слишком много . гипотетического
приходится допускать, следуя Арберу и Паркину; гипотетичными и
никакими ископаемыми находками вплоть до настоящего времени неподг
твержденными являются представления о проантостробиле и о группе
Hemiangiospermae. Подверглась широкой критике интерпретация
эволюции саговниковых, развитая Арб ер ом и Парк иным. И, нако-
нец, совершенно ясно, что в вопросе о прогрессивности насекомоопыления
и примитивности и первобытности ветроопылеиия Арбером и Ларки-
ным допущено определенное одностороннее увлечение.
Тем не менее, разрушая логически представления Веттштейна о
псевдантиевом происхождении цветка и эволюции покрытосемянных от
ныне живущих боковых ветвей голосемянных и нанося тяжелый удар
по представлениям Энглера и Веттштейна, считавшими примитив-
ными покрытосемянными Monochlamydeae, теория Арбера и Парки-
н а сыграла крупнейшую положительную роль в развитии учения о фи-
логении и системе покрытосемянных и явилась существенным вкладом со
стороны палеоботаники в дело, начатое Галлиром и Бесси.
Система К. Меца
С. Metz’s System
192S
Карл М е ц (и его школа, т. н. кёнигсбергская школа) построил свою
систему на основе применения серодиагностического метода.
В двадцатых годах в г. Кёнигсберге и во многих других местах, глав-
ным образом в Германии, была проделана очень большая работа по при-
менению серодиагностического метода к различным группам, как низ-,
ших, так и высших растений. Одно время казалось, что найден точный
и объективный метод, дающий возможность безошибочно судить о род.-
Обзор новейших систем цветковых растений у
ственных отношениях различных групп растений. Впрочем, довольно скоро
выяснилось, что серодиагностический метод и неточен и допускает су-
бъективные истолкования и увлечение методом стало остывать. В насто-
ящее время его можно рассматривать только как некий подсобный и, при-
том, далеко не самый надежный из подсобных методов систематики. Воз-
можно, в дальнейшем, при прогрессе биохимического познания раститель-
ного тела и при очищении метода от влияния различных побочных про-
дуктов биохимической деятельности растений, серодиагностический метод
возродится опять и сослужит службу систематике.
И хотя на обл@жке органа Меца „Botanisches Archiv“ и был изо-
бражен сухой пень, с одним свежим побегом на нем, долженствующим
изобразить серодиагностическое направление в систематике растений, не-
обходимо констатировать, что к настоящему времени этот побег увял,
если не совсем засох, и омоложение пня идет за счет других, более силь-
ных и жизненных ветвей.
Система К. Меца и Г. Цигенспека
С- Metz’s and Н- Ziegenspeck’s System
1926
Что же дало применение серодиагностического метода? Медом
и Медом совместное Цигенспеком (1926) была предложена система
всего растительного мира, изданная в печати в форме „серодиагности-
ческого (кёнигсбергского) филогенетического древа“. В ряде печатных
высказываний М е ц и его последователи дали обоснование этой серодиа-
гностической системе (табл. 34).
Остановимся на некоторых положениях Меда, изложенных в Т925
году в форме 28 основных принципиальных тезисов, в той их части, ко-
торая касается цветковых растений.
Мед считает, что главный ствол растительного мира с Девона до
верхней Юры развивался по линии Psilotales Selaginellaceae-^ Pinaceae
-^Wielandiellaceae- Эта последняя группа принципиально отлична от
беннеттитовых (тезис 206).
Гетероспора возникала в растительном мире по, крайней мере, 5 раз:
Sphenophyllales, Calamariales, Selaginellaceae, период от евспорангиатных
папоротниковKPteridospermae и Hydropterides. Открытосемян-
норть (Gymnospermae) возникала, по крайней мере, 4 раза: Sigilia-
riospermae, Lepidospermae, Coniferales, Pteridospermae
(тезис 216).
Из (низшего) цветка Selaginellaceae—Pinaceae филогенетически воз-
ник высший цветок Wielandiellaceae—Magnoliaceae- Связь Ament ales
с Coniferales отвергается. Высшие, развившиеся вследствие диффе-
ренциации околоцветника цветка, возникали, по крайней мере, 3 раза;
Bennettitaceae, Wielandiellaceae—Magnoliaceae и Gnetaceae (тезис 24).
8
Системы Меца, Гатчинсона Systems Metz’s, Hutchinson’s
Однодольные представляют боковую ветвь развития двудольного
ствола, отщепившуюся между Magnoliaceae и Menis регтасеае (тезис гб).
Симпеталия полифилетична. Primulale s возникли от Centres pe-
rm ае, Eric ales от С olumni ferae, все же остальные, разделяющи-
еся на з очень неравных ряда (ЕЪ еп al es, С am panulatae, Contor-
tae—Tubiflorae—Rub tales), ведут свое происхождение от вершин
Parietales (тезис 27).
Мысль об неоднократном происхождении различных частностей стро-
ения цветка, резко подчеркнута Мецем, плодотворна и в дальнейшем
детально разрабатывалась систематиками. Но, основное понимание Меца
о происхождении цветковых непосредственно от Pinaceae, через некоторые
ископаемые формы из беннеттитовых, конечно, абсолютно неверно и
представляет определенный шаг назад по сравнению с тем уровнем, ка-
кого достигла систематика к двадцатым годам.
Оценивая в историческом аспекте серодиагностическое „древо“ цве-
тковых растений, нужно отметить, что в целом ряде частностей, т. е. в
вопросах о родственных отношениях различных семейств, Мец предло-
жил правильные и здравые решения, в дальнейшем принятые многими
систематиками.
Но, самое интересное, то, что в общем (отвлекшись от ошибочного
представления о Pinaceae, как исходной группы для цветковых) система
цветковых растений Меца почти полностью совпадает и подтверждает
положения Б е с с и-Г а л л и р а и является отрицанием взглядов Энг-
лера и Веттшейна. Примитивными и исходными типами цветковых
Мец считает Magnoliaceae и Ranunculaceae', Monochlamydeae у него явля-
ются результатом длительного и сложного пути развития.
Печатные высказывания самого М е ц а и его последователей полеми-
чески заострены против взглядов Энглера и Веттштейна и, таким
образом, в лице школы Меца, новая систематика получила определенное
подкрепление в своей борьбе с канонами Энглера и Веттштейна.
В этом определенное историческое значение серодиагностической школы.
Система Дж- Гатчинсона
I. Hutchinson’s System
1926; 1934; 1959 (i960).
1928; 1934. Система Гатчинсона была опубликована в два при-
ема: двудольные в 1926 году и однодольные в 19343 (табл. 31 и 33). По-
добно Галлиру и Бесси Гатчинсон формулирует основные морфо-
логические принципы, на которых основано построение системы. Таких
принципов всего Г атчинсон учитывает 24, распределивши их в 4 груп-
пы: общие принципы, по внешнему облику, по основному строению цвет-
ковых растений и по цветкам и плодам. Принципы Гатчинсона во
3 Перепечатанное издание вышло в 1944 г. (Ред.).
1^1
Обзор новейших систем цветковых растений
9
многом, совпадают с принципами Галлира и di°ta Бесси, что приводит
его также к признанию Ran ales и М а gnolial.es древнейшими исхо-
дными типами цветковых растений.
При этом Гатчинсон особенно подчеркивает признаки „по вне-
шнему облику11. Сюда относятся 4 принципа: i) деревья примитийнее
трав, 2) деревья и кустарники примитивнее лиан, 3) многолетники древ-
нее однолетников и 4) водяные растения более молодые, чем сухопутные,
Так же как эпифиты, сапрофиты и паразиты.
Излагая свои взгляды на филогению и построение системы Г атчин-
с о н, также как до него Бесси и Галли р, все время противопоставляет
свои взгляды взглядам Энглера и Прантля. Им составлена особая срав-
нительная таблица отличий своей системы от системы Энглера и Пра-
нтля (а также Бентама и Гукера), в которой особенно подчеркну-
ты 6 основных положений. Во-первых, подчеркнуто, что в противополо-
жность Энглеру примитивными рассматриваются растения с чашечкой
и венчиком’ (при наличии соответствующих других признаков), а расте-
ния без лепестков и чашелистиков являются более совершенными и моло-
дыми. Далее, представляется логическим выводом из теории, что части
цветка покрытосемянных являются видоизмененными листьями. Порядки
рассматриваются в узком объеме. Monochlamydeae распределены среди Poly-
petalae как редукционные формы. Monocotyledones помещены после Dico-
tyledones, от которых они произошли на ранних стадиях. Кроме того,
подчеркивается, что система автора в противоположность системе Энг-
лера, имеет большую поддержку со стороны палеоботаники.
Таковы те особенности системы, которые считал нужным выдвигать
на первый план сам автор. Рассматривая систему Гатчинсона в исто-
рическом аспекте, мы подчеркнем и выдвинем другую её особенность, так
как все указанные выше, не представляются оригинальными и уже были вы-
сказаны в работах Га л лира и Бесс и.
Характерным для системы Гатчинсона (по крайней мере, для си-
стемы двудольных) является внесение в систему общебиологических при-
знаков наравне с признаками строения цветка. Ведь иГаллир и Бес-
си, не говоря уже о школе Э н г л е р а-В е т тш т е й н а, строили систему ,<•
исключительно на основании строения цветка (и плода), совершенно иг-
норируя вегетативные и общебиологические признаки растений.
В этом отношении Гатчинсон сделал шаг вперед, подчеркнув
всвоих принципах в виде особой группы признаки „по внешнему виду",
но, внедряя эти признаки в свою систему двудольных, он сделал ложный
шаг, чем подверг свою систему справедливой жестокой критике. Дело в
том, что Гатчинсон применил в области биологии такой же односто-
ронний подход к классификации, какой раньше применяли часто, и все-
гда неудачно, при установлении групп растений по одному какому-либо
признаку цветка (например, свободнолепестные и спайнолепестные, одно-
дольные и двудольные в качестве признаков групп). Гатчинсон разде-
лил все двудольные на две основные ветви развития—на ветвь, в которой
то Система Гатчинсона Hutchinson’s System
преобладают древовидные растения (Arborescent Habit Predominant) и
ветвь, в которой преобладают травянистые растения (Herbaceous Habit
Predominant).
Обе эти основные ветви происходят от неизвестных ископа-
емых предков Archichlamydeae- Исходной группой для развития пер-
вой (древовидной) ветви является порядок HP agnoliales, для разви-
тия второй (травянистой) ветви—порядок R anales', от этого последнего
произошли однодольные. Гатчинсон, кроме того, резко отличает еще
две ступени (два уровня) развития. Высшей ступенью являются Metaehla-
mydeae, где сосредоточены как эпигиничные, так и гипогиничные поряд-
ки обеих ветвей. В первой же ступени отличаются группы Polypetalae
(два ряда—у древесных и травянистых) и Apetalae, куда заходят линии
развития и от древесной, и от травянистой ветви.
Построенная таким образом система двудольных Гатчинсона при-
обрела совершенно искусственный вид;’искусственность ее доказывается
хотя бы уже тем, что одинитотже порядок Um Ъ el lift огае Г атч и н-
сон помещает в системе два раза в двух местах—в Polypetalae в древе-
сной линии и в Polypetalae травянистой линии. Это одно уже достаточно
наглядно показывает, что отрыв древесных форм от травянистых, как двух
основных эволюционных линии развития всех двудольных растений, со-
вершенно искусственен. Помещение порядка Rosales, Leguminales,
Р assif lor ales, С иеиг Ъ itales, С actales и некоторых других
только в древесной линии развития не оправдывается фактами, так Как
и в Ros ales п в Leguminales, например, травянистые линии раз-
вития не менее мощны, чем деревянистые. Гатчинсон не решился по-
местить эти (и многие-другие) семейства подобно U mbelliflorae в
обеих частях своей системы одновременно, потому что это привело бы в
сущности к полному крушению всей системы.
К однодольным мерка древесного и травянистого габитуса не при-
менена. Здесь представляет интерес критическая переработка установив-
шихся представлении о порядках и объеме семейств. Гатчинсон раз-
бил, например, сем. Liliaceae на целый ряд семейств, даже перенес из
него род Allium в сем. Amaryllidaceae- Во многих случаях новшества,
введенные Гатчинео ном в систему однодольных вполне рациональны
и были одобрены многими последующими систематиками, но в целом си-
стема однодольных Гатчинсона вряд ли может быть приемлема. Отри-
цая значение гипогинии и эиигинии, как определяющих линии развития
цветка, Г атчинсон отходит от своих учителей Г а л л и р а и Бесси, и
приходит к ряду неправильных положений, как, скажем, с указанным им
выше перенесением Allium в сем. Amaryllidaceae.
В общем система Гатчинсона с отмеченными выше отклонения-
ми, является непосредственным дальнейшим развитием взглядов Г ал ли-
ра-Бесс и на филогению (см. конспект системы).
*Дж- Гатчинсон (J. Hutchinson) 1959 (1960). Во втором издании
своего труда, перепечатанный выпуск которого опубликован в i960 г., Га-
тчинсон внес несущественные изменения. Во-первых, он только резче
обособил группы Liguosae и Herbaeeae, а во-вторых, перераспределил
некоторые семейства сростнолепестных растений (табл. 12 и 32). (М. С.)*.
Ч, э, БЕССИ
СН. Е- BESSEY
1845 — 1915
Обзор новейших систем цветковых растений
II
Система А. Г е Й н ц е
A. Heintze’s System
1927
Август Гейнце в 1927 году предложил новую систему и выска-
зал новые взгляды на филогению цветковых растений. Он считает, что
первичными цветковыми должны быть признаны сапрофитные растения с
мелкими безбелковыми семенами и с гладкой сухой пыльцой. Г е й н ц е счита-
ет также, что признаками примитивных двудольных являются обдиплостемо-
ния, синкарпия, у однодольных также Septaldrtiseu и синкарпный гине-
цей. В соответствии с этими взглядами в начале системы (см. конспект)
двудольных Г е й н п е располагает подкласс Bicornes с порядком Eric ales*
состоящим из семейств Monotropaceae, Clethraceae, Diapensiaceae Erica-
ceae, Epacridaceae и Empetraceae- Далее, следующие подклассы в восхо-
дящем порядке: Sympetalae, Synsepalae, Seobiformes и Monadelphiae- Ряд
однодольных соответственно состоит из 3 подклассов: Burmanniaeformes,
Orchidiformes и Liliiformes. Кроме однодольных и двудольных Гейнце
отмечает еще третий, равноценный им, ряд Pseudodicotyledoneae, куда' от-
носятся порядки: 1) Nутphае ales с семействами: Nymphaeaceae (про-
исходит от Eutomaceae), Saururaceae, Piperaceae, и 2) ад noli ales
с-семействами: Trochodendraceae (происходит от Nymphaeaceae), .Magnoli-
aeeae, Lactoridaceae, Anonaceae, Dtyristicaceae, Canellaceae, Calycanthaceae,
Monimiaceae, Lauraceae и Hernandiaceae-
У казанные здесь и многие другие положения Гейнце весьма не-
основательны и приводятся без каких бы то ни было доказательств; пост-
роены они как бы по принципу „наоборот", т. е. перевернуты вверх дном
многие из положений, считающихся б. или м. общепризнанными (напри-
мер, первичность сапрофитов, синкарпии и т. д.).
Рассуждая о происхождении вообще С о г m о phy t а Гейнце очень
легко допускает существование гипотетических вымерщих форм и на ос-
новании этих, ничем не обоснованных допущений, делает выводы важного
принципиального значения. Так, он говорит, что стоит нам только пред"
положить только что вымершую переходную форму между Pomoideae и
Castanea, которую он называет Archicastanea, и мы легко получим такой
ряд развития: Pomoideae (30oxopHa)->Archicastanea(aHeMoxopHs)->Castanea.
(зоохория). Или другой пример: Гейнце строит ряд Butomus->Nuphar->
Trochodendron4, „нужно только вставить между Butomus и Nuphar выме-
рший род“ и тогда мы получаем следующий ряд развития: Butomus (Sep-
taldrusen)->Arehiunphar (Pollenbliiten)->Nuphar (Staminodialnektarien). Так-
же легко подходит Гейнце и к единицам высшего порядка; так, для связи
Оphioglossaceae с псилофитами, придумывает вымерший класс (или тип)'
Archiophioglossaceae- Далее, Cormophyta выводит из Еисасеае при по-
мощи гипотетического Protocormophyton, здесь же утверждается*
В схеме Гейнце ряд этот начинается родом Pleea из Liliaceae. (Ред.).
12
Системы Гейице, Шелленберга Systems Heintze’s, Shellenberg’s
происхождение спорангия из архегония, вследствие чего по Гейнце
„споры и яйцеклетки кормофитов гомологичные явления и обе гапло-
идны" и т. д. и т. д.
Взгляды Гейнце не были признаны ботаниками-систематиками и не
оказали никакого влияния на дальнейшее развитие филогенетической си-
стематики.
Система Г. Шелленберга
G. Shellenberg’s System
1928
Шелленберг не делает попытки построения своей системы, так
как он в общем согласен с системой кёнигсбергской школы, но дает де-
тальный анализ основных положений систематики покрытосемянных на
основе соавнительно-морфологических данных.
Но Шелленбергу три основных вопроса необходимо разрешить
в области учения о филогении покрытосемянных: i) вопрос о происхо-
ждении Angiospermae, 2) вопрос о взаимосвязях однодольных и дву-
дольных и 3) вопрос о том, какая из групп является первичной—Папа-
les или Monochlamydeae.
Шелленберг считает твердо установленным монофилетическое
происхождение покрытосемянных, так как такой сложный аппарат, как
зародышевый мешок покрытосемянных, никаким образом не мог произо-
йти двумя или несколькими путями. Шелленберг не считает возмож-
ным связывать происхождение AngiospermaenH с беннеттитовыми,
ни с кейгониевыми, так как и те и другие представляют боковые ветви
развития, не имеющие прямого отношения к покрытосемянным растениям.
Шелленберг напоминает, что в области зоологической палеонто-
логии прочно обосновалось положение о том, что первичным группам
животных, в том- числе и предкам зверей, свойственны небольшие разме-
ры тела. Такое же положение следует признать и для растений. По-
этому, Шелленберг' полагает, что предками Angiospermae должны
были быть небольшие растения, ископаемые остатки которых имеют еще
меньше шансов на сохранение, чем обычные формы. Поэтому, мы до сих
пор их и не знаем. Он считает, что искать этих ископаемых предков ну-
жно р Юре, если не в Дейпере или еще ниже.
На вопрос о том, какие группы примитивнее, однодольные или дву-
дольные, Шелленберг отвечает в пользу большей примитивности одно-
дольных; к такому заключению он приходит в результате детальных сра-
внительно-морфологических сопоставлений.
Далее Шелленберг приходит к выводу о несомненной примитивно-
сти Пап ales и вторичности Amentiflorae, что доказывается опять-таки
сравнительно-морфологическими данными.
В заключение Шелленберг указывает на то, что явления редук-
ции у покрытосемянных, как прогрессивный процесс эволюции, должны
' - Обзор новейших систем цветковых растений ХЗ
быть гораздо более учитываемы систематиками, чем это до сих пор дела-
лось.
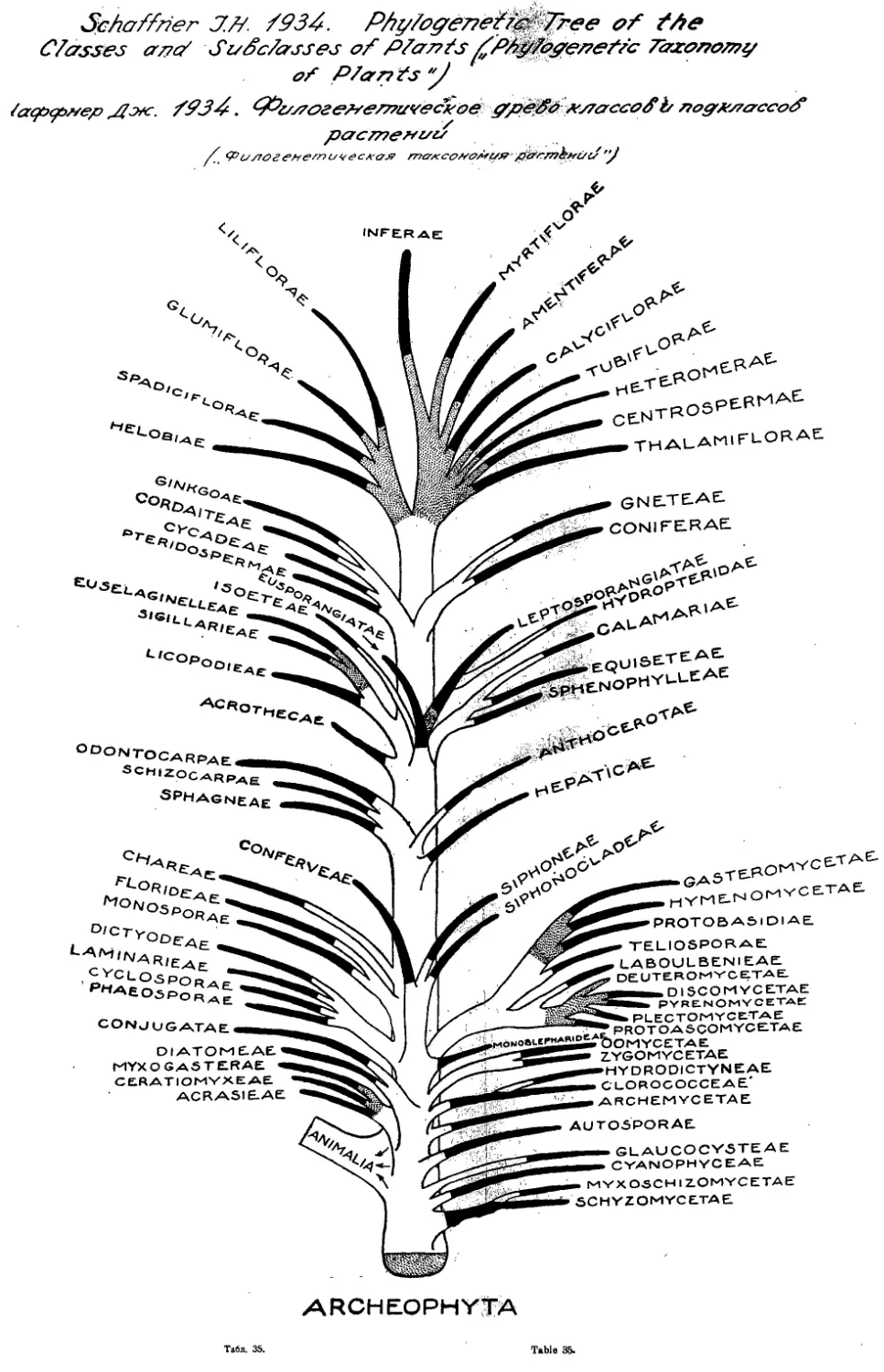
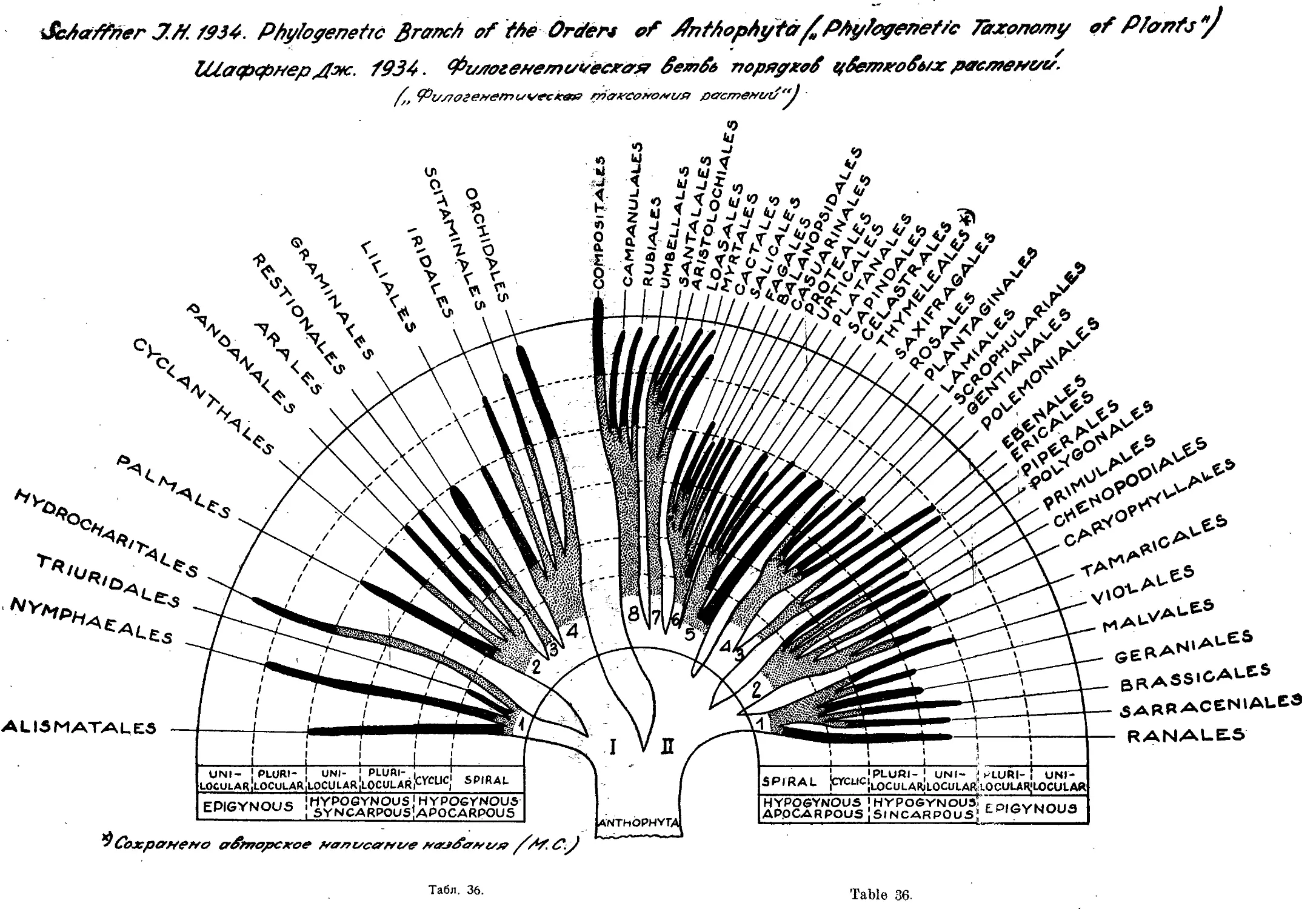
Система Дж. Шаффнера
J. Shaffner’s System
1934
Как последующее развитие идей Бесси представляют также инте-
рес воззрения и система американца Джона Шаффнера5. В течение
ряда лет, начиная с 1905 года, Шаффнер опубликовал целый ряд ста-
тей, посвященных филогении и системе растений, в том числе и покры-
тосемянных. В наиболее законченном виде взгляды Шаффнера были
опубликованы в 1934 году (табл. 35 и 36 и конспект системы).
Шаффнер считает, что.система Бесси открыла новую эру в си-
стематике и только вскользь упоминает о Галлире, указывая, что си-
стема Га л лира имеет много общего с системой Бесси. Такое игнори-
рование творческой роли Галлира в развитии учения о системе вообще
характерно для американских и английских авторов.
Принципы и dicta Шаффнера (всего 31) имеют существенно дру-
гое содержание, чем dicta Бесси, Гатчинсона и правила Галлира-
Это не столько морфологические принципы о первичных и вторичных
признаках, сколько правила, касающиеся построения системы. Интересно
отметить, что Шаффнер придает большое систематическое значение
изучению явления рекапитуляции и в то же время подчеркивает, что
данные генетики, а также явления полиплоидии и неправильности в стро-
ении хромозомного аппарата, имеют малое значение для целей системати-
ки. Здесь же находим заведомо неправильное утверждение о том, что
однодольные филогенетически ниже двудольных и предшествуют им.
Шаффнер устанавливает ю прогрессивных стадий развития расти-
тельного мира; десятой из этих стадий являются покрытосемянные (A n gi-
ospermae). Указывается, что Angiospermae происходят не от ныне
живущих Gymnospermae, а имеют тот же корень происхождения, что
и Gymnospermae — ,от евспорангиатных равноспоровых папоротников-
Подчеркивается, что BennettitalesHe могут рассматриваться как предки
Angiospermae. Также нельзя производить Angiospermae от Са-
ytoniaceae, которые сами представляют большой класс или подкласс прими-
тивных Angiospermae. У Angiospermae закрытые макроспороли-
стики (плодолистики) и новый орган—рыльце. Далее, прослеживаются ос-
новные линии прогрессивной эволюции покрытосемянных от примитивных,
стробилоподобных, апокарпных форм до синкарпных форм с одногнездной
завязью и развитием эпигинии. Подчеркивается, что протопласт Angi-
ospermae обладает громадным комплексом фундаментальных возмож-
ностей (potentialities), так что возможности изменения протоплазмы ста-
• Правильнее: Шаффнер. (Ред.).
14
Система Шаффнера Schaffner’s System
новятся практически необозримыми. Phylum Anthophyta разделен
Шаффнером на два класса: Monocotylae и Dicotylae; причем Monoco-
tylae предшествуют Dicotylae. Среди Monocotylae отмечаются 4 подкласса:
Helobiae, Spadiciflorae, Glnmiflorae u Liliiflorae; среди Dicotylae 8 под-
классов: Thalamiflorae, Centrospermae, Heteromerae, Tubiflorae, Calycif-
lorae, Amentiferae, Myrtiflorae и Inferae. Эти подклассы изображены в
виде конечных ветвей на приложенном филогенетическом дереве всего
растительного мира (табл. 35).
Детализируя филогенетическое дерево цветковых растений до поря-
дков, Шаффнер располагает эти порядки в виде отростков от общего
подклассового корня, достигающих той или иной степени развития в
пределах шести последовательных ступеней или стадий прогрессивной
эволюции. Ступени эти следующие: I) гипогиничный апокарпный со спи-
ральным расположением цветок, 2) гипогиничный апокарпный цикличес-
кий цветок, з) гипогиничный синкарпный с многогнездной завязью цве-
ток, 4) гипогиничный синкарпный с одногнездной завязью пветок, 5)
эпигиничный цветок с многогнездной завязью и 6) эпигнничный цветок
с одногнездной завязью. Очень наглядно все эти отношения показаны на
схеме филогенетического дерева, изображенного в виде куста на фоне
шести кругов, отражающих указанные шесть последовательных ступеней
развития цветковых; ветви, изображающие отдельные порядки, нарисова-
ны разной толщины, соответственно количественному значению их среди
цветковых растений, длина же их доводится до того из шести кругов,
до какой из шести перечисленных выше стадий развития достигает эво-
люция данного порядка. Только один из порядков (а именно С от р о si-
tales} выходит далее за пределы шестого круга (шестой ступени разви-
тия), немногие [12] порядки достигают совершенства шестой ступени,
очень мало [6] порядков достигают пятой ступени и громадное большин-
ство—четвертой (гипогиничные синкарпные цветки с одногнездной завя-
зью). Порядок N ут pH а е ales поставлен Шаффнером среди одно-
дольных.
В своей системе Шаффнер продолжает и разрабатывает взгляды
Б е с с и, придавая, как видно, вслед за Бесси, очень большое значение
положению в цветке завязи. Ценным, в системе Шаффнера является
то, что основные группы (подклассы) цветковых устанавливаются им по
комплексу признаков, как и намеченные им ступени эволюции цветка,
построенные на основании ряда основных признаков цветка, а не на ос-
новании какого-либо одного признака, как это принято у многих систе-
матиков. Но, все же и система Шаффнера, несомненно, страдает одно-
сторонностью, так как кроме строения цветка не приняты во внимание
никакие другие признаки. Кроме того, совершенно ложен, конечно,
взгляд на однодольные, как на группу, предшествующую двудольным,* да и
само разделение цветковых на классы одно-и двудольных. Способ графи-
ческого изображения системы, принятый Шаффнером, очень удобен и
нагляден.
Обзор новейших систем цветковых растении 15
Несмотря на все свои достоинства, большого влияния на системати-
ков взгляды Шаффнера не оказали и непосредственных продолжате-
лей он не имеет.
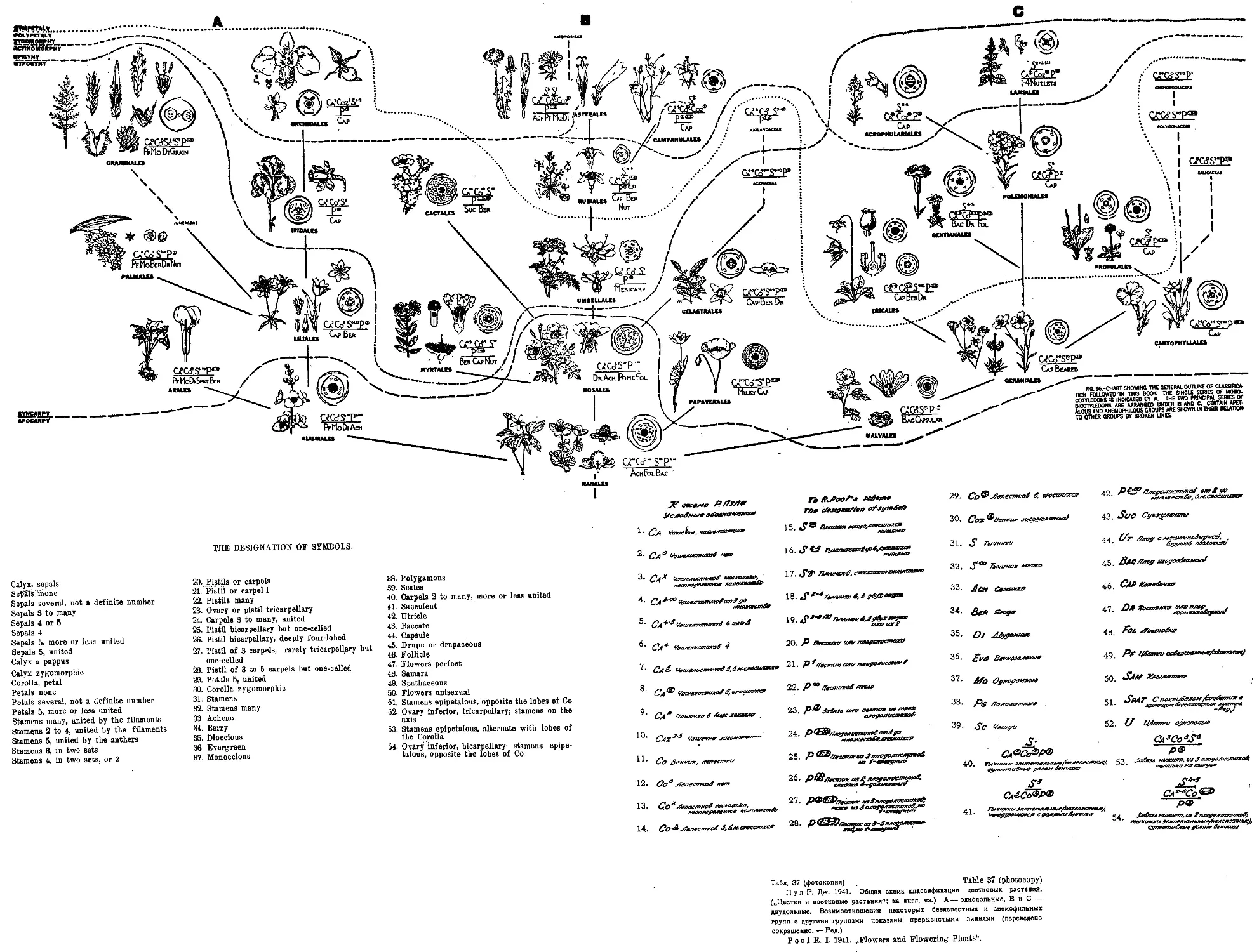
Система Р. Пула
R. Pool’s System
1941
Как признает сам автор, предлагаемая им система является дальней-
шим развитием и усовершенствованием системы Бесси, учеником и по-
следователем которого Р. Пул и является. В основу своей системы Пул
кладет те же принципы, которыми руководствовался и Бесси (табл. 37;
м, также табл. 9 и ю).
Он считает, что группа, от которой произошли цветковые растения
на сегодняшний день представляют собой загадку („...is still very much
f an enigma“; стр. 148). Рассматривая вопрос о Bennettitales Пул
указывает на то, что было время, когда эта ископаемая группа, казалось,
заполняла пропасть между голосемянными и цветковыми растениями. Но, в
настоящее время можно только говорить о том, что вымершие предки
цветковых растений были возможно несколько похожи на Bennetti-
tales.
Пул придает большое значение серодиагностическому анализу, под-
черкивая, что этот метод твердо установил, что цветковые растения про-
изошли от предков, подобных ныне живущим Coniferales-
Пул признает, что наиболее примитивными из ныне живущих ра-
стений являются древесные Magnoliaceae и травянистые Ran ales в пол
ном согласии с Бесси.
Пул считает, что двудольные являются первым появившимся типом
цветковых растений и что однодольные произошли от примитивных Ra-
nales в качестве последующего ответвления.
Далее, Пул излагает свои основные принципы (dicta), подобные
dicta Б е с с и. Они разделены на две группы: вегетативные признаки и
признаки строения цветка.
В первом отделе за примитивные типы считаются растения с колла-
теральным пучком, древесные породы и лианы примитивнее травянистых
растений; простые и неветвистые стебли примитивнее разветвленных; про-
стые листья примитивны, сложные вторичны; мутовчатое и супротивное
расположение вторичны; вечнозеленые листья примитивнее опадающих;
листья с сетчатой нерватурой примитивнее дугонервных листьев. Много-
летники примитивнее двулетников и однолетников.
Во втором отделе примитивными признаками считаются: одиночные
цветки, спиральное расположение частей цветка, неопределенное число
частей цветка; лепестные растения примитивнее безлепестных; свободно*
лепестные примитивнее сростнолепестных, актиноморфные примитивнее
зигоморфных; верхняя завязь примитивнее нижней; апокарпия примитивнее
Система Пула Pool’s System
синкарпии; обоеполые цветки примитивнее однополых; цветки со многими
свободными тычинками примитивнее цветков с немногими сросшимися тй>
чинками; однодольные растения примитивнее двудольных; многосемянные
плоды примитивнее односемянных; семена белковые примитивнее семян
безбелковых; семена с маленьким зародышем примитивнее семян с хорошо
развитым зародышем.
Как видим, эти dicta действительно повторяют ошибочные положе-
ния Бесси и Гатчинсона, нового автор ничего не добавляет; как и
у Бесси, некоторые из этих принципов являются спорными, как,напри-
мер, принятие за первичную форму листа цветковых растений простого
листа, в то время, как есть много оснований думать, что первичным ти
пом листа цветковых растений был лист перистый; многочисленность и
мелкость семян в некоторых случаях может быть не только примитивным,
но и вторичным специализированным явлением (Orobanche и др.).
Далее Пул останавливается на критике основных положений Эн-
глера и Веттштейна, особенно подчеркивая ошибочность бифилети-
ческого взгляда Энглера на происхождение цветковых растений ина
положение групп Apetalae и Amentiflorae в начале системы. Защищая по-
ложение о вторичности Apetalae и Amentiflorae (П у л намеренно избегает
термина Monochlamydeae) автор даже впадает в другую крайность и ут-
верждает,. что энтомофилия вообще является более примитивным призна-
ком и, что анемофилия развилась также на базе энтомофилии. Нет ни-
какого сомнения в том, что анемофилия была первым по времени спосо-
бом опыления цветковых растений и, что энтомофилия появилась позже;
но, с появлением энтомофилии анемофилия не переставала эволюциони-
ровать дальше и к настоящему времени параллельно с энтомофилией дала
вторичные, высоко специализированные типы цветковых растений.
Основным доводом в пользу вторичности Amentiflorae по Пулу
является тот факт, что плодолистики у них всегда сросшиеся и никогда
не наблюдается апокарпия.
Далее Пул ставит в вину Энглеру то, что он обращал мало вни-
мания такому важнейшему признаку, как эпигиния. Этот признак был
выдвинут и поднят на должную высоту в системах Бесси, Гатчинсо-
на и Пула.
Далее подчеркивается, что система Бесси в Америке и системы
Галлира и Гатчинсона в Европе выдержали экзамен времени и на-
шли достаточные подтверждения в ископаемых остатках и в серодиагно-
стическом методе.
Графическое изображение системы цветковых растений у П у л а от-1
личается большой наглядностью, так как на схеме для каждого порядка
(указаны только узловые порядки) дается изображение типичного расте-
ния, диаграммы и формулы цветков. Система, поэтому, получается очень
наглядной и легко обозреваемой.
Но, и в этой системе отсутствует представление об уровнях развития
(как, впрочем, и у Бесси), что снижает ее значение, Правда, на схемё
проведено четыре основных линий, разделяющих каждый раз растения
Обзор новейших систем цветковых растений
17
более примитивные и более вторичные типы, а именно линии, отделяющие
апокарпию от синкарпии, далее линии, отделяющие гипогинию от эпиги-
нии, актиноморфию от зигоморфии и, наконец, полипеталию от симпета-
лии. Но, эти четыре уровня, резко выделенные на схеме Пула, являю-
тся уровнями отдельных признаков, а не уровнем общего развития того
или иного признака; другими словами, на схеме даны только материалы
для построения уроввей, общие же выводы из этого материала не сде-
ланы. ,
Пул в кратком описании своей схемы подчеркивает следующие по-
ложения. Во-первых, установленные на схеме три линии развития цвет-
ковых растений: однодольные и две серии двудольных. Слева на схеме
под литером А изображен^ развитие группы однодольных через Alis-
т ale s от Ran ales, справа под литером С ветвь двудольных, также
начинающаяся от Ranales, идущая через Malvales и Geraniales,
и. приводящая к высшим типам Scrophulariales и Lamiales. Глав-
ная, как ее называет II у л, ветвь (литер В) двудольных занимает на схе-
ме центральное положение, и, отходя также от Ra па les, через Rosales,
в конце концов приводит к таким вторичным типам, как Campanula-
les и Asterales-
Из всего сказанного мы видим, что система Пула не представляет
какого-либо .нового шага в учении о филогении и системе цветковых ра-
стений. Она является несколько видоизмененной системой Бесси; глав-
ное ее достоинство заключается в большой наглядности и легкой обоз-
римости. Главной заслугой Пула является то, что он сумел популяризо-
вать идей Бесси, изложить их в книге, написанной легким, доступным
массовому читателю языком. Книга Пула построена в форме учебного по-
собия; она содержит кроме системы и классификации цветковых расте-
ний также главы о морфологии и анатомии растений, по истории си-
стемы (глава XXVI „От Линнея до Энглера и Бесси"), а также специаль-
ную главу, посвященную гербаризации.
Неудивительно, что она выдержала несколько изданий и широко ра-
спространилась среди ботаников и любителей. Не будучи сам новатором,
Пул сумел хорошо пропагандировать идей новой, послеэнглеровской си-
стематики.
Система Г. Лама
Н. Lam’s System
1948; 1950
Г. Лам рассматривает вопрос таксономии Cormophyta, в частно-
сти покрытосемянных, в ряде своих работ (A new system of the Cormophy-
ta, 1948; Stachyospory and phyllospory as factors in the natural system of the
Cormophyta, 1950 и др.). В первой работе он опубликовал систему Cormo-
phyta в целом, подвергнув известные до него таксономические груп-
пы, по его же выражению, небольшим, но существенным изменениям. В
построении своей системы Лам исходит из понятия о стахиоспории и
филлоспории. Идею о стахиоспории и филлоспории (а также основу
2. А. А. Гроссгейм н М. Ф. Сахокна
< l8 Система Лама Lam’s System
Терминов), как отмечает сам Л а м, он заимствовал у выдающегося ин-
дийского ботаника В. Sahni, выделившего в 1921 г. среди ролосем*н-
яаых две группы: Stachyo sperm а е и Phyllospermae, в .зависи-
мости от того, на какой части растения образуются семена: у растений
группы Staehyospermae они возникают на осевой части и к ней от-
носятся Ginkffoales а С onif er ales, у Phyllospermae семена
развиваются на листьях, как у Pteridospermae и Cycadopiyta.
Как утверждает Лам, это явление свойственно всей группе Сог-
mophyta, а не только Gymnospermae. Поэтому, он предложил по-
нятия о стахиоспории и филлоспории и различал стахиоспоровые
и филлоспоровые растения. Между последними он видит более резкую
разницу, чем между однодольными и двудольными. В их различии наи-
большее значение имеет положение макроспорангия, чем микроспорангия.
Во-вторых, в установлении стахиоспории и филлоспории Лам счи-
тает наиболее существенным характер плацентации: у стахиоспоровых она
вначале центральная и семяпочки обычно ортотропны; у филлоспоровых
она маргинальная, а семяпочки часто анатропны. Таким образом, Лам, при-
верженец телломной теории Циммермана, под стахиоспорией подразу-
мевает явление возникновения спорангиев (изо-микро-или мегаспорангиев)
на оси, обычно, в верхушечной части фертильных телломов; система ве-
твей, окончания которых представляют эти телломы, вначале была дихо-
томией (изотомной), во, впоследствии, в результате эволюционного
развития, трансформировалась в моноподиальную или симподиальную си-
стему. Следы древней дихотомии обнаруживаются преимущественно в ты-
чинках. Хотя у последних спорангии и не развиваются на листьях, однако,
листья могут выполнять в отношении спорангия защитную функцию в
виде стерильных органов—стегофиллов. У стахиоспоровых эволюция на-
правлена на сокращение числа бесплодных телломов.
У филлоспоровых растений спорангий возникает на листе—эвспоро-
филле. Черты древней дихотомии в этом случае сказываются в жилко-
вании. Защита репродуктивного органа (спорангия) осуществляется раз-
личными листовыми образованиями (обверткой, купулой и пр.). У фил-
лоспоровых—склонность к увеличению числа бесплодных телломов.- В- при-
лагаемой схеме (табл. 13) дана система Cormophyta по Ламу (в правой
ее части) с сопоставлением с системой Веттштейна (помещенной в ле-
вой ее части).
Как явствует из данной схемы к стахиоспоровым автор относит сле-
дующие группы (буква S—обозначает стахиоспорию,. Р—филлослорию):
Bryopsida (название группы предложено Лам ом и она включает: а.
Hepaticae и b. Musci);Psilopsida (содержащая: a. Nematopbyta-
les?; b. Psilophytales и c. Psilotales); Lycopsida (включавшая: a. Pse-
udolycopsida; b. Lepidophytales и Lepidospermales; c. Pleuromeiales; d. Ly-
copsidales; e. Selaginellales и f. Isoetales); Spheno psida (объединяющая:
a. Protosphenopsida; b- Pseudoborniales; c. Sphenophyllales; d. Cheirost-
robales; e. Tristachyales и f. Equisetales); только 9 Bennettital.es;
Обзор новейших систем цветковых растений
С on if е Г о ps i d а—название группы также введено Ламоми вообще
она объединяет: a. Ginkgoales; b. Cordaitales и с. Coniferales.
Но стахиоспоровыми не являются растения с вторичной филлоспо-
рией в кладоспорофиллах (Podocarpus, Pinaceae, Araucariaeeae} и <? хвой-
ные; к стахиоспоровым принадлежат также и Prota ngiospermae,
включающие: a. Chlamydospermales (Gnetales) й b. Verticillatae (Casna-
rina).
Филлоспоровыми автор считает: Pteropsida, исключая Caytoni-
ales? (этот таксон вообще включает: a. Filicales; b. Hydropteridales; с.
Noeggerathiales; d. Caytoniales и е. Pteridospermales); Cycadopsida
(наименование для группы предложено также Ланом): таксономический
состав Cy cadopsida по Ламу таков; a. Cycadales; b- Nilssoriiales и с.
Bennettitales; в данном случае йз нее исключаются ? Bennettitales, отне-
сенные, как указано выше, к стахиоспоровым); Coniferae (без Та-
xns). Лам делает оговорку, что филлоспория предпочтительно развива-
ется у пола, примерами чего по его системе служат Bennettitales, Co-
niferae и Cordaitales?
С эволюционной точки зрения Лам учитывает также характер ра-
звития листового органа, в связи с чем в группах Ц. Palae.ocormop-
h yta и III.-М е s о с о гm о р hy t а он различает подгруппы с микрофил-
лией (Р. microphylia и М- microphylla) и подгруппы с макрофиллией или
мегафиллией (Р. macrophylla и М. macrophylla). Микрофиллия сопря-
жена, по Ламу, с стахиоспорией, тогда как макрофиллия—особенность
филлос поровых.
Что же касается A n giosретшае, то эта группа, как очевидно из
схемы, совмещает оба признака—стахиаспорию и филлоспорию. Поэтому,
Лам отличает стахиоспоровые и филлоспоровые покрытосемянные; харак-
тер покрытосемянности у них различный.
Для морфологической диагностики стахиоспоровых покрытосемян-
ных Лам выдвигает такие признаки: i) спорангий у обоих полов разви-
«ается на главной оси; 2) мегаспорангий защищен бесплодными листовы-
ми образованиями—стегофиллами (ложными плодолистиками—псегдоспо-
рОфиллами); ?) плацентация, главным образом, центральная (колончатая)
или базальная (иная плацентация может быть вторичной); 4) семяпочки
чаще ортотропные; 5) завязь преимущественно верхняя; 6) обдиплостемо-
ния; тычиночные нити нередко раздвоены вильчато и в этом Лам усмат-
ривает признак древней дихотомии; 7) цветки первично однополые, редко
двуполые вторично; 8) периант первично простой (редко двойной) или
вовсе не развит, с циклическим расположением долей; 9) жилкование
листа без следов дихотомии. Он приводит и некоторые другие признаки.
В цитируемых работах Лам не дает полной филогенетической сис-
темы цветковых, доведенной до отдельных порядков, ограничиваясь выде-
лением групп по признакам стахиоспории и филлоспории.
Так, среди стахиоспоровых цветковых Лам намечает три группы (в
понимании порядков придерживаясь системы Веттштейна): I. группа од-
нодольных с порядками Н el оЪ гае, S padici florae и Рa ndanales-
20
Система Лама
Lam’s System
Травянистые Н elobiae он связывает родством с последующими группа-
ми; 2. группа травянистых двудольных, включающая Polygonales и
Centrospermae, а также Plumbaginales, Р rimulales и, пред-
положительно, С onv olv ulale s и Ebenales', 3. группа, главным об-
разом, деревянистых двудольных—одноиокровных; в общем в нее влкюче-
ны порядки: F ад ales, Myricales, Salic ales, J и gland ales,
U rticales, Piperales и Euphor biales.
Морфологические особенности филлоспоровых покрытосемянных, в
отличие от стахиоспоровых, Лам усматривает в следующих признаках:
г) мегаспорангий скрыт в настоящих плодолистиках (евспорофиллах, ев-
карпеллах), т. е., образована типичная ангиоспермия; 2) плацентация
маргинальная или почти маргинальная; 3) семяпочки чаще анатроп-
ные; 4) завязь обычно нижняя; 5) тычинки чередующиеся с лепе-
стками и их нити без следов древней дихотомии; 6) цветок рассма-
тривается как специализированный побег; в связи с этим части его рас-
положены первично ациклически, вторично б. м. циклично; он первично
двуполый, раздельнополовость может быть вторичной; 7) древнедихото-
мические признаки сохранились в вегетативных частях—в жилковании
листа.
К филлоспоровым Лам относит; г) из однодольных—Lili flor ае
и их производные, например, Enant io bl ast а е, Су per ales, Glu-
miflorae, S cit amine a e и Gynandrae", 2) из двудольных—P oly car-
pi cae и их производные—Rhoeadales, E 0 sales, условно G utti fe-
ral es, C ontort a e и T ub iflor ае; предположительно P a r i e t a I e s, Ilu-
biales, C ucurbitales и Campanulales и, с большим сомнением,
P rot eales, Sant al ales и H am a m eli d a les. Следовательно, фи ллос-
поровые составляют большую часть цветковых. Важно отметить здесь же, что
стахиоспорию и филлоспорию Лам рассматривает как признаки „древнего
филогенетического раздвоения". Этим он склоняется к бифилетическому
происхождению цветковых. Вместе с тем, с точки зрения эволюционного
совершенства, филлоспорию Лам ставит на более высокую ступень, чем
стахиоспорию. Таким образом, Monochlamydeae, в системе Лама, как в
системах Энглера и Веттштейна, оказываются древней группой—
положение, противоречащее более общепринятому на сегодня взгляду о
вторичности ее „архаических признаков". Из других частных вопро-
сов в разбираемой системе нужно заметить вопрос о родстве Н elob iae,
с одной стороны, и Centrospermae и Р oly д onale s,—с другой.
Напр., Лам энергично настаивает на родстве между Najas и Polygona-
сеае по сходству завязи; по трехмерному цветку Н elobiae сближает с
Рolyдonales', даже по сходству раструба Polygonaceae с лигулой Ро-
tamogeton он устанавливает родство этих двух таксонов. Приводимые им
аргументы мало убедительны для решения вопроса о родстве упомянутых
таксонов. Вызывают возражения и некоторые другие связи. Филогенети-
ческие взаимосвязи между стахиоспоровыми и филлоспоровыми цветко-
выми показаны на „филогенетико-типологической схеме" Лама (табл. 14).
Обзор новейщих систем цветковых растений 2i
Само отнесение растений тех или иных таксонов к группе стахио-
споровых или филлоспоровых связано с большими условностями или
просто с затруднениями. К примеру говоря, для этой пели привлекаю-
тся весьма спорные в смысле архаичности морфологические признаки,
вроде раздвоенности или склонности к ней тычиночных нитей. Не боль-
ше ли основания считать эти признаки вторичными (хотя бы у Lenina,
пожалуй и у Betulaceae, Е и phorb iales и др.).?
Как одну из особенностей системы Лама следует отметить, что он
выключает из покрытосемянных порядок Verticillatae (0 asuari-
nales}, поместив его в Protangiospermae наряду с Chlamydos-
permales (Gnetinae); даже в отношении рода Exocarpus он склоня-
ется к аналогичному решению. Включая Casnarina, наряду с Chlamy-
dos per males, в Protangiospermae, Лам категорически настаи-
вает на филогенетическом родстве Casnarina с родом Ephedra, с другой
стороны, Лам устанавливает филогенетическую связь Verticillatae
с Monochlamydeae, считая обе группы стахиоспоровыми.
По-видимому, в какой-то степени малой точностью критерия объя-
сняется то, что немало семейств оказалось в неопределенном положении
в смысле их принадлежности к группе стахиоспоровых или -филлоспо-
ровых.
Сам Л а м в заключении своей второй работы признает нечеткость
расчленения на стахиоспоровые и филлоспоровые в некоторых группах, и,
что это обстоятельство ограничивает применение его концепции для целей
классификации. Добавлю от себя, что понятия о стахиоспории и фил-
лоспории безусловно заслуживают внимания и под углом зрения этих пре-
дставлений и в дальнейшем нужно вести сравнительно-морфогенетичес-
кий анализ цветковых растений. Исследования Лама в этой области не-
сомненно имеют большую ценность. Немало в его высказываниях оригиналь-
ного. Но, вместе с тем, фактический материал, используемый для этой
концепции, не дает прочного основания для создания истинно филоге-
нетической системы. Система же Лама, к сожалению, не лишена в из-
вестной мере искусственности и немало в ней неясного. С терминологи-
ческой точки зрения система Лама обращает на себя внимание тем, что-
ее автор отказался от таких общепринятых наименований, как Gymno-
spermae, Spermathophyta и Anthophyta, поскольку он находит,
что группы, которым они присвоены, неестественны. Это мнение Лама
не лишено основания. Касаясь времени происхождения покрытосемян-
ных Л а м относит его, по крайней мере, к средней Юре, склоняясь к то-
му, чтобы отодвинуть это время ближе к верхнему Девону (см. геохроно-
логическую систему Cormophyta на табл. 15). Сторонников более ран-
него происхождения Angiospermae, чем общепринятое время—средний
Мел—постепенно становится больше, но принимаемая Лам ом датировка
.слишком гипотетична.
ц Систем# Шоо Soo’s System
Система Р. Шоо
R. Soo’s System
’953
Предковой грудцой современных цветковых растений Шоо также
считает Ро1уамр picas, происшедшие из.Рr o.angiospe.fr® ае. (Табл-
16; см. также конспект). При этом, развитие их по мнению Шоо дол-
жно было осуществляться по шести отдельным эволюционным линиям,,
четыре из которых завершаются порядками Sjmpetalae. В этом заключа-
ется одна из особенностей системы Шоо. Каждую линию образуют, как
подчеркивает автор» группы растений, достигшие идентичную степень эво-
люции. Первая, лини» приводит к узловому порядку Rosales и она
предположительно связана через Н amamelidales и, Т,егеЪiп tho-
les С Monochjamydeae. Наиболее древней боковой ветвью этой линии
является порядок Aristolochiales, образующий самостоятельную ли-
нию (хотя и застывшую) в системах ряда авторов; более непосредственно
связан с Poly car picae один из примитивных порядков—Piperales-
Вторая линия ведет через HPalv ales к Geraniales и далее к
венцу линий—порядку спайнолепестных—Perso п at ае, боковая линия
Т rieoccae предположительно связывает ее опять с Monochiamydeae.
Узловым порядком четвертой линии развития признан С entr о s рег-
тае- Кац вполне справедливо отмечает автор разбираемой системы, эта
линия, включающая шесть групп, весьма сложна по Составу звеньев и,
безусловно, полифилетического происхождения. В частности, наиболее
сложен вопрос о происхождении Monochlanndeae. Последнюю группу
Шоо рассматривает как регрессивную производную группу от древес-
ных Polycar picae* Редукционная линия (стрелки, обращенные
книзу) по Шоо проходит от 0 entrospermae к U rii cales (сем.
Ulmaceae), а затем к Fa gales] в то же время Шоо намечает и другой,
путь редукпии: через Hamamelidales к Fagales. Так или иначе,
Шоо выводит безлепестные древесные из примитивных древесных Pol#*
car picae (ИР ад noli ales). В этом вопросе он расходитсяс Бушем и
Гроссгеймом. Но, как и в системе Буш, эта же линия в схеме Шоо
завершена прогрессивными порядками спайнолепестных— Р rimulales и
Plumbaginal es-
Пятую эволюционную линию, исходящую также непосредственно из
Poly car pic ае, составляют однодольные. Она приводит через Н elo-
biae, с одной стороны к L i I i г f I or ае и, далее, к редукционным поряд-
кам растений с упрощенным строением цветка— С у per a les и Glumi-
florae, а с другой, к таким более развитым зигоморфным порядкам,
как Gynandrae.
Наконец, шестая филогенетическая линия, включающая fi р a dici-
florae и заканчивающаяся порядком Р an dan ales, лишь предполо-
жительно связана непосредственно с Polycarpicae- Шоо сторонник
монофилетического, но полигонного происхождения покрытосемянных. В
вопросе же происхождения цветка он категорически придерживается стро-
билярной теории.
К сожалению, в конспекте („Таблица' III*) таксонов и графичес-
кой филогенетической схеме автором допущены досадные пропуски и
несоответствия (см. примечание в конспекте).
Обзор новейшикеистеицтетковыхраетений 2j
С истец а однодольных И. Кимуры
. Y. Kimura’s System of Monocotyledons
1^0
. К»м у p a рассматривает только систему однодольных, придерживаясь
•».see?«©строении в основном систе4ня«Гатч»ж<о>н а, ио вносит в. иее<
ряд измекевий, как он отмечает, в соответствии с концепциями ряда та*
ксономистов(„Systeme et phylogenie des monocetylddones").
В основу таксономического деления однодольных Кимура кладет
ряд морфологических признаков. Из них важнейшими он считает следу?
юздие 4 признака: i) апокарпию и синкарнию; 2) положение завязи (вер-
хняя или нижняя) и характер плацентации (осевая или париэтальная);
париэтальную плацентацию автор рассматривает как производную от более
примитивной осевой; з) характер околовветника (его дифференцирован-
ность на чашечку или венчик или полное неразвитие), двойной около-
цветник признан более древним, чем не дифференцированный и 4); природу?
белка, т. е. эндосперма; автор предполагает, что крахмалистый тип при-
митивнее мучнистого.
Помимо этих „решающих" признаков автор учитывает и второсте-
пенные (,подтверждающие") признаки, как, например: характер стебля,
в частности степень одревеснения, некоторые морфологические черты ли-
ста, строение интегументов семяпочек и др.
Исходя из анализа однодольных по этим морфологическим призна-
кам Кимура предлагает различать в них 2 группы, 7 порядков и 28
классов, объединяющих 64 семейства (см. табл. 17 и конспект). Обе
группы—I Ароеаграе и II Sy near рае,—как это явствует из йх названий,
автор устанавливает по признакам апокарпии и синкарпии. Первая груп-
па, наиболее примитивная, представлена лишь одним классом—(А.) Н е-
lobiae- Вторая группа подразделена на подгруппы: (11—i) Subsyncar-
рае и (II—2) Соепосаг рае- У представителей Subsynear рае гине-
цей, по пояснению автора, как правило, синкарпный, но часто „модифивиро-
ванно-апокарпный"; под последним автор подразумевает гинецей апокар-
пный в одной части или сращенный слегка (Как у р. Gloriosa из лиле-
йных) или же б. м. разделенный вдоль нектарных швов (как у Alliwsr
из того же семейства, у Hypbaene из пальмовых, тогда как у многих ро-
дов пальм гинецей синкарпный). Подгруппа Subsyncarpae объеди-
няет 2 класса: (В.) Lili florae и (С-) S padici f lovae- В эволюцион-
ной линии она занимает промежуточное положение между примитивны-
ми Н elobiae и наиболее совершенной подгруппой С о eno саг рае-
Остальные 4 класса однодольных составляют подгруппу Соепосаграе-
(D-) Nudiflorae, (Е-) Siccifloraei (Е-) Calyciferae и (&•)
Е pigynae-
Свою систему Кимура иллюстрирует двумя схемами. Первая схема
(табл. 17 в этой книге и fig. I на стр. 140 в подлиннике)—главная, и,
представляет собою филогенетическое древо с показом взаимоотношений-
24
Система Кимуры Kimura’s System
между порядками. Кругами обозначены современные растения, а точками
в центре их—предки данных порядков, по выражению автора „proto-—
растение" („protoplante"). Филогенетически линии связыватОт именно точ-
ки, т. е. „proto—растения", под которыми автор подразумевает также и
и вымершие растения, если даже они и неизвестны в ископаемом состо-
янии. Этим автор подчеркивает, на наш взгляд, то правильное положе-
ние, что не современные растения лежат в основе происхождения тех или
иных таксонов, принадлежащих ныне к другим порядкам, а именно пред-
ковые формы следует считать исходными в их эволюпии. В этом смысле
черные точки, по выражению автора, являются предупреждением от та-
кого неправильного понимания. Например, в этой схеме в круге A.lis-
matales (круг и порядок № i) черная точка обозначает Р г oto-Ali-
sm at ales, точно также в круге Ну dr о ch а г it ales (круг и порядок
№ 2) черная точка соответствует Prot о-Н ydrocharitales, а связываю-
щая обе точки линия указывает, что Р г о t о-Al ism a ta I es явились исход-
ными в возникновении Prot о-Hydro ch а г it ales; связующих линий
может быть несколько в случае, если от „proto—растения" происходят
несколько различных порядков. Само собою разумеется, что от Proto-
Alismatales и Pro to-Hy dr о с ha rit al es произошли современные
представители этих порядков.
> Эти связи более детально изображены во второй схеме (табл. 18 в
этой книге и fig. 3 на стр. 151 в подлиннике), в которой „proto-x“ обоз-
начает вымершие формы, неизвестные в ископаемом состоянии (Proto-
Angiospermae).
Как и многие филогенетики (в том числе Гатчинсон, Бесси,
Веттштейн и др.) Кимура выводит один из узловых порядководно-
дольных Liliflorae из Pr ot о-Н elobiae, противоположность Гал-
лиру, связывавшему происхождение всех однодольных с Liliflorae.
Наиболее тесное родство между однодольными и двудольными К и-
мура, как и ряд других авторов, видит в филогенетической близости
между Helo Ь i а е из однодольных и Polycarpicae из двудольных. Из
отдельных особенностей системы Кимуры отмечу, например, то, что по-
рядки Ро a I es [включающий сем. Poaceae ( — Gramineae'f\ и Су perales,
относимые им к классу Sicciflorae, он рассматривает в группе порядков
с синкарпным гинецеем, т. е. в группе Syncarpae, к тому же он счи-
тает, что плацентация у них париэтальная. Энглер же считая, что у зла-
ков, принадлежащих в его системе к порядку Glumiflo гае, гинецей
апокарпный, на этом основании помещал их почти непосредственно
после Н elobiae-
Следует отметить и то, что непосредственными предками Liliflo-
rae Кимура считает Н elobiae, точнее Р roto-H elobiae- Некото-
рые авторы отрицают подобное взаимоотношение между данными класса-
ми на том основании, что Нelobiae содержит меньшее число видов.
Кимура прав, утверждая в своем возражении сторонникам последнего
взгляда, что нельзя признавать более древней только группу с большим
числом родов и видов.
1844 — 1930
Обзор новейших систем цветковых растений
2$
Точно также нельзя в основном не согласиться с Кимура, как и
с Гр о с сге й м о м, в том, что неправильно представление Гатчинсо-
на о различном происхождении травянистых и деревянистых растений,
тем более в пределах одного порядка. Кимура однодольные рассматри-
вает как гомогенную и монофилетичную группу, происшедшую от дву-
дольных. В этом вопросе Кимура подтверждает точку зрения многих
предшественников. Свою систему К и м у р а оценивает как наиболее син-
тетичную по сравнению с другими системами. В последних, по его мне-
нию, противопоставляются специализированные признаки, этим затумани-
ваются истинные связи между таксонами и такие системы, по его выра-
жению, более аналитичны, чем синтетичны. В принципе это правильно,
но нет оснований все остальные системы считать таковыми. Ряд систем
не менее синтетичен, чем система Кимуры, например, система Гросс-
г-ейма и некоторые другие.
Система двудольных А. Кронквиста
A. Cronquist’s System of Dicotyledons
1957
Наиболее древней и предковой группой среди двудольных Крон-
кв и ст признает традиционную группу Ranales- Но, согласно его фи-
логенетической схеме столь же предковой группой для другой „полови-
ны® двудольных,, наряду с Ran ales, оказывается порядок D Ulen ta-
les- Это одна из главных особенностей разбираемой системы (см. табл. 19
и конспект), Последний порядок Кронквист считает происшедшим от
Ranales, различая их по. признаку последовательности в развитии ты-
чинок: центростремительное развитие у Ranales и центробежное у Dil-
leniales- В этом отношении, следовательно, Ranales более примитивен.
Отмечу некоторые другие особенности данной системы. Главная, как бы
центральная, эволюционная линия от Ranales приводит к порядку
R о sales, занимающему как и в ряде систем других авторов, узловое
положение. Кстати, следует отметить, что порядок Rosales, как и мно-
гие другие порядки, по семейственному составу трактуется здесь широко. В
непосредственное родство с Rosales ставится его производное—поря-
док U rticales, довольно искусственный по своему содержанию, т. к.
включает Balano psidales, F ад ales, Leitner iales, Myric al es
и Urticales s. str. Дальнейшим развитием U rticales, по этой схе-
ме, является C asuarinales, который правильнее, по моему, рассматри-
вать в аспекте более древних связей. Не исключает возражения разъеди-
нение родственных порядков: С orn ales и Garry ales, с одной сто-
роны, и Umbellates, с другой, исходя, по-видимому, в основном из
анатомических различий. Между прочим, в С ornales Кронквист
включил и семейство R.hizo phoraceae (основываясь на эмбриологических
признаках), обычно относимое к М у г tales-
2 6 Система Кронквиста Gronquist’s Syste»
Подтверждая схемой мнение многих филогенетиков о неносредствен-
ной связи Н aloragales с .Sos ales, он относит к нему (правда с со-
мнением) семейство С dllitriehaceae, рассматриваемое другими, как
об этом напоминает сам Крон к вист, среди симпетальныхпорядков. В
отличие от систем некоторых авторов (Буша, Пула) Крон к в ист не
сторонник непосредственного или близкого родства Мal vales с Llera,
п tales, помещая первый в „диллениевой" группе порядков и непосред*
ствеино связывая его с узловым порядком Guttif erales, а второй—в
„раналйенойвыводя его из узлового порядка S a pindales; подтвер-
ждение родства последних он находит и в анатомическом строении древе-
сины, исследованной Heims ch’ ом (1942). Боковой ветвью 8 арёп&а-
les Крон к вист, в согласии с Tippo, рассматривает и Jugland а-
les', в этом его система резко расходится с взглядами ряда авторов, в
частности А. Л. Тахтаджяна. Между системами этих двух авторов
много общего, хотя немало между ними и существенных отличий. С тем
же узловым порядком Sapin dales Кронквист связывает Polyga-
lales в виде боковой ветви, введя в его состав семейство Balsaminaceae,
в противоположность некоторым систематикам, указывающим на родство
этого семейства с Geraniaceae-
Неясными представляются Кронквисту взаимоотношения поряд-
ков Lin ale st С elastrales (с ветвью Е и pho rb i al es) и Santala-
l-es между собою и с узловым порядком 8 a pindale s, но в происхо-
ждении их от Rosales он не сомневается.
В „диллениевой** группе одним из узловых порядков, как и следо-
вало ожидать, выдвинут порядок G aryaphyllales- Впрочем, Кронк-
вист допускает непосредственное происхождение его от Ran ales, по-
лагая, что аналогичное с D illenial е s центробежное развитие тычинок
у него может оказаться лишь параллелизмом, а не признаком иепосред-
ственной взаимосвязи. Другим узловым порядком, филогенетически свя-
зывающим большее число порядков, является Guttiferales* Мнение
Крон к вист а, наряду с мнением других систематиков, о самостоятель-
ности порядка С a ct al es, обладающего в отличие от Caryophy Ha-
le в, нижней завязью и париэтальной плацентацией, должно быть поддер-
жано, в противоположность взгляду Веттштейна и др. Кронквист
также согласен с трактовкой авторов, выделяющих Plum bagin асеае
в особый порядок, в не включает его в Primulales, как поступает,
например, Пул.
Вопреки мнению большинства филогенетиков, Кронквист уста-
навливает особый порядок Lecythidales, с одним семейством Lecy-
thidaceae, выделяя последнее из порядка М yrtales, к которому его и
относили многие. Он диагностирует его очередным листорасположением,
центробежным развитием тычинок, анатомическими признаками древесины
ц эмбриологическими особенностями.
Происхождение порядков, группирующихся вокруг G еп tianales,
Кронквист считает неясным, хотя предполагает их развитие о £ Lina-
les, G elastrales и 8apin dales-
Обзор вовейшихснетем цветковыхрастений 27
Кронк в it с?, помимо всего этого, высказывается вполне опреде-
ленно по поводу таксономического значения и положения в\ филогенети-
ческой системе некоторых семейств (Lennoaceae и др.), получивших прдзг
нание раннее в системах отдельных авторов.
Система Л. Амберже
L. Emberger’s System
i960
Из систем, предложенных в последнее время, пожалуй, наиболее
полифилетична система Амберже (Les v6getaux vascujaires). (См. табл.
20 и конспект).
В своем воззрении о полифилетичности покрытосемянных он исхо-
дит, в частности, из разнообразия последних. По его выражению, каж-
дый большой порядок Prephanerogamae породил Q у m п о в ре г m а е,
которые в свою очередь могли дать начало различным A ng i о s p.erm а е.
Например, как он отмечает, от нескольких потомков Beinnettitales и
других голосемянных (G i п к у о a les, Oycadales) могли произойти те
илд иные группы покрытосемянных, к которым систематически наиболее
близки именно голосемянные. При этом Амберже считает предками
покрытосемянных древние голосемянные, а не современные. Если предками
покрытосемянных являются голосемянные, то таковыми, конечно, нужно
считать именно древних представителей.
Наиболее древней группой цветковых по Амберже следует приз-
нать двудольные, в частности Apetalae и Dialipetalae; одни из них, по
его мнению, не эволюционировали, а от других в дальнейшем произошли
Gamopetalae. Вместе с тем, он допускает, и другой путь возникновения
последних: от общего предка с Dialipetalae. Однодольные А м бе рже
производит от двудольных, в частности, от групп, родственных к Poly?
а гpicae и Piperales', однодольные, по ею утверждению, также поли-
филетичны, но менее, чем двудольные. Однако, Дм бе рже теоретически
не исключает происхождения однодольных непосредственно от Gymno-
s perm а е-
Между прочим, возникновение цветковых Амберже датирует
второй половиной Юры.
Angiospermae Амберже подразделяет на 5 филогенетических
групп, возникших независимо от различных представителей голосемянных.
Эти группы обозначены на схеме римскими цифрами (см. схему в табл. 20;
знак?обозначает неизвестные прямые предки).
Не без основания отказывается Амберже от общепринятого до
веданного времени (да и теперь разделяемого некоторыми полностью иди
частично) деления на Apetalae, Dialipetalae и Gamopetalae.
Приведу в самых общих чертах характеристику этих пяти групп.
Группа I включает единственный, систематически очень изолирован-
ный порядок С asuar inales, сходный с Gymnospermae по ряду
28
Система Амберже Emberger’s System
признаков. Порядок этот одними авторами выводится непосредственно из
раналиевых, другие филогенетически связывают его с Rosales или Н а-
т am el id ales-
П-ая группа включает 2 филы, связанные общим стволом происхож-
дения. Одну филу образует порядок Proteales, другую — порядки
8 antalales и Olacales (многие авторы объединяют последние в один
порядок 8 ant alale s)- Эти порядки одни филогенетики связывают с
узловым порядком Rosales непосредственно или посредством того или
иного порядка, другие же сближают их с сборной группой „Ат enti-
florae*-
III группа составлена двумя сериями порядков: „краиивоцветных“
(„уртикальных") и „сережкоцветных" (Amentiflorae)-
В первую серию включены порядки: U г ticales. Polygon al es,
Centrosperm ales, Primal ales и Plumbaginale s', во вторую—
Fagales, Balano psidales, 8 alio ale s, Myricales, Leitner i-
ales и В a tidales-
Из последней серии исключены порядки Ju gland ales, Julia-
n iales и Gar г у ales, относимые большинством авторов к Amenti-
florae, к которым наиболее близок Juglandales- От Amentiflo-
гае эти порядки Амберже отличает сложными листьями (у Juglan-
dales и Julianiales), вторичной древесиной (J ulianiarles), нижней
завязью (Jug land ales и Garr у ales), чередованием тычинок с покрово-
листиками у Garry ales и Juli an iales и однопокровной семяпочкой.
Наиболее древним и предковым порядком всей группы Амберже
считает U rticales- У ряда порядков группы он находит признаки,
свойственные голосемянным, как, например, промежуток времени между
опылением и оплодотворением, анатомическое строение семяпочек и др.
Между прочим, к Centrosperm ales Амберже относит также
и семейство Thelygonaceae(=Cynoerambaceae), а в U rticales поме-
щено семейство Rhoipteleaceae, сближаемое отдельными авторами с Jug-
landales (напр., Кронквист, 1957)- В „уртикальной" части схема
Амберже совпадает со схемой Буша.
IV группа, по автору, составлена из 8 фил, хотя фил, связанных
непосредственно с гипотетическими предковыми голосемянными независи-
мо друг от друга—-7.
I —ая фила J uglandales — Julianiales включает два близко
родственных порядка: Juglandales и Julianiales, сходных, в тоже
время, с Amentiflorae по ряду признаков.
2-ая фила Terebinthales—R ub iales содержит 7 порядков,
объединенных общим филогенетическим стволом.
Порядки эти следующие: 1) Terebinthales—включает 17 основ-
ных семейств и допольнительно 7 семейств с неясным систематическим
положением; по Амберже связан с седьмой (и восьмой) филой; 2)
Umbelliflorae—с 5 семействами, в том числе Alangiaceae и Ryssa-
сеае, относимые некоторыми авторами то к Cornales, то к Myrta-
les', 3) с предыдущим порядком Амберже, как и Веттштеин (см.
Оброз новейших систем цветковых растений
29
его схему), совершенно правильно связывает тесно порядок G ат г у al es',
вопрос о филогенетическом положении последнего до сего времени вы-
зывает разногласия, поскольку, как отмечает и Амберже, одни поме-
щают его в сборную группу „Amentif er ае“, другие сближают с раз-
личными порядками, такими, как Т erebinthales, Celastrales,Rha-
mnales и Н am am el id ale s', 4) Rubiales—c 6 основными семей-
ствами и одним дополнительным неестественным сем. Loganiaceae, связы-
ваемым автором с Rubiaceae посредством Cinchoneae- Сюда же отнесено
сем. Galyceraceae, помещаемое другими авторами то рядом с Gompositae,
то сближаемое с семействами С am р an ul ales других авторов; 5) Ge-
la st г ales—с 9 семействами, распределяемыми автором в несколько „гомо-
генных семейственных групп11, а именно: а) группа Celastraceae—Icacina-
сеае, б) группа Staphyleaceae (близка к Т erebinthales), с) группа Pe-
ri pt erygiaceae и Erythropalaceae, д) группа Salvadoraceae, сближаемая
другими с Oleaceae (чему противоречит по А м б е р ж е двупокровность се-
мяпочек) или Rhamnales, 6) Rhamnales—с 3 семействами fRhamna-
ceae, Vitaceae и Leeaceae) и 7) Li ди str ales—с одним семейством Ole-
aceae, родственным С elastr ales по строению околоцветника, но сход-
ным, по автору, с Rubiales и 0 ontortae-
3-ью филу Cont ortae—Т ubiflor ае составляют 2 порядка—
С ontortae и Т ubiflor а е- Оба тесно взаимосвязаны общностью про-
исхождения. Порядок Gontortae включает 4 семейства: Gentianaceae,
Jtfenyanthaceae, Аросупасеае и Asclepiadaceae', из этого порядка Амбер-
же исключает сем. Loganiaceae, отнесенное им к Rubiales (правда,
условно), и сем. Buddleiaceae, рассматриваемое им в порядке Tub i flo-
rae- Все шесть упомянутых семейств, а также сем. Oleaceae (отнесенное
Амберже к особому порядку Ligustr ales), Кронквист (г957)
объединяет в один порядок под названием G entianales- Второй по-
рядок Т ubiflor ае, в противоположность первому, является очень круп-
ным, включающим 26 основных семейств и одно дополнительное, мало-
изученное (Columelliaceae), хотя положение и сем. Lentibulariaceae пред-
ставляется Амберже неясным; он же отмечает недостаточную изучен-
ность сем. Humbertiaceae, но сближает его с Convolvulaceae и Solana-
сеае", в составе этого же порядка фигурирует и сем. Plantaginaceae, вы-
деляемое многими филогенетиками в особый порядок Plantag i nates-
Сюда же помещено и сем. Hoplestigmataceae, связываемое некоторыми
авторами с Eb enale s- Амберже считает, что родство входящих в по-
рядок Т ubiflor ае семейств подтверждается серологической реакцией.
4*ая фила Р an dales представлена одним порядком Pan dales,
монотипным в отношении всех подчиненных таксонов вплоть до вида.
У сем. Раndaceae Амберже находит признаки, сближающие его с
G er ani ale s, Т er eb in that е s, С elastr ales и Rhamnales-
5-ая фила С neorales включает также один порядок С neorales
с одним монотипным семейством (Спеогасеае) и 3 видами. Как и подчер-
кивает автор, филогенетические связи Спеогасеае недостаточно выяснены:
jo Система Абыерже Emberger’s System
морфологически оно сходно с Geraniales, а серологически—-с М di-
vale s (Coluntnif er ае)-
6-ая фила Eb enales представлена также одним порядком Е b e-
lt ales, объединяющим 6 семейств: Ebenaeeae, Styracaceae, Symplocaceae,
Sapotaceae, Sarcospermaeeae и Lissocar paceae- Из них Lissocarpaceae с
неясным систематическим положением. Объем этого порядка в данной си-
стеме вполне совпадает с его объемом, принятым в системе Т а х т а д ж. ян а
(1959). Наиболее узко он трактуется Кронк ви стом (1957).
7-ая фила Geraniales (G ruinales) содержит два тесно свя-
занных общим предком порядка: Geraniales и М alv dies- В состав
первого порядка, включающего ю семейств, не входит, вопреки взгляду ряда
авторов, сем. Balsaminaceae, отнесенное Амберже к Тereb inthales-
В этом порядке Амберже рассматривает и такие семейства, которые
отдельными филогенетиками распределены в порядках Р oly д alales,
Linales и др. Сам Амберже признает филогенетическую неоднород-
ность Geraniales в намеченном им объеме. Так, например, ясно род-
ственную группу образуют семейства Geraniaceae, Tropaedlaceae и Oxali-
daceae', обособленное положение занимает, по Амберже, семейство
Limnanthaceae, более гамопетальное, чем предыдущие и, поэтому, дальше
продвинувшееся по пути эволюции; особую группу составляют Linaceae,
Lepidobotryaceae, Ilwmiriaceae и Erylhroxylaceae-, также обособлены как
от перечисленных групп, так и друг от друга, семейства Malpighiaceae и
Dygophyllaceae-
Второй порядок разбираемой филы — Мalvales (Golumnife-
г а е) —объединяет ю семейств, также не образующих монолитную группу.
Если Malvaceae, Bombaceae, Sterculiaceae и Tiliaceae настолько близко
родственны между собою, что допускается их слияние в одно семейство, с
чем согласиться, конечно, нельзя, то другие семейства проявляют филоге-
нетическую отдаленность. В частности, обособленно стоит в системе Elaeo-
саграсеае, хотя Амберже сближает его с Tiliaceae, а положение се-
мейств Chlaenaceae, Scytopetalaceae и Gonystylaceae в данном порядке, по
признанию самою Амберже, вообще спорно. Во-всяком случае, принятое
Амберже содержание порядка, полностью совпадающее с взглядом Вет-
т ш е й н а на объем последнего, более естественно, чем толкование Бесси,
странным образом включившего в него такие семейства, как Ulmaceae,
Moraceae, Urticaceae и др.
8-ая фила Tricoccae включает один порядок Tricoccae с 4
семействами: Euphorbiaceae, Baphniphyllaceae, Dicha petdlaceae и Lopho-
pyxidaceae- Те или иные филогенетики к этому же порядку (имену-
емому иногда Е и pho rb iales) относят некоторые другие семейства. На-
пример, Вихасеае (Веттштейн, Шоо, Кронквист, Тахтаджян),
Aextoxicaceae и Didieraceae (Кронквист), помещенные в системе А м-
б е р ж е в порядок Т er eb inthal es', Callitrichaceae (Веттштейн, Шоо
и др.), принадлежащее по Амберже к порядку М yrt ales и, даже,
Pandaceae (Кронквист), рассматриваемое Амберже, как уже отме-
чено выше, в качестве особой филы. Конечно, „фила Tricoccae* Ам-
1863 — 1931
Обзор иэвейщш систем,цветковых растений
б ер же едва-ли заслуживает обособления как фила, т. к. она, безус-
ловно, представляет собою органическую часть общей эволюционной ли-
нии Geraniales — Mali? ales- В графической.схеме Амберже в су-
щность она так и показана. Да и сам Амберже допускает возмож-
ность объединения данной филы с филой Geraniales—Мalrales. Еще
Бесс-и (1915) семейства,входящие в филу Tricoccae, отнес к порядку
Geraniales- Но, следует заметить, что многие порядки в системе Бе с-
си разнородны по составу. Прав Амберже, считая семейство Euphor-
Ыаееае гетерогенным и даже настолько, что вычлененные из него новые
семейства могут быть более родственными другим порядкам, чем Trico-
ccae.
Наконец, V-ая, наиболее крупная и разнообразная группа, расчле-
нена на 7 фил. Из них 3 филы— Н el оЪ i ае, Spadiciflorae и Li-
lii florae—составляют однодольные, которые по Амберже филогене-
тически связаны с Polycarpicae общим филогенетическим стволом.
В i-ой филе автор выделяет 2 порядка: Р о I year р i сае и Aristolo-
chiales- Порядок Р oly с ar picae, рассматриваемый Амберже, как и
большинством филогенетиков, одним из наиболее древних, включает 29 се-
мейств. Последние он распределяет в группы по наличию секреторных
клеток в паренхиме листа. Этот порядок в трактовке Амберже очень
неоднороден и трактуется им в основном согласно системе В еттште-
йна (отчасти системе Шоо, 195?)- В то же время, системы упоминаемых
авторов в сопоставляемой части существенно отличаются; например, Ве-
ттштейн включил в Р oly с ar pi с ае порядок Arist olochiales,
выделенный в системе Амберже; различны в обоих системах место и
содержание порядка Н am am elid ales, часть семейств которого Ам-
берже отнес к Р oly car picae', имеются и другие отличия в понима-
нии Рolycarpic ае этими авторами. Данную группу правильнее рас-
членить на отдельные, более естественные порядки, как поступили, напри-
мер, из авторов последнего времени Кронквист (1957) и ДР-
Порядок Aristolochiales, как и в системах многих авторов, со-
держит семейства Aristolochiace ie, Rafflesiaceae (охватывающее Cytinaceae
и Mitrastemonaceae) и Hydnoraceae-
2-ая фила Нelob iae (Fluv iales) состоит из трех порядков:
Alismatales, Р о t ат о g et о nales и, условно, Triuridales- Как
и большинство авторов, из однодольных именно Н elob iae считает Ам-
бер же наиболее близким к Polycarpicae- Alismatales с 6 семей-
ствами (i. Butomaceae, 2. Hydrocharitaceae, 3- Alismataceae, 4- Junca-
ginaceae, 5- Scheuchzeriaceae и 6. Aponogetonaceae) Амберже считает
очень монолитной группой.
Из 7 семейств Р ot am og etonales (1. Potamogetonaceae, 2. Ruppi-
aceae,^. Lilaeaceae, 4. Zosteraceae, 5- Posidoniaceae, 6. Zanichelliaceae и 7.
Najadaceae) семейства Potamogetonaceae и Zosteraceae связывают порядок с
другой группой однодольных — с Spadiciflorae', несколько обособ-
ленное место отводит автор Zannicheliaceae и Najadaceae.
32
Система Амберже Emberger’s System
Неясно положение Т riurid ales, несущего по автору черты Не-
lob i а е, Lilii florae, Ranunculaceae « Rosaceae-
В з ью филу Piperales—8рa di ci fl or ае автором включены
порядок Piper ales и группа порядков (надпорядок) 8 р a dici flor ае
в составе AralestPandanalesPalmalesn С у el a nth al es- Амбер-
же филогенетически связывает Piperales общим стволом а 8 padici-
florae, сближая его, с другой стороны, с Pol ycarpicae и Н elo-
biae- Эта точка зрения в основном совпадает с мнением ряда его пред-
шественников, например, Бесси, Буша, Шоо, Гроссгейма, Крон-
квиста, в известной мере Тахта ж ян а и др. Хотя, в более частных
моментах этого вопроса взгляды их не вполне сходны. Иной позиции при-
держиваются Веттштейн, Энглер и др., которые рассматривают
Pi per ales в непосредственном родстве с Monochlamydeae. Семействен-
ное содержание порядков филы таково: Pi per ales—Piperaceae, Sauru-
raceae и Chloranthaceae', Ar ales — Arace'ie и Lemmaceae', P and ana-
les—Pandanaceae, Sporganiaceae и Typhaceae; оба последних порядка
взаимосвязаны общим филогенетическим стволом; Palm ales—Palmaceae,
Phytelephasiaceae и Nipaceae; Cyclanthales—Cyclanthaceae', два пос-
ледних порядка также связаны друг с другом непосредственным общим
предком. Морфологическое сходство Pal males с Gramin ales Ам-
берже объясняет, вопреки взгляду многих авторов, не прямым родст-
вом между ними, а общностью их происхождения, в общем сближая их
с Ро 1у с а г р ic а е- Эволюционное развитие древнего порядка Palma-
les, по мнению Амберже, как и некоторых других авторов, вырази-
лось в сокращении цветкового аппарата; на одной линии оно заверши-
лось семейством Nipaeede, являющимся крайней формой сокращения;
другая линия этого развития привела к семейству Cyclanihaceae, выделя-
емому Амберже, в согласии с системами многих предшественников, в
самостоятельный порядок С yclanthales, но тесно связанный с Pal-
males’, отдельные авторы объединяют оба порядка в один порядок,
иногда со включением и некоторых других семейств.
В системе Амберже, как и почти во всех предшествовавших сис-
темах, в качестве одной из наиболее узловых групп среди однодольных
выделена 4-ая фила Lilii flor ае- Автор справедливо считает ее более
древней, чем об этом свидетельствуют ископаемые представители. Он ука-
зывает на наличие у растений этой филы архаических черт, свойственных
двудольным, и, даже, голосемянным; таковы; боковая семядоля (у Dioscorea-
ceaen Commelinaceae), трахеиды (Agave, Dracaena, Cordyline, lucca, Amaryl-
lis), черешковые листья с сетчатым жилкованием (Dioscoreaceae), дихото-
мия у некоторых AmaryUidaceae, многочисленность тычинок (Smilacoideae)-
В ней автор различает 5 групп порядков; 1) Gramin ales—С о mm eli-
п ale s; порядок Gramin ales представлен одним семейством Gramina-
ceae, a Comm el in ales Сили Enantioblasta e)—с семействами Comme-
linaceae, Mayacaceae-. Xyridaceae, Eriocaulaceae, Centrolepidaceae и Restio-
naceae’, некоторые авторы предпочитают последние три семейства, или по
крайней мере два из них, обособить как отдельные порядки, что пред-
Обзор новейших систем цветковых растений 33
ставляется более правильным [напр., Кимура (1956) отличает порядки
Еrioc aulales и Resti on ales, подчинив последнему Centrolepida-
<5еае]; 2) June ales— Су p erales', June ales составляют семейства
Juncaceae, Thurniaceae, Flagellariaceae и Rapateaceae, рассматриваемые
Амберже как тесно родственные между собою; последним двум семей-
ствам отдельные авторы дают иное филогенетическое толкование; так,
например, Кимура (1956) семейство Flagellariaceae, вместе с семейст-
вами Centrolepidaceae и Restionaceae, относит к порядку Restion ales,
(этого же мнения и Тахтаджян, 1939), семейство Rapateaceae—кХу-
rid a les (последнее Тахтаджян включает в С ommelin ales)', С у-
р er ales представлен только семейством Сурегасеае, объединение кото-
рого с Poaceae (Gramineae) в общий порядок Glumiflor ае, принятое
рядом филогенетиков, едва-ли правильно; 3) Bromeliales с семейст-
вом Bromeliaceae', обособление этого семейства в большей степени отра-
жает его филогенетическую природу, чем объединение с другими семей-
ствами в один порядок, как поступили. Бесси (1915), Шоо (1953) и
некоторые другие; 4) Р ontederiales с семейством Pontederiaceae', вы-
деление этого семейства в отдельный порядок не вполне оправдано; более
правильным следует признать, согласно большинству авторов, его включе-
ние в Liliales', выделение его из Liliales Амберже аргументирует
налйчием мучнистого белка в семенах растений данного семейства; нако-
нец, группа 5) Lil г ales — Dioscoreales наиболее богата семейст-
вами, число которых равно 23; из них наименьшими признаками однодоль-
ных обладают представители Dioscoreaceae, с другой стороны, многие из
этой группы являются весьма современными; группа эта расчленена на 5
порядков: Liliales с ю, Dioscoreales—с 2, S cit aminales—с
6 (хотя Бесси, Энглер, Веттщтейн, Гроссгейм, Шоо и др.
объединяют семейства Musaceae, Strelitsiaceae и Lowiaceae в одно, под
названием Musaceae, в виду их сильной близости), Bur man ni ales—с
3 и Orchidales—с 2 семействами; при этом последние 3 порядка
Амберже эволюционно выводит непосредственно из порядков LiHa-
les и Dioscoreales, независимыми друг от друга линиями; именно эти
3 порядка следует признать, по справедливому утверждению автора, на-
иболее современными потомками Liliales — Dioscoreales- Между
прочим, автор допускает выделение из состава Liliales особого порядка
Rox burghiales с семействами Roxburghiaceae (=Stemonaceae) и Peter-
manniaceae-
В 5-0Й филе—Parietales—Rhoeadales—Synanther ales—
одним из родоначальных порядков Амберже выдвигает порядок Р a rie-
tales, связанный общими предками с Р oly с ar pi с ае и с другими выше
описанными филами. В этой части система Амберже, в известной мере,
перекликается с системами Буша, Гроссгейма, Пу л ле и некоторых
других.
В филе автор отличает 5 порядком; Parietales, Rhoeadales,
С ucurbit ales, Е г ic ales и Synantherales (8 ynanther ае)- Се-
мейства порядка Pariеtales (принятого здесь в общем в энглеровскОм
3. А. А. Гроссгейм и М. Ф. Сахокиа
34
Система Амберже Emberger’s System
толковании) числом 35, разделены им на две категории. Категория А—с
26. основными семействами, это—„типичные Р arietales*', последние, в
свою очередь, разбиты на 3 подгруппы: 1) Dilleniaceae—Dipterocarpaceae
с 12 семействами; соответствует, например, Guttif erales системы Вет-
тштейна и его последователей, 2) Flacourtiaceae—Cochlospermaceae—
из 6 семейств; частично соответствует Р ari etales Веттштейна,
Bixales Кронквиста или Р ar i etal е s Ъ) Cistineae Шоо и 3) Та-
maricaceae—Caricaceae с 8 семействами; соответствует подпорядкам Та-
maricineae, Papayineae и, частично, Flacourtiineae порядка
Р arietales системы Энглера, части Gutti fer ales Бесси, частич-
но Bixales Кронквиста, Tamaricineae и, отчасти, С istine а е
Шоо; в системе Тахтаджяна семейства данной подруппы размещены
в нескольких порядках. Категория В—с 9 семействами (Lacistemaceae,
Loasaceae, Datiscaceae, Begoniaceae, Droseraceae, Ancistrocladaceae- Quiina-
ceae и Fouquieriaceae), происшедшими, по мнению автора, от семейств
„типичных Р ar iet al е su-
Порядки Rhoeadales (с 7 семействами) и Ericales (с 6 се-
мействами), принятые в энглеровском понимании, рассматриваются авто-
ром, как и рядом предшественников, тесно связанными с Р arietales
общими непосредственными предками, а взаимнопараллельными ппоизвод-
ными от Parietales отмечены порядки Sy п anther a I е s и Cucur-
bitales- Эга точка зрения согласуется с мнением большинства филоге-
нетиков и она представляется более правильной, но с ней расходится по
зиция Тахтаджяна (1959), который выводит С am panulales, Aste-
rales (Synantherales по системе Амберже) наряду с параллель-
ными ему эволюционно линиями Polemoniales —S его phulariales—
Lamiales (Tubifloraes смысле Амберже) и Gentianales (С on-
tor tae по Амберже)—Rubial еs из общего предка. А что касается
Cucurbitales, соответствующего части Р as si fl о г ales в понимании
Тахтаджяна, то последний связывает его с совершенно другой ветвью
развития —с С is tales', однако, как показано было выше, порядки Ru-
Ъ iales, С ontortae и Tubiflorae в системе Амберже помещены
во 2 ой и в з-ей филах IV-ой группы, возникшей из Gymnospermae
независимо от V-ой группы, чему в корне противоречит воззрение Тахтад-
жяна. Остальные порядки (в основных своих частях) анализируемой пя-
той филы данной группы в системах Амберже и Тахтаджяна в
общем имеют „париэтальную“ основу.
Неясно положение 6-ой филы с порядком Sarraceniales, хотя в
общем оно созвучно с его положением в системах ряда авторов,
в которых он сближается с Pol year picae, но тут вызывает возра-
жение то, что Амберже устанавливает это родство с Р oly car picae
также через Saxifragaceae (наряду с Droseraceae'). Столь же неясна фи-
логенетическая взаимосвязь составляющих этот порядок семейств: Sarra-
ceniaceae, Nepenthaceae и Cephalotaceae-
7-ая фила — Rosales—JU yrt ales—охватывает 3 порядка: Rosa-
les, М у г tales и Thymelaeales- Порядок Дояа^еяАмберже трак-
Обзор новейших систем цветковых растений
35
тует, какБесстг, Энглер и некоторые другие, весьма широко, вклю-
чая в него вполне заслуживающие порядкового обособления порядки
Н amamelidales и Leguminosales (F ab al е s); даже семейство
Saxifragaceae, с семействами ближайшего с ним родства, следовало от-
членить в качестве самостоятельного порядка, согласно ряду новейших
систем.
Все три порядка данной филы Амберже тесно связывает друг с
другом общим филогенетическим корнем, справедливо разделяя более об-
щепринятую точку зрения о том, что Rosales являются предками Муг-
tales и Thy т el а е ale s- Автор категорически высказывается против
слияния последних двух порядков, допускаемого некоторыми филогене-
тиками.
Однако, правильнее связывать П amamelidales с. особой эволю-
ционной линией, исходящей непосредственно из Р oly car picae и при-
водящей через него к Monochlamvdeae, как это выражено, например, в
некоторых новейших системах (Гроссгейм, Шоо, Тахтаджян). Сам
Амберже в какой-то мере приближает Н amamelidales к Monoch-
lamydeae, говоря о родстве его с Т erebinthales, хотя, с другой сто-
роны, подчеркивает его же связь с Rosales на основе серологических
данных.
Следует отметить и то, что положение Thymelaeales в этой
системе Амберже признает спорным, указывая на возможность его от-
несения „в круг Rhamnales", принадлежащий ко 2-ой филе IV-ой
группы.
Система Хр Я Гоби
Khr. J. Gobi’s System
1916
В 1916 году появилась статья проф. X р. Гоби, „Обозрение систе-
мы растений", занявшая весь XXX выпуск „Ботанических записок". Го-
б и дает схему всей системы растительного мира, но мы остановимся толь-
ко на цветковых растениях (табл. 38 и 59).
Как указывает сам автор, одной из особенностей его системы пок-
рытосемяннных растений явилось то, „что при группировке семянных ра-
стений, т. е. всего Отдела Vi, Spermapbyta, были, между прочим,
впервые приняты во внимание, помимо морфологических данных, также
и проверочные данные над родственными отношениями между растениями
названной категории, добытые путем серодиагностического метода приви-
вок" (стр. IX предисловия.—ГМ. С.). Здесь подразумеваются первые ра-
боты Меца цо применению серодиагносгического метода в систематике,
опубликованные в 1913 году; эта работа широко использована Гоби в
своем построении системы цветковых растений.
Гоби считает, что серодиагностический метод дал окончатетьный
ответ на два важных вопроса систематики цветковых растений: i) о про-
исхождении односемянодольных и 2) о происхождении группы однопо-
кровных.
36 Система Гоби Gobi’s System
Однодольные всегда принимались до этого как особый класс, равно-
ценный и равноправный с двудольными. „Такой взгляд", пишет автор,
„принимался всеми обычно молча, как аксиома, без всяких попыток к
какому либо доказательству". Точно также и вся группа однопокровных,
как обладающая более простым, строением своих цветков, обыкновенно
считалась первичной в смысле своего происхождения"... „И все эти кра-
сивые фантазии или хитросплетения вдруг рухнули"! говорит автор (стр.
X предисловия.—М. С.). Оказывается, что односемянодольные вовсе не
параллельный самостоятельный класс, а только боковая, правда, мощно
развитая ветвь главного ствола двудольных.
Что касается Однопокровных, то они являются лишь упрощенными
вторичными разветвлениями другой из ветвей, отходящей от того же
главного ствола двудольных, Как и первая ветвь, но только несколько
выше нее. Однопокровные являются сборной группой „состоящей из раз-
ных порядков, являющихся лишь в разной степени более или менее сил-
но упрощенными вторичными разветвлениями нескольких больших ветвей,
отходящих -от одного общего им всем главного ствола Двудольных".
Эти мысли, высказанные Г о б и, хотя и навеяны новыми в то время
достижениями серодиагностической школы, были им воплощены в ориги-
нально построеннную систему, в которой целый ряд отношений, еще не
подвергшийся серодиагностической проверке, был самостоятельно изобра-
жен Гоби.
Для своего времени система Гоби явилась крупным шагом вперед,
она идейно заострена против устаревших концепции Энглера и Ветт-
штейна и в этом отношении является более выдержанной, чем си-
стема Н. И. Кузнецова.
Как уже сказано, Г о б и в своей системе отказывается от установле-
ния групп однодольных и однопокровных; это новшество является поло-
жительной чертой его системы. Такими же положительными чертами явля-
ются строго монофилетическое построение системы и первый намек на
учение об уровне развития цветковых растений. Хотя это учение особен-
но детально и глубоко разработано было Н. И. Кузнецовым, но не-
который намек на него мы находим и в системе Гоби, а именно на его
схеме горизонтальной чертой, проведенной через весь чертеж, отделены
„вершинные" группы системы: сюда отошли, во-первых, Synandrae (Си-
curbitaceae, Campanulaceae, Compositae), затем Tubiflorae к Conto-
rtae и, наконец, Li gustr ales вместе c IJ mbelliflorae и Rubi-
ales- Указанные семейства н порядки очень неравноценны в смысле сво-
его филогенетического значения; кроме того, такие „вершины", как Gу-
п and гае, Glumiflorae и др., остались в системе далеко внизу под
этой чертой, так что принцип уровня развития показан на схеме Гоби
в еще очень несовершенной зачаточной форме.
Из частностей системы Гоби некоторый интерес представляет види-
мо особый порядок J unci flor ае, давшего начало, .с одной стороны,
линии через L i li i florae, с другой стороны, приведшей к GI и т i f I о-
гае и FarinosaC' Такое выделение Junciflorae в качестве узло-
ХР. Я. ГОБИ KHR. J. GOBI
1847 — 1919
Обзор оовейщих систем цветковых растений 37
вого порядка филогении известной части покрытосемянных представляет
особенность системы Гоби; это выделение впрочем никем из последую-
щих авторов признано не было.
Интересен также отрыв в системе сем'. Salicaceae, составляющего со-
держание особого порядка Saliciflorae, от других „однопокровных"
и сближение его с порядком Р a riel ales посредством сем. Татаггсасеае,
этот взгляд, как известно, еше недавно защищался многими систематиками.
При всех достоинствах системы Гоби, сделавшего определенный шаг
вперед по пути установления филогенетических отношений между раз-
личными группами цветковых растений, все же нужно признать, что во-
прос о происхождении этих растений разрешен им неудовлетворительно.
Ствол двудольных по Гоби связан с сем. Pinaceae класса Хвойных, а еще
ниже с сем. Selaginellaceae из класса двужгутиковых Плауновых. Сем.
Bennettitaceae на схеме помещено в виде бокового ряда, ведущего начало
из тех же Lyco podinae и заканчивающегося вслепую.
Во времена Гоби уже были опубликованы системы Галлира и
Бесси, в 1914 году был опубликован первый вариант системы Н. И.
Кузнецова; во всех этих системах происхождение покрытосемянных
трактуется значительно иначе и ближе к истине. Гоби ни разу не упо-
минает об названных аяторах; остается впечатление, что он их просто не
знает.
В общем все же нужно считать, что система Гоби, при всех своих
недостатках, представляла определенный шаг вперед в учении о системах
покрытосемянных растений. И нужно только пожалеть, что она была не-
заслуженно позабыта и полностью игнорировалась последующими бота-
никами при своих построениях.
Система Н. И. Кузнецова
N. I. Kuznetzov’s System
1914; 1922; 1936.
Система цветковых растений Кузнецова была опубликована впер-
вые в 1914 году в erq известной книге „Введение в систематику цвет-
ковых растений", где дана схема в виде родословного дерева с детализа-
цией до порядков. В 1922 году опубликована схема без изменения (табл.
40), повторенная в 1936 году; схема эта детализирована до семейств (т.
е., перечислены семейства, составляющие каждый из порядков, но без ука-
зания филогенетических отношений между семействами). В 1922 году
появилась в свет специальная статья, посвященная обоснованию предла-
гаемой системы. В этой статье Кузнецов резко подчеркивает две ос-
новные особенности предлагаемой им системы цветковых растений: г) при-
знание полифилетического происхождения цветковых растений и 2) отказ
от деления цветковых на однодольные и двудольные.
Со своей стороны подчеркнем третью и самую ценную особенность
системы Кузнецова—3) установление последовательных стадий (уров-
38 Система Кузнецова Kuznetzov’s System
ней) филогенетического совершенства цветковых растений на основе эво-
люции плана строения цветка.
Кузнецов высказывается весьма энергично в защиту полифилети-
ческого происхождения цветковых. Он говорит, (1922): чя считаю представ-
ление о монофилетическом происхождении цветковых растений заблуж-
дением, вредно отзывающимся на наших филогенетических построениях,
и, на основании своих исследований, прихожу к выводу о полифилетиче-
ском развитии цветковых (покрытосемянных) растений" (стр. 192).
Всего Кузнецов отличает 6 ветвей покрытосемянных, происшед-
ших от различных предков и, по-видимому, в различное время. Эти ветви
следующие: I) ряд V erticill a iae, 2) ряд Р roteales—Santala-
les, 3) ряд Salicale s, 4) ряд, напоминающий Myricales, очень ра-
зветвленный, приводящий к Urticales, С entros рвгтае и Polygo-
nales, 5) ряд Pi ре г ales—S padici florae и, наконец, наиболее об-
ширный 6) ряд, начинающийся Anonales—Banales и заканчивающий-
ся наиболее совершенными порядками цветковых растений. Первые пять-
рядов произошли от каких-то неизвестных нам вымерших Protogym-
nospermae, шестой ряд произошел от Bennettitales.
Первые пять рядов составляют класс Protoanthophyta Мо-
по chlamy dea; по мнению Кузнецова эти ряды в своем развитии
не перешагнули стадию первичноцветковых (Protoanthophyta) и среди
них вовсе нет представителей настоящих цветковых растений (Euantho-
phyta).
Шестая, самая обширная группа покрытосемянных представлена как
первично цветковыми (ProtoanthophytaPolycarp i с а), так и насто-
ящими цветковыми (Euanthophyta). К Protoanthophyta Po-
lycarpica относятся 7 порядков.
Euanthophyta представлены двумя ступенями (уровнями)1 разви-
тия и двумя линиями развития. Кузнецов считает, что первой, менее
совершенной в эволюционном отношении ступенью развития настоящих
цветковых являются пятикруговые цветки, более совершенной—четырех-
круговые цветки. Одна из линий развития пошла по пути создания трех-
мерных цветков (Pentacyclica trimera), другие по пути создания пяти-
мерных цветков (Pentamera). Кузнецов считает, что первая линия в
своем развитии не поднялась на высоту второй (четырехкруговой) ступени
развития и представлена только одним подклассом (Pentacyclica trimera)*,
вторая линия привела к образованию более совершенных четырехкруго-
вых цветков и, поэтому, она состоит из двух подклассов (Pentacyclica
pentamera и Tetracyclica pentamera).
Таким образом, Кузнецов считает наиболее примитивными груп-
пами цветковых растений одновременно и olycarpicae п все Мопо-
chlamydeae. В этом коренная ошибка Кузнецова, приведшая его с не-
избежностью к принятию ложного положения о полифилетическом про-
исхождении цветковых растений.
В историческом аспекте эта ошибка Кузнецова объясняется темг
что ему, выросшему и воспитавшемуся на основе взглядов Энглера и
Обзор новёйших систем цветковых растений
39
Веттштейна, трудно било отрешиться от этих воззрений и целиком
перейти в лагерь галллировской концепции. В результате получилась из-
вестная половинчатость, стремление примирить непримиримое, признавая
для одних групп растений концепцию Веттштейна, для других — кон-
цепцйю Галлира (Бесси Н. И. Кузнецов, по-видимому, не знал
вовсе). Вот это и является слабым местом системы Кузнецова; искус-
ственность первых пяти рядов, происходящих от совершенно неизвестных
ископаемых предков, совершенно очевидна. Все эти пять рядов причис-
лены Кузнецовым к классу Р г о t о a n t h о р h у t а, хотя сюда же от-
носится порядок Centrospermae, высшие представители которого са-
мим Кузнецовым не могут рассматривавсяиначе, как настоящие цвет-
ковые растения (Alsinaceae, Siltnaceae и др.).
Но, насколько ошибочна эта часть филогенетических построений
К узнецова, принять которую мы не можем, настолько интересны» и
новы его построения, касающиеся громадного большинства цветковых ра-
стений, связанных в своем происхождении с Bennettitales-
Гал л иру чужда идея о ступенях (уровнях) развития цветковых, «у
Бесси эта идея выражена очень нечетко, в»то же время Кузнецов
именно эту идею кладет во главу угла своей филогенетической системы.
Большим шагом вперед явилось то, что разделение покрытосемянных про-
исходит не 'на основании какого-либо односторонне взятого признака,
а на основе общего плана строения цветка, т. е. на основе комплекса
признаков. Один из его главных тезисов—эволюционное развитие от цвет-
ка с неурегулированными отношениями между органами и неопределенно-
го их числа, что связано также с поликарпией и апокарпией (Proto-
anthophyta), к цветку с фиксированными отношениями небольшого
числа органов и с синкар.пным гинецеем (Euan t ho ph у t а). Развитие
Euanthophyta идет по пути дальнейших редукционных явлений в
цветке, что приводит от ступени Pentacyclica к более высокой сту-
пени Tetracyclic а-
Это учение о ступенях (уровнях) эволюционного развития, четко и
убедительно показанное в системе Кузнецова, является.тем новым вкла-
дом в систематику, который внесен Кузнецовым по сравнению с Гал-
лиром и Бесси.
Ступени развития предполагают само собой отражение в системе
принципа времени; первая ступень рассматривается Кузнецовым как
древнейшая, вторая, как промежуточная и третья как новейшая.
Кроме того, необходимо отметить, еще одну особенность системы
Кузнецова, выгодно отличающую ее от зарубежных систем. Ни Гал-
лир, ни Бесси в своих построениях не отражают географического мо-
мента, географических территорий, как плацдарма, на котором происхо-
дит филогения. Таким образом, их построения как бы повисают в воз-
духе, будучи всецело оторваны от арены, на которой развивается расти-
тельный мир. Кузнецов вводит географический принцип в свою сис-
тему; правда, принцип этот в системе Кузнецова играет пассивную
роль, но важно то, что он наличествует в ней. Для всех семейств у
40
Система Козо-Полянского Kozo-Poliansky’s System
Ку з н е ц о в а подчеркнуто их происхождение в широком обобщенном
виде; отмечены типы тропические, антарктические и палеарктические.
В общем система Кузнецова, сделавши соответствующую оговор-
ку насчет полифилетизма, представляет крупное явление в развитии уче»
ния о системах растительного, мира. Она была высоко оценена ботаниками
Советского Союза, но, к сожалению, осталась почти неизвестной ботаникам
зарубежных стран.
Система Б. М. Козо-Полянского
В. М.* Kozo-Poliansky’s System
1922; <947.
В 1922 году была опубликована система цветковых растений Козо-
Полянского (табл. 21 и 41). Систему Козо-Полянского можно
шазвять морфогенетической по преимуществу, так как она построена в ос-
новном в результате глубокого анализа морфогенетических признаков и
целиком основана на них. Автор подвергает детальному просмотру и обсуж-
дению вопрос происхождения основных органов растений—стебля, листа,
: корня, органов размножения и приходит в ряде случаев к оригинальным
.решениям, не принятым в обычных морфологических схемах. Так, автор
•считает, что пластинка листа однодольных есть диафлебий (сросшаяся
.пара прилистников), а язычек—редуцированный лист (эуфиллом), чем под-
тверждается вторичное происхождение листа однодольных от листа типа
„двудольных.
К оз о-По л я иски Д делает попытку реконструкции пращура выс-
4рцх растений и считает, что из ныне живущих водорослей ближе всего
(к теоретической реконструкции стоят флоридеи. Что среди зелены?,. во-
дорослей жили формы, подобные флоридеям, показывает существование
Coleochaete.
„Представляется вероятным, что флоридеи, колеохеты и высшие р.
р. (растения.—Ред.). суть ветви одного корня, так как они—как-бы вари-
ации одного основного типа" говорит автор (стр. 43).
Более близкая к покрытосемянным группа „предпокрытосемянные"
также гипотетична. „Структура покрытосемянных повторяется только у
загадочного, не имеющего места в системе, каменноугольного р. Noegge-
rathia" (стр. 51).
Согласно системе Козо-Полянского класс Angiospermae
(покрытосемянные) относится к Ш-му типу растений Diodotomieae,
который характеризуется следующим образом: „расчленение на трофосому
и гаметосому, по крайней мере по отношению к женским гаметосомам,
не выражено, но может быть, это результат редукции (все тело гамето-
сомы, кроме половых органов, утрачено* (стр. 49).
В 1947 г. Коз о-П о л я н ск и й, еще раз возвращаясь к проблеме си-
стемы цветковых растений, выводит Angiospermae непосредственно
из псилофитов, делая оговорку, что „intermedia ignota"6. (См. конспект
системы).
6 Т. е., „промежуточные неизвестны". (Ред).
Обзор новейших систем цветковых растений
41
Подобно Г а л л и р у и Бесси Коз о-П о ляне ки й сто^т на точке
зрения примитивности многоплодниковых. Значительная часть книги Ко-
з о-П о л я н с к ого посвящена обоснованию этого положения и развер-
нутой критике положений Энглера и Веттттейна. Также детально
обсуждается вопрос о взаимоотношениях одно-и двудольных цветковых
растений; автор приходит к выводу, что „Monocotyledones есть группа
антифилогеническая и естественная; не ветвь, а ступень развития ангио-
сперм, преемственно связанная с примитивными Dicotyledones и возник-
шая в связи с гидро-и геофилией" (стр. 131).
Взвешивая и оценивая все данные морфогении Козо-Полян-
с к и й выдвигает на первый план, как важнейшие и определяющие всю
систему покрытосемянных, признаки карпелляции и плацентации.
Покрытосемянность возникла не одним, а двумя путями; i) путем
смыкания отдельных плодолистиков, сначала расположенных апокарпно,
а затем синкарпно (закрытая карпелляция), 2) путем срастания друг с
другом незамкнутых плодолистиков (открытая карпелляция). „Вообще же,
если ангиоспермия возникла 2 путями, то и система ангиосперм не может
быть монофилетичной"—замечает автор (стр. 137).
Плацентация (семерасположение) может быть и в том и другом
случае или диффузной или упорядоченной (центрально-угловой,
париэтальной,' базальной, колумнарной н т. д.)- Первый случай является
сравнитепьно редким; он первичен, в то время как все остальные случаи
вторичны.
„В основу классификации ангиосперм кладутся свойства гинецея"—
говорит автор (стр. 139). Генетическая классификация гинецеев представ-
ляется им в следующем виде (стр. 140):
Плодолистики закрытые
Рассеянное семерасположение
апокарпия
синкарпия
Краевое семерасположение
апокарпия
синкарпия
. Перегородки целы (центрально-угловая плацентация)
семерасположение без сдвигов
семерасположение ложио-ламинальное
* Перегородки выродились (ложная паракарпия)
Семерасположение колончатое
Семерасположение дониое*’
7 В рукописи А. А. Гроссгейма этот раздел цитируемой части кииги К о-
зо-Полянс кого, вероятно случайно, был пропущен. (Ред.).
42
Система Козо-Паланского Kozo-Poliansky’s System
Плодолистики открытые
Рассеянное семерасположение
Краевое семерасположение
Перегородок нет (типовая паракарпия)
‘ Все швы с семяпочками
Часть швов лишена семяпочек
В полости гинецея перегородки
Перегородки от впячнвания швов
Перегородки от паренхимных новообразований.
Перегородок нет вследствие нх вырождения (колончатая плацен-
тация).
Козо-Полянский дает развернутое графическое изображение
своей системы в виде генеалогического дерева семейств покрытосемянных
(всего 281 семейство). (Табл. 41). Меньшая часть семейств (три ветви)
имеет закрытую карпелляцию (Clausocarpellatae). Ббльшая часть (пять вет-
вей)—открытую (Apertocarpellatae). Семейства не сгруппированы в поряд-
ки; это тоже особенность системы Коз о-П о л ян с к ого, . резко отли-
чающая ее от других систем. В системе особенными обозначениями выде-
лены семейства с первичной диффузной плацентацией (9 семейств), в
разделе Clausocarpellatae выделены семейства с апокарпным гинецеем, а
также семейства с первичным отсутствием сосудов. Хотя в тексте автор
ничего об этом не говорит,—на схеме резко бросается в глаза выделение
некоторых семейств в качестве узловых, дающих начало целому ряду вет-
вей.
Такими семействами средн Clausocarpellatae являются: Tetracentraceae,
Rutaceae, Magnoliaceae, Menis регтасеае; среди Apertocarpellatae: Xyrida-
ceae, Monodoraceae, Flacourtiaceae (дает 19 ветвей), Clusiaceae-
Объем книги не позволил автору дать мотивированное объяснение
взаимного расположения семейств на филогенетической схеме; в основ-
ном оно подчинено тем закономерностям карпелляции и плацентации,
о которых сказано выше. Очень жаль, что в последующие годы автор
не опубликовал нн одной работы, которая освещала бы в деталях его
систему. При изучении филогенетической схемы возникает масса вопро-
сов и недоумений, так как зачастую семейства, которые принято обычно
считать близкими, отрываются автором очень далеко друг от друга (нап-
ример, Umbelliferae и Cornaceae, Rubiaceae и V alerianaceae и многие
другие) и, наоборот, сближаются семейства, обычно рассматриваемые, как
далекие— Umbelliferae и Compositae, Leguminosae и Proteaceae и мно-
гие другие). Семейство Ranunculaceae представлено в виде филогенети-
ческого тупика, в то время как весьма специализированное сем. Menis-
регтасеае фигурирует в качестве крупного филогенетического узла. Ав-
тор говорит, что его схема „заключает ответ (решение) 281 отдельных
филогенических проблем11. Без детальной мотивировки схема эта ско-
рее заключает не ответ, а ряд вопросов.
Н. И. КУЗНЕЦОВ
N- L KUZNETZOV
1864 — 1932
Обзор новейших систем цветковых растений 43
Необходимо отметить, что автор, хотя он и отрицает это (см. выше
цитату), на самом деле стоит на позиции монофилетического происхож-
дения покрытосемянных. Все покрытосемянные произошли от Р г о-А п-
gios perm ае, но путь развития нх с самого начала пошел или по закры-
то-карпеллярному или открыто-карпеллярному типу.
Система Коз о-П олянского представляет крупное явление в ра-
звитии учения о филогении покрытосемянных растений.1 Ее основная цен-
ность—углубленный морфогенетический анализ, позволивший по новому
подойти к решению целого ряда вопросов. Основная идея о двойном
пути карпелляции покрытосемянных, хотя и является дискуссионной,
оригинальна и самобытна. Весьма плодотворно также разработана авто-
ром филогенетическая классификация плацентации. Автор во многом
отошел от проторенных путей и прочно осевших традиций и по новому
взглянул на ряд основных вопросов морфогеиии покрытосемянных (нап-
ример, в вопросе гипогинни и эпигннии, спайнолистности и раздельнолис-
тности околоцветника, происхождении листа и т. д.).
Но, в то же время эта насыщенность системы К о з о-П о л я и с к о-
го морфогенией, примат строения гннецея (см. выше цитату) в вопросе
филогении и системы покрытосемянных, представляют и основную сла-
бость системы. Система К о з о-П о л я н с к о г о односторонне „морфоло-
гична**; все остальные методы и данные филогенетической систематики
(географический, биологический, физиологический и т. д.) обойдены ав-
тором и не нашли отражения в системе.- Эта ее односторонность, „гип-
ноз гинецея** и является слабым местом системы Козо-По ля некого,
по сравнению с другими системами и в первую очередь по сравнению с
системой Кузнецова. Вследствие этого система К о эо-11 о л я н с ко г о
не отражает стадий (ступеней) развития цветковых растений, что так
резко подчеркнуто в системе Кузнецова.
Система К о з о-П о л я и с к ого вызвала при своем появлении рез-
кую и в общем несправедливую критику в Советском Союзе и почти, не
нашла откликов за границей. Такое положение никак нельзя считать
правильным, так как система Коз о-П олянского представляет крупней-
шее оригинальное явление в области систематики растений. Сам автор
в 1945 г. в одной из статей своих („Происхождение цветка Cruciferae в
тератологическом отношении**) замечает, что его гипотеза двойственного
пути происхождения покрытосемянности „не привлекла к себе внимания,-
хотя она плодотворна для филогении и вполне допустима**.
Система Н. А. Буша
N. A. Bush’s System
1924; 1940.
Система, принятая Бушем во втором издании его учебника „Об-
щий курс ботаники, морфологии и систематики растений**, вышедшего в
1924 году, несомненно, создана под большим влиянием системы Кузне-
цова (табл. 42). Буш, каки Кузнецов, выступает здесь, как сторон-
ник полифелитического происхождения цветковых растений, но, правда,
44 Система Буша Bush’s System
отличает уже только две ветви развития. Одна берет начало от Bennetti-
tinae, через гипотетическую, устанавливаемую им группу Protanonales
к порядку Anonales, далее к порядку Pan ales, откуда и развились
пять очень неравноценных ветвей развития основной массы цветковых
растений, другая берет начало от Pteridospermae и через Cordaitinae
непосредственно ведет к гипотетической, устанавливаемой автором группе
Protomonochlamydeae; эта последняя группа дает четыре неравноценных
ветви развития, самая длинная из которых приводит к порядку Centro-
spermae.
Таким образом, эта система Буша, как и система Кузнецова,
пытается примирить взгляды Галлира со взглядами Энглера и в этой
части своей представляет такую же неудачу, особенно ошибочно прямое
выведение Anonales из Sennettitales и прямое выведение Mono-
chlamydeae из Cordaitinae; ничем не обосновано понадобившееся для
этой цели введение гипотетических групп Protanonales и Protomo-
nochlamydeae.
Ценным в системе является отказ автора от расчленения цветковых
растений на одно—и двудольные, раздельно—и сростнолепестные; линии
развития и взаимные отношения порядков в основной массе цветковых
растений также намечены Бушем для своего времени достаточно пра-
вильно.
В заключительной части учебника „Курс систематики высших ра-
стений*, изданного в 1940 году, Буш вновь дает схему системы цветко-
вых растений (табл. 22). Эта система, доведенная до порядков, основана
на принципах Г а л л ир а-Б е с си, но все же в ней еще сохранились не
которые пережитки идей Энглера и Веттштейна. Буш произво-
дит все цветковые из группы Polycarpicae', другими словами, он от-
казался от бифилетизма своей первой системы 1924 года. Система гра-
фически изображена в виде филогенетического дерева; кроме группы
Polycar picae, которая состоит согласно текста из двух порядков
(Anonales и Pan ales), на схеме изображено 43 порядка цветковых
растений; некоторые порядки понимаются очень широко, например, сем.
Palmae включается в порядок 8padiciflorae- В книге Буша рас-
смотрено всего 149 семейств цветковых растений (из более 300) и схема,
построенная в основном на базе этих семейств, не является всеобъемлю-
щей, а до некоторой степени выборочной схемой цветковых растений.
Б у ш не признает деления цветковых на одно—и двудольные, иа
свободно—и сростнолепестные, солидаризируясь в этом отношении со
взглядами Кузнецова. В этом несомненно ценность системы Буша.
Всего Буш отмечает 6 неравноценных по числу порядков и общему
систематическому уровню линий (ветвей) развития цветковых растений.
Линии эти следующие; i) сравнительно мало развита линия, ведущая че-
рез Р i per ales и Р andanales и включающая пальмы и Sp a die ifl о-
гае. Сильно развита линия на базе Не1оЪ гае, дающая две основные
ветви, одна из которых заканчивается Glumiflor ае, другая — Jficro-
spermae. Наиболее сильно развита линия, узловым порядком которой
’ Обзор новейших систем цветковых растений 45
является Rosales', крайними членами развития этого узла являются Муг-
tales, Tubi florae, Eupho rb iales, Rub iales, Leguminosae- 4-я
линия ведет развитие в общем на базе Parietales и доходит до высоты
развития С ampanulatae- Пятая линия очень своеобразна; здесь нет
узловых порядков, через Мyrieales она, с одной стороны, доходит до
J uli a ni ales, а с другой—через U rticales и С entros регтае дает
две вершины: Plumb а д inales и Р rimulales- Шестая линия заклю-
чает единственный порядок—V er ti с 1'll at ае-
Уже из этого краткого обзора видна полная неправильность трак-
товки пятой линии развития, которая целиком построена на взглядах
Энглера о происхождении С entrosperтае и его производных из
Monochlamydeae-
Таким образом, мы видим, что даже в 1940 году такой крупный
систематик, как Буш, не сумел полностью освободиться от традиций
школы Энглера и Веттштейна и отдал им дань в своей системе, тем
самым значительно ее обезценивая (ср. с табл. 23).
Во всяком случае, система Буша 1940 года представляет собой опреде-
ленный шаг вперед по пути упорядочения и дальнейшего оформления прин-
ципов Г а л л и р а-Б е с с и. Многие частные решения Буша вполне пра-
вильны и стоят на высоте современных знаний, например, выделение по-
рядка V erticillat ае в особую самостоятельную ветвь развития, взаи-
мное расположение порядков в наиболее богато расчлененной третьей
линии развития (через Rosales) и некоторые другие. На графической
схеме Буша довольно наглядно показана высота развития, достигаемая
различными порядками и довольно четко вырисовывающая порядки узло-
вого значения.
Система П. М- Жуковского
Р. М. Zhukovsky’s System
1938, 1940.
!
В первом издании своего учебника „Ботаника" Жуковский
полностью сохраняет систему Энглера, хотя и признает, что „она
и является по ряду звеньев несколько устарелой". В тексте упоминаются
системы Кузнецова и Козо - По л я н с к ого.
Во втором издании своего учебника Жуковский принимает си-
стему Бесси в обработке Пула с некоторыми изменениями, а именно:
„Так, например, вслед за многоплодниками излагаются однопокровные
(монохламидные) растения, т. е. та группа растений, которую Ветт-
штейн, Кузнецов и другие считают первичной. Соответственно этому
во втором изданий этой книги изъяты описания семейств некоторых моно-
хламидных из тех порядков, в которые они ^помещены в системе Бесси,
и перенесены в порядки системы В еттштей на". Эти изменения очень
характерны и доказывают насколько живучими являются взгляды Энгле-
ра и Веттштейна и как трудно, даже наиболее передовым ученым на-
шего времени, изжить их до конца. С указанными выше изменениями
46 Системы Жуковского.... Тахтаджяна Zhukovsky’s. Takhtajan’s Systems
система, принятая Жуковским в 1940 году носит, половинчатый ха-
рактер и не может считаться шагом вперед в вопросе построения новой
системы цветковых растений.
Система Б- А- Келлера
В. А. К е I ler’s System
1935
Во втором издании своего курса „Ботаника" акад. Б. А. Келлер
приводит схему Р. Пула с некоторыми незначительными изменениями и
сокращениями. Об отечественных системах в курсе упоминаний нет
(табл. 43).
Система М- И. Годен к и на
М. I- Golеnkin’s System
1937
В своем учебнике „Курс высших растений'1 проф. М. И. Голен-
кин принимает в общих подразделениях покрытосемянных систему Энг-
лера, отличая два класса: Dicotyledones и Monocotyledones и разбивая
класс двудольных на два подкласса: Choripetalae (раздельнолепестные) и
Sympetalae (спайнолепестные). В расположении порядков и семейств по-
крытосемянных у Голе н кин а явно проступает тенденция, не чуждая,
как мы видели, и Кузнецову, как бы примирить противоположные
взгляды Энглера-Веттштейна и Г а л л и р а-Б е с с и. Так, первым
порядком в системе двудольных стоит Poly с ar pie ае (Галлир), и в
тоже время третьим—Piperales-, четвертым—S л и (я? aZ е s, пятым—
Urticiflorae (Энглер). Однодольные Голенкин считает производ-
ными от двудольных и помешает их в конце системы.
В таком виде, какова изложена в учебнике, система Голенкина
не имеет никаких принципиально новых черт и расположение порядков
и семейств в ней является очень мало убедительным. Сторонников и про-
должателей эта система среди систематиков не получила.
Следует отметить, что в обзоре систем, помещенном 1 оленкиным,
совершенно не упоминается такой реформатор системы, как Бесс и, нет
ни одного слова об отечественных, задолго перед этим опубликованных
оригинальных системах Кузнецова и Ко з о-По л я н с ко г о.
Система А. Л. Тахтаджяна
A. L. Takhtajan’s System
1942; 1959.
Начиная с 1941 года Тахтаджян опубликовал ряд работ, в
которых он разрабатывает вопросы, связанные с эволюцией плацен-
тации высших растений. На базе филогенетических выводов, сделан-
ных им на основе изучения явлений плацентации в 1942 году Тах-
таджян предложил свою систему цветковых растений, изображенную
Обзор новейших систем цветковых растений
47
графически в виде „схемы филогенетических взаимоотношений порядков
Angiospermae" (табл. 24).
Исходным типом плацентации для покрытосемянных был маргиналь-
ный, : связанный с перисто-тройственным или многократно-тройствен-
ным прототипом листа. Вследствие „веббинга" и редукции семяза-
чатки из краевого перешли в поверхностное и получилась „рассеянная"
(ламинальная или диффузная) плацентация, являющаяся, таким образом,
вторичной. „Предок покрытосемянных должен был иметь маргинальную
плацентацию" (1941: 149). В этом выводе Тахтаджян резко расходится
с К о з о-П о л я н с к и м (1922), которым высказана была идея о примити-
вности диффузной плацентации.
Тахтаджян различает три типа ценокарпного гинецея: синкарпный
(многогнездный с центрально-угловой плацентацией), паракарпный (одно-
гнездный с париэтальной плацентацией), который может быть выведен из
синкарпного и лизикарпный (одногнездный с колончатой плацентацией),
которая также возникает из синкарпного путем вырождения перегородок.
Таким образом, схема эволюции гинецея может быт представлена
в следующем виде:
паракарпный лизикарпный
синкарпный
t
апокарпный
И в этом коренном вопросе Тахтаджян резко расходится с представле-
ниями школы Козо-Полянского. Мы видели, что согласно Козо-
Полянскому ценокарпия покрытосемянных двойственна, идет по двул/
направлениям: срастания уже замкнутых плодолистиков и срастания еще
не замкнутых плодолистиков, что схематически может быть представлено
следующим образом:
лизикарпный
f
синкарпный
t
апокарпный паракарпный
Другими словами, К о з о-П о л я н с к и й считает паракарпию первичным
аперто-карпеллярным типом. Тахтаджян считает ее вторичным, про-
изводным от синкарпии (путем расклеивания и разворачивания плодоли-
стиков) и этим самым, по-видимому, считает исключением самую возмож-
ность возникновения ценокарпии путем срастания открытых плодолисти-
ков. Вследствие этого, система Тахтаджяна явственно монофилетич-
на, в то время как у Козо-Полянского в системе проявляется
определенная двойственность (но все же не бифилетичность).
48 Сйстема Тахтаджяна Takhtajan’s System
Соответственно изложенному эволюция плацентации представляется
Тахтаджяну в следующим виде.
Гипотетический тип цветковых имеет ламинальную плацентацию,
происшедшую из маргинальной плацентации прототипа. Этот ламиналь-
ный тип выражен у примитивных цветковых двумя вариантами: лами-
нально-латеральным и ламинально-медианным, при чем второй является
явственно производным от первого; основная линия развития плацентапии
цветковых растений шла не по линии ламинальной, а по линии сутураль-
иой плацентации; центральноугловая плацентация проявлятся уже у апо-
карпных гинецеев и достигает своего высшего развития у синкарпных,
где можно отличать ряд ее вариантов; от этой центральноугловой пла-
центации синкарпного гинецея идут две линии развития: х) с одной сто-
роны развивается париэтальная плацентация паракарпного гинепея, по
сути дела тоже сутуральная, но швы здесь вследствие расхождения от
расклеивания плодолистиков приобретают париэтальное положение и 2)
колончатая плацентация лизикарпного гинепея (тоже по существу сутураль-
ная, так как центральная колонка в основном состоит из остатков швов);
редукционные явления во всех рассмотренных случаях приводят к значи-
тельному уменьшению числа плодолистиков, а также числа семязачатков
в завязи.
Исходя из изложенных идей Тахтаджян считает наиболее при-
митивной группой цветковых порядок Ranales и в нем семейства Lar-
dizabalaceae, Nymphaeaceae, Rutomaceae и Hydrocharitaceae, особенно под-
черкивая архаичность сем. Lardizabalaceae-
На графической схеме, представляющей систему покрытосемянных,
изображено 54 порядка в их филогенетических взаимоотношениях, ос-
нованных в основном на строении гинецея и характера плапентапии. В
этой схеме, как и в системе К о з о-П о ля н с ко г о, нет никаких следов
пережитков идей Энглера и Веттштейна.
Примитивной группой цветковых являются Ranales давшие в ви-
де основных ветвей следующие: i) слепо заканчивающуюся небольшую
ветвь Ar is tol о ch i ales, 2) ветвь через Н ата т eli d al es (с боковым
слепым положением на ней Rosales, 3) ветвь через Th eales—самую
разветвленную и богатую порядками, 4) небольшую слепую Р i per ales
и 5) ветвь через Nymphaeales, весьма богатую и разветвленную, со-
ответствующую в общем однодольным прежних систематиков.
Некоторые частности этой системы представляются мало обоснован*
ными и резко расходящимися с обычными систематическими представле-
ниям. Это касается прежде всего положения порядка Rosales, считаю-
щегося слепой ветвью без продолжения; более правильно считать этот
порядок не конечным, а узловым, давшим в дальнейшем богатое филоге-
нетическое рссчленение; это настолько прочно доказано, что во всех но-
вейших системах порядок Rosales рассматривается именно так. С дру-
гой стороны, непонятно и, по нашему мнению, совершенно не обос-
новано, придание порядку The ales крупного узлового значения;
по существу этот порядок Theales играет в схеме Тахтаджяна ту
Обзор новейших систем цветковых растений 49
же роль, как порядок Но sales в схемах других систематиков; в тексте
нет объяснения этому положению. Опять же возбуждает большое сом-
нение порядок Нamamelidales в качестве узлового для целых четырех
порядков, при чем, во всякое случае, выведение из Н amamelidales
порядков F ад ales и U rticales явно неправильно и не соответст-
вует известным на сегодняшний день систематическим данным об этих
порядках. Узловое положение, не вызывающее особых возражений, в
в схеме Тахтаджяна занимают порядки Liliales, С elastr ales,
Geraniales-
Оригинально задуманная и построенная схема Тахтаджяна пред-
ставляет определенный вклад в науку о системе цветковых растений.
Проведение через всю- систему единого морфогенического принципа,
полное изъятие энглер - веттштейновских влияний, придает системе
цельность и стройность. Но, в то же время, о системе Тахтаджяна
нужно сказать тоже, что было сказано о системе Козо-Полянско-
го—она является системой, построенной на одном положении, т. е. она
односторонняя и не может претендовать на универсальность. Примат пла-
центации, о котором говорит система Тахтаджяна, конечно,—слишком
упрощенное понимание филогении цветковых; филогения цветковых—го-
раздо более многогранная проблема. Кроме того и сама плацентация еще
недостаточно изучена у многих групп и толкование ее во многих случаях
является спорным (например, у Cactaceae по К о з о-П о л я н с ко м у —
первичная ламинальная,по Тахта джя ну—вторичная псевдо-ламиналь-
ная и т. д. ) Это не позволяет с полной убежденностью защищать
то или иное положение некоторых порядков в системе. Заслугой автора
является привлечение внимания к одному из важнейших филогенетичес-
ких признаков и оригинальная разработка некоторых вопросов с ним свя-
занных.
*По сравнению с системой, опубликованной в 1954 г-> в последний
вариантчсвоей системы, изданной в 1959 г. („Die Evolution der Angiosper-
meu“), А. Л. Тахтаджян внес более или менее существенные измене-
ния (табл. 25). Прежде всего, он расчленил прежний порядок Мадпо-
liales, выделив из его состава порядки Laurales, Illiciales и
Trochodendrales- Первый порядок, объединяющий 8 семейств, обра-
зует в новом варианте особую эволюционную линию от основного пред-
коАого М ад noli ales, правда завершающуюся вслепую; второй порядок
Illicial es, в составе 2 семейств, в начале же ответвляется от „рана-
лиевой" линии развития и тоже заканчивается вслепую; третий порядок
из 2 семейств, также в виде слепой ветки, связан с линией развития
„сережкоцветных“. Вторая, новая, эволюционная линия, непосредственно
исходящая из М адп о На les— это линия, приводящая к порядку Nelu-
тЪ on ales с семейством Nelumb опасеае, выделенному из порядка
Nymphaeales- Кроме того, в новом варианте, на центральной линии
развития, отходящей от ДГagnoliales, в начале же возникает узловой
порядок Dilleniales, по существу ставший исходным для четырех глав-
ных ветвей с такими крупными узловыми порядками, как The ales,
4. А. А. Гроссгейм и М. Ф. Сахокиа
so
Система Гроссгейма Grossheim’s System
C istales, Rosales и Cunoniales, при чем последний охватывает
ряд семейств, входивших в предыдущем варианте системы в состав Ro-
sales- Таким образом, в новом варианте системы упомянутым порядкам
придано более естественное содержание. Относительно второстепенные
изменения в варианте 1959 г. вызваны, главным образом, увеличением
числа порядков—82 против 54 в прежнем (19^4 г.) варианте (ср. табл.
24 и 25)*. (М. С.).
Система А. А. Гроссгейма
A. A- Grossheim’s System
194S
Система, опубликованная в 1945 году А. Л. Гроссгеймом, ис-
ходит из основных положений школы Г а л л и р а-Б е с с и. В ней сделана
попытка отобразить в системе те биологические предпосылки, которые
определяют в конечном счете то или иное развитие генеративных орга-
нов, а также отобразить основные ступени развития цветковых растений.
Автор отличает в качестве ступеней развития три прогрессивные
стадии: простоцветковые (Protanthophyta), срелнецветковые (Mesa-
nthophyta), и высшие цветковые (Hypsanthophyta), а также одну рег-
рессивную стадию—опрошенные цветковые (Opisthaifthophyta). Ука-
занные ступени устанавливаются не по одному какому-либо морфологиче-
скому признаку, а по совокупности признаков генеративных и, отчасти,
вегетативных органов. На схеме филогенетического дерева эти ступени
находят графическое отображение в виде концентрических кругов, прин-
имающих направление эволюции от центра изображения к его периферии
(табл. 26).
Далее автор рассматривает основные пути или линии развития цве-
тковых растений и устанавливает прогрессивную линию развития насеко-
моопыления и совершенно параллельную и равнозначную ей в биологичес-
ком отношении прогрессивную линию развития ветроопыляемости.
Третья линия биологического развития цветковых растений, приво-
дящая в итоге к регрессивным формам генеративных органов—гидрофилия.
Также регрессивна четвертая основная биологическая линия развития—
переход к гетеротрофному питанию. Различной штриховкой кружков, оз-
начающих порядки, на схеме показана роль в системе цветковых расте-
ний каждого из намеченных путей биологической эволюции.
Автор отказывается нацело от устарелого понятия однодольных и
двудольных, раздельнолепестных и сростнолепестных, однопокровных и
вообще от всех построений, основанных на примере одного какого-либо
признака, и отличает одиннадцать основных стволов развития цветковых
растений, давая им соответствующие названия. 7 из этих стволов берут
начало через порядок Ranales, 3—через порядок Anonales- Сам по
себе ствол I Batrachiophyta, заключающий порядки Ranales и Anona-
les, занимает на схеме центральноузловое, исходное для всех остальных
стволов и ветвей положение; система монофилетична. От ствола Batra-
chiophyta • произошли Дледуюшие стволы: II Hamamelidophyta—короткий
Обзор новейших систем цветковых растений
51
ствол ветроопыляемых растений, III Melophyta—наиболее систематически
расчлененный ствол цветковых растений с преобладанием насекомоопыляе-
мых, IV Teichospermatophyta—также обширный ствол исключительно насе-
комоопыляемых растений, V Proteiophyta—не сильно расчлененный ствол,
для которого характерны явления перехода к гетеротрофии, VI Centrospe-
rmatophyta—богато расчлененный ствол, с преобладанием насекомоопыления,
VII Ebenophyta—короткий, не расчлененный ствол, VIII Krinophyta—богато
расчлененный ствол, давший мощные и высокоспециализированные вет-
ви, как насекомоопыляемых, так и ветроопыляемых растений.
Порядок Anonales дал начало развитию трех стволов: IX Spa-
thophyta—довольно широко расчлененного ствола со своеобразным типом
приспособления к насекомоопнляемости, а также к ветроопыляемости,
X—Itephyta—короткий своеобразный ствол с преобладанием специализи-
рованных ветроопыляемых и XI Kerkidophyta—также короткий очень
своеобразный ствол, высокоспециализированный.
Некоторое представление о явлениях эволюции во времени дает вза-
имное расположение порядков на схеме по направлению от центра схемы
к ее периферии. Географический принцип также находит свое отражение
на схеме, правда, в такой же пассивной форме, каки в системе К у з н е-
ц о в а.
Масштабы развития, т. е. количественное выражение эволюции, отра-
жено графически тем, что кружки на схеме, изображающие порядки,
даны различного диаметра в зависимости от количества видов, содержа-
щихся в порядке. t
В графической схеме (табл. 26) размещено всего 67 порядков цвет- ,
ковых растений; к схеме приложен список стволов, ветвей, порядков и
семейств покрытосемянных растений, так что система доведена до семей-
ства. Объем семейств и порядков принят в основном по П у л л е; всего
в списке приведено 303 семейства, против 388 семейств Гарольда Мол-
д ен к е (1944)-
Система Гроссгейма не вносит чего-либо принципиально нового
в вопросы филогении цветковых растений, но представляет попытку пред-
ставления системы цветковых растений на Бесси-Галлировской основе с
учетом всей многогранности проблемы. В системе нашли отражение био-
логические предпосылки эволюции цветковых растений, подчеркнуты эта-
пы развития, выделены узловые (не специализированные) и специализиро-
ванные порядки. Во многих своих положениях система стоит на уровне
наших современных систематических познаний и представляет некоторый
их синтез.
Конечно, и в ней остается ряд дискуссионных вопросов, как, напри-
мер, нужно ли Ebenophyta считать особым стволом, действительно ли
Itephyta связаны с Anonales, правильно ли происхождение Fagales
и других родственных порядков от Anae ar di al es, правильно ли поло-
жение в системе порядка U rticales и т. д. и т. д. Но, всё это—воп-
росы дальнейшей разработки системы, основные принципы которой можно
считать в общем достаточно обоснованными.
52 Система Комарницкого и Ураиова Komarnitzky’s a. A. Uranov’s System
Система Н. А. Комарницкого и. А- А- У района
N- A. Komarnitzky’s and A- A* Uranov’s System
, 1951; 1962
Система Комарницкого иУранова представляет собою частич-
ное видоизменение системы Н. А. Бутан изложена в главе „Обзор'
эволюции покрытосеменных растений" во втором томе учебника „Бота-
ника", составленного коллективом авторов [Л. И. Курс а новым, Н. А.
Ком а рницк им, К. И. Мейером, В. Ф. Раздорским и А. А.
Урановым (1951).] Сами авторы о своей схеме говорят, что она в основ-
ном соответствует филогенетическим представлениям Буша, „но принята
здесь с рядом существенных изменений в частностях и в ином граф-
ическом изображении" (табл. 27).
Авторы, как и многие их предшественники, считают, что от древне-
йших предков покрытосемянных обособился ряд линий эволюционного
развития. Из намеченных в схеме линий, основной линией, исходной для
большинства порядков покрытосемянных, является линия многоплоднико-
вых (Р о lyca г pic ае)- От последней возникло несколько ветвей система-
тических групп. Из них упомяну те, в отношении филогенеза которых ав-
торы высказывают иную точку зрения, чем Буш. Так, например, они
выводят из многоплодниковых, как самостоятельную,—ветвь центросемян-
ных (С entr os регтае), связанную с многоплодниковыми посредством
порядка Phytolaccales и в дальнейшем эволюционировавшую в по-
рядок сростнолепестных—Р rimulales- Мнение это более приемлемо,
чем мнение Буша, рассматривающего С entro s.permae, и,'связанного
с ним Р rimulales, конечными производными от Monochlamydeae. Сen-
tros регтае—безусловно древняя и филогенетически обособленная
группа, заслуживающая изображения самостоятельной линией. В этой
связи замечу, что древность С ent г о s per тае вполне убедительно дока-
зана М. М. Ильиным. Такую же особую филогенетическую ветвь об-
разует в схеме порядок G ut t if е г ales, дальнейшее развитие которой за-
вершается порядком Eric ales- В системе Буш эти порядки также
представлены в виде производных, а именно, от Parietales- Парал-
лельной независимой ветвью развилась от Р oly с ar pi с ае, согласно дан-
ной схеме, и ветвь Р ar i et ale s, давшая в начальном этапе своего ста-
новления боковую ветвь Rho е ad ales, а в дальнейшей своей эволюции
приведшая к порядкам $ ynanther ales и С ucurbitales, независи-
мым друг от друга.
Как и в системе Буша, наиболее крупная и узловая ветвь Poly-
car picae — это Rosales, четвертая ветвь, исходная для большинства
представителей двудольных, в свою очередь развившая несколько групп
порядков, прошедших самостоятельные пути эволюции.
Отдельной короткой ветвью от Р oly с ar pi с ае изображен поря-
док Р i р в г ales-
Наконец, последнюю особую ветвь, исходящую также от Poly car-
pic ае, составляют порядки однодольных. И-здесь, как в большинстве си-
стем, узловое место отведено порядку лилиецветных (Liliiflorae), из-
Обзор новейших систем цветковых растений
53
которых филогенетически выводится (через Е п anti obi ast ае), в отли-
чие от системы Б у ша, даже порядок Glumiflor ае- В системе Буша про-
исхождение последнего связывается (также через Enantioblastae)
непосредственно с Н elobiae', эта трактовка, мне думается, более близка
к истине.
Остальными, сравнительно мелкими линиями, не получившими даль-
нейшего развития или развившимися слабо, являются следующие: линия
однопокровных, разветвления которой увенчаны порядками Jugland ales'
Р oly д on al es, U rticales и. Fa gales (не образовавших в противо-
положность системе Буша, как отмечено выше, узловой порядок Cent-
er os регтае с производными Р rimulales и Plumbaginales)', в
самом начале от нее отщепилась линия порядка Sal г с ales, линия Ver-
ticillatae (как и в схеме Буша), и, кроме того, линия Sant alales.-
В учебнике „Систематика растений** (1962), составленном в соавтор-
стве с Н. А. Комарнипким и Л. В. Кудряшовым, А. А. Ура-
нов несколько изменил упомянутую выше схему 1951 г. и снабдил ее
-более пространным пояснительным текстом (глава „Обзор эволюции по-
крытосеменных растений**). В этом новом варианте филогенетической схе-
мы покрытосемянных (табл. 28). в отличпе от предыдущего варианта, во-
1-х, отделены от порядков зонтикоцветных (U mb elli florae) И маре-
ноцветных (Rubiales) порядки терпентинных—Terebinthales (пос-
ледний расширен включением в него фисташниковых—А па car diales
и сапиндовых—S a pin dales), бересклетоцветных—С elas trales и кру-
шиноцветных—R h а т па les, в виде самостоятельной ветви, которая свя-
зана в анцестральной части с ветвью порядков геранецветных (Gerania-
les), мальвоцветных (Jtf alv ales) и молочаецветных (Е и ph orb tales)',
обе ветви исходят из розоцветных (Rosales) общим основанием. Во-
2-х, порядок гречихоцветных (Р oly д on al es) выключен из состава
Monochlamydeae и поставлен в родство с центросемянными (С emtr os-
perm а е), в состав которых помешено семейство Phytolaccaceae, в
прежней схеме фигурировавшее в особом порядке Phytolaccales', по-
следнее семейство не без основания рассматривается как предковое в си-
стеме Centr os регтае- В отношении филогенетического положения
порядка Н amamelidales Уранов справедливо высказывается, в согла-
сии с некоторыми авторами, за родство этого порядка с Monochlamydeae, а
не o' Rosales, как многие филогенетики. Поэтому, в схеме он помещен в
основании ветви однопокровных. Кстати, отмечу, что не лишено основа-
ния мнение Уранова о самостоятельном происхождении Monochlamy-
deae от древнейших предков покрытосемянных^ независимо от Р oly car-
pic ае- Monochlamydeae—безусловно, древняя группа. У р а н о в склонен
считать однопокровные даже особым классом (in lift). Вместе с тем, он
.разделяет точку зрения ряда авторов о редукционном развитии цветка
этой группы.
54
Система Куприяновой Kuprianova’s System
В связи с происхождением покрытосемянных Ура нов высказывает
вполне допустимое предположение о неустойчивой энтомофильности древ-
нейших предков покрытосемянных, давших начало двум линиям: одной
(главной) энтомофильной, развившейся через современных Р oly сагргсае
во многие производные от последних, и, второй —линий с неустойчивой
энтомофилией, с ответвлениями, как энтомофильных (Proteales, S ali-
cales, энтомофильные Н а т am е I id ales), так и анемофильных поряд-
ков (ряд однопокровных).
В общем следует отметить, что изменения, внесенные К омар н пи-
ки м и Урановым в систему Буша, представляют определенный ин-
терес. ' ' ,
Система однодольных Л. А, Куприяновой
L. A. Kuprianova’s System of Monocotyledons
1954
Филогенетическое исследование Л. А. Куприяновой касается
лишь класса однодольных, изученного ею с палинологической точки зре-
ния. Палинологические данные в последнее время всё шире используются,
как за рубежом, так и в Советском Союзе, для разрешения вопросов
филогении покрытосемянных. Хотя, в этом направлении предстоит сделать
еще очень многое. Говоря о значении морфологического строения пыль-
цевых зерен для установления филогенетических взаимосвязей между се-
мействами и родами, Куприянова категорически высказывается за то,;
что это может быть „с большей легкостью11 достигнуто именно на основании
признаков пыльцы, чем на основании внешних признаков, которыми обы-
чно только н руководствуются систематики. Применение палинологи-
ческого метода, безусловно, окажется плодотворным и его (как и другие
методы) нужно всемерно поощрять, но не следует его переоценивать, недо-
оценивай значения „внешних признаков11, т. е. собственно морфологии, ко-
торой всегда будет принадлежать главенствующая роль в проблеме филоге-
нии растений. Но, как бы ни было, каждое новое палинологическое ис-
следование, безспорно, представляет большой интерес, поскольку в пелом
изучение вопросов филогении растений примет более комплексный ха-
рактер.
Куприянова намечает следующие этапы морфологической эволю-
ции пыльцевых зерен. Наиболее архаичным морфологическим типом среди
пыльцы однодольных она считает зерна с трехщелевым отверстием; такие
зерна характерны для представителей тропической и субтропической
флор южного полушария. Щелевые зерна имеются и у низших предста-
вителей двудольных, таких, как магнолиевых. Последующим структур-
ным типом зерен с точки зрения эволюции является бороздный тип, даль-
нейшее эволюционное развитие которого получило выражение в поровых
зернах. Но, последнее развитие протекало у обоих классов различными,
путями, приведшими в случае однодольных, преимущественно, к одно-
поровой пыльце, а двудольных—трех-многопоровой, При этом, автор уста-
Б. М. КОЗО-ПОЛЯНСКИЙ в. М. KOZO-POLIANSKY
1890 — 1957
Обзор новейших систем цветкових растений
55
навливает существенную закономерность в отношении структурного разно-
образия пыльцы: наибольшее разнообразие последней наблюдается у тех
представителей, как однодольных, так и двудольных, которые располо-
жены на низких филогенетических уровнях (соответственно у Liliaceae
и Palmae, и, у Р oly с ar pi с ае). На основании палинологического изуче-
ния однодольных Куприянова предлагает схему филогенетического
родства основных семейств этого класса (табл. 29). В ней наглядно по-
казаны три линии развития, исходящие из Proangiospermae- Одна линия
идет от Liliaceae, и у семейств, расположенных по ней, преобладает бо-
роздный тин крупной пыльцы с богатой орнаментацией или пыльца пред-
ставлена скоплениями в виде поллиниев (Orchidaceae)- Эволюция пред-
ставителей этой группы семейств протекала в направлении энтомофилии.
Группа лилиецветных в большинстве систем предшествующих, а также
отдельных последующих авторов, рассматривается как узловое филогене-
тическое звено, исходное для большей части однодольных.
Вторая линия эволюции начинается от Palmae, и, семейства, относя-
щиеся к ней, характеризуются, главным образом, однопоровой мелкой
пыльцой с слабо выраженной орнаментацией.
Обе линии развития — „лилиецветная11 и „пальмовая11—исходят от
общего предк-а. При чем, обе начальные группы—Palmae и 'Liliaceae—
Курприянова считает наиболее древними среди однодольных, и не
менее древними, чем Р oly с ar pi с ае-
Несколько обособленное место отведено автором системы семействам
Агасеае и Lemnaceae, близким к пальмовой группе семейств. Пыльца их
представителей отличается от пыльцы пальмовых и лилейных отсутствием
зерен трехщелевого типа, а от лилейных еще—наличием сетчатой скульптуры
С округлыми ячейками и сплошными стенками. По признаку однопоровой
пыльцы Lemnaceae проявляет родство с семействами „пальмовой линии11,
а именно с Pandanaceae, S parganiaceae, Gentrolepidaceae и др. Третью
линию развития составляют семейства порядка Н elob iae в объеме,
принятом Веттштейном или порядков Н в I о b i а е и Tri иг id ales
в понимании Энглера и Гроссгейма. Наиболее существенной чертой
этой группы следует считать, по автору, наличие особого типа зерен со
многими порами по всей поверхности и бугорчатыми мембранами. Вместе
с тем, для нее характерно отсутствие первичных типов зерен (трехщелевого
вовсе, нет, а однобороздный очень редок)—признак производности их
пыльцы.
Эта линия развивалась по пути приспособления к обитанию в вод-
ной среде и она проявляет непосредственное родство с некоторыми при-
митивными двудольными (с Ranunculaceae и, вероятно, с Nymphaeaceae).
Одним из важных результатов исследования данного автора является более
точное познание отдельных систематических групп, в частности, выделение
особого семейства Limnocharitaceae, включающего роды Limnocharis, Те-
nagocharis и Hydrodeis, рассматривавшиеся систематиками в составе се-
мейства Butomaceae. Новое семейство помещено автором в группе Helo-
biae и оно по характеру пыльцы, отличающейся скульптурной экзиной,
56
Закючение Conclusion
сближается с Hydrocharitaceae, тогда как, собственно Butomaceae (с ро-
дом Butomus) расположено в группе семейств линий от Liliaeeae-
В заключении Куприянова, на основе собственных палинологи-
ческих исследований, соглашается с мнением ряда предшествующих ав-
торов и также делает вывод об искусственности выделения особого класса
однодольных, представляющих филогенетически весьма разнородную груп-
пу растений. В этой связи заслуживает внимания высказывание А. И.
Толмачева, который придерживается этого же взгляда и предлагает
схему, иллюстрирующую в общих чертах параллелизм в развитии групп
однодольных, подчеркивая именно параллелизм развития и отрицая в дан-
ном случае конвергенцию (сходство в строении);
Nymphaeaceae
Protoauthoppyta (или Polycarpicae)/ ;
\Н elobiae
Protoanthophyta -> L i li i fl о r a e;
Protoanthophyta (или Polycarpicae) ->Pi perale S padiciflo-
rae- [См. примечание на стр. 278 — 279 в кн.: Н. А. Буш, Систематика
«ысших растений. (1959)].
Данные Ку при яновои, подтверждая в основных чертах системы
Кузнецова и, в особенности, Буша в части конечнцх ступеней эволю-
ционного развития однодольных, существенно иначе освещают вопрос пе-
рвоначальных этапов становления однодольных. Выведение же однодоль-
ных из многоплодниковых, именно через Нelobiae, безусловно, более,
чем спорно. Это явствует и из сравнительно морфологического анализа
„внешних признаков".
Несогласие в отношении данной системы Куприяновой выс-
казывает Kimura (1956), считающий, что ее точка зрения не подтве-
рждается палинологическими данными Эрдтмана, с которыми якобы
согласуется именно его система.
* *
*
Заключен и е—С onclusion
Из сделанного краткого обзора ясно, что на сегодняшний день нет
ни одной разработанной системы цветковых растений, которая бы явля-
лась общепризнанной и соответствовала бы уровню знаний,' накопленных
в области филогении растений.
Это критическое положение гораздо резче выражено на Западе, чем
в Советском Союзе. Новейшие системы, предложенные на западе [Пул-
ле, 1938 (см. табл. 30), Г. Молденке, 1944] представляют собой опре-
деленную деградацию—шаг назад по сравнению с классическими концеп-
циями Гал лира и Бесси. Явно (Пу л ле) или бессознательно (Мол-
денке)8 авторы скатываются к давно отжившим воззрениям Энглера
и Веттштейна и стараются вдохнуть живой дух в мертвую, отжив-
шую форму. Более прогрессивные системы Гатчинсона и Пула в сво-
8 Критический разбор системы Молденке см. в рецензии А- А. Г росс-
гейм а, опубликованной в БЖ, 1943 г., №3, р. р. 384—386. (Рёд.).
Обзор новейших систем цветковых растений 57
их последних вариантах (Пул, 1941; Гатчинсон, 1946) ничем по су-
ществу не отличаются от первоначальных концеппий и по ряду причин,
изложенных выше, также не могут претендовать на всеобщее признание.
Оба автора со времени опубликования первых своих вариантов (Гат-
чине он, 1926; Пул, 1928) как бы прошли мимо всего того громадного
количества фактов, которые были добыты за этот период морфогенией
и смежными науками, они ничего не позабыли и ничему не научились.
На сегодняшний день обе эти системы отстали от современного состояния
науки о филогении растений и их переопубликование в почти неизменен-
ном виде только еще раз доказывает прежнее состояние учения о фило-
гении и системе цветковых растений на Западе.
Системы, предложенные в Советском Союзе также не могут претен-
довать на общепризнанность и универсальность, но сами по себе они во
многих отношениях глубже разработаны, оригинальнее и в общем итоге
вносят больше для построения будущей системы растений.
Оставляя в стороне половинчатые, мало оригинальные системы Го-
ленкина, Буша и Жуковского, мы должны отметить крупное до-
стижение в системе Кузнецова, ярко отобразившей представление об
уровнях (ступенях) развития цветковых растений на базе обобщенного
анализа строения цветка. Необходимо подчеркнуть большую оригиналь-
ность и самобытность построений Козо-Полянского, развившего уче-
ние о двойственности карпелляции покрытосемянных. Необходимо отме-
тить также оригинально построенную систему Тахтаджяна на базе
разработанной им теории филогении плацентации покрытосемянных. После-
дняя из предложенных в Советском Союзе систем, система Гроссгейма,
является попыткой построения синтетической системы, с учетом по воз-
можности всех сторон многогранного процесса эволюции, с подчеркива-
нием в первую очередь биологических предпосылок эволюции.
Хотя, повторяем, и все эти системы также далеки еще от универ-
сальной системы будущего, но в обшем они стоят на более высоком науч-
ном уровне и это обстоятельство дает уверенность в том, что плодотвор-
ное развитие и дальнейшая разработка проблем филогении и системы мо-
жет быть и в дальнейшем обеспечена ботаниками Советского Союза.
Какие же из основных проблем филогении цветковых растений мо-
жно считать на сегодняшний день прочно установленными и общепризна-
нными?
Мы считаем, что таких проблем две. Установлено монофилетическое
происхождение цветковых растений и установлено еванциевое происхож-
дение цветка. Конечно, прямых доказательств и достоверных, не подле-
жащих критике фактов ни по тому, ни по другому вопросу мы не имеем,
да по сути дела и не будем никогда иметь, но вся сумма относящихся
сюда фактов и явлений, имеющих не прямое, а посредственное отношение
к вопросу, позволяет ботаникам притти к единообразным выводам.
Можно сказать, что громадное большинство систематиков стоит на
той точке зрения, что такое явление, как единообразно построенный
у всех без исключения цветковых растений половой аппарат, не может
иметь двойственного или многократного происхождения. Это то явление,
58
Заключение Conclusion
которое прочно устанавливает монофилетическое происхождейие цветко-
вых растений и иначе не^может быть рассматриваемо. Мы считаем, что
монофилетическое решение вопроса о происхождении цветковых расте-
ний обш.епризнано.
С другой стороны, многочисленные псевданциевые теории происхо-
ждения цветка, начиная с теории Веттштейна, не выдержали серьез-
ной научной критики. Правда, еванциевая теория также не может опе-
реться на неоспоримые палеоботанические факты, но все относящиеся
сюда факты морфогении цветка говорят за то, что она является единст-
венно правильной и, можно сказать, на сегодняшний день в сущности
общепринятой.
К сожалению,, вопрос о примитивном типе покрытосемянных ее мо-
жет считаться на сегодняшний день решенным. Он решен односторонне и
негативно, т. е. в том смысле, что громадным большинством современных
систематиков отвергнута концепция Энглера и Веттштейна о при-
митивности „однопокровных". Такое воззрение на однопокровные, как
на вторичную группу, можно считать по существу ныне установленным.
* В настоящее время совершенно бесспорно, что „однопокровные"
(Monochlamydeae) филогенетически весьма разнородная группа и, поэтому,
искусственная. Что же касается ее вторичности, то несомненно, что вто-
ричны лишь некоторые ее представители, являющиеся филогенетически
более поздними образованиями (наир., J ugland ales)-
Наиболее правильно считать эту группу полифилетичной, связав от-
дельные . „однопокровные" с различными ветвями филогенетического
древа и рассматривать их образованиями различных времен. Гроссгейм,
в общем, правильно поступил, распределив „однопокровные" по различ-
ным стволам и ветвям; в частности, совершенно верно отнесение Myri-
calia и р г tic alia к особым стволам. Но, все же, филогенетическая
сущность группы полностью не раскрыта. На мой взгляд, остается искус-
ственной ветвь AAyricalia, не вполне естественна и ветвь U rticalia,
так как в ней сомнительно положение Phoipteleaceae, более близкого к
Juglandaceae и т. д. Ur tic alia несомненно один из более древних
представителей среди Monochlamydeae* (М. С.).
Но, другая сторона вопроса—что считать за примитив среди цветко-
вых растений,—все же не может считаться окончательно решенной. Прав-
да, и здесь можно отметить, что громадное большинство современных
систематиков согласно признают примитивной группой порядки Лапа-
les и Anonales (Р oly с ar р ic ае), признавая, по-видимому, у покры-
тосемянных только один путь карпелляции, а именно клаузокарпелляцию.
Но, воззрения Козо-Полянского о двух путях карпелляции ни в
каком случае не могут считатсья опровергнутыми; соответственно взгля-
дам Коз о-П оля иск ого должны существовать и другие примитивные
типы покрытосемянных с апертокарпелляиией. Сам К о з о-П о л я н с к и й
в качестве таких исходных для апертокарпеллярной линии развития ти-
Обзор новейших систем цветковых растений
59
пов выдвигает семейства Nymphaeaceae, Taccaceae, Ufonodoraceae и Xyri-
daceae-
Вопрос этот требует дальнейшие; исследований, уточнений и двля-
ется на сегодняшний день, во всяком случае, дискуссионным, а не решен-
ным окончательно.
* В последнее время, ряд филогенетиков, как отечественных, -гак
и зарубежных, более категорически высказывается против стробилярной
или эвантиевой теории происхождения цветковых растений. В частности,
ими отвергается раналиевый тип растений, как основа филогенетической
системы. Из отечественных филогенетиков против этой теории возражают
М. М. Ильин (1951; i960), Я. И. Проханов (i960), М. В. Клоков
(i960) и др.
В частности, Проханов рассматривает стробил кац вторичное об-
разование.
Вопросу о принципиальной основе построения филогенетической си-
стемы цветковых растений Ильин посвятил специальную статью (1951)1
в которой им изложена также критика эвантиевой теории. Поэтому,
приведу некоторые главные его положения. По мнению Ильина, во-
I, невозможно допустить развитие цветка раналиевых из стробила бен-
неттитовых,. а во-2, 'в основу системы цветковых должен быть постав-
лен не один какой-либо порядок, а тем более семейство, а несколько.
В данном вопросе Ильин исходит из того соображения, что образование
новых таксонов осуществляется параллельными филами, хотя из,,... одно-
го, тесно связанного общностью происхождения филетического корня...“
Далее, по этому поводу он говорит:„... биоморфогенез в процессе фило-
генеза совершается монофилетически, но путем политопности“. Но, это
скорее полифилия, т. е. Ильин, по сути явления, придерживается
взгляда о полифилетическом возникновении цветковых, хотя полифилию
различные авторы понимают по разному.
Относительно также весьма дискуссионного вопроса о предках по-
крытосемянных растений Ильин определенно придерживается того взгля-
да, что они произошли от вымерших типов современых гнеталиевых или
оболочкосемянных (С h I am i d 0 s p e rm a e), считая их филогенетически
наиболее близкими к Angiospermae. Эту близость Ильин видит в
том, что у Chlamidospermae имеются настоящие сосуды (каковых
нет у голосемянных); листья их, в частности у Gnetum, очень сходны с
листьями покрытосемянных; у оболочкосемяных иногда развивается при-
митивный околоцветник, а главное, семяпочка нередко целиком скрыта в
плодолистике, а верхушечное образование ее внутреннего интегумента яв-
но сходно со столбиком и рыльцем покрытосемянных. По существу
Ильин высказывается за псевдантиевую теорию Веттштейна о цветке.
Ильин даже полагает, что предки Angiospermae могли дожить до
нынешней эпохи, сохранившись в современной флоре. Подтверждение-
своему предположению он усматривает в факте существования в совреме
иной флоре Ginkgo и Metasequoia, живших еще на заре возникновения
семянных растений. Аналогичного взгляда придерживается и М. В. К л о-
60 Заключение Conclusion
ков. А. И. Толмачев отвергает подобный взгляд (1960)' Действитель-
но, с этим мнением Ильина едва ли можно согласиться; ныне живущие
растения, как-бы архаичны они ни были, не могут быть признаны непо-
средственными предками покрытосемянных.
Филогенетическое родство покрытосемянных с гнеталиевыми несом-
ненно, но остается спорным вопрос о форме этого родства: нуАно ли
рассматривать покрытосемянные хотя бы непосредственными „потом-
ками® гнеталиевых или их роднит общий предок.
Относительно дискуссии вокруг „раналиевой теории" следует заме-
тить, что ее критика располагает на сегодня немалыми вескими аргумен-
тами и на современном уровне знаний о филогенезе растений эта теория
не может быть признана удовлетворительной. Поэтому, ее, по-видимому,
нужно считать пройденным этапом в филогенетическом изучении покры-
осемянных*. (М. С.).
Также продолжает оставаться нерешенным и возбуждающим диску-
ссии другой основной вопрос филогении покрытосемянных—положение в
системе „однодольных® и отношение их к „двудольным".
Для автора настоящей статьи вопрос этот представляется достаточно
решенный путем, полного отказа от разграничения эти-х двух групп, но
такая Точка зрения не может считаться общепризнанной.
В сущности, авторы западно-европейских систем в большинстве слу-
чаев и не отказывались от признания этих групп и не связывали этот
вопрос с антиэнглеровским направлением в систематике. Уже Галлир,
сначала отрекшийся от однодольных, в последнем варианте своей системы
{1912) восстановил эту группу. Гатчинсон с самого начала стоял на
точке признания группы однодольных, также и Пулле и Молденке;
-только Бесси и Пул более илы менее прочно отступают от общеприня-
того деления цветковых растений на одно й двудольные.
Нужно отметить, что почти все авторы советских, систем покрыто-
семянных (Н. И. Кузнецов, Н. А. Буш, Б. М. К о з о-П о л я иск и й,
А. Л. Тахтаджян, А. А. Гроссгейм) стоят на точке зрения непри-
знания старого деления на одно- и двудольные растения. Но, здесь же
следует отметить, что Коз о-П олянс кий в 1947 году отошел от своих
прежних взглядов на этот вопрос и восстанавливает одно- и двудольные
в своей системе, правда, без опубликованной аргументации этого измене-
ния.
При обсуждении этого вопроса полезно припомнить предложение
Гейнце рассматривать кроме одно-п двудольных еще третий, равно-
ценный им ряд псевдодвудольных (Pseudodicotyledoneae), в общем соот-
ветствующий обычно принимаемой группе Р olycar ргсае-
Во всяком случае, вопрос о целесообразности и правильности вы-
деления крупных групп покрытосемянных на основании строения заро-
дыша должен быть подвергнут дальнейшим дополнительным исследова-
ниям и обсуждению.
' ' - /' . V ' f-- '
• Обзор новейших систем цветковых растений 61
Резюмируя все сказанное выше, можно сказать, что перед системати
ками цветковых растений на сегодняшний день стоит в первую очередь
решение следующих капитальных проблем.
I. Окончательное решение вопроса об одном или о двух первич-
ных типах цветковых растений, т. е. решение вопроса о том, является ли
паракарпный гинецей первичным (апертокарпеллярным) или вторичным
(возникшим в результате расхождения и расклейвания клаузокарпелляр-
рного гинецея). В зависимости от того или иного решения этого вопроса
находится то или иное из возможных построений всей системы цветко-
вых растений.
2. Окончательное решение вопроса о том, нужно или не нужно от-
личать среди цветковых два подкласса: одно-и двудольных растений. Воп-
рос о взаимоотношении между этими двумя группами (параллельное разви-
тие из в основном двудольной основы) можно считать решенным. По
мнению многих систематиков, а также и автора настоящих строк, вопрос
о, ненужности расчленения одно-и двудольных также может считаться
предрешенным, но необходимо устранить, последние неясности в этом во-
просе.
*3а таксономическую самостоятельность Monocoty ledoneae высказы-
ваются и новейшие исследователи. Из современных советских филогене-
тиков эту т.очку зрения защищают А. Л. Тахтаджян (1959), Я- И.
Проханов (i960), М. М. Ильин (i960), А- Я. Вага (i960), А. А.
Уранов (1962) и др.
Более того, некоторые авторы (в том числе Проханов и Ильин)-
считают, что- однодольные и двудольные нозникли независимо друг от
друга от общего предка. Проханов, посвятивший этому вопросу специ-
альное исследование, наиболее существенными отличительными признаками
однодольных от двудольных считает „отсутствие камбия.и кольцевой хара-
ктер прикрепления листьев, а в отношении семедолей не столько их число
(одна), сколько верхушечное положение на оси зародаща“. Семейства одно-
дольных из Н elobiae он квалифицирует как непримитивные, габитуаль-
ное же сходство с родственными Ranales (особенно с Nymphaeaceae) он
связывает с конвергенцией, обусловленной экологической средой, а не
филогенетической близостью. К первичным однодольным Проханов
относит „псевдантовые“ Spadiciflorae (в особенности сем. Palmae,
Cyclanthaceae и Агасеае)- У представителей последних, в частности в сем.
Palmae, отмечает Проханов, сочетаются „псевдантового типа" клубочки
однодольных цветков (подсем. Ceroxyloideae) с их нормальным обоеполым
прототипом (подсем. CoryphoideaeY От „псевдантовой" группы Проха-
нов производит Иelobiae и „эвантовую группу трехчленно-пятицикли-
ческих" в порядке Liliiflorae. Рассматривая однодольные филогенети-
чески более ранней группой, чем двудольные, Проханов ставит первые
в начале системы покрытосемянных.
<62
Заключение Conclusion
Точка зрения о независимом происхождении однодольных и дву-
дольных от общего предка, представляется более правильной, чем иные
трактовки их филогенеза*9. (М. С.).
3. Окончательное решение вопроса о всех возможных путях возни-
кновения эпигинии. В зависимости от точного решения этого вопроса эпи-
гиничные цветки не станут рассматриваться как какая-то однородная
группа и получат в системе разнообразное, соответствующее характеру
происхождения эпигинии размещение.
4. Изучение закономерностей проявления разновозрастности призна-
ков (закона корреспондирующих стадий развития или гетерохронии).
5. Установление наиболее вероятного положения цветковых среди
остальных ныне живущих и известных ископаемых растений, а также це-
леустремленные поиски ископаемых растений—действительных предков
цветковых.
При решении этих основных проблем, также как и всяких других
более частных вопросов филогении и системы цветковых растений, не-
обходимо исходить из следующих методических положений.
I. До сих пор все без исключения системы цветковых растений по-
строены на базе строения цветка; если и были попытки внесения в си-
стему иных критериев, то они являлись робкими и оставались все-же как
бы на.заднем плане системы. Между тем, эволюция цветковых растений
охватывает не только генеративные органы, но все тело растения целиком
со всеми органами, тканями и функциями. Система должна строиться,
таким образом, на базе всех морфологических проявлений и с охватом
всех жизненных функций растений, на базе целостного организма, а не
только его репродуктивных органов. Конечно, между вегетативными и
-репродуктивными органами существуют известные корелляции, но и сами
корелляпии и степень п формы кореллятивности еще слишком слабо изу-
чены, чтобы можно было в этом вопросе б, или м. достоверно опираться
на известные закономерности.
Такой постановкой вопроса о системе предполагается, прежде всего,
насыщение системы биологическими данными, изучение и выявление би-
ологических предпосылок эволюции и влияния биологических факторов
на эволюцию всего организма в целом.
Это один из важнейших принципиальных вопросов построения бу-
дущей системы цветковых растений, так как, именно биологические пред-
посылки определяют и направляют общий и частный характер эволюцион-
ного процесса.
2. Отсюда второе принципиально важное требование к работе над
филогенией и системой будущего: изучение и выявление различных ти-
пов эволюции, другими словами разработка вопроса о сравнительной эво-
* Для вопроса о филогении однодольных значительный интерес представляют
тезисы доклада Я. И. П р о х а н о в а—„Класс однодольные, его таксономическая обо-
снованность и положение в системе сосудистых растений11. 1964. Второе Московское
совещание по филогении растений (см- библиографию). (Ред.).
Обзор новейших систем цветковых растений
63
люции, аналогично тому, как мы изучаем и разрабатываем учение о срав-
нительной морфологии (морфогении), сравнительной эмбриологии и т. д.
В этом, отношени гораздо глубже и дальше ушли зоологи система-
тики.
Разнохарактерность эволюционного процесса и основные его катего-
рии установлены в классических трудах академика С е в е р ц о в а и разраба-
тываются дальше академиком Шмальгаузеном и его школой. Из совре-
менных советских ботаников систематиков только немногие разрабатыва-
ют вопросы филогении цветковых растений под этим углом зрения (Тах-
таджян и др).
Ботаникам необходимо освоить зоологический подход к филогейии
и разрабатывать учение о сравнительной эволюции на базе растительного
мира; нет сомнений, что это не будет слепым подражанием зоологам и
ботаникам предстоит выявить новые и особые закономерности, специфи-
чески свойственные эволюции растительного мира.
Само собой разумеется, что эти различные направления эволюции,
обусловленные биологическими предпосылками, должны найти также
свое отражение в системе и ее графическом изображении, что также
представляет собой область творческого труда и новых достижений.
3- Из признания организма в пелом, как объекта филогенетической
системы, явствует также с очевидной ясностью необходимость комплекс-
ного подхода к этому вопросу. Еще Галлир и многие другие система-
тики после него подчеркивали необходимость привлечения к делу фило-
генетического построения целого ряда подсобных методов, кроме основно-
го сравнительно-морфогенетического. Но, одно дело подчеркивать и дек-
ларировать, другое—претворять и осуществлять. И нужно сказать, что на-
стоящего претворения в науку о филогении, так называемых, подсобных
методов, мы до сих пор не видим.
После сравнительно-морфологического наиболее разработанным мож-
но считать метод сравнительно-анатомический; трудами Жеффри, Бер-
ри, Александрова и его школы, накоплен громадный материал, до-
вольно полно освоенный на сегодняшний день филогенетиками. Больше
того, построены даже целые филогенетические системы или фрагменты их
(Жеффри), основанные специально на анатомических данных. Основным
пороком всех этих систем и построений является их односторонность и
полная неувязка с остальными данными филогении.
Громадное количество фактов филогенетического значения накоп-
лено в последнее время эмбриологией, но здесь прямо можно сказать, что
факты эти в учении о филогении пока что почти не освоены. Также
обстоит дело с фактами цитологическими. Большое значение придается
филогенетическим данным, добываемым новой, находящейся в стадии
усиленного и успешного развития, наукой—палинологией и здесь даже
наблюдается, оцять-таки, известное одностороннее увлечение филогенети-
ков палинологическими данными.
Прочно утвердились в качестве важнейших подсобных методов фи-
логенетической систематики—методы географический и фитопалеон-
64
Заключение Conclusion
тологический; здесь не место вдаваться в обсуждение достоинств и поро-
ков этих методов, скажем только, что их применение было и продолжает
быть чрезвычайно плодотворным для развития учения о филогении ра-
стений и построения системы.
Генетический метод -широко использовывали в систематике особенно-
культурных растений и были даже попытки полной подмены систематики
генетикой. Это крайнее увлечение методом можно считать в настоящее
время изжитым, что нисколько не снижает ценности и важности приме-
нения генетического метода, в качестве подсобного, к филогенетическим
целям. Примером .односторонне построенной на базе примата генетики
филогенетической схемы, может служить схема Эдгара Андерсона,
изображающая число хромозом и систематические отличия Gymnos-
permae (1937)-
*13 1934 г- Андерсон (Е. Anderson, „Origin of the Angiosperms")
высказал гипотезу о гибридогенном возникновений покрытосемянных в
результате скрещивания между прародительскими типами хвойных . или
гинкговых, с одной стороны,' и гнетовых, с другой, и аллополиплоидий
(б. м. сходные идей высказывались раннее и другими). Нод прародитель-
скими типами Андерсон подразумевал вымершие группы (в частности,
семейства, роды), сходные с современными формами по числу хромозом.
Например, по его гипотезе, в результате полиплоидного гибридогенеза,
как следствие скрещивания между Oycadales (n=i2) и Gnetales
(п=7), образовались семейство Magnoliaceae и родственные ему семей-
ства (п=19)-
В СССР аналогичного взгляда на происхождение покрытосемянных
придерживался М. Г. Попов, который еще в самых ранних своих рабо-
тах, а особенно в монографиях по отдельным родам., написанных исключи-
чительно талантливо, решающее значение в эволюции таксонов растений и
даже отдельных флор, придавал гибридизационным процессам. Наконец,
происхождение вообще покрытосемянных, в последних своих работах
(1954, а также в книге „Основы флорогенетики", изданной посмертно в
1963 г.), Попов объясняет этими- же процессами. А именно, он счита-
ет, что покрытосемянные возникли как следствие гибридизации между
гнетовыми и беннеттитовыми в меловой период, только однажды и мо-
нотонно. 'Гаким образом, гибридогенез он объявляет „главным механизмом
формообразования". По его мнению первичным центром возникновения
покрытосемянных должна была быть тропическая зона, а первоначальной
жизненной формой—тропическое вечнозеленое дерево. При этом, по пред-
ставлениям Попова, современные семейственные типы развились „как веер
расщепления", т. е. одновременно появились многие семейства. В этом
явлёнии он мыслит две основные эволюционные линий; ксерофильную и
криофильную; побочными линиями были: гидрофильная, эпифитная, пара-
зитарная и др.
Далее Попов утверждает, что еще у примитивных наземных расте-
ний установились основные морфологические структуры. Так, среди пте-
ридофитов он выделяет 3 структурных типа: х) филлофитный (листовой),
Обзор новейших систем цветковых растений 65
2) артрофитный (стеблевой) и 3) микрофильный (мелколистный). При
чем, отдельные классы характеризовались определенной структурой: папо-
ротники были филлофитами, каламиты—артрофитами, плауны—микрофил-
лами. Следует отметить, что выделение микрофильного структурного типа
нарушает логическое построение данной классификации. Но, уже позже,
в возникновении голосемянных и покрытосемянных, по П о п о в у, имело
место лишь смешение ранее существовавших морфологических типов и
прогресс высших растений выражался, как он полагает, не „в усовер-
шенствований структур, а в убыстрений темпов роста11. Автор категори-
чески утверждает, что это смешение морфологических типов было след-
ствием именно отдаленной гибридизации „сперва между филлофитными и
артрофитными птеридоспермами, а потом между гнетовыми, субартрофит-
ными и беннеттитовыми (субфиллофитными) голосеменными, что и при-
вело в конце концов к образованию покрытосеменного типа с очень сме-
шанной структурой4'. При этом у гибридов, как отмечает Попов, появля-
лся гетерозис.
Относительно эволюционного развития жизненных форм цветковых
растений Попов, как и многие авторы, придерживается взгляда на ре-
дукционное направление эволюции цветковых вообще, в частности и это-
го процесса; последний протекал согласно Попову по схеме: веч-
нозеленое дерево-> листопадное' дерево-> лиана (кустарник)-^многолет-
няя трава-> однолетник. Грант (Grant, i960) же полагает, что пре-
вращение . дерева в траву может осуществляться непосредственно, а не
постепенно, через стадии кустарника или полукустарника. Такую после-
довательность, намечаемую Поповым как единственнную, справед-
ливо отвергает, например, М. М. Ильин, который считает, что в других
случаях процесс может иметь обратное направление: от травянистых к
деревьям (И л ь и н, 1951). Такого же мнения придерживается Я-И. Про-
ханов (i960), полагая, что развитие, наир., С entro s регтае шло от
травянистого типа к древовидному.
Как и отмечает Попов, упомянутые морфологические типы, вод-
них случаях, все представлены в одном естественном семействе, напр., в
сем. Linaceae, включающем все биоморфы, начиная с деревьев и кончая
травами, в том числе и однолетние травы, или же они распределяются в
двух или нескольких родственных семействах (напр, представители Mag-
nolia сеае являются деревьями, кустарниками или лианами, а сем. Напипси-
laceae—лианами и травами, в том числе однолетними). Попов признает
также редукционную эволюцию цветка и плода; она выражается в сокраще-
нии числа его частей и членов последних., т. е., в олигомеризации. Кроме
того, уменьшался размер цветка; вследствие этого возникало соцветие.1
Как утверждает Попов, эти преобразования усиливали признак одно-
половости, проявившийся еще у таких древних семейств, как Casuarina-
ceae, Lauraceae и др. .
По мнению Попов-а гибридизация была повторной и многосторо- -
нней, поскольку имели место вторичные, а затем и последующие скре*-
щивания „между сделавшимися константными первичными семейственни-
5. А. А. Гроссгейм и М. Ф. Сахокиа
66
Заключение Conclusion
ми типами, потом третичные скрещивания между вторичными, опять
ставшими константными родами (например, кустарниковыми) и т. д.
Весьма своеобразен взгляд Попова на вопрос о возрасте семейств
цветковых растений и их системе. Им он не придает филогенетичес-
кого значения, поскольку не рассматривает их как эволюционные эта-
пы. Он отличает лишь первичные (их большинство и представлены дере-
вьями) и вторичные (меньшая часть) семейства, включающие кустарники
и травы. Поэтому, эволюционная последовательность некоторых семейств
ему представляется в следующем виде; Magnoliaceae -> Ranunculaceae', Ru-
Ыасеае—> Caprifoliaceae->Valerianaceae, т. e., по его убеждению, это
„соответствует последовательности их появления на Земле". Исходя из
флорогенетических соображений Попов (1962) утверждает, что „...не су-
ществует различий в возрасте у большинства семейств покрытосеменных,
что они все одновозрастны". Из признаваемых 400 семейств первичными,
среднемелового возраста, он считает 350; остальные он относит к вторич-
ным, большинство которых датирует палеогеном, а меньщую часть (Cista-
ceae, Ripsacaceae)—неогеном. В свете этих воззрений Попов (1962)
категорически отрицает большую древность Ran ales по сравнению с
Gompositae- Напротив, он говорит, что „Ranunculaceae с географической
точки зрения, безусловно, моложе Compositae“- Далее: „...абсолютно
неприемлема точка зрения филогенетиков о том, что Roraginaceae как
стростнолепестное семейство моложе, чем любое раздельнолепестное, на-
пример, многоплодниковое Аппопасеае или монохламидное С asuarinaceae* •
Ясно, что этим Попов отрицает „раналиевую" основу большинства со-
временных филогенетических систем. Систему естественных семейств П о-
пов мыслит только как морфологическую, но не как филогенетическую,
рассматривая таксоны лишь как морфологические категории. Им намечены
четыре подкласса двудольных и один подкласс однодольных: i) Poly car-
pic ае, 2) Sync ar pea е, 3) S у m p e t aleae, 4) Monochlamydeae и 5)
Monocotyledones-- Но, поскольку для Попова эти таксоны являются
лишь морфологическими объединениями, он допускает их любое произ-
вольное взаиморасположение. , Поэтому, на его взгляд, базразлично, бу-
дут ли однодольные помещены перед двудольными или после них; он
также не придает значения тому, будет ли соответствующая эволюцион-
ная линия в системе обозначена вначале порядком Eric ales, а на вер-
шине порядком Synantherales, как в системе Энглера или эти по-
рядки будут размещены в соответствии с системой А. П. Де-Каи дол-
ля, предпочитаемой Поповым.
Естественное скрещивание, бесспорно, имеет большое значение для
формообразовательного процесса, но, едва ли оно является столь решаю-
щим в филогенезе, каким оно представляется сторонникам гибридогенного
происхождения покрытосемянных*. (М. С.).
Громадное «количество фактов, накопленных в последнее время био-
химией растений, еще далеко не полностью освоены филогенетической
систематикой, но и в этой области можно отметить попытки односторон-
него использования этих фактов для целей филогенетических построений.
Обзор новейших систем цветковых растений
67
Из всего сказанного следует, что в настоящее время существует оп-
ределенный разрыв между филогенетическими построениями систематиков
и суммой фактов, накопленных смежными дисциплинами. Данные смеж-
ных дисциплин не освоены еще синтетически для целей построения фи-
логенетической системы, почему все, даже самые новые из предложенных
филогенетических построений являются по существу б. или м. односторо-
нними и тем самым неполноценными. Будущая система цветковых расте- ,
ний должна стать подлинно синтетической; вся многогранность проявле-
ния эволюционного процесса в его морфологическом, биологическом, ге-
ографическом и историческом проявлениях, должна быть отражена в си-
стеме органически и гармонично. От дифференциальных, односторонних
систем настоящего—к интегральной синтетической системе будущего—та-
ков путь, неизбежный путь дальнейшего развития науки о филогении
растений.
4. Во всех системах, даже самых новейших, эволюционный процесс
рассматривается как процесс качественно однородный. В лучшем случае,
в некоторых системах сделаны б. или м. удачные попытки отобразить
представление о различных ступенях (уровнях) этого процесса. Показ в
системе стадий эволюционного процесса представляет крупный шаг впе-
ред в разработке учения о системе, но это только первый шаг к реше-
нию более общего вопроса—о показе качественных различий эволюцион-
ного процесса.
В системе необходимо отразить и дифференцировать две основные
линии эволюции—одну, которую можно назвать генеральной линией,
идущую на базе неспециализированных форм (более примитивных типов),
и другую—идущую под знаком специализации. Эта последняя линия так-
же неоднородна: с одной стороны, специализация, приводящая к высоко-
специализированным упрощенным формам, как прогрессивная линия эво-
люции, и другая, идущая в крайних пограничных условиях среды (пара-
зитизм, гидрофилия), приводящая к упрошенным деградированным фор-
мам. Качество эволюционного процесса во всех этих случаях имеет опре-
деленные биологические предпосылки. Для растительного мира первая,
генеральная линия, идет на базе „мезофильного автотрофного наземного
растения, лишенного способностей к сапрофитному или паразитному об-
разу жизни, ксерофильных приспособлений, способности лазить, виться
и пр. Среди покрытосемянных растений эти особенности чаше всего наб-
людаются у лесных и кустарниковых мезофитов11 (Тахтаджян, 1947:
109). По мере изменения средних условий среды, в которой протекает
эволюция, по мере того, как развитие растений переносится в среды одно-
сторонне акцентированные, т. е., главным образом, аридные и более гуми-
дные, в ореофильные условия или в условия . засоленных почв и т. д.,
генеральный тип идиоадаптивной эволюции сменяется более односторон-
ним, специализированным типом прогрессивной эволюции. И, наконец,
68
Заключение Conclusion
при попадании в крайние условия существования, для нветковых расте-
ний являющихся уже пограничными с зонами небытия, эволюция прини-
мает формы специализации регрессивной, дегенеративной.
Значение этих основных типов в общем ходе эволюции совершенно
различно; если генеральная линия неспециализированной эволюции явля-
ется линией сохранения наибольшей эволюционной пластичности и по-
тенцией дальнейшей эволюции, то специализированные линии эволюции
такую пластичность, в основном, в значительной мере теряют, а ряд ветвей
деградирующей эволюции являются полностью слепыми. При более де-
тальном анализе характера эволюционного процесса возможно выделение
еще и других его типов.
Важнейшей органической частью всякой новой системы цветковых
растений должно явиться четкое разграничение в ней не только эволю-
ционных уровней, но и типов эволюции и, в первую очередь, выделение
неспециализированной генеральной линии эволюции. Отсюда важность и
необходимость выделения неспециализированных групп растений, примити-
вных рас и условных групп и правильная расстановка их в системе.
Далее, совершенно необходимо учесть в системе и тот тип эволю-
ционного процесса, который у зоологов получил название „педогенеза®'
(эволюция на основе неотений, филэмбриогенез), на базе которого проис-
ходит как бы омоложение специализированных, даже, в некоторой сте-
пени деградированных групп. Возможность педогенеза обеспечивает не-
прерывность эволюционного процесса. Так как на основе педогенеза мо-
жет происходить эволюция крупных групп, то учет этого типа эволюци-,
онногд процесса особенно необходим при построении системы.
Правда, выявление групп растений, возникших в результате педоге-
неза представляет значительные трудности и еще слишком мало известно
нам достоверных фактов из этой области, но это не может снять поста-
новки самого вопроса в его целом, если даже на сегодняшний день мы
можем отметить в системе только разрозненные и немногочисленные фак-
ты педогенеза—их все же нужно отметить и подчеркнуть. ;
у. Еще одно важное качественное явление процесса эволюции долж-
но также найти свое отражение при построении будущей системы расте-
ний. Это закон ускорения эволюции, впервые сформулированный Эн-
гельсом в „Диалектике природы®.
6. Важнейшей органической частью всякой филогенетической сис-
темы является насыщение ее фактором времени. При этом фактор вре-
мени должен быть показан в системе как ее органическая часть, сочета-
емая со всеми другими факторами и явлениями, а не должен быть вы-
пячен односторонне. В таких односторонне построенных системах, в ко-
торых фактор времени показан как доминирующий (Циммерман и
др.), филогенетические связи или вовсе не показываются или показываются
в очень упрошенном виде. • ,
Такие односторонние системы не являются полноценными и могут слу-
жить только материалом для более совершенных построений.
н. А. БУШ N. A. BUSH
1869 — 1941
Обзор новейших систем цветковых растений 69
Известным приближением к отражению принципа времени в систе-
мах является установление уровней или стадий развития эволюционного
процесса. Изображение системы в форме филогенетического дерева во-
обще подразумевает принцип времени и дает его показ, но, время здесь
как правило, понимается очень условно. Задача состоит в том, чтобы ко-
нстантировать время эволюционных явлений, отразить в системе не время
вообще, а определенные геологические эпохи, к которым приурочено то или
иное явление эволюционного процесса, возникновение той или иной
ветви филогенетического дерева.
Задача эта сложная н трудная, поскольку мы имеем в настоящее
время только фрагментарные данные о точной датировке тех или иных
эволюционных процессов. Эти данные более достоверны для крупных
групп растительного мира и становятся все менее 'достоверными для таких
групп, как порядки, семейства, роды и виды.
Тем не менее, несмотря на трудность проблемы, она должна быть
поставлена во весь рост, так как без того или иного решения этой проб-
лемы филогенетическая система растений не может быть построена.
Вопрос о темпах развития в различные геологические эпохи различ-
ных систематических групп является частным случаем общей картины ра-
звития во времени цветковых растений и должен рассматриваться и ре-
шаться в тесной связи с вопросом о времени развития вообще.
Также чрезвычайно важной и в то же время трудной является
проблема о территориях развития, о пространственном выражении эволю-
ционного процесса. В настоящее время мы изучаем на поверхности земно-
го шара только конечные стадии размещения систематических групп,
фиксируем один исторический „момент" этого непрерывно меняющегося
процесса. Для полного понимания филогенетических отношений необхо-
димо установление территорий возникновения той или иной группы и
всех перемещений ее по поверхности Земли с Момента возникновения до
наших дней.
В противоположность тому, что мы имеем в проблеме времени, здесь
как раз более достоверные сведения мы имеем для групп низшего систе-
матического значения: наиболее достоверные данные о географии видов
и внутривидовых групп, затем менее достоверные данные о родах, семей-
ствах, порядках. Вот почему, наибольшее количество филогенетических
построений с учетом географического принпипа мы имеем в области сис-
тематики видов и родов. Сделаны многочисленные построения, связыва-
ющие течение эволюционного процесса с территориями развития; некото-
рые из этих построений очень удачны и наглядны (например, Лам, Ша-
паренко и др.), но все они, как правило, являются типичными односто-
ронними' построениями, в лучшем случае сочетаемые с принципом времени.
Гармоничное отображение процесса географизма эволюции—задача
будущей филогенетической систематики.
Более просто и легко решается проблема показа масштабов разви-
тия, хотя и здесь есть еще ряд не вполне ясных вопросов. Количествен-
70
Заключение Conclusion
ное выражение процессов эволюции—такая же органическая грань фило-
генетической системы, как и все другие, одного значения и одинаковой
важности. В большинстве построенных систем, однако, эта характерная
особенность эволюционного процесса пренебрегается вовсе (Галлир,
Кузнецов, К о з о-П о л я н с к и и, Тахтаджян и многие другие)
или находит недостаточно четкое отображение (Бесси, Пулле и не-
которые другие).
Количественное выражение эволюционного процесса говорит нам о
степени приспособления той или иной группы к условиям существования,
выявляет группы, угасающие в современной природной обстановке, наме-
чает группы молодые, только начавшие свое эволюционное развитие. По-
каз масштаба эволюционных явлений имеет громадное значение для пони-
мания истинных филогенетических отношений и является совершенно
необходимой принадлежностью всякой филогенетической системы.
Принято, обычно показывать количественные отношения в виде круж-
ков различного диаметра, в более редких случаях в форме других фи-
гур различной площади (например, Бесси). Представляется наиболее це-
лесообразным отойти от традиционной формы кружка и перейти к эл-
липсовидной или овальной фигуре, вытянутой в одну (или в несколько)
сторону, при чем эта вытянутая часть должна показывать направление
или основные тенденции развития данной группы.
7. Совершенно не находит отражения в системах то эволюционное
развитие, какое лает культурная флора. По характеру своему и масшта-
бам проявления культурная флора занимает в настоящее время громадные
территории на земном шаре и играет огромную роль в общей экономике
природы.
Благодаря сознательной деятельности человека, направленной по ли-
нии нового формообразования, культурная флора в настоящее время вклю-
чает колоссальное количество новообразований в форме мелких система-
тических единиц, возникших в историческое время и продолжающих не-
прерывно увеличиваться.
у Этот процесс культурного формообразования протекает под влиянием
качественно отличных типов эволюции (направленная эволюция), ,резко
отличающихся от эволюционных проявлений в дикой природе. Совер-
шенно иными являются также темпы новообразования в культурной фло-
ре, во много раз превосходящие темпы эволюции в дикой природе. Резко
повышены также масштабы формообразования. В системе цветковых ра-
стений будущего должны найти отражение и эти процессы. Быть может,
в качестве предварительной работы, нужйо было бы заняться построением
отдельной филогенетической системы культурных растений, параллельно
системе дикорастущей флоры, а затем сочетать обе эти системы в единую,
тгк как в мире, на поверхности земного шара, существуют оба эти процесса
эволюции и оторвать их друг от друга нельзя.
8. Необходима также углубленная работа еще над одним вопросом—
ографическом изображении системы. Мы имеем целый ряд попыток
Обзор новейших систем цветковых растений
71
различного решения этого вопроса, сводки по которому можно найти в
работах Лама и Шапаренко. В основном авторы графических схем
придерживаются принципа родословного (филогенетического) дерева,
предложенного в классической форме Д а р в и н о м в его схеме дивер-
генции, детализируя и усложняя эту схему различными моментами. Наи-
более резко отступают от этой схемы авторы схем с односторонним пока-
зом принципа времени, у которых схема иногда теряет самое существен-
ное—филогенетические связи между систематическими группами (Уорд,
отчасти Циммерман и др.).
Весьма интересны попытки перехода от плоскостного к трехмерному
показу схемы. Здесь мы пока находимся в области именно попыток, пер-
вых исканий и еще далеки от универсального разрешения этого вопроса.
Очень интересна в этом отношении попытка зоолога Северцова, пред-
ложившего изобразить различные типы эволюционного процесса в различных
плоскостях, при чем в пределах каждой плоскости им принимается пер-
вичная схема дарвиновской дивергенции. Попытка Лама на Западе и
Шапаренко в Советском Союзе сочетать в трехмерной схеме показ
общей эволюции группы с временем и современной географией ее, так-
же представляют большой интерес и достаточно удачное решение неко-
торых частных случаев. Менее удачны сферические системы Лама.
Вопрос о трехмерном изображении системы растений является, во
всяком елучае, очень перспективным и требует дальнейшей разработки.
9. Из всего вышесказанного нам представляется совершенно ясно
такой общий научно-организационный вывод; построение филогенетичес-
кой системы цветковых растений, соответствующей современному уровню
наших знаний и претендующей на общепризнанность, не может быть
выполнено в индивидуальном порядке. Сумма фактов, подлежащих интер-
претации и обобщению, настолько велика, что физически не может быть
освоена отдельным лицом. Только при условии целеустремленной коллек-
тивной работы ряда специалистов по отдельным отраслям знания, работы,
координируемой и направляемой организационным научным центром, воз-
можно плодотворное решение основных проблем филогении и построе-
ние новой системы цветковых растений. При таком характере работы, роль
отдельных, высокоодаренных индивидуальностей, конечно, не будет каким
либо образом стеснена или снижена; больше того, только при такой сов-
местной работе высокоодаренные специалисты могут поднять всю рабо-
ту на должную высосу и наложить печать индивидуальности на то пли
иное решение ряда узловых вопросов.
Роль научно-организационного центра в основном—общее направление
работы, 'выявление первоочередных задач, узких мест, привлечение внима-
ния специалистов к тем или иным группам растений, суммирование до-
сти жений.
72
Заключение Conclusion
Совершенно очевидно, что коллективная работа над проблемой фило-
гении и системой, руководимая научно-организационным центром государ-
ственного масштаба, возможна только у нас в Советском Союзе.
Долг и обязанность советских ботаников организовать эту работу
на должной высоте и нет сомнения, что она пойдет быстрыми и плодо-
творными темпами на благо и дальнейшее процветание нашей советской
науки..
Баку.
20. II. 1948.
Приложение
Supplement I
АКАДЕМИЯ НАУК СССР
т. XIII, № 3
СОВЕТСКАЯ БОТАНИКА
1943
К ВОПРОСУ О ГРАФИЧЕСКОМ ИЗОБРАЖЕНИИ СИСТЕМЫ
ЦВЕТКОВЫХ РАСТЕНИЙ
А. А. Гроссгейм
(Ботанический институт Азербайджанской Академии Наук)
1. ВВЕДЕНИЕ
Большинство современных систематиков стоит на точке зрения мо-
нофилетического происхождения цветковых растений.
Правда, некоторые крупные систематики защищали (Н. И. Кузне-
цов) или защищают (А. Пулле)', полифилетизм и бифилетизм. Вспом-
ним слова Н. И. Кузнецова: «Мне кажется, что нет ничего пагубнее для
истинного понимания цветковых растений и их органа цветка, как пред-
положение, что все цветковые растения возникли из одного корня, от од-
ного исходного типа. Это предположение заставляет нас строить самые
невероятные филогенетические схемы». Несмотря на такие категориче-
ские высказывания, все же мы не можем стать сейчас на точку зрения
полифилетизма. Полифилетизм также нисколько не предохраняет от са-
мых невероятных филогенетических схем; он только отодвигает в сто-
рону трудности, вместо того, чтобы их преодолевать. Некоторые особен-
ности цветковых растений по самой природе своей не могут быть поня-
ты с полифилетической точки зрения.
На самом деле, если почти для всех признаков цветковых растений
можно теоретически допускать возможность полифилетического проис-
хождения, то один признак, объединяющий всю эту обширную группу
растений, а именно — двойное оплодотворение и все процессы, связан-
ные с развитием женского гаметофита и первых стадий зародыша,
никоим образом нельзя толковать как признак, происшедший полифи-
летически. Два или несколько раз от различных предков, двойное оп-
лодотворение не могло произойти; весь ансамбль явлений, связанный с
двойным оплодотворением, мог появиться только на основе одной
какой-либо группы голосемянных растений и только один раз в опреде-
ленное время. Несмотря на необычайное расхождение линий развития
цветковых растений и несмотря на наличие резко отличающихся друг
от друга порядков и даже отдельных ветвей развития, все же необходи-
мо признать, что эти расхождения вторичны, что они являются резуль-
1 A. Pulle. Remarks on the System of the Spermatophytes. — Mededeelingen
von het Botanisch Museum on Herbarium von de Rihksuniverstete, Utrecht, № 43
(1938), 1—17.
76 Гроссгейм—К вопросу о графическом изображении системы...
татом эволюции уже в пределах единой группы цветковых растений, а
не являются первичными, исходными стадиями. Попытка последних
лет Пулле установить дифилетизм цтетковых растений (разные корни
одно- и двудольных) явно неудачна и противоречит всей сумме зна-
ний наших о систематической ценности и .взаимосвязях этих двух групп
растений.
Мы считаем также доказанным эуанциевое происхождение цвет-
ковых растений. Это положение является общепринятым всеми систе-
матиками сегодняшнего дня, и по существу никому серьезно не придет
в голову защищать в настоящее время теорию происхождения цветка
из соцветия. Даже автор наиболее обоснованной псеуданниевой теории,
Веттштейн, в последних изданиях своей систематики цветковых расте-
ний существенно изменил схему происхождения женского цветка в сто-
рону приближения к эуанциальным воззрениям. Из признания эуанцие-
вого происхождения цветковых растений с неизбежностью следует вывод,
что обоеполый цветок является древним и первичным типом, а разнопо-
лость во всех случаях и во всех своих формах является вторичным ре-
зультатом эволюционного процесса, а не его началом.
Из всех эуанциевых теорий наиболее отвечающей сумме совреме -
ных систематических знаний является стробилярная теория. Мы не зна-
ем непосредственных предков цветковых растений, но наиболее убеди-
тельным является в настоящий момент воззрение, выдвинутое Галлиром,
о происхождении цветковых от беннеттитоподобных предков, воззрение,
выдержавшее уже почти полувековой экзамен. Исходя из этих воззре-
ний, первичными цветковыми растениями из общего состава современ-
ного цветкового населения земного шара нужно считать порядки
Двоп; ' - Попытки Пулле возродить веттштейновские воз-
зрения и начинать свои ряды от . •. . хотя бы и в кавычках,
является определенным шагом назад.
При построении всякой филогенетической системы цветковых ра-
стений, на каких бы основных положениях она ни базировалась, необ-
ходимо осветить и как-то отобразить в самой системе следующие кате-
гории эволюционного процесса: !) ступени (или стадии) развития;
2) пути (или линии) развития—вопрос, тесно связанный с вопросом о
причинах и факторах развития; 3) ряды (или ветви) развития; 4) время
(и темпы) развития; 5) место (территория) развития й 6) масштабы раз-
вития.
2. О СТУПЕНЯХ (СТАДИЯХ) РАЗВИТИЯ
«Как лучшую и почти единственную мерку усовершенствования мы
можем считать специализацию, или дифференцировку частей или орга-
нов, предназначенных для различных отправлений, так как вследствие
подобного разделения труда всякое телесное и душевное отправление
становится совершеннее»2. Это положение Ч. Дарвина в применении к
эволюции цветковых растений показывает, что главной формой усовер-
шенствования цветка является та или иная степень специализации опы-
ления.
Переход от низших ступеней развития цветка к высшим во всех
группах и ветвях развития цветковых растений, как это показывает вся
сумма известных нам фактов, совершается в определенной законо-
мерности, которая может быть выражена следующим образом.
2 Ч. Дарвин, Изменения животных и растений. М—Л., 1941.
Обзор новейших систем цветковых растений 77
Прежде всего необходимо отметить развитие, идущее на базе ос-
новной жизненной линии растительного мира — автотрофной (прогрес-
сивная эволюция) от уклоняющихся путей развития гигрофильного и
гетеротрофного (регрессивная эволюция). Каждый из этих путей при-
водит к различным результатам. В первой линии мы можем различать
три основные ступени или эволюционные стадии в развитии цветка от
примитивных форм, приводящих к совершенным вторичным: первично-
простой цветок сменяется усложненным цветком, дальнейшее развитие
которого приводит к более совершенному упрощенному цветку.
Ступень (или стадия) первая (первичная) содер-
жит исходные формы с еще слабым, еще мало специализированным
взаимодействием цветка и среды. Характерными чертами ее являются:
1) неопределенность строения цветка, выражающаяся в нефиксирован-
ном и притом обычно достаточно большом числе членов цветка, этот
признак является основным для первой ступени развития цветка и оп-
ределяющим все остальные; 2) апокарпия, а также отсутствие явлений
срастания в области андроцея и околоцветника; 3) первично-простой
околоцветник, не дифференцированный на внешний .и внутренний кру-
ги. Характерна общая слабая специализация отдельных органов цвет-
ка.
Вторая ступень (или стадия) развития цветка
(осложненная) характеризуется следующими основными призна-
ками: 1) определенностью строения цветка при фиксированном, но; еще
большом числе членов каждого круга и при большом числе кругов
(обычно о); 2) как правило, синкарпным гинецеем; 3) как правило,
верхней завязью; 4) обычно двойным околоцветником, чаще свободно-
лепестным. Характерна большая специализация цветка и всех его ор-
ганов.
Третья ступень (или стадия) развития цветка
(с й е ц и а л и з и р о в а и н о-у п р ощ ейв а я) характеризуется: 1)'
строгой определенностью строения цветка при точно фиксированном и
притом уменьшенном числе членов каждого круга и при уменьшенном
числе кругов (обычно 4); 2) всегда синкарпным гинецеем; 3) как пра-
вило, нижней завязью; 4) двойным околоцветником, который, вследст-
вие поббчной специализации одного из кругов, часто становится упро-
щенным (из одного круга) или вовсе отсутствует. Характерна высокая
специализация цветка и всех его органов.
' Упрощение строения цветка на третьей ступени его развития явля-
ется в эволюционном смысле совершенством по сравнению с первично-
простыми цветками.
Это упрощение касается главным образом количественных отноше-
ний (уменьшение числа кругов, уменьшение числа членов круга до од-
ного, как, например, орхидных и т. п.). В то же время цветки этой сту-
пени являются, несомненно, более сложными и стоящими на более высо-
кой ступени развития; это явствует, уже из того, что для получения од-
ного и того же эффекта (опыление) мы находим здесь меньшее число
органов, чем у цветков более низших ступеней. Такое строение являет-
ся безусловно более высокой стадией развития; эволюция растительного
мира шла и идет путем постепенного подъема всей организации на выс-
шую ступень, что приводит к более «экономному» построению более со-
вершенно-специализированных органов, т. е. к количественно
упрощенным формам.
Таков основной нормальный (т. е. господствующий в те геологиче-
ские эпохи, к которым приурочено развитие цветковых растений) путь
развития. Но, приспособляясь к особым условиям среды, различные
78 Гроссгейм — К вопросу о графическом изображении системы...
группы цветковых растений уклоняются от этого основного нормально-,
го пути эволюции и дают ветви уклоняющегося регрессивного разви-
тия. Эти ветви развиваются в тех случаях, когда растение входит во
взаимосвязи с крайними и резкими изменениями среды. Наиболее край-
ним из этих изменений является переход к сапрофитизму и паразитизму
и в связи с этим—переход от автотрофного к гетеротрофному питанию.
Менее глубокие уклонения организации происходят при переходе цветко-
вых растений к водному образу жизни и к другой крайности — жизни в
предельно засушливых условиях; в последнем случае приспособлен-
ность растения к среде в общем происходит в рамках нормальных для
цветкового растения жизненных процессов, но одно из приспособлений
— суккуленгность—является уже уклоняющимся от обычных физиоло-
гических норм путем, так как оно связано с несколько иным режимом
дыхания. Еще одна крайность среды — повышенная засоленность поч-
вы — также создает особый уклоняющийся тип развития, приводящий
в основном также к суккулентным фермам; физиологические особенно-
сти галофитов, несмотря на большое внимание, уделенное им ботани-
ками, до сих пор не могут считаться выясненными.
Во всех этих случаях уклоняющиеся пути развития приводят к ук-
лоняющимся формам обычно редуцированного, более простого строе-
ния, чем исходные формы. В области цветка это большее упрощение
происходит здесь не вследствие дальнейшей специализации цветка, а
вследствие явлений потери, снятия и редукции его отдельных органов.
Таким образом, этот тип развития резко разнится от .нормального про-
грессивного типа; он является регрессивным, качественно уп-
рощенным, и в отличие от первого типа лучше применять к нему
термин не «упрощенный», а «опрощенный».
Это качественное опрощение всей организации растений приводит
у паразитов к потере способности ассимиляции, к потере листьев; у вод-
ных форм к потере листостебельного строения ( L^mnaceae, Podoste-
шасеае). у суккулентов — к опрощению процесса дыхания. Все эти из-
менения являются соответственными среде, организация является при-
способленной и жизненной в своих условиях среды, но. на общем фоне
эволюции цветковых растений она является определенным шагом в сто-
рону.
Строение цветка у всех этих уклоняющихся типов развития имеет
много общего, и в цветке у них мы найдем более или менее далеко про-
двинувшиеся явления опрощения строения. Эти цветки (на крайних
ступенях опрощения) напоминают внешне упрощенные цветки третьей,
ступени нормального пути развития, но не могут быть причислены к'
этой ступени; не образуют они также особой четвертой ступени, а яв-
ляются боковой, уклоняющейся ступенью, выпадающей из ряда разви-
тия трех ступеней нормальных цветковых растений. ,
Характерными признаками цветков этой боко-
вой (опрощенной) ступени развития являются: 1)
потеря отдельных органов или кругов цветка, могущая заходить очень
далеко — до полной потери околоцветника; 2) редукция внутренних
органов цветка. Наиболее характерными являются отсутствие в цветке
специализации, связанной с нормальными способами опыления (насе-
комоопыление, ветроопыление), и появление специализации, связанной
с уклоняющимися способами опыления (самоопыление, водоопыление).
Цветки третьей ступени развития и уклоняющиеся цветки боковой
ступени имеют иногда внешнее морфологическое сходство; и те и дру-
гие могут иметь некоторое морфологическое сходство с цветками первой
Обзор новейших ейстем цветковых растений
79
ступени, но и те и другие всегда являются вторичными формами.
Эта вторичная «простота», которую первые систематики принима-
ли, как правило, за первичную, еще и до сих пор является тем камнем
преткновения, о который разбиваются многие системы цветковых ра-
стений. Самым ярким примером этого непонимания диалектики разви-
тия цветка и его взаимодействия являются системы Веттштейна и Энг-
лера с их ошибочным представлением о «простоте» цветка Monochlamy-
de?e И хотя мы знаем сейчас, что цветок Monochlamydeae не про-
стой, а упрощенный, все же такова сила гипнотизирующих традиций,
что только немногие систематики делают из этого нужные выводы при
построении системы. Необходимо подчеркнуть прозорливость Галлира,
который, твердо стоя на монофилетической точке зрения на происхож-
дение цветковых растений, должен был пригнать Monochlamydeae
группой вторичной, но, ошибочно считая самую эту группу однородной,
настоящего места в системе для нее он не мог найти. Гетчисон3, как из-
вестно, отказался в своей системе от высказывания по вопросу фило-
гении Monochlamydeae Не далее как в 1938 г. Пулле вновь говорит
об ,mentiflorae и, хотя в тексте своей работы указывает на полифи-
летичность Monochlamydeae, при графическом изображении системы все
же помещает порядки Monochlamydeae в очертаниях общего круга и
весь этот круг располагает в начале рядов двудольных.
Не один какой-либо признак характеризует принадлежность цветка
к той или- иной ступени развития, а совокупность их, почему и нельзя
применять к этим ступеням названия односторонние и притом подчерк-
нуть основанные на одной какой-либо характерной черте.
Так, цветки первых ступеней обычно являются раздельнолепестны-
ми, высших ступеней стростнолепестными; но применять названия «раз-
дельнолепестные» или «сростнолепестные» для обозначения крупных
систематических групп — нельзя. Лучше всего для обозначения ступе-
ней исходить из обобщенных названий, отражающих самый характер и
направление процесса. Поэтому для основных ступеней развития цвет-
ковых растений можно принять следующие термины: простоцветковые
(Protanthophyta), среднецветковые (Mesanthophyta), высшие цвет-
ковые (Hypsanthophytal и опрощенные цветковые lOpisthanthophyta).
Из ныне живущих форм на первой ступени развития стоят 5 поряд-
ков растений: -.noriales, Ranales, Phytolaccales, Helobiae и Triurida-
les и, частично, Rosales- Все остальные порядкгт стоят на второй
и третьей ступенях развития; особенно богата порядками средняя сту-
пень: к ней относятся 33 порядка целиком и 4 частично; к третьей сту-
пени развития относятся 17 порядков целиком и 2 частично; к уклоняю-
щейся боковой ступени развития относятся 5 порядков целиком и на-
которые порядки второй и третьей ступеней частично.
Устанавливая эти общие закономерности ступеней развития цвет-
ковых растений, не нужно ни на минуту забывать, что эволюция охва-
тывает все систематические категории и что в пределах каждого поряд-
ка, каждого семейства, рода и вида также идет непрерывная эволюция.
Поэтому в пределах совершенно естественных порядков, объединяю-
щих формы, тесно связанные общностью происхождения, эволюционные
явления могут пройти все стадии развития, не выходя из рамок по-
рядка, как такового. Порядок Ranales по своим центральным формам
3 По-видимому, опечатка, — должно быть «Гетчинсон» или как пишет автор з
публикуемом «Обзоре..,» — Гатчинсон. (Ред.).
80
Гроссгейм—К вопросу о графическом изображении системы...
должен быть поставлен в системе в пределах первой ступени развития;
в то же время некоторые семейства этого вполне естественного порядка
поднялись в своем развитии до второй ступени; некоторые отдельные
роды — в пределах первично-простого сем.:’anunculaceae — также до-
стигают высот второй ступени (Delphinium, Coasolida, Aconitopsis и др.). В
пределах вполне естественного порядка Helobiae, начинающегося явст-
венно примитивными формами, наблюдаются даже вторичные формы
уклоняющейся ступени развития опрощенного типа.
Порядки второй ступени развития более однородны, но и среди нйх
находятся отдельные порядки, обладающие семействами, достигающи-
ми высшей ступени развития.
Наиболее однородны и выдержаны в систематическом отношении
вторичные порядки третьей ступени развития.
3. О ПУТЯХ (ИЛИ ЛИНИЯХ) РАЗВИТИЯ
(Биологические предпосылки эволюции)
Выше были перечислены те категории взаимосвязей растений и сре-
ды, которые определяют эволюцию- цветка. Каждая из этих категорий
имеет свои характерные особенности, но во всех случаях эти особенно-
сти подчиняются общей эволюционной закономерности: от простых
форм к усложненным и далее к упрощенным или опрощенным.
Линия насекомоопыления, несомненно, самая мощная", включающая
в свою орбиту наибольшее число цветковых растений. Начинаясь ’:гш-
митивным неспециализированным типом насекомоопыления. который в
настоящее время лучше всего можно наблюдать у Anonales — Ranales,
приводит к совершенным специализированным формам. Характер-
ной особенностью этой линии развития является глубокое упрощение
плодущих частей цветка — андроцея и гинецея, наряду с совершенными
формами специализации бесплодных частей цветка. Кроме того, формы
развития насекомоопыляемых цветков различны в зависимости от того,
какие насекомые служат опылителями. Упрощенный цветок Microspe-
тша>- и упрощенный цветок Spathiflorae или Campanula!es раз-
личны по форме, потому что Microspermae выработались во взаимодей-
ствии с хоботными насекомыми (отчасти птицами), a Spathiflorae вы-
работались как группа, специализированная к опылению двукрылыми.
У Microspermae находим сильное упрощение строения плодущей ча-
сти цветка: нижняя завязь, малое число плодолистиков, образующих
одно гнездо, одночленный андроцей, срастание андроцея и гинецея.
Все это — черты глубокого упрощения строения цветка, выносящие
Microspermae на высшую ступень развития цветковых. Одновременно!
покровы цветка развиваются здесь в формах, быть может, наиболее
специализированных, чем у каких-либо других цветковых растений,, у
Spathiflorae плодущие части цветка также чрезвычайно упрощены;
при сохранении верхней завязи мы находим здесь однополость, редук-
цию числа членов андроцея и гинецея. Но, одновременно редуцируется и
околоцветник, так как специфика опыления двукрылыми не требует
развития специализированных форм околоцветника: функции привле-
чения насекомых специализируются не в. околоцветнике, а в прицветни-
ках, образующих покрывало, действующее не только на зрение, ко и на
обоняние насекомых. Как видим, формы приспособления иные, чем у
Microspermae, но конечный результат приводит оба порядка к одной
Обзор иовейшик' систем цветковых растений.
81
и то»;же упрощенной стадии развития цветковых растений. У Rubiales
Campanulales взаимодействие цветка с короткохоботковыми насеко-
мыми приводит к положению среднему между Spathiflorae > Microsper-
mae; упрощение цветка идет очень далеко: нижняя завязь, редук-
ция, числа плодолистиков, одногнездность завязи, один круг гинецея с
малым числом'членов и т. д. Образование компактных соцветий приво-
дит к редукции чашечки и, сравнительно слабой специализации венчи-
ка, Обеспеченность опыления достигается здесь путем образования ком-
пактных соцветий, что напоминает отношения у Spathiflorae, Тии стро-
ения.цветка. Rubiales и Campanulales также нужно признать достигшим
третьей'высшей стадии развития-; как и у Microspermac Spathiflorae.
хотя цветок их все же стоит несколько ниже, не достигая такого край-’
него совершенства специализации, как у первых двух.
К насекомоопыляемым цветковым относятся 22 порядка; к частич-
но насекомоопыляемым — 11 порядков, из них 3 порядка (Microspermae,
Spathiflorae и Rubiales) полностью находятся на последней ста-
дии развития, Umbelliflorae и Campanulales* достигают третьей ста-
дии развития частично.
Второй линией взаимосвязей растений со средой, непосредственно
вызывающей, эволюцию цветка в определенном направлении, является
ветроопыляемостщ Среди современных цветковых растений очень мало
форм, с примитивными приспособлениями для ветроопыления (напри-
мер, порядок Hamamelidales). Почти все современные ветроопыляеь
мые растения стоят уже на очень высокой ступени развития; так, в
третьей наивысшей стадии развития насчитывается 14 • порядков ветро'-
опыляемых, на средней стадии — 1 целиком и 5 частично. Ветроопыле*
ние, как форма взаимосвязей растений и среды, возникло раньше насе>
комоопыления; по происхождению эта форма взаимосвязи является
более древним типом, но она не сменилась и не вытеснена нацело на-
секомоопылением, а продолжала и продолжает развиваться параллель-
но, рядом с насекОмоопылением. поднимаясь на все более совершенные
: ступени. Так как ветроопыление возникло раньше, то прошло больше
времени для выработки наиболее совершенных способов. Поэтому
очень совершенные приспособления к ветроопылению могли возникать
уже в отдаленнейшие геологические периоды. Нет ничего удивительно-
го в том, что уже в меловом периоде и в начале третичного существо-
вали цветки упрощенного типа высоко и специализированно приспособ-
ленные к ветроопылению. В то же самое время насекомоопыление
должно было находиться еще на низких стадиях развития. Поэтому
обычный довод, приводимый в пользу примитивности, скажем, Мопо-
chlamydeae, а именно, что они геологически очень древни, а потому их
тип цветка нужно считать примитивным, не выдерживает критики. Гео-
логически они, конёчйо, очень древни, но это не мешало им уже в древ-
ние геологические эпохи получить вторичный упрощенный тип иветка.
Обычно систематиками принимается положение* что вообще ветро-
опыление является первичным примитивным типом, а насекомоопыле;-
ние вторичным и более совершенным. Несмотря на то, что в некоторых
случаях (Glumiflorae, Cyperales) определенно доказана вторичность
ветроопыления, общее положение о примитивности ветроопыления ос-
тается как бы в силе и утверждается даже в новейших курсах система-
тики цветковых растений (например, Н. А. Буш, 1940 г.). Помимо ссы-
лок на палеонтологические факты, о которых только что сказано, вто-
6. А. А- Гроссгейм и М. Ф. Сахокиа
82 Гроссгейм—К вопросу о графическом изображении цветковых растений
рым соображением, которое выдвигается против признания эволюци-
онной равноценности анемо- и энтомофилии, является следующее: при
анемофилии производится громадное количество пыльцы («драгоцен-
ного материала»), и большая часть ее погибает даром; это неэкономно,
а, следовательно, несовершенно и эволюционно более примитивно. Это
рассуждение не соответствует фактам и имеет порочную особенность
антропоморфизировать объективные явления природы («неэконом-
иость»), Прежде всего, если говорить о количестве пыльцы в цветках
(а не в соцветиях и не на отдельных особях) ветроопылителей, то вряд-
ли можно утверждать, что отдельный цветок ветроопылителя обяза-
тельно приносит больше пыльцы, чем цветок насекомоопылителя; здесь
могут быть и обратные отношения. Соцветие ветроопылителя (напри-
мер, сережка ореха, дуба, березы и т. д.) в сумме приносит также пыль-
цы не больше, чем соцветие насекомоопылителя (Compositae, Dipsaca-
сеае). И если взять отношение числа пыльцевых зерен к числу завя-
завшихся и развившихся семян, то вряд ли эти отношения дадут какое-
либо существенно^ преимущество насекомоопылителям. Может ка-
заться, что ветроопылители производят массу лишней пыльцы потому,
что они «пылят», и пыльцу эту мы видим в воздухе, вне цветка и ра-
стения. Насекомоопылители не пылят, и скопления их пыльцы вне цвет-
ка и растения мы не видим. А между тем, если бы, например, подсол-
нечник пылил и поле, им засеянное, выбрасывало бы пыльцу в воздух,
то, несомненно, в воздухе подучились бы ^учи пыльцы, и мы сокрушен-
но говорили бы о том, сколько драгоценного материала расходуется
даром. С другой стороны, если считать большое количество пыльцы у
ветроопылителей пороком, то и у иасекомоопылителей также найдется
свой порок — своя «неэкономия» и «нерациональность». Мы говорим о
связанности некоторых иасекомоопылителей с известными видами на-
секомых. Отсутствие насекомого определяет в этих случаях невозмож-
ность опыления и в результате может дать вымирание растения (из об-
щеизвестных фактов напомним отношения между клевером и шмеля-
ми, юкками и бабочками, первые опыты плодоводства в Тасмании и
др.). Это создает необеспеченность и неустойчивость существования ви-
да, а, следовательно, является «несовершенством». Однако никто не ста-
нет оспаривать, что подобные тесные связи с насекомыми являются
вторичными, а не первоначальными.
По поводу общего характера всякого биологического развития Фр.
Энгельс говорит в «Диалектике природы»: «Суть же дела, в том, что
каждый прогресс в органическом развитии является в то же время и
регрессом, ибо он фиксирует одностороннее развитие и исключает воз-
'можность развития во многих направлениях»4. Специализированное при-
способление к насекомоопььДению является таким жё односторонним
развитием, как и приспособление к Ветроопылению.
Больше всего пыльцы вырабатывают цветки первой и второй сту-
пени развития, независимо от их насекомо-или ветроопыляемости, т. е.
на той стадии развития, когда еще, нет высокоспециализированных
приспособлений для переноса пыльцы; на третьей же стадии развития
оба типа цветков вырабатывают, примерно, одинаковое количество
пыльцы.
Обычно принято считать первичными ветроопылителями группу
так называемых однопокровных - Monochlamydeae.
По строению цветка прежде всего приходится признать, что эта
группа гетерогенна, отдельные порядки ее имеют разное происхожде-
4 Фо. Э и г е л ь с. Диалектика природы. Изд. 6-е, 1934.
Обзор новейших систем цветковых растений 83
ние и должны быть размещены в различных отделах системы. Морфо-
логия цветка у всех Monochlamydeae явно—вторична, даже с точки
зрения морфологических канонов Энглера, и нужно только удивлять-
ся, как сам автор 35 «морфологических правил» закрывал глаза и про-
ходил мимо морфологических фактов, на примере Monochlamydeae
лучше всяких других подтверждающих его же правила о морфологиче-
ской вторичности. Однополость большинства Monochlamydeae, тже
одна указывает на их вторичность. Развитие нижней завязи у целого
ряда Monochlamydeae малое число кругов цветка (андроцей всегда
однокруговой), малое число членов каждого круга, а в особенности
упрощенный околоцветник, упрощение которого доходит до его полно-
потери (Piperales, Salicales); все это явно вторичные признаки вы-
сокой организации. Расположение тычинок против листочков около-
цветника, на примитивности которого так настаивал Энглер, а впослед-
ствии и Н. И. Кузнецов, может рассматриваться только как явление
вторичное по сравнению с чередованием тычинок с листочками около-
цветника.
Из числа более существенных доводов в пользу примитивности
ветроопыляемых Monochlamydeae выдвигаются факты палеонтологи-
ческие, затем наличие у Monochlamydeae халазогамии и (у некото-
рых) большого числа макроспор в семяпочке. О значении ископаемых
свидетельств сказано выше. Что касается вопроса о халазогамии, то
естественно ‘усомниться в значении этого признака, как первичного.
Мы не знаем, точно, каков был путь пыльцевой трубки у Pteridosper
mae и у Bennettitales, но строение их семяпочки с наличием пыль-
цевой камеры скорее заставляет предположить у них порогамию, а не
халазогамию. У ныне живущих Cycadales наблюдается порогамия, а
не халазогамия. Кроме того, мезогамия открыта и у очень высоко стоя-
щих в системе растений, например у Cucurbitaceae и совершенно
прав Н.А. Буш5, когда он говорит, что, «это обстоятельство, заставляет
сомневаться в примитивности не только мезогамии, но и халазогамии».
Думается, что заключение о первичности халазогамии было сделано и
утверждалось не из анализа особенностей самого процесса халазога-
мии, а обратным путем — от предполагавшегося древнего возраста
растений — халазогамиков. Так как растения, у которых была обна-
ружена халазогамия, общепринято считались в то время древними, то
и эта их особенность стала рассматриваться как древний признак.
Это суждение от обратного наложило печать на все дальнейшие пред-
ставления о халазогамии. Уж одно то, что халазогамия (и мезогамия)
известна у некоторых совершенно не связанных между собой система-
тически групп растений, лишает ее вообще систематического значе-
ния и, заставляет рассматривать как вторичное изменение, которое
может возникнуть от неизвестных нам пока взаимосвязей на различ*
пых ступенях развития цветковых растений.
Указывают также, что примитивность Monochlamydeae и, в част-
ности, Casuarinales подтверждается тем, что у них образуется боль-
шое число макроспор в семяпочке. Интересно, привести по этому по-
воду мнение Н. А. Буша, Который пишет: «В этом отношении казуа-
рины отстали даже от папоротникообразных»6. Такое рассуждение не-
5 Н. А. Буш, Курс систематики. 1940.
6 Ibid, р- 324.
84 Гроссгейм—К вопросу о графическом изображении цветковых растений
верно. Как раз у разноспоровых папоротникообразных вырабатывает-
ся не неопределенное, а вполне определенное и притом небольшое
число макроспор. Такого типа разноспоровых растений, у которых, как
правило, было бы неопределенное и притом большое число макроспор,
мы не знаем, — в природе их нет. Быть может, суфествовали когда-
либо вымершие формы с такими признаками, но судить об этом мы не
можем. Таким образом, сравнить с несуществующим в природе, мно-
гомакросноровым типом ни Casuarinales, ни другие Monochlamydeae
нельзя, а вследствие этого и вея аргументация об их отсталости по
сравнению с папоротникообразными повисает в воздухе и становился
чистой .спекуляцией., С гораздо большим правом мы можем.! рассмат-
ривать многомакроспоровость Monochlamydeae, как уклоняющийся
тип развития, возникший в этой группе как новообразование, а не, яв-
ляющийся филогенетическим пережитком,
В качестве доказательства древности Casuarinales приводится
иногда еще тот аргумент, что этот порядок свойствен Австралийской
флористической области, вообще, говоря, изобилующей древними ти-
пами.
Несомненно, что издревле изолированное положение Австралии,
способствовало сохранению древних форм. Но при этом забывают, что
в то же время продолжительное пребывание в условиях изоляции неиз-
бежно должно было способствовать выработке особых специализиро-
ванных форм, особых форм эволюционных изменений, отличных от тех
форм, 'в которых шла эволюция на материке. Очень многие формы Ав-
стралии являются именно -такими специализированными в условиях
изоляции типами, а, следовательно, вторичными с большим числом
признаков не только не древних, но очень молодых по времени, проис-
хождения и в то же время резко отличных от тех, какие вырабатыва-
лись на материке.
Это развитие в условиях изоляции коснулось целого ряда се-
мейств: и древнего сем. Monimi-aceae (Anonales), и вторичного сем.
Goodeniaceae (Campanulales) и многих других, в том числе и Casua-
rinales, большинство черт строения которых являются явно вторич-
ными. Аргументировать от Австралии необходимо с большой осторож-
ностью.
Обобщая сказанное, укажем, что по сравнению с насекомоопыляе-
мрстью ветроопыляемость приводит к верхним ступеням развития тем
же основным путем — упрощением внутренних, плодущих органов
цветка, но специализации околоцветника здесь не наблюдается, нао-
борот, наблюдается его редукция до полного исчезновения на высших
ступенях развития.
Третья линия взаимодействий растений со средой, отражающаяся
на эволюции цветка, связана со вторичным переходом цветковых ра-
стений к водному образу жизни. Это направление развития связано с
глубокими изменениями во всем организме растений, хотя и не отра-
жается на форме питания, так как все водяные растения остаются ав-
тотрофными. Но морфологические изменения всего тела растения
очень значительны и в крайних своих выражениях приводят к потере
листостебельного строения (Lemnaceae, Podostemaceae), Для гидро-
фильных растений вместе с тем характерна резкая опрощенность цвет-
ка, идущая не по линии специализации, а по линии редукции. Только
у примитивных и более древних водяных растений из порядка T’rnales
"(Nymphaeaceae) иветой является насекомоопыляемым, стоящим на
второй ступени развития. Но уже в пределах этого семейства явствен-
Обзор новейших систем'цветковых растений 85
но‘па-мечается линия редукции цветка (Brasenia и др.). В пределах
этого же примитивного порядка вырабатывается сем. Ceratoohyllaceae.
по крайнему опрощению цветка занимающее третью ступень
развития. В Порядке Helobiae первичные формы цветка водяных ра-
стений (Alismataceae, Butomaceae) проходят быструю эволюцию и
Поднимаются на вторую (Hydrocharitaceae) и на третью опрощен-
ную ступень развития (Potamogetonaceae, Aponogetonaceae). Все
другие порядки и отдельные семейства вОдяных растений являются
производными групп, стоящих на более высокой стадии развития —
на Второй или даже на третьей (например, Leninaceae в порядке Spa-
thiflorae.)
Опрощенные формы водяных растений переходят к ветроопыле-
1 нию, еще чаще к самоопылению и гораздо реже к настоящей гидрофи-
лии. Наиболее обычным и резко выраженным способом является са-
моопыление. В результате развития в этом направлении вырабатыва-
ется высокоопрощенный цветок, но по сравнению с упрощенным цвет-
ком насекомо- и ветроопылителей здесь опрощение идет главным об-
разом по пути редукции и в меньшей степени специализации, так как
для обеспечения самоопыления высокоспециализированные приспо-
собления не нужны. Как и при ветроопылении околоцветник может
быть утерян вовсе, число членов андроцея и гинецея сводится до мини-
мума.
Линйя. гидрофилии на общем > фоне цветковых растений очень не-
< велика. Только 4 Порядка можно назвать целиком приспособившими-
ся к‘водному'образу жизни; в остальных случаях эта линия развития
йМеёт выражение только внутри ‘какого-либо порядка одним или не-
сколькими семействами; таких порядков можно указать 4—5.
Нельзя пройти еще мймо одного очень важного типа взаймодей-
стНия растения со средой, также получившего известное отражение в
эволюции цветка, — это появление сапрофитизма и паразитизма среди
цветковых растений, переход йх от авТОтрЬфного питания к гетеро-
трофному. Специфического типа опыления у паразитов не вырабаты-
вается; в основном у них наблюдается насекомоопыление. Но извест-
ное Опрощение цветка, связанное с гетеротрофным образом жизни,
Дбйзкно быть отмечено. Так. как вся линия развития гетеротрофного
цвЙРКойого рйсТРнИи НредсТавляет сббой лишь опрощение всех его ор-
ганов, приводящее к безлистным и бескорнёвым формам, то в связи
с эТйм общим регрессивным развитием и как часть ёго наблюдается и
некоторое опрощение цветка.
Этот тип эволюций, наряду с гидрофилией, должен считаться ре-
грессивным, понижающим, а не повышающим организацию цветковых
растений: «Члены весьма Высокой группы могут даже приспособлять-
ся, *— ’и, невидимому, это чаёго слуЦййось, — к более Простой Ъки^йй,
и в WOm случйе естественный подбор станет клониться к тому, чтобы
упростить или понизить организацию, так как сложный механизм для
простых действий будет не только -бесполезен, но даже невыгоден»7.
Другие изменения общей биологии цветкового растения мало или
вовсе не отражаются на строении цветка.
Суккулентность — своеобразное изменение растений, связанное о
изменением физиологических его свойств, — на развитии цветка не от-
ражается. Линии упрощения у этой Группы растений нет; высшие фор-
мы-цветка под влиянием суккулейтности не появляются. Цветок Ьс-
7 Ч. Дарвин. Изменения животных и растений. М.-Л., 1941.
86 Гроссгейм—К вопросу о графическом изображении цветковых растений
тается на той же ступени, на какой суккулентность застала исходные
несуккулентные формы. Это может быть и на средней ступени разви-
тия (Cactaceae, Stapelia и др.) и на высшей (Euphorbia).
Влияние другого биологического изменения цветковых растений,
происходящего под влиянием галофитизма, на развитие цветка оста-
ется неясным ввиду неизученности самих биологических свойств га-
лофитов. У многих галофитов наблюдается сильно упрощенный цве-
ток ("Batidales, Chenopodiaceae), но является ли это результа-
том перехода этих растений к ветроопылению или вызвано какими-то
специфическими свойствами галофитного организма как такового, —
этот вопрос сейчас не может быть разрешен.
Все рассмотренные здесь линии развития цветка имеют свои оп-
ределенные биологические предпосылки и являются результатом взаи-
модействия растения и среды, при чем в качестве ведущего звена вы-
ступает то одна, то другая форма взаимосвязи.
При этом в одних случаях изменение и эволюция цветка зависят
от биологических взаимосвязей растения со средой в области генера-
тивных органов как таковых (энтомофилия и анемофилия), в другом
случае эволюция цветка является отражением (вполне или частично)
биологических взаимосвязей растения со средой в области вегетатив-
ной его жизни (гидрофилия, гетеротрофность).
В первом случае мы имеем основную прогрессивную линию эволю-
ции цветковых растений, во вторам случае — несвязанное между со-
бой регрессивное развитие различных групп цветковых растений. Фр.
Энгельс, подчеркивает в «Диалектике природы» положение о том, что
«приспособление к паразитической жизни всегда регресс»8. Таким же
регрессом является и приспособление по самому существу своему су-
хопутных цветковых растений к водному образу жизни. Таким обра-
зом, все наблюдаемые в природе процессы эволюции цветка имеют
свои биологические предпосылки, каковые и должны найти свое отра-
жение при построении системы цветковых растений.
4. О РЯДАХ (ВЕТВЯХ) РАЗВИТИЯ
Вся группа цветковых растений в целом имеет монофилетическое
происхождение; но, в пределах самой группы развитие шло различными
путями, и можно отличить по крайней мере И основных стволов, или
рядов, этого развития. Все они имеют корни или непосредственно в
примитивной группе Batrachiophyta или имеют общие с нею корни;
одни ветви заканчиваются быстро небольшими группами ныне живу-
щих растений; другие, наоборот, весьма многоформенны, имеют слож-
ную систему, рамификаций и в современной флоре играют значитель-
ную роль. Большинство рядов прошло уже все стадии развития цвет-
ковых растений; немногие мы застаем на второй стадии. Три ствола,
исходя из Batrachiophyta имеют более тесные отношения к порядку
Anonoles. и общее с ним происхождение; остальные 7 имеют общее
происхождение с порядком Ranales
Деление всех цветковых растений на одно- и двудольные должно
быть оставлено уже по одному тому, что один ствол однодольных
(Krinophyta)'имеет общность происхождения Ranales, ^другой (Spa-
thophyta) — с Anonales; две ветви, имеющие различное происхожде-
8 Фр. Энгельс. Диалектика природы. Изд. 6-е, 1934.
Обзор новейших систем цветковых растений 87
ние не могут объединяться в один класс. Искусственность деле-
ния на одно- и двудольные растения давно уже установлена в ряде си-
стематических исследований, и рациональность оставления этих клас-
сов в системе давно подвергнута сомнению. Многие крупные система-
тики отказались от этого деления, и потому тем более неожиданной и
странной является попытка Пулле в 1937 г. восстановить опять эти
классы, да еще считать их двумя основными рядами развития цветко-
вых растений.
Также необходимо совершенно отказаться от группы «однопокров-
ные — Monochlamydeae». Это еще более искусственная, чисто фор-
мальная группа, чем классы одно- и двудольных. Порядки, входящие
в Monochlamydeae имеют совершенно различное происхождение ч
должны быть расположены в системе в различных местах; два из этих
порядков (Casuarinales u Salicales) представляют собой особые
основные стволы развития цветковых растений, другие — занимают
место в других стволах развития. Одни из порядков Monochlamydeae
связаны общностью происхождения с Anonales, другие — с Ranales.
Мы считаем, что самые термины одно- и двудольные, а также одно-
покровные не должны совсем фигурировать в современной системати-
ке и могут употребляться только в качестве исторических воспомина-
ний, сыгравших в свое время определенную историческую роль в раз-
витии ботанической науки, но в настоящий момент отживших.
Ствол. I- Batrachiophyta
Центральный ствол цветковых растений — Batrachiophyta,
почти соответствующий группе Polycarpicae наряду с сохранивши-
мися примитивными формами, далеко ушел по пути развития, и неко-
торые его семейства поднимаются на вторую или даже на третью сту-
пень развития. Мы имеем здесь группу, которая при сохранении древ-
них черт строения цветка вовсе не является группой угасающей или
вымирающей, но группой, хорошо приспособившейся к современным
геологическим условиям, во многих своих частях, переживающей в'на-
стоящее время усиленное видообразование (Ranunculaceae в странах
Средиземья, в горах умеренной зоны, Lauraceae в субтропиках, Апопа-
сеае в тропиках. Monimiaceae в южном полушарии и т. д.). Поэто-
му и. количественно Batrachiophyta представлены значительным чис-
лом видов, и оба порядка их, и Ranales и Anonales, стоят в ряду бо-
лее крупных порядков цветковых растений.
Связывая все остальные ветви развития цветковых растений с
Batrachiophyta, , остановимся сначала на тех из них, которые ведут
своё начало от Ranales или имеют с ними общих предков.
Ствол II. Hamamelidophyta
Короткая ветвь с небольшим числом ныне живущих представите-
лей.Эта ветвь не дает дальнейшего продолжения и заканчивается в
основном на второй стадии развития цветковых растений. Правда, сем.
Platanaceae. несомненно родственное сем. Hamamelidaceae и от него
происходящее, по строению цветка может быть отнесено уже к третьей
ступени развития, но основное ядро всего ряда — сем. Hamamelidaceae
— целиком еще находится на второй ступени. В цветках Hamamelida-
с-
88 Гроссгейм—К вопросу о графическом изображении s цветковых . растений
сеае, наряду с.известными усложнениями строения, имеется еще шнхио
черт чисто примитивных (например/строение тычинок). Преобладаю-
щим способом опыления у них является несовершенное ветровое, и в
этом направлении шло развитие всего ствола, заканчивающееся более ,
совершенно ветроопыляемыми Platanaceae.
Ствол IIL Melopbyta
Этот ствол является самым большим и самым многоформенным из
всех рядов развития цветковых растений. Начинаясь порядком Rosales
в пределах которого происходит быстрый переход с низшей стадии,раз-
вития на среднюю ступень, ствол этот дает много ветвей, сосредоточен- <
ных главным образом в пределах второй ступени развития; здесь сосре-
доточены и формы раздельнолепестные, и сростнолепестные, и формы
с верхней, и формы с нижней завязью; сюда относятся один из наи-
более богатых и насыщенных :видами’порядков цветковых, такие, как
Leguminosales с огромным сем. Leguminosae, затем порядки Tubiflorae,
Myrtales. Некоторые порядки достигают высоты развития третьей
ступени: одни, как Rubiales, по линии насекомоопыления, другие
(Tricoccae, Fagales, Myricales, Juglandales, Julianiales, Garryales, Le-
itneriales) по линии ветроопыления, третьи Podostemales, Caliitricha-
les, Hippuridales, по линии самоопыления, как результат приспособле-
ния к водному образу жизни. В некоторых порядках возникает переход
к гетеротрофии.
Если взаимные связи большинства порядков этого крупного ствола
для нас ясны, то этого нельзя сказать об одной принятой нами здесь
в соответствии со взглядами Таллина ветви, а именно о связях
Anacardiales Fagales. Этот вопрос следует рассматривать пока как от-
крытый, так как родственные отношения этих двух групп могут быть
оспариваемы. Многие данные говорят За то, что действительно Fagales
являются упрощенными формами именно Anacardiales, но достаточ-
но .веских доказательств этого .пока привести нельзя. Возможно, что
Myricalia придется в дальнейшем рассматривать как особый само-
стоятельный ствол развития (12-й по счету) и вести их непосредствен-
но от раналиеподобных предков. Одно несомненно: связыватьМупсаНа
с Urticales нельзя никак, и нам представляется, что сближение
этих двух групп, до сих пор практиковавшееся систематиками, есть ре-
зультат, быть может, недоразумения. Как от трехчленных с нижней за-
вязью Myricalia перейти к 4—5*членным с верхней завязью Urticales?
Мы не зйаем среда цветковых ни одного случая эволюции от ниж-
ней завязи к верхней. Считаю, что происхождение Myricalia > Urticales
совершенно .различно, и в системе они должны занимать совершен-
но различные места. • . 1
Ствол IV Teichospermatophyta
Также богат, хотя и не в такой степени, как Melophyta, расчле-
ненный ствол, развитие которого, шло почти исключительно в одном на-
правлении — по линии насекомоопыления; только один порядок, со-
стоящий всего лишь Из одного вида Batidales.отчленился от Rhoeadales
в результате приспособления к ветроопылению. 'Основная ветвь
Teichospermatophyta заканчивается обширным порядком'Camp antild les,
ряд семейств которого находится еще на’второй ступени развития,
Обзор новейших систем цветковых растений 89
но-некоторые семейства, в том числе и семейство самое обильное г вида- -
ми из всех цветковых растений — Compositae — переходит уже на
высиую с!адию развития9.
Ствол V- Proteiophyta
Сравнительно небольшой ствол, состоящий всего лишь из 8 поряд-
ков: первоначальные связи с Ranales утеряны и не совсем ясны. Со-
временные представители начинаются уже с форм, далеко ушедших по
пути развития. Интересно, что тенденция к гетеротрофному образу жиз-
_ни проявляется, как в порядках Santalales, так и Proteales, чтобы
найти свое высшее выражение в замечательном и своеобразном поряд-
ке ' Balanophorales, целиком гетеротрофном, с сильно опрощенным
цветком.
Ствол VI. Centrospermatophyta
Богато расчлененный, своеобразный ствол развития цветковых ра-
стевий. Начинается он примитивными формами порядка Phytolaccales,
а именно сем. Phytolaccaceae,, которое находится еще на первой сту-
пени развития. Происхождение всех остальных Centrospermatophyta
ст Phytolaccaceae может считаться достаточно доказанным. Мы счи-
таем, что от ветроопыляемого типа Centrospermatophyta могли прои-
' зойти Urticales (а не от Urticales е Centrospermae, как считали
Веттштейн, Энглер, Н. И. Кузнецов и др.). Своеобразный в своих выс-
ших -морфологических достижениях порядок Polygonales имеет корни
непосредственно в порядке Phytolaccales. Cactales, есть, несомнен-
но, дальнейшее развитие типа Aizoaceae, a Primulales и Plumbaginales
— .дальнейшее развитие типа Caryophyllaceae. Но если взаим-
ные отношения между порядками ствола Centrospermatophyta, более
или-менее-ясны, то связь Phytolaccales с Ranales не может быть до-
казана непосредственно, так как промежуточные звенья здесь, очевид-
но, утеряны. Во всяком случае примитивное сем. Phytolaccaceae все-
го больше Запоминает некоторые формы Ranales.
Ствол VII- Ebenophyta
Небольшой ствол цветковых растений, состоящий всего из одного
порядка, заключающего в себе К семейств. Имея непосредственные кор-
ни в ‘‘Ranales,-группа эта в своем' развитии поднимается до второй" ста v
дни. и не дает дальнейшего продолжения.
Ствол VIII- Krinophyta
Богато расчлененный ствол, с одной стороны, представленный при-
митивными группами, стоящими на первой ступени развития и имею-
щими непосредственные связи ctRanales, с другой стороны, поднимаю-
щийся до крайних высот третьей стадии развития цветковых растений
как »по «линии насекомоопыления, так ш по линии ветроопыления.
9 Написание названия ствола в статье .даио. двоякое: Teichospermatophyta
И Teichiosperipatophyta Оставляю написание,,принятое в,рукописи «Обзора»"(ред.).
90 Гроссгейм—К вопросу о графическом изображении цветковых растений
Примитивный в своих основных формах порядок Helobiae, начи-
нающий этот ствол, в своих пределах дает несколько линий быстрой
эволюции к опрощенным в связи с водным образом жизни типам. От
него же отчленяется маленький гетеротрофный порядок Triuridaies,
в своем развитии не выходящий за пределы первой ступени.
Порядок Liliiflorae, исходящий из примитивного типа Helobiae,
развивается уже на второй ступени и представляет прекрасный пример
связи с насекомыми на фоне не очень специализированных приспособ-
лений. От этого порядка непосредственно нужно производить Microspe-
rmae — наиболее специализированный к опылению насекомыми, при
глубоком упрощении плодущих органов цветка, порядок цветковых ра-
стений. Этот порядок целиком находится уже на высшей ступени раз-
вития и чрезвычайно богат видами. Единственное его сем. Orchidaceae
по количеству видов стоит почти наравне с двумя другими крупнейши-
ми семействами цветковых растений — с Compositae Leguminosae,
Небольшое уклонение Liliiflorae. в сторону ветроопыления, "пред-
ставленное сем. Juncaceae, дало другое совершенное производное
Liliiflorae — ветроопыляемый порядок Cyperales, весьма богатый
видами и переходящий в третью ступень развития. От Liliiflorae
отчленяется также очень близкий порядок Farinosae (Enantioblastae),
в котором линия ветроопыления получила преимущественное
развитие перед насекомоопылением. От насекомоопыляемой линии
Farinosae отчленились своеобразный порядок Scitamineae остав-
шийся 'на второй стадии развития, а от ветроопыляемой линии обшир-
ный и чрезвычайно жизненный в современных условиях существования
порядок Glumiflorae, поднявшийся на высшую ступень развития.
Ствол IX. Spathophyta
Развитие этого ствола связано с типом Anonales. Можно считать
доказанным, что небольшое сем. Lactoridaceae, имеющее в составе сво-
ем всего лишь один вид и близкое к Magnoliaceae, показывает нам, ка-
ким путем и каким образом от типа Anonales, в результате упрощения
цветка мог возникнуть тип Piperales, а следовательно, и другие, свя-
занные с ним порядки ствола Spathophyta. Связи эти, впрочем, теря-
ются в глубине времен, и можно предполагать только существование
общих вымерших предков всех этих порядков. Один из порядков, раз-
вивавшийся по линии насекомоопыления — Principes — дошел до вто-
рой стадии развития, остальные поднялись на высшую стадию: Spathi-
florae в результате своеобразного приспособления к опылению'при по-
мощи двукрылых насекомых, Piperales Pandanales — в результате
специализированного приспособления к ветроспылению. Интересно от-
метить здесь появление также очень опрощенных и вторичных типов,
приспособившихся к водному образу жизни, как среди Spathiflorae,
так и среди Pandanales.
Ствол X. Itephyta
Относительно небольшая группа ветроопыляемых растений с край-
ней степенью упрощения цветка, представляющая, несомненно, особый
ствол развития, не связанный с другими цветковыми растениями и име-
ющий начало, быть может, среди аноноподобных предков.
Обзор новейших систем цветковых растений 91
Ствол XI. Kerkidophyta
Так же изолированно стоящий ствол, не имеющий ясных родствен-
ных связей с современными цветковыми растениями и несущий ряд вто-
ричных признаков высокой специализации, как в вегетативных частях
^приспособление к ксерофитному образу жизни), так и в области цвет-
ка, весьма специализированно приспособленного к опылению ветром.
Развилась эта ветвь в условиях изоляции в Австралийской области.
Происхождение. Kerkidophyta так же неясно, как и Itephyta; скорее
всего нужно предположить для них какие:либо аноноподобные предки.
5. О времени (и темпах) развития
В идеале система цветковых растений должна быть построена так,
чтобы отобразить последовательность во времени эволюционных стадий
в каждом стволе и ветви. К сожалению, сделать это в настоящее время
не представляется возможным, и, быть может, только в очень отдален-
ном будущем можно будет приблизиться к ответу на вопрос о времени
происхождения той или иной группы цветковых растений. Та последо-
вательность происхождения, какую мы изображаем сейчас на схеме,
дает только самое отдаленное представление о развитии процесса эво-
люции во времени. Если мы считаем, что, скажем Campanulales прои-
зошла в результате длинного пути развития от Ranales, то, несомнен-
но тип Ranales должен был появиться раньше типа Campanulales.
Но фиксировать точно геологическую эпоху появления ни Ranales, ни
Campanulales мы не можем. Кроме того, процесс эволюции и видооб-
разования в каждой группе есть процесс безостановочный, и в группе
Ranales в настоящий момент идут такие же интенсивные процессы
эволюции й видообразования, как и в группе Campanulales. Эта непре-
рывность процесса эволюции, безостановочная и бесконечная изменчи-
вость в любой группе растений, является всеобщей, и всю систему
цветковых растений мы должны представлять в постоянном, непрекра-
щающемся движении. Благодаря этому, вопрос о времени появления
тоц, или иной группы еще более осложняется. Единственно геология
могла бы дать ответ на этот вопрос, если бы геологическая летопись бы-
ла непрерывной. Но, летопись, эта фрагментарна и навсегда останется
таковой. Из древнейших времен,--с самой зари возникновения цветко-
вых растений, мы уже имеем остатки высокоспециализированных
форм, для создания которых уже нужно было продолжительное время
(Salicales, Platanus, Nelumbium и ряд др.). Если сопоставить все
имеющиеся в настоящее время ископаемые находки цветковых расте-
ний и расположить их по геологическим периодам, то последователь-
ность этих находок не будет соответствовать нашим современным пред-
ставлениям о ходе во времени эволюции цветковых растений. Причи-
ной такого несоответствия является не только фрагментарность геоло-
гической летописи; очевидно, здесь нужно поставить еще один вопрос,
без освещения которого вопросы развития цветковых растений не мо-
гут быть обсуждаемы; это — вопрос о темпах развития, о быстроте, с ко-
торой происходили и происходят процессы эволюции в разных группах.
И совершенно ясно, что темпы развития в различные геологические эпо-
хи.ц в пределах различных групп не были одинаковыми. Мы знаем, что
цветковые растения появились на земном шаре сразу и в большом раз-
нообразии представителей; следовательно, на самой заре возникнове-
ния цветковых растений их эволюция становления на более совершен-
92 Гроссгейм—К вопросу о графическом изображении цветковых растений
ные фазы развития должна была совершаться быстрыми темпами; по-
этому среди древнейших известных ископаемых цветковых растений мы
находили уже типы высокого совершенства, специализированно -приспо-
собленные к -ветроопылению, наряду с примитивными насекомоонылне-
мыми типами. Другие (правда, немногие) группы развивались медлен-
ными Темпами и дошли на примитивной стадии цветка до наших дней.
Ranales, Anonales, Helobiae, в своих примитивных представителях
являются древнейшими типами цветковых растений, и палеонтологиче-
ские остатки их мы также знаем с древнейших времен: Но, общий темп-
развития этих групп, был, несомненно, более медленный, что и позво-
лило им дожить до наших дней не только во вторичных, производных,
но-и в примитивных, коренных, формах.
. Все эти вопросы о времени (и о темпах) развития, перед трудно-
стями которых настолько спасовал Пулле, что совсем сложил оружие
и постарался тщательно очистить свою систему от «принципа времени»
вообще, являются чрезвычайно важными, но в настоящий момент мы
можем главным образом их ставить, но не решать. Ответ на эти-вопро-
сы — дело систематики будущего
6. О территории развития
Еще одну категории» явлений должна отобразить система цветко-
вых растений — территорию возникновения систематических групп.
Здесь мы находимся перед рядом неразрешенных и не- могущих быть
разрешёнными в настоящий момент вопросов.
Несомненно, что формообразование, развиваясь на земном Шаре
повсюду и повсеместно, для различных систематических групп могло
быть локализировано, иногда на территории, достаточно обширной, а
иногда и на очень узкой. Здесь нужно прежде всего отличать два воп-
роса: вопрос о территории возникновения систематической группы и
вопрос о территории распространения этой группы; оба вопроса Одина-
ково важны для систематика, но ответ на первый вопрос приближает
к пониманию систематических взаимосвязей между различными груп-
пами растений, а, следовательно, является для Построения системы
'важнейшим. Территории современного распространения и территории
возникновения систематической группы только в редких случаях’могут
совпадать, и знание современного распространения растений Не Может
служить путеводным звеном при решении вопроса о Территории возник-
новения. ^Фрагментарные показания геологической летописи *в некото-
рых случаях дают возможность более или менее достоверно судить о
территориях происхождения, но эти свидетельства ёще скудны, разроз-
нейны, и выводы, сделанные на их:основании, всегда сомнительны. Мы
хорошо знаем территорию развития тургайской (аркто-третичной) фло-
ры и -знаем" характерный флористйчё'ский состав этой флоры. Но зна-
чит ли это, что ее характерные представители возникли на месте? 'Ко-
нечно, нет; самые характерные представители Тургайской‘флоры-МОРЛИ
возникнуть в существенно иной, Древней "флористической области,’от-
туда мигрировать в тургайскую и здесь уже пережить период вторич-
ного расцвета. Сосредоточенность большого числа4 современных-ТГОрИд-
ков под тропиками также не говорит Об их обязательном территори-
альном происхождении В районах1 современных тропиков.
Но все же не нужно совершенно исключать из рассмотрения севре»
менное распространение растений по земному шару при"решении воп-
роса о территории возникновения. Все Же в некоторых случаях совре-
менное распространение служит до Некоторой степени отображением
Обзор.навейшвд систем цветковых растений ; У о
территории возникновения. Трудно допустить*, чтобы обособленные си-
стематические группы» имеющие сейчас распространение в южном по-
лушарии» (Австралия, южная Африка, Антарктика), возникли на терри-
тории/,Палеарктической области. Также трудно допустить южномате-
риковое происхождение целого ряда порядков, характеризующих сей-
час страны северного полушария. Поэтому, если мы попытаемся ввести
в систему, цветковых растений, отображение их современного географиз-
ма,- т® это», будет к пользе, а не к вреду, и мы все. же получим некоторый*
намек на размещение эволюционных процессов не только во времени,
но и в пространстве. Грубо и приближенно принимая только три кате-
гории/территориального размещения растений по земному шару, мы и
попытались это сделать на прилагаемой-.схеме. Вопрос же об истинных
территориях возникновения и последовательного хода развития различ-
ных;, систем этических групп, наравне с вопросом о времени возникнове-
ния и темпах развития, — дело систематики будущего.
7. О масштабах развития
Различные эволюционные явления имели и имеют различное коли-
чественное выражение. Одни явления представляются редкими и коли-
чественно представлены немногими видами, семействами, порядками.
Другие явления, наоборот, обычны и имеют весьма высокое количест-
венное выражение или повсюду по всей 1ерритории земного шара(Сот-
positae, Legaminosae) или более локализированно в определенных
областях (например, Orchidaceae в тропиках). Игнорировать вопро-
сы количественного выражения эволюционных явлений в систематике
нельзя, потому что количественное выражение л данном случае есть
следствие качественных показателей эволюционного процесса, а имен-
но — степени приспособленности и взаимосвязей этого явления со сре-
дой. Более приспособленное взаимодействие растения со средой дает
большие количественные показатели и наоборот. Общее число цветко-
вых растений на земном шаре выражает их высокую степень приспо-
собленности к условиям современной геологической эпохи; на этом фо-
не различные порядки, семейства и роды также показывают различную
степень приспособленного взаимодействия. Систематические группы с
малыми количественными показателями или являются группами древ-
ними, угасающими на фоне современных геологических условий,' или
молодыми, возникающими, еще не успевшими получить достаточное ко-
личественное выражение. В системе цветковых растений должны быть
отражены количественные выражения в пределах различных система-
тических групп, что даст нам представление о масштабах эволюции в
различных группах, а, следовательно, о степени совершенства взаимо-
связей этих групп с окружающей средой современной геологической
эпохи. Тарой количественный учет эволюционных явлений, который мо-
жет быть графически легко выражен, должен рассматриваться, конеч-
но, как отражение какого-то одного момента на фоне постоянно дви-
жущейся эволюции: поэтому представление о связи масштаба явления
с его взаимоприспособлением со средой должно рассматриваться как
относительное и условное, так как мы имеем здесь дело с отражением
самого процесса становления, а’ не его окончательных результатов. По-
этому чрезвычайно важно было бы проследить для каждой систематиче-
ской группы ее количественные изменения, в прошлые геологические
эпохи; у одних групп при этом количественные показатели выявились
бы, как, увеличивающиеся, у других — как уменьшающиеся. Для целей
понимания эволюции растений важны нестолько абсолютные количест-
94 Гроссгейм—К вопросу о графическом изображении цветковых растений
венные показатели, сколько самое направление изменения количествен-
ного выражения. Мы более твердо знаем характер количественных из-
менений только для крупных отделов растительного мира, в том чис-
ле знаем,, что общий путь количественного изменения цветковых расте-
ний для нашей геологической эпохи есть путь увеличения масштабов
эволюционных процессов в этом отделе. Тенденции же количественных
изменений в пределах отдельных систематических групп цветковых для
нас в общем мало известны; с большой осторожностью нужно прини-
мать данные фитопалеонтологии в этом случае, так как их фрагментар-
ность может привести к серьезным ошибкам. При решении этого вопро-
са, я думаю, правильнее базироваться на анализе самого характера
процесса видообразования в пределах данной группы и на ее совре-
менной географии.
Несмотря на все свое относительное значение, элемент количест-
венный, дающий представление о масштабах проявления эволюцион-
ных процессов, в настоящий момент должен найти свое выражение при
построении системы цветковых растений.
По последним подсчетам Бовера число известных до сих пор ныне
живущих видов цветковых растений на земном шаре исчисляется в
182.00010. Истинное число видов, вероятно, приближается к 200.000, так
как еще далеко не все виды цветковых растений известны и описаны.
В настоящее время отличают немного более 300 семейств цветковых
растений, которые составляют содержание принятых нами здесь 67 по-
рядков (против 44 «серий» Энглера). Наиболее выдержанное ограниче-
ние порядков и семейственный состав порядков находим у Пулле
(1937).
8. О НЕКОТОРЫХ ПРИЕМАХ ГРАФИЧЕСКОГО ИЗОБРАЖЕНИЯ
СИСТЕМЫ
Система должна изображаться, согласно давно уже высказанным
взглядам Дарвина, в виде филогенетического дерева, и всякие другие
попытки графического представления системы («динамическая систе-
ма» Гайяты, «система без принципа времени» Пулле и др.) являются
неправильными и противоречащими самому понятию об эволюции. Но
изображать филогенетическое дерево можно двояко: в профиль (обыч-
ный прием) или в виде проекции кроны этого дерева на плоскость; в
этом последнем случае основные стволы и ветви системы располагают-
ся в виде лучей, а в центре'круга может быть помещена та основная
группа цветковых, которую мы считаем наиболее примитивной и от ко-
торой (или через которую, что правильнее) представляем себе проис-
хождение всех остальных групп. Эта группа является, так сказать, ос-
новой филогенетического дерева. Проведя две окружности, мы очерчи-
ваем три последовательно сменяющие друг друга пространства, которые
принимаем, соответствующими трем ступеням развития цветковых ра-
стений, и порядки размещаем соответственно тому, на какой ступени
развития они находятся. При этом некоторые порядки приходится изо-
бражать на границе между двумя кругами. Порядки изображены в ви-
де кругов различной величины. Показ в системе порядков в их относи-
тельной величине придает системе большую наглядность и дает пред-
ставление о роли каждого порядка в составе современной, флоры зем-
50 Beauverd. Excursion botanique en Grece Bull, de la Soc- Bot- de Gdneve
1938.—M. M. Ильин. О количестве видов растений на земном шаре. Сов. ботаника,оЦ
4-5, 1938.
Обзор новейших систем цветковых растений
95
ного шара. Величина порядка при этом определялась нами не по чис-
лу семейств, их составляющих, как сделано у Пулле, а по числу видов
относящихся к порядку, так как только такой подход дает правильное
представление о роли порядка в составе флоры. Представляется для
простоты, что виды порядка собраны в шар, радиус которого вычислен
по известным формулам; на схеме изображены диски, являющиеся про-
екцией таких воображаемых шаров на плоскость. Диски зачерчены
различным образом, соответственно основным линиям развития цветка
цветковых растений; белыми оставлены порядки насекомоопыляемые,
косыми штрихами даны порядки ветроопыляемые; перекрещивающими-
ся штрихами — порядки, приспособившиеся к водному образу жизни,
в основном самоопыляющиеся, и, наконец, черным цветом отмечены по-
рядки, перешедшие к гетеротрофному питанию. Само собой разумеется,
что в некоторых случаях один и тот же диск нужно заштриховать раз-
личным способом. (Табл. 26.—Ред.).
Таким образом этот способ изображения дает нам возможность по-
казать на схеме расположение как прогрессивного типа эволюции, при-
водящей к специализированным упрощенным типам, так и регрессив-
ного, ведущего к опрощенным формам.
Общее направление эволюции на схеме — от центра к периферии.
Показать на схеме направление эволюции регрессирующих типов (гид-
рофильных и гетеротрофных) от периферии к центру было бы непра-
вильно, так как здесь не возвращение к исходному типу (эволюция не-
обратима), а. своеобразное уклоняющееся развитие на более низком
уровне, ведущее к самостоятельным, своеобразным типам вторичного
происхождения. Наиболее правильным графическим приемом в данном
случае был бы показ линий разреза обоих направлений эволюции в
двух различных плоскостях, чего нельзя изобразить на плоскостном
чертеже и для чего нужно было бы построить трехмерную модель (та-
кие модели нужны для музеев и школ).
Названия порядков подчеркнуты различными линиями: пункти-
ром — порядки, имеющие в основном тропическое распространение;
непрерывной чертой — порядки, преимущественно свойственные Го-
ларктике, и прерывистой линией — порядки, свойственные в основном
странам южного полушария. И здесь также у многих порядков харак-
тер подчеркивания получается двойной, а иногда и тройной.
Порядок следования стволов, принятый на схеме, необязателен;
стволы и ветви сгруппированы таким образом, чтобы идущие от аноно-
подобных предков разместились в одной части схемы, идущие от рана-
лиеподобных предков — в другой; но уже в пределах каждого из этих
секторов схемы порядок чередования стволов и ветвей, в виду их рав-
ноценности и параллельности развития, может быть, конечно, изме-
нен, и они могут быть расположены и иначе, чем это сделано на схеме.
Прилагая при этом список .семейств цветковых растений, располо-
женных с указанием линий развития, ступени развития, порядков и ос-
новных ветвей развития, мы воздерживаемся от общей сквозной нуме-
рации семейств и порядков, подчеркивая этим прерывистость этого
списка. Стволы развития и их ветви могут быть расположены и иначе,
чем это сделано мною здесь.
При определении объема порядков и отчасти семейств мы, главным
образом, придерживались Пулле, у которого эта сторона дела разрабо-
вана наиболее удачно, но в то же время нами допущен ряд отступле-
ний. Так, я считаю совершенно необходимым различать порядки
Anonales и Ranales,, а не объединять их вместе, выделять Leguminosae
96 Гроссгейм—К вопросу о графическом> изображении > цветковых растений
из Resales; в качестве самостоятельного, порядка.. Выделяются мною-
также в виде самостоятельных порядков упрощенные и опрощенные*«ти-
ны Myrtles—Halorhagidales. Centrospermatophyta—Theligonales, а так-
же первичный тип Phytolaccales. Объем семейств во „многих случаях
принят также иной. Так, я считаю излишним выделение, целого, ряда
мелких-семейств, принятых Пулле.
9. СПИСОК СТВОЛОВ, ВЕТВЕЙ, ПОРЯДКОВ И СЕМЕЙСТВ
ПОКРЫТОСЕМЯННЫХ РАСТЕНИЙ
(Разрядкой в скобках указаны ступени и определяющие биологические предпосылки
развития)
Отдел Spermatophyta—Покрытосемянные
Ствол I. Batrachiophyta
Порядок 1. Anonales—Аноноцветные (ступень Protanthophyta)
Семейства: j. Magnoliaceag, 2. Calycanthaceae, 3. Ladoridaceae, 4.
Anonaceae, 5- Myristicaceae, (>. Gomortegaceae, д- Monimiaceae, 8. Laura-
ceae, 9. H ernandiaceae, 10. Trochodendraceae (Bce 10 семейств—п pи м ити в-
ноенасекомо опыление).
Порядок 2. Ranales—Лютикоцветные (ступень Protanthophyta)
Семейства: j. Ranunculaceae, 2. Berberidaceae, 3. Lardizabalaceae, 4.
Menis permaceae (t-4 семейства—и а с e к о м о о п ы л е н ие), 5, Nymphaeaceae,
6. Ceratophyllaceae (5—6 семейства—гидрофилия).
Ствол п. Hamameiidophyta
Порядок 1- Hamamelidales—Гамамелиецветные (ступень Mesanthophyta-
Семейства: 1. Нamamelidaceae, 2. Platanaceae (1 и 2 семейства—в е)
троопылен ие).
Ствол in Melophyta
Ветвь 1. Rosales—Розанообразные
Порядок 1 • Rosales - Розаноцветные .(ступень Mesanthophyta).
Семейства: 1. (Jrassulaceae, 2. Cephdlotaceae, 3. Rosaceae, 4. Saxifraga-
ceae, 5. Pittosporaceae- 6. Enmelliaceae, 7. Byblidaceae, 8. Cunoniaceae,
9. JBruniaceae, Ю. Eoridulaceae, 11- Connaraceae- (L—Неем, и а сек омоопы-
ление), 12. Myrothamnaceae, 13. Crossosomataceae (12 и 13 семейства—в e-
троопыление).
Ветвь 2. Leguminosalia—Б о б о во об р азн ы е
Порядок I. Leguminosales—Бобовоцветные (с т у п е н ь Mesanthophyta),
Семейство 1. Leguminosae (н а с е к о м о о п ы л е н и е).
Ветвь 3. М a] v al ia—Мальвообразные
Порядок 1. Pandales—Пандоцветные (ступень Mesanthophyta).
Семейство 1. Pandaceae—Пандовые (н а с е к о м о о п ы л е н и е)-
Порядок 2. Malvales—Мальвоцветные (ступень Mesanthophyta)
Семейства: 1. Elaeocarpaceae, 2. (jhlaenaeeaet 3. Tiliaceae, 4. Malvaceae,
Обзор новейших систем цветковых растении 97
5. ВотЪасеае, 6. Triplochitonaceae, 7. Sterculiaceae, 8. Scytopetdlaceae,
9. Gonystylaceae (все 9 семейств- н асекомоопыление).
Порядок 3. Geraniales—Гераниецветные (ступень Mesanthophyta)
Семейства: 1. Цпасеае, 2. Humiriaceae, 3. Oxalidaceae, 4. Geraniaceae,
5. Limnanthaceae, 6. Tropaeolaceae, 7. Erythroxylaceae, 8. Malpighiaceae,
9. Zygophyllaceae, 10. (Jneoraceae (все 10 семейств—н асекомоопыление)
Порядок 4- Trjcoccae—Трехорешковые (ступень Hypsan th’o phyta).
Семейства: 1. Dichapetalaceae11, 2. Euphorbiaceae, 3. Buxaceae (Bce
3 семейства—ветроопыление).
Ветвь 4. Myrtalia — Миртообразные
Порядок 1. Myrtales—Миртоцветйые (ступень Mesanthophyta)
Семейства: 1. Репаеасеае, 2. Geissolomataceae, 3. Oliniaceae, 4. Thyme-
laeaceae, 5. Elaeagnaceae, 6. Lythraceae, 7. Hetero pyxidaceae, 8. Sonnera-
tiaceae (1—8 семейства-н а с e к о м о о п ы л е н и е), 9. Crypteroniaceae, (ветро-
опыление), ю. Alangiaceae, П. Nyssaceae, 12. Rhisophoraceae, 12. Le-
cythidaceae, 14. Combretaceae, 15. Myrtaceae, 16. Punicaceae, 17. MelasGj-
maceae12, 18. Onagraceae, 19. Hydrocaryaceae (10—18 семейства—н асекомо-
опыление).
Порядок 2. Halorhagidales-Урутецветные (ступень Hypsanthophyta).
Семейства: 1. Н alorhagidaceae13 14, 2. Gunneraceae (°ба семейства—в ет-
роопылен не).
Порядок 3. Thelygonales—Телигоноцветные (ступень Hypsanthophyta)
Семейство 1. Thelygonaceae (ветроопыление).
Ветвь 5. Anacardialia—Су м ах о о б р аз н ы е
Порядок 1 • Anacardiales — Сумахоцветные (ступень Mesanthophvt а
Семейства: 1. Rutaceae, 2. Simarubaceae, 3- Burseraceae, 4. .ceae,
5. T remandr aceae, 6. Polygalaceae, 7. Trigoniaceae, 8. Vochysiaceae, 9. Ana-
cardiaceae, Ю. Sapindaceae, 11. Aceraceae, 12. Hippocastanaceae, 13. Sa-
biaceae, 14. JMLelianthaceae, 15. Balsaminaceae (1 15 семейства—насекомо-
опыление), 16. Aextoxicaceae, 17. Cordiariaceae, 18. Bidiereaceae (16—18
семейства—в етроопыление).
Порядок 2- Bhamnales— Крушиноцветные (ступень Mesanthophyta)
Семейства: 1. Rhamnaceae, 2. Vitaceae (оба семейства—н ас ек ом о опы-
ление).
Порядок 3. Celastrales—Бересклетоцветные (ступень Mesanthophyta)
Семейства: 1, Q apillaceaeu, 2. Рenta phyllaceae15, 3. Corynocarpaceae,
4. Aqiiifoliaceae, 5. G el aHr aceae, 6. Salvadoraceae, 7. Staphyleaceae, 8. Hi-
ppocrateaeeae, 9. Stackhousiaceae, 10. Icacinaceae- <1—10 семейства—на
с екомо on ы л ение).
Порядок 4. Umbelliflorae—Зонтичноцветные (ступень Mesanthophyta)
Семейства: 1. Araliaceae, 2. Согпасеае, 3. Umbelliferae (все 3 семе-
йства—н асекомоопыление).
Порядок 5. Rubiales—Мареноцветные (ступень Hypsanthophyta)
11 Syn. Chailletiaceae 1,Melastomataceae. 13 Halorrhagaceae-
14 Вероятно, Cyrillaceae16 Pentap hylaeaceae (Pentaphyllacaceae). (РеД.).
7. А. А. Гроссгейм и M- Ф. Сахокиа
98 Гроссгейм—К вопросу о' графическом изображении цветковых растений
Семейства: 1. С aprif oliaceae, 2. РмЫасеае, 3. V alet ianaceae, Dipsa-
caceae, 5. Adoxaceae (Bce 4 семейства—н асекомооп мление).
Ветвь 6- Arisfolochiali а—К ирказонообразные
Порядок 1. Aristolochia'es —т!ирказоиоцзегные (ступень Mesa nt ho-
phy ta)
Семейства: 1. Aristolochiaceae (насекомоопыление), 2. Raffiesta-
ceae, 3. Hydnoraceae (2 « 3 семейства-гетеротрофия).
Ветвь 7. Ребоз(ета1|а Подостемообразные
Порядок 1. Podostemalss - Подостемоцветные (ступень Opisthantho-
h у ta)
Семейство 1. Podostemaceae™ (гидрофилия)..
Порядок 2. Hydrostachyales—Гидростахиецветные (ступень Opisthan-
t h о р h у t а)
Семейство 1. Hydrostachyaceae (гидрофилия).
Ветвь 8. М у г ic ai ia—В о с к о в н и ц е о б р аз и ы е
Порядок 1. Fagales — Буноцветные (ступе нь Hyp santhophyta)
Семейства: 1. Betvlaceae, Fagaceae (°ба семейства-в е т р о о п ы л е-
н и е).
Порядок 2. Myrioales—Восковницецветные (ступень Hypsanthophyta)
Семейство 1. Myricaccae (ветроопыленве).
Порядок 3. Balanopsidaks—^аланопсздецветные (ступень Hypsantho-
phyta)
Семейство 1. Balanopsidaceae (ветроопыление).
Порядок 4. Leitneriales—Лейтнериецветные (ступень Hypsanthophy-
ta)
Семейство!, Leitnerioceae (ветроопыленые).
Порядок 5. Garryales—ГаррИецветные (ступень Hypsanthophyta)
Семейство 1. Garryaceae (в етроо п ы л ение).
Порядок 6. Juglandales —Орехоцветные (ступень Hypsanthophyta)
Семейство 1. Juglandatcae (ветроопыление)
Порядок 7. Julianiales —Юлианиецветные (ступень Hypsanthophyta)
Семейство 1. Julianiaceae (ветроопыление).
Ветвь 9. Т u b i f I о г a I iа—Трубкоцв етнообразные
Порядок 1. Ligustraies—Масличноцзетные (ступень Mesanthophyta)
Семейство 1. Olcaceae (насекомоопы лени е).
Порядок 2. Contortae—оарзннкоцветные (ступень Mesanthophyta)
Семейства: 1. Desfordaineaceae, 2. Loganiaeeae, 3. Gentianaceae, 4.
Apocynaceae, 5. Asclepiadaoeae (sce 5 семейств —н ас e к о моопыл ение).
Порядок 3. Tubiflorae— (рубкоцаетные ,с ту пень Mesanthophyta)
Семейства: 1. Convolvuiaceae, 2. Polemoniaceae, 3. Hydrophyllaceae,
4. Borragirtaceae, 5.. Nolanaceae, 6. Solanaceae. 1- Scrophulariceae, 8. Len-
t.ibulariaceae, 9. Gesneriaceae, Ю. Bignoniaceae, 11. Pedaliaeeae, 12. Marty-
niaceae, 13. Aeanthaceae, 14. Verbenaceae, 15. Labiatae, 16. Globulariaceae,
17. Phrymaceae, 18. Alyoporaceae, 19. Columelliaceae-. I1—19 семейства—н a-
секомоопыление), 20. Cuscutaceae, L• Lennoaceaei 22. Qrobanchaceae
(20—22 семейства—г e т e p о т p о ф и я).
16 Podostemonaceae (Ред.).
Обзор новейших систем цветковых растений , 99
Порядок 4. Plantaginales—Подорожникоцветные (ступень Me san th о-
ph у t а)
Семейство 1. Plantaginaceo.e (ветроопыление).
Порядок 5. Hippuridales—Водяные сосенки (ступень Opisthantho-
р h у t а)
Семейство 1. Hippuridaceae (гидрофилия).
Порядок 6. Caiiitrichales—Водяные звездочки (cTyneHbOpisthantho-
р h у t а)
Семейство 1. (j aUitrichaceae (гидрофилия). . .
Ствол ГС Teichospermatophyta
Ветвь 1. R h о е a d a I i а—М а к о о б р аз н ы е
Порядок 1. BhoeadJes Макоцветные (ступень Mesanthophyta)
Семейства: 1. Papateraceae, 2. Cruciferae 3. Tovariaceae, 4. Cappari-
daceae, 5. Resedaceae, 6. JUoringaceae, 1- Bretschneideraceae (Bce 7 семе-
йств—н а секом oo п ы ленив).
Порядок 2. Ratidales— Батиецветные (ступень Hypsanthophyta)
Семейство, 1. Batidaceae. (ветроопыление).
Ветвь 2- Campanula!! а— Колокольчикообразные
Порядок 1. Parietales—Стенкоположные (с т у пен ь М е sa n t h р phy t а)
Семейства: 1. Cistaceae, 2. Bixaceae, 3. Cochlospermaceae, 4. Tamari-
caceae, 5. Fouguieriaceae, 6. Koeberiiniaceae- Ч- Frankeniaceae, 8. Viola-
ceae, 9. Canellaceae, 10. Flacourtiaceae- 11. Stachyuraceae, 12, Turneraceae,
13. Malesherbiaceae, 14. Passifloraceae, 15. Achariaceae, 16. Caricaceae,
1?. Loasaceae, 18. Begoniaceae, 19. Ancistvocladaceae (1—19 семейства—н a-
секомоопыление), 20. Lacistemaccae, 21. Datiscaceae (20—21 семейства—
ветроопыление), 22.' Elatinaceae (гидрофилия)
Порядок 2. Cucurbitales Тыквоцветные (ступень М е s a n t h о р h у t а)
Семейство 1. Cucurbitaceae (н а с е к о м о о п ы л е н и е).
Порядок 3. Campanulales—Колокольчикоцветные (ступень Mesantho-
р h у t а—Н у р s a n t h о р h у t а).
Семейства: 1, Ganipanulaceae. 2. Goodeniaceae, 3. Bruniaceae, 4. Sty-
lidiaceae, 5. G alyceraceae, 6. Compositae (BCe 6 семейств— н а с e к о м о о п ы-
ление).
Ветвь 3. Guttiferalia Зверобоеобразные
Порядок 1. Guttiferales—Зберзбоецветные (ступень Mesanthophyta)
Семейства: 1. ВШепгасеов, 2. Ochnaceae, 3. Mcdusagynac&ae, 4. Euc-
rypJiiaceae, 5. Caryocaraceae, 6. Afarcgraviaceae, "I- Quiinaceae, 8. Thea-
ceae, 9. Guttiferae, Ю. Dipterocarpaceae (BCe семейств—и а с e к о мо о п bi-
ле н и е).
Порядок 2- Diapensiales — Диапензиецветные (ступень Mesantho-
phyta).
Семейство 1. Biapensiaceae (насекомоопыление).
Порядок 3. Ericales-Верескоцветные (ступень Mesanthophyta)
Семейства: 1. (Jlethraceae, 2. Pirolaceae, 3. Ericaceae, 4. Еpacridaceae,
5. Empetraceae (все 6 семейств—н асекомоопыление).
100 Гроссгейм—К вопросу о графическом изображении цветковых растений
Ветвь 4. S а г г а с е n i al i а—С аррацениеобразные
Порядок 1. Sarraceniales—Саррацениецветные (ступень Mesantho-
phyta).
Семейства: 1. Sarraceniaceae, 2. Nepenthaceae, 3. Droseraceae (все 3
семейства—н асекомоопыление).
Ствол V- Proteiophyta
Порядок 1. Proteales—Протейноцветные (ступень Mesanthophyta)
Семейство 1. Proteaceae (насекомоопыление).
Порядок 2- Santalales—Санталоцветные (ступень Mesanthophyta).
Семейства: 1. Olacdceae, 2. Opiliaceae, 3. Ohtohnemaceae14, 4. Grubbi-
aceae, 5. Santalaceae С1—5 семейства — н а с е к о м о о п ы л е н н е), 6. Myzoden-
draceae, 7. Loranthaceae, 8. Cynomoriaceae С6—8 семейства — г и д р о <р и-
лия).
Порядок 3. Balanophorales—Баланофороцветные (ступень Opisthan-
t h о р h у t а).
Семейство 1. Balanophoraceae (гетеротрофия).
Ствол VI. Centrospermatophyta
Ветвь 1. Centrospermaliа—Центросемянообразные
Порядок 1. Phytolaccales—Фитолаккоцветные (ступень Protantho-
р h у t а).
Семейства: 1. Phytolaccaceae, 2. GyrOstemaceaew, 3. kchatocarpaceae
(все 3 семейства—н асекомоопыление).
Порядок 2. Centrospermae—Центросемянные (ступень Mesantho-
phyta).
Семейства: 1. Nyctaginaceae, 2. Aizoaceae, 3. Portulacaceae, 4. Basel-
laceae, 5. Bysphaniaceae, 6. Cary о phyllaceae (!—6 семейства—н а с e к о m o o-
пыление), 7. Chenopodiaceae, 8. Amaranfhaceae (7-8 семейства—вeтрo-
o п ы ле ни e).
Ветвь 2- Urticalia-Крапив о образные
Порядок 1. Urticales—Крапивоцветные (ступень Hypsanthophyta)
Семейства: у. МоГасеае (в етроопыление и насекомоопыление),
2. СаппаЪасеае, 3. Urticaceae, 4. Ulmaceae, 5. Eucommiaceae, 6. Bhoipte-
leaceae (2—8 семейства—в етроопыление).
Ветвь 3. Polygonali а—Г речихообразные
Порядок 1. Polygonales—Гречихоцветные (ступень Mesanthophyta)
Семейство 1. Polygonacede -(насекомоопыление и в.етроо п ы л е-
н и е).
Ветвь 4. Gaeta lia—Кактусообразные
Порядок 1. Cactales—Кактусоцветные (ступень Mesanthophyta)
Семейство 1. Cactaceoe (насекомоопыление).
Ветвь 5. Р г i m u I a I i а— Первоцветообразные * 13
” Octoknematac eae. (Ред.).
13 Gyrostemonaceae. (Ред).
Обзор новейших систем цветковых растений 101
Порядок 1 • Plumbaginales—Свинчаткоцветные (ступень Mesantho-
р h у t а).
Семейство 1. Plwmbaginaceae (насекомоопыление).
Порядок 2. Primulales—Первоцветные (ступень Mesanthophyta)
Семейства: 1. Tbeophrastaceae, 2. Myrsinaceae, 3. Primulaceae (все 3
•семейства—н асекомоопыление).
Ствол viz. Ebenophyta
Порядок 1. Ebenales—Эбеноцветные IетУ пень м е s а п 111 оphytа)
Семейства: 1. Sapotaceae, 2. Ebenaceae, 3. Symplocaceae, 4. Styraca-
ceae, 5. Lissocarpaceae (все 3 семейств—н асекомоопыление).
Ствол Viir Krinophyta
Ветвь 1. Li I i i f I о г ali а—Л и л и eo б p аз н ы е
Порядок 1. Helobiae—Болотникоцветные (ступень Protanthophyta)
Семейства: 1. Alismataceae, 2. Butomaceae (1-2семейства—насекомо-
о п ы л е н и е), 3. Hydrocharitaceae (и асекомоопыление и гидрофилия),
4. Scheuchzeriaceae, 5. Potamogetonaceae, 6. Aponog etonaceae, 1- Najada-
ceae С4—7 семейства—гидрофилия).
Порядок 2. Triuridales—Триуридоцветные (ступень Protanthophyta)
Семейство 1. Triur idaccae (гетеротрофия).
Порядок 3. Liliiflorae—Лилиецветные (ступень Mesanthophyta)
Семейства: 1. Liliaceae, 2. Нaemodoraceae, 3. Stemonaceae- 4. Ата-
ryUidaceae, 5. Veiloziaceae, Taccaceae, 7. Bioscoreaceae, 8. Iridaceae,
9. Burma,nniaceae, 10. (Jorsiaceae C1—10 семейства—н а с e к ом о о п ы л е н и е),
1. Jwncaceae (в е т р о о п ы л е н и е).
Порядок 4. Microspermae—Мелкосемянные (ступень Hypsantho-
phyta).
Семейство 1. Orchidaceae (насекомоопыление).
Ветвь 2. Су per ali а—0 сок о образные
Порядок 1. Cyperdles—Осокоцветные (ступень Hypsanthophyta)
Семейство 1, Сурегасеае (ветроопыление).
Ветвь 3. GI u m i f I о r a I i а—П ленчатоцветнообразные
Порядок 1. Farinosae—Мучнистые (ступень Mesanthophyta)
Семейства: 1. Bromeliaceae, 2. Сommelinaceae, 3. Pontederiaeeae, 4.
С yanastraceae™ •, 5. Philydraceae (1—5 с е м е й ст в а—н асекомоопыление),
6. Bapateaceae, В Thurniaceae, 8. Eriocaulaceae, 9. Xyridaceae, Ю. Jfay-
caceae, 11- Centrolepidaceae, 12. Flagellariaceae, 13. Bestionaceae 13
семейства—в етроопыление).
Порядок 2, Scitaminales—Бананоцветные (ступень Mesa nthop.hyta)
Семейства: 1. Zingiberaceae, 2. Musaceae, 3. Marantaceae, 4. Canna-
ceae (Bce 4 семейства— насекомоопыление). 19
19 Syn. Tecophilaeaceae (Ред.).
102 Гроссгейм—К вопросу о графическом изображении цветковых растений
Порядок 3- Glumiflorae—Пленчатоцветные (ступень Hypsanthophyta?-
Семейство 1. Gramineae (ветроопыление).
Ствол IX. Spathophyta
Ветвь 1. 8 р at h i f I о г a I i а—П о ч атко о б р аз н ы е
Порядок 1. Principes “ Пальмоцветные (ступень М е s a n th о р h у t а)
Семейства: 1. ра1тае, 2. Сyclanthaceac (°ба семейства—н а с е к о м о о-
пыление).
Порядок 2. Spathiflorae—Початкоцветные (ступень Hypsanthophyta)
Семейства: 1. Агасеае (насекомоопыление), 2. Ъетпасеае (гидро-
филия)
Порядок 3. Pandanales—Панданоцветные (с т у п е н ь Hypsanthophyta)
Семейства: 1. Pandunaceie, 2. S'parganiaceac, 3. Typhaceac (все 3 се"
мейства— ветроопыление).
Ветвь 2'- Piperalia—Перечнообразные
Порядок 1. Piperales—Перечноцветные (ступень Hypsanthophyta)
Семейства: 1. Saururaceae, 2. Piperaceae-, 3. Cliloranthaceae, 4. Laci~
stemonaceae29 (все 4 семейства—в етроопыление).
Ствол х Itephyta
Порядок 1. Salicales—Мвоцветные (cryneHbHypsanthophyta)
Семейство 1. Sdlicaceae (ветроопыление).
Ствол xi Kerkidophyta
Порядок 1. Casuarinales—Казуариноцветные (ступень Hypsantho-
phyta)
Семейство ]. С asuarinaceae (ветроопыление).
Баку
15 февраля 1943 г.
20 Lacistemaceae (Ред.).
Приложение II
Supplement II
КОНСПЕКТЫ (ОБЗОРЫ) ФИЛОГЕНЕТИЧЕСКИХ СИСТЕМ
Conspectus (reviews) of the phylogenetic Systems
ВЕТТШТЕЙН P. 1935.
Обзор главных систематических групп подотдела
покрытосемянных.
(„Руководство по систематике растений", т. II.).
I
WETTSTEIN К- 1935.
tibersicht der systematischen Hauptgruppen der Unterabteilung
der Angiospermen-
'(„Handbuch der systematisc’nen Botanik", II. B.).
IX. Stamm. CORHOPHYTA1
II. Abteilung. ANTHOPHYTA
1. Unterabteilung. GYMNOSPEBMAE
1. Klasse. Pteridospermae (Cycadofilicinae)2
2. „ Cycadinae
3. „ Bennettitinae
4. „ Cordaitinae
5. „ Ginkgoinae
6. „ Coniferae
7. „ Gnetinae
2. Unterabteilung. ANGlOSPERMAE
1. Klasse. Dicotyledones
1. Unterklasse. Choripetalae
1. Entwicklungsstufe.3 A. M о n о c h 1 a m у de a e
1 Главные таксоны, принятые автором в данном обзоре: Stamm (лат- Phylum) —
ствол, фила, колено; Abteilung (лат. Division) — отдел; Unterabteilung (лат. Subdi-
vision)— подотдел; Klasse ;лат. Classis) — класс; Unterklasse (лат. Subclassis) —
подкласс; Reihe (лат. Ordo) — порядок; Familie (Fam.) (лат. Familia) — семейство.
2 Представлен только ископаемыми растениями.
s Ступень развития. - (Ред.).
104
Конспект системы Веттштейна Conspectus of the Wettstein’s System
1. Reihe. Verticillatae
1. Fatnilie: Casuarinaceae
2. Reihe. F ag ale s
1. Fatnilie: Betulaceae
2. „ Fagaceae
3. Reihe. Myricales
1. Fatnilie: Myricaceae
4. Reihe. Balanopsidales
1. Fatnilie: Balanopsidaceae
5. Reihe. Leitneriales
1. Fatnilie: Leitneriaceae
6. Reihe. J uglandales
1. Fatnilie: Julianlaceae
2. „ Juglandaceae
7. Reihe. G art yules*
1. Fatnilie: Garryaceae
8. Reihe. Salicales
1. Fatnilie: Salicaceae
9. Reihe. Batidales
1. Fatnilie: Batidaceae
10. Reihe. Urticales
1. Fatnilie: Могасеае
2. К Cannabaceae
3. 99 Ulmaceae
4. п Eucommiaceae
5. 99 Rhoipteleaceae
6. 99 Urticaceae
11. Reihe. Piperales
1. Fatnilie: Piperaceae
2. 99 Saururaceae
3. 99 Chlor anthaceae
4. 99 Lacistetnonaceae
12. Reihe. P roteales
1. Fatnilie: Proteaceae
13. Reihe. S a n t al al e s
1. Fatnilie: Santalaceae
2. 99 Grubbiaceae
3. 99 Opiliaceae
4. 99 Octoknemataceae
5. 99 Olacaceae
6. 99 Myzodendraceae
7. 99 Loranthaceae
8. 99 Balanophoraceae
9. 99 Cynotnoriaceae
в конце этого конспекта. (Ред.)-
4 См. примечание
105
Обзор новейших систем цветковых растений
14. Reihe. Polygonales 1. Familie: Polygonaceae
15. Reihe. Centrospermae 1. Familie: Chenopodiaceae 2. „ Amarantaceae 3. „ Phytolaccaceae 4. „ Thelygonaceae 5. „ Nyctaginaceae 6. n Aizoaceae 7. „ Cactaceae 8. „ Portulacaceae 9. „ Basellaceae 10. „ Caryophyllaceae
16. Reihe. T ricoccae 1. Familie: Euphorbiaceae 2. „ Daphniphyllaceae 3. „ , Dichapetalaceae 4. „ Buxaceae 5. „ Callitrichaceae
17. Reihe. Hamamelidales
1. Familie: H amamelidaceae 2. „ Cercidiphyllaceae 3. „ Euptel(a)eaceae 4. „ Platanaceae
2- Entwicklungsstufe B.Dialypetaleae
18. Reihe. Polycarpicae
1. Familie: Magnoliaceae
2. 99 T rochodendraceae
3. Lactoridaceae
4. Himantandraceae
5. Eupomatiaceae
6. 9) Anonaceae
7. 99 Myristicaceae
8. jj Canellaceae
9. 99 Aristolochiaceae
10. 99 Rafflesiaceae
11. 99 Hydnoraceae
12. 99 Calycanthaceae
13. 99 Gomortegaceae
14. 99 Monimiaceae
15. 99 Lauraceae
16. 99 Hernandiaceae
17. 99 Menispermaceae
18. 99 . Lardizabalaceae
19. 99 Ranunculaceae
20. 99 Berberidaceae
21. 99 Nymphaeaceae
22. 99 Ceratophyllaceae
23. 99 Nepenthaceae
24. 99 Cephalotaceae
25. 99 Sarraceniaceae
106 Конспект системы Веттштейна Conspectus of the Wettstein's System
19. Reihe. Rhoeadales
1. Familie: 2. „ 3. 4. 5. 6. „ Papaveraceae Tovariaceae Capparidaceae -Cruciferae Resedaceae Moringaceae
20. Reihe. P ar i e tales
1. Familie: 2. „ 3. „ 4. 5. 6. 7. „ 8. 9. „ 10. 11. „ 12. „ 13. „ 14. „ 15. „ 16. „ 17. „ 18. „ 19. „ 20. Cislaceae B.ixaceae Cochlospermaceae T amaricaceae Fouquieriaceae Frankeniaceae Elatinaceae Droseraceae Violaceae Flacourtiaceae Stachyuraceae Turneraceae Malesherbiaceae Passif lor aceae Achariaceae Caricaceae Loasaceae Begoniaceae Datiscaceae Ancistrocladaceae
21. Reihe. Guttife rales
1. Familie: Dilleniaceae
2. Actinidiaceae
3. 99 Ochnaceae
4. 99 Strasburgerlaceae
5. 99 Eucryphiaceae
6. 99 Caryocaraceae
7. 99 Marcgraviaceae
8. 99 Quiinaceae
9. 99 Theaceae
10. 99 Guttiferae
11. 99 Dipterocarpaceae
22. Reihe. Rosales
1. Familie: Crassulaceae
2. 99 , Saxifragaceae
3. 91 Cunoniaceae
4. 99 Brunelliaceae
5. 99 Myrolhamnaceae
6. 99 Pittosporaceae
7. 99 Byblidaceae
8. 99 Roridulaceae
9. 99 Bruniaceae
Обзор новейших систем цветковых растений
107
10. „ Podostemonaceae
ц. „ Hydrostachyaceae
/2. „ Rosaceae
lg. „ Crossosomataceae
14. „ Chrysobalanaceae
15, ,, Connaraceae
15, „ Mimosaceae
17, „ Papilionaceae
23. Reihe. M yr t a I e s
1. Fatnilie: Penaeaceae
2. 9? Geissolomataceae
3. Oliniaceae
4. 99 Thumpiaeaceae
5. 99 Elaeagnaceae
6. 99 Lythraceae
7. 99 Heteropyxidaceae
8. 99 Sonneratiaceae
9. 99 Rhizophoraceae
10. 99 Alangiaceae
11. 99 Nyssaceae
12. 99 Lecythidaceae
13. 99 Combretaceae
14. 99 Myrtaceae
15. 99 Punicaceae
16. 99 Melastomataceae
17. 99 Oenotheraceae
18. 99 Halorrhagidaceae
19. 99 Gunneraceae
20. 99 Hippuridaceae
24. Reihe. Columnifer ae
1. Fatnilie: Malvaceae
2. Botnbacaceae
3. n ! ТШс''еае
4. 99 Sterculiaceae
5. 99 Elaeocarpaceae
6. 99 Chlaenaceae
7. 99 Gonystylaceae
8. 99 S cytopetalaceae
25. Reihe. Gruinales
1. Fatnilie: Linaceae
2. Hutniriaceae
3. 99 Oxalidaceae
4. 99 Geraniaceae
5. 99 Limnanthaceae
6. 99 T ropaeolaceae
7. 99 Erythroxylaceae
8. 99 Malpighiaceae.
9. 99 Zygophyllaceae
10. 99 Cneoraceae
108 Конспект системы Веттштейна Conspectus of the Wettstein’s System
26. Reihe. T er eb i nth ales5
1. Familie: Rutaceae
2. 99 Simarubaceae
3. 99 Burseraceae
4. D Meliaceae
5. 99 Tremandraceae
6. 99 Polygalaceae
7. 99 Xanthophyllaceae
8. 99 T rigoniaceae
9. 99 Vochysiaceae
10. 99 Anacardiaceae
11. 99 Sapindaceae
12. Akaniaceae
13. Aextoxicaceae
14. 99 Aceraceae
15. 99 Hippocastanaceae
16. 99 Coriariaceae
17. 99 Cyrillaceae
18. 99 Pentaphylacaceae
19. 99 Sabiaceae
20. 99 Melianthaceae
21. n Corynocarpaceae
22. Balsaminaceae
27. Reihe. C el as 11 -ales
1. Familie: Aquifoliaceae
2. Celastraceae
3. 99 - Salvadoraceae
4. 99 Staphyleaceae
5. 99 Hip poor ate aceae
6. 99 Stackhousiaceae
7. 99 I cacinaceae
28. Reihe. Rhamnales
1. Familie: Rhlamnaceae
2. „ Vitaceae
29. Reihe. Umbelliflorae
1. Familie: Cornaceae
2. „ 3. „! Araliaceae Umbelliferae
2. Unterklasse. Sympetalae
1. Reihe. Plumbagin ales
1. Familie: Plumbaginaceae
* К порядкам 25 и 26 близок порядок Pandales с сем. Pandaceae, не вклю-
ченный автором в общую нумерацию, хотя и приведен им в тексте на стр. 816. (Ред.).
Обзор новейших систем цветковых растении
109
2. Reihe. Primulales
1. Familie: Fheophrastaceae
2. „ Primulaceae
3. „ : Myrsinaceae
3. Reihe. Bicornes
1. Familie: Clethraceae
2. 99 Pirolaceae
3. 99 ' Ericaceae
4. 99 Empetraceae
5. 99 Epacridaceae
6. 99 Diapensiaceae
4. Reiche. D io s p у r ale s
1. Familie: Ebenaceae
2. Я Hoplestigmataceae
3. 99 Styracaceae
4. 9? Symplocaceae
5. 99 Sapotaceae
5. Reihe. T ub if I о r ae
1. Familie: Convolvulaceae
2. 59 Cuscutaceae
3. ?9 Polemoniaceae
4. 99 Hydrophyllaceae
5. 99 Lennoaceae
6. 99 Boraginaceae
7. 99 Nolanaceae
8. 99 Solanaceae
9. 99 Scrophulariaceae
10. 99 Lentibulariaceae
11. 99 Orobanchaceae
12. 99 Gesneriaceae
13. 99 Bignoniaceae
14. 99 Pedaliaceae
15. 99 Martyniaceae
16. 99 Acanthaceae
17. 99 Verbenaceae
18. 99 Labiatae
19. 99 Tetrachondraceae
20. 99 Globutariaceae
21. 99 Phrymaceae
22. 99 Myoporaceae
23. 99 Plantaginaceae
24. 99 Columelliaceae
6. Reihe. С о nt о r t ae
1. Familie: Loganiaceae
2. 99 Buddleiaceae
3. 59 Gentianaceae
4. 99 Menyanthaceae
110
Конспект системы Веттштейна Conspectus of the Wettstein’s System
5. „ A pocynaceae
6. „ , Asclepiadaceae
7. Reihe. Ligustrales
1. Fatnilie: Oleaceae
8. Reihe. Rubiales
1. Fatnilie: Rubiaceae
2. „ Caprifoliaceae
< 3. „ : Adoxaceae
4. „ Valerianaceae
5. „ Dipsacaceae
6. „ Calyceraceae
9. Reihe. Cucurbitales
1. Fatnilie: Cucurbitaceae
10. Reihe. Synandrae
1. Fatnilie: Campanulaceae
2. 99 Lobeliaceae
3. 5? . Goodeniaceae
4. 99 Stylidiaceae
5. 99 Brunoniaceae
6. 9? Compositae
II. Klasse. M n n о с о t у I e d о n e s
1. Reihe. Helobiae
1. Fatnilie: Alismataceae
2. „ Butotnaceae
3. „ H ydrocharitaceae
4. „ Scheuchzeriaceae
5. „ Aponogetonaceae
6. „ Potamogetonaceae
7. „ Najadaceae
8. „ Triuridaceae
2. Reihe. L i I i i f I o-r a e
1. Fatnilie: Liliaceae
2. 99 Stemonaceae
3. 99 Cyanastraceae
4. 99 Pontederiaceae
5. 99 Haemodor aceae
6. 99 Philydraceae
7. 99 Amar у Hid aceae
8. 99 Velloziaceae
9. 99 Iridaceae
10. 99 Juncaceae
11. 99 Flagellariaceae
12. 99 Rapateaceae
13. Thiirniaceae
14. 99 Brotneliaceae
Обзор новейших систем цветковых растений,
111
15. „ Dioscoreaceae
16. Taccaceae
17. Burmanniaceae
Reihe. Е п а п t i 0 b I a star
1. Familie: Commelinaceae
2. „ Mayacaceae
3. Xyridaceae
4. Eriocaulaceae
5. Centrolepidaceae
6. 0 Restionaceae
. Reihe. С у р е г ales
1. Familie: Cyperaceae
Reihe. G 1 и mi florae
1. Familie: Gramineae
Reihe. Sei tarn i n e a e
1. Familie: Musaceae
2. Zingiberaceae
3. Cannaceae
4. Marantaceae
Reihe. Gynandrae
1. Familie: Orchidaceae
Reihe. Spadiciflorae
1. Familie: Palmae
2. 0 Cyclanthaceae
3.' Araceae Lemnaceae
, Reihe. Pandanales
1. Familie: Pandanaceae
2. „ Sparganiaceae
3. „ Typhaceae
Примечание. Ветт н: т е й н, специально касаясь (р. 858) по-
ложения семейства Garryaceae, единственного представителя порядка
GarryaleSi высказывается в пользу его большей морфологической близос-
ти к порядку Umbelliflorae и считает, поэтому, более правильным помес-
тить его в конце Choripetalae, вслед за Umbelliflorae под номером 30.
Однако, в филогенетическом обзоре порядков (р. юбб) и схематическом
изображении их филогенетических взаимосвязей (р. 1068) Веттштейн
помещает (под номером 7) Garryales уже в группе Monochlamydeae» меж-
ду порядками Juglandales и Salicales. правда, с оговоркой; „положение
неясное11, и с вопросительным знаком, но тут же подчеркивает, что этот
112
Конспект системы Веттштейна Conspectus of the Wettstein’s System
порядок стоит на той же высоте морфологического развития, как. и
смежные порядки. В публикуемом в этой книге „Обзоре главных систе-
/матических групп подотдела Angiospermae" Веттштейна, порядку
Garryales мною отведено место в соответствии с последним его взглядом,,
хотя, по моему мнению, правильнее поставить его рядом с Umbelliflorae-
(Ред.).
БЕССИ Ч. Э. 1915.
Таксономия цветковых растений
(„Филогенетическая таксономия цветковых растений*1; на англ, яз.)
BESSEY CH. Е. 1915.
Taxonomy of Flowering Plants
(„The phylogenetic Taxonomy of Flowering Plants11)
PHYLUM6 XIV. ANTHOPHYTA
Class 32. Alternifoliae (Monocotyledoneae)
Sub-C la s s Alternifol iae— S t r 0 b ilo ide ae
Order Ali s m at a les
Family 1. Alismataceae
99 2. Butomaceae
99 3. T rhiridaceae
99 4. S cheuchzeriaceae
99 5. Typhaceae
99 6. Sparganiaceae
99 7. Pandanaceae
99 8. A ponogetonaceae
99 9. Potamogetonaceae
Order Liliales
Family 10. Liliaceae
99 11. Stemonaceae
n 12. Pontederiaceae
99 13. Cyanastraceae
99 14. Philydraceae
99 15. Commelinaceae
99 16. Xyridaceae
99 17. Mayacaceae
W 18. Juncaceae
99 19. Eriocaulonaceae
99 20. Thurniaceae
99 21. Rapateaceae
99 22. Najadaceae
* Phylum — ствол, фила, филум, филия, отдел. Class — класс. Sub-Class —
подкласс. Superorder — надпорядок. Order — порядок. Family — семейство (Ред.).
8. А. А. Гроссгейм и М. Ф. Сахокиа
114 Конспект системы Бесси Conspectus of the BeSSeyS System
Order A r ales
Family 23. Cyclanthaceae
„ 24. Araceae
„ 25. Lemnaceae
Order Palmales
Family 26. Palmaceae
Order G r ami n al e s
Family 27. Restionaceae
„ 28. Centrolepidiac.eae
„ 29. Flagellariaceae
„ 30. Cyperaceae
„ 31. Poaceae
Sub-Class Alternifoliae —
Cotyloideae
Order Hydra les
Family 32. V allisneriaceae
Order Ir idal e s
Family 33. A maryllidaceae
99 34. Haemodoraceae
>9 35. Iridaceae
n 36. V elloziaceae
99 37. Taccaceae
99 38. Dioscoreaceae
99 39. Bromeliaceae
99 40. Musaceae
99 41. Zingiberaceae
99 42. Cannaceae
99 43. Marantaceae
Order Orchidales
Family 44. Burmanniaceae
99 45. Orchidaceae
Class 33. Oppositifoliae (Dicotyledoneae)
Sub-Class Oppositifoliae—Str obiloideae
Super-Order Strobiloideae—Apopetalae—Polycarpellatae
Order Ran al e s
Family 46. Magnoliaceae
„ 47. Calycanthaceae .
„ 48. Monimiaceae
„ 49. Cercidiphyllaceae
„ 50. Trochodendraceae
„ 51. Leitneriaceae
„ 52. Anonaceae
„ 53. Lactoridaceae
„ 54. Gomortegaceae
Обзор! новейших систем цветковых растений
Family 55. Myristicaceae
п 56. Saururaceae
99 57. Piperaceae
99 58. Lacistemaceae
99 59. Chloranthaceae
99 60. Ranunculaceae
99 61. Lardizabalaceae
99 62. Rerberidaceae
99 63. Menispermaceae
99 64. Lauraceae
99 65. Nelumbaceae
99 66. Cabombaceae
99 67. Ceratophyllaceae
99 68. Dilleniaceae
99 69. Winteranaceae
Order M al v al e s
Family 70. Sterculiaceae
99 71. Malvaceae
99 ' ' 72. Bombacaceae
99 73. Scytopetalaceae
99 74. Chlaenaceae
99 75. Gonystylaceae
• 99 76. Tiliaceae
99 77. Elaeocarpaceae
99 78. Balanopsidaceae
99 79. Ulmaceae
99 80. Moraceae
99 81. Urticaceae
Order Sarraceniales
Family 82. Sarraceniaceae
n 83. Nepenthaceae
Order Geraniales
Farhily 84. Geraniaceae
99 85. Oxalidaceae
99 86. Tropaeolaceae
99 87. Balsaminaceae
99 88. Limnanthaceae
99 89. Linaceae
99 90. Humiriaceae
99 91. Eryihroxylaceae
99 92. Zygophyllaceae
99 93. Cneoraceae
99' 94. Rutaceae
99 95. Simarubaceae
99 96. Burseraceae
99 97. Meliaceae
99 98. Malpighiaceae
99 99. T rigoniaceae
99 100. Vochysiaceae
99 101. Polygalaceae
ll6
Конспект системы Бесси Conspectus of the BeSSey’s System
Family 19 99 99 102. Tremandraceae 103. Dichapetalaceae 104. Euphorbiaceae 105. Callitrichaceae
Order Guttiferales
Family 106. Theaceaq
99 107. Cistaceae
99 108. Guttiferaceae
99 109. Eucryphiaceae
99 110. Ochnaceae
99 111. Dipterocarpaceae
99 112. Caryocaraceae
99 113. Quiinaceae
99 114. Marcgraviaceae
99 115. Flacourtiaceae
99 116. Bixaceae
99 117. Cochlospermaceae
99 118. Violaceae
99 119. Malesherbiaceae
99 120. T timer aceae
99 121. Passif lor aceae
99! 122. Achariaceae
99 .123. Caricaceae
99 ' 124. Stachyuraceae
99 125. Koeberliniaceae
Order Rh о e a d a I e s
Family 99 126. Papaveraceae 127. Tovariaceae
99 128. Nymphaeaceae
99 129. Moringaceae
99 130, Resedaceae
99 131. Capparidaceae
99 132. Brassicaceae
Order Caryophyllales
Family 133 Caryophyllaceae
99 134. Elatinaceae
99 135. Portulacaceae
99 136. Aizoaceae
99 137. Frankeniaceae
99 138. T amaricaceae
99 139. Salicaceae ।
99 . 140. Podostemonaceae
99 141. Hydrostachydaceae
99 142. Phytolaccaceae
99 143. Basellaceae
99 144. Amaranthaceae
99 145. Chenopodiaceae
99 146. Polygonacede
99 147. Nyctaginaceae
Обзор воэейших систем цветковых ^растений
Family 148. Cynocramb aceae
„ 149. Batidaceae
Super-Order Strobiloideae—Sympetalae—Polycarpellatae
Order E b e nates
Family 99 150. Sapotaceae 151. Ebenaceae
99 152. Symplocaceae
9 153. Styracaceae
99 154. Fouquieriaceae
Order Ericales
Family 155. Clethraceae
99 156. Ericaceae
99 157. Epacridaceae
99 158. Diapensiaceae
99 159. Pirolaceae
160. Lennoaceae
Order Primula les
Family 161. Primulaceae
99 162. Plantaginaceae
99 163. Plumbaginaceae
99 164. Myrsinaceae
99 165. Theophrastaceae
Super-Order Strobiloideae—Sympetalae —Dicarpellatae
Order Family 99 99 99 99 99 Q e nti an al e s 166. Oleaceae 167. Salvadoraceae 168. Loganiaceae 169. Gentianaceae 170. Apocynaceae 171. Asclepiadaceae
Order Family 99 99 99 99 99 Polemoniales 172. Polemoniaceae 173. Convolvulaceae 1/4. Hydrophyllaceae 175. Borraginaceae 176. Nolanaceae 177. Solanaceae
Order Family 99 99 99 •99 Scrophulariales 178. Scrophulariaceae 179. Bignoniaceae 180. P'edaliaceae 181. Martyniaceae 182. Orobanchaceae
iiS
Конспект системы Бесси Conspectus of the Bessey s System
Family
99
99
99
99
183. Gesner(i)aceae
184. Columeltiaceae
185. Lentibillariaceae
186. Globulariaceae
187. Acanihaceae
Order L am i al e s
Family 188. Myoporaceae
„ 189. Phrymaceae
„ 190. Verbenaceae
„ 191. Lamiaceae
S u b-C lass Oppositifo 1 ia e—C otyloideae
Super-Order Cotyloideae—Apopetalae
Order Rosales
Family 99 192. Rosaceae 193. Malaceae
99 194. Prunaceae
99 , 195. Crossosomataceae
99 196- Connaraceae
99 197. Mimosaceae
99 198. Cassiaceae
99 199. Fabaceae
99 200. Saxifragaceae
99 201. Hydrangeaceae
99 202. Grossulariaceae
99 203. Crassulaceae
99 204. Droseraceae
99 205. Cephalotaceae
99 ' 206. Pittosporaceae
99 f 207. Brunelliaceae
99 208. Cunoniaceae
99 209. Myrothamnaceae
99 210. Bruniaceae
99 21L Hamamelidaceae
99 212. Casuarinaceae
99 213. Eucommiaceae
99 214. Platanaceae
Order M yr tales
Family t . 215. Lythraceae
99 216. Sonneratiaceae
99 217. Punicaceae
99 218. Lecythidaceae
99 219. Melastomataceae
99 220. Myrtaceae
99 221. Combretaceae
99 222. Rhizophoraceae
99 223. Oenotheraceae
99 224. Halorrhagidaceae
99 225. Hippuridaceae
0бзор новейших ситем цветковых растений
119
II
Family 226 Cynomoriaceae
„ 2'27. Aristolochiaceae
„ 228. Rafflesiaceae
„ 229. Hydnornceae
Order Loasales
Family 230. Loasaceae
„ 231. Cucurbitaceae
„ 232. Begoniaceae
„ 233. 'Datiscaceae
„ 234. Ancistrocladaceae
Order С a c t a I e s
Family 235. Cactaceae
Order Cel astг alеs
Family 236. Rhamnaceae
99 237. Vitaceae
ft 238. Celastraceae
до 239. Buxaceae
99 240. AqudfOliaceae
ДО 241. Cyrillaceae
99 242. Pentaphylacaceae
99 243. Corynocarpaceae
99 244. Hippocrateaceae
99 245. Stackhousiaceae
ft 246. Staphyleaceae
99 247. Geissolomataceae
99 248. Penaeaceae
99 249. Oliniaceae
99 250. Thymelaeaceae
99 251. Hernandiaceae
ДО 252. Elaeagnaceae
ДО 253. M yzodendraceae
ДО 254. Santalaceae
99 255. Opiliaceae
,99 256. Grubbiaceae
ДО 257. Olacaceae
ДО 258. Loranthaceae
ДО 259. Balanophoraceae
Order S ap ind al e s
Family 260. Sapirtdaceae
„ 261. Hippocastanaceae
„ 262. Aceraceae
„ 263. Sabiaceae
„ 264. Icacinaceae
„ 265. Melianthaceae
„ 266. Empetraceae
„ 267. Coriariaceae
„ 268. Anacardiaceae
„ 269. Jtiglandaceae
„ 270. Betulaceae
120 Конспект системы Бесси Conspect'is of the Bessey's Bysteisi
Family 99 99 271. Fagaceae 272. Myricaceae 273. Julian(i)aceae 274. Proteaceae
Order U mb e 11 al e s
Family
275. Araliaceae
276. Apiaceae
277. Cornaceae
Super-Order Cotyloideae—Sympetalae
Order Rub i ales
Family 278. Rubiaceae
„ 279. Caprifoliaceae
„ 280. Adoxaceae
„ 281. Valerianaceae
„ 282. Dipsacaceae
Order Campanulales
Family 283. Campanulaceae
„ 284. Goodeniaceae ’
„ 285. Stylidiaceae
„ 286. Calyceraceae
Order Asterales
Family
5?
99
99
99
99
99
99
99
99
99
1287. Helianthaceae
288. Atnbrosiaceae
289. Heleniaceae
290. Arctotidaceae
291. Calendulaceae
292. Inulaceae
293. Asteraceae
294. Vernoniaceae
295. Eupatoriaceae
296. Anthemidacetae
297. Senecionidaceae.
298. Carduaceae
299. Mutisiaceae
300. Lactucaceae
ГАТЧИНСОН ДЖ. i?6c>.
Последовательность порядков и семейств
(цветковых растений)
(„Семейства цветковых растений", тт. I и П, на англ, яз.)
HUTCHINSON J. I960.
(„The families of flowering plants", vol. vol. I a. II)
SEQUENCE OF ORDERS AND FAMILIES 7
Dicotyledones
Division I. Lignosae
1. Magnoliales 4. Dilleni ales
1. Magnoliaceae 2. Ilticiaceae 3. Winteraceae 4. Canellac'eae 5. Schizanaraceae 6. Himantandraceae ' 7. Lactoridaceae 8. Trochodendraceae 9. Cercidiphyllaceae 19. Dilleniaceae 20. Connaraceae 21. Crossosomataceae 22. Brunelliaceae 5. Coriariales 23. Coriariaceae 6. Rosales 1
2. A n no n ale s 10. Annonaceae 11. Eupomatiaceae 24. Rosaceae 25. Dichapetalaceae 26. Calycanthaceae 7. Leguminales i
3. Laurates 12. Monimiaceae 13. Austrobaileyaceae 14. Trimeniaceae 15. Lauraceae 16. Gomortegaceae 17. Hernandiaceae 18. Myristicaceae 27. Caesalpiniaceae 28. Mimosaceae 29. Papilionaceae 8. Cunoniales 30. Pterostemonaceae 31. Cunoniaceae 32. Philadelphaceae
’'A cross -line indicates the climax of a group.
122 Конспект системы Гатчинсона Conspectus of the Hutchinson’s System
33. Hydrangeaceae 34. Grossulariaceae 35. Oliniaceae 36. Greyiaceae 37. Escalloniaceae 38. Baueraceae 39. Crypteroniaceae 18. Casuarinales 67. Casuarinaceae 19. Urticales 68. Ulmaceae 69. Cannabiaceae 70. Moraceae
9. Styracales 40. Lissocarpaceae 41. Styracaceae 42. Symplocaceae 71. Urticaceae 72. Barbeyaceae 73. Eucommiaceae 20. В i x al e s
10. Araliales 43. Cornaceae 44. Alangiaceae 45. Garryaceae 46. Nyssaceae 47. Araliaceae 48. Caprifoliaceae 74. Bixaceae 75. Cistaceae 76. Flacourtiaceae 77. Cochlospermaceae 78. Hoplestigmataceae 79. Achatocarpaceae 80. Lacistemaceae i 21. T h у m el ae at e s
11. H am a me I i d al e s 49. Tetracentraceae 50. Hamamelidaceae 51. Myrothamnaceae 52. Platanaceae 53. Stachyuraceae 54. Buxaceae 55. Daphniphyllaceae 56. Bruniaceae 81. Gonystytaceae f 82. Aquilariaceae 83. Geissolomataceae 84. Penaeaceae 85. Thymelaeaceae 86. Nyctaginaceae 22. Proteales 87. Proteaceae
12. S ali c al e s 57. Salicaceae 23. Pittosporales 88. Pittosporaceae
13., Leitneriales 58. Leitneriaceae 89. Byblidaceae 90. Stegnospermaceae 91. Vivianiaceae
14. M у r i c ale s 59. Myricaceae 92. Tremandraceae 24. C ap p ar id al e s
15. Balanopsidales 60. Balanopsidaceae 93. Capparidaceae . 94. Moringaceae
16. Fag ales 61. Betulaceae 62. Fagaceae 63. Corylaceae 95. Tovariaceae 25. T am a r i.c ale s 96. Frankeniaceae 97. Tamaricaceae
17. J ugland ales 64. Rhoipteleaceae 65. Juglandaceae 66. Picrodendraceae 98. Fouquieriaceae 26. Violate s , 99. Violaceae
Обзор новейших систем цветковых растений
123-
27. Polygal ales
100. Polygalaceae
101. Krameriaceae
102. Trigoniaceae
103. Vochysiaceae
28. L 0 as al e s
104. Turneraceae
105. Loasaceae
29. Passiflorales
106. Malesherbiaceae
107. Passifloraceae
108. Achariaceae
30. Cucurbitales
109. Cucurbitaceae
110. Begoniaceae
111. Datiscaceae
112. Caricaceae
31. С a c t a le s ,
113. Cactaceae
32. T i I i a I e s
114. Dirachmaceae
115. Scytopetalaceae
116. Tiliaceae
117. Sterculiaceae
118. Peridiscaceae
119. Bombacaceae
33. M alv al e s
120. Malvaceae
34. Malpighiales
121. Ixonanthaceae
122. Malpighiaceae
123. Humiriaceae
124. Linaceae
125. Irvingiaceae
126. Huacede
127. Ledocarpaceae
128. Erythroxylaceae
129. Ctenolophonaceae
130. Lepidobotryaceae
131. Balanitaceae
132. Zygophyllaceae
35. Euphorbiales
133. Euphorbiaceae
36. T h e al e s
134. Bonnetiaceae
135. Theaceae
136. Saurauiaceae
137. Actinidiaceae
138. Pellicieraceae
139. Pefitaphylacaceae
140. Tetrameristaceae
141. Marcgraviaceae
142. Caryocaraceae
143. Medusagynaceae
/
37. О chnale s
144. Strasburgeriaceae
145. Ochnaceae
146. Rhodolaenaceae
147. Sphaerosepalaceae
148. Dipterocarpaceae
149. Ancistrocladaceae
38. Ericales
150. Clethraceae
151. Pyrolaceae
152. Ericaceae
153. Epacridaceae
154. Diapensiaceae
155. Monotropaceae
156. Lennoaceae
157. Vacciniaceae
39. Guttiferales
158. Hypericaceae
159. Clusiaceae
160. Eucryphiaceae
161. Quiinaceae
40. M у r t a I e s
162. Myrtaceae
163. Lecythidaceae
164. Rhizophoraceae
165. Sonneratiaceae
166. Punicaceae
167. Combretaceae
168. Melastomataceae
41. Celastrales
169. Pandaceae
170. Aquifoliaceae
171. Salvadpraceae
172. Koeberliniaceae
173. Cneoraceae
174. Cardiopteridaceae
175. Cyrillaceae
176. Icacinaceae
124
Конспект системы Гатчинсона
Jonspectus of the Hutchinson’s System
177. Empetraceae 178. Aextoxicaceae J79. Pentadiplandraceae 180. Celastraceae 181. Corynocarpaceae 182. Stackhousiaceae , 183. Goupiaceae 184. Hippocrateaceae 185. Erythropalaceae 186. Capusiaceae 187. Scyphostegiaceae 49. Sapindates 214. Melianthaceae 215. Sapindaceae 216. Podoaceae 217. Sabiaceae 218. Anacardiacede 219. Aceraceae 220. Hippocastanaceae 221. Staphyleaceae 222. Akaniaceae 223. Julianiaceae
42. Olacales 188. Olacaceae t 189. Opiliaceae 190. Octoknemaceae 191. Aptandraceae ,192. Dipentodontaceae 193. Medusandraceae 224. Didiereaceae 50. Loganiales 225. Potaliaceae 226. Loganiaceae 227. Buddleiaceae 228. Antoniaceae 229. Spigeliaceae
43. S a n t a I a I e s 230. Strychnaceae 231. Oleaceae
194. Loranthaceae 195. Grubbiaceae 196. Santalaceae 197. Myzodendraceae 198. Balanophoraceae 51. Apocynales 232. Placesрегтасеае 233. Apocynaceae 234. Periplocaceae
44. Rhamnales 199. Heteropyxidaceae 200. Elaeagnaceae 201. Rhamnaceae 202. Vitaceae 235. Asclepiadaceae 52. R и b i a I e s 236. Dialypetalanthaceae 237. Rubiaceae
45. Myrsinales ’ 203. Myrsinaceae 204. Theophrastaceae 205. Aegicerataceae 53. Bignon iales 238. Cobaeaceae 239. Bignoniaceae 240. Pedaliaceae 241. Martyniaceae
46. Ebenales 206. Ebenaceae 207. Sapotaceae 208. Sarcospermaceae 47~Rtital e s 54. Verbenales 242. Ehretiaceae 243. Verbenaceae 244. Stilbeaceae 245. Chloanthaceae 246. Phrymaceae
209. Rutaceae 210. Simaroubaceae 211. Burseraceae 212. Averrhoaceae Division II. Herbaceae 55. Ranales . 247. Paeoniaceae 248. Helleboraceae
48. Me Hale s 213. Meliaceae 249. Rantmculaceae 250. Nymphaeaceae 25,1. Podophyllaceae
Обзор новейших систем цветковых растении
125.
252. Ceratophyllaceae
253. Cabombaceae
56. Berberidales
254. Sargentodoxaceae
255. Lardizabalaceae
256. Menis pertnaceae
257. Nandinaceae
258. ’Circaeasteraceae
259. Berberidaceae
57. Aristolochiales
260. Aristolochiaceae
261. Hydnoraceae
262. Cytinaceae
263. Nepenthaceae
58. Piperales
264. Piperaceae
265. Saururaceae
266. Chloranthaceae
59. R h о e ad a I e s
267. Pappveraceae
268. Fumariaceae
60. Cruciales
269. Cruciferae
61. Resedales
270. Resedaceae
62. Caryophyllales
271. Elatinaceae
272. Molluginaceae
273. Caryophyllaceae
274. Ficoidaceae
275. Portulacaceae
63. Pol yg on ales
276. Polygonaceae
277. Illecebraceae
64. Chenopodiales
278. Barbeuiaceae
279. Phytolaccaceae
280. Gyrostemonaceae
281. Agdestidaceae
282. Petiveriaceae
283. Chenopodiaceae
284. Amaranthaceae
285. Cynocrambaceae
286. Batidaceae
287. Baselldceae
65. Ly t h r al e s
288. Lythraceae
289. Onagraceae
290. Trapaceae
291. Haloragidaceae
292. Callitrichaceae
66. Gentianales
293. Gentianaceae
294. Menyanthaceae
67. P rimulal e s
295. Primulaceae
296. Ptumbaginaceae
68. P I an t a g i na le s
297. Plantaginaceae
69. Sax if ragales
298. Crassulaceae
299. Cephalotaceae
300. Saxifragaceae
301. Eremosynaceae
302. Vahliaceae
303. Francoaceae
304. Donati'aceae
305. Parnassiaceae
306. Adoxaceae
70. S ar r ac e ni ale s
307. Droseraceae'
308. Sarraceniaceae
71. Podostemales
309. Podostemaceae
310. Hydrostachyaceae
72. U mb ellale s
311. Umbelliferae
73. Valerianales
312. Valerianaceae
313. Dipsacaceae
314. Calyceraceae
74. Campan ales
315. Campanulaceae
316. Lobeliaceae
j26 Конспект системы Гатчинсона Conspectus of the Hutchinson’s System
75. Goodeniales 317. Goodeniaceae 318. Brunoniaceae 319. Stylidiaceae 84. A I i s m a t a I e s 345. Alismataceae 346. Scheuchzeriaceae 347. Petrosaviaceae
76. Asterales 320. Compositae 85. Tr i ur id ales 348. Triuridaceae
4= sft # # '£ if:
77. Solanales 321. Solanaceae 322. Convolvulaceae 323. Nolanaceae 86. J uncagina les 349. Juncaginaceae 350. Lilaeaceae
78. Person ales 324. Scrophulariaceae 325. Acanthaceae 326. Gesneriaceae , 327. Orobanchaceae 328. Lentibulariaceae 329. Columelliaceae (Heterostylaceae) 351. Posidoniaceae Л A 87. Aponogetonales 352. Aponogetonaceae 353. Zosteraceae
79. Geraniales 330. Gerantaceae 331. Limnanthaceae 332. Oxalidaceae 333. Tropaeolaceae. 334. Balsaminaceae Л # * * 88. Potamogetonales 354. Potamogetonaceae 355. Ruppiaceae 89. Najadales
80. P ole mo ni a I e s 356. Zannichelliaceae
335. Polemoniaceae 336. Hydrophyllaceae 337. Cuscutaceae 357. Najadaceae % & <: &
81. Boraginales 338. Boraginaceae 90. C 0 mm el inales 358. Commelinaceae 359. Cartonemataceae
82. Lam iales 360. Flagellariaceae
339. Myoporaceae 340. Selaginaceae 341. Globulariaceae 342. Labiatae 361. Mayacaceae 91. Xyridales 362. Xyridaceae 363. Rapateaceae
Monocotyledones® Division I. Calyciferae 92. Er iocaulales
83. В ut 0 m al e s 343. Butomaceae 344. Hydrocharitaceae 364. Eriocaulaceae 93. В romeliales 365. Bromeliaceae ,
8 Asterisks * indicates a local climax.
Обзор новейших систем цветковых растений .127
94. Z i п g i b e r a I e s 366. Masaceae 367. Strelitziaceae 368. Lowiaceae 369. Zingiberaceae 370. Cannaceae 371. Marantaceae 390. Dioscoredceae 3$ 102. A gav ales 391. Xanthorrhoeaceae 392. Agavaceae * « * * *
Division II. Corolliferae 95. Liliales 103. P al m die s 393- Palmae
372. Ltltaceae 373. Tecophilaeaccue 374. Trilliaceae 375. Pontederiaceae 376. Stnilacaceae 377. Ruscaceae * * * * * 96. A Istroemeriales 378. Alstroemeriaceae 379. Petermanniaceae 380. Philesiaceae $ 97. Ar ale s 381. Araceae 382. Lemnaceae 3$ * , * * * 98. T у p h al e s 383. Sparganiaceae 384. Typhaceae * * 104‘. Pandanales 394. Pandanaceae 105. С у с I a n i h ales 395. Cyclanthaceae 106. Haemodorales 396. Haemodoraceae 397. Hypoxidaceae 398. Velloziaceae 399. Apostasiaceae 400. Taccaceae 401. Philydraceae 107. В ur mannial e s 402. Burmanniaceae 403. Thismiaceae 404. Corsiacea? * -Ж- * * 108. Orchidales 405. Or chidace ae *
99. Amar у ll id ale s 385. Amaryllidaceae * * * » Ч1 100. Iridales 386. Iridaceae * Division III. G 1 u m i f 1 о г a e 109. J uncalss . 406. Juncaceae 407. Thurniaceae 408. Centrolepidaceae 409. Restionaceae 110. Cyperales i \ \
101. Dioscoreales. 387. Stenomeridaceae 388. Trichopodaceae 389. Roxburghiaceae 410. Cyperaceae * * * * 111. G r a min ale s 411. Gramineae *
ГЕЙНЦЕ А. 1927.
Филогенетическая система Cormophyta
(„Филогения Cormophyta"; на шведском яз. с нем. рез.)9
A. Heintze’s phylogenetic system of the Cormophyta. 1927.
[„The phylogeny of the Cormophyta*4; in Swedish („Cormofyternas fylogeni**)
with Germ, summ ]
CORMOPHYTA
Charophyta Coniferae -> Ang ospermae
t t
Bryophyta Psilotophyta
t f
Psilophyta ->Ophioglossophyta -> Articulatae
/ Ф
Lycopodiophyta Maratiiophyta
ANGIOSPEBMAE
I- Dicotyledoneae
Unterkl.10 Bicornes
E r i c a I e s
Jdiapensiaceae Empetraceae
Monotropaceae -> Ericaceae
I * I
Clethraceae E pacridaceae
Unterkl. Sympetalae
C ontortae -> Gesneri ales
Tetrasperm ae Rub tales
8 В настоящем виде этот конспект составлен мною путем объединения рассе-
янных в тексте книги А. Г ‘е й н ц е схем по отдельным таксонам. (Ред.).
18 Подкласс. (Ред ).
Обзор новейших систем цветковых растений
12g
Сontortae
Gentianaceae Loganiaceae
4 z
Apocynaceae / -> Asclepiadaceae
Gesneriale s
Lentibulariaceae Rhinanthaceae
t. z
Gesneriaceae ——> Scrophulariaceae
Ф . 4 '
Rignoniaceae Solanaeeae
4
HAartyniaceae-----> Pedaliaceae
RиЪiales
Caprifoliaceae -> V alerianaceae
t 4
Rubiaceae Dipsacaceae
Campanulaceae -> Compositae
4
Goodeniaceae -> Stylidiaceae
Tetras permae
Oleaceae Verbenaceae -> Labiatae
Rorraginaceae -> Convolvulaceae -> HydrophyTlaceae
Polemoniaceae -> Plantaginaceae Acanthaceae
Unterkl. Synsepalae
H yr tiflor ae M elast o-matales -> Arist olo chiales
Elaeagnaceae
Thymelaeaceae
Lythraceae
Rhinophoraceae
Proteaceae' Hydrocaryaceae
Oenotheraceae / Lecythidaceae
t z
Sonneratiaceae-----> Hlyrtaceae
4 ' \
Gombretaceae HAelastomataceae
4 4
H alorrhagaceae Punicaceae
^Aristolochiales
Aristolochiaceae -> Rafflesiaceae
9. А.. А. Гроссгейм и M. Ф. Сахокиа
IJO Конспект системы Гейнце
Conspectus of the Heintze’s System
Unterkl. Scobiformes
Sarraceni ales
t
Theales Amentif er ae
t 7
Rosales -----am am elid ales
I \
Paeon iales Datiscales
Rosales
Triuridaceae
Rosaceae -> Umbelliferae Loasaceae
t . • /
Crassulaceae -> Saxifragaceae -> Eittosporaceae
Cunoniaceae
4
M yrothamnaceae
Tremandraceae Bruniaceae
Theales
Salicaceae C aryocaraceae Actinidiaceae
t t 7
Farn'assiaceae <-Theaceae —Marcgraviaceae
/ I
Guttiferae Ochnaceae-----> Dilleniaceae
Malesherbiaceae -> Passifloraceae--> Tumeraceoie
7 I
Pangiaceae Achariaceae---> C ucurbitaceae
Sarraceniales
Droseraceae -> Sarraceniaceae
I
Nepenthaceae
Datiscales
Datiscaceae -> Begoniaceae
Paeoniales
Paeoniaceae -> Berberidaceae -> Menisрегтасеае
4- 4
Ranunculaceae Lardizabalaceae
Hamamelidales
Hamamelidaceae -> Platanaceae
, Обзор новейших систем цветковых растений ijl
Am е nt i f er а е
С asuarinaceae
Fagaceae JHyricaceae
Betulaceae -> Juglandaeeae
Unterkl. Monadelphiae
P oly g alales -> Violates -> Rhoeadales
I \
Gnetales Al el i ales -> Flacourtiales
k
S a pin d ale s<-0 x al г dale s-*M al pig hi ales
Ф I \
U r tic ales T r i с о c c'a e C olumnif erae
О x al id ales Centrospermae-* P rimulales
T am ar I c ales 0 puntiales ЕЪ en ales
k
S antalales
Polygalales
Polygalaceae
Gnetales
(Bredemeyera)
I
Architumboa _> Gue turn
I
Tumboa _> Ephedra
Viо I al es
Violaceae -> Palsaminaceae
Rhoeadales
Fumarioideae -» Hypecooideae -> Papaxeroideae
I
Cruciferde -» Capparidaceae -> Resedaceae
JU el i al e s
ALeliaceae -> Dichapetalaceae Zygophyllaceae
I
Rutaceae -* Simarubaceae -> Vitaceae
Anacardiaceae <- Rurseraceae Rhamnaceae
’ . I
Dipterocarpaceae
^32
Конспект системы Гейнце
Conspectus of the Heintze’s System
Flacourtiales
Lacistemaceae <- Flacourtiaceae -> Eucry phiaceae
Oxalid ales
Hydrostachyaceae Podostemonaceae
t
Erythroxylaceae <- Oxalidaceae
/ I
Connor aceae Geraniaceae
Humiriaceae
t
Linaceae
I
Caricaceae
Leguminosae Tropaeolaceae Limnanfhaceae
C olumnif er a e
Sterculiaceae -> Bombacaceae
I \
Tiliaceae Malvaceae
I \
Bixaceae Cistaceae
I
0 oddos per maceae
Tricoccae
Euphorbiaceae
Malpighiales
M alpighiaceae
Sapindales
Sapindaceae -* Hippocastanaceae
I \
Hippocrateaceae Aceraceae
I
Celastraceae -> Icacinaceae
i I
Staphyleaceae Aquifoliaceae
Urticales
Ulmaceae -> Moraceae -> Urticaceae
Centrospermae
С ary о phyllaceae -> Fortulacaceae
Amaranthaceae -* Polygonaceae
Chenopodiaceae Phytolaccaceae
Nyctaginaceae Aigoaceae
• Обзор новейших систем цветковых {растений
133
Оpuntiales
Cactaceae
Т am aricales
Tamaricaceae -> Frankeniaceae
Prim a Iales
Theophrastaceae Plumbaginaceae
I Z
Pfimulaceae -> Myrsinaceae
E b e n al e s
Styracaceae -> Ebenaceae
I X
Sapotaceae fiymplocaceae
S ant aisles
Olacaceae -> Santalaceae -> Loranthaceae
i X
Hyzodendraceae Balanophoraceae
II- M о no с о t у J e do n e ae
Burmannjaeformes -> Liliiformes
I
Orchidiformes
Uaterkl. Burmaaaiaeformes
Burmanniales: Archiburmanniaceae -> Burmanniaceae
Uaterkl. Orchidiformes
Archapostasiaceae -> Orchidaceae
Musaceae Marantaceae
I t
Zingiberaceae Cannaceae
Uaterkl. Liliiformes
Carnosae -> Farinosae
Helobiae
C ar nos ae
Philydraceae V elloziaceae
Liliaceae -> Colchicaceae -> Iridaceae
Convallariaceae -> Araceae -> Lemnaceae
I IX
Palmae Pandanaceae Cyclanthaceae
I J4 Конспект системы Гейнце Conspectus о! the Heintze’s System
Farinosae
Cyperaceae -> Gramineae
t
Archijuncaceae -> Juncaceae —> Sparganiaceae
I \
Pontederiaceae Bromeliaceae Typhaceae
Eriqcaulaceae <- Bapateaceae -> Xyrtdaceae
Bestionaceae Commelinaceae JHayacaceae
Centrolepidaceae
H el obi ae
Aponogetonaceae Hydrocharitaceae
t /
Butomaceae Alismataceae
I
S cheuchzeriaceae
I
Potamogeionaceae -> Najadaceae
III. Pseudo di cotyledoneae
N у m ph a e al e$ -> Jf a g noli ales
Nymphaeales
Nympkaeaceae -> Sawruraceae -> Piperaceae
M a g n о I i*a I e s
Trochodendraceae
Galycanthaceae <- Magnoliaceae Anonaceae
4- jr
Monimiaceae Lactoridaceae JHyristicaceae
I I
Lawraceae -> Hernandiaceae Ganellaceae
ШАФФНЕР ДЖ. 1934.
Филогенетическая таксономия растений
J. И. SCHAFFNER. 1934.
Pnylogenetic taxonomy of plants
Перечень отделов и подотделов растений
Synopsis of the plants phyla and subphyla
TH ALLOPHYTA
Phylum I—VIII12
META—ТНА^НОРНТТА .:' ,t <
Phylum IX. Bryophyta /
» X- Ptenophyta
„ XI. Galamophyta
, Xll. Lepidophyta.
» XIII. Cycadophfta
„ XIV- Strobflophyta
„ XV- Anthophyta
ПЕРЕЧЕНЬ КЛАССОВ И ПОДКЛАССОВ, НЕКОТОРЫЙ ОТДЕЛОВ
SYNOPSIS OF THE CLASSES AND SUBCLASSES OF ТЙЕ SEVERAL РЙП.А
PHYLUM STROBILOPHYTA’2
1 Coniferae ;
- Gneteae ’ - 'ь > '
PHYLUM ANTHOPHYTA ,
Monocotylae
Subclass H el oh iae. '
в Spadiciflorae *
„ Glumiflorae
„ Liliif 1 orae?
11 Перечня phylum-ов этой группы йе привожу. (Ред.). - \
“ Предшествующие рЪу1швгЫ_опуи1еш>1 мною. (Ред.). J -•
136
Конспект системы Шаффнара Conspectus of the Schaffner’s System
Dicotylae
Subclass Thalamif lorae
„ Centrosperm a e
„ Heteromerae
„ Tnbiflorae
. Calyciflorae
„ Amentiferae
n Myrtiflorae
я Inferae
КРАТКИЙ ПЕРЕЧЕНЬ ПОРЯДКОВ ПОДКЛАССОВ ЦВЕТКОВЫХ РАСТЕНИЙ
BRIEF SYNOPSIS OF THE ORDERS' OF THE SUBCLASSES
OF ANTHOPHYTA
Helobiae
Alism at ales, Num phaeales, T riuridales, H ydrochari-
tales. , .
S p a d i c i f 1 о r a e
Palmales, С у cl ant-hale s, Pandanales, Ar ales-
Glumiflorae
Restionales, G r aminajes-
Liliiflofrae
Liliales, I rid ales, 8 citaminales, 0 r chidales-
T halamif lorae
Hanales, 8 arr aceniales, Bras-Si calcs, Geraniales,
Malv ales, V iolales-
C entrospermae ' "
T am ar i c a I es, Caryophyllales, C heno podiales, Primu-
lales, Polygon ales, Piper-a les-
Heteromerae
Ericales, Ebenales- ;
Tnbiflorae '
Polemoniales, Gentianales, Scrophulariales, Lami-
ales, Plantaginales-
Обзор новейших систем цветковых растений 137
Calyciflorae
Rosales, Saxif ragales, Thy mel(a)e ales, C elastrales, Sa-
pindales-
Amentiferae
Platanales, U rticales, Parietales, C asuarinales, Ba-
lan 0 ps id ales, F agates, Salicales-
M у r t i f 1 о г a e
Cactales, liprtales, Loasales, Aristolochiales, San-
talales-
Inferae
U mb ellales, Rubiale^, Campanulales, Compositales-
1
ШОО Р. 1953.
СИСТЕМА СОВРЕМЕННЫХ SPERMATOPHYTA
SOO R. 1953.
Systems spermatophytorum vivorum
(„Die modernca Gnmdsatze der Phylogenie im
neuen System der Bltitenpflanzen“)
Tabe He III
PHYLUM XIII. GYMNOSPER- MOPHYTA 0 r dines Farniliae
Classis 1. Pteridospermopsida 1- Pteridosper- m a e 4* 2. С у c a dal es 1. Cycadaceae 2. Zammceae
11. Chiamidospermo- psida 1. Be nnettitale s-f* 2. Gnetales 3. Welwitschi- ales Gnetdceae W elwitschiaceae
III. Coniferopsida 1. C or da it ales + 2. G inkg0 al es 3. C 0 n i f e r a I e s Ginkgoaeeae 1. Araucariaceae 2. Podocarpaceae 3. Taxaceae 4. Cephalotaxaceae 5. Pinaceae^ 6. Taxodiaceae T. Cupressaceae1^
4. E ph edr ales Ephedraceae
PHYLUM XIV. ANGIOSPEBMOPHYTA
Classis 1. Dicotyledoneae
Series A. Polycarpicae—Rubiales
1. P oly c ar pic a e
1. A- M a gnoliales
a) M a gn oliin ea e Winteraceae1*
2. JJegeneriaceae
14 Subfam.: Pinoideae et Abietoideae.
15 Subfam.: Thujoideae.Cupressoideae. Juuiperoideae. ,
16 lucl. Trochodendraceae et Tetraijentraceae, excl. Eupteleaceae-
Обзор новейших систем цветковых растений '
3. Magnoliaceaeir 4. Himantandraceae 5. Eupomatiaceae 6. Annonaceae 7. Calycanthaceae 8. Eupteleaceae 9. Cercidiphyllaceae
b) Laurineae 10. Lauraceae (incl. Hemandi- aceae) 11. Gomortegaceae 12. Monitniaceae 13. Mynsticaceae 14. Canellacece
c) Berbertdineae 15. Menispermaceae 16. Lardizabalaceae 17. Berberidaceae
I. B) Ranales 1. Ranunculaceae1* 2. Nymphaeaceae19 3. Ceratophyllaceae
2. Aristolo chiales 1. Aristolochiaceae 2. Rafflesiaceae 3. Hydnordceae
3. Piperales 1. Piperaceae 2. Chloranthaceae 3. Saururaceae 4. Lactoridaceae
4. H am am e I id a I e s 1. Hamamelidaceae 2. Platanaceae 3. Myrothamnaceae 4. Bruhiaceae ' 5. Stachyuraceae 6. Eucommiaceae
5. Rosales
a) Rosineae 1. Rosaceae20 2. 'Crossosomataceae 3. Connaraceae v 4. Crassulaceae
b)Saxifragineae 5. Saxifragaceae21 6. Cunoniaceae (incl. Brunellia- ceae ) 7_. Byblidaceae (incl. Roridu- laceae)
и Ind. Illiciaceae et Schizandraceae.
18 Subfam.: Paeonioideae, Helleboroideae, Anemonoideae.
18 Subfam.: Nelumboideae, Nypmhaeoideae, Cabomboideae.
“ Subfam.: Spirafeoideae, Pomoideae, Rosoideae, Prunoideae, Chrysobalanoideae-
(reap.—Chrysobalanaceae).
81 Subfam.: Saxifragoideae, Hydrangeoideae, Ribesioideae-
140
Конспект системы Шоо
Conspectus of the Soo’s System
8. Pittosporaceae 9. Podostemonaceae 10. Hydrostachyaceae
6. Le gumin о s ae 1. Mimosaceae 2. Caesalpiniaceae 3. Papillonaceae
7. Myrtiflorae a) Thymelaeineae 1. Thymelaeaceae (incl. Gony- stilaceae} 2. Penaeaceae (incl. Geis- solomataceae) 3. Oliniaceae
b) My r tin e a e 4. Elaeagnaceae 5. Lythraceae 6. Heteropyxidaceae
7. Rhizophoraceae 8. Sonneratidceae 9. Crypteroniac&ae 10. Alangiaceae 11. Nyssaceae 12. Myrtaceae 13. Punicaceae 14. Melastomataceae 15. Lecythidaceae 16. Combretaceae 17. Gnagraceae 18. Trapaceae 19j Haloragaceae (incl. Gunne- raceae)
c) Hippuridineae .8. Terebinth ales 20. Hippuridaceae
a) Ratine ae 1. Rutaceae 2. Simarubaceae 3. Burseraceae 4. Meliaceae 5. Malpighiaceae
6. Tremandraceae 7. Vochysiaceae (inPl. Trigo- niaceae) 8. Polygalaceae (incl. Xan- thophyllaceae )
,b) Sapindineae 9. Coriariaceae 10. Anacardiaceae 11. Julianiaceae 12. Aceraceae 13. Aextoxicaceae 14. Hippocastanaceae 15. Sapindaceae
Обзор иалейших систем цветковых растений 141
- 16. Akaniaceae 17. Sabiaceae 18. Mellanthaceae 19. Didiereaceae 20. Balsaminaceae
9. Celastrales 1. Aquifoliaceae 2. Celastraceae 3. Salvadoraceae 4. Staphyleaceae 5. Hippocrateaceae 6. Stackhousiaceae 7. Icacinaceae 8. Cyrillaceae 9. Pentaphyllacaceae 10. Corynocarpaceae 11. Pandaceae
10. Rhamnales 1. Rhamnaceae 2. Vitaceae
11. U mb elliflorae 1. Araliaceae 2. Cornaceae 3. Garryaceae 4. Umbelliferae23
12. Rubiales 1. Rubiaceae2i 2. Caprifoliaceae 3. Adoxaceae 4. Valerianaceae 5. Dipsacaceae
Series B. Malvales—Tubiflorae * 13. M alv ale s 1. Tiliaceae 2. Malvaceae 3. Bombacaceae 4. Sterculiaceae 5. Elaeocarpaceae 6. Chlaenaceae 7. Scytopetalaceae
14. Geranlales 1. Linaceae (incl. Humiriaceae) 2. Oxalidaceae 3. Geraniaceae 4. Limnanthaceae 5. Tropaeolaceae 6. Erythroxylaceae 7. Zygophyllaceae 8. Cneoraceae
15. T r i co с c a e 1. Euphorbiaceae (incl. Daph- niphyllaceae)25
28 Subfam.: Hydrocotyloideae, Saniculoideae, Apioideae.
24 Subfam.: Cinchonoideae, Coffeoideae.
26 Subfam.: Phyllantoideae et Crotonoideae.
I-P
Конспект системы Шоо
Conspectus of the Soo’s System
2. Dichapetalaceae
3. Buxaceae
4. Callitrichaceae
16. Ligustrales
17. С о nt or t a e
18. T ub if I о r a e
i
Oleaceaex
19. Personatae
19a. Plantaginales
1. Gentianaceae27
2. Apocynaceae
3. Asclepiadaceae23
4. Loganiaceae (incl. Buddlei-
aceae)
1. Convolvulaceae29
2. Polemoniaceae
3. Lennoaceae
4. Hydrophyllaceae
5. Boraginaceae
6. Verbenaceae (incl. Phryma-
ceae)
7. Myoporaceae
8. Labiatae
1. Nolanaceae
2. Solanaceae
3. Scrophulariaceae30
4. Bignoniaceae
5. Martyniaceae
6. Pedaliaceae
7. Lentibulariaceae
8. Globulariaceae
9. Acanthaceae
10. Orobanchaceae (incl. Ae-
ginetiaceae)
11. Gesneriaceae33
12. Columelliaceae
Plantaginaceae
Series G- Rhoeadales—Synandrae (—Ericales)
20. Rhoea dales
a) Rhoeadine.ae
b) Capparidineae
c) Resedineae
1. Papaveraceae32
2. Tovariaceae
3. Moringaceae (incl.
schneideraceae?)
4. Cruciferae
5. Capparidaceae
6. Resedaceae
Bret-
21. Sdrraceniales
1. Sarraceniaceae
2 e Subfam.: Oleoideae et Jasminoideae.
21 Subfam.: Gentianoideae et Menyanthoideae.
23 Subfam.: Cynanehoideae et Periplocoideae.
28 Subfam.: Convoivuloideae et Cuseutoideae-
30 Subfam.: Pseudosolanoideae, A.ntirrhino>deae, Rhinanthoideae.
31 Subfam-: Gyrtandroideae et Gesnenoideae.
32 Subfam.: Hypecooideae, Papaveroid eae, Fumarioideae.
Обзор новейших систем цветковых растений. 143 2. Nepenthaceae 3. Cephalotaceae 4. Droseraceae
22. Parietales a) Tamaricineae 1. Tamaricaceae 2. Fouquieriaceae 3. Frankeniaceae
b) Cistineae 4. Cistaceae '• 5. Bixaceae (incl. Cochlosper-- jnaceae) 6. Violaceae 7. Flacourtiaceae (incl. Laci- stemaceae) 8. Turneraceae 9. Malesherbiaceae 10. Passifloraceae 11. Achariaceae 12. Caricaceae
• 13. Loasaceae 14. Begoniaceae 15. Datiscaceae 1 16. Elatinaceae
23. Cucurbitales Cucurbitaceae
24. Cam pan и I ales* 1. Campanulaceae 2. Lobeliaceae33 6. Marcgraviaceae 7. Quiinaceae 8. Theaceae 9. Hypericaceae 10. Dipterocarpaceae 11. Ancistrocladaceae
27. E r i с a I e s 1. Pyrolaceae^ 2. Clethraceae 3. Ericaceae?3
4. Epacridaceae 5. Empetraceae 6. Diapensiaceae
Series D. Ceutrospermae—Monochlamydeae
D 1.2<S. S antalales
a) Santalineae
1. Santalaceae
2. Grubbiaceae * ***
3. Myzodendraceae
*** Далее в подлиннике пропуск. Кроме того, конспект не вполне соответствует
схеме (напр., в схеме пропущены Proteales, Myricales; в конспекте нет Asterales;
имеют место и другие несоответствия). (Ред.).
34 Subfam.: Pyroloideae et Monotropoideae.
38 Subfam.: Rhododendroideae, Arbutoideae, Ericoideae.
144
Конспект системы Шоо
Conspectus of the Soo’s System
4. Opiliaceae
5. Olacaceae (incl. Octoknema-
taceae)
b) L о r anthin e ae 6. Loranthaceae33
c) Balanophorineae7. Balanophoraceae
d)Cynomorineae
iynomoryaceae
D 2. 29. Proteales
30. Centres per mae
a) Phytolaccineae
Proteaceae
b) Portulacineae
1. Phytolaccaceae (incl. Acha-
tocarpaceae)
Gyrostemonaceae
Nyctaginaceae
Thelygonaceae
Aizoaceae
Portulacaceae
7. Basellaceae
2.
3.
4.
5.
6.
c) Caryophyllineae
d) Chenopodineae
8. Caryophyllaceae87
9. Chenopodiaceae
10. Batidaceae
11. Amaranthaceae
31. С a c tales
Cactaceae
32. P r imul al es
1. Theophrastaceae
2. Myrsinaceae
3. Primulaceae
33. Plumbaginales
Plumbaginaceae
D 3. 34. Ebenales
1.
D 4. 35. Polygon ales
Sapotaceae
2. Hoplestigmataceae
Ebenaceae
Lissocarpaceae
?. Diclidantheraceae
5. Styracaceae
6. Symplocaceae
Polygonaceae
3.
4.
36. Urticales
1. Moraceae* 37 38 39
2. Cannabinaceae
3. Urticaceae
4. Ulmaceae^
5. Rhoioteleaceae
37. Fagales
1. Betulaceae
2. Fa^gaceae
38. Juglandales
Juglandaceae
39 Subfam.: Loranthoideae et Viscoideae.
37 Subfam.: Silenoideae, Alsinoideae, Paronychioideae.
38 Subfam.: Moroideae et Artocarpoideae.
39 Subfam.: Ulmoideae et Celtidqideae.
Обзор новейших систем цветковых растений
145
39. Myricales a) Lett пег i ales b) Balanopsidales Myricaceae Leitneriaceae Balanopsidaceae
D 5. 40. S al i c al e s D 6. 41. Verticillatae Salicaceae Casuarinaceae
Classis 11. Monocotyledoneae
Series E. Helobiae—Graminales
42. H e l о о i a e 1. Alismataceae 2. Butomaceae 3. Hydrocharitaceae 4. Zosteraceae 5. Aponogetonaceae 6. Najadaceae 7. J uncaginaceae 8. Lilaeaceae
42a) Triuridales T riuridaceae
43. Liliiflorae a) Liliineae 1. Liliaceaew 2. Stemonaceae 3. Haemodoraceae * A_r.‘.'~ryllidaceaev- 5. Vellosiaceae 6. Dioscoreaceae 7. Taccaceae
b) I ridineae c) Juncineae 8. Iridaceae ' 9. Juncaceae 10. Flagellariaceae
44. Scitamineae 1. Musaceaevi 2. Zingiberaceae 3. Cannaceae 4. Maranthaceae
45. Gynandrae 1. Burmanniaceae 2. Orchidaceaei2,
46. Cyperales Cyperaceae^ "!
47. Farinosae 1. Bromeliaceae 2. Thurniaceae 3. Rapateaceae 4. Cotnmelinaceae 5. Eriocaulaceae 6. Xyridaceae
40 Subfam.: Melanthioideae, Asphodeloideae, Allioideae, Lilioideae, Aspara-
goideae, Dracaenoidoae, Ophiopogonoideae, Smilacoideao.
4t Subfam.: Amaryllidoideae, Agavoideae, Hypoxydoideae.
42 Subfam.: Strelitzioideae et Musoideae.
42 Subfam.: Pleouaudrae et Mouandrae.
44 Subfam.: Scirpoideae et Garicoideae.
10. А. А. Гроссгейм и M. Ф. Сахокиа
146 Конспект системы Шоо L, Conspectus of the Soo s Sysienj
7. Mayacaceae 8. Centroiepidaceae 9. Restionaceae 10. Cynastraceae 11. Pontederiaceae 12. Philydraceae
48. Gram inales Gratnineae
Series F- Spadiciflorae—Pandanales
49. Spadiciflorae 1. Palmae46 2. Cyclanthaceae 3. Araceae46 4. Lemnaceae
50. P andanales 1. Pandanaceaet 2. Sparganiaceae 3. Typhaceae
48 Subfam.: Coryphoideae, Borassoideae, Lepidocaryoideae, Ceroxyloideae,
Phytelephantoideae, Nipoideae.
46 Subfam.: Pothoideae. Monsteroideae, Calloideae, Lasioideao, Philodendro-
deae, Colocasioideae, Aroideae, Pistioideae.
—«•г
КИхМУРА II. 1956.
Систематический список групп, классов, порядков
и семейств однодольных с указанием их филогении
(„Система и филогения однодольных"; на фр. яз-J
(„Systeme et phylog£nie des Monocotyledones")
KIMURA Y. 1956.
Synopsis systematic of the groups, classes, orders and families
of Monocotyledons with indication their phylcgeny
(„System and phylogeny of Monocotyledons'; in Fr.).
I. Apocarpae
A. Helobiae ( Proto-Polycarpicie) -
1- Alismatales — Butomaceae, Limnocharitaceae, Alismataceae-
2. Hydro ch ar it ales ( <- 1) — Hydrocharitaceae, Thalassiaceae,
Halophilaceae, Vallisneriaceae-
3. Scheuchgeriales ( <- 1) — Triglochinaceae (Juncaginaceae),
S cheuchzeriace ae . Apon ogetonaceae-
4. Potambgetonales ( <- 3) — Potamogetonaceae, Zannichellia-
ceae, Zosteraceae-
5. Najadales ( <- 4) — Lilaeaceae, Najadaceae-
6. Tr iur id al es (<- 3) — Triuridaceae-
II. S у n с a r p a e
II—1. Subsyncarpae
B. Liliflorae ( - A)
7. L Hi a les (<- 3) — Petrosaviaceae, Liliaceae, Stemonaceae, Xan
thorrhoeaceae, Trilliaceae, Pontederiaceae, Smilacaceae (подразу
мевая Ruscaceae), Philesiaceae, Agavaceae (подразумевая роды
Yucca, Agave, etc-).
C. Spadiciflorae (<- B) .
8. Ar ec ales (<- 7) — Arecaceae (Palmae).
9. P and an al es (<- 8) — Pandanaceae-
10. Cy clanthales (*- 8) — Cgclanthaceae.
II—2. Coenocarpae
D. Nudiflorae (*- B)
11- Ar ales — Araceae, Lemnaceae-
148 Конспект системы Кимуры Conspectus of the Kimura<s System
12. T у ph ales — Sparganiaceae, Ту phaceae.
E. Sisciflorae (—B)
13. Eriocaulales ( *- 7) — Eriocaulaceae-
14. JRestionales ( *-7) — Bestionaceae, Flagellariaceae, Centra
lepidaceae-
15. J un c ales (*-14) — Juncaceae, Thurniaceae-
16. Poales (*-14) — Poaceae (Gramineae).
17. С у per ales (*-15) -— Cyperaceae.
F. Calyciferae ( * В)
18. С о mm el in ales ( *-7 ) — Commelinaceae-
19. Xyr idales (*-18) — Mayacaceae, Xyridaceae, Bapateaceae-
20. Philydrales (*-18) — Philydraceae-
21. Brom eli ales (*-18) — Bromeliaceae-
22. Z in gib er ales (*-21) — Strelitziaceae, Lowiaceae, Hlvsaceae,
Zingiberaceae, Cannaceae, Marantaceae-
G. Epigynae ( — В)
23. Bioscore ales ( *- 7 ) — Bios cor eaceae, Stenomeridaceae, Tri-
chopodaceae, Petermanniaceae-
24. Amaryll idales (*-7) — AmarylUdaceae (включая Alstroe-
meriaceae, Hypoxidaceae), Haemodoraceae, Vellogiaceae, Teco-
philaeaceae-
25. I r idales (*-24).— Iridaceae-
26. Burmanniales (*-24) — Burmanniaceae-
27. 0 r chidale s (*-24) — Orchidaceae-
28. T accales (*-24) — Taccaceae-
Заглавные буквы обозначают названия классов, цифры—наименова-
ние порядков. Знак ( *- х) указывает на происхождение класса или по-
рядка от „proto—х“; так, например, 13. Eriocardodes (*-------7) происходит
от Proto-Liliales; Т- Liliales (*--3) происходит от Proto-Scheuchgeria-
les... и т. д.
КРОНКВИСТ А. 1957.
Предположительная линейная последовательность
порядков и семейств двудольных
(„Очерк новой системы семейств и порядков двудольных11)
CRONQUIST А., 1957.
Suggested linear sequence of dicotyledons orders and families
(„Outline of a new system of families and orders of Dicotyledons")
I. Ranales
1. Magnoliaceae
2.' Cercidiphyllaceae
3. Winteraceae
4. Trochodendraceae
5. Tetracentraceae
6. Himantandraceae
7. Degeneriaceae
8. Austroballeyaceae
9. Annonaceae
10. Schizandraceae
11. Illiciaceae
12. Eupteliaceaei7
13. Ranunculaceae
14. Nymphaeaceae
15. Ceratophyllaceae
II. Dilleniales
16. Dilleniaceae
17. Paeoniaceae
18. Crossosomataceae
III. Berberidales
19. Berberidaceae
20. Circaeasteraceae
21. Lardizabalaceae
22. Menispermaceae
23. Coriariaceae
24. Corynocarpaceae
25. Sabiaceae
IV. A r i s t о l о c h i a I es
26. Aristolochi.aceae
V. L aur ale s
27. Amborellaceae
28. Monitniaceae (incl. Tri-
menia).
29. Lactoridaceae
30. Gomortegaceae
31. Lauraceae
32. Hernandiaceae
33. Calycanthaceae
34. Eupomatiaceae
35. Myristicaceae
VI. ,C an ell ale s
36. Canellaceae
VII. Piperales 1 ,
37. Saururaceae
38. Piperaceae
39. Chlor ant haceae
VIII. Rosales
40. Connaraceae
41. Cunoniaceae
42. Hatnamelidaceae
43. Myrothamnaceae
44. Hydrangeaceae 41
41 Должно быть: Eitpteleaceae. (Ред.).
150
Конспект системы Кронквиста Conspectus of the Cronquist s System
Es- 86. Eucommiaceae
8i. Balanopsidaceae
XIV. Casuarinales
88. Casuarinaceae
kV. Sapindales
89. Staphyleaceae
90. hielianthaceae
91. Greyiaceae
92. Akaniaceae
93. Bretschneideraceae
94. Sapindaceae
95. Hippocastanaceae
96. Aceraceae
97. Anacardiaceae
98. Burseraceae
99. Simar(o)ubaceae
100. Cneoraceae
101. Rutaceae
102. Meliaceae
103. Zygophyllaceae ~
XVI. Geraniales
104. Oxalidaceae
105. Geraniaceae
106. Limnanthaceae
107. Tropaeolaceae
XVII. Umbellales
108. Araliaceae
109. Umbelltferae
- XVIII. Juglandales
110. Picrodendraceae
111. Rhoipteleaceae
112. Julianiaceae
113. Juglandaceae
XIX. Polygalales
114. Malpighiaceae
115. Trigoniaceae
116. Vochysiaceae
117. Balsaminaceae
118. Stackhousiaceae
119. Tremandraceae
120. Xanthophyllaceae’
121. Polygalaceae
XX. L i n a I e s
122. Humiriaceae
123. Erythroxylaceae
45. Grossulariaceae (incl.
calloniaceoe)
46. Pittosporaceae
47. Bruniaceae
48. Crassulaceae
49. Cephalotaceae
50. Saxifragaceae
51. Rosaceae
52. Leguminosae
53. Krameriaceae
IX. P rot eale s
54. Elaeagnaceae
55. Proteaceae
X. M у r t a I e s
56. Heteropyxidaceae
57. Sonneratiaceae
58. Geissolomataceae
59. Crypteroniaceae
60. Lythraceae
61.. Penaeaceae
62. Thymelaeaceae
63. Hydrocaryaceae .
64. Dialypetalanthaceae
65. Myrtaceae
66. Punicaceae
67. Oliniaceae
68. Onagraceae
69. Melastomataceae
70. Combretaceae
XI. Haloragales
71. Gunneraceae
72. Haloragaceae
73. Hippuridaceae
74. Callitrichaceae
XII. Podostemonal.es
75. ^>odostem.o"a,'eae
76. Hydrostachyaceae
XIII. Urticales
77. Leitneriaceae (inci.
dameleH)
riaianaceae
79. Urticaceae
80. Mnrica-eae
81. Moraceae
82. Ulmaceae
83. Barbeyaceae
8-1. Betulaceae
85. F.agaceae
Обзор новейших систем цветковых растений
151
124. Linaceae
XXI. Celastrales
125. Rhamnaceae
126. Vitaceae
127. Celastraceae
128. Hippocrateaceae
129. Salvadoraceae
130. Dich-netaloreae
131. Aquifoliaceae
132. Icacinaceae
XXII. E и pho r b i ale s
133. Buxaceae
134. Euphorbiaceae
135. Daphniphyllaceqe
136, Aextoxicaceae
137. Pandaceae
138. Didiereaceae
XXIII. Santalales
139. Olacaceae
140. Opiliaceae
141. Santalaceae\
142. Myzodendraceae
143. Loranthaceae -
144. Mitrastemonaceae
145. Hydnoraceae
146. Rafflesiaceae
147. Balanophoraceae
XXIV. Cornales
148. Rhizophoraceae
149. Cornaceae
150. Alangiaceae
151. Nyssaceae
152. Grubbiaceae
XXV. Garry ales
153. Garryaceae
XXVI. Caryophyllales
154. Phytolac'aceae
155. Nyctaginaceae
156. Aizoaceae
157. Portulacaceae
158. iBasellaceae
159. Caryophyllaceae
160. Chenopodiaceae
161. Amaranthaceae
162. Polygonaceae
163. Thelygonaceae
XXVII. В a t i d a le s
164. Batidacece
XXVIII. C act ales
165. Cactaceae
XXIX. Plum baginales
166. Plumb aginaceae
XXX. Guttiferales
167. Caryocaraceae
168. Eucryphiaceae
169. Quiinaceae
170. Elatinaceae
171, Medusaginaceae
172. Guttiferae
173. Hypericaceae
174. Ochrtaceae
175. Chlaenaceae
176. Dipterocarpaceae
177. Theaceae
178. Cyrillaceae
179. Stachyuraceae
180. Actinidiaceae
181. Marcgraviaceae
XXXI. Lecythidales
182. Lecytkidaceae
XXXII. Mai vales
183. Elaeocarpaceae
184. Tiliaceae
185. Buettneriaceae
186. Sterculiaceae
187. Scytopetalaceae
188. Bombacaceae
189. Malvaceae
XXXIII. Rhoeadales
190. Tovart'aceae
191. Papaveraceae
192. Fumariaceae
193. Moringaceae
194. Capparidaceae
195. Cruciferae
196. Resedaceae
XXXIV. Sarraceni ales
197. Byblidaceae
198. Droseraceae
199. Sarraceniaceae
200. - Nepenthaceae
152
Конспект системы Кронквиста Conspectus of the Cronquist’s System
XXXV. Bixales
201. Dioncophyllaceae
202. Flacouriiaceae
203. Bixaceae
204. Cistaceae
205. Violaceae
206. Tamaricaceae
207. Frankeniaceae
208. Samydaceqe
209. Turneraceae
210. Loasaceae
211. Passifloraceae
212. Malesherbiaceae
213. Fouquieriaceae
214., Hoplestigmataceae
215. Caricaceae
216. Achariaceae
217. Begonlaceae
218. Datiscaceae
219. Cucurbitaceae
220. Ancistrocladaceae
XXXVI. Salic ales
221. Salicaceae
XXXVII. E r i с a I e »
222. Clethraceae
223. Ericaceae (sens, lat.)
224. Epacridaceae
225. Diapensiaceae
226. Empetraceae
XXXVIII. Eb envies
227. Styracaceae
228. Ebenaceae
229. Sapolaceae
XXXIX. Prim ulales
230. Theophrastaceae
231. Myrsinaceae
232. Primulaceae
XL. G e nt i an a I e s
233. Buddleiaceae
234. Oleaceae
235. Menyanthaceae
236. Gentianaceae
237. Loganiaceae
238. Apocynaceae
239. Asclepiadaceae
XLI. Plantaginales
240. Plantaginaceae
XLII. Polemoniales
241. Polemoniaceae
242. Hydrophyllaceae
243. Solanaceae
244. Convolvulaceae
245. Cuscutaceae
246. Lennoaceae
XLIII. S crophulariales
247. Scrophulariaceae
248. Myoporaceae
249. Selaginaceae
250. Gesneriaceae
251. Columelliaceae
252. Orobanchaceae
253. Bignoniaceae
254. Acanthaceae
255. Pedaliaceae
256. Lentibulariaceae
XLIV. La mi al e s
257. Boraginaceae
258. Verbenaceae
259. Phrymaceae
260. Labiatae
XLV. Campanulales .
261. Campanulaceae
262. Goodeniaceae
263. Stylidiaceae
264. Calyceraceae
XLVI. Rubiales
265. Rubiaceae
266. Caprifoliaceae
267. Adoxaceae
268. Valerianaceae
269. Dipsac(ac)eae
XLVII. Asterales
270. Compositae
АМБЕРЖЕ Л. i960.
Конспект главных систематических груип
покрытосемянных растений
(„Сосудистые растения", т. II, ч. 1 и 2; на фр. яз.). f
EMBERGER L. i960.
•Conspectus of the principal systematical groups of Angiosperms
(„Vascular plants", v. Il, parts l'a- 2; in Fr.)
(„Les vegetaux vasculaires". T. II, f. 1 et 2)
, GroupeP8
Phylum Casuarinae
Ordre 1.Casuarinales
Fam. Casuarinaceae
G г о u p e II,
Phylum 1. Olacales — Proteales
Ordre 1. Olacales
Fam. 1. Olacaceae ;
„ 2. Opiliaceae
(Pentadiplandraceae)
( Medusandr aceae?)
Ordre 2. S ant al al es
Fam. 1. Grubbiaceae
„ 2. Octoknemataceae
„ 3. Santalaceae
„ 4. Loranthaceae
„ 5. My zodendr aceae
„ 6. Balanophoraceae
„ 7. Cynomoriaceae
Ordre 3. Proteales
Fam. 1. Proteaceae
48 Названия таксонов здесь даны, как в подлиннике, на франц, яз. Привожу их
обозначения на русск. и латинск. яз.: groupe—группа (по лат.: superphylum или
synphylum); phylum—фила, ветвь (по лат.: phylum); ordre—порядок (по лат.: ordo);
fam-, familie—семейство (по лат.; familia). Полная нумерация таксонов произведена
мною; в подлиннике она дана лишь частично (Ред.).
154 Конспект системы . Амберже Conspectus of the Emberger’s System
G г o u p e III.
phylum 1. Urticales—«Amentiflorae»—Centro-
spermales.
„Amentiflorae1-
Ordre 1. F aga les
Fam. 1. Betulaceae
„ 2. Fagaceae
\
Ordre 2. Balanopsidales
Fam. 1. Balanopsidaceae
Ordre 3. Salt cates
Fam. 1. Salicaceoe.
Ordre 4. Myricales
Fam. 1. Myricaceae
Ordre 5. Leiineriales
Fam. 1. Leitneriaceae
„ 2. Didymelaceae
Ordre 6. Bat idales
Fam. 1. Batidaceae
Ordre 7. V rticales
Fam. 1. Vlmaceae
„ 2. Moraceae
„ . 3. Urticaceae
4. Eucommiaceae
5. Rhoipteleaceae
6. Scyphostegiaceae
Ordre 8. Polygon ales
Fam. 1. Polygonaceae
Ordre 9. Centrospermales
Fam. 1. Chenopodiaceae
„ 2 Amaranthaceae
„ 3. Phytolaccaceae
„ 4. Sphenocleaceae
„ 5. Gyrostemonaceae
„ 6. Achatocarpaceae 1
„ 7. Nyctaginaceae
, , 8. Portulacaceae
„ 9. Basellaceae
„ 10. Dysphaniaceae .
49 T. e. дополнительно; это семейства с неясным филогенетическим положе- '
нием. (Ред.).
Доп:49 ”
Обзор новейших систем цветковых растений
155
„ 11. Caryophyllaceae
„ 12. Aizaaceae (Ficoidaceac'
13. Cactaceae
Доп.; 14. Thelygonaceae (Cynocrambaceae)
•)
Ordre 10. Plumbaginales
Fam. 1. Plumbaginaceae
Ordre 11. P r i mu I al e s
Fam. 1. Theophrastaceae
„ 2. Myrsinaceae
„ 3. Primulaceae
G г о u p e IV
Phylum I. Juglandales — Julianiales
Ordre 1.Juglandales
Fam. 1. Juglandaceae
Ordre 2. Julianiales
Fam. 1. Julianiaceae
Phylum II. Terebinthales — Rubiales
Ordre 1. Terebinthales
Fam. 1. Rutaceae
„ 2. Simarubaceae
„ 3. Burseraceae
4. Meliaceae
„ 5. Polygalaceae
„ 6. Trigoniaceae
„ 7. Vochysiaceae
„ 8. Anacardiaceae
„ 9. Sapindaceae
„ 10. Aceraceae
„ 11. Hippocastanaceae
„ 12. Buxaceae
„ 13. 'Aextoxicaceae
„ 14. Akaniaceae
15. Coriariacei e
„ 16. Cyrillaceae
„ Г> '. Pentaphylacaceae
n . „ 18. Diclidantheraceae
/J'on” „ 19. Balsaminaceae
20. Corynocarpaceae
21. Didiereaceae
„ 22. Tremandraceae
„ 23. Sabiacdde
„ 24. Melianthaceae
Ordre 2. U m be 11 if I о r ae-
Fam. 1. Araliaceae
„ 2. Umbelliferae
156
Конспект системы Амберже Conspectus of the Emberger’s System
„ 3. Cornaceae
„ 4. Alangiaceae
„ 8. Nyssaceae
Ordre 3. Garry ales
Fam. 1. Garryaceae
Ordre 4. Rubiales
Fam. 1. Rubiaceae
„ 2. Caprifoliaceae
„ 3. Valerianaceae
„ 4. Dipsacaceae
„ 5. Adoxaceae
„ 6. Calyceraceae
Доп.: „ 7. Loganiaceae
Ordre 5.Celastrales
Fam. 1. Celastraceae
„ 2. Stackhousiaceae
„ 3. Hippocrateaceae
„ 4. Icacinaceae
„ 5. Aquifoliaceae
„ 6. Salvadoraceae -
„ 7. Staphyleaceae
„ 8. Peripterygiaceae
9. Erythropalaceae
Ordre 6. Rh amn al e s
Fam. 1. Rhamnaceae
„ 2. Vitaceae
„ 3. Leeaceae
Ordre 7. Ligustrales
Fam. 1. Oleaceae
Phylum III. Contortales — Tubiflorae
Ordre 1. С о nt о r t ae
Fam. 1. Gentianaceae
„ ’ 2. Menyanthaceae
„ 3. Apocynaceae
„ 4. Asclepiadaceae
Ordre 2. Tubiflorae
Fam. 1. Convolvulaceae
„ 2. Humbertiaceae
„ 3. Polemoniaceae
„ 4. Nolanaceae
„ 5. Solanaceae
„ 6. Buddleiaceae
„ 7. Hydrophyllaceae
8. Lennoaceae
9. Hoplestigmataceae
Обзср новейших систем цветковых растений
157
J? 10. Boraginaceae
5? 11. Scrbphulariaceae 12. Selaginaceae
13. Globulariaceae
14. Orobanchaceae
15. Gesneriaceae 16. Bignoniaceae 17. Pedaliaceae
18. Martyniaceae
» 19. Lentibulariaceae 20. Myoporaceae
21. Acanthaceae
22. Verbenaceae
23. Phrymaceae
24. Labiatae (Lamiaceae) 25. Tetrachondraceae
Доп:. 26. Plantaginaceae 27. Columelliaceae
Phylum IV. Pandales
Ordre 1. Р а п d al е s
Fam. 1. Pandaceae
Phylum V. Cneorales
Ordre 1. Cneorales
Fam. 1. Cneoraceae
Phylum VI. Ebenales
Ordre 1. Ebenales
Fam. 1. Ebenaceae
J, Доп.: 2. Styracaceae 3. Symplocaceae 4. Sapotaceae 5. Sarcospermaceae 6. Lissocarpaceae
Phylum VII. Geraniales (Gruinales)-—Malvales
(Columniferae)
Ordre 1. Geraniales (Gruinales)
Fam. 1. Zygophyllaceae
„ 2. Geraniaceae
„ 3. Tropaeolaceae
„ 4. Oxalidaceae
„ 5. Limnanthaceae
„ 6. Linaceae
„ 7. Lepidobotryaceae
„ 8. Humiriaceae
„ 9. Erythroxylaceae
„ 10. Malpighiaceae
f
.158, Конспект системы Амберже Conspectus of the Emberger's System
Ordre 2. M al v al e s (Columniferae)
Fam. 1. Tiliaceae
„ 2. Elaeocarpaceae
„ 3. Malvaceae
„ 4. Bombaceae
„ 5. Sterculiaceae
„ 6. Chlaenaceae
„ 7. Scytopetalaceae
„ 8. Gonystylaceae
Phylum VIII. Tricoccae
Ordre 1. Tricoccae
Fam. 1. Euphorbiaceae
„ 2. Daphniphyllaceae ,
„ 3. Dichapetalaceae
„ 4. Lophopyxidaceae
G г о u p e V.
Phylum I. Polycarpicae—Aristolochiales
Ordre 1. Polycarpicae
Fam. 1. Magnoliaceae
„ 2. Winteraceae
„ 3. Schizandraceae
„ 4. Degeneriaceae
„ 5. Lactoridaceae
„ 6. Annonaceae
„ 7. Eupomatiaceae
„ 8. Lauraceae
„ 9. Myristicaceae
„ 10. Hernandiaceae
11. Gomortegaceae
„ 12. Monimiaceae
„ 13. Austrobai.leyaceae
„ 14. Menispermaceae
„ 15. Lardizabalaceae
„ 16. Sargentadoxaceae
„ 17. Berberidaceae
„ 18. Ranuticulaceae
„ 19. Circaeastraceae
„ 20. Cabombaceae •
„ 21. Nymphaeaceae
„ 22. Ceratophyllaceae
„ 23. Himantandraceae
„ 24. Tetracentraceae
„ 25. Trochodendraceae
„ 26. Canellaceae
„ 27. Calycanthaceae
„ 28. Eupteleaceae
„ 29. Cercidiphyllaceae
1
Обзор новейших систем цветковых растений
159
Ordre 2. Aristolochiales
Fam. 1. Aristolochiaceae
„ 2. Rafflesiaceae (incl. Cytinaceae et Mitra-
stemonaceae)
„ 3. Hydnoraceae
Phylum II. Helobiae
Ordre 1. A I i s m at ale s
Fam. 1. Butomaceae
„ 2. Hydrocharitaceae
„ 3. Alismataceae
„ 4. Juncaginaceae
« 5. Scheuchzeriaceae
„ 6. Aponogetonaceae
Ordre 2. Potamogetonales
Fam. 1. Potamogetonaceae
„ 2. Ruppiaceae
„ 3. Lilaeaceae
„ 4. Zosteraceae
„ 5. Posidoniaceae
„ 6. Zanichelliaceae
„ 7. Najadaceae
Ordre 3. Triuridales
Fam. Triuridaceae
Phylum III. Piperales—Spadiciflorae (Arales—Panda-
nales— Palmales—Cyclanthales)
Ordre 1. Piperales
Fam. 1. Piperaceae
„ 2. Saururaceae
„ 3. Chloranthaceae
Ordre 2. Arales
Fam. 1. Araceae
„ 2. Lemnaceae
Ordre 3. Pandanales
Fam. 1. Pandanaceae
„ 2. Sparganiaceae
„ 3. Typhaceae
Ordre 4. P alm al e s
Fam. 1. Palmaceae
„ 2. Phy tele phasiacae
„ 3. Nipaceae
160
Конспект системы Амберже Conspectus of the Emberger’s System
Ordre 5. Сус Ianthales
Fam. 1. Cyclanthaceae
Phylum IV. Liliiflorae
A. Groupe d’ordres 60 Commelinales -Graminales
Ordre 1. Commeli n'a les (E n a n t i. ob-
last a e)
Fam. 1. Commelinaceae
„ 2. Mayacaceae
„ 3. Xyridaceae
„ 4. Eriocaulaceae
„ 5. Centrolepidaceae
„ 6. Restionaceae
Ordre 2. Graminales
Fam. 1. Graminaceae (Gramineae)
B- Gr upe d’ordres Juncales Cyperales
Ordre 3. Juncales
Fam. 1. Juncaceae
„ 2. Flagellariaceae
„ 3. Rapateaceae
„ 4. Thurniaceae
Ordre 4. Cyperales
Fam. 1. Cyperaceae
Ordre 5. Bromeliales
Fam. 1. Bromeliaceae
Ordre 6. Ponteder iales
Fam. 1. Pontederiaceae
C. Groupe d’ordres Liliales—Dioscoreales
Ordre 7. Liliales
Fam. 1. Liliaceae
„ 2. Amaryllidaceae
„ 3. Haemodoraceae
„ 4. Velloziaceae
„ 5. Cyanastraceae
„ 6. Philydraceae
„ 7. Iridaceae
„ 8. Geosiridaceae
„ 9. Roxburghiaceae (Stemonaceae)
„ 10. Petermanniaceae
Ordre 8. Doisco reales
Fam. 1. Dioscoreaceae
,, 2. Taccaceae
60 Groupe d’ordres—группа порядков или надпорядок (по лат. superordo)- (Ред.).
Обзор новейших систем цветковых растений
i6l
Ordre 9. S с it ат in ales
Fam. 1. Musaceae
„ 2. Strelitziaceae
„ 3. Lowiaceae
„4. Zingiberaceae
„ 5. Cannacede
,, 6. Marantaceae 4
Ordre lO.Burmannialss
Fam. 1. Burmanniaceae _ '
„ 2. Thismiaceae
„ 3. Corslaceae
Ordre 11. О r chid ale s
Fam. 1. Apostasiaceae
„ 2. Orchidaceae
Phylum V. Parietales-Rhoeadales-Synantherales
Ordre l.Parie tales
А. Типичные Parietales
Fam. 1. Dilleniaceae
2. Actinidiaceae
„ 3. Sauraujaceae
„ 4. Eucryphiaceae
„ 5. Medusagytiaceae
„ 6. Ochnaceae (incl. Luxemburgiaceae)
„ 7. Strasburgeriaceae
„ 8. Caryocaryaceae (=Rhizobolaceae)
„ 9. Marcgraviaceae
10. Theaceae (Ternstroemiaceae)
„ 11. Guttiferae (incl. Hypericaceae)
„ 12. Dipterocarpaceae
„ 13. Fracourtiaceae (incl. Samydaceae}
„ 14. Oioncophyllaceae (~triphyophyllaceae)
„ 15. Violaceae
„ 16. Cistaceae
„ 17. Bixaceae
„ 18. Cochlospermaceae
„ 19. Tamaricaceae
„ 20- Frankeniaceae
21. Elatinaceae
„ 22. Turneraceae
» 23. Malesherbiaceae
„ 24. Passifloraceae
„ 25. Achariaceae
„ 26. Caricaceae
В. Происшедшие от группы предыдущих семейств
„ 27. Stachyuraceae .
„ 28. Lacistemaceae
„ 29. Loasaceae
„ 30. Datiscaceae
„ 31. Begoniaceae
11. А. А. ГроСсгейм и M. Ф. Сахокиа
l61 Конспект системы Амберже Conspectus of the Embergers System
с. Семейства, связанные c Parietales
„ 32. Droseraceae
„ 33. Ancistrocladaceae
„ 34. Quiinaceae
„ 35. Fouquieriaceae (Fouquieraceae)
Ordre. 2. R ho e a d a I e s
Fam. 1. Papaveraceae
„ 2^ Tovariceae
„ 3. Capparidaceae
„ 4. Cruciferae
„ 5. Resedaceae
„ 6. Moringaceae
„ 7. Bretschneideraceae
Ordre З.С и cur bit ales
Fam. 1. Cucurbltaceae
Ordre 4. E ri c ale s
Fam. 1. Clethraceae
„ 2. Pyrolaceae
„ 3. Ericaceae
„ 4. Epacridaceae
„ 5. Empetraceae
Доп.: „ 3. Diapensiaceae
Ordre 5. Synantherales (Syrian-
t h e r ae)
Fam. 1. Campanulaceae
„ 2. Lobeliaceae
„ 3. Goodeniaceae
„ 4. Brunoniaceae
„ 5. Stylidiaceae (Candolledceae)
„ 6. Compositae (incl. Ambrosiaceae)
Phylum VI. Sarraceniales
Ordre 1. Sarraceniales
Fam. 1. Sarraceniaceae
„ 2. Nepenthaceae
„ 3. Cephalotaceae
Phylum VII. Rosales — Myrtales
Ordre f. Rosales
А. Семейства, близкие к типичным В о s al вs
(древние JR о sales?)
Fam. 1. Hafnamelidaceae
„ 2. Platanaceae
„ 3. Bruniaceae
„ 4. Myrothamnaceae
Обзор новейших систем цветковых растений 163
В. Типичные Rosales
1. Группа семейств Saxifragacede—Rosaceae
п 99 5. Crassulaceae 6. Saxifragaceae 7. Pittosporacea& 8. Greyaceae
п 9. Cunoniaceae
м 10. Brunelliaceae
11. Podostemonaceae
12. Hydrostachyaceae 13. Rosaceae 14- Chrysobalanaceae
15. Conparaceae 2. Группа семейств Leguminosae
W 16. Mimosaceae 17. Caesalpiniaceae 18. Papilionaceae С. Семейства, связанные c Rosales
?> 19. Crossosomataceae 20. Byblidaceae
21. Roridulaceae
Ordre 2.Myrtales
Fam. 1. Lythraceae
99 99 5? 99 2. Crypteroniaceae 3. Heteropyxidaceae 4. Sonneratiaceae (=Blattiaceae) 5. Punicaceae
99 99 99 99 6» Rhizophoraceae 7. Lecythidaceae 8. Combretaceae 9. Myrtaceae
99 10. Melastomaceae 11. Onagraceae (=Oenotheraceae) 12, Hydrocaryaceae (=Trapaceae)
V 99 13. Halorrhagidaceae (=Halorrhagaceae) 14. Gunneraceae
99 15. Hippurldaceae 16. Callitrichaceae
99 17. Dialypetalanthaceae
Ordre 3. Thymelaeales
Fam. I. Penaeaceae
99 99 99 99 2. Geissolomataceae 3. Ollniaceae 4. Thymelaeaceae 5. Elaeagnaceae
Конспект новой системы растительного царства
Б. М. Козо-Полянского- 1947.
(Б. М. Козо-Полянский—О новой системе растений. Доклады АН СССРГ
нов. сер., 1947,LVI,3) 1
KOZO-POLIANSKY В. 1947
Conspectus systematis п о Vi г е- g'n i v e g e t a b i 1 i s
A 1. Subregnum SCHIZOPHYTA. — Ex materia anorganica51
В 1. Phylum ARCHEOPHYTA (s. Schaffner). — Ex materia anorganica.
В 2. „ SCHIZOMYCETES. — Ex Archeophyta et Schizophyceae?
В 3. „ SCHIZOPHYCEAE. — Ex Archeophyta.
A 2. Subregnum MYCOPHYTA (incl. Myxophyta-Phytomyxinae). — Ex
Phycophyta.
A3. „ NOMOPHYTA.— Ex ScHizophyta.
В 1. Synphylum PHYCOPHYTA.— Ex Schizophyta.
В 2. „ CORMOPHYT,A.'—Ex. Phycophyta (Pro — Rhodophy-
C. 1. Phylum PROCORMOPHYTA^—Ex Phycophyta.
D 1.. Subphylum Psilophyt a. — Ex Phycophyta.
F 1. Classis Psilophytinae. — Ex. Phycophyta.
F 2. „ Psilotinae. — Ёх Psilophytinae.
D 2. Subphylum Anthocerota. Ex Psilophytinae.
F 1. Classis Anthocerotinae.— Ex Psilophytinae.
C 2. Phylum BRYOPHYTA. — Ex Psilophytinae.
D 1. Classis Sphagnobrya (incl. Atidfeaeales).—Ex Psilophytinae.
D 2. „ Eubrya. — Ex Sphagnobrya.
D 3. „ Hepaticae. — Ex Sphagnobrya.
C 3. Phylum PTER1DOANTHOPHYTA. — Ex Psilophytinae.
D 1; Subphylum Pteridophyta. — Ex Psilophytinae.
E 1. Superclassis Lycopsida. ~ Ex Psilophytinae.
F 1. Classis Iso'itinae (inch Lepidodendrales, Selaginel-
tales, Lepidospermates, Pleuromejales).—
Ex Psilophytinae.
F 2. „ Lycopodinae. — Ex Psilophytinae
E 2. Superclassis P t e r o p sid a. — Ex Psilophytinae.
F 1. Classis Eufilicinae. — Ex Psilophytinae
F 2. „ Leptofilicinae. — Ex Eufilicinae.
E 3. Superclassis Sphenopsida. — Ex Psilophytinae.
F 1. Classis Hyeniinae. — Ex Psilophytinae.
F 2. .. Sphenophylltnae. — Ex Hyeniinae.
F 3. „ Equisetinae. — Ex Hyeniinae. '
F 4. Salviniinae. — Ex Hyeniinae.
“ Здесь (и в других подобных примечаниях) указываются исходные группы.
Обзор новейших систем цветковых растений
165
F 5. Classis? Tingiinae. — Ex Hyeniinae?
F 6. „ АгоШпае. — Ex Tingiinae?
D 2. Subphylum G у m n о s p er m ae. — Ex Psilophytinae.
E 1. Superclassis С у с о d о p h у t a. — Ex Psilophytinae
F 1. Classis Pteridosperminae. — Ex Psilophytinae.
F 2. „ Caytoniinae. — Ex Pteridosperminae.
F 3. „ Cordaitinae. — Ex Psilophytinae.
F 4. ,, Ginkgoinae. — Ex Cordaitinae.
F 5. „ Cycadinae. — Ex Pteridosperminae.
F 6. Bennettitinae. — Ex Cycadinae?
E 2. Superclassis Ephedrophyta. — Ex Cordaitinae?
F 1. Classis Ephedrinae. — Ex Cordaitinae?
E 3. Superclassis Coniferophyta. — Ex Cordaitinae.
F 1. Classis Abietinae. — Ex Cordaitinae.
D 3. Subphylum? A n g i о s p e r m a e. — Ex Psilophytinae?
E 1. Superclassis Proterogenae. —Ex Psilophytinae? In-
termedia ignota.
E 2. „ Metagenae. — Ex Proterogenae.
F, 1. ''Classis Dicotyledoneae (incl. [inter Santalales-
inovuleae] Gnetaceae s. st. et. Tumboaceae).-—
। Ex Proterogenae.
F 2, „ Monocotyledoneae, — Ex Proterogenae.
Приложение III
Supplement III
ЛИТЕРАТУРА - LITERATURE
(Материалы к библиографии по филогении цветковых растений)1
(Materials to the bibliography of phylogeny of flowering plants)
Авдеев В. Д. 1956. Черты эволюции околоцветника покрытосеменных в семействе-
лютиковых. Учен. Зап. Ульяновск. Пед. ин-та, 9.
Александров В. Г. и Добротворская А. В. 1958. О метаморфозе частей цвет-
ка и морфологической природе тычинки (материалы к физиологии цветка по-
крытосеменных растений). «Проблемы Ботаники», III.
А н е л и Н. А. 1944. Материалы о взаимосвязи между филогенией древесных двудоль-
ных н их регенеративной способностью. Сообщ. АН Груз. ССР, V, 9.
Ан ел и Н. А. 1950. Текстура проводящей системы в растениях семейства буковых
(на груз. яз.). Тезисы доклада. IV Научная сессия Зугдидского Гос. Учительско-
го института им. А. Церетели. 26—28. II. 1950. Зугдиди.
Айе ли Н. А. 1952. О систематической принадлежности китайского лимонника и ил-
лнциума на основании анатомического анализа. Сообщения Акад, наук Груз.
ССР, XIII, 4.
А не ли Н. А. 1952. Текстура проводящей системы и некоторые вопросы систематики-
сем. миртовых (на груз. яз. с русск. рез.). Вести. Тбил. бот. сада, 60.
Ан ели Н. А. 1953. Материалы об анатомическом родстве древесных бобовых расте-
ний. Вести. Тбил. бот. сада, 61.
А н е л и Н. А. 1956. Теория «листовых следов» и филогенез покрытосеменных расте-
ний. Тезисы доклада 31.Х.1956 (иа груз. яз.). Первая научная сессия. Кутаис-
ский Сельскохоз. Ин-т.
Анели Н. А. 1956. Об анатомическом родстве растений лавровых и некоторых дру-
гих семейств (на груз. яз. и русск. рез.). Вести. Тбил. ботан. сада, 63.
Айе л и Н. А. 1956. К вопросу новой системы покрытосеменных растений. Тезисы до-
клада. 29.III.1956. Кутаисское Отделение Москов. Общ. Испыт. Прир., Кутаис-
ский Сельскохоз. Ин-т.
Анели Н. А. 1956—1958. К вопросу новой системы покрытосеменных растений. Кута-
исск. отд. МОИП. Кутаисск. Сел.-Хоз. Инет.
Анели Н. А. 1960. Материалы к филогении семейства рутовых. Сборник трудов Тби-
лисского н.-н. химико-фармацевтического института, IX.
Анели Н. А. 1961. Анатомия проводящей системы побега и систематика растений
(Каулифолиарная прово; ящая система древесных двудольных растений). Авто-
реферат докторской диссертации. Институт фармакохимии. Тбилиси.
Базилевская Н. А. 1959. Проблема вида в трудах советских ботаников. Сб. «Ан-
налы биологии», I. М.
Баранов П. А. 1952. Возникновение и развитие растительного мира. М., Воен. изд.
Берг Л. С. 1949. Происхождение наземных растений. Природа, 2.
Берг Р. Л. 1956. Стандартизирующий отбор в эволюции цветка. Б. Ж., 3.
Берг Р. Л. 1958. Дальнейшие исследования по стабилизирующему отбору в эволюций
цветка. Б. Ж., 1.
1 Составил М. Ф. Сахокиа. В списке приведены лишь основные труды общего ха-
рактера, а также некоторые труды, посвященные частным вопросам филогении или фи-
логенетическому изучению отдельных таксономических групп растений. (М. С.).
Обзор новейших систем цветковых растений
Благовещенский А. В. 1925. К вопросу о направленности процесса эволюции.
Бюлл. Ср.-Аз. Гос. Ун., X.
Благовещенский А. В. 1935. Биохимические основы эволюции организмов.
СОРЕНА (Соц. реконструкция и наука. — Ред.), 5.
Благовещенский А. В. 1939. Биохимическая эволюция растений в связи с из-
менением качества ферментов. Усп. совр. биологии, XI.
Благовещенский А. В. 1950. Биохимические основы эволюционного процесса у
растений. М.-Л.
Благовещенский А. В. 1950. Биохимические основы эволюционного процесса у
растений. Изд. АН СССР. М.-Л.
Благовещенский А. В. 1960. Биохимическая эволюция покрытосеменных рас-
тений. Б. Ж., 4.
Благовещенский А. В. 1962. Эволюция белковых комплексов семян и эволю-
ция цветковых растений. Изв. АН СССР, сер. биол., 6.
Бреславец Л. П. 1964. Значение полиплоидии для филогении. Второе Московское
Совещание по дилогении растений. 20—25 марта 1964. ^Тезисы докладов
МОИН. М.
БувН. А. 1924, Общий курс ботаники. Морфология и систематика растении. 2 изд.
М.-Л.
Буш Н. А. 1940. Курс систематики высших растений. Изд. 1. Изд.., Лениигр. Гос.
Унив.
Буш Н. А. 1944. Курс систематики высших растений. Изд. 2. Гос. Уч.-Пед. изд. Мин.
Просвещ. РСФСР. М.
Вага А. Я. 1952. Филема органического мира. Б. Ж., 5.
Васильев В. Н. 1964. Темпы эволюциич Второе Московское Совещание по фило-
гени растений. 20—25 марта. 1964. Тезисы докладов МОИП. М.
Васильченко И. Т. 1936. О значении морфологии прорастания семян для сис-
тематики растений и истории их происхождения. Тр. Бот. Ии-та АН СССР,
сер. I, 3.
Васильченко И. Т. 1938. О значении морфологии прорастания для филогенети-
Ческой систематики цветковых растений. Сов. Бот., 3.
Васильченко И. Т. 1946. К вопросу об эволюционном значении морфологических
есобенностей прорастания цветковых растений. Сб. научи, работ. Бот Ин-та АН
СССР.
Васильченко И. Т. 1948. Неотения у цветковых растений. Тр. Ботан. Инет. АН
СССР, сер. 1, 149, вып. X.
Васильченко И. Т. 1961. О соотношении онтогенеза и филогенеза у высших рас-
тений. Б. Ж., 12.
Вахрамеев В. А. 1947. Роль геологической обстановки в развитии и распростра-
нении покрытосеменных флор в меловое время. Бюлл. МОИП, отд. геол ХХП
6.
В е т тш т е й н Р. 1912. Руководство по систематике растений. М. (см.: Wettstein R.,
1935. Handbuch der systematischen Botanil).
Вет т штейн Р. 1930. Филогения растений (В кн.: Человек и природа. Основы ес-
тествознания в общедоступном изложении. Т. 5), Л.
Виноградов И. С. 1947. Новый принцип систематики покрытосеменных. Бюлл.
МОИП, отд. биол. LII, 5.
Виноградов И. С. 1956. К вопросу о предках покрытосеменных. Уч. зап. Сев,-
Осетниск. Гос. Пед. Ин-та, 20.
Виноградов И. С. 1957. О классах однодольных и двудольных. Уч. зап. Сев.-Осет.
Гос. Пед. Ин-та, 21,2.
Виноградов И. С. 1958. Сокращенное изложение системы покрытосеменных -'Com-
pendium Systematic Angiospermarum). «Проблемы Ботаники», III, Изд. АН СССР.
Литература Literature
, , i ''
Воронин H. С. 1964. Первичные Корневы^ структуры и филогенез высшик расте-
ний. Второе Московское Совещание по филогении растений. 20—25 марта 1964.
Тезисы докладов. МОИП. М.
Гайдуков Н. 1925. О филогенетичеекой системе скрытосеменных в связи с серо-
диагностикой, комплиКациями и конвергенциями. Зап. Белорусок, Гос. Ин-та
Сельск. и Лесн. Хоз., 8.
Г ерасимов а-Н а в а ш и н а Е. Н. 1954. Развитие зародышевого мешка, двойное
оплодотворение и вопрос о происхождении покрытосеменных. Б. Ж., 5.
Г е р а с и м о в а-Н а в а ш и н а Е. Н. 1955. Двойное оплодотворение покрытосеменных,
его природа и происхождение. (Докторская диссертация). Л.
Гоби Хр. (X. Я.) 1916. Обозрение системы растений. Ботанические Записки, изда-
ваемые при Ботаническом Саде Импер. Унив. в Петрограде, в. XXX. Также от-
дельным изд. 1916. Петроград.
Гоби Хр. (X. Я.) 1921. Генетическая классификация плодов. Зап. Лабор.. Семено-
вод., 4.
Голдовский А. М. 1948. Очерки биохимических изменений организмов в ходе
эволюции. Жури, общей биол., IX, 3.
ГоленкииМ. И. 1927. Победители в борьбе за существование Изд. Асе. н.-и. ин-та
при физ.-мат. факультете I Моск. Гос. Унив. М.
Голенкин М. И. 1929. Методы систематики покрытосеменных. Изд. Асе. н.-и. ин-
та при I МГУ, II.
Го лен кн н М. И. 1937. Курск высших растений. Бномедгиз. М.-Л.
Голубев В, Н. 1959. О морфогенезе деревянистых растений и путях морфологи-
ческой эволюции ст деревьев к травам. Бюлл. МОИП, биол.,.64, 5.
Го рож а икин И. Н. 1897. Лекции по морфологии и систематике архегониальиых
растений. I. Biyophyta. Печеночники и мхи. Учен. Зап. Москв. Унив. Отд.
Ест.-ист.
Горожан кин И. Н. 1900. Лекции по морфологии и систематике архегониальиых
растений. II. Pteridophyta. Папоротникообразные. Студ. изд. Москва.
Горожан к-ин И. Н. 1904. Лекции по морфологии и систематики архегоииальных
растений. III. Pteridophyta и Archisperniae. Изд. Общ. Взаимопомощи студ.
естеств. при Моск. Унив Москва.
Григорьев В. Р. 1960. Замечания по поводу метода анализа морфологического
строя. Б. Ж. XLV, 8.
Гроссгейм А. А. 1945. К вопросу о графическом изображении системы цветковых
растений. Сов. Бот., XIII, 3.
Гроссгейм А. А. 1948. Проблемы филогении растений. Б. Ж., 1.
Гроссгейм А. А. 1948. К вопросу о графическом изображении характера эволю-
ции на филогенетических схемах. Б. Ж., 4.
Грушвдгцкий И. В. 1961. Об этапе органогенеза семенных растений, утраченном
в ходе эволюции. Сборн. «Морфогенез растений», 2. Изд. Моек. Гос. Ун-та.
ГрушвицкнйИ. В. 1962. Роль недарзвития зародыша в эволюции цветковых ра-
стений. Комаровские чтения, XIV. АН СССР. М.-Л.
Давиташвили Л. Ш. 1941. Курс палеонтологии. М.-Л.
Дейнега В, А. 1903. Материалы по истории развития листа и заложению в нем
сосудистых пучков. Учеи. Зап. Моск.*ун. Отд. ест.-ист., 18,
Джапаридзе Л. И. 1963; 1965. Пол у растений. I; II. Изд.. Акад. Наук Груз. ССР.
Иист. ботан. Тбилиси. '
Жадов с кий А. Э. 1925. Типы развития зародышевых мешков у покрытосеменных
растений. Журн. Русск. Бот. Общ., '3—4.
Жадове кии А. Э. 1925. Типы развития пыльцы и их филогенетическое значение
Тр. по прикл. бот., ген. и сел., 14, 5.
Жуковский П. М. 1938. Ботаника, Изд. 2-ое. Сельхозгиз. М.
Зажурило К. К. 1934. Проблема рэдословиого дерева цветковых растений в совре-
менном освещении. СБ, 1.
Обзор новейших систем цветковых растений 169
Зажу4>влоК- К. 1936. Следы эволюции плодов в их анатомическом строении. Тру-
ды Воронежского Гос. Унив., 9, 1.
Зажурило К. К. и Кузнецова Е. К. 1939. Природа диффузной плацентации.
Тр. Воронеж. Гос. Ун-та, 10, 5.
ЗамЯгТнин Б. Н. 1951. Основы морфологической инатерпретацин нижней завязи.
Автореферат кандидатской диссертации. Бот. Инет. АН СССР. Л.
Замя.тнин Б. Н. 1951. О нижней завязи осевого происхождения. Б. Ж., 1.
З е р.о.в Д. К. 1955. Основные направления развития растительного мира. Бот. журн.
АН УССР, XII, 2. . '
Иванов С. Л. 1926. Основной биохимический закон эволюции веществ в организ-
мах. Тр. по прикл. ботаи., ген. и сел., XVI, 3.
Иванов С. Л. 1926. Эволюция вещества в растительном мире. Изв. АН СССР, XX.
5—6.
Иванов С. Л. 1936. Эволюция растительного мира с биохимической точки зрения,
Бюлл. МОИП, биол., нов. с, XV, 6.
Ильин М. М. 1950. Поликамбальность и эволюция. «Проблемы ботаники». I. М.-Л.
Ильин М. М. 1951. Основные принципиальные положения к построению новой сис-
темы растений. Б. Ж., 2.
Ильин М. М. 1951. Против идеализма в филогении растений. Б. Ж., 6.
Ильин М. М. 1953. Филогенез покрытосеменных с позиции мичуринской биологии.
Б. Ж., 1.
Ильин М. fyf. 1960. Проблема филогении и филогенеза. Хроника пятого Совещание
по филогении растений Всес. Бот. Общ. в марте 1958 г. АН СССР. Л.
Ильин М. М. 1963. Полиплоидия, видообразование и миграция. Мат. по нет. фл. И
раст. СССР, IV. ,
Каден Н. Н. 1947. Генетическая классификация плодов. Вести. Моск, уиив., 12.
Келлер Б. А. 1933. Материалистическая диалектика индивидуального и филогене-
тического развития растений. Природа, 3—4.
Келлер Б.; А. 1955. Ботаника. 2-ое изд. Сельхозгиз. М.-Л.
Келлер Б. А. 1948. Основы эволюции растений. Изд. Акад Наук СССР. М.-Л.
К рз о-П о л я н с к и й Б. М. 1919. Почему победили цветковые растения. Журн. Русск.
Бот. Общ., 8.
Ком-Палянский Б. М. 1921. О новой схеме родословия Angiospermae. Дневн.
I Веер. Съезда Русск. Бот., 2.
К о з о-П о л я н с к и й Б. М.1921. О происхождении покрытосемянности. Диев. I
Веер. Съезда Русск. Бот., 2.
Козо-Полянский Б. М. 1922. Введение в филогенеческую систематику высших
растений. Воронеж.
К о 3 0-П О л я н с к и й Б. М. 1923. О систематифеском положении сем. Cosipositae.
Жури. Русск. Бот. Общ., VIII.
К о з о-По л я н с к и Й Б. М. 1923—24. О применении анатомического метода^в систе-
( матице покрытосеменных. Бюлл. МОИП, 32, 3—4.
К о з о-П о л я н с к и й Б. М. 1925. Ближайшие перспективы в филогенетической сис-
тематике Angicspermae. Тр. Гос. Тимиряз. и.-и. ин-та, сер. I, отд. эксперим.
эвол., 1, Вологда.
К о эо-Пб л я н с к и й Б. М. 1927—28. Происхождение высших растений. I—II. Во-
логда — Москва.
К о з о-П о л я и с к и й Б. М. 1928. Предки цветковых растений. Изд. Гос. ТимирЯз.
н.-и. ин-та. М.
К о з о-П о л я н с к и й Б. М. 1936. Происхождение цветка. БСЭ, 60. 4
КоздаПол ян с кий Б. М. 1936. О природе цветка Angiospermaf Тр. Воронеж.
Гос. Ун-та, IX, I.
К р з о-Пол я н с к и й Б. М. 1936. Филогения (и филогенез) покрытосеменных-расте-
ний. БСЭ, 57.
К О 3 О-П олянский Б. М. 1936. Уродства у растений. БСЭ; 56.
170 Литература Literature
К о з о-П о л я н с к и й Б. М. 1937. Основной биогенетический закон с ботанической
точки зрения. Воронеж.
К о з о-П о л я н с к и й Б. М. 1937. Тератология цветка и новые вопросы его теории.
I. Сов. Бот., 6.
К о з о-П о л я н с к и й Б. М. 1939. Тератология цветка и новые вопросы его теории.
Бюлл. МОИП, 5—6.
Козо-Полянский Б. М. 1939. Васкуляризация аномальных венчиков у Oenothe-
ra И Campanula. Тератология цветка и новые вопросы его теории. Бюлл. МОИП,
отд. биол., 48.
Козо-Полянский Б. М. 1940. Законы филогенеза растений и дарвинизм. Сб.
«Растение И Среда». Тр. Лаб. ввол. экол., I, АН СССР. М.-Л.
Козо-Полянский Б. М. 1941. Новые успехи морфоботаники. Успехи совр. биол.
XIV, 3.
Козо-Полянский Б. М. 1945. Происхождение цветка Cruciferae в тератоло-
гическом освещении. Тератология цветка и новые вопросы его теории, III.
Б. Ж., 1.
Козо-Полянский Б. М. 1945. Новые успехи поллинистики и проблема эволюции-
высших растений. Усп. совр. биол., XIX, 2.
Козо-Полянский Б. М. 1946. Происхождение рыльца. Усп. современ. биол. 21.
Козо-Полянский Б. М. 1947. О новой системе растений. Докл. АН СССР, нов.
сер., т. LVI, 3.
Козо-Полянский Б. М., 1948. О филогенетическом, значении апомиксиса. Б. Ж., 1.
Козо-Полянский Б. М. 1949. К модернизации системы растительного мира. Тр.
Ворон. Гос. Унив., XV.
К оХз о-П о л я и с к и й Б. М. 1949. Способы изображения эволюционной системы рас-
о Т*"* о
тении. Ь. ш., i.
Козо-Полянский Б-. М. 1950. Значение различных методов в систематике расте-
ний, преимущественно Anthophyta. «Проблемы Ботаники», I.
Козо-Полянский Б. М. 1950. О систематическом положении Gnetaceae. Бюлл.
МОИП, 3.
Козо-Полянский Б. М. 1950. Происхождение цветка и типа цветковых растений.
Бюлл. МОИП, отд. биол., LV, 4.
Козо-Полянский Б. М. 1951. Против идеализма в морфологии растений. Б. Ж., 2.
Козо-Полянский Б. М. 1955. Происхождение покрытосеменных (цветковых) рас-
тений (в связи с проблемой их системы). Тр. Воронеж. Гос. Унив. XLII, 1.
Козо-Полянский Б. М. 1956. Филогения высших растений в СССР. (Обзор
принципов, направлений и достижений). Тр. Воронежск. Ун-та, 36.
Колесников В. П. 1947. Филогенез и сингенез. Докл. АН СССР, 58, 8.
Комарницкий Н. А., - Кудряшов Л. В. и Уранов А. А., 1962. Система-
тика растений. М.
Комаров В. Л. 1936. Происхождение растений. Изд. 5-е. М.-Л.
Кречетович Л. М., 1950. Цветок покрытосеменных растений. Бюлл. МОИП. LV,.
4.
Кречетович Л. М. 1952. Вопросы эволюции растительного мнра. М. ’
Кречетович Л. М. 1958. Новое в проблеме происхождения покрытосеменных. «Про-
блемы Ботаники», III.
Кр и ш т о ф о в и ч А. Н. 1928. Огкрытие в Азии древнейших покрытосеменных. При-
рода, 12.
Криштофович А. Н. 1931. Филогения растений. Природа, 6.
Криштофович А. Н. 1946. Эволюция растительного покрова в геологическом про-
шлом и ее основные факторы. Мат. но ист. фл. И раст-тц СССР, 2. \
Криштофович А. Н. 1950. Эволюция растений по данным палеоботаники. «Про-
блемы Ботаники», I. Изд. АН СССР. М.-Л.
Криштофович А. Н. 1957. Палеоботаника. 4-ое изд. Л.
Обзор новейших систем цветковых растений
17Д
Кудряшов Л. В. 1964. Происхождение односемядольности и значение эмбриоло-
гических признаков для построения системы. Второе Московское Совещание по
филогении растений. 20—25 марта 1964. Тезисы докладов, МОИП, М.
Кудряшов Л. В. 1964. Происхождение односемядольности (на примере Helobiae)
Б. Ж., 4.
Кузнецов Н. И. 1914; 1936. Введение в систематику цветковых растений, Изд.
1-ое н 2-ое. Л.
Кузнецов Н. И. 1920. Схема филогенетического родства цветковых (антофитных)
растений. Изд. тип. Таврич. Губ. Зем., на 1 листе газетного формата. Симферо- _
ПОЛЬ. ‘
Кузнецов Н. И. 1921. Естественная система цветковых растений и дальнейшие
задачи ее разработки. Дневник I Веер. Съезда Русск. Бот., 5.
Кузнецов Н. И. 1922. Основы, методы и задачи естественной филогенетической
системы цветковых растений. Изд. Главн. Бот. Сада РСФСР, XXI, 3.
Кузнецов Н. И. 1924. Политопное происхождение растительного мира. Журн. Он
Агрономии, XXII, (1921—1923).
Куприянова Л. А. 1948. Морфология пыльцы однодольных растений (материа-
лы к филогении класса). Тр. ботаи. ийст. АН СССР, I, в 7.
Куприянова Л. А. 1954. Палинологические данные к филогении класса однодоль-
ных растений. «Вопросы ботаники», I. АН СССР. М.-Л. См. также в Докл. АН
СССР, 1954, XCVIII, 2.
Куприянова Л. А. 1962. Палиоиологические данные к систематике порядков'
Frgales и Urticales. Докл. советских палинологов к I-й международной палино-
лог. КОнф.
Курсанов Л. И., Комарницкий Н. А., Мейер К. И., Раздорский
В. Ф., Уранов А. А., 1951. Ботаника. II (стр. стр. 185—447). Учпедгиз. М.
Ле йсле Ф. Ф., 1959. Данные экспериментальной морфологии и вопрос о природе
цветка. Совещ. по морфоген, раст., II.
Малышев С. И. 1960. Роль пчелиных в истории развития покрытосеменных рас-
тений. Опыление сель.-хоз. раст. пчелами, 3.
Мейер К. И. 1953. Эволюция мужского гаметофита покрытосеменных растений.
Бюлл. МОИП, отд. биол., 5. '
Мейер К. И. 1955. К вопросу о морфологической природе стебля высших растений.
Бюлл. МОИП, отд. биол. 6.,
Мейер К. И. 1958. Морфогения высших растений. Изд-во Моск, ун-та М
Меликян А. П. 1964. Применение данных анатомии спермодермы к системе и фи-
логеннии нимфейных. Вторсе Московское Совещание по филогении растений. 20—
25 марта 1964. Тезисы докладов. МОИП, М.
Мережковский К. С. 1910. Конспективный курс общей ботаники. Казань.
Мечников И. И. 1876. Очерк вопроса о происхождении видов. Вестник Европы
Периздано в 1943 г. в сборнике статей И. И. Мечникова «О дарвинизме».
Михайлов О. Ф. 1957. К вопросу о филогенетическом значении явлений регенера-
ции у растений. Уч. Зап. Тартуск., Унив., 46.
Модилевский Я. С. 1953. Эмбриология покрытосеменных растений, Изд. АН
УССР. Ин-т Ботаники. Киев.
Модилевский Я. С. 1953. К вопросу о происхождении зародышевого мешка по-
крытосеменных растений, Б. Ж., 1.
Монюшко В. А. 1937. Половые формы цветковых растений и закономерности в их
географии и происхождении. Тр. по прикл. бот., ген. и сел., сер., I, № 2.
Мчедлишвили П. А., 1960. К вопросу о проблеме происхождения покрытосемен-
ных растений. «Проблемы филогении и филогенеза». Хроника пятого совещания
по филогении растений Всес. Ботанического Общества в марте 1958 г. Изд АН
СССР. Л.
IJ2
Литература Literature
Мчедлишвили П. А., 1961, Палеобиологические основы проблемы'происхождения
покрытосеменных растений, Тр. Института Палеобиологии Акад. Наук Груз.
ССР» VI; Тбилиси.
П ен е в И. 1955. Произход на покрытосемеиите растения. Природа и анади.е, 8, 2.
София.
Первухина Н. В. 1955. О биологическом значении завязи покрытосеменных и
факторах, обусловивших ее возникновение. Б. Ж., 20.
Первухина Н. В. 1955. Теломная теория и. ее роль в развитии, взглядов на цве-
ток покрытосеменных. Б. Ж„ 6.
Первухина Н. В. 1957. Стробильиая теория происхождения цветка и ее критика.
Тр. Бот. Инет. АН СССР, сер. VII. Морфол. и анат. раст., IV.
Первухина Н. В. 1957. Роль теломноц теории в развитии взглядов иа цветок по-
крытосеменных. Тр. бот. инет. АН СССР, сер. VII. Морфол. и анат. раст., IV.
Первухина Н. В. 1962. Природа нижней завязи, зонтичных и некоторые вопросы
«теории цветка». Тр. Бот. Инет. АН СССР, сер. VIII (Морфол и аиат. раст.), 5.
Первухина Н. В. и М. Д. Иоффе. 1962. Морфология цветка троходендроиа.
(Материалы к филогении-покрытосеменных). Б. Ж., 12.
Поддубна я-А р н о л ь д и В. А. 1930. Эмбриологический метод в систематике по-
крытосеменных растений. Изв. Ассоц. н.-н. ин-т., физ.-мат. I МГУ, III, 1-А.
По д д убн.ая-А рноль д и В. А. 1951. Значение эмбриологических исследований
высших растений для систематики. Успехи совр. биологии, ХлХП, 3(6).
П о д ду б и а я-А р н о л ь д и В. А. 1958. Значение эмбриологических исследований
для построения филогенетической системы покрытосеменных растений. «Проб-
лемы Ботаники», III.
По д д у б и а я-А р н о л ь д и В. А., 1962. Значение эмбриологических исследований
для систематики и филогений иакрыурсемеииых. растений. Юбил. Съезд, чехо-
сл. бот- (1912—1962). 1—8. VII. 1962; Чехосл. Бот. Общ. Прага (ротоприцт).
П о д д у б н а я-А р н о л ь д и В. А. 1964. Общая эмбриология покрытосеменных рас-
тений, м.
Попов М. Г. 1927. Географо-морфологический метод систематики и гибридизацион-
ные процессы в природе. Тр. по прикд. брт- геиет и селекц., XVH, I,
Попов М. Г, 1954. Система по крытое емениЫх растений в. связи с проблемой их эво-
люции. Б. Ж., 6.
Проханов Я. И. 1960. Таксономическое положение однодольных и Проблема их про-
исхождения. Проблемы* филогении и филогенеза. Хроника пятого совещания пр
филогении растений Всес. Бот- Общ-, в марте 1958 г. Л.
Проханов Я. И. 1964. Класс однодольные, его таксономическая обоснованность „
положение в системе сосудистых растений. Второе Московское Совещание по
филогении растений. 20—25 марта 1964. Тезисы докладов'МОИП М.
Проханов Я. И. 1964. Математическая.теория эволюции покрытосемянных (пр
Роберт*), Второе Московское Совещание по филогении растений, 20—25 марта
1964. Тезисы докладов. МО'1П. М.
Раздорский В. Ф. 1949. Анатолия растений (Часть седьмая. Филомррфогенез
растительных структур). М.
Раздорский В. Ф., 1955. Архитектоника растений. М. f
Розанова М. А. 1940. Основы филогенетической систематики высших растений. Уч.
Зап. Лен. Гос. Ун., сер. б. и., 13.
Розанов И. Д,- 1945. Эволюция заррдыдыцрго мешка цветковых растений (Док-
торская диссерт.). Ташкент.
Р ом а н о д И Д- 1960. Параллелизм и эволюции зародышевого мешка покрытосемен-
ных. Вопр. эвол. биотеогр., гецет. и селекц.
Руденко Ф; Е. 1959. Значение мужского гаметофита для систематики покрытосе-
менных (Angiospermae). Б. Ж., 10. ;
Обзор нбвпйпгИхсистёмЦвёТКОвых растений
Савченко М. И. 1957. О природе плодолистика растений. (К истории вопроса о
природе плодолистиков). Тр. Бот. инет. АН СССР, сер. VII {Морфология и
анат, раст.), 4.
Северцов А. Н. 1921. Этюды по теории эволюции. Берлин. ГИЗ.
Северцов А, Н. 1934. Главные направления эволюционного процесса 2-ое изд. М.
Северцов А. Н. 1939. Морфологические закономерности эволюции. Изд. АН
СССР. М.-Л.
Северцов А. Н. 1945. Общие вопросы эволюции. Собр. соч. 3 изд. АН СССР.
М;-л.
С е н я и и и о в а-К о р ч а г и и а М. В. 1959. О победе цветковых с позиции гелиогео-
фйзикй. Изд. Леиингр. Гос. У нив.
С е р е б р я к о в‘И. Г. 1952. Морфология вегетативных органов высШИх растений. М.
Серебряков И. Г. 1954. Биолого-морфологический и филогенетический анализ жиз-
ненных форм покрытосеменных. Ученые записки Моск, городского педагогиче-
ского ин-та им. В. П. Потемкина, 37.
Серебряков И. Г. 19551 Основные направления эволюции жизненных форм у по-
крытосеменных растений. Бюлл. МОИП. LX, 3.
Симпсон Дж. Г. 1948. Темпы и формы эволюции. М.
Скотт Д. Г. 1927. Эволюция растительного мира. Ред. Л. М. Кречетович. М.-Л.
Смирнова Е. С. 1958. К проблеме взаимоотношения однодольных и двудольных
растений. Докл. Москв. с.-х. акад. им. Тимирязева, 39.
Смирнова Е. С. 1959. К познанию рода Podophyllum в связи с проблемой род-
ства однодольных и двудольных растений. Докл. Моск. с.-х. акад. им. Тимиря-
зева.^.
Смирнова Е. С. 1959. О дифференциации покрытосеменных растений на двудольные
н однодольные. Докл. Моск. с.-х. акад. им. Тимирязева, 48.
Смирнова Е. С. 1960. К познанию родства однодольных и двудольных растений
Изв. Тимирязевск. с.-х. акад., 4.
Смирнова Е. С. 1960. Анализ филогенетической системы цветковых растений.
А. А. Гроссгейма. Докл. Моск. с.-х. акад. им. Тимирязева, 53.
Смирнова Е. С. 1961. К морфогенезу семян покрытосеменных растений. Докл.
ТСХА, 72.
Старостин Б. А. 1964. Происхождение и природа цветка (исторический обзор).
Второе Московское Совещание Ио филогении растений. 20—25 марта 1964. Те-
зисы докладов. МОИП. М.
Старостин Б. А. 1965. Системы растений в России в XVIII — начале XIX вв.
Вопросы ист. естествози. и техники, 19.
Стеббинс Дж. Л. 1957. О Гибридном происхождении покрытосеменных. Б. Ж., 10.
Сушкин П. 1915. Обратим ли процесс эволюции? «Новые идеи в биологии», 8.
Съюорд А. Ч. 1936. Века н растения. М.-Л.
Тамамшян С. Г. 1951. Руминация эндосперма-у покрытосеменных. Б. Ж., 5. -
ТамамшянС. Г. 1958. Sanlalales в системе Spermatophyla. «Проблемы Ботаники»,
Ш.
Тамамшян С. Г. 1964. Опыт построения филогенетической системы двудольных на
примере Umbellales и родственных порядков. Второе Московское Совещание
по филогении растений. 20—25 Марта 1964. Тезисы докладов. МОИП. М.
Тахгадняи А. Л. 1941. О примитивном типе плацентации у Angiospermae. И ЗВ.
Арм. Фил. АН СССР, 1(6).
Тахтаджяи А. Л. 1941. Эволюция плацентации у высших растений. Изв. Арм.
Фил. АН СССР. 8(13).
Тахтаджяи А. Л. 1942. Структурные типы гйиецея и плацентация семёзачатков.
‘ Изв. Арм. Фнл. АН СССР. 3—4, (17—18).
Тахтаджяи А. Л. 1943. Соотношения онтогенеза и филогенеза у высших расте-
ний. Тр. Ерев. Гос. Ун., 22.
174 Литература Literature /
Такта джян А, Л. 1943. Эволюция плацентации и филогения высших растений. Те-
зисы докторской диссертации. Ереванск. Гос. Уи-т. Ереван.
ТахтаДжяи А. Л. 1945. Опыт применения теории филэмбриогеиезак объяснению
происхождения зародыша однодольных. Докл. АН Арм. ССР, 3(2).
Тахтаджян А. Л. 1946. Об эволюционной гетерохронии признаков. Докл. АН
ССР, 5(3).
Тахтаджян А. Л. 1947. О принципах .методах и символах филогенетических по-
строений в ботанике. Бюлл. МОИП, отд. биол., LII, 5.
Тахтаджян А. Л. 1948. Морфологическая эволюция покрытосеменных Изд.
МОИП. М.
Тахтаджян А. Л. 1950. Теория филэмбриогенеза А. Н. Северцова и эволюцион-
ная морфология растений. «Проблемы ботан.», I, АН СССР. М.-Л.
Тахтаджян А. Л. 1950. Филогенетические основы системы высших растений,
Б. Ж., 2.
т ахтаджян А. Л. 1951. Пути приспособительной эволюции растений. Б. Ж., 3.
т а х т а д ж я н А. Л. 1952. Теломиая теория и «Новая морфология», Б. Ж., j.
т ахтаджян А. Л. 1954. Вопросы эволюционной морфологии растений. Изд. Ле-
ииигр. Гос. Ун., Л.
Тахтаджян А. Л. 1954; 1961. Происхождение покрытосеменных растений. Изд
1-ое и 2-ое. М.
Тахтаджян А. Л. 1954. Некоторые проблемы эволюционной морфологии покрыто-
семенных. «Вопросы ботаники», II. АН СССР. М.-Л.
Тахтаджян А. Л. 1957. Прямое приспособление или естественный отбор. Б. Ж., 4.
Тахтаджян А. Л. 1957. Экологическая эволюция покрытосеменных и июблема
происхождения умеренной флоры Евразии. Тезисы докл. на делег. съезде Всес.
Бот. Общ., III.
Тихомиров В. Н. 1958. Тератология и проблема происхождения завязи зонтич-
ных. Науч. докл. Высш. Шк. Биол. науки, 3.
Тихомиров В. Н. 1961. Морфогенез плода в семействе Umbelliferae- Морфоге-
нез растений, 2.
Тихомиров В. Н. 1964. Происхождение покрытосеменных растений в связи с про-
блемой построения их филогенетической системы. Второе Московское Совещание
по филогении растений. 20—25 марта 1964. Тезисы докладов. МОИП. М.
Тринклер Ю. Г. 1962. Основные типы развития покрытосеменных растений. Б.
Ж., 1.
Троицкая О. В., 1935. Систематика растений. ОГИЗ. М.-Л.
Ту таю к В. X. 1964. Некоторые закономерности изменчивости цветка в связи с
вопросом о природе его органов. Второе Московское Совещание по филогении
растений. 20—25 марта 1964. Тезисы докладов. МОИП. М.
Толмачев А. И. 1951. О приложении учения А. Н. Северцова об ароморфозе к
исследованию филогении растений. Б. Ж., 3.
Фонт В. 1962. Комплекс призгаков — строение таксонов — естественная система рас-
тений. Юбил. Съезд чехосл. бот. (1912—1962). 1—-8. VII. 1962. Чехосл. Бот.
Общ. Прага (ротоприит).
Хохлов С. С. 1946. Бесполосемениые растения. Уч. Зап. Сарат. ун., 16, 1.
Хохлов С. С. 1946. Исторические предпосылки и эволюционное значение апомикси-
са покрытосеменных. Докл. АН СССР, 52, 9.
Хохлов С. С. 1949. Перспективы эволюции высших растений. Уч. Зап. Сарат. Гос.
Пед. Ин-та, II. '
Хохлов С. С. 1950. Перспективы эволюции высших растений. Саратов.
Хохряков А. П. 1964. Относительная изменчивость и скорость эволюции веге-
тативных и генеративных органов у высших растений. Второе Московское Со-
вещание по филогении растений. 20—25 марта 1964. Тезисы докладов. МОИП. М.
Хржановскии В. Г. 1958, К вопросу о соматической эволюции у покрытосеменных
Докл. Моск. с.-х. акад. им. Тимирязева, 36.
Обзор новейших систем цветковых растений I75
Храановский В. Г. 1959. К развитию метода флорогеиетического анализа. Докл.
Тимнрязевск. с.-х. акад., 46.
Шапаренко К. К. 1939. Эволюция филогенетических схем. Б. Ж., 5—6.
Шмалыаузен И. И. 1940. Пути и закономерности эволюционного процесса. Изд.
АН СССР. М.-Л.
Штемпель Б. М. 1959. К вопросу о местообитании первых покрытосеменных рас-
тении. Ь. m., 1.
Юзепчук С. В. 1946. Апомиксис и эволюция. Тез. научн. сессии Ленингр. ун.
Яковлев М. С. 1946. Однодольность в свете данных эмбриологии. Сов. Бот., 6.
Яковлев М. С. 1958. Принципы выделения основных эмбриональных типов и их
значение для филогении покрытосеменных. «Проблемы ботаники», III.
Яковлев М. С. 1960. Эмбриогенез и его значение для филогении растений. М.-Л.
Я ц е и к о-Х м е л е в с к и й А. А. 1958. Происхождение покрытосеменных по-данным
внутренней морфологии их вегетативных органов. Б. Ж., 3.
Дополнение — Supplement
Имс А. 1964. Морфология цветковых растений. М.
1962. Первое Московское совещание по филогении растений, 22—25 апреля
1962 г.
1960. Сборник «Проблемы филогении и филогенеза». Хроника пятого совещания по
филогении растений Всес. Бот. Общ. в марте 1958 г. (Конспекты докладов к'
прения по ним. — Ред.). Л.
1960. Проблемы филогений растений (по материалам Первого Моск. сов. по
фил. раст. 22—25 апр. 1962). Сб. тр. М.
*
* *
A b b е Е. С. 1935. Studies in the phylogeny of the Betulaceae. I. Floral and inflores
cence anatomy and morphology. Bot. Gaz., 97.
Abel O. 1929. Palaobiologie und Stammesgeschichte. lena.
A 1 e x n a t W. 1922. Sero-diagnostische Untersuchungen fiber die Verwandtschaftsver-
haltnisse innerhalb der Sympetalen. Bot. Archiv, I.
Anderson E. 1934. Origin of the angiosperms. Nature, 133, 3360.
Anderson E. Stebbins G. L. 1954. Hybridization as an .evolutionary stimulus.
Evolution, D, 8.
Andreanszky G. 1952. Uber die Rolle der limnischen Lebensweise in der phylo-
genie der Pflanzen. Acta biol. Acad, scient. hung., 3, 1.
Ankermann F. 1927. Die Phylogenie der Monokotylen. Bot. Archiv, XIX, 1—2.
Arber A. 1925. Monocotyledons. Morphological study. Cambridge.
A r b e r A. 1928. The tree-habit in angiosperms: its origin and meaning. New Phyt., 27,
Arber A, 1933. Floral anatomy and its morphological interpretation. New"Phyt..
XXXII, 3.
Arber A. 1937. The interpretation of the flower: a study of some aspects of morpho-
logical thought. Biol. Rev., 12.
Arber A. 1950. The natural philosophy of plant form.
A r b e r E. A. N. 1899. Relationships of the indefinite inflorescences- Journ. Bot,, 37,
Arber E. A. N. and Parkin J. 1907. On the origin of angiosperms. The Journ, of
the Linn. Soc. Bot., XXVIII.
Arber E. A. N. and Parkin J. 1908. Study on the evolution- of the angiosperms.
Ann, Bot., XXII.
A s a m a K- 1960. Evolution of the leaf forms through the ages explained by the suc-
cessive retardation and neoteny, Sci. Reports Tohoku Univ., Ser. 2, Spec. vol. (4).
Axelrod D. J. 1952. A theory of Angiosperms evolution. Evolution, VI, 1.
Axelrod D. J. I960- The evolution of flowering plants. Evolution. After Darwin,
I. Chicago.
I7*> Литература Literature
Axelrod D. J. 1961. How old are the angiosperms? Americ. Journ. ol Sc., 259.
Babcock E. B, 1938. Phylogeny in the light of genetics and cytology. Current sci.,
special number.
В a i 1 e у I. W. 1912. The evolutionaiy history of the foliar ray in the wood of the
Dicotyledons and its phylogenetic significance. Ann. Bbt., vol. XXVI, no. CHI.
Bailey J. W. 1944. The development of vessels in angiosperms and its significance
in morphological research. Amer. Journ. Bot, 31, 7.
Bailey J. W. 1949. Origin of the angiosperms: the need foi a broadenen outlook.
Journ. Arnold. Arbor., 30, 1.
Bailey J. W. 1953. Evolution of the tracheary tissue of land plants. Amer. Journ.
Bot., 40, i.
Bailey J. W. 1957. The potentialities and .limitations of wood anatomy in the study
of the phylogeny and classification of angiosperms. Jcurn. Am. Arbor., 38, 3.
Bailey J. W. and Sinnott E. W. 1914. Investigations on the phylogeny of angio-
sperms. I—Amer. Journ. Bot., 1; II-Bot. Gaz., LVIII, July, 1; Ш—Amer. Journ,
Bot., 1; IV—Ann. Bot., 28; :
,. 1915. Ead. cit.: V—Amer Journ. Bot., II.
„ 1915. Changes in the fruit type of angiosperms, coincident with the development
of the herbaceous habit. Report Bot. S'oc. Amer, in Science, 41.
Bailey I. W. and Swamy B. G. L. 1951- The conduplicate carpel of dicotyledons
and its initial trends of specialization. Amer. Journ- Bot., 38, 5.
Baker H. G. 1963. Evolutionary mechanisms in pollination biology. Science. 139,
3558. '
Bancroft H. 1930 The arborescent habit in angiosperms. A review. New Phytol.,
29. . '
Bancroft H. 1935. A review of researches concerning floral morphology, Bot. Rev., 1.
Barg'hoorn E. S. 1941 The ontogenetic development and phylogenetic specializa-
tion of rays in the xylem of dicotyledons. III. The elimination of rays. Bulk
, Torrey Bot. Club, 68.
Baum H. 1948. Die Verbreitung der postgenitale Verwachsung im Gynczeum und
ihre Bedeutung fur die typologische Betrachtung der coenokarpen Gynozeums.
Osterr. Bot. Ztschr., 95.
Baum H. 1949. Der einheitliche Bauplan der Angiospermengynozeen und die Homo-
logie ihrer fertilen Abschnitte, Osterr. Bot. Ztschr., 96.
Baum H. 1949. Zur Frage des schi ittweisen Uberganges vorit apokarpen ziim coeno-
karpen Gynozeum. Osterr. Bot. Ztschr. 96.
Baum H. 1950. Unifaziale und subunifaziale Strukturen im Bereich der Bliitenhiille
und ihre Verwendbarkeit fur die Homologisierung der Kelch-und Krdriblatter.
Osterr. Bot. Ztschr., 97.
Baum H. 1950. Lassen sich in der Ontogenese der Karpelle und Staubblatter noch
Anklange an ihre phylogenetische Entwicklung aut Telomen feststellen? Osterr.
Bot. Ztschr.. 97.
Bauman-Bodenheim M. G. 1946. Myodocarpus und die Phylogenie der Umbe-
lliferen-Frucht. Ombelliferenstudien. I. Ber. Schweiz. Bot. Ges., 56.
В a u m a n-B odenheim.M. G., 1.954. Analyse phylogehetischer Entwicklungsorga-
ne bei Angiospermen. Ber. Schweiz. Bot. .Ges., 64.
В a u m-L einfellner H. 1947^ Die, Peltation der Staubblatter und die Phylogenie
der Angiospermen. Phyton, 5. , ’
Beauverie J. and D!u rand M. 1930./L ancienete et la phylogenie des plantes
a fleurs. Rev. Gen. Sci. Pur. App;,'46.,
Becquerel P. 1954. La theorie du Tfelome basfee sur les Rhynia ri’est pas conforme
a la constitution et a revolution des plantes vasculaires. VHI-me Congr. Interma-
tion. de Bot. Paris.
Becqere! P. 1954, L’ontogenie des Fougeres actuelles, demontre-t-elle qu elles
orit fete constituets de telomes modifies an cours • de leur evolution. Compt. R.
Acad. Sc., Paris, 288, no. 1.
Обзор новейших систем цветковых растений. 177
Becquerel Р., 1954. L’ontogenie des Phanerogames, etablie par 1'anatomie dina-
mique nous montre-t-elle qu’elles sont constituees par un agencement de telomes
modifies au cours de la phylogenie? Bull. Museum Nat. d’Hist. Nature., Paris, 26.
Benson M., 1904. The origin of flowering plants. New Phytol., III.
Bentham G. and Hooker J. D. 1894. Evolution and classification. Cont. Bot.
Dept. Univ. Nebr. N. S'. VII.
Berry E. W. 1920 (1918). A sketch of the origin and evolution of floras. Ann. Rep.,
' Smithsonian Inst.
Berry E. W. 1923. Tree ancestors. Baltymore.
Berry E. W. 1945. The beginnings and history of land plants. Johs Hopkins Univ.,
Studies in Geol. 14.
Berry E. W. 1945. The origin of land plants. Baltimore.
Bertalanffy L. von. 1950. An outline of general system theory. Brit. Journ. Philos.
Sci., 1 (2).
Bertrand P. 1937. Isolement preccce de tous les grands groupes de vegetaux vas-
culaires. Bull. Soc., Bot. France, 84, 9/10.
Bertrand P. 1947. Les vegetaux vasculaires. Paris.
В e s s e у С. E. (1893) 1894. Evolution and classification. Proc. Amer. Assoc- Advanc.
Sciences, 42.
Bessey С. E. 1895. The point of divergence of monocotyledons and dicotyledons.
Bot. Gaz., 22.
Bessey С. E. 1897. The phylogeny and taxonomy of angiosperms. Bot. Gaz., XXIV.
Bessey С. E. 1907. Synopsis of plant phyla. Nebresca, University Studies, VII, 4
Bessey С. E. 1909. The phyletic idea in taxonomy. Science, N. S., 29. *
Bessey С. E/1910. The phyla, classes and orders plants. Trans. Amer. Mier. Sot., 29.
Bessey С. E. 1915. Revis, of some plant phyla. Nebrasca Univers. Studies. XIV, 1.
Bessey С. E. 1915. The phylogenetic taxonomy of flowering plants. Annals of the
Missouri Botanical Garden II, 1—2.
В e s s e у С. E. 1916. The origin of the Anthophyta. Michigan Ac. Sc. Ann. Rep., XVII.
В e w s J. W. 1927. Studies in the ecological evolution of the angiosperms. New Phytol,
-XXVI, 1 a. 2.
Blagowestschenski A. W. 1955- Die biochemischen Grundlagen des Evolu-
tionsprozesses der Pflanzen. Akademie-Verlag, Berlin.
Bly th A. 1958. Origin of primary extraxylary stem fibers in dicotyledons. Univ.
Calif. Publ. Bot., 30 (2).
В ocher T. W. 1951. Studies on morphological progrression and evolution in the
vegetable kingdom. Kobenhavn. (Det. k. Danske videnskabernes selskab. Bio-
logiske meddelelser. Bd. 18, no. 13).
Bonne G. 1928. Recherches sur le pedicelle et la ileur des Rosacees- Paris.
Boughey A. S. 1959. Origin and dispersal of flowering plants. Nature, 184 4694.
В о u r e a u E. 1948. L’ontogenie vasculaire des plantules et la phyogenie des pha-
nerogames. Rev. scient, ann. 86, fasc. II, N 3299.
В our e au E. 1952. L’evolution des vegetaux et 1’anatomie des plantules. Annee biol.,
Ser. 3, t. 28, fasc. 7—8.
Boureau E. 1952. L’evolution des vegetaux et 1’anatomie des plantules. Colloque in
ternat. du centre Nation, d. 1, Recher. Sci., 41. Evolution et phylogenie chez
les vegetaux.
Bouygues H. 1918. Structure et developpement de certaines formes vasculaires
anormales du petiole des Dicotyledon.es. Actes Soc. Linn. Bordeaux, 57.
Bower F. O. 1908; reprint 1959. The origin of a land flora. London.
Bower F. O. 1935. Size and form in plants with special reference to the primary
conducting tracts. London.
Boyd L, 1932, Evolution in the monocotyledoncus seedling. Trans, a. Proc. Bot. Soc,
Edinb., 30, 4.
12. А. А. Гросгейм и M. Ф. Сахокиа
178
Литература Literaiure
Braun A. 1874. Bemerknugen uber Plazentenbildung. Verh. Bot. Ver Prov. Bran-
denburg., VIII, Sitzungsber., 30, Jan. '
Breitenfeld Ch. 1957. Genetische Untersuchungen zur Phylogenie und Systema-
tik der intermedium—und labile Gersten. Z. Pflanzenziicht., 38, 3.
Bremekamp С. E. B. 1939. Phylogenetic interpretations and genetic concepts in
taxonomy. Chron. Bet 5, 4—6.
Brown R. 1840. On the relative position of the divisions of stigma and parietal pla-
centae in the compound ovarium of plants. Misc. Bot. Works of Robert Brown, 1.
Brown W. H. 1938. The bearing of nectaries on the phylogeny of filowering plants
Proc. Amer. Phil. Sbc., 79.
Browne J. M. P. 1935. Some viws on the morphology and phylogeny of the leafy
vascular sporophyte. Bot. Rev., 1.
Bugnon P. 1921. L’origine phylogenique des plantes vasculaires... Bull. Soc. Linn.
Norm. 7 ser., -i.
В u v a t R. 1952. Structure evolution et fonctionnement du meristeme apical c.e quel-
ques Dicotyledones. Ann. Sci. Bot. Nat., XI, 13.
Buxbaum F. 1951. Grundlagen und Methoden einer Erneuerung der Systematic
der hoheren Pflanzen. Wien.
C a i n A. I, 1962, The evolution of taxonomic principles. «Mierobial Classific». Cam-
bridge Univ. Press.
C a 1 e s t a n i V. 1933. Le origine e la classificazione della angiosperme. Archivio
Bot., 9.
Camp W. H. 1947. Distribution patterns in modern plants and problems of ancient
dispersals. Ecological Monographs, 17.
Campbell D. H. 1929. The phylogeny of the angiosperms. Bull. Torrey. Bot.
Club., 55.
Campbell D. H. 1930. T'ne phylogeny of monocotyledons. Ann. Bott., XLIV, CLXXIV.
Campbell D. H. 1940. The evolution of the land plants (Embryophyta) California.,
Camp o-D u p 1 a n M. van. 1946. Observations Temploi des grains de pollen en phy-
logenie. (Extr. du Bull. Soc. histoire natur. Toulouse, t. 81). Toulouse. (Trav.
Lab, forest. Toulouse, t. 1, vol. 4, art. 12).
Canright J. E. 1952. The compartive morphology and relationships of the Magno-
liaceae. I. Trends of specialization in the stamens. Amer. Journ. Bot., 39, 7.
Canright J. E. 1953. The comparative morphology and relationships of the Magno-
liaceae. II. Significance of the pollen. Phytomorphology, 3 3.
Canright J. E. 1955. The comparative morphology and relationships of the Mag-
noliaceae. IV. Wood and nodal anatomy, Journ. Arnold Arb., 36.
Canright J. E. 1960. The comparative morphology and relationships of the Ma-
gnoliaceae, III. Carpels. Amer. Journ. Bot., 47, 2.
Carpentier A. 1948. Les dicotyledones homoxylees et l’origine des angiospermes.
Bull. Soc. bot, Nord, France, I, 1.
C e 1 а к о v s к у L. J. 1896, 1900, Ueber den phylogenetischen. Entwicklungsgang
der Bliite und den Ursprung der Blumenkrone. I und II. Zitzber. d. konigl. bohm.
Geseil. d. Wiss.
C h a FR L. 1937. 'The phylogenetic value of certain anatomical features of Dicotyle-
donous woods. Ann. Bot., N, S,. 1,
Chalk L. 1962. Wood anatomy. Adv. Sci., 18 (75).
C h a 1 о n e r W. G. 1960. The origin of vascular plants. Sci. Progr,, 48, 191,
Chauveaud G. 1911. L’appareil conductuer des plantes vasculaires et les phases
principales du son evolution. Ann. Sci. Nat. Bot., 13.
Cheadle V. I. 1948. Observations on the phloem in the Monocotyledoneae. II. Addi-
tional data on the occurence and phylogenetic specialization in structure of the
sieve tubes in the metaphloem. Amer. Journ, Bot,, 35, 2.
Cheadle V. I. 1953, Independent origin of vessels in the monocotyledons and dico-
tyledons. Phytomorphology, 3, 1.
Обзор новейших систем цветковых растений
179
Cheadle V. I. and Whitford N. В. 1941, Observations on the philoem in the
Monocotyledoneae. I- The occurence and phylogenetic specialization in structures
of the sieve tubes in the metaphloem. Amer. Journ. Bot, 28. 8.
C h о d a t R. 1926. La placentation et les enchainements des plantes vasculaires. С. K-
Soc. Phys. Hist. Natur. Gen., XLIII,
Chrysler M. A. 1915. Origin of herbaceous angiosperms. Bot. Gaz., 60, 3.
Clements F. E. and E. S. 1928. Flower families and ancestors. H. W. Wilson Com-
pany, New York,
Clements F. E., Martin E. V. and Long F, L. 1950, Adaptation and origin in
the plant world, Waltham.
Clifford H. T. 1961. Floral evolution in the family Gramineae. Evolution, 15, 4.
Constance L. 1955. The systematics of the angiosperms, In «А Century of pro-
gress in the Natural Sciences, 1853—1953». California Academy of Sciences, San
Francisko.
Cook O. F. 1907. Origin and evolution of angiosperms. Proc. Wash. Acad, of Sc„ IX,
Cook O. F., 1916. Morphology and evolution of leaves. Journ. Wash. Acad., Sci., 6.
Cope E. D. 1904. The primary factors of evolution. Chicago,
Cope E. D. 1957. Forecast of a system of the dicotyledons, Madrono. 14.
Co peland H. F. 1940. The phylogeny of the angiosperms. Madrono. 5.
Corner E. J. H. 1946. Centrifugal stamens. Journ. Arnold Arb., 27.
Corner E, J, H, 1949. The durian theory of the origin of the modern tree. Ann, Bot,
N- S., XIII, 52.
Corner E. J. H. 1953. The durian theory extended—I. Phytcmorphology, 3, 4.
Corner E. J. H. 1954. The durian theory extended—II. The arillate fruit and the
compound leaf. Phytomorphology, 3, 4.
С о r n e r E. J. H. 1954. The durian theory extended—III. Pachycauly and megaspermy—
conclusion. Phytomorpholcgy, 4, (3, 4).
Coulter J. M. 1903. The phylogeny of angiosperms. Dec. Publ. Univ, of Chicago.
First Ser., X.
Coulter'!. M. 1914. The evolution of sex in plants. Univ, of Chicago. Sc. Ser. 1.
Coulter J. M. 1915, The origin of monocotyledony in the grass. Annals, of the
Missouri Botanical Garden, II, 1—2.
С о u 11 e r J. M., Chamberlain C. J., L a n d W. I. G., 1914. The origin of mo-
nocotyledony. Bot. Gaz., 57.
Cowles H. C. 1929. Ecological evolution of angiosperms. Bot. Gaz. 87, 2.
Crete P. 1951. Repartition et interet phylogenetique des albumens a formations haus-
toriales chez les angiosperms et plus particulierement chez les gamopetales. Ann.
Sci. Nat. Bot., П-e Serie, 12.
Crete P. 1964. L’embryogenie et son role dans les essais de classification phylogenetique.
Phytomorphology, 14, 1.
С г о i z a t L. 1943. The concept of inflorescence. Bull. Tcrrey Bot. Club, 70.
Croizat L. 1947. Trochodendron, T’etracentron and their meaning in phyogeny.
Bull. Torrey Bot. Club, 70.
Cronquist A. 1955. Phylogeny and taxonomy of the Compositae. Amer. Midi. Na-
tur., 53, 2.
Cronquist A. 1957. Outline of a new system of families and orders of dicotyle-
dons. Bull, du Jard. Bot. de 1’Etat, Bruxelles, 27 (I).
Cronquist A. 1960. The divisions and classes of plants. Bot. Rev., 26, 4,
Ct о w W. B. 1926. Phylogeny and the natural system. Jcurn. Genetica, XVII.
Cuenod A. 1932, Hypotehese relative a la place des Aionocotyledones dans la classi-
fication naturelie. Bull. Soc. Bot. France, 79.
Gutter E. G. 1957. Studies of morphogenesis in the Numphaeaceae. L, 11. Phytomor-
phology, 7, 1.
180
Литература Literature
Daniker A. U. 1954. Evolution und Epharmose. Aarau, Komm.-Verl Sauerlander.
(Mitf. Bot. Museum Univ. Zurich 200. Ser. Bot. 44). Aus den Verhandl. Schweiz.
Naturforsch. Ges., 134. Jahresversammlung.
D a n i к er A.U. 1959. Was ist eine Blfite? Betracbungen fiber das Wesen u. die Organiza-
tion der Bliiten. Ein Beitrag zur Phylogenie der Angiospermen. Vierteljahrsschr.
Naturforsch. Ges- Jg. 104. Schlussheft.
De Haan H. R. M. 1920. Contribution to the knowledge of the morphological value
and phylogeny of ovule and its integuments. Rev. Trav. Bot. Need. 17.
Delevoryas Th. 1962. Morphology and evolution of fossil plants. New Yorx.
Diels L. 1916. Kaferblumen bei den Ranales und ihre Bedeutung ffir die Phylogenie
der Angiospermen. Ber. Dtsch. Bot. Ges., 34.
Diels L. 1920. Die Methoden der Phytographie und der Systematik der Pflanzen. Ab-
derhalden’s Handb. d. bicl. Arbeitsmethod.
Domin K- 1931. Phylogenetic evolution of the phyllome. Amer. Journ. Bot., 18.
Dornin K- 1944. Soustava rostlin bunecnych s hlediska vyvoje a pribuzenskychvztahu.
Systema plantarum cellularium phylogenetice et ex affinitatis vinculo, tractatum.
Praha.
Douglas G. E. 1944. ТИё inferior ovary. Bot. Rev., 10.
Dykyj-Sajiertova D. 1958. Atmungspigmente, und Phylogenie der Pflanzen.
Preslia, 30, 3.
tames A, J. 1911. On the origin of the herbaceous type in the angiosperms, Ann, Bot,
Vol. XXV, no. XCVII.
Eames A. J. 1929. The role of flower anatomy in the determination of angiosperm,
phylogeny. Proc. Intern. Congress Plant Sci., 1.
E a m e s-A. J. 1931, The vascular anatomy of the flower with refutation of the theory
of carpel polymorphism. Amei’. Journ. Bot., 18.
Eames A, J. 1949. Can the angiosperms be divided into the Phyllosporae and Stachyo-
sporae? Amer. Journ. Bot., 36, 10.
Eames A. J, 1951, Again: «The new' morphology»- New Phyt.. 50.
E b e r E., 1934. Karpellenbau und Plezentations verhaltnisse in der Reihe der Helobiae.
Flora, 27.
E с к a r d t T. 1937. Untersuchungen uber Morphologie, Entwicklungsgeschichte und
systematische Bedeutung des pseudomcnomeren Gynoeciums. Nova Acta Leoo.
Carol. (N. F.), 5.
Eckardt T. 1938. Das pseudomonomere Gynoeceum. Chron. Bot., 4.
Eckardt T. 1941. Kritische Untersuchungen uber das primare Dickenwachstum bei
Monokotylen, mit Ausblick auf dessen Verhaltnis zur sekundaren Verdickung.
Bot. Archiv, 42.
Eckardt T. 1957. Vergleichende Studie fiber die morphologischen Beziehungen
zwischen Fruchtblatt, Samenanlage und Blfitenachse bei einigen Angiospermeii,
zugleich als kritische Beleuchtung der «New Morphology». Neue Hefte z. Morph., 3.
E g 1 e r F. E. 1943. The fructus and the fruit. Chron. Bet., 7.
E hr end о f e r F. 1959, Zusammenhange zwischen Sippenstruktur, Lebensraum
und Phylogenie bei Formenkreisen der Angiospermen. Naturwiss. Rundschau. 12.9.
Elliot J. H- 1933. Growth and differentiation of the vascular system during the-
leaf development in the dicotyledons. Proc, Leeds phil. and. lit, Soc. d.. Sc.
Sect/ 2.
Emberger L. 1944. Les plants fossiles dans leurs rapports avec les vegetaux vi-
vants. Elements de paleobotanique et de morphologie comparee. Massen. Paris.
Emberger L. 1950. La valeur morphologique et l’origine de la fleur. Ann. Biol. Ser.
3, t. 26, f. 6.
Emberger L. 1950. Les Prephanerogames Ann. Sci. Nat. (Bot.). 11 ser., 10.
Emberger L. 1960. Les vegetaux vasculaires. In: Chadefaud M. et Emberger L.,
Traite de Botanique (Systematique). T. II. Fasc. I; II.
Обзор новейших систем цветковых растений l8l
Engler А. 1954. 1964. Syllabus der Pflanzefamilien. Zwolfte Auflage. В, I. u. II,
Berlin. (Herausg. v. H. Melchior..).
Engler A. und G i 1 g E., 1924. Syllabus der Pflanzenfamillien. Neunte und zehnte
Auflage. Berlin.
Engler A. und Prantl H. 1902. Die naturlichen Pflanzenfamilien, 1. Leipzig.
Erdtman G. 1952, Pollen morphology and plant taxonomy. Angiosperms. Stockholm.
Almqvist a. Wiksell.
Erdtman G. 1960. Pollen.wals and angiosperm phylogeny. Bot. Not., 113 (1).
Ernst A. 1908. Zur Phylogenie des Embryosackes der. Angiospermen.. Ber. Dtsch.
Bot. Ges., 26.
Esau K- 1954. Primary vascular differentiation in plants. Biol. Revs. Cambr. Philos.
Soc., 29, 1.
F a-g e г 1 i n d F. 1947. Strobilus und Bliite von Gnetum und die Moglichkeit aus ihrer
Struktur den Bliitenbau der Angiospermen zu deuten, Ark, for Bot., 32, A, 8,
Fagerlind F. 1958. Is the gynoecium of the angiosperms built up in accordance
with the phyllosporous or the stachyospcrous scheme? Svensk bot. Tidskr., 52.
F a h n A. 1953. The topography of the nectary in the flower and its phylogenetic trend.
- Phytomorphology, 3.
Ferre Y. de. 1952. La signification des formes de jeunesse dans 1’evolution. Annee
biol. Ser. 3, t. 28, fasc- 7—8.
Florin R. 1952. Evolution et classification des Gymnospcrmes. In «Evolution pt Phy-
logenie chez les vegetaux». Paris-
Foster A. S. 1939. Problems of structure growth and evolution in the shoot apex
of seed, plants. Bot. Rev., 5, 8.
Foster A. S. and Gifford E. M., Jr. 1959. Comparative morphology of vascular
plants. W. H. Freeman a. Co. New York.
Franz С. E. 1928. Die Brauchbarkeit der Serodiagnostik (spez. f. Monocotyled.).
Berlin. Diss.
Frost F. H. 1930, Specialization in secondary xylem of dicotyledons. I. Origin of
vessel. Bot. Gaz., 89.
Frost F. H. 1930. Specialization in secondary xylem of dicotyledons. II. Evolution
of end walls of vessel segment,. Bot. Gaz., 90.
Frost F. H. 1931. Specialization in secondary xylem of dicotyledons. III. Specializa-
tion of lateral wall of vessel segment. Bot. Gaz., 91.
Gaidukov N. 1927. Uber Konvergenzen und Komplikationen der Algen und der
Angiospermen. Bot. Archiv, XVII, 1—2.
Gaudichaud Ch. 1841. Recherches generales sur 1’organographie la physiologie
et I’organogenie des vegetaux.
Gaussen H. 1952. L’evolution pseudocyclique. Colloque internal, du Centre Nation,
d. 1. Recher. Sci. 41. In «Evol. et phylogenie chez les vegetaux».
Gaussen' H. 1958. La reaction de Vincent et Maule et la phylogenie des plantes
superieures. Extr. Compt. rend... Acad, sci., 247. Paris.
Gaussen H. 1958, L’origine des Angiosperms- Compt, Rend, Acad, Sci., 247, 2.
Georgescu C. und T u t u n a r u V. Mannliche Katzchen des Nussbaumes (Jug-
lans regia L.) auf einjahrigen Trieben und ithre phylogenetische Bedeutung.
Rev. biol., 8. 3.
Gibbs R. D. 1958. Chemical evolution in plants. Journ. Linn. Soc. (Bot.). 56.
Gilibert S'. G. 1940. Evolutionary significance of ring porosity in woody angio-
sperm. Bot. Gaz., 102.
Gliicke H, 1925. Kritische Bemerkungen fiber d. phylogenetische Herkunft der Mono-
cotylen. Flora, Festschr. f. Goebel. 118—119.
Good R. 1956. Features of evolution in the flowering plants. London. (См. рефератив-
ную статью: Благовещенский А. В., Новая попытка установить главные черты
эволюции покрытосеменных. Бюлл. Гл. Бот. Сада АН СССР, 1959. 34.).
182
Литература Literature
G о t h а п W. 1924. Palaeobiologische Betrachtungen uber die fossile Pflanzenwel:.
Berlin.
Goth an W. 1951. Die Entwicklung der Pflanzenwelt im Lauf der Erdgeschichte.
Urania, 2.
Grant V. 1949. Pollination systems as isolating mechanisms in angiosperms. Evo-
lution, 3.
Grant V. 1950. The protection of the ovules in flowering plants. Evolution, 4.
Grant V. 1959. Origins of argiospermous plants. Evolution, 13 (2).
Gregoire V. 1938. La morphogenese et 1’autonomie morphologique de 1’appareil flo-
ra!. I. Le carpelle. La Cellule, 47.
Gregory W. C. 1941. Phylogenetic and cytological studies in the Ranunculaceae..
Trans. Amer. Phil. Soc. Il, 31.
G umin ski S. 1960. Biocbemiczne aspekty ewoluoji roslin. Wiadom. bot., 4, 2.
(на польск. яз.).
Gumppenberg О. v. 1924. Beitrage zur Entwicklungsgeschichte der Blumenblat-
ter mil besonderer Ber.iicksichtigung der Nervatur. Bet. Archiv, VII.
Gundersen A. 1939. Flower buds and phylogeny of dicotyledons. Bull. Torrey
Bot. Club, 66.
Gundersen A. 1950. Families of dicotyledons. Waltham, M-ass.. U .S. A., Chronica
Botanica (A new series of plant science books, vol. 25).
Gundersen A. 1954. World families for angiosperms. Bull. Torrey Bot. Club., В, 1, 3.
Gundersen A. and Hastings G. T. 1944. Interdependence in plant and ani-
mal evolution- Scient. Monthly, 59.
Gusule.ac M. 1939. Der genetische Standpu.nkt in der Taxonomie der Friichte. Bu-
letinui Faculataati de Stiinte din Cernaaut, 12.
Gusuleac M. 1939. Zur Prazisierung der Nomenklatur der Friichte und der Prin-
zipien. ernes natiirlichen Fruchtsystems. Bull. Faculataati de Stiinte din Cernaaut,
12. '
Gusuleac M. 1949. Consideratiuni asupra drirectiei evolutiei la Spermatophyta.
(Plante cu flori). Natura (Romin), 2.
Gusuleac M., Anghel Gh„ Tarnavschi I., Stefureac Tr. I960. Caile
evolutiei in regnul vegetal. Darvinismul si probl. evolutiei in biol. Din materiale-
le consfatuirilor din cadrul Acad. R. P. R. Bukuresti. (на рум. яз.) (Пути эво-
люции в растительном мире).
Guttenberg Н. von. 1960. Griindzuge der. Histogenese hoheren Pflanzen. I. Die
Angiospermen. Berlin.
Guttenberg H. von und Mitarbeitern. 1957. Embryologische und histoge-
netische Untersuchungen an Monokotyledonen. Bot- Studien, 7.
Haan H. R. M. de 1920. Contribution to the knowledge of the morphological value-
and phylogeny of ovule and its integuments, Rev. Trav. Bot. Neerl., 17.
H a g e г u p C. 1934—36. Zur Abstammung einiger Angiospermen durch Gnetales und,
Coniferales. Kgl. Danske Vidensk. Seisk. Biol. Meddel, 11, (4). 13 (6), 14 (4),
Hagerup C. 1938- On the origin of some angiosperms through the Gnetales and the*
Coniferae, III, The gynoecium of Salix cinerea. Ibid., 15, 2.
Hagerup C. 1939. On the origin of some angiosperms through the Gnetales und'
the Coniferae. IV. The gynoecium of Personatae. Ibid., 14.
Haldane J. B. S. 1932. The causes of evolution. New York, and London.
Hall B. A. 1956. Problems and methods in floral anatomy. Phytomorphology, 6.
H a 1 1 i e r H. 1901. Beitrage zur Moiphogenie der Sporophylle und des Trophophylls
in Beziehung zur Phylogenie der Kormophyten. Jahrb. d. Handb. Wissensch.
Anst, XIX.
Hal Her H. 1901. Uber die Verwandtschaftsverhaltnisse der Tubifloren und Ebenalen,
den polyphyletischen Ursprung der Sympetalen und Apetalen und die Anordnung
der Angiospermen iiberhaupt. Vorstudien zum Entwurf eines Stammbaums der
Bliitenpflanzen. Abh. d. naturwiss. Vereins in Hamburg. 16, 2.
Обзор новейших систем цветковых растений
18}
Н а 11 i е г Н. 1902. Uber die Atorphogenie, Phylogenie und den Generatienswechsel der
Achsenpflanzen. Ber. Dsch. Bot. Ges., 20, 8.
Hallier H. 1902. Beitrage zur Morphogenic der Sporophylle und des Trophophylls
in Beziehung zur Phylogenie der Kormophyten. Mitt. Bot. Inst. Hamburg. <
Hallier 1903. Vorlaufiger Entwurf des naturlichen (phylogenetischen) Systems der
Bliitenpflanzen. Bull. Herb. Boiss., II, 3.
Hallier H. 1905. Provisional scheme of the natural (phylogenetic) system of flo-
wering plants. New Phytol. 4, 7.
Hallier H. 1905. Neue Schlaglichter auf d. natiirl. System, d. Dicotyl.
Hallier 1905. Ein zweiter Entwurf des natiirlichen (phylogenetischen) Systems der
Bliitenpflanzen. (Vorlaufige Mitteilung.). Ber. Dtsch. Bot. Ges., В. XXIH.
Halier H. 1908. On the origin of angiosperms. Bot. Gaz., 45.
H a 1 1 i e r H. 1912. L'origin et le systeme phyletique des Angiospermes exposee a
1’aide de leur arbre genealogique. Archives. Neerlandaises des Sciences. Exactes
et Naturelies. Ser. Ill, B. (Sci. Nat.) T, 1, Livr. 3,
Ha 1.1 re г H. 1914. Stammbaum d. Pflanzenreichs. Reinhardt. Von Nebenfleck zum
Menschen. Bd. II—IV. Taf. IX. 3 A.
Hallier H, 1921. Zur morphologischen Deutung der Diskusgebilde in der Dikoty-
lenblfite. Rijks Herb. Leiden, 41.
Hallock F. A. 1930. The relationship of Garrya. The development of flowers and
seeds of Garrya and its bearing on the phylogenetic position of the genus. Ann-
Bot,, XLIV.
H a m m e n .L. van der. 1948. Traces of ancient dichotomies in the angiosperms. Blu-
mea, 6.
Hani M. 1935. . Vergleichende und entwickhmgsgeschichtkche Untersuchungen fiber
Morphologic und Anatomie der Griffel und Griffelaste. Beih. Bot. Zentralbl.,
54, Abt. A., H. i/2.
Hannah M. 1916. A comparative account of epigyny in certain monocotyledons and
dicotyledons. Trans. Amer. Mier. Soc., 35.
H arris T. M. 1932 (1935), The ancestry of angiosperms. C. R. 2, Congr. Strot. Carb, 1,
Haskell G. 1954. Pleiocotyly and differentiation within angiosperms. Phytomorpho-
logy, 4, 1, 2.
Hay at a B. 1921. The natural classification of plants according to the dynamic sys-
tem. leones plantarum Formos.,X.
Hayata B. 1928, The relation between the succession and participation theories and
their bearings upon the natural system. Proceed. Third. Panpacific Science Con-
gress Tokyo, 1926. vol. II.
Hayata B. 1931. Uber das «Dynamische System der Pflanzen». Ber. Dtsch. Bot.
Ges., XLIX, H. 6.
Heberer G. 1913; 1954—1957. Die Evolution der Organismen. Jena 2. Afifl, Stuttgart.
Heil H. 1950. Entwicklungsgeschichte des Pflanzenreiches. 2. Ausk. Berlin.
Heim et alt. 1952, Evolution et Phylogenie chez les vegetaux. Paris. Coll. Int, Centre
Nat, Rech, Sci., 41.
Heintze A. 1927. Cormofyternas fylogeni (Phylogenie de? Cormophyten). Lund.
Hennig W. 1949. Zur Klarung einiger Begriffe der phylogenetischen Systematik.
Forsch. und Fortschritte, Jg. 25, Hf. 11—12.
Hen slow G. 1890. On the vascular system of floral organs and their importance
in the interpretation of the morphology of flowers. Jour. Linn. Soc. Bot. Lon-
don. 28.
Henslow G. 1911. The origin of monocotyie.dons from dicotyledons, through self-
adaptation to a moist of aquatic habit, Ann. Bot., 26.
H e s 1 о p-H a г r i s о n J. 1958. The unisexual flower—a reply to criticism. Phyto-
morphology, 8; (1, 2);
184 Литература Literature
Hess R. W. 1950. Classification of wood parenchyma in dicotyledons. Trop, Woods, 96,
Hili A, W, 1908, The origin of monocotyledons, Ann. Bot., v, XXII, No, XXXVII.
H j el m qu i s t H. 1948. Studies on the floral morphology and phyogeny of the Amen-
tiferae. Lund. Bot. Notiser, Suppl., II, ].
Hocquette M. 1949. Interpretation de la fleur des angiospermes. Biologisch Jaar-
book, 15, 38.
Hoelfgen F. 1922. Sero-diagnostische Untersuchungen fiber die Verwandtschaftsver-
haltnisse innerhalb des Coktmniferen-Artes der Dikotylen. Bot, Archiv, 1.
H о r n e A. S. 1914. A contribution to the study of the evolution of the flower, with
special reference to the Hamamelidaceae, Caprifoliaceae and Cornaceae. Trans-
Linn. London. 8.
Norwood A. R. 1912. The past history of monocotyledons with remarks on their
origin. Scottish. Bol, Rev. 1,
Hruby Karel. 1955. Anatomiske znaky pri taxonomickem Lodnaceni Preslia, 27, 4.
Huber B. 1949. Zur Phylogenie des Jahrringbaues der Rinde. Svensk Bot. Tidskr.,
43, 2—3.
Hu H’s’e’n-H s u 1950. A polyphyletic system of classification of angiosperms. Science
Record (Peking), 3, 2—4.
Hu HsenHsu 1954. The evolutionciy process. In. J, Huxley, A, G, Hardy and
E. B, Ford (ed. by) «Evolution as a process».. London.
H u i s g e n F. 1873. Untersuchungen fiber die Entwicklung der Placenten. Diss. Bonn.
Hunt K. W. 1937. A study of the style and stigma with reference to the nature the
carpel. Amer. Journ. Bot., 24.
Hutchinson J. 1923, 1924, 1925, 1927. Contributions, towards a. phylogenetic
Gasification of flowering plants. I—VI, Kew. Bull., 2, 7; 2, 4; 4; 3.
Hutchinson J. 1929. The phylogeny of flowering plants. Proc. Int, Cong, Pl, Sc„ 1,
Hutchinson J. 1926, 1934, 1959. The families of flowering plants. Vol. I. Dico-
tyledons. First, ed. 1923, (repr. 1944). Vol II Monocotyledons, First ed, 1934.
(repr. 1944). London, Sec. ed, 1959. (repr. 1960),Vol, I, II, Oxford,
Huxley J. S. (ed,) 1940. (1949). The new systematics. Oxford,
Huxley J. S. 1942. Evolution. The modern synthesis. New York-London.
J a a g O, 1949. Entwicklungsgeschichte und Fortpflanzung. Fortschr, Bot., 12.
Jaeger P. 1950. Nectaires floraux et phylogenese. Annee Biol., Ill, 54.
Janchen E. 1932. Entwurf eines Stammbaumes der Blfitenpflanzen nach R. v. Wet-
tstein. Osterr, Bot. Ztschr., 81,
Janchen E. 1950. Die Herkunft der Angiospermen-Bliite und die systematische S’tel-
lung der Apetalen. Osterr. Bot. Ztschr., 97, 2.
Jeffrey C. 1962. The origin and differentiation of the archegoniate land-plants.
Bot. Not. 115, 4.
Jeffrrey E. C. 1903. The structure and development of the stem in the Pteridophyta
and. Gymnosperms. Phil, Trans, Roy, See.. 195 B.
Jeffrey E. C. 1921, Transitional herbaceous dicotyledons. Ann. Bot., 35.
J e f f г e у E. C. 1921, Physiological and morphological correlation in heibaceous an-
giosperms. Bot. Gaz., 71.
Jeffrey E. C. 1925. The origin of parenchyme in geological time. Proc- Nat. Ac.
Sci. Usa.. II.
Jeffrey E. C. 1930. The anatomy of woody plants. Chicago.
Johansen D. A. 1945. A critical survey of the present status of plant embryo-
logy. Bot. Rev., II.
Johansen D. A. 1950. Plant embryology: embryogeny of the Spermatophyta. Chro-
nica Botanica Co., Waltham, Mass.
Johansen D. A. 1951, A modern classification of the plant kingdom, Chronica, Bo-
tanica. 12, 4—6. Biolog;a, 2.
Johnson A. M. 1931, Taxonomy of the flowering plants. New York.
Обзор новейших систем цветковых растений. l8.g
Johnson А’.. А. 1953. Relationship in the Magnoliaceae as determined by the pre-
cipitin reaction. Bull. Torrey Bot. Club, 80, 4.
Joshi A .C. 1935. Criticism of Dr. Thomas’s recent hypothesis on the nature of the
angiospermous carpel. Journ. Botany. British and Foreign, 874, LXXIII.
J о s h i A. C. 1943. Origin of the tiimerous Perianth. Nature, Decembre, II.
Joshi A. C. 1947. Floral histogenesis and carpel morphology. Journ. Indian. Bot.
Soc., 26. 5—6.
Joshi A- C. 1947. The morphology of the gynaeceum. Presidental Address Bot. Sect.,
Indian Sci. Congr. 34th Session. Part II. Dehli.
Just Th. 1918. Gymnosperms and the origin of the angiosperms. Bot. Gaz., 110.
Just Th. 1952. Origine et evolution de la fleur L’Annee Biologique 28, 5—6..
К a p 1 a n J.. Beck J. 1960. TYojrozmerove rodokmene rastlin. Preslia, 32, 1 (на сло-
вацк. яз. с нем. рез.).
Karpowicz W. 1954. Pochodzenie i ewolucja kwiatu. Biol, w szkole 7, 5,
Karsten G. 1918, 1923. Zur Phylogenie der Angiospermen. Ztschr. f. d. Bot. Jahrg.
X., H. 7.. а также Lehrb. d. Bot. f. Hochsch., 16. Aufl.
Kaussmann B. 1941. Vergleichende Untersuchungen fiber die Blattnatur der
Kelch.—Blumen-und Staubblatter. Bot. Archiv., 42.
Kimura Y. 1953. The system and phylogeny of plants. Jap. Journ. Bot., 28, (на
японск. яз., с рез. на англ. яз.).
Kimura Y. 1956. Systeme et phylogenie de= Monocctyledones. Not. System., XV, 2.
Museum Nation. d’Hist. Natur.
К о h z K. 1923. Sero-diagnostische Untersuchungen uber die Verwandtschaften inner-
halb des Rosales-Artes der Dikotylen. Bot. Archiv, III.
Kostyniuk M. 1960. O. mektdrych cechach, procesu ewoluji utrudnijacych ustalenie
pokreivienstwa roznych grup systemu sv.iata roslirmego. Wiadom, bot., 4, 2.
(на польск. яз.) (О некоторых особенностях процесса эволюции, затрудняющих
установление родства разных груцп системы растительного мира).
К о u р г i а п о v a L. А., 1954. Sur la phylogenie des Monocotyledones (d’apres les
donnees palynologiques) Essais de Botanique. I. Acad, de Sciences de I’URRS.
Mosc.-Leningr.
К о z o- Poliansky В. M. 1925. Uber die Anwendung der anthanatormschen Methode
in der Systematik der Angiospermen. Bull. Soc. Natur. Moscoti, n. ser.. XXXII,
3—4.
Krause K. 1923. Eine neue. Form des natiirlichen. System. Naturw., II.
Krause! R. 1949. Einige Fragcn zur Stammesgeschichte der Pflanzen. Vortrag...
Frankfurt am Main, Kramer. (Aufsatze und Reden der Senckenbergischen natur-
forsch. Ges.).
Krausel R. 1956. Zur Geschichte der Angiospermen. Bot. Mag., Tokyo, 69.
Kugler H. 1955. Einftihrung in die Bliitenokologie. Jena.
К u m a z a w a M. 1938. On the ovular structure in the Ranunculaceae and Berberida-
ceae. Jap. Journ. Bot., 14.
К u m a z a v a M. 1938. Systematic and phylogenetic consideration of the Ranuncu-
laceae and Berberidaceae. Bot. Mag., 52.
Kuprianova L. A. 1962. Pollen morphology and phylogeny of Mcnochlamydeae.
International Conference on Palynology. The University of Arizona. Tucson
Kuprianova L. A. 1963. On a Hitherto Undescibed Family belonging to the Amen-
tiferae. Taxon, 1,
Lam H. J. 1936, Phylogenetic symbols, past and present. Acta Biotheoretica, ser. A, 11,
L a m H. J. 1938. Studies in phylogeny. Blumea, 3.
L a m H. J. 1948. A new system of the Cormophyta. Blumea, VI., 1.
I am H. J. 1948. Classification and the new morphology. Acta Biotheoretica, VIII.
jg£ Литература Literature
,Lam H. J. 1950. Stachyospory and Phyllospory as factors in the natural system of the
Cormophyta. Svensk Bot. Tidskr., 44. 4.
L a m H. J. 1950. Morfclo'gia, evolution у filogenia en el reino vegetal. Rev, JJniv,
Madrid, 8.
L a m H. J. 1952. L’evolution des plantes vasculaires. In «Evolution et Phylogenie chez
les vegetaux». Paris,
Lam H. J. 1959. Some fundamental considerations on the New Morphology. Trans, a
Proc. Bot. Soc. Edinb., 38.
Lam H. J, 1959, Taxonomy general principles and angiosperms. «Vistas in botany»,
(ed. by Turill).
Lam H. J, 1961. Reflections on angiosperm phylogeny. II, Facts and theories, Proc, Ko-
ninkl. Nederl. Akad. Wet. Amsterd.. C, 64, 3.
Lange L. 1924. Sero-diagnostische Untersuchungen fiber die Verwandtschaften, in-
nerhalb der Pfianzengruppe der Ranales. Bot. Archiv, V.
Lawrence G. H. M. 1951; 1955. Taxonomy of vascular pl,ants, New York,
Lebegue A. 1952. Recherches embryogeniques sur quclques dicotyledones dtalype-
tales. Ann. Sci. Nat. (Bot.), 13.
Leinf ell ner W. 1950. Der Bauplan des synkarpen Gynozeums. Osterr. Bot. Ztschr.,
97.
L e inf e liner W. 1951. Die U-formige Plazenta als der Plazentatior.'typus der An
giosperms. Osterr. Bot. Ztschr., 98.
Leinf ellner W. 1963. Das Perigon der Liliaceen ist staminaler Herkunft. Osterr.
Bot. Ztschr.. 110, 4.
Lemesle R- 1955. Contribution a I’etude de quelques families de decotyledon.es consi-
derees comme primitives. Phytomorphology, 5 (1).
Leonhardt R, 1949, 1951, Phylogenetisch-systematische Betrachtungen, T. Betra-
chtungen zur Systematik der Compositen. Osterr. Bot, Ztschr. B. 96. 3—4; B.
98, 1—2.
Leonhardt R. 1951. Phylogenetisch-systematische Betrachtungen. II. Gcdanken zur
systematischen Stellung bzw. Gliederung einiger Familien der Choripetalen.
Osterr. Bot. Ztschr., 98.
Leonhardt R. 1959. Phylogenetisch-systematische Betrachtungen. Osterr. Bot.
Ztschr.. Bd. 106, 5.
L e p p i к E. E. Uber die Evolution der Bliite. Sendbrief aus dem Institut fur Gartne-
rische Botanik und Pflanzenscbutz der Staatlichen Lehr-und Forschungsanstalt ftir
Gartenbau in Weihenstephan. Oktober.
L e p p i к E. E. 1955. Some veiwpoints on the origin and evolution of flowering plants.
Acta Biotheoretica, 9, II.
L e. p p 1 к E. E. 1963. Fossil evidence of floral evolution. Llcydia, 26, 2.
L e p p i к E. E. 1956, The form and function of numeral patterns in flowers, Amer.
Journ. Bot., 43, 7.
Leppik E. E. 1957. A new system for classification of flower types, Taxon, VI.
L e p p i к E. E. 1957. Evolutionary relationship between entomophilous plants and
anthophilous insects. Evolution, XI, (4).
Leppik E. E. 1960. Early evolution of flower types. Lloydia. 23, 3.
L e г о у J. F. 1951. La theorie genc-ralisee des carpelles sporcphylles et la fleur
des Juglandales. Comptes Rendiis. 20,
Leroy J. F. 1954. La fleur angiospermienne du point du vue morphologique. Huitieme.
Congres International de Botanique. Sect. Morphologie et Anatomie. Paris.
Lewis D, 1942. The evolution of sex in flowering plants. Biol. Rev. Cambridge Phil.
Soc. 17, (1).
L i H. L. 1955. Classification and phylogeny of Nymphaeaceae and allied families. Amer,.
Midi. Nat., 54, 1.
Li H. L. I960. A theory on the ancestry of angiosperms, Acta Biotheoret, 13, 4,
Обзор новейших систем цветковых растений
187
L i g n i е г О. 1908. Essai sur revolution morphologique du regne vegetal. Assoc.
Franc. 1’Avance. Sci. Compt. Rend. 37 session, Clermont-Ferrand.
L indinger L. 1910. Bemerkungen zur Phylogenie der Monokotylen. Naturwiss, Wo
chenschr., nouv. Ser., 9.
Lindley J. 1930. An introduction to the natural system of botany. London.
Lindley J. 1936. A natural system of botany. 2nd. ed. London.
L i p p m a a Th. 1937, E, V. Tartu iilikooh botaanikaaia siistemaatiiised ja tamegeo-
grafilised kogud. Tartu. (Les collections systematiques et phytogeographiques du-
jardin botaniques de I’universite estonienne a. Tartu, рез.). Acta Instituti et
Horti Botanici Universitatis. Taituensis. V, 3—4.
Lots у J. P. 1906—1908. Vorlesungen fiber Deszendenztheorien mit besonderer Bcrii-
cksichtigung der bctanischen Seite der Frage. Jena.
L о t s у J. P. 1907—1911, 1913. Vorlesungen fiber Stammesgeschichte. Jena
Lots у J. P, 1907, 1909, 1911. Vortrage uber, botanische Stammesgeschichte, Cormophy-
tazzoidiogamia. I, II, III, Jena.
Lyon H. L. 1901. The phylogeny of the Cotyledon. Postelsia, p. 57.
McLean R. C. and W. R. I v i m e y-C о о k. 1956. Textbook of theoretical botany.
2, London.
McNair. J. B. 1934. The. evolutionary status of plant families in relation to some
chemical properties. Amer. Journ. Botany, 21.
McNair, J. B. 1935. Angiosperm phylogeny on a chemical basis. Bull. Torrey Bot.
Club.. 62.
McNair J. B, 1945. Some comparisations of chemical ontogeny with chemical phylogeny
in vascular plants. Lloydia, 8, 3.
M a с r f a г 1 a n e J. M. 1933, The evolution and distribution of flowering plants.
Noll Printing Comp.
M a e k a w a F. 1952, 1956. Topo-morphological investigations on the relation between
stem and leaves and their bearing on the phylogenetic systematics of vascular
plants. Part 1. J. Fac. sci. Univ. Tokyo, Sect, 3, vol. 6. (1952), Part 2, ib, (1956),
M a e k a w a F. 1960. A new consideration on the starting place of Angiosperms. Jap.
Journ. Bot.. 35, 4 (на японск. яз с рез. на англ.-' яз.).
Maekawa F. 1960. A new attempt in phylogenetic classification oi plant kingdom.
J. Fac. Sci. Univ. Tokyo, Sect, 3, 7. 9—14.
Magdefrau K. 1954. Die Geschichte der Pflanzen. In «Evolution der Organismen».
2. Lieferung.
M a g г о u J. 1950. Sur. 1’origine des plantes vasculaires. Comptes rendus Lebdomadai-
res des Seances de l’Academie des Sciences, t. 230, 3.
Ma jumdar G. P. 1956. Carpel morphology. J. Asiatic Soc. Bengal, 22.
Majumdar G. P. 1957. The shoot of higher plants: its morphology and phylogeny.
Journ. Asiatic See. Letters a. Sci., 23. .
M a 1 1 i g s о n F. 1922. Sero-diagnostische Untersuchungen uber die Verwandtschaften
innerhalb des Centrospermen-Artes des Pflanzenreichs- Bot, Archiv, I,
Mangenot G. 1952. L’evolution de 1’ovle, du pistil et du fruit, Ann. Biol., 28,
Markgraf F. 1954. Uber neuere Pflanzensystem. Ber. d. deutsch. botan. Gesellsch,,
LXII, Generalsammlungs-Heft.
Martens P. 1951. Les prephanerogams et le probleme de la graine. La Cellule, 54.
Mason H. L. 1957. The concept of the flower and the theory of homology. Madro^nO"
14. 3.
M a 11 f e 1 d J. 1938. Das morphologische Wesen und die phylogenetische Bedeutung
der Blumenblatter. Ber, Deutsch, Bot, Ges. 5.
Alatthews J. R. 1941. Floral morphology and its bearing on the classification of
angiosperms, Trans, and Proc, Bot, Soc. Edinb., XXXIII. 2,
Mauritzon J. 1939. Die Bedeutung der embryologischen Forschung fur das natiir-
О
liche System der Pflanzen. Lunds. Univ. Arsskr., N. F. Avd. II, 35, 15,
Обзор новейших систем цветковых растений 187
L i g n i e r O. 1908. Essai sur revolution morphologique du regne vegetal. Assoc.
Franc. 1’Avance. Sci. Compt. Rend. 37 session, Clermont-Ferrand.
L indinger L. 1910. Bemerkungen zur Fhylogenie der Monokotylen. Naturwiss, Wo
chenschr., nouv. Ser., 9.
Lindley J. 1930. An introduction to the natural system of botany. London.
Lindley J. 1936. A natural system of botany. 2nd. ed. London.
Lippmaa Th. 1937, E, V. Tartu iilikook botaanikaaia siistemaatiiised ja tamegeo-
grafilised kogud. Tartu. (Les collections systematiques et phytogeographiques du-
jardin botaniques de I’universite estonienne a. Tartu, рез.). Acta Insiituti et
Horti Botanici Universitatis. Tattuensis. V, 3—4.
L о t s у J. P. 1906—1908. Vorlesungen fiber Deszendenztheorien mit besonderer Berfi-
cksichtigung der botanischen Seite der Frage. Jena.
L о t s у J. P. 1907—1911. 1913. Vorlesungen liber Stammesgeschichte. Jena
Lotsy J. P, 1907, 1909, 1911. Vortrage fiber, botanische Stammesgeschichte, Cormophy-
tazzoidiogamia. I, II, III, Jena.
Lyon H. L. 1901. The phylogeny of the Cotyledon. Postelsia, p. 57.
McLean R. C. and W. R. Ivimey-Cook. 1956. Textbook of theoretical botany.
2, London. ,
McNair. J. B. 1934. The evolutionary status of plant families in relation to some
chemical properties. Amer. Journ. Botany, 21.
M cNair, J. B. 1935. Angiosperm phylogeny on a chemical basis. Bull. Torrey Bot.
Club.. 62.
McNair J. B, 1945. Some compensations of chemical ontogeny with chemical phylogeny
in vascular plants. Lloydia, 8, 3.
M a c r f a г 1 a n e J. M. 1933, The evolution and distribution of flowering plants.
Noll Printing Comp.
M a e k a w a F. 1952, 1956. Topo-morphological investigations on the relation between
stem and leaves and their bearing on the phylogenetic systematics of vascular
plants. Part 1. J. Fac. sci. Univ. Tokyo, Sect, 3, vol. 6. (1952), Part 2, ib, (1956),
M a ek a w a F. 1960. A new consideration on the starting place of Angiosperms. Jap.
Journ. Bot.. 35, 4 (на японск. яз с рез. на англ.' яз.).
М а е k a w a F. 1960. A new attempt in phylogenetic classification oi plant kingdom.
J. Fac. Sci. Univ. Tokyo, Sect, 3, 7. 9—14.
Magdefrau K. 1954. Die Geschichte der Pflanzen. In «Evolution der Organismen».
2. Lieferung.
M a g г о u J- 1950. Sur. 1’origine des plantes vasculaires. Comptes rendus Lebdomadai-
res des Seances de I’Academie des Sciences, t. 230, 3.
Majumdar G. P. 1956. Carpel morphology. J. Asiatic Soc. Bengal, 22.
M a j u m d a r G. P. 1957. The shoot of higher plants: its morphology and phylogeny.
Journ. Asiatic See- Letters a. Sci., 23. .
M a 1 1 i g s о n F. 1922. Sero-diagnostische Untersuchungen fiber die Verwandtschaften
innerhalb des Centrospermen-Artes des Pflanzenreichs. Bot, Archiv, I,
MangenotG. 1952. L’evolution de 1’cvle, du pistil et du fruit, Ann. Biol., 28.
M а г к g r a f F. 1954. Uber neuere Pflanzensystem. Ber. d. deutsch. botan. Gesellsch,,
LXII, Generalsammlungs-Heft.
Martens P. 1951. Les prephanerogams et le probleme de la graine. La Cellule, 54.
Mason H. L. 1957. The concept of the flower and the theory of homology. Madro^na
14. 3.
M a t t f e 1 d J. 1938. Das morphologische Wesen und die phylogenetische Bedeutung
der Blumenblatter. Ber. Deutsch, Bot, Ges. 5.
Matthews J. R. 1941. Floral morphology and its bearing on the classification of
angiosperms, Trans, and Proc, Bot, Soc. Edinb., XXXIII. 2,
Mauritzon J. 1939. Die Bedeutung der enibryologischen Forschung ffir das natfir-
О
liche System der Pflanzen. Lunds. Univ. Arsskr., N. F. Avd. II, 35, 15,
188
Литература Literature
Meeuse A. D. J. 1961. The Pentoxylales and the origin of the Monocotyledons. Proc.
Koninkl. Akad.. Wet., C, 64, 4..
M e e u s e A. D. J. 1963. From ovule to ovary: a contribution to the phylogenq of the
megasporangium, Acta, Biotheoretica, vol, XVI, pers III—IV.
Melville E. 1950, A new theory of the angiosperm flower. Nature, 188.
M e r C. 1922. Anleitung zu sero-diagnostischen Untersuchungen fur Botaniker. Bot.
Archiv, I.
Metcalfe C. R. 1960, 1961. Anatomy of the monocotyledons. I. Gramineae, 2, Pal-
mae. Oxford,
Metcalfe C, R, and Chalk L. 1950, Anatomy of the dicotyledons. 1, 2, Oxford.
M e u s e 1 H. 1951. Die Bedeutung der Wuchsform ftir die Entwicklung des naiuriichen
Systems der Pflanzen. Repert. Sp. Nov, Regni Veg., 54.
Mez C, 1924. Serum Reaktionen zur Feststellung von, Verwandtschaftsverhaltnissen
im Pflanzenreich. In Abderhalden’s «Handbuch der Biologischen Arbeitsmetho-
den>, Abt. 11, Teil 1. Berlin-Wien.
M e z C. 1925, Drei Vortrage liber d. Stammesgeschichte d. Pflanzenwelt. Natur-
wissenschaft und Landwirtschaft, 4.
M e z C. 1925. Syllabus 2, ed.
M e z C. 1926. Die Bedeutung der experimentellen Systematik fur die stammesgeschi-
chtliche Forschung, Leopoldina, II.
M e z C. 1926. Die Theorien der Stammesgeschichte Schrift. Konigsberg. Geseisch. Na-
turwiss. Klasse III, 4.
Mez C. 1926. Die Theorien der Phyloge.netik. Bot. Archiv, XVI, H. 1—6.
M e z C, 1926, Die Bedeutung der Sero-Diagnostik ftir die stammesge’schichtliche For-
schung. Bot. Archiv, XVI, 1—6.
M e z C. und Gohlke K, 1914. Physiologisch-systematische Untersuchungen fiber die
Verwandtschaften der Angiospermen. Beitr. zur Biol. d. Pflanzen, XII.
M e z C. und Ziegenspeck H. 1926. Zur Theorie der Sero-Diagnostik. Bot. Archiv,
XII, 1—2.
M e z C. und Ziegenspeck H. 1926. Der Konigsberger serodiagnostische Stamm-
baum. Bot. Archiv, XIII, 5—6.
Mirov N. T. 1963. Chemistry and plant taxonomy. Lloydia. 26, 3.
M о 1 d e n к e H. N. A. 1944. A preliminary classification on the plant kingdom to
families. New York. Bot. Garden February, 11.
Moritz O. 1928, Zur Kritik der Phytoserologie, Biolog, Zentralbl., III. (См. также
рецензию C. Mez’a в Bot. Archiv., 1929, В- 26. H. 3 — 4).
N a g e 1 i C. von. 1884. Mechanisch-physiologische Theorie der Abstammungslehre.
Mfinchen u. Leipzig,
Nelson E. 1954. Gesetzmassigkeiten der Gestaltwandlung im Bliitenbereich, ihre Be-
deutung fur das Problem der Evolution. Publ, mit Unterstiitzung des Schweize-
rischen Nationalfonds zur Forderung der wissenschaftlichen Forschung, Cherne-
ux-Montreux, Nelson, XII.
Nelson E. 1957, Gesetzmassigkeiten der Gestaltwandung im Bliitenbereich, ihre Be-
deutung ffir das Problem der Evolution. Evolution, 11, 1.
Nemejc F. 1950. The natural systematic of plants in the light of the present palae-
ontological documents. Pracha. (Sbornik Narodn. Musea VI, 3).
N eme j c F. 1956. On the problem of the origin and phylogenetic development of
angiosperms. Natodniho Musea v Prase Sbornik. Acta Musei Nationalis Prague,
12. B.2 —3.
Neumayer H. 1924. Die Geschichte der Bliite usw. AbhandL Zool.-Bct. Gesellsch.,
Wien, XIV.
Newman J. V. 1936. Ontogeny of the angiospermic carpel. Nature, 137.
Newman J. V, 1949, The place of ferns and seed plants in classification. Tlhe New
Zealand Sc. Congr. Transact. Roy Soc. N. Z., 77, 7, Novem.
Обзор новейших систем цветковых растений
189-
Nitz’schke I. 1914. Beitrage zur Phylogenie der Monocotyledonen, gegriindet auf
der Embryosackentwicklung apocarper Nymphaeaceen und Helobien. Beitr. zur.
Biol. d. Pfl., XII B.
Norris Th. 1941. Torus anatomy and nectary characteristic as phylogenetic criteria
in the Rhoeadales. Amer. Journ. Bot., 28.
N ovak F. A. 1954. System Angiosperm. Preslia, 26, 4.
О z e n d a P. 1946. La nature morphologique du carpelle. Rev. Scientif., 84. 7.
Ozenda P. 1949. Recherches sur les Dicotyledones apocarpiques. Contribution a letu-
de des Angiosperms dites primitives. Publ. Lab. Ecole Super., ser. Biol., II.
Ozenda P. 1952, Remarques sur quelques interpretation de Ketamine. Phytomorpho-
logy, 2, 4.
Pankow H. 1962. Histogenetische Studien an den Bliiten eimger Phanerogamen, Jena,
Parkin J. 1914. The evolution of the inflorescence. Journ. Linn. Soc,, 42,
Parkin J, 1923. The strobilus theory of angiospermous descent. Proc, Linn, Soc, Lon-
don, 153.
Parkin J. 1924. Stomata and phylogeny. Ann. Bot,, 38,
Parkin J, 1925, The phylogenetic classification of flowering plants, Nature, 11.
Parkin J. 1926. The phylogeny of flowering plants. Nature, v. 177, no. 2944.
Parkin J. 1926, (1927). The evolution and classification of flowering plants. Botan.
Soc. and Exchange Club of the British Isles.
Parkin J. 1926. Comments on the theory of solid carpel and carpel polymorphism.
New Phytol., 25, 3.
Parkin J. 1936. The classical flower and some modern views. Proc. VI Int Bot,
Congr., Amsterdam, 1.
Parkin J. 1945, The classification of flowering plants. North-West Nat., 20.
Parkin J 1948. The stipule corisidered phylogenetically. North-West Nat., 23.
Parkin J. 1951. The protrusion of the connective beyond the anther and ots bearing
on the evolution of the stamen. Phytomorphology, 1, 1.
Parkin J. 1952. The unisexual flower: a criticism. Phytomorphology, 2, 1.
Parkin J. 1953. The durian theory, a criticism. Phytomorphology, 3.
Parkin J. 1955. A plea for a simpler gynoecium. Phytomorphology, 5, 1.
Parkin J. 1957. The unisexual flower again—a criticism. Phytomorphology, 7,1.
Periasamy K. 1962. The ruminate endosperm: development and types of ruminati-
on. Plant embryology. A symposium (Dehli).
Petersen A, E. 1953. A comparison of the secondary xylem elements of certain
species of the Amentiferae and Ranales. Bull. Torrey Bot, Club, 80, 5,
P h i 1 i p s о n W. R. 1953. The relationships of the Compositae particularly as illustra-
ted by the morphology of the inflorescence in the Rubiales a,id the Campamila-
tae. Phytomorphology, 3, 4.
P i j 1 van der L. 1955. Sarcotesta, aril, pulpa and the evolution of the angiosperm
fruit. Proceed, Kon, Neberl, Akad, Wetensch., Amsterd. C, 58, 2; 58, 3.
Pi j 1 van der L. I960-, Ecological aspects of flower evolution. 1, Phyletic evolution.
Evolution, XIV, 4.
Pijl van der L. 1961. Ecological aspects of flower evolution. II. Zoophilous flower
classes. Evolution, XV, 1.
Pilger R. 1922. Die Stamme d. Pflanzenreihs, 2 A,
Plante fol L, 1949, L’ontogenie de la fleur. Fondements d’une theorie floral nou-
velle. Paris.
P 1 a n t e f о 1 L. 1949. A new theory of phyllotaxis. Nature, 163, 4139,
Podpera J. 1938. Jak se vyvinovala myslenka progressu ve studiu fylogenetickeho
vyvoje rise rostlinne. Brno. „Zvlastni otisk z rocenky Masarykovy’mniv. v Brne-’
za r. 1936 — 1937“.
Pool R. J. 1941. Flowers and flowering plants. Sec, ed, McGraw-Hill book Company,
Jnc, New York, a. London.
19°
Литература
Literature
Popov M. G. and Stebbins G. L. 1958, Comments of the origin and phylogeny
of the angiosperms,-Popov M, G, Essay on the origin of angiosperms. — Stebbins
G, L. On the hybrid origin of the angiosperms. — Evolution, 12 2.
Porsch O. 1907. Versuch einer phylogenetischen Erklarung des Embryosackes und
doppelten Befruchtung der Angiospermen. Jena.
Porter C. L. 1959. Taxonomy cf flowering plants, San-Francisco-London.
Potonie H. 1912. Grundlinien der Pflanzen-Morphologie im Lichte der Palaeontolo-
gie, Jena,
Pot о n i e R, 1952, Paleogeographie, sociogenese des plantes et evolution Annee biol.
Ser, 3, t. 28, fasc. 7—8,
Priestley J. H. and Scott L. I, 1933, Phyllotaxis in the dicotyledons from the
standpoint of developmental anatomy. Biol. Rev. 8.
P u 1 1 e A. A. 1937. Remarks on the system of the spermatophytes. Mededeelongen van
het Botanisch Museum en Herbarium van de Rijksuniversteit te Utrecht, 43.
P u 1 1 e A. A. 1938. The classification of the spermatophytes, Chron. Bot,, 4,
P u 11 e A. A. 1952. Compendium van de Terminologie, Nomenclatuur en Systematien
der Zaadplanten. 2, Aufl, Utrecht (1, Aufl, 1938; 2. Aiifl. 1950).
Puri V, 1945. Studies in floral anatomy, III, On the origin and orientation of placen-
tal strands. Proc. Nat. Acad. Sci. (India), 15.
Puri V. 1950. Placentation in angiosperms. Proc. VII Int Bet, Cong,, Stockholm,
Puri V. 1951, The role of floral anotomy in the solution of morphological problems,
Bot. Rev., 17, 7,
Puri V. 1952. Floral anatomy and inferior ovary, Phytomorphclogy, 2, 2—3.
F u r i V, 1952. Placentation in angiosperms. Bot. Rev., 18, 9,
Puri V. 1954. On the relation between leaf and stem. Bull. Bot. Soc. Bengal, 8,
Puri V, 1959. On the structure and evolution of angiosperm carpel. Proc. 9th Internal,
bot. Congr. (Montreal),
P u r i V. 1961. The classical concept of angiosperm carpel: a reassessment. Journ. Ind.
Bot. Soc., XV, 4,
Re c hi n ger К. H. 1956. Probleme und Fortschritte der Systematik der Biiitenpflan-
zen. (B kh.: Congres international de betanique 8-me. Palis. 1954, Comptes ren-
dus...Sect. 3—6).
Raeder F. 1924. Sero-diagnostische Untersuchungen fiber strittige Verwandtschafts-
verhaltnisse innerhalb der Dikotylen, Bot. Archiv, VII,
R e m a n e A. 1952, Die Grundlagen des natiirlichen Systems der vergleichenden Ana-
tomie und der Phylogenetik, Leipzig.
Rendle A. 1893. The origin of monocotyledons plants. Natural Sc., Ill,
R e i n d 1 e A. B. 1903. The origin of the perianth in seed plants. New Phytol., 2.
R e n d 1 e A. B. 1925. The origin of the perianth in seed plants, Cambridge.
Rendle A. B.'1953; 1956, The classification of flowering plants. Vcl, I, Gymnosperms
and Monocotyledons. Vol, II, Dicotyledons. Sec. ed. Cambrige [1. ed, 1904,
1926, (1938); 2. ed. I. 1930].
Reuter K, 1926. Die Phylogenie der Parietales. Bot. Archiv, 16, 1—6,
R e z s 6 S. 1949. A zarvatermok feljcdestorteneti rendszerenek iij kiseriete. A new
sketch of the phylogenetic system of the angiospermous flowering plants. Acta
Geobot. Hung., 6, 2,
Robertson C. 1904. The structure of the flowers and the mode of pollination of
the primitive angiosperms. Bot, Gaz., 37,
Robertson C. 1917, Flowers and insects. XX. Evolution of entomophilous flowers.
Bot, Gaz,, 58.
Rothmaler W. 1948. Uber das natiirliche System der Organismen, Biol. Central-
blatt, 67.
Rothmaler W. 1951/52, Die Gymnospermen und der Unsprung der Angiospermen.
Wiss. Zeitschr. Martin-Luther-Universitat. Halle-Wittenberg. I, II, 4.
Обзор новейших систем цветковых растений 191
Ruff О. 1930, Zur Phylogenie des Columniferen-Artcs der Dikotylen. Bot. Archiv,
31, 1-2.
Rutishauser A. 1959. Pseudogcmous reproduction ano evolution. Recent advances
in botany., 1, (Toronto).
Rutishauser A. I960- Untersuchungen fiber die Evolution pseudogamer Arten. Eer.
Schweiz, bot. Ges., 70.
S a h n i B. 1925. The ontogeny of vascular plants and the theory of recapitulation.
Journ. Ind. Bot Soc., 4
Sakisaka M. and S inoto J, 1930. Critical consideration the phyogenetic system
of classification of plants. I. On the diagnostic character of Phyllum and Sybphyl-
lum. Bot. Mag. Tokyo, 44,
Salisbury E. J. 192Б Floral construction in the Helobiales Ann. Bot,, 40,
S ar gant E, 1903, A theory of the origin of monocotyledons, founded on the struc-
ture of their seedlings, Ann, Bot. XVII,
Sargant E. 1904. The evolution of monocotyledons. Bot, Gaz., 37,
Sargant E. 1908. The reconstruction of a race of primitive angiosperms. Ann. Bot,
v, XXII, no. XXXVI.
Saunders E. R. 1931. Illustrations of carpel polymorphism. VII. New Phytol,, 30,
Saunders E, R, 1936, Some morphological problems presented by the flowers in
Nymphaeaceae. Journ. Bot. bond., 74,
Saunders E. R, 1937, '939. Floral morphology, a new outlook with special referen-
ce to the gynoecium. 1—2, Cambridge,
S ch a eppi H. 1939. Vergleichend-morphologische Untersuchungen an den Staubblat-
tern der Monocotylen, Nova Acta Leopoldina, N, F., 6,
Schaffner J. H. 1905—1922, The classification of plants. Ohio Nat.. 5, 6, 9, 11,
13, 14, Ohio Journ, Sci., 22, \
Schaffner J H. 1924 Principles of plant taxonomy. Ohio Journ. Sci., 24—31,
Schaffner J. H. 1928—1933. Studies in determinate evolution, I—VII. Ohio Journ.
Sci.. 28—33.
Schaffner J. H, 1934. Phylogenetic taxonomy of plants. The Quarterly Review of
Biology, 9. 2.
Schaffner J. H. 1937. The fundamental nature of the flower. Bull, Torrey, Bot,
Club., 65,
Schaffner J. H. 1937. Spiral systems in the vascular plants. Bull. Torrey Bot.
Club, 65.
Schellenberg G. 1928. Beitrage zu einem phylogenetischen System der Bliiten-
pflanzen. Hanz Schinz.—Festschrift. Beiblatt zur Vierteljahrsschrift J. Naturforsch.
Ges. in Ziiich, 73.
Schleiden M. J. 1837. Einige Blicke auf die Entwicklungsgeschichte des vegetabi-
lischen Organismus bei den Phanerogarr.en. Arch, Bwl. Naturgeschichte, 3, 1.
Schmidt E. W. 1917, Bau und Funktion der Siebrohre der Angiospermen. Jena.
S'chnarf K. 1931. Vergleichende Embryolcgie der Angiospermen. Berlin.
Schnarf K. 1933. Die Bedeutung der embryologischen Forscbung fur das naturliche
System der Pflanzen, Biologia Generalis, 9, H, 2,
Schnarf K. 1936, Contemporary understanding of etnbryosac development among
angiosperms. Bot. Rev., 2,
Schnarf K, 1942. Archegonium und Archegontheorie. Biol. Genet., 16,
SchouteJ C 1935. On corolla aestivation and phyllotaxis of floral phylloms. Ver.
Akad. Wet., Amst. Afd. Natuurkunde, Sect. II, 34, 4.
Schrodinger R. 1931. Phylogenetische Ansichten fiber Scheiden und Stipularbin-
dungen, Verhandl, Zcol.—Bot. Ges. Wien. 69.
Schiirhoff P. N. 1919. Zur Phylogenie des angiospermen Ebmryosackes. Ber.
Dtsch, Bot, Ges., XXXVII.
Schiirhoff P. N. 1926, Zur Pleiophylie der Sympetalen auf Grund ihrer Haplonten-
“ntwicklung, Rep, Spes. Nov. Reg. Veg., Beih., 41,
I<J2 Литература Literature
— - • — —-•
Schiirhoff P.N. 1928. Uber die Entwicklung des Eiapparates der Angiospermen.
Ber. Dtsch, Bot. Ges., 46, 8.
Scott D. H. 1909. An introduction to structural botany. Part I. Flowering plants. 7sh ed.
Scott D, H, 1924. Extinct plant and problems of evolution, London.
Scott R, A. 1922. Origin of the seed plants. Aberystwythe Studies. IV Phases of
modern science.
Scott R. A., Borghoorn. E, S., and Leopold E. B.. 1960. How old are the
angiosperms? Amer. Journ. Sci., 258 A,
Senn G. 1904. Die Grundlagen des Hallierschen Angiospermensystems. Beih. Bot,
Zentralbl. Bd. XVIII, H. 1.
Seward A. C. 1933. Plant life through the ages. Cambridge, (В русском перево-
де см.: Съюорд А. Ч. 1936).
S i 11 о п Н. В. 1944. Developmental morphology of vascular plants. New Phytol., 43, 2,
Simpson G. G. 1944. The tempo and mode in evolution, New York. (В русск. пере-
воде см.: Симпсон Дж. Г. 1948).
Simpson G. G. 1953. The major features of evolution. New York.
S i n n о 11 E. W. 1916. The evolution of herbs. Science, 44.
Sinnott E, W, 1922. The significance of the «folliar ray», in the evolution of he'ba-
ceous angiosperms. Ann. Bot., 36
S i n n о t t E. W. 1946, Botany: principles and problems. 4th ed.
Sinnott E. W. and В a i 1 e v I. W. 1914, 1915. (See Bailey I. W, and Sinnott E.W,),
Small J. 1919. The origin and development of the Compositae, New Phyt, Reprint.
N, II. London. (New Phytol., 18),
Smith G. H. 1926, 1928. Vascular anatomy of Ranalian flowers. Bot, Gaz., LXXXII,
1, LXXXV, 1.
Smith R. B, 1950. The roll of mechanics in the evolution of the herbaceous plant
stem. Bot, Gaz, 111, 3.
Solereder H, 1950. Systematische Anatomie der Dicotyledonen.
Sod R. 1949. A zarvatermok fejlodestoitenei rendszerenek uj kiserlete. A new sketch
of the phylogenetic system of the angiospermous flowering plants. Acta Geobot.
Hung., VI, 2,
S о d R. 1953. Die modernen Grundsatze der Phylogenie im neuen System tier Bliiten-
pflanzen. Act. Biol. Acad, Scient, Hung., 4.
Sod — Javorka 1951. A magyar novenyvilag kezikonyve (Handbuch der ungarl-
schen Pflanzenwelt), I—II, Budapest,
Sou ege s R. 1939, Embryogenie et classification «Actualites scient. et industr.», 781.,
Exposes d’embryol. et physiol, vegetales, VIII,
Soueges R. 1954. L’origine du cone vegetatif de la tige et la question de la «ter-
minalite» du cotyledon des Monocotyledones. Ann. Sci, Nat, (Bot.), 15.
S p о г n e K. R. 1948. Correlation and classification in dicotyledons. Proc. Linn. Soc.,
London, 160th session.
S p о r n e K. R. 1949. A new approach to the problem of the primitive Hower, New
Phytol. 48, 2.
S p о r n e K- R, 1954. Statistics and the evolution of dicotyledons. Evolution, 8.
S p о г n e К R- 1954. A note on nuclear endosperm as a primitive character among
dicotyledons. Phytomorphology. 4, 3, 4.
S p о г n e K. R. 1956. The phylogenetic classification of the angiosperms. Biol. Rev.
Cambridge philos. Soc., 31.
Sporne K, R- 1959. On the phylogenetic classification of plants. Amer. Journ.
Bot., 46, 5.
Sprague T. A. 1925. The classification of dicotyledons. I. Evolutionary progression.
Journ. Bot., 63.
Обзор новейших систем цветковых растений. ] 93
Sprague Т. А. 1940, 1945. Taxonomic botany, with special reference to the angio-
sperms. In J- Huxley (ed. by), New Systematics.
Sprague T. A. (1959), 1960, The evolution of the flower. Proc. C. N. F. C„ 32, 3,
Sprotte K. 1940. Untersuchungen fiber Wachstum und Nervatur der Fruchtblatte.
Bot. Archiv, 40.
Starr K. 1937. A critique of plant serology. The Quarterly Review of Biology. vol.
12, no. no. 1, 2. a. 3: (См. также рецензию Ehrle в Bot. Archiv, 1938, B. 39, H. 3.).
Stauffer H. U. 1959. Was ist eine Bliite? Betrachtungen fiber das Wesen und die
organisation der Bliiten. Ein Beitrag zur Phylogenie der Angiospermen. Viertel-
jahrsschr. Naturforsch. Ges. Ziirich, 104, Schlussheft.
Stebbins G. L. 1950. Variation and evolution in plants. Columbia biol. Ser., 16.
Columbia Univ. Press.
Stebbins G. L, 1951. Natural selection and the differentiation of angiosperm fa-
milies. Evolution, 5, 4.
Stebbins G. L. 1953. Plant phylogeny and evolution. Evolution, 7.
Stebbins G. L. 1958. On the hybrid origin of the angiosperms. Evolution, 12, 2.
S t e r n W. I. 1954. Comparative anatomy of xylem and phylogeny of Lauraceae. Trop.
Woods, 100.
St 0 Iley E. 1925. Zur Kritik der Kcnigsberger. Serodiagnostik. Jahresber: d- Nieder-
sachs Geol, Verein., XIX.
SuessenguthK. 1921. Beitrage zur Kenntnis des systematischen Anschlusses
der Monocotylen. Beih. Bot. Zentralbl., XXXVIII, Abt. II.
S u esse n gu th K- und Merxmiiller H. 1952. Uber die Herkunft der Angio-
spermen. Phyton, 4, f. 1—3.
S w a m у B. G. L. and Padmanabhan D. 1962, (1963). A reconnaissance of angio-
sperm embryogenesis. Journ. Indian. Bot, Soc., 41, 3.
S z a f e r W. 1959. Rodowody drzew W swietle ewolucji. Szczecin. (Szczecinskie low-
wo naukowe. Wydzial nauk przyrodniczorolniczych. T. I. Zesz. 2. Pubis. Inst,
bot Univ. Jagellonicae Cracoviensis et Acad. Sci. Polonae 20, 12.
Takhtajan A. 1953. Phylogenetic principles of the system of higher plants. Bot.
Rev., 19, 1.
Takhtajan A. L. 1954. Quelques probleines de la morphologie evolutive des angio-
spermes. Essais de Botanique, II. Acad de Sciences de 1’URSS. Mosc.-Leningr.
Takhtajan A. L. 1957. Pochodzenie okrytonasiennych. Acta. Soc, Bot. Polon,'26, ly
Takhtajan A. L. 1958. Origins of angiospermous plants. Ed. by G. L. Stebbins.
Stechert-Hafner Service Agency. New York.
Takhtajan A. L. 1959. Die Evolution der Angiospermen. Jtna.
Takhtajan A. L. 1964. The taxa of the higher plants above the rank of order..
Taxon, XIII, 5.
T a n s I e у A. G. 1920. The Evolution of plants. New Phytol., 19.
I h 0 m a s E. N. 1907. A theory of the double leaf-trace founded on seedling structure..
New Phytol., 6.
' homas H. H. 1931. The early evlution of the angiosperms. Ann. Bot., 45.
Thomas H. H. 1934. The nature and origin of the stigma. A contribution towards,
a new morphological interpretation of the angiosperm flower. New Phytol.,
XXXIII, 3.
I ho.m a s H. H. 1936. Paleobotany and the origin of the angiosperms. Bot. Rev., II, 8..
Thomas H. H. 1952. The old morphology and the new. Proc, Linn, Soc, Bot. Londo n. 145,
Thomas H. H. 1947. The history of plant form. The Advancement of Science, 4. 15.
Thompson M. L. (J. McL.). 1927. Origin cf the seed habit in plants. Trans. Roy..
Sbc„ Canada,^3. /
Thompson M. L. 1934. Studies in advancing sterility, VII. The state of flowering:
known as angiospermy... Univ. Liverpool, Hartley Bot. Lab. Pub., 72,
Thompson. W; P. ,1918- Independent evolution of vessels in Gnetales and angios-
perms. Bot. Gaz., 65.
13. А. А. Гроссгейм и M. Ф. Сахокиа,
I94
Литература Literature
Thompson W. P. 1923. The relationships of the different types of angiospermic
vessels. Ann. Bot, 37.
Thorne R. F. 1958. Some guiding principles of angiosperm phylogeny. Brittona, 10, 2.
Thorne R. F. 1963. Some problems and guiding principles of angiosperm phyloge-
ny. Amer. Naturalist. 97, 896.
T i p p о О. 1942. A modern classification of the plant kingdom. Chroti. Bot., 7.
T i p p о О. 1946. The role of wood anatomy in phylogeny. The Amer. Midland Natura-
list., 36, 2.
I i s c h 1 e г G. 1928. Ober die Verwendung der. Chromosomenzahl fur phylogene-
tische Probleme bei den Angiospermen. Biol. Zentralbl., 48.
Trapp A. 1956. Zur Morphologie und Entwicklungsgeschichte der Staubblatter sym-
petaier Bliiten. Bot. Studien, Heft 5.
Troll W. 1927. Zur Frage der Herkunft der Blumenblatter. Flora, N. F„ 22 (d. ganz.
Reihe 122. B.).
Troll W. 1928. Zur Auffassung des parakarpen Gynaeceums und des coenokarpen
Gynaceums tiberhaupt. Planta, 6.
Troll W. 1928. Organisation und Gestalt im Bereich der Bliite. Monogr. wiss. Bot.,
I. Berlin.
Tro 1 1 W. 1931, 1932, 1933. Beitrage zur Morphologie des Gynaeceums und des coen-
carpen Gynaeceums. I. Ober das Gynaeceum der Hydrocharitazeen.
Troll W. 1935, 1937, 1938, 1939. Vergleichende Morphologie der hdheren Pflanzen.
Berlin.
Troll W. 1939. Die morphologische Natur der Karpelle. Chron. Bot,, 3.
II. Ober das Gynaeceum von Limnocharis. III. Ober das Gynaeceum von Nigella und
emiger anderer Helleboreen. IV. Ober das Gynaeceum der Nymphaeaceen. Planta, 14,
17, u. 21.
Tur rill W. B. 1942. Taxonowy and phylogeny. Bot. Rev., VII.
Tyler A. A. 1897. The nature and origin of stipules. Ann. N, Y. Acad, Sci., 10.
II nr uh M., 1939. Die morphologische Bedeutung des Karpells. Ein kritischer Sammel-
bericht. Beitr. z. Biol, der Pfl., 26.
Unruh M. 1941. Blattnervatur und Karpellnervatur. Kleiner Beitrag zur morphologis-
chen Deutung des Karpells. Beitr. z. Biol, der, Pfl., 27-
V a g a A. 1946. Eostaimede siisteemi kujunimis kaik ja proeguseed probleemid. Eesti
NSV Tartu rukliku Olikooli, Botanika—alased food. I, (на эст. яз. с русск
рез.). Acta Botanica Univ. Tartuensis. (Вага A. 1946. Развитие системы споро-
вых растений и насущные проблемы их систематики. Бог. Тр. Тартуского
Гос. Унив., 1).
Vaga А. 1960. Korgemate taimede jagamisest hoimkondadeks (О разделении выс-
ших растений на типы). Tartu Olikooli toimetised. Уч. Зап. Тартуск. Унив., 93
(нд эст. яз., рез. русск. н нем.).
Van der Hammen L. См. H a mme n L. van der.
A an der W у k B. W. and J. E. C a n r i g h t. 1956. The anatomy and relationships
of the Annonaceae. Trop. Woods. 104.
V an Tieghem Ph. 1871. Recherches sur la structure du pistil et sur 1’anatomie com-
paree de la fleur. Mem. Savants etrangers a 1’Inst., II, 21.
Vogel S. 1954. Bliitenbiologische Typer, als Elemente der Sippengliederung. Bot. Stu-
dien, Heft 1.
\ uiliemin P. 1923. Classification des Monocotyledones, Comptes Rend. Acad. Sci.,
166.
Walter H. 1952. Grundlagen des Pflanzensystems. Stuttgart.
Walter L. 1963. Das Perigon der Liliaceen ist staminaler Herkunft. Osterr. Bot.
Ztschr., 110, 4.
Walters S. M. 1961. The shaping of angiosperm taxonomy. New Phytol., 60, 1.
V' a 1 t о n J. 1952. L’evolution de teguments et de la protectrice du sporange. Annee
Biol., Ser. 3, 26
Обзор новейших систем цветковых растений. 195
Walton J. 1953. Evolution of the ovule in Pteridosperms. Nature, P2, 4375.
W a r d 1 a w C. W. 1952. Phylogeny and morphogenesis. London.
W a r m i n g-M 6 b i u s 1929. Systematische Botanik. Berlin.
Wee vers Th. 1943. The relation between taxonomy and chemistry of plants. Blu-
mea, 5.
Wernhani H. F. 1913. Floral evolution. New Phytol., 10—IL
Werth E. 1949. Zum Begriff der Hummelblumen. Nat. Gesell. Augsburg, 2.
Wetmore R. H. 1943. Leaf-stem relationships in the vascular plants. Torreya, 43.
Wettstein R. 1907. Die Entwicklung der Blute der angiospermen Pflanzen aus
derjenigen der Gymnospermen. Das Wissen ftir Alle, Jahrg., 45.
Wettstein R. 1909. Was verstehen wir unter monophyetischer und polyphyletischer
Abstammung. Verhandl. d. Zoologisch-Botanischen Gesellschaft (in Wien), 59
(Diskussionsabend am 18 November 1908, veranstaliet von O. Abel.).
Wettstein R. 1910- Der gegenwartige Stand der Kenntnisse von der Staminesges-
chichte der Pflanzen. «Himmel und Erde», 23, 2. Berlin.
Wettstein R. 1914. Phylogenie der Pflanzen. In «Die Kuiiur der Gegenwart», III'.
Teil, IV. Abt., 4 Bd.
Wettstein R. 1925. Die Bedeutung der sero-diagnostischen Methode fiir die phy-
logenetisch-systematische Forschung. Zeitschr. f. indukt. Abslammungs-und Ve-
rerbungslehre, 36, 3/4.
Wettstein R. 1931. Probleme der botanischen Phylogenie. Foischungen und For-
tschritte, 7, 1.
Wettstein R. 1935; 1962. Handbuch der systematischen Botanik. II. В, (I. B. 1933),
Aufl. Leipzig u. Wien. Repr. 1962 by A. Asher Co. Amsterdam. (1, Aufl.
1901; 2. Aufl. 1911; 3. Aufl, 1924.). [„Руководство по систематике растений*.
Русский перевод этого труда (со 2-го немецкого издания) опубликован в 1903—
1912 г.г.: т. 1, Низшие растения — в 1903 г.; т. И, ч. 1, Мхи — Открытосемен-
ные — в 1905 г. и т. II, ч. II, Скрытосеменные — в 1912 г.].
Whitehouse Н. L. К. 1950. Multiple-allelomorph incompatibility of pollen and
style in the evolution of the angiosperms. Ann. Bot., XIV, 54.
Whitehouse H. L. K. 1960. Origin of angiosperms. Nature, 188,
Wieland G. R., 1918; The origin of dicotyls. Science, 48.
Wieland G. R. 1929. Antiquity of angiosperms. Proc. Int. Congr. of Plant Sc.
Wilson C, L. 1937. The phylogeny of the stamen. Amer. Journ. Bot. XXIV.
Wilson C. L. 1941. The evolution of the stamen. Chron. Bot., 6, 11.
Wilson C. L. 1942. The telome theory and the origin of the stamen. Amer. Journ., Bot.
29, '
W' i 1 s о n C. L. 1953. The telome theory. Bot. Rev., 19, 7.
Winkler H. 1939. Versuch ernes «natiirlichen» Systems der Fruchte. Beitr. z. Biol,
d. Pfl., 26, 2.
Winkler FI. 1940. Zur Einigung und Weiterfhhrung in der Frage des Fruchtsystenis.
Beitr. z, Biol. Pfl., 27, 1.
Winkler H. 1941. Verstehen wir das Gyndzeum der Angiospermen schon? Beitr. z.
Biol, d. Pfl., 27. , '
W i t h n e r C. L 1936. The evolution of pollen grains.-Bot. ReV., 2.
Worsdell W. C. 1903. The'origin of the perianth of flowers. New Phytol., 2.
W 0 r s d e 11 W. C. 1904. The structure and morphology of the «ovule», Ann, Bot,, 18,
Worsdell W. C. 1907. The origin of the flower. Sc. Progr., II.
W о r s e c k E. 1922. Sero-diagnostische Untersuhungen fiber die Verwandtschafts
verhaltnisse der Monokotyledonen. Bot. Archiv, II.
Wright S. 1949. Population structure in evolution. Proc. Ainer. Phil. Soc., 93.
Wunderlich R. 1959. Zur Frage der Phylogenie der Endospermtypen bei den An-
giospermen. Osterr. Bot. Ztschr. 106, 3—4.
Ziegenspeck H. 1925. Die Stel?rtheorie und der serologische Stammbaum. Bot
Archiv, X.
Литература Literature
Т9б
Ziegenspeck H. 1925. Der serologische Stammbaum des Pflenzenteichs und Pny-
topalaeontologie. Bot. Archiv, IX, 1—.3.
Ziegenspeck H. 1927. Die systematische Bedeutung der Haptoidrgenerationen ver-
glichen mit den Ergebnissen der Serodiagnostik. Bot. Archiv, XVII.
Ziegenspeck H. 1927. Die systematische Beaeutung der Haploidgenerationen ver-
glichen mit den Ergebnissen der Sero-Diagnostik. Bot. Archiv, XVII, 1—2.
Ziegenspeck H. 1938. Die Phylogenie der Glumiflorae. Bot. Archiv., 39.
Zimmermann W. 1930; 1959. Die Phylogenie der Pflanzen. 1 und 2 Aufl. Jena.
Stuttgart.
Zimmermann W. 1930. Der Baum in seinem phylogenetischen 'Werden. Ber. de-
utsch. Bot. Gesellschaft, XLVIII.
Zimmermann W. 1931. Arbeitswejse der botan. Phylogenetik. Abderhalden’s Han-
dbuch der bfolog. Arbeitsmethod., Abt. IX, Teii. 3, Heft 6.
Zimmermann W. 1935. Die Phylogenie der Angiospermen BEittenstande. Beih. Bot.
Centr., 53, Abt. A. I,
Zimmermann W. 1935. Die Phylogehetische Herkunft der gegensiandigen Blatt-
stellung. Jahrb. Wiss. Bot., 81.
Zimmermann W. 1938. Die Telcmtheorie. Biologie. 7. Jahrg.
Zimmermann E. 1938. «Phylogenie» in Verdoorn’s «Manual of Pteridology».
- Martinus Nijhoff, Netherlands-
Zimmermann W. 1943. Die Methoden der Phylogenetik. In: Heberer. Die Evolu-
. tion der Organismen. Jena.
Zimmermann W. 1944. Griindsatze der Phylogenetik (Evolutionslehre), Tubingen.
Zimmermann W. 1945. Hauptergebnisse der «Telomtheorie»-. Tiibingen.
Zimmermann W. 1948. Phylogenie der Pflanzen. Naturforsehung und Medizin
in Deutschland 1939—46.
Zim-mermann W. 1948. Stammesgeschichtliche Entwicklung unserer Heilpflanzen.
Siidd. Apotheker-lZeitung, 14.
Zimmermann \V. 1949. Die Geschichte der Pflanzen. Stuttgart.
Zimmermann W. 1952. Main results of the «telome theory». The Paleobotanist, 1.
(Birbal Sahni memorial wolume).
Zimmermann W. 1953. Evolution. Die Geschichte ihrer Probleme lind Erkenn-
tnisse. Munchen.
Zimmermann W. 1955. Methoden der Phylogenetik. Heberer’s «Evolution der Or-
ganismen». 2 Aufl. Stuttgart.
Zimmermann W. 1955. Phylogenie des Archegoniaten — Gener ationswechsels.
Feddes Report. Spec. Nov. Regni Veget., 58, 1—3.
Zimmermann W. 1956. On the phylogeny of the steie. Bot, Mag., Tokyo, vol.
69, N 820- 821.
Zimmermann W. 1956. Mikro—und Makroevolution.
Zimmermann W. <957. Phylogenie der Bliite. Phyton, 7, 1—3.
Zimmermann W. 1958. Wie der Generationswechsel der Archegoniaten und der
Phanerogamen entstanden ist. Planta, 51, 4.
Zimmermann W. 1959. Geschichte und Methode der Evolutionsforschung. Arbeit-
stagung zu Fragen der Evolution. (20—24. X ). Jena,
Юй 4. X. 1954. К вопросу морфологической эволюции древесины и филогении
растений. Acta Botanica Sinica, 3, 2.
Дополнение — Supplement I
Ar be r A. 1918—1919. The «law of loss» in evolution. Proc. Linn. Soc. London, 131,
Bailey I. W. 1951. The use and abuse of anatomical data in the study of phylogeny
and classification. Phytomorph., 1.
Bancroft N. 1914. A review of literature concerning the evolution of monoco-
tyledons. New Phytol., 13.
Обзор новейших систем цветковых растений.
Buchholz J. Т. 1919. Studies concerning the evolutionary status of polycotyledony.
Amer, Journ. Bot., 6.
D a t i n C. L. 1909. La morphologie de la germination avec la phylogente. Rev.
Gen. Bot., 21.
Daumann E. 1931.. Zur Phylogenie des Diskusbtldungen: Beitrage zur Kenntnis
der Nektarien, Beih. Bot. Centalbi., 48.
Dor f E. 1955. Plants and the geological time scale. Geel. Soc. Amer.Spec, Paper, 62,
Douglas G. E. 1957, The inferior ovary. Bot. Rev., 23.
Earns A. J. 1961. Morphology of the angiosperms. New York, Toronto, London.
E m b er g er L. 1964. L’inflotescence vue sous Tangle uynamique et phylogenetiqfte.
Bull, de la Soc. Bot. de France. Colloque de morphologie.
Esau K. 1943. Origin and development of the primary vascular tissues. Bot. Rev., 9.
Fagerlind F. 1944. Die tetrasporische Angiospermen — Embrvosack find dessen
Bedeutung fflr das Verstandnis der Entwicklungsmechanik und Phyloge-
nie der Embryosacks. Arkiv Bot., 31 A.
Henslow G. 1893. A theoretical origin of endogens from exogens, through self-adap-
tation to an aquatic habitat. Journ. Linn. Soc. Bot. London, 29.
H i r m e г M. 1935. Die Pteridospermae, insbesondere die Caytoniales und die Enlwi-
cklung der Angiospermae. Proc. Zcsda Int. Bot. Congr. Amsterdam, II.
J о s h i A. C. 1938. Systematic distribution of the Fritillaria — type of embryosac and
the mono-or polyphyletic origin of the angiosperms. Chron. Bot., 4.
Joshi A. C. 1947. Carpel histogenesis and carpel morphology. Journ. Indian Bot, Soc.,
26.
Just T. 1957. Fifty years of paleobotany- Am. Journ. Bot., 44.
К о z o-P о 1 i a n s k у В. M. 1936. On some «third» conceptions in floral morphology.
New Phytol., 35, 5.
L i H. L. 1957. The evolutionary significance of the endosperm and its bearing on the
origin of the angiospeffls. Journ. Wash. Acad. Sci., 47.
Lignier O. 1908. Le fruit des Bennettitees ct 1’ascendance des angiospermes. Mem.
Bot.-Soc, France, 13.
Manum S v c i ft. 1963- Le dekkroete blorr.sterpiantenes oppririnelse. Biyttia, 21, 2,
Nilsson H, 1941. Die Horaologie des angiospermen Embryosackes. Bot. Not.
Parkin J. 1922. The strobilus theory. Proc. Linn. Soc. London, 135.
Parkin J. 1933, The classical carpel and recent attacks. Rpt. Bot, Exchange Club,
Abroath, 1933-
Pchl F. 1928. Der einfaltige Pollen, seine Verbreitung und phylogenetische Bedeu-
tung. Beih, Bot. Centralbl., 45.
P о p h a m R. A. 1952. Developmental plant anatomy Columbus. Ohio,
Pers ch O. 1913. Die Abstammung der Monokotylen und die Bliitennektarieft Ber-
Dtsch. Bot. Qes., 31.
Schwarze C. 1914 Vergleichende entwicklungsgeschichtiiche und histclcgische Un-
tersuchungen reduzierter Staubblatter. Jahr. Wi^s. Bot., 54,
Sifton H. B. 1944. Developmental morphology of vascular plants. New Phytot., 43.
Solereder H. und Meyer F. J. 1928; 1929; 1930. Systematisthe Anatomic der
Monokotyledonen. Berlin.
Soo R. 1961. The present aspect of the evolutionary history of Telomophyta. (Some
ideas concerning the origin of mosses, a new system of Gytnnospermae and a
modern phytogenetical tree of Angiospemae). Ann. Univ. Scient, Budapest. Sec.
biol., 4,
Stadtler G. 1925. Uber Reduktionserscheinungen im Bau der Antherenwand von
AngiOspermen-Bliiteft. Flora, N. F., 116.
Suessenguth K. 1939. Neue Ziele der Botanik. Miihchen-Berlin.
Suessenguth K- i960. The flora of Australia as an index of the antiquity of an-
giosperms. Pacific Sci., 4.
T i p p о О. 1942. A modern classification of the plant kingdom. Chron. Bot., 7.
Ijj8 Литература Literature.
Wodehouse R. P. 1928. The phylogenetic value of pollen-grain characters. Ann. Bot.,
42.
Wodehouse R. P. 1936. Evolution of pollen grains. Bot. Rev., 2,
Zimmermann W 1957. Geschichte der Pflanzen. Stuttgart.
1940. A discussion on phylogeny and taxonomy. Arranged at the request of the Asso-
ciation for the study of systematics. Proc. Linnean soc. London, 152, sesson, p.. 3.
1952. Colloque international sur 1’evolution et la phylogenie chez les vegetaux organise
par le Centre national de la recherche scientifique au Museum national 1’histoire
naturelie du 15 au 20 mai 1952 (Paris). Annee biol., Ser. 3, t. 28. fasc. 5—6,
1952, Evolution et phylogenie chez les vegetaux, Paris. (Collog. internal. Centre nat.
rech. scient., 41).
1963. Symposium on principles and methodes of phylogeny. Philadelphia, pa 27 th Dec.
1962. Amer. Naturalist, 97, 986.
Дополнение—Supplement II
Andrews 1947. Ancient plants and the world they lived. Othaca.
Barkley. 1939. Keys to the phyla of organisms. Missoula, Mont.
Barkley. 1948. Lista de los Ordenes у Familias de las Anthophyta con ejemblos ge-
nericos seleccionados. Revist. Fak. Nac. Agronomia, Colombia, 8.
Barkley. 1949. Un esbozo de clasificacion de los organismos. Revist. Fac. Nac. Agro-
nomia de Medellin. Colombia, 10, 34.
'Benson, 1957. Plant classification. Boston.
Benson. 1962. Plant taxonomy. Methods a. principles. New York-
Breme'kamp С. E. B. 1959. Some comments on the doctrine of evolution. Proc. Kon,
Ned. Acad. Wetens. Amsterdam. C. 62.
Chester K- S. 1937. A critique of plant seroiogy. Quart. Rev. Biol., 12.
Clausen. 1951. Stages in the evolution of plant species. Ithaka. N. Y.
Cock er 11 T. D. A. 1935. The origin of the higher flower plants. Science, 11,-81.
Copeland H. F. 1957- Forecast of a system of the dicotyledons. Madrono 14.
CronquistA. 1965. The status of the general system of classification of flowering
plants. Ann. of the Miss. Bot. Garden, LIT, 3.
Davis and Heywood. 1963. Principles of angiosperm taxonomy; Edinburgh—London.
Erdtman G. 1948. Did dicotyt, plants evist in early jurassic time? Geol. Forening, 1.
Faegri 1937. Some fundamental problems of taxonomy and phylogenetics. Bot. Rev., 3.
Hennig W. 1949. Grundziige einer Theorie der phylogenetischen Systematik. Berlin.
H e s 1 о p-H a г г i s о n, 1955. New concepts in flowering plant taxonomy. London.
! L a w r e n с e С. H. M. 1955. An introduction to plant taxonomy. New York.
Mann. 1952, Systematics of flowering plants. New York—London.
. M а г e s q u e 11 e H. J. 1961 (1962). Sur la filiation des inflorescences. Bull. Soc, Bot.
France, Mem., 117—119.
.Melville E. 1962—63. New theory of angiosperm flower. I—II. Kew. Bull. 16; 17.
Nakai. 1943. Ordines, familiae etc.
Rodriguez 1950. A graphic representation of Bessey’s taxonomic system. Madrono 10.
Sch w arz. 1955. Entstehung der Bliite und system. Stellung der Phanerogamen.’Mitt.
Thur. Bot. Ges., 1.
S w a m у В. G. L. and Parameswaran N. 1963. The helobiae endosperm. Biol.
Rev, Cambridge Philos- Soc., 38. 1.
Thompson J. McL. 1934. Comments on recent statements regarding the nature ori-
gin of the angiospermic stigma. New Phytolog., 33.
Thomson. 1953. Zur Entstehung und Ausbreitung der Angiospermen im Mesophyticum.
Palaont. Zeitschr., 27.
T u r r i 1 1. 1950. Modern trends in the classification of plants. Advanc. of Sci., 26.
Zimmermann W. 1963. Gibt es ausser dem phylogen. System «naturliche». Systeme
der Organismen. Biol. Zebl., 82.
Приложение IV
Supplement IV
Графические схемы филогенетических систем
44 таблицы
Graphical schemes of the phylogenetic systems
44 tables
Hans Hallier,. Arbre genealogique des Angiospermes. I. Proterogenes.
С. Podophylleae
(Paeonieae)
----3. Dilleniaceae
b. Lardizabaleae
23. Amarantaceae 22. Chenopodiaceae 27. Polygonaceae 26. Nyctaginaceae
20. Portulacaceae — 25. PJumbaginaceae 24. Caryophyllaceae
19. Cactaceae 21. Didieraceae
III. Caryophyllinae — 18. Aizoaceae
17. Phytolaccaceae 16. Crassulaceae
10. Halorrbagidaceae 11. Tbelygonaceae 12. Cephalotaceae
9. Podostemaceae • 8. Circaeastraceae II. Nepentbales — 13. Nepenthaceae 14. Parnassiaceae — b. Sarracenieae
6. Nymphaeaceae — 5. Papaveraceae 4. Ranunculactae 7. Ceratophyllaceae 15. Droseraceae a. Pa> nassteae
V. Aristolochiales — 29. Aristolochiaceae - - 3’0. Kafflesiaceae — 31. Hydnoraceae <
2. Menispermaceae 32. Balanophoraceae
IV. Piperinae — 28. Piperaceae
d. berberidopsideae
e. Berberideae
Табл. 1 (фотокопия). Table 1 (photocopy).
Галлир Г. 1912. Генеалогическое древо покрытосемянных. 1. Р г 0-
terogenae. („Происхождение и филетическая система покрыто-
семянных. на фр. яз.)
Hallier Н. 1912. Genealogical tree of Angiosperms. I. Prote-
r о g e n a e. („L'origine et le systeme phyletique des Angiospermes...")
Hans IIaxliek. Arbre genealogique des Angiospermes. 11. Anonophylee
c. Podophylleae
(Pae onia}
Q
X. Columniferae —
157. Bombacaceae
158. Malvaceae — (Neuradeae)
53. Sterculiaceae — (Triplochiton)
i (Bixa)
об. Tiliaceae 1 ' , ,
1 (Cochlospermnm)
55. Elaeocarpaceae
54. Dipterocarpaceae
«3
jprobei'berideae
b£
IX. LAmpanalinae— g
о
CQ
Hydropltylleae
Lennoeae
? 47. Loasaceae
? 48. Campanulaceae —
49. Goodeniaceae
50. Candolleaceae
51. Calyceraceae
52. Conipositae
cd
33. Anonaceae
34. Myristicaeeae
36. Canellaceae
3j. Magnoliaceae — 37. Lactoridaceae
43. Coriariaceae
VI. Anonales
VII. Hamamelinae— g —
tn
I VIII. Uuibelliflorae —
44.
45.
Cornaceae
Umbeiliferae
(Araliaceae)
a —-
Я
38. Calyeanthaceae
39. Monimiaceae — 40. Chloranthacea.
41. Lauraeeae
*
b. La>dizabate<,e
Табл. 2 (фотокопия). Table 2 (photocopy).
Га л ли p Г. 1912. Генеалогическое древо покрытосемянных. II. Ano-
nophyla в. („Происхождение и филетическая система покрытосемян-
ных..?1; на фр. яз.)
Hallier Н. 1912. Genealogical tree of Angiosperms. IL Ano-
n о p h у 1 a e. („L‘origine et le systeme phyletique des Angiosperms../)
Hans Halubr. Arbre genealogique des Angiospermes. HI. Rhodophyles.
la. Proberberideae
e
Podophyll* (Paeonia] XVI. Rosales — 79 XIV. Terebinthinai
у lae 3. Dilleniacea' в — 70. Rutaceae —
.ardizabaleae ttt. Rhodoph' J XV. Protelnae — XIII. Aescnlinae — XII. Grnlnales — 78. Proteaceae 69 Sapindaceae 68. Meliaotbaceae 67 Leguminosae 66. Connaraceae 62 Oxalidaceae — 65. Zygopbyllaceae
XI. Crncialee — 59 Cappandace.ae —
2 XVII. Rhamnalee —| 87. Ampelidaceae 86. Rbamnaceae
83. Saxifragaceae — ? 85. Stackbousiaceae
82. Staphyleaceae
84. Bruniaceae
81. Cunoniaceae
80. Brunelliaceae
71. Cneoraceae
72. Meliaceae 75. Aceraceae a- 0'^'»^
73. Simarubaceae b. Myrtceae
74. Terebintbaceae — ? — 76. Amentaceae — c. Cori/leri»
• d. Casuaiiiteae
11. Urticaceae „ .
e. Beluleae
a Litnmmtheae
z. 64 Balsaminaceae— b. Tcopaeoleae
63. Geraniaceae —
c. Uobaininene
Pe ganuin
60 CriKif'erae
61 Resedacear
Табл. 3 (фотокопия). Table 3 (photocopy).
Г а л л и p Г. 1912. Генеалогическое древо покрытосемянных. III. Rho-
dophylae. („Происхождение и филетическая система покрытосемян-
ных.. .“ на фр. яз.).
Н а 11 i е г Н. 1912. Genealogical tree of Angiosperms. III. R h о d o-
phуlae. („L'origine et le systeme pbyldtique des Angiosperms-.").
Hans IIallieu, Arbre genealogique des Angiospermes IV Ochnigenes 1
I 147 Theophrastaceae
XXV. Primulinae —| Myrsinaceae — 149, Primulaceae
>. Lardiztibaleae .----1---. . c- Podophylleat
+ Ochnigenae 3. Dilleniaceae (Paeonia)
XXIV. Bicorues — 140. Clethraceae — a Saui-aujeae —
101. Maregraviaceae
100. Ternstroemiaceae
99. Pentaphylacaceae
98. Symplocaveae
97. Chlaenaecae
96. Rhopalwarpaceae
93. Octoknemaceae
195. Aquifoliaceae
ЧН Celastraceae (+ Uippccrateaceae)
b. Clethreae
c. Poriduleae
141. Pirolaceae
142. Ericaceae
143 Cyrillaceae
144. Empetraceae
145. Epacridattae
146. Diapensiaceae
133 Rhizophoraceae «
131. Cary< caraceae « 136. Combretaceae
135. Levythidaceae ~ 3“ 137. Lythraceae
g 138. Myrtaceae — 139. Mclastomaceae
130. Penaeaccae
XXII. Baphnales - 129 (Gonysiy/rcH Thymeheaceae— 131. Oliniaceae
132. Elaeagnaveae
i’angteae
109. Euphorbiaceae
Cusea>-ieae
( 110. Salicaceae
’ Idesteae
P'li-uijsieae — 111. Passifluraccae —
XXL Polygalinae
119. Violaceae
?120. Onagraceac
121. Vochysiaceae
122. Trigoniaceae
123. Dichapetalaceae
124. Malpighiaceae
125 Pulygalaceae
92. Quiinuceae
103. Eui ryphiacf-ae
102. Guttifcrae
Turneraceae
Malcsherbiaceae
Papayaceae
113.
114.
XX. Pepouiferae
115 Aehariaceae
?126. Kraineraceae
?12m. Salvadoracv-j'
116 Gucurbitaceae
117. Begoniaceae
11У Datiscueeae
Табл. 4 (фотокопия). Table 4 (photocopy),
Галли p Г. 1912. Генеалогическое древо покрытосемянных. IV.
Ochnigenae 1. („Происхождение и филетическая система покрыто-
семянных"; на фр. яз.)
Hallier Н. 1912. Genealogical tree of Angiosperms IV. Ochni-
genae 1. („L'origine et le systeme phyletique des Angiospermes.
Hans Hallier. Arbre genealogique des Angiospermes. V. Ocbnigenes 2
151. Olacaceae
150. Ebenaceae
157. Sapotaceae
153. Styracaceae
152. Alangiaceae
8
Icacineae— g —
©
154. Santalaceae —
156. Loraotbaceae
Opilieae
155. Myzodendraceae
И
И
158. Pittosporaceae
159. Polemoniaceae
160. Convolvulaceae ®
161. Apocynaceae (+ Asclepiadaceae) — 162. Gentianaceae « |
я §
I 174 "R.uHiAf'Pu.p
163. Loganiaceae — XXIX. Rubiacinae —j ’ p ., .. § $
° I 175. Caprifoliaceae — .£-
> °
164. Solanaceae — ? 165. Myoporaceae
1167. Gesneraceae
168. Oleaceae
169. Bignoniaceae
170. Pedaliaceae
171. Acanthaceae
172. Verbenaceae—173 Labiatae
Табл. 5 (фотокопия). Table 5 (photocopy).
Галлир Г. 1912. Генеалогическое древо покрытосемянных. У.
Ochnigenae 2. („Происхождение и филетическая система покрыто-
семянных"; на фр. яз.).
Hallier Н. 1912. Genealogical tree ef Angiosperms. V. Ochni-
genae 2. .(„L‘origine et In systeme phyletique des Angiosperms .“)
Hans Племен Arbre genealogique des Angiospermes. VI. Monocotyledones
Veratreae
4
DracnenoKi XXXVI. Heleblae—j Tofieldieae XXXV. Cyperales XXXIV. Spadlelfloraa ? 210. 211. Na 212. Al — Prioridaceae jadaceae ismaceae (4 fi'iiomctte.) 208. Juncaceae — 20'. 2)6. Araceae — 207. 205. Typhaceae 204. Pandanaceae 203. Palmae 202. Су c 1 an t haceae я' . Cyperaceae Lemnaceae
г s i" Atphtxidoidt 1 XXXIII. Enantloblastae — 191. Flagellanaceae 192. Gramineae 193. Restiaceae 194. Centrolepidaceae 195 Mayacaceae 196. Xyridaceae 197. Eriocanlaceae 198. Rapateaceae 199. Commelinaceae 200. Pontederaceae 201. Philydraceae
XXXII. Ensatee
184. Agaveceae
185. Bromeliaceat
180. Haemodoracea.
187. Velloziaceae
188. Iridaceae
189. Scitamineae
190. Orchidaceae (+ Corsteaa'
® Lilwdeae — AUmdeae — 180 Amaryllidarear
179 Alstroemeraceae
g . Asparayoideae
I 183 Burmanniaeeaa
5 ' XXXI. Artorrhizae — 182. Taccaceae
? 181. Dioecoreacea*
5 Slemoneae
Luturiageae — Smilaceae
Табл. 6 (фотокопия). Table 6 (photocopy)
Гал лир Г. 1912. Генеалогическое древо покрытосемянных. VI. М 0-
nocotyledoneae. („Происхождение н филетическая система по-
крытосемянных..." на фр.).
Н а 11 i е г Н, 1912. Genealogical tree of Angiosperms. VI- Mon o-
cotyledoneae. <„L‘origine et le systeme phyl6tique des Angio-
sperms...").
fajteirp /". 79/2. Фъ^гогел-ет^есгсгя сс/стема' ц£еттео6б/.х растенш/
/Ую /14. 4.бушу, uj XHc/et/^ype cue/ncffcr/m/xt/ бй/сшс/з: ticrc/T/ewc/ ". 494&J
H./fa//ter'j pte/topene/te system of //ou'-erfng' p/crnfs. 79/2
f Si/jh w Л, frvrr. the „ Toe coarse of sysTemerrtc of OiffOer pfonfs :' T94 0.J
Табл. 7.
Table 7
jBessey CA.C./^PS. CP/arf to sAo/v refaftonsA/p of Me pratov of the AntopAyte
P?eto//ons/>/p is tPa'/соГео' 6г/ pas/Aon: toe areas are approximate/^/ pnport/omrt
to toe пггтбег aP spec/es to toe orpers
toe poy/epene/re /охаеагау ар р/ег^ег/пр p ton to “/
fieccis У. /9/f. Схеме, изоб/оажа’юща'.я АзаыеосАел/ poppg'pof г/femxoft/x
fej/fepo/ xosto/ypof npi/^pest/menoeo про/горг/г/о/гаг/олго/ растенг/г/
paoevee/nSf/ fi/po/f,f noppffxxr
f’t/POsetoemwec/eo-je /nexecorroert/jr iffamKoOs/x paemenvv "J
Табл. 8.
Table 8.
РооГ R.J. /94/. Diagram to s/tou, the sui-cfasses, super-onferj.ana enters of flattering
plants anal toelr relat/anstops acconalng to toe last arrangement 5g Desseg.
fllotrers amt flowering Plants “J
V U E О ©
COTYLOIDEAE
О СОТу,
О«4е<еч
5YMPETALAK
STROBILOIDEAE
ASTERALES
$ YM PET ALAE-DICARPELLATAE
campanulales
COTYLOIDEAE
ORCHiOALES
IRIDALES
HYDRALE5
RUBIALES
LAMIALES
SCROPHUL ARIALES
6ENTIANALES
I
POLEMONIALES
SYMPETALAE-POLVCARPELLATAC
APOPETALAE
J/yn P Ож. /94/., C'xf/AfO, изображающая поа^яасса/, надпорядхс/
/группы порядкоб-ff.C.j и пор ярки ц^еткобыа растемиг/ с/г/х бзаижосбязи,
согласно последнее/ схелге Dessey.
l/бетки и ибеткобые растения
Табл. 9. Table 9-
Филогенетическая система цветковых растений Бесси в изложении
Пула.
Pool R. J. /941 Ot/lhne to show the three principal series of flowering plants,
A, the monocotyledons. 3 and C, the dicotyledons. TOe apetalous and anemophilous groups
are designated frozen l/nes Rearranged from Besseg according to Clements.
/ f/ovcrj ar/taf f/OH'ffr//??’ "J
GRAMINALE3
I raaweem
orcmidales I
OOOSEFOOTS
walnuts W LAM1ALES |
। MINTS ।
PHLOXES I
I
I
WILLOWS
fl/ул P Dw.flri OdufOR схелга трех главных ердпп цбетхоАых
растении : А -однодольные, В и С - дЗддоллные безлепестные i/ewwm-
шильные группы оОознаиены прерыбистыпи линиями.
Ц6&ПК1/
цбетхебые растения "J
V Pcaceae V Лсег&сесге
г) C^tpertgeeae SJ С/>ег>оро<з'дасео'&
Jtentrtre&ov Ро/уфоп&сееге
Sc/tccceo'e
Табл. 10. ТаЫв 10-
Филогенетическая система цветковых растений Бесси в изложений
Пула согласно Клементсу.
drier £~. Ma/?c7 Par*fs? У. /907.
TPife of dngfosperms Tte/oPonj/i/pj.
0n the or/grin of Arfffospewj " Г/te Joi/rio. offfe //mean Soc/efp'.
dpdep ff. и JTopxzr-/ D.W 7907. ЗТ/об/лщо ^зос/т/оотплои/енис/
похрытоеемдюшх / О лрос/схожаемг/г/ /?олрб//77осел?яшрз/.х:.")
Табл. 11.
Table 11.
f/ir/cAtnson J /960 /./near se^t/ence of t/>e orders of Angiosperms
^7Zre /amfZres 0/ ТТм'еггпр pZanfj " / Dicofyfecforj/
£/cefy/ea/onej
— 7erZ>enaZes
-*£ f^rmr/raZcs
p Pvt/oZej
г/ресупа/ел
-** Z^07Z»/<?Zjtf
6az>z/?c7aZej
AfeZZaZej
— PuZaZej
E/TZ/er/crZ&j.
ZZy/rj/naZej
/?f?07r?r)&/ej --
\S<rr?faZa-Zej
GZacafej
—CeZas fraZes—
-~*-Afi/rZa7ej
j—* Gis/ff/eraZcj
T-fr/caZej
0cf?r?aZej
\^-77jeaZes
—*£~,-Apf/Pr7/C'/raS-»—
—* 77er/pf^fi/a/ej
~^-Л/(77кС7/^
—Г/7/aIes^- —
-— C&cfa/ej
-* Ci/c (ypA/fa-fej
Po-J-J-fZ/ppa/pj
7£9<ZS67/pJ
P<97p£/fp/a7e4
Угя/а/еи
Т&тту&г/ссг/'р.}
C/rpprpr/a'c/fej.
Р/ ftajpara/ej
^’Prafea/es
\—7T/yf?7<?p7ea7ej
'S/xafes - —
r-^&r/ScaPes
~^Co’scf£7r/f7a7es
—Jug/Za-po'/T/ps
— F'a’gra,7ej
—Sp/pf/ops/oP'/ej
— /7 1/r7cpZej
— Z e7/perfp7ej
^Jp7/cp7&j
L— TfomaMefatrfer.^
-^^rp/7a7es
•S7^rpcp/ej
-*£&/?&/? fa/es
L ер//7т?/р>ср7ез
~^^osa7es----------
Се>г7аг/<77<?<
— £>t/7enf/77es ——
Z(fc/rp7es
Л nf)/>r>cf7cj
^rcofy/ea/onej
Merbaceae
Afonocofy/ecfones
LamraZes *
Gcrap/fr/aZej*—
Pc7emor>/aZes —
Gerarnfa/ej-— -
Person&Zej — —
Go/anaZei -
.—Xr/e/’^rZs’j'
Go&c/erU&Zej
-Campana/ej *-
—Z'aZer/ar/aZrpj
-*-Z/m6eZZa7eJ -----
—Poabs/emaZej
-* Jarro’ceru&Zej
Sax/frapaZej — •
-P/an ZagzpaZes
-* Pr/m//Za-Ze~s
-*6enp7<7f7<77<?J
-~Zyf/rcrZej
C/rerrojocn/fr/ej
PeZyponaZes
L- Cavyep/fyZZaZej —>
—Яезес/а/ел
—Crac/aZ&j
- £/>oea'c/a'7ej
P/peraZes
/rsj/cZacZ/fcr/ej
Зег/ег/с/а/ел
-fiana/ej-----------
Gram i naZe^s -—
C(/pera7f?j ——
Juncates --------
EGrc/f&aZe^
Sprmarr/a, 'C'j
Zfae/r/co'cra/ef^
-^Ct/cZar/ZT/aZes
-» Ptrffa'antrZej
-* PcrZmpZes
-* r/p'pir'pZej
^Уг/a'a’Zej-------
-* ^maryZ/Zafc/Z^ <
•* Fyp/a/ej
draZef
-* ZZf&JcareaZtrj
-* /iZsZrc^tarfaZej
r/Zafas---------
ZZr/gr/eraZes
Srcm eZfaZ&j
£f/ffcaaZaZe-j
XyrZz/aZffj-^-
-^Comrrf^Zfna/ej —1
AZayaafaZej --------
PC’Za'mape/prxp'Zej-^—X
dpawaeztmaZep -—!
Juncap>/7?aZes^-----j
7У/L/r/c/aZeJ ----J
4 ZZsrr/aZaSef------1
—S t/foznaZes
fft/poffteZ/caZ proangr/ojpermj
T&zTWt/zree* 4.Ж /960. Линейная тме/гедюбмпеяыестлб псря^ю/ ^r^zosper/ncre
^С/лю/стбсг убеткоАблх pwcmm'i/ бг/^ароныс/
Табл. 12.
Table 12.
Old (Wettstein 1935)
New
A Archegoniatae
1. EOCORMOPHYTA
I. BRYOPHYTA
S 1 ) 1. E. gametophytea 1. Bryopsida
II. PTERIDOPHYTA
2. E. sporophytea
1. Psilophytinae ' t S 1 1 --- -.........) 2. Psilopsida
(ine) Psilotalesi
II PALAEOCORMOPHYTA
2. Lyeopodiinae ............ S 1 ) 1. P. microphylla 3. Lycopsida
3. Psilotinae — r —. S 1 —" ) 4. Sphenopsida
4. Artieulatae ~*-*~ 2. P. macrophylla
5. Filicinae 1 • ..— P (somewhat S?) — .— -..........у 5. Pteropsida
( (inel. Pteridospermales)
B. Anthophyta
III. GYMNOSPERMAE III. ME80C0RM0PHYTA
1. Pteridospermae (incl. Caytoniales)_X 1. M. macrophylla
2. Cycadinae
3. Bennettitinae
4. Cordaitinae
5. Ginkgoinae
6. Coniferae
P (+ Si 1 " ) 6. Cycadopsida
2. M. mierophylla
8 (+ P)----- у 7. Coniferopsida
IV. NEOCORMOPHYTA
7. Gnetinae
IV. ANGIOSPERMAE
1. Dieotyledones
A. Choripetalae
a. Monochlamydeae
b. Dialypetaleae
B. Sympetalae ——
2. Monocotyledones —
S-----
9.
P
b. A. phyllosporae
Angiospennae
a. A. stachyosporae
S(+ P’)
8.’ Protangiospermae
(Chlamydospermales,
Verticillatae)
Табл. 13 (фотокопия). Table 13 (photocopy).
Схема филогенетической системы Cormophyta Лам». 1948.
(„Новая система Cormophyta11; на англ, яз.)
The scheme of the phylogenetic system of Cormophyta by
Lam. 1948. („А new system of the Cormophyta11)
II. J. Lan
526
PHYLOGENETIC-TYPOLOGICAL SYSTEM
OF THE
ANGIOSPERMAE
PHYLL. STACH
V.M
POLYCARPICAE.ETC
W.M. WOODY MONO-
CHLAMYDEAE .
H.M. HERBACEOUS DITTO
(• CARYOPHYLLACEAE)
HEL. HELOBIAE (•ALISMAC-BUTOMAC.).
SPADICIFLORAE. PANDANALES
big. 2.
Табл. 14 (фотокопия). Table 14 (photocopy).
Лам X. Дж. 1950. Филогенетическо-типологическая система Angio-
spermae. („Stachyospory and phyllospory as factors In the natu-
ral system of the Cormophyta").
(„Стахиоспория и филлоспория как признаки в естественной системе
Cormophyta"; на англ. яз.).
WYQPSIOA lmop$ lycops
I
QUATERNARY
TERTIARY
CRETACEOUS
JURASSIC
TRIASSIC
PERMIAN
CARBONIFEROUS
DEVONIAN
SILURIAN
CAMBRIAN
comiferI cvcadopsida IprotangosrI angiospermae
PTEROPSIDA
MAO DS
L S I
HSM
TYPOLOGY
phm
D
N
M
c
G.
C.
О
G
E
N
IX s. stachyosporae
p. phylldsporae
M. MONOCOTYLEDONEAE
D. DICOTYLEDONEAE
51 C CONIFERALES
G. GINKVD
CO. COROAITALE5
T. TR1CHOPITYS
P, PITYALES
PT. PTERIDO-
SPERMALES
N
8 bennettitales
P. PENTOXYLALES
FULLY PHYLLDSPOROUS
.......CONNECTIONS
ЗШ C. CYCADALES
NILSSONIALES
MIXEO
CONN
H. HEPATICAE
S. SPHAGNUM
MUSCI
N. NAIADITA
H 5. hepaticites
П P. PSILOTUM
_____T. TMEStPTERIS
L. LYCOPODIUM
S. 5ELAGINELLA
I. ISOETES
P PLEUROMEIA
8. SARAGWANATHIA-
EQUISETUM
NOEGGERATHIALES
STATIC
/ HIGHER
I THALLOPHYTA
FULLY STACHYOSPOROUS
-------- CONNECTIONS
3E.EE. FHJCES EUSPORANGIATAE
F.L. FILICES LEPTO5PORANGIATAE
MA
. О
OS
M.
s.
A
c.
PF,
E
MARATTIALES
OPHIDGLOSSALES
OStlUNOALES
MARSILIA
SALVINIA
AZOLLA
CAYTONIALES
PROTQFILICES
EPHEDRA
VELVITSCHIA
GNETUM
CASUaRINA
PHYLOGENETIC -TYPOLOGICAL
SYSTEM OF THE
CORMOPHYTA
V
У
Табл. 15 (фотокопия). Table 15 (photocopy).
Л а м X. Дж. La m H. J. 1950. Филогенетическо-типологическая
система Cormophyta. („Стахиоспория и филлоспории как признаки
в естественной системе Cormophyta"; на англ. из.). („Stachyospory
and phyllospory as factors in the natural system of the Cormo-
phyta“).
EBENALES
PERSONATAE
TUBIFLORAE
ASTERALES
CAMPANULATAE
CYPERALES GLUMIFLORAE PANDANALES
PLUMBAGINALES
PPIMULALES
CONTORTAE
UGUSTRALES
CACTALES
CENTROSPERMAE
POLYGON ALES
URTICALES <---------
SANTALALES
FAGALES HAMAMELIOALES
I
JUGLANOA^ES '
SALICALES
♦
VERTICILLATAE
RUBIALES
CUCURBITALES ERICALES GYNANDRAE
RHAMNALES UMBELUFLORAE
t ч
CELASTRALES i
GUTT/FERAE SCITAMINEAE FARINOSAE
TRICOCCAE GERANIALES TEREBINTHALES MYRTIFLORAE
т । t I I + PARIETALES
LEGUMINOSAE
ROSALES
SARRACENIALES
LILIIFLORAE
SPALXCIFLORAE
IV.
MALVALES
RHOEADALES
HELOBIAE
II.
I.
ARISTOLOCHIALES
IIL
PIPERALES
VI.
V
P
0
L
Y
C
A
R
P
I
C
A
E
GNE^INAE
PR0AN6I0SPERMAE
Table 16 (photocopy).
Табл. 16 (фотокопия).
Ш о о P. 1953. Родословное древо Angiospermae („Современ-
ные принципы филогении в новой системе цветковых растений", на нем.
яз. с русск. и англ. рез.).
S о о* R. 1953. Genealogical tree of Angiosperms. („Modern prin-
ciples of phylogeny in the new system of flowering plants11; in
Germ, with Engl, and Russ, summ.)
't'mtsra' У, 7956. Schema' ab spsf&ve ef ate Ar рАр7арет?ге
ate\$ /Waaaaafy/eafo/yas
Sys feme ef /о6у7<уете a^es Afom»e»f&/eoimes "J
.Kt/rtypcr &, /$56 Cxew ci/ca?e/w>/ а <ри/?агемгг/ фнадаямшх
/ CwcrT?e/bf<7 сг QDls./*0<5€>*fU0 <lJ
в coorr>gpp,c^eye^ ~o~epV того ггсе пог>Г9*~^Лр~о-г<”'*»">
/ co*p<7us,e**oj 6 *-о*с'У*,*г”* pcr+^ois смсгтн*/**/
Табл. 17.
Table 17.
Kimura У. 1956. Scheme of the system and phylogeny of Mo-
nocotyledons. („System and phylogeny of Monocotyledons’*: in Fr.)
См. „Пояснения к некоторым схемам11.
11
Табл. 18 (фотокопия). Table 18 (photocopy).
Кимура И. Kimura У. 1956. а—схема, изображающая отношение
между методом автора и классическим методом; в—схема, показывающая
связь главных таксономических групп: А—Gymnospermae, В —
Dicotyledoneae, С—Мonocotyledoneae. Proto-x—вы-
мершие растения, неизвестные в ископаемом состоянии. („Система и
филогения однодольных11; на фр. яз.).
а—scheme showing the relationship between authors and clas-
sical methods; b—scheme showing the relationship of general taxo-
nomic groups: A G у m n о s p e r m a e, B—D icotyledoneae, 0—
M onocotyledoneae. Proto-x—extinct plants, not known in
fossil state. („System and phylogenesis of Monocotyledons11: in Fr.).
Табл. 19 (фотокопия).
SAKgACj
МРША.
г
St. SALICAL
38EBENAL
36. ЫХ
VERCAL.
RHOtD
KALVAL.
Table 19 (photocopy).
Кронквист A. 1957. Схема взаимосвязей порядков двудольных.
(,,Очерк новой системы семейств и порядков двудольных11; на англ. яз.).
Cronquist А. 1957. The scheme of relationships of orders of
Dicotyledons; („Outline of new system of families and orders of
Dicotyledons4*).
1LECYTM
[FUicin4as
|Lycopodm4es
[ Articles
P AYANT PU EVOLUER
p. ayant гоСуоыюГ PSILOPH YTINtES
P EVOLUEES OU FIGEES
DANS LEUXS FORMES
PRtPHANtROGAMES
P. EVOLUEES OU FIGEES
DANS LEURS FORMES
G AY ANT PU EVOLUER
j Amenttfiores]
I
lOmbeltifferes
iPnmuiales] [Plumbagmaies]
T^rSbin-
thales
Juglandales
JubaniaJet
Centro^] I
spermales Гг, .*-------,
-----------1 I Polygonalesl
GYMNOSPERMES
G EVOLUtES OU ROBES
DANS LEUXS FORMES
CHLAMYDOSPERMES
[Ephadrales ] jWalwitchiales] [Gnetalesj
Ш
| Pandale» [
- »[ ANGIOSPERMES tM>
[Thyme-1
| laeales |
[MyrtalesJ
[ Encales]
[Synamherees]
HfLOBlfES ________ v
Alismatales TRhoeada/es]
Potamoge 1------------1
tonales
ISPADICIFLORES
[Pandanales ] [Palmales] [CyclanthaleT]
Pane-
tales
Cneorales] ,—--------.
---------[Tncoques]
Ariatolo-
chialas
Polycarplgues]
L. I L11 FLOR ES
[juncalas] jCyperales |
Bromellal— | [ Pontedefialee |
[ Burrnanniales}[Qrchidatei] [Sotamlnaies |
Fig. 12. — Lea lignees d' Xngiospermr* At lenrs relation'- ph\!ogeneti<]ues (*n ’ remarquera le polyphyletisme. la dispersion des « ganiopetales » et la position
des MonocotyUdones (Original)
Табл. 20 (фотокопия). Table 20 (photocopy).
Схема филогенетической системы покрытосемянных Амберже Л.
I960. („Сосудистые растения", т. II, ч. 1; на фр. языке).
Phylogenetic system of Angiosperms by EmbergerL. 1960.
(„The vascular plants11, t. II, f. 1.; in Fr). („Les veg6taux vasculai-
res,“ t. II, f. 1)
PRINCIPES DE LA SYSTEMATIQVE
Табл. 21 (фотокопия). Table 21 (photocopy).
Козо-По лян ский Б. М. 1922. Родословие семейетв диодей с
указанием отношений к ним томий и дигамей. Л—вымершие типы. („Вве-
дение в филогеническую систематику высших растений").
Kozo-Poliansky В. М. 1922. The pedigree of the families
D i od eae with indication of relationships with families T о mi ea e
and Di gam eae. ф—extinct types. („Introduction to the phylo-
genetical systematic of higher plants11; in Russian).
£>yw 77 97 7940 Схема система/ цветка растемл/
GfoffH'ftor#e Mrcrpspermae
Pi/P/a/rs
Сатпратн/РгГае Pfombagvrur/es Pr/miPa/es
£cp/,Or^ej i/n>6e7fff/orae
fr/cafes
P/arrfagina^.
РгдизГга/ез
бегепга/ез
t/rftcafas
/eea/as
ТаРгР/агсге
( ) Сол/сгОау
Ce/rfrooperrnae
0/У&?0<
Рар/ралаРез
Cypera/es
tP77/<?7770p>J
Po/yc arp/ca-e
Зар/ттоОг/аз
СеЬз^Гр/^З P^£7777pa/A5
ела77 rfa^/asfae
Pana'a/rates
Sctfanwe&p
P}7ir/fosv?
Pe/drae
L е$ц//77/У70.м7е
RsHwsPej
SaS/ea/ns
Сисс/гб/Га/ез
Л7.97 /940 77>e scheme of si/sfem of f/wervnp р/ап^г:
/„ Tfo coi/rse of зг/оГетоЛс of fy/ier pProf о "/
Табл. 22.
Table 22
буш б.Л /959. Схема системы цбетхобых растении
/'Систематика
биса/az растении /
Bus/) Н.Л. /959. 77)е scheme of the system of f/owerwy р/amts. {.Bhe sys/emaf/c of /муВег p/an/s "}
Табл. 23.
Table 23.
,4./7. 7942.
^£тярум77урне>/е 777ъ/т7г>/ су туус^с^е^у^с^су^
7акЬГау<гп 4. Z. 7942 The scheme of the рЛу/одеггеСгс reCatronshtps of the orders of frtgmjpermae
ffe scrocr'ijror/ ti/pes of ^r/r>oec//jm ano' the p/o'centaftan "J
Табл. 24.
Table 24
»
an, Evolution dtr
Asterales
Magnobales
Abb. 43
Table 25 (photocopy)
Табл. 25 (фотокопия)
Тахтаджян А. Л. 1959. Схема филогенетических взаимоотношений
порядков Angiospermae. („Эволюция покрытосемянных11; на нем. яз.)
Takhtajan A. L. 1959. Scheme of the phylogenetic relation-
ships of the orders of Angiosperms. („Evolution of Angiosperms11;
in Germ.) („Die Evolution der Angiospermen11».
Схема фи/тогенетичесхего дре/а yfemnofo/z
РССТРСНСС /проекция кроны/
fort росу о eparptsvecxorr изображении сггстеиог yferrofwr растении"
Cofemcrasr fomawura, f&4SlmJt//l,H,3j
Campanulaleg
fiat t dales
Ru6rales
\idntglgfas
Tricoccae
Pr/mt/lales
bi<4® Qalanophorates
frrcgles.
Cuttiferg^\
Urticales
Polqgonales
Uelohrqe
PippuriaWes
Cyperales
\<r lumiflorae
Tarrnosae
\-------
•fcrfaftrneqe
Principe!
peaga/es
Casuarina/ef/f
Ргрегдй.
Sjtqfhtffo.
nales
I fit/florae
ffg/rafes
osa/es
Umbelliflorae
Julianrales ।
7Uql&ri#qfes
leitneriales
fialcgngpsidaleS-
faqales
6eran/afar
.Podostema-
7/ydrosfadfyales
'Parretales I
rrrocenrales
Wi/ricales
'hamпа/.
Angcprafales
Cerrtrosper/wge
Cacto/es
Callffrfcltale.
Tuh/lo№+r-'^
ffljl/alnamelfdafes
Hqlqrhagfcfqh
flhytolaccaiei
Pli/mlaa/nales
ffrprqspermae
7. 3pympePHOU круг—топь/ nepfau ступени разбитии.
2. Средний круг — типы fimapot/ ступени развития.
3. S ггешнии круг — типл/ rnpembetl ступени ргазбитир.
4 Рропиавсхие типы 7. О Насегеомоопыа/кгелгз/е типы.
f. —. Роа/ггр/етииеские типы.й.@ Ветроопыпяеегые типы.
6.--------ft'нтсгрктииеские типы.9,^ l~ugpoqbum>Htae типы.
Ю. W Гетеротрофные типа/.
The scheme of the phylogene/tcal tree of flotrerinp pfants/projec/fon of the crorenj.
(Composed (y f.^.fross/ter'm. 7P45J.
f„ To the question of the-graphical representation of the system of flowering pfonts'^
Табл. 26.
1. Interior ring—types of the first stage of development.
2. Middle ring—types of the second stage of development.
3. External ring—types of the third stage of development.
4. Tropical types.
5. Holarctic types.
Table 26.
6. Antarctic types-
7. Entomopinions types.
8. Anemophilous types.
9. Hydrophilous types.
10. Heterotrophic types.
Схем аг филогенеза покрытосеменных fпо/7.4.бг/ищ, с изменениями)
Аиз книги: преф /r.U.Xypcawotf, Н.А Комогрпицкиг/, проер. АГ.М Меггер, предо. З.гр.Раздррсхггг/,
А. А УрпноА - ^етаиг/зеа'. /7. Сг/сгпемсгтспгсг рсгстепг/гг. 793/J
Табл. 27. Table ’27.
Angiosperms phylogeny scheme after N. A. Bush, with shanges
(from the book: prof. L. 1. Kursanov, N. A. Komarnitzky,
prof. К. I. Meyer, prof. V. F. Rasdorsky and A. A. U.ranov—
,Botany. II. Plants systematic11. 1951; in Russ ).
Табл. 28. Table 28-
(См. „Пояснения к некоторым схемам'1)
(See „Explanationsto some schemes**)
Филогенетическая схема класса однодольных (по данным палинологии).
Polycarpicae । » Kanunculaceae
Табл. 29 (фотокопия). Table 29 (photocopy).
Куприянова Л. А. 1954. („Палинологические данные к филогении
класса однодольных растений*1.)
Kuprianova L. А. 1954. Phylogenetic scheme of the class of
Monocotyledons (from the palynological data). („Palynological data
to the phylogeny of the class of Monocotyledons1*; in Russ. andFr.)
Fig. 15. — Classification des Angiospermes (d’apr^s Pull»).
Табл. 30 (фотокопия). Table 30 (photocopy).
Классификация покрытосемянных по А. Пулле (из книги Л. Ам-
берже—„Сосудистые растения“, т. II, ч. 1; на фр. яз.)
The classification of Angiosperms after A. P u 11 e (from the
book: L. Emberger—„The vascular plants11, t. II, f. 1.; in Fr.). („Les
vegetaux vasculaires11, t. II, f. 1).
ПОЯСНЕНИЯ К НЕКОТОРЫМ СХЕМАМ
EXPLANATIONS ТО SOME SCHEMES
Табл. 17. Table 17
К схеме Кимуры
1. Апокарпия.
2. Субсинкарпия.
3. Синкарпия.
4. Париэтальная плацентация.
5. Осевая плацентация.
6. Завязь верхняя.
7. Завязь нижняя.
8. Осевая плацентация.
9. Париэтальная плацентация.
10. Белок крахмалистый (имеется в виду эндосперм, называемый неудачно
также белком. — Ред.).
11. Белок не крахмалистый.
12. Апокарпия.
13. Синкарпия.
Табл. 28 (фотокопия с дополнением нумерации таксонов). Table 28 (photocopy).
Комарницкий Н. А. и Уранов А. А. 1962. Схема филогенеза покрыто-
семянных (по Н. А. Бушу с изменениями). (Н. А. Комарницкий, Л. В. Кудря-
шов и А. А Уранов, „Систематика растений").
Поправка к таксону № 18: должно быть „Орхидные". См. ниже пояснение к схе-
ме. (Ред.).
Komarnitzky N. A. and U ran от А. А. 1962. Angiosperms phylogeny
scheme after N- A. Bush, with some changes. (N. A. Komarnitzky, L. V.
Kudriashov and A. A. U r a n о v, „Plants systematic11; in Russ.)
К схеме H. А. Комарницкого и А. А. Уранова. 1962. (Табл. 28).
То Komarnitzky's and Uranov's scheme. 1962. (Table 28).
1. Protoanthophyta. 2. Verticillatae. 26. Umbelliflorae. 27. Rhamnales.
3. Hamam elidales. 28. Celastrales-
4. Juglandales. 29. Terebinthales.
5- Fagales. 30. Euphorbiales.
6. Urtieales. 31. Malvales.
7. Salicales. 32. Geraniales.
8- Proteales. 33. Myrtales.
9. Polycarpicae. 34. Leguminosae.
10. Rosales. 35. Synantherales (Synandrales).
11. Piperales. 36. Cucurbitales.
12. Helobiae. 37. Parietales.
13. Liliiflorae. 38. Rhoeadales.
14. Enantioblastae. 39. Ericales.
15. Cyperales. 40. Guttiferale s-
16. Graminales. 41. Primulales.
17. Spadiciflorae. 42. Plumbaginales.
18. Orchidales. 43. Centrospermae.
19. Scitaminales. 44. Polygonales.
20. Scrophulariales. 45. Dialipetalae.
21. Verbenales. 46. Monocotyledoneae.
22. Polemoniales. 47. Sympetalae (Metachlamydeae).
23. Gentianales (Contortae). 48. Dicotyledoneae.
24. Ligustrales. 25. Rubiales. 49. Monochlamydeae.
Александр Альфонсовнч Гроссгейм
ОБЗОР НОВЕЙШИХ СИСТЕМ ЦВЕТКОВЫХ РАСТЕНИИ
Напечатано по постановлению
Ред.-нзд. совета АН Грузинской ССР
* •
Редактор издательства С. К. Чичуа
Техредактор А. Р. Т о д у а
Корректор Н- К. Цхвиравашвили
Подписано к печати 28-12.1965 г. Печатных л. 28,87;
уч.-нзд. л. 24,40. УЭ 02277. Тираж. 1000 экз. Заказ 1556.
. Цена 2 руб. 25 коп.
go3m3Qg3(rara4» „8g(j6ogAgio“, oiio^obo, dgA^oEbjob j. № 8
Издательство «Мецннереба», Тбилиси, ул. Дзержинского № 8
go3<n30g3(m<n&d „3gQ3og6giob“ b(Jo3io, 0>&0(™оЬо( g. £р&о<1оЬ J. № 3/5
Типография Издательства «Мецииеребэд», Тбилиси, ул. Г. Табидэе 3/5'
Главнейшие опечатки
Errata
Стр. Page Табл. Table Строка Line Напечатано Printed Должно быть Should be
Сверху From the top Снизу From the bottom
2 24 ЛИНИИ ЛИНИЙ
11 6 Archiunphar Archinuphar
17 13 идей идеи
20 20 Liliflorae Liliiflorae
33 3 порядком порядков
44 13 Sennettitales Bennettitales
50 17 Молденке Мольденке
53 19 геранецветных гераниецветных
55 . 21 Курприянова Куприянова
56 6 и 8 Молденке Мольденке
56 13 Protoanthoppyta Protoanthophyta
60 19 Молденке Мольденке
97 26 4. Meliaceae
98 9 Opisthanthohyta Opisthanthophyta
114 8 Centrolepidiaceae Centrolepid[i]aceae
124 о 210. Slmaroubaceae 210. Simar[o]ubaceae
128 8 Ang ospermae Angiospermae
134 4 Pnylogenetic Phylogenetic
150 16 Simar(o)ubaceae Simar[o]ubaceae
153 X. груип групп
158 11 Circaeast raceae Circaeast(eiraceae
159 2 Phytelephaslacac Phytelepha$iaeeae
160 14 Gr upe Groupe
165 4 Cycodophyta Cycadophyta
169 31-'-32 1919... Журн. Русск. Бот. Общ., 8. 1949,... БЖ., 3.—
21 7 семейетв семейств
28 Explanationsto Explanations to
'39 2 family families
К книге: Гроссгейм и
Сахокиа „Обзор новейших систем цветковых растений".
1966.
Hutchinson J. /944. The pht/fopeneftc afr'agrram of ftowering ptants
{i The fam/tiej
of fion/er/ng p/ants ff
ffonacofy/ea'on.s
АЙА1Е8
CTCUNTHALES
CWfRALES
(DICOTYLEDONS)
//i/fcfi'nson У /960 £)/crgra/n о/шг/пд //в profari/e р/?у/одепу op/ re7a/7or?sf7ps of ffe orafers of dngfospermj
77?e fcrmf/feo of f7osiferir>g р7ап7з 6 /. D7co7y7&c7or?sJ
Casuartnates
Polygafates
daptntfates
Araffates Yeates
[Urficates
л . - , Vfofates
Bofanopstctetes
Me/rates I
XJ Pt/6 fates
7amarfcates\ ^ufa^es
Apocyna/es
ferbenates
•Sa/fcates
Sfyra^afes^
Cvnanrates,
f.egomfnates
Jagfa/xfates
beifnerfates
ffamamp/'aates \ p/tfosporate
Pagates
£teicates
7ogahfafes -——^Sfg/fe/tiatej f Senates
Ajtrte^tes^^ '\p7aeatesl ^amnates^^'fa^J Goodente/es
| apparfofaes | [ teaApfgPtefes^^T^f^es
Crates
Asferates
, PPeates
Posates /
Passf77ora7es
Cucuri/"fatesf
, \ /.Loasates
Ifaterfanates
Tfffates
Oac fates
P/anfag/nates
Persona/es
Sotenafes
Campanates
Soraginates
Prfmufates
'ч fyfArafes . .
41 ' Umbe/fafes
Cf,enopoatetes 6pnffa„fffes
lamiates
/ ^^'Bi.xafesezZf^
farfafas Pfteenfotes PAy/nefaeates—^ooteates
\ /nnonates
Polygonafes -^——rCarybphyf/ates Saxffragates
Piperafes,
7at/rates
ArfstetecMates>Berierf^
Pofemoniates
Geraniafes
P/ioeacfafes
Peseofates
Cruc fates
Pacfosfemon ates
'Sarracenfates
'Magnofrates
donates—
—To Monecotyteatenes
LiffnasaefaotfanHirtWty »vuotfg J
icotylecfone
fterbaceae /fvn&fmentatly herbaceous)
ffypoffietica7 proangtosperms
Ягтч t/Mccw Дж. /060. Схема', г/забра'жа’тагг^а’т?' &зм<ожмдА? трг/угогемг/та ы Зза'г/мосбя.зг/ порядка# flng/oopes'/ncre
^Ceprei/cmfo' tfSemfcofdAx рсготеяг/с/". f. glfggooobfbfej
Табл. 32.
Table 32.
Hutchinson J. /926. Dzayran? showzny the prefabfe course of erah/tг on of the Dicofytec/ons
Phe famzZies of PfoH/erzny P/ants. _Z. Dicoty/ecZozzs “J
Табл. 33.
Table 33-
ApocynaZes
Сбело-Zes CrZcaZes
“ Polypetalae ”
fparZ)
Aharzinate
Celastrales
GattiAerates
\Afa/pifyhea/es
Theales
Afyrtates
Cacta/es ?
CucuriiZaZes
PassrfZoraZea
Tarnarteafes
Gar/ar/a/es
, ^№ncf/es
Afprs/naZeZ\ /
Lo^aniates
SZi/raccrZes
Umbelliflorae
fparZ)
ZiuifaZes
S&nta/afes
Ofaca/es
SapincZaZes^
Afe/rafes
f?utates
Malvales
leguminosae
iHamanwZ/JaZes
AsferaZes
ZWrZes
METACHLAMYDEAE
JupZanaZaZes
^PapZiorZrctZes
“Apetalae
'Casuarrnates
(frffcafes/partf
fiagtates
X Afyrzcates
Aettoerfafas
GarrpaZes
SaZicaZes
Prateafes
77?yn?etaeafes
Cherzopoa/iateA
C/rticafes
fpart)
Poafostemonates
SoZcrnaZes
-------------BoraqrZnaZej
CampaneZes
Atefyganates
Lyth rates
PZar>Z<r$zi7?aZes
Caryophyllales
Umbelliflorae
fporrf)
OoZemonraZes
GerzAranafes
Prt/7fatates
Polypetalae" Geraniales
(pai-ZJ
Ar/sZoZocZ>iaZes
Rosales
Bix.aZes
Tiliales p
f Силепга^ез
PiZZosporaZes
Zaur#Zes
Dilleniales
Ma^noliales
AriorescenZ A/ai/t PreoZomZncrnf
PfperaZes
Berbert&rZes
fporZ)
Ranales
PoZppcr/aZej
SamrcenZaZes
Saxifrayales
Rhoeadales
ZoZZe/мсв^А
J
BerierrtZaZes
Perbaceotzs ftabft PredoznZnanf
ARCHICHLAMYDEAE---
Crue/afc
СарраггШ^
i»;
f 6
4
Tczzrot/pzcozz Doze.
/926 Схема, изображающая боз/гожныН хоу зболгоции 2>6удоо7рмых,
/Cezzeczcznfo' nfezzzz'ofzzx гмтстенс/е/". J", pfz/qo/zbzzbze.)
Mez unaf Ziegenspeck, sero/cgijcSer S/crmm&atzn?.
i/^racaafe
- - . SerarprAffceae
Sofanace>a\
poS<,„ac.^ «г СоргНоУ&сегге
^r^ena^-
. j ^l^aSerraptr,
fympec
Stxp&Cae
•Compos Here
Мест sere
rf. ,.
AscSepSa
JtSpjara'pj&e
SfeJ St о
фриАмп
CeraSppSerij _
PA
//тп/е77//^>
fjr&Srcrt
lacyfAs
'oaaniocfae
Vw*c
d/SSw.
h°
ceae
niril/7ei/df
Pfafanore
'pAatw^
O’^aeeap
(yOiPrr/p^ffC
'aceae
f/r&cea
7'ujo^s/j
rrefneae
Terra rt/rr>
Wf
CaatAea
4^
PAycomgceios
ScArzaea
/es
Croton
RSri/Sarra
‘nl/c/a'e'/pra
Ai>l/s
2
3
$. Pset/cfo Sj up#
6. limacrcm
? SaroS/za’^a/ac-^erc*
6 i^/P f&na'r e?7cr
9 Ifaettonfofa
z
rrM
AffS
hrreere
Atbc^&ie^ pi
tfaaTtria
JPrtr7farrr
rehtartt/m
AAdrfata
Spn&^nt/m
JficciocQ'rpuj
SHcc7a
Ce^PeeAaefe
CAo7opA a ra
Lorал 7A
Mterfen
3cAfiopA\7fefr»
%: 1
irroonema
rtorrer/Sa
rtnepae
Ttffofr
^r^cyp,fiereyCf,^
X
ItS/pnl
raepae
Co11
« erass*"
ParSf/cptzfi
C^rrcowi
Afyop
O/eacecxp
2>ajrar7Sra' f
'Camlpa7?i//ae<rap
£rybn>a .
/Г,рр.сга^аг.\ c^j/r^
А'/УЪ'геае>
Comacfforp
siOCauiat
Bami
OrcAroSea,
Ъпдгтбега
^farept^cesf
J) irjcorpffc
^УроХгит Om>c,/nr gennto!Pti
^BSopena'rf
M/CraTaM,
JtoMt
PrtaSarta
Mars'St a
Ро/ууа/ес,
Smyt^/rac.
CanereoreS
Z>tp7prec аграсРл
Уг'о/ас,
r><. ,. Tr 'S/ae
^u''
{,,nae.Zv
St/jrarea
лЛи/^Мгссае r
pnefra'
'4r*qf+r/4
v/fap
17/iman,
D7o0n
fiff/oSom/o Gp
series
mra^,
'i
'/toaophuceafes
CSdt^rrtrm
SaAcrna;
Afetervfrtt
t — бьшершие
оригинале нрлаеано ,, (Saryacac"(peg)
S^ey и Т/иеенспегс. 79Z6.
пае ecre \
Sesn&pcate'
ffSgrrgffr'ac eae
Afyrf7nac ear
vl^
Ро/егл
oaor'erecpep t
^a/os>«/>7>0'Sc
В,АЛе,а, VHrcseeoe
///тясе \oe
rps/roe.
S^artopst
Sair'eeoe
A^srerc
Jur,vt.^,p'V>-’r<re r>
eroobp'tj^р^егг.
[fPAy/tog'fos.stsf» S
jtsSefo-ry/on f
PelyfricAum
fAtseercs ,
Gevrgta
Сиргал/слх
Cterrertes
^0<°SiaJei
>nv ^CSadbpAera
Autotroph?
Qakterien .
z/artnO'ceae
aSts/trceere
A&etarh
Pussu/a
aStneae
S' $/./ JPofyporus
arorSrr pAa/7aJ
7 liceprra'on utr
MorcAeHa
. --.- . s\*M;‘,r'i\pezfzo
2>iatc/neae yTyfrrfa
CArtsso tb artacSafes
diayws Йхбя&ал
байт. 1926. Zusammengeste/7f
son Mez und Ziegenspeck
f Sotanisches Src/iir, ХШ, /-2
Т^ёнигсбергехле серо - ^г/агноетогхесаае рор'ослобиое дребо.
Schc/fPier jj/. /934. P/iyfoaenef/e Tree <?/ /Ле
C <** » . £ * 7- r ~ ''-
растении
/" P^cs/Teee’fr'e/'nc/^ecA'&.f? ггмгкс-ея'&Лге/я /Q&ernbffc/c/ "J
inferae
CQ
°O|
ACRA5IE.AE
GNE.TE.AE.
CONIFERAE
SLAUCOCYSTEAE
CYANOPHYCEAE
myxoschhomycetae
» 5CHYZOMYCETAE
diatomeae
MYXOSA5TERAE
-«a»:,
AG,^r. 'SO
^«Gil
°°°~rocAROAe
s₽hagneae
No5-=oR2i
CENTROSPERMAE
th AL AM I FLORAE
0АЭТе.во^СЕТЛЕ
PROTOBASiDIAE
teliosporae
LABOULBENIEAE
DEUTEROMYCETAE.
DISCOMYCETAE
plectomycetae
„=:га^^“’Хтйому“ТАе
ZYGOMYCETAE
HYDRODICTYNEAE
CLOROCOCCEAE’
ARCHEMYCETAE
AUTO5PORAE
archeophyta
Табл. 35.
Table 35.
Partner 7.H 7934. Pfykyenefrc Branch of fte Gr/fen <?f AfMaptyfert/„ PfyPyenefsc
Шаффнер Дж. 7934. Фшюгенета4ёсрб’р бехДа порярхеб цбелжобб/х распгенпп.
Р„ да-ст^^’'"!
«О
Табл, 36.
ТаЫе 36.
«LYFETALY
SEGgMiMFHY
ftCrnTOMORpilY
BPFQYNlf
«YPOGYNY
PALMAUS
SYNCARRY
APOCARPY
(ЖСИ1МШ
Сар
маичыюл»
CAMPANULA LEI
Cra?~S"ePe
RUBIALES
ттдш
Д'
senttanale*
RRIMULALE1
UMBELLALES
CELASTftALES
CA^S^pg*
CARYORHYUJUXS
MYRTALEB
MALVAtt*
4
па «.-CHART SHOWING THE GENERAL OUTUNE OF CLASSIC*
TON FOLLOWED'IN THIS BOOK. THE SINGLE SERIES OF M0B0«
COTYLEDONS IS INDICATED BY A. THE TWO PRINCIPAL SERIES Of
DICOTYLEDONS ARE ARRANGED UNDER В AND ft CERTAIN APET-
ALOUS AND ANEMOPHILOUS GROUPS ARE SHOWN IN THEIR RELATION
TO OTHER GROUPS fff BROKEN UNES
СГСо°*$шР
AchRdlBac
HVNutixts
LAMiAU*
GfG&rS’P0
FHIo DiGrain
GRAMINALES
UL1ALEB ®Eft
UWP°
Cap Beaked
GERANIALS
RfMoDi Spat Вея
ARALEI
1‘*
QgCo?^
Cap
KROPKULAIKALEl
RANALE*
AcHpFMd5(J«™«
гошаотдш
CoSP1
CgG?S“pg»
* @©
GfCo’S^P»
PfMoBcrDrNut
CaCo*5 r
DrAch Rome Fol
ROSALES
mloDiAcN
AUBMALSB
THE DESIGNATION OF SYMBOLS-
CaXZ S P0
Cap Ber Dr
CPC^S^P01
CapBerDa
V G£C£ST®"
® Milky Cap
PAPAVEftALEB
Calyx, sepals
Sepals’mohe
Sepals several, not a definite number
Sepals 3 to many
Sepals 4 or 5
Sepals 4
Sepals 5, more or less united
Sepals 5, united
Calyx a pappus
Calyx zygomorphic
Corolla, petal
Petals none
Petals several, not a definite number
Petals 5, more or less united
Stamens many, united by the filaments
Stamens 2 to 4, united by the filaments
Stamens 5, united by the anthers
Stamens 6, in two sets
Stamens 4, in two sets, or 2
20. Pistils or carpels
21. Pistil or carpel 1
22. Pistils many
23. Ovary or pistil tricarpellary
24. Carpels 3 to many, united
25. Pistil bicarpellary but one-celied
26. Pistil bicarpellary, deeply four-lobed
27, Pistil of 3 carpels, rarely tricarpellary but
one-celled
28. Pistil of 3 to 5 carpels but one-celled
29. Petals 5, united
30, Corolla zygomorphic
31. Stamens
32. Stamens many
33 Achene
34. Berry
35. Dioecious
36. Evergreen
37. Monoecious
38- Polygamous
39- Scales
40. Carpels 2 to many, more or less united
41. Succulent
42. Utricle
43. Baccate
44- Capsule
45. Drupe or drupaceous
46- Follicle
47. Flowers perfect
48. Samara
49. Spathaceous
50. Flowers unisexual
51. Stamens epipetalous, opposite the lobes of Co
52. Ovary inferior, tricarpellary; stamens on the
axis
53. Stamens epipetalous, alternate with lobes of
the Corolla
54. Ovary inferior, btcarneHary-. stamens epipe-
talous, opposite the lobes of Co
Я' Окема ftffy/fe ГЛ/ efa&goattan <п*аут£о&
i ’ £/4 Vataaiaea, >лаюАягеяиааг 15, *5*® Вавяешжаами^сяаеггпка» Muautaao
£/4 poo 1 6. *5* а/аиаохотедоА^рроеапохР utamUMt/
3* CaX Чспае/гиапааюА явсхояаяъ ' хаао«осш^о 17. дЛГ 7a/«ur&iv5,сроеюахаюмоапою/
Ca^'°° go мжояемяЛа 18. £ *** fbtvwe* дбдхячдо»
$ ' Ca Vauopvomcoo# 4 tat»S 19. Крлроа4,./д&гм>д*я
* Ca * йго-илетмл,/ 4$ 20. P Паеямлпа taw тгадаактакя
Ca£ ^tappt/cmt/xoS SiSatoPOcatootto 21. fl Местам tarv тю^мисгяаа f
Ca® VaaopacawpoSS,cpoctaapQt 22. p°° flacmt/Kv6 wrote
9 P Ca^ Vercat'vxer f Лде xarano 23, P ® JpAkm oaemax to троя агодм&еяохоб
Caz^'^ Vpippyxo лмоооряярр^ 24, PQriV/faegM&eBKM»femJgo мможааоА^оаааюея
11 ’ Co SenvtsK, pvnecmiea 25. P 2аяа^ягомпто& •» /-рхк/уныа
12* Co ° УГрросрчооА 2Ь. P^PfffaanntasfwfOffafftvmcaco^ u>iao4-joia»mav
13. Co Х-тЯ/пвс/пкоб мкжар&ео, 'оорррдереюаор axipt/voemte . P&$iP/tgcnwt tofnMjwmmoxoi; •еже toSnaogoPtKnwxoA,ao
14. Co"& JZtnocmpoS 5,&M.CPoeuftsxcP 28. fl(ЙГЛ^ЯяйвфуwЛ*J’/wqpxw»» ASAtfjp >^мдомг
29. Co® JfenocmxoS Si eooeantxar 42. PQ2? /hroffcur&emvxoS am2 go afOOMceemSb', <Ао.а>оегаияаа
30. Coz ®Serrvu* .заеаа&аоръп/ 43. Sl/C
31. S П/vatfxa 44 _ £/r /Zvog e AtetaovKoSogaotf, t SagymoO ooo^otooa
32. S*30 ZtoWHov fiwoaa 45. jaegaoStaaaoaf
33. Pdf еояаята 46, CAP f&oad&our
34. SgA JZeeffo 41. Ся XOcmtoaeo ияв пмд^
35. £)i Абудоомь» 48. Ям, ^ТиотоЛко
36. Stt'fwoaeawo 49. t^вяoyftжSf^яvoя^oo'pЛo»tu^^
37. Mb Offftognaatfrata 50. *SAAf Хяблтаюао
38. P<$ /Голилаагмаа 51 «Ja47* С похрааЛмомpvq&mton ‘ кромиааа^Атеааа^хма рюрюм. -Ъд.;
39. •Sc CA<t>C^>Pt> 52. (Р t{Sem*a однопоа&а Са3Сс^ рв>
40. fh/vuMKc^ ллипотегрогглго/Ьглепееяи№л$/, 53 Jadraa p&sreMW,t'jJpoagoxvep»utta4 egnoomtrftoo уамгг* бено&ю ' 'wvi/trttrwrnotycr S1 -S'4'1 OM&® C^Co<3>
41 ; РФ 54. -Р*4км /таеат, ю Рпмдаяютахо^} CgnpooiaAw» даяяа» Ааачоюа
Табл, 37 (фотокопия) Table 37 (photocopy)
Пул P. Дж. 1941. Общая схема классификации цветковых растений.
(„Цветки и цветковые растения"; на англ, яз.) А—однодольные, В и С —
двудольные. Взаимоотношения некоторых безлепестных и анемофильных
групп с другими группами показаны прерывистыми линиями (переведено
сокращенно. — Ред.)
Pool В. I. 1941. „Flowers and Flowering Plants11.
Synandrae 9(H)
Compositae
Campanulaosae
Coniortae 6(11)
Piperales (10)
Juglaudi Horae (8)
Oynandrae 7(111)
Rubiales 8(H)
Ligustrales tlmbollilorae (23)
<=zFraxina!fcs) <(Ц) -r
T I
Tnbiflorae 6(11)
______T _
Cucurbitaceae
Parietal es (13)
Urticiflorae (11)
-л Ulrnaceae
Gattiferales-—-> (=Theineae)(lo)
Moraceae
Tamaricaceae
Rhoeadales (12)
Papaveraceae
Resedaoeae
: SaurilinCe4^
f 1'ipcraccae
Gruinales (17)
T
Columniferae
p=>Mal vales) (16)
Diospy rales 4(H)
| . Ericales
О Cactaceae
> Loaaaceae
Papa^aceae
Passifloraceae
Lncalea \
(s=Bicornes) 2(11) Primuliflorae \
3(11) \
Violaceae
Phytolaeacea©
Scituniineac 8(111)
Lardizabalaoeae
Myricaceae
Junglandaceae 'x'jf
Caryophyllace'ae
q Amarantaoeae
Portulncaceac ^*****«j
Qhenopod iace ae
i Salicaoeae
Sa kiflvrae (14)
Polygonifloraa (7)
Polygonaceae
Plumbagmales l(II)<——Centrospermae (6)
---------------*----------------'
Liliifllorae 6{III)
Glumiflorae 5(111) ! Farinosae 4(111)
Juncifl orae 3(111)
Spadieiflorae 2(111)
Alismaceae
О
Hydrobiae Gobi
(= Helobiae auct.) 1(1П)
-1
Celastrales (24)
Gupuliferae (9)
Fagaceae x I
Betulacoae
\
Aesculinalee I
(=sSapindales (23)1
Rhanmales T
(j= Ampelidales (25) । |
\ Rutales
(= Terebinthales (22)
Cruciferae
Capparidaceae
Aizoaceae
Rosaceae
Ranales (4)
Rosiflorae (IS)
. Saxifrage Qceae
Crassulaceae
Leguminosae
Myrtiflorae (21)
Myrtaceae
О
Oenotheraceae
------О
Loranthiflorae (20)
j Proteal'es (10)
Объяснение таблицы rv
Таблица эта составлена по филогенетической схемЪ родства семействъ,
данной въ -работЪ г. Меца и его сотрудниковъ (см. прим-йч. на стр. 27). Семейства
послуживпля имъ точками отправлен!» для прнвивочныхъ реакц)Й впрыскиваний
обозначены крести к омъ (X); маленькие нулики (О) же стоять у семействъ
только реагировавших?», но не бившихъ точкам и отправления для нааванныхъ реак*
Ц1й. Bcrfe проведенный на таблиц^ оплошны я лпнш принадлежать названным?» ав-
торфгь, все же остальное, какъ-то: все обведенное пункт» ромъ, равно какъ и
вс% лиши, проведенный черточками—автору итнхъ строкъ.
Bob порядки» поставленные надъ проведенною вверху горизонтальной
прерывистой чертой, состоять ивъ семействъ съ цветками 4-хъ цнклическлмп,
представляющими собою высшую ступень развпт1я цветка всЬхъ цвЪтковыхъ
растешй т. е. состоящего только ивъ 4-членныхъ круговъ. из?» койхъ самый на-
ружный и слъдузопца за нимъ приходятся на околоцнФ.тникъ, а оба сродин-
«ыхъ—по одному на андроцей и гинецей'. „ .
Ср. также согласованную съ этой графической таблицей IV схему взажм-
«аго родства порядковъ семействъ Anglo spermae, указанную на шгЬд. таблиц11 V.
Цифры, стояния въ скобкахъ на этой графич. таблиц-Ь IV, соствЪтствують тремъ
иродольнымъ рядамъ (I, П, III) названной схемы таблицы V.
Nymphaeaceae
Calycauthaceae
Anonaceae
Ranuuculaceae
Berberidnceae
I, Menjspermaceae
->Nepeuth*lea (5)
->Anstolochialee (3)
Polycarpicae (1)
•Hamamelidales (2)
(Anglos
Magnoliaceae
Taxaceae
9—
Сем. Cycadaceae
(Gymno
Sperma
Pierido
(Polyciliata)
Киассъ Fiticmae
permae)
-^=-Gnetaceae
__________i______?
Pinaceae
——Сем, Bennetittaceae
spermae)
phytu
phy ta
(Biciliata)
Классъ Lyeopodmae
Selaginellaee&e
Табл. 38 (фотокопия). Table 38 (photocopy).
Гоби X. Я. 1916. Филогенетическая система
растений („Обоврение системы растений").
Gobi К hr. 1916. Phylogenetic system of
plants. („Review of the system of plants11; in
Russ, and Fr.)
Coniferae
Explication de la planche IV.
Cette planche est dresses suivant le schema phylog^netiqne de parents dee
families publie dans I’ouvraga de M. Mez et ses collaborateure (voir p. 27) Les
- families n6sign6es par une petite croix (X) leur out service comme point de depart
a la reaction par la methode mentionnfie a page 27. Lea petiteg ce role’s (O) ddeignent
les families ijui ifont quereagies n'ayant рая serviea comme point de depart Д
Le reaction ci-dessus mentionnee. ‘Toutes les lignes continues appartiennbnt i ces
auteurs, tandis que tout ce qui est pointille aibsi que tout ce qui est tracd en
traits d’nniou est й moi.
Tons les ordres places au dessus de la IJgne -horizontale entrecoupSe, com-
prennent des families avec des fieurs t£tracycliques qui prdsentent le plus bant
degre d’evolution de la fleur en general (ohea toutes lee plantes' dites. Anthophy-
tes), c’est-a-dire que toute semblabie fleur a’est composie qua de quatre cercles
on verticilles floraux. doDt deux—les dIub exierieun&oiistitQetit -I’envAlpnrin da !
tleur (penanthe) et les deux autres coneutuent ran—I’anorocee, rautre—legynecee.
Voir aussi la ptenche suivante V, ой ев concordance avec cette planche
graphique eet indiqu^ le schema de parent^ rficiproque dea differents ordres de
families d’Angiospermes: les chifires plac^es en parentheses derriere chaque ordre
de cette planche IV, correspondent aux trole rangdee longitudinaiea (С U, III),
indiquees dans le schema.
Таблица V.
Схема взаимнаго родства различныхъ порядвовъ семействъ Angiospermae (Скрытос’Ьмяняыхъ).
Schema de parente reciproque de different* ordres de families d'Angiospermes.
Такъ назыв. Monocotyledeneae Dicotyledoneae (Двус'Ьмянодольныя). ' (Односъмянодольныя).
" . (I) Choripetalae (Archichlamydeae -f- Apetalae) (Свободнолепестный -|- Безлепестный). (II) (HD Synpetalae (Сростнолепестный).
Di—(s. hetero) chlamydeae. Доу—(а. разно)покровныя. Monochlamydeae. Однопокровныя.
jPolyoarpicae Bartl. (partim)
pg* MAgnoiidieB j Ц/ | 1, amamelidales (2) Д, Aristolochiales (3) Ranales (4) 1 1 Nepenthales (&) Uentrosperma 1 | e (6)' ‘ » Polygoniflorae (7) Juglandiflorae (8) l^upuliferae (9) Piperales (10) Urti ciflorae (11) Saliciflorae (14) > Ilydiubiae (—Ilelobiae j,s. Fluviales auct.) (1) ,, Spadiciflorae (2) Junciflorae (3) II. Iharinosae (s. Enantioblastae (4) Plumbaginales (1) Г.?!£тИ1ог“ (5) 6 Lilnflorae (6) 1 Gynandrae (7) Scit.amineae=Arillatae) (8) 1 Ericales (=Bicornes) (2)
L. ;
Rhoeadales (12) i’arietalee (13)
VjTU1U1 ttlUB 1 A IJLUl-i-lUeitJy Columniferae (16) t Primuliflorae (3) Diospyrales (4) Tubiflorae (&) 1 ^oiitortae (6) gynandrae (9) Ligustrales(=Fraxinales) (7) Rubiales (8)
Proteales (19) iuoranthiflorae (20) les (23)
1 • ) Myrtiflorae (21) ; lutales(=Terebinthales) (22) 1 1 Aesculinales (= Sapiudt
, Rhamnales (Ampelidales) (25) Trw Пл1|1ПАГС>Л / : - -- I
Табл. 39 (фотокопия). Table 39. (photocopy).
Г о б и X. Я. 1916. („Обозрение системы растений11).
Gobi К hr. 1916. The scheme of the relationships of different orders
of Angiosperms family. („Review of the system of plants; in Russ,
and in Fr.)
Прилож. к журн. „Известия Глаин. Батан. Сада" Т. XXI, еып. 3. 1922 г
Схема филогенетического родства цветковых (антофитных) растений по И. И. КУЗНЕЦОВУ.
*
I
| SyMMriie-"—
Compositae.
Euanthophyta
Tetracyclica pentamera.
| O/psacales. |
Dipaacaceae.
Valerianaceae.
| Cwtirtie. |
Asclepiadaceae.
Apocynaceae.
Gentianaceae.
Loganiaceae.
THE DESIGNATION OF SYMBOLS
Объяснение знаков:
I
ggplcal types
retie types
bafetic types are not underlined
e moet ancient types
(^Intermediate types
J newest types
Sthe most important families are desi-
r the names of orders
2
3
4
6
.... Типы тропические.
.......- Типы антарктические.
Палеоарктические типы не подчеркнуты.
Древнейшие типы заключены в рамки.
Типы промежуточные заключены в р&мки.
Новейшие типы заключены в рамку.
Примечание. Семейства под названиями поряд-
ков обозначенны лишь важнейшие.
I
0!
Protoanthophyta
Monochlatnydea.
I"” SwtalBlH. ]
Cynonioriaceae.
Balanoyhoraceae.
Lorantbaceae.
Santalaceae.
A
| Proteales"
, Proteaceae.
| YerticdliHe. |
Casuarinaceae.
Euanlhophyta
Pentacyclica
Trimera.
| timlflKu. |
Graniineae.
A
| EMBtiiilashe. |
Rpstionaceae,
Eriocaulaceae.
Xyridaceae.
CnmmeTmaceae.
| Cefltrispermae. |
Sik-naceae.
Paroiticliiaceae.
Alsinaceae.
Portulacaieat!.
Cactaceap.
Aizoacpae.
Nyctagiuaceae.
Phytolaccaceae.
Aiaarantacese.
Chenopodiaceae.
I 6умж1гае. |
Orchidaccae.
A
| Saitamineae. |
Marantaceae.
Caniiaceae.
Zingiberaceae.
Musareae.
| Cyperales. |
Cyperaceao.
Juncaceae.
LHIitlwae . |
Iridaceae.
Dioscoreaceae,
AmaryUidaceae.
Bromeliaceae.
I.iliareae.
Э@ [5]e
| Polygenales. |
Polygonaceae.
I Urticales |
ItiTicacpae.'
Caniiabaceae.
Moraceae.
Ulnisceae.
| Spadiciflerae. |
Lemnaceae
Araceae.
Typtadeae.
Sparganiaceae.
PandaiincHtie.
Vahnae.
-------A
| lalanopaidales. |
Balanopsidaceae.
| SlIlHlM.
Salicaceae.
| Fagales. 7 [
Fagacwe. I W1"ialllt
Betulaceae. . .. .
Juhamaceae.
| Juglandales |
Juglnndaceac.
A
[ fiptites. |
Chloraiitliaeeae,
Sauruft^ae.
Piperaceae,
| |
Stylidiaceae.
Goodeniaceae.
Lobeliaceae.
Campanulaceae.
| Cecwbitaies. |
Cucurbitaueae.
| CamlwhlM. |
CuscuUceae.
ConvolvuUceae.
&
| PassHlirales |
Datiscaceae.
Begoniaceae-
Loa?acrsip,
Uaricaceac
Passifioraceae..
A
| Pirlltales.
Vjolaceae.
Droseraceae.
Elatinaceae.
Frankeniaceae
'Camaricaceae.
Cistaceae.
| Helobiae ]
Najadaceae.
Potamogetonaceae.
Juucaginaceae.
Hydrocharitaceae.
Butomaceae.
AiismaUcpae.
|~ Hepwlhalei/ |
Sarraceniaceae.
Cephalotaceae.
X»p»»lj*aoeaa.
| LIlUtnKS |
Oleaceae.
| PlwabagiMles. |
Plmubaginaceae.
A
| Oiospyrates. |
Sapotaceae.
Styrace^e,
Ebenaceae.
A
| hHwIalM. j
Myrsfaaceae.
Primulaceae.
A
| litirw. |
Diapenaiaceae.
E^acridaceae.
Ericaceae.
Pirolaceae.
Clethracpae
| ArHUlKliliUs. |
Hydnoraceae.
Rafflesiaceae?
Aristolochiaceae.
| Ifaeilifw» |
Globulariaceae.
Labiatae.
Verbenaceae.
Boraginaceae.
| ftblaies. |
Caprifeiiaceae.
Uuoiaceae.
| UmbelUflerae. |
Cornaceae.
Umbelliferae.
Araliaeeae.
| Persinitir |
Plantaginaceae.
Acanthaceae.
Bignoniaceae-
Geeneriaceae.
Orobancbaceae.
Lentibulariaceae.
Scropbulariaceae
Solanaceae.
| Tcliifltrae~ |
Hydrophyllaceae.
Polemoniaceae.
*
Staphyleaceae.
Ceiastraceae,
Aquifoliaceae.
I tniMlK- I
Balaanunaceae.
Geraniaceae.
Linaceae.
Oxalidaceae.
A
| Trictcae- |.
Buxaceae.
Dicbapetalaceae.'
Eupborbiaceae.
| Sapindaies. |
Polygalaceae.
Hippocaetanaceae.
Aceraceae.
Sapindareae.
| Twebiitiwltt.|
Anacardiaceae.
Zygophyllaceae.
V itaceae.
Rhamnaceae.
| llyriales. |
Halorrbagidaceae.
Oenotheraceae,
Punicaceae.
Myrtaceae.
Rbiaophoraceaa.
Lythraceae, ~
A
| Thymlisalet |
Elaeagnaceae.
i liyroeiaeaceae.
I IqwHisit |
Papilionarese.
Mimosaceae.
[ CilwHiifirae.
Bombacaceae.
Malvaceae.
StercuHaceae-
Tiliaceae.
| BattifOTltt. |
DipterocaTpaceae.
Guttiferae.
Hypericaceae.
'fbeaceae.
Dilleniac-aae.
| Saxilrqiltt. |
8axifr*gaceae.
Graesulaceae,'
I tlMla.
Amygdalareae.
Pomaceae.
Rosaceae.
Euanthophyta
Pentaciclica
Pentamera.
| Miudaltt. |
Moringaceae.
Rcsedaceae.
Capparidaceae.
Cruciferae.
Papaveraceae.
| MiwiMliailes:
Platanaceae.
Hamajuslidateae,
| Цва1м. |
Ceratopbyllaccae.
NymphaeaceSfc;
Ranunculaceae.
Berberidaceae.
Menisparmaceae.
A
| <йота!«. |
Lauraceae.
Calycantbaceae.
Anon&ceae.
Lactoridaceae.
T rocii od'end'r aceae.
Magnoliaceae.
| Myricales |
Myricaceae.
Табл. 40. (фотокопия) Table 40. (photocopy)
Кузнецов H. И. 1922. („Основы, методы и задачи естествен-
ной филогенетической системы цветковых растений’4),
Kuznetzov N. 1. 1922. The scheme of the’ phylogenetic
relationships of flowering (antophytic) plants. („Principles, met-,
hods and problems of the natural phylogenetic system of flowe-
ring plants"; in Russ).
Protoanthophyta
Polycarptca.
Издание 3-е (переработанное).
ПЕТРОГРАД.
Типография Главного-Ботанического Сада.
1922.
iB I SNOHl
Oroiahch
Lau'atae
Nolah
ESN«’o----------Q
BORKA9IH KYOROPHYLL
SAX4FRA9
Apertocarpellatae
8А1АНОАИОЯ
Lehho
Eric
Clausocarpellatae.
В их
uphorri
9ooo£ni
9eht!ah
AIERIAH
EPACoio
Acahth
9lOBULARi
MYOPOR
CROPHUIARI
Solan
AFRtFOLI
yay и
PiTTOSPOS
SC AllOK I
EStD
Cork
Ribesi
YTOPETAL
HereНТК
CoMBRET
Balsa min
RlfHCO
ASUARIII
Hippunio
Y0RAH9E
ТАБЛИЦА V,
PlUMSASiH
Basell
PoRTULAt
Nyctm’ih
Amaranth
Caryokhyll OSouiam
CRASSUl OLUHASI
CvRIll
Stapkhf
Meiastom
[CYTHiO
ОКНЕA ATI
ОЬТНК
Paomykth-
KoEBEBliHI
SUHNER
Halorrhamo
Cam tin
Cappamo
Tovar!
Gr-
ille
OiCHAPtTAl
НАИН
ilREMAHDR
9riy
ALIC
OTHDR13I
CORVHOCARF
URANTHI
Hunipi
ItHltBULAAi
Pri’mul
Trap
ehother
Myzooehor
Sahtal
tOAAHTH
PoiEMONI
FoUflliEAl
Stachyur
UMARI
Tamaric
Restioh
Cypea
Sterculi
MalV XTscut
Quiin
llAEOCAH?
Campakul
Cohvolvui
PA PAY
OCHN
Nitrari
Caci
Plaiystimoh
Ci st
В i x
CuNONl
ALESHE-RBI
Marcsravi
Turmer
^KiSSEM»
Z* LoAS
О-----—Q :----------
CuCURSIT Achari q.
Be&ohi
eraki
&UTTIFERAE
Ahcistroclao
IPTEROCARP
Chi A eh
RuijOPHOA
PolY9Al
VoCHYSt
Sapino
Malpi&h!
CRUCIFERAE
YYIH
lACISTEMOH
Franken!
Papaver
JMyrSih
iHEHOPOOi
lAZURIAS
BcRSEAio
Phytoiacc
Balahopsio
ABl
Saurur
Cohark
Crossosomot
Fas
Sarraceni
Haemopor
Cehtrolepio
Eriocaul
CITAMINEAE
Bromeu
CoMME|jn
Г1А9Е11АА1
Typh
Cyahast
Irid
MAAYLllO
UPHI0PO90N
(D . , _________
DASYLIAiONyjTj
Najao
Ar(s.st)
OPohteoer
Sapot
Ebeh
•'"''St yr AC
Phytocre n
Droser
Par mass!
riYONOR
ORafflesi
HeANANDI
Cassythz
PiFER
-polY&oXDiOSCORE
Mowodor Caheii Houttuyhi
ОМеХАп) Qj^j pRAMiHEAE
X^/rtPANDAH *" dfHYTEUPHANT!
OOseuin ijCyclAHTH / CIHy.orochar'it
MehISperm
qPiatah
JllLEHI
Rakuhcvl
ROTE
LESUMIHOSAE
omobt« MohImi
Prim
^ламоёП
Calycahth
Eupomati
Anoh
Myr’istic
Cheor
tlMHAKTH
CHIARI
Emmoi
Zympkyh.
Rut
PilOCARP
Empleur
Cuspaa!
Stakhousi
HlFPOCRATE
Anacaroi
Mvric
U EILAND
CAlliTRKH
IMARUR
PlEURlSAHtH
ICACiH
Celastr
FlaCOURt!
Erythrosperm
Eucryfh!
iRISTOlOCHi
Uli ah!
Pist
Barclay
6)Ceratophvll
№iumbon
BuYOM
Caroms
PrO-An&I os PE R M AE.
Табл. 41 (фотокопия). Table 41 (photocopy).
Козо-Полянский В. M., 1922. Родословие семейств ангиосперм
(„Введение в филогеническую систематику высших растений41).
К о z о - Р о li а пв к у В. М. 1922. The pedigree of the families of
Angiosperms („Introduction to the phylogenetic al systematic of higher
plants"; in Buss.)
—— типы с апокарпией; types with the apocarpy;
1 | типы с первичным отсутствием сосудов (дримисовая структура дре-
весины); types with primarily absent vessels (D rim у s-type
structure of wood),*
ф типы с диффузной плацентацией; types with the diffuse placentation.
Окончания семейственных названий „aceae44 отброшены. The
endings of family-names Maceae“ are discarded.
Compositae
EAYN9)
ELAEA9N SltisSOlOM
>|Trochomhor[
|CEACi»iPHYl|
Eupteu
Chlorahtk
Ta! uric
Betul
Dipsac
Lasoici
AlYCER
Umbelliferae
Rub 1
Hamamelio
UKCA^iN
Анне mi
JApohcseton
Amsmat
luCOMMi
Mok Q"
Penae Thymela
Asclipiao
Apoctn
Tetrasepal.
AoxauRji
Zoster
Cymoooce
PoTAMOSETOH
Tetrohci
Myrothamh
I Tetkacentr I
Puitofes
l/mielPf&r&e
M CampowPrtoe
Щ Aficrospermeze
Сел fees легтеге
ftontoft'er/er/es
Зегр/леГет/ез
Conforfa'e
Prrfygerperfes
Scffem/riire
IPfrcerfes
Ртлтерег/ез
Cppenrles
Зег-ег-лгег/ез
^ЛМет/гет/ез
Tbirfforoe
Parreto/es
PPoeercftr/es
&fomif7orere
РРегтлег/ез
Розо/es
Pa no les
foorra/es
Profanona/es
CmPgotoere
P/errcfophyfer po/pce/rerto
Jcr/ranrerfes
Spat&cr/fonsre I
• T Я8 Уегрг/еглеРг/ез
foyer/es
Сггсо/ез
trip florae Pepera/ej
Не/о&ппе
Mw/cales
Ser Pea les
Perfrcrf/a/ae
'rofomonocplamyefae
Cbro/ar/rPae
‘rmtre
J}rpep/>e/ter
/fenrrettrftnae
Gycao'enae
Gne/inae
Conr/erere
leprcfpppeemere
PrerfarchegeWfftoe
Pter/atopAffto 6/cr7ifffff
P/>ereo&fa/ceae
Phoe/opPe/fer
СРеггерРе//<У
ScprzopAy/er
Mp-KopPt/fo
Mp-KopPyPr
<Mbtmbpe>eStoto
sore orrtoe
Не/егоРалРеге
И ЛРолоРелРеге
M УрерРеглелеелрге
Ж Рзесзлреге
В| pferPometre
Ж Рег/Репеге
||ММЯ^^
p/crepe Her to
Bus h N. /. /924. 77>е scheme oh
the system of plants
f„7he general course of botany "J
Табл. 42.
Table 42.
Soests?ep 5.J7. / 93 S.
Система роретпбеншрг аткяш/еяш/ т/ерр/рррбрг cepfez/cmS рол'рб/т^рселгет/р'б/^: р&стпетуг/г/
по Xffi/ге awepifхамского zfomo'^z/xa' Р./7ргя„£/6егл/ z/ zrfen?X0&/&pa'eznoxojpJf7.£29a./'<: хехо/порбочг/ г/зАгехел’&'упгг/ zscoxpereyex&jnws/.
ОДНОДОЛЬНЫЕ
СпдЙнОЛЕПЕСТНОСТЬ
рАЗДЁЛьнблЁпЁСТНОСТЬ
2
ДВУДОЛЬНЫЕ
3
ORCHIDACEAE
CUCURBITAGE АЕ COMPOSITAE
Зигоморфия
Акгиноморфмя
^•А)С<5^А($).(2).(2)л G(6-3)
________ । 3CR0PHULARIACLAE____________________________________
гбоиа-анбжГ ________ ' .<0RRA6IKA0EAt К<5)0(5) АббЙ)"
^я^гд>жесАМРд7иГАСЦЕ\ | К°РО6О^А f----------------------
КоСб-эАбб<а
4 ОРЕШКА
LABIATAE
К(Б)С(5>Д 4-26(2)
VALLI5NERIACEAE !
ВАЛИСНЕРИЯ,
ЭЛ 09ея. лягушатник AMARYLLIPAGEAE
Кд С з Азчг G(T) У1арцисс, Агавы
PALM ACEAE
ПАЛЬМЫ
Рз-*з Аэ-гз-эб(з)
Завязь нижняя
Завязь верхняя
ARACEAE
Ро Aa-io G(r~ °0)
Синкарпия ___
Апокарпия
ЗАРАЗИХА
IRIDACEAE
SOLANACEAE
LILIACEAE
Рэ«-эАз«з6(3>
«года
Ро Az-7 Go-z)
THEACEAE
Рз*з Аа*з6(б)
КОРОБОЧКА
VIOLACEAE
Фиалка
KsCsAs6(s)
суреяасеае!
Pa*a-oAwGt3-z)l
JUNCAGEAE
’СРГГИЙН
асеНасеае
На см------
К 5-а 9 5' А । о -a G(g)
• - KMMNACEAE
ИРУШинд
'lK5-4C54Ap«Gff-ZJ
Рз.зАз.зВД) /6RAMINEA?|
“ЖХГ’ /
. СARYO PHYLLACEAE
• Ка-411м(ЯСн-оА<1*8ч6еа
коровочка,орех
ifew,
I I Водяной ОРБХ
I К-СчАе.яйм!
Рз.дАэбй)
КОРОБОЧКА
Рз*зАг-| G(J)
КСРО6ОЧКА
KbCsAcoGcs-B)
КОРОБОЧКА
GHENOPOPIACEAE
Ks-oCeAs-l Gcs-Ъ
ОРЕХ
CAGTACEAE
Кактусы
КооСоо Аообд>
Я гбдА—
PRIMULACEA&
ПЕРВОЦВЕТ
R(s) 05 As G(S)
КОроВ&чкА
TYPHACEAE
Рогоз
VITACEA& орешник)
ВИНОГРАД Х-Кэ-вРоАз-4О^)1-
К S-4C5-4A5-A G(2-fe) ХНо^Унмк А J
Ягода
RANUNCULACE
IPERACEAt
К(ЯС<5)А56(г),
Нд'р&ЬОЧКА----
IСБОPH. OPFUJКМ,ЛИСТОВ-
КА, ягода ’
MAGNOLIACEAE
SPARGANIACEAE р3.з-3Д,е?0
ЕЖеголовкА — дмзос J
POTAMOGETONACEAE РоД.-я~б7~9
тдтст ---------- —13------
ALI SM ACEAE Рз*з или КзСзАв-соббш Кз-оо С о - 5*<
СУСАК, ЧАСТУХД , КОР060ЧКА, ' --—-н-.-.т
СТРЕЛОЛИСТ СБОРН ОРЕШКИ
CAPRIFOLIACEAE
Жимолость, КАЛИМА
6'УЗинА , ЛИННЕЯ
RU&IACEAE
Хинное, кофейное
ДЕРЕВО, МАРЕНА,
лодмАренник ,
КОЛОКОЛЬЧИК \
м . X . _ /4tsCa>AsG(a)\
Ks~*C(S‘4)As:46(3'2)z КЬроБОчкд \
Ягода, хортям кА,/
ДРОБНЫЕ ОрСШКИ/
KS-4C(5-4)A54«G«)'
кОРО&ОчКА, ЯГОДА,
КОСТЯНКА, ДРОБИ.
ОРЕШКИ
UMBELLIFERAE..........
'convolvulaceae
ВЬЮНОК, БАТАТ
Гинецей
СИНКАРПНЫЙ
Зигоморфия
Завязь нижняя
I j MYRTACEAE
• I МКРТ,ЭБКАЛМП
I 1K5-4C5-4A00G
•Sync агро из дуооесУ
~ZудрагогрЬс/
Paary snfer/or
Ks Cs As 6(2) 4
Х\вуеЕМ«кк.А
ARALIACEAE
ЛЛЮ1Ц, ЖЕНЬ-ЫЕНЬ
Ka-sCsAsGG7»)
6R0SSULARIACEAE
•мородимА,к₽ь(Я<обник
K5-4Cs.4Ae-4G<2) Ж
ЯГПЯА .
FAGACEAE' -. j ERICACEAE
6УК. дуб-‘ * |Вереск,Брусника
KAWTAH ’ •• клюква
V^°Ag>So'Fi
рех коробочка ягода.
^т®::|косгянкд
О Р EUJMM к, Г РАБ : •
Кч-оС_оА2-ю6(2) ’• J
орех
KtslCtslAsGo;
КОрОЬОЧКА
К(5) C(S)AsG(ZJ
КОРОБОЧКА,
ЯГОДА
KhqCwAa-j G<2)
КОРО БОЧКА, орех,
APQCYNACEAE
КЕН ДЫ Ре». ОЛЕАНДР
)\(5-4)C(B-4)A5-2G(2)
КОРОБОЧКА, ягода.
POLEMONIACEAE
СИНЮХА, ФЛОКС
SALICAGEAE \
ИВЫ, ОСИНА,
ТОПОЛЬ
Ко Со A frioGtz-o^
КОрОБОЧКА —
POLYGONACEAE
ГРЕЧИХА. РЕВЕНЬ.
Щ АВЕ/1Ь
кльСо As-sGa-з)
орел
OLeAGEAE
СИРЕНЬ,ЯСЕНЬ,
GENTIANACEAE
K(»-4>C(S-4)A54G<ZI
LINACEAE
ЛЁЯ
Ks Gs As6(8-2)
КОрОбоЧКА
BAL5AMINA0EAE
,НЕ ТРОНЬ МЕНЯ**
KcaCmAsGia |
КОРОБОЧКА
годл . У .—— —-х
-------Г PAPILIONACEAEa
ROSACEAE К(5)с"л--г--~^
К* £s A,? .ftir '
С 50»н, ОРСШкП , ш
..—-I.AUCTOOKM, w
Кэ Сб или PqqAoqGoo
сбоям листовки
6ERANIACEAE
Ро Аз-з-i G с-1 '
Я г ода, костянк а)
l.?R5ohh^ Ня-оСоАчбса CRUCIFERAE
ЧА’ костянка.орех
URTlCACEAE H^GoAs-aGi
КРАПИВА, рАМИ OpEX, КОСТЯH
^J-MACEAE Кч-вСоАч-абт
ВЯЗ, КАРАРДЧ орех, КОСТ Д НК А
MALVACEAE KsCsA«.G(s-o°)
ХЛОП ЧАТНИК, КОРОБОЧКА,
КВНАФ, МАЛЬВА Д РО&Н.ОРгшкы
К 2*2 См А<.т*6(£)
СТРУМЕК,
ст©учсчбк,ОрЕХ
PAPAVERAOEAE
МАК
К* 02*2 А <ю G (£-со.)
KsCs-o A<o6(s-31
КОРОБОЧКА.СБОРН.
«чп trial ьг ед 1
RUTACEAE
ЛИМ ОН, АПЕЛЬСИН
МАНДАРИН
К5-з Cs-BA.o-feGcs-s)
Коробочка, ягода.
Табл. 43.
Keller В. А- 1935. The system of relationships of some families
of Angiosperms. After the book of American botanist R. Pool „Flo-
Table 43.
wers and flowering plants14,1929, with some changes and abbreviations.
(„Botanic; in Russ )
fynmurmeito 7? 7935. Та&г/уег р<2ус/2?5еуяг>/.х coomszouzowzer r?opj7ysre>5 Лтфгозреглга'е
no ct/cmertzmwre ро'сгпгея'г/г/ rz>.
Z>zcoty/eotones
Т/окоео/у/еаРлез
Gt/mna-
зрегтае
5for/pefafore
Sympe/afo'e
ZZopac/faTPz/iT'ea'e змаТуре/а/еоге
{‘'erS/c/ZZcrZov/T/
РоуоЗез /3/
Л/гргсоЗез /3/
ЗоЗггпорзкга/ее
TV
Ze/toerP>7ej/5J
Ji/pfo'na'o'Zes /3/
*7/r Sneer Zes //0j
—»- PoZpponcrZes //4/
'''—* *~/epZeoepee~20O'e //3/—
''*-*• ZZaTna'mffZ/o'o'Zffs /77/
’ PZi/mioyPrerZes
Pr-rmr/ZerZej /3/
РАоеасГоЗее//5/
। Pern/eZa'Zey/70/ -
\3(r/?7fera7e.s/7//.
/PoserZes /22}
<77уг7<уЗез/73/
, J, e/cz/rZrZerZej /9/
| Syrra-mfro'e /Ze/?
—— Bfcornes /3/
,, IZZeZo iZ^e /2J
; |Z zZ/S/Zerere /7/
я! \S£'ea'0Zro7Ze'sSee/3/
nt*-Ppp0y3P>s /4J
^'8Zun?7/Zoro'e /Р/
'ZSScr/o’mnryeo'e’ /sj
'гб^тппгГга'е /7/
Узрега'Ге2/е7ое<з'е/&/
OoZe/m/yr/erere/fy
•Z>rospyra3es /7)
Zt/i/fZorere/3/
/гуг/з7гог7ез /7/
< Pro-fecrZee /72/
* ЗапГаЗаЗез/ГЗ/
Л. х#х7’ —. r^orr/ora-e^y
^807^70^ 1
^PPaTsy^er/es/^sp
'*~ZZmde7ZZ/Zer0e
779/
tPi/ira'Zeo /8)
l Зупете/геге /7а/
? GarryerZes/7/
? S^ZZeer/es /&}
? SoZro'a'Zes /7/
? P/peroZes ///
___ G^oorrervaem ппесоегмемгр/е coomvou/euc^rР/’Озпгегуо'ет /ерагюггб/е eoomrrourerrczxr рггслоем/т
Pes/nps 27e z/nc/trfrZaJZe reZoZnens/rrps afpZemZp Pesypyj ZPeproioZ/e reZoZronsZyrps e/pZanSJ
к/е&--г7е7г> P. 7935. 7/e 77yiZs ofZZepAyZpyene/Zc reZa/ZemsZrps e/ZZe on/ers of PngrZaspermo’e
Табл. 44.
Tadle 44.