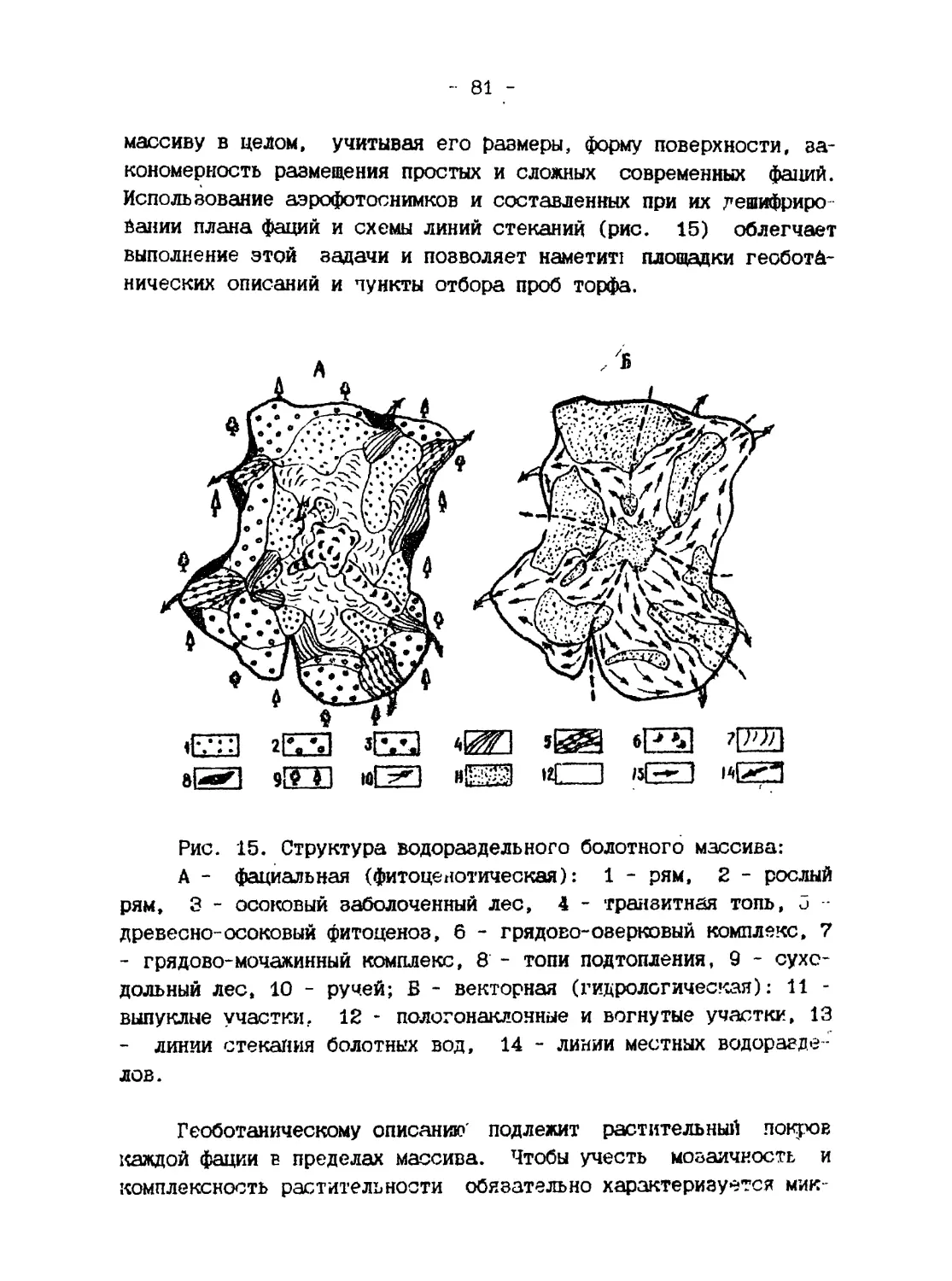
/































































































Теги: растения биология ботаника флора физиология растений учебное пособие геоботаника издательство томск
Год: 1995




Текст
Государственный комитет Российской Федерации
по высшему образованию
ТОМСКИЙ ГОСУДАРСТВЕННЫЙ УНИВЕРСИТЕТ
ВВЕДЕНИЕ К ИЗУЧЕНИЮ РАСТИТЕЛЬНОГО ПОКРОВА
ЛЕСНОЙ ЗОНЫ ЗАПАДНОЙ СИБИРИ
(учебное пособие для полевой практики)
Томск 1995
осударственный комитет Российской Федерации
по высшему образованию
ТОМСКИЙ ГОСУДАРСТВЕННЫЙ УНИВЕРСИТЕТ
ВВЕДЕНИЕ К ИЗУЧЕНИЮ РАСТИТЕЛЬНОГО ПОКРОВА
ЛЕСНОЙ ЗОНЫ ЗАПАДНОЙ СИБИРИ
(учебное пособие для полевой практики)
Томск - 1995
РАССМОТРЕНО И УТВЕРЖДЕНО методи-?ской комиссией
биолого-почвенного факультета
Протокол от /7 1994 г. N Л $
Председатель комиссии Н. С. Москвитина
В учебном пособии дается краткая характеристика, расти-
тельного покрова лесной зоны Западной Сибири, обзор таксонов
разного ранга, описание методики флористических и геоботани-
ческих исследований.
Пособие в полной мере отражает программу полевой практики
по ботанике студентов-биологов 2 курса. Оно позволяет студен-
ту, в случае необходимости (заочное отделение), самостоятельно
пройти учебную практику.
Настоящее учебное пособие может быть полезно также сту-
дентам других естественных факультетов сибирских вузов (геог-
рафам, геологам), а также специалистам по охране природы и
ресурсоведам, которые сталкиваются с необходимостью описания
растительного покрова.
СОСТАВИТЕЛИ: доктор биологических наук A. R Положий
доктор биологических наук Е. П. Прокопьев
кандидат биологических наук Ю. А. Львов
ОБЩИЕ’ОСОБЕННОСТИ РАСТИТЕЛЬНОГО ПОКРОВА
ЛЕСНОЙ ЗОНЫ ЗАПАДНОЙ СИБИРИ
Растительность лесной зоны Западно-Сибирской равнины в
том виде, как мы си наблюдаем сейчас явление исторически
очень молодое. Эта территория лишь 1Г 10.5 тысяч лет назад
освободилась от покрова ледников и предледниковых вод. Около
6-4.5 тысяч лет назад первичные еловые, березовые и сосновые
леса и редколесья уступили место таежным сообществам, а болот-
ный процесс получип широкое развитие. Флора лесной зоны Запад-
ной Сибири сформировалась мигрантами из соседних территорий,
здесь не успели сформироваться эндемичные виды.
По структуре растительного покрова лесная зона Западной
Сибири представляет ботанико-географический феномен - здесь
зональная лесная растительность в большинстве природных райо-
нов оттеснена болотами с водораздельных плато на склоны водо-
разделов и дренированные части речных долин. Интразопальная
растительность - торфяные болота, заболоченные леса, долгопо-
емные торфянистые луга - составляют более половины площади
этой страны. Если исключить сельскохозяйственные земли, то на
долю незаболоченных лесов приходится около трети этой террито-
рии.
Зональным типом растительности в пределах лесной области
в Западной Сибири является равнинная полидоминантная тайга -
темнохвойные леса с доминированием в южных районах пихты си-
бирской (Abies siblrica), а в северных - кедра сибирского
•Pious siblrica) с нередким участием ели (Picea obovata). В
большинстве лесных сообществ обыкновенно присутствие берез
(Betula pendula и Betula pubescens) и осины (Populus tremula).
Этот набор древесных пород слагает эдификаторную синузию рав-
нинной тайги.
На песчаных равнинах - высоких террасах рек, днищах лож-
бин древнего стока, поверхностях предледниковых равнин - расп-
ространены сосновые леса из Pinus siblrica, нередко с
присутствием лиственницы сибирской (Larix siblrica). В поймах
ручьев и небольших рек, где благоприятнее почвенные и ат-
мосферные условия, обычно доминирование переходит к ели. Комп-
лекс природных угодий, характерный для лесной полосы Западной
Сибири слагается из сочетания таежных (или заменивших их мел-
колиственных) лесов, сосновых боров и сфагновых болот.
- 4 -
Таежные сообщества Западной Сибири характеризуются опре-
деленными различиями состава, прослеживающимися как в направ-
лении с запада на восток от Урала к Енисею, так и в широтном
направлении - с юга на север. В Зауралье вплоть до р. Иртыш в
сообщества тайги входят некоторые элементы европейских лесов:
липа (TUia cordata), представители лесного широкотравья. Нап-
ротив, на востоке Западной Сибири обычны в лесах растения,
проникающие ив Алтайского и Средне-Сибирского регионов: жимо-
лость алтайская (Lonicera altaica), ольховник (Duschekia
f rut i ^osa).
Еще более выражены меридиональные различия, позволяющие в
таежной зоне выделить подзоны южной, средней и северной тайги.
Для южной части зоны тайги характерно преобладание коренных и
восстанавливающих лесов с господством пихты (кедрово-пихтовых,
елово-пихтовых и пихтовых), к северу увеличивается доля кедро-
вых (пихтово-кедровых, елово-пихтово-кедровых и чистых кедро-
вых), нередко заболоченных лесов.
В более редкостойных, чем северные, таежных и сосновых
лесах на крайнем юге лесной области под пологом деревьев
господствующая роль принадлежит таежному крупнотравью с боль-
шим участием папоротников (остистого, страусопера, женского,
орляка). Севернее преобладающими становятся сообщества с
лесным мелкотравьем - осокой большехвостой (Carex macroura),
кисличкой (Oxalis acetosella), седмичником (Trientalis
еuropea), плаунами (Licopodium annotinum, L. clavatum), хвощем
лесным (Equisetum syivaticum). Здесь мелкотравные мозаичные
пятна в таежном фитоценозе сочетаются с зеленомошными, образо-
ванными дерниной лесных ’’зеленых” мхов Pleurosium schreberi,
Hylocomium splendens, Rhytidiadelphus triquetrus, Ptiliurn и
другие. В пределах средней тайги зеленомошные синузии стано-
вятся основными.
В Западной Сибири участки коренных лесов, не тронутых про
мышленными вырубками или пожарами, сравнительно редки и неве-
лики по. плонрди. На большей части территории они заменены про-
изводными лесами. На Западно- Сибирской равнине восстановление
леса после сведения древостоя, за исключением только некоторых
типов сосновых лесов, проходит путем чередования пород: на
месте коренных темнохвойных лесов возникают производные с
господством березы и осины. Возобновление темнохвойных пород
5
под пологом мелколиственных лесов идет значительно быстрее в
средней тайге и затягивается на многие десятилетие в южной
тайге.
На южной кромке тайги - на границе с лесостепью и в обжи-
тых сельскохозяйственных районах - от пожаров (особенно силь-
ных в 1915 1916 гг. ) и хозяйственной деятельности человека ко-
ренная растительность почти полностью исчезла. Полоса южных
вторичных березовых и осиновых крупнотравных лесов настолько
хорошо выражена, что многими географами рассматривается как
самостоятельная подзона подтайги или подзона мелколиственных
лесов. О коренном типе растительности здесь напоминают только
отдельные массивы темнохвойных лесов, в том числе охраняемые
населением припоселковые кедровники, да появляющийся местами
подрост пихты.
Местообитания болотной растительности в Западной Сибири
сосредоточены как в понижениях орорельефа, так и на плоских
мало дренированных пространствах широких речных террас и водо-
раздельных плато. Наиболее крупные болотные массивы, точнее -
болотные системы, расположены именно на основных водоразделах
равнины - Обь Иртышском (Большое Васюганское болото),
Обь-Енисейском (болото Комарное, Лотары и пр.). Языки этих бо-
лотных систем проникают, по водоразделам второго порядка, сли-
ваются с терассными болотами, оставляя под лесом только узкие
полосы перегиба склонов.
По типу рельефа Западно-Сибирская равнина - сложно устро-
енная территория: здесь есть невысокие возвышенности - валы,
"материки”, увалы, есть и низменности, как правило, обширные
днища бывших водных потоков и озер. Многие низменности отлича-
ются крайне высокой заболоченностью. Например, Тавда-Кон-
динская низменность в левобережье Иртыша заболочена почти пол-
ностью; более 85 X плошади занято болотами и озерами в Сур-
гутском Полесье - низменности, примыкающие к широтному отрезку
течения Оби; крайне заболоченны террасы р. Васюган и т. д.
Также как и леса, болота Западной Сибири характеризуются
отчетливой поясностью растительного покрова. На юге лесной об-
ласти преобладают низинные (эвтрофные) осоковые, дре-
весно-тростниковые и моховые гипновые болота с плоской поверх-
ностью. По долинам рек эти типы болот проникают далеко на се-
вер, вплоть до границы средней тайги. Торфяной пласт водораз-
. - б -
дельных низинных болот, как правило, с ре немощный - около 2 м
глубиной. Эти болота относительно молодые, возрастом 3-5 тысяч
лет. Но в долинах рек встречаются глубокозалежные торфяники,
сложенные осоково-гипновыми торфами, мощность слоя которых
достигает 6-7 м, да; з 10 м.
Широтная полоса низинных болот проходит по Обь-Иртышскому
водоразделу и его южному склону. На северных склонах водораз-
дела господство переходит к сфагновым переходным и верховым
болотам, в которых низинные травяные и древесные торфа обнару-
живаются только в донных горизонтах залежи. Сфагновые мхи от-
личаются большой влагоудерживающей способностью, благодаря че-
му образованные ими болота, особенно верховые, приобретают вы-
пуклую форму поверхности. Избыточная влага стекает с их скло-
нов, создавая вокруг болотного массива обширные пояса заболо-
ченных лесов и вызывая к жизни новые очаги заболачивания.
01’ приречных таежных увалов до центральных болотных
частей водоразделов прослеживается последовательное изменение
растительности. Таежные фитопенозы сменяются осоковыми к ти
сфагновыми заболоченными лесами, переходящими в крупнососновое
сфагновое болото (рослый рям) и, наконец, в сосново-кустарнич-
ково-сфагновое болото (рям). На водоразделе этот ряд завершают
болотные комплексы - грядо-мочажинные и грядо-озерковые, в ко-
торых безлесные топи разделяются на отдельные мочажины невысо-
кими моховыми грядами, вытянутыми поперек уклона болота и по-
росшими кустарничками и карликовой сосной. В поймах рек, под
обрывом надпойменной террасы, обычны кочковатые осоково-дре-
весные и/Ьфагново-древесные болота - согры.
“•^ри развитии болотного процесса в лесной области Западной
Сибири происходит не только нарастание толщи торфа, но и
расширение границ болотного массива, . образование новых очагов
и площадей заболачивания. В пределах южной тайги агрессия бо-
лот в периоды сухих лет снижается, небольшие по площади масси-
вы даже сокращают свои размеры. Но в средне^ и северной тайге
этот процесс прогрессирует: ежегодно болотами поглощается
десятки тысяч гектаров лесных земель.
В лесной зоне Западной Сибири развито два класса луговой
растительности настоящие луга, возникшие на месте лесов в ре-
зультате хозяйственной деятельности человека, и долгопоемные
болотистые и торфянистые луга в поймах рек в местах длительно-
7
го стояния полой воды. Площади первичных долгопоемных лугов,
незначительные на южной границе лесной зоны, увеличиваются
вниз по течению рек и уже в пределах средней тайги составляют
основную часть пойменных угодий.
Кроме перечисленных типов |>астительности по увалам речных
террас Иртыш, Оби, Томи, Чулыма в пределы лесной области про-
никают степные сообщества типчаковые и ковыльные фитоцено-
зы, а на северной границе области, также в качестве экстразо-
нального эффекта, на ф тяжелых глинистых и торфянистых почвах
развиты участки тундровой растительности.
КРАТКИЙ ОБЗОР ОТДЕЛОВ ВЫСШИХ РАСТЕНИЙ
В настоящее время еще нет единой, общепринятой системы
высших растений. Наиболее широко используется система, предло-
женная в 60-х годах текущего столетия А. Л Тахтаджяном, А. Г.
Кронквистом и В. Циммерманом» отражающая современные представ-
ления о филогенезе высших растений. Основные таксоны этой
системы приняты в соответствии с Международным кодексом бота-
нической номенклатуры (19S0), Авторами внесены изменения в но-
менклатуру таксонов в целях их принципиальной выдержанности (в
основу наименований вышестоящих рангов - семейств, порядков,
классов - положены названия типичных родов).
Схема системы высших растений по А. Л. Тахтаджяну, А. Г.
Кронквисту и В. Циммерману (высшие таксоны).
Regnum Vegetabile - царство растений
Subregnum Embryobionta - подцарство зародышевых растений
Divisio Rhyniophyta - риниофиты (псилофиты)
Divisio Bryophyta - мохообразные
Classis Marchat lops Ida
Classis Bryopsida
Divisio Psilotophyta - псилотовые
Divisio Lycopodiophyta - плауновидные
Classis Lyoopodiopsida
Classis Isoetcpsida
Divisio Equisetophyta - хвощевидные
Classis Hyeniopsida
Classis Sphenophyllopsida
- 8 -
Classis Equisetopsida
Divisio Polypodiophyta - папоротниковидные
Classis Polipodiopsida
Diуisio Pinophyta - голосеменные
Subdivisio Cvcadicae
Classis Lyginopteridopsida
Classis Cycadopsida
Classis Bennetitopsida
Subdivisio Pinicae
Classis Ginkgopsida
Classis Pinopsida
Subdivisio Gneticae
Classis Gnetopsida
Divisio Magno1iophyta - покрытосеменные
Classis Magnoliopsida
Classis Liliopsida
Отдел Rhyniophyta - риниофиты, псилофиты
Древние сухопутные растения. Известны только по ископае-
мым остаткам. Существовали от силура (около 360 млн. лет тому
назад) до середины девона. Описано несколько родов, из которых
главными являются Rhinia, Psilophyton, Asteroxylon,
Protohyenia. Исходная группа в эволюции сосудистых растений.
Отдел Bryophyta - мохообразные (печеночники)
Древние сухопутные растения,.появившиеся предположительно
в силуре или начале девона, но ископаемые остатки их обнаруже-
ны начиная с карбона. Мохообразные представляют’ собой особый
ствол эволюции высших растений. Самой характерной особенностью
мохообразных является преобладание в их жизненном цикле гаме-
тофита (само растение). Спорофит у мохообразных играет подчи-
ненную роль, являясь паразитом или полупаразитом (спорогоний)
на гаметофите. Мохообразные не имеют корней, функцию последних
выполнят ризоиды.
Класс Maruiatlopsida - маршанциевые. Представители этого
класса имеют тело не расчлененное на стебель и листья (маршан-
- 9 -
ция) или имеют дорзивентральные стебли, несущие листья на
спинной и брюшной сторонах (юнгермании). Примером слоевищных
печеночников может служить широко распространенная в Западной
Сибири Marchantia polymorpha. Ее пластинчатые гаметофиты имеют
5-20 см flJt , они двудомные, одни несут архегонии, другие анте-
ридии. Спорогонии состоят из коробочки на очень короткой нож-
ке, внедряющейся гаусториям в ткинь гаметофита.
Класс Bryopsida - листостебельные мхи. Имеют радиальные
стебли, густо покрытые листьями.
Подкласс Bryidae - зеленые мхи. Самая большая группа ныне
живущих мхов. Зеленые мхи широко распространены в сырых лесах,
на лугах, на болотах. Архегонии и антеридии образуются группа-
ми (пс’ювые почки) на верхушках побегов. Окружающие их листья
утрачивают зеленую окраску и образуют защитный покров (перихе-
ций). Спорогоний имеет коробочку и собственную удлиненную нож-
ку, заканчивающуюся гаусторием, внедряющимся в ткань гаметофи-
та (рис. 1). Коробочка на верхушке несет калиптру (колпачек),
представляющий собой оторвавщуюся верхнюю часть архегония. В
верхней части коробочки имеется кольцо из неравномерно утол-
щенных клеток. По кольцу открывается крышечка. Центральную
часть коробочки (урночки) занимает колонка, образующая в верх-
ней части пластинку-эпифрагму. Под крышечкой имеются гигроско-
пичные зубчики, образующие перистом (рис. 2). После сбрасыва-
ния крышечки, если в окружающей среде много влаги, зубчики пе-
ристома завернуты внутрь и прижимают эпифрагму, что удерживает
споры в урночке. В сухую погоду зубчики перистома и эпифрагма
высыхают и споры высыпаются. Спора прорастает нитчатой прото-
немой, на которой закладываются почки, образующие веточки га-
метофита.
Методика определения зеленых мхов. Собранные для опреде-
ления зеленые мхи должны иметь не только побеги, образующие
дернинки, половые почки и спорогонии. Рассматривается характер
дернинки, форма и размер листочков; под микроскопом рассматри-
вается пластинка листа с целю выяснить форму клеток, наличие
жилки, ушек, сосочковидных выростов и других особенностей. Вы-
ясняется расположение архегониев ("бокоплодные” и "вероплод-
- lu -
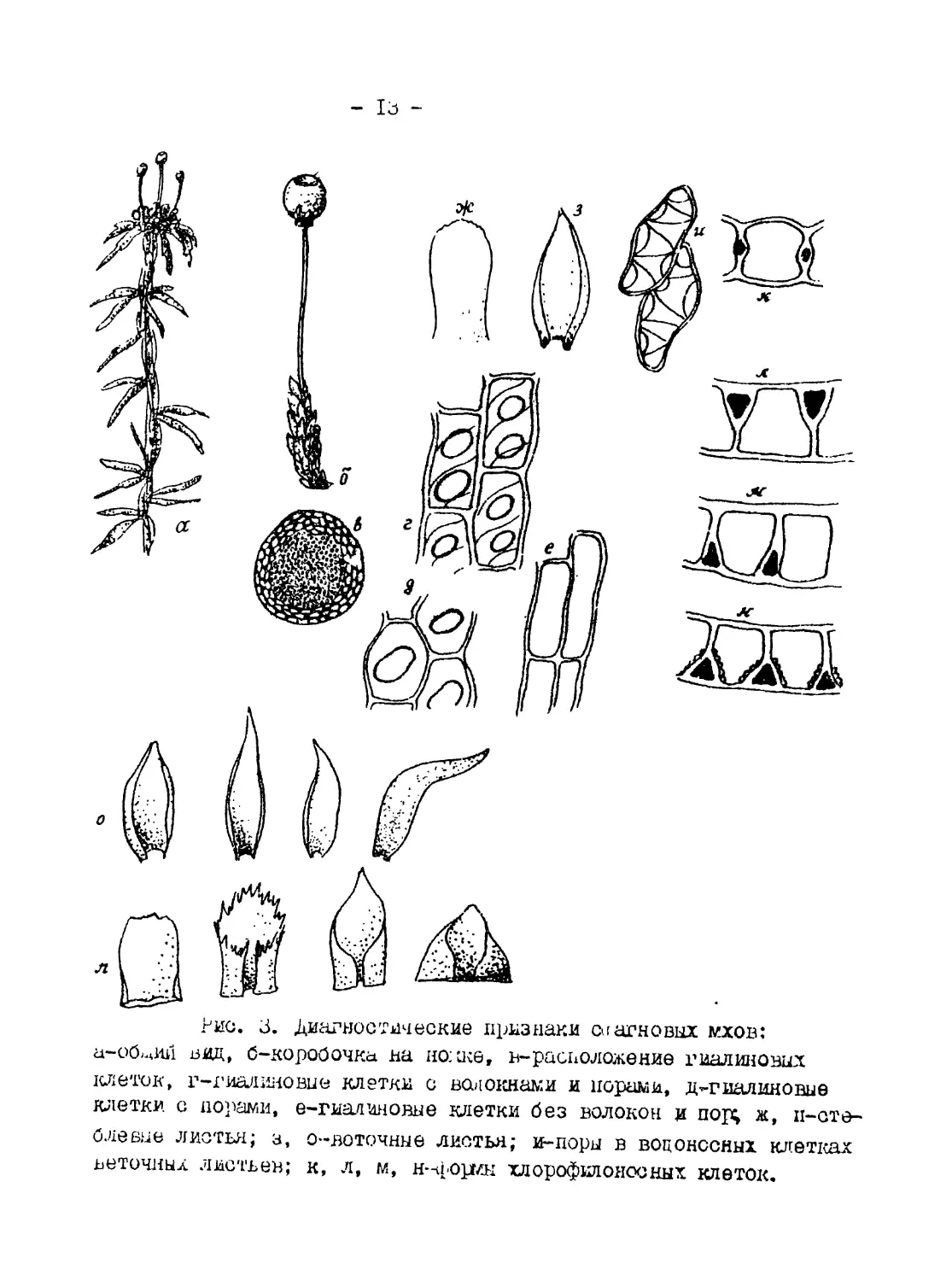
Рис. I. Спорогон зеленого мха:
Рис. 2* Формы перистома
и -
ные”), антеридиев, форма коробочки, наличие и форма колпачка,
положение коробочки по отношению к ножке (склонены я, согну-
тая, горбатая), форма шейки (удлиненная, укороченная, вздутая
и т.д.) (рис. 1), форма зубчиков перистома по краю урночки
(рис. 2). Все эти признаки являются диагностическими для опре-
деления зеленых мхов.
Подкласс Sphagnidae - сфагновые мхи. Сфагновые мхи отли-
чаются от зеленых беловатой, бурой или красноватой окраской.
Стебли их ветвистые, в нижней части ветви более длинные, в
верхней - укороченные, скученные в головку. Взрослые особи
ризоидов не имеют. Сфагновые мхи в стебле и листьях имеют во-
доносные (гиалиновые) клетки. Это мертвые клетки, имеющие спи-
ральные утолшенйя и обычно заполненные водой.
Аухегонии образуются на верхушках коротких побегов, анте-
ридии - на ножках в пазухах листьев. Спорогоний состоит из ко-
робочки и очень короткой ножки. При созревании спорогония из
ткани гаметофита вырастает так называемая ложноножка
(pseudoseta). Коробочка имеет широкую колонку, над которой
сводообразно располагается спорангий. До созревания спор ко-
лонка и спорангий со спорами находятся в полости коробочки.
При созревании спор коробочка открывается крышечкой. Перистом
отсутствует. Из споры вырастает пластинчатая протонема, на ко-
торой развиваются почки, образующие побеги гаметофита.
Подкласс Sphagnidae включает одно семейство Sphagnaceae с
одним родом Sphagnum, который насчитывает более 330 видов.
Методика определения сфагновых мхов. Для, определения
сфагновых мхов с помощью препаровальной иглы от мокрой дернин-
ки отделяют стеблевые •и веточные листья и путем легкого
соскабливания снимают наружный слой ткани стебля (гиалодермы).
Листья и отдельные слои гиалодермы помещают в воду на предмет-
ные стекла, прикрывая их покровными стеклами. Нередко для оп-
ределения требуется приготовить препарат поперечного среза
листа. Для этого делаются тонкие срезы бритвой с ’’головки”
(верхушки побегов) сухого мха.
Диагностическими признаками служат форма стеблевых и ве-
точных листьев, количество и расположение пор в гиалиновых
12 -
клетка;;, форма хлорофиллоносных клеток (прямоугольные, треу-
гольные, веретеновидные, эллиптические и др.), расположение
хлорофиллоносных и гиалиновых клеток и др. На рис. 3 в качест-
ве примера приводятся диагностические признаки двух видов
Sphagnum. Для каждого вида следует составить оригинальное
описание, сделать с объекта зарисовки основных диагностических
признаков, определить вид по определителю. Следует также ука-
зать местонахождение и местообитание каждого вида.
Отдел Lycopodiophyta - плауновидные
Спорофит характеризуется мелколистностью; листья не те-
ломного происхождения, а являются выростами на оси (эмерген-
ции). Спорофилы, как • правило, собраны в колоски. Спорангии
располагаются по одному в пазухах листьев. Гаметофит большей
частью ведет подземный образ жизни, обоеполый. Вегетативное
размножение развито слабо.
В прошлые геологические эпохи существовали как травя-
нистые, так и деревянистые фор:.ы плаунов. Мощные деревянистые
формы (лепидодендроны, сигиллярии) известны по ископаемым
остаткам из отложений карбона. До настоящего времени сохрани-
лись только травянистые формы. Встречающиеся в Западной Сибири
плауновые ‘относятся к . классу LycopodlopsIda, семейству
Lycopodiaceae.
Семейство Lycopodiaceae - плауновые. Равноспоровые формы.
Виды плауновых обитают в равнинных и горных темнохвойных и
сосновых лесах, в арктических тундрах, встречаются в высоко-
горьях. На равнинах Западной’ Сибири распространены виды
Lycopodium и Diphasiastrum. Ниже приводится ключ для определе-
ния плаунов:
1. Листья очередные узколанцетовидные или линейные не
срастающиеся со стеблем......................................2
- Листья супротивные, чешуевидные, сросшиеся со стеблем
Diphasiastrum complanatum (L.) Rothm. -
Дифазиаструм уплощенный
2. Стробилы (колоски) собраны по 2 (редко по 3-4 или оди-
ночные) на длинных ножках. Листья линейные, мелкие (4-6 мм
- Id -
Рис, J. Диагностические признаки са аг но вых мхов:
а-оймий вид, б-коробочка на no:a<et в-расположение гиалиновых
клеток, г-гиалииовие клетки с волокнами и норами, д ^-гиалиновые
клетки с порами, е-гиалиновые клетки без волокон и пор; ж, п-сте-
0ле вне листья; з, о-воточные листья; и-поры в водоносных клетках
веточных листьев; к, л, м, н--формы хлорофклоносных клеток.
14 -
дл.) на верхушке с остевидным придатком
Lycopodium clavatum L. -
Плаун булавовидный
* Стробилы (колоски) одиночные, сидячие. Листья узколан-
цетные более крупные (4.5-7 мм дл.), острые
Lycopodium annotlnum L. -
Плаун гоовалый
Отдел Equisetophyta - хвощевидные
Спорофит имеет членистое строение. От узлов отходя! мел-
кие, но в отличии от плауновых, теломного происхождения
листья. Своим происхождением хвощевидные связаны с риниофита-
ми. В прошлые геологические эпохи, главным образом в карбоне,
существовали деревянистые формы хвощей (каламиты).
В современной флоре этот отдел представлен одним родом
Equisetum. Виды Equlsetum - травянистые равноспоровые расте-
ния. Спорофилы у них имеют вид восьмигранной пластинки ка
центральной ножке, по краю пластинки с внутренней стороны раз-
виваются мешковидные спорангии. Споры кроме экдоспория и эк-
зоспория имеют эписпорий (периний), состоящий из двух спираль-
но скрученных лент, которые раскручиваются и служат прспособ-
лением для распространения спор. Гаметофиты наземные, пластин-
чатой формы, раздельнополые. Размножаются главным образом ве-
гетативно с помощью корневища, в меньшей степени спорами.
Распространены хвощи в лесах, на лесных и пойменных лу-
гах, на болотах. Некоторые виды приспособились к существованию
на обрабатываемых почвах и являются сорняками. Важнейшие диаг-
ностические признаки видов хвоща: имеются или отсутствуют от-
личия, между вегетативными .и спороносными побегами, степень
ветвистости стебля, его анатомические особенности (количество
воздушных полостей), морфология влагалищ (сросшиеся, редуциро-
ванные листья) и др. Ниже приводится ключ длг’ определения ви-
дов хвоща:
1. Спороносные побеги незе окне, образуются весной. Позд-
нее они отмирают и от корневища образуются зеленые, вегетатив-
ные побеги илг спороносные побеги после разбрасывания спор
сохраняются и развивают мутовки зеленых веточек.............2
спороносные и неспороносные побеги зеленые, развиваются
- 15 -
одновременно.. Л.............................................4
2. Спороносные побеги после созревания спор у/ядают, на
смену им развиваются вегетативные побеги с 6-10 ребрами и бо-
ковыми , вверх направленными простыми ветвями
Equlsetum arvense L. - Хвощ полевой
- Спороносные побеги после образования спор развивают му-
товки боковых зеленых пооегов................................3
3. Боковые побеги ветвистые. Влагалища крупные (15-35 мм
дл.) Е. sylvatlcum L. - X. лесной
- Боковые в^тви простые, горизонтальные или книзу изогну-
тые. Влагалища мельче (10-17 мм дл.)
Е. pratense Ehrh. - X. луговой
4. Стебли невысокие <10-20 см выс.) и тонкие (0.5-1.О мм
толщ.) с 6 острыми ребрами. Влагалища трехзубчатые
Е. sclrpoides ichaux. - X. камышковый
- Более крупные растения с другими признаками...........5
5. Стебли жесткие, неветвистые, зимующие
Е. hl emale L. - X. зимующий
- Стебли мягкие, незимующие.............................6
6. Стебли высокие (до 150 см выс.), толстые (2.5-6.О мм
толщ.) с 9-20 ребрами, на срезе с одной крупной полостью
. Е. limosum L. - X. топяной
- Стебли 15-50 см выс. и 1.5-2.5 мм толщ, с 5-8 (редко
12) ребрами; на срезе с одной небольшой и несколькими по ок-
ружности лопастями
Е. palustre L. - X. болотный
Отдел Polypodlophyta - папоротниковидные
Спорофит•макрофильного вида (листья крупные, вайеобравные
имеют теломное происхождение); стебель развит слабо,. чаще в
виде корневища. Спороносные листья обычно не отличаются от ве-
гетативных. Спорангии собраны в синангии или сорусы. Гаметофит
наземный, более или менее мясистый или в виде тонкой пластин-
ки. Папоротниковидные связаны своим происхождением так же как
и плауновидные и хвощевидные с риниофитами. Филогенетический
расцвет этой группы споровых растений относится также к карбо-
ну, когда существовали главным образом деревянистые и отчасти
травянистые формы. В современной флоре попоротниковидные
16 -
представлены гораздо богаче, чем плауны хвощи, они относятся
к 3 классам. В Западной Сибири распространены представители 2
классов - Ophioglossopsida (ужовниковые) и PolypodlopsIda (па-
поротники) .
Наиболее древгчми представителями папоротниковидных во
флорах умеренных широт в настоящее время являются ужовниковые.
Это небольшие травянистые растения, имеющие мясистые корни,
короткое корневище и один лист, подразделенные на вегетативную
и спороносную части, гаметофит мясистый, подземный (роды
Ophiogiossum, Botrychium). Встречаются они редко в темнохвой-
ных лесах.
Большинство ныне живущих папоротниковидных относятся к
подклассу Polypodiidae (настоящие папоротники) класса
PolvpodiopsIda. Это большей частью крупные травянистые расте-
ния с вайеобразными листьями, отходящими от корневища. Споран-
гии собраны в сорусы, споры все одинаковые. Заростки пластин-
чатые, обоеполые. Размножаются вегетативным способом и с по-
мощью спор.
Основными диагностическими признаками при определении
настоящих папоротников являются: наличие или отсутствие диффе-
ренциации на спорофиллы и трофиллы, форма листовой пластинки,
степень ее рассеченности, форма сорусов, их расположение (рис.
4), форма покрывальцу и некоторые др.
Современные разноспоровые папоротники относятся к двум
подклассам класса PolypodlopsIda - Salviniidae и Marslleidae.
В Сибири встречается один род Salvinia. Виды сальвинии - не-
большие водные растения. От узлов отходят по 2 плавающих на
воде листа с цельной пластинкой.й один погруженный, рассеченный
на лопасти лист. Макро- и микроспорангии у них собраны в споро-
карпии (образования, гомологичные сорусам). Гаметофиты как
мужской, так и женский очень редуцированы и существуют крат-
ковременно. Ниже приведен ключ для определения наиболее расп-
ространенных видов настоящих папоротников в ’омской области:
1. Спороносные листья буро-коричневые, они окружены зеле-
ными удлиненно-эллиптическими ' злее крупными (50-150 см дл«),
дважды перистыми неспороносными листьями
Matteuccia struthiopterls (L.) Tod.-
Разнолистник чернокоренный. черная сарана
- Все листья зеленые, спороносные, не отличаются от
- 17 -
Гис. формы и расположение сорусов на вайях папоротников:
a-Polypodium, b-tteridium aquilinum,
c-Dryopreris filix-maa, d~Cyatopterie sudetica.
Рис. 5. Строение женской и мужской линек хвойных в разрезе:
а-желская шинка, б^-кроющан чешуя, б^-семенная чешуя, б^-семя
в-мужскан шишка.
- 18 -
неспороносных........................;.........................2
2. Листья широко треугольные..................... 3
- Листья удлиненно-эллиптические» ланцетовидные или про-
долговато-яйцевидные.......................<.................. 5
3. Листья одиночные, жесткие, на длинных ребристых череш-
ках. Края листа загнуты книзу и прикрывают располагающиеся
здесь сорусы
Pterldium aqullinum (L.) Kuhn -
Орляк обыкновенный
- Листья мягкие, сорусы располагаются с нижней стороны
листа по жилкам............................................. .4
4. Пластинка листьев у основания 15-35 см шир. и почти
такой же длины с нижней стороны негусто покрыта короткими, же-
лезистыми, длинными, простыми беловатыми волосками. Черешок
покрыт черно-бурыми пленками. Покрывальце сорусов продолгова-
тое
Athyrlum filix-femina (L.) Roth -
Кочедыжник городчатый
- Пластинка листьев у основания 6-15 см шир. и немного
меньшей длины (5-13 см), голая. Черешок листа лишь при основа-
нии с бурыми пленками. Сорусы без покрывалец
Gymnocarpium dryopteris (L.) Newm. -
Голокучник трехравдельный
5. Небольшие скальные растения. Листовые пластинки ланце-
товидные 4-12 см дл. на коротких красноватых черенках.' Нижняя
сторона листовой пластинки и черешок кроме тонких волосков
покрыты узкими буроватыми пленками. Сорусы округлые, . позднее
сливающиеся между собой
Woodsla Uvensis (L.) R. Вг. -
Вудсия обыкновенная
- Более’ крупные лесные папоротники с другими признака-
ми.............................................................6
6. Листья дважды-перистые
Dryopteris flllx-mas (L.) Schott -
Щитовник мужской
- Взрослые листья в нижней части всегда трижды пе-
ристые.........................................................7
7. Пластинка листьев продолговато-яйцевидная или яйцевид-
но- ланцетная. Самые нижние вторичные доли на наружной стороне
- 19 -
у нижних первичных долей длиннее всех остальных вторичных до-
лей. Сорусы крупные, покрывальца прикрывают лишь середину его
Dryopteris carthuslana (Vlll.) Н. Р. Fuchs -
Щитовник шартский
- Пластинка листьев продолговато-эллиптическая. Вторичные
доли резко не отличаются по длине. Сорусы продолговатые. Пок-
рывальца полностью закрывает сорусы
Athyrium fillx-femlna (L.) Roth -
Щитовник женский
Отдел Plnophyta - сосновидные (Gymnospermae Голосеменные)
Разнообразные по морфологическим признакам деревянистые,
редко кустарничковые растения . размножаются с помощью семян.
Вегетативное размножение развито слабо.
В современной флере Сибири этот обширный отдел высших
растений представлен только двумя семействами класса Plnopslda
(Plnaceae и Cupressaceae) и одним родом класса Gnetopslda
(Ephedra).
Основную роль в современной флоре Сибири из голосеменных
растений играют представители семейства Plnaceae.
Семейство Plnaceae - сосновые. Деревья и кустарники.
Большинство представителей относится к вечнозеленым растениям
(листья существуют от 3 до 7 лет). Характерными морфологичес-
кими признаками сосновых являются моноподиальное ветвление и
мелкие игольчатые (хвоя) или чешуевидные листья, располагающи-
еся по спирали. Для анатомического отроения ствола характерна
редукция сердцевины, мощное развитие вторичной ксилемы, хорошо
выраженные годичные кольца. Ксилема состоит из трахеид и дре-
весной паренхимы (либриформ и сосуды не образуются). ;
Микростробилы собраны группами, мелкие, состоят из оси,
несущей'чешуевидные микроспорофилы, окрашенные часто в желтый
или красноватый цвет. С нижней стороны микроспорофил несет по
2 микроспорангия с микроспорами. Микроспоры у большинства
представителей с 2 воздушными пузырями. Женские шишки всегда
много крупнее микростробилов, одиночные, состоят из оси, несу-
щей кроющие и семенные чешуи, впоследствии срастающиеся (рис.
5). На верхней стороне семенной чешуи развиваются по 2 семя-
- 20 -
зачатка. Сосновые по ископаемым остаткам гзвестны, начиная с
нижнего мела, предположительно они существовали начиная с юры.
Представители сосновых в современный период играют важную
роль в растительном покрове, образуют различные типы хвойных
лесов.
В Томской области распространены виды 4 родов семейства
•- основых.
Abies - пихта. Корка гладкая, нередко образуются смолян-
ине желваки. Побеги только удлиненные, хвоя короткая, мягкая,
с нижней стороны хвоинки имеют две светлые полоски. Шишки
располагаются вертикально, узкие с пленчатыми чешуями. Созре-
вают шишки в один год и распадаются на чешуи.
Picea - ель. Побеги удлиненные и укороченные. Удлиненные
побеги несут редуцированные пленчатые листья, укороченные -
острые, игольчатые листья (хвою). Шишки узкие, висячие, созре-
вают в один год и опадают целыми.
Pinus - сосна. Корка золотистая или темная, трещиноватая.
Побеги удлиненные и укороченные. Укороченные побеги несут по °
или 5 игольчатых листьев. Шишки разных размеров, при созрева-
нии не распадаются на чешуи.
Larix - лиственница. Хвоя мягкая, нежная,, ежегодно
сбрасывающаяся. На удлиненных побегах хвоинки располагаются
спирально, на укороченных -пучками. Шишки мелкие, созревают в
один год, после рассеяния семян остаются на дереве по несколь-
ку лет.
Семейство Cupressaceae - кипа исовые. В СибИри представ-
лено одним родом Juniperus. Это кустарники с шиловидными, соб-
ранными в мутовки, или чешуевидными листьями. В Западной Сибири
встречается один вид. Женские шишки мелкие, чешуи их более или
менее срастаются между собой и при созревании семян становятся
сочными, в связи с чем шишка имеет ягодообразный вид.
Ключ для. определения хвойных в Томской области:
1. Кустарник или небольшое :еревце (до 4 м выс.) с линей-
но- шиловидными листьями и черными с сизым налетом ягодсобран-
ными женскими нгшками
Juniperus communis L. -
Можжевельник обыкновенный
- 21 -
- Крупные деревья с линейными игольчатыми листьями и де-
ревянистыми чешуями женских шишек...........................2
2. Кроме удлиненных побегов имеются укороченные, которые
в основном и несут зеленые листья...........................3
- Укороченные побеги отсутствуют. Удлиненные побеги пок-
рыты довольно короткими листьями.
3. Хвоя мягкая, короткая, опадающая к концу каждого веге-
тационного периода. Женские шишки шаровидно-яйцевидные 2-4 см
дл., чешуи их яйцевидно-шаровидные, светло-бурые, не опадают
после высыпания ^емян
Larix slbirica Ledeb. -
Лиственница сибирская
-Хвоя плотная, колючая, многолетняя и только на укорочен-
ных побегах................................................. 4
4 Листья собраны по 2 на укороченных побегах. Женские
шишки при созревании сгмян почти округлые, около 5 см дл. Се-
мена мелкие с летучками
Pinus sylvestris L. -
Сосна обыкновенная
- Листья собраны по 5 на укороченных побегах. Женские
шишки продолговато-яйцевидные, 6-13 см дл. Семена крупные
(10-14 мм дл.), темно-бурые, с каменистой оболочкой, без ле-
тучки
Pinus slbirica Du Tour -
Кедр сибирский
5. Листья плоские, тупые, с нижней стороны с 2-мя белыми
полосками воскового налета. Женские шишки прямостоячие, после
высыпания семян распадающиеся на чешуи
Abies slbirica Ledeb. -
* Пихта сибирская
- Листья острые, колючие. Женские шишки висячие, опадаю-
щие вместе с семенами
Picea obovata Ledeb. -
Ель сибирская
Отдел Magnoliophyta (Angiospermae) - цветковые или
покрытосеменные
Крупнейший отдел семенных растений, виды которых опреде-
22 -
ляют состав флоры и характер растительности на всех континен-
тах. Основным признаком покрытосеменных является образование у
них цветка. У большинства видов цветки обоеполые, имеют гине-
цей и андроцей, но значительная часть покрытосеменных имеет
раздельнополые цвет! Абсолютное большинство цветков имеет
околоцветник.
С образованием цветка связаны такие морфологические осо-
бенности, как покрытосемянность и образование плодов.
У покрытосеменных растений полностью редуцирован мужской
заросток (при прорастании пыльцы не образуется ни одной прота-
лиаиьной клетки), а также редуцирована ткань женского заростка
и не образуются архегонии. В связи с редукцией женского за-
ростка эндосперм семени у них имеет другое происхождение (у
голосеменных эндоспермом служит ткань женского заростка, у
покрытосеменшях эндосперм образуется в результате деления эк-
ваториального ядра зародышевого мешка после его оплодотворе-
ния). Плод представляет собой видоизмененный после оплодотво-
рения цветок. Структуру его определяет в основном тип гинецеи.
Осевые органы покрытосеменных, в отличие,от голосеменных, име-
ют сосуды и хорошо развитую механическую ткань (либриформ).
От всех других высших растений покрытосеменные отличаются
разнообразием жизненных форм и огромным разнообразием морфо-
логичексой структуры вегетативных и генеративных органов.
Отличительной особенностью цветковых растений является их
высокая экологическая пластичность, способность осваивать са-
мые различные местообитания. Наряду с высокоэффективным семен-
ным размножением у многих покрытосеменных хорошо выражено ве-
гетативное размножение.
В типе цветковых растений различают два класса:
Magnoliopslda - двудольные, Llliopsida - однодольные.
Двудольные и однодольные растения различаются по ряду
морфологических признаков (табл. 1).
Морфологические признаки наиболее четко выражены между
высоко эволюционно продвинутыми таксонами двудольных и одно-
дольных. У примитивных двудольных и однодольных морфологи-
ческие отличия проявляются не столь резко. Например, у лютико-
вых встречаются трехчленные цветки, у лилейных - четырехчлен-
ные. '
Единой общепринятой системы цветковых до сих пор не су-
Таблица 1
Морфологические различия двудольных и однодольних растений
Признаки
Двудольные
Однодольные
Количество семя-
долей у зародьиа
Тип цветка
Корневая система
Жилкование листьев
Дна*! омическое
строение стебля
две
из большого и неопре-
деленного количества
членов, пятичленный,
четырехчленный
стержневого типа
перистое или сетчатое
приводящие пучки
открытые, за счет
камбия происходит
рост в толщину
одна
трехчленный
мочковатого типа
параллель нонервное
или дугонервное
проводящие пучки
закрытые, вторичный
рост стебля невоз-
можен
ществует. Наиболее детально разработанной и широко используе-
мой является система, разработанная А.Л.Тахтаджяном.
Основные таксоны цветковых растений, представленные вс
флоре Томской области (по системе А.Л.Тахтаджяна):
Класс Magnoliopsida - двудольные
Подкласс Magnoliidae
Пор. Nymphaeales Подмасс Ranun Сем. cull dae Nymphaeaceae ’ ? кувшинковые
Пор. Ranunculaies Сем. Ranunculaceae лютиковые
Пор. Papaverales Сем. Сем. Papaveraceae • Furnari aceae - * маковые, дымянковые
Пор. Paeoniales Сем. Paeoni aceae - пионовые
Подкласс HamameН id idae
Пер. .Betulales Сем. Betulaceae - березовые
Подкласс Сагуophyliidae
Пор. CaryophyHales Сем. Caryophyllaceae - гвоздичные
Сем. Chenopodiaceae - маревые
- 24 -
Пор. Polygonales Подкласс Di Ле Сем. ‘niidae Polygonacea< - гречишные
Пор. Theales Сем. Hypericaceae ~ зверобойные
Пор. Violales Сем. Vlolaceae - Фиалковые
Пор. Capparales Сем. Brassicaceae - крестоцветные
Пор. Sallcales Сем. Salicaceae - ивовые
Пор. Erleales Сем. Ericaceae - вересковые
Сем. Vacctniaceae - брусничные
Сем. Pirolaceae - грушанковые
Пор. Prlmulales Сем. Primulaceae - примуловые
Пор. Maivales Сем. Tiliaceae - липовые
Сем. Malvaceae - мальвовые
Пор. Urticales Сем. Urticaceae - крапивные
Сем. Cannabaceae - коноплевые
Пор. Euphorbiales Сем. Euphorbiaceae - молочайные
Подкласс Rostdae
Пор. Sax ifragales Сем. Grossulariaceae - крыжовниковые
Сем. Crassulaceae - толстянковые
Сем. Saxtfragaceae - камнеломковые
Пор. Rosales Сем. Rosaceae - розоцветные
Пор. Fabales Сем. Fabaceae - бобовые
Пор. Nepentales Сем. Droseraceae - росянковые
Пор. Myrtales Сем. Lythraceae - дербенниковые
Сем. Onagraceae - онагриковые
Сем. Тгарасеае - водяные орехи ,
Пор. Gerantales Сем. Llnaceae - льновые
Сем. Oxalidaceae кисдичковые
Сем. Geraniaceae - гераниевые
Пор. Polygal ales Сем. Polygalaceae - истодовые
Пер. Cornales Сем. Aplaceap - зонтичные
Пор. Rhamnales Сем. Rhamnaceae - крушинные
Подкласс Lamiidae
Пор. Sentianales Сем. Gentianaoeae - горечавковые
Сем. Menyanthaceae - вахтовые
Сем. 1 -biaceae - мареновые
Пор. Роlemontales Сем. Polemonlaceae - синюховые
Сом. Convolvulaceae - вьюнковые
Сем. Boragtnaceae - бурачниковые
Пор. Scrophulari ales Сем. Soianaceae - пасленовые
Сем. Scrophulariaceae - норичника
25 -
Пор. Lamiales Сем. Lamiaceae - губоцветные
Подкласс Asteridae
Пор. Campanulales Сем. Campanulaceae - колокольчиковые
Пор. Asterales Сем. Asteraceae - сложноцветные
Класс Llliopsida - однодольные
Подкласс Alismidae
Пор. Allsmatales Сем. Butomaceae - сусаковые
Сем. Alismataceae - частуховые
Пор. Najadales Сем. Potamogetonaceae - рдестовые
Подкласс Liliidae
Пор. Liliales Сем. Liliaceae - лилейные
Пор. Iridales Сем. Iridaceae - ирисовые
Пор. Orchidales Сем. Orchidaceae - орхидные
Пор. Juncal es Сем. Juncaceae - ситниковые
Пор. Cyperales Сем. Сурегасеае - осоковые
Пор. Poaies Сем. Подкласс Arecidae Роасеае - злаковые
Пор. Typhales Сем. Typhaceae - рогозовые
Систематика цветковых растений основывается главным обра-
зом на морфологических особенностях их генеративных органов -
цветков и плодов. Гербарный материал* собранный без цветков и
плодов, не представляет ценности, поскольку не дает возмож-
ности точно установить систематическую принадлежность растений.
Наиболее трудными для систематики являются крупные и
естественные семейства, виды которых имеют довольно постоянное
строение цветков. К числу таких семейств относятся березовые,
ивовые, крестоцветные, бобовые, зонтичные, норичниковые, гу-
боцветные, сложноцветные, осоковые, злаковые и некоторые дру-
гие.
При определении видов берез основными диагностическими
признаками являются: жизненная форма (дерево, кустарник), мор-
фологические особенности плодов й чешуи женских сережек (рис.
б).
При определении видов ив важнейшими признаками служат:
жизненная форма, морфологические особенности листьев (форма,
консистенция, жилкование, характер опушения), количество тычи-
нок, размер сережек.
У крестоцветных, имеющих довольно устойчивое строение
цветка, диагностика основывается главным образом на морфологи-
- 26 -
Рже. 6. Плоды крестоцветных:
a-Brassica, b-Lepidiua, c-Capsella, d-Draba,
Рио. 7 Плоды ж чешу ж женских соцветии у березы:
а-женское соцветие, б-прицветная чешуя женского цветка, в-плод.
ческих особенностях плодов (соотношение длины и ширины струч-
ка, его форма, характер опушения, способ открывания, анатоми-
ческое строение околоплодника). Большое значение имеет харак-
тер опушения всего растения (рис. 7).
Систематика бобовых в значительной степени основывается
на морфологии плодов(форма, количество перегородок, опушение,
способ открывания), а также структуре листьев.
К числу важнейших диагностических признаков зонтичных от-
носятся морфологические и анатомические особенности их плодов
(вислоплодника), наличие или отсутствие общих (у основания
сложного зонтика) и частных (у основания зонтиков) оберток
(рис. 8).
Норичниковые и губоцветные, имеющие сходное строение
цветков, хорошо отличаются по строению плодов - коробочка у
норичниковых и дробный плод (карцеруля) у губоцветных. Для оп-
ределения родов и видсз в этих семействах кроме деталей строе-
ния цветка широко используются такие признаки, как характер
опушения, морфологические особенности листьев, а у губоцветных
кроме того - строение эфирно-масличных железок.
Основными систематическими признаками сложноцветных явля-
ются: тип соцветия (одиночные, корзинки, сложные соцветия из
корзинок), типы цветков и их сочетания в корзинках (все труб-
чатые, все язычковые, в центре трубчатые, по краю ложноязычко-
вые, воронковидные или нитевидные и др.)» их окраска; строение
оси соцветия (’’цветоложе”); количество и расположение листоч-
ков обертки; форма и придатки семянок.
В семействе осоковых некоторые роды имеют обоеполые цвет-
ки, виды этих родов определяются бее особых затруднений (рис.
у). Наиболее крупным и трудным в систематическом отношении яв-
ляется род Сагех, виды которого имеют довсльий однотипные раз-
дельнополые цветки (рис. 10). Основными диагностическими приз-
наками видов осоки служат: структура колосков (обоеполые,
расположение в них мужских и женских цветков, раздельнополые);
количество рылец; форма, жилкование и окраска мешочков.
В отличие от осоковых абсолютное большинство злаковых име-
ют обоеполые цветки (рис. 10). Важнейшими диагностическими
признаками родов и видов злаков служат: тип соцветия, ко-
личетво цветков в колосках (многоцветковые, одноцветковые),
количество и строение колосковых и цветковых чешуи (наличие
- 28 -
Рис. 8. Строение цветка и соцветий у зонтичных:
а-гинецеи, б-цветок; В, Г, д-ооцветна iegopodlua, Bupleurum, Eryngium,
Рис. 9. Структура обоеполых цветков у осоковых:
a-Sclrpus, Ъ-ВгуарИогшв *
- 2$
мае. 1и* Строение цветков и колосков у осок и злаков:
I - осоки: а-мужскои цветок» б-жанскми цветок, в-обоеполыи
колосок, г*женскии и мужской колоски; II - *яаки: а-колосок
б-формы цветкошл чешуи, в-формы колосковых чешуи*
30 -
ости, место их отхождения, жилкование, опупение и др.); размер
и форма листового язычка, ширина и характер поверхности листа;
строение подземных органов.
МЕТОДИКА ФЛОРИСТИЧЁСКИХ ИССЛЕДОВАНИЙ
Совокупность всех видов растений, населяющих определенный
физико-географический или административный район, называется
флорой.
Флора определенных физико-географических регионов
представляет собой исторически сложившийся комплекс видов,
например, флора Алтая, флора арктической Сибири и др. Флора
административных районов может быть выделена только
искусственно с прикладными целями, т.е. с целью выявления
практически ценных видов, нуждающихся в особой охране и др.
Основной задачей флористических исследований является вы-
явление видового состава растений в целом. Однако проводить
одновременно изучение видового состава $сех групп низших и
высших растений очень сложно, и это требует участия специа-
листов различного профиля. Флористическими исследованиями при-
нято называть изучение видового состава высших растений (не
редко не включая мхи). Низшие же растения изучаются специально
и большей частью по отдельным группам: грибы, водоросли, ли-
шайники.
В задачи флористических исследований кроме познания видо-
вого состава в целом входит детальное изучение распространения
каждого вида, условий их обитания, роли в сложении раститель-
ного покрова. Вхл&ачи флористических исследований входит так-
же выявление хозяйственно.ценных видов (дикорастущих пищевых,
кормовых, технических, лекарственных), вредных и ядовитых
растений, а также редких, сокращающих распространение и исче-
зающих видов.
Существуют два основных метода флористических исследова-
ний; 1. Маршрутный метод и 2. М год конкретных флор.
После предварительного ознакомления с растительным покро-
вом изучаемого района на карту наносятся маршруты, которые
дслжпы пройти исследователи в целях возможно полного ознаком-
- 31 -
ления с видовым составом.
При маршрутных исследованиях проводится гербаризация
растений. Для гербаризации некрупные травянистые растения вы-
капываются и закладываются в гербарные листы (из бумаги, хоро-
шо впитывающей влагу). Крупные травянистые растения закладыва-
ются по частям в разные листы. У деревьев и крупных кустарни-
ков для гербаризации срезают побеги. В гербарий закладываются
растения как правило с генеративными органами(сорусами, стро-
билами, цветками, плодами). При сборе гербария необходимо пом-
нить об основнь:': диагностических признаках таксонов и стре-
миться собрать материал так, чтобы он был пригоден для точного
определения вида (см. характеристику отделов и семейств).
Гербарий следует собирать в умеренном количестве и ни в
коем случае не допускать сбора всех встреченных экземпляров ре-
дких видов. Для закладки гербария в полевых условиях использу-
ются специальные ботанические папки. Растения при закладке т,ер-
бария должны быть тщательно расправлены и по возможности слегка
спрессованы. Одновременно с закладкой растений в каждый лист
вкладывается этикетка, которая заполняется на месте сбора. На
этикетке записывается географический пункт, описывается место-
обитание, указывается дата сбора и фамилия собравшего. Особое
внимание следует уделять характеристике местообитания. Напри-
мер, если растение собрано в лесу, нужно точно написать, какой
это лес, в какой его части вид обитает (по опушкам, на разре-
женных местах и т.д.), относится к числу доминантов или встре-
чается изредка, или единично и т.д. Неправильно написать “На
лугу". Нужно также сказать о местоположении луга, рельефе, о
степени увлажнения, по возможности • о доминирующих видах и о
роли в этом сообществе того вида, к которому составляется эти-
кетка. Гербарий должен быть не только правильно собран и эти-
кетирован, но и хорошо высушен. Для этой цели сразу после про-
ведения однодневного маршрута гербарий закладывается в метали-
ческие сетки для сушки. При этом только очень влажные (водные,
болотные растения) перекладываются в другие, сухие листы бума-
ги. Все остальные растения оставляют в тех же листах, в кото-
рых они были заложены, но перекладывают специальными сукнами
или бумажными “матрасиками". Перекладывать гербарий сухими
сукнами или матрасиками необходимо ежедневно, при этом нужно
раскладывать каждый лист, чтобы растения охватило воздухом.
- 32 -
При высушивании повышенно сочных цветков i эльзуются прокладка-
ми из ваты или мягкой бумаги. После того, как гербарий значи-
тельно подсохнет; его досушивают в тех же прессах, но без
прокладок. Сушить лучше всего на свежем воздухе, только при
необходимости в поитпении. Высушенный гербарий перекладывают
вместе с этикетками в сухую бумагу и перевязывают шпагатом в
пачки по 30-40 листов.
Одновременно со сбором гербарных материалов составляются
флористические списки. Флористические списки представляют со-
бой часть геоботанического описания (см. методику геоботаш-
ческого описания). Целью таких списков является детальный учет
распространения вида и исследования условий его обитания. Ши-
роко распространенные виды нет необходимости закладывать в гер-
барий из каждого местообитания, но учесть все местообитания
необходимо. В том же случае, если в разных местообитаниях вид
изменяет свои признаки (внутривидовая изменчивость), образцы
нужно закладывать в гербарий.
ДЛЯ ТОГО, ЧТОбЫ ДОСТаТОЧНО ПОЛНО ВЫЯВИТЬ ВИДОВОЙ СОСТс
распространение видов, их местообитание., необходимо проводить
маршрутные флористические исследования в одном районе в тече-
нии 2-3 лет, охватив весь вегетационный период.
Наряду с маршрутным методом в настоящее время все шине
используется метод конкретных флор. Автор этого метода
А.И.Толмачев считает, что на обширных пространствах флора мо-
жет быть только условно однородной. Состав флоры существенно
изменяется в зависимости от дифференцированности физико-геог-
рафических условий. Действительно, однородной может быть толь-
ко флора малых территориальных единиц, в пределах которых виды
сочетаются в зависимости от конкретных условий местообитания,
при повторении которых повторяется состав и структура флоры.
Такую вполне однородную флору и называют конкретной флорой.
Площадь конкретных флор различна в разных физико-географи-
ческих условиях, в среднем она может колебаться от 100 до 500
кв. км.
При выделении конкретных доор исследователь проводит ре-
когносцировочное исследование, с помощью которого выделяет ми-
нимальный учас ок, на котором в полной мере представлена фло-
ра, однородная на более обширной площади. Этрт участок деталь-
но изучается в качестве конкретной флоры. Далее при определен-
33 -
ных изменениях'экологических условий и видового состава выде-
ляется новый участок конкретной флоры.
При изучении отдельных горных хребтов на юге Сибири
исследователями выделялось по 4-6 конкретных флор. На участ-
ках конкретных флор собирают материалы для сравнительного ана-
лиза флор как в пределах изучаемой территории, так и по срав-
нению с другими регионами. Метод конкретных флор обеспечивает
высокую точность флористических исследований. На основе поле-
вых флористических исследований и обработки собранных материа-
лов, составляют конспекты, или ’’флоры”.
Более глубоким теоретическим исследованием является срав-
нительный анализ флор. Учитывается богатство флоры (общее ко-
личество видов), структура (соотношение основных таксонов в ее
составе), соотношение экологических групп и некоторые другие
показатели. Эти данные используются при решении сложных воп-
росов, связанных с познанием генезиса современных флор.
При флористических исследования всегда уделяется внимание
выявлению хозяйственно ценных видов, их распространению, оби-
лию. Практически ценные виды подразделяются по характеру их
приименения: дикорастущие пищевые, кормовые, лекарственные,
технические. Учитывая практическую ценность, распространение и
обилие видов, определяется хозяйственная ценность изучаемой
Флоры и могут быть рекомендованы пути ее использования.
Одной из важных задач флористических исследований явля-
ется выявление на изучаемой территории редких и исчезающих
видов. Детально описываются условия местообитания, в которых
встречаются ред^е виды (растительное сообщество, рельеф, соп-
ровождающие виды и т.д.). Выясняются причины, неблагоприятные
для редких видов, вызывающие сокращение их. распространения.
Все выявленные- местонахождения редких видов наносятся на схе-
матическую физико-географическую карту обследуемого района.
Флористические,исследования не могут быть резко отграни-
чены от геоботанических, те и другие направлены на разносторо-
нне изучение естественного растительного покрова.
МЕТОДИКА ГЕОБОТАНИЧЕСКИХ ИССЛЕДОВАНИЙ
Даже беглое знакомство с природой показывает, что виды
растений в большинстве случаев произрастают не изолированно
34 -
ДРУГ от друга, а совместно, образуя более ..ли менее сомкнутые
группировки. При этом по территории виды растений распределены
не равномерно и не случайно, а в соответствии с их экологи-
ческими потребностями локализуются по определенным местообита-
ниям. В результате в каждом конкретном местообитании формиру-
ется группировка совместно произрастающих растений, которая
характеризуется определенным видовым составом и строением. Та-
кая группировка растений называется растительным сообществом,
или фитоценозом.
Известно,/что любая даже сравнительно небольшая террито-
рия, и тем более крупный географический район, никогда не бы-
вают однородными по условиям существования растений. Они ха-
рактеризуются более или менее сложным рельефом, неоднородными
горными породами, разнообразными почвами и т.д. В результате
данная территория расчленяется на массу самых разнообразных
местообитаний, и в каждом из них формируется особый фитоце-
ноз. Так вот вся совокупность фитоценозов той или иной терри-
тории, а также всей Земли в целом образует их растительный
покров, или растительность.
К понятию растительного покрова и фитоценоза можно подой-
ти и с иных позиций. Известно, что к настоящему времени расте-
ния освоили и плотно заселили на Земле все пригодные для них
местообитания, образовав более или менее сомкнутый слой расти-
тельной массы, который покрывает значительные пространства су-
ши, прерываясь лишь на участках безжизненных жарких, холодных
или соленых пустынь и на месте некоторых антропогенных соо-
ружений. Этот слой сомкнутой растительной массы и представляет
собой растительный покров.
Растительный покров Земли в целом и отдельных ее участков
неоднороден на всем занимаемом им пространстве, что обусловле-
но прежде всего неоднородностью на этом пространстве физи-
ко-географических факторов: климата, рельефа, почв и т.д. К
наиболее заметным неоднородностям относятся такие формы расти-
тельного покрова, как например, леса, луга, болота, тундры и
т.д. Эти формы растительного по' эова оказываются также неодно-
роднымр внутри себя и подразделяются на более мелкие структу-
ры. И самой меткой структурой является фитоценоз, который
представляет однородный по видовому составу, строению и усло-
виям обитания участок растительного пежрова.
35 -
По происхождению фитоценозы подразделяются на естествен-
ные и искусственные.
Естественные фитоценозы - это исторически сложившиеся
группировки растений, сформировавшиеся в результате естествен-
ного расселения и распределения по местообитаниям различных
видов. Примерами естественных фитоценозов являются однородные
участки естественного леса, естественного луга, болота и т.д.
Искусственные фитоценозы, называемые агрофитоценозами, созданы
человеком и обычно существуют благодаря поддержке с его сторо-
ны. Примерами агрофитоценозов являются однородные участки
посевов хлебных злаков, сада, огорода и т.д.
Титоценозы можно классифицировать, то есть объединять по
сходству каких-либо признаков в таксономические категории раз-
ных рангов. Основными единицами классификации фитоценозов счи-
таются ассоциация, формация и тип растительности, из которых*
первая является низшим, а последняя - высшим таксоном. Ассоци-
ация объединяет фитоценозы, сходные по видовому составу, коли-
чественным соотношения видов, структуре и условиям местообита-
ния. Например, все однотипные фитоценозы соснового леса с на-
почвенным покровом из брусники (Vaccinium vitls-ldaea L.) и
зеленых мхов, связанные с однородными подзолистыми песчаными
почвами;, относятся к одной ассоциации. Наименование ассоциации
дается по названию преобладающих в ней растений по следующему
принципу:' 1. Сосняк черничный - так называется ассоциация
сосновых лесов с господством в напочвенном покрове черники
(Vaccinium myrtillus L.), 2. Кедровник чернично-зеленомошный -
так называется ассоциация кедровых лесов с господством в на-
почвенном покрове черники и зеленых мхов.
Название.ассоциации можно формировать также по типу слож-
ного прилагательного, , например, мятликоьо-косторово-пырейная
ассоциация, типчаково-ковыльная ассоциация и т.д. При таком
способе, применяемом в основном для наименования травяных
сообществ, на последнее место ставится наиболее обильный или
наиболее важный компонент ассоциации.
Формация объединяет ассоциации, в которых в главном ярусе
доминирует один и тот же вид растений. По этому виду и дается
название формации. Например, все ассоциации с господством в
древостое сосны обыкновенной (Plnus sylvestris L.) образуют
36 -
сосновую формацию, или формацию сосняков. Аналогично выделяют
формации кедровников (Pinus sibirica Мауг.), пихтарников
(Ables sibirica Ledeb.), пырейных (Agropyron repens R.B.) лу-
гов, ковыльных (Stipa joannis Gel.) степей и т.д.
Формации объединяются в типы растительности по принадлеж-
ности господствующих в главном ярусе видов к одной и той же
крупной жизненной форме типа: дерево, кустарник, травянистый
мезофильный многолетник и т.д. Выделяют такие, например, типы
растительности, как леса, кустарниковые сообщества, луга, сте-
пи.
Весь круг вопросов, связанных с изучением разных сторон
фитоценозов: их структуры, классификации, распределения в
пространстве и т.д., - решает одна из ботанических наук, полу-
чившая название геоботаники, или фитоценологии. Поэтому все
исследования, направленные на изучение фитоценозов, называют
геоботаническими исследованиями.
Методика геоботанических исследований включает обширный
арсенал приемов, позволяющих получать разностороннюю информацию
о фитоценозах. Однако при маршрутных герботанических исследо-
ваниях, с которыми студенты как раз и знакомятся на учебной
практике, приходится выполнять прежде всего следующие виды ра-
бот: составлять геоботанические описания, прокладывать геобо-
танические профили, составлять карту растительности обследуе-
мого района. Ниже приводится краткое описание методики выпол-
нения перечисленных работ.
Методика составления геоботанических описаний .
Под геоботаническим описанием понимают характеристику,
составляемую исследователем в полевых,условиях тому или иному
конкретному фитоценозу. Конкретные фитоценозы нередко занимают
более или менее крупные площади, и описать фитоценоз на всей
этой площади не представляется возможным, да в этом и нет не-
обходимости. Геоботанические описания составляются на ограни-’
ченных участка;: фитоценозов, на так называемых "пробных площа-
дях", которые закладываются либо в типичных местах фитоцено-
зов, либо по маршруту черев равные расстояния, но с обязатель-
ным выполнением следующего требования - пробная площадь должна
быть однородной (или однотипной) на всем протяжении.
Для описания разных типов растительных сообществ применя-
ют пробные площади различных размеров: от 4-100 кв. м в тра-
вянистых фитоценозах до 1000-2500 кв. м в лесных сообществах.
Установлено, что пробная площадь таких размеров дает вполне
удовлетворительную характеристику всему фитоценозу, в пределах
которого она закладывается. Форма пробной площади может быть
квадратной или прямоугольной. Для удобства работы пробная пло-
щадь ’’отбивается” в пространстве: стороны ее промеряются ру-
леткой или шагами, а по углам устанавливаются вешки или колыш-
ки.
При описании пробной площади учитываются нс только приз-
наки самого фитоценоза, но и основные условия физико-географи-
ческой среды, в которых формируется данный фитоценоз. Чтобы
придать описанию пробной площади определенную систематичность
и последовательность и не пропустить какого-либо существенного
признака, обычно польз.'ются заранее заготовленным бланком, ку-
да и заносится по определенным рубрикам вся необходимая инфор-
мация. Применяют различные формы бланков в зависимости от то-
го. какой тип растительности описывается и какая ставится
конкретная задача при планировании геоботанических исследова-
ний. Однако при составлении геоботанического описания по блан-
ку любой формы оно должно содержать следующую информацию:
1. Номер описания.
2. Дата составления описания.
3. Фамилия автора описания.
4. Размер пробной площади и примерная площадь описываемо-
го фитоценоза.
5. Географическое положение пробной площади - область,
район, направление и расстояние от ближайшего населенного
пункта.
6. Геоморфологические условия, в которых сформировался
описываемый фитоценоз. Указывается положение фитоценоза в мак-
ро- и мезорельефе, приводится краткая характеристика микро- и
нанорельефа. При этом к макрорельефу относятся крупные формы
рельефа, определяющие общий облик большого участка земной по-
верхности: горные хребты, плоскогорья, низменности, крупные
речные долины и т.д. К мезорельефу относятся мелкие -формы
рельефа, высота которых измеряется метрами или десятками мет-
ров’, а протяженность сотнями метров или километрами, например,
- 38 -
небольшие долины, балки, отроги хребтов, речные террасы. К
микрорельефу относятся еще более мелкие формы высотой несколь-
ко метров и протяженностью в несколько десятков или сотен мет-
ров, например, песчаные бугры, пойменные гривы и межгривные
ложбины, гряды верховых болот. И наконец, к нанорельефу от-
носятся самые мелкие формы рельефа, высота и протяженность ко-
торых измеряется дециметрами. Примерами нанорельефа являются
муравьиные и осоковые кочки, прикомлевые повышения в лесу, мо-
ховые кочки и мочажины на сфагновых болотах и т.д.
7. Условия увлажнения. Указывается тип увлажнения (ат-
мосферное, грунтовое, натечное, смешанное), степень увлажнения
(избыточное, нормальное, недостаточное) и характер проточности
влаги (застойная, затрудненно-проточная, проточная).
8. Почвенные условия. Закладывается и описывается почвен-
ный разрез или, в крайнем случае, по прикопке определяется
почвенная разность и название ее записывается в бланк.
9. Окружение описываемого фитоценоза другими растительны-
ми сообществами, пожарищами, дорогами, постройками и т.д. Н
обходимо не только перечислить все элементы ландшафта, окружа-
ющие фитоценоз, но и у1сазать, на каком расстоянии они распола-
гаются от пробной площади.
10. Название фитоценоза и ассоциации, которое составля-
ется по указанным выше принципам. Из приведенного выше опреде-
ления ассоциации следует, что ассоциации представляют собой
тип сходных (относимых к ней) фитоценозов, в виде которых она
и существует в природе. Следовательно, каждый из этих фитоце-
нозов является конкретным предста .теЛем ассоциации как типа и
несет все ее основные признаки и ‘свойства. Поэтому названия
фитоценозов совпадают с названиями ассоциации, к которой они
относятся. Например, название “сосняк.брусничный** следует дать
каждому встреченному в обследованном районе конкретному участ-
ку соснового леса с напочвенным покровом из брусники
(Vaccinlum vitis-ldaea) и ассоциации, в состав которой входят
данные фитоценозы.
11. ВертигАйьное строение . адземной части фитоценоза,
Основной структурой вертикального строения фитоценоза является
ярус, под которым понимается более или менее четко отграни-
ченный слой растительной массы, образованный растениями од-
чзи жизненной формы - деревьями, кустарниками, травами и т.д.
Ярусы Формируются в результате подбора видов в фитоценозе,
способных произрастать совместно, используя разные горизонты
среды обитания. Это обеспечивает более полное использование
фитоценозом его местообитания, В разных фитоценозах отмечается
разное количество ярусов: от одного в травянистых сообществах
до 3-5 в лесах. Счет ярусов ведется сверху вниз, так что самый
высокий ярус считается первым, а самый низкий - последним. От^
дельный ярус может быть образован одним или несколькими, не-
редко многими•видами растений. При этом преобладающие в ярусах
виды называются доминантами. После расчленения надземной части
фитоценоза на ярусы для каждого из них указывают высоту,
густоту (.плотность), характер горизонтального сложения (равно-
мерное или неравномерное) и доминанты.
Густота или плотность стояния растений в фитоценозе оце-
нивается показателями ’’проективного покрытия”, под которыми
понимается степень покрытия почвы надземными частями растений
в результате их проекции на поверхность почвы. При учете
густоты надземной массы всего фитоценоза говорят об “общем
проективном покрытии”, а при оценке густоты отдельного яруса -
об ’’ярусном проективном покрытии”. Проективное покрытие обычно
выражается в процентах от поверхности пробной площади: 100 %
проективного покрытия означает, что поверхность почвы сплошь
покрыта проекциями растений, а ?0 % проективного покрытия оз-
начает, что 30 % поверхности почвы остается непокрытой надзем-
ными частями растений и просматривается при взгляде сверху.
Имеется немало различных приемов и приспособлений, обеспечива-
ющих достаточно высокую точность учета проективного покрытия.
Однако все они являются довольно трудоемкими. Поэтому при
маршрутных исследованиях проективное покрытие оценивается
обычно глазомерно и при определенном опыте исследователя гла-
зомерная оценка дает вполне удовлетворительные результаты.
12. Видовой (флористический) состав фитоценоза выявляется
путем тщательного осмотра пробной площади и составления списка
зафиксированных на ней видов. Список должен включать: во-пер-
вых, не только нормально развитые цветущие или плодоносящие
растения, но таюке и растения, находящиеся в вегетативном
состоянии, хотя бы даже в виде всходов или сильно угнетенных
взрослых особей; и, во-вторых, не только цветковые растения,
но и напочвенные и зпифитные мхи, лишайники, а также (хотя бы
40 -
шляпочные) грибы. Для облегчения работы список видов составля-
ется по ярусам, а в пределах каждого яруса выявление видов ве-
дется, начиная с наиболее обильных и хорошо^ заметных видов и
постепенно переходя к менее заметным видам. Если при этом
встречаются виды неизвестные или нетвердо известные, то в
списке они обозначаются соответствующими порядковыми номерами
и затем закладываются в гербарий, на этикетках которого указы-
вается номер описания и порядковый номер вида в списке. После
определения гербария Флористический список дополняется и уточ-
няется, то есть вместо соответствующего номера в нем простав-
ляются названия растений. В списке указываются только ла-
тинские названия видов.
Попутно с выявлением видового состава фитоценоза и оформ-
лений флористического списка проводится оценка количественно-
го участия каждого вида в сложении фитоценоза, а также указы-
вается xapaffrep распределения видов по площади фитоценоза и их
фенологическое состояние.
13. Количественное участие вида в сложении фитоценоз а
можно оценить разными показателями: его обилием (числом особей
на единицу площади), весом и видовым проективным покрытием
(покрытием, создаваемым всеми особями данного вида). При марш-
рутных исследованиях обычно используют показатели обилия и ви-
дового проективного покрытия, применяя наиболее простые глазо-
мерные способы их определения.
Обилие видов на пробной площади чаще всего оценивается по
так называемой ’’шкале Друде", которая включает следующие 7
градаций (классов) обилия, имеющих латинское наименование и
записывающихся в сокращенном виде!
Social is - Зое - абсолютное господство, вида в фито- ценозе пр числу особей, участие
других видов незначительно;
Сорlosus -- Cop 3 - особей вида очень много, но абсо- лютного господств^, нет, более или менее значительное участие прини- маю. и другие виды:
- Cop 2 особей вида много;
- Cop 1 - особей вида довольно много;
Spar sus Sp особей ЯНЫ по вила мало, они редко рассе- пробиой площади;
- 41 -
SolItar- Sol - особей вида очень мало, они еди-
нично отмечаются на пробной площа-
ди;
Unicum - Un - встречена единственная особь вида
на пробной площади.
Видовое проективное покрытие оценивается в баллах, каждо-
му из которых соответствует определенный диапазон (класс) зна-
чений проективного покрытия. Применяется несколько различных
шкал баллов проективных покрытий; из них самой простой и удоб-
ной является следующая 5-ти балльная шкала:
4- - вид представлен единичными экземплярами и соз-
дает проективное покрытие меньше 1 %;
1 балл - от 1 до 5 % покрытия;
2 балла - от б до 15 X покрытия;
2 баяла - от 16 до 25 % покрытия;
4 балла - от 26 до 50 Z покрытия;
5 баллов - свыше 50 % покрытия.
Градации приведенных шкал надежно определяются глазомерно
при определенном навыке исследователя. В то же время эти шкалы
являются достаточно дробными и получаемые с их помощью оценки
количественного участия видов в фитоценозе позволяют удовлет-
ворительно решать многие геоботанические вопросы. Причем оби-
лие и видовое проективное покрытие отражают собой разные
аспекты количественного участия видов, поэтому рекомендуется
использовать оба эти показателя одновременно.
14. Распределение растений по площади фитоценоза может
быть более или менее равномерным (диффузным) или явно неравно-
мерным (групповым). Характер распределения растений зависит, с
одной стороны, ст формы роста того или иного вида, с другой
стороны, от неоднородности микроусловий местообитания в преде-
лах фитоценоза, вообще, и пробной площади, в частности. Разли-
чают следующие Форш роста, которые в бланке описания обозна-
чают сокращенно начальными буквами:
I. Каждая особь вида представлена одним надземным побегом
-- "О”;
2. Каждая особь несет несколько или много надземных побе-
гов от одной общей подземной части:
а) стебли в небольшом числе растут кустом или пучком
от одного корневища или корня - "пи",
- 42 -
б) многочисленные побеги образует плотную дерновину
или подушку - ”д",
в) побеги растут более или менее рыхлой зарослью
(латкой) - "л".
Вследствии неоднородности местообитания по площади фито-
ценоза виды независимо от формы роста могут образовать различ-
ные скопления, именуемые пятнами, куртинами или группами и
сокращенно обозначаемые в бланке ч описания соответственно:
ипт”, "к", "гр",
15. Фенологическое состояние видов характеризуется учетом
сезонной фазы развития (фенофазы), в которой находится г мо-
мент описания тот или иной вид растения. Ниже приводится пере-
чень основных фенофаз и их условные обозначения:
- - вегетация до начала цветения,
) - зацветание (и бутонизация),
о - полное цветение
( - отцветание,
+ - созревание плодов,
# - плоды зрелые,
« - вегетация поле осыпания плодов.
Поскольку каждый вид в фитоценозе (и на пробной площади)
почти всегда представлен не одной, а многими особями, которые
в момент описания проходят разные стадии развития, постольку
фенологическое состояние вида оценивается по той фенофазе, в
которой находится большинство его особей.
16. Влияние человека и животных: хозяйственное значение и
использование фитоценоза; краткие предварительные рекомендации,
по рациональному использованию и охране.
Содержащаяся в геоботанических описаниях информация поз-
воляет разрабатывать классификацию . фитоценозов, выявлять
состав и,структуру ассоциаций и других таксонов, решать воп-
росы рационального использования и охраны растительности и
т.д. Однако для решения всех этих (и ряда других) задач боль-
шое значение имеют также данные о распределении фитоценозов в
пространстве и о связи их с физико-географическими факторами
(рельефом, почвами и т.д.). Эти дополнительные сведения пору-
чают в результате проложения геоботанических профилей и геобо-
танического карт ирсвания.
- 43 -
Проложение геоботанического профиля
Под геоботаническим профилем понимают ряд конкретных фи-
тоценозов, последовательно сменяющих друг друга в зависимости
от смены экологических условий при движении по прямой от ка-
кой-то начальной точки к конечной. Следовательно, геоботани-
ческий профиль отражает собой конкретный экологический ряд фи-
тоценозов. Геоботанический профиль вычерчивается в определен-
ном масштабе на бумаге, что позволяет наглядно отравить связь
фитоценозов с устоьиями среды и облегчает выявление основных
закономерностей этой связи. При проложении геоботанических
профилей учитывается прежде всего связь фитоценозов с релье-
фом. Дело в том, что рельеф является одним из важных косвенно
действующих экологических факторов. Именно поэтому*с изменени-
ем рельефа изменяются все основные условия существования расте-
ний - влажность, аэрация, температура, содержание элементов
питания и т.д. К тому же рельеф относится к легкоучитываемьы
даже при маршрутных исследованиях факторам. Кроме рельефа при
проложении геоботанического профиля могут учитываться характер
почв и горных пород, глубина грунтовых вод и некоторые другие
Факторы.
При закладке геоботанического профиля очень важно выбрать
правильное место проложения профиля: его направление, начало и
конец. Закладывать профиль рекомендуется так, чтобы он вкрест
пересекал все основные формы рельефа: в равнинных районах про-
фильный ход лучше вести от уреза реки вкрест речных террас и
дальше к вершине водораздела, а в горах - вкрест направления
горных хребтов. Точное направление профиля задается компасом
или бусолью, а начальная точка его привязывается к хорошо за-
метному постоянному ориентиру на местности с тем, чтобы при
необходимости профиль легко было отыскать в будущем.
Существуют два основных способа прокладки геоботанических
профилей: глазомерный и инструментальный - различающихся по
трудоемкости работ и точности получаемцх результатов. Суть
простого, но зато и неточного глазомерного геоботанического
профилирования заключается в том, что протяженность фитоцено-
зов по ходу профиля промеряют шагами, а изменение рельефа учи-
тывают на глаз. При более точном, инструментальном геоботани-
ческом профилировании . протяженность фитоценоза по профилю из-
44
меряется мерной лентой или рулеткой, а игленение рельефа спе-
циальным прибором - нивелиром. Инструментальное геоботани-
ческое профилирование состоит из следующих операций.
После выбора и направления профиля приступают к подготов-
ке профильной линии. Прежде всего профильную линию расчищают
от деревьев, кустарников и высоких трав, которые могут мешать
гивелированию, и фиксируют линию установкой на ней вешек. За-
тем профильная линия разделяется вбиваемыми в землю колышками
(пикетами) на равные отрезки протяженностью 100-300 м. Эти пи-
кеты, называемые основными, нумеруются в направлении от на-
чальной (нулевой) точки к конечной (0 пикет, 1 пикет, 2 пикет,
3 пикет и т.д.). Кроме основных пикетов на профильной линии
устанавливаются на всех точках перегиба рельефа и пересечения
профилем границ фитоценозов дополнительные пикеты, которые
обозначаются следующим образом: 1) 5+35 - это значит, что до-
полнительный пикет расположен по ходу профиля дальше 5 пикета
на 35 м: 2) 15+10 - дальше 15 пикета на 10 м и т.д. И, нако-
нец, между основными пикетами в середине вбиваются еще колья
без номеров или устанавливаются флажки,. которыми отмечаются
точки постановки нивелира, называемые "станциями". На этом за-
канчивается подготовка профильной линии.
Затем приступают к нивелированию. Суть нивелирования зак-
лючается в том, что с помощью нивелира, устанавливаемого на
треноге последовательно на всех‘станциях, и двух разграфленных
сантиметровыми делениями реек, устанавливаемых у соседних пи-
кетов,, определяется степень повышения (или понижения) мест-
ности у каждого последующего пикета по сравнению с предыдущим
пикетом. Результаты измерений записываются в специальный жур-
нал.
Пс мере продвижения с нивелиром.по ходу профиля произво-
дится выделение фитоценозов, установление их границ не только
по профильной линии, но и на некотором расстоянии от нее по
обе стороны, и картирование растительности i.a миллиметровой
бумаге в определенном масштабе. Нужно помнить, что геоботани-
ческий профиль - это не линия, а полоса шириной 5-250 м в за-
висимости от масштаба работ. Вот на этой полосе и картируется
растительность, или, как говорят, составляется арбис. Кроме
того по ходу профиля составляются геоботанические описания
различных фитоценозов, собирается гербарий, для каждого фпто-
- 45 -
ценоза с той или иной степенью детальности оцениваются почвен-
ные условия, а при необходимости учитываются и некоторые дру-
гие данные по растительности и среде обитания.
Затем все полевые материалы обрабатываются» систематизи-
руются и геоботанический профиль оформляется графически в такой
последовательности: 1) сначала на миллиметровой бумаге на-
носится С в заранее выбранном горизонтальном и вертикальном
масштабе) рельеф с указанием высоты переломных точек; 2) затем
на линию профиля наносятся условные обозначения растительных
сообществ и их границ; 3) и, наконец, внизу профиля на специ-
альных сопряженных с профильной линией полосах изображаются
(красками, штриховкой, условными обозначениями или цифровыми
индексами) сведения о почвах, горных породах, уровне грунтовых
вод и т.д., а также помещается абрисная карта растительности
(рис. 11).
Составление геоботанической карты
Под геоботанической картой понимают такую карту, на кото-
рой обозначены в определенном масштабе контуры ассоциаций,
групп ассоциаций, формаций и других единиц растительности или
контуры разнообразных сочетаний этих единиц. Кроме того, на
геоботанической карте в зависимости от целей ее составления
могут отражаться и другие различные сведения: некоторые осо-
бенности среды (рельеф, почвы и т.п.), хозяйственное использо-
вание растительности, продуктивность растительных сообществ,
степень влияния на растительность человека и т.д. Весь процесс
составления геоботанической карты от начала и до конца называ-
ется геоботаническим картированием. Он подразделяется на два
основных этапа: 1. Полевая геоботаническая съемка, в результа-
те которой составляется предварительный полевой вариант (ма-
кет) карты. 2. Камеральная обработка материалов и составление
окончательного варианта (оригинала карты).
Полевая геоботаническая съемка выполняется на картографи-
ческой основе определенного масштаба, в качестве которой могут
использоваться: топографические карты, материалы аэрофотосъем-
ки, а также планы лесо- и землеустройства. На эту основу и на-
носятся в полевых условиях геоботанические контуры. Различают
два основных метода полевой съемки: площадный и маршрутный.
46 -
Рис. И. Геоботанический профиль через поперечник поймы
Иртыша.
Подтипы растительности: а * луговые степи, б - остепнен-
ные луга, в - настоящие луга, г - сырые и болотистые луга, д
- минеральные торфяные болота, е - прибрежно-водная и водная
растительность. Ассоциации: 1 - узколистномятликово- кострецо-
вая. 2 - заменяющеполынно-типчаковая, 3 - кострецово-пырейная,
4 - двуряднооссково-пырейная, 5 - частухово-пузырчатоосоковая,
6 - ситниково-двуряднооссково-пырейная, 7 - аирово-телорезовая
и аироьо-пузырчатоосоково-тростниковая, 8 - частухово-аиро-
во-пузырчатоосоковая, 9 - ситниког'-двурядноосоково-пырейная,
ситниковая и ситниково-солеросовая,- 10 - ситниково- гусинолап-
чатково-побегообразующеполевицевая, И - тростянковая.
Почвы: I - пойменная луговая слоистая супесчаная, II -
пойменная луговая остепняющаяся слоистая мелкопесчаная, Ш -
пойменная луговая слоистая супесчаная и пойменная луговая сло-
истая легкосуглинистая, IV - пойменная влажнолуговая среднесуг-
линистая. У - пойменная лугово-болотная тяжелосуглинистач, VI -
пойменная влажнолуговая слабосолс гчаковатая среднесуглинистая,
VII - пойменная лугово-болотная слабосолончаковатая тяжелосуг
линистая, УШ - пойменные влажнолуговые средне- и сильносолоча-
коьатые тяжелосуглинистые и пойменные влажнолуговые солончаки,
IX пойменная вдажнолуговая среднесолончаковагая среднесугли-
пистал. X пойменная торфянисто-иловато-болотная слаббсолонча-
X 1_ _ ло.именная. /го^фянрбо^ютн^^ ..
- 47 -
1. Площадная съемка осуществляется путем обхода и осмотра
всей картируемой территории. Во время этого обхода выделяются
все единицы растительного покрова, которые могут быть показаны
в заданном масштабе, и устанавливаются их границы. Для облег-
чения работ вся картируемая территория разбивается пикетами
(вешками или колышками) на сеть мелких площадок следующим об-
разом: 1. Сначала провешиваются с помощью компаса или бусоли
так называемые базисные линии, определяющие внешние границы
участка. 2. Затем на базисной линии через равные расстояния
устанавливаются пикеты. 3. И, наконец, от каждого пикета на
базисных линиях под углом 90 градусов визируются поперечные
ходы, которые также разбиваются пикетами, устанавливаемыми че-
рез те же равные расстояния. Такая же пикетная сеть изобража-
ется в соответствующем масштабе и на топооснове.
Расстояние между пикетами зависит прежде всего от масшта-
ба съемки. Так, например, при геоботанической съемке в масшта-
бе 1 : 25000 расстояние между пикетами должно составлять 25 м,
при съемке в масштабе 1 ; 50000 - 50 м и т.д. При разбивке пи-
кетажной сети очень важно предусмотреть такую ориентацию ее
линий, чтобы они пересекали все основные элементы рельефа в
пределах картируемого участка. Это позволит полнее выявить
пространственную структуру растительного покрова.
В результате разбивки пикетажной сети обследуемая терри-
тория расчленяется на сеть мелких квадратов, внутри которых и
ведется картирование растительности. Осуществляется это следу-
ющим способом: геоботаник продвигается по всем поперечным хо-
дам от пикета к пикету (рис. 12); по мере движения он выявляет
единицы растительности, измеряет их протяженность, и наносит на
топооснову. При этом протяженность выделяемых единиц расти-
тельности по Линии пикетов измеряется мерной лентой, рулеткой
или шагами, а протяженность и очертания их контуров внутри
квадратов между проходимым в данный момент и двумя соседними
ходами оцениваются глазомерно.
В ходе полевой съемки помимо картирования единиц расти-
тельности (или их пространственных комбинаций) производится
также краткая характеристика каждого выделяемого на карте кон-
тура, в нужных местах составляются геоботанические описания
фитоценозов, собирается гербарий и т.д. Все эти материалы за-
тем используются при изготовлении оригинала геоботанической
карты в камеральный период.
- 48 -
Рис. 12. Схема рабочих ходов при глазомерной площадной
съемке: АБ и АВ - базисные линии.
Площадная съемка является довольно трудоемким методом ге-
оботанического картирования. Поэтому она применяется лишь при
выборочном крупномасштабном (1 : 25000 и крупнее) картировании
небольших территорий, когда ставится задача получения деталь-
ной карты, на которой отражаются низшие единицы растительности
- ассоциации или группы ассоциаций.
2. При маршрутной съемке выявление картируемых единиц
растительности и установление границ между ними осуществляется
путем непосредственных наблюдений в природе только вдоль марш-
рутов, в промежутках же между ними границы картируемых единиц
растительности устанавливаются косвенным путем. Маршрутная
съемка применяется в основном при составлении геоботанических
карт среднего масштаба (1 : 50000 - 1*: 100000).
При маршрутной съемке качество ее в значительной степени
зависит от направления и способов проложения маршрутов, а так-
же от густоты маршрутной сети. Маршрута должны распределяться
таким образом, чтобы в пределах картируемой территории с наи-
большей полнотой выявить все типы растительных сообществ,
установить пространственное расположение, форму и размеры кон-
туров и границы между ними. 'Для этого лучше всего маршруты
прокладывать вкрест простирания основных элементов рельефа,
49,-
так чтобы они проходили от наиболее высоких точек водоразделов
до самых низких отметок речных долин и пересекали все формы
рельефа.
По своим очертаниям маршруты могут быть различными. На
практике чаще всего применяют два способа проложения маршру-
тов: способ параллельных пересечений и способ петель. При
съемке способом параллельных пересечений маршруты прокладыва-
ются линейно через равные расстояния, различные в зависимости
от масштаба картирования, и равномерно покрывают всю площадь.
При этом направления рабочих ходов могут быть приблизительно
параллельными и не всегда абсолютно прямыми. При прокладывании
маршрутов по способу петель территория расчленяется на отдель-
ные участки (секторы) применительно к природным особенностям
местности. Затем из центра каждого сектора совершаются после-
довательные петлеобразные обходы (или объезды) территории в
радиальных направлениях. Первый способ проложения маршрутов
применяется обычно при картировании территорий со слабо расч-
ленненым рельефом и относительно несложна растительным покро-
вом, а второй - при геоботанической съемке сильно пересеченной
местности. Иногда приходится сочетать оба способа приложения
маршрутов.
Густота маршрутной сети определяется нельм рядом обстоя-
тельств: качеством картографической основы, масштабом съемки,
сложностью всего ландшафта в целом и его растительного покрова
и т.д. Этот вопрос приходится решать особо в каждом конкретном
случае, учитывая следующие придержки, наиболее часто применяе-
мые в практике геоботанического карти* звания:
Масштаб Среднее расстояние между
рабочими ходами (в км)
1:25000 0.25 - 1.0
1:50000 1.0 - 3.0
1:100000 3.0 - 5.0
1:200000 5.0 - 7.0
Сам процесс маршрутной геоботанической съемки выполняется
в такой последовательности: 1. Сначала производится реког-
носцировочный обход (или объезд) и осмотр вей территории, под-
лежащей ооследованию; 2. Затем на топооснову наносится сеть
рабочих ходов (маршрутов); 3. И, наконец, проводится последо-
вательная отработка маршрутов.
50 -
При этом по мере продвижения ни маршруту пешком, на лоша-
ди иди на автомашине геоботаник ь дет глазомерное картирование
единиц растительности в предел.?-; видимой полосы, при необходи-
мости совершая небольшие дополнительные отходы от маршрута. По
ходу собираются также и другие полевые материалы. Границы кар-
тируемых единиц растительности между соседними маршрутами
устанавливаются в лагере экспедиции методом интерполяции
(’’вставки”), которая осуществляется с учетом всех имеющихся в
распоряжении геоботаника сведений: полевых наблюдений, картог-
рафических материалов по почвам, геоморфологии, геологии, ли-
тературных данных и т.д.
В последние десятилетия широкое применение получила марш-
рутная съемка с использованием аэрофотоснимков, которая осу-
ществляется методом так- называемого маршрутного дешифрирова-
ния. Суть метода заключается в том, что по ходу маршрутов вы-
полняется полевое дешифрирование растительности по аэросним-
кам, т.е. выявление связи определенных типов растительности с
характером изображения их на аэрофотоснимке. И затем выявлен-
ные признаки дешифрирования экстраполируются на всю картируе-
мую территорию. В результате на аэрофотоснимках вычерчиваются
геоботанические контуры.
Итогом полевой геоботанической съемки любого масштаба яв-
ляется полевой вариант (макет) геоботанической карты.
В камеральный период обрабатываются и систематизируются
все полевые материалы (почвенные, геоботанические, флористи-
ческие и т.д.); уточняется легенда, т.е. система единиц расти-
тельности, отображенных на геоботанической карте; уточняются
границы контуров карты и составляется авторский макет карты.
При*этом авторский макет содержит уже всю нагрузку геоботани-
ческой карты: единицы растительности обычно раскрашиваются
разными цветами, а вся остальная дополнительная информация от-
ражайся разнообразной штриховкой, условными обозначениями или
цифровыми индексами (рис. 13).
С авторского макета картографами-чертежниками изготовля-
ется в виде копии оригинал геоботанической карты, который за-
тем может быть размножен тем или иным способом.
•51
Рис. 13. Фрагмент карты растительности поймы Иртыша.
Подтипы растительности: а - настоящие степи, б - луговые
степи, в - остепненные луга, г - настоящи луга.. Ассоциации: 1
- Караганове-типчаково-перистоковыльная, 2 - разнотравно-тип-
чаково-перистоковыльная , 3 - типчаково-уаколистномятликово-
кострецовая, 4 - разнотравно-узколистномятликово- кострецовая,
5 - луговомятликово-пырейно-луговолисохвостовая.
ЭКСКУРСИИ
Экскурсии в леса
Лесами называются растительные сообщества, в которых
основную, формирующую роль играют древесные породы.
Различают хвойные и лиственные леса. Хвойные леса наи-
большее распространение имеют в Сибири. Хвойные леса в свою
очеоедь подразделяются на светлохвэйные и темнохвойные.
Древостой светлохвойных лесов состоит из сосны обыкновен-
ной (Fliius syivectris I..') или лиственницы <Larix sloirica
Ledeb.). иногда с небольшой примесью других дрезесных порол.
г’-то светлые, обычно сухие лес<\ в травяном покрове которых
значительное участие принимают растения, приспособленные к су-
ществованию при недостатке паяги (ксерофиты). Светлохвойные
леса приурочены, в основном, к бедным песчаным и каменистым
почвам. Лиственничные леса значительно дальше, чем сосновые,
продвигаются к ^веру, лиственница хорошо приспосабливается к
существованию на почвах с высоким уровнем в чной мерзлоты.
Темнохвойные леса образуют ель (Picea obovata Ledeb.),
пихта (Abies slbirica Ledeb.) и кедр(Р1пиз slbirica Мауг.).
Древостой темнохвойных лесов отличается большой сомкнутостью
крон, под пологом их хорошо развит подлесок из различных
кустарников. Травяной покров характеризуется распространением
гигрофитов-тенелюбов (Oxalis acetosella L., Trientalis
europea L.), лесных папоротников, хвощей. Обильно развит мохо-
вой покров. Почвы более богатые и влажные. Густые темнохвойные
леса или смешанные леса с преобладанием темнохвойных пород на-
зывают тайгой.
Лиственные леса подразделяются также на 2 группы: широко-
лиственные и мелколиственные. Широколиственные леса распрост-
ранены в европейской части СССР и на Дальнем Востоке, в Сибири
совершенно не встречаются, мелколиственные леса формируются
видами березы (Betula pendula Ehrh., В. pubescens Ehrh.) и
осиной (Populus tremula L.). Травяной покров в этих лесах бо-
гатый и разнообразный. Почвы довольно богатые. Березовые и
осиновые леса в большинстве случаев являются вторичными, раз-
вивающимися на месте сведенных вырубок или пожарах хвойных.
Часто встречаются смешанные хвойно-лиственные леса.
Леса занимают обширнейшее пространство на территории Си-
бири, располагаясь сплошной широкой полосой, ограниченной с
севера боной полярных тундр, а с юга - зоной степей. Они под-
нимаются высоко; в горы, образуя лесной пояс.
^есная растительность имеет исключительно большое и раз-
нообразное хозяйственное значение. Не говоря уже о значении
древесных пород, доставляющих древесину, имеющую самое широкое
и разнообразное хозяйственное использование, в состав кустар-
ничкового и травяного ярусов входит большое количество высоко-
ценных пищевых, лекарственных и других полезных растений.
Основной классификационной единицей считается тип леса.
Тип леса определяется по сочетанию лесообразующей породы, под-
53
леска, мохово-травяного покрова, а также с учетом режима влаж-
ности, .характера почвы и других показателей. Например, основ-
ными типами сосновых лесов являются следующие: 1. Сорняк-бело-
мошник. Подлесок и травяной ярус отсутствуют, почва покрыта
лишайниками (главным образом из рода Cladonia). Занимает сухие
песчаняые холмы. 3. Сосняк-зеленомошник. Подлесок развит сла-
бо. Характеризуется обилием полукустарничков (брусника, черни-
ка). Встречаются ксерофиты. Значительного обилия достигают зе-
леные (гипновые) мхи. Почвы хорошо дренированные. 3.
Сосняк травяной. Имеет довольно хорошо развитый подлесок и
обильный, богатый по видовому составу травяной покров. Моховой
покров развит слабо. Почвы умеренно-увлажненные. 4.
Сосняк-долгомошник. Подлесок и травяной покров развиты незна-
чительно. Обильно развивается моховой покров из Polytrlchum
commune Hedw. Почвы в стадии начала заболачивания. 5. Сосняк
сфагновый. Подлесок и травяной покров почти исчезают, обильно
развиваются сфагновые мхи. Почвы заболоченные.
При изучении лесов пользуются методом описания пробных
площадей (размером не менее 1000 кв. м). Описание травяного
покрова леса можно проводить на• площадках 100 кв. м. г тисание
леса проводится, в основном, по той же схеме, что и других
растительных группировок (см. методика геоботанических иссле-
дований), но имеет свои специфические особенности. В лесах, по
сравнению с другими растительными группировками, наиболее хо-
рошо выражена ярусность^ Различают следующие яруса: 1. Дре-
востой, 2. Подлесок (кустарники), 3. ..«Травяной покров. 4. Мохо-
во-лишайниковый покров.
Характеристика древостоя (1 ярус)
Для количественного соотношения пород в древостое подсчи-
тывают, какое количество экземпляров каждой породы приходится
на 10 деревьев, т.е. определяют процентное соотношение пород и
оставляют формулу леса. Например, * 7Е2К1П+Б. Значит: 70 %
составляет ёль, 20 7. - кедр, 10 X - пихта и единично встреча-
ется береза. ЮС - древостой исключительно из сосны.
Далее определяется сомкнутость крон. Для этого, зайдя в
глуоь леса, нудно посмотреть вверх м определить, какую плошадь
занимают кроны о-'Г'-вьев п? ' равнению с иехду
- 54 -
просветами. Сомкнутость крон выражается баллами от 0.1 до 1.
Если кроны настолько соприкасаются между собой, что между ними
не остается просветов - сомкнутость крон = 1, т.е. 100 X.
Практически это почти не встречается. В густых лесах сомкну-
тость крон чаще всего 0.7-0.8; в разрежениях - 0.5-0.6. Для
хозяйственной у рактеристики древостоя определяют прежде всего
количество стволов на 1 гектар. Это делг.тся путем подсчета
стволов на пробной площадке 2500 кв. м и пересчета на гектар.
Далее необходимо определить среднюю высоту деревьев. Су-
ществует несколько способов определения высоты деревьев.
Простейшим из них является глазомерный способ: нужно отойти от
измеряемого дерева на несколько метров, т.е. на такое расстоя-
ние, откуда наиболее хорошо виден весь ствол, и мысленно отк-
ладывать от основания к верхушке по 2 метра. При некотором на-
выке этот метод дает довольно хорошие результаты.
Диаметр стволов определяют обычно на высоте 1.5 м (на
высоте груди). Диаметр можно определить по длине окружностигпо
формуле , где - диаметр ствола, - длина окруж-
ности, - 3.14. Для быстрого определения диаметра стволов в
полевых условиях можно длину окружности делить округленно на
3. Промеров делается несколько (у более мощных и тонких ство-
лов) и определяется среднее.Следующий показатель качества дре-
востоя - возраст деревьев. Возраст деревьев определяется на
спилах или свежих пнях путем подсчета годичных колец. Различа-
ют классы возраста:
1 кл. - до 20 лет
2 кл. - от 20 до 40 лет
3 кл. - от 40 до 60 дет
4 кл. - от 60 до 80 лет и т.д.
Чтобы закончить характеристику древостоя, необходимо
очень внимателзно осмотреть стволы, ветви, хвою и листья с
целью выяснения пораженности деревьев грибами и вредителями.
Пораженные болезнями части следует собрать. Плодовые тела тру-
товиков собираются с небольшими кусками норки. В дневник за-
писывается количество больных и поврежденных деревьев на 2500
кг. м. Следует собрать лишайники, встречающиеся на ветвях и
стволах деревьев.
При изучении древостоя необходимо закрепить знания по
систематике древесных пород: к каким типам растений и се-
- 65 -
мействам относятся лесообразуювше породы, хорошо усвоить мор-
фологические отличия видов хвойных по окраске и характеру по-
верхности (гладкая, трещиноватая) стволов, общему виду кроны,
расположенно и форме хвои, форме и расположению шишек и др.
признакам.
Далее следует переходить к характеристике возобновления
леса.
На пробной площадке подсчитывается количество экземпляров
подроста каждой породы и переводится на гектар, С помощью сан-
тиметра определяется высота подроста. Составляется таблица
подроста:
Порода Количество экземпляров Высота Возраст
на 1 гектар в см (по кол. мут.)
Закончив исследования древостоя, можно переходить к
описанию подлеска, а также травяного и мохово-лишайникового
покрова.
Для характеристики подлеска составляется список всех
кустарников с обозначением обилия по шкале Друде. Есл* преоб-
ладают высокие кустарники -определяется сомкнутость крон.
Травяной ярус подразделяется на несколько подъярусов.
Флористический список удобнее составлять по подъярусам, обоз-
начая обилие каждого вида и фазу развития. При описании травя-
ного покрова следует также обращать внимание, не встречаются
ли больные растения. Заболевания у растений чаще всего вызыва-
ют паразитические грибы. Поражения встречаются обычно на веге-
тативных органах в виде налетов, подушечек, пятен. Обнаружен-
ные больные растения необходимо заложить в гербарий для опре-
деления болезни.
Мохово-лишайниковый покров. Определяется глазомерно сте-
пень покрытия почвы мхами зеденымй и сфагновыми, а также ли-
шайниками. Все мхи собираются в гербарные листы для определе-
ния. Лишайники также можно закладывать в гербарные листы.или
собирать в коробочки или пакеты. Кроме того, следует обратить
внимание на распространении грибов - микоризаторов и сапрофит-
ных грибов на почве и собрать их в пакеты для определения.
56 -
Экскурсия на луга
К лугам относятся расти!-ельные сообщества, образованные в
основном многолетними мезофильными, то есть приспособленными к
средним условиям увлажнения, травами, растущими в течении все-
го вегетационного периода без летнего перерыва.
По происхождению луга подразделяются прежде всего на
естественные (природные) и искусственные (сеяные). Естествен-
ные луга бывают первичными и вторичными. Первичные естествен-
ные луга возникают в наиболее благоприятных для луговых трав
климатических и почвенных условиях, например, на длительно за-
ливаемых поймах рек, на морских побережьях, в некоторых высо-
когорьях. Вторичные природные луга формируются за счет естест-
венного заселения луговыми травами вырубок, гарей, осушенных
болот и т.д.
По положению в рельефе различают луга материковые, пой-
менные и горные. Материковые луга располагаются на равнинных
междуречьях. Они подразделяются на суходольные луга, занимаю'
щие повышенные участки, увлажняемые только атмосферными осад-
ками; и низинные луга, приуроченные к понижениям рельефа и ув-
лажняемые не только атмосферными осадками, но и близко распо-
ложенными к поверхности почвы грунтовыми водами. Пойменные лу-
га формируются на заливаемых полыми водами днищах речных до-
лин, называемых речньми поймами. По положенно над урезом воды
в реке пойменные луга разделяются на луга низкого, среднего и
высокого уровня. Горные луга наиболее характерны для высоко-
горных районов с влажным и достаточно мягким климатом, где они
нередко образуют особые пояса растительности (альпийский и су-
бальпийский) .
По видовому составу и структуре луга, как тип раститель-
ности, расчленяются на более мелкие единицы (таксоны). Прежде
всег^ луговой тип растительности подразделяется на три класса
формаций: 1. Настоящие луга, образованные типичными мезофита-
ми, 2. Остепненные луга, объединяющие фитоценозы с господством
засухоустойчивых мезофитов, называемых "ксеромезофитами", 3.
Болотистые луга, образованные в основном относительно влаголю-
бивыми мегэфитами, называемыми "гидромеэофитами". Нетрудно
догадаться, что эти классы луговых формаций обусловлены разной
степенью увлажнения местообитаний, относящихся к ним фитоценр-
- 57
зов. Классы формаций луговой растительности далее делятся на
формации и ассоциации (или типы лугов).
Луга на земном шаре занимают огромную площадь, половина
которой (около 100 млн. га) приходится на долю бывшего Советс-
кого Союза.
В Томской области, расположенной в пределах лесной зоны,
луга имеют ограниченное распространение по сравнению с лесами
и болотами, занимая лишь около 3 Z всей территории области.
Вместе с тем общая площадь, покрытая луговьми фитоценозами,
выражается внушительной цифрой, приближающейся к 1 млн. га.
При этом основные массивы лугов Томской области являются пой-
менными. большей частью первичными естественными лугами, фор-
мирование которых связано с высокой и продолжительной поемг
ностью, а также с господством в Обской пойме тяжелых (гли-
нистых и суглинистых) почв. Эти луга характеризуются порази-
тельным однообразием и бедностью их видового состава и очень
широким распространением заболоченных типов, что несомненно
связано с продолжительным затоплением и переувлажнением поймы.
На основных площадках пойменных лугов Томской области до-
минантами являются канареечник (Phalaroides arundinacea (L.)
Rauschert), осока изящная (Carex acuta I.), мятлик болотный
(Роа palustris L.) и некоторые другие гидромезофиты. И только
на самых высоких гривах отмечаются настоящие луга с доминиро-
ванием в травостое типичных мезофитов: пырея ползучего
(Elytrigia repens (L.) Nevski), полевицы белой (Agrostis alba
L.), лисохвоста лугового (A^pecurus pratensis L.) и т.д.
Среди материковых лугов, распре храненных в основном в
южной части Томской области, встречаются как суходольные, так
и низинные типы, относящиеся в основном к вторичным естествен-
ным лугам. Суходольные луга представлены главным образом
классом формаций настоящих лугов. Травостой в них образуют ти-
пичные мезофильные злаки: пыреГ ползучий, мятлик луговой (Роа
pratensis L.), овсяница луговая (Festuca pratensis Huds.), ежа
^борная (Dactylis glomerate L.), вейник тупоколосковый
(Calamagrostis obtusata Trin.), а также многочисленные
представители двудольных. На низинных материковых лугах, от-
носящихся к классу формаций болотистых лугов, доминируют гид-
ромезофильные злаки и ocoiai: вейник Лангсдорфа (Oalamagros* is
langsdorffii (Link) TiinJ. тучка дернистая (L'erchcjjripcja
58 -
cespitosa CL-') Beauv.), осока деркистая (Carex cespitosa L.) и
некоторые другие виды. На низинных лугах обычно формируются
кочки. В последнее время в хозяйствах Томской области расп-
ространены посевы ценных в кормовом отношении многолетних
трав, прежде всего тимофеевки луговой (Phleum pratense L.) и
клевера розового (Trifolium hybridum L.).
При обследовании лугов, так же как и любого другого типа
растительности, составляются геоботанические описания, закла-
дываются геоботанические профили, ведется картирование лугов,
собирается и изучается гербарий луговой флоры и т.д., и все
это выполняется с помощью описанных выше методов. Однако луга
отличаются от других типов растительности по видовому составу,
структуре, характеру использования их человеком, что обязывает
при их изучении несколько видоизменять общую методику геобота-
нических исследований и обращать внимание на некоторые специ-
фические для них особенности.
Прежде всего следует учитывать то, что состав и, в осо-
бенности, структура лугов подвержены сильным изменениям в те-
чении всего вегетационного периода. Поэтому обследование лугов
должно проводиться в сжатые сроки (1-1.5 месяца) и приурочи-
ваться ко времени максимального развития их травостоя.
В связи с тем, что луговые фитоценоэы образованы сравни-
тельно некрупными, травянистыми видами растений, при составле-
нии геоботанических описаний закладываются пробные площади не-
больших размеров, не превышающие 100 кв. м (10-100 кв.м).
Луговые растения очень чутко реагируют на все изменения
среды их обитания, что требует сопровождать обследование лугов
тщательным изучением микро- и мезорельефа, влияющего на
обеспеченность луговых растений влагой, теплом, элементами ми-
нерального питания и другими факторами их существования. В
особенности этс требование необходимо соблюдать при обследова-
нии поименных лугов. Дело в том, что основные факторы поймен-
ных местообитаний: поемность (глубина и продолжительность‘за-
топления полыми водами), айлювиальнссть (мощность и
дисперсность отлагаемых пойменных осадков), степень и характер
увлажнения и т.д. - теснейшим образом связаны с особенностями
микро- и мезорельефа поймы и прежде всего с положением каждого
конкретного участка над уровнем реки и на поперечном профиле
поймы. Чем ниже участок, тем на более продолжительное время он
- 59 -
заливается полыми водами, тем он сильнее увлажнен, тем больше
на негр откладывается аллювиальных наносов (наилка) и тем этот
наилок дисперснее и плодороднее. Чем ближе к реке расположен
участок поймы, тем лучше он дренируется (освобождается от из-
лишней почвенно-грунтовой влаги, благодаря ее оттоку в русло
реки), тем больше га ием откладывается аллювиальных наносов и
тем они грубее и менее плодороднее. Закономерные изменения в
пойме всех перечисленных и некоторых других факторов местооби-
тания несомненно оказывают большое влияние на состав, структу-
ру, продуктивность и другие признаки поименных лугов.
Поэтому обследование поденных лугов должно обязательно
сопровождаться проложением инструментальных профилей через по-
перечник поймы, а также по возможности точной привязкой каждо-
го геоботанического описания на высотном и поперечном профилях
поймы. Инструментальный геоботанический профиль можно привя-
зать к ближайшему водомерному посту.и , используя его данные,
определить режим залдаания ножми водами лугов, расположенных
на разных высотных уровнях. Данные по режиму затопления, до-
полненные сведениями о положении каждого конкретного фитоцено-
за на поперечном профиле nofa&i, а также сведениями выборочного
заложения и анализа почвенных разрезов, позволяют установить
связь различных типов лугов с основными факторами пойменных
местообитаний: поемноетью, увлажнением, аллювиальностью, пло-
дородием почв и т.д., - учет которых оказывает большую помощь
при решении целого ряда вопросов, связанных с изучением, раци-
ональным использованием и охраной пойменных лугов.
Луга являются ценными кормовыми угодьями, используемыми в
качестве сенокосов и пастбищ. Поэтому при их обследовании
большое внимание уделяется оценке кормового достоинства тра-
востоя, определению его продуктивности и характеристике куль-
тур- технического состояния луга.
Кормовое качество луговых уастений зависит, во-первых, от
их химического состава, т.е. от содержания в них протеина,
челка, жиров, клетчатки, витаминов и других веществ, и во-вто-
рых, от поедаемости растений животными, которая связана с та-
кими их признаками, как вкус, запах, мягкость или твердость,
наличие или отсутствие колющих или режущих ротовую полость жи-
вотных эпидермапьных образований на покровах растений и т.д.
Понятно, что перечисленные признаки, а следовательно, и корме-
60
ьос качество изменяются от вида к ".иду. Тем не менее можно вы-
делить более или менее сходные в этом отношении растения, ко-
торые объединяются в так называемые хозяйственно-ботанические
группы. Принято выделять следующие хозяйственно-ботанические
группы: злаки, осоки, бобовые, разнотравье, ядовитые растения.
Если на лугу имеются кустарники и мхи, то они выделяются в
особые группы.
Ясно, что названные группы объединяют виды, не вполне
сходные по кормовому качеству. Но тем не менее каждая хо-
зяйственно-ботаническая группа может быть оценена в этом плане
особо. Группа ядовитых растений вредна на лугах, так как эти
растения вызывают отравление животных. Мхи практически не пое-
даются домашними животными и поэтому не имеют кормового значе-
ния. Кустарники являются в основном грубым, плохо поедаемым
кормом или не поедаются совсем. Разнотравье составлено различ-
ными по кормовому качеству видами, большинство из которых, од-
нако, имеют невысокую ценность. Поэтому в больших количествах
на лугах разнотравье нежелательно. Вместе с тем небольшая при-
месь разнотравья оказывается полезной, так как многие предста-
вители данной группы в небольшой дозе выполняют роль пряных
растений, улучшая вкусовые качества корма.
Группа осок (и других представителей семейства осоковых)
по содержанию питательных веществ относятся к корму среднего
достоинства. К сожалению, многие осоки содержат мало фосфора,
кальция, перевариваемого белка и некоторых других полезных ве-
ществ; в осоках много нежелательного кремнезема, а также име-
ются неприятно пахнущие эфирные масла, снижающие их поедае-
мость. К тому же многие осоки являются жесткими растениями с
острыми шипами по краям листьев, сильно раздражающими сли-
зистые оболочки роговой полости и пищеварительного тракта жи-
вотных. Все э о вместе взятое и обусловливает в целом невысо-
кое достоинство осокового корма, качество которого резко сни-
жается с возрастом осок.
Луговые злаки являются ценными в кормовом отношении
растениями. Они характеризуются сравнительно высоким содержа-
нием питательных веществ и большинство из них поедается отлич
но, хороши или, по крайне мере, удовлетворительно. Кроме того,
многие злаки хорошо отрастают после скашивания и стравливания,
образуя значительную отавную массу й тем самым увеличивая про-
- 61,-
дуктивность травостоя. Указанные особенности злаков и опреде-
ляют их большую роль: так например, в среднем по бывшему ССОР
злаки составляют около 50 X кормового рациона скота. Большое
число видов злаков введено в культуру и возделывается в ка-
честве кормовых растений на значительных иющадях. Из наших
сибирских злаков, доминирующих на лугах, особую ценность
представляют костер безостый (Bromopsis inermis (leyss)
Holub.), пырей ползучий, мятлики луговой и узколистный (Роа
angustifolia L.), полевица белая, канареечник и некоторые дру-
гие.
Группа бобовых растений по кормовому достоинству занимает
первое место среди всех остальных хозяйственно-ботанических
групп. Бобовые содержат повышенное количество хорошо перевари-
ваемых и легко усваиваемых в организме животных питательных
веществ и витаминов. К тому же большинство видов бобовых ха-
рактеризуется хорошей или удовлетворительной поедаемостью жи-
вотными, которая по сравнению с другими хозяйственно-ботани-
ческими группами мало изменяется с возрастом растений. К сожа-
лению, бобовые бывают обильными далеко не на всех естественных
лугах. Поэтому очень важной является задача обогащения естест-
венных лугов ценными бобовыми травами и с здания, искусственных
чисто бобовых или смешанных злаково-бобовых лугов. К ценным в
кормовом отношении бобовым травам, произрастающим на Запад-
но-Сибирских лугах, относятся: клевера луговой (Trifolium
pratense L.), средний (Trifolium medium L.), ползучий
(Trifolium repens L.), пятилистный (Trifolium lupinaster L.),
люцерна серповидная (Medicago falcate. L.), горошек мышиный
(Vicia cracca L.) и другие.
Кормовое качество лугового травостоя и его продуктивность
(урожайность) оцениваются при составлении геоботанического
описания луга. Для этой цели в пределах пробной площади разме-
ром в 100 кв. м закладывается 1С так называемых учетных площа-
док в 0.25-1.0 кв. м каждая. Для получения объективных резуль-
татов учетные площадки лучше всего закладывать регулярным
способом, суть которого заключается в том, что в пределах
пробной площади перпендикулярно к одной из ее сторон намеча-
ются несколько (2-5) прямых линий (ходов), расположенных на
одинаковом расстоянии друг от пруга, и по этим линиям через
равные интервалы закладываются учетные площадки.
На каждой учетной площадке травостой срезается ножницами
- 62 -
для стрижки осей или острым ножом на определенной высоте от
поверхности почвы: на сенокосах - на высоте 7-8 см, на пастби-
щах - на высоте 2-4 см. Полученная с каждой учетной площадки
масса, называемая укосом, взвешивается в сыром состоянии на
бытовых весах с точностью 5-10 г. Затем отбирается "средняя
проба", предназначаемая для определения коэффициента усушки и
качественного состава травостоя. Для этого укосы аккуратно
укладываются комлями в одну сторону на большой клеенке или
брезенте и из разных мест получившегося слоя травы берется по
одинаковой горсти с таким расчетом, чтобы вес всей отобранной
массы (т.е. средней пробы) составлял 1-1.5 кг. Средняя проба
тут же взвешивается в сыром состоянии на бытовых весах. Все
результаты взвешивания записываются в специальные графы блан-
ка, а средняя проба вместе с этикеткой, на которой отмечается
номер описания, помешается в марлевый мешочек или заворачива-
ется в бумагу и направляется на разбор и сушку.
В условиях полевой лаборатории средняя проба разбирается
по хозяйтвенно-ботаническим группам и высушивается до воздуш-
но-сухого состояния. После этого каждая фракция средней пробы,
соответствующая определенной хозяйственно-ботанической группе,
взвешивается на аптекарских вегах с точностью до 0.5 г и путем
сложения навесок всех фракций вычисляется вес средней пробы в
сухом состоянии. Затем определяется содержание в процентах
массы каждой хозяйственно-ботанической группы от веса средней
пробы. Эти данные и будут характеризовать качественный состав
корма обследованного лугового фитоценоза. Для более точного
определения качества корма производится отбор образцов на хи-
мический анализ корма.
При оценке продуктивности лугов определяется количество
на гектар сырой (на пастбищах) или ухой (на сенокосах) массы
травостоя. Для вычисления сырой массы используются данные
взвевг зания укосов с десяти учетных площадок, заложенных в
пределах пробной площади. Для этого находят средний вес одного
укоса из десяти и полученную массу пересчитывают на гектар.
Например, на десяти 0.25 кв. м учетных площадках получены
укосы следующих навесок в граммах: 125, 340, 253, 189, 290,
401, 183, 238, 152, 145. Находим средний вес укоса с учетной
площадки:
125*340+253+189+290+401+183+238+152+145/10 = 2416/10 *-241.6 Г
63 -
Пересчитываем вес сырой массы на гектар ив следующей про-
порции:.
0.25 кв. м - 241.6 г
1 га (10000 кв. м) - х г.
241.6 X 10000 / 0.25 = 2416000 X 4 -
9664000 г » Э664 кг « 96.6 ц/га.
Для нахождения урожая сухой массы вычисляется »соэффициент
усушки, который получают в результате деления сырого веса
средней пробы на сухой ее вес. Пользуясь коэффициентом усушки
находят сухой вес каждого укоса с 10 учетных площадок, а даль-
ше ведут вычисления также, как при определении урожая сырой
массы: сначала находят средний сухой вес одного укоса, а затем
по нему определяют вес всей кормовой массы с гектара.
Определенная таким способом продуктивность луга оказыва-
ется несколько завышенной по сравнению с так называемой '‘хо-
зяйственной продуктивностью", т.е. тем урожаем корма, который
получают в действительности колхозы и совхозы. Дело в том, что
часть урожая луговых трав теряется в процессе сенокошения,
уборки сена, транспортировки его к животноводческий фермам, а
также в процессе стравливания травостоя на пастбищах. Поэтому
для нахождения хозяйственной продуктивности луга необходимо
урожай массы, полученный методом учетных площадок, уменьшить:
1) на 5-10 X для злаковых лугов, 2) на 15-20 X для элако-
во-разнотравных лугов, 3) на 20-25 X для разнотравных и круп-
ноосоковых лугов и 4) на 25-30 X для крупноравнотравных лугов.
Результаты расчетов продуктивности и состава хозяйствен-
но-ботанических групп заносятся в специальные графы бланка
описания.
При наличии геоботанической карты данные по продуктив-
ности лугов и кормовому качеству их травостоев позволяют вы-
числить запасы кормов на определенной луговой площади или в
целом в хозяйстве и определить поголовье скота, которое мсжно
содержать за счет этих кормов.
Под культур-техническим состоянием понимается состояние
луга, оцениваемое с точки зрения его возможного использования
в качестве кормового угодь^ (сенокоса или пастбища). Культур-
техническое состояние луга характеризует наличие или отсутст-
вие на нем деревьев, татарников, пней, разнообразных кочек,
сбоин и троп, засоренности ядовитыми травами и т.д. При харак-
64 -
л. рис гике культур -технического состояния луга не только фикси-
руется наличие или отсутствие всех перечисленных (и некоторых
других) факторов, ухудшающих качество луга, но и производится
количественная оценка отмеченных на данном лугу факторов. Для
всех из них в пределах пробной площади глазомерно определяется
процент покрытия площади, а для деревьев, кустарников, пней и
кочек кроме того определяются средний диаметр и высота, а так-
же количество экземпляров. Последний показатель пересчитывает-
ся затем с пробной площади на гектар.
Все показатели культуртехнического состояния записываются
в специальные графы бланка и учитываются при разработке мероп-
риятий по рациональному использованию, улучшению и охране лу-
гов. Причем предварительно эти мероприятия намечаются уже в
поле при описании каждого конкретного лугового фитоценоза. И
затем в камеральных условиях они уточняются и представляются в
виде обобщенной системы или картограммы, на которых каждый тип
или контур луга получает свою характеристик в этом плане.
В связи с интенсивным развитием животноводства в настоя
щее время очень сильно возрастает пастбищная нагрузка на луга,
нередко приводящая к их ухудшению, а иногда и к полной дегра-
дации лугового травостоя. Поэтому проблема рационального
использования, охраны и улучшения лугов приобретает все боль-
шую остроту. Научные эксперименты и опыт некоторых хозяйств
показывают, что применение рациональной системы использования
в сочетании с недорогостоящими мерами ухода и поверхностного
улучшения (удобрения, подсева ценных кормовых трав, борьбы с
сорняками, удаление кустарников и кочек и т.д.) позволяет по-
вышать продуктивность большинства лугов в 2-5 раз и улучшать
качество их травостоев. Луга с сильно деградированным тра-
востоем или с особо неблагоприятными условиями местообитаний
нуждаются в коренном улучшении, суть которого заключается в
создании искусственных культурных сенокосов и пастбищ на месте
естественного луга.
При этом все мероприятия по рациональному использованию,
охране и улучшению лугов должны разрабатываться и выполняться
на строго научной основе.
При изучении лугов большое внимание уделяется анализу
жизненных форм и экологического состава луговых растений, т.к.
эти вопросы помогают решать целый ряд теоретических вопросов
- 65 -
луговедения и учитываются при разработке мероприятий по рацио-
нальному использование, охране и улучшению луговой раститель-
ности.
Луговые фитоценозы чрезвычайно разнообразны по видовому
составу. Только на лугах бывшего СССР прог,растает около 40b0
вадов трав> относящихся к 582 родам и 76 семействам. Кроме то-
го, здесь отмечается около 50 видов кустарников, представлен-
ных в основном видами ив; несколько десятков видов мхов„ преи-
мущественно гипновых; и несколько десятков древесных пород.
Вместе с тем на лугах чаще всего доминируют представители двух
семейств: злаковых (Роасеае) и осоковых (Сурегасеае).
Среди травянистых луговых растений абсолютно преобладают
многолетники, представленные разнообразными жизненными форма-
ми, среди которых главными являются: корневищные, рыхлокусто-
вые, плотнокустовые, корнеотпрысковые, стержнекорневые, кисте-
корневые, луковичные и клубневые растения. При этом основные
“строители" луговых травостоев - злаки и осоки - относятся к
трем первым жизненным формам.
Практически все луговые травы способны размножаться семе-
нами. В эту группу входят однолетники и из многолетников -
стержнекорнеыве, кистекорневые, рыхло- и ^лотнокустовые расте-
ния. Однако большинство луговых трав способны размножаться не
только семенами, но и специальными органами вегетативного
размножения. Этой способностью обладает корневищные, корнеотп-
рысковые, луковичные, клубневые и некоторые другие растения.
Подавляющее большинство луговых трав являются полными ав-
тотрофами, т.е. самостоятельно обеспечивают себя необходимыми
для них энергией и веществами. Небольшая группа луговых трав в
основном самостоятельно обеспечивает себя веществами и энерги-
ей, но в некоторой степени в этом плане зависит также и от
других организмов. Сюда относятся полупаразиты и симбиотрофные
растения, т.е. растения, вступающие в симбиоз с грибами или
бактериями. И, наконец, третью, еще меньшую по составу, группу
образуют луговые травы-паразиты, получающие необходимые для их
жизнедеятельности элементы питания у других растений, на кото-
рых они паразитируют.
Луга формируются на средне увлажненных почвах, которые
вместе с тем могут сильно различаться по богатству \содержанию
- 66 -
минерального питания) и по засоленности, варьируя от
бедных до богатых и от незасоленных до догэльно сильно засо-
ленных. Поэтому луговые растения, являясь по отношению к ув-
лажнению в подавляющем большинстве мезофитами, оказываются
исключительно разнообразными по отношению к другим экологи-
ческим факторам и относятся к различным экологическим группам.
Среди луговых трав отмечаются олиготрофы (растения, нормально
развивающиеся на бедных почвах), мезотрофы (растения, нуждаю-
щиеся в умеренно богатых почвах), эвтрофы (растения, требующие
богатых почв), гликофиты (растения, нормально развивающиеся на
незаселенных почвах), галофиты (растения, мирящиеся с засолен-
ными почвами) и другие экологические группы.
Анализ жизненных форм и экологии луговых растений выпол-
няется в камеральный период при обработке собранного в экспе-
диции гербария. При этом прежде всего учитываются сведения о
жизненных формах и экологии луговых растений, содержащиеся во
‘’Флоре Западной Сибири" (и в других "флорах"). Однако эти све-
дения, в особенности по экологии, обычно оказываются недоста-
точными и поэтому приходится выполнять специальные работы,
позволяющие оценить экологию луговых растений объективно и бо-
лее детально. Большую помощь в этом плане оказывает составле-
ние местных экологических формул луговых растений по методу
стандартных экологических шкал Л.Г.Раменского.
Экскурсии на болота
Строение и развитие болотных ландшафтов. Болотом называ-
ется избыточно увлажненный участок земной поверхности, который
характеризуется следующими особенностями: 1. Бескислородностью
почвенной влаги, 2. Специфическими почвенными процессами, в
том числе иаког; 1ением торфа, 3. Особой болотной раститель-
ностью. Избыточная увлажненность поверхности и отсутствие
кислорода в воде препятствуют полному разрушению и минерализа-
ции отмерших растительных остатков, они превращаются в торф.
По мере накопления слоя торфа меняется рельеф поверхности бо-
лотного участка, изменяется гидрологическое состояние болота и
поступление питательных веществ в корнеобитаемый слой. Как
следствие этих причин меняется растительный покров болота, а
вновь откладывающийся торф состоит уже из остатков новых
- 67,-
растений. Поэтому торфяное тело болота образовано напластова-
нием разных торфов, различающихся составом растительных остат-
ков.
Все эти особенности болота позволяют рассматривать его
как географический ландшафт с особым геологическим строением,
особой гидрологией, особым растительным и животным миром, осо-
бым типом круговорота веществ и даже местным климатом. Ясно,
что большие площади подобных ландшафтов или их многочислен-
ность оказывают влияние, и иногда очень большое, на всю при-
родную обстановку региона.
Для образования болота нужно, чтобы в течении более или
менее длительного времени поверхность почвы была увлажнена
бескислородной водой; безразлично, каким образом застоялась
вода на поверхности почвы и каким образом она лишалась свобод-
ного кислорода. Застой воды может быть вызван выходом на по-
верхность грунтовых вод, он может произойти благодаря образо-
ванию в почве водоупорных горизонтов, задерживающих избыточную
атмосферную влагу. Болото может возникнуть и на месте озера,
когда оно заполнится осадками и станет очень мелким. Различают
следующие типы воды, питающей болота: 1) атмосферная 2) по-
верхностно-сточная, 3) почвенная и 4) гру товая (рис. 14).
Атмосферная влага насыщена кислородом, но при застаивании
ее на поверхности почвы кислород расходуется на дыхание живот-
ных и растений, на окисление минералов, но более всего потреб-
ляется грибами и микроорганизмами при разрушении отмершего ор-
ганического вещества. Чем толще слой растительного опада на
поверхности почвы, тем быстрее здес з возникают анаэробные
условия. Грунтовые и, частично, почвенные воды лишены кислоро-
да или содержат его в малом количестве. При выходе их на по-
верхность сразу же создаются подходящие для заболачивания
условия. Похожая ситуация возникает также тогда, когда болот-
ные воды увлажняют соседнюю незаболоченную территорию.
Высоко в горах, у подножья ледников и снежников почва
тостоянно увлажнена талой водой, насыщенной свободным кислоро-
дом. Здесь формируются нивальные сообщества растений, аль-
пийские лужайки, в составе которых нет ни одного болотного
растения. Но и здесь можно встретить места, где застаивается
вода или почва увлажнена грунтовыми водами. В этих местах об-
разуются очень своеобразные горные болота. Проточные ьоды, в
- 68 -
г-эм числе речная вода, насыщена ки< лородом и не вызывает забо-
лачивания. Но по берегам наших рек и проток часто образуются
осоковые заросли с обилием болотных растений, даже на крутых
склонах водоема. Как правило, в том месте просачиваются грун-
товые воды. Они-то, а не речная вода, создают заболачивание
почвы и обеспечивают существование болотной растительности.
Застой воды происходит в понижениях рельефа, в его отри-
цательных элементах. Избыточное увлажнение здесь может созда-
ваться совместным воздействием различных по своему происхожде-
нию вод, различающихся также насыщенностью минеральными соля-
ми. Наименее минерализована атмосферная - дождевая, снеговая -
влага, падающая прямо на поверхность болота. Но если эта влага
стечет по склону в болото, то уже успеет насытиться легкораст-
воримыми химическими соединениями. Наиболее насыщена минераль-
ными солями грунтовая вода, поэтому торфяные толщи болот грун-
тового питания обычно обогащены солями кальция, натрия, желе-
за, фосфора и других элементов.
Нередко на ранних этапах развития болота его питают все
типы вод (рис. 14). Болотная вода оказывается насыщенной мине-
ральными солями, и растительный покров образуют растения, тре
бовательные к богатому минеральному питанию, относящиеся к
экологической группе эвтрофов. Это многие виды осок, зеленые
мхи, кустарники и деревья и многие виды травянистых растений.
По мере нарастания слоя торфа поверхность болота оказывается
выше уровня грунтовой воды, и растения-эвтрофы уступают свое
место мезотрофам, нуждающимся в меньших дозах минерального пи-
тания. Однако нередко микрорельеф болота к этому времени ста-
новится очень неровным, на его повышениях поселяются расте-
ния -олиготрофы, требующие минимального почвенного питания, а в
понижениях могут существовать эвтрофы. В соответствующей про-
порции впоследствии остатки тех и других будут законсервирова-
ны в лЭрфе.
При дальнейшем нарастании торфяной толщи центральная
часть болота выходит из под влияния сначала почвенной, а затем
и поверхностно-сточной воды. Увлажнение осуществляется только
атмосферной влагой, и в растительном покрове остаются только
растения-олиготрофы, среди которых господствуют сфагновые мхи.
Анатомо-морфологическое строение этих мхов позволяет им удер-
живать в своем теле до 30 и более раз воды, чем весят они сами
69 -
Рис. 14. Схема строения верхового выпуклого котловин-
ного болотного массива.
Типы вод, питающие болота: А - атмосферные, ПС - поверх-
ностносточные, П - почвенные, Г - грунтовые; виды торфа, сла-
гающие торфяную залежь: 1 - фускум-торф, 2 - ангустифоли-
ум-торф, 3 - сфагновый топяной, 4 - аейхцериево-сфагновый, б -
древесный, б - минеральное ложе болота.
в сухом состоянии. Такая способность мхов позволяет болоту на-
растать выше окружающих его берегов, оно становится выпуклым.
Теперь уже не в болотную впадину стекает вода с суходольных
берегов, а наоборот, болотная вода растекается по ровным
участкам суходолов и создаются с .агоприятные условия для раз-
растания болота вширь/
Особенно привлекают внимание наблюдателей выпуклые болота
в степи, где они небольших размеров. Можно находиться на одном
берегу болота и не видеть человека, стоящего в сотне метров,
но с другой его стороны: середина болота наросла выше роста
человека. В лесной зоне выпуклые болота обычно ведшей по раз-
мерам и иногда запол; пот целиком междуречье. Они тогда стано-
вятся плоско-выпуклыми. В относительно плоской центральной
- 70 -
-их из-за избытка воды формир.огся системы гряд и топких
мочажин и даже системы вторичках болотных озерков. Все же и
здесь можно найти наиболее принанятую часть массива, тогда с
той и другой стороны болота будут видны далекие кромки леса»
иногда находящиеся ниже уровня глав наблюдателя.
Таким образом, путь развития болотного массива определя-
ется постепенным снижением минерализации болотной воды и уве-
личивающейся олиготрофностью болотной растительности. Низинное
болото с богатым питанием и эвтрофной растительностью по мере
развития сменяется переходным болотом с мевотрофной или
микстотрофной (смешанной) растительностью/ наконец, наступает
верховая стадия развития и растительный покров становится оли-
готрофным. Поскольку в верховой стадии развития трофические
свойства болотно/ воды остаются более или менее постоянными,
то главным условием развития болотного массива становится ди-
намика болотной воды» в связи с чем усложняется рельеф поверх-
ности, развивается специфическая гидрографическая сеть, форми-
руются болотные комплексы.
Понятно, что не обязательно болото должно пройти этот
полный путь развития. В условиях постоянного богатого вод-
но-минерального питания оно мс .ет на очень долгий срок задер-
жаться на низинной стадии развития, а многометровая толща тор-
фа быть сложенной одним и тем же видом торфе;. Напротив, на
песчаных террасах и равнинах, например, на севере Томской об-
ласти, болотная вода может оказаться такой бедной минеральными
солями, что болотный процесс сразу начинается с поселения на
песчаном грунте олиготрофных растений, т.е. с верховой стадии
развития. В окрестностях поселка Неготка на террасах реки Тым
мы наблюдали несколько болотных массивов один другого мельче и
моложе. Если в самом старом на дне залегают слои переходного
торфа, то в самых молодых придонный горизонт образован верхо-
вым : )рфом. Можно предположить, что за время существования
старого болота произошло обеднение почв и грунтов террасы реки
Тым.
Следовательно, термины “низинное'*, “переходное" и "верхо-
вое" болото не означают расположение болота на высоких или
низких уровнях рельефа, как это иногда считают, а характеризу-
ют состояние минерального питания болотной растительности -
богатое, среднее и бедное. На самом высоком уровне Западно-Си-
- 71 -
бирской равнины на Обь-Иртышском водоразделе залегает обширное
Большое Васюганское болото. Оно низинное. А ниже его по
абсолютным уровням на склонах этого же водораздела развиты пе-
реходные и верховые болота. Различать эти типы болот можно по
составу и строению растительного покрова, по ботаническому
составу торфа, но используются также критерии химического
состава.
Низинные торфа обычно отличаются сильной разрушенностью
растительных тканей, т.е. высокой степенью разложения; соот-
ветственно, зольность их также выше. Напротив, верховые торфа
содержат малоразрушенные остатки растений, а бесструктурного
органического вещества - торфяного гумуса в них мало. Они от-
личаются слабым разложением и малой зольностью. Принято счи-
тать, что низинным является торф с зольностью от 8 до 20 X и
более, переходным с зольностью 4-8 X, верховым - 2-4 %. Это
так Называемая естественная зольность, получающаяся при разру-
шении и минерализации растительных тканей в процессе их отор-
фовывания. Нередко торф бывает загрязнен минеральным осадком,
выпадающим из просачивающихся сквозь него почвенно-грунтовых
вод, что увеличивает зольность. Например, зольность г.т тнового
торфа пойменных болот Оби достигает 57.7 Верховой торф Заг-
рязнен очень слабо, только тем, что оседает из атмосферы.
Различия в химическом составе болотной воды, в степени
увлажненности болотных почв и в динамике увлажненности приво-
дит к большому разнообразию растительных сообществ и видов
торфа низинных болот. На ог ’их болотах развит мощный ярус де-
ревьев, густой и высокий, на других древостой разрежен и обра-
зован низкорослыми деревцами, на третьих деревья растут только
по отдельным уплотнении* грядам - веретьям; есть болота осоко-
вые, кустарниковые, травяные разных типов, моховые гипновые и
моховые сфагновые и пр. Верховые болота по сравнению с ними
очень однообразны как по состаь/ растительности, так и по
составу торфов, и понятно почему: источник их водно-минераль-
ного питания однороден по своему химическому составу. Переход-
ные и смешанные (микстотрофиые) болота занимают по разнообра-
зию типов промежуточное положение между верховыми и низинными.
Условия произрастания болотных растений. Болотные расте-
ния отличаются рядом специфических особенностей анатомо-морфо-
72 -
логического строения, которые определяются приспособленностью
к условиям среды, в первую очередь к особенностям торфа как
почьы. Естественная влажность торфа очень велика - 90-98 X .
Поэтому у многих болотных раси ий отчетливо выражены призна-
ки, свойственные водным растениям - гидрофитам. В стебле и
корневищах болг ных трав сильно развиты воздушные полости,
позволяющие не только снабжать кислородом корневища и корни,
но даже поддерживать растения на плаву. Однако не вся болотная
вода доступна растениям, значительная часть ее связана с
растительными тканями или входит в состав торфяных коллоидов.
Этот ’'мертвый запас" воды можно определить по точке увядания
растений.
Попробуем вырастить растения в сосудах, наполненных
песком, глиной и торфом.‘ Прекратим полив, и как только расте-
ния начнут увядать, определим влажность почвы, В песчаной поч-
ве оставшаяся вода составит 1-3 X полной влагоемкости, в гли-
нистой - 20-25 %, а в торфе - 42-49.7 X» Иначе говоря, почти
половина воды в болотной почве недоступна растениям. Несмотря
на избыточное увлажнение почвы, болотные растения испытывают
недостаток доступной воды. Поэтому у них наряду с гидроморфны-
ми признаками отчетливо выражены признаки ксероморфные,
свойственные растениям засушливых местообитаний: мощные пок-
ровные ткани, часто с восковым налетом или масличными железка-
ми, узкие, свернутые литья у осок и злаков, мелкие жесткие
листья у кустарничков и т.д.
Из-за плохой обеспеченности кислородом болотных и торфя-
ных почв вырабатываются особые формы роста растений,
несвойственные другим местообитаниям: травянистые растения -
злаки и осоки - образуют в болотных условиях кочки, деревья
развивают горизонтально распростертые корневые системы с дос-
ковилными корнями. Эти приспособления способствуют поднятию
узлоь кущения иди корневых шеек деревьев выше уровня болотной
воды. На приствольных повьвиениях деревьев накапливается перег-
ной и создаются благоприятные условия для поселения многих лу-
говых и лесных растений. Поэтому древесные болота - согры от-
личаются очень пестрый мозаичным покровом растительности: на
приствольных повышениях одни наборы видов, на осоковых и мохо-
вых кочках - другие, а между ними, в понижениях - третьи.
Накопление торфа в разных типах болот происходит с разной
- 73 -
интенсивностью. Здесь имеет,значение жизненная форма доминан-
той. Бескорневые растения - мхи беспрерывно нарастают своей
верхушкой, а нижняя часть стебля вместе с листьями тмирает и
превращается в торф. В моховых болотах происходит послойное
накопление торфа с поверхности, в таких болотах можно выделить
и датировать слои тс эфа иногда с точностью до нескольких лет.
В торфе болот Красноярского края удалось выделить слой торфа с
отложениями 1908 г., в котором сохранились частички вещества,
осевшие на поверхность болот при взрыве Тунгуского метеорита.
Это? слой уже оказался на глубине 33-42 см.
При таком нарастании торфа происходит погребение в нем
корневых систем и узлов кущения растений; многие болотные виды
по этой причине не могут существовать на моховых болотах,
удерживаются виды, способные образовывать придаточные корни на
стебле. У некоторых травянистых растений болот, у кустарничков
- видов ив, багульника, подбела и пр. - появляются мутовки
придаточных корней все вше и выше пр стеблю, в то время как
нижняя часть стебля постепенно отмирает. Подсчитывая эти му-
товки, например, у росянки, можно выявить и величину годового
прироста моховой дернины.
На осоковых болотах нарастание торфяной толщи и отложение
отмерших остатков растений происходит иначе. Корни осок внед-
ряются в уже отложенные слои торфа и там отмирают. Надземные
части растений успевают разрушиться на поверхности залежи и
образуют здесь тонкий слой минерализованного наилка. Нараста-
ние толщи осокового торфа происходит путем внедрения свежих
остатков внутрь торфяного пласта, стратификация торфа здесь
менее отчетлива и в каждом слое торфа могут содержаться остат-
ки разного возраста. Похожие процессы накопления торфа про-
исходят и в древесных болотах. В этих немоховых типах болот
погребение растений происходит слабее, чем на моховых болотах,
соответственно и флора этих болот богаче.
Торф отличается плохой теплопроводностью, он прогревается
медленнее, чем минеральный грунт. В естественных условиях
верхние рыхлые слои торфа и моховая дернина образуют природный
тепловой экран, препятствующий как промерзанию, Tai: к оттаива-
нию торфа. На юге лесной зоны промерзание болот преисходит
только в декабре, а под толстым слоем снега многие болота
вовсе не промерзают. В степи и лесостепи, где снега мело, бо-
лота промерзают сильнее, чем в лесной гене.
/ - 74 -
Гам, торф промерз, отдаивание его происходит также
крайне медленно. Под моховой дерниной, прогреваемой до 30-40
градусов, до середины июля ; . лироте реки Кети может сохра-
няться слой мерзлого торфа. Окол' поселка Средний Васюган сох-
ранились остатки древних мерзлых’ торфяных бугров. Под слоем
талого торфа мощностью 1 здесь покоится реликтовая мерзлота,
Таксим образом, граница вечной мерзлоты в торфяниках проходит
на сотни километров южнее, чем в минеральном грунте. Медленное
оттаивание верхних горизонтов торфа весной существенно сокра-
щает длительность периода вегетации растений. На осушенных
торфяниках зерновые культуры часто не успевают полностью соз-
реть .
Химический состав различных торфов очень неоднороден,
причем многие доступные растениям соединения находятся в мини-
муме. Как правило, даже в низинных торфах ощущается недостаток
того или иного соединения, создающего плодородие почвы, неред-
ко при избытке каких-либо других соединений. В пойменных тор-
фяниках юга Томской области обыкновенны прослои извести и пят-
на солей фосфора (вивианиты), нередко наблюдается избыточное
содержание железа и натрия. Многие химические соединения нахо-
дятся в торфе в недоступном для растений состоянии и высвобож-
даются только при сильном разрушении органической компоненты
торфа. Таким образом, не только физические, но и химические
свойства торфяных почв определяют состав болотных сообществ.
Флора болот и растения-торфообразователи. Несмотря на
жесткие экологические условия, позволяющие произрастать на бо-
лотах только особо приспособленным видам растений, флора болот
довольно богата, б Томской области на болотах встречается бо-
лее 200 видов высших растений, примерно четвертая часть флоры
области. Часть этих видов заходит в болота из лесных и луговых
место битаний, другие виды занимают в болотных условиях особые
экологические ниши и встречаются редко, наконец, относительно
немногочисленная группа растений составляет основу болотной
растительности, эти виды являются доминантами болотных фитоце-
нозов. Особо велика роль тех растений, остатки которых откла-
дываются в виде торфа; своей жизнедеятельностью они преломляют
факторы среды.
- 75
Анализ торфа позволяет уточнить фитоценотическую роль тех
или иных видов растений в болотных сообществах. Например, при
исследовании древесно-сфагнового болота на первый взгляд, по
аналогии с лесом, кажется, что эдификаторами сообщества явля-
ются деревья. Однако анализ торфа и структуры фитоценоза пока-
зывают, что не деревья, а сфагновые мхи трансформируют здесь
среду обитания, откладывают основную массу торфяного волокна.
Деревья имеют хотя и важное, но все же второстепенное значе-
ние.
Список растений-торфообразователей не велик. В Западной
Сибири ведущее место среди них занимают сфагновые мхи. Только
15 X запасов торфа ,в Томской области приходится на долю гипно-
вых, осоковых и древесных торфов, а 85 °' составляют виды сфаг-
нового торфа. В группе олиготрофных сфагнов наиболее распрост-
ранены сфагн бурый (Sphagnum fuscum Klingtr.), сфагн магел-
ланский (S. magellanicum Brid.), * сфагн узколистный (S.
angustifolium С.Jens.), сфагн большой (S. majiis С.Jens.),
сфагн балтийский (S. baiticum С.Jens). В группе мезоолиготроф-
ных сфагнов наиболее обыкновенны сфагн обманчивый (S. f al lax
Klingtr.), сфагн извилистый (S. flexuosum Dozy e Molk сфагн
папиллозный (S. papillosum Lindb.), сфагн оттопыренный (S.
squarrosum Crane.). Требовательные к богатому минеральному пи-
танию сфагн тупой (S. obtusum Watnst.), сфагн бахромчатый (S.
flmbrlatum Wils.), сфагн береговой (S. riparlum Angst.) произ-
растают в местообитаниях переходных и низинных болот. В забо-
лоченных лесах обыкновенны 'Фагн Гиргензона (S. girgensohnil
Russ.), сфагн Вульфа (S. wulfianum birg.). Всего в болотных
местообитаниях лесной зоны Западной Сибири встречается более
30 видов сфагновых мхов;
Способность сфагнов удержит тгь в своем теле значительные
количества воды в сочетании с очень широким распространением
этих растений и образованных ими сообществ приобретает значе-
ние существенного географического фавора, изменяющего физи-
ко-географическую обстановку. Осушение болот уменьшает запас
тепла в грунте и приводит т' понижению температуры воздуха зи-
мой. Летом, наоборот, вследствие уменьшения теплопотока в
грунте и снижения испарения температуры возрастут. Не будь
сфагновых болот, граница вечной мерзлоты в Западной Сибири
проходила бы примерно по линии Томск-Тобольск.
76 -
Друган группа болотных растений - гипновые мхи - принад^
лежит'к различным родам и семействам класса зеленых мхов. В
отличие от сфагнов, населяющ? < и переходные болота, эти мхи
преимущественно произрастают на низинных болотах; они хорошо
переносят избыточное насыщение болотных .вод солями кальция и,
видимо, натрия. Обычны в болотных фитоценозах лесной зоны За-
падной Сибири примерно 20-25 видов гипнов, t из них наиболее
распространены следующие виды: Drepanocladus vernicosus
Warnst., Dr. aduncus Warnst., Dr. fluitans Warnst. CallRrgon
stramineum Kindb., C. giganteum Kindb., C. cordifolium Kintfb.,
Tomenthypnum nitens Loeske., Aulacomnium palustre Schwaegr.,
Meesia triquetre Angstr., Paludella squarrosa Brid., Helodium
blendowii Warnst.
Наиболее распространены гипновые болота в подзоне южной
тайги, где в долинах рек, на низких террасах и поймах созда-
ются условия богатого водно-минерального питания грунтовыми и
поверхностно-сточными водами. На границе лесной и лесостепной
зон гипновые болота размещаются даже на водораздельных прост-
ранствах, образуя широкую полосу, протянувшуюся черев всю За-
падную Сибирь.
Осоки являются обязательным компонентом большинства бо-
лотных фитоценозов. На переходных и низинных болотах лесной
воны Западной Сибири наибольшее распространение получила осока
волосистоплодная (Carex lasiocarpa Ehrh.), на этих же типах
болот обычны осока вздутая (С. vesicarla L.), осока струноко-
ренная (С. chordorrhiza Ehrh.), осока вздутоносая (С.
rhynchophyza С.А.Меу.). На верховых болотах обычны осока топя-
ная (С. lirnosa L.) и осока круглая (С. globularis L.). В
местах с переменным режимом увлажнения ;И в поймах рек большие
площади занимают кочкообразующие гиды: осока дернистая (С.
cespitosa L.), осока омская (С. omskiana Meinsh.), осока изящ-
ная (п. graciris L. = С. acuta (L.) Curt.), осока водная (С.
aquatilis Wahl.).
Осоковая кочка представляет собой переплетение косо вверх
направленных и прижатых друг к другу коротких 1сорневищ и при-
даточных корней; благодаря ее образованию узлы кущения вы-
носятся вверх: чем выше уровень затопления почвы вешними или
дождевыми водами, тем выше кочки. На кочках осок создаются
условия для поселения различных менее влаголюбивых растений, в
том числе и кустарников - таволги иволистной (Spirea
salicifolia L.), кустарниковых видов ив (Salix pentandra L.,
S, rosmarinifolia L. (= S. siblrica Pall.), S. larponum L.)
поселяются карликовые березки и береза пушистая. В поймах рек
поэтому, распространены не столько осоковые, сколько дре-
весно-кустарниково-с зоковые болота. Торфяной пласт таких болот
маломощен; торф насыщен минеральным осадком. Большинство видов
осок торфяных болот не образуют кочки, узлы кущения их не за-
ливаются водой на продолжительный срок; при пересыщении болота
водой торф взбухает и растительный ковер с узлами кущения осок
все равно оказывается на поверхности. Многие другие травя-
нистые растения могут также быть строителями растительного
покрова болот. Виды пушицы (Eriophorum polystachyon L. - Е>
vaginatum L., E. gracile Koch.), чейхцерия (Scheuchzeria
palustris L.), очеретник (Rhynchospora alba Vahl.) распростра-
нены на верховых и переходных сфагновых болотах, причем пушицы
могут откладывать довольно мощные пласты торфа. Особенно бога-
ты травянистыми растениями низинные болота. На них встречаются
папоротник (Thelypteris palustris Schott.) и хвощи (Equisetum
fluviatile L., Equisetum palustre L.), сабельник ^omarum
palustre L.), белокрыльник (Calla p^.ustris L.)> вахта
(Menyanthes trifoliate L.), триостренник болотный (Triglochin
palustris L.), некоторые злаки (Calamagrostls langsdorffii
(Link) Trin., Calamagrostls neglecta (Ehrh.) Gaertn., Mey. at
Scherb., TH set uni sibiricum Rupr.), несколько видов обходных и
пр. Небольшие по площади участки с доминированием вахты или
хвощей,; реже сабельника или тростника довольно обыкновенны в
поймах и на террасах западносибирских рек.
На верховых и переходных болотах обязательно присутствуют
болотные кустарнички - багульник (Ledum palustre L.), подбел
(Andromeda poll fol la L.), болотный мирт (Chamaedaphne
calycuiata (L.) Moench.) й виды клюквы (Oxtcoccus microcarpus
Turcz. ex Rupr. и 0. palustris Fers.). Также обычны здесь
’асскомоядные растения росянки (Drosera rotundifolta L.,. D.
anglica Huds.) и морошка (Rubus chamaemorus L.).
Деревья входят в состав растительного покрова многих ти-
пов болот. На. верховых болотах обыкновенны низкорослые (2-5 м
высотой) Формы сосны, на переходных болотах сосны и берега пу-
шистая представлены более рослыми экземплярами а на низинных
’ 78 -
болотах развиваются разнообразные по составу древостои - кед-
ровые, березовые, сосновые, полидоминантные темнохвойные, сме-
шанные. Обычно высота деревьев здесь также не велика - 8-15 м,
но в условиях богатого проточного питания на мощном слое торфа
могут возникать рослые древостои из хорошо сформированных кед-
ров и сосен.
Наибольшей экологической амплитудой обладает сосна: она
растет и на верховых, и на переходных, и на низинных болотах,
но только в последнем случае она становится здификатором
растительного покрова и торф состоит из разложенной древесины.
На юге лесной зоны обыкновенны древесные пойменные болота -
согры, древостой которых состоит из смеси пород: кедра, сосны,
березы пушистой, лиственницы, иногда встречаются пихта и оси-
на. Микрорельеф таких болот сложный; положительные элементы
его составляют приствольные повышения с основой из корневых
систем деревьев, в понижениях между ними стоит вода или растут
гидрофиты. Деревья обычно растут группами по нескольку ство-
лов, приствольные повышения оказываются довольно обширными, а
высота их поверхности над дном понижений нередко достигает
1-1.5 м. Растительный покров corp отличается сложным мозаичным
строением.
Типы болот и болотных массивов. В большинстве болотных
сообществ более или менее равное участие в строении раститель-
ного покрова и в отложении торфа принимают растения разных
жизненных форм: мхи, травы, кустарнички и кустарники, деревья.
Растительный покров болот; оказывается полидоминантным, а
структура его сложной. Это связано с тем, что роль верхних
ярусов растительности (древесного и кустарникового) в боль-
шинстве типов болот ослаблена, и на господствующее положение
выдвит^ются растения нижних ярусов. С вертикальной сложностью
боло^ой растительности сочетается его сильная горизонтальная
расчлененность: ярко выраженный микрорельеф способствует пят-
нистому, мозаичному строению растительного покрова. Очень
часто мозаичность развивается в комплексность, когда наблюда-
ется закономерное чередование фитоценозов гряд - повышений с
мхами, кустарничками и деревьями и фитоценозов мочажин - топ-
ких понижений, населенных гидрофитами.
По этой причине принято в качестве типов болот выделять
- 79 -
однородные по строению и по составу растительности участки бо-
лот, в пределах которых откладывается однородный торф. Такие
участки или болотные фации бывают простые или сложнее, комп-
лексные. Фация включает как растительный покров участка, так и
отложенный им торф. Геологическим фундаментом современной фа-
ции служит пласт торфа, сложенный остатками растений предыду-
щей по времени фации.
Болотный массив представляет собой вакономерно сложенную
систему болотных фаций. В одном случае эта система может быть
очень простой и образованной одной-двумя фациями, например,
молодое осоковое болото в котловине мезорельефа. В других слу-
чаях болотный массив может быть очень сложным, состоять из
многих, резко отличных друг от друга фаций, например, развитые
водораздельные массивы. Нередко болотные массивы сливаются
между собой и образуют систему болотных массивов. Такими слож-
ными*системами массивов являются большинство болот Западной
Сибири.
В настоящее время принято классифицировать болотные
массивы или по ^преимущественному типу питания современного
растительного покрова (верховые, переходные, низинные) или по
местоположению в рельефе. В последнем случае различают водо-
раздельные массивы, расположенные на междуречьях, террасные и
пойменные массивы и пр. Можно классифицировать их по типу впа-
дины, которую они занимают: массивы котловинные, ложбинные,
притеррасных понижений поймы и т.д. Обычно, характеризуя бо-
лотный массив, используют все эти классификации в одном назва-
нии, указывая и особенности растительного покрова наиболее
развитой фации. Например, террасное котловинное сосно-
во-кустарничково1 сфагновое верховое болото или пойменное при-
террасное береэово-осоковое низинное болото.
Заболоченные земли. Немалые площади в Сибири занимают за-
болоченные земли. Они развиты там, где все условия болотного
процесса напицо, а торф не накапливается или еще не успел.от-
ложиться. В поймах сибирских рек очень широко развиты боло-
тистые и торфянистые ’’луга . Здесь идет большую часть года ин-
тенсивный болотный процесс, но в половодье эти ’’луга” залива-
ются речной водой, богатой кислородом, а осенью обсыхают и
проветриваются. Отмершее ‘ растительное волокно у с п е ь ае т раз р v -
- 80 -
шиться или смешаться с речным аллювием. В результате перемен-
ного увлажнения эти болотные ландшафты могут существовать
очень долгое время без видимою развития или изменения.
Еще большие площади занимают заболоченные леса. Расти-
тельный покров их содержит много болотных видов растений, поч-
ва с признакам7’ болотного процесса (поверхностное и глубинное
оглеение, оруденение); от лесного ландшафта нередко сохраня-
ется только один компонент - древостой.’ Многие из этих заболо-
ченных лесов являются начальной стадией заболачивания, другие
не имеют тенденции превращаться в болото и долгое время могут
находиться в состоянии осокового, болотнотравного или сфагно-
вого леса. Почва этих лесов к осени обсыхает с поверхности и
растительный опад минерализуется. При малейшем изменении усло-
вий среды в сторону, благоприятную для болотного процесса, та-
кие ладшафты трансформируются в короткий срок в торфяные боло-
та.
Если торфяные болота занимают около половины площади
лесной зоны Западной Сибири, то заболоченные земли отнимают от
доступных хозяйственному использованию угодий еще 10-15 X.
Исследованы они очень слабо, что нередко приводит к ошибкам в
их хозяйственном использовании. Травянистые болотные сообщест-
ва используются под сенокосы, причем применение тяжелых сеноу-
борочных машин усугубляет болотный процесс, а сообщества с
развитым древостоем рассматриваются как леса и подлежат выруб-
ке. Часто это приводит к необратимым вспышкам заболачивания,
на месте вырубок восстанавливается древостой уже болотного об-
лика, не пригодный к хозяйственному использованию.
Методы изучения болот. Значительную долю полевых исследо-
ваний болота составляет его характеристика как. сложной ланд-
шафтной системы, находящейся во взаимодействии с окружающими
ландшафтами. Е^ерховое болото в степи может быть окружено со-
лончаками, а в лесной зоне - зеленомошным сосновым лесом, и
хотя растительный покров того и другого массива может быть
схож, различия проявятся в строении торфяной залежи. Эти мас-
сивы будут различны по ходу развития. Поэтому следует подробно
описать окружение болотного массива.
Торфяное болото, особенно крупное, сложено многими фито-
ценозами и их объединениями. Необходимо дать характеристику
81 -
массиву в целом, учитывая его размеры, форму поверхности, за-
кономерность размещения простых и сложных современных фаций.
Использование аэрофотоснимков и составленных при их дешифриро
йании плана фаций и схемы линий стеканий (рис. 15) облегчает
выполнение этой задачи и позволяет наметит! площадки геобота-
нических описаний и пункты отбора проб торфа.
Рис. 15. Структура водораздельного болотного массива:
А - фациальная (фитоценотическая): 1 - рям, 2 - рослый
рям, 3 - осоковый заболоченный лес, 4 - транзитная топь, 3
древесно-осоковый фитоценоз, 6 - грядово-озерковый комплекс, 7
- грядово-мочажинный комплекс, 8 - топи подтопления, 9 - сухо-
дольный лес, 10 - ручей; В - векторная (гидрологическая): 11 -
выпуклые участки,. 12 - пологонаклонные и вогнутые участки, 13
- линии стекания болотных вод, 14 - линии местных водоразде-
лов.
Геоботаническому описанию' подлежит растительный покров
!саждой фации в пределах массива. Чтобы учесть мозаичность и
комплексность растительности обязательно характеризуется мик
- 82 -
рорельеф: форма, размеры и особенности размещения кочек и
гряд, морфология и увлажненность межкочечных понижений и моча-
жин. Составление списка растений производится с расчленением
растительности на ярусы (древесный, травя но-кустарничковый и
лишайниково-моховой) и применительно к элементам микрорельефа.
Это позволяет выявить не только все фитоценозы, входящие в
состав комплексной болотной фации, но и фрагменты фитоценозов
в виде мелких мозаичных пятен. Дополнением к описанию служит
зарисовка плана и профиля болотного участка (рис. 16). Таким
образом, выявляется не только состав и обилие растений фитоце-
ноза, но учитываются их жизненные формы и закономерности раз-
мещения в пространстве.
Рис. 16. Вертикальная структура сосново-кустарничко-
во- сфагнового фитоценоза - ряма.
Виды растений: С - сосна, ВК - вересковые кустарнички
(багульник, Кассандра, подбел), П - пушица. Виды торфа на рис.
14.
- 83 -
Для описания растительности болот требуются знания не
только сосудистых растений, но также мхов, которые необходимо
тщательно гербаризировать, поскольку определение мне их из них
возможно только под микроскопом. Руководствами служат опреде-
лители печеночных и листостебельных мхов.
При исследован! < торфяных отложений производится промер
их глубин и в необходимых точках ручным буром отбираются об-
разцы торфа в виде сплошной колонки от поверхности залежи до
минерального дна. Длина закрывающейся ложки бура 50 см, но
точность исследования повышается, если содержимое ложки расч-
ленить на образчики длиной 10 и 5 см. Образцы торфа подверга-
ются различным специальна анализам. Для выявления ботани-
ческого состава торфяное волокно отмывается на сите и затем
под микроскопом определяются остатки растений с точностью до
вида и оценивается их относительное количество в образце.
Пособиями для ботанического анализа служат специальные опреде-
лители.
Результаты этих анализов могут быть представлены в графи-
ческом изображении (рис. 17), позволяющем проследить сукцессию
болотной растительности в пункте сбора образцов. Серия таких
стратиграфических колонок, отобранных по линии,, пересекающей
болото поперек, дает возможность построить стратиграфический
профиль торфяной залежи и выяснить ход развития болотного
массива.
Экскурсии на водсэш
Водная среда является самой древней средой обитания
растений. Наукой установлено, что именно с водной средой свя-
зано зарождение жизни на нашей планете, в том числе и зарожде-
ние первых растительных организгэв, выход которых на сушу осу-
ществился много позже и уже на достаточно высоком уровне эво-
*тоции растительного мира. Однако и поныне большая часть водое-
мов на Земле (морей и внутренних водоемов) заселена много-
численными видами водных растении, имеющих различные размеры
(от микроскопических до довольно крупных), и относящихся к раз-
ным систематическим группам: водорослям, высшим споровым и
цветковым растениям.
84 -
Рис. 17. Послойные показатели ботанического состава (А),
степени разложения (Б) и стратиграфия торфяной залежи
(В) верхового болота:
Виды растений (А) и отлагаемые ими виды торфа (В): 1 -
сосна, 2 - пушица, 3 - сфагнум фускум, 4 - сфагнум ангустифо-
лиум, 5 - сфагнум магелланикум, 6 - вересковые кустарнички, 7
- сосново-сфагновый торф, 8 - пушицево-сфагновый торф, 9 -
сосново-пушицевый торф.
Во время прохождения полевой практики на втором курсе
стуце 1ты имеют возможность познакомиться с растительным миром
внутренних (континентальных) водоемов. При этом обращается
внимание прежде всего на водные растения, относящиеся к группе
так называемых ’’макрофитов”, т.е. более или менее крупных
растений, различимых невооруженным глазом и представленных в
основном высшими цветковыми растениями. С j микроскопическими
водными растениями студенты знакомятся в соответствии с прог-
раммой на первом курсе обучения.
- 85 -
В связи с тем, что макрофитная водная растительность име-
ет травянистый характер,, при ее изучении применяются в об-
щем- то те же методы и приемы, что и при изучении лугов и дру-
гих травянистых типов растительности. Так же, как и на лугах,
выделяются формации и ассоциации водной растительности, выяв-
ляется флористически! состав фитоценозов, определяется обилие
и видовое проективное покрытие, учитывается густота (общее
проективное покрытие) травостоя, его максимальная высота и
расчленение на ярусы и т.д. При этом используется та же, что
и для лугов, форма бланка геоботанического описания.
Вместе с тем при описании фитоценозов водных макрофитов
используются некоторые особые приемы и учитываются особые фак-
торы местообитания. При описании водных фитоценозов пробная
площадь обычно не фиксируется в пространстве. В связи с по-
ясным распределением водных фитоценозов пробная площадь чаще
всего имеет лентовидную форму. Сбор и учет водных растений
осуществляется с помощью особых приспособлений: варослечерпате-
лей, драг, скребков и т.д., - позволяющих добывать растения из
воды. При описании водных фитоценозов учитываются такие факто-
ры среды, как степень прибойности берега, проточность и глуби-
на водоема, эсарактер дна, прозрачность и гяиический состав во-
ды, газовый режим и некоторые другие.
При изучении водной растительности большое значение при-
дается учету экологии и жизненных форм водных растений, на что
обращается внимание и во время экскурсии на водоемы, и в пери-
од обработга гербария водных растений.
Вода является высокоспецифичной средой обитания, резко
отличающейся от наземных условий существования растений. При-
чем особенности водной среды находят четкое отражение в целом
ряде специфических анатомо-морфологических, биологических,
экологических и физиологических признаков водных растений.
Для большинства водных растений характерно значительное
увеличение поверхности тела в сравнении с его массой, что об-
легчает поглощение необходимых количеств кислорода и доугих
газов, которых в воде содержится меньше, чем в воздухе. Это
достигается развитием у водных растений длинных и тонких
листьев й стеблей, сильным расчленением листовых пластинок на
тонкие нитевидные дольки и образованием воздухоносных полостей
и больших межклетников.
- 86 -
Большая плотность водной среды обуславливает слабое раз-
витие механической ткани в стеблях и листьях водных растений.
При этом элементы механической ткани располагаются в стеблях
ближе к центру, что придает им большую гибкость и предохраняет
от повреждения подвижностью водйой среды.
Высокая обеспеченность водных растений водой, которую они
могут впитывать с растворенными в ней минеральными солями всей
поверхностью тела, обусловливает слабое развитие эпидермиса и
проводящей системы, а также значительную редукцию корневой
системы, которая совсем лишена корневых волосков и поэтому не
выполняет функции питания. Даже у водных растений, ведущих
прикрепленный образ жизни и нередко обладающих достаточно раз-
витой корневой системой, последняя выполняет, главным образом,
функцию прикрепления их ко дну водоема, а не функцию питания.
.Недостаток света в водной среде вызывает у многих водных
растений образование хлорофилла в самой наружной ткани - в
эпидермисе. Эта особенность водных растений обеспечивает луч-
шее использование света в процессе фотосинтеза.
Для многих водных растений характерна разнолистность, или
так называемая ’’гетерофилия’*, суть которой заключается в том,
что подводные, плавающие и воздушные листья на одном и том же
растении различаются как по внешнему, так и по внутреннему
строение. Подводные листья расчленены на мелкие доли и не име-
ют устьиц, в то время как воздушные и плавающие на поверхности
воды листья являются цельными или слабо расчлененными и несут
устьица - первые на обеих сторонах листовой пластинки, а вто-
рые только на верхней стороне.
Все водные растения рассматриваются в качестве одной
крупной экологической группы, называемой гидрофитами. В преде-
лах группы различают две экологические подгруппы; 1) гидатофи-
ты - растения, годностью или большей частью погруженные в воду
и 2) г^рогидрофиты - растения, погруженные в воду только своей
нижней частью. Таким образом, гидатофиты имеют более тесную
связь с водной средой обитания, чем аэрогидрофиты, поэтому
прежде всего для них характерны рассмотренные выше особенности
строения водных растений.
Гидатофиты представлены несколькими жизненными формами.
Прежде всего выделяют подводные гидатофиты и гидатофиты с пла-
вающими на поверхности воды листьями.
- 87 -
Подводные гидатофиты полностью погружены в воду, проникая
на различную глубину. При этом максимальная глубина подводных
гидатофитов, относящихся к группе цветковых рас тент, обычно
не превьвиает 10-12 м и очень редко достигает значений 20-30 м.
У подводных гидатофитов очень своеобразно протекает генератив-
ная фаза развития, а именно, у глубоководных форм опыление,
цветение и образование семян происходит в водной толще; а у
мелководных форм цветоносные побеги обычно выставляются на по-
верхность воды, поэтому опыление у них осуществляется насеко-
мыми или ветром и развитие семян происходит в воздушной среде.
Среди подводных гидатофитов различают растения свободноплаваю-
щие, т.е. обитающие в воде во взвешенном состоянии, и расте-
ния, прикрепленные своими корневыми системами ко дну водоема.
К первой жизненной форме относятся, например, виды пузырчаток
(Utricularia vulgaris L., U. minor L.), а ко второй жизненной
форме - уруть колосистая (MyriophyHum spicatum L.), рдест
сквознолистный (Potamogeton perfoliatus L.), роголистник обык-
новенный (Ceratophyllum demersum L.), элодея (Elodea
canadensis Michx.) и т.д.
У гидатофитов с плавающими листьями все части потужены в
воду, кроме листовых пластинок, которые плавают на поверхности
воды или несколько приподнимаются над ней. Кроме того у них и
цветки обычно приподнимаются над поверхностью воды, хотя с
наступлением ночи у некоторых видов Ли могут закрываться и
опускаться под воду. Данная группа гидатофитов представлена
также двумя жизненными формами: прикрепленной и свсбодноплава-
щей. К первой жизненной форме относится, например, кубышка
желтая (Nuphar lutea Smith.), кувшинки белая (Nymphaea Candida
Presl.) и маленькая (N. tetragona Georgi.), а ко второй жиз-
ненной форме - водокрас (Hydrocharis morsus-ranae L.), болот-
ноцветник (Nymphoides peltata 0. Kuntze), ряска маленькая
(Lemna minor L.) и т.д.
Аэрогидрофиты, ведущие полунагруженный образ жизни и оби-
тающие в прибрежных частях более или менее глубоких водоемов,
на мелководных водоемах и на сильно обводненных болотах, осва-
ивают три среды жизни: воздушную, водную и почвенную, - каждая
из которых имеет для них важное значение. Из почвы аэрогидро-
фиты получают основную часть минерального питания и влаги, из
атмосферы - солнечную энергию, кислород, углекислоту, а из
- 88 -
водной среды - частично и влагу, и минеральные соли, и кисло-
род.
Обитание на контакте трех пред жизни обусловливает соче-
тание в структуре аэрогидрофитов анатомо-морфологических приз-
наков, характерных как для сухопутных растений, так и для ти-
пичных водных растений (гидатофитов).В этом отношении аэрогид-
рофиты представляют собой тип растений, промежуточный между
водными и сухопутными растениями.
С гидатофитами их сближает прежде всего наличие в теле
воздухоносных полостей, имеющих обычно вид мелких пор. Но
иногда, например, у камыша озерного (Scirpus lacustris L.),
поры бывают довольно крупными и хорошо различимыми невооружен-
ным глазом на поперечном срезе или разломе стебля. Наличие
воздухоносных полостей в теле аэрогидрофитов, несомненно, сви-
детельствует о затруднительном снабжении их воздухом. Его
мало содержится и в переувлажненной почве, и в воде, в которой
находятся нижние части растений. В таких условиях растение
сможет обеспечить нормальный газообмен всех своих тканей толь
ко в том случае, если будет запасать атмосферный воздух в сво-
ем теле и передавать его из частей, сообщающихся с атмосферой,
к частям растения, находящимся в водной или почвенной среде.
С сухопутными растениями азрогидрофитов сближает наличие
у них более или менее мощной корневой системы, выполняющей не
только функции прикрепления, но и функцию питания; а также до-
вольно сильное развитие проводящей и механической тканей. Эти
признаки строения аэрогидрофитов говорят о том, что, во-пер-
вых, они добывают воду и элементы минерального питания в
основном по способу сухопутных растений и, во-вторых, аэрогид-
рофиты нуждаются в защите от вредного воздействия (прежде все-
го иссушающего воздействия) атмосферы.
Из нашей (лоры к аэрогидрофитам относятся многие крупные
болотные и прибрежно-водные осоки (Carex aquatilts Wahl., С.
vesicaria L.), тростник (Phragmites australis Trin. ex
Steud.), тростянка (Scolochloa festucacea Link.), виды камыша
(Scirpus lacustris, S. /tabernaemontani Gmelin и др.), стрела-
лист (Sagittaria sagittifolia L.), частуха (Alisma
plantago-aquatica L.), виды рогоза (Туpha latifolia L., T.
angustifolia L.), сусак зонтичный (Butomus umbellatus L.) и
т.д.
- 89 -
Типы континентальных водоемов, к которым относятся озера,
пруды, .реки, ручьи, разнообразны по многим признакам. Они раз-
личаются своими размерами, формой, глубиной, подвижгостью во-
ды, температурным и газовым режимами, составом минеральных и
органических примесей в воде, прозрачностью, рельефом дна, се-
зонными колебаниями уровня воды и т.д. Поэтому в разных типах
водоемов различным оказывается видовой состав гидрофитов и на-
бор образуемых ими водных фитоценозов. В этом легко убедиться
во время экскурсии, осмотрев, например, проточный, полупроточ-
ный и совершенно непроточный водоемы.
Вместе с тем водная растительность оказывается неоднород-
ной также и в пределах каждого отдельного водоема, и мы можем
наблюдать последовательную смену фитоценозов в направлении от
берегов водоема к его центру.Это обстоятельство объясняется
прежде всего тем, что при движении от берегов водоема к его
центру обычно наблюдается увеличение глубины воды и, следова-
тельно, изменяются условия существования водных растений
(условия освещения, температуры и т.д.). В результате прибреж-
но-водные аэрогидрофиты с увеличением глубины последовательно
замещаются разными жизненными формами гидатофитов и образуемые
ими фитоценозы располагаются в виде поясов, сменяющих друг
друга от берегов водоема к его центру.
В достаточно глубоких непроточных водоемах со слабым ук-
лоном их дна от берегов к центру обычно формируются следующие
пояса растительных сообществ:
1. Водоем обычно бывает окружен поясом прибрежных боло-
тистых лугов из канареечника, осоки изпцной, мятлика болотного
и некоторых других гидромезофитов. Иногда пояс болотистых лу-
гов замещается прибрежными зарослями кустарниковых ив (Salix
rosmarinifolla L., S. cinerea L., S. wlmlnalis L. и др.), смо-
родины (Ribes nigrum L.), черемухи (Partus avium Mill.).
2. В прибрежной части водоема до глубины 2-2.5 м формиру-
ется пояс сообществ из аэрогидрофитов: тростника, камышей, ро-
гозов, сусака зонтичного, частухи подорожниковой, тростянки,
манника водного (Glyceria maxima Holmb.), водолюба болотного
(Eleocharis palustris Roem., et Schult.), некоторых осок и т.д.
Обычно эти сообщества характеризуются более или менее густым
травостоем пятнистого сложения. Условия увлажнения в этом по-
псе могут резко меняться в связи с колебанием уровня воды в
- 90 -
водоеме. Нередко во второй половине лета пояс аэрогидрофильных
фитоценозов обсыхает совсем.
3. С увеличением глубины при дальнейшем продвижении от
берега пояс аэрогидрофильных фитэценозов сменяется поясом фи-
тоценозов, образованных прикрепленными гидатофитами с плаваю-
щими на поверхности воды листьями, такими, например, как ку-
бышка, кувшинки, гречиха земноводная (Polygonum amphlbium L.),
рдест плавающий (Potamogeton natans L.) и т.д. В зависимости
от прозрачности воды этот пояс располагается в пределах глубин
от 2-2.5 м до 4 м. В пределах данного пояса обычно присутству-
ют также свободноплавающие гидатофиты: вод окрас, болотиоцвет-
ник, пузырчатка и др.
4. При дальнейшем углублении водоема к его центру вплоть
до 5 м располагается пояс фитоценозов, образованных подводными
прикрепленными и свободноплавающими гидатофитами: рдестами
(Potamogeton perfoHatus L., Р. crispus L.), урутью, элодеей,
полушником озерные (Isoetes lacustris L.) и др.
5. При глубине 5-6 м предыдущий пояс замещается поясом
сообщества нэ подводных мхов (Fontlnalls, Scorpidium,
Cal 11 ergon), донных макрофитных водорослей (Chara и др.) и не
которых свободноплавающих пдатофитов из группы цветковых
растений.
б. При глубине более б м высшие растения уме встречаются
редко и не формируют сомкнутых фитоценозоь, а растительный
компонент водных экосистем образован в основном микроскопи-
ческим фитопланктоном и фитобентоссм.
Таковы закономерности пространственного размещения водных
растительных сообществ. Вместе с тем в каждом конкретном водо-
еме в зависимости от его особенностей набор и степень развития
поясов водных фитоценозов могут быть самши равнообразными.
Поэтому при обследовании растительности каждого водоема жела-
тельно составлять хотя бы схематичную геоботаническую карту,
на что необхоцню обратить внимание во время экскурсии на во-
доемы.
Кроме того, при обследовании водной растительности нужно
учитывать, что картина распределения фитоценозов в пределах
каждого конкретного водоема не остается постоянной во времени,
а постепенно изменяется. Дело в том, что вследствии непрекра-
щающегося накопления на дне водоема отмирающих растений и хи-
- 91
вотных (в том числе и микроскопических) происходит постепенное
обмеление водоема. В результате гидрофиты, обитающие в перифе-
рических частях водоема, и образуемые ими пояса фитоценозов
последовательно смещаются к его центру. Этот процесс может
зайти так далеко, что водоем полностью исчезнет и на его месте
образуется болото, хэс или низинный луг. Таким образом, обсле-
дуя растительность того или иного водоема, нужно обращать вни-
мание на то, в какой стадии зарастания находится данный водо-
ем, что поможет объяснить характер его растительности.
На территории нашей страны - имеется большое количество
разнообразных континентальных водоемов, например, рек у нас
насчитывается около 200 тысяч, а озер - более 300 тысяч. Поэ-
тому ресурсы водных растений в России, несомненно огромны, хотя
до сих пор их запасы не определены даже приблизительно. Уста-
новлено, однако, что в водоемах бывшего СССР произрастает око-
ло 260 видов только цветковых растений. Кроме того, здесь оби-
тает большое число видов водорослей и некоторые представители
мохообразных.
Водная растительность имеет важное значение в жизни жи-
вотных - обитателей водоемов и в жизни человека. Прежде всего
водные растения являются единственнмии согидателями первичного
органического вещества в водоемах. Именно за счет этого ве-
щества, передаваемого по цепям питания, существуют все водные
и ряд земноводных животных: рыбы, моллюбки, ракообразные, во-
доплавающие птицы, лягушки.
Среди водных растений имеются съедобные для человека ви-
ды. Съедобны, например, семена водяного ореха (Тгара nat vis
L.), корневища сусака зонтичного, молодые стебли рогоза. Во
флоре водных растений немало лекарственных видов: 'кубышка,
кувшинки, аир болотный (Acorus calamus L.) и т.д. Многие вод-
ные растения имеют техническое значение. Так, например, осоки,
камыши и тростник дают сырье длг бумажной промышленности; ро-
гоз, камыши и тростник служат строительным материалом; камыши
используются в качестве изоляционного материала и т.д.
Учитывая важное значение водной растительности, необходи-
мо относиться к ней бережно, охранять ее и соблюдать режим ра-
ционального использования. Для увеличения численности полезных
водных растений в настоящее время нередко принимают различные
меры, способствующие интенсивному возобновлению этих растений.
92 -
Так, например, для возобновления водных растений, служа-
щих кормом для рыб, успешно применяется специальная агротехни-
ка.
Однако водная растительность имеет не только положитель-
ное, но и отрицательное значение. Водные растения, например,
способствуют заилению оросительных и осушительных каналов и
прудов. А обильное разрастание в водоемах водорослей может
привести к резкому снижению в воде содержания кислорода, а
следовательно, и к замору рыбы. Понятно, что отрицательное
значение водных растений приходится учитывать и применять
соответствующие меры. Например, для очищения каналов и других
водоемов от обильно разросшихся гидрофитов применяются герби-
циды или разводят в данных водоемах растительноядные виды рыб.
Совершенно ясно, что рациональное использование, охрана и
улучшение водной растительности (так же, как и растительности
любого другого типа) требует тщательно продуманной и научно
обоснованной системы мероприятий.
- 93 -
СПИСОК РЕДКИХ ВИДОВ ВЫСШИХ* РАСТЕНИЙ, НУЖДАЮЩИХСЯ
В ОСОБОЙ ОХРАНЕ
Виды, включенные в ’’Красную книгу СССР” - подлежат госу-
дарственной охране на территории всей страны. Гербаризация их
запрещена.
Башмачек крупноцветковый - Cypripedium macranthon Sw.
Башмачек настоящий или желтый - Cypripedium calceolus L.
Бруннера сибирская - Brunnera siblrica Stev.
Водяной орех, или чилим - Тгара natans L.
Ятрышник шлемоносный - Orchis militaris L.
Виды высших растений, нуждающиеся в местной охране Запад-
ной Сибири. Гербаризация допускается в минимальных количествах.
Башмачек пятнистый - Cypripedium guttatum Sw.
Воронец колосистый - Actaea spicata L.
Гнездовка клобучковая - Neottiahtbe cucullata (L.)
Schlechter
Гроздовник виргинский - Botrychium virginianum (L.) Sw.
Гроздовник ланцетный - Botrychium lanceolatum (S.G.^nel.)
Angstг.
Гроздовник многораздельный - Botrychium multifidum
(S.G.Gmel.) Rupr.
Гусинник зернистый - Gagea granulosa Turcz.
Дремлик ржавый - Eplpactis atrorubens (Hoffm. ex Bernh.)
Schult.
Дягиль аптечный - Angelica archangelica L
Калипсо луковичный - Calypso bulbosa (L.) Oakes
Кандык сибирский - Erythronium siblrlcum (Fisch, et Mey.)
Kryl.
Клевер темнокаштановый - Trifolium spadiceum L.
Колокольчик рапунцелевицный - Campanula rapuncuioides L.
Колокольчик алтайский - Campanula altalca Ledeb.
Красоднев желтый - Hemerocalis f lava L.
Кувшинка белая - Nymphaea Candida J. et C.Presl.
Огонек азиатский - Troliius aslaticus L.
Лилия-саранка - Lilium martagon L.
Любка двулистная - Platanthera bi folla (L.) Rich.
Медуница мягчайшая - Pulmonaria mollis K. Wolff
Пион-марьин корень - Paeonla anomala L.
- 94 -
Примула крупночашечковая - Pi iялПа macrocalyx Bunge
Примула Палласа - Primula oallassii L^hm.
Синеголовник плосколистный Eryngium planum L.
Стародубка (горицвет) сибирская - Adonis apennina L.
Ужовник обыкновенный - Ophioglossum vulgatum L.
Фиалка рсзсеченнолистная - Viola dissecta Ledeb.
Хохлатка прицветничковая - Corydalis braeteat a (Steph.)
Pers.
Черемуха обыкновенная - Padus avium Mill.
Чистотел большой - Che1idoniurn majus L.
Щитовник мужской - Dryopteris filix-mas (L.) Schott
- 95 - .
ТРЕБОВАНИЯ, ПРЕДЪЯВЛЯЕМЫЕ ПРИ СДАЧЕ ЗАЧЕТА
. ПО ПОЛЕВОЙ ПРАКТИКЕ ПО СИСТЕМАТИКЕ ВЫСШИХ
РАСТЕНИЙ С ОСНОВАМИ ГЕОБОТАНИКИ
Полевая практика слагается из работ, выполняемых под ру-
ководством преподават.ля и работ, выполняемых студентами са-
мостоятельно, При приеме зачета преподавателем контролируется
качество выполнения всего комплекса работ, знания и навыки
студентов, полученные в результате их выполнения.
При сдаче зачета необходимо представить индивидуально:
1. Дневник всех проведенных экскурсий.
2. Чегракь, в которую записывались самостоятельно определяемые
виды с характеристикой их основных диагностических признаков,
сопровождаемые оригинальными схематичным рисунками, и условий
местообитания. В конце тетради должен быть приложен список оп-
ределенных растений (по системе А.Л.Тахтаджяна), список ред-
ких, нуждающихся в особой охране растений и список хозяйствен-
но ценных видов в районе практики.
3. Краткий физико-географический очерк района практики с
использованном собственных материалов для характеристики *шоры
и растительности.
Представить от группы 3-5 человек: гербарий (системати-
зированный; хорошего качества и правильно этикетированный.
Растении должны быть распределены по семействам и уложены в
пачки, пригодные для транспортировки гербария.
При подведении итогов практики преподаватель принимает
гербарий от студентов, совместно занимаю дася сбором, проверя-
ет качество гербария, правильность определения и этикетирова-
ния. Индивидуально от каждого студента принимается дневник,
тетрадь дли записи определенных растений, физико-географи-
ческий очнрк района практики. Одновременно преподаватель про-
водит индивидуальный опрос студентов с целью проверки их зна-
ния осн<ип1ых семейств (не менее 150 видов), структуры типичных
в :•айон<’ Фитиценозов, редких видов, нуждающихся в особой охра-
не, а также методик изучения растительного покрова.
СОДЕРЖАНИЕ
Общие особенности растительного покрова лесной зоны
Западной Сибири.......................................... 3
Краткий обзор отделов высших растений.................... 7
Методика флористических исследований.................... 30
Методика геоботанических исследований................... 33
Экскурсии............................................... 51
Экскурсии в леса................................... Ь1
Экскурсии на луга................................. 56
Экскурсии на болота............................... 06
Экскурсии на водоемы.............................. S3
Слисок редких видов высших растений» нуждающихся
в особой охране видов................................... 93
Требования» предъявляемые при сдаче зачета по нолевой
практике по систематике высших растений с основами
’еоботаники............................................. 95
Заказ 96 Тираж 200 экз.
РИО ТГУ , Томск, 29, Никитина,4.