/









































































































































































































































































































































































































































































































































































Похожие




Текст
HEINRICH WALTER
DIE VEGETATION
DER ERDE
in oko-physiologischer
Betrachtung
*
Bd I.
Die tropischen
und
subtropischen Zonen
*
VEB Gustav Fischer Verlag
Jena 1964
Г. ВАЛЬТЕР
РАСТИТЕЛЬНОСТЬ
ЗЕМНОГО ШАРА
Эколого-физиологическая
характеристика
*
Тропические и субтропические зоны
*
Перевод с немецкого
Ю, Я. Ретеюма и И. М. Спич кина
Под редакцией П. Б. Виппера
Издательство «Прогресс»
Москва 1968
Из предисловия автора к первому изданию
Во время многочисленных экспедиций автор имел возможность ознако-
миться со всеми растительными и флористическими царствами (рис. 1), но
человеческая жизнь слишком коротка, чтобы тщательно исследовать все
увиденное. Благодаря современным транспортным средствам земной шар
уже не кажется столь огромным. Именно поэтому возникла настоятельная
необходимость стремиться не только ко все более глубокому анализу, но
и к широкому обобщению имеющихся данных.
Монография «Растительность земного шара» является непосредствен-
ным продолжением книги «Einfiihrung in die Phytologie» («Введение в фито-
логию»), изданной в Штутгарте. Поэтому в тексте автор постоянно ссылает-
ся на материалы, изложенные в этой работе (в ссылках: «Phytologie»).
Много забот доставили автору вопросы номенклатуры. В работе
использованы как данные более старых, так и новых публикаций. Между
тем названия растений часто изменялись. В связи с этим большие трудно-
сти представляли поиски синонимов упомянутых в тексте видов и установ-
ление новейших, официально принятых названий. Поэтому в тексте один
и тот же вид может фигурировать под разными или под устаревшими
названиями.
Г. Вальтер
Штутгарт-Хоэнхейм, 1961 г.
Предисловие автора ко второму изданию
Первое издание книги разошлось уже через год. Читатели встретили
ее с большим интересом. Тем не менее во второе издание мы сочли необхо-
димым внести кое-какие дополнения и сделать некоторую перестановку
материала. В частности, более детально рассмотрен во введении фактор
конкуренции.
Во втором издании, как и в предыдущем, автор поставил перед собой
задачу дать не описание растительности, а сводку экологических, точнее,
эколого-физиологических данных. Начало экспериментальным экологичес-
ким исследованиям было положено в первой половине нашего столетия изу-
чением водного баланса растений; поэтому ученые обратились прежде
всего к аридным районам, в результате последние оказались особенно
детально исследованными.. Важную роль сыграло и то, обстоятельство,
что в крайних условиях существования легче выявить экологические
различия у растений. Этим объясняется и особое внимание, уделяемое
описанию аридных районов в данной книге. Такая установка автора
понятна, особенно если учесть резкие различия в экологии растений
отдельных пустынных’районов, что требует самостоятельного рассмотре-
ния каждого из них.
До настоящего времени при проведении эколого-физиологических
исследований очень мало внимания уделялось тропическим районам
с более или менее продолжительным летним сезоном дождей. Естественно,
что описание таких областей в дащной книге Не могло быть подробным.
Материалов исследований влажных тропиков достаточно много, однако
данных по экспериментальной экологии совершенно недостаточно. Важ-
нейшее значение здесь имеет свет и, следовательно, фотосинтез, для изу-
чения которого мы еще не располагаем простым, пригодным для полевых
исследований методом измерения. Также слабо изучены в тропиках вопро-
сы продуктивности и конкуренции.
Автору высказывались замечания об отсутствии подробного списка
литературы. Однако это сделано вполне сознательно, ибо мы поставили
перед собой задачу дать лишь такое, число работ, которое читатель дей-
ствительно сможет прочитать. Труды, на которые автор ссылается в тексте,
даны в подстрочных примечаниях. В этих работах содержатся более под-
робные указания на соответствующую литературу.
• Автор рассчитывает довести до конца начатую им работу по составле-
нию второго тома. Из числа монографий, посвященных описанию расти-
тельности, одновременно с настоящей книгой выходит в свет труд «Die
Vegetation von Nordamerika und Mittelamerika», редактирование которого
любезно взял на себя профессор-доктор Р. Кнапп.
Пользуюсь случаем принести всем моим сотрудникам глубокую бла-
годарность.
. Г. Вальтер
Штутгарт-Хоэнхейм, 1963 г.
Глава 1
ВВЕДЕНИЕ
РАСТИТЕЛЬНОСТЬ И РАСТИТЕЛЬНЫЕ СООБЩЕСТВА
При изучении растительности объектами исследования являются
не отдельные растения, а растительные сообщества, состоящие из
большего или меньшего числа видов и в совокупности образующие
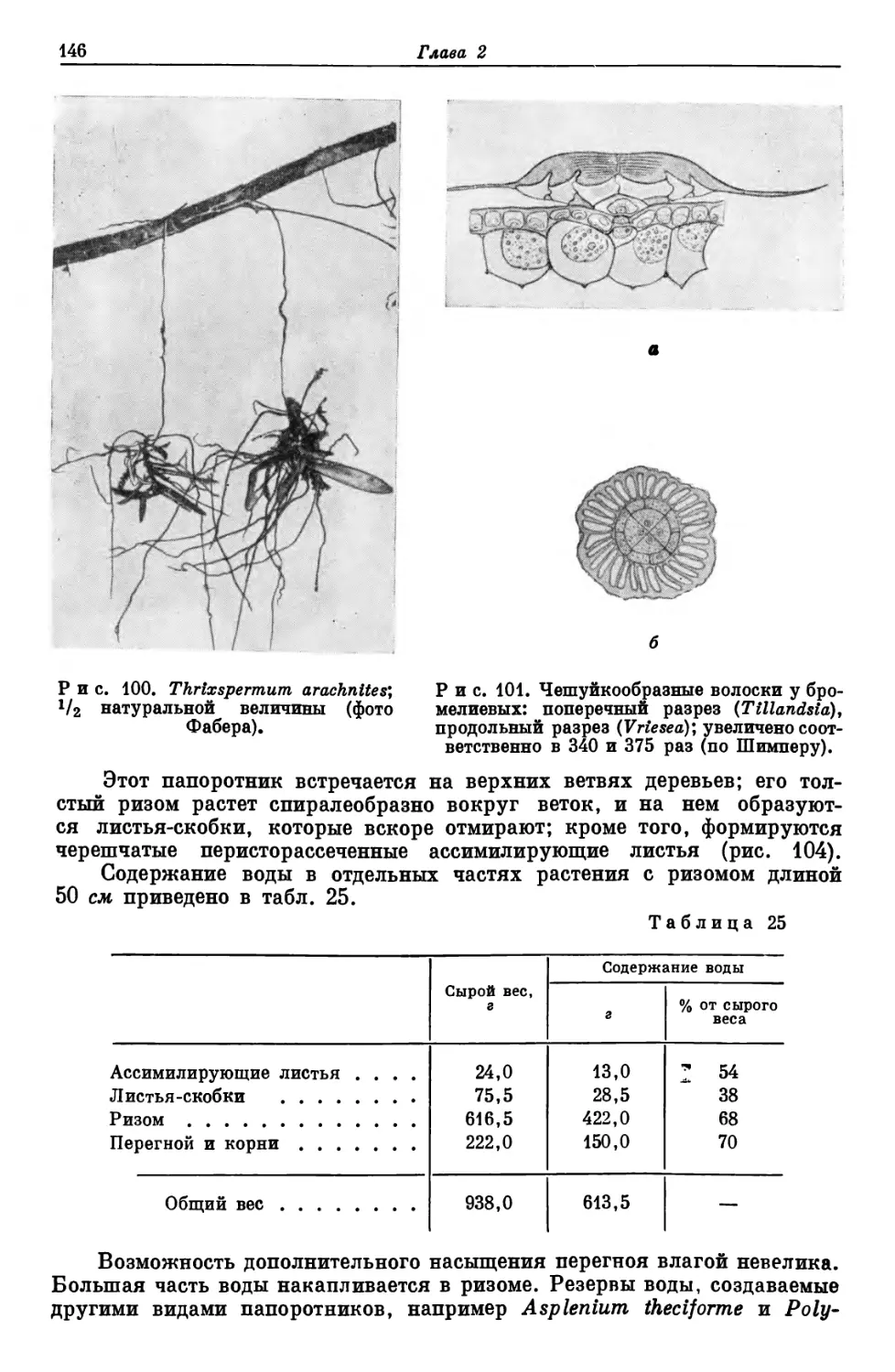
растительный покров, или растительность, данной территории. Естествен-
но возникает вопрос, что же, собственно, представляют собой раститель-
ные сообщества. Ответить на него не так просто. Пожалуй, трудно найти
в ботанической литературе другую проблему, вызвавшую столь оживлен-
ные дискуссии. В этой книге мы не ставим перед собой задачи дать исто-
рический обзор различных мнений, высказанных по этому поводу в разное
время. Разбору теоретических вопросов здесь также не место; мы огра-
ничимся лишь характеристикой растительного покрова земного шара,
изложив прежде наши собственные взгляды, которые предопределяют
подход к изучению растительности. Просматривая соответствующую лите-
ратуру (см. «Phytologie», Bd IV, 1), можно установить существование
двух противоположных точек зрения в вопросе о растительных сооб-
ществах Ч
1. Сторонники первой из них рассматривают растительные сообщества
в качестве фитоценологических единиц и поэтому называют науку о расти-
тельных сообществах «фитосоциологией», или «фитоценологией». В частно-
стях воззрения отдельных ученых могут быть, правда, весьма различными.
Например, если одни рассматривают растительное сообщество как обра-
зование, сходное с организмом, другие видят в нем более сложный ком-
плекс, состоящий из структурных частей фитоценоза более низкого поряд-
ка — синузий.
Некоторые исследователи усматривают наличие четких границ между
отдельными группировками растений, другие же, напротив, считают, что
границы эти неясные и что между различными сообществами существуют
переходные зоны. Фитоценологи ставят перед собой задачу выделить
отдельные сообщества, описать их и объединить в динамическую или
строго иерархическую систему. Прообразом последней являются подраз-
деления, применяемые в систематике растений. Однако в вопросе о прин-
ципах создания такой системы разные ботанические школы к единому
мнению еще не пришли, что вызывало, между прочим, иногда совершенно
бесполезные дискуссии1 2. Сторонниками этой точки зрения являются
1 М. Е. D. Р о о г е. The method of successive approximation in descriptive ecology,
«Advanc. Ecol. Res.», I, 1962, p. 35—68.
2 Точка зрения геоботаников СССР на основные закономерности, свойственные
растительным сообществам, или фитоценозам, в сжатой форме изложена в статье
Е. М. Л а в р е н к о, Основные закономерности растительных сообществ и пути их
изучения, Полевая геоботаника, т. I, изд. АН СССР, М.—Л., 1959.— Прим. ред.
12
Глава 1
Клементс, Уивер, Тенсли, Сукачев, Алехин, Дюрье, Липмаа, Браун-
Бланке, Тюксен, Шарфеттер, Шенников и др. х.
2. Представители другого направления усматривают в отдельных
растениях (индивидуумах) единственно конкретное в природе. В зависи-
мости от местообитания растения объединяются в известные растительные
единства, которые не имеют четких границ. В соответствии с этой точкой
зрения растительный покров представляет собой континуум, иными сло-
вами, он состоит из постоянно изменяющихся сочетаний, с трудом или
вообще не поддающихся разграничению. Этих взглядов придерживаются
Раменский, Негри, Глисон, Кейн, Мейсен, Кёртис, Браун, Уиттеккер,
Пур и др. Математический анализ подтверждает, скорее, именно эту
последнюю точку зрения 1 2.
По мнению автора, реальной действительности больше соответствует
среднее между этими обоими направлениями. Едва ли можно отрицать,
что в природе при сходных условиях местообитания постоянно встречают-
ся одни и те же сочетания видов, мало или почти не отличающиеся одно
от другого, то есть выделяются определенные сообщества. В пределах
небольшого района с различными условиями местообитания их легко
разграничить. Если же растительное сообщество занимает большую пло-
щадь с постепенно изменяющимися климатическими условиями, можно
обнаружить, как исчезают из сообщества одни виды и появляются другие,
пока наконец не возникнет совершенно новая комбинация видов. При
сравнении крайних вариантов легко выделить два разных растительных
сообщества. Если же рассматривать весь ряд, провести четкие границы
между сообществами трудно. В Центральной и Южной Европе, где наибо-
лее ярко выражено фитоценологическое направление, естественные расти-
тельные сообщества, не испытавшие влияния человека, уже не существуют.
Что же касается искусственно созданных человеком лесных, луговых
сообществ и группировок сорняков, то они всегда имеют резко очер-
ченные границы. Сообщества, более близкие к естественным, например
болотные, встречаются в Центральной Европе редко, на большом рас-
стоянии одно от другого, поэтому и в этом случае их границы хорошо
выражены.
Напротив, в районах, заселенных лишь в недавнее время, и причем
очень слабо, например в Юго-Западной Африке, границы между сообще-
ствами весьма расплывчаты 3, что обусловлено исчезновением или появле-
нием отдельных видов. Нельзя предполагать, что при медленном измене-
нии климата одно растительное сообщество внезапно сменяется другим;
обычно переход осуществляется постепенно. Поэтому трудно отрицать
наличие полной взаимонезависимости отдельных видов растений даже
внутри сообщества. Следовательно, как справедливо подчеркивает Эллен-
берг (см. «Phytologie», Bd IV, 1), последней основной классификационной
1 Мы не приводим здесь многочисленных работ по данному вопросу, тем более
что их часто цитируют, например в книге S. А. С a i n, G. М. de Oliveira С a s-
t г о, Manuel of vegetation analysis, New York, 1959 пли R. H. Whittaker, Classi-
fication of natural communites, «The Biol. Review», 28, № 1, 1962 (включает 80 стра-
ниц библиографии); см. также «Phytologie», Bd IV, 1.
2 D. W. Goodall, Objectiv methods for the classification of vegetation, «Austr.
J. Bot.», 9, 1961, p. 162—196; The continuum and individualistic association, «Vegeta-
tion», 11, 1963, p. 297—316.
3 H. Walter, E. Walter, Einige allgemeine Ergebnisse unserer Forschungs-
reise nach Siidwestafrika 1952/53: Das Gesetz der relativen Standortskonstanz; das.
Wesen der Pflanzengemeinschaften, «Вег. Dtsch. Bot. Ges.», 66, 1953, S. 228—236.
Введение
13
Рис. 2. Изменение флористического состава растительного покрова,
обусловленное постепенным изменением климата (пояснения см. в тексте).
•единицей растительных сообществ являются не ассоциации, а виды
растений х.
Если же климат изменяется достаточно быстро, например на горном
склоне, и каждое растительное сообщество образовано одним доминиру-
ющим видом, переход от одного сообщества к другому может осущест-
вляться сравнительно резко. Мы наблюдаем это в горах Европы, где,
например, буковый лес при движении вверх по склону сменяется еловым
лесом. Несмотря на то что климатические условия в горах вдоль склона
изменяются весьма плавно, смена одного сообщества другим происходит
в известной мере скачкообразно. Это объясняется тем, что специфика
растительных сообществ определяется доминирующими видами, и речь
идет, скорее, о смене одних доминирующих видов другими. В горах тропи-
ческих областей и южного полушария, где развиты полидоминантные
леса, выделение вертикальной поясности из-за плавности переходов край-
не затруднено.
Прямая А на рис. 2 характеризует изменение флористического состава
растительного покрова при постепенном изменении климата, когда речь
идет об идеальном континууме. В этом случае различить определенные
растительные сообщества не удается. Другой возможный крайний случай
представлен кривой Г. Растительные сообщества резко разграничены;
изменение растительного покрова происходит скачкообразно, переходные
1 Геоботаники в СССР, как и большинство исследователей за рубежом, за основ-
ную единицу классификации растительных сообществ принимают растительную ассо-
циацию.— Прим. ред.
14
Глава 1
зоны между ними отсутствуют. Однако и этот случай в природе не наблю-
дается. Действительным условиям больше соответствуют кривые В и Б.
На кривой В растительные сообщества еще отчетливо выделяются, но
между ними вклиниваются более или менее широкие переходные полосы.
На рисунке эти полосы занимают небольшое пространство, но они могут
быть и больше площади, занимаемой сообществом. В данном случае пред-
полагается, что флористический состав сообщества в пределах района
своего распространения остается постоянным, даже если климат в изве-
стной мере изменяется. Однако это предположение, по-видимому, далеко
не всегда справедливо, если только климатические влияния не отступают
на задний план вследствие того, что сообщества развиваются в крайних
эдафических условиях местообитания или связаны в своем развитии
с деятельностью человека (сообщества галофитов и сообщества на дюнах,
сообщества луговых и сорных растений). В общем, однако, любое измене-
ние климата отражается и на флористическом составе сообщества. Поэтому
действительным условиям, вероятно, ближе всего отвечает кривая Б.
В зависимости от степени реакции растительных сообществ на изменение
климата и ширины переходных полос выделение определенных сообществ
в каждом отдельном случае либо возможно, либо связано с очень больши-
ми трудностями.
Подчеркивая таким образом взаимонезависимость видов в раститель-
ном сообществе, все же следует признать, что состав ненарушенного расти-
тельного сообщества не является случайным, а обнаруживает известные
закономерные черты.
Под растительным сообществом мы понима-
ем встречающееся в природе более или менее
постоянное сочетание определенных видов
растений, находящихся в экологическом равно-
весии как между собой, так и с окружающей сре-
дой1.
Это равновесие обусловливается:
1. Межвидовой конкуренцией, причем степень конкурентной спо-
собности отдельных видов зависит от их морфолого-физиологических
особенностей и соответствующих условий среды.
2. Известной экологической зависимостью одних видов от других,
что, собственно, и создает возможность их существования в данном расти-
тельном сообществе.
3. Присутствием комплиментарных видов, которые в соответствующих
условиях местообитания дополняют один другого во времени или в про-
странстве или по крайней мере не препятствуют произрастанию других
видов.
Поясним эту мысль на нескольких примерах.
В лиственных лесах Центральной Европы наблюдается интенсивная
конкуренция между основными древесными породами, слагающими древо-
стой,—буком (Fagus silvatica), дубом (Quercus robur и Q. petraea), грабом
(Carpinus betulus). Бук, отличающийся значительной теневыносливостью,
в благоприятных для его роста местообитаниях является наиболее конку-
1 Это определение не распространяется на открытые сообщества, формирующиеся
при заселении растениями новых, свободных территорий. [Вслед за В. Н. Сукачевым
геоботаники СССР под фитоценозом, или растительным сообществом, понимают всякую
конкретную растительность на известном пространстве, однородную по составу, сину-
зиальноп структуре, сложению и характеру взаимодействия между растениями и между
ними и средой.— Ред. ]
Введение
рентоспособной породой, поэтому в горных районах, особенно на извест-
ковых почвах, этот вид господствует в составе лесов. С увеличением тепла
и засушливости климата или, в районах атлантического побережья,
с увеличением влажности на маломощных почвах место бука занимает
дуб. Аналогичная картина наблюдается в затопляемых в течение длитель-
ного времени поймах. Дуб часто растет с примесью граба, широко распро-
странившегося в Центральноей Европе главным образом благодаря воз-
действию человека. Он очень быстро восстанавливается пневой порослью
и поэтому в прошлом широко использовался при ведении низко- и средне-
ствольного хозяйства. Граб более теневынослив, чем дуб, и замещает бук,
как только последний с увеличением континентальности климата достигает
восточной границы своего распространения. Следовательно, в зависимости
от климатических и эдафических условий в лиственных лесах господствует
та или иная древесная порода, либо все три вида развиваются совместно,
образуя смешанные леса.
Аналогичные условия конкуренции существуют и между растениями
напочвенного покрова в лесу, только здесь наряду с другими факторами
среды решающее значение приобретает свет. Условия освещенности почвы
в свою очередь определяются характером древесного полога. Таким обра-
зом, растения напочвенного покрова, хотя и конкурируют между собой,
но в целом зависят от древесного полога, поскольку последний создает
микроклимат — необходимую предпосылку для развития этих растений.
Только в тени лесные растения защищены от конкуренции светолюбивых
видов. Древесный полог не только влияет на освещенность почвы в лесу,
но и на динамику влаги и минеральных веществ в почве. Одновременно
он является гумусообразователем благодаря большому количеству ежегод-
ного опада; гумус создает условия для развития растений, особенно сапро-
фитов, например Neottia nidus-avis в буковом лесу. С другой стороны,
лесная подстилка препятствует развитию мохового покрова.
В еще более тесной зависимости от своих растений-хозяев находятся,
естественно, паразитирующие виды: Lathraea squamaria, виды Melampy-
гит, омелы (Viscum) и др.
Не всегда, однако, верхний ярус занимает господствующее положение
по отношению к нижнему. На верховом болоте, например, господствующая
роль принадлежит сфагновому покрову. Наряду с мхами здесь могут
удержаться только виды, обладающие (на бедном питательными вещества-
ми субстрате) приростом, превышающим ежегодное нарастание мха.
Во влажном океаническом климате мхи растут настолько быстро, что
сосна (Pinus silvestris) не в состоянии с ними конкурировать. В условиях
более континентального климата развитие сфагнума угнетено и верховые
болота все больше и больше зарастают лесом. В конце концов здесь фор-
мируются сфагновые сосняки.
Острая конкуренция существует, однако, не между всеми видами,
входящими в сообщество. Так, комплиментарные виды взаимно дополняют
ДРУГ друга, развиваясь, например, в разное время. В Центральной Европе
во влажных лиственных лесах весной, когда деревья стоят еще без листьев,
первым зацветает весенний чистяк (Ranunculus ficaria). Почти одновре-
менно появляются хохлатки (Corydalis) и Anemone nemorosa. Значительно
позже, когда весенний чистяк уже исчезает, начинают развиваться Мег-
curialis perennis и Asperula odorata, а еще позднее Milium effusum или
Sanicula europaea.
Такую же примечательную последовательность смены аспектов легко
заметить на тучных лугах Центральной Европы. Весной здесь сменяются
16
Глава 1
последовательно Bellis perennis, Тaraxacum officinale, Cardamine pratensis,
Ranunculus acer, Lychnis flos-cuculi, Tragopogon pratensis, Chrysanthemum
leucanthemum, Anthriscus silvestris, Crepis biennis и т. д. Аспект определяет-
ся прежде всего временем цветения, когда большинство Дидов достигает
кульминации в развитии, и характеризуется максимальным покрытием
почвы. В то время как виды, появившиеся раньше, плодоносят, виды,
развившиеся позже, только еще начинают цвести. Растения нижних подъя-
русов должны успеть развиться прежде, чем вырастут виды, образующие
верхний полог травостоя. Такую же характерную смену аспектов можно
обнаружить и в степных районах Восточной Европы, в североамерикан-
ских прериях или саваннах субтропиков. Можно было бы привести
еще множество других примеров.
Во всех этих случаях речь шла о комплиментарных видах, обособлен-
ных во времени своего развития. Однако такая обособленность может
осуществляться и в пространстве. Проявляется она в надземной ярусности
растительности. В лесу различают верхний и нижний древесные ярусы,
кустарниковый ярус, верхний, средний и нижний травяные ярусы
и, наконец, ярус мхов, едва возвышающихся над поверхностью почвы.
В девственном тропическом лесу обычно бывают выражены все перечислен-
ные ярусы, отсутствуют иногда только самые нижние ярусы при условии
недостаточной освещенности почвы.
На пространствах, где развивается только травяной покров, например
на лугах, также выделяются ярусы [подъярусы — в представлении боль-
шинства геоботаников СССР.— Ред.] в зависимости от высоты травостоя.
Помимо надземной ярусности, наблюдается также подземная про-
странственная обособленность корневых систем. В лесу более резко выра-
жена надземная ярусность, в других же сообществах более отчетливым
может быть подземное расчленение; например, на песчаных дюнах Верхне-
Рейнской низменности в типе сообщества Koeleria glauca — Jurinaea мхи
и лишайники распространены только па поверхности песка; корни много-
численных однолетних видов, развивающихся весной, проникают на глу-
бину 5—15 см, большинства многолетних растений — на глубину 20—
60 см и только корни стержнекорневых видов, цветущих осенью, достигают
глубины свыше 150 см. То же можно сказать о степях центральной Анато-
лии и американских прериях (см. «Phytologie», Bd III, 1, S. 273 и след.).
Эта обособленность (в пространстве и во времени) способствует фор-
мированию сообществ с предельно насыщенной надземной и подземной
ярусностью, в которых образующие его виды при данных условиях место-
обитания максимально используют вегетационный период.
Каждое находящееся в равновесии с местообитанием растительное
сообщество увеличивает в пределах своего ареала продуктивность до
известных максимальных величин, соответствующих ресурсам местооби-
тания. Сообщество, так сказать, насыщено видами и отдельными индиви-
дуумами. Это относится даже к сообществам, развивающимся в трещинах
скал или в пустынях.
Отсюда понятно, почему в ненарушенные растительные сообщества
с таким трудом проникают адвентивные виды.
Следовательно, растительное сообщество представляет собой систему,
теснейшим образом связанную с окружающей средой и неотделимую от
нее. Растительность, если не рассматривать ее с чисто описательной точки
зрения, познается экологическими методами. Лишь установив причинные
экологические взаимосвязи, можно глубоко понять растительный покров.
Следует учитывать, что экологическое равновесие — не статическая,
Введение
17
а динамическая система. В пределах каждого сообщества происходит акку-
муляция и трансформация вещества; одни индивидуумы погибают, другие
возникают. Это равновесие не является постоянным, так как условия
местообитания из года в год изменяются. Засушливые годы чередуются
с дождливыми; погода стоит то более теплая, то более холодная. Соответ-
ственно этому изменяется конкурентная способность видов; однако состав
сообщества в целом соответствует неким средним условиям. Стихийные
бедствия могут вызвать сильные опустошения, и обычно проходит немало
времени, пока вновь в сообществе не установится равновесие. В девствен-
ном тропическом лесу такие стадии смен растительных сообществ выделяют-
ся весьма отчетливо, например вдоль путей перемещения тропических
циклонов.
Изменения условий местообитания в определенном направлении при-
водят к перестройке всего сообщества. Если эти изменения продолжаются
длительное время (вследствие понижения уровня грунтовых вод при под-
нятии местности или зарастании водоема либо из-за увеличения мощности
почвенного покрова в результате аккумуляции продуктов выветривания)
и ведут к направленному изменению видового состава сообщества, то
в таком случае наблюдаются смены, или сукцессии, растительных
сообществ (ср. стр. 19).
Основной причиной возникновения производных растительных сооб-
ществ является человек, деятельность ^оторого нарушает равновесие
в сообществах. Постоянное вмешательство человека в ход смены одних
растительных сообществ другими при интенсивном хозяйстве препятству-
ет восстановлению естественного равновесия в сообществах.
Если вмешательство человека постоянно и регулярно, то в этом
случае может установиться равновесие, обусловленное
антропогенным влиянием, иными словами, могут сфор-
мироваться более или менее устойчивые производные растительные сооб-
щества. При интенсивном ведении хозяйства весь растительный покров
практически состоит из таких сообществ, например леса, луга и пашни,
болотные сосновые леса, пастбища, заросли сорняков или воднйх расте-
ний в эвтрофных водоемах Центральной Европы и т. д. Различна лишь
степень изменений, обусловленных влиянием человека (см. «Phytologie»,
Bd IV, 2).
Даже в менее обжитых областях других материков естественный
растительный покров в той или иной мере нарушен вырубкой и выпасом
скота, палами в травяных саваннах, а также подсечно-огневым земледе-
лием в лесных районах.
Территории с истинно девственной растительностью резко сокра-
щаются.
Нужно еще и еще раз отметить, сколь важно — именно для решения
чисто практических вопросов — создавать заповедники не только в живо-
писных, привлекающих туристов районах. К сожалению, опыт показыва-
ет, что общественность и правительства в этих еще девственных уголках
Земли замечают упущение слишком поздно, когда исправить уже ниче-
го нельзя.
Из сказанного о сущности растительных сообществ следует, что их
состав тесно взаимосвязан с местообитанием; под последним с экологи-
ческой точки зрения мы понимаем совокупность физических и химиче-
ских' факторов окружающей среды, воздействующих на растение (см.
«Phytologie», Bd III, 1). Фитоценологи, несомненно, правы, считая, что
видовой состав растительных сообществ лучше всего характеризует
2-1394
18
Глава 1
местообитание, особенно если принять во внимание количественные
соотношения. Это справедливо прежде всего по той причине, что расти-
тельные сообщества в течение продолжительного времени отражают некие
средние условия местообитания, тогда как непосредственная оценка
факторов местообитания характеризует лишь их текущее состояние.
Многолетние наблюдения обычно провести не удается. Однако расти-
тельный покров может служить индикатором местообитания лишь в том
случае, если известны условия местообитания каждого отдельного расти-
тельного сообщества, для чего нужен детальный экологический анализ.
Там, где различия условий местообитания выражены резко, четко
выделяются и разные сообщества, если же условия среды изменяются
постепенно, разграничение сообществ связано с большими трудностями.
Получить представление о характере расчленения растительного
покрова в последнем случае можно лишь при условии, если удается выде-
лить определенное число типов растительных сообществ. Типы расти-
тельных сообществ в нашем представлении объединяют сообщества,
постоянно встречающиеся при известных, часто повторяющихся в одном
районе условиях местообитания Ч Они характеризуются определенным
видовым составом, количественными соотношениями отдельных видов,
а также определенной структурой сообщества, то есть ярусностью. Соот-
ветствующая методика подробно освещена Элленбергом («Phytologie»,
Bd IV, 1, S. 16-66).
Любая типизация, не обоснованная объективными критериями, всег-
да будет несколько субъективной. Удачная типизация требует хорошего
знания района исследований, большою опыта и известного чувства меры.
В зависимости от поставленной цели можно более широко или более узко
трактовать типы растительных сообществ. В Альпах, Средиземноморской
области и Центральной Европе Браун-Бланке с большим мастерством
описал типы сообществ, понимая их сравнительно широко; его искус-
ством объясняется успех работы.
Одновременно Браун-Бланке разработал методику описания и изу-
чения типов растительных сообществ, основанную, правда, только на
количественной оценке; однако она позволяет без особого труда выделять
отдельные растительные сообщества с точностью, достаточной для прак-
тических целей.
С практической точки зрения эта методика не вызывает возражений,
и широкое применение ее весьма желательно. Большинство критиков
отмечают лишь известную догматичность во взглядах сторонников этой
школы, например в представлениях о типах как о неделимых единицах
более высокого порядка, или в попытке создать строго иерархическую
систему, основанную на таксономических принципах.
С экологической точки зрения в построении растительного сообщества
основная роль принадлежит доминирующим видам, или доминантам.
Удаление их приводит к разрушению сообщества. Для характеристики
того или иного сообщества можно использовать также определенные
индикаторные виды, которые часто связаны с особыми условиями место-
обитания и характеризуются узкой экологической амплитудой. Градмап
называет их «ведущими растениями» (Leitpflanzen), Браун-Бланке —
1 Большинство советских геоботаников рассматривают растительную ассоциа-
цию (тип фитоценоза пли тип растительного сообщества) как объедппенпе фитоцено-
зов, сходных по своей синузпальной структуре, с одинаковым составом доминант во
всех основных синузиях, с одинаковыми взаимоотношениями между растениями
и растениями и средой.— Прим. ред.
Введение
19
«характерными видами» (Charakterarten), а Тюксен в последнее время
предложил термин «определяющие виды» (Kennarten). Коль скоро послед-
ние не являются одновременно и доминантами, значение их в построении
сообщества и его экологии невелико. Однако опи могут служить хорошими
индикаторами, хотя далеко не повсеместно.
Лишь учитывая то обстоятельство, что леса Центральной Европы
подвергаются интенсивному лесохозяйственному воздействию и особенно
нарушению состава древостоя, можно понять, почему фитоценологи при
характеристике этих лесов придают столь большое значение характерным
видам травяного покрова. Наоборот, в странах, где практикуется пастьба
скота в лесу, больше всего нарушен травостой. Следовательно, в есте-
ственных условиях главную роль при классификации растительных
сообществ следует отводить господствующему древесному ярусу; травя-
ной и моховой покровы будут иметь значение лишь при выделении более
мелких классификационных единиц.
При изучении растительного покрова конкретного участка необходи-
мо выделять сначала крупные классификационные единицы, а затем, по
мере их изучения, переходить к более мелким. Проблема «с чего начи-
нать» всецело зависит от исследуемого объекта, иными словами, от того
пространства, которое избрано в качестве исходного. В нашем случае
это весь земной шар, так что начинать нужно с выделения самых крупных
единиц. В других случаях это может бткггь материк, например Северная
Америка, или территория меньших размеров, скажем Апеннинский полу-
остров, либо только горный хребет, как Шварцвальд. Иногда же речь идет
о совсем небольших площадях — об участке леса, болота или луга. Всег-
да следует идти от общего к частному. Лишь в этом случае можно полу-
чить представление о растительности конкретного пространства. Сколь
дробным должно быть при этом расчленение растительного покрова,
зависит от цели, которую исследователь поставил перед собой. Так, для
решения практических вопросов лесного хозяйства и луговодства целесо-
образно подразделять растительный покров на очень мелкие единицы;
какой-либо определенной низшей классификационной единицы при этом
не существует Е
Учение о растительном покрове — это не только раздел геоботаники,
но и пограничная с географией область знания1 2 . Поэтому при исследо-
вании растительности, если хотят получить удовлетворительное пред-
ставление о растительном покрове определенного района, важно учиты-
вать и географические факторы. Решению поставленной задачи мало
поможет как составление списка типичных растительных сообществ,
в последнее время практикуемое фитоценологами, так и перечень всех
встречающихся в районе видов.
СУКЦЕССИИ, КЛИМАКС И ЗОН АЛЪНА И РАСТ ИТЕЛЬНОСТЬ
Выше уже отмечалось, что растительные сообщества находятся
в динамическом равновесии с окружающей их средой. При определенных
условиях эта взаимосвязь может, однако, изменяться в известном напра-
влении: одни виды будут исчезать, другие появляться. Со временем
формируется совершенно новое растительное сообщество. Такую после-
1 См. примечание, стр. 13.— Прим. ред.
2 Ср. J. Schmithiise n, Allgemeine Vegetationsgeographie, Berlin, 1959.
[Имеется русский перевод второго издания этой книги: И. Ш м и т х ю з е н, Общая
география растительности, Изд-во «Прогресс», М., 1966.— Ред. ]
2*
20
Глава 1
довательную смену различных сообществ в одних и тех же условиях
произрастания называют сукцессиями. Если при этом смены
стимулируются жизнедеятельностью самой растительности, говорят об
автогенных сукцессиях, если же основной причиной является
изменение условий среды, речь идет об аллогенных сукцессиях х.
Учение о сукцессиях было разработано в Северной Америке Каулсом.
Развитое Клементсом, оно было положено им в основу динамического
подхода к изучению растительности. Клементс исходил из предположения,
что пионерные сообщества на свободных от растений участках воды,
скал, песка или засоленных почв представляют начальные стадии так
называемых сукцессионных серий, конечной стадией которых является
растительное сообщество климакса. Последний, по Клементсу, зависит
исключительно от климатических условий данного района. За исключени-
ем этого, характерного для определенной территории климакса, йсе
другие растительные сообщества, согласно Клементсу, следует рассма-
тривать лишь как временные стадии различных серий. Сукцессию расти-
тельного сообщества, развивающегося в водной среде, Клементс называет
гидросерией, на каменистых почвах — ксеросерией, на засоленных поч-
вах — галосерией.
Первичные сукцессии, по Клементсу, являются преимущественно
автогенными. Тенсли и Браун-Бланке также использовали в своих работах
этот метод динамического подхода к изучению растительности, правда,
последний автор во втором издании своей работы (Braun-Blanquet,
Pflanzensoziologie, 2 Aufl.) уже с известными ограничениями.
Однако исторические данные не подтверждают справедливости изло-
женных взглядов, ибо заселение земной поверхности началось уже
несколько сот миллионов лет назад. Дальнейшая эволюция была связана
с развитием новых форм и групп растений (папоротникообразные, голо-
семенные, покрытосеменные) и изменением климатических условий.
Широкое заселение обнаженных субстратов в северном полушарии, покры-
тых в четвертичном периоде ледниками, началось только после таяния
льда. Однако изменения растительности в послеледниковый период отнюдь
не представляют собой автогенных первичных сукцессий; они обуслов-
лены миграциями различных видов из их ледниковых убежищ и колеба-
ниями климата (см. «Phytologie», Bd III, 2).
Определенные стадии первичных сукцессий можно наблюдать даже
в настоящее время, например там, где сохранялись в ненарушенном виде
отложения отдельных фаз, скажем при заилении озер. Устанавли-
ваются они также в районах распространения дюн или засоленных почв
при сравнении со старыми картами 1 2. Эти стадии могут быть выделены
и в горах при длительных наблюдениях за зарастанием осыпей. Однако
все эти процессы имеют ограниченное, местное значение и протекают
чрезвычайно медленно. В большинстве случаев они, по-видимому, явля-
ются не автогенными, а аллогенными сукцессиями. Заиление озера про-
исходит относительно быстро только там, где, как, например, в устье
рек, наблюдается интенсивная аккумуляция осадков. На песчаных побе-
режьях морей сомкнутый растительный покров образуется только в уда-
1 Вслед за В. Н. Сукачевым большинство геоботаников СССР различают сле-
дующие формы динамики растительного покрова (сукцессии): а) сингенетические сук-
цессии; б) эндоэкогенетические сукцессии; в) экзоэкогенетпческие сукцессии.—
Прим. ред.
2 Ср., например, V. J. Chapman, Studies in salt marsh ecology, IX Changes
in salt marsh vegetation at Scolt Head Island, «J. Ecol.», 47, 1959, p. 619—639.
Введение
21
ленных от берега местах, где ослабевает сила ветра и менее интенсивно
перемещается песок. Аналогично этому зарастание осыпей возможно
только в случае сокращения поступлений обломочного материала. Таким
образом, во всех упомянутых случаях сначала происходит изменение усло-
вий местообитания, а затем уже развитие растительного покрова в новых
условиях среды.
Учение о сукцессиях обычно основывается на предположении о воз-
можности по взаимному расположению растительных сообществ в про-
странстве судить об их последовательности во времени. Однако такой
вывод допустим только при наличии прямых доказательств о последова-
тельности смен растительных сообществ.
Вокруг озера всегда наблюдается циклическое размещение расти-
тельных сообществ: непосредственно у воды располагаются гидрофитные
сообщества, сменяющиеся более мезофитными вплоть до лесных сооб-
ществ. Однако о сукцессионных сериях речь может идти только в том
случае, если при изучении почвенных профилей в них будут обнаружены
озерные осадки. Обычно же такая цикличность расположения раститель-
ных сообществ, часто очень устойчивая в пространстве и во времени,
отражает лишь изменения экологических условий, связанные с разным
уровнем залегания грунтовых вод.
Доказать наличие сукцессий в ксеро- и галосериях едва ли пред-
ставляется возможным, поскольку несомненные признаки первых стадий
этого процесса могут быть обнаружены крайне редко. Поэтому во всех
этих случаях следовало бы говорить только об экологических рядах,
связанных с изменением одного из факторов местообитания (увеличение
мощности слоя почвы на каменистых участках, уменьшение засолен-
ности почвы и т. д.).
Значительно более важную роль играют вторичные сукцес-
сии. Они всегда наблюдаются там, где растительный покров внезапно
полностью или частично уничтожается и развитие растительности начи-
нается на более или менее обнаженной почве, например после лесного
пожара, ветровала, оползня, половодья и т. д. Аналогичные явления про-
исходят и под влиянием воздействий человека на среду или после пре-
кращения ранее имевших известное значение влияний, будь то на осу-
шенном болоте, на отделенных дамбами от моря засоленных болотистых
низинах, на пашнях или на не скашиваемых больше лугах. Такие сук-
цессии обычно протекают быстро, смена растительности прослеживается
из года в год. При этом безразлично, произошло ли нарушение раститель-
ности под влиянием естественных причин или человека. Последнее
в настоящее время является наиболее частой причиной нарушений даже
в слабо населенных местностях (например, выжигание травяного
покрова и т. д.).
Возражения вызывает не только учение о сукцессиях в его крайнем
выражении, но и представление о климаксе, как о конечной стадии сукцес-
сионных серий, определяемой только климатом. Дело в том, что расти-
тельность взаимосвязана с местообитанием, а основные факторы место-
обитания (температура, свет, водный фактор, химические и механические
факторы) в свою очередь зависят как от климата, так и от рельефа и почвы.
При резко расчлененном рельефе и в условиях, где развиваются крайние
варианты почв, влияние климата, характерного для соответствующего
природного района, проявляется не столь сильно. Зависимость расти-
тельности от климата поэтому хорошо выражена лишь на равнинных тер-
риториях, где растения используют только атмосферные осадки, и на
22
Глава 1
почвах среднего механического состава, где не наблюдается ни застоя
влаги, ни ее быстрой инфильтрации. Кроме того, почва должна быть не
слишком бедна и не слишком богата питательными веществами. При
таких условиях в каждой климатической зоне развивается определенная,
соответствующая ей растительность, которая называется зональной.
Это понятие применяется при характеристике крупных единиц расти-
тельного покрова, которые в естественных условиях, не испытавших вли-
яния человека, покрывают большую часть земной поверхности. Посте-
пенное, обусловленное климатом изменение зональной растительности
особенно легко заметить, пересекая материк на самолете, если при этом
путь пролегает не над культурным ландшафтом. Бросается в глаза, что
в крайних условиях обитания (песчаные дюны, солончаки, болота, участки
скал), местами занимающих обширные площади, характер растительности
не соответствует облику зональной растительности данной территории.
В этом случае растительный покров в большей мере обусловлен почвен-
ными условиями, нежели климатом, соответствующим данной зоне, и речь
уже идет об азональной растительности. Правда, послед-
няя все же в известной мере зависит от климата. Например, растительные
сообщества дюн умеренной, субтропической и тропической зон, внешне,
казалось бы, сходные, в флористическом отношении весьма различны.
Это относится также к сообществам галофитов или водных растений.
Нередки случаи, когда зональная растительность встречается
за пределами своей зоны, на местообитаниях, особенно для нее благоприят-
ных; так, лес проникает по влажным долинам рек глубоко в степную
зону, а степная растительность распространяется пятнами на сухих
и теплых местообитаниях в пределах лесной зоны; здесь также речь
идет о сообществах заключительных стадий сукцессионного процесса,
которые, однако, не соответствуют зональной растительности и представ-
ляют уже собой так называемую экстразона л ьную расти-
тельность. Для существования последней необходимо, чтобы изме-
нения климатических условий компенсировались бы большим или мень-
шим постоянством условий местообитания. Речь идет о законе
относительного постоянства местообитания,
к которому мы еще вернемся в гл. 7 (ср. «Phytologie», Bd III, 2, S. 41 )E
Если, например, развитые на обширных равнинах Северной Европы
зональные хвойные леса распространены на границе арктической обла-
сти только на теплых, солнечных южных склонах или в более южных
широтах лишь на холодных северных склонах либо в глубоких ущельях,
то это означает, что температурные условия на всех трех позициях
сравнительно сходны. Точно так же восточноевропейские степи в Цен-
тральной Европе встречаются экстразонально только на солнечных
склонах с маломощными, обычно водопроницаемыми карбонатными почва-
ми. Большее по сравнению с Восточной Европой количество осадков
компенсируется здесь повышенным испарением и меньшим запасом влаги
в почве; это свидетельствует о сходстве условий увлажнения в Централь-
ной Европе и в области распространения сухих восточноевропейских
степей.
Понятие «зональная растительность» более или менее соответствует
понятию «климакс» в узком значении этого термина, особенно если в пос-
ледний не включать неприемлемое в его крайнем выражении представление
1 В геоботанической литературе СССР эта закономерность известна как «пра-
вило предварения» (В. В. Алехин), хотя ему и не придается абсолютного значения.—
Прим. ред.
Введение
23
о сукцессиях. Все попытки сохранить представления о климаксе при помо-
щи теории поликлимакса или таких определений, как «группа климакса»
или «рой климакса», не могут считаться удовлетворительными, так как
они все основываются на концепции первичных сукцессий. Вполне оправ-
дан известный динамический подход к изучению растительности. При
этом важно лишь не терять чувства реальности и не допускать умозри-
тельных выводов.
С большими трудностями сопряжено выявление зональной расти-
тельности в районах, где растительный покров в течение столетий и тыся-
челетий изменялся под воздействием человека. Если никаких следов
естественной растительности на данной территории не сохранилось, сле-
дует обратиться к историческим материалам. Однако и в этом случае
может быть выявлен только общий харатер зональной растительности.
Например, зональной растительностью Центральной Европы были
смешанные лиственные леса с господством бука (Fagus silvaticay В районах
с хорошо выраженным океаническим климатом бук уступил место дубу,
а в областях с менее обильными осадками, по-видимому, был вытес-
нен грабом.
Понятие «зональная растительность» следовало бы использовать
только при описании обширных территорий, например для подразделения
естественной растительности целых материков. Только в этом случае
четко выделятся климатические факторы, а различия, обусловленные
характером почв, отступят на второй план. Однако при более детальных
исследованиях нельзя не учитывать эдафических условий и экспозицию.
В горах климат с высотой изменяется гораздо быстрее, чем на равни-
нах. Поэтому роль климата при расчленении растительного покрова
проявляется в горах на более коротких расстояниях и значительно более
резко, чем на равнинах.
Выделяемые при подъеме в горы подразделения растительного
покрова, соответствующие зональной растительности на равнинах, мы
называем «вертикальными поясами». Растительный покров каждого пояса
следовало бы изучать только в плакорных условиях, поскольку на скло-
нах экспозиция и наклон местности влияют не только на температуру,
но и на гидратурные соотношения, как это наблюдается и в случае экстра-
зональной растительности. Выполнить это требование часто очень трудно,
особенно в горах с резко расчлененным рельефом и крутыми склонами.
В этих условиях следует сравнивать положение границ растительности
на склонах северной и южной экспозиции; среднее между двумя значе-
ниями обычно будет соответствовать пределу на плакорных поверхностях.
Можно также придерживаться данных, получаемых при выделении верти-
кальных поясов отдельно для каждого склона. Последнее обязательно
в том случае, когда по гребню хребта проходит климатическая граница,
что лишает возможности сравнивать растительность на противополож-
ных склонах х.
КОНКУРЕНТНЫЕ ВЗАИМООТНОШЕНИЯ
Выше была дана характеристика конкуренции как очень важного
фактора, определяющего формирование растительных сообществ. На это
еще указывали Клементс, Уивер и Хэнсон (1929 г.). Однако до сего вре-
1 Н. Walter, Klimax und zonale Vegetation. Angewandte Pflanzensoziologie,
Festschr. Aichinger. I, 1954, S. 144—150.
24
Глава 1
мени конкуренции не придавалось должного значения ни в экологии,
ни в учении о растительном покрове.
Конкуренция наблюдается всюду, где на ограниченном пространстве
два или несколько растений оспаривают друг у друга свет, почвенную
влагу и питание. Взаимное влияние, оказываемое конкурентами, имеет
чисто физико-химический характер1. Играет ли в есте-
ственных условиях какую-нибудь роль также выделение особых веществ,
пока не установлено. У гетеротрофных микроорганизмов симбиоз (позво-
ляющий им получать готовое органическое вещество), несомненно, имеет
известное значение. Напротив, автотрофные растения приспособлены
к непосредственному усвоению углерода из углекислоты. Поэтому
аллелопатические взаимовлияния растений, вероятно, проявляются
в выделении ими определенных ингибиторов, которые действуют даже
в весьма малых концентрациях. Возможность такого воздействия пока
удалось доказать лишь лабораторными опытами, главным образом при
изучении растений, никогда не растущих совместно в природе. Речь идет
о корневых выделениях или о веществах, диффундирующих из отмерших
частей растения либо получаемых путем экстракции 1 2. Поскольку между
корневыми системами различных видов, входящих в состав раститель-
ного сообщества, существует тесный контакт, такое непосредственное
воздействие вполне возможно. Правда, Мартин показал, что неповрежден-
ные корни почти ничего не выделяют, поврежденные же, несомненно,
выделяют какие-то вещества. Однако в почве наблюдается быстрый распад
этих веществ (в данном случае речь шла о скополетине). Альгрен и Аамод
установили, что луговые виды в смешанных посевах дают меньшее коли-
чество сухой массы, чем в чистых, и объясняют это токсическим влиянием
корневых выделений; однако Дональд не подтверждает их выводов.
Он выращивал почти все те же виды в 27 различных вариантах, однако взаим-
ной инактивации ни в одном случае не установил 3. Некоторые вещества
попадают в почву при разложении опада; кроме того, в жаркие безветрен-
ные дни в приземном слое воздуха в сомкнутых группировках растений
может накапливаться значительное количество эфирных масел. Тем
не менее прямых доказательств влияния таких веществ на взаимоотношения
между растениями в сообществах еще не получено и вопрос пока при-
ходится оставлять открытым.
1 F. Е. Clements, J. Е. Weaver, Н. С. Hanson, Plant competition,
«Publ. Carnegie Inst, of Washington», № 398, 1929; J. E. Weaver, F. E. Cle-
ments, Plant Ecologie, 1938; E. Schmid, Kausale Vegetationsforschung, Ber.
Geobot. -Forschungsinstitut Riibel f. 1943, 1944; P. В о у s e n-J e n s e n, Causal
Plant Ecology, «Danske Vidensk. Selskab. Biol. Medd.», 21, № 3, 1949; H. Ellen-
b e r g, IJber einige Fortschritte der kausalen Vegetationskunde, «Vegetatio», 5—6,
1954, S. 199—211.
2 R. Knapp, Experimentelle Soziologie der hoheren Pflanzen, Stuttgart, 1955;
G. Grummet, Die gegenseitige Beeinflussung hbherer Pflanzen. Allelopatie, Jena,
1955; B.Rademacher, Gegenseitige Beeinflussung hoherer Pflanzen, Hub. d. Pflan-
zenphys., 11, 1959, S. 655—706.
3 H. L. A h 1 g r e n, 0. S. A a m о d t, Harmful root interactions as a possible
explanation for effects noted between various species of grasses and legumes, «J. Amer.
Soc. Agron.», 31, 1939, p. 982; С. M. D о n a 1 d. Competition between pasture species,
with reference to the hypothesis of harmful root interactions, I, «Comm. Sc. Ind. Res.»,
19, 1946, p. 32—37; P. Martin, Die A.bgabe von organischen Verbindungen, insbe-
sondere von Scopoletin aus den Keimwurzeln des-Hafers, «Z. Bbt.», 45, 1957, S. 475—
506; H. Borner, Excretion of organic compounds from higher plants and its role
in the soil-sicknes problem, «Bot. Rev.», 26, 1960, p. 393—424; F. К о 1 b, Experimen-
telle Untersuchungen zur gegenseitigen Beeinflussung von Kulturpflanzen und Unkrau-
tern, Diss. Hohenheim, 1961.
Введение
25>
Так, например, отсутствие возобновления у Araucaria cunninghamii
при наличии в подросте Backhousia в лесах южного Квинсленда объяс-
няется токсическим действием дигидроангустиона, являющегося главной
составной частью масла, содержащегося в листьях Backhousia. При
концентрации в субстрате, превышающей 1%, это соединение в лабора-
торных опытах препятствовало прорастанию семян Araucaria. Полевые
же опыты показали, что добавление в подстилку листьев Backhousia
не только не задерживало прорастание семян Araucaria, но явно стимулиро-
вало его, хотя концентрация дигидроангустиона составляла 1—2%
Борьба за свет, влагу и питательные вещества проявляется тем рез-
че, чем большее число экземпляров растений встречается на ограниченном
участке и чем крупнее сами экземпляры. Всходы могут расти очень густо,
не мешая друг другу. С увеличением размеров растений конкуренция
между ними все более обостряется. Экземпляры, растущие быстрее
и достигающие большей высоты, раскрывают листья на полном свету.
Они вырабатывают больше органических веществ, чем растения, разви-
вающиеся в их тени. Благодаря этому завоеванное ими преимущество
все время увеличивается. Корневая система высокорослых растений
отличается более быстрым развитием, в связи с чем эти растения лучше
снабжаются водой и минеральными питательными веществами. Угнетен-
ные экземпляры все более отстают в своем развитии и при недостатке
питания погибают, часто не успев зацвести и принести плоды.
В борьбе за свет участвуют только растения, входящие в состав одного
яруса. На ранних стадиях развития подрост древесных пород может кон-
курировать с травостоем; нередко последний угнетает подрост деревьев.
Однако подросшим молодым деревьям, выдержавшим борьбу, затенение
уже не. угрожает. Деревья занимают теперь господствующее положение,
и травам приходится расти либо в тени деревьев, либо они погибают.
Нижние ярусы растительности больше всего страдают от недостатка све-
та. В случае несомкнутости древесного и кустарникового ярусов травя-
ной покров вступает с ними в борьбу за воду и питательные вещества
в почве. В саваннах, например, господствующее положение принадлежит
травяному покрову, а древесным растениям приходится довольствоваться
тем количеством воды, которое остается не использованным травостоем
в конце дождливого периода (ср. гл. 6, стр. 289). Мхи {Sphagnum),
способствующие заболачиванию лесов, могут даже вызвать отмирание
деревьев путем повышения влажности почвы, что в свою очередь препят-
ствует дыханию корней древесных пород.
Следует различать два типа конкуренции: 1) между особями одного
вида, так называемую внутривидовую конкуренцию,
и 2) между особями разных видов — межвидовую конкурен-
цию. Эффект в том и другом случае различный. В сельском и лесном
хозяйствах внутривидовая конкуренция играет большую роль в однови-
довых культурах. При чрезмерно большом числе одновременно подраста-
ющих на пашне растений отдельные их экземпляры настолько страдают
от острой конкуренции, что урожай резко падает,. При широкорядном
посеве или посадке, когда конкуренция почти отсутствует, каждое расте-
ние развивается хорошо, но число растений на единицу площади невели-
1 J. R. Cannon, N. Н. Corbett, К. Р. Н ay dock, J. G. Т г а с к е у,
L. J. Webb, An investigation of the effect of the Dehydroangustione present in the
leaf litter of Backhousia angustifolia on the germination of Araucaria cunninghamii —
an experimental approach to a problem in rain-forest ecology, «Austr. J. Bot.», 10, 1962,
p. 119-128.
26
Глава 1
ко; поэтому максимального урожая получить не удается. Кроме того,
возникает опасность зарастания посевов сорняками. Для каждой посев-
ной культуры в определенных климатических условиях эмпирически
установлены нормы посева и посадки, соблюдение которых позволяет
•снимать наиболее высокие урожаи. Лесовод также создает вначале густые
посадки, так как это способствует очищению стволов от сучьев, а затем
проводит своевременные рубки ухода, тем самым не допуская ослабления
насаждения в целом вследствие конкуренции со стороны отдельных
экземпляров.
Чистые заросли одного вида встречаются и в естественных условиях,
например на гарях в лесах бореальной зоны при наличии хорошего
обсеменения сосны, или на засоленных почвах илистых участков морского
побережья, где появляются тысячи всходов солероса (Salicornia herbaced),
или в западинах пустынь, куда после дождя смываются семена одного
вида и где они затем прорастают, или, наконец, во влажных долинных
лесах, где часто встречаются крупные пятна Impatiens noli-tangere.
Однако эти природные заросли никогда не бывают столь чистыми, как
искусственные, и в них вскоре начинается процесс самоизреживания:
наиболее сильные особи растут, цветут и плодоносят, а слабые погибают.
Следовательно, внутривидовая конкуренция способствует сохранению
вида, причем густые заросли даже защищают вид от конкуренции
со стороны других видов.
При создании защитных насаждений в степях Восточной Европы
наблюдалось следующее. При одиночном посеве желудей травяной покров
заглушает молодые всходы дуба, поскольку они не выдерживают конку-
ренции с травостоем. При посеве же нескольких желудей в гнездо молодой
подрост дуба растет в сомкнутых группах. Расположенные на внешней
стороне гнезда экземпляры вступают в конкуренцию с окружающим
гнездо травяным покровом и подавляются им. Расположенный же в сере-
дине гнезда экземпляр менее страдает от конкуренции с травостоем. Он
растет, поднимается над травяным покровом, развивает мощную корне-
вую систему и со временем превращается в дерево Ч
Совершенно по-иному проявляется межвидовая конкуренция. Для
отдельных видов это настоящая борьба за существование. Вид, выходя-
щий из нее победителем, обильно плодоносит и возобновляется. Угнетен-
ный вид, напротив, либо погибает, либо остается бесплодным, либо плодо-
носит очень слабо, так что из года в год число особей этого вида убывает.
Правда, в длительное время существующих растительных сообществах
со временем во взаимоотношениях между отдельными видами устанавли-
вается известное равновесие. В этом случае несколько господствующих
видов, находящихся в определенных соотношениях, могут участвовать
в построении соответствующего яруса; ими могут быть, например, либо
несколько видов древесных пород в лиственном лесу, либо виды листвен-
ных и хвойных пород в смешанном лесу, либо злаки с видами разнотравья
на лугу или в степи.
Ожесточенная конкуренция между этими видами проявляется, с одной
стороны, в том, что малейшее изменение условий местообитания влечет
за собой изменение количественных соотношений между ними, а с дру-
гой — в том, что одиночно растущие экземпляры растений развиваются
лучше всего на грядках, тогда как в пределах растительного сообщества
1 Е. Г. Орленко, О взапмоотношениях дуба в густых культурах, «Доклады
АН СССР», 102, № 4, 1955, стр. 841-844.
Введение
27
они страдают от недостатка питания. Это еще раз подтверждает тот факт,
что растительное сообщество максимально использует все материальные
ресурсы местообитания, не оставляя свободного про-
странства для видов, чуждых данному сообще-
ств у. Отсюда понятна чрезвычайная стабильность ненарушенных
растительных сообществ.
Факторы местообитания имеют решающее значение в конкурентных
взаимоотношениях между растениями; в зависимости от характера сово-
купного влияния этих факторов и их интенсивности они могут способ-
ствовать росту либо одного, либо другого вида. Засушливые периоды
задерживают развитие видов, требующих обильного увлажнения,
в большей степени, чем засухоустойчивых; заморозки отрицательно вли-
яют на чувствительные к низким температурам вечнозеленые виды, что
часто приводит к их гибели и замене более морозостойкими. Изменение
факторов местообитания в естественных растительных сообществах редко
вызывает непосредственную гибель какого-либо вида; обычно это про-
исходит косвенным путем: путем снижения конкурентной способности
одного вида по сравнению с другими.
Определенный фактор, например поздние заморозки, может по-раз-
ному влиять на определенный вид, поскольку степень его выраженности
зависит также от конкуренции других видов. В одном случае ослаблен-
ный заморозками вид может не устоять в борьбе с сильным конкурентом
и погибает, в другом — он, хотя и повреждается заморозками, однако
выживает. Отсюда ясно, с какой осторожностью нужно подходить к выяв-
лению прямых связей между областью распространения вида и определен-
ными факторами местообитания, тем более что эти связи почти всегда
бывают косвенными.
Это положение иллюстрируется результатами экспериментальных
исследований, проведенных в Хоэнхейме по изучению изменений уровня
грунтовых вод («Phytologie», Bd III, 1, S. 20; Bd IV, 1, S. 115 и след.).
Alopecurus pratensis, Arrhenatherum elatius и Bromus erectus господ-
ствуют в составе луговых сообществ Центральной Европы; первый домини-
рует на влажных лугах, второй — на умеренно увлажненных и третий —
на сухих. Если же их культивировать в специальных экотронах, где
можно контролировать уровень грунтовых вод в пределах от 0 до 150 см
(рис. 3), то в чистых культурах все три вида лучше всего растут при
среднем уровне стояния грунтовых вод. Только смешанные посевы могут
выявить различия в экологии этих видов (рис. 4). Arrhenatherum, обладая
наиболее высокой конкурентной способностью, лучше всего развивается
при среднем стоянии уровня грунтовых вод и вытесняет Alopecurus на
влажные, a Bromus, особенно на почвах, бедных азотом,— на самые
сухие местообитания Ч Со временем опытные участки начинают зарастать
сорняками, семена которых распределяются по поверхности площади
совершенно случайно. В результате конкуренции с упомянутыми видами
злаков и между собой одни виды сорных растений — в соответствии
с характером их распространения на лугах — сохраняются только на
влажной части опытных участков, другие — на умеренно влажной или
сухой 1 2. Характер распространения некоторых видов сорных растений
не связан с положением уровня грунтовых вод.
1 Н. Е ] 1 е nberg, Uber einige Fortschritte der kausalen Vegetationskunde,
«Vegetation», 5/6, 1954, S. 199—211.
2 H. Li e t h, Konkurrenz und Zuwanderung von Wiesenpflanzen, Zschr. f. Acker-
*u. Pflanzenbau, 106, 1958, S. 205—223.
28
Глава 1
Приток
Смешанный посей
Arrhenatherum elatius
Alopecurus pratensis
Dactylis glomerate
Bromus erectus
Bromus erectus
£
ё-
Dactylis glomerata
Alopecurus pratensis
Arrhenatherum elatius
-j
С
В
Смешанный посей
1
I
iiiiiiHiiHiiiHiiimiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiminiiiiiiiunniMiniinniiiniiuimnimiiHHiihiiiHrniiHni
Рис. 3. Продуктивность луговых злаков при постоянном уровне грун-
товых вод.
А — вид экотрона сбоку; Б — вид сверху; на нижнем рисунке приведена схема
опыта, проводившегося в 1953 г. в Хоэнхейме.
Также косвенно влияет на растения и кислотность почвы; поэтому
не удивительно, что вид, встречающийся при наличии острой конкурен-
ции на почвах с узким пределом колебаний pH, при отсутствии конкурен-
ции теряет свои качества индикатора и слабо реагирует на изменение
почвенных условий х.
Таким образом, у растений следует различать физиологический и эко-
логический оптимумы произрастания, первый может быть выявлен при
отсутствии конкуренции, второй связан с наличием конкурентных взаимо-
отношений 1 2. Экологический оптимум наблюдается в условиях место-
обитания, где вид отличается высокой встречаемостью и быстрым ростом.
Экологический оптимум обусловлен не только местообитанием, но и под-
1Н. Ellenberg, Kausale Pflanzensoziologie auf physiologischer Grundlage,
Ber. Dtsch. Bot. Ges., 63, 1950, S. 24—31.
2 В геоботанической литературе СССР физиологическому оптимуму Г. Вальтера
соответствует представление об экологическом оптимуме, а экологическому Г. Валь-
тера — фитоценотический, или биоценотический, оптимум.— Прим, ред.
Введение
29
час, как было показано выше, наличием определенных конкурентов; эко-
логический оптимум может сильно изменяться в зависимости от состава
сообитателей-конкурентов. Так, Hypochoerts radicata развивается в Цен-
тральной Европе только на определенных типах пастбищ. Этот вид был
занесен в Новую Зеландию и, видимо, встретив там меньшую конкурен-
цию со стороны видов местной флоры, распространился почти во всех
группировках растений, вплоть до верхней границы произрастания
деревьев в горах.
Соотношения экологического и физиологического оптимумов про-
израстания растений при различной степени напряженности конкуренции
иллюстируются следующей схемой (рис. 5).
Область распространения вида, растущего в чистых культурах и не
испытывающего конкуренции, в природных условиях сравнительно широ-
ка. Лучше всего растение развивается в условиях физиологического опти-
мума произрастания (рис. 5, Л). При развитии в смешанных культурах
область распространения вида резко сокращается. Высокая встречае-
мость, или, иными словами, экологический оптимум вида в зависимости
от состава конкурентов и степени напряженности конкуренции, смещается
в ту или другую сторону от физиологического оптимума (рис. 5, Б и В).
Вид может характеризоваться даже двумя экологическими оптимумами
(рис. 5, Г и Д). Лишь в случае большей конкурентной способности по
сравнению с другими видами данный вид приобретает господствующее
положение и его экологический и физиологический оптимумы совпадают
(рис. 5, Е); однако область его распространения остается несколько
суженной.
Проиллюстрируем на примере, какие ошибки возможны при оценке
конкурентных взаимоотношений. В штате Миссури (США) Quercus palust-
Р и с. 4. Продуктивность луговых злаков в зависимости от глубины
уровня грунтовых вод.
.1 — в чистых культурах; 2 — в смешанных посевах (Alopecurus pratensis — при
высоком и Hromus erectus — при низком уровне грунтовых вод).
30
Глава 1
ris образует древостои по берегам рек, водоемов и на болотах. На сухих
же возвышенных местообитаниях господствует Quercus rubra. Салливан
и Левитт высказали в связи с этим предположение, что Quercus rubra
отличается большей засухоустойчивостью и легче переносит недостаток
влаги Е Для проверки этого положения с физиологической точки зрения
были изучены молодые дубки обоих видов, выращенные в условиях
нормального и недостаточного увлажнения. Вопреки ожиданию оба вида
Д
Физиологический оптимум
Е Эколого-физиологический
оптимум
Р и с. 5. Основные типы соотношений физиологического и экологического
оптпмумов произрастания растений (Н. Walter, Standortslehre, Stut-
tgart, 1960).
Д — при отсутствии конкуренции; Б — Е — в смешанных культурах. На ордина-
те — продуктивность; на абсциссе — изменение режимов физиологически действую-
щих факторов.
оказались в одинаковой степени засухоустойчивыми. Единственное раз-
личие, как отмечают авторы, состоит в том, что Quercus palustris при обиль-
ном увлажнении растет быстрее, чем Quercus rubra. По нашему мнению,
именно это обстоятельство и объясняет особенности распространения
обоих видов. На влажных местообитаниях Quercus palustris отличается
большей конкурентной мощностью и вытесняет Quercus rubra на более
сухие местообитания, на которых конкурентные взаимоотношения этих
двух видов складываются в пользу последнего. Решающим моментом
в данных условиях местообитания оказывается не засухоустойчивость,
а большая продуктивность.
Конкурентная способность вида — весьма сложное явление. Она
определяется совокупностью морфологических и физиологических свойств
вида, и в частности скоростью прорастания и скоростью роста, характером
строения побегов и корневой системы, ритмом развития и продолжитель-
ностью жизни, способностью к возобновлению и размножению, колпчест-
1 С. Y. Sullivan, J. Levit t, Drought tolerance and avoidance in two
species of oak, «Physiol. Plant.», 12, 1959, p. 299—305.
Введение
31
вом продуцируемых семян и способом их распространения, а также
требованиями к различным факторам местообитания, например темпера-
туре, влажности, свету, химическим факторам и т. д. (ср. «Phytologie»,
Bd IV, 1, S. 120). В следующих разделах мы попытаемся проанализиро-
вать некоторые из этих свойств.
Особую роль у растений, относящихся к одной жизненной форме,
часто играет продуктивность1, поэтому рассмотрим последнюю
несколько пристальнее. Наш обзор мы начнем с продуктивности отдель-
ных растений и перейдем затем к продуктивности целых насаждений.
ПРОДУКТИВНОСТЬ И ПОКАЗАТЕЛЬ ОБЛИСТВЛЕННОСТ И
В процессе фотосинтеза в растениях образуются органические веще-
ства, которые они используют для построения своего организма. При
дыхании, напротив, органические вещества расходуются.
Поэтому физиологи обычно считают, что растения продуцируют тем
больше вещества, чем интенсивнее листья ассимилируют СО2 на единицу
их площади. Такое представление справедливо лишь для какого-либо
определенного момента, но оно не может быть использовано при оценке
продуктивности растения в течение всего вегетационного периода. Послед-
няя зависит в значительной мере от баланса ассимилятов в растении,
иными словами, от характера использования самим растением образовав-
шихся ассимилятов («Phytologie», Bd I, S. 170 и след.; Bd III, 1, S. 399
и след.). Чем больше доля ассимилятов, используемых растением для
построения новых листьев, и чем меньше их доля, расходуемая только на
дыхание, тем больше в конечном итоге будет накоплено сухого вещества.
При этом обнаруживаются известные различия в зависимости от того,
имеем ли мы дело с отдельно стоящими растениями или насаждениями
и посевами 1 2.
Небольшая продуктивность одиночного растения (например, злака)
сразу после прорастания определяется незначительной листовой поверх-
ностью. С увеличением последней возрастает количество накопленного
органического вещества, достигающее максимума непосредственно перед
цветением, пока нижние листья еще не пожелтели и органы размножения
не использовали в процессе дыхания значительной части ассимилятов.
При обильном удобрении азотом (N) продуктивность, то есть урожай,
будет больше. Такеда доказал, что величина урожая зависит главным
образом от площади листьев. Восьмикратный рост урожая достигается
при пятикратном увеличении листовой поверхности, тогда как интенсив-
ность фотосинтеза возрастает при этом всего в полтора раза. Содержание
азота в листьях и соответственно интенсивность их дыхания увеличивают-
ся при этом в шесть раз.
Листья отдельно стоящего растения получают достаточно света.
Ассимиляция листьями СО о с повышением освещенности увеличивается
сначала быстро, затем медленнее и часто достигает максимума при 50%
полного солнечного света. По-иному этОт процесс протекает в чистых
посевах. По мере роста растений и увеличения листовой поверхности
нижние листья в посевах все больше затеняются верхними. Если верхние
1 Продуктивность — количество органического вещества, производимое за еди-
ницу времени.— Прим, перев.
2 Т. Takeda, Studies on the photosynthesis and production of dry matter in
community of rice plants, «Jap. J. Bot.», 17, 1961, p. 403—437.
32
Глава 1
листья при 50% полной дневной освещенности достигают максимальной
интенсивности ассимиляции, нижние листья, значительно менее осве-
щенные, ассимилируют в это же время гораздо слабее. Поэтому продук-
тивность отдельного растения в посевах увеличивается прямо пропорцио-
нально освещенности и достигает наибольших значений при максималь-
ной яркости дневного света. В посевах накопление органической массы
также зависит от площади листьев, только зависимость эта более сложная.
Величина листовой поверхности в посевах и насаждениях выражается
показателем облиствленности (ПО), под которым понимают отношение
общей листовой поверхности к покрываемой ею площади почвы. При
величине показателя менее 2 взаимное затенение листьев не имеет суще-
ственного значения. При увеличении же ПО освещенность вблизи почвы
резко падает и фотосинтез нижних листьев протекает при все более небла-
гоприятных условиях. Одновременно с этим пропорционально увеличению
листовой массы возрастает интенсивность дыхания. Поэтому при опти-
мальных значениях ПО продуктивность будет наибольшей. Чем интенсив-
нее дневной свет, тем выше оптимальное значение ПО. По данным Такеда,
рис характеризуется следующими оптимальными значениями ПО на
стадии колошения:
Радиация, кал/дм2 • сутки 100 200 300 400—500
ПО 3,2 5,8 7,8 около 10
Если ПО при 100 кал!дм2 • сутки равен 5,5, в растении преобладают
потери вещества на дыхание и продуктивность приобретает отрицательный
характер. Следствием этого является пожелтение нижних листьев. Вели-
чина ПО зависит также от расположения листьев. Листья в вертикальной
плоскости пропускают больше света, чем находящиеся в горизонтальной;
поэтому при вычислении величины ПО следовало бы учитывать только
проекцию листьев на горизонтальную плоскость; однако это часто трудно
осуществимо. Значения ПО у риса относительно велики, поскольку его
листья отнюдь не горизонтальны. У многих растений, например у листвен-
ных древесных пород, верхние, сильно освещенные листья ориентированы
ребром к солнцу, тогда как нижние, плохо освещенные, располагаются
строго горизонтально. Брохэм 1 исследовал в Новой Зеландии изменение
ПО в посевах Trifolium repens. В соответствии с колебаниями осве-
щенности в течение года ПО повышается с 0,7 в зимние месяцы до 3,25
в летние. В результате появления новых и отмирания старых листьев
постоянно поддерживается известное равновесие между освещенностью
и ПО, причем в любое время года поверхности почвы достигает незначи-
тельное количество света. Листья этого стелющегося по земле клевера
вначале развиваются в условиях небольшой яркости света. Обладая
более длинными листовыми черешками, новые листья, появляющиеся
летом, располагаются над старыми и таким образом оказываются на
полном свету. ПО связан с количеством листьев и размерами их пла-
стинок.
У растений травяного покрова с хорошо развитыми стеблями молодые
листья всегда развертываются на полном свету, то есть летом в условиях
большей освещенности, чем весной. К концу вегетационного периода они
приобретают все более и более ксероморфный облик, как это отметил
1 R. W. Brougham, The leaf growth of Trijolium repens as in influenced by
seasonal changes in the light environment, «J. Ecol.», 50, 1962, p. 449—459.
Введение
33
Япп 1 у Filipendula ulmaria, и получают все меньше света, так как их
затеняют более молодые, расположенные выше листья.
В аридных районах размеры листовой поверхности зависят от влаго-
обеспеченности растений; в засушливый период площадь листьев в этих
условиях обычно сильно редуцирована (ср. гл. 14, стр. 529).
Деревья с густой кроной имеют два типа листьев: световые и теневые.
То же, по данным Сэки, наблюдается и у растений травяного покрова 1 2.
Световые листья используют более интенсивный свет, тогда как теневые,
приспособленные к малой освещенности, характеризуются слабым дыхани-
ем. У деревьев с ажурной кроной, например у берез, теневые листья отсут-
ствуют. Следует учитывать, что в более высоких широтах средняя суточная
Р п с. 6. Продуктивность тица сообщества Phragmites — Sanguisorba tenuifolia в зависи-
мости от освещенности.
1 — Phragmites', 2 — Sanguisorba tenuifolia', 3 — сопутствующие виды; 4 — пожелтевшие листья;
<5 — освещенность в % полного дневного света.
освещенность меняется в течение вегетационного периода. В северном
полушарии она возрастает до конца июня, а осенью снова уменьшается.
В связи с этим ПО летом достигает оптимальных, а осенью еще более
высоких значений, в результате чего начинается прогрессирующее
пожелтение нижних листьев.
Значительно более сложны условия в многоярусных и многовидовых
сообществах, где совместно обитают светолюбивые и теневыносливые
растения. Последние, будучи приспособленными к небольшой освещен-
ности, используют тем меньше света, чем больше у них ассимилирующих
органов (например, папоротники и мхи).
1 R. Y а р р, Spiraea Ulmaria L. and its bearing on the problem of xeromorphy
in marsh plants, «Ann. Bot.», 26, 1912, p. 815—870.
2 T. S a e k i, Variation of photosynthetic activity with aging of leaves and total
photosynthesis in a plant community, «Bot. Mag.», Tokyo, 72, 1959, p. 404—408.
3-1394
34
Глава 1
ПО таких сообществ слагается из листовых поверхностей видов,
с разной экологией; его следует указывать по возможности отдельно
для каждого яруса.
В качестве примера можно привести, по данным Монси и его сотруд-
ников \ тип сообщества Phragmites — Sanguisorba tenuifolia, развитый в
Японии (рис. 6). На рисунке даны соотношения сырого веса ассимилиру-
ющих частей растений (слева) и неассимилирующих (справа) на различной
высоте от поверхности почвы (через каждые 20 см). На основании сырого
веса можно вычислить площадь листьев, зная, что у Phragmites 1 г сырого
веса (содержание воды 68%) соответствует 37—44 см 1 2 листовой поверх-
ности, а у Sanguisorba tenuifolia — 64—80 см2 (при содержании воды 68%).
Sanguisorba — это осеннее растение. При благоприятных условиях осве-
щения его листья остаются зелеными до середины осени — времени цве-
тения этого вида. Однако в зарослях Phragmites этот вид развивается
только весной, до тех пор пока он не начинает затеняться Phragmites.
Уже летом условия освещения становятся столь неблагоприятными, что
Sanguisorba, не успев зацвести, исчезает из травостоя (рис. 6).
КОНКУРЕНТНАЯ СПОСОБНОСТЬ
И ПОКАЗАТЕЛЬ УСПЕШНОГО РОСТА
Конкурентная способность отдельных видов в разных условиях среды
и по отношению к другим видам сильно различается. Поэтому при эколо-
гических исследованиях конкурентную способность видов желательно
оценивать в количественном отношении. Однако обычно это не представ-
ляется возможным, поскольку число морфологофизиологических свойств
видов, влияющих на интенсивность конкуренции, очень велико.
Необходимой предпосылкой конкурентной способности является про-
дуктивность. Чем больше веществ накапливает растение, тем лучше оно
развивается, обильнее плодоносит и теоретически должно дать больше
или более крупных размеров семян. Стало быть, более высокая продуктив-
ность означает обычно и большую конкурентную мощность. При этом
речь идет не об абсолютном количестве накопленного вещества, а об
относительном, в сравнении с массой, аккумулированной конкурентами.
Это отношение массы сухого вещества, накопленного каждой особью
(МА) вида А, к сухому веществу, аккумулированному особями (Мв) вида В,
мы будем называть показателем успешного роста (УР) (Wiichsigkeit)
вида А. Следовательно, УРа/в = Мд : Мв или УРв/а = Мв : МА.
В определенных условиях среды у растений, относящихся к одной жизнен-
ной форме, показатель успешного роста дает возможность сравнительно
хорошо судить о степени конкурентной способности видов 2. При
сравнении, например, конкурентной способности однолетних растений,
скажем Sinapis alba, Avena sativa, Vicia sativa и Triticum aestivum, выра-
женных в различных комбинациях при одинаковом числе особей на еди-
ницу площади (по 100 экземпляров на 1 м2), выясняется, что Sinapis
обладает наивысшим показателем успешного роста, значительно превос-
1 М. М о n s i, Т. S а е k i, Uber den Lichtfaktor in den Pflanzengesellschaften
und seine Bedeutung fiir die Stoffproduktion, «Jap. J. Bot.», 14, 1953, S. 22—52.
2R. Bornkamm, Zur quantitativen Bestimmung von Konkurrenzkraft und
Wettbewerbsspannung, «Вег. Dtsch. Bot. Ges.», 74, 1961, S. 75—83; Zur Lichtkonkurrenz
von Ackerunkrautern, «Flora», 151, 1961, S. 126—143; Erscheinungen der Konkurrenz
zwischen hoheren Pflanzen und ihre begriffliche Fassung, «Вег. Geobot. Inst. Riibel»,
34, 1963, S. 83—107.
Введение
35
ходя в этом отношении все другие виды; далее следует Avena, затем Vicia
и Triticum, причем разница между двумя последними точно не установ-
лена. Зная соотношение показателя успешного роста Triticum по сравнению
с Sinapis и Triticum по сравнению с Vicia, можно вычислить показатель
успешного роста Vicia по сравнению с Sinapis, ибо УРт/s = УРт/v • УРу/s
или УРу/s —• УРт/s • УРу/т. Отдельная особь Sinapis на эксперименталь-
ном участке площадью 1 м1 2 оказывается угнетенной в чистых посевах
и, наоборот, развивается лучше всего, когда остальные 99 особей отно-
сятся к числу более слабых конкурентов. Напротив, экземпляры наиме-
нее конкурентоспособного вида Triticum достигают наиболее крупных
размеров при чистой культуре. В смешанных же культурах при наличии
мощных конкурентов рост Triticum задерживается. Следовательно,
внутривидовая конкуренция протекает более напряженно, чем межвидо-
вая, лишь в тех случаях, когда в ней участвуют виды, обладающие боль-
шой конкурентной мощностью; в остальных случаях наблюдается обрат-
ная картина.
Показатель успешного роста может быть использован для оценки
конкурентной способности только в случае, когда растения характери-
зуются одинаковой скоростью роста и освещенность не оказывает сущест-
венного влияния на конкуренцию. Если же одно растение отличается
более быстрым ростом и затеняет другое ? то первое всегда будет характе-
ризоваться большей конкурентной мощностью.
Иваки изучал экспериментальным путем конкурентные взаимоотно-
шения у Fagopyrum esculentum и Phaseolus viridissimus В чистых посевах
продуктивность обоих видов была почти одинаковой. Доля листьев
в общем весе растения в абсолютно сухом состоянии составляла у Phaseo-
lus 50—60%, а у Fagopyrum — 20—60%, что свидетельствует об извест-
ном преимуществе первого вида. Однако Fagopyrum растет почти в два
раза быстрее; его листья оказываются в более благоприятных условиях
освещения и ассимилируют интенсивнее. Поэтому в смешанных посевах
листьев Phaseolus достигает лишь 16—35% полного дневного света, в связи
с чем интенсивность ассимиляции резко падает. Если же посеять Fagopy-
rum на 3—13 суток позже Phaseolus, то это благоприятно отразится на
развитии последнего. Однако и в этих условиях Fagopyrum развивается
нормально, быстро перерастает Phaseolus и получает достаточно света.
Следовательно, конкурентная способность
в значительной степени зависит от скорости
роста в высоту, что является характерным видо-
вым признаком, и лишь косвенно связана с про-
дуктивностью растения. Ведущая роль принадлежит ско-
рости роста и во внутривидовой конкуренции, например при густых
посевах подсолнечника (Helianthus annuus). Всходы, появляющиеся из
более крупных семян, растут быстрее, поскольку они лучше освещены 2;
соответственно они накапливают больше сухого вещества; таким обра-
зом, они в нарастающем темпе обгоняют угнетенные растения. В неблаго-
приятных условиях освещения угнетенные растения характеризуются
низкой интенсивностью ассимиляции; однако интенсивность их дыхания,
на что они расходуют основную часть ассимилятов, остается на том же
1 Н. Iwaki, Ecological studies of interspecific competition in plant community, J,
«Jap. J. Bot.», 17, 1959, p. 120—138.
2 S. К u г о i w a, Intraspecific competition in artificial sunflower community,
«Bot. Mag.», Tokyo. 73, 1960, p. 300—309.
3*
36
Глава 1
уровне. У быстрорастущих особей количество накопленных веществ
за вычетом затраченных на дыхание достигает приблизительно 4896
общего количества вещества, накопленного растением, а у сильно угне-
тенных — не превышает 12%. При ухудшении условий освещения это
соотношение приобретает отрицательный характер и растение быстро
гибнет.
Конкурентная мощность отдельных видов всегда зависит от условий
внешней среды, причем надземные части растений конкурируют за свет,
тогда как корни — за влагу и питательные вещества. Однако четко разгра-
ничить эти функции не представляется возможным, поскольку развитие
надземных и подземных органов у растения взаимосвязано.
Влияние света на конкурентные взаимоотношения у Sinapis alba,
Agrostemrna githago, Bromus secalinus и Anagallis arvensis var. azurea изуча-
лось Борнкамом Ч Освещенность составляла 100, 60, 30 и 15% полного
солнечного света. В каждый сосуд Мичерлиха помещалось по 24 экземпля-
ра одного вида в чистых посевах и 2 вида по 12 экземпляров, 3 по 8 или
4 по 6 — в смешанных.
Продуктивность (накопление сухого вещества во всех вариантах,
вместе взятых) и освещенность находятся в логарифмической зависимости.
На долю Sinapis приходится большая часть (80%) накопленного сухого
вещества, а на долю Anagallis — наименьшая (1—2%). В чистых посевах
снижение освещенности с 100 до 60% приводит лишь к небольшому сокра-
щению продуктивности у всех упомянутых видов, тогда как уменьшение
освещенности еще на 15% сокращает продуктивность на 1/4—1/3. Только
у Bromus продуктивность уменьшается в этом случае приблизительно на
50%. Следовательно, этот вид наименее светолюбив. Значительно сложнее
взаимосвязи в смешанных посевах. Anagallis — вид, обладающий наимень-
шей конкурентной мощностью, оказывается в более благоприятных усло-
виях при уменьшении освещенности, что отрицательно влияет на раз-
витие конкурентов; в смешанных посевах двух видов количество накоп-
ленного каждой особью Anagallis органического вещества не зависит от
освещенности, в посевах из трех видов (при 60 и 30% полного солнечного
света) каждый экземпляр Anagallis накапливает уже в четыре раза боль-
ше, чем при полном освещении, а в посевах из четырех видов — даже
в семь раз.
Продуктивность Bromus в посевах из четырех видов при 30% полного
света увеличивается в два раза. Отсюда следует, что конкурентная способ-
ность Anagallis в отличие от других видов повышается с уменьшением
освещенности. Так, например, показатель успешного роста этого вида
по сравнению с Sinapis при 15% полного солнечного света увеличивается
в пять раз по сравнению с Agrostemrna — в 11 и по сравнению с Bromus
{при 30% полного света) — в три-четыре раза. Такая же взаимосвязь
установлена у Bromus с Sinapis и Agrostemrna, тогда как конкурентные
взаимоотношения последних двух видов не зависят от величины освещен-
ности. Не следует, однако, думать, что здесь всегда речь идет о прямом
влиянии света. При недостатке света у сообитателей-конкурентов ухуд-
шается развитие не только побегов, но и в значительно большей степени —
корневой системы. С уменьшением освещенности увеличивается отношение
числа побегов к корням. Следовательно, при недостатке света корневая
конкуренция резко ослабевает, что создает более благоприятные условия
для менее сильных конкурентов.
1 R. Bomkamm, op. cit.
Введение
37
Р и с. 7. Oxalis acetosella в еловом лесу на окопанной площадке, где отсутствует
конкуренция корней (слева), и в естественных условиях (справа) (по Карпову,
уменьшено в 2 раза).
Характер этих взаимосвязей в естественных условиях отражен в рабо-
тах Карпова, проводившего свои исследования в еловых лесах южной
тайги (Piceetum myrtillosum) х. Условия освещения в еловом лесу (при
сомкнутости крон 80%), где велась работа, весьма неблагоприятны.
В составе нижних ярусов распространены преимущественно черника
(Vaccinium myrtillus) и мхи. Кислица (Oxalis acetosella} встречается изред-
ка. Обычно считалось, что под пологом леса травяной покров плохо раз-
вивается из-за недостатка света, однако выяснилось, что в основном это
зависит от конкуренции со стороны корней ели. Если ее исключить, про-
изведя обрезку корней деревьев по периферии опытной площадки (1 м1 2)
на глубину 50 см, растения травяного яруса начинают развиваться
пышнее, хотя условия освещения не изменились. На опытной площадке
прежде всего быстро распространяется Oxalis, подавляющая развитие
мхов. Через несколько лет здесь появляется даже малина (Rubus idaeus),
которая также быстро захватывает площадь. Несмотря на то что интен-
сивность ассимиляции СО2 (в расчете на единицу листовой поверхности)
у Oxalis на контрольных и опытных площадках одинакова, количество
органического вещества, накопленного отдельными экземплярами Oxalis,
на окопанных участках увеличилось в 10 раз. Наряду с этим увеличились
размеры растений и площадь их листовой поверхности (рис. 7). Аналогич-
ные результаты, хотя и не столь ярко выраженные, получены при окопке
площадок с Мajanthemum bifolium и Trientalis еигораеа. В верхних (20 см)
1 В. Г. Карпов, Некоторые вопросы фитоценологии еловых лесов в экспе-
риментальном освещении, «АН СССР, Сообщения лаборатории лесоведения», вып. 6,
1962; О влиянии конкуренции корней деревьев на ассимиляционную деятельность
таежных трав в еловых лесах, «Доклады АН СССР», 140, № 5, 1961; Опыт использо-
вания Р32 для изучения соревнования между корнями деревьев и подроста в лесах
южной тайги, «Доклады АН СССР», 146, № 3, 1962; Некоторые итоги эксперименталь-
ного изучения состава и строения нижних ярусов ельника-черничника, «Проблемы
ботаники», 6, М.—Л., 1962.
38
Глава 1
горизонтах почвы сосредоточено 90 % корней ели и растений травяного
покрова. В условиях очень влажного бореального климата конкуренция
за почвенную влагу не имеет существенного значения, поскольку грунто-
вые воды залегают на небольшой глубине и вполне доступны для расте-
ний. Содержание влаги в почве на контрольных и опытных площадках
в этих условиях практически одинаково и всегда значительно выше коэф-
фициента завядания. В этих лесах конкуренция между растениями идет
в основном за усвояемые формы азота (N). Растения на контрольных пло-
щадках обнаруживают явные признаки азотного голодания, что выра-
жается, в частности, в желтовато-зеленой окраске листьев. Напротив,
на опытных площадках, где отсутствует корневая конкуренция, листья
отличаются темно-зеленым цветом. Содержание питательных веществ
в листьях также свидетельствует о низкой (2,06%) концентрации N
в тканях листа Oxalis при наличии конкуренции с корнями деревьев
и высокой (3,18%) — на опытных площадках, где конкуренция снята из-за
острого недостатка N, который обычно испытывают таежные травы в ело-
вых лесах; в связи с этим площадь их листовой поверхности по сравнению
с неассимилирующими органами достигает небольших размеров. Поэтому
продукты ассимиляции листьев могут обеспечить только дыхание всего
растения. В результате не наблюдается существенного накопления орга-
нических веществ, что и определяет слабый рост этих растений. При
отсутствии контакта с корнями деревьев содержание N в почве и потреб-
ление азота резко улучшается, что способствует увеличению площади
листьев; это благоприятствует накоплению органических веществ, что
в свою очередь способствует увеличению листовой поверхности. Решающее
значение конкуренции за усвояемые формы азота доказывается также
тем, что на опытных площадках, где устранено не только соревнование
между корневыми системами, но и удалены травяной и моховой покровы,
со временем появляются нитрофильные виды или виды, обычно развитые
на богатых лесных почвах.
Аналогичные результаты дали опыты по пересадке под полог елового
леса трав (Aegopodium podagraria, Ajuga reptans и т. д.), развивающихся
на почвах, богатых азотом. Правда, следует учитывать, что обрубленные,
но оставшиеся в почве опытной площадки корни деревьев со временем
отмирают, разлагаются и являются источником пополнения почв пита-
тельными веществами, и в частности азотом. Таким образом, бедность
флористического состава травяного покрова в еловых лесах южной тайги
обусловлена не недостатком света, а конкуренцией за усвояемые формы
азота с корневыми системами деревьев. Мхи влияния этой конкуренции
не испытывают. Они получают питательные вещества с водой, капающей
с деревьев, или из опада хвои и пышно развиваются в еловом лесу.
Конкуренция со стороны корней взрослых деревьев угнетает также
молодой подрост. Поэтому всходы ели развиваются в березовом лесу
крайне медленно, хотя и получают достаточно света. Если исключить
конкуренцию, обрубив вокруг опытной площадки с всходами ели корни
берез, всходы начинают быстро расти. То, что и в этом случае решающее
значение принадлежит конкуренции за питательные вещества, подтвер-
ждается следующим опытом Карпова. В почву контрольных и опытных
площадок равномерно вносился меченый фосфор (Р32). Через несколько
дней после этого измерялась радиоактивность хвои всходов ели. У расте-
ний на опытных площадках она оказалась в 5—6 раз выше, чем у всхо-
дов на контрольных участках. Какого-либо влияния ингибиторов или
выделений корней деревьев установлено не было.
Введение
39
Исследования, проведенные на южных черноземах степной зоны,
показали, что в этих условиях конкуренция за влагу с корнями взрослых
деревьев оказывается для подроста значительно более пагубной, чем
соревнование за минеральные вещества.
ИНТЕНСИВНОСТЬ КОНКУРЕНЦИИ И ЖИЗНЕННЫЕ ФОРМЫ
Выше рассматривались конкурентные взаимоотношения между отдель-
ными особями в период их развития и их отношения к продуктивности.
В случае же межвидовой конкуренции большое значение имеет то обстоя-
тельство, что отдельные виды обычно развиваются и успешно размно-
жаются на одном и том же месте в течение многих поколений. Конку-
рентная способность вида зависит также от продолжительности жизни
одного поколения. Недолговечные виды начинают свое развитие каждый
раз из семени, однолетние виды, например, ежегодно. Долгоживущие
виды, напротив, в течение длительного времени угнетают своих конку-
рентов, прежде чем они погибнут и начнут возобновляться. В общем
конкурентная мощность с возрастом Особи увеличивается. Ниже приво-
дится краткая характеристика основных жизненных форм, различаю-
щихся по форме роста и долговечности их ассимиляционных органов.
Однолетние виды (рис. 8, 7)
Развитие этих видов, начинающееся из семени, всегда заканчивается
в течение одного года; часто оно длится всего несколько недель. При раз-
витии проростка (77) и формировании стебля и первых ассимилирующих
листьев и корней расходуется приблизительно половина содержащегося
в семени сухого вещества. Уже через несколько дней в процессе
фотосинтеза образуются ассимиляты; чистая продукция, или нетто-про-
дуктивность (Пн), выражается в количестве накопленных веществ за
вычетом затраченных на дыхание. Накопленные вещества расходуются
на формирование новых органов и прирост (Д). В дальнейшем листовая
поверхность и нетто-продуктивность увеличиваются до тех пор, пока
тем или иным растением не будет создан общий запас сухого вещества (5).
После этого растение начинает отмирать; некоторое количество содер-
жащихся в отмирающих частях растения веществ переходит в созреваю-
щие, обычно мелкие, но многочисленные (т) семена. Этим цикл развития
одного поколения завершается. Семена переживают неблагоприятное
время года и на следующий год прорастают (т семена), однако часть
из них погибает, поэтому число выросших растений (пг) меньше количе-
ства семян. Еще через год вырастает уже (т')2 растений и т. д. Если одно-
летние растения не испытывают конкуренции, они размножаются очень
быстро. На ограниченной же площади произрастания в густой заросли
чрезвычайно сильно сказывается влияние внутривидовой конкуренции;
в результате этого у угнетенных особей отношение между ассимиляцией
и дыханием становится все более неблагоприятным; растения не цветут
и не плодоносят; в конце концов вся заросль отмирает. Это наблюдалось,
например, в северо-западных штатах Северной Америки, где на участках
с чрезмерным выпасом скота Bromus tectorum образует чистые травостои.
Двулетние виды (рис. 8, II)
У двулетних видов развитие протекает вначале так же, как и у одно-
летних, с той лишь разницей, что у двулетних стебель в первый год
ие образуется; они развивают только прикорневую розетку листьев, сильно
I. Однолетние Виды (теросриты)
[л<2 гл /Л) :
Семя
3f
X
X
К>
+Цветки
m Семена
т'Л
П.Двулетние виды (гемикриптофиты-терофиты)
Год 1
Розетка
Семя
(Л<2 [л
•ГК д-j
к к
L :
I! Запасающие корни
[Л
\\>АС
> к
Год 2
7П
и
К)
+Lift емки-
т Семена
т'П
Ш. Многолетние
\л-& [Л
Семя
.К
виды (геофиты или гемикриптофиты)
Год 1
(Л<^ ।
7Г*\
(К
Запасающие
органы
С
К
X
Год п
^\СН
к н
Запасающие
органы
1К
\л
X
Год 2
Л<^
С~Н~
К
Год(п + 1)
Ш. Многолетние
(вечнозеленые) Виды
Год 1 Год 2-п
(Л^-~ .
Запасающие и т.д..
органы Год3(п^
Л^г |
*A< С+++ М ''Запасающие и т.д.
К Л ; органы
+Цветки-} т Семена
—•-т'П
(как год 1)
Год(п+1)
(Л
Е с —-
К >
.+Щ}етки-*т Семена
л
С ит.д.
К
-+т'П
(как год 1)
Z. Листопадные деревья и кустарники (фанерофиты или хамефиты)
Год 1
(Л^^г ; Почки
^\с^^—\С(Н)
к-------
Год(п+1)
аг*
Семя
Семя
ГЛ
Л\С + +
X к
к
ит.д.
-+к [к
П«
^с+++Е^с—> 'с(н)
К—+ \К
м-Цветки-*\т Семена
-*к lx
Год 2-п
Л
\Л [Л*^ж1 Почки
\С(Н)
',К
2Л
С ит.д.
-+т'П (как год 1)
и т. д.—
Год (3-п)
И. Вечнозеленые деревья и кустарники (фанерофиты или хамефиты)
(развитие, кик и выше, однако в конце года листья не опадают и период покоя не выражен)
Год п
-+Л
->С
ФК
Год (п-1)
[Л-
...Z с-
к-
Рис. 8. Продуктивность
Л — листья;
как J/
Год (п+1]
[Л >
ЕС -------------
К >
,+иЛетки—т Семена
т'П (как год 1)
растений различных жизненных форм.
2 пР°Ростоь\ Л листья; С — стебель; К — корень; Н — запасные вещества;
р ™ . ^етто-продуктивность; д — прирост; т — семена; 2 — общий запас сухого веще-
ства, вертикальный пунктир — начало неблагоприятного сезона; у — отмирание’
++Н вторичный прирост; сплошная вертикальная черта — граница года ’
пунктир — начало
Введение
41
разрастающуюся в течение года, и в таком виде перезимовывают. Нако-
пленные запасные вещества (Н) аккумулируются в подземных частях
в количестве, гораздо большем, чем их образуется в семенах. Таким
образом, второй год растение встречает уже подготовленным, к тому же
розеточные листья, если они не пострадали от морозов, тотчас начинают
ассимилировать. В конечном итоге вырастает крупное цветущее растение,
способное принести много семян. Стало быть, конкурентная способность
двулетних видов на втором году жизни намного выше, чем у однолетних
растений, хотя они и размножаются не так быстро, как последние.
Многолетние виды (рис. 8, III)
В первые годы развитие многолетних видов напоминает развитие
двулетних на первом году жизни; различие состоит лишь в том, что все
надземные органы обычно отмирают и накопленные вещества (Н) откла-
дываются в запасающих органах (корневищах, клубнях, луковицах).
Только последние переживают неблагоприятное время года, частично
сохраняется также корневая система. Количество запасных веществ из
года в год возрастает; улучшаются условия для развития новых побегов;
увеличиваются размеры растений, и через несколько лет они начинают
цвести и плодоносить. Одновременно происходит формирование новых
запасающих органов; на следующий год растения вновь образуют побеги,
а из семян вырастают новые особи. Наряду с этим многолетние виды
размножаются также вегетативным путем; например, образуется несколь-
ко луковиц, причем из каждой развиваются отдельные побеги. Благодаря
большой продолжительности жизни многолетние растения обладают боль-
шей конкурентной мощностью по сравнению с одно- и двулетними видами
и вытесняют последние с их мест произрастания. При помощи семян
многолетние растения в дальнейшем распространяются на соседние
участки.
Многолетние вечнозеленые виды (рис. 8, IV)
Распространены в условиях, где вегетация возможна во все времена
года, например во влажных тропиках. В связи с этим не возникает необ-
ходимости образования запасающих органов. Растение не отмирает еже-
годно и продолжает расти год за годом без значительных перерывов.
Со временем оно начинает цвести и плодоносить. Только некоторые моно-
карпические виды (например, агавы) цветут всего один раз и после этого
отмирают.
Листопадные деревья и кустарники (рис. 8, V)>
В первый год развиваются так же, как многолетние виды (рис. 8, III),
с той лишь разницей, что в конце вегетационного периода отмирают
не все надземные части растения; опадает только листва, а сохраняется
одревесневший стебель с почками, который одновременно служит вмести-
лищем запасных питательных веществ. На построение стебля расходуется
большая часть накопленного сухого вещества, поэтому запасов органи-
ческого вещества для увеличения листовой поверхности остается мало.
В связи с этим в первые годы площадь листьев, а следовательно, и годо-
вая продукция сухого вещества у древесных растений весьма невелика;
у бука она составляет всего 1,5 г. Однако со временем преимущество
деревьев по сравнению с кустарниками увеличивается, поскольку ежегод-
ное развитие начинается из покоящихся почек, расположенных на ство-
лах, поднимающихся из года в год все выше над поверхностью почвы.
42
Глава 1
Хамефиты, или невысокие кустарники и полукустарники, обычно
не превышают высоты 50 см и поэтому в условиях холодной зимы защи-
щены снегом; они распространены также в засушливых областях. Нано-
фанерофиты, или кустарники, достигают высоты 4 м, тогда как некото-
рые древесные породы в отдельных случаях возвышаются более чем
на 100 м {Eucalyptus regnans, Sequoia sempervirens).
Критическим периодом развития у древесных пород являются пер-
вые годы, когда из-за медленного роста они выдерживают конкуренцию
со стороны представителей других ярусов. Крупные семена (Castanea,
Quercus, Fagus) благоприятствуют более быстрому росту всходов г. Кон-
курентная способность подроста древесных пород по сравнению с видами
травяного покрова из года в год увеличивается; подрост продолжает
расти в высоту и наконец обгоняет по высоте своих конкурентов, в свою
очередь начав их затенять. После смыкания крон древесная порода начи-
нает господствовать в сообществе. Достигнутая на этой стадии опти-
мальная величина облиствленности в последующем уже существенно
не изменяется. Таким образом, с увеличением высоты соотношение между
ассимиляцией и дыханием должно было бы приобретать отрицательное
значение, что выразилось бы в сокращении количества накапливаемого
сухого вещества. Однако этого не происходит, поскольку в результате
постоянной редукции нижних ветвей и омертвления древесины в стволах
и корнях наблюдается выравнивание баланса накопления органического
вещества. Вследствие прироста в высоту и по диаметру в стволах увели-
чивается масса заболони, состоящей из деятельных элементов; это вновь
приводит к нарушению баланса накапливаемого вещества. В дальней-
шем с увеличением возраста дерева баланс накопления органического
вещества приобретает отрицательный характер; величина нетто-продук-
тивности оказывается недостаточной для формирования большой листо-
вой поверхности; у стареющего дерева крона редеет, отмирают сначала
отдельные ветви, а затем и весь ствол.
Этой стадии светолюбивые растения, отличающиеся меньшей листо-
вой массой, достигают раньше, чем теневыносливые. Начало этой стадии
зависит также от степени интенсивности ассимиляции и дыхания тех
или иных древесных пород.
Вечнозеленые деревья и кустарники (рис. 8, VI)
Различия с предыдущей группой невелики. Листья иногда сохраняют-
ся в течение нескольких лет; деревья сбрасывают их не одновременно.
При благоприятных в течение всего года температурных условиях
и достаточном увлажнении фотосинтезирующая деятельность листьев
не прерывается. При наличии же ежегодных более или менее длительных
периодов покоя, обусловленных засухой или холодом, вечнозеленые
листья растений приобретают ксероморфную структуру, несмотря на это,
они вырабатывают не меньше органического вещества, чем листья преды-
дущего типа. В природе растения с обоими типами листьев встречаются
даже в одинаковых климатических условиях. В аридных областях Афри-
ки распространены листопадные акации, а в Австралии, в таких же
условиях,— акации с ксероморфными вечнозелеными филлодиями. В саван-
нах и тропических сухих лесах Африки и Южной Америки встречаются
1 Высота однолетнего всхода дуба пропорциональна размеру желудя. См.
Р. G. J а г v i s, The effect of acorn size and provenance on the growth of seedlings of
sessile oak, «Quart. J. Forestry», 57, 1963, p. 11—19.
Введение
43
листопадные древесные породы, а в Австралии — вечнозеленые эвкалип-
ты. Арктическую границу леса образуют листопадные березы, вечнозе-
леные ели и сбрасывающие свою хвою лиственницы. В Альпах выше
всего поднимаются в горы лиственница (Larix decidua) и вечнозеленый
кедр европейский (Pinas cembra), а из кустарников — листопадная Alnus
viridis и вечнозеленая Pinas montana.
Конкурентную способность растений, относящихся к различным
жизненным формам, легче всего изучать на пашне. Здесь сначала доми-
нируют однолетние растения, вскоре вытесняемые двулетними; за ними
следуют многолетние виды травяного покрова, к которым примешиваются
быстро растущие древесные породы и кустарники. В конечном итоге
начинают господствовать древесные породы, а среди них — теневыносли-
вые виды. Поэтому маломощные в конкурентном отношении однолетники
(терофиты) сохраняются в качестве зональной растительности лишь там,
где климатические условия неблагоприятны для произрастания растений
других жизненных форм, например в пустынях, где осадки выпадают
лишь эпизодически. Они развиваются в огромных количествах в полу-
пустынях и пустынях, где ежегодная длительная засуха задерживает
развитие многолетних растений, однако во влажное, хотя и очень непро-
должительное время года в почве накапливается достаточно влаги для
формирования более густого растительного покрова. Климатических
условий, особенно благоприятных для двулетних видов, не существует.
Они развиваются в местах, где растительный покров не уничтожается
ежегодно человеком. Двулетние виды иногда доминируют в засушливых
областях, где деревья почти не растут из-за периодического недостатка
влаги, как, например, в степях.
В благоприятных климатических и почвенных условиях господствуют
деревья. Если человек не уничтожал бы их, то большая часть земной
поверхности и в настоящее время была бы покрыта вечнозелеными или
листопадными лесами. На площадях, где лес сведен, создаются условия,
благоприятные для развития растений, относящихся к другим жизнен-
ным формам (например, на лугах и пашнях). Напротив, в ненарушенных
лесных районах растения этих жизненных форм играют лишь подчинен-
ную роль и господствуют только в местообитаниях, не благоприятных
для произрастания деревьев (скалы, болота и т. д.).
ОБМЕН ВЕЩЕСТВОМ И ЭНЕРГИЕЙ В ЭКОСИСТЕМАХ
Естественный растительный покров находится во взаимодействии
со средой. Природные единства, возникающие благодаря этим взаимо-
связям, называются экосистемами; они состоят из трех основных взаимо-
действующих компонентов Ч
1. Физико-химические факторы атмосферы и почвы.
2. Продуценты — автотрофные организмы, которые создают органи-
ческие вещества и при этом накапливают солнечную энергию частично
в виде химической энергии.
3. Консументы — гетеротрофные организмы, к которым относятся
животные и микроорганизмы; благодаря им органические соединения
в конечном итоге снова превращаются в минеральные.
1 Вместо понятия «экосистема» в ботанической литературе СССР применяется
понятие «биогеоценоз», предложенное В. Н. Сукачевым. Подробную характеристику
основных понятий биогеоценологии см. в книге «Основы лесной биогеоценологии»
под редакцией В. Н. Сукачева и Н. В Дылпса, изд. «Наука», М., 1964.— Прим, ред.
44
Глава 1
В конце концов задача экологии, собственно, и состоит в выяснении
всех деталей превращения вещества и энергии в различных экосисте-
мах Ч От этой конечной цели мы еще очень далеки и знакомы лишь с отдель-
ными звеньями этого процесса. В качестве примера рассмотрим цифровые
данные (в расчете на 1 га), полученные Дювиньо при изучении западно-
европейского смешанного лиственного леса.
Количество солнечной энергии, поступающее в течение года на 1 га
леса, составляет 9-109 ккал. Большая часть ее превращается в тепло
и уходит на нагрев воздуха. Приблизительно V3 расходуется растениями
на транспирацию, то есть на перевод воды из жидкого состояния в газо-
образное. На фотосинтез приходится только 1% поступившей солнечной
энергии.
Половина накопленной в результате ассимиляции органической
массы тотчас расходуется на дыхание растений. Остаток (нетто-продук-
тивность) накопленного сухого вещества составляет приблизительно
12 т/га. Из этого количества около 4 т/га приходится на листья деревьев,
5 — на древесину и примерно 2 т!га — на корни. Остальное, 1 т!гаг
продуцирует подрост.
Площадь листьев составляет приблизительно 4,5 га/га. Таким обра-
зом, показатель облиствленности равен 4,5. Листья и напочвенный покров
ежегодно отмирают и накапливаются в виде опада. Микроорганизмы
в течение нескольких лет разлагают или гумифицируют его.
В естественных условиях в лесах, где запас древесины и количество
гумуса в почве сохраняются на определенном уровне, ежегодно должно
накапливаться столько же вещества, сколько его в течение года разла-
гается, в нашем примере, следовательно, 12 т/га. Большая часть накоп-
ленного вещества разлагается непосредственно микроорганизмами (грибы,
бактерии) при участии также почвенной фауны. Лишь незначительная
часть накопленного вещества потребляется растительноядными живот-
ными, многие из которых в свою очередь поедаются хищниками. Трупы
хищников в конце концов также попадают в почву и там разлагаются.
О незначительной роли высших животных в обмене веществом в лесах
Центральной Европы свидетельствуют данные о валовых запасах орга-
нического вещества (в расчете на 1 га) в 120-летнем древостое.
Надземная масса
Продуценты, т/га Консументы, кг/га
Листья 4 Птицы ............................ 1,3
Ветви 30 Крупные млекопитающие 2,2
Стволы 240 Мелкие млекопитающие . 5,0
Подрост.......... 1 Хищники...................... Не установлено
Итого 275 Итого.............................. 8,5
Кроме того, беспозвоночные . . . Количество не
известно
Подземная масса
Корни . Несколько тонн на 1 га
Гумус........ 1—70 т/га
Почвенная фауна 1 т/га (в том числе дождевые черви около 0,2т/га)
Почвенная флора 0,3 т/га (бактерии — 95-106 на 1 г почвы
актиномицеты—36 • 106 и грибы—1 • 106 на 1 г почвы)
1 Р. Duvigneaud et al., L’ecosysteme. L’ecologie, science moderne de
synthese, pt 5, Bruxelles, 1962; J.D. Ovingto n, Quantitative ecology and the wood-
land ecosystem concept, «Advanc. Ecol, Res.», I, 1962, p. 103—192.
Введение
45
Хотя приведенные цифры весьма приблизительны, было бы большим
достижением, если бы удалось получить аналогичные данные для зональ-
ных растительных сообществ земного шара, однако от этой цели мы еще
очень далеки.
В дальнейшем при характеристике зональной растительности будут
приводиться имеющиеся в распоряжении автора данные о запасах орга-
нического вещества. В основном же придется довольствоваться анализом
взаимосвязей между растительностью и средой, уделяя основное внима-
ние причинной стороне этих взаимосвязей.
НЕКОТОРЫЕ СООБРАЖЕНИЯ О ВОДНОМ РЕЖИМЕ РАСТЕНИЙ
Из различных факторов среды особенно большое влияние на жизне-
деятельность растений оказывают температура и водный режим. Однако
характер воздействия этих факторов на растения принципиально разли-
чен. Растения не обладают почти никакими свойствами, которые могли бы
смягчать воздействие температуры. Они не имеют собственной темпера-
туры и не способны активно препятствовать ни ее повышению, ни пони-
жению. Поэтому температура самих растений существенно не отличается
от температуры окружающей среды. При сильной инсоляции температура
листьев достигает несколько более высоких значений, чем температура
окружающего воздуха; при интенсивной транспирации большое количе-
ство тепла (при 20° С на 1 г воды расходуется 582 кал) расходуется на
переход воды из жидкого состояния в газообразное. Вследствие этого
температура листьев понижается и ее величины могут быть ниже темпе-
ратуры окружающего воздуха; это чисто физический процесс. Таким
образом, измеряя температуру воздуха, мы приблизительно определяем
температуру растения и прежде всего протоплазмы, в которой протекают
все жизненные процессы.
Связь водного режима с жизнедеятельностью растений совершенно
иная. В отличие от низших растений высшие растения в значительно
меньшей степени зависят от водного режима окружающей среды; поэтому
определение влажности воздуха или почвы непосредственно около самого
растения не дает нам возможности судить о том, как протекают жизнен-
ные процессы в самих растениях.
Для понимания этого явления рассмотрим с физиологической точки
зрения, каким образом растение может существовать вне водной среды
Жизнь зародилась в воде, и до настоящего времени она связана с нали-
чием воды. Жизнедеятельность живого вещества, протоплазмы, наблю-
дается только при ее насыщении водой. При высыхании протоплазма
обычно переходит в латентное состояние; в этом случае процесс обмена
веществом в протоплазме не поддается учету.
К жизни на суше удалось приспособиться только одной группе расте-
ний, развитие которой связано с пресной водой 1 2.
1 Н. Walter, Die physiologischen Voraussetzungen fur den iJbergang der auto-
trophen Pflanzen vom Leben im Wasser zum Landleben, «Zeitschr. f. Pflanzenphysiolo-
gie», 56, 1967, S. 170—185.
К. К r e e b, Thermodynamische Betrachtungen zum Wasserhaushalt der Pflanze,
«Zeitschr. f. Pflanzenphysiologie», 56, 1967, S. 186—202.
2 Многое свидетельствует о том, что моря первоначально представляли собой
пресноводные акватории, поскольку соли, содержащиеся в морской воде, являются
той единственно не превратившейся в осадок частью поступающих в море продуктов
выветривания и выбросов при извержениях вулканов. Таким образом накопление
NaCl (происхождение С1, видимо, связано с вулканйческими газами) в морях связано
46
Глава 1
Начальный этап этого процесса связан с одноклеточными водоросля-
ми, такими, как Chlamydomonas и Chlorella\ в дальнейшем процесс идет
через зеленые водоросли (Ghloropliyceae) и Gharaceae к живущим уже
вне водной среды Cormophyta, то есть мхам (Bryophytd), папоротникам
(Pteridophyta) и голосеменным и покрытосеменным растениям. Правда,
некоторые представители низших зеленых водорослей (Protococcales,
Trentepholla и др., рис. 9) живут не в воде, однако относить их к числу
Пойкилогидрические растения
Гомойогидрические
Рис. 9. Схематическое изображение приспособления растений к жизни на
суше.
I. Переход от низших водорослей, обладающих клетками без вакуолей, к пойкилогидри-
ческим наземным водорослям. II. Образование вакуолей у Ghlorophyceae и Gharaceae,
живущих в воде. III. Переход к гомойогидрическим Cormophyta; развитие мхов связано
с высокой влажностью воздуха, однако в засушливых условиях они вторично возвра-
щаются к пойкилогидрическому образу жизни (ср. подвергшиеся сильному плазмолизу
клетки в молярном растворе сахарозы у мхов из влажных местообитаний и клетки, под-
вергшиеся слабому плазмолизу у мхов из сухих местообитаний). У папоротников также
встречаются пойкилогидрические формы, у голосеменных их нет, у покрытосемен-
ных— являются редким исключением (Myrothamnus) (см. текст).
наземных растений нельзя, поскольку они находятся в состоянии актив-
ной жизнедеятельности только тогда, когда они смочены водой или нахо-
дятся в насыщенном водяным паром воздухе. В противном случае они
высыхают, что, однако, не приводит к их гибели. Эта способность высы-
хать присуща протопластам всех растений. Достаточно вспомнить,
например, разносимые ветром одноклеточные организмы, споры мхов
и папоротников или зародыши в семенах высших растений. При этом
речь всегда идет о клетках, не имеющих вакуолей, объем протоплазмы
с историческим развитием земного шара. Если это так, то происхождение современных
морских водорослей следует вести от пресноводных водорослей. Лишь в этом случае
становится понятным, почему обе главнейшие составные части морской соли — натрий
и хлор — вообще не имеют никакого значения в жизни растений.
Введение
47
которых при высыхании уменьшается сравнительно мало. В таких слу-
чаях субмикроскопическая структура протоплазмы остается ненарушен-
ной и при набухании ее жизнедеятельность вновь восстанавливается.
Организмы, обладающие способностью существовать вне водной среды,
гидратура протоплазмы которых определяется степенью увлажнения
окружающей среды, мы называем пойкилогидрическими растениями.
Большинство этих растений приурочено к относительно влажным
местообитаниям, однако они встречаются и в пустынях.
Фитомасса пойкилогидрических растений невелика, поэтому они
не играют существенной роли в сложении растительного покрова. Посколь-
ку освещаемая солнцем ассимилирующая поверхность у них весьма
незначительна, они не могут продуцировать много органического веще-
ства, тем более что эти виды большую часть года находятся в латентном
состоянии.
В эволюционном отношении очень рано началось развитие в водной
среде водорослей с нитчатыми талломами, обладающими относительно
крупными клетками с вакуолями и небольшим количеством протоплаз-
мы типа Spirogyra, Cladophora или Vaucheria (рис. 9, внизу).
Наличие вакуолей при небольшой массе протоплазмы (то есть при
небольшом расходе связанного азота, содержание которого в автотроф-
ных растениях обычно крайне невелико) способствовало формированию
большей поверхности, освещаемой солнцем, что в свою очередь привело
к увеличению количества продуцируемого органического вещества.
Наряду с этим клетка с вакуолью благодаря тургору приобретает проч-
ность. Поэтому в процессе исторического развития выявилось преимуще-
ство этих водорослей по сравнению с водорослями, в клетках которых
нет вакуолей. Правда, наличие вакуолей привело к тому, что, высохнув,
протоплазма в клетках этих растений потердла способность при набуха-
нии восстанавливать свою жизнедеятельность. Дело в том, что слой
протоплазмы, выстилающий оболочку клетки изнутри, при высыхании
испытывает столь сильное механическое воздействие со стороны умень-
шающейся вакуоли и клеточной стенки, что субмикроскопическая струк-
тура нарушается и клетка отмирает.
Развитие вакуолей в клетках привело к важным последствиям. Про-
топлазма этих живущих в воде водорослей, обладающих клетками с вакуо-
лями, существует не только в среде, окруженной водой, но и внутри
клетки граничит с водной средой, клеточным соком; гидратура прото-
плазмы, как мы увидим ниже, определяется теперь уже не внешней,
а внутренней средой.
Развитие вакуолей в клетках растений позволило им переселиться
на сушу по-иному, чем это произошло у пойкилогидрических видов;
этот новый путь оказался более успешным.
Перейдем к рассмотрению конечного члена эволюционного ряда —
к цветковым растениям, у которых на схеме (рис. 9, справа) схематично
изображены три основных органа: корни, стебли и листья.
Цветковые растения — это многоклеточные организмы, у которых
корневая система располагается в среде, содержащей воду, то есть почве,
тогда~как стебли развиваются в условиях относительно сухого воздуха.
Высыханию наземных органов препятствует плохо пропускающая воду
кутикула. Если бы у растений сформировалась покровная ткань, не про-
пускающая воду, но проницаемая для СО2, потерь воды не наблюдалось бы
вообще. Кутикула затрудняет не только водо-, но и газообмен. Поэтому
у органов наземных растений, ассимилирующих СО2, сформировались
48
Глава 1
устьица Ч Внутренние части растений большей частью заполнены жидкой
средой, находящейся в вакуоме, под которым подразумевается сово-
купность всех вакуолей. Благодаря этой внутренней среде — клеточному
соку вакуома, растения приобрели способность сохранять гидратуру
протоплазмы даже в органах, существующих не в водной, а в сухой воз-
душной среде.
Клеточный сок вакуома можно в какой-то степени сравнивать с жидко-
стью, содержащейся в теле животных, однако клеточный сок содержится
в бесчисленном множестве изолированных друг от друга клеток, а частич-
но в специализированных клетках с различным по составу клеточным
соком. Среднее осмотическое давление клеточного сока можно определить,
выделив его из отмерших клеток целых органов.
Физиологию водного режима, сложный механизм функционирования
устьиц и других приспособлений для поглощения, передвижения и испа-
рения воды можно понять, только учитывая, что все они прежде всего
служат для ассимиляции листьями СО2, необходимого для фотосинтеза,
причем водный баланс должен при этом сохраняться на постоянном
уровне при возможно низкой концентрации клеточного сока. Этим путем
поддерживается высокая степень гидратуры и активности протоплазмы.
Гомойогидрические растения остались в известной мере «водными расте-
ниями», однако с внутренней водной средой, концентрация которой почти
не зависит от условий окружающей воздушной среды. Для оценки вод-
ного режима растений поэтому большее значение имеет водная среда
протоплазмы, чем влажность воздуха. Клетки образуют для протоплазмы
своего рода климатическую камеру, определяющую степень ее влажности.
Этим определяется значение влажности внутри камеры, тогда как влаж-
ность вне ее пределов не играет никакой роли. Именно поэтому в раз-
личных климатических условиях сначала всегда надо определять осмо-
тическое давление клеточного сока. Это дает возможность установить,
при каких условиях гидратуры наблюдается жизнедеятельность прото-
плазмы и как влияет на растения засушливость окружающей среды.
Для доказательства того, что гидратура, а следовательно, и набухание,
или гидратация, протоплазмы зависит только от осмотической концентра-
ции клеточного сока, то есть от величины осмотического давления, прове-
дем следующий простой термодинамический опыт (рис. 10).
1. В камеру Пфеффера с полупроницаемой мембраной поместим набу-
хающее тело и все поставим в воду. После достижения равновесия тело
набухнет до максимального уровня.
2. При давлении на тело 14 атм часть воды будет выжата из тела;
это будет продолжаться до тех пор, пока давление набухания не урав-
новесит внешнего давления, то есть не достигнет 14 атм.
3. Заменим часть тела раствором, осмотическое давление которого
должно равняться 14 атм, причем раствор отделим от тела полупрони-
цаемой перепонкой. Равновесие сохранится. Полученная модель дает
представление о насыщенной водой клетке, обладающей сосущей силой,
равной 0, и тургором, равным 14 атм, что соответствует осмотическому
давлению раствора. Набухающее тело, соответствующее слою протоплаз-
мы, выстилающей оболочку клетки изнутри, не набухло до максималь-
1 У цветковых растений, не ассимилирующих СО2 , устьиц почти нет. Интенсив-
ность транспирации у них невелика (R. Hellferich, «Uber die Transpiration der
Bliiten», 32, 1942, S. 493, 516). Кислород, необходимый для дыхания, может из-за
высокого парциального давления воздуха диффундировать сквозь кутикулу. У гетеро-
трофных цветковых растений устьица также редуцированы.
Введение
49
ного уровня, хотя абсолютная упругость пара в клетке вследствие давле-
ния груза на раствор равно этой же величине в воде.
4. Степень набухания не изменится, если мы уменьшим тургор
и соответственно увеличим концентрацию наружного раствора.
5. Степень набухания не изменится и в том случае, если тургор
будет снят совершенно, а концентрация наружного раствора будет соот-
ветствовать концентрации внутреннего раствора.
Во всех этих случаях степень гидратации набухающего тела (про-
топлазмы) останется прежней, пока не изменится концентрация внутрен-
него раствора (клеточного
сока). Набухание не дости-
гает максимального уровня.
Можно привести и пря-
мое доказательство того,
что протоплазма в насы-
щенных водой клетках не
достигает предела набуха-
ния. Для этого растворим
нежные стенки клеток ги-
фов грибов ферментами из
желудочного сока виноград-
ной улитки х. В этом слу-
чае объем обнаженной про-
топлазмы резко сокращает-
ся, а в чистой воде она силь-
но набухает и лопается.
Только в 12,5%-ном раство-
ре сахарозы с осмотическим
давлением, равным 10 атм,
шарики протоплазмы сохра-
няют свою жизнедеятель-
ность (см. пятый вариант
опыта); в протоплазме обра-
зуются вакуоли, и из слоя
протоплазмы, содержащего
ядро, вырастают новые
гифы.
Теперь рассмотрим во-
прос о том, как можно
определить степень гидра-
тации протоплазмы. Обычно
считается, что коллоиды
настолько сильно связывают
воду, что гидратация при
колебаниях влажности из-
меняется мало. Это непра-
ВИЛЬНО.
Р и с. 10. Термодинамический опыт, показываю-
щий, что протоплазма, находящаяся в насыщен-
ных водой клетках, не достигает максимальной
степени набухания.
Кривая, отражающая зависимость гидратации неживых набухающих
тел от относительной влажности, всегда имеет S-образную форму (рис. 11).
Если принять, что максимальное набухание тела происходит при 100%-ной
1 Ch. S I г u п с k, Uber die Entstehung und Reversion enzymatisch erzeugter
Protoplasten, «Biol. Rundschau (Jena)», 3, 1965, S. 242—244.
50
Глава 1
влажности, то при постепенном уменьшении влажности тело отдает боль-
шую часть поглощенной воды. Лишь при влажности воздуха около 96%,
что в растворах соответствует осмотическому давлению в 55 атм (или
понижению точки замерзания на 4,5°), на кривой намечается изгиб.
Это означает, что остаточная влага удерживается сравнительно сильно,
однако говорить о «связанной влаге» в данном случае нельзя г. Установ-
лено, что у лишенных вакуолей клеток, у которых можно измерить объем
протоплазмы, например у карпоспор Lemanea, кривая набухания живой
протоплазмы имеет такую же форму; последняя почти полностью соот-
ветствует кривой набухания раствора, образованного из равных частей
нуклеина, казеина и желатина (рис. 12) 1 2. Физиологические пределы
возможного набухания протоплазмы у гомойогидрических растений наблю-
даются при относительной влажности от 94,7 до 96%, что соответствует
осмотическому давлению, равному 4—55 атм. Более высокие значения
осмотического давления очень редко наблюдались у негалофитов, обычно
эти величины граничат с летальными.
Объем протоплазмы в клетках, обладающих вакуолями, измерить
нельзя. Поэтому невозможно и непосредственное определение изменений
степени гидратации; это можно выяснить только косвенным путем,
поскольку существуют совершенно определенные соотношения между
степенью гидратации и водопоглащающей способностью, которые выра-
жаются в относительной влажности (рис. 12). При колебаниях влажности
в пределах 100—96% эта зависимость имеет почти линейное выражение.
G другой стороны, существует также зависимость между относительной
влажностью и осмотическим давлением (потенциальным осмотическим
давлением л*) раствора:
л*0= —3067 log pIpQ атм,
где p/pQ — относительная упругость пара в термодинамическом смысле,
то есть при изотермических или изобарических условиях. Осмотическое
давление клеточного сока, определяющее степень гидратации протоплаз-
мы, дает нам представление о степени ее набухания, выражаемой при
помощи относительной влажности. Знак — (минус) означает, что суще-
ствует обратная зависимость, иными словами при повышении осмотиче-
ского давления понижается относительная влажность и наоборот. Степень
набухания протоплазмы, определяемую путем измерения относительной
влажности, мы предложили обозначать термином «гидратура», причем
окончание «атура» означает состояние активности (так же как темпера-
тура означает термодинамический параметр состояния тела). Эта вели-
чина безразмерна и не зависит от условий внешней среды; она опреде-
ляется только осмотической концентрацией раствора или состоянием
набухания тела.
Гидратура соответствует следующей термодинамической величине:
а = относительная активность воды (в %).
В гл. 7 (стр. 320—328) эти вопросы рассматриваются более подробно.
Для того чтобы можно было судить об условиях водного режима,
в которых растение существует в естественных условиях, нужно в первую
очередь выяснить, каким образом растение поддерживает наиболее высо-
кий из возможных уровней гидратуры протоплазмы. Об этом можно
1 Это относится также к раствору сахарозы (рис. 11, кривая 1).
2 Н. Walter, Plasma- und Membranquellung bei Plasmalyse, «Jb. wiss. Bot.»,
62, 1923, S. 145—322.
Рис. 11. Набухание нуклеина (2) в зависимости от относительной
упругости пара (по Катцу). Для сравнения приведена кривая (1),
отражающая процесс растворения сахарозы (содержание воды на 1 г
сухого вещества вычислено таким же путем).
Рис. 12. Набухание протоплазмы и неживых тел в зависимости от
относительной влажности воздуха, осмотического давления пли
понижения точки замерзания.
52
Глава 1
судить на основании осмотического давления клеточного сока. Чем выше
давление, тем ниже уровень гидратуры протоплазмы и тем более небла-
гоприятным оказывается водный режим. Учитывая это, у одних растений
устьица находятся в открытом состоянии, с тем чтобы усилить ассими-
ляцию СО2, необходимую для фотосинтеза. Другие растения не ассими-
лируют СО2 и устьица у них находятся в закрытом состоянии, поэтому
у этих растений наблюдается низкое осмотическое давление и высо-
кая гидратура протоплазмы.
В связи с необходимостью поддерживать на постоянном уровне вод-
ный режим и интенсивный газообмен у растений выработались сложные
приспособления, механизм которых связан с передвижением в них воды.
Взаимозависимость между поглощением, передвижением и отдачей
воды определяет степень концентрации клеточного сока, которая должна
оставаться на возможно более низком уровне; это определяет высокий
уровень гидратуры протоплазмы и тем самым способствует ее активности.
Транспирацию следует рассматривать прежде всего как неизбежное зло,
которого растение не может избежать из-за газообмена, осуществляемого
листьями. Передвижение минеральных веществ от корней к листьям
не требует такого большого расхода воды, для этого вполне достаточно
было бы лишь слабой гуттации. Отсюда вытекает также, что интенсивное
накопление вещества всегда связано с большими потерями воды. Быстро-
растущие растения всегда отличаются высокой интенсивностью транспи-
рации.
Все вышеизложенное характеризует лишь растения, растущие на
незасоленных почвах. Если в почве содержатся легко растворимые соли,
например NaCl, растения поглощают некоторое количество этих солей,
и они накапливаются в клеточном соке листьев. У солеустойчивых расте-
ний, называемых галофитами, NaCl способствует набуханию протоплаз-
мы, поэтому более высокое осмотическое давление не понижает ее гидра-
туры. Галофиты растут очень хорошо и при осмотическом давлении,
превышающем 50 атм; некоторое количество соли даже способствует
их росту. В этом случае, помимо величины осмотического давления,
следует еще определять содержание солей (ср. гл. 7, стр. 328—331).
Самые примитивные из Cormophyta — мхи — плохо защищены от
высыхания и поэтому растут лучше всего в условиях, где влажность
воздуха держится на постоянно высоком уровне. Если же они развивают-
ся в засушливых условиях местообитания, значит, они вторично верну-
лись к пойкилогидрическому образу жизни. У этих ксерофитных видов
мхов развиваются маленькие клетки без вакуолей, вследствие чего прото-
плазма, как у некоторых примитивных групп водорослей, может пере-
носить высыхание (рис. 9).
К жизни на суше лучше приспособились папоротники, но и у них
водопроводящая система еще далеко пе совершенна. Поэтому они и раз-
виваются преимущественно во влажных местообитаниях; несмотря на это,
осмотическое давление у них отличается относительно высокими значе-
ниями (см. стр. 132, 151 — 152). Этим объясняется, почему в период пер-
ми — карбона, то есть во времена широкого распространения папорот-
никообразных, сухопутные растения были приурочены почти исклю-
чительно к заболоченным местностям, тогда как в засушливых условиях
растения почти не встречались х. Папоротники, распространенные в на-
1 Н. Е rh а г t, Temoins pedogenetiques de 1’epoque, Permo-Carbonifere, «Comp.
R. Seanc. Soc. Biogeogr.», 335—337, 1962, p. 21—53.
Введение
53
стоящее время в аридных областях (см. рис. на стр. 359), как и мхи, вто-
рично вернулись к пойкилогидрическому образу жизни. Эти растения
обладают крупными клетками, в клеточном соке которых содержатся
различные вещества (например, дубильные вещества — флороглюцин),
затвердевающие при обезвоживании; поэтому объем клеток при высыха-
нии мало изменяется. В других случаях оболочки клеток легко деформи-
руются, и листья при высыхании свертываются, однако и в этом случае
протоплазма сохраняет свою жизнедеятельность. Среди голосеменных
пойкилогидрические растения не встречаются, поскольку у них сосу-
дистая система образована только трахеидами. Эти растения для под-
держания соответствующего уровня гидратуры протоплазмы должны
испарять как можно меньше воды и поэтому отличаются относительно
ксероморфным строением.
Среди покрытосеменных пойкилогидрические растения встречаются
в качестве исключения. Один из этих видов, Myrothamnus flabelltfolia
{Rosales) распространен в Южной Африке (см. гл. 9, стр. 400).
Различные экологические группы засухоустойчивых растений рас-
сматриваются в гл. 7 (стр. 320—328).
КЛИМАТИЧЕСКИЕ ЗОНЫ ЗЕМНОГО ШАРА
Взаимосвязи между климатом и растительным покровом самые тес-
ные. Растительность есть в известной мере отражение соответствующего
климата, поэтому ее можно использовать в качестве косвенного показа-
теля климата для выделения отдельных климатических областей. Не
пытаясь здесь излагать основы климатологии, ниже мы остановимся лишь
на важнейших особенностях климатов х.
Климатом мы называем совокупность погод-
ных условий в их среднем годовом и суточном
ходе. Для эколога важны прежде всего факторы, оказывающие наиболь-
шее влияние на развитие растений, — температура и гидратура.
Так, для ботаника температура в СС не служит показателем коли-
чества тепла, а является величиной, отражающей тепловое состояние
воздуха. Последнее оказывает большее влияние на распространение
растений, чем количество тепла, которое используется в процессе
обмена веществом и энергией.
Это же относится и к фактору воды, поскольку для растений более
важна не абсолютная влажность, а состояние воды, которое мы называем
гидратурой; мерилом гидратуры является относительная упругость пара,
измеряемая в процентах гидратуры (% hy), причем относительная упру-
гость пара чистой воды равна 100% гидратуры. Гидратура растения
зависит не только от прихода воды, но и от ее расхода, то есть от водного
баланса. Этот вопрос подробно рассматривается в гл. 7.
Для правильной оценки влияния водного фактора на растение необ-
ходимо наряду с количеством осадков учитывать также суммарное испа-
рение.
Для распределения тепловых условий на Земле характерны следую-
щие закономерности:
1 W. Кбрреп, Grundri|3 der Klimakunde, Berlin und Leipzig, 1931. [Имеется
русский перевод: В. К ё п п е н, Основы климатологии (климаты земного шара),
Учпедгиз, М., 1938.] Б. П. Алисов, Климатические области зарубежных стран,
1950; Б. П. А л и с о в, О. А. Дроздов, Е. С. Рубинштейн, Курс климато-
логии, ч. I и II, Л., 1952.
54
Глава 1
1. Средняя годовая температура воздуха понижается от экватора
к полюсам, так как количество солнечной радиации, приходящееся на
единицу поверхности, убывает в этих направлениях из-за уменьшения
угла падения солнечных лучей.
2. Температурные различия времен года от экватора к полюсам,
напротив, возрастают в соответствии с сезонными изменениями прихо-
дящей радиации.
Сравнив средние показатели температуры для отдельных широт
северного и южного полушарий, легко заметить, что уменьшение средних
годовых значений температуры в южном полушарии к югу происходит
быстрее, тогда как сезонные различия увеличиваются не столь резко.
Объясняется это меньшей площадью суши в южном полушарии и смягчаю-
щим влиянием океанов.
3. В силу большей подвижности воды и ее высокой теплоемкости
сезонные, суточные и годовые амплитуды температуры над морем меньше,
а над сушей больше. Поэтому с удалением от морского побережья климат
становится все более и более континентальным и смена времен года про-
исходит несколько раньше.
4. Зимой вода, покрытая толстым слоем льда, охлаждается так же,
как суша. Снежный покров на льду вследствие сильного излучения уве-
личивает охлаждение воды еще больше, и, наоборот, весной снег и лед
задерживают ее прогревание.
5. Ветры и морские течения в зависимости от своего направления
оказывают влияние на распределение температуры; с полюсов приносятся
холодные массы воздуха или воды, с экватора — теплые. Над субтропи-
ческими западными побережьями континентов, омываемыми холодными
морскими течениями — Калифорнийским, Перуанским, Канарским и Бен-
гальским,— образуются туманы, напротив, теплые течения — Куро-Сиво,
Игольное, Бразильское и Гольфстрим,— проходящие вдоль восточных
берегов континентов, обусловливают здесь частые грозы и проливные
дожди. В местах встречи теплых течений с холодными водными массами
высоких широт, например у Ньюфаундлендской банки и в Охотском
море, почти круглый год стоит туман. >
Ветры, дующие с моря, выравнивают температуру над сушей, иЛна
приближается к температуре над поверхностью воды. В умереншхй^зоне
господствуют западные ветры, которые определяют более мягкйи климат
в западных частях материков. 7
6. Горы, преграждающие путь ветрам, могут изменить местные тем-
пературные условия в ту или другую сторону.
7. В горах с увеличением высоты средние температуры понижаются,
а экстремальные возрастают; однако в долинах и на вершинах возможны
временные инверсии температуры.
Значительно сложнее на поверхности земли складываются гидра-
турные соотношения (условия увлажнения).
Распределение осадков зависит от общей циркуляции атмосферы.
Под влиянием сезонных областей низкого или высокого давления ветры
над сушей отклоняются от своего первоначального направления, образуя
так называемые муссоны.
Не зная закономерностей общей циркуляции атмосферы, нельзя
понять характер распределения осадков. Последнее же весьма существен-
но, поскольку в тропических и субтропических зонах времена года опре-
деляются не столько ходом температуры, сколько чередованием дождли-
вых и засушливых периодов.
Введение
55
Рис. 13. Воздушные течения в период равноденствия
(по Гебауэру).
Рассмотрим прежде всего условия атмосферной циркуляции для
весны или осени (рис. 13). В это время солнце стоит на экваторе в зените,
что обусловливает здесь сильный прогрев воздушных масс и их адиаба-
тический подъем, приводящий к падению атмосферного давления у по-
верхности земли. Поднимающийся воздух охлаждается, содержащиеся
в нем водяные пары конденсируются, и вода выпадает на землю в виде
зенитальных дождей. Благодаря теплоте конденсации поднимающиеся
воздушные массы остаются более теплыми, чем окружающий их воздух.
На больших высотах поднявшиеся на экваторе воздушные массы растекают-
ся к северу и югу и приблизительно на 40° широты снова опускаются
на землю. Нисходящий воздух нагревается и при этом становится очень
сухим. Вследствие этого осадков здесь мало, атмосферное давление у зем-
ной поверхности всегда высокое, погода обычно безветренная.
Из субтропических зон высокого давления воздух вновь устремляется
к экваториальной зоне низкого давления. Вследствие вращения Земли
ветры отклоняются в северном полушарии вправо, а в южном — влево,
образуя соответственно северо-восточный и юго-восточный пассаты.
Вблизи экватора располагается зона затишья со слабыми переменными
ветрами. В более высоких широтах (50—60°) господствуют западные
ветры, особенно устойчивые в южном полушарии. В полярных областях
вновь преобладает восточный перенос. Здесь, на границе между холодным
воздухом полярных широт и более теплыми воздушными массами низких
широт, непрерывно возникают мощные вихревые движения, которые
в северном полушарии формируются главным образом около Исландии
и в районе Алеутских островов. Циклоны, постепенно заполняясь, пере-
мещаются с запада на восток, иногда отклоняясь к югу Ч В качестве
явления, сопровождающего циклоны, следует отметить фронтальные
1 В южном полушарии, например южнее Австралии, это главным образом пере-
мещающиеся антициклоны, разделенные ложбинообразными областями низкого дав-
ления.
56
Глава 1
дожди, образующиеся на стыке холодных и теплых воздушных масс.
В отличие от зенитальных дождей, идущих только в теплое время года,
циклонические осадки не связаны с каким-либо определенным сезоном,
а выпадают лишь при пересечении данного района циклоном. Поскольку
высота стояния солнца в течение года меняется (летом в северном полу-
шарии оно находится в зените над северным тропиком, а зимой, то есть
летом южного полушария,— над южным тропиком), происходит смещение
системы атмосферной циркуляции (рис. 14). В связи с перемещением
тропических фронтов летом северного полушария на север, а летом южно-
го полушария на юг сдвигается и тропическая зона затишья. Так как
атмосферная циркуляция в южном полушарии развивается интенсивнее,
чем в северном, зона затишья перемещается главным образом между О
и 10° с. ш. Пути движения циклонов также обнаруживают соответствую-
щие уклонения: в северном полушарии зимой циклоны проникают на юг
почти до 40° с. ш., летом же их пути располагаются севернее.
Выделяют следующие главные климатические зоны:
I. Экваториальная зона — приблизительно между 10° с. ш. и 10° ю. ш.
с двумя дождливыми периодами (соответственно положению солнца
в зените), прерывающимися лишь на короткое время. Годовое количество
осадков и влажность весьма велики, месячные температуры различаются
незначительно.
Зима
Р и с. 14. Распределение областей осадков по сезонам (Н. Walter, Stan-
dortslehre, Stuttgart, 1960).
1 — зенитальные дожди; 2 — циклонические осадки. Количество осадков пропорцио-
нально длине штрихов.
II. Тропическая зона —севернее и южнее первой, примерно до 30°
широты. Характеризуется летним дождливым периодом (совпадающим
со стоянием солнца в зените или несколько запаздывающим) и засушли-
вым периодом в течение прохладного времени года. Годовое количество
Введение
57
осадков уменьшается с удалением от экватора; морозы чрезвычайно
редки, влажность воздуха низкая.
III. Зона сухих субтропиков — вблизи конских широт1. Сумма
осадков и влажность воздуха незначительны, однако годовые и суточные
колебания температуры вследствие сухости воздуха и сильного излуче-
ния велики. Наблюдаются ночные заморозки.
IV. Переходная зона с сезоном зимних дождей. Летом она совпадает
с зоной высокого давления и нисходящих сухих воздушных масс, зимой
же ее пересекают циклоны. Морозы бывают чаще, чем в предыдущей
зоне, но собственно холодное время года в этой зоне отсутствует; лето
жаркое. Этот климат характерен для Средиземноморья, Калифорнии,
Южной и юго-западной Австралии, Капской области и юго-запада Южной
Америки.
V—VIII. Умеренная зона с циклоническими дождями, возможными
в любое время года. Годовое количество осадков сокращается с удале-
нием от океана, причем все более выделяется летний максимум осадков.
Одновременно увеличиваются годовые колебания температуры: лето в пре-
делах материков становится более жарким, а зима — более холодной.
Эту зону, охватывающую столь различные для растительности кли-
матические условия, следует подразделить на ряд подзон.
V. Подзона теплого умеренного климата, где собственно зимний
период почти не выделяется и все времена года более или менее влажные.
VI. Подзона типичного умеренного климата с холодной, но не слиш-
ком продолжительной зимой, например в Центральной Европе, или с почти
безморозной зимой, но с очень прохладным летом (резко выраженный
океанический климат).
^VII. Подзона аридного умеренного климата континентального типа,
с резкими температурными контрастами, небольшой суммой осадков
и незначительной влажностью воздуха.
VIII. Подзона бореального, или холодного, умеренного климата,
характерного для обширной зоны, охватывающей Северную Америку
и северную часть Евразии. Лето прохладное и влажное, холодная зима
длится более половины года.
IX. Арктическая (антарктическая) климатическая зона. Незначи-
тельное количество осадков, преимущественно выпадающих в виде снега
и приблизительно равномерно распределяющихся в течение года. Летом
наблюдается полярный день; лето короткое и прохладное, влажность
воздуха обычно высокая. Зона переходит в собственно полярную область,
в которой произрастание растений невозможно.
Указанная зональность климатов осложняется муссонной деятель-
ностью, развивающейся прежде всего на южной и восточной окраинах
Азии; самого большого материка земного шара. Здесь между 10 и 23° с. ш.,
от Восточной Африки до Филиппинских островов, в летние месяцы гос-
подствуют юго-западные и юго-восточные муссоны; низкое давление над
Азиатским материком и высокое давление, наблюдающееся в это время
(зима в- южном полушарии) над Южной Африкой и Австралией, еще
более усиливает их деятельность. Северо-восточный пассат совершенно
не проявляется летом и дует только зимой (при противоположном распре-
делении давления над материками) в виде муссона, проникающего на юг
1 Это широты около 30—35° над океанами; точнее,— области во внутренних
частях субтропических океанических антициклонов со слабыми ветрами и частыми
штилями.— Прим, ред.
58
Глава 1
за экватор. Такого рода муссонные ветры, правда менее резко выражен-
ные, чем в Индийском океане, формируются также в северной Австралии,
у западного побережья Африки (Либерия, Сьерра-Леоне), в Гвинейском
заливе (Камерун), а также на восточном побережье Северной Америки.
На западном побережье Европы они усложняются воздействием циклонов.
Повсеместно, где ветры при переходе с моря на сушу встречают
на своем пути горы, на наветренной их стороне идут обильные дожди,
связанные с подъемом воздушных масс; на подветренной же стороне
стоит относительно сухая погода. Ветры, постоянно дующие в одном
направлении, например пассаты, обусловливают очень резкие климати-
ческие различия на подветренной и наветренной сторонах гор.
Влага, выпадающая на сушу в виде атмосферных осадков, частично
испаряется и вновь возвращается на землю в виде дождя, а частично
стекает обратно в море. Общее количество этой воды на всем земном
шаре определяется в 37 тыс. км3 в год. Примерно в два раза больше испа-
ряется влаги с суши и приблизительно в восемь раз — с поверхности
морей. Трехкратное количество выпадает в виде осадков на сушу и семи-
кратное — на море. Такова общая картина круговорота воды на земле.
Совокупное влияние всех перечисленных факторов с учетом местных
особенностей обусловливает распределение осадков, иллюстрируемое
рис. 15. Обильные осадки выпадают прежде всего там, где на пути насы-
щенного водяным паром ветра встают горы, вызывающие подъем воздуш-
ных масс. Это характерно для побережий Гвианы и Бразилии (от Сан-
Роки до Кабу-Фриу), для восточных берегов Мадагаскара и Австралии,
юго-восточной Новой Гвинеи, где дует пассат. Такие же условия харак-
теризуют юго-западный муссон (в теплое время года) в Либерии, Сьерра-
Леоне на Малабарском берегу, в провинции Аракан, в Тенассериме,
южный муссон — на юго-восточных побережьях Китая, Японии и Кам-
чатки; северо-западный муссон (в холодное время года) — на северо-
западе Японии; северо-восточный муссон — на восточных берегах Тайваня,
Филиппинских островах и Индокитае; северо-западный муссон — в во-
сточной части Малайского архипелага и северный муссон — на северо-
восточном побережье Новой Гвинеи. Орографические осадки наблюдаются
также на наветренной стороне гор океанических островов, стоящих на
пути пассатов или муссонов. При сильных ветрах с моря даже слабо
наклоненные обширные поверхности могут оказать тормозящее влияние
и вызвать подъем воздушных масс на большую высоту. Именно орогра-
фическим эффектом объясняется обилие дождей на равнинах Амазонки,
нижней Брахмапутры и в окаймляющих их горах. Как правило, горы
всегда более влажны по сравнению со своим окружением и часто окутаны
облаками. Только на больших высотах снова становится заметным сокра-
щение количества осадков. Высочайшие, нередко покрытые снегом вер-
шины обычно прорывают облачный покров (пик Тенерифе, гора Каме-
рун, Килиманджаро и др.), представляя необыкновенно живописную
картину в лучах заходящего солнца.
Для оценки климата в целом важны и другие климатические харак-
теристики. Однако их прямое экологическое значение не столь велико,
так как проявляются они косвенно, частично обусловливая температуру
и увлажнение. Можно было бы произвести еще более дробное деление
перечисленных выше основных климатических зон, однако сделать это
далеко не просто. Даже при наличии очень точных карт отдельных кли-
матических элементов для целых материков или обширных областей,
например карт средних годовых температур, январских и июльских
Рис. 15. Карта годовых осадков (Н. Walter, Standortslehre, Stuttgart. 1960).
60
Глава 1
значений, амплитуд температуры, длительности засух, распределения
осадков по временам года, показателей аридности или суммарного испа-
рения ит. д., весьма трудно установить особенности климата конкретного
участка или местности.
Поэтому уже давно предпринимались попытки определить общий
климат (Gesamtklima) отдельных областей, не только выделяя при этом
крупные климатические зоны, но и учитывая особенности климата,
обусловленные размещением морей и континентов и воздействием ветров.
Наиболее детально разработанной климатической классификацией,
основанной на учете режима температуры и осадков и их сезонного рас-
пределения, до настоящего времени остается классификация Кёппена.
Большинство географических работ базируется именно на ней, хотя
в последнее время нередки ссылки на классификацию климатов, предло-
женную Торнтвейтом. В настоящей книге выбран иной путь; тип климата
характеризуется не определенными значениями отдельных его элементов,
например температурой, осадками, суммарным испарением, длительно-
стью засушливых периодов и т. д., а соответствующими климадиаграм-
мами, к рассмотрению которых мы и перейдем.
ГРАФИЧЕСКОЕ ИЗОБРАЖЕНИЕ КЛИМАТА
С ПОМОЩЬЮ КЛИМ А ДИАГРАММ
«Погодой» называют совокупность значений метеорологических эле-
ментов в данный момент. Средний ход погоды в течение года определяет
климат. Следовательно, для установления типа климата необходимо
произвести два интегрирования. Однако ни в одном из учебников климато-
логии не указана методика этих вычислений. Не помогают и самые под-
робные климатические таблицы. Сводные колонки цифровых данных
слишком громоздки, что затрудняет их сравнение. Правильное представ-
ление о климате можно составить, лишь хорошо зная сезонный ход пого-
ды, иными словами, если точно известны длительность и интенсивность
влажного и засушливого времен года или теплого и холодного сезонов.
Таким образом, возникла необходимость разработки нового подсобного
для экологии метода — метода графического изображения климата,
позволяющего с одного взгляда выявить характерные особенности того
или иного типа климата, его различия или сходство с климатами других
местностей.
Указанным требованиям отвечают к л и м а д и а г р а м м ы, на
которых отчетливо видны сезонные изменения климатических условий.
При составлении климадиаграмм используют предложенное Госсеном 1
соотношение между средней месячной температурой и количеством осад-
ков, как 1 : 2. Иными словами, 10° С соответствуют 20 мм осадков. При
таком масштабе (Т : Ос = 1 : 2) засушливая часть года характеризуется
кривой осадков, расположенной ниже температурной кривой. Сравнение
этих двух кривых полезно, поскольку температурная кривая, как уже
отмечалось, может служить показателем годового хода испарения. Она
отражает расход влаги, а кривая осадков — ее приход; обе кривые дают
представление о характере водного баланса данной территории. Непо-
средственно измеренные величины испарения имеются лишь для немногих
1 F. Bagnouls, Н. Gaussen, Saison seche et Indice Xerothermique, Docum.
Cartes Product. Veget. Serie General, t. Ill, vol. 1, art. VIII, 1953, p. 1—48; H. G a u s-
s e n, Experession des milieux par des formules ecologiques; leur representation carto-
graphique, «Colloques Intern, du Centre Nat. Recherche Scient.», 59, 1955, p. 257—269.
Введение
61
станций, поэтому в большинстве климатических формул для характери-
стики условий увлажнения используется соотношение между темпера-
турой и осадками. Величина этого соотношения, определяющая засуш-
ливые и влажные условия, принимаемая различными авторами, не отли-
чается сколько-нибудь заметно от соотношения 1:2, предложенного
Госсеном. К тому же следует учитывать, что резкой границы между засуш-
ливым и влажным временами года не существует. Поэтому любое приня-
тое соотношение в известной мере произвольно, важно лишь, чтобы пра-
вильно были отражены истинные условия.
Однако, помимо изображаемых на диаграмме периода засухи и таких
важных для развития растений элементов климата, как годовой ход тем-
пературы и осадков, не следует забывать и о других не менее существен-
ных для растения факторах, как, например, длительность и режим холод-
ного времени года, для характеристики которых одних лишь средних
месячных данных недостаточно. Морозы наблюдаются и в те месяцы,
средняя температура которых превышает 0°; ранние или поздние, а также
совершенно случайные заморозки имеют для растений особенно большое
значение как фактор отбора. Следовательно, на климадиаграмме должны
найти отражение и эти явления. С другой стороны, климадиаграмму
не следует перегружать подробностями, иначе она утратит свою нагляд-
ность, особенно если климадиаграмма применяется для характеристики
обширных территорий. На практике вполне себя оправдал способ изоб-
ражения, показанный на рис. 16 *.
В дальнейшем оказалось, что графическое изображение засушливого
периода, по Госсену, может быть применено лишь для характеристики
тех условий, где в определенное время года количество выпавших осадков
резко сокращается; однако этот метод теряет свою наглядность при харак-
теристике климатических условий в районах с более равномерным распре-
делением осадков в течение года. Например, на климадиаграмме Одессы
(рис. 16) период засухи почти не выражен. Тем не менее лето в этом степ-
ном районе достаточно сухое, хотя и не столь засушливое, как в Анато-
лии и Средиземноморье. В подобных случаях следует различать засуш-
ливый и полу засушливый периоды, последний можно изобразить графи-
чески; желая при этом подчеркнуть значение испарения, берут соотношение
между температурой и осадками 1 : 3, что соответствует, по Селянинову,
степной области. При этом масштаб для температуры оставляют преж-
ним, считая одно деление в 10° С равным 30 мм осадков. Данная кривая
осадков (прерывистая линия) располагается несколько ниже, чем кривая
осадков при соотношении 1 : 2. Площадь, заключенная между этой кривой
и расположенной над ней кривой температуры, отражает продолжитель-
ность (по горизонтали) и интенсивность (по вертикали) полузасушливого
периода. Диаграмма показывает, что в районе Одессы сухой период
наблюдается только весной и поздним летом. Полузасушливый же период,
напротив, характерен для всего лета.
Во влажных тропиках месячная сумма осадков достигает в некото-
рых местностях нескольких сотен миллиметров. Сохраняя и в этом случае
обычный масштаб, мы получили бы слишком громоздкие диаграммы.
Поэтому целесообразно уменьшить масштаб для осадков, превышающих
100 мм в месяц, в 10 раз и закрашивать соответствующую площадь черным
цветом (рис. 16, Дуала). Черная окраска, таким образом, характеризует
так называемое переувлажненное время года с месячной суммой осадков
1 Н. Walter, Н. L i е t h, Klimadiagramm — Weltatlas, Jena, 1960; 1964.
62
Глава 1
Хоэнхейн (402 м)
. [50-40]
Р и с. 16. Примеры климадиаграмм.
а — станция; Ъ — высота над уровнем моря; с —
число лет наблюдений за температурой (первая
цифра) н осадками (вторая); d — средняя годовая
температура; е — средняя годовая сумма осадков;
/ — средний суточный минимум самого холодного
месяца; g — абсолютный минимум; /г — средний
суточный максимум самого теплого месяца; i — аб-
солютный максимум; j — средняя суточная ампли-
туда температуры; k — кривая средних месячных
температур (1 деление=10°); I — кривая средних
месячных осадков (соотношение 10° С = 20 лии);
т — засушливый период; п — влажное время года;
о — среднее месячное количество осадков, превыша-
ющее 100 мм (масштаб сокращен до 1/ю); Р — кри-
вая осадков (соотношение 10° С=30 мм), площадь
с горизонтальной штриховкой над ней — полуза-
сушливый период; д — месяцы со средним суточ-
ным минимумом температуры ниже 0°; г—месяцы
с абсолютным минимумом температуры ниже 0°; s —
число дней со средней суточной температурой вы-
ше 0°, пли средняя продолжительность безмороз-
ного периода в днях.
более 100мм. Такое большое количество
влаги существенной роли для растений
не играет, так как основная часть ее
расходуется на поверхностный сток.
На диаграммах, характеризующих
влажные области, не выделяются ни по-
лузасушливые, ни засушливые периоды:
кривая температуры всегда располага-
ется ниже верхней (при соотношении
1 : 2) и ниже нижней (при соотноше-
нии 1 :3) кривых осадков (рис. 16, Хо-
энхейм). Областям с достаточным увлаж-
нением присущи совершенно различные
типы климата, например климаты ле-
сов (лиственных и хвойных), климат
лесотундры, тундры и большей части
полярных районов.
Изучение соответствующих диа-
грамм Евразии показало, что на основа-
нии анализа только режима осадков
нельзя провести более дробного рас-
членения территории. В условиях влаж-
ного климата, характеризующегося до-
статочным увлажнением в течение всего
года, значительно большее значение
имеют температурные условия, опреде-
ляющие возможность произрастания
здесь лиственных или хвойных лесов
или смену леса тундрой. В качестве
критерия для такого расчленения нель-
зя использовать ни среднюю годовую
температуру, ни средние или абсолют-
ные минимумы температуры. Более
удачным критерием оказалась продол-
жительность вегетационного периода.
Правда, вегетационный период —
термин довольно неопределенный и тре-
Введение
63
бует более точной интерпретации. Это может быть и безморозный
период, и время года со средней суточной температурой более 5, 10 или
15° С. В качестве показателя при районировании лучше всего исполь-
зовать период со средней суточной температурой более 10° С. Его обычно
определяют по кривым годового хода температуры воздуха. Для выде-
ления зоны субарктических лиственнично-кедровых лесов имеет значение
продолжительность периода со средней суточной температурой ниже
—10° С. Такие леса встречаются в очень холодных континентальных
областях с суровыми зимами, поэтому на графиках, характеризующих
Таблица 1
Название станций Температура возду- ха, °C Название станций Температура воз- духа, *°С
средний минимум абсолют- ный ми- нимум средний минимум абсолют- ный ми- нимум
Лесная зона Степная зона
Киев . -8,8 -30,0 Николаев -6,8 -28,7
Тула . -14,3 -38,4 Мелитополь -7,5 -30,0
Лесостепная зона Умань Харьков Воронеж -9,5 —12,2 —14,1 -33,0 -36,9 -36,5 Маргаритовка Николаевское Оренбург Полупустыня -9,3 -17,1 -19,1 —33,0 -41,0 —41,7
Тамбов . -15,3 —38,7 Бердянск -6,3 -26,5
Пенза . —16,2 —41,4 Астрахань -10,6 -29,8
Ульяновск —17,2 -39,6 Волгоград -13,2 -34,6
Камышин -15,1 —37,2
эти климатические условия, необходимо указывать также продолжитель-
ность сезона с температурой выше —10° С. Примеры соответствующих
диаграмм приведены на рис. 40—43.
При анализе климадиаграмм следует избегать одностороннего под-
хода. Их преимущество состоит именно в том, что они помогают пред-
ставить себе климат как одно целое. Это особенно важно при экологи-
ческих исследованиях, требующих оценки по совокупному воздействию
различных элементов климата, а не по одному какому-либо признаку,
как, например, по длительности засушливого периода.
Для составления климадиаграмм используют средние многолетние
данные, причем число лет наблюдений указывают на графике под назва-
нием станции. Мы получаем, таким образом, тип климата, от средних
значений которого в отдельные годы возможны резкие отклонения.
Отдельные сезоны бывают то холоднее, то теплее «нормы», то они слиш-
ком влажны, то чрезмерно засушливы. Однако развитие растительного
покрова определяется не средними условиями, а действительными, особен-
но когда дело идет о крайних условиях существования.
Климадиаграммы используются для характеристики отдельных типов
климата и их связи с растительностью по следующим соображениям:
1. Естественный растительный покров, находящийся в динамиче-
ском равновесии с окружающей его средой, формировался в течение
многих столетий. На протяжении этого времени состав его испытывал
64
Глава 1
°C нм
°C мм
°C нм
Р и с. 17. Климатограмма’Хоэнхейма (под Штутгартом) ла 50 лет Холодные зимы—
Weltatlas.
изменения около каких-то средних значений, соответствовавших средним
климатическим условиям.
2. Между многолетними средними и экстремальными значениями
в пределах данной климатической области существует известная корре-
ляционная связь. Так, например, при частой повторяемости низких
температур средние минимумы также оказываются низкими; чрезвычайно
засушливым годам соответствуют пониженные годовые суммы осадков.
В табл. 1 приведены средние суточные минимумы температуры самого
холодного месяца и абсолютные минимумы для ряда станций Восточной
Введение
65
1928/29, 1941/42 и др,; сухие летние периоды — 1921, 1949 гг, («Klimadiagramm-
1960»).
Европы. За некоторым исключением, в пределах отдельных расти-
тельных зон заметна довольно высокая корреляция между показа-
телями.
При уменьшении средней многолетней годовой суммы осадков увели-
чивается отношение между количеством осадков самого влажного и самого
сухого годов и засушливые годы становятся более резко выраженными.
По Кёппену, это соотношение максимального годового количества осад-
ков (Мах) к минимальному (Min), полученное за 30—50-летний период
наблюдений для всего земного шара, равно:
66
Глава 1
Таблица 2
Средняя годовая сумма осадков, 200—400 400-600 600-800 1000-1300 Более 1600
Отношение Max/Min 4,4 3,3 2,9 2,4 2,1
Таким образом, то обстоятельство, что при использовании средних
значений учитываются также экстремальные, означает, что средние
величины могут быть применены для оценки климата в экологическом
аспекте, за исключением, правда, некоторых редких случаев, о чем будет
сказано ниже (гл, 12),
Однако такого рода данные применимы только при изучении естест-
венной растительности. Для сельскохозяйственных и лесохозяйственных
культур более существенное значение имеют погодные условия конкрет-
ного года, ибо от погоды зависят величина урожая, а также размеры ущер-
ба, причиняемого стихийными бедствиями многолетним культурам.
Разводить какую-либо древесную породу невыгодно, если она поги-
бает в результате неблагоприятных условий погоды раз в 50 лет или
сильно повреждается каждые 10 лет. Крестьяне или садоводы также
могут мириться лишь со случайными неурожаями. Им важно иметь пред-
ставление о том, как часто повторяются годы с крайне неблагоприятными
условиями погоды; статистические данные, полученные путем вычисления
многолетних средних, большой помощи здесь не окажут, И в этом случае
важно знать погоду отдельного года, иными словами, — совокупное влия-
ние важнейших метеорологических факторов. Представление о них можно
получить по климатограмме, которая по сравнению с климадиаграммоп
более полно отражает климатические условия того или иного района.
Составление климатограмм аналогично построению климадиаграмм:
в виде сплошной линии наносят средние месячные температуры и осадки
за все годы наблюдения и дополняют их другими данными. Следовательно,
климатограмма состоит из непрерывной полосы климадиаграмм за отдель-
ные годы. Приведем в качестве примера климатограмму Хоэнхейма близ
Штутгарта, района с влажным центральноевропейским климатом, за
1907—1956 гг, и климатограмму Анкары, расположенной в засушливых
степях Центральной Анатолии, за 1936—1953 гг. (рис. 17 и 18).
Введение
67
Выделяющиеся на климатограммах экстремально холодные или
экстремально сухие годы следует учитывать при культуре тех или иных
растений.
Приводить климатограммы большого числа станций невозможно, да
и нет необходимости, поскольку они нужны лишь для специальных иссле-
дований. Материалы по интересующему району лучше собирать на месте
и, основываясь на этих данных, самим составлять соответствующие кли-
матограммы.
Получить общее и наглядное представление о различных типах кли-
мата того или иного района помогают карты климадиаграмм, или, по
выражению Тролля, климакартограммы. Климадиаграммы отдельных
Анкары за 1936—1953 гг.
станций наносят на географическую карту. В этом случае участки со сход-
ными климадиаграммами выделяются очень четко и это дает возможность
легко провести границы между районами с определенными типами кли-
мата. При этом нетрудно заметить, что климатические границы отличают-
ся постепенными переходами (исключение представляют горные местно-
сти), в связи с чем между выделенными районами располагаются широкие
переходные зоны, отсутствующие на картах с одними только климати-
ческими границами. Последние получаются путем очень грубой интерпо-
ляции, тогда как на климакартограмме непосредственно видны пункты,
в которых произведены измерения, причем эти измерения приводятся
на соответствующей диаграмме без изменений. Таким образом, предлагае-
мый метод отличается рядом преимуществ и чрезвычайно прост. В настоя-
щее время климадиаграммы всех станций земного шара собраны в атлас
климадиаграмм
Естественно возникает вопрос, пригодны ли описанные выше клима-
диаграммы и климатограммы для характеристики всех климатических
областей земного шара? Климатические индексы и формулы удобны для
характеристики одного определенного района, но они совершенно не
годятся при сравнительном климатическом обзоре разных областей. Что
же касается климадиаграмм, то здесь положение иное, поскольку их
составление не основано на каких-либо предпосылках или предположени-
ях; требуется лишь графически изобразить данные, полученные на метео-
1 Н, Walter, Н. L i е t h, op, cit,
68
Глава 1
25
Р и с. 19—21. Тип
климата I. Экватори-
альный влажный кли-
мат в Колумбии, Ка-
меруне и Австралии
(переходный к ти-
пу П).
Рис. 22—24. Тип
климата II. Тропиче-
ский климат с летни-
ми дождями в Брази-
лии, Южной Африке
(возможны уже замо-
розки) и Австралии.
Р и с. 25—27. Тип
климата III. Аридный
субтропический пу-
стынный климат в Пе-
ру, Юго-Западной
Африке и Аравии.
Кейптаун (12я) tiffM
30
Рис. 28—30. Тип
климата IV. Среди-
земноморский кли-
мат в Чили, Южной
Африке и Португа-
лии.
28 29
Рис. 31—33. Тип
климата V. Умеренно
теплый и влажный
климат в Уругвае,
Южной Африке и се-
верной Анатолии.
31
32
33
Введение
69
Рис. 34—36. Тип
климата VI. Умерен-
ный климат в Норве-
гии, Чили (очень
влажный, мягкие зи-
мы, но прохладное
лето) и США (холод-
ные зимы, но жаркое
лето).
35
36
Рис. 37—39. Тип
климата VII. Арид-
ный умеренный кли-
мат в Центральной
Азии (резко конти-
нентальный), Арген-
тине (с меньшими го-
довыми амплитудами)
и США.
Сармьенто (288м) 10,7*135
37 38 39
Рис. 40—42. Тип
климата VIII. Бо-
реальный холодный
климат в Сибири (рез-
ко континентальный),
центре Европейской
части СССР и Щве-
ции (с меньшими го-
довыми амплитуда-
ми).
Рис. 43—45. Тип
климата IX. Аркти-
ческий климат на се-
вере СССР (континен-
тальный), в Норвегии
(влажный) и Арген-
тине (морской).
Ушцая (8 м) 5^79
йо
45
Рис. 46—48. Тип
климата X. Горные
климаты различных
зон: Цейлон (зона I),
горы Ливана (зона IV)
и Чили (зона III).
48
70
Глава 1
рологических станциях. Выбор масштаба зависит только от поставленных
целей. Использованные нами масштабы для графиков температуры и осад-
ков вполне применимы для всех климатических областей земного шара,
что легко подтвердить несколькими примерами из различных районов.
Основные климатические зоны (I—IX) перечислены на стр. 56. Этот
список следует дополнить типом X, охватывающим горные климаты раз-
личных зон, причем типом X (I) мы будем называть горный климат эква-
ториальной зоны, например климат приэкваториальных Лнд, типом X
(IV) — горный климат западных побережий субтропической зоны, напри-
мер в горах Ливана, типом X (VI) —горный климат в пределах типичной
умеренной зоны, скажем в Альпах.
На каждой из серии рисунков (19—48) приведены три возможных
варианта диаграмм перечисленных типов климата, причем автор старался
подобрать примеры, наиболее характерные для разных континентов.
ПОЧВЕННЫЕ ЗОНЫ ЗЕМНОГО ШАРА
Изучение взаимосвязей между климатом и растительностью показы-
вает, что границы климатических областей совпадают с границами, очер-
ченными растительным покровом, лишь в общих чертах: границы послед-
них более извилисты, чем контуры первых. Объяснение находим в том,
что характер растительности определяется не только климатом, но и поч-
вой, что особенно ярко проявляется в полосе контакта различных зон
растительности. Например, на климадиаграммах степной зоны Восточной
Европы отчетливо выделяется сухой период наряду с полузасушливым
сезоном, продолжающимся все лето; на климадиаграммах лесостепной
зоны обнаруживается только полузасушливый период, в лесной же зоне
отсутствует и последний. Границы между перечисленными выше климати-
ческими зонами вытянуты в виде прямых линий, простирающихся с ЗЮЗ
на BGB 4, тогда как границы зон растительности имеют иную конфигу-
рацию. Последние часто сильно изогнуты; это объясняется тем, что леса
по пескам проникают далеко к югу, а степи, развитые на лёссах, внедря-
ются в виде языков и островов в лесную зону. Дело в том, что травы
благодаря интенсивной корневой системе и особенностям водного режима
особенно хорошо растут на мелкозернистых почвах; древесные же породы,
обладающие экстенсивной корневой системой и экономно расходующие
воду, более приспособлены к крупнозернистой и каменистой почве, о чем
речь еще будет идти ниже (см. гл. 6, стр. 279). Генезис почв и процесс
формирования зональных типов почв рассматриваются в книге «Phytolo-
gie», Bd III, I, S. 435—475 1 2. Здесь мы коротко остановимся лишь на
особенностях распространения различных типов почв.
Тесные взаимосвязи между климатом, почвой и растительностью
иллюстируются следующей схемой:
Климат
I; U
Материнская порода —> Почва Растительность — Флора
Климат определенным образом воздействует не только на раститель-
ность, но и на почву, при этом главную роль играют гидратура и темпера-
1 Н. Walter, Die Klimadiagramme der Waldsteppen und Steppengebiete in
Osteuropa, «Stuttgarter Geogr. Stud.», 69, (Lautensach — Festschrift), 1957, S. 253—263.
2 См. также W. К u b i e n a, Entwicklungslehre des Bodens, Wien, 1948;
R. G a n s s e n, Bodengeographie, Stuttgart, 1957. [Имеется русский перевод: Р.Ганс-
c e н, География почв, M., ИЛ, 1961.— Ред.]
Схема распределения основных типов почв п характерные районы нх ра
сухо и холодно
преимущественно эпизодические н периодические осадки н большие
амплитуды температуры
увлажнение уменьшается
как правило, осадки распредел
зонах еще достаточно больно
репных н тропических — нези
увлажнение увеличивается—
господ-
ствующий
фактор поч-
вообразо-
вания
дополнительное увлаж-
нение в аридном кли-
мате
климат и естественная растительность, преимущественно зональны
полярная
местами ледниковые покровы или обломочные породы почти без растительности н по1
вечная мерзлота (крайний север), местами полигональные почвы (Шницбе
субполяр-
ная
более сухие тундровые почвы, местами еще вечная мерзлота
(Восточная Сибирь, северная Канада)
более влажные тундровые i
(Западная Сибирь)
высокогор-
ная
высокогорные пустынные п тунд-
ровые почвы
(Тибет)
степи
горные черноземы
(сухие горные район i,i СССР)
степи
горно-луговые почвы
(СССР п др.)
высокогорная лесная о
в
умеренно-
прохладная
бессточных пониже-
ниях: галофиты, со-
лончаки, солонцы
(юго-восток СССР,
западная Канада)
полынные степи
серые
вы
(запад
СССР)
и бурые поч-
полупустынь
США, юг
умеренно-
теплая до
субтропи-
ческой
почвы галерейных
лесов
(у аллогенных рек,
например у Нигера)
почвы оазисов
(Сахара)
разреженная ксеро-
фильная раститель-
ность,
пустынные почвы
(Сахара)
отчасти подвижные
дюны без раститель-
ности и почв
тропиче-
ская
ннзкотравпые степи каштановые почвы (юг СССР, запад США) высокотравные степи черноземы (СССР, США) 1 лиственные п хвойные леса серые лесные почвы (СССР, западная Канада)
прерии почвы прерий (США)
заросли кустарников, дубовые леса (дуб пушистый) почвы сухого леса, средиземноморские буроземы (Южная Европа, Северная Африка)
степи южные каштановые почвы (юг США, Судан) степи черноземовидные почвы (регуры, тирсы н др.) (Африка, Индия)
травяные саванны «оземленные» красные суглин- ки и красные почвы (пограничные области в тропи- ках) древесные саванны, сухие леса переменного увлажнения красные суглинки (пограничные области в тропи- ках), возможна латеритизация
хвойные
подзолы
(Сев. Евр
Сев. Аме]
вечиозел!
(постоя
эквато]
сне), ч
Африка
сухо и жарко
Таблица 3
(вития (но Ганссену)
)ны более равномерно; в холодных е амплитуды температуры, в уме- 14 нт е льны е влажно и холодно
почвы дополнительное увлаж- нение во влажном климате горные породы и кли- мат (почвы на изве- стняках н доломи- тах) рельеф п климат (поч- вы склонов)
[в (Антарктика), рген) при сползании почвы на склоне — местами гирляндовые (ополз- невые) почвы
ючвы, местами болотные почвы 1
С верхняя граница леса, низкорос- лые кустарники альпийские гумусовые почвы (Альпы, Анды) ласть, подзолы (Альпы) влажные леса, паст- бища альпийские поймен- ные почвы (Альпы) высокогорная лесная область подзолы, деградиро- ванные рендзины (Альпы) местами на крутых склонах скелетные почвы («ранкеры» н др.)
леса опа, северная часть энкн) s' s' искусст- s' венные пустоши s' н хвойные леса s' вторичные подзолы s' (в области буроземов) Xх листопадные лиственные леса буроземы (Центральная н Западная Европа) влажные луга ваттовые и марше- вые почвы (побережье Северно- го моря) пойменные леса, ни- зинные болота пойменные, болот- ные и глеевые почвы (Центральная Евро- па н др.) буковые леса рендзины (Центральная Европа и др.) теплолюбивые леса, терра-фуска (Центральная н Юж- ная Европа)
/ субтро- / ппческие леса / субтропические / подзолистые почвы / (юго-восток США, / северо-восточная Австралия) речные н морские ман- гровы субтропические и тропические ватто- вые почвы (тропическая Афри- ка) аллювиальные леса, тропические пой- менные п болотные почвы (Амазония) заросли кустарников терра-роса — среди- земноморские крас- ные почвы (С р ед и з емпомо рь е) почвы на корковых известняках, ксеро- рендзпны (жаркие сухие обла- сти, например юж- ная Испания, Север- ная Африка) почвенные ряды, или «катены», возникшие в результате эрозии (но Мильну и Фаге- леру) верхняя часть склонов: элювий, крупный ма- териал средняя часть склонов: коллювий, нередко с почвами, обуслов- ленными климатом нижняя часть склонов: аллювий, мелкий ма- териал; пески, тон- кие пески; глины в понижениях (тропи- ческие н субтропи- ческие, умеренно- влажные и семиарпд- ные области)
пые тропические дождевые леса ино-влажные) шальные бурые суглинки (по Куби- астично оподзоленные (Центральная ), возможна латеритизация
влажно и жарко
Введение
71
турный режим. В свою очередь растительность и почва влияют на климат,
хотя это влияние значительно меньше и ограничено приземным слоем
воздуха. В результате взаимодействия почвы, климата и растительности
возникают определенные природные единства, которые называются
экосистемами. Однако, несмотря на такую взаимозависимость, и почва
и растительность так же, как и их свойства, могут рассматриваться отдель-
но. Растительный покров зависит от флористического состава, тогда как
почва — от материнской породы. Молодые, недавно сформировавшиеся
почвы отражают свойства материнской породы, но по мере развития почв
влияние подстилающей породы ослабевает и все более выделяются харак-
терные особенности, обусловленные климатом и растительностью.
Несмотря на непрекращающееся влияние материнской породы на
почву и тем самым на растительность, зональность, обусловленная клима-
том, почвой и растительностью, имеет много общих черт. Резкое различие
существует между типами почв влажных и аридных областей; в почвен-
ном профиле первых наблюдается нисходящее движение воды, способ-
ствующее некоторому выщелачиванию верхних горизонтов, тогда как
в почвах аридных районов процесс выщелачивания отсутствует. Напротив,
здесь вследствие интенсивного испарения с поверхности почвы и обуслов-
ленного этим подъема воды по профилю в определенных горизонтах или
на поверхности может произойти накопление растворимых соединений,
образующихся в процессе выветривания.i С другой стороны, во влажных
областях растительность всегда отличается большей пышностью, чем
в аридных. Поэтому в почву первых поступает больше органических
веществ и накопление гумуса, особенно в условиях низких температур,
протекает интенсивнее.
Характер процессов выветривания, дальнейшее изменение продуктов
последних, а также формирование и преобразование гумусовых веществ
весьма различны при высокой и низкой температурах. Поэтому почво-
образовательный процесс в тропиках, в умеренной и арктической зонах
характеризуется специфическими особенностями. Ограничимся в настоя-
щей работе лишь ссылкой на схему распределения основных типов почв
земного шара, составленную Ганссеном, с учетом зависимости их от
климата (см. табл. 3). Более подробно типы почв будут рассмотрены при
характеристике отдельных зон растительности.
ОСНОВЫ ТРЕХМЕРНОГО РАСЧЛЕНЕНИЯ
РАСТИТЕЛЬНОСТИ ЗЕМНОГО ШАРА
Крупным климатическим и почвенным зонам, прослеживаемым от
Северного полюса к экватору и от последнего к Южному полюсу, соответ-
ствуют различные зоны растительности.
Массивы суши занимают в северном полушарии большую плошадь,
чем в южном, поэтому распределение климатических зон по обеим сто-
ронам экватора асимметричное. Южное полушарие не является зеркаль-
ным отражением северного; здесь вследствие большого объема водных
масс климат отличается более океаническим характером, отсутствует
обширная зона пустынь, совершенно не выражена умеренная зона.
Например, граница распространения пальм пересекает остров Южный
(Новая Зеландия); лесная растительность острова нередко имеет почти
субтропический характер, тогда как лишь несколько южнее, на архипела-
ге Окленд, уже проходит полярная граница леса. Нигде в другом районе
земного шара она не располагается так близко к субтропикам. Узкая
72
Глава 1
субантарктическая зона в противоположность широкому холодному
умеренному поясу северного полушария, а также в отличие от субаркти-
ческой зоны характеризуется совершенно иными условиями. Наоборот,
полярная зона в Антарктике более резко выражена, чем в Арктике, однако
в нашем обзоре это не играет роли, так как растительный покров здесь
отсутствует.
Асимметричному распределению климатов в обоих полушариях соот-
ветствует аналогичная асимметрия зон растительно-
сти. Поэтому условия, наблюдаемые в северном полушарии, нельзя
автоматически переносить на южную половину земного шара. Это необхо-
димо особенно подчеркнуть в связи с тем, что изучение растительности
исторически началось в северном полушарии. Ботаники невольно пыта-
лись перенести изученные там закономерности на южное полушарие,
что нередко приводило к неправильным представлениям.
То же самое можно сказать о высотных поясах в горах. Распростра-
ненное мнение, что пояса повторяют в горах последовательное чередование
зон растительности, прослеживаемое при движении с юга на север,
приемлемо лишь для северной умеренной зоны и то с известными ограни-
чениями. В остальных областях земного шара условия распределения
растительности в горах значительно сложнее.
Даже в Центральной Европе климат альпийского пояса нельзя счи-
тать идентичным климату Арктики. Общей характерной особенностью
климата этих областей является только короткий вегета-
ционный период. Однако в Арктике суточные контрасты в этот
сезон более сглажены вследствие того, что продолжительность дня здесь-
длиннее, а солнце в известное время года вообще не заходит; кроме того,
днем в Арктике не очень тепло, а ночью не слишком холодно. Напротив,
в альпийском поясе продолжительность дня намного меньше, днем наблю-
дается более интенсивная солнечная радиация, а ночью значительное
излучение. Этим обусловлены резкие суточные колебания, хотя средняя
месячная температура мало отличается от средней месячной температуры
Арктики. Количество осадков в Арктике меньше, чем в Альпах, однако
благодаря равнинному рельефу в тундре наблюдается заболачивание
обширных пространств, в горах же болотистые участки встречаются
лишь в относительно небольших котловинах.
Климат тропических гор не сравним ни с климатом Арктики, ни
с климатом гор Центральной Европы А. Тропики характеризуются высо-
кой постоянной температурой в течение всего года. Разность средних
температур самого теплого и самого холодного месяцев не превышает
нескольких градусов. Следовательно, здесь в равнинных и в горных обла-
стях нельзя на основании показателей температуры выделить отдельные
сезоны. Отличие состоит лишь в том, что в горах с высотой наблюдается
уменьшение средней месячной температуры и одновременно сглаживают-
ся суточные колебания температуры; однако выше пояса облаков суточ-
ная амплитуда температур вновь резко увеличивается. Несмотря на то>
что на высоте 4000 м ежедневно в течение года возможно падение темпе-
ратуры ниже 0°, средняя годовая температура может превысить 5°. Ноч-
ные заморозки здесь едва сковывают поверхность почвы, а выпадающий
иногда снег днем всегда тает. Постоянный снежный покров сохраняется
лишь на высоте около 5500 л, если температура воздуха днем не под-
нимается выше нуля.
1 С. Troll, Der jahreszeitliche Ablauf des N aturgeschehens in den verschiedenem
К1 imagiirteln der Erde, «Studium Generale», 8, 1955, S. 113—133.
й сл К
Рис. 49. Асимметричное распределение растительности в северном и южном полушариях на «идеальном» континенте, по
Троллю.
I. Тропические зоны: 1 — экваториальный дождевой лес; 2 — тропические дождевые леса, развитые в области паееатных орографических осад-
ков; з — тропические саванны (влажные саванны) и листопадные леса; 4 — тропические колючие степи и колючие леса.
II. Внетропические зоны северного полушария: 5 — еухие пустыни; 6 — холодные внутриматериковые пустыни; 7 — субтропические зимне-
зеленые степи; 8 — зона зимних дождей с жарким летом; 9 — зона травяных степей е холодной зимой; 10 — зона влажных муссонных и лав-
ровых лесов с жарким летом; 11 -- летнезелсиыс леса; 12 — океанические летнезеленые листопадные и лавровые леса; 13 — бореальные хвой-
ные леса; 14 — бореальные березовые леса; 15 — субарктические тундры; 16 — арктические холодные пустыни.
III. Внетропические зоны южного полушария: 17 — прибрежные пустыни; 18 — пустыни, развитые в пределах пояеа «гаруа»; 19 — зона зимних
дождей; 20 — субтропические колючие степи (Карру, Монте); 21 — субтропические злаковиики; 22 — субтропические дождевые леса; 23 — уме-
ренно холодные дождевые леса; 24 — степи Патагонии и Повой Зеландии; 25 — субантарктические туееоковые луга и болота; 26 — антаркти-
ческая область материкового льда.
74
Глава 1
Климату Арктики с резко выраженными сезонными изменениями
и небольшими колебаниями температуры в течение суток противоположен
климат тропических гор с почти постоянной в течение года средней
суточной температурой. Переходный тип климата характерен для горных
областей средних широт (см. «Phytologie», Bd III, I, S. 65).
Из сказанного выше следует, что при изучении растительности зем-
ного шара нужно учитывать не только ее размещение в направлении
с севера на юг и с запада на восток, но и ее вертикальную пояс-
ность, то есть рассматривать растительность в трех измерениях. Эту
мысль особенно подчеркивал Тролль и иллюстрировал ее замечательным
описанием растительности тропических гор *.
Распределение климатических и соответственно растительных зон
на «идеальном» континенте в горизонтальной плоскости показано на
рис. 49, а размещение их по вертикали, то есть в третьем измерении,
отражено на меридиональном профиле (рис. 50).
На профиле отчетливо видно асимметричное размещение раститель-
ности по обеим сторонам экватора. Легко заметить, что в северном полу-
шарии горная субальпийская и альпийская растительность по направле-
нию к югу поднимается все выше в горы, постепенно выклиниваясь
и замещаясь совершенно иным типом растительности. Климатические
условия в горах тропической зоны в известном отношении сходны с усло-
виями более низко расположенных поясов в горах южного полушария.
Следовательно, и в характере растительности здесь можно найти мно-
го общего.
Резко выраженный прохладный океанический климат субантаркти-
ческих островов, как и климат тропического высокогорья, отличается
незначительными колебаниями температуры в течение года (Кергелен —
6,5, западная Патагония —4,5, острова Макуори —3,5°). Правда,
суточные амплитуды температуры здесь также очень малы, наибольшие
различия средних часовых температур в течение года достигают всего
4,9°. Если температура близка к 0°, то тогда число дней со сменой отте-
пелей и морозов исключительно велико и промерзание почвы не происхо-
дит. Субантарктика и тропические Анды отличаются большой флори-
стической общностью, обусловленной непосредственной территориальной
связью, отсутствующей между горными системами тропических районов
Африки. Речь идет преимущественно о подушковидных растениях, расту-
щих в заболоченных местообитаниях. Около 64,1% всех подушковидных
растений произрастает в Андах Южной Америки и в субантарктике, из
них 50,5% —в Южной Америке, включая Огненную Землю, и 13,6% —
на субантарктических островах и в Новой Зеландии 1 2. Для этих областей
характерны также следующие формы роста: «туссоковые злаки»— жестко-
листные высокие плотнокустовые злаки различных родов, своеобразные
розеточные деревья, как, например, эспелетии южноамериканских
парамос, древовидные представители рода Senecio — обитатели восточно-
африканских высокогорий, высокие лобелии и некоторые виды люпина,
или «кергеленская капуста», Pringlea antiscorbutica (рис. 123—125,
132, 140).
1 С. Troll, Der asymmetrische Aufbau der Vegetationszonen und Vegetations-
stufen auf der Nord-und Siidhalbkugel, «Berichte Geobot. Forschungsinst. Rubel, f.
1947», 1948, S. 46—83; C. Troll, Die tropischen Gebirge, «Bonner Geogr. Abb.»,
25, 1959.
2 W. Rauh, Der polsterformige Wuchs, «Nova Acta Leopoldina», 7, № 49, 1939.
Введение
75
Отсутствие общих флористических или физиономических черт, харак-
терное для растительности альпийского пояса гор тропиков и раститель-
ности северного полушария, свойственно также растительности верхней
части лесного пояса северного полушария и высокогорным тропическим
лесам. Аналоги последних можно встретить лишь значительно южнее, на
более низких высотных уровнях. Отметим столь характерные роды, как
Podocarpus, Araucaria и Nothofagus или различные древовидные папорот-
ники, представителей семейства Proteaceae или род Gunnera, встречающий-
ся даже в горных лесах пояса облаков Колумбии и Коста-Рики. Из числа
эпифитов нежные Hymenophyllaceae произрастают, за редким исклю-
чением, только во влажных лесах южного полушария.
Южн. Бразилия,
Юго - Бост.
Африка,
Юго-Восточная Тропические Юга-Вост.
Шпиц- сиропа Азия Анды Австралия
Берген ' --------' '---‘--------------------*--------' 'Зап. Патагония Антарктика
Высота, Гималаи НпЙля Ирландия
Р п с. 50. Схема вертикальной поясности растительности земного шара от Арктики
до Антарктики (по Троллю).
При составлении схемы приняты во внимание только постоянно влажные климаты, исключе-
нием являются снеговая линия п область пуны. Одинаковые условные знаки свидетельствуют
о родственных связях растительности, развитой в тропических горах и в более высоких широ-
тах. Во влажных областях отчетливо выделяется асимметрия высотных поясов.
Характерно, что эти леса даже в высоких широтах остаются вечно-
зелеными; лишь очень небольшое число видов рода Nothofagus сбрасывает
листву, но и последние нельзя сравнивать со столь примечательным для
северного полушария типом листопадных деревьев. С экологической точки
зрения вечнозеленые древесные породы с лавровидными листьями, встре-
чающиеся в составе лесов вплоть до южной оконечности Африки, еще
мало исследованы.
В тропических горах южного полушария эти вечнозеленые леса
с еще достаточно хорошо выраженным субтропическим характером
поднимаются непосредственно до верхней границы леса.
Приведенная на рис. 50 схема вертикальной поясности растительно-
сти, составленная Троллем, отражает, как подчеркивает ее автор, лишь
условия, свойственные областям с влажным климатом. Штоккер 4, ана-
лизируя эту схему, отмечает, что профиль, составленный Троллем,
в северном полушарии пересекает области с умеренно влажным климатом
(приблизительно от северной Норвегии до Альп), а в южном полушарии
соответствует наиболее типичным условиям постоянно влажного климата.
Штоккер считает, что если бы этот профиль растительности пересекал бы
1О. Stocker, Das dreidimensionale Schema der Vegetationsverteilung auf
der Erde, «Вег. Dtsch. Bot. Ges.», 76, 1963, S. 168—178.
76
Глава 1
в северном полушарии области с более влажными климатическими усло-
виями и захватывал бы, скажем, Исландию и Ирландию, то асимметрия
профиля в северном и южном полушариях исчезла бы. Следует, однакот
учитывать, что в результате оледенения растительность северо-западных
областей Западной Европы была сильно нарушена и резко отличается от
растительности тихоокеанской части Северной Америки с аналогичным
ярко выраженным океаническим климатом.
Значение такого рода схем не следует переоценивать. Им всегда
присуща некоторая произвольность: схемы будут отличаться в зависимо-
сти от того, на каких примерах они построены и какие из выбранных
примеров можно считать типичными. Флористические различия северного
и южного полушарий столь велики, что не представляется возможным
проводить параллелизацию отдельных поясов растительности.
На Международном ботаническом конгрессе, состоявшемся в 1954 г.
в Париже, оживленно обсуждался вопрос о том, является ли горная
растительность специфической для соответствующей климатической зоны,
и далее — имеется ли аналогия между вертикальными поясами и расти-
тельными зонами более высоких широт, например, соответствует ли пояс
еловых лесов в Альпах зоне хвойных лесов, альпийский пояс — арктиче-
ской зоне и т. д. К сожалению, всеобъемлющего ответа на этот вопрос
дать нельзя.
Схема, приведенная на рис. 50, по-видимому, свидетельствует в поль-
зу второго представления, однако лишь в том случае, если в горах
с понижением температуры резко увеличивается количество осадков, как
это наблюдается в областях с влажным климатом. В аридных районах
положение совершенно иное, что видно на примере средиземноморской
зоны. В горах, где не только зимой, но и летом существует полоса с высо-
кой влажностью и облачностью, над поясом со средиземноморской расти-
тельностью располагается пояс с типичной центральноевропейской расти-
тельностью, например в Апеннинах — пояс буковых лесов. Там же, где
сухой воздух летом достигает даже субальпийских высот, как, например,
в горах Сьерра-Невада на юге Испании, вертикальная поясность расти-
тельности не имеет аналогов в севернее расположенных зонах. Это отно-
сится, например, к поясу субальпийских подушковидных растений.
В горах Тавр и Атлас распространен пояс, образованный кедром
или можжевельником; он также не имеет аналогов в севернее располо-
женных зонах. В горах пустынь влажность с высотой обычно увеличивает-
ся очень незначительно; это определяет слабую выраженность поясно-
сти, которая в этих условиях зависит только от температуры.
На вершинах, возвышающихся над облаками, количество осадков,
как будет показано ниже, резко сокращается; в связи с этим здесь форми-
руется своеобразный, подобный пустынному климат, в условиях которого
в экваториальной зоне сезоны не выделяются. Аналоги этому на более
низких высотных уровнях нигде на земном шаре не наблюдаются (ср. гл. 4,
стр. 207—219).
Итак, на поднятый на Ботаническом конгрессе вопрос однозначного
ответа дать нельзя. Поясность растительности в горах в отдельных слу-
чаях весьма напоминает последовательность широтного расположе-
ния растительных зон на равнине, в других же — никаких общих черт
с последней не обнаруживает. Поэтому при описании отдельных климати-
ческих зон одновременно будет приводиться и характеристика поясов
горных систем, расположенных в пределах той или иной зоны, или же-
будут даны ссылки на аналогии с другими растительными зонами.
Глава 2
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС1
ОБЩИЙ ОБЗОР
Исторически сложилось так, что изучение растительности началось
в северной умеренной зоне. Поэтому мы привыкли рассматривать
растительность других климатических зон, исходя из закономерно-
стей, свойственных растительному покрову северного полушария. В дей-
ствительности же в умеренной зоне в результате приспособления к мест-
ным климатическим условиям с резко выраженными временами года,
с теплым летом и относительно холодной зимой сформировался совершенно
особый тип растительности.
Развитие цветковых растений, несомненно, началось в условиях
тропиков. Тропический лес еще в третичном периоде покрывал большую
часть северного полушария. Эта формация возникла в особо благоприят-
ных для развития растений условиях. Возникновение других формаций
связано с распадом более сложных сообществ на более простые и с умень-
шением видового состава. Дальнейшая эволюция пошла по различным
направлениям; одни виды приспособились к недостаточному увлажнению,
другие — к неблагоприятным температурным условиям.
Таким образом, обзор растительности земного шара целесообразно
начать именно с тропического дождевого леса, а затем перейти к описа-
нию других формаций, развитых в менее благоприятных условиях.
Только такой подход позволит понять особенности растительного покрова
умеренной зоны.
Тропический дождевой лес распространен главным образом в пределах
экваториальной климатической зоны, простирающейся приблизительно
между 10° с. ш. и 10° ю. ш. и характеризующейся небольшими колебани-
ями температуры в течение года. Годовая сумма осадков чрезвычайно
велика и местами достигает нескольких метров. Осадки распределяются
достаточно равномерно в течение всего года, хотя в наиболее типичных
условиях (поскольку солнце дважды находится в зените) выделяются два
максимума. Однако охарактеризованные выше условия свойственны не
всей зоне тропиков. Благодаря дующим в определенные сезоны муссонам
восточная часть Экваториальной Африки, восточные районы Цейлона,
южная оконечность Индостана и некоторые области северо-западной
части Южной Америки отличаются сравнительно большой сухостью
(рис. 15). С другой стороны, на востоке Центральной Америки, на Боль-
1 Приведенные в этом разделе цитаты взяты из книг: Schimper- Faber, Pflan-
zengeographie auf physiologischer Grundlage, Jena, 1935; Richards, The
Tropical Rain Forest, 1952; В iinn i ng, In den Waldern Nordsumatras, Bonn, 1947.
Представления, соответствующие современному уровню знаний в области экологии
и фитоценологии тропического дождевого леса, очень хорошо изложены в монографии
одного из наиболее крупных специалистов по тропикам П. У. Ричардса. [Имеется
русский перевод: П. Ричардс, Тропический дождевой лес, ИЛ, М., 1961.— Ред, ]
78
Глава 2
I Севервый^т роп и к
ших и Малых Антильских
островах, на западном побе-
режье Индостана, в Бирме
и Китае влажный тропиче-
ский климат характерен для
пространств вплоть до Север-
ного тропика, а местами и
севернее. Влажно и жарко
также в горах восточной
окраины Южной Америки
и восточной Австралии. От-
сюда понятно, почему в пре-
делах названных областей
распределение осадков не
всегда соответствует эквато-
риальному типу; здесь часто
выделяется только один,
правда весьма длительный
период дождей, сменяющийся
кратковременным периодом
засухи *.
Районы распространения
тропического дождевого леса
приурочены к этим климати-
ческим условиям (рис. 51),
Наиболее обширный массив
сплошного распространения
лесов может быть выделен
в Южной Америке (бассейн
Амазонки до восточного скло-
на Анд), второй по величи-
не—в Индо-Малайской обла-
сти и несколько меньший
по размерам — в Западной
Африке (от побережья Гви-
нейского залива до бассейна
реки Конго), Помимо этих
трех основных массивов, тро-
пические дождевые леса рас-
1 Как показали исследова-
ния автора (UNESCO, Humid
Tropics Res. Progr. 106 В), сухой
период, продолжающийся один
месяц, почти не отражается па
растительности; двухмесячный
период засухи пе влияет на тро-
пический дождевой лес Индии
в том случае, если годовое коли-
чество осадков достигает по мень-
шей мере 2250 мм, а количество
за четыре месяца — 3500 мм
(рис. 182), При этом период за-
сухи должен совпадать с холод-
ным временем года.
Тропический дождевой лес
79
пространены отдельными участками на влажных наветренных склонах
горных хребтов, простирающихся между Северным и Южным тропиками
(восточная Бразилия, северо-восток Австралии, Индии и т. д.), на менее
крупных горных массивах, например в Восточной Африке на склонах
больших вулканов (Килиманджаро, Меру и др.) и гор Усамбара и Улугу-
ру. С увеличением высоты местности появляются уже горные дождевые
леса, заметно отличающиеся от лесов, занимающих более низкие высот-
ные уровни. Вообще следует подчеркнуть, что структура тропического
дождевого леса и его флористический состав настолько своеобразны в раз-
личных областях, что какое-либо обобщение не всегда уместно.
Ровный годовой ход метеорологических элементов, характеризую-
щий климат экваториальной зоны,— явление, часто лишь кажущееся,
связанное с тем, что обычно исследователи ограничиваются изучением
средних месячных данных, забывая, что в тропиках мы имеем дело не
с сезонными изменениями климата, а с суточными (ср. стр. 80 и след.);
колебания температуры и влажности воздуха в течение суток здесь зна-
чительно более резкие, чем амплитуды средних месячных показателей.
В еще большей мере это относится к высокогорным районам. Тот, кто
бывал во влажных тропиках, не забудет прелести веющего прохладой
раннего утра, когда перед самым восходом солнца в лесу просыпается
жизнь, раздается многоголосый хор птиц, пробиваются первые лучи
солнца и капельки росы на сочных листьях, словно жемчужины, заго-
раются всеми цветами радуги. Воздух свеж и прохладен. Но как только
солнце поднимается над горизонтом, начинаешь испытывать тягостное
чувство, невольно ищешь тени и, уж во всяком случае, надеваешь тропи-
ческий шлем. Постепенно стихает громкое щебетание птиц; воздух стано-
вится все более душным. Небо заволакивается тяжелыми облаками, и вот
уже разразилась гроза. Но дождь быстро проходит, снова светит солнце.
Кругом все мокро. Мимолетная прохлада вскоре сменяется угнетающей
духотой, впечатление такое, что находишься в оранжерее. Лишь к вечеру
начинает тянуть легкий свежий ветерок, а после захода солнца снова все
оживает. Мягкие ночи в тропиках великолепны. На яркий свет лампы
слетаются мириады причудливых насекомых — лакомая добыча для
изящных гекконов. Но свет привлекает и истинных мучителей рода
человеческого — москитов, единственное спасение от которых противо-
москитный полог. В помещении и без того душно, однако приходится
забираться под сетку. Часами беспокойно ворочаешься и, измучившись,
наконец засыпаешь. Так, почти без всяких изменений проходят день
и ночь вне зависимости от времени года. Для человека, жившего в умерен-
ной зоне, длительное пребывание во влажных тропиках превращается
в настоящее мучение.
Эта одна из причин наших скудных знаний в области экологии тропи-
ческого дождевого леса. Существует много хороших путевых описаний,
но почти нет подробных тщательно проведенных многолетних экологи-
ческих наблюдений. Объясняется это просто: подобного рода исследования
возможны лишь на соответствующих станциях и требуют длительного
пребывания в тропиках. На одной из таких станций, в Ботаническом
саду в Богоре (Ява), располагающей прекрасным лабораторным обору-
дованием, в течение нескольких лет работали Трейб и Фабер. Однако
станция находится в пределах культурного ландшафта, а заповедник
Тьибодас расположен сравнительно далеко от нее, выше в горах, что
затрудняет проведение там экологических исследований. Весьма удачно
было выбрано место для ныне ликвидированной опытной биологической
80
Глава 2
станции Амани в горах Усамбара (Восточная Африка), которая находи-
лась на высоте 900 м посреди обширного девственного леса 1 и где автор
в 1934 г. проводил в течение полугода экологические исследования. На
подобных опытных станциях в тропиках ученые в основном работают над
проблемами прикладной ботаники.
Например, в Адиоподуме близ Абиджана (Берег Слоновой Кости,
Западная Африка) уже несколько лет существует Институт исследования
тропиков (Institut d’Enseignement et de Recherches Tropicales).
КЛИМАТ ВЛАЖНЫХ ТРОПИКОВ
Представление о средних многолетних климатических условиях
влажных тропиков дают климадиаграммы расположенных здесь станций.
Некоторые из них приведены на рис. 52—56, а также на рис. 19—21.
Годовое количество осадков в тропиках достигает местами весьма
больших величин. На вершине одного из Гавайских островов — Кауаи —
оно составляет 12 500 мм, у юго-западного подножия вулкана Камерун —
10 500 мм. В городе Черрапунджи, расположенном в горах Кхаси (Ассам)
и отличающемся более прохладным климатом, выпадает 11 630 мм. Но
Рис, 52—54. Климадиаграммы станций, расположенных в области распростране-
ния тропического дождевого леса (бассейн Конго, Южная Америка, Цейлон) («КИ-
madiagramm-Weltatlas», 1960).
даже при таком обилии осадков возможны засушливые периоды. Так,
в том же Черрапунджи в декабре и январе выпадает соответственно
всего 10 и 20 мм осадков.
Однако на приведенных климадиаграммах слабо выделяется важный
показатель тропического климата: характер суточных изменений различ-
ных метеорологических элементов, выражающийся в регулярной повторя-
емости в течение суток их экстремальных значений (ср. экстремальные
значения температуры на рис. 16 для Ду алы).
Рассмотрим в качестве примера климатические условия в Богоре на
Яве, описываемые Штоккером 1 2.
Средние месячные температуры колеблются в пределах 1° (24,3°
в феврале и 25,3°—в октябре), годовое количество осадков составляет
1 R, Е, Moreau, A synecological study of Usambara, Tanganyika Territory,
«J, Ecol.», 23, 1935, p, 1—43; Some ecological data for closed evergreen forest in tropical
Africa, «J, Linn, Soc, (Zool,)», 39, 1935, p, 285—293,
2 O, Stocker, Ein Beitrag zur TranspirationsgroBe im javanischen Regenwald,
«Jb. wiss. Bot.», 81, 1935, S. 464—496.
1. Соцветие Spathodea campanulata; эта древесная порода, достигающая небольшой
высоты, широко распространена в дождевых лесах Уганды.
2. Тропический вечнозеленый дождевой лес в окрестностях Кэрнса (Северный Квин-
сленд). Влажный участок леса, где в составе древостоя преобладает пальма Ltcualla
muelleri.
3. Эвкалиптовая саванна в центральной Австралии около Матаранки (между Алис-
Спрингсом и Дарвином). В травяном покрове преобладает Heteropogon contortus.
Тропический дождевой лес
81
Рис. 55—56. Климадиаграммы
станций, расположенных на Но-
вой Гвинее и островах Фиджи.
4370 мм\ в январе, самом
влажном месяце, выпадает
450, в самом сухом, авгу-
сте,—230 мм. Такое посто-
янство погоды — явление
только кажущееся. Суточный
ход метеорологических эле-
ментов варьирует в весьма
широких пределах.
В солнечные ноябрьские
дни температура в течение
дня может изменяться от 23,4°
в 6 час. до 32,4° в 14 час.,
то есть в пределах 9°. Даже
в период дождей суточные
амплитуды достигают 6—7°,
правда, в пасмурные дни
6—1394
Рис. 57. Ход относительной влажности воздуха в полузасушливый период в Богоре 3—9 октября (по Фаберу).
82
Глава 2
они иногда сокращаются до 2°. Колебания дневной температуры
обусловливают (см. рис. 57) изменения влажности воздуха в пределах
до 25%, от 40 до 100%; только в очень дождливые дни влажность не
падает ниже 90% (рис. 58). При такой высокой температуре дефицит
влажности может достигать весьма больших значений; так, при темпера-
туре 32,3° и относительной влажности 50% он равен 18,3 мм — вели-
чина, характерная для степных областей. Правда, такая сухость воз-
Р и с. 58. Суточный ход метеорологических элементов в Богоре’в дожд-
ливый период (Stocker, Grundl. einer naturgemafi. Gewachshaus-
kultur, Verlag Eugen Ulmer, 1949).
и наблюдается между 8 и 14 часами. Ночью воздух близок к состоянию насы-
щения, что приводит к выпадению росы. Напротив, в аридных районах
повышенная сухость воздуха иногда сохраняется неделями, и даже ночью
влажность столь мала, что, несмотря на значительно более высокую
температуру, роса не образуется.
Резкая выраженность в тропиках суточных колебаний проявляется
в том, что средняя месячная суточная амплитуда температуры в Богрре
достигает 6,9—10,2°; она соответствует амплитуде в Вене (4,7—10,2°), но
значительно превосходит суточную амплитуду, наблюдаемую в Гамбурге
(3,5—7,6°). В области распространения первичного леса бассейна Конго,
Тропический дождевой лес
83
по данным Бернара величина среднего суточного дефицита влажности
равна 13 мм. Бернар считает, что дефицит влажности здесь выше, чем
в северной Франции в июле. При средней месячной амплитуде температу-
ры в 1—2° средние суточные колебания ее достигают 10—12° (абсолют-
ный максимум равен 36°, абсолютный минимум —18°).
По мере подъема в горы увеличиваются облачность, влажность воз-
духа и уменьшается амплитуда температуры. Наряду с понижением сред-
ней годовой температуры с высотой суточные колебания остаются резко
выраженными (см. гл. 4, ср. также рис. 54 с рис. 46).
С высотой увеличивается также разность между суточными макси-
мумами и минимумами температуры, причем более важное значение имеет
снижение минимальной температуры. Изменение видового состава леса
в горах происходит весьма постепенно. Одни виды распространены
только в предгорьях, другие встречаются в верхних частях гор. Четко
выраженная вертикальная поясность отсутствует. Уменьшается число
лиан, орхидей, с другой стороны, значительно чаще встречаются папорот-
ники из семейства Hymenophyllaceae. Бореальные роды (Quercus и Pinus)
распространены только в северном полушарии.
Выше пояса облаков, где почти всегда стелется туман, количество
осадков резко сокращается. Леса постепенно изреживаются, их структура
упрощается, а чисто тропическая флора замещается горной флорой.
Обычно различают предгорный, горный и субальпийский тропический
дождевой лес.
Граница распространения древесной растительности резко обрывает-
ся лишь там, где минимальная суточная температура понижается до
отрицательных значений. Поскольку здесь отсутствует четкое разделение
на сезоны, такое понижение температуры наблюдается почти ежедневно,
и количество дней с отрицательными температурами на определенной
высоте внезапно возрастает. Вечнозеленый тропический горный лес в этих
условиях уступает место альпийской растительности. Такая смена про-
исходит на склонах гор, поднимающихся над поясом облаков, поэтому
тропический альпийский пояс характеризуется не только большим коли-
чеством дней с отрицательными температурами, но и относительной
бедностью осадками. Резкие колебания температуры в течение суток
влияют на влажность воздуха. Так, по наблюдениям Фабера, влажность
воздуха на вершине Пангаронго (Ява) в течение 18 сентября колебалась
в пределах 8—100%.
Вопреки широко распространенному мнению солнечная радиация,
поступающая на поверхность, расположенную перпендикулярно солнеч-
ным лучам, во влажных тропиках меньше, чем в большинстве других
климатических зон, и даже меньше, чем в умеренной. Из-за большой
влажности воздуха в тропиках, несмотря на высокое стояние солнца,
поверхности почвы достигает лишь незначительная часть радиации
(«Phytologie», Bd III, 1, S. 31). Цвет неба обычно голубовато-серый,
а в воздухе почти постоянно присутствует дымка, что может исказить
правильность визуальных наблюдений в тропиках и требует для объек-
тивной оценки условий местообитания применения измерительных
приборов.
То, что радиация в тропиках, несмотря на все сказанное, все же дей-
ствует на человека угнетающе, объясняется другими причинами. Испаре-
1 Е. Bernard, Le climat ecologique de la Cuvette Centrale Congolaise, Bru-
xelles, 1945, «Ref. in Ecology», 30, 1949, p. 265.
84
Глава 2
ние с поверхности тела человека, обладающего собственной температурой,
зависит от абсолютной влажности воздуха, которая определяет отрица-
тельный градиент упругости водяного пара между поверхностью тела
и окружающим воздухом. Холодный воздух содержит лишь небольшое
количество водяного пара, и поэтому даже при его насыщении он кажет-
ся человеку сухим. Если же температура окружающей среды приближает-
ся к температуре тела человека и к тому же влажность воздуха еще доста-
точно высока, градиент упругости водяного пара у поверхности его тела
оказывается весьма малым. Все тело покрывается потом, но с его поверх-
ности почти ничего не испаряется, и охлаждающего влияния испарения
не ощущаешь. Если к тому же человек находится под прямыми лучами
солнца, возможен перегрев организма и, как следствие этого, тепловой
удар. Только ветер приносит облегчение, охлаждая тело. Оценка интен-
сивности радиации в тропиках весьма и весьма субъективна — об этом
свидетельствует опыт автора. Так, при восхождении на вершину Кили-
манджаро постоянно чувствовалось увеличение интенсивности солнечной
радиации, однако на высоте более 4000 м вследствие низкой температуры
воздуха жгучие лучи солнца были даже приятны.
Ни в коем случае нельзя пытаться судить об условиях среды расте-
ний по своим собственным ощущениям. Влажные тропики вовсе не явля-
ются для растений, не обладающих собственной температурой, столь уж
влажными, как это обычно предполагают, во всяком случае для растений,
листья которых нагреваются прямыми лучами солнца. Поэтому не удиви-
тельно, что листья древесных пород, постоянно освещенных солнцем,
нередко отличаются ксероморфной структурой.
Важное значение для развития растений имеет продолжительность
дня, которая в тропиках мало изменяется в течение всего года. Даже
на широте Северного и Южного тропиков самый длинный день продол-
жается всего 13,5, а самый короткий — 10,5 часов. Солнце заходит почти
перпендикулярно горизонту, благодаря чему продолжительность сумерек
здесь короче, чем в более высоких широтах, где солнце пересекает гори-
Р и с. 59. Средняя продолжительность солнечного сияния (в часах)
в Сан-Хосе (Коста-Рика, 9°56' с. ш.; 1135 м над ур. моря) (по
Ханн-Зюрингу).
зонт под острым углом. Обычно все же кратковременность сумерек пре-
увеличивают. В ясные дни с момента погружения солнца за линию гори-
зонта и до того времени, когда на открытой местности уже нельзя читать,
проходит, по нашим наблюдениям, 30 минут, в Южной Европе под 41° с. ш.
Тропический дождевой лес
85
мм/час
Рис. 60. Суточный ход испарения в Джа-
карте (по Брааку).
18 апреля эта величина соста-
вила 40 минут и 22 апреля
севернее Гавра в проливе Ла-
Манш — 48 минут. Только зна-
чительно севернее продолжи-
тельность сумерек увеличивает-
ся. Растения длинного дня
в тропиках обычно не цветут.
Салат, например, можно выра-
щивать в течение всего года,
однако соцветий он не образует.
До полудня в тропиках,
как правило, светит солнце,
дожди идут в основном во вто-
рой половине дня (рис. 59).
Величина испарения, находя-
щаяся в зависимости от сол-
нечной радиации, в тропиках
приблизительно такая же, как
на широте Центральной Евро-
пы летом в пасмурные дни,
и характеризуется весьма четко
выраженным суточным ходом
(рис. 60).
Большинство областей рас-
пространения тропического дож-
девого леса находится в зоне
активной циклонической дея-
тельности. Проходящие ураганы
наносят лесу большой ущерб:
ветер обламывает множество
крупных ветвей, срывает листву с деревьев, ломает тонкие стволы второго,
яруса древостоя, иногда вырывает с корнем большие деревья. Окна,
образовавшиеся в лесу после ураганов, вскоре зарастают главным обра-
зом лианами. Многочисленные стволы поваленных деревьев увеличивают,
опасность возникновения лесного пожара. Опасность возрастает, если,
поблизости расположены плантации, например сахарного тростника,
при очистке которых огонь может перекинуться в лес. Пожары вызывают
дополнительные изменения флористического состава. Вместо сильно пов-
реждаемой огнем ротанговой пальмы {Calamus) в восточной Австралии
широко распространяются лианы {Rubus и Flagellaria), а на свежих гарях
в северном Квинсленде часто появляются обильные всходы некоторых
видов Acacia Ч
При частых ураганах известное равновесие между сообществом и кли-
матом не устанавливается, в связи с чем тропический дождевой лес от-
личается гетерогенной структурой.
МИКРОКЛИМАТ ТРОПИЧЕСКОГО ДОЖДЕВОГО ЛЕСА
Описанные выше климатические условия влияют только на растения,
произрастающие на полном солнечном свету, то есть на растения откры-'
1 L. J. Webb, Cyclones as an ecological factor in tropical lowland rainforest»
(NorthQueensland), «Austr. J. Bot.», 6, 1958, p. 220—228.
86
Глава 2
тых культурных ландшафтов и на полог крон дождевого леса. Внутри
леса господствует совершенно иной климат, более ровный по сравнению
с климатом окружающей территории (рис. 61).
Под пологом леса количество солнечной радиации уменьшается.
В отличие от лиственных лесов умеренной зоны количество света, прони-
кающее в глубь леса, в течение всего года почти не меняется, так как здесь
отсутствует листопад, связанный с определенным сезоном.
Поскольку полог крон в тропическом дождевом лесу обычно не вполне
сомкнут, в глубь леса попадает сравнительно много света, правда уже
значительно ослабленного из-за обилия лиан, эпифитов и деревьев ниж-
них ярусов. Впечатление о небольшой освещенности усиливается из-за
того, что путешественнику приходится пробираться под пологом из почти
смыкающегося подлеска и травяного покрова, достигающих нескольких
метров высоты. Непосредственно поверхность почвы освещена очень слабо
и собственно напочвенный покров, как правило, отсутствует.
В дождевом лесу Восточной Африки близ Амани — восточная часть
гор Усамбара (5° ю. ш., 900 м над уровнем моря), где годовое количество
осадков равно 2000 мм, поверхности почвы достигает лишь около 0,1%
дневного света. Цветковые растения встречаются по большей части в усло-
виях, где освещенность достигает 1% полного дневного света (в пасмур-
ные дни). В солнечные дни определить потребность в свете того или иного
растения очень трудно из-за обилия световых бликов, постоянно меняю-
щих свое положение и на некоторое время резко увеличивающих интен-
сивность света при попадании на лист растения.
Орт, изучавший первичный лес на склонах гор Вирунга в Централь-
ной Африке, также приходит к выводу, что минимум освещенности, необ-
ходимый для развития теневыносливых растений в этих лесах, приблизи-
тельно такой же, как в лиственных лесах Европы, а именно 0,5%. В лесах
южной Нигерии освещенность поверхности почвы также составляет
0,5—1%, увеличиваясь в пределах световых бликов иногда в пять раз;
однако общая площадь бликов невелика. Аналогичные условия наблю-
даются в районе Сантарема, Бразилия
В горных лесах, покрывающих склоны Килиманджаро, на высоте
около 2400 м из-за отсутствия мощно развитого травяного покрова почвы
достигает больше света, поэтому напочвенный покров образован здесь
ярко-зеленым ковром из Selaginella.
Однако приведенные факты вряд ли следует считать общим явлением.
В изобилующих осадками (4500—6000 мм) диптерокарповых лесах север-
ной части Суматры условия освещения, судя по путевым описаниям Бюн-
нинга, значительно более неблагоприятны. Кроны деревьев настолько
сомкнуты, что почти не пропускают света, и поэтому в лесу в течение всего
дня царит «серый сумрак». Измеренная освещенность на поверхности
почвы достигает всего 0,1% полного дневного света — это условия, благо-
приятные лишь для слабого развития протонемы мхов. При освещенности,
достигающей 0,2%, развиваются Selaginella, а также лиственные и пече-
ночные мхи; Hymenophyllaceae и представители других семейств папорот-
1 R. Orth, Zur Kenntnis des Lichtklimas der Tropen und Subtropen sowie des
tropischen Urwaldes, «Gerlands Beitr. zur Geophysik», 55, 1939, S. 52; G. C. Evans,
Ecological studies in the rain forest of Southern Nigeria, «I. Ecol.», 27, 1939, p. 436—
482; G. G. Evans, An area survey method of investigating the distribution of light
intensity in woodlands, with particular reference to sunflecks, «I. Ecol.», 44, 1956,
p. 391—428; P. S. A s h t о n, Light intensity measurements in rain forest near Santarem,
«Brasil. J. Ecol.», 46, 1958, p. 65—70.
Тропический дождевой лес
87
Рис. 61. Суточный ход температуры (верхняя кривая) и относительной
влажности (нижняя кривая) на высоте 1 м над поверхностью почвы в густом
лесу близ Тьибодаса в полузасушливый период (по Фаберу).
ников, а также плауны довольствуются 0,25—О,5?6 полного света. Мхи
-обычно поселяются не на почве, а несколько выше, на мертвых стволах
и досковидных корнях. Столь низкая освещенность не препятствует
развитию даже бегонии, Impatiens, видов семейств Rubiaceae, Commeli-
пасеае и Zingiberaceae.
Более благоприятные условия освещения наблюдались в дождевом
лесу западной Явы близ Тьибодаса (1500 л^). Фабер пишет об этом сле-
дующее: «Изучение светового режима (Lichtklima) в тропическом дожде-
вом лесу показало, что освещенность в нем сильно колеблется; только в
травяном покрове под сомкнутым пологом леса свет распределяется более
или менее равномерно. Большие колебания интенсивности света обуслов-
лены главным образом прерывистостью верхнего яруса древостоя и кожис-
той блестящей поверхностью листьев. Наиболее резкие переходы от све-
та к тени наблюдаются в нижних ярусах леса; в верхних ярусах контрас-
ты смягчаются. Отсюда видно, что освещенность изменяется в направле-
нии, противоположном изменению влажности воздуха и температуры» Ч
В наиболее сомкнутых лесах северной части Суматры влажность
приземного слоя воздуха близка к насыщению, что исключает какие-либо
колебания температуры. В травяном покрове дождевого леса Тьибодаса,
по данным Фабера, средняя минимальная суточная амплитуда темпера-
туры наиболее влажного месяца равна 1,4°, а средняя максимальная
самого сухого месяца —3,2° (рис. 61). На высоте 18 м эти значения уве-
личиваются соответственно до 7,2 и 12,4°.
В качестве еще одного примера можно привести среднюю суточную
амплитуду температуры в первичном лесу на различных высотных уров-
нях (табл. 4) (по Ричардсу).
Суточный ход влажности воздуха на различной высоте над поверх-
ностью почвы приведен на рис. 62; уже на высоте 18 м влажность даже
1 Schimper-Faber, op. cit., S. 344.
88
Глава 2
Рис. 62. Суточный ход относительной влажности воздуха на уровне 1, 5 и 18 м
над поверхностью почвы в дождевом лесу близ Тьибодаса в дождливый период
(по Фаберу).
в сезон дождей часто понижается до 60—40%. Еще более суровые условия г
по-видимому, наблюдаются в пологе крон, где основное количество сол-
нечной радиации поглощается и преобразуется в тепло. Ход метеороло-
гических элементов здесь более ровный, чем в защищенной от солнечных
лучей метеорологической будке. По Ивенсу, максимальный дефицит
влажности в Нигерии не превышал 10 мм, колебания абсолютной влаж-
ности редко достигали 2—4 мм.
Таблица 4
Сухой сезон (по Элли) Диптерокарповый лес (Филиппины), по Брауну
Высота, 0,2 17 26 подле- сок 18
Средняя суточная ампли- туда, °C 1,1—2,9 6,8-7,3 8,6—11,8 5,7 6,0
В табл. 5 приводятся, по данным Брауна, средние максимальные
и минимальные температуры и влажность воздуха в верхних частях крон
Таблица 5
Высота над уров- нем моря, м Характер растительности Температура, °C Влажность, %
средний максимум средний минимум средняя мини- мальная за сутки
80 Вторичный лес 38,2 24,0 82,2 68,6
300 Диптерокарповый дождевой лес 32,4 20,0 — —
450 Диптерокарповый дождевой лес 31,3 19,6 87,9 75,5
740 Дождевой лес предгорий 30,4 18,1 — —
1050 Горный моховой лес 28,1 17,3 Обычно 100 91
Тропический дождевой лес
89
высоких деревьев на горе Маквилинг (Филиппины) на разной высоте над
уровнем моря (цитируется по Ричардсу).
Фабер, подводя итог своим исследованиям, пишет: «В тропическом
дождевом лесу климат отличается относительно равномерным ходом
метеорологических элементов только в самом нижнем ярусе сомкнутого
леса; в верхних ярусах дождевой лес характеризуется сравнительно рез-
кими различиями в степени выраженности отдельных метеорологических
элементов» U
Автор производил в Амани (Восточная Африка) измерения испаре-
ния эвапорометром Пиша (зеленая бумага). На поверхности почвы и на
уровне 1 м нередко в течение нескольких дней вообще не удавалось изме-
рить испарение из-за его малой величины. В очень сухие и ясные дни
испарение в местообитаниях, где развиваются заростки мараттиевых
папоротников, в сухой период достигало за 13 из 70 дней наблюдений
1 мл! сутки; максимальная суточная величина составила 1,7 мл, а наи-
большая за час — около 0,1 мл. На более открытых участках и несколько
выше над поверхностью почвы интенсивность испарения увеличивалась
вдвое.
Напротив, на высоте 70 см на открытом месте над уровнем травяного
покрова в течение суток нередко испарялось от 6 до 10 мл, причем ночью
испарения не происходило. Максимальное испарение за 1 час достигало
1,3 мл — величина, наблюдаемая в умеренных широтах в жаркие солнеч-
ные дни летом.
Большая влажность в первичном дождевом лесу даже после несколь-
ких бездождных дней связана, по нашим наблюдениям, с чрезвычайно
сильной росой, образующейся на кронах деревьев. Стекая вниз, роса
обильно смачивает растения нижних ярусов. Во влажных тропиках роса
выпадает каждую ночь.
На прогалине в первичном лесу близ Амани в течение 40 бездождных
ночей измерялось количество выпавшей росы. Максимальное количество
за ночь составило 0,26 мм, в 14 случаях оно превысило 0,15 мм, в 12 — коле-
балось от 0,15 до 0,1 мм, в 11 случаях было менее 0,1 мм и лишь в течение
трех ночей количество росы было неизмеримо мало (измерения произво-
дились по методу Лейка).
Слой выпавшей росы в 0,1 мм в пересчете на 1 л£1 2, составляет около
100 мл; этого количества вполне достаточно, чтобы вода легко стекала
с обычно несмачиваемых листьев. На более сильно охлаждающиеся листья
в кронах деревьев, вероятно, выпадает значительно больше росы.
Данных о содержании СО2 в тропическом дождевом лесу очень мало.
Учитывая большую продуктивность органического вещества и быстрое
разложение всех органических остатков, в дождевых лесах выделение
углекислоты из почвы должно быть исключительно высоким. Как извест-
но, дыхание почвы в естественных условиях определяется интенсивностью
фотосинтеза растений 2. Поэтому следует предполагать, что содержание
СО2 в приземном слое воздуха будет выше нормы. Очень большие (в деся-
тикратном размере) величины, приводимые Мак-Лином, в исследованиях
Ивенса 3 в Нигерии не получили подтверждения. По данным последнего,,
максимальные концентрации СО2 в утреннее время (до 10 часов) не пре-
1 Schimper-Faber, op. cit., S. 346.
2 H. Walter, W. Haber, Uber die Intensitat der Bodenatmung mit Bemer-
kungen zu den Lundegardbschen Werten, «Вег. Dtscb. Bot. Ges.», 70, 1957, S. 275—282-
3 G. G. Evans, op. cit.
90
Глава 2
вышали более чем в 2 раза норму. Наблюдения Штоккера 1 в Тьибодасе
также показали, что содержание СО2у самой поверхности почвы ненамного
превышало нормальную концентрацию. На плантациях какао (Берег
Слоновой Кости) Леме1 2 установил, что утром в 1 л воздуха содержится
0,7 мг СО2, а после полудня — 0,4 мг. С высотой концентрация СО2
вследствие турбулентности быстро выравнивается. По сообщению Элли,
в дождевых лесах Панамы скорость ветра на уровне 25 м над поверхностью
почвы приблизительно в 10 раз больше, чем у поверхности почвы, а у вер-
шин деревьев еще больше 3. Деревья-гиганты (эмергенты) поэтому испы-
тывают на себе всю силу тропических ураганов; часто происходят вет-
ровалы.
Выше были рассмотрены микроклиматические условия, существую-
щие внутри сомкнутого дождевого леса. Но как только в пределах сплош-
ного лесного массива образуются открытые пространства, сглаживающее
воздействие леса прекращается. В настоящее время обширные площади
леса выкорчеваны частью местным населением, частью плантаторами.
Поэтому небезынтересно изучить лесорастительные условия на таких
площадях. Тщательные экологические исследования провел Киркпатрик 4
на кофейной плантации близ Найроби, в Восточной Африке (1° ю. ш.,
1722 м над уровнем моря).
Хотя указанный район расположен в тропиках, осадков здесь выпа-
дает сравнительно мало (1062 мм), однако роса наблюдается каждую ночь.
Кофейные деревья были высажены рядами; в 20-летнем возрасте они
достигали высоты около 2 м. Для сравнения использовались данные бли-
жайшей метеорологической станции, расположенной в 100 м от плантации.
Температура измерялась в междурядьях на высоте 1,3 м. Суточные коле-
бания температуры, по данным метеостанции, в облачные дни составляли
5—8°, в ясные 12—16°. На плантации амплитуды в облачные дни были
на 10—25%, а в ясные — на 25—50% больше (максимальная амплитуда
22,4°).
В отдельные ясные дни на плантации в полдень температура была
На 3—4° выше (максимум 5°), а ночью — на 2—3° ниже (максимум 4—5°),
чем на метеостанции. При наличии облачности, не закрывающей солнце,
разность в полдень достигала 6,2°. Чем облачнее был день, тем меньше
-становилась разность температур, однако не меньше 1°. Только после полу-
денного дождя иногда отмечалась более низкая температура. При сплош-
ной ночной облачности показания термометров на станции и плантации
были одинаковыми.
В насаждении максимальные температуры были на 0,3—5,9° выше,
минимальные — на 0,0—4,2° ниже. Ветер мало изменял эти соотношения.
Наиболее низкие температуры были отмечены не на почве, а на поверх-
ности крон. По сравнению с открытой местностью поверхность почвы
в междурядьях нагревается сильнее, на глубине 5 см эта разница состав-
ляет 5—8°; непосредственно под растениями почва нагревается слабее
из-за меньшей освещенности солнцем. Большое значение имеет темпера-
1 О. Stocker, Assimilation und Atmung westjavanischer Tropenbaume,
-«Planta», 24, 1935, S. 402—445.
2 G. L e m e e, Recherches eco-physiologiques sur le Cacaoyer. «Rev. gen. Bot»,
63, 1956, p. 41—94.
3 R. C. Mac Lean, Studies in the ecology of tropical rain-forest, «J. Ecol.»,
7, 1919, p. 15—54, 121—172; W. G. Allee, Measurement of environmental factors
in tropical rain-forest of Panama, «Ecology», 7, 1926, p. 273.
4 T. W. Kirkpatrick, Studies on the ecology of coffee plantations in East
Africa (в рукописи).
Тропический дождевой лес
91
тура листьев. В ясные дни она была на 10—15° выше температуры воз-
духа. Наивысшая измеренная температура листьев 46°. Малейшее дви-
жение воздуха понижает температуру на несколько градусов; облако,
закрывшее солнце, может снизить температуру листьев на 10°. Даже если
облака закрывают солнце в течение более чем одного часа, температура
листьев все еще превышает температуру воздуха на 3—5°.
В суточном ходе абсолютной влажности заметны два максимума,
в 8 час. и 16 час. 30 мин., минимальные значения наблюдаются перед
восходом солнца. Относительная влажность уменьшается одновременно
с повышением температуры вплоть до дневного максимума. В посадках
она всего только на 3—4% выше, чем в междурядьях.
Ветер на плантации ощущается только тогда, когда его скорость за
пределами посадок достигает 3 м/сек.
Испарение на широкорядной плантации (3 X 3 Л() в междурядьях
на 15—18% выше, чем в метеорологической будке, а в насаждениях —
на 30% ниже; на узкорядной плантации (2,4 X 2,4 испарение в между-
рядьях на 15—20% выше, а в насаждениях так же, как и в первом слу-
чае, на 30—50% меньше.
Освещенность почвы в междурядьях составляет 49% полного днев-
ного света, в густо облиствленных насаждениях на уровне 1 м ниже вер-
шин — только 1,2%, в сильно подстриженных посадках, в 30 см от вер-
шины,— 26%.
На плантациях, затененных Grevillea или Albizzia, деревьев кофе, рас-
положенных под кроной Grevillea, достигает в солнечные дни около 32—
49% дневного света, а в пасмурные — только 15—20%. Количество света,
проникающего под полог Albizzia, увеличивается соответственно до 50
или 26—33%. Под деревьями с густой кроной листья у кофейных деревьев
крупнее, толще и темнее. Средняя температура здесь приблизительно
на 1° выше, чем на слабо затененных участках посадок; только в полдень
она немного ниже и максимума достигает несколько позже.
Если испарение на незатененной плантации принять за 100%, то на
затененной площади оно составит:
В пасмурные [дни В солнечные дни В очень сухие дни
60-65% 75% 90%
Ветрозащитные насаждения повышают дневную температуру и не-
сколько понижают ночную. Относительная влажность в этих условиях
днем ниже. Экстремальные значения метеорологических показателей
усиливаются на плантациях, расположенных в долинах. На восточных
склонах, соответственно ходу солнца в тропиках, температура выше до
полудня, на западных — после полудня. Северные и южные склоны на
экваторе характеризуются меньшей разностью температур. На склонах
западной экспозиции роса испаряется только^ через 2,5 часа после
восхода солнца, на восточных — через 1 — 1,5 часа.
ПОЧВЫ ТРОПИЧЕСКОГО ДОЖДЕВОГО ЛЕСА*
При изучении почв тропического дождевого леса следует учитывать,
что в отличие от почв умеренной зоны это очень древние образования.
Более молодыми могут быть в тропиках только почвы, сформировавшиеся
1 G. W. Robinson, Soils, London. 1939. R. G a n s s e n., Bodengeographie,
Stuttgart, 1957. (Имеется русский перевод: P. Ганссен, География почв, ИЛ. М.,
1962 — Ped.); Р. V a g е 1 е г, Grundriss der tropischen und subtropischen Bodenkunde,
Berlin, 1938.
92
Глава 2
на вулканических отложениях. Ледниковый период вообще не оказал
какого-либо влияния на растительность и почвы экваториальной зоны.
В отдельных районах Африки в геологическом прошлом имело место чере-
дование влажных и сухих периодов, однако почвенный покров никогда не'
уничтожался полностью. Поэтому здесь вполне вероятны почвы, образо-
вавшиеся в третичный период.
Кора выветривания в тропиках необычайно мощная, местами толщина
ее превышает 20 м; под покровом дождевого леса, даже на сравнительно
крутых склонах эрозия почв не наблюдается и продукты выветривания
остаются на месте своего образования х.
Высокая температура, большая влажность и повышенная концен-
трация СО2 способствуют интенсивному выветриванию силикатов мате-
ринских пород, выщелачиванию оснований и кремнезема; остаточные-
продукты представлены главным образом А12О3 и Fe2O3. Этот процесс
принято называть латеритизацией. Вследствие быстрого разло-
жения органического вещества под пологом дождевого леса гумус не
накапливается. Вода в ручьях иногда приобретает светло-коричневую
окраску — признак того, что в ней содержатся соединения гумуса. Мине-
рализация вод очень небольшая; электропроводность соответствует элек-
тропроводности дистиллированной воды.
В качестве характерного примера можно привести описание почвы,
сделанное Мильном, в первичном тропическом лесу в окрестностях Амани
(Восточная Африка); из работы этого автора заимствованы приводимые
ниже цифры.
Почва даже на крутых склонах представлена мощной толщей красно-
вато-коричневого латеритизованного суглинка, залегающего на гнейсах,,
гранулитах и пегматитах. Модуль SiO2/(Al2O3 + Fe2O3) равен 0,68—0,75..
Подстилка в лесу отсутствует. Поверхность, правда, покрыта опавшими
листьями и ветками, но достаточно их слегка разгрести, как обнажается
почва. В разложении опада большое участие принимают термиты;
несмотря на это, их редко удается заметить, поскольку они не сооружают
термитников в лесу. Невелико также число дождевых червей. Главная
роль в процессах разложения, по-видимому, принадлежит грибам. Поч-
венные горизонты выражены нечетко: в профиле отсутствуют латеритные
конкреции или стяжения. Промывной режим почв обусловливает их
сильную выщелоченность. Реакция почвы кислая (pH = 5,3—4,6). Опад,,
образованный листьями и остатками веток и коры, очень кислый (pH -
= 4,05 — 3,55). В пределах почвенного профиля до глубины 2 м значе-
ния pH практически не изменяются (pH = 4,95 — 5,00). Гумусовый гори-
зонт отсутствует, хотя количество органического вещества местами дости-
гает 2,5—4% (цифровые данные получены методом мокрого сжигания).
Органические вещества находятся в почве в виде растворимых соединений,
что обусловливает в лучшем случае чуть более темную окраску верхних
нескольких сантиметров почвы. Поверхность почвы покрыта гумусом,,
отличающимся сравнительно постоянным отношением углерода к азоту,
что видно из табл. 6 1 2.
Механический состав почвы приведен в табл. 7.
1 П. Ричардс в своей монографии высказывает диаметрально противоположный
взгляд на интенсивность эрозионных процессов под пологом леса.— Прим. ред.
2 Аналогичные значения для неоподзоленных (см. примечание на стр. 97) почв;
в бассейне Амазонки получены Клинге (Н. К 1 i n g е, Beitrage zur Kenntnis der tro-
pischen Boden, «Z. Pflanzenern., Diing. u. Bod.», 97, 1962, S. 106—118.
Тропический дождевой лес
93
Таблица 6
Содержание углерода и азота в почвах Амани
(восточная часть гор Усамбара)
Глубина, см Содержание, % C/N
С N
0-8 4,55 0,364 12,5
10-26 2,85 0,215 13,4
31-47 1,55 0,117 13,2
52-63 1,07 0,089 12,0
70-86 1,09 0,084 13,0
94-104 1,08 0,074 14,2
Таблица 7
Механический состав почвы на старой кофейной плантации Квамкоро близ Амани,
созданной в тропическом дождевом лесу. Высота местности 1000 м над уровнем
моря; годовое количество осадков 2000—2200 мм; наклон поверхности 1:3.
Почва—легкий суглинок; карбонаты выщелочены
Глубина, СЛ1 Окраска почвы Гравий % Грубозер- нистый песок, % Тонко-зер- нистый песок, % Пылеватая фракция, % Илистая фракция, % емкость жание воды [ушно-сухом [НИИ
размер частиц, мм о cd о м. ш §3°
2 1 0,2 0,02 0,002 <0,002 И 5- д о О о О Д о
0-13 Светло-коричневая, блеклая 14,1 24,4 20,5 11,15 39,0 32,4 3,0
15-31 Светло-коричневая 31,5 22,6 19,8 11,1 42,7 33,3 3,2
31-47 Оранжево-коричневая 21,9 20,3 18,8 11,6 45,3 32,7 3,8
78—94 Красновато-коричневая 22,2 24,2 24,7 13,0 35,1 27,0 3,45
125-157 Оранжево-коричневая 16,3 28,9 24,7 14,5 30,1 25,7 3,05
157-188 Пурпурно-коричневая 23,4 32,7 22,7 15,9 29,7 27,7 2,6
220-250 Блеклая лиловая 22,0 32,5 22,8 18,0 25,2 30,5 2,4
Глубже 2,5 м преобладает выветренный гнейс, обломки которого встречаются
и выше по разрезу,
Влажность почвы, определенная в двух повторностях, достигает
в лесу на глубине 40 см 26,8—27,2%,
Питательных веществ в почве обычно очень немного: наличие СаСО3
не установлено, N присутствует только в виде органических соединений
(см, табл, 6). Аммиачный азот отсутствует, обнаружены только следы
нитратного азота; так же незначительно содержание фосфора, Количество
растворимого в разбавленной H2SO4 (pH = 3) фосфора составляло (в %):
На поверхности почвы . .
Несколько глубже
В подпочве
18.10-6
11-12.10-6
ю.ю-6
В районе горы Килиманджаро, напротив, молодые почвы на вулка-
нических отложениях оказались в 5—25 раз богаче фосфором.
94
Глава 2
Из приведенных данных видно, что почвы первичных тропических
лесов бедны питательными веществами, необходимыми растениям. Тем
не менее на таких почвах растет роскошный тропический дождевой лес.
При этом следует учитывать, что климат здесь чрезвычайно влажный;
мы же привыкли к тому, что в таких условиях в умеренной зоне леса
быстро деградируют, уступая место верещатникам. Дальнейшее выветри-
вание горных пород не увеличивает притока питательных веществ, посколь-
ку корни растений из-за большой мощности почвы не достигают коренной
породы. Наиболее богата питательными веществами верхняя часть поч-
венного профиля. Обильные осадки обусловливают нисходящее движение
влаги в почве. Глубина проникновения корней, особенно древесных пород,,
до сих пор почти не изучалась. Судя по вывороченным деревьям, корне-
вая система отличается поверхностным расположением. По мнению Бюн-
нинга, максимальная глубина проникновения корней достигает 75 см.
По данным Элленберга, в лесах вдоль Амазонки (Перу) эта величина не
превышает 10—20 см', по-видимому, эти цифры сильно занижены. Устой-
чивость деревьев в тропических лесах достигается при помощи досковид-
ных корней.
Противоречие между бедностью почвы питательными веществами,
с одной стороны, и богатством растительности, с другой, только кажу-
щееся Ч Естественное тропическое лесное сообщество, не испытывавшее
вмешательства человека, представляет вместе с почвой систему, нахо-
дящуюся в динамическом равновесии. Питательные вещества почвы погло-
щаются растительностью и возвращаются обратно в почву вместе с отмер-
шими частями растений. Устанавливается непрерывный обмен веществом.
Чем интенсивнее в данных климатических условиях протекает в почве
процесс минерализации растительных останков, тем быстрее содержащие-
ся в них питательные вещества могут быть вновь использованы живыми
растениями. Поэтому содержание питательных веществ в почве практи-
чески может быть равно нулю. Все основное количество питательных
веществ, необходимое для развития столь пышной растительности, нахо-
дится в самих живых растениях и меньшая часть — в еще не разложив-
шихся остатках растений. В условиях более холодного климата минера-
лизация замедлена; это приводит к накоплению гумуса, количество
которого увеличивается с понижением температуры. В этом случае запас
питательных веществ сосредоточивается частью в живых растениях, частью
в перегное почвы. Доля этого запаса в растительной массе так же, как
и сама масса, в условиях влажного климата тем меньше, чем ниже темпе-
ратуры. Поэтому во влажных арктических или горных областях наблю-
дается кажущееся несоответствие между разреженной растительностью
и мощными гумусными почвами.
Разобраться в условиях почвообразования во влажных тропиках
можно, лишь учитывая этот круговорот веществ. В естественных условиях
потерь питательных веществ не происходит. Корни представляют своего
рода фильтр; они поглощают все вещества, образующиеся в процессе раз-
ложения, ине поглощают только золи органических веществ, о чем можно
судить по окраске родниковых вод. В Амани эту воду использовали
вместо дистиллированной, причем электропроводность ее была даже ниже,
чем у последней. Поскольку питательные вещества извне не поступают, за
исключением, может быть, только азота (связывание N в результате
1 Н.Walter, Nahrstoffgehalt des Bodens und natiirliche Waldbestande, «Silva»,
24, 1936, S. 201—205; 209—213; H. Walter, Zur Frage nach dem Endzustand der
Entwicklung von Waldgesellschaften, «Der Naturforscher», 13, 1936, S. 151—155. ’
Тропический дождевой лес
95
электрических разрядов во время грозы или жизнедеятельности микро-
организмов), то при нарастающем выщелачивании почв тропический лес
постепенно должен был бы деградировать. В действительности же тропи-
ческие леса в течение тысячелетий не изменились и только в последнее
время целые лесные массивы сведены человеком.
Уничтоженные леса, по-видимому, не смогут больше восстановиться,
ибо после корчевки леса и сжигания пней и порубочных остатков проис-
ходит быстрая минерализация питательных веществ и их постепенное
вымывание из почвы осадками. Лишь небольшое количество их исполь-
зуют культурные растения и сорняки. Поэтому тропические почвы на
вырубках в дождевых лесах неплодородны, что вынуждает местное насе-
ление заниматься подсечно-огневым земледелием. После нескольких лет
обработки почва оказывается непригодной для возделывания и приходит-
ся раскорчевывать новые участки леса. На заброшенных же участках
обычно развивается вторичный лес (рис. 63). Но он вряд ли достигнет
богатства первичного леса, поскольку запас питательных веществ в почве
сильно сократился. При неоднократной обработке одних и тех же участков
под сельскохозяйственные культуры они часто зарастают орляком {Pte-
ridium aquilinum), а при регулярном сжигании травостоя — аланг-алан-
гом (Imperatd).
Приведенная выше характеристика африканского первичного тро-
пического леса во всех деталях совпадает с описанными Зиоли 1 лесами
в бассейне Амазонки. По его наблюдениям, плантации, разбитые на выкор-
чеванных участках тропического леса, из-за прогрессирующего истоще-
ния почвы приходится забрасывать самое большее через три года. Спустя
некоторое время здесь вырастает вторичный лес (капуэйра). Через 10 лет
его можно опять вырубать, чтобы в течение последующих 1—2 лет соби-
рать урожай. «Однако прежний прекрасный высокоствольный лес не
восстановится и через десятилетия; как следствие деятельности земле-
дельцев вместо цветущего культурного ландшафта на амазонской терра-
фирме на обширных площадях развивается скудная кустарникоподобная
капуэйра». Элленберг, напротив, после кратковременного пребывания
в перуанской части бассейна Амазонки пришел к выводу, что вторичный
лес восстанавливает плодородие почвы 1 2. Он высказал предположение, что
во вторичных лесах, где растет много видов из семейства бобовых, идет
быстрое накопление азота в почве. Для более определенных выводов
требуется тщательное изучение этой проблемы.
Видимо, уместно задать вопрос, как первоначально образовался запас
питательных веществ, сосредоточенный в растительной массе первичного
тропического леса, в частности запас столь необходимого растениям фос-
фора. Возможный ответ: исходный запас питательных веществ образовал-
ся в то время, когда мощность коры выветривания была не столь велика
и корни растений непосредственно соприкасались с коренной породой.
В тропиках этот период первоначального накопления следует искать
в весьма далеком геологическом прошлом.
Не все почвы зоны влажных тропических лесов относятся к типу лате-
ритных краснозёмов, формирующихся главным образом на силикатных
породах и известняках. В последнем случае в условиях большой влаж-
1 Н. S i о 1 i, Die Fruchtbarkeit der Urwaldboden des brasilianischen Amazo-
nasgebietes und ihre Bedeutung fur die zukiinftige Nutzung, «Staden-Jb.», 5, 1958,
S. 23—36.
2 H. Ellenberg, Typen tropischer Urwalder in Peru, «Schweiz. Zschr. Forst-
wesen», № 3, 1959, S. 169—187.
36
Глава 2
ности карбонаты интенсивно вымываются и сохраняются лишь глинистые
включения; в результате почвы вскоре приобретают кислую реакцию.
Лишь на обнажениях известняков растительность приобретает специфи-
ческий характер. Из-за хорошей водопроницаемости последних на них раз-
виваются характерные для более сухих тропиков полувечнозеленые леса х.
Рис. 63. Вторичный (производный) лес на побережье север-
ного Камеруна. В центре и справа Elaeis guinensis. Подлесок
образован видами семейств Rubiaceae, Acanthaceae и др.;
особенно характерна Musanga smithii с зонтикообразными
листьями (фото Фабера).
На песчаниках и на очень кислых, богатых кремнеземом изверженных
породах, как и на аллювиальных песках, красноземы обычно не форми-
руются. Эти почвы отличаются наличием элювиальных горизонтов и долж-
1 Лёчерт (W. Lotschert) описал это явление в Сан-Сальвадоре. См. также ссылку
на его работу о Кубе (гл. 3, стр. 187).
Тропический дождевой лес
97
ны быть отнесены к подзолам («Phytologie», Bd 111, 1, S. 454) x. Ричардс
отмечает, что этот тип почв широко распространен не только в умеренной
климатической зоне, но и во влажных тропиках. Приводимый ниже поч-
венный разрез описан в одном из районов Калимантана (Борнео) со сред-
ней годовой температурой 26° и суммой осадков за год 3000 мм (причем
в течение любого месяца выпадает не менее 100 мм осадков). Раститель-
ность резко отличается от типичного тропического дождевого леса. Это
так называемый паданг, подобие зарослей или редколесья, являющий-
ся в этой области, видимо, эдафическим климаксом. Вероятно, сама
материнская порода столь бедна питательными веществами, что расти-
тельность при выветривании последней не могла накопить большого запаса
этих веществ. Почва характеризуется данными, приведенными в табл. 8.
Таблица 8
Профиль тропического подзола низменности
(по Хардону, цитируется по Ричардсу)
Мно-
го
2,7
Ао
А,
А 2
В2
0-10
10-25
25-40
40-70
70-100
Черный полуразложивший-
ся органический материал
Рыхлый серовато-черный
кварцевый песок со скоп-
лениями гумуса
Рыхлый серовато-белый
кварцевый песок
Сцементированный слой,
темно-коричневый очень
плотный кварцевый песок
Рыхлый светло-коричневый
кварцевый песок
5,2
95,6
94,0
86,9
92,1
1,6
0,6
7,2
4,6
7,17 3,9 0,022
8,64 6,1 0,032
0,42 3,9 0,029
0,31 4,6 0,035
I Фракции гравия (> 2 мм) и мелкозема (0,005 — 0,05 льи) опущены.
В другом почвенном разрезе из восточного Калимантана под 20-сан-
тиметровым слоем грубого гумуса (pH = 2,8) выделяется мощный элю-
виальный горизонт (pH = 6,1), расположенный над иллювиальным гори-
зонтом (В) желто-коричневого цвета с pH = 5,4. Тропические подзоли-
стые почвы обычно развиваются или на аллювиальных песках, или на
песчаниках. Аналогичные подзолы встречаются в Таиланде и на островах
Индонезии. На оподзоленных песчаных почвах в Британской Гвиане рас-
1 Некоторые современные, преимущественно американские, исследователи
рассматривают тропические почвы как подзолистые, подразумевая под подзолообра-
зованием выщелачивание или удаление глинистого материала из почвы.— Прим. ред.
1394
98
Глава 2
пространены саванны, заросли кустарников, образованные видами Humi-
ria и леса из видов рода Ерегиа. Эти пески являются продуктами разру-
шения гранитов; пески отложились в мелководном бассейне в третичное
время. В бассейне Амазонки также широко распространены подзолистые
почвы, аналогичные гвианским Ч На них растут леса кампина, высотой
до 25 ж, образованные главным образом представителями семейства
Leguminoseae (Ерегиа и др.) и отличающиеся почти полным отсутствием
подлеска 1 2. Черный цвет воды реки Риу-Негру связан с высоким содер-
жанием в ней золей сырого гумуса. Аналогичные почвы распространены
также на острове Мафия (Восточная Африка). Они покрыты верескоподоб-
ными пустошами, в составе которых преобладают виды Philippia (иа
семейства Ericaceae).
В тропиках, помимо подзолов, встречаются также переувлажненные
почвы, появление которых, как и в умеренной зоне, обусловлено высоким
стоянием грунтовых вод. Речь идет о слабогумусных болотных почвах
или о торфянистых болотных почвах. Первые развиваются в эвтрофных
условиях на серых или желтоватых глинах и имеют много общего с глее-
выми почвами умеренной зоны; слой гумуса очень тонок; аэрация почвы
плохая. Торфянистые почвы распространены менее широко; это почвы
тропических лесных болот (прежде всего это относится к почвам омброген-
ного типа, топогенные же болота обычно покрыты травяной раститель-
ностью 3). Sphagnum встречается только на болотах возвышенностей.
Мощность торфа может превышать 7 м. Из омброгенных болот вытекают
«темноводные» потоки с кислой реакцией, pH которых равен 3,0. Это
олиготрофные болота; зольность торфа невелика. Они соответствуют
верховым болотам умеренной зоны и обусловлены, по-видимому, не толь-
ко обилием осадков, но и бедностью подстилающих пород питательными:
веществами. Топогенные болота, как и европейские низинные болота, обыч-
но эвтрофные.
Однако не все тропические почвы бедны питательными веществами,
и прежде всего почвы, развитые на молодых вулканических отложениях.
Непрерывное поступление питательных веществ происходит здесь в про-
цессе выветривания подстилающих пород после сведения леса. Такие-
территории являются поэтому важнейшими районами поселения в тро-
пиках, часто с весьма плотным сельским населением.
Обработка малоплодородных тропических почв может вызвать их
интенсивную эрозию, вследствие чего на поверхность снова выступают
коренные горные породы. Таким образом, весь процесс накопления
питательных веществ мог бы начаться сначала, если бы прекратилось
вмешательство человека. Конечно, процесс этот протекал бы крайне-
медленно, даже при весьма интенсивном выветривании, характерном для
тропиков. Первичный тропический лес таким путем вполне мог бы восста-
новиться.
Разнообразие типов почв в тропиках находит свое отражение в со-
ставе растительности: типичный дождевой лес преимущественно разви-
вается на латеритных красноземах, на других почвах встречаются рас-
1 Н. S i о 1 i, Н. К 1 i n g е, Uber Gewasser und Boden des brasilianischen Ama-
zonasgebietes, «Die Erde», 92, 1961, S. 205—219; H. К 1 i n g e, Podzol soils in the-
Amazon Basin, «J. Soil Sci.», 16, 1965, p. 95 — 103.
2 M. Takeuchi, The structur of the Amazonian vegetation IV, «J. Fac. Sci.
Univ. Tokyo III», 8, 1962, p. 279—288.
3 Почвообразование омброгенного типа связано с обильными осадками (от греч.
арРро — дождь), развитие топогенных болот обусловлено главным образом релье-
фом. — Прим, перев.
Тропический дождевой лес
99
тительные сообщества иного состава. Во влажных тропиках взаимосвязи
между почвой и растительностью также очень тесные, но выявляются
они нередко лишь при внимательном изучении флористического состава.
Важнейшими для растительности свойствами почвы являются, по-ви-
димому, водопроницаемость, аэрация и мощность; степень кислотности
и практически полное отсутствие извести, по всем данным, имеют мень-
шее значение для растительности. Встречаются ли в тропиках подлинно
базифильные растения — вопрос спорный. Диптерокарповые некоторыми
исследователями рассматриваются как ацидофильные, но это утверждение
признается не всеми. В Бирме и Таиланде диптерокарповые леса якобы
произрастают на известняках, а один из видов, Нореа ferrea, даже пред-
почитает селиться на обнаженных карбонатных породах. Все эти вопро-
сы, в том числе и проблема значения алюминия для растений, еще слишком
мало изучены; приводить же недоказанные предположения и детально
рассматривать необоснованные догадки не имеет смысла.
Подводя итоги, следует еще раз подчеркнуть, что богатство тропиче-
ского дождевого леса отнюдь не является признаком большого плодоро-
дия почвы, как это предполагали первые поселенцы; количество живой
растительной массы зависит прежде всего от влажности климата. Это спра-
ведливо не только для тропиков, но и для всех естественных природных
областей, не испытывающих воздействия человека. Поскольку богатство
растительности на сильно выветренных зрелых: почвах обратно пропор-
ционально запасу питательных веществ в них, культурные почвы во влаж-
ных областях деградируют очень быстро и вторичная растительность,
развитая на них, чрезвычайно скудна.
СТРУКТУРА
ТРОПИЧЕСКОГО ДОЖДЕВОГО ЛЕСА
Ниже мы попытаемся дать общее представление о тропическом дож-
девом лесе, но задача эта не проста, ибо в тропиках, видимо, развито так
же много различных типов леса, как, скажем, в умеренной зоне, а может
быть, и больше. Они еще слишком мало изучены, чтобы их можно было
выделять. Поэтому приводимой ниже характеристике дождевого леса не
следует придавать абсолютного значения. Глубокому познанию тропиче-
ского леса мы обязаны прежде всего Ричардсу Ч
Высота деревьев в тропическом дождевом лесу обычно не превышает
50—55 л, но может быть и выше (вплоть до 60 л«). Число древесных
пород, встречающихся в составе древостоя, как правило, велико; часто
на площади всего 1 га произрастает до 40 видов, иногда даже свыше 100,
однако в отдельных случаях не исключена и известная однородность дре-
востоя. Богатством видового состава отличается также травяно-кустар-
никовый ярус, не говоря уже о большом числе лиан и эпифитов.
Тем не менее внешний облик тропических дождевых лесов на всем зем-
ном шаре почти одинаков.
Стволы у деревьев прямые и необычайно стройные. Они начинают
ветвиться очень близко к вершине; кроны небольшие. В нижних частях
ствола часто развиваются досковидные корни. Кора тонкая и гладкая,
листья обычно крупные, кожистые, темно-зеленые цельнокрайние. Напо-
минают они скорее листья лавровишни (Prunus laurocerasus), чем листья
1 Р. W. Richards, The tropical rain-forest, 1952. [Имеется русский перевод:
П. У. Ричардс, Тропический дождевой лес, М., ИЛ, 1961.— Ред.]
100
Глава 2
Рис. 64. Профильная диаграмма дождевого леса; река
Морабалли, Британская Гвиана (по Дэвису и Ричардсу).
Изображены полоса леса длиной 41 и шириной 7,6 At и все деревья
высотой более 4,6 м.
лавра (Laurus nobilis). Только у древесных пород из семейства Legumi-
noseae листья частично перистые, но и они преимущественно довольно круп-
ные. Листва темная, и цветки резко не выделяются. Обычно различают
три или два яруса деревьев, иногда ярусность выражена недостаточно
четко. Пальмы Pandanus и драконовы деревья не являются типичными
древесными породами дождевого леса; они чаще встречаются в периодиче-
ски затапливаемых лесах, развитых на прибрежных равнинах. Подрост
в лесу не столь непроходим, как об этом нередко приходится читать. Много
неприятностей доставляет скользкая глинистая почва. Непроходимую
зеленую стену создают только опушки леса.
Примечательной особенностью, главным образом горных лесов,
является обилие эпифитов и лиан, свисающих, словно толстые канаты,
с деревьев. Эпифитов и лиан тем больше, чем выше влажность воздуха;
лучше всего они развиваются в менее сомкнутых лесах. В общем же тро-
пический дождевой лес, несмотря на богатство растительных форм, про-
изводит однообразное впечатление, тем более что его облик в течение
года существенно не изменяется. Душный спертый воздух в лесу угнетает,
чувствуешь себя словно в темнице, тишина лишь изредка нарушается
ревом обезьян или криком птицы. Находясь в этом перенасыщенном рас-
тительностью лесу, путнику начинает казаться, что он попал чуть ли не
в мертвое царство.
Со структурой тропического дождевого леса лучше всего можно
познакомиться при помощи вертикальных проекций или, как их назва-
ли Дэвис и Ричардс *, профильных диаграмм.
На рис. 64 представлена профильная диаграмма первичного смешан-
ного леса Британской Гвианы, на которой изображены только деревья
1 Т. A. W. Davis, Р. W. Richards (1933—1934), The vegetation of Mora-
balli Creek, British Guiana; an ecological study of a limited area of Tropical Rain-Forest.
Parts I, II, «J. Ecol.», 21, p. 350—384; 22, p. 106—155.
Тропический дождевой лес
101
высотой более 4,6 ж, образующие три яруса. Средняя высота деревьев
верхнего яруса 35 ж; этот ярус образован видами семейств Lecythiadaceae,
Lauraceae и Araliaceae с зонтиковидными кронами. Средний ярус (высо-
той приблизительно 20 м) состоит из древесных пород, относящихся пре-
имущественно к другим семействам. Оба яруса не сильно сомкнуты и не
всегда хорошо выражены. Наиболее высокие деревья (свыше 42 м) стоят
совершенно обособленно.
Сомкнутый нижний ярус, в среднем высотой несколько более 10 м,
представлен подростом и взрослыми древесными породами из семейств
Апопасеае и Violaceae, отличающимися удлиненной формой крон. Все
более низкие деревья переплетены между собой лианами, которые лишь
редко поднимаются на самые высокие деревья. Под пологом древесного
яруса развиваются кустарники, высокие травы и напочвенный покров,
образованный всходами деревьев, низкими травами, папоротниками
и Selaginella\ ярус мхов отсутствует.
На второй профильной диаграмме (рис. 65) представлен диптерокар-
повый лес, развитый на острове Калимантан (Борнео). Здесь также выра-
жены три яруса высотой около 35, 18 и 8 ж. Верхний, образованный
несколькими видами семейства Dipterocarpaceae слабосомкнутый ярус
резко ограничен от нижеследующего яруса; остальные два яруса плохо
выражены. Они образованы представителями самых различных семейств.
Еще ниже расположены ярус кустарников высотой в среднем до 4 ж и тра-
вяной ярус, достигающий 1—2 м высоты. В последнем очень много всхо-
дов древесных пород (184 всхода деревьев на 10 ж2 и 135 экземпляров видов
травяного покрова). Сомкнутость этого яруса не равномерна и не очень
высока. Моховой покров также отсутствует.
।___________1----------1-----------
н 15 10 5 0
Рис. 65. Профильная диаграмма смешанного диптерокарпового леса; гора Дыолит,
Борнео (по Ричардсу).
Изображена полоса леса длиной 61 и шириной 7,6 м; показаны только деревья высотой более
7,6 м.
102
Глава 2
Третья профильная диаграмма (рис. 66) составлена в первичном
дождевом лесу Западной Африки, в районе, характеризующемся менее
равномерным распределением осадков. Средняя высота деревьев в трех
ярусах достигает соответственно 42, 27 и менее 10 м. Только нижний
древесный ярус полностью сомкнут и перевит лианами. Ярусы кустарни-
ков и трав развиты слабо. Не исключено, что отдельные крупные деревья
в лесу вырублены.
Помимо полидоминантных тропических дождевых лесов, распростра-
нены также леса, в которых доминирует только одна какая-либо древес-
ная порода; на рис. 67 представлена профильная диаграмма такого моно-
доминантного леса из Mora excelsa, развитого на острове Тринидад. Верх-
ний сомкнутый ярус напоминает буковый лес Центральной Европы. Оба
нижних яруса выражены менее четко. По своей структуре с этими лесами
сходны леса из Ерегиа в Гвиане и леса Dacryodes-Sloanea Малых Антиль-
ских островов. В Африке и Азии также известны монодоминантные лесные
сообщества. Ярусы кустарников и трав в лесах такого типа иногда пол-
ностью отсутствуют.
Подробно описаны тропические дождевые леса бассейна Амазонки,
расположенные в 42kjh на север-северо-восток от Манауса Ч На возвышен-
1 М. Takeuchi, The structure of the amazonian vegetation II. Tropical rain-
forest. Fac. Sci. Univ. Tokyo, Sect. 3, 8, 1961, p. 1—26.
Рис. 66. Профильная диаграмма дождевого леса; резервированный лес Шаша. Ниге-
к рия (по Ричардсу).
Изображены полоса леса длиной 61 и шириной 7,6 м и деревья высотой более 4,6 .и.Условные
обозначения: АЬ — Pausinystalia или Corynanthe spec.; Ak — Ako отЪа (не определено); ЕЬ — Ca-
searia bride lioide s', Ek — Lophira procera; Ep — Rinorea spec. (cf. dentata)', Er — Picralima
umbellata", Es — Diospyros confertiflora" Ip — Strombosia spec.; It — Strombosia pustulata ;
<Od —Sc&ttellia kamerunensis’, Om — Rinorea spec. (cf. oblongi folia)', Op — Xylopia guintasii',
Os —Diospyros insculpta; Те — Casearia spec.; Y — Parinarium spec. (cf. excelsa).
Тропический дождевой лес
103
Рис. 67. Профильная диаграмма вечнозеленого леса, образованного Mora excelsa
(М); остров Тринидад (по Бирду).
Изображена полоса леса длиной 61 и шириной 7,6 м.
ных участках (60 м над уровнем моря) деревья достигают высоты 30 м.
В этих условиях растут виды семейства Leguminosae и Sapotaceae; несколь-
ко ниже по склонам преобладают виды Lecythidaceae, сменяющиеся у под-
ножия холмов видами семейства Могасеае. Выделяются 3 яруса деревьев.
Для нижнего яруса характерны густые заросли бесствольных пальм.
В понижениях (40 м над уровнем моря), напротив, степень сомкнутос-
ти крон обычно меньше, ярусность не выделяется, деревья отличаются
меньшими размерами. Бросается в глаза обилие эпифитов из семейств
Агасеае, Orchidaceae, Bromeliaceae, также часто встречаются и эпифит-
ные папоротники. Для всех сообществ типичны деревья с досковидными
корнями.
Этих немногих примеров достаточно, чтобы показать, сколь различ-
ной может быть структура первичного тропического леса. Однородным
юн может показаться лишь при беглом осмотре.
Если считать, что тропический дождевой лес в течение тысячелетий
развивался как единое целое, то это можно себе представить только в пре-
делах более или менее крупных лесных массивов. На небольших же пло-
щадях происходит постоянная замена перестойных деревьев более моло-
дыми, причем временно даже может измениться флористический состав.
О возрасте деревьев, не образующих годичных колец, никаких досто-
верных сведений нет. На основе наблюдений за годичным приростом в неко-
торых случаях удалось определить возраст деревьев, достигающий при-
близительно 200—250 лет.
Падая, лесные великаны образуют в пологе леса окна. Более крупные
из них в первую очередь заполняют виды, доминирующие в составе вто-
ричного леса, например Cecropia — в Южной Америке, Musanga,, или
Schizolobium (рис. 68)—в Африке и Macaranga —в Малайе. В дальней-
шем эти виды вытесняются подростом древесных пород верхнего яруса.
104
Глава 2
Потребность в свете у растений различна. Некоторые из них светолюбивы;
они образуют всходы и вегетируют только на достаточно больших прога-
линах. Другие виды могут расти в нижнем ярусе древостоя, то есть они
начинают свое развитие в глубокой тени.
Количество семян, всходов и молодняка на разных стадиях у разных
видов весьма различно. У некоторых видов деревьев самых верхних яру-
сов иногда совершенно отсутствует подрост или он встречается на очень
небольших по площади участках. Только в монодоминантных лесах гос-
подствующая порода хорошо обеспечена подростом, представленным во
всех ярусах. Прорастание семян обычно происходит сразу, однако иногда
они обладают некоторым периодом покоя. В условиях недостаточной
освещенности почвы в тропическом лесу
, лишь некоторым всходам удается вы-
держать борьбу за существование в те-
чение первых лет жизни. Ричардс
отмечает, что в лесах Британской
Гвианы повсюду встречались всходы
некоторых видов пальм, растущих во
взрослом состоянии только на боло-
те; в лес же плоды этих растений были
занесены птицами. Таким образом,
появление молодого подроста в лесу
не является доказательством начав-
шейся смены, как это утверждают
многие ботаники.
Скорость роста молодых деревьев
целиком зависит от условий окру-
жающей среды. В глубокой тени еже-
годный прирост оказывается совсем
ничтожным. Например, в диптерокар-
повом лесу у молодых деревьев диа-
метром до 5 см прирост стволов в тол-
щину составляет за год менее 0,7 мм,
у более крупных деревьев — почти
1 см. Помимо недостатка света* боль-
шое значения для развития подроста
имеет корневая конкуренция. При
благоприятных же условиях, напри-
мер на лесных прогалинах, деревья
растут гораздо быстрее. Так, у Flin-
dersia в северо-восточном Квинсленде
на вырубке прирост в толщину дости-
гал 2,5 см в год, в то время как в лес-
ной чаще прирост не удалось даже
измерить.
Отсутствие самовозобновления под
пологом древесных пород верхне-
го яруса Обревилль на основе ис-
следований в дождевых лесах Берега
Слоновой Кости объяснил при по-
мощи теории так называемого мозаич-
Рис. 68. Schizolobium excelsum (фото ного возобновления. Он высказал
Тройба). предположение, что в первичном трот
Тропический дождевой, лес
105:
Рис. 69. Многочисленные термитники на вырубке в первичном тропическом лесу;
бассейн Конго близ Янгамби (0° 49' с. ш. 24^ 29' в. д.) (по Ахтниху,} неопублико-
ванные материалы).
пическом лесу любой вид, доминирующий на данном участке, не является
постоянным ни в пространстве, ни во времени; иными словами, на неболь-
ших участках происходит смена одних доминант другими, причем возоб-
новление разных видов про-
исходит циклично. Нечто по-
добное можно наблюдать на
очень ограниченном простран-
стве в сообществах лугов Цен-
тральной Европы. И здесь от-
дельные виды меняют места
своего произрастания как
в пространстве, так и во вре-
мени Ч
Как, вероятно, правильно
замечает Ричардс, это пред-
ставление в известных случа-
ях подтверждается, но универ-
сальным, пригодным для всех
случаев, считать его нельзя.
То, что справедливо в одних
типах дождевого леса, не обя-
зательно может быть прило-
жено к другим.
1 Н. L i е t h, op. cit., S. 35.
Рис. 70. Rafflesia patina', холопа-
разит, паразитирующий на корнях
деревьев; остров Ну-Камбанган,
Ява (фото Фишера).
106
Глава 2
Рис. 71. Различная форма досковидных корней. В середине — Ficus variegata\1
на заднем плане, слева — Ficus cordifolia, справа — Shorea balangaran var. binnen-
dijkii (фото Фабера).
Перестойные деревья при падении образуют окна в пологе леса;
•если же они разрушаются изнутри термитами или грибами, то они разва-
ливаются, не задевая окружающих деревьев. Необычайно много тер-
митов в первичных тропических лесах бассейна Конго. На рис. 69 изобра-
жен участок вырубки, усеянный термитниками.
Около 70% всех видов растений, встречающихся в тропическом
дождевом лесу, относится к фанерофитам — деревьям и кустарникам,
господствующим не только в численном выражении, но и по своей массе.
Помимо фанерофитов, в тропических дождевых лесах представлены и дру-
гие жизненные формы с весьма своеобразной экологией, они часто обра-
зуют оригинальные синузии.
В тропическом дождевом лесу выделяются следующие жизненные
•формы:
1. Деревья и кустарники.
2. Травы.
3. Лианы.
4. Полу эпифиты.
5. Эпифиты.
Гетеротрофные виды — сапрофиты и паразиты — в тропическом дож-
девом лесу играют подчиненную роль; среди последних встречаются
очень интересные виды, например Rafflesia (рис. 70); широко распро-
странены также виды семейства Loranthaceae.
Тропический дождевой лес
107
ЭКОЛОГИЯ ДРЕВЕСНЫХ РАСТЕНИЙ
ТРОПИЧЕСКОГО ДОЖДЕВОГО ЛЕСА
Деревья тропического леса часто отличаются известными особенно-
стями, характерными для многих, но не для всех видов. Выше уже отме-
чалось, что стволы у деревьев гладкие и стройные; кора обычно очень
светлая и тонкая. Корка, как правило, не образуется, так как стволы
практически никогда не попадают под прямые лучи солнца и не испыты-
вают иссушающего воздействия воздуха. Характерно, что одна из древес-
ных пород африканского дождевого леса, Lophtra procera, образует очень
тонкую корку, тогда как у L. alata, вида, чрезвычайно похожего по своим
таксономическим особенностям на предыдущий, но распространенного
в саваннах, корка очень толстая и испещрена глубокими бороздами
(Ричардс). Стволы с шипами, столь характерные для растительности
аридных областей, в дождевом лесу встречаются очень редко. Шиповатые
стволы, например, в Африке имеет Cylicomorpha parvtflora.
Весьма характерная особенность многих большей частью круп-
ных деревьев — досковидные корни (рис. 71 и 72). Они придают
деревьям с поверхностной корневой системой большую устойчивость.
Досковидные выросты образуются от корней, отходящих от основания
ствола, у которых камбий в первые годы функционирует нормаль-
но, и лишь спустя некоторое время начинается отложение вторичной
древесины только по верх- 1
нему краю корня. У взрос-
лого дерева досковидные
корни достигают высоты
до 9 м и на такое же рас-
стояние радиально расхо-
дятся от ствола, постепен-
но уменьшаясь в размерах.
Толщина их колеблется
от нескольких сантиметров
до внушительной величи-
ны. Образование досковид-
ных корней, по-видимому,
связано с избыточным
увлажнением почвы. Воз-
можно, что деятельность
камбия вследствие плохой
аэрации подавляется на
нижней стороне корней.
Досковидные корни обра-
зуются даже у некоторых
европейских древесных по-
род на влажных местооби-
таниях, например у Ulmus
ejfusci в пойменных лесах
или у Populus italica.
Ходульные корни у де-
ревьев тропического леса
встречаются реже — обыч-
Рис. 72. Крупные досковидные корни у Dracon-
tomelum mangiferum.
но только у видов нижнего
яруса древостоя (рис. 73).
108
Глава 2
Почки деревьев дождевого леса в общем защищены очень слабо;
часто эту функцию выполняют только волоски, слизистые выделения
или особым образом развившиеся прилистники и листовые черешки.
Молодая листва на концах растущих побегов часто выглядит слабой
и висит, как увядшая (Schiittellaub) (рис. 74 и 75). При быстром росте
Рис. 73. Ходульные корни у Iriartea orbignyana\
дождевой лес у подножия Кордильер (высота 880•,и)
в восточной Боливии (фото Герцога).
молодых побегов формирование механической ткани часто отстает, поэто-
му молодая листва в первое время бывает вялой. Концы побегов — вна-
чале белые или ярко окрашенные антоцианом в красный цвет — лишь
много позже, по мере приобретения зеленой окраски, становятся жест-
кими. В условиях менее влажного климата развитие молодых побегов
обычно протекает медленнее и идет параллельно с дифференциацией
ткани. Эта вялая молодая листва очень легко засыхает и поэтому наблю-
дается только в условиях высокой влажности. Несколько сходное явле-
ние отмечается в умеренной зоне у молодых светло-зеленых листочков:
бука.
Форма и размер листьев мало варьируют; деревья в подавляющем
большинстве случаев имеют большие цельные листья, нередко с острием,
образующим кончик-капельницу. Из 41 вида, относящихся к 20 семей-
ствам, в лиственном лесу на Цейлоне (Бейкер, Ричардс) 90% имели листья
с кончиками-капельницами. Йодобную же форму имели листья тропиче-
ского дождевого леса на острове Калимантан и в Нигерии. Деревья с рас-
сеченными листьями представляют исключение. В общем размер листьев
убывает с высотой деревьев; листья при этом становятся более кожистымиг
а их кончик-капельница более коротким.
Тропический дождевой лес
109
Рис. 74. Молодая листва на концах расту-
щих побегов у Maniltoa gemmipara (фото
Фабера).
Рис. 75. Крупный план побега
у Maniltoa gemmipara, изобра-
женного на рис. 74.
Большие различия в размерах листьев выявляются при сравнении
листьев одного и того же вида у молодых экземпляров, развитых в ниж-
нем ярусе, с листьями особей, расположенных в верхнем ярусе, или при
сопоставлении листьев на одном дереве — на нижних затененных и на
освещенных ветках.
В дождевом лесу близ Амани (Восточная Африка) автор на одном
к том же дереве обнаружил следующие соотношения (табл. 9).
Таблица 9
Название растений Соотношение длины и ширины листа, сти Отношение
у самого маленького листа у самого большого листа
Myrianthus arboreus 16X7 48 х 19 1 :8
Anthocleista orientalis 22 X Ю 162x38 1 : 28
110
Глава 2
Таблица 1(>
Породы верхнего яруса древостоя Величина осмотического давления, атм
Allanblackia stuhlmannii с кожистыми листьями средней вели- чины Световые листья высокого дерева (нижние ветви) . 16,4
Наиболее старые листья другого дерева 16,5
Молодые листья того же дерева, уже закончившие свой рост 14,7
Листья среднего размера на дереве средней высоты 15,2
Теневые листья молодого дерева .... 13,3
Опавшие, по еще свежие листья с того же дерева 16,3
Piptadenia buchanani Листья молодого дерева, растущего на относительно освещен- ном месте . . . . . . 13,7
Листья 3-метрового дерева, уже закончившие рост 11,1
Молодые, очень нежные листья того же дерева . 8,8
Berlinia schejfieri с топкими кожистыми листьями, похожими на листья бука Старые листья .... 9,7
Побеги со светлыми вялыми листьями 7,1
Рarmarium hoistii Листья с ветки, сломленной сильным ветром 13,&
Albizzia fastigiata. Темно-зеленые листья молодого дерева, уже закончившие свой рост 12,1
Молодые, поникшие светло-зеленые листочки 10,7
Paxiodendron usambarense Крупные кожистые, но не очень толстые листья . . . 13,8
Нижеперечисленные виды встречаются чаще в нижнем ярусе древостоя или должны рассматриваться как породы, через которые идет смена вторичных лесов па первичные Anthocleista orientalis (средняя жилка листа вырезана) Листья большого дерева, уже закончившие свой рост 8,2
Молодые листья того же дерева 8,1
Очень большой лист молодого дерева ... ... 7,7
Листья, еще не завершившие свой рост, на опушке леса 10,2
Macaranga usambariensis Старый лист без средней жилки 9,4
Листья, уже закончившие свой рост, без средней жилки . 9,0
Очень молодые листочки, не более 10 см длиной . 6,1
Polyscias polybotrya Листья, уже закончившие свой рост, с чешуйкамп на нижней стороне ... 9,5
Конус нарастания и прилистники, обильно покрытые чешуй- ками 7,2
Whitjieldia longifolia кустарниковидпой формы Кожистые, как у Aucuba, листья 17,9
Cylicomorpha parvijlora со стволом, покрытым шипами Частично затененные листья 8,0*
Тропический дождевой лес
111
У Polyscias polybotrya на взрослом экземпляре, развитом в верхнем
ярусе, длина листьев составляла 55—95 см, а у молодого деревца того же
вида, расположенного в пределах кустарникового яруса,—170 см.
Эти различия в размерах листьев отражают микроклиматические
особенности, существующие в верхнем и нижнем ярусах древостоя и соот-
ветственно в древесном и кустарниковом ярусах.
Высокая влажность воздуха и хорошая влагообеспеченность деревьев
дождевого леса создают благоприятные гидратурные условия (ср. гл. 7,
стр. 320), выражающиеся в низком осмотическом давлении у листьев.
С октября по декабрь автор изучал в дождевом лесу окрестностей
Амани (Восточная Африка) осмотическое давление у листьев; результаты
приведены в табл. 10. Эти измерения, правда, не коснулись листьев, раз-
витых на вершинах самых высоких деревьев, однако, учитывая резуль-
таты аналогичных определений у деревьев умеренной зоны \ можно пред-
полагать, что величина осмотического давления у них ненамного больше,,
чем у освещаемых солнцем листьев нижних, доступных изучению ветвей.
Таблица И
Древесные породы Величина осмотического- давления, a?n.w
Ficus elastica
Листья, уже закончившие свой рост . 10,8
(Для сравнения листья того же вида из оранжереи в Штутгарте) 9,8
Молодые окончания воздушных корней, толщиной 2 мм . . . Ficus nitida (= retusa), очень высокое дерево 7,7
Листья нижних ветвей, маленькие и жесткие 14,3
Artocarpus integrifolia Листья на пневой поросли Cinnamomum ceylanicum 10,9
Старые листья, кожистые, с сильным запахом . 9,7
Молодые листья, совершенно вялые, ярко-красные Ilex paraguajensis 6,6
Жесткие листья, уже закончившие свой рост . . 13,5
Молодые мягкие, недавно распустившиеся листья . 11,2
В табл. 11 приводятся величины осмотического давления у листьев
древесных пород, интродуцированных в районе Амани.
Автор провел также 45 определений величины осмотического давле-
ния у кофейного дерева. Каких-либо существенных различий в гидратуре
у Coffea arabica, С. liberica и С. robusta не установлено (табл. 12).
Швейтцер на основании изучения обширного клопового материала 1 2
показал, что на Яве кофейное дерево цветет лишь при определенной
величине осмотического давления 3. В условиях слишком влажного кли-
мата нормального цветения не происходит.
1 Н. Walter, Die Hydratur der Pflanze, Jena, 1931.
2 Клон — потомство одного растения, полученное путем вегетативного размно-
жения. — Прим, перев.
3 J. Schweitzer, Handl. 7. Ned. Ind. Naturwat. Congress, 1936, S. 436
(автор не имел возможности ознакомиться с оригиналом).
112
Глава 2
Таблица 12
Осмотическое давление у некоторых видов Coffea1, атм
Название растений У молодых экземпляров Оптимальная величина Максималь- ная величина
Coffea arabica 10,4—12,0 14,4-15,5 18,9
Coffea liberica 9,7 15,8 19,5
Coffea robusta 8.8 13,5-16,6 20,6
1 У теневых листьев величина оптимального давления была несколько ниже:
12,4 — 13 атм.
Эти выводы были подтверждены исследованиями Альвима 1, Почки
у Coffea открываются только в момент некоторого временного понижения
гидратуры растений. Одну из опытных площадок на кофейной плантации
в Перу еженедельно обильно поливали, так что растения не испытывали
недостатка во влаге; в результате ни одна почка не раскрылась и растения
не зацвели. Полив второй площадки начался лишь тогда, когда влаж-
ность почвы достигла величины, близкой к точке завядания. Спустя
10—И дней все почки тронулись в рост и деревья зацвели. Однако при-
чинная связь между уменьшением гидратуры и началом цветения точно
еще не установлена. Поскольку Coffea — растение короткого дня, про-
должительность освещения не оказывает влияния на заложение почек.
Во время необычайно сильной засухи в 1935 г. Швейтцер имел воз-
можность определить максимальную величину осмотического давления
у различных сортов Coffea, Оказалось, что при повышении осмотического
давления интенсивность ассимиляции снижается, но листья, даже несколь-
ко пострадавшие от засухи, через 24 часа после дождя вновь начинают
ассимилировать.
Величина осмотического давления у древесных пород, произрастаю-
щих во влажных тропиках, ниже, чем у видов деревьев умеренной зоны 1 2.
С равномерностью увлажнения в тропических дождевых лесах связана
незначительная величина сосущей силы клеток. Соответствующие изме-
нения были произведены Блумом 3 на Яве. Более точные результаты,
по-видимому, дает метод, использованный Леме при изучении кустарников
в первичных тропических лесах Берега Слоновой Кости 4. Полученные
им значения колеблются в пределах 3—8 атм, У растений открытых мест
сосущая сила клеток резко возрастает; на расчищенных участках она
превышает 20 атм. Еще выше эти значения (до 10—25 атм) в севернее
1 De Т. Р. А 1 v i m, Moisture stress as a requirement for flowering of coffee.
«Science», 132, 1960, p. 354.
2 Жесткость листьев, на которую обращает внимание Фабер, ссылаясь на иссле-
дования Шталя, обусловлена накоплением в них кремния. Это относится прежде всего
к листьям многих лиан, отличающихся интенсивным током влаги. Кремневая кислота,
вымываясь из почвы, доставляется к листу транспирационным током, где и отлагается
в его эпидермисе.
3 G. Blum, Osmotische Untersuchungen auf Java, «Вег. Schweiz. Bot. Ges.»,
42, 1933, S. 550-680.
4 G. L e m e e, La tension de suction foliaire, critere eco-physiologique, des condi-
tions hydriques dans la strate arbustive des groupements vegetaux en Cote d’Ivoire,
-«Naturalia Monspeliensis, Ser. Bot.», 8, 1956, p. 125—140.
Тропический дождевой лес
ИЗ
расположенных областях, где распространены саванны, однако к данным
о величине сосущей силы следует относиться критически *.
Вопросам транспирации в тропиках начали уделять внимание
с 1892 г., когда Габерландт, основываясь на материалах своих исследо-
ваний в Бейтензорге (Богоре) и Граце, высказал предположение, что
интенсивность транспирации у растений влажных тропиков приблизи-
тельно в 2—3 раза ниже, чем в Центральной Европе 1 2. В 1935 г. Штоккер
сопоставил сведения, содержащиеся в ранее вышедшей литературе 3 4,
и провел самостоятельные исследования; методом быстрого взвешивания
он исследовал суточный ход транспирации у трех видов, не свойственных
собственно дождевым лесам. Он установил, что эти виды характеризуются
сравнительно небольшой интенсивностью транспирации и что в полдень
происходит частичное замыкание устьиц, однако он не обнаружил ника-
ких признаков нарушения водного баланса.
В районе Амани автор исследовал транспирацию у некоторых древес-
ных пород; в качестве объекта были использованы виды с крупными
листьями или срезанные ветки; изучение велось при помощи точных тех-
нических весов (грузоподъемностью 10 кг). При сопоставлении транспи-
рации свежих растений с испарением, равным 0,48 мл!час (стандартная
величина), оказалось, что семь исследованных видов деревьев в Амани
транспирируют 2,6—22 мг/г-мин^.
Таблица 13
Интенсивность транспирации у некоторых тропических древесных пород,
мг/г-мин
Виды нижнего и верхнего ярусов древостоя1
Macaranga usambarica (58,6) . . . 22,0
Albizzia fastigiata (62,1) . . . . 14,5
Piptadenia buchanani (45,5) 9,0
Authocleista orientalis (82,1) 8,0
Allanblackia stuhlmannii (71,3) . . . 3,2
Tabernaemontana holstii (70,1) . . 3,0
Myrianthus arboreus (62,7) . 2,6
Ветки и отдельные листья видов Coffea и Thea 2
Coffea robusta (54—66), ветки, среднее из пяти опытов..................3,3 (3,0—4,0)
Coffea robusta, отдельный лист........................................7,3
Coffea liberica (52—69), ветки........................................3,0
Coffea arabica, отдельные листья, среднее из 10 опытов.................7,6 (5,5—12,8)
Thea sinensis, ветки, в среднем........................................1,8 (0,9—2,6)
Thea sinensis (67—72), отдельные листья, в среднем.....................1,5 (0,7—2,7)
1 В скобках приведено’содержание воды в листьях в процентах на сырой вес.
2 Транспирация отдельных листьев определялась с помощью переносных весов Бунге.
У чайного куста получены те же значения интенсивности транспирации отдельных листьев,
что и при исследовании целых веток в пересчете на 1 г сырого веса листьев; у видов Coffea
величина транспирации была в два раза выше.
1 Н. Walter, Zur Klarung des spezifischen Wasserzustandes im Plasma und
in der Zellwand bei der hoheren Pflanze und seine Bestimmung, «Вег. Dtsch. Bot. Ges.»,
76, 1963, S. 40—71.
2G. Haberlandt, Anatomisch-physiologische Untersuchungen liber das
tropische Laubblatt, «Sitz. ber. Akad. Wiss. Wien», 51, 1892, S. 785; E. G i 11 a y,
Vergleichende Studien liber die Starke der Transpiration in den Tropen, «Jb. wiss. Bot.»,
30, 1897, S. 615—644; 32, 1898, S. 477—502; 34, 1900, S. 405—424; K. Holtermann,
Uber den EinfluB des Klimas auf den Ban des Pflanzengewebes. Anat.-physiol. Unter-
suchungen in den Tropen, Leipzig, 1907.
3 O. Stocker., Ein Beitrag zur TranspirationsgroBe im javanischen Regenwald,
«Jb. wiss. Bot.», 81, 1935, S. 464—496.
4 Величины, полученные методом Штоккера, при пересчете составили
бы 5—9 мг!г*мин.
114
Глава 2
Другие культивируемые древесные породы
Artocarpus integrijolia (74) . 5,2
Citrus aurantium (59) ... .4,1
Bambusa vulgaris (49).............................................................. .5,8
Acacia saligna (филлодии) (31) . 5,3
Eucalyptus citriodora (42—48) . 3,4
Casuarina equisetifolia (54) . . .1,8
Интенсивность транспирации, как мы видим, колеблется весьма
значительно, причем приблизительно в тех же пределах, как у растений,
исследованных Пизеком и Картельери в районе Инсбрука при той же
упругости пара в окружающей атмосфере (ср. «Phytologie», Bd III, 1,
S. 310). Относительно низкие величины транспирации или понижение
транспирации в полуденные часы в тропиках становятся понятными, если
принять во внимание перегрев освещенных солнцем листьев.
Выше было показано, что в ясные дни температура листьев кофей-
ного дерева на 10—15° превышает температуру окружающего воздуха.
Такие избыточные температуры 1 тем опаснее для растения, чем выше
температура воздуха, так как у поверхности листа градиент давления
водяного пара возрастает более сильно, чем градиент температуры.
Предположим, что при относительной влажности воздуха, равной
50% , температура воздуха в одном случае будет 10, а в другом 30°, причем
в обоих случаях при сильной инсоляции избыточная температура испаряю-
щей поверхности будет равна 10°, то есть температура самой испаряющей
поверхности в одном случае достигнет 20, а в другом 40°. Тогда при дав-
лении водяного пара, равном полному насыщению, у испаряющей поверх-
ности возникнут градиенты давления, приведенные в табл. 14.
Таблица 14
Давление водяного пара воздуха при £ = 10° (50% от 9,2 мм) То же у испаряющей поверхности (£ = 20°) . 4,6 мм 17,5 мм
Разность 12,9 мм
Давление водяного пара воздуха при £ = 30° (50% от 31,8 мм) . . То же у испаряющей поверхности (£ = 40°) . 15,9 мм 55,3 мм
Разность 39,4 мм
Эти данные показывают, что при одной и той же избыточной темпера-
туре и прочих равных условиях испарение во' втором случае было бы
в три раза интенсивнее. Для защиты от чрезмерной потери воды в листьях
растений имеются особые приспособления, например толстая кутикула,
и способные к замыканию устьица. Упругость пара у поверхности листа
и транспирация при закрытых устьицах сильно снижаются, что, однако,
приводит к перегреву листьев, поскольку не происходит охлаждающего
влияния испарения воды. Для деревьев с освещаемой солнцем кроной
1 Избыточная температура в данном случае — температура поверхности листьев»
превышающая температуру окружающего воздуха.— Прим. ред.
Тропический дождевой лес
115
сильная потеря воды, особенно в полуденное время, могла бы стать при-
чиной гибели. Следовательно, деревьям тропического дождевого леса
в течение нескольких часов приходится испытывать недостаток влаги,
по-видимому не менее жестокий, чем в аридных областях.
Именно этими условиями следует объяснить часто встречающуюся
ксероморфную структуру листьев, благодаря которой растения получают
Рис. 76. Пышное цветение (каулифлория) Dysoxy-
lum caulostachyum, Новая Гвинея (фото Фабера).
возможность постоянно поддерживать стабильный водный баланс с высо-
кой гидратурой (низкое осмотическое давление), несмотря на временами
большой дефицит влажности листьев.
При изучении режима увлажнения в тропическом дождевом лесу
нельзя руководствоваться кривой годового хода осадков, составленной
по многолетним средним месячным данным. Короткие сухие периоды
в отдельные годы бывают во всех экваториальных областях. Но так как,
с одной стороны, расход воды всей транспирирующей массой листьев
в дождевом лесу очень велик, а, с другой — деревья из-за своей относитель-
но неглубокой корневой системы используют лишь незначительную часть
запаса почвенной влаги, они часто испытывают поэтому регулярно повто-
ряющийся недостаток воды. Стеногидрические древесные породы тропи-
116
Глава 2
ческого леса могут переносить эти неблагоприятные условия только путем
сокращения потерь воды почти до нуля.
К аналогичным выводам приходит Коутиньо на основе собственных
тщательных исследований на опытной станции Алтту-да-Серра, располо-
ложенной между Сантусом и Сан-Паулу на восточном побережье Брази-
лии 4. Дождевой лес развивается там при годовой сумме осадков 3600 мм.
Однако июнь или июль иногда бывают бедны осадками, а в некоторые
годы была отмечена даже засуха, продолжавшаяся до 60 дней. В этих
условиях благодаря ксероморфной структуре эпидермиса листьев кути-
кулярная транспирация после замыкания устьиц может сократиться
до 0,3%. Вообще же интенсивность транспирации, не достигающая
6 мг!дм2 • мин, не велика.
Не удивительно поэтому, что даже в наиболее влажных тропических
районах ставят опыты по искусственному дождеванию древесных культур
с целью защиты их от пагубного влияния кратковременных засух. Дело
в том, что важнейшие тропические культуры, такие, как каучуковое
дерево (Hevea brasiliensis), дерево какао (Theobroma cacao) и кофейное дере-
во (виды Coffea), являются стеногидрическими видами. Они отличаются
низким осмотическим давлением и невысокой транспирацией.
Coffea закрывает устьица сразу же после того, как лучи солнца начи-
нают падать на листья. Поэтому кофейные плантации обычно заклады-
вают в полутени других древесных пород. Ассимиляция СО2 в облачные
дни приблизительно в два раза выше, чем в солнечные 2; ее максимальная
величина составляет 4,5, средняя —1,9 мг СО2/дм2-час.
Аналогичная характеристика может быть дана деревьям какао 3.
Температура незатененных, попавших под прямые лучи солнца листьев
уже через 2 минуты поднимается с 28 до 48°. Транспирация увеличивается
в 3—5 раз, и при дефиците влажности листьев всего в 4,5% устьица закры-
ваются в течение 6 минут. Фотосинтез прекращается. Дерево какао гораз-
до в большей степени нуждается в почвенной влаге, чем во влаге воздуха.
При сокращении содержания воды в почве на 2/3 от полевой влагоемкости
сильно понижаются как транспирация, так и интенсивность фотосинтеза.
При искусственном орошении какао можно культивировать даже в пусты-
не Перу, где оно даст очень хороший урожай, так как в этих условиях
дерево не подвержено грибковым заболеваниям. Одно дерево расходует
в сутки около 40 л воды, в год — приблизительно 14 тыс. л, а вся планта-
ция — ориентировочно 900 мм. Что касается фотосинтеза, деревья какао
ведут себя так же, как теневыносливые растения 4. Очень чувствительны
к сухому воздуху молодые, еще нежные листочки. Это свидетельствует
о том, что во влажных тропиках жесткая структура уже закончивших
свой рост деятельных листьев растений, находящихся под прямы-
ми лучами солнца, — необходимое приспособление к местным усло-
виям.
Сведения об измерении ассимиляции СО2 у других видов растений
очень скудны. Все имеющиеся данные приведены в работах, опубликован-
1 L. М. Coutinho, Contribuicao ао conhecimento da ecologia da mata pluvial
tropical, Fac. Fil., Univ, de Sao-Paulo, Bollet. № 257, 1962.
2 F. J. Nutman, Studies of the physiology of coffea arabica L. Photosynthesis
of coffee leaves under natural conditions, «Ann. Bot.», N.S. I, 1937, p. 353—367.
3 P. de T. A 1 v i m, Water requirements of Cacao. FAO Techn. Cacao Meeting,
Acera (Ghana), 1959.
4 G. Lemee. Recherches eco-physiologiques sur le Cacaoyer. «Rev. gen. Bot.»,
63, 1956, p. 41-94.
Тропический дождевой лес
117
ных более 25 лет назад U Применявшиеся в то время методы были менее
точны; кроме того, исследования проводились со срезанными листьями
и при искусственном освещении. У древесных пород собственно дождевого
леса к настоящему времени характер ассимиляции изучен (Штоккером)
только у Stelechocarpus burahol (Anonaceae), вида, обладающего круп-
ными, длинными, жестеподобными листьями с кончиком-капельницей.
Остальные изученные виды — это либо древесные породы сухих тропиков,
либо травяные или культурные растения (см. табл. 15).
Таблица 15
Максимальная интенсивность ассимиляции у различных видов
Название растений Положение листьев по отношению к свету Интенсив- ность ас- симиляции, мг/дм2- час Автор
Stelechocarpus burahol Световые 6,3 Штоккер
Stelechocarpus burahol Теневые 2,6 »
Cassia fistula . Световые 11,0 »
Cassia fistula .... Теневые 8,6 »
Calophyllum inophyllum . Световые 7,3 »
Abutilon darwini . » 7,5 Дастур
Sparmannia africana » 7,5 »
Cocos nucifera » 0,9 Мак-Лин
Musa textilis . . . » 1,0 »
Сахарный тростник . » 5,0 »
Cassia timorensis » 19,2 Гилтей
Cedrela serrulata » 9,0 »
Acalypha tricolor . . . » 7,0 »
Helianthus annuus . » 8,0 »
Nicotiana rustica » 9,0 »
Интенсивность ассимиляции в тропиках отнюдь не выше, чем в дру-
гих климатических областях. Не установлены также какие-либо другие
особенности у тропических растений. Дыхание, несмотря на более высо-
кую температуру, существенно не отличается от дыхания в умеренной
зоне. У многих древесных пород в тропиках листья расположены верти-
кально, в связи с чем они хорошо отражают солнечный свет. При высоком
стоянии солнца пластинка листа получает только ослабленный свет, что,
как мы показали выше, в этих условиях более благоприятно для растений.
По Штоккеру, максимальная ассимиляция у Stelechocarpus наблюдалась
1 Е. Gil tay, Uber die vegetabilische Stoffbildung in den Tropen and Mittel-
europa, «Ann. Jard. Bot. Buitenzorg», 15, 1898, S. 43. F. T. Me Lean, Field studies
of the carbon dioxide absorption of coconut leaves, «Ann. Bot.», 34, 1920, p. 267;
R. H. D a s t u r, The relation between water content and photosynthesis, «Ann. Bot.»,
39, 1925, p. 769; H. von Guttenberg, Beitrage zur Kenntnis der Laubblattassi-
milation in den Tropen, «Ann. lard. bot. Buitenzorg», 41, 1931, S. 115; 0. S t о c k e r,
Assimilation und Atmung westjavanischer Tropenbaume, «Planta», 24, 1935, S. 402—445.
Помимо перечисленных работ, имеются описания исследований, произведенных
в оранжереях. W. Hartenberg, Der Wasser- und Kohlensaurehaushalt tropischer
Regenwaldpflanzen in sommerlicher Gewachshauskultur, «Jb. wiss. bot.», 85, 1937,
S. 641—697; G. Sander-Viebahn, Der winterliche Kohlensaurehaushalt tropi-
scher Gewachshauspflanzen, «Beitr. Z. Biol. d. Pfl.», 37, 1962, S. 13—53.
118
Глава 2
Р и с. 77. Часть ствола Dysoxylum (см. рис. 76),
сфотографированная двумя месяцами позже.
Частично раскрывшиеся плоды висят на корот-
ких безлистных побегах (фото Фабера).
при относительно небольшой
освещенности. Во всяком слу-
чае, все виды дождевого леса,
по крайней мере в стадии под-
роста, должны приспособиться
к развитию в условиях неболь-
шой освещенности.
Накопление органического
вещества в первичном тропи-
ческом лесу зависит не столько
от интенсивности ассимиляции
на единицу листовой площади,
сколько от величины всей ли-
стовой поверхности, приходя-
щейся на 1 га. Последняя здесь
вряд ли намного больше, чем
в лиственных лесах умеренной
зоны. Правда, следует учесть,
что листья в тропиках ассими-
лируют СО2 беспрерывно в те-
чение всего года. Годовое коли-
чество накопленного органи-
ческого вещества составляет
здесь, по данным Фагелера,
100 т!га\ однако даже при на-
личии оптимальных условий
эта цифра явно завышена. Хотя
до настоящего времени более
точные данные еще не полу-
чены, к оценке продуктивности
теперь подходят более осто-
рожно Ч Запасы древесины
в лесных культурах достигают
в тропиках приблизительно 13 т/га, то есть они в два с половиной раза
больше, чем в Центральной Европе (ср. стр. 44). Общий запас сухого
вещества, по-видимому, колеблется в пределах 30—35 т!га. Такие же
количества, видимо, накапливаются и в естественном тропическом дожде-
вом лесу; наоборот, прирост древесины здесь значительно меньше, чем
в посадках, и примерно соответствует приросту в лиственных лесах
Центральной Европы.
Основная масса накопленного органического вещества, попадая
в почву, разлагается, в связи с этим накопления гумуса в тропиках не
происходит. Измерение интенсивности дыхания почвы могло бы послу-
жить известной отправной точкой для оценки действительных величин
накопленного органического вещества. Однако никаких исследований
в этом направлении пока не проводилось. Для этих целей можно было бы
использовать простой метод, предложенный Вальтером и Хабером1 2.
1 J. Week, Moglichkeiten und Grenzen der Zuwachssteigerung bewirtschafteter
Forstflachen in verschiedenen Klimazonen, «Scientia (Como)», 57, 1963, S. 1—4.
2 H. W a 1 t e r, Eine einfache Methode zur okologischen Erfassung des CO2-Faktors
am Standort, «Вег. Dtsch. Bot. Ges»., 65, 1952, S. 175—182; W. Haber., Okologische
Untersuchungen der Bodenatmung, «Flora», 146, 1958, S. 109—157.
Тропически й дождевой лес
119
Одной из особенностей
древесных пород дождевого
леса является каулиф-
лория, то есть образова-
ние цветков, а затем и плодов
непосредственно на стволах.
Пазушные почки в течение
нескольких лет остаются
спящими и только спустя
некоторое время прорывают
кору, после чего начинают
развиваться цветки. В уме-
ренной зоне это явление
известно только у субсреди-
земноморского вида Cercis
siliquastriun, иудина дерева,
и средиземноморского Сега-
ionta siliqua (рожковое дере-
во). В тропическом дождевом
лесу каулифлория распро-
странена очень широко.
Наиболее известный при-
мер —какао, дерево ниж-
него яруса древостоя дож-
девых лесов Америки. Во
всей тропической зоне на-
считывается около 1000 ви-
дов, обладающих каулифло-
Рис. 78. Каулифлория у Couroupita guianensis’,
плоды образуются на безлистных побегах; Бота-
нический сад в Богоре (фото Фабера).
рией.
Только в Африке, по
данным Милдбрэда х, встре-
чается 278 видов. Он раз-
личает несколько типов каулифлории в зависимости от места образо-
вания цветков: 1) на безлистных ветвях, 2) только на главном стволе,
3) у основания ствола, 4) на длинных побегах с листовыми чешуйками,
прикрепленными к стволу или к нижней части ветвей, и 5) на длинных
плетеобразных ветвях, лежащих на поверхности почвы. Некоторые
примеры каулифлории показаны на рис. 76—78.
О причинах каулифлории существует множество предположений,
но большинство из них весьма неубедительно, так, например, Клебс 1 2
считает, что она обусловлена избыточным накоплением в стволе ассими-
лятов. Интересен тот факт, что каулифлория наблюдается почти исклю-
чительно у видов нижнего яруса древостоя, часто относящихся к хиро-
птерогамным видам, то есть видам, опыляемым летучими мышами 3; среди
этих древесных пород нередко распространена хироптерохория — рас-
пространение семян летучими собаками, питающимися плодами этих
1 J.Mildbraed, Wissenschaftl. Ergebnisse der II. deutschen Zentral-Afrika-
Expedition 1910 bis 1911, Leipzig, 1922.
2 G. К 1 e b s, Uber die Rhythmik in der Entwicklung der Pflanzen, «Sitz. ber.
Heidelb. Akad. d. Wiss., math.-nat. Kl.», Abh. 23, 1911.
3O. Porsch, Ein neuer Ту pus Fledermausblume, «Biologia Generalis», 15,
1941, S. 283—294; Van der L. P i j 1, De bloem gezien tegen de achterground van de
bloembiologie, «Vakblad voor Biologen», 47, № 6, 1957; S. V о g e 1, Fledermausblumen
an Siidamerika, «Osterr. Bot. Zschr.», 104, 1958, S. 491—530.
120
Глава 2
видов деревьев. Непривычные для европейца свойства тропических пло-
дов, их непривлекательную окраску, часто неприятные запах и вкус,
к которому нужно привыкнуть, и, наконец, необыкновенное положение
на дереве можно объяснить лишь приспособлением к распространению
летучими собаками, которые лакомятся этими плодами только ночью *.
Рис. 79. Цветочные почки Spathodea campanulata.
Вверху справа — распустившийся цветок, 1/2 натураль-
ной величины (фото Фабера).
Такой способ распространения в Европе не известен. Многие тропические
виды содержат в плодах крупные семена, дающие проростку достаточное
количество питательных веществ, необходимое для того, чтобы в условиях
слабой освещенности близ поверхности почвы поднять листья на высоту,
где становится возможной ассимиляция ими СО2.
Внимание исследователей привлекает еще одна особенность некото-
рых тропических древесных пород — мешочки, наполненные водой,
в которых незадолго перед распусканием цветка находятся погруженные
в воду, выделяемую гидатодами, цветочные почки. Примером может
служить красивое растение Spathodea campanulata и др. (рис. 79, 80).
1 L. van der Р i j 1, The dispersal of plants by bats (Chiropterochory), «Acta Bot.
Neerland», 6,1957, p. 291—315.
Тропический дождевой лес
I2f
Соответствующая литература обобщена Фабе-
ром, однако ни одна работа не объясняет эколо-
гического значения этого явления. Пашер отме-
чает, что такие же наполненные водой мешочки
встречаются у некоторых нетропических видов
пасленовых (Solanaceae) х. У видов семейства
Bromeliaceae соцветие так же часто развивается
в воде, скапливающейся после дождя в ворон-
ках, образующихся из тесно соприкасающихся
в розетке листьев.
Наконец, следует еще остановиться на
явлениях периодичности в тропическом дожде-
вом лесу1 2. В целом, учитывая постоянство
климатических условий в течение года, перио-
дичность здесь не выражена. Лес всегда одина-
ково зеленый. В любое время года цветут какие-
нибудь древесные породы. Цветки часто не-
обыкновенно красивы, но в массе листвы они
почти не заметны. Гораздо больше бросаются
в глаза нередко светлые или ярко-красные моло-
дые листья (рис. 74 и 75), что свидетельствует
о смене листвы. Отдельные деревья, временно
стоящие без листьев, мало заметны на общем
Рис. 80. Наполненный
водой мешочек у Spatho-
dea campanulatа в разрезе
(фото Фабера).
фоне леса.
Известно, что на Новых Гебридах времена года столь плохо выраже-
ны, что местные жители никогда не исчисляют свой возраст в годах,
однако полевые культуры высаживают через строго определенные про-
межутки времени: ямс — когда цветет Erythrina indica, а батат — когда
созревают плоды Alphitonia zizyphoides.
Таким образом, отдельные виды деревьев характеризуются весьма
отчетливо выраженной периодичностью роста и развития. Однако фазы
развития у разных видов не совпадают, более того, часто эти фазы не син-
хронны даже у отдельных особей одного и того же вида. Встречаются дре-
весные породы, у которых листва опадает полностью и дерево стоит неко-
торое время обнаженным (рис. 81). Другие теряют старые листья лишь
после начала развертывания молодых листьев или даже несколько позже.
Древесные породы, отличающиеся непрерывным ростом, неизвестны;
отсутствие периодичности может наблюдаться разве что только у молодых
деревьев. Однако встречаются породы, у которых новые листья на отдель-
ных ветвях появляются не одновременно (рис. 82). Особенно заметно это
у дерева манго (Man gif era indica): на фоне темно-зеленой кроны со ста-
рыми листьями просвечивают светлые ветви с молодой листвой. Нередко
можно увидеть рядом два дерева одного вида — одно в полном зеленом
уборе, другое с обнаженными ветвями (рис. 81).
Виды, регулярно сменяющие свою листву, обнаруживают периодичность
роста, равную в одних случаях 12, а в других —3—4месяцам3. В некото-
рых случаях продолжительность этих периодов равна 9 и даже 32 месяцам.
1 A. Pascher. Uber Wasserkelche von Datura und Arisodus... «Flora», 148.
1960, S. 517—528.
2 Обширный список работ, посвященных этой теме, приведен в книге Шимпера-
Фабера (Schimper - Faber, op. cit., S. 401—402).
3 R. E. Holt turn, Evolutionary trends in an equatorial climate, «Sympos.
Soc. Exp. Biology», № VII, 1953.
122
Глава 2
Пытаясь объяснить закономерную периодичность жизнедеятельно-
сти растений, следует прежде всего предположить, что некоторые виды
весьма чутко реагируют на малейшие сезонные колебания климата, будь
то незначительное изменение продолжительности дня, возможно оказы-
вающее влияние на образование цветков, или небольшое отклонение от
нормы температуры или количества осадков.
Рис. 81. Dipterocarpus incanus на вырубке в первичном тропиче-
ском лесу на Андаманских островах. Часть деревьев облиствлена,
часть в безлистном состоянии (фото Робертсона).
Во всех случаях речь идет об автономной периодичности, которая
может соответствовать периодичности внешних условий. Становится
понятным, почему одни и те же виды в различных климатических условиях
тропиков с более или менее выраженной периодичностью ведут себя по-раз-
ному. Например, тик (Tectona grandis) в культурах на западной Яве,
где осадки распределены исключительно равномерно, никогда не лишается
полностью всех листьев, а в условиях периодически сухого климата вое-
Тропический дождевой лес
123
точной Явы в засушливый период сбрасывает всю листву. В тропических
районах восточной Австралии, где хорошо выражено прохладное время
года, все деревья манго образуют побеги одновременно. Легкость, с кото-
рой древесные породы приспосабливаются к периодичности внешних усло-
вий, у отдельных видов, даже у особей одного вида, выражена не одина-
ково; это дало Бюннингу основания предположить, что речь идет о раз-
личных биотипах.
Листопадные древесные породы, то есть породы, хотя бы кратковре-
менно остающиеся совершенно без листвы, по-видимому, можно встретить
Рис. 82. Cassia fistula с листьями, распустившимися
лишь на некоторых ветвях. Видны длинные черные
плоды (фото Фабера).
в любом районе тропиков. Однако они не характерны для постоянно влаж-
ных тропиков и господствуют лишь в условиях тропического климата
•с четко выраженным сухим периодом.
Очень интересны опыты по акклиматизации в тропиках листопадных
древесных пород умеренной зоны. На Цейлоне на высоте 1800 м были
высажены плодовые деревья, черешчатый дуб и пирамидальный тополь;
Ботанический сад в Тьибодасе на Яве, расположенный на высоте 1500 м
124
Глава 2
Рис. 83. Отдельная
ветвь Coffea robusta,
цветущая и плодоно-
сящая одновременно;
Амани, Восточная Аф-
рика (фото Э. Валь-
тер).
получил саженцы листопадных пород из Европы, Северной Америки
и умеренной зоны Азии. За деревцем бука (Fagus silvatica), высаженным
на вершине вулкана Пангаранго на высоте 3020 м, в течение четырех
лет велись регулярные наблюдения, показавшие, как пишет Фабер, что*
развитие бука происходило неравномерно, как бы толчками; «ветви
реагируют на сезонные колебания климата по-разному. В сухое время
года на дереве имеются как безлистные, так и облиствленные ветви.
Даже в очень сухом 1914 г. наблюдались обе категории ветвей — ого-
ленные и с распустившимися листьями. Ни о каком непрерывном росте
почек пока не может быть и речи» (Schimper-Faber, S.387).
Совершенно аналогично вели себя древесные породы, посаженные
в Ботаническом саду Тьибодаса. Правда, вначале длительное время еще
сохранялась периодичность развития листвы как влияние сезонности кли-
мата их родины; постепенно, однако, ритмика смены листьев перестала
зависеть от чередования сезонов, и с этого момента началось разновременное
облиствление отдельных побегов. Шимпер, изучавший эти посадки в декабре
и январе, сообщает, что у Liriodendron tuliptfera nQuercus pedunculata одни
ветви находились в зимнем, другие — в весеннем, а третьи — в летнем
состоянии. У яблонь и груш, плохо растущих в условиях тропиче-
ского климата, на одном стволе можно было отметить одновременно
все фазы развития, свойственные этим видам в течение всех четырех вре-
мен года. Amygdalus communis стоял в полном цвету. Аналогичные дан-
ные получены и на Цейлоне.
При анализе приведенных выше материалов следует учитывать, что
тропики отличаются коротким днем. Тем не менее растения длинного дня,
произрастающие на больших высотах над уровнем моря, могут начать
цвести под влиянием низких температур. Поэтому плантации Pyrethrum
в Кении закладывают на высоте 2000—3000 м, где температура не подни-
мается выше 15—16°. Primula veris хорошо растет в Индонезии на абсолют-
ной высоте 2400 м, где цветет и обильно плодоносит; до высоты 1400 м
этот вид в горах только вегетирует А.
На острове Реюньон растут персиковые деревья, выращенные из семян,
полученных из Европы. Вначале они ежегодно теряли листья, затем перио-
1 С. G. G. J. Van S t eenis, Die Gebirgsflora der mallsischen Tropen, «Endea-
vour», 21, 1962, S. 183—194.
Тропический дождевой лес
125
ды покоя все более сокращались, и, наконец, деревья вовсе перестали
сбрасывать листву.
Эти данные подтверждают мнение об эндогенной природе наблю-
даемой у растений ритмичности развития листьев, которая при ярко выра-
женной климатической периодичности только следует последней и поэтому
кажется ею обусловленной.
Выше уже упоминалось, что и цветение характеризуется известной
периодичностью. Правда, период цветения может длиться необычайно
долго, так что в любое время года число цветущих экземпляров растений
превышает число нецветущих. Цветение начинается также не одновремен-
но, поэтому рядом с цветущим деревом можно увидеть деревья уже со
зрелыми плодами или деревья с ветвями, цветущими и плодоносящими
одновременно (рис. 83). Бывают моменты и массового цветения (рис. 84).
В короткий период цветения перед глазами наблюдателя развертывается
необыкновенное зрелище: на площади в несколько квадратных километров
в течение одного дня внезапно расцветают все особи одного вида. Ярким
примером массового цветения может служить эпифитная орхидея Den-
drobium crumenatum на Яве (рис. 85).
У этой орхидеи, цветущей всего в течение одного или нескольких
дней, вполне развитые цветочные почки находятся в состоянии покоя.
Они распускаются под влиянием внешнего раздражения, каковым, по-ви-
димому, является внезапное охлаждение, воздуха после сильной грозы.
Fagraea fragrans, по данным Хольттума, в Сингапуре начинает цвести
всегда через 4 месяца после окончания периода дождей, при условии, что
вслед за этим периодом внезапно наступает засуха.
Некоторые растения начинают цвести через определенные, всегда
одинаковые промежутки времени. Два вида рода Нореа и четыре Shorea
Рис. 84. Массовое цветение Strobilanthes cernuus в дождевом лесу; западная Ява
(фото Фабера).
126
Глава 2
цветут через каждые 6 лет. Strobilanthes cernuus, по некоторым данным,
цветет раз в 7 лет, но это не всегда так. Различные виды бамбука и некото-
рые пальмы, как, например, Corypha umbraculifera, цветут через опреде-
ленное количество лет, причем цветение одновременно охватывает обшир-
ную площадь. У южнобразильских бамбуков этот интервал длится 13 лет,
а у Bambusa arundinacea на западном побережье Индостана — даже 30 —
32 года. После цветения происходит массовое отмирание растений.
Рис. 85. Эпифитная орхидея Dendrobium crumenatum;
все экземпляры этого вида расцветают на большой
площади в течение одного дня (фото Фабера).
В этом автор убедился, находясь в северной Австралии южнее Дарви-
на. В теплый период 1951/52 г. осадки не выпадали, а в более прохладный
сухой период 1952 г. сразу зацвели все бамбуковые заросли в долинах
рек и ручьев этого района. В сухой период 1952/53 г., по сообщению Арнд-
та, все растения погибли. Однако семена дали всходы, и в 1958 г. мы уже
любовались молодыми бамбуковыми рощами. В нижнем течении рек
в долинах, всегда достаточно увлажненных, влияние сухого периода не
сказывается столь резко. Заросли бамбука здесь не цветут, но и не поги-
бают. Таким образом, в данном случае цветение было вызвано неблаго-
Тропический дождевой лес
127
приятными условиями увлажнения. Иные варианты требуют еще выяс-
нения. Периодичность цветения можно было бы объяснить, если бы внеш-
ние факторы зависели от солярного ритма. Длительные промежутки меж-
ду периодами цветения могут быть вызваны необходимостью накопления
растением большого запаса питательных веществ для образования цветков.
В связи с этим Фабер отмечает, что листва на цветущих ветвях часто опа-
дает. У некоторых листопадных древесных пород цветущие ветви покры-
ваются листвой позднее, чем ветви, которые только вегетируют. «Некото-
рые древесные породы до цветения являются вечнозелеными, позднее же,
непосредственно перед периодом цветения, они сбрасывают листву. Это
характерно, например, для Schtzolobium exelsum, во всяком случае
на Яве» *.
ЭКОЛОГИЯ ДРУГИХ ГРУПП ЖИЗНЕННЫХ ФОРМ
ДОЖДЕВОГО ЛЕСА
Кустарники
В тропическом дождевом лесу доминирует древостой. Наряду с этим
в лесу развиваются подчиненные древостою ярусы. В кустарниковом яру-
се собственно кустарники, если не считать подроста древесных пород,
играют небольшую роль. Напротив, в нем очень широко представлены,
высокие травы. Эти виды так же, как и Ьолее низкие травы, получают
лишь столько света, сколько его пропускает древостой. Роль корневой
конкуренции пока точно не установлена. О соревновании за воду в усло-
виях насыщенного влагой приземного слоя воздуха, по-видимому, гово-
рить не приходится. Более вероятной представляется конкуренция за
питательные вещества. Этим вопросом занимался, в частности, Костер,
изучавший корневые системы растений в тропиках (см. Ричардс, Тро-
пический дождевой лес). Он установил, что под пологом определенных
видов древесных пород, пропускающих относительно много света, под-
рост отсутствовал, хотя под другими видами, создававшими густую тень,
он был хорошо развит. Костер объясняет эти различия корневой конку-
ренцией подроста и деревьев за кислород, содержание которого во влаж-
ной тропической почве очень ограниченно. Однако, скорее, причину сле-
дует искать в слишком высокой концентрации СО2 в приземном слое
воздуха либо в недостатке в почве нитратов или аммония. Соответствую-
щих исследований во влажном дождевом лесу пока не проведено.
Травы
Видов травяного покрова в дождевом лесу меньше, чем древесных
пород. Это связано, видимо, с недостатком света близ поверхности почвы,
что обусловливает естественный отбор. Характерная особенность травя-
ного покрова дождевого леса в отличие от умеренной зоны состоит в том,
что надземные органы тропических трав живут неопределенно долго;
причину этого следует искать в однородности климатических условии в тро-
пиках в течение всего года. Поэтому виды травяного покрова подчас вырас-
тают до 6 л и часто достигают уровня яруса кустарников, а иногда дажо
и нижнего яруса древостоя, как, например, бананы и крупные экземпля-
ры Scitamineae. Нередко у трав развиваются ризомы или клубни. Правда,.
1 S chim per - F a b er, op. cit., S. 398.
128
Глава 2
последние служат не столько для накопления запасных питательных
веществ, сколько органами вегетативного размножения. Цветение трав
еще менее, чем цветение древесных растений, связано с определенным вре-
менем года, однако оно часто обусловливает периодичность развития,
например у Amor phophallus, у которого сначала развиваются цветки
и лишь позднее — листья. В состав травяного покрова включаются также
многочисленные папоротники и виды Selaginella.
Рис. 86. Напочвенный покров на осветленном участке
в дождевом лесу в Тьибодасе. В центре — крупноли-
стная Begonia robusta, виды Cyrtandra, Elatostema acu-
minatum и др. (фото Фабера).
Размеры и форма листьев трав весьма разнообразны (рис. 86). Замеча-
тельна пестрота их окраски, причем листья могут быть белыми и красными,
отличаться металлическим блеском и даже бархатистой поверхностью.
Однако у большинства видов листья нежные и тонкие, реже сочные, мяси-
стые, как, например, у бегоний.
Наиболее характерной экологической особенностью трав в тропическом
дождевом лесу, видимо, является их приспособленность к существова-
нию в условиях слабой освещенности. Причем известны виды, развиваю-
щиеся только в глубокой тени, и виды, произрастающие в более благо-
Тропический дождевой лес
129
приятных условиях освещения и достигающие на свету максимального
развития. В наиболее затененных участках под пологом леса растения
размножаются в основном вегетативным путем.
Тенелюбивые растения, как показал Фабер в опытах с оранжерей-
ными культурами, транспирируют очень слабо Однако гуттация рас-
тений, особенно ночью, обычно столь интенсивна, что они этим путем
отдают значительно больше влаги, чем путем испарения. В Амани автору
приходилось наблюдать, как тотчас после захода солнца с кончиков листьев
Colocasia начинали непрерывно стекать маленькие капли воды. Под влия-
нием прямого солнечного луча тенелюбивые растения часто закрывают
свои устьица; некоторые же растения, например виды Impatiens, даже
при кратковременном пребывании на солнце завядают.
Как чутко реагируют растения на малейшее уменьшение увлажне-
ния, показывают измерения скорости роста. Незначительное понижение
влажности воздуха в лесу днем немедленно влечет за собой задержку
роста. В этих условиях растение ночью растет быстрее, чем днем 1 2.
Штоккер исследовал в Богоре транспирацию некоторых тенелюби-
вых растений, пользуясь методом быстрого взвешивания. Величина транс-
пирации, как и испарение, оказалась небольшой. В Амани автор изу-
чал транспирацию крупнолистных видов трав — бананов, Colocasia,
Ravenala и др.
В приводимых ниже таблицах цифры в скобках означают содержание
воды в транспирирующих органах в процентах на сырой вес; интенсив-
ность транспирации (мг/г-мин) сопоставляется с испарением, равным
0,48 мл!час (предполагается наличие пропорционального соотношения
между испарением и транспирацией).
Таблица 16
Интенсивность транспирации тенелюбивых видов
по наблюдениям в Амани
Название растений
Интенсив-
ность транспи-
рации,
мг/г • jhuh
Impatiens valleriana (92)
Costus subbiflorus (85)........
Pleomele (Dracaena) papahu (68)
Begonia zimmermannii (92) 1
1,8
1,2
0,2
0,023
1 В течение 24 часов интенсивность транспирации этого
вида достигла только 7 мг/г.
У теневыносливых видов интенсивность транспирации оказалась вы-
ше (табл. 17).
1 F. С. von Faber, Physiologische Fragmente aus einem tropischen Urwald,
«Jb. wiss. Bot.», 56, 1915, S. 197—220.
2 A. M. Smith, On the application of the theory of limiting factors to measu-
rements and observations of growth in Ceylon, «Ann. Roy. Bot. Gard. Peradeniva»,
3, 1906.
9-1394
130
Глава 2
Рис. 87. Ravenala madagascariensis в окрестностях Сингапура.
Слева и справа — молодые экземпляры этого же вида. Слева —
пальма Areca catechu.
Таблица 17
Название растений, условия опыта
Банан (Musdfparadisiaca) (83,8), солнечный день...................
Банан (Musa paradisiaca) (83,8), пасмурный день...................
Дерево путешественников (рис. 87) 'l старый лист..................
(Ravenala madagascariensis) J молодой лист................
Mar ant a arundinacea1 (74,5) 'l на солнце........................
Maranta arundinacea (74,5) J в тени...............................
Таро (Colocasia antiquorum) (81,4)................................
Маниок (Manihot utilissima).......................................
Сахарный тростник (Saccharum of ficinarum) (71)...................
Дынное дерево (Carica papaya), слегка завядшие листья . . . .
Кардамон (Elettaria cardamomum) (73,5)............................
Интенсивность
транспирации,
мг/г • мин
3,3-3,4
8,0
3,7
1,0
4,3
9,0
12,0
7,4
2,8
2,8
1,6
1 Этот вид разводится в тропиках для получения крахмала-арроурута, добываемого из:
корневищ; арроурут вследствие.легкой усвояемости применяется для кормления детей и боль-
ных. — Прим. ред.
Данные об интенсивности транспирации у некоторых видов папо-
ротников приводятся в табл. 18.
Следует особенно отметить, что при опытах в тени транспирация
(сопоставленная с таким же испарением) у папоротников, бананов и Maran-
ta была приблизительно в 2—3 раза больше, чем на солнце, то есть и у этих
видов устьица, по-видимому, частично закрываются на солнце. У Maranta
листья на солнце даже несколько свертывались.
Для сравнения ниже приводятся величины транспирации у некото-
рых других видов (табл. 19).
Тропический дождевой лес
131
Таблица 18
Название растений, условия опыта Интенсивность транспирации, лгг/г • мин
Cyathea usambariensis (47,5) Пасмурный день, в условиях освещения . Пасмурный день, в тени Солнечный день, в условиях освещения . . Солнечный день, в тени Marattia jraxinea (77), быстро завядает и высыхает . Pteridium aquilinum (70) Asplenium nidus (82), эпифит, в течение дня остается свежим . 7,6 16,0 5,4 15,0 4,8 3,3 0,32 Таблица 19
Название растений Интенсивность транспирации, мг/г • мин
Phoenix humilis (55) . . Carludovica palmata (66) Cycas circinalis (71,8) Encephalartos hildenbrandtii (54,3) Monstera deliciosa (79,3) Ананас {Ananas sativa) (85) . . Agave sisalana и др., а также 1 Fourcroya gigantea J в₽емя экспозиции до 4,5 часов . . 3,4 4,2 5,1 3,3 3,7 0,035 0,004-0,025
Приведенные выше цифры —данные лишь небольшого числа опытов.
Тем не менее они показывают, что в общем интенсивность транспирации
как у гигроморфных, так и ксероморфных видов очень низка (Colocasia,
по-видимому, представляет исключение). Суккуленты, как правило,
испаряют чрезвычайно мало влаги.
При столь незначительных величинах транспирации и большой влаж-
ности условий местообитания гидратура остается на одном уровне. И дей-
ствительно, величина осмотического давления у видов травяного покрова
еще ниже, чем у древесных пород.
Полученные автором при исследованиях в Амани (см. табл. 20 и 21)
величины осмотического давления не превышают 10 атм, часто не дости-
гая даже 5 атм, за исключением Brillantaisia madagascariensis, осмоти-
ческое давление у которой составило 15,6 атм (на конце стелющегося
побега), и Pleomele {Dracaena) papahu, у которой оно равно 10.5 атм
(одиночный экземпляр, растущий на прогалине в лесу).
Из приведенных данных видно, что у папоротников, несмотря на
большое количество очень нежных листьев (например, у Adiantum),
осмотическое давление выше, чем у трав.
Данных об интенсивности фотосинтеза у видов травяного покрова
тропического дождевого леса пока не получено. Поскольку травы растут
в условиях слабого освещения, они накапливают известное количество
вещества; по сравнению с сырым весом растения это количество не очень
велико, так как основной составной частью травы является вода (часто
132
Глава 2
Таблица 20
Осмотическое давление у некоторых видов травяного покрова
Название растений Величина осмотического давления, атм
Cylicomorpha parviflora, с млечным соком 8,0
Piper umbellatus, большие листья, в тени 8,9
Piper capensis, очень нежные листья . 6,6
Clinogyne ugandensis ... 8,5
Calathea zebrina, в тени . 8,2
Calathea zebrina, на солнце 8,9
Afframomum или Kaempferia . 7,5
Lobelia kumuriae . . 7,3
Costus subbiflorus 5,7
Fleurya lanceolata (Urticaceae) 8,0
Fleurya lanceolata, стелющийся побег на почве 6,0
Coix lacrimans, открытый заболоченный участок . 8,6
Synadenium (Euphorbia) carinatum .... 5,6
Notenia amaniensis, сложноцветное, внешне похожее на представителя
семейства Crassulaceae 5,6
Impatiens parviflora, свежее растение 4,3-4,7
Impatiens parviflora, завядшее растение 5,4
Streptocarpus ovata 3,6
Streptocarpus ovata, утолщенное основание стебля 4,8
Lissochilus, на солнце в траве . 4,4
Таблица 21
Осмотическое давление у неэпифитных папоротников
Название растений Величина осмотического давления, атм
Cyathea usambariensis (древовидный папоротник), на солнце Cyathea usambariensis (древовидный папоротник), в тени То же растение, молодые еще не развернувшиеся листья Marattia fraxinea, средние листья с синангиями . . ... Microlepis speluncae, у ручья . . Pteris biaurita . . Lonchitis hirsuta Asplenium cicutarium, листья co спорангиями Asplenium unitum Adiantum capillus veneris To же в Аризоне (оазис в пустыне) Pellaea viridis Pellaea viridis, молодое растение ... То же, пожелтевшее и на вид высохшее растение Selaginella spec Pteridium aquilinum, на солнце 10,5 7,5 4,6-5,4 10,7 9,7 8,6 9,0—11,9 8,9-9,1 7,7 11,3 13,9 13,3 10,5 11,0—11,2 10,6 16,2
Тропический дождевой лес
133
на 80 и даже 90%). Положительный баланс накопления вещества объяс-
няется очень слабым дыханием трав. Последнее чрезвычайно характерно
для Aspidistra; этот вид в комнатных условиях хорошо себя чувствует
даже в самом темном углу. Узамбарская фиалка (Saintpaulia usambarien-
sis), которую автор наблюдал в Восточной Африке в глубокой тени леса,
как комнатная культура весьма не требовательна к свету.
В качестве количественного эколого-морфологического признака
часто используют число и размеры устьиц. Поэтому мы приводим ниже
соответствующие данные, полученные в оранжерее в Штутгарте. Число
устьиц у тенелюбивых растений обычно не достигает 100 на 1 мм2, одна-
ко и у тропических светолюбов оно также не очень велико (ср. «Phyto-
logie», Bd III, 1, S. 322 и след.).
Таблица 22
Число устьиц на 1 мм2 нижней поверхности листа 1
Tradescanlia zebrina 15-20 Coffea arabica . . 148 (23)
Begonia hybrida 24 Phoenix canariensis . . 89/91 (22)
Aspidistra elatior 28 Musa sapientium 194 (30)
Cycas circinalis . . 36 (66) Saccharum officinarum 133/60 (40)
Ananas saliva . 42 (27) Bambusa arundinacea 233 (20)
Asplenium nidus 45 (42) Alspphila australis 266 (33)
Monslera deliciosa . . . 46 Citrus auranlium 293 (22)
Encephalarlos lehmannii . . 52 (79) Carica papaya . . 370 (20)
Colocasia odorala . . 52/6 (36) Acacia melanoxylon
Colocasia cochleata 91/6 (35) Филлодии 130/125 (25)
Coslus afer .... 95 Перистые листочки 238/174 (21)
Elellaria cardamomum . . . 100/4 (27) Acacia lalifolia 295/268 (36)
Angiopteris ihysmanniana 111 (45) Manihot utilissima 400 (23)
Ficus elaslica 100-130 (18)
1 В знаменателе — число устьиц на верхней стороне листа; в скобках — длина устьиц
в микронах.
Лианы
Если древостой влияет на травяной покров лишь косвенно, то лианы
и две следующие за ними группы — полуэпифиты и эпифиты — в меха-
ническом отношении обнаруживают прямую зависимость от деревьев,
которые используются ими как опора или место прикрепления.
Обычно всходы лиан нормально растут на почве, однако вскоре их
стебель очень быстро удлиняется без образования при этом достаточно
прочной механической ткани. Чтобы подняться над землей, лианам при-
ходится прикрепляться к другим растениям, причем способы прикрепле-
ния весьма различны.
1. Взбирающиеся или опирающиеся лианы —
обычно кустарники, которые врастают своими ветвями между сучьев
других растений и таким образом находят себе опору. Соскальзыванию
препятствуют колючки, шипы или крючки. Особенно примечательны
в этом отношении «лазящие» пальмы, например Calamus, Тонкий, прочный,
покрытый колючками ствол этой пальмы называют «ротангом». Рахис
их листьев удлинен в виде гибкого, снабженного крючкообразными
шипами жгута. Этим жгутом молодой побег прикрепляется к дереву-
134
Глава 2
Рис. 88. Лианы (Агасеае), лазящие при помощи придаточных корней, в дождевом
лесу Мексики (Трионфо, Грихальва). В середине — Sarcinanthus utilis с двурас-
сеченными листьями. Рядом видны эпифитные Bromeliaceae и папоротники (фото
Карстена)
опоре и постепенно взбирается до его вершины. Когда ротанг прекращает
свой рост, более старые, лишенные листьев побеги соскальзывают и падают
на землю, свиваясь в петли. Тройб измерил часть ствола ротанга; оказа-
лось, что его длина достигает 240 м (ср. данные Фабера). Пальмы-ротанги
встречаются главным образом в Индокитае; они известны и на северо-
востоке Австралии. Колючие, толщиной с руку, лианы из рода Rubus
широко распространены в Новой Зеландии.
2. Лианы, лазящие при помощи придаточ-
ных корней, образуют тонкие придаточные корни, которыми расте-
ния цепляются за кору деревьев, или более толстые, длинные корни,
которыми лианы, словно обручем, охватывают свою опору (например,
лиана из рода Vanilla). К этой же группе лиан относятся виды Freycinetia,
часто обвивающие деревья до самой вершины, и многие виды семейства
Агасеае (рис. 88).
3. Вьющиеся лианы имеют быстро растущую вершину
стебля с длинными междоузлиями; развитие листьев на последних внача-
ле задерживается, как и у этиолированных растений. При помощи враща-
тельных движений стебель обвивает какую-либо вертикальную опору
и плотно к ней прижимается. Со временем он начинает деревенеть и утол-
щается. Таких лиан в тропиках необычайно много. По способу лазания
они сходны с глициниями, которые разводятся в Центральной Европе
(Wistaria sinensis), и видами Aristolochia.
Тропический дождевой лес
135
4. Усиконос-
н ы е лианы облада-
ют особыми, чувстви-
тельными к малейшему
прикосновению органа-
ми, — усиками, которые
также со временем мо-
гут одревеснеть (рис. 89).
В первичном тропи-
ческом лесу широко рас-
пространены древесные
лианы. Число их видов
огромно. Словно тол-
стые переплетающиеся
канаты, обвивают они
стволы от почвы до са-
мой кроны, свисают
с вершин деревьев, сте-
лются по земле и сно-
ва взбираются на дру-
гие стволы (рис. 90).
Длина лиан может пре-
вышать 70 м. По данным
Шенка1, более 90% всех
видов лиан распростра-
нено только в тропиках.
В Вест-Индии около 8%
общего числа видов
представлено древесны-
ми лианами.
Для достижения ос-
вещенной солнцем кро-
ны деревьев лианам не
нужно накапливать осо-
бенно много вещества.
Правда, в молодости им
приходится довольство-
ваться тем небольшим
количеством света, ко-
торое проникает до поч-
вы. Отсюда понятно,
почему лазящие расте-
ния особенно часто
встречаются на более
1 Н. Schenck, Beit-
rage zur Biologie und Ana-
tomic der Lianen, «Bot.
Mitt. Тгореп», H. 4, 1892;
H. 5, 1893.
Рис. 90. Покрытые мхами
лианы в горном дождевом
лесу западной Явы (фото
Фабера).
Рис. 89. Старая сильно утолщенная ветвь усиконос-
ной лианы Dalbergia variabilis] 2/3 натуральной вели-
чины (по Шенку).
136
Глава 2
Рис. 91. Поперечный разрез
ствола лианы Anisosperma passif-
lora\ увеличено в 3,2 раза (по
Шенку).
Рис. 92. Поперечный разрез
ствола лианы Dalechampia ficifo-
На (по Шенку).
разреженных участках леса. По берегам рек и на опушке леса лианы
образуют совершенно непроходимые заросли. Они доставляют много
трудностей при разбивке плантаций. Достигнув вершины дерева и ока-
завшись на сильном свету, лианы образуют столь густую раскидистую
крону, что дерево-опора нередко погибает. В открытой местности лианы
часто приобретают форму низкого кустарника. Обычно же главная ось
у лианы не ветвится. С возрастом она одревесневает и сильно утолщает-
ся, однако сохраняет при этом гибкость; последнее связано с тем, что
ксилема у главного ствола состоит из изолированных друг от друга сосу-
дисто-волокнистых пучков (рис. 91, 92). Снабжение со временем сильно
развивающейся кроны водой происходит по очень длинному и тонкому
стволу. Оно возможно лишь благодаря почти полному отсутствию со-
противления току жидкости в сосудах, очень больших и к тому же
лишенных поперечных перегородок. Перерубив одним ударом топора
ствол у его основания, у некоторых видов лиан можно получить из среза
около стакана чистой питьевой воды. Автор измерил в Амани осмотическое
давление этого клеточного сока; у неизвестного вида лиан оно оказалось
равным 0,3 атм, а у Ampeloctssusvolkensit— 0,7 атм. По Геснеру, электро-
проводимость воды, находящейся в сосудистой системе лиан, прибли-
зительно в 20—100 раз выше, чем электропроводимость воды в почве1.
Водяные нити в этих сосудах могут легко порваться при чересчур
большой сосущей силе листьев; это в свою очередь может нарушить ток
воды и снабжение водой кроны лиан. Именно этим обстоятельством, воз-
можно объясняется распространение древесных лиан почти исключительно
в тропиках, где сосущая сила листьев практически очень мала. По дан-
ным Коутиньо (см. сноску на стр. 116), интенсивность транспирации
у трех изученных видов лиан была ниже, чем у деревьев. У многих лиан
имеются очень резко выраженные различия между теневыми и солнечными
листьями или между первыми и последующими по времени появления
листьями. Морфология и анатомия листьев весьма разнообразны.
Величина осмотического давления, определенная автором в Амани,
сходна как у лиан, так и у деревьев (табл. 23).
1 F. Gessner, Untersuchungen uber den GefaBsaft tropischer Lianen, «Planta»,
64, 1965, S. 186—190.
Тропический дождевой лес
13Т
Таблица 23
Осмотическое давление у некоторых видов лиан
Название растений Величина осмотического давления, атм
Smilax kraussiana, жесткие листья с колючками 12,9
Culcasia spec. (Агасеае) . 12,2—12,3
Cucurbitaceae .... 10,0
Adenia schweinjurthii, нежные теневые листья 7,7
Monstera deliciosa, в посадках 6,7-7,5
Philodendron spec., то же . 6,3-6,7
Ampelocissus volkensii, молодые листья . 5,8
В построении полога среднего и нижнего ярусов древостоя тропиче-
ского дождевого леса лианы играют весьма существенную роль, поскольку
они создают дополнительное затенение почвы. В искусственных насажде-
ниях они относятся к сорнякам и уничтожаются путем обрубки вершин.
Если лианы свисают непосредственно с ветвей дерева, это признак тогог
что дерево-опора давно погибло.
I
Полу эпифиты ;
Полуэпифиты — переходная группа между лианами и [эпифитами.
Они начинают свое развитие либо в качестве лиан, либо эпифитов. Многие
виды семейства Агасеае вначале развиваются как обычные лианы, но затем
нижняя часть их ствола отмирает, непосредственная связь растения с поч-
вой утрачивается, и оно становится эпифитом. О величине транспирации
Monstera можно судить по данным табл. 19.
Обратное развитие наблюдается у некоторых эпифитов; они прорас-
тают на дереве и вскоре образуют длинные воздушные корни, которые
достигают земли. Вслед за этим растение начинает получать воду и пита-
тельные вещества обычным путем из почвы.
К этой группе относятся некоторые виды семейства Агасеае, амери-
канская Coussapoa fagifolia и малайский Pyrus granulosa', последний раз-
вивается то как полуэпифит, то как обычное дерево.
Наиболее примечательную группу полуэпифитов образуют «растения-
удушители». В учебниках обычно упоминаются только фикусы-удуши-
тели (виды Ficus), но в Новой Зеландии встречаются аналогичные виды
и из других семейств. Это Metrosideros robusta (Myrtaceae), прорастающий
почти всегда на Dacrydium и в конце концов полностью заглушающий
своего «хозяина», Griselinia littoralis, G. lucida (Cornaceae), Nothopanax
simplex, N. colensoi (Araliaceae) и др. Виды последних двух родов могут
развиваться и как обычные деревья. Не редкость также, когда удушите ли
поселяются один на другом.
«Удушители», например фикусы, прорастают как эпифиты в развили-
нах крупных ветвей. Проросток кустится, и у него образуются воздушные
корни, одни из которых прилегают к стволу «дерева-хозяина», а другие
свободно свисают. Опускающиеся корни ветвятся и анастомозируют,
пока наконец не достигнут почвы. Постепенно окрепнув, растение раз-
вивает пышную крону, корни его, утолщаясь сжимают все сильнее ствол
дерева-опоры и механически препятствуют росту последнего; в конечном
138
Глава 2
итоге «дерево-хозяин» погибает. С этого момента растение-удушитель
развивается как совершенно независимое дерево. Такие фикусы нередко
становятся самыми крупными деревьями тропического леса; стоя под
ними, трудно поверить, что они начали развиваться как эпифиты высоко
на дереве в развилках ветвей. К этой же группе растений относятся Ficus
retusa и F. bengalensis. Их воздушные корни, превратившиеся в столбо-
видные подпорки, создают впечатление густого древостоя (рис. 93). Наи-
более крупный известный экземпляр «фикуса-удушителя» при высоте
всего 26 м занимает площадь более 2 га\ окружность кроны достигает
530, а ее диаметр — 170 л*.
Величина осмотического давления у видов Ficus в Амани редко пре-
вышала 10—11 атм (см. табл. 24).
Таблица 24
Осмотическое давление у разных видов Ficus
Название растений Величина осмоти- ческого давления, атм
Ficus ulugurensis, в виде эпифита, однако уже укоренившегося в почве Ficus subcalcarata, лишь один корень достиг почвы Ficus rhynchospora^ обычное дерево Ficus rhynchospora, листья, освещенные солнцем . . . 9,6 10,7 10,6 11,8
Эпифиты
Остановимся прежде всего на характеристике сосудистых эпифитов,
то есть эпифитных папоротникообразных и цветковых растений. Они
не укореняются в почве и с экологической точки зрения представляют
собой, вероятно, наиболее интересную группу среди растений тропиче-
ского дождевого леса; в других лесных формациях они почти не встре-
чаются
Эпифиты используют другие растения только в качестве опоры.
Прорастая на ветвях деревьев, высоко над поверхностью почвы, они ока-
зываются в условиях лучшего освещения, однако испытывают трудности
в снабжении водой и питательными веществами. Благоприятную для
своей жизнедеятельности гидратуру эпифиты способны поддерживать
лишь во влажном тропическом лесу. Это удается только благодаря
некоторым приспособительным особенностям корней и побегов.
Что касается происхождения эпифитов, то часть из них произошла
от наземных растений, характеризующихся утолщенными листьями
(бегонии, Piperaceae, орхидеи) и слабой транспирацией, другие — от
растений, произраставших в засушливых условиях (эпифитные кактусы,
например виды Rhipsalis, и, возможно, представители семейства Вгоше-
liaceae). Растения, развивающиеся на скалах, также могут стать эпи-
фитами, поскольку и в данном случае налицо недостаток почвы и опас-
ность высыхания. И, наоборот, некоторые эпифиты, если поблизости нет
деревьев, часто поселяются на скалах.
1 Ряд своеобразных форм, относящихся к другим экологическим группам рас-
тений, прекрасно описал Бюннинг: Е. Banning, Der tropische Regenwald, «Ver-
standliche Wissenschaft», 56, 1956.
Тропический дождевой лес
139
Рис. 93. Мощные воздушные корни Ficus retusa, превратившиеся в столбовидные
подпорки. На ветвях много эпифитов, особенно Poly podium verrucosum; Ботаниче-
ский сад в Богоре (фото Фабера).
Эпифиты — прекрасный индикатор некоторых микроклиматических
условий, особенно — большой влажности воздуха. На свою опору они
почти не оказывают воздействия, и поэтому до сих пор не ясно, существуют
ли какие-либо определенные взаимосвязи между эпифитом и растением-
хозяином. Считают, что многие эпифиты поселяются только на живых
деревьях, предпочитая часто лишь определенные виды, причем некото-
рую роль, возможно, играют свойства корь^, и в частности непрочность ее
у отмерших ветвей. С другой стороны, большие скопления эпифитов на
годном дереве могут вызвать его механическую перегрузку (рис. 94).
Постоянно увлажняя кору, эпифиты, вероятно, содействуют поселению
на дереве-опоре паразитирующих на них грибов. На крупных ветвях или
в развилках ветвей нередко накапливается перегной. Общее количество
этого кислого «эпифитного гумуса», по оценке Клинге 4, достигает несколь-
ких тонн на 1 га. Скопляющийся мелкозем содержит много органических
остатков и минеральных частиц (в Сальвадоре — принесенные с вулка-
ническим пеплом и пылью), и даже обломков раковин улиток. Эпифиты
способны также фиксировать азот, получая его с атмосферной влагой,
которая в тропиках, по-видимому, более богата нитратами, или в виде
ионов аммония из воды, капающей с листьев деревьев и содержащей
в 3 раза больше NH3, чем дождевая вода. Таким образом, в тропическом
лесу формируется новый, постоянно влажный биотоп, благоприятный
даже для существования древесных лягушек.
1 Н. Kling е, Uber Epiphy tenhumus aus El Salvador (Zentralamerika) I und
III, «Pedobiologia», 2, 1962, S. 1—8; 1963, S. 102—107.
Рис. 94. Прогалина в дождевом лесу близ Амани (восточная часть гор Усамбара);
на деревьях эпифиты Asplenium nidus и лианы (фото Фабера).
Тропический дождевой лес
141
В тропических лесах на деревьях высоко над почвой в скоплениях
детрита вокруг эпифитов или в полостях клубней Myrmecodia и Hydno-
phytum (рис. 95, 96), а также в мешковидных листьях дисхидий поселяют-
ся многие виды муравьев. Некоторые виды строят на деревьях настоящие
гнезда, в которых прорастают семена эпифитов; издали кажется, что эти
гнезда сплетены из живых растений. Муравьи предпочитают некоторые
виды Bromeliaceae, Gesneriaceae, Агасеае, Solanaceae, Ficus myrmecophila,
Peperomia, Phyllocactus и др. Уле наблюдал в бассейне Амазонки настоя-
щие муравьиные «цветники», словно декоративные цветочные горшочки,
свисавшие с ветвей деревьев *. Скопления воды в воронках, образующих-
ся из тесно соприкасающихся в розетке листьев, у некоторых видов Brome-
liaceae служат местом, где разводятся личинки москитов и различных
водных насекомых.
Во внетропических областях распространены почти исключительно
бессосудистые эпифиты; во влажных лиственных лесах Еврдпы из папо-
ротников произрастает лишь Polypodium vulgare, обычны же мхи, лишай-
ники и некоторые водоросли. Все это пойкилогидрические виды, хорошо
переносящие полное высыхание. В тропиках, наоборот, исключительно
велико число гомойогидрических эпифитных сосудистых растений.
В составе семейства Orchidaceae преобладают эпифитные растения;
число же растущих на почве невелико. Виды семейства Bromeliaceae
пр ед став л ены пр еиму ще ст-
венно эпифитами и расте-
ниями, обитающими на
скалах 1 2.
То, что большинство
эпифитов в дождевом лесу
предпочитает расти на де-
ревьях, а не на почве, объяс-
няется их большим светолю-
бием и крайне скудным осве-
щением поверхности почвы
в сомкнутом лесу. В более
разреженном лесу эпифиты
не выдерживают конкурен-
ции с быстро растущими
наземными растениями. Эпи-
фиты можно подразделить
на две, не столь уж резко
отличающиеся друг от дру-
га группы:
1. Теневые эпи-
фиты. Поселяются на ниж-
1 Е. U 1 е, Blumengarten
der Ameisen am Amazonenstrome,
«Veg. Bilder», 3, H. 1, 1905.
2 Они распространены толь-
ко в Америке. Лишь один вид,
растущий на почве, Pitcairnia
feliciana Harms et Mildbr., из се-
мейства Liliaceae, встречается
в Западной Африке (Hutchin-
son, The families of flowering
plants, VII, 1959, p. 577).
Рис. 95. Клубень Myrmecodia tuberosa (фото
Фабера).
142
Глава 2
них частях стволов деревьев и часто располагаются в развилках крупных
ветвей. В основном представлены папоротниками.
2. Солнечные эпифиты. Растут в верхней части крон дере-
вьев, нередко спускаясь до первой развилки ветвей. Чем тоньше веткиг
тем меньше размеры эпифитов. К солнечным эпифитам относится большое
число видов орхидей, оплетающих своими воздушными корнями ветви
растения-опоры. Крохотная орхидея в Амани вместе с соцветием едва
достигала в высоту нескольких сантиметров. Однако среди орхидей извест-
ны и очень крупные экземпляры (рис. 97).
При подразделении эпифитов на две эти группы более важным кри-
терием, видимо, следует считать водный режим, а не условия освещения.
В верхней части крон деревьев влажность воздуха уменьшается, и поэто-
му развитым здесь эпифитам приходится существовать между периодами
выпадения осадков в более суровых засушливых условиях, чем эпифит-
ным растениям, распространенным в нижних частях стволов. Эпифит-
ные орхидеи нередко встречаются даже в очень сухих саванновых
лесах.
Рис. 96. Продольный разрез клубня Myrmecodia tuberosa\ хорошо видны внутрен-
ние полости; !/2 натуральной величины (фото Фабера).
К неблагоприятным условиям увлажнения эпифиты приспосабли-
ваются по-разному. Особенно важное значение имеет накопление в местах
произрастания перегноя, который образуется из отмерших частей расте-
ний. Asplenium nidus (рис. 98) накапливает детрит между листьями; кор-
ни и медленно разлагающиеся жилки отмерших листьев скрепляют его.
У Platycerium (рис. 99), Drynaria и других видов формируются особые
листья-скобки, прилегающие к стволу дерева-опоры и быстро отмираю-
щие; однако даже мертвыми они продолжают удерживать скопившуюся
почву. Образованию перегноя способствуют муравьи, по-видимому снаб-
жающие эпифиты азотом и другими питательными веществами. Сущест-
Тропический дождевой лес
14$
вуют ли в этих условиях азотные бактерии, не установлено. Реакция
перегноя кислая
Шпаннер установил, что Myrmecodia лучше усваивает аммонийный
азот, чем нитратный 1 2.
Эпифиты, которые не в состоянии накапливать между корнями почвуг
должны обладать приспособлениями, позволяющими им быстро собирать
Рис. 97. Эпифитная орхидея Grammatophyllum specio-
sum\ гнездовой эпифит, обладающий отрицательным гео-
тропизмом (фото Фабера).
дождевую воду или росу при одновременном сокращении транспирацион-
ных расходов.
Широко известны выполняющие эту роль клетки веламена, распола-
гающиеся с наружной стороны воздушных корней орхидей (рис. 100),
и чешуйкообразные волоски на листьях бромелиевых (рис. 101); тесно
соприкасающиеся в розетке листья этих растений образуют воронки,
1 Y. В о у е г, Contribution а Г etude de I’ecophysiologie de deux fougeres epiphy-
tes: Platycerium stemaria et P. angolense, «Ann. Sci. Natur., Bot. (Paris)», Ser. 12, 5r
1964, p. 87—228.
2 L. Spanner, Untersuchungen uber den Warme- und Wasserhaushalt von
Myrmecodia und Hydnophytum, «Jb. wiss. Bot.», 88, 1939, S. 243—283.
144
Глава 2
Рис. 98. Эпифитный папоротник Asplenium nidus в дождевом лесу
западной Явы (фото Фабера).
в которых скапливается дождевая вода (рис. 102) Ч Значение кувшино-
и мешковидных листьев Dischidia (рис. 103), в которые врастают корни,
не выяснено. Эпифитные виды Nepenthes используют воду, накапливаю-
щуюся в их кувшинах, для регулирования своего водного баланса.
Клубни и суккулентные побеги многих орхидей, а также Rhipsalis,
суккулентные листья бромелиевых и утолщенные ризомы папоротников
позволяют создавать довольно значительные запасы влаги.
Приводимые ниже четыре примера характеризуют особенности тене-
вых и солнечных эпифитов, исследованных автором в Амани.
1. Теневой эпифит —Asplenium nidus (см. также табл. 18). «Гнездо»
диаметром 2,5 м состояло из 24 уже закончивших свой рост (длина самого
большого листа 104 см) и трех молодых листьев. В сухой период отдельные
части растения весили (в г):
Листья......................... 958
Ризомы с корнями и прилипши-
ми частицами почвы........... 5099
Всего
6057
1 Более подробно с эколого-физиологической точки зрения были исследованы
два вида Tillandsia, распространенные в засушливых районах тропиков. Они всасы-
вают лишь капающую воду, которая сначала удерживается капиллярными силами
волосков на чешуйках, а затем медленно впитывается живой тканью. Осмотическое
давление равно 6—8 атм, то есть оно столь же низко, как и у суккулентов; интенсив-
ность транспирации также весьма невелика, однако растения переносят сильный
недостаток влаги. Ср. R. В i е b 1 е, Zum Wasserhaushalt von Tillandsia recurvata L.
und Tillardsia usneoides auf Puerto Rico, «Protoplasma», 58, 1964, S. 345—368.
Тропический дождевой лес
145
За 20 дней величина испарения воды папоротником, который поме-
стили на защищенной от дождя веранде дома, стоявшего на опушке тропи-
ческого леса, составила 1287 г. Растение при этом не погибло, хотя и выгля-
дело несколько увядшим; Следует заметить, что в лесу не бывает столь
длительного сухого периода, да и воздух там значительно влажнее.
Рис. 99. Гнездовой эпифит Platycerium willinckii
с листьями-скобками в Ботаническом саду Богора
(фото Хусманса).
Влажность сильно гумусированной почвы под двумя другими гнездо-
выми папоротниками составляла (в % от сухого веса):
I II
23/XI 1934 г. (сухой период) 152 96
7/XII 1934 г. (дождливый
период) 220 317
Следовательно, в перегное, на котором развиваются папоротники,
всегда содержится достаточный запас воды, благодаря которому расте-
ния, видимо, никогда не страдают от недостатка влаги.
2. Солнечный эпифит —Drynaria laurentii.
146
Глава 2
Рис. 100. Thrixspermum arachnites}
V2 натуральной величины (фото
Фабера).
Рис. 101. Чешуйкообразные волоски у бро-
мелиевых: поперечный разрез (Tillandsia),
продольный разрез (Vriesea)} увеличено соот-
ветственно в 340 и 375 раз (по Шимперу).
Этот папоротник встречается на верхних ветвях деревьев; его тол-
стый ризом растет спиралеобразно вокруг веток, и на нем образуют-
ся листья-скобки, которые вскоре отмирают; кроме того, формируются
черешчатые перисторассеченные ассимилирующие листья (рис. 104).
Содержание воды в отдельных частях растения с ризомом длиной
50 см приведено в табл. 25.
Таблица 25
Сырой вес, г Содержание воды
г % от сырого веса
Ассимилирующие листья .... 24,0 13,0 Z 54
Листья-скобки 75,5 28,5 38
Ризом 616,5 422,0 68
Перегной и корни 222,0 150,0 70
Общий вес 938,0 613,5 —
Возможность дополнительного насыщения перегноя влагой невелика.
Большая часть воды накапливается в ризоме. Резервы воды, создаваемые
другими видами папоротников, например Asplenium theciforme и Poly-
Тропический дождевой лес
147
podium lanceolatum, еще меньше, что приводит к свертыванию их листьев
в сухой период. Два последних вида—это уже пойкилогидрические расте-
ния; они особенно характерны для семейства Hymenophyllaceae. По харак-
теру своей жизнедеятельности эти папоротники, листья которых образо-
ваны только одним слоем клеток, напоминают уже мхи. Они распростра-
нены главным образом в горных тропических лесах. Физиология папорот-
ников изучена Геснером и Хэртелем, правда не в лесу, а в оранжерее
Р и^Сж 102. Эпифиты — Nidularium eleutheropetalurn (Brome-
liaceae) и Hillia ulei, развитые на Japandariba spruceana\
Перу, бассейн Амазонки, окрестностьюримагуаса (фото Уле).
Мюнхена *. Со своей стороны Коутиньо (см. сноску на стр. 116) устано-
вил, что в дождевом лесу Бразилии Hymenophyllum polyanthos переносит
дефицит влаги, достигающий 94 %.
1 F. Gessner, Die Assimilation der Hymenophyllaceen, «Protoplasma», 34,
1940, S. 102—116; Wasserhaushalt der Epiphyten und Lianen. Handbuch der Pflan-
zenphysiologie, Bd III, 1956, S. 915—950; О. H art el, Physiologische Studien an
Hymenophyllaceen, II, «Protoplasma», 34, 1940, S 489—514.
148
Глава 2
3. Солнечные эпифиты — орхидеи, не образующие больших скоп-
лений перегноя.
У клубневых Bolbophyllum определялся сырой вес разных частей
растения. Результаты взвешивания двух растений (в %) приведены
в табл. 26.
Таблица 26
Листья Клубни Ризомы Корни
Растение I 46 40 8 6
Растение II .... 60 30 5,5 4,5
Содержание воды в листьях составляет 92% их сырого веса, а в клуб-
не — 93%. Следовательно, и листья, и клубни служат растению храни-
лищем воды. Корни при кратковременном
Рис. 103. Dischidia imbricata
с мешковидными листьями, края
которых плотно прилегают к
опоре (фото Фабера).
увлажнении могут поглотить влагу в коли-
честве 60% своего веса.
При взвешивании крохотного растеньи-
ца с пятью листьями и 10 клубеньками ока-
залось, что само растение весило 1,1 (а,
листья —0,16, клубеньки—0,87 и корни —
0,07 а.
У Angraecum на долю корней прихо-
дится лишь 5% сырого [веса; в побеге
и суккулентных листьях содержится 92%
воды.
Некоторые орхидные обладают сильно
развитой корневой системой, достигающей
70% общего веса растения (при веламене,
наполненном воздухом); при увлажнении
в течение 15 минут корневая система погло-
щает воду в количестве 30—170% собствен-
ного веса.
У эпифитных орхидей транспирируют
все органы растения, даже корни. Поэтому
транспирацию следует изучать, используя
для этого целое растение, осторожно отде-
лив его от опоры. Такого рода девятиднев-
ный опыт был проведен с тремя растениями,
подвешенными на защищенном от дождя
месте веранды.
Уменьшение содержания воды в расте-
ниях представлено на рис. 105. Кривые
несколько отличаются между собой; так,
кривые для бесклубневых орхидей имеют
вид кривых усыхания. Эти растения по
истечении 9 дней потеряли 20—25% перво-
начального веса, но нисколько при этом
не пострадали. Первые признаки завя-
дания появились лишь через 10—12 дней.
Однако в естественных условиях орхидеям
не приходится столь долго обходиться без
Тропический дождевой лес
149
воды, так как здесь их по меньшей мере ежедневно смачивает
роса.
Клубневые виды в течение первых 12 дней равномерно теряли за сут-
ки около 2,5% воды.
При погружении одних только воздушных корней орхидей на 1 час
в воду сырой вес растения увеличивался на 15%, это свидетельствует
о том, что растение способно очень быстро восполнить потерянную влагу.
Орхидеи, распространенные в тропических областях с четко выражен-
ным бездождным периодом продолжительностью около месяца, теряют
на это неблагоприятное время свои листья; остается только толстый
или почти клубневидный побег, практически не испаряющий влаги.
С началом дождей на нем появляются новые листья.
4. Эпифитные кактусы. Эта группа растений представлена в Восточ-
ной Африке Rhipsalis cassytha. Аналогичный вышеописанному опыт по
определению транспирации был проведен с крупным экземпляром этого
Рис. 104. Эпифит Drynaria laurentii, закрывающий толстую
ветвь дерева; Гдождевой лес в окрестностях Амани, Восточная
Африка (фото Э. Вальтер).
растения. Спустя 28 дней после начала наблюдений кактус потерял 28%
своего первоначального веса и обнаружил явные признаки засыхания.
В отличие от кактусов, развитых в аридных областях, Rhipsalis не пере-
носит длительного сухого периода.
Измерения интенсивности транспирации у тропических Codonanfhe
gracilis и Hypocyrta radicans (Gesneriaceae), произведенные Коутиньо мето-
дом быстрого взвешивания, показали, что максимальные величины у этих
видов не превышают 1—2 мг/дм2-мин. У орхидеи Maxiliaria picta и бро-
мелиевых Vriesea altodaserrae и V. inflata интенсивность транспирации
оказалась столь малой, что иногда ее вовсе не удавалось измерить (ср.
с интенсивностью транспирации ананаса, представленной в табл. 19).
Коутиньо недавно установил «эффект Соссюра» у различных видов
эпифитов; он доказал, что ночью они поглощают СО2 из воздуха, а днем
ассимилируют его (ср. стр. 362). При этом содержание кислоты в кле-
точном соке ночью повышается, а pH понижается. Это установлено
у эпифитных бромелиевых Aechmea pectinata и Neore gelia concentrica,
150
Глава 2
Рис. 105. Динамика сырого веса
эпифитных орхидей, размещенных
под навесом.
Для опыта были взяты целые растения
с воздушными корнями. Из четырех видов
орхидей только у Bolbophyllum развиты
клубни. Растения не цвели, поэтому
нельзя было определить их видовую
принадлежность.
а также у растущих на почве: Bilbergia
атоепа, Quesnelta testudo и Cannistrum
cyathiforme. Такой же эффект обнару-
жен у орхидеи Epidandrum ellipticum'
у эпифитов Maxillaria ptcta (Orchida-
сеае) и Vriesia inflata (Bromeliaceae)
и у всех видов Gesneriaceae фотосинтез
протекает обычным путем. «Эффект Со-
ссюра» отмечается только у суккулен-
тов, у которых не наблюдается резких
различий между ассимилирующей па-
ренхимой и водопроводящей тканью 4.
В общем следует отметить, что орхи-
деи и Rhipsalis являются не ярко выра-
женными суккулентами, примерно та-
кими, как европейские виды Sedum
и Sempervtvum. Этим условиям соответ-
ствует и гидратура, о чем свидетель-
ствуют низкие значения осмотического
давления, приведенные в табл. 27.
У папоротников величина осмоти-
ческого давления оказалась выше, чем
у цветковых растений, что иллюстри-
руют табл. 28—30.
Вент 1 2, изучая сообщества эпи-
фитов дождевого леса в Тьибодасе
на Яве, установил отчетливую зависи-
мость их развития от условий освеще-
ния и количества накопленного пере-
гноя.
Свойства древесной коры также
играют известную роль, причем Вент
предполагает, что значение имеют не
только ее физические свойства, но
и химический состав, а также состав
стекающей по стволу воды; например,
развитие Castanopsis, вероятно, зависит от дубильных веществ, содержа-
щихся в коре. По сообщению Ричардса, Utricularia schimperi в Бри-
танской Гвиане растет только на толстых подушках печеночников в бо-
роздах стока дождевой воды по стволам деревьев.
Семена эпифитов обладают характерными приспособлениями, свя-
занными со способами их распространения. Семена эпифитов каким-то
образом должны попасть на деревья. Поэтому они либо мелкие, как пыль
(Orchidaceae), либо обладают летучками и легко разносятся ветром
(Asclepiadaceae, Gesneriaceae и др.). Некоторое значение имеет и перенос
1 L. М. Coutinho, Untersuchungen fiber die Lage des Lichtkompensation-
spunktes einiger Pflanzen zu verschiedenen Tageszeiten mit besonderer Beriicksichtigung
des «de-Saussure-Effektes» bei Sukkulenten, Beitr. zur Phytologie (Walter — Festschr.),
Stuttgart, 1964, S. 101—108; Algunas informa^oes sobre a capacidade ritmica diaria da
fixa^ao e acumula^ao de CO2 no escuro em epffitas e erbaceas terrestre da mata pluvial,
Bolet. № 294, Fac. Fil. Cienc. e Letras da Univ. Sao Paulo, Botan. 21, 1965.
2 F. W. W e n t, Soziologie der Epiphyten eines tropischen Urwaldes, «Ann. Jard.
Bot. Buitenz.», 50, 1940, S. 1—98.
Тропический дождевой лес
151
Таблица 27
Название растений Величина осмоти- ческого давления, атм
т, f листья ... .... Клуоневые орхидеи клубни Орхидеи (бесклубневые) с длинными fl., воздушными корнями, но без утол- <] II f щенного стебля 1 III Rhipsalis cassytha д , . л f листья Asclepiadaceae, каучуконос КЛубни 3,1 2,0 4,3 6,1 6,6 5,2—5,6 10,8 3,8 Таблица 28
Название растений
Величина осмоти-
ческого давления,
атм
Asplenium nidus, молодые, еще пе развернувшиеся
листья ......................................
Asplenium nidus, листья, уже закончившие свой
*’ рост....................
Asplenium nidus, старые листья . . . J .
6,1
10,2—10,9
11,1—11,7
Таблица 29
Drynaria laurentii; растение I растет в более
сухих условиях, чем растение II
Величина осмотического давления, атм
ассимилирую- щие листья скобочные листья ризомы
I 12,6 7,0 5,7
II 10,2 5,7 4,8
Таблица 30
Папоротники, переносящие высыхание; величина
осмотического давления приведена для свежих листьев
Название растений Величина осмоти- ческого давления, атм
Vittaria lineata (или isoetifolia), на солнце 15,7
Vittaria lineata (или гзоёН/оИа), в тени 10,6
Polypodium phymatodes, на солнце . . 15,0
Polypodium phymatodes, в тени 11,6
Polypodium phymatodes, тонкий ризом 10,4
Oleandra africana . . 9,4
Asplenium sinuatum . . 9,0
Asplenium macrophyllum 11,7—12,7
Asplenium protensum, увядшие листья 11,5
Asplenium theciferum, свернутые листья 33,3
Lycopodium cernuum, облиствленные побеги .... 11,6
Lycopodium cernuum, главный стебель 8,4
152
Глава 2
Рис. 107. Лист, покрытый
многочисленными эпифильны-
ми мхами, лишайниками и водо-
рослями, в натуральную вели-
чину (фото Фабера).
Рис. 106. Эпифитный мох Barbella javanica
в дождевом лесу Явы (фото Фабера).
семян с помощью птиц (Cactaceae, Bromeliaceae); птицы способствуют
расселению Tillandsia usneoides, используя ее вегетативные части для
постройки своих гнезд. Важную роль в распространении семян эпифитов,
видимо, следует отвести также муравьям. Семейства, чьи представители
обладают тяжелыми семенами, эпифитов не имеют.
Споры папоротников без труда попадают на деревья, где разви-
ваются их проталлии Ч
Наряду с перечисленными сосудистыми эпифитами в тропиках рас-
пространены и бессосудистые — многочисленные мхи и водоросли, обыч-
1 Заростки — половое поколение папоротникообразных; маленькие пластинки^
на которых развиваются антеридии и архегонии.— Прим, перев.
Тропический дождевой лес
153
но находящиеся в тесной ассоциации с сосудистыми эпифитами. Наряду
с мхами, сплошь покрывающими ветви деревьев, интерес представляют
и мхи, свисающие в виде нитей; повисая в воздухе, они достигают иногда
метра в длину (рис. 106). Лишайники, обладающие очень медленным
ростом, встречаются в небольшом количестве. Их можно обнаружить
только на открытых солнечных местообитаниях вместе с устойчивыми
к засухе мхами, главным образом вблизи верхней границы древесной рас-
тительности.
Особую группу образуют эпифиллы, развивающиеся на поверх-
ности живых листьев, где они живут продолжительное время (рис. 107).
Поскольку эпифиллы не успевают размножиться за кратковременный
период жизни листа, они чаще встречаются на очень влажных местооби-
таниях, например вблизи лесных ручьев, на берегах которых всегда
обильна молодая поросль. Не безразличны эпифиллы также к условиям
Рис. 108. Эпифитная орхидея Т aeniophyllum zollingeri; на
рисунке показаны зеленые ассимилирующие воздушные
корни (фото Фабера).
освещения и степени увлажнения поверхности листа. Первыми заселяют
листья лишайники и водоросли, среди которых пионерами являются
Trentepohlia и Phycopeltis, вытесняемые позднее печеночниками.
Среди эпифильных водорослей и лишайников различают виды, про-
никающие в ткань листа, и виды, проникающие только в кутикулу,
но не в эпидермис *. В последнее время в условиях этого биотопа обна-
ружены азотофиксирующие бактерии (Azotobacter, Beijerinckia), а также
актиномицеты, дрожжи и грибы 1 2.
1 Н. Fitting., Uber die Beziehungen zwischen den epiphyllen Flechten und
den von ihnen bewohnten Blattern, «Ann. Jard. bot. Buitenz». Suppl., 3, 1910, S. 505—518.
2 J. Ruinen, The Phylosphere I. An ecological neglected milieu, «Plant and
Soil», 15, 1961, p. 81—109.
154
Глава 2
^Изучая водный режим эпифильных водорослей, мхов и лишайников,
Реннер 1 наблюдал у эпифильного мха (Ephemeropsis tjibodensis) интерес-
ное явление. Протонема мха развивается в виде очень мощных длинных
нитей, однако листостебельные побеги, образующие впоследствии поло-
вые органы, отличаются столь малыми размерами, что создается впечат-
Р и с. 109. Холопаразит Balanophora elongata в горном
лесу западной Явы. В середине — мужское соцветие; 1/3
натуральной величины (фото Фабера).
ление, будто коробочка сидит непосредственно на протонеме. Следова-
тельно, в этом случае речь идет о явлении, аналогичном тому, что наблю-
дается у сосудистого эпифита Taeniophyllum. У этой орхидеи развиты
только плоские зеленые ассимилирующие корни, из которых непосред-
ственно вырастает побег с цветками и несколькими маленькими чешуй-
кообразными листьями (рис. 108).
Сапрофиты и паразиты
Гетеротрофные высшие растения в тропическом дождевом лесу мало-
заметны. Крупные растения встречаются очень редко, основную массу
1 О. Renner, Zur Kenntnis des Wasserhaushalts javanischer Kleinepiphyten,
«Planta», 18, 1933, S. 215—287.
Тропический дождевой лес
155
составляют очень маленькие растения. Из числа однодольных сапрофиты
представлены семействами Burmanniaceae, Liliaceae, Orchidaceae и Triu-
ridaceae; из двудольных — семействами Gentianaceae и Polygalaceae.
Их можно увидеть, естественно, только в местах скопления органических
остатков, на валеже, в заполненных опадом листьев понижениях рельефа
или в пространствах между досковидными корнями. Они предпочитают
глубокую тень и постоянно влажную почву. Они могут вовсе отсутство-
вать на больших пространствах, а если и встречаются, то обычно неболь-
шими группами.
Удивительно, что высшие растения в качестве паразитов редко встре-
чаются в тропическом дождевом лесу. Очень мало здесь корневых парази-
тов; все они являются холопаразитами. Они представлены лишь двумя
семействами: Balanophoraceae (рис. 109) и Rafflesiaceae с родами Mycetan-
the и Rafflesia. Виды Rafflesta встречаются только в Индонезии и парази-
тируют на корнях Vitaceae. Как известно, вегетативное тело Rafflesia
редуцировано вплоть до клеточных нитей, пронизывающих корни расте-
ния-хозяина. Только перед цветением на корнях, выступающих над
поверхностью почвы, или у комля ствола развивается мощная толща
ткани, из которой появляются огромные цветки (рис. 70). Цветок Raf-
flesia arnoldi, распространенной на Суматре, отличается ярко-красным
или коричневато-красным цветом, иногда превышает 1 м в диаметре
и издает сильный трупный запах. Виды Cuscuta, которые в тропиках
Вест-Индии паразитируют и на деревьях, проникая до самой кроны,
а также Cassytha, относящаяся к семейству Lauraceae, не характерны
для дождевого леса. Более широко распространены эпифитные полупара-
зиты, принадлежащие к семейству Loranthaceae. Они образуют гигант-
ские шары, заметные в кронах деревьев-хозяев благодаря яркой окраске
цветков.
Обращают на себя внимание гиперпаразиты. Так, один из видов
Loranthus паразитирует на другом виде своего собственного семейства1.
Известны даже примеры гиперпаразитизма второй степени, когда парази!
поселяется на паразите паразита и только последний — на дереве-хозяине.
Заключение
Тропический дождевой лес не испытал столь строгого есте-
ственного отбора, как растительность других климатических зон.
Во влажных тропиках есть все условия для существования самых
различных экологических групп растений, начиная от ярко вы-
раженных гигроморфных форм и кончая ксерофитами или сук-
кулентами. Вместе с тем тропический дождевой лес — это источник совре-
менного разнообразия экологических форм. Именно его флора в процессе
многовековой эволюции пополняла растительный мир новыми жизнен-
ными формами, приспособившимися впоследствии к крайним условиям
существования — к климату с четкой периодичностью времен года,
с холодными или засушливыми периодами.
Вряд ли удастся когда-нибудь точно установить, приспособились
ли вторично некоторые из выходцев из тропических лесов к дождевому
лесу (например, бромелиевые) с тем, чтобы здесь в качестве древесных
эпифитов поселиться в крайних условиях местообитания, или же все эти
формы возникли в самом дождевом лесу.
1М. Koernicke, Biologische Studien an Loranthaceae, «Ann. Jard. bot.
Buitenz.», Suppl., 3, 1910, S. 665—697.
Глава 3
ДРУГИЕ ТИПЫ РАСТИТЕЛЬНОСТИ
ВЛАЖНЫХ ТРОПИКОВ
БОЛОТНАЯ И ВОДНАЯ РАСТИТЕЛЬНОСТЬ
Обилие осадков и относительно слабое испарение в экваториальной
зоне создают известный избыток воды, которая, скапливаясь
в понижениях рельефа, способствует образованию здесь болот
и озер. На затопляемых длительное время местообитаниях лесная рас-
тительность обычно не развивается. Злаковники возникают всюду, где
во впадинах собирается достаточно воды, чтобы почва была переувлаж-
нена даже в период засухи. Болота занимают в тропической Африке
огромные площади. Описание наиболее крупных болот приводится ниже.
При перелете из Ливингстона в Найроби трасса полета проходила
над болотами Луканга, западнее Брокен-Хилла, и далее над озерами
Камполомбо и Бангвеоло, где в 1873 г. умер Д. Ливингстон (исследова-
тель Африки, открывший водопады Виктория). Оба озера окружены обшир-
ными болотами общей протяженностью, вероятно, около 150 км. В север-
ной части болота поперек простиранию озера протягивается гряда, обыч-
но встречающаяся в грядово-мочажинных болотах. Западная часть озера
Бангвеоло почти полностью отделена песчаной косой от остального озера.
На этой косе было много хижин, а берега озера, окаймленные болотами
и широким поясом плавающих водных растений, вряд ли были доступны.
Заболоченная территория вокруг озера Мверу и западнее него была необи-
таема. К северо-западу от озера Руква и к югу от озера Эяси,
болота занимают большие пространства. Озера Эяси, Маньяра и
Натрон, а также некоторые другие озера приурочены к восточноафри-
канскому грабену; они расположены в относительно засушливой области,
отличающейся сильным испарением. Озера лишены стока; в них накап-
ливается сода.
Чрезвычайно многочисленны болота вокруг озера Виктория; не менее
многочисленны они и в долинах, расположенных на север от озера, уже
в Уганде. Здесь распространены заросли папируса и группы болотной
пальмы — Phoenix reclinata\ реже встречается пальма Raphia. Заболачи-
вание этой территории усилилось после подъема уровня озера Виктория,
когда на Ниле была сооружена плотина Джиния. Об этом свидетельствуют
заболоченные лесные массивы и стволы погибших деревьев.
По данным Васаво 4, в Уганде болота распространены на площади
12 800 км2, что составляет 6% всей территории. Они покрыты сомкнутыми
зарослями Cyperus papyrus (при pH = 5,5 —6,4), которые пропускают
так мало света, что препятствуют развитию всходов. Между этими зарос-
лями и поясом, образованным Phoenix reclinata, развиваются чистые зарос-
ли из Miscanthidium*, на кислых почвах (pH = 4,6 — 5,5) в этих зарослях
встречается даже Sphagnum.
1 D. Р. S. W a s a w о, Some problems of Uganda swamps, IUCN Publ., new ser. № 4,
1964, p. 196—204.
Другие типы растительности влажных тропиков
157
Наибольшие по площади болота образованы Нилом. На расстоянии
80 км от озера Виктория Нил протекает через озеро Кьога; окружающие
озеро болота простираются, вероятно, на расстояние более 200 км. Еще
дальше к северу Нил протекает в узкой долине и ниже по течению по водо-
падам Мерчинсон низвергается в Центральноафриканский грабен. После
пересечения северной оконечности озера Альберт Нил начинает имено-
ваться Альберт-Нил. Неподалеку от границы с Суданом, выше Нимуле,
горный хребет перегораживает Нил, который течет здесь по обширным
заболоченным территориям. По другую сторону хребта (под названием
Белый Нил) река впадает в огромную, расположенную на высоте около
400 м котловину, которую Нил частично затапливает. Вместе с левым
притоком —Бар-эль-Газаль —Нил протекает здесь, вероятно, по наи-
более обширным болотам («сэдд»), площадь которых, по данным Меншин-
га, достигает 150 тыс. кл^21. Максимальная протяженность этой заболо-
ченной площади с севера на юг и с востока на запад составляет более
600 км. Автору довелось наблюдать эти болота с самолета во время павод-
ка. Почти вся площадь болот, за исключением небольших участков,
была покрыта водой — солнце отражалось в больших зеленых, часто
правильной округлой формы поверхностях. Это были пространства,
покрытые плавающими водными растениями, или сплавины, образованные
Vossia cusptdata или Суperus papyrus, и, кроме того, «плавающие газоны»
из Eichhomia и Pistia. Эти болота прерываются многочисленными озерами
с открытой водной поверхностью, связанными между собой сетью меанд-
рирующих протоков.
На островках среди болот расположены отдельные хижины и поля.
Здесь распространена пальмовая саванна, в составе которой господствуют
Hyphaene thebaica и Borassus aethiopica или Acacia и Balanites (на глинистых
почвах). В период засухи уровень воды понижается и значительная часть
затопленной площади снова появляется над водой. На ней развивается
злаковый покров, образованный Hyparrhenta rufa или Setaria incrassata.
На более влажных участках преобладают Echinochloa stagnina и Е. руга-
midalis с примесью Phragmites communis и Veteveria nigritana 1 2.
В результате интенсивного испарения болота Сэдд снижают расход
воды в Ниле почти вдвое. Строительство отводного канала позволило бы
не только осушить эти огромные пространства болот, но и удвоить водо-
носность Белого Нила. Кроме того, из этих болот в Нил проникает завезен-
ный из Южной Америки водяной гиацинт — Eichhomia crassipes, который
не только мешает судоходству, но и накапливается у плотины выше Харту-
ма в таких огромных количествах, что реку можно свободно перейти
по этому своеобразному мосту 3.
Если к этим болотам добавить площадь заболоченных пространств
1 Н. Mensching, Nordafrika in «Die Grosse Illustrierte Liinderkunde»,
Bd II, 1963.
2 См. карту растительности в работе М. N. Harrison, J. К. Jackson,
Ecological classification of the vegetation of the Sudan, «Forest. Bull.», № 2, Khar-
toum, 1958.
3 Борьба c Eichhomia крайне трудна. Несмотря на то что в 1956—1957 гг. для
уничтожения эйххорнии в Конго было израсходовано 50 млн. бельгийских франков,
ежедневно мимо Леопольдвиля по реке Конго за час проплывает 150 т эйххорнии
(по Лебрену, из работы: Н. W i 1 d, Harmful aquatic plants in Africa, «Kirkia», 2, 1961,
p. 1—66).
Местами такой же вред приносит завезенная из Южной Америки Salvinia auri-
culata. Это растение в водохранилище Кариба (Родезия) покрывает водную поверх-
ность слоем толщиной 25 см, на этом субстрате в дальнейшем образуются заросли
Vossia и Scirpus cubensis.
158
Глава 3
в южной Катанге и в бассейне Конго, то можно себе представить, сколь
велики площади, занимаемые сообществами водных растений, по сравне-
нию с остальным растительным покровом тропической Африки.
Злаковники, развитые в понижениях, где почвы периодически пере-
увлажняются, часто незаметно переходят в сообщества болотных растений.
Совершенно иной растительный покров развивается там, где количество
осадков, выпадающих в период дождей, меньше величины испаряемости,
особенно в течение длительного периода засухи. Тектонические движе-
ния, связанные с образованием грабена, создали множество бессточных
областей, где накапливались отложения, возникающие при выветривании
основных пород. Отсутствие более поздних морских отложений и отдален-
ность моря не способствуют концентрации хлоридов. Поэтому здесь наблю-
дается накопление натрия. В связи с этим большинство бессточных озер
в сухом восточноафриканском грабене, как уже упоминалось, являются
горькосолеными (натриевыми).
На берегах горькосоленого озера Накуру (Кения) наблюдаются следу-
ющие пояса растительности. В воде растут Scirpus sp., далее Juncellus
и Sporobolus spicatus\ несколько выше растет Cynodon dactylon. За преде-
лами пояса засоленных почв — Cyperus rotundus и Pennisetum clandesti-
пит, тычиночные нити которого достигают длины 6 см. Озеро окружает
заболоченный лес из Acacia xanthophloea, отличающийся желтыми ство-
лами. Тысячи фламинго населяют эти натриевые озера.
Злаковники, распространенные в понижениях, нередко постепенно
переходят в сообщества, развитые на засоленных почвах. Кроме вышеупо-
мянутых видов, здесь встречаются также Sporobolus robustus, Diplachne
fusca и др. Наиболее низкие участки депрессий, пересыхая, покрываются
белыми выцветами солей; растения здесь не растут. Эти засоленные почвы
пока еще достаточно подробно не изучены.
На затопляемых длительное время местообитаниях вместо лесной
растительности буйно разрастаются сообщества водных и болотных
растений, широко распространенные на равнинных территориях. Сообще-
ства водных растений тропиков флористически отличаются от аналогичной
растительности умеренной зоны, однако эти различия не столь велики,
как у сухопутных сообществ. Водные растения-космополиты часто встре-
чаются и в тропиках Ч
Зарастающие водоемы могут быть расчленены на три пояса:
а) пояс погруженных и свободноплавающих растений,
б) пояс растений с плавающими листьями и
в) пояс болотных растений.
а) В этом поясе доминируют Hydrilla verticillata, Ceratophyllum demersum,
виды Myriophyllum, Patamogeton и др. Особую роль играют заросли
свободно плавающей Eichhornia, которая была занесена из Нового Света
в водоемы Старого Света. Быстро приспособившись к новым условиям, это
растение превратилось в отдельных районах в настоящий бич для населе-
ния, как это наблюдается, например, в области Сэдд (верховья Нила),
где оно мешает судоходству и затрудняет лов рыбы1 2.
В этом же поясе развиваются Lemna, Azolla, Salvinia, а в Новом Свете
1 F. Ruttner, Die Wasservegetation einiger Seen Niederlandisch-Indiens,
«Veg. Bilder», 25, H. 7. 1940.
2 См. M. J. Chadwick, Some observations of the ecology of Eichhornia
crassipes Solms., «8. Ann. Rep. Hydrobiol. Res. Univ, of Khartoum», 23—28, 1960
и T. A. G a y, Ecological studies of Eichhornia crassipes Solms in the Sudan, «J. Ecol.»,
48, 1960, p. 183—191.
Другие типы растительности влажных тропиков
159»
Р и с» 110. Заросли Papyrus по заболоченному берегу озера^Банг-
веоло (фото Розена).
Jussiena repens, б) К растениям с плавающими листьями относятся разви-
тые в следующем поясе виды Nymphaea, в том числе распространенный
в Африке египетский лотос (N. lotus) и голубой лотос (N. coerulea.) Здесь
же встречаются виды Limnanthemum и Trapa, Euryale, а в бассейнее Ама-
зонки — знаменитая Victoria regia. У виктории большие тонко перфори-
рованные плавающие листья с загнутыми кверху краями. Дождевая вода,
попадающая на листья, стекает по многочисленным стоматодам. Для боль-
шинства тропических болотных растений, например Nelumbium, характер-
на несмачиваемость их листьев благодаря особому веществу, выделяю-
щемуся из сосочков на нижней стороне листьев Ч
1 F. Gessner, Hydrobotanik, Bd I, II, Berlin, 1955; 1959. Ср. Bd II, S. 32,
co многими интересными экологическими данными о тропических водных растениях.
160
Глава 3
Необычайно своеобразны растения из семейства Podostemonaceae,
порядок розоцветных. Они встречаются в тропиках только в быстротеку-
щих водах и в пенящихся потоках водопадов. Растения отличаются удиви-
тельными вегетативными образованиями, которые имеют вид слоевища
или коры и этим напоминают водоросли или мхи. Экология этих растений
впервые исследована Геснером и Хаммером в районе водопадов Карони,
главного притока реки Ориноко 4. У распространенных здесь четырех
видов цветки образуются около возвышающегося над водой места прикрепле-
ния растения и опыляются насекомыми. Вегетативные органы погружены
в воду; у этих растений нет ни проводящей ткани, ни приспособлений,
предохраняющих их от испарения, поэтому они быстро засыхают при
малейшем спаде воды. Осмотическое давление клеток при плазмолизе
составляет 6,6 атм. В стоячей воде растения гибнут; лучше всего они
развиваются в тропической «черной воде», то есть воде, окрашенной золя-
ми гумуса в коричневый цвет и содержащей небольшое количество СО2
и электролитов. Поэтому для изучения фотосинтеза необходимо обогащать
эту воду углекислотой. В местах развития Podostemonaceae пена водопа-
дов и стремнин постоянно поглощает СО2 из воздуха. Podostemonaceae —
типичные светолюбивые растения, только один вид этого семейства распро-
странен в тенистых ручьях.
Podostemonaceae очень крепко прикрепляются к подводным скалам
и в таком состоянии из-за резких колебаний уровня воды существуют
весьма непродолжительное время (не дольше двух месяцев). Однако смена
поколений следует непрерывно и притом достаточно быстро. Как только
очень мелкие, длиной 0,04 мм, семена попадают на периодически обсыхаю-
щие части скалы, внешний слой оболочки семени быстро, буквально
меньше, чем за минуту, набухает и эта желатиноподобная, похожая на
пектин масса скрепляет семя со скалистым основанием настолько прочно,
что смыть его не удается даже сильной струей из шланга, в) Болотные
растения представлены видами: Typha, Cladium, Phragmites, Nelumbium,
Sagittaria, Pistia, Limnophyton, а также многочисленными Сурегасеае,
из которых Cyperus papyrus (рис. 110), достигающий нескольких метров
в высоту, образует в Африке гигантские заросли (болота Сэдд) 1 2. Очень
характерны для тропических водоемов плавающие острова, образованные
прежде всего растениями, которые укореняются на берегу водоема и чьи
плавающие стебли на несколько метров вдаются в открытую водную
поверхность. Это Ipomoea reptans, Jussiena repens, Polygonum barbatum,
а также вида семейства Gramineae и Сурегасеае. Растения, развивающие-
ся на этих плавающих островах и увлажняемые лишь дождевой водой,
отличаются очень кислой реакцией. На подобного рода сплавинах находят
даже Sphagnum с Rhynchospora, Eriocaulon, Xyris, а из насекомоядных
наряду с Utricularia в Индо-Малайской области — также различные виды
Nepenthes. Линд и Виссер, проводившие исследования на озере Виктория,
в районе Энтеббе, выделили несколько поясов болотной растительности
1 F. Gessner, L. Hammer, Okologisch-physiologische Untersuchungen an
den Podostemonaceen des Caroni, «Int. Revue ges. Hydrobiol.», 47, 1962, S. 497—541.
2 Мигахид изучил в области Сэдд почвенные условия, условия увлажнения,
экологию и транспирацию Cyperus papyrus, Vossia cuspidata, Pistia stratiotes'.
A. M. M i gah i d, Report on a botanical excursion to the Sudd Region, 1, II, Cairo
Univ. Press., 1948; Velocity of water current and its relation to swamp vegetation in the
«Sudd» region of the upper Nile, Cairo Univ. Press, 1952; Further observations on
the flow and loss of water in the «Sudd» swamps, Cairo Univ. Press, 1952; A. №. M i-
gahid, A. A. Abd el Rahman, M el Shaffei ali, An ecological study
of a swampy island in the upper Nile, Arabic Congress, 1955, S. 677—718.
Другие типы растительности влажных тропиков
161
Коренной п
берег 1------------------------Далина реки----------------------------------
(terra firme)
с бысокостболь- Галерейный
ным лесом лес
(ретичные Уровень
породы половодья
Меженный
уровень
Рис. 111. Поперечный профиль долины Амазонки в ее нижнем течении (пример
«светловодного потока») (по Сиоли).
Современные
намытые почвы
в зависимости от высоты стояния воды. Здесь по краю внешнего пояса
часто встречаются сплавины с зарослями папируса х.
В прибрежном поясе, где формируются болота озерного происхожде-
ния, поселяются древовидные однодольные, среди которых преобладают
виды Pandanus, Raphia, Calamus и др., Д в Африке, например, Phoenix
reclinata. Приспособленные к плохо аэрируемым почвам, эти виды пальм
на почвах избыточного или переменного увлажнения являются более
конкурентоспособными, чем большинство двудольных 1 2. Именно поэтому
пальмы растут по берегам рек в составе периодически затопляемых лесов,
сходных по своей экологии с пойменными лесами Центральной Европы.
Характер распределения растительности в долине нижнего течения Ама-
зонки представлен на рис. 111. Здесь различают три типа рек.
1. «Agua Ьгапса», «светловодные потоки»—очень мутные, несущие
большое количество взвешенных частиц, с водой, окрашенной в глинисто-
желтый цвет. Берут начало в Андах.
2. «Agua preta», «черноводные потоки»— с прозрачной водой кофейно-
бурого цвета из-за высокого содержания гумусовых коллоидов. Верховья
таких рек, вроде Риу-Негру, теряются в обширных болотных лесах.
3. Реки с прозрачной желто-зеленой водой, берущие свое начало
в районах выровненного рельефа, например на плато Мату-Гросу.
Наиболее интенсивная аккумуляция осадков происходит в реках
«Agua Ьгапса» 3, плоские долины которых имеют ширину от нескольких
метров до 100 км. При подъеме уровня воды, который начинается при-
близительно в конце года, пойма затапливается; течение реки с удалением
от русла замедляется, и взвешенные частицы оседают. Окаймляющий
берега реки галерейный лес играет роль своеобразного сита; здесь про-
исходит наиболее интенсивное осадконакопление и образуются возвышаю-
щиеся над урезом воды береговые валы, полого понижающиеся к затоплен-
ной пойме—«варзеа» (varzea). Галерейный лес по мере удаления от русла
1 Е. М. Lind, S. А. V i s s е г, A study of a swamp at the north end of Lake
Viktoria, «J. Ecol.», 50, 1962, p. 599—613.
2 Особенно благоприятные условия для разведения кокосовых пальм на песча-
ных морских побережьях тропиков, видимо, связаны с близким залеганием уровня
грунтовых вод, постоянный сток которых направлен к морю.
3 Н. S i о 1 i, Sedimentation im Amazonasgebiet, «Geol. Rundschau», 45, 1947,
S. 608—635.
11 — 1394
162
Глава 3
становится все более низкорослым и наконец сменяется злаковыми болота-
ми—«campos de varzea». Самые низкие участки поймы заняты мелковод-
ными озерами нередко огромных размеров. За озерами, в сторону от
реки, поднимается поросший лесом коренной берег — «terra firme», сложен-
ный из осадков плиоценового озера, существовавшего в бассейне Амазон-
ки. Нижние части склона коренного берега покрыты болотным лесом
«igapo», затопленным иногда в течение нескольких месяцев, поскольку
в половодье вода часто поднимается на 15 м выше меженного уровня Ч
В составе лесов «игапо», образованных небольшим числом видов, домини-
рует Eugenia inundata, сбрасывающая листву на ветвях, покрытых водой.
Вечнозеленые виды образуют остальную часть древостоя. Кустарники
в лесу местами достигают высоты 5 м. В более глубоководных местах
развивается водная растительность, представленная Polygonum hispidum,
Jussiena, Neptunia, Victoria regia и злаками Paspalum repens, а также
Oryza perennis. Сообщество плавающей водной растительности образуют
Eichhomia azurea, Phyllanthus fluitans, Ceratopterispteridoides с участками
Pistia, Salvinia, Azolla, Lemna, Ricciocarpus и др. 1 2.
В реках с прозрачной водой осадки, преимущественно песок, откла-
дываются уже в верхнем течении. В «agua preta» седиментация также
незначительна.
Затопляемые леса широко распространены и в бассейне Конго, но
сведения о них очень скудны.
В устьях рек, где сказывается влияние приливов и отливов, затопляе-
мые леса постепенно сменяются мангровами, к описанию которых мы
еще вернемся.
Помимо лесов, характеризующихся сильными колебаниями уровня
грунтовых вод, в тропиках на низменностях широко распространены
настоящие постоянно влажные болотные леса, похожие на заболоченные
леса Центральной Европы. В случае кислой реакции среды здесь может
начаться процесс торфообразования, что дает нам основание говорить
о лесах, развивающихся на торфяных болотах. Физиномически они напо-
минают дождевые леса, однако их видовой состав беднее, они не так
роскошны и более разрежены, благодаря чему под древесным пологом
могут развиваться болотные растения 3.
Бюннинг описал леса такого типа на восточном побережье северной
части Суматры 4. Корни древесных пород расположены в поверхностном
слое слабокислой (pH = 3,0) почвы. Частое явление — досковидные и хо-
дульные корни. У многих видов встречаются также воздушные и коленча-
тые корни. Более обильно, чем в дождевых лесах, представлен бамбук.
Мощность залежи торфа иногда превышает несколько метров 5,
Характерная особенность этих лесов — так называемые метловидные
корни, свойственные определенным видам деревьев. Они образуются на
стволе на высоте одного или нескольких метров от уровня почвы и отходят
от ствола перпендикулярно его поверхности, достигая 0,5—1 м длины
и больше. Предположение Бюннинга о роли этих корней как поставщиков
1 F, Gessner, Hydrobotanik, Bd II, Berlin, 1959, S. 245—249.
2 M, Takeuchi, The structure of the Amazonian vegetation, VI, «J, Fac. Sci,
Univ,», Tokyo, III, 8, 1962, p, 297—304,
3 J. A, R, Anderson, The ecology and forest types of peat swamp forests of
Sarawak and Bruneil in relation to their silviculture, Edinburgh, 1964 (тезисы дис-
сертации).
4 E. Biinning, In den Waldern Nordsumatras, Bonn, 1947.
5 Ср. H. Str aka, Uber Moore und Torf auf Madagaskar und den Maskarenen,.
«Erdkunde», 14, 1960, S. 81—98.
Другие типы растительности влажных тропиков
163
воды в период высыхания поверхности торфяной почвы вряд ли верно.
Метловидные корни могут использовать только атмосферную влагу, одна-
ко во время выпадения осадков, намокает и почва, и поэтому неглубоко
расположенные в почве корни будут в этом случае получать достаточно
воды. Адсорбция в значительных количествах водяного пара воздуха или
росы также маловероятна.
Возникает вопрос, могут ли в определенных условиях влажных
тропиков формироваться естественные травяные ландшафты? Вероятно,
нет; исключение составляют заросли злаков на сплавинах по берегам
зарастающих озер. Развитию древесной растительности препятствует
лишь периодическая засуха, в течение которой почва совершенно пересы-
хает. Однако во влажных тропиках таких условий не существует. Естест-
венные пожары в дождевом лесу не возникают даже при частых грозах.
Только на участках, где человек вырубает и выжигает лес, почва вскоре
покрывается злаковым дерном. При неоднократном выжигании этих
участков формируются обширные травяные ландшафты даже в таких влаж-
ных районах, как Суматра, где, например, вокруг озера Тоба в округе
Батак широко распространены злаковники, образованные аланг-алангом
(Imperata arundinacea) и Saccharum spontaneum. Правда, здесь, в цен-
тральной части острова, отделенной от моря горными хребтами, иногда
наблюдается засушливый период.
МАНГРОВАЯ РАСТИТЕЛЬНОСТЬ
На ботаника, впервые попавшего в тропическую область со стороны
моря, наиболее сильное впечатление производят мангровы, зеленой стеной
встающие из морских вод близ побережья. Во время прилива над водой
возвышаются лишь кроны деревьев, а при отливе обнажаются их своеобраз-
ные дыхательные корни. Лишь за поясом мангровой растительности можно
заметить стройные силуэты кокосовых пальм. Мангровые леса пред-
ставляют совершенно своеобразный мир с особыми, только ему присущими
формами растительной и животной жизни.
Мангровы развиваются лишь в местах, защищенных от сокрушитель-
ных ударов волн коралловыми рифами или прибрежными островами. Ман-
гровы приурочены к местообитаниям, периодически затопляемым морем;
мангры совершенно не переносят заморозков. Такие условия наблюдаются
не только в экваториальной зоне, но и далеко за ее пределами. Поэтому
мангровы, лучше всего развитые во влажных тропиках, встречаются и за
пределами тропиков, отличаясь в этом случае более бедным видовым соста-
вом. Северной границей их распространения являются Бермудские остро-
ва (32° с. ш.), северная оконечность Красного моря (залив Акаба, 30° с. ш.)
и юг Японии (32° с. ш.). В южном полушарии мангровы заходят за Дурбан
в Южной Африке (33° ю. ш.), окаймляют Австралию (38° ю. ш.) и прони-
кают на север Новой Зеландии. Мангровы распространены даже на
острове Чатам, расположенном восточнее Новой Зеландии (44° ю. ш.).
Подобно всем видам прибрежных сообществ, мангры распространены
очень широко и не ограничены экваториальной зоной. В флористическом
отношении область Индийского и западной части Тихого океанов отличает-
ся богатством видового состава (табл. 31) по сравнению с областью Атланти-
ческого океана. Хотя роды растений в обеих областях совпадают, их видо-
вой состав совершенно различен. Из числа видов восточных мангров
все 20, приведенные в табл. 31, произрастают на Яве, в Новой Гвинее и на
Филиппинских островах; в Восточной Африке известно 14, в северной
И*
164
Глава 3
Австралии ;гакже лишь 14, в Китае и на Тайване — 4, а в Новой Зелан-
дии — только один вид.
Т а б л л да 31
Распределение видов восточной мангровы
(но Foxworthy, 1910, с некоторыми изменениями;
цитируется по Шимпер — Фаберу; 1
Вост. Африка Мадагаскар Цейлон Индия Бирма Спам и Кохинхина Малакка Суматра св Д ЗС Борнео (Калимантан) Нов. Гвинея Австралия Филиппины Формоза Ю/кп. Япония Япония Китай |
Rhlzophora mucronate д_ _Д 4- 4- д- j _ 4 4 - 1- г д_ 0
Rhlzophora conjugata . . 0 4- i + д- 4 + 4 Д 0 4 0 0 0 0
Brugulera gymnorrhiza д- + + Д- + 1- 0 д- 0 0
Brugulera erlopetala . . -- 0 + 0 -f- -г -4- 1- • - д Ч- 0 0 Ч-
Brugulera caryophylloldes 0 0 + + 4- 4- 4 4- 0 г 0 4- 0 0 0 0
Brugulera parvlflora 0 0 0 + 4- 4- _д 4- -4- t- - д 0 0 0 0
Cerlops candolleana ч- д + + 4- + 4- 1- 0 0 0 0
Ceriops rorburghlana -- 0 Чд 4- _1_ 4- - - - • - 0 0 0 0 0
Sonneratla aclda + 0 4- + -4- т _L. Д- Д 0 4 0 0 0 0
Sonneratla alba . 0 “I -Д Д- 4- 4- + 4- 1- X 0 д_ 0 0 0 0
Xylocarpus mo I и ccen s i s 0 Ч- + -- jl -Д - --L- 0 4 0 0
Xylocarpus granatum Д- 0 0 0 -4 -- 0 0 4 4- ч- 0 -Д 0 0
Liunnitzera litorea 0 0 0 д- и 0 Т Д- -- г- 4 • - L 0 +- 0 0
Lumnltzera racemosa. — г + 4~ 4- 0 0 0 4 0 ! - 4 1_ 1- 0 0
Aegiceras cornlciilatuni 0 0 + д- 4- 4- д- Д- х Д Г ч- 0 0 0 +
Aegiceras floridum 0 0 0 0 0 0 0 0 0 4 0 ч- 0 1- 0 0
Avicennia officinalis . . Scyphl phora hydro phyl- -д д- -г + -L ч- Д- -- д д J. - i- 0 -д
lacea .... 0 0 —L- + -г- 4- 1- -X X —- 4 0 -д 0 0
Acanthus illclfolius "Г 0 д_ + Д- + 4- 4 • - — 4 0 4 0 4
Nipa f rut leans 0 0 ~4 0 -4 4- д- 4- 4 4 - ~4 0 0 0 0
Общее число видов 12 7 <3 17 18 17 17 20 I17 20 14 20 4 10 1 4
1 Крестики означают шличие, ноли—отсутствие соответствующих видов. В таблице при-
ведены старые названия видов.
Амплитуда прилива на разных побережьях не одинакова, но даже на
одном и том же берегу она обнаруживает периодичность в зависимости
от фазы луны. Различают средний уровень приливов и средний уровень
отливов. В полнолуние и новолуние соответственно уровень приливов
поднимается выше (сизигийный прилив), а уровень отливов падает ниже
среднего уровня. В первую и последнюю четверти Луны возникают квад-
ратурные приливы с наименьшей амплитудой. Наконец, дважды в год,
в весеннее и осеннее равноденствие, приливы (так называемые равно-
денственные) достигают наибольшей высоты. Мангровая растительность
Другие типы растительности влажных тропиков
165
распространена во всей зоне этих приливных явлений до границы наибо-
лее высоких уровней приливов. Лишь у нижней границы отливов можно
встретить участки, лишенные растительности (рис. 112, 113). Древесные
породы, слагающие мангровы, являются настоящими галофитами. Неко-
торые из них, правда, могут расти и в пресной воде, но лучше развивают-
ся все же в соленой. Подобно всем облигатным галофитам, они накапли-
вают в клетках много поваренной соли (рис. 123) и отличаются суккулент-
ностью своих листьев.
Расчленение мангровых зарослей на пояса зависит от трех различ-
ных факторов, определяющих конкурентную способность отдельных видов:
1) частоты и продолжительности затопления;
2) характера субстрата: песчаные или илистые отложения;
3) в устьевых участках рек — от соотношения примеси пресной воды
к соленой морской. Концентрация смеси пресной и соленой воды
подвержена колебаниям, зависящим от амплитуды приливов,
направления ветров и водоносности реки.
Перечисленные факторы обусловливают поясность мангровых сооб-
ществ, причем в каждом поясе доминирует либо один, либо два, а иногда
и несколько видов. Поясность мангровы зависит от конкурентной способ-
ности слагающих ее видов; последняя в свою очередь связана с условиями
местообитания. Чем богаче видовой состав мангровых зарослей, тем слож-
нее конкурентные взаимоотношения и тем труднее выявление закономер-
в
т
0,50
3,00
0,50
2,00
0,50
1,00
0,50
0,00
ЗА5м
_2г92м
Сизигийный прилив
2,26м
3,51 н Полоса песка,
лишенная
—растительности
_______
Квадратурный прилив
Пояс
Sooner а На
Рис. 112. Сезонное изменение высоты прилива и поясность мангровых зарослей
у Танги, Восточная Африка (по Вальтеру и Штейнеру).
Зачернена зона, затапливаемая только во время равноденственных приливов. Внизу показаны
фазы Луны и месяцы.
166
Глава 3
.Рис. 113. Внешний пояс мангровы во время
отлива у Танги. Чистые заросли Sonneratia alba;
видны дыхательные корни и полоса, лишенная
растительности (по Вальтеру и Штейнеру).
Рис. 114. Мангровы (заштриховано) у берегов
Восточной Африки близ Танги, развитые под
защитой коралловых островов и рифов (по Валь-
теру и Штейнеру).
ностей поясного расчленения
мангровы. Отсутствие отдель-
ных конкурентов может при-
вести к изменению в последо-
вательности размещения ви-
дов по поясам; поэтому дан-
ные, собранные в одном
районе, нельзя автоматически
переносить на другой флори-
стически отличный район. Не
менее важны также различия
в количестве осадков, опре-
деляющие степень засоленно-
сти изредка затопляемых, то
есть наиболее высоко распо-
ложенных участков манг-
ровы.
Прежде чем перейти к
рассмотрению сложной систе-
мы поясности мангровых со-
обществ, познакомимся с эко-
логией относительно более
простых и хорошо изученных
мангров, окаймляющих побе-
режье Восточной Африки
(рис. 114) 4.
Названные прибрежные
мангровы в отличие от сооб-
ществ, развитых в устьях рек,
растут под защитой коралло-
вых островов вдали от воздей-
ствия речных вод и испыты-
вают влияние только одной
морской воды с осмотическим
давлением около 24 атм.
Поясное распределение манг-
ровы иллюстрируют рис .115 —
118. Bruguiera не образует
здесь самостоятельного пояса,
а растет совместно с Rhizo-
phora и Ceriops. Только вдоль
русел рек этот вид проникает
до внутреннего края мангро-
вы с солоноватой водой,где
появляются уже отдельные
экземпляры папоротника Ac-
rostichum аигеит.
1 Н. W а 11 е г, М. Stei-
ner, Die Okologie der Ostafrika-
nischen Mangroven, «Zschr. Bot.»,
30, 1936, S. 65—193.
Другие типы растительности влажных тропиков
167
Рис. 115. Поясность прибрежной мангровы в Восточной Африке (по Вальтеру
и Штейнеру).
На коралловых рифах мангры растут в виде одиночных экземпляров
или группами (рис. 119, 120). Конкуренция между отдельными расте-
ниями здесь отсутствует, и поясность поэтому не выражена.
Климат в районе Танги (5° ю. ш.) относительно сухой для данной
широты (см. рис. 121). На климадиаграмме очень четко выделяются два
сезона дождей. Средняя месячная влажность воздуха в 14 час. колеблется
от 68 до 76%. Испарение в сухой период за сутки равно 9,2—12 мл; испа-
рение за 1 час в полуденное время превышает 1 мл, максимальное достига-
ет 1,25 мл!час. Такая величина испарения характерна, между прочим,
для некоторых районов Центральной Европы в жаркие летние дни. Хотя
испарение собственно мангровы в два раза меньше, все же следует счи-
таться с тем, что во время отливов происходит некоторое увеличение
концентрации почвенного раствора. В действительности оказалось, что
содержание солей в почве повышается при удалении от моря, причем
в зарослях Avicennia осмотическое давление почвенного раствора дости-
гает 40 атм; вместе с этим увеличивается и осмотическое давление
в листьях растений. Разность осмотического давления почвенного раство-
ра и осмотического давления в листьях составляет 8—9 атм, и лишь
у Avicennia равно около 20 атм (рис. 122). Установлено, что содержание
хлоридов в клеточном соке растений в основном равно концентрации поч-
венного раствора и различие состоит лишь в других осмотически активных
веществах (сахарах, органических кислотах и т. д.; см. рис. 123). Важно
отметить, что эта разность в 8—9 атм приблизительно соответствует вели-
чине осмотического давления у деревьев дождевого леса (см. стр. 110 —
111). Следовательно, мангры, несмотря на высокую концентрацию солей
в морской воде, десугируют воду не хуже, чем деревья, растущие на неза-
соленных тропических почвах; Avicennia имеет в этом отношении, пожалуй,
даже некоторые преимущества. Этот вид мангров наиболее далеко прони-
кает за пределы тропиков, вплоть до прибрежной пустыни залива
Акаба.
Водный баланс мангров хорошо уравновешен. Листья у них типично
суккулентные, в них хорошо развита водоносная ткань (рис. 124). Транс-
пирируют мангры очень слабо. При интенсивности испарения 0,53 —
0,62 мл!час транспирация различных видов составила 1,8—2,6 мг на
1 г сырого веса в 1 мин, или 6,1—8,8 мг!дм2 > мин; только Avicennia
и Loranthus (паразитирующий на Lumnitzera') расходуют воды во время
транспирации в два раза больше. Таким образом, мангры по сравнению
с древесными видами тропического девственного леса транспирируют гораз-
до слабее. Потери воды у них на единицу сырого веса меньше, чем у Sedum
maximum; лишь Avicennia по режиму испарения близка к последнему.
Даже при столь невысокой интенсивности транспирации в листьях мангров
началось бы быстрое отложение солей, если бы корни постоянно поглоща-
ли морскую воду. В жизнедеятельности мангров зна-
168
Глава 3
Рис. 116. Пояс Sonneratia\ в отличие от рис. 113
изображен во время прилива (по Вальтеру и
Штейнеру).
Рис. 117. Ходульные корни молодого экземп-
ляра Rhizophora mucronata; на заднем плане —
Ceriopscandolleana, окрестности Танги (по Валь-
теру и Штейнеру).
чительно большее
значение имеет не
водный режим, а со-
левой баланс.
Различают соленакопля-
ющие мангры и солевыделя-
ющие мангры, принадлежа-
щие к родам Avicennia, Aegi-
ceras и Aegialitis. В условиях
сухого климата Восточной
Африки нижняя сторона ли-
стьев солевыделяющей Avi-
cennia часто бывает покрыта
кристаллами NaCl (рис. 125
и 126). Кристаллы соли, об-
ладая высокой гигроскопич-
ностью, за ночь поглощают
влагу из воздуха и раство-
ряются. Но это отнюдь не
означает, что у растений
имеет место листовая адсорб-
ция атмосферной воды. Во
влажном климате поваренная
соль не выкристаллизовыва-
ется, она просто смывается
дождем.
Ш оландер и его сотруд-
ники, работающие в северном
Квинсленде, исследовали
процесс образования и уда-
ления соли из листьев манг-
ров и связали его с деятель-
ностью особых железок Ч Этот
процесс характеризуется от-
четливым суточным ходом,
особенно у Aegialitis, а также
у Aegiceras; максимальная ин-
тенсивность процесса наблю-
дается в полдень. При затене-
нии листьев выделение соли
прекращается. Концентрация
раствора, в виде отдельных
капелек выступающего из
железок, у Aegialitis состав-
ляет 1,8—4,9%, у Aegiceras
0,9—2,9% и у Avicennia —
1Р. F. Scholander,
Н.Т. Hammel, W. G а г е у,
The salt balance in mangroves,
«Plant physiol.», 37, 1962, p. 722—
729; Sap pressure in vascular
plants, «Science», 148, 1965, p.
339—346.
Другие типы растительности влажных тропиков 169
Рис. 118. Заросли Avicennia marina, образующие внутренний пояс
мангровы, и полоса, лишенная растительности; хорошо видны дыхательные
корни, похожие на молодые стебли спаржи; Нианиани у Танги, Восточная
Африка (по Вальтеру и Штейнеру).
I
Рис. 119. Рифовые мангровы на абразионной террасе кораллового
острова Уленге у побережья Восточной Африки. У Avicennia на обна-
женных коралловых скалах образуется большое число воздушных корней,
что характерно и для других видов мангровы (по Вальтеру и Штейнеру).
4,1% , тем самым в два раза превышая в некоторых случаях соленость мор-
ской воды. Выделившиеся соли состоят на 90% из NaCl и на 4% из КС1,
что приблизительно соответствует химическому составу морской воды.
Количество выделившейся за день (9 часов) соли достигает 0,2—3,5 мг
на 10 см2 листовой поверхности, что значительно меньше количества транс-
пирируемой влаги. Это говорит о способности корней всасывать раствор
с небольшим содержанием солей. Действительно, данные исследований
показали, что вода в сосудах растения представляет собой 0,2—0,5%
170
Глава 3
Рис. 120. Rhizophora mucronata на
обнаженных скалах кораллового ост-
рова Уленге (фото Э. Вальтер).
нескольких атмосфер. Поэтому
раствор хлорида, осмотическое давле-
ние которого повышается еще на1— 2
атм только присутствием других солей.
У соленакопляющих видов мангро-
вы концентрация соли в сосудах прибли-
зительно в 10 раз меньше. Регулиро-
вание концентрации соли в растении
осуществляется либо путем сбрасывания
старых листьев (вместе с ними происхо-
дит и элиминация некоторого количества
соли), либо путем частичного переноса
солей в молодые растущие листья. Дело
в том, что концентрация солей в молодых
листьях в период их развития увеличи-
вается (в сравнении с низкой транс-
пирацией) относительно быстро. Незна-
чительная концентрация солей в тра-
хеях и трахеидах не связана с высо-
ким сосущим действием листьев и непро-
ницаемостью клеток корней для хлори-
дов. В таком случае корневое давление
в них должно было бы быть равным
20 атм. Однако Шоландер и его сотруд-
ники обнаружили лишь незначительное
отрицательное давление, не превышавшее
можно предположить, что корни играют
роль своего рода опреснителей: будучи окруженными соленой морской
водой, они тем не менее направляют в сосуды растения почти чистую
воду. Необходимую для этого энергию растение получает в процессе
дыхания, осуществляемого в почве, лишенной О2, дыхательными кор-
нями.
У слабо транспирирующих проростков живородящих видов мангровы
осмотическое давление очень низкое, оно не превышает 13—18 атм, не
высоко и содержание хлоридов 4. Объясняется это низкой концентрацией
солей в трахеях и трахеидах, не испытывающих сильного корневого
давления. Поскольку непосредственная связь между сосудами материн-
ского растения и сосудами проростка отсутствует, в переносе питательных
веществ, по-видимому, должны участвовать клетки, выполняющие функ-
цию железок и расположенные в семядоле и в окружающем ее слое клеток.
Этим вопросом занимался Кипп-Голлер 1 2 и в последнее время Панье 3,
проводивший более детальные исследования на живом материале с исполь-
зованием Rhizophora mangle. По его данным, осмотическое давление
в околоплоднике достигало 30 атм, а в семядоле проростка — 24,4 атм\
концентрация же клеточного сока в тканях, окружающих проросток,
равнялась 16,4 атм. Доля хлоридов при относительно высоком содержа-
нии сахаров также оказалась очень низкой. Гистохимический анализ
этих тканей и семядоли свидетельствует о более высоком содержании
1 Н. Walter, М. Steiner, op. cit.
2 A. Kipp-Goller, Lber Ban und Entwicklung der viviparen Mangroven-
keimlinge, «Zschr. Bot.», 35, 1939/40, S. 1—40.
3 F. Pannier, Estudio fisiologico sobre la viviparia de Rhizophora mangle L.f
«Acta Cient. Venezolana», № 6, 13, 1962, p. 184—197.
Другие типы растительности влажных тропиков
171
в них фосфатазы, меченого фосфора (Р32) и о большей интенсивности про-
цесса дыхания и т. д. Все это указывает на активную деятельность желе-
зок, играющих важную роль в снабжении живородившегося проростка
питательными веществами.
У крупного проростка неживородящей Avicennia осмотическое давле-
ние достигает таких же величин, как у материнского растения, однако
хлориды в нем практически отсутствуют.
Вскоре после укоренения величина осмотического давления и содер-
жание хлоридов в проростках уже ничем не отличаются от материнских
растений. Рисунки 127—129 знакомят читателя с проростками некоторых
мангров. Дыхание проростков мангров подробно изучал Чэпмен, не
установивший, правда, какой-либо специфики 4.
В зарослях мангровы засушливого побережья Восточной Африки
внимание привлекает лишенная растительности песчаная полоса, окай-
мляющая внутренний край мангровы. Эта полоса дважды в год на несколь-
ко дней затапливается во время равноденственных сизигийных приливов.
После отлива почва высыхает и вместе с этим увеличивается концентрация
солей в ней 1 2. В остальное время года осадки опресняют песчаный грунт.
При таких условиях, по-видимому, здесь не могут произрастать ни галофи-
ты, ни вообще какие-либо растения (рис. 115, 118 и 130).
Таким образом, выявленные закономерности в поясности восточно-
африканской мангровы, несомненно, свйзаны с солевым режимом почвы,
определяющим в свою очередь конкурентную способность отдельных
видов. В других климатических условиях при ином флористическом соста-
ве может возникнуть другой тип поясности, что будет определяться уже
иными факторами.
Так, например, южнее, на побережье Мозамбика, Sonneratia уже
более не встречается, a Avicennia образует здесь не только внутренний,
но — на песчаных почвах — и внешний край мангровых зарослей. Юж-
ная граница распространения Bruguiera, Rhizophora и Avicennia проходит
около 33° ю. ш.
Каждая растительная зона характеризуется своей, только ей при-
сущей фауной. Однако Макнэ и Калк предполагают, что зависимость
здесь лишь косвенная и что решающее значение и для животных, и для
растений имеют глубина залегания грунтовых вод, длительность затопле-
ния и степень засоленности субстрата 3 4.
Из всех видов, свойственных мангровам, Avicennia дальше всех
проникает в аридные районы и отличается наибольшей холодоустойчиво-
стью. Avicennia marina var. resintfera —единственный представитель
мангров, еще встречающийся на острове Северный Новой Зеландии, где
на широте Окленда он уже подвержен влиянию заморозков. По данным
Чэпмена и Рональдсона 4, проводивших исследования в окрестностях
1 V. J. Chapman, Respiration studies of mangrove seedings, I, II, «Bull.
Marine Sc. Gulf. Carribb.», 12, 1962, p. 137—167, p. 245—263.
2 Харрис, исследовавший на Ямайке концентрацию солей морской воды, сохра-
нившейся в углублении песчаной полосы, установил, что осмотическое давление рас-
твора было равно 69 атм.
3 W. М а с n а е, М. К а 1 k, The ecology of the mangrove swamps at Ithaca Island,
Mozambique, «J. Ecol.», 50, 1962, p. 19—34; W. M a c n a e, Mangrove swamps in South
Africa, «J. Ecol.», 51, 1963, p. 1—25.
4V. J. Chapman, J. W. Ronaldson, The mangrove and salt-marsh flats
of the Auckland isthmus, «New Zeal. D.S.I.R. Bull.», 125, 1958.
Рис. 121. Климадиаграмма
Амани (горы Усамбара, в 50
Танги (побережье Восточной Африки) и
км от моря) («Klimadiagramm-Weltatlas»,
1960).
Рис. 122. Наибольшие и наименьшие величины
осмотического давления у разных видов манг-
ровы и осмотическое давление почвенного рас-
твора (Н. Walter, Standortslehre, 2. Aufl.,
1960).
□ а ffltf
Рис. 123. Величина осмотического давления (а)
и содержание хлоридов (б) и других осмотически
активных веществ (в) у мангров Восточной
Африки (Н. Walter, Standortslehre, 2. Aufl.,
1960).
1 — Sonneratia alba', 2 — Rhizophora mucronata’, 3 — Ce-
riops candolleana', 4 — Avicennia marina’, 5 — Bruguiera
gymnorrhiza’, 6 — Lumnitzera racemosa\ 7 — Xylocarpus-
obovatus.
Другие типы растительности влажных тропиков
173
Р п с. 124. Поперечные разрезы листьев различных
мангров (по Вальтеру и Штейнеру).
а - Sonneratia alba\ б — Rhizophora mucronata', в — Ceriops
eandolleana', г — Bruguiera gymnorrhiza; d — Avicennia marina',
e - - Lumnitzera racemosa\ ж — Loranthus sp.; з — Suaeda
monoica.
Окленда, у южной границы распространения мангров, Avicennia уже не
может расти при температуре —2° С, повторяющейся раз в 5—10 лет.
Avicennia образует только внешний пояс на илистых маршах и по направ-
174
Глава 3
лению в глубь страны сменяется
видами, свойственными засоленным
маршам: Salicornia australis, Juncus
maritimus, Samolus, Triglochin, Aster
и др. Несомненно, что Avicennia, луч-
ше всего развивающаяся на песчаном
грунте, очень плохо переносит сла-
бо аэрируемые илистые почвы. Если
вдоль стоковых борозд на ваттах 1
этот вид еще достигает высоты 2 —3 ле,
Рис. 125. Железка, выделяющая соль, то на ваттах его высота уменьшается
на листе у A vicennia marina (по Фаберу). д0 30 см. Интересно отметить, что
проростки Avicennia, длительное вре-
мя затапливаемые приливами, обладают способностью к интрамоле-
кулярному [анаэробному—Перев.] дыханию.
Познакомимся теперь с результатами исследований в некоторых
других районах распространения мангровы. Лёчерт приводит в своей
статье1 2, посвященной растительности Сальвадора, профиль мангровы
в береговой зоне (рис. 131). Детального анализа условий засоленности
почвы он не производил. Тем-
пературные условия и режим
испарения аналогичны во-
сточноафриканским. Величи-
на испарения, по данным
Лёчерта, составляла 0,8
мл!час', в Центральной же
Европе фиксируемые макси-
мальные значения лишь ред-
ко достигали 0,9 —1,25 мл/час.
Лишенный растительности
пояс у внутреннего края здесь
отсутствует, непосредственно
же у кромки воды растет Rhi-
zophora mangle. Это наблюда-
ется в тех случаях, когда в
составе мангровы отсутствует
Sonneratia, или же в мангро-
вых, развитых В устьях рек. Р и с. 126. Листья Avicennia marina, покрытые
По Панье 3, в Венесуэле, в кристаллами соли (фото Фабера).
окрестностях Каракаса, Rhi-
zophora развивается лучше всего в солоноватых водах устьев рек (см.
также рис. 132). В опытных условиях Rhizophora, выращивавшаяся в сосу-
де с пресной (75%) и морской водой (25%), развивалась также хорошо,
как и в природе. Воздушно-сухой вес листьев при такой концентрации был
в 5,6 раза, а корней — в 3,4 раза больше, чем в морской воде (в естествен-
ных условиях). Осмотическое давление в клетках, к сожалению, не опреде-
лялось. Этим данным противоречат результаты, полученные Штерном
1 Ватты — прибрежные участки мелкого моря, затопляемые во время прилива
и обнажающиеся при отливе.— Прим. ред.
2 W. Lotschert, Vegetation und Standortsklima in El Salvador, «Botan.
Studien», H. 10, Jena, 1959.
3 F. P a n n i e r, El efecto de distintas concentraciones salinas sobre el desarrollo
de Riiophora mangle L., «Acta Cient, Venezolana», 10, 1959, p. 68—78.
Другие типы растительности влажных тропиков
175
и Фойгтом, согласно которым
проростки Rhizophora mangle
лучше всего растут в 100%
морской воде1. Поясность и сте-
пень засоленности почв в рай-
оне устья реки Сьерра-Леоне
исследовал Гледхилл 1 2 3 4.
На побережье Камеруна
близ Дуалы Rhizophora mangle
также растет в слабосоленой
воде (общее осмотическое дав-
ление равно 5,1 атм, а давле-
ние, обусловленное хлоридами,
—4,4атм). Соответственно осмо-
тическое давление в листьях,
достигающее 19,8—24,5 атм,
следует считать сравнительно
низким, однако содержание хло-
ридов в клеточном соке (осмо-
тическое давление равно 10,8 —
14,6 атм) оказалось значитель-
но более высоким, чем в поч-
венном растворе. Следователь-
но, Rhizophora, как облигатный
галофит, накапливает соль
в своем клеточном соке. Даже
у экземпляров, выращенных
ОПЫТНЫМ путем В пресной воде, Р и с. 127. Проростки Bruguiera parviflora^
содержание хлоридов составля- висящие на материнском дереве (фото Фа-
ло 41%, а осмотическое дав ле- бера).
ние равнялось 4,7 атм.
Осмотическое давление мангров исследовали также Харрис на Ямай-
ке, Сен-Гупта вблизи Калькутты и Бхаруча в окрестностях Бомбея 3.
Собранные данные подтверждают наши выводы.
Обусловленное муссонными дождями сильное опреснение воды близ
Бомбея вызывает и соответствующее падение осмотического давления.
Необычайно сложны условия в Индо-Малайской области, где мангро-
вы исследованы Троллем и Драгендорфом и подробно описаны Бюннин-
гом 4. И здесь внешний край мангровы образует Sonneratia alba, зарос-
ли которой простираются, однако, дальше в открытую воду. Avicennia
marina, видимо, развивается только на песчаной почве и проникает дале-
1 W. L. Stern, G. К. Voigt, Effect of salt concentration on growth of red
mangrove in cultures, «Bot, Gaz.». 121, 1959, p. 36—39.
2 D. Gledhill, The effect of the Aberdeen Greek mangrove swamp, «J. Ecol.»,
51, 1963, p. 693-703.
3 J. A. Harris, The physico-chemical properties of plant saps in relation to
phytogeography, Minneapolis, 1932; J. Sen-Gupta, Die osmotischen Werte bei
einigen Pflanzen in Bengal (Indien), «Вег. Dtsch. Bot. Ges.», 53, 1939, S. 783—795;
F. R. Bharucha, B. S. Navalkar, Studies in the ecology of mangrove, «Journ.
Univ. Bombey», 9, 1942, p. 93—100.
4 W. Troll, O. Dragendorff, Uber die Luftwurzeln von Sonneratia und
ihre biologische Bedeutung, «Planta», 13, 1931, S. 311—473; E. Biinning, In den
Naldern Nordsumatras, Bonn, 1947.
176
Глава 3
Р и с. 128. Срезанная ветвь живо-
родящей Rhizophora mucronata с про-
ростком, мангровы около Танги
(фото Э. Вальтер).
ко вверх по рекам, что подтверждается
и наблюдениями автора на востоке
Адстралии. Rhizophora представлена не-
сколькими видами и образует сомкнутые
леса, где деревья имеют высоту 20—30 м
(до 35 —40 л) и где почвы достигает лишь
1—5% полного дневного света. Вместе
с Rhizophora растут и другие виды, напри-
мер Sonneratia acida, для развития кото-
рой, по Бюннингу, «необходима высокая
кислотность почвы». Эти соображения
требуют проверки в условиях экспери-
мента, поскольку здесь может наблю-
даться расхождение между экологиче-
ским оптимумом, связанным с вышеопи-
санными местообитаниями, и физиологи-
ческим оптимумом, который может
определяться совершенно иными значе-
ниями pH почвы. Поясность мангровой
растительности, как уже отмечалось, обу-
словлена не столько непосредственно
факторами местообитания, сколько кос-
венно степенью интенсивности конкурен-
ции. Иной, по сравению с Восточной
Африкой, экологический ареал отдель-
ных видов в Индо-Малайской области
обусловлен тем, что эта оптимальная
для развития мангровой растительности
область характеризуется чрезвычайным разнообразием флористического
состава и климатических условий (см. табл. 31 и рис. 134—136), тогда как
в Восточной Африке прибрежныэ мангровы находятся близ крайних
пределов своего развития.
Мангры отличаются своеобразными дыхательными корнями, или
пневматофорами. Еще Румфуис (1660 г.) обратил на них внимание, но
лишь Гебель (1886 г.), изучив анатомию пневматофор, смог назвать их
органами дыхания. Однако правомерность этого вывода не удалось дока-
зать ни Вестермайеру (1900 г.), ни Троллю и Драгендорфу (1931 г.).
Последние считали на основе собственных тщательно выполненных морфо-
логических исследований, что у Sonneratia на подземной части пневмато-
фор близ поверхности почвы образуются тонкие поглощающие корни. По
мере намыва почвы пневматофоры растут вверх, образуя активно действу-
ющие корни на все более высоких уровнях, однако на постоянной высоте
под поверхностью почвы. Значение пневматофор в жизнедеятельности
мангров удалось установить экспериментальным путем лишь Шоландеру
и его сотрудникам х. Оказалось, что корневые чечевички пронизаны мно-
жеством настолько мелких отверстий, что они пропускают только воздух,
но не воду. Во время прилива, когда пневматофоры полностью покрывают-
ся водой, кислород, содержащийся в межклетниках, расходуется на дыха-
ние; создается пониженное давление, так как углекислота, легко раство-
ряющаяся в воде, улетучивается. Как только при отливе корни появляют-
1 Р. F. S ch о 1 a n d er, L. van Dam, S. I. Scholander, Gas exchange
in the roots of mangroves, «Am. J. Bot.», 42, 1955, p. 92—98.
ЮЗ
Рис. 129. Проростки Rhizophora conjugata при отливе; между ними вид-
ны коленчато изогнутые корни Bruguiera gymnorrhiza] мангровы на
южной Яве (фото Фабера).
Рис. 130. Лишенная растительности полоса у внутреннего края
мангровы, затапливаемая лишь дважды ггод, Ндуми под Тангой
(по Вальтеру и Штейнеру).
Культуры Cocos nucifera г-Молодые культуры
и fheobroma Cacao Cocos nucifera
Avicennia и
Layuncularia
Layuncularia iC8
racemosa
\ Нйзш, цр. прилива 1
о. Эспириту -Санто
4м „ Л . г . -
2 м Древни и oepeeooouj
Ом ------------------
Avicennia nitida
I Layuncularia Rhizophora
I I / mangle
Conocarpus
erectra
Rhizophora
manyle
Зйпера^Сш^ан-дель-Госо о. Тортуеа
1км
1900м
Pithecollobium
| dulce \
“ icereus
1280м 260m
oaccns
subylooosa
Эль-Триушро
(суша)
Рис. 131. Схематический профиль мангровы на побережье Сальвадора (по Лёчерту).
178
Глава 3
Рис. 132. Rhizophora mangle в устье Арройо-Соледад на тихоокеанском побережье
Нижней Калифорнии (фото Гейма).
Рис. 133. Внешний пояс мангровы на северном побережье Явы
при отливе. Впереди — Avicennia marina, на заднем плане —
(более высокие деревья) A. officinalis (фото Фабера).
ся над водой, давление выравнивается и корни начинают засасывать
воздух. Таким образом, в пневматофорах происходит периодическое
изменение содержания кислорода, синхронное ритму приливов и отливов.
Другие типы растительности влажных тропиков
179
Рис. 134. Заросли Bruguiera gymnorrhiza на Андаманских островах (фото
Робертсона).
При отливе содержание кислорода почти равно 20%, а во время прилива
падает до 10% и ниже г.
Рис. 135 воспроизводит своеобразный облик дыхательных корней
Xylocarpus и Bruguiera. Примерно такого же типа корни развиваются
у Rhizophora. У Rhizophora функцию дыхательных корней выполняют
выступающие из воды части ходульных корней. В илистой почве на них
образуются пучки пористых корешков, межклетники которых связаны
с межклетниками ходульных корней и через чечевички.последних — с окру-
жающим воздухом.
То, что формирование дыхательных корней непосредственно не
обусловлено кольматажем, подтверждается их нормальным развитием
на обнаженных коралловых скалах, где не происходит аккумуляции
мелкоземистого материала (рис. 119, 120).
Читателя, интересующегося более подробным описанием поясности
мангровы на разных континентах, мы отсылаем к специальным моногра-
фиям. В заключение приведем лишь описание мангровы, развитой в устье
реки, данное Шимпером (рис. 136).
«Похожий на лагуну залив в южной части Явы с трех сторон охвачен
отлогими берегами суши и лишь на юге отделен от Индийского океана
холмистым островом Камбанган. Несколько рек лениво текут к заливу,
1 F. Gessner, op. cit., Bd. II, S. 168—185.
180
Глава 3
едва врезаясь в пойму. Их незначительные уклоны позволяют приливной
волне подниматься далеко вверх по течению. Дробясь на несколько рука-
вов, многие реки формируют обширные дельты, которые во время прилива
затапливаются морем, а в период отлива несколько возвышаются над его
уровнем. Пожалуй, трудно найти лучшие условия для развития мангро-
вой растительности, и действительно, она отличается здесь необычайно
пышным развитием.
Рис. 135. Дыхательные корни мангров. На переднем плане — змеевидно
изогнутые корни Xylocarpus moluccensis, между ними и на заднем плане справа —
коленчато изогнутые пневматофоры Bruguiera (фото Фабера).
Путешественнику, плывущему на каноэ вдоль берега бухты или по
одной из многочисленных речных проток, беспрерывно открываются все
новые пейзажи. Если на открытых крутых побережьях, за малым исклю-
чением, только Rhizophora mucronata способна выдержать удары волн
и размножаться, то на пологих берегах бухты, где прибой слабо выражен,
условия для существования разных видов приблизительно одинаковы,
и в борьбе за пространство побеждает то один, то другой вид. Лодка сколь-
зит то вдоль густых зарослей ризофор, то вдруг попадает в тень серебри-
сто-серой, похожей на иву Avicennia marina var. alba. Тусклая зелень
доминирующей местами Sonneratia acida сменяется уходящей в воду
узкой живой изгородью из Nipa fruticans. Там и сям бросаются в глаза
причудливые очертания Xylocarpus granatum, в маленькой изящной кроне
которого блестят крупные, величиной с голову человека, коричневато-
желтые плоды; на зеленом фоне выступают кусты Aegiceras corniculatum,
увешанные белоснежными цветками и изогнутыми, словно рог, плодами.
Оба вида Bruguiera (Br. gymnorrhiza, Br. parviflora}, редко развивающиеся
во внешнем (по отношению к морю) поясе, часто встречаются во внутрен-
них частях мангровых зарослей, где кроны Br. gymnorrhiza высоко подни-
маются над другими деревьями; на стволах Bruguiera уже издалека видны
Другие типы растительности влажных тропиков
181
большие, вздутые стебли Hydnophytum montanum; менее заметны невысо-
кие с невзрачными цветками экземпляры Br. parviflora.
Отлив открывает взору путешественника причудливую картину лаби-
ринта ходульных корней Rhizophora или «газона» из корней Avicennia
и Sonneratia со снующими рыбками (особенно интересны своеобразные
виды Periophthahnus) и крабами. На других участках илистого дна обна-
жается змеевидно изогнутые корни Xylocarpus granatum.
Рис. 136. Мангровы Явы. На переднем плане— Nipa fruticans,
за ней Avicennia и Sonneratia (фото Фабера).
Приливная волна скрывает на некоторое время весь этот хаос корней,
даже нижние ветви ризофор и оказываются под водой. Через тол-
щу воды просвечивают листья молодых экземпляров Rhizophora mucronata.
Эпифиты в мангровах встречаются относительно редко, во внешнем
по отношению к морю поясе и на Rhizophora mucronata они обычно полно-
стью отсутствуют. Очевидно, их развитию не благоприятствует соленый
субстрат. В глубоких заливах и во внутренних частях обширной мангровы,
куда не проникают соленые брызги, эпифиты появляются чаще: например,
виды Platycerium, Hydnophytum montanum, отдельные экземпляры Poly-
podium и один вид Loranthus. Наряду с ними всегда встречаются лишай-
ники, но мхов, совершенно не переносящих соли, здесь нет (Schimper—
Faber, S. 568-571).
РАСТИТЕЛЬНОСТЬ ПЕСЧАНЫХ БЕРЕГОВ
На всех открытых морских берегах мангровая растительность отсут-
ствует. Так, мангровы не характерны почти для всего западного побере-
жья Африки, за исключением лишь устьев крупных рек и лагун. Сильный
прибой формирует здесь более или менее широкий отлогий песчаный берег,
всхолмленный иногда дюнами. Здесь распространена специфическая,
182
Глава 3
характерная для тропической зоны, растительность с соответствующей
поясностью. Рассмотрим в качестве примера типичные условия индо-
тихоокеанской области.
Полоса песчаного берега, граничащая с морем и перемываемая штор-
мовыми волнами, обычно лишена растительности. Непосредственно за ней
следует первый пояс, представленный типом сообщества Ipomoea pes сар-
гае — Canavalia (рис. 137). Длинные стелющиеся побеги этого красиво
цветущего вида Ipomoea так же хорошо, как и Canavalta из семейства
бобовых, закрепляют песчаный грунт. В этом поясе распространены
и Sesuvium portulacastrum (рис. 138), некоторые злаки, например Sporobo-
lus virginicus, а в восточной части области — своеобразный Spinifex
(рис. 139) и др. Виды, свойственные этому поясу, еще испытывают на себе
воздействие соленой воды. Sesuvium относится к галофитам; по данным
Бюннинга, листья Ipomoea pes саргае усеяны солевыделяющими желез-
ками, что следовало бы, однако, проверить. Ipomoea растет преимуществен-
но на песчаных дюнах уже за пределами полосы засоленных почв, однако
песок здесь содержит некоторое количество NaCl Ч Анализ клеточного
сока в листьях растений, распространенных на песчаной береговой полосе
острова близ Танги, проведенный Вальтером и Штейнером, дал следую-
щие результаты:
Дата Общее осмотическое давление, arn.vi Давление, обусловленное NaCl, атм Содержание хлоридов, %
I pomoea pes caprae
9/XI I 12,7 1 7’6 I 60
10,7 1 2,5 23
Hibiscus tiliaceus
(кустарник высотой 2 лг, растет несколько дальше от берега)
25/XI | 25,5 | 5,2 [ 20
Sesuvium portulacastrum
(типичный галофит, растет ближе к кромке воды
и временами затапливается)
11/X 27,5 21,4 78
25/XI (после сухого периода) 32,3 19,0 59
Колебания этих величин в зависимости от засоленности и сухости
почвы очень велики.
В качестве примера приведем определения осмотического давления
у Sesuvium, по Харрису 2.
Ямайка, морское побережье 43,0 п 52,6 атм
Ямайка, пустыня, в составе
которой преобладают какту-
сы и акации 19,5 и 49,9 атм
1 Н. Walter, М. Steiner, op. cit., S. 190.
1 J. A. Harris, op. cit.
Другие типы растительности влажных тропиков
183
Флорида (15 анализов) . . от 11,6 до 34,2 атм
Содержание хлоридов в
двух анализах .... 41 и 44%
Гавайи (4 анализа) . .16,5 атм (у пруда), 26,1 ,
33,5 и 40,6 атм (коралловый берег) .
Вслед за видами травяного покрова на тропических побережьях
появляются кустарники (рис, 139), Обширные пространства дюн в тро-
пиках не известны, так как формирующиеся обычно за поясом низкорос-
лой растительности редколесья Barringtonia (рис. 140) закрепляют песок
и препятствуют его перевеванию. Здесь встречаются также следующие
виды деревьев: Calophyllum inophyllum, Terminalia catappa, Hibiscus
tiliaceus, Tournefortia argentea, Pandanus tectorius (рис, 141), Casuarina
equisetifolia. Древостои, образованные последним видом, часто занимают
обширные площади. Плоды этих растений хорошо плавают в воде (рис, 142),
и поэтому они встречаются на всех морских побережьях тропиков. Дре-
весные растения часто перевиты паразитирующей Cassytha (рис. 143).
На берегах Атлантического океана Ipomoea pes саргае распространена
наряду с другими, не свойственными предыдущей области видами травяно-
го покрова и древесными породами, образующими сомкнутые или разре-
женные древостои высотой до 20 м или более. На берегах Вест-Индии
широко распространен кустарник Coccolpba unifera, а также Hippomane
Рис. 137. Ipomoea pes саргае на отлогом песчаном морском берегу на южном
побережье Явы. На переднем плане — прибитые прибоем плоды и семена расте-
ний, развитых на морских побережьях (фото Фабера).
Рис. 138. Sesuvium portulacastrum — широко распространенное в тропиках растение,
развитое на песчаных морских берегах. Коралловый остров в Батавском заливе (фото
Фабера).
Рис. 139. Заросли кустарников на берегу кораллового острова в Батавском заливе.
Здесь распространены: Pemphis acidula, Desmodium umbellatum, Clerodendron inerme,
Premna integrifolia^Colubrina asiatica, Scaevola frutescens, Caesalpinia crista, Guettardia
speciosa и др. На переднем плане — Spinifex litoreus (фото Фабера).
Другие типы растительности влажных тропиков
185^
Рис. 140. Заросли Barringtonia speciosa на южном побережье западной Явы (фото-
Блея).
mancinella, Chrysobalanus icaco и др. Лишь немногие виды, например
Terminalia catappa (Combretaceae) и Thespesia populnea (Malvaceae),
являются пантропическими.
Большинство видов этого удаленного от моря пояса, по-видимому
относится к факультативным галофитам, поскольку засоленность почвы
здесь уже сравнительно невелика. Гудинг (цитируется по Ричардсу}
приводит следующие данные (табл. 32).
, Т а б л и ц а ч 32
Концентрация
солей в почвен-
ной воде, %
Средняя
обводненность
песка, %
Полоса пионерной растительности (50 м от берега
моря)............................................
Полоса Ipomoea pes caprae (100 м от берега) . . .
Полоса Coccoloba (150 м от берега)...............
5,3
1,8
0,4
0,26
0,48
1,65
Величина осмотического давления клеточного сока, обусловленная
хлоридами, у большинства растений еще весьма значительна. У кокосовой
пальмы давление клеточного сока листьев, обусловленное хлоридами,,
достигает 40% при общем осмотическом давлении, равном 15 атм (Валь-
тер и Штейнер). Кокосовая пальма может развиваться и на незасоленных
местообитаниях в 50—100 км от берега моря, однако в этих условиях она
186
Глава 3
Рис. 141. Заросли Pandanus tectorius
на берегу небольшого кораллового ост-
рова в Батавском заливе (фото Фабера).
Рис. 142. Плоды прибрежных растений,
приспособленные к плаванию в воде.
Слева вверху — Cerbera manghas, вверху
справа — Terminalia catappa (внизу по-
перечный разрез) и внизу слева — Ca-
lophyllum inophyllum (по Шимперу).
Рис. 143. Cassytha filiformis (Lau-
гасеае)’— широко распространенный па-
разит, опутывающий густой сетью своих
красно-желтых нитей ветви кустарни-
ков и трав (фото Фабера).
не встречается большими насаж-
дениями, поскольку отличается
низким урожаем. Мы видели
обильно плодоносящую пальму
даже на одном из островов Нила
близ Асуана. Правда, в условиях
этого резко выраженного пустын-
ного климата она требовала
еженедельного полива.
Другие типы растительности влажных тропиков 187
За пределами береговой полосы распространены леса, характерные
для местных климатических условий; однако вблизи моря они обычно
вырублены или нарушены человеком.
Уничтожение растительности на тропических побережьях может быть
обусловлено и естественными причинами, например периодическими
ураганами, или тайфунами. Правда, об особенно большом ущербе гово-
рить не приходится, так как лес быстро восстанавливается» Значительно
тяжелее урон, который наносят вызванные ураганами штормовые
волны, не только уничтожающие растительность, но и смывающие
почву Ч
В Бразилии на мысе Кабу-Фриу (восточнее Рио-де-Жанейро) на
песчаных участках береговой полосы, так называемых рестингах, с их
характерным мозаичным распределением растительности, в настоящее
время проводятся очень детальные микроклиматические и экологические
исследования 1 2.
РАСТИТЕЛЬНОСТЬ СУХИХ МЕСТООБИ ТАНИЙ
Во влажных тропиках сухие местообитания — редкое явление.
Быстро высыхает после каждого дождя лишь обнаженная поверхность
скал. Несмотря на это, лес сплошь покрывает каменистые склоны, на
которых даже скалистые утесы едва заметны. Конечно, на таких место-
обитаниях могут встречаться виды, обычно отсутствующие в лесу. В пер-
вичном лесу в горах Усамбара (Восточная Африка) на скалах часто нахо-
дили ксероморфный саговник — Encephalartos.
Поверхность, смачиваемая только во время дождя, пригодна для
поселения эпифитов так же, как и стволы деревьев и ветви. Поэтому
непосредственно на обнаженных скалах могут поселяться многие эпифиты,
лишь бы здесь было достаточно света и в первую очередь виды, предпо-
читающие кислый субстрат, в лесах полуострова Малакка, например,
Nepenthes и Rhododendron. При этом они предпочитают лесные прогалины
и вершины гор, где условия освещения более благоприятны. Так, на
скалистом гребне самой высокой возвышенности в резервации Бинна-
Бурра (Квинсленд) автор однажды обнаружил большую куртину Asple-
nium nidus.
Особенно сухими являются скалы, сложенные пористыми известня-
ками. Поэтому на таких местообитаниях поселяются растения, отличаю-
щиеся от окружающего растительного покрова. Это показано в работе
Лёчерта, посвященной изучению карстовых явлений на останцах «моготе»
западной Кубы 3. Даже эти особенно сухие местообитания в тропиках
1 J. D. Sauer, Effect of recent tropical cyclones on the coastal vegetation of
Mauritius, «J. Ecol.», 50, 1962, p. 275—290; D. I. В 1 u m e n s t о c k, F. R. Pos-
her g, C. G. J о h n s о n, The re-survey of typhoon effects on Jaluit Atoll in the Marshall
Islands, «Nature», 189, 1961, p. 618—620.
2 L. D a u, Microclimas das Restingas do Sudeste do Brasil, «Arquiv. Mus. Nat.»,
50, 1960, p. 79—134; W. P. Ormond, Ecologia das Restingas do Sudeste do Brasil,
«Arquiv, Mus. Nat.», 50, 1960, p 185—236; Ср. также P. Dansereau, Zonation et
succession sur la restinga de Rio de Janeiro, «Rev. Canad. de Biol. (Montreal)», 6, 1947,
p. 448-477.
3 W. L 6 t sc h er t, Die Ubereinstimmung von geologischer Unterlage und
“Vegetation in der Sierra de los Organos (Westcuba), «Вег. Dsch. Bot. Ges.», 71, 1958,
S, 55—70.
188
Глава 3
Р и с. 144. Поясность растительности на останцах,
сложенных известняками («моготе»), в западной
части Кубы (по Лёчерту).
покрыты непроходимыми за-
рослями первичной расти-
тельности, поясность кото-
рой схематически показана
на рис. 144. Округлую вер-
шину занимает своеобразный,
похожий на пальму вид се-
мейства Rutaceae {Spathelia
brittonii). Несколько нижег
на недоступных участках ра-
стет ВотЪах emarginatum, на
перегибах склона — Gaussia
princeps'SL Thrinax microcarpa,
а на гладких стенах скал —
Agave tubulata, Anthurium
venosum и Philodendron lace-
rum. Philodendron scandens
образует длинные до 10 м по-
беги, свисающие над сталак-
титами. Здесь встречается
также эндемичный саговник
Mier осу cas calocoma.
С сухостью местообитаний связаны бутылкообразная форма стволов
деревьев, в древесине которых накапливаются большие запасы воды
{ВотЪах и эндемичный вид пальмы Gaussia), обилие суккулентов {Agave,
Рис. 145. Vaccinlum vartnglifoltum, растение-пионер в кратере вулкана Пап ан-
даян, западная Ява (фото Хойманса).
Другие типы растительности влажных тропиков
189
Leptocereus, Selinicereus, Rhipsalis) и экстенсивный характер корневой систе-
темы большинства видов. Реакция субстрата щелочная (pH = 7,7 и более).
Совершенно иная растительность развивается на Кубе на кислых
песчаных почвах (pH = 5,0—5,5), где распространены дубовые и сосновые
леса с сопутствующими видами тропического происхождения. Зависимость
растительности от подстилающих пород редко где выражена столь четко,
как это наблюдается на западе Кубы.
Сукцессии, начинающиеся на обнаженных скалах, в тропиках можно
наблюдать на вулканических породах после извержения вулкана (рис. 145).
Протекают они сравнительно быстро и уже через 100—150 лет иногда
достигают стадии леса. После извержения в 1883 г. лишенный раститель-
ности остров Кракатау, несмотря на свое изолированное положение, через
50 лет был покрыт лесом, очень сильно напоминающим обычный вторич-
ный лес.
ТРОПИЧЕСКИЙ КУЛЬТУРНЫЙ ЛАНДШАФТ
Область вечнозеленого тропического дождевого леса претерпела боль-
шие изменения под влиянием человека. В густонаселенных районах тро-
пиков, например на Яве, не сохранилось ни одного не потревоженного
человеком участка леса. Исключение составляют лишь некоторые
труднодоступные возвышенности.
Поэтому Ричардс в конце своей книги «Тропический дождевой лес»
призывает последовать примеру Национального парка Альберт в Африке
и создать в разных уголках континентов, особенно на низменностях,
обширные природные заповедники девственного леса. Лишь такая мера
может служить гарантией сохранения лесов, необходимых для научных
исследований и изучения вопросов большой практической важности.
Это особенно необходимо в связи с тем, что тропический дождевой лес
даже после однократной вырубки вследствие бедности почвы больше уже
не восстанавливается. На его месте развивается вторичный лес, нередко
даже более густой и непроходимый, чем первичный.
Вследствие нерациональной выборочной рубки, при которой выбира-
ются только самые ценные породы, уже в настоящее время большие мас-
сивы девственного леса сильно изменены и обесценены. Еще более непо-
правимый ущерб наносит практикуемой местным населением подсечно-
огневое земледелие. Для того чтобы иметь возможность возделывать
полевые культуры, аборигены вырубают определенный участок леса, за
исключением лишь самых крупных трудно поддающихся валке деревьев.
Срубленные стволы и ветви сжигают, почву с золой перекапывают и затем
возделывают маис, суходольный рис, ямс и маниоку. Через несколько лет,
поскольку почва очень бедна питательными веществами и совсем не удо-
бряется, урожай резко снижается. Тогда вырубают новый участок леса,
а старую пашню забрасывают, и она зарастает малоценными породами.
По истечении длительного времени на месте заброшенной пашни разви-
вается производный лес, отличающийся отсутствием равновесия между
сообществом и местообитанием (рис. 63).
Эти леса более разрежены, и поэтому в них разрастаются много-
численные лианы.
Древесные породы вторичного леса обладают быстрым ростом и за
три года могут достичь высоты 12 м. Это прежде всего связано с благо-
приятными условиями освещения. Однако они, как правило, недолговеч-
ны, и поэтому вторичный лес всегда отличается меньшей высотой, чем
190
Глава 3
первичный. Виды собственно первичного леса появляются только на
более поздних стадиях сукцессии. Их всходы нуждаются в известном зате-
нении, иначе они заглушаются быстрорастущими породами.
Во многих областях площади, покрытые первичным лесом, уже были
однажды заняты под культурами; теперь для расширения возделываемых
площадей человек вынужден вырубать и вторичный лес. Чем выше плот-
ность населения, тем чаще приходится распахивать одну и ту же площадь,
тем меньше времени остается для восстановления леса и тем заметнее
деградация почвы. В конце концов на этих участках развиваются заросли,
в составе которых часто доминируют Ptertdium или Gleichenia. Следова-
тельно, подобный метод ведения хозяйства представляет собой самый
хищнический способ обработки земли. Он таит в себе постоянно расту-
щую угрозу, поскольку плотность населения повсеместно быстро увеличи-
вается. Немало способствовали сокращению площади первичных лесов
и современные плантации. Благодаря высокопроизводительным машинам
намного увеличились раскорчеванные площади, в связи с чем огромные
массивы леса обречены на безвозвратную гибель Ч
В последние годы рациональное хозяйство в первичных лесах ведется
уже в большинстве тропических стран (тиковые, эвкалиптовые леса и др.)1 2.
Здесь обычно закладывают плантации каучука, масличной пальмы, хин-
ного дерева, какао, кофе, чая, сахарного тростника и бананов. Место
вырубленных прибрежных зарослей всюду занято огромными рощами
кокосовых пальм, перемежаемыми заселенными участками.
При повторных палах на залежах могут возникнуть открытые зла-
ковые сообщества, в которых сохраняются лишь устойчивые к пожарам
виды деревьев, например в Индо-Малайской области Тectona grandis,
Pinus merkusit или Melaleuca leucadendron.
Co временем в тропиках формируется культурный ландшафт, так же
сильно отличающийся от девственных лесов, как современные ландшафты
в Центральной Европе отличаются от первобытных природных форма-
ций. На чужестранца этот ландшафт производит впечатление более девст-
венного образования, чем первичный лес. Угодья аборигенов состоят из
мозаично расположенных небольших участков леса, групп деревьев
и более обширных, нередко поросших травой открытых пространств,
между которыми разбросаны возделываемые земли. Ландшафт напоминает
саванну; однако это впечатление обманчиво, поскольку речь идет о все
том же культурном ландшафте или производной влажной саванне, в соста-
ве которой доминируют виды, чуждые девственному лесу. Повторными
вырубками человек препятствует развитию здесь древостоя. Травяной
покров девственного леса, приспособленный к развитию в условиях
значительного затенения и поэтому обладающий небольшой интенсивно-
стью роста, не может использовать обилие света на выкорчеванных пло-
щадях. Эти виды не выдерживают конкуренции со светолюбивыми зла-
ками, развивающимися в тропиках в период летних дождей; последние
максимально используют короткий вегетационный период и отличаются
очень быстрым ростом в благоприятных условиях освещения. Если же
корчевание сопровождается регулярными палами, то в особенно благо-
1 II. Н е s s m с г, Der kombinierte landw. und forstwirtschaftliche Anbau. I. Tro-
pisches Afrika, Stuttgart, 1966.
2 H. Lamprecht, Tropenwalder und tropische Waldwirtschaft, «Beitr. Zsclir.
Schweizer Forstverein», Zurich, 1961; Europaischer Waldbau und Waldbau in den Tro-
pen, «DerForst-und Holzwirt», 19, № 20, 1964; Cber Waldbau in tropischen Entwicklungs-
landern, «Schweizer Zschr. f. Forstwesen», № 4, 1964, S. 211—227.
Другие типы растительности влажных тропиков
19Г
приятном положении оказываются прежде всего виды, почки возобновле-
ния которых располагаются на уровне поверхности почвы и не поврежда-
ются пожаром (ср. стр. 277). Устойчивы против огня древесные породы
с толстой корой, а также пальмы, не имеющие чувствительного к высоким
температурам камбия. Поэтому пальмы в таких производных сообществах
встречаются значительно чаще. К тому же они очень декоративны, и поэто-
му их обычно высаживают на плантациях и в городах. При отсутствии со-
обитателей-конкурентов пальмы хорошо растут на самых различных
местообитаниях. Поскольку путешественники обычно видят только эти
культурные ландшафты, широко распространилось мнение, что пальмы —
это наиболее характерное растение тропиков.
В заселенных районах тропиков, как и в умеренной зоне, повсеместно
распространены быстрорастущие сорные травы и кустарники. Наиболее
часто встречающиеся на островах Малайского архипелага виды Eupatorium
pallescens, Ageratum conyzoides и Lantana camara занесены сюда из тропи-
ческой Америки. На островах Фиджи коренные леса сменились обшир-
ными пространствами, покрытыми Psidium guajava, Lantana, Metastoma
malabaricum и др.
Изучение вторичных лесов и различных производных от них сооб-
ществ с целью сохранения в будущем производительности тропических
почв является и во влажных тропиках важной задачей, ибо дело идет
о борьбе с хищнической обработкой земли и деградацией почв. Это имеет
огромное значение для всего человечества, так как именно области влаж-
ных тропиков являются тем еще не использованным резервом, который
сможет прокормить быстрорастущее население земного шара. Используя
рациональные приемы ведения хозяйства и достаточно удобряя почву,,
в благоприятном климате тропиков можно получать самые высокие
в мире урожаи в расчете на единицу возделываемой площади. Непремен-
ным условием, однако, должно быть разумное соотношение между исполь-
зованием земли и ее охраной.
К сожалению, вплоть до настоящего времени во всех облесенных
и слабо заселенных местностях самой распространенной формой земле-
пользования остается уже упомянутое выше подсечное земледелие
При достаточно большой общей территории, где применяется под-
сечное земледелие, и небольшой площади возделываемой земли может
возникнуть состояние известного длительного равновесия. Переход на
новые земли вызывается следующими тремя факторами: 1) истощением
почвы, 2) зарастанием сорняками и 3) появлением вредителей 1 2. Истоще-
ние очень кислых, бедных питательными веществами почв, например
белых песчаных почв в Британской Гвиане, является главной причиной,
обусловливающей необходимость обработки новых участков; на молодых
почвах, развитых на вулканических породах, это явление никогда не
наблюдается. В процессе зарастания возделываемых участков сорняками
большую роль играют злаки. Их роль в этом процессе особенно важна
в областях, где развиты редкостойные леса с хорошо выраженным травя-
ным ярусом. На малоплодородных почвах труд, затрачиваемый на про-
палывание, не окупается полученным урожаем. Выгоднее раскорчевать
1 Местные обозначения этой системы в различных областях мира: taungya (Бир-
ма), chena (Цейлон), ladang (Малайский архипелаг), kaingin (Филиппины), milpa
(Центральная Америка) chitimene (Северная Родезия).
2Н. Popenoe, The pre-industrial cultivator in the tropics, IUCN Publ.
new ser., № 4, 1964, p. 66—73; J. P h i 1 1 i p s, Shifting cultivation, ibidem, p. 210—220.
В этой статье помещен расширенный список литературы.
192 Глава 3
новую площадь под культуры, чем бороться с сорняками на старой.
В Юго-Восточной Азии особенно злостным сорняком является Imperata
cylindrica. Со временем на небольших возделываемых участках появляют-
ся различные вредители, например птицы, грызуны и насекомые, уничто-
жающие большую часть урожая. Поэтому целесообразно непрерывно
забрасывать одни и обрабатывать другие участки. Если же при увеличе-
нии плотности населения системой подсечного земледелия перестают
пользоваться, необходим немедленный переход на современные методы
обработки почвы, что должно сопровождаться уничтожением сорняков,
удобрением почвы, борьбой с вредителями и т. д. Следует устранить опас-
ность почвенной эрозии.
Глава 4
ДОЖДЕВЫЕ ЛЕСА
ВБЛИЗИ ВЕРХНЕЙ ГРАНИЦЫ РАСПРОСТРАНЕНИЯ
И ВЕРТИКАЛЬНАЯ ПОЯСНОСТЬ В ГОРАХ
ТРОПИЧЕСКОЙ ЗОНЫ
ЗАКОНОМЕРНОСТ И ВЕРТИКАЛЬНОЙ ПОЯСНОСТИ
И ЗОНАЛЬНОГО РАСПРЕДЕЛЕНИЯ РАСТИТЕЛЬНОСТИ
В ТРОПИКАХ
Тропические дождевые леса лучше всего развиваются на нижних
частях склонов гор. Климат здесь теплый, характеризуется ровным
ходом температуры и достаточно влажный. Однако дренаж почв
тем не менее хороший. На равнине же влажность воздуха в период между
дождями может сильно снижаться, а обилие осадков и замедленный поверх-
ностный сток обусловливают развитие почв застойного увлажнения.
С уменьшением высоты меняется и внешний облик растительности.
Дождевой лес нижних частей склонов уступает место дождевому
лесу низменностей, характеризующемуся меньшим количеством
лиан и эпифитов, а также присутствием пальм, хорошо переносящих
неравномерное увлажнение почвы и периодическое затопление; в еще
более влажных условиях местообитания появляются виды Pandanus.
В настоящее время дождевые леса на низменностях почти повсеместно
сведены и замещены плантациями сахарного тростника, риса или других
полезных растений.
Выше предгорной полосы количество осадков резко возрастает,
и одновременно понижается средняя годовая температура. Наибольшая
влажность и наименьшие колебания суточной температуры наблюдаются
в поясе облаков, где развиты горные тропические дожде-
вые леса1. Физиономически они мало отличаются от типичного дожде-
вого леса, однако в их составе появляется и ряд новых видов деревьев.
Кроме того, увеличивается число древовидных папоротников, распростра-
ненных в предгорьях, преимущественно в глубоких ущельях. Необычайно
обильны лианы и эпифиты; среди последних с подъемом в горы начинают
преобладать папоротники и плауны. Особенно широко распространены
Hymenophyllaceae и мхи; на лесных прогалинах почва обычно покрыта
ковром из Selaginella. В наиболее влажных условиях виды Bambusa
часто образуют непроходимые заросли.
Еще выше в горах, на высотах 2500—3000 м, количество осадков вновь
сокращается; средняя суточная температура снижается, а суточные коле-
бания температуры становятся более заметными; минимальная темпера-
тура постепенно приближается к нулю. На этом рубеже происходит
и существенное изменение характера леса. Листва вечнозеленых деревьев
принимает более ксероморфный облик. Из состава эпифитов исчезают орхи-
деи. Кроны деревьев увешаны лишайниками из рода Usnea. Это пояс
субальпийского тропического дождевого леса.
Выше этого пояса температура уже падает ниже нуля. Днем здесь еще
тепло, и эти условия не изменяются в течение всего года; количество
1 Н. Lamprecht, Estudios selviculturales en los bosques del valle de la Mucuy
cerca de Medina, Univ, da los Andes, Fac. de Ingen. Foredtal. Merida (Venez.), 1954,
198
Глава 4
Рис. 148. Лес, образованный Beilschmiedia tawa с примесью субтро-
пических элементов на острове Северный (Новая Зеландия). Вулканичес-
кое плато на высоте 600 м (фото Кокайна).
и интенсивной вулканической деятельности на острове Северный, продол-
жавшейся до недавнего геологического прошлого.
В качестве пионеров при заселении освободившихся территорий
выступали прежде всего виды Podocarpaceae. Последние затем постепенно
вытеснялись субтропическим лиственным лесом; важную роль в этом
процессе в качестве растения-удушителя, развивавшегося на Dacrydium,
играл представитель семейства Myrtaceae Metrosideros robusta. В настоя-
щее время виды Podocarpaceae часто образуют сообщества с примесью
субтропических элементов, и только в горах они частично еще сохранили
первоначальный девственный облик. Однако и здесь более конкурентно-
способными, по-видимому, оказываются виды Nothofagus, что наводит
на мысль об их все более широком распространении в будущем х. Отсюда
понятно, что не современные взаимоотношения растений, а лишь истори-
ческие данные могут вскрыть специфические особенности распределения
растительного покрова Новой Зеландии.
1 R. G. Robbins, Tho podocarp broadleaf forest of New Zealand, Trans. Roy.
Soc, N, Z., «Bot.», 1, 1962, p. 33—75.
Дождевые леса вблизи верхней границы распространения
199
По своему характеру и фло-
ристическому составу новозе-
ландские леса Nothofagus ана-
логичны западнопатагонским,
развитым в южном Чили; опи-
сание последних выходит за пре-
делы настоящей книги. В юж-
ном полушарии субтропические
вечнозеленые леса распростра-
нены также на юго-востоке Аф-
рики, на восточном побережье
южной Бразилии и на восточном
склоне Анд и районе Ту кумана.
Удивительно, что субтропиче-
ские вечнозеленые леса выше
в горах (начиная с высоты
1200 ж) резко сменяются летне-
зелеными лесами1, родствен-
ные связи которых надо искать
в северном полушарии (Alnus,
Juglans, Sambucus, Prunus, Ber-
beris). Это же явление наблю-
дается еще только в горах суб-
тропиков северного полушария
(Мексика, южный склон Гима-
лаев, Юго-Восточная Азия, юж-
ная Япония).
Рис. 149. Лес, образованный Nothofagus fusca
в районе фьордов острова Южный (Новая
Зеландия); в напочвенном покрове Polysti-
chum vestitum. Физиономически этот лес на-
поминает буковый лес Центральной Европы,
однако листья у Nothofagus вечнозеленые
и гораздо мельче, чем у бука (фото Рейда).
здесь они характеризуются присутствием
Растительность этих райо-
нов прекрасно описана, однако
здесь не проводилось экологи-
ческих исследований.
Совершенно своеобразные
условия наблюдаются в веч-
нозеленых умеренных дожде-
вых лесах на юге Австралии и
на близлежащей Тасмании. И
вечнозеленого N othofagus cunninghamii, реликтовые местообитания которого
обнаружены даже на границе Квинсленда и Нового Южного Уэльса 1 2.
Однако он господствует только в самых влажных районах западной
Тасмании, где растет совместно с некоторыми другими представителями
антарктической флоры. На юге штата Виктория и в Центральной Тасма-
нии Nothofagus образует смешанные древостои с видами Eucalyptus,
типичными представителями флоры Австралии. В условиях влажного
климата эвкалипты достигают огромных размеров и намного возвышаются
над деревьями Nothofagus. Стройные стволы Eucalyptus regnans имеют
1 К. Hueck, Waldbaume und Waldtypen aus Nordwest — Argentinien, Berlin,
1954; K. Hueck, Pflanzengeographisch-forstwirtschaftliche Probleme aus dem Nord-
westen Argentiniens, «Zschr. i. Weltforstwirtschaft», 17, 1954, S. 219—225.
2 В Тасмании известен также листопадный вид Nothofagus*, он встречается
в горах на влажных местообитаниях, например на водоносных крупнокаменистых
осыпях в долинах рек вблизи границы древесной растительности. Это Nothofagus
gunnii, мелколистный кустарник высотой 4—5 м с красивой желтой окраской листвы.
На влажном западе Тасмании этот вид растет также по склонам гор.
196
Глава 4
Физиономически субтропические дождевые леса почти не отличаются
от тропических х. Крупнолистные виды сменяются древесными породами
с листьями средней величины. Несколько реже встречаются лианы и эпи-
фиты. Рассматриваемые тропические и субтропические дождевые леса
образованы видами не австралийской флористической подобласти, а Индо-
Малайской и, таким образом, относятся к палеотропической области.
Изменение состава леса становится заметным лишь с появлением в древо-
стое хвойных (Araucaria bidwillii, A. cunninghamii) и увеличением коли-
чества древовидных папоротников (рис. 146). В лесах северной и суб-
тропической частей Новой Зеландии доминирует каури (Agathis, рис. 147),
тогда как на севере Квинсленда она встречается лишь в виде отдельных
экземпляров. На Калимантане каури распространена в горах. Agathis
дает ценную смолу, которую обычно находят в почве в виде крупных полу-
окаменелых комков; для облегчения ее поисков леса постепенно уничто-
жались и в результате частых пожаров деградировали в бесплодные
пустоши; остатки лесов Agathis сохранились лишь на небольших участ-
ках. Широко распространены в Новой Зеландии и характерные для тро-
пических субальпийских лесов виды Podocarpus*, здесь встречаются также
хвойные Dacrydium, Phyllocladus и многочисленные виды, свойственные
Рис. 146. Субтропический лес в горах Баниа, юго-восточный Квинсленд,
в составе которого встречается Araucaria bidwillii. На опушке леса, вдоль
ручья растет папоротник Dicksonia antarctica (фото Фриера).
1 L. J. W е b Ь. A physiognomic classification of Australian rain forest, «J. Ecol.»,
47, 1959, p. 551—570; G. N. В a u r, Nature and distribution of rain-forests in New South
Wales, «Austr. J. Bot.», 5, 1957, p. 190—233.
Дождевые леса вблизи верхней границы распространения
197
Рис. 14.7. Лес Вайпуа в северной части Новой Зеландии. Толстые стволы — Aga-
this australis, тонкие — Beilschmiedia tarairi (фото Кокайна).
Меланезии (рис. 148). Несмотря на то что в Новой Зеландии господствует
почти умеренный климат \ леса сохраняют субтропический облик
и отличаются обилием эпифитов и лиан (Metrosideros, Freycinetia)\ на
севере встречаются даже пальмы. 1
В умеренном дождевом лесу общий облик резко меняется. В его
составе появляются представители антарктической флоры, и, в частности
род Nothofagus, характеризующийся вечнозелеными листьями небольших
размеров. Эти леса отличаются монодоминантностью (рис. 149) или господ-
ством небольшого числа видов.
Субтропический характер лесов Новой Зеландии и произрастание
здесь N othofagus — явление, достаточно любопытное, ибо объяснить его
местными климатическими условиями не удается. В распространении
лесов Nothofagus нельзя усмотреть какой-либо закономерности: островное
положение на вулканическом плато острова Северный, более широкое
развитие по направлению к югу, полное отсутствие в некоторых районах
острова Южный. Подобное мозаичное чередование субтропических лесов
и лесов, образованных южным буком в Новой Зеландии, возможно, объяс-
няется отсутствием равновесия между современным растительным покро-
вом архипелага и его климатом. Состояние равновесия не могло быть
достигнуто в условиях значительного оледенения на острове Южный
1 J. W. Dawson, The New Zealand lowland Podocarp forest. Is it subtropical?
«Tuatara», 9, 1962, p. 98—116.
198
Глава 4
Рис. 148. Лес, образованный Beilschmiedia tawa с примесью субтро-
пических элементов на острове Северный (Новая Зеландия). Вулканичес-
кое плато на высоте 600 м (фото Кокайна).
и интенсивной вулканической деятельности на острове Северный, продол-
жавшейся до недавнего геологического прошлого.
В качестве пионеров при заселении освободившихся территорий
выступали прежде всего виды Podocarpaceae. Последние затем постепенно
вытеснялись субтропическим лиственным лесом; важную роль в этом
процессе в качестве растения-удушителя, развивавшегося на Dacrydium,
играл представитель семейства Myrtaceae Metrosideros robusta. В настоя-
щее время виды Podocarpaceae часто образуют сообщества с примесью
субтропических элементов, и только в горах они частично еще сохранили
первоначальный девственный облик. Однако и здесь более конкурентно-
способными, по-видимому, оказываются виды Nothofagus, что наводит
на мысль об их все более широком распространении в будущем х. Отсюда
понятно, что не современные взаимоотношения растений, а лишь истори-
ческие данные могут вскрыть специфические особенности распределения
растительного покрова Новой Зеландии.
1 R. G. Robbins, The podocarp broadleaf forest of New Zealand, Trans. Roy.
Soc, N, Z., «Bot.», 1, 1962, p. 33—75.
Дождевые леса вблизи верхней границы распространения
199
По своему характеру и фло-
ристическому составу новозе-
ландские леса Nothofagus ана-
логичны западнопатагонским,
развитым в южном Чили; опи-
сание последних выходит за пре-
делы настоящей книги. В юж-
ном полушарии субтропические
вечнозеленые леса распростра-
нены также на юго-востоке Аф-
рики, на восточном побережье
южной Бразилии и на восточном
склоне Анд и районе Тукумана.
Удивительно, что субтропиче-
ские вечнозеленые леса выше
в горах (начиная с высоты
1200 ж) резко сменяются летне-
зелеными лесамих, родствен-
ные связи которых надо искать
в северном полушарии (Alnus,
Juglans, Sambucus, Prunus, Ber-
beris). Это же явление наблю-
дается еще только в горах суб-
тропиков северного полушария
(Мексика, южный склон Гима-
лаев, Юго-Восточная Азия, юж-
ная Япония).
Р и с. 149. Лес, образованный Nothofagus fusca
в районе фьордов острова Южный (Новая
Зеландия); в напочвенном покрове Polysti-
chum vestitum. Физиономически этот лес на-
поминает буковый лес Центральной Европы,
однако листья у Nothofagus вечнозеленые
и гораздо мельче, чем у бука (фото Рейда).
Растительность этих райо-
нов прекрасно описана, однако
здесь не проводилось экологи-
ческих исследований.
Совершенно своеобразные
условия наблюдаются в веч-
нозеленых умеренных дожде-
вых лесах на юге Австралии и
на близлежащей Тасмании. И здесь они характеризуются присутствием
вечнозеленого Nothofagus cunninghamii, реликтовые местообитания которого
обнаружены даже на границе Квинсленда и Нового Южного Уэльса 1 2.
Однако он господствует только в самых влажных районах западной
Тасмании, где растет совместно с некоторыми другими представителями
антарктической флоры. На юге штата Виктория и в Центральной Тасма-
нии Nothofagus образует смешанные древостои с видами Eucalyptus,
типичными представителями флоры Австралии. В условиях влажного
климата эвкалипты достигают огромных размеров и намного возвышаются
над деревьями Nothofagus. Стройные стволы Eucalyptus regnans имеют
1 К. Hueck, Waldbaume und Waldtypen aus Nordwest — Argentinian, Berlin,
1954; K. Hueck, Pflanzengeographisch-forstwirtschaftliche Probleme aus dem Nord-
westen Argentiniens, «Zschr. f. Weltforstwirtschaft», 17, 1954, S. 219—225.
2 В Тасмании известен также листопадный вид Nothofagus*, он встречается
в горах на влажных местообитаниях, например на водоносных крупнокаменистых
осыпях в долинах рек вблизи границы древесной растительности. Это Nothofagus
gunnii, мелколистный кустарник высотой 4—5 м с красивой желтой окраской листвы.
На влажном западе Тасмании этот вид растет также по склонам гор.
200
Глава 4
среднюю высоту 75 м, однако известны также экземпляры высотой до
97 м, Достоверны сведения, что длина некоторых срубленных деревьев
достигала 110 м, приведенная же в работе Мюллера цифра 145 м осталась
пока неподтвержденной. Такой же высоты достигают Eucalyptus gigan-
tea, Е, obliqua.
Экология лесов, образованных видами Eucalyptus и Nothofagus,
исследована в Тасмании достаточно детально. Эти леса возникают всегда
после лесных пожаров. Под пологом Nothofagus Eucalyptus не возобновляет-
ся. Всходы Eucalyptus и Nothofagus появляются одновременно на свежих
гарях, однако Eucalyptus, отличающийся более быстрым ростом, вскоре
опережает южный бук и образует верхний ярус.
Состав указанных лесов зависит от того, как часто повторяются
лесные пожары. Гилберт 1 установил следующие закономерности форми-
рования этих лесов.
1) Если на данной территории пожары не возникают в течение по
меньшей мере 350—400 лет (максимальный возраст эквалиптов), на ней
развиваются только леса из Nothofagus с примесью Atherosperma moschata
и ярусом из древовидного папоротника Dicksonia antarctica (рис. 150).
Высота деревьев Nothofagus не превышает 40 м; максимальный возраст
450 лет. Atherosperma обычно ниже, и ее предельный возраст 250 лет.
Dicksonia иногда достигает высоты 3 м и растет даже при очень слабой
освещенности (около 1% полного солнечного света). Из эпифитов встре-
чаются только папоротники, многие представители семейства Hymeno-
phyllaceae и мхи. Чем беднее субстрат, тем разнообразнее видовой состав
сообщества; на очень бедных почвах появляются также Eucalyptus simon-
dsii, а из хвойных — Phyllocladus,
2) Если интервалы между пожарами не превышают 350 лет, то форми-
руется смешанный лес. Над ярусом из Nothofagus и Atherosperma возвы-
шается ярус Eucalyptus высотой 75 м (до 90 м), образованный видами
Е. regnans, Е, gigantea или Е, obliqua, кроны которых, однако, не образу-
ют сомкнутого полога (рис. 151). Одновозрастность верхнего яруса этих
лесов свидетельствует о возобновлении гарей непосредственно после лес-
ных пожаров. Огонь не повреждает твердую оболочку семян и даже спо-
собствует их прорастанию. Поскольку подрост эвкалипта растет быстрее,
чем Nothofagus, и не испытывает недостатка в свете, он образует пер-
вый ярус.
3) Если лесные пожары повторяются 1—2 раза в столетие, Nothofa-
gus вытесняется из нижнего яруса древостоя следующими видами деревь-
ев и кустарников: Pomaderris, Olearia, Acacia,
4) При повторяемости пожаров каждые 10—20 лет возникает чистый
эвкалиптовый лес, из состава которого, однако, выпадает сильнее повреж-
даемый огнем Е, regnans. На малоплодородных почвах развивается
Е, simondsii,
5) В результате очень частых пожаров возникают изреженные
саванноподобные древостои или, на более бедных почвах, заросли кустар-
ников из видов Myrtaceae и Proteaceae. При большой влажности суб-
страта обширные площади занимают заросли осоки Mesomelaena sphaero-
cephala с примесью видов семейства Restionaceae. над которыми возвы-
шаются редкие кусты Leptospermum, Melaleuca, Banksia.
1 J. М. G i 1 b е г t, Forest succession in the Florentine Valley, Tasmania, «Proc.
Roy. Soc. Tasmania», 93, 1959, p. 129—151; T. M. Cunningham, The natural
regeneration of Eucalyptus regnans, Bull. № 1, School of Forestry, Melbourne, 1960.
и срутьг
50.
-150
Р и с. 150. Профильная диаграмма умеренного дождевого леса, не тронутого пожа-
рами, Тасмания (по Гильберту).
Рис. 151. Профильная диаграмма смешанного умеренного дождевого леса; по-
жары здесь повторяются каждые 200—300 лет (по Гильберту).
Р п с. 152. Эпифиты из семейств Psilotaceae и Lycopodiaceae, распрост-
раненные на деревьях в тропическом дождевом лесу на склоне горы
Геде, Ява.
I — Psilotum comptanaturrr, 2 — Lycopodium nummulariifoliurrr, 3 — L. phtegmarina
(а, в, c — в натуральную величину)»
Дождевые леса вблизи верхней границы распространения
203.
В погоне за кенгуру-валлабие, предпочитающим открытые простран-
ства, местные жители еще до появления белых поселенцев пускали так
часто палы (Mesomelaena сгорает даже на корню), что это способствовало
изменению облика некогда облесенных территорий.
ГОРНЫЙ ДОЖДЕВОЙ ЛЕС НА ЯВЕ
На Яве дождевые леса низменностей в настоящее время полностью
сведены, поэтому все исследования первичных лесов проводятся в поясе
горного дождевого леса в окрестностях Тьибодаса, на высоте 1500 м на
Рис. 153. Altingia excelsa (разамала) в дождевом лесу Тьи-
бодаса (фото Фабера).
склоне вулкана Геде, где расположен ботанический сад в Богоре. Только
здесь еще можно найти почти нетронутый первичный тропический лес.
Он очень подробно изучен, а благодаря работам Коордерса (1918—1923) 1
1 Schimper-Faber, Pflanzengeographie auf physiologischer Grundlage,
3 Aufl., Jena, 1935.
204
Глава 4
Рис. 154. Дождевой лес на берегу озера Телага-Варна, расположенного в кратере.
В центре, у берега,— дикий банан (Musa cf. zebrina) (фото Фабера).
хорошо известен и его флористический состав. В этих лесах развито мно-
жество эпифитных- орхидей и лиан, древовидных папоротников, эпифит-
пых плаунов (рис. 152) и видов семейства Hymenophyllaceae, а также мхов.
По данным Коордерса (1914 г.), из 575 встречающихся здесь цветко-
вых растений к деревьям относится 165 (29%) видов, к кустарникам и тра-
вам 350 (61%) видов и к лазающим растениям 60 (10%) видов.
Хотя влажность на этой высоте чрезвычайно велика (ср. стр. 193),.
дождевой лес еще отличается необыкновенной пышностью и изобилует
высокими (до 60 Jt) деревьями, например Altingia excelsa (рис. 153).
Создается впечатление предельной насыщенности пространства, что дало
основание Юнгхуну говорить о «horror vacui» [боязнь пустоты.— Перев.\.
Просветы между стволами деревьев заполнены лианами (рис. 90), стволы
и ветви увешаны эпифитами (рис. 102). Среди них много небычайно красиво
цветущих видов, например Medinilla, Rhododendron javanicunr, послед-
ний выше границы леса растет и на почве. Под пологом леса из высоко-
рослых травянистых видов развиты дикие бананы (виды Musa, рис. 154),
а на почве — различные бегонии (рис. 86).
Мейджер 1 произвел недавно в Тьибодасе на высоте 1500 м перечет
совершенно нетронутого участка леса на площади 1 га, где были учтены
все деревья диаметром более 10 см (на уровне груди). На пробной площади
оказалось 333 вида явнобрачных и папоротников; из них древесных
пород — 78, кустарников — 40, лиан — 30, лазающих растений — 10,
1 W. М е i j е г, Plantsociological analysis of montane rain forest near Tjibodast
Westjava, «Acta Bot. Neerlandica», 8, 1959, p. 277—291.
Дождевые леса вблизи верхней границы распространения 205
эпифитов — около 100 и 73 вида трав. Только у 59 древесных пород диа-
метр ствола превышал 10 см‘, из числа последних у 284 экземпляров был
измерен диаметр ствола. Результаты измерений приведены в табл. 33.
Таблица 33
Распределение 284 стволов по ступеням толщины
Ступени, см . 10-20 21-30 31—40 41—50 51-60 61-70 71-80 >80
Число стволов, шт. 121 44 39 21 16 20 20 3
У 19 видов деревьев (из них 13 представлены подростом) диаметр
ствола был меньше 10 см. Число стволов у половины видов не превышало
одного, а господствовавший в составе древостоя вид был представлен
33 деревьями. Максимальная высота деревьев достигала 60 м. В верхнем
ярусе древостоя доминировали Altingia excelsa и Castanea javanica, Schima
wallichii\ реже встречались Quercus induta, Q. pseudomoluccana и одиночно
Engelhardtia spicata. Под пологом древостоя отмечено небольшое число
всходов всех древесных пород. Восстановление леса происходит только
в просветах крон. В крупных окнах полога появляются виды вторичного
леса. Максимальный возраст деревьев, по-видимому, не превышает 200—
250 лет, средний же возраст у деревьев на пробной площади около 130 лет.
Ежегодный отпад составляет 0,77—0,83^1.
Средняя температура в районе опытной площади равна 17,7° (мини-
мальная 12, максимальная 27°), влажность колеблется.в пределах 80—
90%, а годовая сумма осадков составляет 3400 мм\ продолжительность
сухого периода не превышает трех недель. Гумусовый горизонт вулкани-
ческих почв, на которых развиты эти леса, достигает 20—30 см‘, на глуби-
не 70—80 см залегает коренная порода. О высоком плодородии почвы
свидетельствуют развитые здесь Cyrtandra, Elatostema, Begonia, Strobilan-
thes. На бедных тропических лесных почвах эти виды либо вообще не
растут, либо встречаются значительно реже.
Примечательно, что в горных лесах Индо-Малайской области рас-
пространены семейства (Fagaceae) и роды {Rhododendron, Vaccinium, Pinus),
родина которых расположена значительно севернее. Некоторые из них,
например Androsace, Primula, Gentiana, семейство Ranunculaceae, встре-
чаются на относительно большой высоте (выше 2000—2500 м) Е Пред-
ставители Ranunculaceae распространены также в наиболее высоких горах
Австралии и в Новой Зеландии; Rhododendron в Австралии произрастает
только на севере Квинсленда.
Заморозки на почве (до —10°) на высоте 2000 м наблюдаются только
на открытых участках с травянистой растительностью, тогда как в лесу
они не были отмечены. Высокогорные плато, покрытые после вырубки леса
травянистой растительностью, представляют собой настоящие морозо-
бойные ямы. Лес в этих условиях не возобновляется, ибо чувствительные
к заморозкам всходы древесных пород погибают при первом же значи-
тельном понижении температуры.
Наиболее высокие точки на Яве — вершины молодых вулканов —
находятся в пределах вертикальной границы распространения леса, поэто-
му выше расположенные пояса здесь не выражены. Правда, сами вершины
1 C.G.G.J. van S teenis, Die Gebirgsflora der malesischen Tropen, «Endea-
vour», 21, 1962, S. 183—194.
Рис. 155. Средняя вершина вулкана Лаву (3340 л«), восточная Ява. По краю кра-
тера растут Vaccinium varingiifolium, Anaphalis и др., на дне кратера мелковод-
ное озеро, окаймленное зарослями Festuca nubigena — Tripogon exiguus.
-4
Рис. 156. Альпийская растительность на вершине вул-
кана Пангранго (3020 ж), западная Ява. Бело-серые
кустарники — Anaphalis javanica; на заднем плане по
краю кратера — криволесье, образованное Vaccinium varin-
giifolium, в центре — участок, покрытый злаками, осо-
ками, мхами и лишайниками (фото Фабера).
Дождевые леса вблизи верхней границы распространения
207
безлесны (рис. 155), но это объясняется уже эдафическими причинами.
На слаборазвитых вулканических почвах первыми поселяются альпий-
ские виды. К числу последних относится яванский эдельвейс, Anaphalis
javanica (рис. 156, 157), за развитием которого на вершине Пангранго
(3000 м) в течение многих лет наблюдал Лёувен. Всходы этого вида только
на тринадцатый год достигли высоты 20 см. По данным Лёувена, растения
при толщине стебля 15 см имеют возраст по меньшей мере 100 лет (ср.
данные, приведенные в работе Шимпера — Фабера).
В табл. 34 приводится характеристика вертикальной поясности
растительности в горах Индонезии, по наблюдениям Стиниса (цитирует-
ся по Троллю х).
Таблица 34
Вертикальная поясность в горах Индонезии
Вертикальные пояса Высота, м Характер растительности
Нивальный пояс >4500 Снеговая граница _ Вечные снега
Альпийский пояс 4000-4500 Граница древесной растительности Каменистая пустыня с мхами, ли- шайниками и небольшим количе- ством явнобрачных, главным обра- 1 зом злаков и осок
3600-4000 Низкорослые кустарники (одиночно или группами), хвойные
Субальпийский пояс Граница леса
2400—3600 Густой низкорослый лес с отдель- ными более высокими деревьями г часто покрытыми мхом, хвойные
Горный пояс 1500-2400 Сомкнутый высокоствольный лес; ниже 2000 м постепенно умень- шается количество мхов
Пояс предгорий 1000-1500 Сомкнутый высокоствольный лес;
мхов мало
АЛЬПИЙСКАЯ РАСТИТЕЛЬНОСТЬ ТРОПИ ЧЕСКИХ АНД
Далеко в пределы зоны вечных снегов вздымают свои вершины тропи-
ческие Анды. Развитая здесь альпийская растительность известна под
названием «парамос». Высокогорных опорных пунктов для проведения
экологических исследований здесь нет; только в Андах южного Перу
в течение многих лет проводились наблюдения за температурой воздуха
(ср. «Phytologie», Bd III, 1). На высоте несколько ниже 4000 м в тече-
ние всего года бывают дни, когда наблюдается отрицательная температура.
На рис. 158 изображена климадиаграмма станции Уанкапо (3353 м,
12° ю. ш.). Здесь уже отчетливо выражены дождливый и сухой периоды,
хотя средняя месячная температура колеблется в незначительных пре-
делах (8,9—13,0°).
1 С. Т г о 1 1, Die tropischen Gebirge, «Bonner Geogr. Abhandlungen», H. 25, 1959.
2
Рис. 157. Растения, обычно встречающиеся на вершинах вулканов западной Явы
(фото Фабера).
1 — Rhododendron retusum (1/б натуральной величины); 2 — Gaultheria fragrantissima, цветет
и плодоносит (3/4 натуральной величины); 3 — Anaphalis javanica, цветет (1/5 натуральной
величины); 4 — Gaultheria nummularioides (1/б натуральной величины).
Дождевые леса вблизи верхней границы распространения
209
Тролль (1959 г.) приводит в своей работе данные Эспинозы (табл. 35)
о температурных условиях альпийского пояса Эквадора.
Таблица 35
Температура воздуха на различной высоте над уровнем
моря в пределах альпийского пояса Эквадора
Станция Высота над уровнем моря, м Период наблюдений Среднемесячная температура, °C Крайние значения, °C Суточная амплитуда, °C
макси- маль- ная мини- маль- ная макси- мум мини- мум макси- мальная мини- мальная
Котопахи 3600 VI. 1930 г.— 7,5 5,4 17,3 —1,5 14,9 2,2
IX. 1931 г. (П) (VII) (XII) (VIII) (VIII. (VIII.
1931 г.) 1930 г.)
Крус-Лома 3950 V.-X. 6,8 5,9 14,0 1,5 10,2 2,6
1931 г. (V) (IX) (X) (VIII) (X) (X)
Гомессиат I 4450 VI.-X. 4,5 2,1 12,7 —1,8 9,2 1,1
1931 г. (VI) (VIII) (VI) (VIII) (IX) (X)
Г омессиат 4720 VII.-X. 0,9 0,7 3,5 —2,0 4,3 0,6
II 1931 г. (VII) (IX) (IX) (VII)
°C
21,9
В последнее время экологические исследования в перуанских Высо-
ких Андах (8—15° ю. ш.) проводил Хирш г. Он выделил:
1) пояс подушковидных растений (4200—5100 м);
2) пояс пуны (4000—4500 м);
3) растительность сухих глубоких долин.
1. Пояс подушковидных растений развит в наиболее влажных усло-
виях, по периферии горных массивов. Сообщества подушковидных расте-
ний образованы своеобразными растениями-подушками, морфологии
которых посвящена работа Рау 1 2. В их составе господствуют виды
Plantago, Pychophyllum, Azorella (рис. 159) и Werneria. Для климатиче-
ских условий этого пояса характерно большое число дней с отрицатель-
ной температурой, однако, несмотря
на это, настоящего периода покоя
у растений здесь не наблюдается. Тем-
пература поверхности почвы быстро
повышается в утренние часы и в тече-
ние дня остается на достигнутом уров-
не; после 16 часов она начинает па-
дать. Приблизительно такой же ход
температуры воздуха наблюдается
и вокруг растений-подушек. Соответ-
ственно влажность приземного слоя
воздуха днем понижается почти до
60%, однако дефицит влажности
вследствие низкой температуры неве-
лик (менее 5 мм).
1 G. Н i г s с h, Zur Klimatologie und
Transpiration an Vegetationsgrenzen, «Beitr.
Biol. d. Pflanzen», 33, 1957, S. 371—422.
2W. Rauh, Der polsterformige Wuchs,
«Nova Acta Leopoldina», 7, № 49, 1939.
Уанкайо (3353н)
20
-15,4
-1,4
11,2*845
нм
1100
-80
60
40
20
Рис. 158. Климадиаграмма Уанкайо
(3353 м); из-за отсутствия данных по
абсолютному минимуму температуры
воздуха трудно судить о возможности
заморозков в теплое время года (ок-
тябрь — март).
210
Глава 4
В качестве объектов изучения были выбраны Plantago rigida, образу-
ющий плотные низкие (50 см) подушки (листья не особенно жесткие
с устьицами, расположенными на верхней стороне листовой пластинки)
и Pycnophyllum spec. (Caryohpyllaceae), образующий круглые напомина-
ющие куртины мха подушки диаметром до 1 м.
Рис. 159. Альпийская растительность на высоте 4800 м около
Чальяпаты, Восточные Кордильеры (Боливия). Плотные подушки
Azorella diapensioides, между ними кусты Polylepis и дернины
Stipa ichu (фото Тролля).
Максимальная интенсивность транспирации Plantago в полдень
достигала 800 мг!дм2-час, а у Pycnophyllum едва составляла половину этой
величины. Хирш на основе анализа кривой хода транспирации приходит
к выводу о сокращении интенсивности транспирации в первой половине
дня. Однако, вероятно, речь идет всего лишь о разбросе значений, обу-
словленном небольшим числом измерений. Уменьшение интенсивности
транспирации в первой половине дня вообще не объяснимо, поскольку
в полдень ее интенсивность почти в четыре раза превышает утреннюю
величину.
2. Пояс пуны развит во внутреннем нагорье Анд (рис. 160). Осадков
в этой части Анд выпадает меньше, период Покоя у растений обусловлен
сухим временем года. Несомкнутый покров образован господствующими
здесь кустовыми злаками — Calamagrostis, Festuca (рис. 161) и Stipa,
среди которых в период дождей развиваются более низкорослые виды
травяного покрова. Особенно примечательны суккулентные кактусы.
Быстро высыхающий с поверхности гумусовый горизонт имеет мощность
до 1 м. Днем температура поверхности почвы может достигать 38°. Влаж-
ность воздуха к полудню понижается до 28% при дефиците насыщения,
равном почти 10 мм. Испарение, к сожалению, не измерялось. Ночью
условия здесь очень сходны с наблюдаемыми в поясе подушковидных
растений.
Интенсивность транспирации определялась только у кактусов:
подушковидного Tephrocactus floccosus и шаровидного (диаметром 20 см)
Дождевые леса вблизи верхней границы распространения
211
Oroya borchersii. Tephrocactus покрыт плотным волосяным войлоком
и колючками, неопушенный Oroya снабжен многочисленными колючками.
Днем растения сильно нагреваются (температура поверхности растения
на 24° выше окружающей), причем Tephrocactus сильнее, чем Oroya.
Однако, по-видимому, из-за опушения интенсивность транспирации перво-
го вида достигала менее 150 мг!дм2 • час, тогда как у второго она была
в два раза выше. В общем же величина транспирации, как это обычно
наблюдается у кактусов, очень невелика.
3. Растительность сухих долин развивается только в глубоких каме-
нистых ущельях. Из-за небольшой абсолютной высоты температура
всегда остается здесь выше нуля, а у поверхности почвы достигает даже
49°. Влажность воздуха в приземном слое понижается с 100% ночью до
17% в полдень, при дефиците, равном 39 мм.
Интенсивность транспирации изучалась у трех видов: низкорослой
Acacia, Tillandsla paleacea и Opuntia subulata. Результаты получены
совершенно разные.
Максимальная интенсивность транспирации Acacia превышала
700 мг!дм* • час, резко сокращаясь к полудню в крайних условиях место-
обитания до 100 мг!дм2 • час; интенсивность транспирации Tillandsia
была в два раза меньше и днем не понижалась столь резко, как упреды-
Р и с. 160. Ландшафт пуны на высоте 3850 м; нагорье Боливии в верховьях Десагуа-
деро. В широких межгорных долинах на отдельных участках распространены заросли
тола (это преимущественно смолистые эрикоидного типа кустарнички из семейства
сложноцветных — Lepidophyllum, Baccharis, Mikromeria и др.) и дернины — Festuca,
Stipa, Calamagrostis и др. (фото Тролля).
212
Глава 4
Рис. 161. Festuca orthophylla («iru-ichu»); боливийская пуна, высота 4200 м-
На заднем плайе — вулкан Сахма (6520 л<) (фото Тролля).
Рис. 102. Густоопушенная свечеподобная Lobelia telekii и древовидный Seneciv
keniodendron, развитые на склоне горы Кении (4100 м), Восточная Африка
(С. Troll, 1959).
дущего вида. Что касается Opuntia, то, как и предполагалось, этот вид
отличался лишь слабым испарением с поверхности. Поверхностное рас-
положение корней Tillandsia и Opuntia не позволяет растениям исполь-
Дождевые леса вблизи верхней границы распространения
213
зовать почвенную влагу, но Tillandsta, несомненно, пополняет свой запас
воды ночью при обильной росе с помощью чешуйковидных волосков. Aca-
cia, напротив, укореняется достаточно глубоко.
Рис. 163. Парамос близ Боготы (Колумбия) на вы-
соте 3500 м. Древовидная Espeletia grandiflora среди
зарослей Calamagrostis effusa (фото Тролля).
Еще более влажными, чем пояс подушковидных, являются «парамос»,
развитые в пределах альпийского пояса и распространенные в высоко-
горьях Центральной и Южной Америки от 10° с. ш. до приблизительно
8° (15°) ю. ш. Наиболее северные парамос, развитые в Коста-Рике, иссле-
.довал Вебер х. По его данным, постоянно влажный прохладный климат
в этих условиях способствует интенсивному накоплению гумуса, кислот-
ность которого достигает 3,8—4,0. В парамос, так же как и в альпийском
поясе Новой Зеландии, распространены высокие кустовые злаки, назы-
ваемые местным населением «туесок», что аналогично немецкому «Вй-
schel-Graser» [дерновинные злаки.— Ред.]. Однако такое название может
создать ложное представление об облике растения, поскольку «дернины»
достигают высоты 0,5—1 м при таком же диаметре. Скорее они напомина-
1 Н. Weber, Die Paramos von Costa Rica und ihre pflanzengeographische Ver-
kettung mit den Hochanden Siidamerika, «Akad. d. Wiss., Mainz, Math.-Naturw. KI.»,
№ 3, 1958.
214
Глава 4
ют крупные кочки осоки. Очень жесткие листья не опадают и после пожел-
тения. Поэтому луга из туссоковых злаков никогда не имеют цвета свежей
зелени и нередко кажутся совершенно желтыми. Альпийские туссоковые
луга образованы видами Festuca, Deschampsia, Danthonia, Stipa, Calama-
groslis и Andropogon. Также характерны для горных альпийских поясов
Рис. 164. Lupinus alopecuroides', парамос на склонах
Чимборасо (Эквадор,высота 4800л*) (фото Вебера).
экваториальной зоны своеобразные древовидные формы, представленные
в Африке гигантскими крестовниками (рис. 162), в Южной Америке —
древовидными эспелетиями (рис. 163), в Индо-Малайской области —
Anaphalis (рис. 156); все это роды, относящиеся к семейству сложноцвет-
ных. Кроме того, здесь встречаются древовидные папоротники и све-
чеподобные лобелии и Лунины (рис. 162 и 164).
С удалением от экватора изменяется климат, а вместе с ним и характер
горной растительности. Более заметно выделяются времена года, возраста-
ют амплитуды температуры, наибольшие значения которой могут
достигать 50° (табл. 36). Более резко выражены сухие периоды, сокращает-
ся сезон летних дождей. Соответственно растительность принимает все
более ксероморфный облик; в конце концов появляются пустыни с харак-
терными для них суккулентами или засоленными почвами. Эта последо-
вательность прекрасно прослеживается в Андах Южной Америки, где
более ксерофитные альпийские формации называются в отличие от «пара-
мос» «пуна» (рис. 165, 166).
Р п с. 165. Схематический поперечный профиль Высоких Анд [в районе экватора (Л) и около 16—18° ю. ш. на высоте
озера Титикака (В) (С. Troll, 1959).
1 — полувечпозелспый дождевой лее; 2 — тропический дождевой лес; з — тропический вечнозеленый горный лес пояса «tierra tem-
plada»; 4 — высокогорный лее и лес пояса облаков «tierra ITia»; 5 — парамос; 6 — заросли мезофитпых кустарников и рощи Polylepis,
7 — заросли колючих и суккулснтпых кустарников в глубоких долинах; 8 — заросли колючих и суккулентных кустарников в нагорных
долинах; 9 — пустыня; 10 — влажная пуна; 11 — сухая и колючекустарниковая пуна; 12 — саванны жарких низменностей; 13 — сне-
говая линия; 14 — пояс туманов и облаков.
216
Глава 4
Таблица 36
Температура воздуха в Андах в пределах пояса пуны
(Troll, Die tropischen Gebirge, «Bonner. Geogr. Abhandl.», H. 25, 1959)
Зтанция и ее зстоположение Высота над уровнем мо- ря, м Среднегодо- вая темпе- ратура, °C Годовая амплитуда, °C Крайние значения, °C Выборочные суточные амплитуды, °C
M( макси- мум мини- мум ампли- туда макси- мум мини- мум
1 Ф Кито, 0°14' 2850 12,6 0,4 24 3 21 14,8 (VIII) 11,4 (IV)
s « И S Куско, 13°27' 3380 10,0 3,6 23 — 2 25 16,3 (VII) 10,9(11)
И Л 2 ° Ла-Пас, 16°30' 3690 9,3 4,6 24,2 -2,7 26,9 15,8 (VIII) 12,1 (I)
4 ss i ф и co Оруро, 17°38' Ла-Кьяка, 3706 8,3 6,7 23,5 -10,1 33,6 17,2 (VIII) 9,6(1)
® 1 L 22°10' 3458 9,5 9,6 30,7 -18,0 48,7 26,6 (VIII) 18,7(1)
CO I г Винкокая 4380 1,9 7,1 19 -22 41 24,1 (VIII) 14,4(11)
Я J г-*! 1 к 1 Мина-Агилар 1 Трес-Крусес 4600 3,1 11,0 16,3 -14,2 30,5 — —
Элленберг высказал мнение, что перуанские парамос представляют
собой лишь стадию деградации естественных лесов, обусловленную вмеша-
тельством человека \
Подтверждением его вывода может служить рис. 50, где (между 10'
и 30° ю. ш.) нанесена верхняя граница распространения Polylepis. Виды
этого рода розоцветных имеют древовидную форму и образуют заросли,
напоминающие криволесье в Альпах; в отличие от других древесных пород
они не страдают от заморозков. Отдельные рощи Polylepis встречаются
в горах до высоты 4500 ж, причем обычно вне зависимости от истинной
границы распространения древесной растительности 1 2. В противополож-
ность Альпам в Андах нижняя часть альпийского пояса частично была
заселена уже несколько тысяч лет назад, и Polylepis явился единственным
источником топлива для населения. Древесина его используется даже
на местных рудниках, а медно-красная кора применяется для дубления
кожи. Таким образом, вполне возможно, что встречающиеся сейчас рощи
представляют собой остатки некогда более широко распространенных
лесов, особенно если учесть весьма небольшой прирост на этих высотах.
Средняя годовая температура здесь равна 6,9—2,8°; принимая во внима-
ние ежедневные заморозки, ее нельзя назвать высокой. Осадки не превы-
шают 600—800 мм в год. Если рассматривать верхний предел распро-
странения Polylepis (на рис. 50) как границу произрастания древесной
растительности, то асимметричная форма этой границы в северном и юж-
ном полушариях исчезнет и сохранится только заранее предполагаемое
понижение уровня этой границы во влажной экваториальной области,
где развиты настоящие парамос.
Альпийский пояс в экваториальной области южного полушария,,
который ни в климатическом, ни в физиономическом отношениях не
1 Н. Ellenberg, Wald oder Steppe? Die natiirliche Pflanzendecke der Anden
Perns I u. II, «Die Umschau in Wiss. u. Techn.», 1958, S. 645—648, 679—681.
2 К. H u e c k, Der Polylepis-Wald in den venezolanischen Anden, eine Parallele-
zum mitteleuropaischen Latschen-Wald, Angew. Pflanzensoz. (Klagenfurt), H. 17, 1962r
S. 57—76.
Дождевые леса вблизи верхней границы распространения
217
соответствует альпийскому поясу Альп, Тролль (1959 г.) предложил
называть «парамос», использовав для этого южноамериканское название.
Под понятием «альпийский» в настоящее время понимают только высотный
пояс, расположенный между снеговой линией и границей древесной
растительности, причем этот пояс имеет свои отличительные особенности
во всех климатических зонах. Значительные трудности представляет
только вопрос о соподчинении пояса кустарников, который расположен
выше лесного пояса; этот пояс иногда четко отграничен, иногда же обра-
зует лишь постепенный переход от верхней границы леса, высота которого
вверх по склону постепенно уменьшается (Новая Зеландия). Переход же
к выше расположенным туссоковым лугам весьма отчетлив. В таких
случаях надо проявлять известную гибкость, поскольку нельзя верти-
кальную поясность разных горных систем уложить в одну систему. Тем
Рис. 166. Распространение пуны в Центральных Андах близ озера Титикака (С. Troll,
1959).
1 — пояс сухой и засоленной пуны; 2 — пояс влажной пуны; з — пампасы, развитые на пло7
скогорьях Атакамы; 4 — вечнозеленые дождевые и высокогорные леса восточного склона Анд»
5 — летнезеленые, выше в горах вечнозеленые, тропическо-субтропические леса (тукумано-
боливийский лес); 6 — острова саванн в горах (средняя часть пояса горных лесов); 7 — пояс
мезофитных кустарников на западном склоне Анд (пуна); 8 — заросли кустарников в долинах
восточного склона Анд; 9 — долины, по которым дуют восточные компенсационные ветры; 10 —
основная климатическая граница в Андах; западная граница осадков и туманов, возникающих
при подъеме воздушных масс, приходящих с востока, и вечновлажной растительности; 11 — вос-
точная граница пустошей, образованных Lepidophyllurrr, 12 — основные горные хребты и вос-
точная окраина Анд; 13 — восточная граница пуны; 14 — вулканы.
Верти-
каль-
ные
пояса
Малайзия
(по Стинису)
Восточная Аурика
(по Хэдбергу)
6000м'
5800-
5600-
5400-
5200-
5000м - .53
Колумбия
(по Куатрекасасу)
4600-
4400-
42OO--
‘000М-
3800-
3600-
3400-
3200-
3000м-
2800- j,
2600-
2400-
2200-'
2000/1-
1800-
ГоРи?п
4400-s
4100-
3500
270lr
----2400------
2300-
.2000
сз
F3T3
Горный
пояс
^lepls1
'81Л Пуна
О „ О О о о
Сежа
ВГЛ^-3500^-
1800
Перу-Воливия
(по Вебербаузру
и Траллю)
Мексика
(по Троллю и др.)
Восточные
Гималаи
(по ШВейнфурту)
Гавайи
(по Мак-Коши,
Иелеру и ап.)
Ланд-
шафтный
пояс
4700
о
Senecw-
Lobelia -
Alchemilla
/ 4800-
J rculc/tium I ТА41
—±^4500^-&j
-BBl-(Polylepis) — gi
Espeletia S'1
.t$<T
food
Хбайный лес
(Pinus - Abies)
3200
^o^-3000^^
ГШ -
1200-
1000м -
800-
600 -
Пояс
предгорий
Tierra
II7TII
1800----
Tierra
templada
/ООО-
Tierra
caliente
3/00
Низкоросло
1000-
2500
°—^00^
'Смешанный
ООО
лес
ООО
(Pinus-
uuercus-
°Alnus)
templada
— 800—
Tierra
caliente
сл
'—СЛ--------?44-00—
Saussurea sacra
^ВГД^-МО^-
юросли рододендрона
о о Хооино- о о
5 рододендроновый^
rt л Л
------3700----
Argyroxlphium
sandraicense
-1500-
1000
лес
----600-----
Низкогорный
пояс
° °o °oT-o
g Высокогорный о
° О о О о °
Горный
дождевой
лес
-сл- 1 -вгд- 2
-ВГЛ- 3 llllllllllll *
P и c. 167. Схема вертикальной поясностп тропических гор (С. Troll, 1959).
1 — снеговая линия (сл); 2 — верхняя граница отдельных деревьев (вгд)-, з — верхняя граница леса (вгл)\ 4 — пояс бамбука;
5 — пояс Ericaceae; 6 — пустынный пояс высокогорий; 7 — вечные снега; 8 — Hypericum leschenaultii (Малайзия), Н. leucopty-
chodes (Восточная Африка), Н. laricifolium) Анды).
Дождевые леса вблизи верхней границы распространения
219
Рис. 168. Верхняя граница леса; Попокатепетль, высота около 4000 м; здесь раз-
виты редкостойные древостои Pinus hartwegii; на переднем плане — Senecio calcarius,
вдали видна покрытая снегом вершина (фото Пурпуса).
не менее схема (рис. 167), предложенная Троллем в его великолепной
работе, посвященной сравнительному описанию растительности тропиче-
ских гор, представляет удобную отправную точку. На схеме, а также на
рис. 50 видно, сколь отличны условия |в тропических горах северного
полушария от условий в южной половине земного шара. Севернее эквато-
ра леса насыщены бореальными элементами: в Центральной Америке
и Азии они образованы видами родов Pinus, Quercus, а в Азии, кроме
того, многочисленными видами Lithocarpus, и в том числе голарктическими
родами Rhododendron и Vaccinium, из которых первый проникает через
горы Юго-Восточной Азии и Индо-Малайскую область в Австралию
(северный Квинсленд).
Тролль отмечает, что верхняя граница леса, образованная в Мексике
Pinus hartwegii, совершенно своеобразна и аналогична тому, что наблю-
дается обычно в местах контакта сосновых лесов с прерией. При подъеме
в горы древостой изреживается (рис. 168).
ВЕРТИКАЛЬНАЯ ПОЯСНОСТЬ КИЛИМАНДЖАРО
Вулканы Восточной Африки, в том числе Килиманджаро (6000 м),
занимают среди гор тропиков особое место в силу своего изолирован-
ного положения на окружающей равнине и отсутствия прямой связи
220
Глава 4
с горными системами, расположенными далее на север или юг. В январе
1935 г. автор проводил экологические исследования в верхней части аль-
пийского пояса Килиманджаро. К тому времени район исследований был
уже хорошо изучен: оценка климатических условий дана Клуте; расти-
тельность подробно описана Фолькенсом (1897 г.), а позднее — Тоблером;
детальная фитоценологическая характеристика проведена в последнее
время Селтом и главным образом Клётцли, составившим схему вертикаль-
ной поясности вдоль маршрута: от приюта Бисмарка через приюты Петер-
са и Кибо до вершины х.
Килиманджаро (рис. 169) — сложный стратовулкан с тремя эруптив-
ными центрами и многочисленными паразитическими кратерами. Древней-
ший эруптивный центр — Шира (3906 ж), представляющий в настоящее
время почти ровное плато. Одну из двух вершин вулкана Мавензи
(5150 3f) образует лавовая пробка главного жерла, окруженная сильно
размытым чехлом туфов. Ее возникновение относят к раннему плиоцену,
тогда как вторая вершина —Кибо (5900 м, ранее считалось 6010 3f) с сохра-
нившимся кратером датируется средним плейстоценом. С вершины берут
начало многочисленные ледники; в 1898 г. ледники спускались по юго-
западному склону до высоты 4000 м (по северо-восточному —до 5700 ж),
но в 1930 г. — уже только до 4500 в дальнейшем они отступили еще
больше (рис. 173). Следы ледниковой деятельности плювиального периода
прослеживаются до высоты 3600 м.
Вертикальная поясность растительности схематически изображена
на рис. 169. Юго-западный склон, открытый влиянию юго-западного
муссона, очень влажный, северо-восточный же, расположенный в ветро-
вой тени, значительно более сух (здесь развиты леса, очень похожие на
распространенные в западной части гор Усамбара, рис. 178). Этим обус-
ловлено постепенное выклинивание пояса дождевого леса к востоку,
а также более низкое положение ледниковых языков на юго-западной
стороне вулкана.
Основание вулкана возвышается над равниной, сложенной гнейсами
и простирающейся на высоте 750 м над уровнем моря. До высоты 1100 м
на пологих склонах горы развивается разреженная саванна, называемая
«садовая саванна», поскольку внешне она напоминает плодовый сад
с невысокими, равномерно разбросанными по травяному покрову дере-
вьями. Далее следует полоса культур — посадки кофейных деревьев
и несколько выше по склону — преимущественно плантации банана. Час-
тично эти культуры занимают уже ранее раскорчеванные лесные площади.
И лишь начиная с высоты около 1800 м появляется девственный лес. Он
покрывает нижний крутой уступ и на юго-западном склоне простирается
до высоты 3100 м. Это типичный высокогорный тропический лес пояса
облаков, в котором широко представлены эпифиты {Lycopodium) и папо-
ротники, особенно Hymenophyllaceae и эпифитные мхи (рис. 170, 171) 1 2.
Виды Selaginella покрывают почву сплошным ковром.
Субальпийский лес образован Olea chrysophylla, Ilex mills, Juniperus-
1 F. К 1 u t e, Ergebnisse der Forschungen am Kilimandscharo, 1912, Berlin, 1920;
G. V о 1 k e n s, Der Kilimandscharo, Berlin, 1897; G. T о b 1 e r-W о 1 f f, Fr. Tob-
ler, Vegetationsbilder vom Kilimandscharo, «Veget. Bilder», 12, H. 2,3, 1914; G. S a 1 t,
A contribution to the ecology of upper Kilimanjaro, «J. Ecol.», 42, 1954, p. 375—423;
F.Klotzli, Zur Pflanzensoziologie des Siidhanges der alpinen Stufe des Kilimandscha-
ro, «Вег. Geobot. Forsch.— Inst. Rubel fur 1957», 1958, S. 33—58.
2 R. В i e b 1, Austrocknungsresistenz tropischer Urwaldmoose auf Puerto Rico,
«Protoplasma», 59, 1964, S. 277—297.
Рис. 169. Схема вертикальной поясности Килиманджаро (с данными о годовом
количестве осадков в мм).
Рис. 170. Высокогорный
тропический лес пояса обла-
ков на склоне Килиманджа-
ро (высота около 2500м). На
переднем плане— Agauria
saliclfolia^ за ними — Scheff-
lera volkensii и Paxlodendron
usumbarense, сплошь покры-
тые эпифитными мхами,
Hymenophyllaceae и други-
ми видами папоротников.
На почве Selaginella, Impa-
tienS) Viola abyssinica и др.
(фото Тоблера).
222
Глава 4
procera, Podocarpus, milanjianus и Hagenia abyssinica. По открытым участ-
кам канделябровидные Euphorbiaceae поднимаются до высоты 2130 м,
a Cynodon dactylon образует здесь плотную дернину.
Вблизи верхней границы леса (рис. 172) часто встречаются Podocarpus
и Hagenia (сплошь покрытые Usnea). Обращают на себя внимание древо-
видная Crotalaria и красивый, с крупными цветками вид Hypericum,
достигающий здесь высоты 6 м. Выше начинается альпийский пояс, в пре-
делы которого длинными языками по ущельям поднимается лес. Переход
между лесным и альпийским поясами образуют заросли кустарников
(Ericaceae). Кусты Erica arborea и Philippia excelsa достигают высоты 6 м
и более, скрывая под собой двухметровые экземпляры Pteridium aquilinum
и густой покров Lycopodium clavatum.
Собственно альпийский пояс прослеживается почти до седловины
между обеими вершинами — Кибо и Мавензи. На этих высотах разви-
вается крайне скудный растительный покров (рис. 173). По данным Клётц-
ли, уже с высоты 4000 м начинается холодная пустыня, где встречаются
лишь отдельные растения-подушки. Последних представителей расти-
тельного покрова — Koeleria gracilis и Hilichrysum newii — он нашел
несколько выше отметки 5000 м.
В нижней части альпийского пояса распространены типичные туссо-
ковые луга, развивающиеся на торфянистых почвах (рис. 174); это дало
основание Фолькенсу назвать эту часть альпийского пояса поясом торфя-
ников. Клётцли считает, что в составе этих лугов доминируют Exotheca
abyssinica и Agrostis volkenisii (семейство Gramineae). Автора, побывавшего
в этих местах, поразили цветущая Kniphofia и кусты Protea kilimandscha-
rica. Клётцли упоминает еще о развитых в этих условиях Anemone thomso-
ni, Dierama pendulum (семейство Iridaceae) и орхидеи Disa stairsii. Они
растут совместно с видами Helichrysum. На менее влажных участках
разрастаются заросли Adenocarpus mannii и Artemisia afra. По данным
Селта, на сильно заболоченных местах наблюдается Carex monostachya,
растущая совместно с Deschampsia caespitosa var. и Alchemilla argyro-
phylla.
В верхней части альпийского пояса преобладают виды Ericaceae,
достигающие высоты 0,5—1 м. Philippia trimera (рис. 175) и Blaeria
johnstoni. До этой высоты иногда поднимаются Protea и Adenocarpus.
свойственные нижней части альпийского пояса. Весьма своеобразна
Рис. 171. Эпифитные
папоротники. На перед-
нем плане—Begonia теу-
eri-johannis. Первичный
лес на склоне Килиманд-
жаро (фото Э. Вальтер).
Рис. 172. Опушка леса
у приюта Бисмарка. Де-
ревья (Hagenia abyssinica
и Podocarpus) покрыты
лишайниками из рода
Usnea\ слева от здания —
Erica arborea (фото
Э. Вальтер).
Рис. 173. Вершина Ки-
бо с языками многочис-
ленных ледников. Вид с
седловины между Кибо
и Мавензи (фото Э. Валь-
тер).
Рис. 174. Нижняя часть
альпийского пояса (тус-
соковые луга); наиболее
высоко расположенная
роща образована A gauria
salicifolia. Выше по скло-
ну (темное пятно) — верх-
няя часть альпийского
пояса (Ericaceae). Сле-
ва — заснеженная верши-
на Кибо, справа — Ма-
рензи (фото Э. Вальтер).
224
Глава 4
Рис. 175. Кустарнички, развитые в альпийском поясе Килиманджаро.
1 — Euryops dacrydioides (в натуральную величину); 2 — общий вид расте-
ния (уменьшено); 3—4 — растение, ранее называвшееся Ericinella mannii (веро-
ятно, Philippia)\ <5 — то же, уменьшено (рисунок Анхейсера).
достигающая высоты 1 м Lobelia deckenii (рис. 176). На заболоченных
местообитаниях наряду с Carex monostachya хорошо развиваются древо-
видные крестовники (рис. 177), ранее известные под названием Senecio
johnstonii, теперь же подразделяемые на два вида — Senecio kilimanjari
Дождевые леса вблизи верхней границы распространения
225
Рис. 176. Lobelia deckenii и виды Ericaceae в верхней части
альпийского пояса Килиманджаро (фото Э. Вальтер).
(встречается, начиная с высоты 3100—4000 м) и S. cottoni (3700—4500 м).
Оба вида могут достигать высоты 7,5 м. Группу этих необычайно краси-
вых растений, о которой Мейер упоминал еще в 1898 г., Селт через
50 лет увидел почти неизменившейся. Видимо, эти растения достигают
большого возраста (ср. Anaphalis. стр. 207).
Несколько выше 4000 м на поверхность выходят родники. Расти-
тельный покров здесь более разрежен; вместо Ericaceae здесь развиваются
виды Compositae, и в частности Euryops dacrydioides (рис. 175), Helich-
rysum newii и др., к которым присоединяются злаки Festuca abyssinica
и Pentaschistis borussica. На заболоченных участках вокруг выходов род-
ников распространены чистые заросли Euryops и Carex monostachya
с Alchemilla johnstonii. Гигантские крестовники больше не встречаются.
На высоте 4400 м доминируют виды Helichrysum, образующие белесые
подушки, Festuca kilimanjarica и Koeleria gracilis, растущие в виде отдель-
ных экземпляров под защитой обломков скал.
Согласно Клётцли, эти климаксные сообщества (высотные подразде-
ления альпийского пояса) «постепенно переходят один в другой»; у автора
создалось такое же впечатление.
Сходный растительный покров развит и в пределах альпийского пояса
других вулканических массивов Восточной и Центральной Африки 1.
На рис. 179 приведена схема распределения растительности в пределах
альпийского пояса гор Рувензори, составленная Хауманом.
Этот невулканический горный массив отличается чрезвычайно влаж-
ным климатом. На высоте 2200 м годовая сумма осадков, видимо, равна
4000 мм. а вблизи вершины понижается лишь до 1600 мм. Осадки прак-
тически выпадают каждый день, поэтому не удивительно, что здесь широко
распространен Sphagnum.
На сильно увлажненных склонах гор Вирунга, Рувензори, Элгон,
Абердэр, Кения распространен пояс бамбуковых зарослей, слаборазви-
тый на горе Меру и совершенно отсутствующий на Килиманджаро.
1 О. Н е db erg, Vegetation belts of the East African Mountains, «Svensk. Bot.
Tidskr.», 45, 1951, p. 140—202.
226
Глава 4
Рис. 177. Ущелье с зарослями гигант-
ских крестовников на высоте 3800 м
(фото Э. Вальтер),
Рис, 178. Сухой горный лес без лиан и
эпифитов. На прогалине растут древо-
видные лобелии; лес Шуме в западной
части гор Усамбара (фото Э. Вальтер).
Альпийской флоре Африки посвящены работы Хэдберга Она насчи-
тывает около 278 видов, из которых 225, или 81%,— эндемы Африки;
94 вида отмечены только на одной горе; часто встречаются викарирующие
виды. Из неэндемичных видов 7 распространены в бореальных областях,
7 — в Южной Африке, 4 — в Средиземноморье и 2 — в Гималаях. Осталь-
ные виды встречаются в умеренной зоне обоих полушарий или являются
почти космополитами.
После этого краткого обзора можно перейти к данным собственно
экологических исследований.
В тропиках климат отличается небольшими сезонными и относитель-
но большими суточными колебаниями температуры, поэтому ход темпера-
туры выравнивается в почве уже на небольшой глубине, ниже которой
температура в течение года остается постоянной, и в результате отсут-
ствия в тропиках снежного покрова и постоянной теплопроводности
всегда влажной почвы соответствует средней температуре воздуха у поверх-
ности земли. Как показали исследования, проведенные в Амани, средняя
температура воздуха, измеренная в метеорологической будке, равна
постоянной температуре затененной почвы. На открытых солнечных
участках постоянная температура почвы на 2—3° выше (табл. 37).
Из таблицы видно, что в тени постоянная температура почвы устанав-
ливается уже с глубины около 30 см, а на открытых участках — лишь
с глубины 60 см.
1 О, Н е d b е г g, The phytogeographical position of the Afroalpine flora, «Rec,
Advanc. Bot. Toronto», 1961, p. 914—919; Afroalpine vascular plants, a taxonomic revi-
sion, «Symb. Bot. Upsal.», 15/1, Upsala, 1957.
Дождевые леса вблизи верхней границы распространения
227
Т а б л и ц'а 37
Температура почвы у юго-западного подножия Килиманджаро
(плантация кофе на высоте 1180 м, 2 января 1935 г., в 8 час.;
мощная, очень однородная, рыхлая почва на выветренных
вулканических породах)
Открытый солнечный участок Сильно затененный участок под деревом
Глубина, см Температура, °C Глубина, см Температура, °C
5 19,8 6 19,3
16 22,8 14 19,8
44 24,4 23 20,4
52 24,4 27 21,4
60 24,5 39 21,8 ”
68 24,7 л 52 21,8 Постоянная
70 2/ 7 1 Постоянная 70 21 8 температура
87 24* 7 / темпеРатУРа 94 21,8 -
В табл. 38 сопоставляются полученные на разных участках постоян-
ные температуры почвы со средней температурой воздуха на метеостан-
циях (названия станций даны в таблице курсивом, а температура воздуха
выделена полужирным шрифтом).
Таблица 38
Географическая точка или характеристика растительности (Высота над уровнем моря, м Местообитание
в тени, °C на солнце, °C
Седловина Кибо-Мавензп 4400 — 8,0
Пологий склон, покрытый злаковым 'дер-
ном Около 4300 — 8,0
Ущелье (поток холодного воздуха) . . . 3800—4000 5,3 —
Заросли Ericaceae выше ущелья . . 3800—4000 — 8,9
Верхняя граница леса . . 2600 11,2 —
Там же, травяной покров . £600 12,0 —
Там же, открытый участок 2600 — 12,7
Открытый участок в лесу 2200 — 16,5
В глубокой тени у нижней границы леса 1840 16,5 —
Станция Мамба . 1550 17,8 —
Плантация кофе 1180 21,8 24,7
Станция Моши . .... 1150 20,8 —
Горы Усамбара:
Станция Амани 900 19,9 —
Там же в лесу 900 19—20
Па побережье:
Станция Танга . . 28 25,4
Там же, открытый участок 28 — 27,5—28
Климат в восточной части гор Усамбара близ Амани значительно
более влажный и поэтому более прохладный, чем в области развития
саванн в окрестностях Моши. Невысокие значения средней годовой тем-
228
Глава 4
пературы, достигающей на склоне Килиманджаро на высоте 4000 м 5,3°,
возможно, объясняются инверсионным влиянием потока холодного возду-
ха (ночной минимум —11°); с другой стороны, величины (8°), полученные
на высоте 4400 м на освещенных солнцем участках, слишком высоки для
Рис. 179. Схема распределения растительности в зависимости от условий место-
обитания; субнивальный пояс гор Рувензори на высоте более 3600 м (С, Troll, 1959)
1 — Philippia; 2 — Senecio div. spec.; 3 — Helichrysum stuhlmannii; 4 — Carex runssorensis',
5 — Deschampsia и Subularia; 6 — Alchemilla div. spec.; 7 — Sphagnum; 8 — Lobelia wollastonii;
9 — мхи и лишайники.
Дождевые леса вблизи"верхней границы распространения
229
среднегодовой температуры. Последняя на высоте 4400 м, то есть у верх-
него предела распространения растительности, должна равняться +5°,
то есть достигать таких же значений, как и в лесистой центральной части
Швеции. Из этих данных явствует, сколь неблагоприятен на этой высоте
суточный ход метеорологических элементов для произрастания растений;
этим также можно объяснить появление в тропических горах ниже снего-
вой границы широкого пояса высокогорных пустынь, совершенно отсут-
ствующего в горах умеренной зоны.
О температурных условиях альпийского пояса Килиманджаро мож-
но судить по данным Клуте, полученным на высоте 4160 м на затененных
участках с 19 августа по 12 октября 1912 г. (табл. 39).
Таблица 39
Сроки наблю- дений, час. Средняя тем- пература, °C Температура, °C Показания минималь- ных термометров на от- крытом воздухе
в ысота, м °C
7 0,1 ( средний . . -1,8 4030 -7,3
14 5,6 Минимум { ( абсолютный -4,0 4330 -7,9
21 0,5 Максимум 11,5 4340 -8,2
Среднесуточная амплитуда 10,0
Весьма своеобразно распределение атмосферных осадков. Альпий-
ский пояс расположен в тропиках высоко над пологом облаков и поэтому
получает мало осадков. Уже в лесах субальпийского пояса количество
осадков резко сокращается.
Селт сообщает о характере распределения осадков, изучавшихся
в течение пяти лет на более влажном юго-западном склоне Килиманд-
жаро. Максимум осадков выпадает на высоте 2400 м. На высоте 3600 м
в зависимости от экспозиции склона количество осадков варьировало
очень резко. В среднем на трех участках годовая их сумма равнялась
1016, 608 и 174 мм. На высоте 4200 м количество осадков также достигало
174, а на высоте 4800 м — лишь 75 мм в год. Согласно новейшим наблю-
дениями Метеорологической службы Восточной Африки, на седловине
(4400 м) за год выпадает около 300 мм осадков, а у кратера — менее
100 мм (Клётцли).
Правда, в альпийском поясе, видимо, гораздо большую роль играет
конденсационная влага, не поддающаяся непосредственному измерению.
В первой половине дня (в 10—11 час.) облака часто поднимаются до аль-
пийского пояса, влага, осаждаясь на поверхности скал, обильно смачи-
вает их.
Интенсивность солнечной радиации с высотой постоянно возрастает.
За время с 1 по 6 января 1935 г. на высоте 3800 м с помощью двух ртутных
термометров с полушаровыми резервуарами автору удалось получить
следующие величины максимальной интенсивности радиации, выражен-
ной в виде разности температур:
1180 м — 13,4°; 2180 м - 17,2°; 3800 м - 18,2°.
Абсолютные максимумы интенсивности радиации, вероятно, не были
измерены, так как в полдень небо всегда затягивалось облаками1.
1 В безоблачный день 18/XII в Амани (980 м) максимальная разность температур,
измеренных с помощью пары термометров (один из них зачернен, другой покрыт белой
краской), достигала 17,2°.
230
Глава 4
Рис. 180. Связь интенсивности солнечной ра-
диации с высотой солнца на Яве (по Бёреме).
Джакарта расположена на берегу моря; уменьшение
радиации в августе — октябре объясняется большим
содержанием пыли в воздухе в это относительно сухое
время года.
Интенсивная солнечная
радиация в пределах альпий-
ского пояса, отмеченная и на
Яве (рис. 180), обусловливает
сильный нагрев приземного
слоя воздуха. При темпера-
туре воздуха 7,2 в злаковой
дернине температура подня-
лась до 21°, а в подушке Heli-
chrysum — до 17 —29°. На вы-
соте 4000 м (температура воз-
духа 8,2°) в подушке Alche-
Tiiilla температура достигала
18,5°, а в дернине злаков —
даже 41°.
Как только солнце скры-
вается в тумане, температура
быстро падает. Утром мини-
мальная температура на от-
крытом месте достигала —3,5°,
в ущелье под гигантским Se-
necto —11° (рис. 177), в по-
душке Helichrysum —2°, а в
трещине скалы —2,1°.
Резкие колебания темпе-
ратуры повторяются изо дня
в день в течение всего
года. Столь своеобразные
климатические условия не
известны больше нигде на
земном шаре.
Несмотря на низкую температуру воздуха вследствие сильной инсо-
ляции, испарение достигает относительно больших значений.
На высоте 3800 м (6.1.1935 г.) при безоблачном небе были получены
следующие величины испарения в мл/час (табл. 40).
Таблица 40
Сроки наблюдений Высота над поверх- ностью почвы, см Температура, °C
150 30 2
8 — 9 час. 1,1 0,84 0,77 Воздуха 11,4
9 — 10 час. 25 мин. 1,15 1,10 1,00 Поверхности почвы 28, в дер- нине злаков 36,5
10 час. 25 мин. —11 час. 40 мин. 1,12 1,05 1,05 Поверхности почвы 33,5 , в дер- нине злаков 36,5
Седьмого января за период 7 час. 10 мин. — 10 час, 40 мин., пока небо
было безоблачным, испарение достигало 0,66 мл!час, Несмотря на туман
Дождевые леса вблизи верхней границы распространения
231
и периодически выпадавший небольшой дождь, испарение за сутки в этот
и последующие дни достигло следующих величин:
Высота над поверхностью почвы, см Испарение, мл
150................................. 7,7-10,9
30 ................ 6,6-8,9
2.............................. 5,8-8,1
Шестого января в 10 час. 25 мин. еще до появления тумана
было проведено несколько определений интенсивности транспирации
(в мг/г-мин) у разных растений (табл. 41); испарение достигало в это
время 1,05 мл!час.
Таблица 41
Ericaceae
Erica arborea . . .
Philippia cf. trimera
Compositae
2,6 Euryops dacrydioides 1,4
6,1 Helichrysum cf. newii . .3,6
Полученные невысокие значения приблизительно соответствуют
интенсивности транспирации у распространенной в альпийском поясе
Loiseleuria procumbens. 1
В этих крайних климатических условиях растения приобретают ксе-
роморфную структуру. Большинство малакофильных растений покрыто
белым войлочным покровом. У некоторых растений развиваются эрикоид-
ные листья. У видов гигантского Senecio крупные листья по своему строе-
нию очень напоминают Tussilago farfara.
Аналогичные климатические условия, по данным Хэдберга, сущест-
вуют и на горе Кения Ч В долине Телеки на высоте 4200 м с 2 по 9 августа
термограф зарегистрировал следующие максимальные и минимальные
значения температуры:
1) на склоне, 0,5 м над уровнем почвы . . . +9,5 и —1,0°,
2) на дне долины, 1,0 м над уровнем почвы ...+15,0 и —6,5°.
На дне долины были развиты заросли Lobelia keniensis. Растения должны
приспособиться к существованию в этих суровых условиях, отличающихся
большим числом дней в году с отрицательной температурой. По данным
Хэдберга, растущие части растений защищены от сильных колебаний
температуры по-разному. Ночью, например, Lobelia и Senecio закрывают
свои розетки. Кроме того, у Lobelia точка роста погружена в 10-сантимет-
ровый слой воды, который ночью замерзает только частично. Ствол
покрыт старыми листьями или толстым пробковым слоем. У растений-
подушек (например, Festuca pilgeri ssp. supina) черешки отмерших листьев
также не отпадают.
Чередование положительных и отрицательных температур способ-
ствует развитию процессов солифлюкции, которые в свою очередь опреде-
ляют мозаичный характер распределения растений, и в частности Heli-
chrysum citrispinum var. hoehnelli, Carduus keniensis, Festuca sp. и Senecio
majeri-johannis.
1 О. H ed b erg, Ekologisk specialesering in den afroalpine floran,
Svensk Naturvetenskap, 1963, S. 158—170.
Глава 5
ТРОПИЧЕСКИЕ ПОЛУВЕЧНОЗЕЛЕНЫЕ
И ЛИСТОПАДНЫЕ ЛЕСА
СЕЗОННЫЙ ЛИСТОПАД С ЭКОЛОГИЧЕСКОЙ ТОЧКИ ЗРЕНИЯ
Если в тропическом дождевом лесу листопад не связан с определен-
ным временем года, то в полувечнозеленых и тропических листо-
падных лесах это типично сезонное явление, причем, видимо,
опадение листвы здесь также обусловлено сухим периодом, как в уме-
ренной зоне; считается, что оно вызвано наступлением зимы. Однако
в последнем случае эта зависимость чисто случайна, ибо летнезеленые
деревья теряют листву зимой даже в том случае, если их поместить в оран-
жерею или пересадить в местность с очень мягкой, безморозной зимойт
например в Капскую область.
Правда, в этом случае деревья стоят без листьев более короткий
период. С весенним потеплением на деревьях начинают распускаться
молодые листья, одни — раньше, другие — несколько позже. В листо-
падных тропических лесах это явление также не может быть объяснено
так просто, как это может показаться на первый взгляд. В сухие годы
листопад наступает раньше, после сезона обильных дождей — позже.
Однако появление листьев не совпадает с началом периода дождей;
деревья часто цветут до появления листьев, задолго до начала сезона
дождей. Это подтверждается как литературными данными, так и наблю-
дениями автора, изучавшего это явление в полувечнозеленых лесах
Панамы, у листопадных эвкалиптов северной Австралии и у колючей
акации в Юго-Западной Африке. Таким образом, в данном случае решаю-
щим фактором является не гидратура, а повышение температуры.
Можно высказать предположение, что листопадные породы должны
быть распространены и в областях, где выражены два сезона — очень
сухой, жаркий и более прохладный, но дождливый, — и что на деревьях
должны появляться молодые листья в конце сухого сезона. Однако этого
не наблюдается. Очевидно, облиствление у деревьев связано не с пониже-
нием температуры, а, наоборот, повышение температуры дает импульс
к появлению новой листвы после периода покоя. В областях, где осадки
выпадают зимой, распространены вечнозеленые древесные породы, у кото-
рых листва также появляется весной в период повышения температуры
в начале сухого сезона. Листопадные виды (например, платаны в Средизем-
номорье), развитые в таких условиях, на влажных местообитаниях сохра-
няют листву летом и во время засухи.
Известен только один вид, распространенный в пустыне Сонора (север-
ная Мексика, Аризона), кустарник Fouquieria splendens, на котором листья
появляются после каждого сильного дождя и, по-видимому, в любое время
года нередко до шести раз в год (ср. стр. 364 и след.), чтобы затем, после
нескольких недель засухи, снова опасть, следовательно, этот вид не имеет
настоящего периода покоя. Вообще же периодичность облиствления,
видимо, обусловлена температурным фактором.
Тропические полу вечнозеленые и листопадные леса
233
В областях с одним неблагоприятным сезоном приспособление расте-
ний к изменению среды развивалось в двух противоположных направ-
лениях.
1) В одних случаях это выражалось в приобретении листьями ксеро-
морфной структуры, позволявшей им переносить неблагоприятное время
года (жестколистные формы в областях, где осадки выпадают зимой,
Eucalyptus, акации с филлодиями, зимнезеленые лиственные породы, как
Ilex. Hedera и др., а также зимостойкие хвойные).
2) В других это было связано с сокращением срока жизни листьев,
их опадением в неблагоприятное время года, что компенсировалось,
однако, некоторой гигроморфностью их структуры, как это наблюдается
у всех листопадных древесных пород.
Развитие листьев того или иного типа связано с конкурентной спо-
собностью данного вида. Преимущество ксероморфных вечнозеленых
растений состоит в том, что фотосинтетическая деятельность, а следова-
тельно, и процесс накопления вещества может начаться у них в любое
время при наступлении благоприятных условий; зато и расход вещества
на построение листьев у них относительно большой. Листопадные же виды
должны ежегодно обновлять свои листья, однако в силу своей гигроморф-
ной структуры они расходуют на построение листовой поверхности срав-
нительно мало вещества.
В тропических областях с летним сезоном осадков и длительным
засушливым периодом в течение прохладного времени года доминируют
листопадные древесные породы; исключение составляет лишь Австралия,
где распространены вечнозеленые эвкалипты, а у видов Acacia вместо
листьев образуются филлодии. Видимо, соответствующих предпосылок
для формирования опадающей листвы здесь не было. Только в Северной
Австралии среди элементов индо-малайского происхождения встречаются
некоторые листопадные породы: Gyrocarpus. Bauhinia, Brachychiton.
Cochlospermum (Bixaceae) и др.
Представьте себе наше удивление, когда южнее Дарвина мы обнару-
жили на обширной площади листопадные эвкалипты; это было в конце
октября, и деревья только что начали покрываться крупными мягкими
светло-зелеными листьями. В литературе это явление не отмечалось.
Наиболее часто листопадная форма встречалась у Eucalyptus alba. Анало-
гично ведут себя также Ей. grandifolia. Ей. bigalerita, Ей. confertifolia
и Ей. clavigera. У двух последних видов листья более жесткие и утолщен-
ные. При наличии длительных дождей листва опадает лишь частично. Сле-
довательно, речь идет о видах, факультативно сбрасывающих листву, что
характерно и для многих других древесных пород, свойственных полу-
вечнозеленым лесам. Как и последние, они распространены в областях,
изобилующих летними осадками и отличающихся коротким периодом
засухи, или в полосе перехода к вечнозеленому дождевому лесу. Однако
здесь, по-видимому, они обладают конкурентной способностью только
в неблагоприятных условиях местообитания, например на каменистых
гребнях гор или в понижениях рельефа на тяжелых почвах, сильно высы-
хающих в сухое время года. На таких участках вечнозеленые эвкалипты,
свойственные влажным лесам, не выдерживают конкуренции, а виды
вечнозеленых Eucalyptus с резко выраженными ксероморфными листьями,
развитые в засушливых областях, видимо, отстают от листопадных видов
по степени продуктивности органического вещества и тем самым по пока-
зателю успешного роста. Преимущество листопадных пород сказывается
в том, что они создают большую листовую поверхность при минимальных
234
Глава 5
затратах вещества. Большая же листовая поверхность способствует более
интенсивному накоплению органического вещества и более быстрому
росту (см. стр. 31 и след.).
Таким образом, в определенных климатических областях листопад обу-
словливает повышенную конкурентную способность. Однако в некоторых
флористических областях, например в Новой Зеландии, где распростра-
нены только вечнозеленые виды, развитые как на границе леса с засушли-
выми областями (на плато Отаго), так и у верхнего предела распростра-
нения леса, предпосылки листопада вообще отсутствуют. Завезенные
сюда листопадные виды Rosa в короткое время превратились в злостных
сорняков и распространились в пограничной полосе между лесом и зла-
ковником. При попытках лесоразведения на юге острова Южный оказа-
лось, что Acer pseudoplatanus превращается в настоящий сорняк, тогда как
Betula и Larix легко акклиматизируются, так же как и кустарники Sam-
bucus nigra, Ribes grossularia, R. sanguineum и различные виды Salix.
ТРОПИЧЕСКИЕ ПОЛУ ВЕЧНОЗЕЛЕНЫЕ
И ЛИСТОПАДНЫЕ ЛЕСА
В условиях, где засушливый период длится в течение 1—2,5 месяцев,
вечнозеленый дождевой лес может развиваться лишь там, где выпадает
большое количество осадков (2500—3000 мм) (см. цветное фото 2, стр. 80).
Однако и этого срока уже достаточно для возникновения в таких районах
известной периодичности в развитии растений. Она выражается в одно-
временном сбрасывании листвы и появлении новых листьев, в прекра-
щении в засушливое время жизнедеятельности эпифитных орхидей; эти
леса отличаются также менее пышным развитием. Поэтому Бирд называет
их вечнозелеными сезонными лесами *. В этих лесах, распространенных
в бассейне Амазонки, близ Монти-Алегри, штат Пара, Эштон определял
освещенность; его данные согласуются с результатами исследований,
проведенных Эвансом в южной Нигерии 1 2. Переход от типичного дожде-
вого леса к лесу, где выражены сезонные явления, осуществляется
весьма постепенно. Состав и структура леса претерпевают существенные
изменения лишь при увеличении продолжительности засушливого перио-
да или сокращении количества осадков. В этих случаях верхний ярус
древостоя образуют листопадные породы, в то время как в нижних ярусах
сохраняются вечнозеленые виды. Поэтому в нижних ярусах древостоя
эти леса называются полувечнозелеными.
При ухудшении условий гидратуры увеличивается число листопадных
пород. В соответствии с этим различают тропические влажные и сухие
листопадные леса.
Эти леса занимают в Африке почти все пространство южнее Сахары
до рек Купене и Замбези на юге материка, со всех сторон окаймляя эква-
ториальную область распространения вечнозеленого дождевого леса 3.
В Азии упомянутые леса сосредоточены на полуострове Индостан, в Индо-
китае, а также в засушливых областях Индонезии. В Южной Америке
большую часть материка также занимают эти растительные формации.
1 J. S. Beard, Climax vegetation in tropical America, «Ecology», 2$, 1944.
p. 127-158.
2 G. С. E v a n s, Ecological studies in the rain forest of Southern Nigeria, «J. Ecol.»,
27, 1939, p. 436—482.
3 W. Mans hard, Ein Vorschlag zur Gliederung und Benennung von Vegeta-
tionsformationen in Afrika siidlich der Sahara, «Geogr. Taschenb.», 1960/61, S. 454—463.
Тропические полу вечнозеленые и листопадные леса
235
В Австралии, напротив, в соответствующей климатической области листо-
падные виды деревьев почти совершенно отсутствуют; их заменяют здесь
вечнозеленые виды Eucalyptus,
Рис. 181. Климадиаграммы станций, расположенных в вечно-
зеленых (Мармаган), полувечнозеленых (Белгаум), влажных
(Бидар) и сухих (Джайпур) муссонных лесах Индии («Klima-
diagramm-Weltatlas», 1960).
Область развития этих лесов отличается от области распространения
вечнозеленого дождевого леса главным образом четко выраженными сезон-
ными изменениями климата, обусловленными в основном неравномерным
распределением осадков. Известную роль играют также температурные
условия, особенно на значительном удалении от экватора и при увеличе-
нии продолжительности засушливого периода. Эта область еще может
быть отнесена к тропикам, поскольку здесь не бывает заморозков, однако
засушливый период здесь заметно холоднее. В этих условиях выражены
два сезона: теплое время года с большим количеством осадков и сухой
прохладный сезон.
Дожливый и сухой периоды особенно резко выделяются в областях,
где выражены муссоны, например в Индии, обусловливая распространение
здесь сухих и влажных муссонных лесов. Характер
дождей — зенитальный или мусонный — не влияет на развитие расти-
тельного покрова. Лауэр строит свою классификацию тропических лесов
236
Глава 5
Рис. 182. Зависимость развития различных лес-
ных формаций от количества осадков (ордината)
и продолжительности засушливого периода
(абсцисса) в Индии.
I — вечнозеленый и II — полувечнозеленый тропиче-
ский дождевой лес; III — муссонный лес (А — влаж-
ный, Б — сухой); IV — саванна (колючекустарнико-
вая); V — пустыня.
только на основании продол-
жительности периода засухи1-
Однако этого явно недоста-
точно, поскольку для выяв-
ления закономерностей зо-
нального распространения
лесов необходима всесторон-
няя характеристика климата
в целом. В качестве иллю-
страции этого положения при-
водятся соответствующие кли-
мадиаграммы (рис. 181).
В условиях нарастающей
сухости климата количество*
осадков или продолжитель-
ность засушливого периода
не могут быть приняты в ка-
честве критерия, определяю-
щего границы распростране-
ния различных лесных фор-
маций, поскольку продолжи-
тельный период засухи может
компенсироваться большим
количеством осадков.
Н еобхо димо, следов а-
тельно, учитывать как коли-
чество осадков, так и продол-
жительность засушливого пе-
риода. Это явствует хотя бы
из исследований, проведен-
ных по поручению ЮНЕСКО
в Индии 1 2. При классифика-
ции растительности использо-
вана карта лесов, составлен-
ная Чемпионом, в течение
длительного времени прово-
дившим исследования в лесах
Индии 3. Величины осадков
и продолжительность периода
засухи определены по соот-
ветствующим климадиаграм-
мам. Если на ординате отложить количество осадков, а на абсциссе —про-
должительность засушливого периода по данным отдельных станций, то
станции, расположенные в пределах области распространения той или иной
лесной формации, сгруппируются в резко ограниченном контуре (рис. 182).
Если бы единственным критерием классификации лесов была бы продолжи-
тельность засухи, то линии раздела проходили бы строго вертикально. При
учете одних только осадков линии раздела располагались бы, по-види-
1 W. Lauer, Humide und aride Jahreszeiten in Afrika und Siidamerika und ihre
Beziehungen zu den Vegetationsgiirteln; «Bonner Geogr. Abh.», 9, 1952.
2 UNESCO, Humid Tropics Res. Progr. 106B.
3 H. H. Champion, A preliminary survey of forest types of India and Burma,
«Indian Forest Records N.S. Silvicult.», I, New Delhi, 1936.
Тропические полу вечнозеленые и листопадные леса
237
мому, горизонтально. В действительности же кривые проходят под углом,
причем во влажных областях круче, а в сухих — положе.
Таким образом, смена тропического дождевого леса полувечнозеле-
ным лесом и последнего вечнозеленым сухим лесом в значительно большей
степени определяется продолжительностью засушливого периода, чем
годовой суммой осадков. Противоположная зависимость характерна при
смене вечнозеленого сухого леса саваннами и пустынями.
Аналогичная зависимость, видимо, существует и в тропических обла-
стях Африки и Южной Америки, с той лишь разницей, что здесь годовое
количество осадков и продолжительность периода засухи в каждом
отдельном случае будут изменяться в соответствии с особенностями кли-
мата. Проверить высказанные предположения довольно трудно, посколь-
ку растительность в этих районах сильно нарушена человеком и до сих
пор здесь не проведено тщательных геоботанических исследований, ана-
логичных выполненным, например, в Индии.
Поскольку в сомкнутом дождевом лесу человек не может найти всего
необходимого для жизни, первые поселенцы в этих областях прежде всего
начинали борьбу с лесом. Именно этим объясняются огромные масштабы
истребления тропических лесов. Обратный процесс —восстановление
леса — обычно начинается слишком поздно. В Австралии и сегодня еще
среди фермеров господствует мнение о необходимости полного уничтоже-
ния леса: «травинка дороже двух деревьев».
Уничтожение леса и в тропиках начинается с небольших вырубок;
в дальнейшем после зарастания вырубок травой уже не представляет
4-6 6-8 8-ю 10-11 12
м
50-
40’
31-
28-
18-
0-
Вечнозеленый Вечнозеленый
дождевой лес сезонный лес
Число сухих месяцев
количество осадков,
Незначительное
выпадающее регулярно каждый год
Заросли Полупустыня Пустыня
суккулентных с господством
кустарников суккулентов
Неравномерное количество осадков по годам
Яолувечнозеленый Сухой листа- Сухие листопад- Кустарниковая
сезонный лес ладный лес
е г 1 * X Глувина проникновения
• - •'___г " 7 корней
ные заросли полупустыня Пустыня
кустарников
t
Высокотравные саванны Низкотравные саванны
Многолетние культуры вез орошения | с_ орошением ------
Культуры на выжженных и Культуры дождливого
выкорчеванных участках периода
Культуры сырого периода
Рис. 183. Зависимость развития различных лесных формаций в тропических об-
•стях Перу от продолжительности периода засухи и количества осадков (по Эллен-
бергу).
На схеме показаны также пути деградации лесов в саванны после пожаров и пригодность лес-
ных почв для использования под сельскохозяйственные культуры.
большого труда, путем ежегодного выжигания травы, наносить все больше
и больше повреждений древостою до тех пор, пока лес не сменится откры-
тым, напоминающим саванну ландшафтом. Поскольку в области распро-
странения полувечнозеленых и тропических влажных листопадных лесов
238
Глава 5
Рис. 184. Полувечнозелевый тропический лес с господ-
ством Dipterocarpus tuberculatus, Верхняя Бирма. На перед-
нем плане лиана Spatholobus roxburghii (фото Чемпиона).
осадки выпадают более регулярно, чем в области развития сухих листо-
падных лесов, в связи с чем первая область более удобна для развития
земледелия, именно первые два типа леса 1 почти полностью уничтожены.
Видимо, в связи с этим географы применяют термин «влажные саванны».
Напротив, сухие листопадные леса, не столь благоприятные для развития
земледелия, еще сохранились на обширных площадях.
Более детальные экологические исследования в этих лесах до настоя-
щего времени не проводились. Не останавливаясь подробно на их флори-
стическом составе, ограничимся лишь их кратким общим описанием.
На рис. 183 на примере Перу и прилегающих аридных территорий схема-
тически представлены отдельные типы леса. Поскольку описание расти-
тельности аридных районов будет приведено позднее, здесь коротко оста-
новимся лишь на характеристике трех типов леса:
1 Автор трактует понятие «тип леса» совершенно по-иному, чем это принято
в СССР. В некоторых случаях это в известной степени соответствует представлению
о формации, в иных же это отражает взгляды автора на закономерности распределе-
ния климатов и сопутствующей им растительности.— Прим. ред.
Тропические полу вечнозеленые и листопадные леса
239
Рис. 185. Tetrameles nudiflora после листопада; во-
сточная Ява.
а) тропических полувечнозеленых дождевых лесов;
б) тропических влажных листопадных лесов, или влажных муссон-
ных лесов;
в) тропических сухих листопадных лесов, или сухих муссонных лесов.
а) Тропические полувечнозеленые дожде-
вые леса. Близкие по своей структуре к вечнозеленым лесам, они
отличаются от последних лишь несколько меньшим числом видов. Неко-
торые древесные породы образуют чистые древостои. Досковидные корни
и каулифлория наблюдаются реже.
Особенно хорошо развиты тропические полувечнозеленые дождевые
леса в муссонной области Индостана, Бирмы (рис. 184), Таиланда и полу-
острова Малакка *, где они непосредственно примыкают к полосе вечно-
1 Н. Н. Champion, A preliminary survey of forest types of India and Burma.
«Indian Forest Records N.S. Silvicult.», I, New Delhi, 1936; G. S. Puri, Indian
Forest Ecologie, New Delhi, Calcutta, 1960; H. О g a w a, K. Y о d a, T. К i r o, A pre-
liminary survey on the vegetation of Thailand, «Nature life SE Asia», 1, 1961, p. 21 —
157;Тай-ван Труп, Экология и классификация лесной растительности Вьетна-
ма, Л., 1962.
240
Глава 5
Рис. 186. Тиковый (Tectona grandis) лес во влажное время года; восточная Ява.
Почва покрыта Gastrochilus panduratum.
зеленых лесов. В дождливое время года полувечнозеленый лес существенно
не отличается от вечнозеленого и древостой в среднем не различается по
высоте; однако отдельные деревья никогда не достигают тех размеров,
Тропические полу вечно зеленые и листопадные леса
241
Рис, 187. Лес, образованный Pentacme suavis, Верхняя Бирма (фото Оливера).
которые свойственны наиболее крупным деревьям в вечнозеленом лесу;
кроме того, верхний ярус древостоя в полувечнозеленом лесу характери-
зуется большей разреженностью. В полувечнозеленом лесу еще очень
242
Глава 5
много лиан и эпифитов. Различия становятся заметными лишь в период
засухи. Деревья, периодически сбрасывающие всю листву, встречаются
и в вечнозеленом лесу, однако листопад там не связан с каким-либо опре-
деленным временем года. В верхнем ярусе полувечнозеленого леса до 30%
деревьев относится к числу листопадных видов (рис. 185), нижние же яру-
сы включают еще значительное количество вечнозеленых особей; листья
у деревьев преимущественно более мелкие, чем в вечнозеленом дождевом
лесу. Молодые листья обычно появляются за месяц до начала муссонных
дождей, что связано с повышением температуры. Это свидетельствует
о наличии в почве даже к концу засушливого периода достаточного запаса
влаги, необходимого для начала вегетации.
Годовая сумма осадков в описываемой лесной области, как правило,
превышает 1500 мм. В горах, где температура несколько ниже, а влаж-
ность воздуха очень высока (туманы), вечнозеленый горный дождевой
лес может развиваться, несмотря на отсутствие осадков в течение несколь-
ких месяцев. Так, на берегу моря у подножия Западных Гатов в Индии
распространены полувечнозеленые леса, а в горах, где количество осад-
ков местами превышает 5000 мм, преобладают вечнозеленые леса. Послед-
ние в качестве экстразональной растительности встречаются также на
влажных местообитаниях вдоль русел рек.
В Африке, в бассейне Кванго, вечнозеленые и полувечнозеленые леса
простираются южнее 5° ю. ш. Однако между последними и сухими листо-
падными лесами бассейна Замбези вклиниваются обширные простран-
ства злаковников, несмотря на то, что климат здесь достаточно влажный.
Девред 1 считает, что прежде эта область была занята полувечнозелеными
лесами переходного типа. С увеличением сухости климата способность
к восстановлению этих лесов после пожаров и раскорчевания уменьши-
лась и в конечном итоге они были полностью уничтожены человеком.
На их месте распространились злаковники.
Эта область, где развиты леса переходного типа, в Африке, видимо,
не выражена, поскольку обширные массивы тропического вечнозеленого
леса непосредственно граничат с открытыми ландшафтами — антропо-
генными саваннами, являющимися стадией деградации полувечнозеленых
лесов.
Это же может быть сказано и о саваннах Гвинеи, развивающихся
при годовой сумме осадков 1000—1500 мм, они занимают обширные про-
странства в Нигерии, примыкая на юге к дождевым лесам. Эта область
распространения саванн плотно населена, и огонь здесь регулярно выжи-
гает значительные площади. Первоначально здесь был развит полувечно-
зеленый лес с господствующими в верхнем ярусе Abbizzia adianthifolia,
A. zygia и Bombax buonopozense и нижним ярусом, образованным вечно-
зелеными породами. Теперь в производных саваннах встречаются Daniella
с Elaeis или Uapaca 1 2. Хопкинс также отмечает, что в Нигерии при сумме
годовых осадков, достигающей 1232 м, и засухе, продолжающейся 4—
5 месяцев, после пожаров полувечнозеленые леса сменяются вторичными
саваннами 3.
1 R. D е vred, Limite pbytogeographique occidento-meriodionale de la region
guineenne au Kwango, «Bull. Jard, Bot, Bruxelles», 27, 1957, p, 417—431.
2 D, Clayton, Derived savanna in Kabba-Province, Nigeria, «J. Ecol.», 49,
1961, p, 595-604,
3 В, H о p k i n s, Vegetation of the Olokemeji Forest Reserve,Nigeria I, «J, Ecol.»,
50, 1962, p. 559—598.
Тропические полувечнозеленые и листопадные леса 243
Деградация лесов наблюдается даже в заповедниках. Из Националь-
ного парка в районе водопада Мерчисон в Уганде из-за эпидемии сонной
болезни в 1912 г. было эвакуировано все население, что вызвало резкое
увеличение поголовья слонов. Повреждаемые ими деревья засыхали и при
первом же пожаре гибли. Постепенно тропические леса, образованные
Chlorophora excelsa, и полувечнозеленые леса из Terminalia glaucescens
сменились безлесными злаковниками. С 1932 по 1956 г. общая площадь
древостоя сократилась на 55—60%. Сейчас здесь распространяются
только немногочисленные породы: не повреждаемая слонами Hapoptelia
grandis или огнестойкие виды, например Lonchocarpus laxtflorus \
Аналогичные описанным полувечнозеленые дождевые леса встре-
чаются также в Южной Америке 1 2, где они граничат с вечнозелеными
лесами.
б) Листопадные тропические леса (влажные мус-
сонные леса). Влажные листопадные леса известны в Индии под назва-
нием муссонных лесов. Естественных злаковников в Индии нет, поскольку
они все возникли в результате деятельности человека 3. Важнейшие леса
в Индии образованы тиком (Тectona grandis) и салом (Shorea robusta)
с сопутствующими им видами. Тик (рис. 186) играет большую роль и в лес-
ном хозяйстве; он достигает высоты 40 м и вместе с Terminalia tomentosa
и D iospyros melanoxylon образует в центральной части Индии зональные
сообщества, сохранившиеся в настоящее ,время лишь в виде небольших
пятен 4. В сезон дождей эти леса имеют гигрофитный облик; они зеленеют
и в засушливое время года, поскольку листья опадают постепенно. Листья
на деревьях появляются несколько раньше начала дождливого периода,
поэтому леса стоят обнаженными лишь очень недолго. Видовой состав
менее разнообразен, чем в полувечнозеленых лесах, и нередко древостой
образован одним видом. Поскольку леса этого типа более разрежены по
сравнению с полувечнозелеными дождевыми лесами, в них формируется
сомкнутый травяной покров. Лианы и эпифиты еще довольно много-
численны. Большинство растений цветет в сухое время года.
С уменьшением осадков количественно в составе древостоя начинает
преобладать Pentacme suavis (рис. 187); в еще более засушливых условиях
господствуют леса из Terminalia tomentosa, образующие переход к сухим
лесам.
В качестве примера трехмерного распределения растительности
в муссонной области ниже приводится краткое описание растительного
покрова Бирмы 5.
В теплой прибрежной полосе, где количество осадков превышает
1500 мм, распространен полувечнозеленый дождевой лес (высотой 40—
50 м); выделяются два яруса древостоя (Dipterocarpaceae, Leguminosae,
Meliaceae, Sapindaceae и др.), ярус кустарников (Anonaceae, Rubiaceae,
Euphorbiaceae), ярус трав (Агасеае, Zingiberaceae) и большое число лиан
1 Н. К. Buechner, Н. С. Dakins, Vegetation change induced by elephants
and fire, «J. Ecol.», 49, 1961, p. 752—766.
2 F. S. В e a r d, см. стр. 234; ср. К. Hueck, Die Walder Venezuelas, «Beih.
Fortsw. Cbl.», H. 14, 1961.
3 R. O. Whyte, S. V. Venkataram ana n, P. M. Dabadghao, The
grasland of India, 8. Congr. Internet. Botan., Compt. Rend. Sect., 7,8, Paris, 1954,
p. 46-53.
4 К. К. В h a t i a, A mixed Teak forest of Central India, «J. Ecol.», 46, 1958,
p. 43—63.
5 J. Vidal, Conditions ecologiques, groupements vegetaux et flore de Laos,
«Bull. Soc. Bot. Fran^.», Mem., 1959, p. 3—41.
244
Глава 5
и эпифитов (Orchideaceae). На вырубках широко распространен Eupato-
гшт odoratum. На маломощных почвах развиваются листопадные леса,
образованные Pentacme, Shorea, Terminalia tomentosa и Dipterocarpus.
Древесный ярус здесь достигает меньшей высоты и менее сомкнут, напоч-
венный покров хорошо выражен. В переходной полосе развиты леса,
где господствует Tectona. В горах (высота 600—800 м над уровнем моря,
количество осадков 3000 мм) распространены вечнозеленые диптерокар-
повые леса с пальмами и бамбуком. На высотах 1000—2000 м преобла-
дают леса, образованные видами Fagaceae (Castanopsis, Lithocarpus,
Pesania, Quercus) и Lauraceae. Половина видов относится к числу листо-
падных. На высоте более 2000 м деревья покрыты Usnea и мхами, а почва
покровом из Selaginella. Здесь встречаются также хвойные (Pinus тегки-
sii, Р. khasya, Keteleeria), распространению которых способствуют пожары
и выпас скота.
Смену времен года в муссонной области Индии подробно описал Мие-
ра; на равнине верхнего Ганга он выделяет 6 фенологических пери-
одов Ч
1) В марте и апреле температура начинает повышаться и суточная
амплитуда ее достигает 25°. На деревьях появляются молодые листья.
В этот период цветет большинство трав.
2) Май и июнь — самые сухие и жаркие месяцы года. Рост про-
должается лишь на влажных местообитаниях.
3) Основная масса осадков выпадает в июле и августе. Снова стано-
вится прохладнее; в это время наблюдаются минимальные суточные коле-
бания температуры. Это период массового прорастания семян и макси-
мального роста деревьев.
4) В сентябре и октябре ливневые дожди сменяются солнечной пого-
дой. Условия для роста чрезвычайно благоприятны; цветут и плодоносят
однолетние виды.
5) В ноябре и декабре стоит прохладная погода; заканчивается
период роста.
6) В январе и феврале наблюдаются минимальные температуры —
это самое холодное время года и период покоя у растений; напочвенный
покров изреживается.
В результате обильных осадков низменные участки в течение 4—
8 месяцев оказываются затопленными. После спада воды здесь на илистых
почвах быстро развиваются сообщества травянистых растений. Леса
сохранились лишь в очень немногих местах, поскольку равнинные терри-
тории подвергаются интенсивному выпасу скота и покрываются постепен-
но зарослями колючих растений. Более возвышенные участки возделы-
ваются.
Огава с сотрудниками1 2 опубликовал весьма подробное описание
растительности Таиланда, которое иллюстрируется картой растительно-
сти и профилем вертикальной поясности; кроме того, они изучали продук-
тивность растительных сообществ влажных и сухих местообитаний,
а также некоторых вторичных группировок, а именно:
1) Разреженный сухой диптерокарповый лес, расположенный на
высоте 300 м\ редко стоящие деревья достигают высоты 20 м, высота тра-
вяного покрова 20—30 см.
1 R. М i s г a, The status of the plant communities in the upper Gangetic plain,
«J. Ind. Bot. Soc.», 38, 1959, p. 1—7.
2 H. Ogawa, K. Yoda, T. К i r o, op. cit.
Тропические полувечнозеленые и листопадные леса 245
2) Несколько более влажный смешанный листопадный лес; деревья
достигают высоты 20—25 м, травяной покров редкий.
3) Влажный вечнозеленый галерейный лес высотой 30 м, очень
сомкнутый, с лианами, но лишь небольшим количеством эпифитов.
4) Умеренный вечнозеленый лес, произрастающий на высоте 1200 м,
образованный Pinus khasia, Terminalia, Artocarpus, и др. видами, дости-
гающими высоты 20 м. В нижнем ярусе древостоя развиты Symplocos,
Quercus, Castanopsis и др. Множество эпифитных орхидей; лианы отсут-
ствуют.
5) Злаковник из Eragrostis, Andropogon, Agrostis, Setaria и др.
6) Хорошо развитый злаковник высотой до 2,3 м, образованный
Themeda и Arundinaria, на старых термитниках.
7) Густые заросли Arundo donax (высота 300 м, вдоль берегов реки),
похожие на бамбук, побеги которого достигают высоты 7 м.
На семи пробных площадях были подсчитаны запасы растительной
массы (см. табл. 42) в тоннах на 1 га в абсолютно сухом состоянии (римские
цифры в таблице означают запасы массы: I — стволов и ветвей; II —
листьев; III — валовой запас, включая подрост и корни; IV — показа-
тель облиствленности; V — нетто-продуктивность надземных частей рас-
тений, за вычетом годового прироста древесины).
Таблица 42
Место наблюдений I п Ш IV V
1. Сухой диптерокарповый лес 44,7 4,9 65,9 4,3 7,8
2. Смешанный лес 52,6 4,9 77,0 4,2 8,0
3. Галерейный лес 270,0 19,5 377,6 16,6 25,3
4. Умеренный лес . 163,4 14,5 231,4 12,6 18,9
5. Злаковник — — 13,5 — 11,3
6. Злаковник на термитниках . — — 42,1 — 38,5
7. Заросли Arundo donax — — 162,5 23,3 51,9
Упомянутые авторы исследовали также содержание углерода и азота
как в растительной массе, так и в почве. Показатель облиствленности
галерейного леса, по-видимому, завышен. Для сравнения авторы приводят
ПО тропических дождевых лесов (8,5—Берег Слоновой Кости; 9,5—
Нигерия), умеренных лесов северной Японии (менее 7) и южной Японии
(более 7). Чрезвычайно велик ПО зарослей Arundo; это объясняется тем,
что листья у этого вида расположены почти вертикально. Кроме того,
ширина зарослей достигала всего 3—5 м, что создавало для растений
дополнительное боковое освещение. Большая нетто-продуктивность свя-
зана с высоким показателем облиствленности. Нетто-продуктивность
леса, образованного Abies sachalinensis на севере Японии, равна: хвои —
4,5, древесины — 16,8, общей массы — 21,4 т!га (брутто-продуктив-
ность — 47,7 т/га), а вечнозеленого леса из Disylium racemosum на юге
Японии составила: листьев — 11,4, древесины — 9,2, общей массы —
21,6 т/га (брутто-продуктивность — 73,1 mlea}.
Нетто-продуктивность тропических лесов Огава оценивает в 40 т!га,
а брутто-продуктивность, учитывая, что потери на дыхание достигают
60—70 т!га, более чем в 100 т!га (ср. стр. 44).
246
Глава 5
ТРОПИЧЕСКИЕ СУХИЕ ЛИСТОПАДНЫЕ ЛЕСА
И ЗАРОСЛИ КОЛЮЧИХ КУСТАРНИКОВ
Первые называют также сухими муссонными, или саванновыми,
лесами. В этом разделе будут также рассмотрены формации, развитые
в экваториальных районах и распространенные в наиболее засушливых
местообитаниях; они представлены зарослями колючих кустарников,
в составе которых встречаются многочисленные очень интересные сукку-
ленты. Переходная полоса от сухих листопадных лесов к саваннам,
характеризующаяся уменьшением летних осадков по мере удаления от
экватора, будет описана несколько ниже. Тропические сухие листопадные
леса под различными названиями описаны в Южной Америке и глав-
ным образом в Африке (рис. 189). Определение «сухой» очень точно харак-
теризует внешний облик этих лесов в засушливое время года, когда они
стоят обнаженными (рис. 190). Голые ветви не спасают от жгучих лучей
солнца, земля покрыта желтой засохшей травой; в случае недавно про-
шедшего пожара все вокруг бывает покрыто черной угольной пылью
и в воздухе еще долго стоит запах гари. Но уже перед началом лет-
них дождей деревья покрываются светло-зелеными, часто словно покры-
тыми лаком листьями. Многие деревья цветут, но цветки у них обычно не-
взрачны. Изредка можно увидеть деревья, буквально увешанные гроз-
дьями золотистых цветков, например Burkea africana, или виды Егу-
thrina, крупные нежно-розовые цветки которых распускаются до того,
как развернутся листья. Последние напоминают цветущие персико-
вые деревья весной. С началом дождей трогаются в рост и травы, на поч-
ве появляются многочисленные всходы разнотравья и многолетних
злаков.
Сухие леса, развитые в области летних дождей, в зависимости от тех
или иных условий среды отличаются весьма разнообразным флористиче-
ским составом. Они подробно описаны, однако в них до сих пор не прово-
дились эколого-физиологические исследования. В Замбии и Танзании
наиболее широко распространены леса, образованные Brachystegia и Iso-
berlinia или Julbernardta (все — Leguminoseae). Эти леса напоминают раз-
витые в Европе низкоствольные дубравы; деревья достигают высоты 9—
12 м', степень сомкнутости полога — 60—80%. Деревья трогаются в рост
за месяц до начала летнего периода дождей; цветут они в это же время.
Деревья стоят без листьев в течение более прохладного засушливого
периода; продолжительность пребывания деревьев в таком состоянии
зависит от водного режима почв. В этих лесах развит подлесок, однако
травяной покров почти не выражен. Здесь встречаются пойкилогидриче-
ские папоротники Pellaea hastata и Acttniopteris radiata. Подробное описа-
ние этих лесов приводится в работе Берта Ч
Вопреки широко распространенному мнению господствующие в этих
лесах древесные породы очень легко повреждаются огнем. В результате
многократных пожаров лес может быть уничтожен полностью; особенно
опасны палы, пускаемые до начала периода дождей 1 2.
1 В. D. В u г t t, Some East African vegetation communities, «J. Ecol.», 30, 1942,
p. 65—146.
2 C. G. Trapnell, Ecological results of woodland burning experiments in
Northern Rhodesia, «J. Ecol.», 47, 1959, p. 129—168.
Тропические полу вечнозеленые и листопадные леса
247
Р и с. 188. Ареал Adansonia digitata (по Шпрингеру).
Более разреженные леса, где развивается более сомкнутый травяной
покров, образованы Colophospermum торапе (леса мопане), различными
видами Combretaceae, Pterocarpus или видами Acacia, Albizzia, а также
Commiphora. При увеличении сухости на элювиальных почвах происходит
смена сухих лесов зарослями колючих кустарников. Среди них часто
встречается баобаб (Adansonia digitata), который обычно сохраняется при
расчистке лесных участков; благодаря причудливой форме ветвей он
придает своеобразный облик ландшафту (рис. 194). У гигантских экзем-
пляров баобаба высотой более 20 м окружность ствола равна 20 м. Запас
воды в древесине достигает иногда 120 тыс. л. Поэтому баобаб, сбрасы-
вающий в сухое время года листья, может легко переносить продолжи-
тельную засуху. Adansonia digitata встречается на границах лесной обла-
сти и в переходной области к саваннам, развитие которых обусловлено
климатическими причинами (рис. 188). В более влажных областях этот
вид приурочен к сухим местообитаниям, например к обнажениям водо-
проницаемых коралловых известняков на восточноафриканском побе-
режье. Деревья, встречающиеся в селениях, обычно посажены челове-
ком и, следовательно, растут вне пределов своего естественного ареала.
Это, в частности, относится к экземплярам, распространенным на остро-
вах Зеленого мыса, Канарских, на Мадейре и на островах Индийского
Рис. 189. Редкостойный сухой лес в облиствленном состоянии на северо-востоке-
Замбии. На переднем плане — четырехметровый термитник (фото Фриса).
Тропические полу вечно зеленые и листопадные леса
249
океана. На Мадагаскаре и в северо-западной Австралии встречается
несколько родственных видов.
Возраст крупных экземпляров легко переоценить. Ствол одного
дерева, посаженного в Хартуме у Голубого Нила, уже через 30 лет достиг
диаметра почти 1 м. Суорт (Сэлисбери) установил радиоуглеродным мето-
дом, что возраст одного крупного дерева достигал 1000 лет Ч
Деревья в сухих листопадных лесах обычно не превышают высоты 20 м.
В таких лесах мало тени, и они, скорее, напоминают низкоствольные
леса умеренных широт. Известные под названием «миом б о», эти леса
в Восточной Африке покрывают огромные пространства. В их составе
господствуют виды Leguminosae {Brachystegia и Isoberlinia)\ флористи-
ческий состав этих лесов очень разнообразен; они проникают далеко
в северные районы Юго-Западной Африки, где их уже называют
«мопане».
Почки древесных пород, распространенных в сухих листопадных
лесах, хорошо защищены от засухи, а стволы покрыты толстой коркой
(рис. 191), которая предохраняет их от губительного влияния частых
лесных пожаров. Эпифиты отсутствуют; редко встречающиеся здесь лианы
имеют толстые деревянистые стволы (рис. 192, 193).
Вблизи экватора в районах с небольшим количеством осадков, выпа-
дающих в течение двух периодов дождей, например в Восточной Африке,
часто распространены очень густые непроходимые заросли колючих кус-
тарников; помимо различных акаций, выделяющихся своими зонтиковид-
ными кронами, здесь развиты и другие виды колючих кустарников.
Очень хорошо эти заросли развиваются на богатых осадками морских
побережьях, где они произрастают на водопроницаемых коралловых
скалах. Над зарослями этих кустарников возвышаются отдельные экзем-
пляры A dans onia digitata с причудливо изогнутыми сучьями и ветвями.
В сухое время года безлистные ветви баобаба увешаны крупными продол-
говатыми плодами. Эта прибрежная полоса в основном уже раскорчевана,
оставлены лишь баобабы, образующие наиболее характерную черту ланд-
шафта (рис. 194).
В еще более засушливых условиях вблизи экватора развиваются
тропические полупустыни, расположенные в ветровой тени высоких гор.
Климат наветренных и подветренных склонов хребтов может быть совер-
шенно различным, особенно в области пассатов, где направление ветров
постоянно.
Чрезвычайно своеобразные растительные формы можно встретить,
например, в небольшом засушливом районе Восточной Африки, распо-
ложенном между горами Паре и Усамбара (рис. 195).
Точных метеорологических наблюдений здесь не производилось, но,
вероятно, это самое засушливое место вблизи экватора (4—5° ю. ш.).
Средняя годовая температура здесь около 28° (температура почвы на глу-
бине 40 см равна 30,5°); количество осадков колеблется от 100 до 200 мм.
Осадки выпадают в течение двух дождливых сезонов, что благоприят-
ствует росту суккулентов. Вокруг расстилается своеобразная полупустыня,
в составе которой господствуют очень интересные виды суккулентов.
Наряду с видами Sansevieria, образующими отдельные крупные куртины,
1 Эти данные заимствованы из монографии о баобабе — A. Springer, Der
Affenbrotbaum (Adansonia digitata L.), Mskr., 139 S., 1 Karte. В этой работе исполь-
зована обширная литература, приводится ареал и описывается значение баобаба
в жизни местного населения. Работа еще не опубликована; копия рукописи хранится
в Ботаническом институте в Штутгарт — Хоэнхейме.
Рис. 190. Сухой листопадный лес (Boswellia papyrifera) в засушливое время года на
•северном склоне Абиссинского нагорья в районе Дембеласа (фото Швейнфурта).
Рис. 191. Поперечный срез ствола
Sweetia dasycarpa*, эта порода рас-
пространена в сухих лесах Брази-
лии (по Уормингу).
Рис. 192. Лиана Fockea damarana (Ascle-
piadaceae) в сухом лесу, где господствуют
Sclerocarya schweinfurthiana и Gyrocarpus
americana] выходы доломитов, горы Отави,
Юго-Западная Африка (фото Э. Вальтер).
Тропические полу вечнозеленые и листопадные леса
251
стеблевыми суккулентами Euphorbia tirucalli и Caralluma (из рода
Stapelia\ см. рис. 196) здесь встречаются похожая на глыбы серого камня
Pyrenacantha (Icacinaceae) и Adenia globosa, чей ствол похож на большой
зеленый шар, из вершины которого во все стороны расходятся многочис-
Р и с. 193. Лиана (Fockea
damarana), напоминающая
гигантскую змею.
Рис. 194. Баобаб (Adansonia digitata) и масличная пальма (Elaeis guineen-
sis) — остатки прежде развитых здесь лесов; окрестности Бомы, нижнее Конго
(фото Уинтера).
252
Глава 5
Рис. 195. Восточная Африка между Тангой и Килиманджаро.
ленные, изогнутые в виде лука, лишенные листьев, но усаженные шипами
зеленые ветви (рис. 197). Ствол настолько закрыт ветвями, что его совсем
не видно. По кустарникам вьются многочисленные виды Cissus, родствен-
ные виноградной лозе; их суккулентные стебли очень напоминают некото-
рые виды Cereus (семейство Cactaceae). Среди колючих кустарников выде-
ляется необычайно красиво цветущий, но очень ядовитый Adenium
(рис. 198). Один из видов этого рода образует на острове Сокотра деревья
с уродливыми стволами (рис. 199).
Суккуленты встречаются в тропиках и в других районах, особенно на
сухих местообитаниях, например по скалистым склонам. В Африке это
прежде всего канделябровидные молочаи и виды А1оё, а в Америке —
кактусы.
Также широко распространены сухие листопадные леса и в Южной
Америке, например на равнине Гран-Чако. Ниже будет дано описание
только бразильских «campos cerrados» и «каатинги», подробно изу-
ченных Равичером и Ферри1.
«Campos cerrados» развиты в области, где осадки выпадают летом и их
годовое количество превышает 1300 мм, а средняя годовая температура
достигает 21,2°. Необыкновенно мощные (до 20 м) почвы аккумулируют
осадки, выпавшие в течение трех лет, так что с глубины 2 м они остаются
1 F. Rawitscher, The water economy of the vegetation of the «Campos cerra-
dos» in southern Brazil, «J. Ecol.», 36, 1948, p. 237—268; M. G. Ferri, L. M. С о u-
t i n h o, Contribuigao ao conhicimento da ecologia do Cerrado, Bol. № 224, Univers, de
Sao Paulo, 1959, p. 103—150.
Тропические полу вечнозеленые и листопадные леса
253
Рис. 196. Засушли-
вый район Восточной
Африки, расположен-
ный между горами
Паре и Усамоара.
Среди куртин Sanse-
vieria отдельные поч-
ти черные цветки Са-
гаИита, обладающие
очець неприятным за-
пахом (фото Э. Валь-
тер).
Рис. 197. Засушливый район Восточной Африки, расположенный между горами
Паре и Усамбара. Зеленый шарообразный ствол Adenia globosa; передние колючие
ветви удалены. На заднем плане — Acacia spiro carp а, рядом — колючие Euphorbia
сипе ata и др. (фото Винклера).
254
Глава 5
Рис. 198. Среди колючих кустарников цветет Ade-
niuin obesum (Аросупасеае). Красиво цветущие, но очень
ядовитые виды Adenium содержат яд, используемый
для наконечников стрел (фото Бюса).
постоянно влажными в течение всего года. Поэтому все глубоко укоре-
няющиеся древесные растения являются вечнозелеными видами и отли-
чаются круглогодичной, очень интенсивной транспирацией. К растениям
с поверхностно расположенными корнями относятся лишь злаки, высы-
хающие в сухое время года, и листопадные древесные породы с чрезвы-
чайно своеобразными подземными стволами (ксилоподиями). К этой груп-
пе принадлежит также «бесствольная» пальма Acanthococcos. После про-
растания семян ее ствол растет вниз, в почву, из которой выходят только
листья, обладающие отрицательным геотропизмом. Иногда вершина ство-
ла достигает глубины 50—60 см, ствол изгибается на 180° и растет вверх.
Часть ствола, направленная вниз, в дальнейшем отмирает и разлагается.
Эти растения очень устойчивы против пожаров, ежегодно возникающих
в период засухи. Из эпифитов встречаются лишь засухоустойчивые виды
Bromeliaceae и Cactaceae (рис. 200).
Тропические полу вечнозеленые и листопадные леса
255^
Из недавно опублико-
ванного обзора 1 явствует,
что под понятием «campos
cerrados» объединяются со-
вершенно различные типы
растительности: от сомкнутых
лесов до открытых саванн.
Частично это естественные,
природные образования, ча-
стично они возникли под воз-
действием антропогенных
факторов. Саванноподобный
облик ландшафта обычно свя-
зан с влиянием ежегодных па-
лов, которые индейцы пуска-
ли задолго до прихода белых.
Редкостойность этих ле-
сов может быть обусловлена
также и естественными при-
чинами, а именно бедностью
почв питательными вещества-
ми и микроэлементами. Око-
ло Пирассунунга (штат Сан-
Пауло) автор видел остатки
полувечнозеленого дождево-
го леса, распространенного
на почве, развитой на ба-
Рис. 199. A den turn socotranum (Apocynaceae)
в западной части Сокотры; диаметр ствола до-
стигает 2 м. Справа — Cissus subaphylla, сле-
ва — Croton socotranus (фото Коссмата).
зальтах; эти леса в этих ус-
ловиях, видимо, являются
зональным типом раститель-
ности. Редколесные саванны
(Campos cerrados), в составе
которых растет Roplea (Proteaceae), напротив, приурочены к бед-
ным песчаным почвам (сходные условия наблюдаются в Квинсленде,
стр. 261). О бедности этих песчаных по^в фосфором и калием, а также-
и микроэлементами свидетельствуют следующие опыты, проведенные
Вейчи с хлопчатником, кукурузой и соей на опытной станции ISIP (уро-
жай дан в кг!га)\
Хлопчатник Кукуруза Соя
Полное удобрение . . 2460 4860 1200
To же, без азота . . . 1440 3600 —
« » » фосфора . . 270 860 240
« » » калия 70 1300 450
« » » серы .... 1570 4170 680
« » » цинка и бора 870 4080 750
« » » кальция . . 980 3900 1100
Контроль — — 240
1 Simposo sobre do Cerrado, Univers, Sao Paulo, 1963.
256
Глава 5
Эти опыты дают возможность судить о том, что хлопчатник реагирует
сильнее на недостаток цинка и бора, чем на недостаток азота в почве.
Урожай в этом случае достигает примерно х/3 величины, полученной при
внесении полного удобрения. В отличие от хлопчатника кукуруза почти
Рис. 200. Cereus megalanthus на "крупном экземпляре Ficus, Тара-
пото, Перу (фото Уле).
не реагирует на недостаток бора и цинка, а у сои эта реакция хорошо
выражена (урожай достигает примерно 3/5 максимальной величины).
Посевы Panicum maximum также свидетельствуют об остром недостатке
цинка и бора в почве (при полном удобрении урожай достигает 18 ml га,
а без внесения микроэлементов — 2 ml га).
То же самое, вероятно, следует сказать и о «чапарале» в Венесуэле,
редкостойной древесной саванне. Хюк в отличие от Вереши не придает
Рис. 201. Цветущее «бутылочное дерево» Cavanillesia arborea (Bombacaceae); ка-
атинга в окрестностях Тамбури, Бразилия (фото Уле).
258
Глава 5
решающего значения влиянию пожаров \ но он, по всем данным, не прав,
ибо при годовой сумме осадков, равной 1700 мм, и почти трехмесячной
засухе, даже при весьма равномерной в течение года температуре, дости-
гающей 28,2°, здесь должен бы был произрастать полувечнозеленый лес.
В качестве крайне сухого варианта листопадного леса можно рас-
сматривать каатингу в засушливой северо-восточной части Бразилии.
Хюк 1 2 пишет, что каатинга является «листопадным сухим редколесьем,
зеленеющим в период дождей и образованным из низких или средней
высоты деревьев и колючих кустарников, довольствующихся любой, даже
самой плохой почвой. Оно вполне соответствует «thorn forest», или колю-
чему редколесью, других жарких стран. Экологические условия характе-
ризуются прежде всего недостатком влаги в течение многих месяцев,
крайней неравномерностью выпадения осадков, относительно низкой влаж-
ностью воздуха и интенсивным испарением. В центре засушливой области
в Полигоно-да-Сека засуха нередко длится на протяжении нескольких
лет, превращаясь в настоящий бич для населения». В каатинге растут
своеобразные запасающие воду «бутылочные деревья» (рис. 201), много-
численные суккуленты, например кактусы, неэпифитные Bromeliaceae
(рис. 202) и виды семейства Euphorbiaceae, также относящиеся к сукку-
лентам. Столь большое разнообразие суккулентов обусловлено характе-
ром распределения осадков; годовое количество осадков, достигающее
500 мм, выпадает в течение длинного дождливого периода, продолжающего-
ся с октября по декабрь, и короткого, охватывающего май — июнь
(рис. 203). Следовательно, условия здесь приблизительно такие же, как
в дождевой тени африканских гор Паре (ср. стр. 249—252).
Ферри 3, проводивший в каатинге исследования транспирации несук-
кулентных растений, выяснил, что виды, не сбрасывающие листву в период
засухи, закрывают устьица и характеризуются весьма слабой кутикуляр-
ной транспирацией, хотя их листья и не отличаются ксероморфной струк-
турой. Даже в период дождей транспирация растений понижена; таким
образом, их поведение совершенно противоположно тому, что наблюдается
в «campos cerrados».
С особыми условиями мы встречаемся в Австралии в областях, где
осадки выпадают летом. О тропических и субтропических дождевых лесах,
развитых на влажном восточном побережье Австралии и образованных
индо-малайскими элементами, уже упоминалось (стр. 195 и след.). Флори-
стически родственные им реликтовые полувечнозеленые леса встречаются
лишь на очень ограниченных площадях. Один такой лесной массив автору
вместе с д-ром Уэббом удалось обнаружить в 60 км юго-западнее Маунт-
Гарнета. Этот лес отличается следующим составом и структурой:
Древесный ярус (20—25 м) не сомкнут и образован Brachychiton
australe («бутылочные деревья», ср. рис. 204), Gyrocarpus americana, Pleio-
gynium cerasiferum, Polyscias elegans и одним видом Ficus, единственной
вечнозеленой породой в этом лесу.
Кустарниковый ярус (2,5—3 м) очень сомкнут, отличает-
ся богатством видового состава и образован частично из вечнозеленых,.
1 К. Hueck, Verbreitung, Okologie und wirtshaftliche Bedeutung der «Cha"
parrales» in Venezuela, «Вег. Geobot. Inst. Riibeb>, 32, 1961, S. 192—203; V. V a r e-
schi, La estacion biologica de los llanos de la Sociedad Venezolana de Ciencias Natu-
rales у su tarea, «Bol. Soc. Venez. Cienc. Natur.», № 96, 1960, p. 107—134.
2 K. Hueck, Die waldgeographischen Regionen und Unterregionen von Siid-
amerika, «Geogr. Taschenbuch», 1960/61, S. 224—234.
3 M. G. Ferri, Contribuigao ao conhecimento da ecologia do Cerrado e da Caa-
tinga, Univ. Sao Paulo, «Botanica», 12, № 195, 1955.
Рис. 202. Cereus leucostele и Hohenbergia catingae (Bromeliaceae); каатинга близ
Кальдерона, Бразилия (фото Уле).
260
Глава 5
Рис. 203. Климадиаграмма области
развития каатинги, по данным Фер-
ри. Влажное время года короткое,
два сезона дождей благоприятствуют
росту суккулентов. На абсциссе месяцы
с июля по июнь. Кривая температуры
интерполирована.
частично из листопадных видов: Celastrus balocularis (листья как у Ilex),
Strychnos arborea, Carissa ovata, Melia dubia, Alstonia constricta, Maba
humilis, Trema aspera, Ehretia membranifolia, Canthium lucidum, Geijera
salicifolia, Erythroxylon australe, Turraca brownii и многих других видов.
Напочвенный покров почти отсутствует; встречаются
отдельные экземпляры Cheilanthes tenuifolia и небольшие подушки мха.
Лианы, нередко толщиной с руку, или обвивающие одна дру-
гую — Parsonsia, Cissus, Cynanchum, Celastrus,
Эпифиты представлены видами Bolbophyllum и Dendrobium, а
также пойкилогидрическими Polypodiaceae.
Леса, в составе которых господствуют индо-малайские элементы,
произрастают особенно у границ своего распространения, на почвах,
Рис. 204. «Бутылочные деревья» (Brachychiton, Sterculiaceae) на вырубке в Бриге-
лоу-скраб, окрестности Мандаберры, южный Квинсленд (фото Фрайера).
Тропические полу вечнозеленые и листопадные леса
261
развитых на вулканических (базальты) породах (см. цветное фото 2,
стр. 80).
С изменением состава материнских пород и уменьшением количества
осадков густой дождевой лес тотчас сменяется редкостойным эвкалипто-
вым лесом. Граница между дождевым и эвкалиптовым лесом выражена
Рис. 205. Влажный эвкалиптовый лес в юго-восточ-
ном Квинсленде, горы Тамбурин. Пальмы Archontophoe-
nix cunninghamii относятся к числу субтропических
элементов (фото Кёртиса).
очень резко как физиономически, так и флористически. Во влажный эвка-
липтовый лес (рис. 205) проникают лишь немногие индо-малайские эле-
менты. Вообще же эвкалиптовые леса очень существенно отличаются
от полувечнозеленых, влажных листопадных и сухих лесов остальной
части тропиков. За исключением небольшого числа факультативно листо-
падных эвкалиптов (ср. стр. 232), произрастающих только в более влаж-
ных и теплых районах, все остальные эвкалипты, даже встречающиеся
в аридных районах, являются вечнозелеными видами с кожистыми, но
отнюдь не склерофильными листьями. Кроме того, листовые пластинки
262
Глава 5
Рис. 206. Влажный эвкалиптовый лес в юго-восточном Квинсленде. В центре —
Eucalyptus mtcrocorys (фото Фрайера).
у них располагаются вертикально. Поэтому все эвкалиптовые леса, даже
распространенные во влажных областях, кажутся очень светлыми; в свя-
зи с этим повсюду в них развит более или менее высокий и сомкнутый тра-
вяной покров, во всяком случае в областях, где осадки выпадают летом.
Тропические полу вечнозеленые и листопадные леса
263
Рис. 207. Более сухой эвкалиптовый лес в окрестностях Виндзора, Новый Южный
Уэльс. В центре — Eucalyptus micrantha (фото «Dept. Agric. N. S. Wales»).
Виды Eucalyptus хорошо приспособлены к совместному произрастанию
со злаками (ср. стр. 300); в связи с этим между влажным и сухим эвкалип-
товыми лесами и открытыми саваннами с эвкалиптами существуют лишь
небольшие различия (рис. 206—208 и цветное фото 3, стр. 80). Вечнозеле-
264
Глава 5
Рис. 208. Эвкалиптовая саванна; на переднем плане — травяные деревья (Хап-
thorrhoea), южный Квинсленд (фото Бенкрофта).
ные эвкалипты в горах Австралии поднимаются до верхней границы леса
(цветное фото 1, стр. 272), где до шести месяцев в году лежит снег. К сожа-
лению, эколого-физиологические исследования до настоящего времени
не проводилось в эвкалиптовых лесах, развитых в областях, где осадки
выпадают летом.
ТРОПИЧЕСКИЕ ПАРКОВЫЕ ЛАНДШАФТЫ
Сначала познакомимся с природными условиями вдоль трассы, про-
стирающейся от восточноафриканского грабена, где развиты многочис-
ленные содовые озера, через озеро Руква мимо южной оконечности озера
Танганьика и дальше через озера Бангвеоло и болота Луканга (западнее
Брокен-Хилла) к водопадам Виктория на реке Замбези (рис. 209). Эта
трасса фактически проходит по очень плоскому водоразделу между Атлан-
тическим (водосборный бассейн Конго) и Индийским океанами, высота
которого достигает 1100—1400 м над уровнем моря. В этой области летом
идут сильные дожди; они сменяются резко выраженной засухой с несколь-
ко более низкой температурой воздуха.
Влияние смены влажных и засушливых условий в отдельных случаях
усиливается под воздействием рельефа. Даже при очень небольших пере-
падах высоты в период дождей вода движется с возвышенностей в неболь-
шие понижения сплошным потоком. В это время происходит сортировка
материала по механическому составу. На возвышенностях развиваются
элювиальные почвы, из которых выщелочены воднорастворимые соеди-
нения; на склонах — коллювиальные почвы, характеризующиеся наряду
с вымыванием и процессом вмывания, заиления, и, наконец, в понижениях
Р п с. 209. Заболоченные районы (штриховка) в Замбии,
Танзании, Уганде и южном Судане. Черная линия —
трасса полета из Ливингстона в Найроби, далее в Энтеббе
(у озера Виктория) и через Джубу в Хартум.
266
Глава 5
распространены иллювиальные почвы. Вода, сортируя на своем пути
почвенные частицы, откладывает более крупные на склонах, а илистые —
в понижениях.
Таким образом, формируется связанная с рельефом серия почв
(Мильне называет ее «катеной», цепью); на этих почвах развиваются
группировки растений, отражающие смену почвенных условий1.
Элювиальные почвы относительно сухи, коллювиальные обладают
несколько большей влажностью, однако при этом они все же хорошо аэри-
рованы; что касается тяжелых иллювиальных почв, то они достаточно
водонепроницаемы; их верхние горизонты в период дождей перенасыщают-
ся влагой и в таком состоянии отличаются вязкостью и плохой аэрацией,
при спаде воды эти почвы высыхают и становятся твердыми, как камень.
В таких переменно увлажняемых местообитаниях деревья не растут,
а развиваются злаковники. К сожалению, водный и воздушный режимы
этих почв в стационарных условиях не изучались. Нет также данных
и о корневых системах растений. Поэтому при экологической характери-
стике произрастающих здесь растений приходится довольствоваться опи-
саниями путешествий и мелкомасштабными картами растительности,
имеющимися в литературе, посвященной этим районам 1 2.
Крутизна склона и подстилающие горные породы, а также количество
и распределение осадков влияют на формирование почв и на флористиче-
ский состав растительных сообществ. Микрорельеф оказывает влияние
на водный режим и степень аэрации почвы; например, на термитных
постройках, развитых на элювиальных почвах, в лесных массивах появ-
ляются засухоустойчивые стеблесуккулентные канделябровидные моло-
чаи, виды Boscia, Commiphora или баобаб (Adansonia digitata). На иллю-
виальных почвах на развитие растительности термитников оказывают
влияние лучшие условия аэрации, поэтому здесь на фоне окружающих
злаковников формируются сомкнутые древостои. Эта мозаичность расти-
тельного покрова, связанная с чередованием участков, покрытых лесом
и травяным покровом, хорошо заметна с самолета и выражается в виде
смены светлых и темных круглых пятен.
В более влажной части тропической области, где дожди идут летом,
во время относительно короткого дождливого периода часто выпадает
огромное количество осадков до 1000—2000 мм. Обширные равнинные
пространства в течение нескольких месяцев оказываются под водой.
В период же засухи глинистые почвы, свойственные этим территориям,
очень сильно высыхают. Следовательно, речь идет о периодически увлаж-
няемых местообитаниях. В таких условиях лес не может существовать,
но хорошо растут травы, а из древесных растений — пальмы, в Африке,
например Borassus и Hyphaene, встречающиеся вместе с акациями в виде
одиночных экземпляров на фоне злакового покрова (рис. 210). Следова-
тельно, речь идет о природных, обусловленных эдафическими причинами
злаковниках (иногда это может быть пальмовая саванна), развитых в опре-
деленных условиях рельефа. Открытые пространства злаковников с пере-
межающимися островками леса образуют парковый ландшафт. К сожале-
нию, в этих условиях не изучались ни длительность затопления, ни усло-
1 C.G.T. Morrison, А. С. Hoyle, J. F. Hope-Simpson, Tropical
soil-vegetation catenas and mosaics, «J. Ecol.», 36, 1948, p. 1—84.
2 B. D. Burtt, Some East African vegetation communities, «J. Ecol.», 30, 1942,
p. 65—146; J. M. R a t t г а у, H. W i 1 d, Vegetation map of the Federation of Rhodesia
and Nyassaland, «Kirkia», 2, 1961/62, p. 94—104.
Тропические полу вечнозеленые и листопадные леса
267
вия аэрации почвы, ни ее влагосодержание в период засухи, ни строение
корневых систем, ни периодичность развития растительного покрова. Эти
области посещаются обычно только в засушливое время года, так как
в период дождей они непроходимы.
Рис. 210. Пальмы Borassus на фоне злакового покрова; участок,
переувлажненный в дождливый сезон; окрестности Бомы, нижнее
Конго (фото Уинтера).
В качестве примера приведем парковый ландшафт, распространен-
ный в северной части юго-западной Африки (между реками Омурамба
и Овамбо), в области развития сухих листопадных лесов. Это совершенно
равнинная местность, легкодоступная в сухое время года. Перед взором
наблюдателя открываются обширные пространства, покрытые злаков-
ником, с живописно разбросанными по нему небольшими группами
деревьев или островками леса, иногда с одиночными пальмами между
ними. На первый взгляд трудно понять, какие причины обусловливают
чередование лесных и безлесных участков. Однако это стало для нас
268
Глава 5
совершенно ясным тогда, когда мы после нескольких очень сильных дож-
дей оказались в этом районе, где даже на джипе с трудом можно было
передвигаться. Все пространство вокруг было затоплено; над водной
гладью, словно острова, возвышались только группы деревьев. Микро-
рельеф здесь очень слабо выражен, однако даже небольшая разность
высот в 20—50 см может обусловливать чередование лесных и безлесных
пространств. Если период дождей затягивается, то вода, не затопляя
островки леса, расположенные на возвышенных пространствах, начинает
течь медленным широким потоком, причем в различных точках района
нельзя даже точно установить направление стока — на север, к реке
Кунене, или на юг, к солончаку Этоша-Пан. Во всяком случае в сезон
дождей, в течение 4—5 месяцев, почва постоянно насыщена водой, что
губительно действует на древесную растительность. В самых глубоких
впадинах вода стоит еще дольше и в сухой период испаряется очень мед-
ленно. В этих глинистых блюдцах-впадинах, так называемых влеяхг
развивается болотная растительность, оживляемая красиво цветущими
видами Amaryllidaceae. Древесные породы здесь не растут, так как почва
во время засухи совершенно пересыхает; трава желтеет, болотные расте-
ния сохраняются только в самых влажных частях впадин (см. обзорную
карту района на рис. 287, ср. также рис. 211).
Возникает вопрос, каким образом здесь сформировался микрорельеф.
Тролль описал в Восточной Африке парковый ландшафт, названный им
Рис. 211. Парковый ландшафт северо-восточнее солончака Этоша-Пан. На перед-
нем плане — влей, плоская впадина, заполненная водой. Далее — крупный экзем-
пляр Combretum primigenium среди Sclerocarya schweinfurthiana (Anacardiaceae) (фото
Э. Вальтер).
Тропические полу вечно зеленые и листопадные леса
269
«термитной саванной», или «затопляемой саванной». Здесь на фоне зла-
кового покрова выделяются облесенные участки, занимающие при бли-
жайшем рассмотрении старые термитники. По мнению Тролля, заброшен-
ные термитники содержат больше питательных веществ, что и благоприят-
ствует росту деревьев. Более подробно этот вопрос изучал Хессе х; он
пришел к иным выводам, чем Тролль. Термиты используют для своих
построек исключительно материал, добываемый ими из-под почвы, при-
чем этот материал они не перерабатывают. Пока постройки обитаемы,
термиты тщательно следят за ними и препятствуют их зарастанию. На
заброшенных же термитниках, напротив, развивается более пышная рас-
тительность. Так, Agave sisalana в культуре обычно лучше растет на тер-
митниках. Однако объясняется это не высоким содержанием питательных
веществ, а более благоприятными физическими свойствами грунта. Почва
здесь отличается большей мощностью и лучше дренирована. При разбивке
плантаций не следует разравнивать старые термитники, поскольку они
сложены из совершенно неплодородного стерильного грунта.
Замечено также, что при пожарах в сухих лесах огонь обычно щадит
облесенные термитники, тогда как выжигание равнинных участков спо-
собствует развитию густого травостоя. Возникают так называемые «тер-
митные саванны» (ср. рис. 213).
В затапливаемой в период дождей местности, на более приподнятых
и хорошо дренируемых термитниках ]^огут расти древесные породы;
на покрытых же водой участках происходит отложение глинистых час-
тиц; здесь на тяжелой, плохо аэрируемой почве развиваются только
злаки.
В северной части Юго-Западной Африки также во многих случаях
выяснилось, что группы деревьев приурочены к очень плоским, развалив-
шимся старым термитникам. В сухое время года ветер навевает около
них пыль и мелкий песок, расширяя тем самым площадь возвышений.
Таким образом, «термитная саванна» не может рассматриваться в качестве
однородного ландшафта, она состоит из мозаичных сообществ, причем
в одних местообитаниях развивается сухой листопадный лес, а в других —
злаковник. То же можно сказать и о «затопляемых саваннах», если их
‘Существование обусловлено различиями в условиях рельефа. На место-
обитаниях с переменным увлажнением, цомимо травяного покрова, раз-
виваются только пальмы, возможно, благодаря способности образовы-
вать придаточные корни.
Аналогичные условия существуют, по описаниям Бантинга и Ли,
в центральном Судане 1 2. Затопляемые участки с глинистыми почвами
покрыты злаковниками. Разность высот в 10 см уже обусловливает изме-
нение характера растительного покрова. Осадки проникают в почву только
на глубину 30—40 см; запас воды в почве приблизительно равен 100 мм;
при отсутствии осадков он расходуется растениями в течение трех недель.
Однолетние растения в этом случае быстро погибают. Господство однолет-
них злаков и отсутствие многолетних — несомненное следствие сильных
пожаров.
Эдафически обусловленные саванны встречаются также в северной
части Суринама, где выпадает за год 2000—2500 мм осадков. Причиной
1 Р. R. Hesse, A chemical and physical study of the soils of termite mounds in
East Africa. «J. Ecol.», 43, 1955, p. 449—461.
2 A. H. Bunting, J. D. Lea, The soils and vegetation of the Fung, East
Genral Sudan, «J. Ecol.», 50, 1962, p. 528—558.
270
Глава 5
их развития могут быть залегающие на небольшой глубине водонепро-
ницаемые слои каолина, уплотненные горизонты почвы, а также смыв,
верхних слоев почвы вблизи рек, базис эрозии которых понижается.
Эрозия способствует иссушению почвы Ч
Часто наблюдаемая даже в необжитых тропических районах мозаика
ландшафтов не поддается расшифровке с первого взгляда. Лишь после-
многолетних исследований и подробных экологических изысканий удает-
ся выявить взаимосвязи растительного покрова и среды и установить-
причины этих связей, вызванные различными условиями микроклимата,,
рельефом, экспозицией и почвами.
Окончательные же выводы можно будет сделать лишь после создания
здесь опытных станций, рассчитанных на длительный период исследо-
ваний.
ТРОПИЧЕСКИЕ ЗЛАКОВНИКИ
Злаковники, развитие которых обусловлено климатическими при-
чинами, распространены и в тропической лесной области и в лесной обла-
сти умеренной зоны только в горах выше лесного пояса. Тем не менее,
естественные злаковники, как уже было сказано, широко распростра-
нены в тропиках 1 2. Выделяются следующие злаковники:
I. Первичный, или естественный, злаковник, и в частности:
А. Эдафически обусловленный злаковник.
1. В местообитаниях с переменным увлажнением:
а) злаковник, развитый на плоских водоразделах (в Восточной
Африке называется «дамбо»);
б) временно затопляемый при паводках злаковник, распро-
страненный в долинах рек;
в) злаковник, приуроченный к очень сырым в период дождей,
но пересыхающим в сухое время года понижениям (называется
«мбуга»).
2. В сухих местообитаниях с маломощными почвами.
Б. Злаковник, образовавшийся в горах в результате естественных
пожаров.
II. Вторичный, или антропогенный, злаковник, возникновение которого*
связано с воздействием человека (вырубка, палы, пастьба скота).
Пролетая в самолете над Замбией, можно очень отчетливо проследить,
как зарождаются на водоразделах реки. Сначала заметны лишь широкие
соединяющиеся друг с другом травяные полосы. Уклон местности здесь
столь незначителен, что дождевая вода скапливается в этих неглубоких
понижениях и лишь медленно стекает вниз. Ниже по склону в средней
части полос уже обозначается русло реки, однако широкие полосы травя-
ного покрова еще долго покрывают берега рек. При нарастающей крутиз-
не склонов долины рек углубляются, пойма, образованная травяным
покровом, полностью исчезает, и в виде узких блестящих лент реки пере-
секают сплошное море лесов. При отсутствии дальнейшего уклона мест-
ности реки впадают в обширные болота (рис. 212).
1 I. Р. Bakker, Uber den Еinflufl von Klima, jiingerer Sedimentation und
Boden prof ilentwickhmg auf die Savannen Nord-Suiinams, «Erdkunde», 8, 1954, S.
89—112.
2 A. P. G. Michelmore, Observations on tropical grasslands, «J. Ecol.»,.
27, 1939, p. 282—312.
Тропические полу вечно зеленые и листопадные леса
271
Рис. 212, Распространение лесов (точечный пунктир) и зла-
ковнпков в области болот Луканга на территории Замбии
(рисунок сделан по аэрофотоснимку),
Тропические злаковники недавно были подробно изучены Визи-
Фитцджеральдом Ч В их составе доминируют Loudetia simplex, виды
Hyparrhenia и другие виды подсемейств Andropogoneae и Paniceae, Листья
злаков расположены на высоте 50 см’, стебли поднимаются до высоты 1 м\
истинное покрытие почвы достигает здесь 50%, Изредка здесь встречаются
геофиты и несколько видов разнотравья, Границы между этими злаков-
никами (дамбо) и сухими лесами, образованными Brachystegia, проходят
1 D, F. Vesey-Fitzgerald, Central african grasslands, «J, Ecol.», 51,
1963, p. 243—274.
272
Глава 5
очень резко; менее резко злаковники отграничены от древостоев Сот-
bretum, в то время как заросли акации часто весьма постепенно переходят
в открытые злаковники.
Злаковники встречаются также на склонах, в местах, увлажняемых
после дождей выходящими на поверхность грунтовыми водами, благодаря
чему почва здесь дольше остается влажной.
Большие площади занимают злаковники в обширных депрессиях или
в долинах-грабенах, повсюду, где вследствие тектонических движений
русла рек перегораживаются и образуются подпруженные бессточные
бассейны. В таких местах на известняках формируются насыщенные
в период дождей влагой черные почвы («cotton soils»), называемые в Вос-
точной Африке «мбуга». Они распространены в Кении, а также в районе
Серенгети и в степях Массаи; в период дождей они непроходимы в течение
многих месяцев. В сухое время года эти почвы сильно высыхают, уплот-
няются и растрескиваются, что делает передвижение по ним в это время
весьма опасным. Содержание илистой фракции в этих почвах очень высо-
кое; черная окраска почвы связана с наличием в ней темных глинисто-пере-
гнойных комплексов. Несмотря на значительную влажность и вязкость
почв в период дождей, вода проникает в них лишь на небольшую глубину
(правда, точных данных о влажности почвы пока нет). На фоне злакового
покрова встречаются лишь одиночные деревья. Среди них несколько видов
акаций, среди которых наиболее распространена Acacia drepanolobium.
Она достигает высоты 3 м\ у основания обоих прилистников, имеющих
вид колючек, у этой акации образуются крупные шаровидные галлы,
в стенках которых до их затвердения муравьи проделывают ходы; муравьи
используют галлы в качестве гнезд. Если дотронуться до ветки акации,
то муравьи выбегают из галлов, готовые наброситься на нарушителя покоя.
Когда дует ветер, галлы издают особый звук (в связи с этим акация и назы-
вается «поющей»). В феврале, то есть после начала периода дождей, ака-
ция цветет и у нее развиваются нежные перистые листья. Однако боль-
шую часть года акация стоит без листьев.
Галлы образуются также у Acacia formicarum, A. seyal var. fistula,
A. malacocephala (дает гуммиарабик) и др.
Кроме перечисленных акаций, на этих почвах растет нетребователь-
ная к условиям среды Balanites aegyptiaca\ у нее развиваются длинные
зеленые колючки. К юго-востоку от Найроби этот вид образует очень
разреженные древостои, в которых деревья растут на больших, но одина-
ковых расстояниях одно от другого. Возможно, у них развиваются очень
длинные поверхностные корни. Степень покрытия почвы едва превышает
1%. Травостой здесь отличается большой степенью сомкнутости; он обра-
зован главным образом Pennisetum mezianum и Themeda triandra с неболь-
шой примесью Digitaria milanjana и одиночно встречающимися экземпля-
рами Pennisetum stramineum.
Очень часто в понижениях рельефа образуется плотный цементиро-
ванный карбонатами горизонт, иногда сверху перекрытый слоем принесен-
ного водой песка. После дождя верхние слои этой почвы «плывут», однако
в глубину вода не просачивается. В период засухи эти почвы пересыхают,
и поэтому они покрыты лишь редким травяным покровом. Такие пятна
нередко встречаются среди обширных лесных пространств.
Наконец, поймы крупных рек и площади вокруг болот или озер также
покрыты злаковниками. Здесь распространена так называемая термитная
•саванна, где деревья растут на постройках термитов. Леса часто растут
по берегам рек на береговых валах, тогда как затапливаемые площади
1. Верхняя граница от-
дельных деревьев (Euca-
lyptus niphophila) в
Австралийских Альпах.
Видна вершина горы
Косцюшко (2241 лг). На
переднем плане альпий-
ские луга с белой Cel-
misia longifolia (Compo-
sitae) и желтым Ranun-
culus lappaceus.
2. Вид с Холугберга на
высохшие заросли тра-
вяного покрова близ
фермы Грюнау; южная
часть Юго-Западной
Африки. В руслах вре-
менных водотоков — за-
росли кустарников и от-
дельные деревья.
3. Травяной
покров,
Т ропические полу вечнозеленые и листопадные леса
273
позади этих валов покрыты злаковниками. С экологической точки зрения
нецелесообразно называть эти парковые ландшафты саваннами, как это
нередко делают географы.
Лишь немногие древесные породы постоянно и закономерно встре-
чаются среди злаковников. Это прежде всего пальмы Borassus и Hyphaene\
они растут на фоне травяного покрова одиночными экземплярами. Syzy-
gium cordatum чаще встречается на крутых обрывах или термитниках.
Acacia nilotica —это типичный представитель пойменных лесов, перено-
сящий двух-трехмесячное затопление паводковыми водами, однако поги-
бающий в стоячей воде.
С экологической точки зрения значительно труднее объяснить раз-
витие злаковников в высокогорьях. Они встречаются на высоте более
1700 м, несколько ниже или в пределах лесного пояса, образованного
Podocarpus и расположенного на высоте 1800—2200 м\ здесь среди траво-
стоя встречаются одиночные экземпляры невысоких древесных пород:
Protea, Erythrina, Dombeya, Cussonia, Faurea, Myrica и Uapaca (Euphorbia-
ceae). В составе травяного покрова господствует Themeda triandra.
По данным Михельмора, в этих условиях и даже на пологих склонах лес не
развивается из-за плохой аэрации почвы. Он связывает это также с боль-
шим количеством осадков и меньшей величиной испарения в горах. Лес
появляется лишь на крутых каменистых склонах и в глубоко врезанных
долинах, отличающихся лучшим дренажам. Наряду с этим отмечается
производный характер злаковников, обусловленный постоянными пожа-
рами, пастьбой или распашкой. Остатки леса сохранились лишь во влаж-
ных ущельях. Здесь также следовало бы провести экологические иссле-
дования.
Гораздо меньшую площадь занимают злаковники, развитые на мало-
мощных сухих почвах. Они приурочены к скалистым обнажениям, покры-
тым твердой коркой из окислов железа. Вероятно, они являются дальней-
шей стадией деградации производных злаковников, возникших на месте
сведенных лесов. Твердая корка препятствует возобновлению леса. Во
всех населенных местностях широко распространены производные зла-
ковники, возникающие после уничтожения леса путем раскорчевки,
пожаров или пастьбы.
Здесь сохраняются отдельные деревья, в связи с чем возникает саван-
ноподобный ландшафт, однако эти саванны являются антропогенными.
Площади злаковников при повторных пожарах постоянно расширяются.
Даже опушки влажных лесов из года в год повреждаются огнем и усту-
пают место злаковникам. Резкая граница между лесом и этими вторич-
ными злаковниками в большинстве случаев обусловлена пожарами. Часто
встречаются чистые злаковники, где господствует один вид злака. На
месте сведенных дождевых лесов развиваются заросли Pennisetum риг-
ригеит, тогда как на месте более сухих лесов — Imperata cylindrica. При
ежегодных палах злаки развиваются еще лучше, поскольку огонь уничто-
жает мертвую ветошь. Лишь в тех случаях, когда зеленеющие после дождя
злаковники начинают интенсивно стравливаться скотом, ценные в кормо-
вом отношении виды злаков вытесняются малоценными.
Эти вторичные саванны и злаковники в настоящее время распростра-
нены настолько широко, что часто определяют облик ландшафта. Посколь-
ку в большинстве случаев трудно доказать их производный характер,
географы относят их часто к коренным образованиям. Этим и объясняется
выделение зоны коренных «влажных саванн», где господствуют высоко-
рослые злаки и зоны коренных «сухих саванн» с низкорослыми злаками,
274
Глава 5
которые окаймляют зону дождевого леса. Фактически же речь идет
о вторичных группировках, развивающихся на месте определенных кате-
горий лесов. В качестве зональной растительности саванны и злаковники
появляются лишь в условиях, где выпадает значительно меньшее количест-
во осадков (см. стр. 285 и след.).
Правда, напрашивается вопрос, нельзя ли и огонь рассматривать как
естественный фактор. Защитить полностью леса от пожаров практически
невозможно и даже опасно, поскольку при отсутствии палов в них накап-
ливается слишком много сухой подстилки; если в этом случае вследствие
удара молнии или неосторожности местных жителей возникнет пожарт
то действие огня окажется особенно разрушительным.
Этот вопрос наиболее детально изучался в провинции Наталь. Зла-
ковник, образованный Themeda triandra, покрывает здесь все склоны Дра-
коновых гор в пределах лесного пояса, где господствует Podocarpus lati-
folius; леса приурочены к крутым склонам и ущельям. На фоне злакового
покрова растут виды Protea, достигающие высоты 3—4 м; таким образом,
здесь развивается разреженная древесная саванна. Корка толщиной до
2 см хорошо защищает Protea от повреждения огнем; деревья гибнут лишь
при очень сильных пожарах. Только там, где палы пускаются ежегодно
и где в результате этого опад не накапливается, сила огня не бывает боль-
шой. Всходы Protea переносят такие палы; со временем здесь могут
образоваться сомкнутые древостои. В тех случаях, когда пожар распро-
страняется вниз по склонам, огонь также не достигает большой силы.
В настоящее время палы в большинстве случаев пускаются созна-
тельно или возникают по неосторожности; однако Киллик подчеркивает,
что в области Драконовых гор очень часто наблюдались пожары, вызван-
ные ударами молнии. В 1957 г. 17% пожаров было вызвано молнией,
а в остальные годы — 6—13% х. Эти пожары возникали преимущественно
во время первых весенних гроз, когда травостой бывает особенно сухим.
Однако пожары могут возникать также и в период засухи, в частности
когда с обрывов падают обломки твердых кварцитов и, ударяясь друг
о друга, высекают сноп искр. Таким образом, пожары наблюдались
здесь еще до поселения человека, правда не так часто, как сейчас, но все
же достаточно часто, чтобы влиять на формирование растительного покро-
ва. Это же может быть сказано и о пожарах в горных районах тропической
Африки. Иногда палы пускаются сознательно, чтобы предотвратить воз-
никновение естественных пожаров.
Таким образом, в горах распространены естественные первичные зла-
ковники, не связанные в своем происхождении ни с климатическими при-
чинами, ни, вероятно, также и с эдафическими причинами, а обусловлен-
ные естественными пожарами. Этой же точки зрения придерживается
Байер 1 2. Если такие злаковники в течение длительного времени оградить
от огня, то из их состава полностью исчезнет Themeda triandra, поскольку
она вскоре будет вытеснена другими видами злаков. Нельзя, однако, при-
давать слишком большое значение подобным естественным пожарам. По
данным Тальбота, в Уганде палы практически всегда пускаются сознатель-
но с целью предотвратить наступление леса. Во взаимоотношениях леса
и злаковников наряду с климатом, почвой и огнем известную роль играют
1 D. J. В. К i 1 1 i с k, An account of the plant ecology of the Cathedral Peak Area
of the Natal Drakensberg, «Bot. Survey of South Africa», № 34, 1963.
2 A. W. Bayer, The ecology of grasslands. The grasses and pastures of South
Africa, Johannesburg, 1955.
Т ропические полу вечнозеленые и листопадные леса
275
также и дикие животные х. При сильном стравливании травяной покров
частично уничтожается, и это способствует развитию древесных растений.
При недостаточно сильном стравливании, наоборот, начинают преобладать
высокорослые виды злаков. Если домашний скот поедает преимуществен-
но злаки, то под влиянием пастьбы диких животных злаковники дегради-
руют не так сильно, поскольку жирафы поедают листья деревьев, носоро-
ги— листья кустарников, антилопы-гну — лишь сочные зеленые листья
злаков, зебры — более старые, но еще зеленые листья, другие же живот-
ные питаются сухими листьями и колосьями. Кроме того, дикие животные
делятся на кочующие и оседлые виды. Поэтому кормовая емкость естест-
венных пастбищ в Восточной Африке примерно в 2—15 раз выше для
диких животных, чем для домашнего скота. Биомасса одних диких живот-
ных достигает примерно 12—25 т/га, биомасса скота на фермах — около
5—7,5, а у местного скота эти значения снижаются до 2,5—3,5 т!га.
Нельзя, однако, переоценивать роль крупных диких животных в фор-
мировании тропических злаковников. Стада диких животных являются
особенностью Африки. Они полностью отсутствуют в Южной Америке,
однако тем не менее и там в тропических областях, где количество осадков
достигает 500—2500 мм в год, злаковники, образование которых обуслов-
лено эдафическими причинами, покрывают большие пространства. Таким
примером могут служить льяносы в бассейне Ориноко. Именно для харак-
теристики этих ландшафтов впервые был применен термин «саванна».
По Бирду, саванны — это естественная растительность, приуроченная
к развитым почвам на сглаженных формах рельефа; почвы здесь отличаются
неблагоприятным водным режимом: они периодически то пересыхают, то
переувлажняются. Саванны, по Бирду, являются «эдафическим климак-
сом»; пожары в них бывают очень часто, однако они не являются причиной
возникновения саванн и их дальнейшего развития 1 2.
Таким образом, описанные выше топогенные злаковники
с климатической точки зрения распространены в областях, где потенци-
ально должен расти лес.
1 L. М. Т а 1 Ь о t, Savanna, IUCN Publ., new ser. № 4, 1964, p. 88—97.
2 J. S. Beard, Savanna, IUCN Publ., new ser. № 4, 1964, p. 98—103; «Ecol.
Monogr.», 33, 1953, p. 149—215.
Глав а 6
ЗОНАЛЬНЫЕ САВАННЫ
КАК ПЕРЕХОДНАЯ СТУПЕНЬ К АРИДНОЙ ЗОНЕ
ПОНЯТИЕ «САВАННА»
Хотя понятия «саванна» и «степь» были введены для обозначения
определенных типов растительности, в последнее время они при-
меняются географами скорее для обозначения соответствующих
климатических зон. Тролль 1 первоначально предложил называть все
гигрофильные растительные сообщества в тропиках «саваннами», а все
ксерофильные сообщества —«степями», вне зависимости от различий
в характере растительного покрова. Однако, поскольку гигрофит и ксеро-
фит — это относительные и трудноопределимые понятия, Егер 1 2 придал
понятию «саванна» чисто климатическое значение, совершенно не связан-
ное с какой-то конкретной зональной растительностью.
Влажная саванна, по Егеру,— это климатическая зона области тро-
пических летних дождей с засушливым периодом длительностью 2,5—
5 месяцев, сухая саванна — зона, где этот период продолжается 5—
7,5 месяцев, и колючекустарниковая саванна — зона с периодом засухи
длительностью 7,5—10 месяцев. Тролль, согласившись с этим определе-
нием, отмечает, что к саваннам, возникающим под воздействием эдафи-
ческих, биотических или антропогенных факторов, могут быть отнесены
все группировки растительности — от лишенных древесной растительно-
сти злаковников до лесных сообществ включительно 3. Тем самым это
понятие было лишено своего первоначального ботанико-географического
смысла и стало применяться для обозначения климатической зоны, где
распространены различные растительные формации. В этом случае, по
мнению автора, следовало бы пользоваться чисто климатической термино-
логией, говоря, например, о «влажной области тропических летних дож-
дей» и т. д. Если же, напротив, привлекаются ботанические термины, то
они должны применяться для характеристики какой-то определенной
категории растительного покрова. Совершенно необходимо различать
естественные и производные, антропогенные, формации, даже если они
физиономически сходны. В противном случае изучение их в экологиче-
ском аспекте будет невозможно.
Поэтому в настоящей работе термин «степь» (которым так часто непра-
вильно пользовались, особенно исследователи Африки) употребляется
только для обозначения злаковников внетропической зоны. Как известно,
русское слово «степь» применяется для обозначения развитых на юге
Восточной Европы злаковников, аналогичных американским прериям
1 С. Troll, Gedanken und Bemerkungen zur okologischen Pflanzengeographie,
«Geogr. Zschr.», 14, 1935, S. 380.
2 F. Jaeger, Zur Gliederung und Benennung des tropischen Graslandgiirtels,
«Verb. Naturf. Ges. Basel», 56/2, 1945, S. 509-520.
3 C. Troll, Das Pflanzenkleid der Tropen in seiner Abhangigkeit von Klima,
•«Boden und Mensch, Deutscher Geographentag, Frankfurt, 1951», 28, 1952, S. 35—66.
Зональные саванны как переходная ступень к аридной зоне 277
В соответствии со взглядами Шимпера, Друде и других исследовате-
лей саванны представляют естественную однородную зональную расти-
тельность области тропических летних дождей с сомкнутым злаковым
покровом, на фоне которого равномерно распределены древесные расте-
ния — деревья или кустарники. Эта обусловленная климатическими
условиями саванна встречается в Африке, вероятно, только при годовом
количестве осадков менее 600 мм и, скорее всего, соответствует описанной
Егером «колючекустарниковой саванне». Если при небольшом количестве
осадков в пограничных с пустыней областях на песчаных почвах разви-
вается злаковый покров без древесных растений, то целесообразнее всего
говорить о злаковниках (цветные фото 2, 3, стр. 272).
Физиономически очень сходные «саванны» занимают большие площа-
ди также и в более влажных частях Африки и Южной Америки. Сейчас
уже нет никакого сомнения, что речь идет о вторичных растительных сооб-
ществах, возникших из различных описанных выше типов леса. Причи-
ной их возникновения являются палы, искусственно пускаемые каждый
год человеком для выжигания сухой травы. Даже в малонаселенных обла-
стях этим путем можно изменить растительный покров на обширной терри-
тории. При регулярных пожарах в конце концов достигается известное
равновесие — возникает растительный покров, представленный сомкну-
тым травостоем, на фоне которого разбросаны огнестойкие древесные поро-
ды. После прекращения пожаров вновь восстанавливается первоначаль-
ная лесная растительность, остатки которой сохранялись во влажных
ущельях, на незадернованных термитниках или скалистых выступах. На
путешественника подобный ландшафт производит впечатление вполне
естественного образования, такое же, как европейские луга или верещат-
ники; при кратковременном пребывании здесь часто нелегко сделать
вывод о происхождении этого ландшафта \ и только тщательное изучение
позволяет говорить об антропогенном характере этих образований. Вряд
ли можно возражать против применения термина «производные саванны»,
однако последние нужно строго отличать от обусловленных климатом
зональных саванн, занимающих гораздо меньшую площадь.
Производные саванны можно классифицировать в зависимости от
климатических условий: в более влажных областях злаки достигают
большей высоты, чем в засушливых; состав огнестойких древесных пород
также различен. Эти примеры не опровергают производного характера
саванн; нужно только освободиться от представления, будто в слабонасе-
ленных областях Африки и Южной Америки все еще сохранилась нетрону-
тая растительность. Она может быть найдена только после очень долгих
и старательных поисков.
Часто возражают, что пожары возникают также и естественным
путем — например от ударов молнии, и это совершенно верно. Однако
такие пожары возникают гораздо реже и не производят больших разруше-
ний в лесных областях. Лишь после того как под влиянием деятельности
человека (путем многократного выжигания в сочетании с пастьбой) леса
были осветлены и возникли обширные площади злаковников, интенсив-
ность и длительность действия пожаров настолько усилилась, что
естественная растительность исчезла полностью.
1 J.Schmithiisen, Allgemeine Vegetationsgeographie, Berlin, 1959. О южно-
американских саваннах см. R. Goodland, On the savanna vegetation of Galaboze,
Venezuela and Rupununi, Britisch Guiana, «Bol. Soc. Venezol. Cienc. Natur.», № 110,
26, 1966, p. 341—359.
278
Глава 6
* t У т
^-3
4
5
Р ис. 213. Ландшафты, которые Тролль относит
к «саваннам». В действительности же это макро-
мозаика совершенно различных растительных
формаций: чистых злаковников, саванн и боль-
ших или меньших лесных массивов.
1 — галерейные саванны; 2 — овражные саванны!
з — саванны с лесами по береговым валам(«леса бан-
ке»); 4 — галерейно-термитные саванны; 5 — овражно-
термитные саванны; 6 — термитные саванны с лесами
по береговым валам и «термитными островами».
Можно указать еще на
один природный фактор, а
именно на многочисленные
стада антилоп и зебр, которые
производили такое большое
впечатление на первооткры-
вателей в Африке. В эко-
системах, развитых в этих
областях, они, безусловно,
играют известную роль, одна-
ко маловероятно, чтобы они
оказывали существенное вли-
яние на развитие раститель-
ности. Во многих случаях эти
стада были вытеснены в еще
не заселенные человеком обла-
сти. Однако и здесь обилие
дичи бросается в глаза толь-
ко в тех случаях, когда в за-
сушливые годы стада собира-
ются в более обеспеченных
водой районах, где еще сохра-
нилась сочная трава. Дичью
богаты прежде всего полуза-
сушливые области, где рас-
пространены зональные высо-
котравные саванны.
Тем не менее одно обстоя-
тельство бросается в глаза:
в Африке с ее большими стадами диких животных акации всегда покрыты
колючками, тогда как в саваннах Австралии, где обитают только кен-
гуру, никогда не встречающиеся крупными скоплениями, акации и почти
все другие древесные растения лишены их (ср. стр. 291) х.
Парковые ландшафты с экологической точки зрения также не следо-
вало бы называть «саваннами», как это делает Тролль (рис. 213) 1 2, посколь-
ку в этом случае речь идет о мозаично распределенных по площади есте-
ственных экстразональных лесных сообществах, развитых вдоль рек, во
влажных ущельях или на не затопляемых в период дождей возвышениях,
окруженных периодически затопляемыми злаковниками, пальмовыми
саваннами или даже производными саваннами. Слишком широкая трак-
товка понятия «саванна» обесценивает его научное значение.
В производных саваннах развиты почвы, свойственные лесам, из
которых они возникли, однако эти почвы обеднены гумусом, ибо органи-
ческая масса уничтожается при пожарах. Азота и фосфора в них содержит-
ся также меньше, чем в лесных почвах. Сильное переувлажнение в сезон
дождей и сильное иссушение]в засушливый период приводят к образова-
нию на небольшой глубине плотной железистой коры, часто выступаю-
щей в результате эрозии на дневную поверхность. Следовательно, почвы
1 Н. Walter, Die Bedeutung des Grosswilds fiir die Ausbildung der Pflanzen-
decke, «Stuttagart, Beitr. z. Naturk.», № 69, 1961.
2 C. Troll, Savannentypen und das Problem der Primarsavannen, Proc. 7.
Intern. Bot. Congr., Stockholm, 1950.
Зональные саванны как переходная ступень к аридной зоне
279
бедны и могут давать хорошие урожаи только после соответствующего
удобрения.
В этой области хорошо выражены почвенные ряды, или катена —
в смысле Мильне,— то есть закономерная смена почв на склонах гор по
направлению от вершины к основанию. Если на возвышениях обычно
развиты красные почвы, содержащие железо и каолин, то в понижениях
распространены черные, трескающиеся в сухое время года почвы, содер-
жащие монтмориллонит. Последние более плодородны, но требуют тща-
тельной обработки и в условиях примитивного земледелия обычно
не используются. В период дождей они сильно затрудняют движение
транспорта.
Следующие главы посвящены описанию зональных саванн, обуслов-
ленных климатическими причинами и развитых в условиях сухих тропи-
ков, где лес в плакорных условиях уже не может произрастать.
ТРАВЯНАЯ И ДРЕВЕСНАЯ РАСТИТЕЛЬНОСТЬ
В КАЧЕСТВЕ АНТ АГОНИСТОВ
Коренные саванны — это однородные растительные сообщества, где
на фоне злакового покрова распространены отдельные более или менее
далеко отстоящие друг от друга древесные растения (рис. 214). Если дере-
вья разделены расстоянием, примерно в ^—10 раз превышающим их высо-
ту, как это часто наблюдается, например, в центральной части Калахари,
то ландшафт производит оригинальное впечатление. Наблюдателю кажет-
ся, будто он стоит на лесной поляне с одиночными деревьями, окруженной
со всех сторон густым лесом. Однако достичь леса никогда не удается, ибо
поляна перемещается вместе с идущим, а граница леса постоянно отсту-
пает (рис. 215).
В таких саваннах на одной и той же площади распространены
два типа растений, являющиеся большей частью антагонистами.
В умеренной зоне мы встречаем, например, пространства, покрытые или
чистым травяным покровом (степь, прерия) или древесной растительностью
(леса, кустарники). Их взаимопроникновения не происходит, поскольку
«лесостепь» представляет собой мозаичный макрокомплекс, в котором эле-
менты леса и степи в большинстве случаев остаются резко отграничен-
ными.
Деревья и злаки зональной саванны представлены различными эко-
логическими типами. Тем не менее можно указать на некоторые характер-
ные свойства, показательные, с одной стороны, для большинства древес-
ных пород, а с другой — для типичных злаков, предопределяющие раз-
витие того или иного типа растительности Ч При этом имеются в виду
многолетние злаки, исключая колючие и ксерофильные виды, чуждые
зональным саваннам и характерные для иных эколого-физиологиче-
ских условий.
Типичные злаки отличаются в первую очередь спецификой водного
режима. По отношению к теплу различия между древесными породами
и злаками значительно меньше; это выражается в том, что и тех и других
представителей можно найти от тропиков до Арктики. Особенности вод-
ного режима злаков проявляются как в специфике отдачи, так и в погло-
щении воды.
1 Н. Walter, Grasland, Savanne und Busch der ariden Teile Afrikas in ihrer
«kologischen Bedingtheit, «Jb. wiss. Bot.», 87, 1939, S. 750—860.
280
Глава 6
Рис. 214. Древесная
саванна в Юго-Запад-
ной Африке, ферма
Граслаагте близ Оть-
юваронго
(фото Я В альте о).
Р и с. 215. Древесная
саванна с Acacia gi-
raffae; в травостое
Schmidtia quinqueseta,
ферма Дорнфонтейн,
Юго-Западная Афри-
ка (фото Э. Вальтер).
Злаки, как правило, отличаются очень интенсивной транспирацией,
не уменьшающейся даже в период засухи. При недостатке воды происхо-
дит резкое уменьшение гидратуры клеток листьев, то есть быстрое
повышение осмотического давления, ведущее к отмиранию клеток. Отми-
рание начинается с верхушек листьев и распространяется к их основа-
ниям. В течение короткого времени ярко-зеленые участки злаковников
высыхают и приобретают желтый и даже бурый оттенок.
Поскольку почки возобновления у большинства злаков расположены
у основания стебля и защищены от полного иссушения окружающими
их листовыми влагалищами, они остаются живыми в течение многих
месяцев засухи и при благоприятной температуре и достаточном увлаж-
нении вновь образуют побеги.
У чувствительных к холоду субтропических злаков отмирание над-
земной части иногда может быть вызвано заморозками. Однако и в этом
случае почки возобновления, так же как и корневая система, не погибают.
При высыхании растений на корнях этих злаков (Aristida, Eragrostis
и др.) отмирает кора и ее клетки съеживаются; отмерший наружный слой
Bo Bo
5н
Ca w и ®
2(7
P и~с. 216 и 217. Совмещенные вертикальные и горизонтальные проекции растительности
(без злаков)'близ Фойгтсгрунд в Юго-Западной Африке. На рис. 216 изображено русло неболь-
шого временного водотока, окаймленное акациями (Acacia nebrownii-glandulifera), а на
рис. 217 — поперечный профиль долины реки Тсуб, где развита галофильная и псаммо-
фильная растительность; в левой части рисунка — скалистый склон. Влаголюбивые виды
акаций и Albizzia укореняются в трещинах скал, вблизи русел временных водотоков или
достигают своими корнями уровня грунтовых вод (незасоленных).
Ас — Acacia nebrownii (glandulijeraY, Ac. d.— Acacia detinens; Ac. h.— Acacia horrida", Ai — Tetragonia
schenckii', Al — Aloe\ Alb — Albizzia anihelmintica’, Am — Argemone mexicana\ Aq —солоноватые по-
верхностные воды; Asc — Asclepiadaceae; Bo — Boscia foetida; Bl — Blume a gariepina; Ca — Catophra-
ctes alexandri; Ly — Lyciurrr, Sa — Sansevieria; Su — Suaeda monoica', Go — Gomphocarpus fruticosus',
Sa — Salsola aphylla\ Cr — Cryophytum', Ms — Mesembryanthemum; Ps — Psilocaulon\ Rh — Rhigozum
trichotomum.
282
Глава 6
.коры с остатками корневых волосков и застрявшими в них песчинками
образует чехол вокруг живого центрального цилиндра Е
В засушливых областях распространены также крупные злаки, вет-
вящиеся не только у основания, но и на вершине стебля, причем их стебли
и расположенные над поверхностью почвы почки возобновления сохра-
няются в течение периода засухи. В качестве примера можно назвать очень
распространенную в Юго-Западной Африке Aristida uniplumis (рис. 219),
стебли которой только желтеют, но не высыхают полностью. Уже перед
началом отрастания хлорофилл в них восстанавливается и стебли вновь
начинают зеленеть. Это явление пока детально не изучено 1 2.
В состоянии покоя, то есть после отмирания транспирирующих орга-
нов, злаки потребляют лишь очень немного воды, что позволяет им пере-
носить длительные периоды засухи даже при небольшом запасе почвенной
влаги. Наблюдения автора на юге и юго-западе Африки показали, что
даже в конце засушливого сезона незначительных количеств воды, сохра-
нившихся в почве, достаточно для покрытия имевшегося расхода. Чем
выше злаки, тем больше воды они расходуют во время засухи. Поэтому
высокорослые виды встречаются только в саваннах, развитых в областях,
где сухой период не бывает особенно продолжительным.
Если в почве сохраняется некоторый избыток влаги, то злаки трога-
ются в рост еще до начала дождей. Такую картину можно наблюдать,
например, по берегам небольших русел временных водотоков; правда,
в этом случае листья развертываются полностью только после первого
хорошего дождя. Песчаные дюны, покрытые редким растительным покро-
вом, накапливают больше воды, и поэтому расположенные на них отдель-
ные кусты Aristida ciliata зацветают даже до начала периода дождей.
Анализы показали, что влажность почвы на глубине 1 м равнялась
1,25%, а на глубине 1,25 м достигала 2%. Почву можно было сжать
в комок, что свидетельствовало о ее достаточной влажности.
Из-за низких температур начало вегетации у злаков наблюдается
в этих условиях лишь в конце засушливого периода. Тропические злаки
пребывают в состоянии покоя до окончания периода ночных заморозков
и подъема температуры до 20°. Поэтому обильный полив в холодное время
года (август) не смог сократить период покоя у злаков. Напротив, у выко-
панного и перенесенного в теплое и влажное помещение Eragrostis Lehman-
niana зеленые листочки появились уже через 14 дней; у Eustachys paspaloi-
des через 2 недели появились молодые корни, а вскоре после этого и листья.
Вполне возможно, что тропические злаки в холодное время года находят-
ся в состоянии зимнего покоя и развитие возобновляется только после
окончания этого периода.
Сильная транспирация злаков в период их активного роста должна
компенсироваться столь же интенсивным поглощением воды. Поскольку,
однако, злаки всегда образуют сомкнутый травостой, корневая система
каждого экземпляра ограничена лишь небольшим объемом почвы. Поэто-
му интенсивное поглощение влаги растением возможно только при боль-
шой насыщенности этого объема почвы корнями, что обычно наблюдается
в сообществах злаков и при достаточной обводненности почвы. Злаки
1 М. Н enri ci, Structure of the cortex of grass roots in the more arid regions
• of South Africa, Dep. of Agric., «Sci. Bull.», № 85, 1929; A. P. G о о s s e n s, Notes
• on the anatomy of grass roots, «Trans. Roy. Soc. South Africa», 23, pt I, 1935; H. Wal-
ter, op. cit. (рис. 3).
2 M. H e n r i c i, Physiological plant studies in South Africa, I u. II, 11. u. 12
^Report Direct. Veter., Educ. a. Research, Pretoria, 1927.
Зональные саванны как переходная ступень к аридной зоне
283
лучше всего развиваются в области тропических летних дождей, где
преобладают мелкоземистые, обладающие большой влагоемкостью почвы
и где в период вегетации выпадает много осадков («Phytologie», Bd III,
1, S. 284).
Совершенно по-иному реагируют на эти условия древесные растения.
Они в основном отличаются от других экологических форм растений спо-
собностью уже при незначительном дефиците влаги сокращать интенсив-
ность транспирации путем смыкания устьиц. Вечнозеленые жестколист-
ные виды (склерофилы) могут благодаря этому переносить продолжитель-
ные периоды засухи, в то время как другие виды в сухое время года
частично или полностью сбрасывают листья.
Однако и вечнозеленые и листопадные породы тратят во время засу-
хи воду на транспирацию, поэтому необходимо известное, хотя и незначи-
тельное восполнение этих потерь за счет почвенной влаги.
В кустарниковой саванне, развитой в Юго-Западной Африке, где
господствуют Aristida uniplumis в травостое и кустарники Boscia foetida
(Capparidaceae), Catophractes alexandri (Bignoniaceae) и Rhigozum trichoto-
mum (Bignoniaceae), определялась интенсивность транспирации у покоя-
щихся растений в период засухи (в ноябре); взвешивались целые ветви
или небольшие кусты через каждые 30 минут. Интенсивность транспи-
рации составляла у Boscia в среднем 7,8% сырого веса в сутки, у Catoph-
ractes — 6,4% и у Rhigozum — 5%. При незначительной величине
транспирации обрезка ветвей не повлекла за собой снижения интенсив-
ности транспирации, изменения которой в целом следуют за кривой
испарения.
К сожалению, не удалось определить интенсивность транспирации
кустарниковых акаций в безлистном состоянии, поскольку они росли
в обследуемой области только в условиях относительно хорошего обес-
печения водой, например в руслах временных водотоков или в расще-
линах скал, и в ноябре уже цвели. Транспирация цветущих ветвей
достигала 2,0—3,5% сырого веса в час. Поскольку транспирация самих
цветков была незначительной, а листочки были еще очень мелкими, то
относительно высокие общие значения транспирации свидетельствуют об
интенсивной транспирации ветвей; последняя была, бесспорно, больше,
чем у вышеуказанных кустарников, где она достигала 5—8% в сутки.
Акации в качестве элементов зональной растительности встречаются
только в областях с относительно большим количеством осадков, а в более
засушливых — в виде экстразональной растительности лишь в особенно
благоприятных местообитаниях (рис. 216 и 217).
Потеря воды путем транспирации в кустарниковой саванне (где
господствуют виды Catophractes и Rhigozum) на площади 100 м2 составляла
примерно 350 г в сутки. В течение всего периода засухи, то есть восьми
месяцев, этот расход достиг бы 84 л, что соответствует менее чем 1 мм
осадков. В предшествующий год выпало 265 мм осадков. Если даже учесть
весьма приблизительный характер этих вычислений, то все же видно,
сколь мал должен быть запас воды в почве, чтобы в течение всего периода
засухи поддерживать существование растений, находящихся в состоя-
нии покоя.
Несмотря на незначительные величины транспирации, кустарники
в течение засушливого периода вынуждены поглощать известное количест-
во воды из почвы, поскольку их собственных запасов едва ли может хва-
тить на одну неделю. В отличие от злаков кустарники обладают более
экстенсивной корневой системой, которая очень далеко простирается
284
Глава 6
в стороны от ствола 1 и в глубь почвы, но в общем слабо ее пронизывает.
На каменистых лишенных мелкозема почвах, а также в областях, где
осадки выпадают зимой и глубоко промачивают почву, это вполне целесо-
образно. На мелкокомковатых же почвах, наоборот, экстенсивная корне-
вая система кустарников уступает в борьбе за влагу интенсивной корневой
системе злаков. Поэтому кустарники растут со злаками только в местах,
где в период засухи запасы воды в почве превышают количество влаги,
израсходованной злаками.
Исходя из вышеизложенного, можно следующим образом выразить
отношение древесных растений и злаков к влаге.
Древесные растения:
1) нуждаются в большом количестве осадков;
2) эти осадки могут выпадать зимой;
3) запасы воды в почве в период засухи должны быть достаточными,
чтобы обеспечить хотя бы минимальный расход влаги растениями;
4) высокая влагоемкость почвы не обязательна; иными словами,
почва может отличаться крупнокомковатой структурой и даже каменисто-
стью, поскольку экстенсивная корневая система простирается далеко
в горизонтальном и вертикальном направлениях.
Злаки (многолетние, несклерофильные):
1) могут существовать и при меньшем годовом количестве осадков;
2) эти осадки должны, однако, выпадать в период вегетации, то
есть летом;
3) в период покоя растения практически не поглощают воды из почвы;
4) почва должна обладать относительно высокой влагоемкостыо-
и, следовательно, содержать достаточно мелкозема; такая структура поч-
вы позволяет интенсивной корневой системе в период вегетации поглощать
из небольшого объема почвы достаточное количество воды; интенсивная
транспирация у злаков наблюдается только при больших запасах влаги
в почве, при недостатке воды листья злаков быстро высыхают. .
С учетом высказанного становится понятным характер распределения
древесной и травяной растительности в засушливых областях, который
зависит как от климата, так и от почв.
Можно предложить следующую схему:
I. Область зимних дождей:
Склерофильная древесная растительность, злаки угнетены.
II. Область летних дождей:
А. Каменистая почва: преобладает древесная растительность, зла-
ки угнетены.
Б. Более мелкозернистая почва:
а) малое количество осадков (примерно 100—150 мм) — господст-
вуют чистые злаковники, древесные растения отсутствуют;
б) большее количество осадков (примерно 250—500 мм) — саван-
на; отдельные экземпляры древесных растений на фоне злакового по-
крова;
в) большое количество осадков (более 500 мм) — леса, злаки
угнетены.
По мнению Шимпера, можно говорить о климате травяных и древес-
ных формаций. Эколог не может согласиться с мнением К. Тролля, что
коренной ошибкой фитогеографической концепции Шимпера была его
1 У Zizyphus lotus радиальная протяженность корней достигала 20 ле, а у Тата-
rix и Acacia tortilis — даже 50 м (по Каушу). См. также гл. 13, стр. 485.
Зональные саванны как переходная ступень к аридной зоне 285
точка зрения, будто лес и тропический злаковник являются противопо-
ложностью в климатическом отношении. Речь идет о разных вещах.
Когда говорят о зависимости между растительностью и климатом, то при
этом должна приниматься во внимание только зональная расти-
тельность, развитая на равнинных территориях и в средних усло-
виях существования. Появление травяных сообществ в качестве зональной
растительности (то есть не в крайних условиях существования) действи-
тельно связано с совершенно определенным климатом: осадки должны
выпадать в вегетационный период в количествах, недостаточных для раз-
вития леса х. Если же злаковники встречаются вне пределов этой климати-
ческой области, то тогда речь идет об экстразональной, или
азональной, растительности, развитие которой обусловливается
рельефом, крайними вариантами почв или, наконец, вмешательством чело-
века (пожары, пастьба, сенокошение). В этих случаях не следует доволь-
ствоваться только внешним обликом ландшафтов; необходимо выяснять
причинные связи, весьма различные в каждом отдельном случае. Следова-
тельно, эколог не имеет права ставить знак равенства между одним сооб-
ществом злаков и другим; это больше свойственно географам. Поэтому
нужно попытаться выяснить, каким образом в саваннах совместно суще-
ствуют оба антагониста — древесные растения и злаки, поскольку, как
правило, между ними развертывается жесточайшая конкуренция и они
'большей частью взаимно исключают друг друга.
।
КОНКУРЕНТНЫЕ ВЗАИМООТНОШЕНИЯ В САВАННАХ
Во флоре умеренной зоны отсутствуют виды деревьев, способные
произрастать одиночно на фоне травяного покрова. Здесь распростране-
ны только виды, образующие сомкнутые древостои. Именно поэтому между
лесом и степью проходит довольно резкая граница и лишь узкая полоса
кустарников часто вклинивается между ними. Иначе обстоит дело в тро-
пиках, где распространены древесные породы в основном с Зонтиковой
кроной (особенно виды акаций, рис. 218), не образующие древостоев,
достаточно огнестойкие и обладающие способностью порослевого возоб-
новления.
Поэтому между зоной сомкнутого лрса и открытыми травяными про-
странствами (цветные фото 2, 3, стр. 272) существует очень широкая пере-
ходная полоса, где одиночные деревья и кустарники распространены на
фоне травяного покрова. Это саванны, развитые в областях, где осадки
выпадают летом (рис. 219 и цветное фото 3, стр. 80).
Рассмотрим в качестве примера изученные автором смены раститель-
ности в Юго-Западной Африке. Чтобы понять, как будут складываться
конкурентные взаимоотношения, проследим их в условиях постепенного
увеличения количества летних осадков. Распространенная здесь зональ-
ная растительность развита на выровненном рельефе и почвах, поглощаю-
щих всю атмосферную влагу. На рис. 220, а количество осадков составляет
около 100 мм. Выпавшая влага не проникает глубоко в почву и задержи-
1 Хотя годовое количество осадков в «степях» Серенгетти в Восточной Африке
достигает 530—785 мм, однако они выпадают крайне нерегулярно и в течение отдель-
ных месяцев иногда вообще не наблюдается дождей. Здесь также распространены
почвы с прослойками, сцементированными известью. Ср. G. D. Anderson,
L. М. Talbot, Soil factors affecting the distribution of the grasland types on the
Serengetti plains, «J. Ecol.», 53, 1965, p. 33—56.
286
Глава 6
вается в верхних слоях, где располагаются корневые системы низкорос-
лых кустовых злаков. В течение вегетационного периода они с помощью
своей интенсивной корневой системы поглощают всю воду, которая затем
испаряется через листовой аппарат. Исчерпав большую часть запаса водыг
надземные органы злаков, за исключением точек возобновления, отмира-
ют. Оставшаяся в почве влага также расходуется злаками в период,
засухи. Отсутствие в почве достаточного количества не израсходованной:
злаками влаги препятствует поселению здесь каких-либо древесных
растений.
При количестве летних осадков, равном 200 мм (рис. 220 б), почва
увлажняется на большую глубину, что способствует развитию более
высокорослых видов злаков. Однако и они в течение лета поглощают из.
почвы практически всю воду. При дальнейшем увеличении количества
летних осадков (рис. 220, в) злаки уже не могут расходовать всю накоп-
ленную в почве влагу, и поэтому даже в период засухи почва содержит
какой-то резерв воды; таким образом, впервые создаются предпосылки
для развития отдельных древесных растений, представленных вначале'
лишь мелкими кустарниками. С увеличением влажности климата возра-
стает количество не использованной злаками влаги и увеличиваются число-
и размеры древесных растений, развивающихся среди травяного покрова
(рис. 220, а). Таким образом, кустарниковая саванна сменяется древесной?
саванной. Однако во всех этих случаях большей
конкурентной мощностью отличаются злаки.
В засушливые годы многие древесные породы погибают; при наличии же
обильных осадков они пышно развиваются и даже появляются новые
всходы древесных пород. Только в случае резкого увеличения количест-
ва осадков, что создает предпосылки для смыкания древесного полога
и формирования сухих лесов, возникает обратная зависимость. В зате-
Р и с. 218. Acacia benthamii в саванне, провинция Наталь (фото Бьюса).
Зональные саванны как переходная ступень к аридной зоне
28Т
Рис. 219. Древесная саванна, снятая с гряды дюн. Одиночные деревья Аса^
cia giraffae среди цветущей Aristida (Stipagrostis) uniplumis, западная часть
Калахари, ферма Доннерсберг, Юго-Западная Африка (фото Э. Вальтер).
ненных местах под пологом леса злаки отмирают и сохраняются только*
в более освещенных просветах (рис. 220, г).
Для засушливых областей характерна еще одна жизненная форма —
суккуленты (листовые и стеблевые суккуленты). Они обладают
специальными запасающими воду органами, которые наполняются после
дождей и затем длительное время покрывают потребность растений в воде.
Из-за слабой транспирации суккуленты отличаются лишь незначитель-
ным приростом сухого вещества. Поэтому они растут крайне медлен-
но и не способны конкурировать с другими растениями; они встреча-
ются только в крайних условиях существования — чаще всего в пусты-
нях, где другие растительные формы уже не могут развиваться (см. стр. 353
и 429). В гумидных областях суккуленты могут произрастать на очень
маломощных почвах, быстро иссушаемых! после дождя; но даже в тече-
ние этого короткого срока суккуленты успевают запасти воду, кото-
рая затем медленно расходуется ими до выпадения следующего
дождя.
Таким образом, господство отдельных жизненных форм определяется
наряду с климатическими причинами и эдафическими условиями. В лю-
бой климатической зоне смена почв сопровождается сменой раститель-
ного покрова. Это очень наглядно видно в Верхнем Карру (Южная
Африка) (см. стр. 428 и рис. 342).
В некоторых аридных областях, например в Сомали \ внимание*
путешественника привлекает чересполосное распределение растительно-
сти, особенно заметное на аэрофотоснимках. Подобную же картину автор*
наблюдал в центральной Австралии. Полосы кустарниковых акаций
чередуются с полосами, почти лишенными растительности или покрыты-
ми только злаками. Во всех этих случаях были выявлены некоторые
1 С. А. Е. Hodge, Vegetation strips in Somaliland, «J. Ecol.», 50, 1962,
p. 465-474.
288
Глава 6
различия в структуре или механическом составе почвы, что однако, не
всегда является причиной возникновения полос. По большей части полосы
идут поперек малозаметного склона, на котором под влиянием воды и вет-
ра отложился материал различного состава.
Рис, 220, Схематическое изображение смены злаковников (а и б) саваннами (в) и сухи-
ми лесами (г).
Зональные саванны как переходная ступень к аридной зоне
289
ЗАРАСТАНИЕ КУСТАРНИКАМИ — УГРОЗА ЗЕМЛЕДЕЛИЮ
В ЗОНЕ САВАНН
Как уже говорилось выше, в более засушливой части тропиков при
уменьшении количества осадков приблизительно с 600 до 250 мм про-
исходит смена лесной растительности злаковниками; при этом конку-
рентная способность злаков непрерывно возрастает, вследствие чего
древесная саванна сменяется вначале кустарниковой саванной и, наконец,
низкорослой кустарниковой саванной. Наряду с климатом какую-то роль,
безусловно, играет также практикуемое местным населением ежегодное
выжигание травы, а в естественных условиях — пожары, возникающие от
ударов молнии. Лишь в последние годы во вновь освоенных под сельское
хозяйство районах подобные методы ведения хозяйства были запрещены.
Огонь оказывает, особенно на древесные растения, избирательное
действие; в результате в саванне растут только огнестойкие виды или
виды, обладающие способностью возобновляться порослевым путем после
пожара. У многих акаций на корневой шейке на глубине примерно 20 см
от поверхности почвы расположены спящие почки, которым огонь не
может причинить вреда. Температура на различной высоте над поверхно-
стью почвы во время пала в злаковнике достигала следующих величин х:
наивысшая температура 280—560° была зарегистрирована на высоте 50 см\
у самой поверхности почвы возможно кратковременное, на 2—3 минуты,
повышение температуры до 70—100°; на глубине же 2 см влияние огня
уже не сказывается.
Толстокорые древесные породы переносят пожары без всякого ущер-
ба. При частых пожарах в саваннах сохраняются только пирофиты, то
есть устойчивые к огню породы. В Австралии одревесневшие плоды неко-
торых видов (Hakea, Xylomelum, Banksia, Callistemon и др.) семейств
Proteaceae и Myrtaceae (частично) в течение многих лет висят на ветвях
и раскрываются только после пожаров. Однако проходит еще некоторое
время, прежде чем семена упадут на остывшую золу и прорастут.
Запрет на огневую очистку сухих злаковников в ряде сельскохозяй-
ственных районов связан с тем, что на выгоревших площадях первые же
дожди вызывают сильнейшую эрозию почвы и частично уносят питатель-
ные вещества вместе с золой. Кроме того^ слишком частые пожары обу-
словливают отрицательный отбор среди Злаков. Поскольку скот весьма
охотно посещает начинающие постепенно зеленеть выжженные участки,
это ослабляет развитие более ценных видов, тогда как менее ценные со
временем начинают преобладать. Если бы палы не сопровождались выпа-
сом, то это, вероятно, приносило бы меньше вреда.
С прекращением практики выжигания злаковников фермеры связы-
вают зарастание территории кустарниками; это явление в засушливых
тропических областях всех частей света происходит во все более угрожа-
ющих масштабах. В процессе зарастания степень участия колючих кустар-
ников в саваннах увеличивается, и со временем возникают колючекустар-
никовые заросли, не пригодные для пастьбы. Таким образом, этот процесс
угрожает самой основе существования фермеров 1 2.
1 A. Pitot, Н. Masson, Quelques donnees sur la temperature au cours des
feux de brousse aux environs de Dakar, «Bui. Inst. Frang. d’Afrique Noire», 13, 1951,
p. 711—732; N. C. W. Beadle, Soil temperatures during forest fires and their effect
on the survival of vegetation, «J. Ecol.», 28, 1940, p. 180—192.
2 H. Walter, Die Verbuschung, eine Erscheinung der subtropischen Savan-
nengebiete, und ihre okologischen Ursachen, «Vegetatio», 5/6, 1954, S. 6—10.
290
Глава 6
Рис. 221. Многочисленные всхо-
ды Acacia detinens на оголенной,,
вытоптанной скотом территории,
ферма Осомбусатьюру, Юго-За-
падная Африка. На последующих
рисунках 222—224 запечатлены
различные стадии зарастания зла-
ковников колючими кустарни-
ками.
Рис. 222. Смыкание ПологаAca-
cia detinens и Dichrostachys nutans.
На заднем плане — Acacia giraf-
fae у сухого русла, ферма Омам-
бонде-Ост.
Рис. 223. Непроходимые колю-
чие заросли Acacia maras, ферм»
Хазельдене.
При прекращении палов кустарники начинают разрастаться|не повсе-
местно, а лишь на фермах с очень большим количеством скота или вблизи
водопоев, то есть там, где вследствие слишком интенсивной пастьбы много-
летние злаки сильно ослаблены или полностью уничтожены. Возрастаю-
щая изреженность травяного покрова приводит к постепенному сокраще-
нию расхода воды на транспирацию и, как следствие, к увеличению
запасов влаги в почве к концу периода дождей. Это способствует лучшему
развитию древесных растений и их более быстрому возобновлению.
В обычных условиях всходы древесных пород не выдерживают конкурен-
ции с корнями злаков; лишь очень немногим из них удается выжить.
Зональные саванны как переходная ступень к аридной зоне
291
Если же в процессе пастьбы уничтожаются злаки, то это благоприят-
ствует появлению всходов древесных растений и образованию в дальней-
шем непроходимых колючекустарниковых зарослей (рис. 221—224).
Распространению колючих кустарников способствуют также живот-
ные, поедающие при недостатке злаковых пастбищ плоды акаций. Семена
последних проходят кишечный тракт животных, не теряя всхожести,
и хорошо прорастают в их выделениях 4.
Всхожесть семян Acacia giraffae, взятых из зрелых плодов, из экскре-
ментов слонов и некоторых копытных, через 5 недель равнялась 92, 78
и 59%; первые всходы появлялись соответственно через 11, 9 и 5 дней.
Рис. 224. Заключительная стадия зарастания кустарниками
(Dichrostachys nutans) местности близ Цумеба.
Таким образом, энергия прорастания в двух последних случаях была
значительно выше, что для аридных областей имеет большое значение.
Наличие почти у всех древесных растений африканской саванны
колючек, вероятно, можно объяснить многовековым отбором, обуслов-
ленным пастьбой диких животных. В Австралии, где только кенгуру могут
быть отнесены к пастбищным животным, и в Новой Зеландии, где вообще
не было никаких млекопитающих (за исключением нескольких видов
летучих мышей), почти совершенно отсутствуют виды колючих растений
или в крайнем случае встречаются растения с колючими листьями, распро-
страненные, однако, чаще под пологом леса, чем на открытых местах.
Здесь встречаются только колючие злаки, известные под местным назва-
нием «spinifex» (Triodia, Plectrachne).
Если пастьба диких животных оказывает столь сильное избиратель-
ное действие на древесные растения, то спрашивается, почему она не
приводит к вытеснению злаков кустарниками. В этом случае решающую
роль играет совершенно различный характер стравливания травы дикими
животными и домашним скотом. Первые чрезвычайно подвижны и посто-
янно меняют места пастьбы; при этом злаки не объедаются до основания
1 О.A. Leistner, On the dispersal of Acacia giraffae by game, «Koedoe», № 4,
1961, p. 101—104.
292
Глава 6
последовательно одно растение за другим. Стада копытных никогда долго
не задерживаются и у водопоев, где им грозит опасность встречи с хищ-
никами. Происходит именно такое стравливание, какого пытаются
добиться с помощью мелкозагонной пастьбы или пастбищеоборота.
При повсеместно практикуемой на огороженных фермах бессистемной
пастьбе нагрузка на травостой совсем иная. Скот ленив и малоподвижен.
Лишь объев всю траву вблизи водопоя, он начинает искать ее несколько
дальше. В результате вокруг водопоя многолетние травы полностью уни-
чтожаются, а на отдаленных участках фермы остаются вообще нетрону-
тыми. Поэтому заросли кустарников появляются прежде всего вблизи
водопоев и лишь со временем распространяются на остальную территорию.
Рис. 225. Старые заросли A cacia detine ns. Достигнув примерно
50-летнего возраста, кусты отмирают, а в просветах поселяется
Aristida uniplumis. При отсутствии пастьбы вновь восстанавли-
вается кустарниковая саванна, ферма Отьитамби.
Участки фермы, не посещаемые животными, не зарастают кустарниками,
даже если там и прекращено выжигание травы. Введение мелкозагонной
пастьбы позволило бы сохранить травостой и тем самым предотвратить
наступление кустарников
Заросли кустарников придают зональной растительности облик,
свойственный более влажному климату. Напрашивается вопрос, насколь-
ко эта смена необратима. Достаточно длительных опытов пока не про-
ведено, однако некоторые наблюдения говорят о том, что при увеличении
размеров и числа кустов на единицу площади расход воды будет пре-
вышать количество выпадающих осадков. Поэтому в засушливые годы
угнетенные экземпляры кустарников отмирают, заросли изреживаются
и в них укореняются злаки. В очень сомкнутых зарослях кустарники не
1 Н. Walter, О. Н.Vol k, Die Grundlagen der Weidewirtschaft in Siidwestaf-
rika, Stuttgart, 1954.
Затопляемые саванны в результате пастьбы зарастают кустарниками вследствие
уплотнения почвы, ускорения поверхностного стока воды и сокращения периода
затопления. См. по этому вопросу Н. Wilhelmy, Das grofle Pantanal im Mato
Grosso, «Deutsch. Geographentag Wurzburg 1957», Seite 45—71.
Зональные саванны как переходная ступень к аридной зоне
293
возобновляются, вероятно, из-за сильного затенения всходов (рис. 225).
Достигнув через несколько десятилетий предельного возраста, кустарники
отмирают и сменяются уже внедрившимся травостоем, который быстро
смыкает свой полог. В его составе остаются лишь одиночные более моло-
дые кусты. Конечно, ни один фермер не станет ждать так долго и будет
стремиться возможно быстрее уничтожить эти заросли. Как этого добить-
ся — зависит в основном от затрат, которые он может себе позволить.
Одна лишь огневая очистка кустарников не дает должного результата
(рис. 226).
Зарастание злаковников кустарниками представляет большую опас-
ность для слабонаселенных, но освоенных под животноводство территорий
Рис. 226. Начало формирования сомкнутых зарослей Acacia giraf-
fae после выжигания кустарников, ферма Омамбонде-Ост.
и менее опасно для областей, где преобладает местное население. На ферме
обычно живет лишь одна семья, располагающая небольшим количеством
рабочих рук. Расход топлива в домашнем хозяйстве очень невелик. В тузем-
ных же деревнях, тесно группирующихся вокруг источников воды,
потребляют много хвороста для приготовления пищи, используя для этого
также и колючие кустарники. Поэтому, хотя здесь и не наблюдается
зарастания местности кустарниками, однако при чрезмерном стравлива-
нии может возникнуть опасность опустынивания территории, что сопро-
вождается сильнейшей эрозией почвы. Этот процесс с экономической точки
зрения еще более неблагоприятен, чем вышеописанный.
РА СТИТЕЛЬНОСТЬ,
ОБУСЛОВЛЕННАЯ ЭДАФИ ЧЕСКИ МИ ПРИЧИНАМИ,
В ЗОНЕ САВАНН И ЗЛАКОВНИКОВ
Зональная растительность в засушливых тропических областях раз-
вита только на сравнительно мощных песчаных почвах в равнинных
условиях. Суглинистые или глинистые почвы при медленно идущем про-
цессе химического выветривания формируются лишь в понижениях релье-
294
Глава 6
Рис. 227» Проекция типичного участка растительного покрова в Юго-За-
падной Африке» На горизонтальной проекции не нанесены злаки» Крести-
ками отмечены отмершие растения (из Walter, Volk, Grundlagen der
Weidewirtschaft in SW-Afrika, Verlag Eugen Ulmer, Stuttgart, 1954)'
фа. В более влажных частях зоны саванн эти периодически увлажняемые
местообитания заняты чистыми злаковниками, на фоне которых разброса-
ны одиночные пальмы. Развитые здесь черные почвы тяжелого механиче-
ского состава в сухом состоянии покрываются глубокими трещинами,
а после сильных дождей становятся вязкими, кашеобразными. Однако
в генетическом отношении они очень далеки от черноземов; черную же
окраску им придают сильно набухающие при увлажнении устойчивые
структурные отдельности. В засушливых частях зоны саванн в большин-
стве случаев наблюдается слабое засоление периодически увлажненных
почв. На этих солонцеватых почвах в Юго-Западной Африке растут виды
кустарниковых Salsola. Встречаются также и суккулентные формы
Mesembrianthemum.
Очень часто в области распространения саванн, там, где на поверх-
ность выходит выветрившаяся порода, встречаются местообитания, лишен-
ные почвенного покрова. Чаще это явление наблюдается на горных скло-
нах. В таких местах злаки сменяются разреженными зарослями кустарни-
ков, степень покрытия которых снижается пропорционально уменьшению
количества осадков. Эти группировки образуют постепенный переход
к пустыне, на границе которой часто распространены суккуленты или
своеобразные древесные породы, запасающие в стволах большое количест-
во воды (см. стр. 394)» Саванноподобные сообщества распространены также
на маломощных песчаных почвах и в климатических условиях, где при
большей мощности песчаных почв должен был бы развиться злаковник 4.
В качестве примера можно привести распределение растительного
покрова в южной части Юго-Западной Африки у Мариенталя, где в сред-
нем выпадает 185 мм осадков. Представление о характере растительности
может дать рис. 227. Степень покрытия почвы достигает 40%; запас сухого
вещества, накопленного злаками и однолетниками, составляет 400 кг! га.
Господствует Aristida uniplumis, достигающая высоты 60 см. Маломощ-
1 Н» Walter, Grasland, Savanne und Busch der ariden Teile Afrikas in ihrer
okologischen Bedingtheit, «Jb» wiss» Bot»», 87, 1939, S» 750—860»
Зональные саванны как переходная ступень к аридной зоне
295
пая почва подстилается песчаником раз-
личной степени трещиноватости. Часть
выпавших осадков накапливается в пес-
чаной почве и без остатка используется
злаками, часть же просачивается вглубь
и скапливается в трещинах породы,
оставаясь недоступной для злаков. Эта
влага используется экстенсивной кор-
невой системой кустарников.
В этом случае, следовательно, не
существует никакой непосредственной
конкуренции между злаками и кустар-
никами, скорее, они дополняют друг
друга в отношении использования поч-
венной влаги. Напротив, между тремя
Рис. 228. Почвенный разрез и ха-
рактер распространения корней
Rhigozum (вверху) и Catophractes
(внизу). Точками обозначена срав-
нительно мелкозернистая почва,
пунктиром—оборванные корни (из
W а 11 е г, Vo 1 k, Grundlagen der Wei-
dewirtschaft in SW-Afrika, Verlag
Eugen Ulmer, Stuttgart, 1954).
встречающимися здесь видами кустар-
ников — Boscia foetida, Catophractes ale-
xandri и Rhigozum trichotomum — сущест-
вует острая конкуренция. Boscia достига-
ет наиболее крупных размеров и расходу-
ет наибольшее количество воды. Catoph-
ractes занимает промежуточное положе-
ние, a Rhigozum., наименьшее по размерам, одновременно и менее требова-
тельно к условиям среды. Благоприятные для развития кустарников
местообитания связаны с подстилающими породами, в трещинах которых
скапливается достаточно воды. Семена кустарников равномерно распре-
деляются по площади; однако выживают только всходы, укоренившиеся
в водоносных трещинах, не занятых корнями других растений. Часто
всходы всех трех видов укореняются в одном месте, однако для взрослых
экземпляров запасов воды уже не хватает и Boscia вытесняет два других
вида. Их отмершие остатки часто встречаются под кустами Boscia. В усло-
виях, где воды не хватает для развития Boscia, лучше всего растет Catoph-
ractes, тогда как Rhigozum вынужден довольствоваться наименее благо-
приятными для развития остальных видов местообитаниями, где песок
подстилается тонкозернистым песчаником. Если эти площади достаточно
обширны, то на них из древесных растений растет только Rhigozum
(рис. 228). Более благоприятны местообитания вдоль русел временных
водотоков, где эпизодический сток пополняет время от времени запас
поступающей в трещины воды. Здесь по берегам потоков пышно разви-
ваются Catophractes или Boscia, а по дну встречаются густые заросли
одного вида акации (рис. 216).
При изменении почвенных условий, например при увеличении мощно-
сти песчаного покрова, кустарники полностью вытесняются злаками.
Если же, напротив, мощность верхнего песчаного слоя уменьшается или он
совсем исчезает, то злаки уступают место кустарникам. Анализ почвен-
ных условий помогает понять характер пространственного распределения
злаков и кустарников.
Характер растительного покрова меняется также в зависимости от
угла наклона пластов. При наклоне пластов, благоприятном для выхода
инфильтрационной воды на поверхность, развиваются сомкнутые древо-
стои. В противном случае эти местообитания отличаются сухостью и на
них растут только суккуленты, например виды Aloe (рис. 217).
296
Глава 6
ЗОНАЛЬНОЕ РАСПРЕДЕЛЕНИЕ РАСТИТЕЛЬНОСТИ
ПРИ СНИЖЕНИИ КОЛИЧЕСТВА ОСАДКОВ
В СУБТРОПИЧЕСКИХ ОБЛАСТЯХ
В Юго-Западной Африке по мере уменьшения летних осадков наблю-
дается следующая смена растительного покрова:
Тропический
сухой листо-
падный лес
(рис. 229)
Саванна
(рис. 219)
Злаковник —>
(цветные фото
1—3, стр. 272)
Пустыня
причем различные типы саванн, существование которых обусловлено
климатом, развиваются при годовом количестве осадков от 500 до 250 мм.
Можно предполагать, что аналогичное распределение растительного
покрова наблюдается и в области летних дождей южнее Сахары, однако,
как показывают исследования Смита 4, здесь дело обстоит иначе; выпадают
злаковники, а саванны проникают в области, где годовые осадки достигают
даже 900 мм. Количество осадков колеблется в Судане от нуля у Вади-
Хальфа (на границе с Египтом) до 1400 мм и более в областях, гранича-
щих с Конго. Рельеф ровный, почвы на северо-западе песчаные, на юго-
востоке глинистые. При движении с севера на юг, по мере увеличения
количества осадков, наблюдаются следующие смены растительного
покрова (рис. 230):
1. Пустыня (desert) —менее 50 мм осадков в год.
2. Акациевая полупустыня (Acacia-desert-scrub).
3. Низкотравная акациевая саванна (Acacia-shortgras country).
4. Высокотравная акациевая саванна (Acacia-tallgras country).
5. Тропический сухой листопадный лес (mixed deciduous fireswept
forest).
1. В пустыне травяной покров (называемый по местному «gezzu»)
развивается только после эпизодических дождей, привлекая кочевников
с их стадами верблюдов. В остальное время глинистые почвы лишены
растительности. На песчаных почвах к югу от изогиеты 50 мм встречается
Acacia flava (A. ehrenbergiand). По руслам временных водотоков этот вид
заходит еще дальше на север, где растет вместе с Capparis decidua, Maerua
crassifolia и Leptadenia spartium.
2. Акациевая полупустыня развивается на песчаных почвах при
количестве осадков 50—250 мм и на глинистых — при осадках до 400 мм.
Здесь доминирует Acacia tortilis, и наряду с ней, кроме вышеназванных
видов, встречаются также Acacia raddiana и Boscia senegalensis. Вдоль
русел временных водотоков появляется Acacia seyal, а в более широких
долинах —A. mellifera. На песчаных дюнах, подстилаемых глиной,
растет Salvadora persica, а на обширных площадях, покрытых песками,—
Panicum turgidum и виды Суperus. На берегах Нила встречаются Acacia
albida, A. seyal, Zizyphus spina-christi, Balanites aegyptiaca, а также пальма-
дум Hyphaene thebaica.
3. В низкотравной акациевой саванне на глинистых почвах господ-
ствует Acacia mellifera, растущая здесь совместно с Boscia и Cadaba rotun-
difolia, а на песчаных почвах Acacia Senegal (дающая гумиарабик) с видами
1 J. S m i t h, Distribution of tree species in the Sudan in relation to rainfall and.
soil texture, Khartoum, 1949.
Зональные саванны как переходная ступень к аридной зоне
2S7
Рис. 229. Тропический сухой листопадный лес (Copaifera торапе) в северной
части Юго-Западной Африки; на переднем 1 плане — река Угаб с галерейным
лесом, образованным Acacia albtda, Euclea pseudebenus, Tamarix (фото Э. Вальтер).
Albizzia. Эти саванны развиваются на песчаных почвах при количестве
осадков, достигающем 250—400 мм, и на глинистых — при 400—600 мм.
На затопляемых в период дождей участках встречаются чистые древостои
Hyphaene. Травостой образован однолетними видами, используется под
выпас и ежегодно выжигается. Acacia melltfera часто образует одновозраст-
ные древостои, которые затем одновременно отмирают и погибают от огня.
На их месте возникают заросли дерновинных злаков с Cadaba.
4. Даже в высокотравной акациевой саванне, где злаки достигают
высоты 1,5-2,5 м, доминируют однолетние виды. Сильные пожары проходят
по всей территории; их останавливают только реки или иногда лишенные
травяного покрова площади, имеющие | местное название «mahal», на
которых всходы появляются после одного из ранних дождей, а затем засы-
хают. Чистые злаковники развиваются на площадях, затопляемых в период
дождей; лишь на термитниках растут деревья (стр. 279). В составе
этих саванн господствует Acacia seyal, образующая чистые древостои или
смешанные cBalanites aegyptiaca. Обширность области, занятой этими саван-
нами и протягивающейся вплоть до годовой изогиеты 900 мм, обусловлена
частыми пожарами. На каменистых возвышенностях со слаборазвитым
травяным покровом, не затрагиваемых огнем, встречаются виды, свой-
ственные сухим листопадным лесам, как, например, Lonchocarpus, Stereo-
spermum, Sterculia, Anogeissus, Boswellia, Ficus, и даже бамбук (Oxyte-
nanthera) *.
Область распространения этих саванн характеризуется сменой лишен-
ных травостоя площадей —«mahal», обусловленных неравномерным выпа-
дением осадков, высокотравными акациевыми саваннами. Лишенные тра-
вяного покрова участки быстро зарастают кустарниками; на них формиру-
1 В области распространения этих саванн находятся также большие болота
Сэдд Верхнего Нила.
298
Глава 6
тотся одновозрастные заросли акаций (см. стр. 290). После их отмира-
ния временно образуется открытый злаковник с Balanites. Перечисленные
стадии сукцессии могут наблюдаться одновременно на соседних участках.
5. Только при годовом количестве осадков свыше 900 мм развитие
травяного покрова подавляется древесной растительностью и появляются
сухие, большей частью смешанные леса, образованные широколистными
Рис. 230. Распределение почв, растительности и годового количества
осадков в Судане (по Смиту, с некоторыми изменениями).
а — количество осадков, мм; б — песчаные почвы; в — железистые песчаники;
г — глинистые почвы. 1 — пустыня; 2 — акациевая полупустыня; 3 — низко-
травная акациевая саванна; 4 —высокотравная акациевая саванна; 5 — тропи-
ческий сухой листопадный лес. Кружками обозначены облесенные горные массивы.
породами. Чистые древостои создают только Isoberlinia doka и Anogeissus
schimperi. Травостой образован многолетними видами, которые начинают
вегетировать сразу же после первых дождей, когда к ним присоединяются
луковичные и клубневые растения. Многие распространенные здесь роды
древесных встречаются также и в Южной Африке (Sclerocarya, Вигкеа,
Pterocarpus, Strychnos и др.), другие же представлены в Индии родствен-
ными видами (Tamarindus, Mimusops, Dalbergia).
Зональные саванны как переходная ступень к аридной зоне
299
В благоприятных условиях увлажнения встречаются реликтовые
участки дождевого леса, образованные Chlorophora excelsa, Ceiba pentandra
и др., причем в составе яруса кустарников нередко присутствует Cof-
jea robusta.
При рассмотрении вышеописанных смен растительного покрова бро-
сается в глаза, что даже на песчаных почвах близ границы с пустыней
акации играют большую роль, чем злаки, и что в области развития саванн
многолетние злаки отсутствуют вовсе. Напрашивается вывод о вторич-
ности группировок акаций, которые, скорее, следует рассматривать
как стадии деградации, обусловленные антропогенными факторами.
В Юго-Западной Африке многолетние злаки после длительного чрезмерно-
го стравливания также исчезают полностью, в результате чего изменяются
конкурентные взаимоотношения в саваннах. Однолетние злаки, прораста-
ющие только после первых дождей и образующие ежегодно заново не
только надземную часть, но и корневую систему, не являются столь
сильными конкурентами для кустарников, как многолетние. Последние,
быстро развиваясь, истощают почти весь запас воды, содержащийся
в корнеобитаемом слое.
Области, расположенные южнее Сахары, в течение многих столетий
неразумно использовались кочевниками для пастьбы верблюдов, коз
и овец. Даже при незначительной плотности населения нельзя найти тер-
риторию, не пострадавшую от неумеренной пастьбы. К этому еще добав-
ляется обычай ежегодно выжигать траву, в результате чего сильно стра-
дают многолетние злаки, тогда как лежащие на поверхности почвы семена
однолетников сохраняются, поскольку они вряд ли подвергаются дейст-
вию температуры выше 100° (см. стр. 289). Изменяется не только состав
травостоя. Верблюды и козы не щадят и древесные растения. К этому
добавляется большая потребность местного населения в топливе и древес-
ном угле. Даже в очень отдаленных областях кустарники безжалостно
вырубаются и приходится прибегать к защитным мерам против подобной
хищнической рубки *. В результате этого на обширных пространствах
развивается производный растительный покров, образованный отдель-
ными колючими кустарниками и однолетниками; он мало чем напоминает
исходную растительность. Единственный многолетний злак, растущий
в пограничных с пустыней районах, Panicum turgidum встречается очень
редко. Не удивительно, что в этих условиях на охраняемых участках
кустарники развиваются лучше; однако в целом исходный растительный
покров не восстанавливается.
Проникновение высокотравных саванн в гумидные области точно так
же может быть следствием ежегодных сильных пожаров. Смит, вероятно,
был прав, придавая этим саваннам значение пирогенного климакса и отри-
цая, таким образом, их зональный характер, обусловленный климатом.
В ранее приведенных примерах почвы отличались более или менее
мелкозернистой структурой (супесчано-суглинистой). На каменистых
же почвах злаки деградируют, и тогда в условиях Юго-Западной Африки
наблюдаются следующие смены растительного покрова:
Сухой Заросли сухих Кустарничко-
листопадный —> листопадных —> вал полупу- —> Пустыня
лес кустарников стыня
1 R. Halwagy, The impact of man on semi-desert vegetation in Sudan,
«J. Ecol.», 50, 1962, p. 263—273. См. также: Ch. R osetti, Observations sur la vege-
tation au Mali Oriental, FAO, № UNSF/DL/ES/4, Roma, 1962.
300
Глава 6
В области, где осадки выпадают зимой, как было показано выше,
господствуют древесные растения,, тогда как виды травяного покрова
практически отсутствуют. С уменьшением количества осадков понижает-
ся высота древесных растений, сменяющихся кустарничковой полу-
пустыней, а затем пустыней. В этом случае злаки выпадают и возникает
следующий ряд:
Жестколист-
ный лес
Заросли же-
стколистных
кустарников
Кустарничко-
вая полупу- —>
стыня
Пустыня
Если осадки выпадают в течение двух сезонов или выпадают нерегу-
лярно, то роль суккулентных растений возрастает (цветное фото 1,.
стр. 36) и возникает следующий ряд:
Заросли колючих
кустарников (с сук-
кулентами)
Полупустыня с гос-
подством суккулен- —>
тов
Пустыня
Обязательным условием всех этих смен является наличие соответ-
ствующих жизненных форм в описываемых природных областях. В Австра-
лии, например, полностью отсутствуют суккуленты, так же как и листо-
падные кустарники, свойственные саваннам. У эвкалиптов развиваются
кожистые вечнозеленые листья, а у акаций — жесткие вечнозеленые*
сплющенные или цилиндрической формы филлодии. Своеобразны также*
нигде больше не встречающиеся злаки, представленные колючими подуш-
ками. Это обычно полусферические подушки диаметром более метра,
которые у одного вида с возрастом увеличиваются, а у других распада-
ются, поскольку они нарастают радиально только с северной стороны.
Эти злаки относятся к родам Triodia и Plectrachne. Кроме того, в Австра-
лии между областями зимних и летних дождей нет, как это наблюдается
в Африке, пустынь; здесь располагается область, где в среднем выпадает
100 мм осадков, по-видимому очень равномерно распределяющихся
в течение года; в некоторые годы вообще не случается ни одного дождя,
и тогда первые осадки могут выпасть в любое время года. Для этих
областей характерны полусуккулентные кустарнички (Chenopodiaceae).
Злаки, образующие колючие подушки, распространены в бедных осадка-
ми областях, где явно преобладают летние дожди, но отчасти они выпадают
еще и зимой. Таким образом, области зимних (а) и летних (б) осадков свя-
заны с полупустыней весьма сложными переходами, которые схемати-
чески можно представить следующим образом (см. также цветные фото Зг
стр. 80 и 1, 2, стр. 432).
а) Эвкалипты-]-жестко- —> Эвкалипты-[-кустар-
листные кустарники нпчкп ->
Заросли Acacia апеига-*
б) Эвкалиптовая саван- / Кустарники-(-колючие
Кустарничковая (Che-
nopodiaceae) полупу-
стыня
на
злаки
Если профиль провести не через центральную, а через западную-
часть Австралии, то из схемы выпадают заросли кустарничковых солянок
(salt bush), вместо которых в наиболее засушливых областях ряда (б)
развиваются чистые заросли злаков, образующих колючие подушки,
а в наиболее бедных осадками областях ряда (а) — чистые заросли Acacia
апеига (мульга-скраб) с похожими на сосновые иглы филлодиями. В пере-
ходной полосе обе формации мозаично вклиниваются одна в другую.
Зональные саванны как переходная ступень к аридной зоне
301
Таким образом, видно, сколь разнообразным может быть характер
растительного покрова в зависимости от распределения осадков и флори-
стического состава. Для каждой области характерны свои особенности.
С экологической точки зрения, безусловно, нельзя ставить знак равенства
между эвкалиптами австралийских саванн и листопадными кустарниками
африканских и южноамериканских саванн. Первые гораздо лучше при-
способлены к сосуществованию со злаками в богатых осадками областях
летних доящей, поскольку дают мало тени. Поэтому древостои, похожие
на саванны, встречаются даже в областях с довольно большим количест-
вом годовых осадков (см. стр. 261 и след.).
Элленберг в Перу выделяет следующие смены растительности
(рис. 183):
а) Регулярно выпадающее небольшое количество осадков:
Сухой листо- Заросли суккулент- Полупустыня
падный лес —> ных кустар- —> с господством —> Пустыня
ников суккулентов
б) Количество осадков сильно колеблется из года в год:
Сухой листо- Заросли сухих Кустарнико-
падный лес —> листопадных —> вая полупу- —> Пустыня
кустарников стыня
Итак, после беглого ознакомления с растительностью аридных обла-
стей перейдем к более детальной ее характеристике.
Глава 7
РАСТИТЕЛЬНОСТЬ
СУБТРОПИЧЕСКИХ АРИДНЫХ ОБЛАСТЕЙ
ОБЩАЯ ХАРАКТЕРИСТИКА АРИДНЫХ ОБЛАСТЕЙ
В аридных областях растения вследствие малого количества осадкой
и сильного испарения испытывают недостаток воды в течение боль-
шей части года. Слаборазвитый растительный покров приспособлен
здесь к перенесению неблагоприятных условий увлажнения.
Аридные области занимают примерно 35% земной поверхности, при-
чем располагаются симметрично по обе стороны тропической зоны
(рис. 231). Они приурочены к субтропической зоне северного и южного
полушарий, а в Азии и Северной Америке заходят также далеко в умерен-
ную зону. При попытке найти критерий для точного разграничения арид-
ных областей от гумидных исследователи всегда сталкиваются с извест-
ными трудностями.
В большинстве случаев для этого не могут быть использованы данные
о количестве осадков, так как при равной величине осадков в субтропи-
ках будет господствовать климат пустыни, а в Арктике, напротив, очень
влажный климат. Эту границу большей частью проводят там, где величина
испарения с открытой водной поверхности за год превышает годовое коли-
чество осадков. До сих пор еще не выработано способа измерения испаре-
ния, дающего достоверные значения, поэтому приходится пользоваться
косвенными признаками:
1. Бессточные озера могут существовать только в тех случаях, когда
испарение превышает осадки. Во влажных областях, где испарение с вод-
ной поверхности меньше выпадающих на эту поверхность осадков, каждое
понижение рельефа, не имеющее подземного стока, со временем пере-
полнится водой и возникнет поверхностный сток. Не следует, однако,
думать, что в аридных областях у озер не бывает стока. Если, например,
водосбор озера достаточно велик, то впадающие в него реки несут избыточ-
ное количество воды и в этом случае, несмотря на сильное испарение,
озеро все же будет иметь хотя бы периодический сток.
2. Еще одним признаком аридных областей является наличие в них
периодически пересыхающих водотоков; они не возникают в тех случаях,
когда истоки реки находятся вне пределов аридной зоны, как, например,
у Нила, Оранжевой реки, Колорадо, Волги и т. д. С другой стороны,
также и в богатых осадками областях в засушливое лето может наблюдать-
ся временное прекращение поверхностного стока, как это часто происхо-
дит, например, в Средиземноморье.
3. Наконец, в аридных областях широко распространены засоленные
почвы, что связано с недостаточным количеством осадков, необходимых
для вымывания образующихся при выветривании солей. Во влажных
областях существуют лишь весьма ограниченные площади засоленных
земель, например вокруг минеральных источников, выносящих на
Рис. 231. Аридные области земного шара (по UNESCO Program for Arid Lands).
a — крайне засушливые; б — засушливые; в — полузасушливые.
1 — пустыня Сонора; 2 — пустыня туманов „Намиб; 3 — чилийско-перуанская прибрежная пустыня; 4 — Карру:
5 —засушливые области Центрально11 Австралии; 6 — Сахара с египетско-аравийской пустыней.
б
-'304
Глава 7
поверхность легкорастворимые соединения из определенных прослоек
горных пород, или же на морском побережье, где сказывается влияние
морской воды.
Однако всех этих признаков еще недостаточно для четкого разграни-
чения аридных и гумидных областей, поскольку на земле существуют
лишь несколько аридных территорий, в которых бездождный период не
приурочен к строго определенному времени года. С другой стороны, суще-
ствуют также области с влажным климатом, где хорошо выражены более
или менее длительные периоды засухи, в связи с чем здесь господствуют
тропические сухие леса или саванны, о которых уже говорилось.
Следовательно, и в засушливых областях необходимо учитывать смену
сезонов, и поэтому можно вновь воспользоваться климадиаграммами, на
которых очень четко выделяются аридные области (заштрихованная пло-
щадь значительно меньше площади, покрытой пунктиром). Однако
к этому вопросу не следует подходить слишком упрощенно, поскольку
положение контура, покрытого пунктиром, или расчленение его на две
части, имеет существенное значение при характеристике аридных условий
той или иной области.
Объективной границы между аридной и гумидной областями вообще
не существует; каждая попытка такого разграничения в какой-то мере
произвольна и производится только из соображений целесообразности.
Переход от одних климатических условий к другим всегда осуществляется
постепенно, если, конечно, климатический рубеж не связан с высоким
горным хребтом.
Существует несколько различных типов аридного климата, которые
также выделяются более или менее произвольно. Эту мысль поясняют
следующие климатические диаграммы.
1. Аридные области, где осадки выпадают зимой и засушливый период
выражен летом (рис. 232—234).
2. Аридные области, где осадки выпадают летом и засушливый
период выражен зимой (рис. 235—237).
3. Аридные области с двумя периодами осадков или без ясно выражен-
ного сезона дождей (рис. 238—240).
4. Крайне засушливые аридные области, где осадки выпадают
эпизодически или осадков вообще нет (рис. 241—243).
Обе последние диаграммы (побережье Юго-Западной Африки и Чили)
характеризуют, собственно, еще один тип пустынь, а именно пустыни
туманов. Здесь не выпадает осадков, но очень часто бывают туманы, влаги
которых, однако, недостаточно для того, чтобы в этих крайне суровых
условиях изменить характер пустыни, почти лишенной растительности.
Туман оказывает влияние всего лишь на гребнях и на склонах гор, где
он конденсируется при движении воздуха; в низинах он лишь повышает
влажность воздуха и вызывает снижение температуры (см. гл. 9 и 10).
Об аридных областях умеренной и холодной зон, развитых при
других температурных условиях, речь пойдет уже во втором томе.
Крайне засушливые области называют пустыней, но даже и это
понятие не имеет четкого определения 4. Только в очень немногих пусты-
нях из-за крайней сухости растительность отсутствует полностью (за
исключением оазисов) или же после редких дождей развивается лишь на
короткое время (эфемеры).
1 Н. Schiffers, Die Sahara, Stuttgart, 1950; A. Gabriel, Die Wiisten
•der Erde und ihre Erforschung, «Verstandl. Wiss.. Naturw. Abt.», 76, 1961.
1. Прибрежная почти лишенная растительности часть пустыни Намиб (Юго-Запад-
ная Африка). На заднем плане — горы Россинг.
2. Четыре экземпляра Lithops на каменистой осыпи (ферма Стенкампспан близ Апинг-
тона, ЮАР).
Растительность субтропических аридных областей
305
Очень часто при определении «пустынь» решающее значение приобре-
тают соображения экономического порядка. Под пустыней тогда понимают
область, в которой оседлое население не может без орошения использовать
земли под сельское хозяйство. Дальше всего в пустыню вторгается экстен-
сивное скотоводство, в то время как богарное земледелие прекращается
уже в менее аридных условиях климата. Часто совершенно необитаемые
«пустыни», например в центральной Австралии или в Калахари, вовсе
не являются самыми засушливыми частями континента. В данном случае
решающими являются не климатические, а другие факторы. Известную
роль могут играть состояние средств сообщения или же песчаные дюны,
затрудняющие доступ в эти области и создающие видимость их непригод-
ности для пастбищного хозяйства; такое же значение могут иметь отсут-
ствие пригодной питьевой воды или же очень проницаемые известковые
почвы, препятствующие развитию здесь какой-либо растительности.
Арика (5м) 19^°;
Рис. 232—243. Климадиаграммы пустынных областей («Klimadiagramm-Weltatlas»,
1960).
232—234 — области, где осадки выпадают зимой; 23 5—237 — области, где осадки выпадают
летом; 238—240 — области, где осадки выпадают в любое время года или где выражены два
дождливых сезона; 241—243 — области, где осадки выпадают эпизодически.
243
306
Глава 7
При определении «пустыни» очень многое зависит от возможностей
сравнения. Для жителя влажных областей Северной Америки юго-запад-
ная часть США, граничащая с Мексикой, уже является «пустыней Сонора».
Египтянин же, знакомый с крайними условиями пустыни, не относит
прибрежную полосу своей страны к пустыне, хотя она значительно засуш-
ливее и беднее растительностью, чем пустыня Сонора.
Поэтому не следует удивляться, что словом «пустыня» часто обозна-
чают весьма различные ландшафты. Во всяком случае, в этой главе будут
описаны лишь пустыни, существование которых обусловлено климатом,
то есть аридные области, которые характеризуются сравнительно неболь-
шой сомкнутостью растительного покрова; однако даже эти пустыни
столь сильно отличаются друг от друга, что их необходимо рассматривать
раздельно. Общие для всех пустынь данные будут приведены ниже, тогда
как их отличительные особенности изложены в гл. 8—14.
ВЛАГООБЕСПЕ ЧЕННОСТЬ РАСТЕНИЙ АРИДНЫХ ОБЛАСТЕЙ
HP И СНИЖЕНИИ СОМКНУТОСТИ РАСТИТЕЛЬНОГО ПОКРОВА
И НЕРАВНОМЕРНОМ РАСПРЕДЕЛЕНИИ ВОДЫ В ПОЧВЕ
Если задать вопрос, в чем выражается большое сходство ландшафтов
аридных областей во всех частях земного шара, то в первую очередь
следовало бы указать на два явления: уменьшение значения растительного
покрова и увеличение роли горных пород в формировании общего облика
ландшафта. Если обычно растительность покрывает поверхность земного
шара и защищает ее от эрозии, то в аридных областях наблюдается иная
картина. В пустыне выветривание наряду с эрозией способствует форми-
рованию резко выраженных форм рельефа, которые часто, несмотря на
равнинные условия, напоминают ландшафт высокогорья. Только учи-
тывая незначительную сомкнутость растительного покрова в аридных
областях, становится понятным, почему, несмотря на очень малое количе-
ство осадков, растения здесь достаточно обеспечены водой 4.
Возможность существования растений в очень бедных осадками пусты-
нях обычно вызывает удивление и склонность приписывать этим расте-
ниям совершенно особые эколого-физиологические свойства. Однако
более тщательные исследования свидетельствуют о наличии лишь некото-
рых особенностей. На местах произрастания растений в пустыне, в преде-
лах площади, охваченной их корнями, всегда имеется вода, конечно, в том
случае, если речь идет о растениях с развитой корневой системой, а не
о растениях с водозапасающими органами и тканями. Нужно, в частности,
учитывать, что осадки, выпадающие в количестве нескольких миллимет-
ров, соответствуют нескольким литрам воды, поступающим на 1 м1 2
поверхности почвы; это количество воды всегда следует сопоставлять
с растительной массой (на той же площади в 1 л2), что, однако, до настоя-
щего времени делалось редко.
Установить зависимость степени сомкнутости растительного покрова
от количества осадков можно только при условии неизменности других
факторов, например температуры или почвенных условий. Кроме того,
сравнению подлежат только сходные группировки растений.
При наличии этих предпосылок можно показать, что растительная
масса, а в связи с этими транспирирующая поверхность уменьшаются про-
1 По данным Лавренко (см. стр. 467), на это указывали также Свешникова
и Заленский в своей работе «Водный режим аридных областей Средней Азии и Казах-
стана», 1956. Автору не удалось познакомиться с этой работой.
Растительностъ субтропических аридных областей
307
порционально количеству осад-
ков. Автору удалось доказать
это впервые в Юго-Западной Аф-
рике при сравнении различных
не затронутых пастьбой злаков-
ников, развитых на песчаных
почвах В этой области осадки
выпадают только летом и их ко-
личество убывает по направле-
нию от более влажного северо-
востока страны к побережью,
от более чем 500 мм практи-
чески до нуля (рис. 244). Одно-
временно уменьшаются высота
травостоя и степень сомкну-
тости растительного покрова;
при количестве осадков менее
100 мм в год злаковники сменя-
ются пустыней. Зависимость
между количеством осадков и на-
коплением сухого вещества над-
земными органами представлена
на рис. 245, на котором хорошо
видно, как с уменьшением осад-
ков пропорционально падает и
продуктивность растительности.
Поскольку сравнение велось
только со злаками, можно выска-
Р и с. 244. Годовое количество осадков в Юго-
Западной Африке (в мм) (по Г. Вальтеру).
1 — точки, где брались образцы для анализа кор-
мовых растений; 2 — пробные площади, где опре-
делялась продуктивность травяного покрова.
зать предположение, что полученная закономерность характеризует не
только запасы сухого вещества, но и площадь транспирирующей поверхно-
сти злаков, иными словами, площадь этой поверхности уменьшается пропор-
ционально снижению количества осадков. Это означает, что на едини-
цу площади транспирирующей поверхности во
влажных и аридных областях приходится оди-
наковое количество осадков; иначе говоря,
отдельные растения снабжаются водой в арид-
ных областях ненамного хуже, чем в областях
достаточного увлажнения1 2.
Автор в дальнейшем показал, что эта закономерность действительна
и в отношении других типов растительности; это подтвердилось, напри-
мер, в Австралии, при изучении эвкалиптовых лесов. Здесь определялись
не только количество ежегодно образующейся массы листьев (листовой
опад), но и непосредственно площадь их поверхности. При проведении
1 Н. Walter, Grasland, Savanne und Busch der arideren Teile Afrikas in ihrer
okologischen Bedingtheit, «Jb. wiss. Bot.», 87, 1939, S. 750—860.
2 Ухудшение гидратуры растений может возникнуть в связи с тем, что при
постоянном уровне снабжения водой потенциальное испарение возрастает с умень-
шением количества осадков. Однако это верно только в масштабах года. Непосред-
ственно в период дождей, который соответствует вегетационному периоду, различия
в величинах потенциального испарения не очень велики, даже наоборот — при умень-
шении количества осадков вегетационный период в целом сокращается. Поэтому
злаки, развитые во влажных областях и отличающиеся крупными размерами, сме-
няются в засушливых областях другими видами, быстрее заканчивающими свое раз-
витие.
308
Глава 7
таких исследований необходимо
проводить раздельное сравне-
ние участков первичного леса
с подлеском и лесных культур
без подлеска, отличающихся
благодаря этому большей мас-
сой листьев.
Указанная закономерность
изображена на рис. 246; мак-
симальные значения количест-
ва листового опада и площадь
поверхности опавших листьев
были приняты в обоих случаях
за 100. Если не считать неко-
торых отклонений, неизбежных
при подобных определениях,
то в целом пропорциональный
характер зависимости выра-
Р и с. 245. Зависимость продуктивности зла- жен довольно отчетливо; в дан
ковников от количества осадков в Юго-За- ном случае это проявляется
падной Африке (по Вальтеру). при значительно больших ко-
личествах осадков — примерно
от 500 до 1500 мм и более; кроме того, эта закономерность в отличие
от предыдущего случая выявлена в области зимних дождей А.
Таким образом, расход воды на единицу площади
листовой поверхности во влажных и аридных
областях остается приблизительно одинако-
в ы м. Эта закономерность распространяется только на растения, погло-
щающие воду посредством корневой системы. В общем в аридных областях
при уменьшении площади листовой поверхности развивается более мощ-
ная, неглубоко залегающая корневая система. С увеличением засушливо-
сти климата соотношение между надземными и подземными частями расте-
ний все больше сдвигается в пользу последних. Исключение в этом
отношении составляют лишь суккуленты, которые нельзя сравнивать
с другими растениями.
В крайне засушливых областях с количеством осадков меньше 100 мм
следует обратить внимание на неравномерное распределение воды в почве,
что отражается на характере растительного покрова. Отсутствие сомкну-
того растительного покрова обусловливает здесь сильный поверхностный
сток. Поэтому вне зависимости от количества выпавших осадков на возвы-
шениях почва суше, чем на дне русел временных водотоков или в пониже-
ниях. В связи с этим растительность также неравномерно распределена
по площади и концентрируется там, где запасы воды в почве больше; в этих
местах растения также обеспечены водой, как и во влажных областях
(рис. 247). Если принять, например, что при годовом количестве осадков,
равном 25 мм, в результате поверхностного стока в понижение стекает
40% выпавших осадков с площади, в 50 раз большей, чем сама депрессия,
1 Подобную же закономерность установил Уэру в Тунисе. По его данным, уро-
жай с 1 га оливковых насаждений снижается параллельно уменьшению количества
осадков; причем урожай с одного дерева не изменяется, тогда как число деревьев
на 1 га уменьшается пропорционально количеству осадков. Н. N. le Houerou,
Ecologie, phytosociologie et productivite de 1’olivier en Tunisie meridionale, «Bull.
Service Carte Phytogeogr.», ser. B, t. IV, Fasc. 1, 1959, p. 7—72.
Растительность субтропических аридных областей
309
то окажется, что в распоряжение растений, обитающих в этой впадине,
поступает количество воды, соответствующее 500 мм осадков.
Изменение характера растительности особенно хорошо прослежи-
вается с самолета. Если в богатых осадками областях растительность
равномерно распределяется по всей площади и в понижениях отмечается
Рис. 246. Зависимость количества листового опада и площади
поверхности листьев от количества осадков в эвкалиптовых
лесах Австралии (данные заимствованы из О. W. Loneiagan,
Forest Dep. of W. Australia).
1 и 3 — первичный лес; 2 и 4 — лесные культуры.
лишь увеличение ее сомкнутости, то в крайне засушливых областях наблю-
дается прерывистый характер распространения растительности 4, иными
словами, происходит оголение все большей территории и концентрация
растительности в долинах и понижениях 1 2. Поскольку вода здесь просачи-
1 Th. М о п о d, Modes «contractes» et «diffus» de la vegetation saharienne, Biolo-
gy of deserts, London, 1954, p. 35—44.
При бороздной эрозии эта растительность распространена в виде полос в лож-
бинах, тогда как при потоках воды, захватывающих большие площади, она более
равномерно распределена по всей территории.
В Сомали, в областях, где выпадает 150 мм осадков, можно было наблюдать
параллельные полосы, заросшие A ndropogon, направленные вдоль склона. Наличие
между этими полосами лишенных растительности площадей обусловлено пастьбой,
образованием корки на поверхности почвы и более сильным стоком. Влажность почвы
под полосами растительности достигала 10%, а на оголенных площадях между ними —
6—7% (по С. F. Hemming, Vegetation areas in Somaliland, «J. Ecol.», 53, 1965,
p. 57—67; ср. также стр. 462).
2 Нечто сходное наблюдается в переходной зоне между лесом и степью. С уве-
личением засушливости лесная площадь расчленяется все больше и лес концентри-
руется в долинах. Таким образом, и здесь речь идет о «концентрации» лесной расти-
тельности.
310
Глава 7
вается довольно глубоко и не испаряется, корни растений проникают очень
глубоко в почву (см. рис. 372 — 373, 384). Таким образом, видно, что
глубина проникновения корней не всегда связана со снижением количества
осадков.
Рис. 247. Переход от равномерного распределения растительного
покрова (7, 2) к прерывистому (3, 4) при уменьшении количества
осадков в крайне засушливых областях.
Размещение культур в местах, лучше всего обеспеченных влагой, при-
менялось в сельскохозяйственной практике с давних времен. В пустыне
Неджеб (Негев) была открыта древняя оросительная система, служившая
в прежние времена для сбора поверхностных вод, поступающих после
выпадения осадков с вышерасположенных площадей, и отвода воды на
нижележащие поля. В последнее время некоторые из этих оросительных
систем были реконструированы и с их помощью удалось даже при незна-
чительном количестве годовых осадков собрать определенный урожай1.
Размер поверхностного стока в этом случае достигал 20—40% общего
количества выпавших осадков.
На побережье Египта подобные системы, сохранившиеся со времен
Римской империи, использовались для полива виноградников. Еще и сей-
час при количестве осадков, равном всего лишь 100 мм. здесь можно
выращивать маслины, направив к оливковым садам поверхностный сток
с окружающих каменистых склонов. Водосборная площадь при этом
должна быть по меньшей мере в 4 раза больше возделываемой, а потери
на инфильтрацию — минимальными. При соблюдении этих условий
земледелие становится здесь вполне возможным.
1 М. Evenari, L. Schana, М. Т a dm о г, Y. Aharon i, Ancient Agri-
culture in the Negev, «Science», 133, 1961, p. 979—996, см. также: «Runoff-Farming
Report», Rehovot, 1963.
Растительность субтропических аридных областей
311
Неравномерное распределение влаги в пустыне заметно не только
в условиях пересеченной местности, но и на почти идеальной равнине.
В последнем случае вода не концентрируется в отдельных водотоках,
а движется широким потоком на больших площадях. Для этого достаточно
незаметного для глаза уклона, равного 1 : 2000. Этот поток движется
медленно, в связи с чем вода просачивается в почву. Чем дольше пласт
воды покрывает поверхность почвы, тем больше воды просачивается
вглубь. Поэтому степень сомкнутости растительного покрова увеличивает-
ся по направлению движения потока воды. Только в наиболее низкой
части водосборной площади условия снова меняются, так как вода в зави-
симости от степени интенсивности дождя продолжает или течь дальше,
пока вся не впитается в почву, или же ее сток прекращается еще раньше.
В нижней части водосборной площади происходит накопление принесен-
ных водой нерастворимых соединений, в результате чего начинается
засоление почвы. Наиболее мощные потоки воды достигают бессточных
котловин, где соли отлагаются на поверхности почвы. При этом образу-
ются настоящие солончаки (цветное фото 1, стр. 368).
При близком залегании водопроницаемых известняков поверхност-
ный сток обычно полностью отсутствует. Такие территории почти совер-
шенно лишены растительности, как, например, обширная равнина Нал-
ларбор на юго-западе Австралии. Только в небольших долинах, заполнен-
ных мелкоземом, могут существовать отдельные древесные растения. На
песчаных территориях поверхностный сток также слабо выражен, зато,
как будет показано ниже, песчаная почва в аридных местностях отличается
более благоприятными условиями увлажнения, чем в гумидных.
Приведенные выше соображения относятся в первую очередь к много-
летним растениям, поскольку только они в состоянии приспособиться
к суровым климатическим условиям пустынь. В неблагоприятные перио-
ды они или теряют большую часть своей надземной массы, или полностью
отмирают. В годы с большим количеством осадков многолетники пышно
разрастаются и их всходы появляются на новых площадях. Наряду с ни-
ми во всех аридных областях распространены еще эфемеры, развитие
которых связано с количеством осадков текущего года. Они обусловли-
вают так называемую «буферность» растительности, выражающуюся
в известной зависимости между увеличением количества осадков и нара-
станием растительной массы. В благоприятные по количеству выпавших
осадков годы многолетние виды не в состоянии использовать всей имею-
щейся в их распоряжении воды. Это помогают им сделать эфемеры. Они
прорастают тысячами и сплошным ковром — зеленым во время вегетации
и пестрым во время цветения — покрывают всю пустыню. Они значи-
тельно меньше многолетников, не приурочены к определенным местообита-
ниям и растут повсюду, где почва длительное время остается влажной,
поэтому в засушливые периоды эфемеры появляются только в пониже-
ниях рельефа. Если почва увлажнена незначительно, то осмотическое
давление у однолетников быстро растет и развивается много низкорослых
растений, быстро начинающих цвести, но приносящих лишь немного зре-
лых плодов и семян. Если же сезон дождей затягивается, то в благо-
приятных условиях гидратуры растения сильно разрастаются и к концу
вегетационного периода образуют большое число цветков и плодов,
пополняя запас семян в почве пустыни. Обилие эфемеров зависит, таким
образом, от количества осадков (цветное фото 1, стр 432). Количество
осадков определяет также флористический состав эфемеров и, в частности,
по следующим причинам:
312
Глава 7
1. Некоторые однолетние виды прорастают только в очень узких
температурных пределах. Поэтому прорастание одних видов и состояние
покоя других зависят от температурных условий, наблюдаемых после
выпадения дождя1.
2. Эфемеры отличаются различной быстротой роста и потребностью
в воде. Среди них встречаются гигроморфные виды с нежными листьями,
которые быстро растут, но требуют хорошего увлажнения почвы и раз-
виваются в условиях не слишком сильного испарения. Другие отличают-
ся более ксероморфным строением, часто густо опушены или имеют
облик суккулентов. Они гораздо лучше переносят короткий засушливый
период во время дождливого сезона, развивают более мощную корневую
систему, но зато растут гораздо медленнее.
3. Во время короткого благоприятного для роста эфемеров периода
конкуренция среди них выражена так же сильно, как и в любом другом
растительном сообществе. При большом количестве осадков господствуют
гигроморфные и быстрорастущие виды, если же осадки выпадают нерегу-
лярно и верхние слои почвы в периоды между дождями иссушаются,
то тогда доминируют более устойчивые к недостатку влаги ксероморфные
виды.
Поэтому в отличие от многолетних растений состав эфемеров сильно
меняется по годам. Когда в 1934 г. в Юго-Западной Африке после трех
очень засушливых лет начался небывалый по количеству осадков сезон,
продолжавшийся несколько месяцев, фермеры утверждали, что появились
совершенно новые, неизвестные им виды. Тщательная проверка этих сооб-
щений показала, что речь шла о видах, которые развиваются во влажных
местообитаниях и поэтому в обычные годы малозаметны, поскольку они
цветут только под защитой скал или на дне русел временных водотоков;
таким растением, например, является сложноцветное Nidorella. Благо-
даря крайне обильным осадкам и переувлажнению почвы в составе эфеме-
ров начала господствовать Nidorella, желтые цветки которой были вид-
ны повсюду.
У геофитов, обладающих известным сходством с эфемерами, благода-
ря их очень короткому периоду вегетации и замиранию жизнедеятельности
при наступлении засухи наблюдается хорошо выраженная зависимость
между количеством осадков и развитием растений. Геофиты — в боль-
шинстве случаев луковичные и клубневые растения, запасающие органы
которых обычно находятся глубоко в почве. По данным местных жителей,
при небольшом количестве осадков указанные геофиты якобы вообще не
развиваются в течение многих лет, а затем, после обильных дождей,
в отдельные годы очень пышно цветут. Автор не мог проверить этих дан-
ных, поскольку это возможно только путем многолетних наблюдений,
однако они кажутся вполне правдоподобными. В период засухи почва,
где находятся клубни и луковицы, полностью высыхает. Только при нали-
чии обильных осадков, глубоко промачивающих почву, вновь образовав-
шиеся корни начинают поглощать влагу в количествах, обеспечивающих
формирование цветоносов и листьев. При отсутствии этих условий
луковицы и клубни, защищенные слоем почвы от высыхания, пребывают
в состоянии покоя в течение нескольких лет.
В отдельные годы геофиты трогаются в рост еще до начала
периода дождей. Это свидетельствует о наличии какого-то, пусть даже
1 F. W. Went, Ecology of desert plants: I and II, «Ecology», 29, 1948, p. 242—
353; 30, 1949, p. 1—13, 26—38.
Растительность субтропических аридных областей 313
незначительного запаса воды, оставшегося в глубоких слоях почвы после
предшествующих, богатых осадками лет. Это наблюдается, как правило,
на площадях, сильно стравленных скотом. Возможно также, что в луко-
вицах или клубнях некоторых геофитов накапливается столько воды,
сколько необходимо для образования цветоносов без дополнительного
поглощения воды извне.
Что касается конкуренции между многолетниками и эфемерами,
то она вряд ли имеет место. Как уже указывалось, многолетники слабо
пронизывают корнями верхние слои почвы и поэтому не в состоянии
использовать всю воду, накапливающуюся в почве во время обильных
дождей. Кроме того, многолетние виды образуют очень разреженный
травостой и вряд ли могут сильно затенять эфемеры. Напротив, разрежен-
ный полог многолетних растений оказывает даже благоприятное влияние
на эфемеры, поскольку в его тени уменьшается испарение с поверхности
почвы и нежные растения хорошо предохраняются от воздействия сол-
нечных лучей. Аналогичная картина наблюдается и после окончания пери-
ода дождей, когда на открытых площадях покров эфемеров уже высох,
а под защитой древесного полога он еще сохраняется некоторое время.
Все то, что говорилось об эфемерах, развитых в пустынях, где осадки
выпадают более или менее регулярно, справедливо и для пустынь с эпизо-
дическими осадками. Только здесь эфемеры появляются не ежегодно,
а лишь в годы или в областях с исключительно обильными осадками. Так,
например, западная часть пустыни Намиб в Юго-Западной Африке обыч-
но почти лишена растительности, но в 1934 г., когда там выпало более
100 мм осадков, пустыня покрылась многоцветным ковром мезембриан-
темумов; автор видел их даже в 1935 г., хотя и в меньших количествах;
это связано с большими запасами воды, накопленными суккулентами, дав-
шими возможность последним не только просуществовать больше года
без осадков, но даже цвести.
ЗАВИСИМОСТЬ ВОДНОГО РЕЖИМА РАСТЕНИЙ
В ЗАСУШЛИВЫХ ОБЛАСТЯХ ОТ ПОЧВЕННЫХ УСЛОВИЙ
Из всей просачивающейся в почву воды, количество которой зависит
не только от суммы выпавших осадков, нр, как было показано, и от нали-
чия стока, только часть может быть исйользована растениями. Чтобы
пояснить эту мысль, воспользуемся схемой, изображенной на рис. 248.
Осадки, выпадающие на поверхность почвы, количество которых ино-
гда увеличивается за счет поверхностного стока с вышележащих площа-
дей, частично просачиваются в почву, а частично тотчас же испаряются
или стекают. Просочившаяся вода удерживается частицами почвы в виде
пленочной влаги. Избытка влаги, достигающей грунтовых вод в форме
гравитационной влаги, в засушливых областях обычно не бывает. Почва
периодически увлажняется лишь до определенной глубины; ниже располо-
жена «мертвая зона», не содержащая доступной растениям влаги.
Поверхностный сток поступает по сухим руслам в долины. После
очень сильных дождей образуется мощный поток воды, который в при-
брежных пустынях может достичь моря, в других же случаях заканчивает-
ся в бессточных депрессиях. После небольших дождей вода быстро про-
сачивается в русло рек, которые питают грунтовые воды. При высоком
уровне грунтовых вод вода обычно испаряется с поверхности почвы.
Таким образом, грунтовые воды вне речных долин питаются главным
образом за счет воды, просачивающейся через трещины, и периодического
314
Глава 7
стока по руслам пересыхающих рек. Грунтовые воды могут также вообще
отсутствовать.
Существование растительности главным образом зависит от количе-
ства пленочной влаги в почве. Она наиболее доступна для растений,
причем ее запас в почве зависит не только от количества осадков и частоты
их выпадения, но в значительной степени и от структуры почвы.
На величину поверхностного стока или притока воды наряду с укло-
ном оказывает влияние также и водопроницаемость почвы. В общем мак-
Р и с. 248. Круговорот воды в аридных областях (из Walter, Volk, Die Grund-
lagen der Weidewirtschaft in SW-Afrika, Verlag Eugen Ulmer, Stuttgart, 1954).
симальный поверхностный сток наблюдается на глинистых почвах, на
песчаных почвах он бывает незначительным, а на рыхлых каменистых
осыпях полностью отсутствует. Чем больше поверхностный сток, тем
меньше воды просачивается в почву. Следовательно, с этим связано
увеличение сухости глинистых почв. Конечно, сток с поверхности скал
также очень велик, однако он уменьшается благодаря просачиванию
воды по трещинам. Смит 1 указывает на то, что в засушливых областях
поверхностный сток сильно возрастает благодаря уплотнению поверхно-
сти обнаженной почвы под действием копыт животных, давления авто-
мобильных шин или ударов дождевых капель. Это же может быть сказа-
но о периодически затопляемых почвах, особенно если они отличаются
более тяжелым механическим составом. На пойменных террасах речных
долин растительность поселяется с трудом, поскольку после каждого
дождя потоки воды уносят с собой семена.
Не только поверхностной сток, но и количество просочившейся
в почву рыхлосвязанной воды зависит от типа и механического состава
почвы. Следует особенно подчеркнуть, что основные, важные для жизне-
деятельности растений показатели водного режима в аридных областях
существенно отличаются от таковых во влажных областях (см. «Phytolo-
gie», Bd III, 1, S. 191 и след. 1 2). Глинистые почвы, развитые во влажных
областях и обладающие высокой влагоемкостью, характеризуются
наибольшей влажностью, тогда как песчаные почвы и особенно камени-
стые значительно суше; в аридных же областях соотношение обратное,
поскольку почвы здесь никогда не бывают полностью увлажнены. Влага
накапливается в этих условиях только в верхних горизонтах, на глубине,
1 J. Smith, op. cit.
2 Н. Walter, Die Wasserveihaltnisse an verschiedenen Standorten in humi-
den und ariden Gebieten, «Beih. Bot. Zbh», 49 (Erg. Bd), 1932, S. 495—514.
Растительность субтропических аридных областей
315
до которой просачивается вода в период дождей. Предположим, что выпа-
ло 50 мм осадков. Это количество воды увлажнит лишь верхние 10 см
глинистой почвы, 50 см песчаной почвы и гораздо глубже просочится
в каменистую почву, где вода удерживается только мелкоземом, скаплива-
ющимся в трещинах скал.
После дождя влага начинает испаряться. На глинистой почве, кото-
рая вскоре растрескивается, верхние 5 см быстро высыхают, в результате
чего теряется 50% выпавших осадков. Поверхность песчаной почвы также
Глинистая Песчаная
Рис. 249. Глубина промачивания почв разного механического состава при
одинаковом количестве осадков («Phytologie», III, 1, 2. Aufl., Verlag Eugen
Ulmer Stuttgart, 1960).
/ — / — нижняя граница промачиваемого осадками слоя почвы; v—v — нижняя граница
пересыхающего вследствие испарения слоя почвы. Глинистая почва теряет половину
просочившейся воды, песчаная — 1/ю часть, а каменистая — практически ничего.
Каменистая
высыхает, однако нити капиллярно подвешенной влаги быстро обрыва-
ются, и таким образом более 90% осадков сохраняется в почве. На каме-
нисто-скальных грунтах испарение практически отсутствует, в связи
с чем они отличаются оптимальной аккумулирующей способностью
(рис. 249). Поэтому в аридных областях в сходных климатических усло-
виях на скалах распространена древесная растительность, а на песках —
травяная, в то время как глинистые почвы почти совершенно лишены
растительности. Этот вывод автора, полученный им при изучении аридных
территорий запада Северной Америки, был в дальнейшем подтвержден
работами Хиллеля и Тедмора *, проводившимися в пустыне Неджеб.
По данным этих авторов, при одинаковом годовом количестве осадков
величина доступной растениям воды достигает 35 мм на лёссах, примерно
50 мм на скалистых участках (где много осадков теряется за счет поверх-
ностного стока), 90 мм на песчаных почвах и, наконец, 250—500 мм
в сухих долинах (получающих дополнительный приток воды).
Если в скалах имеются трещины, то фильтрующаяся вода может
достичь по ним грунтовых вод. Поэтому скапливающийся в расселинах
мелкозем увлажнен от поверхности почвы до уровня грунтовых вод, что
позволяет корням растений, используя трещины, проникать до грунтовых
вод; в связи с этим растения обеспечены водой на протяжении всего года.
В этих условиях распространены виды, с глубоко идущими стержне-
выми корнями. Однако такой способ укоренения встречается очень редко,
поскольку в целом глубина проникновения корней зависит от мощности
увлажненного горизонта, который в засушливых областях в общем очень
невелик (см. стр. 308).
1 D. Hillel, N. Та dm or, Water regime and vegetation in the Central Negev
Highlands of Israel, «Ecology», 43, 1962, p. 33—41.
316
Глава 7
Интересный пример использования подземных вод при облесении
пустынь автор наблюдал близ Басры в Месопотамии. Здесь на глубине
15 м сосредоточены неисчерпаемые запасы грунтовых вод, пополняемые
фильтрующимися через гравий водами Евфрата и Тигра. При годовом
количестве осадков, равном в этих условиях 120 мм, увлажняются лишь
верхние горизонты почвы. Расположенный ниже слой сухой почвы пре-
пятствует проникновению корней до уровня грунтовых вод, в связи с чем
после зимних дождей развиваются лишь эфемеры. Местное население при
помощи колодцев поднимает на поверхность грунтовые воды и использу-
ет их для полива овощных культур (лука, томатов, тыквы) на небольших
площадях. При высоких летних температурах (до 50°) приходится до
восьми раз в день подавать воду в поливные борозды. Вследствие сильно-
го испарения вскоре начинается засоление почвы, поэтому овощи удается
выращивать только в течение одного года. Между овощными культурами
производят посадки тамарикса (Tamarix articulata), которые быстро уко-
реняются. Поскольку почва (вследствие обильного полива) хорошо
увлажнена вплоть до уровня грунтовых вод, корни тамарикса продол-
жают и после прекращения полива расти до тех пор, пока не достигнут
грунтовых вод. Теперь дальнейшее существование деревьев обеспечено.
Их вырубают каждые 25 лет, но они вновь возобновляются пневой
порослью. Так, в пустыне на месте брошенных орошаемых земель выра-
стает лес.
Если наряду с влиянием климата, воздействием рельефа учитывать
также особенности почвы, ее структуру и влагосодержание, то придется
признать, что в аридных областях условия местообитания бесконечно
разнообразны. Поэтому нельзя безоговорочно согласиться с мнением
Штоккера, что формы приспособления у растений отличаются большим
разнообразием, чем местообитания. Такое утверждение будет оправдан-
ным лишь при очень схематическом объединении различных местообита-
ний в несколько обобщенных категорий и одновременной попытке рас-
сматривать каждую незначительную специфическую реакцию у растений,
например разницу в интенсивности транспирации как отличительную
особенность. Эта точка зрения может быть верной постольку, поскольку
на ограниченных, кажущихся нам однородными местообитаниях встре-
чаются различные экологические типы растений; в этом случае большей
частью невозможно установить, действительно ли почва отличается одно-
родностью или это только кажущееся явление. К этому надо добавить, что
каждый вид обладает определенным экологическим ареалом, поэтому
одни и те же экотипы могут встречаться в различных местообитаниях.
Поскольку видовая численность и тем более число экотипов строго ограни-
чены, а местообитания отличаются бесконечными градациями, то скорее
можно даже согласиться с точкой зрения, противоположной взглядам
Штоккера.
ЗАКОН ОТНОСИТЕЛЬНОГО ПОСТОЯНСТВА МЕСТООБИТАНИЙ
И СМЕНЫ БИОТОПОВ
При изучении растительности Юго-Западной Африки в глаза прежде
всего бросается обширность экологических ареалов многих древесных
пород Ч Здесь встречаются виды, которые распространены от границ
1 Н. u. Е. Walter, Einige allgemeine Ergebnisse unserer Forschungsreise nach
Siidwestafrika, 1952/53: Das Gesetz der relativen Standortskonstanz; das Wesen der
Pflanzengemeinschaften, «Вег. Dtsch. Bot. Ges.», 66, 1953, S. 228—236.
Растительность субтропических аридных областей
317
с пустыней, где количество осадков не превышает 100 мм, до древесной
саванны. Конечно, их биотопы отличаются в разных частях ареала. Если
в засушливых областях эти виды произрастают только по дну сухих русел
или в расщелинах скал, то в более влажных частях ареала они приурочены
к пескам, а в богатых осадками областях растут на сухих склонах,
поскольку на плакорных пространствах они не в состоянии конкуриро-
вать с другими, более быстрорастущими видами. Таким образом, про-
исходит хорошо выраженная смена биотопов, подчиняющаяся следующей
закономерности: сувеличением сухости климата воз-
растает влажность биотопов, на ко то р ы х раз-
виваются растения, иными словами, меньшее количество
осадков в условиях засушливого климата компенсируется большей влаж-
ностью почвы, и, таким образом, водный режим различных местообитаний
остается более или менее постоянным.
Если в качестве критерия принять влажность почвы С учетом харак-
тера и структуры почвы, рельефа и величины поверхностного стока,
то выявляется следующая последовательная смена биотопов от наиболее
засушливых к наиболее влажным.
1. Склоны с влагонепроницаемыми почвами и сильным поверхност-
ным стоком.
2. Затопляемые террасы с поверхностно уплотненными почвами.
3. Плакорные пространства с тяжелыми почвами в условиях интен-
сивного испарения. I
4. Плакорные территории, покрытые хорошо дренированными
песками.
5. Песчаные дюны (закрепленные).
6. Осыпи на склонах и каменистые почвы, где испарение
незначительно.
7. Скалы с трещинами, хорошо аккумулирующими воду.
8. Увлажненные участки близ выхода грунтовых вод у подно-
жия склонов.
9. Русла временных водотоков, эпизодический сток в которых попол-
няет запасы влаги в почве.
10. Сухие долины с постоянным уровнем грунтовых вод, залегаю-
щим на небольшой глубине.
Если проследить распространение отдельных видов от влажных
областей к самым засушливым, то смена биотопов будет происходить
в вышеупомянутой последовательности. Конечно, отдельные виды будут
отличаться потребностью в воде и строением корневой системы.
Редко два вида последовательно встречаются в совершенно одинако-
вых местообитаниях. Одни предпочитают скалы, другие — пески. Неко-
торые древесные породы, распространенные, как правило, в более
влажных условиях, можно вдруг встретить в засушливых областях на
покрытой трещинами скале или у подножия гранитного склона, другие
же, напротив, поселяются только в долинах с близко расположенными
грунтовыми водами, где они переносят временное затопление и выдержи-
вают натиск бурных вод. Общая тенденция, однако, хорошо выражена
и поэтому позволяет сформулировать закон относительного
постоянства местообитаний и смены биотопов
(см. «Phytologie», Bd III, 2, S. 41 и след.).
При изменении климата в пределах ареала
какого-либо вида в определенном направлении
происходит смена биотопов этим видом, что
1 лава /
компенсирует изменения климата; таким обра-
зом, условия местообитания в различных био-
топах остаются более или менее одинаковыми.
Этот закон характеризует не только изменения условий гидратуры
в аридных областях, но может быть применен также и при оценке дру-
гих факторов, определяемых изменениями климата (см. стр. 22).
Особенно яркие примеры, подтверждающие правильность этого зако-
на, можно найти у Смита 4. Смит выделяет в Судане 14 различных биото-
пов, отличающихся по степени влагообеспеченности; это позволяет расте-
ниям существовать в различных климатических условиях. Отдельные виды
могут расти при следующей амплитуде годовых осадков:
Acacia seyal . .
Khaya senegalensis
Prosopis ajricana .
Tamarindus indica
Dalbergia melanoxylon
370—800 мм
400—1050 »
480-1100 »
500-1200 »
360-1200 »
При этом, конечно, в раличных климатических областях эти виды
встречаются на разных биотопах. Например, Acacia tortilis может расти
при количестве осадков, равном
50 мм — по дну русел временных водотоков в пустыне,
150 » —близ Хартума на песчаных почвах,
300 » — в провинции Кассала на глинистых почвах,
500 » — на сухих склонах холмов Бутана.
Hyphaene thebaica встречается при количестве осадков, равном
100 мм — по руслам временных водотоков и вдоль рек,
600 » —на красных суглинках,
750 » —на возвышениях, сложенных суглинками,
900 » —на тяжелых глинистых почвах.
Sterculia setigera при количестве осадков, равном 1200 мм, растет
в плакорных условиях; при сокращении количества осадков до 300 мм
встречается только на скалистых возвышениях. Albizzia aylmeri на гли-
нистых почвах развивается при 800 мм осадков, а на песчаных дюнах
довольствуется всего 300 мм осадков.
Смит приводит очень много примеров, свидетельствующих о более
благоприятных условиях на песчаных почвах по сравнению с глинистыми.
Выбранный им в качестве объекта исследования Судан особенно удобен
для таких сравнений, поскольку его западная часть занята обширными
песчаными равнинами, тогда как плакорные территории на востоке стра-
ны покрыты глинистыми отложениями.
В общем можно сделать вывод, что для видов, произрастающих на
песчаном субстрате, требуется лишь 2/3 осадков, необходимых для разви-
тия тех же видов, но распространенных на глинистых почвах. Так,
например, основная область распространения Acacia Senegal на песках
ограничена изогиетой 400 мм, а на глинистых почвах — 600 мм.
Об отношении разных видов к воде можно судить по их распростра-
нению на плакорных почвах, находящихся в равных условиях увлажне-
ния. В Судане господствующие в составе саванн виды акации сменяют друг
1 J. S m i t h, Distribution of tree species in the Sudan in relation to rainfall and.
soil texture, Khartoum, 1949.
Растительность субтропических аридных областей
319
друга при увеличении количества осадков следующим образом: Acacia
flava, A. orfota, A. tortilis, A. raddiana, A. mellifera, A. fistula, A. Senegal,
A. seyal, A. drepanolobium, A. campylacantha, A. sieberiana, A. albida,
A. hebecladoides, A. abyssinica. Конечно, в условиях, где выпадает одина-
ковое количество осадков, разные виды могут расти на одной территории,
Рис. 250. Распространение различных древесных пород в Судане в зави-
симости от механического состава почвы и годового количества осадков
(по Смиту, с некоторыми изменениями).
1 — Prosopis africana] 2 — Khaya senegalensis; 3 — Acacia seyal] 4 — A. melliiera.
Прямая линия отделяет виды акаций от видов, свойственных тропическим сухим
лесам.
однако в этом случае они встречаются в разных биотопах: одни приуро-
чены к засушливым местообитаниям, другие развиты на более влажных.
Поскольку увеличение содержания илистой фракции в почвах аридных
областей неблагоприятно отражается на водном режиме почвы, в распро-
странении разных видов выявляется ясно выраженная зависимость между
количеством осадков и содержанием илистой фракции в почве (рис. 250).
По Уэру (см. примечание на стр. 308), в Тунисе максимальный урожай
с одного дерева Olea на песчаных почвах получают при годовом количе-
стве осадков, равном 230 мм, а на тяжелых почвах — при количестве
осадков не менее 500 мм. В очень засушливых условиях климата культуры
хорошо растут только на легких почвах; во влажном климате песчаные
почвы слишком быстро выщелачиваются.
Чем больше почва содержит глинистых частиц, тем больше воды тре-
буется растениям и, следовательно, тем больше должно быть количество
выпавших осадков. Условия развития древесных растений на глинистом
субстрате становятся более благоприятными, когда вокруг них накапли-
320
Глава 7
вается приносимый ветром песок и образуются дюны, на вершинах кото-
рых в конце концов поселяются кустарники (рис. 366 и 369).
Гребни неподвижных дюн представляют особенно благоприятные
местообитания для древесной растительности. Такая картина наблюдается,
например, на юге пустыни Калахари, где на территории 117 000 км1 2
ряды дюн высотой 5—51 м и длиной до 3 км располагаются параллельно,
на расстоянии примерно 230 м одна от другой, в направлении с северо-
запада на юго-восток. Песок на гребнях дюн отличается большей крупно-
зернистостью, чем в понижениях, поэтому на вершинах дюн может произ-
растать целый ряд древесных пород: Albizzia anthelminthica, Terminalia
sericea. Acacia giraffae, Boscia albitrunca, и эндемичная, отличающаяся
малой конкурентной способностью Acacia haematoxylon Ч Albizzia и Ter-
minalia, как более мезофильные виды, распространены в более богатых
осадками северных областях.
ВАЖНЕЙШИЕ ЭКОЛОГИЧЕСКИЕ ГРУППЫ РАСТЕНИЙ
НЕЗАСОЛЕННЫХ НОЧВ АРИДНЫХ ОБЛАСТЕЙ
И ИХ ГИДРАТУРА
В жизнедеятельности растений аридных областей основное значение
имеет вода. При проведении экологических исследований недостаточно
в общих чертах ознакомиться с водным режимом; необходимо установить
его воздействие на отдельные растения. Это достигается путем определе-
ния гидратуры самих растений, или, точнее, степени насыщения водой
живой протоплазмы.
Поскольку эти термины мало что говорят геоботаникам и , как пока-
зывают запросы, даже некоторым экологам, ниже приводится краткое
объяснение этих понятий 2.
Под гидратурой мы понимаем в самом общем смысле степень насыще-
ния влагой какого-либо тела, которая выражается через относительную
упругость пара в процентах.
Относительная упругость пара — это отношение упругости водяного
пара р над поверхностью тела (раствора или набухающего тела) при дан-
ной температуре и давлении к упругости водяного пара р0 над чистой
водой при тех же условиях. Вода должна находиться в жидком состоя-
нии, поскольку при замерзании лед и любой раствор имеют одинаковую
упругость насыщения; это связано с тем, что растворенные в воде соли
понижают упругость водяного пара по сравнению с его упругостью над
чистой водой. Гидратура, или степень насыщения воздуха водой, выра-
жается относительной влажностью, то есть отношением, выраженным
в процентах, фактического содержания водяных паров (в г/м3), или фак-
тической упругости водяного пара (в мм pm. cm. или в мб) в воздухе, соот-
ветственно к содержанию водяных паров в воздухе для состояния насы-
щения или к упругости насыщающего водяного пара при той же темпера-
туре воздуха.
Между гидратурой, измеренной в процентах (сокращенно % Ьу),
и потенциальным давлением набухания, или потенциальным осмотиче-
1 О. A. Leistner, The plant ecology of the Southern Kalahari, Dissert, Stellen-
bosch, 1964.
2 Cm. «Phytologie», Bd I, 1950, S. 72—78, 326—356; Bd III, 1, 1960, S. 207—256.
См. также сноску на стр. 332.
Н. Walter, Zur Klarung des spezifischen Wasserzustandes im Plasma und
in der Zellwand bei den hoheren Pflanzen und seine Bestimmung, «Вег. Dtsch. Bot.
Ges.», 76, 1963, S. 40—71.
Растительность субтропических аридных областей
321
ским давлением, которое мы для краткости называем просто осмоти-
ческим давлением,
зависимость:
WT =
существует следующая математическая
10007?- Т -s . р
-----м-----In — ,
М ро
где WT — потенциальное давление набухания, или осмотическое давле-
ние в атм при абсолютной температуре Т\ R — газовая постоянная, рав-
ная 0,08207; Т — абсолютная температура (например, температуре 20° С
соответствует 293°); s — удельный вес воды (при 20° С = 0,9982); М —
молекулярный вес воды (равный 18) и —-------относительная упругость
Ро
пара в %, которую мы называем гидратурой (hy). Знак минус означает,
что существует обратная зависимость, то есть что осмотическое давление
возрастает с уменьшением гидратуры.
Если в формулу подставить константы, то при температуре 20° С,
получаем
1У20 = -1333,5 In 4- = — 3067 log .
Ро Ро
По этой формуле определена зависимость между гидратурой и осмо-
тическим давлением при температуре 20° (табл. 43).
Таблица 43
hy, % IV, атм hy, % W, атм | hy, % IV, атм
100 0 95 68,4 85 217
99,5 6,7 94 82,5 80 298
99 13,4 93 96,7 75 384
98,5 20,1 92 111 70 476
98 26,9 91 125 60 681
97,5 33,8 90 140 50 924
97 40,6 89 155 40 1221
96,5 47,5 88 171 30 1606
96 54,5 87 186 20 2146
95,5 61,4 86 201 10 3065
Как известно, протоплазма во взрослой растительной клетке отделена
от внешних частей клетки и со стороны наполненной клеточным соком
вакуоли полупроницаемыми перепонками. Вся клетка как единое целое
окружена эластичной целлюлозной оболочкой. Для характеристики осмо-
тического давления применимо уравнение (все значения в атм)
S = W-P,
где S — сосущая сила клетки, определяющая водопоглощающую (всасы-
вающую) способность клетки; W — осмотическое давление клеточного
сока и Р — тургор. Поскольку живая протоплазма отделена от клеточ-
ного сока полупроницаемой перепонкой, она должна находиться в состоя-
нии равновесия (или иметь ту же гидратуру) с клеточным соком.
Как известно, живая протоплазма, если в ней не происходит никаких
физико-химических изменений, обнаруживает сходную зависимость сте-
21-1394
322
Глава 7
пени набухания от гидратуры, как л мертвая Ч Поэтому любое повышение
осмотического давления (снижение гидратуры) должно одновременно
повлечь за собой уменьшение набухания (снижение степени насыщения
протоплазмы водой). Такое уменьшение набухания протоплазмы не может
не отразиться на характере происходящих в ней физиологических про-
цессов. Таким образом, величина осмотического давления может служить
показателем подобных изменений, и поэтому ее определению мы придаем
особое значение. Осмотическое давление является показателем гидратуры
протоплазмы растения и тем самым — состояния его тургесцентности 1 2.
Определить осмотическое давление криоскопическим методом очень
несложно. Выдвигавшиеся же против этого метода возражения были про-
верены и признаны неосновательными3.
При нарушении водного режима растения, когда потери влаги путем
транспирации временно превышают количество воды, поглощаемой кор-
нями, содержание воды в растении снижается.
Поскольку большая часть воды, если не говорить о проростках, содер-
жится в вакуолях, снижение содержания воды в растении должно про-
явиться не только в ослаблении напряжения клеточной стенки вследствие
потери тургора, но и в повышении концентрации клеточного сока, то есть
в повышении осмотического давления. С этим связано также снижение
степени набухания протоплазмы4.
Повышение осмотического давления и снижение тем самым гидратуры
протоплазмы является первым поддающимся точному
определению и физиологически важным след-
ствием нарушения водного режима. При его быс-
тром уменьшении клетки вновь обретают тургесцентность и растение воз-
вращается в прежнее состояние. Если же потери воды слишком велики,
повышение осмотического давления и снижение гидратуры протоплазмы
могут вызвать необратимые повреждения, в результате чего клетка отми-
рает. Предельную величину, при которой наступает гибель многих кле-
ток, называют максимальным осмотическим давле-
нием.
Однако, если нелетальное нарушение водного режима повторяется
слишком часто или продолжается длительное время, в протоплазме воз-
никают регулирующие процессы, вызывающие активное повышение осмо-
тического давления благодаря новообразованию осмотических веществ.
Протоплазма приобретает при этом другие физиологические свойства,
что отражается также на морфологическом строении органов и их физио-
логических свойствах (ксероморфное строение, большая устойчивость).
Возникающая при недостатке воды цепь реакций, насколько мы можем
1 Н. Walter, Protoplasma und Membranquellung bei Plasmolyse, «Jb. wiss.
Bot.», 62, 1923, S. 145—213. См. также H. Walter, op. cit., 1963.
2 Конечно, набухание протоплазмы зависит еще и от ряда других факторов,
например от электростатического заряда, на который влияет адсорбция ионов, и т. д.
Такие изменения набухания также будут оказывать физиологическое действие. Имеют
ли они значение при экологических исследованиях водного режима в природе, не выяс-
нено. В отношении же галофитов, о которых сказано несколько ниже, их значение
несомненно.
3 Н. Walter, О. Weissmann, t)ber die Gefrierpunkte und osmotischen
Werte lebender und toter pflanzlicher Gewebe, «Jb. wiss. Bot.», 82, 1935, S. 273—310.
4 О том, не сказывается ли на ходе физиологических процессов в протоплазме
уже одно уменьшение напряжения клеточной стенки, нет пока никаких данных.
Напротив, уменьшение набухания протоплазмы, как показали многочисленные опы-
ты, влияет на все жизненные процессы (см. «Phytologie», Bd I, S. 346 и след.; Bd III,
1, S. 221—231).
Растительность субтропических аридных областей
323
ее проследить, не пускаясь
щий вид:
в необоснованные гипотезы, имеет следую-
Нарушение водного
режима
Снижение тургора и
повышение осмоти-
ческого давления
Уменьшение
гидратуры
протоплазмы
Регулирующие про- 1
—> цессы и изменение —>
структуры протоплазмы
Изменение физиоло-
. гических свойств
клеток
Изменение мор-
► фологического
строения органов
При сравнении, например, теневых и световых листьев, взятых с одно-
го дерева, но выросших при различных условиях гидратуры, нетрудно
установить, что осмотическое давление у освещенного листа в состоянии
насыщения водой и полной тургесцентности всегда выше, чем у теневого,
В соответствии с этим освещенный солнцем лист обладает ксероморфным
строением и физиологически лучше приспособлен к недостатку воды.
Эти различия не могут быть выявлены посредством определения степени
оводненности или сосущей силы клеток, а только через величину осмо-
тического давления.
Те же закономерности выявляются при сравнительном изучении
какого-либо растения весной при благоприятных условиях гидратуры
и засушливым летом, когда оно чаще страдает от недостатка воды. Листья,
распустившиеся летом, с морфологической и физиологической точек зрения
резко отличаются от весенних листьев. Их осмотическое давление даже
при полной тургесцентности, то есть полном насыщении водой, более
высокое. Сосущая сила в этом случае, напротив, равна нулю. Лишь содер-
жание воды на 1 г сухого вещества даже в состоянии насыщенности будет
у летних листьев ниже. Соответствующие примеры приведены в следую-
щих разделах.
Все вышеизложенное должно объяснить, п о ч
и и
ода
о я
а в
знание
движения воды в растении, а для понимания в ц е л о м
растений.
Принимая гидратуру растений в качестве основы для выделения эко-
логических групп растений, прежде всего различают следующие две
основные их группы:
А. Пойкилогидрические низшие растения: гидратура этих растений
и окружающей среды очень близки по своим значениям; как только гидра-
тура опускается несколько ниже 100%, их жизнедеятельность переходит
в латентное состояние.
Б. Гомойогидрические высшие растения; они сохраняют даже в край-
них условиях существования в пустынях очень высокую гидратуру про-
топлазмы; их органы не переносят иссушения.
Обе группы этих растений встречаются в аридных областях. Первая
представлена водорослями, и прежде всего Cyanophyceae, затем лишай-
никами, гораздо реже мхами, частично печеночниками, например ксеро-
фильными Ricciaceae, рядом папоротников, некоторыми видами Sela-
ginella и одиночными видами покрытосеменных, например Myrothamnus,
н
в
т
Д
н
л
растительности
ется лимитирующим
приводятся
Последнее служит показателем
аридных
я
о
н
в
л я
Д
а
иные об
ему при и з у ч е-
областей, где
фактором, п о с-
осмотическом
гидратуры растения,
н
е
которой необходимо отнюдь не для простого объяснения пере-
экологии
и
и.
324
Глава 7
изображенным на рис. 312—315 (см. «Phytologie», Bd 111, 1, S. 209
и след. х.
Наличие этих растений в пустынях свидетельствует о существовании
хотя бы кратковременных, влажных сезонов, в течение которых растения
растут и размножаются. После каждого дождя ранее совершенно сухие
растения через короткий промежуток времени приобретают ярко-зеленую
окраску и возобновляют свой рост (рис. 271) 1 2. Интенсивность их фото-
синтеза пока еще глубоко не изучалась.
Обычно для развития пойкилогидрических растений наиболее благо-
приятным является прохладное время года с высокой влажностью воздуха
даже при незначительном количестве осадков. В некоторых прибрежных
пустынях в этот сезон часто наблюдаются туманы, что особенно благо-
приятствует развитию лишайников. В крайних условиях существова-
ния — в пустынях, где воздух в течение длительного времени остается
очень сухим, эта группа растений отсутствует.
На протяжении большей части года жизнедеятельность пойкило-
гидрических растений в пустынях затухает. Они полностью высыхают
и на поверхности почвы подвергаются воздействию высоких температур,
достигающих 60° и выше. Они развиваются преимущественно в затенен-
ных местах под защитой обломков скал, где температура не бывает столь
высокой и где влага после дождей сохраняется дольше. Некоторые водо-
росли поселяются под прозрачными обломками кварца 3. Данных о засу-
хоустойчивости растений аридных областей и их жаростойкости, особенно
в сухом состоянии, не получено. Однако известно, что скальные лишай-
ники в Европе отличаются крайней выносливостью (см. «Phytologie»,
Bd 111, 1, S. 50). Наличие лишайников в пустынях свидетельствует об их
хорошей приспособленности к местным условиям.
Более сложна экология гомойогидрических высших растений. Их
способность поддерживать уровень гидратуры в крайне засушливых
условиях пустыни чуть ниже 100% представляет собой поразительное
явление, которое уже давно привлекало внимание экологов.
Подробный анализ этого вопроса проведен в другой работе автора
(«Phytologie», Bd 111, 1), поэтому здесь будут изложены лишь наиболее
существенные выводы.
Прежде всего необходимо еще раз подчеркнуть, что водный режим
растений может рассматриваться только сопряженно с их продуктив-
ностью, поскольку, например, сокращение потерь воды путем замыкания
устьиц и понижения интенсивности транспирации влечет за собой прекра-
щение ассимиляции СО2, а тем самым и фотосинтеза.
С одной стороны, растения аридных областей должны активно асси-
милировать СО2, не снижая уровня гидратуры, а с другой стороны,
в засушливые периоды они должны почти полностью прекращать отдачу
1 Найденное недавно в Западной Африке на гранитных останцах растение
из семейства Сурегасеае — Trilepis pilosa хорошо переносит полное иссушение, лишь
слегка при этом желтея. При опускании в воду, содержание влаги в нем повышается
с 8 до 56% и листья на свету приобретают зеленую окраску уже в течение суток.
Укоренившиеся растения после полива начинают зеленеть от основания стеблей
к верху. В сухом состоянии растение остается живым по меньшей мере в течение года.
См. D. J. Hambier, A poikilohydrous, poikilochlorophyllous angiosperm from
Africa, «Nature» (London), 191, 1961, p. 1415, 1416.
2 H. R. Oppenheimer, A. H. Halevy, Anabiosis von Ceterach off ici-
narum Lam. et D., «Bull. Res. Coun. Israel», D. 11, 1962, p. 127—147 (с обширной
библиографией).
3 Об «оконных водорослях», стр. 381.
Растительность субтропических аридных областей
325
воды, даже если в результате этого происходит снижение интенсивности
фотосинтеза или полное его прекращение. К этим требованиям совершен-
но по-разному приспособились отдельные группы гомойогидрических
растений.
1. Эфемеры развиваются в течение очень короткого периода
(наступающего регулярно или наблюдаемого лишь эпизодически в арид-
ных областях), когда почва содержит достаточное количество влаги,
позволяющее растениям даже при активном газообмене поддерживать
высокий уровень гидратуры:
а) у однолетников (терофитов) в течение продолжительных
неблагоприятных для развития периодов сохраняются только семена, а
б) у геофитов подземные запасающие органы.
С экологической точки зрения и по условиям гидратуры эти эфемеры
близки к терофитам и геофитам, развитым на солнечных местообитаниях
в умеренной зоне. Они отличаются лишь очень интенсивным ростом,
2. У суккулентов (негалофитных видов) вода накапливает-
ся в водоносных тканях, развитых в листьях, стеблях или подземных
органах; запасы воды пополняются в благоприятные периоды, когда
почва увлажнена х. На время длительной засухи растения почти гермети-
чески изолируются от окружающей среды (как воздуха, так и почвы)
и крайне экономно расходуют накопленную влагу. Вследствие затруд-
ненного газообмена эти растения накапливают очень незначительное коли-
чество вещества и медленно растут. Обмен СО2 у этих растений отклоняет-
ся от нормального (накопление органических кислот при дыхании и их
разложение в процессе фотосинтеза). Суккуленты широко распростране-
ны в аридных областях, где выражены два коротких периода дождей
в году. Они полностью отсутствуют в Австралии, не считая редко встре-
чающейся Sarcostemma australis.
3. Ксерофиты в периоды засухи продолжают существовать
в более или менее активном состоянии; в этот период газообмен у них
продолжается, хотя временами он заметно сокращается. Для покрытия
расходов на транспирацию и сохранение нормального уровня гидратуры
они нуждаются в постоянном поступлении воды из почвы. Как указыва-
лось на стр. 308 и далее, ксерофиты в аридных областях достаточно обеспе-
чены влагой (при расчете на площадь транспирирующей поверхности).
Однако в отдельных случаях они испытывают затруднения в обеспечении
влагой.
Количество осадков, поступающее в аридных областях на единицу
поверхности почвы, сравнительно невелико, поэтому у ксерофитов раз-
вивается очень мощная, широко разветвленная корневая система. Только
у этой группы растений подземная масса превосходит надземную, чему
способствует также более раннее завершение роста надземных органов
по сравнению с корневой системой. К тому же ксерофиты отличаются
более низкими значениями гидратуры.
1 Суккуленты с подземными органами, запасающими воду (корни, корневища,
клубни), гораздо более многочисленны в аридных областях, чем это обычно думают
(«Phytologie», Bd III, S. 264—265). На небольшой площади в Южной Африке встре-
чаются: много видов бобовых, Euphorbia (7 видов), Fockea edulis и др. Pachypodium
succulentum, Р. bispinosum, Pelargonium spp., Mesembryanthemum spp., Raphionacme
zeyheri^ Ceropegia ampliata, Ipomoea argyroides, Kedrostis spp., Chamarea spp., Othonna
auriculifolia, Senecio spp., Doria carnosa, D. eriocarpa. Cp. R. A. Dyer, The vegeta-
tion of the Divisions Albany and Bathurst, «Bot. Survey South Africa», № 17, Preto-
ria, 1937.
326
Глава 7
При возникновении дефицита влаги, связанного со снижением гидра-
туры, или повышении осмотического давления у малакофильных ксеро-
фитов (обладающих мягкими листьями) происходит резкое сокращение
площади транспирирующей поверхности.
При этом в большинстве случаев крупные мезоморфные листья сме-
няются менее крупными ксероморфными, а при дальнейшем увеличении
водного дефицита растение сбрасывает почти все листья; это дает возмож-
ность растениям сохранить гидратуру почек (рис. 286) на необходимом
уровне.
Оршан и Цанд 1 определяли площадь транспирирующей поверхности,
сокращающуюся в период засухи у растений, распространенных в пустыне
Неджеб {Anabasis articulata, Zygophyllum dumosum, Noea macrantha,
Haloxylon articulatum, Artemisia herba-alba, A. monosperma) и у малако-
фильных видов, развитых в восточной части Средиземноморья (Poterium
spinosum, Phlomis viscosa, Teucrium polium, Thymus capitatus). У растений
пустыни эта поверхность уменьшилась на 96—72%, а у средиземномор-
ских видов на 49—25%. У Artemisia monosperma, пустынного растения,
встречающегося на песчаных дюнах, поверхность сократилась на 55%.
Таким образом, сокращение площади транспирирующей поверхности
идет параллельно нарастанию засушливости климата.
Усклерофильных ксерофитов с жесткими листьями
или с безлистными прутьевидными побегами, а также у растений с филло-
диями, филлокладиями или кладодиями редукция транспирирующей
поверхности происходит только в редких случаях, когда отмирают целые
части растения. Склерофилы в критические периоды сокращают интен-
сивность транспирации и при почти полном отсутствии газообмена сводят
до минимума потери воды. Играет ли при этом какую-нибудь роль большая
сосущая сила корней, связанная с повышенным осмотическим давлением
в клетках, до настоящего времени в естественных условиях еще не удалось
доказать1 2. Однако некоторые особенности ксероморфной структуры
листьев свидетельствуют о возможности такого явления.
Обе выделенные выше группы ксерофитов разграничены недостаточно
четко. Между суккулентами и ксерофитами встречаются и промежуточ-
ные формы. Ксерофиты также сильно различаются по интенсивности
транспирации и фотосинтеза.
К этим двум группам ксерофитов нужно добавить еще одну группу,
чаще встречающуюся в засушливых областях умеренной зоны; это стено-
гидрические ксерофиты, которые при недостатке воды сразу закрывают
устьица, вследствие чего прекращается фотосинтез. При длительной засухе
растения начинают голодать, однако листья не высыхают, а только жел-
теют; осмотическое давление у них не увеличивается, а часто даже сни-
жается. К их числу относятся многие однодольные геофиты, виды Euphorbia
и др. Типичным представителем этой группы, распространенным в пустыне
Сонора, является Fouquieria splendens (гл. 8).
4) В каждой аридной области всегда встречаются местообитания, где
в течение всего года растения обеспечены достаточным количеством воды.
1 G. О г sh a n, G. Z a n d, Seasonal body reduction of certain desert halfshrubs,
«Bull. Res. Counc. Israel», D, II, 1962, p. 35—62.
2 Некоторые данные, подтверждающие этот вывод, имеются в работах:
К. К г е е b, Zur Frage des negativen Turgors bei mediterranen Hartlaubpflanzen unter
natiirlichen Bedingungen, «Planta», 56, 1961, S. 479—489; Untersuchungen zum Wasser-
haushalt der Pflanzen unter extrem ariden Bedingungen, «Planta», 59, 1963, S. 442—458.
Растительность субтропических аридных областей
327
Под этим подразумеваются не только открытые водоемы и выходы источ-
ников, но также и внешне засушливые местообитания, которые, однако,
отличаются наличием увлажненных горизонтов или грунтовых вод
на незначительной глубине.
В этих условиях в пустынях могут произрастать даже чуждые им
растения. Они часто проникают в пустыню из соседних более влажных
областей. Большей частью эти растения отличаются мезоморфным строе-
нием листовых органов и усиленной транспирацией, интенсивность кото-
рой даже в период засухи не уменьшается или сокращается лишь очень
незначительно. Как это удалось доказать Ланге, часто именно большая
интенсивность транспирации и позволяет им существовать в аридных
условиях Поскольку эти виды принадлежат к родам, основная часть
ареала которых захватывает тропические области, где осадки выпадают
летом и где высокие температуры не совпадают с периодом засухи, здесь
не было предпосылок для выработки жаростойкости у растений. Жаро-
стойкость, например, тыквенных растений, свойственных пустынным
областям, или видов Abutilon столь мала (они переносят температуру лишь
немногим выше 40°), что только охлаждающее действие транспирации
сохраняет температуру листьев на уровне, выше которого начинается
их отмирание. Уменьшение интенсивности транспирации при недостаточ-
ной обеспеченности водой влечет за собой быстрое повышение температуры
листьев и гибель растения от перегрева. У типичных растений пустынь,
интенсивность транспирации которых сильно уменьшается в период
засухи, температура листьев поднимается даже выше 50°, что, однако,
не влияет на их жизнедеятельность. Естественный отбор у пустынных
растений шел именно в этом направлении.
В зависимости от условий гидратуры выделяются стеногидри-
рические виды, отличающиеся небольшими различиями между
оптимальным и максимальным осмотическим давлением, и эвригид-
рические — с большим диапазоном между этими величинами. Одна-
ко при определении засухоустойчивости гораздо более важное значение
имеет их гидростабильность или гидролабильность,
что связано или с быстрым повышением осмотического давления при
наступлении засухи, или со способностью достаточно долго поддерживать
неизменный уровень гидратуры. Эти свойства связаны с различными мор-
фолого-физиологическими особенностями растений, например характером
и строением транспирирующих органов и корневой системы, способами
регулирования отдачи и поглощения воды, а также эффективностью водо-
проводящей системы. Очень важное значение имеет также протяжен-
ность корней; однако в засушливых областях, где почвы обычно очень
уплотнены, это может быть установлено лишь с большим трудом.
Эфемеры, как правило, представлены стеногидрическими, гидро-
лабильными видами, обладающими лишь незначительной засухоустой-
чивостью.
К суккулентам относятся стеногидрические, но крайне гидростабиль-
ные и поэтому часто особенно засухоустойчивые виды.
Ксерофиты — это по большей части хорошо выраженные эвригидри-
ческие виды; причем малакофильные виды выделяются известной гидро-
лабильностью, а склерофильным, напротив, свойственна гидростабиль-
1 О. L. Lange, Untersuchungen iiber Warmehaushalt und Hitzeresistenzs
mauretanischer Wiisten- und Savannenpflanzen, «Flora», 147, 1959, S. 595—651.
328
Глава 7
ность. Поэтому первые относительно засухоустойчивы, тогда как вторые
отличаются большой засухоустойчивостью х.
Выше была дана схематическая характеристика основных групп
пустынных растений. Поскольку; однако, каждая из пустынь отличается
своей спецификой и, кроме того, характеризуется своими особыми эколо-
гическими группами растений, их описание будет приведено в дальней-
шем при более детальной характеристике каждой из пустынных областей.
Однако прежде следует остановиться еще на одном, очень важном для
аридных условий явлении — на засолении почв.
ЗАСОЛЕНИЕ ПОЧВ И ГАЛОФИТЫ
Наряду с условиями водного режима на характер растительного
покрова аридных областей оказывает влияние засоление почвы. В извест-
ной степени это обусловливается климатическими причинами. В бессточ-
ных областях не происходит выноса легкорастворимых солей, и поэтому
в результате непрерывного испарения соли накапливаются в почве. Одна-
ко это не может рассматриваться как чисто климатическое явление,
поскольку в одинаковых климатических условиях в одном случае наблю-
дается сильное засоление почв, а в других оно полностью отсутствует.
По обычным представлениям, легкорастворимые соли накапливаются
исключительно в процессе выветривания горных пород, что, однако, невер-
но, а вся проблема много сложнее.
В почвах аридных областей основное значение имеют соли NaCl
и MgCl2. Частично здесь присутствуют также легкорастворимые суль-
фаты — Na2SO4 и MgSO4. Однако сульфаты чаще встречаются в форме
CaSO4, то есть гипса, который, однако, при настоящем засолении не играет
никакой роли вследствие своей незначительной растворимости. Натрий
содержится в больших количествах почти во всех горных породах, подвер-
гающихся выветриванию. Совершенно иначе обстоит дело с хлором, встре-
чающимся только в составе немногих, очень редких минералов. Поэтому
при выветривании кристаллических горных пород NaCl никогда не накап-
ливается в сколько-нибудь значительных количествах. В связи с этим
в аридных областях в местах выхода кристаллических пород засоленные
почвы практически не встречаются. Засоление хорошо выражено в облас-
тях, сложенных осадочными породами морского происхождения (юрскими,
меловыми, третичными). Здесь часто наблюдаются выцветы солей на гра-
ницах контакта различных пластов выветривающихся горных пород.
Осадочные породы морского происхождения содержат растворен-
ные в морской воде соли, главным образом NaCl. При выветривании этих
пород морская соль выносится на поверхность, растворяется дождевой
водой, переносится в бессточные котловины и здесь накапливается.
Таким образом, в аридных областях дождевые воды перемещают лег-
корастворимые соли с более высоких мест в более пониженные. Поэтому
возвышенные части рельефа редко подвергаются засолению, тогда как
отрицательные формы почти всегда засолены. Соли накапливаются в основ-
ном в местах испарения капиллярной влаги (рис. 251). Отложение солей
происходит также в условиях прекращения поверхностного стока, если,
1 С экологической точки зрения вряд ли целесообразно изучать засухоустойчи-
вость растений на срезанных побегах или листьях или в горшечных культурах, где
корневая система ограничена небольшим объемом почвы. О засухоустойчивости мож-
но судить только непосредственно в естественных условиях, поскольку отдача
и поглощение воды взаимосвязаны и взаимообусловлены.
Растительность субтропических аридных областей
329
Рдело реки
с пресной ВодВй Поименные поверхностно увлажненные почВы $
4 1*1
!________L.............ul.I-
Речная терраса
Рис. 251. Накопление солей в долине реки Свакоп, пустыня Намиб, Юго-Западная
Африка («Phytologie», III, 1, 2 Aufl., 1960).
Сплошные стрелки указывают направление и скорость движения грунтовых вод, пунктирные —
испарение. Концентрация солей повышается у бортов долины; выцветы солей (6) появляются
у подножия террасы, там где прекращается ток воды.
конечно, увеличение концентрации раствора не приведет еще раньше
к кристаллизации солей. Поэтому соли на пути движения воды выпадают
в соответствии со степенью их растворимости: сначала СаСО3, затем
CaSO4 и, наконец, NaCl вместе с другими легкорастворимыми соедине-
ниями. При высоком уровне грунтовых вод в аридных областях капилляр-
ное поднятие влаги обеспечивает постоянную увлажненность поверхно-
сти почвы; в этом случае вследствие постоянного испарения поднимающей-
ся по капиллярам влаги на поверхности почвы накапливается особенно
много солей и они образуют корку. В аридных областях все влажные
местообитания сильно засолены, даже если грунтовые воды или источники
содержат сравнительно немного легкорартворимых солей.
Возвышенные участки подвергаются засолению только в очень засуш-
ливых условиях, например в пустынях Египта, поскольку эпизодические
осадки не могут растворить соли и обеспечить их вынос.
Накопление значительных количеств солей может быть осуществлено
только путем переноса их водой, поэтому засоление обычно наблюдается
в областях, пограничных с пустыней, где еще выпадает достаточное коли-
чество осадков. В крайне сухих пустынях, где нет воды, необходимой для
переноса солей, засоление почти не выражено; кроме того, в самих горных
породах содержится незначительное количество солей.
При поездке через совершенно безжизненную южную часть Ливий-
ской пустыни, от Асьюта в долине Нила до оазиса Эль-Харга, где не выпа-
дает осадков, на протяжении более 200 км, автор не заметил никаких
признаков засоления. Непосредственно же в самом оазисе, где грунтовые
воды залегают очень близко, сразу же появляются выцветы солей.
Однако засоление не всегда бывает обусловлено переносом солей
водой; часто это связано с деятельностью ветра. Во время сильного при-
боя на морских побережьях в субтропических областях вода разбрызги-
вается на мелкие капельки, которые успевают испариться, а остающаяся
соленая пыль уносится ветром в глубь материка и там осаждается на поч-
ву. В пустыне Намиб капли прибрежного тумана содержат соль; граница
его продвижения в сторону суши является одновременно пределом рас-
пространения засоленных почв.
Это явление очень детально изучалось в Западной Австралии. Соглас-
но Тиклу х, содержание хлористого натрия в дождевой воде оказалось
равным: на побережье 15—50 мг!л, а внутри страны 4—20 мг!л. Выпавшая
во влажных прибрежных областях соль поверхностным стоком вновь
уносится в море, и поэтому здесь не наблюдается засоления. Перемещаю-
1 L. J. Н. Т е a k 1 е, The salt (sodium chloride) content of rain water, «J. Dep.
Agric. West Australia», 14, 1937, p. 115—133.
330
Глава 7
щаяся таким путем соль называется «циклической». В засушливых внут-
ренних областях, где отсутствует сток, накопление солей неизбежно.
В 1933—1936 гг. на западном побережье Австралии близ Мерредина,
в 250 км к востоку от Перта, и на южном побережье у Селмон-Гамса,
в 100 км севернее Эсперанса, определялось содержание солей в дождевой
воде. Количество солей, поступавших в почву, составляло в среднем
за год: в Мерредине — 18 кг!га, а в Селмон-Гамсе — 30 кг!га. С этими дан-
ными хорошо согласуется большая засоленность почв у Селмон-Гамса.
Дополнительные данные о содержании солей в осадках 1936 г. были
получены с 4 береговых и 5 внутриматериковых станций, где, кроме того,
определялось ежегодное поступление солей в почву (табл. 44).
Таблица 44
Береговые станции Среднее со- держание со- лей в дожде- вой воде, мг /л Количество солей, посту- пающих в почву, кг/га Внутримате- риковые станции Среднее со- держание со- лей в дожде- вой воде, мг/л Количество солей, посту- пающих в почву, кг/га
Перт ... 27,2 340 Кулгарди . . 19,8 40
Джералдтон . 47,3 194 Кью . . . 15,8 41
Эсперанс 34,1 290 Мандивинди 5,8 11
Кордон . . 13,5 43 Уилуна . . . 6,6 11
Роулина 10,4 10
Как и можно было ожидать, количество солей, перенесенных дожде-
выми осадками к внутриматериковым станциям, в некоторых случаях
удаленным от моря более чем на 600 км, оказалось гораздо меньшим, чем
на побережье; вследствие господства западных ветров преобладал пере-
нос солей с западного берега. Однако не следует думать, что вся соль,
приносимая осадками, имеет морское происхождение. Уже на очень незна-
чительном удалении от берега вследствие крайней засушливости климата
образуются небольшие засоленные впадины, остающиеся сухими в тече-
всего лета. С их поверхности, покрытой соляной коркой, в сухое время
года ветер может сдувать соляную пыль. Такое «опыление» солью
играет заметную роль в образовании солонцовых почв в степных областях
СССР. Здесь источником соли является сильно пересыхающий летом
Сиваш.
Наконец, засоленные почвы встречаются на прибрежных равнинах,
образовавшихся вследствие поднятия морского дна, например на Каспий-
ской низменности, или на месте некогда обширных, а ныне почти пересох-
ших водоемов, как, например, вокруг Большого Соленого озера в штате
Юта или близ озера Туз в Анатолии. Соли могут также накапливаться
на орошаемых, но недостаточно дренированных почвах. Вполне вероятно,
что именно таким образом произошло засоление обширных пространств
в Месопотамии. Вокруг Ура, окруженного много тысячелетий назад цве-
тущими сельскохозяйственными угодьями (о чем свидетельствуют древ-
ние оросительные сооружения), сегодня простирается лишь солончако-
вая пустыня \
Таким образом, видно, что в почвах засушливых областей накопление
солей происходит по-разному.
1 К. К г е е b, Okologische Grundlagen der Bewasserungskulturen in den Subtro-
pen, Stuttgart, 1964.
Растительность субтропических аридных областей
331
Растения засоленных почв называют галофитами. Поскольку
соли накапливаются главным образом в понижениях, куда стекает также
и вода, почвы здесь представлены в основном влажными солончаками
(см. цветное фото 1, стр. 368). Развитые в этих условиях гигрогало-
фит ы не испытывают затруднений в потреблении влаги, поскольку повы-
шенная концентрация почвенного раствора уравновешивается соответ-
ствующей концентрацией клеточного сока и высоким осмотическим давле-
нием (см. «Phytologie», Bd III, 1, S. 477—491). Ксерогалофиты
растут на менее засоленных, но зато периодически сильно высыхающих
почвах, где высокая концентрация почвенного раствора сочетается с недо-
статком воды.
Водоудерживающая сила почвы слагается из сорбционных сил твер-
дой части почвы, зависящих от механического состава почвы и ее влаж-
ности, и осмотического давления, обусловленного концентрацией почвен-
ного раствора. Количество поглощенной растением воды определяется,
следовательно, суммой этих двух величин. Эта теория, пожалуй, неверна.
Ксерогалофиты, например, поглощают легкорастворимые соли из почвен-
ного раствора и накапливают их в клеточном соке (см. стр. 452), уравно-
вешивая тем самым осмотическое давление почвенного раствора; таким
образом, сохраняется лишь действие сорбционных сил, обусловленных
физическими свойствами почвы. Так, например, при изучении всхожести
семян удалось выяснить, что их прорастание тормозилось лишь сорб-
ционными силами почвы, а осмотическое' давление, несмотря на то, что
оно было в 100 раз больше, не оказывало никакого действия Ч
1 N. Colli s-G eorge, J. Е. Sands, Comparison of the effects of the physi-
cal and chemical components of soil water energy on seed germination, «Austr. J. Agric.
Res.», 13, 1962, p. 575—584.
Глава 8
ПУСТЫНЯ СОНОРА
УСЛОВИЯ СРЕДЫ
Изучение экологии растений пустынь началось в 1903 г. после создания
Лаборатории по изучению пустынь института Карнеги в Вашинг-
тоне (Desert Laboratory of the Carnegie Institution of Washington)
в Таксоне (Аризона), то есть на севере области, простирающейся от мек-
сиканского штата Сонора до склонов плоскогорья Колорадо в США.
На протяжении почти полувека сотрудники лаборатории изучали эколо-
гию растений сначала под руководством Д. Мак-Дугала, а затем Ф. Шрива.
В 1929—1930 гг. автор в качестве стипендиата имел возможность рабо-
тать в лаборатории, и приобретенные здесь знания определили направле-
ние его дальнейшей деятельности х.
Обобщающая работа Шрива «Растительность пустыни Сонора» вышла
в свет только после его смерти в 1950. В этом же году в Таксоне была
ликвидирована и Лаборатория по изучению пустынь. Поэтому нам кажет-
ся уместным начать описание растительности аридных областей с обзора
фундаментальных экологических исследований, проведенных в Таксоне.
Пустыня Сонора простирается от 35 до 23° с. ш. Таксон расположен
в ее северо-восточной части (32° с. ш.) на высоте 728 м над уровнем моря
(рис. 252). О климатических условиях можно судить по климадиаграммам
(рис. 253—256). Средняя годовая температура в Таксоне составляет 19,5°.
Средняя месячная минимальная температура в течение года выше нуля,
однако в зимние месяцы наблюдаются ночные заморозки (особенно в «озе-
рах холодного воздуха»).
Количество осадков колеблется от 56 мм на западе, у нижнего Коло-
радо, до 400 мм и более у восточной границы гор. В окрестностях Таксона
выпадает в среднем 293 мм. Следовательно, речь идет скорее о полупусты-
не, хотя эту наиболее засушливую часть США и называют «пустыней».
Особенно интересно распределение осадков по сезонам. В то время
как на западе дожди выпадают в основном зимой, в Таксоне наблюдаются
два дождливых периода: один — с декабря по март, а другой — с июля
по сентябрь. Дальше к югу, в Мексике, осадки выпадают преимуществен-
но летом. Таким образом, в Таксоне выделяются четыре времени года:
1) зимний дожливый период (к тому же еще и холодный); 2) засушливый
период начала лета с наивысшими температурами в июне (средняя макси-
мальная 37,1°); 3) летний дождливый период с отдельными сильными гро-
зовыми ливнями и 4) засушливый период начала осени (табл. 45).
В табл. 45 приведены лишь средние значения количества осадков;
в отдельные годы могут наблюдаться сильные отклонения. Шрив графиче-
1Н. Walter, Die Hydratur der Pflanze und ihre physiologisch-okologische
Bedeutung, Jena, 1931. Все приведенные в этом разделе данные о гидратуре (осмоти-
ческом давлении) заимствованы из указанной работы.
Рис. 252. А — пустыня Сонора (выполнена штрихами с изолиниями годовых осадков, мм); Б — распро-
странение Carnegiea и Pachycereus в пустыне Сонора (по Шриву, с некоторыми изменениями). Это викариру-
ющие виды, ареалы которых соприкасаются только у побережья Калифорнийского залива.
334
Глава 8
Рис. 253—256. Климадиаграммы станций, расположенных на севере пустыни
Сонора.
ски показал изменчивость продолжительности засушливых периодов
в отдельные годы. Засушливым он считал период, когда на протяжении
не менее чем 30 дней, количество выпавших осадков не превышало 3,5 мм,
Таблица 45
Осадки
%
Зимний дождливый период (декабрь — март) .... 92 31
Засушливый период начала лета (апрель — июнь) 22 7
Летний дождливый период (июль — сентябрь) .... 144 49
Засушливый период начала осени (октябрь —ноябрь) 37 13
поскольку менее интенсивные дожди не оказывают влияния на раститель-
ность (рис. 257, взят из работы Шраца) 4. В 1929 г. в Аризоне годовой
ход и количество выпавших осадков были близки к норме, тогда как
в 1930 г. настоящего засушливого периода в начале лета не было, а засуш-
ливый период в начале осени был очень кратким. Данные об испарении
в более прохладное время года представлены на рис. 259. Большей частью
они колеблются от 1,0 до 1,2 мл!час\ аналогичные величины в середине
лета наблюдаются в центре Европы.
Как и во всех пустынных областях, в Аризоне преобладает механиче-
ское выветривание горных пород. Скалы, нагревающиеся на солнце до
70° С, могут охлаждаться после внезапно выпавших дождей до 20°. Нерав-
номерное сжатие отдельных минералов способствует разрушению горной
породы. Образовавшийся в результате этого песок, поднимаемый вихрями
1 Е. Schratz, Vergleichende Untersuchungen iiber den Wasserhaushalt von
Pflanzen im Trockengebiet des siidlichen Arisona, «Jb. wiss. Bot.», 74, 1931, S. 153—290.
лета период осени
Рис. 257. Распределение и продолжительность засушливых периодов в Таксоне
за 1908—1929 гг. (по Шриву).
Рис. 258. Погодные условия в районе Лаборатории по изучению пустынь (Таксон)
в 1929—1930 гг.
Вверху; 1 средняя максимальная и средняя минимальная температура; за педелю; 2 — сред-
няя месячная температура; з — месячное количество осадков.
Внизу; сумма осадков за трое суток.
336
Глава 8
ветра, усиливает дальнейший процесс эолового выветривания. Правда,
песчаные дюны образуются вследствие этого лишь у северной оконечности
Калифорнийского залива, западнее Юма, уже вне пределов пустыни
Сонора Ч
В окрестностях Таксона развита кактусовая пустыня. Значительная
часть поверхности (25%) занята небольшими возвышенностями, сложен-
ными вулканическими породами или гранитами, обломки которых покры-
вают все склоны и после сильных ливней сносятся потоками воды. Круп-
ные глыбы остаются в верхней части склона, а более мелкие смываются
внпз по склону. Таким образом, на каждом склоне образуется ряд раз-
личных по механическому составу пустынных почв, начиная от крупно-
каменистых осыпей вблизи вершин, сменяемых несколько ниже камени-
стыми и щебнистыми осыпями, и кончая тончайшими илистыми почвами
в долинах. Параллельно изменяется также и крутизна склонов. Вершины
и более крутые склоны покрыты осыпями из крупных обломков; у основа-
ния осыпей склоны резко выполаживаются и переходят в гладкие, как
стол, лишь слегка наклоненные поверхности конусов выноса (bajada)
с развитой на них хрящевато-песчаной почвой. Уклон составляет около
4—12 м на 1 км, или в среднем 0,008. Переход к сложенным аллювием
долинам достаточно резок. Песчаное русло реки глубоко и круто врезано
в пойму. У реки Санта-Крус, притока Хилы, впадающей близ Юмы
в Колорадо (см. рис. 252), подрусловой поток не иссякает на протяжении
всего года.
Выровненные поверхности конусов выноса занимают около половины
всей площади пустыни. Они перерезаны многочисленными более или
менее глубокими руслами временных водотоков. После ливней, прино-
сящих часто за 24 часа более 50 мм осадков, обычно сухие русла на корот-
кое время, часто лишь на несколько часов, превращаются в пенящиеся
горные потоки. Огромные массы воды заполняют русло реки и надолго
затопляют окружающую равнину, откладывая на пойме тончайшие фрак-
ции ила и приносимые со склонов растительные остатки. Поэтому почвы
пустыни очень бедны гумусом и не подразделены на горизонты. Однако
в целом почвы на конусах выноса не столь уж каменисты, как это обычно
считают. Более или менее крупные обломки концентрируются лишь на
поверхности, образуя своеобразную «мостовую пустыни», под которой
накапливается много мелкозема.
Почвы отличаются сухостью и хорошей аэрацией; однако после ред-
ких, но сильных дождей днища долин и особенно бессточные котловины
могут в течение длительного срока оставаться затопленными. Ухудшение
воздушного режима почвы губительно отражается на древесных расте-
ниях. Нередки случаи, когда после продолжительного затопления отми-
рали даже 30—50-летние деревья.
Во всех бессточных понижениях, где вода застаивается, медленно
испаряясь при этом, начинается накопление солей. Однако в пустыне
Сонора засоленные почвы редки, поскольку при выветривании в основном
распространенных здесь кристаллических пород освобождается мало хло-
ристого натрия. Только в долинах, где иногда накапливается гумус,
наблюдается образование «черных щелочных почв» (по классификации
американских авторов). Значительно шире распространены почвы, у кото-
рых на небольшой глубине образуется горизонт, цементированный карбо-
1 Р. J. Rempel, The crescentic dunes of the Salt on Sea and their relation to
the vegetation, «Ecology», 17, 1936, p. 347—358.
Г~
Заросли кактусов, образованные Helianthocereus pasacana, близ Тилькары на севере Аргентины.
Пустыня Сонора
337
патами («hardpan», или «caliche» 1). Мощность этого слоя колеблется от
нескольких миллиметров до 10 см и более; по твердости этот горизонт
напоминает бетон. При смыве верхних горизонтов почвы этот твердый,
напоминающий брекчию слой выходит на дневную поверхность. Он
Даты наблюдений
Рис. 259. Максимальное испарение в дни' наблюдений зимой 1929/30 г.
в районе Лаборатории по изучению пустынь (Таксон, Аризона).
Рис. 260. Вид из Лаборатории по изучению пустынь. Вдали — горы с обшир-
ными конусами выноса. На переднем плане — русла временных водотоков.
Растительный покров образован Prosopis velutina (русла временных водотоков),
Larrea tridentata (конуса выноса), Carnegiea gigantea, Fouquieria (склоны) и
повсеместно встречающимися опунциями (фото Фабера).
1 F. S h г е v е, Т. D. М а 1 1 е г у, The relation of caliche to desert plants, «Soil.
Sci.», 35, 1933, p. 99-112.
338
Глава 8
большей частью непроницаем для воды и препятствует проникновению
корней в более глубокие слои почвы. Предпосылкой для его возникнове-
ния является определенное содержание извести в горных породах. При
выветривании гранитов карбонатный горизонт не образуется. Конкреции
карбонатов появляются уже в полузасушливых областях, например
в черноземах (см. «Phytologie», Bd III, 1, S. 338). Они образуются обычно
у средней границы промачивания почвы дождем. Чем суше климат, тем
ближе к поверхности отлагаются карбонаты. Уплотненные карбонатные
слои возможны и на меньшей глубине — там, где испаряется поступающая
по капиллярам вода
РА СТИТЕЛЬНОСТЬ
Характер растительного покрова тесно связан с вышеописанными
условиями среды 1 2. Здесь можно выделить следующие типы сообществ:
а) Тип сообщества Carnegiea —Encelia (рис. 261), развитый на воз-
вышенностях и крутых склонах в условиях хорошей влагообеспеченно-
ности.
Осадки здесь легко проникают в почву. Скопляющийся между облом-
ками скал и камнями мелкозем хорошо удерживает влагу, которая слабо
испаряется и беспрепятственно используется растениями. Инфильтрация
воды происходит медленно и продолжается в течение нескольких недель
после каждого дождя. Шрив показал, что в этих условиях при осадках,
равных или менее 2 мм. влажность почвы не повышается. Осадки,
достигающие 3 мм. могут оказывать влияние только при условии выпаде-
ния на влажную почву 3. В засушливый период влажность почвы сни-
жается, однако на глубине 30 см влажность почвы не бывает ниже 9%
в мае и 10% в ноябре. Количество недоступной для растений воды не опре-
делялось; о достаточных запасах воды в почве в этих условиях свидетель-
ствует то, что осмотическое давление у Larrea в засушливый период почти
не повышалось (см. рис. 284).
На скалах и каменистых почвах хорошо развиваются кустарники;
помимо двух упомянутых, здесь господствуют следующие виды:
Cercidium microphyHum (рис. 262)
Fouquieria splendens (рис. 281)
Acacia constricta
Celtis pallida
Opuntia versicolor и др.
Ferocactus wislizenii
Jatropha cardiophylla
Prosopis juliflora var. velutina
Ephedra tr if urea
Heteropogon contortus
Hibiscus coulteri
Lippia wrightii
Selaginella arizonica и, кроме-
того, пять видов папорот-
ников.
1 В Испании сцементированная известью корка на поверхности почвы обра-
зуется при годовом количестве осадков, достигающем 100—500 мм (оптимальное коли -
чество осадков — 150—250 льп). Это обусловлено высокой температурой и длительным
засушливым периодом, поскольку при равномерном распределении осадков в течение
года возникают только известковые конкреции. Корка образуется или на поверхности
почвы, или в виде прослоек в самой почве в тех ее горизонтах, где наблюдается испа-
рение влаги. См. Е. Rutte, Kalkkrusten in Spanien, «Neues Jb. f. Geol u. Palii-
ontol», 106, 1958, S. 52-138.
2 F. S h г e v e, Vegetation of the Sonoran Desert, I, Carnegie Inst, of Washington.
Publ. 591, 1951; V. M. Spalding, Distribution and movements of desert plants.
Carnegie Inst, of Wash., Publ. 113, 1909.
3 F. Shreve, Rainfall as a determinant of soil moisture, «Plant World», 17,
1914, p. 8.
Пустыня Сонора
339
Рис. 261. Общий характер растительного покрова пустыни Сонора, образованного
Carnegiea gigantea, Ferocactus wislizenii (рядом с фигурой человека) и Opuntia fulgida.
На переднем плане — кустарнички Encelia farinosa\ крупные кусты справа —
Olneya tesota. Долина Пима, гора Санта-Каталина близ Таксона (фото из архива Лабо-
ратории по изучению пустынь).
Larrea в этих условиях встречается в виде одиночных экземпляров,
a Prosopts более широко распространен в поймах рек. В целом в Аризоне
в подобных условиях произрастает более 300 видов. Carnegiea встречает-
ся лишь там, где продолжительность заморозков не превышает 24 часов 4;
это обычно наблюдается на южных склонах, где при безоблачной погоде
ночью бывают заморозки, а днем температура всегда поднимается выше
нуля. В этих наиболее благоприятных для развития местообитаниях
Larrea образует особенно густые заросли; зимой осмотическое давление
у этого вида достигает наименьших значений именно в подобных усло-
виях (рис. 263).
б) Тип сообщества Larrea — Franseria, распространенный на почти
выровненных конусах выноса.
У подножия крупнокаменистых осыпей, на каменистых почвах
в составе вышеупомянутого типа сообщества появляется Larrea. Ниже по
1 F. S h г е v е, The influence of low temperatures on the distribution of the giant
cactus, «Plant World», 14, 1911, p. 136; W. V. T u г n a g e, A. L. Hinckley,
Freezing weather in relation to plant distribution in the Sonoran Desert, «Ecol, Monogr.»,
8, 1938, p. 529—550.
Рис. 262. «Кактусовый лес» из Cercidium microphyllum и Carnegiea gigantea
близ Таксона. Среди них несколько кустов Acacia constricta, а на переднем плане —
Opuntia julgida*
Разница высот ,м
Рис. 263. Зависимость осмотического давления у Carnegiea gigantea (сопостави-
мые данные) от условий произрастания (из Н. Walter, Hydratur der Pflanze,
1931).
Пустыня Сонора
341
склону развиты хрящевато-песчаные почвы, у которых на небольшой
глубине часто формируется уплотненный карбонатный горизонт. На этих
крайне сухих местообитаниях из числа названных видов встречается
только Larrea tridentata, обычно в виде угнетенных экземпляров, теряю-
щих в засушливый период почти все листья. Вместе с Larrea здесь растут
также несколько видов Franseria (Compositae) и много разнообразных
опунций, отличающихся формой стеблей (рис. 264).
в) Тип сообщества Prosopis velutina, развитый на аллювиальных
пылеватых суглинках.
Частые разливы рек обусловливают глубокое промачивание почвы,
почти до уровня грунтовых вод. Prosopis обладает разветвленной поверх-
ностной корневой системой (простирающейся радиально на 20 м) и одно-
временно глубоко проникающими в почву стержневыми корнями. В этих
условиях развивается редкостойный лес с почти соприкасающимися зон-
тиковидными кронами деревьев. Высота деревьев достигает 15—18 ж,
а диаметр стволов — 1,2—1,5 м (рис. 265). Поскольку эти леса на протяже-
нии длительного времени были единственным источником топлива, они
в настоящее время почти полностью уничтожены. Местами еще встречают-
ся сомкнутые порослевые древостои, образованные Prosopis с участием
Acacia greggii, Celtis pallida, Condelia lycioides, видами Lycium, а также
лианами (Clematis drummondii, Vitis arizonica и т. д.)—большей частью
видами, стрежневые корни которых достигают грунтовых вод. Обычно
же эти площади распаханы и заняты хлопчатником, люцерной и ячменем;
они поливаются при помощи насосов, подающих воду из колодцев. Почвы
здесь очень мощные и плодородные. В понижениях, где после разлива
рек вода длительное время испаряется, могут накапливаться соли. Здесь
поселяются галофиты — Atriplex и даже Suaeda moquini. Зимой в ясные
ночи воздух здесь сильно охлаждается. Шрив провел сравнение минималь-
ных температур на холме Тумамок, где расположена Лаборатория
по изучению пустынь, и у подножия холма в пойме. У подножия холма тем-
пература в ясные ночи могла быть на 13° ниже, а в дождливые дни эта
разница составляла всего лишь 0,2° Поэтому зимой очень часто наблю-
даются ночные заморозки. Однолетники, вегетационный период которых
на влажной почве мог бы отличаться большей продолжительностью,
здесь из-за слабой морозоустойчивости отмирают уже в конце октября,
a Prosopis регулярно сбрасывают листья зимой, хотя экземпляры, разви-
тые на склонах, остаются часто облиствленными.
г) Тип сообщества Populus — Salix, развитый в поймах, непосред-
ственно по берегам рек.
Сухие русла рек врезаны в пойму на глубину 1—2 м, а в исключи-
тельных случаях даже на 12—15 м. Углубление ложа происходило,
по-видимому, лишь в течение последних 75 лет, когда вследствие усилен-
ной пастьбы растительный покров изреживался все больше и больше,
что вызывало усиление поверхностного стока и эрозии почвы.
Пастьба повсеместно и притом очень существенно изменила весь
облик растительности. В результате почти полностью исчезли одни виды,
а вместо них сильно распространились некоторые виды кактусов, и преж-
де всего опунции, не поедаемые скотом. У некоторых опунций, отличаю-
щихся цилиндрической формой и снабженных длинными колючками,
отдельные выросты отваливаются при малейшем прикосновении, как,
например, у Opuntia fulgida (cholla). Они могут зацепиться за шерсть
1 F. Shreve, Cold air drainage, «Plant World», 15, 1912, p, 110—115.
342
Глава 8
Рис. 264. Larrea tridentata (крупные темные кусты) и Franseria dumosa (опушен-
ные кустарнички) близ Пайлот-Ноб, в пустыне Мохаве (фото из архива Лабора-
тории по изучению пустынь).
Р и с. 265. Мескитовый' лес (Prosopis) в пойме пересыхающей реки Саладо,
Южная Калифорния (фото Гейма).
животных, которые переносят их на большие расстояния; при падении на
землю они быстро укореняются.
Ложа рек по большей части лишены растительности из-за подвижно-
сти субстрата, смещаемого ежегодно паводками. В зависимости от ско-
Пустыня Сонора
343
Рис. 266. Cercidium torreyanum на дне сухого русла реки близ Таксона (фото из
архива Лаборатории по изучению пустынь).
рости течения потока русло может быть каменистым, галечниковым или
песчаным. По берегам распространены участки пойменного леса; грунто-
вая вода залегает здесь на небольшой глубине. Виды, слагающие эти леса,
очень близки к породам, развитым в листопадных лиственных лесах уме-
ренной зоны. Это виды Populus и Salix, Fraxinus velutina var. toumeyi
или Sambucus mexicana, напоминающая Sambucus nigra.
Полые воды, постепенно размывая( корни деревьев, вырывают их
и, протащив немного, оставляют ниже йо течению.
На перемываемых паводковыми водами песчаных участках, располо-
женных вблизи русла реки, распространены по большей части быстро-
растущие кустарники —Baccharis и Hymenoclea, не страдающие от час-
тичного заноса их песком.
д) Растительность русел временных водотоков (уошей, арройо).
Эти русла начинаются в верхней части склона в виде небольших
борозд, которые, сливаясь, превращаются в обширные овраги. Послед-
ние, все больше углубляясь и расширяясь, часто образуют небольшие
каньоны или постепенно переходят в широкие и неглубоко врезанные сухие
русла временных водотоков. В истоках этих русел, где вода после
дождя устремляется вниз с большой силой, почва покрыта крупными
обломками. В низовьях скорость потока уменьшается и здесь отклады-
ваются галька и песок.
Водный режим по берегам русел временных водотоков всегда болеее
благоприятен и почва лучше и глубже увлажнена. В нижнем течении
могут даже образовываться скопления грунтовых вод. В связи с этим
виды, распространенные в основном на склонах возвышенностей, разви-
344
Глава 8
ваются здесь пышнее, имеют более крупные листья и часто сохраняют
листву даже в периоды засухи. Из уже упоминавшихся видов чаще всего
здесь встречаются Celtis pallida и Lycium, наряду с ними в этих условиях
встречаются и новые виды. Вблизи устьев распространены Cercidium tor-
reyanum (рис. 266) и виды, развивающиеся на аллювиальных почвах, —
Prosopis и Acacia,
В глубоких каньонах на состав растительного покрова влияют потоки
стекающего по склонам холодного воздуха, и поэтому здесь отсутствуют
легко повреждаемые заморозками виды. По данным Шрива, температура
на дне долины в 20 час. на 5° ниже, чем на 31 м выше по склону. Однако
мощность слоя холодного воздуха не превышает 18 м от дна каньона. Эти
неблагоприятные температурные условия и одновременно благоприятная
гидратура дают возможность развиваться в каньонах представителям
выше расположенных горных поясов, например вечнозеленым дубам.
Нередко здесь встречается также и Platanus wrightii,
е) Вертикальная поясность на высоких горах.
К востоку от Таксона мощные горные хребты Санта-Каталина и
Санта-Рита поднимаются на высоту до 3000 м, В горах выпадает
большое количество осадков, и зимой они длительное время покрыты
снегом.
Рис. 267. Склоны горы Санта-Каталина близ Таксона на высоте 1500 м. Нижняя
граница распространения Quercus oblongifolia и Q. arizonica (темные кусты). Впе-
реди справа — Nolina microcarpa, в середине — Dasylirion wheeleri и Yucca schottit
(фото из архива Лаборатории по изучению пустынь).
Пустыня Сонора
345
Рис. 268. Каменистая пустыня с Pinus edulis var. monophylla. На заднем плане —
Большой Каньон (Колорадо).
На высоте 1000 м характер растительного покрова существенно
меняется, несмотря на то, что отдельные представители пустынной флоры
проникают по благоприятным местобитаниям до высоты 1500 и даже
2000 м. Крупные кактусы исчезают полностью; здесь развивается саванна,
образованная Prosopis, в которой между одиночно растущими кустами Pro-
sopis распространены дерновинные злаки. Наряду с ними здесь встречают-
:346
Глава 8
ся различные виды листовых суккулентов — Agave и Yucca, а также Dasy-
lirion и Nolina (рис. 267). На высоте около 1300 м появляются отдельные
экземпляры, а выше 1500 м сомкнутые группы вечнозеленых дубов, обра-
зующие пояс дубовых лесов. Это сомкнутый склерофильный лес высотой
15—20 м, образованный видами Quercus oblongifolia, Q. emoryi, Q. hypo-
leuca, реже Q. reticulata, а также Arbutus arizonica, Arctostaphylos pungens
и др.
В этих почти еще не затронутых деятельностью человека лесах можно
получить представление о девственном облике жестколистных лесов
Средиземноморья.
На высоте около 2000 м вечнозеленые дубовые леса сменяются сосно-
выми лесами из Pinus ponderosa ssp. scopulorum, к которой выше присоеди-
няется Pinus strobiformis.
Далее, на высоте 2400 м, на северном склоне распространены пихто-
вые леса из Abies concolor с примесью Pseudotsuga taxifolia. По южным
-склонам сосна поднимается до наиболее высоких вершин.
В этих климатических условиях выпадает пояс листопадных листвен-
ных лесов. Лишь на более крутом северном склоне горы Болди автор
видел зимой безлистные заросли дуба.
На плоскогорье Колорадо, расположенном на высоте около 2100 м
на севере Аризоны, распространен пояс сосновых лесов из Pinusponderosa',
в нижней части пояса часто развиваются саванноподобные группировки
Рис. 269. Кустарничковая полупустыня на спуске в Большой Каньон, примерно
на высоте 1100 м. Заросли Coleogyne ramosissima,rOpuntia polyacantha, Atriplex canescens
и Ephedra viridis (фото Мак-Дугала).
Пустыня Сонора
347
из Juniperus utahensis и J uniperus monosperma. Здесь встречается также
Pinus edulis var. monophylla с одиночной хвоей в пучках.
Это плоскогорье прорезано крупнейшей в мире речной долиной —
Большим Каньоном. Его длина достигает 350 км, а глубина —800 —
1800 м. Ширина верхнего края каньона местами превышает 10 км, его
склоны образованы фантастическими террасами, сложенными осадоч-
ными породами; по нижней части каньона в кристаллическом ложе текут
мутные воды реки Колорадо, образующие многочисленные водовороты.
В Большом Каньоне Колорадо отчетливо выражена вертикальная
поясность растительности, которая, однако, не соответствует зональным
сменам, характерным для южной Аризоны. На покрытых подвижными
осыпями склонах обычно не развивается сомкнутый растительный покров.
В верхней части склонов еще встречаются отдельные деревья сосны и мож-
жевельника (рис. 268). На высоте 1200 м распространена Artemisia tri-
dentata, образующая вместе с Coleogyne ramosissima (Rosaceae), Atriplex
canescens, Ephedra viridis, Opuntia polyacantha и другими видами сообще-
ства, развитые на границе с кустарничковой полупустыней (рис. 269).
Верхнюю границу леса в Аризоне можно наблюдать лишь на вершине
Сан-Франциско, поднимающейся на Колорадском плато почти до высоты
4000 м. Здесь выше пояса Abies — Pseudotsuga распространен пояс еловых
лесов wzPicea engelmannii, образующей верхнюю границу леса. На южных
склонах у верхнего предела леса встречается еще один низкорослый вид
Abies. I
В ходе этого краткого обзора лесной растительности мы, однако,
отвлеклись от характеристики аридных областей, поэтому читателям,
интересующимся поясностью в горах, рекомендуем превосходные работы
Ф. Шрива х.
ЭКОЛОГИЧЕСКИЕ ГРУППЫ РАСТЕНИЙ ПУСТЫНИ СОНОРА
Как было показано выше, количество осадков в аридных областях
сильно колеблется от года к году. Та же закономерность присуща и Ари-
зоне 1 2. Избыток воды во влажные годы используется многочисленными
эфемерами (однолетниками и геофитами), не развивающимися в засушли-
вые годы (см. стр. 311).
Эфемеры
Как уже указывалось, в Аризоне выражено два дождливых сезона:
1) зимний дождливый период со средними суточными температурами
около 10—15° (причем ночной минимум может быть даже ниже нуля)
и с осадками, выпадающими в виде моросящих затяжных дождей;
2) летний дожливый период со средними суточными температурами
около 30° (максимальными выше 40°) и осадками, выпадающими большей
частью в виде сильных ливней.
Два дождливых сезона определяют развитие эфемеров зимой и летом.
Однако сомнительно, чтобы один и тот же вид мог встречаться и зимой
и летом.
1 F. S h г е v е, The vegetation of a desert mountain range as conditioned by cli-
matic factors, Carnegie Inst, of Washington, Publ. 217, 1915; Conditions indirectly affe-
cting vertical distribution on desert mountains, «Ecology», 3, 1922, p. 269—274.
2 R. R. Humphrey, A detailed study of desert rainfall, «Ecology», 14, 1933,
p. 31—34.
348
Глава 8
Причина столь резкого разграничения связана с несовпадением опти-
мальных температур, необходимых для прорастания эфемеров. Как уда-
лось показать Шриву, зимние однолетники трогаются в рост при темпе-
ратуре около 15—18°, а летние —около 27—32° Почти все виды быстро
прорастают также при температуре, отклоняющейся от оптимальной не
Рис. 270. Зависимость высоты растений и осмотического давления
от условий местообитания у Solarium elaeagnifolium (из Н. W a J t е г,
Die Hydratur der Pflanze, 1931).
более чем на 3—5°. Эфемеры прорастают очень быстро; это необходимо
в связи с тем, что их проростки должны успеть укорениться за корот-
кий период, во время которого поверхностные слои почвы еще увлаж-
нены.
Развитие эфемеров тесно связано с внешними условиями. Если вскоре
после прорастания наступает засуха, то всходы погибают или достигают
карликовых размеров (высоты 1 см)\ несмотря на это, растения цветут
и плодоносят. Зато при более благоприятных условиях растения дости-
гают высоты 20—30 см и обильно цветут. Высота некоторых летних эфе-
меров равняется 2—3 м.
В период зимних дождей летние эфемеры не прорастают; точно так
же летние дожди не прерывают покоя зимних эфемеров. В западной части
1 F. W. W е n t, Ecology of desert plants: I and II, «Ecology», 29, 1948, p. 242—
353; 30, 1949, p. 1—13, 26—38.
Пустыня Сонора
349
•области, где летом осадки выпадают лишь изредка, летние эфемеры почти
не встречаются. Однако ареал их простирается далеко в глубь Техаса,
где осадки выпадают только летом. Продолжительность жизни однолет-
ников редко превышает 6—8 недель, однако в благоприятных местооби-
таниях, например по склонам берегов оросительных каналов, они могут
переносить также засуху и превращаться в многолетников. Это, в частно-
сти, свойственно некоторым занесенным сюда сорнякам, поскольку в обыч-
ных для них условиях они развиваются как однолетники. Приспособлен-
ные к низким температурам зимние эфемеры являются элементами голарк-
тической флоры. Они представлены знакомыми нам родами и семействами;
кроме того, среди них часто встречаются сорные растения Европы, напри-
мер Е г odium, Sonchus asper, Hordeum murinum, Malva silvestris и т. д.
В противоположность этому летние эфемеры являются элементами
неотропической области; среди развитых здесь сорняков встречаются
виды, широко распространенные в субтропиках, например Tribulus
terr estr is.
В октябре 1929 г., по наблюдениям автора, сложились очень благо-
приятные условия для развития летних эфемеров, так как перед этим
с 28 июня по 24 сентября выпало 200 мм осадков, а последний дождь
принес около 60 мм. Летний дождливый период отличался почти тропиче-
скими условиями: высокой относительной влажностью и высокой темпера-
турой. Однако температура поверхности влажной почвы не превышала
обычных значений. Поскольку глубин^ проникновения корней эфемеров
в среднем не достигает 15 см, их период вегетации заканчивается одно-
временно с высыханием верхних слоев почвы. Крупнокомковатая почва
на склонах гор после прекращения дождей высыхает на эту глубину
примерно за 3 недели, мелкокомковатая почва, развитая в затопля-
емых паводковыми водами речных долинах,— примерно за 6 недель.
Бездождная погода, установившаяся с 24 сентября до 31 декабря, позво-
лила автору детально проследить за последовательным отмиранием
эфемеров.
Уже к 15 октября большая часть видов, отличающихся наиболее неж-
ными листьями (Boerhavia, Mollugo, Datura), погибла; злаки (Bouteloa,
Panicaceae) высохли. Начали желтеть Amaranthus palmeri, Euphorbia
heterophyНа, и лишь отдельные экземпляры, растущие в тени кустарников,
сохраняли ярко-зеленую окраску. Напротив, виды с суккулентными
листьями (Trianthema portulacastrum, Allionia incarnata) или несколько
опушенные (Kallstroemia, Tribulus, Cladothrix lanuginosa, Solanum elaeag-
nifolium) существовали до середины ноября. По данным Кэннона А, корни
Kallstroemia проникают на глубину 22 см. Очень сочная Martynia (осмо-
тическое давление 9,4 атм), так же как и другие виды, приуроченные
к пойме, погибли после ночного заморозка 31 октября.
Гидратура у всех названных видов достигает значений, свойственных
широколистным мезофитам (табл. 46).
Суточные колебания осмотического давления незначительны. Лишь
в период засухи давление постоянно повышалось до максимальных
(летальных) значений, при которых растения отмирали.
Зависимость высоты растений и гидратуры от условий местообитания
наглядно показана на рис. 270. У Solanum elaeagnifolium развиты колючки
ц ксероморфные опушенные листья; исследованные экземпляры росли на
1 W. А. С a n п о n, The root habits of desert plants, Carnegie Inst, of Washing-
ton, Publ. 131, 1911.
350
Глава 8
Таблица 46
Осмотическое давление летних эфемеров (в атм)
Оптимальное Макси- мальное
Euphorbia heterophylla 13-14 17
Kallstroemia grandiflora 15,4 18,4
Amaranthus palmeri 14,5 17,2
Tribulus terrestris 13,2 21,7
Cucurbita digitata . . 13,4 —
Cladothrix lanuginosa 18,9 34,8
Allionia incarnata <20 25
Trianthema portulacastrum 18,4 30,9
затопляемых участках в небольшой лощине. В центре понижения, где*
почва была лучше увлажнена А, растения достигали более крупных раз-
меров, у краев же, на сухой почве, они не превышали нескольких сан-
тиметров.
Зимний дождливый сезон начался 31 декабря после 97-дневной засу-
хи, во время которой не было ни одного дождя. С января по март выпало
127 мм осадков, обусловивших хорошее развитие зимних эфемеров.
В окрестностях Таксона распространены 44 летних и 109 зимних
видов эфемеров. Дальше к западу, в пустыне Мохаве, в области, где осадки
Таблица 47
Осмотическое давление у зимних эфемеров (в атм)
Оптимальное Макси- мальное
Erodium cicutarium Sonchus asper . . Hordeum murinum Malva silvestris Marrubium vulgare Brodiaea spec. Sicyos spec. Rumex hymenosepala Delphinium scaposum Anemone sphenophylla Verbena ciliata . . Pentstemon wrightii Sphaeralcea pedata Erodium texanum Senecio douglasii Plantago ignota . . . 9—10 10,6 12,6 15,2 15,4 7,4 8-9 9,2 10,5 14,3 14,4 14,2 17,1 13,4 10,8 11,5 >14,7
Streptanthus arizonicus 14,8 >20,2
Parietaria obtusa . . . Phacelia tanacetifolia 12,5 16,2 >14,2
1 F, Shr е v е, Rainfall, runoff and soil moisture under desert conditions, «Ann.
Assoc. Amer. Georg.», 24, 1934, p. 131 —156.
Пустыня Сонора
351
Рис. 271. Пойкилогидрические виды Cheilanthes lindheimeri и Selaginella ari-
zonica. Пустыня Сонора близ Таксона (фото Э. Вальтер).
выпадают зимой, число зимних эфемеров возрастает почти до 500. 73 вида*
встречаются в обеих областях. .
Поскольку дефицит влажности воздуха в прохладные зимние месяцы-
намного ниже, чем летом, зимние эфемеры отличаются более нежной
и гигроморфной структурой. Многие из них, например Delphinium sca-
posum и Anemone sphenophylla, растут на защищенных от прямых лучей
солнца северных склонах. Наличие в их составе европейских сорных
растений свидетельствует о сходстве климатических условий периода
зимних дождей с климатическими условиями Центральной Европы
в конце весны — начале лета; это сходство отражается также и на гидра-
туре растений (табл. 47).
Эфемеры не приурочены к определенным местообитаниям и почвам.
Они растут там, где достаточно влаги для их развития. Обычно сильные
дожди хорошо промачивают любую почву, а слабые увлажняют лишь
русла временных водотоков и почвы низин. Если дождевая вода сносит
семена в понижения, то там часто формируются одновидовые заросли.
По данным Шрива, на 1 м2 влажной глинистой почвы встречается до 1200'
всходов Plantago. В тени кустарников, где почва дольше остается влаж-
ной, покров эфемеров более сомкнут. На песчаной почве они растут на
больших расстояниях друг от друга. После сильных мартовских дождей
в апреле цветущая эшшольция придала обширным площадям в западной
части пустыни желтый аспект, а в пустыне Мохаве огромная территория
была покрыта низкорослыми белыми цветущими экземплярами Oenothera
и фиолетовыми Abronia.
Примерно в это же время в связи с повышением температуры начи-
нают прорастать и многолетние виды, причем они по большей части сначала
цветут и лишь затем у них начинают формироваться вегетативные органы.
Пойкилогидрические папоротникообразные растения
Эта небольшая, приуроченная к скалам группа папоротников раз-
вивается аналогично эфемерам с той лишь разницей, что их листья не*
отмирают в засушливый сезон. С наступлением засухи они высыхают,.
Глава 8
однако, смоченные, они через короткий промежуток времени вновь начи-
нают зеленеть. Во влажном воздухе листья распускаются очень медлен-
но 4. Следовательно, они ведут себя’подобно мхам, однако листья папорот-
никообразных имеют ксероморфный облик. Они бывают или жесткими
(Pellaea mucronata), или сильно опушенными, или покрыты чешуйками
(виды Notholaena и Cheilanthes, Selaginella arizonica). Виды с очень неж-
ными листьями, как, например Adiantum cap Ulus veneris, встречаются
только в тени у источников. Cheilanthes wrightii, также обладающий неж-
ными листьями, растет лишь во влажных местах, никогда не освещаемых
солнцем. Впрочем, даже ксероморфные виды всегда растут с северной
стороны скал. Возможно, это обусловлено тем, что только в этих условиях
их заростки развиваются нормально. Во время засухи папоротники имеют
столь невзрачный облик, что их с трудом можно найти. Осмотическое
давление у папоротников можно определить только на свежих экземпля-
рах (табл. 48).
Таблица 48
Осмотическое давление у пойкилогидрических
видов (в атм)
Gymnopteris hispida 13,4
Pellaea mucronata 14,1—15,4
Notholaena hookerl . . 14,6—15,8
Notholaena sinuata var. integerrima 15,4
Notholaena aschenborniana 13,4—16,1
Cheilanthes wrightii 15,9—16,0
Cheilanthes jendlert . . . . 17,7—18,3
Cheilanthes lindheimeri (рис. 271) . 24,1
Selaginella arizonica (рис. 271) . 13,6—16,7
Автор изучал эту группу растений также в Карру (ЮАР).
Общие замечания о многолетних видах
Пустыня Сонора не отличается крайне засушливыми условиями
климата. Поэтому здесь развивается (даже если учитывать только много-
летние виды) еще относительно сомкнутый растительный покров. Степень
покрытия почвы не превышает 30%, а часто она даже меньше 10%. Высо-
та большинства видов достигает лишь 0,5—1,5 л; однако встречаются
и гигантские кактусы высотой 11 м, а иногда даже до 16—18 м. Высокие
деревья растут только в галерейных лесах, окаймляющих берега сухих
потоков с постоянным уровнем грунтовых вод.
Из-за подземного смыкания корневых систем даже у очень редко
стоящих экземпляров Larrea tridentata, развитых в плакорных условиях
на сухих песчаных почвах, другие многолетние виды не могут поселиться
на кажущихся свободными пространствах между кустами. На скалах
все благоприятные для поселения участки, связанные с трещинами,
также заняты многолетними растениями. Поэтому подрост, для которого
не остается свободного пространства, почти совершенно отсутствует.
Древовидный кустарник Cercidium microphyПит достигает возраста 300 —
400 лет, и лишь после гибели старого экземпляра подрост может занять
1 Е. S. Spalding, Absorption of atmospheric water by desert shrubs, «Bull.
Torrey Bot. Club», 33, 1906, p. 367; Absorption of water by leaves, «Bot. Gaz.», 41,
1906, p. 262.
Пустыня Сонора
353
его место. Шрив изучал на протяжении многих лет возобновление этого
вида, отличающегося обильным плодоношением на пробной площадке 4.
В течение 8 лет наблюдений на площади 557 м1 2, было обнаружено 923 всхо-
да, однако через 5 лет выжило лишь 4, поскольку большинство погибло
в первый же год (см. «Phytologie», Bd III, 1, S. 271). Только в случаях,
когда растения достигают таких размеров, при которых они могут терять
в крайне засушливые годы при недостатке воды часть своих надземных
органов, а в благоприятные годы восстанавливать их за счет вторичного
отрастания, их дальнейшее существование бывает обеспечено. Поэтому
у большинства пустынных кустарников обычно имеется очень много отмер-
ших и лишь несколько живых ветвей, свидетельствующих о предшество-
вавших засушливых годах.
Суккуленты <
В большей части пустыни Сонора, которую называют также «кактусо-
вой пустыней», обильно встречающиеся суккуленты представлены много-
численными видами Cactaceae (цветное фото 1, стр. 336). В Южной Кали-
форнии распространен только один суккулентный вид из семейства
Euphorbiaceae Pedilanthus macrocarpus, тогда как в Африке молочайные
отличаются большим разнообразием суккулентных форм.
Кактусовые характеризуются весьма! разнообразной формой:
а) колоннообразной, ветвящейся с середины стебля (Carnegiea.
Pachycereus) или уже у основания (Lophocereus. Lemaireocereus) (рис. 272);
Рис. 272. Lemaireocereus thurberi (в середине);[и Fouquieria splendens у пере-
вала Пику, пустыня Сонора, Мексика_(фото Э. Вальтер).
1 F. Shreve, Establishment behavior of the Palo Verde, «Plant World», 14 f
1911, p. 289; The establishment of desert perennials, «J. Ecol.», 5, 1917, p. 210.
354
Глава 8
Рис. 273. Корневая система Ferocactus wisli-
zenii (по Кэннону, с некоторыми изменения-
ми) (из H. Walter, Standortslehre, 1960).
Вверху — корни простираются в горизонтальном
направлении на глубине всего 2 с.н;
внизу — план корневой системы того же экзем-
пляра.
б) шаровидной (Ferocactus);
в) подушковидной, возникающей благодаря многократному повтор-
ному ветвлению (Echinocereus, Mammillaria);
г) цилиндрические опунции
(Qpuntia fulgida, О. acanthocarpa
и более мелкие О. bigelovii,
О. arbuscula и др.);
д) плоские опунции (много-
численные виды).
Экология кактусов подроб-
но изучалась в Лаборатории по
изучению пустынь, однако эти
исследования пока не обобщены.
Все кактусы, за исключе-
нием Penicereus, укореняюще-
гося при помощи стержневого
корня в руслах временных водо-
токов, обладают крайне поверх-
ностной, далеко распространяю-
щейся в радиальном направле-
нии корневой системой, распо-
ложенной на глубине всего
нескольких сантиметров (рис.
273). Следовательно, кактусы
могут поглощать воду, накап-
ливая ее в тканях стебля и
листьев только после дождя,
пока верхние слои почвы еще увлажнены. В период засухи окончания
всасывающих корней отмирают и растение вынуждено обходиться только
собственными запасами воды.
Кауш изучал развитие корней у кактусов в оранжерейной культу-
ре Ч Первый полив растений был произведен лишь спустя 6 месяцев после
начала опыта. У одного вида опунций уже через 8 часов начали форми-
роваться всасывающие корни; через 24 часа они были отмечены у 50%
кактусов, а через 72 часа — у 80% растений.
Крупные экземпляры Carnegiea запасают до 2000—3000 л воды и могут
больше года обходиться без новых поступлений влаги 1 2. На отмершем,
стебле часто сохраняются живые ответвления, которые могут цвести
в следующем году. Однако они никогда не укореняются, даже если их
закопать в почву. Ferocactus wislizenii используется индейцами в качестве
источника питьевой воды в пустыне. Они срубают верхушку и выдалбли-
вают деревянным пестиком углубление, где накапливается клеточный сок,
который можно вычерпывать пригоршнями и пить. Правда, на основании
собственного опыта автор считает вкус этого сока ужасным, однако для
человека, гибнущего от жажды, вкус уже не имеет никакого значения.
1 W. К a u s h, Der EinfluB von edaphischen und klimatischen Faktoren auf
die Ausbildung des Wurzelwerkes der Pflanzen unter besonderer Beriicksichtigung eini-
ger algerischer Wlistenpflanzen, Habilitationsschrift, Darmstadt, 1959.
2 D. T. MacDougal, The water balance of desert plants, «Ann. Bot.», 26,
1912, p. 71; Variations of the water balance, Carnegie Inst, of Washington, Publ. 141,
1910; D.T. MacD ougal, E.S. Spalding, The water balance of succulent plants,
Carnegie Inst, of Washington, Publ. 141, 1910, D. T. M a c D о u g a 1, E. R. Long, I. G.
Brown, End results of desiccation and respiration in succulent plants, «Physiol-
Res.», 1, 1915, p. 289.
Пустыня Сонора
355
Опыты по изучению транспирации кактусов в естественных условиях
проводились главным образом Мак-Дугалом.
a) Carnegiea gigantea, см. рис. 261 и 262.
Этот гигантский кактус достигает высоты 10—12 лг; иногда встречают-
ся и более высокие экземпляры. Диаметр стебля 30—40 см. У растений
высотой 5—8 м образуется 5—6 отходящих наподобие канделябров и рас-
тущих кверху боковых ответвлений. Вторичное ветвление является исклю-
чением и наблюдается лишь у очень старых экземпляров.
Корни Carnegiea углубляются в почву не более, чем на 1 м, тогда как
в горизонтальном направлении они простираются близ поверхности
почвы на расстояние до 30 м. Цилиндрический стебель, образованный
анастомозирующими друг с другом одревесневшими тканями, представля-
ет прочный каркас. Один из видов дятлов часто устраивает свои гнезда
в стеблях кактусов, где они обволакиваются тканями каллюса, которые
постепенно пробковеют. Белые цветки раскрываются в мае, преимущест-
венно вблизи вершины и на юго-западной стороне стеблей; опыляются они
пчелами. Сочные плоды кактуса поедаются птицами; их собирают и индей-
цы племени папаго. Старый экземпляр кактуса приносит до 200 плодов;
в каждом из них содержится около 1000 семян. Семена распространяются
при помощи животных и муравьев. Плоды созревают за 2—4 недели до на-
чала периода летних дождей; семена прорастают летом, однако к осени
сохраняются лишь единичные всходы. Через 2—3 года они достигают
высоты всего нескольких миллиметров, а через 10 лет — 1,5-2,0 см. В воз-
расте примерно 30—50 лет растения не превышают высоты 1 м. Лишь
после того как растения достигнут высоты 2—3 м, дальнейший прирост
составляет примерно 10 см в год. Возраст наиболее старых экземпляров,
вероятно, равен 150—200 годам.
Carnegiea распространена в пустыне Сонора к востоку от реки Коло-
радо и на восточном побережье Калифорнийского залива (рис. 252, Б).
В горах этот вид проникает до высоты 1200—1500 лг, особенно широко
он распространен в окрестностях Таксона. Вес старых экземпляров дости-
гает 2000—4000 кг, в отдельных случаях — 6000—7000 кг. В среднем в рас-
тениях содержится 85—91 % воды. Меньше всего воды содержится в ниж-
ней части ствола, заполненной в основном одревесневшими тканями цент-
рального цилиндра.
- По данным Мак-Дугала, сразу после срезки в ноябре интенсивность
транспирации растений в лабораторных условиях достигла 2—3 мг воды
на 1 г сырого веса в сутки. После этого растения оставляли под открытым
небом. К весне (февраль — март) интенсивность транспирации снизилась
до 1,5 мг на 1 г сырого веса в сутки, а через год (декабрь — май) — до 1 мг
в сутки. У взрослого экземпляра интенсивность транспирации составляла
всего 1/1700—1/9300 сырого веса. Интенсивность транспирации всходов
достигала 1/12 сырого веса в сутки, а у более взрослых растений доходила
до 1/46. Взрослые экземпляры погибали от потери воды, превышавшей
23% сырого веса, а у всходов эта величина составляла около 54%
и наступала через 100—120 дней.
Обусловленное потерей воды изменение объема осуществляется у как-
тусов благодаря механизму действия ребер, сходному с действием кузнеч-
ных мехов Ч Ребра сближаются без всяких нарушений механических тка-
ней, находящихся в наружных слоях коры. При поглощении растением
1 Е. S. Spalding, Mechanical adjustment of the Sahuaro (Cereus giganteus)
to varying quantities of stored water, «Bull. Torrey Bot. Club», 32, 1905, p. 57.
356
Глава 8
влаги ребра расходятся и объем стебля снова увеличивается. Амплитуда
растяжения на южной стороне больше, однако на северной стороне растя-
жение после дождя сохраняется гораздо дольше. Максимальное расстоя-
ние, на которое раздвигаются ребра, составляет между двумя ребрами
35%, или около 2,5 см. Мак-Дугал и Сполдинг (1910) проследили годовую
амплитуду толщины стеблей. Минимум наблюдался в периоды засухи
в начале и конце лета. После каждого дождя толщина стебля сразу начи-
нает увеличиваться; это продолжается до тех пор, пока не будет достигнут
предел набухания. Амплитуда и скорость набухания индивидуальны
у каждого растения. Высокие температуры благоприятствуют набуха-
нию, низкие — замедляют его. Кактус растет главным образом летом.
Цветение начинается после периода зимних дождей; цветки раскрываются
при восходе солнца, в течение дня. Интенсивность их транспирации очень
незначительна.
Осмотическое давление свежего клеточного сока, по данным Мак-Ду-
гала, равно 6—8 атм и 9,8 атм — после потери 23,3 % воды. Однако следует
учитывать, что осмотическое давление клеточного сока в разных тканях
хотя и незначительно, но различается. В тканях ребер, содержащих хло-
рофилл, давление примерно на 1—2 атм выше, чем в паренхиме ребер,
тогда как давление сока в паренхиме коры и сердцевине на 1,0—1,5 атм
ниже. Меристема у вершины стебля отличается несколько более высоким
давлением, поскольку здесь не развиты запасающие воду ткани. Кроме
того, на южной стороне стебля, где ребра расположены ближе друг к дру-
гу, давление клеточного сока примерно на 2 атм выше, чем на северной
стороне. Лишь после выпадения дождя давление выравнивается или даже
наблюдается небольшой сдвиг в обратном направлении; это связано с тем,
что на южной стороне, где сильнее развиты проводящие воду ткани, запа-
сы воды восстанавливаются быстрее (табл. 49).
Таблица 49
Осмотическое давление (в атм) у Carnegiea в период осенней засухи и сезон
зимних дождей
(осадки выпали 24 января и 19 марта)
Паренхима ребер (на высоте 1,5 м) 24/Х 10/XI 5/1 24/1 (образцы взяты с четырех растений) 14/11 19/III 27/III
Южная сторона . . — 6,0 7,2 5,9 4,7 5,7 5,9 5,3 5,2 4,1
Западная сторона 5,5 5,6 — — — —
Северная сторона — 5,1 5,3 5,9 5,2 6,0 6,0 4,7 5,8 —
У отломанного ответвления стебля ио истечении 4 месяцев осмотиче-
ское давление в паренхиме ребер повысилось до 9,3—10,5 атм, а у про-
лежавшего в течение 9 месяцев соответствующие значения достигали 10,7
(северная сторона) и 13,6 атм (южная сторона). Однако при высыхании
осмотическое давление у кактусов повышается не всегда.
Если растения высыхают медленно, что сопровождается пожелтением,
наблюдается даже некоторое снижение осмотического давления вследствие
потери осмотически активных веществ в процессе дыхания.
Зависимость осмотического давления от высоты взятия образца изу-
чалась на экземпляре кактуса высотой 1,5 м (табл. 50).
б) Ferocactus (Echinocactus) wislizenii.
Этот кактус редко достигает высоты свыше 1 м (рис. 274) и приобре-
тает более шаровидную форму, чем предыдущий вид; в экологическом
Пустыня Сонора
357
Таблица 50
Зависимость осмотического давления у Carnegiea от высоты взятия образца
Расстояние от вершины, см Диаметр стебля, С31 Осмотическое давление в паренхиме, атм
коры (южная сторона) сердцевины коры (северная сторона)
9 23 6,3 5,6 6,1
30 29 6,4 6,2 5,8
60 30 — 6,0 —
90 25 — 6,3 —
120 23 6,1 — 5,7
Конус нарастания = 10,0 атм.
отношении он напоминает Camegiea. Интенсивность транспирации у этого
вида, пожалуй, еще ниже, чем у Carnegiea. Она колеблется в зависимости
от степени насыщения водой в пределах 0,7—5,0 мг на 1 г сырого веса
в сутки.
Мак-Дугал в течение нескольких лет изучал опытный экземпляр как-
туса, имевшего вес 49,39 кг и площадь поверхности 15 600 см2\ экземпляр
содержал 45 кг воды. В течение опыта вес кактуса изменялся следующим
образом (табл. 51).
Таблица 51
Сырой вес, г
Исходные данные 5 ноября 1908 г. 17 010
Без воды до мая 1909 г 11370
Укорененный в почве до октября 1909 г. . 20370
Без воды в лаборатории до мая 1910 г. . 13 335
По данным Мак-Дугала, осмотическое давление у Ferocactus достига-
ло 3—5 атм\ после сильной потери воды — 7 атм, а для экземпляров,
пожелтевших после потери 36,2% воды,— всего 2,2 атм.
Из приведенных данных видно, что процесс высыхания у кактусов
очень сложен (Мак-Дугал, Лонг, Браун); выделяются три фазы.
1. Преобладание потери воды, возрастание концентрации сахаров,
увеличение содержания кислот.
11. Сокращение количества сахаров; содержание воды (на единицу
сырого веса), несмотря на потери, остается постоянным; снижение содер-
жания кислот.
Ill. Начало быстрого гидролиза и пожелтения.
Среднесуточные потери веса у опытного экземпляра Ferocactus (весом
37,6 кг), в течение шести лет находившегося в затененном помещении,
достигали следующих значений:
Год наблюдений 1-й 2-й 3-й 4-й 5-й 6-й В среднем
Потеря веса, г 10,5 5,4 4,0 4,4 3,2 2,9 5,0
.358
Глава 8
Р и с, 274, Небольшой экземпляр Ferocactus wislizenii (фото Э. Вальтер).
Рис, 275, Ferocactus wislizenii, экземпляр, изображенный на рис, 274, Харак-
терна эксцентричность центрального цилиндра и различная форма ребер (см, также
рис, 276).
По истечении шести лет запасы воды не изменились и по-прежнему
равнялись 94%, тогда как содержание гидролизуемых углеводов в запа-
сающих тканях снизилось с 26 до 11%, а в наружних тканях коры —
с 32,5 до 23,3%. Концентрация клеточного сока оказалась более низкой,
чем перед началом опыта. К концу опыта растение потеряло 30% своего
Пустыня Сонора
359
первоначального веса при неизменном содержании воды; следовательно,
30% сухого вещества было израсходовано в процессе дыхания.
Автор тщательно изучил различия в концентрации клеточного сока
на разных частях опытного среза растения, изобразив полученные данные
в виде изоосмотических линий (рис. 275 и 276).
В
Рис. 276. Величина осмотического давления (в атм) на поперечном
срезеFerocactus wislizenii (из Н. W al t е г, Die Hydraturder Pflanze, 1931).
Асимметрия в распределении на срезе проводящих пучков и в ширине
ребер полностью совпадает с неравномерным распределением гидратуры,
а также с различной интенсивностью роста на юго-западной и северо-
восточной сторонах стебля. Более медленный рост юго-западной стороны
обусловливает эксцентричность стеблей кактусов. Потеря равновесия
у очень старых экземпляров кактусов приводит к их падению
и гибели.
Конус нарастания отличается у Ferocactus наибольшим осмотическим
давлением, которое достигает здесь 6,9 атм, тогда как в паренхиме сердце-
вины оно не превышает 4,4 атм, а в паренхиме коры — 5,3—5,6 атм.
Величины осмотического давления у вырванного с корнем и в течение
9 месяцев пролежавшего на боку экземпляра кактуса приведены на
рис. 277. Расположение цветочных почек зависит от распределения гид-
ратуры на поперечном срезе. Это особенно четко выражено у Carnegiea
и Pachycereus. На северо-восточной стороне (где осмотическое давление
наименьшее) цветочные почки обычно не закладываются. Цветки раскры-
ваются сначала на юго-западной стороне, где осмотическое давление
360
Глава 8
41см
Рис. 277. Величины осмотического дав-
ления (в атм) на продольном срезе Feroca-
clus wislizenii через 9 месяцев после его
выкопки.
достигает наибольших значений,
в) Опунции.
Эта группа растений, и сре-
ди них плоские опунции, обла-
дает наибольшим ареалом. Холо-
достойкие виды, развитые в низ-
котравных прериях, достигают на
севере границы Канады. В пу-
стыне Сонора они представлены
многими мало отличающимися
друг от друга видами, часто с
крупными кладодиями.
Максимальная интенсивность
транспирации у опунции в по-
левых условиях достигает 5 мг
на 1 а сырого веса в сутки; однако
эти растения способны в неблаго-
приятное время настолько сильно
ограничивать транспирацию, что, по-видимому, могут в течение 2—3 лет
обходиться без дополнительного потребления воды. При этом более
молодые кладодии перехватывают воду из старых кладодий, которые
медленно желтеют и отмирают. Первые способны даже вновь укореняться,
На рис. 278 представлено падение веса во время высыхания опунций на
протяжении трехмесячного опыта в полевых условиях и в лаборатории.
На интенсивность транспирации влияет также экспозиция кладодий
(Споэр) (табл. 52).
Пустыня Сонора
361
Таблица 52
Потери влаги в процессе транспирации у Opuntia Ыакеапа
при различной экспозиции кладодий (в %)
Экспозиция В тени
Ю-С B-3
Уменьшение веса за период 28/11 — 5/IV
(34 дня)..........................18,5 16,3
Уменьшение веса за период 15/V — 28/VI
(44 дня) 24,7 26,7
5,8
23,3
Кладодии, освещенные солнцем, нагреваются очень сильно; в частно-
сти, сильный нагрев отмечен при ориентации на восток — запад в 14 час.,
а при ориентации на север — запад — в 11 час.; в 12.30 наблюдался мини-
мальный нагрев, а максимум наступал около 16 час. Максимальные
температуры в естественных условиях достигали 53° в июне и 55°
в июле. ।
Наряду с водным режимом Споэр изучал также и углеводный обмен 1 2.
Кактусы отличаются большим содержанием полисахаридов; последние
при гидролизе (с 1%-ной НС1) дают 34,1 % d-глюкозы и 65,9% 1-ксилозы.
В течение первых четырех часов процесс испарения идет у них так же,
как и у чистой воды. Поэтому трудно себе представить, чтобы они могли
способствовать ограничению транспирации. Слабая интенсивность транс-
пирации кактусов связана с малым числом устьиц и непроницаемостью
эпидермиса и наружных тканей коры, особенно при сильном обезвожи-
вании последних, вызванном их высыханием.
Молодые побеги относительно богаты полисахаридами. Содержа-
ние пентозанов при потере влаги в процессе испарения повышается, а
моносахаридов снижается; это определяет сезонные колебания их содер-
жания.
При высыхании в результате окисления углеводов в процессе дыха-
ния образуется вода, поэтому вес падает быстрее, чем запасы воды в рас-
тении.
Опытный экземпляр Opuntia phaeacantha потерял за 189 дней 60%
первоначального веса, тогда как содержание воды снизилось всего лишь
с 84,75 до 72,68%. Образование воды при дыхании, наблюдаемое также
у животных организмов, распространенных в аридных биотопах, помогает
многим из них долгое время обходиться без воды.
Ливингстон в 1907 г. установил, что у кактусов относительная транс-
пирация достигает максимальных значений ночью и минимальных днем 2;
что же касается абсолютной транспирации, то ее величина ночью также
очень значительна и не уступает дневным значениям.
1 Н. A. S р о е h г, The carbohydrate economy of cacti, Carnegie Inst, of Washing-
ton, Publ. 287, 1919; Photochemische Vorgange bei der diurnalen Entsauerung der Sukku-
lenten, «Bioch. Zschr.», 57, 1913, S. 95.
2 В. E. Livingston, Relative transpiration in cacti, «Plant World», 10»
1907, p. 110.
362
Глава 8
Шрив подтвердила это иным путем1. Она доказала, что побеги осо-
бенно у цилиндрических опунций периодически меняют свое положение.
У Opuntia versicolor побеги медленно поникают в период засухи и быстро
выпрямляются после дождя. Наряду с этим у них выражены суточные
колебания, связанные с изменениями тургора: днем — выпрямление,
а ночью — поникание. Наблюдения за водным балансом этого вида пока-
зали, что ночью преобладает отдача воды и тургесцентность уменьшается.
Днем происходит обратный процесс. Измерения длины и ширины растений
с помощью ауксографа свидетельствуют об уменьшении размеров как-
тусов ночью и увеличении их днем. Это означает, что устьица у кактусов
открываются ночью; в это же время в основном осуществляется газообмен,
и в частности поглощение СО2. Следовательно, углекислота сначала свя-
зывается в растении, а затем днем под действием солнечного света в про-
цессе фотосинтеза восстанавливается и превращается в углеводы.
По Споэру (1913), у Opuntia versicolor содержание кислот в клеточном
соке достигает наибольших значений при восходе солнца и наименьших
к 17 час. Для нейтрализации 1 мл сока утром требовалось 2,45 мл 0,1 нор-
мального раствора КОН, а в послеполуденные часы — всего лишь 0,31 мл.
Углекислота, поглощенная ночью, накапливается, видимо, в форме орга-
нических кислот 2. Это явление, свойственное кактусам, напоминает пове-
дение грызунов, распространенных в пустынях; эти грызуны во избежание
потерь воды выходят из своих нор на поиски пищи только ночью и пере-
варивают ее днем во время сна в норах.
Таким образом, кактусы служат прекрасной иллюстрацией неодно-
кратно высказываемого положения, что при проведении экологических
исследований необходимо рассматривать все процессы в совокупности
и во взаимной связи. Водный режим оказывается, например, теснейшим
•образом связанным со всем обменом веществ.
Значения гидратуры у опунций зависят от экспозиции их кладодий.
Осмотическое давление,
атм
в 7 час. в 14 час.
Opuntia engelmanii Экспозиция север —юг . . 11,0 11,7
Экспозиция восток —запад 10,6 10,3
Opuntia phaeacantha (экспозиция восток-запад, продольный разрез) Солнечная сторона . . . 15,5
Теневая сторона ... . .... 14,1
Opuntia versicolor (поперечный разрез цилинд- рического побега) Верхушка 15,7
Низ .... 15,2
1 Е. В. Shreve, An investigation of the causes of autonomic movements in
succulent plants, «Plant World», 18, 1915, p. 297, 331; An analysis of the causes of varia-
tions in the transpiring power of cacti, «Physiol. Res.», 2, 1916, p. 73.
2 Этот «эффект Соссюра» подробно описывается в работе Нюрнберга; там же упо-
минаются виды суккулентов, у которых проявляется этот эффект. См. Е. L. Nuern-
berg к, Endogener Rhythmus und CCVStoffwechsel bei Pflanzen mit diurnalem Siiurer-
hythmus, «Planta», 56, 1961, S. 28—70.
Пустыня Сонора
363
Автору не удалось установить
с достаточной достоверностью нали-
чие суточных колебаний осмотическо-
го давления. У взаимосвязанных
побегов осмотическое давление раз-
личалось лишь незначительно. При
высыхании опунции давление вначале
повышается, затем оно слегка снижа-
ется и растения желтеют. Наблюде-
ния показали, что давление повы-
шалось до 8 января, а затем, с на-
чалом периода дождей, вновь начало
падать (табл. 53).
Наиболее низкое осмотическое
давление (6,7 атм) было отмечено
у молодых побегов, наивысшее
(19,4 атм)—у цилиндрической Орип-
.tia arbuscula, отличающейся слабо
выраженной суккулентностью.
г) Другие виды кактусов.
Другие виды кактусов с эко-
логической точки зрения подробно
не изучены. Развитие Pachycereus
pringlei (рис. 279), самого крупного
кактуса, распространенного в север-
ной Мексике, а также других видов
колоннообразных кактусов ничем не
-отличается от развития Carnegiea.
Рис. 279. Pachycereus pringlei близ
Пуэрто-Либертада, штат Нью-Мексико
(фото Э. Вальтер).
Таблица 53
ОсмотическоеУдавление у опунций в засушливый
период начала осени и в сезон зимних дождей
(в атм)
Октябрь , Начало j января Март
Opuntia toumeyi 8,8 15,4-15,8 8,4
Opuntia Ыакеапа 8,8 15,1 10,0
Opuntia versicolor 7,4 16,6-17,8 11,9
В марте после обильных осадков в северной Мексике и на побережье
Калифорнийского залива при изучении осмотического давления были
получены следующие значения (в атм):
Pachycereus pringlei
Carnegiea gigantea
Lemaireocereus thurberi
Lophocereus schottii
I. Паренхима ребер —4,2
хлорофиллоносная ткань —
8,8
11. Паренхима ребер — 5,4—6,2
Паренхима ребер—4,1
Паренхима коры —4,4
Паренхима коры —4,7—4,9
364
Глава 8
Приводим также некоторые величины, полученные при изучении
небольших колоннообразных, а также шаровидных кактусов:
Echinocereus jendleri
Echinocereus polyacanthus
Echinocereus rigidissimus .
Mammillaria grahami
24 января .
. 8,1 (24.1)
. 3,7 (7.Ill)
5,4 (7.Ill)
5,8 (наружные слои)
. 4,8 (внутренние слои)
Некоторые характерные виды кустарников
Кактусы — наиболее хорошо изученная группа растений пустыни
Сонора; однако и среди кустарников также можно назвать несколько
характерных видов, и в частности Fouquieria splendens, представителя
небольшого семейства Fouquieriaceae, Cercidium (Parkinsonia) microphyIla
из семейства Caesalpiniaceae и Larrea tridentata (Covillea glutinosa) из семей-
ства Zygophyllaceae.
a) Fouquieria splendens,
В южной части пустыни Сонора Fouquieria splendens замещается дру-
гими видами. Это эндемичный род, развитый только в пустыне Сонора
(рис. 280); в горах виды этого рода выходят за пределы пустыни, подни-
маясь до высоты 1800 м (2100 м), Fouquieria — очень своеобразное с эко-
логической точки зрения растение. Побеги у Fouquieria тонкие, достигают
длины 3—4 м (10 3i); их число доходит до 25; они отходят от корневой шей-
ки под углом более 45° (рис. 281). Удлиненные побеги развиваются только
в период летних дождей. Они несут нежные листья, пластинки которых
скоро опадают, а листовые черешки превращаются в колючки. В их пазу-
хах развиваются укороченные побеги с пучками нежных листьев длиной
1,5—4,0 см и шириной 0,5—1,5 см; они появляются в любое время года
после интенсивных дождей, но вскоре желтеют, как только пересохнет
почва. Поэтому новые листья развиваются иногда по 5—6 раз в год. Устьи-
ца равномерно распределены на обеих сторонах листьев; их число дости-
гает 160 на 1 см1 2, Деятельность устьиц изучал Лотфилд Ч В конце периода
зимних дождей распускаются соцветия, напоминающие пламенеющие
ярко-красные кисти.
Поскольку Fouquieria облиствлена только в периоды, когда почва
увлажнена, а ветви защищены пробковым слоем от потерь воды, этот вид
по характеру водного режима может быть скорее отнесен к мезофитам.
Этот вопрос подробно изучался Кэнноном и Ллойдом 2.
О скорости развития листьев можно судить по следующим данным:
после дождя, выпавшего 11 июля, размеры почек заметно увеличились;
13 июля они достигли длины 7—8 мм; 14 июля уже распустились листья
длиной 1,5 см, В период засухи через 5 дней после полива кусты начинали
зеленеть. Ллойд обвязал одну из веток тряпкой, которую периодически
увлажняли. Через 8 дней на этой ветке распустились листья длиной 2 см.
Вода при этом, по-видимому, поглощалась почками и листовыми рубцами.
1 J. V. Lotfield, The behavior of stomata, Carnegie Inst, of Washington,
Publ. 314, 1921.
2 W. A. Cannon, On the transpiration of Fouquieria splendens, «Bull. Torrey
Bot. Club», 32, 1905, p. 397. F. E. Lloyd, The artificial induction of leaf formation
in Ocotillo, «Plant World», 9, 1906, p. 56; The relation of transpiration and stomatai
movements to the water-content of the leaves in Fouquieria splendens, «Plant World»,
15, 1912, p. 1.
Р и с. 280. А — ареал эндемичного рода F ouquieria\ Б — ареал Larrea iridentata в пустыне Сонора (по
Ф. Шриву, с некоторыми изменениями).
1 — Fouquieria splendens', 2 — F. peninsularis; 3 — F. macdougalii.
366
Глава 8
Рис. 281. Fouquieria splendens (в центре), Larrea
tridentata (слева), Opuntia arborescens (между ни-
ми) и Carnegiea gigantea (на заднем плане слева).
Склон конуса выноса близ Таксона (фото Апхофа).
Транспирация у Fou-
quieria отличается большой
интенсивностью: максималь-
ные значения достигают
8,25 мг!мин-дм2. Ночью ра-
стения поглощают больше
воды, чем отдают, а днем на-
оборот. Содержание воды
в листьях достигает макси-
мума при восходе солнца;
однако эта величина колеб-
лется всегда в пределах 70—
74% сырого веса. Суточные
колебания осмотического дав-
ления не установлены. Дав-
ление у распускающихся мо-
лодых листьев равно 10,4лапл4,
а затем быстро повышается
до 14,4 атм. У листьев, за-
кончивших свой рост, осмо-
тическое давление клеточ-
ного сока достигает 17,2—
19,6 атм. При недостатке
воды листья не высыхают,
а лишь желтеют, причем ос-
мотическое давление сначала
повышается до 21,4 атм,
а затем постепенно снижает-
ся до 16 атм и, наконец,—
до 11,7 атм. При пожелте-
нии наблюдается отток основ-
ных питательных веществ иа
листьев в стебли, что, вероят-
но, имеет определенное значе-
ние при частом отмирании листьев. Пожелтение, видимо, начинается сразу
после нарушения фотосинтезирующего механизма вследствие потерь воды
и известного уменьшения гидратуры, возможно вызывающего закрытие-
устьиц. Этот процесс очень сходно протекает у Jatropha cordifolia, кустар-
ника из семейства Euphorbiaceae, у которого листья напоминают листья
березы. Осмотическое давление у этого вида при пожелтении листьев сни-
жалось с 17,6 до 11,8 атм. Представитель семейства Fouquieriaceae —
Idria columnaris — встречается на Калифорнийском полуострове; кроме-
того, этот вид был еще обнаружен к востоку от залива близ Либертада.
У этой древесной породы, достигающей высоты 6—8 м, стволы имеют
бутылкообразную форму (рис. 282); нижняя, утолщенная часть ствола
служит резервуаром, где накапливается вода. Диаметр ствола равен
50—60 см (рр 75 см). Губчатое строение тканей способствует быстрому
изменению объема при поглощении или отдаче воды растением. От ствола
почти горизонтально отходят тонкие ветви; на них развиваются листья,
отличающиеся известной ксероморфностью структуры; осмотическое дав-
ление у них достигает 17,8—19,1 атм, в то время как осмотическое давле-
ние в утолщенных стволах равно 12,6 атм- Вода накапливается также
в стволах встречающихся вместе с Idria видов Bursera (Elaphrium) из.
Пустыня Сонора
367
семейства Burseraceae. Осмотическое давление в листьях последних
достигало 18,5 атм, а в тканях ствола — 11 атм.
б) Cercidium microphyllum.
У этого вида зеленый ствол и зеленые ветви (рис. 262) выполняют
роль фотосинтезирующих органов. Перистые листья состоят из 3—5 пар
жестких круглых листочков диаметром менее 1 мм. У молодых растений
площадь поверхности листочков в 4 раза больше. Зимой они часто окра-
шены в красный цвет и отличаются меньшей засухоустойчивостью
Рис. 282. Молодой цветущий экземпляр Idria columnaris (высотой 2 ,u);
после прошедших зимних дождей большая часть листьев опала. Сле-
ва — облиствленный экземпляр Рachycormus discolor (фото Гейма).
Листья появляются только после обильных летних или зимних дож-
дей и редко остаются на ветвях дольше чем 6—10 недель. Затем они жел-
теют и опадают, тогда как черешки еще несколько недель держатся на
368
Глава 8
ветвях. Даже у полностью облиствленных растений общая листовая
поверхность всегда меньше поверхности стеблей. Это кустарникоподобное
дерево достигает высоты 5—6 м(8—10 .и). Цветение начинается в апре-
ле — мае после зимних дождей. Семена прорастают только на следующий
год в период летних дождей.
Эти растения приурочены к скалам и крупным осыпям; их корни
используют скопляющуюся в трещинах скал влагу. В засушливые годы
большая часть побегов отмирает, и тогда Cercidium возобновляется порос-
лью от пня. Анатомические особенности Cercidium, его транспирация,
водный режим и влияние окружающей температуры изучала Шрив 1.
Величина осмотического давления у ксероморфных листочков отно-
сительно высока и незначительно колеблется с ноября по февраль в пре-
делах 26,6—31,5 атм. В сухих местообитаниях давление может достигать
даже 34,7—36,8 атм, что близко к значениям, отмеченным у ксерофильных
видов. Аналогичные величины отмечены также у близких видов — Cercidium
(Parkinsonia) torreyanum (23,6—27,1 атм) и Parkinsonia aculeata (у моло-
дых деревьев — 15,5 атм, у взрослых—26,5 атм) (рис. 283).
в) Larrea (Covillea) tridentata.
У этого вида листья покрыты лаком и сильно пахнут креозотом после
каждого дождя. Larrea распространен и за пределами пустыни Сонора
(рис. 280, Б), встречается он также в пустыне Мохаве и в отдельных
частях пустыни Большого Бассейна. Близкие виды широко распростра-
нены в засушливых областях Аргентины 1 2.
Рис. 283. Древовидная Parkinsonia aculeata и Olneya tesota (сзади); впе-
реди— кусты Encelia farinosa, Питикито, Сев. Мексика (фото Э. Вальтер).
1 Е. В.Shreve, The daily march of transpiration in a desert perennial, Carnegie
Inst, of Washington, Publ. 194, 1914.
2 J. Morrell o, Estudios botanicos en las regiones aridas de la Argentina, I
u. II, «Revista Arg. NW-Argentine», 1, 1955, p. 301—370, 385—524 (Tucuman); III,
2, 1956, p. 79-152.
I
1. Солончак севернее Калгурли. На переднем плане — цветущий Carpobrotus (Mesem-
bryanthemum); вдали— более засоленная полоса с Arthrocnemum и Salicornia; на зад-
нем плане — соленое озеро.
2. Triodia basedowii и цветущие экземпляры Накеа multilineata на песках во внутри
континентальной части Западной Австралии близ Уилуны.
Пустыня Сонора
369
В и с. 284. Годовой ход осмотического давления у Larrea tridentata. Окрестности Лабо-
ратории по изучению пустынь в Таксоне. В благоприятных условиях развита ассо-
циация Carnegiea — Encelia.
Условия среды: 1 — благоприятные; 2 — сочень сухие; 3 — поливные.
Larrea отличается незначительной требовательностью и исключи-
тельной приспособляемостью к окружающей среде. Larrea может больше
года обходиться без воды, но не менее хорошо себя чувствует и в местности
с годовым количеством осадков 500 мм, лишь соответственно меняется
габитус растения х.
Этот кустарник начинает ветвиться уже от корневой шейки; в даль-
нейшем ветви продолжают многократно ветвиться во всех направлениях,
причем нижние ветви распластываются почти горизонтально чуть выше
поверхности почвы (рис. 260 и 264). В благоприятных условиях Larrea
достигает высоты нескольких метров и образует до ста ветвей, покрытых
многочисленными листьями длиной до 28 мм. В неблагоприятных услови-
ях куст имеет небольшие размеры, а листья не превышают в длину 4 мм',
последние при увеличении сухости частично опадают. Однако даже почти
безлистные ветви долгое время остаются живыми и после дождя на них
вновь распускаются листья. Благодаря потере части ветвей и сокращению
площади листовой поверхности, а также сильному ограничению расхода
воды Larrea способен существовать даже в очень засушливых условиях,
где другие виды погибают. При улучшении условий увлажнения в расте-
ниях начинает накапливаться вещество и в результате этого увеличивает-
ся прирост. После длительного периода засухи при первых же дождях
на кустах появляются многочисленные мелкие желтоватые цветки.
Даже при благоприятных условиях увлажнения осмотическое давле-
ние у Larrea не бывает ниже 28—30 атм. Только у молодых листьев на
растущих побегах концентрация клеточного сока достигает 22—24 атм.
При недостатке влаги и нарушении водного баланса осмотическое давление
1 V. М. Spalding, The Creosote Bush (Covillea tridentata) in its relation to
water supply, «Bot. Gaz.», 38, 1904, p. 122; E. H. Runyon, The organisation of the
creosote bush with respect to drought, «Ecology», 15, 1934, p. 128—138.
370
Глава 8
возрастает до 55 атм и выше; при этом листья не опадают, а лишь слегка
буреют и свертываются. Это повышение давления, конечно, нельзя рас-
сматривать как приспособление; оно является всего лишь следствием дефи-
цита влаги. Засухоустойчивость именно и проявляется в том, что растения
выдерживают это снижение гидратуры. Годовой ход осмотического давле-
ния и его зависимость от условий местообитания представлены на рис. 284.
г) Другие экологические типы кустарников.
Приведенными тремя примерами далеко не исчерпывается разнообра-
зие экологических особенностей склерофильных растений. Собственно,
можно сказать, что почти каждый вид кустарника представляет самостоя-
тельный экологический тип.
Simmondsia chinensis (calif ornica) — низкий кустарник с тонкими
ветвями, но с очень толстыми суккулентными листьями, напоминающими
семядоли, но только жесткими. В противоположность суккулентам осмо-
тическое давление у него очень высокое — от 34 до 40 атм. После трех-
месячной засухи давление поднялось до 49,1 атм. У молодых листьев
концентрация сока меньше, однако она также достаточно велика и дости-
гает 20,8 атм. Свежесрезанные ветви теряли в первый же час 8,5—11,0%
сырого веса, во второй час — еще 3,2—3,6%, а в последующие часы —
всего 0,6—0,9%. Гидростабильность этого вида связана с его способностью
сильно ограничивать интенсивность транспирации.
Несмотря на это, описываемый вид не встречается в окрестностях
Таксона в крайне засушливых условиях, тогда как у северной границы
пустыни Сонора, около плотины Рузвельта, этот кустарник сплошь покры-
вает склоны даже южной экспозиции.
Mortonia scabrella — небольшой кустарник, распространенный к юго-
востоку от Таксона и приуроченный только к выходам известняков. Изда-
ли растение имеет серый оттенок; оно покрыто тесно расположенными,
почти чешуевидными мелкими жесткими листьями, сохраняющимися
в течение 6—7 лет. Клетки эпидермиса отличаются крупными размерами
и очень толстыми наружными стенками. Устьица мало углублены, пали-
садная ткань характеризуется многослойностью.
В лаборатории ветки Mortonia уже через 5 дней высыхают до воз-
душно-сухого состояния и содержат около 20% воды. Осмотическое
давление после дождя достигало всего лишь 23,2—25,8 атм. Анатоми-
ческое строение вряд ли создает возможности для больших колебаний
водного режима растений.
Mortonia, вероятно, обитает только на известняках, содержащих
достаточное количество доступной для растений влаги; правда, соот-
ветствующих исследований в этом отношении не было проведено.
Acacia — род, особенно широко распространенный в тропических
областях, где осадки выпадают летом; некоторые виды этого рода встре-
чаются и в пустыне Сонора. В зимние месяцы растения полностью или
лишь частично сбрасывают свои тонкие перистые листья. Акация растет
только при наличии в почве влаги, которую она успешно извлекает при
помощи своей широко разветвленной корневой системы.
В нормальных условиях осмотическое давление у листьев равно
примерно 26 атм, однако при недостатке воды оно может повышаться
до 40 атм и больше. Без постоянного притока воды листья вряд ли долго
могут существовать.
Мескит — Prosopis juliflora ssp. velutina — вид, характерный для
следующего высотного пояса; в пустыне Сонора он встречается в пой-
Пустыня Сонора
371
менных лесах, занимающих обширные затопляемые террасы (рис. 265).
У этой древесной породы чрезвычайно сильно развита корневая система,
при помощи которой она даже в периоды засухи достаточно хорошо
обеспечивает себя водой. Мескит, растущий по берегам русел временных
водотоков или, реже, в расщелинах скал, также, вероятно, в этих усло-
виях всегда приурочен к наиболее обеспеченным влагой местообита-
ниям. Поэтому не удивительно, что у этих экземпляров мескита в пери-
оды засухи осмотическое давление существенно не повышается. В сред-
нем давление равно 25—28 атм. После месячной засухи оно возрастает
до 30—32 атм, а еще через месяц —до 34—35 атм.
После первых заморозков, которые особенно ощутимы в долинах,
листья опадают. На склонах они сохраняются в течение всей зимы.
В засушливых условиях концентрация клеточного сока повышается
до 41,1 атм. Листья, развивающиеся в условиях плохой влагообеспе-
ченности, отличаются мелкими размерами и более ксероморфной струк-
турой; это явление будет подробнее рассмотрено при характеристике
Encelia farinosa. В северной Мексике у молодых листьев, распускаю-
щихся на молодых ветвях, давление в конце марта составило 19,9 атм.
Листопадные деревья и кустарники (представители голарктической
флоры) также проникают в пустыни, однако здесь они приурочены
к очень влажным местообитаниям, например берегам пересыхающих
рек, в которых в течение всего года не иссякает подрусловой поток, или
к местам выходов источников у подножия горных склонов. К этой группе
принадлежат виды Salix и Populus, а также растения, упомянутые на
стр. 343.
Эти виды как по габитусу, так и по гидратуре не отличаются от
видов, распространенных в умеренной зоне. Осмотическое давление
у них равно примерно 19—20 атм.
У молодых листьев рода Vitis концентрация клеточного сока дости-
гает 9,6 атм. У европейской виноградной лозы Vitis vinifera осмоти-
ческое давление равно 10,5 атм.
Полукустарники с мягкими опушенными листьями
(малакофилъные растения)
Encelia farinosa — полукустарник из семейства сложноцветных, высо-
той 60—80 см (рис. 285), с ломкими ветвями, диаметром 5—10 мм, и череш-
чатыми листьями 3—7 см длиной. Ветви и листья густо опушены, так
что растение издали кажется белым. Желтые соцветия на длинных цвето-
носах возвышаются на 10—15 см над облиствленными побегами; они
цветут дважды в год — во время обоих дождливых периодов. В Аризоне
этот вид растет главным образом на склонах гор.
Транспирация у Encelia farinosa на хорошо увлажненных почвах
относительно интенсивна *. Особенно интересна зависимость между
гидратурой и морфологическим строением листьев. С интенсивной транс-
пирацией связаны значительные суточные колебания осмотического
давления ( в среднем 4,5 атм).
У растений, произрастающих на более влажных северных склонах,
осмотическое давление достигает 22—23 атм; крупные и слабо опушен-
ные гигроморфные листья имеют здесь зеленоватый оттенок. В более
1 Е. В. Shreve, Seasonal changes in the water relations of desert plants, «Eco-
logy», 4, 1923, p. 226; Factors governing seasonal changes in traspiration of Encelia
farinosa, «Bot. Gaz.», 77, 1924, p. 432.
472
Глава 8
Рис. 285. Полукустарник Encelia farinosa; гора Тумамок близ
Таксона, Аризона (фото Э. Вальтер).
Рис. 286. Гигроморфные, мезоморфные и ксероморфные листья
у Encelia farinosa, в середине — почки (фото Э. Вальтер).
засушливых условиях осмотическое давление равно 28 атм\ здесь раз-
виваются мезоморфные, то есть более густо опушенные, беловатые
и несколько меньшего размера листья. Такие же листья развиваются
и на северном склоне, но лишь после того как концентрация клеточного
сока в течение засушливого периода повысится до 28 атм. Если засуха
затягивается, осмотическое давление у растений повышается, размеры
Пустыня Сонора
373
листьев уменьшаются и их структура становится более ксероморфной;
гигроморфные листья отмирают и опадают при осмотическом давлении,
равном 36—38 атм. Ксероморфные листья растение теряет лишь при
величинах давления, достигающих 40 атм и более. В период длительной
засухи к концу декабря давление достигает 55 атм. Растение к этому
времени полностью лишается листьев, и лишь вокруг почек сохраняют-
ся мелкие очень густо опушенные листочки (рис. 286). После сильных
дождей в марте листья начинают энергично распускаться и концентра-
ция клеточного сока в молодых гигроморфных листьях вновь восстанав-
ливается до 22—23 атм. Таким образом, Encelia присущи качества
типичного малакофильного ксерофита, то есть у него очень велики коле-
бания осмотического давления, связанные с относительно поздним сокра-
щением интенсивности транспирации и зависящим от этого сильным
дефицитом влаги; кроме того, с этим связано сокращение транспириру-
ющей поверхности сначала путем образования все более мелких ксеро-
морфных листьев и затем за счет постепенной потери листьев.
Развитие листьев и колебания осмотического давления осущест-
вляются аналогично и у двух видов Franseria. Franseria dumosa распрост-
ранена шире и, кроме того, вероятно, более засухоустойчива, чем Encelia.
В противоположность им Lippia ivrightii из семейства губоцветных
отличается меньшей засухоустойчивостью, она приурочена к защищен-
ным местообитаниям на северных скалистых склонах. Ее листья слабо
опушены, а осмотическое давление меньше, чем у предыдущих видов
(15—31 атм).
ЗАКЛЮЧЕНИЕ
Представители различных экологических групп растений, разви-
тые в пустыне Сонора, не могут быть объединены в подразделения, пред-
ложенные Шимпером и Максимовым, при классификации ксерофитов.
Незначительная интенсивность транспирации в период засухи свойствен-
на главным образом суккулентам, а также Cercidium и видам, которые
способны переносить еще больший дефицит влаги, как, например, Lar-
rea и Encelia.
Особенно важным свойством растений является их способность
поддерживать определенный уровень гидратуры; стеногидрические виды
не переносят слишком сильного повышения осмотического давления,
тогда как эвригидрические, напротив, менее чувствительны к его коле-
баниям. Между этими крайними вариантами наблюдаются постепенные
переходы.
Не следует, однако, забывать, что критическая фаза развития расте-
ний, ограничивающая их естественное распространение в аридных обла-
стях,— это первые годы жизни. Всходы очень чувствительны к неблаго-
приятным условиям среды и поэтому продолжают развиваться лишь
в случае, если благоприятные годы с короткими периодами засухи непре-
рывно следуют один за другим. Подобные условия наблюдаются лишь
в пустынных областях, не отличающихся ярко выраженной засушливо-
стью. Таким образом, способность вида переносить чрезвычайно суровые
условия во взрослом состоянии не может способствовать расширению аре-
ала этого вида, если он погибает в первые критические годы своей жизни.
С другой стороны, этот вид при известном уходе, например периодиче-
ском орошении, может существовать в культурах. Поэтому естествен-
ный ареал каждого вида всегда намного меньше площади его распро-
странения в культуре.
Глава 9
ПУСТЫНЯ ТУМАНОВ НАМИБ
РЕЖИМ ОСАДКОВ
В качестве второго типа пустынь выбрана крайне засушливая область
на западном побережье Африки, где дожди выпадают редко, но
отмечается большое число дней с туманами. Эта область — пустыня
Намиб — простирается узкой полосой (шириной около 100 км) вдоль
берега Атлантического океана, омываемого холодным Бенгуэльским
течением; она тянется от Анголы на севере (южнее Мосамедиша, при-
мерно от 15° ю. ш.) за Оранжевую реку и Малый Намаленд, вклиниваясь
в западную часть Капской провинции (примерно до 33° ю. ш.). В южной
части пустыни (до бухты Людериц) сказывается влияние зимних дождей,
тогда как в средней и северной частях — тропических летних дождей,
конечно, в случае если осадки вообще выпадают.
С экологической точки зрения лучше всего изучена средняя часть
пустыни Намиб близ Свакопмунда Ч
Количество осадков в 50% лет составляет менее 10 мм; в отдельные
годы их с трудом можно измерить. Очень редко количество осадков
достигает 30 мм. Однако в 1934 г. за два месяца (март — апрель) выпало
почти 150 мм осадков, причем в марте их количество составило ИЗ мм,
а в апреле — 27 мм. Влияние этих осадков сказалось и в следующем
году и привело к развитию пышного растительного покрова. Во время
следующих научных экспедиций (в 1938 и 1953 гг.) здесь наблюдался
обычный, почти лишенный растительности ландшафт. Единственными
постоянно несущими воду реками в этой области являются Оранжевая
на юге и Кунене — на севере. В основном же здесь преобладают сухие
русла периодических водотоков.
Пустыню Намиб можно разделить на несколько частей (рис. 287):
1) южная часть пустыни Намиб включает территорию южнее реки Оран-
жевой, достигая на севере бухты Людериц; 2) средняя часть — прости-
рается до мыса Кросс; 3) северная, наиболее слабо изученная часть,—
к северу от мыса Кросс. В средней части нас интересует лишь область,
расположенная севернее сухого русла Куизеб, поскольку южная часть
пустыни Намиб представляет собой обширную, лишенную раститель-
ности область подвижных дюн (рис. 287). Кайзер 1 2, проводивший геологи-
ческое обследование Алмазной пустыни южнее бухты Людериц, описал
также и механизм образования дюн. Песок, морского происхождения
и возникший при выветривании местных пород, перемещается преоблада-
1 Н. Walter, Die okologischen Verhaltnisse in der Namib-Nebelwiiste (Siid-
westafrika) unter Auswertung der Aufzeichnungen des Dr. G. Boss (Svakopmund), «Jb.
wiss. Bot.», 84, 1936, S. 58—222. Здесь же приведена библиография. Недавно близ
Гобабиса, примерно в 115 км к югу-востоку от Уолфиш-Бея, непосредственно у сухого
русла Куизеб, создан исследовательский институт.
2 Е. К aiser, Dip Diamantwiiste Siidwestafrikas, Bd II, Berlin. 1926.
Пустыня пгуманов Намиб
Ж
Рис. 287. Юго-Западная Африка. Пустыня Намиб, расположенная в прибрежной
полосе шириной около 100 км.
1 — возвышенные участки внутреннего плато (1500—2000 м над ур. моря); 2 — горные
поднятия (выше 2000 jw); з — долины пересыхающих рек.
ющими здесь южными ветрами к северу. Только при половодье по руслу
Куизеб песок выносится обратно в море. Однако, поскольку воды Куизе-
^ба лишь очень редко достигают моря (в прошлом столетии примерно
376
Глава 9
8 раз), дюны могут пересекать устье Куизеба у Уолфиш-Бея и задер-
живаются лишь перед правобережьем реки Свакоп Ч Севернее Свакопа
начинаются мелко- и крупнокам'енистые пустыни, лишь местами всхол-
мленные невысокими дюнами. Эта область и была, собственно, объектом
нашего изучения (см. цветное фото 1, стр. 304).
Пустыня Намиб довольно равномерно повышается в восточном
направлении от берега моря до подножия внутреннего плато; уклон
составляет 10 м на 1 км до высоты 1000 м (рис. 288). Далее плато в отдель-
ных местах возвышается крутыми уступами, превышая иногда высоту
2000 м. К востоку плато постепенно понижается, переходя в обшир-
ную песчаную пустыню Калахари, западную часть которой называют
Зандфельд.
Средняя часть пустыни Намиб располагается в области юго-восточ-
ных ветров. Влажные воздушные массы, идущие с Индийского океана,
пересекая континент, отдают влагу в Натале и Трансваале и достигают
Атлантического океана чрезвычайно горячими и сухими. Это происходит
в холодное время года и только в случаях, когда в глубине материка
наблюдается устойчивое высокое давление, а над морем в это время
проходят циклоны. Над сильно нагретой территорией пустыни часто
образуется местный барический минимум, который затягивает холодный
и влажный морской воздух с юго-запада, поскольку около Свакопмунда
он проходит над холодным морским течением с температурой пример-
но 15°. Ночью на материк в юго-западном потоке движется туман, посто-
янно стоящий над холодным морским течением; туман рассеивается
Область подьена Внутреннее плато Зандуельд
М
2000-
100 км
1000-
внешний внутренний
Намиб
Кальтешузен
Лжикалсуопар
Отьимоингве
Нью-Хьюсис Хозварте
УВиндхукЗеис
Оитвлей Годадис
Пас
0-
100-
Свакопмунд
200-
300-
400-
500-
Среднегодовое количество осадков
мм
Рис. 288. Схематический профиль через Юго-Западную Африку примерно но
22,5° ю. ш. и среднегодовое количество осадков, уменьшающееся с востока на запад;
в пустыне Намиб осадки почти не выпадают (по Н. Walter, 1936).
1 Лишенные растительного покрова дюны сравнительно богаты насекомыми,
например интересными псаммофильными и бескрылыми жуками (Tenebrionidae),
которые питаются принесенными ветром частицами гумуса, скапливающимися на под-
ветренных склонах дюн. Здесь песок может содержать до 36,3% органического веще-
ства. См. также С. К о с h, Some aspects ol abundant life in the vegetationless sand of
the Namib Desert dunes, «J. of S. W. Afr. Sci. Soc.» (Windhoek), 15, 1961, p. 76—92.
Пустыня туманов Намиб
377
только днем, когда почва пустыни прогревается. Туманы редко про-
никают в глубь материка более чем на 50 км. Постоянное взаимодействие
между восточными и западными ветрами, то есть между сухим воздухом
пустыни, нагретым до температуры около 40°, и морским влажным возду-
хом с небольшими температурными колебаниями, характерно для пусты-
ни Намиб. Несмотря на близость холодного морского течения, которое,
казалось, должно было бы смягчить климат пустыни, здесь сохраняются
крайне суровые условия для развития растительности (рис. 289). Чтобы
понять это, необходимо подробнее рассмотреть процесс образования
тумана. Поскольку влияние тумана и росы в данных условиях трудно
разделить, мы будем рассматривать их совместно, называя «оседанием
тумана».
По данным Гюлланда, в Свакопмунде в 1903 г. было всего 94 дня
с туманами, а в 1905 г. число их достигло 215 *. В 1927/28 г. отмечено
143 дня с туманами со следующим распределением по месяцам:
VII VIII IX X XI XII I II III IV V VI За год
10 15 22 10 12 8 7 10 13 14 10 12 143
На станциях, расположенных в 35 км от побережья в горах Рос-
синг, наблюдения проводились уже в течение 3 лет 1 2:
1913 г. 1914 г. 1915 г.
VII VIII IX X XI XII I II Ш IV V VI 1 VII VIII IX X XI XII I II
8 15 14 8 2 — — 28 23 7 10 12 9 24 30 30 25 7 25 12
Это означает, что с июля 1913 г. по июнь 1914 г. было в общей слож-
ности 127 дней с туманами, а в календарном 1914 г. —даже 204 дня.
В южной части пустыни Намиб число дней с туманами также не
меньше. По материалам Вайбеля, в 30 км севернее бухты Людериц
в Анихабе с июля 1913 г. по июнь 1914 г. наблюдалось 295 случаев выпа-
дения росы со следующим распределением по месяцам:
VII VIII IX X XI XII I II III IV V VI
30 27 22 27 17 27 27 1 | 22 28 24 27 17
За время своей многолетней преподавательской деятельности в Сва-
копмунде Г. Босс провел многочисленные экологические исследования
и любезно предоставил автору материалы своих наблюдений3. Из 180 дней,
1 Giilland, Das Klima von Swakopmund, «Mitt, deutsch. Schutzgeb.», 20,
1907, S. 131.
2 L. W a i b e 1, Winterregen in Deutsch-Sudwestafrika, Hamburg, Univ. Abb-
Auslandsk., 9. Reihe, C. «Naturw.», 4, 1922.
3 H. Walter, op. cit.
378
Глава 9
Р и с. 289. Климадиаграмма Свакопмунда.
в течение которых Босс измерял
испарение в 1934 и 1935 гг.,
134 дня были с туманами. По-
вторяемость туманов является не
менее важным показателем, чем
количество воды, выпадающее на
поверхности почвы при тумане
или росе. Мы опускаем описание
методики этих измерений и при-
водим здесь лишь общие выводы.
Без точных измерений можно легко переоценить количество осад-
ков, приносимых туманом. Оно достаточно велико, и ночью в Свакоп-
мунде вода непрерывно капает с крыш; домашние хозяйки собирают
эту влагу, поскольку водопроводная вода очень жестка. Если прихо-
дится ночевать в пустыне, то утром одежда бывает совершенно
мокрой.
Все проводившиеся измерения показали, что приносимые отдель-
ным туманом осадки составляют самое большее 0,7 мм, а в большинстве
случаев они меньше 0,1 мм. Это совпадает с результатами измерений
количества осадков, выпадающих в виде росы в самых различных клима-
тических областях.
При среднем числе дней с туманами около 200 общее количество
осадков, приносимых туманами за год, будет составлять примерно 40 —
50 мм. Если бы эти 40 мм выпадали в виде одного или немногих ливней,
то они имели бы для растительности огромное значение. Однако в дан-
ном случае их влияние, по крайней мере в равнинных условиях, ничтож-
но, поскольку на песчаных почвах роса в самых благоприятных случаях
проникает вглубь едва на 2,0—3,5 см, а на глинистых почвах —всего
лишь примерно на 1—2 см. Как только туман рассеется и солнце про-
греет почву, эта влага немедленно испарится. Подобные условия пре-
пятствуют развитию здесь цветковых растений. Иначе обстоит дело,
когда осадки в виде тумана выпадают на скале, стекают с нее, как с кры-
ши дома, и проникают в трещины скал, где вода защищена от испарения.
Здесь уже могут поселяться растения, поскольку 0,1 мм осадков при
стоке с площади 5 м2 дают не менее 0,5 л воды.
ЭКОЛОГИЧЕСКОЕ ЗНАЧЕНИЕ ТУМАНА
Неоднократно поднимался вопрос, встречаются ли растения, которые
способны своими листьями непосредственно поглощать влагу туманов
в количестве, достаточном для их существования. В таком случае листья
этих растений должны обладать способностью быстро поглощать воду
и наряду с этим препятствовать ее быстрой отдаче, что осуществляется,
например, с помощью чешуеобразных волосков у бромелиевых (рис. 101)
или веламена, покрывающего воздушные корни у эпифитных орхидей.
Однако ни у одного из видов, произрастающих в пустыне Намиб, не
было обнаружено каких-либо приспособлений подобного рода.
Само собой разумеется, что при смачивании листьев поглощается
известное количество воды, конечно, если у листьев не развита особенно
толстая кутикула или водонепроницаемый восковой налет. При длитель-
ном увлажнении росой увядшие растения вновь обретают тургесцент-
ность, однако это еще не означает, что они существуют за счет влаги,
приносимой туманом или росой.
Пустыня туманов Намиб
37S
Рис. 290. Местообитания растений на скалах
пустыни Намиб, увлажняемые осадками, при-
носимыми туманом.
К числу таких растений
можно отнести лишь те, ко-
торые, не извлекая воду при
помощи корней, довольству-
ются лишь увлажнением над-
земных органов туманом или
росой. Марлот 1 приводит
примеры таких приспособле-
ний, развитых у растений,
в области Карру. При этом
он различает способность
растений поглощать влагу
при помощи волосков, на-
бухающих клеток эпидерми-
са, прилистников, отлича-
ющихся большей гигроско-
пичностью, и воздушных кор-
ней, расположенных в пазу-
хах листьев. Однако признания возможности поглощения воды этими орга-
нами само по себе еще недостаточно; необходимо доказать, что растения,
поглощая таким образом влагу, способны существовать в естест-
венных условиях; таких доказательств пока еще не получено.
Воздушные корни Cotyledon cristata и набухающие клетки эпидермиса
у Crassula decipiens, вероятно, выполняют такую же функцию, как
и чешуеобразные волоски у бромелиевых.
Именно в пустыне Намиб, где при частых туманах лишь в исклю-
чительных случаях можно учесть количество выпавших осадков, хорошо
выражены условия, необходимые для формирования таких приспособле-
ний у растений, и тем не менее растения, существующие за счет влаги
туманов, здесь не встречаются. Равнинные прибрежные части пустыни
Намиб совершенно лишены растительности, за исключением местообита-
ний с близким уровнем грунтовых вод. Лишь по направлению в глубь
материка, где уменьшается число дней с туманами, но зато возрастает
количество дождевых осадков, начинают появляться отдельные ра-
стения.
Иначе обстоит дело на скалистых склонах, где влага, приносимая
туманом, конденсируясь, стекает в трещины (рис. 290). Здесь растения
поселяются даже при очень небольшом количестве осадков. Они раз-
виваются в холодный, изобилующий туманами период, а не летом, когда
дожди выпадают крайне редко. Среди этих растений особенно много
суккулентов. Однако они поглощают воду обычным путем — при помо-
щи корней. Они могут с таким же успехом расти и в других природных
областях, где, правда, нет туманов, но почва увлажняется редко выпа-
дающими дождями. Поэтому, хотя в этом случае и можно говорить
о растительности, развитию которой в этих условиях способствуют тума-
ны, будет совершенно неверным выделять их как специфические
растения, существование которых зависит от туманов. Растения
развиваются при определенном количестве воды в почве, и им совершенно
безразлично, поступает ли эта вода в виде дождей, тумана, росы или
же путем полива.
1 R. Marloth, Das Kapland. Wiss. Ergeb. d. Deutschen Tiefsee-Exped., Bd II,
'Teil 3, Jena, 1908.
380
Глава 9
Само собой разумеется, что при недостатке влаги продолжительный
туман или роса улучшают гидратуру растения или путем увлажнения
поверхности листьев, или уменьшения интенсивности облучения, или
резкого сокращения интенсивности транспирации; тем самым увеличи-
вается продолжительность жизни травянистых растений. Насколько
велико значение этих факторов в каждом отдельном случае, оценить
более точно крайне трудно.
Изучая этот вопрос, Босс в естественных условиях в окрестностях
гор Россинг провел ряд наблюдений на растениях, корни которых были
помещены в резиновые камеры, чем исключалась возможность поглоще-
ния воды корнями. Ночью температура воздуха снизилась до 6,5°
и наблюдалось сильное образование росы и тумана. Босс изучал два
вида семейства Acanthaceae, которые, как считалось, существуют за
счет влаги туманов, и один колючий суккулент из семейства Asclepia-
daceae. В течение суток вес растений (в г) изменялся следующим обра-
зом (табл. 54).
Таблица 54
Pe.tatidium variable Monee lima deserticola Hoodia currori
17 июля, 19 час. 157,3 30,2 198
18 » , 7 час. 166,0 35,5 200
9 час. 30 мин. 135,5 — —
19 час. 123,7 20,0 192,3
Из таблицы видно, что несмотря на то, что растения поглощали
значительное количество воды ночью, днем они быстро высыхали.
У Hoodia, видимо, наблюдалось лишь набухание колючек и эпидермиса.
Насколько быстро в пустыне Намиб в течение дня сменяются влаж-
ные и засушливые условия погоды, можно судить по данным Босса.
В сентябре у восточной границы пустыни Намиб близ Шпицкопье вече-
ром господствовал восточный ветер, в 19 час. относительная влажность
при температуре воздуха 17° была равна 14% (упругость водяного пара —
2 мм рт. ст.). При смене направления ветра на юго-западное упругость
пара повысилась к 21 час. до 4,4 мм рт. ст., к 22 час. — до 8,2 мм; при
температуре воздуха 9,5° относительная влажность воздуха поднялась
до 94%, и вскоре температура упала до точки росы. Ночью минимальная
температура опустилась до 2°; наблюдалась сильная роса. На следую-
щее утро снова подул восточный ветер, и уже около 7 час. упругость
пара вновь снизилась до 2,3 мм рт. ст., а относительная влажность
при температуре воздуха 8° составила всего 20%.
При столь резких суточных атмосферных колебаниях туман не
оказывает никакого влияния. Иначе обстоит дело во влажных областях,
где рост растений зависит от измеримого количества осадков. В этих
условиях неизмеримые осадки в виде тумана и росы создают лишь допол-
нительное увлажнение.
Подчас бывает трудно определить разницу между туманом и дож-
дем, когда речь идет о моросящем тумане или моросящем дожде. Это
относится, в частности, к туманам на Столовой горе, к оазису туманов
Эрковит на побережье Красного моря в Судане или к Гаруа на юге Перу
(см. стр. 415).
Пустыня туманов Намиб
381
Во всех этих случаях влажный насыщенный водяными парами
воздух поднимается вверх по крутому склону; происходит конденсация
влаги, количество которой так значительно, что его можно определить
при помощи обычного дождемера. Количество выпавших осадков резко
возрастает, если при ветре моросящий туман встречается с вертикальным
препятствием, на котором конденсируется влага («Phytologie», Bd III,
1, S. 155 и след.).
Вышеприведенные соображения не относятся к лишайникам —
растениям, действительно существующим за счет влаги туманов и не
зависящим от почвенной влаги. На окраине пустыни Намиб, где туманы
особенно часты, они покрывают фактически все плохо поддающиеся
Рис. 291. «Оконные» водоросли (по Фогелю, с некоторыми изменениями).
Зависимость развития водорослей от условий освещения под обломками кварца (слой
водорослей зачернен). На больших обломках, затрудняющих проникновение света,
водоросли развиваются только по краям (слева и справа), зато на маленьких облом-
ках они покрывают всю нижнюю сторону кварца (середина).
выветриванию скалы и обломки породы, тогда как во внутренних райо-
нах, где возрастает количество осадков, но уменьшается число дней
с туманами, они практически исчезают.
Фогель приводит характеристику своеобразных лишайников южного
Намиба % способных использовать влагу туманов. Их слоевища, разме-
ром всего в несколько миллиметров, погружены в почву; на нижней
стороне слоевищ наблюдаются хорошо развитые ризины. Водоросли
расположены на нижней стороне слоевищ и защищены толстым, частич-
но пигментированным верхним корковым слоем от слишком сильного
действия света. Фогель совершенно обоснованно сравнивает эти лишай-
ники с часто встречающимися в той же пустыне Намиб «оконными» расте-
ниями из рода MesembryanthemuTTi, как, например, Lithops. Возможно,
еще более своеобразны «оконные» водоросли, использующие в качестве
«окон» прозрачные обломки кварца, на нижней стороне которых, то
есть уже ниже поверхности почвы, они образуют слой медно-зеленого
цвета 1 2. Сквозь прозрачные куски кварца проникает достаточное коли-
чество света; наряду с этим лишайники используют влагу туманов, сте-
кающую под камни и сохраняющуюся дольше под их защитой (рис. 291).
1 S. Vogel, «Niedere FensterpfJanzen» in der siidafrikanischen Wiiste, «Beitr.
Biol. Pfl.», 31, 1955, S. 45-135.
2 Об этом же упоминает Бидл (стр. 443) при описании солянковой пустыни
в Австралии; по данным автора, такие водоросли встречаются в центральной части
пустыни Намиб и в пустыне между Каиром и Суэцем. Это наблюдается также в пу-
стыне Неджеб (устное сообщение Эвенари).
В Неваде под кусками кварца развиваются Protosiphon cinnamomeus п Anacystes
montana, а также азотфиксирующпе виды Nostoc и др. В этих водорослях содержится
много азота (7—8% сухого абс. веса). В 1 км от места проведения взрывов ядер-
1ных установок никаких следов водорослей не было найдено, поскольку они, видимо,
382
Глава 9
РАСТЕНИЯ ПУСТЫНИ НАМИБ И ИХ ЭКОЛОГИЯ
В зависимости от условий среды* 1 автор различает
А) растительность равнин:
а) прибрежной части пустыни Намиб,
б) внутриконтинентальной части пустыни Намиб;
Б) растительность скал:
а) на мраморе (известковом шпате), на склонах гор, сложенных
кварцитами и диабазами,
б) на горных хребтах, сложенных гнейсами и гранитом;
В) растительность русел временных водотоков и сухих долин:
а) русла потоков со слабосолоноватой водой,
б) русла потоков с солоноватой водой,
в) устья сухих долин;
Г) переходная зона, образованная Welwitschia.
Растения равнин
В прибрежной части пустыни Намиб господствует щебнистая пусты-
ня. Около 50% поверхности почвы здесь как бы сцементировано камня-
ми; этот слой защищает от выдувания лежащий ниже богатый слюдой
пылеватый песок. Незначительные количества песка, уносимые ветром,
откладываются в виде песчаных кос с подветренной стороны препят-
ствий. Уже на глубине 1—5 см залегает горизонт, цементированный
гипсом и известью до твердости камня; осадки обычно проникают до
этого уровня. Лишь в местах, где благодаря выраженному микрорельефу
почва лучше промачивается, этот уплотненный слой располагается
гораздо глубже. Пестрые листоватые и корковые (накипные) лишайники
сплошь покрывают разбросанные по поверхности камни.
После сильных дождей, прошедших в 1934 г., почва внезапно покры-
лась многочисленными растениями. В среднем степень покрытия почвы
составила 20%, а в небольших понижениях —50 и даже 90%.
В составе покрова преобладали Mesembryanthemum salicornioides,
Hydrodea bossiana (рис. 292), Drosanthemum paxianuirr, кроме того, встре-
чались плотные подушки Aizoon dinteri и Zygophyllum simplex.
В 1935 г. около половины растений погибло. Возможно, что осадки,
приносимые туманом, несколько продлевают жизнь растений, однако
ненадолго. В живых остались лишь галофитные суккуленты, отличаю-
щиеся крайне слабой транспирацией. Все базальные части Hydrodea
отмерли и высохли, тогда как апикальные еще продолжали существо-
вать за счет накопленных резервов влаги, поскольку о каком-либо непо-
средственном поглощении воды не могло быть и речи.
погибли под воздействием высокой температуры. См. L. М. Shields, F. Dr о-
u е t, Distribution of terrestrial algae within the Nevada test site, «Am. J. Bot.», 49,
1962, p. 547—555.
Согласно Фоллманну (G. F о 1 1 m a n n, Fensterflechten in der Atacamawiiste,
«Die Naturwissensch.», 52, 1965, S. 434, 435), влажность водорослей через 6 часов после
их смачивания росой при температуре почвы, равной 62°, все еще достигала 44%.
1 Фотографии и список растений, собранных в пустыне Намиб близ Гобабиса,
приведены в работе W. G i е s s, Some notes on the vegetation of the Namib Desert,
Sci. Pap. Namib Des. Res. Station № 3, 1962. Наполненное в 1963 г. водой сухое
русло Куизеба (в 11-й раз с 1830 г.) запечатлено на рисунках в работе С. Кос h,
Der Kreis, 2/3, Windhoek, 1963.
Пустыня туманов Намиб
Рис. 292. Hydrodea bossiana — однолетний ярко выражен-
ный суккулент из рода Mesembryanthemum. Цветки желтого
цвета (фото Динтера)
Анатомическое строение Mesembryanthemum salicornioides изучал Цем-
ке, работавший в Высшей сельскохозяйственной школе Штутгарт-Хоэн-
хейма Ч Междоузлия у этих видов достигают толщины карандаша и дли-
ны 1 см. У супротивных удлиненно цилиндрических суккулентных
листьев развивается пузырчатый эпидермис; листья недолговечны и вско-
ре опадают (рис. 293). В этом случае функции листа (фотосинтез) выпол-
няют мясистые, часто красноватые более молодые части стебля, поверх-
ность которых так же густо покрыта пузырчатыми клетками (рис. 294).
1 Е. Zemke, Anatomische Untersuchungen an Pflanzen der Namibwiiste, «Flo-
ra», N. F., 33, 1939, S. 365-416.
384
Глава 9
Хотя эти клетки плотно прижаты друг к другу, между ними все же
остаются свободные пространства, в которых располагаются устьица,
до 54 на 1 мм2 (рис. 295 и 296)..
Р и с. 293. Mesembry-
anthemuYn, salicornioi-
des. Часть стебля с
увядшими листьями.
Р и с. 294. Поперечный срез старого стебля М. salicor-
nioides. Справа пузырчатый эпидермис, К — опробковев-
шая эндодерма (с пробковой тканью), до которой отми-
рает кора (по Цемке).
Рис. 296. Пузырчатый эпидермис М. sali-
cornioides и устьица в плане (по Цемке).
Р и с. 295. Пузырчатый эпидермис и
устьица М. salicornioides па попереч-
ном срезе (по Цемке).
Пустыня туманов Намиб
385
Рис. 297. Цветущие мезембриантемумы вскоре после дождя в пустыне Намиб
между Аусом и Людериц (фото Ветштейна).
Тонкие стенки пузырчатых клеток покрыты едва заметным слоем
кутикулы, что, однако, не увеличивает интенсивности кутикулярной
транспирации. Еще более крупные пузырчатые клетки развиваются на
широких листьях Cryophyturn (1,2—2,0 мм). Кутикула на них настоль-
ко тонка, что обнаружить ее можно лишь с большим трудом. Клеточный
сок пузырчатых клеток, судя по реакции с AgNO3, видимо, содержит
больше хлоридов, чем остальные клетки. Число устьиц достигает 45 —
50 на 1 мм2.
Вода, содержащаяся в тканях более старых частей стебля Mesembry-
anthemum salicornioides, со временем расходуется, и тогда эти ткани
отмирают. Клетки эндодермы опробковевают, а под ней формируется
пробковая ткань (рис. 294). Одновременно сильно одревесневают клетки
центрального цилиндра, в результате чего базальные части растений
оказываются образованными одними лишь тонкими одревесневшими
междоузлиями.
В почвах прибрежной части Намиба присутствуют легкорастворимые
соли. Пылеватый песок содержит 0,274% NaCl и 3,34% сульфатов (пре-
имущественно гипса) в расчете на сухой вес. Цементированный карбо-
натами горизонт содержит всего лишь 0,1% NaCl. После дождей влаж-
ность почвы, вероятно, бывает очень высокой.
Хотя процессы засоления здесь слабо выражены, у этих растений,
как и у всех облигатных галофитов, наблюдается повышенное содер-
жание в клеточном соке хлоридов. Осмотическое давление достигает
следующих величин (табл. 55; в скобках указано число взятых образ-
цов).
У галофитов не отмечается никакой определенной зависимости
между величиной осмотического давления, содержанием солей и при-
ростом у растений. Два экземпляра Zygophyllum, отличавшиеся величи-
ной осмотического давления (16,3 и 33,5 атм), развивались одинаково
386
Глава 9
хорошо. Только при наиболее высоких значениях концентрации клеточ-
ного сока растения начинают отмирать.
Таблица 55
Название растений Осмотическое давление, Содержание хлоридов, %
Hydrodea (2) 30,5-46,9 70—80
Mesembryanthemum (4) 28,6-31,1 30—40
Aizoon (8) 20,9-52,8 22-52
Zygophyllum simplex (5) 16,3-53,2 66—95
Вышеупомянутые виды — это эфемеры, развивающиеся только после
выпадения осадков. Основная часть их ареала, особенно у мезембрианте-
мумов, расположена в южной части пустыни Намиб, где они красочно
цветут после первого хорошего зимнего дождя (рис. 297).
Наряду с этими эфемерами в 1935 г. встречались также некоторые
многолетние виды, обычно приуроченные к увлажненным понижениям
рельефа или устьям сухих русел временных водотоков. Это Zygophyllum
stapffii и Arthraerua leubnitziae.
После обильных осадков, выпавших в 1934 г., они распространились
также и на равнинных территориях, однако уже в 1935 г. находились
в угнетенном состоянии и в последующие годы исчезли.
У суккулентных величиной с большую монету листьев Zygophyllum
stapffii мезофилл состоит из тонкостенных клеток, служащих как для
накопления воды, так и для фотосинтеза. Наружная стенка клеток эпи-
дермиса утолщена, однако кутикула слабо развита. Устьица очень
немногочисленны (примерно 10 штук на 1 мм2). Осмотическое давление
у растений (на основании 15 наблюдений) достигает 27,5—55,6 атм\
однако уже при величинах давления более 40 атм растения развиваются
очень плохо. Содержание хлоридов достигает высоких значений —
70—80%. Особенно сильно от недостатка воды страдает Arthraerua (Ama-
rantaceae), достигающая более крупных размеров только на участкахг
остающихся влажными в течение продолжительного времени. Это расте-
ние не является суккулентным; листья у него полностью редуцированы
а разветвленные побеги напоминают Equisetum limosum (рис. 298 и 299).
Осмотическое давление клеточного сока колеблется (13 образцов) в пре-
делах 24,2—33,2 атм. По данным Босса, у отдельных растений наблю-
далось и более высокое давление (40 атм и выше, а максимум 51,5 атм).
Однако эти данные были получены в 1932 г., отличавшемся длительным
периодом засухи. Содержание хлоридов достигало всего 9—48%; вели-
чина по сравнению с суккулентными галофитами очень небольшая. В про-
тивоположность суккулентным галофитам Arthraerua, наряду с хлорида-
ми накапливает много сульфатов и поэтому относится к числу несукку-
лентных галофитов, накопителей сульфатов. Как известно, сульфаты
уменьшают набухание коллоидов («Phytologie», Bd III, 1, S. 487).
Arthraerua хорошо задерживает переносимый ветром песок и всегда
растет на рассеянных по равнине дюнах, достигающих высоты около 1 м
(рис. 299). Надземные органы растений частично оказываются погребен-
ными песком. В отличие от Zygophyllum stapffii, вокруг которого ветер
Рис. 298. Arthraerua leubnitziae (или близкий вид) из семейства Amarantaceae (фото
Диптера).
Рис. 299. Дюны, заросшие Arthraerua leubnitziae, в прибрежной части пустыни Намиб, севернее нижнего тече-
ния Куизеба. Слева — Euphorbia cervicornis.
Пустыня туманов Намиб
389
также насыпает дюны, у Arthraerua не образуются придаточные корни.
У обоих видов развиваются стержневые корни, уходящие глубоко в грунт
и достигающие влажных горизонтов почвы. Последние всегда хорошо
выражены в низовьях крупных сухих русел пересыхающих водотоков.
Согласно Боссу, интенсивность транспирации у названных видов
примерно в 10 раз выше, чем у мезембриантемумов, а дефицит влаги
небольшой (около 10%). Таким образом, во влажных условиях эти виды
отличаются уравновешенным водным балансом, тогда как в засушливых
они развиваются очень плохо.
Не совсем ясны причины засоления прибрежной части пустыни
Намиб. Богатые солями морские отложения в пустыне Намиб не встре-
чаются; обширные же залежи соли, наблюдаемые, например, у мыса
Кросс, частично возникли на месте отделенных от моря лагун. По-види-
мому, засоление равнинных территорий следует отнести, скорее, за счет
туманов. Очень сильный прибой у побережья Юго-Западной Африки
распыляет морскую воду на мельчайшие капли, которые вместе с тума-
ном ветер гонит в сторону материка. По данным Кайзера, во влаге,
приносимой туманом, содержится 244 мг/л ионов хлора, что соответ-
ствует 0,4 г поваренной соли. Если взять это количество за основу наших
расчетов и принять, что годовая сумма осадков достигает 50 мм, что
соответствует 50 л1м\ то на 1 м2, в течение года 1 будет отлагаться 20 г
NaCl. Это немалая величина, тем более что при незначительном количе-
стве осадков поверхностный сток удаляет лишь очень небольшую часть
выпавших солей.
Признав этот эоловый путь переноса солей как реальность, можно
объяснить, почему почва засолена только в прибрежной части пустыни
Намиб, в полосе влияния туманов, в то время как внутренние терри-
теории не обнаруживают никаких признаков засоления. Граница прохо-
дит примерно в 50 км от побережья.
Внутриконтинентальная часть пустыни Намиб отличается совер-
шенно иным растительным покровом. Галофиты, подобные Zygophyllum
stapffii и Arthraerua, здесь отсутствуют и вместо них развиваются злаки —
различные виды Aristida. По направлению в глубь материка увеличи-
вается степень сомкнутости покрова, образованного дерновинными
злаками; полностью сомкнутый злаковый покров развивается на возвы-
шенных территориях (рис. 300 и цветные фото 1—3, стр. 272). Песчаная
почва отличается щелочной реакцией (pH = 7,3—9,0); в ней обнаруже-
Аы лишь следы растворимых солей. В 1935 г. автор наблюдал отмершие
кусты злаков, встречавшиеся также и на навеянных ветром песчаных
гребнях в прибрежной части пустыни Намиб. Эти злаки могли появиться
здесь только после дождей, прошедших в 1934 г., но высохли на следую-
щий же год. Осмотическое давление у злаков невысокое (у Aristida \hoch-
stetteriana —16,6 атм)-, при высыхании почвы высыхают и листья. У мно-
голетних злаков вокруг корней образуется чехол из песчинок (см.
стр. 282). У сопутствующих злакам видов разнотравья (Compositae,
Acanthaceae), относящихся к числу мягколистных ксерофитов, осмоти-
ческое давление повышается только при увядании. Суккулентные листья
развиваются лишь у Sesuvium digynum (Aizoaceae). Однако низкие зна-
чения осмотического давления (14,6—18,8 атм, при отмирании —
27,7 атм) и небольшое содержание хлоридов (2—15%) свидетельствуют
1 См. также G. Boss. Niederschlagsmenge und Salzgehalt des Nebelwassers an
der Kiiste Deutsch-Siidwestafrikas, Bioklim, H. 1, 1941.
Рис. 300. Aristida brevifol а на песках внутриконтинентальной части пустыни
Намиб, близ Кубуба.
Рис. 301. Acacia giraffae и A. horrida в долине с близко залегающими грунто-
выми водами; внутриконтинентальная часть пустыни Намиб, близ Кубуба.
Пустыня туманов Намиб
391
о том, что этот вид не может быть отнесен к числу настоящих галофитов.
В благоприятных условиях местообитания, например у подножия
обнажений гранита, с которых стекает дождевая вода, или в местах
скопления грунтовых вод под руслами пересыхающих водотоков, уже
появляются деревья и кустарники, свойственные саваннам и сухим
лесам (рис. 301). Низкое осмотическое давление, которым, по данным
Босса, отличаются листья Terminalia porphyrocarpa (23 атм), свидетель-
ствует о благоприятном водном режиме.
Растения на скалах
На скалах в пустыне формируются относительно благоприятные
условия для произрастания растений, поскольку здесь даже небольшие
осадки стекают в трещины, где влага защищена от испарения и где созда-
ется известный запас воды. В прибрежной части пустынй Намиб точно
так же только на скалах и в местах с близким уровнем грунтовых вод
создаются условия, где могут существовать многолетние растения. Осо-
бенно благоприятны условия для роста растений на трудно под-
дающихся выветриванию обнажениях кварца, диабаза, а также мрамора
(кристаллические известняки). Частые, но очень незначительные осадки,
приносимые туманом, способствуют развитию на скалах многочислен-
ных лишайников, а также цветковых растений, среди которых преобла-
дают суккуленты, отличающиеся небольшими размерами:
1. Стеблевые суккуленты Trichocaulon dinteri (рис. 302) и Т. pedi-
cellatum, Hoodia currori, Euphorbia brachiata.
2. Листовые суккуленты —виды Lithops (цветное фото 2, стр. 304).
Hereroa bossii (Mesembryanthemum), Anacampseros albissima, Aloe aspe-
rifolia, Cotyledon orbiculata.
3. Стеблевые суккуленты (временами облиствленные) — Pelargoni-
um roessingense, Sarcocaulon marlothii, Othonna protecta, Senecio (Kleinia)
longiflorus, Adenia (Echinithamnus) pechuelli (рис. 303).
Кроме этих видов, в местах скопления мелкозема, в 1935 г. встре-
чались некоторые виды разнотравья, а также одиночные экземпляры
галофитов.
Рис. 302. Trichocaulon dinteri в рас-
селинах скал; горы Витпорт в пустыне
Намиб (фото Э. Вальтер).
Рис. 303. Adenia (Echinothamnus) pe-
chuelli (Passifloraceae) в трещинах скал;
горы Витпорт в пустыне Намиб (фото
Э. Вальтер).
П1мм
Р и с. 304. Lithops ruschiorum (по
Цемке).
а — эпидермис молодых листьев с утол-
щенным слоем целлюлозы на стенках кле-
ток и кутикула; б — эпидермис старых
листьев, слой целлюлозы на стенках кле-
ток инкрустирован кристаллами; пункти-
ром обозначены кутикулярные слои;
в — эпидермис молодого листа с устьи-
цем. В середине каждой клетки видно
шаровидное утолщение кутикулы.
OjriM
Рис. 305. Hereroa bossii (по Цемке).
а — поперечный срез листа с клетками, содержащими дубильные вещества (г); б - идио-
бласт. содержащий дубильные вещества при большем увеличении; pl — периферийный и г/ --
центральные проводящие пучки.
Пустыня туманов Намиб
393-
Сходный состав, но еще более богатый суккулентами обнаружен на
скалах близ бухты Людериц. Здесь уже преобладают мезембриантемумы.
Анатомическое строение этих своеобразных суккулентов изучалось неод-
нократно; Цемке приводит в своей работе полный список литературы
по этому вопросу Ч
«Оконные» растения изучал также Шмуккер 1 2. На рис. 304 изобра-
жено строение эпидермиса молодых и старых листьев этих растений.
Устьиц у них или очень немного (от 3 до 15 штук на 1 мм 2), или они
отсутствуют вовсе. У Негегоа устьица более многочисленны (23 на 1 мм2\
и к тому же этот вид отличается другим строением листа (рис. 305).
Стеблевые суккуленты характеризуются небольшой интенсивностью
транспирации, причем это относится и к растениям третьей группы,
когда они находятся в безлистном состоянии. У Sarcocaulon стебли
покрыты пробковым слоем, клетки которого наполнены смолой. Этот
покров из смолы сохраняется на отмерших растениях и, если его под-
жечь, горит, как коптящая свеча (отсюда и местное название растения —
«свеча бушменов»). Листовые суккуленты также отличаются незначи-
тельной транспирацией. К результатам соответствующих исследованийг
проведенных Форесмитом в Карру, мы еще вернемся.
О строении эпидермиса других суккулентов можно получить пред-
ставление по рис. 306 и табл. 56.
Таблица 56
Название растений Толщина слоя кутикулы, К Толщина целлюлоз- ного слоя наружной стенки клет- ки, ц Устьица
число на 1 Л1.«2 размер (длинах ши рииа), ц
Trichocaulon dinteri . 2,5-3,5 2,5 24 26 X 15
Hoodia currori . . . 6 2,5 19 35 X 19
Caralluma lateritia 1 ... 3,5 4-6 17 30 х 15
Tavaresia grandiflora 2 2 17 29 х 12
Euphorbia dinteri . 6-8,5 3,5 40 28 X 15
Euphorbia lignosa . 6-7 1 22 34 X 13
Euphorbia spec. 1 3-6 2-3 23 32 X 14
1 Встречается в более влажной северо-восточной
части стран л! близ Цумсба.
Из листовых суккулентов в пустыне Намиб встречаются невысокие
Aloe asperifolia, тогда как крупные A. rubrolutea распространены пре-
имущественно во внутренних областях материка, a Sansevieria — на севе-
ро-востоке Юго-Западной Африки; в более засушливых условиях они
растут лишь под защитой скал.
Число устьиц и их размеры приведены в табл. 57.
При сильной инсоляции температура поверхности листьев у сук-
кулентов на 10—15° выше окружающей.
Как и следовало ожидать, эти растения отличаются небольшим
осмотическим давлением (табл. 58).
1 Е. Zemke, op. cit. Автор предоставил в распоряжение Цемке собранный
им материал.
2 Th. Schmucker, Okologie der Fensterblatter, «Planta», 13, 1931, S. 1 — 17.
394
Глава 9
У суккулентных растений в отжатом соке обнаружены хлориды, хотя
и в очень небольшой концентрации. Исключением являются мезембриан-
темумы (Hereroa, Lithops), поскольку их осмотическое давление дости-
гает 14,5—16,6 атм, а содержание хлоридов составляет примерно 25%,
что свидетельствует об их явно галофитном характере.
Таблица 57
Название растений Устьица
число на 1 At At 2 | размер (длинах ши- рина), ц,
верхняя сторона нижняя сторона
Aloe asperifolia . 25 24 59x43
А1оё rubrolutea . . 41 51 54 X 26
Sansevieria cylindrica 39 53 х 26
Senecio longiflorus 1 25
1 В пустыне Намиб всегда в безлистном состоянии.
Совершенно иным видовым составом отличаются растительные груп-
пировки, развитые на обнажениях гранитов, гнейсов или сланцев. Эти
Таблица 58
Осмотическое давление у растений, развитых на скалах
в пустыне Намиб (в атм)
Название растений По Боссу по Вальтеру
Trichocaulon dinteri . . 5,3-5,7 —
Trichocaulon pedicellatum 6,6—8,6 —
Hoodia currori ... 5,8-6,4 9,4
Cotyledon orbiculata 2,4 (?) 7,0
Aloe asperifolia 6,6 6,5-8,0
Senecio longiflorus ... 8,5—10,8 7,2-9,2
Adenia pechuelli (стебли) . 5,2—8,0 —
Adenia pechuelli (листья) . 12,5 —
Pelargonium rossingense . 8,8 8,4
(цветет)
Sarcocaulon marlothii . — 9,7—12,3
Euphorbia brachiaia . — 11,8
Othonna protecta 21,1 и 21,4 —
породы подвергаются гораздо более сильному механическому выветри-
ванию, в результате которого образуются большие россыпи, на которых
мелкие суккуленты не могут удержаться. Здесь встречаются только
большие колоннообразные, ветвящиеся у основания экземпляры Euphor-
bia dinteri и канделяброобразные А1оё dichotoma. Последний вид особен-
но характерен для более удаленных от побережья опустыненных тер-
риторий Юго-Западной Африки (рис. 307 и 308); здесь обширные пло-
щади покрыты крупными высотой более 1 м кустами молочая с тонкими
Пустыня, туманов Намиб
395
цилиндрическими побегами (рис. 308); наряду с ними появляются также
галофиты. Детальные экологические исследования в этих условиях не
проводились.
Р и с. 306. Эпидермис у различ-
ных суккулентов (по Цемке).
а — Euphorbia dinteri’, б — Euphor-
bia spec, (близ Цумсба); в — Ноо-
dia currori (Asclcpiaclaccac); г —
Senecio longiflorus (Goinpositac),
c* — Aloe asperifolia', e — Sansevi-
eria cylindrica; ж — Caralluma
lateritia (Asclcpiadaccac). Кутику-
ла зачернела.
На скалах у восточной границы пустыни Намиб встречаются очень
своеобразные древесные формы с толстыми суккулентными стволами,
напоминающие Idria и виды семейства Burseraceae, распространенные
в пустыне Сонора С Частично они так?ке относятся к семейству Burse-
raceae, как, например, Commiphora glaucescens, С. oblanceolata\ здесь
растут также поразительно похожая на них Euphorbia guerichii, Cissus
cramerianus и Moringa ovaliifolia; на юге, в бассейне реки Оранжевая,
распространен Рachypodium namaquanum. Те же древесные формы встре-
чаются и во внутренних частях страны на известняках и доломитах гор
Отави (рис. 309 и 310).
В областях, граничащих с пустыней, распространены уже пойкило-
гидрические папоротники, как, например, виды Actiniopteris, Notholaena
и другие (рис. 311), а из цветковых растений Myrothamnus flabellifolia
(рис. 312). Этот кустарник достигает высоты 50 см и внешне напоминает
сухую метлу. Он разрастается на каменистых склонах. Достаточно неболь-
шого дождя, и уже спустя полчаса куст покрывается мелкими блестящими
1 Ср. также сходные формы, распространенные па острове Сокотра; см.
G. В. Р о р о v, The vegetation of Socotra «J. Linnean Soc. bond. Bot.», 55, 1957,
p. 706—720.
396
Глава 9
зелеными листочками; клетки последних даже спустя год после пере-
возки в сухом состоянии все еще обладали способностью плазмолиза.
Ветви этого кустарника не укореняются; супротивные листья не обладают
ксероморфной структурой (рис. 31*3). Крупные толстые клетки эпидермиса,
сменяются мелкими, более тонкими, причем это наблюдается и на верх-
ней, и на нижней стороне листа. При высыхании под воздействием сил
сцепления листья складываются (рис. 314 и 315). При смачивании вес
и площадь поверхности листьев увеличиваются на 62%. Анатомическое-
строение не дает возможности объяснить способность этих растений пере-
Р и с. 307. Aloe di-
chotoma близ Кет-
мансхопа, Юго-За-
падная Африка:
(фото Э. Вальтер).
Рис. 308 Пустыня близ Гаруба в Юго-Западной Африке на высоте примерно 700
над ур. моря. На заднем плане — отдельные экземпляры Aloe dichotoma; на перед-
нем — полушаровидные кусты Euphorbia cf. gummifera (фото Марлота).
Рис. 309. Cissus juttae (с суккулент-
ным стволом) на известняках. Ферма
Граслагте близ Отыоваронго, Юго-
Западная Африка (фото Э. Вальтер).
Рис. 310. Р achy podium, lealii (gigan-
teum), Apocynaceae, за ним— Commi-
phora berberidifolia в горах на облом-
ках доломита. Ферма Гейдельберг
близ Цумеба, Юго-Западная Африка
(фото Э. Вальтер).
Р и с. 311. Pellaea hastata — пойкилогидрический папоротник среди облом-
ков горной породы. Ферма Фойгтланд, Юго-Западная Африка (фото
Э. Вальтер).
Рис. 312. Myrothamnus flabellifolia в сухом состоянии, Науклюфт, Штоль-
ценфельз (фото Э. Вальтер).
Пустыня туманов Намиб
399
Рис. 314. Листья Myrothamnus flabellifolia (по
Цемке).
а — сухие; б — насыщенные влагой.
Рис. 315. Поперечный срез
листьев, изображенных на
рис. 314.
а — сухие; б — насыщенные
влагой.
носить полное высыхание. Число устьиц, расположенных большей частью
в бороздках, составляет на верхней стороне листа 64 на 1 мм2, а на ниж-
ней — 78.
В этой области в местах выхода грунтовых вод растут виды Ficus,
например стелющийся по отвесным скалам F. guerichiana, а между облом-
ками скал часто встречается Sterculia guerichii — древесный вид с круп-
ными нежными листьями.
В небольших углублениях, возникающих при выветривании гранита,
встречается оригинальное водное растение. Здесь вода сохраняется всего
лишь несколько дней после дождя. Почва покрыта слоем песка мощностью
1—2 см, густо пронизанного корнями Chamaegigas intrepidus. В период
400
Глава 9
-засухи живыми остаются только корни и сморщенные совершенно сухие
клубнеобразные подводные листья. После увлажнения листья быстро
Рис. 316. Цветущий Chamaegigas intrepidus в небольшом
углублении в обломках гранита. Ферма Оконге, Юго-
Западная Африка (фото Э. Вальтер).
набухают, начинают зеленеть, и спустя несколько часов их размеры уве-
-личиваются в 10 раз. Еще через несколько дней развиваются плавающие
листья и цветки (рис. 316), а прежде чем вода полностью испарится,
успевают созреть и семена 4.
Растения, в руслах временных водотоков
и сухих долинах
Как это вообще характерно для засушливых областей, в пустыне
Намиб русла временных водотоков и долины пересыхающих рек являются
наиболее благоприятными местообитаниями для развития растений. При
заполнении русла водой песок речного ложа промачивается на большую
глубину и сохраняет влагу на протяжении многих лет даже в поверх-
ностных слоях.
Положение верховьев сухой долины и характер горных пород, встре-
чающихся в ее водосборном бассейне, определяют степень минерализа-
ции воды. При быстром стекании воды по поверхности скал лишь первая
паводковая волна оказывается насыщенной солями. Если же вода лишь
медленно просачивается через слой песка или щебня, то большая часть
ее испаряется, а остаток все более обогащается солями. На рис. 317 схема-
тически представлен процесс отложения солей в сухих долинах.
На дне долины вода выходит из-под слоя песка на поверхность. Песок
хорошо увлажнен; на нем растут галофиты. Далее вода течет по поверх-
ности вниз по руслу. При этом она испаряется, течение ее замедляется,
а содержание солей возрастает. В конце концов соли начинают выпадать
1 Н. Heil, Chamaegigas intrepidus Dtr., eine neue Auferstehungspflanze, «Beih.
Bot. Zbl.», 1, 41, 1925, S. 41—50.
Пустыня туманов Намиб
401
сначала лишь по краям потока, где образуют слабый налет, а затем отла-
гаются в виде сплошной соляной корки. Ниже этой границы в местах, где
прекращается ток воды, русло остается сухим. Зона выпадения солей
постепенно, по мере уменьшения притока воды, перемещается вверх
по руслу, причем граница выпадения солей может меняться в течение
суток. Ночью, в связи с меньшим испарением, вода течет дальше по доли-
не, днем испарение возрастает и путь воды по поверхности сокращается.
Вследствие этого образуется множество рукавов с отложениями солей —
своего рода дельта. Среди соляных пятен возвышается несколько отмер-
ших кустов Tamarix, что свидетельствует о протекавшем здесь еще совсем
недавно быстром и полноводном потоке с гораздо меньшей концентра-
цией солей.
Рассмотренный здесь процесс засоления равным образом повторяется
и в пределах более обширных понижений во всех пустынных областях.
То же явление наблюдается и в крупных долинах пересыхающих рек.
Даже если подрусловые воды этих рек вовсе не выходят на поверхность,
верхние горизонты почвы в низовьях оказываются все же несколько
увлажненными.
Постоянный поток грунтовых вод или линзы подземных вод наблю-
даются лишь в очень обширных сухих долинах. В небольших руслах вре-
менных водотоков и в верхнем течении сухих долин в почве сохраняется
лишь пленочная влага. Однако этого уже достаточно для существования
растений. При больших запасах воды в прчве длительные периоды засухи
могут переносить даже более крупные растения. Здесь также устанавли-
вается известное соотношение между размером транспирирующей поверх-
ности и запасами воды. В мелких сухих руслах с пресноводными пото-
ками воды встречаются Citrullus ecirrhosus и несколько видов разнотравья,
широко распространенных обычно в период дождей (виды Tribulus и (Лео-
те). При увеличении запасов воды в почве здесь поселяются, кроме того,
отдельные виды кустарников, например Bauhinia pechuelii, Parkinsonia
africana, Commiphora dulcis и, возможно, также некоторые виды акаций.
При избыточном увлажнении грунтовыми водами развиваются Rhus
lancea и Salvadora persica, а также виды деревьев — Euclea pseudebenus,
Acacia giraffae и A. albida. Последние по берегам пересыхающих рек
встречаются в составе настоящих галерейных лесов в виде крупностволь-
ных деревьев.
По сухим руслам распространены также заносные растения (Nico-
tiana glauca и Ar genome mexicana), а также сорные виды (Ricinus, Datura
metel). На песчаных дюнах нижнего течения Куизеба с неглубоким зале-
ганием грунтовых вод развивается своеобразный безлистный, покрытый
зелеными шипами кустарник Acanthosicyos horrida (Cucurbitaceae). Его
плоды охотно употребляет в пищу коренное население, а семена продают
в качестве заменителя миндаля (рис. 318). Многие виды разнотравья
распространяются по руслам пересыхающих водотоков из восточных
частей страны в глубь пустыни. Число устьиц у этих видов гораздо боль-
ше, чем у суккулентных видов (табл. 59).
Сравнительно большое число устьиц свидетельствует об интенсивном
газообмене у этих видов при условии хорошего обеспечения водой.
Если в воде содержится много растворимых солей, появляют-
ся кусты Tamarix, а также Lycium tetrandrum. Если же в устьях пе-
ресыхающих рек образуются обширные периодически затапливаемые
площади, а грунтовые воды располагаются близко к поверхности
почвы, эта территория начинает заселяться типичными галофитами
в следующей последовательности по мере увеличения влажности и степе-
ни засоления: Zygophyllum stapffii, Cryophytum barkleanum, Arthraerua
leubnitziae, Salsola, Suaeda fruticosa, Arthrocnemum glaucum. Бессточные
дефляционные котловины \ куда после дождей, хотя бы даже и очень
Внутрипочденныи
сток под слоен
песка
Выход 0оды на
поверхность
Прекращение стока
и испарение Воды;
соляная корка
Замедление течения;
по краям потока налеты
соли
Tamarix и Zygophyllum stapfii
Отмершие кусты
Tamarix
Р и с. 317. Схема отложения солей по руслу временного водотока ниже зоны проса-
чивания в условиях пустыни Намиб (по Н. Walter, 1936).
Рис. 318. Песчаные дюны, заросшие Acanthosicyos horrida, в нижнем течении Куи-
зеба вблизи Уолфиш-Бея. На сухой растрескавшейся почве пятнами растет склеро-
фильный злак Eragrostis spinosa.
редких, сносятся соли с окружающих возвышений и где происходит
интенсивное накопление солей, полностью лишены растительности.
Наличие соляной корки на поверхности почвы еще ничего не говорит
о содержании солей в грунтовых водах. При стоянии грунтовых вод на
глубине менее 1 м поверхность почвы будет постоянно увлажняться вслед-
1 Е. Kaiser, op. cit.
Пустыня туманов Намиб
403
ствие капиллярного поднятия воды; последняя, непрерывно испаряясь,
оставляет на почве кристаллы соли, которые в конце концов образуют на
поверхности белую корку. Если в почве содержатся органические веще-
ства, то образуется сода и корка приобретает черный оттенок. В местах
выхода слабосоленых грунтовых вод на засоленных болотах растут
Phragmites, Diplachne paucinervis, Sporobolus pungens, Juncellus (Cy perus)
laevigatus и др.
Таблица 59
Число устьиц на 1 мм2 на верхней
(первая цифра) п на нижней (вторая цифра)
стороне листьев
Lyperia litoralis 56- 56
Justicia arenicola . 119-110
Petalidium variabile 193-183
Helichrysum roseo-niveum 94—126
Tripteris arborescens 136-132
Gazania varians . 64—150
Растения, развитые в руслах временных водотоков и в сухих долинах,
укореняются не особенно глубоко, проникая лишь до уровня влажных
горизонтов песка. Даже в долинах крупных пересыхающих рек корни
деревьев углубляются обычно не более чем на 1—2 м, но зато распростра-
няются далеко в горизонтальном направлении. Осмотическое давление
изучалось у экземпляра Citrullus, веретеновидный корень которого раз-
ветвлялся на глубине 15 см, а более тонкие корни углублялись в почву
под небольшим углом. Влажность песчаной почвы достигала на глубине
40 см 1,3% ; в воде были обнаружены следы хлоридов и нитратов; сульфаты
отсутствовали. Осмотическое давление почвенного раствора не превышало
1 атм. Концентрация клеточного сока в листьях Citrullus достигала
13,6 атм, а в целых побегах — 10,8 атм. Содержание хлоридов в боль-
шинстве случаев не превышало 20%. Осмотическое давление у остальных
видов разнотравья, развитых в сухих руслах, где не было отмечено при-
знаков засоления, также колебалось в пределах 12—18 атм и лишь
в исключительных случаях превышало 20 атм\ у суккулентных кустар-
ников оно составляло около 13 атм, а у остальных — 20—22 атт. Зна-
чительно более высокое осмотическое давление (35—50 атм) отмечено
у Lyperia litoralis (Scrophulariaceae), причем у этого вида на долю одних
только хлоридов приходится 23—26 атм. Это еще один пример, свидетель-
ствующий о способности галофитов накапливать хлориды даже в незасо-
ленных местообитаниях. Zygophyllum stapffii и Arthraerua также растут
в незасоленных сухих руслах, однако они встречаются чаще и развивают-
ся лучше в рядом расположенных засоленных долинах.
Срезанные ветви растений, характерных для незасоленных сухих
долин, очень быстро увядают, что говорит об их интенсивной транспи-
рации. Однако небольшие величины осмотического давления свидетель-
ствуют о благоприятном для них водном режиме. На примере древесных
растений видно, что их развитие не связано с наличием большого
количества осадков, выпавших в 1934 г., поскольку они смогли перенести
предшествующие периоды засухи в 1930—1933 гг. С другой стороны, виды
травяного покрова вновь исчезают при длительном отсутствии дождей
и истощении запасов влаги в небольших руслах временных водотоков.
404
Глава 9
Солевой режим в крупных пересыхающих долинах довольно сложен.
Верховья реки £вакоп расположены в области развития саванн. Серия
проб грунтовых вод, взятых в различных местах вдоль течения Свакопа,
свидетельствует о постепенном увеличении минерализации первоначаль-
но пресной воды. В табл. 60 приводится содержание хлоридов и сульфатов
в граммах на 1 л воды.
Таблица 60
Место взятия проб
верховья (Окаханджа) среднее течение (Нудис) устье (Свакопмунд)
Хлориды (NaCl) 0,008 0,23 2,28
Сульфаты (Na2SO4) 0,012 0,05 0,39
Увеличение концентрации солей объясняется двумя причинами.
1. Высоким уровнем грунтовых вод во многих местах русел пересы-
хающих долин, что способствует увлажнению почвы вплоть до поверхно-
сти и, как следствие этого, — сильному испарению воды. Это явление
наблюдалось в нижнем течении Свакопа у Палмхорста, где близ порога
грунтовые воды выходят на поверхность. Каждое утро вода течет здесь
довольно широким потоком, который на протяжении дня постепенно
ослабевает, так что вечером русло уже можно пересечь, не замочив ног.
К следующему утру течение восстанавливается полностью. Таким обра-
зом, испарение в дневные часы превышает приток воды, а это в свою оче-
редь вызывает соответствующее повышение концентрации солей (см.
также рис. 251).
2. Следующей причиной увеличения концентрации солей является
поступление соленых вод из русел боковых притоков. Выпадающий
в пустыне Намиб дождь смывает всю скопившуюся на возвышениях
соль в русла водотоков, которые переносят ее в Свакоп. Поэтому первые
ливневые потоки несут особенно сильно минерализованную воду, тогда
как при более продолжительных дождях содержание солей в ней падает.
В период обильных дождей, выпавших в 1934 г., сухой остаток вод под
рекой Свакоп у Свакопмунда снизился с 2,27 до 0,49 г!л. Поскольку,
однако, удельный вес солоноватой и соленой воды различен, смешения
вод не происходит. Поэтому на некоторой глубине от поверхности почвы
рядом могут течь потоки, отличающиеся разной степенью засоления. Этим
же объясняются сильные колебания солености вод в колодцах, располо-
женных недалеко один от другого.
В устье Свакопа грунтовые воды расположены на глубине 62 см\
почва увлажнена до самой поверхности. Вокруг отдельных растений ветер
насыпает небольшие песчаные дюны. Наносная почва, отложенная павод-
ком в 1934 г., содержит много органического вещества; в более глубоких
слоях почвы образуется сероводород, и она приобретает черную окраску
(благодаря сульфиду железа). Корневая система растений не проникает
глубже 10—30 см.
Здесь развиваются в основном галофиты и солевыносливые виды
(табл. 61).
Из числа упомянутых видов тамарикс относится к сульфо-галофитам,
поскольку наряду с хлоридами он в отличие от суккулента Suaeda содер-
жит в клеточном соке очень много сульфатов.
Пустыня туманов Намиб
405'
Таблица 61
Название растений Осмотическое давление, атм Содержание хлори- дов в клеточном соке растений, %
Кустарники Tamarix articulata . 12,8 75 (рассоление)
Lycium tetrandrum 19,6 60
Suaeda jruticosa .......... 26,5 50
Суккулентные галофиты Arthrocnemum glaucum . ... . Злаки и осоки 28,0 75
Diplachne paucinervis 23,3 55 (рассоление)
Phragmites communis var 15,1 35
Juncellus (Cyperus) laevigatus1 . . . 13,8 60
Разнотравье Heliotropium curassavicum 12,1 60
Nesaea fleckii 16,7 40
1 На побережье Флориды, по данным Гарриса, давление достигает 15,0 атм,
а содержание хлоридов 70%.
На поверхности почвы вследствие испарения накапливаются соли.
Осмотическое давление почвенного раствора на глубине 1 см составило
75 атм, но уже на уровне 30 см — всего 3,6 атм. Таким образом, расте-
ния вполне обеспечены влагой. Семена, видимо, могут прорастать только
в совершенно определенные периоды, когда снижается содержание солей
в верхних слоях почвы.
Сходная растительность развивается на морских побережьях, где
песок перемешан с разлагающимися водорослями. Здесь к перечислен-
ным выше видам присоединяются также ярко выраженные нитрофилы,
как, например, Atriplex muralis, Datura metel, Nicotiana glauca.
На влажной засоленной почве часто встречается Cryophyturn barkleya-
num (род Mesembryanthemum) с крупными листьями, покрытыми пузыр-
чатыми клетками эпидермиса размером 1 мм, напоминающими кристаллы.
Осмотическое давление составляло у них 22,8 и 26,2 атм, а содержа-
ние хлоридов —40 и 65%. У Arthrocnemum развиты безлистные ассими-
лирующие стебли, напоминающие Salicornia. Толщина кутикулы равна
4,5 |1, а число устьиц — 81 на 1 мм2.
Меньшей суккулентностыо отличаются виды солянок; ксероморфный
облик и сильное опушение свойственны Salsola zeyheri. Во внутриконти-
нентальных частях страны солянки развиваются на временно затопляе-
мых, но большей частью очень сухих территориях, почвы которых напо-
минают такыры, характерные для глинистых пустынь юга СССР
Осмотическое давление у этих видов достигало 48,3 и 61,9 атм при
содержании хлоридов, равном 40%. Они также накапливают много суль-
фатов. Приведенные значения характерны для галофитов; это понятно,
поскольку в 1935 г. еще преобладали благоприятные условия увлажнения.
В 1932 г., по данным Босса, осмотическое давление у изучаемых
растений было значительно выше (табл. 62).
1 R. Ganssen, Siidwest-Afrika, Boden und Bodenkultur, Berlin, 1963.
406
Глава 9
Таблица 62
Название растений Осмотическое давление, атм
Suaeda fruticosa Salsola zeyheri . . . Arthrocnemum glaucum Cryophytum barkleyanum . Heliotropium curassavlcum . до 44,3 » 74,0 » 33,0 » 40,8 » 32,4
Еще более высоким давлением отличаются эти виды, по данным Кил-
лиана и Фореля, в Бени-Униф (Алжирская Сахара) и, по материалам
Адриани, в Монпелье (юг Франции) 1 (табл. 63).
Таблица 63
Название растений Осмотическое давление, атм
Беии-Униф Монпелье
Suaeda fruticosa 26,2—51,7 ДО 80
Arthrocnemum glaucum 41,2—49,8 40—100
Ознакомление с анатомо-морфологическими особенностями различ-
ных групп растений, развитых в пустыне Намиб, дает возможность сде-
лать вывод, что разнообразие экологических форм, определяемое много-
образием условий среды в одной климатической области, часто бывает
гораздо большим, чем в сходных условиях произрастания, но приурочен-
ных к разным климатическим областям.
Экология Welwitschia mirabilis
Наиболее известным растением, распространенным в пустыне Намиб,
является Welwitschia mirabilis. Этот вид в систематическом отношении
резко отличается от других голосеменных1 2. У него развиваются два листа,
расположенные над подсемядольным коленом; растения растут очень долго,
причем могут достигать возраста 100 лет и более (рис. 319 и 320). Вель-
вичия — эндемик пустыни Намиб, однако и в пределах пустыни этот вид
встречается лишь в рамках узкой полосы, расположенной примерно
в 50 км от берега и простирающейся с перерывами от Хоуп-Майн между
Куизебом и Свакопом до Мосамедиша на юге Анголы.
У мужского экземпляра, диаметр ствола которого достигает 18 —
27 см, свекловидное подсемядольное колено возвышается над поверх-
ностью почвы примерно на 20 см и на 30 см углубляется в нее. На глубине
30 см его диаметр резко сокращается; здесь от корневой шейки отходит
стержневой корень толщиной 1 см. На глубине 60 см толщина корня все
еще не меняется.
1 N. J. Adrian i, Recherches sur la synecologie de quelques associations halo-
philes mediterraneenes, Sigma, № 32, 1934; Ch. Killian, L. Faure 1, Observa-
tions sur la pression osmotique des vegetaux desertiques et subdesertiques de I’Algerie,
«Bui. Soc. Bot. Franc.», 80, 1933, p. 775; La pression osmotique des vegetaux du Sud
Algerien, «Am. Physiol.», 12, 1936, p. 5.
2 Fr. Markgraf, Welwitschiaceae. In Engler-Prantl: Natiirliche Pflanzen-
familien, 2. Aufl., 13, 1926, S. 419—429.
Пустыня туманов Намиб
407
Рис. 319. Мужской цветущий экземпляр Welwitschia mirabilis.
Диаметр седлоообразнрй вершины растения — 80 см,
пустыня Намиб, в 20 км от Свакопмунда.
По данным Босса, корни вельвичии не проникают в почву глубже
1—1,5 м. Кроме этого, ветвящегося в более глубоких слоях почвы стерж-
невого корня, у вельвичии развиваются еще боковые корни, отходящие
от корневой шейки на глубине 7 —10 см и обильно ветвящиеся около
поверхности почвы.
Таким образом, у вельвичии корни, располагаются двумя ярусами.
Поверхностные корни поглощают влагу' из верхних горизонтов почвы,
смачиваемых небольшим количеством выпадающих здесь осадков, тогда
как стержневой корень служит для десукции почвенной влаги из более
глубоких слоев; однако он не достигает уровня грунтовых вод. Развитие
вельвичии не связано с грунтовыми водами. Точно так же вряд ли может
идти речь о поглощении влаги туманов или росы, поскольку вельвичия
растет на границе между внутренней и прибрежной частями Намиб,
куда туманы вторгаются лишь очень редко. Зато в этой части пустыни,
вероятно, чаще выпадает незначительное количество летних осадков.
Вельвичия обычно встречается на возвышенных плато на границе
внутреннего Намиба, причем собственно не на самом плато, начинающем
зеленеть после каждого хорошего дождя, а в верховьях сухих русел вре-
менных водотоков или пересыхающих долин, представленных здесь широ-
кими, неглубокими, едва заметными рытвинами. После сильных дождей
эти углубления собирают воду со всех вышерасположенных участков,
и поэтому почва в них увлажняется на глубину до 1,5 м. Почвенная влага
сохраняется в течение многих лет и используется растениями, распо-
ложенными далеко друг от друга в этих широких понижениях. Расстоя-
ния между отдельными растениями обычно превышают 20 м.
408
Глава 9
Наряду с вельвичией в 1935 г. здесь встречалось много отмерших
дернин злаков и одиночных экземпляров Arthraerua и Zygophyllum stapf-
fii, а также некоторых других видов (негалофитов). Ближе к низовьям
сухих русел, где почва лучше увлажнена, появляется растительность,
свойственная пересыхающим долинам и представленная Bauhinia, Раг-
kinsonia, Salvadora, а при наличии засоления — Tamarix.
При раскопке корней вельвичии были взяты образцы почвы; анали-
тические данные приведены в табл. 64.
Таблица 64
Горизонт, С At Почва Состав на 100 г абсолютно сухой пэчвы, г
вода NaCl сульфаты NaNO3
0-5 Крупный щебень 0,19 0,004 Нет 0,010
40 » » 2,3 Нет Нет 0,002
60 Сцементирован- ный слой 5,9 Нет Гипс 0,005
На глубине 60 см под верхними сухими горизонтами почвы распола-
гался очень влажный слой крупного песка. В другом разрезе на глубине
55—60 см в зоне роста корней почва также была влажной и содержала
0,008 г NaCl.
Вельвичия может расти и на склонах гор, покрытых крупными облом-
ками. Так, в одном случае вельвичия была обнаружена на высоте 10 м,
над сухим руслом, а в другом — даже на 100 м выше подошвы склона.
В трещинах выветрившейся пегматитовой жилы также был найден экзем-
пляр вельвичии. Однако эти местообитания не типичны для вельвичии;
растения здесь растут в виде одиночных экземпляров и развиваются плохо.
Рис. 320. Женский цветущий экземпляр Welwitschia mirabilis. Концы ли-
стьев засыпаны песком, пустыня Намиб.
Пустыня туманов Намиб
409
Возможно, они используют здесь влагу, стекающую по стенкам тре-
щин в результате конденсации тумана. В подсемядольном колене вельви-
чии может накапливаться известное количество воды, несмотря на отсут-
ствие специальных тканей, служащих для этой цели.
При ухудшении водного режима листья вельвичии высыхают. Сохра-
няется только меристема; ее клетки начинают расти при благоприятных
условиях влагообеспеченности. У опытного экземпляра спустя 3 недели
после выпадения 16,5 мм осадков прирост листа в длину составил 10 см.
Босс считает, что годичный прирост листьев достигает 30—40 см. У экзем-
пляров, пересаженных в Свакопмунд, ежегодный прирост равнялся 15 см.
В 1934 г. листья у вельвичии достигали необыкновенно крупных
размеров, а соцветия были поражены грибами (возможно, Aspergillus
niger). Вельвичия — двудомное растение; опыляется Odontopus sexpun-
ctatus\ возобновляется очень плохо; дает много пустых семян, здоровые
семена прорастают через 8—10 дней. Несмотря на это, Босс наблюдал
среди взрослых растений всего лишь 5 проростков. Возможно, это зависит
от засоления и быстрого иссушения верхних горизонтов почвы. Животные
поедают листья вельвичии лишь в случае крайней нужды.
Об анатомическом строении вельвичии можно получить представле-
ние по рис. 22, приведенному в книге Г. Вальтера (см. сноску, стр. 374) Ч
Лист отличается крайне ксероморфной (склерофильной) структурой.
Склеренхима и крупные склереиды, набыщенные мелкими кристаллами
щавелевокислого кальция, придают листу большую жесткость. Кутикула
отличается большой толщиной. Число устьиц составляет 98 на 1 мм1 2
на верхней стороне листа и 114 — на нижней.
Транспирация характеризуется повышенной интенсивностью при откры-
тых устьицах и резко сокращается при недостатке воды. В критические
периоды, кроме того,вследствие высыхания апикальных частей листа проис-
ходит сокращение листовой поверхности. Срезанные листья начинают увя-
дать. Испаряя 1,55 мл!час в полуденное время, отрезок листа (поверх-
ности срезов были покрыты парафином) за 80 минут терял 16,5% сырого
веса. Кэннон при помощи других методов (кобальтовая бумага) устано-
вил, что величина транспирации на единицу поверхности у вельвичии
примерно в 10 раз больше, чем у суккулентов, и соответствует транспира-
ции жестколистных растений 2.
Строение листьев не дает оснований к выводу о возможности погло-
щения поверхностью листьев влаги туманов или росы, о чем неоднократ-
но упоминалось в литературе. Поверхность листьев не смачивается, и во
время утреннего тумана листья покрыты отдельными капельками воды.
Только срез ствола всасывает воду, как губка. Еще предстоит выяснить,
имеет ли это явление какое-либо экологическое значение. С жестколист-
ными растениями вельвичию объединяет относительное постоянство вели-
чины осмотического давления, лишь незначительно меняющейся в за-
сушливые и дожливые годы.
По данным Босса, осмотическое давление достигает у вельвичии сле-
дующих значений (в атм) (табл. 65).
В табл. 66 приведены данные автора о содержании клеточного сока
вельвичии.
1 См. также R. J. Rodin, Leaf anatomy of Welwitschia, I und II, «J. Bot.»,
45, 1958, p. 90—103. У Родина и у Цемке перечислена более старая литература.
2 W. A. Cannon, General and physiological features of the vegetation of the
more arid portions of southern Africa, Carnegie Inst, of Wash., Publ. № 354, 1924.
410
Глава 9
Из материалов, приведенных в таблице, видно, что в клеточном соке
вельвичии содержится около 20—25% хлоридов; это свидетельствует
о том, что вельвичии не может бь1ть причислена к галофитам,хотя и может
Таблица 65
Старое
растение
Молодое
растение
Засушливый 1932 г.
Дождливый 1934 г.
39,6
34,8
35,4
34,9
расти на слабо засоленных почвах. В клеточном соке содержится также
довольно много сульфатов (4—5%). Содержание хлоридов в клеточном
соке галофитов, росших рядом с вельвичией, составляло: у сульфо-гало-
фита Arthraerua —35% и у хлоридо-галофита Zygophyllum —81—95%.
Таблица 66
Осмоти- ческое давление, атм Хлориды Моноса- хариды, мг/мл Дисаха- риды, мг/мл Концен- трация сахаров, атлг
п/100 атм %
17/11 Область распро- странения вельви- чии Основание листа 32,3 15,9 6,6 20 20,6 7,4 3,1
Верхушка листа 33,5 18,3 7,7 23 31,0 9,7 4,6
17/Ш Дорога в Витпорт у Зебровых гор Весь лист . . . 34,0 20,4 8,5 25 36,9 6,9 5,2
То же у другого растения 33,4 17,5 7,3 22 37,0 2,3 5,0
Возраст крупных экземпляров вельвичии может превышать 100 лет.
Возраст одного растения с полуразложившимся стволом (диаметром около
1,5 м) и одним живым листом на одной стороне ствола, определенный
радиоуглеродным методом (С14), достигал 500—600 лет 4.
1 Н. Н е г г е, The age of Welwitschia bainesii (Hook, f.) Carr: C14-research, «J. S.
Afr. Bot.», 27, 1961, p. 139.
Глава 10
ЧИЛИЙСКО-ПЕРУАНСКАЯ ПУСТЫНЯ
С ОАЗИСАМИ ТУМАНОВ
TJМАИ КАК ИСТОЧНИК ВЛАГИ
Подобно тому как холодное Бенгельское течение создает особые
климатические условия у берегов Юго-Западной Африки, Перуан-
ское течение обусловливает особенности климата пустыни, развитой
на побережьях севера Чили и юга Перу. Температура воды Перуанского
течения несколько выше, чем Бенгельского, и составляет зимой около
17°, а летом около 24°. Отличия выражаются и в том, что Анды защищают
побережье от влияния восточных пассатов, а горы высотой 500—800 м
близко подступают к самому берегу (рис. 333). В Юго-Западной же Афри-
ке пустыня Намиб характеризуется лишь незначительным поднятием
в глубь материка (с уклоном всего около 1 : 100). Более высокие горные
хребты, как, например, горы Бранденберг, Эронго, плоскогорье Комас,
горы Науклюфт и Царис, расположены далеко от берега, уже вне зоны
влияния юго-западных ветров и прибрежных туманов. Лишь несколько
не очень высоких хребтов, как, например, горы Россинг, подходят к берегу
на расстояние до 30 км (см. цветное фото 1, стр. 304).
Это орографическое своеобразие Перу определяет сильное воздействие
берегового тумана, известного под названием «гаруа»; под влиянием тума-
на на крутых склонах прибрежных гор, «ломас», в зимние месяцы с час-
тыми туманами в пустыне, обычно лишенной растительности, возникают
настоящие оазисы туманов, где развивается особый тип эфемеровой пус-
тыни, так называемая растительность «лома».
Прибрежная пустыня простирается примерно от 6 до 27°30' ю. ш.
и там, где наблюдается зимний максимум осадков, переходит в полупусты-
ню и в область развития жестколистной растительности. К северу, наобо-
рот, возрастает количество летних осадков и пустыня сменяется сначала
зарослями сухих кустарников, а затем листопадными и, наконец, полу-
вечнозелеными тропическими лесами. Характеристика прибрежных
пустынь дана в работах Рейхе (1907) и Вебербауэра (1911—1945), а также
Бернингера (1925), Тролля (1930—1956), X. и. М. Кёпке (1953—1954),
Шмитхюзена (1956) и Рау (1951, 1958). Общее описание растительного
покрова приводит Кёпке \ Более подробные экологические исследования
в последнее время провел Элленберг (1959); он тщательно изучил расти-
тельность «лома» и осадки, не поддающиеся измерению обычными мето-
дами 1 2.
1 Н. W. К ое реке, Synokologische Studien an der Westkiiste der peruanischen
Ande, «Bonner Geogr. Abh,», H. 29, 1961.
2 H. E lie n b er g, Uber den Wasserhaushalt tropischer Nebeloasen in der Kiis-
tenwiiste Perus, «Вег. Geobot. Forsch.-Inst. Rubel fiir 1958», 7, 1959, S, 47—74.
412
Глава 10
Данные о климатических условиях прибрежной равнины приведены
в табл. 67 и на климадиаграммах (рис. 322—324).
Р и с. 321. Юго-западная часть Перу. На карте показано расположение метео-
рологических станций. Измерения количества осадков, приносимых тума-
нами, проводились близ Лачая.
322
324
Рис. 322—324. Климадиаграммы прибрежной пустыни юго-запада Перу и севера
Чили (Антофагаста). Данные об осадках отклоняются от указанных в табл. 67, что
всегда бывает в засушливых областях при сопоставлении разных периодов наблю-
дений.
Чилийско-перуанская пустыня с оазисами туманов
413
Таблица 67
Количество осадков на побережье Перу (в лмг)
(по Элленбергу)
Станции Лето Зима Лето За
береговые материковые I ”1 Ш IV V VI VII VIII IX X XII XII ГОД
Чиклайо (31 м) 1 1 1 1 1 1 1 0 0 0 0 0 0 1 0 5
Трухильо (26 м) 0 0 1 0 0 0 0 0 0 0 0 0 3
Парамонга (15 м) 1 0 0 0 6 1 1 3 2 0 0 0 15
Лима (137 м) 1 0 1 0 3 5 6 7 6 2 1 1 34
Каньете (36 м) 0 1 1 0 9 3 4 3 3 1 1 12 40
Писко (6 м) 0 0 0 0 0 0 0 1 1 0 0 0 2
Пуэрто-Ломас 0 0 0 0 1 0 0 0 0 2 0 0 3
(10 м) Такна (457 л<) 3 2 0 2 4 3 3 7 8 6 3 2 43
Из этих данных видно, что чилийско-перуанское побережье, как
и пустыня Намиб, практически лишено осадков. Поэтому развитие рас-
тений в столь суровых условиях обусловлено исключительно наличием
грунтовых вод; в частности растения поселяются вблизи устьев рек, спа-
дающих с Анд. В лагунах встречаются заросли галофитов, однако мангро-
вы в области влияния холодного Перуанского течения совершенно отсут-
ствуют.
Процесс возникновения туманов в этой части побережья напоминает
таковой в пустыне Намиб. Непосредственно на берегу туман образуется
довольно редко и держится только ночью. Пустыня здесь лишена расти-
тельности. Лишь на подвижных дюнах встречаются полосы или куртины,
образованные Tillandsia, вокруг которых иногда накапливается песок
в виде холмиков высотой до 1 м. Эти растения, о которых, в частности,
упоминает также Бернингер при описании растительности в Чили, где
они развиты на высоте 800—1200 м (рис. 325), часто непосредственно
лежат на почве *. Корни у них слаборазвиты и не используются ни для
закрепления растений, ни для поглощения воды. Вода поглощается
листьями с помощью чешуевидных волосков, типичных для бромелиевых;
это наблюдается только после крайне редких дождей или после конденса-
ции тумана во влажные ночи и в утренние часы зимой (южного полуша-
рия). Интенсивность транспирации у Tillandsia крайне незначительна,
хотя поверхность листьев даже при сильном тумане нагревается до 30°.
Водный режим этих истинных «растений туманов» недавно был под-
робно изучен П. де Т. Альвимом и Андрэ Узеда. Они любезно предоста-
вили автору свои еще не опубликованные данные.
В качестве опытного растения была выбрана Tillandsia straminea
(рис. 326). Осмотическое давление в клетках листьев оказалось равно
7—8 атм. Если листья держать 24 часа в эксикаторе при относительной
влажности воздуха от 80 до 100%, то лишь при почти максимальном
насыщении воздуха парами наблюдается очень небольшое увеличение
веса листьев; в этом случае, видимо, нельзя также полностью исключить
конденсацию пара. Из этого следует, что тилландсия поглощает воду
при помощи листьев только при их увлажнении.
’О. Berninger, Extreme Ausbildung einer Nebelvegetation in der nord-
chilenischen Waste, «Zschr. Ges. Erdkunde», № 9—10, 1925.
414
Глава 10
Интенсивность транспирации у Tillandsia определяется в естествен-
ных условиях довольно легко. Растения, общий вес которых достигает
около 3 кг. были уложены на пластмассовую пленку и оставлены на месте.
Растения взвешивались в 9 час., когда роса уже испарялась, и в 17 час.
Исследования проводились в июл’е при особо благоприятных погодных
условиях, поскольку туман и роса в это холодное время года наблюдаются
особенно часто. Средняя температура равнялась 14°, а средняя упругость
водяного пара —12 мм рт. ст. Результаты представлены на рис. 327;
отмечается увеличение веса в течение ночи и его снижение днем. В сред-
нем суточный прирост сырого веса в течение июля составил 0,17%. Таким
образом, рост даже в это время года происходит чрезвычайно медленно.
В неблагоприятное же время года он, вероятно, полностью прекращается.
Во всяком случае, опыты показали, что тилландсии существуют только
за счет влаги туманов или росы, которую они поглощают при помощи
упомянутых чешуевидных волосков. Это и дает им возможность расти
в пустыне, где единственным источником влаги являются туманы. Тйл-
ландсии, пожалуй, единственно известные до настоящего времени среди
покрытосеменных настоящие «растения туманов». В пустыне Намиб
и в Южной Африке не удалось обнаружить ничего аналогичного. Конеч-
но, необходимо тщательно изучить эти растения с физиологической точки
зрения. Поскольку растения обладают действующими корнями, не исклю-
чена возможность поглощения водн также из почвы, сильно увлажненной
туманом или росой (см. стр. 379).
В нескольких километрах от берега морской воздух наталкивается
на стену гор; поднимаясь по склону, он охлаждается, благодаря чему
Рис. 325. Tillandsia werdermannii. растущая в полосе туманов на севере Чили, близ
Такны, образует небольшие куртины на сухой песчаной почве.
Чилийско-перуанская пустыня с оазисами туманов
415-
Рис. 326. Полосы, образованные Tillandsia latifolia, Т. purpurea и Т. straminea у под-
ножия Морро Солар, южнее Лимы. На заднем плане — море (фото Элленберга).
возрастает вероятность образования здесь туманов (рис. 328). На высоте
примерно 600 ж, где восходящий холодный воздух встречается с более
теплым воздухом, образуется густой туман — г а р у а, который в тече-
ние нескольких месяцев может возникать ежедневно, даже не рассеиваясь
после полудня, что обусловливает высокую влажность воздуха. К тому
же условия для образования капелек тумана здесь, как подчеркивает
Элленберг, особенно благоприятны. Это определяется следующими обстоя-
тельствами:
1. На побережье постоянно господствует легкий ветер от южного
до юго-западного направлений, скорость которого хотя и редко превы-
шает 4 м/сек, однако достаточна, чтобы; непрерывно приносить новые
массы тумана.
2. Крутые склоны прибрежных гор способствуют вынужденному
подъему и охлаждению воздуха, что в свою очередь приводит к почти
непрекращающемуся процессу конденсации находящейся в нем влаги
и образованию туманов,
3. Температура воды Перуанского течения значительно выше, чем
Бенгельского. Вследствие этого содержание водяного пара в воздухе
здесь выше, что при равной степени охлаждения способствует образова-
нию большего количества влаги.
Конденсация влаги туманов происходит на твердых препятствиях,
мимо которых движется туман. На большую поверхность выпадает
и больше влаги из тумана. Поэтому дождемером можно измерить лишь
часть воды. На поверхности почвы конденсируется сравнительно мало
влаги, но зато на растениях — намного больше, причем количество влаги,
естественно, должно возрастать с увеличением размеров растений. Поэто-
му на поверхности деревьев конденсируется особенно много воды; она
416
Глава 10
капает с них непрерывно. Без движения воздуха количество влаги,
возникшее в процессе конденсации, было бы очень небольшим. Количе-
ство воды в 1 м3 тумана колеблется очень сильно (от 0,01 до 5,0 г), состав-
ляя в среднем 0,4—0,8 г. Если примерно 0,5 г будет задержано кронами
деревьев, то при скорости ветра 1 м/сек (то есть 3,6 км/час) это будет соот-
ветствовать 1,8 л воды, или почти 2 мм осадков в час. Так как число дней
с туманом достигает за зиму 120 и туман ежедневно держится в течение
многих часов, то в результате конденсации под пологом деревьев может
накапливаться значительное количество влаги.
Рис. 327. Водный баланс тилландсии в прибрежной пустыне Перу
(по Альвиму и Узеда).
А —> вес в 9 час. после выпадения росы; Б — вес в 17 час. (подробности
в тексте).
Результаты измерений (табл. 68), проведенных в 1944—1954 гг.,
свидетельствуют об очень большом количестве годовых осадков под поло-
гом деревьев (в скобках указаны максимальные и минимальные значе-
ния).
Таблица 68
Среднее количество осадков (в мм)
Открытый участок Под пологом Casuarina Под пологом Eucalyptus
168(121-219) 488 (262-819) 676 (252-1240)
Во всех трех случаях наибольшее количество осадков, сконденси-
ровавшихся из тумана, было зарегистрировано в 1948 и 1949 гг., но зато
годы с минимальным количеством осадков совпадали в редких случаях.
Чилийско-перуанская пустыня с оазисами туманов
417
ЮЗ
>20'С
СВ
Р и с. 328. Схематический профиль берегового склона в Перу с данными
о частоте туманов (гаруа) и распределении растительности в течение зимы
южного полушария (по Эллепбергу).
Рис. 329. Влажность почвы (в %) под редкостойным эвкалиптовым на-
саждением около Лачая в 1957 г. Туманы в 1957 г. начали образовываться
только в июне (по Элленбергу).
Зимой с появлением туманов влажность почвы под деревьями быстро
повышается до глубины 1,5 м (рис. 329). Наблюдения показали, что запас
влаги в почве даже больше количества выпавших осадков. Распростра-
нение корневых систем растений связано с распределением влаги в почве
(рис. 330). При неглубоком промачивании корни травянистых растений
проникают на глубину до 50 см, тогда как при более сильном промачи-
вании корни деревьев проникают на значительно большую глубину
и на протяжении года в почве на глубине 1 м и больше образуется даже
некоторый запас воды. Испарение и тем самым транспирация вряд ли
могут быть измерены в туманные дни, однако в умеренно облачные дни
их величина уже заметно возрастает (максимальное испарение 0,45 мл/час),
а в засушливый период достигает больших значений (при температуре
воздуха, равной почти 28°, испарение превышает 0,7 мл1час).
Из этого следует, что гидратура в течение зимних месяцев благо-
приятна для развития растительности. Однако это наблюдается только
на определенной высоте над уровнем моря; ниже условия для развития
растительности менее благоприятны.
418
Глава 10
РАСТИТЕЛЬНОСТЬ ЛОМА
За прибрежной полосой, где распространены тилландсии, еще на
несколько километров вглубь простирается пустыня, а затем местность
начинает постепенно повьмпаться, Из растений первыми появляются тай-
нобрачные, «Это в большинстве случаев синие водоросли (Nostoc commune
и др.), покрывающие не более 30% поверхности почвы; в отличие от тил-
ландсий они не растут на подвижных песках, В холмистой местности
на огромных площадях, например южнее Лимы у Пачакамак, на песках
развиваются кустистые лишайники (виды Cladonia); этот ландшафт напо-
минает дюны Северо-Западной Европы, покрытые лишайниками; в отли-
чие от последних лишайники развиваются на субстратах, характеризую-
щихся нейтральной или щелочной реакцией. Здесь же встречаются облом-
ки породы, покрытые пестрым покровом из корковых лишайников; это
наблюдается, например, в окрестностях Моро Солар, южнее Лимы» К
Выше по склону, где туманы приносят больше влаги, место тайно-
брачных занимают эфемеры. Из однолетников здесь часто встречаются
Осадки, мм
1944~1954гг.}В среднем за год
крайние значения
Засушливый 1957г.
Запасы влаги 8 почве в 1957г.,мм ~60
Почва, лишенная
растительности
168
121-219
79
~ 140
блага,
накопившаяся
в предыдущие
годы
Глубина
промачивания
почвы в 1957г,
-WIUADIL
Почвенный
просриль
Глубина проникно-
вения корней
Растения
полосы туманов
Содержание
влаги в почве
Рис. 330. Количество осадков и влажность песчаной почвы в эфемеровой пусты не*
(«лома») близ Лачая (по Элленбергу).
1Н, Ellenberg, op. cit., S. 51,
Чилийско-перуанская пустыня с оазисами туманов
419
Рис. 331. Сомкнутый растительный покров, развитый в полосе туманов (Euphorbia
lactiflua, Cereus, Echinocactus и др.) близ Тальталя (фото Мортенсена и Бернингера).
эндемичные виды Nolana, однако злаки пока еще не появляются. Только
еще выше в горах количество злаков начинает увеличиваться, травяной
покров смыкается и достигает высоты 60 см. В его составе встречаются
покрытая жгучими волосками Loasa urens. Кроме того, Nicotiana paniculata,
виды Chenopodium, Malvaceae, Caryophyllaceae, Galinsoga, Urtica и евро-
пейские сорняки (Sonchus oleraceus, Stachys arvensis, Erodium cicutarium,
Stellaria media и др.). Здесь распространены также геофиты, например
виды Iridaceae, Solanum, Oxalis и др. У Stenomesson coccineum во время
туманной зимы развиваются только листая, а оранжево-красные цветки
распускаются летом.
Древесные породы в настоящее время в этих условиях почти пол-
ностью отсутствуют, однако в культуре здесь хорошо растут вечнозеле-
ные Casuarina и Eucalyptus, удовлетворительно переносящие длительный
засушливый период. Местные древесные породы встречаются только
в виде одиночных экземпляров на каменистых почвах. Это Acacia тасга-
cantha, Carica candicans и Caesalpinia tinctoria, на ветвях которой посе-
ляются эпифитные мхи, лишайники и даже миниатюрная полусукку-
лентная Peperomia crystallina. Заросли кустарников, по вполне обоснован-
ному предположению Элленберга, можно рассматривать как остатки
прежних лесов.
Сходная растительность, развитая в полосе туманов, отличающаяся,
однако, большим числом суккулентных форм, распространена на севере
Чили (рис. 331).
Обычно безлюдная область туманов преображается зимой, когда
с высыхающих в зимние месяцы пастбищ сюда с Анд спускаются кочев-
ники со своими стадами. На протяжении веков они вырубали местные леса
420
Глава 10
на топливо, причем для этой цели использовались даже корни. Это привело
к полному уничтожению лесов, восстановлению которых препятствовала
пастьба скота. Животные не трогали только ядовитых кустов Croton,
колючей Acacia macracantha и кактусов. Истреблению лесов способство-
вала, вероятно, также большая потребность прибрежных поселений в дре-
весном угле.
Сохранившийся небольшой лесной массив близ Атикипа может дать
представление об исходном растительном покрове. В его составе господ-
ствует преимущественно один вечнозеленый вид Eugenia, отличающийся
очень жесткими листьями; кривые стволы этой древесной породы дости-
гают высоты 5—8 м и образуют плотно сомкнутый полог. Здесь встречает-
ся также Capparis. В тени древесного полога растет несколько видов
гигроморфных папоротников
На высоте около 700 м плотность облаков уменьшается, а на уровне
800—1000 м проходит их верхняя граница и сопряженная с ними граница
растительности «лома». У верхнего предела распространения этого пояса
в южном Перу, в частности в местах скопления облаков, появляются круп-
ные пятна тилландсии, вокруг которых, как и на побережье, подчас обра-
зуются небольшие песчаные дюны (см. рис. 325 и 328). На сухих подвет-
ренных северных и восточных склонах поселяются лишайники и кактусы.
За полосой прибрежных гор, куда туманы уже не проникают, вплоть до
главной цепи Анд простирается лишенная растительности пустыня.
Наиболее ярко выраженным вариантом ее является пустыня Атакама
на севере Чили (рис. 332).
Однако даже и в этой пустыне встречаются растительные организмы.
Автотрофные растения представлены здесь почвенными водорослями
с сильно разветвленными слоевищами: Schizothrix atacamensis на более
низких, a Calothrix desertica — на более возвышенных участках; две формы
Cyanidium развиваются в трещинах диоритов на побережье Чили 1 2.
Элленберг установил, что степень развития растений зависит от интен-
сивности конденсации влаги туманов. Туман в этом случае заменяет
дождевые осадки; за исключением тилландсии растущие здесь растения
не имеют никаких особых приспособлений для поглощения влаги туманов.
Среди однолетников встречаются и гигроморфные и мезоморфные виды,
а среди многолетников, вынужденных переносить длительные периоды
засухи, — склерофильные формы. Все эти виды поглощают воду из почвы
при помощи корней. Поэтому можно говорить об «оазисах туманов», но
следует избегать термина «растения туманов», иначе к их числу пришлось
бы отнести также Sonchus arvensis, S tacky s arvensis, Erodium cicutarium
1 Такие оазисы туманов встречаются также на западном склоне прибрежной
части Кордильер в центральном Чили (здесь выпадает 250 мм осадков) и близ Эрка-
вита на Красном море (218 мм). Более подробные данные приведены в работах:
J. К ummer ow, V. Matte, F. Schlegel, Zum Problem der Nebelwalder an
der zentralchilenischen Kiiste, «Вег. Dtsch. Bot. Ges.», 74, 1961, S. 135—145; M. Kas-
sas, Certain aspects of landform effects on plant water resources, «Bull. Soc. Geogr.
Egypte», 33, 1960, p. 45—52. См. также G. Foilman n, Eine dornbewohnende
Flechtengesellschaft der zentralchilenischen Sukkulentenformationen mit kennzeichender
Chrysothrix noli-tangere Mont., «Вег. Dtsch. Bot. Ges.», 73, 1960, S. 449—462. В этой
области в течение 10—11 месяцев продолжается засушливый период, однако туманы
наблюдаются в течение всего года. Этот же автор описывает распространенные на севе-
ре Чили (18—30° ю. ш.) на высоте 0—1600 м сходные сообщества, в составе которых
господствует Anaptychia intricata («Вег. Dtsch. Bot. Ges.», 74, 1961, p. 495—510).
Здесь встречаются также Т rentepohlia и на кактусах тилландсии.
2 G. Н. Schwabe, Zur autotrophen Vegetation in ariden Boden IV, «Osterr.
Bot. Z.», 107, 1960, S. 281—309.
Чилийско-перуанская пустыня с оазисами туманов
421
Рис. 332. Пустыня Атакама на севере Чили (примерно 27° ю. ш.). Растения ветре-
чаются только в руслах временных водотоков (фото Мортенсена и Бернингера.)
и другие виды европейских сорняков, являющиеся компонентами расти-
тельности «лома».
Схематический профиль растительности от побережья до вершин Анд
приведен на рис. 333.
Засушливое высокогорное плато Альтиплано, расположенное
к востоку от главного хребта, называют пуна. Пуна простирается до гра-
ницы с Аргентиной, где и была описана Кабрерой *. В этой области близ
тропика Козерога на высотах 3300—4300 м условия чрезвычайно суровы.*
Средняя температура равна 8—10°, минимальная —18°, максимальная
30°. Заморозки бывают во все времена года; снеговая линия проходит
на высоте 5500 м. Осадки выпадают летом (декабрь — март); их количе-
ство достигает 103—324 мм в год. Количество осадков уменьшается к югу;
они выпадают очень неравномерно. В наиболее засушливой западной
части пуны встречаются выходы солей. Восточная окраина более увлаж-
нена и частично окультурена. Здесь возделываются Vicia faba, Zea mays,
Medicago sativa, а также Solanum andigenum и Chenopodium quinoa. Из
числа заносных сорняков здесь встречаются Meli lotus indicus, Rumex
crispus, Convolvulus arvensis, Brassica campestris.
Из высших растений к сложноцветным относится 23% всех видов,
к злакам —13% и к бобовым —7%. Растительный покров сухой пуны
большей частью образован Fabiana densa (Solanaceae), Psila boliviensis
(Compositae) и Adesmia horridiuscula (Leguminosae). Это полупустыня,
где господствуют невысокие кустарники; степень покрытия не превы-
шает 10%. Несколько выше по профилю появляются Prosopis ferox,
колоннообразный кактус Oreocereus и древовидный Polylepis tomentella.
1 A. L. G а b г е г а, VI La vegetation de la Puna Argentina, «Revista de Invest.
Agricolas», 11, № 4, 1958.
422
Глава 10
Рис. 333. Поперечный профиль Анд на широте Лимы от побережья до области Вы-
соких Анд (по Кёпке из С. Troll, 1959).
1 — море и устья рек; 2 — пустыни; з — поля гуано на островах; 4 — полоса густого тумана
(гаруа); -5 — растительность «лома» на холмах прибрежной полосы; в — пустыни с покровом
из Tillandsia; 7 — прибрежный лес; 8 — субнивальный пояс растений-подушск; 9 — злаков-
ники пуны; 10— злаковая степь с редкими кустарниками; 11 — скалистые склоны, покрытые
видами бромелиевых; 12 — редкостойный вечнозеленый лес; 13 — кустарниковая степь; 14 —
склоны долин, покрытые летнезеленым травостоем; 15 — заросли Саггса и Jalropha\ 16 — за-
росли колоннообразных кактусов; 17 — ледники и фирновые поля.
Местами преобладают дерновинные злаки —Festuca scirpifolia или Реп-
nisetum chilense, Дерновинные злаки доминируют также в наиболее высо-
ких частях Анд, выше 4300 м {Festuca orthophylla, F, chrysophylla, Роа
gymantha и др.). Кроме того здесь распространены растения-подушки —
Azorella yareta (Umbelliferae), Pycnophyllum (Caryophyllaceae), Opuntia
nigrispina, а на засоленных почвах Anthobryum triandrum (Frankeniaceae),
Salicornia pulvinata. Галофиты встречаются в разных сочетаниях; Spo-
robolus rigens чаще всего растет совместно с видами Atriplex, a Lycium
humile с Distichlis humilis.
Глава 11
ПУСТЫНЯ КАРРУ
ОБЩАЯ ХАРАКТЕРИСТИКА
Пустыня Карру 1 расположена в Южной Африке между прибрежной
возвышенностью с резко выраженным сезоном зимних дождей
и внутренним нагорьем Южной Африки, где преобладают летние
дожди Каппу, как и пустыня Сонора, расположенная близ Таксона,
является скорее полупустыней, чем пустыней. От моря она отделена гор-
ными хребтами Звартберг и Лангенберг, простирающимися параллельно
южному берегу и превышающими 2000 м. Территория между этими гора-
ми и крутым склоном нагорья и представляет собой собственно плато
Большое Карру (рис. 334), распадающееся на многочисленные отдельные
котловины. Годовое количество осадков в этой области обычно не превы-
шает 200 мм. Дожди идут преимущественно осенью. К востоку возрастает
количество летних осадков. Температуры высокие. На севере к Большому
Рис. 334. Южная Африка. Область Карру. Заштрихована территория распростра-
нения растительности Капской области, где осадки выпадают зимой.
1 На языке готтентотов karroo означает «засушливая местность».
424
Глава 11
Карру примыкает нагорье Верхнего Карру, простирающееся за Оранже-
вую реку, где оно углубляется в южную часть Оранжевой Республики.
Вблизи северной границы Карру, в заповеднике «Veld Reserve Fauresmith»,
возглавляемом М. Хенричи, автор проводил экологические исследова-
ния х. Распространенные здесь столовые горы разделены отдельными кот-
ловинами, сток которых направлен в Оранжевую реку.
Вследствие значительной высоты (1100—1300 3i) зимы в Верхнем
Карру холодные. Ночью часто бывают заморозки. Территория занята
исключительно пастбищами, перемежаемыми лишь очень небольшими
площадями пашни. Только чуть южнее Блумфонтейна начинается область
возделывания кукурузы. Три климадиаграммы дают представление о кли-
мате отдельных частей Карру (рис. 335—337).
Поскольку Карру расположено между областью, где осадки выпадают
зимой, и областью с преобладанием летних осадков (аналогично пустыне
Сонора), эта пустыня характеризуется исключительным разнообразием
суккулентов. Наряду с суккулентами некоторую роль играют и кустар-
нички (преимущественно сложноцветные), особенно ценимые как корм для
овец. В растительном покрове Малого Карру, расположенном южнее гор
Звартберг (рис. 338), уже появляется примесь капских элементов, свой-
ственных области зимних дождей. В расположенном севернее гор Зварт-
берг Большом Карру, напротив, преобладают суккуленты (рис. 339);
наряду с ними здесь широко распространены геофиты (Iridaceae, Amaryl-
lidaceae, Liliaceae и клубневые Oxalis), Обильные весенние дожди иногда
превращают обширные территории в течение 1 —2 месяцев в настоящее
море цветов. Из кустарничков в Верхнем Карру встречаются Pentzia,
Pteronia, Eriocephalus, Chrysocoma, Euryops, Diplopappus, Tripteris и дру-
гие, образующие «велд» с господством сложноцветных; кроме того, здесь
распространены виды Lycium, Zygophyllum и несколько видов Mesembry-
anthemum с суккулентными листьями, но одревесневшими побегами.
В Большом Карру растут также и более крупные древесные породы,
например Olea verrucosa, Euclea, Osyris, Ehretia, виды Rhus (Rhus lancea), не-
которые виды акации или Salix capensis и Royena (Diospyros). Однако эти
виды встречаются только в местах, хорошо обеспеченных влагой.
Для Большого Карру типичны следующие суккуленты:
1) Стеблевые суккуленты — виды Euphorbia (Е. таи-
retanica и др.), виды Asclepiadaceae: Stapelia, Caralluma, Hoodia, Tricho-
caulon и, кроме того, Kleinia (Senecio), Sarcocaulon, Pelargonium (оба
последних обладают мезоморфными листьями, сбрасываемыми в период
засухи) и Cotyledon fascicularis, образующий переход к следующей группе.
Рис. 335—337. Климадиаграммы области Карру («Klimadiagramm-Weltatlas», 1960).
1 Н. Walter, Grasland, Savanne und Busch der ariden Teile Afrikas m ihrer
okologischen Bedingtheit, «Jb. wiss. Bot.», 87, 1939, S. 750—860. В этой работе при-
водятся описываемые ниже опыты. F F
Пустыня Карру
425
Рис. 338. Малое Карру близ Ледисмита.1 Здесь широко распространены
Carissaferoxfpodonaea thunbergii, Euclea undulata, Mesembryanthemum spp.,
Cotyledon wallichii, Elytropappus rhinocerbtis и др. Развитие земледелия
возможно лишь при орошении (фото Марлота).
2) Листовые суккуленты, вроде видов Cotyledon и Crassula,
Portulacaria и Anacampseros (Portulacaceae), лилейные, например виды
A loe, Haworthia, Gasteria, Apicra, и внешне похожие на камни Titanopsis,
Lithops (цветное фото 2, стр. 304), Argyroderma, Didymaotus и неко-
торые виды Crassula.
3) Корневые суккуленты с подземными запасающими
органами, например Pachypodium bispinosum, виды Asparagus, Fockea
и др. У относящихся к стеблевым суккулентам низкорослых видов Euphor-
bia часто развиваются крупные по размерам подземные запасающие воду
органы. '
Эти суккуленты проникают в Малый Намакваленд и в южную часть
Юго-Западной Африки на расстояние, где еще сказывается влияние зим-
них дождей (рис. 340).
В Карру распространен также вид, относящийся к семейству Diosco-
reaceae — Testudinaria elephantipes с шишковатым, покрытым пробковым
покровом стволом, и два вида Encephalartos (Cycadaceae). Кроме того,
здесь встречаются некоторые пойкилогидрические папоротники и некото-
рые однолетники. В отдельных бессточных котловинах в пустыне Карру
распространены засоленные почвы, однако крупные соленые озера или
обширные соляные отложения здесь отсутствуют, поскольку в этой обла-
сти не встречаются молодые морские осадочные породы. На засоленных
почвах развиваются галофиты, например виды Tamarix и Salsola, Atri-
plex halimus и различные виды Mesembryanthemum. Подробное описание
растительности Южной Африки приводит Марлот 4.
1 R. М а г 1 о t h, Das Kapland, Wiss. Ergeb. Deutsch. Tiefsee-Exp., Bd II»
Jena, 1908.
Рис. 339. Большое Карру близ Лейнгсберга. Покров образован суккулентами Euphor-
bia mauretanica (преобладает), различными видами Mesembryanthemum, Cotyledon
orbiculata и Rhigozum trichotomum (фото Бруннталера).
Рис. 340. Карру близ Штикфонтейна (Малый Намакваленд). Преобладают
виды Mesembryanthemum и Galenia africana. На переднем плане — Cheiri-
doрsis spec, (род Mesembryanthemum) (фото Ветштейна).
Пустыня Карру
427
РАСТИТЕЛЬНОСТЬ ВЕРХНЕГО КАРРУ
В районе Фаурсмита намечается переход к злаковникам Верхнего
Велда; здесь уже ясно выражен сезон летних дождей.
Из данных климадиаграмм (рис. 337) видно, что уже в Фаурсмите
осадки выпадают преимущественно летом, а зимние месяцы (июнь —
сентябрь) отличаются засушливостью. Среднегодовое количество осадков
достигает 412 мм. В июне — июле возможны заморозки, правда большей
частью только ночные.
В областях, где происходит стык различных типов растительного
покрова, характер размещения растительности определяется почвенными
।
Рис. 341. Ландшафт в окрестностях Фаурсмита, Южная Африка.
Травостой уничтожен в результате пастьбы; начало овражной эро-
зии (фото Э. Вальтер).
условиями. И действительно, в Верхнем Карру в непосредственной бли-
зости друг от друга развиваются различные типы растительности. Эта
область называется также Broken Veld, поскольку злаковники здесь не
развиваются на крупных площадях, а расчленяются на отдельные полосы
участками с другим растительным покровом, приуроченным к столовым
возвышенностям (рис. 341). Влияние почвенных условий на распределе-
ние растительности иллюстрируется схемой, представленной на рис. 342.
Столовые возвышенности с едва заметным уклоном сложены долери-
тами; участки с небольшим количеством мелкозема встречаются только
в трещинах скал. На этих площадях развивается типичная для Карру
растительность, в составе которой преобладают суккуленты. Крутые
склоны возвышенностей покрыты крупнокаменистыми осыпями; на них
ранее преобладала вечнозеленая Olea verrucosa. В настоящее время древо-
стои ее в окрестностях Фаурсмита сильно изрежены рубками. Основной
фон образуют кустарники: Rhus erosa, Euclea ovata var. undulata, Osyris
compressa (abyssinica), Ehretia hottentotica (Boraginaceae), Royena micro-
phylla, R. cuneifolia, Chilianthus arboreus (Logan iaceae). Своеобразная
Cussonia spicata с огромными лопастными листьями кажется в этом окруже-
нии совершенно чужеродным элементом. Здесь же растут более мелкие
428
Глава 11
кустарники, кустарнички и некоторые виды травяного покрова. Под
защитой скал часто поселяются пойкилогидрические папоротники
{Pellaea hastata, Ceterach cordata, Cheilanthes hirta). Среди них встречается
и один вид Riccia, талломы которого покрывают площадь размером
с ладонь и свертываются при высыхании.
Низкорослый флористически бедный лес лучше развивается на южных
склонах (более прохладных в условиях южного полушария). Как извест-
но, участки с сильно расчлененным рельефом в засушливых областях
относительно хорошо обеспечены водой и поэтому на них лучше всего
растут древесные растения. О хорошей влагообеспеченности этих участков
свидетельствует и то, что злак Themeda triandra, распространенный
севернее на больших площадях, встречается здесь лишь в трещинах скал.
На более сухих и крутых северных склонах часто куртинами растут круп-
ные алоэ; здесь проходит граница их ареала, о чем свидетельствуют ноч-
ные заморозки, повреждающие эти растения.
Ближе к подножию крутизна склонов уменьшается, а песчаная почва
сменяется суглинистой. Здесь гораздо суше, и на этих площадях разви-
ваются низкорослые злаки, например Eragrostis lehmanniana, а ближе
к средней части долины — Sporobolus ludwigii. Степень покрытия состав-
ляет около 30%. Кустарнички и виды разнотравья встречаются только
изредка.
Врезанное в аллювиальную равнину русло реки по крутым берегам
окаймлено остатками галерейного леса. В его составе ранее, вероятно,
встречались акации, вырубленные впоследствии на топливо. Сейчас здесь
распространены только кустарники: Rhus lancea, Zizyphus mucronata,
Lycium hirsutum, Royena decidua 1 и на более влажных участках Melian-
thus comosus и Salix capensis.
Грунтовые воды под сухим в течение большей части года руслом реки
способствуют лишь развитию Gomphocarpus fruticosus и сорных расте-
ний— Nicotiana glauca, Argemone mexicana, Tagetes minuta и многих
других. При паводках эти растения заносятся песком, однако появляются
снова и снова. Там, где за береговым валом образуется широкое плохо
дренируемое понижение, начинается засоление почвы. Здесь появляются
виды Salsola и Suaeda atramenttfera, виды рода Mesembryanthemum, а также
Psilocaulon, интересный Nanathus vittatus и Lithops salicola (рис. 343 и 344).
I Суккуленты
Ложе реки с подрусловым
потоком грунтовых вод
Рис. 342. Влияние почвенных условий на распределение растительности близ Фаур-
смита (Н. Walter, 1939).
I__столовые возвышенности, сложенные долеритами, с небольшими участками мелкозема,
где господствуют суккуленты; II — крупнокаменистые осыпи по склонам, где могут расти дре-
весные породы с экстенсивной корневой системой; III — мелкозернистая наносная почва, силь-
но увлажняемая в период дождей, доминируют злаки; IV — галерейный лес, образованный
древесными породами, отличающимися глубокой стержнекорневой системой, достигающей уров-
ня грунтовых вод.
1 Royena, по современным представлениям, относится к роду Diospyros.
Пустыня Карру
429
На влажных засоленных местах растет злак Sporobolus tenellus. Засоленные
участки окружены кустарниками, свойственными области Карру, Pentzia
incana, Pteronia glauca, Eriocephalus glaber, E. spinescens, Tripteris pachyp-
teris, своеобразным Thesium hystrix, а также колючим Mesembryanthemum
cf. spinosum. Распространенная здесь тяжелая глинистая почва уже слиш-
ком суха для развития злаков. Как показали наблюдения, уровень грун-
товых вод в засоленных почвах в засушливый период находился на глу-
бине 2,0—2,5 м. Осмотическое давление почвенного раствора равнялось
0,3 атм, что свидетельствовало об отсутствии засоления на этих глубинах.
На лишенном растительности участке почва растрескалась на полиго-
нальные отдельности; в верхней части профиля величина pH достигала 7,4,
а на глубине 10 см —8,4. Следовательно, здесь был развит такыр.
ЭКОЛОГИ ЧЕСКИ Е ИССЛЕДОВАНИЯ
Они проводились автором в течение засушливого периода в 1937 г.,
когда на протяжении 6 месяцев в Фаурсмите не выпало ни одного дождя.
Столь длительная засуха благоприятна для проведения исследований,
поскольку экологическая специфика растений засушливых областей опре-
деляется не интенсивностью транспирации в благоприятное время года,
а этим показателем в сухое время года, потому что потери воды не должны
приводить к снижению уровня гидратуры растений до значений, грани-
чащих с летальными. ’
Суккуленты и галофиты
Эта группа растений особенно характерна для области Карру. Стеб-
лесуккулентные молочаи реагируют на недостаток влаги так же, как
кактусы, и могут существовать, не поглощая воды в течение многих меся-
цев. Аналогично ведут себя и виды алоэ, цветущие именно в период засухи.
Несмотря на большую поверхность, интенсивность транспирации соцве-
тий алоэ очень невелика. В один из дней наблюдений интенсивность транс-
пирации этого вида составила 8,5—13,5 мг на 1 г сырого веса, а после ноч-
ного заморозка, повредившего растения, она оказалась вдвое больше. Осмо-
тическое давление нектара,
обильно выделяемого нектар-
никами, равнялось 29,0 атм.
Более подробно транс-
пирация изучалась у мел-
ких суккулентов. Интенсив-
ность транспирации у них
оказалась столь незначитель-
ной, что взвешивания про-
водились раз в несколько
дней. Было установлено, что
в промежутке между пер-
Таблица 69
Название растений Интенсивность транспирации, % от исходного веса
Kleinia radicans 3,3
Massonia (с клубеньками) 1,7
Crassula platyphylla . . . 0,9
Mesembryanthemum unidens 1,0
Mesembryanthemum hamatum 1,0
Bergeranthus glenensis . 0,5
вым и вторым днем проис-
ходит уменьшение интенсивности транспирации примерно на V2—*/з
первоначальной величины, затем она приобретает постоянный характер
и составляет в сутки около 1% исходного веса (табл. 69).
Интересно, что во время опыта мезембриантемум (срезанные побеги)
продолжал цвести как ни в чем не бывало, причем все время раскрыва-
лись новые цветочные почки.
Рис. 343—344. Низина со слабозасоленной глинистой почвой. Ферма
Роземари близ Фаурсмита, Южная Африка. На рис. 343 — пятно Lithops sali-
cola, видны «оконные» листья; на рис. 344 — Nananthus vittatus, видны только
верхушки листьев.
Пустыня Карру
43f
Снижение интенсивности транспирации в первый день свидетельст-
вует о все еще продолжающемся поглощении растениями некоторого
количества воды из почвы и их способности легко сокращать интенсив-
ность транспирации после срезания. Если засуха затягивается, в растении
происходит перераспределение влаги и ее отток из старых листьев в более
молодые. В результате старые листья высыхают и отпадают.
Значительно подробнее изучена экология Lipthops salicola (рис. 343
и 345).
Влажность тяжелой глинистой почвы в пределах слоя, где развиты
корни Lithops, достигает всего 4,5—5,4%. Почва отличается щелочной
Рис. 345. Корневые системы Lithop? (слева) и Nananthus (спра-
ва) (см. также рис. 343 и 344).
реакцией (pH = 8,6—8,8). На глубине 30 см отмечено некоторое накопле-
ние Na2CO3 и MgCO3; NaCl накапливался в верхних 15 см в количестве
до 1%, сульфаты и нитраты, напротив, во много меньших количествах.
У этих суккулентов всасывающие корни развиваются только
во влажное время года, когда концентрация солей невелика. В период
засухи они существуют за счет накопленных запасов влаги. Осмотическое
давление достигало у них в это время 19,2 атм', у растений, находивших-
ся в лаборатории в сухом состоянии в течение 10 дней, оно колебалось
от 20,4 до 21,2 атм, а у одного поврежденного экземпляра равнялось
24,1 атм. Содержание хлоридов, как и у всех представителей рода
Mesembryanthemum, оказалось высоким и превышало 40%.
Для оценки водного режима были проведены следующие опыты.
1. Все растение (Lithops) целиком с мелкими корнями было извлечено
из почвы, помещено в бюксу и обернуто ватой, наружу выступали лишь
так называемые «оконные листья»; таким образом, транспирация у этих
видов происходила в условиях, близких к природным. Потеря веса
за 16 дней не составила даже 0,2 г (4,5% исходного веса).
2. Неповрежденные базальные части завядших растений опуска-
лись в воду; через 15 дней прибавка составляла 15% исходного веса,
однако растения по-прежнему оставались в завядшем состоянии. При
небольших же надрезах эпидермиса через 24 часа растение поглощало
количество воды, равное 30% исходного веса. Новые всасывающие корни
у растений образовались только через 3 недели.
3. Если с водой соприкасается только «оконная» поверхность расте-
ний, то они поглощают в сутки столь мало воды, что эта величина почти
не поддается измерению. Однако она очень быстро увеличивается, если
сделать несколько надрезов эпидермиса. Если на одном корне разви-
ваются два побега, то достаточно, чтобы один из них поглощал воду через
432
Глава 11
надрезы; у второго в этом случае также увеличится степень насыщения
клеток водой. Из этого видно, что не может быть и речи о непосредствен-
ной адсорбции влаги тумана или росы «оконной» поверхностью ра-
стений.
После дождя у растений сначала образуются новые корни, и только
после этого они начинают всасывать влагу.
Аналогично реагирует на эти условия Nananthus, у которого лишь
верхушки многочисленных листьев выступают над поверхностью почвы
(рис. 344 и 345). В лабораторных условиях у Nananthus (при ежеднев-
ном смачивании) уже через девять дней образуются новые корни, после
чего растения начинают энергично поглощать воду.
Кроме того, изучалась экология Pachypodium succulentum, облада-
ющего суккулентными корнями. Безлистные колючие побеги высотой
20—30 см возвышаются над подземным клубнем, над которым прости-
рается поверхностная корневая система. В клубне содержалось 266 г
воды (950% сухого веса). Осмотическое давление клеточного сока состав-
ляло 4,2 атм. Таким образом, воды в клубне вполне достаточно для
длительного снабжения лишенного листьев маленького побега влагой,
необходимой для обеспечения очень слабой транспирации. По данным
Марлота, вес клубня одного старого экземпляра Pachypodium достигал
7 кг при весе безлистного побега, равном 68 г. Так же, как и у Pachypo-
dium, у видов Asparagus, сохраняющих зеленую окраску в период засухи,
за сутки интенсивность транспирации достигает всего лишь 7,5 мг на
1 г сырого веса, несмотря на значительный запас воды в их подземных
клубнях или сочных корнях.
В связи с этим становится понятным, почему у этих растений уровень
гидратуры в течение длительного периода засухи существенно не сни-
жается. За исключением представителей рода Mesembryanthemum, осмо-
тическое давление у остальных видов менее 10 атм (табл. 70).
Таблица 70
Осмотическое давление клеточного сока у суккулентов,
распространенных в Карру (в атм)
Liliaceae Euphorbiaceae
Aloe grandidentata (листья) ... 5,0 Aloe grandidentata (цветки) 7,9 А1оё hereroensis 6,8 Haworthia tessellata (свежее расте- ние) 4,1 Haworthia tessellata (увядшее расте- ние) 6,3 Euphorbia cf. pulvinata 8,5
Compositae
Kleinia radicans . . 8,3 Senecio longiflorus 9,5
Crassulaceae Portulacaceae
Cotyledon decussata .7,6 Crassula platyphylla 7,7 Crassula harveyi ........ 9,6 Anacampseros ustulata 15,3
1. Море цветов после дождя в области распространения Acacia апеига (мульга). Жел-
тые цветки — Waitzea аигеа, белые — Helipterum (оба вида сложноцветные), окрест-
ности Уилуны во внутриконтинентальной части Западной Австралии.
2. Солянковая пустыня на севере Южной Австралии близ заповедника Кунаморе.
Преобладают Atriplex vesicaria с Kochia sedifolia. На заднем плане — невысокие
деревья вблизи сухого русла временного водотока.
Пустыня Карру
433
Из всех упомянутых в таблице видов только Anacampseros отличает-
ся более высокой концентрацией клеточного сока, но у этого вида
не развиваются суккулентные побеги и только корни обладают мясистой
консистенцией. Crassula, Kleinia и один вид Euphorbia могут встречаться
на слабо засоленных почвах, однако осмотическое давление у этих
растений едва ли достигает больших величин (примерно 3—4 атм).
В противоположность упомянутым суккулентам виды Mesembryan-
themum как на незасоленных, так и слегка засоленных почвах всегда
обладают осмотическим давлением, превышающим 10 атм (табл. 71).
Таблица 71
Осмотическое давление у растений на незасоленных
или слегка засоленных (2 последних образца) почвах
(прочерк — не определялось)
Название растений Осмотическое давление, атм Содержание хлоридов, %
Mesembryanthemum saxicolum 16,4-17,4
Bergeranthus glenensis .... 16,5-17,6 —
Stomatium erminium 17,7—21,0 —
Mesembryanthemum unidens 18,2-20,6 38
Mesembryanthemum ferox . . 19,1 —
Mesembryanthemum hamatum . . . . 23,4 —
Mesembryanthemum cf. spinosum . . 27,6 —
Mallephora mollis 29,4 48
Lithops salicola 19,2—24,1 43
Nananthus vittatus 18,5-25,7 42
Большие значения осмотического
обусловлены содержанием хлоридов,
на незасоленных почвах. Это дает
основание относить виды Mesemb-
ryanthemum к типичным галофитам.
Содержание сульфатов достигало
у них 1,0—3,6%, и только у Mal-
lephora оно равнялось 11,0%. На
типичных солончаках эти величи-
ны значительно выше, но у видов
Mesembryanthemum они вряд ли мень-
ше, чем у Chenopodiaceae, расту-
щих в тех же условиях (табл. 72).
Виды Chenopodiaceae отличаются очень высоким
давления в значительной степени
даже если растения развиваются
Таблица 72
Название растений Осмотическое давление, jvt
Psilocaulon (Mesembryan- themum) Salsola spp Suaeda atramentifera 41,3—48,1 50,7-74,9 54,8-66,7
осмотическим
давлением даже на незасоленных почвах, что ясно видно при сравнении
данных табл. 72 с данными табл. 73.
Salsola, а также Atriplex 1 обладают незначительной суккулентно-
стью и относятся к сульфогалофитам. Они могут расти и в более сухих
местообитаниях (ксерогалофиты), тогда как Suaeda всегда приурочена
к сырым засоленным почвам.
1 Atriplex nummularis занесен из Австралии.
434
Глава 11
Таблица 73
Осмотическое давление у растений на почти незасоленных почвах
Название растений Осмотическое давление, атм Содержание хлоридов, % Содержание сульфатов, %
Salsola spp 51,7-65,3 23 17
Suaeda atramentifera 35,9-47,2 — —
Atriplex nummularia 51,8-57,1 29 16
Пойкилогидрические папоротники
Представители этой группы в период засухи находятся в воздушно-
сухом состоянии. Поэтому вес растений, если их не увлажнять, не
изменяется. В лучшем случае эти растения, как и все другие гигроскопи-
ческие материалы, прибавляют в весе на несколько десятых грамма
в более прохладные и влажные дни и снова теряют их в жаркие и сухие.
Влажность молодых воздушно-сухих, но, безусловно, живых листьев
достигает 6,7—8,0% сухого веса. На слегка увлажненной почве удалось
найти полусухой экземпляр Cheilanthes hirta, у этого экземпляра содер-
жание воды в листьях составляло 95% сухого веса, а осмотическое
давление клеточного сока равнялось 20,5 атм.
Если дать возможность сухим растениям впитать влагу, слегка
обсушить фильтровальной бумагой, а затем проследить за их транспи-
рацией, то выяснится, что кривая, характеризующая транспирацию
в строго контролируемых лабораторных условиях, в точке перехода от
испарения капиллярной влаги с влажной поверхности к действитель-
ной транспирации через устьица не нарушает свой плавный ход. Следо-
вательно, папоротники практически столь же незначительно сокращают
интенсивность своей транспирации, как и мхи. Высыхание растений
начинается в момент исчезновения в почве доступной для растений влаги.
Высохшие корни в первое время после увлажнения совершенно неспо-
собны поглощать воду; эту роль выполняют, как и у суккулентов, вновь
образовавшиеся всасывающие корни х.
Водный режим кустарничков
Транспирацию кустарничков, развитых в Карру, изучал Хенричи 1 2.
Проанализируем водный режим растений в сухой период года. В качестве
объекта были взяты растения, которые в течение 5—10 лет выращивались
на опытных площадках в условиях, близких к естественным (без оро-
шения).
Часто посаженные кустарнички в августе — сентябре находились
еще в состоянии покоя. Они стояли без листьев и были покрыты лишь
мелкими почками. Осмотическое давление определялось лишь у Hermanni а
candidissima*, оно оказалось равным 37,3 атм. Напротив, у кустарничков,
корни которых развивались в большем объеме почвы, например у экзем-
1 Н. R. Oppenheimer, А. Н. Halevy, Anabiosis von Geterach officina-
nm Lam. et D., «Bull. Res. Conn. Israel.», D, 11, 1962, p. 127—147.
2 M.Henrici, The transpiration of Karroo bushes, «Sc. Bull. S. Africa», № 185,
1940; The transpiration of large Karroo bushes, «Sfr. . A J. of Sc.», 37, 1940, p. 156—163.
Пустыня Карру
435
пляров, встречавшихся вдоль дорог, уже распустились маленькие зеле-
ные листочки. Осмотическое давление у них было примерно 22—
30 атм.
Проведенные опыты свидетельствуют о различиях в ходе транспира-
ции у высохших растений, находящихся в состоянии покоя, и у расте-
ний, уже слегка тронувшихся в рост. При ежечасном взвешивании
довольно крупных побегов одного из кустарничков оказалось, что у расте-
ний, находящихся в состоянии покоя, в течение дня интенсивность
транспирации практически не менялась и ее величина была очень
небольшой. У растений, слегка тронувшихся в рост, наиболее интенсивная
транспирация наблюдалась утром, а затем ее интенсивность все больше
и больше снижалась. Из этого следует, что растения в состоянии покоя
почти не испаряют воды, тогда как у растений, тронувшихся в рост,
наблюдается иная картина. Рост последних начинается сразу после
начала поглощения из почвы воды. Влажность почвы на участках, где
кустарнички посажены менее густо, была заметно выше, однако наивыс-
шей она оказалась на площадках, находящихся под паром. Отдельные
встречавшиеся здесь экземпляры Salvia cleistogama, не испытывавшие
никакой корневой конкуренции, прекрасно росли и цвели. Они внешне
напоминают Salvia pratensis и относятся, следовательно, к мезоморфным
малакофильным видам. Осмотическое давление у них достигало 15,2—
18,7 атм, влажность листьев — 410% сухого веса; кроме того, они
отличались очень интенсивной транспирацией. Одно из растений, взятых
для анализа, завяло уже через 20 минут после того, как его срезали.
Поскольку укоренившиеся растения в естественных условиях не увядали
даже в полуденные часы, следует предположить, что их корни погло-
щали из почвы не менее 60 г в час. В радиальном направлении корни
этого растения простирались на расстояние 40 см, а в глубину почвы
более чем на 25 см, Влажность почвы на глубине 10 см составляла 10,5—
12,7%.
Таким образом, при небольшой сомкнутости растительного покрова
даже в засушливое время в распоряжении растений остается еще доста-
точно влаги.
Влияние недостатка влаги на водный режим
жестколистных растений
Хотя жестколистные деревья и кустарники в засушливые периоды
года преимущественно не теряют листьев, интенсивность их транспирации
настолько мала, что она не может быть установлена путем взвешивания
через короткие промежутки времени. Для этой цели приходится исполь-
зовать крупные ветви, а время экспозиции продлевать до 1 часа. В тече-
ние дня кривая интенсивности транспирации не снижается непрерывно,
как это наблюдается при высыхании растений, а изменяется параллельно
кривой испарения, то есть в первой половине дня растет, а затем падает
(рис. 346), из чего следует, что у некоторых склерофильных растений усть-
ица остаются закрытыми.
О гидратуре этих растений можно судить по следующим значениям*
осмотического давления (табл. 74).
Из таблицы видно, что эти величины, всегда более высокие у скле^
рофильных растений, вовсе не достигают особенно высоких значений^
Таким образом, растения способны даже в условиях плохой обеспечен-^
ности водой поддерживать свой водный баланс на постоянном уровне^
28*
436
Глава 11
Т archonanthus обладает
Таблица 74
Название^растений Осмотическое давление, атм
Olea verrucosa 37,4
Chilianthus arboreus . . . 33,3
Euclea undulata 29,8
Osyris compressa 34,7-35,8
Tarchonanthus minor . . . 29,0
Rhus erosa ... 27,5
Rhus ciliata 24,6
Rhus burchellii 36,8
Rhus burchellii (в свежем состоя-
нии) 29,2
Viscum capense 1 30,7
Viscum rotundijolium* 51,7
1 На свежем Rhus burchellii.
2 На очень сухом Ehretia hottentotica.
относительно мягкими, силь-
но опушенными листьями, у
Osyris, напротив, развиты
слегка суккулентные листья;
однако оба они не являются
галофитами (содержание хло-
ридов достигает только 14%,
а сульфатов — 15%). Совер-
шенно иной внешний облик
имеет Cussonia spicata (Ага-
liaceae); на вершине сук-
кулентного стебля у этого
растения развивается пучок
очень крупных листьев, каж-
дый из которых весит более
20 г. Листья (общий вес 44 г)
испаряли за сутки только
1 г воды. Cussonia должна
быть отнесена к особой груп-
пе суккулентов.
расположена восточнее Ист-
Основная часть ареала этого вида
Лондона, где период засухи менее выражен. Несомненно, что Cussonia
благодаря накопленному запасу влаги хорошо переносит периоды за-
сухи. Однако это возможно только при десукции влаги из поч-
вы. Это может быть отнесено ко всем склерофильным растениям.
Суточный расход влаги на
Рис. 346. Дневной ход транспирации (в г1час)
жестколистных растений в период засухи (28 ав-
густа, заповедник Велд, Фаурсмит, ЮАР).
транспирацию жестколистны-
ми растениями достигает при-
близительно 1/22 мм, или 9 мм
за шестимесячный период за-
сухи. Это немного, но все же
в 10 раз больше величины,
определенной для кустарни-
ковых саванн, образованных
Catophractes и Rhigozum (см.
стр. 283). В сухих злаковни-
ках, развитых в этой же об-
ласти, суточный расход вла-
ги путем транспирации сос-
тавил 1/20 мм, то есть при-
мерно 7 мм в течение перио-
да засухи. В корнеобитаемом
слое почвы, по-видимому,
всегда имеется достаточный
запас доступной для расте-
ний влаги.
Таким образом, расте-
а — испарение, мл/час; б — Rhus erosa; в — Osyris
compressa; г — Rhus ciliata. Погода 28 августа в за-
поведнике Велд; штиль, безоблачное небо, температу-
ра по сухому и смоченному термометру в 9 час. была
соответственно: 15,4 и 9,0°; в 13 час.— 22,5 и 12,4°;
в 16 час.— 22,0 и 10,5°; в 18 час.— 19,2 и 19,0°; мини-
мальная температура накануне ночью — 2°, в следую-
щую ночь 4-1°.
ния, относящиеся к различ-
ным экологическим группам,
в разных почвенных усло-
виях по-разному переносят
неблагоприятное засушливое
время года.
Глава 12
ЗАСУШЛИВЫЕ ОБЛАСТИ ЦЕНТРАЛЬНОЙ АВСТРАЛИИ
СРАВНЕНИЕ АВСТРАЛИИ С АФРИКОЙ
Для того чтобы сопоставить расположение Австралии и Африки
по широте, следует мысленно сдвинуть Австралию на 100° долготы
на запад; в этом случае окажется, что Австралия расположится
на территории Южной Африки, включая Мадагаскар, если бы последний
простирался на 14° дальше к югу (рис. 347). Если наложить карты обоих
континентов друг на друга без изменения широтного положения, то
территория Западной Австралии совпадет с Южно-Африканской Респуб-
ликой, южной половиной Юго-Западной Африки, Ботсваной и Юж-
ной Родезией. Северную территорию Австралии можно сравнить с Мозам-
биком, а восточный Квинсленд — с Мадагаскаром. Сходство климатиче-
ских условий видно из климадиаграмм, изображенных на рис. 348—353.
Западная Австралия не столь засушлива, как африканская пустыня
Намиб, поскольку у австралийского берега нет аналога холодному
Бенгельскому течению. Центральной Австралии в Южной Африке
соответствует Калахари (часто называемая неправильно пустыней). Это
только «жаждущая страна», где песчаный покров поглощает все выпав-
шие осадки. С климатической точки зрения Калахари, наоборот, отли-
чается большей влажностью по сравнению с расположенными дальше
к западу областями Юго-Западной Африки; соответственно и раститель-
ный покров Калахари также более сомкнут.
Во внутренних областях Австралии отсутствуют пустыни, сущест-
вование которых обусловлено климатом. Однако площади засушливых
территорий в Австралии в связи с большей протяженностью материка
в широтном направлении более обширны, чем в Южной Африке.
Если тем не менее на географических картах Австралии к северу
от озера Эйр располагается область, которая называется «пустыней
Симпсон», а западнее от нее — еще большая по площади «пустыня Гиб-
сона», то это объясняется расхождениями в определении понятия пусты-
ни. Обе эти области Австралии, так же как и Калахари, отличаются
отсутствием поверхностного стока. Частично они покрыты песками,
частично же здесь развиты каменистые почвы.
На юге расположена равнина Налларбор («nulla arbor»— означает
«лишенная деревьев») протяженностью более 700 км с запада на восток
и свыше 300 км с севера на юг. Эта равнина представляет собой карсто-
вое плато, сложенное третичными известнякамих. На севере нередко
встречаются также выходы меловых пород, а на юге обнажения миоце-
новых отложений образуют обрыв в сторону моря высотой 60 м. Неболь-
шое количество выпадающих здесь осадков сразу же просачивается
1 R, S, A d am s о n, Т, G, В, О s b о г n е, On the ecology of the Ooldea district,
«Transact. Roy. Soc. S. Australia», 46, 1922, p. 539—564.
438
Глава 12
Рис. 347. Сопоставление положения Австралии и Африки по широте.
в глубь 250-метровой толщи известняков, частично перекрытых сверху
слоем красноватой почвы мощностью 30 см. Здесь распространены раз-
личные карстовые образования; провальные воронки часто ведут в пеще-
ры. Как и в других «пустынях» Австралии, отсутствие поверхностных
грунтовых вод препятствует заселению этих областей. Поэтому Тейлор
при описании Австралии определяет пустыню как область с незначи-
тельным количеством осадков и бедной специализированной флорой
и фауной, которые не могут быть использованы оседлыми животновода-
ми, даже если они обосновались в смежных с пустыней областях уже
50 лет назад \ Этим он хотел подчеркнуть также и фактор времени, име-
ющий определенное значение при освоении недавно заселенных областей.
В Южной Африке и на Мадагаскаре, на восточных склонах гор,
расположенных на восточном побережье, в отличие от засушливых
внутренних районов выпадает очень много осадков. Это же наблюдается
и в Австралии, однако горы здесь значительно ниже. Несмотря на это,
сравнение восточного Квинсленда с восточным побережьем Мадагаскара
(рис. 350 и 351) и восточной части Нового Южного Уэльса с областью
побережья, лежащей перед Драконовыми горами, вполне возможно.
Что же касается штата Виктория и особенно Тасмании, то они располо-
жены настолько далеко к югу, что не имеют в Африке аналогов.
ОСАДКИ В ЦЕНТРАЛЬНОЙ АВСТРАЛИИ
Область, где в течение года выпадает менее 250 мм осадков, про-
стирается от западного побережья Австралии (мыс Карнарвон и Онслоу)
«плоть до южного побережья (Юкла). На востоке ее граница проходит
около Уилканнии и Брокен-Хилла. Площадь равнины Налларбор и обла-
сти, расположенной от Порт-Огаста, где выпадает 200 мм осадков в год,
1 G. Taylor, Australia, 6. Aufl., London, 1951. Количество осадков в этих
«пустынях» колеблется от 150 до 500 мм.
352
353
Рис. 348—353. Климадиаграммы Австралии (рис. 348, 350 и 352) и Африки
(рис. 349, 351 и 353) («Klimadiagramm-Weltatlas», 1960),
440
Глава 12
Рис. 354. Аридные области Австралии с годовым количеством осадков менее 250 мм,
около 200 мм и менее 150 мм в год.
отличается уже гораздо меньшими размерами, ее северная граница
проходит южнее Алис-Спрингса.
Очень незначительна площадь области, где выпадает менее 150 мм
осадков в год. В ее пределах расположены лишь станции Шарлотт-Уотерс,
Вильям-Крик, Кук и Мулка. В районе последней, в 50 км к востоку от
восточного берега большого озера Эйр, зарегистрировано минимальное
в пределах Австралии среднегодовое количество осадков, равное
101 мм (рис. 354).
При оценке экологических условий аридных областей наряду с общим
количеством осадков существенное значение имеет также и их распре-
деление во времени. В этом отношении климат Австралии имеет одну
неповторимую особенность.
Вернемся опять к Африке; здесь на севере Сахары расположены
пустынные области, где незначительное количество осадков выпадает
только зимой, например, в Каире (30° с. ш.); в центральной части Сахары
осадки практически отсутствуют полностью, а южнее около 20° с. ш.
они выпадают редко и то только в летние месяцы. Сравнительно неболь-
шие размеры Австралийского континента сглаживают контрасты в рас-
пределении осадков по временам года. Осадки в центральной Австралии
связаны как с тропическими циклонами, пересекающими 30° ю. ш. при
своем движении к югу \ так и антарктическими депрессиями, которые
1 J. Gentill i, Die Klimate Australiens, «Z. Ges. Erdkunde 1955», Berlin,
S. 206—238.
1935
99
Рис. 355. Годовой ход осадков за период 1918—1950 гг. в Мулке (50 км\
восточнее озера Эйр).
442
Глава 12
могут приносить осадки даже в северную часть этой аридной зоны.
Поэтому на юге осадки выпадают преимущественно зимой, а в северной
части центральной Австралии — главным образом летом. Между этими
областями расположены станции, которые, судя по средним многолет-
ним данным, отличаются ровным ходом осадков в течение года. Однако
на самом деле здесь бывают годы, когда почти не выпадает осадков,
и годы с периодами сильных дождей, причем осадки могут выпадать
в любое время года.
Анализ режима осадков в самом засушливом месте континента
(станция Мулка) за 1918—1950 гг. (рис. 355) показал, что количество
осадков здесь очень сильно колеблется в отдельные годы. Например,
в 1929 г. выпало 18 мм, что составило 18% средней многолетней величины,
а в 1920 г.—298 мм, или почти 300%. Отношение между количеством
осадков в наиболее засушливые и наиболее дождливые годы равно при-
мерно 1 : 16. Как видно из рис. 356, равномерного колебания годовых
сумм осадков около средней величины, равной 101 мм, не происходит.
Примерно на 19 засушливых лет приходится всего 14 влажных. Наиболее
часто наблюдаются засушливые годы с количеством осадков от 40
до 60 мм.
Неравномерно распределение осадков и в течение года (рис. 355).
С октября по март и в июне количество осадков за месяц составляет
более 50 мм. В июле — сентябре и апреле — мае количество осадков
очень редко превышает 25 мм.
Растения, распространенные в этой области, должны быть приспособ-
лены к продолжительным (более года) периодам засухи и в то же время
к использованию осадков, которые могут выпасть в любое время года.
Это определяет развитие в таких условиях своеобразной кустарниковой
полупустыни, в составе которой господствуют виды семейства Che-
nopodiaceae.
В пустыне Намиб, в пограничных между пустыней и саванной обла-
стях, где летом выпадает 100—200 мм осадков, на песчаных почвах
развиваются чистые злаковники. В Австралии они отсутствуют, если
не считать зарослей склерофильной Triodia, о которых говорится ниже.
Чистые злаковники, распространенные на больших площадях, встре-
чаются в Австралии только в области летних дождей, где за год выпадает
250—750 мм осадков. В составе травяного покрова здесь доминируют
виды Astrebla (рис. 357). Злаковники, как правило, приурочены к отло-
жениям мелового периода (мергелям) или к аллювиальным почвам (бас-
сейны рек Уоррего и Балонн во внутреннем Квинсленде). Эти тяжелые
почвы в период дождей перенасыщаются влагой и сильно высыхают
в сухое время года, когда в них образуются трещины метровой глубины.
Эверист приводит следующее описание почвенного разреза:
0—15 см—серая или серо-бурая рыхлая глина;
15—75 см—серая уплотненная крупнокомковатая глина;
75—125 см—желто-бурая глина с выцветами гипса;
ниже—выветрившаяся материнская порода.
Таким образом, существование здесь злаковников обусловлено эда-
фическими причинами; это доказывается также тем, что на рядом распо-
ложенных участках, но уже на почвах другого типа развиваются эвкалип-
товые саванны или заросли кустарниковых акаций.
У видов Astrebla период покоя приходится на зимние месяцы (апрель—
сентябрь). При поездке автора по этой области в 1958 г. начало дождли-
Засушливые области центральной Австралии
443
вого периода запоздало и вся территория вплоть до начала декабря
выглядела совершенно безжизненной. После дождя злаки быстро отра-
стают и в течение двух недель заканчивают развитие вегетативных органов,
а через 6 недель после начала роста уже созревают семена. По данным
Эвериста, вместе с Astrebla здесь встречаются некоторые многолетние
виды разнотравья (Boerhavia, Sida, Psoralea и др.), однолетники (Iseilema,
Dactyloctenium) и представители семейства маревых (SalsolaJ^ali, Атагап-
thus и др.), а после спорадических зимних дождей также Lepidium, Helipte-
гит и др. и, кроме того, некоторые многолетние злаки: DigitarianDichan-
thium Ч 4
Степень покрытия почвы дерновинами Astrebla часто не превышает
4%. Корневища у злаков в период засухи хорошо защищены плотными
сухими листовыми влагалищами. На длительно затопляемых участках
Astrebla вытесняется Marsilia hirsuta.
РА СТИТЕЛЪНОСТЪ
Почвы и растительность засушливых областей западной части Ново-
го Южного Уэльса подробно описал Бидлу 1 2. Здесь, как и вообще в наи-
более бедных осадками частях Австралии, где дожди могут выпадать
в любое время года, развита солянковая полупустыня (цветное фото 2,
стр. 432).
На большей части территории господствуют виды Atriplex и Kochia,
причем наиболее широко распространены Atriplex vesicaria (saltbush)
и Kochia sedifolia (bluebush). Оба кустарничка достигают высоты 30—
60 см. На западе Австралии солянковые полупустыни развиты близ
Калгурли; здесь заросли Kochia и Atriplex мозаично распределены среди
разреженных эвкалиптовых лесов, занимая обычно небольшие пониже-
ния рельефа. Частично эти виды заходят также под полог эвкалиптовых
древостоев.
Заросли Atriplex — Kochia распространены также на равнинах
Налларбор, в местах, где на известняках лежит тонкий слой красной
глинистой почвы. Здесь Kochia, очевидно, встречается чаще, чем Atriplex.
После дождя здесь появляются некоторые однолетники, вроде Salsola kali
и пр. В небольших понижениях рельефа встречаются одиночные экзем-
пляры невысокого кустарника Pittosporum phillyraeoides.
Маревая полупустыня занимает наибольшие площади в северной
части штата Южная Австралия, откуда она простирается на восток до
Брокен-Хилла в Новом Южном Уэльсе.
Kochia sedifolia и Atriplex vesicaria образуют как смешанные, так
и чистые заросли, причем Kochia преобладает на почвах, богатых изве-
стью. Характер современного почвенного покрова в этой области связан
с климатическими условиями прошлого, которые Джессап (устное сооб-
щение) представляет следующим образом.
Климат Южной Австралии в третичном периоде был значительно
прохладнее и, как это подтверждают ископаемые остатки, здесь повсе-
местно был распространен Nothofagus. Кроме того, согласно результатам
палеомагнетических исследований, Австралия находилась тогда в более
1 D. Davidson, The Mitchell Grass association of the Longreach district,
«Univ, of Queensland. Papers», 3, Brisbane, 1954, p. 45—59.
2 N. C. W. В e a d 1 e, The vegetation and pastures of Western New South Walex,
Sydney, 1948.
444
Глава 12
высоких широтах. Затем началось потепление, продолжавшееся в плей-
стоцене Ч
В течение послеледникового, периода климат менялся от более
засушливого к более влажному. Современный климат, по мнению Джес-
сапа, на севере южной части Австралии гораздо суше по сравнению
с предшествующими периодами.
Все это доказывается следующим:
1. Равнина Налларбор в настоящее время лишена почвенного покро-
ва; последний ранее, при более аридных условиях климата, был развеян
и перемещен в восточном направлении. В расположенных далее на восток
Рис. 356. Колебания годовых осадков в Мулке (см. рис. 355 и 381).
областях, где наблюдаются выходы гранитов и кислых пород, часто
встречаются известковые отложения эолового происхождения в незави-
симости от их приуроченности к той или иной высоте над уровнем
моря.
2. В каменистой пустыне (gibber plains) встречаются небольшие по-
нижения, свободные от обломков породы и в связи с этим заросшие Atrip-
lex ragodioides, A. nummularia. Их возникновение можно связать с изме-
нениями климата; в аридных условиях ветер навевает вокруг кустов
небольшие песчаные дюны. Образующийся после сильного дождя
поверхностный поток откладывает между песчаными холмами ил и камни.
Мелкозем выдувается, а камни покрывают почву сплошным крепко
цементированным слоем. С увеличением аридности климата кустарники
отмирают и песок развевается ветром. На месте небольших дюн обра-
зуются понижения, в которых после изменения климата в сторону боль-
шей влажности начинает собираться вода, постепенно они вновь зара-
стают кустарниками. При вторичном иссушении климата описанный
процесс повторяется. Это отчетливо прослеживается в строении почвен-
ного профиля, в котором каменистый субстрат чередуется с песчаными
прослойками, тогда как между ними на лишенных камней участках
встречаются островки растительности.
3. С наступлением засушливого периода котловинные выходы солей
1 См. также N. Т. Burbidge, The phytogeography of the Australian region.
«Austr. J. Bot.», 8, 1960, p. 75—212.
Засушливые области центральной Австралии
445
подвергаются развеванию; в результате вокруг них образуются дюны,
вытянутые в направлении наиболее сильных (но не преобладающих)
ветров. Так, в Западной Австралии, где господствуют западные ветры,
но большей силой отличаются восточные ветры, дюны всегда располага-
ются с западной стороны котловин. Дюны сложены из очень рыхлых
слоев гипса, поверхность которых после дождя легко уплотняется,
и чередующихся с ними прослоек песка; это свидетельствует об измене-
ниях климата. Легкорастворимые соли выдуваются и уносятся ветром;
осаждаясь в виде пыли, они способствуют слабому засолению соседних
участков. О происхождении солей в этой области трудно сказать что-
нибудь определенное, потому что вокруг озера Эйр (настоящий солончак)
расположены меловые, а дальше к югу — третичные осадочные породы.
Присутствие солей в дождевой воде может быть объяснено наличием
в воздухе соляной пыли местного происхождения.
Поскольку Atriplex vesicaria и Kochia sedifolia охотно поедаются
овцами, покров солянковой полупустыни местами сильно изрежен. На
северо-востоке Южной Австралии вследствие чрезмерной пастьбы уничто-
жено, вероятно, 9/10 растительного покрова полупустыни.
На остальной части этой аридной области господствуют Acacia
апеига, известная как «мульга», и виды Triodia, обычно называемые
«спинифекс».
Акации сменяют доминирующие в Австралии эвкалипты (этот род
представлен 600 видами) в условиях, где климат становится слишком
сухим для развития последних. Эвкалипты не связаны с каким-либо
определенным типом климата. Они растут в областях, где осадки выпа-
дают как зимой, так и летом; наряду с этим они встречаются и в усло-
виях влажного климата на востоке страны. Правда, здесь на базальтах
они вытесняются тропически-субтропическим дождевым лесом (см.
стр. 260). Некоторые виды эвкалиптов тесно связаны с определенными
местообитаниями, однако в основном они крайне нетребовательны
к почвенным условиям. Высота эвкалиптов различна.
Если во влажных областях нередко можно встретить деревья высо-
той более 75 м (Eucalyptus diverstcolor — в Западной Австралии, Е. reg-
nans — в Виктории и на Тасмании), то с уменьшением количества осад-
ков они становятся более низкорослыми и, наконец, на границе с обла-
стью зимних дождей образуют лишь заросли кустарниковых эвкалиптов
(см. рис. 357). Речь идет о кустах, отдельные побеги которых появляют-
ся из большого одревесневшего подземного клубня (lignotuber). Граница
проникновения эвкалиптов в аридную область связана со степенью
выраженности влажного времени года, что хорошо прослеживается на
климадиаграммах. В собственно аридных областях распространен древо-
видный Eucalyptus camaldulensis (rostrata), всегда связанный с пересыхаю-
щими долинами, где близко залегают грунтовые воды.
Кроме Acacia апеига (мульга), в засушливой области встречаются
и многие другие виды акаций. Вместе с видами Casuartna они проникают
по песчаным дюнам в солянковую полупустыню. Здесь растут только
вечнозеленые акации с хорошо развитыми филлодиями. Как правило,
в наиболее засушливых областях у акаций преобладают тонкие цилин-
дрические филлодии, а в менее засушливых — более широкие и пло-
ские. У Acacia апеига развиваются цилиндрические и плоские филлодии.
Обе формы акации могут расти рядом, однако у «влажной» границы
ареала в Западной Австралии и в Квинсленде встречается только форма
с плоскими филлодиями.
446
Глава 12
В той части области распространения Acacia апеига, где наряду
с зимними дождями осадки выпадают также и летом, этот вид вытесняется
на песчаных почвах видами злаков из рода Triodia и несколько реже —
видами Plectrachne, образующими колючие подушковидные дерновины
(цветное фото 2, стр. 368); очень жесткие свернутые листья этих растений
оканчиваются колючим острием. В Астралин их обычно называют спини-
фекс (не следует путать с родом Spinifex, виды которого распространены
на морских побережьях, рис. 139).
На западе, по направлению от залива Шарк и мыса Карнарвон
в глубь материка, в бассейне реки Мурчинсон, где осадки преимуществен-
но выпадают летом (в среднем 250—300 мм) и причем весьма эпизодиче-
ски, на больших площадях господствуют злаковники, образованные
главным образом склерофильным злаком Triodia pungens. Этот же вид
встречается и в центральных районах Австралии, где выпадает больше
осадков; здесь он приурочен к сухим каменистым склонам.
Таким образом, склерофильные злаки с экологической точки зре-
ния заметно отличаются от обычных злаков (см. стр. 279 и след.). Прав-
да, они также лучше всего развиваются в области, где осадки выпадают
летом и на песчаных почвах, однако, подобно склерофильным древесным
Рис. 357. Карта растительности Австралии (обобщено по многим источникам).
I — тропически-субтропический дождевой лес с индо-малайскими флористическими элемента-
ми; 2 — влажный эвкалиптовый лес; з — сухой эвкалиптовый лес; 4 — злаковники, образо-
ванные Astrebla; 5 — разреженные заросли кустарниковых эвкалиптов (мали-скраб); 6 — заросли
Acacia апеига и др. (мульга); 7 — солянковая полупустыня, образованная видами A triplex
и Kochia; 8 — сообщества злаков с колючими листьями (Triodia); 9 — необитаемые области,
называемые «пустынями»; 10 — альпийские луга (только на юго-востоке, в Австралийских
Альпах).
Засушливые области центральной Австралии
447
растениям, растут и на каменистых почвах. Склерофильная структура
листьев дает им возможность переносить очень длительные периоды
засухи и расти в областях, где осадки летом выпадают лишь эпизодиче-
ски. Это же может быть сказано и о склерофильных злаках, распростра-
ненных в Юго-Западной Африке, с одной стороны, на прибрежных дюнах
и на юге Калахари, а с другой — в южной части страны, на местообита-
ниях, обеспеченных атмосферными осадками (Eragrostis spinosa, Е. суре-
roides, Aristida namaquensis, A. brevifolid). В этих условиях, при наличии
осадков, растения хорошо обеспечены влагой. К склерофильным зла-
кам, вероятно, может быть отнесен и распространенный в Северной
Африке ковыль — Stipa tenacissima.
На севере Австралии, где летом дожди идут более регулярно, появ-
ляются типичные злаки с мягкими листьями, высыхающими в период
засухи. Это главным образом виды родов Andropogon и Panicum и занос-
ный злак Cenchrus ciliaris.
Почвы засушливых областей Западной Австралии характеризуются
наличием уплотненных горизонтов (hardpan). Обычно этот горизонт
цементирован карбонатами, однако в Западной Австралии, где извест-
няки почти не встречаются, они заменяются силикатами. Окиси кремния
в этих почвах весьма подвижны, в связи с чем уже на глубине 10—15 см
образуется твердый слой различной мощности, отличающийся пластин-
чатой структурой. На таком субстрате преимущественно и растет Acacia
апеига в сочетании с разными видами кустарников и полукустарников
и особенно многочисленными видами Eremophila (Муорогасеае). Осталь-
ная лишенная растительности поверхность почвы обычно обнажена,
и только после достаточно сильного дождя она покрывается эфемерами:
после зимних дождей преимущественно сложноцветными, видами родов
Helipterum, Helichrysum, Waitzia, Schoenia и др., а после летних дож-
дей — злаками и видами Trichinium (Amaranthaceae). В блюдцеобразных
понижениях в благоприятные годы, как это было, например, в 1958 г.,
образуется густой ковер цветущих эфемеров (цветное фото 1, стр. 432).
В Западной Австралии широко распространены котловины с выходами
солей (около 200 крупных и бесчисленное множество мелких). Это остатки
прежней речной системы, развитой в более влажный период плейстоцена.
В последующем реки были перепружены во многих местах песчаными
перемычками. В настоящее время лишь после очень сильных дождей
котловины превращаются в частично соединяющиеся между собой озера
(рис. 358). Галофиты вокруг этих котловин распространены концентри-
ческими кругами, расположение которых усложняется гипсовыми дюнами
(цветное фото 1, стр. 368).
В заключение приведем описание пустыни Симпсон по Кроккеру Ч
Пустыня включает площади песков, расположенные к северу от озера
Эйр (ранее они назывались «пустыня Арунт»), и обширные территории
с каменистыми или щебнистыми почвами. Песчаная пустыня простирается
примерно на 480 км при ширине около 400 км. Развитые здесь песчаные
валы достигают длины около 250 км и высоты 7,5—30,0 м; они расположе-
ны в 250—500 м один от другого и вытянуты в направлении господствую-
щих ветров, с юго-востока на северо-запад. Подвижными являются лишь
гребни дюн; восточные склоны дюн более круты, чем западные, что,
1 R. L. Crocker, The Simpson Desert Expedition 1939. The soils and vegeta-
tion of the Simpson Desert and its borders, «Trans. Roy. Soc. South-Australia», 70,
1946, p. 2.
448
Глава 12
Они отсутствуют во влажных прибрежных областях и на равнине Налларбор, сло-
женной известняками.
вероятно, обусловлено преобладанием западных ветров, На севере эти
гряды переходят в обширную песчаную равнину, среди которой встре-
чаются понижения, сложенные глинами или с выходами солей.
Пустыня Симпсон отличается крайне выраженной засушливостью
и в перечне больших песчаных пустынь земного шара стоит на пятом месте
по степени сухости, Еще большей засушливостью отличаются следующие
пустыни (перечислены в последовательности убывания степени засушливо-
сти): Центральная Сахара, включая Ливийскую пустыню, Такла-Макан
(южный Синьцзян), Руб-эль-Хали (Саудовская Аравия) и северо-западная
Сахара. Менее суровы условия в пустынях Кара-Кум (Туркменская
ССР). Чиуауа (Мексика) и Тар. или Синд (Индия).
Дожди здесь выпадают крайне нерегулярно, годовое количество осад-
ков колеблется от 100 до 250 мм при среднем значении около 150—200 мм.
Кислотность песка, слагающего дюны, достигает 6,5—7,3; в меж-
дюнных понижениях значение pH несколько больше — 6,4—8,8. Пески,
слагающие дюны, почти не содержат извести и совершенно лишены
растворимых солей.
Засушливые области центральной Австралии
449
Пески перекрывают бурую пустынную почву с хорошо выраженной
каменистой прослойкой. Нередко эти почвы выходят на дневную поверх-
ность. Их кислотность достигает 7,6—8,7, а содержание солей — 1,3—
1,5%. Дренируемые понижения рельефа засолены значительно меньше.
Серые или серо-бурые аллювиальные почвы содержат около 50% гли-
ны (pH = 8,1).
Гребни дюн весьма подвижны; на них большей частью растут единич-
ные экземпляры Zygochloa paradoxa совместно с многолетниками Ptilotus
latifolius, Sida corrugata var. pendunculata. На склонах дюн растут
Crotalaria cunninghamii и C. dissitiflora. После дождей развиваются одно-
летники — Salsola kali и различные виды злаков {Plagiosetum refractum,
Eriachne aristided), Trichodesma zeylanicum и др.; еще ниже по склону
появляются Blennodia pterosperma и Goodenia cycloptera.
Основной фон и .здесь образует Triodia basedowii. Он покрывает
нижние части склонов, междюнные понижения и песчаные равнины
в северной части пустыни. Вместе с ним растут отдельные виды разно-
травья и кустарники — Накеа leucoptera, Grevillea stenobotrya, G. junci-
folia, Eremophila longifolia, E. latrobei, а также представители семейства
Chenopodiaceae — Rhagodia spinescens и Euchylaena tomentosa. Семейство
Euphorbiaceae представлено Euphorbia wheeleri n Adriana hookeri. Расту-
щая на склонах дюн Triodia закрепляет песок. Этими же свойствами
обладают некоторые акации и Dodonaea. После зимних дождей здесь
появляются Helipterum, Schoenia rosea и др. Таким образом, судя по
развитым здесь группировкам растений, растительность песков в
пустыне Симпсон относится к лишь ненамного менее аридному типу, чем
таковая на песчаных дюнах Западной Австралии.
На севере пустыни, на песках, распространена группировка: Triodia
basedowii — Grevillea juncifolia — Eucalyptus pachyphylla. На участках,
где почва покрыта плотной сцементированной корой, появляется траво-
стой, образованный Astrebla pectinata, Atriplex и видами Bassia,
На каменистых почвах преобладают Chenopodiaceae, а на скалистых
возвышениях, то есть в наиболее благоприятных местообитаниях,—
Acacia cambagei, A. tetragonophylla, Cassia desolata, различные виды
Eremophila и т. д.
На временно затопляемых почвах, по берегам пересыхающих потоков
с постоянным током грунтовых вод, растут Eucalyptus coolabah и появля-
ются заросли Atriplex nummularia или Chenopodium auricomum, а также
Muehlenbeckia cunninghamii.
На равнинах, где после дождей почва насыщается влагой, приносимой
реками, видовое разнообразие увеличивается.
На развитых вокруг озера Эйр дюнах (содержащих гипс), рас-
пространена колючая Nitraria schoberi (Zygophyllaceae). На гребнях дюн
встречается Zygochloa paradoxa. Из кустарников здесь представлен
только Накеа leucoptera, а из однолетников — многочисленная Sal-
sola kali.
Таким образом, растительность этой наиболее засушливой «пустын-
ной области» Австралии отличается отнюдь не столь уж бедным видо-
вым составом, как это может показаться на первый взгляд. В ее составе
отмечено 350 видов, относящихся к 50 семействам. Из них 76 видов
характерны для областей песчаных территорий. Пески и растущие на них
жесткие не поедаемые скотом злаки представляют собой основное в ре
пятствие для заселения этих областей.
450
Глава 12
В описываемых аридных областях Бидл и Чан смогли обнаружить
на тяжелых, а также каменистых почвах, покрытых редким раститель-
ным покровом, Azotobacter, Clostridium и сине-зеленые водоросли (Nostoc,
Anabaena) 4. В почвах всегда содержатся нитраты. Так же, как в пусты-
не Намиб, водоросли особенно хорошо развиваются под прозрачными
обломками кварца, торчащими из почвы. Местами почва на 1/6 площади
покрыта, особенно на участках проявления плоскостной эрозии, круп-
ным, диаметром 0,5—3,5 см, щебнем, нижняя поверхность которого
наполовину обросла водорослями. Водоросли покрывают, таким обра-
зом, около 1/12 всей поверхности почвы. Бидл и Чан предполагают, что
эти водоросли могут фиксировать азот.
ЭКОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ
Солянковая полупустыня
В 1925 г. Осборн в полупустыне, образованной Atriplex и Kochia
(северо-восток Южной Австралии), на сильно пострадавшей от пастьбы
территории, в пределах обширной огороженной площади создал лабо-
раторию, где он вместе с Вудом и группой сотрудников проводил эколо-
гические исследования 1 2.
Опытная станция расположена близ пункта Кунамор, на террито-
рии овцеводческого хозяйства площадью 300 тыс. га, примерно в 60 км
севернее железнодорожной станции Юнта по пути к Брокен-Хиллу. За
год здесь выпадает в среднем несколько больше 200 мм.
Здесь возникла типичная полупустыня, образованная A triplex и Kochia
(цветное фото 2, стр. 432); суглинистая почва характеризуется большим
содержанием извести в горизонте В; степень кислотности почвы дости-
гает 7,7, содержание хлоридов —0,1%. Встречающиеся здесь песчаные
дюны содержат конкреции СаСО3; кроме того, распространены пески
(мощностью 150 см}, перекрывающие глины, которые содержат желези-
стые оолиты и гипсовый горизонт на глубине 90 см\ и те, и другие пески
не содержат ни хлоридов, ни сульфатов. На песчаных дюнах растут
Acacia апеига, A. burkittii, Eremophila sturtii, Stipa nitida (однолетник),
а на песчаной равнине — Casuarina lepidophloia, под пологом которой
развивается Bassia uniflora. На илистых почвах встречаются виды Hetero-
dendron и Муорогит (древовидные формы), а также кустарниковидные
Eremophila и Cassia.
Atriplex vesicaria —кустарничек, живущий 12 лет; как и у многих
видов семейства Chenopodiaceae, у него развиты слегка суккулентные
листья, опадающие при продолжительной засухе. Корневая система
1 N. С . W. Beadle, Y. Т. Tchan, Nitrogen economy in semi-arid plant
communities, I u. II, «Proc. Linn. Soc. N. S. Wales», 80, 1955, p. 62—70, 97—104.
2 T. G. В. О s о b r n e, Introduction and general description of the Koonamore
Reserve for the study of the saltbush flora, «Trans. Roy. Soc. South Australia», 49,1925,
p. 290; T. G. B. Osborne, J. G. W о о d, On some halophytic and non halophytic
plant communities in arid South Australia, «Trans. Roy. Soc. South Australia», 47,
1923, p. 388—399; On the growth and reaction to grazing of the perennial saltbush Atrip-
lex vesicaria, «Proc. Roy. Soc. N. S. Wales», 57, 1932, p. 377; J. G. Wood, On the
climate and vegetation of the Koonamore Vegetation Reserve to 1931, «Proc. Roy. Soc.
N. S. Wales», 60, 1935, p. 392; Regeneration of the vegetation on the Koonamore Vege-
tation Reserve 1926—1936, «Trans. Roy. Soc. South Australia», 60, 1936; The physio-
logy of xerophytism in australian plants, «J. Ecol.», 22, 1934, p. 69—87; The selective
absorption of chlorine ions and the absorption of water by the leaves in the genus Atri-
plex, «Austr. J. Exp. Biol. a. Medic. Sci.», 11, 1925, p. 45—56.
Засушливые области центральной Австралии
451
расположена очень неглубоко, примерно на глубине 10—20 см; на этом
уровне, расположенном выше плотного карбонатного слоя, корни про-
стираются далеко в горизонтальном направлении. Поэтому отдельные
кусты растут довольно далеко один от другого п покрывают 10—25%
поверхности, а в очень засушливых условиях —еще .меныце.
Kochia в противоположность Atriplex живет, вероятна, дольше. Глу-
боко идущая корневая система, проникающая по трещина^ уплотненного
слоя карбонатных конкреций на 3—4 м в почву и. простирающаяся
настолько же в горизонтальном направлении, обусловливает появление
этого вида в местах, где почва промачивается значительно глубже. Kochia
приурочена к карбонатным почвам и более засухоустойчива, чем A triplex
vesicaria. В качестве пионера на стравленных скотом и выбитых площадях
поселяется Bassia patenticuspis (Chenopodiaceae); корни этого вида про-
никают в глубину до 2 м.
По поводу возможности причисления Atriplex vesicaria к галофитам
существуют разные мнения. Вуд отвечает на этот вопрос отрицательно;
он считает, что хотя Atriplex и встречается на засоленных почвах, одна-
ко обычно в этих почвах содержание хлоридов не превышает пример-
но 0,1%. По мнению автора, при решении этого вопроса основное внима-
ние должно быть уделено не почве, а самим растениям. Так, в частности,
было установлено, что в листьях Atriplex всегда содержится много солей,
главным образом хлоридов, даже если в почве обнаружены лишь их
следы Ч
Согласно Вуду, 90% сырого веса Atriplex vesicaria приходится на
воду; высока доля хлоридов. На основании данных, приведенных
в табл. 75, видно, что между содержанием солей в почве и растении нет
определенной зависимости; однако с увеличением количества солей
в почве увеличивается и их содержание в растениях. Содержание солей
Таблица 75
Содержание солей в % сухого веса
В почве . . 0,05 0,07 0,12 0,68
В растении . • 14,1 23,5 15,3 32,9
у Atriplex повышается в сухой период. Данные анализа листьев Atriplex
nummularia, собранного в Ботаническом саду Аделаиды на незасоленной
почве, приведены в табл. 76.
Таблица 76
Растворимые всего . соли, С1-1 СаО К+1 Na+C Сульфаты
25,0 7,78 2,20 6,20 6,00 Следы
1 N. С. W. В е a d 1 е, R. D. В. W h а 1 1 е у, J. В. G i b s о n, Studies in halo-
phytes II. Analytical data on the mineral constituents of three species of Atriplex and
their accompanying soils in Australia, «Ecology», 38, 1957, p. 340—344.
452
Глава 12
Высокому содержанию солей соответствует и величина осмотиче-
ского давления, достигающего у Atriplex vesicaria 40 атм, а у Л. paludo-
sa —65 атм, у A. vesicaria осмотическое давление, определенное дру-
гим методом, равно 50 атм, а у A. spongiosa — 55 атм.
Способность Atriplex nummularia накапливать соли была выявлена
Эшби и Бидлом в опытах с водными культурами1. Они сравнивали осмо-
тическое давление отжатого сока с осмотическим давлением питательных
растворов и установили, что осмотическое давление сока примерно на
16 атм превышает эту величину у растворов (табл. 77).
Таблица 77
Питательный раствор Осмотическое давление, атм Разница, атм
питательного раствора отжатого сока
Без добавления NaCl 0,3 16,0 15,6
50 мг/л NaCl 1,7 17,6 15,9
200 » » 7,9 24,0 16,1
400 » » 14,7 31,6 16,9
600 » » 23,3 41,5 18,2
Растения развивались при подкормке солями значительно лучше
йричем добавление NaCl давало больший эффект, чем внесение Na2SO4.
Эти данные свидетельствуют о том, что виды Atriplex являются
типичными галофитами, а именно ксерогалофитами, которые растут на
почвах, содержащих незначительные количества солей (при пересчете
на сухой вес); поскольку, однако, влажность этих почв часто бывает
очень незначительной, концентрация солей в них может достигать боль-
ших величин.
Приведенный пример показывает, что поверхностное изучение место-
обитаний не дает еще оснований к выделению различных экологических
групп растений.
У листьев Atriplex и Chenopodium развиты пузыревидные волоски.
Выяснено, что у Atriplex vesicaria они выделяют соли. Концентрация
солей в клеточном соке волосков очень высока. Пузыревидные волоски
существуют очень недолго и быстро высыхают, однако при этом постоян-
но образуются все новые волоски. Посредством отмерших волосков
некоторое количество солей удаляется из листьев. Поскольку в сухую
погоду высохшая соль остается на поверхности листьев, определить содер-
жание солей и осмотическое давление (криоскопическим методом) можно
только с известной ошибкой.
Бергер-Ландефельдт так же считает, что соли выделяются при помо-
щи пузыревидных волосков (см. стр. 492). Ранее высказанное предполо-
жение Вуда о том, что хлориды содержатся главным образом в сосудисто-
волокнистых пучках и в хлоренхиме листа вокруг жилок, а в волосках
встречаются лишь в незначительных количествах, должно, таким обра-
зом, приниматься с поправками. Увеличение веса листьев у Atriplex
1 W. С. A s h Ь у, N. С. W. В е a d 1 е, Studies in halophytes, III. Salinity factors
in the growth of Australian saltbushes, «Ecology», 38, 1957, p. 344—352.
Засушливые области центральной Австралии
453
во влажной атмосфере в отличие от эвкалиптов или акаций связано
с солью, сохраняющейся на поверхности листьев. Речь в этом случае
идет, вероятно, не о поглощении воды листьями, а о ее адсорбции соля-
ми, находящимися на поверхности листьев (см. стр. 175).
В одинаковых условиях (срезанные ветви, погруженные в воду)
интенсивность транспирации у видов семейства Chenopodiaceae в 4—7 раз
меньше, чем у австралийских склерофитов.
У видов семейства Chenopodiaceae устьица равномерно распределены
на верхней и нижней поверхности листьев; число устьиц на 1 мм2 приве-
дено в табл. 78.
Таблица 78
Виды семейства Chenopodiaceae Число устьиц Iia 1 AtAt2 Склерофиты (для сравнения) Число устьиц на 1 мл12
Atriplex vesicaria . . . 240 Acacia апеига . . 224
Atriplex stipitata . . 256 Acacia (8 видов) 116-384
Kochia sedifolia 100 Cassia sturtii .... 220
Salsola kali . . 112 Er emophila (5 видов) 164-249
Bassia (7 видов) . 46-104 Eucalyptus (5 видов) 224-240
растений, по данным Вуда, приве-
Таблица 79
Число устьиц у жестколистных
дено в табл. 79.
Название семейства
Число устьиц на
1 мм2 (верхняя+
4- нижняя сторона
листьев)
Proteaceae
Leguminosae
Myrtaceae .
Epacridaceae (только
нижняя сторона лис-
тьев)
145+19,0
277 + 21,7
307 + 32,0
302+5,0
Было выяснено, что площадь поверхности отдельных устьиц обратно
пропорциональна их числу; таким образом, общая площадь поверхности
устьиц вне зависимости от их числа остается более или менее постоян-
ной. Эта закономерность, вероятно, не всегда выражена.
У других видов семейства Chenopodiaceae хлоридов содержится
меньше, чем у A triplex, а количество сульфатов не достигает 0,5% сухого
веса. Осмотическое давление достигает у этих видов следующих вели-
чин (табл. 80).
Для проверки этих ранее полученных данных 8 и 9 октября 1958 г.,
во время затянувшегося периода дождей, в заповеднике Кунамор и в его
ближайших окрестностях была проведена серия анализов. Листья после
взвешивания в сыром состоянии высушивались; затем определялся их
сухой вес и содержание воды. Высушенные листья использовались для
определения осмотического давления и содержания хлоридов; все ана-
лизы проводились в Штутгарт-Хоэнхейме.
454
Глава 12
Таблица 80
Название растений Осмотическое давление, am.и Название растений Осмотическое давление, атм
Kochia sedifolia 43 Kochia pyramidata . 47
Kochia planifolia 35 Kochia tomentosa ... 38
Kochia triptera 47 Bassla obliquicuspis 47
Для сравнения: Acacia aneura . 68 Zygophyllum fruticulosum 10
Acacia burkittii . 55 Zygophyllum prismato-
Eremophila scopartia 50 thecwn .... 7
Myoporum platy car pum 43 Lycium dustrale 8
К сухой массе листьев добавлялась вода до тех пор, пока не был
достигнут исходный сырой вес листьев; затем вся масса выдерживалась
в хорошо закрытом сосуде в течение 45 минут при 90°. Вслед за этим
под давлением 100 атм выжимался сок; в заключение определялось
осмотическое давление и содержание хлоридов в соке. Опыты, проведен-
ные в Штутгарте с различными растениями, показали, что ошибка при
определении осмотического давления вышеуказанным методом не пре-
вышала 1 атм. Например, осмотическое давление у плюща, измеренное
криоскопическим методом, равнялось 16,5 атм (среднее из 5 анализов),
а давление, определенное вышеописанным методом (также среднее из
5 анализов), достигло 16,1 атм Ч
Поскольку перед отбором образцов в заповеднике Кунамор долгое
время шли дожди, глубоко промочившие почву, полученные величины
отражали не крайние, а скорее оптимальные условия. Кроме того, сле-
дует учитывать, что дождь смыл соли, отложившиеся на поверхности
листьев Atriplex. Результаты представлены в табл. 81—83, где осмоти-
ческое давление (W) выражено в атм, давление, обусловленное NaCl,—
также в атм (С1), а осмотическое давление, обусловленное хлори-
дами,— в %.
Эти значения соответствуют данным, приведенным в таблицах на
стр. 451—452. Небольшие величины давления у пятого образца связаны
со снижением концентрации клеточного сока после дождя. Величина
осмотического давления и содержание хлоридов у Arthrocnemum приве-
дены в табл. 87 (образец 2).
Высокое осмотическое давление, обусловленное хлоридами, позво-
ляет рассматривать Atriplex vesicaria в качестве настоящего галофита.
Листья у этого вида отличаются суккулентностыо, а содержание воды
достигает 230—270% сухого веса.
Величины осмотического давления у Kochia (табл. 82) в два раза
меньше отдельных значений, приведенных в табл. 80. Однако они досто-
верны, поскольку результаты автора совпадают с данными Гарриса,
полученными им во время исследований в штате Юта. Содержание хлори-
1 По этому вопросу см. R. Т h г е n, «Zschr. Bot.», 26, 1934, S. 461; Н. К no-
de 1, «J b. wiss. Bot.», 87, 1939, S. 558. В среднем ошибка не превышает 0,4 атм. Этот
метод очень удобен, если в распоряжении экспедиции нет оборудованной лаборатории.
Требуется лишь умение взвешивать п высушивать образцы, причем более крупные
образцы можно взвешивать на простых аптечных весах с роговыми чашками.
Засушливые области центральной Австралии
455
Таблица 81
Осмотическое давление у Atriplex vesicaria при возрастающем (с первого
по шестой образец) засолении почвы
лъ образца Условия произрастания W С1 %
1 Заросли кустарниковых эвкалиптов на слабозасоленных песчаных дю- нах . ... 37,4 22,6 59
2 Под Casuarina на песках 53,5 32,0 60
3 Заросли Atriplex с примесью Муоро- гит 55,4 42,4 76
4 Ассоциация Atriplex-Kochia .... 65,4 45,2 69
5 Солончак в окружении ярко-зелено- го Arthrocnemum 48,6 37,5 77
6 Солончак Уэй, Западная Австралия (3. IX. 1958 г.) . . 70,1 58,9 84
дов у Kochia низкое, а осмотическое давление, обусловленное хлоридами,
отличается значительными колебаниями. Поэтому виды Kochia следует,
вероятно, относить к факультативным галофитам. Если осмотическое
давление, обусловленное хлоридами, достигает у Atriplex (образец 4)
45 атм, то у Kochia в тех же условиях местообитания оно составляет
всего лишь 13,7 атм\ в другом случае у Atriplex (образец 2) —давление
32 атм, а у Kochia (образец 5) — всего 10,2 атм. В растениях Kochia
sedifolia и Atriplex vesicaria содержится одинаковое количество воды.
Таблица 82
Осмотическое давление у видов Kochia в заповеднике Кунамор
№ образца Название растений и условия произрастания W С1 %
1 Kochia sedifolia, чистые заросли близ Кунамора ’ 25,0 5,4 21
2 К. sedifolia, там же (повторный анализ) 24,6 4,6 19
3 К. sedifolia рядом с Atriplex (табл. 81, образец 4) 33,7 13,7 41
4 К. sedifolia, Калгурли, Западная Австралия (28/IX 1958 г.) . . . . 26,3 3,3 13
5 К. lanosa рядом с Atriplex (табл. 81, образец 2) 22,3 10,2 46
6 К. spp., солончак Уэй (3/1X 1958 г.) 26,7 15,0 56
7 К. georgeii, солончак Уэй, на скале 25,4 4,3 17
В табл. 83 приведены величины осмотического давления у других
типичных для этой области растений.
Перечисленные в табл. 83 виды, вероятно, также не являются гало-
фитами. Правда, содержание солей в тонких побегах Casuarina lepido-
phloia с редуцированными листьями оказалось необычно высоким. Следо-
456
Глава 12
Таблица 83
Осмотическое давление у различных видов в заповеднике Кунамор
№ образца Название растений и условия произрастания W С1 %
1 Casuarina lepidophloia среди зарос-
лей Atriplex 25,8 8,5 33
2 То же на песчаной дюне, старые
деревья 25,1 8,5 34
3 То же, молодая корневая поросль 16,8 6,5 38
4 Heterodendron oleaefolium среди за-
рослей Atriplex Около
27,5 3,0 11
5 То же, вершина песчаной дюны . . Около
24,2 1,1 5
6 Муорогит platycarpum рядом с
Atriplex (табл. 81, образец 3) 33,5 6,7 20
7 Acacia апеига, песчаная дюна, рядом
с Atriplex Около
23,0 5,0 22
8 Acacia burkittii, песчаная дюна 24,1 6,4 26
9 Eremophila sturtii рядом с Acacia
апеига 26,1 3,6 14
10 Eucalyptus oleosa на незасоленных
песчаных дюнах 28,2 5,1 18
вательно, этот вид должен по крайней мере обладать солевыносливостыо,
что относится, по-видимому, и к Casuarina equisetifolia, часто встречаю-
щейся в культуре на побережьях морей. В одном из образцов содержа-
ние воды у этого вида составляло 104% сухого веса, у Heterodendron —
80—90%, Муорогит —154, Acacia апеига —87, A. burkittii — 108,
Eremophila —129 и у Eucalyptus —80%. Поскольку у Acacia апеига
и Heterodendron удалось отжать лишь немного сока, полученные значе-
ния нельзя рассматривать как абсолютно точные.
Кроме этих видов, осмотическое давление определялось у двух
свежих, зеленых, однолетних, явно суккулентных галофитов.
Цветущий и плодоносящий экземпляр Zygophyllum iodocarpum рос
по краю заросли Atriplex; осмотическое давление у него достигало
19,8 атм, а содержание хлоридов — 16,1 атм, или 81%. У Babagia
acroptera, росшей в слабозасоленной долине, соответствующие значения
были равны 20,8 атм, 8,7 атм (С1) и 41%. Содержание воды составляло
у Zygophyllum и у Babbagia более 600% сухого веса.
Условия в солянковой полупустыне Австралии напоминают засуш-
ливые области юго-западных штатов в США (рис. 359). Здесь на плоско-
горьях Пейнтед-Дезерт севернее Большого каньона Колорадо часто
растет только Atriplex confertifolia. Распределение осадков сходно с на-
блюдаемым в солянковой полупустыне Австралии.
Гаррис определил осмотическое давление и частично содержание
хлоридов у этого вида, а также у Atriplex canescens, встречающегося на
Засушливые области центральной Австралии
457
засоленных участках в пустыне Сонора Автор дополнительно к этому
вычислил величину осмотического давления, обусловленного хлоридами
в %; все эти данные приведены в табл. 84.
Таблица 84
Осмотическое давление клеточного сока и содержание
хлоридов у двух видов Atriplex (по Гаррису)
Atriplex canescens Atriplex confertifolia
W Cl % W Cl %
21,7 8,4 39 24,8 4,5 18
24,5 9,6 39 28,3 12,1 43
27,7 6,5 23 38,6 36,2 94 !
30,9 10 32 38,7 16,6 43
36,0 9,7 27 75,2 28,7 38
38,3 16 42 82,9 55 66
39,7 28,5 72 99,4 40,6 41
40,3 13,7 34 104,2 61,2 59
46,8 17,7 38 154,5 76 49
49,3 42,7 87 ! 156,2 93 59
107,7 51 47 202,5 80 39
Из приведенных в табл. 84 данных видно, что виды Atriplex также-
являются типичными галофитами, накапливающими много NaCl в кле-
точном соке. Содержание сульфатов относительно невелико. Из трех
анализов выяснилось, что у A. canescens при незначительном содержании
хлоридов сульфатов содержится примерно вдвое меньше, а при высоком
содержании хлоридов количество сульфатов составляет примернс
часть. То, что между величиной осмотического давления и содержа-
нием хлоридов нет никакой четкой зависимости, не удивительно, по-
скольку образцы брались на различной степени засоленных и увлаж-
ненных почвах. Водный и солевой режим тесно связаны друг с другом; от
них зависит величина осмотического давления и содержание хлоридов.
У Atriplex canescens (75% образцов) осмотическое давление не превы-
шало 40 атм. Отмеченное в одном случае исключительно высокое давле-
ние (более 100 атм) не может быть без дальнейшей проверки признано
достоверным.
Atriplex confertifolia встречается в значительно более сухих и засо-
ленных местообитаниях. В этих условиях у 20% образцов осмотическое
давление не превышало 40 атм, у почти 50% образцов оно было выше
100 атм, а у некоторых даже превышало 200 атм. Однако эти величины
нельзя считать абсолютно достоверными, поскольку после продолжи-
тельного засушливого периода поверхность листьев могла быть покрыта
солью, содержавшейся в отмерших волосках.
Как известно, у австралийских видов Kochia, в соке, отжатом и&
листьев, содержится немного солей. По данным Гарриса, в США у Kochia
vestita наблюдается та же картина. У этих видов не развиваются волоски,
1 J. A. Harris, The physico-chemical properties of plant saps in relation to.-
phytogeography, Minneapolis, 1932.
458
Глава 12
Рис. 359. Atriplex canescens (на переднем плане) у озера Солтон-Си в Кали-
форнии (фото Мак-Дугала).
выделяющие соль. Осмотическое давление (29 образцов) у Kochia vestita,
развитого в штате Юта, оказалось равным 18,9—54,6 атм, а содержание
хлоридов — 9—39%.
Таким образом, создается впечатление, что растения, распростра-
ненные в штате Юта и на севере Аризоны, экологически сходны с вида-
ми солянковой полупустыни, растущими в Австралии; к тому же клима-
тические условия этих территорий очень похожи; различия сводятся
к тому, что зимой в Австралии почти не бывает морозов, тогда как в соот-
ветствующих областях США зимы могут быть довольно холодными.
Заросли колючих злаков (спинифекс)
и заросли Acacia апеига (мулъга)
в центральной Австралии
Экология своеобразных склерофильных колючих злаков Triodia
и Plectrachne изучена еще очень слабо.
Один из видов рода Triodia (10 видов), Triodia basedowii, распро-
странен на песчаных почвах в наиболее засушливых частях Австралии,
тогда как Triodia pungens встречается дальше к северу от Западной
Австралии до Квинсленда, а в центральной Австралии покрывает также
каменистые осыпи, нередко образуя чистые травостои.
Центральная часть больших дерновин Т. basedowii, достигающих
высоты 50 см, довольно скоро отмирает; дальнейший рост продолжается
только на северной стороне дерновин. Поэтому круглая дерновина сна-
чала превращается в кольцо, а затем в открытую к югу подкову, которая
в конце концов распадается на отдельные части (цветное фото 2,
•стр. 368). Степень покрытия почвы составляет до 40%. У Т. pungens
дерновины, имеющие форму подушки, сохраняются значительно дольше;
дерновины достигают высоты 2 м и диаметра 4 м.
Засушливые области центральиой Австралии
459
Междоузлия этих злаков отличаются своими размерами: короткие
междоузлия чередуются с длинными. После дождей начинается интен-
сивный рост корней; они проникают в почву на глубину 3 м и очень
густо пронизывают верхние 100 см почвы Ч
У Т. pungens, как и у Ammophila, листья свернуты; они содержат
мало хлорофиллоносной ткани и много склеренхимы, образующей
несколько слоев под эпидермисом. В эпидермисе имеются клетки, выде-
ляющие смолу, которая покрывает поверхность листьев. Такие смоля-
ные пленки встречаются также у развитых в этой области видов родов
Eremophila (Муорогасеае), Dodonaea, Olearia (Compositae) и др. Устьица
всегда расположены как на внешней, так и на внутренней сторонах.
При свертывании листа устьица на внутренней стороне листа оказы-
ваются закрытыми. При сжимании клеток паренхимы закрываются
и устьица, расположенные на наружной стороне листа; тем самым расте-
ние сокращает потери воды до едва измеримого минимума.
Таким образом, эти склерофильные злаки отличаются ярко выражен-
ной ксероморфной структурой, которая позволяет им переносить даже
очень сильную засуху. У обычных злаков лист очень быстро высыхает
при малейшем недостатке влаги.
Прирост молодых листьев Triodia (при хорошей влагообеспеченности
и высокой температуре) достигает 3,5 мм в сутки. Кормовое значение
Triodia basedowii невелико, тогда как молодые листья Т. pungens охотно
поедаются скотом. Как уже упоминалось, севернее реки Мерчисон
в Западной Австралии распространены чистые злаковники, вытесняющие
здесь заросли Acacia апеига, которые чередуются с участками,
покрытыми Triodia basedowii на песках. Границы между отдельными
сообществами выражены очень нечетко. Увеличение годовой суммы
осадков на 25—30 мм может привести к смене одного сообщества другим.
Даже незначительные различия почвенных условий или изменение
направления дождевых потоков могут влиять на степень взаимопроник-
новения различных растительных группировок. Большое влияние ока-
зывают пастьба скота, а также вызываемые человеком пожары. На редкий
покров огонь не оказывает существенного влияния. Хорошо горят лишь
травостои, образованные Triodia, причем образуются такие же густые
черные клубы дыма, как в случае если горит смола.
Близ Алис-Спрингса после удара моЦнии возник пожар, распростра-
нившийся более чем на 50 км. Спустя 18 месяцев на выгоревшей площади
появилось много всходов Triodia, Aristida arenaria, нескольких видов
разнотравья и многочисленных эфемеров. Появление последних связано
с отсутствием корневой конкуренции с Triodia, поскольку обычно Triodia
сама расходует всю имеющуюся в наличии воду. После длительной
засухи листья желтеют, но после первого же дождя вновь начинают
зеленеть. Ниже приводится краткое описание травостоя, образован-
ного Triodia и развитого на красной песчаной почве.
Степень покрытия почвы —50%; дерновины —в виде подушек
высотой 50 см', высота одиночных кустов — до 2 м.
3 Triodia basedowii
1 Plectrachne schinzii и одиночно встречающиеся экземпляры Cassia spec.,
1 N. Т. В u г b i d g е, Morphology and anatomy of the Western Australian species
of Triodia R., Br. I u. II, «Trans. Roy. Soc. South Australia», 69, 1945, p. 303—308;
70, 1946, p. 221—229; Ecological notes on the vegetation of 80 Mile Beach («J. Roy.
Soc. West. Australia», 28, p. 157—164) and on the De Grey-Coongan Area (ibid., 29,
p. 151 — 161), 1941 — 1942.
460
Глава 12
Eucaplyptus gamophylla, Накеа spec., Grevillea juncifolia, Acacia
aneura.
Определение осмотического давления у этих склерофильных злаков
проводилось близ Алис-Спрингса под руководством Р. Слатьера 4. По-ви-
димому, эти злаки отличаются гидростабильностью, а их клетки обла-
дают большой сосущей силой даже при небольшом дефиците воды.
Основным видом, распространенным в аридных областях Австралии,
несомненно, является Acacia aneura, заросли которой местные жители
называют м у л ь г а. Этот вид доминирует на огромных территориях,
которые с самолета выглядят, как безбрежное серо-бурое море, преры-
ваемое лишь пятнами солончаков, грядами песчаных дюн и сухими рус-
лами рек.
Внешний облик Acacia aneura очень разнообразен. Филлодии у ака-
ции могут быть плоскими и цилиндрическими; иногда у стволов акации
высотой 4—6 м ветви простираются почти горизонтально, иногда же
ствол начинает многократно ветвиться от основания. Площади, занятые
древостоями, и степень их сомкнутости часто изменяются в зависимости
от условий влагообеспеченности. Иногда, в небольших понижениях,
кусты образуют почти непроходимую чащу; через нее, однако, можно
пробраться, поскольку у мульги в противоположность африканским
акациям стсутствуют колючки. Филлодии охотно поедаются овцами.
У Acacia aneura очень хорошо развита корневая система, глубоко*
проникающая в трещины уплотненных слоев почвы. На мощных почвах
основная масса корней развивается в пределах верхних 2 м. Корни,,
простирающиеся в горизонтальном направлении на небольшой глубине,,
придают акации устойчивость, остальная часть сильно ветвящихся корней
снабжает растения водой. У акации очень твердая древесина. Благодаря
выделению смолы кажется, что филлодии покрыты шеллаком. Содержа-
ние сухого вещества в эфирной вытяжке может достигать 4—15%; его
количество возрастает с увеличением сухости местообитаний. Соцветия,
напоминающие желтый колос, достигают длины 2—5 см. Поскольку
осадки выпадают нерегулярно, начало цветения у акации не связано
с определенным временем года; растения зацветают всегда после выпав-
шего дождя. Плоды и семена созревают лишь после сильных дождей.
Acacia aneura не выносит никакого засоления; в остальном обладает
широким экологическим диапазоном. В сухих местообитаниях она часто
образует разреженные чистые древостои высотой 1,8—3,0 м, обычно без
подлеска, иногда смешанные древостои с видами Acacia, Eremophila,
Cassia и др. Многочисленные эфемеры появляются только после хоро-
ших дождей, и тогда деревья растут среди моря цветов.
В зависимости от интенсивности осадков и времени года наблю-
дается смена аспектов, причем часто на значительных площадях господ-
ствует один вид (цветное фото 1, стр. 432).
Поскольку семена Acacia aneura перед прорастанием набухают
в течение нескольких недель, этот вид возобновляется лишь в очень
влажные годы. Семена A. aneura, по-видимому, очень долго сохраняют
свою всхожесть, как и большинство семян бобовых с твердой оболочкой.
К сожалению, точных данных о всхожести семян этого вида пока не
получено. Деревья растут примерно до тридцати лет. Очень часто на
Acacia паразитируют Loranthus.
1 R. О. S 1 a t у е r2 Annual Progress Report for 1957 —1958, Climatology Unit
CSIRO.
Засушливые области центральной Австралии 461
Слатьер изучал водный режим Acacia апеига близ Алис-Спрингса *.
В этой области выпадает в среднем около 250 мм осадков, причем 75%
приходится на 6 летних месяцев. Как и во всех других засушливых
областях, осадки здесь выпадают крайне нерегулярно. Часто бывают
засухи, длящиеся более трех месяцев. Наблюдения велись в течение
6 месяцев; образцы для наблюдений отбирались еженедельно. Определя-
лось относительное содержание воды в филлодиях, осмотическое давле-
ние и сосущая сила клеток. Слатьер применял разработанный им грави-
метрический метод определения сосущей силы клеток. Этот метод
в Хоэнхейме проверял Крееб; выяснилось, что он дает достаточно надеж-
ные результаты. Если не учитывать потери на дыхание, то полученные
значения сосущей силы оказываются несколько заниженными1 2.
Результаты исследования Слатьер представил в виде кривых, отра-
жавших зависимость между относительным содержанием воды и сосу-
щей силой, а также осмотическим давлением. Величины осмотического
давления были, по-видимому, вычислены, а не измерены. При сосущей
силе, достигавшей 130 атм. филлодии не повреждались, а величина
сосущей силы была больше осмотического давления, что могло бы сви-
детельствовать о наличии отрицательного тургора. Весьма ксероморфная
структура филлодий, по-видимому, свидетельствует о реальности этого
допущения. Однако окончательно об этом судить можно лишь после
публикации более подробных данных.
Автор взял несколько образцов близ Алис-Спрингса 23 и 24 октября
1958 г. после продолжительной засухи. Осмотическое давление и содер-
жание хлоридов определялось методом, описанным на стр. 456; получен-
ные результаты приведены в табл. 85.
Таблица 85
Осмотическое давление у Acacia апеига
(в 40 км севернее Алис-Спрингса)
№ образца Характеристика растений W G1 %
1 Старое дерево с иглообразными филлодиями 25 4,5 18
2 Маленький куст с широкими серы- ми филлодиями 27 4,8 18
3 Молодой куст со свежими зелеными филлодиями 26 3,5 13
По-видимому, этот вид отличается ярко выраженной гидростабиль-
ностью (ср. табл. 83, образец 7); вряд ли у него можно обнаружить коле-
бания гидратуры. Влажность в трех образцах достигала 58, 65
и 80% сухого веса. Однако эти величины ничего не говорят о дефиците
влаги, и разницу между ними скорее следует отнести за счет неодинако-
вого развития механических тканей и клеточных стенок.
1 R. О. Slatyer, Aspects of the tissue water relationships of an important
arid zone species (Acacia апеига F. Muell.) in comparison with two mesophytes, «Bull.
Res. Coun. Israel», D. 8, 1960, p. 159—168.
2 К. К r e e b, Uber die gravimetrische Methode zur Bestimmung der Saugspannung
und das Problem des negativen Turgors, «Planta», 55, 1960, S. 274—282; К. К r e e b,
M. О n a 1, Dasselbe II, Mitteilung, «Planta», 56, 1961, S. 409—415.
462
Глава 12
Особенности строения листьев Triodia basedowii не представляют
возможности взять образцы; в связи с этим исследовались образцы с трех
кустарников, распространенных среди травостоя Triodia на песках
в 100 км севернее Алис-Спрингса; поскольку у Acacia kempeana удалось
отжать лишь немного сока, полученные значения (табл. 86), как
и у A. aneura, нельзя рассматривать как абсолютно точные.
Таблица 86
Осмотическое давление кустарников, растущих среди покрова, образованного
Triodia
Да образца Название растений W С1 %
1 Eucalyptus gamophylla, фертильный молодой экземпляр с суккулепт- ными листьями . . . 17,0 5,0 29
2 Grevillea juncea с жесткими цилин- дрическими листьями 18,0 1,1 6
3 Acacia kempeana с широкими фил- лодиями 24,5 6,8 28
Несмотря на длительные периоды засухи, осмотическое давление
у этих видов, как и у Acacia aneura, оказалось невысоким. Вполне оправ-
дано невысокое содержание хлоридЬв; это связано с тем, что в централь-
ной Австралии почти не встречаются засоленные почвы, несмотря на то,
что эта область практически лишена стока. Влажность составляла
у Eucalyptus 95%, у Grevillea— 53% и у Acacia—80% сухого веса.
Севернее Алис-Спрингса Слатьер наблюдал чередование поперек
склона полос густых зарослей Acacia aneura с зарослями злаков. Оказа-
лось, что уклон (в среднем равный 1 : 500) на протяжении склона выра-
жен по-разному. В пределах зарослей акации он соответствует 1 : 600,
а на более крутых участках, покрытых зарослями злаков, —1 : 400. Вслед-
ствие этого дождевая вода слегка застаивается в зарослях акации,
которые в результате! получают несколько больше влаги, чем простран-
ства, покрытые в верхней части склона зарослями Eragrostis eriopoda,
а в нижней, более эродируемой, почти совершенно лишенные расти-
тельности.
Вследствие корневой конкуренции подлесок не развивается; лишь
под пологом Acacia aneura иногда в этих условиях встречаются чахлые
кусты Eremophila или пойкилогидрический папоротник Cheilanthes tenui-
folius. Под акацией заметны приствольные повышения (около 4 см),
связанные с поверхностным залеганием корней и накоплением опада
вокруг стволов. Вследствие этого текущие по почве дождевые потоки
обходят стволы акаций, но зато примерно 50% осадков, задерживаемых
кроной, стекает по стволу. ПоэтОлМу вода распределяется в почве очень
неравномерно. Пастьба скота препятствует зарастанию этих пространств
(см. стр. 460), поскольку молодые, лишенные колючек кусты акации
охотно поедаются животными.
Вероятно, из всех древесных пород, распространенных в Австралии,
Acacia aneura занимает наибольшие площади. Ареал этого вида про-
стирается с запада через всю центральную Австралию и на востоке далеко
заходит в Квинсленд. На восточной границе у Сент-Джорджа автор
Засушливые области центральной Австралии 463
наблюдал сомкнутые почти чистые древостои до 10 м. В холмистой
местности возвышения всегда заняты зарослями акации, а пониже-
ния рельефа эвкалиптовой саванной (Eucaplyptus melanophloia или
Е. populnea).
В этой же области на тяжелых глинистых светло-серых слегка солон-
цеватых почвах распространен еще один вид акаций, образующий круп-
ные заросли, — Acacia harpophylla (бригелоу-скраб). Стволы акации могут
достигать высоты 10—12 м.
Почвы, на которых развивается Acacia h., очень своеобразны.
Вследствие попеременного высыхания и увлажнения почвы здесь наблю-
даются такие же явления, какие происходят в Арктике при замерзании
грунта и последующем его оттаивании. Эти почвы (gilgay) растрескивают-
ся на многоугольные отдельности, которые напоминают полигональные
арктические почвы. Многоугольные в плане площадки представляют со-
бой впадины (gilgay или «melon holes») глубиной до 1 м и диаметром 4—5 м.
На разделяющих их узких грядах растут акации. Во впадинах раститель-
ность не развивается, поскольку в период дождей (годовое количество
осадков равно 600 мм) они заполняются водой. Эти впадины возникают
следующим образом. В период засухи в почве образуются трещины
глубиной до 1 м, куда ветер или вода приносят мелкозем. При увлажне-
нии мелкозем набухает и выступает над поверхностью почвы, обра-
зуя узкие невысокие гряды, между которыми формируются впадины.
У Acacia harpophylla формируется широко разветвленная корневая
система, расположенная близко (на глубине 5—10 см) к поверхности
почвы. От этих горизонтально расположенных корней в глубь почвы
отходят якорные корни; кроме того, на них образуется поросль, способ-
ствующая загущению и возобновлению древостоя. Эта поросль затрудняет
расчистку зарослей акации, и поэтому ее приходится уничтожать хими-
ческими средствами, используя для этой цели самолеты
Влажные местообитания в центральной Австралии
В бессточных понижениях засушливых областей происходит
накопление солей (см. стр. 329 и след.), приводящее к образованию
солончаков или соленых озер. Почва в этих понижениях всегда увлаж-
нена, даже если на ее поверхности! образуется соляная корка.
Влажность почвы и содержание солей снижаются от середины впадины
к ее краям. Соответственно этому растительный покров располагается
концентрическими кругами. Такие соленые озера встречаются прежде
всего в засушливых внутренних областях штата Западная Австралия
(рис. 358) и в северной части штата Южная Австралия. Их размеры
колеблются от совсем небольших понижений до таких обширных впадин,
как, например, озеро Эйр. Окружающая эти впадины растительность
образована гигрогалофитами; среди них в Австралии, как и всюду,
господствуют виды семейства Chenopodiaceae. В качестве примера выбра-
но небольшое соленое озеро в окрестностях Уилуны; 1 сентября 1958 г.
оно частично еще было заполнено водой. Здесь ясно выражена смена
следующих поясов растительности:
Первый пояс (окаймляет водную поверхность) шириной 30 м недавно
был покрыт водой, поэтому старые затопленные кусты Arthrocnemum
отмерли; появляются новые всходы.
1 Автор знакомился с этой областью под руководством С. Эвериста Брисбен).
464
Глава 12
Второй пояс (пояс Arthrocnemum — Salicornia) шириной 80 м. В ниж-
ней части (40 лг) пояса эти сочные солянки покрывают только около
3% поверхности почвы, тогда как в верхней части (40 лг), где к ним при-
соединяются другие виды (Atriplex inf lata, Kochia glomerata, Bassia
и Frankonia), степень покрытия почвы достигает уже 10%.
Третий пояс — пояс кустарничков — шириной около 1 км\ изредка
встречаются виды Bassia, Scaevola spinosa и Cratystylis (Compositae);
сомкнутый покров образует однолетний злак Aristida arenaria, у которого
в момент наблюдений уже созрели семена и налицо были признаки
отмирания.
Четвертый пояс — незасоленная площадь, покрытая зеленым ковром
однолетников, среди которых встречаются изредка Kochia pyramidata
и виды Eremophila.
Более сложны условия, когда солончак граничит с крутой гипсовой
дюной. Солончак Уэй, к югу от Уилуны, представляет собой настоящий
лабиринт засоленных котловин, гипсовых и песчаных дюн; в последний
раз он заполнялся водой в 30-х годах. В настоящее время дно этого огром-
ного понижения покрыто довольно сомкнутыми зарослями Arthrocnemum
и Salicornia. У края понижения был обнаружен один экземпляр Kochia.
Кроме того, у подножия гипсовой дюны росли отдельные экземпляры
Frankenia, Plagianthus holmsii (Malvaceae) и крупнолистный суккулент
Zygophyllum. Метром выше на гипсовой дюне среди густой заросли,
образованной Melaleuca, встречались отдельные экземпляры Casuarina.
Еще выше вместе с казуариной росла Acacia longifolia, а на самой
дюне росли Eucalyptus lesouefii, Atriplex vesicaria, A. paludosa, Lycium
и мелколистный Zygophyllum.
В табл. 87 приводятся величины осмотического давления у образ-
цов, собранных 3 сентября 1958 г. во впадине Уэй Ч
Таким образом, чуть выше уровня солончака, на гипсовых дюнах
уже могут расти виды, отличающиеся низким содержанием хлоридов,
тогда как настоящие галофиты, даже если они растут на песчаной дюне,
обнаруживают очень высокое содержание хлоридов (образцы 9 и 10).
Второго сентября 1958 г. для анализа были взяты растения, расту-
щие на засоленных почвах в районе Уилуны. Наряду с Acacia апеига
и Triodia basedowii здесь распространены различные виды Eremophila
(Муорогасеае). Они часто растут в крайне сухих местообитаниях, их
блестящие, покрытые лаком листья на ощупь кажутся совершенно сухи-
ми, хотя фактически влажность листьев составляет 120% сухого
веса. Величины осмотического давления у растений, распространенных на
незасоленных почвах, приведены в табл. 88.
Из данных этой таблицы видно, что Eremophila не отличается ни
высоким осмотическим давлением, ни повышенным содержанием хлори-
дов. Эвкалипты, приуроченные в аридных областях к сухим долинам,
как и виды, распространенные во влажных прибрежных областях (W =
13—16 атм, G1 = 1,5—5 атм), отличаются невысокой концентрацией
клеточного сока. Только Bassia, обладающий суккулентными листьями
и относящийся к числу галофитов, даже на незасоленных почвах накап-
1 Автор смог познакомиться с этим районом, присоединившись к экспедиции
Топографического отдела Организации научных и промышленных исследований Бри-
танского содружества наций (CSIRO), руководимой Дж. Мабуттом. Ботанические
исследования в экспедиции вел Н. Шпек. Для проведения подробных экологических
исследований не хватило времени; собранные образцы были в дальнейшем исследо-
ваны Е. Бихлером в Штутгарт-Хоэнхейме.
1. Вади-Хофф близ Хельвана (ОАР). Ландшафт каменистой пустыни (гамада); на
переднем плане — русло высохшего водотока. Полосы растительности выражены
только вдоль песчаного русла.
2. Щебнистая пустыня (серир) вдоль шоссе Каир — Суэц (ОАР). В руслах временных
водотоков на отложениях песка (светлые полосы) встречаются отдельные растения.
Более темная поверхность, покрытая щебнем, лишена растительности.
Засушливые области центральной Австралии
465
Таблица 87
Осмотическое давление у растений, развитых на солончаке Уэй;
виды перечислены в последовательности, связанной с высотой местообитаний
над уровнем солончака
№ образца Название растений п условия произрастания W С1 %
1 Arthrocnemum, солончак . ... 53,6 36,5 68
2 Arthrocnemum из Купа мора (для сравнения) 48,7 36,5 75
о Kochia spec., пограничная полоса 26,7 15,0 56
4 Zygophyllum (крупнолистный), верх- няя часть засоленного пояса . . 16,4 13,1 80
5 Plagianthus holmsii, пнжняя гипсо- вая терраса 32,5 13,2 41
6 Casuarina, дерево, растущее ниже всех ... 17,8 4,9 27
7 Murtaceae, там же 27,7 5,4 19
8 Eucalyptus lesouefii, на песчаной дюне 23,9 3,6 15
9 Zygophyllum, мелколистный .... 13,7 11,0 80
10 Atriplex vesicaria, на песчаной дюне 70,1 58,9 84
ливает относительно много хлоридов. Его влажность достигает почти
470% сухого веса.
В целом можно сказать, что виды, распространенные в засушливых
областях Австралии и относящиеся к числу негалофитов, также обла-
дают способностью поддерживать высокий уровень гидратуры (низкое
Таблица 88
Осмотическое давление у растений, развитых на незасоленных почвах;
окрестности Уплуны
№ образца Название растений и условия произрастания W (1 %
1 Eremophila jreisii, очень сухо . 15,4 1,3 10
2 Eucalyptus oleosa, сухая долина . . 22,8 1,4 6,1
3 Bassia spec. (Chenopodoiaceae) . . . 29,7 12,9 44
осмотическое давление) даже в течение длительных периодов засухи; это
связано с тем, что они лучше обеспечены влагой, чем это предполагается
в большинстве случаев (см. стр. 306 и след.); кроме того, эти виды могут
при недостатке влаги очень резко сокращать газообмен и тем самым
транспирацию, сохраняя таким путем водный баланс на постоянном,
уровне. Растения, развитые в крайне засушливых природных областях,
где засуха бывает выражена в течение длительного времени, могут испы-
тывать нарастающий дефицит влаги в тканях. Для предотвращения этого
в растениях прекращается процесс накопления органического вещества.
Наблюдается ли у растений в период засухи ассимиляция углекислоты,
выделяемой при дыхании, или же происходит потеря органического
вещества — пока еще точно не установлено.
466
Глава 12
Область вокруг Алис-Спрингса, расположенная почти в самом центре
Австралии, у южного тропика, > отличается еще одной особенностью.
В общем очень монотонный равнинный характер местности нарушается
здесь несколькими горными хребтами (например, горы Макдоннелл), что
способствует обогащению флористического состава растительности. Горы
сложены докембрийскими кварцитами и песчаниками, вздымающимися
на высоту 1200 м (наивысшая точка — 1460 м) над равниной (650 м над
уровнем моря).
Если на равнине (см. стр. 459) в зависимости от почвенных условий
чередуются заросли Acacia апеига и колючих злаков {Triodia basedowii)t
то в горах наблюдается совершенно иная картина. На крутых скалистых
склонах растут лишь отдельные низкорослые Callitris glauca, В рассели-
нах скал встречаются Macrozamia macdonnellii (Cycadaceae) и Capparis
mitchellii, Лишь на более пологих нижних частях склонов появляется
покров, образованный Triodia pungens; еще ниже по склону к этому виду
присоединяется Acacia апеига.
Эту область прорезает несколько сухих русел рек (creeks). Река Тодд
принадлежит к бассейну реки Финке, впадающей в озеро Эйр, однако ее
воды большей частью испаряются или просачиваются в грунт, прежде
чем успевают достичь озера. На песчаных отмелях в сухих руслах рас-
тет Nicotiana velutina; по краю сухого русла встречается Eucalyptus са-
maldulensis, а на нижней террасе распространены Накеа lorea и Н, inter-
media, Напочвенный покров образован злаком Themeda australia, свой-
ственным более увлажненным областям на востоке Австралии.
На широких сложенных глинами речных террасах часто встречается
Atriplex nummularia, ближе к их краю растут многочисленные кусты
Eremophila, В сухих руслах небольших рек Eucalyptus camaldulensis
замещается Е, рариапа; ослепительно белая кора последнего хорошо
видна даже ночью. Вместе с последним видом растут Е, dichromophloia^
Acacia estrophiolata, виды Melaleuca и др. Одиночные экземпляры пере-
численных видов встречаются и на скалистых участках. В совсем мелких
ложбинках стока произрастает Citrullus ecirrhosus; на затопляемых пло-
щадях распространена Atalaya hemiglauca, размножающаяся корневой
порослью; большие площади часто бывают покрыты зарослями Acacia
estrophiolata и Ventilago viminalis, а в периодически затопляемых пониже-
ниях разрастаются Themeda australis, Digitaria brownii и крупная Aristida
nitidula. Если грунтовые потоки пересекаются выходами кварцитов, то
создается подпор и в этом месте образуются воронки, вокруг которых
встречаются реликты, как, например, Ficus platypoda, Dodonea viscosar
лиана Marsdenia australis; на скалах Sarcostemma australis, а у воды —
Ricinus и Hibiscus,
Большой известностью пользуется долина реки Палмер близ Херманс-
берга; здесь, в самом сердце Австралии, растет пальма Livistona murryae, не-
смотря на то, что граница ареала этого рода проходит чуть южнее Дарвина.
По устному сообщению Чиппендейла, там же распространены Typha
angustifolia, Phragmites karka, Najas major, Xerotes dura, Cryptandra spa-
thulata (Lauraceae), Rulingia magniflora (Sterculiaceae), Baeckea polystoma,
Hibbertia glaberrina, Teucrium grandiusculum; в других убежищах релик-
тов изредка встречаются папоротники: Adiantum hispidulum, Nephrolepis
cordifolia, Cyclosorus gongylodes и Lindsaea ensifolia.
Около водоемов или в тенистых влажных расселинах скал в пусты-
нях в большинстве случаев распространены реликты, отражающие
иные, ранее существовавшие климатические условия.
Глава 13
ПУСТЫНЯ САХАРА
ОБЩАЯ ХАРАКТЕРИСТИКА
Крупнейшая пустыня земного шара — Сахара расположена в Север-
ной Африке, от Атлантического побережья на западе до долины
Нила на востоке (рис. 360). Полоса пустынь простирается еще
дальше на восток по направлению к Красному морю, захватывает Аравию
и доходит вплоть до пустыни Синд (Тар) на северо-западе Индии х. В севе-
ро-западной части ареала пустыни граничат со средиземноморской
областью жестколистных деревьев и кустарников, а в восточной части
Средиземного моря полоса пустынь доходит до побережья и далее к востоку
тянется в глубь Сирии и Месопотамию На северной стороне высо-
ких ирано-среднеазиатских гор полоса пустынь также широка
и простирается вплоть до пустыни Гоби. Однако в области ирано-
туранских и монгольских пустынь климатические условия и в пер-
вую очередь холодные зимы определяют совершенно иной характер
растительного покрова. Поэтому краткая характеристика этих пустынь
будет дана при обзоре растительности умеренных областей 1 2.
Северная граница субтропических пустынь совпадает с границей
промышленной культуры финиковой пальмы, поскольку она может куль-
тивироваться лишь в областях с достаточно высокой температурой, при
большой сухости воздуха и полном отсутствии холодных периодов;
только в этих условиях финиковая пальма дает товарную продукцию,
пригодную для экспорта. Граница возделывания финиковой пальмы
совпадает на западе с южным склоном Анти-Атласа и Сахарского Атласа.
По направлению к востоку рощи финиковой пальмы иногда достигают
побережья.
Гораздо труднее провести южную границу пустынь, поскольку
здесь переход осуществляется постепенно; южнее Сахары расположен
судано-синдский пояс, простирающийся через южный Иран до полуостро-
ва Индостан 3. Переходная область (с количеством осадков 100—200 мм)
между южной Сахарой и областью развития саванн в Судане называется
зоной Сахель 4. Здесь осадки выпадают летом. В сахельской зоне господ-
ствуют злаковники с разбросанными по ним немногочисленнымидеревь-
1 Н. Schiffers, Die Sahara, Stuttgart, 1950. «Сахара» на арабском языке
означает «желтый» или «красноватый», а также «пустынная равнина».
2 Характеристика основных флористических элементов пустынь приведена
в работе: Е. М. Л а в р е н к о, Основные черты ботанической географии пустынь
Евразии и Северной Африки, АН СССР, Комаровские чтения, вып. 15 (6/ХП
1960 г.), М.-Л., 1962.
3 М. Z о h а г у, On the geobotanical structure of Iran, «Bull. Res. Counc. Israel»,
Sect. D. 11 (Suppl.) Jerusalem, 1963.
4 «Sahel» означает «берег»; иными словами, эта зона является берегом обширных
пустынь, расположенных на севере.
30*
468
Глава 13
Ш|1 ЕЖ12 1ПШПППП13 Сх%714 П7715
Рис. 360, Сахара (по Шифферсу, несколько изменено).
1 — горные системы; 2 — пески; з — впадины, лежащие ниже уровня моря; 4 — уступы
высокогорных плато; 5 — источники.
ми. Южная граница Сахары проходит примерно по 19° с. ш. в западной
части Африки и 18° — в восточной. Сахельская зона расположена между
20 и 16° с. ш.
Протяженность Сахары от Атлантического океана до Красного моря
превышает 6000 км, а с севера на юг она простирается на расстояние
более 1800 км. Таким образом, площадь Сахары достигает примерно
9 млн. км2. Следовательно, Сахара больше Австралии и лишь немного
меньше Европы. В Сахаре выделяется несколько бессточных впадин,
отрезанных от моря складчатыми горами Атласа и ливийским, а также
нубийским поднятиями.
За исключением отдельных более высоких горных массивов, бес-
сточные впадины представляют собой выровненные поверхности, сложен-
ные осадками молодого возраста. Наиболее низкие участки этих впадин
часто заполнены песком и дюнами, однако вопреки широко распростра-
ненному мнению песчаные пустыни занимают всего 20% общей площади
Сахары. На более высоких возвышенностях на поверхности наблюдаются
выходы кристаллических пород. Некоторые возвышения, подобно
горам Ахаггар, достигают высоты 3000 м и даже выше, как, например,
нагорье Тибести. Широко распространены каменистые пустыни, поверх-
ность которых покрыта корой выветривания (гамады); они пересекаются
сухими руслами временных водотоков (вади) х. Неглубокие депрессии
выполнены отложениями плейстоцена; здесь развиты щебнистые (per,
серир) пустыни или пустыни, покрытые грядовыми песками (эрг). У север-
ной границы Сахары в бессточных впадинах происходит отложение
илистых частиц и одновременно, в результате сильного испарения,—
накопление солей. Постепенно образуются обширные солончаки и глини-
стые впадины (шотты, себха). Среди бескрайней пустыни рассеяны оазисы,
существование которых связано с выходами грунтовых вод. Все без
исключения оазисы освоены человеком (рис. 361).
1 См. W. Meckelein, Forschungen in der zentralen Sahara, Braunschweig, 1959.
Пустыня Сахара
469
По режиму осадков в Сахаре можно выделить три зоны: северную,
центральную и южную. В северной зоне осадки выпадают зимой и коли-
чество их не превышает 200 мм за год. К югу количество осадков посте-
пенно уменьшается, и в центральной зоне они выпадают лишь эпизоди-
чески. Поэтому эта область отличается крайней бедностью видового
состава. По данным Киллиана х, в окрестностях Феццана, в центральной
части Сахары, встречаются лишь следующие виды: Zygophyllum simplex.
Z. album, Fagonia bruguieri, Traganum nudatum, Cornulaca monacantha.
Alhagi camelorum и Aristida pungens. Многолетники растут лишь там,
где после эпизодических дождей накапливается вода и почва остается
достаточно влажной даже при отсутствии осадков в течение многих лет.
В высоких горах центральной Сахары вряд ли выпадает большее
количество осадков. Огромные пространства, например на юге Ливийской
пустыни, практически совершенно лишены осадков. На протяжении
более 200 км пути автор не встретил здесь ни одного растения.
Еще дальше к югу количество осадков снова медленно возрастает,
однако здесь они выпадают летом, что иллюстрируется климадиаграммами
на рис. 362—364. Такое распределение осадков обусловливает различ-
ный характер растительного покрова на юге и севере Сахары 2. Через
1 Ch. Killian, Observations sur la biologie des quelques plantes fezzanaisses,
«Publ. Inst. Rech. Sahar», 4, 1945, p. 1. I
2 P. О z e n d a, Flore du Sahara septentrional et central, Centre Nat. de la Rech.
Scient., 1958. В этой работе интерес представляет также введение, стр. 9—96;
Р. Q и ё z е 1, La vegetation du Sahara, Geobotanica Selecta, Bd 2, Stuttgart, 1965.
Рис. 361. Оазис Сивана севере Ливийской пустыни. Роща финиковых пальм в
Агурми. На переднем плане — песчаная пустыня с отдельными полузасыпан-
ными песком финиковыми пальмами, на заднем плане — засоленная впадина
(себха) (фото Рюбеля и Улингера).
470
Глава 13
Рис. 362—364. Климадиаграммы северной, центральной и южной Сахары.
центральную Сахару проходит граница между голарктической и палео-
тропической флористическими областями. В составе флоры преобладают
сахаро-аравийские элементы, явно связанные с голарктикой. На севере
здесь ощутимо влияние Средиземноморья, выражающееся в распростра-
нении видов Astragalus, Reseda, Plantago, Salsola и других, проникающих
до центральной Сахары, в то время как на юге суданские элементы палео-
тропической флоры не заходят слишком далеко на север. К суданским
элементам относятся виды родов Indigofera, Tephrosia, Hibiscus, Cleome,
Acacia, Eragrostis, Cyperus и др.
Сахаро-аравийские виды могут быть разделены на две группы:
одни обладают очень большим ареалом, тогда как распространение других
ограничено только Африкой; следовательно, последние являются уже
собственно сахарским элементом. Среди последней группы одни виды
чаще встречаются на западе, а другие — на востоке Сахары. Здесь рас-
пространены также виды с крайне ограниченным ареалом: например, Fredo-
На (Anabasis) aretioides, подушковидное растение, встречающееся в Сахаре
к югу от Орана. В целом эндемизм в Сахаре выражен очень отчетливо.
Правда, в большинстве случаев речь идет о викариантах средиземно-
морских видов или о микроэндемиках. Даже если не учитывать последних,
по данным Озенды, в Сахаре встречаются 162 эндемика, что составляет
примерно 25% общего числа распространенных здесь видов. Это свидетель-
ствует о том, что пустыня способствует изоляции таксонов.
В Сахаре преобладают семейства: Chenopodiaceae (17 родов), Crucife-
гае (31 род), число видов которых, однако, уменьшается к югу, и Zygoph-
yllaceae (7 родов).
Пустыни вовсе не должны отличаться бедностью флористического
состава. Например, флора Карру или пустыни Сонора очень богата,
особенно суккулентами. Однако этого нельзя сказать о Сахаре; в флори-
стическом отношении она в десятки раз беднее равновеликой по площади
Южной Европы. До настоящего времени в центральной Сахаре, по дан-
ным Озенды, обнаружено 450 видов высших и 75 видов низших растений,
а в нагорье Тибести — соответственно 350 и 45 видов. Бросается в глаза
бедность мхами и лишайниками. Пойкилогидрические папоротники
отсутствуют в Сахаре полностью. В этом, в частности, проявляется
большая сухость данной пустыни.
Только северная окраина и Атлантическое побережье Сахары, где
часто наблюдаются туманы, более богаты лишайниками. Здесь растут
также и настоящие суккуленты (Euphorbia, Senecio). В горах Ахаггар
встречаются два вида Caralluma. Если в тропиках на площади 10 тыс. км2
можно обнаружить примерно 3—4 тыс. видов, в Европе — 1—2 тыс., то
в Сахаре — не более 150 видов.
Пустыня Сахара
471
Флористический состав и пути проникновения средиземноморских
элементов далеко в пределы центральной Сахары могут быть поняты
только с учетом генезиса флоры. В плейстоцене в Сахаре выпадало зна-
чительно больше осадков. Об этом, в частности, свидетельствуют глубоко
эродированные долины и речные террасы, которые могли возникнуть
только в период выпадения повышенного количества осадков. О большей
влажности климата в недалеком прошлом можно судить также по наход-
кам костей жирафов, слонов и носорогов, а также по наскальным рисун-
кам, найденным в ныне совершенно необитаемых областях. В неолитиче-
ских стоянках обнаружена пыльца Pinus halepensis, Quercus ilex, Callit-
ris, Cupressus, Juniperus. Эти средиземноморские виды ныне полностью
отсутствуют в Сахаре. Лишь реликтовые экземпляры этих видов встре-
чаются изредка в горах центральной Сахары. Приведенные факты сви-
детельствуют о том, что южная граница средиземноморской области
после плейстоцена сильно отодвинулась к северу.
Северная граница тропической области, видимо, наоборот, или
вообще не изменилась или изменилась значительно меньше. Это и обу-
словило возникновение крупнейшей пустынной области земного шара.
ПОЧВЫ И РАСТИТЕЛЬНОСТЬ
Поскольку в пустыне не происходит накопления гумуса, здесь
распространены примитивные почвы — продукты выветривания горных
пород. В пустыне преобладает физическое выветривание, хотя не исклю-
чено и химическое выветривание, связанное с воздействием различных
солей и кислот, содержащихся в воде. Из-за отсутствия перегноя почвы
пустынь практически являются безжизненными. Правда, Киллиан
и Фэйе 1 установили, что все пустынные почвы отличаются активной
жизнедеятельностью микроорганизмов и даже интенсивным выделе-
нием СО2, однако эти данные требуют подтверждения, так как степень
интенсивности дыхания почвы не может превосходить степень активности
накопления органического вещества растительным покровом 1 2. Посколь-
ку на больших пространствах растительность вообще не развита, то
вполне естественно, что в почвах должны отсутствовать как газообмен,
лак и деятельность микроорганизмов. Исключение представляют лишь
места, где скапливается органическое вещество, например в плотных
подушках некоторых растений или под отдельными кустами в виде
листового опада. Здесь при благоприятных условиях увлажнения могут
развиваться микроорганизмы, и в частности нитрифицирующие бакте-
рии; именно с их деятельностью связано относительно высокое содержа-
ние нитратов в пустынных почвах 3.
Свойства пустынных почв определяются прежде всего горными
породами, на которых они формируются. Наиболее действенное влияние
в пустыне оказывает ветер, а вода в настоящее время играет меньшую
роль. Под воздействием ветра образуются либо эродированные площади,
либо аккумулятивные равнины.
1 Ch. Killian, D. Felier, Recherches sur les phenomenes microbiologiques
des sols sahariens, «Ann. Inst. Pasteur», 55, 1935, p. 573.
2 H. Walter, W. Haber, Uber die Intensitat der Bodenatmung usw., «Вег.
Dtscb. Bot. Ges.», 70, 1957, S. 275—282.
3H. Vargues, Etude microbiologique de quelques sols sahariens en relation
avec la presence d’Anabasis aretioides Coss, et Moq. Trav. Lab. Beni-Ounif № 30,
Proc. Intern. Symp. on desert research, Jerusalem, 1952.
472
Глава 13
Каменистая пустыня, или гамада
В результате ветровой эрозии с возвышенных участков плоскогорий
сносятся все продукты выветривания, а под воздействием дефляции
обтачиваются все обнажения и выходы горных пород. На поверхности
накапливаются обломки породы величиной примерно с кулак и образуется
своеобразная пустынная корка. Возникает каменистая пустыня, или
гамада 1 (рис. 365).
Гамады формируются в результате разрушения осадочных пород
мелового и третичного возраста, занимающих наибольшие площади
в Сахаре. Поскольку речь идет об осадочных породах морского происхо-
ждения, в них всегда содержатся соли. Под пустынной корой залегает
более или менее мощный красновато-бурый пылеватый слой, возникший
в результате выветривания известняков и мергеля; его выдуванию пре-
пятствует корка, покрывающая поверхность, образующаяся в результате
выпадения солей. Атмосферные осадки так незначительны, что они не
могут вымыть эти соли; в результате химических процессов, обуслов-
ленных конденсацией влаги в холодные ночи и ее испарением с поверхно-
сти породы днем, на утесах, обломках и щебне образуется черно-бурый
пустынный загар, состоящий из окислов железа, марганца и кремнезема.
Последний придает каменистой пустыне мрачный облик (цветное фото 1,
стр. 464).
Рис. 365. Каменистая пустыня (гамада) близ Хелуана (ОАР). Обнаженные
в результате ветровой эрозии выходы горных пород и осыпп. В сухой долине
временного потока (вади) встречаются отдельные растения (фото Штоккера)
1 Hamada (араб.) — отмирать.
Пустыня Сахара
473
Поверхность гамады местами глубоко рассекается каньоноподоб-
ными долинами, крутые склоны которых покрыты каменистыми
осыпями.
Вследствие незначительных запасов влаги в почве и часто довольна
высокого содержания солей в каменистой пустыне растения обычно не-
развиваются. Только в областях со слабо выраженными периодами
дождей в расселинах скал можно встретить отдельные экземпляры
растений.
Щебнистая пустыня (рег, серир)
При выветривании гетерогенной материнской породы, например*
конгломератов, легче поддающаяся разрушению цементирующая масса
уносится ветром, а на поверхности накапливаются щебень или галька.
В этом случае говорят об автохтонных щебнистых пустынях. Аллохтон-
ные пустыни возникают в местах, где скапливаются мощные аллювиаль-
ные отложения, из которых ветром вынесены наиболее мелкие частицы.
Такие наносные равнины, в частности, были образованы Нилом на севере
Египта в плювиальный период.
Щебнистые пустыни, распространенные между Каиром и Суэцем,
образовались в результате выветривания флювиальных отложений оли-
гоцена или плио-плейстоцена (цветная вклейка 2, стр. 464).
Под находящимся на поверхности слоем щебня в виде тонкой порош-
ковидной массы накапливается гипс. На несколько большей глубине
(около 50 см) залегает цементированный гипсом горизонт, сложенный
из прослоек песка и гальки. Существуют два возможных варианта возник-
новения подобных уплотненных горизонтов.
1. Просачивающаяся в грунт дождевая вода увлекает с собой в рас-
творенном виде CaSO4, который выкристаллизовывается у нижней грани-
цы промачивания почвы. В результате испарения устанавливается восхо-
дящий ток воды и вместе с ней вверх вновь поднимается часть растворен-
ного гипса. Вода, испаряясь, оставляет на поверхности почвы рыхлый
слой гипса.
2. Слой кристаллического гипса возник ранее при более влажных
климатических условиях, в то время как слой рыхлого гипса представляет
собой недавнее образование.
Щебнистая пустыня, как правило, слабо засолена, но тем не менее
содержание хлоридов в гипсоносных слоях может достигать 1 %
и более.
Слой щебня на поверхности часто бывает настолько плотным, что
корни с трудом проникают сквозь него. В связи с этим в щебнистых
пустынях, особенно в более засушливых центральных частях Сахары,
растения не встречаются, В более влажных условиях южноалжирской
Сахары, по данным Озенда, распространены разреженные зарослиHalo-
xylon (Clienopodiaceae), среди которых встречаются несколько сопутству-
ющих саксаулу растений. Поверхность этих пустынь выровнена или
образована плоскими холмами, разделенными между собой более или
менее широкими песчаными долинами, которые вливаются в обширные
песчаные равнины. Растительный покров на этих холмах крайне скуден
и развивается преимущественно по понижениям на северных склонах
холмов, В местах скопления песка, и в долинах пересыхающих рек, и па
равнинах, покрытых песком, возникают более благоприятные условия,
для развития многолетних видов.
474 Глава 13
На песках растут виды Aristida, а также геофиты и некоторые одно-
летники. На суглинистых почвах появляются колючие Cornulaca (Cheno-
podiaceae), Randonia (Resedaceae) или Zygophyllum album и Hyoscyamus
muticus.
Песчаная пустыня, или эрг (арег)
Песок, сносимый ветром с возвышений, аккумулируется в пониже-
ниях рельефа, где отлагается с подветренной стороны мелких препят-
ствий или перед любым уступом рельефа. Мощность этих песчаных отло-
жений часто незначительна. Наряду с этими участками встречаются огром-
ные пространства грядовых песков, называемые местными жителями
«эрг». Эти песчаные пустыни алжирской Сахары занимают площадь,
равную территории Франции. При устойчивом направлении ветра
образуются серповидные дюны, или барханы, пологие с наветренной
стороны и крутые с подветренной. При частой смене ветров дюны не
двигаются и только их гребень постоянно перевевается в зависимости от
направления ветра.
Песок не всегда состоит из зерен кварца, иногда он образуется
в результате выветривания известняков. В отдельных случаях зерна
песка бывают покрыты пленкой окиси железа, которая в наиболее сухих
и жарких областях придает песку красноватый оттенок, и тогда вся
местность кажется окрашенной в ярко-красный цвет. В более влажных
условиях, например вблизи побережья, пески имеют желто-бурую
окраску.
В аридных областях в местах накопления песков возникают благо-
приятные условия для скопления влаги. Так, в песках Большого Запад-
Р и с. 366. Песчаная пустыня Мармарика (на севере Ливии) с небольшими дюнами,
покрытыми полукустарниками (фото Гелингера).
Пустыня Сахара
Mb
ного эрга близ Бени-Аббеса, несмотря на ничтожное количество осадков,
в более глубоких слоях накапливается влага. Данные об эпизодических
дождях, давших более 5 мм осадков за 1946—1950 гг., приведены
в табл. 89 (из F. Pierr е, Proc. Montp. Symp., 1955).
Т а б л и ц а 89
Бедность осадками сочетается с высокими температурами воздуха; так, в июле максимум равен 43,3° и минимум 27,7°, а в январе соответ- ственно 18,8 и 3,1°. Соответственно велику и испаряемость, возрастающая по мере удаления от побе- режья Средиземного моря Таблица 90 (табл. 90). Испаряемость за год в северной и центральной На движущихся дюнах, Сахаре обоазованных неппепывно
перемещающимся и поэтому Место наблюдения совершенно сухим песком, ра- д Удаленность от побережья, к.и Испаряе- мость, мм
’стения, конечно, не могут развиваться. Однако на менее о подвижных песках встреча- с * & ются растения, корни кото- т/.ин рых достигают влажных ело- „ к тт Бени-Аббес ев. Наиболее характерным 1 1 Адрар . из распространенных здесь видов является Aristida pun- На берегу 300 425 575 850 1383 2916 4052 4809 5033
gens с жесткими колючими
листьями. Вместе с ней растут Tamarix articulata, Ephedra alata, Retama
retain. Genista saharae и Calligonum comosum, а также эндем северо-запад-
ной Сахары С. azel. Из видов травяного покрова можно назвать Cyperus
eonglomeratus, Moltkia ciliata и эндемичный злак Danthonia fragilis.
Многолетние виды, создающие известное препятствие для ветра, часто
растут на образующихся вокруг них небольших дюнах (рис. 366). При
помощи широко разветвленной корневой системы они поглощают в течение
всего года достаточное количество воды из влажных слоев песка, поэтому
у них периодичность развития выражена недостаточно ясно.
Сухие долины временных потоков, или вади
В пустыне наряду с ветром вода также играет определенную, хотя
и меньшую роль. Многочисленные эрозионные образования — сухие
большую часть года долины временных потоков (вади) — нарушают
476
Глава 13
монотонность рельефа каменистых и щебнистых пустынь, Дождевые
потоки стекают в долины с окружающих возвышений и отлагают на дне
русла песок и щебень, Вода при этом вымывает соли и сравнительно
глубоко увлажняет аллювиальные почвы, Эти местообитания особенно
благоприятны для произрастания растений, Чем больше водосборный
бассейн сухих долин, тем большее количество воды протекает в них,
Нередко в вади возникает подрусловой поток грунтовых вод, и тогда по
краям долины и на оползневых склонах, даже в самом центре Сахары
вырастают небольшие рощи, В средней части долины и на ее склонах
растения не встречаются; это связано с перемещением почвы при паводках
(рис, 367 и цветное фото 1, стр, 464),
Обычно в сухих долинах растут несколько видов акаций и злак
Panicum turgidunr, вместе с ними встречаются Zizyphus, Maerua, Balani-
tes, Pennisetum dichotomum, Zilla (рис, 368), Ephedra, безлистное зонтичное
Pituranthos и многие другие. На слабо засоленной почве вместо перечис-
ленных видов появляются Tamarix articulata (Т, aphylld) с Zygophyllum
или Salsola с другими видами маревых, На юге к ним присоединяются
Calotropis, Aerva и Ipomoea — элементы тропической флоры,
Впадины с засоленной глинистой почвой
(дайа, себха, или шотты)
Вода, стекающая по сухим долинам, несет с собой глинистые частицы;
они откладываются в местах просачивания воды в грунт, или в бессточных
понижениях,
При наличии в этих понижениях внутрипочвенного стока здесь фор-
мируются незасоленные почвы, хорошо увлажненные на всю глубину
профиля, В этих условиях встречаются Pistacia, Zizyphus, а также более
мелкие многолетние виды и эфемеры,
После дождя в небольших незасоленных впадинах особенно обильно
развиваются однолетние виды (терофиты), Лемэ 1 выделил близ Ревуаль-
Бени-Унифа три типа сообществ:
1, Во впадинах с глинистой почвой —тип сообщества Althaea ludwi-
gii — Trigonella anguina.
2, В песчаных руслах потоков — тип сообщества Lotononis dichotoma.
3, На песчаных наносах — тип сообщества Asphodelus pendulinus.
Степень покрытия почвы терофитами может достигать 5—20% (до
50%); корни у них проникают на глубину всего 20 см. Корни терофитов
очень быстро углубляются в почву, поскольку верхние слои последней
быстро высыхают; так, в одном из пунктов наблюдений после выпадения
8 мм осадков спустя 20 дней верхние 10 см уже не содержали доступной
для растений влаги,
В бессточных понижениях, где вода беспрерывно испаряется, проис-
ходит сильное накопление солей, В результате образуются солончаки,
часто покрытые белыми выцветами солей, Местное население называет их
«себха», или «шотты» (рис. 369),
В Сахаре засоленные почвы играют лишь незначительную роль,
поскольку количество осадков здесь так мало, что соли не перемещаются
и не аккумулируются в понижениях. Солончаки распространены в основ-
ном в переходной полосе между пустыней и побережьем Средиземного
1 G. Le m е е, Les associations a therophytes des depressions sableuses et limo-
neuses non salees et des rocailles aux environs de Beni Ounif, «Vegetatio», 4, 195.‘k
p. 137-154.
Рис. 367. Широкая сухая долина временного потока (вади) в египетско-ара-
вийской пустыне. Впереди слева — гамада, в центре — кусты Zilla spinosa и
Zygophyllum coccineum в сухом русле, сложенном песком и галькой (фото
Штоккера).
Р и с. 368. Молодой экземпляр Zilla spinosa в цвету (фото Штоккера).
478
Глава 13
Рис. 369. Впадина с засоленной глинистой почвой в Вади-эн-Натрун. На
небольших песчаных дюнах Zygophyllum album (фото Штоккера).
моря и вдоль него, где сложились особо благоприятные для их образова-
ния условия: наличие пород с высоким содержанием солей и очень высо-
кий уровень грунтовых вод. Преобладают хлористые соединения (натрия
и магния), встречаются также и сульфаты. В некоторых случаях разви-
ваются карбонатно-известковые солончаки.
В пустыне всюду, где грунтовые воды стоят близко и поверхность поч-
вы бывает постоянно влажной, образуется корка. В ее составе известь,
гипс или хлориды и сульфаты в зависимости от содержания солей в грун-
товых водах. Белый налет солей часто наблюдается и в междюнных пони-
жениях.
Состав растительности на этих влажных и засоленных почвах меняет-
ся в зависимости от содержания солей и влажности почвы. На более сухих
почвах распространены виды Salsola, Traganum nudatum и Zygophyllum
album, на более увлажненных — виды тамарикса, на слабозасоленных
почвах —Atriplex halimus, а на влажных и сильнозасоленных —виды
Suaeda, Frankenia и другие. Наибольшей солеустойчивостью, видимо,
обладают Halocnemum strobilaceum и виды Saltcornia.
Иные условия наблюдаются в Ливийской пустыне (описанной Шток-
кером *) во впадине Вади-эн-Натрун. Эта впадина расположена к запад-
северо-западу от Каира и представляет собой понижение шириной 5—
20 км и длиной 160 км\ оно расположено на 23 м ниже уровня моря. В этой
низине насчитывается 16 крупных и мелких соленых озер, питаемых филь-
трующимися в грунт водами Нила. По пути нильская вода растворяет
соли, содержащиеся в соленосных горизонтах горных пород, и в какой-то-
1 О. Stocker, Das Wadi Natrun, «Veget. Bilder», 18, H. 1, (1927/28).
Пустыня Сахара
479
степени минерализуется. Наивысший уровень воды в озерах отмечается
в январе, примерно на 2—3 месяца позже паводка на Ниле, причем раз-
ница в уровнях составляет всего лишь около 30 см. В местах выхода грун-
товых вод развивается болотная растительность, образованная Typha
latifalia, Phragmites communis, Eragrostis bipinnata и Juncus acutus. В этих
условиях иногда наблюдается поверхностное засоление почвы, однако
корни растений проникают в более глубокие влажные слои почвы, где
концентрация солей незначительна. Так, по данным Штоккера, на глубине
30 см под экземпляром Juncus содержание солей в грунтовой воде дости-
гало всего лишь 0,9%.
В бессточных озерах сильное испарение содействует накоплению
солей, выпадающих на поверхности в период засухи. Наряду с поваренной
солью (NaCl) здесь образуются также сульфат натрия (Na2SO4) и сода
(Na2CO3).
Периодически высыхающие и покрытые соляной коркой глинистые
почвы совершенно лишены какой бы то ни было растительности (рис. 369).
Это типичные себхи. При уменьшении степени засоленности здесь
поселяются наиболее солеустойчивые галофиты. На пятнах незасоленного
песка, отложенного на этих пространствах, возникают более благоприят-
ные условия для прорастания семян; вокруг растущих здесь растений со
временем ветер навевает маленькие дюны; на них встречаются Zygophyl-
lum album, Z. simplex, Alhagi maurorum и виды тамарикса.
Co времени исследований Штоккера эта область сильно изменилась.
Автор посетил ее в 1960 г. Грунтовый поток, идущий от Нила, в настоя-
щее время перехвачен перед его впадением в озера и используется для
орошения возделываемых земель. Вода содержит около 0,03% раство-
ренных солей. Вади-эн-Натрун превратился в густонаселенный оазис,
население которого наряду с сельским хозяйством занимается кустарным
промыслом. Поэтому естественная растительность здесь вряд ли сохра-
нилась.
Оазисы
В местах выхода на поверхность слабоминерализованных подземных
вод даже в пустынях могут расти некоторые гидрофильные растения.
Если же в этих условиях образуется постоянное зеркало воды, то в водо-
емах развиваются широко распространенные водные растения.
Наряду с водорослями здесь появляются Potamogeton, Ceratophyllum,
Utricularia, Lemna. Вдоль берегов в водоемах появляются заросли, в со-
ставе которых Typha, Phragmites, Scirpus и Erianthus, переходящие на
берегу в заросли Tamarix gallica, и Т. nilotica.
Естественная растительность оазисов была образована пальмой-дум
(Hyphaene thebaica), Acacia albida и другими видами акаций, а также вида-
ми Маегиа и Capparis, Calotropis procera, Citrullus colocynthis и пр.
В наше время эти группировки полностью вытеснены финиковой паль-
мой (Phoenix dactylijera) (рис. 361) и другими культурными растениями:
ячменем (Hordeum) и бобами (Vicia faba), Ficus sycomorus и Albizzia lebbek,
вероятно, также интродуцированы человеком. В оазисы занесены и мно-
гие злостные сорняки и рудеральные растения. Даже при небольшом коли-
честве солей, содержащихся в воде, в условиях сильного испарения в оази-
сах может начаться засоление почвы, которое возможно устранить только
путем дренажа и длительного промывания почвы. Поэтому хорошо дрени-
руемые почвы более пригодны для культур, чем глинистые и большей
частью засоленные почвы. Особое значение при этом имеет качество воды.
480
Глава 13
Какие же культуры можно выращивать в оазисах при наличии хоро-
шей воды и дренированных почв? Ответ на этот вопрос можно получить
на основе наблюдений в оазисах на юге Ливийской пустыни — Эль-Харга
(количество осадков за 20 лет равно нулю) и Барис, а также в Асуане на
Ниле (в среднем 3 мм осадков в год). Здесь выращиваются без затенения
пшеница, ячмень, бобы, клевер александрийский (зимой), люцерна, хлоп-
чатник и клещевина.
В зимние месяцы со средней температурой 12—15° можно также выра-
щивать почти все овощи. Больший интерес представляют многолетние
культуры, выживающие при средних летних температурах выше 30°.
Наиболее широко в оазисах культивируется финиковая пальма;
основные породы ветрозащитных полос: Casuarina, Prosopis, Dalbergia
и Eucalyptus camaldulensis (rostrata), встречающийся также в крайне за-
сушливых районах Австралии в местах близкого залегания грунтовых
вод. На участках, окаймленных ветрозащитными полосами, выращиваются
оливы, цитрусовые, виноград, абрикосы, миндаль, гуайава и инжир.
Еще лучших результатов можно достичь при выращивании культур
в тени финиковых пальм. В результате затенения и защиты от ветра под
пологом пальм формируется особый микроклимат, который способствует
развитию цитрусовых, граната, папайи и даже бананов. Открытые план-
тации бананов автор видел и в Иврихо (Иорданская низменность), однако
там бананы высаживались в глубокие траншеи, так что над поверхностью
почвы поднимались лишь верхушки листьев. При выращивании чувстви-
тельных к высоким температурам культур необходимо создавать сомкну-
тые насаждения с более мягким микроклиматом. Это легче осуществить
в центре большого оазиса, чем на его границе с пустыней. При соблюдении
этого условия и при наличии обильного орошения (зимой раз в 2 недели,
а летом еженедельно) даже в крайне засушливых условиях пустыни
можно выращивать почти все тропические растения; об этом свидетель-
ствует опыт Ботанического сада, расположенного на острове Китченер,—
одном из островов Нила близ Асуана. Лишенная растительности пустыня
простирается здесь непосредственно до самого берега реки. На острове,
по указанию Китченера, были посажены древесные породы, интродуци-
рованные главным образом из Индии; сейчас они образуют высокий
тенистый лес. Этот опыт имеет очень важное значение, поэтому ниже при-
водится полный перечень всех тропических видов деревьев, большинство
которых цветет и плодоносит.
Из этого списка видно, что практически все растения, выращиваемые
в Индии, могут расти также и в оазисах Сахары. Из тропических культур
здесь встречаются даже кокосовая и масличная пальмы, хлебное дерево,
манго, гвоздичное дерево, маниок, бананы, кофе и другие
Список растений,
выращиваемых на острове Китченер близ Асуана
(прямым шрифтом выделены культурные растения)
Gymnospermae: Cephalotaxus drupacea, Cycas.
Monocotyledones:
Musaceae: Strelitzia angusta.
1 В перуанской прибрежной пустыне в условиях орошения можно выращивать
даже какао (устное сообщение Альвима). Культуры какао дают здесь высокие урожаи,
поскольку в условиях сухого воздуха они не поражаются болезнями.
1. Солончак близ Рас-эль-Хикмы на побережье Средиземного моря (ОАР): Salicornia
fruticosa, Zygophyllum album, Reaumuria muricata, Aeluropus litoralis.
2. Полупустыня с покровом из Artemisia herba alba близ Рас-эль-Хикмы, на побережье
Средиземного моря (ОАР).
Пустыня Сахара
481
Palmae: Areca lutescens, Arenga saccharifera, Borassus flabelliformis, Calamus
indicus, Caryota urens, Cocos nucifera, C. ronianzoffiana, Cryptostegia mada-
gascariensis, Elaeis guinensis, Kentia balmoriana, Latania borbonica, Livis-
tona chinensis, Raphis flabelliformis, Roystonia (Oreodoxa) regia, Sabal pal-
metto, Thrinax barbadensis, Th. elegans.
Zingiberaceae: Alpinia nutans.
Dicotyledones:
A c a n t h a c e a e: Sanchezia nobilis.
Anacardiaceae: Anacardium occidentale, Mangifera indica, Spondias dul-
cis, Sp. mangifera.
A p ocynaceae: Alstonia scholar is, Beaumontia grandiflora, Carissa grandi-
flora, Tabernaemontana coronaria, Thevetia nereifolia, Trachelospermum fas-
miuoides.
Aquifoliaceae: Ilex paraguajensis.
Araliaceae: Aralia papyrifera, Brassaia actinophylla.
Bignoniaceae: Bignonia magnifera, B. purpurea, Kigelia pinnata, Mark-
hamia (Dolichandrone') platycalyx, Spathodea nilotica, Tabebuia chrysotricha,
T. grandiflora, T. guafacana, T. pentaphylla.
Bombacaceae: Bombax malabaricum, Eriodendron sp., Pachira excelsa.
Boraginaceae: Cordia alba, C. heterophylla, C. myxa.
Caesalpiniaceae: Bauhinia heterophylla, Cassia artemisioides, C. fistula,
C. nodosa, C. siamea, Saraca indica, Tamarindus indica.
Capparidaceae: Capparis zeylanica.
С а г i с a c e a e:| Carica papaya.
Combretaceae: Terminalia arjuna, T. catappa.
Convolvulaceae: Ipomoea batatas.
Ebenaceae: Diospyros ebenus.
E u p h о г b i a c e a e: Acalypha marginata, Aleurites moluccensis, Antidesma
buuius, Hura crepitans, Manihot utilissima.
Flacourtiaceae: Flacourtia cataphracta, Fl. indica, Xylosma longifolia.
G u t t i f e г a e: Garcinia dulcis.
Lauraceae: Cinnamomum camphora.
Loganiaceae: Buddleia asiatica.
M a 1 v a c e a e: Chorisia speciosa.
M eliaceae: Amoora rohituca, Cedrela toona, Khaya senegalensis, Melia aze-
darach, Swietenia mahagoni, Sir. microphylla.
M i m о s a c e a e: Acacia catechu, A. sundra, Adenanthera pavonina.
M о г a c e a e: Arthrocarpus incisa, A. integrifolia, Ficus nitida, F. macrophylla,
F. misorlusis, F. pandurata, F. platypoda, F. repens, F. sycomorus, F. vogelii,
Madura aurantiaca.
M у rtacea e: Callistemon lanceolatum, Eugenia gambolana, Eu. javanica, Eu.
rosea, Eu. uniflora, Feijoa sellowiana, Melaleuca armillaria, M. styphelioides.
N yctaginaceae: Bougainvillea precatorius.
Oleaceae: J asminum sambac.
Papilionaceae: Butea frondosa, Dalbergia sericea, Indigofera pulchella, Ma-
chaerium tipu, Sophora secundiflora.
Pittosporaceae: Hymenosporum flavum.
Proteaceae: Stenocarpus stenaptera, Grevillea robusta.
Punicaceae: Punica granatum.
Rosacea e: Eryobotrya japonica.
R u b i a c e a e: Catesbaea spinosa. Coffea arabica, Txora, parviflora, Randia dume-
torum.
482
Глава 13
Rut а с е а е: Murraya exotica.
S an t al асе ае: Santalum album.
Sapindaceae: Dodonaea visco sa, Nephelium longana, N. tomentosa.
Sapotaceae: Bumelia sp., Chrysophyllum cainito9 Mimusops caffra.
Sterculiaceae: Sterculia foetida, St. lurida.
Verbenaceae: Citharexylum quadrangulare, Clerodendron siphonanthus^
Tectona grandis.
К БИОЛОГИИ РАСТЕНИЙ САХАРЫ
В связи с крайней засушливостью Сахара отличается не только бед-
ностью флористического состава, но и меньшим числом экологических
групп растений по сравнению с другими пустынными областями. Поэтому
исследователь, изучавший только Сахару, как правило, не может соста-
вить правильное представление о разнообразии экологических групп рас-
тений, распространенных в аридной зоне.
Многочисленные и разнообразные многолетние суккуленты, развитые
в американских и южноафриканских пустынях, полностью отсутствуют
в Сахаре. Это же можно сказать и о пойкилогидрических видах, и в част-
ности о папоротниках и мхах. Даже лишайники и почвенные водоросли
встречаются здесь редко. В Сахаре распространены суккулентные гало-
фиты (Chenopodiaceae, Zygophyllaceae).
Наиболее широко представлены в Сахаре однолетники, то есть эфе-
меры, использующие для своего развития эпизодические дожди и отли-
Р и с. 370. Подушки Fredolia (Anabasis) aretioides (Chenopodiaceae) в щебнистой
пустыне близ Бен-Сирега у северной границы алжирской Сахары. Между
ними — Limoniastrum feei (Plumbaginaceae), а на заднем плане — в сухой
долине Pistacia therebinthus var. atlantica (фото Гейма).
Пустыня Сахара
483
чающиеся крайне коротким периодом вегетации; некоторые виды быстро
прорастают и плодоносят уже через 8—15 дней. Они не обладают какими-
либо особенными физиологическими свойствами; их легко выращивать
летом в Центральной Европе.
Рис, 371, Корневые системы растений алжирской Сахары близ Ревуаль-Бени-Упифа.
Черным показан песчаник (по Каушу),
После сильного дождя пустынная почва покрывается зеленым ков-
ром однолетников; несмотря на свою кратковременность, они тем не
менее имеют большое значение для пастбищного хозяйства,
Некоторые однолетники, например галофитные виды Mesembryan-
themum, отличаются суккулентностыо; благодаря накопленным запасам
влаги живут даже на полностью высохшей почве до шести месяцев,
Здесь встречаются также и геофиты; они столь же внезапно, как
и эфемеры, появляются после дождя, и после короткого периода вегетации
их почки возобновления в течение длительного периода остаются в почве
в состоянии покоя, Они чаще встречаются в пограничных с пустыней обла-
стях, К их числу относятся некоторые луковичные растения из семейства
Liliaceae, Pancratium trianthum (Amaryllidaceae), а также некоторые
корневищные растения: Ferula и др,
Многолетние виды, то есть собственно ксерофиты, встречаются изред-
ка в сравнительно хорошо увлажняемых местообитаниях, Надземные
органы у них слабо развиты по сравнению с корневой системой; корни
развиваются в пределах увлажненного слоя почвы; очень хорошо выраже-
на поверхностная корневая система, При наличии на известной глубине
постоянно влажного слоя почвы или близкого уровня грунтовых вод,
у растений образуется глубоко идущий стержневой корень, Корневые
системы некоторых видов, распространенных в Сахаре, изображены на
рис, 371-373,
В этих условиях у растений хорошо выражена редукция листовой
поверхности, При недостатке воды листья опадают или не образуют-
ся вовсе, как это наблюдается, например, у прутьевидных и колючих
кустарников (Zilla, Launaea—Zollikoferia, Retama, Pituranthos и др J.
В критические периоды интенсивность транспирации у этих растений
сокращается до минимума; таким образом, несмотря на незначительные
запасы воды в почве, гидратура у этих видов сохраняется в течение дол-
31*
484
Глава 13
гих периодов засухи на неизменном уровне. При наличии постоянно
увлажненных слоев почвы или близко залегающих грунтовых вод расте-
ния не испытывают затруднений в снабжении водой. В этих местообита-
ниях растут даже деревья. По данным Озенда, здесь распространены
следующие древовидные растения: Ephedra alata, Cupressus dupreziana,
Phoenix dactylifera, Hyphaene thebaica, Populus euphratica, Ficus salicifolia,
Maerua crassifolia, Acacia raddiana, A. seyal, A. arabica, A. albida, Cassia
lanceolata, C. obovata, Balanites aegyptiaca, Pistacia atlantica, Tamarix
(6—10 видов), Periploca laevigata, Calotropis procera, Salvadora persica,
Olea laperrini. К их числу следует также добавить несколько кустарников,
достигающих подчас значительных размеров. В этих условиях встре-
чаются также гигроморфные виды, например Citrullus colocynthis и другие;
эти виды происходят из тропиков и поэтому производят здесь впечатление
инородных элементов. Листья у них не перегреваются только благодаря
большой интенсивности транспирации L Особую группу образуют ксеро-
и гигрогалофиты. Их число, безусловно, больше, чем полагают (содержа-
Р и с. 372. Корневая система ZMa macroptera в понижении
с глинистой почвой близ Ревуаль-Бени-Унпфа (по Каушу).
ние солей в клеточном соке растений Сахары пока не изучалось). К этой
группе, вероятно, относится большинство видов семейств Chenopodiaceae,
Aizoaceae, Zygophyllaceae, Plumbaginaceae и др. Они накапливают соли
в клеточном соке, даже если растут на малозасоленных песках.
1 О. L. Lange, Untersuchungen iiber Warmehaushalt und Hitzeresistenz maure-
tanischer Wiisten und Savannenpflanzen, «Flora», 147, 1959, S. 595—651.
Пустыня Сахара
485
В северо-западной Сахаре распространено оригинальное растение
Fredolia (Anabasis) aretioides, образующее крупные, высотой 50—60 сл,
полушаровидные плотные подушки (рис. 370). Растения такого типа чаще
встречаются в антарктической флористической области, и в частности
в высокогорных районах Южной Америки и Новой Зеландии.
Р и с. 373. Корневая система Fredolia (Anabasis) aretioi-
des\ щебнистая пустыня близ Ревуаль-Бени-Унифа (по
Каушу).
Большую роль в жизни растений пустыни играет также ветер. Очень
многие виды опыляются при помощи ветра, например злаки, виды Che-
nopodiaceae и др. Условия опыления у других растений еще достаточно
подробно не изучались. Ветер оказывает особенно сильное влияние на
распространение семян и плодов. Лишь эпизоохорные растения встре-
чаются в Сахаре чаще, чем анемохоры; эндозоохоры отсутствуют здесь
полностью; это связано с бедностью Сахары животными и особенно пти-
цами. С экологической точки зрения интересна иерихонская роза (Anasta-
tica hierochuntica), ветви которой в период засухи при созревании плодов
загибаются вверх и внутрь; все растение превращается в шар, легко
отламывающийся и катящийся по земле; только после дождя набухаю-
щие ветви распрямляются, разбрасывая вокруг семена.
ЭКОЛОГИ ЧЕСКИЕ ИССЛЕДОВАНИЯ
В АЛЖИРСКОЙ САХАРЕ И В ЛИВИИ
Экспериментальные экологические исследования в северо-западной
части Сахары были начаты Фиттингом; в 1911 г. он опубликовал резуль-
таты своих исследований, проведенных в Бискре — области, где в сред-
нем выпадает 299 мм осадков К Несмотря на то что в дальнейшем не все
1 Н. F i t t i n g, Die Wasserversorgung und die osmotischen Druckverhaltnisse
der Wiistenpflanzen, «Zschr. Bot.», 3, 1911, S. 209—275.
486
Глава 13
полученные им данные были подтверждены, его работа послужила
толчком для проведения других многочисленных работ в этом направлении.
Значение осмотического давления для растений пустыни впервые
было показано Фиттингом. Он определял осмотическое давление плазмо-
литическим методом при помощи растворов KNO3 и NaCl. Высокие зна-
чения осмотического давления у пустынных растений Фиттинг рассмат-
ривал как полезное приспособление, позволяющее им извлекать воду
даже из почти сухой почвы.
В дальнейшем Киллиан, изучавший те же растения, однако в более
неблагоприятное время года и в более засушливых условиях (Ревуаль-
Бени-Унифе, где выпадает всего 100 мм осадков), показал, что при помо-
щи плазмолитического метода получают завышенные показания Ч В своей
работе Киллиан пользовался криоскопическим методом; сравнительные
данные приведены в табл. 91.
Из этой таблицы видно, что у некоторых галофитов (Atriplex, Statice,
Limoniastrum и Frankenia pulverulenta) величины осмотического давления
совпадают, в остальных же случаях величины, полученные плазмолитиче-
ским методом, завышены.
В настоящее время известно, что в условиях пустыни, где встречаются
многолетние растения с глубоко развитой корневой системой, водный
режим почв далеко не столь неблагоприятен, как это считалось раньше
(см. стр. 306 и след.).
Кроме того, нельзя рассматривать повышение осмотического давления
в качестве целесообразного приспособления к недостатку воды; напротив,
оно свидетельствует о нарушении водного баланса, даже если растение
в результате одновременного повышения сосущей силы может вследствие
этого извлекать из почвы несколько больше воды. Сильное снижение
гидратуры, во всяком случае, не может сказываться благоприятно на
жизнедеятельности растений. Вероятно, никому не придет в голову рас-
сматривать снижение веса у голодающего человека как целесообразное
приспособление, даже если благодаря этому и улучшится степень исполь-
зования имеющегося в наличии недостаточного количества пищи. Влаж-
ность любой почвы, и прежде всего песчаной почвы, в пустыне так неве-
лика, что даже сильное повышение осмотического давления не может
помочь растению в получении дополнительных количеств влаги. В послед-
нее время было выяснено, что при хорошей влагообеспеченности осмоти-
ческое давление у растений пустыни, не считая галофитов, достигает
величин лишь немного больших, чем у растений влажных областей. Расте-
ния, растущие в наиболее суровых условиях пустыни, обладают настолько
выраженным постоянством осмотического давления, что даже при очень
длительной засухе у них отмечается лишь незначительное повышение
концентрации клеточного сока.
Как уже было сказано, у этих растений в критические периоды
интенсивность транспирации сильно снижается, и поэтому они доволь-
ствуются незначительными запасами воды в почве. Кроме того, у этих
растений имеется мощная корневая система (см. стр. 489).
О водоудерживающей силе пустынных почв пока еще не получено
достаточно данных. Можно предполагать, что у почв, отличающихся круп-
нозернистой структурой, она невелика. На первый взгляд может показать-
1 Ch. Killian, L. Faure 1, Observations sur la pression osmotique des vege-
taux desertiques et subdesertiques de I’Algerie, «Bull. Soc. Bot. Franc.», 80, 1933,
p. 775; La pression osmotique des vegetaux du Sud Algerien, «Ann. Physiol.», 12,
1936, p. 5.
Пустыня Сахара
Таблица 91
Величина осмотического давления у растений в Сахаре, атм
Название растений Бискра (по Фиттингу, плаз- молитический метод, при- близительно) Ревуаль-Бени-Униф (по Кил- лиану, криоскопический метод)
Citrullus colocynthis И 5,5-7,7
Erodium pulverulentum 15-21 9,9
Erodium pulverulentum 27—53 (пустыня) —
Asteriscus pygmaeus , . Atriplex halimus 15—18 (обработанная пло- щадь) 36-53 12,5 35,5—70 (октябрь)
Atriplex halimus 18—24 (дюны) —
Atriplex halimus 42—53 (галечниковая пус- —
Fagonia glutinosa . тыня) 70—100 и выше 9,7—17,3
Statice delicatula 30—53 (засоленное болото) 20,7—48,6 (октябрь)
Peganum harmala . 42-70 14,9—34,4 (октябрь)
Neriurn oleander . 21—24 12,4—22,3 (январь)
Capparis spinosa 53-70 18,4—21,3 (ноябрь)
•Capparis spinosa 27—42 (обработанная пло- —
Phoenix dactylifera щадь) 42—53 (засоленное болото) 16,8—19,5 (обработанная
Phoenix dactylifera . . 36—42 (Уэд-Бпскра) площадь) 28,9—32,6 (дюны)
Limoniastrum guyonianum 53—70 (засоленное болото) 49,6—53,4 (засоленное бо-
Traganum nudatum 70 (пустыня) 100 и выше лото) 20,2—30,7 (дюны)
Traganum nudatum — 22,9—29,4 (засоленное бо-
Suaeda vermiculata Свыше 100 (засоленное бо- лото) 24,9—27,4 (дюны)
Suaeda vermiculata лото) Свыше 100 (пустыня) 22,9—29,4 (засоленное бо-
Frankenia pulverulenta 21-30 лото) 40,6
Frankenia pulverulenta 27—53 (засоленное болото) —
Frankenia thymifolia Свыше 100 (пустыня) 37,1
ся, что очень высокие значения 1 водоудерживающей силы почвы, полу-
ченные Хардером (1930 г.) в Ревуаль-Бени-Унифе, противоречат этому;
однако, во-первых, образцы были взяты Хардером вне почвенной толщи,
где распространены корни растений, а во-вторых, именно в пустынных
почвах вода крайне неравномерно распределена по профилю; поэтому
полученные значения ничего не говорят о фактической величине водо-
удерживающей силы той части почвы, где расположены активные корни
растений. Кроме того, для определения водоудерживающей силы приме-
нялся капиллярный метод; однако значения, полученные этим методом
на сухих почвах, завышены.
1 R. Harder, Uber den Wasser- und Salzgehalt und die Saugkrafte einiger
Wiistenboden Beni Unifs (Algerien), «Jb. wiss. Bot.», 72, 1930, S. 665—699.
488
Глава 13
Особенно важно выяснить, продолжается ли ассимиляция у растений
пустынь во время длительных периодов засухи? В этом отношении боль-
шой интерес представляют опыты Хардера, Фильцера и Лоренца U Они
определяли интенсивность ассимиляции СО2 у живых растении непосред-
ственно в природе, накрывая растения целлофановыми мешками и про-
тягивая через мешки воздух, В результате сильного нагрева накрытых
мешками растении и небольшой скорости воздушного потока интенсив-
ность фотосинтеза у опытных растений была, безусловно, значительно
ниже, чем в естественных условиях. Тем не менее оказалось, что все рас-
тения даже после почти годичной засухи продолжали ассимилировать
или, во всяком случае, не находились в состоянии покоя.
Интенсивная ассимиляция в августе наблюдалась только у Haloxylon
articulatum, в связи с этим он мог цвести и плодоносить. Значительно
меньше величина ассимиляции была у Limoniasirum feei. Интенсивность
ассимиляции у обоих видов после выпадения осадков в сентябре остава-
лась на прежнем уровне, В противоположность этому незначительная
интенсивность ассимиляции у Fredolia aretioides резко повысилась после
дождя.
Еще большие различия до и после дождя были отмечены у Zolliko-
feria arborescens; в период засухи она начинала желтеть и отмирала, однако
после выпадения осадков (20 мм) интенсивность ассимиляции резко
повысилась,
К сожалению, при проведении этих исследований не определялась
гидратура растений. Вполне вероятно, что у первых двух видов она
существенно не изменилась, а у последних повысилась, Haloxylon и Ыто-
niastrum могут быть отнесены к числу галофитов, накапливающих соли.
Основное значение для аккумуляции вещества в условиях пустыни
всегда имеет водный режим. Водный режим определяет большие различия
в размерах и в ритме развития отдельных экземпляров одного и того же
вида, поскольку часть из них растет в благоприятных, а часть в небла-
гоприятных условиях среды,
В 1953 г, Штоккер в этом же районе Сахары изучал микроклимат,
водный режим, фотосинтез, а также дыхание растений. Результаты рабо-
ты пока обобщены лишь в виде предварительного сообщения 1 2,
В этой работе приведены кривые, характеризующие ход транспира-
ции и ассимиляции в апреле — мае у Zilla macroptera (из влажных и сухих
местообитаний), Zizyphus lotus, Fredolia (Anabasis) aretioides, Retama retam,
Citrullus colocynthis, Capparis spinosa, Phragmites communis и Frankenia
thymoides. Все растения отличались большой интенсивностью транспира-
ции, не снижавшейся даже в полуденные часы. Совершенно по-иному
выглядит дневной ход ассимиляции. За исключением Zilla (из влажных
условий) и Phragmites, у которых в течение всего дня наблюдалось
накопление органического вещества, у всех других растений интен-
сивность ассимиляции уже утром резко снижалась и к полудню расход
вещества часто преобладал над приходом. Создается впечатление, что эти
данные не вполне соответствуют действительности: апрель и май — это
наиболее благоприятный период для развития растений пустыни; в это
время у них отмечается интенсивный рост, а интенсивная транспирация
1 R. Harder, Р. Filzer, A. Lorenz, Uber Versuche zur Bestimmung
der Kohlensaureassimilation immergruner Wiistenpflanzen wiihrend der Trockenzeit
in Beni Unif (algerische Sahara), «Jb. wiss. Bot.», 75, 1932, S. 44—194.
2 0. Stocker, Der Wasser- und Assimilationshaushalt siidalgerischer Wiistcn-
pflanzen, «Вег, Dtsch. Bot. Ges.», 67, 1954, S. 289—299.
Пустыня Сахара
489’
свидетельствует о хорошей влагообеспеченности и об открытых устьицах.
Если растения даже и в это время года не отличаются энергичной асси-
миляцией, то когда же еще они могут накапливать необходимые им орга-
нические вещества?
Бергер-Ландефельдт (цит. на стр. 491), проводивший близ Большого
Сирта опыты по определению ассимиляции у галофитов и получивший
сходные результаты, комментировал их следующим образом: «Во всяком
случае, нужно полагать, что все эти наблюдавшиеся нарушения нор-
мального хода ассимиляции являются следствием неестественных условий
опыта», понимая под нарушением хода ассимиляции выделепие СО2
в полуденное время.
На влияние перегрева и обусловленные этим ошибки при определении
ассимиляции даже в условиях мягкого климата южных районов ФРГ
недавно указал Босиан Штоккер и Фивег, напротив, считают, что
поскольку их наблюдения продолжаются всего одну минуту, то «точность
опыта значительно больше инструментальных ошибок» 1 2. Из-за отсут-
ствия параллельно проведенных наблюдений невозможно судить о вели-
чине ошибок в проведенных опытах. Не исключено, что в пустыне в полу-
денные часы перегрев может наступить менее, чем за одну минуту.
Сотрудник Штоккера Кауш занимался изучением корневых систем
растений в Ревуаль-Бени-Унифе 3. В этих условиях растения встречаются
только в руслах временных водотоков и в сухих долинах или в бессточ-
ных котловинах и более значительных понижениях. Грунт образован
преимущественно песчаником, частично растрескавшимся или перекры-
тым щебнем, песком или глиной. В почве на некоторой глубине выражен
цементированный карбонатами горизонт. Как подчеркивает Кауш,
создается впечатление, что здесь существуют очаги инфильтрации влаги
в почву. Ими могут быть, в частности, трещины в породах, к которым
в основном приурочены растения (рис. 371). На рис. 372 и 373 изображены
корневые системы пустынных растений. Часть корней располагается на
небольшой глубине (10—40 сл); эти корни поглощают влагу из верхних
увлажненных слоев почвы после относительно слабых дождей; другая
часть корней проникает довольно глубоко в почву или в трещины породы,
где почва долгое время остается влажной, а запасы воды все время попол-
няются. Таким образом, в этих условиях глубина проникновения корней
связана с глубиной промачивания почвы 4. Следует заметить, что глубоко
расположенные влажные слои почвы не имеют ничего общего с грунто-
выми водами.
Водный баланс многолетних злаков близ Ревуаль-Бени-Унифа изу-
чал Лемэ 5. Эти злаки в общем хорошо обеспечены водой. Корни, покры-
тые песчаным чехлом (см. стр. 281), далеко простираются в горизонталь-
1 G. В о s i а п, Zum Problem des Kiivettenklimas, «Вег. Dtsch. Bot. Ges», 72,
1959, S. 391—397.
2 0. Stocker, G. H.Viehweg, Die Darmstadter Apparatur zur Momentan-
messung der Photosynthese unter okologischen Bedingungen, «Вег. Dtsch. Bot. Ges.»,
73. 1960, S. 198—207.
3 W. К a u s c h, Der Einfluss von edaphischen und klimatischen Faktoren auf
die Ausbildung des Wurzelwerkes der Pflanzen unter besonderer Beriicksichtigung
einiger algerischer Wiistenpflanzen, Habilitationsschrift, Darmstadt, 1959.
4 Данные о максимальной протяженности корневых систем в пустынных обла-
стях приведены в работе М. Z о h а г у, On hydroecological relations of the Near East
Desert Vegetation,^ Proc. Madrid Symp., Unesco Arid Zone Res., 16, 1961, p. 199—212.
5 G. L e m e e, L’economie de 1’eau chez quelques graminees vivaces du Sahara
septentrional, «Vegetatio», 5/6, 1954, p. 534—541.
490
Глава 13
ном направлении и сильно углубляются в почву. Интенсивность транспи-
рации соответствует величине транспирации злаков в Центральной Европе;
если же сравнивать по отношению к испарению, то в этих условиях
интенсивность транспирации достигает меньших величин. Интенсивность
транспирации сильно сокращается при свертывании листьев злаков, что
наблюдается, в частности, при определенном дефиците влаги (см. табл. 92).
Таблица 92
Уменьшение интенсивности транспирации при свертывании листьев
у злаков
Название растений Сокращение интенсивности транспирации, % Дефицит влаги, при котором начинается свертывание листьев, %
Lygeum spartum . 70 8-20
Aristida pungens 70 6-24
Stipa parvijlora 75 18-40
Andropogon laniger
верхние листья 30 40
нижние листья . 60 90
Andropogon annulatus . 83 50 верхние, 90 нижние
Cynodon dactylon . . 50 30 верхние, 80 нижние
Pennisetum dichotomum Листья высыхают 18-38
Aeluropus litoralis Листья высыхают 80
Поскольку растения хорошо обеспечены влагой, величина сосущей
•силы не превышает 6—15 атм\ это же относится и к осмотическому давле-
нию; исключением являются: Lygeum (слабо выраженный галофит)
и Aeluropus (типичный галофит) (табл. 93).
Таблица 93
Осмотическое давление у пустынных
злаков (атм)
Aristida pungens
Aristida obtusa .
Andropogon laniger
Lygeum spartum
Aeluropus litoralis
9,6-21,3
18,2
6,4-13,0
23,6-26,6
25,2-55,3
Два последних вида относятся к числу галофитов, которые распро-
странены по периферии пустыни. Здесь в условиях более влажного кли-
мата выпавшие осадки стекают в понижения, где затем испаряются;
в этих понижениях образуются солончаки с типичными галофитами.
Озенда описал растительность таких солончаков на плоскогорьях
южного Алжира Киллиан в своей работе указал на индикаторное
1 Р. О z е n d a, Observations sur la vegetation d’une region semiaride: Les Hauts-
Plateaux du Sud-Algerois, «Bull. Soc. Hist. Natur. Afrique du N.», 45, 1954, p. 134—169.
Пустыня Сахара
491
значение отдельных галофитов и описал биологию одного из гало-
фитов — Frankenia pulverulenta \
Краткий обзор растительных сообществ, приуроченных к солонча-
кам, составлен Дюбуи и Симмоно 1 2.
Особенно обширные солончаки встречаются вдоль побережья Сре-
диземного моря в Ливии. Наибольший из них — Тауорга — занимает
площадь около 5000 км2 на западе
500ft
Рис. 374. Поперечный срез листа
Atriplex mollis с пузыревидными волос-
ками (по Бергеру-Ландефельдту).
Большого Сирта; экология произра-
стающих здесь растений изучена
Бергером-Ландефельдтом3. Средне-
годовое количество осадков, по дан-
ным трех станций, расположенных на
северо-западе, северо-востоке и севере
солончака, составляло 60, 127 и
132 мм. Почва в течение всего года
Рис. 375. Срез листа Nitraria retusa.
Пунктиром показаны клетки, содержа-
щие дубильные вещества (по Бергеру-
Ландефельдту).
остается постоянно влажной, тогда как концентрация солей в почвенном
растворе подвержена сильным колебаниям. В дождливый период
зимой и весной, вследствие притока больших масс воды концентрация
солей достигает незначительной величины. В это время появляются
всходы растений. Затем на протяжении лета концентрация солей
вследствие испарения непрерывно повышается и на поверхности
1 Ch. Killian, Sols et plantes indicatrices dans les parties non irriguees des
oasis de Figuig et de Beni-Ounif, «Bull. Soc. Hist. Nat. Afr. du N.», 32, 1941; Observa-
tions sur la biologic d’un halophyte saharien, Frankenia pulverulenta, «Trav. Inst.
Rech. Sahar», 7, 1951; La vegetation autour du Chott Hodua, indicatrice des possibilites
culturales, et son millieu edaphyque, «Ann. Inst. Agric. Algerie», 7, 1953, p. 5.
2 A. D u b u i s, P. S i m о n n e a u, Les unites phytosociologiques des terrains
salins de terrains salins de 1’Quest Algerien, «Trav. d. Sect. Pedol. et Agrologie», Bull.
№ 3, 1957, p. 1—20.
3 U. Berge r-L andefeldt, Beitriige zur Okologie der Pflanzen nordafri-
kanischer Salzpfannen, «Vegetatio», 7, 1957, S. 169—206; 9, 1959, S. 1—47.
492
Глава 13
Рис. 376. Поперечный срез междоузлия стебля Arthrocnemum glaucum (по
Бергеру-Ландефельдту).
почвы образуется соляная корка; однако в области распространения
корней осмотическое давление почвенного раствора не превышает 34 атм.
Осмотическое давление у растений достигало высоких значений: у Nit-
raria retusa —35,4 атм, у Salicornia fruticosa — 69,9—72,4 (давление,
обусловленное хлоридами,— 50,9—54,5 атм). Необычно высоким ока-
залось давление у Atriplex mollis (123 — 156 атм, осмотическое дав-
ление, обусловленное хлоридами,— 103—132 атм). Однако Бергер-
Ландефельдт совершенно обоснованно сомневается в правильности полу-
ченных значений, поскольку в отжатый из листьев сок могли попасть
соли, накопившиеся в высохших пузыревидных волосках, оставшихся на
поверхности листьев (см, стр, 452), На листьях развивается огромное
количество пузыревидных волосков (рис. 374). На рис. 375 видны клетки
Nitraria retusa, заполненные дубильными веществами, и крупные идио-
бласты; последние при завядании листьев быстро теряют тургор, однако
в случае притока воды мгновенно набухают снова. В палисадной ткани
междоузлий Arthrocnemum glaucum встречаются крупные склереиды,
придающие прочность растениям (рис. 376). Nitraria, Atriplex, Salicornia
и Arthrocnemum теряют в течение суток в процессе транспирации немного
воды. Эти виды наиболее широко распространены среди многолетников.
Наряду с ними встречаются также и другие виды Chenopodiaceae и Plum-
baginaceae, а весной — еще целый ряд однолетних видов.
Пустыня Сахара
493
РАСТИТЕЛЬНОСТЬ НАГОРЬЯ Т И БЕСТ И
Это изолированное нагорье расположено в центральной части восточ-
ной Сахары; оно находится несколько южнее северного тропика, возвы-
шаясь над окружающей равниной до высоты 3400 м, Обзор флоры и рас-
тительности приводится в работе Р. Мэра и Т. Моно Ч
В горах Тибести распространено 396 видов; 40 из них — низшие рас-
тения. В расположенном далее к западу массиве Ахаггар до настоящего
времени было собрано 350 видов. При большом сходстве флористического
состава обоих районов флора Тибести все же отличается некоторыми
особенностями:
1) незначительным числом северных (средиземноморских) элементов;
2) увеличением степени участия южных (сахельских) элементов (45%
но сравнению с 8% в Ахаггаре);
3) большим числом восточных (сахаро-аравийских или судано-синд-
•ских) элементов.
Степень участия тех или иных элементов в строении покрова меняет-
ся в зависимости от высоты над уровнем моря.
Массив Тибести сложен кристаллическими породами, скрытыми под
покровом песчаников, на платообразной поверхности которых возвы-
шаются вулканические конусы.
Климат массива напоминает климат Сахары. Метеорологические
наблюдения проводились здесь в общей сложности только 31 месяц в тече-
ние 4 лет. Средняя температура в Бардаи (1000 м) равна 25,1°, средний
максимум 37,8° и средний минимум 2,8°, абсолютный минимум
составил —2°. На высоте более 2000 м температура может опускаться
ниже —10°, но после ночных заморозков, достигающих —12°, температура
днем повышается до +14—19°. Более скудны данные об осадках. Из
имеющихся отдельных наблюдений следует, что в мае выпадает наиболь-
шее количество осадков. Так, например, в мае 1934 г. в трех понижениях
было зарегистрировано 35, 52 и 455 мм (из них 450 мм выпало за 5 суток).
Наблюдаются и годы почти без осадков. Что касается больших высот,
то, судя по характеру растительного покрова, там должно выпадать боль-
ше осадков. Вблизи горных вершин развита сильная облачность. Здесь
встречаются многочисленные выходы подземных вод; некоторые источ-
ники имеют постоянный излив воды. Однако, несмотря на это, в целом
нагорье Тибести напоминает пустыню.
Моно выделяет, так же как и в Ахаггаре, следующие высотные пояса:
Сахаро-средиземноморский
Сахаро-тропический
верхний
нижний
ниже
2400-3400 м
1700-2400 .н
1700 м
В пределах сахаро-тропического пояса развита очень сухая саванна,
образованная Acacia и Panicum; эта саванна простирается вокруг
горного массива. В горах в составе этих саванн встречаются реликты,
проникшие сюда с юга. В частности, на юго-западном склоне нагорья
в горных ущельях широко распространены сахельские виды Acacia laeta,
A, stenocarpa, Boscia salicifolia, Carissa edulis, Rhus villosa, Cordia gharaf,
Ehretia obtusifolia и даже пальма дум — Hyphaene thebaica. В сухих доли-
1 R. М a i г е, Th. Monod, Etudes sur la flore et la vegetation du Tibesti, «Mem.
Inst. FranQ., Afr. Noire», № 8, Paris, 1950.
494
Глава 13
нах у подножия гор и на песках растут чуждые для Сахары виды: (Леоте,
Cassia, Crotalaria, Buchea, Geigeria, Cenchrus biflorus и другие. Лишь зна-
чительно выше в горах появляются средиземноморские элементы, рас-
пространенные также и в северной Сахаре: Diplotaxis acris, Rumex, Ero-
dium malacoides, Astragalus, Globularia alypum, Lavandula, Salvia, Ephedra
altissima.
Для нижней части средиземноморского пояса характерны: Helianthe-
тит lippii, Н. ellipticum, Artemisia judaica, Teucrium polium, Myrtus
nivellei, Argyrolobium abyssinicum, Echinops bovei, Cuscuta planiflora,
Andropogon hirtus, а для верхней части средиземноморского пояса —
Ballota hirsuta, Sisymbrium reboudianum, Nepeta tibestica, Silene spp.,
Oxalis corniculata, Malva parviflora, M. rotundifolia, Cheilanthes pteridioides,
Senecio coronopifolius, Bromus rubens, Stipa tibestica и многие другие. Это
уже виды, которые растут в более влажных условиях климата.
Некоторые виды являются реликтами более влажных климатических
периодов, как, например, распространенные вокруг источников Ficus
salicifolia и Adiantum capillus veneris. В своей работе Мэр и Моно приво-
дят полный список всех собранных растений с указанием для каждого
образца географического местоположения и высоты над уровнем моря.
Глава 14
ЕГИПЕТСКО-АРАВИИСКАЯ ПУСТЫНЯ,
ПУСТЫНИ СИНАЙ И НЕДЖЕБ
УСЛОВИЯ СРЕДЫ И РАСТИТЕЛЬНОСТЬ
Зта пустынная область расположена на восточной окраине Сахары
(рис. 377); условия среды здесь сходны с условиями в Сахаре.
Близость этой пустынной области к Нилу позволила еще в конце
XIX века Фолькенсу провести в ней экологические наблюдения и дать
анатомическое описание некоторых растений пустыни Ч В частности,
он высказал предположение, что опушение листьев может иметь значение
для обеспечения растений влагой за счет росы. Очень подробные иссле-
дования транспирации и водного режима растений, а также водного
и солевого режимов почвы провел в дальнейшем Штоккер 1 2.
Однако стационарные исследования были начаты лишь в последнее
десятилетие, после того как был создан опорный пункт Каирского уни-
верситета (34-й километр шоссе Каир —Суэц).
А. Мигахид и А. Абд эль Рахман изучали в основном водный режим
растений, тогда как М. Кассас больше внимания уделял общей харак-
теристике условий среды.
Каир расположен на 30° с. ш. Здесь выпадает крайне незначительное
количество осадков в период с октября по май, а чаще с ноября по март
идут редкие и слабые дожди (см. рис. 378). В среднем за период с 1941
по 1950 г. выпало 21,3мм в год, причем в отдельные годы количество осад-
ков колеблется от 6,9 до 42,6 мм. Это количество столь невелико, что
на климадиаграмме (метеостанция Хельван близ Каира) невозможно
выделить более влажный период (рис. 379). В течение года осадки выпа-
дают не чаще пяти-шести раз; бездождные периоды длятся большей частью
9—10 месяцев. Как и во всех засушливых областях, осадки выпадают
здесь крайне неравномерно. На основе многолетних наблюдений (1906—
1965 гг.) получена средняя величина, равная 27 мм, с возможными откло-
нениями до 15,2 мм в ту или иную сторону. Вероятность выпадения годо-
вых осадков (рис. 381) очень близка к тому, что наблюдается на станции
Мулка в Австралии (рис. 356).
Засушливые годы с количеством осадков ниже нормы повторяются
гораздо чаще, чем влажные годы, хотя в некоторые из них и выпадает
сравнительно много осадков. Среднемесячная максимальная температура
наиболее теплого месяца достигает 35°, а абсолютный максимум превы-
шает 40°. Зато в зимние месяцы температура может падать до 3°, и в такие
холодные ночи обычно выпадает роса. С другой стороны, в жаркие дни
относительная влажность воздуха опускается иногда до 4%; в наиболее
теплые месяцы испарение достигает 22 лм/100 м2, а в самые холодные —
1 G. V о Ike ns, Die Flora der Agyptisch-Arabischen Wiiste, Berlin, 1887.
2O. Stocker, Der Wasserhaushalt agyptischer Wiisten- und Salzpflanzen,.
Jena, 1928.
Глава 14
Рис. 377. Восточная Сахара и прилегающие к ней пустынные области.
1 — пески; 2 — впадины, расположенные ниже уровня моря.
около 8 лгл/100 м1 2. Этому благоприятствуют, кроме того, устойчивые
и сильные ветры, скорость которых достигает 5—10 м/сек. Различия в на-
греве склонов южной и северной экспозиции, согласно наблюдениям
Кассаса и Имама \ достигают наибольших величин при наиболее низкой
высоте стояния солнца, то есть в январе. Это важно в связи с тем, что зимой
ъегетирует большинство растений. В это время года почва после обильных
осадков покрывается несколько более плотным ковром эфемеров.
Почвенные условия здесь столь же неблагоприятны, как и климат.
В гамаде юго-восточнее Каира близ Хельвана на поверхность выходят
желто-серые богатые солями эоценовые известняки и мергель. Восточнее,
вдоль шоссе на Суэц, в почвах щебнистой пустыни содержится много
гипса. Лишь в широких долинах, где скапливается песок, создаются более
благоприятные условия для развития растительности.
1 М. К assas, М. Imam, Climate and microclimate in the Cairo desert, «Bull.
Soc. Geogr. Egypte», 30, 1957, p. 25—52.
Египетско-аравийская пустыня, пустыни Синай и Неджеб
497
Рис. 378. Годовой ход осадков] в Каире в 1941 —
1950 гг.
Рис. 379—380. Климадиа-
граммы Хельвана, расположен-
ного в пустыне близ Каира,
Рис. 381. Колебания годовых осадков близ Каира в 1906—
1955 гг.
В пустынных областях растительный покров сильно нарушен чело-
веком, поскольку он используется для выпаса овец, коз, ослов и верблю-
дов. В результате при стравливании и без того скудной растительности
498
Глава 14
почти полностью исчезли лучшие кормовые растения (в частности, зла-
ки), а остались лишь немногочисленные ядовитые или сильно опушенные
и колючие растения. Кроме того, следует учитывать, что все древесные
растения используются кочевниками на топливо.
Представление о естественной растительности могут дать лишь запо-
ведные территории, однако в египетско-аравийской пустыне их нет.
Характеристика растительности пустыни, которая будет приведена
ниже, заимствована из работ Штоккера, Дэвиса, Кассаса; кроме того,,
она составлена на основании путевых впечатлений автора в 1960 г.
Растительность каменистой пустыни
Из-за высокой концентрации солей и сухости почвы растительный
покров в гамаде большей частью не развивается. Только в трещинах
скал растут некоторые хасмофиты: Erodium glaucophyllum, Reaumuria
hirtella, Helianthemum kahiricum, Fagonia mollis и др. Нередко здесь
встречаются также лишайники (Lecanora, Lecidea, Physcia, Verrucaria,.
Endocarpon, Ramalina, Usnea).
Выпадающие зимой осадки стекают в каменистой пустыне по едва
заметным желобкам стока, заполненным рыхлыми продуктами выветри-
вания и соединяющимся в более крупные промоины глубиной обычно не
более 2 лг, в этих временно увлажняемых руслах происходит вымывание
солей, и поэтому в них уже встречаются отдельные растения, обычно
эфемеры: Anastatica hierochuntica, Diplotaxis acris (рис. 382) и D. harra,
Zygophyllum simplex, Trigonella stellata, Plantago ovata и др., а также
многолетники, зеленеющие лишь зимой: Asteriscus graveolens, Iphiona
mucronata, Farsetia aegyptiaca, Erodium arborescens (рис. 383) и, кроме
того, Echinops spinosus, Calligonum comosum, Reaumuria hirtella и
Ephedra.
Если вода, текущая по долинам временных водотоков, падает с усту-
пов плато, то образуются ущелья с вымоинами по дну и отвесными сте-
нами. По дну ущелья на влажной почве между обломками породы растет
Rumex vesicaria, а в трещинах по склонам ущелья укореняется Capparis
spinosa.
Вокруг невысыхающей воронки глубиной 4 м, образованной водо-
падом, падающим с высоты 50 м, встречаются два реликта более влаж-
ных периодов — Ficus pseudosycomorus и Adiantum capillus veneris. Послед-
ний распространен повсеместно в засушливых субтропиках, в таких же
постоянно увлажненных местообитаниях.
1 О. S tocke г. op. cit., а также Die agyptisch-arabische Wiiste, «Veget. Bil-
dcr», 17, H. 5/6, 1926; P. H. D a v i s, The vegetation of the deserts near Cairo, «J.
Ecol.», 41, 1953, p. 157—173; M. Kassa s, Landforms and plant cover in the Egyptian
desert, «Bull. Soc. Geogr. Egypte», 26, 1953, p. 193—205; M. Kassas, M. Imam,
Habitat and plant communities in the Egvptian desert, I —V, «J. Ecol.», 40. 1952.
p. 342—351; 41, 1953, p. 248—256; 42, 1954, p. 424 — 441; 47, 1959, p. 289—310:
M. Kassas, M. A. Z ahr an, Studies on the ecology of the Red Sea coastal land,
I. «Bull. Soc. Geogr. d’Egypte», 35, 1962, p. 129—175; M. Kassas, M. S. E l-A b у a d,
On the phytosociology of the desert vegetation of Egypt, «Ann. of Arid Zone», 1. 1962,
p. 54—83; M. Kassa s, M. S. E l-A b у a d, W. A. G i r i s, The limestone plateau,
«J. Ecol.», 52, 1964, p. 107—119.
Сведения об отдельных видах можно также найти в вышедших к настоящему
времени томах «Flora of Egypt», составленной V. Tackholm и М. Drar. Этими же авто-
рами составлен краткий определитель «Students Flora of Egypt».
Рис. 382. Слева — Anastatica hierochuntica (иерихонская роза) в свежем состоя-
нии; в середине — два свернувшихся экземпляра, справа — Diplotaxis \acris.
Дождевая рытвина в гамаде близ Хельвана (фото Штоккера).
Рис. 383. Erodium arborescens в небольшом вади близ Хельвана.
500
Глава 14
Растгппелъиость сухих долин временных потоков
Несколько более сомкнут растительный покров в крупных промои-
нах, дно которых покрыто щебнем; под большим уклоном они впадают
в широкие заполненные песком сухие долины временных потоков или так
называемые вади (рис. 367 и цветное фото 1, стр. 464).
Кассас выделяет в окрестностях Хельвана следующие типы сообществ:
1. Stachietum aegyptiacae; развит на обнажениях кристаллического
известняка по дну сухих долин; в трещинах скал растут хасмофиты. Пре-
обладает Stachys aegyptiaca, достигающий большого возраста (корни диа-
метром до 5 см). Характерны также Limonium (Statice) pruinosa, Fagonia
mollis, F. kahirina и Erodium glaucophyllum. Иногда здесь доминируют
Asteriscus graveolens и Reaumuria hirtella.
2. Zygophylleto-Anabasietum; распространен на каменисто-щебнистых
почвах мощностью около 50 см; содержание мелкозема в этих почвах не
превышает 30%.
У обоих доминирующих видов Zygophyllum coccineum и Anabasis
setifera хотя и развиты суккулентные листья и стебли, однако они должны
быть, скорее, отнесены к числу галофитов, чем суккулентов; иными сло-
вами, они не запасают воду и, вероятно, накапливают хлориды. Повсе-
местное распространение Zygophyllum объясняется его несъедобностью.
Наряду с этими видами здесь встречается также много многолетников
и эфемеров.
3. Zilletum spinosae; развит на мощных почвах, влажность которых
на глубине 60—70 см всегда достигает 3%, содержание солей — около
0,2—0,8%, органических веществ — менее 1 %. Поскольку Zygophyllum
coccineum, судя по годичным кольцам, не достигает возраста старше 5 лет,
он вытесняется в этих условиях кустарником Zilla spinosa (рис. 368),
имеющим высоту 150 см при диаметре куста до 2 м. Почвы образованы из
прослоек грубого аллювия, чередующихся с илистыми прослойками,—
это связано с изменением скорости течения паводковых вод. Между двумя
последними ассоциациями наблюдаются различные переходы.
4. Злаковник; развит на песчано-илистых почвах мощностью более
100 см. В составе злаковника господствуют Panicum turgidum и Pennisetum
dichotomum; из них первый стравливается скотом сильнее. Степень покры-
тия почвы может достигать местами 80%. Число сопутствующих видов
здесь гораздо меньше.
5. Заросли кустарников; развиты на незаливаемых паводками терра-
сах. В их составе преобладают Nitraria retusa (tridentata), Lycium arabi-
cum, Atriplex halimus и виды Tamarix. Кустарники достигают высоты
1—2 м; Tamarix articulata — это дерево высотой до 10 м. Крупные мас-
сивы встречаются редко из-за интенсивной вырубки на топливо. Заросли
Atriplex сильно повреждены скотом. Более сомкнутые галерейные леса
развиваются только при наличии постоянного зеркала грунтовых вод,
расположенного на глубине 15 м от поверхности, а вблизи побережья —
всего 2 м.
Растительность щебнистой пустыни
Вдоль шоссе Каир — Суэц, между возвышенностью Мокаттам (300 м)
на западе и горами Атаква (700 м) на востоке, простирается обширная
всхолмленная щебнистая пустыня, поверхность которой покрыта темной
зоркой, так называемым пустынным загаром. Пологие склоны холмов
Египетско-аравийская пустыня, пустыни Синай и Неджеб
501
прорезаны небольшими руслами временных водотоков, заполненными
песком; в связи с этим они отличаются более светлым оттенком. У подно-
жия склонов эти русла сливаются и образуют обширные долины или замк-
нутые бассейны. Засоленность песка небольшая, и в местах его скопления
развивается довольно сомкнутый растительный покров. Здесь распро-
странены следующие типы растительных сообществ: 1) тип сообщества,
где доминирует Haloxylon salicornicum] степень покрытия почвы менее
5%; 2) травостои, в составе которых преобладают Panicum turgidum,
и иногда Lasiurus hirsutus, растущий в небольших ложбинах стока; 3) тип
сообществ, где господствует Zilla spinosa. Общее представление об этом
ландшафте дает цветное фото 2, стр. 464.
В лучше увлажняемых руслах потоков наряду с Panicum turgidum
встречаются крупные кусты Retama retain и даже небольшие деревья
Acacia raddiana или A. tortilis. Последние, однако, уже почти все выруб-
лены.
Кассас приводит в своей работе списки растений, собранных на
пробных площадках, с указанием константности и встречаемости отдель-
ных видов. Мы же ограничимся простым перечислением видов, растущих
вблизи опорного пункта Ботанического сада Каирского университета
(табл. 94).
Таблица 94
Многолетники (в порядке уменьшения степени обилия)
Haloxylon salicornicum Citrullus colocynthis Hyoscyamus muticus Zilla spinosa Pituranthos tortuosus Retama retam Crotalaria aegyptiaca Pennisetum dichotomum Panicum turgidum Farsetia aegyptiaca Heliotropium luteum Odontospermum graveolens Convolvulus lanatus Echinops spinosus Fagonia arabica Artemisia judaica Artemisia monosperma Daemia cordata Cleome arabica Pancratium sickenbergeri Halogeton alopecuroides Astragalus forskalei Linaria aegyptiaca Gypsophila capillaris
Двулетники или однолетники
Euphorbia cornuta Centaurea aegyptiaca
Однолетники (в порядке уменьшения степени обилия)
Mesembryanthemum jorskalei Trigonella stellata Schismus calycinus Ifloga -fontanesii Plantago ovata Anthemis melampodina Matthiola livida Zygophyllum simplex Malva parviflora Erodium pulverulentum Tribulus alatus
Однолетники развиваются в течение короткого благоприятного вре-
мени года, подобно зимним эфемерам, распространенным в Аризоне;
многолетники же переживают длительный период засухи. Отсутствие сре-
ди них типичных суккулентов {Haloxylon обладает слабо выраженной
502
Глава 14
суккулентностыо) говорит об их способности поглощать воду в течение
всего года. Мы вернемся к этому вопросу в следующем разделе.
В приведенном выше списке отсутствует Zygophyllum coccineum, осо-
бенно часто встречающийся в вади близ Хельвана. Однако он появляется
на 101-м км шоссе Каир — Суэц; Haloxylon salicornicum, напротив, здесь
не встречается, равно как и в сухих руслах временных потоков. Видимо,
флористические различия определяются содержанием кальция в почве:
Zygophyllum coccineum приурочен к известнякам, a Haloxylon их избегает.
В вади у 101-го км степень покрытия почвы растительным покровом
достигала 20%. Наряду с отдельными экземплярами Acacia tortilis и Lyci-
ит arabicum здесь были широко распространены следующие многолет-
ники: Zygophyllum coccineum и Launea (Zollikojeria) spinosa, кроме того,
встречались Retama retam, Zilla spinosa, Panicum turgidum, Echinops
spinosus и Farsetia aegyptiaca.
Вызывает удивление богатство флористического состава раститель-
ного покрова пустынь, способного существовать при среднем количестве
осадков около 25 мм в год. Правда, при этом не следует забывать о незна-
чительной сомкнутости покрова, степень покрытия которого в среднем
не превышает 1 %. Поэтому отдельные растения могут быть относительно
хорошо обеспечены влагой (см. стр. 306 и след.).
ЭКОЛОГИ ЧЕСКИЕ ИССЛЕДОВАНИЯ,
ПРОВЕДЕННОЕ БЛИЗ КАИРА
Экология растений пустыни изучалась сотрудниками Ботанического
института Каирского университета в щебнистой пустыне вдоль шоссе
Каир — Суэц (см. стр. 500) и вблизи Хельвана в Вади-Хофф; последний
достигает длины 15 км, глубоко врезан в поверхность каменистой пустыни
и впадает в долину Нила примерно в 20 км к югу от Каира (рис. 365, 367,
цветное фото 1, стр. 464).
Исследования, проведенные вдоль шоссе Каир — Сущу
Равнинная территория местами пересекается широкими долинами
и понижениями, к которым главным образом и приурочена раститель-
ность. В этих депрессиях степень покрытия почвы многолетниками не
превышает 10%; здесь обычно доминирует Haloxylon. Рыхлый грубозер-
нистый песок хорошо промачивается; пленочная влага относительно легко
усваивается растениями. Недоступная для растений влага не превышает
1%.
Содержание влаги в почве. Наблюдения показали, что
верхние 25 см почвы почти регулярно промачиваются зимними осадками,
тогда как летом они полностью высыхают. Начиная с глубины 50 см и до
глубины 100 см и более, расположен слой почвы, который на протяжении
всего года всегда содержит какое-то количество доступной для растений
влаги, пополняемое в отдельные годы за счет воды, просачивающейся
после особенно сильных дождей. Некоторую роль при этом играет и вода,
стекающая после ливней в понижения с окружающих возвышений. Слой
1 А. М. М i g a h i d, A. A. A b d el Rahman, Studies in the water economy
of Egyptian desert plants. I. Desert climate and its relation to vegetation, «Bull. Inst.
Desert Egypte», 3, 1953, p. 5—24; II. Soil water conditions and their relation to vege-
tation, 3, 1953, p. 25—27; III. Observations on the drought resistance of desert plants,
3, 1953, p. 39—83.
Египетско-аравийская пустыня, пустыни Синай и Неджеб
503
почвы, расположенный на глубине 25—50 см, иногда промачивается после
сильных дождей, однако в бедные осадками годы он высыхает полностью,
В этих случаях нередко на протяжении многих лет между верхним перио-
дически увлажняемым и нижним по-
стоянно влажным слоями существует
совершенно сухой горизонт почвы,
В качестве примера можно при-
вести данные о влажности почвы, по
материалам Мигахида и Абд эль Рах-
мана (табл, 95), Работы проводились
восточнее Каира в 1949/50 г. Опре-
делялась влажность песчаной почвы;
недоступная растениям влага равна
0,8%, Последний дождь был отмечен
в ноябре 1948 г,: выпало 0,7 мм осад-
ков, Осадки, выпавшие в течение зимы
1948/49 г,, в противоположность бо-
лее влажной зиме 1949/50 г, увлаж-
нили лишь верхние слои почвы, Ниже
25 см в феврале 1949 г, располагал-
ся сухой горизонт почвы. Некоторые
небольшие отклонения в значениях
влажности авторы связывают с кон-
денсацией водяных паров воздуха
в почве. Однако этот процесс пока
еще изучен слабо.
Грубозернистая песчаная почва
в результате испарения с поверх-
ности высыхает на глубину не более
30 см. Ниже уровня 50 см распола-
гается постоянно влажный горизонт;
крайне скудная растительность не
в состоянии исчерпать всю накоплен-
ную в этих горизонтах влагу, к тому
же пополняемую в годы с обильны-
ми осадками, как это наблюдалось,
например, в 1950 г.
Ночное охлаждение почвы вы-
зывает выпадение на ее поверхности
росы. Поскольку, однако, роса днем
быстро испаряется и в период засухи
ее роль в увеличении влажности
почвы, даже рано утром, совершенно
ничтожна, очевидно, что этот процесс
конденсации влаги не может способ-
ствовать улучшению снабжения во-
дой растений (см, также стр, 506),
Корневая система и
развитие растений. Корни
пустынных однолетников развивают-
ся на глубину до 21 см, а в радиальном
направлении до 35 см. В связи с этим
развитие растений зависит от обес-
Р и с. 384. Корневая система Pituran-
thos tortuosus (по Каушу).
Влажность песчаной почвы (в процентах на абсолютно сухую навеску)
Таблица 95
Год . 1949 1950
Дата выпадения осад- ков . 7/II 10/П 13/V 23/XI 19/XII 26/1 — 17/III — — —
Количество осадков, мм . 2,9 0,4 — 0,2 — — — — — 6,1 6,9 9,6 — 9,2 — —
Дата взятия образцов 17/11 17/Ш 17/IV 5/V 23/VI 12/VII — 19/IX — 25/XI 22/XII — 19/11 23/1II 10/IV 6/V 8/VI
Образцы взяты с глу- бины, с м:
5 2,7 1,5 0,6 0,4 0,3 0,2 — 0,4 — 5,5 2,7 — 1,7 2,8 0,9 0,5 0,3
10 2,0 2,0 1,1 0,5 0,3 0,5 — 0,7 — 2,9 4,3 — 1,7 3,3 2,7 0,7 0,4
25 1,0 1,2 1,6 2,1 1,0 0,8 — 0,8 1,0 1,2 — 8,4 2,5 3,5 3,9 1,3
50 3,2 3,3 3,3 3,3 3,2 3,3 — 2,6 — 2,9 2,7 — 2,9 6,0 6,4 6,4 5,2
75 2,9 2,7 2,7 2,8 2,7 2,7 — 3,5 — 2,7 3,4 — 2,4 2,7 3,2 — 4,0
Глава 14
Египетско-аравийская пустыня, пустыни Синай и Неджеб
505
печенности влагой верхних горизонтов почвы, увлажняемых только
в дождливый период. Семена однолетников быстро прорастают после пер-
вых же дождей. Если осадков выпало мало, развивается только часть
видов, достигающих карликовых размеров. Например, в 1949 г. из видов,
перечисленных на стр. 501, были отмечены только Trigonella, Malva
и Erodium, в то время как в 1950 г. все 11 видов пышно развивались. Как
только верхние слои почвы пересыхают, однолетники отмирают, посколь-
ку их корни не достигают постоянно увлажненных слоев почвы; только
у Zygophyllum simplex иногда развивается более глубокая корневая систе-
ма. Тогда этот вид продолжает существовать и в течение периода засухи.
У многолетних растений, наоборот, корни достигают нижних постоян-
но влажных горизонтов. Обычно стержневой корень, ветвящийся в более
глубоких слоях почвы, проникает у большинства видов до глубины 130 см,
а в руслах пересыхающих потоков еще глубже (рис. 384). В горизонталь-
ном направлении корни простираю! зя на расстояние 220 см у Haloxylon
и на 440 см у Hyoscyamus. Как и вез te, у злаков развивается интенсивная
корневая система, тогда как у других видов — экстенсивная.
Семена многолетников прорастав т лишь через некоторое время после
того, как их смочит дождь, и то только в том случае, если дожди идут доста-
точно долго и вымывают тормозящие рост вещества из оболочки плодов
или семян. Наряду с этим необходимо промачивание почвы на всю
глубину профиля. В противном случае всходы погибают вместе с однолет-
никами, как только начинается засуха.
Надземные части многолетников по сравнению с корневой системой
развиты очень слабо. Поэтому листья относительно хорошо обеспечены
влагой, несмотря на незначительную влажность почвы. Интенсивность
транспирации достигает больших величин у Citrullus. Максимальная
интенсивность транспирации, равная 8,5 мг!г-мин, наблюдается в ноябре
в 10 час. утра, после чего ее интенсивность резко снижается. Транспира-
ция Haloxylon salicornicum отличается гораздо меньшей интенсивностью:
максимум, отмеченный между 7 и 10 час. утра, составлял всего 5 мг!г-мин\
минимальная интенсивность равнялась около 1 мг1г-мин.
Наименьшая интенсивность транспирации отмечепа у видов Mesem-
bryanthemum. Даже в утренние часы растение расходовало всего 4 % общего
количества воды, содержащейся в растении, а позже — в 5 раз меньше.
Столь незначительный расход воды позволяет этому однолетнику существо-
вать еще некоторое время после того, как высохнут верхние слои почвы.
Приведенные примеры показывают, что, несмотря на крайне засуш-
ливые условия пустыни, водный баланс распространенных здесь много-
летников сохраняется на протяжении всего года на более или менее посто-
янном уровне. Это подтверждают также относительно низкие значения
осмотического давления. Только у Haloxylon, который, скорее, является
галофитом и накапливает соли в клеточном соке, величины осмотического
давления достигают больших значений; они колеблются от 30,4 атм
в январе до 43,2 атм в августе. Колебания осмотического давления в тече-
ние года сходны с изменениями величин испарения. В табл. 96 приведены
крайние значения осмотического давления у некоторых видов, распро-
страненных в этих условиях.
Эти данные свидетельствуют о том, что даже в этих крайне засушли-
вых условиях пустыни осмотическое давление у растений не достигает
больших величин. Несмотря на это, в литературе до настоящего времени
сохраняется точка зрения, что пустынные растения отличаются высоким
осмотическим давлением. Далее, из приведенных материалов видно, что
506
Глава 14
Таблица 96
Наименьшие и наибольшие величины
осмотического давления (в атм)
Hyoscyamus muticus . 18,7—26,9
Pituranthos tortuosus 15,4—22,9
Zilla spinosa . . . 16,7-18,7
Citrullus colocynthis . 11,1—13,6
Echinops spinosus . 15,9-18,3
Heliotropium luteum . . 16,8-17,0
Retama retam . 24,1-25,5
Artemisia sp. 15,2
однолетники развиваются в пустыне лишь там, где влага содержится
в верхних горизонтах почвы в легкодоступной для растений форме. Для
существования несуккулентных видов необходимо наличие в почве неко-
торого количества влаги даже в период, когда у них сокращается расход
влаги на транспирацию. Когда запасы воды в почве становятся ничтож-
ными, растение переходит в состояние покоя. В процессе перехода в период
покоя растение постепенно высыхает, что можно проследить по постепен-
ному повышению концентрации клеточного сока. Менее обеспеченные
водой отдельные органы растений при этом начинают отмирать. Если
этот процесс не будет приостановлен, то растение в конце концов погибнет.
Многолетние виды приспособлены к перенесению длительных засух
и достигают возраста 100 лет и более; они встречаются в пустыне только
в местах, где никогда полностью не иссякает запас воды в горизонтах
почвы, пронизанных корнями. Если дожди выпадают лишь эпизодически
и нерегулярно, как, например, в центральной Сахаре, то здесь практиче-
ски могут встречаться лишь эфемеры, семена которых, возможно, более
десяти лет сохраняются в почве, не теряя всхожести.
В этих условиях эфемеры развиваются в образованных дождевыми
потоками бороздах и мелких понижениях, где почва лучше увлажнена
и куда вода смывает семена этих растений.
Значение стока в этих пустынях автор мог наблюдать во время поезд-
ки через Сирийскую пустыню из Багдада в Иерусалим в марте 1955 г.
На фоне почти безжизненной пустыни отчетливо выделялись две зеленые
полоски, простиравшиеся с обеих сторон асфальтированного шоссе.
Незадолго до этого прошел дождь, и вся вода с шоссе стекала на обо-
чины; влажность почвы здесь увеличилась примерно в 5—10 раз; это
и обусловило возможность развития растений.
Абд эль Рахман и эль Хадиди приводят следующие данные о конден-
сации влаги в почвах пустыни близ Каира. На глубине 5 см авторам не
удалось установить никаких колебаний влажности почвы в течение ночи.
Эти изменения отмечены только на поверхности почвы. Максимум наблю-
дается обычно утром в 6—7 час., а минимум — вскоре после полудня
Максимальные суточные колебания этих значений достигают лишь вели-
чины немногих долей процента, причем влажность почвы остается в это
время ниже величины влажности завядания. Лишь утром 27 января 1956 г.
влажность почвы достигла 0,99%, однако днем эта величина быстро упала
до 0,1% и менее. Из этого следует, что роса и гигроскопическая влага не
имеют никакого значения для растений, распространенных в этой пустыне.
Круговорот воды в растительных сообщест-
вах. Данные о расходе воды в растительных сообществах на площади
Египетско-аравийская пустыня, пустыни Синай и Неджеб 507
100 м2 позволяют судить о том, какая часть осадков используется расте-
ниями на транспирацию и сколько накапливается сухого вещества.
Изучением этих вопросов занимались Абд эль Рахман и эль Хадиди
в тех же условиях в 1955/56 г.1. В зимние месяцы выпало в общей слож-
ности 23,4 мм осадков (рис. 385); в результате запасы влаги увеличи-
лись только в верхних 25 см почвы, что способствовало развитию эфе-
меров. Многолетники должны были использовать запасы влаги, накоп-
ленные в более глубоких слоях почвы в предыдущие богатые осадками
годы. Иногда выпадала роса; эта влага способствовала повышению влаж-
ности лишь верхних 2 см почвы, однако и в этом слое влажность почвы
составляла всего 0,16—0,91%. Эта величина столь ничтожна, что ее мож-
но не принимать в расчет. Урожай зеленой массы растений на площади
размером 100 м2 достигал следующих величин (в кг) (табл. 97).
Таблица 97
Panicum turgidum . 1,960
Haloxylon salicornicum . . 1,245
Mesembryanthemum jorskalei . 9,718
Erodium pulverulentum 0,070
Schismus calycinus 0,046
Остальные, не перечисленные в таблице виды были представлены
одиночными экземплярами.
Таким образом Mesembryanthemum накапливает около 75% зеленой
массы. У отдельных видов периодически изучался ход транспирации
в течение суток; на основе этого вычислялся общий расход влаги на транс-
пирацию всем сообществом.
Все сообщество (на площади 100 м2) расходовало в течение одного часа
11 — 12 ноября следующее количество воды: максимум, равный 1,02 кг,
наблюдался в 14 час., а минимум, равный 0,5 кг,— около 20 час. 25—
27 апреля эти величины достигали соответственно 1,38 кг в 13 час. и 0,31 кг
в 24 час. Кривая расхода влаги соответствовала примерно ходу испарения.
Суточный расход воды перед началом дождей в октябре (когда еще
не были развиты эфемеры) составлял 16,0 кг на площади 100 м2 (рис. 386).
В ноябре и декабре эта величина снизилась в результате возросшей влаж-
ности воздуха; эфемеры успели в это время лишь частично развиться.
В январе — феврале сомкнутый растительный покров расходовал 18,05 кг
воды; эта величина снизилась по мере отмирания эфемеров до 11,75 кг.
Максимальная интенсивность транспирации наблюдалась у эфемеров.
Так, днем максимальная интенсивность транспирации составляла у Schis-
mus 599 мг!г-час, у Erodium — 325, у Panicum и Haloxylon — лишь немно-
го более 100, тогда как наименьшая интенсивность транспирации наблю-
далась у видов Mesembryanthemum — всего 28 мг!г-час. Она снижается
у этого вида по мере развития растений с 41 мг у проростков до 19 мг!г-час
у взрослого растения. Одновременно падает и содержание воды в самом
растении.
Хотя Mesembryanthemum и накапливает 75% общей массы, однако он
расходует всего 37% общего количества воды. Это примерно равно коли-
честву воды, расходуемому Panicum, и вдвое превышает эту величину
у Haloxylon. Эфемеры проросли после первого дождя, выпавшего 9/10 ноя-
1 A. A. Abd el Rahman, М. N. el Hadid у, Observations on the water
output of the desert vegetation along Suez Road, «Egypt. J. Bot.», 1, 1958, p. 19—38.
508
Глава 14
Р п с. 385. Климадиаграмма Капра,
зима 1955/56 г.
Р и с. 386. Расход воды на транспи-
рацию растительным покровом пу-
стыни на площади 100 м2\ участок
расположен вдоль шоссе Каир —
Суэц; зима 1955/56 г. (по Абд эль
Рахману и Хадпди).
1 — расход воды эфемерами; 2 — рас-
ход воды многолетниками.
бря. По отношению ко всей массе растений они расходовали следующее
количество воды: в декабре — 5,4%, январе — 40,5, феврале — 44,1,
марте — 39,2 и в мае — 9,7% (ср. с рис. 386). Без учета многолетников,
поглощающих влагу из более глубоких почвенных горизонтов, расход
воды эфемерами (в мм) достигал следующих значений, приведенных
в табл. 98.
Т а б л и ц а 98 В
Месяц XII I II ш I V V
Расход воды 0,2 2,1 2,3 1,6 0,8 0,3
В общей сложности расход эфемерами достигал примерно 7,3 мм\
иными словами, 32% выпавших осадков (то есть 23,4 мм) было израсхо-
довано эфемерами на транспирацию за время их развития.
На ровную поверхность площадью 100 м2, в период дождей выпало
2340 л осадков. Эфемеры накопили 9834 кг зеленой массы, или 518г сухого-
Египетско-аравийская пустыня, пустыни Синай и Неджеб
509
вещества. Если учесть, что на эту площадь не поступало дополнительной
влаги (путем стока), то окажется, что только 0,4% осадков было накоплено
в тканях растений.
Поскольку эфемеры (на площади 100 м2) в целом израсходовали на
транспирацию 730 кг воды и накопили 518 г сухого вещества, продуктив-
ность транспирации составила 518 : 730 = 0,71; иными словами, на каж-
дый килограмм израсходованной на транспирацию воды эфемеры накап-
ливали 0,7 г сухого вещества. Коэффициент транспирации равен 1409 —
величина, очень большая по сравнению со значениями, наблюдаемыми
в Центральной Европе (400—600).
В Юго-Западной Африке, согласно расчетам автора, каждые 100 мм
осадков дают возможность растительному покрову накопить около
Рис. 387. Изменения сырого и сухого веса эфемеров на площади
100 ле2; зима 1955/56 г. (по Абд эль Рахману и Хадиди).
1000 кг!га сухого вещества. Если выпадет 23,4 мм осадков, то будет накоп-
лено 234 кг!га, или 2,34 кг на 100 м2. В Египетской пустыне количество
накопленного сухого вещества не превышало 0,52 кг, что составляет менее
0,25 величины, наблюдавшейся в Юго-Западной Африке. Разница связана
с тем, что при незначительном количестве осадков их основная часть
расходуется на испарение. Кроме того, растительный покров в Юго-
Западной Африке (чистый злаковник) и в окрестностях Каира, где пре-
обладают суккулентные виды Mesembryanthemum, резко отличен и поэто-
му такое сравнение не может быть проведено.
Динамика накопления массы у эфемеров представлена на рис. 387.
Конкурентные взаимоотношения. С целью изу-
чения развития растительности на протяжении года Абд эль Рахман про-
вел картирование двух пробных площадей размером 100 м2 в январе,
феврале и июне 1952 г.1. На площадях было отмечено 8 многолетников,
2 двулетника и 7 однолетников. Степень покрытия почвы многолетними
видами составляла немногим более 10%; в их составе преобладал Panicum
1 A. A. A b d el Rahman, Studies in the water economy of Egyptian desert
plants. IV. Establishment and Competition, «Bull. Inst, Desert Egypte», 3, 1953,
_p. 84-93.
510
Глава 14
turgidum. Однолетники, очень многочисленные в феврале, в июле, напро-
тив, бесследно исчезли.
Оказалось, что нежные однолетники могут погибнуть уже после
первой песчаной бури. По данным Абд эль Рахмана и эль Хадиди, после
песчаной бури, продолжавшейся с 27 по 30 января 1956 г. (скорость
ветра более 15 м/сек), примерно половина всех растений была уничто-
жена: некоторые были вырваны с корнем, другие высушены или повре-
ждены перемещавшимся песком. Степень покрытия уменьшилась на
31% х. Многолетние виды, наоборот, оказались ветроустойчивыми. У мно-
голетников на первой пробной площадке в июле сохранился один всход
Salvia aegyptiaca и два Haloxylon, а на второй площадке — только один
всход Centaurea aegyptiaca. Несмотря на наличие свободных пространств,
растения здесь возобновляются плохо, и поэтому у многолетников отсут-
ствует корневая конкуренция. Следовательно, вопреки нашим прежним
высказываниям в этих условиях остается достаточно свободного про-
странства для развития новых растений. Правда, автор указывает, что
пастьба скота на пробных площадках обусловливает изреженность покро-
ва; пастьба оказывает особенно сильное влияние на покров в крайних
условиях существования растений в засушливых областях. Этот довод,
однако, не исключает высказанной выше мысли, что в крайних условиях
существования растения возобновляются естественным путем столь сла-
бо, что не могут обеспечить развитие растительного покрова на участках,,
покрытых песками (рис. 388).
Исследования,
проведенные в Нади-Хофф близ Хелъвана
Годовое количество осадков в этой области составляет в среднем
31 мм', после выпадения 10 мм здесь начинается поверхностный сток
воды по лишенному растительности плато, сложенному известняками.
Вода концентрируется сначала в мелких промоинах, которые, сливаясь
друг с другом, постепенно углубляются и наконец переходят в широкие
обычно сухие русла, лишь в исключительно дождливые годы на короткое
время заполняющиеся водой. Легко просачивающиеся в песок осадки
создают в почве некоторый запас влаги, обусловливающий здесь развитие
растительности; сомкнутость покрова возрастает от неглубоких дождевых
борозд, где встречаются одиночные экземпляры растений, до днищ вади,
где иногда распространены даже заросли Acacia и Tamarix. Степень
сомкнутости растительного покрова была бы еще больше, если бы здесь
не паслись верблюды и деревья не вырубались на топливо. Водный режим
развитых здесь растений на протяжении одного года изучал Батануни 1 2.
На схематическом профиле, проведенном через Вади-Хофф (рис. 389),
можно выделить 6 экотопов: 1) плато с неглубокими дождевыми борозда-
ми; 2) уступ скалы, у подножия которой растения развиваются весь
день в тени, за исключением раннего утра; 3) собственно русло вади;
4) первая пойменная терраса; 5) вторая (надпойменная) терраса и 6) склон
1 A. A. A b d cl R a h m а и, е 1 М. N. Н a d i d у, Some observations on the
effect of wind on the desert vegetation along Suez Road, «Bull. Soc. Geogr. Egypte», 32,
1959, p. 207-216.
2 К. H. Batanouny, Water economy of desert plants in Wadi Hoff, Ph. D.-
thesis, Cairo University, 1963*; A. A. A b d el Rahman, К. H. Batanouny,.
The water output of the desert vegetation in the different microhabitats of..., «J. Ecol.»r
53, 1965, p. 139-145.
Египетско-аравийская пустыня, пустыни Синай и Неджеб
511
гамады, покрытый осы-
пями. Почвы слабораз-
виты: преобладают кру-
пные обломки породы
и щебень, среди которых
встречаются прослойки
крупнозернистого, реже
мелкозернистого песка
и глины. Только во 2 и 4
экотонах на долю или-
стой фракции прихо-
дится более 10%. Вла-
гоемкость почвы невысо-
ка, и влажность завя-
дания равна 1 % или
даже менее. В экотонах
1—4 почва не засолена;
в экотопе 5 содержание
солей в почве достигает
1—2,5%, а в экотопе 6—
даже более 5%.
Влажность
почвы. Влажность
м2 4
верхнего 50-сантимет-
рового слоя определя-
лась один раз в месяц.
Верхние горизонты поч-
вы бывают увлажнены
И
I
4
5
8
10
Panicum
о Centaurea
Panicum
Panicum
Haloxylon
Panicum
О Centaurea
Q Haloxylon
Рис. 388. Распределение многолетних растений на
площади 100 м2; степень сомкнутости покрова сравни-
тельно велика (шоссе Каир — Суэц) (по Абд эль Рах-
ману и Хадиди).
только в период зимних дождей, а летом почва здесь находится
в воздушно-сухом состоянии. Однако в целом в почве всегда содержит-
ся какое-то количество доступной для растений влаги (табл. 99)~
Степень покрытия почвы приведена в табл. 99 не случайно, поскольку
запас влаги в период засухи зависит не только от количества просочив-
шейся в почву воды, но также и от количества израсходованной расте-
ниями влаги. Многолетние растения не могут расти в перемещающемся
во время паводков грунте вади (3), а сильно засоленные почвы на скло-
м О 2 4 6 8 10 П А 20 30 40 50 60 70 80 90 ЮО НОм
Рис. 389. Поперечный профиль через Вади — Хофф (ио Батануни, несколько изменено).
1 — 6 — экотопы с соответствующими почвенными разрезами (слева — глубина, справа — общее количество солей, содержащихся в горизонтах 0—25
и 25 — 50 см, выраженное в % от веса сухой почвы). Горизонтальный масштаб правой половины рисунка уменьшен на 1/5. Вертикальный масштаб
изображен вверху слева.
1. Посадки Ricinus communis на песчаных дюнах (север Синайского полуострова).
2. Роща финиковых пальм на песчаном берегу Средиземного моря близ Эль-Ариша
(север Синайского полуострова).
Египетско-аравийская пустыня, пустыни Синай и Неджеб 513
нах (6) вообще не пригодны для развития растительности (см. также
работу Штоккера).1 Существенное значение имеет то обстоятельство, что
в год исследований, когда выпало всего 19,5 мм осадков, в почве всегда
имелась доступная для растений влага; видимо, корни проникают на глу-
бину более 50 см в трещины материнской породы. У эфемеров корни про-
никают неглубоко в почву; однако этот недостаток компенсируется
непродолжительностью периода вегетации, во время которого верхние
слои почвы достаточно хорошо увлажнены. В большом количестве эфеме-
ры развиваются только в очень влажные годы.
Круговорот влаги у растений. У наиболее широко распро-
страненных многолетников — Pennisetum dichotomum, Zilla spinosa и Zygo-
phyllum coccineum регулярно один раз в месяц определялся суточный ход
транспирации. Наблюдения подтвердили ранее выявленную закономер-
ность, что растения и в период засухи способны поглощать воду из почвы;
в это время в течение непродолжительного срока днем устьица у них
остаются открытыми, в связи с чем может осуществляться процесс фото-
синтеза. С увеличением сухости воздуха растение сокращает расход
воды; это выражается во временном снижении интенсивности транспира-
ции около полудня; максимальная интенсивность транспирации наблю-
дается в 10 час. утра. Летом интенсивность транспирации выше, чем
зимой. При поливе интенсивность транспирации летом у Pennisetum
и у Zilla увеличивается примерно в два раза; это признак того, что сокра-
щение транспирации произошло вследствие затруднений при поглощении
воды. У Zygophyllum развиты суккулентные листья, поэтому интенсив-
ность транспирации у него при пересчете на 1 г сырого веса гораздо ниже,
чем у предыдущих видов. Летом интенсивность транспирации у Zygophyl-
lum, несмотря на гораздо более сильное испарение, достигает таких же
значений, как и зимой. Такая же незначительная интенсивность транс-
пирации наблюдается и у однолетнего вида Zygophyllum simplex', этот вид
накапливает так много воды в листьях, что иногда может перенести
период засухи. Наибольшей интенсивностью транспирации отличаются
тонколистные эфемеры (максимум 14—25 мг!г • мин), однако и у мягко-
листных Stachys aegyptiaca, Artemisia fragrantissima и Diplotaxis harra
установлены аналогичные величины. Сокращению расхода воды в период
засухи способствуют не только закрытые устьица; это достигается и путем
сбрасывания листьев у растений или путем образования летних более
мелких листьев или колючек, то есть путем уменьшения транспирирую-
щей поверхности, как это наблюдается, например, у Zilla, Pennisetum,
Lavandula coronopifolia, Iphiona, Diplotaxis harra, Pituranthos tortuosus,
Stachys aegyptiaca и др.
Батануни определял в марте и июне величину расхода воды расти-
тельным покровом, развитым в различных местообитаниях на площади
100 м2 (табл. 100).
На основании этих величин можно определить примерный расход
воды растительным покровом в год в различных условиях среды; так,
в экотопах 1 и 2 (рис. 389) эта величина оказалась равной 180 мм, в эко-
топе 4 — около 400 мм и в экотопе 5 — около 30 мм. Конечно, к этим
величинам нужно добавить также расход воды эфемерами в зимние меся-
цы, а также испарение; величина расхода воды эфемерами колеблется
из года в год в зависимости от количества осадков. Небольшой по сравне-
нию с растительной массой расход воды в экотопе 2 связан с господством
1 О. Stocker, op. cit.
514
Глава 14
Таблица 100
Вес растительной массы и расход воды на площади 100 м2
Экотоп Месяц Сырой вес многолетни- ков, кг Расход воды, кг
1. Плато . . Март Июнь 15,2 15,3 44,8 51,0
2. Подножие скального уступа Март Июнь 47,4 42,8 45,9 48,7
3. Первая терраса Март Июнь 46,8 44,3 114,2 120,0
4. Вторая терраса Март Июнь 6,1 5,5 8,3 8,0
Таблица 101
Наименьшие (первый столбец) и наибольшие (второй столбец)
величины осмотического давления у растений, распространенных
в Вади-Хофф (атм), В скобках указаны месяцы
(по Батануни)
1. Многолетники Farsetia aegyptiaca 18,2 (I) 23,0 (X)
Achillea fragantissima . 17,0 (I) 22,7 (X)
Lycium arabicum 19,1 (III) 21,3 (XI)
Asteriscus graveolens 16,3 (II) 19,0 (IX)
Stachys aegyptiaca 13,8 (IV) 19,3 (XI)
Zilla spinosa:
Экотоп 1 (плато) 14,2 (II) 16,6 (IX)
» 2 (подножие уступа) 12,1 (II) 14,4 (IX)
» 4 (первая терраса) 14,2 (II) 14,9 (XI)
Iphiona mucronata:
Экотоп 1 (плато) 13,1 (HI) 23,3 (XII)
» 2 (подножие уступа) 11,6 (II) 16,0 (XII)
» 4 (первая терраса) . 13,6 (IV) 17,8 (XI)
Pennisetum dichotomum 11,2 (III) 14,7 (XI I>
Ochradenus baccatus . 19,7
Diplotaxis harra 22,0
Retama retam . . . 14,9 18,2
Pituranthos tortuosus . 13,5 17,0
Heliotropium arabianense 14,3 15,1
Pergularia tomentosa 16,3
Fagonia kahirina . . . 12,2
Lavandula coronopifolia 11,2
Scrophularia deserti 12,1
Centaurea aegyptiaca (двулетник) 13,1 14,4
Египетско-аравийская пустыня, пустыни Синай и Неджеб
515
2. Эфемеры
Diplotaxis acris —13,6; Trigonella stellata — 13,2; Plantago ovata — 11,6.
3. Галофиты Nitraria retusa 25,3 (I) 33,0 (VII)
Zygophyllum simplex (однолетник) Zygophyllum coccineum 28,9
Экотон 1 (плато) 29,1 (IV) 35,6 (XI)
» 2 (подножие уступа) 20,6 (II) 29,3 (X)
» 4 (первая терраса) . . 23,6 (IV) 32,1 (X)
здесь Zygophyllum coccineum (отличается слабой интенсивностью транспи-
рации) и большим затенением. Таким образом, в целом, в этой крайне
засушливой пустыне растения отнюдь не так уж плохо обеспечены влагой.
Осмотическое давление. Об относительно благоприят-
ных условиях влагообеспечения свидетельствует также осмотическое
давление у встречающихся здесь растений; за исключением галофитов,
величины осмотического давления относительно низки и мало меняются
в течение года (табл. 101).
Осмотическое давление, обусловленное хлоридами, не определялось.
Однако у видов Zygophyllum оно всегда очень высокое (см. стр. 386 и 465).
Что касается Nitraria, то этот вид обычно приурочен к засоленным место-
обитаниям (см. стр. 449 и 492).
Суточные колебания осмотического давления весьма незначительны,
особенно у Pennisetum’, у Zilla и Farsetia они составляли около 3 атм, что
свидетельствует об уравновешенности водного баланса. После полива
осмотическое давление у Pennisetum снижалось на 2,3 атм, а у Zilla
только на 1,7 атм\ это свидетельствует о том, что дефицит влажности
даже летом не бывает большим. На основании приведенных данных
можно сделать вывод, что способность пустынных растений переносить
завядание вовсе не связана с какими-то особыми свойствами протоплазмы.
Преодолению засухи, скорее, способствует сильное развитие корневой
системы и сокращение транспирирующей поверхности у этих растений.
РАСТИТЕЛЬНОСТЬ ПОБЕРЕЖЬЯ СРЕДИЗЕМНОГО МОРЯ
В ЕГИПТЕ
В направлении от Каира к Александрии, по мере увеличения коли-
чества осадков, увеличивается и сомкнутость растительного покрова.
В марте 1960 г. автор совершил поездку по соединяющей оба города
автостраде. Непосредственно за Каиром простиралась почти лишенная
растительности пустыня. Только у края асфальтового покрытия шоссе
(эффект местного стока) росли Aristida plumosa, A, pungens^ Panicum
turgidum, Zygophyllum coccineum и одиночно некоторые другие виды.
В одном понижении вокруг Aristida pungens образовались небольшие
дюны высотой 60 см. В середине пути, сначала небольшими зелеными
пятнами, развитыми только в понижениях, которые далее к северу
все более расширялись, появляется Mesembryanthemum forskalei совместно
с Artemisia monosperma.
На 112 км от Александрии (примерно в 84 км от побережья) было
проведено краткое описание растительности; степень покрытия почвы —
516
Глава 14
5%; господствуют Artemisia monosperma и Pituranthos tortuosus и несколько
реже встречается Panicum turgidum. Кроме того, отмечены эфемеры —
Mesembryanthemum forskalei, Trigonella, Ifloga, Erodium.
Вскоре начинает встречаться Thymelaea hirsuta, а на 88 км, пример-
но в 60 км от побережья, уже распространена полупустыня, образованная
Artemisia monosperma и Thymelaea hirsuta*, степень покрытия почвы здесь
достигает почти 10%.
Еще дальше к северу появляются Plantago albicans, Gymnocarpos
decander, Zygophyllum album и Asphodelus, из числа геофитов Allium
и Muscari, а также эфемеры — Matthiola, Salvia lanigera, Malva и т. д.
У 53 км начинаются поля ячменя; кочевники сеют его сначала в пони-
жениях, а ближе к морю — повсеместно. В благоприятные годы посевы
дают зерно, а в плохие отводятся под выпас.
На 29 км шоссе проходит вдоль озера Марьют, за которым виднеют-
ся уже прибрежные дюны. Близлежащие площади покрыты ковром цвету-
щих эфемеров, в составе которых преобладает Chrysanthemum coronarium.
Прибрежная полоса к западу от озера Марыот отличается неоднород-
ным рельефом. Непосредственно перед кромкой берега тянется полоса
дюн шириной 0,5—2 км*, дюны сложены крупнозернистым известковым
песком, отличающимся псевдооолитовой структурой; песок частично
закреплен растениями. Затем следует депрессия, наиболее глубокие
части которой заняты влажными солончаками, покрытыми зарослями
галофитов. По направлению в глубь материка эта депрессия сменяется
скалистыми гребнями, сложенными известняками плейстоценового воз-
раста, за пределами которых начинается пустынное Ливийское плоско-
горье, образованное миоценовыми известняками. Скалистые уступы
возвышаются не более чем на 35 м над уровнем моря.
Растительность этой части побережья Египта описана Тадросом Ч
Обширные пространства покрыты Thymelaea hirsuta, растущей совмест-
но с Zygophyllum alba и Asphodelus microcarpus. Растительный покров
деградировал здесь в результате многовековой пастьбы. В составе покро-
ва господствуют виды, не поедаемые скотом. После второй мировой войны
в этой области наблюдалось любопытное явление: на больших площадях
разрослась Kochia indica. Первое сообщение о находках этого однолет-
ника было опубликовано Драром. Согласно Шишини и Тодей, всхо-
жесть его семян лишь редко сохраняется больше года. Драц изучал кор-
мовое значение этого вида 1 2. В первые годы после войны Kochia охотно
поедалась скотом. Поскольку Kochia indica распространилась на этой
территории после войны и в районе военных действий, местное население
считало, что этот вид занесен сюда войсками. Исследования, однако,
показали, что семена этого растения, распространенного в северо-запад-
1 Т. М. Т a d г о s, An ecological survey of the semi-arid coastal strip of the western
desert of Egypt, «Bull. Inst. Dcisert Egypte», 6, 1956, p. 26—56; T. M. T a d г о s,
В. A. M. A t t a, Further contributions to the study of the sociology and ecology of the
halophilous plant communities of Mareotis (Egypt.), «Vegetatio», 8, 1958, p. 137—160;
The plant communities of the barley fields and uncultivated desert areas of Mareotis
(Egypt.), «Vegetatio», 8, 1958, p. 161—175.
2 M. D r ar, A report on Kochia indica Wight in Egypt, «Bull. Inst. Desert Egyp-
te», 2, 1952, p. 54—58; E. D. H. el Shishin y, D. Thoday, Inhabition of germi-
nation in Kochia indica, «J. Exp. Bot.», 4, 1953, p. 10—22; Effect of temperature and
desiccation during storage on germination and keeping quality of Kochia indica seeds,
«J. Exp. Bot.», 4, 1953, p. 403—406; O. D r a z, Some desert plants and their uses in ani-
mal feeding, Publ. Inst. Desert, № 2, 1954, p. 1 — 95; D. Thoday, T. M. T a d г о s,
E. D. H. el S h i s h i n y, Kochia indica Wight and its dispersal in Egypt, «Bull. Inst.
Desert Egypte», 6, 1956, p. 57—66.
Египетско-аравийская пустыня, пустыни Синай и Неджеб 517
ной Индии от Дели до Инда, были присланы в 1945 г. из Института сель-
скохозяйственных исследований Уэйт (Аделаида, Австралия) в Египет.
Обилие семян, приносимых этим растением, и шаровидная форма,
которую это растение приобретает после созревания семян, позволили
Kochia очень быстро распространиться в последующие влажные годы
(иногда выпадало более 200 мм осадков), тем более что сначала в районах,
занятых войсками, скот не выпасался. Однако в дальнейшем вследствие
сильного стравливания и незначительного количества осадков в течение
ряда лет кохия постепенно стала исчезать. Семена этого однолетника
прорастают только поздней весной, а развивается он в основном летом,
когда ощущается недостаток в свежем корме для скота. Семена кохии,
как уже было сказано, сохраняют всхожесть только в течение одного года;
если этот год будет засушливым, то запас семян в почве может погибнуть.
Кроме того, при сильном стравливании у растений вряд ли вообще обра-
зуются семена.
Экологические исследования проводились в 225 км западнее Алек-
сандрии, близ мыса Рас-эль-Хикма, выдающегося более чем на 2 км
в море; западный берег мыса крутой и скалистый, восточный — отлогий,
сложен песками. Ближайшая метеорологическая станция расположена
в Мерса-Матрухе (см. климадиаграмму на рис. 380). По сравнению с Хель-
ваном близ Каира (рис. 379) климатические условия на побережье
отличаются меньшей засушливостью. В Мерса-Матрухе наибольшее
количество осадков выпадает в декабре и январе. Среднегодовое коли-
чество осадков достигает 158 мм (крайние значения 49 и 275 мм)\ это
в 6 раз больше, чем близ Каира. Летом здесь температура ниже, а отно-
сительная влажность воздуха значительно выше, чем близ Каира; в июле
максимальные значения достигают 82%. С июля 1955 г. по июнь 1956 г.
на мысе Рас-эль-Хикма метеонаблюдения проводил Абд эль Гавад Айяд Ч
С ноября по март здесь выпало 159,6 мм осадков (табл. 102).
Таблица 102
Количество осадков, выпавших зимой
1955/56 г. близ Рас-эль-Хикмы (мм)
XI XII I II ill В сумме
87,2 47,2 11,6 1,8 11,8 159,6
Абсолютный максимум температуры равен 41°, а абсолютный мини-
мум + 5°. Суточная амплитуда температуры на побережье незначительна
и составляет зимой 6°, а летом еще меньше. Это обусловливает очень
высокую относительную влажность воздуха (82% в июле, а в февра-
ле 61%). Лишь в исключительных случаях, когда дует хамсин, влаж-
ность может снижаться на 10%.
Испаряемость (за год) примерно в два раза меньше, чем в пустыне:
Александрия Хельван Вади-Х альфа
1825 ,u.u 3830 мм 5660 мм
1 Abd el Gaw ad M. Ay у ad, An ecological study of Ras El Hikma-District,
Thesis, Cairo University, 1957,
518
Глава 14
Очень часто в течение года (в среднем 102 раза) выпадает роса; наибо-
лее часто это наблюдается в бездождные периоды в мае и июне.
Количество выпавшей в течение года росы (по дням) на мысе Рас-эль-
Хикма приведено в табл. 103.
Общее количество росы, выпавшей в 1955/56 г., равнялось 11,0 мм;
однако в течение любого месяца это количество не превышало 3 мм.
Только однажды выпало 0,35 мм, 7 раз — 0,2 мм, чаще же всего коли-
чество выпавшей росы не превышало 0,1 мм.
В 60 км к западу от Александрии, близ Бург-эль-Араба, и в 5 км
от побережья Средиземного моря Арвидссон и Хельстрем измеряли коли-
чество выпавшей росы х; эти наблюдения проводились в наименее благо-
приятные для этого месяцы — с декабря по февраль. Исследования про-
водились при помощи воронок, изготовленных из дерева, алюминия
и пластмассы. Наибольшее количество росы, которое стекло по стенкам
воронки площадью 1 м1 2, составило 400 см3, или 0,4 мм. Наибольшее
количество росы было отмечено в деревянных воронках; однако эти
величины были все же несколько ниже наблюдавшихся при измерении
росы по методу Дувдевани. Количество росы на листьях Asphodelus
microcarpus, Olea еигораеа и Hordeum vulgare достигало всего 1/5—1/4 вели-
чины, наблюдавшейся в деревянных воронках.
В одном из сообщений Арвидссон указывал, что он не обнаружил
существенных различий в количестве росы, выпавшей на растениях
в Бург-эль-Арабе и на острове Оланд (Швеция) 2. Максимальное коли-
чество росы на одном растении достигало 0,1 мм, а на отдельном листе —
0,2 мм. Особенно обильная роса была на Chenopodium album, Ch. murale
и на Beta. Число ночей, когда в Бург-эль-Арабе выпадала роса, равно 130;
минимальное количество росы выпадало в апреле — мае и в августе —
октябре. В Рас-эль-Хикме минимальное количество росы практически
выпадало в течение всей зимы, с ноября по март.
В почвах, распространенных в этой части Египта, начиная с глу-
бины 50 см, всегда содержится доступная для растений влага. В преоб-
ладающих здесь песчаных почвах количество пленочной влаги не пре-
вышает 13%, а влажность завядания составляет около 1%. Верхние
слои почвы насыщаются влагой только в период дождей. На глубине
50 см влажность почвы всегда была более 3,4%, на глубине 75 см —
выше 4,1% и на глубине 100 см — более 4%.
С 7 ноября по 10 апреля почва по всему профилю была хорошо увлаж-
нена. Роса редко увлажняет почву на глубину до 5 см; даже очень силь-
ная роса в период засухи не может увеличить влажность почвы выше
величины коэффициента завядания. В большинстве случаев роса про-
мачивает лишь верхние 2 см почвы, причем эта влага быстро испаряется
после восхода солнца; после 10 час. утра в почве больше не остается
доступной растениям влаги. Об этом свидетельствуют следующие наблю-
дения за влажностью почвы, проводившиеся в течение трех дней в апре-
ле, мае и июне, после выпадения росы (табл. 104).
Представление о том, что на глубине 2 см у растений всасывающие
корни утром поглощают воду, а в остальное время суток окружены
высохшей почвой, очень маловероятно, поэтому весьма сомнительно, чтобы
1 I. Аг vidsson, В. Hellstrom, «Ass. Internal. d’Hydrologie Publ.»,
36, 1955.
2 I. Arvidsson, Plants as dew collectors, «Assembl. Gener. Toronto, 1957-
Compt. Rend, et Rapp.», 2, 1958, p. 481—484.
Т а б л и ц а ЮЗ
количество росы (по дням), выпавшей в 1955/56 г. на мысе Рас-эль-Хикма (в jx = 0,001 jwjw)1
1 Измерения проводились по методу Дувдеванп: использовалась горизонтально расположенная пластинка, количество определялось с учетом
числа и размера капель.
Египетско-аравийская пустыня, пустыни Синай и Неджеб
520
Глава 14
Таблица 104
Испарение росы и влажность почвы на глубине 0, 2 и 5 см (в % от веса
сухой почвы)
Срок на- блюде- ний, час. 1956 г.
16—17 апреля 20—21 мая 27—28 июня
0 см | 2 см 5 см 0 см 2 см 5 см 0 см 2 см | 5 слг
14 0,42 0,76 0,96 0,38 0,45 0,55 0,26 0,34 0,42
5 3,84 2,02 1,38 4,08 1,27 0,14 — — —
6 3,68 1,86 1,35 3,89 1,27 1,23 2,91 1,26 0,88
7 1,86 1,85 1,32 2,30 1,03 0,93 2,50 0,87 0,76
8 1,49 1,83 1,29 1,40 1,05 0,73 1,34 0,75 0,75
10 0,82 1,51 1,27 0,73 1,03 0,76 0,43 0,69 0,67
12 0,53 1,16 1,25 0,37 0,77 0,75 0,32 0,65 0,65
14 0,55 0,87 1,00 0,29 0,76 0,74 0,29 0,42 0,48
роса могла влиять на развитие растений х. Может быть, конденсация
влаги оказывает какое-то влияние на развитие эфемеров, поскольку
последние уже в апреле, когда в верхних 25 см почвы остается слишком
мало влаги, начинают засыхать. Смачивание листьев росой в утренние
часы и увлажнение верхних 2 см почвы могут только несколько замедлить
процесс отмирания эфемеров, который наблюдается в апреле и мае.
Конденсация влаги в более глубоких слоях песчаных почв столь незна-
чительна, что в периоды засухи она никогда не достигает величин влаж-
ности завядания; поэтому она также не может иметь никакого значения
для растений, В связи с большим количеством осадков, которые выпадают
зимой, растительность в окрестностях Рас-эль-Хикмы гораздо богаче,
чем в окрестностях Каира, и прежде всего здесь значительно больше
многолетников. Состав растительных сообществ и степень покрытия
почвы в первую очередь зависят от характера почвы 1 2,
Автор выделяет здесь несколько типов растительных сообществ,
развитых в различных условиях местообитания:
1, На каменистых почвах (степень покрытия почвы
5—10%; pH = 8,1; карбонаты кальция — 25%; гумус и соли — следы)
встречаются 3:
5 Gymnocarpos decander
5 Thymelaea hirsuta
5 Globularia arabica
4 Pituranthos tortuosus
4 Echinops spinosus
4 Dactylis hispanica
3 Helichrysum conglobatum
2 Helianthemum kahiricum.
На обращенных к морю утесах, выработанных в оолитовых извест-
няках, степень покрытия почвы растениями не превышает 1%. Растения
1 По данным Массона («Bull, Inst, Franc, Afrique Noire», 10, 1948, p, 1—181),
в прибрежной пустыне близ Дакара, где часто наблюдаются туманы, поверхностные
корни Polycarpea nivea и Sporobolus spicatus поглощают влагу росы из верхнего слоя
почвы мощностью 1 см, однако одной только этой влаги для существования этих
растений недостаточно (см, стр, 378 и след,),
2 М, М i g a h i d, A, A, A b d el R ahm a n, M, el S h a f e i, M, A, H am-
rn о uda, Types of habitat and vegetation at Ras El Hikma, «Bull, Inst, Desert Egyp-
te», 5, № 2, 1955; M, Abd el Gawad Ayyad, An ecological study of Ras El
Hikma-District, Thesis, Cairo University, 1957,
3 Встречаемость в баллах от 1 до 5.— Прим. ред.
Египетско-аравийская пустыня, пустыни Синай и Неджеб
521
встречаются здесь лишь в расселинах и вымоинах (Statice pruinosa^
Frankenia revoluta, Inula crithmoides, Atriplex portulacoides) или растут
на песке, навеянном у подножия скал,— Agropyrum junceum, Euphorbia
paralias, Nitraria retusa.
2. В аллювиальных впадинах с мощными суглинисты-
ми почвами (pH = 7,4 — 8,2; карбонаты кальция — 15—20%; гумус —
0,6/6, соли — следы; растительность отличается большей сомкнутостью
благодаря стоку воды со склонов впадин) распространены:
5 Artemisia herba alba
4 Asphodelus microcarpus
4 Noaea mucronata
3 Plantago albicans
3 Allium roseum
2 Lycium europaeum
2 Salvia lanigera
2 Stipa lagascae
2 Anabasis articulata
2 Matthiola humilis
и другие эфемеры.
На почвах, содержащих гипс (pH = 8,0—8,5) и растворимые соли
(0,34%), расчет жестколистный злак Lygeum spartum.
3. На перемещающихся песчаных дюнах (pH = 8,0—8,7;.
карбонаты кальция — 30—35%; растворимые соли — 0,2%) распро-
странены злаки: Ammophila arenaria и Agropyrum junceum’, на более ста-
рых закрепленных дюнах число видов увеличивается:
5 Ammophila arenaria
5 Echinops spinosus
3 Echium sericeum
3 Agropyrum junceum
2 Ononis vaginalis
2 Silene succulenta
2 Launea sp.
2 Cyperus capitatus
1 Centaurea pumila
1 Crucianella maritima
1 Juncus acutus
1 Pituranthos tortuosus.
На менее мощных песчаных толщах, помимо перечисленных видов,,
встречаются также растения, свойственные другим местообитаниям.
4. На засоленных почвах с грунтовыми водами, располо-
женными на глубине 80—200 см (см. цветное фото 1, стр. 480) (pH =
=8,3—8,7; карбонаты кальция — 20—30%; гумус — 0,25%; растворимые
соли — 2—5%, в том числе хлориды 0,5—3%, местами белые выцветы
солей) растут:
5 Arthrocnemum glaucum
4 Aeluropus lagopoides
3 Zygophyllum album
3 Frankenia revoluta
3 Suaeda pruinosa
1 Atriplex sp.
Весной, кроме этих видов, здесь встречается еще эфемер — Sphenopus
divaricatus. Иногда в этих условиях развивается Halocnemum strobilaceum,
а на выровненных засоленных участках — Suaeda fruticosa и Salsola
tetrandra. В засоленных микропонижениях, где в почве содержание
растворимых солей не превышает 1%, наряду с Suaeda fruticosa растут
также Mesembryanthemum crystallinum и М. nodiflorumг.
Влажные песчаные почвы на побережье моря покрыты Aeluropus
lagopoides, Frankenia revoluta, Nitraria retusa, а также растениями, свой-
ственными песчаным дюнам.
5. На болотах распространены заросли Phragmites communis.
1 Более подробные данные о различных группировках, распространенных на засо-
ленных почвах, и о почвах, развитых к западу от Александрии, приводят Тадрос
и Альта (см. сноску на стр. 516).
522
Глава 14
Таблица 105
Величина осмотического давления (W) и осмотического давления,
обусловленного хлоридами (С1), у растений в окрестностях
Бург-эль-Араба, атм
W Cl
1. Виды, развитые на слабо засо- ленных почвах Thymelaea hirsuta на скалистом гребне 34 14
Pituranthos tortuosus на песчаной дюне . 25 20
Ammophila arenaria на песчаной дюне ... 40 21
Acacia saligna (посажена) на песчаной дюне 19 12
2. Виды, встречающиеся в культу- ре Olea еигораеа (без полива) 59 9
Olea еигораеа (без полива) 52 5
Olea еигораеа (с поливом) . . 33 3,5
Olea еигораеа (с поливом) . . 22 7
Amygdalus vulgaris в саду 39 14,6
Zizuphus jujuba в саду 30 18
Ceratonia siliqua в саду 31 9
Phoenix dactylifera на кладбище . 26 13
Pinus cf. halepensis близ кладбища . 65 33
Schinus terebinthifolius на кладбище 30 20
Acacia saligna недалеко от мечети 15 7
3. Галофиты, распространенные на засоленных почвах Suaeda fruticosa близ артезианского колодца (поч- ва слабо засолена) 40 33,5
Salicornia fruticosa на засоленном болоте (слабое засоление) 79 67
Salicornia fruticosa на засоленном болоте (слабое засоление) 85 66
Salicornia herbacea (почва сильно засолена) ИЗ 102
Suaeda sp. (почва сильно засолена) ... 130 100
Halocnemum strobilaceum 116 102
Часть территории в Рас-эль-Хикме огорожена; на ней пастьба скота
запрещена. Динамика растительного покрова изучалась на 23 пробных
площадках размером 1 м1 2 каждая. За 4 года произошли следующие
изменения Ч
1) На каменистых почвах степень покрытия почвы возросла с 2 до
13,5%, в основном за счет видов, распространенных ниже по профилю.
2) В песчаных понижениях степень покрытия почвы повысилась
с 15,1 до 20 %. Особенно возросли обилие охотно поедаемого скотом
1 A. A. Abd el Rahman, М. A. Hammond a, Vegetation development
n a fenced area at Ras El Hikma in four years, «U.A.R.J. Bot.», 3, 1960, p. 1—11.
Египетско-аравийская пустыня, пустыни Синай и Неджеб 523
Ononis vaginalis и степень покрытия почвы этим видом (с 0,2 до 10,6%),
количество экземпляров за 4 года увеличилось с 113 до 778.
3) С гребней дюн исчез Echinops spinosus', зато увеличилось обилие
Helianthemum ellipticum и Crucianella maritima\ появились новые в этих
условиях виды: Echium sericeum, Zygophyllum album, Ononis vaginalis,
что, видимо, связано с прекращением вытаптывания растений животными.
4) За пределами огороженного участка растения, поедаемые скотом
{Echium sericeum, Ononis vaginalis, Lotus creticus и Echiochilon frutico-
sum), развивались значительно хуже, чем непоедаемые.
Судя по характеру растительности, в окрестностях Рас-эль-Хикмы
на глубоких суглинистых почвах развита полынная {Artemisia herba
alba) полупустыня, занимающая значительно более обширные простран-
ства в Северо-Западной Африке, а также в Палестине и Сирии (см. цвет-
ное фото 2, стр. 480 и стр. 541 и 545).
Креб 1 изучал осмотическое давление и содержание хлоридов (в кон-
це периода засухи с 27 августа по И сентября) у растений, распростра-
ненных в окрестностях станции, расположенной близ Бург-эль-Араба
в 7 км от берега моря (табл. 105). Осмотическое давление определялось
по методу, описанному на стр. 454 (при температуре 31°).
Когда изучаются виды, выделяющие соли, никогда нельзя быть
уверенным, не попадет ли в изучаемый образец уже выделенная расте-
ниями соль. Поэтому полученные величины недостаточно достоверны.
Например, осмотическое давление у Limoniastrum monopetalum равно
70 атм, а давление, обусловленное хлоридами,—37 атм, у Tamarix
articulata на засоленном болоте соответствующие величины достигают
49 и 26 атм, а на кладбище — 120 и 97 атм.
Следует обратить внимание на высокое содержание хлоридов у нега-
лофитных видов; исключением является только Olea.
ПАЛЕОКЛИМАТ ЕГИПТА
\
Исторические данные свидетельствуют о том, что в римскую эпоху
Средиземноморское побережье Ливии и Египта было относительно густо
заселено. Это подтверждают и многочисленные развалины древних
городов. Не говорит ли это о том, что климат в прошлом был более благо-
приятным, чем в настоящее время?
Эту проблему изучал Бутцер; он провел анализ геологических,
геоморфологических, исторических и археологических материалов 1 2.
За последние 300 тыс. лет климат на территории Северо-Восточной
Африки неоднократно изменялся. Вполне вероятно, что в палеолите
существовали более благоприятные климатические условия, причем
влажные и прохладные периоды неоднократно сменялись засушливыми
и жаркими. При годовом количестве осадков около 200 мм и несколько
более низкой среднегодовой температуре в условиях современной пусты-
ни могла произрастать растительность, сходная с современной расти-
тельностью Киренаики. В пустыне в то время еще жил человек; об этом
свидетельствуют многочисленные следы материальной культуры, обна-
1 К. К г е е b, Untersuchungcn zum Wasserhaushalt der Pflanzen unter extrem
ariden Bedingungen, «Planta», 59, 1963, S. 442—458.
2 K. W. В u t z e r, Studien zum vor- und friihgeschichtlichen Landschaftswandej
der Sahara. III. Die Naturlandschaft Agyptens wiihrend der Vorgeschichte und der dyna-
stischen Zeit: «Abh. Math.-Naturw. KI., Akad. d. Wiss. Mainz, Jahrg. 1959», № 2,
S. 44—122. Здесь приведена библиография по излагаемому вопросу.
524
Глава 14
ружейные даже в центральной части Ливийской песчаной пустыни. Уже
в эпоху палеолита количество выпадавших здесь осадков уменьшилось;
в связи с этим местное население переселилось из пустыни ближе к Нилу,
долина которого в то время не была еще столь глубоко врезана. В связи
с изменением гидрологического режима Нила в долине реки началось
отложение ила, приносимого летними паводками с Абиссинского нагорья.
По Бутцеру, аридные климатические условия существуют здесь пример-
но 18 тыс. лет. В те времена в галерейных, пойменных и заболоченных
лесах еще водилась разнообразная дичь. Местные жители питались также
рыбой и устрицами.
На дальнейшее изменение конфигурации долины Нила повлияло
повышение уровня Средиземного моря; в результате таяния континен-
тальных льдов уровень моря повысился более чем на 50 м; примерно
в 3—4 тысячелетии до н. э. уровень достиг наивысшей отметки; в это
время он на 4 м превышал современный. В долине Нила речная эрозия
сменилась аккумуляцией рыхлого материала. Мощность речных наносов
выше Каира составляет в среднем 9,7 м, а между Куэ и Асуаном — 6,7 м.
Примерно 60% ила отложилось в период до 3000 лет до н. э. В последую-
щие годы уровень моря неоднократно колебался и за 2500—1500 лет
до н. э. располагался на 2 м выше, а за 400 лет до н. э. на 2,5 м ниже
современного; далее к 100 г. до н. э. он повысился на 50 см и только-
в третьем столетии н. э. достиг современной отметки.
Климат в эпоху неолита, вероятно, также был несколько более
влажным, чем в настоящее время; в это время в пустыне, в долинах пере-
сыхающих рек, возможно, еще существовали разреженные древостои.
По Бутцеру, субплювиальный период продолжался с 5000 до 2350 г. до н. э.
Только начиная с эпохи шестой династии фараонов здесь господ-
ствуют сходные с современными засушливые условия. Со времени суще-
ствования Древнего царства до примерно 500 г. до н. э. климат оставался
крайне засушливым. Уже на рельефах периода Древнего царства отсут-
ствуют изображения слонов, носорогов и жирафов, а силуэты львов
и гривастых баранов появляются все реже. В период Среднего и Нового
царств в сценах охоты уже не изображается степная растительность. На
рельефах Нового царства еще встречаются рисунки представителей болот-
ной фауны — бегемотов, крокодилов, первобытных быков, различных
видов птиц, а также рыб.
Период с 500 г. до н. э. до 300 г. н. э. характеризуется высокими
паводками. В результате дюны, окаймляющие долину Нила в западной
части Среднего Египта, были покрыты илом. Это создало довольно благо-
приятные условия для развития земледелия в долине Нила, несмотря
Таблица 106
Период (годы н. э.) Показатели изменений климата
300-800 Меньшее число наводнений, интенсивное накопление песка
800-1200 1200-1450 1450-1700 Вновь частые и сильные паводки Засушливые условия, образование дюн Сильные наводнения, но местами сохраняются засушли- вые условия
После 1700 Образование современных дюн в Среднем Египте
Египетско-аравийская пустыня, пустыни Синай и Неджеб
525
на то, что климат не изменился (если не считать некоторых незначи-
тельных колебаний климата, представленных в табл. 106).
В общем нет никаких достоверных данных, свидетельствующих
о том, что после окончания субплювиального периода в течение эпохи
неолита (продолжавшейся еще во время постройки пирамид) климат
претерпел существенные изменения. В прошлом количество осадков было
не больше, чем в настоящее время. Нельзя недооценивать хозяйственную
деятельность человека в течение последних двух тысячелетий и прежде
всего ущерб, нанесенный пастьбой скота, особенно ощутимый там, где
растительность плохо возобновляется в период длительной летней засу-
хи (см. стр. 310).
ХАРАКТЕРИСТИКА ЭКОЛОГИЧЕСКИX УСЛОВИЙ
НА СИНАЙСКОМ ПОЛУОСТРОВЕ
Синайский полуостров простирается к северу от Суэцкого канала
до Рафы или Газы. На юге он ограничен Суэцким заливом и заливом
Акаба. На Синайском полуострове выделяются: а) узкая прибрежная
полоса, переходящая на юге в б) плато Эт-Тих, и в) высокогорная область,
занимающая южную треть полуострова (рис. 377).
Прибрежная полоса на севере Синая резко отличается от западной
части побережья Египта. Здесь на больших площадях распространены
высокие дюны (высота до 40—50 ж), которые, однако, сравнительно слабо
подвижны. Правда, асфальтированное шоссе, идущее из Эль-Кантары
(у Суэцкого канала) в Эль-Ариш, местами слегка заносится песком, но
его очистка, так же как и защита железнодорожной линии, не представ-
ляет больших трудностей.
По сравнению с остальной частью побережья Средиземного моря на
побережье Синайского цолуострова выпадает наименьшее количество
осадков; на побережье между Порт-Саидом и Эль-Аришем выпадает
менее 100 мм осадков. Далее к северо-востоку количество осадков быстро
возрастает (Эль-Ариш — 100 мм, Рафа — около 200 мм, Газа — около
400 мм). Поскольку перевеваемые ветром дюны почти полностью лишены
растительности, примерно 40% выпавших осадков накапливается в более
глубоких слоях песка или питает подземные воды. Поэтому под слоем
песка по направлению к морю движется йоток грунтовых вод (по расчетам
на 1 км берега ежедневно в море поступает 50—164 м3 воды \ Очень
часто этот водоносный горизонт пресной воды вблизи берега располагает-
ся над слоем более тяжелой соленой воды, поступающей в результате
инфильтрации из моря. В междюнных понижениях уровень грунтовых
вод расположен на глубине всего нескольких метров от поверхности,
что позволяет выращивать здесь финиковые пальмы. Для этой цели
используют стеблевые отпрыски, которые закапывают в песок так, чтобы
их нижний конец соприкасался с влажной почвой, а на поверхности
оставались бы лишь связанные в пучок концы листьев. После образова-
ния придаточных корней начинается рост побегов. При высоком уровне
грунтовых вод поверхность почвы постоянно увлажняется благодаря
капиллярному поднятию воды; в результате испарения воды на поверх-
ности почвы накапливаются соли. В этих случаях в междюнных пони-
жениях развивается галофитная растительность, в составе которой в наи-
1 G. L. Paver, J. N. J о г d a n, Report on reconnaissance of hydrological and
geophysical observations in North Sinai coastal area of Egypt, Publ. Inst. Desert Egyp-
te, № 7, Mataria, 1956.
526
Глава 14
более глубоких частях депрессий встречаются Arthrocnemum, Halocnemum,.
Salicornia, Juncus acutus, Suaeda, несколько выше — Nitraria, Tamarix,
Limonium и, наконец, там, где влияние засоления уже не сказывается, —
Zygophyllum и Thymelaea, Часто. здесь образуются настоящие соле-
ные озера.
На берегу моря финиковые пальмы обычно растут настолько близко
к полосе прибоя, что в шторм волны докатываются по песку до основания
стволов. Создается впечатление, что деревья растут в море. В действитель-
ности же корни деревьев расположены в пресноводном водоносном гори-
зонте (рис. 390, см. также цветное фото 2, стр. 512). Если бы дело обстоя-
ло иначе, то можно было бы поливать пальмовые рощи морской водой.
Дюнный песок после дождя содержит некоторое количество пленоч-
ной влаги, которое из-за грубозернистости песка очень невелико. Однако
местное население умело использует эту воду для выращивания одно-
летней клещевины. Весной, когда песок еще сохраняет влажность после
зимних дождей, на обращенных к морю склонах дюн высевают семена
клещевины. Корни растений постепенно углубляются во влажную почву
и поэтому непрерывно поглощают влагу вплоть до периода, когда созре-
вают семена, даже если после посева больше не выпадет осадков. Обяза-
тельным условием является, конечно, малая подвижность песка; в случае,
если песок не перевевается, корни растений не обнажаются. По наблюде-
ниям автора, клещевина в этих условиях хорошо плодоносит (см. цветную
вклейку I, стр. 512). Вдоль железнодорожного полотна для защиты
от песчаных заносов сажают Acacia saligna и Eucalyptus', они также растут
без всякого полива.
На побережье в устье вади Эль-Ариша расположен наиболее круп-
ный населенный пункт полуострова — Эль-Ариш. Площадь водосборного
бассейна составляет 22 500 км2, или почти половину всей Синайской
области. На южной части водораздела этой речной системы выпадает,
вероятно, не более 25 мм осадков, поэтому русло вади лишь очень редко
наполняется водой. В 1948 г. через плотину Руафа (в 50 км от побережья)
прошел 21 млн. м3 воды, а в 1951—3 млн. м3; в последующие годы русло
в створе плотины оставалось сухим. Тем не менее в низовьях вади наблю-
дается приток грунтовых вод (частично из района распространения дюн);
это дает возможность вести в Эль-Арише, хотя и в небольших масшта-
бах, поливное земледелие. В садах выращивают яблони, груши, гуайяву
(Psidium guajava), инжир, масличную пальму и цитрусовые, а в самом
русле зимой — зерновые культуры. Одиночными группами по краям
вади растет Tamarix,
По направлению в глубь материка песчаные дюны переходят в гамаду.
Степень покрытия почвы растительным покровом не превышает здесь
10%; в его составе встречаются: Zygophyllum dumosum, Thymelaea hirsuta,
Artemisia monosperma и Haloxylon salicornicum, а на легких песчаных
почвах — Panicum turgidum.
Монотонность гамады нарушают одиночные столовые горы, дости-
гающие высоты 1087 м. По данным Кассаса, побывавшего на столовой
горе Рас-эль-Ахмар, расположенной в 20 км от побережья (абсолютная
высота 256 м, высота над уровнем пустыни — 100 м), поверхность скал
на вершине покрыта лишайниками (проективное покрытие достигает
90%), что свидетельствует о том, что здесь часто выпадает роса или наблю-
даются туманы. Здесь распространены упомянутые выше цветковые
растения. В верхней части склонов горы преобладает Anabasis articulata,
а на песчаных участках у подножия наряду с Panicum turgidum растет
Египетско-аравийская пустыня, пустыни Синай и Неджеб
Рис. 390. Схема распределения грунтовых вод на побережье моря близ Эль-Ариша
и культур финиковой пальмы и клещевины.
1 — поток пресных грунтовых вод; 2 — поток соленых грунтовых вод, поступаю-
щих из моря. В первой междюнной котловине видны заросли галофитов.
также Aristida scoparia. Столовая гора Гебель-эль-Магхара (высота 750 м)г
сложенная из юрских пород, расположена примерно в 100 км к юго-
западу от Эль-Ариша. На станции Эль-Хамма, у юго-восточной подошвы
горы, выпадает (обычно в январе и феврале) 90 мм осадков. Було соста-
вил список видов, распространенных на гребне горы протяженностью
45 км и шириной 20 км х. Среди них встречаются Caralluma sinaicar
Sedum cf. viguieri, J uniperus phoenicea, Ephedra alata, виды Astragalus,
Colutea, Iris sisyrinchium, многочисленные лилейные и др. Здесь растет
также папоротник Notholaena vellea', кроме того, здесь встречается доволь-
но много средиземноморских видов: Lavandula pubescens, Teucrium polium,
Thymus bovei, Globularia arabica и др. Таким образом, на скалах создают-
ся благоприятные для существования растений условия. Количество осад-
ков, выпадающее на вершинах гор, вероятно, также несколько больше.
Центральную часть Синайского полуострова занимает плато Эт-Тих,
расположенное на высоте (в среднем) 700 м над уровнем моря.
Количество осадков на станции Нехль (высота 400 м) составляет всего*
26 мм. Состав развитой здесь растительности мало отличается от состава
растительности египетско-аравийской пустыни, распространенной на за-
паде, и пустыни Неджеб — на востоке от Синайского полуострова.
Зохари, дважды пересекший эту территорию, составил краткий
обзор растительности Синайского полуострова1 2. Каменистая пустыня
почти лишена растительности. На скоплениях песка, в понижениях,
руслах временных водотоков, в вади встречаются группировки, состав
которых сильно варьирует в зависимости от мощности слоя песка, рель-
ефа местности, трещиноватости пород и т. д. Обычно здесь встречаются
Anabasis articulata, Zygophyllum dumosum, Zilla spinosa, Reaumuria hir-
tella, Noaea mucronata, Panicum turgidum, Haloxylon salicornicum (мес-
тами доминирует). На закрепленных или малоподвижных песках рас-
пространены Aristida scoparia и A. plumosa, Artemisia monosperma,
Retama retam, Panicum turgidum. В пересыхающих руслах долин,
впадающих в Суэцкий залив, наряду с акацией широко распространены
Ephedra alata и Tamarix mannifera, выделяющий при укусах насекомых
сладкий засыхающий сок, так называемую манну пустыни 3.
1 L. В о u 1 о s, Flora of Gebel El-Maghara, North Sinai, Agric. Ext. Dep., Herbar.
Sect., Cairo, 1960. Тот же автор в 1959 г. опубликовал список растений, собранных
близ Газы. Р. Range, Die Flora der Isthmuswiiste «Ges. Pal.-Forsch.», 7, Berlin, 1921.
2 M. Z о h a r y, Vegetational transects through the desert of Sinai, «Palestine
J. Bot.», Jerus, Ser. 3, 1944, p. 57, 78.
3 Сведения, что «манна» в Синайской пустыне — это скопление приносимых
ветром стеблей Lecanora esculenta, по сообщению V. Taekholm, не соответствуют дей-
ствительности.
528
Глава 14
При неглубоком уровне засоленных грунтовых вод типична
Nitraria retusa, вокруг которой ветер навевает песок. Наряду с Nitraria
здесь распространены Tamarix articulata и Т. tetragyna, а также виды
Zygophyllum. На засоленных влажных почвах растет Suaeda vermi-
culata.
Южная часть Синайского полуострова образована горами, сложен-
ными эруптивными породами (большей частью гранитами); четыре верши-
ны превышают высоту 2 тыс. л; из них Гебель-Муса достигает высоты
2285 м, а наивысшая точка — пик Катерин — 2641 м. При перелете из
Суэца в Акабу с самолета хорошо видны эти вершины, высоко вздымаю-
щиеся над облаками.
В последнее время растительность Синайского полуострова, кроме
Зохари, изучали также Мигахид и др.
Горный массив образует подобие треугольника с вершиной, обра-
щенной на юг. На севере он отделен от плато Эт-Тих песчаной равниной,
подстилаемой нубийскими песчаниками. На станции Тор, в южной части
Суэцкого залива, за год выпадает 13 мм осадков, а испаряемость дости-
гает 3500 мм. В горах зимой снег лежит очень долго. Среднегодовое
количество осадков в горах пока не установлено. Зохари на основании
характера растительного покрова считает, что здесь выпадает 300 мм,
а метеорологи — не более 50 мм. Во всяком случае, редкие дожди на
короткий срок превращают реки в стремительные потоки. Осадков
здесь выпадает достаточно, чтобы питать источники у подножия гор.
Близ одного из них расположен известный монастырь св. Екатерины.
Во влажных ущельях встречаются кустарники и изредка деревья; среди
них —Ephedra alata, Crataegus sinaica, Moringa peregrina, Ficus pseudo-
sycomorus, F. carica var. rupestris, Cupressus sempervirens. На заболоченной
почве и вокруг заполненных водой ям растут Scirpus holoschoenus, Juncus
arabicus, Veronica anagallis, Equisetum ramosissimum, Mentha microphylla,
Origanum syriacum и даже Adiantum capillus veneris. В орошаемых садах
наряду с финиковыми пальмами, маслинами и гранатами выращивают
также миндаль, персики, сливы, яблони, груши и виноград.
В горах выражена поясность растительности.
На высоте около 100G м господствуют Artemisia Judaica и Zilla spi-
nosa', несколько выше появляется Artemisia herba alba, сопутствуемая
целым рядом ирано-туранских видов, хорошо переносящих низкие зимние
температуры. Лишь на высоте 1600 м флористический состав резко
меняется, причем в составе покрова появляются также и средиземномор-
ские виды. На этой высоте доминируют Phlomis аигеа и Pyrethrum santoli-
noides\ на высоте 2000 м Phlomis вытесняется Artemisia herba alba. Вместе
с полынью здесь встречаются Роа sinaica, Nepeta septemcrenata и др.
Изолированное положение Синайских гор способствует развитию
эндемизма; здесь распространены следующие эндемы: Galium sinaicum,
Anarrhinum pubescens, Phlomis aurea, Thymus decussatus, Nepeta septem-
crenata, Otostegia sinaitica, O. kaiseri, Convolvulus spicatus, Centaurium,
malzacianum, Primula boveana, Rosa arabica, Cotoneaster orbicularis, Arabi-
dopsis kneuckeri, Dianthus sinaicus, Silene leucophylla, S. schimperiana
и др. Это, вероятно, реликты средиземноморского происхождения.
1 А. М. М i g a h i d, M. el Shafei All, A. A. Abd el Rahman,
M. A. H a m m о u d a, Ecological observations in Western and Southern Sinai, «Bull.
Soc. Geogr. Egypte», 32, 1959, p. 165—206.
Египетско-аравийская пустыня, пустыни Синай и Неджеб
529
ЭКОЛОГИ ЧЕСКИ Е ИССЛЕДОВАНИЯ,
ПРОВОДИВШИЕСЯ В ПУСТЫНЕ НЕДЖЕБ
Северная граница пустыни Неджеб проходит между Газой, располо-
женной на берегу Средиземного моря, и Мертвым морем; на юг эта пусты-
ня простирается вплоть до залива Акаба. Осадки выпадают здесь с октя-
бря по апрель; на севере пустыни в среднем выпадает 150 мм, а на юге —
20 мм 1. Средняя температура самого холодного месяца (январь) дости-
гает на севере пустыни 11,5°, а самого теплого (августа) —27,3°. Сред-
няя относительная влажность воздуха колеблется от 37% (в июле) до 55%
(в январе). Роса выпадает в течение 163 дней в году, преимущественно
в летние месяцы. Туманы бывают очень редко. Таким образом, климат
на севере пустыни сохраняет некоторые средиземноморские черты; осталь-
ная часть территории напоминает север Синайского полуострова (рис. 391).
На плоскогорьях развита почти лишенная растительности гамада, где
изредка встречается Reaumu-
ria palaestina. На каменистых
склонах вади, достигающих
высоты 2—3 м, преобладает
Zygophyllum dumosum. На
террасах вади растет Haloxy-
lon articulatum, а по руслу
Retama retam (рис. 392).
Экология растений каме-
нистой пустыни, где преобла-
дает Zygophyllum dumosum,
изучалась в период летней
засухи в августе 1961 г. в 30 км
южнее Беершебы1 2. Среднее
количество осадков состав-
ляет здесь 80 мм.
На пробной площадке
размером 100 м2 встречались
следующие растения: 39 эк-
земпляров Zygophyllum dumo-
sum , 8 — Reaumuria palaestina,
2 —Atractylis serratuloides и
258 однолетних экземпляров
Salsola inermis. Данные о рас-
ходе на транспирацию всем
растительным сообществом,
за исключением расхода влаги
Reaumuria и Atractylis, при-
водятся в табл. 107.
P ii c. 391. Карта растительности пустыни Пед-
жеб (по Зохарп).
1 — Retametea; 2 — Anabasidctca (Zygophyllelalia);
3 — Arteinisietea; •/ — Anabasidetca (Anabasic'.etalia);
5 — Haloxylolea; 6 — Suaecletea.;
1 D. Z о h а г у, Vegetation
map of the Central and Southern
Negev., «Palestine J. Dot.», Jerus,
Ser. 6, 1953, p. 27—36.
2 D. Z о h a r y, G. О rsh a n,
The Zygophylletum duinosi and its
hydroccology in the Negev of Israel,
«Vegetatio», 5/6, 1954, p. 341—350.
530
Глава 14
Рис. 392. Схема распределения растительности по поперечному
профилю вади (по Зохари и Оршанскому).
Если дневную продолжительность транспирации принять равной
10 часам, то расход влаги на транспирацию растительным сообществом
достигнет 1,1 кг в день, или 30 л в месяц.
Таблица 107
Расход влаги на транспирацию в период летней засухи
(август 1951 г.)
Название растений Средняя ин- тенсивность транспира- ции, мг/г • час Вес всей листовой массы, г Расход влаги, г/час
Zygophyllum dumosum 72 875,5 60,4
Salsola inermis . . 192 256,4 49,3
Всего 1131,9 109,7
Хотя прорастание Salsola inermis (однолетник) после дождей про-
исходит в течение ноября — февраля, развитие ее, однако, начинается
только в марте и кончается в сентябре. Вместо крупных зимних листьев
летом появляются небольшие по размерам листья. Летние однолетники
распространены только там, где растительный покров нарушен и где
в связи с этим в почве сохраняется влага; при наличии более сомкнутого
покрова они после прорастания не выдерживают конкуренции за почвен-
ную влагу с зимними однолетниками, а летом подавляются многолетни-
ками. На площадях, где развивается только Salsola inermis, из года в год
увеличивается сомкнутость травостоя; это происходит до тех пор, пока
растения не начнут ослабевать, а затем погибать вследствие внутри-
видовой конкуренции (см. стр. 39) Ч Однолетники в этой области, где
осадки выпадают зимой, представлены только рудеральными растениями.
Определение расхода воды в зарослях Zygophyllum было повторено
весной 1952 г. Кроме того же числа экземпляров Zygophyllum, на площадке
встречалось значительно больше эфемеров; в связи с этим увеличился
и расход воды (табл. 108).
Zygophyllum сбрасывает летом свои перистые листья и сохраняет
только суккулентные черешки; в связи с этим летом у Zygophyllum пло-
щадь транспирирующей поверхности сокращается по сравнению с весен-
ним периодом более чем в 7 раз. Кроме того, весной интенсивность транс-
пирации (при расчете на 1 г сырого веса) увеличивается в 2,75 раза по
сравнению с летними величинами; в результате только один Zygophyllum
1 М. N е g b i, М. Е v е n а г i, The means of survival of some desert summer annu-
als, «Proc. Madrid Symp., Unesco Arid Zone Res.», 16, 1961, p. 249—259.
Египетско-аравийская пустыня, пустыни Синай и Неджеб
531
расходует в марте в 20 раз больше воды, чем в августе. Добавив к этим
величинам количество воды, израсходованной однолетниками, получим,
что в марте растительный покров расходует на транспирацию в 30 раз
больше влаги, чем в августе. Расход воды весной составляет в день 32,4 кг,
а в месяц — около 1000 л.
Таблица 108
Расход влаги на транспирацию весной (31 марта) 1952 г.
Название растений Средняя ин- тенсивность транспира- ции, мг/г • час Вес всей листовой массы, г Расход влаги, г!час
Zygophyllum dumosum 198 6448,7 1276,8
Эфемеры 297 6610,0 1963,2
Всего 13 058,7 3240,0
На основании определения влажности почвы выяснилось, что весной
даже верхние слои почвы хорошо увлажнены, тогда как летом (с учетом
засоленности почвы) доступная для растений влага находится только
в более глубоких горизонтах
почвы (начиная с глубины
60 см). На основании расчет-
ных данных, суммарное испа-
рение в зимние месяцы до-
стигает примерно 6400 л,
а летом —только около 300 л,
что хорошо согласуется с вы-
шеприведенными величинами
расхода влаги на транспира-
цию. Расход влаги на транс-
пирацию за год составляет
60—70 мм, причем в засу-
шливый летний период эта
величина не превышает 2 мм.
Поскольку в почве содер-
жатся легкорастворимые соли,
I
Таблица 109
Величина осмотического давления у растений,
развитых в гамаде в районе Беершебы
Название растений Осмотическое давление, атм
23/VIII 1951 г. 31/III 1952 г.
Zygophyllum dumosum 72,5 40,0
Salsola Inermls . Gymnarrhena micran- 116,8 59,5
tha Нет данных 39,5
упомянутые выше виды дол-
жны быть отнесены к числу ксерогалофитов. Об этом свидетельствуют
также и высокие значения осмотического давления этих растений
(табл. 109).
Содержание хлоридов в клеточном соке не определялось.
Растения могут развиваться в крайне неблагоприятных условиях
существования в гамаде только благодаря способности резко сокращать
расход влаги в летние месяцы. Это достигается путем
1) значительного уменьшения растительной массы, связанного с отми-
ранием эфемеров;
2) сильной редукции транспирирующей поверхности листьев, осу-
ществляемой при сбрасывании перистых листьев у Zygophyllum',
3) сокращения интенсивности транспирации на х/3, что происхо-
дит, несмотря на увеличение испарения с поверхности почвы.
532
Глава 14
В связи с большим количеством осадков, выпадающим на севере
пустыни Неджеб, вади часто бывают заполнены водой. Даже при 10 мм
осадков (при площади водосбора, равной 50 км1 2) расход воды в вади дости-
гает 30 тыс. м^/час. Значение пересыхающих долин рек для хозяйства
очень невелико, если в них не перекрыть сток и не приступить к накоп-
лению воды в почве. Соответствующие опыты были проведены в пустыне
Неджеб Ч Уже в верховьях в долине сооружается каскад плотин, что
облегчает проникновение воды в почву и обеспечивает развитие более
сомкнутого растительного покрова. В крупных долинах сооружают
плотины с верхним сбросом воды. Накопленные воды используют для
разведения поливных культур на защищенных от паводков террасах.
Конечно, эти культуры можно возделывать только в том случае, если вади
будут хотя бы раз в год наполняться водой.
Наиболее крупное вади в восточной части пустыни Неджеб (Вади-
Араба) достигает ширины 5—20 км и является продолжением Иордан-
ского грабена. Вади-Араба впадает в Мертвое море на высоте 396 м ниже-
уровня моря. Водораздел расположен на высоте 240 м выше уровня
моря, в 70 км от залива Акаба. Экологические исследования проводились
на западном склоне Иорданской впадины 2 и на северном берегу Мертвого
моря 3. Эта наиболее глубокая депрессия на земном шаре характеризует-
ся пустынным климатом; годовое количество осадков составляет здесь
65—140 мм] средняя месячная температура в январе равна 9,8°, в июне —
38,1°. В Джерихо средняя годовая температура достигает 23,1°. Все
почвы (за исключением некоторых оазисов) сильно засолены. Выделяются:
1) автогенные, или автоморфные, засоленные почвы, сформировавшиеся
в процессе выветривания соленосных горных пород; наибольшее количе-
ство солей наблюдается в этих почвах на определенной глубине; 2) гидро-
генные, или гидроморфные, засоленные почвы; соли выпадают на поверх-
ности почвы вследствие испарения засоленной грунтовой воды.
У некоторых видов, представленных здесь, определялась величина
осмотического давления. Наиболее низкое осмотическое давление было
обнаружено у глубоко укореняющихся растений, корни которых, однако,
не достигают уровня засоленных грунтовых вод (Prosopis jarcata и Alhagt
maurorum). Наоборот, у всех галофитов осмотическое давление превыша-
ло 25 атм, а иногда поднималось даже выше 100 атм. Выявлена прямая
зависимость между величиной осмотического давления у растений
и концентрацией солей в почвенном растворе.
У галофитов на постоянно влажных почвах колебания осмотического
давления в течение года невелики, чего нельзя сказать о Salsola tetran-
dra (рис. 393).
1 М. Е v о п а г i, D. К о 1 1 е г, Desert agriculture problems and results in Israel,
The future of arid lands, Washington, 1956, p. 390—413.
2 M. E venari. Root condition of certain plants of the wilderness of Judaea,
«J. Linn.. Soc.», 51, 1938, p. 382—388; The physiological anatomy of the transpiratory
organs and the conducting systems of certain plants typical of the wilderness of Judaea.
«J. Linn. Soc.», 51, 1938, p. 389—407; M. E v e n a r i, R. Richter, Physiological-
ecological investigations in the wilderness of Judaea, «J. Linn. Soc.», 51, 1938.
p. 333-381.
3 M. Z о h a r y, A vegetation map of western Palestine, «J. Ecol.». 34, 1947.
p. 1 —19; E. Shmuel i, Ecological investigations in Palestine. II. The water balance
of some plants of the Dead Sea salty soils, «Palestine J. Rot., Jerus. Ser.», 4,, 1948,
p. 117—143; M. Z о h a r y, G. Or s h a n s k y, Structure and ecology of the vegeta-
tion in the Dead Sea region of Palestine, «Palestine J. Rot., Jerus. Ser.», 4, 1949,.
p. 177 — 206,
Египетско-аравийская пустыня, пустыни Синай и Неджеб
533
Несколько южнее, в окре-
стностях Вади-Араба, климат от-
личается еще большей засушли-
востью; годовое количество
осадков снижается с 60 мм на
севере до 20 мм на юге. Темпе-
ратура колеблется от 3 до 45е,
а относительная влажность —
от 20 до 60% . Только в долинах
пересыхающих рек почвы слабо
засолены Ч
Зохари и Оршан проводили
исследования в двух типах сооб-
ществ: 1) Acacia — Anabasis;
встречается в слабоврезанных
вади и руслах временных водо-
токов, пересекающих гамаду в
пределах южного отрезка Вади-
Араба, и 2) Haloxylon persicum;
встречается на песчаных равни-
нах и дюнах 1 2.
Ш 2F Г ЕГ Ж И 2Z X Ж ХЕ I X
Р и с. 393. Годовой ход осмотического давле-
ния у некоторых галофитов, распространенных
в окрестностях Мертвого моря (по Зохари и
Оршанскому).
Аг.— A)Инос нетит glaucum', An.— Anabasis arti-
culala', Si).— Suaeda palaestina', S.t.— Satsota teti'an-
dia; S.v.— Salsola villosa.
1. В почвенном профиле под зарослями акации, начиная уже с глу-
бины 1 ж, обнаружена доступная для растений влага; установлена макси-
мальная влажность почвы на глубине 2 ж, (6,1%), а на глубине 2,5 м она
снижается до 2,4%. Содержание солей в почве незначительно. Корни
акации проникают до глубины 1,5 м. В течение года интенсивность транс-
пирации определялась 8 раз у следующих растений: Acacia spirocarpa,
A. raddiana, Anabasis articulata, Ochradenus baccatus, Lycium arabicum,
Haloxylon salicornicum. Обе акации отличались очень большой интенсив-
ностью транспирации, причем интенсивность транспирации не понижа-
лась даже летом. Среднесуточные значения интенсивности транспирации
колебались в течение года от 500 до 1000 мг!г-час. В августе в полуденные
часы у Acacia spirocarpa интенсивность транспирации превышала даже
2500 мг!г-час. Следовательно, водный баланс сохранялся на постоян-
ном уровне так, же, как и осмотическое давление, величина которого на
протяжении всего года колебалась около115 атм. В условиях, где рас-
пространена акация, почвы лишь слабо засолены.
Интенсивность транспирации у четырех остальных менее высоких
кустарников всегда была значительно меньше (рис. 394). Водный баланс
у сбрасывающего летом листья Ochradenus (Resedaceae) сохранялся на
постоянном уровне, поскольку величина осмотического давления у этого
вида в течение года не превышала 20 атм, тогда как у других видов
отмечалось очень сильное повышение давления до 50 атм (рис. 395).
Lycium также теряет листья летом, что, вероятно, объсняется меньшей
активностью корней. Возможно, что Lycium, Anabasis и Haloxylon отно-
сятся к числу соленакапливающих галофитов, однако пока это еще не
1 R. К а г я с h о n. The hamada in Wadi Araba its properties and possibilities
of afforestation, Ilanoth (Israel). 2, 1953, p. 19—45. Здесь приведены результаты поч-
венных анализов.
2 М. Z о li а г i, Outline of the vegetation of Wadi Araba, «J. Ecol.», 32, 1945,
p. 204—213; Ecological studies in the vegetation of Near Eastern Deserts. [. Envi-
ronment and vegetation classes, «Israel Exploration J.», 2, 1952, p. 201—215;
M. Z oh ary, G. Orschan, Jbid.. П, Wadi Araba «Vegetatio», 7, 1957, p. 15—37.
534
Глава 14
Рис. 394. Колебания средне-
часовых значений интенсивно-
сти транспирации у видов, рас-
тущих в зарослях акации в юж-
ной части Вадп-Араба (по Зо-
харп и Оршану).
As.—Acacia spirocarpa', Ar.— Aca-
cia raddiana', L.— Lycium arabicum',
H.— Haloxylon salicornicum\ O.—
Ochradenus baccalus; An.— Anaba-
sis arliculala.
вполне ясно, поскольку у них не опреде-
лялось содержание хлоридов в клеточном
соке.
2. Haloxylon perslcum развивается в не-
сколько иных условиях влагообеспеченности.
Haloxylon perslcum (Chenopodiaceae) — это
дерево высотой 3—4 м с членистыми гибкими
суккулентными побегами. Haloxylon встреча-
ется вместе с безлистным кустарником Calll-
gonum comosum (Polygonaceae). Это элементы
ирано-туранской флоры; они в основном рас-
пространены в песчаных пустынях Централь-
ной Азии. По данным Васильева, эти виды
в Туркмении отличаются наиболее высокими
из полученных до настоящего времени зна-
чениями интенсивости транспирации *. Однако
на следующий год Кокина установила, что
эти величины были завышены в 2—3 раза.
Кроме того, в этих условиях изучались еще
два вида —Retama retam и Zilla spinosa —
оба они являются элементами сахаро-аравий-
ской флоры.
Как известно, песчаные почвы в пустыне
лучше всего обеспечены влагой. Однако на-
блюдения показали, что песок здесь до глу-
бины 1,2 м не содержал доступной для рас-
тений влаги. Только в марте влажность
почвы на глубине 30 см составляла 2,4%. Корни всех четырех изученных
видов проникают в почву очень глубоко и черпают влагу из глубоких
постоянно увлажненных слоев почвы, в связи с чем и отличаются высо-
кой интенсивностью транспирации (при пересчете на сырой вес). Так,
интенсивность транспирации у Haloxylon perslcum достигает в июне
700 мг!г-час. Летом эти виды сбрасывают листья, в связи с чем интен-
Таблица ПО
Интенсивность транспирации у растений, растущих в зарослях Haloxylon
persicunv в мг< час 1
Дата Ha loxylon persicum Calligonum comosum Relama relam Zilla spinosa
стебель листья
F w F w F w F w F 1 w
24/111 396 456 792 402 — — — 954 1794
27/IV 654 978 522 738 510 852 606 774 870 996
20/VI 696 — 462 — 642 — 432 — — —
15/VIII 558 882 270 570 372 996 324 618 — —
16/IX 444 750 300 462 234 1038 294 546 — —
28/X 318 516 192 282 210 420 384 552 — —
23/1 84 120 138 222 186 408 192 180 450 564
1 F — в пересчете на 1 г сырого веса, W — в пересчете па 1 г воды.
1 J. М. Vassiljev, ЁЬег den Wasserhaushalt der Pflanzen der Sandwuste
im sudostlichen Karakum, «Planta», 14, 1931, S. 225—309.
О-----------О 1
▲----------А 2
о----------□ 3
Рис. 395. Годовой ход осмотического давления у неко-
торых видов, растущих в зарослях акации в южной
части Вади-Араба (по Зохари и Оршану).
1 — Acacia raddiana', 2 — Anabasis articutata; 3 — Lycium
arabicum; 4 — Ochradenus baccatus; 5 — Hatoxyton saticornicum.
Рис. 396. Годовой ход осмотического давления у ви-
дов, растущих в зарослях Haloxylon persicum (по Зохари
и Оршану).
1 — Hatoxyton persicum', 2 — Retama retam', 3 — Zitta spinosa
4 — Cattigonum comosum.
536
Глава 14
сивность транспирации у них несколько снижается (табл. 110); летом
ассимиляция осуществляется зелеными стеблями.
Осмотическое давление у Zilla й Calligonum немного больше 10 атм.
У Retama осмотическое давление в августе повышается до 30 атм',
Эфемера
Lycium Zygophyllum
Retama
Noaea
Haloxylon persicun.
Рис. 397. Морфологические типы растений засушли-
вых областей Ближнего Востока, отличающиеся по ха-
рактеру водного режима (по Зохари).
Косой штриховкой показаны ассимилирующие части расте-
нии, отмирающие при недостатке влаги; белым — части расте-
ний, сохраняющиеся :период засухи; точками — неассимили-
рующие части растений.
у Haloxylon весной эта величина составляет около 30 атм, а затем непре-
рывно увеличивается и достигает 56 атм', как ни странно, но это наблю-
дается в январе (рис. 396). Вполне возможно, что такое большое осмоти-
ческое давление связано с накоплением солей в более старых частях
тканей (в результате интенсивной транспирации).
Если попытаться составить биологический спектр этой пустынной
области, то окажется, что жизненные формы Раункиера для этой цели
не годятся. В условиях пустыни важно не положение почек возобновления,
а способ перенесения засухи. Поэтому Зохари выделяет несколько групп
(типов) растений, отличающихся по характеру водного режима Чрис. 397):
1 М. Z о h а г у, Hydro-economical types in the vegetation of Near East deserts.
Sympos. Biol. And Product, of hot and cold deserts, London, 1952, p. 56—67.
Египетско-аравийская пустыня, пустыни Синай и Неджеб
537
1. а) Эфемеры, заканчивающие свое развитие в течение трех зимних
месяцев и отмирающие уже в марте (тип Filago, очень много видов);
б) однолетники, криптофиты и гемикриптофиты, живущие до июня (тип
Launaea, очень много видов); в) однолетние растения, прорастающие
лишь весной (март); летом они теряют крупные листья, но сохраняют
мелкие чешуевидные зеленые листья; эти растения живут вплоть до
зимы (виды Salsola).
2. Зимнезеленые кустарники, сбрасывающие летом все листья
(Lycium arabicum, Anagyris foetida и др.).
3. Кустарнички, сильно редуцирующие в период засухи транспириру-
ющую поверхность листьев; к их числу относятся Reaumuria palaestina,
Salsola villosa, Suaeda palaestina, S. asphaltica, Artemisia monosperma,
Zygophyllum dumosum и др.
4. Вечнозеленые прутьевидные кустарники, которые зимой теряют
свои мелкие листья; в течение всего года они сохраняют зеленые ассими-
лирующие побеги; из них лишь часть отмирает летом, как у Retama retam,
Calligonum comosum и др.
5. Безлистные ксероморфные виды, теряющие летом часть своих
зеленых ломких побегов, как, например, Ephedra.
6. Мягколистные ксерофиты, теряющие летом нижние серые гиг-
роморфные листья и сохраняющие лишь мелкие ксероморфные, напри-
мер Artemisia herba alba.
7. Кустарнички, у которых в пазухах листьев на молодых побегах
образуются небольшие почковидные укороченные побеги; после сбрасыва-
ния листьев они ассимилируют СО2 в течение всего лета (Reaumuria).
8. Колючие кустарнички, образующие в начале лета колючие боко-
вые побеги с мелкими листочками; последние отмирают в периоды засу-
хи (Noaea).
9. Безлистные галофиты с членистыми побегами; летом зеленая кора
прошлогодних побегов у них отмирает и опадает в виде колец, как
у Anabasis articulatum, Haloxylon articulatum и др.
10. Сходны с предыдущей группой; сбрасывают летом также и часть
старых побегов (Haloxylon persicum).
Наконец, нужно упомянуть деревья, которые сбрасывают старые
листья только после того, как летом будут полностью сформированы
новые листья. К их числу относятся Acacia и Tamarix, развитие которых
связано с близким уровнем грунтовых вод.
Таким образом, растения всех выделенных типов во время летней
засухи в условиях недостатка влаги редуцируют транспирирующую
поверхность (см. стр. 326). Эта редукция, вероятно, связана с достиже-
нием в периферийных частях органов летальных пределов гидратуры.
У эфемеров отмирают все надземные органы; остаются только семена
или подземные части.
Более простую классификацию предложил в 1953 г. Оршан Ч Он
различает следующие группы (типы) растений:
I. Уходящие от засухи f 1. Терофиты
I 2. Виды, втягивающие побеги
II. Переносящие засуху f 1. Сбрасывающие побеги
1 2. Сбрасывающие листья.
1 G. О г s h a n, Note on the application of Raunkiaer’s system of life forms in
arid regions, «Pal. J. Bot.», Jerus, Ser. 6, 1953, p. 120—122.
35-1 39 4
538
Глава 14
С усилением сухости климата возрастает удельный вес видов, отно-
сящихся к первой подгруппе. Это особенно заметно при сопоставлении
процентного соотношения видов разных подгрупп, представленных в той
или иной провинции, с возрастающей засушливостью климата (табл. 111).
Таблица 111
Биологический спектр доминантой в отдельных провинциях
Палестины (в %)
Название провинций Число ВИДОВ I И
1 2 1 2
терофи- ты втягива- ющие побеги сбрасы- вающие побеги сбрасыва- ющие листья
Средиземноморская 90 30 37 17 16
Ирано-туранская 70 47 30 19 4
Сахаро-аравийская 71 60 13 27 0
Расход воды в летнее время наряду с редукцией транспирирующей
поверхности регулируется снижением интенсивности транспирации на 1 г
сырого веса транспирирующих органов, причем настолько значительным,
что интенсивность транспирации летом, несмотря на значительно боль-
шую величину испарения, чаще всего оказывается гораздо меньше
интенсивности транспирации весной (рис. 398). Особенно ярко это про-
является у видов Artemisia, Noaea и Salsola villosa. Исключение составля-
ют лишь акации, круглогодично связанные с грунтовыми водами, а также
Haloxylon persicum и Atriplex halimus, которые также зависят от запасов
влаги в почве.
Несмотря на редукцию транспирирующей поверхности и снижение
интенсивности транспирации, гидратура растений летом ухудшается.
Осмотическое давление летом всегда несколько увеличивается. Это
повышение может быть очень небольшим у одних видов, и прежде всего
у Acacia tortilis и?Artemisia monosperma, и очень значительным у других,
например у Zygophyllum dumosum или Salsola tetrandra. Последние два
вида являются галофитами; повышение осмотического давления у них
не всегда связано с ухудшением условий роста растений. Однако
в большинстве случаев очень высокие значения осмотического давления
(выше 50 атм) уже связаны с сильным снижением жизнедеятельности
растений.
Более подробно 1 проводилось изучение эколого-физиологических
особенностей Tamarix aphylla (articulata). Этот вид повсеместно распро-
странен в Палестине, где встречается на незасоленных местообитаниях,
отличающихся высоким уровнем грунтовых вод. Несмотря на это, этот
вид переносит засоленность почвы. Молодые экземпляры встречаются
на незасоленных песках (дюнах), выделяют белые кристаллы СаСО3,
а старые растения, корни которых достигают засоленного суглинистого
слоя, подстилающего песок, наоборот, выделяют главным образом кри-
1 Y. W a i s е 1, Ecological studies in Tamarix aphylla (L.). Karst. II. The water
economy FYTON (Argentina), 15, 1960, p. 19—28. III. The salt economy, «Plant and
Soil», 13, 1961, p. 356-364.
Египетско-аравийская пустыня, пустыни Синай и Неджеб
539
сталлы NaCl. При культуре
в воде рост тамарикса задер-
живается уже при наличии
0,1 и 0,2 М (0,6-1,2%) NaCl,
а при наличии 0,3 М в раст-
ворах рост у Tamarix пре-
кращался, хотя растения еще
продолжали существовать; в
растворах, содержащих 0,4
и 0,5 М NaCl, растения поги-
бали. При различном содер-
жании NaCl в питательном
растворе количество выделя-
емых Са и К всегда остается
одинаковым, выделение же
натрия и хлора, напротив,
возрастает до тех пор, пока
концентрация NaCl в пита-
тельном растворе не достигнет
0,2 М\ при концентрации
NaCl в питательном растворе,
равной 0,3—0,4 М и обу-
словливающей отсутствие
прироста и незначительную
интенсивность транспирации,
выделяются меньшие количе-
ства NaCl. При хорошей
влагообеспеченности интен-
сивность транспирации у Ta-
marix достигает больших зна-
чений и характеризуется
одновершинной кривой, рост
которой наблюдается около
полудня. При недостатке
влаги кривая, характеризую-
щая ход транспирации, сна-
чала отличается двухвершин-
ностыо, а в дальнейшем
0 250 500 1000 мг/г-час
--------1--------1 ।
-------------1 Tamarix articulata
—3 Reaumuria palaestina
Lycium arabicum
3 Artemisia monosperma
—1 Retama retam
Zilla spinosa
3 Retama duriaei
• Nitraria retusa
Salsola outrani
Zygophyllum dumosum
Haloxylon salicornicum
Ochradenus baccatus
—=з Calligonum comosum
— J Salsola villas a
1 Salsola inermis
__--------==1 Haloxylon articulaium
----- J Noaea mucronata
1 Arthrocnemum glaucum
Artemisia herba alba
^=> Anabasis articulata
Acacia raddiana
-----------------! Acacia
spirocarjja
Haloxylon persicum
Atriplex halimus
________i________।________________i________________
Рис. 398. Интенсивность транспирации весной
(незалитые строки) п летом (залитые строки) у
видов, распространенных в пустынных областях
Палестины (по Зохари).
сохраняется только один подъем кривой перед полуднем; таким образом
интенсивность транспирации днем начинает сокращаться довольно рано.
Несмотря на то что выделяющаяся на листьях соль растворяется
влагой, поступающей из влажного воздуха, поглощения воды растениями
этим путем не наблюдается.
ПЕРЕХОДНАЯ ПОЛОСА К ОБЛАСТИ РАЗВИТИЯ
СРЕДИЗЕМНОМОРСКОЙ ЖЕСТКОЛИСТНОЙ РАСТИТЕЛЬНОСТИ
На Синайском полуострове и тем более в северной части пустыни
Неджеб можно проследить переход к области средиземноморской расти-
тельности. К сахаро-аравийским флористическим элементам здесь
присоединяются ирано-туранские и изредка средиземноморские элементы.
По направлению на северо-восток в пределах Сирийской пустыни, вскоре
начинают преобладать ирано-туранские элементы. Они характерны для
540
Глава 14
Рис. 399. Флористические подразделения Ближнего Востока (М. Z oh агу,
On the geobotanical structure of Iran, «Bull. Res. Counc. Israel», Sect. D, 11
(Suppl.), Jerusalem, 1963. При годовом количестве осадков, равном 350 лш, на
юго-востоке Пакистана (в пустыне Синд или Тар) встречаются типичные па-
леотропические (суданские) рода: Euphorbia, Acacia, Commiphora, Salvadora,
Grewia, Crotalaria, Aerva, Callotropis, Eleusine, Panicum, Cumbopogon и др.
См. R. L. W г i g h t, Arid Zone (UNESCO), 25. Sept. 1964).
1 — Средиземноморская провинция; 2 — Ирано-туранская провинция; 3 — Сахаро-
аравийская провинция. В пустыне Синд, по данным Зохари, встречаются суданские
флористические элементы, относящиеся к палеотропической области. Поэтому пра-
вильнее выделять Судапо-синдскую и Сахаро-аравийскую провинции.
пустынных областей с низкими зимними температурами. С продвижени-
ем на север и более высоко в горах уже ясно выделяется холодное время
года; здесь исчезает финиковая пальма (древесная порода, характерная
для Сахары). На западе Палестины по мере приближения к берегу Среди-
земного моря и с увеличением количества зимних осадков начинают
преобладать средиземноморские элементы. При количестве осадков,
превышающем 400—500 лиц характер ландшафта меняется —ксерофиль-
ные кустарники и кустарнички сменяются склерофильными древесными
растениями, а последние — вечнозелеными древесными породами.
Сахаро-аравийская, ирано-туранская и средиземноморская провин-
ции не могут быть резко разграничены, поэтому на рис. 399 приводятся
лишь схематичные границы этих выделов Ч Такая расплывчатость границ
объясняется тем, что между средиземноморской лесной зоной 1 2 и сахаро-
1 М. Z о h а г у, Ecological studies on the vegetation of the Near Eastern Deserts.
I. Environment and vegetation classes, «Israel Explor. Journ.», 2, 1952, p. 201—215.
2 В работах, использованных Г. Вальтером при составлении этого раздела
и самим автором монографии, понятие «зона» применяется как для обозначения зоны,
так и подзоны, полосы, подобласти, провинции или пояса растительности, а пояс,
в свою очередь иногда соответствует растительной зоне или полосе. Во избежание
путаницы в подписях под рисунками и в этом разделе главы дается дословный пере-
вод текста. — Прим. ред.
Египетско-аравийская пустыня, пустыни Синай и Недсмсеб
541
аравийской или ирано-туранской пустынными зонами вклинивается ряд
промежуточных зон, например зона полупустыни, где господствуют виды
Artemisia, степная зона с преобладанием злаков и переходная зона, где
доминируют древесные виды. Схема растительных зон восточной части
Средиземноморья и карта растительности Палестины, составленные
Бойко 4, дают представление о многообразии типов растительности
на территории Палестины (рис. 400, 401).
Наряду с вышеназванными элементами по Иорданской впадине
далеко на север проникают также судано-синдские элементы (примерно
39 видов) и прежде всего различные виды акации, а также Eragrostis
bipinnata, которая образует на юго-восточном берегу Мертвого моря,
в районе выхода источников, заросли высотой до 3 м, покрывающие
площади в несколько квадратных километров, и ряд других видов.
В древостоях, образованных Zizyphus spina christi, Balanites aegyptiaca,
встречаются также Callotropis procera, Solanum incanum, Acacia tortilis
и др. К югу от Мертвого моря распространены, кроме того, Salvadora
persica, Moringa aptera, Cordia gharaf, Maerua crasstfolia. Изредка в релик-
товых местообитаниях встречаются чистые древостои Acacia albida.
В верхнем течении Иордана 2, в болотах Хула, распространен даже
тропический реликт — Cyperus papyrus. Эти болота расположены на
высоте 100 м над уровнем моря и отличаются высокой температурой
воды —25—35° в сентябре и 12—17° в декабре. Реакция воды слегка
щелочная (pH = 6,95—7,65).
В Палестине проведено очень много геоботанических исследований.
Список работ по этому вопросу опубликован Опенгеймером 3. Карты
растительности и почв составил Зохари 4.
В зоне полупустыни, где в год выпадает не менее 150 мм осадков,
на тяжелых лёссовых почвах развивается Artemisia herba alba, в то время
как на песчаных почвах в основном распространен Zizyphus lotus. Вокруг
кустов накапливается песок, поэтому Zizyphus по большей части распо-
лагается на вершинах небольших песчаных дюн. Оба вида распростране-
ны от Марокко до Ирана, и таким образом могут быть отнесены к числу
мавритано-иранских или мавритано-туранских элементов.
При увеличении количества осадков полупустыня сменяется злако-
выми степями; в Северо-Западной Африке в их составе господствует
альфа — Stipa tenacissima, а в Передней Азии господство переходит к дру-
гим высокорослым видам ковыля (Stipa lagascae, St. szovitziana) и Aristida.
Степи в Передней Азии почти полностью уничтожены в результате пасть-
бы и замещены полупустыней, в составе которой преобладает Artemisia
herba alba', таким образом, полупустыня распространилась далеко за пре-
делы своих естественных границ. Еще больше разнотравья и геофитов,
выдерживающих более низкие зимние температуры, встречается в степях,
1 Н. Boyko, A new plant-geographical subdivision of Israel (as an example
for Southwest Asia), «Vegetatio», 5/6, 1954, p. 309—318.
2 H. R. Oppenheimer, An account of the Huleh Swamps, «Pal. J. Bot.»,
Reh. Ser., 2, 1938, p. 34—39; M. Z о h a r y, G. О r s h a n s k y, The vegetation of
the Huleh plain, «Pal. J. Bot.», Jerus. Ser., 4, 1947, p. 90—104.
3 H. R. Oppenheimer, Geobotanical research in palestine vegetation, «Vege-
tatio», 3, 1952, p. 301—320.
4 M. Zohary, A vegetation map of western Palestine, «J. Ecol.», 34, 1947,
p. 1 — 19; A geobotanical soil map of western Palestine, «Pal. J. Bot.», Jerus Ser., 4,
1947, p. 24—35; N. Feinbrun, M. Zohary, A geobotanical survey of Transjor-
dan, «Bui. Res. Council of Israel, Sect. Botany», 5, 1955, p. 5—35; R. Gradman n,
Die Steppen des Morgenlandes, «Geogr. Abhandl.», 3, Reihe, H. 6, 1934.
542
Глава 14
образующих переход к степям, распространенным в центральной Анато-
лии. Однако и от них в результате пастьбы и пахоты вряд ли что-то
осталось. На большей части площади они замещены полупустыней,
в составе которой преобладает Artemisia fragrans Ч
Влажнотропический пояс у»
Рис. 400. Схема растительных зон восточной части Средиземноморья
(по Бойко).
Условные обозначения те же, что и на рис. 401. Судано-деканский пояс называется
сейчас судано-синдсннм поясом.
Поскольку в этой переходной полосе не проводилось экологических
исследований, было бы интересно более подробно изучить экологию
Artemisia herba alba и некоторых злаков.
Между открытыми степными пространствами и сомкнутыми древо-
стоями из жестколистных пород вклиниваются участки с разреженным
древесным пологом, образованным отдельно стоящими деревьями или
кустарниками, под которыми в прошлом, видимо, был развит злаковый
покров. Эти саванноподобные ландшафты нигде не сохранились в перво-
начальном виде; сейчас встречаются лишь их жалкие остатки. Можно
выделить следующие обусловленные климатом типы (рис. 401):
1. Термофильный тип, образованный Pistacia mutica\ этот вид вместе
с распространенной в Северо-Западной Африке Р. atlantica относится
к числу мавритано-иранских элементов. В настоящее время встречаются
лишь отдельные экземпляры этих древесных пород; вместо развитого
в прошлом злакового покрова широко распространилась Artemisia
herba alba.
2. Преимущественно вблизи моря распространены полувечнозеле-
ные дубы, близкие к Quercus aegilops (в Палестине — Q. ithaburensis,
в Анатолии —Q. macrolepis). Они появляются при количестве осадков
свыше 400 мм в год. При меньшем количестве осадков возникают
заросли кустарников, в составе которых встречаются летнезеленые виды
из родов Amygdalus, Rhamnus и Crataegus.
3. В континентальных областях с холодными зимами наряду с летне-
зеленым дубом (Q. infectoria и др.) встречается также древовидный можже-
вельник (Juniperus excelsa., J. foetidissimd).
1 H. Walter, Das Problem der Zentralanatolischen Steppe, «Die Naturwissen-
schaften», 43, 1956, S. 97—102.
Рис. 401. Карта растительности Палестины (по Бойко).
Средиземноморская лесная зона:
1 — типичная; 2 — полувлажная; <? — полузасушлпвая.
Аридная (переходная) зона кустарников:
4 — анатолийско-иранская; J —мавритано-ирапская.
Степная зона:
€ — анатолийская; 7 — ирано-туранская; 8 — мавритано-иранская (тер-
мофильная).
Пустынная зона:
9 — центральноазиатская, или ирано-туранская; 10 — сахаро-аравийская.
С у дано-синдская зона:
11 — заросли Cyperus papyrus (болота Хула); 12 — изолированные место-
нахождения в районе Мертвого моря.
544
Глава 14
Оппенгеймер 1 изучал водный режим Quercus ithaburensis, распро-
страненного на обнажениях известняков. Этот дуб отличается чрезвычайно
интенсивной транспирацией в июне и сентябре после пятимесячного»
периода засухи (табл. 112).
Таблица 112
Интенсивность транспирации Quercus ithciburensls,
в мг/Ъ'Час
Сроки наблюдений, час. 28 июня Сроки наблюдений, час. 21 сен- тября
15.27 925 8.30 1000
16.35 1095 11.00 500
При заходе солнца 490 После полудня 650
Приведенные данные свидетельствуют о том, что водный баланс
у этого вида сохраняется на постоянном уровне. Последнее подтверждает-
ся также наблюдениями за осмотическим давлением; в частности, восьмого
октября, еще до начала периода дождей, осмотическое давление дости-
гало только 20,3 атм.
Возможность большого расхода воды у Quercus ithaburensis зависит
от крупного размера сосудов, значительно более широких, чем у вечно-
зеленого Q. coccifera var. calliprinos. К тому же большие расстояния между
деревьями способствуют повышению запасов почвенной влаги.
Весьма своеобразны литофиты, изучением которых занимались
Зохари и Оршанский 1 2. Они распространены в средиземноморской обла-
сти, где зимой выпадает 400—700 мм осадков, однако растут там в наибо-
лее засушливых местообитаниях и поэтому будут рассматриваться в этом
разделе книги.
В отличие от хасмофитов корни литофитов распространяются не по-
трещинам скал, а непосредственно растворяя породу3. Поэтому они
встречаются на доломитах, кристаллических известняках, литографских
сланцах и на известковых туфах. Корни всходов проникают в породу
только в тех случаях, когда семена прорастают в небольших углублениях
или в выемках на поверхности породы, то есть в местах, где однород-
ность ее структуры уже несколько нарушена водой. В дальнейшем корни
продвигаются по менее плотным участкам породы, часто меняя направ-
ление роста, и достигают длины до 50 см. Кроме этих корней, у растений
развиваются еще тонкие корешки, проникающие в небольшие заполнен-
ные мелкоземом углубления, расположенные на поверхности скал.
В течение очень жаркого и бездождного лета изучалась экология
Varthemia iphionoides (Gompositae) и Podonosma syriacum (Boraginaceae).
Это невысокие кустарнички. У Varthemia более крупные листья развивают-
ся с ноября по июль, а мелкие летние листья — с июня по декабрь;
цветет с сентября по октябрь. У Podonosma зимние листья сохраняются
1 Н. R. Oppenheimer, The water, turn-over of the Valonea oak, «Pal. J.
Bot.», Reh. Ser., 7, 1949, p. 177—179.
2 M. Zohary, G. Orshansky, Ecological studies on lithophytes, «Pal. J.
Bot.», Jerus. Ser., 5, 1951, p. 119—128; M. L i t a v, Mycorrhizal association in dwarf
shrub species growing in soft cretaceous rocks, «J. Ecol.», 53, 1965, p. 147—153.
3 H. R. Oppenheimer, Penetration active des racines de buissons mediter-
ranees dans les roches calcaires, «Bull. Res. Counc. Israel», 5D, 1956, p. 219—222.
Египетско-аравийская пустыня, пустыни Синай и Неджеб 545’
с декабря до конца августа, а летние — с июня до февраля. Период
цветения растянут с февраля до конца августа. Интенсивность транспира-
ции у обоих видов летом и зимой достигает одинаковых величин и состав-
ляет 1000—1400 мг/г*час, что намного превышает интенсивность транс-
пирации растущих в этих же условиях Poterium spinosum (644 мг/г^час)
или Cistus salviifolius (336 мг/г ♦ час), Один экземпляр Varthemia расхо-
дует в течение суток 60—120 г воды, или 10—20 кг в течение лета. Несмо-
тря на это, водный баланс у этих видов, как и у относящегося к той же
группе растений Stachys palaestina, сохраняется на постоянном уровне;
осмотическое давление летом у Varthemia равно 23—29 атм, у Podosma —
16—19 атм и у Stachys— 15 атм. Высокая влагоемкость пород (8—22%),
на которых развиваются растения, способствует улучшению их водного
режима. Водопроницаемость породы также велика: за одну минуту
просачивается 1 мл воды. Даже в разгар лета влажность породы, на
которой росли растения, достигала 2,5%, Растения поглощают влагу,
поступающую по капиллярам, и расходуют при этом запас воды, содер-
жащийся менее чем в 2 ж3 породы; этим определяется степень сомкну-
тости растительного покрова, появляющегося на этих породах,
В восточной части Средиземноморской области (на территории Пале-
стины и в Сирии) между зоной пустыни и в прошлом облесенной среди-
земноморской зоной расположена область полупустыни или степей.
Аналогичная картина наблюдается и в Северо-Западной Африке, Однако
здесь высокие горы Атласа (более 4000 м над уровнем моря) определяют
большое разнообразие климатических условий. Увеличение количества
осадков по направлению к Атлантическому океану и при подъеме в горы,
большая сухость на подветренных склонах гор и снижение температуры
при подъеме в верхние пояса гор обусловливают формирование здесь
разнообразного растительного покрова, издавна привлекавшего внима-
ние ботаников. Поскольку в горах Атласа широко распространены леса,
характеристика этих условий будет дана при описании растительности
Средиземноморья, Сейчас можно ограничиться упоминанием, что и в этой
области, особенно на высокогорном плато между Телль-Атласом и Сахар-
ским Атласом, на больших площадях возникли полупустыни, образованные
Artemisia herba alba, и степи, в составе которых господствуют Stipa
tenacissima или Lygeum spartum.
Количество осадков в Средиземноморской области составляет около
250 мм', они распределены в течение года довольно неравномерно. Сред-
няя температура наиболее теплого месяца достигает 28°; зимы холодные;
снегопады бывают редко; число дней с отрицательными температурами
равно 48, Подробное описание растительности и почв приводится в рабо-
тах Озенда, Дюбуи и Симонно *,
Artemisia herba alba распространена преимущественно в понижениях
на незасоленных пылеватых суглинках. Наряду с господствующей здесь
полынью в этих понижениях встречается много однолетников и очень
редко — геофиты.
По данным Дюбуи и Симонно, степень покрытия почвы достигает
35—70%; высота растений —более 25 см. Характеристика почв из двух
разрезов глубиной до 70 см приводится в табл, ИЗ, в %,
1 Р, О z е n d a, Observations sur la vegetation d’une region semiaride: Les Hauts-
Plateaux du Sud-Aglerois, «Bull, Soc, d’Hist, Nat, de 1’Afrique du Nord», 45, 1954,
p, 134—169;^ A, Dubuis, P, Simonneau, Contribution a Г etude de la vegeta-
tion de la region d’ATn Skrouna, Serv, Etudes Sci, Gouv, Gener, de I’Algerie, 1954.
546
Глава 14
Таблица 113
Почвен- ный разрез pH Крупный песок Мелкий песок Пыль Глина Карбона- ты каль- ция NaCl
I 8,8 0 29,8 53 15,7 13 0,037
II 8,7 4,8 44,3 26,9 22,7 12,8 0,033
Почвы могут отличаться и более легким механическим составом
(табл. 114, в %), однако состав сообщества при этом не меняется, а умень-
шается лишь степень покрытия почвы (30—40%); высота травостоя
в этом случае не превышает 10—15 см.
Таблица 114
Горизонт, см pH Хрящ Крупный песок Мелкий песок Пыль 1 Глина 1 Карбона- ты каль- ция NaCl
0—15 8,4 8,4 0 76,7 13,8 8,3 30,1 0,01
J 5—40 8,7 0 0 42,3 52,8 4,1 75,8 0,07
40-100 8,8 0 0 66,8 13,7 18,8 73,5 0,11
С увеличением засоленности почвы наряду с полынью начинают
встречаться галофиты: Atriplex mauretanica и Suaeda fruticosa. При уве-
личении засоления, особенно часто наблюдаемого на высокогорном плато,
развиваются чистые заросли галофитов.
В противоположность полыни Stipa tenacissima распространена пре-
имущественно по возвышениям и склонам, где имеются более или менее
каменистые почвы или верхние горизонты насыщены обломками породы.
Обширные площади, покрытые Stipa tenacissima, простираются к востоку
от Хомса (Ливия); здесь производится сбор альфы, используемой при
приготовлении бумаги; это важная статья экспорта.
Lygeum spartum предпочитает, с одной стороны, хорошо аэрируемые
почвы более легкого механического состава, а с другой — засоленные.
Lygeum часто встречается по внешнему краю зарослей галофитов.
Озенда указывает, что эти виды хотя и образуют чистые заросли,
однако чаще встречаются в смешанных сообществах.
К сожалению, экология этих трех видов до сих пор не изучалась.
Полыни относятся к числу мягколистных ксерофитов, а оба упомянутых
злака следует причислить к особому типу склерофильных злаков, о кото-
рых уже шла речь раньше (см. стр. 447). Изучение экологии этих злаков
во влажные периоды и периоды засухи могло бы дать интересные
результаты.
Stipa tenacissima распространена на площадях, покрытых ранее
лесами из вечнозеленого дуба и других древесных пород, растущих уже
в пределах средиземноморской зоны.
Собственно средиземноморская зона1 простирается
вокруг всего Средиземного моря и выклинивается лишь на засушливом
побережье Египта. В пределах этой зоны выделяются две субзоны, отли-
чающиеся океаническим и более континентальным климатом (рис. 400).
1 См. примечание на стр. 540.— Прим. ред.
Египетско-аравийская пустыня, пустыни Синай и Недмсеб 547
К последней относится прибрежная область от Алжира до Киренаики,
восточная Греция и побережье Передней Азии. В пределах этой субзоны
при увеличении влажности климата можно выделить полувлажную
подзону, где господствует Laurus nobilis. Эта подзона образует переход
к субсредиземноморским летнезеленым лиственным лесам. В горах,
расположенных в континентальной части Средиземноморья, встречаются
Curpessus horizontalis (на Кипре, начиная с высоты 700 м) и Cedrus libani
( в горах Тавра, начиная с высоты 1200 м, на Крите — примерно с высо-
ты 900 м), выше расположен пояс колючих подушечников.
В восточной части средиземноморской зоны распространен Quercus
coccifera (на территории Палестины var. calliprinos), вместе с этим видом
дуба часто встречаются другие жестколистные растения: Phillyrea,
Arbutus andrachne, Myrtus, Pistacia lentiscus, Smilax и др. В более теп-
лых местообитаниях господствует Ceratonia siliqua. Из хвойных пород
здесь распространена сосна алеппская (Pinus halepensis), а севернее
и выше в горах — близкий к ней вид Pinus brutia; на песчаных почвах
растет Pinus pinea; рощи, образованные этим видом сосны, сохранились
лишь в очень немногих местах.
Экологическая характеристика средиземноморской зоны, учитывая
•ее почти сплошную облесенность в прошлом, будет приведена в следую-
щем томе при описании растительного покрова умеренной зоны.
ОГЛАВЛЕНИЕ
ИЗ ПРЕДИСЛОВИЯ АВТОРА К ПЕРВОМУ ИЗДАНИЮ
ПРЕДИСЛОВИЕ АВТОРА КО ВТОРОМУ ИЗДАНИЮ
Глава 1
ВВЕДЕНИЕ
РАСТИТЕЛЬНОСТЬ И РАСТИТЕЛЬНЫЕ СООБЩЕСТВА (И). СУКЦЕС-
СИИ, КЛИМАКС И ЗОНАЛЬНАЯ РАСТИТЕЛЬНОСТЬ (19). КОНКУРЕНТ-
НЫЕ ВЗАИМООТНОШЕНИЯ (23). ПРОДУКТИВНОСТЬ И ПОКАЗАТЕЛЬ
ОБЛИСТВЛЕННОСТИ (31). КОНКУРЕНТНАЯ СПОСОБНОСТЬ И ПОКА-
ЗАТЕЛЬ УСПЕШНОГО РОСТА (34). ИНТЕНСИВНОСТЬ КОНКУРЕНЦИИ И
ЖИЗНЕННЫЕ ФОРМЫ (39). ОБМЕН ВЕЩЕСТВОМ И ЭНЕРГИЕЙ В ЭКОСИ-
СТЕМАХ (43). НЕКОТОРЫЕ СООБРАЖЕНИЯ О ВОДНОМ РЕЖИМЕ РАСТЕ-
НИЙ (45). КЛИМАТИЧЕСКИЕ ЗОНЫ ЗЕМНОГО ШАРА (53). ГРАФИЧЕСКОЕ
ИЗОБРАЖЕНИЕ КЛИМАТА С ПОМОЩЬЮ КЛИМАДИАГРАММ (60). ПОЧ-
ВЕННЫЕ ЗОНЫ ЗЕМНОГО ШАРА (70). ОСНОВЫ ТРЕХМЕРНОГО РАСЧЛЕ-
НЕНИЯ РАСТИТЕЛЬНОСТИ ЗЕМНОГО ШАРА (71).
Глава 2
ТРОПИЧЕСКИЙ ДОЖДЕВОЙ ЛЕС
ОБЩИЙ ОБЗОР (77). КЛИМАТ ВЛАЖНЫХ ТРОПЦКОВ (80). МИКРО-
КЛИМАТ ТРОПИЧЕСКОГО ДОЖДЕВОГО ЛЕСА (85). ПОЧВЫ ТРОПИЧЕ-
СКОГО ДОЖДЕВОГО ЛЕСА (91). СТРУКТУРА ТРОПИЧЕСКОГО ДОЖДЕ-
ВОГО ЛЕСА (99). ЭКОЛОГИЯ ДРЕВЕСНЫХ РАСТЕНИЙ ТРОПИЧЕСКОГО
ДОЖДЕВОГО ЛЕСА (107). ЭКОЛОГИЯ ДРУГИХ ГРУПП ЖИЗНЕННЫХ
ФОРМ ДОЖДЕВОГО ЛЕСА (127).
Глава 3
ДРУГИЕ ТИПЫ РАСТИТЕЛЬНОСТИ ВЛАЖНЫХ ТРОПИКОВ
БОЛОТНАЯ И ВОДНАЯ РАСТИТЕЛЬНОСТЬ (156). МАНГРОВАЯ РАСТИ-
ТЕЛЬНОСТЬ (163). РАСТИТЕЛЬНОСТЬ ПЕСЧАНЫХ БЕРЕГОВ (181). РАС-
ТИТЕЛЬНОСТЬ СУХИХ МЕСТООБИТАНИЙ (187). ТРОПИЧЕСКИЙ КУЛЬ-
ТУРНЫЙ ЛАНДШАФТ (189).
Глава 4
ДОЖДЕВЫЕ ЛЕСА ВБЛИЗИ ВЕРХНЕЙ ГРАНИЦЫ РАСПРОСТРАНЕНИЯ
И ВЕРТИКАЛЬНАЯ ПОЯСНОСТЬ В ГОРАХ ТРОПИЧЕСКОЙ ЗОНЫ
ЗАКОНОМЕРНОСТИ ВЕРТИКАЛЬНОЙ ПОЯСНОСТИ II ЗОНАЛЬНОГО
РАСПРЕДЕЛЕНИЯ РАСТИТЕЛЬНОСТИ В ТРОПИКАХ (193). ВЕЧНОЗЕЛЕ-
НЫЕ СУБТРОПИЧЕСКИЕ И УМЕРЕННЫЕ ДОЖДЕВЫЕ ЛЕСА (195). ГОР-
НЫЙ ДОЖДЕВОЙ ЛЕС НА ЯВЕ (203). АЛЬПИЙСКАЯ РАСТИТЕЛЬНОСТЬ
ТРОПИЧЕСКИХ АНД (207). ВЕРТИКАЛЬНАЯ ПОЯСНОСТЬ КИЛИМАН-
ДЖАРО (219).
Глава 5
ТРОПИЧЕСКИЕ ПОЛУВЕЧНОЗЕЛЕНЫЕ И ЛИСТОПАДНЫЕ ЛЕСА
СЕЗОННЫЙ ЛИСТОПАД С ЭКОЛОГИЧЕСКОЙ ТОЧКИ ЗРЕНИЯ (232). ТРО-
ПИЧЕСКИЕ ПОЛУВЕЧНОЗЕЛЕНЫЕ И ЛИСТОПАДНЫЕ ЛЕСА (234). ТРО-
550
Оглавление
ПИЧЕСКИЕ СУХИЕ ЛИСТОПАДНЫЕ ЛЕСА И ЗАРОСЛИ КОЛЮЧИХ
КУСТАРНИКОВ (246). ТРОПИЧЕСКИЕ . ПАРКОВЫЕ ЛАНДШАФТЫ (264).
ТРОПИЧЕСКИЕ ЗЛАКОВНИКИ (270).
Глава 6
ЗОНАЛЬНЫЕ САВАННЫ КАК ПЕРЕХОДНАЯ СТУПЕНЬ
К АРИДНОЙ ЗОНЕ
ПОНЯТИЕ «САВАННА» (276). ТРАВЯНАЯ И ДРЕВЕСНАЯ РАСТИТЕЛЬ-
НОСТЬ В КАЧЕСТВЕ АНТАГОНИСТОВ (279). КОНКУРЕНТНЫЕ ВЗАИМО-
ОТНОШЕНИЯ В САВАННАХ (285). ЗАРАСТАНИЕ КУСТАРНИКАМИ —
УГРОЗА ЗЕМЛЕДЕЛИЮ В ЗОНЕ САВАНН (289). РАСТИТЕЛЬНОСТЬ,
ОБУСЛОВЛЕННАЯ ЭДАФИЧЕСКИМИ ПРИЧИНАМИ, В ЗОНЕ САВАНН
И ЗЛАКОВНИКОВ (293). ЗОНАЛЬНОЕ РАСПРЕДЕЛЕНИЕ РАСТИТЕЛЬНО-
СТИ ПРИ СНИЖЕНИИ КОЛИЧЕСТВА ОСАДКОВ В СУБТРОПИЧЕСКИХ
ОБЛАСТЯХ (296).
Глава 7
РАСТИТЕЛЬНОСТЬ СУБТРОПИЧЕСКИХ АРИДНЫХ ОБЛАСТЕЙ
ОБЩАЯ ХАРАКТЕРИСТИКА АРИДНЫХ ОБЛАСТЕЙ (302). ВЛАГООБЕС-
ПЕЧЕННОСТЬ РАСТЕНИЙ АРИДНЫХ ОБЛАСТЕЙ ПРИ СНИЖЕНИИ СОМ-
КНУТОСТИ РАСТИТЕЛЬНОГО ПОКРОВА И НЕРАВНОМЕРНОМ РАСПРЕ-
ДЕЛЕНИИ ВОДЫ В ПОЧВЕ (306). ЗАВИСИМОСТЬ ВОДНОГО РЕЖИМА
РАСТЕНИЙ В ЗАСУШЛИВЫХ ОБЛАСТЯХ ОТ ПОЧВЕННЫХ УСЛОВИЙ
(313). ЗАКОН ОТНОСИТЕЛЬНОГО ПОСТОЯНСТВА МЕСТООБИТАНИЙ
И СМЕНЫ БИОТОПОВ (316). ВАЖНЕЙШИЕ ЭКОЛОГИЧЕСКИЕ ГРУППЫ
РАСТЕНИЙ НЕЗАСОЛЕННЫХ ПОЧВ АРИДНЫХ ОБЛАСТЕЙ И ИХ ГИД-
РАТУРА (320). ЗАСОЛЕНИЕ ПОЧВ И ГАЛОФИТЫ (328).
Глава 8
ПУСТЫНЯ СОНОРА
УСЛОВИЯ СРЕДЫ (332). РАСТИТЕЛЬНОСТЬ (338). ЭКОЛОГИЧЕСКИЕ.'
ГРУППЫ РАСТЕНИЙ ПУСТЫНИ СОНОРА (347). ЗАКЛЮЧЕНИЕ (373).
Глава 9
ПУСТЫНЯ ТУМАНОВ НАМИБ
РЕЖИМ ОСАДКОВ (374). ЭКОЛОГИЧЕСКОЕ ЗНАЧЕНИЕ ТУМАНА (378).
РАСТЕНИЯ ПУСТЫНИ НАМИБ И ИХ ЭКОЛОГИЯ (382).
Глава 10
ЧИЛИЙСКО-ПЕРУАНСКАЯ ПУСТЫНЯ С ОАЗИСАМИ ТУМАНОВ
ТУМАН КАК ИСТОЧНИК ВЛАГИ (411). РАСТИТЕЛЬНОСТЬ ЛОМА (418)..
Глава 11
ПУСТЫНЯ КАРРУ
ОБЩАЯ ХАРАКТЕРИСТИКА (423). РАСТИТЕЛЬНОСТЬ ВЕРХНЕГО
КАРРУ (427). ЭКОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ (429).
Глава 12
ЗАСУШЛИВЫЕ ОБЛАСТИ ЦЕНТРАЛЬНОЙ АВСТРАЛИИ
СРАВНЕНИЕ АВСТРАЛИИ С АФРИКОЙ (437). ОСАДКИ В ЦЕНТРАЛЬНОЙ
АВСТРАЛИИ (438). РАСТИТЕЛЬНОСТЬ (443). ЭКОЛОГИЧЕСКИЕ ИССЛЕ-
ДОВАНИЯ (450).
Глава 13
ПУСТЫНЯ САХАРА
ОБЩАЯ ХАРАКТЕРИСТИКА (467). ПОЧВЫ И РАСТИТЕЛЬНОСТЬ (471).
К БИОЛОГИИ РАСТЕНИЙ САХАРЫ (482). ЭКОЛОГИЧЕСКИЕ ИССЛЕДО-
ВАНИЯ В АЛЖИРСКОЙ САХАРЕ И ЛИВИИ (485). РАСТИТЕЛЬНОСТЬ
НАГОРЬЯ ТИБЕСТИ (493).
Оглавление
551
Глава 14
ЕГИПЕТСКО-АРАВИЙСКАЯ ПУСТЫНЯ,
ПУСТЫНИ СИНАЙ И НЕДЖЕБ
УСЛОВИЯ СРЕДЫ И РАСТИТЕЛЬНОСТЬ (495). ЭКОЛОГИЧЕСКИЕ ИССЛЕ-
ДОВАНИЯ, ПРОВЕДЕННЫЕ БЛИЗ КАИРА (502). РАСТИТЕЛЬНОСТЬ
ПОБЕРЕЖЬЯ СРЕДИЗЕМНОГО МОРЯ В ЕГИПТЕ (515). ПАЛЕОКЛИМАТ
ЕГИПТА (523). ХАРАКТЕРИСТИКА ЭКОЛОГИЧЕСКИХ УСЛОВИЙ НА СИ-
НАЙСКОМ ПОЛУОСТРОВЕ (535). ЭКОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ,
ПРОВОДИВШИЕСЯ В ПУСТЫНЕ НЕДЖЕБ (529). ПЕРЕХОДНАЯ ПОЛОСА
К ОБЛАСТИ РАЗВИТИЯ СРЕДИЗЕМНОМОРСКОЙ ЖЕСТКОЛИСТНОЙ РАС-
ТИТЕЛЬНОСТИ (539).
Г. Вальтер
РАСТИТЕЛЬНОСТЬ
ЗЕМНОГО ШАРА
Редактор Л. Бабинцева
Художник В. Кузяков
Художественный редактор Л. Шканов
Технический редактор И. Володина
Корректор Г. Артемьева
Сдано в производство 26/Х 1967 г.
Подписано к печати 2/Х 1968 г.
Бумага 70 X 1081/ie- Бум. л. 141/в.
39,6 печ. л. + 9 цв. вкл.
Уч.-изд. л, 46,11. Изд. № 11/6810.
Цена 4 р. 19 к. Зак. 1394
Издательство «Прогресс»
Комитета по печати при Совете Министров
СССР
Москва, Г-21, Зубовский бульвар, 21
Московская типография № 16
Тлавполиграфпрома Комитета по печати при Совете
Министров СССР
Москва, Трехпрудный пер., 9.