/






































































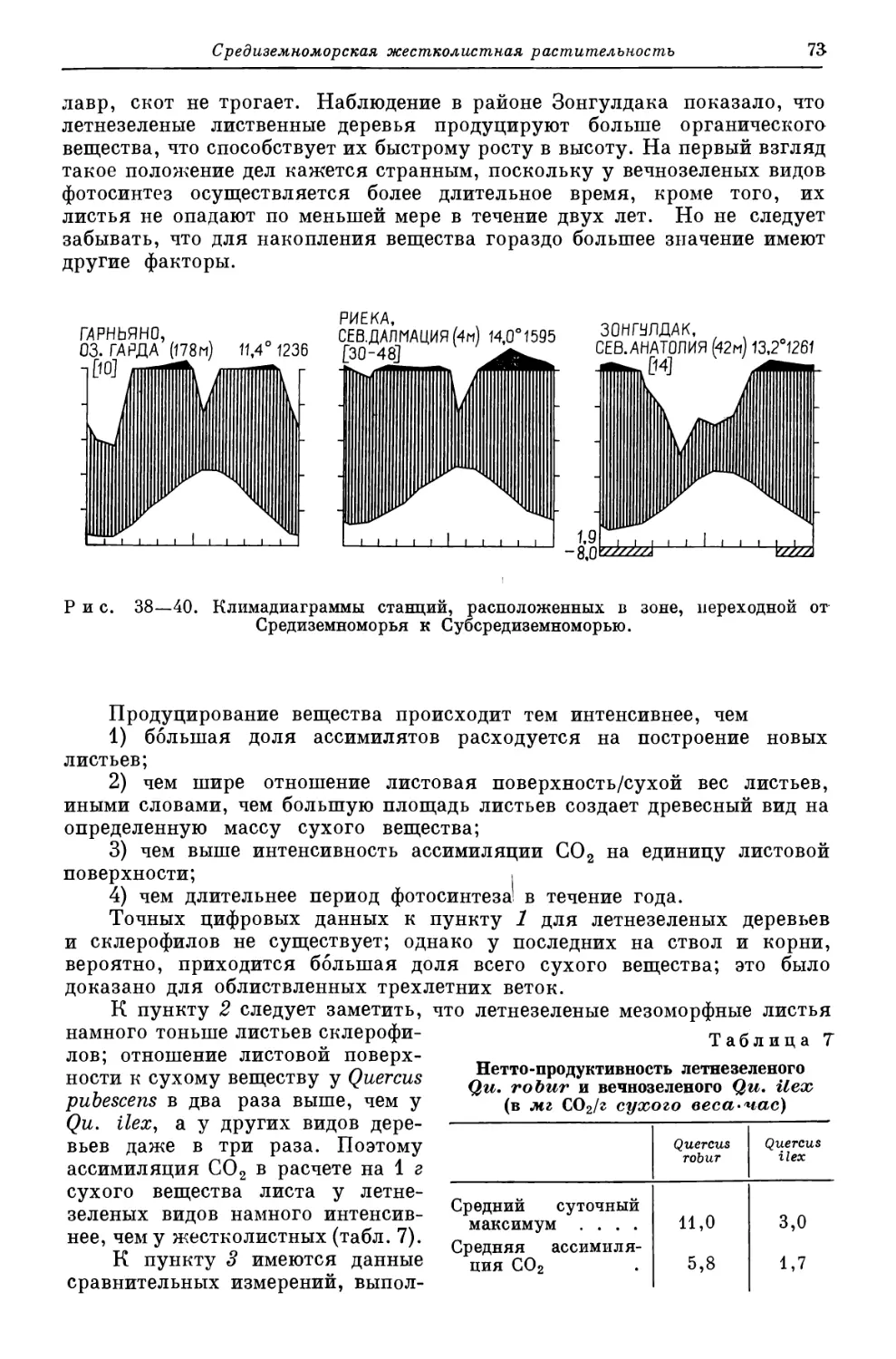




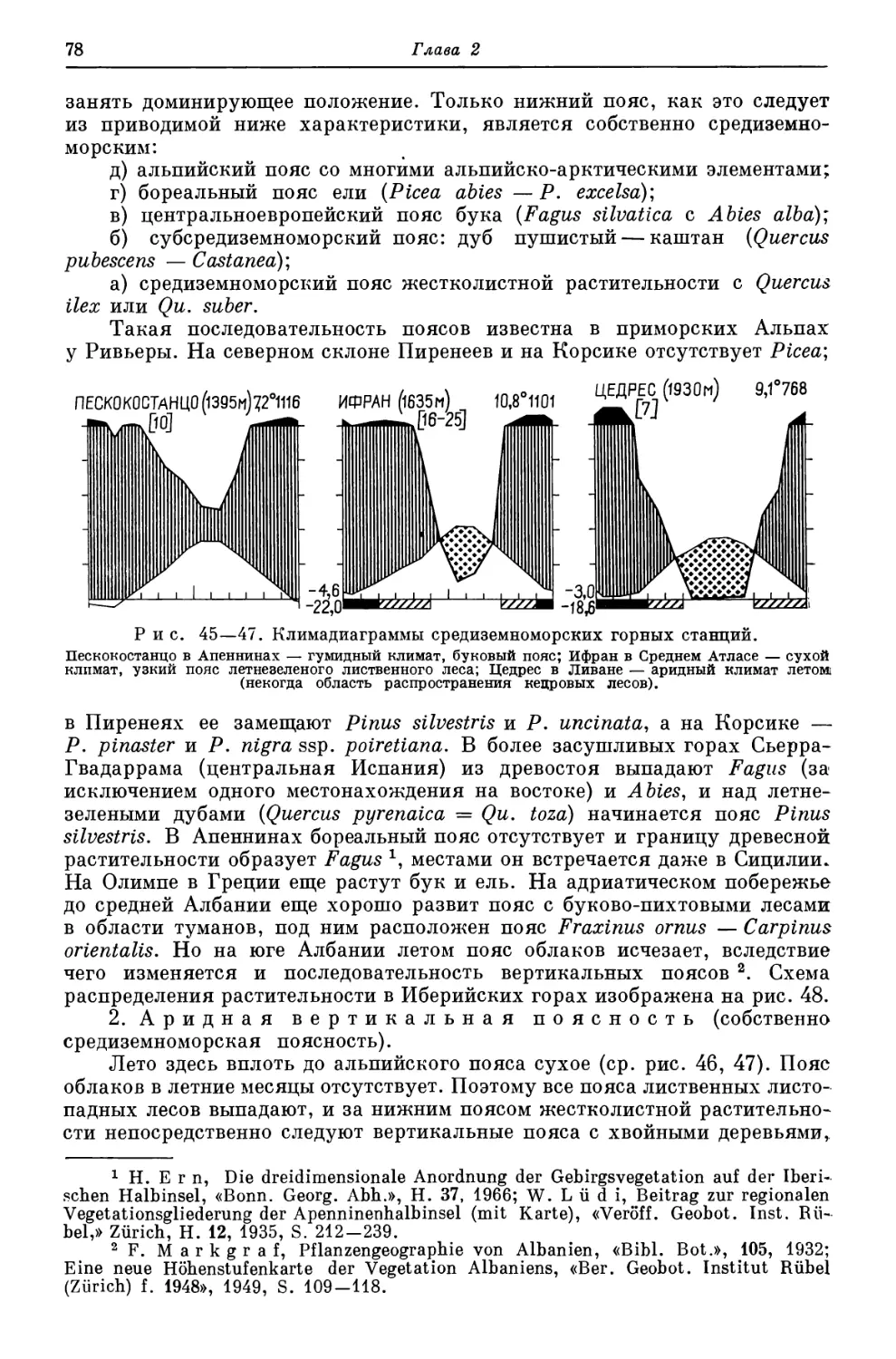





























































































































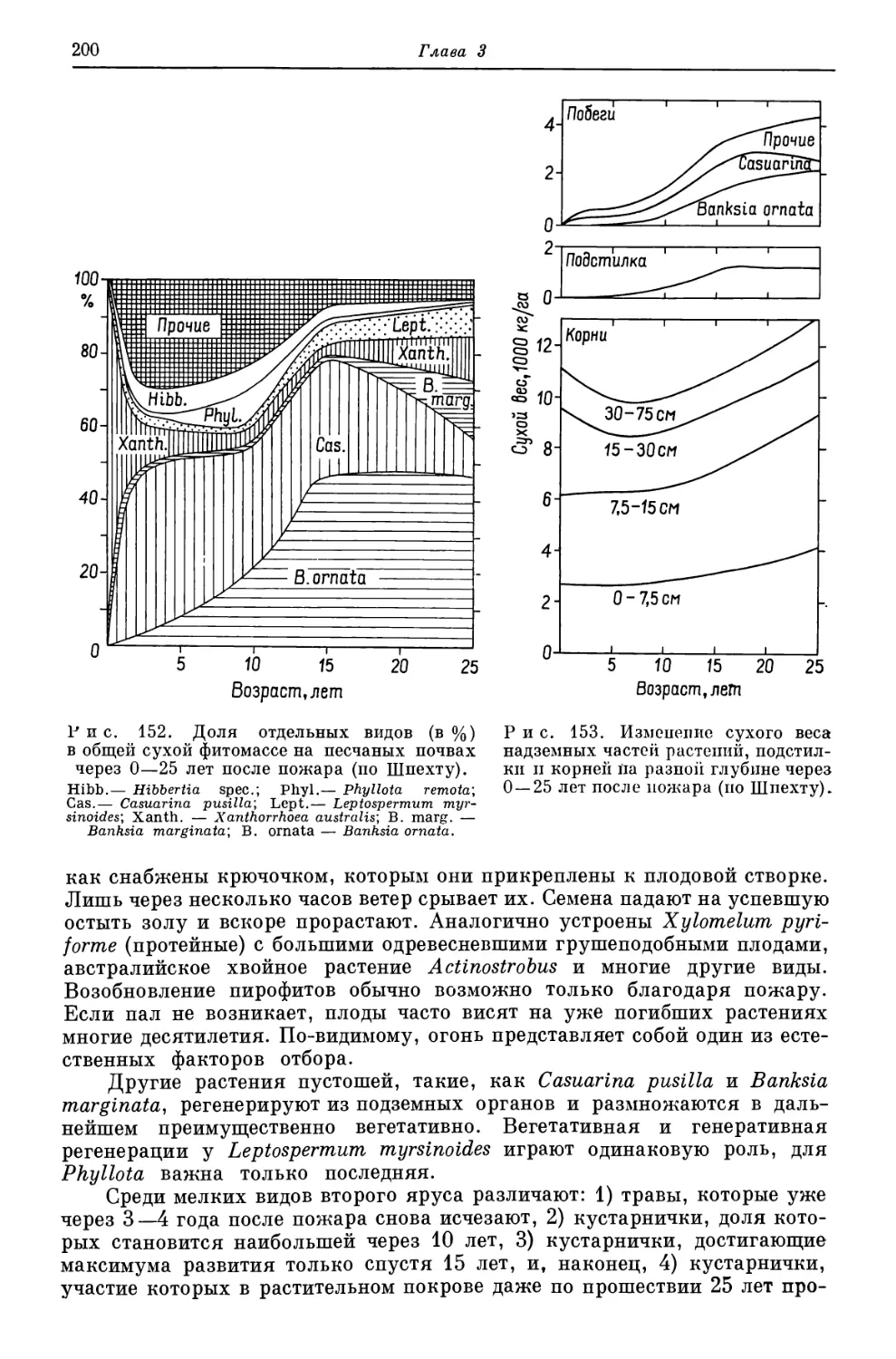





















































































































































































































Похожие




Текст
HEINRICH WALTER
DIE VEGETATION
DERERDE
in oko-physiologischer
Betrachtung
♦
Bd II.
Die gemapigten
und
a/rktischen Zonen
*
VEB Gustav Fischer Verlag
Jena 1968
Г. ВАЛЬТЕР
РАСТИТЕЛЬНОСТЬ
ЗЕМНОГО ШАРА
Эколого-физиологическая
характеристика
♦
пь. II
Теса умеренной зоны
♦
Сокращенный перевод с немецкого
Т. П. Лисовской и Ю. Я, Ретеюла
Под редакцией Т. А. Работнова
Издательство «Прогресс»
Москва 1974
Главы 1 — 3 перевел Ю. Я. Р ете ю м,
главы 4 — 5 перевела Т. П. Лисовская.
Редакция литературы по экономике и географии
© Перевод на русский язык, издательство «Прогресс», 1974 г.
В
20901-333
006(01)-74
Предисловие автора
При написании второго тома [в русском издании тома второй и тре-
тий.— Ред.}, в котором дана обобщающая эколого-физиологическая
характеристика умеренной и арктической зон, автор столкнулся с гораздо
большими трудностями, чем те, которые сопровождали его работу над
первым томом. Если в основу последнего могли быть положены резуль-
таты экспериментальных исследований, правда для некоторых районов
не слишком многочисленных, то при подготовке материалов по умеренной
зоне мы располагали данными исследований хотя и обширных, но обычно
посвященных отдельным, частным проблемам.
Отмечаемая в последнее время общая тенденция, крен в сторону
специализации, неблагоприятно отражается прежде всего на экологии.
Дело в том, что экология с ее постановкой вопроса о причинности явлений
требует синтетического подхода к анализу, а это сопряжено с необходи-
мостью привлечения данных многих смежных отраслей науки.
В настоящей работе автор попытался — в той мере, в какой это ныне
возможно,— творчески обобщить имеющиеся сведения и дать обзор эко-
лого-физиологических взаимосвязей между растительным покровом нашей
планеты и окружающей его средой. Никакая компиляция, требующая преж-
де всего составления очень полного списка литературы, такого обзора
заменить не может х. В США, Японии и Скандинавских странах много-
численные интересные эколого-физиологические исследования, проведен-
ные в последнее время, критически еще не обобщены. Хорошие обзорные
работы имеются только на русском языке. Поскольку в западных странах
они мало известны, мы старались изложить их по возможности полнее.
Некоторые читатели, несомненно, будут в претензии к автору, что районы,
особенно хорошо им известные, описаны вскользь. Но эти сожаления,
быть может, послужат стимулом к тому, чтобы эти читатели сами опубли-
ковали сводные обзоры по таким районам. Автор, правда, посетил многие,
но, конечно, далеко не все страны мира, однако даже самое тщательное
изучение литературы не может заменить собственных наблюдений. Поэто-
му автор особенно признателен руководству факультета агрономии и вете-
ринарии в Буэнос-Айресе и Исследовательского института INTA в Арген-
тине за приглашение, которое позволило ему познакомиться с умеренной
зоной Южной Америки. На рис. 1 показаны районы, которые автор
посетил в 1965—1966 гг.
1 От составления полного списка литературы, занявшего бы несколько сотец
страниц, мы отказались и здесь, как и в т. I; в подстрочных примечаниях приводим
лишь работы, на которые мы непосредственно ссылаемся, а также труды, представляю-
щие собой обзорные работы. [Ссылки на т. I даны по изданию: Г. Вальтер, Расти-
тельность земного шара, М., «Прогресс», 1968.— Ред.}
6
Предисловие автора
Рис. 1. Поездки автора в 1965—1966 гг. в умеренной климатической
зоне Южной Америки.
В настоящее время в биологии преобладает тенденция изучать явле-
ния на молекулярном или электронном уровне; но если в физике наряду
с исследованиями ядра производится изучение Вселенной, то и в биоло-
гии следовало бы стремиться к познанию не только микро-, но и макро-
строения.
«Молекулярной ботанике», относящейся к области физи-
ологии, нам хотелось бы противопоставить «континентальную
геоботанику»—сферу деятельности экологов.
Г. Вальтер
Штутгарт-Хоэнхейм, 1966 г.
Глава 1
ВВЕДЕНИЕ
КРИТИЧЕСКИЕ ЗАМЕЧАНИЯ К МЕТОДАМ НОЛЕВЫХ
ЭКОЛОГО-ФИЗИОЛОГИЧЕСКИХ ИССЛЕДОВАНИЙ
Развитие эколого-физиологического направления исследований, толч-
ком для которого послужили работы Э. Варминга (1895) и А. Шим-
пера (1898), ныне имеет уже более чем полувековую историю. В Цент-
ральной Европе особенно бурное его развитие приходится на период
после первой мировой войны. Почти одновременно это направление иссле-
дований зародилось также в США, в Лаборатории по изучению пустынь
Института Карнеги в Таксоне (штат Аризона), где оно было связано
с именами Д. Мак-Дугала, Ф. Клементса, Дж. Э. Уивера, Ф. Шрива и др.
Для эколого-физиологических исследований под открытым небом
потребовалось разработать особые полевые методы измерений. Они были
проверены прежде всего в умеренной ^оне, описанию которой главным
образом и посвящен данный том. Предпошлем поэтому рассмотрению
частных проблем несколько общих замечаний, касающихся этих методов,
чтобы читатель мог лучше оценить надежность и достоверность получен-
ных результатов.
Работа на естественном местообитании без вспомогательных средств
хорошо оборудованной лаборатории связана с особыми трудностями.
Не представляется, например, возможным использовать стационарную
очень чувствительную аппаратуру. В то же время при постоянно меняю-
щихся условиях внешней среды и неоднородности изучаемого материала
большая точность отдельных измерений обычно не дает никаких преиму-
ществ. Целесообразнее производить большое число измерений на многих
параллельных пробах, с тем чтобы получить средние значения, поскольку
единичные измерения, какими бы точными они ни были, как правило,
мало о чем свидетельствуют. Строго говоря, ни один опыт нельзя воспро-
извести, ибо погодные условия и предыстория объекта каждый раз будут
различными.
Особенно важно проводить исследования в одном и том же месте
и по возможности в течение всего года, дабы выявить годовой ход отдель-
ных процессов или хотя бы установить их сезонное изменение. Нередко
длительное наблюдение растительности обеспечивает выяснение эколого-
физиологических зависимостей и подсказывает момент, когда следует
провести измерения для контроля. Многочисленные, но случайные изме-
рения в период кратковременного пребывания в ранее незнакомом месте
обычно не поддаются интерпретации. Не будет преувеличением сказать,
что при экологических исследованиях предпосылкой успеха является
известное проникновение в жизнь растения, как бы вживание в его «нуж-
ды», умение прочувствовать жизненные условия растения в благоприят-
ное и неблагоприятное для него время года. Исследователь должен быть
непосредственно связан с природой. Чисто лабораторный биолог, не сумев
разобраться в сложной структуре экосистемы, откажется от такой работы
8
Глава 1
как якобы не научной. Разумеется, одного выявления причинных связей
путем наблюдений совершенно недостаточно. Нужно всегда стремитьсяу
насколько это возможно, доказать их с помощью точных исследований.
Эксперимент обычно осуществляет сама природа (засуха, резкое похоло-
дание и т. д.), экологу же остается в нужный момент и подходящим спосо-
бом установить результат этого эксперимента измерениями. Всегда, прав-
да, желателен контроль наблюдений в лаборатории при постоянных
условиях.
Из опыта прошедших десятилетий можно извлечь некоторые уроки.
Применявшиеся вначале весьма примитивные методы исследований с тече-
нием времени все более и более совершенствовались \ тем не менее они
еще и сегодня не удовлетворяют всем требованиям. Каждый метод содер-
жит свой источник ошибок, поэтому точность публикуемых цифр нельзя
переоценивать. Чаще они дают нам только представление о порядке вели-
чин, а нередко лишь отправную точку для качественной оценки 1 2.
1 «Phytologie», Bd. Ill, 1 (Standortslehre); L. S t eu b i n g, Pflanzenokologisches
Praktikum, Methoden und Gerate, Berlin—Hamburg, 1965.
2 В русском переводе опущено описание некоторых наиболее часто применяемых
методов полевых эколого-физиологических исследований, сохранены лишь ссылки
на литературу. Измерение транспирации: I. Н о 1 z, Uber Streuung der Transp irations-
werte bei verschiedenen Blattern einer Pflanze und bei artgleichen Pflanzen eines Bestan-
des, «Stzbr. Osterr. Akad. Wiss., math.-naturw. Kl»., Abt. I, 164, 1955, S. 659-721;
G. E g e r, Untersuchungen zur Methode der Transpirationsbestimmung durch kurzfristige
Wagung abgeschnittener Pflanzenteile, besonders an Wiesenpflanzen, «Flora», 145, 1958,
S. 374-420. Определение дефицита насыщения: H. D. В а г г s, Р. Е. W е a t h е г 1 е у,
A reexamination of the relative turgidity technique for estimating water deficits
in plants, «Austr. J. Biol. Sci.», 15, 1962, p. 413-428. В англо-американской литературе
употребляется выражение «relativ turgidity», («относительная тургеусентность»), что
неверно, поскольку речь идет об измерении относительного содержания воды, а не
о тур горном давлении; R. Bornkamm, Stand ortsb edingungen und Wasserhaushalt
von Trespen-Halbtrockenrasen usw., «Flora», 146, 1958, S. 23-67; H.P olst er, Gasstoff-
wechselphysiologische Untersuchungen an Geholzen zur Frage der Vitalitat der Pflanze,.
«Stb. Deutsch. Ak. Landw.-Wiss.», 13, H. 9, Berlin, 1964; H. R. Oppenheimer,
Zur Kenntnis kritischer Wassersattigungsdefizite in Blattern und ihre Bestimmung, «Plan-
ta», 60, 1963, S. 51—69. Осмоmическое давление: H. W a 1 t e г, О. W e i s m a n n, Uber
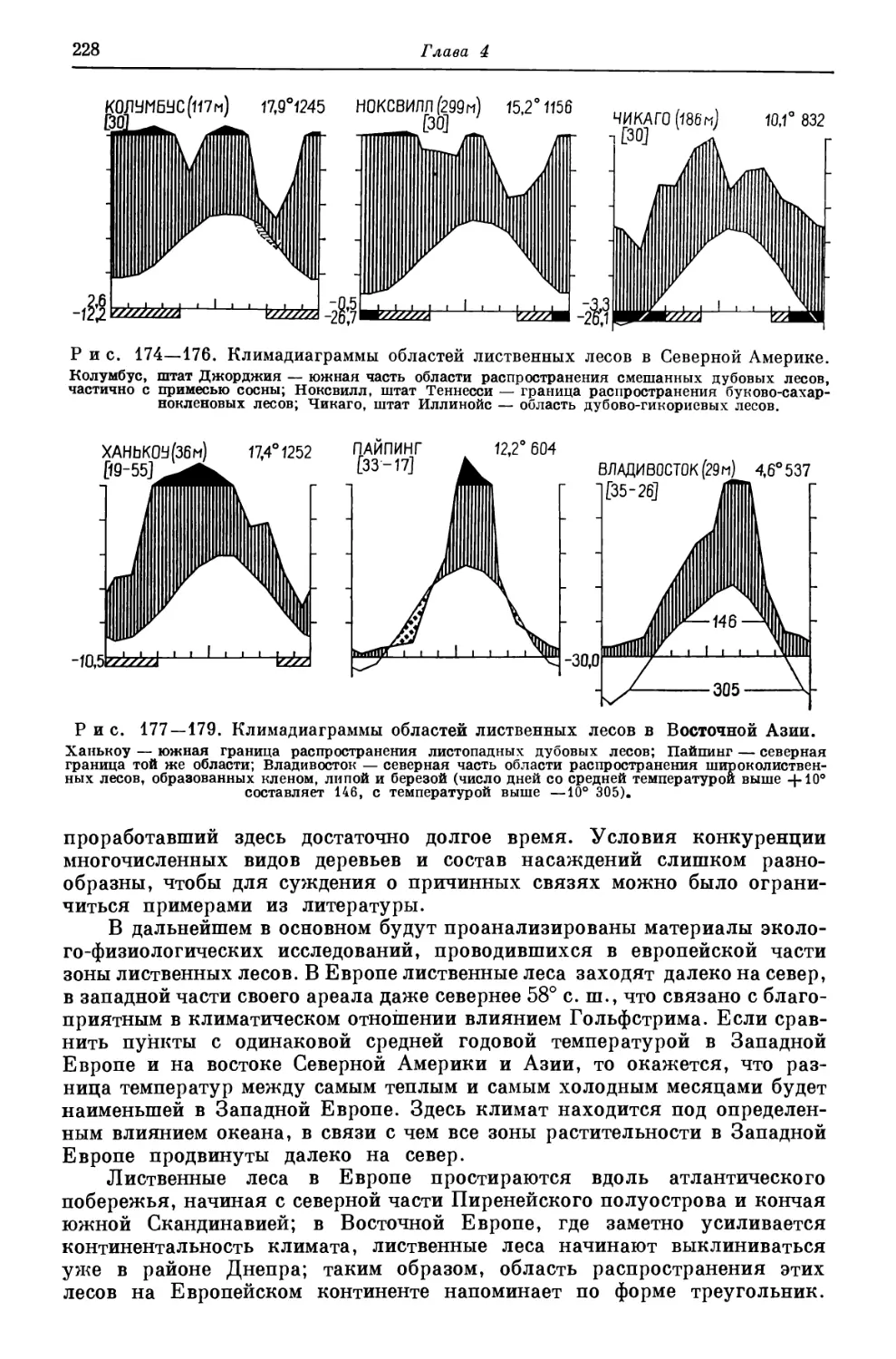
die Gefrierpunkte und osmotischen Werte lebender und.toter pflanzlicher Gewebe,
«Jb. wiss. Bot», 82, 1938, S. 273—310; S. W. В r e c k 1 e, Okologische Untersuchungen
im Korkeichenwald Kataloniens (Nordspanien), Hohenheim, 1966/67 (дисс.). Измерение
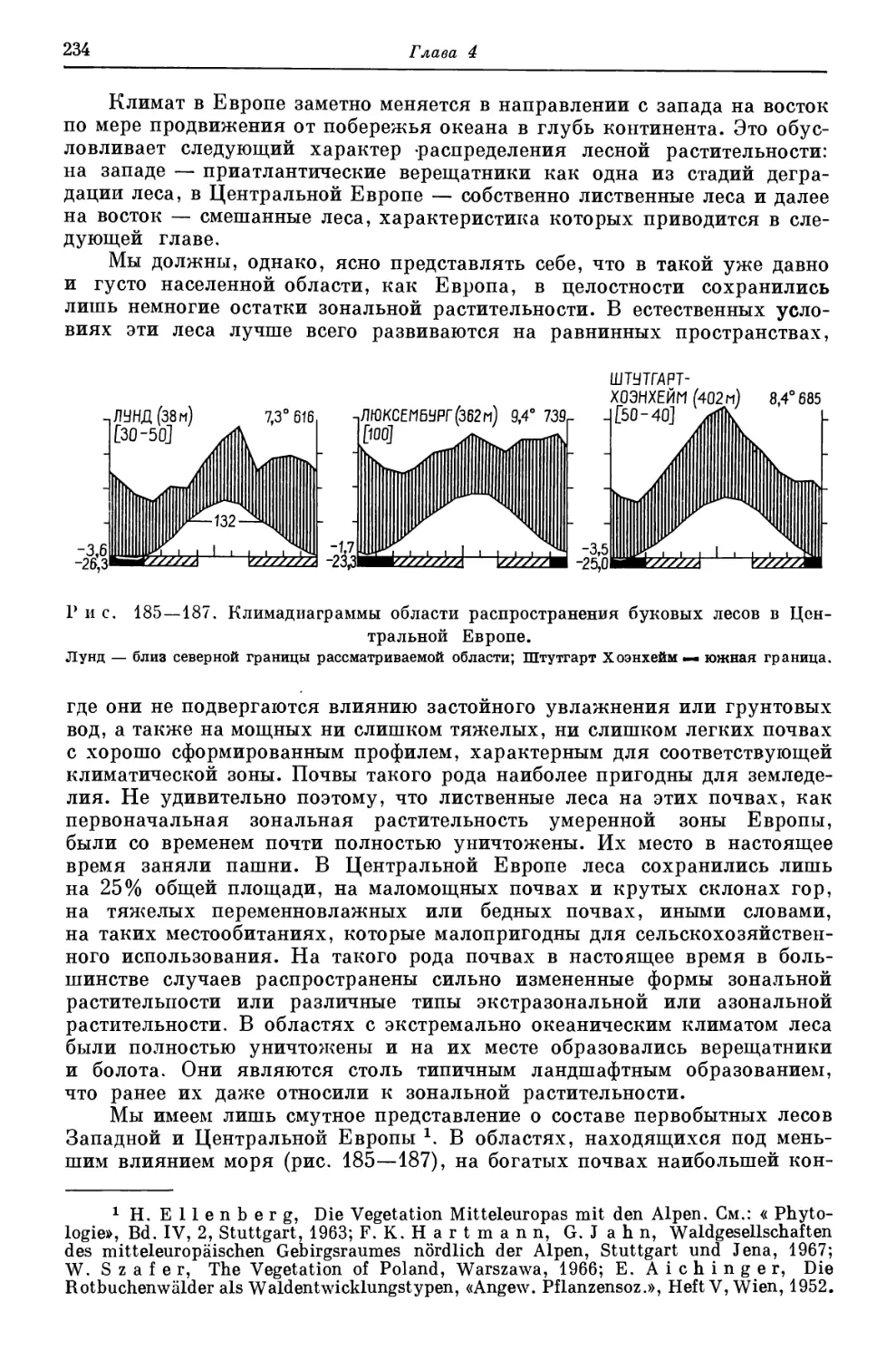
сосущей силы; Н. Rehder, К. К г е е b, Vergleichende Untersuchungen zur Bestim-
mung der Blattsaugspannung mit der gravimetrischen Methode und der Schardakow-
Methode, «Вег. Deutsch. Bot. Ges.» 74, 1961, S. 95—98; A. S c h a f 1 i, Uber die
Eignung der Refraktometer- und der Schardakow-Methode zur Messung osmotischer
ZustandsgroBen, «Protoplasma», 58, 1964, S. 75—95; К. К r e e b, Uber die
gravimetrische Methode zur Bestimmung der Saugspannung und das Problem des
negativen Turgors I und II (mit Onal), «Planta», 55, 1960, S. 274—282; 56, 1961,
S. 409—415; H. D. Barrs, Comparison of water potentials in leaves as measu-
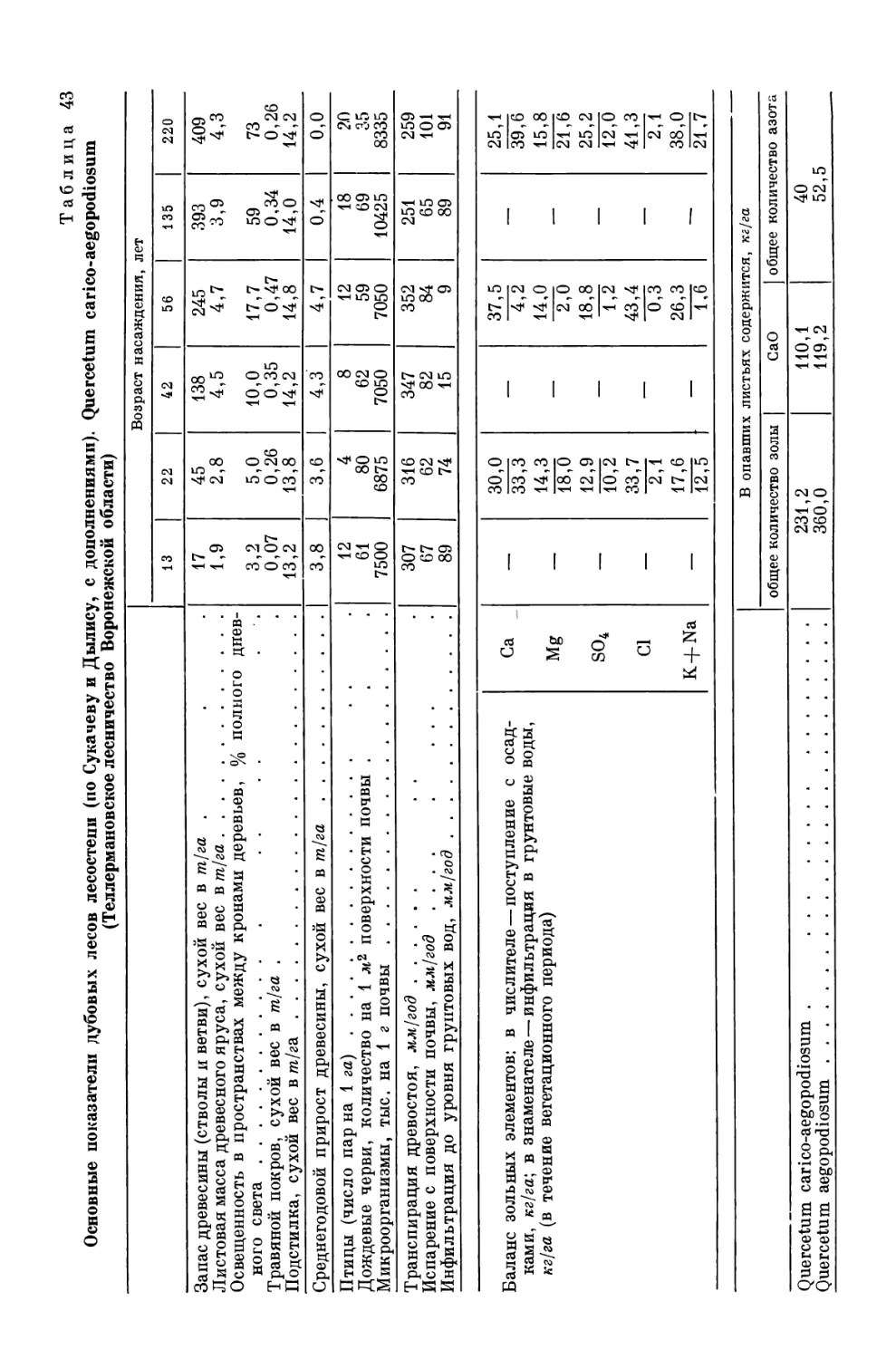
red by two types of thermocouple psychrometer, «Austr. J. Biol. Sci.», 18, 1965, p. 36—52;
D.F. Gaff, D.J.Carr, An examination of the refractometric method for determining
the water potential of plant tissues, «Ann. Bot. N. S.», 28, 1964, p. 351—368; H. W a 11 e r,
Zur Klarung des spezifischen Wasserzustandes im Plasma, «I—III. Ber. Deutsch. Bot.
Ges.», 76, 1963, S. 40—71; 78, 1965, S. 104—114; J. Slaviko v a, Eine okologische Met-
hode zur Wurzelsaugkraftmessung, «Preslia», 35, 1963, S. 241—242; 37, 1966, S. 419—
428; 38, 1966, S. 15—22. Методы определения фотосинтеза: W. Tranquillini,
Uber den EinfluB der Ubertemperaturen der Blatter bei DauereinschluB in Kiivetten auf
die okologische CO2-Assimilationsmessung, «Ber. Deutsch. Bot. Ges.», 67, 1954, S. 191 —
204; G. В о s i a n, Zum Kuvettenklimaproblem, «Flora», 149, 1960, S. 167—188;
F. E. E c k a r d t, Le principe de la soufflerie climatisee, applique a I’etude des echanges
gazeux de la couverture vegetale, «Oecol. Plant»., 1, 1966, p. 369-400; W. Tranquil-
lini, Blattemperatur, Evaporation und Photosynthese bei verschiedener Durchstromung
der Assimilationskiivette, «Вег. Deutsch. Bot. Ges.», 77, 1964, S. 204—218. Содержание
азота: H. Z 6 t t 1, Die Bestimmung der Stickstoffmineralisation im Waldhumus durch
Введение
9
Возможность воспроизведения значений, полученных тем или иным
методом, еще не служит гарантией правильности этих значений. Работа
с числовыми показателями и их детальная интерпретация не могут счи-
таться научным подходом и часто ведут к ошибочным выводам, если
не учитывается величина погрешностей. Поэтому следует пользоваться
не всеми опубликованными в литературе данными либо применять их
с известной осторожностью.
Трудности, которые приходится преодолевать при эколого-физиоло-
гических исследованиях, несомненно, очень велики, однако это не зна-
чит, что перед ними нужно капитулировать и полностью переключаться
на работу в лаборатории.
Задачи экологии иные, нежели задачи физиологии. Детально е зна-
ние каждой отдельной функции растения при строго определенных усло-
виях с одним изменяющимся фактором не означает понимания поведения
растения в целом, поскольку растение находится в непрерывно изменяющей-
ся окружающей среде и в условиях конкуренции с другими растениями.
Эколого-физиологические исследования отличаются от чисто физиоло-
гических главным образом тем, что
а) рассматривают растение как единый организм, отдельные жизнен-
ные функции которого тесно взаимосвязаны;
б) проводятся при постоянно меняющихся условиях окружающей
среды, когда временами может резко активизироваться какой-либо один
экстремальный фактор; i
в) учитывают фактор конкуренции, свидетельствующий о, как пра-
вило, косвенном характере проявления отдельных внешних факторов;
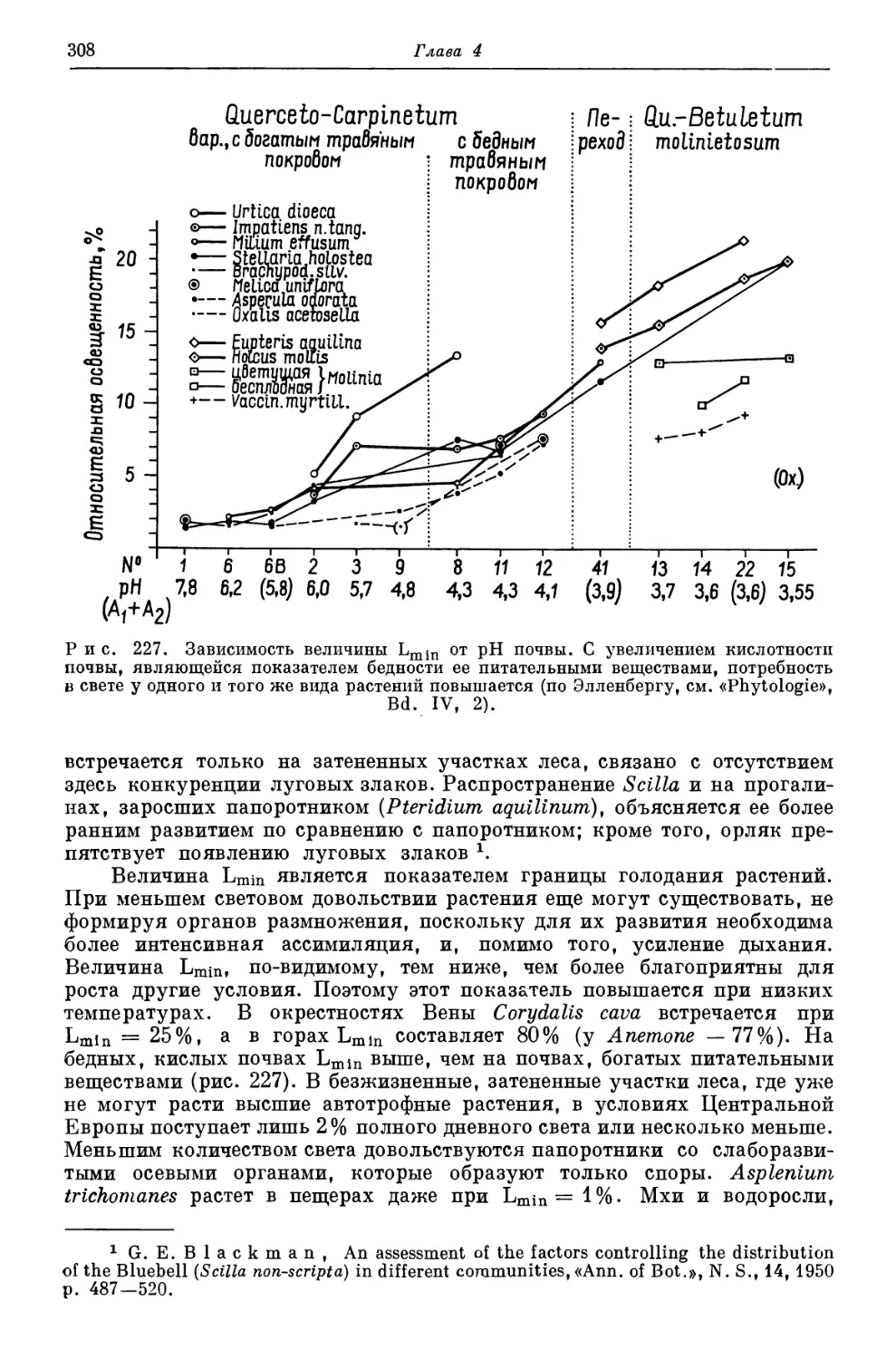
это находит свое отражение в том, что конкурентную способность одних
видов эти факторы увеличивают, а других понижают;
г) основываются на требовании следовать от анализа к синтезу,
иными словами, на необходимости понять то значение, какое имеет для
существования растения в естественных условиях взаимозависимость
различных факторов.
ОБЩАЯ ХАРАКТЕРИСТИКА КСЕРОМОРФНОЙ СТРУКТУРЫ
ТРАНСНИРИРУЮЩИХ ОРГАНОВ РАСТЕНИЙ
Различные экологические группы засухоустойчивых растений описа-
ны в т. I, стр. 320—328. Однако следует упомянуть еще одну группу.
С ней мы встретимся в засушливых областях умеренной зоны. Это группа
стеногидрических ксерофитов, устьица которых при
недостатке воды тотчас закрываются, вследствие чего фотосинтез пре-
кращается. Поэтому при длительной засухе растение голодает, его листья
не засыхают, а лишь желтеют и осмотическое давление не повышается,
а нередко даже понижается. К этой группе принадлежат многие одно-
дольные геофиты, а также травянистые виды Euphorbia и некоторые другие.
Таким образом, можно выделить следующий ряд групп растений
по возрастающей гидростабильности:
den Brutversuch, «Z. Pflzern., Diing. u. Bodenk.», 81, 1958, S. 35—50; Methodische Unter-
suchungen zur Bestimmung der Mineralstickstoffnachlieferung des Waldbodens, «Forstw.
Cbl.», 79, 1960, S. 72; Dynamik der Stickstoffmineralisation im organischen Waldboden-
material. I. Beziehungen zwischen Bruttomineralisation und Nettomineralisation, «Plant
and Soil», 13, 1960, S. 166-182; H. Ellenberg, Stickstoff als Stand ortsfaktor, «Вег.
Deutsch. Bot. Ges.», 77, 1964, S. 82—92.
10
Глава 1
а) малакофильные (с мягкими листьями) ксерофиты,
б) склерофильные ксерофиты,
в) стеногидрические ксерофиты и
г) суккуленты.
Не только в тропиках и субтропиках, но и в умеренной зоне встре-
чаются гумидные и аридные районы, или влажные и сухие местообитания,
а следовательно, и соответствующие формы растений, приспособившихся
к засушливым условиям и отличающихся особой анатомо-морфологиче-
ской структурой, которую мы называем «ксероморфной». Перечислим ряд
признаков, характерных для растений с ксероморфной структурой: мел-
кие листья или отсутствие листьев, сильноразвитая механическая ткань,
слаборазвитые межклетники, многослойная палисадная паренхима, изо-
латеральное строение листьев, у которых отсутствует губчатая паренхима,
толстая кутикула, густое опушение, глубокое расположение устьиц, груп-
пы клеток с крупным внутренним пространством вокруг проводящих пуч-
ков листьев и т. д. (ср. «Phytologie», Bd. Ill, 1). Среди ксероморфных
признаков необходимо, однако, различать видоизмененные приспособле-
ния и наследственно закрепленные признаки. Первая реакция растения
на длительный период определенной сухости окружающей среды выра-
зится в нарушении водного баланса и повышении осмотического давления
клеточного сока. Одновременно снижается гидратура протоплазмы, что
в свою очередь оказывает влияние на меристематические клетки. Видимо,
при этом происходит какое-то физико-химическое изменение протоплазмы
или изменение ее структуры, своего рода отвердевание, сопровож-
даемое усилением засухоустойчивости и резким повышением осмотиче-
ского давления; в результате вновь образующиеся органы приобретают
ксероморфное строение. Мы убедимся в этом, если проследим развитие
солнечных и теневых листьев на одном и том же дереве. Почки, заложен-
ные год назад на освещаемых солнцем ветках, водный режим которых
вследствие интенсивной транспирации был несколько напряженным, в сле-
дующем году дают солнечные листья с более ксероморфной структурой;
при насыщении клеток водой их осмотическое давление будет более высо-
ким, чем осмотическое давление теневых листьев. Совершенно аналогично
поведение видов одного рода, произрастающих на влажных или сухих
местообитаниях. Разница лишь в том, что их ксероморфные признаки
наследственно закреплены в результате отбора соответствующих мутантов.
Совокупность различных признаков ксероморфности никогда не
встречается у одного вида растений. Например, густо опушенным листьям
свойственна жесткость, обусловленная немеханической тканью; листья,
покрытые толстой кутикулой, обладают вместе с тем рыхлым мезофил-
лом и т. д. Эти признаки всегда рассматривались как приспособление
к особым условиям окружающей среды, но так бывает отнюдь не всегда.
Анатомо-морфологическую структуру определяют:
а) систематическое положение вида, то есть особенности генотипа
или наследственность, предопределяющие, каким образом вид может
реагировать на условия внешней среды;
б) условия окружающей среды, к которым вынуждены приспосабли-
ваться растения, чтобы не погибнуть. Однако в зависимости от наследст-
венности, то есть от положения растения в ботанической системе, это
приспособление может быть различным, всегда существуют многие воз-
можности;
в) наконец, известную роль играют также исторические факторы, ибо
они определяют состав флор; флора Австралии, например, отличается
Введение
И
от флоры остальных материков, поэтому при одинаковых с другими
континентами условиях обитания здесь часто можно обнаружить иные
формы; так, в аридных областях Австралии совершенно отсутствуют сук-
куленты (т. I, стр. 443 и след.).
Из сказанного следует, что «чистых» признаков приспособления
не существует, они всегда обусловливаются «характерной структурой»,
определяемой наследственностью, различной у отдельных таксонов,
и вместе с тем они являются признаками внутренней организации расте-
ния. Конвергенции, конечно, встречаются, но в большинстве случаев
формы кажутся конвергентными только на первый взгляд. Опытный
исследователь всегда обнаружит генетически обусловленные различия,
даже если объектом его наблюдений станут лишь вегетативные органы
(например, отличит канделяброобразные кактусы от молочаев). Во многих
случаях ксероморфные признаки обусловлены прежде всего таксономиче-
ски и почти не зависят от окружающей среды. Так, все виды рода Pinus,
распространенного от субарктических районов до тропиков и от гумидных
областей до аридных, обладают одной и той же ксероморфной игольчатой
структурой листьев, внешний вид деревьев также одинаков. Этот факт
служит препятствием для разработки системы жизненных форм: ведь
столь резко выделяющиеся своим обликом пальмы или злаковые типы
относятся к определенным семействам. Нельзя строго разграничивать
между собой признаки приспособления и признаки организации. Этого
не следует забывать при проведении экологических наблюдений. Преобла-
дание ксероморфных видов Melaleuca или Casuarina на постоянно влажных
местообитаниях в Австралии для европейского исследователя кажется
странным. С другой стороны, древесные виды с крупными мезофильными
листьями, которые можно обнаружить в пойменных лесах Европы, неиз-
вестны в Австралии. Существуют ксерофильные и гигрофильные виды
Casuarina, но резко выраженная ксероморфная структура сохраняется
и у тех и у других1.
Выявить признаки приспособления можно, сравнив между собой
виды одного рода, то есть виды с достаточно идентичной наследственностью,
но встречающиеся в различных условиях окружающей среды. После работ
Заленского и Келлера такие исследования проводились многократно
(ср. «Pliytologie», Bd. Ill, 1).
Удалось установить, что у родов Ga\lium и Asperula и многих других
признаками ксероморфности должны считаться следующие количествен-
ные показатели:
1) сокращение площади листовой поверхности,
2) большая плотность жилкования,
3) большее число устьиц на 1 мм2 листовой поверхности,
4) большая толщина наружного эпидермиса или кутикулы.
Вместе с тем выяснилось, что это правило применимо отнюдь не ко
всем родам (не относится оно, например, к Chenopodium) и что перечислен-
ные признаки совсем не обязательно являются специфической реакцией
растения на увеличивавшуюся сухость воздуха. Обычно они лишь прояв-
ления задержки в развитии листьев, которые также обнаруживаются
при недостатке азота, затрудненном его поглощении при низких темпе-
1 Как доказал Бидл, ксероморфность (точнее, пейноморфность) австралийских
растений обусловлена главным образом резким недостатком фосфора в большинстве
типов почв, а не только недостатком воды. См.: N. С. W. Beadle, Soil phosphate and
its role in molding segments of the Australian flora and vegetation, with special reference
to xeromorphy and sclerophylly, «Ecol.», 47, 1966, p. 992 — 1007.
12
Глава 1
ратурах или даже при избытке воды в почве, то есть в условиях недоста-
точной ее аэрации1.
Таким образом, мы сталкиваемся здесь с так называемыми п е й н о -
морфозами, явлениями недостаточности, реакция на которые со сто-
роны представителей различных семейств не обязательно должна быть
одинакова.
Если же речь идет о задержке в развитии органов растения, то в этой
ситуации изучать следует не только вполне сформировавшиеся структуры
на распустившихся листьях, но и их онтогенетическое развитие начиная
с момента появления молодых зачатков листьев на конусе нарастания.
Такого рода наблюдения выполнены Василевской 1 2. Изучая преиму-
щественно листья древесных растений, распространенных в горах Средней
Азии или культивируемых там в садах, Василевская выделила три типа
развития: 1) у мезофильных видов: плодовые деревья, а также род Acer,
Aesculus, Juglans, Catalpa и др.; 2) у более ксерофильных летнезеленых
видов в натуре: Amygdalus, Cerasus microcarpa, Pistacta vera, Zizyphus
jujuba, Ammodendron conollyv, 3) у ксероморфных вечнозеленых растений:
Magnolia grandtflora, Buxus sempervirens, Evonymus japonica, культивируе-
мых в садах и более требовательных к воде. Чем мезофильнее вид, тем более
продолжителен рост листьев, тем больше листовая поверхность и тем
тоньше лист.
У первого типа растений развитие листа продолжается 18—62 дня;
при недостатке влаги рост начинается позже, растение развивается очень
медленно, так что листья остаются мелкими. Наблюдаются незначитель-
ные количественные различия в структуре, они касаются размеров кле-
ток, числа устьиц на 1 мм2 листовой поверхности, густоты жилкования
и т. д. При ранней дифференциации мезофилла число клеточных слоев
сохраняется постоянным. При увеличении слоев палисадной ткани раз-
витие листа может продолжаться около месяца. Виды разного таксономи-
ческого положения могут по-разному реагировать на затруднение в обес-
печении водой.
У второго типа растений рост листьев заканчивается через 10—18 су-
ток. Уже в течение первых дней число слоев паренхимы вследствие
переклинального деления клеток заметно увеличивается. Деление и растя-
жение клеток во всех слоях паренхимы происходят более или менее
одинаково. Благодаря этому формируются однородные, плотно прилегаю-
щие друг к другу клетки паренхимы без крупных межклетников. Харак-
терно, что уже на ранней стадии развития происходит утолщение наруж-
ного эпидермиса. Внутреннее пространство клеток эпидермиса вследствие
набухания клеточной оболочки может уменьшиться. Толщина листа
в течение лета увеличивается. Быстрое развитие листьев позволяет ксеро-
морфным видам использовать благоприятное весеннее время для интен-
сивного продуцирования органического вещества; вместе с тем эти особен-
ности развития обусловливают формирование качественно иной структуры
листьев и повышенную засухоустойчивость, в результате листья сохра-
няются в течение засушливого лета. Вокруг проводящего пучка часто
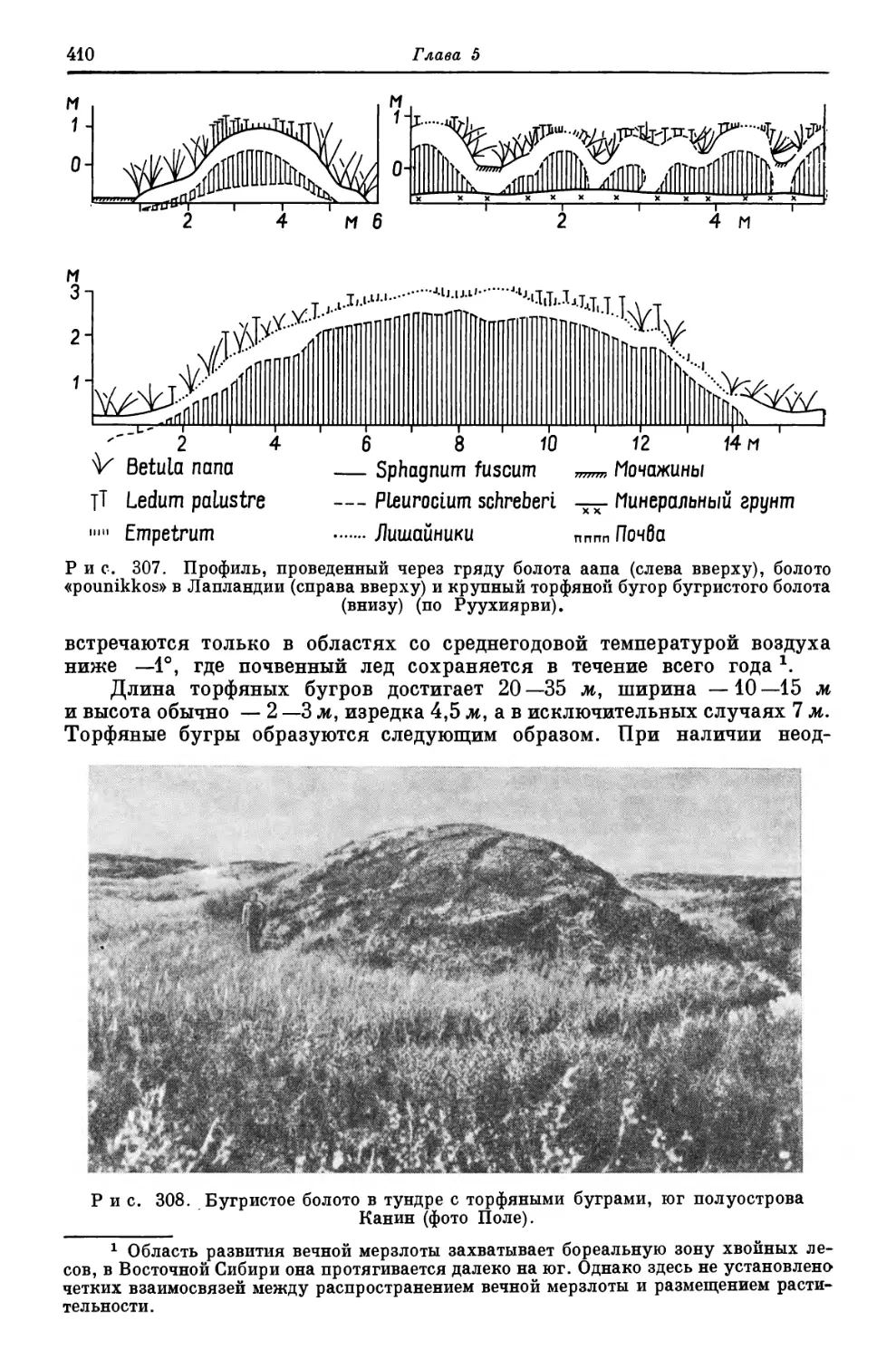
образуются сильно вакуолизированные крупные клетки, представляющие
1 Н. G г е b, Der Einfluss tiefer Temperaturen auf die Wasser-und Stickstoffaufnah-
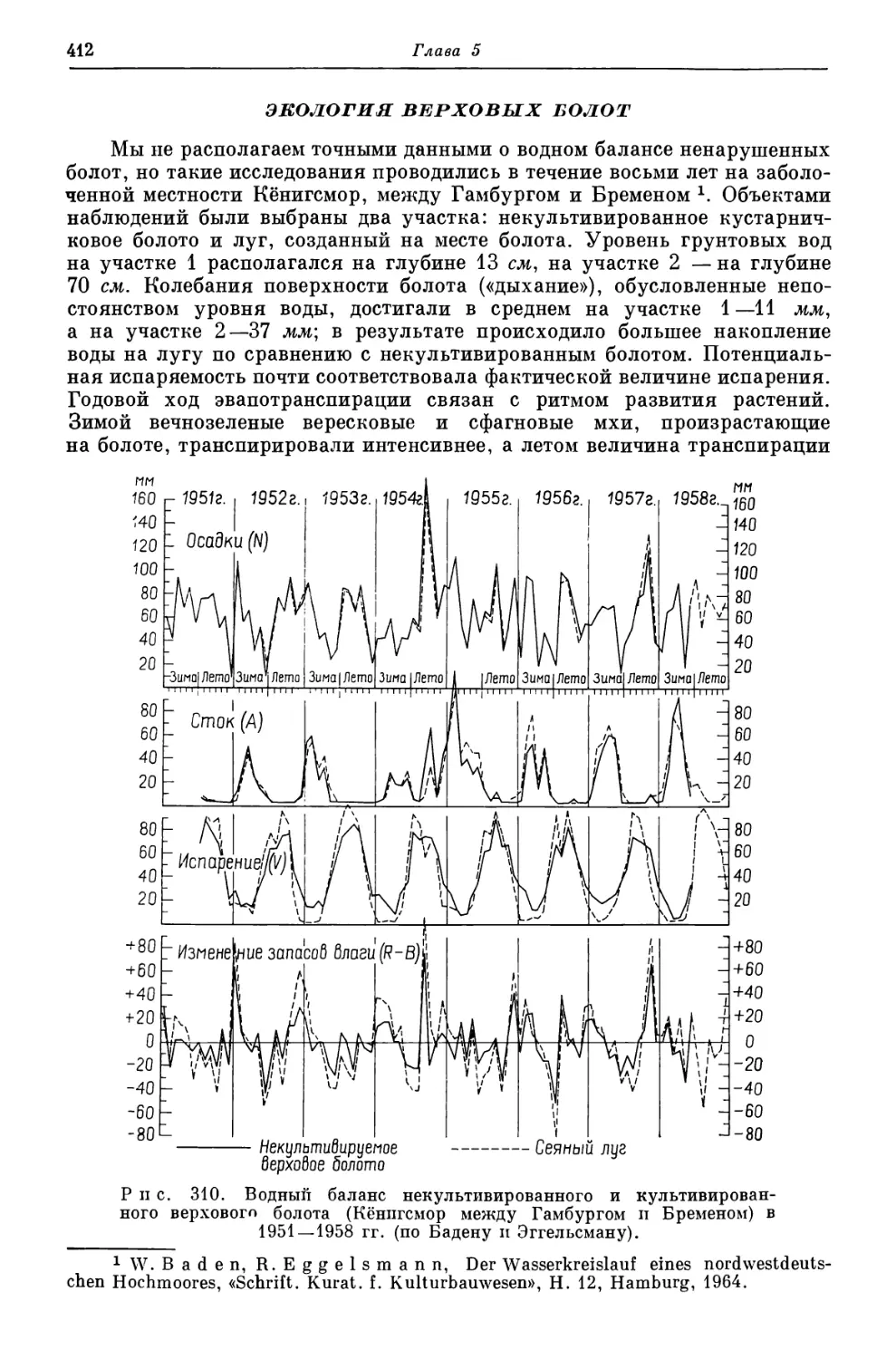
me der Pflanzen und ihre Bedeutung fur das «Xeromorphieproblem», «Planta», 48, 1957,
S. 523—563; L.O.Lundquist, Wasseruberschuss und St ickst off mangel als Ursache
gewisser Strukturanderungen bei Mesophyten, «Sv. bot. Tidskr.», 49, 1955, S. 387—418.
2 В. К. Василевская, Развитие листьев у засухоустойчивых растений,
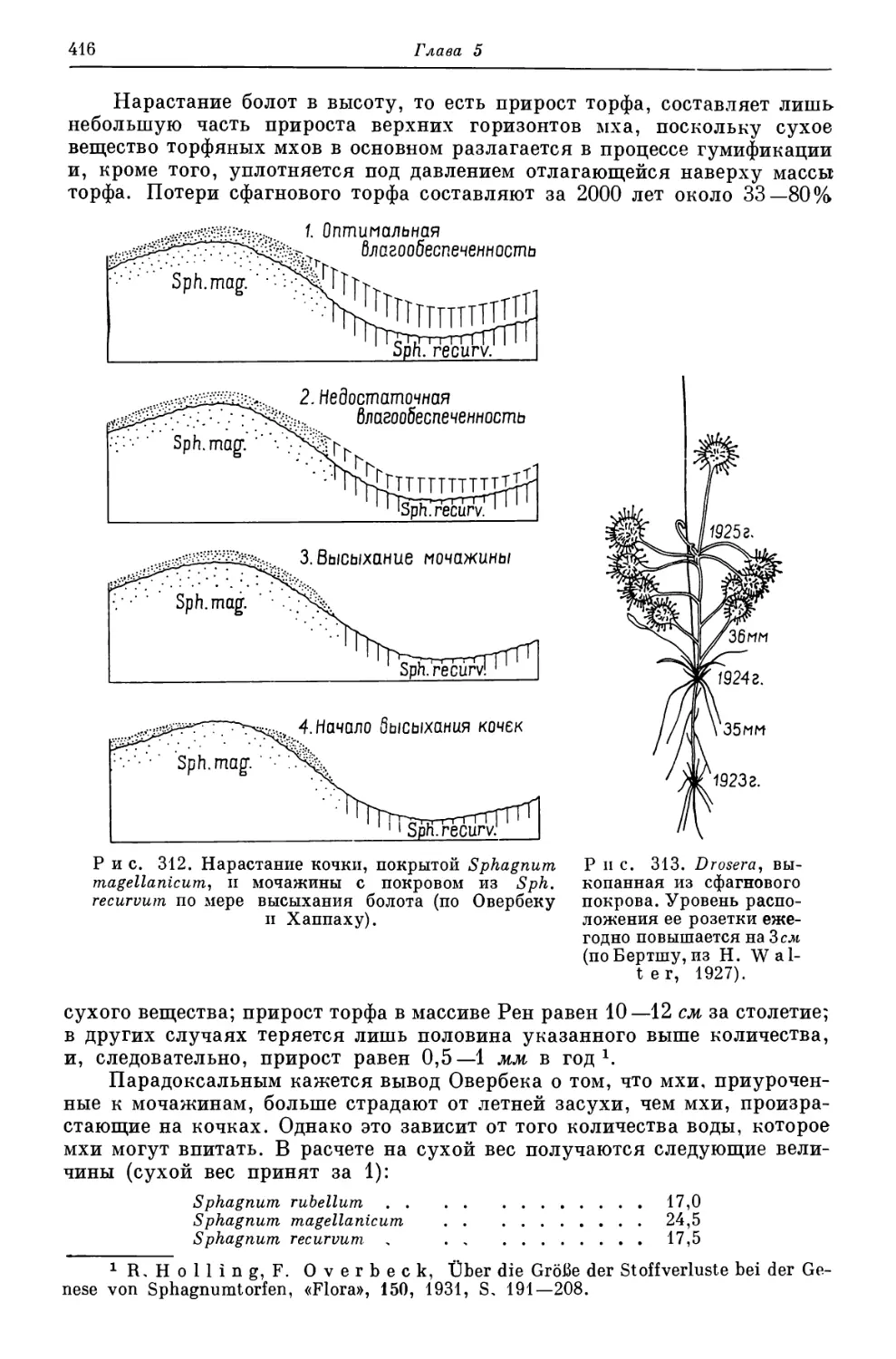
Ашхабад, 1954.
Введение
13
собой своего рода водный резервуар; они помогают восполнить дневной
недостаток влаги в других клетках, благодаря чему в растении длитель-
ное время поддерживается интенсивный газообмен.
Третий тип охватывает ксероморфные вечнозеленые растения, часто
встречающиеся на влажных местообитаниях. Листья у них развиваются
очень медленно и, хотя часто обладают толстым кутинизированным эпидер-
мисом, одновременно характеризуются чрезвычайно рыхлым мезофиллом
с очень крупными межклетниками. Палисадная паренхима нередко слабо
дифференцирована. Не исключено, что толстая кутикула представляет
собой первичный признак. Его можно обнаружить у многих древесных
растений влажных тропиков; он был характерен уже для первых наземных
растений, например для псилофита Rhynia, Вместе с тем число устьиц
на единицу листовой поверхности у ксероморфных вечнозеленых видов
гумидных районов относительно невелико (ср. цикадовые, Monstera,
Phoenix, Ficus, Coffea', т. I, стр. 133).
Ограничимся здесь этими немногими примерами, поскольку сегодня
мы располагаем результатами лишь первых исследований развития ксеро-
морфных органов листьев. К тому же очень велико разнообразие жизнен-
ных форм. С некоторыми другими примерами мы еще познакомимся.
ЭКОТИПЫ И ЭКО КЛ ЫНЫ
I
Основной структурной единицей в систематике, или таксономии,
растений является вид. Эколог также пользуется этой единицей, когда
характеризует изучаемые им в поле растения или когда исследует при-
чинные взаимосвязи растений с окружающей средой. Однако мы знаем,
что виды генетически неоднородны, что они представлены популяциями,
то есть смесью многих различных биотипов. Вид — абстрактное понятие,
устанавливаемое систематиком. На основе обильного материала он объеди-
няет особи, отчетливо отличающиеся от других особей, но характеризую-
щиеся важными, с точки зрения систематика, совпадающими морфологи-
ческими признаками. Такие виды нередко заселяют обширную площадь,
климат которой обычно неоднороден. Кроме того, в пределах этой площади
часто наблюдается приуроченность видов к местообитаниям с разными
почвами или с различными микроклиматическими условиями.
Назовем в качестве примера сосну (Pinus Silvestris'), распространенную
на пространстве от Лапландии до южных гор Испании и в Сибири. Ее мож-
но встретить в бореальных лесах вместе с елью, прежде всего на гарях,
или с примесью дуба в районах лесостепи. Растет она на сухих скалистых
гребнях и на влажных верховых болотах. Изучив сосны разного проис-
хождения, но выращенные в одинаковых условиях окружающей среды,
легко установить их отчетливые экологические различия в отношении
способности к росту, холодо- и засухоустойчивости и проч. Обычно сосны
различаются и морфологически — по длине иголок, форме кроны, строе-
нию ствола и т. д. Однако все это признаки, не имеющие значения для
выделения вида, поскольку они склонны к модификации. Следовательно,
установленные при экологическом исследовании определенные свойства
сосны нельзя относить ко всем остальным особям, принадлежащим виду
Pinus Silvestris, Строго говоря, эти свойства специфичны только для расы
или места происхождения изученного вида.
Первым это отметил Кернер. Он культивировал растения одного
и того же вида, растущие на равнине и в горах и образующие четко раз-
14
Глава 1
личающиеся между собой равнинные и горные формы, под Веной (абсолют-
ная высота 180 м), близ Инсбрука (569 м) и в Тирольских Альпах (2195 м)
и наблюдал за ними в течение 6 лет. Равнинные формы в горных питомниках
хотя и видоизменялись (оставались маленькими, имели более узкие листья,
меньшее количество цветков при более сочной окраске), но достаточно
резко отличались от росших рядом с ними горных форм. Аналогично вели
себя альпийские формы в культуре на равнине рядом с равнинными
формами.
Эти наблюдения позволили Кернеру сделать вывод, что равнинные
и горные формы есть наследственно различающиеся между собой расы
одного и того же вида, признаки которых могут изменяться в пределах
определенных границ.
Правда, Бонье в 1920 г. пришел к противоположному заключению,
а именно что равнинные и горные формы являются лишь модификациями,
вызванными различием условий существования (рис. 2иЗ)х. Однако пред-
ставления Кернера были подтверждены более поздними исследованиями.
Мнение Бонье опровергнуто, как основанное на недостаточно точных
наблюдениях.
Весьма подробно рассмотрел этот вопрос Турессон. Он также устано-
вил, что особи одного и того же вида, происходящие из различных кли-
матических областей или из совершенно несходных друг с другом место-
обитаний, в культуре при одинаковых условиях среды сохраняют в основ-
ном свой первоначальный внешний облик. Он предложил называть эти
наследственно закрепленные расы, наблюдающиеся в природе в специфи-
ческих экологических условиях, экотипами и различал
а) климатические экотипы, например равнинные и горные расы;
б) эдафические экотипы, например расы, связанные с песчаными
почвами открытых пространств или с засоленными лугами;
в) биотические экотипы, формирование которых обусловлено тем или
иным видом их использования. Примеры: Phleum pratense — стелющаяся
форма на пастбищах и прямостоящая форма на сенокосах или двухукосная
раса клевера лугового в Южной Европе и одноукосная морозоустойчи-
вая — в Северной Европе.
Правда, необходимо подчеркнуть, что на экологически различных
местообитаниях наряду с наследственно закрепленными расами сущест-
вуют также ненаследуемые модификации, которые по чисто внешним
признакам неотличимы от экотипов. Так, например, из трех растений
Succisa pratensis, росших на приморском лугу как карликовые формы,
после пересадки в сад два растения сохранили свой карликовый рост,
третье же приняло обычную форму; следовательно, карликовый рост
последней представлял собой лишь модификацию. Известно также, что
наземные формы водных растений являются исключительно модификацией.
Например, Polygonum amphibium образует в воде, на влажной почве
и на сухих песчаных дюнах три совершенно различные ненаследуемые
формы.
В дальнейшем, однако, было установлено, что представление об
экотипе как об особой эколого-генетической единице в большинстве слу-
чаев может быть приемлемо лишь с оговорками. Это убедительно подтвер-
дили продолжавшиеся в течение 30 лет точные исследования, начатые
1 Модификации сахарной свеклы, наблюдающиеся в культуре на низменности
(160 м над уровнем моря) и в высокогорных районах Пиренеев (2860 м), изучал Ласкомб:
G. Lascombes, Les facteurs de la morphologie vegetales a haute altitude, «Oecologia
Plantarum», 1, 1966, p. 275—300.
Рис. 2. Taraxacum officinale: P — выращен на низменности, М — выращен высоко
в горах (в Альпах) (обе 2/5 натуральной величины); М1 = М, нат. величина (по Бонье,
считавшему, что различные жизненные формы растений являются всего лишь моди-
фикациями) »
a
Р|и с. 3. Нeltanthemum chamaecistus (= nummularium) (см. рис. 2): а — форма, харак-
терная для низменности, б — альпийская форма., обе 1/2 нат. величины; форм^ в =
= форме б, нат. величина. t ’
Введение
17
в 1922 г. в Калифорнии Холлом, а после 1932 г. продолженные Клаузеном,
Кеком и Хизи (ср. «Phytologie», Bd. Ill, 1). В качестве примера рассмот-
рим результаты изучения культуры Achillea millefolium s.l.— растения,
которое встречается на обширном пространстве от Станфорда (побережье
Калифорнии) до высокогорных районов Сьерра-Невады (более 3000 м
над уровнем моря) и внешний облик которого весьма разнообразен. Собран-
ные в местах произрастания, Achillea были посеяны в питомнике под Стан-
фордом в 22,5 км от побережья на высоте 30 м над уровнем моря (ср.
«Phytologie», Bd. Ill, 1).
Результаты опытов (форма роста и высота растений) схематически
изображены на рис. 4. Для высоты приведена вариационная кривая;
стрелка указывает среднюю высоту растений.
Были выделены немногие основные типы (ср. римские цифры на рис. 4):
I. Береговой тип развивался интенсивно без периодов покоя,
отличался приземистым ростом и суккулентностью. Эти признаки на есте-
ственном местообитании под влиянием соленого морского ветра выражены
несколько более резко.
II, III, V. П р едгорный тип на его естественном местообита-
нии испытывает летнюю засуху. Растения сильнее опушены и имеют более
серую окраску. Рост их происходит преимущественно в период зимних
дождей, летом растения находятся в состоянии покоя. В Станфорде эти
признаки сохранялись даже при поливу растений летом.
IV. Дол и н н ы й тип распространен па влажных местообитаниях
в Калифорнийской продольной долине; в природных условиях растения
этого типа достигают необычайно большой высоты — до 2 м\ в культуре
они сохранили свой интенсивный рост. Период покоя у растений не наблю-
дался ни в естественном для них местообитании, ни в питомнике, даже
при непрерывном поливе. Такого рода факты, по-видимому, говорят
в пользу понятия «экотип»; все три названных типа отчетливо отличаются
один от другого.
VI—XV. Горный тип ведет себя иначе. Его поведение нельзя
назвать однообразным, по пет и оснований для подразделения его на
отдельные подтипы. Он постепенно изменяется с высотой как в морфоло-
гическом, так и эколого-физиологическом отношениях. В горах осадков
выпадает больше, поэтому растения на нижних частях гор выше, чем
в предгорьях, летний период покоя у нйх менее четко выражен; в усло-
виях мягкой зимы района Станфорда рост таких растений не прекращается.
Растения, развившиеся из семян, собранных с более высоких уровней,
отличались приземистой формой; выходцы из высокогорных районов
в питомнике представляли собой настоящие розеточные растения. Вместе
с тем зимний период покоя даже в культуре под Станфордом всегда был
выражен достаточно отчетливо. Восточный склон Сьерра-Невады (место-
обитания XII—XV) характеризуется значительно более континенталь-
ными климатическими условиями. Зимой здесь очень холодно, летом очень
жарко, осадков выпадает мало. Achillea растет здесь на влажных лугах.
В питомниках под Станфордом эти растения после более короткого зимне-
го периода цвели особенно обильно.
Для сравнения на опытных площадках под Станфордом культивиро-
вали также растения родом с острова Кыска (Алеутские острова, высота
365 м над уровнем моря). Эти растения холодного влажного климата
морфологически были, правда, похожи на растения высокогорного типа,
так как достигали лишь 8 см в высоту, но их период зимнего покоя был
относительно коротким.
2-1248
Береговые горы Калифорнийская Сьерра-Невада
пробольная
дпппип
Сьерра-Невада большой бассейн
Рис. 4. Схематический профиль от Тихого океана до Большого Бассей-
на; показаны места сбора семян Achillea millefolium, культивировав-
шихся затем в Станфорде (изображены среднее растение и вариационная
кривая). Хотя все экземпляры росли в одинаковых условиях, они
в основном сохранили свой первоначальный облик.
Введение
19
Из приведенных данных явствует, что морфологические свойства
не обязательно идут в ногу с эколого-физиологическими. Для горного
типа характерны плавные переходы у наследственно закрепленных рас,
тем более постепенные, чем ближе одно к другому располагались места
взятия проб. Таким образом, можно говорить не о четко обособленных
экотипах, а лишь о непрерывном ряде экотипов, называемом э к о к л и -
ном. Видимо, следует предцолагать, что существование экотипов часто
лишь кажущееся, если число проб слишком мало или если отсутствуют
пробы из переходных зон между разными местообитаниями.
В продолжение опыта экземпляры Achillea различного происхождения
были высажены в горах на высоте 1400 м и у границы древесной расти-
тельности на высоте 3050 м, где вегетационный период составляет соответ-
ственно только 145 и 67 дней. Все равнинные формы у границы произраста-
ния деревьев зимы не перенесли.
На уровне 1400 м предгорным формам был навязан зимний период
покоя, однако они сохранили при этом и свойственный им период летнего
покоя. Через 1—2 года все растения обычно погибали.
Очень интересным оказалось поведение высокогорных форм. У грани-
цы отдельных деревьев период зимнего покоя длится 9 месяцев. Весной
растения быстро пробуждаются, вегетируют, цветут и плодоносят за корот-
кое время. Напротив, на равнине близ Станфорда они находятся в состоя-
нии покоя лишь 3 зимних месяца. Весной они дают очень слабые побеги,
летом несколько крепнут, но рост их всегда остается умеренным.
Напрашиваются следующие выводы.
1. Периодичность развития растений Achillea разного происхождения
приспособлена к климатическим условиям их первичного местообитания.
Такая приспособленность не связана с модификациями, а закреплена
наследственно, ибо она сохраняется и в совершенно иных климатах.
Проще всего объяснить возникновение наследственного приспособления,
если предположить, что внутри популяции, в которой постоянно возни-
кают новые мутанты, отбираются биотипы, сумевшие оптимально приспо-
собиться к данным климатическим условиям. Отбор происходит в отно-
шении как морфологических, так и эколого-физиологических свойств,
причем не обязательно происходит отбор всех морфологических призна-
ков; они могут быть обусловлены также плейотропией или быть связан-
ными с экологическими признаками приспособления.
2. Нельзя говорить об оптимальных климатических условиях или
благоприятных условиях произрастания, действительных для всех мест
происхождения растения; например, для высокогорных рас суровый
высокогорный климат значительно благоприятнее мягкого климата Стан-
форда. У растений в естественном местообитании наблюдается гармониче-
ское сочетание процесса развития с процессами ассимиляции и диссими-
ляции, которое при других климатических условиях нарушается. Холодо-
устойчивость высокогорных форм гораздо выше и более соответствует
климату в естественном местообитании, чем холодоустойчивость равнинных
форм. С другой стороны, предгорные формы существенно более засухо-
устойчивы вследствие того, что их развитие прерывается периодом
летнего покоя.
3. Если климат или условия существования растений непрерывно
изменяются, то будут меняться и наследственно закрепленные морфолого-
экологические признаки приспособления. Следовательно, когда мы гово-
рим об экотипах, то обычно речь идет лишь об отдельных членах, произ-
вольно выхваченных из непрерывного ряда.
2*
20
Глава 1
4. Наряду с наследственно закрепленными признаками приспособле-
ния всегда встречаются и функциональные приспособительные признаки,
представляющие собой всего лишь модификации. Характер и величину
модификаций предсказать нельзя. Внешне они не отличаются от наслед-
ственно закрепленных признаков приспособления и могут быть выявлены
только экспериментальным путем в условиях непрерывно изменяющихся
факторов среды. При скрещивании различных экотипов образуются поли-
гибриды; большинство как морфологических, так и эколого-физиологиче-
ских признаков вызвано многими парами генов. Расщепление Г2-генерации
осуществляется весьма сложным путем; возникает генетически разно-
образный материал, в котором под влиянием окружающей среды проис-
ходит отбор. Решающими для конкурентных взаимоотношений являются
прежде всего все эколого-физиологические свойства, но отнюдь не все
морфологические особенности. Часто последние вообще не оказывают
влияния, но если они регулируются теми же генами, что и экологические
свойства, то экотипы выделяются также морфологически. Разделение
видов на подвиды и вариации, следуя таксономической точке зрения,
может быть экологически целесообразным, но нередко таким не является.
Как показали исследования в Калифорнии, полиплодия часто не имеет
экологического значения.
5. Существуют элементарные виды с ограниченной областью распро-
странения, которые вместе с другими родственными элементарными вида-
ми образуют сборный вид; в этом случае они характеризуются таким же
распространением, как вид с рядами экотипов, составляющих экоклин.
Различие состоит в том, что элементарные виды (эковиды) не скрещи-
ваются, то есть они генетически обособлены, и поэтому обладают четко
ограниченными ареалами. Такие виды называют викарирующими.
Приведенные выше данные исследования экотипов в последнее время
подтверждены многими наблюдениями1.
Проанализируем еще раз факты, относящиеся к сосне (Pinus Silvestris).
Ланглет показал, что лапландская, центральношведская, верхнеитальяп-
ская и испанская сосны по холодоустойчивости и по способности к росту
различаются между собой, причем эти свойства изменяются параллельно
с изменением содержания сухого вещества и сахара в сеянце 1 2. С увеличе-
нием продолжительности вегетации на месте происхождения со 105 дней
на севере до 189 дней на юге содержание сухого вещества в проростке
уменьшается с 40% в Лапландии и 31% в южной Швеции до 29% в Верх-
ней Италии и 27% в горах Испании. Масса сухого вещества в растениях
Швеции была определена на 580 опытных площадках. При этом оказалось,
что она весьма плавно сокращается в направлении с севера на юг; столь
же плавно и в том же направлении изменяются экологически важные
свойства. Следовательно, и в данном случае речь идет не об экотипах,
а об экоклинах.
Эти закономерности, видимо, присущи и ели, таксономически под-
разделяемой на ель европейскую (Picea excelsa = Р. abies) и ель сибирскую
1 Новейшую сводку, в которой эта проблема рассматривается с таксономической
точки зрения, представляет работа: Р. Н. D a vis, V. Н. Н еу wood, Principles of
Angiosperm Taxonomy, Edinburgh und London, 1964. См. также: J. Heslop-Har-
r i s о n, Forty years of genecology, «Advanc. Ecol. Res.», 2, 1964, p. 159—247.
2O.Langle t, Studien uber die physiologische Variabilitat der Kiefer und deren
Zusammenhang mit dem Klima, «Medd. Skogsforsokanst.», 29, Stockholm, 1936,
S. 219—470.
Введение
21
(Picea obovata), которые, однако, связаны между собой в переходной
области промежуточными формами (Р. fennica, Р. medioxima, Р. uralensis).
У бука (Fagus silvatica и Fagus orientalis) также известны промежуточные
формы — балканская (F. moesicd) и крымская (F. taurica). По-видимому,
во всех этих случаях правильнее будет говорить об экоклинах.
Для практики факт существования рядов экотипов имеет исключи-
тельно важное значение. В лесоводстве, в частности в Швеции, где леса
разделены на зоны шириной 200 км по меридиану \ в настоящее время
используется только тот посевной материал, место происхождения кото-
рого соответствует району разведения. В сельском хозяйстве также учи-
тывается место происхождения семян. Однако для эколога эти положения
создают трудности в том плане, что данные, полученные им при изучении
растений данного вида, неприменимы ко всем особям этого вида. Следо-
вательно, нужна большая осторожность при обобщениях. Она нужна
в отношении всех свойств, рассматриваемых как признаки приспособления
к окружающей среде, то есть свойств, с экологической точки зрения
особенно интересных.
В последнее время наблюдается тенденция проводить исследования
эколого-физиологических свойств, используя климатические камеры, что-
бы таким способом установить степень приспособления растения к опреде-
ленным условиям среды. Выводы, сделанные на основании таких опытов,
таят в себе опасность. Дело в том, что даже в самых конструктивно совер-
шенных климатических камерах и фитотронах невозможно точно воссоз-
дать условия окружающей среды. Исследования, проведенные Клаузеном,
Кеком и Хизи с расами Achillea millefolium в пасаденском фитотроне,
показали, что хотя реакции растений на колебания температуры п про-
должительность дневного освещения у разных рас и были различны, но
они не всегда соответствовали ожидаемым. Высокогорные расы, происхо-
дившие из района южпее 40° с. ш., в условиях жаркого дня и теплых ночей
начинали цвести, как это ни странно, только при долготе дпя 24 часа,
лапландской же расе при тех же температурных условиях нужна была
всего 8-часовая длительность дня. Поэтому интерпретация результатов
физиологических исследований при строго контролируемых условиях
и одном переменном факторе также требует большой осмотрительности.
Анализ необычайно осложняется тем, что характер приспособления зави-
сит от взаимодействия всех постоянно Изменяющихся факторов среды.
1 Как недавно было установлено, Fagus silvatica даже на такой небольшой террито-
рии, какую занимает Бельгия, подразделяется на 5 экологических рас. См.: A. Ga-
lon х, La variabilite genecologique du hetre commun (Fagus silvatica L.) en Belgique,
«Trav. Stat. Rech. Eaux et Forets», Ser. A, № 11, 1966.
Глава 2
СРЕДИЗЕМНОМОРСКАЯ ЖЕСТКОЛИСТНАЯ РАСТИТЕЛЬНОСТЬ
РАСТИТЕЛЬНЫЕ ЗОНЫ ЗА ПРЕДЕЛАМИ ТРОПИКОВ
И ПЕРЕХОДНАЯ ОБЛАСТЬ ЗИМНИХ ДОЖДЕЙ
В тропических и субтропических зонах, описанных в т. I, времена
года определяются прежде всего чередованием дождливых и засуш-
ливых периодов; они же служат основой для климатического райони-
рования и для выделения зон растительности. Количество осадков в на-
правлении от экватора к субтропическим широтам непрерывно уменьшает-
ся. При господствующих пассатных ветрах, устойчиво дующих в течение
всего года, на восточных побережьях материков и восточных склонах гор
выпадает больше дождей. Эти общие закономерности нарушаются в опре-
деленных областях земного шара муссонной деятельностью (ср. расти-
тельные зоны 1—3 на рис. 5).
Значительные колебания в годовом ходе температуры наблюдаются
только по мере удаления от экватора. Однако даже в субтропиках в про-
хладное время года температура падает ниже нуля лишь редко и ненадол-
го. Собственно холодного времени года с длительными морозными перио-
дами здесь нет.
Совершенно иные климатические условия характеризуют внетропи-
ческие зоны. Времена года определяет здесь преимущественно годовой
ход температуры, причем наблюдается более или менее продолжительный
холодный сезон с устойчивыми морозами. Осадки же, как правило цикло-
нического происхождения, обычно столь четко по временам года не рас-
пределяются. Преобладающий в средних широтах западный перенос обус-
ловливает тот факт, что на западных окраинах материков, в общем, выпа-
дает больше дождей, чем в восточных, удаленных от моря районах суши.
Соответственно различают океанический, влажный, климат с меньшими
контрастами температуры в течение года и континентальный, сухой,
климат с более резкими изменениями температуры. Лишь на восточных,
прилегающих к океанам побережьях климат вновь становится более
влажным1.
Таким образом, во внетропических областях в отличие от тропиков
не представляется возможным выделить климатические и растительные
зоны, следуя строго широтным поясам. Но в полярных областях циркум-
полярная зональность вновь проступает более резко. Некоторые откло-
нения от общих условий климата и здесь обусловлены наличием горных
хребтов (см. на рис. 5 зоны растительности 4—9).
Все сказанное выше относится только к северному полушарию с его
обширными массивами суши — Евразией и Северной Америкой. В южном
1 Новейшие представления о территориальном изменении климата можно почерп-
нуть из работы: J.Bluthgen, Allgemeine Klimageographie, 2. Aufl., Berlin, 1966
(Bd. II des Lehrb. d. Allg. Geogr.). [Есть русский перевод: И. Б лютген, География
климатов, т. I—II, М., 1972—1973.—Ред.]
Средиземноморская жестколистная растительность
23
полушарии внетропическая область почти не выражена (ср. т. I, рис. 49,
«идеальный» континент х). Лишь южная оконечность Южной Америки,
небольшая часть юго-восточной Австралии, Тасмания и Новая Зеландия
простираются за пределы 35° ю. ш. в сторону полюса. Климат материков
в высоких южных широтах из-за небольшой их площади резко океани-
ческий, с несущественными колебаниями температуры в течение года.
Только в дождевой тени высоких горных хребтов преобладают засушливые
условия.
Поэтому в настоящем томе мы будем рассматривать преимущественно
обширные территории северного полушария и лишь вкратце охарактери-
зуем значительно меньшие по площади массивы суши южного полуша-
рия, тем более что эколого-физиологические исследования здесь почти
не проводились.
Провести границу между умеренной зоной и субтропической не очень
просто. В области влажного климата на восточных окрайнах материков
тропическо-субтропическая зона весьма постепенно переходит в умеренно
теплую (т. I, стр. 195—200). Поэтому разделяющая их граница всегда
будет более или менее произвольной. Иначе обстоит дело в аридных
областях. Здесь между аридной субтропической зоной пустынь и более
влажной умеренной зоной в западных частях материков всегда вклини-
вается весьма своеобразный и экологически интересный переходный пояс;
летом он всецело находится под влиянием субтропической области высо-
кого давления с присущей ей жаркой сухой погодой, зимой же в этом
поясе идут циклонические дожди, типичные для умеренной зоны. Если
лето здесь засушливое, то зима прохладная и влажная, причем в отдель-
ные годы могут даже происходить вторжения холодов.
Облик растительного покрова в переходной области определяют
сообщества жестколистных древесных растений, резко отличающиеся
от несомкнутой растительности сухих субтропиков; в более высоких
широтах они постепенно переходят в леса умеренной зоны (рис. 5, зона 4).
По этой причине мы не будем относить растительную зону с зимними
дождями к субтропикам, а ее рассмотрение в нашей книге предварим
описанию растительного покрова собственно умеренной зоны. Увеличение
континентальности и сухости климата в восточном направлении характер-
но также и для зоны зимних дождей. Соответственно леса из жестколист-
ных деревьев по мере движения на северо-восток сменяются травяной
растительностью степей и, наконец, пустынями, но уже с холодной зимой.
Местами эти пустыни непосредственно граничат с пустынями субтропиков.
За влажной или сухой умеренной зоной в северном полушарии сле-
дует бореальная зона хвойных лесов, которая представляет собой наибо-
лее обширную на Земле, сравнительно однородную область растительно-
сти (рис. 5, зона 8).
В области арктического климата развита уже циркумполярная без-
лесная тундра.
В южном полушарии картина иная. Зона, соответствующая арктиче-
ской тундре, здесь почти полностью отсутствует. Только на самой южной
оконечности Американского континента (Огненная Земля) и на крайних
островах южных морей встречаются безлесные территории, для характе-
ристики которых может быть применен термин «антарктическая тундра».
В Антарктиде растительного покрова нет.
1 С. Troll, Karte der Jahreszeiten-Klimate der Erde, «Erdkunde», 18, 1964,
S. 5-28. '
Рис. 5. Схема растительных зон Земли (упрощено). Эдафическп обусловленные типы растительности (песчаные пустыни Австралии,
обширные заболоченные районы, мангровы и т. д.), а также растительность, измененная деятельностью человека, не учтены.
Тропические и субтропические зопы
1 — вечнозеленые тропические дождевые лееа низменностей и горных склонов (лееа нояеа туманов); 2 — тропичеекпг нолувечпозелепыс п листопадные
леса; 2а — еухие леса, естественные еаванны и заросли колючих кустарников, частично злаковипкп зоны тропиков п субтропиков; з — тропические п суб-
тропические полупустыни и пустыни, простирающиеся в еторопу полюсов только до 30° (35°) е. и ю. ш. (в других местах — растительность, отмеченная
в пункте 7а).
Умеренная и арктическая зоны
4 — переходная область жеетколиетной растительности (е зимними дождями), включая аридные области е зимними дождями; 5 — вечнозеленые листвен-
ные леса умеренно теплой зоны или зон е постоянно влажным прохладным климатом; в — летпсзелспые лиственные лееа умеренной зоны е зимним
периодом покоя (неморальная зона); 7 — злаковпики умеренной зоны (степи, прерии, пампа); 7а — полупустыни п пустыни с холодной зимой (простираются
в сторону полюсов от 30—35° е. и ю. hi.), в Тибете — холодная пустыня; 8 — бореальная зона хвойных лееов северного полушария; О — арктичеекая’зона
тундры северного полушария; 10 — горы с различными поясами растительности, включая альпийский п нивальный пояса.
Средиземноморская жестколистная растительность
25
Не развита в южном полушарии и бореальная зона хвойных лесов.
Здесь могут быть выделены лишь сравнительно небольшие ареалы расти-
тельности умеренной зоны, которые мы вкратце охарактеризуем после
обзора переходной зоны с зимними дождями.
Все это многообразие типов растительности умеренной и арктической
зон еще более увеличится, если принять во внимание горные хребты с их
весьма разнородными вертикальными поясами растительности.
Однако наибольшую трудность представляет тот факт, что вследствие
плотного заселения многих областей человеком сегодня в них почти пол-
ностью уничтожена естественная растительность и повсеместно создан
культурный ландшафт. Поэтому мы нередко будем вынуждены иметь
дело с растительными сообществами, которые испытали в той или иной
мере антропогенное влияние и с экологией которых нам предстоит позна-
комиться несколько ближе, чем это было необходимо при изложении
материалов, вошедших в первый том. Мы, однако, оставляем в стороне
описание собственно культурных формаций.
ОБЩАЯ ХАРАКТЕРИСТИКА РАСТИТЕЛЬНОГО ПОКРОВА
СРЕДИЗЕМНОМОРЬЯ1
Жестколистная растительность занимает в Средиземноморской обла-
сти наибольшую площадь и лучше всёго изучена. Этот тип раститель-
ности опоясывает все Средиземное море, оставляя незанятыми только
Ливию и побережье Египта (рис. 6).
Вместе с тем вряд ли какая-нибудь другая зона растительности
на Земле столь сильно нарушена человеком, как средиземноморская.
Рис. 6. Области зимних дождей на западных побережьях материков.
1 — средиземноморский тип климата; 2 — аридные области, преимущественно с зимними дождями.
1 Подробные сведения о важнейших видах растений Средиземноморья и характе-
ристику его растительного покрова читатель найдет в трехтомной прекрасно иллюстри-
рованной работе: М. R i k 1 i, Das Pflanzenkleid derMittelmeerlander, Bd. 1—111, 2. u.
3. Aufl., Bern, 1943—1948.
26
Глава 2
Р и с. 7. Маквис, образованный Quercus ilex, на горе Монтсеррат (Каталония), высота
S00—1200 м над уровнем моря (фото Г. Вальтера).
Средиземноморье — колыбель человеческой культуры, но одновременно
и убедительное свидетельство изменений природы, вызванных деятель-
ностью человека. Средиземноморский климат особенно благоприятен для
человека. Зима здесь достаточно мягкая, поэтому меры защиты от холода
не нужны. Вместе с тем влаги хватает, чтобы зерновые злаки и другие
однолетние культурные растения росли зимой и созревали ранней весной.
Наступающая затем вскоре летняя засуха не мешает разведению маслины,
инжира, миндаля или винограда, которые дают богатый урожай. Нетре-
бовательные домашние животные, такие, как козы или ослы, даже засуш-
ливым летом находят для себя достаточно корма. Много здесь и родников,
не пересыхающих в течение всего года, так что острый недостаток воды
ощущается редко. В более засушливых окраинных районах уже давно
ведется орошаемое земледелие. Отсюда понятно, почему именно в этой
климатической зоне произошло развитие высоких культур классического
древнего мира.
Лучшие почвы Средиземноморья превращены в настоящее время
в культурные земли, но ощутимо изменена также растительность необ-
рабатываемых территорий в результате порубок, выпаса (главным обра-
зом коз) и пожаров. Особенно резко проявляется воздействие человека
в районах, где рост растений задерживается длительным периодом летней
засухи. Из-за высокой плотности населения потребность в древесине была
здесь всегда столь велика, что естественный прирост вечнозеленых дубовых
лесов, типичных для этой климатической области, не покрывал ее. Со вре-
менем леса были уничтожены полностью, так как возобновления их из-за
выпаса скота не происходило. Начавшаяся в конце концов эрозия охвати-
ла огромные территории и разрушила на них почвенный покров. Перво-
Средиземноморская жестколистная растительность
27
начально плодородные или покрытые лесами земли постепенно превраща-
лись в каменистую пустыню со скудной растительностью. Такие безра-
достные ландшафты ныне занимают большую часть территории Среди-
земноморья. Это означает, что в наших исследованиях мы вынуждены
изучать не естественную зональную растительность, а лишь различные
стадии ее деградации. Наиболее благоприятные для леса местообитания
полностью заняты сельскохозяйственными угодьями. Сохранившиеся
лишь кое-где остатки лесов деградировали и уступили место кустарнико-
вым зарослям — маквису, занимающему непригодные для возделывания
земли с маломощным почвенным покровом (рис. 7).
Остатки вечнозеленых дубовых лесов уцелели только в Среднем
Атласе. Аналогичные леса еще можно найти в нижнем поясе гор Аризоны.
В этих почти еще не затронутых деятельностью человека насаждениях
можно получить представление о девственном облике жестколистных лесов
Средиземноморья. Напротив, в более заселенной юго-западной части
Калифорнии, климат которой близок климату Средиземноморья, первич-
ный вечнозеленый дубовый лес деградировал и его заменил кустарник,
называемый здесь чапарралем: формация растительности, соответ-
ствующая маквису. Аналогичную склерофильную растительность, правда
с совершенно иным флористическим составом, можно еще наблюдать
на юго-западе и юге Австралии, в Южной Африке и среднем Чили (рис. 6).
Экологические исследования склеррфильных типов растений прово-
дились лишь в небольшом .числе районов, главным образом в Средиземно-
морье, что дает нам основание рассмотреть эту область более подробно.
КЛИМАТ СРЕДИЗЕМНОМОРСКОЙ ОБЛАСТИ
ЗИМНИХ ДОЖДЕЙ
Как и в первом томе, для характеристики климатических условий
используем метод графического изображения климата — метод климадиа-
грамм Ч
Рис. 8—10. Климадиаграммы береговых станций Средиземного моря.
Танжер — западное побережье, относительно влажный климат; Мессина — переходная зона;
Бейрут — восточное побережье, лето очень сухое.
При изучении климадиаграмм для береговых станций Средиземного
моря (рис. 8—10) сразу бросается в глаза характерная особенность здешних
климатических условий: мягкая, влажная зима и жаркое, засушливое лето1 2.
1 Для подробного ознакомления с методом климадиаграмм отсылаем читателя
к т. I, рис. 16.— Прим. ред.
2 Н. W а 1 t е г, Н. L i е t h, Klimadiagramm-Weltatlas, VEB Gustav Fischer Ver-
lag, Jena, 1960—1967.
10° 5°з.д. 0° 5°в.д. 10° 15* 20° 25° 30° 35° 40° 45° 50°
Рис. И. Годовое распределение осадков в северной части Средиземноморья (по Фишеру).
Севернее линии а — область с дождями во все времена года; южнее линии Ъ — область с зимними дождями; между линиями а и Ъ — область с дождями в пс-
г риод равноденствий (максимум осадков весной и осенью) (по Шимпер-Фаберу).
Средиземноморская жестколистная растительность
29
Зимой циклоны, перемещающиеся из района Исландии в сторону
Европейского материка, проникают далеко на юг и приносят с собой
в Средиземноморскую область обильные дожди (рис. 11). Летом, напротив,
отрог азорского антициклона распространяется на юг Европы и оттесняет
циклоны к северу. В результате Средиземноморье оказывается в области
сухих субтропиков. На северо-западной его окраине летнюю засуху смяг-
чает близость Атлантического океана, но в более континентальной восточ-
ной части засуха проявляется особенно резко.
Количество осадков зимой значительно варьирует: оно одно на бере-
говой станции, расположенной у подножия горы, и другое на станции,
за которой расстилается равнина. Ривьера у подножия Приморских Альп
или южная Анатолия с круто поднимающимся хребтом Тавр отличаются
обильными осадками. Напротив, на равнинном побережье Египта выпадает
так мало дождей (около 160 мм), что его ландшафты скорее напоминают
пустыню. Кроме того, выделяют средиземноморскую океаническую кли-
матическую зону (от северо-западной Африки до восточного побережья
Адриатического моря) и континентальную (располагающуюся юго-вос-
точнее названной выше линии и простирающуюся до Палестины; ср. т. I,
рис. 400). Близость моря обусловливает сравнительно высокую влажность
воздуха во всей области, отчего летом здесь часто бывает душно.
Ночью воздух охлаждается слабо, поэтому росообразование незначи-
тельно.
Холодного периода зимой в этой1 климатической зоне не бывает,
но лишь на немногих станциях отмечается отсутствие дней с морозом
(абсолютная минимальная температура на самых «теплых» станциях равна:
Гибралтар —1,1°, Танжер —0,1, Алжир +0,2, Филипвиль +0,9, Сира-
кузы 0,0, Триполи +1,2, Газа —1,1°).
Влияние очень холодной европейской зимы иногда распространяется
до северных районов Средиземноморья и сопровождается снегопадами
и длительными заморозками, как это случилось, например, зимой 1928/29,
1956 и 1961/62 гг.1. Природная растительность без вреда переносит воздей-
ствие холода, но культуры (маслины, цитрусовые, пинии, алеппская
сосна) и декоративные растения (пальмы и др.) иногда сильно поврежда-
ются морозом. Поэтому плантации бананов даже в самых южных частях
области (район Аланьи в южной Анатолии, Ливан) могут серьезно постра-
дать в такие зимы.
Климатические условия с зимними дождями прослеживаются в севе-
ро-восточном направлении до северной Анатолии и некоторых районов
Кавказа (несколько смягчены они в южной части Крыма), в Передней
Азии — до северо-западного горного обрамления равнины Индостана
и даже Афганистана (рис. 6). Соответственно ряд средиземноморских
элементов флоры распространен до северо-западного Пакистана.
Особенно многочисленные экологические исследования, с которыми
мы еще познакомимся, проведены на средиземноморской станции SIGMA
в Монпелье (южная Франция), основанной Браун-Бланке. Климадиаграм-
ма (рис. 12) подробно характеризует условия местоположения этой стан-
ции. Средняя температура времен года здесь следующая: зимы 6,7°,
весны 13,4, лета 22,6 и осени 15,2°.
Дожди обычно проливные, что вызывает интенсивную эрозию почв.
Роса выпадает лишь незадолго перед ненастными днями или вскоре после
1 А. у. О. de Bolos, Efectos de los frios de febrero de 1956 sobre la vegetacion,
«Goll. Bot.», Barcelona, 5, 1956, p. 165—171.
30
Глава 2
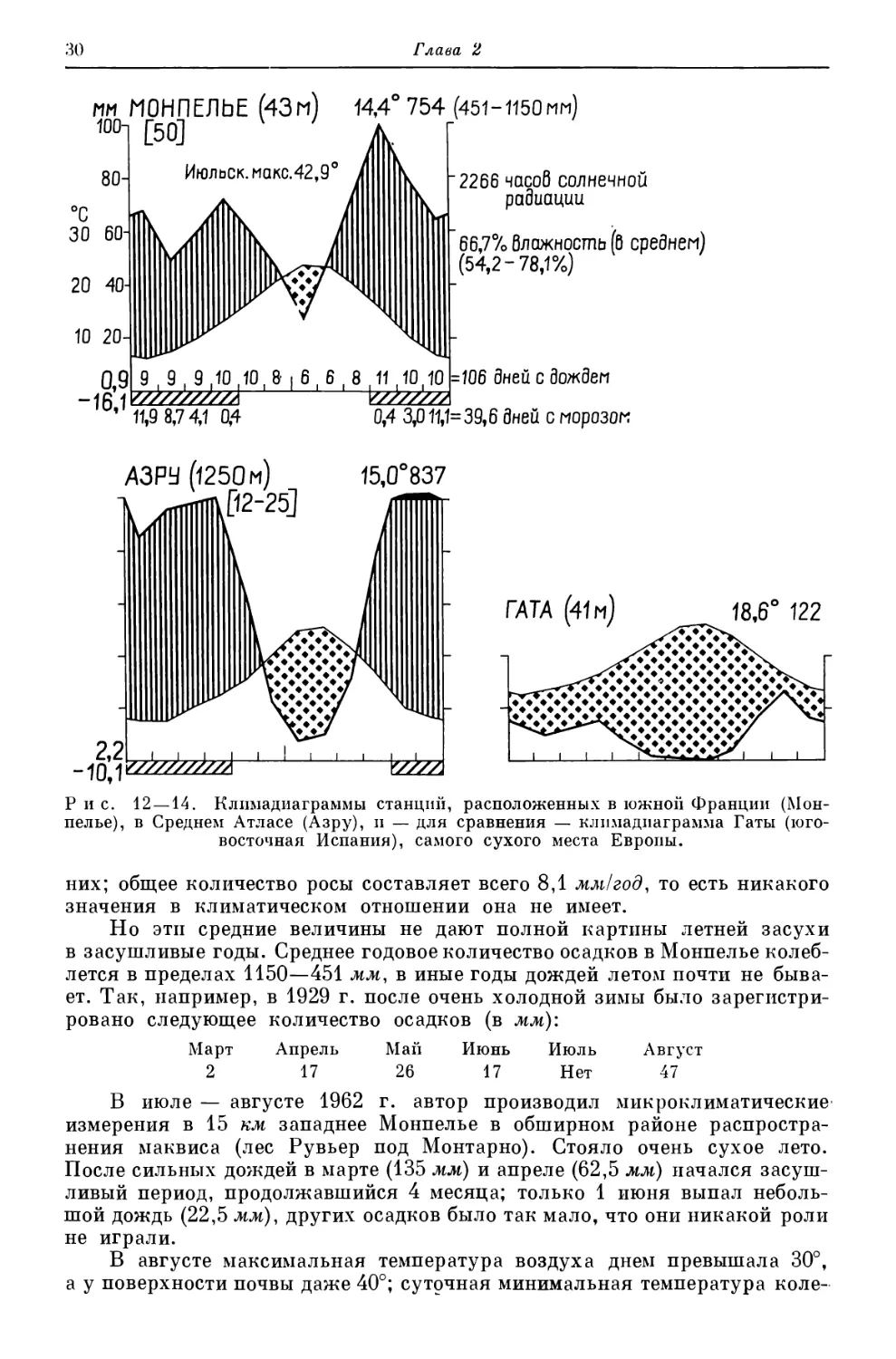
им МОНПЕЛЬЕ (43м) 14,4° 754 (451-1150мм)
100-1
80-
60-
20
40-
10
20-
°C
30
[50] • •
Июльск.макс.42,9
,9 9 9 10 10, & ! 6 , 6 , 8 ,11,10,10
11,9 8,74,1 0,4
"2266 часов солнечной
равиации
66,7% Влажность (б среднем)
(54,2-78,1%)
=106 дней с ЗожЗем
0,4 3,011,1= 39,6 Зней с морозом
0,9
-16,1
Рис. 12 —14. Климадиаграммы станций, расположенных в южной Франции (Мон-
пелье), в Среднем Атласе (Азру), и — для сравнения — клпмадиаграмма Гаты (юго-
восточная Испания), самого сухого места Европы.
них; общее количество росы составляет всего 8,1 мм/год, то есть никакого
значения в климатическом отношении она не имеет.
Но эти средние величины не дают полной картины летней засухи
в засушливые годы. Среднее годовое количество осадков в Монпелье колеб-
лется в пределах 1150—451 мм, в иные годы дождей летом почти не быва-
ет. Так, например, в 1929 г. после очень холодной зимы было зарегистри-
ровано следующее количество осадков (в мм):
Март Апрель Май Июнь Июль Август
2 17 26 17 Нет 47
В июле — августе 1962 г. автор производил микроклиматические
измерения в 15 км западнее Монпелье в обширном районе распростра-
нения маквиса (лес Рувьер под Монтарно). Стояло очень сухое лето.
После сильных дождей в марте (135 мм) и апреле (62,5 мм) начался засуш-
ливый период, продолжавшийся 4 месяца; только 1 июня выпал неболь-
шой дождь (22,5 мм), других осадков было так мало, что они никакой роли
не играли.
В августе максимальная температура воздуха днем превышала 30°,
а у поверхности почвы даже 40°; суточная минимальная температура коле-
Средиземноморская жестколистная растительность
31
балась от 11,0 до 15,8°. Содержание влаги в воздухе составляло 10 а/ле3.
Днем влажность воздуха понижалась до 20% (один раз до 10%), ночью
увеличивалась до 90%. Слабая роса отмечалась только два дня. Дефицит
насыщения составлял 20—25 а/ле3, а у поверхности почвы — 50 г/м3.
Испарение, по измерениям стандартным испарителем Пише (зеленая
бумага) на высоте 1 м (солнечный день) в полдень, часто превышало 1 мл/час
в течение 5—7 часов, а в отдельные дни достигало 1,5—1,6 мл/час. Таким
образом, оно значительно больше, чем в жаркие летние дни в Центральной
Европе \ но существенно меньше, чем в районах пустынь. В тени же, под
кронами Quercus ilex, испарение лишь редко превосходило 0,6 мл/час.
Измерения Браун-Бланке, предпринятые 4 июля 1934 г., также
показали, что максимальное испарение на открытых солнечных участках
равно 1,4 мл/час (в пересчете на зеленую бумагу) и 0,6 мл/час в тени
Quercus ilex, а по измерениям 11 апреля — соответственно 1,5 и 0,3 мл/час.
Брекле (см. сноски на стр. 8) производил широкие микроклиматиче-
ские измерения в течение почти целого года на северо-востоке Испании
у Бланеса (Коста-Брава) и, кроме того, в трех других местах, вплоть
до границы произрастания пробкового дуба на высоте 650 м над уровнем
моря. Ход климатических элементов в этих районах аналогичен таковому
в Монпелье: в июле и августе максимум испарения в полуденные часы
достигает 1,7 мл/час, но обычно не превышает 1,0—1,4 мл/час, в прохлад-
ное время года испарение колеблется в пределах 0,3—0,8 мл/час. Самая
высокая температура воздуха днем в июле и августе может перейти отмет-
ку за 30°, но, как правило, она ниже. Температура освещенных солнцем
листьев пробкового дуба значительно превышает температуру воздуха
(наивысшая измеренная разница температур листьев и воздуха 14°). Однако
в тени это отличие практически незаметно. Поверхность почвы в полдень
часто нагревалась до 50° и выше (максимальная измеренная температура
+ 68°); дневной максимум на глубине 2 см был равен 44° (суточная ампли-
туда 22°), на глубине 10 см — 31° (амплитуда 6°).
Следует учитывать, что Монпелье и Бланес расположены у северной
границы области жестколистной растительности. В южных районах опи-
санные климатические условия летом выражены еще более резко.
ЗОНАЛЬНАЯ РАСТИТЕЛЬНОСТЬ
СРЕДИЗЕМНОМОРСКОЙ ОБЛАСТИ
Первоначально Средиземноморье, несомненно, было покрыто густыми
лесами, как бы странным это сегодня пи казалось при взгляде на голые
скалы с редкой растительностью. Наиболее хорошо развитые леса, естест-
венно, были распространены на мощных почвах, ныне превращенных
в культурные земли. Но и склоны гор были залесены. Человек свел леса,
вызвал к жизни эрозию почв, в результате которой почвенный покров
был разрушен и уничтожен. В наше время существование лесов на горных
склонах здесь уже невозможно, так как слой почвы слишком тонок и не
в состоянии накопить достаточное количество влаги. Даже если сегодня
объявить растительность скал заповедной, первоначальный лес на этих
местообитаниях не восстановится. Он останется кустарникообразным
маквисом. И все же на местообитаниях с благоприятными условиями может
возникать густой тенистый лес, о чем свидетельствуют стройные деревья
1 В Центральной Европе испарение редко превышает в полдень в течение 1 часа
величину, равную 1,0 мл/час.
32
Глава 2
Рис. 15. Ветка Quercus cocci]era.
Quercus ilex, которые иногда встре-
чаются среди маквиса, в парках
или на обочинах улиц. И в прош-
лом лес, конечно, никогда не был
дремучим первобытным лесом и не
достигал высоты буковых лесов
Центральной Европы, но до 20 м
кроны деревьев, видимо, все же
поднимались.
Еще в средние века вечнозеле-
ные дубовые леса подступали к
окраинам Монпелье, но за время
с 1290 по 1790 г. они были выруб-
лены, главным образом на топливо
для стекольных заводов. В на-
стоящее время необрабатываемые
земли представляют собой откры-
тую гаригу, то есть местность,
поросшую невысоким (до 1 м) кустарником с примесью Quercus coccifera
(рис. 15), называемым здесь «гарулией» (отсюда гарига), или различными
видами Cistus, Rosmarinus, Juniperus и др. (рис. 16). Встречаются также
отдельные участки маквиса, то есть островки низкорослого леса с 20-лет-
ним периодом оборота рубки. Лес используется на дрова или для выжи-
гания угля; вырубки в течение нескольких лет служат пастбищем для
овец. Господствующий древесный вид, вечнозеленый Quercus ilex, легко
восстанавливается порослью от пней и уже через 6—7 лет плодоносит,
Рис. 16. Гарига с цветущим Cistus albidus; на переднем плане —большой полусфе-
рический куст Euphorbia dendroides; западная Ривьера (фото Э. Вальтер).
Средиземноморская жестколистная растительность
33
но более крупные деревья дают семена лишь через 10—12 лет. Ныне дро-
ва уже никому не нужны, угольщиков стало мало, выпас овец сократился,
так что хозяйственное значение маквиса убывает. Некоторый доход при-
носит еще только охота на кабанов и зайцев.
Современный маквис дает такое же неправильное представление
о прежних лесах, как на широте Центральной Европы молодая буковая
поросль — о высоком буковом лесе. На основе тщательного изучения
34 небольших участков сохранившегося на юге Франции лесного массива
Браун-Бланке 1 воссоздал структуру и состав естественного леса, назван-
ного им Ouercetum ilicis galloprovincialis. В своей работе он приводит
следующие данные (цифры после названий растений означают их «обилие»
по шкале Браун-Бланке 1 2 * 4).
Древесный ярус — сомкнутый, высота 15—18 м, представлен только Quercus
ilex (5).
Кустарниковый ярус — высота 3—5 м (до 12 ле); Buxus sempervirens (2),
Viburnum tinus (1), Phillyrea media (1), кроме того, единичные экземпляры
(+): Phillyrea angustifolia, Pistacia lentiscus, P. terebinthus, Arbutus unedo,
Rhamnus alaternus и Rosa sempervirens.
Лианы — Smilax aspera (2), Lonicera implexa (1), Clematis flammula (1), Hedera helix
(1), Lonicera etrusca (+)•
Травяной покров — покрывает не более 30% площади: Ruscus aculeatus (2),
Rubia peregrina (1), Asparagus aculeatus (1), Carex distachya (1), единичные
экземпляры (+): Viola scotophylla, Asplenium adiantum-nigrum, Stachys
officinalis, Teucrium chamaedrys и ЁирТгогЫа characias. Встречаются также
некоторые более редкие и сопровождающие видыя
Ярус мхов — слабо развит: Drepanium cupressiforme (1), Eurynchium circinnatum
(1), Scleropodium purum (1), Brachythecium rutabulum (+) и некоторые
другие виды8
Для старых древостоев особенно типичны: Viburnum tinus, Ruscus aculeatus, Carex dis-
tachya и Asplenium adiantum-nigrum, а также Hedera helix.
В этих лесах на ветвях деревьев много различных мхов (Fabronietum с 6 видами) и ли-
шайников (Parmelietum trichoterae с 7 обычно встречающимися и в Цен-
тральной Европе видами Parmelia, а также A naptychia ciliaris, Ramalina
spec, и др.).
Непосредственно поверхность почвы в таком лесу освещена очень
слабо; климат внутри леса более ровный по сравнению с климатом окру-
жающей территории, максимальные температуры приблизительно на 10е
ниже, чем за пределами леса, но влажность воздуха выше; движение воз-
духа даже при мистрале почти не ощущается.
Под пологом леса Quercetum ilicis формируется средиземноморская
красная почва, или терра-росса, причем вне зависимости от типа мате-
ринской породы, которой могут быть известняки или доломиты, мергели,
песчаники, базальты, сланцы, гнейсы, граниты или, наконец, аллювиаль-
ные отложения.
Типичный профиль терра-росса приводит в своей работе Браун-Бланке:
Ао, глубина 1—2 (5) см — подстилка из слабо разложившихся листьев, в ниж-
ней части с мицелием грибов.
1 J. Braun-Blanquet, La Chenaie d’Yeuse mediterraneenne (Quercion
ilicis) «Sigma, Comm.», 45, «Mem. Soc. Sc. nat. Nimes», 5, 1936.
2 В шкале Браун-Бланке приняты следующие обозначения: г — крайне редко,
с очень незначительным покрытием; + — редко, с очень незначительным покрытием;
1 — многочисленные особи, но с небольшим проективным покрытием или довольно
редко, но с большой площадью покрытия; 2 — весьма обильно или занимает не менее
5% площади пробной площадки; 3 — покрытие от 25 до 50% при любом числе особей;
4 — покрытие от 50 до 75% при любом числе особей; 5 — покрытие свыше 75% при
любом числе особей.— Прим. ред.
3-01248
34
Глава 2
Ai, глубина 5—15 (25) см — тонкозернистый, хорошо аэрируемый, богатый гуму-
сом (6—10 до 20%) горизонт, бурого до черно-серого
цвета; содержит тонкие корни растений и обильную
фауну; выщелочен, pH обычно равен 7,1.
Bi, глубина 30—150 см — мелкокомковатый, плотнее, чем горизонт Af, рас-
падается на угловатые отдельности; буро-красного
цвета, бедный гумусом (1—4%); пронизан корнями
кустарников, много дождевых червей; pH = 7,2—8,2.
В2, мощность обычно более 50 см — глинистый, очень пластичный, плотный, лишен
гумуса; на воздухе высыхает с образованием тре-
щин; буро-красный до желтоватого, с черными пят-
нами (окислы марганца); немного корней деревьев
и крупных кустарников; pH = 7,4—8,3.
С — материнская порода.
Мелкоземистый материал в основном (90%) состоит из тонкого песка
и пыли (приблизительно в равных долях), содержание глины незначитель-
но, несколько увеличивается лишь в горизонте В.
Объемный вес почвы: горизонт Ai — 1,04; В4 — 1,48; В2 — 1,64. Содержание-
воздуха составляет (в скобках даны средние значения): Ai — 10,8—24,4% (18,7%);
В4 - 7,2-9,0% (7,9%), В2 - 1,6-5,1%.
В почвах, формирующихся на известняковых породах, нередко на
глубине 1—2 м располагается горизонт кирпично-красного цвета. Посла
смыва верхних горизонтов почвы в результате эрозии эта красная окраска,
столь характерная для культурных почв Средиземноморья, проступает
на поверхности; смытая же красноцветная почва отлагается в понижениях.
В выветрелых известняковых скалах трещины и промытые водой полости
бывают иногда заполнены до глубины 10 м ископаемой красноцветной
почвой.
Сильно уплотненный горизонт В2 хорошо удерживает воду, поэтому
содержание воды в почве вряд ли падает ниже 16 — 20% . Этим объясняется,
почему при слабой глинистости почвы в ней всегда содержится достаточ-
ное количество воды, которое может быть использовано растениями.
Совершенно аналогичный почвенный профиль описал Руша ль в районе Ровинь,.
Истрия. Гранулометрический анализ материала до глубины 2 м дал следующее:
содержание тонкозернистого песка и пыли — 92—95%, глины — только 1,3—3,8%.
Приводимые им значения водного потенциала почвы, несомненно, завышены.
Смена аспектов в лесах Quercetum ilicis начинается вслед за окончанием
периода осенних дождей, когда все снова пробуждается к жизни: про-
растает множество семян, жестколистные виды дают побеги и иногда
цветут второй раз. Однако низкие температуры тормозят рост растений.
В глубине леса морозы редко причиняют вред. Почва никогда не промер-
зает, поэтому семена многих растений прорастают уже зимой. В первые
весенние дни после последних морозов зацветают Euphorbia characias,
в марте — Phillyrea, Rhamnus и Juniperus oxycedrus\ в конце марта (позд-
нее, чем на открытых местах) начинают вегетировать травы: Carex. Viola,
а также Arum italicum и Aristolochia. Время основного цветения — май,
когда цветет и одновременно сбрасывает старые листья Quercus ilex.
В июне раскрываются цветки Clematis, Rosa, Rubus и Lonicera. Плодоносят
травы в июне — июле, древесные растения — осенью. Летняя засуха —
период покоя длительностью 2 — 3 месяца. Совпадение периода самых
высоких температур со временем наибольшей сухости особенно пагубно
отражается на растительности. Большинство всходов погибает уже в пер-
вое лето. На пробной площадке из 151 проростка через год осталось толь-
ко 13, причем из 38 проростков дуба выжили лишь 8, а через 3 года —
Средиземноморская жестколистная растительность
35
только 7. Корни этих проростков в течение года проникли на глубину
60—80 см (до 120). Большое значение имел размер семян. У Viburnum
tinus первое лето пережил только один проросток из 71, причем во второе
лето погиб и он.
Границы распространения этих лесов (в широком понимании тип
Quercion ilicis) показаны на рис. 17. В южной Франции граница проходит
на высоте 250—300 м и лишь местами достигает отметок 500 м над уров-
нем моря; в изобилующей дождями Италии и в Далмации она отделяется
от моря только очень узкой береговой зоной; в Испании эти леса подни-
маются до уровня 500 м и выше (700—800 м). На юге Испании и в Среднем
Атласе тип Quercetum ilicis характеризуется большей примесью Pistacia
lentiscus; наблюдаются также некоторые флористические различия. Здесь
этот тип относится уже к лесам горного пояса: под Малагой он развит
на высоте 700—1300 м, а в горах Атласа — в зоне 900—1800 м над уровнем
моря. Климатические условия в этой зоне, видимо, соответствуют условиям
в Монпелье: климадиаграмма для Азру (1250 м) аналогична климадиаг-
рамме для Монпелье (ср. рис. 12 и 13). В нижнем поясе прежде, вероятно,
преобладали другие виды жестколистных растений, а именно дикая мас-
лина (Olea oleaster) и рожковое дерево (Ceratonia siliqua, рис. 18) Ч Сегод-
ня эти леса (Oleo-Ceratonion) полностью сведены.
Рис. 17. Область естественного распространения лесов, образованных Quercus
ilex (7), растительности типа Oleo-Ceratonion (2) и рощ с Argania (3) в западном Среди-
земноморье (по Браун-Бланке, с изменениями).
В областях с типичным океаническим климатом и на очень бедных
кислых почвах место Quercus ilex занимает пробковый дуб (Qu. suber,
рис. 19). Здесь заготавливают пробку, снимая кору со стволов деревьев
(рис. 20) 1 2. На западе Греции Quercus ilex распространен в более широкой
зоне, протягивающейся до северного Пелопонесса, но в районе Эгейского
моря этот вид встречается только на островах (высокоствольные леса
1 Для Ceratonia, как и для Cercis siliquastrum, характерна каулифлория. В этом
поясе часто встречается также Pinus halepensis (обычно в искусственных насаждениях).
На юго-востоке Испании местами господствует климат пустынь.
2 Ch. Sauvage, Recherches geobotaniques sur les suberaies maroccaines,
«Trav. Inst. Sc. Gherifien», Serie Botan, № 21, Rabat, 1961.
3*
36
Глава 2
Рис. 18. Ceratonia siliqua (рожковое дерево) с плодами; на
переднем плане — Chamaerops humilis\ Испания, севернее
Валенсии (фото Э. Вальтер).
на Эвбее и трех горных отрогах полуострова Халькидика (по Хорвату) х.
В Анатолии редкие экземпляры Qu. ilex можно увидеть лишь в юго-запад-
ной части региона, несколько чаще он встречается на западном побе-
режье Черного моря. Вообще же в этих областях, в восточной Греции
и на анатолийском побережье Средиземного моря, отличающихся более
континентальным климатом, Quercus ilex повсеместно замещается Qu. сос-
cifera. Обычно это кустарник высотой 0,5—1,5 м, но иногда дерево высо-
той до 20 м. Представляет ли он собой экотип, развившийся в результате
частых пожаров, неизвестно, так как соответствующие исследования
не проводились. В Палестине выделяют особую форму кермесового дуба —
1 L. Н о г v a t, Pflanzengeographische Gliederung Siidosteuropas, «Vegetatio»,
5/6, 1954, S. 434—447; R. Knapp, Die Vegetation von Kephallinia, Griechenland,
Koenigstein, 1965; S.-W. Breckle, Okologische Untersuchungen im Korkeichenwald
Kataloniens, Hohenheim, 1966 (дисс.).
С редиземноморская жестколистная растительность
37
Рис. 19. Область распро-
странения пробкового дуба
(Quercus suber) — прерыви-
стая линия; точки — изо-
лированные местонахожде-
ния (по Целлеру).
I
Quereus coccifera var. calliprinos. Кроме него, здесь произрастает летне-
зеленый Qu. ithaburensis.
Леса в восточном Средиземноморье тоже сведены, но Зохари пишет
об обширных территориях, занятых маквисом х. Последний существенно
не отличается от западносредиземноморской растительности, лишь неко-
торые западные элементы замещены восточными, например уже в южной
Анатолии очень часто можно встретить Styrax officinalis. Для наиболее
теплых местообитаний типичны группировки Ceratonia — Olea, в которых
всегда в большом количестве присутствует Pistacia lentiscus.
Рис. 20. Редкий лес, образованный пробковым дубом (Quercus suber), в Марокко
(лес Мамора). С нижней части стволов кора снята (обнаженный ствол яркого буро-
красного цвета) (фото Г. Вальтера).
1 М. Z о h а г у, Plant life of Palestine, «Ghron. Bot.», N. S. № 33, 1962.
38
Глава 2
СТАДИИ ДЕГРАДАЦИИ РАСТИТЕЛЬНОСТИ
И ЭКОЛОГИЯ АССОЦИАЦИИ
BEACHVPODIUM RAMOSUM — PHLOMIS LYCHNITIS
Читатель уже получил представление о первичной растительности
Средиземноморской области, познакомившись с приведенным выше описа-
нием вечнозеленых дубовых лесов. Как уже подчеркивалось, эта расти-
тельность в настоящее время повсюду уничтожена. Густой вечнозеленый
дубовый лес бесполезен для человека. Дичи в нем мало, поэтому охота
имеет небольшое значение. Годится лес только на дрова, но 20-летняя
пневая поросль для этой цели более удобна, чем старые деревья, которые
трудно колоть. Кроме того, прирост древесины с возрастом деревьев,
видимо, быстро уменьшается. Все это послужило причиной тому, что уже
в древние времена первичный лес был замещен низкоствольным лесом,
то есть маквисом.
Маквис — это первая стадия деградации. Высота его около 4 м.
На вырубках сначала развивается растительный покров, состоящий из зла-
ков и разнотравья и используемый для выпаса скота. Стремление рас-
ширить площадь пастбищ привело к дальнейшему уничтожению маквиса.
Особого труда это не требовало, достаточно было поджечь кустарник
во время летней засухи. Когда не хватало дров, то на топливо шли и кор-
ни, поэтому порослевого восстановления не происходило. На низмен-
ностях с мощными почвами, где лес был сведен, успешно развивалось
земледелие, здесь закладывали виноградники, сажали фруктовые деревья.
При чрезмерном длительном выпасе пастбища часто завоевывали
ядовитые или колючие сорняки, борьба с которыми также велась с по-
мощью огня. Сильные ливни смывали со склонов мелкозем, обнажая мате-
ринскую породу.
Превратившаяся в пустыню Средиземноморская область — наиболее
яркий, но и печальный пример того, как человек хищнической эксплуата-
цией земли подрывает основу своего существования. Климат в районах
с выраженным карстом ухудшается. Количество осадков, правда, остается
прежним, но нарушается водный режим всей территории. Разливы рек
и наводнения после дождей сменяются безводием сухих русел и острым
недостатком воды во время засухи. Поэтому обширные территории сегод-
ня покинуты населением. Когда-то густонаселенные и богатые районы
теперь полностью обнищали. Прекращение обработки земли ведет к регене-
рации растительности, но, поскольку почва уничтожена, восстановление
растительности протекает крайне медленно. Лишь в последнее время
в некоторых районах начали разводить лес, но это требует больших затрат.
На всей территории Средиземноморья можно наблюдать стадии дегра-
дации и регенерации в самых различных вариантах. Не вдаваясь в геобо-
танические подробности, рассмотрим эти условия на примере Монпелье,
где они лучше всего изучены.
На рис. 21 и 22 схематически показаны стадии деградации маквиса —
в первом случае развитого на плотных юрских известняках, во втором —
на плейстоценовых слабоизвестковистых, но богатых кремнеземом почвах.
а) Маквис менее однороден и поэтому отличается большим флористи-
ческим разнообразием, чем сомкнутый высокоствольный лес. На вырубках,
возобновляемых каждые 20 лет, в первые годы появляются светолюбивые
травы и кустарнички; также быстро развиваются обычно угнетенные
в сомкнутых древостоях из-за недостатка света кустарники, но вскоре
Средиземноморская жестколистная растительность
39
Рис. 21. Стадии деградации растительного покрова на юрских известняках
(по Браун-Бланке).
1 — Quercetum ilicis; 2 — стадия Qu. coccifera; 3 — Brachypodietum ramosi; 4 — стадия
Euphorbia characias.
P n c. 22. Стадии деградации растительности на плейстоценовых силикатных
почвах (по Браун-Бланке).
11— Quercetum ilicis; 2 — стадия Qu. coccifera', 3 — стадия с Lavandula stoechas\ 4 — ассо-
циация Corynephorus articulatus — Helianthemum guttatum] 5 — стадия C Erica scoparia в по-
нижении с наносной почвой.
их затеняет пневая поросль основных древесных пород. Однако промежу-
ток времени до следующей сплошной рубки столь короток, что кустарники
не исчезают, а продолжают существовать в угнетенном состоянии в густой
тени. Сохраняются они также на обочинах дорог и на участках с мало-
мощным почвенным покровом. Таким образом, маквис представляет собой
пеструю смесь различных стадий смены растительности на вырубках или
местах случайных лесных пожаров.
В то время как флора лесов, развивающаяся на разнотипных почвах,
не обнаруживает существенных различий, флористический состав на
вырубках весьма чутко реагирует на физические и химические свойства
почв. Так, например, ацидофильные виды отличаются от других видов,
так пли иначе приспособленных к жизни на известковых почвах. Правда,
в отдельных случаях не всегда ясно, является ли решающим фактором
значение pH, как таковое, или, что вероятнее, главную роль играют иной
водный режим, неодинаковое содержание питательных веществ и т. д.
Ацидофильными на юге Франции являются многочисленные виды вереско-
Рис. 23. Типичные виды средиземноморской флоры на открытых место-
обитаниях.
2 — Daphne guidium’, 2 — Thymelaea passerina', 3 — Lavandula stoechas\ 4 — Phillyrea
media’, 5 — Erica multiflora’, 6 — Globularia alypum.
Средиземноморская жестколистная растительность
41
вых, например Arbutus unedo, Erica arborea, E. scoparia, кроме того, Cistus
salviaefolius, Lavandula stoechas (рис. 23), Myrtus communis и др.1.
Более или менее связаны с карбонатной почвой Cistus albidus, Erica
multiflora (рис. 23), Rosmarinus officinalis, Lavandula latifolia, Thymus
vulgaris, Asphodelus cerasifer.
Через несколько лет маквис становится очень густым, а разросшиеся
колючие кустарники и вьющееся растение Smi lax aspera делают его совер-
шенно непроходимым. Большинство древесных растений маквиса на юге
Европы имеет форму кустарника; однако многие старые экземпляры
Quercus ilex, а также большинство других видов деревьев отличаются
толстым стволом. В Анталье (южная Анатолия) автору довелось увидеть
огромные деревья, внешне очень похожие на старые дубы, но это была
Pistacia lentiscus. У Arbutus unedo ствол нередко достигает у основания
50 см в диаметре; настоящими высокими деревьями могут стать также
Ceratonia siliqua, Pistacia terebinthus, Phillyrea media, Laurus nobilis, если
дать им возможность свободно расти.
б) При регулярной вырубке через 6—8 лет и при расширении выпаса
виды деревьев постепенно исчезают из растительного покрова и их место
занимают Rosmarinus, Cistus, Erica, Genista scorpius и Quercus coccifera.
Дуб кермесовый скотом не поедается и поэтому быстро захватывает все
новые и новые площади. Его часто выжигают, но он снова возобновляется
и образует плоские широкие кусты высотой до 1,5 м, занимающие иногда
до 50% площади. Это так называемый Cocciferetum, представляющий
здесь, на юге Франции, в области с влажным океаническим климатом,
стадию деградации, а в континентальных районах — зональную расти-
тельность.
в) Между кустами Quercus coccifera при умеренном выпасе хорошо
растет злак Rrachypodium ramosum. При сведении кермесового дуба путем
выжигания формируется оптимальная для выпаса скота стадия — ассо-
циация Rrachypodium ramosum — Phlomis lychnitis, которая геоботани-
чески подробно изучена 1 2.
г) При чрезмерном выпасе уничтожается и это травяное сообщество;
на почти голой почве остаются только виды растений, не поедаемые
скотом.
На кислых кремнистых почвах прогрессирующая деградация проте-
кает аналогично, различие состоит лишь в том, что флористический состав
отдельных стадий иной (рис. 22).
В южной Франции выпас овец в последнее время заметно сокращается.
Не возделываются и менее плодородные пахотные земли или культурные
земли, качество которых ухудшалось вследствие эрозии почв. Многие
деревни покинуты, так как труд, вложенный в сельское хозяйство, уже
не окупается. На брошенных землях происходит демутация, отдельные
стадии которой соответствуют стадиям деградации. Складывается пестрая
мозаика многих растительных сообществ, разнообразие которых усугуб-
1 На богатом осадками побережье Далмации на известняках встречаются Myrtus,
Arbutus, Erica arborea, Cistus salviaefolius. Анализ шести почвенных проб, взятых из го-
ризонта размещения корней этих видов, показал, что мелкоземистый материал не выще-
лочен, а содержит известь. Величина pH (КС1) колеблется в пределах 6,5—7 (в пробе,
взятой под Rosmarinus, рН=6,0).
2 J. Braun-Blanquet, Die Rrachypodium, ramosum-Phlomis lychnitis-
Assoziation der Roterdeboden Siidfrankreichs. Cm.: Festschr. C. Schroter, «Veroff.
Geobot. Inst. Rubel», H. 3, 1925; F. R.Bharucha, Etude ecologique et phytosocio-
logique de Г association a Rrachypodium ramosum et Phlomis lychnitis des Garigues Lan-
guedociennes, «Sigma», № 18, 1932.
Глава 2
Красноземы на плотных известняках
Деградировавшие лиг
и кустарниковое пастбище
Переотложенные каменистые
карбонатные почвы
Пастбище на заброшенных
культурных землях
| СтаЗия теросритов
Brachypodium ramosum (оптимум!) |
/Ассоциация Brach.ram.-Phlomis lychn.) СтаЗия Thymus vulgaris-Brachypodium
aistachyon
пас
Чрезмер
ный быпас и выжигание
мерный выпас Вы
\ £ Чрез
СтаЗия
Brachypodium ramosum
ЧрезмерныйХ^ьтас
Asphodelus cerasifer Стадия Fuphnrhin t _ СтаЗия
СтаЗия геосритов Cynareen “------► Genista scorpius
Стадия
Lavandula
Latif о На
Прекращение\выпаса,
выжи
гание
СтаЗия Cistus
monspeliensis-
С. albidus
Редкоеувыжигание
СтаЗия Quercus coccifera
СтаЗия
Juniperus
oxycedrus
- Brachypodium ramosum
(оптимум!)
Выжигание и выпас
СтаЗия Rosmarinus-Cistus
Quercetum ilicis
(зональная растительность
спустя Злительное время)
P и c. 24. Схема стадий деградаций и реградации растительности в Лангедоке (южная
Франция) на двух различных типах карбонатной почвы в зависимости от характера
и интенсивности использования растительности (по Браун-Бланке, с изменениями).
ляется различием местообитаний. Схема на рис. 24 дает представление
о важнейших сукцессиях в Лангедоке, характер которых от места к месту
-опять-таки неодинаков Ч В результате может сложиться любое число
различных растительных сообществ. Эти открытые формации низких
кустарников или кустарничков часто имеют местные названия: гарига —
во Франции, томилляры — в Испании, фригана — в Греции, бата —
в Палестине. Местами на обширных площадях резко доминируют отдель-
ные виды, такие, как Thymus vulgaris, Rosmarinus officinalis или виды
Lavandula. В восточном Средиземноморье особенно широко распространен
колючий полукустарник Poterium spinosum, нередко сплошь покрывающий
склоны гор (рис. 25). Часто встречается также Juniperus oxycedrus.
1 G. Kuhnholtz-Lordat, L’equilibre agro-sylvo-pastoral est-il irreme-
diablement comrpomis dans la garrique mediterraneenne, «Rev. Forest. Franc.», № 12,
1954, p. 755—764.
Средиземноморская жестколистная растительность
43
Под Монпелье обширные площади занимает ассоциация Brachypodium
ramosum — Phlomis lychnitis. Браун-Бланке и Бхаруха провели деталь-
ные исследования ее состава и экологии, поэтому рассмотрим эту ассо-
циацию подробнее, с тем чтобы выявить ее различие с зональной расти-
тельностью (Quercetum ilicis). Сообщество предпочитает известковые поч-
вы и оптимально развивается на юрских известняках. В течение 6—7 меся-
цев в году здесь производится интенсивный выпас овец.
Почвенный покров маломощный, каменистый и водопроницаемый,
верхний горизонт благодаря гумусу темный, более глубокие — красно-
бурого цвета (pH = 6,8—7,2) х. Мелкозем содержит 0,3—58% СаСО3
и состоит преимущественно из тонкозернистого песка и пыли.
Микроклиматические условия — особенно у поверхности почвы, при
воздействии солнечных лучей — очень суровые.
В растительном покрове доминирует Brachypodium ramosum (занимает
около 50% площади). Вид относится к неизвестному । в Центральной
Европе типу кустарниковидных злаков, которые разветвляются не только
у основания, но и вверху, так как прямостоящие стебли перезимовы-
вают 1 2. Корни растения проникают на глубину до 1 л^, тем не менее в раз-
гар лета листья обычно засыхают. На участках, где в течение 4 лет выпас
скота не производился, продукция сырой массы, по данным Бхаруха,
составила перед наступлением засухи в конце мая 1325 г!м* (13,25 т/га),
Рис. 25. Полусферические кусты Poterium spinosum на побережье Эгейского моря
(Греция) совместно с Thymus и Agropyrum (фото Э. Вальтер).
1 J.Braun-Blanquet, Etudes sur la vegetation mediterraneenne III. Con-
centration en ions H et calcimetrie du sol, «Bull. Soc. Bot. de France», 71, 1924,
p. 639—879.
2 Этот тип широко распространен в тропиках, как и древовидные злаки, напри-
мер виды ВатЬия.
44
Глава 2
Рис. 26. Взаимосвязи
изменений погоды, периода
ассимиляции и времени
цветения по наблюдениям
в окрестностях Монпелье-
(по Бхаруха).
а при выпасе скота — только 375 з/л1 2 (3,75 т/га) Ч Поскольку запасы
влаги в почве невелики, многолетние растения не могут образовать сомкну-
того покрова. Из числа других растений особенно характерен Phlomis
lychnitis. Розетки листьев этого вида засыхают, за исключением
самых молодых листочков. Сопутствующими видами являются Brachypo-
dium distachyon, Thymus vulgaris, Eryngium campestre, Sedum nicaeense.
У геофитов Iris chamaeiris и Asphodelus cerasifer весной распускаются
необычайно красивые цветки; в середине лета растения погибают.
На обычно открытых местообитаниях после первых осенних дождей
прорастают семена многочисленных терофитов (озимые однолетники).
К терофитам относится 43% всех входящих в сообщество видов, 30%
представлены гемикриптофитами и 8% — геофитами. Из числа древесных
растений наряду с Quercus coccifera весьма часто встречается Juniperus
oxycedrus (пастбищный сорняк).
Периодичность развития сообщества показана на рис. 26. На диаграм-
ме видно, что кульминация развития листовых органов приходится
на период с конца марта до начала июня, а высшая фаза цветения — на
конец мая. К концу июня терофиты исчезают, но молочайные и Sedum
еще в июле нормально вегетируют.
Сильное избирательное влияние на флористический состав оказывает
выпас:
а) скот не поедает все виды Euphorbia, виды Cistus, Phlomis lychnitis,
Eryngium nCarlina corymbosa;
1 Сухой вес не определялся, но он, очевидно, не соответствовал бы годовой про-
дукции, так как стебли многолетние.
Средиземноморская жестколистная растительность
45
Рис. 27. Pinus maritima на скалистом побережье итальянской Ривьеры
(фото Э. Вальтер).
б) скот поедает только молодые побеги Quercus coccifera, Juniperus
oxycedrus, Phillyrea angustifolia и скусывает лишь цветки Thymus vulgaris
(зимой также листья), Lavandula latifolia, Echium pustulatum, Psoralea
bituminosa и Asphodelus', у Genista scorpius и Juniperus oxycedrus животные
поедают только плоды;
в) целиком животные съедают Brachy podium, Sedum, Aphyllanthes,
Ononis.
Р и с. 28. Сосна алеппская (Pinus halepensts) близ Дубровника, Далмация. На заднем
плане — маквис с Quercus ilex, Erica verticillata, Pistacia lentiscus, Myrtus communis
(на известняках) и др. (фото Адамович).
Средиземноморская жестколистная растительность
47
Рис. 29. Лес, образованный Pinus brutia, на высоте 1000 м, юго-западная Анатолия.
Справа на переднем плане — Cercis siliquastrum (фото Э. Вальтер).
При чрезмерном выпасе последняя группа растений вытесняется
с пастбища, первая же, напротив, достигает оптимального развития.
В конце концов остаются одни молочаи и колючие сложноцветные. При
пожаре погибают и эти растения, огонь щадит только геофиты с их глубо-
ко расположенными запасающими органами, например Asphodelus.
Таким образом формируются повсеместно распространенные в Средиземно-
морье асфодилловые группировки, воспетые уже в Древней Греции, но пред-
ставляющие собой крайнюю стадию деградации.
Особенно большое значение в Средиземноморской области имеют раз-
личные виды сосны. На западе области это Pinus maritima, предпочитаю-
щая бедные известью почвы (рис. 27), на юге — Pinus halepensis, исполь-
зуемая в последнее время в южной Франции и Италии для лесонасаждения.
Этот вид распространен также в Далмации (рис. 28), Греции и Палестине.
В Анатолии он замещается очень близкой к нему Pinus brutia (рис. 29).
Здесь, в континентальной части Средиземноморской области сосны
играют еще большую роль. В горах они поднимаются выше зоны вечно-
зеленых дубов. В Анатолии, например, где лесопосадки практически
не производились, Pinus brutia распространена на южном склоне Тавра
до высоты 1000 м и в средней части склона формирует чистые древостои,
иными словами, сосновые леса принадлежат здесь к нижнему вертикаль-
ному поясу растительности. На некоторых участках побережья леса
спускаются даже к самому морю; возникает вопрос, не относится ли здесь
сосна к нижнему средиземноморскому поясу, то есть к зональной расти-
тельности (см. стр. 80, примечание 2).
При более внимательном осмотре этих лесов под древесным ярусом
можно обнаружить сомкнутый низкий маквис, росту которого препят-
48
Глава 2
Р и с. 30. Лес из Pinus pinea близ Пизы (Италия), на опушке леса —
Cistus salviaefolius с белыми цветками. В подлеске Quercus ilex (фото
Э. Вальтер).
ствует постоянная вырубка его на дрова. Под пологом маквиса семена
Pinus brutia не прорастают х, следовательно, возобновление ее не проис-
ходит. Поэтому вполне вероятно, что регенерация сосны наступает лишь
1 Вероятно, автор имеет в виду, что проростки (или молодые растения) погибают
под покровом маквиса.— Прим. ред.
Средиземноморская жестколистная растительность
49
после лесных пожаров, чем, видимо, объясняется и однородность древо-
стоя. Таким образом, встречающиеся в самом нижнем поясе гор сосновые
леса следует рассматривать как вторичные, возникшие после пожаров.
Поскольку лесные пожары в Анатолии случаются часто, становится понят-
ным столь широкое распространение этих лесов. Частично их развитие
на малоплодородных почвах (например, на серпентинитах) 1 обусловлено
также эдафическим фактором. Лишь выше в горах, где из-за низкой
температуры зимой вечнозеленая растительность существовать уже не
может, сосны образуют естественный вертикальный пояс.
Очень трудно сказать что-либо определенное об естественном распро-
странении современного характерного для Средиземноморья дерева —
пинии (Pinus pined). Принадлежит она, несомненно, к нижнему поясу,
так как очень чувствительна к морозам, и, по-видимому, росла всегда
только на бедных приморских песках у побережья. Лес в южной Анато-
лии, между Аланьей и Антальей, очевидно, естественный. С еще большей
долей вероятности это можно сказать о лесах, расположенных, например,
в районах Равенны и Пизы (рис. 30).
Сосны (иногда также кипарисы) являются видами, используемыми
при лесопосадках, ибо они растут быстрее жестколистных деревьев и дают
деловую древесину. Поэтому сосна и кипарис за последнее время все
более доминируют в картине ландшафта. В окрестностях Монпелье все
леса, состоящие из Pinus halepensis, посажены начиная с конца XVIII в.,
на первой известной карте растительности эти площади еще обозначены
словом «пустошь». Сосна особенно хорошо развивается в сообществе,
образуемом Rosmarinus officinalis, Juniperus phoenicea, Staechelina dubia,
Erica multiflora, Lavandula latifolia и Aphyllanthes monspeliensis.
В последние годы в Средиземноморье на более теплых местообита-
ниях при лесоразведении все чаще используют австралийские виды эвка-
липта, которые пригодны для изготовления бумаги. Речь идет о Eucalyp-
tus camaldulensis (rostrata) и Ей. gomphocephdla, в Марокко также о Ей. tere-
ticornis, Ей. rudis, Ей. viminalis. Для добычи масла возделывают Ей. citrio-
dora, а танина — Ей. sideroxylon1 2. Лесопосадки при орошении дают
большую массу древесины.
ЭКОЛОГО-ФИЗИОЛОГИ ЧЕСКИ А ИССЛЕДОВАНИЯ
ВОДНОГО РЕЖИМА И ФОТОСИНТЕЗА РАСТЕНИЙ
В СРЕДИЗЕМНОМОРСКОЙ ОБЛАСТИ3
Вода как экологический фактор
и влагообеспеченностъ растений
Количество осадков, выпадающих в Средиземноморье зимой, отно-
сительно велико. Летом, во время засухи, верхние горизонты почвы
быстро высыхают, однако глубокие ее слои, иногда до уровня грунтовых
вод, накапливают достаточно влаги. Поэтому влагообеспеченность расте-
ний в значительной мере зависит от мощности почвенного слоя, глубины
1 W. Krause, Pflanzendecke und Standort auf Serpentin, «Hb. d. Pflanzen-
physiol.», IV, 1958, S. 755 ff.
2 K. Hueck, Reisebeobachtungen 1960 uber die Aufforstungen in Israel, «Forstw.
Cbl.», 79, 1960, S. 257—269; L’Eucalyptus au Maroc, Ministere de 1’Agriculture, Rabat,
1960; J. G e n t i 11 i, Il fattore termico nell’ecologia degli eucalitti, «Centro Sperimet.
Agric. e Forest.», Publ. 4, Roma, 1960, p. 53—119.
3 A. P i s e k, Immergriine Pflanzen, Hb. d. Pflanzenphysiol., Bd. V, 1960, S. 415—
459 (с подробной библиографией).
4-01248
50
Глава 2
проникновения корней, величины испаряющей поверхности (индекса
листовой поверхности) и интенсивности транспирации. При описании
аридных районов мы отметилй, что водоотдача всегда соответствует
возможной влагообеспеченности (см. т. I, стр. 306—320).
Маломощные каменистые почвы на возвышенностях и мощные аллю-
виальные почвы в долинах — очевидный результат процессов эрозии.
Запасов влаги в почве долин и на равнинных участках достаточно для
выращивания винограда, миндаля, инжира, маслины и т. д. Эти много-
летние культуры без полива переносят летнюю засуху и хорошо плодоно-
носят. Важно лишь, чтобы густота насаждений соответствовала запасам
влаги в почве.
Жестколистная растительность в настоящее время распространена
лишь на местообитаниях с маломощным почвенным покровом. На участ-
ках, подстилаемых сильно трещиноватыми скальными породами, на неко-
торой глубине могут накопиться значительные запасы влаги, на площа-
дях же, где материнская порода представлена монолитной скалой, аккуму-
ляция воды невелика. В зависимости от характера почвы и глубины
проникновения корней влагообеспеченность двух рядом стоящих деревьев
бывает совершенно различной, соответственно неодинаковым окажется
и их развитие. Там, где растет большое дерево, можно не сомневаться,
что влаги в почве достаточно, там же, где стоит чахлое деревце, воды
явно не хватает. Брекле проследил, что корни Quercus ilex и Qu. suber
проникают в гранитную скалу по трещинам на глубину 10—12 м.
Длительность жизни листьев у отдельных видов жестколистных
растений также различна. У Quercus ilex двухлетние листья обычно сохра-
няются, реже сохраняются трехлетние, четырехлетние встречаются как
исключение. Угнетенные в росте деревья на теневой стороне часто побегов
не дают и имеют только однолетние листья. Arbutus unedo, а также Vibur-
num tinus обычно часть двухлетних листьев теряют. Phillyrea angustifolia
может сохранять все трехлетние листья, напротив, у Pistacia lentiscus
и Erica multiflora они встречаются редко. Очень старые листья бывают
у Buxus sempervirens. Ilex aquifolium, Smilax aspera*, у Ruscus нередко наб-
людаются многолетние филлокладии.
В течение летнего засушливого периода запас влаги в почве постепен-
но расходуется, соответственно сокращается влагообеспеченность расте-
ния. Каким образом растения реагируют на недостаток влаги: сокращают
транспирацию, закрывая устьица и тем самым замедляя фотосинтез, или
же снижают уровень гидратуры (повышение осмотического давления;
см. т. I, стр. 50)? В пустынях на всех древесных растениях можно заметить
множество сухих ветвей, отмерших в годы сильной засухи. В Средиземно-
морье это явление не наблюдается, следовательно, здесь недостаток влаги
большого ущерба древесным растениям не наносит х.
Анализ кривых годового хода осмотического давления у растений
одного и того же местообитания позволяет судить о том, в какой мере
отдельные виды поддерживают уровень гидратуры протоплазмы в тече-
ние года. Такого рода исследования проводились зимой 1929/30 г. под
Монпелье в маквисе. Опытную площадку подстилали скальные извест-
няки, пронизанные трещинами и промоинами 1 2.
1 В 1962 г. на юге Франции на кустах Quercus ilex видны были отдельные отмер-
шие ветки. Но это следствие поражения жуком Choerebus, личинки которого прогры-
зают ходы в древесине, отчего ветки засыхают.
2J.Braun-Blanquet, Н. Walter, Zur Okologie der Mediterranpflan-
zen, «Jb. wiss. Bot.», 74, 1931, S. 697—748.
Средиземноморская жестколистная растительность
51
30 о ,0 ,
20 — ч Т —О— 78=^ 6
Л"' V- “°—J
1 J 4 л с ) duercus ilex „солнечный лист
IU • 1 Quercus ilex, теневой лист л л
□ л UL. UUUUlJGl U
и и т и И W и X 31 31Г I п ш
Рис. 31. Кривые изменения осмотического давления в течение года
у склеро- и малакофильных видов (по Браун-Бланке и Вальтеру).
В качестве примера приведем данные экологических исследований
каменного дуба {Quercus ilex). Аналогичные результаты дало изучение
Qu. coccifera и Pistacia lentiscus (рис. 31).
Заметного повышения осмотического давления у Quercus ilex в летние
месяцы не наблюдалось. Наибольшее его значение отмечено в апреле
1929 г. как следствие холодной зимы, когда была зафиксирована мини-
мальная температура —12,4°. Дней с морозом было в 4,5 раза больше,
чем зимой 1930 г. (абсолютный минимум —1,1°), но осадков втрое меньше.
Соответственно величина осмотического давления зимой 1929/30 г. была
меньше. В 1929 г. засуха продолжалась до августа. Ко времени появления
побегов в мае осмотическое давление понизилось.
4*
52
Глава 2
Совершенно иная картина наблюдалась на том же местообитании
у Viburnum tinus и Cistus albidus, а также у Thymus vulgaris и Phlomis
lychnitis. Это все малакофильнйе виды с мягкими опушенными листьями,
дающие побеги в апреле и затем, но менее обильно, в октябре, после нача-
ла осенних дождей. В мае осмотическое давление у этих видов резко
поднялось —до 40 атм (рис. 31); в условиях хорошей влагообеспечен-
ности осмотическое давление незначительно (12—17 атм).
Таким образом, при условиях, характерных для местообитания под
Монпелье, склерофильные виды выделялись известной гидростабильностью,
а малокофильным, напротив, была свойственна гидролабильность. Виды
Arbutus unedo, Lonicera implexa и Juniperus oxycedrus занимают проме-
жуточное положение; у теневыносливых Ruscus aculeatus, Rubia
peregrina и Smilax aspera величина осмотического давления составляла ме-
нее 20 атм, то есть растения обладали значительной засухоустойчиво-
стью.
Приведенные данные справедливы прежде всего для изученного
местообитания в 1929 г. Для определения возможных колебаний осмоти-
ческого давления автор проанализировал 180 проб, взятых в районе Мон-
пелье в августе 1962 г. после четырехмесячной засухи. Пробы отбирались
с самых различных местообитаний.
На местообитаниях с достаточной влагообеспеченностью значения
осмотического давления у жестколистных растений были невелики, и рас-
тения, несмотря на засуху, выглядели свежими. Недостаток влаги на
очень сухих местообитаниях —например, бровка каменоломни, где почва
высыхала не только сверху, но и с боков, или холм из кварцевой гальки
под Бассан-Виасом, почва которого содержала очень мало мелкозема,—
проявлялся уже во внешнем виде растений; вместе с тем у них было
заметно повышено осмотическое давление.
В табл. 1 сопоставлены некоторые данные.
Таблица 1
Осмотическое давление у склерофилов на влажных
и сухих местообитаниях (в атм)1
Название растений Местообитания Наибольшие значе- ния по другим авторам 2
относительно влажные сухие по Ру- шалю (Р.) по Гут- тенбергу и Буру (Г.)
Quercus ilex . . 21—23 31,5—39,4 30,3 32,6
Pistacia lentiscus .... 22-25 31,9 29,1 28,5
Phillyrea media 29,7 56,1 з 60,0 60,2
Olea europaea .... 22,2—23,5 46,9 60,6 51,7
Arbutus unedo . . . 21,3—25,3 42,3 40,6 32,2
Rhamnus alaternus — 45,7 — 37,0
1 Е. Rouschal, Zur Okologie der Macchien I. Der sommerliclie Wasserhaus-
halt der Maccliienpflanzen, «Jb. wiss. Bot.», 87, 1939, S. 436—523; Bsitrage zum
Warmehaushalt der Macchien, «Osterr. Bot. Z.», 87, 1938, S. 43; H. Gutten-
berg, H. Bulir, Stu dim uber die Assimilation und Atmung mediterraner Mac-
chienpflanzen wahrend der Regen- und Trockenzeit, «Planta», 24, 1935, S. 163—265.
2 Для Myrtus communis Гуттенберг приводит следующие значения осмотиче-
ского давления; в апреле (Бриони) — 33,4 атм, в сентябре (Корсика) — 53,4 атм.
3 Ph. angustifolia.
Средиземноморская жестколистная растительность
53
Виды с мягкими листьями выглядели угнетенными, только на отно-
сительно влажных местообитаниях и в тени они были олиственны. На сол-
нечных участках, напротив, наблюдалось обильное плодоношение, но све-
жими оставались только кончики побегов с мелкими, неразвитыми листи-
ками. У Viburnum tinus даже в тени листья казались увядшими. Значе-
ния осмотического давления для этих видов приведены в табл. 2.
Таблица 2
Осмотическое давление у малакофильных видов
на влажных и сухих местообитаниях (в атм)
Местообитания Наибольшие значе- ния по другим авторам
Название растений относительно влажные сухие по Ру- шалю по Гут- тенбергу и Буру
Viburnum tinus1 Cistus albidus . . Виды Cistus . . Phlomis lychnitis . ... Lavandula latifolia . . Rosmarinus officinalis . . . 1 Занимает промежуточное видами. Табл. 3 содержит резуль у видов Spartium и Buxus (с у теневыносливых Laurus и 30,6 26,7—27,8 21,0—25,0 21,0 21,6 29,5 положение i ►таты опре; 1ТНОСЯЩИХС Heder а, а 1 47,0 36,8-38,3 36,6 32,5 ! 35,2 между склеро- з;еления oci я уже к су также у х£ 43,2 37,3 и мал ан иотическ бсредизе юйных г 42,2 офильными ого давления мноморским), [ вересковых.
Таблица 3
Осмотическое давление у различных видов растений
на относительно влажных и сухих местообитаниях (в атм)
Название растений Местообитания Наибольшие значе- ния по другим авторам
относительно влажные | сухие по Ру- ша лю по Гут- тенбергу и Буру
Spartium junceum Buxus sempervirens 13,9 (P. 14,1) 24,8 35,7 43,0-48,5 27,2
Laurus nobilis Hedera helix . . Pinus hale pensis Pinus maritima Pinus salzmanni Juniperus oxycedrus . . Juniperus phoenicea . Erica arborea . 17,7 (июнь) 14,0—15,7 15,1 17,7 16,4 20,8 26,1 21,3 не исслед. 40,6 30,1 28,3
Erica cinerea — 48,2 по Жиру, 72,8
Erica multiflora Erica scoparia . Calluna vulgaris 20,5 23,8 35,8 по Жиру, 24,1
54
Глава 2
Анализ табличных данных позволяет сделать следующие выводы.
У типичных склерофилов, растущих на относительно влажных место-
обитаниях, даже к концу засушливого периода осмотическое давление,
всегда колеблющееся в пределах 21 —25 атм. не повышается; исключение
составляет Phillyrea, Напротив, на очень сухих местообитаниях иногда
фиксируется значительный рост давления, заметно отражающийся на
состоянии растений.
У малакофильных видов осмотическое давление весной обычно рав-
но 15 атм. в августе даже на относительно влажных почвах оно воз-
растает, тогда как на сухих, неблагоприятных местообитаниях это увели-
чение ничтожно мало; одновременно у растений резко сокращается листо-
вая поверхность. Несомненно, что эти виды значительно более чувстви-
тельны к засухе. Экология теневыносливых жестколистных видов изучена
слабо, в литературе обычно приводятся лишь низкие максимальные зна-
чения осмотического давления, например для Ruscus aculeatus — 16,7 атм
(Р.), для Smtlax aspera — 20,8 (Р.) и 20,5 (Г.), для Asparagus acutifolius —
23,3 атм (Р.). Лишь у лавра, который, правда, встречается и на открытых
участках, величина осмотического давления больше (см. табл. 3). Хвойные
ведут себя так, как склерофилы Ч
Не совсем еще ясно, каким образом склерофилы поддерживают свой
водный баланс на постоянном уровне во время засухи. При хорошей
влагообеспеченности они интенсивно транспирируют, что подтверждают
опыты Бергена (1904 г.) с отрезанными и поставленными в воду ветками 1 2.
Гуттенберг, применивший тот же метод, установил, что летом транспира-
ция вследствие более высокого потенциального испарения резко вырастает,
причем виды Cistus отличаются особенно большой водоотдачей 3. Но тогда
возникает вопрос, как ведут себя виды в естественных условиях: сохра-
няется ли устойчивость их водного баланса при а) неограниченной транс-
пирации, б) затрудненной или в) максимально уменьшенной транспира-
ции? Дело в том, что вслед за сокращением транспирации снижается
интенсивность ассимиляции СО2. Следовательно, полностью закрытые
устьица означают состояние голодания.
Ответ на вопрос могут дать только опыты по измерению транспира-
ции. Такие исследования предпринял Рушаль на острове Бриони (север-
ная часть Адриатического моря). Измерения производились поздним
летом незадолго до наступления дождливого периода и в первые дни
после его окончания.
Из анализа построенных Рушалем кривых суточной транспирации
явствует, что при хорошей влагообеспеченности, прежде всего до полудня,
интенсивность транспирации достигает очень высоких значений. Если
же растение испытывает недостаток влаги, транспирация сокращается
(табл. 4).
Эти данные показывают, что при недостатке воды величина транспи-
рации сокращается до долей ее нормального значения. Тем не менее
1 По Жиру, атлантическая незасухоустойчивая Erica cinerea на одинаково сухих
местообитаниях характеризуется бурным ростом, тогда как у средиземноморской Erica
multiflora интенсивность роста очень мала.
2 J. Y. Bergen, Transpiration of sun leaves and shade leaves of Olea europaea
and other broad-leaved evergreens, «Bot. Gaz.». 38, 1904, p. 285 ff.; J. Y. Bergen,
Relative Transpiration of old and new leaves of Myrtus type, «Bot. Gaz.», 38, 1904, p.
p. 446 ff.
3 H. Guttenberg, Studien fiber das Verhalten des immergriinen Laubblattes
der Mediterranflora zu verschiedenen Jahreszeiten, «Planta», 4, 1927, S. 726—779.
Средиземноморская жестколистная растительность
55
Таблица 4
Интенсивность транспирации у некоторых
склерофильных видов (в % от содержания воды)
за 1 час (средние значения)
Название растений Влажная почва Сухая почва
Quercus ilex ...... 92 (11,7)1 15 (1,3)
Pistacia lentiscus .... 88 (18,3) — —
Phillyrea media .... 102(13,9) 32 (2,5)
Olea europaea 72 (12,8) 21 (3,4)
Arbutus unedo ... 48 (6,2) Ю(2,2)
Rhamnus alaternus . . . 45 (10,7) — —
1 В скобках приведено значение транспирации
в мг/дм2 .мин.
л в этом случае в течение суток несколько раз происходит обмен всей
содержащейся в листьях воды, что возможно только при известном погло-
щении влаги растением. Отсюда ясно, что почва, как и следовало ожидать,
содержит достаточное количество влаги, доступной для растений, и что
их корневая система проникает до влагосодержащего горизонта. В обна-
жениях в бортах каменоломен корни Quercus ilex прослежены до глубины
10 м, а корни Quercus coccifera — до 5 м. Корйевая система Phillyrea angusti-
folia и Pistacia lentiscus проникает столь же глубоко.
Приведенные значения интенсивности транспирации подтверждаются
измерениями, предпринятыми автором в августе 1962 г. в маквисе Мон-
тарно под Монпелье. При расчете на сырой вес для Quercus ilex и Arbutus
unedo получены значения 3—6 мг1г*мин. Аналогичные величины опреде-
лены для Erica arborea и Е. scoparia. При опытах использовались не толь-
ко торзионные весы и мелкие верхушки веток, но и прецизионные весы
и крупные ветви (весом до 300—500 г). Ветки быстро доставляли с опыт-
ной площадки в помещение, где находились весы, их взвешивали, выно-
сили наружу на 30 минут и снова взвешивали (потери веса составляли 10 —
15 г). Вычисление производилось на 1 г сырого веса листьев. Это соответ-
ствовало разнице между исходным весом и весом веток без листьев после
опыта, с учетом количества испарившейся воды, которая в основном
поступала из веток. Отмечено хорошее совпадение данных, полученных
обоими методами.
Совершенно иная величина транспирации установлена у кустарника
Quercus ilex на очень сухом местообитании: она в 2—4 раза меньше при-
веденных выше значений и приближается к границе точности измерения.
Кустарник низкий, и листьев на нем меньше, чем у того же вида на влаж-
ном местообитании.
Многочисленные сравнительные измерения транспирации Quercus ilex
и Qu. suber произвел Брекле (см. стр. 36), работавший в районе Бланеса.
В мае — июне молодые листья транспирировали значительно слабее, чем
старые, но к августу интенсивность транспирации тех и других сравнялась.
У Qu. ilex летом испарение сокращается более резко: уже после 8—9 часов
утра кривая суточной транспирации круто идет вниз и остается на этом
уровне до вечера. Напротив, у Qu. suber четко выделяются два дневных
максимума. Соответственно у пробкового дуба наблюдается менее выров-
ненный водный баланс, он несколько менее гидростабилен и характери-
56
Глава 2
Таблица 5
Максимальные скорости сокодвижения
в стволе при хорошей влагообеспеченности
ранним летом (I) и в разгар летней
засухи (II) (в см/час)
Название растений I II
Olea еигораеа 268 72
Arbutus unedo 150 40
Laurus nobilis 306 121
Spartium junceum 108 72
Fraxinus ornus 778 105
зуется четко выраженным пиком осмотического давления летом. Не уди-
вительно поэтому, что Qu. suber предпочитает районы с океаническим
климатом. Здесь он, особенно на слабокарбонатных почвах, более жизнеспо-
собен, чем дуб каменный (Qu. ilex), так как благодаря более высокой про-
дуктивности отличается быстрым ростом и, следовательно, большей
конкурентной мощностью.
Таким образом, ксероморфное строение склерофилов позволяет им при
необходимости полностью задерживать водоотдачу и тем самым предотвра-
щать на длительное время засыхание листьев. Деревья с мезоморфной
листвой не в состоянии этого делать, так как кутикулярная транспирация
у них выше. При остром недо-
статке воды, когда испарение
в течение продолжительного вре-
мени превышает поглощение влаги
растением, его листья засыхают.
Величина сублетального дефицита
влаги, который листья еще спо-
собны переносить, менее важна,
чем уровень, до которого может
быть снижена транспирация и ко-
торый уже означает затрудненную
степень поглощения воды расте-
ниями.
У срезанных вместе с листья-
ми веток после высушивания по-
глощение влаги полностью пре-
кращается. Листья на ветках Quer-
cus pubescens и Sorbus torminalis, разложенных на открытом воздухе, уже
через 4 часа засыхают, для листьев же Quercus ilex и Heder a helix на это
требовалось много дней. Arbutus и Ilex aquifolium отдают воду также
очень медленно, но уже через 2 суток наблюдается некроз, на листьях
появляются бурые или черные пятна.
Показателем затрудненного поглощения воды растениями во время
летней засухи может служить также величина скорости сокодвижения,
определенная Рушалем 1 (табл. 5).
В условиях водного дефицита жестколистные кустарники уменьшают
потребление воды почти в четыре раза, кустарник Spartium с безлистными
прутьевидными ветвями и, следовательно, малой транспирирующей пло-
щадью — только на 1/3, а листопадный Fraxinus — более чем в семь раз.
Скорость сокодвижения в корнях Olea, равная 148 и 52,3 см/час, соответ-
ствует сокращению водопотребления приблизительно в три раза.
Таким образом, экологическое значение склерофильной структуры
проявляется в том, что она позволяет растениям — при хорошей влагообес-
печенности, открытых устьицах и благодаря их большому числу 1 2—под-
держивать интенсивный газообмен и эффективно продуцировать органи-
ческое вещество; при недостатке влаги часть устьиц закрывается, то есть
транспирация приводится в соответствие с поступлением воды из корней,
и, наконец, в периоды острого дефицита влаги транспирация при закры-
1 Е. Rouschal, Die Geschwindigkeit des Transpirationsstromes in Macchien-
Geholzen, «Stz.-Вег. Akad. Wiss. Wien, math.-nat. KI.», I, 146, 1937, S. 119.
2 По Брекле, согласно измерениям в Португалии, Испании и южной Франции,
у Quercus ilex и Qu. suber число устьиц на 1 мм2 составляет 414—541.
Средиземноморская жестколистная растительность
57
тых устьицах практически прекращается, что дает возможность растению
пережить засуху до начала дождей без потери листвы.
Большая жесткость листьев уменьшает их перемещение из одного
положения в другое при сильном ветре. Возникает эффект, сходный с дей-
ствием кузнечных мехов. Следствием его является отсутствие вынужден-
ного газообмена,
Градман проводил исследования устьиц склерофилов в районе Неапо-
ля, Обнаружив, что устьица даже в августе были немного открыты, он
указал на тот факт, что склерофилы летом активны (исключение состав-
ляют растения в крайних условиях обитания), хотя и не в такой степени,
как в благоприятное время года (апрель — июнь) г. Отсюда можно заклю-
чить, что фотосинтез также есть процесс непрерывный, никогда полностью
не прекращающийся.
Правда, газоаналитические измерения, проведенные Гуттенбергом
и Буром поздним летом на Корсике, как будто показали незначительность
у растений чистой ассимиляции; исследователи даже считали, что расте-
ния в период засухи «работают на износ», расходуют запасные вещества
и находятся «на грани возможности существования». Но, судя по облику
растений, этого нельзя сказать для периода летней засухи. Если учесть
источники ошибок метода, использованного этими авторами, то выделение
листьями СО2 в полдень, скорее, следует рассматривать как признак силь-
ного их перегрева в кювете при опыте, В климатических камерах, широко-
применяемых в последнее время, всегда выявляется положительный баланс
ассимиляции (Эккард, устное сообщение).
Одной из характернейших черт растений — обитателей районов с зим-
ними дождями является резкое преобладание у них жесткой листвы.
Ее развитие наблюдается у видов, принадлежащих различным семейст-
вам, например у видов буковых, маслинных, бобовых, вересковых, лилей-
ных и почти у всех протейных, а также рестионовых Южной Австралии
и Южной Африки, Столь длинный перечень свидетельствует о том, что
анатомо-морфологическое строение растений имеет экологическое значе-
ние; этим вопросом уже давно занимались многие исследователи * 2,
В частностях склерофильные признаки растений различны. Так,
наряду с обычно мелкими или даже эрикоидными листьями встречаются
и крупные листья, как, например, у Arbutus или Protea, Однако необычай-
но толстая внешняя стенка эпидермиса у последних обеспечивает постоян-
ство формы листа. Особенно примечательны волокна склеренхимы, или
склереиды (ср, Olea, Phillyrea, Hakea и др,), которые препятствуют изме-
нению длины листа или разделят нижнюю и верхнюю стороны эпидермиса.
Межклетники редуцированы, поэтому и клетки паренхимы при напряже-
ниях сцепления не могут сжаться; более того, даже клетки палисадной
ткани часто бывают соединены друг с другом по бокам, и тогда их разде-
ляют только поясовидные межклетники.
ХН,Gradman n, Die Windschutzeinrichtungen an den Spaltoffnungen der Pflan-
zen, «Jb, wiss, Bot,», 62, 1923, S, 449—527,
2 A, Tschirch, Uber einige Beziehungen des anatomischen Banes der Assimi-
lationsorgane an Klima und Standort mit spezieller Beriicksichtigung des Spaltoffnungs-
apparates, «Linnaea», 43, 1882; G, Haberland t, Physiologische Pflanzenanato-
mie, Leipzig, 1918; H, Guttenberg, Anatomisch-physiologische Untersuchungen
uber das immergriine Laubblatt der Mediterranflora, «Englers Bot, Jb.», 38, 1907, S, 383—
443; F, F i r b a s, Uber die Ausbildung des Leitungssystems und das Verhalten der
Spaltoffnungen im Friihjahr bei Pflanzen des Mediterrangebietes und der Tunesischen
Steppen und Wiisten, «Beih, Bot, Cbl, I», 48, 1931, S, 451—465,
58
Глава 2
Возникает вопрос, не служат ли все эти приспособления одной цели:
создать отрицательные значения тургорного давления и большую сосу-
щую силу, возможно превышающую 100 атм.
Эту проблему весьма подробно изучал Крееб, усовершенствовавший
разработанный Слатьером метод определения сосущей силы клеток Ч
В лабораторных условиях ему удалось в подсыхающих листьях Buxus
выявить отрицательный тургор, однако проверка опытов в поле на скле-
рофилах в северо-восточной Испании (Коста-Брава) и — при еще более
экстремальных условиях — на севере Египта показала, что хотя в отдель-
ных случаях, возможно, и возникает слабое отрицательное тургорное
давление, но придавать ему экологическое значение вряд ли сле-
дует.
Данные, приводимые Гривом для склерофилов юго-западной Австра-
лии, требуют проверки, так как использованный им метод, по-видимому,
не дает абсолютно правильных значений 1 2.
Но даже если отрицательные значения тургорного давления и не
возникают, склерофильная структура имеет тем не менее важное значе-
ние, так как увеличивает гидростабильность вида. Термодинамическая
модель, изображенная на рис. 10 в первом томе, показывает, что в камере
Пфеффера с жесткими стенками возникновение сосущей силы (увеличение
концентрации наружного раствора, варианты 4 и 5) компенсируется пони-
жением тургорного давления, причем осмотическое давление клеточного
сока и степень гидратации набухающего тела (протоплазмы) не изменя-
ются. Если стенки клеток упругие, увеличение сосущей силы и снижение
тургора обусловливают снятие напряжения со стенок клетки, и она отдает
воду; концентрация клеточного сока повышается, соответственно прото-
плазма сжимается. Это наблюдается у мягколистных растений в период
обычной летней засухи, когда их осмотическое давление быстро
растет.
Напротив, у склерофилов уменьшения объема не происходит, то есть
они ведут себя как камера Пфеффера. Не наблюдается также заметного
повышения осмотического давления и сжатия протоплазмы. Снижение
тургора не вызывает завядания листьев, так как последние обладают
жестким строением. Лишь при сильной засухе и очень большой величине
сосущей силы листья немного деформируются, объем клеток несколько
-сокращается и осмотическое давление возрастает (ср. табл. 1).
Жестколистные виды — вечнозеленые и переносят только кратко-
временные легкие заморозки; гидростабильность они сохраняют лишь
при умеренной засушливости и поэтому не растут в пустынях; жестко-
листные виды распространены преимущественно в областях с зимними
дождями, где наблюдаются мягкие, хотя и не совсем безморозные, зимы,
а период летней засухи выражен весьма резко. Важно одно: чтобы в поч-
ве еще сохранялось некоторое количество влаги, достаточное для поддер-
жания сильно уменьшившейся транспирации.
1 К. К г е е b, Zur Frage des negativen Turgors bei mediterranen Hartlaubpflanzen
unter naturlichen Bedingungen, «Planta», 56, 1961, S. 479—489.
2 В. J. Grieve, Negative turgorpressures in sclerophyll plants, «Austral. J.
Sci.», 23, 1961, p. 376—377. Грив применил метод Шардакова, которым в Монпелье
в благоприятные времена года пользовался и Редер, но получил значения сосущей
силы, значительно превосходящие величину осмотического давления. Редер считает
их завышенными и предлагает рассматривать только как относительные (Н. R е h d е г,
Saugkraftmessungen an mediterranen Immergrunen mit der Schardakow-Methode, «Вег.
Deutsch. Bot. Ges.», 74, 1961, S. 84—92).
Средиземноморская жестколистная растительность
59
У жестколистных растений устьица большей частью глубоко погру-
жены в ткань листа. Иногда листья имеют устьица, которые открываются
не непосредственно наружу, а в узкие канальцы. Такое строение настоль-
ко своеобразно, что невольно напрашивается мысль о его экологическом
значении. Градман (см., стр. 57) с помощью опытов на моделях попытался
доказать, что при ветре эта особенность строения в большей степени сокра-
щает транспирацию, чем поглощение СО2, что означало бы более интен-
сивное использование воды для продуцирования вещества. Правда,
не ясно, можно ли переносить полученные на моделях данные на устьица
с их микроскопическими размерами. Однако в последнее время Лархеру 1
удалось доказать, что отношение A/W (где А — ассимилированное при
фотосинтезе количество СО2, a W — одновременно тем же листом транспи-
рированное количество воды) у склерофильных листьев не остается
постоянным (при условии, что часть устьиц закрывается), а характери-
зуется максимумом, при котором вода используется оптимально. Прибор
«Uras» позволил одновременно измерять поглощение СО2 и водоотдачу,
благодаря чему возросла точность определения величины А/W. Наиболь-
шее сопротивление, которое вынуждена преодолевать молекула воды при
диффузии из межклетников наружу, обусловлено устьицами, но это
положение не обязательно по отношению к молекулам СО2, диффундирую-
щим к хлоропластам. Для них наибольшим, видимо, является сопротив-
ление, возникающее при прохождении молекул СО2 сквозь плазмалемму
или при их диффузии через узкие межклетники между клетками палисад-
ной ткани. Если это так, то частичное закрытие устьиц должно сильнее
отразиться на диффузии воды, чем на диффузии СО2, иными словами,
•отношение А/W есть величина переменная. Действительно, Лархер уста-
новил, что, когда устьица срезанных листьев дуба начинают закрываться,
величина А/W возрастает до определенного оптимального значения,
и, лишь когда устьица почти совсем смыкаются, она вновь уменьшается.
Рост отношения А/W более заметен для Quercus ilex, чем для Qu. pubes-
cens. У растений Olea еигораеа, посаженных в горшки, при высыхании поч-
вы в течение первых трех дней происходило сужение отношения, затем
последовало его увеличение; оптимум был достигнут лишь на четвертый
день опыта. Следовательно, вполне возможно, что величина А/W дости-
гает оптимального уровня при частично закрытых во время летней засухи
устьицах. Однако для полного решения вопроса нужны дополнительные
исследования непосредственно в природе k У Olea устьица хотя и не углуб-
лены в ткань листа, но зато они плотно окружены защитными волосками,
что, очевидно, играет аналогичную роль. Таким образом, не исключено,
что глубокое расположение устьиц в большей мере задерживает диффузию
молекул Н2О, чем молекул СО2.
Все описанные исследования проведены в северной части Средиземно-
морской климатической области. Как же отражается засуха на поведении
растений, произрастающих в ее южных пределах? На севере Африки
соответствующая зона Quercus ilex относится к горному поясу, в пределах
которого выпадают сравнительно обильные осадки и где условия, по-види-
мому, ненамного засушливее, чем на севере области. Эколого-физиологи-
1 W. L а г с h е г, Transpiration and photosynthesis of detached leaves and shoots
•of Quercus pubescens and Quercus ilex during dessication under standard conditions, «Bull.
Res. Counc. Israel», 80, 1960, p. 213—224; W. L a r c h e r, Orientierende Untersuchun-
gen uber das Verhaltnis von CO2-Aufnahme zur Transpiration bei fortschreitender Boden-
trocknung, «Planta», 60, 1963, S. 339—343.
60
Глава 2
ческое изучение склерофилов в Северной Африке не проводилось, но
имеются данные опытов, выполненных в Палестине \
Исследовались виды в отсутствии орошения на плантации под Иеру-
салимом. У Olea при засухе транспирация резко падала, однако осмоти-
ческое давление было в пределах 22,4—27,0 атм. Аналогичная картина
наблюдалась у Pinus halepensis, осмотическое давление которой оставалось
относительно высоким (22,3—30,1 атм). У Arbutus andrachne (31,1 атм)
и Ceratonia siliqua (19,1—20,6 атм) при закрытых устьицах транспирация
полностью прекращалась. Следовательно, Ceratonia —хорошо выражен-
ный гидростабильный вид (оптимальные значения осмотического давления
около 14 атм), но летом растение находится в состоянии покоя. Laurus
и Rosmarinus без полива сильно страдают от засухи.
Другая серия опытов осуществлена у подножия горы Кармель, где
маквис занимает северный склон (уклон 20°). Вместо западносредиземно-
морского Quercus ilex господствует Quercus coccifera var. calliprinos, Laurus
встречается как примесь. Микроклиматические условия в разгар лета
аналогичны условиям под Монпелье. Интенсивность транспирации опре-
делялась методом быстрого взвешивания срезанных листьев в марте и апре-
ле (благоприятное время года), а также в июне и сентябре (период засухи).
Опыты подтвердили приведенные выше результаты: реакции на среду
у Quercus calliprinos мало чем отличаются от поведения Quercus ilex. Летом
транспирация в расчете на 1 г сырого веса вследствие того, что устьица
полузакрыты, резко сокращается до 2—2,5 (6) мг!г*мин в отличие от
4—6(10) мг!г*мин весной; тем не менее осмотическое давление возрастает
с 20,9 атм всего до 25,7—27,7 атм (экстремальное значение 31 атм).
У Phillyrea media устьица летом открыты и транспирация приблизи-
тельно столь же интенсивна, как весной. Вместе с тем гидратура сущест-
венно уменьшается (осмотическое давление достигает 55 атм), что, однако,
не наносит ущерба растению.
Laurus nobilis здесь, у крайней южной границы его распространения,
менее устойчив. Уже в июне устьица едва открыты. Транспирация весьма
слабая. В сентябре устьица обычно полностью закрыты, транспирация
столь мала, что измерить ее почти невозможно. Но и в этих условиях
осмотическое давление поднималось до 36,8 атм*, такое значение для этого
вида, вероятно, является максимальным.
На том же местообитании растет листопадная Pistacia palaestina (близ-
кая к Р. terebinthus). Вид отличается относительно высокой интенсив-
ностью транспирации летом, хотя устьица открыты мало. Оппенгеймер
предполагает, что кутикулярная транспирация его мезоморфных листьев
весьма значительная, поэтому сокращение транспирации по этой причине
невозможно. Тем не менее водный баланс у этого вида сохранялся на
постоянном уровне, поскольку величина осмотического давления не пре-
вышала 24,7 атм.
Следует, однако, учитывать, что величина
транспирации сама по себе еще не дает пра-
вильного представления [о всем процессе о т д а -
1 Н. R. Oppenheimer, Zur Kenntnis der hochsommerlichen Wasserbilanz
mediterraner Geholze, « Ber. Deutsch. Bot. Ges.», 50 a, 1932, S. 185—245; Studies on the
water balance of unirrigated woody plants, «Palestine J. Bot. Rehov.», Ser. 6, 1947,
p. 63—76; Summer drought and water balance of plants growing in the Near East,
«J. Ecology», 39, 1951, p. 356—362; An experimental study on ecological relationship and
water expenses of mediterranean forest vegetation, «Palestine J. Bot., Rehov.», Ser. 8,
1953, p. 103—124.
Средиземноморская жестколистная растительность
61
ч и воды растениями во внешнюю среду, так как
последняя зависит от площади всей транспи-
рирующей поверхности. У дерева с большой интенсивностью
транспирации, но с маленькой общей листовой поверхностью водный
баланс будет более выровненным, чем у дерева со слабой транспирацией,
но с очень большой листовой поверхностью.
Перейдем теперь к экологии малакофильных видов. От жесткоуст-
роенных листьев листья этих видов отличаются тем, что их объем при воз-
никновении напряжений сцепления легко уменьшается. Особенности
строения листа допускают резкие колебания содержания воды в клеточ-
ной ткани. Именно это свойство типично для мягколистных видов. В усло-
виях хорошей влагообеспеченности они обнаруживают очень большую
интенсивность транспирации. При недостатке воды листья завядают,
а осмотическое давление повышается (рис. 31—32). Невыровненный вод-
ный баланс Рушаль обнаружил у Cistus villosus, а также у Teucrium polium,
Т. chamaedrys, Thymus vulgaris и Satureja montana. Неустойчивость вод-
ного режима отчасти объясняется неглубокой корневой системой; по
Браун-Бланке, глубина проникновения корней не превышает 20—30 см.
Однако не исключено, что нежные мочковатые корни уходят на глубину,
в два раза большую. Верхние почвенные горизонты летом высыхают
столь сильно, что в августе в Монтарно мы не обнаружили никакого
«дыхания» почвы. Это означает, что жизнь во всех содержащих гумус
горизонтах в этот период замирает. Если обычно осмотическое давление
у малакофильных видов колеблется в пределах 10—15 атм, то летом,
даже на местообитаниях с благоприятными условиями, оно возрастает
до 20—30 атм, а с неблагоприятными — до 40 атм. Одновременно проис-
ходит резкое сокращение листовой поверхности, подобное описанному
автором в Аризоне у Encelia (т. I, стр. 371—373). Крупные листья засы-
хают и опадают, поздним летом на растениях видны только сухие сопло-
дия и еще живые почки на концах побегов. В отличие от Cistus у Thymus
и Rosmarinus общая листовая поверхность из-за небольшого размера самих
листьев с самого начала не очень велика. У некоторых видов Cistus,
например у Cistus monspeliensis, С. laurifolius и С. ladaniferus, транспира-
ция листьев вследствие смолистого налета на них сильно понижена. Редук-
ция листовой поверхности в этом случае происходит в более ограничен-
ных масштабах. Срезанные побеги Cistus monspeliensis с лакированными,
очень клейкими листьями высыхают крайне медленно, даже после лежа-
ния на ярком солнце в течение двух дней они выглядят почти свежими.
Поскольку эта группа малакофильных растений переносит период
засухи только благодаря сильной редукции своей поверхности, она,
по-видимому, менее приспособлена к данным климатическим условиям,
нежели склерофилы, конкурентная борьба с которыми оказывается
мягколистным растениям не под силу. Поэтому малакофильные виды
встречаются исключительно на местообитаниях, вторично измененных
пожаром или вырубкой.
Во время длительной засухи склерофильные растения затененных мест
продолжают существовать только в более мягких условиях, связанных с
затенением. В менее засушливом климате они хорошо развиваются и на
солнечных местообитаниях; примером может служить лавр во влажном
средиземноморском районе северной Анатолии или на северных скло-
нах гор.
Отметим еще вкратце поведение некоторых других экологических
типов растений. Зимой влаги всегда более чем достаточно, поэтому на всех
62
Глава 2
открытых участках осенью прорастают семена бесчисленных терофитов,
которые весной цветут и плодоносят. Высота их нередко не превышает
нескольких сантиметров, а корни пронизывают слой толщиной всего 1,5 —
3,5 см (более крупные растения — до 15 см). К концу весны, когда верхние
слои почвы высыхают, развитие растений заканчивается. Осмотическое
давление у терофитов всегда очень низкое Ч
Аналогичная картина наблюдается у геофитов (Allium, Asphodelus,
Narcissus, Iris, Orchidaceae, Arum); отличие лишь в том, что их листья
очень слабо транспирируют и сравнительно долго не опадают после начала
засухи. Ксероморфным строением обладает Aphyllanthes monspeliensis.
У Iris chamaeiris, верхняя половина листьев которых была совершенно
сухая, величина осмотического давления составила 11,4—12,5 атм. Мно-
гие травянистые растения переносят весь засушливый период, не повышая
своего осмотического давления, а лишь несколько сокращая листовую
поверхность; листья часто не засыхают, а лишь желтеют, например
у Euphorbia characias (рис. 32). У ксероморфного Eryngium campestre
с увядшей наполовину листвой осмотическое давление в конце августа
составляло 22,2 атм, у Dorycnium suffruticosum (в неблагоприятных усло-
виях обитания сбросил все листья) — 23,2 атм, а у Bupleurum rigidum
(цветущий экземпляр, но со слегка завядшими прикорневыми листья-
ми) — 23,9 атм. Для поздноцветущих трав весьма характерно, что в нача-
ле цветения сильно транспирирующие листья желтеют, а затем засыхают;
запасы вещества поступают в стебли с соцветиями, которые несут только
маленькие верхушечные листья и потому расходуют мало воды. Следова-
тельно, основную массу вещества эти виды продуцируют незадолго до
наступления собственно засушливого периода. Растения этой группы боль-
шей частью имеют глубоко уходящий в почву, нередко утолщенный стерж-
невой корень (например, у Eryngium).
Суккуленты в периоды засухи также продолжают свою жизнедеятель-
ность. При возникновении дефицита влаги вода поступает из старых листь-
ев к молодым, поэтому в живых частях растения заметного повышения
осмотического давления не происходит. Так, для Sedum nicaeense в авгу-
сте характерны значения 6,2—8,8 атм, для Sedum altissimum в апреле —
7,6, а в июне — 8,7 атм. Поверхностная корневая система не позволяет
суккулентам восполнить израсходованное количество воды за счет почвен-
ной влаги. Пример суккулентного терофита — Umbilicus pendulinus; вели-
чина его осмотического давления повышается с 5,4—6,7 атм весной до
13,3 атм в июне, перед отмиранием растения.
В областях с зимними дождями естественные злаковники, типич-
ные для районов с летними дождями, отсутствуют. Только среди терофи-
тов имеется много видов злаков, однако многолетние злаки встречаются
редко и лишь на некоторых, измененных человеком местообитаниях. На
участках, используемых как пастбища, из злаков доминирует Brachypo-
dium ramosum. Побеги возникают весной и с приходом осенних дождей.
В целом начало роста средиземноморских видов весной зависит от глубины
проникновения их корней: чем ближе к поверхности располагается корне-
вая система, чем быстрее прогреваются верхние слои почвы, тем раньше
появляются побеги. У склерофилов с глубоко проникающими корнями,
а также у Viburnum tinus побеги возникают лишь в конце мая. У Brachypo-
dium, как и у всех типичных злаков, регуляция водоотдачи не происхо-
1 W. S i m о n i s, Beobachtungen zur Okologie einiger Therophyten in zwei medi-
terranen Pflanzengesellschaften, «Vegetatio», 5/6, 1954, S. 553—561.
20
15
10
20
15
10
-----Euphorbia nicaeensis
_----Euphorbia characiqs^
— Urospermum dalechampii >> .
— Ononis minutissima /
Рис. 32. Кривые изменения осмотического давления в течение года у раз-
личных видов трав: стеногидрических ксерофитов (виды Euphorbia), сукку-
лентов (Sedum), геофитов (Asphodelus), злаков (Brachypodium) и др. Пунк-
тир — месяцы 1931 г., когда Бхаруха не нашел свежих листьев (по Бха-
руха, кривые с буквами Бр.-В. — поданным Браун-Бланке и Вальтера).
<64
Глава 2
дит. При уменьшении влажности почвы первыми засыхают нижние листья,
затем кончики верхних, несколько позже их листовые пластинки и, нако-
нец, влагалище листа; стебли, как правило, остаются живыми и только
при очень сильной засухе частично отмирают. Злаковый покров летом
выглядит безжизненно сухим, но осенью он вновь зеленеет и возобновляет
свой рост. На рис. 32 приведен годовой ход осмотического давления
у Brachypodium. Характерен быстрый рост давления ранним летом, при-
чем нужно учитывать, что из-за небольшой массы листьев пробы приходи-
лось брать каждый раз на другом месте.
Большое число кривых годового хода осмотического давления, в том числе для
нескольких местообитаний, составил Брекле (см. стр. 36). Данные охватывают все лет-
ние месяцы, отсутствуют наблюдения только для периода с середины сентября до нача-
ла ноября. Исследовались:
1) жестколистные виды Quercus ilex, Qu. suber, Qu. coccifera, Qu. pubescens, Qu. aegi-
lops (Ботанический сад), Arbutus unedo, Phillyrea angustifolia, Myrtus communis,
Pistacia lentiscus, Coriaria myrtifolia, Pinus pinea, Daphne gnidium (листья желтеют);
2) некоторые виды с отчетливыми максимумами осмотического давления летом:
Rosmarinus officinalis, Erica scoparia, E. arborea, Ulex parviflorus, Sarothamnus scoparius,
Calycotome spinosa, Spartium junceum, Cistus salviaefolius (до 50 атм), C. monspeliensis
(свыше 80 атм), Lavandula stoechas (до 65 атм), Helichrysum stoechas (до 50 атм).
Lonicera implexa, Psoralea bituminosa, Rubia peregrina.
Интересно, что у Cytinus hypocistis, паразитирующего на Cistus, давление не пре-
вышало 8—10 атм', видимые части растения-паразита тотчас отмирают, как только
величина осмотического давления у растения-хозяина превысит 20 атм, что происходит
уже в мае. Еще более узкий диапазон колебания давления (8—10 атм) отмечен у орхи-
деи-сапрофита Limodorum abortivum; уже при давлении 12 атм она наполовину засыха-
ет. Оба вида транспирируют очень слабо; гетеротрофным видам интенсивный газообмен
не нужен. У однолетней Vicia dumetorum давление колеблется в пределах 10—15 атм;
если оно в мае быстро повышается, растения погибают.
Пойкилогидрические сосудистые растения в Средиземноморье боль-
шой роли не играют. Деление года на постоянно влажный и засушливый
периоды неблагоприятно для них. Они более конкурентоспособны там,
где в течение одного сезона короткие влажные периоды часто сменяются
сухими. В средиземноморском климате такие условия свойственны только
скалистым местообитаниям с маломощным почвенным покровом, который
во влажный сезон успевает быстро высохнуть между двумя дождями.
Поэтому излюбленные места поселения Ceterach officinarum или Notho-
laena maranthae — это трещины скал и щели стен. Оппенгеймер и Халеви
в обзорной статье, посвященной экологии Ceterach officinarum, отмечают,
что заростки этого вида способны к высыханию, но полностью при этом
не погибают Ч Единственные европейские виды геснериевых — Ramondia
и Haberlea — являются пойкилогидрическими цветковыми растениями
окраинных районов Средиземноморской области 1 2. Ramondia pyrenaica
растет, например, в густом лесу на скале у станции канатной дороги на
Монсеррат в северо-восточной Испании. На несколько более влажных
северных склонах можно заметить маленькую Selaginella denticulata. Для
углублений в скалах, наполняющихся зимой водой, а летом быстро высы-
хающих, характерны низкорослые виды Isoetes, которым иногда сопут-
ствуют Marsilia и Pilularia. Экология их пока еще не изучена.
1 Н. R. Oppenheimer, А. Н. Halev у, Anabiosis von Ceterach officinarum
La. et D., «Bull. Res. Counc., Israel» D, 11, 1962, p. 127—147.
2 H. Walter, Die Hydratur der Pflanze, 1931; L. К a p p e n, Der EinfluB des
Wassergehaltes auf die Widerstandsfahigkeit von Pflanzen gegenuber hohen und tiefen
Temperaturen, untersucht an Blattern einiger Fame und von Ramondia myconi, «Flora»,
Abt. B, 156, 1966, S. 427—445.
Средиземноморская жестколистная растительность
65
Почва в пересыхающих летом небольших водоемах с глинистым дном
иногда сохраняет некоторую влажность. В результате здесь развивается
довольно пестрый растительный покров. В качестве примера на рис. 33 пока-
зан профиль и растительность одного такого высыхающего летом водое-
ма недалеко от Монпелье. Влажность почвы уменьшается от центра впади-
ны к периферии, что отчетливо проявляется в развитии видов растений.
У Potentilla reptans на сухом местообитании (2) отмечено заметное сниже-
ние гидратуры (повышение осмотического давления почти на 1 атм)*,
соответственно развиваются и в четыре раза меньшие по площади и более
Рис. 33. Профиль высохшего в августе водоема под Монпелье (лес Рувьер).
1 — сухая, лишенная растительности почва в средней части водоема, с трещинами усыха-
ния; 2 — относительно сухое местообитание; 3 — несколько более влажное местообитание
Potentilla reptans (листья растения в 1/3 нат. величины и их осмотическое давление); 4 и
5 — травостой Holoschoenus romanus, высота которого к краевой части снижается вдвое,
причем осмотическое давление повышается только с 15,0 до 15,2 атм.
ксероморфные листья. У болотного растения Holoschoenus romanus увели-
чивающаяся сухость почвы также сдерживает рост, но величина осмоти-
ческого давления почти не изменяется. Такое кажущееся противоречие
типично, как будет показано и объяснено р гл. 6, т. III, для болотных
растений. В описываемом районе, сложенном мергелями, на отдельных
уступах эродированного склона растет множество экземпляров Molinia
coerulea. Высота растений вместе с метелками достигала 60 см, в августе
должно было начаться их цветение. Осмотическое давление составляло
12,5 атм, стало быть, в почве было еще много влаги, хотя поверхность ее
выглядела сухой и пыльной.
Тепло как экологический фактор
и температурные условия
У средиземноморских видов важную роль, помимо водного, играет
также тепловой баланс.
Инсоляция летом вызывает перегрев листьев в сравнении с темпера-
турой воздуха. Препятствовать перегреванию растения может только
охлаждающее влияние транспирации. Но поскольку у склерофилов
интенсивность транспирации сильно сокращается именно летом, было бы
5-01248
66
Глава 2
Рис. 34. Суточный ход
температуры листа Quercus
ilex: обычного (Л) и смазан-
ного вазелином (С); 2 сен-
тября 1960 г., Коста-Брава,
Испания (по О. и Р. Ланге).
L — температура воздуха вблизи
листа; U — температура влаж-
ного диска эвапориметра; F —
относительная влажность воз-
духа вблизи листа; S — интен-
сивность солнечного излучения.
интересно выяснить, до какого уровня может подниматься температура
листьев и как это повышение отражается на жизнедеятельности растений \
На рис. 34 приведены кривые суточного хода температуры воздуха
и температуры транспирирующего листа Quercus ilex, нетранспирирующего
листа (смазанного вазелином или отрезанного), влажной фильтровальной
бумаги, кривые влажности воздуха вблизи листа, а также интенсивности
инсоляции. Измерения произведены Ланге на северо-востоке Испании
с И августа по 3 сентября, то есть в период, когда транспирация среди-
земноморских видов растений сильно сокращается. Максимальное значе-
ние ее интенсивности у Quercus suber составило 10 мг!дм1 2 -мин, у Rham-
nus —5 мг/дм2•мин, обычно же оно было намного меньше. При столь
слабой транспирации охлаждающего его влияния отметить не удалось.
Максимальные же температуры листьев были существенно ниже границы
устойчивости, иными словами, ниже температуры, которая при воздейст-
вии в течение 30 минут повреждает 50% листьев (табл. 6).
У проб, взятых на высоте 900—1000 м, граница устойчивости была на
2,5—4° ниже, однако и максимальные температуры в этих местах не были
столь высокими. Следовательно, средиземноморские виды от жары не
страдают. Авторы этих исследований задают вопрос, не может ли высокая
1 O.-L. Lange, R. Lange, Untersuchungen uber Blattemperaturen, Transpi-
ration und Hitzeresistenz an Pflanzen mediterraner Standorte (Costa Brava, Spanien),
«Flora», 153, 1963, S. 387—425; Die Hitzeresistenz einiger mediterraner Pflanzen in Ab-
hangigkeit von der Hohenlage ihrer Standorte, «Flora», 152, 1962, S. 707 — 710.
Средиземноморская жестколистная растительность
67
Таблица 6
Температура листьев и границы устойчивости у склерофилов
летом (по Ланге)
Название растений Превышение температуры листьев по сравнению с температу- рой воздуха, °C Максим, температура листьев, °C Граница устойчи- вости, °C
Quercus ilex 15,6 43,6 54
Quercus suber . . 12,2 43,9 53
Rhamnus alaternus 15,0 41,6 52
Pistacia lentiscus 10,1 36,9 —
Myrtus communis 17,6 44,6 52
Lonicera Unplexa 18,4 47,7 52
Cistus albidus . 10,6 43,9 47
Cistus salviaefolius . 10,5 42,3 49,5
Rosmarinus officinalis — — 48
Lavandula stoechas .... — — 47
Smilax aspera . 17,5 45,6 54
Ruscus aculeatus — — 55,5
I
температура листьев отрицательно сказываться на фотосинтезе в полуден-
ное время, как это предполагали Гуттенберг и Бур. Л архер установил, что
компенсационная точка у Quercus ilex обычно равна 10 тыс. лк. У сильно
перегретых листьев освещенность всегда намного выше; однако известно,
что компенсационная точка весьма изменчива. При опытах в климатиче-
ских камерах до настоящего времени выделение СО2 растениями днем
не отмечено.
Совершенно иная картина наблюдается у травянистых растений
(Н eliotr opium europaeum, Convolvulus althaeoides, Psoralea bituminosa) или
у орошаемых культур (Solanum melongea, Cucumis melo, Citrullus vulgaris).
Интенсивность их транспирации очень велика — иногда она превышает
30 (50) мг!дм2*мин. Вследствие охлаждающего действия транспирации
температура листьев по сравнению с температурой нетранспирирующих
листьев ниже на 5,7° (Psoralea), 8,5° (Н eliotr opium), 13,9° (Convolvulus)
и 15,7° (Solanum). Благодаря этому температура листьев не достигает 40°,
что очень важно, поскольку граница устойчивости этих видов равна 44 —
47°. Следовательно, резкое уменьшение интенсивности транспирации
могло бы повлечь за собой гибель растения от перегрева. Правда, не
ясно, возможно ли вообще столь сильное сокращение интенсивности
транспирации у этих видов и не повреждается ли растение еще до момента
перегрева вследствие слишком большой потери воды. У Psoralea осмотиче-
ское давление в июле быстро возрастает с 10 до 35 атм, после чего листья
обычно засыхают. У растений в состоянии сильного завядания граница
устойчивости повышается, так что не известно, идет ли речь о тепловом
повреждении или о повреждении в результате обезвоживания.
Реальность возникновения тепловых повреждений в естественных, то
есть не испытывающих влияния человека, условиях до настоящего време-
ни точно не доказана. Гораздо чаще, особенно в Средиземноморье, повре-
ждения происходят вследствие слишком низкой температуры, правда,
ущерб в основном причиняется культурным и декоративным растениям,
а также посадкам леса. Итак, мы подошли к вопросу о морозоустойчивости.
5*
68
Глава 2
Nerium oleander 1953/54г.
—
Laurus nobilis 1952/53a 1953/542.
4»
Arbutus unedo —
Viburnum tinus — 1 — Зима Апрель-пай । Лето
Olea europaea Ы
—
Pinus pinea = ZZZZZ
Quercus ilex
Chamaerops humilis —
Trachycarpus forimei
Cupressus sempervirens ZZZZ2
Cedrus atlantica мм
Cedrus deodara 1= Piifia, №
нпы
Citrus trif., побеги —
-2 ~4 -6 -8 -10 -12 -14 -16 -18 -20 -22
Морозоустойчивость
Рис. 35. Морозоустойчивость созревших' листьев растений Средиземноморья в раз-
ное время года. Левый конец прямой указывает температуру, при которой проявляют-
ся повреждения, причиненные морозом, правый — температуру, при которой лист
погибает; вертикальным штрихом отмечена величина повреждения, равная 15%.
У некоторых видов, таких, как Myrtus и Cupressus, сначала замерзают жилки листьев
(по Лархеру).
Лархер 1 исследовал морозоустойчивость склерофилов у северной
границы их распространения. Срезанные ветки медленно охлаждались
в холодильной камере, выдерживались в ней при минимальной температу-
ре в течение трех часов, затем им давали медленно оттаять. Результаты
опыта иллюстирует рис. 35. Зимний покой у склерофилов не установлен.
Повреждения листьев, обусловленные морозом, объясняются образовани-
ем льда. Зимой морозоустойчивость растений выше, так как образование
льда происходит лишь при более низкой температуре. Возможно, что уве-
личение морозоустойчивости есть следствие более высокого содержания
сахара. Известно, что с наступлением холодов крахмал в клетках исчезает
и осмотическое давление повышается (см. гл. 6). После холодного периода
фотосинтез сокращается, однако для Olea достаточно двух дней без
мороза, чтобы способность ее к ассимиляции полностью восстановилась.
При прогрессирующем образовании льда растения погибают.
Из рис. 35 явствует, что вечнозеленые склерофилы могут переносить
сравнительно низкие температуры; Ceratonia особенно чувствительна
к ним, Quercus ilex наиболее морозоустойчив. Таким образом, северная
граница естественного распространения жестколистных лесов, видимо,
определяется не столько низкой температурой, сколько конкурентными
1 W. L archer, Kalteresistenz mediterraner Immergruner und ihre BeeinfluBbar-
keit, «Planta», 44, 1954, S. 607—635; Winterfrostschaden in den Parks und Garten von
Arco und Riva am Gardasee, «Veroff. Museum Ferdinand. Innsbruck», 43, 1963. S. 153 —
199.
Средиземноморская жестколистная растительность
69
взаимоотношениями с летнезелеными лиственными деревьями. И тем не
менее слишком холодные зимы в Центральной Европе не позволяют выра-
щивать склерофилы даже в садах, где конкуренция отсутствует. В Ан-
глии с ее мягкими зимами Quercus ilex хорошо растет еще в районе Хар-
вича (52° с.ш.). Cistus albidus, если его укрывать на зиму, выдерживает
условия под Гейдельбергом. Cistus laurifolius более устойчив и встречается
даже в окраинных районах центральной Анатолии; старый куст его, без
укрытия зимой, рос в Хоэнхейме под Штутгартом, пока его не срубили при
строительных работах. В ботаническом саду Гейдельберга без укры-
тия много лет стоял Ruscus aculeatus, но замерз в холодную зиму 1928/29 г.,
причем осмотическое давление поднялось до 25—35 атм. Однако морозо-
устойчивость в той степени, какая существует у хвойных деревьев умерен-
ной зоны (см. гл. 5), у склерофилов не вырабатывается:
Экология культурных растений
Завершим наш обзор экологической характеристикой многолетних
культурных растений Средиземноморья. Рассмотрим виды, которые пере-
носят засушливое время без полива и притом обильно плодоносят. Это
склерофильная маслина (рис. 36) и листопадные инжир, миндаль и вино-
Р и с. 36. Olea еигораеа на
берегу озера Гарда. Пора-
жение растения грибом
Polyporus robustus, разру-
шающим древесину, объяс-
няет своеобразную форму
его стволов. Больную дре-
весину вырезают, из-за чего
образуются глубокие щели,
ствол при этом нередко рас-
щепляется (фото Фабера).
70
Глава 2
Рис. 37. Годовой ход транспирации листьев апельсинов
(в г на 1 г сырого веса) на солнце и в тени (Яффа); кривая
показывает изменение осмотического давления в течение года
(по Оппенгеймеру и Менделю).
град. Для их возделывания в горных местностях на склонах устраивают
террасы, создают наиболее благоприятные условия для роста растений:
формируют по возможности мощный слой почвы, высаживают растения
на большом расстоянии одно от другого, удаляют сорняки.
Глубоко проникающая корневая система позволяет этим деревьям
эффективно использовать почвенную влагу, накопленную в период зим-
них дождей. Оппенгеймер (см. стр. 60) показал, что миндаль и инжир
летом интенсивно транспирируют и все же характеризуются низким осмо-
тическим давлением и малым водным дефицитом.
У хорошо плодоносящей виноградной лозы в августе под Монпелье
после очень длительной засухи осмотическое давление, несмотря на сла-
бое завядание листьев, составляло только 13,5 атм, а у неплодоносящего
инжира с пожелтевшими из-за сухости листьями — 16,7 атм. На неблаго-
приятных почвах, например на террасированных склонах, урожай в срав-
нении с затратами труда непропорционально мал. Поэтому там, где
представляется возможность более легкого заработка, плантации этих
культур забрасывают. На равнине урожай с 1 га оливковых насаждений
колеблется параллельно изменению количества осадков (см. т. I, сноску
на стр. 308). В более сухих районах Туниса, под Сфаксом, на площади
в 1 га высаживают только 16 деревьев.
Теплые районы Средиземноморья представляют собой важнейшую
область разведения цитрусовых: апельсинов, лимонов, мандаринов и пум-
мело. Родина этих культур —вероятно, тропические области Юго-
Восточной Азии с летними дождями х. В теплое время года цитрусовые
потребляют много воды, поэтому в Средиземноморье они плодоносят толь-
ко при искусственном орошении. Прохладный сезон на родине цитрусо-
вых — засушливый период. Поэтому они в известной мере засухоустой-
чивы. Но в климате Юго-Восточной Азии зима для них — одновременно
1 Н. R. Oppenheimer, Adaptation des agrumes a la secheresse, IV. Congres
Intemat. de Г Agrumiculture Mediterr., Tel Aviv; H. R. Oppenheimer, K.Men-
d e 1, Agric. Rs. Station Rehovot, «Bull. 25», 1930.
Средиземноморская жестколистная растительность
71
период покоя. Возможно, этим объясняется, почему цитрусовые в усло-
виях влажной средиземноморской зимы не обнаруживают активности.
Даже в Палестине их осмотическое давление отчетливо возрастает зимой,
как только температура почвы падает ниже 16° (рис. 37).
Умеренную засуху цитрусовые переносят без ущерба для себя; в этот
период растения крайне экономно расходуют накопленную влагу Ч
КОНКУРЕНТНЫЕ ВЗАИМООТНОШЕНИЯ
МЕЖДУ ВЕЧНОЗЕЛЕНЫМИ ЖЕСТКОЛИСТНЫМИ
И ЛЕТНЕЗЕЛЕНЫМИ ЛИСТВЕННЫМИ ДЕРЕВЬЯМИ
Средиземноморская зона жестколистной растительности в северном
направлении переходит в зону летнезеленых лиственных лесов; в этом
же направлении уменьшается летняя засуха и возрастает суровость зим.
Но и в пределах собственно средиземноморской зоны встречаются древес-
ные виды, сбрасывающие листву на зимний период. Особенно широко
распространены Pistacia terebinthus (в Палестине Р. palaestina), как лиана
Clematis flammula, на востоке Средиземноморья — также Styrax officinalis.
С экологических позиций исследованы Pistacia terebinthus, или Р. pa-
laestina. О последней мы уже писали. Кривая осмотического давления для
Р. terebinthus повторяет ход давления для Р. lentiscus с той лишь разницей,
что у Р. terebinthus величина его, как это вообще свойственно летнезеленым
видам, на 3 атм, а у молодых листочков даже на 7 атм ниже; максимум
-составлял в июле 23,5 атм, а в августе —23,7, в другое время —18,0 —
20,9 атм (рис. 31). Рушаль поздним летом получил значение 21,0—24,9 атм.
Цифры свидетельствуют о стабильности водного баланса. У Р. terebinthus
транспирация, обычно весьма интенсивная, в засушливый период заметно
сокращается, но и тогда растение отдает воду энергичнее, нежели это
делают склерофилы. Это подтверждается наблюдениями автора под Мон-
тарно. Данные об относительной общей листовой поверхности отсут-
ствуют.
Вместе с Р. terebinthus на острове Бриони (Истрия) растет Fraxinus
ornus, ясень манный, водный баланс которого менее уравновешен; расте-
ние транспирирует в два раза слабее, но тем не менее обнаруживает рост
осмотического давления с 23,2 до 42,1 атм. Типично субсредиземномор-
ский вид, он, однако, встречается в Далмации даже под Дубровником.
Лишь несколько севернее Монпелье в маквисе появляются некоторые
субсредиземноморские элементы; вечнозеленые Buxus sempervirens и Ilex
aquifolium и листопадные Quercus pubescens, Sorbus domestica и S. tormi-
nalis.
На одном и том же местообитании эти летнезеленые виды транспири-
руют интенсивнее, чем склерофилы. Они обладают свойством лишь в не-
значительной мере сокращать водоотдачу; листья на срезанных ветках, как
уже упоминалось, на солнце очень быстро вянут. Тем не менее водный
1 R. К arsch on, D. Heth, The water balance of a plantation of Eucalyptus
camaldulensts Dehn, Rehovot, 1967.
По Картону п Хету, посадки эвкалипта на береговой равнине Палестины расхо-
довали за вегетационный период весь запас воды в почве. За год здесь выпадает 640 мм
осадков, кроны деревьев перехватывают 96 мм, сток составляет 83 мм. Потенциальная
испаряемость равна 1460 мм!год, в январе — 1,6 мм!сутки, в июле — 6,5 мм!сутки.
Отношение транспирация/потенциальная испаряемость для эвкалиптов зимой состав-
ляло 0,84, летом — только 0,1, то есть летом транспирация резко сокращалась. С 1 га
ллощади получали 11,1 м? древесины. На открытом участке сток достигал 320 мм.
72
Глава 2
баланс у этих растений даже в августе сохраняется на постоянном уровне.
По измерениям автора, величина осмотического давления обычно не пре-
вышает 20—22 атм; на самом сухом местообитании концентрация клеточ-
ного сока составила 27,8 атм. При одном измерении у Sorbus domestica
она равнялась 22,6 атм.
Можно предположить, что при менее благоприятных условиях водный
баланс у растений нарушается и летнезеленые деревья утрачивают конку-
рентную способность. В переходной зоне севернее Монпелье легко заме-
тить, что во всех понижениях рельефа с мощными почвами Quercus pubescens
начинает доминировать над Quercus ilex. Дуб пушистый (Quercus pubescens)
растет здесь быстрее, если только вследствие постоянного стравливания
скотом его рост не сдерживается искусственно. На участках с тонким
почвенным покровом этот вид отсутствует. Не встречаются здесь и виды
Sorbus, которые никогда не образуют плотного древостоя, а обычно раз-
бросаны по местности в виде единичных экземпляров. Quercus ilex на
очень сухих местообитаниях (крутые южные склоны, известняковые греб-
ни) заходит далеко на север, поскольку на таких местообитаниях, несмо-
тря на большее количество атмосферных осадков, преобладает период
длительной засухи. В соответствии с законом относительного постоянства
местообитаний (т. I, стр. 316 и след.) увеличение количества осадков ком-
пенсируется большей величиной поверхностного стока и меньшей влаго-
емкостью почвы. Еще дальше на север (до Рейна у Базеля) проникает
Buxus sempervirens.
Особенно резко проявляется конкуренция между вечнозелеными
и летнезелеными древесными породами во влажных климатических райо-
нах со средиземноморским типом сезонного распределения осадков, на-
пример в северной Анатолии, частично в северной Испании, на наиболее
влажных участках Адриатического побережья, в частности под Котором
и Опатьей, а также в окрестностях озера Гарда х. Здесь даже летом выпа-
дает столько осадков, что могут произрастать летнезеленые лиственные
деревья (разные виды Quercus, Castanea, Carpinus, Tillia, Acer)', с другой
стороны, зимы настолько мягкие, что склерофилы не вымерзают (рис. 38 —
40). Из последних в этих влажных районах широко представлен прежде
всего лавр (Laurus nobilis), не отсутствуют полностью и другие склеро-
филы.
В северной Анатолии к перечисленным видам примешиваются Rhodo-
dendron ponticum и Prunus laurocerasus. Исследования автора в этом районе
позволили сделать вывод, что летнезеленые лиственные деревья в конку-
ренции с вечнозелеными склерофилами одерживают верх. Они растут
в высоту намного быстрее и, не будь вмешательства человека, вскоре обра-
зовали бы верхний ярус леса 1 2.
Высокие стволы листопадных деревьев здесь вырубают, а пневую
поросль охотно поедает пасущийся скот; склерофилы же, и в особенности
1 Бейг (I. В eug, «Вег. Deutsch. Bot. Ges.», 78, 1965), на основе пыльцевых ана-
лизов установил, что Quercus ilex мигрировал в район озера Гарда уже в позднелед-
никовое время, исчез в период кратковременного наступания ледников (8100—7700 лет
до н. э.). а затем снова проник сюда вместе с сосной. Термические условия не позво-
ляли дубу распространяться шире, поэтому он сохранился лишь экстразонально. Леса
состояли преимущественно из Quercus pubescens. Ostrya carpinifolia и Fraxinus ornus
впервые появились в 800—600 гг. до н. э., когда леса превратились в низкоствольные
насаждения. Olea в средние века была известна только в культуре.
2 Н. Walter, Die heutige okologische Problemstellung und der Wettbewerb
zwischen mediterraner Hartlaubvegetation und den sommergrunen Laubwaldern, «Вег.
Deutsch. Bot. Ges.», 69, 1956, S. 263—273.
Средиземноморская жестколистная растительность
73
лавр, скот не трогает. Наблюдение в районе Зонгулдака показало, что
летнезеленые лиственные деревья продуцируют больше органического
вещества, что способствует их быстрому росту в высоту. На первый взгляд
такое положение дел кажется странным, поскольку у вечнозеленых видов
фотосинтез осуществляется более длительное время, кроме того, их
листья не опадают по меньшей мере в течение двух лет. Но не следует
забывать, что для накопления вещества гораздо большее значение имеют
другие факторы.
Рис. 38—40. Климадиаграммы станций, расположенных в зоне, переходной от
Средиземноморья к Субсредиземноморью.
Продуцирование вещества происходит тем интенсивнее, чем
1) большая доля ассимилятов расходуется на построение новых
листьев;
2) чем шире отношение листовая поверхность/сухой вес листьев,
иными словами, чем большую площадь листьев создает древесный вид на
определенную массу сухого вещества;
3) чем выше интенсивность ассимиляции СО2 на единицу листовой
поверхности; ।
4) чем длительнее период фотосинтеза' в течение года.
Точных цифровых данных к пункту 1 для летнезеленых деревьев
и склерофилов не существует; однако у последних на ствол и корни,
вероятно, приходится большая доля всего сухого вещества; это было
доказано для облиствленных трех летних веток.
К пункту 2 следует заметить, что летнезеленые мезоморфные листья
намного тоньше листьев склерофи-
лов; отношение листовой поверх-
ности к сухому веществу у Quercus
pubescens в два раза выше, чем у
Qu. ilex, а у других видов дере-
вьев даже в три раза. Поэтому
ассимиляция СО2 в расчете на 1 г
сухого вещества листа у летне-
зеленых видов намного интенсив-
нее, чем у жестколистных (табл. 7).
К пункту 3 имеются данные
сравнительных измерений, выпол-
Таблица 7
Нетто -про ду ктивность летне зеленого
Qu. robur и вечнозеленого Qu. ibex
(в мг СО2/г сухого веса > час)
Quercus robur Quercus ilex
Средний суточный максимум .... Средняя ассимиля- ция СО2 11,0 5,8 3,0 1,7
74
Глава 2
Quercus pubescens
„ Quercus ilex
11 ---------------—
ST
10 -
9 -
со
8
7
б
5
4
3
2
1
О
1
2
Е
3
5
4
3
? А
2
1
о
2
Е
-16 -12 -8-404 8 12 16 20 24 28 32 36 40 44 48 52 56 60
Температура листьев, °C
PJnc. 41. Зависимость нетто-продуктивности и дыхания побегов от
температуры, начало образования льда в листьях, морозо- и жароустой-
чивость летнезеленых (Quercus pubescens) и вечнозеленых (Qu. ilex) дубов
(по Лархеру).
а) Период опыта: март — ноябрь; 1 — прирост этого года летом; 2 — конец сен-
тября 1958 г. после кратковременной засухи; з — ноябрь 1959 г.; 4 — прирост прош-
лого года. Жирная прерывистая линия: июль — сентябрь; тонкая прерывистая
линия: март.
б) Опыт в течение зимы. Прирост Quercus ilex этого года; 1 — безморозный период,
декабрь 1958 г. и январь 1960 г.; 2 — морозный период, январь 1959 г.; з — опыты
по снижению устойчивости. Пунктирные кривые — состояние растений летом. Усло-
вия опыта: освещенность — 10 к.ак; содержание СО2—0,033—0,035 объемных про-
центов; влагообеспеченность оптимальная: у Qu. pubescens дефицит насыщения
4—7%, у Qu. ilex дефицит насыщения 4—9%.
пенных Лархером \ Опыты производились со срезанными и поставленными
в воду ветками в лаборатории при освещенности 10 тыс. лк и разной темпе-
ратуре (рис. 41). Из рисунка явствует, что ассимиляция в расчете на сухой
1 W. L archer, Jahresgang des Assimilations- und Respirationsvermogens von
Olea europaea L., ssp. sativa Hoff., et Link., Quercus ilex L. and Quercus pubescens Willd.
-aus dem nordlichen Gardaseegebiet, «Planta», 56, 1961, S. 575—606; Zur Assimilations-
bkologie der immergrunen Olea europaea und Quercus ilex und der sommergriinen Quercus
pubescens im nordlichen Gardaseegebiet, «Planta», 56, 1961, S. 607—617.
Средиземноморская жестколистная растительность
75
Рис. 42. Изменение продуктивности вечнозеленых и летне-
зеленых дубов в течение года (по Лархеру). Общая продук-
тивность у Quercus pubescens при постоянной хорошей влаго-
обеспеченности всегда выше.
вес у Quercus pubescens приблизительно в 2,5 раза больше, чем ассимиля-
ция листьев Qu. ilex летом; однако при расчете на листовую поверхность
различия сглаживаются. ।
Что касается пункта 4, то в этом отношении склерофилы имеют пре-
имущество. Способность листьев Quercus ilex к ассимиляции зимой, прав-
да, несколько ниже, чем летом, особенно после морозных периодов, но это
все же имеет свое значение (рис. 42).
Рис. 43. Liquidambar orientalis в пойменном лесу; реликтовое местообитание в юго-
западной Анатолии (фото Э. Вальтер).
76
Глава 2
Рис. 44. Цветущий Nerium oleander на галечниках речного русла; Испания,
северная Андалузия (фото Г. Вальтера).
На основании приведенных данных и с учетом действительных темпе-
ратур у озера Гарда Лархер вычислил величину накопления вещества
(в г на 1 г веса веток) однолетними побегами Qu. ilex, Olea еигораеа и, для
сравнения, побегами Quercus pubescens. Для вечнозеленых видов он полу-
чил значения 17,9 и 14,1, а для летнезеленого дуба в условиях постоянного
влагообеспечения — 22,9. Таким образом, в районах Средиземноморья,
достаточно влажных и в летнее время, вечнозеленые виды, несмотря на
более длительный период возможного фотосинтеза, накапливают меньшую
массу вещества. Следует заметить, что при этом не учтена большая доля
древесины, приходящаяся в общем сухом весе склерофилов на стволы
и корни.
Эти расчеты подтверждают наблюдение автора, что при хорошей
влагообеспеченности в летние месяцы летнезеленые лиственные деревья
успешно конкурируют со склерофилами.
В типично средиземноморской климатической области с длительной
летней засухой летнезеленые виды также вытесняют вечнозеленые виды
на всех достаточно влажных местообитаниях, например в поймах рек.
Мы встречаем здесь лиственные деревья, такие, как виды Populus, Ulmus
campestris или виды Alnus, прежде всего Platanus orientalis, а на юго-запа-
де Анатолии третичный реликтовый вид Liquidambar orientalis (рис. 43).
Но как только в небольших долинах грунтовые воды временно иссякают,
летнезеленые растения сменяет вечнозеленый вид Nerium oleander, рас-
пространение которого, правда, ограничено южной частью Средиземно-
морской области, где сильных зимних заморозков уже не бывает (рис. 44).
В горах количество осадков летом возрастает с высотой; происходит
и существенное изменение характера растительного покрова. И здесь
Средиземноморская жестколистная растительность
77
вечнозеленая жестколистная растительность быстро вытесняется летне-
зелеными лиственными деревьями. Мы подходим к вопросу о вертикаль-
ной поясности Средиземноморья, отличающейся большим разнообразием.
ВЕРТИКАЛЕН АН < ПОЯСНОСТЬ
В СРЕДИЗЕМНОМОРСКОЙ ЗОНЕ
В горах с увеличением высоты средняя годовая температура пони-
жается и одновременно сокращается вегетационный период. Аналогичную
картину мы наблюдаем на низменности, передвигаясь с юга на север.
Если климат в обоих случаях гумидный, то есть вода не является ограни-
чивающим фактором, то и характер изменения растительности в том
и другом случае будет сходным. Это правило обычно формулируют сле-
дующим образом: последовательность вертикальных
поясов в горах представляет собой сокращен-
ное повторение зон растительности, сменяю-
щих одна другую в направлении с юга на север.
Однако это правило достоверно далеко не всегда. Для Западной Евро-
пы оно в общем справедливо, но и здесь в частностях встречаются несо-
ответствия, например у пояса пихты (Abies) в горах нет зонального анало-
га; пояс Alnus viridis — Pinus montana частично заменяется зоной Betula —
Pinus Silvestris. В климатическом отношении вертикальные пояса и зоны
также не в полной мере созвучны друг с другом. Продолжительность дня
на севере летом значительно больше, стало быть, и температурные раз-
личия дня и ночи меньше, чем в горах. Зимой, напротив, в горах дни
длиннее, а инсоляция бывает столь интенсивной, что растительный покров,
если он не находится под снегом, заметно нагревается.
Аналогия исчезает полностью, как только появляется разница в сте-
пени обеспеченности водой. Вследствие этого именно в Средиземноморье,
образующем переходную область от аридной субтропической зоны к гу-
мидной умеренной, имеется особенно большое разнообразие в последова-
тельности вертикальных поясов, тем более что в последних известную
роль часто играют реликтовые виды, которые севернее отсутствуют (Cedrus,
Cupressus, различные виды Pinus, Abies и Juniperus). Мы не можем оста-
навливаться здесь на всех особенностях в|ертикальной поясности отдель-
ных горных систем, ограничимся описанием только основных типов Ч
Если горный гребень представляет собой климатическую границу, то
последовательность вертикальных поясов этого хребта на южном и север-
ном или западном и восточном склонах будет различной.
Автор выделяет два главных типа средиземноморской вертикальной
поясности.
1. Гумидная вертикальная поясность. Характе-
ризуется резким увеличением количества осадков с высотой и одновремен-
но сокращением периода летней засухи, которая в поясе облаков и тумана
вообще не наступает (рис. 45). Это дает возможность центральноевропей-
ским, а на большей высоте также бореальным и альпийским элементам
1 Общий обзор с двумя меридиональными профилями, относящимися к западно-
средиземноморской (южная Франция — Марокко) и восточносредиземноморской
(Крым — Палестина) областям, читатель найдет в работе Кюммеля: К. К u m m е 1,
Die Stellung Sudfrankreichs und der Krim im west- und ostmediterranen Vegetationsprofil,
Bonn, 1949. Согласно последним данным, некоторые варианты вертикальной поясности
’требуют уточнения.
78
Глава 2
занять доминирующее положение. Только нижний пояс, как это следует
из приводимой ниже характеристики, является собственно средиземно-
морским:
д) альпийский пояс со многими альпийско-арктическими элементами;
г) бореальный пояс ели (Picea abies —Р. excelsa);
в) центральноевропейский пояс бука (Fagus silvatica с Abies alba);
б) субсредиземноморский пояс: дуб пушистый — каштан (Quercus
pubescens — Castanea);
а) средиземноморский пояс жестколистной растительности с Quercus
ilex или Qu. suber.
Такая последовательность поясов известна в приморских Альпах
у Ривьеры. На северном склоне Пиренеев и на Корсике отсутствует Picea;
Рис. 45—47. Климадиаграммы средиземноморских горных станций.
Пескокостанцо в Апеннинах — гумидный климат, буковый пояс; Ифран в Среднем Атласе — сухой
климат, узкий пояс летнезеленого лиственного леса; Цедрес в Ливане — аридный климат летом1
(некогда область распространения кедровых лесов).
в Пиренеях ее замещают Pinus silvestris и Р. uncinata, а на Корсике —
Р. pinaster и Р. nigra ssp. poiretiana. В более засушливых горах Сьерра-
Гвадаррама (центральная Испания) из древостоя выпадают Fagus (за
исключением одного местонахождения на востоке) и Abies, и над летне-
зелеными дубами (Quercus pyrenaica = Qu. toza) начинается пояс Pinus
silvestris. В Апеннинах бореальный пояс отсутствует и границу древесной
растительности образует Fagus х, местами он встречается даже в Сицилии.
На Олимпе в Греции еще растут бук и ель. На адриатическом побережье
до средней Албании еще хорошо развит пояс с буково-пихтовыми лесами
в области туманов, под ним расположен пояс Fraxinus ornus — Carpinus
orientalis. Но на юге Албании летом пояс облаков исчезает, вследствие
чего изменяется и последовательность вертикальных поясов 1 2. Схема
распределения растительности в Иберийских горах изображена на рис. 48.
2. Аридная вертикальная поясность (собственно
средиземноморская поясность).
Лето здесь вплоть до альпийского пояса сухое (ср. рис. 46, 47). Пояс
облаков в летние месяцы отсутствует. Поэтому все пояса лиственных листо-
падных лесов выпадают, и за нижним поясом жестколистной растительно-
сти непосредственно следуют вертикальные пояса с хвойными деревьями,
1 Н. Е г n, Die dreidimensionale Anordnung der Gebirgsvegetation auf der Iberi-
schen Halbinsel, «Bonn. Georg. Abb.», H. 37, 1966; W. L u d i, Beitrag zur regionalen
Vegetationsgliederung der Apenninenhalbinsel (mit Karte), «Veroff. Geobot. Inst. Rii-
bel,» Zurich, H. 12, 1935, S. 212-239.
2F. M arkgr af, Pflanzengeographie von Albanien, «Bibl. Bot.», 105, 1932;
Eine neue Hohenstufenkarte der Vegetation Albanians, «Вег. Geobot. Institut Rubel
(Zurich) f. 1948», 1949, S. 109-118.
Рис. 48. Вертикальная поясность растительности в горах Пиренейского полуострова, сложенных кристаллическими горными поро-
дами, по профилю с СЗ на ЮВ (по Эрну).
1 — листопадный дубовый лес (Qu. robur, Qu. petraea)', 2 — лес c Qu. ругепагса', 3 — лес с каменным дубом (Qu. ilex)', 4 — буковый лес (Fagus silvatica)',
Г) — березовый лес (Betula verrucosa)', 6 — сосновый лес (Pinus silvestris)', 1 — смешанный лиственный лес с Quercus, Tilia, Acer', 8 — горный лес Сьерра-
Невады (Sorbus, Prunus и др.); 9 — высокогорные луга (травы и злаки); 10 — вересково-монокевельниковая кустарничковая пустошь с Vactinium t Calluna,
Juniperus; 11 — дроко-можжсвсльниковая кустарничковая пустошь с Cytisus, Genista, Erica', 12 — средиземноморский колючеподушечный кустарничковый
цояс; 13 — сухие луга с Festuca (см. рис. 57).
SO
Глава 2
прежде всего с видами Pinus, но также и с видами A bies. Если горные хреб-
ты^служат климатической границей, то на наветренной стороне форми-
руется гумидная поясность, а в ветровой тени — аридная. Такая картина
наблюдается, например, на Корсике \
Аридную вертикальную поясность в типичном развитии можно видеть
на востоке Средиземноморской области, в частности в южной Анатолии 1 2.
На южном склоне гор Тавр, ведущем к Средиземному морю и отлича-
ющемся сравнительно обильными зимними осадками, четко выделяется
следующая последовательность вертикальных поясов:
е) альпийский пояс, в нижней части с зарослями колючеподушечников;
д) у границы древесной растительности Juniperus excelsa и J. joetidissima',
г) субальпийский пояс с Cedrus libanotica3 и Abies cilicica (на влажных
участках) или видами Juniperus (на более сухих местообитаниях);
в) горный пояс с Pinus nigra ssp. pallasiana (слабо развит);
б) верхний средиземноморский пояс с Pinus brutia,
а) собственно средиземноморский пояс жестколистной растительности
(ср. стр. 47) с Ceratonia, развитый на теплых приморских низмен-
ностях.
В изобилующем осадками юго-восточном секторе Анатолии, в горах
Аманус, развит еще пояс облаков с Fagus orientalis, в котором летняя за-
суха не проявляется.
На северном склоне Тавра зимой дождей мало, поэтому нижние сре-
диземноморские пояса отсутствуют, например у киликийского Аладага 4.
Это высокое нагорье поднимается над степью, достигающей 1500 м абсо-
лютной высоты, до отметок 4000 м. В деградированной степной раститель-
ности доминируют Artemisia caucasica, A. fragrans, Achillea santolina и др.
Следующий пояс был представлен сухим лесом, ныне полностью выруб-
ленным. Изредка встречаются отдельные экземпляры Juniperus excelsa,
J. drupacea, J. foetidissima рядом с дубами (Quercus libani) и осиной. Еще
выше располагается пояс пихты с Abies cilicica, нижняя граница которого
местами опускается до 1600—1700 м, а верхняя (на отметках 2400 м)
намечена отдельными группами деревьев (но не отдельными экземпляра-
ми). Но и эти позиции большей частью заняты зарослями колючеподушеч-
ников, хотя и относящихся к субальпийскому поясу, но часто занимаю-
щих место лесов на значительно более низких участках горного склона
(рис. 49). Это характерно также и для других горных систем, например
для Атласа.
Над ярусом колючеподушечников, выше отметки 2700 м, начинается
собственно альпийский высокогорный пояс, образуемый Seslerietum апа-
tolicae с большим числом цветущих растений (виды родов Draba, Aethione-
та, Lamium, Androsace, Taraxacum, Crepis, Ranunculus, Veronica, Coryda-
lis и сравнительно редко крупный Cirsium ellenbergii). Верхняя граница
пояса колючеподушечников ограничивается низкой температурой. До-
1 Н. Ellenberg, «Е igenbiirt ige» und «fremdbiirtige» Vegetationsstufung auf
Korsika. Cm.: Beitr. z. Phytol. (Waiter-Festschrift), «Arb. Landw. Hochsch. Hohenheim»,
30, Stuttgart, 1964, S. 145—154.
2H. Walter, Die Vegetationsgliederung Anatoliens, «Flora», 143, 1956,
S. 295—326.
3 H. M а у e r, M. S e v i m, Die Libanonzeder, «Jb. Ver. Schutz Alpenfl.», Miin-
chen, 1958, S. 1—20.
4 H. M. Schiechtl, R.Stern, Studien uber Entwaldung im Kilikischen Ala
Dag, «Вег. Nat .-Med. Ver. Innsbruck», 53 (Festschrift Gams), 1963, S. 173—192;
H.M. Schiechtl, R. Stern, E. H. W eiss, In anatolischen Gebirgen, Klagen-
furt, 1965.
Средиземноморская жестколистная растительность
81
минирующими являются здесь Onobrychis cornuta и Astragalus angusttfo-
lius, виды же колючеподушечников встречаются реже, причем на более
высоких уровнях, где ветры сдувают снег и растения должны быть морозо-
устойчивее.
Вертикальная поясность в горах Троодос на Кипре 1 аналогична по-
ясности на южном склоне Тавра, но выше зоны с Pinus brutia здесь вкли-
нивается пояс полувечнозеленых Quercus alnifolia и Qu. infectoria, в суб-
альпийском поясе появляется Cedruslibanotica ssp. brevifolia. На северном
склоне острова над узким поясом жестколистной растительности развит
пояс с Cupressus sempervirens, спускающийся до отметки 1000 м. Этот вид,
15 20 25 30 кп
Рис. 49. Вертикальная поясность растительности в киликийском Аладаге,
северная часть Тавра (по Шихтлю и Штерну).
Между прерывистыми линиями мог бы существовать лес, но вследствие вырубки и вы-
паса он в нижней части склонов замещен степью с Artemisia, а в верхней—зарослями
колючеподушечников.
на естественных местообитаниях отличающийся горизонтальным располо-
жением ветвей, встречается также в некоторых районах южной Анато-
лии, на Крите (рис. 50) и в Киренаике 2.
Пояс с Cedrus libanotica выражен такз|ке в горах Ливана 3, где он,
включая деградированную растительность, занимает площадь около
3 тыс. га на высоте 1400—1800 м, преимущественно на северных и запад-
ных склонах. На более влажных местообитаниях появляется также Abies
cilicica. Сумма осадков составляет 800—1500 мм (ср. рис. 47, Цедрес).
Дожди идут зимой, но и летом после полудня горы часто окутаны обла-
ками. Из сопутствующих видов следует назвать Berberis cretica. Cotoneas-
ter nummularia, Juniperux oxycedrus, Quercus calliprinos, Qu. infectoria,
Rosa glutinosa и травы Rubia aucheri. Geranium libanoticum. Все это виды
открытых местообитаний и сорняки пастбищ.
Весьма сложна вертикальная поясность в Атласских горах, отдель-
ные районы которых характеризуются резкими различиями климатических
1 Сообщение М. Пура.
2 М. Zohar у, G. Orsha n, An outline of the geobotany of Crete, «Israel
J. Bot.», 14 (Supplement), 1966; H.N. Le Nouerou, Les resources pastorales et four-
rageres en Libye, FAO-Bericht, 1964.
3 E. W. Beals, The remnant cedar forests of Lebanon, «J. Ecol.», 53, 1965,
p. 679—694.
6-01248
82
Глава 2
Рис. 50. Cupressus sempervirens на юге Крита. Слева — культивируемый
пирамидальный кипарис (фото Марьяни).
%
условий. Типичный средиземноморский пояс жестколистной растительно-
сти, образованный Quercus ilex, представляет собой уже настоящий гор-
ный пояс, располагающийся над поясом с Tetraclinis articulata (= Calltt-
Средиземноморская жестколистная растительность 83
ris quadrivalvis) 1 —Juniperus phoenicea. На влажных северных склонах
Среднего Атласа за поясом с Quercus ilex следует пояс летнезеленого
Quercus lusitanica ssp. faginea с примесью Acer monspessulanum, по стволам
которого от почвы до самой кроны взбирается плющ Hedera helix. Выше
растут леса из Cedrus atlantica, в составе которых часто присутствует
Ilex aquifolium (рис. 51, 52) 1 2. Далее следует пояс с Juniperus thurifera,
образующий границу лесной растительности. В значительно более засуш-
ливом Высоком Атласе этот последний непосредственно примыкает
к средиземноморскому поясу с Quercus ilex. Лесная растительность всюду
сильно деградирована. Схематические профили на рис. 533 дают пред-
ставление о расположении и последовательности вертикальных поясов.
Аналогичную картину мы видим в Алжире и Тунисе, с той лишь раз-
ницей, что к востоку климат становится суше. Но и здесь всегда более
увлажнены северные склоны гор 4. На побережье еще развита истинно
средиземноморская растительность типа Oleo-Ceratonion с Pistacia lentis-
cus. Местами растет высокоствольный Quercus coccifera. В районах с наибо-
лее обильными осадками Quercus suber занимает лишь незначительную
площадь. На тощих почвах распространены леса, образованные Pinus
halepensis\ однако на большинстве местообитаний древостои из Р. hale^
pensis деградированы и представляют собой гаригу с видами Rosmarinus
и Erica. Несколько выше в горах в качестве подлеска в сосновых лесах
появляется Quercus ilex, который с подъемом в горы начинает все больше
доминировать.
В более теплых климатических условиях развивается зона с ви-
дами Tetraclinis articulata (« туя») и Juniperus phoenicea. И для этой
зоны характерно чередование разных стадий деградации гариги.
Р и с. 51. Районы распро-
странения Cedrus atlantica
в Марокко (заштрихованы).
Ареал простирается до вос-
точной границы Алжира (по
де Вильду, с изменениями).
1 В Европе этот вид встречается только на сухом юго-востоке Испании под Кар-
тахеной.
2 J.J.F.E. de Wilde, Cedrus atlantica Manetti in Marokko, «Medel. Bot. Tuin.
Belmonte Arbor. (Wageningen)», 5, 1961, S. 93—103.
3 L. Emberger, Aper^ue general zur la Vegetation du Maroc (mit Vegetations-
karte 1:1 5000 000). Cm.: «Veroff. Geobot. Inst. Riibel», Zurich, 1939, S. 40—157;
W. R a u h, Vegetationsstudien im Hohen Atlas und dessen Vorland, «Sitzber. Heidelb.
Akad. Wiss.», Jg. 1952. 1.
4 H.Gaussen, Verne t, «Bull. Serv. Carte Phytogeogr.», Serie A, 3, Paris, 1958.
6*
84
Глава 2
Рис. 52. Пояс с Cedrus atlantlca в алжирском Атласе (на высоте 1550 м над уров-
нем моря).
Там, где количество осадков резко уменьшается, древесные по-
роды в конце концов исчезают и их место занимает средиземно-
морская зона полупустынь, образованная Stipa tenacissima, Lygeum
spartum и Artemisia herba-alba. С увеличением сухости климата зона
полупустынь переходит в пустыню.
Лё Уэру в работе, посвященной Тунису, весьма подробно описал
растительные сообщества этой зоны, попутно рассмотрев также их цен-
ность как пастбищ Ч Первичная растительность в результате чрезмерного
выпаса и распашки повсюду сильно изменена. Предложены меры для
улучшения этих мест.
Чрезвычайно засушливы горные склоны грабена Иордана; только на
западных склонах осадков выпадает достаточно, чтобы на них можно было
различить средиземноморские пояса растительности; восточные склоны
пустынны (ср. рис. 54, 55) 1 2.
Летняя засуха отражается на характере аридной средиземноморской
поясности вплоть до самых высоких вершин гор. Ее воздействие обуслов-
1H.N.Le Houerou, Recherches ecologiques et floristiques sur la vegetation
de la Tunisie meridionale, 1 et 11 Univ. d’Alger, Inst. Rech. Sahar., Memoire № 6, 1959;
Les paturages naturels de la Tunisie aride et desertique, Unit, de Devel., Tunis, 1962.
2 B.Kasapligil, A report on the vegetation profiles of the forest and grazing
lands in Jordan, Econ. Planing Division, Amman, 1965; Plants of Jordan with notes on
their ecology and economic uses, Forestry Department, Amman, 1956.
Средиземноморская жестколистная растительность
85
ливает и иной флористический состав средиземноморского альпийского
пояса по сравнению с Альпами с их многими аркто-альпийскими элемен-
тами, требующими для своего развития постоянно влажного климата.
В условиях сильной летней засухи такие в большинстве случаев эндемич-
ные элементы встречаются в горах Средиземноморья только там, где поч-
ва из-за таяния снега летом остается постоянно влажной (рис. 56). Обычно
же доминируют растения сухих травянистых группировок, и в особенности
колючие подушечники, весьма типичные для альпийского пояса гор Сьер-
ра-Невада на юге Испании, гор Анатолии и Высокого Атласа (рис. 57).
Колючеподушечниковые растения относятся к самым разным семействам
и в вегетативном состоянии различаются с трудом; следовательно, это
прекрасный пример конвергенции, которой следует придавать определен-
ное экологическое значение. Для гор Атласа характерны: Alyssum (Ptilotri-
chum) spinosum (крестоцветные), Arenaria pungens (гвоздичные), Erinacea
pungens, Cytisus balansae (бобовые), Bupleurum spinosum (зонтичные),
Рис. 53. Вертикальная поясность Высокого Атласа (по Рау).
1 — Zizyphus lotus’, 2 — Acacia gummifera', 3 — Chamaerops humilis’, 4 — Tetraclinis (CallitrisY, 5 —
Juniperus phoenicea', 6 — Quercus ilex’, 7 — Juniperus thurifera; 8 — пояс колючеподушечников;
9 — Argania spinosa’, 10 — суккулентные молочайные; 11 — Buxus sempervirens', 12 — Launea
(Zollikoferia) spinosa’, 13 — Phoenix dactylifera; 14 — Stipa tenacissima’, 15 — Artemisia herba-alba\
16 — Cedrus atlantica’, 17 — Populus.
Рис. 54. Карта
грабена Иордана и
Мертвого моря с
годовыми изогие-
тами (в мм).
Цифрами 1—6 обозна-
чены профили, изо-
браженные на рис. 55.
Рис. 55. Верти-
кальная поясность
растительности на
склонах грабена
Иордана и в горах
южнее Мертвого
моря (по Казипли-
гилю, с изменения-
ми).
1 — маквис; 2 —
Quercus и ' Pistacia;
3 — Ceratonia; 4 —
плодовые культуры;
5 — Pinus halepensis;
6 — Cupressus; 7 —
Juniperus; 8 — степь;
9 — Zizyphus; 10 —
пустыня Ретама;
11 — Acacia; 12 —
пустыня с Haloxylon;
13 — пустынные гале-
рейные леса; 14 —
каменистая пустыня;
15 — понижения с на-
носными почвами.
Р и с. 56. Некоторые гигрофильные виды альпийского пояса гор Сьерра-
Невада.
1 — Linaria nevadensis; 2 — L. glacialis; 3 — Viola nevadensis; 4 — Artemisia nevadensis;
5 — Dianthut brachyanthus; 6 — Galium pyrenaicum; 7 — Ranunculus acetosellaefolius",
8 — Plantago nivalis (нат. величина).
88
Глава 2
Рис. 57. Верхний альпийский пояс гор Сьерра-Невада с Festuca indigesta. Лестнице-
образное расположение полос растительности — следствие процесса солифлюкции
(фото Г. Вальтера).
Bufonia murbeckii (гвоздичные). В Передней Азии формацию растений-
подушек образуют преимущественно виды Astragalus и Acantholimon
(свинчатковые) (рис. 58) х.
Эколого-физиологические исследования этих любопытных форм расте-
ний до настоящего времени не проводились. Кезель 1 2 предполагает, что
при интенсивной инсоляции в горах Средиземноморья, где летом небо
безоблачное и стоит сильная засуха, дефицит насыщения воздуха днем
должен быть очень большим. Плотная структура подушек, видимо (изме-
рения не производились), способствует значительному повышению влаж-
ности воздуха внутри них. Благодаря этому транспирация листьев и
сильно одревесневших побегов могла бы понизиться. Побеги, вырастаю-
щие за пределы подушки, засыхают.
В плейстоцене горы Средиземноморской области были покрыты лишь
небольшими ледниками, поэтому на изолированных участках здесь сохра-
нились многочисленные третичные реликты, например пятиигольчатая
сосна (Pinus рейсе) в Албании, Picea omorica в Иллирии, Abies pinsapo
в южной Испании, А. тагоссапа в Рифе, А. numidica в Алжире и др. 3.
1 Н. Gams, Die Tragacantha-Igelheiden der Gebirge um das Kaspische, Schwarze
und Mittellandische Meer, «Veroff. Geobot. Inst. Rubel, Zurich», 31, 1954, S. 217—243.
2 P. Quezel, Peuplement vegetal des hautes montagnes de I’Afrique du Nord,
Encyclop. Biogeogr. et Ecolog., X, Paris, 1957.
3 H. Mayer, Abies pinsapo Boiss. in der Betischen Kordillere Siidspaniens,
«Forstl. СЫ.», 81, 1962, S. 48—62; M. M. J a n k о v i 6, Betrachtungen fiber die gegensei-
tigen Beziehungen der MoIika-(Pinus рейсе) und Panzerkiefer (Pinus heldreichii) usw.,
«Bull. Instit. Botan. Univ. Beograd», 1 (V), 1960, S. 141—180.
Средиземноморская жестколистная растительность
89
Рис. 58. Подушки Astragalus и Acantholimon на высоте 2000 м над уровнем моря;
Эрджиясдаг, Анатолия (фото Цедербауэра).
Особенно интересны в этом отношении Канарские острова. Они подоб-
ны музею, экспонаты которого позволяют воссоздать картину эволюции
растительного мира соседнего Африканского континента начиная с мело-
вого периода. «Экспозицию» музея образуют не только остатки флоры,
ныне известной в Южной Африке, но и тропические элементы, реликты
лесной флоры субтропиков северного полушария, а также элементы
средиземноморской зоны (Quercus ilex) или дроково-эрикоидных пустошей1.
Нашлось здесь место и представителям флоры Сахары и Альп.
Все эти элементы в разное время мигрировали из Африки на острова
и сохранились здесь на подходящих местообитаниях, тогда как на мате-
рике, в районах, соседних с островами, они полностью были вытеснены
последующими волнами растительности. Лишь в результате заселения
Канарских островов человеком эта сложная мозаика растительности была
сильно нарушена. Большие площади земли были превращены в сельско-
хозяйственные угодья или пустыри, причем средиземноморский элемент
стал доминирующим. Из 1389 видов 505 — эндемики; 33% родственны
средиземноморским видам.
Отсюда понятно, почему растительный мир Канарских островов издав-
на интересовал ботаников 1 2. Однако мы вынуждены ограничиться в нашем
1 Н.Meuse 1, Die Reliktvegetation der Kanarischen Inseln in ihren Beziehungen
zur siid- und mitteleuropaischen Flora. Cm.: Gers ch, Gesammelte Vortr. mod.
Probl. Abstammungsl. I., Jena, 1965.
2 H. C h r i s t, Vegetation und Flora der Canarischen Inseln, «Bot. Jb.», 6, 1885r
S. 458; H. Schenck, Beitrage zur Kenntnis der Vegetation der Canarischen Inseln,
«Wiss. Erg. deutsch. Tiefsee-Exp. «Valdivia», 2, 1, 1907, S. 225; M. H i r m e r, Beitrage
zur Kenntnis der Geholz-Formationen auf Teneriffa, «Veg. Bilder», 16, H. 8, 1924/26;
so
Глава 2
обзоре лишь кратким описанием вертикальных поясов растительности,
прослеживаемых на вулканическом острове Тенерифе от уровня моря до
вершины пика Тейде (3707 м). Из климадиаграмм, составленных для раз-
ных высот (рис. 59—61), видно, что климат здесь средиземноморско-арид-
ный, черты которого особенно ярко проступают на нижних и верхних
частях склонов; на последних безморозным является только период с мая
по сентябрь. Особенность острова, расположенного в зоне северных пасса-
тов, состоит в том, что поднимающиеся по северному склону воздушные
Рис. 59—61. Климадиаграммы острова Тенерифе.
'Санта-Крус — на уровне моря, очень сухой климат; Ла-Лагуна — нижняя граница пояса облаков;
Исанья —• верхняя граница лесной растительности.
массы вызывают образование гряды облаков, обычно развитой на высоте
500—1700 м. Поэтому растительность на этих высотах постоянно испыты-
вает увлажняющее влияние тумана, который оседает на деревья (неизме-
ряемые осадки) и обусловливает известную влажность почвы даже в тече-
ние летних месяцев, когда осадков выпадает мало (ср. т. I, стр. 412 и след.).
В этих условиях на северном склоне острова развит влажный лавровый
лес, в котором присутствуют многие элементы субтропического дождевого
леса (рис. 63). Естественный растительный покров острова в наше время
сохранился лишь на отдельных участках. Все же в общих чертах еще
можно составить карту растительности и выделить последовательность
вертикальных поясов (рис. 62, 63).
а) Зона пустыни в низменных районах южной оконечности
острова, лежащей в ветровой тени. Местами здесь преобладает Launea (Zol-
likoferid) spinosa; встречается также Zygophyllum fontanesii, а на восточных
островах при аналогичных условиях также Suaeda vermiculata. Все это
сахаро-аравийские элементы.
б) Полупустыня с суккулентами на нижних частях
склонов распространена значительно шире, на влажных же северных скло-
нах она простирается всего до высоты 400 м, а на сухих южных — до
800 м. На молодой сухой почве, сформировавшейся на лаве, доминирует
бело-серая столбчатая кактусообразная Euphorbia canariensis, на почвах
с большим количеством мелкозема — Ей. regis-jubae с Kletnia (Senecio)
neriifolia. Каменистые участки на верхних и потому уже более влажных
уровнях занимает богатая видами суккулентная растительность с пред-
c. Burchard, Beitrage zur Okologie und Biologie der Kanarenpflanzen, «Bibl. Bot.»,
98, 1929; K.M agdefrau, Die Moosvegetation der Lorbeerwalder auf Teneriffa, «Flo-
ra», 137, 1944, S. 125—138; E. S c h m i d, Beitrage zur Flora und Vegetation der Kana-
rischen Inseln, «Вег. Geobot. Inst. Rubel f. 1953», 1954, S. 28—53; A. D a v у de
V i r v i 1 1 e, Contribution a Г etude de I’endemisme vegetal dans I’Archipel des Cana-
ries, «Rev. Gen. Bot.,» 68, 1961, p. 201—213; E.Oberdorfer, Pflanzensoziologische
Studien auf Teneriffa und Gomera, «Beitrage naturk. Forsch. SW-Deutschl.», 24, 1965,
S. 47—104.
Рис. 62. Карта растительности острова Тенерифе (по Себальо
и Ортуньо, с изменениями Обердорфера).
1 — полупустыня с Zygophyllum и Launea (Zollihoferia)’, 2 — сообщества
суккулентов Kleinia, Euphorbia и замещающие сообщества; 3 — лавровые
леса с Erica и замещающими сообществами; 4 — леса с Pinus canariensis,
а также кустарники из Cistus и Cytisus, с замещающими сообществами;
5 — умеренная горная полупустыня с Spartocytisus', 6 — каменистые осыпи,
поросшие Viola cheiranthifolia; 7 — холодная горная пустыня с криптогам-
ными растениями. А— В — линия профиля, изображенного на рис. 63.
Тейде
близ И кода Эль-МеЭано
Рис. 63. Профиль острова Тенерифе с ССЗ на ЮЮВ (см. рис. 62)
с вертикальными поясами растительности (по Себальо и Ортуныо, из
Обердорфера).
Z — пустыня с Zygophyllum и Launea (Zollihoferia).
92
Глава 2
ставителями рода Aeonium (Sempervivum), видами Dendrosonchus и др.
Кроме того, известен ряд интересных эндемичных видов (Boraginaceae,
Labiatae, Rubiaceae, Compositae). Пришлыми видами являются Opuntia
ficus-indica, агавы и, вероятно, Cryophytum (Mesembryanthemum) crlstal-
linum. К числу туземных в этом поясе принадлежат драконовое дерево
(Dracaena draco), а также Канарский финик (Phoenix canariensis), плоды
которого несъедобны; его часто разводят в Средиземноморской области
как декоративное растение. Естественным местообитанием финика, по-ви-
димому, служат долины с водотоками.
Полупустыне с суккулентами также свойственны африканские черты,
она напоминает засушливые районы атлантического побережья Марокко*
и местами даже область Карру.
На Канарских островах, как и повсюду, эта аридная форма расти-
тельности широко распространилась за счет лесов, сведенных человеком.
Вся стекающая вода здесь тщательно собирается и используется для
полива обширных плантаций бананов. Их разбивают на низменных участ-
ках, не подверженных заморозкам. Климат позволяет выращивать также
ананасы, сахарный тростник и хлопчатник. Заложены прекрасные сады.
На неорошаемые же площади теперь поступает меньше воды, поэтому
они стали суше.
в) Пояс лавровых лесов занимает на северной стороне
острова умеренно теплую, изобилующую туманами зону на высоте более
400 ж, простирающуюся местами до отметки 1200 м над уровнем моря.
Туман, перехватываемый кронами деревьев, постоянно и в достаточной
мере увлажняет почву. Крупные массивы первичных лавровых лесов до
наших дней практически не сохранились. Судя по уцелевшим остаткам,
в их составе, помимо лавра (Laurus canariensis), очень близкого к среди-
земноморскому виду Laurus nobilis, принимал участие ряд древесных
пород, явно связанных с тропическими или умеренно субтропическими
вечнозелеными дождевыми лесами главным образом Южной Африки;
здесь растут лавровые Ocotea foetens, Apollonius canariensis и Persea indica,
чайные Visnea mocanera или Ilex platyphylla и Z. canariensis. Влияние
южной атлантико—средиземноморской области выражается в распро-
странении видов Prunus lusitanica и Smilax. В целом лавровые леса
Канарских островов, по-видимому, близки к третичным лесам Европы
и представляют собой своего рода реликт.
Там, где лес сведен и где туман уже не оказывает столь сильного
увлажняющего влияния, все резче проступают черты средиземноморского
климата с периодом летней засухи (ср. рис. 60). В этом поясе развивается
пустошь с невысокими редко стоящими деревьями, образуемая Erica
arborea и Myrica faya и многими средиземноморскими элементами, или же
наблюдаются различные стадии деградации (ср. данные Обердорфера).
На плантациях взамен тропических возделываются средиземноморские
растения: лимон, инжир, виноград, реже маслина, каштан съедобный
и лещина, косточковые и семечковые плодовые деревья; в садах растут
камелии, араукарии, кипарисы, эвкалипты, дуб пробковый, Spartium^
Arundo donax и др.
г) Пояс сосновых лесов лишь с трудом можно отграничить
от пояса лавровых лесов. Трехигольчатая Pinus canariensis встречается
в последнем на всех сухих местообитаниях, на скалистых гребнях и т. д.;
лишь на высоте 1200—2000 м, где туман оседает менее часто, она образует
свой пояс. Ближайший родственник ее — Pinus longifolia, растущая в Ги-
малаях; близкие формы известны из третичных отложений Европы. В ка-
Средиземноморская жестколистная растительность
93
честве представителей подлеска сосновых лесов назовем виды Cistus,
Cytisus и Adenocarpus.
Из культурных растений распространены виноград, грецкий орех
и миндаль. Картофельные поля из-за большой сухости воздуха приходит-
ся тщательно засыпать пемзовой дресвой, чтобы предотвратить испарение
с поверхности почвы.
д) Горная пустыня резко отделяется от расположенного ниже
ее лесного пояса и начинается там, где приблизительно на высоте 2000 м
проходит верхний предел развития пассатной облачности. Таким образом,
верхняя граница леса является здесь границей засушливости (ср. т. I,
стр. 219 и рис. 168). Доминирующий вид растений на этих высотах —
дрок с белыми цветками, «ретама» (Spartocytisus supranubium); следует
упомянуть также эндемиков Sisymbrium bourgaeanum и Cheiranthus scopa-
rius с сине-фиолетовыми цветками. Необычайно своеобразен растущий
в более влажных впадинах Echium bourgaeanum] во время цветения (см.
цветное фото 2) он выбрасывает красноватые соцветия длиной несколько
метров и напоминает тогда густоопушенные свечеподобные растения тро-
пических высокогорий (т. I, рис. 162 и 164).
е) Альпийские ковры на каменистых осыпях
развиты на высотах более 2600 м. В условиях низких температур и преж-
де всего характерной для этих мест большой сухости воздуха выживают
только многолетние ползучие растения с серебристыми листочками; это
эндемичные Viola cheiranthifolia и Silene nocteolens. И в этом отношении
мы наблюдаем условия, аналогичные условиям в тропических высоких
горах. На высотах 3100—3200 м (вершина пика Тейде) цветковые растения
уже не встречаются. Шенк упоминает только некоторые сине-зеленые
водоросли (Scytonemd), мхи Weissia verticillata и Frullania nervosa и раз-
личные виды Cladonia.
СУБСРЕДИЗЕМНОМОРСКАЯ ЗОНА
К северу от собственно средиземноморской зоны длительность и ин-
тенсивность летней засухи начинают постепенно убывать, а зимы становят-
ся холоднее (рис. 64—66). Изменения претерпевает и растительность:
жестколистные растения, как мы видели, завещаются летнезелеными лист-
венными видами. Это прежде всего разные виды дуба: дуб пушистый (Quercus
pubescens), на востоке также Qu. cerris, в северной Испании Qu. pyrenaica
(toza) и др. В западной части ареала виды дуба растут совместно с Acer
monspessulanum, A. opalus, Sorbus domestica, S. torminalis, Prunus mahaleb,
Cornus mas, Amelanchier ovalis и др., на побережье Адриатического моря —
с Fraxinus ornus, С arpinus orientalis, Ostrya carpinifolia, Acer obtusatum,
Cercis siliquastrum. Распространены также некоторые сорта груш с узкими
листьями с белым опушением; на западе это Pirus amygdalijormis, на
востоке — Р. elaeagrifolia, кроме того, Staphyllea pinnata, Rhus cotinus
и др. Более требователен к теплу Celtis australis Ч
Большинство вечнозеленых видов исчезает. Дальше к северу распро-
странен только Buxus sempervirens] Rubia peregrina встречается даже на
севере Франции. Некоторые травянистые виды и геофиты Средиземномор-
ской области проникают в субсредиземноморскую зону (Dorycnium suffru-
1 В. Huber, Im Orneto-Ostryon des mittleren Eisack- und oberen Etschtales,
•«Mitt. Deutsch. Dendrolog. Ges.», № 62, 1962.
94
Глава 2
Рис. 64—66. Климадиаграммы станций, расположенных в субсредиземноморской
зоне.
Баланс на Роне — период засухи отсутствует, выделяется только сухое время (заштриховано);
Мостар — в Герцеговине, Югославия; Ушак, Анатолия — длительный период летней засухи, но
холодная зима (область распространения Pinus pallasiana).
ticosum, виды Ophrys), но обычно и они замещаются здесь другими видами,
которые в свою очередь известны на теплых местообитаниях даже в Цен-
тральной Европе. Мы познакомимся там с ними как с «ксеротермическими
элементами».
В результате многократных рубок и особенно усиленного выпаса
скота первоначальные леса субсредиземноморской зоны на большей
части территории деградировали и замещены летнезеленым низкостволь-
ным лесом или кустарником — аналогом маквиса; на Балканском полу-
острове такой тип растительности известен как ш и б л я к. Интенсив-
ный выпас коз может стать причиной полного исчезновения растительно-
сти — остается голая, безжизненная скала, как это случалось раньше
в карстовых районах Далмации или в Черногории. Однако достаточно
было прекратить пастьбу коз, как в течение всего нескольких лет мест-
ность покрывалась зеленью подраставшего кустарника.
Много дискуссий вызвал вопрос о том, относить ли описываемую нами
зону к собственно средиземноморской зоне или рассматривать ее как
принадлежащую к центральноевропейской зоне лиственных лесов. В горах
с различным характером вертикальной поясности ответить на этот вопрос
особенно сложно Ч Судя по тому, что древесные породы представлены
летнезелеными видами, экологически субсредиземноморская зона, несом-
ненно, ближе к центральноевропейской зоне лиственных лесов. К среди-
земноморской зоне следовало бы относить только типичную вечнозеленую
жестколистную растительность, а в горах — соответствующие пояса.
Однако в резко континентальном климате или на сухих местообитаниях
летнезеленые лиственные деревья занимают в этой переходной зоне весьма
подчиненное положение и замещаются разными подвидами сосны примор-
1 Е. Oberdorfer, Gliederung und Umgrenzung der Mittelmeervegetation auf
der Balkanhalbinsel, «Вег. Geobot. Inst. Rubel (Zurich) f. 1947», 1948, S. 84—111;
C. R e g e 1, Uber die Grenze zwischen Muttelmeergebiet und Mittelleuropa in Griechen-
land, «Вег. Deutsch. Bot. Ges.,» 58, 1940, S. 155—165; Zur Frage der Grenze zwischen dem
Mittelmeergebiet und Mitteleuropa in Griechenland, «Вег. Geobot. Inst. Rubel f. 1946»,
1947, S. 15—22; F. Markgraf, Die Siidgrenze mitteleuropaischer Vegetation auf der
Balkanhalbinsel, «Вег. Deutsch. Bot. Ges.», 60, 1942, S. 118—127; Zur Abgrenzung der
Mittelmeervegetation, «Vegetatio», 3, 1952, S. 324—325; I. H о r v a t, Die Grenze der
mediterranen und mitteleuropaischen Vegetation in Siidosteuropa im Lichte neuer pflan-
zensoziologischer Forschungen, «Вег. Deutsch. Bot. Ges.», 75, 1962, S, 91—104.
Средиземноморская жестколистная растительность
95
ской (Pinus pinaster) и сосной черной (Pinus nigra). Подвид последней
ssp. salzmanni распространен в западной половине \ тогда как ssp. pal-
lasiana можно считать типичным для континентальной части субсредизем-
номорской зоны. Но поскольку в этой области обширные низменности
отсутствуют, речь должна идти скорее о вертикальном поясе (рис. 66).
Представляется наиболее целесообразным рассматривать субсреди-
земноморскую зону как переходную. Центральноевропейская зона начи-
нается лишь там, где максимум осадков приходится на летние месяцы.
Более или менее детальные эколого-физиологические исследования
в Субсредиземноморской области не проводились, имеются лишь неко-
торые данные для полосы, переходной к центральноевропейской зоне
лиственных лесов. Субсредиземноморский Quercus pubescens в Централь-
ной Европе замещается видами Qu. petraea или Qu. robur, а субсредиземно-
морская Pinus nigra на сходных местообитаниях — Pinus silvestris. В пере-
ходной зоне эти виды местами растут близко один к другому, например
оба вида сосны; в этом случае Р. nigra приурочена к более сухим место-
обитаниям, чем Р. silvestris. В Восточных Альпах (Караванке) известны
даже местообитания, где оба вида образуют смешанные древостои, в кото-
рых Бейзель произвел сравнительные измерения транспирации1 2. Он
нашел, что однолетняя хвоя сосны обоих видов всегда транспирирует сла-
бее, чем 2—3-летняя. При сопоставлении же одновозрастной хвои одно-
временно полученные кривые суточного хода транспирации, несмотря на
сильный разброс значений, свидетельствуют о меньшей интенсивности
транспирации у Р. nigra. Поскольку значения транспирации вычислялись
в миллиграммах на 1 г сырого веса в минуту, полученный результат
может быть объяснен меньшей площадью более жесткой хвои Р. nigra.
Кроме того, устьица у этого вида расположены несколько глубже, однако
число их на 1 мм2 поверхности листа на 10% больше, чем у Р. silvestris.
Величина интенсивности транспирации хвои еще не позволяет судить
о расходе воды растением в целом, так как последний в большей мере
зависит от размера всей траспирирующей поверхности. Поэтому был
определен общий сырой вес всей хвои у деревьев в возрасте приблизитель-
но 10 лет, вычислена водоотдача за сутки и — с учетом местоположения —
с 1 м2 поверхности почвы. Получены следующие значения интенсивности
транспирации поздним летом: для Р. nigra — 2,4 мм/сутки, а для Р. sil-
vestris — 2,9; таким образом, отношение составляет 100 : 121. Вероят-
ность больших ошибок при вычислении заставляет, однако, сомневаться
в достоверности этой пропорции. Но даже если результат верный, он еще
не означает, что Р. nigra на сухих местообитаниях более конкурентоспо-
собна, чем Р. silvestris. Транспирация не является жизненно важной
функцией растений, а представляет собой прежде всего процесс, сопут-
1 J. Braun-Blanquet, La foret de Pinus salzmanni de Saint Guilhem-le-
Desert, «Sigma»/ Comm. № 133, «Collectanea Bot.», 4, Fasc. Ill, Barcelona, 1955,
p. 435-489.
2 D.Beysel, Vergleichende Transp ir at ionsmessungen an Pinus nigra Arnold und
Pinus silvestris L., «Вег. Deutsch. Bot. Ges.», 73, 1960, S. 429—444.
Измерения осмотического давления у видов растений шибляка позволили уста-
новить отчетливый годовой ход его со следующими максимальными значениями в авгу-
сте: Quercus cerris — 35 атм (наименьшие колебания), Qu. conferta — 43 атм, Qu.
petraea — 46 атм (вблизи границы распространения). У видов травяного покрова мак-
симальная величина давления достигала: у Chrysanthemum corymbosum — 27 атм,
Lathyrus niger — 26, Galium aristatum — 30, Stellaria holostea — 28 и у Cytisus austria-
cus — 35 атм. Ср.: M. Kojic, M. J ankovic, Uber die Hydraturverhaltnisse der
thermophilen Waldgesellschaft von Quercus conferta und Quercus cerris auf der Arada bei
Belgrad, «Вег. Deutsch. Bot. Ges.», 80, 1967, S. 71—79.
96
Глава 2
ствующий фотосинтезу. Ассимиляция же осуществляется только при
открытых устьицах. Способность к конкуренции зависит от количества
продуцируемого сухого вещества и прироста в высоту. Следовательно,
измерения транспирации должны были сопровождаться определением
величины продуктивности в условиях оптимального и слабого влаго-
обеспечения. Из собственного опыта автору известно, что при облесении
грубообломочной, а значит, водопроницаемой осыпи в Шварцвальде
3-летние Р. nigra развивались значительно лучше, чем Р. silvestris,
и быстро перерастали их. Бейзел подчеркивает также, что осмотическое
давление у однолетней хвои Р. nigra, составлявшее 19,5—21 атм, было
ниже, чем у хвои Р. silvestris (20,4—27,5 атм)9, это свидетельствует о менее
напряженном водном балансе у Р. nigra.
Однако выводы, основанные на единичных измерениях, часто бывают
неправильными. Следует опираться только на кривые осмотического
давления, характеризующие весь период вегетации. Решающее значение
для способности растения переносить засуху в большинстве случаев имеет
протяженность корней, особенно в сравнении с массой надземных побегов.
Можно предположить, что именно этот фактор обусловливает превосход-
ство Р. nigra при заселении местообитаний с неблагоприятными усло-
виями.
Субсредиземноморская область в условиях океанического климата
•служит переходной зоной к западноевропейским областям лесов и вере-
щатников, в Центральной Европе — к буковым лесам, а на Балканском
полуострове — к более сухим типам лиственных лесов лесостепи (ср.
стр. 233). Средиземноморские элементы можно обнаружить в пределах
этой области еще во Фракии, а на западном побережье Черного моря —
на пространстве приблизительно до Варны.
Необычайно интересный полностью изолированный форпост среди-
-земноморско-субсредиземноморской растительности расположен на Юж-
ном берегу Крыма. На участке побережья протяженностью 50 км (от
мыса Форос до Алушты) климат имеет отчетливый средиземноморский
характер: более 30% осадков выпадают зимой, средняя температура зим-
них месяцев почти равна 5°. Но поскольку средний годовой минимум
температуры колеблется от —8 до —10° (абсолютный минимум от —11 до
—16°), характер растительности здесь скорее субсредиземноморский.
Представлена она преимущественно древесными видами Quercus pubescens
и Juniperusexcelsa (рис. 67), но на подходящих местообитаниях встречается
также ряд главным образом восточносредиземноморских элементов, как-
то: Arbutus andrachne, Juniperus oxycedrus, Pistacia mutica, Rhamnus
alaternus, Rhus coriaria, Jasminum fruticans, Vitex agnuscastus, Pinus pithy-
usa var. stankevici (близкая к P. halepensis), Cistus tauricus, Ruscus pontica,
Euphorbia biglandulosa, Psoralea bituminosa, Lasiagrostis bromoides и др.1.
Горный пояс образует лес из Pinus pallasiana, в котором 32,6% видов
принадлежат средиземноморскому элементу в широком понимании тер-
мина и 29,6 — бореальному. Растительность характеризуется отчетливо
выраженным периодом зимнего покоя и летним главным периодом веге-
тации. Напротив, в нижнем поясе, образуемом Juniperus и Quercus, отме-
чаются два максимума развития — весной, а также осенью с некоторым
ослаблением его летом, причем зима не является периодом абсолютного
покоя. В этом поясе 45% видов представлены средиземноморскими и толь-
1 Н. И. Р у б ц о в, Л. В. М а ха ев а, М. С. Ш а л ы т, И. Н. К о т о в а,
Природа Крыма. Растительный мир, Симферополь, изд-во «Крым», изд. 2-е, 1966.
Средиземноморская жестколистная растительность
97
Рис. 67. Южное побережье Крыма. Juniperus excelsa и средиземноморские элементы
(фото Клепинина).
ко 11,2% бореальными элементами1. Средиземноморская растительность
Крыма после некоторого перерыва прослеживается и на побережье Кав-
каза, между Новороссийском и Туапсе; южнее климат становится столь
влажным, что растительный покров приобретает облик, типичный уже
для Колхиды.
СРЕДИЗЕМНОМОРСКАЯ ОБЛАСТЬ СТЕНЕЙ АНАТОЛИИ
Там, где на востоке Средиземноморской области годовая сумма осад-
ков падает ниже 400 мм и вегетационный период вследствие длительной
летней засухи и продолжительного холодного времени года сокращается
до 4 месяцев, древесная растительность уступает место степи. В пределах
Передней Азии она раскинулась в центральной Анатолии, а также на
территории стран так называемого «плодородного полумесяца», то есть
по горным склонам, обрамляющим междуречье Евфрата и Тигра с северо-
запада, севера и востока. В древности это был один из крупнейших цент-
ров цивилизации.
В полосе, пограничной с областью степей, некогда были распростране-
ны летнезеленые рощи с древовидными видами Juniperus, от которых
вследствие тысячелетних вырубок и пастьбы в наше время сохранились
лишь жалкие остатки. С увеличением сухости климата древесная расти-
1 Н.Б.Белянин а, Ритм сезонного развития растений и растительных сооб-
ществ южного склона Крымских гор, «Бюлл. Моск, о-ва испытателей природы», отд.
биологии, т. 67, вып. 5, 1962, стр. 90—104; А. А. Корчагин, О возможности
облесения крымской Яйлы, «Вести. Ленингр. университета», № 18, серия «Геология
и география», вып. 3, 1960, стр. 97—109.
7-01248
Р п с. 68. Обзорная карта Анатолии с центральноанатолийской областью степей (по Вальтеру).
Высота местности передана толщиной линий.
Средиземноморская жестколистная растительность
99
тельность замещается средиземноморскими злаковниками, сегодня также
полностью деградировавшими. Более подробно проводилось изучение
степных растительных формаций Анатолийского плоскогорья, в центре
которого лежит большое бессточное соленое озеро Туз. С экологической
точки зрения центральноанатолийская степь особенно интересна.
Анатолия — горная страна (рис. 68). Большая часть ее территории
первоначально была покрыта лесами континентально-субсредиземномор-
ского типа \ На средних высотах с холодными временами года (см. рис. 66)
доминирующим древесным видом является Pinus nigra ssp. pallasiana.
Центральный район Анатолии представляет собой нагорье, с трех сторон
окруженное хребтами. На западе сплошная горная преграда отсутствует,
но и здесь отдельно стоящие горные массивы препятствуют проникнове-
нию в глубь континента влажных воздушных масс со стороны моря.
Климат нагорья поэтому чрезвычайно сухой, как это видно на климадиа-
грамме для Анкары (рис. 69). Летняя засуха продолжается с начала июня
до середины октября. Годовая сумма осадков — 341 мм’, основная их мас-
са выпадает зимой, но и в мае заметен некоторый максимум. Последний
связан с частыми грозами, вызванными подъемом еще влажных воздуш-
ных масс над уже согревшимся нагорьем1 2. Холодное время года длится
с декабря до марта, но оно часто прерывается оттепелью.
Климатические условия, и прежде всего сухость, неблагоприятны
для формирования лесной растительности; в этих условиях успешно могут
развиваться только степные сообщества3. Но центральная Анатолия—-
не ровное плато; обширные ландшафты котловин чередуются здесь с не-
сколько более влажными горными хребтами или потухшими вулканами 4.
Сохранившиеся на определенных местообитаниях небольшие рощи,
общипанные скотом кустарники из дуба и одиночные деревья свидетель-
ствуют о том, что на возвышенных частях рельефа или в глубоких ущельях
острова леса прежде были распространены значительно шире. В настоя-
щее время остатки лесов удерживаются лишь на 7—8% общей площади.
По Узлу, ранее леса занимали 50—55%,
но эти данные, видимо, существенно пре-
увеличены 5.
Между субсредиземноморской лесной
зоной и зоной степей в качестве переходного
типа растительности распространены за-
росли кустарников, образованные Junipe-
rus oxycedrus, Cistus laurifolius, Quercus pu-
bescens, Colutea cilicica, Pirus elaer^rifolia,
видами Crataegus и Amygdalus. В ка-
честве восточных, иранских, элементов
к ним примешиваются Juniperus excelsa
и J. foetidissima.
Рис. 69. Климадиаграмма
Анкары, станции, расположенной
у северной границы центрально-
анатолийской степи.
1 Н. Walter, Vegetationsgliederung Anatoliens, «Flora», 143, 1956,
S. 295-326.
2 Местное население называет этот дождливый период «киркикинди», что означает
«40 послеполудней» (с грозовыми дождями).
3 Н.Walter, Das Problem der zentralanatolischen Steppe, «Die Naturwiss.», 43,
1956, S. 97-102.
4 H. Louis, Das naturliche Pflanzenkleid Anatoliens, Stuttgart, 1939.
5 S. U s 1 u, Untersuchungen zum anthropogenen Charakter der zentralanatolischen
Steppe, «Giessener Abh. z. Agrar-u. Wirtsch. Forsch.», 12, 1960.
7*
100
Глава 2
Нижняя граница горных лесов со степной зоной проходит, по Лиу,
на севере области степей на высоте 1100 л, на юге, у подножия Тавра,—
на высоте 1400—1500 м. Большая часть собственно степи в настоящее
время занята пашней; возделывается озимая пшеница с применением
методов сухого земледелия (с черным паром через год). Остальная пло-
щадь используется как пастбище.
Площадь под посевами пшеницы с 1928 по 1960 г. утроилась, вместе с тем вдвое
увеличилось и поголовье скота. Пшеницу высевают в октябре после первых дождей;
к началу засухи, в июне, она созревает. Урожай полностью зависит от количества
Р и с. 70. Годы сильной задержки роста сосны (узкие годичные кольца)
между 1801 и 1900 гг. На оси ординат показан дефицит прироста древесины,
превышающий 50% (по Кристиансен-Венигеру).
зимних и весенних осадков. Поскольку их сумма в Анатолии, как и во всех аридных
районах, изменяется от года к году, хорошие урожаи сменяются полным неурожаем.
По данным Кристиансен-Венигера, за период с 1928 по 1961 г. было 8 среднеурожай-
ных лет, 7 лет урожайных, 4 года высокоурожайных, 6 лет собирали низкий, 4 года —
плохой урожай; за это же время было 5 недородов, из них 2 катастрофических (в 1928
и 1945 гг.) г. Количество осадков лимитирует также прирост древесины в окраинных
районах степной зоны. Засушливые годы находят отражение в узких годичных кольцах
в поперечном разрезе стволов сосен. Кристиансен-Венпгер вместе с Гаснером провел
дендроклиматические исследования в Анатолии. Результаты представлены в виде
диаграммы замедления роста сосны, росшей в 55 км северо-западнее Анкары на абсо-
лютной высоте 850 м (рис. 70). Все годы XIX в. характеризовались уменьшением роста
в толщину более чем на 50%. Голодные 1874 и 1875 гг., а также засушливые 1898
и 1899 гг. отмечены очень узкими годичными кольцами. Какая-либо периодичност]»
засушливых периодов не выявляется, поэтому предсказать засуху не представляется
возможным. Неосуществимость такого прогноза подтверждают также измерения годич-
ных колец деревьев, которые автор предпринял в Юго-Западной Африке.
Нераспаханные площади служат в центральной Анатолии естествен-
ным пастбищем. Но и здесь состав растительного покрова вследствие
чрезмерного выпаса скота, прежде всего коз, сильно изменен. В худшем
случае исчезают все многолетние виды, за исключением колючих или
ядовитых, сохраняются только однолетние сорняки. Весной цветущие
сорняки образуют пестрый красочный ковер, который, однако, быстро
пропадает. В засушливое время почва голая. В годы засухи часто про-
исходит большой падеж скота.
1 F. Christiansen -W ей ige г, Gefahrdung Auatoliens durch Тгоскеи-
jahreund Diirrekatastrophen, «Z. f. Ausland. Landwirtsch.», 1964, S. 133 — 147; G. G a s s-
ner, F. Christiansen -Weniger, Dendroklimatische Untersuchungen uber
die Jahresringentwicklung der Kieferii in Anatolien, «Nova Acta Leopoldina», N. F.,
Bd. 12, № 80.
Средиземноморская жестколистная растительность
101
Отдаленные от жилых мест районы используются для выпаса овец.
Здесь растут Artemisia fragrans с Роа bulbosa и многие терофиты \ а также
колючие астрагалы, виды Noea и Eryngium. На каменистых почвах в го-
рах распространены только колючие виды, которые не трогают даже
козы. Возникшая стадия деградации представлена сообществом колючих
подушечников с видами Astragalus (Т ragacanthd) и Acantholimon (свин-
чатковые), характерными и для некоторых высокогорных районов Ирана.
Широкое распространение Artemisia fragrans, встречающейся вокруг соле-
ных озер на умеренно засоленных почвах вместе с Camphorosma, также
есть следствие слишком интенсивной пастьбы. Это полукустарник с очень
жесткими одревесневшими нижними частями побегов; овцы скусывают
только верхние травянистые части, Роа bulbosa также устойчив к выпасу
и размножается при помощи многочисленных луковичек. Но все это
виды уже многократно измененного растительного покрова степей. Пред-
ставление о том, как выглядела естественная степная растительность, мы
получили, посетив огороженный, защищенный от скота водосбор водо-
хранилища, снабжающего Анкару водой.
Здесь состав растительности совершенно иной. На мощных почвах
развивается травянистая степь. Жизнь пробуждается уже в феврале —
марте, когда зацветают геофиты — виды Crocus, Muscari, Gagea, Colchi-
cum и др, (позднее такжеOrnithogalum). Наиболее обильное цветение про-
исходит в мае. Растительный покров кажется в это время сомкнутым, но
у оснований растений еще остаются свободные пространства. Из злаков
преобладают ковы л и (Stipa pulcherrima, S. lagascae} и Bromus tomentellus,
а также Festuca ovina var, vallesiaca, виды Phleum и Melica. Весьма много-
численно многолетнее разнотравье, относящееся к родам Potentilla, San-
guisorba, Melilotus, Hypericum, Eryngium, Thymus, Galium, Scabiosa,
Phlomis, Onosma, Centaurea и др. Astragalus atropurpureus не принадлежит
к растениям с колючими листьями, Vinca herbacea же относится к числу
летнезеленых видов. Фенологические наблюдения проведены Марк-
графом 1 2,
Таким образом, эта заповедная степь уже напоминает травянистые
степи Восточной Европы, только здесь шире представлены виды, харак-
терные для Средиземноморья, Севернее береговой линии Черного моря
летняя засуха несколько ослабевает, зимд же становится намного холод-
нее и длительнее. Нов обоих случаях растения имеют возможность веге-
тировать в течение 4 благоприятных месяцев: в центральной Анатолии это
март — июнь, в Восточной Европе — апрель — июль, в степях северной
Месопотамии с более короткими зимами, но особенно продолжительной
засухой — февраль, март, апрель, май. Мнение о том, что в естественных
условиях центральноанатолийская степь представляла бы собой травя-
нистую степь с обилием злаков, подтверждают многочисленные островки
степей. Их находят преимущественно в распаханных областях, где выпас
скота не производится. Они занимают межевые полосы и участки вдоль
дорог,
О былом существовании травянистой степи свидетельствует и совре-
менный почвенный профиль, полностью соответствующий профилю обыч-
ного чернозема (см, гл, 7); отличие состоит лишь в том, что верхние гори-
1 Н. В i г a n d, Erste Ergebnisse der Vegetations-Untersuchungen in der zentrala-
natolischen Steppe 1, «Bot. Jb.», 79, 1960, S. 255—296.
2 F. Markgraf, Die jahreszeitliche Entwicklung einer Steppenflache bei An-
kara, «Вег. Geobot. Institut Riibel, Zurich», 32, 1961, S. 236—244.
102
Глава 2
зонты менее обогащены гумусом и выглядят скорее бурыми, чем черными.
Под действием соляной кислоты вскипает уже поверхность почвы (pH =
= 7,2—9,1). На глубине 70 см в. виде налета карбонатов («псевдомице-
лий» русских исследователей) встречаются выделения извести, а ниже
(до 150 см) ее конкреции. Поглощающая способность почвы до глубины
1,5 м приблизительно соответствует годовому количеству осадков. На-
блюдались также остатки прежних ходов степных грызунов («кротовин»
русских почвоведов). Экологию распространенных ныне в степи видов,
иными словами, видов деградировавшего растительного покрова, деталь-
но изучал Биранд1. Исследования производились им в 1936 г., отличав-
шемся более или менее обычными климатическими условиями (рис. 71).
Максимальное испарение в жаркие месяцы в полдень достигает 2—3 мл!час
(эвапорометр Пише, зеленая бумага). Почва редко промерзает более чем
на 5 см (в декабре 1936 г. — на 30 см). В год исследований еще в мае почва
по всему профилю была хорошо увлажнена, в июне высохли верхние 20 см,
в июле верхние 80 см; содержание воды на глубине 120 см даже в августе
уменьшилось лишь незначительно.
Соответственно величина осмотического давления геофитов, период
развития которых приходится на весну, не превышала 6—9 атм, хотя
корневая система их не проникает глубже 4—8 см.
Весенние эфемеры, исчезающие уже в июне, также, несмотря на не-
глубокое (20 см) укоренение, отличаются низким осмотическим давлени-
ем —10—15 атм, а у засыхающих растений — до 20 атм. Частично это
виды, в умеренной зоне Центральной Европы являющиеся сорняками:
Adonis flammeus, Lepidium draba, Reseda lutea, Amaranthus retrojlexus,
Senecio vernalis, а помимо того, виды Hypecoum, Glaucium, Cerinthe, Micro-
meria и др. Иначе ведут себя летние растения, вегетирующие до июля или
августа, некоторые из них — в течение всего лета и осени. Чем меньше
глубина проникновения их корней, тем раньше они перестают вегетиро-
вать, а некоторые экземпляры погибают совсем. При благоприятных усло-
виях осмотическое давление у весенних эфемеров едва превышает давле-
ние, отмечаемое у весенних видов, однако при недостатке влаги оно подни-
мается до 30 атм. Листья у этих видов мягкие, обычно сильно опушенные,
при плохом водообеспечении засыхают снизу вверх, что обусловливает
непрерывное уменьшение транспирирующей поверхности. В первые дни
своего развития растения энергично расходуют влагу, поэтому суточные
колебания осмотического давления у них весьма резкие. Постепенно вод-
ный дефицит возрастает, высокие значения осмотического давления на-
блюдаются уже в утренние часы, интенсивность транспирации сокращает-
ся, колебания осмотического давления в течение суток сглаживаются.
Это продолжается до тех пор, пока осмотическое давление не достигает
максимальных значений, после чего надземные части растения засыхают.
В июне отмирают виды Achillea santolina, Delphinium orientale, Ranunculus
illyricus, Ajuda chia, Anchusa italica, Linaria corifolia, виды Moltkia,
а также степные злаки Stipa pulcherrima и Festuca sulcata. В июле засыха-
ют Coronilla varia, виды Linum, Teucrium и Salvia, Phlomis herba-venti,
Onosma aucheriana, очень характерная Artemisia jragrans и известная
гармала (Peganum harmala), растущая вблизи поселений и мест отдыха
караванов,1 так как ее не едят даже верблюды.
1 Н. A. Birand, Untersuchungen zur Wasserokologie der Steppenpflanzen bei
Ankara, «Jb. wiss. Bot.», 87, 1938, S. 93—172; Untersuchungen uber die Wurzelsysteme
der Steppenpflanzen, «Gommun. Fac. Sci. Univ. Ankara», 3, 1952, S. 219—235.
Средиземноморская жестколистная растительность
103
Рис. 71. Погодные условия в Анкаре в 1936 г. (по Биранду).
Кривые температуры показывают для каждого месяца (сверху вниз): абсолютный максимум, сред-
ний суточный максимум, среднюю температуру за месяц, средний суточный минимум и абсолют-
ный минимум.
У свежих листьев Artemisa осмотическое давление равно 12—15 атм,
у сухих оно в два раза выше. Осмотическое давление у свежих растений
Peganum составляет 16—19 атм, при засыхании оно увеличивается до
28 атм. Отдельные корни этих видов иногда проникают в почву на глуби-
ну более 1,2 м; во время засухи они снабжают водой едва транспирирую-
щие, но живые листовые почки надземных побегов. Листья Peganum
и, несомненно, Artemisia в свежем состоянии отличаются очень большой
интенсивностью транспирации. В первое время, пока верхние горизонты
почвы еще сохраняют высокую влажность, водоотдача компенсируется
всасыванием влаги корнями. В благоприятные годы или на более влажных
местообитаниях все перечисленные виды длительное время остаются жи_
104
Глава 2
выми. После сильного осеннего дождя у них могут даже развиться новые
сосущие корни, и растение еще раз дает побеги.
У всех видов, поздно развивающихся и цветущих во время самой
сильной засухи, имеется глубокопроникающий стержневой корень, кото-
рый разветвляется лишь на очень большой глубине, в постоянно влажных
слоях почвы. Благодаря этому растения относительно хорошо обеспечены
влагой. Осмотическое давление у них во время засухи обычно не достига-
ет 15 атм, хотя транспирация может быть очень интенсивной. К этой
группе относятся виды Eryngium campestre, Scolymus hispanicus, Euphorbia
tinctoria, E. aleppica, E. myrsinites, 2 вида Echinophora, а также колюче-
подушечные растения (Acantholimon и многие виды астрагала).
Особое положение занимает верблюжья колючка — колючее расте-
ние из семейства бобовых (Alhagi camelorum). Ее канатоподобные, ветвя-
щиеся и образующие корневые побеги стержневые корни уходят на много
метров в глубину и достигают уровня грунтовых вод. Глубина проникно-
вения корней 17-месячного растения может достигать 4,5 м, а 30-месячно-
го — 7,65 м. Транспирация этого вида характеризуется очень сильной
интенсивностью, правда, общая транспирирующая поверхность его невели-
ка, поскольку летом на нем образуется больше зеленых колючек, чем
листьев. Верблюжья колючка растет обычно небольшими группами (воз-
никшими из корневых отпрысков), которые поздним летом выделяются
на общем сухом фоне своим зеленым цветом.
Хорошая влагообеспеченность перечисленных выше видов проявляет-
ся также в том, что колебания осмотического давления в течение суток
даже у интенсивно транспирирующих растений невелики или совсем от-
сутствуют.
Биранд изучал водную экологию древесных растений у границы степ-
ной зоны. Они также обладают мощной и глубоко проникающей корне-
вой системой; несмотря на это, у них удается заметить некоторый дефицит
влаги во время засухи: транспирация уменьшается, осмотическое давле-
ние возрастает с 16—19 атм весной до 29—38 атм в августе и сентябре,
суточные колебания его сокращаются. В засушливые годы листья начи-
нают сохнуть уже в конце лета, а в годы очень сильной засухи отмирают
целые ветви. Это явление известно во всех аридных районах (ср. т. I,
стр. 353).
Анатомическое строение листьев засухоустойчивых видов и древесных
растений характеризуется отсутствием губчатой паренхимы и часто
развитием водоносной ткани в средней части мезофилла, особенно усилен-
ным вокруг проводящих пучков.
Человек очень рано заселил окраинные зоны средиземноморских
степных районов, так как нашел здесь наиболее благоприятные условия
для возделывания зерновых культур. Древнейшие следы хлебопашества
обнаружены в странах уже упоминавшегося «плодородного полумесяца»
(Иерихон, Бейда, Жармо). В Анатолии центр царства хеттов располагался
у северной краевой зоны, в области стыка леса и степи. Степь распахивали
под зерновые культуры, лес давал древесину и служил местом охоты. Но
степь позволяла также развивать животноводство. В своем первоначаль-
ном составе богатая разнотравьем злаковая степь являла собой прекрасное
пастбище. Поголовье скота в то время было еще столь невелико, что выпас
не вызвал деградации растительного покрова. Таким образом, степь
предоставляла человеку, располагавшему только примитивными орудия-
ми труда, наилучшие условия для существования, более благоприятные,
чем он мог найти в лесу. Отсюда ясно, почему именно в этой полосе возни-
Средиземноморская жестколистная растительность
105
кли первые крупные поселения, явившиеся основой дальнейшего развития
культуры.
Вместе с тем такой ход событий привел к тому, что растительный по-
кров именно в этих районах очень рано и необратимо был изменен чело-
веком. Вырубки и выпас скота со временем вытеснили лес и способство-
вали расширению ареала степей во всех степных районах мира. Ныне
степи занимают большие площади, чем это соответствует климатическим
условиям. Чрезмерная эксплуатация обусловила дальнейшую деградацию
степей в сторону полупустыни, а в наиболее засушливых районах — в
сторону антропогенной, то есть связанной с деятельностью человека, пу-
стыни.
Процесс деградации можно наблюдать сегодня на всей территории
Передней Азии, и прежде всего там, где на поверхность выходят легко
поддающиеся эрозии мягкие третичные толщи горных пород. Происхо-
дит развитие бедлендов, на которых растительный покров часто вообще
отсутствует.
Глава 3
ОБЛАСТИ С ЗИМНИМИ ДОЖДЯМИ
И СРЕДИЗЕМНОМОРСКИМ КЛИМАТОМ ЗА ПРЕДЕЛАМИ ЕВРАЗИИ
И ПРИМЫКАЮЩИЕ К НИМ УМЕРЕННЫЕ ЗОНЫ
КАЛИФОРНИЙСКАЯ ЗОНА
ЖЕСТКОЛИСТНОЙ РАСТИТЕЛЬНОСТИ
Область с зимними дождями, мягкой зимой и жарким сухим летом,
соответствующую средиземноморскому климату Европы, в Северной
Америке можно найти только на западной окраине континента. Она
занимает сравнительно узкую полосу тихоокеанского побережья и про-
стирается от Ванкувера на юго-западе Канады до Нижней Калифор-
нии (см, рис, 6) \ Сухость климата увеличивается здесь с севера
на юг и одновременно в направлении от побережья, характеризуемого
частыми туманами, на восток, до подножия высочайших горных хребтов
(Каскадные горы, Сьерра-Невада, Сан-Бернардино),
Однако истинно средиземно морским климатом обла-
дают только три области: полоса побережья от Монтерея до Лос-Анджеле-
са (несколько севернее города) и примыкающие к ней глубинные районы
до гор Сан-Бернардино, Калифорнийская продольная долина севернее
Сакраменто и, наконец, территория вдоль западного склона Сьерра-
Невады,
Севернее области с истинно средиземноморским климатом климати-
ческие условия имеют субсредиземноморский (с относи-
тельно коротким засушливым периодом), а на побережье —даже влаж-
ный средиземноморский характер. Южнее же этой
области, а также в Калифорнийской продольной долине, к югу от Сакра-
менто, климат обретает черты, свойственные сухому Средизем-
номорью, или становится даже похожим на климат полупустынь.
Приведем в качестве примера несколько климадиаграмм (рис, 72—77),
Рис, 72—74. Климадиаграммы области жестколистной растительности Калифорнии,
•Сан-Франциско — побережье с туманами, низкие летние температуры; Пасадена — истинно среди-
земноморский климат; Сан-Диего — юг Калифорнии, аридный средиземноморский климат.
1 UNESCO-FAO, Bioclimatic map of the mediterranean zone with explanatory
notes, «Arid Zone Research», XXI, 1963.
Области средиземноморского климата за пределами Евразии
107
Рис. 75—77. Климадиаграммы станций, расположенных за пределами зоны жестко-
листной растительности.
Ванкувер — у границы Канады; Йосемитский национальный парк — горы (хвойные леса); Лас-
Вегас — пустыня восточнее Долины Смерти. I
Климат области обусловливает развитие склерофильной раститель-
ности, во многих отношениях сходной с маквисом Средиземноморья. Эту
формацию называют здесь «чапарраль» (рис. 78) (от испанского «сйарагга» —
кустарниковидный дуб) х. Все преобладающие виды (всего их 44) —
вечнозеленые.
Рис. 78. Чапарраль в окрестностях Сан-Франциско.
1 W. S. Cooper, The broad-sklerophyll vegetation of California, Carnegie Inst,
of Washington, Publ. № 319, 1922; R. К n a p p, Die Vegetation von Nord- und Mittel-
amerika, См.: H. W a 1 t e r, Vegetationsmonogr. d. einz. Grofiraume, Bd. I, Stuttgart
und Jena, 1965.
OJ
P и c. 79. Некоторые виды растений области жестколистной растительности
Калифорнии.
1 — Arctostaphylos tomentoscr, 2 — Adenostoma fasciculatum', 3 — Cecmothus cuneatus’, 4 —
C. papillosus\ 5 — Pickeringia montana\ 6 — Quercus dumosa (пат. величина).
Области средиземноморского климата за пределами Евразии
109
Рис. 80. Распространение Adenostoma fasciculatum, Sequ-
oiadendron ( = Sequoia) gigantea и Sequoia sempervirens в Ка-
лифорнии.
Кустарник обычно достигает высоты 1,5—2 м (рис. 78). Наиболее распространена
похожая по внешнему виду на Erica Adenostoma jasciculatum (высотой 0,3—4,0 м) из се-
мейства розоцветных, с цветками, напоминающими цветки Spiraea (рис. 79); район ее
произрастания (рис. 80) совпадает с ареалом типичной формации чапарраль. Листья
у Adenostoma маленькие, линейные; вид часто образует чистые естественные насаждения.
Из других видов следует назвать: кустарниковидные дубы (Quercus dumosa и др.),
Dendromecon rigidum (кустарнпковпдное растение из семейства маковых), розоцветные
Heteromeles arbutifolia, Cercocarpus betulaefolius, Prunus ilicifolia, различные виды Rhus,
относящиеся к сумаховым. Rhamnus crocea (с листьями, едва отличимыми от листьев
Prunus ilicifolia) и 25 различных видов Ceanothus (семейство крушиновых), из которых
С. cuneatus и С. velutinus нередко резко доминируют, особенно на старых гарях. Ряд
видов этого семейства сбрасывает листву. Характерным, но редко образующим заросли
кустарником является Garrya elliptica (из семейства Garryасеае, близкого к Согпасеае).
Особенно широко представлено семейство вересковых, прежде всего 18 кустарниковид-
ными видами Arctostaphylos, из которых важнейший —A. tomentosa; A. manzanita
и А . glauca — викарпрующпе виды, причем первый встречается севернее, а второй
южнее Сан-Франциско. Ареалы некоторых эндемичных видов весьма ограничены. Виды
Dendromecon, Diplacus glutinosus (Scrophulaceae) напоминают средиземноморские виды
Cistus', сложноцветное Ericameria arborescens похоже на виды Cytisus.
Отсутствие в чапаррале деревьев (рис. 78) не есть признак деградации,
а объясняется небольшим количеством осадков (в среднем 500 мм). Форма-
ция чапарраль обладает очень большой устойчивостью и в области основ-
ного своего распространения доминирует на всех склонах вне зависимо-
сти от экспозиции. Какие-либо сведения о том, что первоначальный
характер растительности был иной, отсутствуют. Анатомически и морфо-
110
Глава 3
логически растения, очевидно, очень хорошо приспособлены к специфике
сухого климата с зимними дождями. Древесные виды появляются в Кали-
форнии лишь севернее, где выпадает больше осадков и климат несколько
прохладнее, то есть в области, переходной к субсредиземноморскому
климату.
Типичными древесными видами этих сильно разреженных и деградировавших
жестоколистных лесов являются: Quercus agrifolia, Qu. chrysolepis, Qu. engelmanniif
Qu. wislizenii, Pasania densiflora, Castanopsis chrysophylla, Myrica californica, Umbellu-
laria calif arnica (лавровые) и Arbutus menziesii.
Более умеренный климат рассматриваемой области проявляется в том,
что здесь, как и в Средиземноморье, к вечнозеленым видам присоединяют-
ся листопадные. В собственно жестколистном лесу с 13 доминирующими
видами, помимо названных выше 9 вечнозеленых видов, это еще 4 листо-
падных вида: Quercus keloggii, Qu. lobata, Acer macrophyllum и Aesculus
californica. В горах по мере увеличения высоты в составе древостоя появ-
ляются хвойные деревья, прежде всего дугласия (Pseudotsuga menziesii =
taxifolid). Под верхним древесным ярусом в виде подлеска развит Чапар-
раль, причем в этом ярусе уже около 2/5 всех видов — листопадные. Час-
тично это те же семейства, которые в субсредиземноморской области Сре-
диземноморья играют известную роль; например, наряду с листопадным
дубом представлены виды: Corylus, Amelanchier, Prunus, Cercis, Acer,
а также Aesculus и Ceanothus. Но лес Калифорнии значительно богаче видами
и содержит большее число эндемиков по сравнению с лесами Средиземно-
морской области.
Под влиянием деятельности человека, особенно из-за многократного
выжигания, жестколистный лес, подобно тому как это произошло в Среди-
земноморье, местами был вытеснен; в результате чапарраль распростра-
нился дальше к северу и вверх по склонам гор.
С другой стороны, в краевой зоне Калифорнийской продольной доли-
ны, где проходит граница засушливости, чапарраль оттеснен злаковника-
ми, а в южных прибрежных районах — пустошью с кустарничком Artemi-
sia californica. Искусственные палы, если они пускаются человеком не
слишком часто, виды, образующие чапарраль, переносят без всякого
ущерба; благодаря молодой, возникающей после обжига поросли кустар-
ник становится только гуще. Регулярные же пожары для них губительны.
В 1870-х годах местные жители промышляли тем, что выкапывали широко-
разветвленные корни жестколистного кустарника и продавали их в горо-
дах на топливо. Такая форма использования кустарниковых зарослей
была равносильна их полному уничтожению. Еще большие разрушения
произвела раскорчевка территории под плантации фруктовых деревьев.
В настоящее время в Калифорнийской продольной долине с ее обширными
орошаемыми угодьями, там, где еще сохранились клочки невозделанной
земли, можно найти только однолетние злаки, как Avena fatua, A. barbata,
Bromus mollis, В. rigidus и др., или травы Medicago hispida, Erodium cicuta-
rium, Hypochoeris glabra и др. Согласно Клементсу, первоначально здесь
была степь с преобладанием Stipa, но Купер и ее считает вторичной и пред-
полагает, что чапарраль — за исключением наиболее сухих районов —
ранее был распространен шире, но затем из-за постоянного применения
палов в целях расширения пастбищных угодий был вытеснен.
В самой южной и наиболее сухой части области зимних дождей, то
есть южнее Лос-Анджелеса, кустарничковую гаригу следует считать
зональной растительностью. Доминирующим видом здесь является Arte-
misia californica. Эта гарига приблизительно соответствует полупустыне
Области средиземноморского климата за пределами Евразии
ш
Средиземноморья с A. herba-alba. Кроме того, представлены полукустар-
ники Salvia и Eriogonum (семейство гречишных) с беловато опушенными
листьями, а также некоторые кустарники с безлистными прутьевидными
ветками (например, Lotus scoparius), напоминающие виды Spartocy-
stisus Средиземноморья Ч
В силу горного характера местности различные типы растительности
очень сложным образом проникают друг в друга, причем на влажных
местообитаниях развиваются даже хвойные леса. Последовательность
размещения всех важнейших типов растительности схематически показана
Рис. 81. Профиль рас-
тительности на нижних
частях склонов гор Сан-
Габриэль в Южной Ка-
лифорнии (R. Knapp,
Die Vegetation Nord-und
Mittelamerikas, 1965).
1— шалфейно-полынная рас-
тительность (Salvia mellife-
ra — Eriogonum fascicula-
tum — Artemisia californica)',
2 — чапарраль c Adenostoma,
Ceanothus crassifolius, Quercus
dumosa', 3 — лес из Quercus
chrysolepis\ 4 — лес, образо-
ванный Pseudotsuga macrocar-
pa\ 5 — калифорнийский
лавр (Umbellularia californi-
ca); 6 — летнезеленый лес
c Acer macrophyllum.
на рис. 81. Характерно, что и здесь во влажных ущельях летнезеленые
лиственные деревья, например Acer macrophyllum, видимо, приобретают
большую конкурентную мощность по сравнению с жестколистными видами
при условии, что летом они могут использовать близкозалегающие
грунтовые воды.
Чем дальше к северу, тем прохладнее становятся зимы и тем менее
отчетливо выражен летний засушливый период. Эти перемены особенно
ощутимы в той области побережья, где благодаря близости холодного Кали-
форнийского морского течения в летние месяцы часто образуются густые
туманы. В Сан-Франциско температура в июле и в туманную погоду лишь
ненамного выше, чем в теплые дни в январе.
В соответствии с изменением климата, проявляющимся и в горах по
мере увеличения высоты, претерпевает существенные изменения и расти-
тельность: хвойные леса в большей мере теснят жестколистную раститель-
ность (ср. стр. 123).
Список всех видов, встречающихся в чапаррале, приведен в работе
Купера.
После этой краткой общей характеристики растительности перейдем
к рассмотрению экологических проблем.
1 Отсутствие злаков в чапаррале и гариге частично объясняют токсическим дей-
ствием особых веществ, выделяемых растением и обнаруженных, в частности, у Adeno-
stoma и Salvia. Однако не ясно, могли бы злаки вообще конкурировать в естественных
травостоях. Ср. работу: W. Н. М u 1 1 е г, С. Н. М и 1 1 е г, «Bull. Torrey Bot. Club»,
91, 1964; 93, 1966; W.H. Muller, С. H. Muller, B. L. Haines, « Science»,
143, 1964; 144, 1965. Доказана реальность выделения различных терпенов, которые,
будучи смыты росой, якобы повреждают проростки злаков. Однако корневая система
Salvia и Artemisia не исследовалась, так что не исключено, что истинной причиной
образования лишенных злаков полос вокруг травостоя Salvia-Artemisia является кор-
невая конкуренция. Ср. также: С. Н. М u 1 1 е г, R. d е 1 Moral, Soil toxicity induced
by terpenes from Salvia leucophylla, «Bull. Torrey Bot. Club», 93, 1966, p. 130 — 137.
112
Глава 3
Корневая система жестколистных видов, в том числе и видов, распро-
страненных в Калифорнии, проникает очень глубоко в грунт, поскольку
летом верхние 100 см почвы часто не-содержат влаги, доступной для расте-
ний. Тем не менее древесные породы и в это время года получают доста-
точное количество воды; это явствует из того, что даже летом деревья,
у которых была срублена или сгорела крона, продолжают еще в течение
3 недель давать поросль. Из-за сокращения транспирирующей поверхности
того небольшого количества воды, которое всасывается корнями, уже
достаточно, чтобы почки раскрылись. Летом запас влаги в почве под
жестколистными лесами больше, чем под чапарралем.
Осенние дожди почти не вызывают роста древесных пород, поскольку
требуется много времени, прежде чем вода проникнет в глубокие, прони-
занные корнями горизонты почвы; верхние же плоскостелющиеся корни
после летней засухи утрачивают активность. Так, например, осенью после
первого сильного дождя 31 октября повышение содержания влаги в почве
на глубине 50 см было отмечено лишь 22 ноября, а на глубине 100 см —
только 5 декабря. Однако к тому времени температура так низко упала,
что интенсивный рост побегов уже был невозможен. Апогей развития
растительности приходится на апрель, когда влагообеспеченность доста-
точно хорошая, а температура воздуха быстро повышается. Подробное
изучение биологии 68 растений чапарраля, принадлежавших 18 видам,
показало, что можно выделить три типа корневой системы Ч Корни пер-
вого типа, включающего важнейшие виды чапарраля {Adenostoma fascicula-
tum, Arctostaphylos glandulosa, Ceanothus leucodermis, Photinia arbuttfolia,
Quercus chrysolepis и Qu. dumosa), развиваются больше в глубину, чем
в горизонтальном направлении. Максимальная измеренная глубина
(с учетом местообитаний в глубоких расселинах скал) составила 4—8,5 м.
В стороны корни все же расходились дальше, чем простирались побеги.
Корни второго типа ветвятся больше по горизонтали (радиус 3—6,5 м),
но мало проникают в глубину (1,4—2,6 м). К третьему типу относятся
преимущественно полукустарники, чьи корни длиной 0,9—1,8 м расхо-
дятся радиально и углубляются лишь на 0,6—1,5 м. Особое положение
занимает суккулент Yucca whipplei (рис. 82); ее корни уходят только на
75 см в глубину, но простираются в стороны на расстояние до 4 м. Ис-
следования проводились на площадках, где в течение 16—72 лет не было
пожара.
У всех видов имеется микориза, у видов Ceanothus на корнях обра-
зуются клубеньки; вопрос о том, связывают ли они азот, пока точно не
решен.
Листья растений чапарраля овальные, ланцето- или игловидные
(Adenostomd). Интересна конвергенция Prunus ilicifolia и Rhamnus crocea,
колючие листья которых почти неразличимы. Опушение листьев типично
для 48,7% видов, у двух видов верхняя часть листьев покрыта лаковым
налетом. У Ceanothus, как и у Nerium, на листьях можно наблюдать опу-
шенные углубления, а в них выступающие устьица. Для определения
интенсивности транспирации важно знание не столько размера влаго-
отдачи на единицу листовой поверхности, сколько величины всей транс-
пирирующей поверхности листа, о которой, однако, точные данные отсут-
ствуют, В работе Купера приведены поперечные срезы листьев 15 видов
и отмечено, что толщина кутикулы в среднем составляет 5,6 мкм, на
1Н. Н ellmers, J, S. Horton, G. J uhren, J, O’Keefe, Root sys-
tems of some chaparral plants in Southern California, «Ecology», 36, 1955, p. 667—67Я.
Р и с. 82. Отдельные красиво цветущие экземпляры Yucca whipplei в чапаррале,
образованном Arctostaphylos manzanita, Prunus ilicifolia, Adenostoma fasciculatum, Cea-
nothus cuneatus, Cercocarpus ledifolius и др.; горы Сан-Бернардино в Калифорнии (фото
Упхофа).
114
Глава 3
верхней стороне листа — до 10 мкм. Клетки, содержащие дубильные веще-
ства, в листьях разных видов растений чапарраля наблюдаются так же
часто, как в листьях растений Средиземноморской области.
Старые листья вечнозеленых видов начинают ассимилировать СО2
еще весной, пока не разовьются молодые листья. В июне, когда фотосинтез
начинают осуществлять молодые листочки, они опадают. Таким образом,
процесс накопления сухого вещества у этих видов может продолжаться
в течение всего года, если только они не страдают от недостатка воды.
Напротив, Aesculus, как листопадный вид, сбрасывает листья уже летом,
причем тем раньше, чем засушливее лето. Благодаря этому отдача воды
этим древесным видом резко уменьшается, но вместе с тем сокращается
период фотосинтеза.
Большую роль в области развития чапарраля играют пожары. Де-
тальное исследование сукцессии на выгоревших площадях произведено
на южном склоне гор Сан-Бернардино, где изучено сообщество Adenostoma-
Ceanothus crassifolius1. Виды Adenostoma, Arctostaphylos glandulosa, Ceano-
thus leucodermls и Quercus dumosa после прекращения пожара дают обиль-
ные побеги. Напротив, Ceanothus crassifolius и Arctostaphylos glauca поги-
бают; однако уже на следующий год многие семена этих видов всходят,
так что их доля в сообществе не уменьшается.
Поскольку растительный покров в первые годы после пожара сильно
разрежен, первые же дожди вызывают на выжженных участках сильную
эрозию почв, сдерживаемую, однако, вскоре появляющимися однолетни-
ми и многолетними травянистыми видами. Однолетнее растение Phacelia
brachyloba уже на следующий год после пожара может распространиться
по всей площади. Случайно занесенная сюда Brassica nigra покрывает
наибольшую площадь на второй, a Bromus rubens и В. tectorum — на тре-
тий год после прекращения пала. Через пять лет однолетники обычно
исчезают, травянистые многолетние виды сохраняются несколько дольше.
Интересно поведение луковичных растений, которые в Калифорнии
в отличие от Южной Африки занимают подчиненное положение. Сохранив-
шиеся в почве луковицы зацветают на следующий год после пожара и да-
ют много всходов. В последующие годы растения не цветут; возникшие
из семян особи растут медленно, причем виды Brodiaea и Calochortus
образуют только один лист. В Южной Африке также отмечено, что луко-
вичные растения зацветают лишь после пожара.
Пожары, повторяющиеся каждые 12 лет, состава формации чапарраля
не изменяют. Если формация длительное время не подвергается действию
огня, то в ней появляются виды жестколистного леса: Prunus ilicifolia
и Rhamnus сгосеа. Если через два года после пожара кустарник снова
охватывает пал, то все всходы кустарниковых видов погибают; в результа-
те древесные растения постепенно вытесняются из состава растительного
покрова. Полукустарники, такие, как Artemisia и Salvia, после пожара
размножаются семенами и переносят частые палы лучше, чем раститель-
ность чапарраля. Большинство пожаров устраивается местным населени-
ем умышленно, с целью получения хороших пастбищных угодий. Но
1 J.S.Norton, С. J. Kraebel, Development of vegetation after fire in the
chamise chapparal of Southern California, «Ecology», 36, 1955, p. 244—262.
По Комареку, пожары в Калифорнии следует рассматривать как естественный
фактор. Только в июле 1965 г. во многих лесных округах неоднократно (до 4—5 случа-
ев) возникали пожары, вызванные ударом молнии, в некоторых округах было отмечено
до 28 пожаров. Повторялись они и в августе (см.: Н. W а 1 t е г, Das Feuer als natiir-
licher klimatischer Faktor).
Области средиземноморского климата за пределам Евразиии
115
систематически проведенные опыты показали, что слишком частое выжи-
гание растительности влечет за собой отрицательные последствия, так
как на выгоревших площадях увеличивается поверхностный сток воды
после дождя и вместе с тем развивается сильная эрозия, вызывающая
необратимые изменения почвы х. Содержание влаги в почве на гарях
к концу лета в верхних 50 см оказалось ниже, на большей же глубине
выше, чем на площадях, не затронутых пожаром. Это и понятно, посколь-
ку развивающиеся на пожарищах однолетние растения обладают поверх-
ностной корневой системой, тогда как древесные растения на контрольных
площадках добывают воду из более глубоких горизонтов почвы.
При необходимости удаления кустарниковой растительности чапар-
раля можно поступить проще — достаточно высеять траву тотчас после
контролируемого пожара 1 2. Проростки кустарников в результате конку-
ренции со стороны трав столь сильно задерживаются в своем развитии,
что их корни в первый год не могут достичь глубоких, более влажных
горизонтов почвы и летом, в засуху, погибают.
На площадях с интенсивным выпасом скота из растительного покрова
вытесняются также многолетние виды и сохраняются только однолетние
травы и злаки, поэтому летом пастбища полностью выгорают 3. Проблемы
пастбищного хозяйства, имеющие уже, скорее, прикладной характер,
достаточно полно освещены в литературе 4 *, поэтому мы позволим себе на
них не останавливаться.
В Калифорнии практика выжигания в 1 сочетании с пастьбой также
получила широкое распространение. Естественная растительность оказа-
лась столь сильно нарушенной, что ныне здесь преобладают лишь раз-
личные стадии ее деградации, за исключением, разумеется, тех пло-
щадей, которые не заняты сельскохозяйственными угодьями или поселе-
ниями.
Очень интересна наблюдаемая на морском побережье сукцессия на
песчаных дюнах, в которой злаки не принимают участия. Начинается
она с суккулентных видов Abronia (семейство никтагиновых) и Mesembry-
anthemum chilense, к которым примешиваются Convolvulus soldanella
и Ambrosia chamissonis=Franseria bipinnatifida б. Затем следуют полу-
кустарники Eriogonum parvifolium (гречишные), Lupinus chamissonis,
а также сложноцветные Ericameria ericoides и Eriophyllum stoechadifolium.
Дальше в глубь суши простирается уже чапйрраль с Adenostoma и Arcto-
staphylos, за которым может следовать лес, образованный Quercus agrifolia.
На полуострове Монтерей растет эндемичный для этого района вид сосны
Pinus radiata (рис. 83). В Северной Америке Р. radiata заселяет только этот
весьма ограниченный по площади район, но на других материках, напри-
мер в Австралии, а также в Новой Зеландии и южном Чили после
иптродуцирования она превратилась в важнейший J объект лесного
1 A.W.Sampson, Effect of chaparral burning on soil erosion and on soil-moi-
sture relations, «Ecology», 25, 1944, p. 169—191.
2 A. M. S c h u 11 z, J. L. L a u n c h b a u g h, H. H. В i s w e 11, Relation-
ship between grass density and bush seeding survival, «Ecology», 36, 1955, p. 226—244.
3 J.R. В entley ,M. W. Talbot, Annual-plant vegetation of the California,
foothills as related to range management, «Ecology», 29, 1948, p. 72—79.
4 Z. N a v e h, Mediterranean grassland in California and Israel, «I. Range Mana-
gem.», 13, № 6, 1960.
6 А. К о h 1 e r, Ambrosia chamissonis (Less) Greene, ein Neophyt der chilenischem
Pazifikkiiste, «Вег. Deutsch. Bot. Ges.», 79,1966. Этот занесенный в центральные районы.
Чили вид встречается там в сходных климатических условиях и выступает каЙ важней-
ший фактор образования дюн. ; 'V'iLH
8*-
116
Глава 3
Рис. 83. Pinus radiata на полуострове Мон-
терей, Калифорния. Этот вид, отличающийся
очень быстрым ростом, в настоящее время
очень широко распространен в среднем Чилп,
Южной Африке, юго-западной Австралии
и Новой Зеландии (фото Фабера).
хозяйства, поскольку дает
больший прирост древесины,
чем любая другая туземная или
экзотическая хвойная порода.
Обширные территории были за-
сажены в этих районах Pinus
radiata. Ее древостои резко
изменили картину ландшафта.
В названных странах виды
Cupressus macrocarpa (рис. 84)
и С. goveniana («кипарис Мон-
терея»), естественным местопро-
израстанием которого является
только обрывистое побережье
или песчаный пляж полуострова
Монтерей, стали излюбленной
породой при насаждении ветро-
защитных полос даже в клима-
тических условиях, отличаю-
щихся от климата их родины.
Оба вида оказались легкопри-
спосабливающимися, чего от
них, судя по весьма ограничен-
ному району распространения,
нельзя было ожидать.
ЭНСИНАЛЬНАЯ
РА СТ И ТЕЛЬНОС ТБ
В АРИЗОНЕ
Область Южной Калифор-
нии с сухим средиземномор-
ским климатом в направлении
к юго-востоку и в соответствии
с уменьшением суммы осадков
переходит сначала в полупусты-
ню и затем в настоящую пустыню
(Долина Смерти, пустыня Мохаве, пустыня Колорадо). Еще дальше к вос-
току, в Аризоне, начинается пустыня Сонора, подробно описанная в т. I.
Интересно распределение осадков по сезонам. Помимо умеренно влажного
периода зимних дождей, здесь наблюдается второй дождливый период,
приходящийся на лето. На обширных площадях доминирует Larrea tri-
dentata, вероятно идентичная L. divaricata, занимающей еще большие тер-
ритории на востоке Аргентины. Однако в горах Аризоны над пустыней
с Larrea или кактусами и узкой переходной зоной, представленной саван-
ной с Prosopis, развит вертикальный пояс жестколистной растительности
(рис. 85, ср. также рис. 260 в т. I), соответствующий чапарралю. Отличие
в том, что здесь шире представлены различные виды вечнозеленых дубов
(рис. 86), благодаря чему эту растительность называют «энсинальной»
(от испанского «encina»—вечнозеленый дуб).
Окрестности Таксона, массив Санта-Каталина (2790 м), 50 лет назад,
когда растительный покров почти еще не был нарушен человеком, были
Рис. 84. Cupressus macrocarpa, эндемичный вид калифорнийского побережья. На
других материках, в районах без заморозков, С. тасгосагра часто используется при
насаждении ветрозащитных полос (фото Фабера).
Рис. 85. Распространение
чапарраля в Калифорнии (1)
и энсиналя в Аризоне (2),
а также первичных злаковни-
ков и кустарничковой расти-
тельности в Калифорний-
ской продольной долине (5)
(R. Knapp, Die Vegetation
Nord-und Mittelamerikas, 1965).
118
Глава 3
Рис. 86. Очень редкий энсииальный древостой с Quercus oblongifolia и Qu. emoryi,
Аризона (фото Упхофа).
детально исследованы Шривом х. В период с конца лета 1929 г. до весны
1930 г. автору также удалось провести здесь некоторые исследования.
Вертикальную поясность гор Санта-Каталина иллюстрирует рис. 87.
Рис. 87. Схематический профиль поясов растительности в горах
Санта-Каталина, Аризона (по Шриву).
В горах плювиометрические станции отсутствуют, но можно допу-
стить, что режим осадков здесь, вероятно, соответствует их годовому ходу
в Таксоне, лежащему у подножия (см. т. I, рис. 257). Около 51% осад-
1 F. S h г е v е, The vegetation of the desert montain range as conditioned by clima-
tic factors, Carnegie Inst, of Washington, Publ. № 217, 1915.
Области средиземноморского климата за пределами Евразии
119
ков выпадает в виде сильных грозовых ливней в летнее дождливое время,
31 % — зимой. В Таксоне годовая сумма осадков равна в среднем 293 мм,
но в горах с высотой она увеличивается.
В 1911—1912 гг., когда Шрив проводил свои исследования, в горах
измерялись только летние осадки. С высоты 900 м над уровнем моря до
отметки 2400 м сумма дождевых осадков возрастает в четыре раза. Сухой
период перед началом лета с увеличением абсолютной высоты имеет все
меньшее значение, так как он во все большей степени совпадает с зимним
периодом покоя. Если в пустыне этот период длится в среднем 15 недель,
то на высоте 1800 м он сокращается уже до 11, а на высоте 2400 л, где снег
лежит длительное время, —до 6 недель.
Вечнозеленые дубы зимой сохраняют листву и сбрасывают ее только
в апреле, когда молодые листья уже полностью развились. Наиболее ин-
тенсивный вегетативный рост дубов происходит во время летних дождей,
в июле—августе. Хотя ход климатических элементов в пределах энсиналь-
ного пояса несколько иной, чем в типичных районах с зимними дождями,
характер растительности и здесь и там очень близок, сходен и ее флори-
стический состав.
Шрив различал нижний и верхний энсинальные пояса.
а) Нижний энсиналъный пояс
Характерная его черта та, что произрастающие здесь дубы, как
и в чапаррале, имеют кустарниковидную форму и образуют редкий
древостой. Речь идет об отчетливо различаемом поясе, переходном от
верхнего пояса пустынь к собственно лесному поясу. Последний начина-
ется на северном склоне на высоте 1200 л, а на южном — на высоте 1450 м.
Главными представителями являются: Quercus oblongifolia, Qu. arizonica (напоми-
нает Qu. ilex), Juniperus pachyphloea, Arctostaphylos pungens и Vauquelinia californica
(семейство розоцветных). На прогалинах, помимо различных видов злаков, можно
найти ряд листовых суккулентов, как Yucca macrocarpa, Agave palmeri, A. schottii,
Dasylirion wheeleri и Nolina microcarpa.
В каньонах первые дубы встречаются уже на высоте 823 л, то есть
значительно ниже, чем в собственно лесном поясе. В постоянно влажных
ущельях этого пояса растут, что весьма характерно (см. стр. 72, среди-
земноморская растительность), листопадные виды Populus spec., Fraxinus
toumeyi, Juglans major, Sapindus drummondii, Platanus wrightii, Vitis arizo-
nica, а также вечнозеленый Cupressus arizonica. Иными словами, здесь
встречаются те же роды, что и в Средиземноморской области. Из вереско-
вых место Arbutus, который типичен лишь для более высокого пояса,
пояса сосновых лесов, занимает кустарниковидный Arctostaphylos.
б) Верхний энсиналъный пояс
Quercus oblongifolia в этом поясе отсутствует, Qu. arizonica представлен
только в нижней его части; замещаются эти виды Qu. reticulata, а в самой
верхней части пояса — Qu. hypoleuca. Доминирует во всем поясе Qu.
emoryi, листья которого напоминают листву Qu. coccifera, обитателя восточ-
носредиземноморского района. Помимо Juniperus pachyphloea, встречается
также Pinus cembroides. Часто наблюдаются кустарники Arctostaphylos
pungens и Garrya wrightii. На высоте 1680—1860 м уже распространены
120
Глава 3
плотные древостои высотой до 10 м. Напочвенный покров в этих лесах
очень редкий. На слабокарбонатных почвах произрастают Pteridium aquili-
пит var. pubescens, ряд пойкилогйдрических папоротников и Selaginella
rupicola, последняя предпочитает здесь открытые места, тогда как ниже
она держится в тени скал. Осадков в этом поясе выпадает больше, испаре-
ние, как показали измерения, меньше, следовательно, засуха выражена
слабее, чем в нижнем энсинальном поясе. Подробный список встречающих-
ся видов приведен в работе Шрива.
На высоте 1800 м на северном склоне и 1900 м на южном верхний эн-
синальный пояс переходит в пояс сосны, в котором доминирует Pinus
ponderosa var. scopulorum, образующая мощные, высокие стволы. Quercus
emoryi, Arctostaphylos и Garrya, а также Pinus cembroides исчезают, но Qu.
reticulata и особенно Qu. hypoleuca еще проникают в сосновый лес, где
образуют кустарниковый ярус. Последний вид на уровне 2000—2500 м
представлен даже деревьями высотой 12 м, их стволы достигают 1,2 м
в обхвате. Кроме того, появляется Arbutus arizonica, поднимающаяся до
высоты 2400 м. Ceanothus fendleri, как представитель столь широко распро-
страненного в чапаррале рода, образует в ярусе кустарников густые
заросли при условии, что дубовый подлесок отсутствует. В долинах ручьев
количество листопадных древесных видов возрастает. Помимо Platanus,
начиная с высоты 2000 м, можно встретить Qu. submollis, Alnus acuminatat
Juglans rupestris, Salix taxvfolia, Prunus virens и Rosa fendleri.
Пояс летнезеленых лиственных лесов полностью отсутствует, так
как период засухи проявляется здесь еще слишком резко. Только на
крутом северном склоне возвышенности Олд-Белди в соседних горах
Санта-Рита известен лес, образованный летнезеленым видом Quercus
gamblei.
Над поясом сосен должен бы следовать пояс с Pseudotsuga и Abies
concolor, но, поскольку горы Санта-Каталина слишком низки, эти краси-
вые леса распространены только на северном склоне, где поднимаются
на высоту 2250—2700 м. Зимой они долго покрыты снегом, поэтому
вечнозеленые энсинальные виды здесь совершенно отсутствуют. Вместо
них встречаются листопадные виды, такие, как Qu. submollis и Populus
tremuloides. На вершине горы Сан-Франциско (северная Аризона), на
высоте свыше 3000 м, над поясом пихты еще развит пояс с Picea engelman-
nii, образующий границу лесной растительности с альпийским поясом.
Безморозный период на высоте 1500 м длится около 210 дней, а на
уровне 2400 м — только 120 дней. Но в энсинальном поясе холодного
времени года не бывает, заморозки случаются лишь по ночам, днем из-за
сильной инсоляции всегда тепло. Только в поясе сосновых лесов наблю-
даются дни с морозом, то есть даже в полуденные часы температура здесь
не поднимается выше 0°.
Разность высот от нижней границы распространения вида до самых
высоких мест произрастания может достигать для отдельных видов 1500 м.
Такая гигантская амплитуда возможна лишь благодаря смене биотоповг
осуществляющейся согласно закону относительного постоянства место-
обитаний (ср. т. I, стр. 316). Так как у нижней границы вода находится
в минимуме, виды сначала появляются только во влажных долинах
ручьев или в каньонах, а несколько выше —на северных склонах; на
оптимальной высоте экспозиция роли не играет, но ближе к верхней гра-
нице решающим фактором становится температура, поэтому растения
предпочитают здесь более благоприятные южные склоны. Самые высокие
местонахождения располагаются на узких гребнях гор, приблизительно
Области средиземноморского климата за пределами Евразии
121
на 150—200 м выше местообитаний в долинах. Аналогичная закономер-
ность наблюдается в Альпах. Для тех условий можно предполагать, что
горные хребты представляют собой местообитания, защищенные от воз-
действия человека. В незаселенных горах Аризоны антропогенный фактор
значения не имеет. Следовательно, решающим оказываются естественные
причины — защищенность растений в долинах от потоков холодного
воздуха.
В Таксоне автор имел возможность изучить зависимость между вели-
чиной осмотического давления у энсинальных видов и хвойной расти-
тельности лесного пояса и распространенностью этих видов. Если границы
распространения какого-либо вида определяются внешними условиями,
а не конкуренцией, то величина осмотического давления оказывается наибо-
лее низкой там, где условия роста вида оптимальные. Границы распро-
странения можно изучать и в пределах очень небольшого пространства.
Рис. 88. Схема распространения Quercus oblongifolia в пред-
горьях Санта-Рита (Аризона) и величина осмотического давления
в листьях на различных местообитаниях.
Тем самым появляется возможность одновременно брать хорошие, сравни-
мые пробы и сопоставлять величины осмотического давления на различных
участках в пределах ареала вида.
В качестве примера на рис. 88 приведены данные для Quercus оЫоп-
gtfolia, образующего нижнюю границу энсинального пояса с разрежен-
ной растительностью полупустынь; вид преимущественно встречается на
северных склонах и лишь редко переходит на северо-восточные и северо-
западные стороны гор. В пограничной зоне древостой разреживается;
деревья, особенно на возвышенностях с маломощным почвенным покро-
вом, принимают кустарникообразную форму. На рис. 88 видно, что раз-
ность осмотических давлений у растений на наиболее неблагоприятных
(30,9 атм) и наиболее благоприятных (22 атм) местообитаниях превышает
8 атм\ для жестколистных видов такую разность следует рассматривать
как очень значительную. Многочисленные измерения показали, что
значения осмотического давления у различных видов вечнозеленого дуба
в зимние месяцы колебались в пределах 19,0—32,9 атм, у Arctostaphylos —
в пределах 24,2—48,8 атм, и лишь немногие измерения давления у Arbu-
tus дали величину от 20,7 до 25,8 атм. Если учесть воздействие сильных
ночных заморозков в этом поясе, то на основании наблюдений в Средизем-
номорской области можно считать, что полученные значения соответ-
ствуют ожидаемым.
Две следующие серии исследований проведены в северной Аризоне на
вершине Сан-Франциско, в пределах верхних лесных поясов (рис. 89, 90).
122
Глава 3
склонах пика Сан-Франциско и величина осмотического
давления в пробах, взятых на разной высоте.
Рис. 90. Схема распространения Pinus aristata на
склонах пика Сан-Франциско (Аризона) и величина
осмотического давления в пробах, взятых на разной
высоте.
На схемах отчетливо заметно, что Picea engelmannii предпочитает
северные, a Pinus aristata —южные склоны. У верхней границы распро-
странения вида, обращенной к альпийскому поясу, встречаются угнетен-
ные формы; их осмотическое давление
лением экземпляров на оптимальных
Таблица 8
Осмотическое давление
у Pinus aristata летом
(вершина Пайкс-Пик, Скалистые горы)
Осмотическое
Местообитание давление,
атм
У границы древесной ра-
стительности, высота 3560 л, измененные вет- ром деревья 21,7
В поясе сосновых лесов,
высота 2120 м, редко стоящие деревья .... 21,3
по сравнению с осмотическим дав-
местообитаниях резко повышено.
Отбор проб производился в де-
кабре, то есть зимой, когда на
границе с альпийским поясом
условия окружающей среды очень
суровые. Летом условия более
благоприятны. При измерениях,
выполненных 16 июля на горе
Пайкс-Пик (Скалистые горы, Ко-
лорадо), такие различия не уста-
новлены (табл. 8).
Следует, однако, заметить,
что точное определение зависимо-
сти между состоянием гидратуры
у растений и районом их распро-
Области средиземноморского климата за пределами Евразии
123
странения станет возможным только тогда, когда мы будем располагать
кривыми годового хода осмотического давления у растений, полученными
на различных местах произрастания (ср. гл. 5) \
ХВОЙНЫЕ ЛЕСА ТИХООКЕАНСКОГО ПОБЕРЕЖЬЯ
СЕВЕРНОЙ АМЕРИКИ1 2
Калифорнийская область жестколистной растительности на севере
переходит в зону хвойных лесов, простирающуюся через территорию
Британской Колумбии на север, в пределы Аляски. Зимой в этой зоне
также идут дожди, и чем дальше на север, тем их больше; в этом же на-
правлении уменьшается воздействие засухи. Соответствующий аналог
этим хвойным лесам, образующим непосредственный переход от средизем-
номорской к бореальной зоне, в Средиземноморье отсутствует. В запад-
ной части последнего субсредиземноморская зона переходит в зону лист-
венных лесов с максимумом осадков летом. Условия произрастания хвой-
ных лесов тихоокеанского побережья Северной Америки напоминают,
скорее всего, вертикальные пояса в восточном Средиземноморье, где
с увеличением осадков и понижением температуры за поясом жестколист-
ной растительности следуют различные пояса хвойных пород с видами
Pinus, Abies, Cedrus и Juniperus.
На побережье Калифорнии хвойные деревья встречаются уже север-
нее залива Монтерей, а далее к северу их преобладание становится все бо-
лее заметным, чему благоприятствуют густые туманы летом. Туман часто
конденсируется в кронах деревьев, растущих на опушках леса, и вода
каплями стекает вниз по стволам, обильно увлажняя почву; однако по
мере удаления от кромки леса количество влаги в почве, конденсируемое
из тумана, быстро уменьшается. Влагомеры, установленные под деревья-
ми, позволили определить следующее количество стекающей воды летом
в бездождный период (в мм) 3:
Опушка леса —> В глубь леса
1500 435 225 180 46
На некотором расстоянии от моря, куда туманы проникают лишь ред-
ко, леса уже не получают конденсационной Ьоды, но тем не менее и в этой
полосе летнюю засуху смягчают часто образующаяся облачность и большая
влажность воздуха.
На западе Северной Америки широко распространены хвойные породы:
Pseudotsuga menziesii (дуглассия), Abies concolor, Pinus ponderosa и P. con-
torta (рис. 91); центром распространения Tsuga heterophylla и Thuja plicata,
а также Picea sitchensis является северная часть Тихоокеанского побережья,
но эти виды проникают и в Калифорнию. Напротив, Sequoia sempervirens,
Sequoiadendron gigantea, Libocedrus decurrens, Pinus lambertiana, Abies
magnifica и другие виды встречаются только в Калифорнии или заходят
лишь ненамного севернее.
1 Н. Walter, Die Hydratur der Pflanze und ihre physiologisch-okologische Be-
deutung, Jena, 1931.
2 R. Knapp, Die Vegetation von Nord- und Mittelamerika, 1965; A. W. К u-
c h 1 e r, Potential natural vegetation of the conterminous United States, «Amer. Geog-
raph. Soc.», Spec. Publ. № 36, New York, 1964.
3 G.T.O b er lan d er, Summer fog precipitation on the San Francisco Peninsula,
«Ecology», 37, 1956, p. 851—852.
124
Глава 3
Рис. 91. Pinus contorta var. murrayana в каньоне реки Йеллоустон, Скалистые горы-
На полосе побережья от залива Монтерей до северной границы Кали-
форнии среди хвойных пород доминирует Sequoia sempervirens (см. рис. 80).
Ее стройные стволы нередко достигают в высоту 100 м. Sequoia образует
густые чистые древостои почти без подлеска, но массивы первичных,
девственных лесов в наше время почти не встречаются. В деградированных
в результате вырубки лесах иногда формируется подлесок из видов чапар-
раля; последний на подходящих местообитаниях даже замещает лес-
Области средиземноморского климата за пределами Евразии
125
Рис. 92. Хвойные леса, образованные A hies procerty,, в Каскадных горах (фото Фабера).
Севернее и выше в горах секвойя растет совместно с Pseudotsuga menziesii
var. viridis.
Sequoia sempervirens дает прекрасную деловую древесину, поэтому лес
на больших площадях вырублен. Плодоношение и возобновление у этого
вида характеризуются некоторым своеобразием. На взрослых деревьях
часто шишек не бывает. В известной мере напряженный водный режим,
по-видимому, определяет особенности цветения и плодоношения вида.
Даже при густом посеве проростки не переживают первого года в том
-случае, если почва в хороших насаждениях покрыта лесной подстилкой;
проростки, избежавшие нападения насекомых или грибов, погибают из-за
корневой гнили. Выживают они только на сырых почвах, например в ме-
стах отложения наилка в речных долинах ^ли там, где удален гумусный
горизонт х. Рост и прирост древесины протекают весьма интенсивно. За
30 лет ствол может достичь высоты 24 м и диаметра 40 см. Поросль дают
даже старые деревья, хорошо развивается и поросль от пня. Деревья
растут до 500-летнего возраста, а затем начинают засыхать. Самому ста-
рому дереву приписывается возраст 1800 лет. Родственный вид Sequoiaden-
dron gigantea, мамонтово дерево, реликтовый ареал которого находится
в горах Сьерра-Невада на высоте 1400—2500 м (см. рис. 80), достигает
возраста 4000 лет; максимальная высота его равна 88 м при диаметре
-ствола у основания свыше 4 м.
Севернее 42° с. ш. леса из секвойи на побережье сменяются хвойны-
ми лесами с Picea sitchensis, Thuja plicata, Tsuga heterophylla, в которых
встречаются также Abies grandis и Pseudotsuga menziesii.
При движении в глубь суши, на пространстве от залива Монтерей до
Орегона, в составе вечнозеленых дубовых лесов (образованы Quercus
1 D. W. Mulder, J. Н. Hansen, Observation on cone bearing of Sequoia
sempervirens, «Calif. Forestry and Forest Products», № 26, 1961; Biotic factors in natural
regeneration of Sequoia sempervirens, Vortrag Internet. Union Forest Res., Wien, 1961.
126
Глава 3
Рис. 93. Pinus ponderosa на вершине Сан-Франциско на высоте около 2500 м над
уровнем моря (фото Лаборатории по изучению пустынь, Таксон).
chrysolepis, Qu. wislizenii с Arbutus menziesii, Castanopsis, Lithocarpus,
Umbellularia и др.) в северном направлении появляется все большая при-
месь Pseudotsuga menziesii, пока и эти леса не сменятся лесами из Pseudo-
tsuga menziesii с Thuja plicata и Tsuga heterophylla, причем последние за-
паднее Каскадных гор простираются до Канады (рис. 92).
Сходная смена жестколистной растительности хвойными породами
наблюдается в Южной и средней Калифорнии на западном склоне Сьерра-
Невады на уровне 1000—1700 м, где сумма осадков с высотой увеличи-
вается, а температура понижается. В этом случае формируются разно-
образные смешанные древостои многочисленных видов хвойных, таких,
как Abies concolor, Libocedrus decurrens, Pinus lambertiana, P. ponderosa,
Pseudotsuga macrocarpa, P. menziesii и др. У нижней границы распростра-
нения этих лесов в подлеске еще встречаются жестколистные виды: Quer-
cus, Arctostaphylos и Ceanothus. Смешанный состав лесов в значительной
мере обусловлен пожарами, возникающими во время летней засухи. Часто
повторяющиеся пожары препятствуют накоплению опада, не нанося,
однако, большого ущерба древостою. Лесные пожары, несомненно, можно
рассматривать как естественный фактор, благодаря которому в природе
происходит отбор устойчивых к огню видов. В противоположность этому
современная борьба с лесными пожарами ведет к тому, что из-за большого
скопления горючего материала древостой при пожаре полностью уничто-
жается.
Области средиземноморского климата за пределами Евразии
127
Естественное возобновление этих хвойных лесов происходит без
труда г. Сосны развиваются преимущественно на пожарищах или на про-
галинах, образовавшихся в результате падения старых стволов. В север-
ной Калифорнии сходные, богатые видами хвойные леса встречаются
и в более низменной части Калифорнийской продольной долины; просле-
живаются они вплоть до юго-западных районов Орегона. Высоко в горах
их сменяют леса из Abies magnifica, а на сухих горных склонах — леса,
состоящие почти исключительно из Pinus ponderosa (рис. 93). Во всей
западной части материка последние очень широко распространены там,
где климат столь сухой, что обладающие большей конкурентной мощ-
ностью древесные виды (в том числе и Abies concolor) из древостоя выпада-
ют. В первичном состоянии этот лес может обладать весьма различной
структурой. Обычно в нем выделяются 2 яруса. Верхний древесный ярус
образуют деревья, возраст которых достигает 250—500 лет. Нижний
древесный ярус сильно угнетен. При гибели деревьев верхнего яруса
в пологе леса образуются окна, которые тотчас же заполняются видами
второго яруса. В Калифорнии в составе лесов из Р. ponderosa появляется
Pinus jeffreyi, близкая к Р. ponderosa.
Пояс Pinus ponderosa вместе с развитым над ним поясом из Pseudotsuga
menziesii встречается в горах более сухой западной части Северной Амери-
ки на пространстве от южной Аризоны до Британской Колумбии. На запа-
де Канады большую роль играет прежде всего Tsuga heterophylla, а в суб-
альпийском поясе— Ts. mertensiana 1 2. Лрса, раскинувшиеся севернее,
входят уже в бореальную зону.
СЕВЕРОАМЕРИКАНСКАЯ АРИДНАЯ ХОЛОДНАЯ ОБЛАСТЬ
ЗИМНИХ ДОЖДЕЙ
За линией Каскадных гор и Сьерра-Невады калифорнийская область
зимних дождей простирается еще дальше на восток, захватывает плато
Большой Бассейн и доходит почти до Скалистых гор. Но климат плато уже
УИННЕМАККА (1306м) 8,4°214
п[бд-б1]Неба-4-^х к
-8,3
-28.9
Юта
СОЛТ-ЛЕЙК-СИТИ (1300м) 10,6°414
[19-57]----
Рис. 94. Климадиаграммы области полупустыни с Artemisia tridentata: Рино, Уин-
немакка, Солт-Лейк-Сити (зона, переходная к злаковникам).
настолько холодный и осадков на более низменных и теплых позициях
выпадает так мало (рис. 94), что собственно жестколистная растительность
здесь отсутствует; в лучшем случае она встречается лишь местами.
Поднимающиеся над высокой возвышенной равниной горные хребты,
особенно их западные, открытые ветру склоны, получают больше осадков,
благодаря чему на них кое-где развиваются леса (рис. 95).
1 D.Mulder, Struktur, Verjtingung und Wuchspotential einiger Kalifornischer
Waldtypen, «Schrift. Forstl. Fak. Gottingen», 33, 1965, S. 115—128.
2 V. J. Kr a jina, Bioclimatic zonPs; and classification of British Columbia,
«Ecol. West. N.-Am.», Vancouver, 1, 1965, p., 1—17.
128
Глава 3
Рис. 95. Зависимость количества осадков (вверху) от характера рельефа (внизу) на
примере широтного профиля западной части Северной Америки приблизительно по
38° с. ш. (см. «Phytologie», Bd. Ill, 1).
Мы встречаемся здесь с вертикальной поясностью, напоминающей
последовательность вертикальных поясов в южной Аризоне; отсутствует
здесь только энсиналъный пояс, то есть пояс вечнозеленых дубов, для
которых зима с небольшим количеством осадков оказывается слишком
холодной. Его замещает пояс полыни, который в качестве зональной расти-
тельности занимает большую часть возвышенной равнины на высотах
1200—1800 м. Речь идет о полупустыне, в составе которой преобладает
Artemisia tridentata (рис. 96). Покров из Artemisia развивается преимуще-
ственно на тяжелых почвах пониженных участков рельефа, а на возвышен-
ностях с каменистыми почвами нередко мозаично чередуется с сообщест-
вами «пинион». Последние представляют собой очень разреженные, напоми-
нающие саванну древостои низкорослых хвойных («пигмейные хвойные»)
с сомкнутостью крон 40% и менее и максимальной высотой 9 м (на сухих
местообитаниях только 2 м). Насаждения пинион образуют два вида сос-
ны — Pinus edulis и своеобразная Pinus monophylla с отдельно стоящими
круглыми иголками, а также ряд древовидных видов Juniperus (J. uta-
hensis, J. monosperma, J. scopulorum). Местами наблюдается зарастание
территории кустарником Juniperus monosperma как следствие слишком
интенсивного выпаса (ср. т. I, стр. 289). Там, где скот не пасется, развив-
шиеся из семян растения Juniperus вследствие конкуренции со стороны
злаков растут плохо и при пожаре гибнут х.
Вертикальная поясность схематически показана на рис. 97: пояс
креозотового куста представляет собой полупустыню на самых низких
участках района (ниже 1000 м) с почти единственным видом Larrea tri-
dentata ( = Covillea glutinosd) из семейства парнолистных. В качестве
зональной растительности она проникает только до 37° с. ш., где сме-
няется полупустыней с Artemisia tridentata^ зима здесь уже очень суровая
(см. рис. 94).
В качестве промежуточного пояса местами развита кустарниковая
пустыня, в южной части штата Юта — с господствующим видом Coleogyne
1 Т. N. J ohnsen, One seed Juniper invasion of Northern Arizona grasslands,
«Ecol. Monogr.», 32, 1962, p. 187—207.
Области средиземноморского климата за пределами Евразии
129
Рис. 96я Полупустыня с Artemisia tridentata в долине Туэле, штат Юта.
На заднем плане — Juniperus utahensis (по Кирнею).
ramosissima (семейство розоцветных), а южнее — как аризонский чапар-
раль с вечнозелеными дубами Quercus turbinella и Qu. fendleri. На крайнем
юго-западе штата Юта пояс соприкасается с областью распространения
Рис. 97. Схема высотной по-
ясности растительности в горах
северной/Аризоны и штата Юта.
оригинального растения юкки (Clistoyucca brevifolia) (см. цветное фото 5),
За поясом полынной полупустыни и низких хвойных (пинион) следует
пояс Pinus ponderosa, о котором мы уже говорили. Но там, где для Pinus
ponderosa условия среды оказываются неблагоприятными, над поясом
9-01248
130
Глава 3
пинион снова появляются представители вечнозеленого чапарраля —
низкие кустарники Ceanothus velutinus, виды Arctostaphylos, Cercocarpus
и Pachystima, правда, последние уже совместно со многими листопадными
видами: Quercus gambelii, Acer grandidentata, Physocarpus malvaceus (семей-
ство розоцветных), а также виды Amelanchier, Symphoricarpus, Rhus,
Purshia (семейство розоцветных) и др. На схеме этот промежуточный пояс
назван предгорным поясом кустарников.
На более высоких хребтах выше располагаются пояса хвойных пород
с Pseudotsuga menziesii N&T.glauca и Abies concolor и, наконец, пояс Picea
engelmannii, образующий границу древесной растительности и альпийского
региона. Возможны местные отклонения от этой общей схемы 1.
Полынной растительности следует придавать особое значение, так
как она занимает обширнейшие, служащие пастбищами пространства
между хребтами Калифорнии и Скалистыми горами. Но кормовая цен-
ность Artemisia tridentata невелика; поэтому интенсивный и нерациональ-
ный выпас скота благоприятствует ей. В результате за счет прежних зла-
ковников полупустыня с господством Artemisia распространилась далеко
за пределы своих естественных границ 1 2.
Вид Artemisia — это полукустарник высотой около 1,5—2 м, дости-
гающий возраста 25—50 лет. Диаметр ствола у корневой шейки 7—15 см,
корни проникают в глубину на 3 м, но наряду со стержневым корнем имеет-
ся много боковых, развитых в верхних горизонтах почвы. В нормальных
зарослях на 100 м2 площади приходится до 75 особей. Частыми спут-
никами полыни являются виды Chrysothamnus (семейство сложно-
цветных).
В поясе полынной полупустыни, где годовое количество осадков
составляет 150—200 мм, в понижениях рельефа нередко можно видеть
обширные участки, занятые галофитами, и даже соленые озера. Обычно
это бедные видами сообщества на засоленных почвах, в которых преобла-
дают определенные виды семейства маревых: Allenroljea occidentalis,
Sarcobatus vermiculatus, виды Salicornia, Suaeda, Atriplex и Kochia, а также
галофильные злаки (виды Distichlis, Sporobolus, Spartind).
Можно было бы думать, что при столь широком распространении по-
лынной полупустыни по крайней мере экология и физиология господ-
ствующих видов исследованы достаточно подробно. В действительности
это не так. Большинство работ посвящено проблемам выпаса, причем
проблемам скорее местного значения. Но все же Харрис определил вели-
чину осмотического давления у важнейших видов растений этой области 3.
Только для одной Artemisia tridentata произведено 208 измерений. Речь
идет о типично малакофильном виде с мягкими, сильно опушенными,
приятно пахнущими листьями. Для этой группы ксерофитов характерно,
что при хорошем обеспечении растений влагой значения осмотического
давления у них весьма низкие (10—15 атм). Однако при недостатке воды
в почве эти значения очень быстро возрастают. Поэтому наиболее часто
приводимые Харрисом величины колеблются от 20 до 35 атм', максималь-
ное измеренное им осмотическое давление составляло 73 атм. У растений
с таким высоким осмотическим давлением листья обычно уже засохшие
1 А.М. Woodbury, Distribution of pigmy conifers in Utah and Northeastern
Arizona,«Ecology», 28, 1947, p. 113—126.
2 J. H. Robertson, Responses of range i1 asses to different intensities of com-
petition with sagebrush (Artemisia tridentata Nutt.), «Ecology», 28, 1947, p. 1 — 16.
3 J.A.Harris, The physico-chemical properties of plant sap in relation to phy-
togeography, Minneapolis, 1934.
Области средиземноморского климата за пределами Евразии
131
и опадают; засуху переносят только верхушечные почки, которые при
улучшении влагообеспеченности снова оживают.
Artemisia tridentata развивается на бурых полупустынных почвах,
не содержащих легкорастворимых солей, но характеризующихся наличием
на сравнительно большой глубине карбонатного горизонта. Так как
осадки зимой выпадают в виде снега, вода при его таянии проникает срав-
нительно глубоко в почву, хотя общее количество осадков относительно
невелико; растения, имеющие стержневой корень, в период засухи могут
поглощать воду из этих глубоких почвенных горизонтов.
Для расчленения растительного покрова весьма важен такой пока-
затель, как содержание солей в почве.
Были изучены свойства почвы на местообитаниях разных видов расте-
ний полупустыни, встречающихся в почти чистых естественных насажде-
ниях; как показывают данные табл. 9, местообитания различаются между
собой прежде всего содержанием солей в почвенном профиле х.
Таблица 9
Содержание солей в почве на глубине 15—45 см в северных
полупустынях Северной Америки под различными видами растений,
образующих естественные насаждения
Artemisia tridentata Eurotia lanata Atriplex conferti- folia Atriplex nutallii Sarcobatus vermicu- latus
Общее содержание лег- корастворимых солей, % 0,13 0,19 0,37 0,69 0,82
Электропроводность поч- венной вытяжки (КХ103) 2,8 4,6 8,1 14,4 13,2
Содержание натрия, мг!кг 298 586 995 1693 1865
Для растений концентрация солей в почвенном растворе, из которого
они получают воду, гораздо важнее, чем количество солей, накопленных
почвой. В течение года его концентрация в зависимости от содержания
воды в почве очень сильно колеблется, но в данном случае она не изуча-
лась. Необходимы также детальные исследования характера размещения
корневой системы в почве. Глубина проникновения корней названных
видов превышает 2 м. Виды Atriplex — ксерогалофиты, a Sarcobatus —
гигрогалофит. Весьма обширный район засоленных почв обнаружен в шта-
те Юта вокруг Большого Соленого озера.
ЖЕСТКОЛИСТНАЯ РАСТИ ТЕЛЪНОСТЬ СРЕДНЕГО ЧИЛИ
И СОСЕДНИХ ОБЛАСТЕЙ1 2
Аналогом калифорнийской области жестколистной растительности
северного полушария в южном полушарии примерно на тех же широтах
является среднечилийская область. Начинается она на побережье у
1 D. Н. G a t е s, L. A. S t о d d а г t, С. W. Cook, Soil as a factor influencing
plant distribution on salt-deserts in Utah, «Ecol. Monogr.», 26, 1956, p. 155—175.
2 Краткий обзор растительности Чили читатель найдет в работах: К. Hueck,
Die Walder Siidamerikas, см.: H. W a 1 t er, Vegetationsmonogr. d. einz. Groflraume,
9*
132
Глава 3
31° ю. ш.( а в долинах Главной
цепи Кордильер—у 33° ю. ш.
и простирается к югу вдоль
берега океана до 37° ю. ш.,
а во внутренней продольной
долине — до 38° ю. ш. Это
также типичная область
с зимними дождями, причем
годовое количество осадков
увеличивается с севера на юг
(рис. 98). В отличие от Кали-
форнии Чили, за исключе-
нием его южной части, резко
отделено от внутренних об-
ластей континента высокой
горной цепью Анд.
Севернее 31° ю. ш. коли-
чество осадков сокращается
до 200 мм и менее; здесь раз-
вита колючекустарниковая
пустыня с суккулентами, ко-
торая у Антофагасты пере-
ходит в особо суровую (экст-
ремную) пустыню, практи-
чески лишенную осадков.
Последняя протягивается
вплоть до центральных райо-
нов Перу (см. т. I, стр. 411
и след.). Напротив, южнее
38° ю. ш. сумма осадков резко
возрастает, летняя засуха не
выражена. Жестколистные
леса сменяются сначала лет-
Р и с. 98. Области| растительно-
сти Чили (по Шмитхюзену, с из-
менениями) и климадиаграммы.
Северное Чили: 1 — северные Высокие
Анды; 2 — пояс пустынь; з — область
кустарничков Малого севера и ксеро-
фитная кустарниковая растительность.
Среднее Чили: 4 — область жестколи-
стной растительности; 5 — область
летнезеленого леса. Южное Чили:
6 — вечнозеленые дождевые леса уме-
ренной зоны; 7 — тундровидная расти-
тельность холодной зоны; 8 — субан-
тарктический летнезеленый лес; 9 —
патагонская степь; 10 — южные Анды.
Bd. II, Stuttgart und Jena, 1966; К. R eiche, Grundziige der Pflanzenverbreitung in
Chile, «Die Veget. d. Erde», VIII, Leipzig, 1907; O. Berninger, Wald und offenes
Land in Sud-Chile seit der spanischen Eroberung, «Geogr. Abh.» 3, Reihe 1, Stuttgart,
1929; E. Werdermann, Die Pflanzenwelt Nord- und Mittelchiles, «Veget. Bilder»,
21. Reihe, H. 6/7, 1931; J.Schmithiisen, Die raumliche Ordnung der chilenischen
Vegetation, «Bonn. Geogr. Abh.», H. 17, 1956, S. 1—86; E.Oberdorfer, Pflanzen-
soziologische Studien in Chile, Flora et Vegetatio Mundi, Bd. II, 1960.
Области средиземноморского климата за пределами Евразии
133
незелеными лиственными лесами с Nothofagus obliqua, которые далее к югу
переходят в умеренные вечнозеленые дождевые леса.
Таким образом, территорию Чили, которая занимает прибрежную
полосу шириной 200 км и протягивается от 18 до 57 ° ю. ш., то есть более
чем на 4300 км, можно подразделить на три крупные части, в свою очередь
распадающиеся на более мелкие. Поскольку эколого-физиологические
исследования в Чили практически не проводились, приходится ограни-
читься краткой характеристикой растительности и ее связи с климатом.
Допускаемые нами в дальнейшем параллели с Северной Америкой вполне
оправданны, поскольку Чили во многих отношениях является точной
копией Калифорнии, однако флора здесь совершенно иная.
Вся Северная Америка относится к голоарктическому флористическому царству,
и только в его южных аридных районах чувствуется влияние неотрописа; Чили же
входит в неотропическое царство с большой долей антарктических элементов на край-
нем юге Ч
Чисто неотропической является флора районов пустынь северного Чили, поэтому
обнаруживаются некоторые ее черты, общие с флорой пустынь юго-запада Северной
Америки. Остальные неотропические элементы Чили относятся к флоре влажных тро-
пиков Южной Америки. Но поскольку после образования Анд в третичном периоде
климат Чили сильно изменился, а неотропические элементы оказались полностью изо-
лированными от влажных южноамериканских тропиков, в Чили сформировался очень
резко выраженный эндемизм. Наблюдается странное явление: представители обычно
тропических семейств, таких, как Gesneriaceae, Bignoniaceae, Elaeocarpaceae, Sapota-
ceae, Monimiaceae, Lauraceae, Loganiaceae, Myrtaceae, Flacourtiaceae, Icacinaceae,
Bambuseaen др., в Чили приспособились к средиземноморскому, а на юге материка —
даже к умеренному климату. Наряду с этим в геологический период, когда еще суще-
ствовал мост суши через Антарктику к Австралии и Новой Зеландии, в Южную Амери-
ку с юга мигрировали антарктические элементы, в среднем Чили, однако, почти уже
не встречающиеся. Поэтому не удивительно, что южное Чили имеет очень тесные фло-
ристические связи именно с Новой Зеландией: здесь насчитывается, например,
421 общий род. Из них 28 одновременно встречаются и в Австралии, а 3 — также в Аф-
рике. Напротив, в Южной Америке есть только 8 родов, общих с Австралией, которые
в Новой Зеландии неизвестны, и только один, представленный также и в Африке,
и в Австралии, но не в Новой Зеландии. Сходство с флорой Новой Зеландии в направле-
нии к югу все более усиливается и особенно резко проявляется у антарктических поду-
шечных растений болот1 2.
Эти флористические различия помогают понять, почему в Калифорнии
чапарраль с увеличением количества осадков переходит в хвойный лес,
тогда как в Чили среднечилийская жестколистная растительность сме-
няется вечнозеленым или листопадным смешанным лесом, в котором наря-
ду с тропическими элементами очень важное место занимает прежде всего
антарктический род Nothofagus.
Охарактеризуем вкратце условия среды в аридном северном Чили.
Перуанская особо суровая (экстремная) пустыня (см. т. I, стр. 411 —
422) заходит частично и на территорию Чили. Местность поднимается
здесь двумя уступами от уровня моря до высоких гор. Береговая полоса
в северной половине лишена растительности, только в низинах (чилькас)
встречаются небольшие кусты сложноцветных (Tessarta absinthoides, Вас-
charis petiolata, Pluchea chingoy о); свободные пространства между ними
часто занимает солончаковый злак Dtstichlts. Там, где земля в долинах
рек, стекающих с Анд, не возделана, Schinus molle (семейство сума-
ховых) и бобовые Prosopis fuliflora и Р. tamarugo, а также чанар, Geoffroea
1 Среди них роды Nothofagus, Drimys, Aristotelia, Discaria, Azorella и др. См.:
R. G о о d, A geographical survey of the flora of temperate South Amerika, «Ann. of
Bot.», 47, 1933, p. 691—725.
2 E. J. G о d 1 e y, The botany of Southern Chile in relation to New Zealand and
the Subantarctic, «Proc. Roy. Soc. В», 152, 1960, p. 457—475.
134
Глава 3
Рис. 99. Формация «тола» с различными кустарничками семейства сложноцветных
в пуне на высоте около 3700 м, плоскогорье Альтиплано в Боливии (фото Тролля).
( = Gourliea) decorticans, образуют небольшие массивы галерейных лесов.
На высоте 1900 м встречаются кактусы, особенно хорошо развивающиеся
на уровне 3000 м, где выпадает уже больше осадков. В пуне, на высотах,
превышающих 3500 м, господствует Baccharis tola (рис. 99), а на уровне
4000 м начинается альпийский пояс с подушечными растениями из семей-
ства зонтичных: Laretia compacta и Azorella. Здесь и в северо-восточной
части до 22°ю.ш. примечательно дерево семейства розоцветных Polylepis
incana («кеньоа»).
На береговых хребтах от Икике до Тальталя, то есть приблизительно
в полосе от 20 до 25° ю. ш., так же как в Перу (см. т. I, рис. 331), распро-
странена растительность туманов, представленная цветковыми растения-
ми, многочисленными лишайниками и водорослью Trentepohlia.
Выпадающие изредка в пустыне интенсивные зимние дожди (как,
например, в 1965 г.) определяют развитие красочной, но короткоживущей
растительности эфемеров, состоящей из маленьких однолетников или
геофитов (Alstroemeria, Hippeastrum, Nolana alba, Viola с желтыми цвет-
ками и др.) г.
Южная часть северного Чили, где дожди идут несколько чаще, пред-
ставляет собой уже кустарничковую полупустыню, растительный покров
которой образован преимущественно видами сложноцветных (Haplopap-
1 Характерной особенностью растительности пустыни в области туманов между
Антофагастой и Кокимбо являются «земляные» кактусы, так глубоко погруженные
в землю, что на поверхности они почти незаметны. От утолщенного стержневого корня
на глубине всего 1,5 см отходят боковые корни, задача которых, вероятно, состоит в том,
чтобы поглощать влагу туманов. См.: P.W eisser, Zur Kenntnis der Erdkakteen in
Chile «Вег. Deutsch. Bot. Ges.», 80, 1967, S. 331—338.
Рис. 100. Trichocereus chilensis и различные кустарники западнее Овалье (фото
Мортенсена и Бернингера).
Рис. 101. Puya chilensis (бромелиевые) и цветущие Heliotropium stenophyllunr, южная
часть провинции Кокимбо (фото Скотсберга).
136
Глава 3
pus), к которым южнее присоединяются кустарники и суккуленты, такие,
как колоннообразные кактусы (рис. 100), Риуа (рис. 101) и др., а весной,
после зимних дождей,— многочисленные весенние эфемеры.
Переходной зоной к жестколистной растительности можно считать
заросли колючих кустарников с господствующим видом Acacia caven.
В среднем Чили условия среды осложняются тем, что южнее долины
Аконкагуа (приблизительно на 33° ю. ш.) местность расчленяется в широт-
ном направлении на Береговой хребет, Чилийскую продольную долину
и западный склон Анд. Вновь возникает аналогия с условиями в средней
Калифорнии. Продольная долина располагается в дождевой тени берегово-
го хребта, и поэтому здесь суше. Этим объясняется, почему все границы
растительности в продольной долине резко изгибаются к югу.
На территории среднего Чили с присущим ему средиземноморским
климатом 1 осели в свое время первые испанские колонизаторы. Таким
образом, эта ранее всего заселенная, ныне с наибольшей плотностью насе-
ления часть Чили. Поэтому естественная растительность пострадала здесь
особенно сильно, хотя и не в такой мере, как в Средиземноморье. Возник-
ший культурный ландшафт имеет вполне средиземноморский облик, тем
более что местные поселения напоминают своим видом деревни и города
Испании. В садах и парках мы встречаем те же декоративные растения,
например Phoenix canariensis, Magnolia grandtflora, платаны, шелковицу
и Eryobothria japonica, из кустарников — олеандр и гортензию, Ligustrum
japonicum и Evonymus japonica, у домов — Wistaria и Bougainvillea. Куль-
турные растения, например цитрусовые, маслины, орехи, виноград, пше-
ница и т. д., не говоря уже о сорняках и рудеральных растениях, также
представлены средиземноморскими видами.
Первоначально территория среднего Чили была покрыта низким
жестколистным лесом. Кустарниковая растительность, похожая на
чапарраль, отсутствовала. Преобладают почвы типа красноземов и желто-
земов. Различают ксерофитные леса «литре» (Lithraea caustica, семейство
сумаховых), «киллай» (Quillaja saponaria, розоцветные), «больдо» (Peumus
boldus, Monimiaceae) и более влажные леса, «пеймо» (Cryptocarya rubra)
и «беллото» (Beilschmiedia miersii) (оба вида — семейства лавровых)
(рис. 102) 1 2. Чисто внешне эти леса, достигающие 10—15 м высоты, напо-
минают средиземноморские леса из Quercus ilex. Сходство особенно заметно
у Peumus boldus как в отношении формы и цвета листвы, так и формы кро-
ны. Более похожи на кустарники виды родов Azara (Flacourtiaceae),
Escallonia (камнеломковые), Colliguaja (молочайные), Kageneckia (розо-
цветные), Schinus (сумаховые), Maytenus (бересклетовые), Muehlenbeckia
(гречишные) или ядовитая для скота Oestrum parqui из семейства паслёно-
вых. Колючими кустарниками являются Trevoa trinerva (крушиновые)
и Proustia pyrifolia (сложноцветные); Lucuma valparadisea (сапотовые)
имеет съедобные плоды; вблизи воды растут виды Myrceugenia (семейство
1 На недавно опубликованной ЮНЕСКО карте областей с зимними дождями тем
же условным знаком, что в Чили, обозначены значительно более обширные районы
в Аргентине, главным образом в Патагонии, хотя они отличаются совершенно иным
климатом и другой растительностью. Это может создать неправильное представление
об истинном положении. См.: «Arid Zone Res.», XXI, UNESCO-FAO, 1963.
2 Lithraea caustica (семейство сумаховых), если дотронуться до ее листьев, вызы-
вает у большинства людей сыпь на всем теле и сильную лихорадку, иными словами, она
столь же ядовита, как растущий в Северной Америкег кустарник Rhus toxicodendron
(также сумаховые). Кору Quillaja saponaria — мыльного дерева, — содержащую сапонин,
используют вместо мыла и применяют в медицине. Из листьев Peumus boldus приготов-
ляют чай.
Рис. 102. Некоторые виды чилийской жестколистной растительности.
1 — Quillaja saponaria; 2 — Escallonia arguta; 3 — Kageneckia oblonga; 4 — K. angu-
stifolia; 5 —Colliguaja odorifera; 6 — C. integerrima; 7 — Lithraea caustica; 8 — Satureja
oirgata; 9 — Baccharis rosmarinifolia; 10 — Aristotelia maqui (Elaeocarpaceae).
138
Глава 3
сапотовых) и Fuchsia. Своеобразна единственная пальма Чили — Jubaea
spectabilis. Это эндемик, обитающий в узкоограниченном районе берегового
хребта, прежде всего в долине Окоа. северо-восточнее Вальпараисо. Спут-
ники ее — обычные жестколистные виды. У автора создалось впечатление,
что этот вид, как и большинство пальм, предпочитает влажные местооби-
тания в долинах рек или кажущиеся очень сухими, заполненные обломоч-
ным материалом ложбины на склонах гор.
От жестколистных лесов Чили в северной части области сохранились
лишь небольшие остатки во влажных ущельях или на северных склонах
гор, па обширных же пастбищных угодьях в продольной долине широко
распространены разреженные естественные насаждения колючей зонтико-
образной Acacia caven высотой около 3 м. Ландшафт с зарослями акации
напоминает кустарниковую саванну тропической области с летними
дождями. Но это не настоящая саванна, так как здесь отсутствует сомкну-
тый многолетний злаковый ярус; состав травяного покрова образуют
занесенные из Европы однолетние злаки и другие терофиты. Это сооб-
щество, необычное для области зимних дождей, большинство ботаников
считают стадией деградации жестколистной растительности, обусловлен-
ной воздействием огня. Зоологи же, основываясь на своеобразии фауны,
рассматривают сообщество как естественное. Решить этот вопрос нелегко.
Но если учесть, что в Сантьяго, расположенном в продольной долине,
количество осадков составляет 359 мм при средней годовой температуре
14°, то трудно себе представить, чтобы в таких условиях жестколистный
лес был зональной растительностью. Однако этого количества осадков
достаточно для развития разреженного древостоя Acacia, в котором хоро-
шо развитые корневые системы растений, возможно, смыкались, так что
это могло быть замкнутое сообщество. Избытком же влаги в зимние месяцы
пользуются терофиты, представленные севернее эндемичными видами;
в окрестностях Сантьяго, где дожди выпадают чаще, последние были
вытеснены европейскими видами Ч
Согласно такой интерпретации, на широте Сантьяго можно обнару-
жить следующие варианты естественного деления растительности по
местообитаниям: на равнинных площадях — кустарник из Acacia; на
каменистых сухих северных склонах — экстразональное сообщество сукку-
лентов с Porlieria, Trevoa, Colletia и др.; на более влажных южных склонах
и в ущельях — жестколистная растительность (экстразональная). Можно
предположить, что колючие кустарники акации в результате пастьбы
скота и выжигания расселились далеко за границы района своего прежне-
го распространения. Двигаясь по продольной долине к югу, то есть
в направлении увеличения количества осадков, легко заметить, что жестко-
листная растительность все активнее занимает северные склоны, завоевы-
вая новые и новые площади. Заросли акации становятся все более густыми
(можно говорить чуть ли не о закустаривании территории), и, наконец,
там, где годовая сумма осадков превышает 700 мм, даже на равнинных
участках их начинает вытеснять жестколистная растительность, о чем,
правда, в настоящее время свидетельствуют только небольшие группы
деревьев или отдельно стоящие деревья. Следовательно, вполне вероятно,
что зона колючих кустарников, образующая переход от полупустыни
к области развития жестколистной растительности, в наиболее засушливых
частях продольной долины выдвигается сравнительно далеко на юг. На
1 Аридность этого района подтверждает явление засоления почв, никогда не на-
блюдающееся в районах собственно жестколистной растительности.
Области средиземноморского климата за пределами Евразии
139
побережье же, где сухость даже в разгар лета несколько смягчается часты-
ми туманами, а также на западном склоне Анд условия благоприятны для
существования жестколистной растительности.
Вертикальная поясность в горах северной части зоны жестколистной
растительности (до 34—359 ю. ш.) соответствует аридному средиземно-
морскому типу (ср. стр. 78). Жестколистная растительность проникает
до высоты 1400 ж, где переходит в формацию ксерофитных кустарников
с Valenzuellia trinervia (семейство сапиндовых) и видами Berberts. На высоте
2000 м ее сменяют альпийские сообщества. Поскольку западный склон Анд
очень крутой, в альпийском поясе распространена главным образом расти-
тельность осыпей: желтые Tropaeolum, фиолетовые Schtzanthus (паслено-
вые), а также амариллисовые (Alstroemeria ъ Hip peastrum uniflorum и др.).
На ровных площадях появляются некоторые патагонские элементы
(Mulinum spinosum, Chuquiraga и др.). Необыкновенно красочны кармино-
вые цветки Calandrinia (портулаковые), оранжевые Quinchamalium (санта-
ловые), розовые цветки видов Oxalis, желтые и черно-лиловые кальцеоля-
рии (норичниковые), сверкающие подушки Viviania rosea (гераниевые)
и др.; менее эффектны желтые и белые орхидеи, беловатые лилейные,
голубые и желтые касатиковые; сложноцветное Perezia diverstfolia называ-
ют эдельвейсом Анд. В верхнем поясе мы встречаем характерные плоские
подушки Azorella, Pozoa, Laretia (все из семейства зонтичных) и Anarthro-
phyllum (бобовые).
В заповеднике Алеман на высоте 200Q м еще растут серебристый
тополь, дуб, липа, вяз, вишня, яблоня, Syringa', культивируется кукуру-
за и люцерна. На северных склонах долины Маипу приблизительно на
высоте 1700 м можно видеть остатки пояса хвойных лесов из Austrocedrus
(Libocedrus) chilensis. В южной части пояса жестколистной растительности
в соответствии с типом гумидной вертикальной поясности между поясом
жестколистной растительности и альпийским регионом вклинивается пояс
умеренного дождевого леса южного Чили (см. рис. 108).
Весьма своеобразные климатические условия в зоне побережья Чили
создает холодное морское течение Гумбольдта. Благодаря сравнительно
низкой температуре своих вод оно способствует образованию береговых
туманов. Число дней без тумана в течение всего года здесь сравнительно
невелико. Туман, который обычно держится на определенной высоте,
существенно ослабляет поток солнечной радиации. Но этим его экологи-
ческое воздействие не ограничивается. Влага, приносимая туманом, кон-
денсируется на крутых скалистых склонах горных хребтов и стекает по
поверхности почвы. Благодаря этому почва получает дополнительную,
помимо дождевых осадков, влагу, причем в значительном количестве.
В результате в пределах этого пояса туманов происходит пышное раз-
витие растительности. Сходную картину мы уже видели в Перу(см.т.1,
стр. 411).
В зависимости от макроклимата того или иного участка последствия
влияния приносимой туманами влаги весьма различны (рис. 103).
В экстремпой пустыне под 18° ю. ш. растут виды Tillandsia, растения, кото-
рое существует исключительно за счет влаги туманов (рис. 104) Ч Несколь-
ко южнее, под 22° ю. ш., в поясе туманов встречаются сообщества стебле-
вых суккулентов, которые поглощают влагу из почвы, увлажняемой тума-
ном, при помощи корней (рис. 105). В области полупустынь, как и в Перу,
1 О. Berninger, Extreme Ausbildung der Nebelvegetation in der nordchile-
nischen Waste, «Z. Ges. f. Erdk.», 1925, S. 382.
140
Глава 3
н
2000п
1000-
Пустыня cTillandsia
0 области туманов
Формация
стеблевых
суккулентов
Токопилья Папосо Тальталь
Пояс пустынь
Растительность
„лома"
28е I
Уаско
зо7] '
Фрай-Хорхе
Область
кустарничков
Лес из Кампана
Quillat . п м ,, t
\ I Лес из Nothofaqus
ЖесткоХ Ju obliqua
листный я rcs
лес о J
1 ^Жестколистная
туманоо у в растительность
-----о. о о
n°o°oV°S о
Раститель-
ность района
Ла-Серена
Область жестколист-
ной растительности
Рис. 103. Меридиональный высотный профиль северочилийского побережья с дан-
ными о среднем высотном положении берегового тумана (между пунктирными линиями)
и растительности пояса туманов (косая штриховка). На горе Кампана расположен
самый северный лес из Nothofagus (по Шмитхюзену, с изменениями).
развивается так называемая растительность «лома». Повсеместно распро-
странены многочисленные лишайники, и уже почти у границы зоны жестко-
листной растительности в районе Фрай-Хорхе расположен знаменитый
«лес туманов», о котором так много написано х. Особый интерес к нему
вызван тем, что в климате с малым количеством осадков, в среднем около
150 мм!год, он имеет такой же состав, как леса влажного южного Чили
под Вальдивией, существующие в климатических условиях с годовой
суммой осадков 1000—2500 мм.
Для измерения количества воды, конденсирующейся из тумана,
Куммеров использовал три прибора: туманомер модели «Хоэнпрейсенберг»
конструкции Грунова, который он установил на опушке леса (I), дожде-
мер, установленный под кроной 5-метрового дерева в 8 м от края леса (II),
и обычный дождемер для определения дождевых осадков (III) 1 2. Измере-
ния продолжались 15 недель; показания снимались еженедельно, причем
в первый срок наблюдений (с 13 по 29 октября 1961 г.) постоянно моро-
сило. Результаты приведены в табл. 10.
По данным табл. 10 можно вычислить среднее количество воды за
неделю: для I—32,5 мм, для II — 50,2 мм и для III —0,9 мм.
Если предположить, что количество воды в остальное время года
было таким же (а подобное допущение при регулярности действия туманов
вполне обоснованно), то годовое количество влаги составит: для I — около
1800 мм и для II — приблизительно 2800 мм. Последнее значение, возмож-
но, завышено и, конечно, в известной мере случайно, так как вода в преде-
лах кроны дерева стекает неравномерно.
1 J. Schmithusen, Die raumliche Ordnung der chilenischen Vegetation,
«Bonn. Geogr. Abh.», H. 17, 1956; Munos у Pisano, Estudio de ]a vegetation
у flora de los parques nacionales de Fray Jorge у Talinay, Santiago, 1947: J. Kum-
mer о w, V. Matte, F. S c h 1 e g e 1, Zum Problem der Nebelwalder an der
zentralchilenischen Kuste, «Вег. Deutsch. Bot. Ges.», 74, 1961, S. 135 — 145;
G. Foilman, P. Weisser, Oasis de neblina en el Norte de Chile, «Bol.
Univers, de Chile», № 67, 1966.
2 J. К u m m e г о w, Quantitative Messungen des Nebelniederschlages im Walde
von Fray Jorge an der nordchilenischen Kuste, «Die Naturwissenschaften», 49, 1962,
S. 203—204.
Рис. 104. Tillandsia werder mannii, растущая в полосе туманов на севере Чили, близ
Такны, образует небольшие куртины на сухой песчаной почве (фото Мортенсена
и Бернингера).
Рис. 105. Пустыня в области туманов на побережье Тальталя. На конусе выноса —
стеблесуккулентные кактусы (фото Мортенсена и Бернингера).
142
Глава 3
Таблица 10
Количество воды, образовавшейся в результате конденсации тумана
в районе Фрай-Хорхе в период с 13 октября 1961 г.
по 27 января 1962 г. (в мм)
Дни измерений I II III Дни измерений I II III
29.Х.1961 г. 108,5 239,4 8,8 17.XII.1961 г. 24,5 37,8 0,0
5.XI.1961 г. 18,5 9,7 1,1 24.XII.1961 г. 18,0 34,4 0,0
12.XI.1961 г. 23,9 12,5 2,2 31.XII.1961 г. 27,4 26,4 0,0
19.XI.1961 г. 58,8 178,7 0,0 7.1.1962 г. 34,6 14,8 0,0
26.XI.1961 г. 50,0 97,0 0,0 14.1.1962 г. 24,8 28,7 0,0
3.XII.1961 г. 33,4 28,5 0,0 21.1.1962 г. 14,4 0,0 0,0
10.XII.1961 г. 37,2 34,4 0,0 27.1.1962 г. 27,0 11,0 1,1
В 1962—1965 гг. с целью получения более точной величины конден-
сации за год Куммеров продолжил измерения с помощью свободно стояще-
го туманомера. На рис. 106 отражены месячные количества. Годовые сум-
мы осадков (дождя и влаги тумана) приведены в табл. И.
Таблица 11
Сумма дождевых осадков и осадков,
приносимых туманом, за год (в мм)
Вид осадков 1962 г. 1963 г. 1964 г. 1965 г. 1
Дождь 79 272 59 326
Туман 598 535 860 651
Сумма .... 677 807 919 977
1 Только за январь — сентябрь.
1962 г. был очень сухой, но все же измерения и в этот год показали,
что осадков, приносимых туманом, вполне достаточно для существования
лесной растительности, особенно если учесть, что кроны деревьев наиболее
эффективно конденсируют влагу из тумана, которая затем стекает по ство-
лам и без потерь проникает в почву. Кроме того, растения в период дей-
ствия тумана не транспирируют, следовательно, потери влаги у них не
происходит.
Продолжительность действия тумана на том же местообитании реги-
стрировалась с ноября до октября 1964 г. самописцем увлажнения системы
Вёльфке. Детальное изучение данных измерения показало, что увлажнение
влагой тумана происходило преимущественно с 18 до 9 часов. В течение
20 месяцев только 10,5% суммарной длительности действия тумана падало
1964 г.---
Р и с. 106. Данные измерений в «лесу туманов» района Фрай-Хорхе (по Кум-
мерову, не опубликовано).
Вверху — суммарная (солнечная и рассеянная) радиация, пробелы — пеинтерпретируемые
участки ленты; в середине — длительность увлажнения; внизу — суммы дождевых осадков
и осадков из тумана за месяц.
144
Глава 3
Рис. 107. «Лес туманов» района Фрай-Хорхе (Чили). Преобладающая древесная
порода — Aextoxicum punctatum', на основании стволов — Peperomia coquimbensis',
моховой покров образован Mielichhoferia demissa (фото Фольмана).
на дневные часы — с 9 до 18 часов. Отсюда становится понятным высокое
значение суммарной радиации, измерявшейся актинографом Робича.
В летние месяцы она соответствует суммарной радиации в Гамбурге,
а в зимние месяцы приблизительно в два раза выше. Во всяком случае,
ее достаточно для фотосинтеза деревьев леса.
Важнейшие древесные породы «леса туманов»: Aextoxicum punctatum (Euphor-
biaceae), Myrceugenia correifolia (миртовые), Drimys winteri (магнолиевые, Wintera-
ceae), Griselinia scandens (кизиловые) — дерево-удушитель на Myrceugenia, Fuchsia
lycioides и Raphithamnus spinosus (вербеновые) (рис. 107). Кроме того, встречаются
еще 10 видов, которые одновременно характерны для дождевого леса района Валь-
дивии, удаленного отсюда более чем на 1000 км. Эпифиты представлены двумя видами
Peperomia, Sarmienta repens (геснериевые) и двумя видами папоротников (Hymenophyllum
и Asplenium). Самый старый лес из Aextoxicum очень густой, высотой около 25 м,
диаметр ствола одного из старых деревьев около 1 м. Все ветви густо покрыты мхами и
Usnea. В один из дней января 1966 г. стоял такой густой туман и вода так обильно капа-
ла с деревьев, что Куммеров и автор, осматривавшие лес, промокли до нитки.
«Лес туманов» начинается на западном склоне гор на высоте 400 м
приблизительно в 3 км от моря. Гребень горы возвышается на 500—680 м.
На восточном склоне туман низко не опускается. Температура воздуха
2 января 1966 г. в тумане была 12°, а на солнечном восточном склоне, где
уже начинается полупустыня с высокими колонновидными кактусами,—
25° (см. рис. 100).
Из числа важнейших видов растений следует назвать: Trichocereus chilensis, Flou-
rencia turifera (сложноцветные), Adesmia bedwellii (эндемик), Cassia coquimbensis, Proustia
pungens, Porlierahygrometrica и кустообразный Heliotropium stenophyllum (см. рис. 101).
Области средиземноморского климата за пределами Евразии
145
Adesmia поражена двудомным паразитом Pilostylis berterii (рафлезиевые) х, присоски
•которого пронизывают ветки; на одном растении он образует соцветия одного пола
(заражение, вероятно, происходит на стадии проростка). На Trichocereus паразитирует
широко распространенная во всем районе Phrygilanthus aphyllus (ремнецветниковые)
с красными или желтыми ягодами. Благодаря туману несколько увеличивается влаж-
ность воздуха и на восточном склоне, о чем свидетельствует множество лишайников,
свисающих с колючек канделяброобразных кактусов.
Существование этого леса в столь большом отдалении от его основной
области распространения говорит о том, что занимаемое им местообита-
ние — реликт прежнего, более влажного периода, когда такие леса,
видимо, были развиты вдоль всего побережья. Пока не установлено,
является лес реликтом третичного времени, эпохи оледенения или после-
ледникового периода.
В целом для области жестколистной растительности Чили весьма
характерно, что на всех территориях, где производился выпас скота или
проявлялась иная деятельность человека, эндемичная травйная расти-
тельность почти полностью вытеснена европейско-средиземноморскими
пастбищными и рудеральными растениями. Устойчивые к антропогенному
влиянию виды здесь не развились, поскольку вся область прежде была
облесена. В противоположность этому в Европе, в частности в Средизем-
номорье, в результате длившейся тысячелетиями деятельности человека
из состава различных естественных сообществ произошел отбор таких
спутников культуры, которые на местообитаниях, нарушенных антропо-
генным воздействием, объединились в новыЬ сообщества; ныне эти сооб-
щества на всем земном шаре в соответствующих климатических областях
берут верх в конкуренции с местными видами.
За зоной жестколистной растительности к югу следует зона летнезеле-
ных лесов, образованных Nothofagus obliqua («робле»), а южнее, как
и в более высоком поясе гор,— Nothofagus procera (nervosa) («раули»).
Зона Nothofagus соответствует переходной субсредиземноморской зоне
в Европе с летнезеленым Quercus pubescens, в пределах которой максимум
осадков смещается с зимы на лето. В Чили картина иная. Здесь зимний
максимум выражен также очень резко, но засуха летом не столь сильная
(см. рис. 98, климадиаграмма Темуко). Аналогами следует, скорее, считать
анатолийскиеилиинсубрийские леса (ср. рис. 40), в подлеске которых еще
встречаются многие вечнозеленые виды, что наблюдается и в Чили. Своеоб-
разие состоит лишь в том, что в южном ЧиЛи, характеризующемся боль-
шей суммой осадков, состав леса вновь образуют вечнозеленые породы
деревьев, например вечнозеленый Nothofagus] dombeyi с относительно
мелкими жесткими листьями. Только опыты по культивированию этих
пород деревьев помогли понять, почему в относительно сухом районе
преобладают летнезеленые, а во влажных — вечнозеленые виды.
В продольной долине южная граница распространения жестколистной
растительности проходит приблизительно по 38° ю. ш., а в береговых
хребтах — по 37° ю. ш.; обычно же границу проводят по реке Мауле
(35°30' ю. ш.). Для западного склона Анд границей считают 36 параллель.
Соответственно южная граница летнезеленого леса следует, по Бернингеру,
в продольной долине по 41°30' ю. ш., в береговых хребтах — по 40°, а на
западном склоне Анд — по 39° ю. ш. Но Nothofagus obliqua образует на
береговом хребте — на двух его высоких вершинах, Кампана и Роблес
(2222 ж),— далеко выдвинутый к северу форпост.
1 J. Kummerow, Pilostyles berterii Guill., eine wenig bekannte Rafflesiacee
in Mittelchile, «Z. f. Bot.», 50, 1962, S. 321—337.
10-01248
146
Глава 3
Эти вершины выступают высоко над покровом тумана (см. рис. 103),
так что о его конденсации не может быть и речи. Видимо, здесь выпадает
больше дождевых осадков, но соответствующие измерения отсутствуют,
хотя на одной из вершин расположена передаточная радиостанция.
О большом количестве осадков свидетельствует тот факт, что в долине,
лежащей у подножия гор, растут пальмы Jubaea.
Леса «робле» очень похожи на наиболее хорошо развитые дубовые леса
умеренной зоны Центральной Европы. Деревья в них достигают высоты
35—40 ж. Между 39 и 40° ю. ш. зона этих лесов переходит па аргентинскую
сторону Аид. Высота древостоев «раулп» такая же. Поражают мощные-
стволы деревьев. Ствол одного старого дерева смогли охватить только
13 человек. По своему облику «раули» больше напоминают европейские
буковые леса.
В нижнем древесном ярусе этих лесов появляются вечнозеленые виды:
Persea lingue (лавровые), Eucryphia cordijolia (камнеломковые), Aextoxi-
сит punctatum, с которыми автор познакомился в «лесу туманов» области
Фрай-Хорхе, и Laurelia aromatica (Monimiaceae).
Эколого-физиологические исследования жестколистной раститель-
ности Чили немногочисленны. Анатомо-морфологическое изучение защит-
ных приспособлений против засухи еще в 1894 г. проводил Мейген х.
Он указал на влияние летом пыли, менее отрицательно воздействующей
на растения с гладкими листьями. Подробно описывает признаки приспо-
собления у растений, испытывающих недостаток влаги, Рейхе (см. стр. 130)г
в их числе часто наблюдающуюся водосодержащую ткань в виде круп-
ных клеток эпидермиса. Водным балансом в последнее время первым
заинтересовался Кубицки 1 2, определивший осмотическое давление также-
и у древесных растений южного Чили. У жестколистных растений (Реи-
mus boldus, Cryptocarya, Sophora tetrapterd) оно изменялось от 15 до 22 атм,
у вечнозеленых видов с лавроподобными листьями, переходных к видам
умеренной зоны,— от 10 до 16, реже до 20 атм. Очень невелики колебания
осмотического давления (13—15 атм) у Nothofagus dombeyi, N. nitida,
N. betuloides. Эти виды еще встречаются на высотах более 1000 м. Как
и везде, осмотическое давление у теневых листьев всех древесных пород,
всегда ниже, чем у солнечных. Отчетливую зависимость величины осмоти-
ческого давления от высотного положения местообитания растений уста-
новить не удалось. Измерения у Araucaria araucaria дали величину осмоти-
ческого давления 17—19 атм. При изучении листопадных субантаркти-
ческих видов Nothofagus pumilio и N. antarctica нижний предел оказался
равным 13,5 атм, а верхний лишь редко превышал 18 атм.
Поскольку пробы отбирались и на сухих местообитаниях, а также пос-
ле длительной засухи, можно сделать вывод, что водный баланс растений,,
в общем, уравновешен. Он подтверждается также дополнительными опре-
делениями сосущей силы клеток методом Шардакова, который дает, скорее,
относительные ее значения. В ясные дни наблюдались колебания дневного
хода, ночью же, видимо, достигалась полная влагонасыщенность.
В условиях резко выраженного океанического климата виды лаврово-
го леса по своей гидратуре сравнительно близки к растениям тропическо-
го дождевого леса, прежде всего к формам с мелкой листвой.
1 F, М е i g е n , Biologische Beobachtungen aus der Flora Santiagos in Chile.
Trockenschutzeinrichtungen, «Bot, Jb.», 18, 1894, S, 394—487,
2 К, К u b i t z k i, Zur Kenntnis der osmotischen Zustandsgrofien siidchilenischer
Holzgewachse, «Flora», 155, 1964, S. 101—116.
Области средиземноморского климата за пределами Евразии
147
Интересные экологические наблюдения совершенно иного рода, и не
на эндемичных, а на культурных или занесенных растениях, провел Швабе,
работавший в продольной долине у южной границы зоны жестколистной
растительности, под Мининко (37°40' ю. ш.), на высоте 180 м над уровнем
моря Ч
Это были фенологические и фенометрические исследования, пред-
принятые в связи со своеобразием климата Чили. Как известно, он отли-
чается несоответствием между интенсивной инсоляцией (созвучной поло-
жению района на низкой широте) и относительно низкой температурой
воздуха, обусловленной холодным течением Гумбольдта. Это несоответ-
ствие проявляется в климате побережья и увеличивается по мере движения
к северу. Как следствие, температура воздуха днем по сравнению с низкой
температурой ночью относительно высока, чем, возможно, объясняется
интенсивный рост некоторых завезенных растений (незначительные потери
на дыхание ночью), таких, как Pinus radiata, Acacia melanoxylon, Eucalyp-
tus globulus и Beta.
Перепад температур особенно ощутим вблизи поверхности почвы,
и, вероятно, поэтому происходит очень пышное развитие розеточных
сорняков, например Echium vulgare, в прохладное время года. Кроме того,
ритм климата в Чили совершенно иной, нежели на родине разводимых
здесь культурных растений или занесенных видов.
Под Мининко средняя годовая сумма осадков равна 1300 мм. Но
поскольку зимой южного полушария дожди’ идут с апреля по август,
летняя засуха еще отчетливо выражена. Ход кривой осадков прямо про-
тивоположен ходу кривой температуры, иными словами, период с наимень-
шим количеством осадков совпадает с периодом, характеризующимся
наивысшей температурой, и наоборот.
Прежде, когда лесов здесь было больше, контрасты между отдельными
временами года сглаживались. В открытом же культурном ландшафте
они обостряются и в течение года вызывают в ритме развития не туземных
для Чили растений несколько кризисов или импульсов.
1. Зимний кризис обусловлен избытком осадков при относительно
низкой температуре, сдерживающей рост; период зимнего покоя не насту-
пает. В это время года растения почти не поглощают минеральных пита-
тельных веществ, поэтому последние вымываются из почвы дождем. Почва
истощается.
2. Весенний импульс развития является следствием быстрого повы-
шения температуры в условиях, когда почва еще содержит достаточно
влаги; начинается интенсивное продуцирование вещества и быстрый
рост.
3. Кризис в начале лета резко прерывает этот процесс — наступает
засуха; в середине ноября прирост в высоту почти прекращается, наруше-
ние же роста может вызвать аномалии в морфологическом строении орга-
нов растения (скрючивание листьев, искривление формы, необычные раз-
ветвления побегов и др.).
4. Летняя засуха, усиливаемая сухими южными ветрами, наносит
ущерб различным видам культур и вызывает снижение урожая. Избыток
воды в почве в предшествовавший период обусловливает развитие у расте-
ний поверхностной корневой системы. Но летом верхние горизонты почвы
быстро высыхают. Корни не успевают проникать в более глубокие и влаж-
1 G. Н. Schwabe, Die okologischen Jahreszeiten in Klima von Mininco (Chile),
«Bonner Georg. Abh.», 17, 1956, S. 139—183.
10*
148
Глава 3
ные слои, поэтому растения испытывают затруднения в поглощении воды.
Минеральное питание ослабевает. Так как почвы здесь бедны питательны-
ми веществами, у растений обнаруживаются признаки голодания. Что-
бы избежать этого, в почву приходится вносить удобрения и микроэле-
менты.
5. Проявляющийся поздним летом стимул в развитии обусловлен
началом сезона дождей, когда температура воздуха еще достаточно высока.
Снова возобновляется рост, иногда в этом же году, а не будущей весной.
Открываются цветочные почки; например, у сирени видны одновременно
и семена и цветки, а из соцветий Echlum vulgare вырастают розетки.
В Центральной Европе после редкого сухого лета осенью также можно
наблюдать второе цветение фруктовых деревьев. В Чили же подобные
аномалии бывают каждый год.
Местные растения зоны жестколистной растительности приспособи-
лись к этому ритму климата и развиваются нормально, что нельзя ска-
зать о видах, происходящих из других климатических областей.
Экологические исследования такого рода не только интересны с науч-
ной точки зрения, но и имеют огромное практическое значение. Следовало
бы во всех климатических зонах точнее изучать чужеродные культурные
растения и адвентивные виды с позиций возможных нарушений или ано-
малий в их развитии.
УМЕРЕННАЯ ЗОНА ЮЖНОГО ЧИЛИ
Южнее полосы летнезеленых лесов, образованных Nothofagus. начи-
нается обширная зона вечнозеленых умеренных дождевых лесов южного
Чили (рис. 98 и 108). В ее северной части (до Пуэрто-Монта) еще четко
прослеживаются контуры Чилийской продольной долины, расположенной
между береговыми хребтами и Андами. Но далее к югу она погружается
ниже уровня моря 1. Береговой хребет сначала находит свое продолжение
на большом острове Чилоэ, а затем выступает лишь в виде цепочки остро-
вов, расположенных близ материка. Хребет Анд к югу также понижается,
но, поскольку климат в этом направлении становится холоднее, в горах
образуются фирновые поля и ледники. Южнее 46° ю. ш. уже развито
покровное оледенение.
Южное Чили лежит на пути постоянных западных ветров и поэтому
изобилует дождями. Часть приносимых ветрами осадков выпадает на
восточном склоне Анд, благодаря чему в Аргентине растут аналогичные
чилийским леса, правда образуя очень узкую полосу. Причина в том, что
на широте озера Науэль-Уапи количество осадков с 4000 мм у чилийско-
аргентинской границы убывает на протяжении всего 45 км ro 1000 мм.
определяя таким образом границу лесной растительности.
Минимум осадков на юге Чили также приходится на летние месяцы,
однако дождей в это время года достаточно, чтобы пастбища оставались
зелеными. Период летнего покоя у растений обычно отсутствует. Поэтому
здешний культурный ландшафт напоминает ландшафт Центральной Евро-
пы. В самой северной части зоны при лесопосадках используют Pinus
radiata. в других местах — преимущественно дуглассию (Pseudotsuga
1 После землетрясения 1960 г. район Вальдивии опустился на 1,8 м. поэтому боль-
шие площади культурных земель ныне затоплены. В 1966 г. из воды торчали только
проволочные заборы, разделявшие отдельные землевладения.
Области средиземноморского климата за пределами Евразии
149
м
5000-
о Libocedrus chilensis
4000-
зооо-
Г' rx /л -..Cty
Высокогорная формация
S Podocarpus salignus
• Podocarpus andinus
n Podocarpus nubigenus
x Saxegothea conspicua
T Araucaria araucana
I Fitzroya cupressoides
p Pilgerodendron uviferum
2000-.
1000-г
формация кустарничков:
''Д'"—Полупустыня
с колючими
кустарниками
суккулентами/
30е 32е
N.pumilio+antarctica
Nothofagus
alpina(procera)
N. purnilio
N.antarctica
-1000
34*
36*
P P Р
48°
н
г200О
so s s
40°
Летнезеленые
леса c Nothofagus лес
obliqua
42° 44° 46° u 48° 50°W.uz.
Дождевой. Себеропатагонскии Суоантаркти-
лес дожоебой лес с N.dombeyi ческии оожЗе-
ВальЗибии и N.betuloides ‘ бой лес с
N.betuloides
иласть жестколист-
ной растительности
о
Рис. 108. Меридиональный высотный профиль растительности на западном склоне
чилийских Анд (по Шмитхюзену). 1
menziesit). В искусственных насаждениях еще растут субтропические вечно-
зеленые деревья, даже пальмы, но летнезеленые лиственные виды (Castanea,
летнезеленый дуб, Tilia, Acer, Robinia) все же преобладают. Для винограда
лето уже слишком прохладное и сырое. Из полевых культур возделывают
пшеницу, овес, корнеплоды. Однако основную площадь занимают паст-
бища1. В продольной долине леса большей частью сведены, но в горах
только еще начинают применять подсечное земледелие, поэтому кое-где
на высотах сохранились прекрасные девстьенные леса. Но это не должно
успокаивать, и уже сейчас следует создавать заповедники, иначе будет
поздно.
Лесную зону южного Чили можно подразделить на 3 нечетко отгра-
ниченные между собой подзоны: 1) в а л ьдивийск у ю (до 42—43°ю. ш.),
2) патагонскую (до 47—48° ю. ш.) и 3) на крайнем юге—м а г е л-
л ан скую, или субантарктическую. Видовой состав лесов к югу
беднеет. В самой северной части, до широты острова Чилоэ, известную
роль еще играют элементы тропического происхождения, на юге же явно
преобладают антарктические элементы 1 2. Во влажных районах побережья
распространены болота с покровом из Sphagnum acutifolium, Drosera
uniflora, Pinguicula chilensis, Schizaea fistulosa, а также пустоши с верес-
ковыми и Empetrum rubrum.
Вальдивийский умеренный дождевой лес по пышности вряд ли усту-
пает тропическому дождевому лесу, а по запасам древесины, вероятно,
даже превосходит его. Поражает богатство видового состава, в котором
представлены древесные породы большей частью с вечнозеленой листвой.
Важнейшая древесная порода патагонской зоны — вечнозеленый
Nothofagus dombeyi («коиуэ») наряду с Eucryphia cordifolia («ульмо») и др.
Назовем прежде всего Aextoxicum punctatum, Eucryphia cordifolia, Laurelia aro-
matica, L. serrata, Drimys winteri, Flotovia diacanthoides (сложноцветное), Maytenus boa-
ria, протейные Guevina avellana, Lomatia ferruginea и L. hirsuta, Nothofagus dombeyi,
а также летнезеленые Nothofagus obliqua и N. procera. Из кустарников встречаются
1 E. К 1 a p p, Futterbau und Futterwirtschaft in Chile, «Bonner Georg. Abh.», H.
17, 1956, S. 87—137.
2 L. H a u m a n n, La foret valdivienne et ses limites, «Fac. Cienc. Medicas
de Buenos Aires», № 34, 1916.
150
Глава 3
Рис. 109. Пышный
дождевой лес на юге
острова Чилоэ. В древес-
ном ярусе доминируют
Nothofagus dombeyi и
Eucryphia cordifolia, Dri-
mys winteri (меньших
размеров, на переднем
плане). Из эпифитов —
Fascicularia bicolor (бро-
мелиевые) (фото Скот-
сберга).
Weinmannia trichosperma, Caldcluvia paniculata (оба Cunoniaceae), Raphithamnus cyano-
carpus (вербеновые), Ugni molinae (миртовые), Coriaria ruscifolia и др.; рядом с кустар-
ником, на прогалинах, можно найти бамбук (виды Chusquea), Среди лиан легко заметить
Hortensia scandens, достигающую толщины с руку и выбрасывающую белые шарообраз-
ные соцветия, Mitraria coccinea (геснериевые) с красивыми красными цветками, Boqui-
la trifoliata (лардизобаловые), Dioscorea brachybotrya и др.; отмечены также некоторые
цветковые эпифиты; часто встречаются мхи, лишайники, а также гименофилловые
папоротники. Вполне возможно, вальдивийский лес представляет собой реликт третич-
ного времени, без изменений переживший на этих широтах период оледенения. Броса-
ется в глаза своей оранжевой корой Myrceugeniella apiculata (миртовые), «аррайан»,
занимающий узкую полосу вдоль рек и по берегам озер.
видами (рис. 109). На лесных прогалинах быстро распространяется бамбук
(Chusquea quila или Ch, coleu) с неполыми стеблями высотой 4—5 м, обра-
зующий очень густые заросли. Возобновление леса происходит лишь
вслед за годом массового цветения бамбука, так как после цветения рас-
тения отмирают. Предполагают, что это явление повторяется каждые
10 лет.
Отдельные виды вальдивийского леса проникают далеко на юг: Weinmannia,
Caldcluvia, Laurelia serrata и миртовые; Drimys winteri образует подлесок.
Области средиземноморского климата за пределами Евразии 151
Южнее 48-й параллели Nothofagus dombeyi замещается также вечнозеленым Notho-
fagus betuloides, «гуиндо». Высота деревьев не превышает 18—20 м, к югу она еще боль-
ше снижается; южнее пролива Бигль лес едва достигает в высоту 6—8 м. Число видов
древесных растений в этом направлении также сокращается.
На всех островах южнее 48° ю. ш., лежащих западнее материка, дре-
весная растительность отсутствует. Острова покрыты подушечниковыми
•болотами с Donatio, (Stylidiaceae), Astelia (лилейные), Oreobolus (осоковые)
и Gaimardia (Centrolepidaceae) (сфагнум здесь отсутствует); южная же часть
Огненной Земли и остров Наварино еще заняты лесами: на наветренной
стороне лес образован Nothofagus betuloides, во внутренних районах —
N. pumilio и N. antarctica (см. карту в работе Го дли). Вечнозеленый
N. nitida встречается на острове Чилоэ и в области фьордов, где он
растет преимущественно на заболоченных, плохо дренируемых почвах
{рис. 110). ।
Болота Магеллановой зоны, распространенные также в горах Сан-
Педро (остров Чилоэ) и Пеллада, по флористическому составу очень
близки к растительности субантарктических островов, лежащих южнее
Новой Зеландии. Подушечные растения представлены викарирующими
видами. Для обозначения здешней растительности можно употребить
понятие «антарктическая тундра». Температура в течение всего года колеб-
лется здесь лишь в пределах 6—8°.
Из вышеизложенного следует, что в условиях очень влажного уме-
ренного климата южного полушария четкое разграничение зоны умерен-
ных лесов на подзоны отсутствует. Начиная с теплой умеренной зоны,
чипы леса лишь постепенно переходят один в другой, сходя на нет у антарк-
тической границы леса, причем вечнозеленый тип растительности сохра-
няется на всем протяжении. 1
Рис. 110. Дождевой лес с господством Nothofagus nitida и Weinmannia trichosperma
(слева). В воде — крупная бурая водоросль Macrobystis pyrifera. Область фьордов Чи-
ли, пролив Месье (фото Скотсберга).
152
Глава 3
Рис. 111. Nothofagus pumilio зимой (без листвы, с полупарази-
том Myzodendron punctulatum) и вечнозеленый N. betuloi'des на глав-
ном острове Огненная Земля близ Ушуаи (фото Скотсберга).
Более четко выделяются вертикальные пояса. В верхних поясах гор
снова появляются летнезеленые виды, прежде всего Nothofagus pumilio
(«ленга») (рис. 111). Над границей древесной растительности N. pumilio
образует обширные криволесья (рис. 112). Трудно объяснить причину
распространения здесь другого летнезеленого вида, Nothofagus antarctica
(«ньире»). По-видимому, он занимает лишь окна в лесу и развивается
только там, где другие виды Nothofagus не растут; либо образует криво-
лесье над границей древесной растительности на сырых ровных участках
или в холодных низинах; либо встречается как дерево и в поясе с N. dom-
beyi или даже на окраине степи, а также на неблагоприятных местообита-
Рис. 112. Вид с высоты 2200 м из Валье-де-лас-Ньеблас (долина туманов) на трого-
вую долину Рио-Ренегадо с границей
подветренной стороне северо-западных
и N. antarctica
древесной растительности, образованной на
ветров кустарником из Nothofagus pumilio
(фото Бернингера).
Рис. 113. Pilgerodendron (Libocedrus) uvtferum и Nothofagus betuloides (справа) в об-
ласти фьордов Чили, архипелаг Королевы Аделаиды (фото Скотсберга).
154
Глава 3
ниях (заболоченные почвы, щебнистые осыпи, почвы с близким залеганием
грунтовых вод); либо как растение-пионер поселяется на вулканических
пепловых почвах или древних лавовых потоках.
С видами Nothofagus связаны определенные паразиты. В качестве полупаразитов
выступает ряд видов» Myzodendron, образующих особое семейство Myzodendraceae,
близкое к санталовым. Их развивающиеся на ветвях побеги с длинными волосками
на соплодиях напоминают лишайники (рис. 111). Распространение этого семейства
ограничено территорией Чили и соседней облесенной частью аргентинской Патагонии.
Из других паразитов назовем виды Cyttaria, образующие собственную группу
среди аскомицетов. Их шарообразные, величиной со сливу плодовые тела выходят
из ветвей; мицелий образует огромные деревянистые наросты размером с голову. Име-
ются сведения, что Cyttaria паразитирует также на австрало-новозеландских видах
Nothofagus.
Хвойные деревья важного места в растительном покрове Чили не
занимают в отличие от тихоокеанского побережья Северной Америки, где
они широко распространены.
Pinaceae здесь полностью отсутствуют, но встречаются представители других семейств Ч
Cupressaceae: Austrocedrus (Libocedrus) chilensis, Pilgerodendron uviferum (рис. 113),
Fitzroya cupressoides.
Podocarpaceae: Saxegothea conspicua, Podocarpus nubigenus, P. salignus(=chilina), P. an-
dinus, Dacrydium fonkii.
Araucariaceae: Araucaria araucana (= imbricata) (см. рис. 114).
Все эти виды растут чрезвычайно медленно, хотя некоторые из них достигают
2000-летнего возраста. Поэтому они не выдерживают конкуренции с лиственными
деревьями и довольствуются известными экологическими нишами в различных верти-
кальных поясах и разных зонах растительности, иными словами, растут там, где виды,
обладающие большей конкурентной мощностью, отсутствуют. Древесина их частично
очень ценная, поэтому существует опасность, что они будут вырублены, так как под-
рост практически отсутствует. Большинство видов производит впечатление реликтовых,
подобно Sequoiadendron gigantea в Калифорнии. Saxegothea, напоминающая Taxus,—
монотипный род, близкородственный Microcachrys tetragona, растущему в Тасмании;
так же монотипный род Fitzroya родствен Diselma archeri (Тасмания), которую прежде
относили к роду Fitzroya. Dacrydium fonkii — кустарник высотой несколько дециметров,
встречающийся на сфагновых и осоковых болотах в полосе от 40° ю. ш. до Огненной
Земли. Из других 20 видов Dacrydium D. cupressinum представляет собой в Новой
Зеландии большое дерево.
Ни одна из чилийских хвойных пород при лесопосадках не исполь-
зуется, предпочтение отдается экзотам. Распространение некоторых видов
хвойных иллюстрирует меридиональный профиль западного склона Анд,
составленный Шмитхюзеном (см. рис. 108). Отдельные зоны в Андах
вплоть до 34°30' ю. ш. выступают как вертикальные пояса.
Наибольшее значение из хвойных пород имеют Fitzroya («алерке»),
араукария («пино») и Austrocedrus («ципрес»). Fitzroya в западной Патаго-
нии достигает в высоту 50—60 м при диаметре ствола до 3 м', растет она
до 2000 лет. При осмотре одного такого срубленного дерева оказалось,
что оно выросло на старом, поваленном, но еще хорошо сохранившемся
также 2000-летнем стволе.
Fitzroya встречается как на низменностях с болотами, поросшими
сфагнумом, так и на горных склонах вблизи верхней границы леса. Некогда
обширные насаждения в продольной долине в окрестностях Пуэрто-Монта
теперь вырублены, но культурной почвы здесь не создали. В горах
этот вид распространен шире.
1 J.Schmithusen, Die Nadelholzer in den Waldgesellschaften der siidlichen
Anden, «Vegetatio», 9, 1960, S. 313—327.
Области средиземноморского климата за пределами Евразии
155
Рис. 114. Лес из Araucaria агаисапа, в подлеске Nothofagus pumilio; Кордильера-де-
Науэльбута на высоте 1400 м над уровнем моря (фото Бернингера).
Словно нришелец из далекого геологического прошлого выглядит
Araucaria агаисапа с ее широкими чешуевидными иголками, бесформенны-
ми ветвями и крупными шишками. И действительно, араукарии относятся
к древнейшему роду хвойных и должны рассматриваться как реликтовые.
В Чили А, агаисапа имеет два ареала: один небольшой — в береговом
хребте на высоте 700 м над уровнем моря, другой более обширный —
в Андах, между 37 и 40° ю. ш., на высоте 1600-^1800 м, а на восточном скло-
не на высоте 600—800 м. Из-за интенсивной вырубки ареалы повсеместно
быстро сокращаются. Предельная высота дерёва 35 м. Сухость араукария
переносит сравнительно хорошо.
Еще большей засухоустойчивостью обладает Austrocedrus, играющий
в ландшафте примерно такую же роль, как сосна в центральноевронейских
широтах. Его можно встретить на сухих местообитаниях или на маломощ-
ных ночвах, то есть в условиях, в которых другие лесные деревья разви-
ваются плохо. Поэтому на чилийской стороне Анд он распространен мало,
и только у верхней границы леса зоны жестколистной растительности.
На аргентинском склоне у границы со степями Патагонии Austrocedrus
в качестве зональной растительности сменяет лес из N olhofagus. Эту смену
можно хорошо наблюдать под Барилочем у озера Науэль-Уани, где на
расстоянии около 50 км количество осадков сокращается с 1500 до 500 мм.
С изолинией годовых осадков 1500 мм примерно совпадает граница
лесов, образованных Nothofagus dombeyi.
В этом районе, расположенном вблизи границы засушливости, миртовые деревья
растут только по берегам озер. Известен чистый древостой Myrceugeniella apiculata,
156
Глава 3
занимающий площадь 18 га на полуострове Кетриуэ у озера Науэль-Уапп. В глубокой
тени этого леса в изобилии встречается сапрофит Arachnitis uniflora из семейства Bur-
manniaceae; его невысокий (10—20 см) стебель несет один крупный бледный, своеобраз-
ной формы цветок. Очень часто миртовые слагают на берегах формацию, напоминающую
мангровые и простирающуюся в открытую воду на 10—20 м от берега; корни растений,,
сплетаясь, образуют на поверхности воды плотный, поросший мхами покров, выдержи-
вающий тяжесть взрослого человека; глубина воды под покровом — около 30—40 см.
Непосредственно у кромки воды располагается пояс Myrceugenia exsuca (высота до 4 л*),
за которым следует зона с Myrceuginella apiculata (до 12 м)\ верхнюю часть склона зани-
мают древостои Nothofagus dombeyi (высотой более 25
В зоне с осадками менее 1500 мм развиваются леса с Austrocedrus,
часто образующим обширные чистые древостои. Примечательна гибель
этих лесов на больших площадях, хотя никаких паразитов или поврежде-
ний от пожаров замечено не было. Предполагается, что климат Патагонии
за последнее время стал теплее и суше (сильное отступание ледников), что
и вызвало сокращение площади лесов. Вероятно, одного засушливого
года было достаточно, чтобы леса из Austrocedrus на наиболее сухих место-
обитаниях погибли. Процесс засыхания деревьев особенно заметен в недав-
но созданном национальном парке у озера Пуэло. Если количество осадков
сокращается до 1000 мм и более, лес становится реже, Austrocedrus в нем
уже немного, но преобладают Nothofagus antarctica, Schinus patagonica,
Maytenus boaria, протейные Lomatium hirsutum и Embothrium coccineum
(последняя в декабре маркирует эту зону темно-красными соцветиями),
а также кустарник из Diostea juncea (вид семейства вербеновых, напоми-
нающий Spartium), Discaria serratifolia, Fabiana imbricata (эрикоидное
паслёновое) и мелкие кустарники Berberis и Ribes.
Экстразонально Austrocedrus еще встречается в более влажных районах
Патагонии на скалах северной экспозиции, где растет вместе с Mayte-
nus boaria. Это последние островки южночилийского леса в Аргентине
восточнее линии Анд (о растительности Патагонии см. т. III, гл. 3).
ЖЕСТКОЛИСТНАЯ РАСТИТЕЛЬНОСТЬ
КАНСКОЙ ПРОВИНЦИИ И ЗОНА, ПЕРЕХОДНАЯ
К ОБЛАСТИ КАРРУ1
Область зимних дождей средиземноморского типа занимает в Африке
ее крайнюю юго-западную оконечность и в северном направлении перехо-
дит в полупустыню Карру (см. рис. 6, а также т. I, рис. 334). К востоку
вдоль побережья протягивается умеренно теплая влажная климатическая
зона с вечнозелеными лесами, состоящими преимущественно из тропи-
ческих элементов. В отличие от Южной Америки Африканский материк
в собственно умеренную зону не простирается, поэтому умеренные расти-
тельные зоны здесь полностью отсутствуют. Климатические условия обла-
сти характеризуют климадиаграммы на рис. 115—120.
Жестколистная растительность занимает в Южной Африке очень незна-
чительную площадь. Вместе с тем она отличается столь большим богат-
ством видов и таким своеобразием флористического состава, что есть все
основания выделить Капскую провинцию в самостоятельное флористи-
ческое царство — Capensis. * II,
1 R. Mario th, Das Kapland, Wiss. Ergeb. d. Deutschen Tiefsee-Exp., Bd.
II, Teil III, Jena, 1908; R. S. A d a m s о n, The Vegetation of South Africa, Monogr.
Brit. Emp. Veget., London, 1938.
Области средиземноморского климата за пределами Евразии
157
Рис. 115—117. Климадиаграммы области жестколистной растительности Южной
Африки.
Рис. 118—120. Климадиаграммы районов за пределами] области жестколистной
растительности в Южной Африке.
На небольшом полуострове Кейп, 50 км в длину и менее 10 км в ширину,
можно встретить свыше 2000 различных видов, то есть столько же, сколь-
ко насчитывается во всей Центральной Европе. В заповеднике Джонкерс-
хук под Стелленбосом на площади всего 2000 га было обнаружено
2000 видов.
Для лучшего понимания характера растительности приведем краткий
обзор ее флористического состава х>.
Своеобразие флоры Капской провинции обусловлено распространением следую-
щих семейств и таксонов, встречающихся только на полуострове Кейп или имеющих
здесь свой основной ареал.
*Семейства: Proteaceae, Restionaceae, Penaeaceae, Bruniaceae, Geissolomaceae, Stil-
baceae, Retziaceae, Grubbiaceae.
Трибы: Diosmeae среди Rutaceae.
Роды: Erica (почти 600 видов), Cliffortia (розоцветные, co 108 видами), Muraltia (исто-
довые, 115 видов), Metalasia и Stoebe (сложноцветные), Roridula (2 вида, семейство
Byblidaceae, к которому относятся еще только 2 австралийских вида Byblis).
Все протейные, помимо вида Brabejum stellatifolium, относятся к подсемейству
Persoonioideae. Семейство богато представлено и в Австралии, но другими родами.
В Африке широко распространен самый примитивный род Faurea. Из 100 видов Protea
только немногие встречаются за пределами Капской провинции.
Из семейства Restionaceae один только род Restio представлен в Южной Африке
117 видами (1 на Мадагаскаре, 27 в Австралии).
В раннетретичное время Ericaceae в Капской провинции еще не были распростра-
нены в отличие от Restionaceae. Семейство Eriaceae голарктическое, но тем не менее
ареал Erica arborea простирается далеко за эти районы от Средиземноморья до гор Тро-
пической Африки. В Южной Африке возникли не только многие виды Erica, но и как
1 М. R. Levyns, Migration and origin of the Cape Flora, «Trans. Roy. Soc. S.
Afr.», 37, Part 2, 1964, p. 85-107.
158
Глава 3
редукционный ряд роды Eremia, Acrostemon, Syndesmanthus, Sympieza, Salaxis co мно-
гими видами.
Это действительно умопомрачающее обилие видов в пределах отдельных родов
можно объяснить только историей развития флоры. Доказано, что в раннетретичное
время климат в этих широтах был более влажный. Южную Африку заселяла лесная
флора с вечнозелеными лавроподобнымп листьями. Ее состав формировали генети-
чески различные элементы: 1) древние элементы, такие, как хвойные Podocarpus
и Widdringtonia, 2) южные антарктические лесные элементы, как Cunonia, Brabejum,
Metrosideros, 3) северные элементы: Olea, Celtis, Ilex, Pittosporum, Apodytes. Когда кли-
мат внезапно стал более засушливым, древняя флора в Капской провинции погибла,
сохранились лишь редкие реликты. На ее место мигрировала засухоустойчивая скле-
рофпльная капская флора. Переселение сопровождалось бурным развитием видов
некоторых родов. Аридность климата продолжала возрастать, и в горах Малого
Карру капская флора вынуждена была покинуть наиболее сухие местообитания в кот-
ловинах и перейти на более влажные возвышенности. Освободившиеся местообитания
заняла засухоустойчивая флора Карру. Последняя также очень богата видами. Мы
различаем:
а) мелкие суккуленты, как Mesembryanthemum, ныне разделенные на более
чем 100 мелких родов, Zygophyllaceae, Stapelia, Euphorbia и Crassulaceae;
б) мелкие ксерофильные кустарнички, преимущественно сложноцветные (Ptero-
nia с 60 видами, Eriocephalus с 20 видами, Pentzia и др.);
в) более поздние иммигранты с севера: Euclea, Acacia, Рарреа, Schotia, Rhus,
Buddleia и суккуленты А1оё со многими эндемичными видами, Portulacaria и др.
Растительность Капской провинции в настоящее время состоит из
трех компонентов, не смешивающихся между собой, а мозаикообразно
проникающих друг в друга; компоненты эти следующие:
1) капская растительность, или собственно жестколистная; она засе-
ляет близкие к морю районы на юго-западе, обращенные к морю, более
богатые осадками южные склоны гор, а также все обычно окутанные обла-
ками горные вершины высотой более 1250 м в Малом Карру;
2) вечнозеленая лесная растительность, встречающаяся как реликт
в самых сырых горных ущельях;
3) растительность Карру, занимающая только котловины или горные
склоны в ветровой тени, где количество осадков не превышает 250 мм.
Почвы области жестколистной растительности отличаются исключи-
тельной бедностью. Важнейшие горные породы, которыми сложены горные
хребты Капской провинции, относятся к капской системе, перекрытой
конгломератом Двайка, моренными отложениями пермо-карбонового оле-
денения. Конгломерат Двайка слагает низы системы Карру. Подстилающие
ее породы капской системы представлены песчаником Столовой горы —
очень твердой кварцевой породой мощностью до 1500 м, образующей
почти отвесные склоны. Почвы нагорий — бедные и кислые, грубозер-
нистые и маломощные; содержание гумуса в верхнем горизонте не превы-
шает 6—9,5%. В небольших углублениях гумуса иногда накапливается
чуть больше. Корпи древесных растений проникают по трещинам па глу-
бину 3—4,5 м.
Рассмотрим сначала особенно интересную растительность Столовой
горы и условия ее произрастания Ч Вершина последней представляет
собой относительно ровную поверхность на высоте около 1000 м над уров-
нем моря; к югу возвышенность медленно понижается и переходит в гор-
ный хребет полуострова Кейп. К северу Столовая гора круто обрывается
и отделена от северных и восточных горных гряд обширной песчаной рав-
ниной Кейп-Флетс, созданной морем в недавнее геологическое время.
Климат Столовой горы определяется ее положением на стыке теплых
влажных воздушных масс, которые поступают с Индийского океана
1 R. S. Adamson, The plant communities of Table Mountain, «J. Ecol.», 15,
1927, p. 278-309.
Области средиземноморского климата за пределами Евразии
159
с юго-восточными ветрами, и прохладных воздушных масс, перемещающих-
ся с запада над холодным Бенгельским течением. Поэтому на восточном
склоне горы количество осадков (1600 мм) в два раза превышает их сумму
на западном склоне (800 мм). Влажный воздух, адиабатически поднимаясь
по восточному склону, охлаждается. На высоте 600 м часто достигается
точка росы и образуются облака. При дальнейшем подъеме конденсация
облачной влаги все возрастает; каждый твердый предмет покрывается
капельками воды. На западном склоне воздушные массы опускаются
и нагреваются. Вода снова переходит в парообразное состояние. Но так
как часть ее уже выпала, облако рассеивается на более высоком уровне
(обычно на высоте 900 м над уровнем моря). Таким образом, облака оку-
тывают только верхнюю часть горы, образуя знаменитое «облачное покры-
вало», столь характерное для всех гор Капской провинции. Летом облач-
ность несколько смягчает засуху на вершинах гор. Иногда там даже выпа-
дают дождевые осадки, правда в таком малом количестве, кто измерить
их нельзя. Марлот упоминает, например, о юго-восточном шторме, буше-
вавшем в феврале 1905 г. шесть дней подряд. В Кейптауне же (северо-
западный склон горы) не упало ни капли дождя; на Столовой горе обычным
дождемером было отмечено только 4 мм осадков. Контрольный дождемер,
в который были воткнуты ветки, чтобы конденсирующаяся вода стекала
в мерный сосуд, накопил 152 мм. Плоская вершина горы после шторма
летом выглядела словно зимой: на ровных скалах стояли лужи воды,
ложбины были переполнены до краев, участки, занятые Restionaceae,
представляли собой мокрые болота. Благодаря этим особым условиям
Рис. 121. Древостой Leucadendron argenteum на склонах «Львиной головы» близ
Кейптауна на высоте 300 м. Диаметр самого толстого ствола — 60 см (фото Марлота).
1'60
Глава 3
Рис. 122. Protea grandiflora на южном склоне Столовой горы, высота 200 м над
уровнем моря (фото Марлота).
®о всех горах, не слишком удаленных от побережья, экспозиция склонов
«играет для растительности большую роль, нежели высота их местообита-
ния: южные склоны оказываются влажными, северные, напротив, получа-
ют мало осадков.
Зональной растительностью в Капской провинции следует считать
похожий на маквис кустарник, называемый «финбос». Единственный
древесный вид представлен в нем Leucadendron argenteum (семейство про-
тейных). Это так называемое серебряное дерево достигает высоты
15 м (рис. 121). Его естественный ареал на влажных склонах Столовой
горы на высоте менее 500 м очень невелик. Марлот, заставший Капскую
провинцию еще такой, как она выглядела почти 100 лет назад, с еще пер-
вичной растительностью, называет, кроме того, отдельные виды мощных
деревьев — Olea verrucosa и Gymnosporia laurina (Celastraceae). Он пред-
полагает, что ранее оба вида были значительно шире распространены,
но, скорее всего, это элементы вечнозеленого леса, особенно далеко про-
никшие в Капскую провинцию.
К сравнительно высоким жестколистным растениям следует отнести
также Protea grandiflora. древесный кустарник высотой 5—8 м (до 12
{рис. 122), а также Р. arborea и Р. mellifera (=repens), Leucospermum сопо-
'Сагрит и различные виды Mimetes (рис. 123).
В заповеднике Джонкерсхук под Стелленбосом имеются уже 20-лет-
ние густые заросли Protea nerifolia высотой 4,5 м, а среди них — много-
численные деревца Widdringtonia cupressoides, которые могут достигать
большей высоты. По мнению Вихта (устное сообщение), в этом районе
в качестве климаксного сообщества должен был бы образоваться лес
«с Gymnosporia (=Maytenus), Olea. Olinia и Cunonia. Однако развитие
Области средиземноморского климата за пределами Евразии
161
Рис. 123. Жестколистный кустарник из протейных на восточном склоне Столовой
горы выше Вейнберга на высоте 200 м. Слева — Protea incompta, справа — Leucosper-
тит сопосагрит, на переднем плане—маленький куст цветущей Protea grandtflora,
а рядом слева — крупные листья Haemanthus coccineus (фото Марлота).
в этом направлении нарушается периодически повторяющимися пожа-
рами.
Жестколистный кустарник имеет в среднем высоту 1—4 м (рис. 124).
Верхний его ярус состоит из протейных; пдд их пологом растут виды
Rhus, Passerina, Clitfortia, множество С. ruscifolia с колючими листьями,
местные бобовые — виды Aspalathus и Borbonia, а также Gymnosporia
buxifolia и Asparagus capensis с более острыми шипами. Встречаются много-
численные виды Erica или эрикоидные формы других родов (рис. 125).
В самом нижнем, густом, ярусе распространены полукустарники Pelargo-
nium, Stoebe, Felicia, Aristea, Fagelia и др., злаки и Сурегасеае занимают
резко подчиненное положение; у самой земли растут Haemanthus coccineus,
виды Oxalis, различные геофиты и некоторые папоротники, как Cheilanthes
multifida и Blechnum australe. Терофиты обычно полностью отсутствуют.
Во время летней засухи все травянистые части растений засыхают. На
западном склоне Столовой горы, получающем меньше осадков, кустарник
более редкий и флористический состав его несколько иной. Часто встре-
чается паразит Cassytha capensis (семейство лавровых).
Неизмененные жестколистные естественные насаждения ныне уже не
найти. Важнейший экологический фактор здесь — огонь. Поэтому всегда
речь идет лишь о различных стадиях сукцессии после палов.
11—01248
162
Глава 3
Рис. 124. Жестколист-
ный кустарник на скло-
нах Столовой горы на
высоте 700 м над уров-
нем моря. Protea супа-
roides с крупными цвет-
ками (см. цветное фото 6)
на заднем плане — Вег-
zelia lanuginosa (фото
Марлота).
Protea grandiflora переносит выжигание без ущерба для себя и снова
дает побеги. То же самое можно сказать о Leucadendron. Большинство же
других кустарников погибает и возобновляется только семенами х.
Смена растительности после пожара в общих чертах такова:
1. В первый год преобладают геофиты — виды гладиолуса, Watsonia
rosea (рис. 126) и др. Они соответствуют стадии с Asphodelus в Средиземно-
морье. Многие виды цветут только после воздействия огня, обычно же
геофиты образуют только вегетативные побеги. Появляются однолетники,
но, так как почвы здесь бедные, они не играют заметной роли, как в Среди-
земноморье.
2. Спустя некоторое время развиваются травянистые растения, ча-
стично злаки, например Danthonia macrantha и Andropogon hirtus.
3. На следующей стадии выступают Restionaceae и виды вересковых
вместе с мелкими кустарниками (Rhus, Pelargonium, Anthospermum).
4. Лишь приблизительно через 7 лет снова вырастают древесные виды,
прежде всего протейные. Они могут достигать большого возраста, но одре-
1 P.G.J о г d а п, The influence of a fire in winter on the reproduction of four spe-
cies of the Proteaceae, «Tydskrif. v. Natuurwetenskappe», 5, 1965, p. 27—31.
Р и с. 125. Эрикоидные виды флоры полуострова Кейп.
1 — Ghidia pinifolia (Thymelaeaceae); 2 — Leucadendron levisancus (Proteaceae); 3 — Berzelia abrota-
noides (Bruniaceae); 4 — Diosma succulentum (Rutaceae); 5 — Phylica paniculata (Rhamnaceae); 6 —
Coleonema album (Rutaceae); 7 — Agathosma capitatum (Rutaceae). Нат. величина.
И*
164
Глава 3
Рис. 126. Watsonia rosea на склоне Столовой горы, выгоревшем
в предыдущем году. Высота 300 м над уровнем моря. Молодые
облиственные побеги — корневые побеги Protea grandtflora.
(фото Марлота).
воспевают и цветут мало. Для получения соцветий, идущих в продажу,
рекомендуется сильно обрезать ветки. Периодическое выжигание, несом-
ненно, относится к факторам, накладывающим отпечаток на современную
растительность. Но поскольку зарастание протекает очень медленно
и почва многие годы остается обнаженной, дожди легко ее размывают,
и уже очень скоро развивается прогрессирующая эрозия почв.
Условия существования и характер растительности на плоских вер-
шинах столовых гор иные, нежели описанные выше. В значительной сте-
пени они зависят от мощности почвенного покрова. Протейные также
встречаются, но отнюдь не в таком количестве. Преобладают Restionaceae
и Ericaceae. Но и здесь мы преимущественно наблюдаем различные стадии
сукцессии, вызванной пожаром.
На открытых скальных поверхностях развиваются исключительно
лишайники (Pertusaria lactea, Parmelia conspersa, Umbilicaria rubiginosa 1
и др.), на затененных участках, кроме того, Trentepohlia occulta; в расщели-
нах растут мхи, а на влажном моховом покрове — Utricularia capensis
и Drosera cuneata. В местах с хорошо выраженным песчаным горизонтом
доминируют Restionaceae. На более мощных почвах их сменяют вересковые
1 Этот вид маркирует границу опускания «облачного покрывала» на западном
склоне.
Области средиземноморского климата за пределами Евразии
165
совместно с видами Helichrysum и Helipterum, а также Cliffortia, Репаеа,
Brachysiphon и отдельные протейные. Пустошь с Restionaceae и Ericaceae
преобладает на сухих территориях. Своеобразный папоротник Schizaea
pectinate, — хороший показатель бывшего пожара, так же как и геофиты.
В небольших впадинах образуются заболоченные участки с Restio и мохо-
вым покровом (Campylopus), на котором часто поселяется Drosera trinervia.
В более крупных низинах распространены заросли Elegia (семейство
Restionaceae) высотой в рост человека. Во влажных ложбинах и на хорошо
увлажняемых склонах встречаются Berzelia или сложноцветные Osmitopsis
и Todea barbara (Osmundaceae). На постоянно мокрых склонах растет
сфагнум, а в подушках мха — красивейшая орхидея Капской провинции
Disa uniflora.
Сухие участки между обломками скал являются наиболее благоприят-
ными местообитаниями для развития суккулентов, как Rochea coccinea
(толстянковые), Othonna dentata (сложноцветные), и различных Mesem-
bry anthemum.
Физиономический облик растительности повсюду в Капской провин-
ции приблизительно одинаков, однако богатство флоры обусловливает
столь огромное разнообразие видов, что распознать флористический состав
определенных растительных сообществ почти невозможно. Наряду с груп-
пировками, похожими на маквис, повсеместно на склонах встречаются
напоминающие гаригу заросли кустарников, которые Мар лот называет
холмистой пустошью. Их образуют различнейшие кустарнички (Erica-
ceae, Thymelaeaceae, Rutaceae, низкие Proteaceae, Compositae Leguminosae),
разнотравье и геофиты. Красиво цветущими видами последних Капская
провинция особенно богата. Здесь насчитывается свыше 350 видов, принад-
лежащих семействам Orchidaceae, Haemodoraceae, Iridaceae, Amaryllidaceae
и Liliaceae. Злаки, напротив, полностью отступают на задний план. Наз-
вать следует роды Ehrharta и Pentaschistis. Вид Е. villosa, относящийся
к роду Ehrharta, распространился в юго-западной Австралии.
Весьма примечательно растительное сообщество, приуроченное к бере-
гам ручьев. К нему относится ряд кустарников, обычно ксероморфного
облика: Phylica buxifolia, Psoralea pinnata, Indigofera cytisoides, Podalyria
calyptrata. Poly gala myrtifolia, Athanasia parviflora, Pelargonium cucullatum
и принадлежащая семейству Bruniaceae —\Berzelia lanuginosa с белыми
шарообразными соцветиями. На более влажных восточных склонах Сто-
ловой горы уже упомянутый Brabejum stellatifolium образует насаждения
высотой 5 м.
Необыкновенно своеобразна формация Palmiet, развитая в руслах
мелководных рек. Ее густые заросли образует Prionium serratum (Junca-
сеае), напоминающий карликовую пальму или, скорее, панданус; ствол
его, толщиной с руку, усаженный широкими зубчатыми листьями, весной
оканчивается соцветием-метелкой длиной 1,5 м. Скопления растений силь-
но замедляют течение и способствуют осаждению взвешенных в воде частиц.
Для высокотравных болот характерны два вида Typha\ Cladium maris-
cus и красивая, известная в Европе как калла Zantedeschia aethiopica.
В воде растут виды Potamogeton, Nymphaea и Limnanthemum, в перио-
дически возникающих после дождей водоемах — прежде всего Aponogeton
distachyon и менее часто встречающиеся Oxalis natans^ Crassula natans
и виды Scirpus.
Здесь не место останавливаться на описании растительности засолен-
ных маршей и песчаных дюн на морском побережье. Упомянем только
поселяющийся на прибрежных песках кустарник Calvaria (Sideroxylori)
166
Глава 3
inermis (семейство сапотовых) — тропический элемент с крупными блестя-
щими листьями; иногда он образует на дюнах густые низкие, изогнутые
ветром древостои.
Местность Кейп-Флетс некогда представляла собой обширную терри-
торию, занятую дюнами, которые очень мешали движению транспорта.
Дюны закрепили, высеяв австралийские акации с филлодиями: Acacia
cyanophylla (прежнее название A. saligna) и A. cyclopis х. На тощих песча-
ных почвах эти связывающие азот кустарники оказались настолько конку-
рентоспособными, что полностью вытеснили первичную растительность.
С самолета, подлетающего к Кейптауну, эта обширная песчаная равнина
во время цветения акации кажется ослепительно желтым ковром. Но оба
вида распространились дальше и теперь встречаются уже на всех склонах
Столовой горы, угрожая подавить местную растительность. Еще одним
бедствием являются колючие австралийские Накеа pectinata и Н. acicula-
ris. Напротив, часто высаживаемые эвкалипты (Ей. cladocalyx, Ей. camaldu-
lensis, Ей. diversicolor) широко не распространяются. Большую опасность
представляют разные виды сосны, используемые при лесопосадках и даю-
щие деловую древесину. Прежде высаживали Pinus pinea и Р. maritima
(= pinaster), теперь — преимущественно Pinus radiata, а в сухой местности
также Р. halepensis. На склонах Столовой горы до самой ее вершины встре-
чаются одиночные сосны, выросшие из семян, занесенных ветром. В райо-
не, объявленном заповедником, они будут вырублены. В летнее время
посадкам сосны угрожают пожары.
В составе капской флоры совершенно отсутствуют лиственные породы
с мезо- или гигроморфными листьями. Тем более бросаются в глаза выса-
женные у домов старые дубы (Quercus robur); хорошо растут каштаны,
платаны, а также Albizzia lophanta. По долинам ручьев тянутся рощи
Populus canescens, древесного вида, уже полностью акклиматизировавшего-
ся здесь.
В Капской провинции очень резко проявляется фенологическая
интерцепция. Иными словами, у видов, происходящих из умеренных зон
северного полушария, наблюдается иная последовательность фенологи-
ческих фаз. В Южной Австралии нас поразило, что 16 октября 1958 г.
Robinia pseudacacia уже цвела, а виноград дал побеги. Но на Populus alba
были лишь сережки, а виды Salix только зазеленели. Таким образом,
согласно фенологическому календарю Центральной Европы, Robinia и
виноград опередили в своем развитии другие растения на 2 месяца.
Аналогичную картину автор наблюдал в Кейптауне: 26 сентября 1963 г.
дубы были уже совершенно зеленые, у Castanea vesca развернулись моло-
дые листочки, виноград пустил побеги, a Populus только сбросил сережки.
Своеобразной ксерофитной формой капской растительности является
формация «реностер», служащая переходным звеном к растительности
Карру. Она типична для тех районов области зимних дождей, где осадков,
в общем, выпадает несколько меньше. Это сообщество кустарничков,
в котором абсолютно преобладает серо-зеленый, высотой до 1 м Elytropap-
pus rhinocerotis (сложноцветные). Местами он равномерно покрывает огром-
ные пространства, как это можно видеть, например, в Робертсон-Карру
или в западной части Малого Карру; но обычно образуемый им покров —
это не естественный, а вторичный тип растительности. Elytropappus энер-
гично заселяет заброшенные пашни и гари, куда ветер заносит его семена.
1 Е. R. Roux, History of the introduction of Australien Acacias on the Gape
Flats, «S. Afr. J. Sci.», 57, 1961, p. 99—102.
Области средиземноморского климата за пределами Евразии
167
Его смолистые ветки скот не трогает, а поедает другие виды; такое селек-
тивное стравливание также способствует быстрому распространению
реностера.
Как и у всех вторичных сообществ, сопутствующие растения в рено-
стере в зависимости от истории происхождения весьма разнообразны.
Наряду с другими кустарничками сложноцветных встречаются многие
луковичные и клубневые растения, особенно хорошо развивающиеся после
пожаров. Правда, слишком частые палы способствуют тому, что кустар-
нички реностер вытесняются растением Bobartta spathacea (семейство
касатиковых), обладающим длинными ситниковидными листьями. В соста-
ве реностера встречаются также вересковые, но протейны^ отсутствуют
полностью.
В области собственно жестколистной растительности в сообществе
реностер со временем снова появляются кустарники. Первыми проникают
Cltffortia ruscifolia или Dodonaea и Metalasta, за ними постепенно следуют
другие. В пределах Малого Карру, напротив, примешиваются крупные
суккуленты, как Cotyledon paniculata и Euphorbia mauretanica и характер-
ный для Карру кустарник Euclea undulata.
Плато Карру не является однородной областью ни в климатическом,
ни в фитоценологическом отношении. В тех районах, где дожди идут
не только зимой, но и летом, могли бы произрастать злаки, даже если
общее количество осадков не достигает 300 мм. Но из-за хищнического
в течение столетий выпаса овец злаки оттеснены или полностью исчезли.
Их место заняли кустарничковые сообщества сложноцветных с видами
Pentzia, Pteronia, Pegplettia, Chrysocoma, Eriocephalus и др. или Rhigozum,
Zygophyllum, колючие кустарники Lycium с Salsola zeyheri, наконец,
Рис. 127. Aloe ferox под Риверсдейлом (юго-западная часть Капской провинции).
Длинные сухие стебли — Ornithogalum altissimum (фото Марлота).
168
Глава 3
Рис. 128. Aloe plicatilis (высотой 2,5 м) в пес-
чаниковых скалах под Тулбагом (фото Марлота).
кустарнички Mesembryanthe-
mum с одревесневшими стеб-
лями, но суккулентными
листьями и др. На камени-
стых склонах часто преобла-
дают крупные суккуленты:
Portulacaria afra, Cotyledon
orbiculata или C. pantculata,
Crassula arborea, Euphorbia
mauretanica или виды Aloe
(рис. 127—128). Интересные
суккуленты, которыми сла-
вится Карру, отнюдь не рас-
пространены повсеместно,
а связаны с особыми, часто
ограниченными по площади
условиями мест обитания.
Либо это кварцевые жилы,
обычно выделяющиеся своим
белым цветом на фоне бурых
скал, либо небольшие пло-
щади, покрытые белой квар-
цевой галькой или кварцевой
дресвой. По наблюдениям
автора, водный режим этих
мест особенно неблагоприя-
тен; почти невыветренная по-
рода содержит мало мелко-
зема и не задерживает воду.
На таких местообитаниях
медленно растущие мелкие
суккуленты защищены от конкуренции других видов и поэтому не
вытесняются ими; встречаются они здесь во множестве, причем в необы-
чайном видовом разнообразии.
В настоящее время растительность Карру представляет собой пеструю
смесь различных флористических элементов. Кустарнички сложноцветных,
видимо, являются проникшими с юга элементами жестколистной расти-
тельности ксерофитного строения. Родина большинства суккулентов —
западное побережье Южной Африки (севернее Капской провинции), то
есть область Намакваленд. Прежде участие злаков, несомненно, было
значительно большим. Как, вероятно, справедливо предполагает Акокс,
современный полупустынный характер Карру является следствием про-
должавшегося столетиями выборочного стравливания растительности ско-
том, причем все ценные кормовые растения, прежде всего многолетние
злаки, были уничтожены. Поэтому малопитательные растения, и в первую
очередь несъедобные виды, смогли захватить обширные площади х.
Мы приводим здесь две карты Акокса, свидетельствующие о прогрес-
сирующем изменении растительности Южной Африки (рис. 129). Верх-
няя карта отражает характер растительности, предположительно господ-
ствовавшей около 1400 г. н. э., то есть до появления здесь первых евро-
XJ. Р. Н. А с о с k s, Veld types of South Africa, «Bot. Survey of S. Africa», Me-
moir № 28, Pretoria, 1953.
Рис. 129. Изменение растительности Южной Африки со времени начала колониза-
ции территории европейцами (по Акоксу).
Вверху — незадолго перед началом колонизации, около 1400 г.н.э.; внизу — современное состояние.
1 — пустыня; 2 — собственно Карру с суккулентами; 3 — собственно Карру и карруподобный ку-
старник; 4 — кустарники бушвелд; 5 — кустарники бушвелд и злаковники; в — низкотравные
злаковники: 7 — смешанные злаковники; 8 — высокотравные злаковники; 9 — вечнозеленый дожде-
вой лес: 10 — открытый маквис; 11 — истинный маквис (финбош); 12 — граница аридного Карру.
170
Глава 3
Рис. 130. Саванна с Protea в Драконовых горах на высоте 2000 м (фото Бевса).
пейцев. Ранее заселявшие страну бушмены были столь малочисленны,
что вряд ли могли оказать существенное влияние на растительность.
Карта составлена на основе первых описаний страны и распространения
реликтовых растений. На карте видно, что в то время леса простирались
вдоль влажного восточного побережья почти без перерыва через мыс
Игольный и далее к западу. Карру с суккулентами располагалось только
на западном побережье. Собственно Карру с более густой кустарничковой
растительностью занимало западную часть современной Капской провин-
ции. Жестколистной растительности Акокс отводит лишь очень ограничен-
ную площадь; но мы отнесем к этому типу растительности также западную
часть, названную им «bush-clump veld» (велд 1 с групповым распределени-
ем кустарников).
Карта растительности 1950 г. выглядит иначе. На южном и восточном
побережьях от леса следов почти не осталось. Наибольшая площадь,
занятая им, находится в окрестностях Книсны. Во всей южной части
до Порт-Элизабета место леса заняла жестколистная растительность,
вероятно прежде всего вследствие раскорчевки, вырубки на дрова и пожа-
ров. Форпосты последней сегодня можно встретить в Драконовых горах
(рис. 130) 1 2. Большая часть прежней растительности Карру деградировала,
ее заменила пустыня. Растительность собственно Карру распространилась
далеко за реку Оранжевую. Развитые здесь прежде злаковники уничтоже-
1 Под «велдом» (от голланд. «veld» — поле) в Южной Африке понимаются про-
странства, покрытые травяной растительностью. — Прим, ред.
2 J. М u i г, R. A. D у е г, R. S. A d a m s о n, D. I. В. К i 1 1 i с k, «Bot.
Survey of S. Africa» (Pretoria), № 13, 1929; № 17, 1937; № 18, 1938; № 34, 1963.
Области средиземноморского климата за пределами Евразии
171
ны вследствие слишком интенсивного и селективного стравливания.
Акокс считает, что если землепользование в его современной форме сохра-
нится, то в ближайшие 100 лет вся территория современного Карру пре-
вратится в пустыню, а растительность собственно Карру продвинется
до Вааля и через Лесото далее на восток.
От ранее широко распространенной лесной растительности на Столо-
вой горе во влажных ущельях сохранились лишь единичные реликтовые
местонахождения. Последний крупный лесной массив находится на бога-
том осадками южном побережье, близ Книсны х. Это флористически
обедненный древостой, типичный для субтропических лесов, протягива-
ющихся вдоль влажного восточного побережья Африки через провинцию
Наталь, область Транскей и восточную часть Капской провинции 1 2, Срав-
нительный анализ этих лесных территорий показывает, что около 60 дре-
весных видов встречаются только в Натале и Транскее и полностью отсут-
ствуют на востоке Капской провинции; 130 видов распространены от
Наталя до восточной части Капской провинции, но их нет в Книсне;
ареал 130 видов простирается от Наталя до Книсны; только 25 видов —
эндемики лесов района Книсны,
Но и эти древесные породы являются формами, происшедшими от субтропиче-
ских видов; развились они, вероятно, в результате изоляции лишь в недавнее геоло-
гическое время.
Из видов, эндемичных в Южной Африке, назовем известную спарманию (Spar-
mannia africana), Empleurum serrulatum (Rutaceae), Dodonaea thunbergii (Sapindaceae),
Hartogia capensis (Celastraceae), Botryceras laurind (Anacardiaceae), Virgilia capensis
(Leguminosae), Platylophus trifoliata (Cunoniaceae), Cussonia thyrsiflora (Araliaceae),
Myrsine gilliana (Myrsinaceae), Royena glabra (Ebenaceae), Euclea acutifolia, E. polyand-
ra, E, racemosa (Ebenaceae), Olea exasperata (Oleaceae), Gonioma camassi (Apocynaceae),
Freylinia undulata (Scrophularia), Cluytia polyfolia, C., pubescens, C, rubricaulis (Euphor-
biaceae), Myrica burmannii (Myricaceae).
Обеднение видового состава прослеживается далее на запад, в сторону
Столовой горы, и объясняется, помимо прочего, климатом (возрастание
засушливости) и бедностью почв. Многочисленность сохранившихся по
периферии остатков прежней растительности, по-видимому, свидетель-
ствует о том, что еще в третичное время климат на всей территории Южной
Африки был более влажный и что леса почти без перерывов простирались
до Столовой горы.
Похоже, что некоторые роды представляет собой реликты прежнего,
очень широкого в южном полушарии распространения. Например, Podo-
carpus или Cunonia, помимо Южной Африки, встречаются и в Новой
Каледонии, W iddringtonia — также эндемик, родственный видам Callitris
в Австралии и Новой Каледонии, К листопадным относятся лишь немногие
древесные виды: Kiggelaria (Flacouriaceae), Calodendrum (Rutaceae), Ekeber-
gia (Meliaceae), Plectronia (Rubiaceae), Ficus capensis (Moraceae), Rhus laevi-
gata (Anacardiaceae), Celtis rhamnifolia (Ulmaceae), Heteromorpha arborescens
(Umbelliferae). Но сбрасывают листву эти виды не зимой, а во время
засухи; во влажные годы они могут остаться зелеными. Следовательно,
это также тропическо-субтропические элементы.
Климатические условия района Книсны иллюстрирует климадиаграм-
ма для Джорджа (см. рис. 120). Климат постоянно влажный. Количество
1 J,F,V, Phillips, Forest succession and ecology in the Knysna region, «Bot.
Survey of S, Africa», № 14, 1931; The principal forest types in Knysna region, «S. Afr.
J. Sci.», 25, 1928, p. 188.
2 H. Weimarck, Phytogeographical groups, centres and intervals within the
Cape Flora, «Lunds Univ. Arsskrift», N. F. Avd. 2, 37, № 5, 1941.
172
Глава 3
осадков около 1000 мм (максимум дождей в марте и сентябре). Температу-
ра воздуха никогда не опускается ниже 0°, во всяком случае, в лесу, хотя
над травяной лужайкой приборы .могут в единичных случаях отметить
и ее отрицательные значения. На сплошных вырубках лес восстанавливает-
ся с трудом, так как заморозки на почве убивают молодые побеги. Пожа-
ры от ударов молнии возникают крайне редко; но местные жители поль-
зовались огнем, очевидно, уже до прихода сюда европейцев и тем самым
вызвали известное сокращение площади лесов и способствовали замещению
лесной растительности устойчивыми к огню жестколистными видами.
Глубина проникновения корней деревьев в лесу близ Книсны невели-
ка, не превышает 1 м. Ежегодный прирост в толщину составляет у Podo-
carpus falcatus 3,5 мм', у других древесных видов он также редко бывает
больше 5 мм', только у Olinia cymosa был установлен прирост 15 мм.
Как видим, местные древесные виды растут медленно. Поэтому для
лесопосадок используют древесные породы родом из других стран. Прежде
это была Pinus maritima (= pinaster}, сегодня — Pinus radiata, а также
австралийские Eucalyptus diversicolor и Acacia melanoxylon. Эвкалипт
в возрасте 40 лет, когда он достигает 30 м в высоту и 80 см в диаметре,
считается спелым для рубки. Из местных деревьев ценную древесину для
изготовления мебели дают Ocotea bullata и Podocarpus. Большая часть лесов
ныне состоит из искусственных насаждений. Меньшая часть, где еще мож-
но встретить слонов и обезьян, находится под охраной, однако совсем
недавно и здесь вырубались деревья с наиболее ценной древесиной. Струк-
тура леса, по нашим записям, следующая.
Верхний древесный ярус: Podocarpus falcatus 1 is Р. lattfo-
lius (с чешуеобразной корой).
Нижний древесный ярус: высота около 25 м, постепенно пере-
ходит в ярус кустарников; очень густой и богат видами (до-
минанты отсутствуют): старая пневая поросль Ocotea bullata
(Lauraceae), Trichocladus crinitus (Hamamelidaceae), Platy-
lophus trtfoliatus (Cunoniaceae), Halleria lucida (Scrophulria-
ceae), Gonioma camassi, Olea laurifolia ( = capensis}, Curtisia,
Lachnostylis и др.
Лианы: Secamone (Asclepiadaceae), Rhoiocissus (Vitaceae) и др. (некото-
рые виды отличаются очень толстым стволом).
Эпифиты: Peperomia, Vittaria isoettfolia и другие папоротники.
Ярус травянистых растений отсутствует: почва (подзо-
листая) покрыта подстилкой; слой гумуса; отчетливый под-
золистый горизонт. На влажных участках встречаются дре-
вовидные папоротники (Hemitelia)', растущий на обочинах
дорог Blechnum capense достигает высоты 2 м.
В качестве самых западных форпостов на более влажной восточной
стороне Столовой горы произрастают близкие по составу леса, однако
число древесных пород в них сократилось до 34; в древесном ярусе растет
листопадный вид Celtis. Из Podocarpus представлен только Р. latifolius;
следует еще назвать Ocotea bullata, Curtisia faginea (обе с крупными листья-
ми), Gymnosporia acuminata и др. В нижнем ярусе можно заметить Halleria
lucida, Maurocenia frangularia и др., на влажных местообитаниях — Сипо-
nia capensis, на сухих — Olea, Olinia, Elaeodendron. Из лиан: Secamone
alpini, Scutia myrtina (встречается и как кустарник) и Rhoiocissus capensis',
1 Крупнейший экземпляр Р. falcatus в возрасте около 1500 лет достигал в высоту
40 м (высота ствола 21 м) и 6,5 м в обхвате.
Области средиземноморского климата за пределами Евразии
173
из эпифитов: Peperomia retusa и Polypodium lanceolatum. В ярусе трав выде-
ляются Cenociphium lanceolatum, родственный Carex, и крупная Knoltonia
capensis (Ranunculaceae). На сырых местах растут крупнолистные папо-
ротники, как, например, Hemitelia capensis или Hypolitis sparsisora, а на
тенистых скалах с высачивающейся водой — Hymenophyllum capense,
Н. tunbridgense и Н. marlothii.
Хотя климат этого района умеренно теплый, флористический состав
лесов заставляет вспомнить леса влажных субтропиков. Собственно уме-
ренная зона растительности в Африке вообще отсутствует.
Эколого-физиологические исследования в
области жестколистной растительности Южной Африки практически
не проводились. При огромном богатстве видов усилия ботаников были
направлены прежде всего на изучение флоры. Кноблаух изучал экологи-
ческую анатомию ассимилирующих органов жестколистных видов Южной
Африки L
Он различал:
1) эрикоидные листья с бороздой на нижней стороне, в которой между
волосками расположены выступающие устьица [виды Erica, Stilbe (Ver-
Ьепаесае), Anthospermum (Rubiaceae) и др.];
2) пиноидные листья, развивающиеся у представителей совершенно
различных семейств: Cliffortia falcata (Rosaceae), Psoralea pinnata, Aspa-
lathus (Leguminosae), Diosma (Rutaceae), Lachnaea (Thymelaeaceae), Leucos-
permum caniculatum, Aulax pinifolia, Niveniq, (Proteaceae) и др.;
3) свернутые листья у Royena rugosa (Ebenaceae), Cliffortia spec.
(Rosaceae) и др.;
4) плоские, но жесткие, обычно изотеральные листья, иногда стоящие
вертикально, как у многих видов Protea, Royena lucida, Leucadendron
и Penaeay
5) прутьевидные побеги с редуцированными листьями, например
у Psoralea aphylla (Leguminosae).
Таким образом, наблюдаются частые конвергенции и появление оди-
наковых типов; впрочем, это свойственно и другим областям жестко-
листной растительности. Параллель может быть продолжена. Анатоми-
ческие признаки, встречающиеся у средиземноморских видов, обнаружи-
ваются и здесь, а именно очень толстые наружные стенки эпидермиса
с сильно развитой кутикулой, маленькие межклетники в мезофилле,
многослойная палисадная паренхима, частое развитие клеток, заполнен-
ных бурыми дубильными веществами, углубленные устьица, склереиды
в мезофилле и т. д.
Как мы видели, осмотическое давление у средиземноморских склеро-
филов во время засухи повышается лишь незначительно. Этот вопрос
следовало изучать и у видов Южной Африки. Ван-Стаден каждый месяц
в течение года брал пробы в районе Кейптауна и определял сырой и сухой
вес листьев. Высушенные пробы были высланы в Хоэнхейм под Штутгар-
том для определения величины осмотического давления1 2.
Рассмотрим три типичных примера. Все пробы взяты на северо-
западном склоне (уклон 25°) у подножия гор Готтентоте-Голланд на высоте
80—120 м над уровнем моря. Почва — водопроницаемый песчаный сугли-
1 Е.Knoblauch, Okologische Anatomie der Holzpflanzen der siidafrikanischen
immergriinen Buschregion, Giessen, Tubingen, 1896.
2 H. Walter, J. van Stade n, Uber die Jahreskurven des osmotischen
Wertes bei einigen Hartlaubarten des Kaplandes, « J. South Afr. Bot.», 31, 1965, S. 225—
236.
174
Глава 3
Рис. 131. Кривые содержания воды (в % сухого веса) и осмотического давления (в атм)
у молодых и старых листьев Protea arborea (вверху) и количество атмосферных осадков
вблизи местообитания в период исследований (внизу). Средние значения двух парал-
лельных проб (по Вальтеру и Ван-Стадену).
нок (с обломками пород), залегающий на песчанике Столовой горы. Кустар-
ник высотой 2 м, покрытие 60%. Речь идет о типичном маквисе с протей-
ными, в составе которого доминируют Protea neriifolia. Protea arborea,
Protea repens ( = mellifera) и Leucospermum conocarpum. Ниже яруса кустар-
ников расположен 80-сантиметровый ярус с Restionaceae, из которых была
исследована Elegia stipularts. В 1963—1964 гг. засуха, стоявшая с сентября
по апрель, смягчалась редкими сильными ливнями в ноябре, декабре
и феврале. Влажное время года началось в мае проливными дождями.
На рис. 131 приведены кривые осмотического давления у молодых
и старых листьев Protea arborea, а также кривые содержания в них воды.
На диаграмме видно, что первые представляют более или менее зеркальное
отражение вторых. Осмотическое давление у молодых листьев Protea
до января было ниже, чем у старых листьев; в дальнейшем же разность
не выходила за пределы точности измерений. Лишь после того, как расте-
ние в начале мая, когда температура уже поднялась, дало побеги, давле-
ние у молодых листьев упало до 7 атм. Напротив, содержание воды
в молодых листьях всегда больше, чем в старых. Оно особенно резко уве-
личивается после образования побегов. Это различие следует относить
на счет накопления нерастворимых зольных компонентов в старых листьях
(или окремнения). Сухой вес повышается, что обусловливает кажущееся
понижение содержания воды, отнесенного к сухому весу. Именно в этом
Области средиземноморского климата за пределами Евразии
175
Рис. 132. Содер-
жание воды (вверху)
и осмотическое дав-
ление (внизу) у Leu-
cospermum conocar-
рит (по Вальтеру
и Ван-Стадену).
состоит недостаток метода определения содержания воды. При измерении
осмотического давления не приходится прибегать к относительным еди-
ницам.
Повышение осмотического давления в период засухи — признак
известной напряженности водного баланса. С началом зимних дождей
кривая идет вниз.
В отличие от Protea Leucospermum характеризуется чрезвычайно
уравновешенным водным балансом (рис. 132). Величина осмотического
давления колебалась в пределах 10—15 атм. Содержание воды также из-
менялось незначительно, в пределах от 220 до 255%. Возможно, корневая
система Leucospermum проникает глубже в почву, а быть может, у него
меньше общая листовая поверхность.
В экологическом отношении резко отличается от Leucospermum расте-
ние из семейства Restionaceae — Elegta. Колебания величины его осмоти-
ческого давления и содержания воды в течение года весьма значительны
и зеркально симметричны (рис. 133). Наименьшему содержанию воды,
равному 100%, соответствует наивысшее осмотическое давление —
26,7 атм, а наибольшему содержанию воды — наинизшее осмотическое
давление — 13,3 атм. Произведения обеих величин 26,7 X 100 = 2670
и 13,3 X 220 = 2926 почти одинаковы; это свидетельствует о том, что
изменения осмотического давления обусловлены главным образом разли-
чием в содержании воды. Резкие колебания осмотического давления,
вызванные погодными условиями, несмотря на малую транспирирующуку
поверхность этих прутьевидных растений, объясняются их близко
Рис. 133. Содержание
воды (вверху) и осмоти-
ческое давление (внизу)
у Elegia stipularis (семей-
ство Restionaceae) (по
Вальтеру и Ван-Стаде-
ну).
1.11.631.12.631.1.641.2. 1.3. 1.4. 1.5. 1.6. 1.7. 1.8. 1.9. 1.10.1.11.64-
176
Глава 3
поверхностной корневой системой и высыханием верхних горизонтов
почвы.
Эти три примера характеризуют 3 типа: гидролабильный (Elegia),
гидростабильный (Leucospermum) и промежуточный между ними (Protea).
В общем же, можно сказать, что гидратура у жестколистных растений
Капской провинции колеблется мало. Правда, лето 1963/64 г., когда
проводились исследования, было не слишком сухим. В засушливые годы
амплитуда колебаний, вероятно, будет несколько выше.
ОБЛАСТЬ ЖЕСТКОЛИСТНОЙ РАСТИТЕЛЬНОСТИ
АВСТРАЛИИ 1
Если на глобусе мысленно переместить Австралию по занимаемой
ею широте к востоку, то окажется, что Перт точно совпадает с Кейптау-
ном, а юго-западный выступ Австралии расположится на южной оконеч-
ности Африки (т. I, рис. 347). Почти идентичны климатические условия
Рис. 134—139. Климадиаграммы области с зимними дождями в Австралии.
Керридейл — район влажного леса «керри»; Перт — вблизи леса «джаррах»; Джералдтон — север-
нее границы эвкалиптовых лесов; Аделаида— юг Австралии; Мельбурн и Хобарт (восточная часть
Тасмании) — умеренно теплый климат.
двух районов (т. I, рис. 348, 349). И здесь и там отсутствует область уме-
ренного климата. Климатически аналогичная юго-западной Австралии
область повторяется в Южной Австралии в районе Аделаиды (см. рис. 6).
К югу она продолжается в сторону Виктории, где в районе Мельбурна
к ней примыкает область влажного умеренного теплого климата; близкие
условия характерны также для Тасмании (рис. 134—139) 1 2. К северу
1 L. Diels, Die Pflanzenwelt von West-Australien siidlich des Wendekreises.
Cm.: Engler-Drude, Die Vegetation der Erde, Bd. VII, Leipzig, 1906; J. G. Wood,
The phytogeography of Australia, «Monogr. Biol.», 8, 1959, p. 291—302.
2 J. Gentill i, Die Klimate Australiens, «Die Erde», 1955, S. 206—238.
Anigozanthos humilis (Haemodoracesie), около V2 натуральной величины; болото Кен-
нингтон под Пертом, юго-западная Австралия (фото Э. Вальтер).
Banksia coccinea (протейные,) приблизительно натуральная величина; окрестности
Элбени, юго-западная Австралия (фото Э. Вальтер).
Echium bourgaeanum (бурачни-
ковые) высотой 2—4 м; склон
вулкана Тейде (остров Тенериф),
высота 2000—2300 м над уровнем
моря. На переднем плане спра-
ва — Cheiranthus scoparius, на
заднем плане — Spartocytisus nub-
igenus (фото П. Морица).
Liliuni bulbiferumy Южные Альпы
до абсолютной высоты 2100 м.
Лавинный путь с лиственницами
на фоне темного хвойного леса
(фото П. Амона).
Цветущие виды Rhododendron.,
образующие подлесок в лесах
Вирджинии (Северная Америка).
Yucca (Clistoyucca) brevifolia (вы-
сотой до 10 3t), желтые цветки
сложноцветных, фиолетовые цвет-
ки вербеновых и др.; пустыня
в юго-восточной части Калифор-
нии (фото Дж. Мюнха).
Proiea cynaroides, около !/г нату-
ральной величины; Капская про-
винция в Южной Африке (фото
Сатур).
Eriophorum scheuchzeri (осоковые),
пушица альпийских озер (до вы-
соты 2900 м над уровнем моря)
(фото Г. Кёнига).
Области средиземноморского климата за пределами Евразии
177
область зимних дождей переходит в обширный засушливый район внутрен-
ней Австралии, причем в этом же направлении быстро уменьшается коли-
чество осадков и изменяется характер их распределения по временам года.
Аналогично тому, как это наблюдается в Большом Карру, все больше
проявляется влияние летних дождей (см. т. I, стр. 438 и след.).
Таким образом, область с зимними дождями в Австралии имеет много
общего с Южной Африкой, однако характер растительности, несмотря
на некоторое флористическое сходство, здесь совершенно иной. В Австра-
лии в этом климате преобладают древесные растения, представленные
исключительно различными видами рода Eucalyptus. Сходство с Южной
Африкой создает присутствие общих семейств — Proteaceae и Restionaceae.
Однако все роды австралийских Proteaceae отличны от африканских родов
и относятся преимущественно к подсемейству Grevilleoideae. Листья этих
растений в отличие от протейных Африки обычно узкие, рассеченные или
игловидные (рис. 140). Роды Накеа и Grevillea насчитывают сйыше 100 ви-
дов. В Австралии, так же как в Южной Африке, можно наблюдать взрыв-
ное образование видов в пределах некоторых родов, особенно частое
у Eucalyptus и Acacia. Род Eucalyptus занимает во всей Австралии столь
ведущее положение и характеризуется столь многими биолого-экологи-
ческими особенностями, что мы вынуждены подробно описать его, иначе
обстановка в Австралии не будет понятна.
Род Eucalyptus относится к широко распространенному в тропиках семейству
Myrtaceae, к подсемейству Leptospermoideae, почти исключительно представленному
в австралийской флоре. Своеобразно устроен цветок у эвкалипта: он не имеет ни чашели-
стиков, ни лепестков. Нижнюю часть цветочной почки образует воронковидная тру-
бочка чашечки, причем иногда у верхнего ее края выступают чашелистики в виде четы-
рех зубчиков. Вместо лепестков на трубочке чашечки находится крышечка, оперкулюм,
развившаяся из сросшихся и утолщенных лепестков. Когда цветок распускается, кры-
шечка отпадает и находящиеся под ней многочисленные согнутые тычинки выпрямля-
ются. Тычинки располагаются вокруг диска, служащего нектарником. После опыления
из трубки чашечки формируется коробочка с многочисленными семенами.
Знание строения соцветий, цветочных почек и плодов необходимо для правиль-
ного определения приблизительно 600 видов. Почти все виды распространены
только в Австралии. Лишь ареал 5—6 видов несколько выходит за пределы Австралии,
и только 2 вида в ней не встречаются, а известны в Новой Гвинее и на Филиппин-
ских островах. Все виды представляют собой деревья или кустарники высотой от
нескольких метров до 100 м. Многие виды характеризуются быстрым приростом в вы-
соту п большой массой древесины.
У кустарниковых форм ствол также обычно деревянистый, но представляет собой
крупный подземный одревесневший клубень, лигнотубер. Образование его становится
понятным, если проследить за своеобразным ростом всходов многих эвкалиптов, что
очень важно с экологической точки зрения. У большинства видов, за исключением тех
немногих, которые распространены только в гумпдных районах или на почвах с близ-
ким залеганием грунтовых вод (Eucalyptus diversicolor, Ей. regnans, Ей. grandis, Ей.
camaldulensis), через несколько недель пли месяцев после прорастания в пазухе семя-
долей пли первых двух парах листьев образуется вздутие, в котором откладываются
запасные вещества; это вздутие обладает способностью сформировать новые листовые
почки, если верхняя часть проростка отмирает. Со временем вздутия сливаются, уве-
личиваются в объеме и, как два наплыва на стебле и в верхней части корпя, нарастают
вниз. Так образуется подземный лигнотубер. Если основание проростка засыпать зем-
лей, вздутие продолжает расти кверху. При уничтожении побега в результате лесного
пожара, засухи или стравливания скотом из лигнотубера вырастают все новые и новые
побеги, причем он увеличивается в объеме. Эта особенность роста обусловливает боль-
шую резистентность эвкалиптов и их способность к восстановлению Ч В засушливых
районах деревья не образуются, а происходит формирование кустарниковидного, высо-
той всего несколько метров типа эвкалиптов, называемого «малли»; в этом случае из од-
ного огромного лигнотубера вырастают несколько ветвей одинаковой толщины.
1 М. R. J а с о b s, The growth and regeneration of Eucalyptus, «J. Austral. Inst.
Agric. Sci.», 17, 1951, p. 174—183.
12-01248
Рис. 140. Австралийские протейные.
1 _ Banhsia marginata; 2 — В. serrata; 3 — Dryandra mucronulata; 4 — Banhsia ericifclia;
5 — B. spinulosa. Нат. величина.
Области средиземноморского климата за пределами Евразии
179
Древесина эвкалиптов кольцепоровая, годичные кольца выделяются отчетливо.
Быстрый рост многих видов эвкалипта и продуцирование ими большой массы древесины
не являются следствием особенно интенсивного фотосинтеза на единицу листовой
поверхности, а объясняются способностью быстро создавать большую листовую поверх-
ность. Последняя связана с определенной особенностью роста. Рассматривая побег
эвкалипта, в пазухе каждого молодого листочка можно заметить длинный стебель
с верхушечной почкой. Это уже развивающиеся боковые ветки. На них тоже образуются
(на расстоянии 2—3 см один от другого) листья, пазушные почки которых сразу дают
побеги. Поэтому у эвкалиптов в отличие от других деревьев ежегодно вырастают боко-
вые ветки нескольких порядков. В результате очень быстро формируется крона с об-
ширной листовой поверхностью, способная к продуцированию большой массы сухого
вещества. В северном Трансваале Ей. saligna уже через 6 лет дает крепежный лес,
а через 12 лет — деловую древесину. Период же оборота рубки Pinus caribaea в той
же области составляет 25 лет, то есть он вдвое дольше, древесина же ее намного легче.
В Австралии прирост эвкалиптов обычно сдерживается поражением их насеко-
мыми, которые объедают почки и листья молодых боковых веток. Но и в этом случае
у основания пазушных побегов снова образуются дополнительные почки, тотчас даю-
щие побеги. После лесных пожаров в Австралии также часто наблюдаемся быстрый рост
растений, возникших из семян.
Например, Ей. grandis через 2 года после посадки на гари достиг 12 м в высоту,
так как огонь уничтожил всех насекомых. Но уже на третий год деревья были объедены.
Со временем между степенью поражения деревьев насекомыми и ростом деревьев уста-
навливается известное равновесие.
Листья всех видов Eucalyptus очень похожи, они простые, удлиненно заостренные,
вечнозеленые (ср. т. I, стр. 233), кожистые и снабжены масляными железками. Распо-
ложены листья (у взрослых растений) очередно и вертикально. Поэтому эвкалиптовые
леса кажутся очень светлыми; эвкалипты, даже молодые, не переносят затенения.
В Восточной Австралии на плодородных почвах эркалипты не выдерживают конкурен-
ции с тропическо-субтропическими дождевыми лесами (см. т. I, стр. 261), но господ-
ствуют на тощих почвах, на которых виды индо-малайского дождевого леса расти
не могут. Виды эвкалипта образуют леса на всей территории Австралии от уровня моря
до границы древесной растительности в горах п встречаются как в тропиках с летними
дождями и в гумпдных районах, так и областях с зимними дождями. Эвкалипты исче-
зают в аридной части Австралии, как только годовая сумма осадков снижается до ме-
нее 300 мм в области с зимними дождями и до менее 600 мм в районах с летними дождя-
ми. Однако Ей. camaldulensis (= rostrata) можно найти и в Центральной Австралии, они
приурочены здесь к сухим руслам рек с неглубоким залеганием грунтовых вод.
Отдельные виды рода Eucalyptus экологически приспособились к очень узкоогра-
ниченным условиям мпкроместообитаний как в отношении климата, так и почвы х.
Конечно, существуют и более широко распространенные виды, но они распадаются
на экоклины без видимой морфологической дифференциации. Так, например, при изу-
чении Ей. viminalis, а также Ей. coccifera и Ей. pauciflora оказалось, что степень моро-
зоустойчивости их проростков отчетливо зависит от происхождения семян. В Западной
Австралии отдельные виды иногда образуют чистые насаждения, в Восточной же
Австралии преобладают смешанные древостои. По ранным Приора, при этом возникают
очень сложные взаимосвязи.
Род Eucalyptus распадается на 5 подродов:
Macranthera
Renanthera
Adnata
Corymbosa
Eudesmia
(в него входит половина всех видов)
(менее одной четверти)
(менее одной четверти)
(приблизительно одна десятая)
(около 12 видов, ограниченно распространенных).
Внутри подродов известны гибриды. Однако область их распространения совпа-
дает с ареалами отдельных видов, развивающихся на экологически различных место-
обитаниях. Последние обычно очень четко отграничены. По-видимому, гибриды обла-
дают меньшей конкурентной мощностью и живут менее долго, почему, как правило,
отсутствуют в старых насаждениях. Чаще они наблюдаются на площадях, измененных
деятельностью человека. В смешанных насаждениях, то есть там, где на одном местоо-
битании присутствует несколько видов, эти виды принадлежат к разным подродам
и гибридов не дают.
Интересно, что внутри четырех крупных подродов можно выделить ряды-гомо-
логи как с точки зрения морфологии и анатомии (образование плодоносящих молодых
растений, трехцветковые соцветия, характер образования коры), так и в отношении
1 L.D.Pryor, Species distribution and association in Eucalyptus, «Biography and
Ecology in Australia. Monogr. Biol.», 8, 1959.
12*
180
Глава 3
климата, условий среды, принадлежности к вертикальным поясам и выбора местооби-
таний. В Снежных горах по мере увеличения высоты виды разных подродов параллель-
но сменяют друг друга, причем границы распространения по высоте отдельных подро-
дов не всегда совпадают; в результате формируются разнообразные смешанные насаж-
дения. Очевидно, это относится и к местообитаниям с различными эдафпческимп усло-
виями; благодаря этому возникают разнообразнейшие сочетания видов. Приор предпо-
лагает, что такого рода сообщества на очень бедных почвах могут обладать некоторым
преимуществом в отношении питания растений. Например, виды подрода Renanthera,
возможно, представляют собой облигатные микоризные растения. Но пока это только
гипотеза.
Итак, можно утверждать, что в Австралии разные виды рода Euca-
lyptus практически одни образуют все леса, за исключением отдельных
районов восточного побережья. Эти виды характеризуются такой же
экологической дифференциацией, какую мы наблюдали на других мате-
риках у различнейших родов и видов хвойных и лиственных пород. И тем
не менее приходится констатировать, что леса Австралии слабо диффе-
ренцированы.
Пример Австралии показывает, что разделение растительности по
одному физиономическому признаку недостаточно. Оно должно быть
основано не только на экологических факторах, но и на флористическо-
таксономических данных. На это мы уже указывали при рассмотрении
засушливых районов Австралии в т. I. То же следует сказать и о ее гумид-
ных областях. Физиономически растительность этих районов Австралии
нельзя параллелизовать с растительностью других флористических царств.
Весьма своеобразны и формы цветков австралийских растений (см. цвет-
ное фото 1).
Лес «керри»
На юго-западе Австралии количество осадков со 1500 мм сокращается
в северо-восточном направлении, одновременно усиливается и становится
более продолжительной летняя засуха. Соответственно сменяют друг друга
различные лесообразующие виды Eucalyptus. Во влажной части области
зимних дождей, где годовая сумма осадков превышает 1250 мм, господ-
ствующим видом является «керри» (Ей. diversicolor). Это дерево высотой
до 85 м (в среднем 60—75 м), с очень прямым и стройным стволом
(рис. 141). Степень сомкнутости крон не превышает 65%. Леса светлые.
Ярус кустарников покрывает в среднем 50% площади. В нем часто доми-
нирует Trymalium spathulatum (Rhamnaceae) с широкими тонкими листья-
ми, называемый «орешником», или растет орляк (Pteridium) с 1,5-метро-
выми листьями, покрывающий до 60% площади. Их присутствие придает
лесу вполне гигрофильный облик. Ярус травянистых растений высотой
20 см с многочисленными Bossiaea amplexicaulis покрывает около 40% .
Редко встречаются лианы Clematis и Hardenbergia. Над кустарниковым
ярусом лианы не поднимаются. После лесного пожара первыми появляют-
ся всходы видов Acacia, имеющих «твердые» семена. Среди Acacia пред-
ставлены формы как с перистыми листьями (A. pentadenia, A. pulchella),
так и с филлодиями (A. urophylla и др.). Кроме того, встречается Albizzia
distachya. Изредка можно заметить также ксероморфные формы, например
некоторые виды бобовых или Hakea amplexicaulis с листьями, похожими
на листву Ilex.
Примесь других видов деревьев наблюдается редко. Назовем Ей.
calophylla, распространенный и в следующей лесной зоне, и низкую Casua-
rina decussata. По берегам рек растут мелколистные, вполне ксероморфного
Рис. 141. Лес «керри» (Eucalyptus diversicolor) в Южной Австралии. Деревья дости-
гают высоты 85 м, в подлеске обычно Acacia pulchella.
182
Глава 3
облика виды семейства миртовых, как Agonis flexuosa и A. junipertna,
и узколистное бобовое — Gompholobium. На крайнем юге керри сменяют
некоторые другие высокорослые виды Eucalyptus, распространение кото-
рых, правда, более локализовано.
Почвы под этими влажными средиземноморскими лесами обычно
мощные, глинистые, с почти нейтральной реакцией. Профиль их напоми-
нает разрез бурых лесных почв под буковыми лесами Центральной Евро-
пы. О бедности почв питательными веществами свидетельствует массовое
развитие Pteridium. Встречаются также насекомоядные растения: на
открытых участках нередки Drosera pallida и D. stolontfera, а в заболочен-
ном кустарнике из миртовых на торфе, образованном корнями растений
семейства Restionaceae, было найдено очень редкое растение — Cephalotus
(рис. 142). Его кувшинообразные листья (канны) приспособлены для ловли
насекомых. В одном канне этого вида нашли 18 крупных муравьев.
О развитии Ей. diversicolor можно привести следующие сведения. Цветочные почки
развиваются в течение 2,5 лет. Период основного цветения приходится на декабрь
(время засухи), но, в общем, цветение растягивается на более длительное время. Для
созревания плодов после опыления требуется 1,5 года, так что весь цикл развития до со-
зревания семян продолжается 4 года. Рассевание семян происходит в летние месяцы
на протяжении двух лет. Каждые 4—5 лет деревья плодоносят особенно обильно. Если
осенью выпадает 100 мм дождя, проростки появляются через 3 недели. На возникших
из семян растениях уже в первый год образуются 14 листьев. На почве, удобренной
золой (после лесного пожара), растения в первый год достигают высоты 80 см, на вто-
рой год — уже 3 м. Дальнейший годовой прирост в течение первых 25 лет составляет
1,2—1,5 м, а затем, до 80-летнего возраста, около 30 см. Затем рост прекращается, изме-
няется форма кроны. У молодых деревьев она плотная, удлиненно-коническая, сильно
облиствленная. У старых деревьев крона зонтиковидная. Терминальный побег в них
Рис. 142. Cephalotus follicularis с кувшинообразными (каннами) и обычными листь-
ями (масштаб — круглый карандаш) на влажной болотистой почве под Пембертоном,
юго-западная Австралия (фото Э. Вальтер).
Области средиземноморского климата за пределами Евразии
183
отмирает, но из пазушных почек развиваются новые побеги, поэтому разреженная кро-
на кажется состоящей из отдельных кустиков листьев. Около 50% листьев после обра-
зования новых побегов, то есть через год, опадают, остальные погибают через 2, реже
через 3 года. Листовой опад в первый год теряет 39% веса, остаток медленно разлагает-
ся. Предельный возраст деревьев — 350 лет, но прирост древесины у деревьев в воз-
расте свыше 120 лет очепь мал. При правильном лесопользовании обеспечивают есте-
ственное возобновление: треть деревьев оставляют в качестве семенников, неиспользо-
ванная древесина сжигается на месте рубки, с тем чтобы обогатить почву золой; воз-
никает очень густой подрост; разреживание происходит естественным путем, так как
жердняк применения не находит.
Область леса керри мало заселена. На раскорчеванных участках
устраивают молочные фермы или закладывают плантации яблонь, но при-
рост древесины в лесу столь велик, что, видимо, рациональнее вести
лесное хозяйство Ч Для технических целей древесина эвкалипта менее
пригодна, чем древесина сосны. Опыты посадки Pinus radiata показали,
что продуцирование древесины этим видом приблизительно $ 4 раза боль-
ше, чем в эвкалиптовом лесу, при условии, что почва не испытывает недо-
статка в микроэлементах (в частности, в цинке) (табл. 12). На очень бед-
ных песчаных почвах побережья высаживают также Pinus maritima
(= pinaster). Посадка всех других видов сосны не дала положительных
результатов: величина прироста древесины в куб. метрах на 1 га в год
оказалась в этих насаждениях приблизительно в 10 раз меньше. Древе-
сина эвкалипта обладает более высоким объемным весом.
' Таблица 12
Прирост древесины у различных пород леса
«керри» (по Стьюарту)
Вид
Прирост, в куб. фут.
на 1 акр в год
«Джаррах» (Eucalyptus таг-
ginata) ....................
«Керри» (Eucalyptys diver-
sicolor) .............
Pinus pinaster ....
Pinus radiata . .
15-25
75—100
100—150
300-400
Еще 60 лет назад Дильс, изучавший растительность юго-западной
Австралии, не смог осмотреть район леса керри из-за отсутствия там дорог.
В настоящее время освоение многих районов Западной Австралии только
начинается. Не удивительно поэтому, что среди всех областей с зимними
дождями австралийский район претерпел наименьшие изменения под
влиянием человека.
Лес «джарраос»
Совершенно иной характер присущ лесу «д ж а р р а х» {Eucalyp-
tus marginata, рис. 143), расположенному в несколько более сухой истинно
средиземноморской зоне. Количество осадков здесь 625—1250 мм в год.
Засуха летом резко выражена. Вид образует чистые насаждения лишь
с небольшой примесью Ей. calophylla. Леса джаррах связаны с определен-
1 D. W. R. Stewart, Forestry as a form of land use in the South West of
W. Australia, «I. Austral. Inst. Agric. Sci.», 1958, p. 101—111.
184
Глава 3
Рис. 143. Лес «джаррах» (Eucalyptus marginata) с травянистыми
деревьями (Kingia australis, между ними Xanthorrhoea preissii).
Стволы деревьев почернели после пожара. На переднем плане —
Г. Вальтер (фото Э. Вальтер).
ными, сравнительно бедными питательными веществами почвами, которые
развиты на обширной территории. В прибрежных районах на известковых
почвах джаррах уступает место «туарту» (Ей. gomphocephala), на влажных
почвах — Ей. patens, а в поймах — Ей. rudts.
Лес джаррах намного ниже, светлее и ксерофильнее леса керри.
Ей. marginata — дерево высотой не более 40 м, в среднем около 15—20 м;
предельный его возраст — 200 лет. Сомкнутость лесного полога редко
достигает 50%. Растения, возникшие из семян, образуют лигнотубер
и в течение первых 15 лет практически не подрастают. Продукция древеси-
Области средиземноморского климата за пределами Евразии
185
ны приблизительно в 8 раз меньше, чем у Ей. diverstcolor. Но древесина
очень тяжелая и не поражается термитами. Нижний древесный ярус обра-
зуют Casuarina preisti и виды Banksia (протейные). В ярусе кустарников
много ксероморфных представителей протейных, бобовых и миртовых.
На общем фоне выделяются травянистые деревья — Xanthorrhoea preisti
и Kingia australis. Они весьма огнестойки, пожар только способствует их
расселению. Часто встречается и Macrozamia fraseri (Cycadaceae). Ярус
травянистых растений поражает богатством флористического состава
(Epacridaceae, Goodeniaceae, отдельные орхидеи и т. д.). Особенно инте-
ресно присутствие насекомоядных Drosera, которые в юго-западной
Австралии представлены 47 видами. Одни из них — маленькие розеточные
растения. Их хорошо защищенная верхушечная почка переживает засуху,
располагаясь на поверхности почвы. Уже после первого дождя она дает
побег, засыхающий сразу после цветения. Другие растения пережидают
летнюю засуху в виде подземного клубня и с началом држдей образуют
либо очень красивые большие розетки, либо длинные слабые побеги
с маленькими очередными листьями, которые своими клейкими железисты-
ми волосками прикрепляются к веткам кустарников. Такие растения
поднимаются на высоту до 1 м и бросаются в глаза своими крупными,
нередко яркими цветками. Широкое распространение и частая встречае-
мость видов Drosera служит признаком бедности почв питательными веще-
ствами. Речь идет об очень богатых SiO2 почвах с железистыми конкреция-
ми, называемых латеритами. Типичный) профиль па вершинном плато
хребта Дарлинг следующий:
Верхние 80 см\ сильно сцементированные кремнекислотой железистые конкреции,
вверху размером с грецкий орех, внизу — с горошину; верхние 20 см очень
твердые.
Ниже: выветрившийся гранит желтоватой или местами красноватой окраски.
По-видимому, это ископаемая почва, сформировавшаяся в третичном
периоде при климатических условиях, сходных с современным муссонным
климатом Индии.
Заселение зоны лесов джаррах человеком началось лишь в последнее
время. Леса часто страдают от пожаров, поэтому стволы деревьев кажутся
обугленными.
По направлению в глубь суши за поясбм джаррах следует зона «вандо»
(Ей. redunca ssp. wandoo). В районах с годовым количеством осадков
625—500 мм этот вид образует разреженные леса («вудленд»). В настоящее
время большая часть лесной территории занята фермерскими хозяйствами
в окружении сеяных пастбищ. Большие деревья не корчуют, выжигают
только кустарник и взрыхляют почву. В юго-западной Австралии, где
осадки выпадают зимой, собственно злаковники отсутствуют (ср. т. I,
стр. 300), хотя в растительном покрове злаки встречаются часто. Для
использования на пастбищах они, однако, служить не могут.
Высеваются привозные семена трав, как, например, Lolium rigidum. Но при бед-
ности почв питательными веществами проблема пастбищ была решена лишь благодаря
рациональному удобрению. Потребовалось сильное удобрение почв суперфосфатом.
Кроме того, при высеве трав добавляют семена средиземноморского вида Trifolium
subterraneum, обогащающего почву азотом. Это, правда, однолетний вид, но его плоды
углубляются в почву, и потому каждый год происходит его самосев. Лишь эти меры
позволили поднять производительность овечьих ферм в зоне зимних дождей. Важней-
шее значение для сельского хозяйства Австралии с ее очень древними почвами имеет
также внесение в них микроэлементов.
186
Глава 3
Рис. 144. Редкий эвкалиптовый лес вблизи его аридной границы. Деревья Eucalyp-
tus salmonophloia и Ей. longicornis; на переднем плане — кусты Daviesia (бобовые)
и Grevillea (протейные).
По мере увеличения сухости климата возрастает численность видов
Eucalyptus. К ранее названным добавляются Ей. astringens, Ей. loxophleba,
Ей. salmonophloia, Ей. salubris и многие другие (рис. 144). В области, пере-
ходной к аридной зоне, где сумма осадков приблизительно равна 300 мм,
уже насчитывается около 30 видов эвкалипта. Здесь встречаются и различ-
ные формы «малли» х.
Песчаные пустоши (рис. 145)
В юго-западной Австралии песчаные почвы столь бедны, что на них
не растут даже самые неприхотливые виды эвкалипта. Поэтому обширные
площади заняты здесь пустошью с протейными высотой 0,5—0,75 м. Пусто-
ши распространены на северо-западе области с зимними дождями восточ-
нее Джералдтона и устья реки Мёрчисон (осадки менее 500 мм!год), а так-
же в более влажном районе юго-восточнее гор Стерлинг. Неплодородность
здешних почв служит также препятствием развитию сельского хозяйства.
1 Область, где количество зимних дождей сокращается с 500 до 300 мм, ныне
занята культурными землями — это пояс пшеницы. Здесь снимают самые высокие уро-
жаи, так как в более влажных районах очень велико поражение растений ржавчиной,
а в более сухих местностях устойчивый сбор не гарантирован. Практически возделыва-
ют только пшеницу, ибо при полной механизации полевых работ она требует наимень-
шей затраты рабочей силы; для внесения удобрений и борьбы с сорняками используют
самолеты.
Области средиземноморского климата за пределами Евразии
187
Рис. 145. Типичная австралийская пустошь с протейными. Видны светлые соцветия
Conospermum stoechadis’, небольшие темные деревца — хвойное Actinostrobus arenarius
(фото Э. Вальтер).
В Европе нам кажется естественным, что с самыми бедными песчаны-
ми почвами связана флористически скудная растительность, состоящая
из немногих нетребовательных видов. В Австралии картина обратная.
Песчаные пустоши наиболее богаты видами, причем разнотравье и злаки
почти не представлены. В пойме реки Калган в кустарнике «малли» с
зарослями протейных на 100 ж2 площади насчитали 63 вида кустарников
и кустарничков, а всего 90 видов.
Своеобразие пустошам придают древесные гемикриптофиты, то есть
протейные, ствол которых расположен горизонтально непосредственно
над поверхностью почвы; на определенно^ расстоянии одна от другой из
него вертикально вверх отходят ветки. Из числа таких видов можно
назвать: Banksia prostrata, В. repens, Dryandra prostrata, D. nivea. Очень
низкорослым растением является также Casuarina humilis. На периодически
увлажняемых песках наряду с различными видами Drosera встречается
Utricularia menziesii с подземным клубнем. Ее листья собраны в маленькую
розетку, на которой сидят мелкие ловчие пузырьки. Цветок у растения
очень крупный, ярко-красный, почти не возвышающийся над землей.
В центральной части этой области в период жаркого сухого лета
-образуются небольшие засоленные впадины. Зимой дожди вымывают соли
из верхних горизонтов почвы, и тогда появляются всходы многочисленных
несолевыносливых терофитов, например Hydrocotyle calyculata, и галофитов
Arthrocnemum, Carpobrotus (Mesembryanthemum), Plagianthus (мальвовые).
Вокруг таких засоленных местообитаний наблюдается очень четко выра-
женная «зональность»: у границы с засоленными почвами растут Mela-
leuca cuticularis (миртовые), образующая густой кустарник, и много-
численные виды семейства Centrolepidaceae.
188
Глава 3
Южная Австралия
Климат Южной Австралии очець похож на климат юго-запада матери-
ка, но растительность и условия ее произрастания здесь совершенно дру-
гие. Горы Лофти достигают высоты 670 м. С высотой количество осадков
увеличивается от 450 мм на побережье до 1200 мм на вершине возвышен-
ностей. Почвенный покров также далеко не однороден. Произрастающие
здесь виды эвкалипта отличны от видов, распространенных в юго-восточ-
ной Австралии, к тому же они многочисленнее. Растительность представ-
лена различными типами, начиная с кустарников малли и кончая сухими
эвкалиптовыми лесами из Eucalyptus obliqua и Ей. baxtert, приблизительно
соответствующими лесу джаррах. О растительности Южной Австралии
написано много сводных работ, а также отдельных монографий, осве-
щающих и почвенные условиях. Число видов, занесенных из Южной Афри-
ки или Средиземноморья и ныне в качестве сорняков занимающих, иногда
значительные площади, в Южной Австралии больше, чем на юго-западе
страны, больше здесь и земель под возделываемыми растениями.
Эколого-физиологические исследования
в юго-западной Австралии
Весьма примечательно, что на юго-западе Австралии, несмотря на
значительные территориальные колебания годовой суммы осадков (от 1500
до меиее 500 мм), повсеместно господствует одна и та же жизненная фор-
ма, а именно вечнозеленый (кожистые листья) эвкалиптовый лес.
Почему листья с уменьшением осадков не становятся более ксероморф-
иыми, можно понять, лишь сопоставив количество осадков с индексом
листовой поверхности древесного яруса различных лесных зон. В т. I,
стр. 307, мы уже вкратце отметили, что масса ежегодно образующегося
листового опада и площадь поверхности опавших листьев в расчете на
1 га уменьшаются пропорционально количеству осадков и что, следова-
тельно. влагообеспеченность единицы листовой поверхности в гумидных
и аридных областях приблизительно одинакова. В табл. 13 и 14 приведены
более точные цифры, причем раздельно для первичных лесов и молодых
насаждений, поскольку общая листовая поверхность, а таким образом,
и листовой опад в последних больше. Первичный лес разрежен и отли-
чается более густым подлеском, листовую поверхность которого следова-
ло бы учесть, что, однако, не сделано; в молодых густых насаждениях
какой-либо подлесок отсутствует, так что общая площадь листовой
поверхности слагается из площади поверхности листьев древесного яруса.
Надо учесть, что количество осадков не измерялось непосредственно
на опытных площадках, а получено путем интерполяции данных бли-
1 J. G. W о о d, The vegetation of South Australia, Adelaide, 1936; R.L. Croc-
ker, Soil and vegetation relationships in the lower south-east of South Australia, «Tran-
sact. Roy. Soc. S.-Australia», 68, 1944, p. 144—172; R. L. S p e c h t, R. A. Perry,
Plant ecology of the Mount Lofty Range, ibidem, 72, 1948, p. 91—132; R. L. Spech tr
A reconnaissance survey of the soils and vegetation of County Buckingham, ibidem, 74,
1951, p. 79—107; H. A. M a r t i n, Sklerophyll communities in the Inglewood
District, Mount Lofty Range, South Australia, ibidem, 85, 1961, p. 91—120;
R. L. S p e c h t, R. F. В г о w n e 1 1, P. N. Hewitt, The distribution of Euclyptus-
elaeophorba, ibidem, 85, 1961, p. 155—176; Flora conservation in South Australia,,
ibidem, 85, 1961, p. 177 — 196.
Области средиземноморского климата за пределами Евразии
189
Таблица 13
Первичный лес с подлеском
Количество осадков Листовой опад Листовая поверх- ность
мм относит. кг/га относит. М2/м2 | относит.
1530 100 2400 100 1,114 100
810 53 1280 53 0,57 50
750 49 1500 62 0,66 58
Таблица 14
Молодые насаждения без подлеска
Количество осадков Листовой опад Листовая поверх- ность
мм относит. кг/га относит. М2/м2 относит.
1330 100 2740 100 1,30 100
1290 97 2040 75 1,05 81
560 42 1290 47 1 0,39 30
жайших метеорологических станций и что определение массы листового
опада сопряжено с небольшими ошибками. И тем не менее, несмотря на
некоторые значительные расхождения цифр в таблицах, сопоставимость
их выступает весьма отчетливо. Однако известное нарастание ксероморф-
ности листьев с уменьшением количества осадков, несомненно, наблю-
дается, ибо с увеличением засушливости климата листья летом более дли-
тельное время пребывают в условиях острого дефицита влажности воздуха.
Сравнительные исследования анатомии листьев различных видов эвка-
липта не производились.
В дождливый сезон, в августе 1958 г., автором было отобрано несколь-
ко проб и получены значения осмотического давления, приведенные
в табл. 15 и 16. Попутно определялось содержание хлоридов. Оно введено
в таблицы под рубрикой «содержание NaCl», так как известно, что в
Западной Австралии соль, приносимая с атмосферными осадками, играет
важную роль в образовании засоленных почв (см. т. I, стр. 330).
Из приведенных в таблицах данных видно, что осмотическое давление
у жестколистных растений Австралии в дождливый сезон очень низкое.
Осмотическое давление у протейных соответствует давлению у растений
того же семейства в Южной Африке. Содержание хлорида также невелико.
До настоящего времени отсутствуют данные о годовом ходе величины
осмотического давления у типичных видов. Известна лишь работа Грива,
которая содержит несколько цифр, полученных в засушливый период
и относящихся к различным видам. Пробы были взяты к концу засухи,
причем часть их отобрана в районе Перта (концентрация отжатого сока
определялась методом Баргера), часть же — в горах Лофти в Южной
Австралии; осмотическое давление измерено плазмолитическим методом
(табл. 17).
Как видно, величины осмотического давления на 50% превышают
значения, полученные автором весной для Banksia, Stirlingia и Xanthorr-
190
Глава 3
Таблица 15
Содержание воды (в % сухого веса), осмотическое давление (в атм)
и содержание хлоридов в клеточном соке (в атм)
у некоторых видов леса «керри»
Описание пробы Содер- жание воды Осмо- тичес- кое давле- ние Содер- жание NaCl
Eucalyptus diversicolor Листья цветущих веток, свободно стоящее дере- во высотой 6 м 120 12,8 3,1
Листья нецветущих веток, дерево (5 м) в ред- ком лесу 125 14,8 3,0
Листья молодого дерева (2 л), в тени ... 130 13,4 2,9
Листья кроны, 100 — 150-летнее дерево (60 м) 115 14,2 2,6
Листья с вершины того же дерева . 116 13,4 1,5
Trymalium spathulatum (крушиновые) Куст в лесу, тень, листья гигроморфные . . . 184 12,3 2,4
Куст в лесу, при разреженном распределении 164 12,6 2,0
Куст при полном освещении солнцем 130 16,0 1,5
Agonis flexuosa (миртовые) Листья куста (4 л), берег реки Ли-Фрой . 115 12,7 2,8
Таблица 16
Содержание воды (в % сухого веса), осмотическое давление (в атм)
и содержание хлоридов (в атм) у некоторых видов леса «джаррах»
Описание пробы Содер- жание воды Осмо- тичес- кое давле- ние Содер- жание NaCl
Eucalyptus marginata
Листья с нижних ветвей большого дерева . 106 16,3 5,7
Листья молодых деревьев (1 — 2 м высоты) . 104 13,1 3,0
Casuarina freiseriana молодые побеги . 106 14,5 3,9
Виды Banksia (протейные)
В. menziesii, листья 97 13,3 2,4
В. menziesii, листья 94 14,7 4,7
В. attenuata, листья . 75 15,4 3,6
Виды Накеа (протейные)
Н. prostrata, листья (под Пертом) 118 15,3 2,7
Н. spec., листья (гранитная скала, хр. Дарлинг) 70 19,7 2,7
Stirlingia latifolia (протейные), листья . 142 13,6 1,8
Xanthorrhoea preisii (доля NaCl неточно) 4,7
Листья, очень хрупкие (под Пертом) . 198 9,3
Листья, очень хрупкие (под Пертом) . . 180 10,6 3,1
Листья, старый экземпляр (хр. Дарлинг) . . . 161 14,0 6,3
Macrozamia jraseri (цикадовые), части очень жест- 112 10,9 1,2
ких листьев
Daviesia divaricata (бобовые), листья .... 108 11,5 1,2
Нelichrysum cordatum, очень мягкие листья . . 390 8,1 3,1
Borya nitida (ксероморфная, из лилейных) Молодые листья' растения-подушки (гранитная 170 10,9 1,6
скала, хр. Дарлинг)
Области средиземноморского климата за пределами Евразии
191
Таблица 17
Величина осмотическою давления
(в атм) у некоторых видов
жестколистной растительности
(по Гриву)
Перт (Западная Австралия)
Banksia menziesii . . . . 21
Stirlingia latifolia ... 21
Hibbertia hypericoides . 27
Bossiaea eriocarpa........... 21
Hardenbergia comptoniana . 14
Kennedya prostrata . . 21
Хребет Лофти (Южная Австралия)
Eucalyptus odora . 30
Acacia pygnantha . 25
Hibbertia stricta .... 21
Leptospermum myrsinoides . . 18
Xantorrhoea semiplana 15
hoea. Следовательно, речь идет о возникновении известного дефицита насы-
щения, хотя и не очень резко выраженного. Соответственно этому измере-
ния интенсивности транспирации, произведенные Гривом под Пертом
в разное время года, показали некоторое сокращение транспирации
в период засухи Весной кривые суточного хода интенсивности транспи-
рации характеризовались пиком в полуденное время, летом же этих
пиков было два. Поздним летом кривые были более пологими и лишь
изредка резко поднимались перед полуднем. Листья видов с неглубоко
расположенной корневой системой, таких, как Hibbertia и Bossiaea, в кон-
це лета столь мало содержали влаги, что отжать клеточный сок из них
не удалось; растения крайне экономно расходовали воду. Результаты
наблюдений позволяют, таким образом, говорить об экологическом сход-
стве этих видов с малакофильными губоцветными Средиземноморья.
В летние месяцы только у Eucalyptus marginata интенсивность транспира-
ции при открытых устьицах достигала 7,2 мг!г*мин, по-видимому, благо-
даря глубоко проникающей корневой системе и хорошей обеспеченности
растения водой. Напротив, у Ей. calophylla транспирация резко понизи-
лась. Однако при резком недостатке вод^г эвкалипты также обладают
способностью сильно сокращать интенсивность транспирации, закрывая
устьица и изолируясь тем самым от окружающего пространства. Отноше-
ние кутикулярной транспирации к устьичной составляет у Eucalyptus
1 : 60. Количество устьиц у них приблизительно равно 300 на 1 мм1 2 по-
верхности листа, у протейных — 150 на 1 мм2. Таким образом, эвкалипты
австралийской области зимних дождей в эколого-физиологическом отно-
шении близки к вечнозеленым дубам. Можно ожидать, что летом уменьше-
ние интенсивности транспирации у австралийских склерофилов вслед-
ствие частичного смыкания устьиц обусловливает также сокращение асси-
миляции СО2. Соответствующие измерения, однако, не проводились.
Грив опубликовал несколько данных определения сосущей силы,
выполненного методом Шардакова 2. Эти значения намного превышают
1 В. J.Crieve, Transpiration of Western Australian (Swan Plain) sclerophylls,.
«J. Roy. Soc.», 40, 1956, p. 14—30; К. К r e e b, Transpirationsmessungen an einigen
australischen Immergriinen im Bereich des feuchten Eucalyptuswaldes und der Mulga-
buschsavanne, «Oecol. Plant.», 1, 1966, s. 235—244.
2 B.J.Grieve, Negative turgor pressures in sklerophyll plants, «Austr. J. Sci.»r
23, 1961, p. 375—377.
Ри с. 146. Примеры конвергенции у жестколистных видов.
Я — Pittosporum phillyraeoides (Pittosporaceae); 2 — Acacia linearis; 3 — A. floribunda (Leguminosae);
-4 — Hakea saligna (Proteaceae); 5 — Leptospermum resiniferum; 6 — Metrosideros viminalis (Myrta-
ceae); 7 — Styphclia cunninghami (Epacridaceae). Нат. величина.
Области средиземноморского климата за пределами Евразии
193
величину осмотического давления, что должно было бы свидетельствовать
об отрицательном тургорном давлении, но, скорее всего, определения
сосущей силы были неточными L
Морфологические особенности жестколистных растений, их ксеро-
морфные признаки такие же, как у видов из других областей с зимними
дождями; при этом, однако, нигде не наблюдается так много заметных
конвергенций у представителей различных семейств, как именно на юго-
западе Австралии. Принадлежность многих видов к определенным семей-
ствам часто очень трудно установить, если отсутствуют органы размноже-
ния. Приведем несколько примеров такого рода конвергенции:
1) широкие кожистые листья: у Myrtaceae, Leguminosae (филлодии),
Proteaceae, Pittosporaceae, Epacridaceae (рис. 146);
2) эрикоидные листья: у Myrtaceae, Epacridaceae, Leguminosae и
Goodeniaceae;
3) игловидные листья: у Proteaceae и Leguminosae, у Casuarina —
иглоподобные побеги;
4) афиллия: у Goodeniaceae и Leguminosae;
5) крылатые ассимилирующие стебли: у Leguminosae и Umbelliferae.
Очень хорошо выражены также ксероморфные анатомические призна-
ки (ср. Hakea suaveolens и др.). Известно, что ксероморфность часто пред-
ставляет собой собственно пейноморфоз, то есть, в общем, она отражает
явления недостаточности в обеспечении растений элементами минераль-
ного питания, а не только приспособленность к дефициту влаги. Явления
недостаточности наблюдаются, например, и при азотном голодании. Как
мы уже подчеркивали, почвы Австралии бедны питательными веществами;
особенно неблагоприятны в этом отношении песчаные почвы пустошей.
Поэтому возникает вопрос, не связан ли особенно резко выраженный
ксерофильный характер флоры Австралии со скудостью ее почв. Недоста-
ток одного азота не может быть решающим фактором. Дело в том, что
ксероморфность необычайно четко проявляется у многих австралийских
растений из семейства бобовых, например у 400 видов Acacia секции
Phyllodinae, иногда образующих чрезвычайно твердые филлодии (рис. 147),
хотя у бобовых имеются корневые клубеньки с бактериями, связывающими
азот. Но почвы бедны также фосфором и микроэлементами, и именно недо-
статок фосфора играет в Австралии очень большую роль 1 2. Установлено,
что очень многие ксероморфные виды, и ^прежде всего из семейства про-
тейных, являются весьма нетребовательными к условиям среды расте-
ниями, которые именно на неплодородных почвах обладают большой
конкурентной мощностью. Поэтому не удивительно, что и в условиях
влажного климата Восточной Австралии, прежде всего под Сиднеем,
на бедных песчаных каменистых почвах развита изобилующая представи-
телями семейства протейных растительность пустошей, физиономически
весьма похожая на растительность юго-запада Австралии. Неприхотли-
вость свойственна также растениям, распространенным в районе восточ-
нее Аделаиды в Южной Австралии. Район получил название «90-мильной
пустыни» только по той причине, что песчаные почвы здесь малопригодны
для возделывания и потому до настоящего времени не заселены. Расти-
1 Н. W а 1 t er, Zur Klarung des spezifischen Wasserzustandes im Plasma und in
der Zellwand bei hoheren Pflanzen und seine Bestimmung, «Ber. Deutsch. Bot. Ges.»,
76, 1963, S. 40-71.
2 N. C. W. Beadle, Soil phosphate and the delimitation of plant communities
in Eastern Australia, «Ecology», 43, 1962, p. 281—288.
13—01248
1
Рис. 147. Различные формы филлодий у Acacia.
1 — А. armata; 2 — A. marginata', 3 — A. decipiens; 4 — A. alata. Нат. величина.
Области средиземноморского климата за пределами Евразии
195
тельность района и условия ее произрастания с экологической точки зре-
ния фундаментально исследованы Шпехтом \ поэтому мы рассмотрим их
здесь подробнее.
Песчаные пустоши
Почвы под растительностью песчаных пустошей кислые и очень бедные
азотом, фосфором, нередко калием, а также медью, цинком и молибденом.
В районах с большим количеством осадков развиты подзолы; с увеличени-
ем аридности процесс оподзоливания ослабевает и происходит засоление
почв. В профиле засоленных почв уже отчетливо наблюдается горизонт,
обогащенный растворимыми соединениями. Низкое плодородие почв свя-
зано с бедностью материнской породы. Там же, где она представлена сили-
катами и при выветривании дает глину, развивается не пустошь, а саванна
со злаковым покровом.
Количество осадков в районе, исследованном Шпехтом, составляет
450 мм1год\ 70—75% приходится на зимние месяцы — с мая по ноябрь.
Каждый пятый год — засушливый; засуха длится 7 месяцев, причем
дождей в это время практически не бывает.
Температура почвы колеблется в течение года в следующих пределах:
Глубина, см
7,5
15
30
°C
<1,9—45,0
4,1-36,0
5,8—29,0
Заморозки на почве могут случаться в каждом месяце, но с ноября
по февраль они редки. Часто оседает роса. Пустошь формируется там, где
мощность слоя песка не менее 1,2 м.
Верхние 15—25 см (горизонт А) песчаной почвы окрашены гумусом
в серый цвет. Сплошной слой подстилки на поверхности отсутствует.
Горизонт А2 желтый и лишен гумуса, горизонт А3 характеризуется ржавы-
ми пятнами и залегает непосредственно на засоленной глине горизонта В
со столбчатой структурой. Значения pH в почвенном профиле колеблются
от 5,8 до 6,4. Типичными склерофилами являются Eucalyptus baxteri,
9 видов протейных, 2 вида казуариновых, Xanthorrhoea, бобовые и др.
Основной вегетационный период растений приходится на лето, так как
песчаная почва до января сохраняет влагу; даже в феврале и марте
в горизонте В имеется вода.
Слегка волнистый из-за множества дюн рельеф обусловливает неко-
торое разнообразие растительности, однако четкое отграничение расти-
тельных сообществ не наблюдается.
Исследована корневая система 91 вида. У 25% видов обнаружены
корни, проникающие глубоко в почву, включая горизонт В, а также
корни, расходящиеся в верхних 25 см почвы горизонтально; последние
у некоторых видов дают корневые отпрыски. У мелких многолетних
видов (42%) корневая система приповерхностная, пронизывающая верх-
ние 30 или 60 см. Основной период вегетации этих видов приходится
на весну. Все корневищевые и клубневые растения, а также Stipa semibar-
bata имеют корни только в верхних 30 см почвы. У Xanthorrhoea australis
корневая система очень тонко разветвленная, пронизывает почву радиаль-
1 R. L. S р е с h t и др., Dark Island Heath (Ninety-mile plain), South Australia,
I—VI, «Austral. J. Bot.», 5, 1957, p. 52—114, 137—172; 6, 1958, p. 59—88.
13*
196
Глава 3
Рис. 148. Содержание воды в песчаной почве (в % сухого веса)
под растительностью пустоши в конце периода засухи и перед
началом дождливого сезона (3 февраля) (по Шпехту).
Крестиками показаны места взятия проб. Растения представлены: Banhsia
ornata, Xanthorrhoea australis (Xanth.), Casuarina pusilia (Gas.), Acrotriche
affinis (Ac.), Lepidosperma carphoides (L. ca) и Schoenus tepperi (S.).
но более чем на 1 м и в глубину на 2,5 м и более. Корни Drosera и орхидей
проникают всего на 5—7 см в глубину; это эфемерные виды. Если опреде-
лить сухой вес корней в почве, то оказывается, что основная масса их
расположена в верхних 25 см, причем больше всего корней сосредоточено
на глубине 15 см.
Растительность пустошей часто подвержена пожарам, огонь уничто-
жает надземные части растений и щадит корневые системы, в результате
около 70% всех видов дают побеги из базальных почек или корней. Пита-
тельные вещества растения черпают с помощью корней из горизонта В.
Они накапливаются с опадом в горизонтах Ао и Аь обеспечивая тем
самым питание растений с поверхностной корневой системой.
Очень интересно распределение влаги в почве. Песчаную почву считают
однородной, но не в отношении содержания в ней влаги. Из-за наличия
растительного покрова дождь смачивает поверхность почвы неравномерно:
Области средиземноморского климата за пределами Евразии
197
Рис. 149. Распределение влаги в песчаной почве после дождя, выпавшего на
почти сухую почву (20 марта) (по Шпехту).
Обозначение видов растений то же, что на рис. 148, кроме того, Hibbertia stricta (Hib.)#
Phyllota pleurandroides (Phyl.) и Ait'oloma conostephioides (Ast.).
5,0-10,0%
надземные части растений отводят дождевую воду либо к стволу, либо
к периферии кроны, поэтому с этих участков в почву попадает больше
влаги. В результате возникает весьма сложная картина распределения
воды в почве. Как только дождь прекращается, вода частично просачи-
вается глубже в почву, частично всасывается кончиками корней. Но
поскольку последние распространены также неравномерно, картина
содержания воды в почве становится еще более запутанной. Шпехт в тече-
ние двух лет с периодичностью месяц производил измерения. В большом
числе точек в пределах профиля через растительное сообщество он брал
почвенные пробы на различной глубине и определял содержание воды
в процентах от сухого веса почвы. Точка завядания почвы (сосущая сила
15 атм) составляла: в горизонте At —1%, в горизонте А2 —0,7 и в гли-
нистых слоях —7—17%. Результаты обработки проб приведены на
рис. 148—151. Крестиками обозначены места проб. Рисунки показы-
вают, сколь трудно брать репрезентативные средние пробы для определе-
ния содержания воды даже в песчаных почвах и как малоинформативны
отдельные пробы.
У крупных растений, таких, как Banksia ornata и Xanthorrhoea australis,
дождевая вода стекает по стволу вниз. Поэтому под пологом этих растений
влажность почвы обычно всегда выше, что способствует их росту. Чем
старше становятся растения, тем энергичнее вытесняют они малооблист-
венные виды, которым приходится довольствоваться меньшим количест-
вом воды. Измерения транспирации у крупных растений подтвердили
результаты исследований Грива в юго-западной Австралии. В дождливое
198
Глава 3
14Q-2to%
^2,0-3,07.
>°о°о 3,0-4,07.
[\$$J 4,0-5,07.
Ширина, срутп
5,0-/0,07.
>10%
Рис. 150. Распределение влаги в песчаной почве после очень
влажной зимы (8 июля) (по Шпехту).
Обозначение видов растений то же, что на рис. 148—149, кроме того,
Hibbertia sericea (Hib. ser.), Lepidosperma laterale (L. co.) и Hypolaena
fastigiata (Hyp.).
время года кривые суточного хода транспирации характеризовались
одним пиком (полуденный максимум), в период летней засухи — двумя
пиками, причем по мере высыхания почвы пики все более и более сглажи-
вались. Xanthorrhoea транспирирует особенно слабо. Тем не менее главным
периодом роста остается лето при температуре свыше 18°.
Видовой состав пустоши определяется повторяемостью пожаров.
После пала развивается весьма своеобразная сукцессия. Быстрее всего
дает побеги Xanthorrhoea. Около 90% общей массы надземных частей
приходится в первый год после пожара на этот вид. Его доля в следующие
годы вследствие более интенсивного развития других видов быстро сокра-
щается и, достигнув 5—10%, остается постоянной. Не изменяется и коли-
чество особей, так как Xanthorrhoea цветет лишь после пала, а семена
обычно уничтожаются насекомыми; поэтому даже на гарях всходов этого
вида не наблюдалось. Напротив, Banksia ornata возобновляется после
пожара исключительно в результате появления всходов. Поэтому ее доля
Области средиземноморского климата за пределами Евразии
199
0,5-1,0%
1,0-2,07.
^2,0-3,0%
23 з, 0-4,07.
4,0-5,0%
5,0-10%
>10,0%
Рис. 151. Распределение влаги в песчаной почве перед на-
чалом летней засухи (13 декабря) (по Шпехту).
Обозначение видов растений то же, что на! рис. 148—150, кроме того,
Boronia caerulescens (Вог.), Baeckea еггсаеа (В.).
в надземной растительной массе очень медленно увеличивается, приблизи-
тельно до 50% на 15-й год, а затем остается более или менее неизменной.
Основная масса сухого вещества у 25-летних экземпляров, достигающих
высоты 2,5 м, приходится на одревесневшие соплодия, поскольку плоды
раскрываются лишь после пожаров. Только на площадях, на которых
в течение 50 лет не было пожара, где торчат одни отмершие кусты, наблю-
далось раскрывание плодов и рассеивание отдельных семян, но всходов
было очень мало.
Таким образом, Banksia относится к экологическому типу пиро-
ф и т о в, очень широко распространенному в Австралии и представлен-
ному прежде всего видами протейных. У этих видов плоды на живых
растениях не раскрываются. Если же положить, например, ветку Накеа
platysperma со старыми, одревесневшими, закрытыми плодами на десять
минут в костер, то плоды вскоре лопаются и разделяются на 2 створки,
в которых находятся крылатые семена. Но семена тотчас не выпадают, так
200
Глава 3
Рис. 152. Доля отдельных видов (в %)
в общей сухой фитомассе на песчаных почвах
через 0—25 лет после пожара (по Шпехту).
Hibb.— Hibbertia spec.; Phyl.— Phyllota remoia\
Cas.— Casuarina pusilia', Lept.— Leptospermum myr-
sinoides', Xanth. — Xanthorrhoea australis', B. marg. —
Banksia marginata', B. ornata — Banhsia ornata.
Возрастает
Рис. 153. Изменение сухого веса
надземных частей растений, подстил-
ки и корней па разной глубине через
0 — 25 лет после пожара (по Шпехту).
как снабжены крючочком, которым они прикреплены к плодовой створке.
Лишь через несколько часов ветер срывает их. Семена падают на успевшую
остыть золу и вскоре прорастают. Аналогично устроены Xylomelum pyri-
forme (протейные) с большими одревесневшими грушеподобными плодами,
австралийское хвойное растение Actinostrobus и многие другие виды.
Возобновление пирофитов обычно возможно только благодаря пожару.
Если пал не возникает, плоды часто висят на уже погибших растениях
многие десятилетия. По-видимому, огонь представляет собой один из есте-
ственных факторов отбора.
Другие растения пустошей, такие, как Casuarina pusilia и Banksia
marginata, регенерируют из подземных органов и размножаются в даль-
нейшем преимущественно вегетативно. Вегетативная и генеративная
регенерации у Leptospermum myrsinoides играют одинаковую роль, для
Phyllota важна только последняя.
Среди мелких видов второго яруса различают: 1) травы, которые уже
через 3—4 года после пожара снова исчезают, 2) кустарнички, доля кото-
рых становится наибольшей через 10 лет, 3) кустарнички, достигающие
максимума развития только спустя 15 лет, и, наконец, 4) кустарнички,
участие которых в растительном покрове даже по прошествии 25 лет про-
Области средиземноморского климата за пределами Евразии
201
должает возрастать. Из числа однолетних видов в первый год после пала
наблюдались только два. Рис. 152 отчетливо иллюстрирует разное участие
важнейших видов пустошной растительности в отдельные годы после
прекращения пожара. На рис. 153 показано увеличение: а) сухого веще-
ства надземной растительной массы важнейших видов, б) подстилки
и в) сухого вещества подземных частей, причем следует учесть, что боль-
шая часть последних переносит пожары без всякого ущерба. Спустя 25 лет
масса подземного сухого вещества все еще в три раза больше массы над-
земного. В 50-летнем естественном насаждении на опытной площадке
надземное сухое вещество составило 15 тыс. кг, из которых 12 920 ^отно-
сились к Banksia ornata и лишь 10 кг — к Casuarina. После пожара насчи-
тывалось 36 видов, через 25 лет —только 20 и через 50 лет —лишь 10
(на 6 опытных площадках размером по 5 X 10 ярдов). Наиболее благо-
приятные условия роста наблюдаются в первые годы после пожара, так
как обеспеченность водой вследствие маленькой транспирирующей поверх-
ности в этот период особенно велика и, кроме того, на выжженных уча-
стках зола содержит много питательных веществ. Однако вскоре вступает
в силу конкуренция, и со временем в составе растительности начинают
преобладать немногие виды с крупными растениями, а мелкие постепенно
подавляются. Их погибшие побеги часто видны под крупными кустами,
в то время как между кустами растения остаются живыми. За 25 лет
масса продуцированного вещества составляет (в кг/га)\
Надземные части растений 27 500
Подземные органы 13 500
Итого 31 000
Подстилка 7 500
Всего 38 500
Итак, в среднем за 25 лет продуцируется около 1500 кг!га сухого веще-
ства в год, что с учетом количества осадков и большой продолжительности
вегетационного периода немного. Такая сравнительно невысокая продук-
тивность объясняется недостатком питательных веществ в почве.
Помимо процесса накопления сухого вещества, исследовался также
круговорот важнейших элементов — С, N, Р, К, Na, Са, Mg, Zn, Си и Мп.
В общем, содержание этих элементов в надземных органах растений
до 15-летнего возраста естественного насаждения увеличивается парал-
лельно увеличению массы продуцированного сухого вещества, но затем
остается на одном уровне. Только содержание марганца продолжает резко
уменьшаться, что, вероятно, связано с сокращением листовой массы
Banksia ornata. Со временем питательные вещества концентрируются
в плодах Banksia и в отмерших листьях Xanthorrhoea (около V4—х/3 общего
количества). Масса питательных веществ в подстилке также постоянно
возрастает; особенно заметен этот рост в подземных органах. Извне в расте-
ние поступает только азот, связываемый корневыми клубеньками Phyllota
и микоризой Casuarina. Но в более старых естественных насаждениях оба
вида вытесняются другими видами. Небольшое количество других элемен-
тов растения извлекают из горизонта В. В общем, однако, содержание
питательных веществ в верхних горизонтах почвы в первые годы после
пожара быстро уменьшается вследствие восстановления растительного
покрова, а затем уже почти не изменяется. Прогрессирующее накопление
элементов минерального питания в определенных над- или подземных
органах растений и в неразлагающейся подстилке в течение 50 лет должно
было бы вызвать деградацию растительности. Но до истечения этого срока
202
Глава 3
происходит — обычно благодаря воздействию огня — минерализация
органической массы. В результате сукцессия может начаться снова: одни
виды будут исчезать, другие появляться.
Поскольку описанные выше условия, по-видимому, аналогичны и для
маквисоподобной растительности других областей с зимними дождями
и малоплодородными почвами, мы описали их столь подробно. Питание
растений становится еще более недостаточным, если в результате периоди-
ческой вырубки или выпаса скота из экосистемы постоянно удаляются
питательные вещества. Тогда возникают необратимые явления деградации,
исключающие восстановление первоначальных условий, если только
выветривание материнской породы не вызовет смены состава минераль-
ных элементов. В Средиземноморье почвы, в общем, не столь бедны, как
в Австралии.
Растительность дюн
Небезынтересно сопоставить растительность песчаных пустошей
с сообществами, развитыми на песчаных дюнах у морского побережья.
В последнем случае речь идет об очень молодом почвенном образовании,
причем песок первичных и вторичных дюн еще находится в движении.
Дюны иногда достигают высоты 25 м. Их растительность в Южной и юго-
западной Австралии весьма сходна. Ограничимся здесь описанием района
дюн на побережье под Котсло, недалеко от Перта х.
Выделяется обычная зональность:
1. Песчаный пляже отдельными Cakile maritima и Arctotheca nivea.
Последняя занесена из Южной Африки. Это однолетнее (местами многолетнее) расте-
ние служит причиной образования лишь временных небольших дюн.
2. П е р вичные дюны, низкие, сформировавшиеся благодаря кустам Spi-
nifex hirsutus.
3. Вторичные дюны, с искусственными насаждениями Ammophila аге-
naria и Spinifex longifolius. Степень покрытия — в среднем около 50%, песок еще пере-
мещается, особенно в местах, нарушенных ветром. На защищенных участках наблю-
даются:
а) суккулентные виды, как Tetragonia zeyheri, Carpobrotus (Mesembryanthemum)
aequilaterus, Scaevola crassifolia, Anthericum divaricatum (занесен из Южной Африки);
б) мягколистные виды Calocephalus brownii, Olearia axillaris, Angianthus cunning-
hamii (все Compositae), Pelargonium drummondii',
в) склерофилы, например Myoporum insulare, Acacia cyclopis, а также Scirpus
nodosus и Lepidosperma gladiatum.
4. Третичные дюны, степень покрытия — 100%, песок закреплен. Рас-
тительность представляет собой приблизительно метровый (больший рост ограничивает
ветер) вечнозеленный, богатый видами кустарник (пустошь), в котором, однако, про-
тейные полностью отсутствуют. Назовем здесь Spyridium globulosum
(Rhamnaceae) с паразитирующими на нем Cassytha glabella (лавровые), Calothamnus
quadrifidus (мало), Acacia cyclopis (много), Acanthocarpus preissii (колючая Liliaceae),
Exocarpus cf. spartea (паразит, сандаловые), Scaevola, Threlkeldia diffusa (Chenopodia-
ceae), Rhagodia baccata (то же семейство, с красными ягодами), Templetonia retusa
(Leguminosae), Olearia axillaris (Compositae).
5. Лесочки, образованные хвойной Callitris robusta совместно c Santalum (полу-
паразит).
Совершенно иной состав этой пустоши и полное отсутствие протейных
объясняется большим содержанием извести в песке, а также, вероятно,
влиянием морских солей, заносимых на пляж сильным прибоем. Если
в береговом обрыве обнажаются известняки, то пустошь занимает всю его
1 G. G. S m i t h, A guide to sand dune plants of South-Western Australia, «W. Austr.
Naturalist», 6, № 1, 1957.
Области средиземноморского климата за пределами Евразии
203
поверхность до самой кромки воды; позади пустоши начинается открытый
лес с «туартом» {Eucalyptus gomphocephala), достигающим высоты 20 м.
Степень покрытия почвы в лесу —20%. Верхний ярус кустарников
высотой 6 м покрывает 30% поверхности {Banksia attenuata, В. menziesii и
Casuarina freiseriana). Остальная часть площади занята очень густым,
богатым видами нижним ярусом кустарников (высотой 1 л) с небольшой
примесью злаков и трав. Лианы представлены здесь Hardenbergia comptonii
(семейство бобовых).
Нас интересовало, в какой мере растения дюн обеспечены водой
во время длительной летней засухи и какова экология перечисленных
выше типов растительности. Изучалось содержание воды в листьях, осмо-
тическое давление клеточного сока и концентрация солей в нем, а также
водоотдача срезанных частей растений. Если растения летом страдают
от недостатка воды и имеют летний период покоя, то есть плотно закры-
вают устьица, то их водоотдача после срезания должна, была осуще-
ствляться медленнее, чем у вполне тургосцентных растений.
Пробы были взяты зимой, в условиях хорошей влагообеспеченности;
осмотическое давление определялось сухим методом в Штутгарте. Отбор
проб в конце периода засухи, 16 марта, когда было особенно жарко
и ветрено, производил Смит. Результаты исследований приведены
в табл. 18.
Таблица 18
Содержание воды, осмотическое Давление и давление,
обусловленное хлоридами, у растений дюн
в юго-западной Австралии
Название растений Содержание воды, % сырого веса Величины зимой, в конце августа
осмоти- ческое давление, атм содержание хлоридов
летом | зимой атм | %
Малакофильные
Calocephalus 61 80 12,3 6,6 54
Pelargonium 74 80 6,8 2,7 40
Olearia 62 71 14,0 6,0 43
Склерофильные
Myoporum 81 83 15,8 11,1 67
Acacia 64 63 20,8 10,5 51
Злаки
Spinifex hirsutus 72 70 13,5 7,2 55
Spinifex longifolia 74 70 10,9 5,3 49
Ammophila arenaria Галофиты 69 66 — —
и суккуленты
Atriplex isatidea — 75 37,8 22,7 60
Carpobrotus Z П0беги r l верхушки 86 89 94 15,0 9,2 11,3 6,7 75 73
Arctotheca Z побеги 65 91 9,8 7,5 76
l верхушки — 92 6,6 5,1 77
( листья 88 91 11,9 6,6 55
Tetragonia < верхушки по- l бегов Scaevola Г побеги 1 листья — 93 10,2 3,7 36
77 89 12,5 8,9 72
— 89 17,0 14,1 83
1 2 3 4 5
-------Зима
-------Лето
Рис. 154. Кривые потери воды при высыха-
нии для срезанных побегов растений — обитателей
песчаных дюн на юго-западе Астралин (зимние
данные по Вальтеру, летние — по Смиту).
Области средиземноморского климата за пределами Евразии
205
Из данных таблицы видно, что мягколистные виды летом характери-
зуются известным недостатком влаги, который, однако, не превышает
дефицита, обычно наблюдаемого у этих видов в жаркие дни в полдень.
У склерофильных видов и у злаков дефицит не установлен. Но он обнару-
жен у суккулентов, часть которых имеет поверхностную корневую систему.
Однако и в данном случае дефицит невелик, исключением является только
Arctotheca. Этот однолетний вид переживает лето лишь при условии
не слишком жестокой засухи. Не исключено, что во время отбора проб
этот вид уже отмирал.
В согласии с небольшим дефицитом влаги летом у растений не было
отмечено и какого-либо сокращения транспирации. Потеря в весе у сре-
занных побегов летом и зимой была одинаковой. Летом ход кривой потери
воды при высыхании для всех видов, за исключением Carpobrotus, был
в соответствии с более интенсивным испарением несколько круче. Только
для Carpobrotus кривая оказалась очень пологой, а летом значительно
более пологой, чем Зимой. Это свидетельствует о том, что этот, в общем,
очень слабо транспирирующий суккулентный вид в сухой период в извест-
ной мере сокращал транспирацию (см. рис. 154).
Величину осмотического давления и давление, обусловленное хлори-
дами, удалось определить только на пробах, взятых зимой. Поскольку
летом дефицит воды мал и условия гидратуры благоприятные, можно пред-
полагать, что эти значения в это время года не повышаются. В общем,
значения невелики, несколько выше они|у Acacia и необычайно высоки,
как это типично для галофитов, у Atriplex, в листьях которого всегда
содержится много хлоридов.
Во время шторма и сильного прибоя растения дюн обильно смачи-
ваются распыленной морской водой. Они солеустойчивы и поглощают
сравнительно много соли. Как все виды Mesembryanthemum (см. т. I,
стр. 433), Carpobrotus накапливает в своих тканях хлориды; так же ведут
себя Archotheca и Scaevola. Это дает основание относить все суккулентные
растения пляжной полосы к типичным галофитам. Spinifex hirsitus для
злака необычно сочный, но содержание воды в нем, как это ни странно,
невелико.
Лагуны с засоленными маршами в Южной Австралии встречаются чаще, чем
на юго-западе материка. Здесь в приливно-отливной зоне можно обнаружить последний
пояс мангровы — низкие кусты Avicennia marina var. resinifera. Несколько выше растут
Arthrocnemum arbusculum совместно с Salicornia australis, Kochia oppositifolia n Suaeda
australis. Заросли Arthrocnemum halocnemoides с примесью Frankenia pauciflora и Diphys-
Таблица 19
Содержание хлоридов в клеточном соке некоторых
видов, растущих в солоноватой воде
(Новый Южный Уэльс)
Название растений Содер- жание воды,% сырого веса Осмоти- ческое давле- ние, атм Содержание хлоридов
атилг %
Avicennia marina var. resinijera . . 62 40,5 22,3 55
Acacia bakeri 60 13,5 7,1 53
Araucaria cunninghamii 65 12,8 5,1 40
Cupaniopsis anacardioides 65 12,8 3,7 29
Pyrrosia confluens , , . 85 5,1 1,0 20
206
Глава 3
та australis (Mesembryanthemum) во время прилива уже не затапливаются. Там, где
уровень грунтовых вод лежит ниже, на сухих слабозасоленных (0,3% NaCl) почвах пре-
обладает A triplex paludosum с видами Bassia, служащий уже переходным звеном к ку-
старниковым засоленным пустошам «малли». На более влажных местообитаниях,
напротив, распространены травяные сообщества Sporobolus virginicus с Juncus maritimus
или Distichlis spicata c Cladium и Plantago coronopus, граничащие в глубине суши с раз-
реженным древостоем Casuarina stricta.
Д-р. Уэбб отобрал 3 января 1959 г. несколько проб растений, росших в полосе
солоноватых вод недалеко от устья реки Брунсвик в Новом Южном Уэльсе, и переслал
их автору для исследования. Берег реки здесь густо порос видами Avicennia (мангро-
вы) и типичными негалофитами, как Araucaria, Acacia и Cupaniopsis. Выступавшие
из земли корни деревьев и отмершие ветви покрыты папоротником-эпифитом Pyrrosia
confluens. Интересно было сравнить содержание хлоридов у этих разнородных элемен-
тов. Как следует из табл. 19, A vicennia поглощает очень много соли и характеризуется
высоким осмотическим давлением, хотя и обладает свойством выделять соли, которые
откладываются на поверхности листьев. У других видов доля хлоридов также значи-
тельна, но абсолютное количество их невелико.
УМЕРЕННАЯ ЗОНА И ВЕРТИКАЛЬНАЯ ПОЯСНОСТЬ
РАСТ И ТЕЛЬНОСТ И В АВСТРАЛИИ, ТАСМАНИИ
И НОВОЙ ЗЕЛАНДИИ
У меренные дождевые леса
Все восточное побережье Австралии отличается влажным климатом
в течение всего года. Поэтому в его северной части мы находим тропиче-
ский вечнозеленый дождевой лес, который к югу переходит в субтропиче-
ский (см. т. I, стр. 195 и след.); лишь в южной части штата Виктория
можно встретить умеренно теплый вечнозеленый дождевой лес. Отличается
он от субтропического тем, что в его составе присутствуют антарктические
элементы: нотофагус (Nothofagus cunninghamii), древовидный папоротник
Dicksonia antarctlca и др. Эти виды встречаются в горах Нового Южного
Уэльса. Последние местонахождения известны на юге Квинсленда.
На юге Виктории и в центральной части Тасмании в поясе влажного
дождевого леса нотофагус образует нижний древесный ярус в лесах
из Eucalyptus, Стройные стволы Eucalyptus regnans здесь, во влажном
климате, поднимаются до высоты в среднем 75 м, но известны деревья
высотой 97 м. Длина старых срубленных стволов достигает 110 ранние
сведения Мюллера о высоте деревьев, равной якобы 145 м, проверить уже
не представляется возможным. Примерно такой же высоты достигают
Ей. gigantea и Ей. obliqua.
Умеренные леса Тасмании, образованные видами Eucalyptus и Nothofa-
gus, длительно исследовались с экологической точки зрения. Леса этого
типа всегда развиваются только после лесных пожаров. Возобновления
эвкалипта в глубокой тени под нотофагусом не происходит. Лишь после
того, как пожар уничтожит лес, на выгоревшей площади одновременно
появляются проростки эвкалипта и нотофагуса. Однако более быстро
растущий эвкалипт вскоре опережает соперника и образует верхний дре-
весный ярус.
Состав этих лесов зависит от того, как часто повторяются лесные
пожары. Гилберт установил следующие закономерности1.
1 J.M. Gilbert, Forest succession in the Florentine Valley, Tasmania, «Proc.
Roy. Soc. Tasmania», 93, 1959, p. 129—151; T. M. Cunningham, The natural
regeneration of Eucalyptus regnans, Bull. № 1, School of Forestry, Melbourne, 1960.
Области средиземноморского климата за пределами Евразии
207
1. Если па данной территории пожары не возникают в течение
по меньшей мере 350—400 лет (максимальный возраст эвкалиптов), что
на влажной западной покатости Тасмании обычно, на ней развиваются
только чистые нотофагусовые леса с примесью Atherosperma moschata
(Monimiaceae) и ярусом из древовидного папоротника Dicksonia antarctica
(рис. 155). Высота деревьев Nothofagus не превышает 40 м; максимальный
возраст —450 лет. Atherosperma обычно ниже, и ее предельный возраст
250 лет. Dicksonia иногда поднимается до 3 м и растет даже при 1% осве-
щенности. Из эпифитов встречаются только папоротники (много гимено-
филловых) и мхи. Чем беднее субстрат, тем разнообразнее видовой состав
сообщества; на очень бедных почвах появляются также Eucalyptus simond-
sii и хвойный Phyllocladus.
2. Если лесные пожары повторяются чаще чем один раз в 350 лет,
образуется смешанный лес, в котором над древесным ярусом 1-го типа
поднимается верхний ярус Eucalyptus высотой 75 м (до 90 л^), образован-
ный видами (Ей. regnans, Ей. gigantea и Ей. obliqua), кроны которых,
однако, не образуют сомкнутого полога (рис. 156, 157). Одновозраст-
ность верхнего яруса этих лесов свидетельствует о возобновлении гарей
непосредственно после лесных пожаров. Огонь не повреждает твердую
оболочку семян и даже способствует их прорастанию. Поскольку подрост
эвкалипта растет быстрее, чем нотофагус, и не испытывает недостатка
в свете, он образует первый ярус.
3. Если лесные пожары повторяются 11—2 раза в столетие, то Nothofa-
gus вытесняется из нижнего яруса древостоя следующими видами деревьев
и кустарников: Pomaderris, Olearia и Acacia.
4. При повторении пожаров каждые 10—20 лет возникает чистый
эвкалиптовый лес, из состава которого, однако, выпадает сильнее повреж-
даемый огнем Ей. regnans. На малоплодородных почвах развивается
Ей. simondsii.
5. В результате очень частых лесных пожаров формируются изрежен-
ные саванноподобные древостои или — на более бедных почвах — заросли
кустарника из миртовых и протейных, а на очень влажных площадях —
обширные болота с Mesomelaena (Gymnoschoenus) sphaerocephala (Сурегасеае),
Restionaceae, а также Drosera и Utricularia, над зарослями осоки возвы-
шаются редкие кусты Leptospermum, Melaleuca и Banksia.
В погоне за кенгуру-валлабие, предпочитающим открытые простран-
ства, местные жители еще до прихода белых часто пускали палы (Mesome-
laena сгорает даже на корню). В результате леса на обширной площади
деградировали (рис. 158). Ныне пожары на западе Тасмании случаются
реже и наблюдается новое наступание леса.
Ходьбу во влажном дождевом лесу сильно затрудняет своеобразный A nodopetalum
biglandulosum (Cunoniaceae), называемый здесь «горизонталью». Это маленькое дерево,
ствол которого поднимается прямо до высоты 5—6 м. При дальнейшем росте ствол при-
нимает горизонтальное положение или даже изгибается в направлении к земле. Побеги
также сначала растут вверх, а затем склоняются вниз. В результате на высоте прибли-
зительно 10 м возникает своего рода платформа из тесно переплетенных между собой
стволов и ветвей. Путнику не остается ничего иного, как пробираться по этой плат-
форме, как только на его пути возникает такая «горизонтальная» заросль.
В Тасмании встречается и другой вид нотофагуса — листопадный
N. gunnii, представляющий собой мелколистный кустарник высотой
4—5 м, листва которого осенью приобретает красивую окраску. Растет
он в горах, вблизи границы древесной растительности, на влажных глыбо-
н ФУГИ
-150
Р и с. 155. Профильная диаграмма умеренного дождевого леса, не тронутого
пожарами, Тасмания (по Гильберту).
Древесный ярус из Nothofagus cunninghamii (доминант) и Atherosperma moschata, ярус
кустарников образован древовидным папоротником (Dicksonia antarctica).
Рис. 156. Профильная диаграмма смешанного умеренного дождевого леса,
уничтожавшегося пожаром приблизительно через каждые 200—300 лет
(по Гильберту).
Верхний древесный ярус образован Eucalyptus obliqua, более густой нижний древесный
ярус — Nothofagus cunninghamii и Atherosperma moschata, в ярусе кустарников домини-
рует Dichsonia antarctica.
Рис. 157. Лес из Eucalyptus regnans, нижний древесный ярус состоит из
Nothofagus cunninghamii и Atherosperma moschatum. Ниже располагается почти
сомкнутый ярус древовидного папоротника — Dicksonia antarctica (фото
Э. Вальтер).
Рис. 158. Болото с Mesomelaena sphaerocephala на перевале Кинг-Уильям,
горы Эрроусмит. На переднем плане — цветущий Leptospermum scoparium (фото
Вальтер).
Рис. 159. Схема распределения растительности на территории Национального парка
Маунт-Филд, Тасмания (по Швейнфурту, с изменениями).
1 — сухой эвкалиптовый лес; 2 — влажный эвкалиптовый лес; з — дождевой лес (особенно широ-
ко распространен на изобилующем осадками западном склоне, локальное развитие у водопада
Рассел); 4 — субальпийский лес из Eucalyptus coccifera’, 5 — пояс кустарников; 6 — болота с расте-
ниями-подушками; 7 — горный лес из Athrotaxis; 8 — альпийский пояс жестких растений-поду-
шек; 9 — долеритовые скалы.
Области средиземноморского климата за пределами Евразии 211
Р и с. 160—162. Климадиаграммы Тасмании (см. также рис. 139).
ЗИан — умеренно гумидный климат западной части острова; Спрингс — климат леса из Eucalyp-
tus-Nothofagus} Мина — климат нагорья близ озера Грейт-Лейк, граница древесной растительно-
сти и пояса жестких растений-подушек.
вых осыпях, на изобилующей осадками западной стороне острова,
а также на нижних частях склонов.
Последовательность расположения вертикальных поясов в горах
Тасмании, этого очень интересного с экологической точки зрения острова,
обладающего живописнейшими ландшафтами, иллюстрирует схематиче-
ский профиль через территорию Национального парка Маунт-Филд
(рис. 159) \ Диаграммы (рис. 160—162) знакомят читателя с климатом
острова.
Тасмания расположена уже в области влияния западных ветров,
отчетливо преобладающих прежде всего в зимние месяцы. Поэтому климат
западной стороны острова очень влажный. Здесь выпадает за год 2500 мм
осадков, причем ни в одном месяце их количество не составляет
менее 100 мм. В результате обширные площади заболочены и безлюдны.
На восточной стороне, напротив, климат очень приятный (600 мм осадков
в год, слабая засуха летом, см. рис. 139); вся местность густо заселена.
Первичная растительность представлена саванноподобными сухими эвка-
липтовыми лесами, которые по мере увеличения влажности переходят
во влажные, а затем в дождевые леса. |
Граница древесной растительности, образованная кустарниковид-
ными Eucalyptus coccifera, проходит на высоте 1200 м. Альпийский пояс
характеризуется распространением жестких подушечных растений, изо-
билует антарктическими элементами и интересными эндемичными видами
(рис. 163).
Существенно более важное место занимает умеренный лес в Новой
Зеландии (рис. 164) 1 2. Здесь, на севере острова Северный, распространены
субтропические леса с Agathis, пальмами и многочисленными меланезийско-
тропическими элементами. Флористически они резко отличаются от суб-
1 U. Schweinfurt h, Studien zur Pflanzengeographie von Tasmanien, «Bon-
ner Geogr. Abh.», H. 31, 1962; W.D.J ackson, Vegetation. Cm.: «Atlas of Tasmania»,
p. 30—35 (с картон растительности), Hobart, 1965; W.M.C ur t i s, Forests and flowers
of Mount Wellington, Tasmanian Museum, Hobart, 1966.
2L. Cockayne, The vegetation of New Zealand. Cm.: Engler-
D r u d e, «Die Veget. d. Erde», 7, 1936, новое издание 1958; New Zealand Forestry,
Wellington, 1964; U. Schweinfurth, Neuseeland, «Bonner Geogr. Abh.», H. 36,
1966.
14*
212
Глава 3
тропических лесов Австралии. В Новой Зеландии нет, например, местных
видов эвкалипта или акации. Вечнозеленые дождевые леса с видами
Podocarpus (рис. 165) и Dacrydium cupressinum протянулись почти до южной
оконечности острова Южный, где климат уже отчетливо умеренно влаж-
ный. Но здесь они граничат с типичными умеренными лесами Nothofagus,
которые характеризуются монодоминантностью или господством неболь-
шого числа видов деревьев (рис. 166). В Новой Зеландии известны 5 видов
нотофагуса, отличных от австралийских видов. Как это ни странно,
но распространение лесов субтропического типа и лесов с Nothofagus нельзя
объяснить климатическими причинами. Леса с нотофагусом занимают
островные ареалы даже на вулканическом плато острова Северный. К югу
они встречаются, правда, чаще, но на острове Южный они уже отсут-
ствуют на обширных территориях (например, в Уэстленде) и замещены
субтропическими лесами. Подобное мозаичное чередование лесов разных
типов, возможно, объясняется отсутствием равновесия между современ-
ным растительным покровом архипелага и его климатом. Установлению
такого равновесия препятствовало обширное оледенение острова Южный
в недавнем геологическом прошлом, а также интенсивная вулканическая
деятельность на острове Северный, продолжающаяся до настоящего време-
ни. Так, например, обширные территории острова Северный до 1700-х годов
были покрыты мощной толщей вулканического пепла. В качестве пионеров
при заселении освободившихся территорий выступали прежде всего виды
Podocarpaceae, распространению которых способствуют птицы. Но пред-
ставители этого семейства постепенно вытеснялись субтропическим лист-
Р и с. 163. Заболоченная поверхность нагорья близ озера Грейт-Лейк, Тасмания;
между подушками A brotanella forsterioides видны Astelia alpina, Gleichenia alpina и др.
На заднем плане — лес из Eucalyptus coccifera (фото Э. Вальтер).
Области средиземноморского климата за пределами Евразии
213
венным лесом, в котором важную роль играл Metrosideros robusta (миртовые)
в качестве растения-душителя, развивавшегося на Dacrydium. В настоящее
время виды Podocarpaceae часто образуют сообщества с примесью субтро-
пических элементов, и только в горах эти леса частично еще сохранили
первоначальный, девственный облик. Однако и здесь более конкуренто-
способными, по-видимому, оказываются виды Nothofagus, что наводит
на мысль об их все более широком распространении в будущем. Отсюда
понятно, что лишь исторические данные могут помочь вскрыть специфи-
ческие особенности распределения растительности Новой Зеландии,
но такого рода анализ выходит за рамки нашей задачи х. По своему харак-
Р ис. 164. Первичный умеренный дождевой лес на острове Чатем, расположен-
ном восточнее Новой Зеландии. Лес, прежде занимавший всю площадь острова,
ныне большей частью выкорчеван (фото Кокейна).
1 McKelvey, Forest colonisation after recent volcanity at West Taupo, «The
New Zeal. J. of Forestry», 6, 1953, p. 435—448; R. G. R о b b i n s, The podocarp broad-
leaf forest of New Zealand, «Trans. Roy. Soc. N. Z. Bot.», 1, 1962, p. 33—75; J.T. Hol-
lo way, Forests and climate in the South Island of N. Z., «Trans. Roy. Soc. N. Z.»,
82, 1954, p. 329—410; Symposium: 1. Evidence for ecologically significant changes in cli-
mate during the postglacial period in New Zealand. 2. Vulcanity and vegetation in the
Rotorua district, «Proceed. N. Z. Ecolog. Soc.», 10, 1963, p. 21—72.
214
Глава 3
Рис. 165. Заболоченный лес с Podocarpus dacrydioides, в подлеске—
Phormium tenax, Новая Зеландия, остров Южный (фото Кокейна).
теру, флористическому составу, а также климату леса с Nothofagus, осо-
бенно на юго-западе острова Южный, аналогичны западнопатагонским
лесам, развитым в южном Чили (см. рис. 166).
Особенностью этого очень влажного, незаселенного района фьордов
являются «полосы поваленного леса», которые отмечает Швейнфурт
(рис. 167) х. Они напоминают участки леса, вырванные лавинами, но отли-
1 U. Schweinfurth, Uber eine besondere Form der Hangabtragung im neu-
seelandischen Fjordland, «Z. Geomorph.», N. F. 10, 1966, S. 144—149.
Рис. 166. Лес, образован-
ный Nothofagus jusca, в рай-
оне фьордов острова Юж-
ный (Новая Зеландия); на
почве— Polystichum vestitum
(фото Рейда)9
МИЛФ0РД-СА9НД(бм) 9,9’6337
Рис. 167. Климадиаграммы Милфорд-Саунда (умеренно гу-
мидный климат района фьордов острова Южный, Новая Зелан-
дия) и, для сравнения, Келберна под Веллингтоном (нормаль-
ный климат).
КЕЛВЕРН, ВЕЛЛИНГТОН (127м) 12,3°
216
Глава 3
чаются от последних тем, что располагаются в середине лесного массива
на крутых склонах, достигают 2—6 м в ширину и образуются под влия-
нием силы тяжести. Когда вес подросшего древостоя становится слишком
большим, происходит срыв всего слоя растительности вместе с корнями
и почвой. Обнажается скальный грунт, который спустя некоторое время
вновь заселяют растения, сначала лишайники, мхи и папоротники,
затем кустарники и, наконец, деревья, закрепляющиеся корнями в тре-
щинах скалы. Обвал повторяется, как только вес достигает критической
величины.
Большой урон наносят нотофагусовым лесам Новой Зеландии заве-
зенные туда европейские благородные олени, размножение которых
совершенно не поддается контролю. Олени съедают и вытаптывают моло-
дую поросль нотофагуса х. Уничтожение лесов на крутых склонах в горах
вызывает эрозию почв и катастрофические наводнения.
Вертикальные пояса и альпийская растительность
Австралии, Тасмании и Новой Зеландии
Расположенные на юго-востоке Австралии Снежные горы (2240 м)
поднимают свои вершины высоко над границей древесной растительности
(1700—1950 м). Не удивительно, что на их склонах представлены все вер-
тикальные пояса растительности, включая альпийский пояс. Образуют
их различные виды эвкалипта, сменяющие друг друга по мере подъема
в горы, правда, в картине ландшафта эта смена почти не отражается. Даже
границу произрастания деревьев отмечает кустарниковидный эвкалипт
Ей. niphophila} защищенный в течение почти 6 месяцев толстым снежным
покровом, он легко переносит зимние морозы (рис. 168, 169). Попытки
культивировать этот вид в саду Хоэнхейма, в Центральной Европе, окон-
чились неудачей. Без достаточной снежной защиты эвкалипт каждый год
вымерзал до основания, затем, правда, снова давал побеги, но через
несколько лет погиб.
Рис. 168. Климадиаграммы наиболее высоко расположенных
станций в Снежных горах, юго-восточная Австралия.
Цифры внизу (10, 14) — средняя длительность безморозного периода.
1 НА W а 11 е г, Uber die Bedeutung des GroBwildes fur die Ausbildung der Pflan-
zendecke, «Stuttgarter Beitr. z. Naturk.», № 69, 1961; K. A. W о d z i c k i, Introduced
mammals of New Zealand, DSIR Bull. K? 98, Wellington, 1950.
Области средиземноморского климата за пределами Евразии
217
Р и с. 169. Граница древесной растительности (Eucalyptus niphophild) на ropeJKoc4io-
шко, Новый Южный Уэльс (Австралия); середина лета, но видны пятна снега (фото
Э. Вальтер).
О растительности альпийского пояса, занимающего около 5 тыс. км\
Костин написал подробную монографию1. Он находит здесь много ана-
логий с европейскими Альпами. Правда, флора здесь совершенно инаяг
если не считать нескольких общих родов, таких, как Ranunculus, Caltha,
Veronica, Euphrasia, Luzula, Роа, Carex и др. Вересковые, встречающиеся
в Альпах, замещаются в Австралии видами Epacridaceae. Горы сложены
гранитами и гнейсами. Годовая сумма | осадков равна 2000—2250 мм;
холодный период длится 4—6 месяцев (см;, рис. 168); средняя годовая тем-
пература от +2,3 до +4°; температура самого теплого месяца 6,3—9,7°г
наинизшая —11°.
Для альпийского пояса гор южного полушария очень характерны
альпийские туссоковые луга (альпийские злаковники). Туесок — это
высокие, полусферической формы дерновины, образуемые злаками с жест-
кими листьями. Даже в высохшем состоянии растения сохраняются
долгие годы, вот почему общий фон альпийских (туссоковых) злаковников
всегда имеет желтую окраску.
В Австралии альпийский злаковник, тип сообщества Роа
caespitosa —Danthonia nudiflora, развит только в плоских долинах. Каме-
нистые склоны выше границы древесной растительности покрыты сооб-
ществом кустарничков Epacridaceae, заменяющих кустарнички вересковых
1 А. В.Costin, A study of the ecosystem of the Manoro Region of New South
Wales, Sydney, 1954; The high mountain vegetation of Australia, «Austr. J. Bot.», 5,
1957, p. 173—189.
218
Глава 3
Центральной Европы. Здесь можно встретить виды: Epacridaceae (Epacris,
Leucopogon, Riched), Leguminosae (Oxylobium, Hovea, Bossiaed), Myrtaceae
(Kunzea, Baeckia, Leptospermum, Callistemori), Protaceae (Orites), Thyme-
laeaceae (Pimelea), Labiatae (Prostrantherd), Compositae (Olearid) и Gonife-
ralis (Podocarpus alpinus).
Многие из этих кустарничков имеют шпалерную форму, например
Podocarpus.
Альпийские матты, соответствующие ассоциации Nardetum
с Arnica в Альпах, состоят из сообщества Celmisia longifolia — Роа caespi-
tosa. Вегетационный период длится 3—8 месяцев; там, где он длительнее,
преобладает Роа, при более кратком — Celmisia из семейства сложно-
цветных, с крупными белыми соцветиями. Здесь встречаются 5 видов Ranun-
culus, среди них R. anemoneus, напоминающий Anemone alpina', очень
крупными цветками отличается Euphrasia alsa, тогда как у Gentiana
цветки невзрачные, бледно окрашенные.
Из антарктических элементов можно обнаружить Oreobolus, Uncinia,
Асаепа, Azorella. Общими с Новой Зеландией являются Celmisia и Aciphyl-
la. Из адвентивных видов известны Роа annua, Р. pratensis, Rumex aceto-
sella, Cerastium glomeratum, Hypochoeris radicata, Taraxacum officinale
и др. Формацию скальных трещин представляет тип сообщества
Brachycome nivalis —Danthonia alpicola. Brachyome (семейство сложно-
цветных) напоминает мелкую Chrysanthemum', к ней присоединяются
Cystopteris fragilis var., Blechnum pennamarina, Asplenium flabellifolium,
Cardamine hirsuta, Alchemilla novaehollandiae и др.
Только на редких местообитаниях ниже границы сохранения снежни-
ков встречается растительность снежных долинок:
тип сообщества Plantago muelleri —Montia australasica с антарктическими
элементами Caltha introloba, Abrotanella и Oreobolus. Вегетационный период
не превышает 2—3 месяцев.
На наветренных участках склонов формации Loiseleurietum Альп
соответствует тип сообщества Epacris petrophila — Veronica (Hebe) densifo-
lia, в котором встречаются форма Luzula campestris и желтый Ranunculus
muelleri.
Антарктические подушечники, распространенные в Тасмании и Новой
Зеландии, в Австралии полностью отсутствуют. Выше и ниже границы
древесной растительности развиты болота, причем преобладают болота
переходного типа (тип сообщества Carex gaudichaudiana — Sphagnum crista-
turn), а на склонах и небольших возвышенностях также омброгенные
верховые болота с кочками (pH = 4,5) и мочажинами (pH = 6,8). Кочки
состоят из Sphagnum cristatum и Epacris paludosa, с Е. serpyllifolia и Cal-
listemon sieberi, ниже растут Richea continentis, Restio australis и Astelia
alpina, тогда как мочажины заполнены Polytrichum commune, Breutelia
pendula и Carex gaudichaudiana. В результате интенсивного выпаса скота
растительность разрушается и развивается эрозия почв Ч В настоящее
время эта область приобрела важное экономическое значение благодаря
сооружению водонапорных плотин и гидроэлектростанций.
Значительно сложнее картина распределения растительности в Тас-
мании1 2. На горе Филд-Уэст в дождевом лесу средних поясов господ-
1 А. В. С о s t i n, High mountains catchments in Victoria in relation to land use,
Victoria, 1957; Vegetation of high mountains in Australia in relation to land use. Cm.:
Biography and Ecology in Australia, The Hague, 1959.
2 H. W a 1 t e r, Hohenstufen und apline Vegetation in Australien, auf Tasmanien
und auf Neuseeland, «Ber. Geobot. Inst. Riibel (Ziirich)», 31, 1960, S. 67—71.
г Области средиземноморского климата за пределами Евразии
219
ствуют гигантские эвкалипты, прежде всего виды Ли. obliqua и Ей. regnans,
выше по склону — Ей. regnans и Ей. gigantea. В поясе тумана к ним
присоединяются уже антарктические элементы, как Nothofagus cunningha-
mii и Phyllocladus (хвойные).
Рис. 170. Dracophyllum traversii (Epacridaceae) (слева на переднем и на
заднем плане) на опушке леса с Nothofagus', остров Южный, Новая Зелан-
дия, высота 900 м над уровнем моря (фото Гиббса).
В субальпийском поясе снова представлено несколько видов эвка-
липта (Ей. subcrenulata, Ей. coccifera) и хвойные Athrotaxis cupressoides
и A. selaginoides. Richea scoparia из семейства Epacridaceae вырастает
до высоты 3 м (см. рис. 159).
Граница древесной растительности выражена слабо. Ниже по склону
Ей. coccifera еще имеет древовидную форму, выше его сменяет кустовидный
220
Глава 3
Рис. 171. Верхняя граница леса на восточном склоне Новозеландских Альп
(остров Южный), высота 1200 м над уровнем моря. Резкая граница между лесом
и поясом альпийских злаковников «туесок». С подъемом вверх леса из Nothofagus
fusca сменяются древостоями N. clifforttoides и N. menziesii (фото Симпсона
и Скотт Томсона).
(высотой до 1,5 ле) Nothofagus cunninghamii, затем следует кустарник
из Gaultheria (вересковые) и Bauer а (камнеломковые).
В альпийском поясе на плоскогорьях характерно наклоненное
по ветру криволесье с эндемичными хвойными (Diselma archeri и Phero-
sphaera (Dacrydium) hookeri), а также шпалерный кустарник Microcachrys
tetragona. Между кустами растут Richea scoparia (здесь мелкая), Astelia
alpina, Orites acicularis Celmisia longifolia и др. В заболоченных низинах,
где снег долго не тает, развиваются плотные, сросшиеся в сложную мозаику
подушечники (Abrotanella, Pteridopappus, Donatia и др.). Они принадле-
жат разным семействам, но по вегетативным признакам почти не разли-
чаются. На внешней поверхности подушек поселяются мелкие эпи-
фиты.
Совершенно иная картина открывается перед нами в Новой
Зеландии, где эвкалипты полностью отсутствуют. На острове
Южный, в районе горы Кука (свыше 3700 ле), нивальный пояс на крутых
склонах поднимается до 3300 м. Снеговая линия на влажной западной
стороне острова проходит на высоте 1800 ле, а на восточной — 2400 ле. Для
альпийского пояса типичен туссоковый злаковник с Chionochloa (Dantho-
nia) flavescens; на скалистых местообитаниях выделяются гигантские
подушки Raoulia и Hastia. Субальпийские кустарники прослеживаются
Области средиземноморского климата за пределами Евразии
221
в восточной части острова до 1300 м. Представлены они Podocarpus nivalis,
Phyllocladus alpinus, одревесневшими кустами сложноцветных (Senecio,
Cassinia, Olearia) и др.
У перевала Артур легко прослеживается вертикальная поясность:
лесной пояс состоит на западной стороне из смешанного леса с Podocarpus
{Podocarpus hallii, Р. dacrydioides, Dacrydium cupressinum, Weinmannia,
•Griselinia, Pseudowinter a, Metrosideros lucida и др.). Лес постепенно пере-
ходит в субальпийский кустарник, в составе которого резко выделяются
Fuchsia excorticata совместно с Nothopanax, Olearia, Hebe, Dracophyllum
(рис. 170); над зарослями кустарников возвышаются хвойные Libocedrus
bidwillii и Dacrydium beforme. Важнейшим древесным видом является
Metrosideros lucida (семейство миртовых), корой которого питается заве-
зенный из Австралии опоссум. Гибель деревьев влечет за собой опасность
возникновения эрозии почв. Над кустарником, на высоте 1200 м, снова
расстилаются туссоковые злаковники с преобладанием Chionochloaflavescens
(см. работу Швейнфурта, 1966).
Лесные пояса на восточной стороне острова, закрытой от западных
ветров, имеют совершенно иной облик. Здесь распространены бедные
видами леса с Nothofagus, которые ниже по склонам, в сухом районе Отаго,
сменяются туссоковыми злаковниками, образованными Festuca novazee-
landiae (рис. 171, см. также т. III, гл. 3). Растительность острова Север-
ный мы опишем здесь лишь на примере вертикальной поясности южного
склона вулкана Эгмонт (2500 м).
2400—2500 м
1800-2400 м
1650—1800 м
1500-1650 м
1350-1500 м
1100—1350 м
Ю50—1100 м
1000—1050 м
Около 900 м
Около 800 м
Около 610 м
От вершины до снеговой линии лежат вечные снега.
Обломочные осыпи, лишенные растительности.
Растения на осыпях, степень покрытия — 5%; Роа colensoi, Epilobium,
Helichrysum, Montia.
Альпийские матты, степень покрытия — 50%; кроме вышеназванных:
Luzula, Agrostis muscosa, Ranunculus, Celmisia, Coprosma, Gaultheria,
Forstera, Gunnera, покровы из Racomitrium lanuginosum var. pruino-
sum и Rreutelia elongata, Stereocaulon vesuvianum.
Альпийские злаковники «туесок»: Chionochloa cf. flavescens, Hebe,
Senecio, Cassinia и в качестве растений-пионеров на лавовых полях
связывающие азот Coriaria pteridioides и С. plumosa.
Субальпийский кустарник высотой 1—2 м; Senecio, Olearia, Hebe,
Dracophyllum, Coprosma, Nothopanax colensoi, N. simplex, Griselinia
litoralis. Последние три вида представлены ниже по склону во всех
вертикальных поясах; с уменьшением высоты растения становятся
крупнее.
Переходный лесо-кустарник высотой 3 м. Флористический состав
дополняется видами Podocarpus hallii и Libocedrus.
Субальпийский лес, высота деревьев 4—5 м', тип сообщества Podo-
carpus — Fuchsia — Griselinia.
10-метровый лес из Рodocarpus hallii и Weinmannia', ярус кустарников,
лианы (Rubus cissoides, Rhipogonum), эпифитные папоротники (а также
Lycopodium).
20-метровый лес, Dacrydium, Metrosideros, Weinmannia', много эпифи-
тов (орхидеи, Astelia, папоротники, растения-удушители). Metrosi-
deros robusta дает всходы как растение-удушитель на Dacrydium
cupressinum.
Пышный субтропический лес высотой более 30 м (тип сообщества
Podocarpus ferrugineus—Reilschmiedia tava — Elaeocarpus hookeriana) c 3
ярусами деревьев и ярусом кустарников (древовидные папоротники);
травяной ярус (Astelia, папоротники), лианы и многочисленные эпи-
фиты (Metrosideros diffusa, Senecio kirkii, Pittosporum conifolium, Collo-
spermum, Dendrobium, Earina, Lycopodium, папоротники, а также
Hymenophyllaceae).
222
Глава 3
Рис. 172. Пояс кустарников в горах Маунгатуа (остров Южный. Новая
Зеландия) на высоте 885 м над уровнем моря. В центре Aciphylla (Umbelliferae;
с соцветиями высотой 1,8 м, слева Phorniium colensoi (Liliaceae), на переднем
плане справа — Celmisia coriacea (Compositae). Все виды с пучковидным распо-
ложением листьев (фото Симпсона и Скотт Томсона)в
Особенно характерен тот факт, что какие-либо резкие границы отсут-
ствуют. Лес становится все ниже, постепенно переходит в кустарник,
последний превращается в кустарниковые заросли, и только злаковник
туесок издалека четко выделяется благодаря желтому цвету. Граница
древесной растительности не выражена. Некоторые древесные виды про-
ходят сквозь все вертикальные пояса и в конце концов едва достигают
50 см в высоту.
В районе горы Эгмонт Nothofagus отсутствует, но в виде перемежаю-
щихся островков леса встречается даже на вулканическом плато. Напри-
мер, на еще действующем вулкане Руапеху (2760 м) он образует на высоте
1275 м весьма заметную границу древесной растительности, до которой
поднимается и высокая Cordyline indivisa. Выше снова тянутся 3-метровые
заросли кустарников Nothopanax colensoi, Phyllocladus, Hebe, Olearia,
Senecio, Coprosma, Dracophyllum, Gleichenia и т. д., которые на уровне
1550 м не превышают 10—30 см. Отчетливо выраженный пояс туссоковых
лугов на молодых лавовых обломочных осыпях не развивается. На высоте
1880 м растительный покров уже начинает распадаться; на склонах
появляются первые снежники.
На рис. 172 и 173 изображены некоторые интересные типы растений,
характерные для кустарникового пояса и пояса растений-подушек
острова Южный.
Области средиземноморского климата за пределами Евразии
223
Рис. 173. Альпийский пояс растений-подушек в горах Гумбольдта
(остров Южный, Новая Зеландия) на высоте 1735 м над уровнем моря.
В центре — подушки Phyllachne colensoi (Stylidiaceae) в окружении
Celmisia hectori (Compositae) (фото Симпсона и Скотт Томсона).
ПРОБЛЕМА ПРИСПОСОБЛЕНИЯ РАСТЕНИЙ
К ФАКТОРАМ СРЕДЫ. ОБЩИЕ ЗАМЕЧАНИЯ
Области зимних дождей занимают на земле сравнительно небольшую
площадь. И если тем не менее они здесь столь детально рассмотрены, то это
обосновано следующим.
1. Эти области, для которых характерен средиземноморский тип
климата, представляют собой переходные зоны от субтропических арид-
ных районов к умеренным зонам; поэтому здесь на небольшом расстоянии
224
Глава 3
экологические условия произрастания растений изменяются достаточно
резко. Соответственно одно растительное сообщество внезапно сменяется
другим, и растительный покров в целом приобретает весьма пестрый облик.
2. Речь идет о гомоклиматах, представленных на всех материках.
Климадиаграммы областей с преобладанием зимних осадков столь похожи
одна на другую, как, пожалуй, ни в какой другой зоне. Но поскольку
состав флоры отдельных материков весьма различен, легко понять, что
для определения типа растительности не только окружающая среда,
но и флора имеет решающее значение. Это обстоятельство эколого-физио-
лог должен всегда учитывать. При одинаковых климатических условиях
на одном местообитании развивается эвкалиптовый лес с травянистыми
деревьями (Австралия), на другом — либо вечнозеленый дубовый лес
(Средиземноморье, Калифорния, Аризона), либо низкий кустарник про-
тейных, например в Капской провинции, либо смешанные древостои,
родственные связи которых указывают на влияние влажных тропиков
{Чили).
3. Однако во всех случаях в составе растительного покрова домини-
руют древесные растения. Злаки почти полностью отсутствуют; появ-
ляются они лишь там, где количество осадков для произрастания древес-
ных растений уже недостаточно и где зимы настолько мягкие, что злаки
продолжают вегетировать и в зимний сезон дождей. Но поскольку злаки
эти принадлежат не к тропическим таксонам, а к таксонам умеренной
зоны, они остаются активными даже при относительно низкой темпера-
туре зимой.
4. В растительном покрове средиземноморских областей с зимними
дождями резко доминируют склерофильные, то есть жестколистные,
виды растений. Малакофильные виды (с мягкой листвой) играют второ-
степенную роль. Суккуленты в зоне собственно жестколистной раститель-
ности отсутствуют полностью. Особенно типичны для этой зоны анатомо-
морфологические пейноморфозы, и прежде всего там, где почвы характе-
ризуются не только сухостью летом, но и бедностью питательными веще-
ствами. Прекрасно растут в областях с зимними дождями суккулентные
виды; об этом свидетельствует акклиматизация агав и опунций в Среди-
земноморье. Правда, держатся они только на местообитаниях, естествен-
ный покров которых нарушен человеком. Растут они медленно, требуют
полного солнечного освещения и потому не выдерживают конкуренции
со склерофилами.
5. Характерное морфолого-анатомическое строение склерофильных
видов, сохраняемое ими при одинаковых климатических условиях в совер-
шенно различных флористических царствах (Голарктика, Неотропиче-
ское царство, Капская провинция и Австралийская область), есть признак
того, что эта жизненная форма хорошо приспособлена к условиям внеш-
ней среды, иными словами, что она целесообразна. В век каузальных
исследований отвыкли говорить о целесообразности. Так называемый
телеологический способ рассмотрения считается просто ненаучным, хотя
целесообразность строения и функций живого организма является пред-
посылкой возможности его существования часто во враждебной, то есть
угрожающей жизни, среде.
6. Следовательно, нет никакого противоречия между каузальными
и телеологическими способами познания. Принцип целесообразности
в физике отражен в законе наименьшего действия (принцип Ле Шателье).
Согласно этому правилу, воздействие внешнего фактора на равновесную
-систему вызывает в ней реакции, противодействующие производимому
Области средиземноморского климата за пределами Евразии
225
изменению. Растительный организм — сложная биологическая система,
находящаяся в динамическом равновесии с окружающей средой Ч Если
последняя изменяется, например ухудшается обеспеченность водой,
растение восстанавливает равновесие, сокращая транспирирующую
поверхность и интенсивнее развивая корневую систему; благодаря этому
нарушение водного режима по возможности компенсируется и водный
баланс сохраняется на постоянном уровне.
При недостатке питательных веществ в почве аналогичные пейно-
морфозы, то есть обусловленные дефицитом соответствующих элементов
изменения в морфологическом строении органов, дают растению возмож-
ность закончить свой цикл развития с меньшим количеством питательных
веществ. Если в потомстве таких растений случайно возникают мутации,
приводящие к тем же морфологическим изменениям, то эти мутации
в аридных условиях (или в условиях, когда минеральное питание ослаб-
лено) будут обладать положительной селекционной ценностью, в гумид-
ной же среде или на почвах, богатых питательными веществами,— отри-
цательной; ксероморфные растения, например, растут медленно и с обыч-
ными растениями конкурировать не могут. Ксероморфные признаки,
обусловленные мутациями, наследственно закрепляются. Вместе с тем
распространение мутантов из-за наличия конкуренции будет ограничено
сухими (или бедными минеральными элементами) местообитаниями,
то есть возникнут особые экотипы или даже( новые виды, если дальнейшие
мутации приведут к генетической изоляции. Такого рода пейноморфные
виды, встречающиеся как на сухих местообитаниях, так и в гумидных
условиях (или на сырых почвах неблагоприятных местообитаний), осо-
бенно распространены среди Ericaceae, Proteaceae, Restionaceae. Известны
они и среди вечнозеленых дубов; достаточно вспомнить, например, засухо-
устойчивые Quercus ilex и Qu. suber, связанные во влажной средиземно-
морско-атлантической области с бедными почвами. Весьма характерно
также для многих видов дрока, что они, с одной стороны, способны пере-
носить засуху, а с другой — встречаются в области атлантического
климата на местообитаниях, бедных питательными веществами. Если
сравнить между собой реакции видов различных таксонов, то оказывается,
что не только степень пластичности, но и характер модификаций и мутан-
тов может быть совершенно разным. Таким путем возникли разнообразные
экологические типы, о которых мы уже говорили.
7. Тот, кто признает значение отбора в эволюции организмов, не будет
отрицать целесообразность их устройства. Ведь селекционной ценностью
обладает только мутация, наиболее приспособившаяся к данной окру-
жающей среде; при этом само собой разумеется, что каждый организм
обладает многими таксономически важными признаками, не имеющими
ни положительной, ни отрицательной селекционной ценности. Рацио-
нальность определенного свойства также должна быть доказана экспери-
ментально.
Строение организма может считаться целесообразным в условиях
естественной окружающей среды. И напротив, если организм подвергнуть
воздействию искусственно созданного фактора (например, воздействию
искусственных гормонов роста, используемых как гербициды), то реакция
организма может оказаться совершенно нецелесообразной и даже привести
к его гибели.
1 Вместе с тем это управляемая система, которую можно рассматривать и с кибер-
нетической точки зрения.
15—01248
226
Глава 3
Этими соображениями мы хотели дополнить анализ признаков
приспособления растений к факторам среды, изложенный в данном раз-
деле.
Несколько слов о растительности умеренно гумидной зоны южного
полушария, к которой в следующих разделах мы уже не вернемся. Пло-
щадь этой зоны по сравнению с площадью аналогичной зоны северного
полушария очень мала. Следовательно, генофонд в соответствующих
флористических областях был столь невелик, что развиться могла только
малотипичная умеренная флора. Часто речь здесь идет о представителях
гумидного тропического пояса, приспособившихся к низкой температуре,
тем более что в умеренно гумидной зоне южного полушария с ее более
океаническим климатом резко выраженное холодное время года отсут-
ствует. Встречаются здесь и антарктические элементы, в общем, однако,
малочисленные. Напротив, флора умеренной зоны северного полушария
настолько своеобразна, что ее вполне обоснованно относят к особому
флористическому царству — Голарктике.
Глава 4
ЛИСТВЕННЫЕ ЛЕСА УМЕРЕННОЙ ЗОНЫ
СЕВЕРНОГО ПОЛУШАРИЯ - НЕМОРАЛЬНАЯ ЗОНА
ОБЩАЯ ХАРАКТЕРИСТИКА ЛИСТВЕННЫХ ЛЕСОВ
ВОСТОЧНОЙ ЧАСТИ СЕВЕРНОЙ АМЕРИКИ,
ВОСТОЧНОЙ АЗИИ И ЕВРОПЫ
Прежде чем перейти к более подробной характеристике зоны летне-
зеленых лиственных лесов, распространенных в областях умеренного
климата, необходимо выявить ее местоположение внутри крупных
лесных зон северного полушария.
Бореальные хвойные леса умеренно холодного климата образуют
циркумполярную зону, южная граница которой расположена между 50
и 60° с. ш., а северная лишь в некоторых местах не достигает Полярного
круга; в большинстве же случаев она заходит за Полярный круг
до 70° с. ш. Расположенная южнее зона лиственных лесов не покрывает
сплошь всю территорию собственно умеренной зоны (см. рис. 5). Леса
распространены прежде всего на юго-востоке Североамериканского кон-
тинента и в восточной части Азии. Эти области распространения листвен-
ных лесов отграничены с юга субтропическими влажными лесами Флориды
или Восточного Китая, а с севера — бореальными хвойными лесами;
в переходных зонах преобладают смешанные леса. Представление о кли-
матических условиях дают климадиаграммы расположенных здесь стан-
ций (рис. 174—176 и 177—179). Лиственные леса Северной Америки
и Восточной Азии, являющиеся остатками арктотретичной флоры плио-
цена, отличаются .исключительным разнообразием видового состава х.
Очень красочны цветки Rhododendron в Аллеганских горах (см. цветное
фото 4). В западном направлении при уменьшении количества атмосферных
осадков лиственные леса в Северной Америке и в Восточной Азии перехо-
дят в безлесные злаковники (прерии или степи умеренной зоны).
Пространственная структура лиственных лесов отличается большой
сложностью, поэтому мы характеризуем ее в упрощенной форме (рис. 180
и 183) и, кроме того, иллюстрируем фотографиями (рис. 181—182 и 184).
Автор не проводил длительных экологических исследований в лист-
венных лесах Северной Америки и знает восточноазиатские леса только
по литературным данным. Хотя в Северной Америке и в Азии, а именно
в Японии, выполнено большое число интересных эколого-физиологических
исследований, обобщить их в состоянии только опытный специалист,
1 R. Knapp, Die Vegetation Nord- und Mittelamerikas. См.: H. W a 11 e r, Vegeta-
tionsmonographien der einzelnen GroBraume, Bd. I, Stuttgart und Jena, 1965;
M.M. Grandtner, La vegetation forestiere du Quebec meridional, Quebec, 1966;
Chi-wu Wang, The forests of China, «Cabot. Found. Publ.», № 5, 1961; T. P о c s,
Analyse aire-geographique et ecologique de la flore du Vietnam Nord, Eger (Ungarn),
1965 (указано положение южной границы распространения голарктических лиственных
лесов); Т. Suzuki, The highest vegetation units of Japan and their areas, «Contr.
Biol. Inst., Ooita Univ.», № 68, 1967 (на япон., карта растительности и пояснения
к ней — на англ.); А.И.Толмачев, О флоре острова Сахалин, «АН СССР, Бота-
нический институт, Комаровские чтения», 12, М.— Л., 1959; Д. П. Воробьев,
Растительность Курильских островов, М.—Л., 1963.
15*
228
Глава 4
Рис. 174—176. Климадиаграммы областей лиственных лесов в Северной Америке.
Колумбус, штат Джорджия — южная часть области распространения смешанных дубовых лесов,
частично с примесью сосны; Ноксвилл, штат Теннесси — граница распространения буково-сахар-
нокленовых лесов; Чикаго, штат Иллинойс — область дубово-гикориевых лесов.
Рис. 177—179. Климадиаграммы областей лиственных лесов в Восточной Азии.
Ханькоу — южная граница распространения листопадных дубовых лесов; Пайпинг — северная
граница той же области; Владивосток — северная часть области распространения широколиствен-
ных лесов, образованных кленом, липой и березой (число дней со средней температурой выше 4-10°
составляет 146, с температурой выше —10° 305).
проработавший здесь достаточно долгое время. Условия конкуренции
многочисленных видов деревьев и состав насаждений слишком разно-
образны, чтобы для суждения о причинных связях можно было ограни-
читься примерами из литературы.
В дальнейшем в основном будут проанализированы материалы эколо-
го-физиологических исследований, проводившихся в европейской части
зоны лиственных лесов. В Европе лиственные леса заходят далеко на север,
в западной части своего ареала даже севернее 58° с. ш., что связано с благо-
приятным в климатическом отношении влиянием Гольфстрима. Если срав-
нить пункты с одинаковой средней годовой температурой в Западной
Европе и на востоке Северной Америки и Азии, то окажется, что раз-
ница температур между самым теплым и самым холодным месяцами будет
наименьшей в Западной Европе. Здесь климат находится под определен-
ным влиянием океана, в связи с чем все зоны растительности в Западной
Европе продвинуты далеко на север.
Лиственные леса в Европе простираются вдоль атлантического
побережья, начиная с северной части Пиренейского полуострова и кончая
южной Скандинавией; в Восточной Европе, где заметно усиливается
континентальность климата, лиственные леса начинают выклиниваться
уже в районе Днепра; таким образом, область распространения этих
лесов на Европейском континенте напоминает по форме треугольник.
Рис. 180. Карта растительности восточной части Северной Америки (по Кнаппу,
с дополнениями).
1 — районы, покрытые льдом; 2 — северная тундра; 3 — южная тундра; 4 — лесотундра; 5 —
бореальные хвойные леса; 6 — северные лиственные леса с примесью отдельных видов хвойных
пород; 7 — буково-сахарнокленовые лиственные леса; 8 — смешанные дубовые леса; 9 — дубовые
леса с большой примесью сосны (частично доминирующей); 10 — пойменные и заболоченные леса,
частично субтропического характера; 11 — дубово-гикориевые леса; 12 — северные прерии с осино-
выми колками; 13 — высокозлаковые прерии; 14 — смешанные и низкозлаковые прерии (Великие
равнины).
Рис. 181. Пышный, флористически богатый (свыше 25 видов древесных пород на 0,5 га) лиственный лес
в Северной Каролине (фото министерства земледелия США, Лесная служба).
Рис. 182. Дубово-гикориевый лес (Quercus alba и виды Сагуа) в Национальном парке Шенандоа (фото мини-
стерства земледелия США, Лесная служба).
Рис. 183. Сильно схематизированное зональное расчленение растительности Восточ-
ной Азии (по Вангу).
1 — тропически-субтропический дождевой лес, 2—4—вечнозеленые умеренно теплые леса, образо-
ванные дубом, Schima и видами лавровых (2 — леса с Pinus yunnanensis, ср. рис. 184; 3 — с Pinus
massoniana’, 4 — без сосны); 5 — смешанный меэофитный лес; 6 — лиственные леса из летнезеленых
видов дуба; 7 <— северные широколиственные леса, состоящие из клена, липы и березы (В — по-
следняя доминирует); 8 — горные хвойные леса с преобладанием Abies (A), Picea (Р) и Larix (L);
9 — безлесные районы (степи и пустыни); 10 — альпийская растительность и горные тундры.
На врезке сопоставлены положения Китая, Монголии, США и Мексики по широте.
Рис. 184. Леса нижней части пояса гор (высота 2250 м) севернее Юнняньфу, образо-
ванные Pinus yunnanensis и Quercus griffithii (фото Хандель-Мацетти).
Лиственные леса умеренной зоны северного полушария — неморальная зона 233
Правда, на восток от Днепра зона лиственных лесов еще продолжается
узкой полосой лесостепи, примыкающей на севере к очень широкой зоне
смешанных лесов, северная граница которой тянется от южной Швеции
к устью Невы и далее до нижней Камы и Южного Урала 1 у Свердловска.
Только в пределах этбй зоны и встречаются еще широколиственные
породы, в то время как мелколиственные виды, к которым относятся
береза (Betula), осина (Populus tremula), ольха серая (Alnus incana), а также
рябина (Sorbus aucuparia) и многие виды рода Salix, распространяются
в Европе до северной границы бореальной зоны и проникают даже
в Сибирь. В западной части ареала европейская зона лиственных лесов
ограничивает с севера средиземноморскую зону жестколистных лесов;
между этими двумя зонами располагается субсредиземноморская пере-
ходная зона. В восточной части ареала зона лиственных лесов образует
северную границу восточноевропейских злаковых степей. Эти закономер-
ности представлены в нижеследующей схеме:
Бореальная зона { Березовые леса
Зона хвойных лесов или тайга
Умеренная зона J Приатлантические
L верещатники
t
т Смешанные леса
Централ ьноевро- ф
пейские листвен-
ные леса |
Субсредиземно- J
морская зона
(Леса из лавро-
видных пород)
Каштановые леса
и леса из пушис-
того дуба
Леса из черной
сосны
I----Степи-----
Среднеазиат-
ские пустыни
Полупустыни
Степи-
t
Средиземномор- ( Жестколистная растительность в областях
ская зона < сзимнимидождями
( Запад Восток
Характерной чертой европейской зоны лиственных лесов является
уже упомянутая выше исключительная бедность видового состава, осо-
бенно древесного яруса. За исключением бука (Fagus silvatica) и граба
(Carpinus betulus), а также двух видов дуба (Quercus petraea nQu. robur),
ни одна из лиственных пород не доминирует в лесах. Ясень (Fraxinus
excelsior), клен (Acer pseudoplatanus и A. platanoides), а также липа (ТШа
cordata и Т. platyphyllos) и вяз (Ulmus scabra и U. laevis) встречаются
в лесах часто; остальные виды (Prunus avium, Pirus и Matus) имеют под-
чиненное значение.
Эта флористическая бедность есть следствие оледенения Земли в плей-
стоцене. Продвижению лесной растительности на юг в тот период пре-
пятствовали альпийские ледники и Средиземное море. Большинство
видов лиственных деревьев плиоценовых лесов, отличавшихся в Европе
богатством видового состава, вымерло или не восстановилось в после-
ледниковый период, среди них Juglans, Castanea, Aesculus, Liquidam-
bar и др.
Поскольку основные эколого-физиологические исследования прово-
дились именно в европейских лиственных лесах, мы охарактеризуем евро-
пейский сектор зоны лиственных лесов более подробно, уделив меньшее
внимание другим районам распространения.
1 Точнее, Среднего Урала. — Прим. ред.
234
Глава 4
Климат в Европе заметно меняется в направлении с запада на восток
по мере продвижения от побережья океана в глубь континента. Это обус-
ловливает следующий характер -распределения лесной растительности:
на западе — приатлантические верещатники как одна из стадий дегра-
дации леса, в Центральной Европе — собственно лиственные леса и далее
на восток — смешанные леса, характеристика которых приводится в сле-
дующей главе.
Мы должны, однако, ясно представлять себе, что в такой уже давно
и густо населенной области, как Европа, в целостности сохранились
лишь немногие остатки зональной растительности. В естественных усло-
виях эти леса лучше всего развиваются на равнинных пространствах,
Лунд — близ северной границы рассматриваемой области; Штутгарт Хоэнхейм — южная граница.
где они не подвергаются влиянию застойного увлажнения или грунтовых
вод, а также на мощных ни слишком тяжелых, ни слишком легких почвах
с хорошо сформированным профилем, характерным для соответствующей
климатической зоны. Почвы такого рода наиболее пригодны для земледе-
лия. Не удивительно поэтому, что лиственные леса на этих почвах, как
первоначальная зональная растительность умеренной зоны Европы,
были со временем почти полностью уничтожены. Их место в настоящее
время заняли пашни. В Центральной Европе леса сохранились лишь
на 25% общей площади, на маломощных почвах и крутых склонах гор,
на тяжелых переменновлажных или бедных почвах, иными словами,
на таких местообитаниях, которые малопригодны для сельскохозяйствен-
ного использования. На такого рода почвах в настоящее время в боль-
шинстве случаев распространены сильно измененные формы зональной
растительности или различные типы экстразональной или азональной
растительности. В областях с экстремально океаническим климатом леса
были полностью уничтожены и на их месте образовались верещатники
и болота. Они являются столь типичным ландшафтным образованием,
что ранее их даже относили к зональной растительности.
Мы имеем лишь смутное представление о составе первобытных лесов
Западной и Центральной Европы х. В областях, находящихся под мень-
шим влиянием моря (рис. 185—187), на богатых почвах наибольшей кон-
1 Н. Ellenberg, Die Vegetation Mitteleuropas mit den Alpen. Cm.: « Phyto-
logie», Bd. IV, 2, Stuttgart, 1963; F. K.Hartmann, G. Jahn, Waldgesellschaften
des mitteleuropaischen Gebirgsraumes nordlich der Alpen, Stuttgart und Jena, 1967;
W. S z a f e r, The Vegetation of Poland, Warszawa, 1966; E. A i c h i n g e r, Die
Rotbuchenwalder als Waldentwicklungstypen, «Angew. Pflanzensoz.», Heft V, Wien, 1952.
Лиственные леса умеренной зоны северного полушария — неморалъная зона 235
Рис. 188. Буковый лес
(Fagus silvatica) близ
Людвигсхафена у Боден-
ского озера. Естествен-
ное возобновление под
пологом леса (фото
Фойхта).
курентной способностью обладает бук (Fagys silvatica) (рис. 188). Поэтому
буковые леса должны рассматриваться как зональная растительность;
лишь иногда в чистых буковых лесах наблюдается примесь других лист-
венных пород. Бук теряет свое господствующее положение в приатлан-
тических областях на очень бедных выщелоченных почвах, а также в самых
Рис. 189—191. Климадиаграммы области распространения буковых лесов в горах.
236
Глава 4
засушливых частях Центральной Европы, например на Верхнерейнской
низменности и в междуречье Заале и Эльбы, где распространены темно-
цветные почвы, в Средней Чехии и Южной Моравии. В горных районах
на благоприятных почвах бук достигает оптимального развития
(рис. 189—191). В условиях океанического климата бук нередко образует
верхнюю границу распространения древесной растительности (рис. 192).
Обычно же на больших высотах к буку примешивается пихта, и только
на вершинах среднегорий и в верхней части северного склона Альп пихта
уступает свои позиции ели. Таковы климатические границы произраста-
ния буковых лесов х. Наряду с климатом распространение бука ограни-
Р и с. 192. Буковый лес у верхней границы распространения леса на высоте 1110
над уровнем моря (Баварско-Чешский Лес). Возраст деревьев 140 лет. Под пологом
леса естественное возобновление (фото Фабрициуса).
1 О характере распределения буковых лесов в Юго-Восточной Европе см.;
R. Soo, Die regionalen Fagion-Verbande und Gesellschaften Sudosteuropas, «Stud.
Biol. Hung.», 1, Budapest, 1964.
Лиственные леса умеренной зоны северного полушария — неморальная зона 237
Р ис. 193. Карта распространения лесов Центральной Европы перед нача-
лом нашей эры, составлена на основе данных пыльцевых анализов (по
Фирбасу, упрощено; из работы: Н. Walter, Arealkunde, 1954).
1 — засушливые области без бука (сумма годовых осадков менее 500 мм)\ 2 — сме-
шанные леса с буком на более низких высотах (частично преобладает дуб); 3 — горные
буковые леса; 4 — буковые леса с незначительной примесью сосны; 5 — горные буковые
леса с примесью пихты и ели (д) или (А) субальпийские буковые леса; 6 — области
с преобладанием сосны и с примесью дуба на песчаных почвах; 7 — смешанный дубово-
грабовой лес; 8 — то же с примесью ели, иногда доминирует сосна.
чивает и эдафический фактор: бук не растет на застойно увлажненных
и переувлажненных почвах, на легких песчаных почвах и сухих скалах.
На таких местообитаниях, покрытых в настоящее время лесом, встречаются
другие типы леса как представители экстразональной и азональной
лесной растительности. Нередко последние занимают большие площади,
особенно обширные на почвах с застойным увлажнением или на почвах,
находящихся под воздействием грунтовых вод. На таких почвах разви-
ваются дубово-грабовые леса, которые фитоценологи неправильно при-
нимают за климаксовую формацию. Карта лесов Центральной Европы,
которую составил Фирбас (на основе данных спорово-пыльцевого ана-
лиза) на начало нашей эры, хорошо отражает прошлое распространение
лесов (рис. 193); такое же четкое представление о распределении лесов
дает схематический профиль, составленный Элленбергом (рис. 194);
однако следует учесть, что он переоценивает роль дуба на равнинах.
Вплоть до позднего средневековья человек своей деятельностью способ-
ствовал расширению ареала дуба.
Постоянное вмешательство человека содействует изменению лесов
в определенном направлении, а именно образованию более разреженных
древостоев, то есть редкостойных лесов, зарослей кустарников и верещат-
ников. В результате культурный ландшафт в лесной зоне отражает более
238
Глава 4
1000 км
Рис. 194. Схематические профили через территорию Центральной Европы с указа-
нием вертикальных поясов растительности (по Элленбергу, см. «Phytologie», Bd. IV, 1).
1 — нивальный пояс; 2 — пояс альпийских лугов; з — лиственница; кедр европейский; 4 — ель;
5 — ель и пихта; 6 — сосна (сосна обыкновенная); 7 — пихта, ель, бук; 9 — бук с дубом; 10 —
дуб; 11 — горная сосна (криволесье).
Бук предпочитает океанический климат, хвойные леса — континентальный. ГРЛ — граница
распространения леса; СЛ — снеговая линия.
засушливые условия по сравнению с климатом, характерным для данной
территории.
К востоку от границы распространения бука происходит смена
зональной растительности и начинается область, где господствует граб
(Carptnus betulus); это наблюдается в междуречье Вислы, среднего
Днестра и Днепра, восточнее которого эта древесная порода почти
не встречается (рис. 195). Небольшой реликтовый участок с грабом распо-
ложен около Донца. В свою очередь граб вытесняется дубом черешчатым
(Quercus robur), однако дубовые леса образуют лишь узкую полосу между
степью на юге и смешанными лесами на севере. Дубовые леса достигают
южных отрогов Урала, где в их составе увеличивается роль липы (Tilia
cordata). Нередко в результате деятельности человека дубовые насаждения
сменяются чистыми липовыми лесами, которые широко используются для
пчеловодства; по-видимому, нектарообразование в цветках липы протекает
более интенсивно на востоке, чем на западе.
В средней части Восточной Европы зональная растительность пред-
ставлена смешанными лесами, образованными хвойными
и широколиственными древесными породами. Однако смешанные леса
далеко не однородны. В этой зоне распространены леса, в древесном
ярусе которых совместно растут лиственные и хвойные породы (в боль-
шинстве случаев сосна в верхнем ярусе и дуб в нижнем ярусе), чистые
хвойные леса (в основном из Picea abies) и чистые лиственные леса. Раз-
личные типы леса мозаично чередуются друг с другом. Лиственные леса
приурочены к лучшим, богатым известью почвам, обычно развитым
в западной части ареала или на склонах южных экспозиций; напротив,
хвойные леса преимущественно встречаются на склонах северной экспози-
ции и на равнинных участках с холодными почвами застойного увлаж-
нения.
Лиственные леса умеренной зоны северного полушария — неморальная зона 239
Рис. 195. Восточные границы не-
которых центральноевропейских ви-
дов лиственных деревьев и западные
границы сибирских хвойных пород
(по Вальтеру—Алехину, 1936).
Fa — Fagus silvatica; Са — Carpinus betu-
lus; Qu — Quercus robur; Fr — Fraxinus
excelsior; Ab — Abies sibirica; La — Larix
sibirica; Pi — Pinus sibirica (cembra).
В центре Восточной Европы леса такт^е были в свое время почти пол-
ностью уничтожены человеком. На севере прежде других в пахотные
земли были переведены плодородные почвы, занятые лиственными лесами,
поэтому здесь до наших дней сохранились преимущественно хвойные
леса. На юге в первую очередь вырубались хвойные породы, которые
дают лучшую товарную древесину, вот почему в сохранившихся там лесах
преобладают виды лиственных деревьев. Прежние обширные лесные
площади в настоящее время покрыты лугами; это чаще всего наблюдается
в заболоченных ольховых лесах, где нередко формируются луга с отдель-
ными куртинами деревьев («Laubwiesen»). Бессистемное ведение лес-
ного хозяйства приводит к господству на больших пространствах пород-
пионеров; в этом, в частности, причина широкого распространения почти
чистых березняков на бедных почвах с близким уровнем грунтовых вод.
Подлесок в лиственных лесах этой части зоны (смешанных лесов в меньшей степени,
чем древесный ярус, подвергается крайним воздействиям климата; поэтому по своему
составу он напоминает ярус кустарников лиственных лесов Центральной Европы. Боль-
шинство центральноевропейских видов проникает далеко на север и на восток до Урала,
например кустарники: Corylus avellana, Lonicera xylosteum, Evonymus europaea (на восто-
ке сменяется Evonymus verrucosa), Prunus padus, Salix caprea, Daphne mezereum, Rubus
idaeus и др. Такой же характер распространения свойствен и следующим видам травя-
ного покрова: Asperula odorata, Asarum europaeum, Stellaria holostea, Pulmonaria obscura,
Viola mirabilis, Lathyrus vernus, Aegopodium podagraria, Festuca silvatica, Milium effusum,
Brachypodium silvaticum, Neottia nidus-avis и др.
Восточнее Днепра отсутствуют Acer pseudo plat anus, Prunus avium, Sorbus tormina-
lis, Pulmonaria mollissima, Polygonatum latifolium, Scilla bifolia, Arum maculatum и др.
Вместо вышеуказанных видов появляются новые: Crataegus sibirica, Cornus sibi-
rica, Lonicera tatarica, Bupleurum aureum, Cacalia (Senecio) hastata, Siler trilobium,
Mulgedium macrophyllum и др.
Южная граница распространения леса на контакте со степью выра-
жена недостаточно четко. Здесь развита переходная зона с макромозаич-
ным характером распределения лиственных лесов и северных луговых
степей. Эту зону называют лесостепью (см. т. III, гл. 2).
240
Глава 4
В юго-западной части зоны лиственных лесов Восточной Европы
появляются уже некоторые субсредиземноморские элементы: Quercus
pubescens, Cotinus coggygria и др<; это форпосты шибляка — листопадных
низкоствольных лесов субсредиземноморской зоны \ распространенных
на Балканском полуострове.
Восточноевропейские леса отделены степной зоной от лиственных
лесов северного склона гор Крыма и Кавказа. Здесь на небольшом про-
странстве зональность выражена в обратной последовательности: степь
сменяется лесостепью, затем южнее идут лиственные леса, дубовые
леса, а еще выше при большем количестве атмосферных осадков —
буковые леса, образованные Fagus orientalis. Буковые леса занимают про-
странство от северных склонов Кавказских гор и Закавказья вплоть
до района распространения колхидских лесов, которые приурочены
к треугольнику, образуемому берегом Черного моря от Сухуми до Батуми,
Главным Кавказским хребтом на северо-востоке и горами Закавказья
на юго-востоке. Колхида характеризуется продолжительно влажным,
теплым, безморозным климатом, который обеспечивает возможность
произрастания здесь многочисленных вечнозеленых лиственных древес-
ных растений. Колхида и расположенная на юго-западе Каспийского
моря Ленкорань (колхидо-гирканская область) — исключительно богатая
с флористической точки зрения область с многочисленными эндемами
и отдельными третичными реликтами, как-то: Zelkowa, Pterocarya, Dtospy-
ros lotus, Parrotta persica, Juglans regia, Acer laetum, многочисленные виды
Quercus, Albizzia julibrissln, Ficus hircana, Danae racemosa и др. Здесь мы
вновь сталкиваемся с остатками арктотретичной флоры а.
Колхидские леса, простирающиеся на запад от Колхиды через всю
Малую Азию вдоль южного побережья Черного моря, делятся па соб-
ственно колхидскую, или колхидо-средиземноморскую, нижнюю ступень
и более высокорасположенную колхидо-центральноевропейскую ступень,
особенно хорошо выраженную на отвесном побережье северной части
Малой Азии. В обеих ступенях древесный ярус образован летнезелеными
лиственными породами. В нижней ступени господствуют дуб, каштан
и серебристая липа (Tilta tomentosa); роль бука в этих лесах невелика.
На сухих местообитаниях иногда встречаются элементы средиземномор-
ской флоры: Quercus ilex, Phillyrea media, Arbutus unedo и A. andrachne,
Erica arborea, Pistacia terebtnthus, Rhus coriaria; часто большие площади
занимают Laurus nobilis и Myrtus. В верхней ступени доминирует бук
(Fagus orientalis), и только в более освещенных местах вместе с ним растут
Carpinus, Ostrya, Acer campestre и др. Для обеих ступеней колхидских лесов
характерно наличие вечнозеленого подлеска, представленного невысо-
кими нежестколистными деревьями и кустарниками. В подлеске наиболь-
шее значение имеет Rhododendron pontlcum, Ilex aqul folium, на влажных
местообитаниях — Prunus laurocerasus, Vaccinium arctostaphylos, Buxus
sempervtrens и Daphne pontica. Более резко выраженным средиземномор-
ским характером отличаются Ruscus aculeatus, R. hypoglossum, Smllax
и листопадный Tamus communis. На освещенных пространствах, а нередко
и внутри леса Rhododendron образует непроходимые заросли. У верхней
границы распространения к буку примешивается пихта.
1 I. Н о г v a t, Pflanzengeographische Gliederung Siidosteuropas (mit Vegetations-
karte), «Vegetatio», 5/6, 1954, S. 434—447.
2 A. N о irf a 1 ise, M. H. D j a z i г e i, Contribution a la phytogeographie de
la foret caspienne, «Bull. Soc. Roy. Bot. Belg.», 98, 1965, p. 197—214.
Лиственные леса умеренной зоны северного полушария — неморальная зона 241
? Fagus
f Carpinus
т Tilia I Abies
а Quercus т Pinus Silvestris
ft Castanea т ” nigra
Гис. 196. Вертикальная поясность растительности в горах севернойАнатолии
(по Кюне, с дополнениями).
Верхняя граница заштрихованной поверхности соответствует уровню моря для вычерченного над
ней профиля. По мере удаления от моря возрастает континентальность климата. Бук вытесняется
пихтой, а та в свою очередь — сосной. Северный склон гор более увлажнен, чем южный склон,
обращенный в противоположную от моря сторону.
Леса Колхиды интересны тем, что среди лиственных лесов европейско-
переднеазиатской области они выделяются богатством видового состава,
который при продвижении в северо-западном направлении через Балканы
к Атлантическому океану становится все более бедным. В этом, как уже
упоминалось, проявляется влияние эпохи оледенений. В ледниковый
период ледники Кавказа проникали глубоко в предгорья, однако значи-
тельная часть лесных растений смогла пережить это время в более теплой
Колхиде. Область распространения Rhododendron ponticum в ледниковый
период ограничивалась двумя районами: южной частью Иберийского
полуострова (см. рис. 197) и Колхидой Ч Северная Анатолия отличается
особенно сложными экологическими условиями, поскольку здесь от берега
моря в глубь суши, возвышаясь одна над другой, кулисообразно протяну-
лись три горные цепи. По мере удаления от моря климат в долинах стано-
вится все более континентальным. Обращенные к морю северные склоны
1 На основании межледниковых находок ископаемых остатков растений, обна-
руженных Гёттингом у Инсбрука, можно прийти к выводу, что Rhododendron ponticum
до ледникового периода был распространен по всей Южной Европе.
16-01248
242
Глава 4
гор более увлажнены, чем защищенные от влияния моря южные склоны.
В результате этого от склона к склону меняется последовательность рас-
положения вертикальных ступеней, начиная от средиземноморских и кол-
хидных ступеней и кончая альпийскими. В качестве примера анализа
вертикальной поясности приводится ранее не публиковавшаяся схема
Кюне (рис. 196).
ОБЛАСТЬ ПРИ АТЛАНТИЧЕСКИ X ВЕРЕЩАТНИКОВ
Общие замечания
Уже в западной части средиземноморской области на бедных
известью почвах встречаются верещатникоподобные сообщества, обра-
зованные Erica cinerea, Lavandula stoechas, некоторыми видами семейства
ладанниковых (Cistus crispus, С. ladaniferus), а также многочисленными
лишайниками (виды С Ladonia) и несколькими видами мхов (Poly trichum
juniperinum, Р. piliferum), однако только в горах северо-западнее Гиб-
ралтара, где летом выпадает большее количество осадков, происходит
существенная смена растительного покрова. На открытых пространствах
в лесах из пробкового дуба здесь развиваются настоящие верещатники.
Ладанник вытесняется видами Halimium с желтыми цветками, увеличи-
вается число видов семейства вересковых, появляются многочисленные
виды дрока, в том числе иберийский Pterospartum tridentatum, различные
виды Cytisus (С, candicans, С, triflorus, С, linifolius) и виды Sarothamnus
(S. baeticum, S. welwitschiae, S. catalanicus) и др.
В еще очень теплой и влажной юго-западной части Иберийского
полуострова 1 встречаются реликтовые лавролистные виды, которые,
помимо того, распространены только на Канарских островах, и элементы,
иногда встречающиеся также в Колхиде (Кавказ, Северная Анатолия),
среди них: Quercus lusitanica ssp. canariensis ( = Qu. mirbeckii), Prunus lusi-
tanica (aff. P. laurocerasus), Rhododendron ponticum ssp. baeticum (рис. 197)
и т. д. Здесь же отмечены и интересные насекомоядные: Drosophyllum
lusitanicum (рис. 198) и крошечные Utricularia lusitanica, а также эпифит
Davallia canaricum (семейство многоножковых).
Верещатники в условиях очень выровненного климата с мягкой
зимой и влажным летом распространены вдоль атлантического побережья
Португалии, в северной Испании, западной Франции, а также в Англии,
Ирландии и Шотландии вплоть до Норвегии (рис. 199—204). К северу
все большее число средиземноморских элементов исчезает (Arbutus unedo,
Rubia peregrina, Ceterach officinarum и Umbilicus pendulinus встречаются
вплоть до Ирландии). Они вытесняются иберийскими элементами, среди
которых значительным видовым разнообразием отличаются дроки. Роды
Pterospartum, Sarothamnus, Ulex, Genista и Adenocarpus представлены
в верещатниках большим или очень большим числом видов, так же как
род Erica.
Помимо того, встречаются также виды, область распространения
которых простирается через Иберийский полуостров на север (иберийско-
атлантические), а также флористические элементы, центр распространения
которых расположен севернее (эуатлантические элементы); в Шотландии
и Норвегии к ним присоединяются бореально-атлантические или арктиче-
1 Die Pflanzenwelt Spaniens, I и II (red. W. Liidi), «Veroff. Geobot. Inst. Rubel»,.
H. 31, Zurich, 1956.
Лиственные леса умеренной зоны северного полушария — неморальная зона 243
Рис. 197.^Rhododendron ponticum ssp. baeticum (в центре) в пес-
чаниковых горах на юге Испании. Над Rhododendron — Quercus
suber, рядом — Pteridium aquilinum (фото Э. Вальтер).
ские элементы (Empertum, Vaccinium, Phyllodoce, Cassiope и др.). Особенно
хорошо выражен этот сдвиг ареалов у вересковых и у видов дроков среди
бобовых.
Виды вересковых, распространенные в верещатниках
Эусредиземноморские: Erica arborea и Е. multiflora.
Западносредиземноморские: Erica terminalis.
Средиземноморско-атлантические: Е. cinerea, реже Е. scoparia.
Иберийские элементы, ареал которых ограничен Иберийским полуостровом: Erica lusi-
tanica, Е. mackaiana, Е. umbellata, Е. australis.
16*
244
Глава 4
Иберийско-атлантические элемен-
ты, встречающиеся вплоть до
Ирландии и в некоторых слу-
чаях до Шотландии: Erica
ciliaris, Е. vagans, Е. mediter-
ranean Е. tetralix и цветущая
крупными цветками Daboecia
cantabrica.
Атлантическо-европейские элемен-
L- ты, представленные наиболее
широко распространенным
видом Calluna vulgaris, кото-
рый занимает пространство
от Лазурного берега и Коста-
Брава почти вплоть до Арк-
тики и Урала, минуя лишь
засушливые районы Юго-
Восточной Европы.
Виды дроков, распро-
страненные в вере-
щатниках
Средиземноморские: Sparti-
ит junceum и Calicotome
villosa.
Рис. 198. Drosophyllum lusita-
nicum во влажных расселинах
песчаниковых скал на юге Испа-
нии (фото Э. Вальтер).
Рис. 199—201. Климадиаграммы областей с резко выраженным океаническим кли-
матом.
Биарриц — юго-западная Франция; Корк — западная Ирландия; Берген — Норвегия.
Рис. 202—204. Климадиаграммы областей с менее резко выраженным океаническим
климатом.
Флиссинген — Голландия; Бремен—северо-западная Германия; Эдинбург — восточная Шот-
ландия.
Лиственные леса умеренной зоны северного полушария — неморальная зона 245
Западносредиземноморские виды: Calycotome spinosa, Argyrolobium linnaeanum (= Су-
tisus argenteus), Cytisus sessiliflorus, Genista (Cytisus) purgans, G. scorpius, G. cinerea.
Средиземноморско-атлантические: Ulex parviflorus.
Иберийские элементы: представлены другими многочисленными видами вышеуказан-
ных родов; на Иберийском полуострове находится их генетический центр.
Иберийско-атлантические: Ulex nanus и U. gallii.
Эуатлантические: Ulex europaeus и Genista anglica.
Субатлантические, доходящие до Центральной Европы: Sarothamnus scoparius. Genista
sagittalis. Genista pilosa.
Центральноевропейские: Genista germanica.
Евразиатские, не связанные с близостью океана: Genista tinctoria.
К северу роль видов дроков уменьшается, зато увеличивается значе-
ние видов вересковых. Какими причинами обусловлена столь различная
форма ареалов близкородственных видов? В направлении к средиземно-
морской области ограничивающим фактором является засушливое лето,
в направлении к северу и особенно к востоку — холодная зима, посколь-
ку не только Ulex europaeus, но и Sarothamnus в этих условиях часто
обмерзают. Хотя эти виды вновь возобновляются корневыми отпрысками,
периодическая потеря надземных частей сильно снижает их конкурентную
мощность по сравнению с морозостойкими видами. Даже в Центральной
Европе с ее небольшими морозами и малым количеством снега возможны
повреждения Calluna vulgaris холодами, что связано с высыханием
на морозе (см. стр. 327). Способность вереска переносить сильные морозы
в горах Центральной Европы и в Восточной Европе объясняется его
защищенностью в течение зимы достаточно мощным снежным покровом.
Для большинства вечнозеленых атлантических и атлантическо-
средиземноморских видов в приатлантических областях менее важное
значение имеет влажность лета и гораздо большую роль играет мягкость
зимы. Это хорошо прослеживается в Новой Зеландии, где Sarothamnus
scoparius и Ulex europaeus широко распространены как сорняки на
пастбищах.
Зимы в Новой Зеландии мягкие. Но в районе Отаго на острове Южном
влажность быстро уменьшается по направлению в глубь суши и корен-
ные леса уступают место сухим злаковпикам. На пастбищах, ранее покры-
тых лесом, распространен папоротник-орляк (Pteridium aquilinum ssp.
esculentum); он вытесняется в области, переходной к злаковникам, Sarotha-
mnus. Дальше всего в засушливые райрны проникает Ulex. Характер
распространения этих трех видов в Новой Зеландии противоположен
тому, что наблюдается в Европе, где Ulex растет в прибрежной полосе
Атлантики, Sarothamnus является субатлантическим флористическим
элементом, a Pteridium проникает через Центральную Европу далеко
на восток. Такой характер распределения обусловлен, однако, не умень-
шением влажности, а усилением морозов по мере продвижения на восток.
Ulex часто обмерзает уже на западе Германии, Sarothamnus — в ее цен-
тральных частях, в то время как папоротник-орляк, почки возобновления
которого зимуют в почве, хорошо переносит даже восточноевропейскую
зимнюю стужу.
Океанический климат ярко выражен в Ирландии
Средняя годовая температура колеблется здесь между 9 и 10,5°, средняя темпе-
ратура января достигает 3,5—3,7°, а июля—около 14 — 16°, Разница в средних темпера-
турах самого теплого и самого холодного месяца нигде не превышает 8,5—10,5°, причем
средняя дневная амплитуда в течение всего года составляет приблизительно 6,5°,
1 Die Pflanzenwelt Irlands (red, von, W. Liidi), «Veroff, Geobot, Inst. Riibel»,
H. 25, Zurich, 1952.
246
Глава 4
Иногда здесь наблюдаются слабые морозы. Температура опускается до —10°
только раз в 4—5 лет. Снег лежит на западе острова в среднем около трех-четырех дней
в году, а на востоке — около восьми-десяти дней. Высокая температура (до 25°) сохра-
няется не больше двух-трех дней.
Годовое количество осадков равно 750—1000 мм (в горах до 1500 мм) и распреде-
ляется в течение года очень равномерно. Даже в самые сухие месяцы (апреле — мае)
выпадает свыше 45 мм осадков в виде моросящих затяжных дождей. Среднее годовое
количество осадков отклоняется от нормы не более чем на 25%. С этим связана посто-
янно наблюдаемая высокая влажность воздуха; дефицит насыщения не превышает 2,9 л* б.
При большой облачности продолжительность солнечного сияния снижается до 31%
максимально возможной.
Ровный, без резких колебаний климат Ирландии способствует произ-
растанию различных геоэлементов. Поэтому не удивительно, что на
острове, как уже было упомянуто, встречаются не только средиземномор-
ские элементы, но в наиболее низких местоположениях даже арктические:
Dry as octopetala, Saxifraga oppositifolia, Selaginella selaginoides, Salix
herbacea (на скалах северной экспозиции), Rubus chamaemorus (одно место-
нахождение на высоте 550 м над уровнем моря на склоне северной [экспо-
зиции).
Однако в Ирландии растут также виды влажных субтропиков, как,
например, папоротники Adiantum capillus-veneris, Hymenophyllum tunbrid-
gense, H. peltatum и Osmunda regalis.
Во влажном климате распространение растений в большей степени
зависит от кислотности почв. Многие базифильные виды развиваются
только на почвах, содержащих известь, в том числе и виды, которые
в Центральной Европе отнюдь не приурочены к известнякам.
С другой стороны, благодаря высокой постоянной влажности в запад-
ной части острова ацидофильные виды растут на известняках, даже если
они не изолированы от горной породы слоем кислого торфа.
Верещатники приатлантических областей не являются естественной
растительностью. Как и существование гарриги Средиземноморья, суще-
ствование верещатников обусловлено антропогенными причинами; они
развиваются на площадях, ранее занятых лесом Ч В наше время эти леса
полностью уничтожены, поэтому трудно восстановить их прежний состав.
Древесный ярус слагался в первую очередь из летнезеленых видов дуба:
в субсредиземноморском районе это Quercus pubescens, и Qu. pyrenaica
(toza), севернее — Quercus petraea и Qu. robur, еще севернее возрастает
роль березы (Betula verrucosa, В. pubescens), рябины (Sorbus aucuparia)
и сосны (Pinus silvestris). Последняя не встречается в прибрежной области
с резко выраженным океаническим климатом, что, возможно, связано
с повреждениями, которые наносит сосне солевая пыль. Вероятно, в суб-
средиземноморской области на песчаных почвах Pinus pinaster (= maritima)
всегда играла большую роль, так же как это наблюдается в настоящее
время на юго-западе Франции (Ланды). Атлантическо-средиземноморский
вид Ilex aquifolium часто встречался в этих лесах и доходил вплоть до Нор-
вегии. На бедных известью горных породах растения верещатников,
очевидно, росли в составе подлеска, поэтому они смогли широко рас-
пространиться после уничтожения древесного яруса. Верещатники как
самостоятельная форма растительности первоначально занимали очень
небольшие пространства, преимущественно маломощные или сырые торфя-
нистые почвы, особенно широко в горах выше границы леса.
1Ph. Duchaufour, Recherches ecologiques sur la chenaie atlantique fran-
$aise, «Ann. de Г Ёсо1е Nation, des Eaux et Forets», 11. Fasc. 1, Nancy, 1948.
Лиственные леса умеренной зоны северного полушария — неморалъная зона 247
Совсем иной была лесная растительность на известняковых почвах,
более богатых питательными веществами. Здесь могли расти ясень {Fraxi-
nus excelsior), вяз (Ulmus glabra), ольха {Alnus glutinosa) и лещина
{Corylus avellana). Бук. {Fagus silvatica) и тисс {Taxus baccata) проника-
ют по известнякам далеко в приатлантическую область, несмотря на то
что подстилка бука способствует образованию грубого гумуса. Бук не
достиг Ирландии в послеледниковый период; в настоящее время после
интродукции он хорошо растет на горных породах, богатых известью.
В Англии, только на юго-востоке, бук является туземным растением.
На плодородных почвах леса уничтожены и превращены либо в луга,
интенсивно используемые для выпаса скота, либо в пахотные земли.
В приокеанической области причины уничтожения леса были
следующие: 1) давнее заселение этой территории человеком и 2) бед-
ность почв, которые в условиях постоянно влажного климата сильно
выщелачиваются и обладают тенденцией к образованию Грубого гумуса.
В данном случае здесь происходит то же, что наблюдается и во влажном
тропическом дождевом лесу (ср. т. I, стр. 94 и след.). Ненарушенный дев-
ственный лес, где в процессе круговорота веществ потери питательных
веществ не происходит, может существовать вечно. Однако в результате
вмешательства человека такой лес очень легко деградирует.
В малонаселенных областях с типичным морским климатом еще
до сегодняшнего дня сохранились мощные лесные массивы, например
леса, образованные Nothofagus, в западной части острова Южный Новой
Зеландии или в южном Чили. Можно также упомянуть хвойные леса,
развитые на тихоокеанском побережье Северной Калифорнии и Британ-
ской Колумбии. Напротив, в приатлантической области лиственных лесов
Западной Европы ненарушенные леса не сохранились.
Одна лишь вырубка леса не могла привести к его полному уничтоже-
нию. Причинами явились также выпас, проводившийся здесь издавна,
применение регулярных палов для улучшения пастбищ, а после введения
стойлового содержания скота — использование дернины в качестве под-
стилки в скотных дворах. Грубый гумус после высушивания часто сжигали
на пашнях с целью удобрить почву. Все эти меры препятствовали возоб-
новлению леса и во влажном климате способствовали большим потерям
питательных веществ и увеличению кислотности почвы. В этих условиях
могли сохраниться только малотребовательные к элементам минерального
питания растения верещатников; к тому же при застое воды почва забола-
чивалась. Что формирование верещатников началось под воздействием
человека еще в доисторическое время, убедительно доказано при помощи
пыльцевого анализа торфяников. Иверсен приводит поучительный пример
из южной Ютландии \
В почвенном разрезе под облесенным в настоящее время верещатником на глуби-
не около 1 м обнаружен оторфованный слой грубого гумуса. Судя по данным пыльце-
вого спектра нижней части профиля, здесь первоначально был дубовый лес с липой,
ольхой и лещиной, а также с примесью Ilex aquifolium, Malus silvestris и пионерных
пород Betula и Frangula. В то время были развиты типичные приатлантические листвен-
ные леса с бедным напочвенным покровом {Anemone, Pteridium, Dryopteris и Polypodium);
пыльца злаков почти отсутствовала. Картина резко меняется на глубине около 30 см:
пыльца дуба почти исчезает, а количество пыльцы Calluna, которая до этого почти
не встречалась, увеличивается и достигает 50% . Датировка по С14 свидетельствует, что
смена ландшафтов произошла в начале нашей эры (740 + 100). В этом горизонте обнару-
жена также прослойка древесного угля, пыльца зерновых культур, гречихи и сорняков.
Все это говорит о воздействии человека, который выжег леса и превратил освободив-
1 J. Iversen, Retrogressive vegetational succession in the postglacial,
«J. Ecol.», 52, (Suppl. 1), 1964, p. 59-70.
248
Глава 4
шуюся площадь в пастбища, что подтверждается наличием пыльцы Trifolium repens,
являющегося пастбищным индикатором. Более детальный анализ прослойки древесно-
го угля приводит к выводу, что сразу после пожара распространились папоротник-ор-
ляк и иван-чай (Chamaenerion angustifolium). Только затем наблюдается резкое увеличе-
ние количества пыльцы Calluna.
На других участках современных верещатников оторфованный слой грубого
гумуса был удален и использован на топливо. В результате почва и лес потеряли запас
питательных веществ. На выщелоченных песках, расположенных под слоем грубого
гумуса, лес не восстанавливается.
На основании пыльцевой диаграммы можно судить, что воздействие человека
на природу в приатлантических областях началось уже в каменном веке.
В областях, климат которых носит черты резкого воздействия океана,
возникновение верещатников способствует быстрому подъему уровня
грунтовых вод и образованию «покровных» болот. До сих пор не выяснен
Рис. 205. Водный баланс елового леса под Ленин-
градом (по Кощееву, 1955).
Левая колонка — осадки, правая — эвапотранспирация древос-
тоя (штриховка) и других растений (клетки): 1 — 100-летний
древостой (сомкнутость крон 0,8); 2 — вырубка с Vaccinium
myrtillus', 3 — более старая вырубка с Polytrichum commune
(начало заболачивания); 4 — еще более старая вырубка с по-
кровом из Sphagnum и молодым подростом; 5 — молодняк
5—8-летнего возраста из ели и пионерных пород; 6 — молодой
древостой ели, возраст 12—15 лет (сомкнутость крон 0,9 —1,0);
7 — древостой в возрасте 25—30 лет; 8 — древостой в возрасте
50 лет; 9 — спелый древостой в возрасте 70—80 лет; 10 — дре-
востой в возрасте 85—90 лет.
вопрос, что расходует больше воды: лес, верещатник или болото. На Меж-
дународном ботаническом конгрессе в Эдинбурге (1964) Пенман высказал
точку зрения, что испаряемость с поверхности постоянно увлажненной
почвы не зависит от характера растительного покрова. Количество испа-
рившейся влаги в этом случае соответствует притоку энергии, который
необходим для перевода жидкой воды в состояние пара. Наряду с сол-
нечными лучами энергию растительному покрову поставляет и ветер,
если он теплее, чем транспирирующие органы; в противном случае ветер
уносит тепловую энергию. Наконец, энергия теряется вследствие турбу-
лентности воздуха и утечки тепла в почву. Оба процесса усиливаются под
влиянием нагрева приземного слоя воздуха и поверхности почвы. Пра-
вильная оценка этих факторов, помимо которых имеются и другие, менее
значимые, довольно сложна, особенно при сравнении леса, верещатника
и болота. Исследования, проведенные в зоне очень влажных хвойных
лесов, опровергают предположение, что испаряемость леса при постоян-
ной хорошей влагообеспеченности не больше величины испаряемости
низкорослого растительного покрова.
Лиственные леса умеренной зоны северного полушария — неморалъная зона 249*
Рис. 205 составлен по материалам Кощеева, из которых видно, что-
эвапотранспирация после вырубки леса достигает только около 25%
количества выпавших осадков х. Со временем это приводит к заболачива-
нию и распространению Poly trichum commune и Sphagnum. С ростом молод-
няка возрастает и величина эвапотранспирации, и, когда лес достигнет
возраста 25 лет, она опять будет соответствовать количеству выпавших
осадков. Если же развитию молодняка воспрепятствует вмешательство
человека, как это случилось в приокеанических областях, где распро-
странены верещатники, то это приведет к образованию болот на всех
плохо дренированных почвах. Поэтому в океанических областях развиты)
омброгенные «покровные» болота 1 2, которые на породах, бедных известью,
вне зависимости от рельефа, как одеяло, покрывают обширные простран-
ства. Они особенно типичны в западной Ирландии и Шотландии. Тем
не менее покровные болота нельзя рассматривать в качестве зональной
растительности, расположенной ниже границы леса, поскольку толчок
их образованию дал человек, уничтожив леса и препятствуя их возобно-
влению. Наличие в болотах под слоем торфа остатков древесины свиде-
тельствует о том, что здесь раньше рос лес.
Можно, конечно, считать, что заболачивание является лишь есте-
ственным следствием ухудшения климата, связанного с увеличением,
влажности.
Но это предположение опровергается тем фактом, что на других кон-
тинентах, которые, разумеется, резко отличаются от Европы своим фло
ристическим составом, лес растет и в более влажных условиях, например
в южном полушарии (табл. 20).
Исключением не является и западная Тасмания, где встречаются
болота с Mesomelaena (ср. стр. 207), поскольку известно, что эти болота
раньше постоянно выжигались коренными жителями, а в настоящее
время охотниками за валлаби.
В океанической области наблюдаются постепенные переходы между
сухими верещатниками на хорошо дренированных кислых минеральных
почвах (например, песках), верещатниковыми болотами с начальной
стадией образования торфа и покровными болотами. Чем влажнее
климат, тем заметнее преобладание заболоченных пространств.
При длительном использовании верещатников, возникших на месте
лесов, под выпас происходит отбор растений, и в составе верещатников
начинают преобладать колючие или несъедобные виды: Ulex, Sarothamnus,
Juniperus и т. д. Попытки уничтожить эти растения огнем в большинстве'
случаев не приносят успеха.
Большое влияние на растительность оказывает кролик, которого
ввезли из Франции в Англию и Ирландию; он быстро распространился
и лишь в последние годы был почти полностью уничтожен миксоматозом.
Вполне возможно, что численность кролика со временем опять увеличится.
1 Известно, что содержание влаги в почвенном профиле под лесом всегда меньше,,
чем на открытом месте: в буковом лесу — 68%, в березовом лесу — 74%, в сухое время
года соответственно 50 или 65%. Ср.: V. Т. Aaltonen, Boden und Wald, Berlin,
1948, S. 83—95 (в работе упоминаются опыты Высоцкого в лесостепи п приводятся дан-
ные по величине стока, полученные Бургером в Швейцарии); A. J. Rutter, Die
Wasserverdunstung der Walder, «Endeavour», 26, 1967, S. 39—43. Автор приводит сле-
дующие сведения (Слайбурн, Йоркшир, 1956—1965): лесные культуры (Picea sitchen-
sis) — 801 мм, путь — 456 мм (при орошении) и 416 мм (без орошения).
2 Освальд (Oswald, 1925) назвал эти болота «верховыми болотами, покрывающими
местность» (terrainbedeckendes Hochmoor); английские авторы называют их «одеяль-
ными болотами» (blanket-bog). — Прим. ред.
250
Глава 4
Таблица 20
Сравнительная характеристика климата территорий
с покровными болотами и областей распространения
леса
Раститель- ность Местность Сред- няя годо- вая темпе- рату- ра, °C Годовое количе- ство осадков, мм Средний суточный минимум самого холодного месяца, °C Абсолют- ный ми- нимум темпера- туры, °C
Районы рас- простране- ния по- Блэксод- Пойнт (зап. Ирландия) 9,9 1321 3,9 -6,6
кровных болот Тайри (зап. Шотландия) 8,5 1204 3,4 -6,7
Леса из Nothofagus Мильфорд (Новая Зе- ландия) 9,9 6337 1,1 -5,0
Зеехан (зап. Тасмания) 10,9 2470 3,5 —6,5
Пуэрто-Ай- зен (южное Чили) 8,9 3018 2,2 -7,2
Кобо-Рапер (южное Чи- ли) 8,6 2210 3,3 -2,2
Кролик не только препятствует возобновлению леса и развитию расти-
тельности верещатников, он благоприятствует распространению злаков:
Festuca ovina, Nardus stricta, Sieglingia decumbens, Deschampsia flexuosa x.
Растительность верещатников Шотландии
Верещатники Шотландии хорошо описаны в работе, вышедшей
в последние годы 1 2. Уничтожение леса в Шотландии началось еще в неоли-
те, поэтому в настоящее время здесь не сохранилось естественных лес-
ных насаждений.
В наиболее теплых местообитаниях растут два вида дуба (Quercus
petraea и Qu. robur) и UImus glabra, а на известняках встречается и ясень
(Fraxinus excelsior). Береза (Betula pubescens) распространена всюду.
В горах совместно с березой произрастает особая форма Pinus silvestris.
Кроме этих видов, древесные породы представлены Prunus avium, Р. padus, Malus
silvestris, Corylus avellana, Crataegus monogyna, Prunus spinosa, Viburnum opulus, Sorbus
aucuparia, Populus tremula, а кустарники — различными видами Salix, Juniperus com-
munis.
Травяной покров этих насаждений образует переходы между богатыми разно-
травьем лесами на хороших почвах с Mercurialis perennis, Allium ursinum, Oxalis aceto-
sella, Anemone nemorosa, Primula vulgaris, Endymion (Scilla) non-scriptus и лесами, раз-
витыми на бедных почвах, с Calluna, Vaccinium, Lycopodium annotinum и т. д.
1 A. G. Tansley, The British Islands and their vegetation (2 vols.), Cambrid-
ge, 1949.
2 The Vegetation of Scotland (ed. J. H. Burnett), Edinburgh and London, 1964.
Лиственные леса умеренной зоны северного полушария — неморалъная зона 251
Рис. 206. Ареал кустарничковых верещат-
ников с Calluna в Шотландии (по Вайтлес).
Большие площади, покрытые вереском, обозначены
пунктиром, маленькие — залиты черным цветом.
Calluna vulgaris — основной вид шотландских верещатников. Он
является «национальным цветком» этой страны. О его широком распро-
странении в Шотландии можно судить по рис. 206. Такие верещатники
встречаются на площади от северной Шотландии до Норвегии. Во многих
областях они исчезают, поскольку используются под посадки сосны.
Напротив, в Шотландии они сохранились, так как в течение всего года
на них выпасаются овцы и крупный рогатый скот или же опи служат
охотничьими угодьями. Только
на более i богатых почвах низ-
менностей ведется земледелие.
Верещатники развиваются на
самых бедных почвах. Типич-
ная почва — железистый подзол
с более или менее мощным
горизонтом грубого гумуса (Ао),
серым горизонтом AiT образо-
ванным гумусом с обесцвечен-
ными кварцевыми зернами; не
всегда выраженным оподзолен-
ным горизонтом А2 и желези-
сто-ортштейновым горизонтом
В. Там, где ортштейновый гори-
зонт плотный и мощный, вода не
просачивается в глубь почвы,
что приводит к образованию
глея или даже торфа. В запад-
ных областях, где выпадает
наибольшее количество осадков,
верещатники вытесняются по-
кровными болотами и сохра-
няются лишь на крутых скло-
нах.
В Шотландии почти все
верещатники представляют со-
бой производные сообщества,
возникшие на месте уничтожен- ।
ных лесов. В естественных условиях верещатники встречаются только
на маломощных почвах, не благоприятных для развития деревьев,
и в первую очередь в горах выше границы распространения леса. На
не подверженных обжигу в течение длительного времени верещатниках
появляется подрост березы, рябины (Sorbus aucuparia) и сосны. Верещат-
ники выжигают для стимулирования роста молодых побегов вереска
и для улучшения качества пастбища.
Наряду с Calluna в состав верещатников Шотландии входят следующие виды,
имеющие существенное значение: Erica cinerea, Е. tetralix, Vaccinium myrtillus, V. vitis-
idaea, Empetrum nigrum, Arctostaphylos uva-ursi, а также Deschampsia jlexuosa, Molinia
coerulea, Nardus stricta, Trichophorum caespitosum, виды Eriophorum, Carex binervis,
C. pilulifera и др. Juniperus communis и Ulex europaeus — типичные представители
верещатников, возвышающиеся над Calluna. Заросли Calluna в возрасте 5—8 лет
образуют сомкнутый ярус высотой 40 см (до 60—80 см). Сопровождающие виды форми-
руют второй ярус высотой 10—20 см. Третий ярус высотой 5—10 см образуют Potentilla
erecta и Galium saxatile, но в основном мхи Pleurozium schreberi и Hylocomium splendens.
Поверхность почвы покрывают невысокие мхи: Hypnum cupressiforme, Pohlia nutans
и различные виды ’лишайников.
252
Глава 4
Сквозь ярус, образованный Calluna, проникает примерно 20% днев-
ного света; у поверхности почвы освещенность часто составляет лишь 0,5%.
Кусты Calluna сильно тормозят движение воздуха, поэтому в их зарос-
лях в большинстве случаев господствуют почти полный штиль и высокая
влажность воздуха. При освещении солнцем температура в ярусе Calluna
на 2—5° превышает температуру почвы.
При рациональном выжигании возникают очень густые заросли
Calluna, в которых лишь изредка встречаются мхи или лишайники.
В верхних 10 см почвенного профиля в таких зарослях формируется
плотный корневой войлок, образуемый корнями стелющихся побегов.
Множество одиночных корней проникает до глубины 75 см, образуя
густое сплетение непосредственно над ортштейповым горизонтом. На
песчаных почвах корни в большинстве случаев распределяются более
или менее равномерно до глубины 80 см, в то время как у одиночно расту-
щей Calluna на сырой болотной почве рост корней в глубину ограничи-
вается верхними 5—10 см (до 20 см)', в горизонтальном направлении корни
распространяются на расстояние до 60 см. По сравнению с вереском
корни собственно болотных растений (Eriophorum vaginatuiri) проникают
в таких местах гораздо глубже (до 50 см).
Ежегодный опад Calluna составляет в среднем 421 кг/га. Опад происхо-
дит осенью; опадают не листья, а укороченные побеги. В результате зимойг
когда растения страдают от морозного высыхания, поверхность листьев
редуцируется. В почву поступают не все продукты разложения подстилки;
в первый год в результате разложения теряется от 1,/10 Д° 1/го сухого веще-
ства. Средний запас подстилки достигает 7500 кг!га сухой массы, что-
примерно в 17 раз больше годовой продукции
Флористический состав верещатников различен. Он зависит: 1) от вре-
мени, которое прошло с момента превращения леса в верещатник, 2) от
характера и степени интенсивности выпаса и 3) от частоты пожаров.
Оптимального развития Calluna достигает при 8—15-летних интервалах
между пожарами. При частом выжигании (каждые 6 лет) Calluna плохо
возобновляется генеративным путем; в этом случае в зависимости от сте-
пени влажности местообитания распространяются другие виды: Pteridtum
aquilinum, Vaccinium myrtillus, Nardus stricta, Molinia coerulea. Juncus-
squarrosus и т. д. Таким образом, верещатники являются производным как
факторов местообитания, так и воздействия человека. Эту двойственность
необходимо учитывать при анализе их флористического состава. Различия
в климате также оказывают влияние на формирование верещатников.
В более резко выраженных океанических условиях верещатники характеризуют-
ся присутствием Erica cinerea. Кроме того, здесь распространены Carex binervis, Hype-
ricum pulchrum, Blechnum spicant, Listera cordata, Dactylorchis maculata ssp. ericetorum,.
Rhacomitrium lanuginosum, Plagiothecium undulatum. В северных вариантах верещат-
ников обычны Trientalis europaea, Empetrum nigrum, Vaccinium vitis-idaea n Hylocomium
splendens, a Ulex minor и Cuscuta epithymum здесь отсутствуют, в то же время еще встре-
чается Genista anglica — единственный более южный вид.
Четкий экологический ряд связан с возрастающей влажностью почв
(от подзолистых к глеевым и торфяным почвам) (табл. 21).
Сырые верещатниковые болота в западной Шотландии сменяются
типичными покровными верховыми болотами. Они расположены в мест-
ностях с высотой менее 450 м над уровнем моря, в них доминируют виды
Sphagnum, образующие сплошной покров. Среди мхов встречаются еди-
1 Е. Cormack, С. Н. Gimingham, Litter production by Calluna vulga-
ris (L.), «Hull. J. Ecol.», 52, 1964, p. 285-297.
Лиственные леса умеренной зоны северного полушария — неморальная зона 253
Таблица 21
Изменение флористического состава верещатников
при увеличении влажности почвы
Увеличение влажности
05 u Ф Erica cinerea Calluna vul- Erica tetra- Erica tetra-
g garis lix lix
S Calluna vul- Erica tetra- Molinia coe- Tricho-
Ф garis lix rulea phorum
ф caespito-
E-i sum
U Deschampsia Juncus Nardus stric- Eriophorum
jlexuosa squarrosus ta vagina-
Ф turn
Vaccinium Calluna Myrica gale
Ф И myrtillus vulgaris
о N arthecium Carex echi-
ф tr ossifra- nata
Е Ч gum
О К \
ничные экземпляры Trichophorum caespitosum, Eriophorum angustijolium,
E. vagmatum, Calluna vulgaris, Erica te^tralix, Molinia coerulea, Drosera
rotundijolia, D. anglica, Narthecium ossijragum, Myrica gale и т, д, В моча-
жинах растут Menyanthes trifoliata, Untricularia minor, Carex limosa,
иногда Rhynchospora alba и R. fusca.
Болота этого типа образуются только в областях с количеством дней
с осадками более 235 в год. Самое крупное покровное болото Шотландии
занимает площадь 2500 км1 2 *. Такие болота встречаются в Озерном округе
Англии, в Северном Уэльсе и Дартморе и особенно часто в западной
Ирландии, где Schoenus nigricans играет значительно большую роль,
чем в Шотландии,
Мох Rhacomitrium lanuginosum s,l, покрывает в Северном Уэльсе
большие пространства, чем Sphagnum Г Этот вид господствует на при-
атлантических покровных болотах, а также на скалах в горах с резко
выраженным океаническим климатом, например на вулканических горных
породах вершины Эгмонт (Новая Зеландия) выше границы леса.
Годовой прирост этого мха не превышает 5—15 мм, поэтому он встре-
чается только в условиях, где отсутствует конкуренция со стороны более
быстрорастущих видов, Rhacomitrium впитывает влагу по капиллярам,
расположенным с внешней стороны побега; там, где влажность воздуха
мала, он быстро высыхает и поэтому хорошо развивается только в постоян-
но влажном климате. Оптимальная величина нетто-ассимиляции наблю-
дается при содержании воды, равном 300—400%, но очень сырых место-
обитаний мох избегает. Оптимальная температура нетто-ассимиляции
(определенной по СО*4) равна 13—15°, При температуре ниже 10° или выше
20° интенсивность фотосинтеза незначительна. Эти особенности обусловли-
вают нормальный период роста мха на Британских островах: 1) интенсивный
рост в длину с марта до мая, 2) формирование боковых побегов и развитие
1 J.H, Tallis, A study of the biology and ecology of Rhacomitrium lanuginosum
Brid, Ph, D. Thesis, University College of North Wales, 1957; Growth, reproduction, and
physiology, «J, Ecol.», 47, 1959, p. 325—350; Growth studies on Rhacomitrium lanugino-
sum, «The Bryologist», 67, 1964, p. 417—422,
254
Глава 4
органов размножения с июня до августа, 3) вторичный рост в длину с конца
августа до сентября, 4) созревание спорогониев в течение сентября и начале
октября. Оптимальный ход этого процесса может быть нарушен аномаль-
ными погодными условиями, В Ьухое время года наступает период покоя.
Уже при влажности 95% мох высыхает в течение 2—3 дней, Прн содер-
жании воды, равном 100%, листочки начинают свертываться.
Слабый выпас благоприятствует Calluna и ограничивает распростра-
нение Erica cinerea. При более интенсивном выпасе разрастаются злаки
(Calluna плохо выносит вытаптывание): на более богатых почвах —
Festuca ovina, на очень бедных — Deschampsia flexuosa, а на влажных
почвах — Nardus и Molinia, Мы не приводим характеристику отдельных
ассоциаций. Отметим лишь, что в каждом сообществе происходят цикли-
ческие изменения, обусловленные старением или отмиранием Calluna,
Вереск, если его рост не нарушается пожарами или стравливанием скотомг
достигает своего полного развития в возрасте 15—20 лет, В возрасте
25 лет отдельные побеги начинают отмирать, а число побегов в центральной
части куста снижается, В таких местах вереска поселяются Cladonia
silvatica и другие лишайники. После разложения отмерших побегов и обра-
зования в центральной части куста свободного пространства, на нем
разрастается Arctostaphylos uva-ursi, В подушку, сформированную побе-
гами Arctostaphylos, в дальнейшем проникают молодые растения Calluna,
которые затеняют толокнянку и вызывают со временем ее гибель,
В верещатниках с развитым моховым покровом свободные простран-
ства заполняются Pleurozium schreberi и Hylocomium splendens', среди мхов
растут Vaccinium vitis-idaea и У, myrtillus, пока в конечном итоге оба вида
не вытесняются молодыми растениями Calluna, Такие циклы приво-
дят в старых сообществах верещатников к формированию мозаичной
структуры х.
Охота и экстенсивный выпас овец не являются рациональным способом
использования огромных пространств, занятых верещатниками. Счи-
тается, что одной овце требуется приблизительно 1,2—2,8 га пастбищ.
Продолжительное использование и периодическое выжигание верещат-
ников должно в будущем привести к более сильному оподзоливанию
и обеднению почв. Поэтому прилагаются усилия к повышению эффек-
тивности вересковых пастбищ путем применения удобрений и замены
Calluna злаками; второй путь связан с посадками леса на верещатниках.
Для этой цели используются лиственница, Picea sitchensis и некоторые
другие виды деревьев.
Аналогичные кустарничковые верещатники распространены также
в северной Шотландии выше границы леса на высоте 400—1000 м над
уровнем моря и на центральном плоскогорье на высоте 800—1200 м,
В нижней части этого вертикального пояса Calluna встречается еще
довольно часто, выше доминируют Vaccinium myrtillus и Empetrum her-
maphroditum. Кроме этих видов, здесь растут стелющийся можжевельник
(Juniperus communis ssp. папа), альпийские кустарнички (Arctous alpina,
Loiseleuria procumbens и др.) и альпийские лишайники, а среди мхов
обширные площади покрывает Rhacomitrium lanuginosum. Расчленение
на сообщества, которое в значительной степени зависит от срока появления
и схода снежного покрова, будет более подробно освещено при характе-
ристике арктической зоны или альпийского пояса.
1 A. S. W a 11, Pattern and Process in plant community, «J. Ecol.», 35, 1947,.
p. 1—22.
Лиственные_леса умеренной зоны северного полушария — неморальная зона 255
Верещатники Центральной Европы
Приатлантические верещатники распространены вдоль западных
берегов Европейского континента и простираются от Нидерландов
до западной части Северогерманской низменности, захватывая также
территорию Центральной Европы х. Узкой полосой верещатники тянутся
от Шлезвиг-Гольштейна вдоль побережья Балтийского моря до устья
реки Неман. Особенно широко известны Люнебургские верещатники,
занимавшие наибольшую площадь примерно в 1800 г. (рис. 207). Эти вере-
щатники также являются вторичным образованием. Они возникли на месте
лесов и сохранились лишь благодаря вырубке леса, выпасу овец, пожарам
и сдиранию дернины. При использовании дернины побеги и корни вереска
подрезаются в форме четырехугольников, снимаются мотыгой вместе
с верхним 10-сантиметровым слоем грубого гумуса и используются в скот-
ных дворах в качестве подстилки. Из оставшихся в почве корней Calluna
vulgaris образуются придаточные побеги, и таким образом через 10—12 лет
восстанавливается однородный, сомкнутый верещатник. В настоящее
время дерн снимается в основном во влажных верещатниках, сформиро-
ванных Erica. Лес на верещатниках возобновляется или естественно —
ветер заносит семена сосны и березы (рис.'|208),—-или при помощи
Рис. 207. Люнебургские можжевеловые (Juniperus communis) верещатники. На
переднем плане — обкусанные скотом кустарники.
1 Р. Gra.ebner, Die Heide Norddeutschlands und die sich anschlieBenden Foi-
mationen in biologischer Betrachtung. Cm.: Engler-Drude, «Die Vegetation der Erde»,
5, 1901. В работе описывается состояние верещатников в начале нашего столетия. Сов-
ременные вересковые пустоши кратко охарактеризованы Элленбергом (См.: «Phytolo-
gie», Bd. IV, 2, S. 641—666); P. H e i n e m a n n, Les landes a Calluna du district Picar-
do-Braban^on de Belgique, «Vegetatio», 7, 1956/57, p. 99—147; C. Van den Ber-
gh e n, Etude sur la vegetation des dunes et des landes de la Bretagne, «Vegetatio»,
8,1958/59,p. 193—208; R. Schubert, Die zwergstrauchreichen azidiphilen Pflan-
zen gesellschaften Mitteldeutschlands, «Pflanzensoziologie», Bd. 11, Jena, 1960.
•256
Глава 4
Рис. 208. Верещат-
ник из Calluna с еди-
ничной сосной (Pinus
silvestris), способст-
вующей облесению
верещатников.
посадок сосны. Только в заповеднике у Вильседерберга, где для сохране-
ния экстенсивно используемого культурного ландшафта производится
выпас овец, уцелели обширные пространства верещатников.
На лесных полянах в сухих сосновых лесах западной части бореаль-
ной области встречаются еще участки верещатников; однако лес в этих
-•сосняках растет настолько интенсивно, что подавляет развитие вереска,
который поэтому растет здесь преимущественно в составе подлеска. Вере-
щатники распространены также высоко в горах западной части Централь-
ной Европы (Арденны, Эйфель, Зауэрланд, Вогезы, Шварцвальд). Здесь
проходит граница распространения формации верещатников х. Там, где
^еще развито овцеводство (оно уже почти исчезло), Calluna служит основ-
ным кормом для овец; предпочитающий же более влажные местообитания
атлантический вид Erica tetralix не поедается скотом, что благоприят-
ствует его распространению как пастбищного сорняка. Вытаптывание
отрицательно влияет на кустарнички, которые сменяются злаками
'(Festuca ovina, а в горах — Nardus strictd). Nardus, являющийся показате-
лем бедности почв, не реагирует положительно на удобрение пастбищ
-фосфором и известью. В этом случае он заглушается более быстро расту-
щими и требовательными к условиям среды видами.
Внесение удобрений (N, Р, S, К, Са и Mg) во влажный верещатник
(Molinia coerulea — Erica tetralix) Высокого Венна приводило через
*8 лет к полному исчезновению видов Sphagnum. Изменялись количе-
ственные соотношения видов: типичные для верещатников виды заглуша-
лись злаками и осоками, положительно реагировавшими на удобрения.
Избыток N благоприятно влиял на молинию; однако этот вид не реагировал
на излишек фосфора. Существование верещатника как растительного
'•сообщества прежде всего обусловлено недостатком азота 1 2.
Все кустарники, распространенные в верещатниках, характеризуются
выровненным водным балансом в летние месяцы (низкими величинами
1 Различные гипы верещатников занимают большие площади в Альпах, где они
также являются производными растительными сообществами (ср.: E.Aichinger,
-«Angew. Pflanzensoz.», Н. XII — XIV, Wien, 1956—1957).
2 N. Sougnez, Reactions floristiques d’une lande humide aux tum^res mine-
rales, «Oecologia Plantarum», 1, 1966, p. 219—234.
Лиственные леса умеренной зоны северного полушария — неморалъная зона 257
и незначительными суточными колебаниями осмотического давления).
Почва верещатников достаточно влажная, соотношение побегов и корней
относительно небольшое. Эколого-физиологические исследования были
посвящены в основном изучению транспирации растений верещатни-
ков х. Лист Calluna vulgaris имеет относительно толстую кутикулу,
а устьица расположены в опушенных волосками углублениях. Мезофилл
отличается очень рыхлой структурой. При хорошем водоснабжении
наблюдается довольно интенсивная транспирация, особенно если ее
перевести на единицу поверхности почвы, покрытую растительностью.
При недостатке влаги величина транспирации может быть сильно снижена.
Поэтому Calluna в состоянии расти даже на побережье Средиземного моря
(Лазурный берег). В табл. 22 представлены некоторые показатели,
полученные Лемэ в Эльзасе.
Таблица 22
Интенсивность транспирации (в г/г сырого веса-час)
Название растений Открытые местообитания Затенен- ные место- обитания
сухие влажные
Calluna vulgaris 0,4—4,1 1,1—7,3 2,3-4,5 1,0—1,5
Genista pilosa ... — 1,8-3,7
Teucrium scorodonia . . 3,8—7,3 — 1,8-4,0
Pinus silvestris 1,7—3,9 — —
Oxalis acetosella .... — — 0,9—1,3
Критическим периодом для вечнозеленых видов верещатников
являются зимы, когда температура падает ниже 0° и почва замерзает.
К востоку зимы становятся все более суровыми. Относительно ксеро-
морфные листья Calluna теряют влагу даже зимой, особенно интенсивно
в конце зимы, когда температура воздуха начинает повышаться, а почва
все еще остается холодной. В это время снежный покров, защищающий
растения, уже отсутствует и утратившие закалку побеги вереска ночью
легко замерзают, а днем страдают от повреждений в результате физиологи-
ческой сухости, о чем можно судить по повышению осмотического давле-
ния 1 2. Чувствительные к морозам виды (Erica vagans и Ulex europaeus)
встречаются только в приатлантических областях. Myrica gale меньше
других повреждается заморозками, поскольку она сбрасывает листья
осенью. Calluna, распространенная повсюду в Центральной Европе, очень
часто вымерзает в южной Германии. Однако вереск хорошо переносит
длительный зимний период в западной бореальной области, охватывающей
территорию от Швеции до Северной Двины, что объясняется наличием
1 О.Stocker, Die Transpiration und Wasserokologie nordwestdeutscher Heide-
und Moorpflanzen am Standort, «Z. Bot.», 15, 1923, S. 1—41; G. Breitenspre-
c h e r, Vergleichende Transpirationsmessungen an Pflanzen der Hiddenseer Diinenheide,
«Mitt. Naturw. Ver. Neuvorpommern u. Rugen», 62, 1935, S. 5—92; G. L e m e e, Recher-
ches sur I’economie de 1’eau des sous-abrisseaux xeromorphes de landes, «Ann. Sci. Nat.,
Bot. Ser.», 11, 7, 1947, p. 53—85.
2 R. T h r e n, Jahreszeitliche Schwankungen des osmotischen Wertes verschiedener
okologischer Typen in der Umgebung von Heidelberg, «Z. Bot.», 26, 1934, S. 449—526;
D.Podlech, Untersuchungen zur Okologie atlantischer Pflanzen an ihrer Verbreitungs-
grenze, «Osterr. Bot. Zschr.», 107, 1960, S. 121—152.
17-01248
258
Глава 4
Рис. 209. Ареал Ericc. cinerea (по Бёхеру).
Знаком х обозначено местоположение за пределами ареала, вблизи Бонна и на Лигурийском
побережье.
здесь зимой постоянного, мощного снежного покрова, под которым Calluna
не страдает от последствий высыхания на морозе.
Лётчерт и Хорст провели в Люнебургских верещатниках подробные
микроклиматические наблюдения, исследования почвы (содержание влаги,
величина pH, выделение СО2) и измерение интенсивности транспирации х.
Для приатлантических верещатников характерна олиготрофность.
Мало требовательны к обеспечению питательными веществами виды как
сырых (Myrica) и влажных (Erica tetralix), так и сухих местообитаний.
Myrica растет на почвах с высоким уровнем грунтовых вод, избегает
верховых болот и вытесняется ольхой (Alnus) на почвах, богатых
питательными веществами. Подлех считает, что в Центральной Европе
ареал Ulex и Sarothamnus обусловлен климатом, а ареал Erica tetralix
и Genista anglica — скорее эдафическими факторами (оба вида — кальце-
фобы). Только опыты с исключением конкурентного влияния растений,
поставленные в различных почвенных условиях, могут помочь решению
этого вопроса. По мере удаления от атлантического побережья в глубь
1 W. Lotschert, Beitrage zur Okologie der subatlantischen Zwergstrauchhei-
de NW-Deutschlands, «1, 11 u. HI. Beitr. z. Biol. d. Pfl.», 37, 1962, S. 331—410; 41,
1965, S. 359—366. W. Lotschert, K. Horst, Zur Frage jahreszeitlicher pH-
Schwankungen. 11. Untersuchungen an Heide- und Waldstandorten, «Flora», 152, 1962,
S. 689—701; K. Horst, Klima und Bodenfaktoren in Zwergstrauch- und Waldgesell-
schaften des Naturschutzparks Liineburger Heide, «Natursch. u. Landschaftspfl. in Nie-
dersachen», H. 2, 1964.
Лиственные леса умеренной зоны северного полушария — неморальная зона 259
континента крайне олиготрофные виды и кислые почвы (pH = 4,2)
встречаются все реже и реже.
Ареал Erica cinerea совпадает с границей распространения верещат-
ников, приуроченных к приатлантическим областям (рис. 209). Этот вид
встречается от Португалии до западной Норвегии, а в Центральной
Европе доходит лишь до Бонна; единичное местонахождение этого вида
обнаружено на Лигурийском побережье. Ареал Erica cinerea ограничи-
вается на востоке январской изотермой +2°, а на юге — июльской изо-
термой + 22°. Это означает, что конкурентная способность Erica в течение
морозного зимнего периода и сильной летней засухи резко понижается,
в результате чего Erica cinerea вытесняется другими видами. Ареал Erica
tetralix заходит уже несколько дальше, в субатлантическую область.
Бёхер в работе, посвященной Фарерским островам, приводит харак-
теристику различных типов приатлантических верещатников Ч В ниже-
приведенной схеме дается перечень основных типов верёщатников:
Типы верещатников
(характеризуются господствующими видами)
СЕВЕР
Rhacomitrium
Empetrum — Calluna — Vacc. myrtillus— Hylocomium
Calluna — Erica 'cinerea
ЗАПАД (западная Норвегия, северная Шотландия) ВОСТОК
Calluna — Erica cinerea — Ulex
Calluna — Thymus (Англия, Франция) Calluna Calluna
-------> --------------------------> --------------->
Daboecia (Азор- Erica scoparia — Molinia Genista Antennaria
ские о-ва) -j-виды дроков anglica (Германия)
(юго-западная Франция, Ланды)-------------------------1
Erica vagans— Е. cinerea — Е. scoparia Средиземно-
Е. umbellata-\- многочисленные виды дроков----->морский маквис
(Португалия, северная Испания) (на почвах, не
содержащих
известь)
ЮГ
I
ЛЕС КАК ЭКОСИСТЕМА, ИЛИ БИОГЕОЦЕНОЗ
И ЛЕСНОЙ КЛИМАТ
Лес, как и вообще любое стабильное, длительно существующее сооб-
щество (ср. т. I, стр. 20 и след.), образует совместно со средой единство,
в котором круговорот вещества и превращение энергии находятся в состоя-
нии динамического равновесия. Это единство, образованное органической
природой (растениями, животными, микроорганизмами) и неорганической
средой, или факторами местообитания (тепловым, или температурным,
водным, или гидратурным, световым, химическим и физическим факто-
рами), мы называем экосистемой, или, по В. Н. Сукачеву, для
определенного растительного сообщества, биогеоценозом. Эко-
система — это ограниченное, рассматриваемое как целое гомогенное
1 Т. W. Booher, Studies on the plant-geography of the north-atlantic heath-
formation, «Kgl. Danske Vidensk. Selskab., Biol. Medd.», 15, 1940 p. 1—64.
17*
260
Глава 4
Атмосфера
Солнечная
Осадки
. I Отложение пыли
Обмен ◄—
энергией
(турбулентность,
конвекция)
Растения ) йзбух
[Птицы
Млекопитающие
I Насекомые
Сток
Е
Почва
I
Инфильтрация
Рис. 210. Схема обмена энергией и веществом биогеоценоза, или
экосистемы, с окружающей средой.
пространство биосферы, в пределах которого в течение длительного
периода времени существенно не меняется состав организмов и условия
окружающей среды.
Экосистема, или биогеоценоз, не является замкнутой системой, так
как в нее поступают извне солнечная радиация и атмосферные осадки.
Газообмен (СО2, О2, водяной пар) осуществляется между слоем атмосферы,
входящим в экосистему, и всей атмосферой путем турбулентного пере-
мешивания воздуха или при помощи ветра. В гумидных областях часть
влаги, выпавшей с осадками, проникает в почву и достигает уровня грун-
товых вод или стекает по поверхности почвы. Известная доля энергии,
поступающая посредством инсоляции, уходит из экосистемы путем отра-
жения, излучения или конвекции. Большая же часть инсоляции расхо-
дуется на испарение. В экосистему с пылью привносятся минеральные
частицы; кроме того, в экосистеме происходит обмен органических веществ
с окружающей средой, осуществляемый животными при их миграции
(рис. 210).
Баланс вещества и энергии в экосистеме, как правило, находится
в состоянии равновесия. Лишь в болотах происходит накопление вещества
и энергии в отложениях торфа; аналогичная картина наблюдается также
в пионерных растительных сообществах, которым не свойственно состоя-
ние равновесия, и в сообществах, отражающих последующие стадии
сукцессии. При вмешательстве человека изъятое количество вещества
и энергии должно быть снова восстановлено, в противном случае наблю-
дается деградация растительного покрова.
Ниже приводится характеристика лесных биогеоценозов умеренной
зоны, а именно лиственных лесов. В следующей главе будут рассмотрены
хвойные леса. Многочисленные данные получены при изучении дубовых
лесов (на площади 2000 га) в Теллермановском лесном массиве, располо-
женном на правом берегу Хопра (левый приток Дона) в Воронежской
области (приблизительно на 51,5° с. ш.) Ч Результаты работ опублико-
1 Е.М.Лавренко, Опыт изучения биогеоценозов на примере Теллерманов-
ской дубравы Воронежской области, «Изв. Геогр. об-ва», № 1, 93, 1961, стр. 23—33.
Лиственные леса умеренной зоны северного полушария — неморальная зона 261
ваны в трудах Сукачева, Дылиса и Молчанова на русском языке и поэтому
недостаточно известны на Западе Ч
Исследования позволили возможно более точно охарактеризовать
свойства определенного биогеоценоза, тем самым были осуществлены
многие положения Международной биологической программы. Поэтому
мы заимствуем из вышеупомянутых трудов большое число таблиц,
несколько видоизмененных автором.
Общим руководителем работ был В. Н. Сукачев, руководство на месте
осуществлялось А. А. Молчановым. В исследованиях принимало участие
большее число специалистов, работавших в различных областях знания.
Изучались:
а) микроклимат насаждений путем измерения температуры и радиации
на различной высоте,
б) водный режим насаждения,
в) взаимоотношения почвы и подстилки с напочвенным покровом
и микрофлорой,
г) фитомасса древесного яруса и яруса трав, а также продуктивность
и круговорот веществ,
д) фауна, ее состав и количественные соотношения.
Метеорологические исследования на пробных площадях проводились
на 12 станциях. Кроме того, было установлено 300 дождемеров и соору-
жены многочисленные установки, возвышавшиеся над кронами деревьев.
Инфильтрация влаги в почву и рост корней изучались в траншеях, выко-
панных на глубину до 5 м.
Исследования велись в различных типах леса лесостепной зоны. Гос-
подствующие здесь дубовые леса различаются между собой степенью
увлажненности местообитания. В травяном покрове доминируют Aegopodi-
ит podagraria или Carex pilosa или одновременно оба вида. Дуб {Quercus
robur) растет с примесью ясеня {Fraxinus excelsior) или липы {Tilia corda-
ta); во влажных эрозионных оврагах леса образованы вязом {Ulmus laevis)
с примесью лещины {Corylus avellana). В замкнутых бессточных пониже-
ниях лесостепи встречаются солонцеватые почвы. По мере увеличения
степени солонцеватости к дубу сначала примешивается Acer campestre,
а затем и Evonymus verrucosa; на настоящих солонцах дубовые леса изре-
живаются и в конце концов исчезают.
Аналогичные исследования проводились также в различных типах
лиственных лесов Европы и Северной Америки, но они никогда не велись
с такой степенью детальности в одном лесном массиве. Поэтому ниже
в основном будут приведены результаты работ в дубовых лесах Теллер-
мановского массива.
Радиационные и температурные условия
В учении о факторах местообитания (см. «Phytologie», Bd. Ill) про-
анализированы общие условия теплообмена в приземных слоях атмосферы
и в верхних горизонтах почвы1 2. Остановимся лишь на показателях, харак-
1 В. Н. Сукачев, Н. В. Дылис, Основы лесной биогеоценологии, 1964;
А.А.М ол чанов, Лес и климат, 1961; Гидрологическая роль сосновых лесов на пес-
чаных почвах, 1952.
2 D.M.Gates, Energy exchange in the biosphere, New York, 1962: R. G e i g e r,
Das Klima der bodennahen Luftschicht, 3. Aufl, Braunschweig, 1950. [Есть русский пере-
вод: P. Гейгер, Климат приземного слоя воздуха, М., 1965.— Ред.]
262
Глава 4
терных для лиственного леса. В качестве примера рассмотрим упоминавшие-
ся выше дубовые леса лесостепной зоны. Табл. 23 содержит средние вели-
чины радиации, полученные в течение года в этих лесах; данные характери-
зуют поток солнечной радиации на ровную поверхность и на поверхность
склонов разной экспозиции с углом наклона 30°.
Таблица 23
Суточная радиация при средней облачности (в кал/сл€2)
I н ш IV V VI VII VIII IX X XI XII
Г оризонтал ьная поверхность 20 41 105 196 278 317 267 220 160 79 25 15
Южный склон . . 57 93 163 251 307 330 285 260 230 149 61 46
Западный склон 18 41 94 179 247 287 240 203 151 75 24 14
Восточный склон 20 40 90 173 240 280 237 195 146 74 22 16
Северный склон — — — 11 85 170 220 180 121 48 — —
Как видно из таблицы, в умеренных широтах влияние различий
в экспозиции наиболее резко проявляется весной и осенью. Около 50%
поступающей радиации приходится на световое излучение, которое может
быть использовано в процессе фотосинтеза; эта величина колеблется
в течение дня. Наибольшая величина физиологически активной радиации
наблюдается около полудня, когда солнце находится в зените; вечером ее
значения резко падают. Лишь небольшая часть светового излучения не-
посредственно используется в процессе фотосинтеза. В лесу она составляет
около 3%. Известно, что фотосинтез протекает наиболее интенсивно в крас-
ных лучах солнечного спектра и наименее активно — в зеленых. Поэто-
му изменение спектрального состава света играет в лесу определенную
роль. В табл. 24 приводятся данные, характеризующие спектральный
состав света в дубовом лесу лесостепи в зависимости от стадии развития
листьев.
Таблица 24
Спектральный состав света под кронами деревьев в дубовом лесу
(в % от его состава над кронами) 1
Фенология Длина (в мкм) и цвет волн
0,730 красный 0,615 оранже- вый 0,580 желтый 0,530 зеленый 0,463 синий 0,420 фиолето- вый 0,330 и меньше ультра- фиолето- вый
18. III. Почки за- крыты 63 □7 53 49 48 47 45
15. IV. Почки рас- крываются . . 61 44 39 38 37 38 32
10. V. Молодые листья .... 21 7 7 6 6 6 5
4-25. VI. Взрос- лые листья . . 15 5 5 4 3 3 3
1 Возраст леса 23 года, сомкнутость крон 0,9—1,0.
Лиственные леса умеренной зоны северного полушария — неморалъная зона 263
Из таблицы видно, что под полог леса проникает значительно больше
лучей красной части солнечного спектра, чем лучей синей части Ч
Выделяют прямую солнечную радиацию и рассеянную. Прямая
солнечная радиация проникает под древесный полог в меньшем количе-
стве, чем рассеянная; значительную роль в этом распределении играет сте-
пень сомкнутости крон.
В табл. 25 представлен радиационный режим ряда сообществ дубовых
лесов, развитых на разных почвах.
Таблица 25
Суммарная, рассеянная и отраженная радиация
в ясные дни в различных типах дубовых лесов
Тип леса Радиация над древесным пологом Радиация под кронами деревьев
I п III 1 IV | V
суммарная, кал/см% .мин рассеянная, кал/см2 мин суммар- ная, % от I рассеян- ная, % от II отражен- ная, % от III
Quercetum aegopo- diosum .... 0,69 0,15 2,9 10,0 30,0
Querceto-Tilietum Querceto-Fraxine- 0,82 0,21 4,0 19,5 24,5
tum 0,69 0,22 4,4 18,0 31,2
Querceto-Aceretum campestris . . 0,66 0,17 4,0 10,1 27,1
Quercetum c Evo- nymus 0,75 0,18 7,2 33,2 33,3
Quercetum на со- лонцах 0,64 0,11 8,7 40,4 25,4
Условия освещенности в лесу меняются в пределах древесного полога
и травяного покрова; определенное влияние при этом оказывает возраст
древостоев. Условия роста травяного покрова наименее благоприятны
в молодняках; в очень старых насаждениях травостой развивается значи-
тельно лучше. В табл. 26 приводятся осредненные показатели освещенно-
сти, полученные в солнечные дни; суммарная радиация над кронами дости-
гала в дни наблюдений 62—68 клк.
Травяной покров в молодых насаждениях, куда проникает 0,6%
полного дневного света, отсутствует. Травостой появляется лишь в 22-
летних насаждениях и по мере увеличения возраста леса становится все
более густым, что находит ясное отражение в его продуктивности (см.
табл. 27).
В насаждениях в возрасте 220 лет травяной покров иногда заглушает-
ся кустарниками (ср. показатель освещенности на высоте 1,3 м над по-
верхностью почвы в табл. 26).
Изменение освещенности под пологом деревьев существенно зависит
от наличия или отсутствия яруса кустарников. Это подтверждают наблю-
дения, проведенные в различных сообществах дубовых лесов. В зависи-
1 На поверхности почвы в лесу наблюдается красно-зеленая тень с ярко выражен-
ным максимумом в темно-красной части спектра (около 0,710 мкм) и в более широкой
зеленой части спектра (около 0,550 мкм).
Таблица 26
Освещенность в дубовых лесах различного возраста
Место наблюдения Возраст насаждения, лет
13 22 42 58 135 220
лк % лк % лк % лк % лк % лк | %
Над кронами 64 210 100 62 377 100 64318 100 68 508 100 65 000 100 68 217 100
Между кронами . 1 925 3,2 3 069 5,1 6 394 10,0 12 000 17,7 38 493 59,0 48 659 72,5
На половине высоты древостоя . . 625 1,2 1 660 2,8 3 693 5,8 7 695 10,1 И 563 18,0 14 921 21,0
На х/з высоты древостоя 506 0,8 1 293 2,2 2 893 4,1 3 468 5,1 4 208 6,6 7 167 10,6
На высоте 1,3 м над почвой .... 385 0,6 796 1,3 1393 2,2 2 986 4,1 3 686 5,7 5119 7,5
У поверхности почвы 350 0,6 454 0,8 1038 1,6 1317 1,9 1383 2,1 1 143 1,7
Лиственные леса умеренной зоны северного полушария — неморальная зона 265
мости от сомкнутости и высоты яруса кустарников освещенность про-
странства в пределах 0—10 м очень различна (табл. 28).
Кроны дуба развиваются лучше всего при условии, когда солнечные
не только верхушки деревьев;
Таблица 27
Световое довольствие
на поверхности почвы
и продуктивность травяного
покрова
лучи освещают всю поверхность крон,
в этом случае интенсивность света в про-
странстве между кронами взрослых
деревьев достигает 45—50 клк.
Древесный полог является актив-
ной поверхностью леса, которая погло-
щает и переводит в тепло большую часть
солнечного излучения. Поэтому по-
верхность крон нагревается интенсив-
нее; здесь отмечена наиболее высокая
температура воздуха в течение дня.
Ночью в результате излучения сильнее
всего охлаждается верхняя часть крон.
Это видно из табл. 29, данные которой
характеризуют температуру воздуха
в насаждениях различного возраста.
Такая инверсия температуры не на-
блюдается в древостоях с опавшей лист-
вой; здесь самая высокая температура
днем отмечена у поверхности почвы.
а
Воз- раст насаж- дения, лет Световое довольствие на поверх-* ности почвы, % Сухая масса, кг/га
13 0,6 Нет
22 0,8 120
42 1,6 464
58 1,9 475
135 2,1 Максимальная, но не под- твержденная точными дан- ными
Величина радиации, достигающей поверхности почвы, и температура
почвы зависят также от степени сомкнутости крон.
Таблица 28
Освещенность на различной высоте над поверхностью
почвы при условии, что суммарная радиация
над поверхностью крон равна 65 клк
Высота над поверхностью почвы, м Quercetum aegopodio- sum Ouerceto- Tilietum Querceto- Fraxi- netum Querceto- Aceretum Quercetum c Evo- nymus
10 20 700 21 700 24 700 31400 37 000
1,3 1680 2 350 5120 3 450 6 300
0,5 Поверх- ность поч- 880 1680 ^990 1620 2 650
вы 630 1250 1540 790 680
В табл. 30 приводятся показания температуры почвы, наблюдавшиеся
в июле в сообществе Quercetum aegopodiosum.
От температуры почвы зависит деятельность микроорганизмов, что
в свою очередь определяет скорость разложения подстилки и минерали-
зации органических остатков растений.
В лесу отмечена высокая влажность воздуха; на поверхности почвы
она выше, чем в кронах деревьев (табл. 29). Температура в насаждении
понижается по мере приближения к поверхности почвы; поэтому дефицит
насыщения воздуха в лесу и особенно у поверхности почвы достигает
меньших величин, чем на открытом пространстве. С этим связано
ослабление транспирации у напочвенных растений, имеющих листья
гигроморфной структуры. Скорость ветра в лесу заметно уменьшается
(табл. 31).
266
Глава 4
Таблица 29
Средняя суточная температура и влажность воздуха
в лесу с распустившейся листвой.
Вегетационный сезон 1954 г.
Высота над поверхностью почвы, м Средняя температура, °C Средний максимум, °C Средний минимум, °C Средняя влажность воздуха,%
Дубовый древостой, возраст 6 лет, высота 2,2 м
4,2 17,0 25,0 5,3 74
2,2 17,3 33,4 2,8 77
1,0 16,9 27,9 4,5 86
0,05 15,4 22,9 4,7 94
0,00 15,3 22,0 4,8 98
Дубовый лес, возраст 27 лет, высота 9,2 м
11,2 17,0 25,8 5,1 75
9,2 17,2 33,6 2,6 77
4,0 16,3 26,7 4,4 85
2,0 15,9 24,9 4,4 86
0,05 15,6 22,7 4,7 92
0,00 15,3 21,9 5,8 98
Дубовый лес, возраст 220 лет , высота 31 . и
33,0 17,3 25,9 5,1 76
31,0 17,4 30,8 3,8 78
20,0 17,2 28,9 4,4 81
10,0 16,4 26,8 4,4 86
2,0 16,2 28,9 4,5 85
0,0 15,5 22,9 5,8 98
Таблица 30
Средняя суточная температура почвы
на различной глубине (в °C)
Глубина почвы, см Возраст насаждения, лет
2 22 42 56 220
5 18,7 14,9 16,3 16,1 15,5
10 17,5 13,7 15,4 15,1 14,3
15 16,9 13,0 14,6 14,3 13,8
30 16,5 12,6 14,3 13,9 13,2
50 15,8 11,4 13,4 13,1 12,6
Таблица 31
Скорость ветра в дубовом лесу (высота 25 м) весной
во время распускания листьев и с распустившейся листвой
(15.VI). Скорость ветра над поверхностью крон (2,4 ле/сек)
принята за 100%
Высота над по- верхностью почвы, м 21 апреля 15 мая 21 мая 15 июня
м/сек % м/сек % м/сек % м/сек %
18,0 1,16 48 1,09 45 1,15 48 0,65 27
10,0 0,93 39 0,90 38 0,78 33 0,51 21
5,0 0,83 35 0,63 26 0,52 22 0,31 13
1,5 0,59 25 0.56 23 0,35 15 0,2 9
Лиственные леса умеренной зоны северного полушария — неморалъния зона 267
Ветер, если он теплее воздуха в лесу, повышает температуру воздуха
леса, если холоднее —то выносит тепло из леса. Кроме того, ветер влияет
на обмен водяного пара и СО2.
Круговорот воды
В лесную экосистему влага поступает с осадками, выпадающими на
поверхность древесного полога. Где же находится источник этой влаги?
Связана ли она с испарением с поверхности океана и дальнейшим перено-
сом испарившейся влаги на большие расстояния или эта влага возникает
в результате эвапотранспирации растительного покрова и турбулентности
воздуха над ним? Поскольку скорость ветра на высоте 3 км, где обычно
располагаются облака, достигает 30 км/час, метеорологи придерживаются
точки зрения, что осадки, выпадающие над Восточной Европой, не пре-
вышают 37% от общего количества влаги, содержащейся в воздушных
массах, перемещающихся над этой территорией. Из общего количества
влаги 13% образуется в результате местного круговорота между расти-
тельным покровом и атмосферой, а 87% — вследствие испарений с поверх-
ности морей.
Осадки, выпадающие на поверхность древесного полога, частично
задерживаются листьями деревьев. В дубовых лесах Теллермановского
массива, отличающихся высокой степенью сомкнутости крон, при годовом
количестве осадков, равном 523 мм, полог леса независимо от возраста
насаждения задерживает 49—64 мм (в среднем 60 мм), то есть около 11 —
12% общего количества осадков. Лишь часть влаги просачивается в почву,
а еще меньшее количество проникает в грунтовые воды, причем инфильтра-
ция наблюдается в лесостепи только весной после таяния снега Ч
Влага, поступающая в лес в виде осадков и не достигающая уровня
грунтовых вод, расходуется на поверхностный сток, испарение и транс-
пирацию растений. В табл. 32 представлен водный баланс дубовых на-
саждений в зависимости от степени сомкнутости крон.
Передвижение воды в почве зависит от типа почвы и ее структуры.
Данные табл. 33 характеризуют инфильтрацию атмосферных осадков
в почву в течение трех лет в старых дубовых насаждениях, развитых на
темно-серых лесных суглинках. 1
Круговорот влаги в дубравах определяется густотой леса и нижних
ярусов, снижающих поверхностный сток воды. В изученных дубовых
лесах, развитых в плакорных условиях, поверхностный сток достигает
следующих величин (в % от годового количества осадков):
Quercetum aegopodiosum ..........................1,6
Querceto-Fraxinetum...............................4
Querceto-Aceretum ................................18
Quercetum на солонцах.............................32
1 Галу изучал в Бельгии водный и энергетический балансы трех смешанных лист-
венных насаждений на почвах, богатых известью: на участках северной экспозиции,
юго-юго-восточной экспозиции и на площади, расположенной на высоте 210 м над уров-
нем моря. Он установил, что водный баланс в насаждении, расположенном на склоне
южной экспозиции, в период с мая до середины июля был отрицательным, а средняя
годовая температура на 1—2° выше, чем в насаждении на ровном участке. Это отрази-
лось на флористическом составе лиственного леса, приуроченного к склону южной
экспозиции: среди кустарников обнаружены теплолюбивые элементы — Ligustrum
и Cornus mas (A. G а 1 о u х, Budgets et bilans dans Г ecosysteme foret, Bruxelles, 1963)
268
Глава 4
Таблица 32
Водный баланс (в мм) дубовых насаждений
в возрасте 45—48 лет в зависимости от степени
сомкнутости крон1
Элементы водного баланса Степень сомкнутости крон
1,0 0,85 0,75 0,5
Поверхностный сток 13 13 14 13
Испарение с поверх- ности почвы . . . 77 72 81 96
Испарение с поверх- ности крон . . . 70 48 43 23
Транспирация 421 381 392 379
Эвапотранспирация и сток 581 514 530 511
Инфильтрация на глубину 5 м 8 75 59 78
1 После проведенного в 1954 г. прореживания го-
довое количество осадков в следующие 4 года в среднем
составляло 589 мм. Испарение с поверхности крон равно
величине интерцепции; сток, инфильтрация и испарение
с поверхности почвы измерялись непосредственно, а ве-
личина транспирации получена расчетным путем.
Таблица 33
Инфильтрация воды в почву на глубину 5 ле (в мм)
Год Инфильтрация Возраст насаждения, лет
4 15 25 45 50 138 220
Инфильтрация в почву 347 354 351 338 334 346 340
1953 Вода, задержан- ная почвой . . 246 180 208 288 260 253 239
Инфильтрация до глубины 5 м . . 101 175 143 50 74 93 101
Инфильтрация в почву 171 167 162 164 155 162 154
1954 Вода, задержанная почвой .... 169 145 147 149 149 136 119
Инфильтрация до глубины 5 м 2 22 15 15 6 26 35
Инфильтрация в почву 398 381 374 370 374 384 388
1955 Вода, задержанная почвой .... 289 246 205 283 279 280 259
Инфильтрация до глубины 5 м 109 135 169 187 95 107 129
Лиственные леса умеренной зоны северного полушария — неморалъная зона 269
Основное количество выпавших осадков выносится из леса поверх-
ностным стоком во время таяния снегов, когда почва еще мерзлая. В лесах
с рыхлой подстилкой талые воды легко просачиваются в грунт. Поэтому
на больших площадях величина стока уменьшается по мере увеличения
степени облесенности территории. Табл. 34 иллюстрирует эту зависимость
в лесостепи Воронежской области.
Таблица 34
Зависимость величины поверхностного стока
от степени облесенности лесостепи
При незначительном поверхностном стоке даже на наклонных участках
в лесу процессы эрозии почвы не выражены; напротив, на вырубках
в результате эрозии выносится до 10—15 т/га почвы в год. Влага, сохра-
нившаяся в почве, всасывается корнями р течение вегетационного периода
и восполняет расходы на транспирацию.
БИОМАССА ЛИСТВЕННЫХ ЛЕСОВ УМЕРЕННОЙ ЗОНЫ
Фитомасса леса
В отличие от остальных экосистем лес накапливает в стволах большое
количество органических веществ, которые разлагаются только после
отмирания деревьев. Это особенно ясно выявляется при определении
фитомассы в отдельных зонах растительности по профилю, проведенному
через Восточную Европу от берегов Северного Ледовитого океана до пу-
стынь Центральной Азии (табл. 35).
Из таблицы видно, что фитомасса травяных сообществ не превышает
1,5 т/га. В кустарниковых сообществах фитомасса составляет 3 т/га\
при наличии отдельных деревьев она равна примерно 7 т/га и только
в лесах достигает 100 — 250 т/га. Налицо четкая зависимость фитомассы
от климата. И тем не менее продуктивность растительного покрова зави-
сит прежде всего от наличия древесных пород в пределах той или иной
зоны, их высоты и возраста. Только с этим обстоятельством связано
увеличение фитомассы в наиболее резко выраженных пустынных услови-
ях, где распространены заросли древовидных видов Haloxylon. Однако
было бы неправильным отождествлять фитомассу с живой биомассой расте-
ний, поскольку в лесах основная часть стволовой древесины состоит из
мертвой ткани, которая начинает разлагаться только при заболевании
деревьев или после их отмирания.
В отличие от древесной растительности надземная фитомасса травяных
сообществ образуется в результате прироста одного года, который можно
сравнивать с приростом однолетних побегов древесных пород. Наиболь-
шая фитомасса накапливается не в тропических дождевых лесах, а в лесах
влажной и теплой умеренной зоны, как, например, в юго-восточной
270
Глава 4
Таблица 35
Фитомасса в различных зонах растительности
Зоны растительности Сухой вес, т/га
1. Растительность Арк-
тики
Холодная пустыня с Dryas . . . 0,6
Тундра с видами осоковых . . . 1,2
Тундра с видами Salix 3,0
Тундра с Betula папа 3,2
Лесотундра с березой и елью . . 2. Лесная растительность 7,3
Северная часть зоны еловых лесов
(лшпайниково-моховый тип) Средняя часть зоны еловых лесов 90
(тип Vaccinium myrtillus) . . . Южная часть зоны еловых лесов 130
(тип Oxalis acetosella) Зона лиственных лесов с дубра- 220
вами 260
3. Степная раститель-
ность
Зона северной луговой степи . . Зона типичных ковыльных степей 1,5
с разнотравьем 1,2
Зона типчаково-ковыльных степей 1,0
Зона сухих степей с Artemisia 4. Пустынная раститель- 0,5
ность
Зона полынных полупустынь . . Зона кустарничковых пустынь с 0,7
эфемерами Южная часть зоны пустынь с 1,0
зарослями Haloxylon ... 7,0
Австралии и Тасмании, где эвкалипты достигают высоты 100 м, а нижний
ярус в лесах образован Nothofagus высотой до 40 м\ это же наблюдается
и в лесах Калифорнии с Sequoia sempervirens, достигающей высоты 100 м.
В дождевых лесах южного Чили запасы древесины достигают 4000 м?!га.
Количество фитомассы до сих пор учитывается недостаточно полно,
поскольку из-за трудности определения запасов подземных частей растений
их масса не включается в общую фитомассу. Видимо, наибольшая масса
подземных органов сосредоточена в лесных сообществах, так как здесь
образуется значительное количество корневой древесины.
Соотношение веса отдельных органов дерева в общей фитомассе дубрав
лесостепной зоны представлено в табл. 36.
Таблица 3
Сухой вес отдельных органов деревьев в
дубовых лесах различного бонитета (в т/га)
Бонитет Воз- раст, лет Листья Ветви Стволы Корни Всего
I 55 3,3 26,6 167,0 87,5 284,4
II 52 3,9 23,5 166,1 50,8 244,3
III 55 2,3 19,7 97,6 29,8 149,4
IV 47 2,6 15,0 53,9 39,1 110,6
Лиственные леса умеренной зоны северного полушария — неморалъная зона 271
В табл. 37 приведены величины общей фитомассы лиственных лесов
различного возраста, а также сухой вес надземной части живого травя-
ного покрова.
Таблица 37
Общая фитомасса надземных и подземных частей
древесного яруса, а также надземной части живого
травяного покрова в Quercetum carico-aegopodiosum
(в т/га)
Возраст насаждения, лет
10 20 40 60 140 220
Общая фито- масса . . . 18,9 47,8 142,5 249,3 397,0 413,3
Фитомасса тра- вяного по- крова . . . 0,07 0,265 0,350 0,475 0,340 0,265
По сравнению с общей фитомассой древесного яруса количество фито-
массы травяного покрова с увеличением возраста насаждения повышается
незначительно, достигая максимума в древостоях возрастом 60 лет;
в дальнейшем по мере старения древостоя фитомасса травостоя уменьшает-
ся; видимо, это связано с угнетающим влиянием кустарников на травяной
покров. В лесах других типов максимум живой биомассы травяного
покрова сохраняется до предельного возраста древостоя, тогда как общая
фитомасса древесного яруса быстро уменьшается в результате отмирания
отдельных деревьев. Это положение может быть проиллюстрировано на
примере сосновых лесов (см. табл. 71). В этих лесах по достижении ими
возраста 120 лет прирост древесины прекращается, и поэтому их следует
вырубать.
Индекс листовой поверхности и освещенность
i
Основными органами, производящими органическое вещество в био-
геоценозе, являются зеленые части растений, то есть листья. Поэтому
большое значение имеет правильная оценка листовой массы, или, точнее,
листовой поверхности. У лиственных пород умеренной зоны листья суще-
ствуют только в течение одного вегетационного периода, а затем опадают.
Вместе с опавшими ветками они образуют подстилку. Общее количество
сухого вещества, опавшего с одного дерева в течение его жизни, в 3—4 раза
больше сухого вещества, накопленного в виде фитомассы. На рис. 211
представлена зависимость между ежегодно продуцируемой деревьями
органической массой (приростом) и возрастом дубового леса (по Родину).
Если из брутто-прироста вычесть опад, то есть ежегодно образующуюся
массу подстилки, то мы получим нетто-прирост, который на графике
представлен площадью, расположенной между двумя кривыми. Только
в насаждениях в возрасте 25—35 лет нетто-прирост превосходит массу
подстилки, обычно же он меньше. В среднем вес листа площадью 1 дм2
равен: у лиственных деревьев — 1,5 г (сухой вес), у хвойных — 2 г.
Общая площадь листьев насаждения обычно выражается при помощи
так называемого индекса листовой поверхности (ИЛП), под которым пони-
272
Глава 4
мают отношение общей площади листьев насаждения к покрытой ими
поверхности почвы. Как известно, в благоприятных условиях среды
индекс листовой поверхности зависит от интенсивности дневного света.
Оптимальное значение индекса наблюдается в случае, когда теневые
листья, расположенные в нижней части кроны, имеют положительный
баланс вещества. При недостатке влаги или плохом обеспечении листьев
минеральной пищей (в первую очередь азотом) индекс листовой поверхно-
сти не достигает оптимального значения. Обеспеченность влагой и эле-
тп/га
18-
---1---1---1--1---1---1---1--1---1---1--1---1---г
О 20 40 60 80 100 120
Возраст, лет
Рис. 211. Зависимость годовой
брутто-продуктпвности органической
массы (прироста) и запаса подстил-
ки от возраста дубовых насаждений
(по Родину).
ментами минерального питания явля-
ется фактором, определяющим величину
ИЛП.
В качестве примера можно привести
исследования Крея, проведенные в по-
севах Heltanthus annuus х. Во влажное
лето 1963 г. ИЛП равнялся 9, в сухое
лето при такой же агротехнике ИЛП
достигал лишь 4—5, то есть был в 2 раза
меньше. Листья не только ассимили-
руют СО2, но и испаряют влагу; поэтому
растение приспосабливает свою транс-
пирирующую поверхность к количеству
влаги, поступающей из почвы. Увели-
чение засушливости климата способст-
вует изреживанию лесов. ИЛП восточно-
европейских лиственных лесов равен
4—6, хвойных лесов с многолетней хво-
ей — 9 —12.
Листовая масса в вышеописанных
лиственных лесах лесостепи достигает
в среднем 7,5 т/га, ИЛП равен 4,9 1 2 * * *.
Определенное влияние на величи-
ну ИЛП оказывают также содержа-
ние питательных веществ в почве и по-
ступление азота. Количество питатель-
ных веществ и содержание азота в большинстве случаев снижаются
по мере увеличения степени кислотности почвы. Элленберг обнаружил
четкую связь между ИЛП и величиной pH почвы при изучении лесов
в окрестностях Ганновера, расположенного в атлантической климатической
области Центральной Европы. В этих условиях индекс листовой поверх-
ности снижался с 8—9 при pH = 6,0 во влажных дубово-грабовых лесах
(Querceto-Carpinetum) до 3—4 в дубово-березовых лесах (Querceto-Be-
tuletum) на бедных кислых почвах. Величина ИЛП падает при чрезмерной
влажности и связанной с этим плохой аэрации почвы, а также при недо-
статке питательных веществ (рис. 212).
1 R. К г eh, Untersuchungen uber den Aufbau und dieStoffproduktion vonSonnen-
blumen und Maisbestanden, Hohenheim, 1966 (дисс.).
2 В старых древостоях, образованных Quercus petraea на силикатных почвах в при-
атлантической области Англии, ИЛП равен 4,9, тогда как общий сухой вес листьев,
площадь которых равна 23,6 X Ю6, достигает лишь 3,82 т!га. Общий вес подстилки
равен 3,86 т/га (из них 2,2 т приходится на осенний опад). В подстилке содержится
(в кг)'. 41,0 N; 2,2 Р; 10,5 К; 23,8 Са; 3,9 Mg; 1,7 Na; 117,4 золы; 1960 С и 18,6 X
X Ю6 ккал/га энергии. См.: А. С а г 1 i s 1 е, А. Н. F. Brown, Е. J. W h i t е, Lit-
ter fall, leaf production and the effects of defoliation by Tortrix viridana in a sessile Oak
(Quercus petraea) woodland «J. Ecol.», 54, 1966, p. 65—76.
Лиственные леса умеренной зоны северного полушария — неморальная зона 273
Рис. 212. Зависимость индекса листовой поверхности различных типов лиственных
лесов от средней величины pH почвы (по Элленбергу, см. «Phytologie», Bd. IV, 2).
Корневая система и водный режим
Корневая система, при помощи которой деревья поглощают из почвы
влагу, состоит из двух типов корней: 1) многолетних осевых корней, кото-
рые обеспечивают распространение корневой системы в горизонтальном
и вертикальном направлениях, и 2) ответвляющихся от них многочислен-
ных сосущих корней, окончания которых впитывают воду и минеральные
вещества; окончания сосущих корней существуют недолго и, вероятно,
заново отрастают каждый год. У древесных пород распространена мико-
риза; гифы грибов разрастаются в лесной почве, расширяя площадь,
с которой корни деревьев усваивают зольные вещества и азот. Структура
корневой системы аналогична строению побегов: многолетние ветви обра-
зуют остов, а растущие на них в течение короткого периода листья —
активную поверхность.
Основная масса корней и наибольшее число корневых окончаний
расположены в верхних, насыщенных гумусом слоях почвы, где сосредо-
точены питательные вещества, потребляемые растениями х. У большин-
ства лиственных деревьев развиты также корни, проникающие вертикально
в глубь почвы. В периоды засухи, когда верхние горизонты почвы высы-
хают, эти корни поставляют растению влагу, правда относительно не-
большое ее количество, и предохраняют тем самым деревья от усыхания.
Опытами, проведенными в лесостепи, установлено, что основная часть
влаги впитывается поверхностными корнями; для проверки этого в первом
варианте опыта были удалены горизонтальные корни на глубину 40 см,
а во втором — вертикальные корни; одновременно определялась величина
транспирации древостоя (табл. 38).
Как видно, удаление вертикальных корней почти не влияет на интен-
сивность транспирации. Дальнейшие опыты с искусственным увлажнением
1 Характер распределения п численность корневых окончаний, поглощающих
влагу и питательные вещества, изучались Веллером у яблони (Хоэнхейм, 1965). Ябло-
ня в возрасте 25 лет образует около 21 млн. корневых окончаний.
18-01248
274
Глава 4
Таблица 38
Интенсивность транспирации (мм) в дубовых
насаждениях (по Цельникер)
Насаждение Кон- троль Корни удалены
горизонталь- ные вертикальные
Чистый дубняк Дубняк с при- месью остро- листного 194 155=80% 191=98%
клена . . . 129 81=63% 114=88%
почвы, проведенные в период засухи, подтвердили эти наблюдения. При
увлажнении верхних горизонтов почвы интенсивность транспирации резко
повышалась, увлажнение же более глубоких горизонтов не оказало замет-
ного влияния на транспирацию. Вертикальные корни имеют большое
значение в жизнедеятельности лесов, распространенных в районах с за-
сушливым климатом. Роль этих корней увеличивается в тех случаях, когда
они достигают горизонта грунтовых вод и способствуют тогда длительному
и бесперебойному снабжению деревьев водой.
В общем можно заключить, что корневая система приспособлена
к тому, чтобы обеспечить потребности растения в поглощении [влаги из
почвы. Увеличение расхода воды листьями приводит к более интенсивному
развитию корневой системы1.
При хорошей влагообеспеченности эвапотранспирация лесных древо-
стоев зависит от дефицита влажности, величина которого в свою очередь
обусловливается температурой воздуха. Поэтому интенсивность транспи-
рации любой древесной породы, распространенной в определенных услови-
ях местообитания, находится в прямой зависимости от температуры возду-
ха (коэффициент корреляции, по Иванову и Силиной, равен примерно 0,9).
Максимальная интенсивность транспирации в течение суток в период
полного облиствления леса определяется количеством энергии, поступив-
шим с солнечной радиацией, и вычисляется по формуле: М = 0,6 ^/585,
где М — количество воды в мм, Q —инсоляция в ккал/см2,. При затруднен-
ном водоснабжении транспирация ограничивается тем количеством воды,
которое могут впитать корни из почвы. Если это количество опускается
ниже определенного минимума, древостой отмирает.
На южной границе распространения лиственных лесов Восточной
Европы максимальная интенсивность транспирации достигает в течение
вегетационного периода 100—120 мм.
Транспирация древостоев изучается также методом быстрого взвеши-
вания обрубленных ветвей; используется также метод лизиметров. Полу-
ченные этими методами результаты^резко отличаются друг от друга (ср.
«Phytologie», Bd. Ill, 1), а источники ошибок настолько велики, что нет
смысла их обсуждать. Наиболее правильна точка зрения, что в летний
период в лиственном лесу нет воды, которая в состоянии просочиться до
уровня грунтовых вод. Таким образом, осадки, выпадающие в течение
летних месяцев, вновь возвращаются в атмосферу, в основном путем
1 W. К a u s с h, Н. Е h г i g, Beziehungen zwischen Transpiration und Wurzel-
werk, «Planta», 53, 1959, S. 434—448.
Лиственные леса умеренной зоны северного полушария — неморальная зона 275
транспирации древесным ярусом. Иными словами, средний транспирацион-
ный расход равен количеству осадков в течение вегетационного периода
(в мм):
В Центральной Европе (Fagetum) ....................... 350—400
В Восточной Европе (Querceto-Carpinetum)................. 250—300
В лесостепи (Quercetum)............................... 150—200
В табл. 39 приводится интенсивность транспирации древостоя в посад-
ках лиственных пород (возраст 13—19 лет) в Деркульской степи в 1950 —
1956 гг.
Таблица 39
Листовая масса и транспирация древостоя лиственного леса в степи
в 1950—1956 гг.
Год и условия увлажнения
1950 опти- мальный 1951 опти- мальный 1952 влаж- ный 1953 засуш- ливый 1954 очень засуш- ливый 1955 очень влаж- ный 1956 влаж- ный
Листовая мас- са, т/га . . 5,86 7,31 7,68 5,10 6,69 10,70 8,89
Транспирация, мм 120 135 177 135 116 276 144
Как явствует из таблицы, в среднем за семь лет транспирационный
расход составляет 158 мм. При искусственном орошении лесных посадок
(из тополя) в полупустыне около Волгограда величина транспирации мо-
жет достигать 1000—1650 мм.
Лиственные породы имеют гидростабильные листья; дефицит воды
в листьях деревьев, растущих в лесостепи, не превышает 10—20% сырого
веса, или 20—30% запаса воды. При больших потерях воды листья
начинают преждевременно отмирать. В результате расход воды древостоем
в период засухи снижается до минимума. По данным русских ученых,
коэффициент транспирации у всех древесных пород равен 200—250. По
Польстеру, его значение составляет: у бука — 169, у дуба —344 Ч У мо-
лодых яблонь коэффициент транспирации равен 200—230, при недостатке
фосфора —482 (по Райнкену). Материалов, полученных в древесных
культурах, пока еще недостаточно для определения амплитуды изменения
коэффициента транспирации в зависимости от климата.
Зоомасса в лесу
Продуцентами органического вещества в лесной экосистеме являются
зеленые растения. В экосистеме, находящейся в состоянии равновесия со
средой, величина ежегодно продуцируемого органического вещества соот-
ветствует количеству минерализованной массы. В девственном лесу запас
фитомассы не меняется в течение длительного периода времени. Разложе-
ние или минерализация почти всегда осуществляются микроорганизмами
(бактерии, грибы и протисты), живущими в почве (включая и подстилку).
1 Н. Polster, Die physiologischen Grundlagen der Stofferzeugung im Walde,
Munchen, 1950.
18*
276
Глава 4
Частичное участие принимают и животные организмы. В процессе разло-
жения из растительной субстанции формируется вещество животных.
Животные расходуют значительно больше органического вещества, чем
создают сами. Они часто ускоряют процесс разложения, поскольку их
выделения разлагаются быстрее, чем остатки растений \ В то же время
участие животных усложняет круговорот веществ, так как при этом
возникают новые пищевые цепи. Фитофаги поедаются плотоядными пер-
вого порядка, последние уничтожаются плотоядными второго порядка
и т. д. Наряду с этим возникает еще одна пищевая цепь, начинающаяся
от тех животных, которые потребляют мертвое растительное вещество
(сапрофаги) и которых в свою очередь поедают плотоядные первого поряд-
ка. Эти пищевые цепи, которые могут быть продлены, представлены на
рис. 213.
Трудно разработать классификацию животных, основываясь только
на способах их питания, поскольку, например, птицы могут употреблять
в пищу семена, ягоды и насекомых. Часть сапрофагов питается отмершими
частями растений, другие существуют за счет экскрементов (копрофаги)
или трупов животных (некрофаги). Благодаря подвижности животные
могут выносить органические вещества из биогеоценоза или обогащать
его ими. В качестве примера можно привести косуль, пасущихся на лугах
вне леса, лисиц, ищущих добычу за его пределами, насекомых, улетающих
из леса, и т. д. Животные не только ускоряют разложение органического
вещества, но и распределяют его в горизонтальном и вертикальном направ-
лениях (роющие животные, дождевые черви, муравьи ) 1 2. Подобно тому
как растения образуют в лесу различные ярусы, животные организмы,
и в особенности беспозвоночные, также располагаются в пределах опре-
деленных горизонтов; здесь животные находят подходящие для своей
жизнедеятельности «ниши», однако они не всегда тесно приурочены к ним.
Например, многие гусеницы, питающиеся в кронах деревьев, окуклива-
ются в почве.
Различают 3 группы животных в зависимости от размеров площади,
в пределах которой они обитают:
1) животные, жизненный цикл которых протекает в определенном
биогеоценозе или в его части;
2) животные, эпизодически покидающие биогеоценоз;
3) животные, не связанные с определенным биогеоценозом и переме-
щающиеся на большие расстояния, например многие виды млекопитаю-
щих и птиц. Птицы во время перелетов залетают даже на другие кон-
тиненты.
Надземная фитомасса растений в лесном сообществе в 1000 раз
больше биомассы животных (см. т. I, стр. 44). В дубовых лесах лесостепи
биомасса животных достигает следующих величин (в кг!га сухого веса):
растительноядные млекопитающие (копытные и грызуны) — 2,2, хищ-
ники — 0,1, птицы — 0,2, в целом — 2,5. Количество растительной массы,
поглощаемой млекопитающими, составляет 327 кг!га\ большая часть этого
вещества вновь возвращается вместе с экскрементами животных. В зоне
1 Большое значение имеет размельчение животными отмерших органов растений,
что увеличивает их поверхность и облегчает разложение грибами и бактериями.—
Прим. ред.
2 В ясеневых лесах вес дождевых червей достигает 1500 кг! га. CM.:A.Finckh,
Okologische und bodenkundliche Studien uber die Leistungen der Regenwiirmer fur die
В odenfruchtbarkeit, «Z. Pflzern., Diing. und Bodenk.», 58, 1952, S. 120—145.
Лиственные леса умеренной зоны северного полушария — неморальная зона 277
смешанных лесов Восточной Европы наиболее крупные травоядные жи-
вотные — лоси —потребляют 8% фитомассы подлеска; менее половины
(34%) этого вещества лоси расходуют на дыхание. В Центральной Европе
обычно насчитывается около 20 голов крупной дичи на 1000 га покрытой
лесом площади. В лесах, которые служат охотничьими угодьями, всегда
имеется перенаселенность крупными животными. Только в годы катастро-
фического размножения насекомых-вредителей уничтожается очень зна-
чительная часть фитомассы, вплоть до отмирания деревьев х.
Наземные животные, добывающие корм в лесу, в силу их небольшой
численности вряд ли имеют значение, но они нередко оказывают большое
влияние на структуру леса, так как ограничивают или препятствуют возоб-
новлению части видов. Например, плоды определенных видов деревьев
могут быть полностью уничтожены (орешки бука и желуди грызунами),
а приживанию всходов препятствуют обкусывающие их животные.
При слишком высоком поголовье оленей в смешанных лесах возоб-
новление бука и пихты не происходит; в этом случае выживает только
подрост ели. В Новой Зеландии благородные олени опустошают большие
площади лесов.
Иногда животные содействуют распространению определенных дре-
весных пород: сойки прячут желуди под подстилку, кабаны разрыхляют
почву, а роющие животные перекапывают почву и создают тем самым
благоприятные условия для возникновения всходов.
Растения, расположенные в разных ярусах леса, различаются между
собой способами распространения семян и плодов; каждый вид растений
использует свой способ распространения зачатков, являющийся для них
наилучшим.
1. Растения древесного яруса являются анемохорами (Carpinus, Ulmus,
Acer, Fraxinus, Tilia, Populus, Salix, виды хвойных и т. д.) или синзоохо-
рами (Quercus, Fagus). Распространению их семян способствуют относи-
тельно большая скорость ветра в кронах деревьев, а также животные
(белка, сойки), которые собирают крупные плоды, закапывают в почву
И' затем нередко используют их лишь частично.
2. Кустарники являются эндозоохорами. Их семена разносятся птица-
ми, которые питаются сочными плодами подлеска — ягодами или костян-
ками.
3. Виды травяного яруса — эпизоохоры — распространяются отно-
сительно крупными животными; часть видов самостоятельно разбрасывает
семена. На лесных прогалинах, где движение воздуха более интенсивно,
среди видов травянистых растений часто встречаются анемохоры.
4. Виды трав, распространенные в приземном ярусе, большей частью
мирмекохоры; они имеют семена с элейосомами (Corydalis, Viola, Luzula
и др.) и частично образуют плоды-ягоды.
Почвенная фауна, представленная мелкими животными — сапро-
фагами,— играет гораздо большую роль, чем наземные животные. Био-
масса сапрофагов нередко достигает величины 1 т/га', в лесах, развитых
на почвах, богатых перегноем, основную часть биомассы составляют
дождевые черви. В буковых лесах Дании на муллевых почвах биомасса
почвенной фауны равна 76,8 г/м1 2, а в лесах на почвах с грубым гумусом —
16,6 г/м2. В дубовых лесах лесостепи биомасса достигает 81,2 г/м2. Поч-
1 В лиственных лесах Америки в оптимальные годы насекомые поедают от 4,4 до
11,7% общей площади листьев. См.: J. R. В г а у, Measurement of leaf utilization as an
index of minimum level of primary consumption, ««Oikos», 12, 1961, p. 70—74.
278
Глава 4
Обет
С02 атмосферы
. итомасса „ a Т „ , „»
(надземная)/Фитофаги—►Плотоядные I —* Плотоядные!! со
Растительный опад + Экскременты и трупы животных
Корни — » Сапрофаги —►Плотоядные 1->ЛлотояЗные1)
О „ * 1 я 1 У
Микроорганизмы почвы *< J
биомасса
почвы
Рис. 213. Схема пищевых цепей и круговорота углерода, протекающего
параллельно с процессом превращения энергии, в лесу.
венные организмы потребляют значительное количество питательных
веществ. В дубовых лесах лесостепи деятельность почвенной фауны устра-
нялась добавлением в почву нафталина, что не оказывало влияния на раз-
витие бактерий и грибов. В результате интенсивность разложения под-
стилки снизилась до 9%, в то время как на контрольных площадях эта
величина достигала 55%. Сапрофаги, однако, обладают большой избира-
тельной способностью по отношению к пище. Опавшие листья Fraxinus,
Tilia и Ulmus быстро потребляются ими, листья клена и граба перерабаты-
ваются медленнее, листья же Quercus и Fagus поедаются сапрофагами
неохотно. Только незначительная часть поглощенной пищи разлагается
животными; основная масса выделяется в виде экскрементов, в которых
содержится огромное число микроорганизмов. Помимо того, особенно
в экскрементах дождевых червей, непереваренная пища бывает переме-
шана с минеральными частицами почвы. В результате деятельности поч-
венной фауны реакция почвы становится более благоприятной, почва
обогащается азотом, ускоряется процесс распада, а продукты распада
частично превращаются в гумус. Таким образом, почвенная фауна оказы-
вает большое влияние на формирование почвы.
Рис. 213. схематично отражает взаимоотношения растений и живот-
ных в лесном сообществе. Более подробный анализ многосторонних вза-
имосвязей фауны с окружающей средой выходит за пределы задач этой
работы.
Микроорганизмы в почве
Третьим компонентом биомассы являются микроорганизмы — живая
часть почвы, тесно связанная с корневой системой растений. Определить
живую массу микроорганизмов в почве очень трудно, поэтому точные
показатели этой величины пока неизвестны. По данным Штруггера, масса
бактерий достигает 0,03—0,28% веса почвы. Наряду с ними в почве
встречаются грибы, актиномицеты и протисты. Почвенные микробиологи
для некоторых типов почв принимают следующие соотношения (в ьг/га):
грибы —1000—1500, актиномицеты — до 700 и протисты — от 100 до
Лиственные леса умеренной зоны северного полушария — неморалъная зона 279
Рис. 214. Дыхание почвы весной (слева)
и осенью (справа) в лиственном лесу в окрест-
ностях Рура (по Хаберу).
А — дыхание почвы; В — температура почвы на
глубине 10 см\ С — максимальная и минимальная ।
температуры приземного слоя воздуха; D — облач-
ность (точки разной интенсивности) и осадки (стрел-
ки, направленные вниз). Пунктирные линии пост-
роены по данным, полученным в других насажде-
ниях окрестностей Рура (до 21 мая в сосново-бере-
зовом лесу, с 26-мая — в смешанном лиственном
лесу).
300, По данным других исследователей, доля грибов, актиномицетов
и протестов составляет 5—10% бактериальной массы почвы. Эти цифры
мало что говорят и имеют большее значение для характеристики обра-
батываемых почв. Грубая подстилка разлагается главным образом грибами
и бактериями, и только в дальнейшем к этому процессу подключаются
актиномицеты.
Основуню роль в лесных почвах играют грибы, поскольку они не только разлагают подстилку, но и образуют микоризу и получают часть органиче- ского вещества из корней. Хабер 1 выя- вил хорошую взаимосвязь между дыха- нием почвы и количеством бактерий в нелесных почвах; такая корреляция отсутствует в лесных почвах, где бакте- рии встречаются в меньшем количестве, чем грибы. В табл. 40, заимствованной у Сукачева и Дылиса, приводится содер- жание грибов в различных почвах. Количество (в тыс, на 1 Таблица 40 грибов в почве г сухой почвы)
Почва Обрывки гиф Микроско- пические грибы
Под лесом Окультурен- ная поляна 565 12 27 42
Деятельность микроорганизмов в значительной степени определяется
температурой и гидратурой почвы. Последняя зависит не столько от со-
держания воды, сколько от относительной влажности почвенного воз-
1 W, Н a b е г, Vergleichende Untersuchungen der Bodenbakterienzahlen und der
Bodenatmung in verschiedenen Pflanzenbestanden, «Flora», 147, 1959, S. 1—34.
280
Глава 4
духа. Микроорганизмы активизируются при влажности воздуха в почве
(= гидратуре) более 96%. При высокой влажности и затрудненном газо-
обмене аэробные организмы погибают, а для анаэробных создаются благо-
приятные условия. В кислых почвах нитрификации не происходит. Лесные
растения в этом случае имеют в своем распоряжении азот в форме аммония.
Фиксатор азота — Azotobacter — почти всегда встречается в обрабатывае-
мых почвах и обычно отсутствует в лесных почвах с кислой реакцией гуму-
сового горизонта.
Мы оставляем в стороне проблему жизнедеятельности ризосферы
и микробного населения почвы. Отметим, что основная масса микроорга-
низмов приурочена к почвенным горизонтам, содержащим гумус, посколь-
ку существование гетеротрофных организмов зависит от наличия орга-
нических веществ.
Об активности микроорганизмов в почве судят по «дыханию» почвы,
которое фиксируется по количеству СО2, выделяемому из почвы. Хабер
видоизменил предложенный Вальтером метод абсорбции 1 и провел в тече-
ние 12 часов весной и летом соответствующие непрерывные наблюдения
в лиственном лесу (рис. 214). Поскольку дыхание почвы зависит от темпе-
ратуры и влажности почвы, ход кривых, изображенных на рис. 214,
с трудом поддается анализу. В целом выделяемое почвой количество СО2
определяется массой органического вещества, которое поступает из над-
земных частей растений в корни или в виде подстилки в почву 1 2.
Почвенные условия
В лесах умеренной зоны выделяют подзолистые, лессивированные
почвы, парабуроземы, бурые лесные почвы, темно- и светло-серые лесные
почвы, ранкеры 3, гумусированные карбонатные почвы, глеевые почвы
(возникают при высоком уровне грунтовых вод) и слаборазвитые почвы
местообитаний пионерных пород. Подробная характеристика этих почв
приводится в учебниках по лесному почвоведению.
Почва является важным компонентом биогеоценоза, поскольку в ней
содержатся вода и питательные вещества, за счет которых существует
фитоценоз.
Гумус в верхних горизонтах почвы формируется из подстилки. Свой-
ства гумуса зависят от состава подстилки, климата и материнской породы.
Легкоподвижные фульвокислоты и малоподвижные гуминовые кислоты
являются наиболее значимыми составными частями гумуса. Различные
типы леса характеризуются определенным соотношением между содер-
жанием гуминовых кислот (Сг) и содержанием фульвокислот (Сф). В Вос-
точной Европе отношение повышается при движении с севера на юг;
в подзолистых почвах зоны хвойных лесов оно составляет 0,2, в почвах
зоны смешанных лесов равно 0,5, а в почвах лиственных лесов лесостепи
достигает 0,8. Чем выше это соотношение, тем благоприятнее эдафические
условия для развития растений.
1 Н. Walter, Eine einfache Methode zur okologischen Erfassung des CO2-Fak-
tors am Standort, «Вег. Deutsch. Bot. Ges.», 65, 1952, S. 175—182; W. Haber, Okologi-
sche Untersuchungen der Bodenatmung* «Flora», 146, 1958, S. 109—157.
2 H.Walter, W. Haber, ЁЬег die Intensitat der Bodenatmung usw., «Вег.
Deutsch. Bot. Ges.», 70, 1957, S. 275—282.
3 Почва, состоящая из одного, большей частью кислого, гумусового горизонта,
расположенного непосредственно на горной породе.— Прим. ред.
Лиственные леса умеренной зоны северного полушария — неморальная зона 281
Изменение влажности почвы на разной глубине определяется при
помощи нейтронного зонда; для изучения передвижения воды в почве
в качестве индикатора используется тритий.
В лёссовых парабуроземах под буковым лесом дождевые осадки
после длительного периода засухи прежде всего заполняют пространства
между уменьшившимися в объеме почвенными агрегатами, особенно
интенсивно в тех случаях, когда гумусные агрегаты обладают плохой
смачиваемостью. В глубоких слоях почвы вода перемещается очень мед-
ленно, со скоростью 80—95 см!год, постепенно вытесняя более старый
почвенный раствор. Во время засухи древесные породы поглощают из
почвы всю воду, связанную давлением менее 20 —25 атм. После засушли-
вого лета в период с 30 ноября 1964 г. по 24 ноября 1965 г. баланс воды
в почве слагался из следующих величин (в мм) х:
Осадки (очень влажный год)...................901
Задержание осадков древесным пологом . . . 194
Поверхностный сток .......................... 0
Инфильтрация ...............................707
Удержанная почвой влага ....................160
Инфильтрация глубже 130 см..................152
Эвапотранспирация...........................395
ПРОДУКЦИЯ ВЕЩЕСТВА И ЕГО КРУГОВОРОТ
В БИОГЕОЦЕНОЗЕ ЛИСТВЕННОГО ЛЕСА
В настоящее время трудно проанализировать все цепи материально-
энергетических превращений в лесу, поскольку до сих пор изуча-
лись лишь частные случаи этого сложного процесса. Мёллер 1 2 в Дании пер-
вым определил первичную продуктивность древесного яруса, используя
уравнение, составленное Бойсен-Йенсеном:
Na—R —Bv —Av — WV = Z+S,
где (в mlга)\
Na — нетто-ассимиляция листьев;
R —дыхание (ствола, ветвей и корней);
Bv— осенний листовой опад;
Av — опад отмерших ветвей;
Wv — трудноопределимый опад корней;
Z —прирост древесной массы (надземной и подземной);
S —обычно неучитываемый урожай семян, достигающий в отдель-
ные годы больших величин.
Эти показатели измерялись непосредственно или, если это не пред-
ставлялось возможным, получались путем расчетов.
1 Н.-Р. Blume, К.-О: Miinnich, U. Zimmermann, Das Verhalten
des Wassers in einer LoB-Parabraunerde unter Laubwald, «Z. Pflern. Diing. u. Bodenk.»,
112, 1966, S. 156-168.
2 C.M.Moller, Untersuchungen fiber die Laubmenge, Stoffverlust und Stoffpro-
duktion des Waldes, Kopenhagen, 1945; D.Muller, J.Nielsen, Ein Diagramm
der Stoffproduktion im Buchenwald, «Вег. Schweiz. Bot. Ges.», 64, 1954, S. 487—494.
См. также: E. Gaumann, Der Stoffhaushalt der Buche Fagus silvatica L. im Laufe
eines Jahres, «Вег. Schweiz. Bot. Ges.», 44, 1935; H. Schroder, Ein Versuch, die
Oberflachenentwicklung eines Baumes (Fagus silvatica) zahlenmaBig zu bewerten, «Вег.
Deutsch. Bot. Ges.», 44, 1926, S. 579—583,
282
Глава 4
Определение величины R на спилах стволов выявило, что интенсивное
дыхание свойственно только наружным слоям древесины, прежде всего
Таблица 41
Расход на дыхание древесиной
в течение года (в % от массы
сухого вещества)
Высота древостоя/ м Бук Ясень Ель
5,9 11,0 12,5 8,0
10,9 9,1 10,1 6,6
15,9 7,6 8,0 5,6
20,9 6,3 6,2 4,8
25,9 5,4 5,0 4,2
30,9 4,8 4,2 3,7
камбию. Изучение дыхания на отдельных
древесных кольцах шириной 2,5 см, распо-
ложенных последовательно от внешней сто-
роны ствола к его центру, показало сильное
снижение интенсивности дыхания от первого
кольца ко второму; это снижение проявля-
ется более резко у древесных видов с выра-
женной ядровой древесиной (дуб), чем у видов
деревьев с заболонью (бук). Внутренние ча-
сти ствола практически не влияют на расход
энергии в процессе дыхания, и поэтому эта
величина пропорциональна поверхности
ствола. При старении дерева поверхность
ствола по отношению к массе древесины
уменьшается; в связи с этим расход на
дыхание древесиной, вычисленный в % от массы сухого вещества,
сокращается с увеличением возраста древостоя (табл. 41).
Опад листьев определяется по количеству опавших листьев. Величина
листового опада всегда меньше запаса подстилки древостоя, поскольку
подстилка состоит из опавших листьев и сучьев. Опад отмерших ветвей
рассчитывается сложным методом, исходя из количества ветвей, образую-
щих крону. Опад корней, который не мог быть измерен, приравнивался
к величине опада ветвей. У дуба и сосны необходимо также учитывать
опад коры, у бука и ели он не имеет значения.
Прирост древесины определяется по таксационным таблицам с добав-
лением 18% для учета корневой древесины. Прирост древесины вычисляет-
ся путем умножения величины прироста у основания ствола на прирост
Рис. 215. Ассимиляция и продуцирование органического вещества
в буковом лесу Дании (по Мёллеру и Мюллеру — Нильсену) (см.: «Phy-
tologie», Bd. Ill, 1).
1 — брутто-ассимиляция древесного яруса; 2 — нетто-продуктивность листьев за
вычетом опада листьев; 3 — надземный и подземный прирост древесины.
Лиственные леса умеренной зоны северного полушария — неморалъная зона 283
в высоту. В исследованных насаждениях различного бонитета величина
прироста у основания ствола была приблизительно одинакова; поэтому
запас древесины зависел от прироста в высоту и возрастал пропорцио-
нально последнему.
Величина прироста древостоев с возрастом увеличивается, достигает
максимума и затем снижается, однако кульминация всех видов прироста
не происходит одновременно; сначала наблюдается кульминация прироста
в высоту, несколько позже — прироста по диаметру и в последнюю оче-
редь — прироста древесной массы.
Рубки ухода незначительно изменяют запас древесины, но зато вли-
яют на качество прироста, поскольку в результате прореживания сокра-
щается число стволов, а оставшиеся обладают лучшим приростом.
Рис. 215 схематично иллюстрирует продуктивность букового леса
в Дании.
Таблица 42
Продуктивность (в ml га) тропического дождевого леса (Берег Слоновой Кости)
и букового леса (Дания) (по Мюллеру и Нильсену)
Тип леса I Брутто-про- дуктивность (сумма II и Ш) II Расход на дыхание 1 III Нетто-продуктивность
листья стебли корни продуктивность опада продуктивность древесины
листья стеб- ли 1 2 корни 2 надзем- ная подзем- ная
Тропический дождевой лес 52,5 16,9 18,5 3,7 2,1 1,9 0,4 7,5 1,5
Буковый лес 23,5 4,6 4,5 0,9 2,7 1,0 0,2 8,0 1,6
1 Из-за высоких температур в тропическом дождевом лесу в процессе дыхания расходуется
75% брутто-продукции, в буковом лесу — только 43%.
2 Цифры приблизительные.
Как видно, брутто-фотосинтез ассимилирующих листьев достигает
в буковом лесу в возрасте 40—60 лет 23,5 т!га в год; большая часть
этого вещества, синтезированного листьями, поступает в другие
органы дерева, расходуется на дыхание и на отмирающие части.
Оставшаяся доля, не превышающая 10 т, приходится на прирост древеси-
ны стволов и корней. Годовой прирост древесины стволов бука не превыша-
ет 8 т1га, в среднем 6 т!га. Аналогичные величины характерны и для ели;
из-за меньшего объемного веса древесины ели увеличение ее объема пре-
восходит таковой у бука. Прирост объема ели в среднем составляет 17 лг3,
тогда как у бука он достигает только 11 м3 х. Этим же методом определя-
лась продуктивность вечнозеленого тропического дождевого леса Берега
Слоновой Кости (4° с. ш.); нетто-продуктивность дождевого леса лишь
немного превышает продуктивность букового леса (табл. 42) 2.
Общая поверхность листьев в дождевом лесу невелика. У доминирую-
щих деревьев верхнего яруса развиты только световые листья, а у угне-
1 Воздушно-сухой вес 1 м3 древесины составляет (в кг): у дуба — 760, бука —
720, сосны — 520, ели — 470, тополя — 420.
2 D.Muller, J. Nielsen, Production brute, pertes par respiration et pro-
duction nette dans la foret ombrophile tropicale, «Forstl. Forsogsvaesen in Danmark», 29,
1965, s. 69—160. При рациональном ведении хозяйства лесные культуры в тропиках,
естественно, могут обеспечить значительно более высокие приросты древесины, чем
леса Центральной Европы.
284
Глава 4
тенных и невысоких деревьев — только теневые листья. Индекс листовой
поверхности не превышает 3,2; масса листьев составляет 7,3 т сырого
веса, или 2,5 т сухого веса, на 1 га. Число стволов диаметром > 3 см
равно 832 на 1 га, их запас в абсолютно сухом состоянии достигает 240 ml га,
объем — 421 м\ Индекс листовой поверхности подлеска также невелик
и равен 0,5. В какой мере эти данные имеют значение для других типов
тропических лесов, покажут дальнейшие исследования.
Подробный анализ круговорота веществ провел в смешанном дубовом
лесу в Арденнах Дювиньо, в котором он охватил и другие части биомассы,
а также определил состав золы. Важнейшие результаты этой работы
приведены во введении к т. I.
В последние годы Медвецка-Корнас проводила аналогичные исследо-
вания в буковом лесу (Fagetum carpaticum), расположенном на высоте
410—427 м над уровнем моря на рендзинах в области распространения
известняков юрского периода Ч
Возраст букового леса 70—80 лет, проективное покрытие древесного полога —
94%, травяного яруса — 30%; вегетационный период длится с апреля по октябрь;
подстилка разлагается через 3 года. Показатели продуктивности букового леса;
Фитомасса, кг/га Годовой прирост, ке/га
Древесина Листья Чешуйки, цветки, плоды . . Травяной покров 256 201 2 905 44 7 100 (среднее за 1953-1963 гг.) 2 905 117 46
Итого 259 150 10 168
Продуктивность букового леса сопоставлялась с производительностью свежего
луга (Arrhenatheretum), расположенного там же: надземная фитомасса луга — 3855 кг!га,
годовой прирост — 8738 кг/га. Последний значительно больше фитомассы, посколь-
ку в течение всего вегетационного периода происходит отмирание отдельных частей
растений. Урожай воздушно-сухого сена составлял 5000 кг!га.
Уиттекер исследовал продуктивность лесов Грейт-Смоки-Маунтинс
в Северной Америке 1 2.
В климаксовых лесах (лиственный лес с примесью пихты), по данным Уиттекера,
продуктивность достигает следующих величин:
Надземная фитомасса .......................... 500—610 ml га
Годовой прирост............................ 10 000—12 000 кг/га
В верещатниковых лесах сухих местообитаний и в лесах верхнего пояса годовой
прирост достигает меньших величин и равен 4200—6500 кг/га. Наибольшая продуктив-
ность отмечена в лесах, образованных Picea и расположенных на высоте 1400 м над
уровнем моря. Буково-пихтовые леса, приуроченные к южным склонам, характери-
зуются более высокой продуктивностью, чем те же леса на северных склонах гор. Дре-
востои, где удалены старые деревья, более продуктивны, чем непрореженные.
Годовое количество осадков в этой области достигает 1270—1500 мм, высоко
в горах выпадает до 2000 мм. В долинах среднемесячная температура января равна
4—5°, июля 23—25°.
Наиболее подробно круговорот веществ в дубовом лесу лесостепной
зоны Восточной Европы рассмотрен в работе Сукачева и Дылиса; эти
исследования проведены с учетом возрастного состава насаждений,
1 A. Medwecka-Kornas, Ecosystem studies in a beech forest and meadow
in Ojcow National Park, Krakow, 1967.
2 R. H. Whittaker, Forest dimensions and production in the Great Smoky
Mountains, «Ecol.», 47, 1966, p. 103—121.
Таблица 43
Основные показатели дубовых лесов лесостепи (по Сукачеву и Дылису, с дополнениями). Quercetum carico-aegopodiosum
(Теллермановское лесничество Воронежской области)
Возраст насаждения, лет
13 22 42 56 135 220
Запас древесины (стволы и ветви), сухой вес в т/га . 17 45 138 245 393 409
Листовая масса древесного яруса, сухой вес в т/га ............. 1,9 2,8 4,5 4,7 3,9 4,3
Освещенность в пространствах между кронами деревьев, % полного днев-
ного света . . . . . 3,2 5,0 10,0 17,7 59 73
Травяной покров, сухой вес в т/га . 0,07 0,26 0,35 0,47 0,34 0,26
Подстилка, сухой вес в ттг/га 13,2 13,8 14,2 14,8 14,0 14,2
Среднегодовой прирост древесины, сухой вес в т/га ........... 3,8 3,6 4,3 4,7 0,4 0,0
Птицы (число пар на 1 га) 12 4 8 12 18 20
Дождевые черви, количество на 1 м2 поверхности почвы . 61 80 62 59 69 35
Микроорганизмы, тыс. на 1 г почвы 7500 6875 7050 7050 10425 8335
Транспирация древостоя, мм/год 307 316 347 352 251 259
Испарение с поверхности почвы, мм/год .... . ... 67 62 82 84 65 101
Инфильтрация до уровня грунтовых вод, мм/год 89 74 15 9 89 91
Баланс зольных элементов: в числителе — поступление с осад- ками, кг/га\ в знаменателе — инфильтрация в грунтовые воды, кг/га (в течение вегетационного периода) Са Mg — 30,0 33,3 14,3 18,0 — 37,5 4,2 14,0 2,0 — 25,1 39,6 15,8 21,6
SO4 12,9 18,8 25,2
10,2 1,2 12,0
Cl 33,7 43,4 41,3
2,1 0,3 2,1
17,6 26,3 38,0
K-f-Na — 12,5 — 1,6 —• 21,7
В опавших листьях содержится, кг/га
общее количество золы GaO общее количество азота
Quercetum carico-aegopodiosum . 231,2 110,1 40
Quercetum aegopodiosum 360,0 119,2 52,5
286
Глава 4
круговорота воды, а также баланса некоторых зольных элементов
(табл. 43).
Из сравнения материалов, полученных Дювиньо при изучении сме-
шанных лесов Центральной Европы, и данных табл. 43 видно, что запас
древесины в исследовавшихся лесах почти одинаковый. Масса листьев
в них также одинакова. Она достигает максимума в дубовых лесах
в возрасте 40 лет и остается на том же уровне в насаждениях, возраст
которых более 200 лет. Масса травяного покрова в дубовых молодняках
невелика; она постепенно увеличивается, достигая максимума в зрелых
древостоях, а затем уменьшается в перестойных лесах, несмотря на их
изреженность (это связано с угнетающим влиянием яруса кустарников,
которые развиваются в разреженных дубравах). Интересно, что масса
подстилки остается неизменной вне зависимости от возраста насаждений.
Наибольший прирост древесины отмечен в дубовых лесах возрастом
42—56 лет; он лишь ненамного меньше прироста древесины в лиственных
лесах Центральной Европы. В древостоях, возраст которых свыше 100 лет,
прирост практически прекращается. В старых дубравах в древостое
преобладает масса незеленых частей деревьев, поэтому листовая масса
(количественно больше не меняющаяся) в состоянии покрыть лишь расходы
на дыхание древостоя и ежегодный опад. Нетто-продуктивность старых
дубрав незначительна, поэтому она обеспечивает только ежегодное обнов-
ление листовой поверхности и массы сосущих корней.
Разделы, посвященные участию в круговороте веществ дубрав живот-
ных и микроорганизмов, содержат недостаточное количество материалов.
Расход воды на эвапотранспирацию и на проникновение ее в глубь
почвы составляет в среднем около 450 мм!год и не зависит от возраста
Таблица 44 Баланс питательных веществ в буковом лесу (в кг /га -год)
Эле- мент Поглоще- но корня- ми Израсхо- довано на под- стилку Накоплено в древе- сине
Са 96 82 14
К 15 10 5
Р 13 10 3
N 50 40 10
лесами накапливается только С1“
леса. Эта величина соответствует коли-
честву осадков, выпадающих в лесостеп-
ной зоне. Чем выше транспирация дре-
востоя, тем меньше влаги поступает
в грунтовые воды. Проникновение воды
за пределы возможности ее использова-
ния растениями наблюдается только
в период таяния снега, когда деревья
еще не облиствлены и не транспири-
руют.
Баланс зольных элементов в молод-
няках находится в состоянии равно-
весия. В почве под молодыми дубовыми
В насаждениях в возрасте 56 лет проис-
ходит накопление всех зольных элементов; видимо, они концентрируются
в быстро увеличивающейся массе древесины. В старых насаждениях на-
блюдается потеря Са и в меньшем количестве Mg; одновременно с этим
повышается содержание сульфатов, хлоридов и в меньшей степени щелоч-
ных металлов \
Некоторые сведения о круговороте питательных веществ в буковом
лесу сообщаются в работе Денглера (табл. 44) * 2.
При рубках из леса изымаются питательные вещества в количествах,
указанных в последней колонке табл. 44. Поэтому Са, К и Р должны вновь
ХН. П. Ремезов, Л. Е. Родин, Н. И. Базилевич, Методические
указания к изучению биологического круговорота зольных веществ и азота наземных
растительных сообществ в основных природных зонах умеренного пояса, «Бот. журн.»,
48, 1963, стр. 869-877.
2 A. D engler, Waldbau auf okologischer Grundlage, Berlin, 1930.
Лиственные леса умеренной зоны северного полушария — неморальная зона 287
поступать в почву путем выветривания материнских горных пород. Азот
восполняется микроорганизмами, связывающими N. В связи с большой
потребностью в кальции бук оптимально развивается на известковых
почвах и плохо — на выщелоченных почвах в приатлантических областях.
Бук отрицательно реагирует на удаление подстилки, вместе с которой
в этом случае выносится большая часть кальция и азота, участвующих
в круговороте веществ. Уже Гессельманом было показано, что наибольшее
значение среди питательных веществ, поступающих в растение из почвы,
имеет азот Ч Он потребляется растениями в форме аммония (NH*) или
нитрата (NO“). Для жизнедеятельности растений имеет значение не общее
количество связанного азота в почве и не содержащееся в почве в какой-
то момент абсолютное количество NH* или NO", а постоянное новообразо-
вание этих соединений из органического азота, связанного в гумусе и под-
стилке, иными словами, способность почвы образовывать доступные для
растений формы азота. Превращение органического N в неорганический,
или минерализация азота,— это биологический процесс, осуществляемый
микроорганизмами почвы. Часть минерального азота (нетто-продукция
процесса минерализации) используется растениями, тогда как другая
часть брутто-продукции используется микроорганизмами для построения
их тел и таким образом вновь превращается в недоступные корням выс-
ших растений соединения. При неблагоприятных условиях аэрации,
в частности при очень высокой влажности почвы, часть азота в результате
денитрификации уходит из почвы в форме элементарного азота. В резуль-
тате лабораторных исследований форм гумуса Цётль вычислил соотноше-
ние между минеральным азотом и общим количеством азота, содержащим-
ся в гумусе 1 2. Цётль определил среднюю величину интенсивности минера-
лизации гумуса в 5% в год, что соответствует образованию не менее
31 кг/га NHJ или NOg. В опытах с посевом семян ели на свету присутствие
минерального азота в почве не обнаруживается. Контрольный опыт, без
посева семян в почву, показал, что почти 2/3 минерального азота потребля-
ется всходами ели, тогда как оставшаяся треть используется организма-
ми ризосферы. Корни растений не в состоянии усвоить азот из почвы
в количестве, превышающем величину нетто-минерализации. Органиче-
ские соединения азота для них недоступны.
При анализе круговорота веществ в лесной экосистеме необходимо
знать фактическое количество образующихся в естественных условиях
NH^ и NO3, а также вычислить годовой ход образования этих форм азота
в определенных местообитаниях (Элленберг).
На рис. 216 даны две кривые: одна — для богатого основаниями
бурозема под кленово-ясеневым лесом (Aceri-Fraxinetum), вторая — для
подзолистого бурозема с грубым гумусом под Luzulo-Fagetum. В обоих
случаях пробы брались только в верхних горизонтах почвы. В первом
случае в почве образовалось 0,076 г!л NO3 в год и только 0,022 г/л NH*
(в сумме 0,098 г/л) 3. Во втором случае из-за высокой степени кислотности
1 Н. Н esselman, Studien fiber die Nitratbildung in natiirlichen Boden und
ihre Bedeutung in pflanzenokologischer Hinsicht, «Medd. Stat. Skogsfors. Anst.», 21, 1917,
S. 297—527; H. Hesselman, Die Bedeutung der Stickstoffmobilisierung in der
Rohhumusdecke fur die erste Entwicklung der Kiefern- und Fichtenpflanzen, «Medd.
Stat. Skogsfors. Anst.», 23, 1927, S. 337—432.
2 H. Z 0 t t 1, Die Mineralstickstoff-Anlieferung in Fichten und Kiefernbestanden
Bayerns, «Forstw. Gbl.», 79, 1960, S. 221—236.
3 В нитрофильных лесных сообществах с Urtica dioica (Штутгарт-Хоэнхейм)
степень образования минеральных форм азота была в 3,5—6 раз больше (L. М а г-
kovic-Gospodaric, «Acta Botan. Groatica», XXVI).
288
Глава 4
почв нитрификации не было и в почве образовался только NH* в количе-
стве 0,105 г/л. В теплые летние месяцы наблюдается наиболее интенсивное
образование минеральных форм азота; в отдельных случаях этот процесс
зависит от влажности почвы. Заслуживает быть отмеченным, что общее
количество минерального азота несколько больше в почвах с грубым
гумусом, чем в муллевых буроземах. Это явление характерно только для
Рис. 216. Зависимость годового хода поступления минеральных
форм азота от температуры и влажности почвы в грубом гумусе
подзолистого бурозема (Luzulo-Fagetum) и в мулле бурозема, бога-
того основаниями (Aceri-Fraxinetum), в окрестностях Цюриха
(по Элленбергу).
верхних горизонтов почвы. В более глубоких горизонтах кислых почв
с грубым гумусом происходит резкое снижение количества N, а в богатых
почвах содержание минерального азота сохраняется относительно высо-
ким до подпочвы (рис. 217). Значение этого явления для растений призем-
ного яруса будет рассмотрено дальше. Табл. 45 содержит данные о круго-
вороте азота в искусственных посадках сосны, проведенных в области
лиственных лесов центральной Англии.
Лиственные леса умеренной зоны северного полушария — неморальная зона 289
В процессе разложения подстилки в лиственных лесах важную
роль играют гименомицеты. Сайто установил увеличение содержания
аммонийного азота и глюкозы при заражении их культурой Collybia. Что же
касается микроскопических грибов, то они снижают содержание пита-
тельных веществ в подстилке. Collybia выделяет антибиотики, которые
тормозят развитие микроскопических грибов и бактерий \
j____1___
Мулль
ИзВестпь
20 30
40 N мг/л
Cartel- Fagetum
РенЗзины
От
20-
40-
1111
NOS'''.'-"
Мулль
."Валовая
/ аккумуляция
Суглинок
Дсег1-Ргах1пе1ит
pH Эвтрофный бурозем
(слабо оглеенный)
...." -1
.....'Мовер-мулль
; Melico-Fagetum
; бурозем
I
I Супесь
П 1 1 1 1 1 1 ! 1 1
4 5 6 7 8 pH
Рис. 217. Минерализация азота на различной глубине
в почвах разных типов; август 1961 г., окрестности
Цюриха (по Элленбергу).
Пояснения см. рис. 216 (Aceri-Fraxinetum).
Luzulo-Fagetum
ОпоЗзоленный
бурозем
Наиболее богата азотом подстилка под Robinia pseudacacia, посколь-
ку этот вид деревьев ассимилирует элементарный азот при помощи клу-
беньковых бактерий. .Robinia используется для облесения сухих и теплых
почв, бедных азотом (например, каменистых осыпей). В подстилке под
Robinia условия для образования минеральных форм азота особенно
1 Т. S a i t о, Goactions between litter-decomposing hymenomycetes and their asso-
ciated microorganisms during decomposition of beech litter, «Sci. Rep. Tohoku Univ.,
Ser. IV (Biol.)», 31, 1965, p. 255—273.
19-01248
290
Глава 4
Таблица 45
Круговорот азота в 55-летнем сосновом лесу
в центральной Англии (в кг/га N)1
Содержание в живых растениях:
в деревьях................... 453
в растениях нижних ярусов 40
Содержание в подстилке и гумусе . . . 409
Итого 902
Ежегодное потребление растениями:
деревьями...................... 87,5
растениями нижних ярусов ... 37,5
Итого , 125
Содержание в годовом опаде (листья, вет-
ви и т, д,):
деревьев............................. 64
растений нижних ярусов......... 39
содержание в годовом опаде при
рубке (хворост, корни и т. д.) 13
Итого в подстилке Т16
Остаток от потребления (125—116) ... 9
В ежегодно удаляемой стволовой древеси-
не: ......................... . 3
остаток........................... 6
В годовом слое нарастающей подстилки 5
Годовой прирост экосистемы ......... 11
Если величина прироста превышает раз-
мер погрешности расчетов, то она может быть
обусловлена деятельностью микроорганизмов,
фиксирующих азот.
1 J. D. Ovington, Quantitative ecology and
the woodland ecosystem concept, «Ecol. Res.», 1, 1962,
p. 103—192.
благоприятны. Это доказывается тем, что в зарослях Robinia широко
распространяются рудеральные виды и растения, характерные для выру-
бок: Galium aparine, Chelidonium majus, Urtica dioica, Alliaria officinalis,
Bromus sterilis, Veronica hederifolia, виды Galeopsis, Moehringia trinervia
и др. Подстилка из листьев Rubinia отличается очень узким отношением
С/N, равным 14—17, и очень сильно изменяет биологию почвы Ч
При фотосинтезе солнечная энергия превращается в химическую
энергию образовавшегося органического вещества. При изучении круго-
ворота энергии в экосистеме леса необходимо определить содержание
энергии в органическом веществе, выраженное в калориях, Овингтон
провел такие исследования в Англии в культурах сосны различного
возраста 1 2,
1 Н. Gemeinhardt, Bodenmikrobiologische Beitrage zum Robinienproblem,
«Arch, f. Forstwesen», 8, 1959, S, 1078—1116; 9, 1960, S, 1082—1104,
2 J.D.Ovington, Some aspects of energy flow in plantations of Pinus sylvestris
«L., Ann. of Bot. N, S,)>, 25, 1961, p. 12—20.
Лиственные леса умеренной зоны северного полушария — неморалъная зона 291
ЭКОЛОГО-ФИЗИОЛОГИ ЧЕСКИЕ ИССЛЕДОВАНИЯ
ШИРОКОЛИСТВЕННЫХ ДЕРЕВЬЕВ
Из-за своих размеров деревья являются трудным объектом для экс-
периментальных исследований. Эти трудности усугубляются также неод-
нородностью условий среды, окружающей дерево. На поверхности дре-
весного полога температура достигает других величин по сравнению
с нижней или внутренней частями кроны. Без особых приспособлений
верхние части кроны почти недоступны для экспериментального изучения.
Поэтому рассмотрение дерева в качестве единого целого почти недостижи-
мо, хотя и крайне необходимо при проведении эколого-физиологических
исследований. Это вынуждает ставить опыты с молодыми деревьями.
Рост деревьев — явление, которое можно понять, исходя из конку-
ренции за свет. Чем выше растения и, следовательно, чем больше они
возвышаются над уровнем конкурентов, тем лучше они обеспечены светом,
необходимым для фотосинтеза. Но вместе с тем у более высоких деревьев
возникают затруднения со снабжением крон водой. Высокое корневое
давление, требуемое для поступления воды к вершинам деревьев, и свя-
занное с этим повышение концентрации клеточного сока приводят к ухуд-
шению гидратуры плазмы. Для жизнедеятельности дерева прежде всего
необходимо обеспечить поступление воды к вершинам, несмотря на кон-
куренцию нижерасположенных частей цроны. Это достигается тем, что
в водопроводящей системе, в местах присоединения боковых ветвей к ос-
новным путям движения воды, возникает значительное сопротивление,
тогда как обеспечение водой побегов облегчается в результате увеличения
площади водопроводящей системы1. Это способствует гармоническому
развитию всей кроны, что наблюдается у одиночно стоящих старых
деревьев бука, липы и дуба, у которых все части дерева получают доста-
точное количество света. Поэтому при одинаковой освещенности осмоти-
ческое давление почти не различается у листьев, расположенных в раз-
личных частях кроны. У листьев благородного каштана (Castanea vesca),
произрастающего на опушке леса в окрестностях Гейдельберга, осмоти-
ческое давление повышалось с 16,4 атм на высоте 2,5 м до 18,8 атм на
высоте 22 м, а у ели — с 18,4 атм на 21-й мутовке, считая сверху, до
20,7 атм на самой высокорасположенной мутовке; при этом следует
учесть, что верхние части кроны были лучше освещены, чем нижние 1 2.
Различия в освещенности имеют большое значение для растений. Свето-
вые листья отличаются более высоким осмотическим давлением и более
ксероморфной структурой, чем теневые листья, расположенные на той же
высоте в кроне. Поэтому с эколого-физиологической точки зрения эти
листья неравноценны. Уже в предыдущем году, когда закладываются поч-
ки, можно заранее определить, будут ли листья световыми или теневыми.
Исход определяется не столько условиями освещения, сколько условиями
гидратуры. Поскольку в тканях меристемы отсутствует тургор, гидратура,
а тем самым и набухание плазмы зависят от средней величины сосущей
силы, присущей водопроводящей системе ветвей, на которых закладыва-
ются почки. Освещенные ветви характеризуются более напряженным
водным балансом по сравнению с затененными ветвями, поэтому гидра-
1 I. В. Huber, Die Saftstrome der Pflanzen, «Verstandliche Wissenschaft»,
Bd. 38, 1956.
2 H. Walter, Grundlagen des Pflanzenlebens, «Phytologie», Bd. I, 4. Auf.,
1962, S. 335.
19*
292
Глава 4
турные условия в листьях, заложенных в почках на освещенных ветвях,
менее благоприятны. В результате после распускания листьев следующей
весной размеры клеток остаются небольшими и у листьев появляются
количественные морфологические признаки, характерные для световых
листьев (большее число устьиц и жилок,
Таблица 46
Соотношение между величиной
осмотического давления
и густотой сети жилок
у листьев бука
Fagus silvatica Осмо- тиче- ское давле- ние, атм Густо- та сети жилок, лии/слс2
Световые листья . . . 16,3 954
Теневые листья . . . 11,6 657
меньшие размеры клеток и площади листьев,
большая толщина листьев ит. д.) (см.: «Phyto-
logie», Bd. Ill, 1). В целом более напряженный
водный баланс световых листьев определяет
более высокое осмотическое давление по срав-
нению с теневыми листьями. В табл. 46 сопо-
ставляются величины осмотическою давле-
ния с густотой сети жилок у листьев бука.
В табл. 47 приводятся сравнительные
анатомические признаки световых и теневых
листьев бука, по данным Шрамма х.
Большие количества воды, поступающей
в атмосферу в результате транспирации,
восполняются за счет ее поглощения из
почвы. Поступление и отдача воды очень
тонко регулируются растениями в постоянно
меняющихся условиях окружающей среды. Деревья в отличие от боль-
шинства травянистых растений при затрудненном водоснабжении свое-
временно ограничивают транспирацию, частично смыкая устьица. Поэто-
му в целом суточные колебания осмотического давления листьев деревьев
Таблица 47
Анатомическая структура световых и теневых листьев бука
Тол- щина листа, мкм Число клеток эпи- дермиса на 1 лш2 Число устьиц на 1 лии2 нижней стороны листа Длина жилок, мм/мм%
верхняя сторона нижняя сторона
Световые листья . . . Теневые листья 160 73 1725 938 3000 1125 413 ИЗ 12,20 8,80
относительно невелики даже в крайне неблагоприятных условиях, при
резком сокращении запасов воды в почве, что видно на примере Pirus
elaeagrifolia, произрастающей на границе распространения леса (опре-
деляемой сухостью) в центральной Анатолии (табл. 48).
Из данных таблицы видно, что по мере ухудшения водного баланса,
обусловливающего увеличение осмотического давления в первой полови-
не дня, суточные колебания давления возрастают, но лишь в начале
вегетационного периода; затем они быстро снижаются и окончательно
прекращаются в августе при полностью закрытых устьицах.
Этот процесс протекает по-иному только у деревьев, растущих в по-
стоянно сырых местообитаниях и поэтому никогда^не страдающих от не-
1 См.: К. R u b n е г, Die pflanzengeographischen Grundlagen des Waldbaues,
4. Aufl., Radebeul u. Berlin, 1953.
Лиственные леса умеренной зоны северного полушария — неморальная зона 293
Таблица 48
Суточные колебания осмотического давления
при увеличении засушливости летом у JPirus elaeagri folia
в окрестностях Анкары (по Биранду)
Апрель Май Июнь Июль Август Сен- тябрь
Величина в первую половину дня, атм Суточные колебания, % ОТ предыдущей величины . . . 17,6 19,9 24,2 29,9 34,0 34,0
8,4 10,0 8,1 2,7 0,1 0,0
хватки воды; такой породой является тополь черный {Populus nigra) 1.
В искусственно созданных условиях засухи эти виды деревьев неспособны
регулировать расход воды. Процесс транспирации и ассимиляции СО2 у
этих видов протекает нормально до наступления продолжительной засухи;
затем ассимиляционный аппарат необратимо изменяется, листья начина-
ют желтеть и преждевременно опадают. В отличие от тополя у осины
{Populus tremula), развивающейся в относительно сухих местообитаниях,
в первые же дни засухи предусмотрительно закрываются устьица. В ре-
зультате почти полностью прекращается фотосинтез; однако сохранившая-
ся без повреждений листва при улучшении условий водоснабжения вновь
может возобновить ассимиляцию СО2.
Поэтому гидро лабильный черный тополь, отличающийся большой
продуктивностью, конкурентоспособен только на постоянно влажных
местообитаниях. В местообитаниях, подверженных засухе, более продук-
тивна и засухоустойчива гидростабильная осина, своевременно замыкаю-
щая устьица.
При хорошей влагообеспеченности световые и теневые листья лист-
венных деревьев транспирируют в зависимости от условий испарения, то
есть пропорционально величине испаряемости на солнце или в тени.
Интенсивность транспирации у световых и теневых листьев сокращается
даже при небольшом дефиците насыщения листьев; у более восприимчивых
теневых листьев даже раньше, чем у световых. При помощи метода ин-
фильтрации при этом можно установить процесс закрывания устьиц.
При большом дефиците насыщения наблюдается только кутикулярная
транспирация 1 2. Одинаковая реакция световых и теневых листьев сви-
детельствует о единообразном строении водопроводящей системы у этих
листьев. Любое нарушение водного баланса вызывает повышение сил сцеп-
ления в сосудах, что проявляется в изменении толщины ствола. Измере-
ния диаметра ствола с помощью дендрометра проводились Краузе, Фрид-
рихом и Мак Дугаллом; они установили периодические суточные колеба-
ния, которые накладываются на расширения ствола путем вторичного
прироста в толщину. В течение дня происходит сокращение диаметра,
ночью — расширение. Отклонения могут достигать 0,5 мм. Эти колебания
1 G. Neuwirth, Н. Polster, Wasserverbrauch und Stoffproduktion der
Schwarzpappel und Aspe unter Diirrebelastung, «Arch. f. Forstwesen», 9, 1960, S.789—
2 A.Pisek, E.Gartellieri, Zur Kenntnis des Wasserhausbalts der Pflan-
zen. IV. Baumeund Straucher, «Jb. wiss. Bot.», 88, 1939, S. 22—68.
294
Глава 4
Р и с. 218. Суточный ход скорости транспирационного потока
у трех видов деревьев (по Шуберту, из работы Губера).
Пунктирная линия означает нулевую величину транспирационного потока
в срезанной ветке (при использовании компенсационного метода указывает,
что срез был произведен в одно и то же время).
зеркально отражают изменения температуры воздуха и потенциального
испарения, которые, так же как и транспирация, зависят от инсоляции х.
Кривая изменений диаметра ствола несколько отстает от кривой хода
транспирации; причина — в эластичности системы сосудов. Параллельно
Таблица 49
Максимальная скорость транспирационного
потока (в м/час) (по Губеру)
Кольцепоровые
лиственные де-
ревья . . .
Рассеяннопоро-
вые лиственные
деревья . . .
Хвойные деревья
4-44
1-6
1,2—1,4
Для сравне-
ния: травя-
нистые рас-
тения —
10—60, лианы-
150
с увеличением и снижением интен-
сивности транспирации и сил сцеп-
ления повышается и падает ско-
рость транспирационного потока
в сосудах; ночью она почти не
выражена; утром наступает быст-
рый подъем, достигающий после
полудня при благоприятной погоде
почти стабильной максимальной
величины; вечером наблюдается
снижение (рис. 218). Благодаря
эластичности системы
сосудов транспирационный поток начинается
утром с концов ветвей и затем перемещается по направлению
к основанию ствола; вечером транспирационный поток сначала прекращает-
ся в ветвях, тогда как ночной поток продолжается в комле еще в течение
нескольких часов. Абсолютная скорость транспирационного потока,
естественно, зависит от величины потребляемой воды и от размеров про-
водящей площади; поэтому у кальцепоровых деревьев, образующих только
одно годичное кольцо, эта скорость выше, чем у рассеяннопоровых деревь-
ев. В табл. 49 представлена наибольшая скорость транспирационного
потока у различных деревьев в полдень.
1 I. Impens , J. Schale k, Daily trunk diameter changes of a Pirus Malus
in relation to cyclic environmental factors, «Mededel. Labor. Plantecologie», № 8, Gent.
1964. Резюме в «Arch. f. Meteorol., Geophys. u. Bioklimat.», 13, 1964, S. 400—403;
G. Mitscherlich, W. Mull, E.Kunstle, P. Maurer, Ertragskundlich-
okologische Untersuchungen im Rein- und Mischbestand, «Allg. Forst.- u. Jagdzschr.»,
136, H. 10—12, 1965; 137, H. 1—5, 1966.
Лиственные леса умеренной зоны северного полушария — неморальная зона 295
Водный режим растений необходимо рассматривать во взаимосвязи
с фотосинтезом, поскольку движения устьиц влияют на поглощение СО2.
Для продуцирования органического вещества хорошо освещаемые свето-
вые листья имеют большее значение, чем теневые листья. При напряжен-
ном водном балансе теневые листья, как уже упоминалось, закрывают
устьица, тогда как световые листья продолжают ассимилировать СО2.
При фотосинтезе листьев надо различать брутто-ассимиляцию и нетто-
ассимиляцию:
Брутто-ассимиляция = нетто-ассимиляция — расходы на одновремен-
ное дыхание листьев.
Сначала определяют нетто-ассимиляцию, или «видимую ассимиляцию».
Дыхание изучают, затемняя листья в возможно более сходных условиях.
При этом исходят из предположения, что процесс дыхания листьев про-
текает одинаково и в темноте и на свету; однако это не может быть дока-
зано экспериментальным путем. Затем рассчитывают брутто-ассимиляцию.
При повышении температуры увеличение брутто-ассимиляции идет не так
быстро, как потери на дыхание; поэтому оптимальная температура нетто-
асспмиляции относительно невелика и не превышает 15—20°. В связи
с этим даже небольшое повышение температуры листьев, помещенных для
измерения ассимиляции СО2 в камеру, приводит к ошибочным результа-
там. Необходимо поэтому при количественной интерпретации таких опы-
тов проявлять осторожность. Они больше пригодны для качественной
характеристики процесса.
С экологической точки зрения величина нетто-ассимиляции имеет
особое значение, поскольку дает возможность рассчитать продукцию
органического вещества. Однако нетто-ассимиляция не соответствует
нетто-продуктивности, то есть приросту сухого вещества растения. Для
получения нетто-продуктивности нужно из нетто-ассимиляции всех
листьев вычесть расход на дыхание всех незеленых частей растения
и величину дыхания листьев ночью. При одинаковой нетто-ассимиляции
нетто-продуктивность тем меньше, чем больше масса незеленых частей
растения и чем интенсивнее их дыхание.
Суточный ход нетто-ассимиляции листьев бука в целом аналогичен
изменениям транспирационного режима; это означает, что оба процесса
связаны с состоянием устьичного аппарата. При оптимальной влагообес-
печенности нетто-ассимиляция зависит от интенсивности освещения:
наибольшая величина нетто-ассимиляции отмечена на лучше всего осве-
щенной вершине дерева, наименьшая — в наиболее затененных частях
кроны; у очень высоких деревьев величина нетто-ассимиляции на верши-
нах несколько снижается из-за трудности подачи воды на такую высоту.
Сухость воздуха увеличивает значение фактора воды, фотосинтез тене-
вых листьев снижается, поскольку у них преждевременно закрываются
устьица. Если сухость воздуха усугубляется сухостью почвы, то устьица
и у световых листьев смыкаются, что ограничивает транспирацию. При
таких погодных условиях больше всего вещества продуцируется в пред-
полуденные часы. В пасмурные дни, но при светлой облачности бук про-
дуцирует не меньше, а пожалуй, даже больше вещества, чем в ясные
теплые дни1.
1 Н. Polster, Die physiologischen Grundlagen der Stofferzeugung im Walde,
Munchen, 1950; A. P is ek, W.Tranquillini, Assimilation und Kohlenstoffhaushalt
in der Krone von Fichten (Picea. excelsa Link) und Rotbuchenbaumen (Fagus silvatica L.),
«Flora», 141, 1954, S. 237—270; P. J. Kramer, The role of water in tree growth.
Treegrowh, New York, 1962, p. 171—182; T.P.K oslowski, Photosynthesis, climate
and tree growth. Tree growth, New York, 1962, p. 149—164.
296
Глава 4
Таблица 50
Интенсивность дыхания и точки
компенсации у световых и теневых листьев
(по Бойсен-Йенсену)
Дыхание, jug СОг/^М'Час Точка ком- пенсации, % дневного света
Бук световой лист теневой лист fсвето- 1,о 0,2 1,3 0,3
вой
Ясень 4 лист тене- 1,2 1,4
вой k лист 0,4 0,4
меняется
Конкурентные взаимоотношения деревьев зависят прежде всего:
1) от теневыносливости подроста и 2) от прироста в высоту и возраста,
которого могут достичь отдельные деревья. Поэтому долгоживущие тене-
выносливые виды деревьев в благоприятных условиях местообитания со
временем вытесняют светолюбивые виды деревьев. Последние исчезают,
поскольку они не могут возобновляться под пологом теневыносливых
деревьев. Светолюбивые виды сохраняются лишь время от времени в освет-
ленных местах, возникших под влиянием внешних воздействий (бурелом,
лесной пожар, вырубка), или в местообитаниях, неблагоприятных для
произрастания теневыносливых видов. Зональная растительность поэто-
му всегда образуется наиболее
теневыносливым видом деревьев,
образующим сомкнутый древесный
полог, при условии, что остальные
факторы не снижают его конку-
рентную способность.
Теневые листья теневыносли-
вых деревьев не только иначе
реагируют на водный режим, но
они в состоянии более оптимально
использовать слабое освещение,
чем сильное. Интенсивность дыха-
ния у теневых листьев выражена
слабее, а точка компенсации, та
есть интенсивность света, при
которой брутто-ассимиляция рав-
на расходу на дыхание, распо-
ложена ниже, чем у световых
листьев (табл, 50),
течение вегетационного периода.
Точка компенсации
У бука (Fagus silvatica) в период с 10 мая до 29 июля она постепенно сни-
жается: у световых листьев с 1000 до 300 лк, у теневых — с 350 до 100 лк \
Различие свойств световых и теневых листьев имеет большое значе-
ние, поскольку первые освещены значительно сильнее, чем вторые (ср,
табл, 26), Максимальная интенсивность фотосинтеза даже у световых
листьев достигается примерно при 40% полного дневного света. Однако-
эти листья редко используют всю интенсивность солнечного света, по-
скольку их листовые пластинки обычно расположены под большим углом;
наоборот, теневые листья повернуты перпендикулярно к падающим лучам,
что особенно хорошо видно на деревьях, растущих вблизи опушек леса.
Такое расположение световых листьев способствует проникновению
в кроны деревьев большего количества света; положительный баланс
вещества наступает даже у листьев, расположенных в наиболее затенен-
ных частях кроны, при ИЛП, равном 4—5, При отрицательном (в среднем)
балансе вещества, наблюдаемом при очень слабой освещенности теневых
листьев, они начинают желтеть и опадают.
Световой минимум вида деревьев — это наименьшая интенсивность
света, выраженная в процентах от полного дневного света, или то коли-
чество света, при котором еще могут существовать листья внутри кроны.
При уменьшении светового минимума увеличиваются сомкнутость крон
1 Н, L i е t h, Grenze und Anwendungsmoglichkeiten der kolorimetrischen CO2-
Bestimmung, «Planta», 51, 1958, S, 705—721; Uber den Lichtkompensationspunkt der
Landpflanzen, I. und II., «Planta», 54, 1960, S, 530—576,
в
Лиственные леса умеренной зоны северного полушария — неморалъная зона 297
деревьев и затененность леса. У отдельно стоящих деревьев, находящихся
в более благоприятных условиях, световой минимум иногда бывает мень-
ше, чем в сомкнутом древостое (табл. 51).
Таблиц^ 51
Световой минимум основных видов
лиственных деревьев и некоторых хвойных
(по Визнеру и Гессельману)
Вид лиственных деревьев На от- крытом месте, % в дре- востое, % Хвойные На от- крытом месте, %
Fagus silvatica 1,2 1,6 Picea abies 3,61
Carpinus betulus — 1,8 Pinus Silve-
stris . . 10
Acer platanoides — 1,8 Larix deci- dua . . . 20
Acer campestris 2,3 —
Quercus robur 5,01 4,0
Betula verrucosa 11,0
Populus tremula Sorbus. aucupa- 11,0
ria 12,0
Fraxinus excel-
sior . ... 17,0
1 Данные получены в Швеции, поэтому трудносо-
поставимы с другими величинами.
Световой минимум зависит от размеров минерального питания де-
ревьев. На бедных почвах ИЛП ниже, чем на богатых, световой минимум
выше, а леса более изрежены. По Гессельману, световой минимум лещины
(Corylus avellana), произрастающей на скудных почвах, достигает 5,3—
4,3%, а на богатых почвах он равен 2%. Пока еще не выявлены причин-
ные связи, объясняющие это явление.
Световой минимум теневых листьев вцдов деревьев не является пока-
зателем того, сколь сильное затенение может перенести подрост этих же
видов. Световые листья взрослых деревьев получают много света, а"тепе-
вые в крайних случаях должны только обеспечить дыхание в течение
суток. В отличие от взрослых деревьев у подроста все листья находятся
в неблагоприятных условиях освещения; листья подроста должны не
только покрывать расход на дыхание всего растения, но и накапливать
вещество, необходимое для образования листьев в следующем году; если
это не произойдет, растения погибнут. В окрестностях Берлина, в Эбер-
свальде, в течение прохладного, пасмурного года были определены преде-
лы теневыносливости подроста, который был хорошо обеспечен питатель-
ными веществами и водой и не испытывал корневой конкуренции
(табл. 52) \
1 Н. L у г, G. Н о f f m a n n, W. E ng el, Uber den EinfluB unterschiedlicher
Beschattung auf die Stoffproduktion von Jungpflanzen einiger Waldbaume, II., «Flora»,
155, 1964, S. 305—330; P.Burschel, J.Huss, Die Reaktion von Buchensamlingen
auf Beschattung, «Forstarch», 35, 1964, S. 253; P. Burschel, J. Schmaltz, Die
Bedeutung des Lichtes fur die Entwicklung junger Buchen, «Allg. Forst -und Jagdz.» 136,
1965, S. 193—210.
298
Глава 4
Таблица 52
Минимум светового довольствия подроста
некоторых видов деревьев (в % дневного света)
Вид лиственных деревьев Хвойные
Betula pendula 12—15
Quercus borealis 5
Fraxinus excelsior 2—3
Tilia cordata 1
Pinus silvestris 12
Larix decidua 10—12
Picea abies . . . 3—4
Pseudotsuga taxi-
folia . . — 1
Из данных таблицы видно, что подрост выдерживает примерно такое
же затенение, как и теневые листья взрослых деревьев (у ясеня значи-
тельно в большей степени); видимо, это связано с сокращением величины
дыхания всего растения, С увеличением степени затененности подроста
продукция вещества непрерывно снижается. Особенно заметно уменьшает-
ся в валовой сухой массе доля корней, тогда как прирост в высоту даже
увеличивается, В результате этого подрост быстро поднимается над зате-
няющим ярусом травянистых растений и начинает распускать листья
в лучших условиях освещения. Это же явление наблюдалось и в долине
реки Везер с сеянцами бука, достигшими возраста 1—2 лет. Продукция
ими вещества при освещенности, равной 20% дневного света, достигала
около 50% продукции на открытом месте; при усиливающемся затенении
продукция вещества начинала резко сокращаться, однако даже при осве-
щенности, равной 1 %, вещество все еще продуцировалось в размере 2—
6% масссы на открытом пространстве. Угнетение подроста бука начинается
при степени затененности менее 10% (на богатых почвах при значительно
меньших значениях). Рост подроста бука в высоту прекращается при
освещенности менее 20% полного дневного света. При световом доволь-
ствии 12% размеры листьев в 2 раза превышают таковые у листьев, вырос-
ших при полном дневном свете. Продукция вещества в густых посадках
саженцев бука при полном дневном свете составляет до 8 т!га^ при свето-
вом довольствии 1% —только 250—500 кг! га1.
Наиболее критический период наступает для всходов, когда они
израсходуют запасы, содержащиеся в семенах, У сеянцев дуба это про-
исходит только на втором году жизни, при неблагоприятных условиях
освещения они вскоре погибают. Иногда сеянцы в течение первых лет
жизни не увеличивают свою листовую поверхность, В этом случае отно-
шение продуктивной массы листьев к возрастающей из года в год непро-
дуктивной массе стеблей и корней изменяется в неблагоприятном для раз-
вития сеянцев направлении; так продолжается до тех пор, пока баланс
веществ не утрачивает равновесия, и тогда подрост отмирает.
Древостой хорошо использует поступающие в него солнечные лучи
благодаря ступенчатому распределению листьев в кроне деревьев и раз-
личному положению листьев по отношению к падающим лучам. Фото-
синтез древостоя в условиях наиболее благоприятной влагообеспеченности
повышается параллельно с ростом освещенности до максимальных зна-
чений, наблюдаемых при полном солнечном свете; в отличие от того, что
установлено в физиологических экспериментах с отдельными листьями,
расположенными перпендикулярно падающему свету, в них максимальная
1 См. сноску на стр. 297,
Ачень сухая
Сухая
Умеренно
сухая
Умеренно
сбежая
Сбежая
Умеренно
блажная
Влажная
Умеренно
сырая
Сырая
Очень сырая
Воба
Для леса слишком сухо
(Pinus) Многочисленные светолюбивые беребья и кустарники (Pinus)
QUERCUS PETRAEA, ROBUR или PUBESCENS
OOOOOOOOOOOOOOOOOOOOOOOOOOOOOOOOOO
)оСЬи0ы Quercus, Sorbusjilia
Carpinus betulus
a.PETRAEAo°
О
Q.ROBURg
о
Betula opendula
о
о
о
о
о
о
о
о
Виды Acer,
Fraxlnus excelsior
FAGUS SILVATICA
Bu3bi Quercus
Tilta cordata
Carpinus
betulus
OOOOOOOOOOOOOOOOOOOOOOOOOOOOOOOOOO
Betula pubescens Carpinus betulus Дсег pseudoplatanus
QUERCUS robur FRAXINUS excelsior
Вивы Ulmus
BETULA ALNUS glutinosa
.(Ptnus) RUBESCENS
Fraxinus excelsior
Acer pseudoplatanus
Ulmus scabra
Для леса слишком сыро
Очень Кислая Умеренно
кислая кислая
Слабо
кислая
P и c. 219. Зависимость распространения лесо-
образующих пород Центральной Европы от
влажности почвы и ее реакции (насыщенность
основаниями) как следствие конкурентных вза-
имоотношении. Размер шрифта отражает степень
доминирования породы (по Элленбергу, см.
«Phytologie», Bd. IV. 2).
Рис. 220. Зависимость распространения видов
лиственных деревьев и сосны от влажности почвы
и содержания глинистых частиц (питательных
веществ) в области широколиственных лесов за-
паднее Днепра (по Кожевникову, изменено).
о—5 — очень сухая, сухая, свежая, влажная, сырая,
•заболоченная почва; А — Г — песок, супесь, суглинок, се-
рая лесная почва на лёссе.
Область господства бука
Нейтраль- Щелочная
ная
--------№
--------Остролистный клен и липа
--------Граб
-------- Ясень и яоор
нннн Рябина
-------- Черная ольха
300
Глава 4
1
3 4
5
Очень сухая
Сухая
Умеренно
сухая
Умеренно
свежая
Свежая
Умеренно
влажная
Влажная
Умеренно
сырая
Сырая
Очень сырая
Вова
Populus tremula
£___________r__________________________________ч
porbus aria [dmelanchier avails Rosa pomlfera I
|S.torminalis’Cotoneaster Itegerr. Cornus mas I
l₽runus mahJBerberis vulgaris I
oqOOOO OO(
Sarothamnus
° scoparius
Lonicera periclymenum
о
о
о
< б >
Ilex aquifolium
Rhamnus frangula
Rubus serpens u 9p.
о
'-------
°oooo ooc
Pinus montana prostrat
• Prunus spin.; Cornus sanguin.
! Rosa can. ’ Rhamnus cath.
I Crataegus | Evonymus eur.
i Corylus av.pDaphne mezer.
»Lonlc.xy lost! deer campestre
!<------1-'--------
ISambucus :
j racemos |
! Rubus id. : Sambucus
[Viburnum • nigra
j opulus j
I
^00000^000000000000000000
Viburnum Lantana
Clematis vitalba
Ligustrum vulgare
•Ribes uva crispa
Rubus caestus
Vitis silvestris
(Hu mu Lus rRlbes alpinum
i lupulus / Prunus padus
^Salix aurita.cineria
Myrica gale^Ribes nigrum
• 1
о о о 2
Очень Кислая Умеренно кислая Слабо Кис- Нейтраль- Щелоч-
кислая кислая лая ная ная
Рис. 221. Экологический ареал основных видов кустарников Центральной Европы
(аналогичный изображенному на рис. 219) (по Элленбергу, см. «Phytologie», Bd. IV, 2).
J — теневыносливые виды; 2 — область распространения буковых лесов.
интенсивность фотосинтеза отмечена уже при освещенности, равной
40% дневного света.
Среди всех видов деревьев, распространенных в Центральной Европе,
у бука (Fagus silvatica) развивается наиболее густая крона. Поэтому этот
вид растет в любых благоприятных для него местообитаниях и образует
зональную растительность. К востоку, при увеличении континентальности
климата, бук, как уже было упомянуто, вытесняется грабом (Carpinus
betulus'). Это также теневыносливый вид, который там, где климат стано-
вится слишком сухим для него, может сменяться светолюбивым дубом
(Quercus robur). В приатлантической области Западной Европы с бедными,
кислыми и постоянно влажными почвами для бука благоприятны только
водопроницаемые карбонатные почвы. В этой части Европы, как уже
говорилось, зональные леса образованы дубом (Quercus petraea, Qu. robur)1.
1 В приатлантической области почвы под буковыми лесами очень быстро дегради-
руют, поскольку в них образуется мощный слой грубого гумуса; это явление не наблю-
дается под дубовыми лесами (A. G а 1 о u х, Le hetre et 1а degradation des sols forestiers
loessiques, «Bui. Soc. Roy, Forest. Belgique», 1953, p. 225—235). Стекающие co стволов-
Лиственные леса умеренной зоны северного полушария — неморалъная зона 301
Дальнейшее подразделение лесов в пределах любой климатической
зоны связано с почвенными условиями; решающее значение имеют влаж-
ность и реакция почвы, поскольку последняя свидетельствует о содержа-
нии питательных веществ в ней.
Зависимость конкурентной способности отдельных лиственных пород
•от факторов местообитания выявлена на основе эмпирических наблюдений
работников лесного хозяйства и фитоценологов. Эти результаты, получен-
ные в условиях Центральной Европы, обобщены Элленбергом (рис. 219).
Схема показывает господство бука (Fagus silvatica) в Центральной Европе.
По-иному складывается обстановка в области распространения лиственных
лесов в западной части Восточной Европы (рис. 220). Здесь доминируют
дуб и сосна.
Экологические особенности кустарников рассмотрим очень кратко.
Кустарники не играют существенной роли в сильно затененных зональ-
ных лесах; их значение возрастает на опушках леса, на участках, где
лес искусственно уничтожен, или в наименее пригодных для произраста-
ния деревьев местообитаниях. Все виды кустарников оптимально разви-
ваются в условиях полного дневного освещения. Экологический ареал
кустарников, отражающий их распределение по отдельным местообитани-
ям в результате межвидовой конкуренции, изображен на рис. 221. Как
и у деревьев, в крайние условия существования вытесняются наименее
конкурентоспособные виды, а наиболее конкурентомощные занимают
лучшие местообитания. ।
ЭКОЛОГО-ФИЗИОЛОГИ ЧЕСКИ Е ИССЛЕДОВАНИЯ
ТРАВЯНОГО ПОКРОВА В ЛИСТВЕННЫХ ЛЕСАХ
Древесный ярус в лесах получает полное дневное освещение, а травя-
нистые растения довольствуются светом, который проникает сквозь
кроны деревьев и ярус кустарников, если последний развит. Среди травя-
нистых растений встречается много видов, которые лучше растут в тени
и поэтому не встречаются за пределами леса. В табл. 23—31 приведены
данные, характеризующие изменения условий местообитания в дубовых
лесах около поверхности лесной почвы по сравнению с условиями на
верхней поверхности крон деревьев. В лиственных лесах, образованных
другими видами деревьев, эти показателе сходны, хотя и различаются
в количественном отношении. По сравнению с открытым местом поверх-
ность почвы в лесу освещена значительно меньше, колебания температуры
после облиствления деревьев также менее выражены, а влажность воздуха
и в верхних горизонтах почвы устойчиво высокая; медленное движение
воздуха в лесу способствует увеличению количества водяного пара под
пологом деревьев.
Поэтому с экологической точки зрения виды травяного покрова в лесу
относятся к числу теневыносливых растений и гигрофитов.
Условия освещения в лиственном лесу отличаются большой сложно-
стью х. Визнер изучал освещенность на поверхности почвы в лесу и опре-
дуба осадки обладают нейтральной реакцией, со стволов бука — кислой (pH = 4,0—
—4,5); почва вокруг стволов бука обеднена известью и зарастает мхами, свойственными
кислым почвам (A. G а 1 о u х, Compts. Rend, И. Congr. Un. Internal. Inst. Rech. Forest.
Rome, 1953, Firenze, 1954, p. 343—356).
1 W. Tranquiln i, Das Lichtklima wichtiger Pflanzengesellschaften,
«Hb. d. Pflanzenphysiologie», Bd. V, 1960, S. 304—338.
302
Глава 4
Рис. 222. Интенсивность света, определенная при помощи самопишущего люксмет-
ра, в первой половине солнечного дня и после полудня при переменной облачности
(И мая 1959 г.) в Хоэнхейме (по Литу).
1 — открытая местность; 2 — в тени леса со средней величиной светового довольствия 6%. Пики
на кривой — многочисленные пятна света на поверхности почвы в лесу. Общая высота ординаты
равна 100 тыс. лк.
делил «световое довольствие», иными словами, интенсивность света на
поверхности почвы в процентах от освещенности за пределами леса Ч
В лесу можно различать глубоко затененные места и менее затененные,
а также период хорошей освещенности поверхности почвы перед облист-
влением деревьев и период неблагоприятных условий освещения после
облиствления (Солсбери) 1 2. Световое довольствие легко определить в те
дни, когда небо равномерно покрыто облаками, и значительно труднее
в дни, когда на почве в лесу мелькают многочисленные пятна света
(рис. 222). Световое довольствие на поверхности почвы зависит также от
высоты стояния солнца; эта величина будет отличаться в первой и во
второй половине дня от таковой в полдень; весной она иная, чем в середине
лета, когда солнце расположено высоко на небосклоне. Фотосинтез тра-
вяного покрова зависит, естественно, не от светового довольствия, а от
абсолютной интенсивности света, которая ежедневно меняется. Поэтому
Андерсон 3, изучая световой климат лиственного леса в окрестностях
Кембриджа (Англия), в течение трех лет измерял суммарную радиацию
за сутки на трех различных участках в лесу:
1) в сомкнутом древостое,
2) на небольшой прогалине,
3) на большой поляне, диаметром 20 ж.
Прямая и рассеянная радиация изучалась раздельно. Радиацион-
ный режим в климатических условиях Англии, отличающихся резко
выраженной переменностью облачности, подвержен большим колебаниям.
На протяжении одной недели величина суммарной радиации за сутки
может изменяться 5 раз. Наглядное представление о радиационном режи-
ме можно получить, рассматривая рис. 223. На открытом местообитании
кривые суммарной радиации за месяц симметричны и характеризуются
максимумом с июня по июль. Наоборот, в лиственном лесу при увеличении
1 J. W i е s п е г, Der LichtgenuB der Pflanzen, Leipzig, 1907.
2 E. J. Salisbury, The oak-hornbeam woods of Hertfordshire. «J, Ecol.», 4,
1916, p, 83-117; 6, 1918, p. 14-52.
3 M.C. Anderson, Studies of the Woodland light climate, «J. Ecol.», 52, 1964,
p. 27—41, 643 — 663; H. Schliit er, Licht- und Temperaturmessungen an den Vegeta-
tionszonen einer Lichtung («Lochhieb») im Fichtenforst, «Flora», Abt. B, 156, 1966.
S. 133—154.
Лиственные леса умеренной зоны северного полушария — неморольная зона 303
степени сомкнутости древесного полога максимум все больше и больше
сдвигается к апрелю. При сопоставлении величины месячной суммарной
радиации (в % от радиации за год. поступающей на соответствующие
Р и с. 223. Среднемесячная радиация в окрестностях Кембриджа (Англия) (по
Андерсону, дополнено).
На открытом местообитании: 1 — суммарная радиация, 2 — прямая солнечная радиация, <3 —
рассеянная радиация. В лиственном лесу: 4 — на большой поляне, 5 — в сомкнутом древостое.
а — распускание листьев; б — полное облиствление; в — листопад.
местообитания) видно, насколько мала интенсивность света в лесу
в период между облиствлением и листопадом (рис. 224). Благоприятные
условия освещения до начала облиствления деревьев создают возможность
Рис. 224. Месячная суммарная радиация в % от радиации за год, поступающей на
соответствующие местообитания (тот же эксперимент, что и на рис. 223) (по Андерсону,
дополнено).
1 — открытое местообитание, 2 — большая поляна, 3 — древостой, незадолго до облиствления.
особой экологической группе растений — весенним геофитам — пройти
полный цикл развития, прежде чем появятся конкурирующие виды.
Эти весенние геофиты имеют подземные запасающие органы (корне-
вища, луковицы, клубни), расположенные на небольшой глубине под
рыхлой подстилкой. Небольшая теплоемкость подстилки способствует ее
304
Глава 4
Листья Carpinus
распустились
полностью
Рис. 225. Последовательность распускания листьев (штриховка) и цвете-
ния (залито черным) весенних геофитов до начала облиствления леса Scilla
nutans = Endymion nonscriptus (по Солсбери, из работы: Н. Waite г,
1927).
быстрому нагреву весной, когда солнечные лучи, почти не задерживаемые
древостоем, непосредственно достигают поверхности почвы 1. Уже в апре-
ле температура может достигать 25—30°, поэтому геофиты, у которых
подземные органы расположены близко к поверхности no4Bbi(Ranunculus
jicaria, Scilla bifolia, Corydalis, Anemone и др.), быстро образуют надзем-
ные облиственные побеги и затем цветут. Корни деревьев распространены
в более глубоких и медленнее прогреваемых горизонтах почвы; в связи
с этим деревья покрываются листьями позже, чем виды травяного покро-
ва. Фенологические спектры представлены на рис. 225.
Как происходит продуцирование вещества у этих видов, показал
Коич на примере Leucojum vernum, Galanthus nivalis и Allium, ursinum 1 2.
В Ботаническом саду Хоэнхейма у первых двух видов формирование
надземных органов начинается с 19 марта и продолжается до 12 апреля —
24 мая. Листья распустились полностью к 14 апреля. Площадь листьев
определялась планиметром. Величина продуцированного сухого вещества
вычислялась как разность общего сухого веса растений и сухого веса
луковиц до начала их прорастания. Образцы (5—6), взятые в период
развития растений, позволяли определить величину ассимиляции в г су-
хого веса на 1 л2 листовой поверхности и на протяжении дня.
1 F. F i г b a s, Uber die Bedeutung des thermischen Verhaltens der Laubstreu fur
die Friihjahrsvegetation des sommergriinen Laubwaldes, «Beih. Bot. Zbl.», 44, II, 1928,
S. 179—198; A. G a 1 о u x, Rythmes energetiques et phenophases en chenaie calcaire,
«Bull. Soc. Roy. Bot. Belg.», 99, 1966, p. 189—200. Автор выделяет 10 фенофаз в дубовом
лесу; по его данным, в течение весны и раннего лета в сообществе продуцирование веще-
ства все больше перемещается от травяного покрова в древесный ярус, что связано с ра-
диацией и температурными условиями.
2 М. К о j i с, Uber den Transpirationskoeffizienten der Fruhjahrsgeophyten,
«Flora», 154, 1964, S. 212—214; Der Assimilationshaushalt einiger Zwiebelpflanzen unter
Laboratoriums- und feldmabigen Bedingungen, «Вег. Deutsch. Bot. Ges.», 77, 1964,
S. 313—316; liber die Wasserbilanz und Stoffproduktion bei Fruhjahrsgeophyten, «Вег.
JDeutsch. Bot. Ges.», 79, 1966, S. 188—197.
Лиственные леса умеренной зоны северного полушария — неморальная зона 305
Наряду с этим изучался расход воды на транспирацию в течение
каждых трех дней. Величина транспирации нарастала до 14 апреля и до-
стигала почти 2 г на растение в сутки. По мере пожелтения листьев расход
воды вновь начал снижаться. Общее количество воды, израсходованное
на транспирацию, и общее количество продуцированного вещества дают
возможность вычислить коэффициент транспирации. Осредненные величи-
ны этих показателей для отдельно взятого растения приведены в табл. 53.
Таблица 53
Средние величины ассимиляции и коэффициент
транспирации двух весенних геофитов
Leucojum ver-
num ....
Galanthus niva-
lis
Макси-
маль-
ная
листо-
вая по-
верх-
ность,
см2
Про-
дуци-
рован-
ное
сухое
веще-
ство, г
Средняя
величина
ассимиляции,
г/м2-сутки
Коэф-
фи-
циент
транс-
пира-
ции
2,78 0,138 8,0—13,31
1,82 0,062 7,2—7,8
87
НО
1 He учитывается, поскольку выпадает из общего
порядка величин.
Столь небольшая продукция сухого вещества вначале вызывает
удивление. Однако следует принять во внимание высокое содержание
воды в растениях. Коэффициент транспирации А Шит ursinum равен 103,
а в среднем для всех
трех видов он равен 100.
Это самый низкий из
известных в настоящее
время показателей. Не-
значительная величина
коэффициента транспи-
рации, возможно, свя-
зана с развитием расте-
ний влажной и прох-
ладной весной. Так как
коэффициент транспира-
ции Allium сера даже
летом не превышает 191,
то можно предполо-
жить, что слабая транс-
200-
Коэффициент
транспирации
Рис. 226. Зависимость коэффициентов транспирации
у Allium ursinum и А. сера от температуры и дефици-
та насыщения воздуха (по Кёнигу и Кончу).
пирация свойственна
всем однодольным растениям Ч На рис. 226 представлена зависимость
коэффициента транспирации Allium ursinum, и для сравнения А. сера,
от температуры и дефицита насыщения воздуха.
Развитие весенних геофитов в период до начала облиствления леса
и их пожелтение после распускания листвы могут натолкнуть на мысль, что
1 Этот вывод нельзя отнести ко всем однодольным, поскольку коэффициент тран-
спирации у злаков значительно выше.— Прим. ред.
306
Глава 4
их отмирание обусловлено нехваткой света. Однако это неверно, посколь-
ку в условиях культуры при достаточном количестве света листья геофи-
тов желтеют в то же самое время или даже несколько раньше. Как эта
было показано на примере Ranunculus ficaria, речь идет об эндогенном
ритме развития: после цветения и заполнения запасающих органов
(корневых клубней и почек в пазухах листьев) прочие части растения
отмирают Запасающие органы находятся в состоянии покоя вплоть до»
осени. Поздней осенью и зимой из-за низких температур растения не
развивают надземных побегов, они появляются только ранней весной. На
экспериментальным путем было доказано, что временное воздействие
холода (при температуре от 4-2 до +4°) ускоряет прорастание клубней
после окончания периода покоя.
Эндогенный ритм развития и яровизация низкими температурами
обусловливают раннее развитие растений до распускания листьев на
деревьях; геофиты заполняют в лиственном необлиствленном лесу пустую-
щую экологическую нишу 1 2. Используя метод половинок листьев Саксаг
Мудрак определил величину ассимиляции СО2 у этого вида. При тем-
пературе 5—15° наблюдается наиболее благоприятный баланс. При тем-
пературе 15—25° баланс нередко отрицательный3. Оказалось, что тем-
пература имеет большее значение, чем интенсивность света. Затененные
листья ассимилировали более энергично, чем освещенные; однако освещен-
ные растения развивались значительно быстрее затененных. Фаза разви-
тия растения влияет на фотосинтез листьев: в период цветения фотосинтез
достигает минимальных значений4. Максимальная суточная величина
сухого вещества, продуцированного в процессе ассимиляции, равна, па
Мудраку, 10,6 мг/час*м2 (у Anemone — 12,8 мг). В течение короткого
вегетационного периода растение, у которого площадь листьев всего 10 —
11 см2, производит 250—300 мг вещества. Эти данные полностью совпада-
ют с величинами, полученными при изучении Leucoujum и Galanthus.
Суточный ход транспирации и ассимиляции изучался (каждые 2 —
3 дня) у весенних геофитов в культуре, а у Anemone nemorosa и Arum
maculatum в природных условиях 5. У этих видов устьица слабо регулиру-
ют отдачу воды. В солнечные дни температура листьев значительно выше
температуры окружающей среды; в результате, несмотря на высокую
влажность почвы, водный баланс растений нарушается и интенсивность-
фотосинтеза снижается. Наиболее высокие значения нетто-ассимиляции
1 Такое же явление характерно и для сахарно-кленовых лесов, распространенных
в Квебеке (Канада). В этих лесах весенние геофиты представлены Claytonia caroliniana,
Erythronium americanum, Trillium erectum и др. Два первых вида прекращают свое
развитие до начала облиствления леса и существенного ухудшения освещенности (см.:
Р. Е. Vezina, М. М. Grand ter, Phenological observations of spring geophytes
in Quebec, «Ecol.», 46, 1965, p. 869—872).
2 F. Mudrack, Uber die Assimilationstatigkeit und das Wachstum von Ficaria
verna, «Planta», 23, 1935, S. 71—104; H. A u g s t e n, Die Wirkung tiefer Temperaturen
auf die Entwicklung von Ranunculus ficaria L., «Ber. Deutsch. Bot. Ges.», 70, 1957r
S. 234—244.
3 По Лундегарду, оптимальная температура фотосинтеза у Anemone nemorosa
равна 8—10°. См.: Н. Lundegardh, Klima und Boden in ihrer Wirkung auf das
Pflanzenleben, 5. AufL, Jena, 1957; см. также: E. Lohr, Photosynthese von Anemone ne-
morosa, «Physiol. Plant.», 5, 1952, S. 221—227.
4 M. Spanin g, Die Assimilation einiger Friihjahrs- und Sommerpflanzen im
Verlaufe ihrer Vegetationsperiode, «Jb. wiss. Bot.», 89, 1941, S. 574—614.
5H. Bethke, H. Haas, O. Stocker, Uber den Wasser- und Photosyn-
thesehaushalt einiger Fruhjahrsgeophyten, «Flora», Abt. B, 156, 1965, S. 8—49.
Лиственные леса умеренной зоны северного полушария — неморалъная зона 307
достигают 0,14—0,22 мг СО2/дм2>мин, а продукция вещества, вычислен-
ная в сахарозе, равна 1—1,5 г/м2 -сутки. Это в 6—7 раз меньше, чем при
непосредственном определении ассимиляции по приросту сухого веса,
сходные показатели установлены для зерновых культур. Поэтому
данные, полученные в середине солнечного дня при изучении фото-
синтеза в ассимиляционной камере, вызывают сомнение, поскольку
они могут отражать специфический режим, создаваемый камерой. Мало-
вероятно, чтобы ассимиляция весенних геофитов, развивающихся в тече-
ние короткого вегетационного периода продолжительностью менее двух
месяцев, с относительно короткими днями, была столь незначительной.
На лугах, в условиях полного освещения солнцем они развиваются
нормально: не увядают и не прерывают хода ассимиляции.
После облиствления деревьев условия освещения поверхности лесной
почвы не только ухудшаются, но к тому же становятся неравномерными.
Это отражается на характере распределения лесного травяного покрова.
На более осветленных местах травяной покров очень хорошо развит,
а на затененных участках встречаются лишь немногие растения, которые
часто не плодоносят. Различия в флористическом составе яруса травяни-
стых растений в буковых, дубово-грабовых и чистых дубовых лесах
в первую очередь зависят от различий в условиях освещения.
Визнер изучал световое довольствие отдельных видов в типичных
для них местообитаниях. Он установил, что большинство лесных растений
различаются по максимальному для них световому довольствию (Lmax),
так как они не встречаются при полном дневном освещении, и по мини-
мальному световому довольствию (Lmjn), то есть по наименьшей относи-
тельной освещенности, при которой у них еще выражен положительный
баланс веществ, отчего растения не погибают (табл. 54).
Таблица 54
Lmax и Lmin лесных растений (в %)
Corydalis cava
Anemone пето-
rosa ....
Lamium macu-
latum ....
50—25
40-20
67—12
Alliaria officinalis
Geranium robertia-
num . ...
Cynanchum vince-
toxicum . .
Lathyrus vernus
33—20
Prenanthes purpu-
rea .............
33-9
74-4
67-64, 5
(до 3%
у вегетатив-
ных особей)
10-5
(до 3%
у вегетатив-
ных особей)
Почему лесные растения растут только в тени? Это объясняется раз-
ными причинами. Многие виды отличаются большой гигрофильностью;
они произрастают на постоянно влажных почвах или не выносят высокого
дефицита насыщения воздуха. Местообитания, освещенные полным днев-
ным светом, для них слишком сухи. Во влажном горном климате Corydalis
cava и Anemone nemorosa растут на незатененных местообитаних. В Англии
для Scilla (= Endymion) non-scripta установлено, что оптимальные условия
для ее роста реализуются при полном дневном освещении; то, что она
20*
308
Глава 4
Ле-; Qu.-Betuletum
pexodj molinietosum
с бедным
травяным
покровом
Querceto-Carpinetum
вар., с богатым травяным
покровом
ж ,Л ’-----1----1----1----1----1—
№ 1 6 6В 2 3 9
pH 7,8 6,2 (5,8) 6,0 5,7 4,8
(At+A2)
8 11 12 41 13 14 22 15
4,3 4,3 4,1 (3,9) 3,7 3,6 (3,6) 3,55
Рис. 227. Зависимость величины Lmin от pH почвы. С увеличением кислотности
почвы, являющейся показателем бедности ее питательными веществами, потребность
в свете у одного и того же вида растений повышается (по Элленбергу, см. «Phytologie»,
Bd. IV, 2).
встречается только на затененных участках леса, связано с отсутствием
здесь конкуренции луговых злаков. Распространение Scilla и на прогали-
нах, заросших папоротником (Pteridium aquilinum), объясняется ее более
ранним развитием по сравнению с папоротником; кроме того, орляк пре-
пятствует появлению луговых злаков Ч
Величина Lmm является показателем границы голодания растений.
При меньшем световом довольствии растения еще могут существовать, не
формируя органов размножения, поскольку для их развития необходима
более интенсивная ассимиляция, и, помимо того, усиление дыхания.
Величина Lrain, по-видимому, тем ниже, чем более благоприятны для
роста другие условия. Поэтому этот показатель повышается при низких
температурах. В окрестностях Вены Corydalis cava встречается при
Lmin = 25%, а в горах Lmin составляет 80% (у Anemone —77%). На
бедных, кислых почвах Lmin выше, чем на почвах, богатых питательными
веществами (рис. 227). В безжизненные, затененные участки леса, где уже
не могут расти высшие автотрофные растения, в условиях Центральной
Европы поступает лишь 2% полного дневного света или несколько меньше.
Меньшим количеством света довольствуются папоротники со слаборазви-
тыми осевыми органами, которые образуют только споры. Asplenium
trichomanes растет в пещерах даже при Lmin = !%• Мхи и водоросли,
1 G. Е. Blackman, An assessment of the factors controlling the distribution
of the Bluebell (Scilla non-scripta) in different communities, «Ann. of Bot.», N. S., 14, 1950
p. 487—520.
Лиственные леса умеренной зоны северного полушария — неморальная зона 309
у которых развиты лишь
зеленые ассимилирующие
клетки, встречаются даже
при Lmin менее 0,1% днев-
ного света.
Растения, произра-
стающие в глубокой тени,
в солнечные дни уравно-
вешивают баланс вещества
интенсивным фотосинтезом
в те моменты, когда на них
падают солнечные блики.
Характер изменения осве-
щенности на поверхности
лесной почвы в первой по-
ловине довольно ясного
дня и после полудня при
переменной облачности
представлен на рис, 222,
Даксер 1 исследовал
фотосинтез травянистых
растений лиственных ле-
сов летом при слабом осве-
щении, В этих условиях
устьица целый день оста-
ются открытыми. Опти-
мальная температура нет-
то-ассимиляции—15—18°,
Различают две группы рас-
тений в зависимости от их
приспособленности к сла-
бому освещению.
1, Виды, у которых
после облиствления де-
ревьев снижается интен-
сивность дыхания; в ре-
зультате либо нетто-асси-
миляция достигает поло-
жительных величин, на-
пример у папоротника
Aspidium jilix-mas и под-
роста (Fagus, Fraxinus),
либо предотвращается сни-
жение ассимиляции ниже
точки компенсации (у Оха-
lis или Viola),
2, Виды, у которых
ход ассимиляции не пере-
страивается после обли-
1 Н, D а х е г, Uber die
Assimilationsokologie der Wald-
bodenflora, «Jb, Wiss, Bot,»,
80, 1934, S, 363—420,
Расстояние от опушки леса,м
Рис, 228, Увеличение продуктивности травяного
покрова с повышением светового довольствия при
приближении к опушке леса. Среднее из двух серий
пробных площадей в еловом лесу (вычислено по
данным Даксера), В травяном покрове господствуют
Oxalis acetosella, Senecio fuchsii и Geranium rober-
tianum.
Рис, 229, Суточный ход содержания СО2 в сосно-
вом лесу (среднее за июнь — июль); II бонитет, воз-
раст 70—80 лет, сомкнутость крон — 0,7 (по Кобак),
1 — на поверхности почвы; 2 — на высоте 15 см от поверх-
ности почвы; з — на высоте 1,3 л; 4 — у основания крон;
5 — в кроне; 6— на высоте 2—3 м над поверхностью крон.
310
Глава 4
Полный Знойной свет,лк
сэ о о о о О О ОООООООООСЭ
о о о о о о о о оооооооосэ
О О О О ООО ОООСЭСЭСЭСЭСЭСЭСЭ
т- CXI СО о LO о -V- сП LDOminOLOCO^
т- т- CO-r-CNCNCNCOOJCxjC'J
Освещенность поверхности почвы в лесу, лк
ю о о о □□□ ю оооосэсэсэсэа
с? ю о ю о о о счиооооиосэсэют
04 04 т- с\| т- ’r-CNC4Xj-COCOCQx-
О О OOQO
о о сэсэсэсэ
С\1 ОО COOCCOCN
hepaticc
Апетпопе henatico.
18 апреля
430 630 8 30 1О30 13 15
29 мая
Рис. 230. Опыты для выяснения момента уравновешивания суточного баланса
(УСБ) у различных видов лесных трав.
Суточный баланс уравновешен, когда количество С02, выделенное в процессе дыхания ночью,
вновь ассимилируется днем; в результате в колбе восстанавливается нормальная концентрация С02,
отмеченная предыдущим вечером. Ночью 18 апреля у Anemone hepatica наблюдалось интенсивное
дыхание, однако УСБ наступило уже около 9.30 утра. После периода плохой погоды в ночь на
29 мая величина дыхания оставалась небольшой; УСБ не было достигнуто из-за малой освещенности.
После многих дней хорошей погоды 12 июня интенсивность дыхания ночью вновь увеличилась;
УСБ произошло около 12.30, как и у подроста Fagus. В этот день точка УСБ была достигнута:
у отличающейся слабой интенсивностью дыхания Asperula — около 10.40, у Asarum — около
13.30, у Viola — после 15 часов, тогда как баланс подроста Carpinus и Hedera оставался отрицатель-
ным. Температура измерялась в стеклянных колбах, размещенных на поверхности лесной почвы.
ствления; точка компенсации у них не меняется. Эти виды голодают летом;
они характеризуются отрицательным балансом. Только осенью, после
осветления древесного полога, ассимиляция растений достигает положи-
тельных величин. К этой группе принадлежит много лесных видов, разви-
вающихся летом: Fragaria vesca, Lathyrus vernus и L, montanus, Stellar la ho-
lostea, Hypericum perforatum. Anemone nemorosa (пока листья не погибли)
и вечнозеленая Pirola rotundifolia. При увеличении освещенности ближе
к опушке леса растения развиваются значительно лучше. Это хорошо вид-
но при сопоставлении продуктивности травяного покрова (на 1 л1 2)
и светового довольствия (рис. 228) Ч
Даксер исследовал фотосинтез отдельных листьев. Продуктивность
может быть определена лишь при изучении растения в целом. Такую
работу провел К. Зиб (неопубл.), используя метод Alvik.
Эти опыты могут быть использованы только для получения качествен-
ных показателей. Большей точностью отличаются летние наблюдения,
когда интенсивность дыхания растений травяного покрова в лесу резко
снижается и круговорот С02 достигает небольших значений (рис. 229).
1 По данным Медвецка-Корнас (цит. стр. 284), в буковом лесу на известняках, где
господствуют Asperula odorata п Asarum europaeum, валовая надземная фитомасса тра-
вяного покрова увеличивается с 2 г/м2 сухого вещества в апреле до 4,5 г!м2 в июне,
в затем к поздней осени снижается до 1,5 г!м2. Подземная сухая масса также достигает
наибольших величин в июне; однако она мало колеблется около 3,5 г!м2', одна четвертая
’часть сухого вещества приходится на корни, а остальное — на подземные побеги.
У Asarum количество надземной сухой массы остается почти неизменным в течение года,
у Asperula оно колеблется довольно значительно.
Лиственные леса умеренной зоны северного полушария — неморалъная зона 311
Полный Знебной свет, лк
Освещенность поверхности почвы 8 лесу, лк
С02 б колвах, мг
2 мая 15 мая 4 июня
Рис. 231. Опыты со всходами некоторых видов деревьев, произведенные в течение
трех разных по погодным условиям дней. Обозначения см. рис. 230.
Такой способ приспособления растений к неблагоприятным условиям
освещения летом не выражен у Hedera helix и Fragaria vesca, но зато хоро-
шо прослеживается у Dryopteris filix-mas, Anemone nemorosa, A. hepatica,
Viola silvatica, Asperula odorata, Asarum
europaeum, Convallaria majalis и у сеянцев
Fagus, Carpinus, Quercuspetraea и Abies alba.
Пасмурная погода перед началом облист-
вления леса снижает величину дыхания;
интенсивность дыхания вновь увеличива-
ется при наступлении солнечной погоды
(рис. 230). Создается впечатление, что
снижение интенсивности дыхания связано
с голоданием растений и низким обеспече-
нием углеводами. Рис. 231 —232 иллюст-
рируют некоторые примеры. Ниже приво-
дится обобщение результатов этих иссле-
дований.
Продуктивность лесных травянистых
растений определяется не относительной
освещенностью, а суммой дневного света,
достигающей поверхности лесной почвы.
Поэтому уже перед началом полного обли-
ствления деревьев суточный баланс (реас-
симиляция выделяемого за ночь СО2)
в очень пасмурные дни не уравновешива-
ется. После облиствления древостоя виды,
приспособленные к небольшому освеще-
нию, стабилизируют суточный баланс
только в ясные дни; в дождливую погоду
это явление не наблюдается. Hedera и Fra-
garia не снижают заметно интенсивности
своего дыхания, поэтому летом суточный
Полный дневной свет,лк
N С\1 г-
Освещенность
поверхности почвы
6 лесу,лк
О *“Т-----------1-----1----1----1-----1
Рис. 232. Опыт, проведенный
в пасмурный осенний день, когда
листья с деревьев почти полно-
стью опали. Все три вида дости-
гают точки УСБ около 11 часов,
в том числе Hedera, отличающаяся
слабой интенсивностью дыхания
(ср. состояние 12 июня на рис.
230). Обозначения см. рис. 230.
312
Глава 4
Интен-
Р и с. 233. Компенсационные точки освещения (сплошная линия)
и дыхание в темноте (прерывистая линия) у Allium ursinum и Asarum
europaeum в лиственном лесу (по Литу).
баланс у них редко уравновешивается. Это происходит только осенью,,
после начала листопада (рис. 232). й
Кабен изучал фотосинтез Lamium galeobdolon в течение целого года.
Он установил, что растения летом голодали. Наибольшей интенсивности
фотосинтез достигал осенью; зимой баланс еще сохранялся положитель-
ным х.
Лит изучал способность некоторых лесных теневыносливых растений
приспосабливаться к различным условиям освещения. Он определял это
по изменению компенсационной точки света 1 2. Компенсационная точка
перед началом распускания листьев равна примерно 900 лк, после облист-
вления быстро снижается до 200 лк и затем остается на этом уровне без
изменений. Снижение обусловлено уменьшением интенсивности дыхания
(рис. 233), тогда как фотосинтетическая деятельность протекает без замет-
ных изменений (см.: Даксер, стр. 309). Весной при наступлении поздних
заморозков компенсационная точка повышается иногда в течение не-
скольких дней, однако это не влияет на процесс дыхания (рис. 234).
В этом случае фотосинтетический аппарат повреждается низкой темпера-
турой воздуха.
1 Н. К a b е n, Die Schwankungen der CO2-Assimilation von Lamium galeobdolon.
(L.) Crantz im Laufe eines Jahres, «Biol. Zbl.», 78, 1959, S. 213—231.
2 H. L i e t h, Uber den Lichtkompensationspunkt der Landpflanzen. II, «Planta»,
54, 1960, S. 555—576.
Лиственные леса умеренной зоны северного полушария — неморальная зона 313
Компенсационная точка весенних геофитов изменяется по-иному.
Время облиствления совпадает у них с пожелтением листьев; в соответ-
ствии с этим резко повышается компенсационная точка; повышение это
продолжается до тех пор, пока полностью не прекратится фотосинтез.
Снижение компенсационной точки после облиствления, то есть уменьше-
ние интенсивности дыхания от ранней весны к лету, является, по-види-
мому, в основном эндогенной реакцией растения, поскольку это снижение
наблюдается также у растений, растущих в благоприятных условиях
освещения за пределами леса. У Oxalis acetosella развиваются только
теневые листья; это выражено в меньшей степени, если растения культиви-
ровать при световом довольствии, равном 50% дневного света. Макси-
Р и с. 234. Компенсационные точки освещения у Allium ursinum
в 1958 г. (вверху) и в 1959 г., когда наблюдалось два периода заморозков
(внизу) (по Литу).
1 — световое довольствие, определенное люксметрами, расположенными на расстоя-
нии 6 м друг от друга в травостое Allium; 2 — растения, произрастающие в зате-
ненных местообитаниях; з — растения, пересаженные на открытое место; последние
находились под большим воздействием заморозков и раньше пожелтели.
314
Глава 4
мальная величина фотосинтеза у кислицы при освещенности, достигаю-
щей 8000 лк, почти в 3 раза выше, чем у растений, встречающихся в самых
затененных участках леса. Интенсивность дыхания на единицу поверхно-
сти листьев у первых также выше, чем у растущих в тени. У растений,
культивируемых на свету, клетки палисадной ткани несколько длиннее,
а сырой вес единицы поверхности листьев в 1,5 раза больше Ч
Субатлантический вид Teucrium scorodonia, приуроченный к кислым
лесным почвам, относится к евритопным в отношении условий освещения
видам. Если этот вид культивируется при освещенности в пределах 100 —
12% дневного света, то продуктивность суточной ассимиляции на единицу
поверхности листьев уменьшается соответственно падению освещения.
Поскольку в тени образуется большее число листьев, большая по разме-
рам площадь листьев компенсирует меньшую интенсивность ассимиляции;
в связи с этим продуктивность побега остается примерно одинаковой при
любой освещенности 1 2.
Интенсивность фотосинтеза зависит не только от света, но и от дру-
гих факторов, и в частности от поглощения СО2 листьями. Концентрация
СО2 в приземных слоях воздуха в лесу может повышаться, и особенно
ощутимо в утренние часы, до начала турбулентного перемешивания
воздуха в глубине леса. В результате усиливается фотосинтез растений
почвенного покрова. При крайне низкой величине СО2 в воздухе
ветер препятствует снижению содержания углекислоты в слое воздуха,
непосредственно соприкасающемся с ассимилирующим листом. Погло-
щение СО2 заметно усиливается при малых скоростях ветра (в ин-
тервале между штилем и скоростью ветра, равной 1 м/сек) и достигает
максимума при скорости 3 м/сек, в 2—3 раза превышая в эти часы абсорб-
цию при штиле 3. Ускорение абсорбции СО2 при усилении движения
воздуха можно изучать экспериментальным путем. Сосуд высотой 12 см
с отверстием диаметром 3,5 см, в который вливают 10 мл 0,5 Н КОН,
используют в качестве модели листа, ассимилирующего СО2. Метод
позволяет доказать, что меньшая подвижность воздуха около поверхности
почвы сильнее тормозит абсорбцию СО2 по сравнению со стимулирующим
ее влиянием возможного повышения концентрации СО2. Таким образом,
не только малая интенсивность света на поверхности почвы, но и отсут-
ствие ветра неблагоприятно влияет на фотосинтез 4.
Фотосинтез зависит также от уравновешенности водного баланса.
Влажность воздуха около поверхности лесной почвы относительно высо-
кая, инсоляция невелика, а почва в большинстве случаев достаточно
влажная. Иногда наблюдаются длительные периоды без осадков; в этом
случае водный баланс видов травяного покрова с поверхностно располо-
1 Е. Lohr, Н. С. Hansen, Physiologische Spannweite der Schattenpflanze
Oxalis acetosella, «Planta», 60, 1963, S. 46—50.
2 M.Mousseau, Influence de Г eclairment sur Г assimilation journaliere et an-
nuelle du Teucriuin Scorodonia L. en conditions naturelles, «Oecologia Plantarum (Mont-
pellier»», 1, 1966, p. 103—115.
3H. D en ek e, Uber den EinfluB bewegter Luft auf die Kohlensaureassimilation,
«Jb. wiss. Bot.», 74, 1931, S. 1—32.
4 H. Walter, W. Zimmermann, OkologischeCO2-Absorptionsmessungen
in verschiedenen Pflanzenbestanden, «Z. f. Bot.», 40, 1952, S. 251—268. При изучении
ассимиляции CO2 скорость прохождения воздуха в камере была небольшой, поэтому
полученные величины слишком малы, не говоря уже о других источниках ошибок
(см.: W. Tranquillini, Blattemperatur Evaporation und Photosynthese bei ver-
schiedener Durchstromung der Assimilationskiivette, «Вег. Deutsch. Bot. Ges.», 77, 1964,
S. 204—218).
Лиственные леса умеренной зоны северного полушария — неморалъная зона 315
женной корневой системой нарушается; именно в этот период обостряется
конкуренция за почвенную влагу корней деревьев и связанных с ними
микоризных грибов. На небольших прогалинах с почвами, мало пронизан-
ными корнями деревьев, среди видов травяного покрова встречаются инди-
каторы влажных мест. Элленберг в дубово-грабовом лесу в конце периода
необычно сухого лета наблюдал особо резкое завядание у растений —
индикаторов влажных мест и установил четкую связь между степенью
увядания и давлением почвенной влаги х. Славикова определила макси-
мальные величины всасывающей силы корней у различных видов лесных
трав, помещая растения в сосуд, где почва высыхала до тех пор, пока
растения не начинали увядать. Обнаружились четкие различия у видов,
произрастающих в пойменных лесах, и растений, приуроченных к более
сухим типам леса (табл. 55) 1 2.
Таблица 55
Максимальные величины всасывающей силы корней
у различных видов лесных трав (в атм) (по Славиковой)
Milium effusum 1 6,3 Geum urbanum 29,9
As e rum europaeum . 10,6 Fragaria vesca . 28,1
Lamium galeobdolon 13,8 Carex muricata . . . 28,6
Viola riviniana .... 15,5 Lamium maculatum 28,8
Aegopodium podagraria 15,9 Poa trivial is 36,0
Hepatica nobilis .... 19,1 Cj/nanchum vincetoxi-
cum .... 37,0
Mercurialis perennis . . 19,6 Viscaria vulgaris . 43,7
Pulmonaria officinalis 20,7 Sanguisorba minor . 48,3
Calamintha clinopo-
dium ....... 53,0
1 В левой колонке виды пойменных лесов.
По мере достижения точки завядания листья теряли тургор; поэтому,
определяя осмотическое давление листьев, можно было бы установить
и величину их всасывающей силы, что послужило бы контролем при изу-
чении всасывающей силы корней.
В другом исследовании Славиковой показано, что максимальная
всасывающая сила корней ясеня равна 37—39 атм и, следовательно,
выше, чем у видов трав, произрастающих на том же участке 3. Это дока-
зывает, что корни деревьев в борьбе за влагу отличаются большей кон-
курентной мощностью по сравнению с видами травяного покрова. Поэто-
му вблизи границы распространения буковых лесов, определяемой сухо-
стью (субконтинентальная область Чехии), корни бука используют всю
почвенную влагу, ничего не оставляя травам. В результате в таких лесах
растения нижних ярусов отсутствуют (тип Fagetum nudum). При изоля-
ции корней бука на границах пробных площадок и исключении корневой
1Н. Ellenberg, Uber die Zusammensetzung, Standort und Stoffproduktion
bodenfeuchter Eichen-Buchen-Mischwaldgesellschaften Nordwestdeutschlands, «Mitt,
florist.-soziol. Arb.-gem. Niedersachsen», 5, 1939, S. 3—135.
2 J. Slavikova, Die maximale Wurzelsaugkraft als okologischer Faktor,
«Preslia», 37, Praha, 1965, S. 419—428.
3 J.Slavikova, Wechselbeziehungen der Wurzelsaugkraft bei einigen Kompo-
enten der Eschenphytozdnosen, «Preslia», 38, Praha, 1966, S. 15—22.
316
Глава 4
конкуренции деревьев на этих участках начинают появляться травяни-
стые растения Ч
То же явление автор наблюдал в Анатолии в лесах, образованных
Abies bornmuelleriana и Pinus halepensis, вблизи границы их распростра-
нения, определяемой сухостью. В этих лесах не развиты подлесок и тра-
вяной покров; однако виды травяного покрова появляются тотчас же на
прогалинах, где отсутствует конкуренция корней деревьев. Сухость пре-
пятствует также разложению подстилки, которая в этих лесах обычно
достигает большой мощности. Не меньшее значение имеет и конкуренция
корней деревьев за питательные вещества, и в первую очередь за азот.
В мощном профиле бурозема с хорошо развитым муллевым гумусом обычно
выражено несколько горизонтов, где располагаются корни. Корни трав
приурочены к верхним горизонтам гумуса, корни деревьев простираются
несколько глубже. Травянистые растения получают достаточное количе-
ство питательных веществ, поэтому даже наиболее требовательные виды—
Asperula odorata, Mercurialis perennis, Lathyrus vernus и др. — хорошо
развиваются в этих условиях. На маломощных почвах, где формируется
слой грубого гумуса, корни располагаются больше в верхнем горизонте
почвы. Поскольку в грубом гумусе накапливается достаточное количест-
во N, деревья в обоих случаях растут хорошо, если только их рост не
ограничивается недостатком влаги. Травянистые же растения вынуждены
довольствоваться тем, что не использовано корнями деревьев. В почвах
с грубым гумусом не использованных деревьями питательных веществ
остается очень немного, поэтому в таких лесах произрастают только
малотребовательные виды: Luzula luzuloides, Deschampsia flexuosa, Poten-
tilla sterilis, Melampyrum pratense, Teucrium scorodonia и даже Vaccinium
myrtillus, а на сухих местообитаниях — Calluna vulgaris.
Таким образом, флора напочвенного яруса не может быть точным
индикатором роста древесного яруса, что подтверждается практикой
ведения лесного хозяйства. Часто она дает лишь представление о количе-
стве питательных веществ, которое не используется древесным ярусом.
Подстилка, образованная листьями, оказывает большое влияние на
напочвенный покров в лесу. Опавшие листья осенью ровно покрывают
всю поверхность почвы. Это исключает возможность произрастания мхов.
Поэтому в лиственных лесах мхи растут только на выступающих из почвы
камнях или у оснований стволов деревьев. По этой же причине в лесу
отсутствуют виды с зимующими розетками, в то время как на лесных
полянах они распространены довольно часто (Digitalis purpurea, виды
чертополоха и др.).
Напочвенный покров лиственных лесов Центральной и Восточной
Европы отличается большим разнообразием в зависимости от климата,
почвы и состава древесного яруса; влияние зонального климата в лесу
сказывается довольно слабо. Древесный ярус прямого воздействия на
травяной покров не оказывает. Их взаимоотношения имеют характер
косвенных взаимодействий и проявляются во влиянии на микроклимат
и почву посредством подстилки, химические свойства которой (величина pH,
содержание N) и быстрота разложения различны у разных видов деревьев
(рис. 235). Если на небольшой территории определенные виды травяни-
стых растений встречаются лишь под покровом определенных видов
1 J.Slavikova, EinfluB der Buche (Fagus silvatica) als Edifikator auf die Ent-
wicklung der Krautschicht in Buchenphytozdnosen, «Preslia», 30, Praha, 1958, S. 19—42.
В работах Славиковой используется метод Шардакова, отличающийся большой точно-
стью при определении всасывающей силы корней.
Лиственные леса умеренной зоны северного полушария — неморалъная зона 317
СЛ
pH
Рис. 235. Средняя продолжительность естественного разложения подстилки
из листьев различных видов деревьев в сравнимых условиях. Слева — отношение
С/N в свежей подстилке, справа — pH подстилки (по Шефферу и Ульриху, см.
«Phytologie», Bd. IV, 2).
деревьев, то это свидетельствует о том, что эти растения имеют благо-
приятные условия местообитания под данными видами деревьев. В иных
климатических или эдафических условиях эти виды трав могут быть при-
урочены к другим видам деревьев. При этом следует иметь в виду, что
современные леса Центральной Европы не являются естественными лесами.
Моно- или полидоминантный состав древесного яруса или часто наблюдае-
мая одновозрастность насаждений в значительной мере определяются
работниками лесного хозяйства. В отличие от естественных они более
гомогенные. Очень существенно, что при использовании древесины в этих
лесах не остаются отмершие, медленно гниющие на поверхности почвы
стволы, которые в девственном лесу создают местообитания, резко отли-
чающиеся от соседних. Условия освещения на этих участках более благо-
приятны, разлагающаяся выщелоченная древесина имеет кислую реакцию
и относительно бедна питательными веществами, конкуренция с корнями
деревьев, а в первое время также и с травянистыми растениями отсутству-
ет. На единственном участке девственного леса в Центральной Европе
(если не считать небольшого по площади леса в Чехословакии), который
расположен в Нижней Австрии, на высоте 1000 м распространены
мозаичные взаимопроникающие группировки буково-пихтового леса,
развитого на почвах, богатых питательными веществами, и елового леса
на бедных почвах (см. гл. 5). Условия освещения в этом лесу неоднородны,
поскольку на месте упавших стволов образуются прогалины. Применение
методов ведения лесного хозяйства привело к однообразию и четкому
разграничению новых, вновь созданных человеком насаждений. От есте-
ственной лесная растительность отличается также там, где производится
выпас скота. В результате вытаптывания, уплотнения почвы, отложения
318
Глава 4
2 3
5
Очень
сухая
Сухая
Умеренно
сухая
Умеренно
сбежая
Сбежая
Умеренно
блажная
Влажная
Умеренно
сырая
Сырая
Очень
сырая
ВоЗа
Группа Cladonia
о
Гр. Vaccinium
myrtillus
Гр. Blechnum
О
О
......О...
°°с
Гр. Molln i
I| montana | ^гУзап^етит corymbosum I
рооооо^(Го оооо 15 о ооооооосбооооооооооой
fp.Desch.;
flexuosai
Гр- ;
Pteridiumi
Гр. Anemone'Гр. Galeobdolon
Tp.Ajuga | Гр. Ficaria
Гр. Cephalanthera
Гр. Corydalis
О О О О О O|t
a !i
oooooojooooooooo
Гр. с.
Гр. Fi 11-
,,j remota -
Гр. Carex acutiformis
30000000оооо
Carex pendula
ffr'' i
‘I Fp.TheLy-
'! pteris
Гр. Caltha
о
м
о о о о 1
Л
Очень Кислая Умеренно Слабо Кислая Ней- Щелочная
кислая кислая кислая траль-
ная
Р и с. 236. Примерное распределение в зависимости от влажности и кислотности
почвы экологических групп травянистых растений, мхов и лишайников в лиственных
(Лесах Центральной Европы (по Элленбергу, см. «Phytologie», Bd. IV, 2).
1 — область распространения буковых лесов.
экскрементов и выборочного стравливания травяной покров полностью
изменяется, становится более однородным, характеризуется наличием
многих рудеральных видов и видов — индикаторов пастбищного исполь-
зования. В большинстве случаев в результате уничтожения подроста
происходит изреживание лесов, начинается их задернение, и в конечном
счете они деградируют до лесолугов (парковых лесов). Такие леса господ-
ствовали в Центральной Европе в средние века, а в настоящее время еще
встречаются в районах, где распространены лесные пастбища.
Среди лесных трав имеются виды, одинаково реагирующие на влаж-
ность или реакцию почвы, которая отражает содержание питательных
веществ в ней. Эти виды часто растут вместе и могут служить показателем
определенных условий местообитания на ограниченной территории.
Поэтому их объединяют в эколого-фитоценотические группы.
Элленберг обобщил материалы Шленкера и Шенхара и объединил
виды, встречающиеся в юго-западной части Центральной Европы, в труп-
Лиственные леса умеренной зоны северного полушария — неморалъная зона 319
пы (см. «Phytologie», Bd. IV, 2) Ч Поскольку это обобщение имеет узко-
региональное значение, на рис. 236 приводится только экологическая
схема, отражающая распростране-
ние отдельных групп, названных по
наиболее характерному для них
виду.
Эти группы развиваются в раз-
ное время. В лиственных лесах
Центральной Европы выделяется
следующая смена аспектов:
1) весенний аспект с весенни-
ми геофитами — до начала облист-
вления деревьев;
2) раннелетний аспект с цвету-
щими летнезелеными и вечнозеле-
ными видами разнотравья — сразу
после облиствления;
3) позднелетний аспект, харак-
теризующийся прежде всего цвету-
щими злаками;
2) зимний аспект с голой под-
стилкой из листвы и немногочис-
ленными вечнозелеными листьями
или побегами Oxalts, Asarum,
Sanicula, Stellaria holostea и др.
Эта последовательная смена
аспектов хорошо представлена на
фенологических спектрах, состав-
ленных Элленбергом для богатого
разнотравьем грабового леса Цент-
ральной Европы (рис. 237) и Мед-
вецка-Корнас (ссылка на стр. 284)
для букового леса, развитого на
известняках (в качестве сравнения
и для Arrhenateretum). Более под-
робные сведения об экологических
условиях и структуре лесов Цент-
ральной Европы содержатся в кни-
ге Элленберга (см. «Phytologie»,
Bd. IV, 2).
1 См. также: Н. Schluter, Ein
Beitrag zur Frage okologischer und sozio-
logischer Artengruppen mit einem Beispiel
aus dem Thiiringischen Schiefergebirge,
«Arch. f. Forstwesen», 6, 1957, S.
44-58.
к
к
К
Рис. 237. Фенологический спектр видов
деревьев, кустарников и травянистых ра-
стений во влажных дубово-грабовых ле-
сах (по Элленбергу, см. «Phytologie»,
Bd. IV, 2).
1 — листья текущего года; 2 — перезимовавшие
листья; з — цветки.
Виды Зубово-грабовых лесой
Carpinus betulus
Alnus glutinosa
Tilia cordata
Acer pseudoplatanus
Fagus silvatica
GLuercus robur
Fraxinus excelsior
Ribes grossularia
Sambucus nigra
Lonicera xylosteum
Crataegus spec.
Prunus padus
P. avium
Acer campestre
Corylus aveHana
Humulus lupulus
Evonymus europaeus
Rubus idaeus
R.caesius
Cornus sanguinea
Hedera helix
Лех aquifolium -p-
Ярус трабянистых
Leucoium vernum __
Gagea lutea
Anemone пет. и. ranunc.
Adoxa moschatellina _
Ranunculus ficaria
Corydalis cava
Veronica hederifolia
Arum maculatum
Allium ursinum
Дрейесный ярус
Ярус
(ОесённёзёленыёТда
куста
£
Pulmonaria officinalis I
Primula elation
Mercurialis perennis |
Ranunculus auricomus
Paris quadrifolius i
Ranunculus lanuainosus
Palyganatum murttflorum
Geum urbanum I
Stellaria nemorum |
Geranium robertianum
Vicia sepium
Phuteuma spicatum
MeL.ica uniflora
Galium aparlne
Melandrium dioecum
Poa nemoralis
Aegopodium podagraria
Carex remota
Stachus silvaticus
Dactylis glomerata
Chaerophyllum temulum.
r j p*"a______________r
Lamium maculatum
"Scrophularia nodosa
Crepis paludosa
Urtica dioeca
Jmpatiens noli tangere
Campanula tracheltum
C. latifolia
Elymus europaeus
FiGpendula ulmaria
Festuca gigantea
Circaea liitetiana
Brachypodium silvaticum
Galium silvaticum I
Г~
Athyrium filix femina
Dryopteris filix mas
D.Linnaeana
D.austriaca dilatata
Anemone hepatica
Luzula pilosa
Oxa Its acetosella
Patentilla sterilis
viola silv. u.Riviniana
Glechoma hederaceum
Lamium galeobdolon
Carex silvatica
Stellaria holostea
Veronica mpniana
Asperula odorata
Sanicula europaea
Milium effusum
Deschampsia caespitosa
летнезеленые бибы
К.
ПозЗноцйетущие
Л BBh
к
летнёзеленые бибы
4
Папоротники
Зимнезеленые бибы
Oh
1 2 3 4 5 6 7 8 9 10 11 12
1 Ш2 ШИШ 3
йВВь
320
Глава 4
ПОВЕДЕНИЕ РАСТЕНИЙ ЛИСТВЕННЫХ ЛЕСОВ ЗИМОЙ
Зона лиственных лесов, в отличие от зоны жестколиственных лесов
характеризуется наличием холодного времени года. Продолжительность
холодного периода и экстремальные температуры зимой возрастают при
движении с юга на север и с запада на восток по мере увеличения конти-
нентальности климата. Состав лиственного леса меняется в том же на-
правлении. В основном эти изменения затрагивают древостой и в мень-
шей степени —нижние ярусы леса. При усилении зимних холодов широко-
лиственные породы вытесняются хвойными, свойственными бореальной
зоне хвойных лесов. Трудно выявить отдельные факторы, определяющие
эту смену. При нарастании континентальности климата в юго-восточном
направлении зимы становятся холоднее, а лето более жарким и сухим.
Наряду с этим сокращается и продолжительность вегетационного сезона,
что особенно хорошо проявляется в горах, здесь выражена смена листвен-
ных пород по мере подъема в горы; последние в верхней части лесного
пояса обычно сменяются хвойными породами. Несмотря на возрастающую
континентальность, летом в горах более прохладно и выпадает больше
осадков; поэтому воздействие низких зимних температур на растения
более ощутимо в горах, чем на равнине.
Влияние низких зимних температур на растения очень многосторон-
не. Повреждения, причиняемые холодом, могут оказывать непосредствен-
ное воздействие; в этом случае говорят о морозобойных повре-
ждениях. Повреждения могут происходить при длительных морозных
периодах, в особенности в солнечную погоду, когда надземные части
растений, возвышающиеся над снегом, высыхают; причина в том, что
растения транспирируют даже зимой, в то время как корни не могут
поглощать воду из замерзшей почвы. В этом случае речь идет о м о р о з-
ном высыхании. Оно имеет значение лишь тогда, когда растения
достаточно морозоустойчивы и непосредственно не повреждаются низкими
температурами.
Лиственные породы в отличие от вечнозеленых видов хвойных деревь-
ев сбрасывают перед наступлением зимних холодов наиболее подвержен-
ные низким температурам ассимилирующие органы — листья; на ветвях
остаются почки, защищенные чешуйками от высыхания. Это не
наблюдается у лиственных пород, развитых в теплых климатических
зонах.
Осевые органы также хорошо защищены от высыхания пробкой
и корой. Растения не имеют анатомо-морфологических приспособлений
для защиты от воздействия низких температур и осуществляют это физи-
ологическим путем, повышая сопротивляемость живых клеток холоду.
Этот процесс называют закалкой. Закалка растений осуществляется
поздней осенью при понижении температуры и при первых небольших
заморозках. Это частично эндогенный процесс, однако он происходит
под прямым воздействием низких температур, поэтому растения подготов-
лены к закалке осенью больше, чем весной или летом. В связи с этим
морозоустойчивость растений меняется в течение года. Морозоустойчивость
можно легко определить экспериментальным путем. Отдельные части
растений в течение определенного времени подвергают воздействию
низких температур; после оттаивания устанавливают, произошло или не
произошло повреждение растений. Ход морозоустойчивости важнейших
видов лиственных деревьев в течение зимнего полугодия (октябрь — май)
Рис. 238. Изменение морозоустойчи-
вости двух видов дуба в течение зим-
них месяцев. За морозоустойчивость
принимается температура,при которой
повреждается до 15—20% отдельных
частей растений (по Тиллу).
1 — почки в состоянии покоя; 2 — рас-
пускающиеся почки; 3 — молодые листья.
Р и с. 239. Морозоустойчивость двух ви-
дов клена (по Тиллу). Пояснения см.
рис. 238.
Рис. 241. Морозоустойчивость двух ви-
дов липы (по Тиллу). Пояснения см.
рис. 238. Tilia cordata более морозоустой-
чива и распространяется дальше на север.
Р и с. 240. Морозоустойчивость бука
в холодные зимы 1952/53 и 1953/54 гг.
(по Тиллу). Пояснения см. рис. 238.
21—01248
322
Глава 4
Рис. 242. Морозоустойчивость отдельных тканей осевых
органов двух сортов плодовых деревьев в течение зимних
месяцев (по Лархеру и Эггартеру).
1 — вторичная кора (старая); 2 — вторичная кора (молодая); 3 —
камбий; 4 — древесина. Фенологические данные: К — раскрывание
почек, В1 — начало цветения, М — бутонизация, L — распускание
листьев.
представлен на рис. 238—241 х. Как видно, Quercus robur, ареал которога
проникает дальше на восток и северо-восток континентальной климати-
ческой области, более морозоустойчив по сравнению с Qu. petraea. Вид
Acer pseudoplatanus поднимается в условиях океанического климата выше
в горы по северному склону Альп, чем A. platanoides; однако последний
вид клена распространяется дальше на восток, в область холодного кон-
тинентального климата.
Морозоустойчивость древесных пород в разные годы резко меняется;
это особенно хорошо видно на примере Fagus silvatica. Степень закалки
в значительной мере зависит от хода температуры. Начиная с конца декаб-
ря и в январе при умеренной температуре растения легко теряют закалку.
1 О. Т i 1 1, Uber die Frostharte von Pflanzen sommergriiner Laubwalder, «Flora»,.
143, 1956, S. 499—542; H. J a h n e 1, tjber die Frostresistenz bei Waldbaume, «Arch. L
Forstwesen», 8, 1959, S. 697—725.
Лиственные леса умеренной зоны северного полушария — неморальная зона 323
что наблюдалось в Центральной Европе зимой 1952/53 г. Декабрь и ян-
варь 1955/56 г. были также очень теплыми. После неожиданно низких
температур в феврале 1956 г. растения, частично поте-
рявшие закалку, сильно пострадали от моро-
зов. Наоборот, в исключительно суровую зиму 1928/29 г. в Центральной
Европе в январе были отмечены очень низкие температуры, поэтому расте-
ния сохранили свою закалку и холодный февраль оказал лишь незначи-
тельное влияние.
Как видно, проблема холодоустойчивости очень сложна: эта слож-
ность усугубляется еще тем, что у одного и того же дерева морозоустой-
чивость отдельных тканей различна и может меняться в течение года.
Такая изменчивость особенно подробно изучалась у плодовых деревьев
(рис. 242) Ч Большое значение имеет, какая из тканей повреждена моро-
зом, поскольку от этого зависит возможность нормальной жизнедеятель-
ности растения после частичного повреждения. Особенно опасны повреж-
дения камбия, которые могут наблюдаться теплой весной, когда камбий
преждевременно теряет закалку и замерзает во время поздних замороз-
ков. Низкие зимние температуры сами по себе менее опасны для растений,
чем зимы с аномальным ходом температуры, а именно мягкие зимы с позд-
ними заморозками.
Наиболее опасны поздние заморозки, наступающие после распуска-
ния почек. Нежные молодые листья очень чувствительны к заморозкам
и погибают уже при небольшой отрицательной температуре. В результате
лиственное дерево может в течение одной ночи потерять все листья. Если
же дерево и не погибает, а на нем образуются новые листья из спящих
почек, то это способствует его сильному ослаблению. При повторных
заморозках конкурентная способность соответствующего вида снижается,
и он достигает границы своего распространения. Если вид защитить от
конкуренции, выращивая в ботанических садах, его можно культиви-
ровать далеко за пределами естественного ареала; так, бук растет в Кие-
ве и южной Финляндии.
Интересно проследить за развитием бука в горах, где он образует
верхнюю границу распространения деревьев, то есть в условиях отсут-
ствия конкуренции с другими видами деревьев. Такая ситуация наблюдает-
ся в горах с ярко выраженным океаническим климатом: в Южных Аль-
пах, в Апеннинах, а также на вершине фельдберг в Шварцвальде, где
Мюллер-Штолль проводил экологические исследования 1 2.
В этих условиях, в особенности на южных склонах, почки бука
крупнее и имеют более выпуклую форму. Укороченные побеги отличаются
большим числом междоузлий и большей толщиной. Почки тяжелее всего
в ноябре; в это время они достигают наивысших значений сухого веса
и одновременно характеризуются относительно большой влажностью.
В течение зимы сухой вес почек уменьшается в результате расхода вещест-
ва на дыхание; влажность снижается вплоть до января. Осмотическое
давление, влажность и количество сахара в почках бука в лесу, располо-
женном на высоте 1240 м над уровнем моря, меньше, чем на верхней
границе распространения деревьев на высоте 1435 л. Эти величины меня-
ются в течение года аналогичным образом (табл. 56).
1 W. Larcher, Н. Е g g art er, Anwendung des Triphen yltetrazoliumchlo-
rids zur Beurteilung von Frostschaden in verschiedenen Achsengeweben bei Pirus-Arten
und Jahresgang der Resistenz, «Protoplasma», 51, 1960, S. 595—619.
2 W. R.Muller-Stoll, Beitrage zur Okologie der Waldgrenze am Feldberg
im Schwarzwald, «Angew. Pflanzensoz. (Klagenfurt)», 2, 1954, S. 824—847.
21*
324
Глава 4
Таблица 56
Результаты изучения листовых почек бука в окрестностях горы
Фельдберг (Шварцвальд) (по Мюллеру-Штоллю)
Показатель Высота над уровнем моря, м 1935 г. 1936 г. 1936 г.
15.XII 9.II 2.III 9.V 26.IX 8.XI
Осмотическое давле- ние, атм 1435 1240 22,6 21,8 23,3 22,4 19,2 17,4 5,6 5,5 9,9 11,5 20,1 17,3
Общее количество сахара, атм 1435 1240 9,6 9,1 8,5 7,6 5,3 5,1 1,6 1,3 1,7 2,1 9,5 8,1
Влажность, % сухо- го веса 1435 1240 77,3 71,5 78,7 71,0 84,0 86,6 238 249 135 152 86,9 88,3
Произведение: влаж- ность X осмотиче- ское давление 1435 1240 1740 1560 1830 1590 1610 1505 1330 1370 1335 1750 1740 1530
В сентябре перед листопадом осмотическое давление и количество
сахара в почках относительно низкое, однако влажность остается еще
довольно высокой. После листопада и перехода к зимнему покою осмоти-
ческое давление и содержание сахара резко повышаются, а влажность
почек понижается. В течение зимы эти величины не меняются; только
весной, после активизации деятельности почек, осмотическое давление
и количество сахара в почках уменьшаются, а влажность резко возрастает.
На основании данных Мюллера-Штолля автором дополнительно рас-
считано произведение влажности и осмотического давления. Увеличение
этого показателя в зимние месяцы позволило выявить активное повыше-
ние осмотического давления зимой благодаря образованию сахара, что
всегда связано с закалкой растения. При снижении закалки весной эта
величина опять уменьшается. Различия осмотического давления на
высоте 1435 м и 1240 м очень незначительны; по сравнению с деревьями
в лесу повреждения криволесья бука у верхнего предела распространения
деревьев в результате морозного высыхания не установлены (см. гл. 5).
Несмотря на это, в июне 1936 г. у верхней границы леса молодые листья
оказались сильно поврежденными заморозками (до —3°), так что в конце
июня на деревьях, росших на открытых участках, не осталось почти ни
одного зеленого листа. В течение лета образовались новые облиствленные
побеги, за исключением наиболее высоких местоположений в горах, где
из-за краткости вегетационного периода побеги уже не могли сформиро-
ваться и в большинстве случаев зимой погибли. Таким образом, поздние
заморозки, видимо, являются основной причиной, определяющей верх-
нюю границу произрастания бука в горах. Аналогичные повреждения,
вызванные поздними заморозками, автор наблюдал и в Крыму, где бук
также образует верхнюю границу распространения деревьев на северном
склоне Яйлы. Бук не произрастает на плоскогорье Яйлы; это вообще
характерно для последних форпостов бука в горах, обычно приуроченных
к местообитаниям, защищенным от ветра. Как известно, ветер ограни-
чивает распространение деревьев в горной местности.,
Лиственные леса умеренной зоны северного полушария — неморалъная зона 325
На восточной границе ареала поздние заморозки снижают конкурент-
ную способность бука; в результате он вытесняется грабом.
Переходим к рассмотрению последствий морозного высыхания.
Учитывая, что лиственные деревья сбрасывают листья, можно высказать
предположение, что морозное высыхание («физиологическая сухость»)
не оказывает на них влияния. Это характерно для Центральной Европы.
В Восточной Европе с ее продолжительным зимним периодом и очень
холодной, но часто солнечной зимней погодой величина расхода воды
у безлистных ветвей может быть зимой даже больше, чем у вечнозеленых
видов хвойных Ч
У лиственных деревьев, относящихся к одному роду, величина транс-
пирации зависит от их распространения: виды, ареалы которых прости-
раются дальше на север, транспирируют меньше, чем виды того же рода,
но встречающиеся южнее (ср. Salix, Alnus, Acer). Эти закономерности
представлены в табл. 57.
Таблица 57
Относительная величина транспирации ветвей деревьев
(по опытам Иванова, проведенным в Ленинграде)
(Larix = 1)
Abies sibirica . 0,3 Alnus incana . 1,86
Pinus silvestris 0,6 Alnus glutinosa . 2,59
Pinus cernbra . 0,9 Acer platanoides . 1,45
Picea obovata . 0,8 Acer pseudoplatanus 1,60
Picea abies . . 1,5 Acer campestre . . 2,53
Larix decidua . 1,0 Acer tataricum . . . 2,68
Betula humilis . 0,77 Fraxinus excelsior . 2,10
Betula pubescens 2,09 Fagus silvatica . . . 2,91
Betula verrucosa 2,41 Quercus pedunculata . 3,38
Salix caprea . . . . 0,95 Ulmus montana . . . 3,48
Salix purpurea . . . 1,93 Carpinus betulus . . 4,58
Salix alba 2,97 Tilia platyphyllos . 4,91
Площадь поверхности ветвей хвойных пород (за исключением Larix)
значительно больше, чем поверхность безлистных ветвей лиственных
деревьев; по данным Гордягина, даже если рассчитывать транспирацию
на сырой вес, северные хвойные меньше расходуют воды, чем расположен-
ные в тех же условиях лиственные деревья, сбросившие листву. Это объяс-
няется преимуществом ксероморфной структуры хвои, у которой устьица
полностью закрыты. Только с началом таяния снега транспирация
хвойных видов усиливается, что связано с возвратом их активности
и раскрыванием устьиц.
На основании исследований, проведенных зимой в окрестностях
Казани, удалось установить, что величина транспирации у видов хвойных
и лиственных деревьев, ареал которых простирается далеко на север
(березы, клена остролистного, осины, черемухи и, возможно, липы), при-
мерно одинакова. Расход воды у дуба черешчатого был в четыре раза
больше; такие же величины отмечены у ясеня, сирени и яблони.
1 L. I van of f, Uber die Transpiration der Holzgewachse im Winter, I: u. II.,
«Вег. Deutsch. Bot. Ges.», 42, 1924, S. 44—49, 210-224; A. G о r d i a g i n, Uber die
winterliche Transpiration einiger Holzgewachse Ostrublands, «Beih. Bot. Zbl.», 46, I,
S. 93—118; В. И. Образцова, Зимняя транспирация деревьев и кустарников
в условиях степной зоны УССР, «Физиология растений», т. 3, 5, 1956, стр. 409—413.
326
Глива 4
В мягкие зимы, характеризующиеся чередованием холодных и теп-
лых периодов, у безлистных деревьев и кустарников транспирация дости-
гает относительно высоких значений, например на Украине наблюдались
значения, превосходящие наблюдаемые в севернее расположенных райо-
нах с холодной зимой в 3—1*3 раз. При оттепелях потери воды ветвями
должны были возмещаться водой, поступающей из ствола.
Проблема холодоустойчивости осложняется тем, что определенный
в таксономическом отношении вид деревьев далеко не однороден с эколо-
го-физиологических позиций и расчленяется на экоклины или клинотипы
(см. гл. 1). Особи одного и того же вида, распространенные в пределах
северной и южной частей какой-либо области, непохожи в отношении
морозоустойчивости и повреждений от морозного высыхания, несмотря на
отсутствие морфологических различий, имеющих значение для системати-
ки. Большое значение при проведении эколого-физиологических иссле-
дований имеет место происхождения вида; поэтому нельзя переносить
закономерности, свойственные особям, произрастающим в южной части
области распространения или в долинах, на растения того же вида, расту-
щие на севере ареала или в горах.
Отрицательные температуры зимой влияют и на травяной покров,
несмотря на то что они сильно смягчаются в почве, где расположены под-
земные органы растений, и в снежном покрове, предохраняющем от вы-
мерзания надземные части растений. По данным Тилла, морозоустойчи-
вость характеризуется четким годовым ходом: понижением с марта по
май, незначительностью летом, повышением с октября по декабрь и мак-
симумом в зимние месяцы. Корневища растений в зимний период менее
морозоустойчивы, чем почки или перезимовывающие листья, подвергаю-
щиеся воздействию более низких отрицательных температур (рис. 243)
В пределах естественного ареала морозостойкость растений всегда
достигает такого уровня, который позволяет им в любое время года пере-
носить минимумы, не превышающие обычные годовые отрицательные
значения; поэтому низкие зимние температуры не причиняют вреда расте-
ниям. Поскольку в теплое время года отрицательных температур, как
правило, не бывает, понижение холодостойкости не опасно для растений.
Это снижение холодостойкости необходимо для их жизнедеятельности,
так как морозостойкость постоянно связана с уменьшением активности
плазмы.
Сущность процесса закаливания пока еще изучена недостаточно хоро-
шо. Известно, что закаливание происходит только на свету, что подтвер-
ждается определенным увеличением содержания сахара и изменением
соотношения сахарозы и глюкозы.
Устойчивость к морозу и к высыханию изучалась также у папоротни-
ков как у спорофитов, так и у гаметофитов. Наблюдается постепенный,
различный в зависимости от времени года переход от пойкилогидрических
папоротников, которые могут полностью высыхать, к гомойогидрическим,
которые высыхания не выдерживают. В общем, папоротники более устой-
чивы к высыханию зимой Ч Сильные повреждения, вызванные отрица-
тельными температурами, наблюдаются у видов, которые культивируются
в садах и скверах за пределами их естественного ареала: у Prunus lauro-
1 L.Карре n, Untersuchungen fiber den Jahresablauf der Frost-,Hitze- und Aus-
trocknungsresistenz von Sporophyten einheimischer Polypodiaceen (Filicinae), «Flora», 155,
1964, S. 123—166; Untersuchungen uber die Widerstandsfahigkeit der Gametophyten ein-
heimischer Polypodiaceen gegeniiber Frost, Hitze und Trockenheit, «Flora», 156, 1965,
S. 101-115.
Лиственные леса умеренной зоны северного полушария — неморальная зона 327
Р ис. 243. Годовой ход морозоустойчивости некоторых видов лесных травяни-
стых растений в 1952/53 г. (по Тиллу).
I — корневища; 2 — листья; 3 — молодые распускающиеся листья; 4 — цветки; 5 — цветоч-
ные почки. Корневища видов рода Polygonatum, ареал которых распространяется дальше на
север или выше в горы, обладают меньшей морозоустойчивостью, поскольку они лучше, благо-
даря снежному покрову, защищены от влияния низких температур.
cerasus, Aucuba japonica, видов Cedrus и т. д. На восточных границах
распространения виды с субатлантическим ареалом сильно повреждаются
морозами; в холодные зимы в Шварцвальде у жарновца (Sarothamnus)
отмерзает вся надземная часть. На следующий год он вновь отрастает.
При повторных обмерзаниях в холодные периоды года вид погибает на
форпостах своего ареала. Таким образом, ареал вида начинает сокращать-
ся и вид сохраняется только в условиях более мягкого климата; в более
благоприятные климатические периоды вид вновь распространяется па
новые территории, насколько это позволяет современный культурный
ландшафт.
Изменения границ ареалов происходили в широком масштабе в тече-
ние ледникового и послеледникового периодов; в результате этих перемеще-
ний сформировалась современная сильно обедненная флора Европы;
многие виды или не смогли вернуться в более теплые рефугиумы, или не
имели возможности распространиться за пределы этих убежищ.
При отсутствии снежного покрова надземные органы напочвенного
покрова, сохраняющиеся зимой, повреждаются морозом, но, кроме того,
и страдают от морозного высыхания (физиологической сухости). Растения
испытывают физиологическую сухость не столько во время наиболее
сильных морозов, сколько весной, когда температура воздуха поднимает-
ся выше нуля, а почва еще не оттаяла; в это время корни не в состоянии
восстановить то количество воды, которое расходуется в процессе транс-
пирации. Процесс высыхания растений проявляется в непрерывном
увеличении осмотического давления; после превышения критической
328
Глава 4
величины соответствующие органы растения отмирают. Больше всего от
физиологической сухости страдают освещенные солнцем части растенийт
поскольку они расходуют наибольшее количество воды. Они прежде-
временно теряют морозостойкость, поэтому при последующих сильных
заморозках могут погибнуть от вымерзания. Причину отмирания расте-
ний (повреждение морозом или физиологическая сухость) можно устано-
вить только путем тщательных исследований.
Этой проблемой занимался Трен г. По его данным, в окрестностях
Гейдельберга на фоне в общем мягкой зимы в отдельные дни с сильными
морозами расход воды у покрытых хвоей ветвей ели {Picea abies) и сосны
{Pinus silvestris) существенно не отличался от величины транспирации
у безлистных ветвей бука и дуба. Напротив, у безлистных побегов черни-
ки {Vaccinium myrtillus) даже при расчете на сырой вес величина транс-
пирации была в 3 раза выше, а у жарновца метельчатого {Sarothamnus
scoparius) — почти в 10 раз. У вечнозеленых барвинка {Vinca minor)
и вереска {Calluna vulgaris) транспирация может достигать в 20 раз боль-
ших величин. Наивысшие значения получены у травянистых видов:
Linaria cymbalaria и Parietaria ramijlora, побеги которых на стенах домов
сохраняются живыми до зимы; величина их транспирации была в 30 —
50 раз выше, чем у древесных видов; суккулентнып вид Sedum rejlexum
транспирирует зимой очень слабо, в тени — так же, как сосна, а на
солнце приблизительно в 3—4 раза сильнее. Все упомянутые выше иссле-
дования проводились в одинаковых условиях среды. Растения, располо-
женные под снежным покровом или глубоко в расселинах стен, значи-
тельно меньше расходуют влаги на транспирацию. Во время оттепелей
они могут восполнить дефицит воды. При малоснежных зимах растения
постепенно высыхают, а осмотическое давление у них непрерывно растет.
Конечно, нельзя не учитывать активного повышения осмотического
давления, вызванного увеличением содержания сахара в процессе
закаливания. У плюща это увеличение составляет 7,8 атм’, соответ-
ственно критическое осмотическое давление зимой выше — около
24,6 атм, летом оно не превышает 22 атм. Полувечнозеленые побеги
Teucrium scorodonia и перезимовывающие розетки Digitalis purpurea
зимой 1931/32 г. страдали от сильного морозного высыхания; это
субатлантические элементы, произрастающие недалеко от их восточной
границы распространения. Незадолго до отмирания их осмотическое
давление достигало соответственно 24,5 и 28,8 атм. Устойчивость побегов
Vaccinium myrtillus, Calluna и Sarothamnus при морозном высыхании была
больше. Однако и эти растения, если они росли на бесснежных участках,
страдали ранней весной от физиологической сухости. В отличие от них
у Pinus и Picea никогда не наблюдается критическое осмотическое давле-
ние, даже если они встречаются на освещенных солнцем местообитаниях.
Жизнедеятельность растений изучалась также в окрестностях Гей-
дельберга в очень суровую зиму 1928/29 г. 1 2. В период с 1 января до
7 марта, по данным метеорологической станции, в Гейдельберге было
всего три безморозных дня. Абсолютный минимум 12 февраля 1929 г.
составлял —21°. Повреждение растений наблюдалось только в начале
марта; в это время температура воздуха начала повышаться, а глубина
1 R. Т hr en, J ahreszeitliche Schwankungen des osmotischen Wertes verschiedener
Okotypen usw., «Z. f. Bot.», 26, 1934, S. 449—526.
2 H. Walter, Die Winterschaden an unseren immergriinen Pflanzen wahrend
der Kalteperiode Januar —Marz 1929 und ihre Uhrsache, «Die Naturwissenschaften», 17,
1929, S. 854—859.
Лиственные леса умеренной зоны северного полушария — неморальная зона 329
промерзания почвы дости-
гала свыше 1 м. Зимние
побеги летнезеленых ра-
стений (Urtica dioica, Che-
lidonium mafus, Rumex
acetosella, Teucrium scoro-
donia, Thymus serpyllum)
могут перезимовывать
только под снегом, как
и побеги полузимнезеле-
ных Luzula silvatica, De-
schampsia flexuosa, Vacci-
nium myrtillus и видов
Rubus. У вечнозеленых
видов, распространенных
в приатлантических обла-
стях, в этот период обмер-
зали все надземные части
(Hedera helix, Ilex aquifo-
lium, Helleborus foetidus,
Sarothamnus scoparius).
Центрально европейские
виды повреждались лишь
в наиболее неблагоприят-
ных местообитаниях (Taxus
baccata, Vinca minor, Vis-
сит album). Субсредиземно-
морский Buxus sempervirens
не пострадал, хотя его
осмотическое давление в
результате расхода воды
повышалось до 78 атм.
Вечнозеленые хвойные по-
роды бореальной зоны,
а также Pinus nigra, Р.
strobus, Abies alba и Pseu-
dotsuga хорошо перене-
сли эту зиму. В довольно
холодную зиму 1946/47 г.
у растений не было обна-
Р и с. 244. Промерзание (изотерма 0°) различных
типов почв и их оттаивание весной (по Кройцу, см.
«Phytologie», Bd, III. 1).
ружено почти никаких по-
вреждений; вероятно, это связано с чередованием холодных периодов
и оттепелей. Большое значение имеет глубина промерзания почвы
зимой и то, насколько быстро она оттаивает после окончания холодного
периода. Оба фактора зависят от характера почвы. Сильно гумусирован-
ные почвы, в особенности торфяные, оттаивают очень медленно. Из рис. 244
видно, что богатая гумусом почва на глубине 10 см оттаивает на месяц,
позже, чем базальтовый щебень. Поэтому в Центральной Европе на торфя-
ных почвах редко встречаются вечнозеленые виды, распространенные-
в приатлантических областях; они чаще растут на каменистых осыпях
или песках.
Осмотическое давление (в атм) у листьев видов субсредиземпоморского пояса
образованного Quercus pubescens (по Педротти)1
Таблица 58
Название растений 8.V 15.VI 29.VII 23.VIII 21.IX 18.X 23.X.1963Tr.
Quercus pubescens 13,2 19,5-20,3 20,7—21,2 24,4 19,5 20,7 22,6 (19,2 7. X)
Fraxinus ornus 18,1—19,5 20,9 24,7 27,1 22,3 24,4 24,5(18,5—19,8)
Acer monspessulanum . . . 14,9 16,4 23,4 — 23,9 24,0 23,5
Ostrya carpinifolia . . . 16,3 19,1 20,3 23,5 21,0 18,6 21,4
Pyrus mat us 27,0 25,3 29,7 31,0! 27,5 26,5 25,0*
Prunus mahaleb . 22,2-23,1 24,2-24,5 27,3 25,6 24,3 24.5—27.9 24,1—25,3
Cornus mas 16,4 18,8 22,1 25,8 21,7 21,3 23,7
Ligustrum vulgare, прошлогодние листья, новые листья с июня . . 30,2 29,6 36,0 36,2! 29,4 31,7
Cotinus cogyggria 12,4-12,5 21,9 18,7 21,6 19,5 19,6 19,6 (красные 15,8)
Cyiisus sessilifolius . . 21,6 22,9 25,9 — — 22,2 24,6
Helleborus foetidus 19,6-23,3 12,6-18,5 19,0 23,2 22,7 19,8 21,2(22,9 17. II. 1964 г.)
Lonicera etrusca . . 21,2 22,1 26,0 30,8! 25,0 22,6 —
Cistus inc anus var. villosus . . 14,1—15,5 12,6-13,3 14,5 20,9 15,5 16,6 18,1 (21,6 17. II. 1964 г.)
1 Выделенные шрифтом цифры означают оптимальную величину осмотического давления, цифры в скобках — у сильно пожелтевших листьев. Самые
высокие значения осмотического давления в августе отмечены знаком!; снижение величии в сентябре и октябре объясняется дождливой осенью.
Лиственные леса умеренной зоны северного полушария — неморалъная зона 331
ПОЯС БУКА В ГОРАХ СРЕДИЗЕМНОМОРСКОЙ ЗОНЫ
Горные буковые леса, сходные с центральноевропейскими, проника-
ют далеко в Средиземноморскую область зимних дождей. Например, по
Апеннинам, правда только по вершинам гор, которые летом почти посто-
янно закрыты облаками, снижающими влияние летней засухи, они
доходят вплоть до Сицилии Педротти изучал этот вопрос в районе Бо-
лоньола у Камерино (Италия), расположеном на высоте 1070 м над
уровнем моря. Годовая сумма осадков здесь 1473 мм; минимум отмечен
летом, однако с июня по август выпадают 248 мм, что соответствует коли-
честву, обычному для Центральной Европы. Среднегодовая температура,
равная 8—9°, мало отличается от наблюдаемой в зоне буковых лесов.
В результате большого количества зимних осадков образуется мощный
•снежный покров, который держится несколько менее пяти месяцев.
Поэтому можно полагать, что бук, приуроченный к этому поясу облаков
•средиземноморских гор, в состоянии уравновешивать водный баланс
в относительно сухие летние месяцы.
Таблица 59
Осмотическое давление (в атм) у листьев видов субсредиземноморского пояса,
образованного Quercus pubescens1
Название растений 30.IV 26.V 28.IV i 26.VII 28.IX 7.XI
Quercus pubescens 13,7 22,9—25,7 19,6—20,3 27,8 22,5 24,2
Fraxinus ornus . . Acer monspessula- 18,2—19,0 23,1—25,4 22,5—25,0 33,5! 30,9 22,5—24,9
nurn 17,8 20,0 21,7 30,6! 28.8 19,7
Acer opalus . . 13,8-14,1 19,1 16,4-18,3 26,6 21,8 22,9—23,4
Ostrya carpinifolia 15,9 19,2 16,6 24,7 20,8 23,3
Lonicera etrusca 18,2—19,6 17,0 24,2 29,2 24,5 19,6 (15.III. 1964 г.)
i Продолжение табл. 58.
Наше предположение подтверждает флористический состав напоч-
венного покрова горных буковых лесов, который аналогичен развитому
в этих же лесах Центральной Европы и представлен Anemone ranunculoides,
Corydalis cava, Actaea spicata, Asperula odorata, Dentaria bulbtfera, Sanicula
europaea, Polygonatum multiflorum и т. д. Это доказывается также данны-
ми изучения осмотического давления, которые Педротти провел в Умбрий-
ских Апеннинах в окрестностях Камерипо 1 2. На восточном склоне гор
выражена типичная гумидная последовательность средиземноморских
поясов:
а) горный пояс, образованный Fagus silvatica (от 1000—1700 до
2000 м над уровнем моря);
Ь) субсредиземноморский пояс, сформированный Quercus pubescens
(от 500 до 1000—1200 м над уровнем моря);
1 Е. Oberdorfer, Beitrag zur Kenntnis der Vegetation der Nordapennin,
«Beitr. naturk. Forsch. S-W-Deutschl.», 26, 1967, S. 83—139.
2 F. P e d г о t t i, Contributo alia conoscenza dell’idratazione e della pressione
osmotica nelle specie di tre assozioni forestall delle Marche, «Giorn. Bot. Ital.», 70, 1963,
p. 398-424; 72, 1965, p. 93-113.
Таблица 60
Осмотическое давление (в атм) у видов средиземноморского пояса, образованного Queucus ilex 1
Название растений 15.Ill 30.IV 26.V 28.VI 26.VII 28.IX 7.XI 24.1 1964
Quercus ilex (5 м), старые, С Quercus ilex (5 ле), старые, 10 35,1 32,0 25,5 26,5
34,2 29,7 23,5 17,4 21,9 24,6 23,3 32,2!
Quercus ilex (5 ле), молодые, С Arbutus unedo (2—3 ле), старые 11,8 13,9—16,2 17,8
27,4—29,6 28,6 20,7—22,8 20,4—24,3 24,4
Arbutus unedo (2—3 ле), моло-
дые Viburnum tinus (2—3 ле) ста- 15,8 18,8 22,5 24,7 23,1—23,4 24,0—27,4
рые, С Viburnum tinus (2—3 ле), ста- 23,5-23,8 24,6 23,7 19,2—20,5 24,8
рые, К) Viburnum tinus (2—3 ле), моло- 20,1-23,8 23,4 23,9 22,2
дые, С 20,4-23,2 21,9—22,6 28,7! 21,4-25,2 19,2—20,0 24,6
Phillyrea latifolia (1 ле), старые Rhamnus alaternus —5 ле), 33,9—36,3 Молодые 21,4 33,2 36,1 34,6
старые Rhamnus alaternus (4—5 ле), 31,6-31,7 30,9-32,5 28,7
молодые Pistacia lentiscus, старые . . . 25,5—28,0 23,8—24,4 23,9—24,0 24,0 30,5 31,2 27,4 32,3!
Pistacia lentiscus, молодые 18,6 21,8—21,9 28,8 26,9 24,1 23,0
Pistacia terebinthus ... — 12,8-13,4 19,6—20,8 20,7 23,0 21,4 — —
Smilax aspera, старые .... 26,1-33,7 27,2 30,9 21,8 18,9 19,8 25,5!
Smilax aspera, молодые . . . Asparagus acutifolius, старые 12,6 16,5-17,2 19,4
34,8-35,0 36,8 33,1 31,4 27,2—33,2 27,5
Asparagus acutijolius, молодые Rubia peregrina (c 30. IV моло- 14,7-14,8 13,6-15,5 14,8-23,7 27,8 20,1—20,2!
17,0-18,0
дые) 19,7 11,0-11,9 11,7-12,5 12,3—12,7 14,9 15,9
Ruse us aculeatus, старые . 25,9-29,2 23,6—29,7 23,5-25,7 17,3—22,5 25,2 20,9 21,2-21,3 26,2
Ruse us aculeatus, молодые . . Teucrium flavum (c 26.V моло- 11,7 10,3-10,6 18,5
дые) 19,8—23,0 19,1 13,8 15,9 25,2—25,3! 20,2 17,0
Coronilla emerus ... . . — 16,0 17,4-17,9 21,0 27,0! 20,8 20,8 —
Cyclamen neapolitanum . 10,4—10,7 8,5—8,7 7,7 6,6 Мертвые Мертвые 7,1 12,3
1 Образцы у вечнозеленых видов были взяты со старых (прошлогодних) и с молодых (этого года) листьев. С и Ю означают сенепную и foamvin якепп-
зииии с КЛОНЯ.
Лиственные леса умеренной зоны северного полушария — неморальная зона 333
с) средиземноморский пояс, образованный Quercus ilex в наиболее
низко расположенных местообитаниях (до 500 м над уровнем моря).
У наиболее характерных видов, развитых в упомянутых выше горных
поясах, в 1963 г. примерно раз в месяц брались образцы листьев и опре-
делялось осмотическое давление криоскопическим методом: в поясе бу-
ка — в течепие вегетационного периода с 29 мая по 24 октября; в суб-
средиземноморском поясе — с 8 мая по 23 октября;в средиземноморском
поясе — с 15 марта по 24 января 1964 г. Число образцов было недостаточно
велико для детальной характеристики годового хода осмотического давле-
ния, поэтому полученные результаты представлены в таблицах, в которых
материал приводится раздельно для каждого пояса (табл. 58—60).
В поясе бука летом осмотическое давление заметно не увеличивается
ни у древесных видов, ни у видов трав; таким образом, водный баланс
здесь, как это наблюдается и в Центральной Европе, остается на постоян-
ном уровне. Небольшое постепенное увеличение осмотического давления
к осени следует рассматривать как нормальное явление.
У многих видов субсредиземноморского пояса максимум осмоти-
ческого давления отчетливо выражен в августе. Практически в той или
иной степени он наблюдается у всех видов и возникает под воздействием
летней засухи, поскольку облака в это время года всегда расположены
высоко. Особенно незначителен летний максимум у Cistus, тогда как
в средиземноморском поясе осмотическое давление у него обычно резко
повышается. Падение осмотического давления осенью доказывает, что
временное нарушение водного баланса летом растения переносят без
ущерба. Виды средиземноморского пояса хорошо приспособлены к лет-
ней засухе, которая существенно не нарушает водного баланса у склеро-
филов (см. табл. 60). В этом поясе наиболее высокое осмотическое давле-
ние у растений наблюдается зимой. У малакофильных видов (Teucrium
flavum) и у молодых листьев Viburnum tinus максимумы давления хорошо
выражены; это же отмечается и у Coronilla emerus, которая уже может
быть отнесена к числу субсредиземноморских видов.
Г ла в а 5
ХВОЙНЫЕ ЛЕСА УМЕРЕННОГО ХОЛОДНОГО КЛИМАТА —
БОРЕАЛЬНАЯ ЗОНА
БОРЕАЛЬНО-НЕМОРАЛЬНАЯ ЗОНА СМЕШАННЫХ ЛЕСОВ
Между зонами широколиственных и хвойных лесов расположена
переходная область, в которой чистые лиственные и хвойные леса
мозаично взаимопроникают и в которой наряду с этим развиты
смешанные насаждения. Смешанные леса широко распространены в Се-
верной Америке (рис. 245—247); здесь они проникают в глубь немо-
ральной зоны, а их существование обусловлено эдафическими причинами
Рис. 245—247. Климадиаграммы зоны сметанных и хвойных лесов Северной
Америки.
Монреаль — северная часть зоны лиственных лесов, образованная смешанными лесами; Стнвен-
вилл — зона хвойных лесов; Тимминс — граница смешанных н хвойных лесов. Основное значение
имеет не столько среднегодовая температура, сколько продолжительность периода со средней
суточной температурой выше 10° (на рисунке числа 111 и 117). Виды лиственных деревьев требуют
более 120 дней.
(см. Кнапп, ссылка на стр. 227). Хвойные в этих лесах представлены
различными видами Pinus: Pinus rigida, Р. virginiana, Р. pungens и Juni-
perus virginiana на юго-востоке, a Pinus strobus (рис. 248), Р. resinosa
и Tsuga canadensis на северо-востоке. Из лиственных деревьев встречаются
различные виды рода Quercus и некоторые другие виды *. Смешанные
насаждения приурочены к более бедным почвам, в основном песчаным
или кислым, и к сухим местообитаниям, где конкурентная способность
лиственных деревьев заметно снижается. Распространению смешанных
древостоев способствуют пожары; в большинстве случаев эти леса пред-
ставляют различные стадии послепожарпых сукцессий. Ареал смешанных
лесов Восточной Азии показан па рис. 183 1 2.
1 F. Dansereau, Phytogeographia Laurentiana,«Gontr. Inst. Bot. Montreal».
№ 75, 1959.
2 См. также: S. D a n e r t, S. Gei ler, P. Hanelt, Vcgctationskiindliche“
Studien in Nordostchina (Mandschurei) und der Inneren Mongolei, «Feddes Repert».
Beih. 139, 1960.
Хвойные леса умеренного холодного климата — бореальная зона
33£
Рис. 248. Смешанный лес, образованный Pinus strobus, в Аллеганском нацио-
нальном парке Пенсильвании (фото министерства земледелия США, Лесная
служба).
В европейской области условия развития этих лесов менее сложны
(рис. 249—251). В южной (Скандинавии зона в прошлом чистых листвен-
ных лесов, образованных буком, сменяется бореально-неморалыюй зоной,,
которая простирается до «Limes Norrlandicus», достигая 60° с. ш. В пре-
делах этой зоны доминируют ель (Picea abies) или сосна (Pinus silvestris)^
а наиболее богатых почвах и в более благоприятных в климатическом
отношении местообитаниях встречаются небольшие участки лиственного
леса, образованного в основном дубом (Quercus robur), а на юго-западе —
336
Глава 5
Рис. 249—251. Климадиаграммы европейской зоны смешанных лесов.
Стокгольм — южная граница распространения хвойных лесов; Москва — зона смешанных лесов;
Казань — вблизи границы распространения лиственных лесов и непосредственного перехода сте-
пей в бореальную зону хвойных лесов (континентальный климат).
Quercus petraea и Fagus silvatica; из числа других лиственных деревьев
здесь растут Ulmus glabra, Acer platanoides, Fraxinus excelsior, Tilia cordata
и балтийский эндем Sorbus intermedia \
В Восточной Европе южнее границы распространения ели основной
хвойной породой является Pinus silvestris. Она образует в районе Полесья
(между Киевом и Минском) на моренах и флювиогляциальных отложениях
ледникового периода чистые или смешанные с дубом и другими листвен-
ными породами насаждения. Выделяют многие типы леса, для наимено-
вания которых используются особые названия.
1. Б о р — чистый сосновый лес (Pineta) на бедных флювиогляциаль-
ных песках; в зависимости от степени влажности почвы эти леса характе-
ризуются различным составом нижних ярусов (см. рис. 271).
2. Суборь — дубово-сосновый лес (Querceto-Pineta)f в котором
верхний древесный ярус образован сосной, а нижний — дубом; нижние
ярусы слаборазвиты; почва слабоподзолистая, легкая супесь.
3. Сугрудки — сосново-лиственные леса; по К Леонову — Carpi-
no-Nemoreto-Pineta; под Nemoreta он понимает смешанные дубовые леса
с липой, вязом и кленом.
Граб (Carpinus betulus) образует в сугрудках нижний древесный
ярус. Эти леса встречаются в виде отдельных островов и образуют пере-
ход к чистому лиственному лесу (груд), развитому в Восточной Европе
только на наиболее богатых серых лесных почвах, подстилаемых лёссом.
В чистых лиственных лесах дуб образует верхний древесный ярус,
а граб — нижний (Carpino-Querceta); на востоке Европейской части
СССР граб исчезает, и здесь леса сформированы дубом и липой (Tilieto-
Querceta или Nemoreta).
Взаимосвязь вышеупомянутых типов леса с определенными почвен-
ными условиями показана на рис. 252. Бонитет сосновых, а также дубовых
насаждений возрастает с увеличением богатства почвы питательными
веществами; при этом имеет значение и степень влажности почвы (рис. 253.)
Зависимость распространения отдельных видов деревьев и основных
видов травянистых растений нижних ярусов от условий местообитания
в упомянутых выше лесах представлена на рис. 220 и 254. Эти естествен-
ные леса изменяются различным образом под воздействием человека.
1. Во всех лесах выпасается скот, что препятствует развитию подлес-
ка и способствует распространению сорняков.
1 The Plant Cover of Sweden, Acta Phytogeogr. Suecia 50, Uppsala, 1965.
Хвойные леса умеренного холодного климата — бореальная зона
337
2. Лесные пожары чаще всего бывают в чистых сосновых лесах и
и очень редко в лиственных лесах. Поэтому на месте сосняков особенно
распространены вторичные сообщества, возникшие после пожара.
3. Вырубка нижнего яруса преимущественно влияет на дубово-
сосновые леса, поскольку после удаления дуба формируются чистые
«сосновые леса, и меньше — на другие типы леса.
4. Заготовка древесины особенно сильно изменяет сосново-листвен-
ные леса, так как после вырубки сосны они превращаются в чистые лист-
венные леса.
Рис. 252. Взаимосвязь типов леса и почвенных условий западнее
Днепра (по Погребняку).
А — бор (Pinetum) на песчаной почве; сосна с единичной березой; Б — суборь
(Querceto-Pinetum) Г на супеси; нижний древесный ярус образован дубом; В — суг-
рудок (Carpineto-Querceto-Pinetum) на суглинке; нижний ярус древостоя сформиро-
ван Carpinus betulus\ Г — груд (Carpineto-Quercetum) на серых лесных почвах, под-
стилаемых лёссом; ^верхний древесный ярус образован дубом, нижний — Carpinus',
развит ярус кустарников.
5. В основном раскорчевывают лиственные леса, произрастающие на
самых богатых почвах, и гораздо реже леса других типов; чистые сосно-
вые леса на песках вообще не раскорчевываются, поскольку почвы, на
которых они растут, непригодны для земледелия.
Смешанные сосновые леса раньше были широко распространены на
песчаных почвах центральной Германии. Применением лесохозяйствен-
ных мероприятий они большей частью превращены сегодня в чистые
сосновые насаждения. Даже однородные в прошлом лиственные леса,
такие, как дубово-березовые леса на песчаных почвах в западной части
Центральной Европы и леса Верхнерейнской низменности, в настоящее
время превратились в чистые сосняки с примесью бука в нижнем ярусе.
Смешанные пихтово-буковые леса распространены в верхней половине
пояса бука в горах Центральной Европы. Еще выше в горах к ним при-
мешивается ель. Склоны северной экспозиции обычно покрыты еловыми
лесами, южные склоны — буковыми, а восточные и западные склоны —
смешанными лесами.
Возможность существования смешанных насаждений в естественных
условиях может быть доказана наличием в природе девственного леса.
22-01248
338
Глава 5
Такой лес произрастает на территории Нижней Австрии, в Восточных
Альпах близ Лунца. Благодаря своей недоступности лес Ротвальд никог-
да не вырубался и использовался только
как угодье для охоты. Это типичный
смешанный древостой, образованный
равным числом стволов бука, пихты
и ели. Лес разделен на два участка —
Т) Сосна
Ш Дуб
А_Ольха
-------Pteridium aquilinum
-------Cytisus ruthenicus
-------Vaccinium myrtillus
------- Pirolo secunda
-------Asarum europaeum
------- Sphagnum spec.
-t-n । । Aegopodium podagraria:
Filipendula ulmaria
Рис. 253. Зависимость бонитета (римские
цифры) древостоев в возрасте 100 лет от влаж-
ности (0—5) и типа почвы (Л — Г). Поясне-
ния см. рис. 220 (по Погребняку).
Рис. 254. Зависимость распро-
странения основных видов лесных
растений зоны смешанных лесов
Восточной Европы от условий
местообитания (по Погребняку).
0—5 — увеличение влажности почвы;
А — Г — возрастание количества пита-
тельных веществ в почве (ср. рис. 220).
Ниже приводится описание двух типичных местообитаний Малого девст-
венного леса, который занимает площадь 56 га и более однороден, чем
Большой девственный лес.
Описание девственного леса Ротвалъд
(Нижняя Австрия)
Характеристика местообитания: южный шлейф горы Дюрен-
штейн с небольшим уклоном на запад, высота 1000 ле, годовое количество осадков около*
1700 мм; материнская горная порода — известняк, залегающий на глубине 1—1,5 мг
почва — бурая лесная, чередующаяся с дерново-карбонатной, суглинистая с мощным!
(8 см), сильно гумусированным горизонтом А и тонким слоем подстилки.
Хвойные леса умеренного холодного климата — бореальная зона 339
Структура древостоя: древесный ярус образован несколькими вида-
ми разной высоты, средняя степень сомкнутости крон равна 70—90% . Отдельные старые
пихты достигают высоты 50 м при диаметре на высоте груди 1 м. Средняя высота грех
древесных пород, образующих древостой (пихта, бук, ель), около 35—40 м. Под верх-
ним древесным ярусом развит более разреженный нижний ярус (высотой до 20 м),
сформированный теми же видами. Лишь местами на участках, где отсутствует верхний
ярус, в составе нижнего яруса доминирует бук и разрастаются кустарники. Большое
поголовье благородных оленей повреждает подрост пихты, обкусывая его вершины.
Ель возобновляется на большом числе отмерших поваленных стволов. На ветровале
всходы ели лучше освещены и развиваются на кислом субстрате, в связи с чем подрост
ели часто располагается рядами. Мертвые стволы бука обычно долгое время не падают
и покрыты мощными плодовыми телами трутовика (Fomes fomentarius). Запас древесины
достигает 1000 плотных кубометров на 1 га; из них 30% приходится на лиственные поро-
ды, а 70% — на хвойные. Древесина лиственных деревьев представлена только буком,
поскольку Acer pseudoplatanus и Ulmus montana встречаются в лесу единично. Среди
хвойных преобладает пихта. Древесный ярус неоднороден, бук и пихта часто образуют
куртины. Состав травяного покрова меняется на разных участках леса. Его своеобра-
зие выражается в том, что в нем совместно растут виды, развитые обычно под покровом
лиственного леса, и растения, приуроченные к кислым почвам; последние произра-
стают на слое гумуса, образовавшемся в результате разложения сгнивших стволов
(Vaccinium myrtillus. Lycopodium annotinum, Blechnum spicant и др.). Моховый покров
встречается только на ветровале и на выступающих над поверхностью почвы обломках
известняка. При описании леса мхи не учитывались. Были выбраны два больших уча-
стка площадью 600 м2 каждый. На площади I число видов равно 34, на площади II —
23. Описание древостоя проведено Г. Вальтером и Г. Фрайтагом (обилие по Браун-
Бланке): I 11
Верхний древесный [ ярус
Высота, м 30-50 20—45
Сомкнутость крон, % 70 85
Fagus silvatica 3 4
A bies alba 3 3
Picea abies . . . 1 2
Acer pseudoplatanus + —
Ulmus montana (+) —
Нижний древесный ярус
Высота, м 8-15 5-20
Сомкнутость крон, % 7-10 15
Fagus silvatica 1 2
A bies alba 1 1
Picea abies . . Ярус кустарников 1 —
Высота, м 0,4-1,5 0,3-1,2
Покрытие, % 45 5
Fagus silvatica 3,4 1
Picea abies + 1
Abies alba . . . + —
Acer pseudoplatanus + —
Daphne mezereum Ярус травянистых растений — X
Высота, см 5 — 10 5—10
Покрытие, % . . 50 30
Всходы деревьев: Fagus 1 1
Picea 1 1
A bies + 1
Acer + X
Oxalis acetosella . . 3 2
Cardamine trifolia 2,3 2
Vaccinium myrtillus 1,3 1,2
Asperula odor ata 1,3 2
Lamium galeobdolon 1 X
Lycopodium annotinum + , 3 1,3
Prenanthes purpurea . . . . +
22*
340
Глава 5
I II
Viola silvatica . . + +
Neottia nidus-avis + , 2 +
Veronica officinalis r +
Adenostyles glabra + , 2 +
Ranunculus nemorosa . + —
Cicerbita muralis + —
Dryopteris disjuncta . . + , 3 —
Dryopteris austriaca + —
Dryopteris filix-mas + —
Athyrium filix-femina . . + —
Luzula maxima + , 2 —
Polygonatum verticillatum + , 2 0 —
Fragaria vesca .... —
Calamagrostis varia 0 —
Carex silvatica . . . + —
Epilobium montanum + —
Asplenium viride + —
Solidago virgaurea + —
Senecio fuchsii . . 0 —
Lycopodium selago r —
Rosa pendulina . . r —
Carex digitata ... — + , 2
Dryopteris phegopteris — + .2 (+, 2)
Blechnum spicant . . . —
Luzula pilosa — +
Euphorbia amygdaloides — +
Sanicula europaea — +
Lonicera nigra . . . — r
Если не считать небольшого по площади леса в Чешском Лесу,
Ротвальд является уникальным крупным массивом первобытного леса
в Центральной Европе. Однако не следует думать, что в Ротвальде рас-
пространен единственный тип девственного леса, свойственного зоне
смешанных лесов. Судя по существующим первобытным лесам в умеренных
климатических зонах за пределами Европы, они представлены большим
многообразием типов. Смешанные насаждения чаще встречаются в дев-
ственных, чем в искусственно созданных лесах; но бывают и одновозраст-
ные девственные леса, образованные одним видом деревьев, что наблюда-
ется, в частности, при быстром облесении гарей сосной.
Чаще распространены разновозрастные насаждения, однако внешне
они обычно не отличаются от одновозрастных; это связано с тем, что по
достижении 60 лет прирост древесины резко сокращается и деревья раз-
ных классов возраста (от 60 до 400 лет и более) становятся похожими друг
на друга. Если деревья достигают возраста свыше 2000 лет, тогда доста-
точна замена одного дерева на 1 га в год. Подрост в этих лесах может прак-
тически отсутствовать, несмотря на нормальное возобновление древостоя.
Известны также леса, где деревья представлены самыми различными
классами возраста или распределены мозаично по площади; это наблю-
дается в тех случаях, когда древесные виды возобновляются не на больших
площадях, а лишь в окнах, образовавшихся в результате падения крупных
деревьев; первыми на этих участках появляются растения-пионеры.
Многие виды деревьев не возобновляются под пологом материнского
древостоя; в этом случае он может восстановиться лишь в результате
какой-либо катастрофы. Это периодически наблюдается в древостоях
хвойных пород, где естественным путем после удара молнии возникают
лесные пожары. Поскольку в девственных лесах накапливается огромная
масса мертвой сухой древесины, лесные пожары могут нанести большой
Хвойные леса умеренного холодного климата — бореальная зона
341
ущерб. Имеет также значение массовое размножение насекомых-вредите-
лей после лет с экстремальными климатическими условиями. Перестой-
ные лиственные древостои часто уничтожаются во время бурь. Участки
лиственного леса, пострадавшие от бури, хорошо известны еще со времен
первых поселений в Северной Америке, поскольку образовавшиеся зава-
лы препятствовали дальнейшему проникновению в глубь девственного
леса. В 1872 г. разразилась буря, которая уничтожила лес на площади
длиной свыше 60 км и шириной в среднем более 1 км. На этих обезлесен-
ных площадях в течение столетия наблюдалась смена различных типов
леса, возобновлявшаяся после новых катастроф; в результате крупные
площади первобытных лесов длительное время остаются неоднородными
(см. «Phytologie», Bd. Ill, 1).
ОСНОВНЫЕ ВИДЫ ДЕРЕВЬЕВ
БОРЕАЛЬНЫХ ХВОЙНЫХ ЛЕСОВ
По мере продвижения на север увеличивается значение температур-
ного фактора для растительности. Это определяет не только смену зоны
лиственных лесов зоной смешанных лесов, а затем бореальной зоной
хвойных лесов, но п расчленение зоны хвойных лесов, или, как ее назы-
вают, тайги. В евросибирской области в пределах тайги в направлении
с запада на восток усиливается континентальность климата: зимняя тем-
пература понижается, а летняя повышается; резко увеличивается ампли-
туда температуры, которая достигает в районе полюса холода в Восточ-
ной Сибири (Оймякон) 100° (абсолютный минимум — 70°). Только на
Дальнем Востоке на побережье
Тихого океана выражен океани-
ческий климат. Особенное значе-
ние для лиственных деревьев,
в большей степени, нежели для
хвойных, имеет короткий вегета-
ционный период, обусловленный
недостатком тепла в летние месяцы.
В умеренной зоне, где осадки
в течение года выпадают равномер-
но, по температура выше 10° дер-
жится в среднем менее 120 дней,
вегетационный период для широ-
колиственных деревьев слишком
мал; их конкурентная способность
хвойными породами, образующими чистые хвойные леса или леса с при-
месью мелколиственных пород. Мелколиственные породы широко распро-
странены в океанических областях, например виды берез господствуют
в лесах западной Скандинавии и на Камчатке. Продолжительность периода
от начала распускания листьев до осеннего листопада у мелколиственной
березы (рис. 255), образующей на западе Европы северную границу
леса, в Финляндии в зависимости от широты местности изменяется
от 114 до 145 дней (табл. 61) Ч
” В зоне хвойных лесов, так же как и в зоне лиственных лесов, зональ-
ная растительность образована наиболее теневыносливыми видами.
1 K.Rubner, Die pflanzengeographischen Grundlagen des Waldbaues, 4. AufL,
Radebeul u, Berlin, 1953,
Таблица 61
Продолжительность периода (в днях)
от начала распускания листьев
до листопада у березы на различной
широте в Финляндии (по Кояла)
66—69° с. ш. 114 (3.VI —25.IX)
64—66° с. ш. 118 (29.V-24.IX)
63° с. ш. 130 (23. V-30, IX)
62° с, пт. 140 (18.V-5.X)
61° с. ш. 142 (17.V-6.X)
60° с, ш. 145 (17.V-9.X)
резко снижается, и они вытесняются
.Р и с. 255. Березняк в возрасте 40 лет в юго-западной Финляндии зимой (фото Каян-
дера).
Хвойные леса умеренного холодного климата — бореальная зона
343
Рис. 256. Разновозрастный еловый лес-зеленомошник в северной
Швеции. Возраст самых старых деревьев свыше 250 лет, высота
большого высохшего дерева достигает 22 м (фото Хессельмана).
В евросибирской области это ель (Picea abies и Р. obovatd), сибирская
пихта (Abies sibiricd) и сибирский кедр (Pinus cembra ssp. sibiricd). Они
формируют «темнохвойные леса». Только в условиях, где понижается
конкурентная способность этих видов или где они уничтожены частыми
пожарами, распространены «светлохвойные леса», в составе которых
доминирует сосна. В крайне континентальных условиях Восточной
Сибири господствуют летнезеленые лиственницы (Larix sibirica, L. da-
hurica и др.).
Основной породой европейской части зоны хвойных лесов является
ель: в западной части ареала — Picea abies (рис. 256), в восточной
Р. obovata- в местах контакта развиты переходные формы (Р. fennica,
344
Глава 5
Рис. 257—259. Климадиаграммы европейской бореальной зоны.
Эстерсунд — Швеция; Оулу — Финляндия; Архангельск — берег Белого моря. Указано число
дней со средней температурой выше 4-10°, а на рис. 259 также число дней со средней температурой
выше —10°.
Рис. 260—262. Климадиаграммы Сибири для лет с экстремально холодными зи-
мами.
Иркутск — около южной границы тайги (частично в пределах лесостепи); Николаевск-на-Аму-
ре — крайний восток Сибири (пояснения см. рис. 257—259).
Р. medioxima, Р. uralensis — это экотипы или климатические расы).
В пределах европейского сектора также широко распространена сосна
(Pinus silvestris).
Ель, у которой развита поверхностная корневая система, растет на
подзолистых почвах с постоянно влажным верхним горизонтом почвы.
Влажное, прохладное лето Северной Европы благоприятствует ей. Отно-
сительно короткий вегетационный период и продолжительные холодные
зимы увеличивают конкурентную способность ели по сравнению с листвен-
ными породами. Но это не является обязательным условием для успешного
ее произрастания, о чем свидетельствуют посадки ели в Центральной
Европе на участках, ранее занятых лиственными лесами. Лесоводы при-
меняют меры для предотвращения в насаждениях ели конкуренции с дру-
гими видами. Прирост ели в области распространения лиственных лесов
больше, чем в зоне хвойных лесов.
Только в условиях ярко выраженного океанического климата с мяг-
кой зимой ель повреждается красной гнилью: у ели формируются очень
широкие годичные кольца и сопротивляемость повреждениям
понижается, поэтому нижняя часть ствола легко поражается грибами,
что бывает заметно по образованию вздутий. Резкая континентальность
климата также неблагоприятна для ели. В Западной Сибири ее распро-
странение ограничивают Abies sibirica и Pinus cembra (западная граница
ареала показана на рис. 195). В районе Амура она замещается близко-
родственным видом.
Климадиаграммы евросибирской зоны хвойных лесов дают пред-
ставление о влажных климатических условиях этой области (рис. 257—
Рис. 263. Девственный] лес из
Larix dahurica на западном склоне
Хингана, 800—1300 л над уровнем
моря (фото Матцошима).
Р и с. 264. Larix leptolepis
(= kaempferi) у подножия горы
Нантай (среднее Хондо), высота
1200 м над уровнем моря (фото
Мийоши).
Глава 5
Рис. 265. Лес, образованный Cryptomeria japo-
nica, близ Никко (среднее Хондо), высота 1000 м
над уровнем моря (фото Фабера).
262). Продолжительность
теплого периода летом
определяется числом дней
с температурой выше +10°.
Для обозначения длитель-
ности и интенсивности
зимних холодов использу-
ется число дней с темпе-
ратурой выше —10°. Чем
меньше число таких дней,
тем суровее зимы и конти-
нентальнее климат (рис.
260—262). Ель в Западной
Сибири проникает на север
дальше, чем Abies sibirica
и Pinus sibirica, но зато
последние два вида подни-
маются выше в горы. Кедр
(Pinus cembra ssp. sibirica)
в Сибири образует верх-
нюю границу леса. В горах
Алтая и на Урале на гра-
нице с альпийским поясом
кедр растет в виде стла-
ника (Pinus sibirica v.
humistrata). Эта форма кед-
ра не идентична распрост-
раненному в Восточной
Сибири выше границы леса
виду Pinus pumila, также
формирующему стланики;
Pinus pumila генетически близок Pinus sibirica. Наибольшей холодо-
устойчивостью отличается лиственница. Основными видами являются
Larix sibirica, распространенная в западной части Евразии (вплоть до
северо-восточных областей Европы, рис. 195), и Larix dahurica (восточ-
нее Енисея, рис. 263). В настоящее время выделяют еще несколько мелких
видов лиственницы, встречающихся в основном на востоке Сибири х.
Лиственничные леса в Сибири покрывают 2,5 млн. км2; евросибирская
зона хвойных лесов также занимает значительно большую площадь по
сравнению с остальными растительными зонами земного шара. Большое
число других видов хвойных распространено в Китае и Японии (рис. 264—
265).
В Центральной Европе и Малой Азии сохранились лишь небольшие
естественные участки хвойных лесов, приуроченные к более высоким
горным массивам (Альпы, Пиренеи, Карпаты, северо-восточная Анатолия,
Кавказ). Здесь встречаются Picea abies (= excelsa), Pinus silvestris и P. mon-
.tana; в Центральных Альпах распространены викарирующие виды:
Larix europaea (= decidua) и Pinus cembra s. str. На границе с поясом бука
развиты древостои, образованные Abies alba (= pectinata) (рис. 266).
В нижних частях гор в настоящее время встречаются одновозрастные
чистые культуры хвойных пород.
1 А. Я. Орлов, Хвойные леса Амгунь-Буреинского междуречья, М., 1955.
Хвойные леса умеренного холодного климата — бореальная зона
347
Р ис. 266. Малонару-
шенный лес, образован-
ный Abies alba и Picea
abies, в Шварцвальде.
В напочвенном покрове
Vaccinum myrtillus и мхи
(фото Фойхта).
Основным видом хвойных деревьев бореальной зоны Северной Аме-
рики является белая ель — «white spruce» (Picea glauca). Она доминирует
в лесах от Ньюфаундленда до Берингова пролива х. Larix laricina
и Abies balsamea играют здесь меньшую роль. На бедных почвах растут
Pinus banksiana и Picea mariana («black spruce»). На юго-востоке Северной
Америки, в Аппалачских горах, распространена Picea rubens («red spruce»)
(рис. 267). Среди мелколиственных древесных видов наиболее часто
встречаются Betula papyri]era (на Аляске также В. resinijerd) и Populus
tremuloides (иногда Р. balsamifera). Thuja occidentalis приурочена к восточ-
ной части бореальной зоны. Экология хвойных лесов изучалась в равнин-
ных условиях и в горах. Прежде чем перейти к обсуждению результатов
1 R. К n а р р, Die Vegetation Nord- und Mittelamerikas, Stuttgart u. Jena,{1965;
см. также: J. S. R о w e, Forest regions of Canada, «Dep. North Aff., Forest Branch Bull.»,
123, Ottawa, 1959 (с картой растительности); A. G a 1 о u x, Les forets de I’Amerique
septentrionale temperee, «Pruefst. Groenendaal», Ser. A., № 7, 1951; V. К u j a 1 a,
Waldvegetationsuntersuchungen in Kanada, «Ann. Acad. Sci. Fennica», Ser. A., IV, Biol.,
7, 1945, p. 1—434.
348
Глава 5
этих исследований, необходимо отметить, что климатические условия
бореальной зоны и пояса горных хвойных лесов не являются идентичны-
ми; такой же идентичностью с экологической точки зрения не отличаются
и растения одного и того же вида, распространенные в плакорных усло-
виях и в горах. В этом случае речь идет о различных экотипах или гео-
графических расах.
Рис. 267. Picea rubens в Новой Англии (США) с подростом Abies balsamea (фото
министерства земледелия США, Лесная служба).
Хвойные леса умеренного холодного климата — бореальная зона
349
ТИПЫ ЛЕСА БОРЕАЛЬНОЙ ЛЕСНОЙ ЗОНЫ ЕВРОПЫ
Каяндер выделил типы леса в пределах бореальной зоны хвойных
лесов Финляндии х. Он назвал их по доминирующим видам травяно-кус-
тарничкового покрова.
1. Тип Calluna с СаИйпа vulgaris и лишайниками.
2. Тип Vaccinium с Vaccinium vitis-idaea.
3. Тип Myrtillus с Vaccinium myrtillus (рис. 268).
4. Тип Oxalis с Oxalis acetosella и другими травянистыми растениями.
К одному типу леса с лесоводческой точки зрения он относил все
участки леса, растительность которых в возрасте спелости при нормальной
сомкнутости древесного яруса характеризуется сходным видовым соста-
вом и однородными экологическими условиями. Поэтому к определенному
типу леса следует причислить также участки, отличительные черты кото-
рых могут рассматриваться как временные. Эти отклонения могут быть
обусловлены возрастом древесного яруса или способом пользования
лесом. Тип леса отражает в первую очередь климатические и эдафические
факторы местообитания, которые сохраняются и в том случае, когда учас-
ток полностью лишается растительного покрова, например на вырубках,
где отсутствует ярус деревьев. Ниже приводятся примеры изменений
на вырубках в различных типах леса. i
1. На вырубках сосняка типа Calluna в большинстве случаев исчезают гипновые
мхи и кустистые кладонии. На их месте появляются невысокие бокальчатые кладонии
и более мелкие мхи; вереск (Calluna) становится низкорослым и распределяется в виде
густых групп. Появляется Arctostaphylos и бурно разрастаются злаки Festuca ovina
п Сalamagrostis.
2. На вырубках в типе Vaccinium также почти полностью исчезают очень мощный
моховой покров и Vaccinium myrtillus; разрастаются и цветут злаки Deschampsia
flexuosa и Сalamagrostis, обычно не плодоносящие в старых насаждениях. То же
наблюдается и у A ntennaria dioica и Chamaenerium angustifolium. Часто в больших
количествах появляются Arctostaphylos, Calluna и Rumex acetosella. Если на вырубке
выпасают скот, злаки, за исключением Agrostis, вытесняются.
3. В типе Myrtillus вырубка влияет аналогичным образом. Vaccinium myrtillus,
покрывавший в лесу большую часть поверхности почвы, исчезает и замещается Vacci-
nium vitis-idaea. Злаки Deschampsia flexuosa, Agrostis, Calamagrostis и разнотравье Rubus
saxatilis, Veronica officinalis, Chamaenerium и Galium развиваются более пышно, чем на
вырубке второго типа. Местами появляется Pterid^uni aquilinum, тогда как Majanthe-
тит bifolium и Trientalis еигораеа сильно угнетены. Выпас сказывается сильнее, по-
скольку скот находит здесь больше корма.
4. На вырубках типа Oxalis проявляется влияние более благоприятных почвен-
ных условий. Теневыносливые виды, свойственные старым насаждениям, вытесняются,
а вместо них в типе Oxalis-Majanthemum появляются типичные для лесов неморального
типа виды: Milium effusum, Urtica dioica, Stellaria nemorum, Chaerophyllum silvestre,
Aegopodium podagraria и т. д.; во влажном, богатом папоротниками подтипе распростра-
няются Deschampsia caespitosa, Triticum caninum и многочисленные виды разнотравья:
Angelica silvestris, Filipendula ulmaria, Geum rivale и т. д.
В общем, типы леса легче выделяются в старых насаждениях, чем
если исходить из растительности вырубок.
Сходные типы леса выделил Сукачев в пределах бореальной зоны
хвойных лесов Восточной Европы. Ниже помещена составленная Сукаче-
вым схема еловых лесов (Piceeta), встречающихся на влажных место-
обитаниях 1 2.
1 A. K.Cajander, Uber Waldtypen, Acta for. fennica, Helsinki, 1909; Wesen
und Bedeutung der Waldtypen, «Silva fennica», 15, Helsingfors, 1930, а также:
« Intersylva», 3, 1943. См. также: V. T. Aaltonen, Boden und Wald, Berlin, 1948.
2 См. также: H. W a 1 t e r, Die Vegetation Osteuropas, 2. Aufl., Berlin, 1943.
Р п с. 268. Еловый лес в возрасте 80 лет (подтип Oralis — Myrtillus по Каяндеру)
в южной Финляндии (фото Каяндера).
Рис. 269. 250-летний еловый лес с хорошо развитым моховым покровом в северной
Финляндии (фото Каяндера).
352
Глава 5
Еловые леса (Piceeta, сокращенно Pic.)
(Усиление заболачивания и
торфообразования)
(Увеличение содержания извести,
мягкий гумус)
Верховое -ч— Piceetum Piceetum
болото sphagno-
sum
poly-
tricho-
sum
Pic. typicum
-> Piceetum -> Переходит в лист-
петого-
sum
венные
леса
Pic. herbosum
увлажнения с хоро-
Переходит в пойменные леса (ряд возрастающего
шей аэрацией почвы)
Типичные еловые леса на подзолистых почвах всегда состоят из трех
ярусов: древесного яруса, яруса травянистых растений и напочвенного
покрова (мхов). Эти леса отличаются довольно бедным видовым составом,
что связано с интенсивной конкуренцией корней деревьев и неблагоприят-
ными условиями освещения; на почве развивается мощный моховой
покров (Hylocomium splendens, Pleurozium schrebert, Rhytidiadelphus trt-
queter, пятнами Pttlium crista-castrensis, Dicranum scoparium и D. undula-
tum).
На рис. 269 изображен типичный еловый древостой. На основании
состава травяного покрова выделяют три фации ельников:
1. Piceetum oxalidosum (ельник-кисличник),
2. Piceetum myrtillosum (ельник-черничник),
3. Piceetum vacciniosum (ельник-брусничник).
Piceetum oxalidosum занимает центральное положение в схеме еловых
лесов, составленной только для южной части бореальной зоны; в зоне
смешанных лесов ельники-кисличники могут переходить в Pic. nemorosum,
Р ис. 270. Cornus suecica в цветущем состоянии, Швеция (фото Е. Вальтер).
Хвойные леса умеренного холодного климата — бореальная зона
353
где ель растет с примесью дуба и липы, а в травяном покрове появляются
представители широколиственных лесов (Asperula odorata, Pulmonaria
officinalis, Stellaria holostea, Lamium galeobdolon и т. д.). Ельники-кислич-
ники образуют также переход к Pic. herbosum, в составе которого рас-
пространены виды заболоченных и пойменных лесов. Вторая фация связана
с повышением влажности и содержания грубого гумуса в почвах и при
дальнейшем увеличении застойного увлажнения приводит к образованию
болот. Третья фация с покровом из Vaccinium vitis-idaea характеризуется
образованием грубого гумуса в сухих условиях и является связующим
звеном с сосновыми лесами. К сопутствующим видам относятся Empetrum
nigrum и Ledum palustre. Очень типичны для бореальных хвойных лесов
Linnaea borealis, Trientalis europaea, Circaea alpina, виды орхидных —
Listera cordata и Calypso bulbosa, различные виды грушанок, Lycopodium
annotinum, а в западной части зоны Cornus suecica (рис. 270).
В качестве примера приводится характеристика насаждений, распо-
ложенных в северной части зоны (Архангельская область) и в области, где
хвойные леса граничат с лиственными лесами (окрестности Москвы).
Флористический состав травяно-кустарничкового покрова типичного елового леса
в Архангельской области; цифрами показано обилие видов (+ означает единично,
5 — сплошной покров):
3 Oxalis acetosella
2—3 Vaccinium myrtillus
+ Vaccinium vitis-idaea
1 Linnaea borealis
+ Listera cordata
-f- Calypso bulbosa
1 Lycopodium annotinum
+ Lycopodium clavatum
1 Majanthemum bifolium
1 Trientalis europaea
1 Dryopteris phegopteris
+ Dryopteris linneana
+ Dryopteris spinulosa
+ Atragene sibirica
+ Equisetum silvaticum
+ Deschampsia flexuosa
+ Aconitum excelsum
4" Angelica silvestris
+ Pirola (Moneses) uniflora
+ Pirola media
-f- Pirola minor
Hr Pirola rotundifolia
+ Luzula pilosa
1 Melica nutans
+ Orobus vernus
+ Paris quadrifolia
Pirola (Ramischia') secunda
- j- Veronica officinalis
+ Geranium silvaticum
- f- Cirsium heterophyllum
- j- Convallaria majalis
4- Fragaria vesca
4" Rubus saxatilis
4- Solidago virgaurea
4" Vicia sepium
Менее характерен флористический состав Травяного покрова еловых лесов в ок-
рестностях Москвы. Описание проводилось на площади 100 м2. Были выявлены сле-
дующие виды:
4 Oxalis acetosella
1 Deschampsia caespitosa
+ Melica nutans
+ Poa nemoralis
1 Carex digitata
+ Luzula pilosa
+ Actaea spicata
+ Ajuga rep tans
+ Athyrium filix femina
+ Prunella vulgaris
+ Campanula persicifolia
4- Convallaria majalis
4" Crepis paludosa
Dryopteris felix mas
4" Dryopteris spinulosa
1 Fragaria vesca
-- Galium schultesii
- - Geranium silvaticum
- - Lysimachia vulgaris
- - Majanthemum bifolium
- - Paris quadrifolia
- - Pirola secunda
- - Pirola rotundifolia
1 Rubus saxatilis
1 Solidago virgaurea
1 Stellaria holostea
4“ Vaccinium vitis-idaea
1 Veronica chamaedrys
4- Equisetum pratense
Эта пробная площадь заложена в области распространения смешанных лесов,
недалеко от южной границы ареала ели. В связи с этим в составе травяного покрова
отсутствуют почти все характерные спутники ели; травостой по своему составу напоми-
нает напочвенный покров лиственных лесов.
23-01248
354
Глава 5
Сосновые леса отличаются более широкой экологической амплитудой^
чем еловые:
(Усиление заболачивания)
Сосновые леса (Pineta, сокращенно Pin,)
(на сухих почвах)
.Pin. cladinosum
Верховое
болото
<—Pin,<— Pin, <—
shag- poly-
nosum tricho-
sum
Pinetum
hylocomiosum
(Увеличение содержания
извести)
—> Pin, —> Степные леса (на бога-
nemoro- тых известью почвах)
sum
Pinetum herbosum
(на влажных, хорошо аэрируемых почвах)
Как видно, сосновые леса близки к еловым и образуют много заме-
щающих типов леса. Сосновые леса встречаются на более сухих место-
обитаниях, где условия произрастания для ели неблагоприятны. Они
Рис. 271. Зависимость,
роста сосны на песчаных
почвах от глубины зале-
гания грунтовых вод (по
Погребняку),
С увеличением влажности
почвы изменяется и состав
нижних ярусов. Выделяют:
А, — Pinetum cladinosum
с лишайниковым покровом;
Аг — Pinetum hylocomiosum
с Vaccinium vitis-idaea; Аз —
Pinetum myrtillosum с при-
месью березы (уровень грун-
товых вод на глубине
1—2 м); А4 — Pinetum uli-
gino-ledosum на торфянисто-
глесво-подзолистых почвах;
А5 — верховые болота с еди-
ничными соснами па торфя-
ных почвах.
в основном приурочены к бедным, кислым почвам, на которых в северной
части зоны распространен Pin, cladinosum — сосняк с лишайниковым
покровом с Calluna vulgaris, Vaccinium vitis-idaea, Arctous (=--Arctosta-
phylos) alpina, Lycopodium selago, Antennaria dioica и др, В крайних усло-
виях существования сохраняются только лишайники (виды Cladonia,
Cetraria), Сухостью отличаются и местообитания степных лесов на юге
лесной зоны; эти леса относятся к типу Pin, substepposum с покровом
из Koeleria glauca, Veronica spicata, Jurinea cyanoides, Genista tinctoria,
Cytisus ruthenicus, Dracocephalum ruyschianum и к Pin, stepposum, в кото-
ром растут уже настоящие степные виды: Stipa pennata s,l,, Agropyrum
cristatum, Linum perenne, Linaria genistijolia и т, д. Эти степные леса
приурочены к богатым известью песчаным почвам; в травяном покрове
преобладают псаммофиты: Koeleria glauca, Euphorbia segueriana, Helichry-
sum arenarium, Peucedanum arenarium и др. Иногда встречаются лишай-
ники и мхи (среди них Tortula (Syntrichium) ruralis), доминирующие
на наиболее сухих почвах,
В пределах Pinetum hylocomiosum, аналогично Piceetum typicumy
можно выделить те же три фации:
1) Pin, oxalidosum, 2) Pin, myrtillosum, 3) Pin. vacciniosum.
Хвойные леса умеренного холодного климата — бореальная зона
355
Из перечисленных фаций наиболее типичен для сосновых лесов Pin.
vacciniosum,; Pin. oxalidosum встречается значительно реже; вторая
фация так же, как и в еловых лесах, переходит в болота. Существование
этих фаций, приуроченных к песчаным почвам, зависит от близости уровня
грунтовых вод (рис. 271).
В Восточной Европе сосновые леса развиваются на песчаных почвах следующих
типов:
Тип почвы Растительность
Сухие леса: бор (а и б) и суборь
(в и г):
а) сухие дюнные пески (слабо оподзо-
ленные)
б) подзолистые песчаные почвы
в) подзолистые супесчаные почвы
г) подзолистые супесчано-суглинистые
почвы.
Сырые леса на глеевых почвах
а) подзолистые, оглеенные песчаные
почвы и песчаные глеевые почвы с
с торфом
б) подзолистые, оглеенные супесчаные
почвы и суглинисто-супесчаные гле-
евые почвы с началом торфообразо-
вания
в) подзолистые, оглеенные супесчано-
суглинистые почвы и супесчано-су-
глинистые глеевые почвы с низин-
ным торфом.
Сухой сосновый лес с лишайниковым по-
кровом (Pinetum cladinosum)
Влажный сосновый лес с моховым покро-
вом (Pinetum hylocomiosum)
Влажный, разнотравный дубово-сосно-
вый лес (Querceto-Pinetum pteridio-
sum)
Сосново-лиственные леса на западе с гра-
бом (Carpino-Nemoreto-Pinetum)
Сырой сосновый лес (Pinetum myrtillosum
и Pinetum uligino-ledosum)
! Сырой дубово-сосновый лес (Querceto-
Pinetum myrtillosum и Qu.-P. ledo-
sum)
Сырой сосново-лиственный лес на западе
с грабом (Carpino-Nemoreto-Pinetum
myrtillosum)n богатый папоротниками
сосново-ольховый заболоченный лес
(Nemoreto-Pinetum alnosum).
Pinetum nemorosum — это смешанные насаждения, нижний ярус
которых образован дубом и липой, а подлесок — лещиной (Corylus avel-
land). Ярус травянистых растений почти аналогичен травостою листвен-
ных лесов. В сосновых лесах широко распространены виды грушанок,
особенно Chimaphila umbellata и Pirola chlorantha, из орхидных —Goodyera
repens, а из плаунов — L. complanatum.
Типы сосновых лесов, аналогичные соответствующим типам еловых
лесов, не могут быть включены в состав зональной растительности. Это
производные леса, возникшие после лесных пожаров. Pinus silvestris
легко заселяет гари, растет здесь быстро и поэтому часто образует одно-
возрастные древостои. Под ярусом сосны обычно развивается подрост
ели (рис. 272); в дальнейшем более теневыносливая ель вытесняет сосну,
если только вторичный пожар не уничтожит ее подрост. На гарях под
пологом сосны часто разрастаются Molinia coerulea, Calamagrosits epi-
geios, Pteridium aquilinum; эти виды расположены в последовательности,
отражающей увеличение сухости местообитаний. Лесные пожары в настоя-
щее время в большинстве случаев возникают по вине человека. Старые
статистические данные (табл. 62) дают представление о частоте лесных
пожаров в Восточной Европе, возникающих от различных причин («Phyto-
logie», Bd. Ill, 1).
Пожары, вызванные естественными причинами (от удара молний),
случаются относительно редко. Но уже сама вероятность таких пожаров
свидетельствует о том, что огонь может выступать как естественный
23*
356
Глава 5
Р и с.” 272. Сосновый лес (тип Myrtillus) в возрасте 50 лет с подростом ели, централь-
ная Финляндия (фото Каяндера).
фактор; это необходимо учитывать в малонаселенных районах, где борьба
с лесными пожарами не проводится и где они могут охватывать большие
площади, поскольку в девственных лесах накапливается много мертвой,
легковоспламеняющейся древесины.
Таблица 62
Частота лесных пожаров, обусловленных
различными причинами (в %)
В результате неосторожного обращения с
огнем ................................ 58
От переброски огня с соседних участков . . 30
В результате поджогов................... 10
От ударов молний .... 2
То, что сосновые леса в естественных условиях в пределах бореальной
лесной зоны существуют благодаря лесным пожарам, подтвердилось при
ознакомлении с первобытным лесом в заповеднике Муддус (шведская
Лапландия) (рис. 273) х. В этих не тронутых человеком лесах сосна пред-
ставлена только определенными классами возраста: возраст самого старого
поколения достигает 410 —430 лет, более молодого — около 280 лет,
следующего—125 лет и самого молодого — 50 лет. На стволе очень
старой сосны остались два следа от пожаров. На основании подсчета
годичных колец установлено, что одно повреждение возникло от пожара
1 Экскурсия во времяУП Международного ботанического конгресса в Стокгольме.
Хвойные леса умеренного холодного климата — бореальная зона
357
Рис. 273. Девственный лес в заповеднике Муддус в шведской Лапландии: еловые
и сосновые леса, болота и озера (фото Э. Вальтер).
около 125 лет назад, а более старые — около 400 лет назад. После этих
пожаров появились соответствующие поколения сосны. Остальные два
пожара не повредили ствол сосны.
Таким образом, девственный лес не всегда характеризуется гомоген-
ной структурой, он может быть представлен различными стадиями сук-
цессий.
Вблизи арктической границы распространения леса сукцессии осу-
ществляются очень медленно. В северной Швеции первая стадия березы
длится около 150 лет, следующая стадия сосны продолжается до 500 лет,
и лишь затем наступает заключительна^ стадия ели. Пожары происходят
и во влажной бореальной области, где всегда повторяются летние засухи.
Лесная подстилка высыхает и легко воспламеняется х.
Лесные пожары в хвойных лесах классифицируются следующим
образом:
1) низовые пожары, во время которых гибнет ель, обладающая поверх-
ностной корневой системой, тогда как сосна сохраняется;
2) верховые пожары, обычно повреждающие сосну;
1 Температура измерялась в горящей подстилке сосновых лесов. В среднем наи-
более высокая температура достигает 340° (140—545°). Невысокая температура (< 300°)
связана с небольшой массой воспламеняющегося напочвенного покрова (< 60 г сухого
вещества на 1 ле1 2). Температура повышается по мере увеличения количества горючего
материала, что особенно хорошо выражено в случаях, когда горит большая масса дре-
весины. Максимальная температура при низовых пожарах отмечена на высоте 5 ему
очень высокая температура обычно держится в течение короткого срока (наиболее высо-
кая из измеренных температур — 20 сек). Скорость ветра во время опытов оказывала
небольшое влияние на величину температуры. См.: D. W. Smith, J. Н. Spal-
ding, The temperatures of surface fires in Jack Pine Barren: I and II, «Canad. J. Bot
44, 1904, p. 1285-1298.
358
Глава 5
3) торфяные пожары, при которых торфяные залежи тлеют, но не
горят открытым огнем; эти пожары возникают в заболоченных лесах.
Табл. 63 содержит старые статистические данные о числе различных
типов пожаров в Восточной Европе и размерах площади, охваченной ими.
Таблица 63
Число и площадь лесных пожаров
в Восточной Европе
Тип пожара G 1923/24 до 1928/29 гг. 1938 г.
число площадь, тыс. га площадь, тыс. га
Низовой . . 66 937 3570 4264
Верховой 2 667 488 477
Торфяной .... 1276 43 30
Обширные площади березовых и сосновых лесов в северной части
бореальной лесной зоны связаны с наличием большого числа пожаров
в прошлом. Береза и осина — это породы-пионеры, которые первыми
распространяются на вырубках. Только спустя 20—30 лет на песчаных
почвах появляется сосна, а на влажных суглинистых почвах •— ель.
БИОГЕОЦЕНОЗЫ ХВОЙНЫХ ЛЕСОВ
Хвойные леса отличаются от лиственных в основном тем, что листовая
поверхность у них (за исключением лиственничных лесов) сохраняется
в течение всего года. Поэтому весной перед облиствлением и осенью после
начала листопада в хвойных лесах на поверхности почвы не создается
благоприятных условий освещения, как это наблюдается в лиственных
лесах.
Таблица 64
Средняя температура почвы (в °C) в период
с 15.VI по 15.VII под пологом насаждения
Pinus silvestris — Vaccinium vitis-idaea
в Московской области
Возраст насаждения, лет Глубина почвы, см
5 10 15 30
Разновозрас тный 19,6 18,0 16,8 16,1
33 12,6 12,3 11,9 10,9
65 16,6 16,0 14,8 14,0
150 17,9 16,9 15,6 14,7
Сосна в возрасте 150 лет с густым нижним еловым ярусом 12,8 11,4 10,5 10,0
Хвойные леса умеренного холодного климата — бореальная зона
359
Результаты исследований, проведенных в евросибирской бореальной
зоне, подробно изложены в упомянутой выше работе Сукачева и Дылиса
«Основы лесной биогеоценологии» (1964).
Температура почвы в хвойных лесах зависит от характера древостоя,
его сомкнутости и возраста (табл. 64).
Наиболее низкая температура отмечена под сомкнутым пологом
молодых насаждений. Особенно ощутимо на величину температуры
в сосновом лесу влияет нижний густой еловый ярус.
Большое значение имеет также годовой ход температуры на различной
глубине почвы (табл. 65).
Таблица 65
Температура (в °C) на различной глубине почвы в еловом и березовом лесу;
для сравнения приводятся данные, полученные на безлесном участке
около Мустиала в Финляндии (по Гомену)
Глу- бина, м I II III IV V VI VII VIII IX X XI XII Сред- няя за год
Безлесный участок (площадью 3 га в еловом лесу)
0,5 1,53 1,22 1,01 1,16 4,55 10,36 12,97 12,99 10,37 6,79 3,61 2,10 5,75
1.0 2,34 1,79 1,53 1,44 3,20 7,57 10,20 11,16 9,94 7,51 4,74 3,13 5,40
2,0 3,45 2,78 2,43 2,09 2,63 4,91 Ело 7,26 вый лес 8,72 8,75 7,65 5,84 4,31 5,10
0,5 0,84 0,42 0,23 0,34 0,81 4,49 8,86 9,82 8,35 5,80 3,10 1,54 2,80 3,73
1,0 1,88 1,35 1,04 0,90 1,06 3,32 6,70 8,29 7,78 6,26 4,23 3,81
2,0 3,26 2,62 2,13 1,85 1,81 2,48 4,58 6,25 6,78 6,26 5,25 4,18 3,97
Березовый лес
0,5 0,76 0,23 0,03 0,33 2,11 7,22 10,17 10,74 8,98 6,01 6,48 2,95 1,54 4,29
1,0 2,16 1,61 1,21 1,06 1,85 4,89 7,44 8,69 8,20 4,37 2,98 4,26
2,0 3,73 3,05 2,71 2,42 2,42 4,02 5,85 7,17 7,60 7,05 5,76 4,62 4,71
Температура освещенной солнцем почвы на 2° выше, чем под пологом
елового леса. Светлый березовый лес занимает промежуточное положение.
Определенное влияние на температуру почвы в лесу оказывает под-
стилка. По сравнению с обнаженной почвой почва, покрытая подстилкой,
замерзает на месяц позже; глубина промерзания последней на 40% меньше.
Большую роль играет также сомкнутость древостоя: чем гуще древостой,
тем меньше солнечного света проникает под его полог и тем маломощнее
снежный покров, препятствующий; промерзанию почвы. В северной
Швеции почва в разреженном древостое промерзала на глубину 30 см
и оттаивала в начале июня, в сомкнутом — промерзала на глубину 85 см,
а оттаивала лишь в начале августа, то есть почти в конце вегетационного
периода.
В этой связи важно знать глубину залегания корней ели. В табл. 66
приведены данные о распределении корней по профилю различных почв
в еловых лесах Вологодской области.
Скорость ветра резко снижается даже в разреженных сосновых лесах;
табл. 67 отражает результаты наблюдения в сосновых лесах Московской
области.
Еще сильнее тормозят движение воздуха еловые леса; при скорости
ветра 2,8 м/сек на открытом поле скорость ветра в глубине леса на рас-
стоянии 40 м от опушки леса достигает лишь 1—1,5%.
360
Глава 5
Таблица 66
Распределение массы корней по профилю почвы
в некоторых типах еловых лесов
Тип насаждения и почва Глубина, СЛ1 Количе- ство кор- ней, кг/га
I. Еловый лес с Oxalis 3—10 1780
acetosella и большим 10-20 2700
числом папоротников на 20-30 720
слабоподзолистой почве 30-40 47
40—50 13
II. Еловый лес с Vaccini- 2-8 4180
um myrtillus на сильно- 8-10 1380
подзолистой почве 10—20 300
20-30 01
30-40 0
40-50 0
III. Очень разреженный 0-4 4—11 140 0
еловый лес с Equisetum 4 4 4 R п
silvaticum и Sphagnum 1 1 1 и 4R ОП V П
на торфянисто-подзо- листо-глеевой почве 1U V
Таблица 67
Скорость ветра в сосновых лесах на расстоянии 200 м
от опушки леса (в % от скорости ветра на открытом месте)
Возраст леса и сомкнутость крон Скорость ветра за пределами леса, м/сек
1,2 2,2 2,7 3,5 5,2 6,2 7,0
65 лет, сомкну- тость 0,4 . . . 12 18 20 29 44 61
65 лет, сомкну- тость 0,9 . . . 8 10 12 17 28 39 46
150 лет, сомкну- тость 0,8, ель во втором ярусе 0 2 4 8 15 17 19
Таблица 68
Перехват осадков кронами хвойных лесов (в мм; в среднем за год)
Годовые осадки, мм Возраст насаждения, лет
20 40 60 80 100 120 140 160
Сосновый лес под Архангельском . . 525 55 86 98 120 118 117 116 112
Сосновый лес под Москвой 550 127 150 140 135 120 105 100 97
Еловый лес, Воло- годская область . . 600 106 178 175 170 166 160 158 —
Хвойные леса умеренного холодного климата — бореальная зона
361
Был определен эффект задержания осадков кронами сосновых и ело-
вых древостоев с одинаковой степенью сомкнутости крон, равной 1,0
(табл. 68).
Поверхностный сток в хвойных лесах с мощной подстилкой из хвои
больше, чем в лиственных лесах с рыхлой подстилкой. В условиях равнин-
ного рельефа годовой сток в среднем достигает: в еловых лесах на подзо-
листых почвах 15—26%, в сосновых —около 7% (в лиственных лесах
часто лишь 1,6—4%). В период таяния снега сток в еловых насаждениях
выше, чем в сосновых; минимальное значение стока отмечено в листвен-
ных лесах.
В сосновых лесах изучался расход влаги древесным ярусом на транс-
пирацию (табл. 69).
Таблица 69*
Расход влаги на транспирацию (в мм) в различных типах сосновых лесов
с 15.IV по LX в северной части зоны тайги (I) и в зоне смешанных лесов (II)
(в скобках — суммарное испарение)
Возраст леса, лет Сомкну- тость крон С лишай- никовым и моховым покровом С Vacci- nium vitis-idaea С Vacci- nium myrtillus С большим числом видов раз- нотравья С Poly- trichum Со Sphag- num
I 35 1,0 88 (211)
60 1,0 — 183 197 (332) — — —
160 0,8 115 (250) 168 (310) 176 (340) — — —
270 0,7 — 97 114 (274) — — —
II 13—20 1,0 162 (355) 201(392) 221(430) - (435) - (400) — (385)
30—40 0,8—1,0 171 (356) 250 (405) 273 (461) 218 (463) — (410) - (458)
40 1,0 188 (455) 242 (405) 256 (490) 268 (495) - (455) - (438)
60 0,9-1,0 173 (393) 223 (429) 230 (464) 245 (470) 165 (475) 119 (351)
60 0,7 - (351) 236 (395) - (399) - (451) - (463) -£343)
150 0,5 - (391) - (432) — (484) 2151
150 0,8—1,0 161 (328) 135 (360) 165 (391) — (395) — (365) —
150, второй ярус обра- зован елью 1,0 — 260 (500) — — — —
1 Возраст 80 лет, сомкнутость крон 0,9.
Как видно, количество транспирируемой влаги зависит от бонитета
насаждения. Чем выше бонитет, тем больше масса транспирирующих
листьев, тем выше продуктивность леса. Эта зависимость особенно хорошо
выражена в шестидесятилетием насаждении в области смешанных лесов.
Расход влаги на транспирацию увеличивается по мере нарастания богат-
ства и влажности почв, а затем резко снижается при заболачивании почвы.
Транспирация лесов на севере зоны тайги значительно ниже, чем в более
теплой и более продуктивной зоне смешанных лесов. Возраст насаждения
также влияет на транспирацию; расход влаги на транспирацию достигает
максимума в период кульминации прироста, который для сосны на севере
наступает в 60 лет, а южнее —уже в 30 —40 лет.
Данные по эвапотранспирации еловых лесов приведены в табл. 70.
Эвапотранспирация чистых березовых лесов с покровом из Vaccinium
myrtillus в средней тайге и зоне смешанных лесов достигает тех же величин
или несколько меньших, чем в еловых лесах.
362
Глава 5
х
Таблица 70
Суммарное испарение в еловых лесах в различных зонах
(в мм)
Возраст леса, лет Сомкну- тость крон С Vacci- nium vitis-idaea С Vacci- nium myrtillus С Oxalis acetosella Со Sphag- num
Северная тайга
1,0 | - 1 248 | - 1 | 3701
Средняя тайга
7 1,о — 201 — —
18 1,0 — 387 —
37 1,0 —> 449 —
110 1,0 — 334 — 330
135 1,0 — 323 440 —
Южная тайга
65 1 1,0 1 - | 440 | 500 1 -
Зона смешанных лесов
20 1,0 335 410 521
40 1,0 398 456 521
55 1,0 445 501 530
80 0,9 500 510 518
120 1,0 — — 428
150 0,9 331 390 405
1 Тип леса с Filipendula ulmaria.
Увеличение фитомассы древесного яруса в сосняках с Vaccinium myrtil-
lus характеризуют данные табл. 71.
Средняя продукция сухого органического вещества в лесах Ленин-
градской области почти одинакова в одновозрастных древостоях, образо-
ванных различными видами деревьев (сосной, елью и березой) и соста-
вляет 5,5—5,6 т/га»год. Средний годовой прирост древесины (сухой вес)
Таблица 71
Надземная и подземная фитомасса древесного яруса
и напочвенного покрова в сосновых лесах (сухой вес в т(га)
Возраст леса, лет
20 40 60 80 1 1 100 120 140 160 180 200 220 240 260
Сухой вес древес- ного яруса . . 39 77 141 213 248 270 271 267 266 254 252 248 249
Сухой вес напоч- венного покрова 0,8 2,8 13,1 14,1 14,5 15,5 16,8 17,4 18,7 19,4 19,5 19,5 19,5
в южной тайге равен 5 т!га, в средней тайге —3 т!га. На рис. 274 и 275 го-
довой прирост древесины представлен в виде брутто-продуктивности
сосновых и еловых лесов, а годовой запас подстилки — органической
йассой, поступающей в почву. Прирост надземной фитомассы с увеличе-
Хвойные леса умеренного холодного климата — бореальная зона
363
нием возраста леса и интенсивность накопления опада изображены
на рис. 276; в старых насаждениях запас подстилки в 3—4 раза превышает
таковой в молодняках. Опад не накапливается в лесу, поскольку он все
время разлагается почвенными животными и микроорганизмами; это
происходит до тех пор, пока не наступит равновесие между поступлением
Р и с. 274. Брутто-продукция (годо-
вой прирост) древесного яруса п годовое
поступление подстилки в сосновых лесах
различного возраста. Нетто-продуктпв-
ность представлена между двумя кривыми;
она достигает наибольших значений в на-
саждениях возраста 30—50 лет (по Ро-
дину).
Р и с. 275. То же в еловых лесах. Наи-
более высокая нетто-продуктивность
наблюдается в насаждениях возраста
40—50 лет.
опада и его разложением. В одинаковых почвенных условиях подстилка
в еловом лесу всегда характеризуется кислой реакцией, содержит меньшее
количество бактерий и отличается более низкой интенсивностью «дыхания
гумуса», чем подстилка в буковом лесу \ В еловом лесу наиболее интен-
сивный распад подстилки наблюдается в самом верхнем ее слое (Forna),
тогда как опад бука особенно энергично разлагается в пределах верхней
части превращающегося в модер слоя подстилки. Скорость полной мине-
рализации подстилки может быть вычислена на основе ее дыхания. Срав-
нительные данные по скорости разложения подстилки на различных поч-
вах в окрестностях Гамбурга приведены в табл. 72.
1F. Н. М еуег, Vergleich des mikrobiellenAbbausvon Fichten-undBuchenstreu
auf verschiedenen Bodentypen, «Arch. f. MikrobioL», 35, 1960, S. 340—360.
364
Глава 5
Хвойная подстилка минерализуется медленно, поэтому в верхних
слоях почвы в хвойном лесу содержится относительно много органиче-
ских веществ. В лесах северной части бореальной зоны, где низкие тем-
пературы и большая влажность почвы препятствуют разложению под-
Рис. 276. Зависимость прироста
надземной фитомассы (7) и количе-
ства подстилки (2) от возраста Pinetum
myrtillosum. В старых насаждениях
в возрасте свыше 100 лет фитомасса
не увеличивается, тогда как запас
подстилки продолжает нарастать
(по Молчанову).
стилки, накапливается большое коли-
чество грубого гумуса или образуется
торф. В очень неблагоприятных усло-
виях среды продуктивность древесного
яруса снижается: она может быть даже
меньше продуктивности напочвенного
покрова. В качестве примера могут слу-
жить заболоченные леса Вологодской,
области (табл. 73).
Таблица 72
Время, необходимое для полного
разложения подстилки бука и ели
(число лет)
Тип почвы
Бук
Ель
Бурозем...........
Подзолистый бурозем
Псевдоглеевый под-
зол ..............
Железистый подзол
9,4
10,3
20
40
17
40
121
121
Индекс листовой поверхности в ти-
пичных древостоях хвойных лесов
бореальной зоны выше, чем в листвен-
ных лесах. Хвоя существует не менее
двух лет. Индекс листовой поверхности
в сосновых древостоях зоны смешанных
лесов достигает 9,0 —9,7, в еловых ле-
сах Вологодской области —11,3. Двухлетняя хвоя обычно отличается!
меньшей фотосинтетической активностью, чем однолетняя.
Таблица 73
Годовой прирост сухого вещества в разных ярусах
заболоченных лесов (в кг/га)
Тип леса Древес- ный ярус Ярус кустар- ников Ярус травя- нистых растений Ярус мхов Всего
Ельник болотно-травя- ной 850 8 89 959 1906
Сосняк кустарничково- сфагновый 104 — 15 1661 1780
У хвойных пород развита эктотрофная микориза. Гифы грибов спо-
собствуют значительному расширению поверхности корневой системы)
Хвойные леса умеренного холодного климата — бореальная зона
365
и облегчают тем самым растению процесс поглощения минеральных пита-
тельных веществ, в первую очередь азота. Микориза образуется при
избыточном количестве сахара, который накапливается в корнях хвойных
в результате недостатка азота. При внесении искусственных удобрений
корни начинают потреблять такое ^количество азота, в присутствии кото-
рого весь сахар используется на синтез белка; в этом случае микориза
не развивается х. Микориза обычно формируется на корнях деревьев,
приуроченных к местообитаниям с бедными почвами. Речь идет о настоя-
щем симбиозе, поскольку при наличии микоризы рост деревьев улуч-
шается, а при обрезке корней и прекращении притока продуктов ассими-
ляции грибы не образуют плодовых тел.
По Бьёркману, подъельник (Monotropa hypopitys) — сапрофит, широко
распространенный в еловых лесах, — косвенно паразитирует на ели,
с которой он связан при помощи гифов грибов, образующих микоризу
у Мonotropa и у ели 1 2.
В зоне хвойных лесов развиты типичные подзолистые почвы, а при
высоком уровне грунтовых вод — подзолисто-глеевые почвы. В профиле
этих почв хорошо выражены горизонт грубого гумуса (AJ, подзолистый
горизонт (Ае) и иллювиальный горизонт (В). Как уже упоминалось
(см. wrp. 359), образование минеральных форм азота в слое грубого гумуса
может быть относительно высоким. С увеличением глубины этот процесс
в почве резко сокращается. Поэтому большинство корней деревьев при-
урочено к верхним горизонтам почвы, где располагаются и корни травя-
ного покрова. Здесь ярко выражена корневая конкуренция. На сухих
местообитаниях или в период длительной засухи летом конкуренция
идет за воду, в остальных случаях большей частью за азот.
При обрубке корней деревьев влажность лесной почвы относительно
.мало повышается в течение летних месяцев (табл. 74).
Таблица 74
Запасы влаги в верхних 0—50 см лесной почвы
при наличии и отсутствии конкуренции корней
(в мм выше коэффициента завядания);
южная часть зоны тайги
Тип леса Корни деревьев обрублены Контрольный участок
30.VI | 30. VIII 30.VI | 30. VIII
Ельник кислично-па- поротниковый . . . 120 98 89 57
Разнотравный берез- няк . . 115 88 79 28
1 Этот взгляд, высказанный Бьёркманом, в последнее время оспаривается
<Р. X. Майером. Последний считает, что повышению содержания сахара в корнях способ-
ствуют действующие вещества, которые выделяет гриб. Этот гриб принимает активное
участие в образовании микоризы. Чаще всего микориза встречается на почвах, богатых
питательными веществами. Количество микоризы не уменьшается после внесения азот-
ных и фосфорных удобрений.См.: F. Н. М е у е г, Neue Erkenntnisse iiber das Zusammen-
leben von Pilz und Baum, «Die Umschau», H. 11, Frankfurt a.M.,1964;H. К о b erg,
Ddngung und Mykorrhiza. Ein GefaBversuch. mit Kiefern, «Forstwiss. Zbl.», 85, 1966,
S. 321-384.
2 E. Bjorkman, Uber die Natur der Mykorrhizabildung unter besonderer
BeriicksichtigungTder Waldbaume, «Forstwiss. Zbl.», 75, 1956.
366
Глава 5
Рис. 277. Сухой сосновый лес с покровом из лишайников (заповедник Муддусг
шведская Лапландия). Сосна возобновляется только вокруг отмершего дерева, в центре
снимка (фото Э. Вальтер).
Если в сосновых лесах на маломощных почвах, подстилаемых грани-
тами, деревья летом потребляют всю воду, накопившуюся в верхних
слоях почвы, то травяной покров не развивается (см. стр. 357); в этих
условиях растут только лишайники (Cladonia, Cetraria и др.) и мхи, кото-
рые не расходуют влагу из почвы \ В таких обычно разреженных древо-
стоях по той же причине отсутствует и подрост. Только после отмирания
старых деревьев появляется возможность для развития всходов (рис. 277).
В хвойных лесах конкуренция за минеральные вещества и особенно
за азот выражена более ярко, чем в лиственных лесах.
По данным Ромеля и Тамма, в Швеции внесение минерального азот-
ного удобрения увеличивает прирост еловых древостоев в течение первых
четырех лет; остальные минеральные удобрения без добавления азота
оказывают незначительное влияние. После внесения удобрений содержа-
ние N в хвое повышается, однако только небольшая часть (около 20%)
азота, использованного в качестве удобрения, накапливается в надземных
частях деревьев. Оставшаяся часть азота используется напочвенным
покровом и микроорганизмами 1 2.
1 Мхи (например, Hylocomium splendens) в еловом лесу получают минеральное
питание с водой, стекающей с крон деревьев; в связи с этим под кронами ели мхи раз-
виваются значительно лучше (до 100 г/л2), чем по мере удаления от стволов. См.:
С. О. Tamm, Growth and plant nutrient concentration in Hylocomium proliferum (L)
Lindb. in relation to tree canopy, «Oikos», 2, 1950, p. 60—64; Growth, yield and nutri-
tion in carpets of a forest moss (Hylocomium splendens), «Medd. Stat.Skogsforskn. Inst.»r
43, 1953, p. 1 — 140.
2 С. О. T a m m, Die Nahrstoffaufnahme gediingter Fichten- und Kiefernbestander
«Arch. f. Forstwesen», 12, 1963, S. 211—222; С. О. T a m m, С. G a r b о n n i e r, Plant
nutrients and forest yield, «Stat. Skogsforskn. Inst.», 82, 1961.
Хвойные леса умеренного холодного климата — бореальная зона
367
Из опытов следует, что азот в лесах находится в минимуме для
деревьев 1 и в еще большей мере для травяного покрова или всходов
деревьев.
При изучении содержания питательных веществ в хвое сеянцев ели
или в листьях видов травяного покрова, растущих в условиях конкурен-
ции с корнями деревьев и при исключении ее в результате обрубки корнейг
Рис. 278. Динамика накопления радиоактивного фосфора
в хвое сеянцев ели после внесения удобрений (по Карпову).
1 — березовый лес с покровом из Vaccinium myrtillus на сильноподзоли-
стой почве; 2 — березовый лес с хорошо выраженным травяным покровом
из Aegopodium на слабоподзолистой почве. Верхние кривые — при отсут-
ствии конкуренции с корнями березы, нижние — в условиях конку-
ренции.
установлено, что устранение корневой конкуренции ведет к значитель-
ному увеличению содержания питательных веществ, в особенности азота
(табл. 75).
Таблица 75
Содержание питательных веществ в хвое и листьях растений
травяного покрова в березовом лесу (в % сухого веса) (по Карпову)
Название растений Конкуренция корней деревьев выражена Конкуренция корней деревьев исключена
зола 1 N Р Са зола N 1 р | Са
Сеянцы Picea abies . . 3,69 1,38 0,23 0,11 3,73 1,70 0,26 0,19 j
Oxalis acetosella . . 9,89 2,06 0,57 0,89 9,71 3,18 0,43 1,07
Vaccinium myrtillus . . 4,85 2,19 0,18 0,94 4,84 2,70 0,24 0,95
Calamagrostis langsdorfii . . . 14,26 1,85 0,31 0,11 13,87 2,82 0,31 0,16
Aegopodium podagraria .... 12,52 1,49 0,50 1,40 12,00 3,11 0,58 1,60
Исследования с применением меченого фосфора свидетельствуют,
4то его поглощение сильно затрудняется конкуренцией со стороны корней
деревьев (рис. 278). Эти опыты подтверждают также, что более интенсив-
1 Н. Z б t t 1, Die Mineralstickstoffanlieferung in Fichten-und Kiefernbestiinden
Bayerns, «Forstw. Cbl»., 79, 1960, S. 221—236; Anhiiufung und Umsetzung von Stickstoff
im Waldboden, «Вег. Deutsch. Bot. Ges.», 78,1965, S. 167—180; M. R u n g e, Untersu-
chungen uber die Mineralstickstoff-Nachlieferung an nordwestdeutschen Waldstandorten,
«Flora», 155, 1965, S. 353—386.
Цётль, изучавший круговорот азота в лесах Баварии, выявил, что в угнетенных
древостоях сосны за год образуется минеральных форм азота — 8,8 кг!га, а в хорошо»
развитых древостоях— 77,8 кг!га.
368
Глава 5
ное поглощение N после обрубки корней не может осуществляться за счет
азота, поступающего в почву из разлагающихся обрубленных корней.
Сеянцы деревьев плохо развиваются в старом древостое не столько
из-за недостатка света, сколько во многих случаях из-за недостаточного
обеспечения питательными веществами в результате корневой конкурен-
ции (рис. 279). Это же наблюдается и у сеянцев пихты, которые могут
приспособиться к слабому осве-
Р и с. 279. Рост сеянцев ели в березовом
лесу бореальной зоны (по Карпову).
1 — при отсутствии конкуренции с корнями взрос-
лых деревьев; 2 — в условиях конкуренции.
щению; при этом подавляется
развитие осевого побега и раз-
растаются боковые ветви. В ре-
зультате у сеянца пихты фор-
мируется очень приземистая, но
широкая крона, что способст-
вует поглощению света даже при
его небольшой интенсивности.
Сеянцы сосны после устранения
конкуренции с корнями взрос-
лых растений растут быстрее
только в 2—3 раза, а при
улучшении условий освещения—
в 10 раз.
Опыты по изучению кон-
куренции корней, проведенные
в старом буковом лесу в воз-
расте 190 лет при освещенности
около 20% полного дневного
света, свидетельствуют, что во
влажные годы корни материн-
ского древостоя не оказывают
влияния на подрост бука х.
Проведены следующие 4 серии
экспериментов:
1. Ненарушенный участок: со-
хранена конкуренция с сорняками
и корнями материнского древо-
стоя.
2. Конкуренция сорняков устра-
нена путем неоднократного удаления
их надземных частей; конкуренция
материнского древостоя сохранена.
3. Конкуренция материнского
древостоя устранена; корни деревьев
обрублены на глубину 80 см\ конку-
ренция сорняков сохранена.
4. Устранены все виды кон-
куренции.
Буковый лес развит на мощном псевдоглеевом буроземе, который характеризуется
'Следующими признаками: реакция — сильно кислая; содержание извести — неболь-
шое; обеспеченность калием и фосфорной кислотой — от умеренной до достаточной;
мощность гумусового горизонта — 3—4 см (модер); потери от прокаливания составля-
ют 5,4%.
Корни взрослых деревьев бука проникают на глубину 80 ел, до уплотненного
горизонта. На опытных участках произрастали сорняки: Calamagrostis epigeios, Rubus
idaeus, Agrostis alba и Chamaenerium angustifolium.
1 P. Burschel, J. Schmaltz, Untersuchungen uber die Bedeutung von
Unkraut-und Altholzkonkurrenz fur junge Buchen, «Forstwiss. Zbl.», 84, 1965,
:S. 201—164.
Хвойные леса умеренного холодного климата — бореальная зона
369
Высота саженцев бука в среднем достигала 19 см', на второй год, по данным
измерений, они развивались нормально. На открытой площадке в районе Райнсхардс-
вальда (севернее Касселя), расположенном на высоте 340 м над уровнем моря, в период
наблюдении стояла прохладная и влажная погода без засухи.
В конце второго вегетационного периода влияние конкуренции корней материн-
ского древостоя все еще не было отчетливо выражено, тогда как продуктивность сор-
няков снизилась на 25—30%, что связано с недостатком света.
Эти результаты нельзя обобщать. В каждом отдельном случае следует определить,
какой из факторов оказывает наибольшее влияние. Например, в более северных широ-
тах в сильно разреженных сосновых древостоях девственного леса Муддус (шведская
Лапландия), решающее значение, по-видимому, имеет корневая конкуренция. Неза-
висимо от условий освещения подрост сосны появляется лишь на прогалинах, образую-
щихся после отмирания старых деревьев (см. рис. 277).
Ранее уже упоминалось (т. I, стр. 36 и след.) о характере изменений
состава травяного покрова в бореальной зоне хвойных лесов при исклю-
чении конкуренции с корнями взрослых деревьев. Степень освещенности
около поверхности лесной почвы обычно не имеет существенного значе-
ния. Виды, приуроченные к лесосекам, часто способны выносить
сильное затенение; несмотря на это, они не растут в лесу, поскольку
не выдерживают конкуренции с корнями деревьев за минеральное пита-
ние. Если исключить корневую конкуренцию, то даже при неблагоприят-
ных условиях освещения Deschampsia caespitosa и Dactylis glomerata
развиваются почти нормально и цветут. Только Chamaenerium сохраняется
в этиолированном состоянии и не плодоносит. По данным Карпова, в лес-
ной почве содержится на 1 м1 2 1300—1500 семян видов, произрастающих
на лесосеках {Deschampsia caespitosa, Rubus idaeus, Chamaenerium, Epilo-
bium montanum)1. Эти семена были занесены ветром, птицами и грызунами
или сохранились в почве на местах бывших лесосек. Пока еще не уста-
новлено, каким образом корни деревьев препятствуют их прорастанию.
При обрубке корней деревьев семена прорастают, тогда как удаление
подлеска не влияет на их всхожесть. Результаты посева семян этих видов
в почву березовых лесов отражены в табл. 76.
Содержание азота при отсутствии корневой конкуренции повышалось
в листьях на 60—70%. В засушливые годы известную роль играла и влаж-
ность почвы. На участке II с более богатой почвой корневая конкуренция
деревьев выражена более сильно; после ее исключения растения травяного
покрова развиваются хуже, чем на участке I. Причина в том, что на более
богатых почвах деревья поглощают больше питательных веществ, быстро
растут и образуют более сомкнутый полог; в связи с этим создаются менее
благоприятные условия освещения для напочвенного покрова 2.
Табл. 77 содержит данные о количестве азота, необходимого для
продукции сухого вещества в среднем на 1 га в год 3.
Азот, содержащийся в листьях деревьев, не уходит из круговорота
веществ в насаждении, если только не удаляется подстилка, тогда как
в сельском хозяйстве с урожаем ежегодно отчуждается большое количе-
ство почвенного азота: при возделывании картофеля — 61 кг!га, при
посеве ржи — 52 кг!га. Потеря зольных элементов в лесных посадках
также намного меньше, чем в посевах сельскохозяйственных культур
(табл. 78).
1 В.Г. Карпов, О видовом составе живых семян и запасе'их в почве ельника-
черничника, «Тр. Москв. Общ. испыт. природы., отд. биол.», т. 3, 1960, стр. 131—140.
2 В. Г. Карпов, Новые экспериментальные данные о механизме смен лесных
сообществ в таежной зоне, «Бот. журнал», т. 49, 8, 1964, стр. 1101—1119.
3 L.Tschermak, Waldbau auf pflanzengeographisch-okologischer Grundlage,
Wien, 1950.
24-01248
370
Глава 5
Таблица 76
Продуктивность видов, высеянных в двух типах
березовых лесов при наличии н отсутствии
конкуренции со стороны корней деревьев
(в г сухого веса)
I. Тип сообщества Calamagrostis — Vaccinium
myrtillus;
II. Тип разнотравный с Aegopodium podagraria
Тип Высеянные виды Конкуренция корней
выражена | исключена
I Deschampsia caespi- tosa ....... Agrostis vulgaris Festuca pratensis . . Chamaenerium angus- tifolium 0,036+0,002 0,018+0,003 0,022+0,002 0,420+0,029 7,910+0,205 0,620+0,067 1,812+0,075 8,175+0,273
II Deschampsia caespi- tosa Agrostis vulgaris Festuca pratensis . . Chamaenerium angu- stifolium . . 0,010+0,001 0,007+0,001 0,012+0,002 0,027+0,018 4,820+0,318 0,435+0,054 0,783+0,083 6,821+0,219
Таблица 77
Количество азота (кг/га-год), необходимое
для создания листьев и древесины в лесах
Буковый лес Еловый лес Сосновый лес
Создание листьев 42-44 30—32 28
Создание древе-
сины 9—10 10-13 6
Итого 51-54 40-45 34
Запас фитомассы в хвойных лесах различного типа представлен;
в табл. 79.
Ниже приводятся данные о запасах подстилки в хвойных лесах
Восточной Европы.
Поскольку в пределах зоны хвойных лесов температура понижается
при движении к северу, интенсивность разложения подстилки замед-
ляется в том же направлении; поэтому на поверхности почвы в северной
и средней тайге накапливается около 57—69 т/га опада. В горных сосно-
вых лесах южной -Болгарии запасы подстилки очень невелики и достигают
всего 6,6—8,5 т/га. Масса подстилки существенно зависит от типа лесаг
что видно из следующих данных, полученных при изучении лесов в Бело-
вежской пуще (в т1га)\
Еловые леса с Vaccinium myrtillus . 34,0—52,6
Елово-дубовые леса ............... . . 17,9 — 19,7
Дубовые леса с примесью ели . . . . 11,2 — 12,8
Дубово-буковые леса ................... 8,2—10,0
Хвойные леса умеренного холодного климата — бореальная зона
371
Таблица 78
Количество зольных веществ (в -кг/га < год),
необходимое для построения листьев
и древесины в различных лесах
Калий Фос- форная кисло- та ; Из- весть
Буковый [ листья . 10 И 81
лес \ древесина 5 3 15
Еловый Г листья . 5 6,5 60
лес [ древесина . . 4 1,5 10
Сосновый Г листья . 5 4 19
лсс ( древесина . . 2 1 9
Для сравнения:
картофель 120 36 40
свекла ... . . 184 32 40
луговое сено . . ... 80 30 50
Таблица 79
Запас фитомассы в различных органах деревьев и других растениях
в хвойных лесах (в т/га)
Тип древостоя Воз- раст, лет Хвоя или листья Древе- сина Корни Травяно- моховой покров Итого
Моховая лесотундра, гумусо- во-железистый подзол (Ар- хангельская обл.) (северная тайга) 200 2,0 99,2 29,2 5,2 135,6
Еловый лес-зеленомошнпк с Oxalis, сильноподзоли- стая почва (южная танга) 93 10,0 | 239,5 65,4 1,0 315,6
Папоротниково-разнотравный еловый лес, подзолистая почва (южная тайга) . . 115 15,8 289,6 78,8 0,6 384,8
Пойменный еловый лес, тор- фянисто-перегнойная пес- чайная почва (зона смешан- ных лесов) 45 15,7 223,7 102,4 ? 341,8
Сосновый лес с Vaccinium vi- tis-idaea, подзолистая пес- чаная почва (зона смешан- ных лесов) 95 3,9 181,8 62,8 ? 248,5
Для сравнения: дубовый лес (carico-aegopodiosurn), темно- серая супесчаная почва (Теллермановское лесниче- ство, Воронежская обл.) . . 55 3,5 149,1 85,1 0,4 238,1
Среднегодовое количество опада в различных лесах (в т!га)'.
Лиственичный лес —2,7 Дубовые леса —3,9
Еловые леса —5,0 Для сравнения: субтропические дождевые
Сосновые леса —4,1 леса в Южном Китае—11,6
372
Глава 5
Процесс разложения подстилки в хвойных лесах осуществляется
медленнее, чем в лиственных лесах (см. табл. 72), и протекает по-иному.
В процессе гумификации в почвах лиственных лесов образуется большое
количество гуминовых кислот, тогда как в почвах хвойных лесов пре-
обладают фульвокислоты. Отношение углерода в составе гуминовых
кислот (Сг) к углероду, содержащемуся в фульвокислотах (Сф), повы-
шается по мере продвижения с севера на юг:
Северная и средняя части зоны хвойных лесов 0,2
Южная часть зоны хвойных лесов 0,5
Зона смешанных лесов . . 0,5—0,7
Зона лесостепи 0,8
В зависимости от состава видов деревьев и подлеска наблюдаются
те или иные отклонения от указанных выше закономерностей. Фульво-
кислоты, являющиеся коллоидами гумуса, относительно подвижны и силь-
нее воздействуют на глинистые частицы, разлагая последние; при этом
вымываются Fe2O3, А12О3, а в почве сохраняется только SiO2. Поэтому
фульвокислоты, образующиеся в основном под еловым лесом, способ-
ствуют оподзоливанию почв и образованию эллювиального подзолистого
горизонта (Ае) и иллювиального горизонта В, куда вмываются коллоиды
гумуса, а также Fe2O3 и А12О3. Степень оподзоливания почв уменьшается
параллельно уменьшению содержания фульвокислот при движении
с севера на юг. В почвах полузасушливых областей богатые Са гуминовые
кислоты находятся в неподвижном состоянии; здесь формируется мощный
гумусовый горизонт, характерный для черноземов степной зоны. Этот
гумус отличается большой сорбционной способностью и обусловливает
высокое плодородие почвы. Почвенная фауна и микрофлора, принимающие
участие в образовании разных типов гумуса — сильнокислого грубого
гумуса, насыщенного фульвокислотами, и слабокислого до нейтрального
мулля, насыщенного гуминовыми кислотами, — резко отличаются
по своему составу. В почвах с грубым гумусом преобладают грибы, в поч-
вах с муллем — бактерии; в последних встречается также значительно
больше дождевых червей. Промежуточное положение между вышеупомяну-
тыми типами гумуса занимает модер (см. «Phytologie», Bd. Ill, 1). В пре-
делах бореальной зоны хвойных лесов годовое количество осадков посте-
пенно снижается при движении с юга на север. В том же направлении
возрастает увлажненность почв; кажущийся парадокс разъясняется тем,
что величина испарения при низкой температуре, особенно в течение
прохладного лета, снижается еще быстрее. На ровных площадях в преде-
лах бореальной зоны хвойных лесов уровень грунтовых вод располагается
очень высоко, особенно весной после таяния снега. В связи с этим в зоне
хвойных лесов широко распространены сырые почвы с хорошо выражен-
ным глеевым горизонтом (G). Корни углубляются в эти почвы только
летом, когда происходит временное опускание уровня грунтовых вод;
только в этот период кислород может поступать в пропитанный водой
горизонт G и окислять железо. При этом возникают характерные ржаво-
коричневые или желтые пятна, чаще всего вдоль ходов, образуемых кор-
нями. В зависимости от того, где расположен глеевый горизонт, в гори-
зонтах А, В или глубже, выделяют подзолистые глеевые почвы типов
а, Р или у.
Глеевые почвы типа а — это болотные почвы, на которых сомкнутый
хвойный лес не развивается: 0-глей — это периодически переувлажняе-
мые почвы, характеризующиеся неглубоким распространением корней
Хвойные леса умеренного холодного климата — бореальная зона
373
деревьев. В глеевых почвах у уровень грунтовых вод залегает в среднем
на глубине 1,5—2 корни проникают здесь достаточно глубоко и дости-
гают капиллярной каймы грунтовых вод, благодаря этому они обеспечи-
вают деревья влагой в течение достаточно длительного периода времени.
Эти местообитания наиболее благоприятны для хвойных лесов.
В табл. 80 приводятся данные, характеризующие продукцию и круго-
ворот веществ в двух типах еловых лесов (север бореальной зоны),
в сосновом лесу (северо-восточная часть зоны смешанных лесов) и в вер-
ховом болоте с сосной (Западная Сибирь).
Таблица 80
Продукция и круговорот веществ в бореальной зоне1
Piceetum hylocom. Архангельская обл. Pinetum vaccin. Мордов- ский заповед- ник I бонитет Верховое болото с сосной в Зап. Сибири
I бонитет IV бонитет
Возраст древостоя, лет 200 125 71 100
Фитомасса, т/га 340,8 173,5 279,9 37,0
зеленые части, ml га 21,1 11,5 13,9 15,1
многолетние надземные части, ml га . 234,6 121,3 202,4 17,9
корни, т/га 85,1 40,7 63,6 4,0
Первичная продукции, т/га . . . 7,9 6,0 6,1 3,4
зеленые части, т/га 3,3 4,3 2,4 3,2
многолетние надземные части, т/га . 3,25 1,3 2,8 0,17
Годовой опад, т/га . . . 5,9 4,1 4,7 2,5
опад, в % от фитомассы 1,7 2 2 7
зеленые части, т/га 2,8 2,7 2,1 2,3
многолетние надземные части, ml га . 1,8 0,9 2,2 0,12
корни, ml га 1,3 0,5 0,4 0,08
Истинный прирост, т/га 2,0 1,9 1,4 0,9
Подстилка, ml га . Не опре- делено 25,0 44,8 Не опре- делено
Зола-j-N в фитомассе, кг!га . . . 3966 1460 1878 609
в зеленых частях, кг/га 1 856 392 358 447
в многолетних надземных частях, кг/га . ’ 2239 594 1076 114
в корнях, кг/га 871 474 444 48
для первичной продукции потребляется, кг 1га 193 145 85 109
с годовым опадом возвращается, кг 1га . 147 93 58 73
удерживается приростом, кг/га . ... 46 52 27 27
Азот N в фитомассе, кг 1га для первичной продукции потребляется, 1978 447 664 229
кг/га 75 47 27 40
с годовым опадом возвращается, кг/га 52 29 16 25
Зола в фитомассе, кг/га для первичной продукции потребляется, 1988 1013 1214 380
кг/га 118 98 59 69
с опадом возвращается, кг/га 95 64 42 48
Возвращающиеся с опадом органогены 2, кг/га 64 41 32 21
1 л. Е. Родин, Н. И. Базилевич, Динамика органического вещества и биологиче-
ский круговорот в основных типах растительности, М.-Л., 1965.
2 Под органогенами подразумевается сумма элементов Са, К, Р и S.
374
Глава 5
ПОЯС ХВОЙНЫХ ЛЕСОВ В ГОРАХ
t ЦЕНТРАЛЬНОЙ ЕВРОПЫ
Обширной зоне хвойных лесов, простирающейся на севере евро-
сибирского района, в горах Центральной Европы соответствует узкий
пояс, приуроченный к верхней части горной и субальпийской полосы;
в этом поясе растут те же или близкородственные виды хвойных деревьев \
Сосна (Pinus silvestris) представлена тем же видом, что и на севере зоны
хвойных лесов; однако в горах Центральной Европы встречаются другие
географические расы или экоклины (см. гл. 1). Ареал сосны в Центральной
Европе непосредственно смыкается с североевропейской частью ее ареала;
в настоящее время трудно установить, какие из насаждений сосны воз-
никли естественным путем, а какие созданы искусственно. Сосна и бе-
реза — это виды, которые в послеледниковый период первыми начали
захватывать площади, освобождающиеся после отступления ледника
в Европе; в дальнейшем сосна была вытеснена более конкурентоспособ-
ными видами и сохранилась до настоящего времени только в крайних
условиях местообитания. Более или менее целостная область распро-
странения сосны простирается на запад от Эльбы в направлении к рав-
нине, расположенной в междуречье Рейна и Майна, и через горные массивы
в верховьях Рейна вплоть до Западных и Приморских Альп. Изолирован-
ные местонахождения сосны встречаются в Центральном массиве во Фран-
ции, в Пиренеях, в горах центральной части Испании и Сьерра-Невады
(южная Испания). На Балканском полуострове вплоть до северной Греции
также распространены сосны; обособленные местообитания сосны известны
и в северной Анатолии, и на Кавказе. Сосна не распространена в зоне
степей, однако в устье Днепра отмечен ее форпост, образованный тремя
деревьями (рис. 280). В Альпах реликтовые экземпляры сосны приурочены
к отдельным сухим каменистым участкам, находящимся под влиянием
фёна.
Центральноевропейская пихта (Abies alba) очень резко отличается
от сибирской пихты (Abies sibirica) с экологической точки зрения. В период
оледенения ее рефугиум находился в юго-западной Европе, откуда пихта
в дальнейшем распространилась в Центральную Европу, где она в настоя-
щее время встречается на ограниченной площади, в основном в верхней
части пояса буковых лесов. С запада на восток ареал пихты простирается
от Пиренеев до Карпат и Родоп, а с юга на север от Корсики, Сицилии
и северной Албании до Северо-Германской низменности (рис. 281) 1 2.
На юге к области распространения пихты примыкают ареалы нескольких
подвидов (А. borisii-regis, А. bornmuelleriana) и родственных реликтовых
видов: A. nordmanniana (западный Кавказ), A. cilicica (южная Анатолия),
A. cephalonica (Греция), A. pinsapo (южная Испания, Рифф) и A. numidica
(северное Марокко).
Ель (Picea abies) представлена в'Центральной Европе особыми геогра-
фическими расами того же вида, который встречается и в Северной Европе.
Она распространена в Центральной Европе в двух областях (герцинско-
карпатской и альпийско-центральноевропейской частях ареала), которые
1 К. Rubner, Die pflanzengeographischen Grundlagen des Waldbaues, 4. Aufl.,
Radebeul u. Berlin. 1953; L. Tschermak, Waldbau auf pflanzengeographisch-
•okologischer Grundlage, Wien, 1950.
2 J.Mattfeld, Das Areal der WeiBtanne, «Mitt. d. D. Dendrol. Ges.», 2, 1926;
Pflanzen.areale, 1. Reihe: Die europaischen Afcies-Arten, Karte 14—16, Jena, 1926—1928.
Хвойные леса умеренного холодного климата — бореальная зона
375
Р н с. 280. Ареал сосны (Pinus silvestris) (по Рубнеру и др., см. также рис. 281—285).
отделены друг от друга и от североевропейской части^ареала узкими
полосами (рис. 282). Герцинско-карпатская часть ареала в послеледнико-
вый период была заселена елью из восточного рефугиума, а вторая частя
ареала — из юго-восточного рефугиума (о различиях мест происхождения
см. в работе Рубнера). Ареал ели не доходит до Пиренеев. Обычно он,
образует в горах, главным образом в Северных Альпах и Карпатах, пояс-
расположенный выше пояса бука и пихты, а в континентальных условиях
Центральных Альп заменяет последнюю.
В Центральных Альпах верхнюю границу леса образуют листвен-
ница (Larix decidua) и кедр (Pinus cembra), близкородственные видам,
произрастающим в континентальной Сибири. На верхней границе лесного
пояса в Центральных Альпах лиственница является пионером; в даль-
нейшем она вытесняется кедром. Поэтому лиственница встречается
по руслам лавин и проникает глубоко в горные долины (цветное фото 3).
Оба вида распространены и в Карпатах, а лиственница в виде особой
формы также в Судетах (300—800 м над уровнем моря) и в Польше
(рис. 283). В горах Албании, Греции и Болгарии растет Pinus рейсе; этот
вид. как и Р. cembra, относится к группе сосен, у которых развиваются
пучки с пятью хвоинками в каждом (рис. 284). У Pinus montana —цен-
тральноевропейского вида, приуроченного к субальпийскому поясу
(Пиренеи, Альпы, Карпаты) выше границы леса, — развиваются пучки
с двумя хвоинками. Горная сосна не является видом, родственным кедро-
вому стланику (Р. pumila, 5 хвоинок в пучке); она образует стелющуюся
форму (рис. 285) или растет прямыми стволами (Spirke) и спускается
с гор по верховым болотам далеко в предальпийскую область (см. «Phyto-
logie», Bd. Ill, 2).
Р ис. 281. Ареал (Abies alba) и некоторых других видов пихты.
Р и с. 282. Ареал ели (Picea abies = excelsa) и реликтового вида Picea oinorica.
Хвойные леса умеренного холодного климата — бореальная зона
377
Рис. 283. Ареал
лиственницы европей-
ской (Larix decidua).
Рис. 286 дает представление о высотной поясности в Альпах в зави-
симости от условий климата (океанического на внешних окраинах Альп,
континентального в Центральных Альпах).
Определение границ естественного распространения хвойных пород
в Центральной Европе сопряжено с большими трудностями. С началом
применения рациональных методов ведения лесного хозяйства в этой
области были созданы культуры сосны — путем посева или посадок;
в настоящее время преимущественно культивируется ель, отличающаяся
большой продуктивностью легкой древесины. Искусственно созданные
чистые сосновые и еловые древостои, не испытывающие конкуренции
с лиственными породами, за пределами области естественного распро-
странения в пониженных местообитаниях или на более богатых почвах
Рис. 284. Ареал кед-
ра (Pinus cembra) в
Европе и сосны руме-
лийской (Pinus рейсе)
на Балканах (у этих
видов по 5 хвоинок в
пучке).
378
Глава 5
Рис. 285. Верхняя грани-
ца распространения деревь-
ев в Центральных Альпах,
образованная лиственницей
(Larix decidua) и горной со-
сной (Pinus montana) на вы-
соте около 1700 м над уров-
нем моря, западный Тироль
(фото_Фабера).
Пояса:
Северная окраина Альп
Центральные Альпы
Южная окраина Альп
Я-~
леса
ниваль-
ный
низкогор-г
ный \
преЗгор- г
ный \
альпий-
ский
субаль-
пийский
горный
2
ГГЖТГ i т
Ель >
Пихта ° бук
Ель'
Ду®Г.бук
Дуб пушистый
^уоТраб
осна
ихта
о о о * о О
гДуб—буКоо-з ~оЪ
?V^°I°oX°o~Oo°° ° ° °
° о0Дубы°0Каштань1о°
<££ о°Л°О о° ’о° ’ ° 0 5 о
Рис. 286. Схематический поперечный профиль Альп в направлении с севера на юг
(по Элленбергу).
Характер поясности: в Северных Альпах — гельветская поясность с преобладанием лиственных
лесов, в Центральных Альпах — пеннинская, без лиственных пород, а на юге — инсубрийская,
переходящая в субсредиземноморскую область.
Лиственница
км
3
О
Хвойные леса умеренного холодного климата — бореальная зона
379
дают больший прирост древесины, чем в естественных условиях произ-
растания х.
Структура и состав лесов в Центральной Европе определяются систе-
мой мероприятий, применяемой в лесном хозяйстве, что косвенно влияет
и на характер подлеска. В течение последних столетий лесной ландшафт
в Центральной Европе неоднократно менялся. Господство хвойных лесов
установилось здесь сравнительно недавно.
В средние века большое значение имел выпас свиней в лесах. Вначале это способ-
ствовало развитию дуба, В дальнейшем выпас скота в лесах и неумеренное пользование
древесиной привели к полной деградации лесов и формированию зарослей кустарни-
ков. Даже в горах лес вырубался хищническим образом для нужд стекольных и металлур-
гических заводов. Нехватка топлива и подсечное хозяйство способствовали развитию
порослевых лесов; дубовые низкоствольники использовались для получения корья,
необходимого для кожевенных заводов; такое низкоствольное хозяйство велось вплоть
до первой четверти нашего столетия. Возросшая потребность в товарной древесине
наряду с дровяной обусловила еще в средние века переход к среднествольному хозяй-
ству, которое в настоящее время больше не применяется. При переходе к высокостволь-
ному хозяйству в последнее время наблюдается увеличение степени участия в лесах
видов хвойных. Высокоствольные леса по своей структуре очень сходны с естественными
лесами; однако из-за одновозрастности и монодоминантности они более однообразны,
чем последние; кроме того, высокоствольные леса отличаются более однородным соста-
вом нижних ярусов, поскольку на почве отсутствуют отмершие стволы старых деревьев.
Только в последние годы в Центральной Европе приступают к созданию смешанных
насаждений, с тем чтобы уменьшить кислотность почвы и предотвратить вспышки раз-
множения насекомых-вредителей; наряду с этим прекращены сплошные рубки на боль-
ших площадях, В результате вновь меняется структура древостоя и характер лесного
ландшафта.
ЭЕО ЛОГО-ФИЗ ИО ЛОГИ ЧЕСКИЕ ИССЛЕДОВАНИЯ
ХВОЙНЫХ ПОРОД
Штальфельт подробно изучил водный режим елового леса в Швеции
в 1938—1942 гг3. Кроны деревьев задерживают большую часть атмосфер-
ных осадков, которые не достигают поверхности почвы; в летние месяцы
эта величина составляет 57,2%, а в остальные — 52,3% от количества
осадков, выпадающих на открытом месте; таким образом, кроны деревьев
в еловых лесах Швеции перехватывают больше осадков, чем менее сомк-
нутые еловые леса Восточной Европы (см. табл. 68). Моховой покров
и подстилка задерживают 18,4% осадков в летние месяцы и 9,3% —
в остальные; в результате корни могут использовать только 24 или 38%
ют общего количества выпавших осадков. Это составляет в летние месяцы
90 мм, а в остальные — 202 мм; всего за год — 292 мм. Влага удержи-
вается в основном в верхнем (20 см) слое почвы, развитой на морене;
только 35 мм просачивается ежегодно в более глубокие слои почвы
(70—150 см). Верхние 50 см пронизаны корнями; лишь в исключительных
случаях корни ели проникают на глубину от 80 до 100 см. Уровня грунто-
вых вод влага практически не достигает, поскольку весь ее запас расхо-
дуется на транспирацию елового насаждения в возрасте 40 лет. Величина
транспирации в летние месяцы (с мая по август) достигает 211 мм, на влаж-
ных местообитаниях —378 мм. В последнем случае часть влаги, израсхо-
дованной на транспирацию, потребляется из горизонта грунтовых вод.
1 Е. A i с h i п g е г, 1, Rotfohrenwalder als Waldentwicklungstypen, 2. Fichten-
walder und Fichtenforsten als Waldentwicklungstypen,«Angew. Pfranzensoziol.», H. VI,
VII, Wien, 1952.
2 M. G. S t a 1 f e 1 t, «Kgl. Lantbruksakad. Tidskr.», 83, 1, 1944.
380
Глава 5
Клемм изучал водный баланс в культурах хвойных, определяя содер-
жание воды в стволах с помощью гамма-лучей1. В результате исследова-
ний выяснилось, что Picea и Larix значительно лучше регулируют свой
водный режим в период засухи, чем Pseudotsuga.
Польстер изучал водный режим и фотосинтез в молодняках хвойных
пород и у бука в лесном питомнике (см. стр. 275), а Пизек и Картельери
(см. стр. 293), а также Пизек совместно с Транквиллини — у ели на есте-
ственном местообитании в окрестностях Инсбрука 1 2. Исследовались
интенсивность транспирации и ассимиляции СО2 у световых и теневых
побегов. Связь ассимиляции с транспирацией и шириной устьиц световой
и теневой хвои в ясные и пасмурные летние дни аналогична тому, что
наблюдается у бука (см. стр. 295). Основное отличие в том, что ель является
вечнозеленой породой и продуцирует органическое вещество весной
и осенью, то есть в то время года, когда деревья бука еще не покрылись
листьями или уже сбросили их. Указанный период является наиболее
важным для ели. Поскольку в это время днем уже тепло, но не слишком
жарко, нетто-ассимиляция сохраняется на довольно высоком уровне,
а расход на дыхание в прохладные ночи относительно невелик; в связи
с этим ель продуцирует значительное количество вещества. Поздней
осенью после ночных заморозков ассимиляция СО2 временно приостана-
вливается. Только с наступлением холодного времени года с отрицатель-
ными температурами у ели начинается период длительного покоя и она
не ассимилирует даже в ясные дни. Одновременно резко снижаются
и потери на дыхание; поэтому расход вещества, несмотря на длительный
дефицит в балансе, очень незначителен и его величина уменьшается при»
понижении температур зимой. Одновременно хвоя теряет свою свежую,,
зеленую окраску, и хлоропласты с трудом различаются даже под микро-
скопом.
Ход дневного баланса, изученный Целлером в относительно холодную
зиму 1946/47 г. и в мягкую зиму 1947/48 г., дает представление о процес-
сах ассимиляции и дыхания у ели в районе Штутгарт-Хоэнхейма, где
наблюдения проводились на высоте 400 м над уровнем моря за пределами
ее естественного ареала. В эксперименте использовался колориметриче-
ский метод Олвика (см. «Phytologie», Bd. Ill, 1), наиболее удобный для
определения незначительной ассимиляции СО2, в то время как величины
при интенсивном фотосинтезе вследствие изменяющейся концентрации СО2.
могут иметь только относительное значение3. Как видно из рис. 287,.
на котором представлен ход ассимиляции и дыхания у ветвей ели в холод-
ную зиму 1946/47 г. 23 ноября при температуре от+З до +8°, нетто-асси-
миляция (белые колонки) над нулевой линией заметно превышала вели-
чину дыхания (черные колонки); выделенное ночью в процессе дыхания
количество СО2 после восхода солнца было вновь ассимилировано и уже
через 2 часа суточный баланс был уравновешен. В пасмурный день
14 декабря при температуре немного выше 0° газообмен резко снизился,
поэтому суточный баланс достиг равновесия только спустя 3 часа. После
первых дней с морозом 18 декабря газообмен почти полностью прекра-
щается так же, как и накопление органического вещества. При длительных
1 W. Klemm, Wasserhaushaltsuntersuchungen an Fichte, Douglasie und Larche
wahrend einer Diirreperiode, «Flora», Abt. B., 156, 1966, S. 232—235.
2 A. Pisek, W. Tranquillini, Transpiration und Wasserhaushalt der
Fichte (Picea excelsa) usw., «Physiol. Plant.», 4, 1951, S. 1—27.
3 0.Zeller, Uber Assimilation und Atmung bei tiefen Temperaturen, «Planta»,
39, 1951, S. 500—526.
Хвойные леса умеренного холодного климата — бореальная зона
381
Рис. 287. Ход ассимиляции и дыхания у ветвей ели холодной зимой 1946/47 г.
(по О. Целлеру).
А — амплитуда температуры на открытом месте (тонкая линия) и в экспериментальных^колбах (жир-
ная линия); Б — интенсивность ассимиляции (белые колонки) и интенсивность дыхания (черные
колонки); этим величинам следует придавать относительные значения; В—уравновешивание суточ-
ного баланса (в часах) после восхода солнца (нулевая линия) и интенсивность света во время асси-
миляции вплоть до момента равновесия суточного баланса; степень интенсивности света пред-
ставлена пятью ступенями, обозначенными штриховкой различной густоты (интенсивность сни-
жается по мере сгущения штриховки).
I
отрицательных температурах жизнедеятельность растения затухает (кру-
жок около даты 20 декабря). Внезапное потепление 27 декабря и особенно
15 января способствовало временному усилению газообмена и незначи-
тельному накоплению вещества, которое окончательно прекратилось
21 января. В течение долгого периода с отрицательными температурами
ель находилась в состоянии зимнего покоя, лишь в феврале наблюдались
признаки слабо выраженного газообмена. Фотосинтез у ели возобновился
в марте; несмотря на то что температура в это время превышала 10°,
фотосинтез осуществлялся очень медленно и достиг ноябрьских величин
лишь в апреле.
Этот процесс протекал совсем по-иному в течение очень мягкой зимы
1947/48 г. (рис. 288). Умеренные холода во второй половине февраля
не способствовали установлению полного зимнего покоя, хотя газообмен
в это время достигал небольших значений. Фотосинтез начался сразу же
после потепления и 6 марта при температуре около 10° достиг уже значи-
тельной интенсивности.
382
Глава 5
В области естественного распространения ели с продолжительным
зимним периодом зимний покой может длиться почти полгода. В феврале
у ветвей ели, принесенных в теплую комнату, резко повышалось дыхание;
в результате отношение величины ассимиляции СО2 к расходу на дыхание
Рис. 288. Ход ассимиляции и дыхания у ветвей ели в мягкую
зиму 1947/48 (по О. Целлеру). См. пояснения к рис. 287.
в темноте при температуре от +12 до +16° составляло лишь 4,6, в то время
как на открытом месте при слабом газообмене и температуре —1,5° оно
достигало 27. В период зимнего покоя устьица должны находиться в закры-
том состоянии с тем, чтобы не только прекратился круговорот вещества,
но и максимально сократилась отдача воды (см. табл. 57).
Зимний покой хвойных ярко выражен в северной части бореальной
зоны, характеризующейся длинными ночами и низким солнцестоянием.
Напротив, в горах Центральной Европы, особенно на склонах южной
экспозиции, в ясные дни при сильной инсоляции температура хвои может
подняться выше точки замерзания. Однако этого недостаточно для того,
чтобы прервать зимний покой, поскольку в течение безоблачных ночей
температура сохраняет отрицательные значения.
У вечнозеленых видов лиственных деревьев, произрастающих
в областях с очень мягкой зимой, например у лавровишни (Prunus lauro-
cerasus), полный зимний покой не выражен, и СО2 продолжает выделяться
Хвойные леса умеренного холодного климата — бореальная зона
383
даже на свету. Интенсивность дыхания ветвей, принесенных в теплую
комнату, повышается столь резко, что суточный баланс уравновешивается
с трудом.
Павлетич и Лит определили точку компенсации освещенности у видов-
хвойных после короткого периода с отрицательными температурами
на фоне в общем очень мягкой зимы; они установили, что у Pinus silvestris,
Abies alba, Taxus baccata и Chamaecyparis nutkaensis этот показатель резко-
повышается, что, очевидно, связано с инактивированием фотосинтетиче-
ского аппарата 1. У Picea abies, Tsuga canadensis и Sequoiadendron gigantea,
а также у вечнозеленых лиственных растений (Prunus laurocerasus, Buxus
sempervirens, Hedera helix и Mahonia aquifolium) эта реакция не устано-
влена. После того как ветви этих растений были помещены на 24 часа
в теплое помещение (12—15°), компенсационная точка резко снизилась-
только у Buxus и Prunus.
Вейзе изучал обмен веществ у подроста ели в течение зимнего полу-
годия в период снижения морозостойкости после искусственного промора-
живания1 2. В то время как фотосинтез растений медленно усиливался,
пока не достиг значений, свойственных контрольным, не подвергавшимся
воздействию отрицательных температур экземплярам, интенсивность
дыхания сначала резко увеличилась, а затем постепенно начала снижаться
до величин, которые наблюдались перед промораживанием. Одновременно-
с фотосинтезом повышалась и интенсивность транспирации (устьица
начали раскрываться).
Большое значение имеют эколого-физиологические исследования, про-
веденные в области естественного распространения хвойных пород Инсбрук-
ской школой. Изучались Picea abies и Pinus cembra 3. У последнего вида
была выявлена четкая связь между замерзанием хвои и фотосинтезом.
После того как замерзнет клеточный сок, хвоя остается в переохлажденном
состоянии; когда же начинается замерзание хвои, то оно происходит
медленно, скачкообразно до тех пор, пока вся вода, которая может замерз-
нуть при определенной температуре, не превратится в лед. Количество
льда определяется калориметрическим методом.
В апреле выжатый клеточный сок хвои Pinus cembra замерзает при
температуре —2,4°, а неповрежденная хвоя на ветвях кедра лишь при
температуре —3 или —4°. В данном случае речь не идет о точке замерзания
живой хвои, поскольку при температуре —4° по истечении 12 часов в лед
переходит только 36% общего количества воды, а это свидетельствует
о том, что оставшийся раствор стал на 100/64 более концентрированным.
При наличии линейной связи между концентрацией раствора и точкой
замерзания последняя должна понизиться у клеточного сока до 2,56°;
это соответствует данным, полученным в результате непосредственных
наблюдений. При дальнейшем понижении температуры количество замерз-
шей в хвое воды увеличивается (табл. 81).
1 Z. Pavletic, Н. Lieth, Der Lichtkompensationspunkt einiger immer-
griiner Pflanzen im Winter und Fruhjahr, «Вег. Deutsch. Bot. Ges.», 71, 1958, S. 309 — 314..
2 G. W e i s e, Untersuchung fiber den EinfluB von Kaltebelastungen auf die physio-
logische Aktivitat von Forstgewachsen, «Biol. Zbl.», 80, 1961. S. 137—166.
3 W. Tranquillini, K. Holzer, Uber das Gefrieren und Auftauen von
Coniferennadeln, «Вег. Deutsch. Bot. Ges.», 71, 1958, S. 143—156; A. P i s e k, G. Reh-
n e r, Temperaturminima der Netto-Assimilation von mediterranen und nordisch-al-
pinen Immergriinen, «Вег. Deutsch. Bot. Ges.», 71, 1958, S. 188—193; The nature of the-
temperature optimum and minimum of photosynthesis, «Bull. Res. Counc. of Israel»,
8D, 1960, p. 285—289; Immergriine Pflanzen (einschlieBlich Coniferen), «Hb. d. Pflanzen-
physiol.», Bd. V, 1960, S. 415—459.
384
Глава 5
Таблица 81
Зависимость замерзания воды в хвое Pinus cembra от температуры
Температура хвои, °C
Среднее количество замерз-
шей воды, % общего за-
паса влаги.............. О 36 42
Относительная упругость
пара (гидратура), % . 98,2 96,4 94,7
48
93
51 54 58
91,3 89,7 около
70
Из таблицы видно, что основная масса воды превращается в лед уже
'при температуре —4°, оставшаяся вода замерзает при температуре
ниже —8°. Это не является особенностью живых клеток и не обусловлено
наличием в них «свободной» или «связанной» воды, а наблюдается также
у растворов сахара и других набухающих тел, что хорошо видно при
сопоставлении кривых гидратуры растворов и набухающих тел. Поэтому
замерзает тем большее количество воды, чем выше содержание воды
в ветвях и чем ниже осмотическое давление. Фотосинтез практически
прекращается уже в начальный период замерзания хвои при температуре
юколо —4°; дыхание останавливается только тогда, когда вся легкозамер-
зающая вода превратится в лед, что отмечается при температуре от —6
до —8°. Замерзшая хвоя не погибает даже при температуре —40° при
условии, что она приобрела закалку зимой; после оттаивания фотосинтез
и дыхание протекают у нее нормально. Это же наблюдается и у Picea
abies в отличие от средиземноморских жестколистных растений, которые
не обладают морозоустойчивостью и у которых интенсивное образование
льда означает гибель клеток.
Опыты, проведенные Целлером, свидетельствуют о том, что замерза-
ние хвои у видов хвойных приводит к определенным последствиям. Чем
дольше период, в течение которого хвоя находилась в замерзшем состоянии
и чем ниже в это время была температура, тем длиннее срок (часто много
дней), необходимый для того, чтобы после оттаивания у хвои восстано-
вился нормальный фотосинтез; однако фотосинтез не достигает летних
величин. Закаленные клетки находятся в менее активном состоянии,
чем незакаленные, но первые отличаются большей холодостойкостью
и лучше переносят низкие отрицательные температуры. Морозоустойчи-
вость меняется в течение года. Она возрастает осенью с наступлением
первых ночных заморозков; при этом содержание сахара в клеточном
соке и осмотическое давление быстро повышаются, а соотношение саха-
розы и глюкозы резко падает. Весной в первый более теплый день морозо-
устойчивость снижается, а содержание сахара и осмотическое давление
уменьшаются скачкообразно1. По данным Хебера, при закалке происхо-
1 W. Ulmer, Uber den Jahresgang der Frostharte'einiger immergriiner Arlen der
alpinen Stufe, sowie der Zirbe und Fichte, «Jb. Wiss. Bot.», 84, 1937, S. 553—592;
A. Pisek, Frostharte und Zusammensetzung des Zellsaftes bei Rhododendron ferrugi-,
neum, Pinus cembra und Picea excelsa, «Protoplasma», 39, 1950, S. 129—146; A.Pisek
R. Schiessl, Die TemperaturbeeinfluBbarkeit der Frostharte von Nadelholzern und
Zwergstrauchern an der alpinen Waldgrenze, «Вег. Naturwiss. med. Ver. Innsbruck», 47,
1946, S. 33—52; M. Steiner, Zum Chemismus der osmotischen Jahresschwankungen
-einiger immergriiner Holzgewachse, «Jb. Wiss. Bot.», 78, 1933; S. 564—622; Die Zusam-
Хвойные леса умеренного холодного климата — бореальная зона
385
Рис. 289. Годовой ход морозоустойчивости хвои в возрасте до года у ели (по
Пизеку и Шисслю).
Под морозоустойчивостью подразумевается самая низкая температура, которую растения
могут^ перенести без повреждений в течение 1,5 часа. Ветви закаливались на протяжении
четырех дней под воздействием температур, отмеченных на рисунке; изнеживание прово-
дилось во влажной камере при комнатной температуре.
1—контроль; 2—закалка; 3—режим изнеживания; 4—то же плюс насыщение водой.
дит концентрация сахара в протоплазме, а не в клеточном соке; накопив-
шийся в протоплазме сахар предохраняет систему фосфорилирования
от воздействия низких температур и, как это уже давно предполагалось,
оказывает известное защитное влияние х.
Морозоустойчивость можно повысить искусственным путем, в основ-
ном осенью и весной, воздействуя на растение низкими температурами
(рис. 289). В результате утраты закалки при температуре выше 0° морозо-
устойчивость снижается, особенно сильно поздней зимой, начиная
с декабря. Сопротивляемость ели низким температурам в области естест-
венного произрастания достигает в холодный период года почти —40°,
тогда как Pinus cembra, распространяющийся в горах значительно выше
ели, переносит температуру ниже —40°. В естественных условиях хвойные
деревья непосредственно не повреждаются низкими температурами;
в частности, это не наблюдается в Сибири, где температура опускается
ниже —60°. В период длительного полного зимнего покоя накопленное
органическое вещество не расходуется, поэтому долгую полярную ночь
растения переносят без ущерба.
Опыты, проведенные в Инсбруке, выявили влияние минимальной
температуры на ассимиляционную способность ели. В течение всей зимы
Пизек и Винклер брали в качестве образцов ветви с деревьев, растущих
на высоте 600 и 1840 м над уровнем моря, а затем проверяли их способность
ассимилировать СО2 в лабораторных условиях при температуре 12°
и освещенности, равной 10 тыс. лк (рис. 290) * 1 2. Холодный период на высоте
600 Jf, если исключить временное похолодание в ноябре, начинается лишь
mensetzung des Zellsaftes bei hoheren Pflanzen in ihrer okologischen Bedeutung, «Ergebn.
d. Biol.», 7, 1939, S. 151—254; W. S cheumann, S. В or t i t z, Studien zur Physio-
logic der Frosthartung bei Coniferen, «Biol. Zbl.», 84, 1965, S. 489—500.
1 U. W. Heber, K.A.Santarius, Loss of Adenosin-Triphosphate synthesis
caused by freezing and its relationship to frost hardiness problems, «Plant Physiol.», 39,
1964, p. 712—719.
2 A. Pisek, E. Winkler, Assimilation und Respiration der Fichte in ver-
schiedener Hohenlage und der Zirbe an der Waldgrenze, «Planta», 51, 1958, S. 518—543.
25—01248
386
Глава 5
Рис. 290. Различия в ассимиляционной способности ели, произрастающей
на высоте 600 м и 1840 м около Инсбрука (по Пизеку и Винклеру).
Вверху — суточная минимальная температура воздуха, измеренная вблизи опытных
площадок; внизу —величина дыхания. Заштрихованная поверхность дает представление
о том, насколько выше ассимиляционная способность ели, произрастающей ниже
в горах, по сравнению с елью, растущей у верхней границы распространения деревьев,
на высоте 1840 м.
в конце января и длится до конца февраля; кратковременные отрицатель-
ные температуры наблюдаются еще в начале марта. Только в течение этого
недолгого времени ель, растущая здесь, находится в состоянии зимнего
покоя. Напротив, минимальная температура около Патшеркофеля с сере-
дины ноября и почти до середины апреля не превышает 0°. Зимний покой
в этих условиях длится 5 месяцев; в это время хвоя желтеет и после оттаи-
вания начинает ассимилировать СО2 не сразу. Фотосинтетическая актив-
ность хвои ели, произрастающей на границе распространения деревьев
на высоте 1840 м, на протяжении всего года несколько ниже, чем у хвои
ели, растущей ниже в горах; возможно, это связано с тем, что выше
в горах дыхание при одинаковой температуре отличается большей интен-
сивностью. Дыхание хвои не имеет четкого годового хода; даже зимой
в короткие оттепели хвоя на свету начинает выделять СО2. Заштрихован-
ная поверхность на рис. 290 свидетельствует о снижении ассимиляционной
способности хвои с увеличением высоты над уровнем моря. Ель, приуро-
ченная к верхней границе деревьев, накапливает очень мало органического
вещества. Прирост деревьев в высоту невелик, и ель формирует здесь
узкие годичные кольца.
Граница леса в Северной Европе и в Северных Альпах образована
елью. На севере Европы и по мере подъема в горы лето становится короче
и прохладнее, а зима — длиннее и холоднее. Поэтому часто обсуждается
вопрос, какой из факторов ограничивает дальнейшее продвижение леса,
образующего четкую и узкую границу на крайнем пределе своего рас-
пространения. При приближении к границе леса высокоствольные деревья
встречаются еще довольно часто. Лес, в особенности у полярной границы,
изреживается, затем прирост в высоту деревьев сокращается, они стано-
вятся корявыми и низкорослыми и наконец полностью исчезают
(рис. 291). Эта внезапная потеря способности роста не может быть обусло-
влена чисто климатическими причинами, поскольку все элементы клима-
Хвойные леса умеренного холодного климата — бореальная зона
387
Р и с. 291. Ветровая форма ели у верхней границы распространения деревьев,
массив Шварцвальд. Нормально развиты только покрытые снегом нижние ветви
(фото Кляйна).
та — температура, продолжительность вегетационного периода, мощ-
ность снежного покрова и т. д.— постепенно изменяются при движении
в северном направлении и с увеличением высоты в горах без каких-либо
скачкообразных повышений или снижений этих показателей у границы
распространения леса. Последняя четко выражена даже в условиях
выровненного рельефа. Поэтому граница леса, очевидно, обусловлена
эколого-физиологическими причинами Ч При увеличении крутизны скло-
нов и повышении степени каменистости почвы сплошной лесной массив
у верхней границы произрастания распадается на куртины или отдельные
деревья; при отсутствии воздействия человека это связано с почвенными
условиями.
В Альпах различают верхнюю границу распространения леса и верх-
нюю границу распространения деревьев. Лес, начиная с определенной
высоты над уровнем моря, постепенно изреживается, пока наконец
не остаются отдельные разбросанные по склонам деревья, которые на гра-
нице произрастания деревьев принимают форму криволесья.
1 И. Клаусен (доклад на Ботаническом конгрессе в Эдинбурге в 1964 г.) на основе
наблюдений, проведенных у верхней границы распространения деревьев в горах Кали-
форнии, считает, что низкорослость и корявость Pinus albicaulis, Р. murryana и Tsuga
mertensiana обусловлены генетическими причинами. Эти экземпляры плодоносят и рас-
тут в пределах одного высотного пояса вместе с типичными древовидными формами того
же вида, которые образуют куртины из семян, занесенных ветром из предгорий. Кри-
волесье из Р. albicaulis встречается на высоте 3600 л, где формируются стелющиеся кур-
тины диаметром до 20 м. В отлйчие от американских видов сосны европейские виды —
Pinus cembra и Р. silvestris — не образуют криволесья. Корявые формы ели, приурочен-
ные к верхней границе распространения деревьев, с генетической точки зрения не от-
личаются от древовидных форм, растущих ниже в горах.
388
Глава 5
В результате подробных исследований, проведенных Инсбрукской
станцией по изучению субальпийских лесов, подчиненной Федеративному
лесному опытному институту в Вене, были установлены различия между
теоретической климатической, потенциальной и фак-
тической границами леса1. Климатическая граница леса
проходит на определенной высоте над уровнем моря; в горах она обусло-
влена понижением температуры, связанным с увеличением высоты. Лес
никогда не поднимается в горы до этой границы, так как этому препят-
ствуют местные условия среды, например скалистые обрывы, скопление
щебня, холодные потоки воздуха в долинах (ледниковые ветры) и т. д.
Таким образом, в естественных условиях лес достигает только потен-
циальной границы распространения 1 2. Во всех населенных районах под
влиянием человека или в результате естественных катастроф леса у верх-
ней границы сильно изрежены или уничтожены; сохранившиеся в настоя-
щее время остатки соответствуют фактической границе распространения
леса. Это особенно хорошо выражено при выпасе на альпийских лугах,
когда человек сводит лес для организации горнопастбищного хозяйства
и заинтересован в расширении пастбищ за счет сокращения лесопокрытой
площади 3. Поскольку лес плохо возобновляется и растет здесь у верхней
границы произрастания, ущерб, причиненный человеком, не может быть
ничем компенсирован. На месте прежних лесов в настоящее время на боль-
ших площадях встречаются менее требовательные к условиям среды кустар-
никовидная форма Pinus montana, кустарники (Alnus viridis) или кустар-
нички (виды родов Rhododendron, Vaccinium и т.д.); в естественных условиях
эти виды растут только в пределах узкой полосы, примыкающей к верхней
границе почти сомкнутого леса. Лес распадается на куртины или пред-
ставлен отдельными экземплярами деревьев на каменистых почвах.
Реагируя на непостоянство климатических условий, климатическая
и потенциальная границы леса в горах смещаются то вверх, то вниз.
Продвижение древесных растений в альпийский пояс происходит не сплош-
ным фронтом, поскольку выше границы распространения деревьев выра-
жена микромозаичность различных форм рельефа. Только после смыкания
полога леса условия местообитания становятся более однородными. Слож-
ные условия среды у верхней границы леса затрудняют проведение лесо-
восстановительных работ с целью предотвращения лавин.
Условия среды на верхней границе распространения деревьев в горах
далеко неоднородны. Теневыносливые древесные виды образуют четкую
границу леса (Picea, Fagus), тогда как светолюбивые породы, например
виды Pinus (Р. flexilis, Р. aristata в Скалистых горах), фор-
мируют изреженные древостои. Это особенно характерно для
Р. hartwegii, которая в Мексике растет около альпийской границы рас-
пространения деревьев, где не развит снежный покров и климат отли-
чается относительной сухостью.
В Новой Зеландии выделяют два типа границ. Граница распростране-
ния деревьев, образованная Nothofagus, четко очерчена (в Южной Америке
она в большинстве случаев переходит в широко простирающуюся полосу
1 См. обобщающие работы: Н.М. Schiechtl, Н. Friedel, I. Neuwin-
g е г, М. М о s е г, W. Т г a n q u i 1 1 i n i, N. М a i г, «Allgemeine Forstzeitung»,
77, Folge 6,—Wien, 1966.
2 H. Friedel, Verlauf der alpinen Waldgrenze im Rahmen anliegender Gebirgs-
gelande, «Mitt. Forstl. Bundes-Versuchsanst», H. 75, Wien, 1967.
3 R.S t ern, Der Waldriickgang im Wipptal, «Mitt. Forstl. Bundes-Versuchsanst»,
H. 70, Wien, 1966.
Хвойные леса умеренного холодного климата — бореальная зона
389
криволесья из Nothofagus). В тех же случаях, когда вечнозеленые виды
лиственных деревьев субтропического происхождения достигают альпий-
ского пояса, эта граница не выражена. В этом случае высота деревьев
постепенно уменьшается, и в итоге древесные виды вытесняются видами
Рис. 292. Годовой ход осмотического давления у хвои ели (Picea
engelmannii), произрастающей на различной высоте на пике Пайкс(Коло-
радо), по данным Голдсмита и Смита. Резкое повышение осмотического
давления у деформированных елей, растущих на высоте 3800 м над
уровнем моря, объясняется дефицитом влаги (Н. W а 1 t е г, Die Hyd-
ratur der Pflanze, 1931).
кустарников (Senecio, Olearia, Hebe и др.), которые вместе с альпийским
злаковником туесок образуют мозаикоподобную переходную зону х.
Как было сказано выше, низкая температура не вызывает непосред-
ственной гибели растений от мороза, поскольку предел морозоустойчи-
вости располагается значительно ниже наблюдаемых зимой температур.
Однако отрицательные температуры зимой могут способствовать повреж-
дению растений в результате морозного высыхания (физиологической
сухости). Правда, как уже упоминалось (стр. 325), транспирация хвойных
зимой очень незначительна; однако замерзшие растения не могут воз-
местить израсходованную воду и поэтому при длительных морозах посте-
пенно высыхают 1 2.
Значение физиологической сухости показано в исследованиях, про-
веденных Голдсмитом и Смитом в 1931 г. Эти авторы изучали годовой ход
осмотического давления у хвои ели (Picea engelmannii), выросшей на раз-
личных почвах пика Пайке (Колорадо, США) (рис. 292); верхняя граница
леса и криволесья в этих условиях проходит на высоте 3800 м над уровнем
1 Е. Р. W а г d 1 е, A comparison of alpine timber lines in New Zealand and North
America, «New Zeal. J. Bot», 3, 1965, p. 113—135.
2 Повреждения, нанесенные культурам хвойных около Гамбурга суровой зимой
1962/63 г., по данным Майера, связаны с физиологической сухостью (cm.:F. Н. М е у е г,
Frosttrocknis bei Koniferen, «Вег. Deutsch. Bot. Ges.», 78, 1965, S. 193—202. См. также:
G. M i c h a e 1, Untersuchungen uber den Wasserhaushalt einiger immergriiner Geholze
im Winter unter besonderer Berucksichtigung der Frosttrocknis, Tharandt-Dresden, 1964
(дисс.).
390
Глава 5
моря. На оптимальной высоте (2700 м) величина осмотического давления
почти не колеблется в течение года. Низкие значения, зафиксированные
с июля по август, связаны с началом роста молодых побегов. Небольшое
увеличение осмотического давления в мае на высоте 2400 м вызвано
сухостью, выраженной у нижней границы произрастания ели. Значи-
тельно более заметно повышалось осмотическое давление на границе
леса па высоте 3750 м в апреле — в период, когда дни становятся длиннее
и солнце стоит высоко. В это время на границе криволесья на высоте
3800 м отмечено наиболее высокое значение осмотического давления,
равное 50 атм1. Характерно, что резкое повышение давления наблюдается
не зимой, а весной, когда растения начинают транспирировать при еще
не оттаявшей почве, но уже при повышенной температуре воздуха. Неясно,
чем обусловлено различие в величине осмотического давления, достигаю-
щее^ атм у экземпляров ели, произрастающих на высоте 3750 м и 3800 ж,
и почему деревья внезапно принимают корявую форму. Выяснение этого
вопроса содержится в экологических исследованиях, проведенных на гра-
нице леса в Северных Альпах 1 2. Михаэлис установил, что частая инверсия
температуры в солнечные дни на границе леса зимой ведет к превышению
температуры хвои над температурой воздуха, что способствует расходу
хвоей влаги.
Конечно, при кратковременном оттаивании на солнце вода может
перемещаться из ветвей в хвою 3. Однако у верхней границы распростра-
нения деревьев молодые побеги начинают развиваться поздно, а заморозки
осенью наступают рано, поэтому хвоя не успевает созреть. В связи с этим
покровная ткань,и особенно кутикула,формируются неправильно4. Поэтому
хвоя не может предотвратить расход воды; весной, когда водопроводящая
ткань еще не оттаяла, в хвое наблюдается большой дефицит влаги. Это
отражается на величине осмотического давления, которое достигает
65,7 атм. Вся хвоя, расположенная над снежным покровом, высыхает;
сохраняется только хвоя па ветвях, покрытых снегом. В связи с этим
рост деревьев в высоту прекращается; высота криволесья соответствует
мощности снежного покрова. Таким образом, верхнюю границу
распространения деревьев обусловливают
короткое лето, продолжительность которого
не благоприятствует созреванию хвои, и су-
хость зимой, способствующая высыханию хвои. Необхо-
димо еще установить степень морозоустойчивости несозревшей хвои, кото-
рая может оказаться столь незначительной, что при низких отрицательных
температурах хвоя будет отмирать. В этом случае хвоя должна была бы
опадать уже зимой, а не весной.
1 Н. Walter, Die Hydratur der Pflanze, Jena, 1931.
2 P.Michaelis, Okologische Studien an der Baumgrenze. I. Das Klima und die
Temperatur der Pflanzenorgane, «Ber. Deutsch. Bot. Ges.», 50, 1932, S. 31—42; II. Die
Schichtung der Lufttemperatur, Windgeschwindigkeit und Evaporation fiber einer Schnee-
flache, «Beih. Bot. Zbl.», 52, B, 1934, S. 310—332; III. Uber die winterlichen Tempera-
turen der pflanzlichen Organe, insbesondere der Fichte, ibidem, S. 333—377; IV. Zur Kenn-
tnis des winterlichen Wasserhaushalts; V. Osmotischer Wert und Wassergehalt wahrend
des Winters in den verschiedenen Hohenlagen, «Jb. wiss. Bot.», 80, 1934, S. 169—243,
337—362.
3 W. Larcher, Zur spatwinterlichen Erschwerung der Wasserbilanz von Holz-
pflanzen an der Waldgrenze, «Ber. Natur.-med. Ver., Innsbruck», 53, 1963, S. 125—137.
4 Только через 3 месяца после начала распускания почек кутикула у хвои ели
достигает нужной толщины. См.: О. L. Lange, E.-D. Schulze, Untersuchungen
fiber die Dickenentwicklung der kutikularen Zellwandschichten bei Fichtennadeln, «For-
stwiss. ZbL», 85, 1966, S. 27—38.
Хвойные леса умеренного холодного климата — бореальная зона
391
Закономерности, отмеченные на границе произрастания еловых лесов,
повторяются на пределе распространения Pinus montana, произрастающей
на несколько сот метров выше ели. Сосна выдерживает более низкие
температуры и поэтому по склонам гор поднимается выше ели. Развитие
горной сосны также предрешается взаимным влиянием укороченного
вегетационного периода и высокой физиологической сухости зимой; эти
причины формируют резко выраженную верхнюю границу распростране-
ния сосны, которая характеризуется внезапным повышением осмотического
давления у хвои х.
Причинная обусловленность северной границы леса не изучалась,
однако она, очевидно, определяется теми же факторами. Несмотря на то
что в высоких широтах зимой совершенно не сказывается влияние солнеч-
ной радиации, значение которой также невелико и весной при низком
-стоянии солнца, иссушающее воздействие, аналогичное наблюдаемому в
горах, оказывают в этих условиях сильные полярные вётры.
Подробные исследования с учетом всех факторов были проведены
Инсбрукской исследовательской группой у Обергургля (Тироль) на цен-
тральноальпийской границе леса, образованной кедром (Pinus cembra)
и лиственницей (Larix decidua). Работы проводились в связи с проблемой
разведения леса на лавиноопасных участках. Исследования велись
на склоне 30—35° на высоте 1820—2240 м над уровнем моря. Граница
распространения кедрово-лиственничного леса расположена на высоте
2000—2150 м.
Турнер изучал световой и радиационный климаты 1 2. Большое влия-
ние на показания станции, расположенной в горной долине (в данном
случае на высоте 1940 м над уровнем моря), оказывают окружающие горы,
закрывающие горизонт. Этим обусловлено снижение прихода суммар-
ной радиации, равное 10% в июне и более 60% в декабре (28% в год).
Интенсивность радиации в полдень при ясной погоде в горной долине
на 11 —12% выше, чем на равнине в окрестностях Вены (200 м над уровнем
моря).
Наивысшее значение суммарной радиации за сутки зарегистрировано
в июне — 755 кал!см2, (без горизонтального экранирования эта величина
составляла бы 850 кал/см2).
Рассеянная радиация при безоблачном небе невелика: в течение дня
в июне она равна 60 кал/см2, в декабре — 18 кал!см2. После выпадения
снега в результате отражения со склонов величина рассеянной радиации
может повыситься на 40% . При облачном небе величина рассеянной радиа-
ции очень непостоянна; в среднем в мае она составляет 225 кал/см2,
в декабре — 35 кал!см2 (после выпадения свежего снега — 51 кал/см2).
При переменной облачности вследствие отражательной способности
облаков мгновенная величина излучения может резко повыситься. Макси-
мальные значения, полученные в этих условиях, равны 2,20—2,25 кал/см2,
или 118% солнечной постоянной, то есть интенсивности солнечной радиа-
1 М. Steiner, Winterliches Bioklima und Wasserhaushalt der Pflanzen an der
alpinen Baumgrenze, «Bioklima. Beibl.», H. 2, 1935, S. 57—65.
2 H. Turner, Uber des Licht-und Strahlungsklima einer Hanglage der Otztaler
Alpen bei Obergurgl und seine Auswirkung auf das Mikroklima und auf die Vegetation,
«Arch. f. Meteor. Geophys. u. Bioklim.», Ser.B, 8, 1958, S. 273—325; Maximaltemperatu-
ren oberflachennaher Bodenschichten an der alpinen Waldgrenze, «Wetter und Leben»,
10, 1959, S. 1—11; Die globale Hangbestrahlung als Standortsfaktor bei Aufforstungen
in der subalpinen Stufe, «Mitt. Schweizer. Anst. Forstl. Versuchswesen», 42, H. 3, 1966;
W.Tranquillini, The physiology of plants on high altitudes, «Ann. Rev. of Plant
Physiol.», 15, 1964, p. 345—362.
392
Глава 5
ции вне атмосферы. В результате сильного облучения хвоя кедра пере-
гревается. Температура хвои в безоблачные дни превышает температуру
окружающего воздуха в среднем па 5° (максимально на 10°). Еще значи-
тельнее нагреваются освещенные солнцем верхние слои почвы. Самая
высокая температура хвои составляла 32,5° (температура почвы на глу-
бине 0,2—1,0 см была равна 49,3°). В апреле при сильном воздействии
солнечных лучей и низкой температуре воздуха хвоя подвергается воз-
действию колебаний температуры (суточная амплитуда достигает 34°).
Всходы кедра, растущие на непокрытой растительностью почве, в боль-
шинстве случае погибают от перегрева в первый же солнечный день. При
безветренной погоде и сильной радиации темная поверхность грубо-
гумусной почвы, расположенной на склоне (угол наклона 35°, юго-запад-
ная экспозиция), нагревалась в течение часа до 75—80°! В это же время
на соседнем затененном склоне температура была на 57° ниже и достигала
только 23°. Интенсивность суммарной освещенности в полдень при безоб-
лачном небе в июне составляла в среднем 136 тыс. Х-люкс, а в ноябре —
495 тыс. Х-люкс (1 Хефнер-люкс = 0,9 принятой в международной прак-
тике величины «люкс»). При переменной облачности мгновенная интен-
сивность освещенности может повыситься до 190 тыс. Х-люкс.: Эти дан-
ные характеризуют радиационные условия на альпийской границе леса
в Центральных Альпах; в горах влияние инсоляции выражено значительно
более резко, чем на полярной границе леса.
Увеличение интенсивности радиации на альпийской границе леса
и еще выше в альпийском поясе обусловливает сильный нагрев поверх-
ности почвы и способствует формированию особых микроклиматических
условий. Это приводит к мозаичному распределению местообитаний
и растительных сообществ в зависимости от их расположения, экспозиции
и степени затененности, в частности деревьями. Большую роль в горах
играет ветер, который удаляет нагретый воздух с поверхности почвы;
скорость ветра на высоте 3 м на границе леса, расположенной на высоте
2000 м над уровнем моря, на 40% выше, чем в долине на высоте 1800 м,
а на высоте 2200 м даже на 70% выше. В результате сильной инсоляции
с увеличением высоты в горах почвы по отношению к температуре воз-
духа становятся относительно теплее (абсолютно они, конечно, холоднее).
Поэтому продолжительность вегетационного периода в этих условиях г
видимо, в большей степени ограничивается температурой почвы на глу-
бине 10 см, чем температурой воздуха х.
Особенно важным климатическим фактором является снежный покров.
По данным метеорологической станции, расположенной на высоте 2072 м
над уровнем моря в области Центральных Альп, годовая сумма осадков
составляет здесь 831 мм. Это очень небольшая величина, причем 60%
этого количества выпадает в течение летнего полугодия. В среднем осадки
выпадают каждые 2 дня; периоды без дождей могут длиться 4 дня, в край-
них случаях — 14 дней. Максимальная мощность снежного покрова дости-
гает 103 см; снег лежит в среднем около 174 дней. Среднемесячная тем-
пература воздуха в течение шести месяцев (ноябрь — апрель) за 5 лет
наблюдений была ниже 0°; только в июле и августе температура дости-
гала 10°. Абсолютный максимум составлял 27,6° (июль), а абсолютный
минимум —20,2° (январь); наиболее высокая суточная амплитуда в отдель-
ные месяцы равнялась 12,2—16,7°. Среднегодовая температура в 1957 г.
1 H.Aulitzky, Bodentemperaturverhaltnisse an einer zentralalpinen Hanglage
beiderseits der Waldgrenze I-III, «Arch. f. Meteor., Geophys. u. Bioklim.», 10, 1961,
S. 445—532; 11, 1962, S. 301—376.
Хвойные леса умеренного холодного климата — бореальная зона
393
составляла 1,8°, а температура источников — 4°. В горах на больших
площадях однородный климат не выражен. Рельф, инсоляция, ветер
и степень затененности обусловливают местные микроклиматические
различия; решающую роль здесь играет продолжительность бесснежного
Ветер
-О
/I
Рис. 294. Годовой ход температуры
почвы в 1957 и 1958 гг. на глубине 0—1м
на местообитаниях, покрытых мощным
и незначительным снежным покровом
(по Аулицки).
Р и с. 293. Влияние ветра, солнечной ра-
диации и степени затененности деревьями
на мощность снежного покрова и продол-
жительность бесснежного периода (по
Аулицки). Пояснения см. в тексте.
периода. Рис. 293 иллюстрирует микроклиматические особенности раз-
личных местообитаний. В случаях I и II показано влияние рельефа на отло-
жение снега при различном воздействии ветра. Бесснежный период на осве-
щенных солнцем подветренных склонах может быть непродолжительным,
поскольку здесь образуются снежные наносы, которые поздно оттаивают;
в результате этого зима резко переходит в разгар лета. В случае II южный
склон рано освобождается от снега, однако почва не перегревается вслед-
ствие сильного воздействия ветра, дующего преимущественно в одном
направлении. В случаях III и IV эффект воздействия рельефа усиливается
394
Глава 5
влиянием облесения; наряду с этим появляется новый фактор, связанный
с затенением почвы. Поэтому климатическая граница у верхнего предела
произрастания леса в горах располагается между преимущественно откры-
тыми и подвергнутыми влиянию ветра местообитаниями и территорией,
занятой высокоствольным лесом, где ярко выражены различия в степени
инсоляции.
В верхних частях пояса гор развиты подзолистые почвы; в зависи-
мости от продолжительности бесснежного периода и теплового режима
они варьируют и представлены подзолистым ранкером, железистым под-
золом, железисто-гумусовым подзолом, подзолистым буроземом и т. д.
На рис. 294 сопоставлен ход температуры почвы на покрытом снегом
участке в районе метеорологической станции, расположенной на освещен-
ном солнцем склоне с небольшим углом падения, и температуры почвы
па затененном склоне с маломощным снежным покровом. Почва промер-
зает очень глубоко па почти бесснежных склонах; при наличии мощного
снежного покрова среднемесячная температура понижается только в преде-
лах верхних 25 см и то лишь на десятые доли градуса ниже точки замерзания.
Во время наблюдений экстремальные температуры на поверхности
почвы достигали —17,7° и +80о. При наличии более длительного периода
наблюдений амплитуда температуры предположительно могла бы пре-
высить 100°. Эти данные пригодны для характеристики непокрытой расти-
тельностью почвы. Любой растительный покров, и в частности кустар-
нички, снижает экстремальные величины, поэтому максимальная тем-
пература практически не превышает 20°. Лес особенно сильно влияет
на эти показания.
Исследования проводились на склоне 30° на высоте 1890—1940 м
над уровнем моря в кедрово-лиственничном лесу. Возраст насаждения
60—70лет(с отдельными перестойными деревьями);степень сомкнутости крон
равна 35—40%; развит густой ярус кустарничков; почвы — железистый
подзол с хорошо выраженным слоем подстилки и грубого модера. Про-
должительность солнечного сияния около поверхности почвы в кедровом
лесу очень невелика и составляет на поляне 51% от астрономически
возможной, а в лесу, на наиболее затененном участке, — лишь 3,5%.
Температура поверхности почвы сильно колеблется. При плохом
газообмене почвы даже непродолжительная инсоляция может нагреть
ее поверхность. На прогалинах около освещенной солнцем опушки тем-
пература достигала наиболее высоких значений,но уже па небольшой глуби-
не в почве эти различия сглаживаются. В течение вегетационного периода
температура почвы, начиная с глубины 10 см, постоянно равна 5—10°.
Здесь, около верхней границы леса, в течение зимы в естественных
условиях проводились эколого-физиологические исследования Pinus
cembra (рис. 295) с применением самописцев; при этом изучались не только
условия произрастания взрослых деревьев, но и молодых сеянцев,
поскольку последние определяют возобновление древостоя. Как видим,
речь идет о единственных в своем роде наблюдениях Ч
1 W. Tranquillini, Standortsklima, Wasserbilanz und CO2-Gaswechsel
junger Zirben (Pinus cembra L.) an der alpinen Waldgrenze, «Planta»,F49,1957, S. 612—
661; Die Stoffproduktion der Zirbe (Pinus cembra L.)an der Waldgrenze wahrend eines
Jahres I, II, «Planta», 54, 1959, S. 107—151; Beitrag zur Kausalanalyse des Wettbewerbs
okologisch verschiedener Holzarten, «Вег. Deutsch. Bot. Ges.», 75, 1963, S. 353—364;
Die Abnangigkeit der CO2-Assimilation junger Larchen, Fichten und Zirben von der Luft-
und Bodenfeuchte, «Planta», 60, 1963, S. 70—94; Photosynthesis and dry matter producti-
on of trees at high altitudes. Formation of Wood (CM.:For. Trees, Acad. Press New York,
1964).
Хвойные леса умеренного холодного климата — бореальная зона
395
Рис. 295. Pinus cembra в горах Ортлер (Южный Тироль) на высоте 2200 м
над уровнем моря. На заднем плане — вершина Кёнигс-Шпитце (3857 м).
Водный баланс растений в этих условиях постоянно обеспечен.
Правда, климат в Центральных Альпах, хотя и отличается относительной
сухостью, однако остается еще достаточно влажным. При низкой темпе-
ратуре величина испарения незначительна. Очень редко испарение в пол-
день превышает 1 мл/час (Пише, зеленая бумага). Поэтому почвы постоянно
увлажнены даже на небольших высотах в горах. Только в течение пере-
ходного периода между поздней зимой и ранней весной может идти речь
о физиологической сухости (см. стр. 389); однако она может оказать влия-
396
Глава 5
Минимальная температура хвои В преВыЭущую ночь
Р и с. 296. Суточная вели-
чина нетто-ассимиляции у
не покрытых снегом моло-
дых растений Pinus cembra
в период с 24 октября по 13
ноября, окрестности Обер-
гургля (Тироль) на высоте
2100 м над уровнем моря.
Ночные температуры ниже
—4° резко снижают фото-
синтез на следующий день
(по Транквилини).
ние только на сеянцы, не покрытые’снегом, поскольку запасы воды в их
осевых органах меньше, чем у старых деревьев.
Большое значение для жизнедеятельности растений при коротком
вегетационном сезоне имеет продукция вещества. Поэтому особое внима-
ние уделялось изучению фотосинтеза молодых сеянцев кедра. Летом при
температуре выше 0° в процессе фотосинтеза вещество продуцируется
нормально. Осенью при резком снижении температуры ночью (ниже —4°)
хвоя замерзает; на следующий день, как известно, фотосинтез сокра-
щается даже при благоприятной температуре. Чем чаще это наблюдается,
тем меньше поглощается СО2, пока наконец накопление органического
вещества не прекращается полностью (рис. 296). Это происходит при
отрицательной температуре почвы, когда растения не могут поглощать
воду, а устьица плотно закрыты. Начинается зимний покой, который
не прерывается даже в отдельные теплые дни Ч Если хвоя в это время
покрыта снегом, она остается зеленой. Если же хвоя выставлена
на солнце, она начинает желтеть, хлоропласты группируются вокруг
ядра или в углах складчатой паренхимы, а количество хлорофилла сни-
жается до 1/3 от летней величины.
Из-за сильной инсоляции хвоя редко замерзает даже зимой в солнеч-
ные дни. Во время зимнего периода (159 дней) в течение 42 дней льда
в хвое не было, и только в течение 2 дней она полностью замерзала. Расте-
ния лучше сохраняются под снегом при постоянной температуре ниже 0°.
Влажный снег может способствовать развитию грибов (Lophodermium
pinastri на кедре и Herpotrichia nigra на горной сосне и ели), кото-
рые повреждают молодые сеянцы.
1 По данным последних исследований, подрост Pinus cembra, выращиваемый в теп-
лой оранжерее, также резко снижает фотосинтез в декабре. Минимум отмечен в январе,
после чего опять наблюдается повышение фотосинтеза. Это явление выражено и при
нормальной продолжительности дня и при долготе дня 8 или 12 часов и объясняется,
очевидно, эндогенными причинами. См.: S. В a m Ь е г g, W. Schwarz, W. Tran-
quil 1 i n i, Influence of day length on the photosynthetic capacity of stone pine (Pinus
cembra L.), «Ecology», 48, 1967, p. 264—269.
Хвойные леса умеренного холодного климата — бореальная зона
397
При повышении температуры весной баланс СО2 остается отрицатель-
ным; только постепенно растения начинают фотосинтезировать и накапли-
вать сухое вещество (рис. 297). Фотосинтез наступает тем раньше, чем
более зеленой сохранилась хвоя. Поэтому молодые деревья, как только
они выйдут из-под снега, могут начать ассимилировать СО2, тогда как
старым деревьям для этого необходимо больше времени.
Длительное изучение нетто-фотосинтеза и дыхания незеленых органов
позволило установить годовой ход баланса СО2 у молодых сеянцев кедра
(рис. 298). Наряду с этим годовой баланс был определен непосредственно
методом расчета прироста сухого вещества у целых сеянцев. Эти резуль-
таты не соответствуют друг другу. Данные, полученные первым методом,
оказались в три раза больше полученных вторым путем. Транквилини,
считавший раньше, что органическое вещество выделяется корнями
(микориза), в дальнейшем правильно объяснил отсутствие совпадения
этих данных тем, что при изучении фотосинтеза в сосуде складываются
более благоприятные условия температуры, чем в естественных условиях.
Ход накопления органической массы в течение года в целом соответствует
показателям, представленным на рис. 298. Вегетационный период длится
с середины мая до конца октября. Расход вещества зимой под снегом
•составляет только 6% от величины брутто-фотосинтеза. В целом в течение
года образуется очень мало органического вещества. Вещество продуци-
руется в течение 25 лет. пока деревце не достигает высоты 80 см.
Рис. 297. Суточный ход нет-
то-ассимиляции у Pinus cembra
ранней весной (по Транквили-
ни).
Вверху — подрост кедра; 2 апре-
ля — баланс СО2 все время отрица-
тельный, 15 апреля — положитель-
ный только во второй половине дня,
27 апреля — баланс слабо положи-
тельный, 30 апреля—нормальный и в
мае фотосинтез, очень интенсивный
Внизу — сопоставление выцветшей
зимой ветви старого дерева, имев-
шей 29 апреля отрицательный ба-
ланс вещества, с зеленым молодым
подростом, только что вышедшим
из-под снега и обладавшим 27 апре-
ля положительным балансом.
398
Глава 5
С02(мг) ’
60-
40-
20-
п Г днем—
Дыхание< ночью -
I корней
Брутто-ассимиляция
°C
-Ю
< Расход С02 под снегом /
I п лг
Нетто-прихоЗ
С02 мг/г-сутки
-8
-6
-4
-2
>--2
Среднемесячная температура
воздуха
МетеойуЗка, 2м
У У1 ИГ Ж IX
—4
-6
-8
-10
0
О
Р и с. 298. Годовой ход баланса СО2 у подроста Pinus cembra на границе
распространения деревьев в Обергургле, Тироль (по Транквплини).
В Центральных Альпах верхнюю границу леса вместе с кедром {Pinus'
cembra) образует и лиственница {Larix decidua) с летнезеленой хвоей. Хвоя
начинает распускаться только в середине июня, когда прекращаются
ночные заморозки, и желтеет уже в конце сентября; молодая хвоя лист-
венницы очень страдает от заморозков. Кедр накапливает вещества
в течение 181 дня, тогда как летнезеленая лиственница всего только
на протяжении 107 дней. Поэтому возникает вопрос, каким образом
лиственница может конкурировать с кедром.
Измерения фотосинтеза, проведенные одновременно у обоих видов,
свидетельствуют о том, что лиственница по сравнению с кедром ассими-
лирует летом в 2 раза больше СО2 в пересчете на сухой вес хвои. Это*
объясняется тем, что количество ассимилирующей паренхимы на попереч-
ном разрезе хвои составляет у лиственницы 80%, а у кедра —60%.
Кроме того, у кедра более мощно развиваются эпидермис и клетки склерен-
химы — непродуктивная ткань, которая увеличивает сухой вес хвои.
Таким образом, в этих условиях складывается картина, сходная с той,,
которая наблюдается у вечнозеленого Quercus ilex по сравнению с летне-
зеленым Q. pubescens. Различия сглаживаются, если фотосинтез у кедра
и лиственницы соотнести с одинаковой поверхностью хвои. Для этого*
из накопленного в течение дня количества вещества нужно вычесть расход
на дыхание в течение ночи; величина последнего у лиственницы несколько
выше, однако так же, как у кедра, составляет 8—9% от значений нетто-
ассимиляции. Длятого чтобы вычислить валовую продукцию растения, необ-
ходимо выявить общее количество ассимилирующей хвои. Установленот
что масса хвои у молодых лиственниц почти в 3 раза превышает эту массу
у молодых кедров, а в возрасте 12 лет у лиственниц, растущих на мине-
ральных почвах, даже в 6 раз. Кроме того, поскольку лиственница,
несмотря на короткий вегетационный период, ассимилирует в расчете
на 1 г хвои в год на 47% больше СО2, общая продукция вещества у лист-
Хвойные леса умеренного холодного климата — бореальная зона
399
венницы в возрасте 4 лет будет почти в 4,5 раза больше, чем у кедра того же
возраста, а у лиственницы в возрасте 12 лет даже в 8,5 раза. Только
в возрасте 25 лет у лиственницы оказывается хвои меньше, чем у кедра,
и она отстает в росте от кедра. Таким образом, лиственница лишь в началь-
ный период жизни превосходит кедр и быстрее растет в высоту, что наблю-
дается только на благоприятных для лиственницы минеральных почвах;
на почвах с грубым гумусом ее быстро опережает кедр. В дальнейшем
теневыносливый кедр захватывает все местообитания. Взаимоотношения
лиственницы и кедра напоминают взаимоотношения сосны и ели; раз-
личие выражается в том, что в последнем случае оба вида являются вечно-
зелеными породами.
Необходимо коротко остановиться на влиянии ветра. Прямое воз-
действие ветра относительно невелико; косвенное —проявляется в меха-
нических повреждениях, которые вызывают деформацию кроны деревьев
(флагообразные кроны деревьев). Непосредственно над поверхностью снеж-
ного покрова происходит шлифовка снегом, когда кристаллы льда с огром-
ной силой обрушиваются ветром на ветви деревьев. Поэтому у одиноко
стоящих деревьев ели часто можно наблюдать следующую картину: у них
развиты ветви на нижней части ствола, находящейся зимой под снегом;
затем следует участок ствола без ветвей, выступающий над снегом и откры-
тый воздействию переносимого ветром снега; верхняя часть ствола рас-
положена настолько высоко, что кристаллы льда ее не достигают, поэтому
здесь опять появляются хорошо развитые боковые ветви (см. рис. 291).
Мы не располагаем данными об эколого-физиологических исследова-
ниях у полярной границы леса. Зимы там длиннее, а воздействие инсоля-
ции не проявляется. Поэтому зимний покой, видимо, выражен лучше,
а вещества в этот период расходуется меньше. Возможно, в этих усло-
виях растения больше страдают от физиологической сухости, поскольку
здесь не наблюдается случайного оттаивания ветвей, которое обеспечивает
подачу воды к хвое; зато влияние ветра более ощутимо.
Снежный покров в горах всегда хорошо выражен, тогда как в бореаль-
ной зоне его мощность часто очень невелика. Поэтому от низких зимних
температур страдают даже типичные для этой зоны виды, среди них
черника (Vaccinium myrtillus), которая в малоснежную зиму 1963/64 г.
сильно обмерзала на северном побережь^ Ботнического залива и в резуль-
тате этого не цвела летом Ч Черника не встречается в местообитаниях
с постоянно маломощным снежным покровом: у основания стволов, на коч-
ках склонов южной экспозиции и т. д. (см. стр. 328).
ТИПЫ БОЛОТ И ИХ РАСПРОСТРАНЕНИЕ
В климатическом отношении бореальная зона хвойных лесов харак-
теризуется длинной холодной зимой и относительно прохладным влажным
летом. Величина испарения в этой области значительно меньше годового
количества осадков. Поэтому образуется избыток влаги, который стекает
по поверхности почвы или просачивается в почву до уровня грунтовых вод.
Постоянный нисходящий ток влаги в почве способствует вымыванию
оснований, и в результате образования коллоидов гумуса почва принимает
кислую реакцию.
1 Р. Н a v a s, Pflanzendkologische Untersuchungen im Winter, «Aquilo (Oulu)Ser.
Botan.», 4, 1965, S, 1—36,
400
Глава 5
Большая часть бореальной зоны расположена в равнинных условиях,
поэтому уровень грунтовых вод здесь залегает обычно недалеко от поверх-
ности почвы и в течение преобладающей части года располагается на глу-
бине 50 см. Тем самым создается возможность произрастания на таких
местообитаниях деревьев. Если уровень грунтовых вод периодически
повышается и достигает поверхности почвы, то деревья угнетаются. Леса
изреживаются и в конце концов переходят в болота; последние очень
характерны для бореальной зоны и господствуют на обширных террито-
риях этой зоны. Болота занимают большие площади в равнинных областях
Фенноскандии, в северной части бореальной зоны Восточной Европы.
Территория Западной Сибири (прежде всего в районе Оби и Иртыша)
почти сплошь покрыта болотами, которые занимают пространство вплоть
до русел рек. Кроме того, болота распространены в некоторых частях
Камчатки и Аляски, на Лабрадоре и в областях, расположенных южнее
Гудзонова залива \
Среди экологических рядов еловых и сосновых лесов этой зоны
(стр. 352 и 354) выделяется ряд, отражающий увеличение кислотности
и степени заболоченности почв; его замыкает Piceetum или Pinetum spha-
gnosum. Продолжают ряд верховые болота. Конечный член другого ряда
этих лесов, развивающегося на почвах, богатых питательными вещест-
вами, Piceetum herbosum, в процессе заболачивания переходит в поймен-
ные леса, а последние —в плоские, или низинные, болота, которые при-
урочены к площадям неглубокого залегания минерализованных грунто-
вых вод; эти болота очень широко распространены также в зоне листвен-
ных лесов и даже в степной зоне.
Болото с геологической точки зрения — это участок, где имеется
'Слой торфа мощностью от 20 —30 см. Если слой торфа достигает меньшей
толщины или содержание горючего вещества составляет лишь 15—30%,
то в этом случае говорят о заболоченных почвах 1 2. С геоботанической
точки зрения болота — это растительные сообщества, приуроченные
к местообитаниям с высоким уровнем грунтовых вод вне зависимости
от мощности отложений торфа, на котором они размещены. При высоком
•стоянии грунтовых вод болотные растения укореняются очень неглубоко,
и поэтому их жизнедеятельность определяется только свойствами верх-
них горизонтов торфа. В дальнейшем изложении болота будут рассматри-
ваться только с геоботанической точки зрения без анализа их стратигра-
фии и датировки времени образования отдельных горизонтов с помощью
пыльцевого анализа (см. «Phytologie», Bd. Ill, 2).
В зависимости от источника поступления воды, питающей болото,
выделяют три типа болот:
1. Топогенные болота, связанные с грунтовыми водами;
эти болота встречаются в наиболее глубоких понижениях рельефа, в кот-
ловинах и долинах рек или там, где на поверхность почвы выходят источ-
1 В Восточноевропейско-сибирской области болота занимают 3% общей площа-
ди (с заболоченными почвами — 9%). Самое крупное болото в Васюгане покрывает
площадь, равную 53 700 км2 и, таким образом, превышает территорию Дании. Ней-
штадт по характеру растительного покрова выделяет 10 типов болот: 1) северные релик-
товые болота, 2) собственно верховые болота, 3) злаковые и гипновые низинные болота,
4) центральносибирские болота, 5) болота горных долин, 6) амурские низинные боло-
та, 7) вулканические болота, 8) карельские болота, 9) субтропические болота и 10)
топогенные низинные болота. См.: М. I. Neustadt, Prinzipien der Rayonierung der
2Moore in der UdSSR, «Ann. Acad. Sci. Fenn.», A, № 89, III, 1966, p. 1—29.
2 F. Overbeck, Die Moore Niedersachsens, 2 Aufl., Bremen-Horn, 19£0.
Хвойные леса умеренного холодного климата — бореальная зона
401
ники. К этому типу принадлежат различные по своему характеру низин-
ные болота.
2. Омброгенные болота, которые в отличие от предыдущих
совершенно не связаны с грунтовыми водами. Они существуют исключи-
тельно за счет атмосферных осадков, выпадающих на их поверхность,
и поэтому могут возвышаться над окружающими формами рельефа. Эти
болота называют верховыми; в типичных случаях они отличаются выпук-
лой поверхностью с относительно крутыми склонами.
3. Солигенные болота; также существуют за счет атмосфер-
ных осадков. Поскольку поверхность этих болот вогнутая, они не возвы-
шаются над окружающей их местностью; дополнительно эти болота
питает вода, стекающая со склонов.
Химический состав воды в названных трех типах болот резко разли-
чен. Грунтовая вода топогенных болот может содержать большое коли-
чество растворенных минеральных питательных веществ. Поэтому эти
болота называют эвтрофными, или минеротрофными.
Вода, выпадающая в виде атмосферных осадков и питающая омброгенные
болота, почти не содержит растворимых веществ; эти болота характери-
зуются бедностью питательными веществами; их называют олиго-
трофными, или омбротрофными; здесь растут только мало-
требовательные к содержанию питательных веществ виды растенийх.
Количество веществ, растворенных в дождевой воде, в значительной мере
зависит от степени загрязнения воздуха дымом или пылью. В качестве
примера приводится содержание минеральных веществ в дождевых
осадках, выпадающих в области с малозагрязненным воздухом (табл. 82).
Таблица 82
Содержание минеральных веществ (в кг) в дождевых осадках,
выпадающих в течение одного года на 1 га площади в Финляндии (по Виро)
Орга- ниче- ские Неоргани- ческие SiO2 ГегОзЧ- +А120з Са Mg к Na р S Cl N
27,40 31,70 2,91 1,31 1,99 1,31 2,45 1,83 1,83 5,16 4,76 5,87
В солигенных болотах вода, стекающая со склонов, содержит больше
питательных веществ, чем чистая дождевая вода; поэтому эти болота
занимают промежуточное положение и в большинстве случаев являются
минералотрофными. В табл. 83 приведены данные о химическом составе
воды в верхаловых и низинных болотах.
Органическое вещество в воде верхового болота представлено золями
гумуса. Вода окрашена в бурый цвет (цвет крепкого чая) и имеет кислую
реакцию (pH = 3,7—4,0). Вода низинного болота в зависимости от содер-
жания извести обладает слабокислой (pH = 5,5—6,0) или даже щелочной
1 G. E.Du Rietz, Die Mineralbodenwasserzeigergrenze als Grundlage einer na-
tiir lichen Zweigliedernung der nord- und mitteleuropaischen Moore, «Vegecatio», 5/6,
1954, S. 571—585.
В висячее солигенно-омброгенное болото почвенная влага, насыщенная минераль-
ными веществами, может поступать с верхней части склона; проникая в йолото, вода,
если она бедна растворимыми веществами, теряет свои питательные свойства. В этом
случае нижняя часть болота постепенно переходит в омбротрофное (см.: U. J ens еп.
Die Vegetation des Sonnenberger Moores im Oberharz und ihre okologischea Bedingungen,
«Natursch. u. Ldschaftspfl.», H. 1, Hannover, 1961.
1/2 26-01248
402
Глава 5
Таблица 83
Содержание минеральных солей в воде
верховых н низинных болот в частях
па 106 (по Раманну)
Химическое соединение Верховое болото Низинное болото
К2О .... 2,44 22,85
Na2O . .... 1,98 7,39
СаО 11,70 62,61
MgO 2,08 6,51
FeO . . 9,03 40,08
р2о5 0,95 6,45
SiO2 3,62 10,87
Зола 32,78 164,5
Органическое веще-
ство . . 127,41 288,3
реакцией (pH >7,0). В Скандинавии граница распространения растений,
являющихся индикаторами почвенной минерализованной воды, проходит
там, где в 1 л воды содержится 1 мг Са. На севере, где выпадает избыточное
количество осадков, обычно развиты выщелоченные подзолистые почвы,
поэтому грунтовые воды бедны минеральными солями и содержат золи
гумуса. Поэтому в этих областях трудно провести четкую границу между
верховыми и низинными болотами. Часто выделяют переходные мезо-
трофные болота, занимающие промежуточное положение между верхо-
выми и низинными болотами. Развитие растений зависит не только от коли-
чества минеральных питательных веществ, но и от характера движения
воды. Если корни растений постоянно соприкасаются с обновленной водой
(приточная вода или прибой волн), то это способствует большему потребле-
нию питательных веществ даже из воды, бедной минеральными веще-
ствами; в этих местообитаниях распространяются виды, более требователь-
ные к богатству почв (вдоль водотоков, края вымоин).
В первую очередь остановимся на характеристике омброгенных
болот, которые в зависимости от климатических условий делятся на целый
ряд типов. Типичные безлесные верховые болота (рис. 299, 2)
формируются в областях с океаническим климатом на северо-западе
Центральной Европы и в южной части Фенноскандии. Там, где климат
характеризуется сухостью в летние месяцы, на болотах поселяются
деревья; в этом случае образуются лесные верховые болота (рис. 299, 7),
распространенные в предальпийской области Центральной Европы и в Вос-
точной Европе. Здесь растут сосна (Pinus silvestris, Р. montana) и различ-
ные кустарнички. По мере увеличения океаничности климата верховые
болота становятся более плоскими и почти не выступают над поверх-
ностью окружающего рельефа (рис. 299, 3). В дальнейшем они постепенно
переходят в солигенные болота (рис. 299, 4). В последних
насыщенная минеральными веществами вода, стекающая с окружающих
склонов, часто неравномерно распределяется по площади болота, что
обусловливает мозаичное распределение минеротрофной и омбротрофной
растительности. Такие болота в Финляндии называют а а п а (рис. 299, 5);
их называют также островными или грядово-мочажин-
Хвойные леса умеренного холодного климата — бореальная зона
403
ними болотами1. Они распространены дальше к северу, где уже
сказывается влияние почвенного льда. В областях с ярко выраженным
океаническим климатом, например в западной Шотландии, влажность
столь велика, что болота образуются вне зависимости от рельефа; это
покрывающие местность болота, или покровные
болота, они занимают обширные пространства (рис. 299, б).
Рис. 299. Схематический разрез различных типов омброген-
ных и солигенных болот (по Освальду, из Овербека).
1 — лесное верховое болото; 2 — типичное верховое болото; 3 — плос-
кое верховое болото; 4 — солигенное болото; 5 — болото аапа (ост-
ровное болото); 6 — покровное болото. От 1 к 6 — увеличение оке-
аничности климата (5 — северный тип).
Северному полушарию свойственны следующие закономерности
в размещении типов болот.
Покровные болота распространены в пределах зоны лиственных лесов
с резко выраженным океаническим климатом; верховые болота — в преде-
лах зоны лиственных лесов и южной части бореальной зоны с океаниче-
ским климатом; лесные верховые болота — в слабоконтинентальной части
зоны смешанных лесов; аапа — в центральной и северной частях зоны
хвойных лесов; бугристые болота (особый тип торфяных бугристых болот,
о которых еще будет идти речь) — в лесотундре и прилегающих к ней
областях.
Основные области распространения различных типов болот в Европе
показаны на рис. 300. Их распределение в горах иллюстрирует рис, 301.
Сфагновые болота широко развиты в Западной Сибири на плоских
водоразделах и в области крупных ледниковых озер в бассейне реки Обь 1 2.
1 Н, S j б г s, Oberflachenmuster in den borealen Torfgebieten, «Endeavour», 20»
1941, S. 217—244 (с хорошими фотографиями),
2 N, Katz, Uber die Typen der Moore der westsibirischen Nied erung und ihre
geographische Zonation, «Вег, Deutsch. Bot, Ges.», 48, 1930, S. 13—25,
404
Глава 5
Рис. 300. Основные области распространения различных типов болот в Северной
Европе (по Кацу и Еурола, с дополнениями).
1 — бугристые болота; 2 — аапа; 3 — типичные верховые болота; 4 — покровные болота; 5 —
лесные верховые болота; 6 — горные болота. Белый цвет — южные области, где преобладают топо-
генные болота.
Рис. 301. Приуроченность различных типов болот к горным поясам в Западной Ев-
ропе на примере двух схематических профилей, проведенных с запада на восток п с се-
вера на юг (по Освальду и Овербеку).
При движении с юга на север можно выделить три типа верховых болот,
занимающих ограниченную площадь.
1. Типичные выпуклые верховые болота, встречающиеся вплоть
до зоны лесостепи; обычно представлены сухими кустарничковыми боло-
Хвойные леса умеренного холодного климата — бореальная зона
405
тами (Cassandra, Vaccinium vitis-idaea) с Pinus silvestris. Такие болота часто
расположены среди засоленных почв.
2. Влажные верховые болота с Pinus silvestris, Cassandra и Ledum
в южной и центральной частях бореальной зоны.
3. Кустарничковые болота в северной части бореальной зоны с Pinus
sibirica, образующей придаточные корни над слоем вечной мерзлоты.
В бореальной зоне Сибири, помимо того, широко распространены
огромные плоские верховые болота с мочажинами. В лесотундре встре-
чаются бугристые болота; размеры торфяных бугров по мере продвижения
к Арктике уменьшаются. В Сибири не развита переходная зона, образо-
ванная болотами аапа.
Омброгенные и солигенные болота являются сравнительно молодым
природным образованием с новыми растительными сообществами, кото-
рые, очевидно, возникли лишь в послеледниковый период. В северо-
западной Европе преобладают собственно верховые болота; климат форми-
руется здесь под отепляющим влиянием Гольфстрима.
На омбротрофных болотах развиваются следующие типичные цветко-
вые растения, распространенные от бореальной до субарктической зоны
и малотребовательные к содержанию в почвах питательных веществ ь.
Eriophorum vaginatum, Trichophorum caespitosum, Rhynchospora alba, Carex
limosa, Drosera rotundifolia, D. anglica, Andromeda polifolia, Vaccinium
oxycoccus, V. vitis-idaea, V. uliginosum, Calluna vulgaris, Empetrum nigrum.
Помимо того, на западе чаще встречаются Narthecium ossifragum, Erica
tetralix, Drosera intermedia и Eriophorum angustifolium и только на востоке
Ledum palustre и Chamaedaphne calyculata, а на севере Rubus chamaemorus,
Retula папа и Scheuchzeria palustris.
Особенностью экстремально-омбротрофных покровных болот, расположенных
на западном побережье Ирландии, является произрастание вблизи берега этих болот
Schoenus nigricans — вида, обычно свойственного минеротрофным и часто щелочным
местообитаниям. В прибрежные болота вместе с атмосферными осадками, содержащими
морскую воду в распыленном состоянии, поступает относительно большое количество
электролитов, главным образом Na+ и С1~; несмотря на это, концентрация электроли-
тов в воде прибрежных болот при повышенной влажности климата лишь ненамного
больше, чем у омбротрофных болот в глубине страны. Опыты, проведенные с водными
культурами Schoenus nigricans, свидетельствуют, что этот вид одинаково хорошо растет
как при высоком, так и при низком содержании кальция и при величине pH в пределах
4—8. Растение, однако, очень чувствительно к ионам алюминия. При содержании
алюминия, равном 0,55 мг!л, рост корней замедляется вдвое. Концентрация алюми-
ния в воде болот, расположенных в глубине страны, в которые поступают глинистые
частицы из окружающих почв, значительно выше, чем в прибрежных болотах Ир-
ландии, в течение всего года находящихся под воздействием морских ветров. Развитию
Sch. nigricans благоприятствует отсутствие морозного периода, поэтому этот вид в
Ирландии пребывает в вечнозеленом состоянии и ассимилирует даже зимой 1 2.
Основными растениями верховых болот, способствующими их форми-
рованию, являются сфагновые мхи, то есть виды Sphagnum. Их анатоми-
ческая структура позволяет им впитывать воду как губка в количествах,
в несколько раз превышающих их вес. Верхняя часть мхов растет в высоту,
а нижняя отмирает и оторфовывается; таким путем отлагается торф и осу-
ществляется прирост верхового болота. Основной побег Sphagnum, от кото-
рого ответвляются наклоненные вниз боковые побеги, действует как
1 Полный перечень всех лишайников, мхов и сосудистых растений приведен в ра-
боте: К. Muller, Zur Flora und Vegetation der Hochmoore des nord-westdeutschen
Flachlandes, «Schr. Naturw. Ver. Schlesw.-Holst.», 36, 1965, S. 30—70.
2 J, H. Sparling, The occurence of Schoenus nigricans in blanket bogs, «I.
Ecol.», 55, 1967, p, 1—31.
406
Глава 5
фитиль, и способствует капиллярному подъему воды. Это дает возмож-
ность видам Sphagnum, которые не переносят иссушения, осуществлять
прирост в высоту при помощи окончаний побегов, возвышающихся над
уровнем грунтовых вод. Отдельные виды Sphagnum в этом отношении ведут
себя по-разному (рис. 302): в воде растет только Sph. cuspidatum, в то
время как Sph. magellanicum (= medium), Sph. rubellum, Sph. acutifolium
и чаще встречающийся на востоке Sph. fuscum могут произрастать, рас-
полагаясь до 50 см над уровнем грунтовых вод. Различная экология
Рис. 302. Высота над уровнем грунтовых вод, на которой
могут расти различные виды рода Sphagnum (по К. Мюллеру).
видов Sphagnum способствует образованию кочек и мочажин, характерных
для верховых болот, и формированию тем самым микрорельефа на болотах
(рис. 303).
В верховых болотах выделяют 3 уровня: верхний, образованный
кочками, средний, образованный моховым покровом, и нижний, пред-
ставленный мочажинами. Средний уровень может быть очень хороша
выражен, но иногда он отсутствует полностью.
Мочажинами называют углубления в верховых болотах,
в которых можно увязнуть. Грунтовая вода в мочажинах расположена
очень близко от поверхности; при таянии снега они наполняются водой.
Ржи с.1 303. Микрорельеф двух болот (увеличено в 2 раза): а — относительно ровный,.
б — с резко выраженными кочками и мочажинами (по Овербеку).
Хвойные леса умеренного холодного климата — бореальная зона
407
Рис. 304. Мочажина между кочками Eriophorum vaginatum, озеро Тойфель
под’Берлином (фото Хюка, из Н. Walter, 1927).
В мочажинах произрастают гигрофильные сфагновые мхи и некоторые
виды высших растений (Carex limosa, Scheuchzeria palustris) (рис. 304).
Из мочажин выступают округлые возвышения высотой до 50 см — кочки.
На кочках обычно отчетливо выражена микропоясность раститель-
ности. Чем выше на кочках располагаются мхи над уровнем грунтовых
вод, тем меньше среди них встречается гигрофильных видов рода
Sphagnum', последние в этом случае вытесняются Sph. magellanicum,
Sph. rubellum и Sph. acutifolium. В восточных областях высокие кочки
покрыты Sphagnum fuscum.
Соединяясь, мочажины образуют |постоянно наполненные водой
маленькие или крупные болотные озера. Максимальная глубина их
достигает 3 м, обычно 1,5—2 м; они не имеют твердого дна, поскольку оно
образовано мягким детритом. Болотные озера образуются в засушливые
годы, когда мочажины высыхают, сфагновые мхи отмирают и появляется
большое количество водорослей. В последующие годы на этих участках
моховый покров не нарастает, и мочажина постепенно наполняется водой.
Раньше считали, что кочки и мочажины постоянно сменяют друг друга,
поскольку виды Sphagnum, образующие кочки, со временем развиваются
все медленней, пока прирост в высоту не прекращается полностью. Сфаг-
новые мхи на вершине кочки вытесняются Polytrichum strictum, а послед-
ний при увеличении сухости сменяется лишайниками (виды рода Cladonia,
Cetraria). В результате рост кочки приостанавливается и торф разла-
гается. В конечном счете отмершую кочку перекрывают соседние увлаж-
ненные участки, поэтому бывшие мочажины превращаются в кочки,
а последние — в мочажины. Стратиграфические исследования, проведен-
ные в торфяниках, не всегда подтверждают вывод о закономерной смене
408
Глава 5
торфа мочажин и торфа кочек. Часто торф представлен однородным нара-
стающим вверх покровом из сфагнума \ Избыток воды должен стекать
с верхового болота. Ложбины стока, иногда глубоко врезающиеся в торф,
называются р ю л л е н. Вода, вытекающая по ложбинам из болота,
собирается в обводненной зоне вокруг верхового болота, называемой
л а г г (рис. 305). Поскольку в этой части болота вода соприкасается с мине-
ральной подпочвой, она обогащается питательными веществами; поэтому
здесь распространены мезотрофные виды переходного болота или эутроф-
ные виды низинного болота.
Более крутые и поэтому лучше дренируемые склоны болота
отличаются относительной сухостью и обычно зарастают кустарниками
или даже деревьями.
Рис. 305. Болото Ауг-
стумаль в дельте реки Не-
ман. Типичное верховое
болото с мочажинами, лож-
бинами стока и обводненной
окраиной, занятой переход-
ным болотом (по Веберу).
В Центральной Европе в наши дни не встретишь девственных болот.
Они сильно изменены в результате осушения, разработки торфа или
переведены в сельскохозяйственные угодья и находятся в стадии превра-
щения в верещатники или в культурный ландшафт. Это же можно сказать
и о небольших по размеру болотах, расположенных в области среднегорий
и в предгорьях Альп.
Неизмененные болота сохранились только в Фенноскандии и Восточ-
ной Европе.
Здесь развит особый тип верхового болота — грядово-моча-
жинное верховое болото. При небольшом уклоне поверх-
ности болота кочки растут быстрее в горизонтальном направлении у ниж-
него края и сбоку, чем у верхнего края, где застаивается вода, в связи
с чем он отличается большей крутизной. Кочки принимают удлиненную
форму и простираются в направлении, перпендикулярном падению;
таким путем формируются отдельные параллельные гряды.
Сходная структура наблюдается у распространенных севернее болот
аапа. Грядово-мочажинные верховые болота являются омбротрофными,
включая и углубления между грядами — удлиненные мочажины. В соли-
генных аапа развиты минеротрофные вытянутые мочажины, которые
в Финляндии называются «rimpis», а в Швеции «flarke»; их растительный
1 F. Overbeck, Aufgaben botanisch-geologischer Moorforschung in Nordwest-
deutschland, «Вег. Deutsch. Bot. Ges.», 76, 1963, S. 2—12.
Хвойные леса умеренного холодного климата — бореальная зона 409
покров отличается однородностью (виды Carex и Eriophorum, Menyanthes,
Scheuchzeria, Equisetum fluviatile, Juncus stygius и др.)* Это вязкие и непро-
ходимые углубления; уровень воды в них горизонтальный, даже если
длина углублений достигает нескольких сот метров. Они простираются
поперек уклона участка и отделяются друг от друга высокими валами,
или грядами, покрытыми омбротрофной растительностью; гряды
образуют ступени, подобные террасам на рисовом поле. Уровень мине-
ротрофной воды в мочажинах располагается особенно высоко весной
Рис. 306. Болото аапа в северной
Финляндии (по Еурола иРуухиярви).
Зависимость расположенияг гряд ' (густой
пунктир) и мочажин («rimpis») (белые пят-
на) от наклона местности. В верхней ров-
ной части мочажины широкие, а гряды пре-
рывистые и размещены беспорядочно; в
средней, более крутой части почти сплошь
развиты сухие поверхности (редкий пунк-
тир); только в нижней части со слабым
уклоном встречаются мочажины и хорошо
выраженные взаимосвязанные гряды.
после таяния снега. Вода накапливается между грядами, прорывается
сквозь них в наиболее низких местах и попадает в соседнюю мочажину.
Выпуклый характер гряд в болотах аапа в известной степени обусло-
влен почвенным льдом, который сохраняется под ними почти до конца
лета. Еще большее значение имеет боковое давление льда; зимой слой
льда покрывает мочажины и простирается в горизонтальном направлении.
Солифлюкция не имеет большого значения, поскольку мочажины обра-
зуются только при небольшом уклоне местности (рис. 306). Влияние поч-
венного льда особенно ощутимо в бугристых болотах, расположенных
севернее. В переходной к этим болотам зоне в Финляндии выделяют
«р о u n i к к о s ». Это комплекс кочек, возникших под влиянием вспу-
чивания грунта почвенным льдом (рис. 307).
В бугристых болотах, распространенных в лесотундре (рис. 308),
нередко образуются многолетние торфяные бугры вспучивания. Бугры
27 - 01248
410
Глава 5
V Betula папа — Sphagnum fuscum Мочажины
уТ Ledum palustre — Pleurocium schreberi — Минеральный грунт
Empetrum ....Лишайники пппп Почва
Р и с. 307. Профиль, проведенный через гряду болота аапа (слева вверху), болото
«pounikkos» в Лапландии (справа вверху) и крупный торфяной бугор бугристого болота
(внизу) (по Руухиярви).
встречаются только в областях со среднегодовой температурой воздуха
ниже —1°, где почвенный лед сохраняется в течение всего года х.
Длина торфяных бугров достигает 20—35 м, ширина —10—15 м
и высота обычно — 2 —3 м, изредка 4,5 ле, а в исключительных случаях 7 м.
Торфяные бугры образуются следующим образом. При наличии неод-
Р и с. 308. Бугристое болото в тундре с торфяными буграми, юг полуострова
Канин (фото Поле).
1 Область развития вечной мерзлоты захватывает бореальную зону хвойных ле-
сов, в Восточной Сибири она протягивается далеко на юг. Однако здесь не установлено
четких взаимосвязей между распространением вечной мерзлоты и размещением расти-
тельности.
Хвойные леса умеренного холодного климата — бореальная зона
411
нородного растительного покрова на повышенных местах накапливается
меньше снега, и мерзлота здесь быстрее сковывает торфяную почву,
отчего между слоями торфа образуются прослойки льда. К этим прослой-
кам подтекает вода с сосед-
них незамерзших участков
торфяных почв; возникают
все более мощные прослойки
льда, которые вспучивают
торф, расположенный над
ними. Поскольку летом лед
не тает полностью, вспучен-
ное возвышение частично со-
храняется; в связи с этим
следующей зимой здесь откла-
дывается еще меньший слой
снега, почва замерзает ско-
рее, масса льда из года в год
становится больше, а торфя-
ной бугор все выше (рис. 307).
Летом размеры бугра несколь-
ко уменьшаются; в результате
вокруг него образуется капа-
вообразное углубление, за-
полненное водой. В нем растут
Betulanana, а также различ-
ные виды пушицы: Eriopho-
rum angusttfolium и Е. scheu-
chzeri; последний вид харак-
терен также для альпийских
болот (см. цветное фото 7).
С увеличением высоты бугра
его вершина становится А су-
ше; растительный покров на
вершине бугра исчезает, по-
являются трещины, и разви-
Р и с. 309. Распространение болот в Финляндии
(в % от общей площади территории) (по Ильвес-
сало).
1—1 — граница между областью распространения аапа
и областью бугристых болот; 2—2 — граница между об-
ластью распространения аапа и зоной верховых болот
(по Руухиярви и Еурола).
вается ветровая эрозия. После
того как лед растает, на месте полностью разрушенного торфяного бугра
возникает озерко. Бугры представляют собой полуископаемые образова-
ния, которые, видимо, сформировались в континентальный климатиче-
ский период около 1000 лет до н. э. В настоящее время они находятся
в стадии таяния или эрозионного разрушения.
Аапа и бугристые болота — это основной комплексный тип болот,
развитый в циркумполярной области, тогда как омбротрофные верховые
болота приурочены к областям умеренной зоны с океаническим климатом.
В Фенноскандии подробно изучалось региональное размещение различных
типов болот \ В Финляндии болота занимают большую площадь терри-
тории страны, что связано с особенностями климатических, топографи-
ческих и почвенных условий (рис. 309).
1 R. Ruuhijarvi, Uber die regionale Einteilung der nordfinnischen Moore,
«Ann. Bot. Soc. Zool.-Bot. Fennicae Vanamo», 31, № 1, 1960; S. Eur ol a, Uber die re-
gionale Einteilung der siidfinnischen Moore, ibidem, 33, № 2, 1962; N.M aimer, The
Southern Mires in «The plant cover of Sweden», «Acta Phytog. Suecia», 50, 1965, p. 149—
158; H. S j 6 r s, Northern Mires, ibidem, p. 180—188; A. P e r s s о n, Mountain Mires,
ibidem, p. 249—256.
412
Глава 5
ЭКОЛОГИЯ ВЕРХОВЫХ БОЛОТ
Мы не располагаем точными данными о водном балансе ненарушенных
болот, но такие исследования проводились в течение восьми лет на заболо-
ченной местности Кёнигсмор, между Гамбургом и Бременом х. Объектами
наблюдений были выбраны два участка: некультивированное кустарнич-
ковое болото и луг, созданный на месте болота. Уровень грунтовых вод
на участке 1 располагался на глубине 13 см, на участке 2 —на глубине
70 см. Колебания поверхности болота («дыхание»), обусловленные непо-
стоянством уровня воды, достигали в среднем на участке 1 —11 мм,
а на участке 2—37 мм-, в результате происходило большее накопление
воды на лугу по сравнению с некультивированным болотом. Потенциаль-
ная испаряемость почти соответствовала фактической величине испарения.
Годовой ход эвапотранспирации связан с ритмом развития растений.
Зимой вечнозеленые вересковые и сфагновые мхи, произрастающие
на болоте, транспирировали интенсивнее, а летом величина транспирации
Рис. 310. Водный баланс некультивированного и культивирован-
ного верхового болота (Кёнигсмор между Гамбургом и Бременом) в
1951 — 1958 гг. (по Бадену и Эггельсману).
1 W. Bade n, R.Eggelsmann, Der Wasserkreislauf eines nordwestdeuts-
chen Hochmoores, «Schrift. Kurat. f. Kulturbauwesen», H. 12, Hamburg, 1964.
Хвойные леса умеренного холодного климата — бореальная зона 413
была больше у растений на лугу. Поэтому над лугом относительная влаж-
ность воздуха летом была выше, а колебания температуры меньше. Вод-
ный баланс участков изображен на рис. 310. Количество осадков превы-
шает величину испарения, поэтому сток наблюдается в течение всего
года; все же в летние месяцы он часто бывает незначительным (табл. 84).
^Таблица 84
Средняя величина стока на болоте Кёнигсмор
(по Бадену и Эггельсману)
Элементы баланса Зима Лето Год
Осадки, мм 294 351 645
Сток, % от осад- ков Болото 59 10 32
Луг 65 9 34
Колебания стока на лугу значительно меньше выражены, чем на боло-
те, где при максимальном уровне воды после таяния снега величина стока
в 2—4 раза превышает сток па лугу. Поэтому трудно согласиться с точкой
зрения, что болота являются накопителями и регуляторами стока. Уже
на кустарничковом болоте с низким уровнем грунтовых вод условия менее
благоприятны, чем па лугу, возникшем под воздействием человека.
Нетронутые болота, где грунтовые воды почти достигают поверхности,
имеют еще меньшую возможность аккумулировать влагу.
Субстратом нарастающих верховых болот являются верхушки побегов
сфагновых мхов и их оторфованные отмершие части. Микробиология
верхнего горизонта торфа, на котором растут высшие растения, и прежде
всего многочисленные виды вересковых, интенсивно изучалась Бургефом Ч
Вересковые характеризуются наличием эндотрофной микоризы.
Бургеф выделил большое число грибов-симбионтов вересковых и опреде-
лил два из них: Cladosportum macrocarpurp Preuss и Oidiodendrum griseum
Robak. Микориза способствует расширению площади подземных всасы-
вающих органов вересковых, чем и можно объяснить выраженную нетре-
бовательность этих видов. В клетках корней содержится большое количе-
ство катехина — дубильного вещества, которое привлекает грибы. В свою
очередь гифы грибов поставляют видам вересковых минеральные пита-
тельные вещества. Фиксации атмосферного азота при этом не происходит.
На рис. 311 обобщены результаты исследований, проведенных Бурге-
фом. Дождевая вода (pH = 6,2) попадает на моховый покров и просачи-
вается глубже в торф. Профиль торфа состоит из нескольких горизонтов.
Самый верхний горизонт, зеленого цвета, образован Sphagnum-, его мощ-
ность не превышает нескольких сантиметров; реакция кислая (рН = 3,8!).
На глубине, куда свет уже не проникает, побеги мха отмирают; здесь
развит узкий горизонт серого цвета. Под ним расположен темнеющий
книзу коричневато-черный горизонт, в котором происходит гумификация
торфа. Он простирается до уровня грунтовых вод и еще содержит О2.
В этом слое распространяются тонкие корни вересковых. Глубже они
1 Н. Burgeff, Mikrobiologie des Hochmoores, Stuttgart, 1961.
414
Глава 5
Рис. 311. Профиль торфа типичного верхового болота и харак-
тер биологических процессов, протекающих на различной глубине (по
Бургефу, с изменениями).
отмирают, однако здесь еще встречаются живые подземные части побегов.
В этом же горизонте происходит частичное разложение органических
соединений под воздействием свободно живущих грибов или грибов-
симбионтов.
Ниже уровня грунтовых вод торф приобретает желтую окраску
и содержит много H2S; в анаэробных условиях начинается гниение белков
и восстановление сульфатов, которые (окисленный SO2) поступают в болото
с атмосферными осадками (в Предальпах около озера Хим в размере
10—14 кг!га). Этот слой начинается под кочками с глубины 40 —80 см,
а в мочажинах —с глубины 5—30 см. Здесь могут существовать только
корневища Carex, обладающие крупными межклетникамивеличина
pH = 4,1. В результате анаэробных процессов в более глубоких слоях
торфа образуются газы, которые иногда выделяются на поверхность в вгде
1 Содержание кислорода в проточных грунтовых водах значительно выше, чем
в стоячей воде. В условиях застойного увлажнения Molinia coerulea не растет; живые
корни Molinia могут существовать только при отсутствии H2S и там, где pH выше 1,2.
Напротив, Menyanthes (как и Carex inflata) выдерживает концентрацию H2S, равную
8 мг!л, и pH = 0,98. В этом случае О2 поступает из корней в почву, на поверхности пх
эпидермиса выделяется окись железа, что препятствует проникновению H2S. См.:
W. Armstrong, D. J. Boatman, Some field observations relating the growth
of bog plants to conditions of soil aeration, «I. Ecol.», 55, 1967, p. 101—110.
Хвойные леса умеренного холодного климата — бореальная зона
415
пузырьков. Они содержат СН4, Н2, N2, СО2 и H2S. В местах выхода этих
газов ощущается запах H2S х.
Непрерывный прирост в высоту видов Sphagnum является важным
экологическим фактором, поскольку цветковые растения, развитые среди
мохового покрова, должны суметь приспособиться к нарастанию болота
в высоту. Увеличение в длину прилегающих к поверхности видов Sphag-
num может достигать в мочажинах 30—40 см, в основном в горизонтальном
направлении. Более существенное значение имеет прирост в высоту видов
Sphagnum, образующих на кочках густую подушку. В северо-западной
Германии эта величина у Sphagnum magellanicum (= medium) равна
3,5—11,7 см, у Sph. rubellum —3,3—10,7 см и зависит от степени обеспе-
ченности растения водой в течение вегетационного периода 1 2. Зимой
вертикальный прирост почти не выражен. При отсутствии атмосферных
осадков прирост зависит главным образом от высоты расположения вер-
хушек побегов над уровнем воды в болоте. Поэтому по мере увеличения
высоты кочек он постепенно уменьшается.
В континентальных условиях Восточной Европы мхи растут медлен-
нее (табл. 85).
Таблица 85
Годовой прирост (в см) сфагновых мхов (по Гребенщиковой)
Вид Sphagnum pH Высота над уров- нем грун- товых вод, см Линейный годовой прирост Сред- ний при- рост
juscum ... 4,38 23—37 0,5—0,9 0,7
magellanicum 4,18 14-26 1,1—1,3 1,2
papillosum . . 4,08 9-16 1,0—2,0 1,4
parvifolium 4,17 5—16 2,4—3,5 3,3
dusenii . 3,75 1-4 2,8-5,4 4,2
cuspidatum 3,70 0,5 4,1-10,7 7,1
Годовая продукция сфагновых мхов представлена в табл. 86.
Таблица 86
Продукция сухого вещества (в т/га год)
на участке, покрытом различными видами
Sphagnum (по Овербеку)
Вид Sphagnum Шлезвиг- Гольштейн Массив Рен Восточная Европа
fuscum . . 4,2-8,0 2,0-3,2
rubellum . . . 2,1-2,6 3,7-9,6 —
magellanicum 2,5—3,5 4,1—7,2 2,0-2,7
parvifolium — — 2,7-3,5
dusenii .... 4,9-6,7 11,2—15,0 —
1 Химическое и микробиологическое разложение листьев Juncus squarrosus иссле-
довали в течение двух лет; установлено, что при этом расход сухого вещества за год
достигает 20—30% . См.: Р. М. Latter, J.B.Cragg, The decomposition of Juncus
squarrosus leaves and microbiological changes in the profile of Juncus moor,«J. Ecol.»,
55, 1967, p. 465—482.
2 F. Overbeck, H. Hapach, Uber das Wachstum und den Wassergehalt
einiger Hochmoorsphagnen, «Flora», 144, 1957, S. 335—402.
416
Глава 5
Нарастание болот в высоту, то есть прирост торфа, составляет лишь
небольшую часть прироста верхних горизонтов мха, поскольку сухое
вещество торфяных мхов в основном разлагается в процессе гумификации
и, кроме того, уплотняется под давлением отлагающейся наверху массы
торфа. Потери сфагнового торфа составляют за 2000 лет около 33—80%
f. Оптимальная
Рис. 312. Нарастание кочки, покрытой Sphagnum
magellanicum, и мочажины с покровом из Sph.
recurvum по мере высыхания болота (по Овербеку
и Хаппаху).
Р п с. 313. Drosera, вы-
копанная из сфагнового
покрова. Уровень распо-
ложения ее розетки еже-
годно повышается на Зел
(поБертшу,из Н. Wal-
ter, 1927).
сухого вещества; прирост торфа в массиве Рен равен 10—12 см за столетие;
в других случаях теряется лишь половина указанного выше количества,
и, следовательно, прирост равен 0,5—1 мм в год1.
Парадоксальным кажется вывод Овербека о том, что мхи, приурочен-
ные к мочажинам, больше страдают от летней засухи, чем мхи, произра-
стающие на кочках. Однако это зависит от того количества воды, которое
мхи могут впитать. В расчете на сухой вес получаются следующие вели-
чины (сухой вес принят за 1):
Sphagnum rubellum ............ 17,0
Sphagnum magellanicum .......... 24,5
Sphagnum recurvum , 17,5
1 R, H о 11 i n g, F. Overbeck, Uber die GroBe der Stoffverluste bei der Ge-
nese von Sphagnumtorfen, «Flora», 150, 1931, S, 191—208.
Хвойные леса умеренного холодного климата — бореальная зона
417
Правильнее соотносить вес воды с объемом (100 см3) срезанного
мохового покрова; в этом случае количество воды будет зависеть от густоты
мохового покрова (в г):
Sphagnum rubellum . ,
Sphagnum magellanicum
Sphagnum recurvum
46
19
10
Эти данные свидетельствуют о том, что покров мха в мочажинах
отличается большей разреженностью; величина испарения у них выше,
Р п с. 314. Повышение
уровня зоны кущения
у Carex jusca на кочке,
образованной Sphagnum
(по Бертшу).
чем у мхов, покрывающих кочки; поэтому
можачины высыхают быстрее,чем кочки, в ко-
торых накапливается большее количество
воды. В результате мхи, приуроченные к мо-
Р п с. 315. Побеги Vactinium oxycoccus в сфагно-
вом покрове. Весной молодые побеги растут вер-
тикально сквозь слой сфагнума, переросшего
клюкву в течение предыдущего года; затем побеги
стелются по поверхности мха. Старые побеги со
временем спрессовываются под слоем уплотнен-
ного торфа (по Гроссе-Браукману).
чажинам, быстрее прекращают свой рост в периоды засухи и, отмирая,
раньше окрашиваются в белый цвет, чем мхи, растущие па кочках
(рис. 312).
Нарастание поверхности покрова, образованного Sphagnum, наряду
с олиготрофностью и кислой реакцией торфа верхового болота оказывает
сильное влияние на состав цветковых растений болот. Здесь могут расти
только те виды, корневища которых с расположенными на них почками
возобновления способны ежегодно нарастать на высоту, соответствующую
приросту мха, как это наблюдается у видов Drosera (рис. 313), Eriophorum
и Carex (рис. 314). Виды с горизонтально расположенными побегами также
вынуждены весной сначала расти вертикально вверх, чтобы выйти
Р и с. 316. Vaccinium myrtillus, почти перекрытая покровом из
Sphagnum (по Бертшу).
Рис. 317. Ель в возрасте около 9 лет, ствол которой на глубину 20 см
погребен под оторфованным слоем мха (по Бертшу, из Н. W а 1 t е г,
1927).
Хвойные леса умеренного холодного климата — бореальная зона
419
Рис. 318. Ель, полупогребенная в кочке, образованной
Sphagnum, Тироль, 1200 м над уровнем моря (фото Фабера).
из слоя мха (рис. 315) х. По отдельный годичным побегам этих растений
можно определить скорость прироста мохового покрова. Если корневища
растений с почками возобновления не нарастают в высоту, то растения
постепенно перекрываются мхами, и только верхушки их побегов высту-
пают над поверхностью мохового покрова (рис. 316); цветковые растения
отмирают. Это происходит и с сеянцами деревьев, растущими на болоте
(рис. 317 и 318); сеянцы выживают только при ослабленном приросте мха
(склоны болота, лесные верховые болота в Восточной Европе). Поэтому
естественно, что многие виды, среди них Calluna, Vaccinium uliginosum
(рис. 319) и др., могут расти только на более высоких кочках. Эти виды
сразу же начинают господствовать, как только болото становится суше.
В итоге моховое болото превращается в кустарничковое. На рис. 320
изображена микропоясность растительности двух кочек, прирост в высоту
которых остановился и вершины которых поэтому покрыты Entodon
schreberei или Polytrichum strictum. ( i
1 G. Grosse-Brauckmann, Zur Artenzusammensetzung von Torfen,
«Вег. Deutsch. Bot. Ges.», 76, 1963, S. 22—37.
420
Глава 5
Рис. 319. Кочка верхового болота, прирост в высоту которой остановился.
Вершина кочки покрыта лишайниками {Cladonia rangiferina), среди которых
может существовать Vaccinium uliginosum (фото Фабера).
Клеточные оболочки некоторых видов Sphagnum, особенно у произ-
растающих на кочках, окрашены в красный цвет; эту окраску вызывают
соединения, напоминающие антоциан, но отличающиеся от него по своему
химическому составу. Экспериментальным путем доказано, что основным
фактором, вызывающим такую окраску, является низкая температура,
около 2,5°; наряду с этим определенную роль играют и другие факторы,.
Рис. 320. Микропоясность-
растительности на двух кочках
(по Бертшу).
Вверху: а — Entodon schreberi
с Vaccinium myrtillus и V. uligino-
sum, б — Sphagnum parvifolium с V.
oxycoccus; в — Sph. magellanicum
c Andromeda polifolia или Eriopho-
rum vaginatum.
Внизу: a — Polytrichum strictum
c Calluna vulgaris, 6 — Sph. parvi-
folium с V. vitis-idaea, ниже —
V. oxycoccus; в — то же, что и на
первой кочке.
Хвойные леса умеренного холодного климата — бореальная зона
421
На высоте верхушек
побегов горной сосны g.
0,6-
0,5-
0,4-
0,3-
0,2-.
0,1-
__________J I
Г 22’487. j
I (ветер) j
,_._J i--
Г 23’407.
I На поверхности сфагнового покрова
28’557. ......... I
: 25’437.
Местообитание
: Vaccinium ’
..•33’607.
(на почве),
31’507.
26’407.
-]«,5’707.
15’100%!
-------II
14,5’78%!
Местообитание Plagiothecium undulatum JI
~ i?99% 12,5*98% 12,5’100%; j
“i-------1--------1 1 1 1 A
10 11 12 13 14 15 16
Время,часы
P и с. 321. Величина испарения, измерявшегося при помощи эвапорометра Пише
(зеленая бумага), на гппновом болоте вблизи Лунца в середине сентября, Нижняя
Австрия (по Вальтеру).
ограничивающие рост растений. В зарослях видов Sphagnum, окрашенных
в красный цвет, начинается разложение хлорофилла, что можно рас-
сматривать как повреждение растений. Заросли со свежими побегами,
хорошо обеспеченные водой, не меняют своей окраски \
На верховых болотах отмечены очень большие колебания темпера-
туры, чаще всего после того, как их верхний слой (особенно кочек) высох-
нет и приобретет в связи с этим незначительную теплоемкость. Это наблю-
дается на осушенных верховых болотах, которые при ночном излучении
сильно охлаждаются и поэтому легко подвергаются воздействию поздних
заморозков; днем под влиянием поступающей радиации они значительно
нагреваются. Максимальная температура поверхности болота с сухим
торфом равна 77°, на черном торфе кочек Polytrichum strictum — 69°,
в сухих, еще не поврежденных зарослях, образованных сфагновыми
мхами, — 60°, на сырых — только 43°, а в сфагновых мхах в мочажинах —
не более 36°1 2.
1 Н.Rudolph, Zur Frage der Membranochromie bei Sphagnen I und II, «Flora»,
155, 1964, S. 250—293; «Planta», 64, 1965, S. 178 — 185.
2 H.Schmeidl, Oberflachentemperaturen in Hochmooren, «Wetter und Leben»,
17, 1965, S. 87-97.
422
Глава 5
Наиболее подробные экологические исследования верховых болот
провел Фирбас Ч Экстремальной температуре соответствует также наи-
более интенсивное испарение, которое измерялось эвапорометром Пише
(зеленая бумага); при сильной инсоляции около полудня величина испаре-
ния была очень высокой даже для климатических условий Центральной
Европы (рис. 321): на высоте 2 см над поверхностью почвы в течение одного>
часа испарялось около 0,6 —0,8 мл (максимум 0,87 мл), а на высоте 30 см —
около 1,0 мл (максимум 1,42 мл) 1 2. Зимой почва не промерзает очень глу-
боко, однако лед тает довольно долго, и поэтому развитие растительности
весной сильно запаздывает. Летом почва остается относительно холодной
даже на небольшой глубине. Около полудня в почве наблюдается резкий
перепад температур и разница между температурой на глубине 1 см
и 12,5 см может превышать 20°. Таким образом, местный климат верховых
болот отличается резко выраженной континентальностью: сильным нагре-
вом поверхности при инсоляции; низкими температурами корнеобитаемых
горизонтов почвы; возможностью заморозков в ясные ночи; длительным
сохранением мерзлоты в почве; низкими средними температурами воздуха
и сокращенным вегетационным периодом. Поэтому на верховых болотах
встречаются бореальные флористические элементы и даже ледниковые
реликты. Типичные для верховых болот растения (см. стр. 405), за исклю-
чением насекомоядных Drosera, характеризуются рядом анатомо-морфо-
логических признаков, которые обычно называют «ксероморфными»: отно-
сительно слабое развитие поверхности ассимилирующих органов, толстые
наружные стенки эпидермиса, большая густота устьиц, хорошо развитая
водопроводящая система, но одновременно рыхлый мезофилл. Как
ни странно, эти свойства наиболее ярко выражены у растений на сырых
участках сфагновых болот, а не у растений, приуроченных к сухим кочкам.
У этих растений водный режим сохраняется на постоянном уровне. Осмо-
тическое давление повышается только холодной зимой. Фирбас высказал
предположение, что «ксероморфность» обусловлена не столько трудно-
стями в обеспечении влагой, сколько бедностью субстрата питательными
веществами. Дальнейшие исследования показали, что в первую очередь
имеет значение недостаток азота 3. Наряду с этим было установлено, что
бореальные виды легко поглощают воду из холодных почв, тогда как
потребление азота сопряжено с большими трудностями 4. В связи с этим
следует говорить не о «ксероморфизме», а о явлениях недостаточности,
то есть о пейноморфозе.
Совершенно другой характер свойствен растительности эвтрофных
топогенных плоских и низинных болот. О них речь пойдет в т. III, гл. 2.
1 F. F i г b a s, Untersuchungen uber den Wasserhaushalt der Hochmoorpflanzen,
«Jb. wiss. Bot.», 74, 1931, S. 459—696.
2 H. Walter, Verdunstungsmessungen auf kleinstem Raum in verschiedenen
Pflanzengessellschaften, «Jb. wiss. Bot.», 68, 1928, S. 233—288.
3 К. M о t h es, Ernahrung, Struktur und Transpiration, «Biol. Zbl.», 52, 1932,
S. 193—223; W. R. M ii I I e r-S t о I I, Der EinfluB der Ernahrung auf die Xeromorphie
der Hochmoorpflanzen, «Planta», 35, 1947, S. 225—251; W. S i m о n i s, CO2-Assimila-
tion and Xeromorphie der Hochmoorpflanzen, «Biol. Zbl.», 67, 1948, S. 77—83.
4В.П.Дадыкин, О водном режиме и питании растений на холодных почвах,
«Доклады АН СССР, нов. сер.», 70, 1950, стр. 1073—1076; Н. G г е b, EinfluB tiefer
Temperaturen auf die Wasser-und Stickstoffaufnahme der Pflanzen, «Planta», 48, 1957,.
S. 523—563.
ОГЛАВЛЕНИЕ
ПРЕДИСЛОВИЕ АВТОРА
Глава 1
ВВЕДЕНИЕ
КРИТИЧЕСКИЕ ЗАМЕЧАНИЯ К МЕТОДАМ ПОЛЕВЫХ ЭКОЛОГО-ФИЗИОЛО-
ГИЧЕСКИХ ИССЛЕДОВАНИЙ (7). ОБЩАЯ ХАРАКТЕРИСТИКА КСЕРОМОРФ-
НОЙ СТРУКТУРЫ ТРАНСПИРИРУЮЩИХ ОРГАНОВ РАСТЕНИЙ (9). ЭКОТИПЫ
И ЭКОКЛИНЫ (13).
Глава 2
СРЕДИЗЕМНОМОРСКАЯ ЖЕСТКОЛИСТНАЯ РАСТИТЕЛЬНОСТЬ
РАСТИТЕЛЬНЫЕ ЗОНЫ ЗА ПРЕДЕЛАМИ ТРОПИКОВ И ПЕРЕХОДНАЯ
ОБЛАСТЬ] ЗИМНИХ ДОЖДЕЙ (22). ОБЩАЯ ХАРАКТЕРИСТИКА РАСТИТЕЛЬ-
НОГО ПОКРОВА СРЕДИЗЕМНОМОРЬЯ (25). КЛИМАТ СРЕДИЗЕМНОМОРСКОЙ
ОБЛАСТИ ЗИМНИХ ДОЖДЕЙ (27). ЗОНАЛЬНАЯ РАСТИТЕЛЬНОСТЬ СРЕДИ-
ЗЕМНОМОРСКОЙ ОБЛАСТИ (31). СТАДИЯ ДЕГРАДАЦИИ РАСТИТЕЛЬНОСТИ
И ЭКОЛОГИЯ АССОЦИАЦИИ BRACHYPODIUM RAMOSUM — PHLOMIS
LYCHNITTS (3 8). ЭКОЛОГО-ФИЗИОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ В СРЕДИ-
ЗЕМНОМОРСКОЙ ОБЛАСТИ (49). КОНКУРЕНТНЫЕ ВЗАИМООТНОШЕНИЯ
МЕЖДУ ВЕЧНОЗЕЛЕНЫМИ ЖЕСТКОЛИСТНЫМИ И ЛЕТНЕЗЕЛЕНЫМИ
ЛИСТВЕННЫМИ ДЕРЕВЬЯМИ (71). ВЕРТИКАЛЬНАЯ ПОЯСНОСТЬ В СРЕДИ-
ЗЕМНОМОРСКОЙ ЗОНЕ (77). СУБСРЕДИЗЕМНОМОРСКАЯ ЗОНА (92). СРЕДИ-
ЗЕМНОМОРСКАЯ ОБЛАСТЬ СТЕПЕЙ АНАТОЛИИ (97).
Глава 3
ОБЛАСТИ С ЗИМНИМИ ДОЖДЯМИ И СРЕДИЗЕМНОМОРСКИМ КЛИМАТОМ
ЗА ПРЕДЕЛАМИ ЕВРАЗИИ И ПРИМЫКАЮЩИЕ К НИМ УМЕРЕННЫЕ ЗОНЫ
КАЛИФОРНИЙСКАЯ ЗОНА ЖЕСТКОЛИСТНОЙ РАСТИТЕЛЬНОСТИ (106).
ЭНСИНАЛЬНАЯ РАСТИТЕЛЬНОСТЬ. В АРИЗОНЕ (116). ХВОЙНЫЕ ЛЕСА
ТИХООКЕАНСКОГО ПОБЕРЕЖЬЯ СЕВЕРНОЙ АМЕРИКИ (123). СЕВЕРОАМЕ-
РИКАНСКАЯ АРИДНАЯ ХОЛОДНАЯ ОБЛАСТЬ ЗИМНИХ ДОЖДЕЙ (127).
ЖЕСТКОЛИСТНАЯ РАСТИТЕЛЬНОСТЬ СРЕДНЕГО ЧИЛИ И СОСЕДНИХ
ОБЛАСТЕЙ (131). УМЕРЕННАЯ ЗОНА ЮЖНОГО ЧИЛИ (148). ЖЕСТКОЛИ-
СТНАЯ РАСТИТЕЛЬНОСТЬ КАПСКОЙ’ПРОВИНЦИИ И ЗОНА, ПЕРЕХОДНАЯ
К ОБЛАСТИ КАРРУ (156). ОБЛАСТЬ ЖЕСТКОЛИСТНОЙ РАСТИТЕЛЬНОСТИ
АВСТРАЛИИ (176). УМЕРЕННАЯ ЗОНА И ВЕРТИКАЛЬНАЯ ПОЯСНОСТЬ
РАСТИТЕЛЬНОСТИ В АВСТРАЛИИ, ТАСМАНИИ И НОВОЙ ЗЕЛАНДИИ (206)
ПРОБЛЕМА ПРИСПОСОБЛЕНИЯ РАСТЕНИЙ К ФАКТОРАМ СРЕДЫ. ОБЩИЕ
ЗАМЕЧАНИЯ (222).
Глава 4
ЛИСТВЕННЫЕ ЛЕСА УМЕРЕННОЙ ЗОНЫ СЕВЕРНОГО
ПОЛУШАРИЯ — НЕМОРАЛЬНАЯ ЗОНА
ОБЩАЯ ХАРАКТЕРИСТИКА ЛИСТВЕННЫХ ЛЕСОВ ВОСТОЧНОЙ ЧАСТИ
СЕВЕРНОЙ АМЕРИКИ, ВОСТОЧНОЙ АЗИИ И ЕВРОПЫ (227). ОБЛАСТЬ ПРИ-
АТЛАНТИЧЕСКИХ ВЕРЕЩАТНИКОВ (242). ЛЕС КАК’ЭКОСИСТЕМА, ИЛИ
БИОГЕОЦЕНОЗ, И ЛЕСНОЙ КЛИМАТ (259). БИОМАССА ЛИСТВЕННЫХ ЛЕСОВ
УМЕРЕННОЙ ЗОНЫ (269). ПРОДУКЦИЯ ВЕЩЕСТВА И ЕГО КРУГОВОРОТ
В БИОГЕОЦЕНОЗЕ ЛИСТВЕННОГО ЛЕСА (281). ЭКОЛОГО-ФИЗИОЛОГИЧЕ-
СКИЕ ИССЛЕДОВАНИЯ ШИРОКОЛИСТВЕННЫХ ДЕРЕВЬЕВ (291). ЭКОЛОГО-
ФИЗИОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ ТРАВЯНОГО ПОКРОВА В ЛИСТВЕН-
НЫХ ЛЕСАХ (301). ПОВЕДЕНИЕ РАСТЕНИЙ ЛИСТВЕННЫХ ЛЕСОВ ЗИМОЙ
(320). ПОЯС БУКА В ГОРАХ СРЕДИЗЕМНОМОРСКОЙ ЗОНЫ (331).
Глава 5
ХВОЙНЫЕ ЛЕСА УМЕРЕННОГО ХОЛОДНОГО КЛИМАТА — БОРЕАЛЬНАЯ ЗОНА
БОРЕАЛЬНО-НЕМОРАЛЬНАЯ ЗОНА СМЕШАННЫХ ЛЕСОВ (334). ОСНОВНЫЕ
ВИДЫ ДЕРЕВЬЕВ БОРЕАЛЬНЫХ ХВОЙНЫХ ЛЕСОВ (341). ТИПЫ ЛЕСА
БОРЕАЛЬНОЙ ЛЕСНОЙ ЗОНЫ ЕВРОПЫ (349). БИОГЕОЦЕНОЗЫ ХВОЙНЫХ
ЛЕСОВ (358). ПОЯС ХВОЙНЫХ ЛЕСОВ В ГОРАХ ЦЕНТРАЛЬНОЙ ЕВРОПЫ
(374). ЭКОЛОГО-ФИЗИОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ ХВОЙНЫХ ПОРО Д
(379). ТИПЫ БОЛОТ И ИХ РАСПРОСТРАНЕНИЕ (399). ЭКОЛОГИЯ ВЕРХОВЫХ
БОЛОТ (412).
Г. Вальтер
РАСТИТЕЛЬНОСТЬ
ЗЕМНОГО ШАРА
т. II
Редактор Л. Бабинцева
Мл. редактор М. Бродовская
Художник В. Кузяков
Художественный редактор В. Пузанков
Технический редактор Л. /Барова
Корректор В. Пестова
Сдано в производство 21.9. 1973 г.
Подписано к печати 31.5. 1974 г.
Бумага 70хЮ81/1в-тип. № 1. Бум. л. 131/4
Печ. л. 37,12+0,35 цв, вкл. Уч.-изд. л. 35,25
Изд. № 16950 Цена 2 р. 97 к. Зак. 01248
Издательство «Прогресс» Государственного
комитета Совета Министров СССР по делам издательств,
полиграфии и книжной торговли
Москва, Г-21, Зубовский бульвар, 21
Ордена Трудового Красного знамени
Московская типография № 7 «Искра революции»
Союзполиграфпрома при Государственном комитете
Совета Министров СССР по делам издательств,
полиграфии и книжной торговли
Москва, К-1, Трехпрудный пер., 9