/






































































































































































































































Автор: Андреева Е.Н. Баккал И.Ю. Горшков В.В.
Теги: экология лесное хозяйство
ISBN: 5-7997-0452-5
Год: 2002




Текст
: Ж.
РОССИЙСКАЯ АКАДЕМИЯ НАУК
БОТАНИЧЕСКИЙ ИНСТИТУТ ИМ. В. Л. КОМАРОВА
МЕТОДЫ ИЗУЧЕНИЯ
ЛЕСНЫХ СООБЩЕСТВ
САНКТ-ПЕТЕРБУРГ
2002
Печатается по решению Ученого совета Ботанического института
им. В. Л Комарова РАН, протокол № 13 от 5 ноября 2001 г.
Рецензенты.
доктор биол. наук, профессор В В Никонов,
доктор географ, наук, профессор А С Алексеев
Ответственные редакторы:
доктор биол. наук, профессор В. Т Яр.мишко,
кандидат биол. наук, с. н. с. И В Лянгузова
Методы изучения лесных сообществ. — СПб.: НИИХимии СПбГУ,
М54 2002. — 240 с., Библиогр. 203 назв. Ил. 76. Табл. 16.
Авторы: Е Н Андреева, И. Ю. Баккал, В. В. Горшков, И. В. Лянгузова,
Е А Мазная, В Ю. Нешатаев, В. Ю. Нешатаева, Н. И Ставрова, В Т
Ярмишко, М А Ярмишко.
ISBN 5-7997-0452-5
Коллективная монография является результатом опыта многолетних комплексных
полевых и экспериментальных исследований сотрудников лаборатории экологии раститель-
ных сообществ Ботанического института им В Л Комарова, изучавших структуру и ди-
намику естественных и антропогенно нарушенных лесных сообществ Европейского Се-
вера и Северо-Запада России В книге приведены основные методы, широко используе-
мые в геоботанических и экологических исследованиях, многие из которых модифициро-
ваны или усовершенствованы авторами, а также новые методики Основными принципа-
ми изучения растительных сообществ являются комплексность и количественная оценка
(измерение) их параметров в целом и отдельных компонентов, а также характеристик
местообитаний
Издание предназначено для студентов старших курсов и магистрантов биологичес-
ких, лесохозяйственных и географических факультетов высших учебных заведений, аспи-
рантов и исследователей, специализирующихся в области геоботаники, лесоведения, эко-
логии, охраны окружающей среды
Издание осуществлено при поддержке
Санкт-Петербургского научного центра РАН
ISBN 5-7997-0452-5 © авторы, 2002
ЛР№040815от 22 05 97
Подписано к печати 04 07 2002 Формат бумаги 60X84 1/16 Бумага офсетная
Печать офсетная Объем 15 усл п л Тираж ЮООэкз Заказ 2534
Отпечатано в отделе оперативной полиграфии НИИХ СПбГУ
с оригинал-макета заказчика
198504, Санкт-Петербург, Старый Петергоф, Университетский пр . 26
СОДЕРЖАНИЕ
ВВЕДЕНИЕ..............................................................7
РАЗДЕЛ 1. ОБЩИЕ МЕТОДЫ ИЗУЧЕНИЯ ЛЕСНЫХ СООБЩЕСТВ
И ИХ КОМПОНЕНТОВ...................................................9
1 1. Принципы и методы классификации лесных сообществ.................9
1.2. Рекогносцировочное обследование территории Маршрутные методы
изучения лесных фитоценозов...........................................24
1 3 Подбор и закладка постоянных пробных площадей в лесных сообществах . 33
1.4 Закладка учетных площадок и профилей на постоянных пробных площадях . 37
1.5 Измерение характеристик древесного яруса... ... . . 42
1.6 Измерение характеристик нижних ярусов и полога подроста
в лесных сообществах............................................... 55
1.7 Методика измерения характеристик эпифитного лишайникового покрова
стволов деревьев............................................... . . 63
1.8 Методика описания почв. Краткая классификация лесных почв........68
РАЗДЕЛ 2. СПЕЦИАЛЬНЫЕ МЕТОДЫ ИЗУЧЕНИЯ ЛЕСНЫХ
СООБЩЕСТВ.........................................................76
2.1 Методы изучения биологической продуктивности древесного яруса,
полога подроста и подлеска ...................................... . 76
2.2 Методы изучения биологической продуктивности нижних ярусов
лесных сообществ..................................................... 89
2.3 Оценка запасов растений, имеющих хозяйственное значение..........95
2.4. Методы изучения семенной продуктивности растений...............102
2 5. Методы оценки качества и жизнеспособности семян ............ .. 108
2.6. Методика изучения банка семян в почве............... . . ... 114
2.7. Методы изучения ценопопуляций растений .................. . . 119
2.8. Количественная оценка биоразнообразия......... .... . 127
2.9. Методы изучения мохового покрова ............................. 130
2.10. Методы изучения подземных частей растений ...... . .... 13"
РАЗДЕЛ 3. МЕТОДЫ ИЗУЧЕНИЯ ЛЕСНЫХ СООБЩЕСТВ
В УСЛОВИЯХ НАРУШЕННЫХ МЕСТООБИТАНИЙ....................154
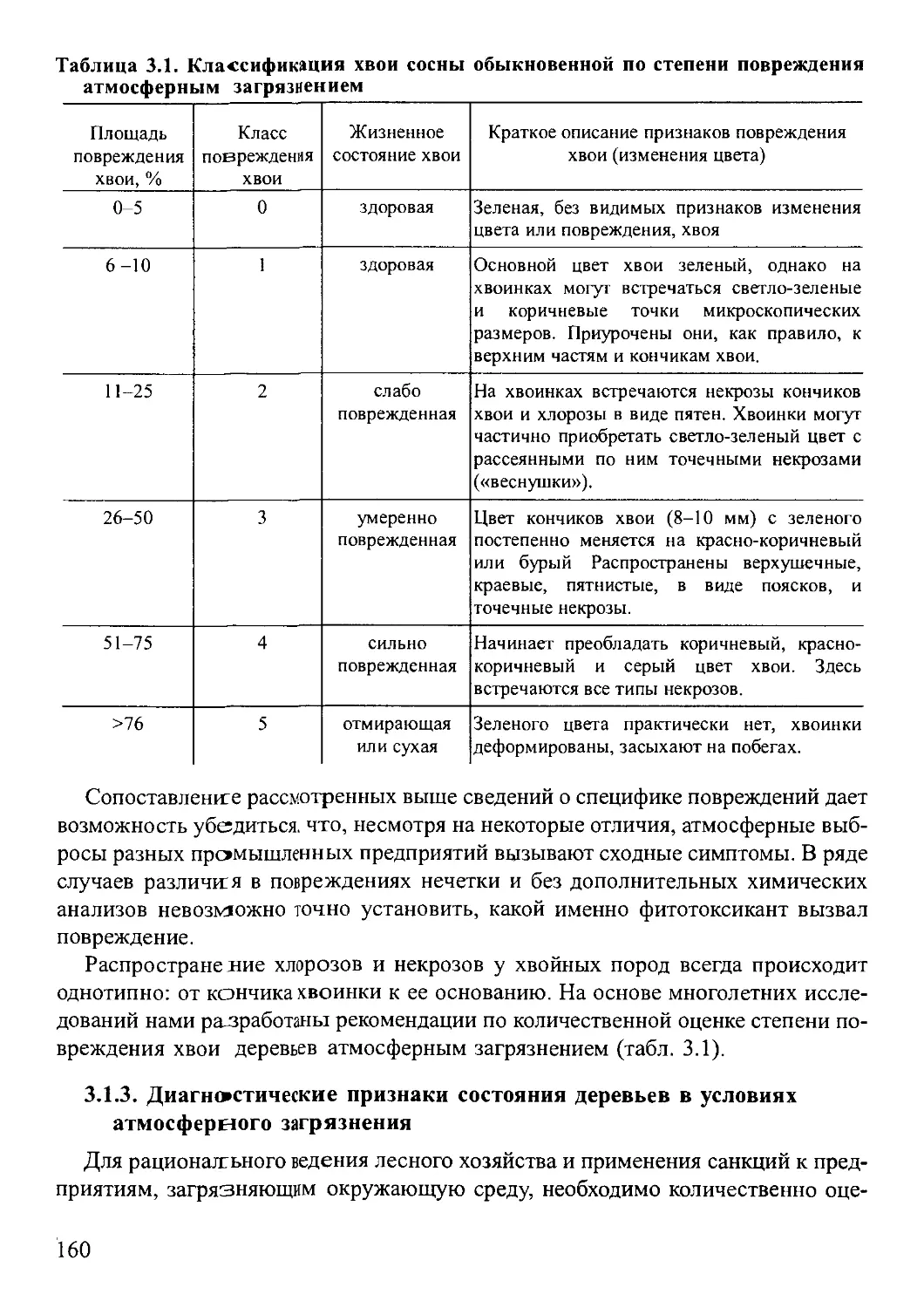
3 1 Диагностика повреждений и оценка жизненного состояния деревьев
и древостоев в условиях промышленного атмосферного загрязнения ... . 154
3 2 Использование эпифитного лишайникового покрова лесных сообществ
для индикации атмосферного загрязнения.............................. 165
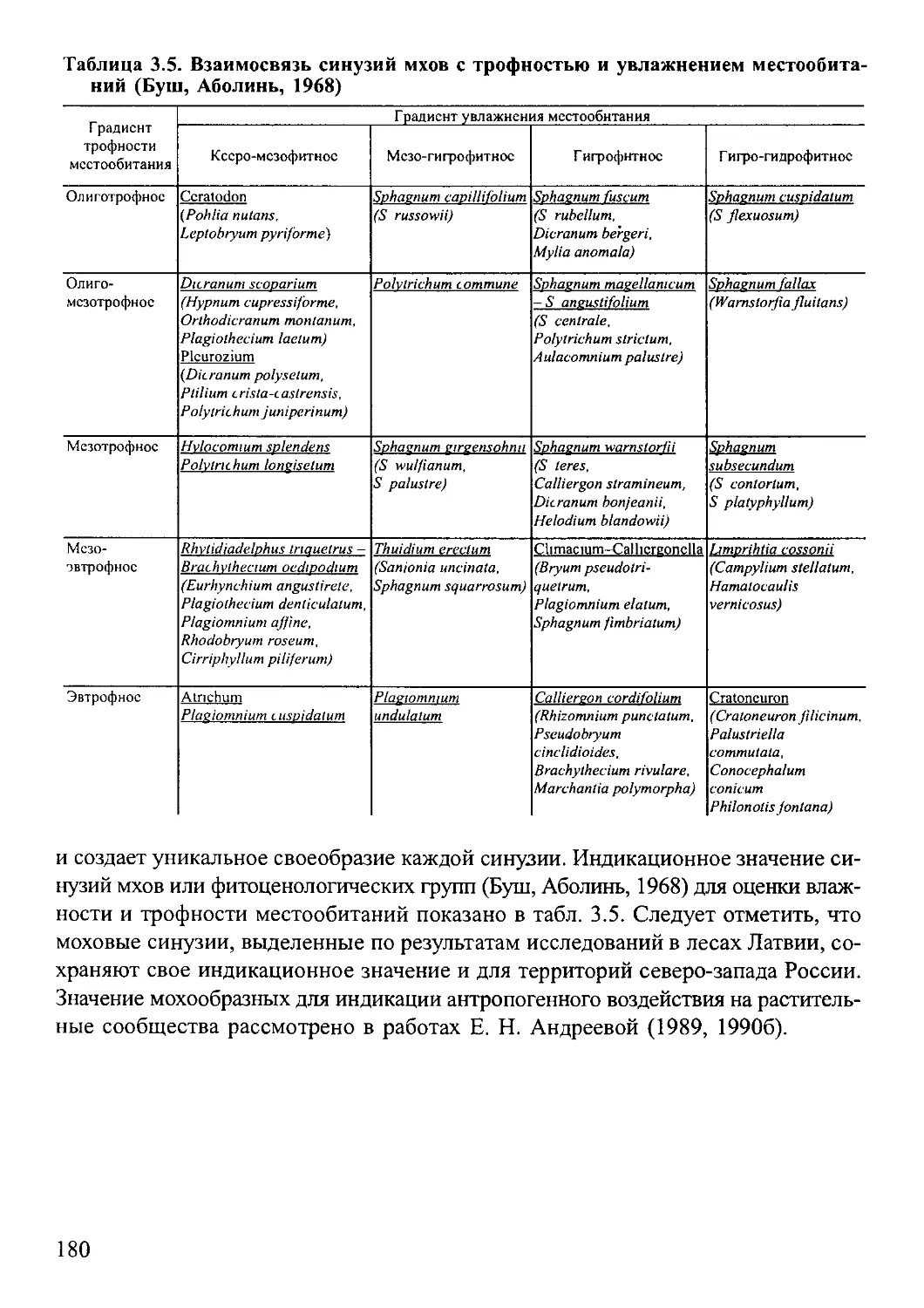
3 3 Использование мхов для индикации характеристик местообитания.170
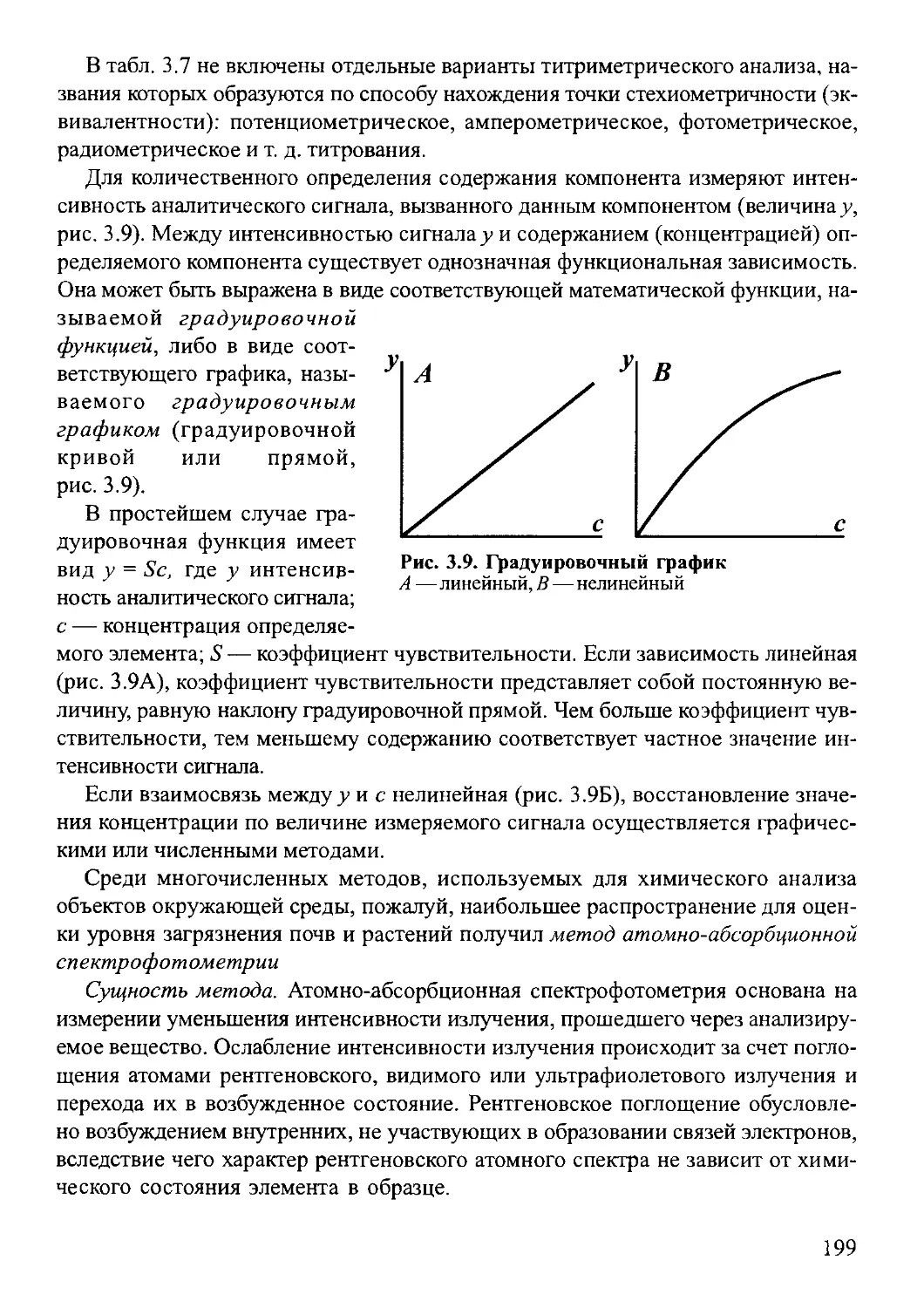
3 4 Химический анализ почв и растений ............. .. . . 181
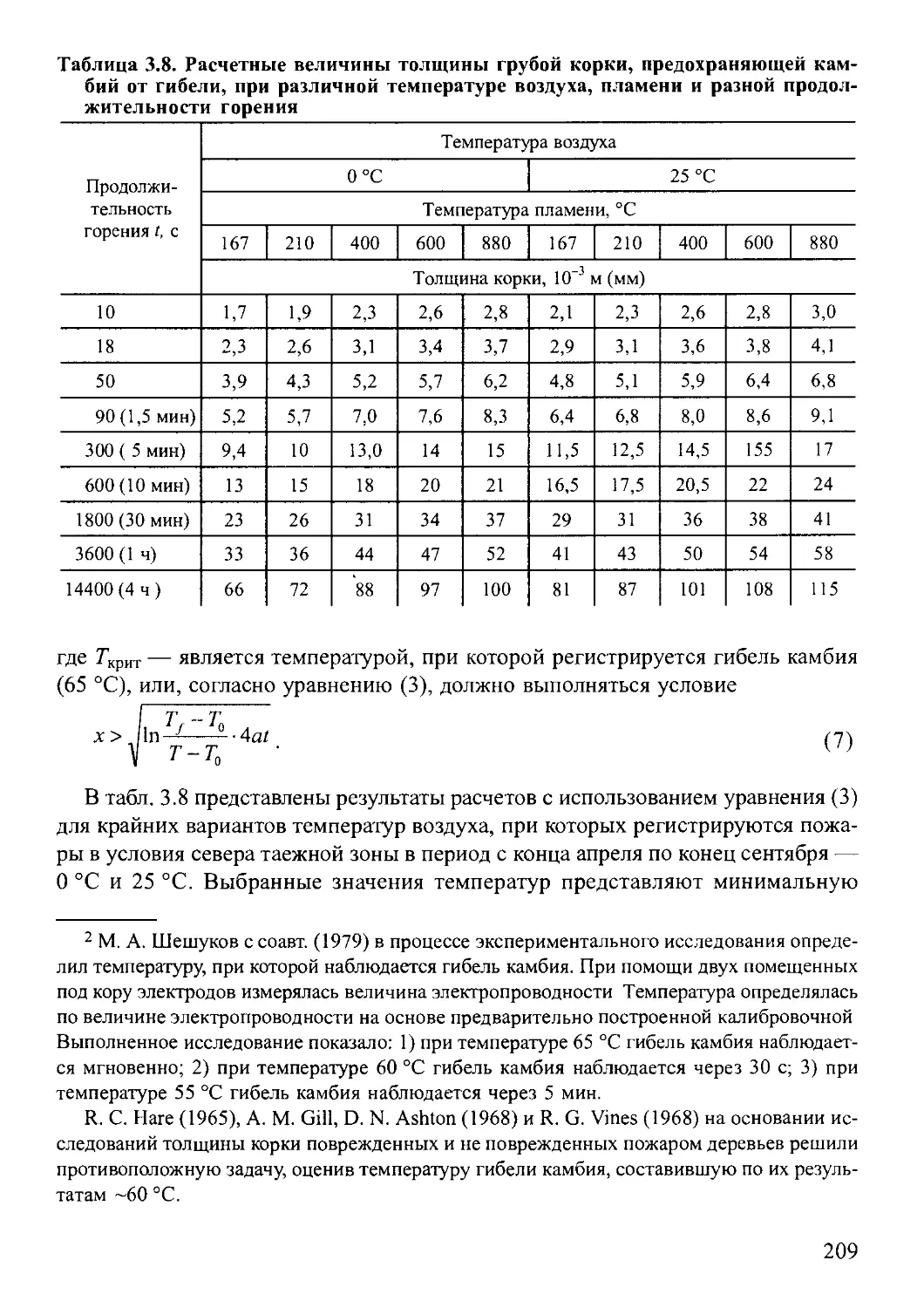
.3 5 Принципы и методы анализа давности и периодичности пожаров..201
РАЗДЕЛ 4. ОСНОВНЫЕ ПРИНЦИПЫ СТАТИСТИЧЕСКОЙ
ОБРАБОТКИ ДАННЫХ.......................................214
ОСНОВНАЯ ЛИТЕРАТУРА..............................................232
METHODS FOR FOREST COMMUNITY STUDY
CONTENTS
INTRODUCTION.................................................................7
PART 1. GENERAL METHODS FOR STUDY
OF FOREST COMMUNITIES AND THEIR COMPONENTS................................9
1.1 . Principles and methods of forest communities classification . . 9
1 2 Reconnaissance inspection of the territory Rout methods for study of forest
phytocoenoses.......................................................... 24
1.3 . Selection and registration of study sites in forest communities...... 33
1.4 . Techniques of sampling plots and profiles positioning within study site ... .... 37
1.5 Estimation of stand characteristics ..................... . . 42
16 Estimation of ground vegetation and regrowth characteristics.......... 55
1 7. Estimation of the epiphytic lichen cover of tree trunks and its habitats ... .63
1.8 . Investigative methods in pedography. Brief soil classification...... 68
PART 2. SPECIAL METHODS FOR STUDY
OF FOREST COMMUNITIES....................................................76
2.1. Estimation of biological production of tree layer, regrowth and shrub layer ... 76
2.2. Estimation of biological production of the ground vegetation... ... 89
2.3. Estimation of stock for commercially important plants................. 95
2.4. Estimation of seed-producing ability of plants....................... 102
2.5. Estimation of quality and vulnerability of seeds......................108
2.6. Estimation of seed banks in soil..................................... 114
2.7. Methods for plant coenopopulations study............................. 119
2.8 Estimation of biodiversity ......................................... 127
2.9. Methods for moss study ............................................ 130
2.10 Estimation of plant underground organs. .........................139
PART 3. METHODS FOR STUDY FOREST COMMUNITIES
IN THE PERTURBED SITES...............................................154
3 1 Injury diagnostics and estimation of tree and stand living status tn polluted areas . . 154
3 2 Using of epiphytic lichen cover of tree trunks for diagnostics of air pollution ... 165
3 3 Using of mosses for sites indication .................................................. 170
3 4 Chemical analysis of soils and plants......................................... 181
3 5 Principles and methods of analysis of fire age and periodicity................ 201
PART 4. BASIC PRINCIPLES OF STATISTICS.....................................................214
REFERENCES...................................................................................232
6
ВВЕДЕНИЕ
Бореальные леса выполняют важнейшие биосферные функции и имеют боль-
шое экономическое значение. На протяжении последнего столетия они испыты-
вают все возрастающие нагрузки в результате интенсивного хозяйственного ос-
воения (различные виды рубок, гидролесомелиорация, побочное пользование и
др.), лесных пожаров и воздействия локального и регионального промышленно-
го загрязнения. Перед современной лесной фитоценологией и экологией стоят
задачи детального исследования особенностей реакции лесных биогеоценозов
на изменение характеристик внешней среды, совершенствования методов изу-
чения их структуры и динамики, биоиндикации промышленного загрязнения, а
также прогнозирования последствий различных антропогенных и природных на-
рушений.
Решение этих задач на современном уровне имеет особое значение в связи с
тем, что продолжающееся разрушение лесов может иметь глобальные катастро-
фические последствия, так как на земном шаре ненарушенные леса являются
одним из основных факторов биотической регуляции среды: поддержания ста-
бильных, пригодных для жизни параметров биосферы.
Предлагаемая монография является результатом опыта многолетних комплек-
сных полевых и экспериментальных исследований коллектива авторов, изучав-
ших структуру и динамику естественных и нарушенных под влиянием антропо-
генных воздействий лесных сообществ Европейского Севера и Северо-Запада
В процессе этих исследований использовались разнообразные методы, существу-
ющие в геоботанике и экологии, создавались их модификации и разрабатыва-
лись новые методики в соответствии с конкретными задачами. При этом основ-
ными принципами исследований были принцип комплексности и принцип коли-
чественной оценки (измерения) параметров исследуемых объектов — раститель-
ных сообществ, их компонентов и характеристик местообитания.
Для получения надежных, репрезентативных данных существенное значение име-
ет достижение правильного представления о разнообразии лесных сообществ на изу-
чаемой территории, их принадлежности к различным стадиям динамических рядов,
что обеспечивает правильный подбор объектов исследования. Поэтому в данной ра-
боте уделено внимание рассмотрению основных принципов и методов классифика-
ции лесных сообществ, а также методов подбора и закладки постоянных и времен-
ных пробных площадей и учетных площадок (разд. 1). Среди методов изучения ком-
понентов лесных сообществ прежде всего рассмотрены основные, базовые, позво-
ляющие получить данные о наиболее общих, фундаментальных характеристиках раз-
личных ярусов фитоценозов, внеярусной растительности и лесных почв
7
Кроме того, в данной монографии представлен ряд специальных методов ис-
следования компонентов лесных сообществ (разд. 2), использование которых
диктуется необходимостью получения детальных характеристик объекта или спе-
циальными задачами.
Некоторые подходы и методики, необходимые при изучении структуры и ди-
намики сообществ, подверженных хроническому воздействию различных вне-
шних факторов нарушения или восстанавливающихся после кратковременных
воздействий, рассмотрены в разд. 3. Авторы стремились обосновать мысль о том,
что оценка степени нарушенности лесных сообществ является необходимым эта-
пом любых стационарных и маршрутных геоботанических и экологических ис-
следований. Практическая значимость такого подхода определяется тем, что ра-
стительность является одним из наиболее чувствительных индикаторов состоя-
ния природных комплексов и биосферы в целом.
Большое значение корректного применения статистических методов обработки
количественных данных, выбора способов представления результатов в научных
работах, а также известные авторам наиболее распространенные ошибки и труд-
ности у начинающих исследователей явились основанием для включения в по-
собие разд. 4, посвященного обзору некоторых важных вопросов использования
методов математической статистики в геоботанических и экологических иссле-
дованиях.
В задачи данного издания входило представление основных методических
принципов и конкретных методов классификации, исследования структуры, ди-
намики и оценки состояния лесных сообществ. Безусловно, ряд важных конк-
ретных методических вопросов не был затронут в связи с ограниченным объе-
мом книги. В первую очередь авторы старались представить те методы изуче-
ния лесных сообществ, апробация или разработка которых осуществлялась ими
лично в течение многих лет. К сожалению, ограниченный объем издания не
позволил представить полный список процитированной литературы, поэтому в
конце книги приводятся только наиболее важные обобщающие работы.
Авторы выражают надежду, что эта книга в определенной мере восполнит по-
стоянно существующий дефицит методической литературы и послужит поводом
для последующих изданий, целью которых будет совершенствование и развитие
предлагаемых методов исследования, а также создание новых методических раз-
работок. Мы заранее выражаем искреннюю признательность всем, кто выска-
жет замечания и пожелания по ее улучшению.
8
РАЗДЕЛ 1. ОБЩИЕ МЕТОДЫ ИЗУЧЕНИЯ
ЛЕСНЫХ СООБЩЕСТВ И ИХ КОМПОНЕНТОВ
1.1. ПРИНЦИПЫ И МЕТОДЫ КЛАССИФИКАЦИИ
ЛЕСНЫХ СООБЩЕСТВ 1
Изучение совокупности любых природных объектов включает стадию накоп-
ления информации и последующее выделение групп объектов, сходных по оп-
ределенному набору признаков, принимаемых как наиболее важные, показатель-
ные. Таким образом, производится выделение типов объектов и при необходи-
мости их дальнейшее последовательное объединение в более крупные группы,
т. е. строится их классификация.
Классификация лесных сообществ необходима для представления о разнооб-
разии, сходстве и различиях лесной растительности различных регионов, для кар-
тографирования лесной растительности, для оценки особенностей влияния фак-
торов местообитания на лесные растительные сообщества, для рационального
ведения лесного хозяйства и выбора стратегии охраны лесной растительности.
Различные направления в классификации лесных сообществ
Растительные сообщества характеризуются рядом признаков, которые прини-
маются в качестве критериев при разработке классификаций. Наиболее важны-
ми из них являются:
• флористический состав — набор видов, слагающих сообщества;
• структурно-ценотические особенности — вертикальное (ярусы, пологи) и
горизонтальное (синузии, микрогруппировки) строение; количественное соотно-
шение популяций видов каждого яруса (доминанты, содоминанты, второстепен-
ные виды); статус (роль) популяций видов в сообществе (эдификаторы, соэди-
фикаторы, ассектаторы);
• эколого-ценотические особенности — приуроченность сообществ к опре-
деленным условиям местообитания (положение в рельефе, гидрологический
режим, свойства почвообразующих пород); количественное соотношение эколо-
гических групп видов в составе сообществ, положение сообществ в экологичес-
ких рядах;
• динамический статус — положение сообществ в сукцессионных рядах
Первые попытки классификации лесов, связанные с лесохозяйственными за-
дачами, известны еще с середины XIX века. В России идея о наличии различ-
ных типов лесорастительных условий появляется уже в конце 40-х годов XIX
века в исследованиях А. Варгаса де Бедемара (1848); позже идея о типах насаж-
дений (типах леса) была ясно сформулирована В. Я. Добровлянским, Д. М. Кор-
1 Авторы В. Ю Нешатаева, Н И Ставрова, В Ю Нешатаев
9
/кит кпм, (' Ф Бергером, Г Ф Морозовым, А. Крюденером как применительно
к иесоводс । ну, i ак и с позиций лесоведения и геоботаники. Г. Ф. Морозов был
первым, кк> последовательно изложил в публикациях основы лесной типологии.
I к> подход к выделению «типов насаждений» основывался на критериях общ-
1ЮС1И условий местообитания: климата, рельефа, почвенно-грунтовых условий,
.1 ипоке свойств лссообразующих пород и влияния человека. Очень важным для
дальнейшего развития лесной типологии было его указание на существование
основных природных Типов и временных типов насаждений (Морозов, 1912,
1930, 1971) Развитие лесной типологии в России, СССР и некоторых зарубеж-
ных cipanax, атакже анализ различных классификационных направлений и школ
нредоавлены в обстоятельных обзорах С. А. Дыренкова, О. Г. Чертова (1975) и
JI II Рысина (1982).
В первой половине XX века в геоботанике сформировались три основных клас-
сификационных направления: эколого-фитоценотическое, наиболее активно
развивавшееся в России и Германии, эколого-флористическое франко-швейцар-
ской школы Браун-Бланке и морфолого-статистическое шведской школы Дю-
Рие (Александрова, 1969; Нешатаев, 2001).
До настоящего времени нет единого подхода к выделению классификацион-
ных единиц лесной растительности. Несомненно, классификация лесных расти-
юльных сообществ должна учитывать все их наиболее существенные признаки:
наличие древесной эдификаторной синузии, характерные черты структуры, ко-
личественное соотношение видов в пределах разных ярусов, положение в дина-
мических рядах, особенности связи с определенными условиями местообита-
ния
На таком подходе основаны принципы эколого-фитоценотического направ-
ления классификации лесных сообществ, которое разрабатывалось и разрабаты-
ваемся представителями Московско-Ленинградской (Санкт-Петербургской) гео-
бшаничсской школы В. Н. Сукачевым (1928, 1931, 1939, 1964), А. А. Корчаги-
ным (1940, 1946), А. А. Ниценко (1961, 1971), А. П. Щенниковым (1962, 1964),
В И. Василевичем (1961, 1985), Н. В. Дылисом (1964), А. И. Уткиным совмес-
пю с II В. Дылисом (1968), Л. П. Рысиным (1971, 1974, 1975), И. X. Блюмента-
лем (1990), В. Ю. Нешатаевым (2001) и др. При выделении основных классифи-
кационных единиц на первый план выдвигаются признаки растительного сооб-
щес шва, хотя значение придается и комплексу факторов местообитания. В каче-
С1не основной таксономической единицы при классификации лесных фитоце-
ноюв В. Н. Сукачев рассматривал тип леса. Сущность своего подхода он отра-
шл в следующем определении: «Тип леса или лесная ассоциация объединяет все
насаждения (лесные сообщества), однородные по видовому составу всех их яру-
сов, по сложению и жизненным свойствам, характеризующиеся биологически
однородными местообитаниями» (Сукачев, 1928). Реализация эколого-фитоце-
1Ю1пческого подхода к классификации лесных сообществ отражена В. Н. Сука-
10
чевым (1939) в широко известной схеме эдафо-фитоценотических рядов, выпоп-
ненной на примере еловых лесов, и в обобщенной схеме типов леса, нрсдс! ви-
ляющей «эдафо-фитоценотические ареалы» ряда основных лссообразующич
пород.
С появлением теории биогеоценологии (Сукачев, 1942, 1947) тип леса опре-
деляется В. Н. Сукачевым и его последователями не как тип лесного фитоцено-
за (синоним лесной ассоциации), а как тип лесного биогеоценоза, т. е системы,
включающей не только растительное сообщество, но и другие как живые, так и
неживые компоненты, и отражающей характер их материально-энергетических
связей. В соответствии с этим в рамках одного типа леса объединяются отдель-
ные лесные биогеоценозы, однородные «по составу древесных пород, по дру-
гим ярусам растительности и фауне, по микробному населению, по климатичес-
ким, почвенным и гидрологическим условиям... по восстановительным процес-
сам и направлениям смен в них» (Сукачев, 1964).
Безусловно, биогеоценотический, системный подход является, с теоретичес-
кой точки зрения, более правильным не только при изучении лесных сообществ,
но и тундровых, степных и др. Однако он очень сложен для применения на прак-
тике; в большинстве случаев в классификациях лесных биогеоценозов при оп-
ределении таксономических единиц с достаточной полнотой учитываются ха-
рактеристики только двух компонентов — растительного сообщества и экотопа,
т. е. те же основные признаки, что и при классификации лесных фитоценозов
Эколого-фитоценотическое направление в классификации, по нашему убеж-
дению, является более целесообразным при изучении лесных сообществ по срав-
нению с эколого-флористическим и морфолого-статистическим, так как отли-
чается легкостью распознавания классификационных единиц в природе, позво-
ляет учитывать при их выделении наиболее существенные признаки лесных со-
обществ: наличие древесной эдификаторной синузии, структуру фитоценозов и
ценотическую роль различных компонентов сообществ (через проективное по-
крытие, встречаемость), условия местообитания.
Следует подчеркнуть, что многие из названных выше исследователей отме-
чали важность учета динамического статуса лесных сообществ при их типо-
логии и классификации.
Динамический подход к классификации лесных сообществ характеризуется
рассмотрением основной типологической единицы — типа леса (или серии ти-
пов леса) как динамической системы. Сущность динамического подхода впер-
вые была сформулирована в работах А. К. Каяндера (Cajander, 1926), С. Я Со-
колова (1929), Б. А. Ивашкевича (1933). Они считали, что при выделении клас-
сификационных единиц должны объединяться лесные сообщества, сменяющие
друг друга в возрастных и восстановительных рядах в тех или иных конкрет-
ных условиях местопроизрастания. Эта идея была развита в работах Б 11 Ко-
лесникова (1956, 1958а, б, 1961, 1974), И. С. Мелехова (1968, 1976, 1988).
1 1
В. Б. Сочавы (1972). В последние десятилетия динамический подход к класси-
фикации лесных сообществ разрабатывался Л. П. Рысиным (1983), В. Н. Федор-
чуком (1979, 1992), В. С. Ипатовым с сотрудниками (1990; 1991; 1992; 1999;
2001) С позиций биогеоценотического подхода Л. П. Рысин (1983) определяет
тип леса как совокупность лесных биогеоценозов (участков леса), объединен-
ных общностью условий местообитания и трансформирующихся в процессе ес-
тественного развития в один и тот же коренной тип лесных биогеоценозов. Это
означает, что тип леса может включать сообщества, образованные разными дре-
весными породами, в тех случаях, когда в процессе развития происходит смена
пород.
Один из вариантов классификации типов леса с учетом динамического аспекта
разработан группой санкт-петербургских (ленинградских) ученых — В. Н. Фе-
дорчуком, С. А. Дыренковым, О. Г. Чертовым и их последователями для целей
лесоведения и лесного хозяйства. Они считают, что при выделении лесотиполо-
гических единиц важное значение имеют признаки условий местообитания лес-
ных сообществ, которые должны учитываться в равной мере с признаками
растительности. На основе указанного подхода выделяются серии, или семей-
ства, типов леса, которые являются основной единицей в данной типологичес-
кой классификации и объединяют все лесные биогеоценозы и производные от
них вырубки и гари в сходных местообитаниях (Чертов, Дыренков, 1972; Ды-
ренков, 1973, 1982; Федорчук, Дыренков, 1975; Федорчук, 1976, 1979, 1992). Типы
леса в пределах семейств выделяются по преобладающей древесной породе и
характеристикам местообитания. Степень сходства местообитаний определяет-
ся исходя из целевого назначения типологической классификации. Обычно в одну
серию типов леса включают лесные биогеоценозы и вырубки, формирующиеся
на месте коренного или длительно производного типов леса.
Ниже будут охарактеризованы особенности подходов к классификации лес-
ных фитоценозов и. биогеоценозов, применяемых санкт-петербургскими геобо-
таниками, лесоведами и экологами и используемых в своей работе авторами дан-
ного методического руководства.
1.1.1. Эколого-фитоценотическая классификация лесных сообществ'
Основные синтаксономические единицы. Основной классификационной
единицей лесной растительности является ассоциация — абстрактное понятие,
объединяющее конкретные лесные фитоценозы, сходные по совокупности при-
знаков. Определение ассоциации, принятое Брюссельским ботаническим конг-
рессом (1910), устанавливает следующие ее важнейшие признаки: флористичес-
кий состав, физиономия (строение, внешний вид) и условия местообитания.
В рамках эколого-фитоценотического направления русской геоботанической
1 Автор В Ю Нешатаева
12
школы принято следующее определение ассоциации: «к одной ассоциации от-
носятся фитоценозы, сходные по господствующим в каждом ярусе видан рас -
тений, находящимся в близких количественных соотношениях, при значитель-
ном сходстве состава сопутствующих видов и их количественных соотноше-
ний» (Шенников, 1964). Иными словами, к одной ассоциации относятся фито-
ценозы сходного состава и строения, отражающего сходство взаимоотноше-
ний между растениями в сходных условиях местообитания Близкие по этим
признакам сообщества имеют общие статические (структурные, физиономичес-
кие) и динамические черты: сходные амплитуды сезонных и разногодичных из-
менений, сходное положение в сукцессионных рядах, сходную реакцию на вне-
шние воздействия (Шенников, 1964).
Таким образом, основой для объединения сообществ в одну ассоциацию яв-
ляется общность видового состава доминантов и характерного ядра сопряжен-
ных видов, встречающихся в фитоценозах, относящихся к данной ассоциации
на всем ареале ее распространения. Следовательно, ассоциация рассматривает-
ся как синтаксономическая единица низшего ранга, имеющая определенный гео-
графический ареал и являющаяся основной единицей для построения иерархи-
ческих классификаций. При детальных исследованиях растительности рекомен-
дуется использовать синтаксоны, подчиненные ассоциации, например субассо-
циации и др. (Резолюция..., 1972).
Субассоциации выделяются по следующим признакам: различия в количе-
ственных соотношениях доминантов и содоминантов, наличия нехарактерных для
ассоциации в целом содоминантов, а также некоторых сопутствующих видов,
особенностей экологических условий местообитания. Следовательно, субассо-
циации — это экологические варианты ассоциации. Различаются субассоциации
климатогенные, эдафогенные, региональные и сукцессионные (Шенников, 1964)
Основной единицей среднего ранга является формация, выделяемая по домини-
рующему виду (видам) господствующей эдификаторной синузии К одной форма-
ции относятся ассоциации, в которых господствующий ярус сложен одним и тем
же доминирующим видом (видами) (Шенников, 1964). При выделении формации
рекомендуется учитывать не только доминирующие виды, но также принимать во
внимание и другие признаки сообществ: общность доминанта климаксовой серии
для серийных сообществ, ботанико-географический и эколого-ценотический ареал
эдификатора, особенности структуры сообществ и т. п. (Резолюция..., 1972).
Основной синтаксономической единицей высшего ранга является тип рас-
тительности, выделяемый на физиономически-экологической основе по пре-
обладающей экобиоморфе (или набору экобиоморф) господствующей эдифика-
торной синузии. К одному типу растительности относят все формации, сооб-
щества которых в господствующем ярусе сложены одной и той же экобиомор-
фой.
Диагностические признаки ассоциаций. При исследовании свойств конк-
13
ретных растительных сообществ (фитоценозов) и ассоциаций можно выделить
две группы признаков — аналитические (частные) и синтетические (общие)
Частные признаки характеризуют отдельные конкретные фитоценозы — видо-
вой состав, количественные соотношения видов, особенности структуры и дав-
ность нарушения (положение в динамическом ряду). Общие признаки характе-
ризуют ассоциацию, их можно установить методом сравнения фитоценозов, от-
носимых к данной ассоциации. Эти признаки в совокупности составляют диаг-
ноз ассоциации, поэтому они называются диагностическими признаками ассо-
циации (Шенников, 1964).
Диагностические признаки ассоциации устанавливают на основе сравнитель-
ного изучения нескольких фитоценозов в сводной таблице геоботанических опи-
саний (табличный анализ). Таблица представляет собой сводку описаний и со-
держит информацию о видовом составе и ярусном строении (строки) и обилии
(проективном покрытии) видов разных ярусов для каждого конкретного фито-
ценоза (столбцы).
По таблице определяют общее количество видов в ассоциации, характеризую-
щее ее флористическую насыщенность и являющееся диагностическим призна-
ком. Таблица дает возможность установить для каждой пары фитоценозов коэф-
фициент флористической общности (в %), для расчета которого можно использо-
вать какой-либо из стандартных показателей (коэффициенты Жаккара, Коула, Бра-
вэПирсопа и др.). На основе данных таблицы можно вычислить средний ко-
эффициент общности флористического состава ассоциации. Чем больше его
значение, тем более однородны по флористическому составу фитоценозы ас-
социации, степень флористической однородности также является диагности-
ческим признаком ассоциации. Кроме того, таблица позволяет охарактеризо-
вать ассоциацию по степени постоянства присутствия в ней тех или других
видов, т. е. установить константность видов — важный диагностический при-
знак ассоциации. Константность определяется по количеству фитоценозов, в
которых встречен данный вид, и выражается в процентах от общего количе-
ства описаний. Обычно различают пять классов константности - I — 1-20 %,
II — 21-40 %, III — 41-60 %, IV — 61-80 %, V — 81-100 %. Проанализиро-
вав количество и состав доминирующих видов господствующего яруса и под-
чиненных ярусов, можно определить, является ли данная ассоциация монодо-
мииаитной или полидоминантной, а также установить особенности ее сложе-
ния и структуры (Нешатаев, 1987). Эти характеристики также являются диаг-
ностическими признаками ассоциации.
Содержащиеся в таблице (или в приложении к ней) сведения об особеннос-
тях местообитания каждого фитоценоза дают возможность составить сводную
характеристику условий местообитания лесных сообществ данной ассоциации,
а 1акжс проанализировать воздействие на сообщества ассоциации антропоген-
ных факторов (например, при сравнении сообществ с различной интенсивнос-
14
тью хозяйственного воздействия).
Таблица является основным документом при выделении и описании ассоци-
ации, позволяющим судить, насколько оправданы трактовка данной ассоциации,
ее объем, варьирование диагностических признаков и т. п. При достаточном ко-
личестве описаний количественные характеристики видов и сообществ ассоци-
ации можно проанализировать статистическими методами. Публикация фитоце-
нотических таблиц позволяет сравнить ассоциации, выделенные разными авто-
рами в различных географических районах, выявить их общие черты и регио-
нальные различия. В геоботанической литературе имеется немало примеров
детального анализа больших массивов описаний с приведением полных фито-
ценотических таблиц и разработкой детальной классификации.
Правила наименования формаций, ассоциаций и субассоциаций. Клас-
сификация растительности, использующая комплекс признаков растительных
сообществ, требует точной, понятной и удобной номенклатуры. Система ла-
тинских наименований растительных ассоциаций и формаций была впервые
предложена в 1896 г. Сернандером и впоследствии развита и дополнена мно-
гими другими геоботаниками. В то же время, отсутствие четко сформулиро-
ванных номенклатурных правил длительное время являлось тормозом для ус-
пешного развития и совершенствования эколого-фитоценотического направ-
ления классификации. Попытка обобщить накопленный опыт и сформулиро-
вать единые правила наименования синтаксонов была предпринята В. Ю Не-
шатаевым, предложившим в 1989 г. Проект Всесоюзного кодекса фитоценоло-
гической номенклатуры. В последующие годы поступило много предложений
по его усовершенствованию и дополнению. В новой редакции Проект Всерос-
сийского кодекса фитоценологической номенклатуры опубликован в 2001 г.
(Нешатаев, 2001).
Название формации образуют из наименования доминирующего вида (или
содоминирующих видов) с прибавлением к корню родового названия суффикса
«-eta». Например: сосняки — Pineta, ельники — Piceeta, березняки — Betuleta,
ольховники — Alneta и т. п. Если необходим видовой эпитет, то его приводят в
родительном падеже (например, Pineta sylvestris, Piceeta abietis, Betuleta
pubescentis). Если название формации образуется из названий двух содоминиру-
ющих видов, то к родовому названию вида, стоящему на первом месте, прибав-
ляют суффикс «-eto», а к родовому названию вида, стоящему на втором месте
— суффикс «-eta». Между названиями видов ставится дефис. Например: елово-
березовые леса — Piceeto-Betuleta, березово-сосновые леса — Betuleto-Pineta,
липово-дубовые леса — Tilieto-Querceta и т. д. Если необходимо использовать
видовые эпитеты при наименовании формаций, оба видовых эпитета приводят-
ся в родительном падеже (например, Piceeto abietis-Betuleta pubescentis, Betuleta
pubescentis-Pineta sylvestris и т. п.).
При наименовании ассоциаций используют бинарные названия, состоящие
15
из существительного, — названия формации, в котором суффикс «-eta» заменя-
ется на суффикс «-etum» — и прилагательного (эпитета), указывающего на до-
минанта (или содоминантов) подчиненного яруса. Например: сосняк лишайни-
ковый — Pinetum cladinosum, ельник-черничник — Piceetum myrtillosum, берез-
няк вейниковый — Betuletum calamagrostidosum. При необходимости можно ука-
зать два содоминирующих вида подчиненных ярусов, тогда к основе латинского
родового названия первого вида добавляется суффикс «-oso», например: сосняк
багульниково-сфагновый — Pinetum ledoso-sphagnosum, сосняк воронично-ба-
гульниковый — Pinetum empetroso-ledosum. Видовые эпитеты доминантов под-
чиненных ярусов при необходимости могут приводиться или быть заключены в
скобки, а при последующем упоминании могут быть опущены. Например: со-
сняк голубично-багульниковый — Pinetum vaccinioso uliginosii — ledosum palustris
или Pinetum vaccinioso (uliginosii) — ledosum (palustris), далее — Pinetum
vaccinioso-ledosum.
Название суб ассоциации, используемой в рамках фитоценотического подхо-
да, состоит из названия ассоциации, к которой она относится, и эпитета, соеди-
ненных сокращенным словом «subass», эпитет может быть образован от назва-
ния какого-либо вида, характеризующего субассоциацию, либо может являться
прилагательным, характеризующим экологические, структурные и другие при-
знаки субассоциации или ее географическое распространение. Например: сосняк
черничный на песках — Pinetum myrtillosum subass. arenosum, ельник сфагно-
вый с брусникой — Piceetum sphagnosum subass. vacciniosum vitis-idaea.
1.1.2. Принципы динамического подхода к классификации лесных
сообществ1
Согласно рассматриваемому подходу, основная классификационная единица
лесной растительности представляет собой абстрактную динамическую систе-
му, состоящую из совокупности элементов — стадий, Последовательно сменяю-
щих друг друга в рядах формирования, восстановления (или разрушения) ос-
новной климаксовой стадии, характеризующейся наиболее полным соответствием
условиям данного экотопа1 2 (местообитания). Климаксовая стадия характеризу-
ется стабильностью всех основных характеристик лесного сообщества при ус-
реднении во времени и пространстве. Это означает, что на фоне постоянно иду-
щих локальных изменений на отдельных участках (микросукцессий) и в отдель-
ные периоды (под влиянием климатических флуктуаций) в среднем для всего
сообщества эти характеристики остаются постоянными. Согласно результатам
1 Автор Н И Ставрова
2 Под экотоном понимается совокупность факторов абиотической среды, не изменя-
ющихся под влиянием сообществ (климат, рельеф, гидрологический режим, свойства поч-
вообразующих пород).
16
исследования восстановительной динамики Европейских бореальных лесов, вре-
мя восстановления и стабилизации основных характеристик сообществ после
полного разрушения составляет примерно 200-250 лет (Корчагин, 1954; Пуш-
кина, 1960; Кулешова и др., 1996; Горшков, 2001), т. е. бореальные лесные сооб-
щества с такой давностью последнего нарушения могут рассматриваться как кли-
максовые или близкие к ним.
Каждая стадия динамического ряда представляет собой тип состояния лесно-
го сообщества в конкретных условиях местообитания, который характеризуется
определенной давностью последнего нарушения, отличается определенным на-
бором лесообразующих видов, определенной видовой, возрастной и вертикаль-
ной структурой древостоя, составом и соотношением видов подчиненных яру-
сов и т. д. Следует признать, что у сторонников динамического подхода к клас-
сификации лесных сообществ нет единого мнения в отношении объема и но-
менклатуры выделяемых стадий и, соответственно, основных классификацион-
ных единиц. Так, Б. П. Колесников считал низшей единицей динамической (ге-
нетической по Б. П. Колесникову) классификации лесной растительности «тип
насаждения», который объединяет «участки леса, принадлежащие к одноимен-
ным стадиям возрастных или восстановительных смен и однородные по комп-
лексу лесорастительных условий, по составу древесных пород, по другим яру-
сам растительности и фауне и по взаимоотношениям растений со средой...» (Ко-
лесников, 1956). В частности, в возрастном цикле кедра корейского, начиная с
его появления под пологом, он выделял 8 стадий — типов насаждений в соот-
ветствии с видовой и возрастной структурой древесного яруса и ролью рассмат-
риваемого поколения кедра в его составе.
К одному типу леса, который является основной классификационной едини-
цей, по Б. П. Колесникову (1958а, б), следует относить типы насаждений, пред-
ставляющие все стадии одного цикла возрастных или коротко восстановитель-
ных [без смены пород] смен, совершающихся в пределах территорий, сходных
по положению и свойствам. По мнению Б. П. Колесникова, в полном научном
названии типа леса должны быть отражены все его диагностические показате-
ли. На практике сторонники динамического подхода придерживаются следую-
щего принципа номенклатуры: название эдификатора (эдификаторов) с указани-
ем доминантов нижних ярусов и условий местообитания.
При наличии длительно восстановительных смен [при смене пород], в преде-
лах определенного типа местообитания выделяется генетический ряд (серия)
типов леса. Б. П. Колесников полагал, что такой подход к классификации лес-
ной растительности позволяет не только проводить инвентаризацию лесных со-
обществ, но и учитывать «ведущие закономерности лесообразовательного про-
цесса». По общности основного доминанта древесного яруса типы леса в клас-
сификации Б. П. Колесникова объединяются в лесные формации.
И. С. Мелехов (1968, 1976, 1988) так же, как Б. П. Колесников, подходил к
17
определению типа леса как динамической системы. Именно И. С. Мелехов ввел
понятие «динамическая типология». Согласно его определению, лесные сообще-
ства, относящиеся к одному типу леса, отличаются общностью морфологии,
происхождения, общими особенностями лесорастительных условий и тенден-
ций развития Одной из наиболее важных характеристик типа леса И. С. Меле-
хов считает его динамичность. В соответствии с этим он выделял этапы (ста-
дии), предшествующие формированию собственно лесного сообщества (типы
гарей, вырубок), этапы формирующегося типа леса и этап сложившегося типа
леса. Если этот процесс происходит без существенных изменений в составе и
продуктивности древостоя, то, согласно И. С. Мелехову, речь идет о формирова-
нии одного типа леса. Если происходит смена доминирующих видов в составе
древостоя и нижних ярусов сообществ, то выделяется ряд типов леса. Например,
на вырубке ельника зеленомошно-черничного формируются последовательно два
типа леса: березняк, затем ельник. Следует отметить, что И. С. Мелехов большое
внимание уделил разработке типизации сообществ, формирующихся на началь-
ных стадиях восстановления лесных сообществ после рубок (Мелехов, 1959).
В классификации лесных сообществ, предлагаемой В. С. Ипатовым с сотруд-
никами (1990; 1991; 1992; 2001), выделяемые классификационные единицы име-
ют несколько иной объем, однако принцип подхода к построению классифика-
ционной схемы тот же. Для обозначения отдельных стадий динамического ряда
В. С. Ипатов предлагает использовать термин, принятый для основной класси-
фикационной единицы в геоботанике — ассоциация1 . При этом к одной ассоци-
ации относятся одинаковые стадии в динамических рядах. В таком понимании
ассоциация соответствует типу насаждения по Б. П. Колесникову. Ассоциа-
ции, объединяемые одним эдификаторным видом, образуют биотопическую си-
стему ассоциаций (т. е. систему ассоциаций с одинаковым характером биотопа,
формируемым одним и тем же эдификатором), которая соответствует типу леса
в классификации Б П. Колесникова Совокупность биотопических систем ассо-
циаций, объединенных одним и тем же экотопическим климаксом, согласно
В. С. Ипатову, представляет собой экотопическую систему ассоциаций (т. е. си-
стему ассоциаций, развивающуюся в условиях одного экотопа) — тип леса (ге-
нетическую серию типов леса по Б. П. Колесникову).
Таким образом, по определению В. С. Ипатова, к одному типу леса относят-
ся лесные сообщества, произрастающие в сходных условиях экотопа и имею-
щие общий климаксовый эдификатор — один вид или комбинацию видов древес-
ных растений (Ипатов, 1990). Как видно из изложенного, Б. П. Колесников и
В. С. Ипатов понимают тип леса в несколько разном объеме, однако оба вариан-
та представленных классификаций отражают принцип динамического подхода.
1 В. С Ипатов выделяет еще более низкую по рангу единицу — «социацию», кото-
рую он рассматривает как «оперативную», используемую только на стадии первичной
сортировки материала для упорядочения больших массивов описаний.
18
Следует отметить, что под сходными условиями экотопа подразумеваются
биологически равноценные сочетания конкретных факторов абиотической сре-
ды, под влиянием которых формируются одинаковые климаксовые типы леса (или
серии типов леса по Б. П. Колесникову). Отдельные факторы конкретных мес-
тообитаний могут при этом несколько различаться, в частности, некоторые ха-
рактеристики почвообразующих пород, на которых формируются сосняк лишай-
никовый или ельник чернично-зеленомошный, могут различаться в разных по
климатическим характеристикам регионах. Могут наблюдаться и различия в со-
ставе эдификаторных видов древесных растений на промежуточных этапах вос-
становления одного и того же климаксового типа леса. Формирование разных
динамических рядов с одинаковым заключительным типом леса в сходных мес-
тообитаниях связано с действием разных по характеру и интенсивности преоб-
разования исходного экотопа нарушающих факторов', рубок, пожаров, массо-
вых ветровалов, а также влиянием случайных причин (наличие или отсутствие
семян данного вида, погодные флуктуации в начальный период восстановления
И т. д.)
Некоторые исследователи не считают оправданным использование в динами-
ческой классификации лесных сообществ, предложенной В. С. Ипатовым, тер-
мина «ассоциация», уже применяемого в определенном смысле и объеме при
классификации растительности. Учитывая замечания, сделанные в начале раз-
дела, с этим можно согласиться лишь отчасти. Следует признать не совсем удач-
ным термин «тип насаждения», особенно применительно к начальным стадиям
восстановления после полного разрушения древесного яруса лесных сообществ
под влиянием естественных или антропогенных катастрофических факторов.
Однако эти терминологические проблемы вполне преодолимы. Например, для
обозначения классификационных единиц разного ранга могут быть использова-
ны термины, предложенные С. Я. Соколовым, Б. П. Колесниковым и И. С. Ме-
леховым: стадия, тип леса и серия типов леса.
Следует отметить, что описанные выше варианты динамической классифика-
ции лесных сообществ по сути принципа, положенного в их основу, не предназ-
начены для классификации культур местных или интродуцированных видов дре-
весных растений. Сообщества на территориях, где характеристики конкретного
местообитания (например, положение базиса эрозии, уровень грунтовых вод, хи-
мические свойства почв) не являются в среднем постоянными на протяжении
достаточно длительного периода (по крайней мере нескольких сотен лет), а под-
вержены постоянному изменению под влиянием природных процессов или че-
ловеческой деятельности, формируют динамический ряд с гипотетической (по-
тенциальной) заключительной стадией, которая может быть реализована при пре-
кращении действия фактора, нарушающего стабильность экотопа.
На основе принципов динамической классификации в пределах типа леса со-
сняка лишайникового могут быть выделены, например, стадии: а) сосняка ли-
19
шайниково-верескового с давностью пожара 20 лет и доминированием в мохо-
во-лишайниковом покрове пионерных видов мхов, бокальчатых и трубчатых форм
лишайников, а в травяно-кустарничковом ярусе — вереска обыкновенного; б)
сосняка чисто лишайникового с давностью пожара 150 лет и доминированием в
мохово-лишайниковом ярусе кустистого лишайника Cladina stellaris, а в травя-
но-кустарничковом — брусники и вороники; с) сосняка мертвопокровно-толок-
нянкового (с погибшим мохово-лишайниковым ярусом и доминированием в тра-
вяно-кустарничковом ярусе толокнянки) в зоне разрушения лесных экосистем
под влиянием промышленного загрязнения и др.
Подводя итог обсуждению принципов динамического подхода к классифика-
ции лесных сообществ, следует еще раз подчеркнуть важность учета не только
особенностей их состава и строения, но и динамического статуса, взаимной связи
в динамических рядах, связи с определенным комплексом факторов местооби-
тания, которые представляют собой фундаментальные характеристики изу-
чаемых объектов
1.1.3. Комбинированный метод выделения и распознавания типов леса'
Типологические классификации лесов создаются для многоцелевого исполь-
зования в лесном хозяйстве и лесоводстве. Назначение типологической класси-
фикации — лаконичное объяснение природы анализируемых объектов по их по-
ложению в системе. Важное значение придается типологической классификации
как инструменту генерации и проверки гипотез о закономерностях изменения в
пространстве и во времени характеристик лесных биогеоценозов. Всякое лесо-
водственное и лесохозяйственное исследование должно начинаться с привязки
исследуемых объектов к имеющейся лесной типологии и заканчиваться указа-
нием типологических единиц, для которых его результаты справедливы.
Рассматриваемые ниже методы выделения и распознавания типов леса разра-
ботаны в Ленинградском (Санкт-Петербургском) НИИ лесного хозяйства
(СПбНИИЛХ) В. Н. Федорчуком и его сотрудниками (Федорчук и др., 1974;
Федорчук, Дыренков, 1975, Федорчук, 1976а,б, 1979, 1992; Федорчук и др., 1981;
Федорчук, Бурневский, 1986; Нешатаев, Федорчук, 1989). Метод выделения ле-
сотипологических единиц СПбНИИЛХ является вариантом комбинированного
метода исследования растительности и ее местообитаний, обоснованного и раз-
работанного Л. Г. Раменским (1938).
Объектами лесотипологической классификации являются лесные биогеоце-
нозы и производные от них биогеоценозы вырубок и гарей. Метод типологичес-
кой классификации заключается в сведении многообразия объектов и их при-
знаков к конечному числу лесотипологических единиц таких, что: 1) они макси-
мально сходны внутри себя по заданным хозяйственно значимым признакам;
Автор В Ю Нешатаев.
20
2) отличаются друг от друга по минимальному числу легко учитываемых при-
знаков, необходимых и достаточных для их распознавания (отбор наиболее ин-
формативной системы показателей).
Для рассматриваемого направления характерно четкое выделение целей клас-
сификации, ранжирование их по важности, определение конкретных способов
использования классификации. Основная цель классификации — прогноз для
каждого конкретного участка лесного биогеоценоза на основе его типологичес-
кой принадлежности всех возможных состояний растительности, в особенности
продуктивности и устойчивости древостоев как в процессе естественной дина-
мики, так и при хозяйственном вмешательстве. Из других наиболее важных за-
дач, для решения которых используется лесная типология, следует назвать вы-
бор главной породы (древесной породы, выращивание которой в данных усло-
виях наиболее экономически выгодно), целесообразного в хозяйственном отно-
шении состава древостоя и способов его формирования, выбор способа мелио-
рации местообитания и оценка его возможной эффективности, выбор способа
лесовосстановления после рубки.
Основной типологической единицей классификации СПНИИЛХ является се-
рия (семейство) типов леса. В одну серию типов леса включают относительно
устойчивый (коренной, длительно производный или условно коренной) тип лес-
ного биогеоценоза и лесные биогеоценозы вырубок и гарей, которые переходят
в него в течение периода лесовыращивания (продолжительность жизни одного
поколения преобладающей древесной породы) при условии действия современ-
ного режима климата и дренажа (в том числе искусственного осушения или под-
топления). Следует иметь в виду, что на месте одного коренного типа леса под
влиянием осушения, подтопления, улучшения водно-физических свойств почв
на месте брошенных сельхозугодий или обеднения почв периодическими пожа-
рами может возникать другой условно коренной тип леса, по которому и следу-
ет выделять серию типов леса, объединяющую потенциальный условно корен-
ной тип и биогеоценозы, рассматриваемые как стадии его формирования.
Типы леса выделяют в пределах серий по преобладающей древесной породе
(Резолюция..., 1973). Серии типов леса примерно соответствуют понятиям «тип
леса» лесотипологов географического направления (Колесников, 1956), «тип
лесорастительных условий», принятый в Латвии, «тип развития леса» Э. Айхин-
гера, «форма местообитания» лесотипологов Германии.
Понятие серии типов леса близко к типу леса в понимании В. С. Ипатова
(2001), но не тождественно ему. Согласно В. С. Ипатову, к одному типу леса
относятся все стадии сукцесии, идущие в направлении климакса. Полагая, что
лишайниковые сосняки на дренированных песках в Южной Карелии сменяются
климаксовыми сосняками зеленомошными, он относит и те, и другие к одному
типу леса (Ипатов и др., 1991). Исследования СПбНИИЛХ на Карельском пере-
шейке также дают основания полагать, что лишайниковые сосняки могут сме-
21
няться зеленомошными и, более того, возможно, существуют на месте еловых
лесов благодаря периодическим пожарам, обедняющим местообитания в резуль-
тате выгорания гумуса и лесной подстилки (Чертов, 1981). Однако в классифи-
кационной системе СПбНИИЛХ выделена самостоятельная лишайниковая серия
типов леса на сильно дренированных песках, поскольку процесс смены лишай-
ников зелеными мхами занимает время, по-видимому, большее, чем период ле-
совыращивания. Таким образом, в классификации СПбНИИЛХ в серию типов
леса объединяют стадии формирования или коренного, или длительнопроизвод-
ного (каким считают сосняк лишайниковый), или условнокоренного (на осушен-
ных местообитаниях, на брошенных залежах) типа, а не обязательно климаксо-
вого (как у В. С. Ипатова) типа леса.
В основу разработки типологической классификации какого-либо региона мо-
гут быть положены комплексные описания пробных площадей, заложенных с
учетом требования репрезентативности. На пробных площадях выполняют так-
сационное описание древесного яруса с инструментальным определением сред-
них характеристик элементов леса (полноты, высоты, диаметра, возраста и др.),
геоботаническое описание, определение мощности верхних почвенных горизон-
тов в 20 прикопках и описание почвенного разреза. Рекомендуется начинать изу-
чение типов леса региона с коренных и близких к ним старовозрастных участ-
ков леса на ненарушенных местообитаниях. Производные биогеоценозы реко-
мендуется изучать после того, как разработана типологическая классификация
коренных, длительнопроизводных и условно коренных биогеоценозов.
При выделении серий типов леса используются такие признаки растительно-
сти и местообитаний, которые не меняются в процессе возрастных и коротко-
восстановительных смен. Показано, что из признаков растительности наиболее
устойчивым в процессе восстановительных смен является наличие групп инди-
каторных видов (Федорчук, Кузнецова, 1995), выделяемых в процессе таблич-
ной сортировки геоботанических описаний методами школы И. Браун-Бланке
(Александрова, 1969; Миркин, 1978; Нешатаев, 1987). Использование индика-.
торных групп снимает трудности классификации и распознавания растительных
сообществ с нечетко выраженными доминантами (например, неморальных и тра-
вяноболотных лесов). В коренных лесах с ясно выраженными доминантами клас-
сификация по индикаторным группам совпадает с классификацией по доминан-
там живого напочвенного покрова. Среди производных мелколиственных насаж-
дений настолько распространена конвергенция нижних ярусов, что по их доми-
нантам трудно судить об исходной и потенциальной растительности (Ниценко,
1971). Дело в том, что многие типы нижних ярусов, выделяемые по преоблада-
ющим видам, развиваются в сериях смен на местах самых разных коренных ас-
социаций, например, вейниковые и орляковые ярусы могут возникать как на ме-
сте черничников, так и кисличников (Ниценко, 1971; Федорчук, Бурневский,
1986).
22
Экологическую однородность и своеобразие фитоценонов (бсзранговых групп
описаний), выделенных с помощью групп дифференцирующих видов, контро-
лируют, используя балльные оценки по таблицам Л. Г. Раменского (Раменский и
др., 1956). В. Н. Федорчук (1976а), показал, что совместное использование ме-
тодов И. Браун-Бланке и Л. Г. Раменского позволяет выделять дискретные, хо-
рошо распознаваемые единицы растительности с помощью групп индикаторных
видов.
Ведущими признаками местообитания для выделения серий типов леса и ти-
пов местообитаний выбраны гранулометрический состав почвы и почвообразу-
ющей породы, степень дренированности почвы, степень аллювиальности и по-
емности, наличие вечной мерзлоты и форма гумуса. Последнее понятие объеди-
няет как подстилку, так и собственно почвенный гумус. В пределах одной био-
климатической провинции эти факторы определяют формирование в каждом типе
местообитаний, как правило, одного коренного типа биогеоценоза, почвы одно-
го подтипа и определенного набора серийных и производных типов биогеоце-
нозов (Чертов, 1981).
Выделение основных типологических единиц, серий типов леса, ведут мето-
дом перекрестной разделительной таблицы (Федорчук, Дыренков, 1975). По стро-
кам таблицы размещают предварительно выделенные группы местообитаний, по
столбцам — фитоценоны. Элементами таблицы являются номера описаний, со-
ответствующих групп местообитаний и фитоценонов. Анализ и перестройка таб-
лицы ведутся таким образом, чтобы добиться более полного соответствия групп
местообитаний и фитоценонов, уменьшить количество получившихся групп опи-
саний при сохранении их однородности по увлажнению и активному богатству
почвы (по шкалам Л. Г. Раменского), по продуктивности (классу бонитета) дре-
востоев, по аналитическим показателям почвы, группам индикаторных видов. В
окончательном варианте таблицы описания, отнесенные к одной серии типов
леса, расположены в одной клетке.
Изучение хозяйственно важных свойств лесотипологических единиц возможно
вести лишь в том случае, когда обеспечено одцозначное определение разными
лицами выделенных лесотипологических единиц. Поэтому выделенные едини-
цы должны быть охарактеризованы относительно небольшим числом четких, про-
стых для распознавания в натуре и устойчивых диагностических признаков, ко-
торые коррелятивно связаны с большим числом других признаков и свойств
выделенных единиц. Для распознавания серий типов леса используются чаще
всего следующие диагностические признаки: наличие или отсутствие растений,
входящих в индикаторные группы видов, доминирование некоторых экологически
показательных видов растений или их групп, мощность подстилки и торфа, сте-
пень разложения торфа, мощность гумусового горизонта, гранулометрический
состав минеральной части почвы, характер подстилающей горной породы (Фе-
дорчук, Дыренков, 1975). Для удобства распознавания типологических единиц
23
разработаны специальные определители и характеризующие таблицы (Федор-
чук и др., 1981; Федорчук, Бурневский, 1986, раздел 5.8).
На основе собранного материала и разработанных типологических классифи-
каций создаются региональные кадастры типов леса (Региональные..., 1990),
содержащие подробную информацию о составе, продуктивности, структуре, по-
чвах и лесохозяйственных особенностях лесов по их типам. Особой проблемой
является сопоставление типологических единиц, выделенных для разных реги-
онов, и разработка обобщенной классификации типов леса для всей страны, а
также обоснование и выделение типологических единиц высокого ранга. Сле-
дует признать, что методические подходы к такому обобщению в лесной типо-
логии разработаны еще недостаточно (Федорчук, 1992).
1.2. РЕКОГНОСЦИРОВОЧНОЕ ОБСЛЕДОВАНИЕ ТЕРРИТОРИИ.
МАРШРУТНЫЕ МЕТОДЫ ИЗУЧЕНИЯ ЛЕСНЫХ ФИТОЦЕНОЗОВ 1
Комплексное изучение лесных сообществ независимо от целей и-задач иссле-
дования необходимо начинать с рекогносцировочного обследования. Рекогнос-
цировка является первым этапом, предваряющим детальные геоботанические
исследования и проведение лесоустроительных работ. Цель рекогносцировочного
обследования состоит в получении общих сведений о характере растительного
покрова территории, на которой предполагается проведение исследований, ус-
тановлении соответствия между имеющимися картографическими данными, ма-
териалами лесоустройства и действительным распределением различных типов
растительных сообществ на исследуемой территории. Обследование лесов мо-
жет производиться как самостоятельный вид работ или в качестве одного из под-
готовительных мероприятий к стационарному или полустационарному их изу-
чению.
В результате рекогносцировочного обследования должны быть получены сле-
дующие сведения:
1) общая ландшафтная характеристика района исследований;
2) состав основных лесообразующих видов, разнообразие и распределение
различных типов лесных сообществ;
3) факторы внешнего нарушения и примерная степень нарушенное™ лес-
ных сообществ;
4) имеющиеся транспортные пути, их состояние и возможность использова-
ния для более детальных маршрутных и стационарных исследований лесной
растительности;
1 Автор В Ю Нешатаева
24
5) схема маршрутных исследований с предварительно намеченными пунк-
тами закладки постоянных и временных пробных площадей.
Типы маршрутных геоботанических исследований
Маршрутные геоботанические исследования лесных сообществ, в зависимо-
сти от целей изучения, подразделяются на рекогносцировочно-маршрутные и
детально-маршрутные. Рекогносцировочно-маршрутные методы применяются
при обследовании лесов обширных, ранее не изученных территорий в целях
проведения инвентаризации растительности, разделения лесных массивов на од-
нородные участки с выделением наиболее распространенных растительных ас-
социаций (или типов леса) и предварительного учета их площадей. При этом
исследуемый район покрывается редкой сетью маршрутов, инструментальные
методы таксации древостоя, как правило, не применяются.
При детально-маршрутных исследованиях, планируемых на основе предва-
рительно собранных и проанализированных литературных и картографических
материалов, а также результатов рекогносцировочного обследования, исследуе-
мый район покрывается частой сетью геоботанических маршрутов, которые
опираются на существующую квартально-визирную сеть лесоустройства. Наря-
ду с глазомерным описанием лесных фитоценозов используют инструменталь-
ные методы таксации древостоев.
Подготовка маршрутных исследований
При планировании маршрутных исследований очень существенным является
подготовительный этап камеральных работ, во время которого геоботаник дол-
жен ознакомиться с существующими литературными источниками, характери-
зующими растительность района будущих исследований, его геологию, геомор-
фологию, климат, почвенный покров. Геоботаник до выезда в поле должен хо-
рошо знать флору района, подлежащего обследованию и, по возможности, био-
логию и экологию основных растений (Программы..., 1932).
Для планируемого района работ необходимо собрать весь существующий кар-
тографический материал в виде топографических карт масштаба 1 : 100 000 либо
1 : 200 000, при детальных исследованиях — 1 : 25 000, аэрофотоснимков, кос-
мических снимков, планов лесонасаждений, специальных карт (почвенных, гео-
логических, геоморфологических, климатических и др.). По возможности гео-
ботаник должен ознакомиться с материалами лесотаксационных описаний, ве-
домостями пробных площадей, лесоустроительными отчетами и т. п., чтобы
иметь наиболее полное представление о степени изученности лесных террито-
рии и в полной мере использовать имеющиеся материалы. Наличие топографи-
ческих карт обязательно при любых маршрутных исследованиях. Если плани-
руется проведение геоботанической съемки в крупном или среднем масштабах
(1 : 1000 — 1 : 100 000), то необходимо подготовить два комплекта топографи-
25
чсских карт: один — съемочного масштаба, другой — более мелкого. Если пер-
вый комплект будет использоваться как рабочая основа для нанесения контуров
растительности, то желательно иметь каждый лист в двух экземплярах. Возможно
использование черно-белых и цветных ксерокопий либо компьютерных распе-
чаток топокарт При приобретении топокарт на район исследований желательно
включить в перечень заказываемых карт также листы, примыкающие к рабочим
картам, поскольку во время исследований может возникнуть необходимость по-
лучить информацию о растительности окружающих территорий (Исаченко, 1999).
Важнейшим источником информации о растительности и ландшафтах изуча-
емых территорий служат аэрофотоснимки (АФС) и космические снимки (КС).
АФС наиболее доступны, их нередко используют в качестве рабочей основы для
геоботанической съемки. АФС обновляются намного чаще, чем топокарты, по
ним можно получить наиболее свежую информацию о состоянии изучаемой тер-
ритории При работе с АФС необходимо помнить, что они имеют искажения мас-
штаба, не позволяющие точно совмещать их с топоосновой, на краях снимка ка-
чество изображения хуже. Один снимок в масштабе 1 : 25 000 покрывает терри-
торию 3,5 х 3,5 км, поэтому для протяженных маршрутов необходимо иметь со-
ответствующий комплект АФС (Исаченко, 1999).
Космические снимки, выполненные в различных спектральных каналах, со-
держат обширную информацию об изучаемом районе. Они покрывают значи-
тельную территорию, поэтому их желательно использовать при средне- и мел-
комасштабной съемке, а также в работах обзорного характера.
Приборы и оборудование для маршрутных исследований
До выезда на полевые работы маршрутный геоботаник-лесовед должен обес-
печить себя специальным оборудованием, необходимым в лесу, в дополнение к
общему набору оборудования для геоботанических исследований. Сюда нужно
отнести компас или буссоль для равнинных условий, гониометр или горный ком-
пас для горных территорий, геодезический высотомер (альтиметр) или барометр-
анероид (для определения высоты над уровнем моря).
В настоящее время для привязки точек по маршруту исследований к геогра-
фической координатной сетке, а также для определения абсолютной высоты ме-
стности все чаще применяют электронное радионавигационное устройство GPS.
Этот прибор вычисляет местоположение пользователя, определяет параметры
движения, отслеживая сигналы, посылаемые спутниками системы GPS. Каждый
из 24 спутников дважды в день огибает Землю по точным орбитам и передает
информацию о своем местоположении. Работа GPS-приемника основана на при-
еме и автоматической обработке радиосигналов от спутников, поэтому для точ-
ного определения местоположения необходима хорошая радиовидимость спут-
ников, находящихся в данный момент над горизонтом, т. е. отсутствие препят-
ствий на пути распространения сигнала от спутника до GPS-приемника. Сигна-
26
лы ослабевают и не проходят сквозь вершины гор и холмов, здания, людей, ме-
талл, сомкнутый лесной полог. Обычно в любой точке Земли над горизонтом
находятся от 4 до 8 спутников GPS. Для определения трехмерных координат
(широты, долготы и высоты над уровнем моря) приемнику необходимо принять
сигналы от 4 спутников. Для определения двумерных координат (широты и дол-
готы) достаточно уловить сигналы от 3 спутников. После того, как GPS-прием-
ник захватит достаточное количество спутников, на его экране отобразятся ме-
стоположение пользователя, направление и скорость его передвижения. На эк-
ране отображаются местоположение пользователя в двух координатах (широта
и долгота) или в трех координатах (широта, долгота и высота над уровнем моря).
Помимо отображения местоположения (географических координат) прибор со-
держит встроенные одометр (измеритель пройденного пути), компас и часы. За-
фиксированные путевые точки можно записывать в память прибора и использо-
вать для прокладки курса на карте, введенной в запоминающее устройство GPS-
приемника. Работая в режиме «Магистраль», прибор с помощью графического
изображения показывает направление на выбранную путевую точку, расстояние
до нее, курс и скорость передвижения. Работая в режиме «Компас», прибор с
помощью графического отображения стрелки компаса показывает направление
на выбранную точку прибытия, а с помощью вращающегося изображения шка-
лы компаса — направление движения. Имеется индикатор отклонения от курса,
который фиксирует все изменения в направлении передвижения.
Специальные приборы и оборудование включают таксационную вилку, рулетку
(20 м, 50 м), складной метр, портновский сантиметр, возрастной бур, полното-
мер Биттерлиха, высотомер (для измерения высоты деревьев), угломер (для оп-
ределения крутизны склона), складную рамку, 8-кратную лупу, почвенный нож,
лопату, топор, пилу. Кроме того, необходимы гербарная папка, гербарный пресс,
газеты или фильтровальная бумага для сбора и сушки гербария, крафт-бумага
(или плотная оберточная бумага) для сбора образцов мхов и лишайников, по-
чвенные мешочки, таксационный планшет, линейка, бланки геоботанических
описаний, полевые дневники. При работе на лесных торфяниках и торфяных
болотах нужен торфяной бур для зондирования торфяной залежи и отбора об-
разцов торфа.
Способы регистрации наблюдений при маршрутных исследованиях
Существует две основные формы регистрации полевых наблюдений: бланки
геоботанических описаний (ведомости, журналы наблюдений, регистрационные
книги) и полевые дневники (блокноты, тетради). Сложные технические спосо-
бы регистрации маршрутных наблюдений (надиктовывание информации на дик-
тофон или запись в портативный компьютер-ноутбук) не получили широкого при-
менения и используются довольно редко, поскольку в полевых условиях они не
всегда удобны в применении.
27
Использование бланков геоботанических описаний при маршрутных иссле-
дованиях имеет целый ряд преимуществ в связи с формализованностью блан-
ков, сведением количественной информации в табличную форму, наличием ко-
лонок и строк для последующей камеральной обработки. Бланк указывает ис-
следователю те позиции, которые обязательно должны быть заполнены (харак-
тер микрорельефа, тип увлажнения, проективное покрытие травяно-кустарнич-
кового и мохово-лишайникового яруса и т. п.). Для специалиста с малым опы-
том работы бланк — своеобразная подсказка, дающая возможность не упустить
ничего существенного (Исаченко, 1999). Важное преимущество бланков — воз-
можность их быстрой сортировки по заданным критериям при последующей ка-
меральной обработке, удобство введения информации в компьютерные базы дан-
ных (Исаченко, 1999)
Очень широко применяется также способ фиксации маршрутных наблюдений
в полевых дневниках, которые дают большую свободу формы ведения записей.
Однако следует помнить, что эффективно обработать материалы полевых днев-
ников могут только сами авторы записей, от чего ценность полевого дневника
как единицы хранения собранной информации существенно меньше, чем у блан-
ков, которые могут быть обработаны достаточно широким кругом лиц.
Подбор и закладка временных пробных площадей
Работу по закладке пробных площадей следует начинать с рекогносцировоч-
ных маршрутов, при этом особое внимание следует обращать на рельеф и гид-
рологические условия местности. Необходимо разобраться в формах рельефа и
сопоставить с ними флористические особенности лесных сообществ, характер
древостоя и подчиненных ярусов, то есть предварительно наметить выделение
типов леса (или растительных ассоциаций).
Следующий этап работ состоит в выборе типичных мест для закладки проб-
ных площадей. Каждую ассоциацию следует характеризовать по описаниям не-
скольких пробных площадей (не менее трех), чтобы иметь представление о ва-
рьировании особенностей каждого типа. Количество пробных площадей долж-
но быть увеличено, если лесные сообщества, относимые к одному типу леса,
разновозрастны. Их необходимо охарактеризовать отдельными пробными пло-
щадями, чтобы иметь представление о возрастных стадиях основных типов леса
(Сукачев и др., 1957).
Размер пробной площади должен быть не меньше площади выявления фито-
ценоза, которая заметно варьирует в зависимости от сложности состава и струк-
туры лесного фитоценоза. Обычно для описания сообществ северных монодо-
минантных лесов достаточна пробная площадь 400-500 кв. м, т. е. 20 х 20 м или
25 х 25 м. Для более сложных хвойных полидоминантных и широколиственных
лесов пробная площадь должна быть не менее 1000 кв. м. Для горных разрежен-
ных лесов и лесотундровых редколесий рекомендуется закладка пробных площа-
28
дей размером 50 х 50 м. Применяемый иногда размер пробной площади в 100 кв. м.
для лесов безусловно недостаточен. Форма пробной площади может быть различ-
ной, но предпочтительнее форма близкая к квадрату (Сукачев и др., 1957).
Так как точный перечет всех деревьев на пробной площади и затем пересчет
их на гектар имеют важное значение, то пробную площадь следует закладывать
по возможности инструментально, точно определить ее размеры, закрепить ее в
натуре при помощи столбов или колышков и привязать ее по топокарте или к
каким-нибудь ориентирам, например к просекам. Если есть план лесонасажде-
ний, пробную площадь необходимо на нем обозначить. При наличии GPS-при-
емника нужно определить трехмерные координаты пробной площади и записать
их в запоминающее устройство прибора, а также в бланк или полевой дневник.
Для закладки временных пробных площадей достаточно иметь три шнура по
20 м каждый, которые предварительно размечаются по 10 м. По краям каждого
шнура и в его середине привязывают петли для колышков. Колышки рекомен-
дуется заготовить заранее, их желательно окрасить в яркий цвет. Длина колыш-
ков — 50-70 см, диаметр — 2-3 см. При закладке временных пробных площа-
дей прямые углы между сторонами устанавливают с помощью буссоли. Направ-
ление сторон пробной площади обычно ориентируют по компасу на севср-юг и
восток-запад (Заугольнова и др., 2000). Особое внимание необходимо обращать
на то, чтобы пробная площадь была вполне типичной для данного местообита-
ния и на всем своем протяжении достаточно однородной по мезорельефу и ус-
ловиям увлажнения. Пробные площади лучше закладывать в местах, достаточ-
но удаленных от дорог, лесосек, гарей и других нарушений естественной лес-
ной растительности, если, конечно, эти нарушения не изучаются специально (Су-
качев и др., 1957). Размещение закладываемых пробных площадей на местнос-
ти может производиться различными способами в зависимости от условий изу-
чаемого района и целей исследования. Наиболее часто используют методы опор-
ных геоботанических профилей, произвольных маршрутов и геометрический.
Метод опорных геоботанических профилей
Маршрутные исследования растительности на обширных территориях с вы-
раженным мега-, макро- или мезорельефом обычно проводят методом опорных
геоботанических профилей. Опорные профили в виде прямых либо ломаных ли-
ний закладывают в типичных ландшафтах изучаемого района перпендикулярно
к направлению простирания основных форм рельефа: от речных пойм до водо-
разделов или от морских побережий до ближайших горных вершин. Линию про-
филя прокладывают инструментально с точной нивелировкой и привязкой по то-
покарте или аэрофотоснимку. Профиль с помощью компаса и 50-метровой ру-
летки (мерной веревки) разбивают на пикеты с регулярным шагом в 50, 100 или
200 м. Точки пикетов отмечают колышками, в них определяют местоположение,
положение в рельефе, высоту над уровнем моря (в метрах), экспозицию (по 8
29
или 16 румбам) и крутизну склона (в градусах). В реперных точках профиля, а
при необходимости и в дополнительных точках, закладывают временные проб-
ные площади и выполняют полные геоботанические описания (Нешатаев и др.,
1994). В зависимости от строения рельефа местности линии профилей могут быть
либо параллельными, либо пересекаться.
Метод опорных профилей наиболее эффективен при проведении геоботани-
ческой съемки для составления крупномасштабных геоботанических карт. В от-
личие от геометрического способа закладки пробных площадей метод профили-
рования обеспечивает высокую репрезентативность выбора типичных местооби-
таний. В горных районах метод опорных профилей с закладкой пробных пло-
щадей через равные промежутки с определением абсолютной высоты над уров-
нем моря (с помощью авиационного высотомера, барометра-анероида или GPS-
приемника) — наиболее удобный способ изучения высотной поясности расти-
тельности (Исаченко, 1999).
Единственным недостатком метода опорных профилей является сложность, а
иногда и невозможность закладывания линейных профилей при высокой степе-
ни расчлененности рельефа и наличии естественных преград, из-за которых точ-
ный азимут профиля бывает трудно выдержать. Кроме того, линейный принцип
прокладывания профилей не всегда дает возможность выявить все разнообразие
растительных сообществ на изучаемой территории.
Геометрический метод размещения пробных площадей
Этот метод наиболее прост в применении. Пробные площади размещают ре-
гулярно в вершинах геометрических фигур (квадратов или прямоугольников), об-
разующих сплошную сеть. Размер стороны фигуры задан изначально. Он может
быть различным в зависимости от целей исследования и степени детализации
изучения растительного покрова и составлять от 100 м до 1-2 км. Точки проб-
ных площадей заранее наносят на топографическую карту или аэроснимок, а за-
тем при помощью компаса и рулетки (или мерного шнура) их привязывают на
местности. Геометрический метод удобен для использования на равнинах с рав-
номерным характером условий увлажнения, с хорошей проходимостью и отсут-
ствием резких естественных границ. Его несомненными достоинствами являют-
ся репрезентативность выборки и удобство последующей статистической обра-
ботки. Широко используется при детальном крупномасштабном картографиро-
вании заповедных территорий. Основной недостаток метода, кроме высокой тру-
доемкости, состоит в том, что при одинаковых расстояниях между пробными
площадями, которые зачастую не совпадают с размерами растительных конту-
ров, существует вероятность в итоге получить выборку описаний с явным пре-
обладанием одних, наиболее распространенных типов сообществ и отсутствием
других, не менее характерных, но реже встречающихся (Исаченко, 1999).
30
Метод произвольных маршрутов
Этот метод дает возможность преодолеть недостатки двух предыдущих мето-
дов. Маршруты и точки закладки пробных площадей выбирают исходя из осо-
бенностей рельефа и строения растительного покрова с учетом информации, со-
держащейся на аэрофотоснимках и космических снимках. Планируется сеть мар-
шрутов, пересекающих изучаемый район в различных направлениях, как вдоль,
так и поперек направлений основных форм рельефа. Точки для закладки проб-
ных площадей следует выбирать в зависимости от разнообразия положений в
рельефе и условий местообитания (Исаченко, 1999). Желательно размещать проб-
ные площади в коренных, условно коренных или длительно производных сооб-
ществах. В отдельных случаях, например при изучении антропогенного воздей-
ствия на растительность, пробные площади закладывают в нарушенных сооб-
ществах, находящихся на разных стадиях восстановительных сукцессий (напри-
мер, различной давности гари, вырубки, залежи, осушенные участки и т. п ).
При использовании метода произвольных маршрутов нужно стремиться к тому, что-
бы количество описаний каждого типа растительных сообществ (но не менее 3 описа-
ний каждого типа) было пропорционально его относительной площади в пределах ис-
следуемой территории и при этом соответствовало заданной точности исследования.
Описание растительных сообществ на пробных площадях
Геоботаническое описание пробной площади необходимо проводить со всей
возможной при маршрутном методе исследований полнотой. В первую очередь
следует детально охарактеризовать древостой и подчиненные ярусы лесного фи-
тоценоза. Большое внимание необходимо уделять характеристике почвенно-грун-
товых условий, лесной подстилки и условий увлажнения.
На пробной площади проводят детальный учет флористического состава фи-
тоценоза по ярусам, перечет деревьев и кустарников. Для каждого вида опреде-
ляют проективное покрытие (в процентах или в баллах), среднюю высоту, фе-
нофазу и жизненность.
Для древесного яруса указывают диаметр и сомкнутость крон, средние и мак-
симальные диаметр и высоту стволов деревьев, средний и максимальный воз-
раст, количество стволов, полноту и бонитет для каждого элемента леса. Состав
пород древостоя определяют отдельно для каждого подъяруса перечислением всех
входящих в него пород и оценивая участие каждой из них по 10-балльной шка-
ле. Далее необходимо отметить степень обеспеченности возобновления древо-
стоя. Следует отдельно учитывать всходы (высотой от 1 до 10 см) и подрост
(более высокие экземпляры). При характеристике всходов и подроста следует
указать их видовой состав, возраст, высоту, количество экземпляров, происхож-
дение (порослевое, семенное), характер распределения по площади, жизненное
состояние. Количество подроста и всходов учитывают либо на всей пробной пло-
31
щади в целом, либо на специальных учетных площадках с последующим пере-
счетом на гектар.
Далее необходимо остановиться на описании подлеска — кустарникового яру-
са. Необходимо отметить общую сомкнутость полога подлеска в десятых долях
единицы, видовой состав образующих его пород, проективное покрытие, мак-
симальную и господствующую высоту каждого вида. Следует также охаракте-
ризовать общее состояние каждой породы, ее жизненность, фенофазу, происхож-
дение и особенности возобновления.
При описании травяно-кустарничкового яруса следует отметить его суммар-
ное проективное покрытие, особенности сложения, сезонный аспект, вертикаль-
ную (подъярусы) и горизонтальную (синузии) структуру, высоту и сложение каж-
дого подъяруса. Далее необходимо установить полный флористический состав
травяно-кустарничкового яруса, отметить проективное покрытие (в процентах),
среднюю высоту, фенофазу и жизненность каждого вида. Определение проек-
тивного покрытия может производиться глазомерно или инструментально (с при-
менением сетки Раменского, рамки Ипатова (Ипатов, 2000) и т. п.) либо на всей
пробной площади в целом, либо на 10-25 учетных площадках размером 1 кв. м,
заложенных в пределах пробной площади регулярным или случайным способом.
Для характеристики мохово-лишайникового яруса необходимо указать его
общее покрытие в процентах (отдельно для напочвенных, эпиксильных и эпи-
литных видов), мощность покрова (в сантиметрах), соотношение мхов и лишай-
ников, видовой состав, проективное покрытие, характер произрастания (отдель-
ными экземплярами или дерновинами) и распределения и жизненность для каж-
дого вида. Отдельно описывают внеярусную растительность (лианы, эпифиты)
и отмечают имеющиеся крупные грибы.
После описания всех компонентов фитоценоза необходимо отметить имею-
щиеся признаки антропогенных воздействий: рубок, пожаров, выпаса скота, гид-
ролесомелиорации, рекреации, атмосферного загрязнения и т. д. Эти факторы в
различной степени влияют на характер сообществ и могут сильно менять их
облик.
В заключение необходимо указать место данной пробной площади в экологи-
ческих рядах, наличие переходов к окружающим фитоценозам, отметить веро-
ятную историю происхождения данного фитоценоза и его хозяйственное значе-
ние. При описании производного лесного сообщества необходимо установить,
какой здесь существовал коренной тип и происходит ли его восстановление.
32
1.3. ПОДБОР И ЗАКЛАДКА ПОСТОЯННЫХ
ПРОБНЫХ ПЛОЩАДЕЙ В ЛЕСНЫХ СООБЩЕСТВАХ '
Создание сети постоянных пробных площадей (ППП) с последующим прове-
дением на них длительных комплексных исследований — хорошо известный и
надежный метод наблюдений, позволяющий получить разностороннюю досто-
верную информацию о состоянии лесных экосистем и их динамике, о взаимоот-
ношениях основных лесообразующих пород на разных этапах их роста и разви-
тия, о реакциях отдельных видов на изменения природных и воздействие антро-
погенных факторов и т. д. Систематические наблюдения за растительными со-
обществами, позволяющие фиксировать происходящие изменения в их составе,
структуре, состоянии и продуктивности должны стать важной составной частью
фундаментальных исследований.
Методические рекомендации разработаны на основе многолетних комплекс-
ных исследований, проводимых коллективом лаборатории экологии раститель-
ных сообществ Ботанического института им. В. Л. Комарова РАН на Европейс-
кой территории Российской Федерации в лесных сообществах в фоновых усло-
виях, где деятельность человека не наблюдается или она минимальна, а также в
сообществах, испытывающих воздействие антропогенных факторов различной
интенсивности (пожары, атмосферное загрязнение, рубки и др.). В основу при-
водимых методик положен принцип количественной оценки (измерения) пара-
метров исследуемых объектов — растительных сообществ и их компонентов.
Выбор места для закладки ППП. Подбор и закладка ППП должны выпол-
няться по общепринятым (ГОСТ 16128-70) и подробно описанным в литературе
методикам (Моисеев, 1971; Рысин и др., 1988; Баккал и др., 1990; Ярмишко, 1997
и др.) Когда целью исследования является изучение естественной динамики лес-
ной растительности, серия постоянных пробных площадей закладывается в ко-
ренных или условно-коренных типах лесных сообществ, являющихся зональны-
ми или региональными эталонами лесов. При этом необходимо, чтобы к участ-
ку леса с постоянными пробными площадями не примыкали объекты мелиора-
тивной сети, значительные по площади вырубки, сельскохозяйственные угодия,
линии электропередачи, проезжие дороги и рекреационные маршруты.
Когда основной целью исследований является оценка восстановления лесных
сообществ после различных внешних воздействий (рубки, пожары, ветровалы и
пр.), ППП следует закладывать в сообществах с разной давностью нарушения в
пределах одного типа леса.
1 Автор В Т Ярмишко
33
При изучении и оценке воздействия промышленного атмосферного загрязне-
ния на лесные экосистемы и их компоненты ППП должны располагаться на раз-
личном удалении от источника эмиссии по направлению преобладающих вет-
ров и в фоновых условиях (контроль) вне зоны влияния загрязнителей. Серия
ППП должна включать сообщества, относящиеся к одному типу леса и имею-
щие примерно одинаковую давность пожара или рубки. Принадлежность лес-
ных сообществ к одному исходному типу леса на участках с нарушенным рас-
тительным покровом определяется по положению в рельефе, механическому
составу почвообразующих пород и морфологии почвенного профиля.
Каждая ППП в своих границах должна быть однородна по характеристикам
растительности и почв.
О степени нарушенное™ участка леса и типе повреждающих факторов мож-
но судить по состоянию различных компонентов сообщества: возрасту древо-
стоя, наличию механических повреждений и пожарных шрамов на стволах де-
ревьев, состоянию ассимиляционных органов растений, состоянию эпифитной
лишайниковой растительности и др.
Размеры и форма ППП. Размер ППП зависит в значительной степени от воз-
раста древостоя и числа стволов на единицу площади; в древостое IV класса
возраста и старше желательно, чтобы было не менее 200 деревьев с диаметром
4-6 см и более. Обычно вполне отвечает этим условиям площадь 0,25 га; при
необходимости она может быть уменьшена до 0,1 или увеличена до 0,5 га. ППП
придается прямоугольная или квадратная форма. Для этого применяется бус-
соль — угломерный инструмент, предназ-
Рис. 1.1. Буссоль
1 — лимб, 2 — коробка; 3 — диоптры;
4 — втулка для крепления
наченныи для непосредственного измере-
ния на местности магнитных азимутов
(рис. 1.1). Ориентация ППП может быть
произвольной, но удобнее, когда стороны
ППП ориентированы по сторонам света.
Отбивка границ пробной площади.
Работу начинают с установки углового
столба, например юго-западного. Рядом
со столбом (или непосредственно на нем)
располагают буссоль и по градусному
кольцу задают направление западной и
южной сторон ППП. Точность проклад-
ки каждой из сторон контролируется при
помощи глазного диоптра буссоли. Для
удобства работы можно использовать
вешки — хорошо различимые с большо-
го расстояния шесты, служащие времен-
ными ориентирами. В указанных направ-
34
Рис. 1.2. Формы и размеры столбов для офор-
мления пробных площадей
1 и 2 — угловые столбы (со щеками для надпи-
сей) для обозначения постоянных пробных пло-
щадей; 3 - пикетажные колышки
лениях натягивают шнуры. На выбранном расстоянии фиксируют положение
северо-западного и юго-восточного угловых столбов ППП. Установив буссоль
на северо-западный угловой столб, проверяют точность закладки западной
стороны и намечают направление северной стороны ППП. Отмерив необхо-
димое расстояние, устанавливают северо-восточный угловой столб. В резуль-
тате получается ППП прямоугольной или квадратной формы с ориентацией
по сторонам света.
Отбивку границ ППП целесообразнее делать весной или осенью, когда на де-
ревьях отсутствуют листья. Угловые пикеты соединяют туго натянутым по пря-
мой линии шнуром. При необходимости вдоль сторон прорубают узкие визиры,
однако размер нарушений должен быть минимальным (как правило, убираются
лишь мешающие визированию боковые ветви). Затем вдоль шнура через каж-
дые 10 м вбивают пикеты (вешки) высотой 1,0-1,5 м.
Разбивка ППП на квадраты. Пикеты на противоположных сторонах проб-
ной площади соединяют туго натянутым по прямой линии шнуром. Визиры при
этом не прорубают. При наличии густого подроста или подлеска шнур можно
поднять на высоту до 1,5 м. Обозначенные шнуром линии размечаются пикета-
ми через каждые 10 (5) м. При уста-
новке пикетов средняя ошибка в оп-
ределении их положения не должна
превышать 0,1 м.
Оформление ППП. Все 4 угла
ППП закрепляют стандартными стол-
бами (рис. 1.2). Надпись делают чер-
ной краской. На столбе указывают но-
мер квартала и пробной площади,
размер ППП, название организации и
год закладки ППП.
Привязка ППП. Привязка
(рис. 1.3) осуществляется без проруб-
ки визиров по отношению к какому-
либо постоянному ориентиру, чаще
— к квартальному столбу. Параметры
привязки заносятся в паспорт ППП.
Паспорт ППП. В паспорт ППП
заносятся общие сведения о пробной
площади. Указывается примерная
площадь лесного массива, тип сооб-
щества и его площадь, номер и раз-
мер пробной площади, дата описания,
фамилии исполнителей. Можно ис-
35
Рис. 1.3. Схема привязки ППП 62
пользовать сложные номера, вклю-
чающие в себя год описания и гео-
графический район. Например: «Л-
08-97», что соответствует пробной
площади № 8 в Лапландском госу-
дарственном биосферном заповед-
нике, заложенной в 1997 году.
Кроме того, необходимо указать
сведения об административном и
географическом положении пробной
площади: район, ближайший насе-
ленный пункт или природный
объект (река, озеро, горный массив)
и более детальные указания, позво-
ляющие при необходимости опреде-
лить положение пробной площади
на географической карте или схеме.
Общие характеристики местообитания. После завершения закладки ППП
необходимо определить следующие характеристики:
1) превышение над локальным базисом эрозии (превышение над элементом
ландшафта с выходящими на поверхность грунтовыми водами — рекой, озером,
болотом), определяется с использованием высотомера-эклиметра на основе из-
мерения расстояний и углов;
2) крутизну склона, измеряется эклиметром;
3) экспозицию склона, определяется при помощи компаса;
4) давность пожара, определяется на основе анализа спилов или кернов (кер-
ны отбираются при помощи возрастного бурава Пресслера) живых деревьев, име-
ющих пожарные повреждения стволов; необходимо обследовать не менее 5 де-
ревьев в радиусе 50-100 м от пробной площади, т. к. повреждения отдельных
деревьев могут быть обусловлены причинами непирогенного характера (напри-
мер, животными, человеком, падением соседних деревьев); в случае совпадения
давности нарушения камбия на 5 или более деревьях, зарегистрированную дав-
ность нарушения можно считать давностью пожара; подробно методика опреде-
ления давности пожара и контроля беспожарных периодов изложена в разд. 3.3.
Описание компонентов растительного сообщества. После закладки проб-
ной площади описание лесного сообщества необходимо проводить в следующей
последовательности:
1) определение характеристик подчиненных компонентов сообщества (тра-
вяно-кустарничкового и мохово-лишайникового ярусов, полога подроста); начи-
нать описание с других компонентов не следует, т. к. при определении их пара-
метров происходит нарушение нижних ярусов (см. разд. 1.6);
36
2) определение характеристик древесного яруса и подлеска (см. разд. 1.5);
3) описание внеярусной растительности (подробнее см. разд. 1 7);
4) описание почвенного разреза и прикопок (см. разд. 1.8).
Геоботанические описания следует делать на специально подготовленных
бланках или в полевом журнале в форме таблицы, где по вертикали указывают-
ся последовательно все измеряемые характеристики местообитания и сообще-
ства, по горизонтали (в шапке) — номера пробных площадей или площадок.
1.4. ЗАКЛАДКА УЧЕТНЫХ ПЛОЩАДОК И ПРОФИЛЕЙ
НА ПОСТОЯННЫХ ПРОБНЫХ ПЛОЩАДЯХ 1
Для измерения различных характеристик нижних ярусов и подогов лесных
сообществ (полога подроста древесных растений, подлеска, травяно-кустарнич-
кового и мохово-лишайникового ярусов), исследования процессов семеношения
древесных растений, а также для проведения различных полевых эксперимен-
тов используются методы постоянных учетных площадок, профилей, трансект
(Иваненко, 1947; Побединский, 1966; Маслаков, 1968; Раменский, 1971; Злобин,
1972; Орлова, 1981; Санников, Санникова, 1985; Пугачевский, 1988; Маслов,
1990). Размер, форма, количество, характер расположения и способ фиксации
учетных площадок в пределах пробной площади могут различаться в зависимо-
сти от целей исследования и особенностей состава и строения изучаемого сооб-
щества. Размер учетных площадок варьирует в очень широких пределах: от «точ-
ки» (метод уколов) до нескольких сотен квадратных метров. В практике геобо-
танических и экологических исследований в большинстве случаев закладыва-
ются учетные площадки квадратной формы размером 0,2 х 0,2 м, 0,25 х 0,25 м,
0,5 х 0,5 м, 1x1 м, 2 х 2 м, 5 х 5 м. Наиболее универсальными и часто исполь-
зуемыми являются площадки размером 1 х 1 м. Значительно реже применяют
площадки прямоугольной формы (0,25 х 0,5 м, 0,5 х 1 м, 2 х 1 м и т. д.). Заклад-
ка круговых учетных площадок, как правило, связана с особой целью исследо-
ваний, в частности с оценкой влияния особей древесного яруса на характерис-
тики полога подроста, напочвенного покрова, лесной подстилки. При этом в от-
личие от описанных выше площадок, используемых для изучения состава и
структуры нижних ярусов сообществ, круговые площадки часто имеют более зна-
чительный размер: их радиус обычно составляет от 2 до 10 и даже 20 м (Санни-
ков, 1992). Крайним вариантом учетной площадки является учетная точка. Ме-
тод, использующий для измерения характеристик сообществ учетную точку, на-
зывается методом уколов. Учет проводится с использованием тонких спиц. Уко-
лы выполняются по одному или спицы закрепляются на специальной раме или
1 Автор Н. И. Ставрова
37
круге. Подсчитывается процент уколов (от общего числа выполненных уколов),
при которых спица касается растений данного вида. Эта величина и характери-
зует проективное покрытие вида.
Существует несколько вариантов расположения учетных площадок на проб-
ной площади, основными являются случайный и типический. При собственно
случайном (рандомизированном) способе заложения площадок положение каж-
дой из них полностью не зависит от положения всех остальных. Например, пло-
щадки закладываются вдоль параллельных профилей, причем как расположение
профилей на пробной площади, так и расположение площадок на них определя-
ются по таблице случайных чисел. Однако такой метод используется сравнитель-
но редко. Как правило, площадки располагают вдоль нескольких параллельных
или взаимно перпендикулярных трансект, расположенных на одинаковом рас-
стоянии друг от друга (5-25 м в зависимости от размеров пробной площади).
При детальном исследовании изменения характеристик нижних ярусов сообществ
в зависимости от положения относительно особей древостоя могут закладываться
серии учетных площадок на трансектах, расположенных по радиусу от стволов
деревьев до межкроновых участков или между деревьями в группах (Лукина,
Никонов, 1996).
Расположение площадок на трансектах определяется по заранее определен-
ному правилу, например по таблице случайных чисел (Злобин, 1972) или через
равные расстояния друг от друга (Санников, 1992; Горшков и др., 1995). Во вто-
ром случае они могут составлять от 2 до 10 м. И тот, и другой способ закладки
в природных растительных сообществах не нарушает рандомизации выборки,
хотя во втором случае положение каждой точки зависит от положения других.
Следует отметить, что наиболее удобной при закладке площадок является ори-
ентация трансект по сторонам света.
При типическом методе закладки места расположения площадок выбираются
субъективно после визуального изучения всей пробной площади, например, опре-
деленное число площадок закладывается в каждой из выделенных визуально пар-
целл. Обычно исследователи ставят такую задачу с целью дать количественную ха-
рактеристику нескольких, наиболее типичных с их точки зрения, участков в преде-
лах растительного сообщества, различающихся по видовому составу нижних яру-
сов, плотности и сомкнутости древесного яруса и т. д. (Ястребов, Познанская, 1993;
Лукина, Никонов, 1996). С позиций строго количественного подхода к определению
характеристик растительного сообщества как целого такой способ является некор-
ректным. Более приемлемым вариантом этого метода является пропорциональный,
при котором число закладываемых площадок пропорционально примерной площа-
ди каждой выделяемой парцеллы, и следовательно, может быть произведен расчет
величин, характеризующих изучаемое растительное сообщество в целом.
Большинство авторов признает метод закладки учетных площадок по регу-
лярному принципу наиболее оптимальным для получения объективных результа-
38
тов (Грейг-Смит, 1967; Василевич, 1969, 1972; Миркин, Розенберг, 1979). Именно
этот метод наиболее широко используется в современных геоботанических ис-
следованиях. Одним из его основных достоинств является получение коррект-
ных оценок не только средних значений различных характеристик, но и их ва-
рьирования в пределах пробной площади.
Учетные площадки на трансектах могут располагаться одиночно или блока-
ми по несколько площадок. Используется также метод «вложенного» располо-
жения учетных площадок, когда в пределах площадки большего размера после-
довательно одна в другой располагают площадки более мелкого размера. После-
дние два способа (блоки и «вложенные» площадки) позволяют провести срав-
нение точности измерения или установить особенности динамики изучаемых ха-
рактеристик в зависимости от использования различной единичной учетной пло-
щади, а также определить площадь выявления той или иной характеристики (на-
пример, числа видов).
Вопрос о количестве учетных площадок, закладываемых в пределах каждой
пробной площади, решается исходя из задач исследования и особенностей стро-
ения нижних ярусов лесного сообщества. Общим правилом является то, что чем
меньше размер учетных площадок, тем большее их количество должно быть за-
ложено для измерения характеристик с заданной точностью. При использова-
нии площадок размером 1 х 1 м наименьшее необходимое их число равно 20.
Такой объем выборки применяется для получения самой общей характеристики
нижних ярусов лесных сообществ (когда их исследование не является основной
целью работы) или в отдельных случаях в лесных сообществах с однородным
сложением нижних ярусов.
Полученные на основе анализа выборки из 20 площадок значения коэффици-
ента вариации исследуемой характеристики (характеристик) могут быть исполь-
зованы для расчета числа учетных площадок, необходимого для определения
среднего значения характеристик с заданной точностью. Расчет производится по
формуле (Зайцев, 1984) N = (tv / г)2, где N— искомый объем выборки, у — ко-
эффициент вариации в процентах, г — допустимая погрешность в процентах,
t— критерий достоверности Стьюдента, определяемый по таблице в соответ-
ствии с выбранным уровнем значимости. Например, согласно приведенной фор-
муле, для определения общего проективного покрытия травяно-кустарничково-
го яруса в северотаежных зеленомошных сосновых лесах (коэффициент вариа-
ции покрытия составляет ~40 %) с погрешностью 10 % при доверительном уров-
не Р= 0,95 (t -2,0) необходимо заложить более 60 площадок размером 1 х 1 м.
Для определения с аналогичной точностью общего проективного покрытия мо-
хово-лишайникового яруса (коэффициент вариации -16 %) достаточно заложить
10 площадок, плотности подроста (коэффициент вариации -200-300 %) — 1500—
3500. В практике геоботанических исследований для последнего случая обычно
ограничиваются закладкой значительно меньшего числа площадок — 100-200,
39
погрешность измерения при этом возрастает до 30-50 %. Однако такой резуль-
тат является приемлемым, так как столь значительное варьирование характерно
только для очень низких значений средних величин характеристик, в частности
проективного покрытия единично встречающихся в сообществе видов, плотно-
сти подроста в ненарушенных лесных сообществах. В этих случаях биологичес-
кий смысл полученного значения характеристики не изменяется с увеличением
погрешности измерения до 30-50 %. Например, плотность подроста, составля-
ющая 1 или 2 тыс. экз. • га 1, практически одинакова в сравнении с плотностью,
характерной для нарушенных сообществ и достигающей 200 тыс. экз. • га-1. Если
цель исследования предусматривает детальное изучение редких видов с проек-
тивным покрытием, составляющим доли процента, или возобновительных про-
цессов в климаксовых сообществах, число учетных площадок увеличивают до
необходимых величин или дополнительно используют другие методы учета (на-
пример, регистрацию наличия или отсутствия вида на линейных трансектах).
В большинстве случаев для получения средних значений основных характе-
ристик с погрешностью 5-10 % требуется заложить на территории пробной пло-
щади 60-200 учетных площадок размером 1 х 1 м (Побединский, 1966; Злобин,
1972; Санников, Санникова, 1985; Пегов, 1992). Если используются площадки
малого размера, например 0,2 х 0,2 м, их число на пробной площади увеличива-
ют, в ряде случаев до 1000-4000 (Маслов, 1990). При использовании точечных
площадок (уколов) для определения характеристик с удовлетворительной точ-
ностью всегда требуется большое число (несколько тысяч) замеров.
Кроме метода учетных площадок для регистрации характеристик нижних яру-
сов лесных сообществ часто используются методы учетных профилей и трансект.
В пределах профилей производится сплошной учет изучаемых объектов, поэто-
му методом профилей изучают характеристики конкретных особей, чаще всего
древесных растений и кустарников. Например, в лесных сообществах учитыва-
ют особи древесных растений высотой более 0,2 м (более 0,5 м) с диаметром на
высоте груди менее 6 см, относящиеся к пологу подроста и нижнему пологу дре-
весного яруса, а также растения, формирующие подлесок (Пугачевский, 1988;
Маслов, 1990). Такие учетные профили имеют, как правило, ширину 1-2 м и
располагаются параллельно друг другу на равном расстоянии (5-15 м) или по
диагоналям пробной площади. Постоянные учетные профили могут использо-
ваться также для измерения характеристик подроста высотой менее 0,2 м (0,5 м),
травяно-кустарничкового и мохово-лишайникового ярусов. В этом случае они
имеют ширину 0,2-1 м и состоят из примыкающих друг к другу прямоугольных
или квадратных учетных площадок (Маслов, 1990).
Линейные трансекты закладываются на территории пробной площади так же,
как профили. Регистрация характеристик на трансектах ведется относительно ли-
нейного отрезка, а не площади, как при использовании площадок или профи-
лей. Трансекты разбиваются на отрезки определенной длины (0,5 м, 1 м и т. д.),
40
в пределах которых регистрируется изучаемая характеристика, например нали-
чие или отсутствие вида, совместная встречаемость пар видов (Демьянов, 1978).
Положение постоянных учетных площадок на пробной площади обычно фик-
сируется четырьмя деревянными или металлическими колышками, расположен-
ными по ее углам, а иногда дополнительно белым синтетическим шпагатом, зак-
репленным по периметру
площадки. Более экономным,
менее трудоемким и в мень-
шей степени нарушающим
напочвенный покров являет-
ся способ фиксации площа-
док или блоков площадок при
помощи одного колышка, рас-
полагаемого всегда в опреде-
ленном, например, северо-за-
падном углу (рис. 1.4). При
таком способе закладки
трансекты, вдоль которых бу-
дут располагаться площадки,
должны всегда быть ориенти-
рованы строго по сторонам
света. Положение учетных
профилей на территории
пробной площади фиксирует-
ся четырьмя колышками и
двумя параллельными лини-
ями белого синтетического
шпагата. Каждая учетная
площадка и каждый профиль
(5-25 блоков на пробную площадь)
Рис. 1.4. Схема размещения площадок для измере-
ния характеристик напочвенного покрова, плот-
ности возбновления и плотности опавших ши-
шек
Залитыми черными кружками обозначено положение
почвенных прикопок. Залитыми черными квадратами
обозначено положение маркировочных колов с номе-
рами. в этих точках проводится измерение сквозисто-
сти полога древесного яруса и измерения относитель-
ной суммы площадей сечений древесного яруса Рас-
стояние между блоками площадок постояно для тер-
ритории пробной площади
нумеруются. Номер площадки пишется краской на боковой стороне определен-
ного колышка (например, северо-западного) или наносится ручкой на малень-
кую бирку из тонкой жести, которая фиксируется на колышке гвоздиком.
Закладку постоянных учетных площадок и профилей при их регулярном раз-
мещении рекомендуется осуществлять следующим образом. Перед началом ра-
боты определяется необходимое количество и соответствующий целям исследо-
вания размер учетных площадок. Рекомендуется, чтобы площадь учета, занима-
емая площадками, составляла не менее 2 % территории пробной площади Ис-
ходя из этих величин определяется число трансект, расстояние между ними и
расстояние между учетными площадками на трансектах. Затем по двум проти-
воположным сторонам пробной площади отмеряются расстояния между трансек-
тами, положение концов трансект фиксируется колышками, а сами трансекты на-
41
мечаются при помощи белого синтетического шпагата. Вдоль каждой трансек-
ты через определенный интервал колышками отмечается положение одного из
углов учетной площадки. Например, если трансекта ориентирована с севера на
юг, то при движении вдоль трансекты в этом направлении через определенный
интервал на ней фиксируется северо-западный угол каждой площадки или бло-
ка площадок (рис. 1.4). Все установленные колышки сразу нумеруются. Анало-
гичным образом фиксируется положение всех площадок на всех трансектах. При
проведении измерений на площадке ее положение намечается при помощи ме-
таллической или деревянной рамки нужного размера и формы. Рамка распола-
гается таким образом, чтобы колышек на трансекте являлся ее северо-западным
углом. Если колышек на трансекте является северо-западным утлом блока из 4
площадок, то после описания первой площадки рамка последовательно перекла-
дывается сначала на восток, затем на юго-восток и юг.
1.5. ИЗМЕРЕНИЕ ХАРАКТЕРИСТИК ДРЕВЕСНОГО ЯРУСА 1
В пределах всей совокупности особей древесных растений в лесном сообще-
стве выделяют основную структурную часть — древесный ярус (древостой).
Основным критерием выделения древесного яруса является критерий высо-
ты особей. В большинстве классических исследований (Третьяков, 1927, 1957;
Сукачев, 1931; Анучин, 1971; Корчагин, 1976) к древесному ярусу относят дре-
весные растения, высота которых составляет более ’А (иногда — более ’/г) вы-
соты наиболее высоких особей древесных растений в составе лесного сообще-
ства. В современных исследованиях (Linder et al., 1997; Engelmark, 1998; Горш-
ков, 2001) к древесному ярусу рекомендуется относить все древесные растения
высотой более 1,3 (или 1,5) м. Древесные растения, высота которых меньше
указанных пределов, относят к пологу подроста
Древесный ярус является главным, ценозообразующим компонентом лесного
сообщества. Характеристики древесного яруса определяются параметрами место-
обитания (климатом, рельефом, свойствами почвообразующих пород), а также
типом, давностью и интенсивностью последнего нарушения. В свою очередь, дре-
весный ярус формирует внутреннюю среду фитоценоза (биотоп), оказывающую
большое влияние на характеристики подчиненных компонентов лесных сообществ.
Древостой представляет собой сложную по составу и структуре систему, сфор-
мированную совокупностью взаимосвязанных в своем происхождении и разви-
тии особей древесных растений. В пределах древесного яруса на основе ряда
критериев могут быть выделены отдельные структурные части (компоненты,
подъярусы, пологи).
1 Автор В Г Ярмишко
42
В большинстве случаев в пределах древесного яруса выделяется перечетная
часть, в состав которой входят деревья с диаметром на высоте груди 6 (4) см и
более (в молодых древостоях — 2 см и более). Эта часть древостоя определяет
его основные биологические и лесохозяйственные параметры; доля общей био-
массы и запаса древесины, приходящаяся на неперечетную часть древостоя, как
правило, незначительна.
В фитоценологических и лесоведческих исследованиях в случае выраженной
вертикальной расчлененности древесного яруса принято выделять в его преде-
лах отдельные подъярусы (иногда называемые I II и т. д. ярусы) и/или пологи.
Подъярусы и пологи выделяются на основе различий древесных растений по
высоте1. В лесоведении выделение подъярусов (пологов) в древостое рекомен-
дуется, только если входящие в них особи древесных растений по высоте отли-
чаются не менее чем на 20-30 % и имеют запас не менее 20-30 % от общего
запаса яруса (Третьяков, 1927, 1957; Анучин, 1971; Поляков, Набатов, 1983).
В бореальных лесах образование четко выраженных подъярусов обусловлено
в основном периодическим воздействием внешних нарушающих факторов, в пер-
вую очередь лесных пожаров средней и высокой интенсивности, приводящих к
гибели всех или значительной части особей в древесном ярусе и формированию
поколений деревьев, различающихся по возрасту, видовому составу и высоте.
При изучении временной динамики характеристик древесного яруса, в зави-
симости от поставленных задач, в его составе могут быть выделены также гос-
подствующий и подчиненный компоненты (сформированные успешно развива-
ющимися и отстающими в росте, угнетенными особями), допожарный и после-
пожарный компоненты и др.
В лесоведении в составе древесного яруса принято выделять структурные
части, называемые элементами леса. Под элементом леса понимается часть дре-
весного яруса (или древесный ярус в целом), состоящая из деревьев одного вида,
формирующих один подъярус и различающихся по возрасту не более чем на
2 класса (т. е. на 20 лет для лиственных и на 40 лет для хвойных древесных ра-
стений).
Оценка параметров древесного яруса выполняется на основе измерения ха-
рактеристик отдельных особей древесных растений, входящих в его состав, и
определения характеристик его структурных и динамических компонентов. Пе-
1 В. Н. Сукачев (Сукачев, 1931; Сукачев, Зонн, 1961) и вслед за ним А. А Корчагин
(1976) и некоторые другие исследователи придерживаются мнения, что подъярусы в
древостое выделяются в том случае, если различие в высоте особей связано с их видо-
вой спецификой (т. е. если в древостое представлены разные виды); если же различие в
высоте обусловлено различным возрастом или степенью угнетенности особей одного
вида, то в древостое могут быть выделены отдельные пологи (обычно 2-3), являющиеся
в отличие от подъярусов динамическими, непостоянными компонентами.
43
Рис. 1.5. Постоянная пробная площадь в со-
сняке чернично-сфагновом в Ленинград-
ской области
ред выполнением измерений харак-
теристик древесного яруса прово-
дят нумерацию деревьев.
Нумерация деревьев. Кора всех
стоящих деревьев диаметром не
менее 6 см слегка зачищается по
периметру на высоте 1,3 м. С юж-
ной стороны зачищается несколько
большая площадка; масляной крас-
кой на уровне 1.3 м от корневой
шейки ставится горизонтальная
черта и над ней — номер дерева
(рис 1.5). Высота цифр — 5 см.
Иногда практикуется нумерация
деревьев с использованием различ-
ных металлических пластинок (би-
рок) с предварительно нанесенны-
ми на них номерами. При раздвое-
нии ствола дерева ниже высоты
1,3 м каждому из двух стволов да-
ется свой номер. Нумерация дере-
вьев проводится по 10-метровым
полосам челночным движением,
начиная с северо-западного угла.
В том же порядке получают номе-
ра квадраты размером 10 х 10 м.
В конце работы производится про-
верка. В том случае, если обнару-
жены пропущенные при перечете деревья, им даются очередные порядковые но-
мера, а в перечетной ведомости как примечание указывается порядковый номер
ближайшего дерева.
Желательно также пронумеровать все сломанные и ветровальные деревья.
Обеим частям сломанных стволов (стоящей и лежащей) дается один номер Если
слом произошел ниже высоты 1,3 м, то нумеруется часть ствола, лежащая на
земле. Черта и номер ставятся на расстоянии 1,3 м от корневой шейки. При этом
сломанные и ветровальные стволы следует отличать от валежа, к которому от-
носятся лежащие разлагающиеся стволы, продавливаемые твердым предметом
и зарастающие мхами и травами.
При повторных наблюдениях нумерация деревьев подновляется. Экземпляры
подроста, диаметр которых достиг 6 см, переводятся в древостой и получают
очередные порядковые номера. Номера деревьев, выпавших из древостоя или
44
перешедших в валеж, другим де-
ревьям не присваиваются.
Измерение длины окружно-
сти (периметра) ствола произ-
водят металлической рулеткой на
уровне горизонтальной черты
(1,3 м) с точностью до 1 см. У со-
прикасающихся стволов допуска-
ется измерение полупериметра
между точками, фиксированны-
ми краской на противоположных
Рис. 1.6. Измерение диаметра ствола с помощью
деревянной мерной вилки
сторонах ствола; полученный результат при этом удваивается. Результат измере-
ния заносится в паспорт ППП с указанием номера дерева и породы (условным
обозначением, принятыми в лесоустройстве). Рассчитанные диаметры округляются
до целых сантиметров. При повторных наблюдениях вновь измеряют периметры
лишь у живых деревьев.
Измерение диаметров стволов деревьев осуществляют мерной вилкой. Она
представляет собой штангенциркуль (рис. 1.6), выполненный из текстолита, дере-
ва или другого материала и состоит из мерной линейки и двух ножек, одна из ко-
торых может свободно передвигаться по линейке. Мерная линейка имеет длину
до 90 см. На широкой стороне линейки по обеим сторонам нанесены шкалы с по-
лусантиметровыми делениями (цифры поставлены через 2 см). В научно-иссле-
довательских целях используются мерные вилки с миллиметровыми делениями.
При этом одной и той же шкалой можно пользоваться как при измерениях диа-
метров отдельных деревьев, так и при перечетах в древостоях.
Диаметры стволов растущих деревьев принято измерять на строго определен-
ной высоте — 1,3 м от шейки корня. Эта высота соответствует уровню груди
среднего человека, поэтому диаметр, измеренный на высоте 1,3 м, называется
диаметром на высоте груди. При измерении диаметров подвижную ножку от-
водят от неподвижной, прикладывают вилку линейкой вплотную к стволу (к чер-
те, помеченной масляной краской) перпендикулярно его оси и прижимают под-
вижную ножку (рис. 1.6). В таком положении снимают отсчет измеряемого ди-
аметра. При этом записывают цифру того деления, которое видно на линейке со
стороны руки, держащей подвижную ножку. Учитывая то, что ствол обычно не
имеет идеальной окружности, диаметр измеряют два раза во взаимно перпенди-
кулярных направлениях и берут среднее из двух показаний, округляя цифру до
ближайшей ступени толщины. Для того чтобы исключить появление системати-
ческих ошибок, к мерной вилке предъявляют следующие требования:
1) ножки в момент измерения диаметра должны быть параллельны между со-
бой и перпендикулярны мерной вилке;
2) подвижная ножка должна свободно передвигаться по мерной линейке;
45
3) деления на шкалах должны быть четкими, хорошо видны;
4) длина ножек должна быть не меньше половины измеряемого диаметра.
В настоящее время имеется ряд оригинальных конструктивных решений мер-
ных вилок, однако они не выпускаются серийно и не получили широко приме-
нения в науке и на производстве.
Определение возраста деревьев. У 20-30 живых деревьев, выбранных про-
порционально распределению по ступеням толщины, с помощью бурава Прес-
слера по керну определяется возраст с точностью до 1 года. В разновозрастных
древостоях число пробуренных деревьев желательно увеличить до 50 шт.
Керн берется буром с восточной стороны ствола на высоте 0,3 м от шейки
корня. После взятия керна отверстие забивается плотной пробкой из древесины
лиственной породы (сухим сучком) или закрывается садовым варом. При нали-
чии стволовой гнили бурав дезинфицируется в растворе любого антисептика.
К полученному возрасту по керну прибавляется возраст на пень, равный в сред-
нем 5 годам. Результат определения возраста заносится в паспорт ППП. При по-
вторных наблюдениях число деревьев с кернами желательно увеличить. Для более
точного определения возраста деревьев часто используют спилы ствола на уровне
корневой шейки.
По кернам и спилам желательно также определить ширину водопроводящей
зоны, патологию ствола и радиальные приросты ствола по десятилетиям или по-
годичную их динамику.
Определение прироста древесины по годичным кольцам. В процессе мно-
голетних исследований в лесных сообществах мы сконструировали два приспо-
собления для изучения и анализа годичных колец деревьев по кернам.
Для зажима и последующей зачистки кернов, отобранных возрастным бура-
вом, используют простое при-
способление (рис. 1.7). Оно со-
стоит из пластин и винтов для
зажима. Образец древесины 1
помещают в паз 2 приспособле-
ния и зажимают пластинами 3 с
помощью винтов 4 таким обра-
зом, что он выступает над повер-
хностью прибора на 1-1,2 мм.
Затем остро заточенным инстру-
ментом (нож, бритва и т. п.) про-
изводят срез выступающей час-
ти образца. Здесь уместно под-
черкнуть, что влажные образцы
древесины легко и хорошо зачи-
щаются, срезы имеют ровную
Рис. 1.7. Приспособление для подготовки дре-
весных образцов (кернов) к подсчету шири-
ны годичных колец
46
без шероховатостей поверхность с
четко различимыми границами го-
дичных колец. В хорошо подго-
товленных кернах легко различа-
ется ранняя и поздняя части го-
дичного кольца.
При длительном хранении кер-
ны теряют свою естественную
влажность, становятся довольно
Рис. 1.8. Прибор-дендрометр для измерения
ширины годичных колец древесных образ-
цов, сконструированный на базе микроско-
па МБС-1
твердыми и подготовка их к ана-
лизу имеет определенные сложно-
сти, поэтому такие образцы перед
зачисткой целесообразно опустить
на 15—20 мин в кипящую воду. По мере подсыхания подготовленную поверхность
образцов рекомендуется смачивать спиртово-глицериновой смесью.
Для измерения ширины годичных колец деревьев мы сконструировали спе-
циальный прибор-дендрометр (рис. 1.8) на базе стереоскопического микроскопа
МБС-1. Он состоит из предметного столика 1, ячейки для образцов (кернов) 2,
микрометрического винта 3, основания 4, нониуса 5, деленного на 100 равных
частей, ручки вращения всей подвижной системы 6, узла крепления к микро-
скопу 7, миллиметровой шкалы 8. Параметры микрометрического винта и но-
ниуса рассчитаны таким образом, что одно деление на нониусе соответствует
0,01 мм.
Методика измерений ширины годичных колец деревьев на этом приборе со-
стоит в следующем. Вначале производится совмещение шкалы микроскопа с об-
разцом древесины, расположенным в ячейке 2 или вместе с приспособлением
(рис. 1.8) на столике 1. Совместив шкалу микроскопа с границей последнего
(внешнего) годичного кольца, производят отсчет на нониусе 5. Затем ручкой 6
приводят в движение нониус и перемещают предметный столик с образцом до
совмещения микрошкалы с границей следующего кольца или его части (грани-
цей между ранней и поздней древесиной) и производят второй отсчет. Разница
в отсчетах и представляет собой ширину годичного кольца или искомой его ча-
сти, выраженную в миллиметрах. Конструкция прибора позволяет производить
измерение ширины годичных колец с точностью до 0,01 мм. При анализе отно-
сительно узких годичных колец (до 1 мм) отсчеты снимаются только на нониу-
се, а при ширине их более 1 мм рекомендуется пользоваться как нониусом 5,
так и миллиметровой шкалой 8.
Вышеописанный прибор можно использовать для измерения ширины годич-
ных колец на образцах в виде кернов и древесных спилов, а также при исследо-
вании физиологически активной части корневых систем и других органов рас-
тений. Простота конструкции и надежность приборов в работе позволяют при-
47
Рис. 1.9. Приборы для измерения высот
деревьев
А — эклиметр, Б — маятниковый высотомер
менять их с одинаковым успехом
как в лабораторных, так и в по-
левых условиях.
Измерение высоты деревьев
проводится с помощью высото-
меров (рис. 1.9). Известными гео-
дезическими инструментами для
измерения высот деревьев явля-
ются эклиметры (рис. 1.9А), ук-
лономеры и оптические высото-
меры (например, зеркальный ре-
ласкоп В. Биттерлиха). Измере-
ние производится с точностью
до 0,5-1 м у тех же деревьев, у
которых определялся возраст.
Для каждой породы на милли-
метровке строится график зави-
симости высоты от диаметра, ко-
торый прикладывается к паспор-
ту ППП. Нанесенные точки по-
зволяют оценить достаточность
числа измеренных высот для всех ступеней толщины и пологов и, соответствен-
но, необходимость дополнительных измерений. Для деревьев максимальных ди-
аметров замеряется не менее 5-10 высот. При повторных учетах измерение вы-
сот необходимо проводить у тех же, но только живых деревьев.
В качестве высотомера можно использовать мерную вилку (рис. 1.10). В не-
подвижной ножке имеется отверстие, в котором укрепляют нить, оканчивающую-
ся отвесом. На подвижной ножке точно напротив точки прикрепления нити с от-
весом ставят черту и принимают ее за нулевое деление. Измерение высот осуще-
ствляется следующим образом Исследователь отходит от дерева на некоторое рас-
стояние так, чтобы хорошо была видна вершина измеряемого дерева. Подвижную
ножку устанавливают в такое положение, чтобы расстояние между ножками в сан-
тиметрах было равно расстоянию от исследователя до дерева в метрах. Далее вдоль
внутренней стороны неподвижной ножки визируют на вершину дерева. При этом
нить с отвесом займет вертикальное положение и на шкале подвижной ножки
укажет деление, соответствующее высоте дерева до глаза наблюдателя.
В равнинной местности, чтобы получить всю высоту дерева, нужно к изме-
ренной части высоты прибавить рост исследователя до глаз. В горной местнос-
ти при измерении высоты необходимо визировать инструмент на вершину и на
основание ствола и брать два отсчета. Если основание ствола расположено ниже
наблюдателя, высота ствола будет равна сумме отсчетов на вершину и на осно-
48
вание ствола, а если
основание ствола
выше наблюдателя,
то высота ствола бу-
дет равна разности
отсчетов на вершину
и на основание.
Точность опреде-
ления высот с помо-
щью мерной вилки
обычно составляет
±5-8 %. Измерение
высот мерной вил-
кой в современных
условиях применяет-
ся в исключитель-
ных случаях, только
при отсутствии бо-
лее совершенных
Рис. 1.10. Схема измерения высоты дерева мерной
вилкой
высотомеров.
Среди оптических высотомеров наиболее распространен зеркальный релас-
коп В. Биттерлиха. Он является универсальным прибором и позволяет измерять
все основные таксационные показатели дерева и насаждения: высоты деревьев,
их диаметр на любой высоте, видовую высоту, длину горизонтальных линий, угол
наклона местности, сумму поперечных сечений деревьев на 1 га.
Реласкоп представляет собой портативный прибор по размерам немного боль-
ше фотоаппарата. В корпусе реласкопа имеется маятник с нанесенными на нем
измерительными шкалами. Изображение этих шкал переносится на измеряемые
объекты зеркальным устройством. По бокам корпуса имеются два окошка из ма-
тового стекла. В верхней части прибора в передней и задней стенках корпуса
имеется глазок для визирования на измеряемые объекты. Для измерения высоты
дерева устанавливают величину базисного расстояния (15, 20, 25 или 30 м), ко-
торое определяется с помощью дальномерного устройства реласкопа.
Определение полноты древостоя полнотомером. Полнотомер (или угловой
шаблон) В. Биттерлиха (рис. 1.11) представляет собой деревянный брусок дли-
ной 1 м, на одном конце которого имеется металлическая прицельная рамка с
вырезом 2 см, служащая предметным диоптром. Отношение ширины диоптра к
длине бруска составляет 2/юо = '/50, поэтому при различной длине бруска может
быть и соответствующая ширина диоптра.
При определении суммы площадей поперечных сечений стволов исследова-
тель становится в насаждении, прикладывает брусок к щеке на уровне глаза и
49
Рис. 1.11. Полнотомер В. Биттерлиха
1 — схема прибора; 2 — определение сумм площадей поперечных сечений
визирует вдоль бруска через диоптр на все деревья, которые попадают в поле
зрения, постепенно поворачиваясь на 360°. Визировать необходимо на ствол на
высоте 1,3 м от шейки корня.
При визировании на деревья исполнитель подсчитывает деревья, полностью
закрывающие просвет диоптра, а деревья, касающиеся внутренних сторон ди-
оптра, считает два за одно. Деревья, вписывающиеся в предметный диоптр и не
касающиеся его внутренних сторон, не учитываются. Количество учтенных де-
ревьев, согласно вышеприведенному правилу, будет составлять сумму площадей
поперечных сечений в кв. м на 1 га.
Для большей портативности полнотомер В. Биттерлиха может быть изготов-
лен в виде пластинки из цветного органического стекла на гибкой базисной це-
почке. Если на пластинке сделать предметный диоптр шириной 1 см, то длина
базисной цепочки составит 0,5 м. В таком исполнении угловой шаблон стано-
вится более портативным.
Сквозистость древесного яруса (доля свободного пространства, образован-
ного просветами между кронами и в кронах деревьев) измеряется в одном из
углов площадки. Величина сквозистости определяется с помощью специально-
го прибора — сквозистомера (Ипатов и др., 1979). Основными элементами скво-
зистомера являются зеркало и расположенная над ним рамка. Сквозистость оп-
ределяют как обратную величину площади проекций крон и стволов деревьев,
попадающих в рамку на плоскости зеркала. Зеркало расчерчено линиями, обра-
зующими прямоугольники, площадь которых составляет 5, 10, 25, 50 % от пло-
щади зеркала, что служит ориентиром при определении сквозистости. При из-
50
мерении сквозистости сквозистомер располагается на расстоянии ~20 см от глаз
с тем, чтобы характеризуемая область небесного свода («угол захвата») состав-
ляла 27° по вертикали и 60° по горизонтали. Вся область небесного свода де-
лится на 4 сектора по основным сторонам света. По вертикали каждый сектор
делится на 3 области: нижнюю (10-37°), среднюю (37-53°) и верхнюю (53-90°).
Для нижней области каждого сектора делают два измерения, для средней и вер-
хней — по одному. Предлагаемая методика является одной из модификаций
метода, предложенного В. С. Ипатовым с сотрудниками (1979).
Определение категорий состояния деревьев. Категории состояния дерева
указываются по 10-балльной шкале (Рысин и др., 1988); для стоящих деревьев
используются баллы от 0 до 5. Баллы 6-0 используются для обозначения буре-
ломных и ветровальных стволов (в том случае, если они занумерованы). Катего-
рии состояния определяются по совокупности признаков: ажурности кроны, при-
росту по высоте, состоянию ветвей, ствола и корней.
0 — здоровые деревья — без внешних признаков ослабления; с густой зеле-
ной кроной, с нормальными для данного возраста и условий местопроизраста-
ния приростами последних лет;
1 — ослабленные деревья — характеризуются слабо ажурной кроной, повреж-
дением до 1/3 фотосинтезирующего аппарата, укороченным приростом в высо-
ту, усыханием отдельных ветвей, повреждением или небольшим местным отми-
ранием ствола, отдельных корневых лап;
2 — сильно ослабленные деревья — с ажурной кроной, с повреждением и усы-
ханием до 2/3 фотосинтезирующего аппарата, с сильно укороченным приростом
или без него, суховершинные, со значительными повреждениями, поражениями
ствола, корневых лап; в ряде случаев наблюдается частичное заселение дерева
стволовыми вредителями при местном типе ослабления;
3 —усыхающие — с сильно изреженной кроной или только отдельными жи-
выми ветвями; с повреждением более 2/3 фотосинтезирующего аппарата, листва
(хвоя) желтеет и осыпается, текущего прироста по высоте нет, по стволу и кор-
ням возможны насечки и единичные свежие поселения стволовых вредителей;
4 — свежий сухостой — деревья, усохшие в текущем году, с желтой или бу-
рой листвой (хвоей) или без нее; по стволу — свежие поселения короедов, воз-
можны личинки 1-3-го возраста. Усыхание хвойных деревьев без заселения ко-
роедами встречается редко, среди лиственных пород — чаще;
5 — старый сухостой — деревья, усохшие в прошлые годы; хвои (листвы)
нет; вершинка, как правило, обломлена; кора и мелкие веточки легко отвалива-
ются; стволовые вредители вылетают или вылетели;
6 — свежий ветровал — деревья, которые в прошлом году относились к катего-
риям 0-3, а в год наблюдений выпали с оголением корневой системы; пожелтевшая
листва (хвоя) местами может сохраняться, текущего прироста нет; по стволу, как
правило, свежие поселения короедов или энтомокомплекса, летных отверстий нет;
51
7 — старый ветровал — лежащие деревья с вывороченными корневыми си-
стемами, по времени — это отпад прошлых лет; кора легко отваливается, ство-
ловые вредители вылетают или вылетели;
8 — свежий бурелом — деревья, которые в прошлом году относились к кате-
гориям 0-3, а в год наблюдений выпали со сломом ствола ниже границы кроны;
пожелтевшая листва (хвоя) местами еще может сохраняться; текущего прироста
нет, по стволу как правило, свежие поселения короедов или энтомокомплекса,
летных отверстий нет;
9 — старый бурелом — деревья со сломанным ниже границы кроны ство-
лом, по времени — это отпад прошлых лет; кора легко отваливается, стволовые
вредители уже вылетели.
С целью повышения объективности оценки состояния живых деревьев жела-
тельно учитывать и такие признаки, как наличие водяных побегов и контрфор-
са, каллюсообразование и т. п.
Картирование деревьев. Для составления плана древостоя измеряются ко-
ординаты отдельных деревьев. За начало координат, как правило, принимается
юго-западный угол ППП. Измерения проводятся при помощи 10-метровой ру-
летки и мерной рейки (шеста) длиной 5,5 м в соответствии со схемой картиро-
вания. На каждом 10-метровом отрезке линий, проходящих через пикеты, для
деревьев, попавших на полосу картирования, измеряются расстояния от начала
рулетки до рейки и от рулетки до ствола, при этом рулетка и рейка должны со-
ставлять прямой угол. К расстоянию от рулетки до ствола сразу прибавляют ра-
диус дерева (получая, таким образом, координату центра ствола), после чего
результаты измерений заносят в специальную таблицу. Оба расстояния даются с
точностью 0,1 м. Окончив картирование, проверяют наличие в таблице всех но-
меров деревьев. В камеральных условиях исходя из таблицы и схемы картиро-
вания координаты деревьев, измеренные относительно 10-метровых отрезков
отдельных линий, переводят в абсолютные координаты относительно утла ППП,
принятого за «0». Для этого к первой координате прибавляют расстояние от на-
чала линии до начала соответствующего 10-метрового отрезка. Ко второй коор-
динате при картировании «вверх» прибавляют расстояние от линии картирова-
ния до самой нижней линии, при картировании «вниз» координату вычитают из
этого расстояния. При повторных наблюдениях дополнительно измеряются ко-
ординаты деревьев из подроста, диаметр которых достиг 6 см.
Для достижения большей точности измерения координат желательно увели-
чить число линий картирования. С этой целью используются линии, проходя-
щие через постоянные колышки трансект для учета подроста и подлеска.
Ярус подлеска (кустарниковый ярус) не относится к числу характерных ком-
понентов ненарушенных или слабо нарушенных бореальных хвойных лесов. В
наибольшей степени он представлен в производных типах лесных сообществ,
развивающихся на стадиях восстановления северных хвойных лесов после раз-
52
личных внешних нарушений, а также на начальных этапах формирования лес-
ной растительности на вновь образовавшихся субстратах естественного и ант-
ропогенного происхождения в процессе первичных сукцессий в поймах рек, на
отложениях ледников, оползней, вулканов, отвалах горных разработок и т. д.
Для измерения характеристик яруса подлеска на постоянных пробных пло-
щадях в лесных сообществах используются методы постоянных учетных пло-
щадок и профилей (разд. 1.4). В пределах пробной площади в зависимости от ее
размера закладывается от 2 до 10 регулярно расположенных профилей шириной
2 м и длиной, соответствующей длине одной из сторон пробной площади. В пре-
делах каждого профиля фиксируются примыкающие друг к другу или располо-
женные через 3-5 м одна от другой квадратные учетные площадки (размером
2 х 2 м). При малой плотности особей и разреженности яруса подлеска исполь-
зуются примыкающие площадки, т. е. учет растений проводится по всей длине
профиля. При хорошо выраженном, имеющем достаточно высокое проективное
покрытие ярусе подлеска измерение всех (или части) его характеристик осуще-
ствляется на расположенных по профилю учетных площадках. При закладке для
описания нижних ярусов сообществ блоков учетных площадок, состоящих из
4-9 площадок размером 1 х 1 м, можно использовать эти же блоки для регист-
рации характеристик яруса подлеска, принимая 1 блок за 1 учетную площадку.
Учет особей высотой менее 0,5 м во всех случаях производится на площадках
для описания нижних ярусов сообществ (разд. 1.6).
На каждой площадке для каждой особи подлеска фиксируется видовая при-
надлежность и категория состояния, определяется сомкнутость яруса. При де-
тальных исследованиях дополнительно определяют возраст каждой особи кус-
тарника, высоту и ширину куста, величину общего проективного покрытия и про-
ективного покрытия каждого вида. Кроме того, для каждой учетной площадки
фиксируется ряд характеристик условий местообитания: относительная сумма
площадей сечений и сквозистость древесного яруса, характеристики верхних
горизонтов почвы (разд. 1.6).
При определении морфометрических параметров и возраста растений яруса под-
леска следует принимать во внимание особенности морфологии кустарников. Со-
гласно определению И. Г. Серебрякова с сотрудниками (1954), кустарники — это
«деревянистые растения, обычно не превышающие 1,5-4,5 м высотой, с главной
осью, функционирующей лишь в начале их онтогенеза, и представляющие собой
во взрослом состоянии систему сменяющих друг друга укореняющихся скелет-
ных осей с коротким жизненным циклом... быстрым спадом годичного прирос-
та... и поэтому незначительной высотой... В отличие от деревьев у кустарников
одновременно функционируют скелетные оси двух-трех и даже четырех поколе-
ний, чем и обусловливается их многоосность». Формирование куста начинается с
развития из семени главной оси, рост которой в высоту постепенно замедляется
и через 7-15 лет (в зависимости от вида) прекращается полностью, после чего
53
главная ось отмирает. Этот процесс сопровождается появлением из спящих почек
у основания главной оси боковых побегов II порядка, на которых, в свою оче-
редь, формируются побеги третьего порядка и т. д. При увеличении возраста ку-
ста более старые оси постепенно отмирают, их заменяют более молодые; одно-
временно на одной материнской оси существуют оси нескольких порядков, что и
способствует созданию формы куста (Серебряков и др., 1954; Корчагин, 1960).
Высота особей яруса подлеска регистрируется как среднее из высоты 3-5 осей
разных порядков. Высота определяется при помощи линейки, мерного шеста или
высотомера.
Ширина куста определяется по двум взаимно перпендикулярным направле-
ниям при помощи рулетки, аналогично измерению ширины крон деревьев.
Возраст куста в большинстве случаев определяется приблизительно. Труд-
ность определения возраста растений яруса подлеска состоит в том, что с тече-
нием времени старые оси кустов, как правило, постепенно отмирают, разруша-
ются и заменяются новыми, возраст каждой из которых значительно меньше
общего возраста куста. У некоторых видов главная ось сохраняется наряду с
боковыми в течение всей жизни. К таким растениям относится широко распро-
страненный вид кустарникового яруса бореальных лесов можжевельник обык-
новенный (Juniperus communis L.), у которого в более сухих местообитаниях осо-
би нередко принимают форму небольшого дерева. В этом случае определение
возраста проводится на поперечном срезе у основания главной оси или на керне
так же, как это делается для древесных растений. Для видов кустарников, у ко-
торых наблюдается отмирание главной оси, определение возраста может быть
выполнено по методу, предложенному И. Г. Серебряковым (1954). Общий воз-
раст особи кустарника (или парциального куста у видов, для которых характер-
но развитие длинных корневищнообразных подземных побегов, дающих начало
новым кустам), определяется по формуле
А = а + (п Ь),
где А — полный возраст куста; а — возраст скелетной оси последнего порядка
(определяется по числу годичных колец у основания периферического побега
или по числу линейных годичных приростов, если они хорошо различимы); п
— наибольшее число порядков скелетных осей куста; b — интервал кущения,
т. е. разность в возрасте пар материнских и дочерних осей куста (устанавливает-
ся на основе измерения у 50 экземпляров данного вида и принимается за посто-
янную для данных условий величину).
Сомкнутость яруса определяется как доля площади проекций кустов (в про-
центах) от площади учетной площадки без учета просветов внутри кустов.
Проективное покрытие отдельных видов и общее проективное покрытие
регистрируется как доля площади проекции всех надземных частей растений дан-
ного вида или яруса в целом (в процентах) от площади учетной площадки.
54
Категория состояния растений яруса подлеска определяется на основе тех
же критериев, что и категория состояния деревьев (см. выше).
1.6. ИЗМЕРЕНИЕ ХАРАКТЕРИСТИК НИЖНИХ ЯРУСОВ
И ПОЛОГА ПОДРОСТА В ЛЕСНЫХ СООБЩЕСТВАХ 1
Основным методом сбора данных при детальном исследовании характерис-
тик травяно-кустарничкового и мохово-лишайникового ярусов и полога подрос-
та древесных растений на постоянных пробных площадях в лесных сообществах
служит метод постоянных учетных площадок. Наиболее удобными для прове-
дения комплексных исследований нижних ярусов сообществ являются площад-
ки размером 1 х 1 м. Методика закладки учетных площадок подробно изложена
в разд. 1.4.
На каждой площадке проводится измерение всех основных характеристик по-
лога возобновления (подроста) древесных растений, мохово-лишайникового и
травяно-кустарничкового ярусов. Кроме того, для каждой площадки или блока
площадок фиксируется ряд характеристик условий местообитания: относитель-
ная сумма площадей сечений и сквозистость древесного яруса, характеристики
верхних горизонтов почв.
1.6.1. Изучение характеристик возобновления древесных растений
Критерии выделения полога подроста
Для выделения полога подроста в бореальных лесных сообществах в отече-
ственных и зарубежных исследованиях и методических руководствах предлага-
ется использовать следующие критерии: величину диаметра на высоте 1,3 м, вы-
соту и возраст особей. При этом разные авторы определяют разные количествен-
ные границы одних и тех же параметров и используют разные параметры, в за-
висимости от целей исследования и особенностей исследуемых лесных сооб-
ществ (табл. 1.1).
Отечественные исследователи при выделении единой категории — подроста,
наиболее часто используют критерий соотношения со средней высотой I, наи-
более высокого яруса древостоя. К подросту при этом относят особи высотой
менее 0,25 или 0,5 высоты I яруса древостоя (Анучин, 1952, 1971; Нестеров, 1954;
Корчагин, 1956, 1976; Тихонов, Набатов, 1995). Часто используется также кри-
терий величины диаметра на высоте 1,3 м: особи древесных растений с диамет-
ром на высоте груди менее 6 (иногда 3-5) см считают подростом, остальные особи
относят к древесному ярусу (Зябченко, 1974, 1984; Молчанов, 1974; Казимиров
1 Авторы И Ю Баккал, В. В Горшков, Н И Ставрова
55
Таблица 1.1. Критерии выделения полога подроста в бореальных лесах
Категория особей Параметры Автор
высота, м диаметр (1,3 м), см возраст, лет
Подрост < 0,5 Н I яр — — Анучин, 1952
Подрост <0,5 HI яр — > 1 Нестеров, 1954
Подрост < 0,25 Н I яр — — Корчагин, 1956, 1976
«Всходы» <0,1 — — Сукачев и др , 1957
Подрост 0,1 <<Н 1яр — —
Подрост — <6,0 — Зябченко, 1984, Дыренков, 1984
Самосев — <2,0 — Молчанов, 1974
Подрост — 2,0< <6,0 —
Подрост — <3,0 — Казимиров и др, 1977
Подрост мл генер — — < 10 Факторы регуляции ,1983
Подрост ст генер — <6,0 > 10
Самосев — __ <2(10)" ГОСТ 18487-87
Подрост <0,25 Н I яр — >2(10)
Подрост <0,25 HI яр2) < 6.01’ <404) Тихонов, Набатов, 1995
Самосев — — 2-5 Федорчук и др , 1998
Подрост — <6,0 >5
Seedlings — <2,5 — Dix, Svan, 1971
Saplings — 2,5-10,0 —
Seedlings < 1,3 (1,37) — — Sykes, Horrill, 1971; Engelmark, 1998, Youngblood, 1995, Linder et al, 1997
Seedlings < 0,3 м — —
Transgressives > 0,3 м < 1,0 — Purchase, La Roi, 1982
Saplings — 1,0-5,0 —
Seedlings < 1,3 — — Gagnon, Bradfield,
Saplings > 1,3 < 10 — 1986
Seedlings + saplings — — <40 Steijlen et al, 1994
Примечания: О — цифра в скобках означает вариант значения; 2 3) — общий критерий,
3) — критерий для сообществ с одновозрастным старым древостоем; 4) — критерий для
сообществ с разновозрастным древостоем
56
и др., 1977; Факторы регуляции..., 1983; Дыренков, 1984; Федорчук и др., 1998)
В составе возобновления, как правило, выделяют 3 качественные категории
особей: 1) всходы — особи возрастом до 1 (2) лет; 2) самосев (подрост младших
генераций) — особи в возрасте от 2 (3) до 5 (10) лет или все особи с диаметром
на высоте 1,3 м менее 2 см; и 3) собственно подрост (подрост старших генера-
ций) — особи возрастом более 5(10) лет или с диаметром на высоте груди от 2
до 6 см.
В значительном числе зарубежных работ к возобновлению относят все особи
древесных растений высотой менее 1,3 (1,37; 1,5) м, остальная часть популяции
рассматривается как древесный ярус (Sykes, Horrill, 1971; Engelmark et al., 1993;
Youngblood, 1995; Linder et al., 1997).
Другим часто используемым критерием является величина диаметра на вы-
соте 1,3 м — менее 5 (или менее 10) см (Dix, Svan, 1971; Purchase, La Roi, 1983;
Gagnon, Bradfield, 1987). Возобновление подразделяется обычно следующим
образом: 1) подрост (seedlings), включающий особи с диаметром на высоте 1,3 м
менее 1 (в других случаях менее 2,5) см или особи с высотой менее 1,3 м; 2)
молодые деревья (saplings) с высотой более 1,3 м или диаметром на высоте гру-
ди более 1 (2-2,5) см, но менее 10 см. Как в отечественных, так и в зарубежных
исследованиях для выделения полога возобновления иногда в качестве основ-
ного используется критерий возраста. В этом случае, как правило, к подросту
относят все особи в возрасте менее 40 лет (Steijlen et al., 1995; Тихонов, Наба-
тов, 1995).
Применение разных методик приводит к различию получаемых результатов
(как по подросту, так и по древесному ярусу) и затрудняет сопоставление дан-
ных разных исследований. Если отнесение особей к древесному ярусу выполня-
ется на основе величины диаметра на высоте груди, а выделение полога подро-
ста основано на критерии высоты или возраста, возможен неучет части особей
древесных растений при описании их популяций в пределах сообществ. Прежде
всего это относится к лесным сообществам с разновозрастными популяциями
древесных растений и непрерывным распределением особей по высоте и диа-
метру. Только в лесах с резко дискретным распределением особей древесных ра-
стений по высоте и диаметру (например, при высоте древостоя 12-20 м и высо-
те особей в пологе подроста, не превышающей 1 м) данные по численности и
основным параметрам полога подроста, полученные при его выделении разны-
ми методами, могут практически не различаться. Это явление характерно для
лесов с одновозрастным высокополнотным древостоем.
В соответствии с этим наиболее универсальным и корректным методом вы-
деления полога возобновления следует признать использование абсолютной вы-
соты особей. Удобной и широко применяемой является высота 1,3 м (или 1,4;
1,5 м): к возобновлению относят все особи древесных растений высотой мень-
ше указанных значений. При этом в составе древесного яруса (совокупность
57
особей высотой более 1,3 м) может быть при необходимости особо выделена
перечетная часть, состоящая из деревьев с диаметром 6 см и более на высоте
1,3 м, а в составе возобновления — всходы и подрост.
Измерение характеристик возобновления
На каждой площадке регистрируются все особи древесных растений, относя-
щиеся к пологу возобновления (особи старше 1 года, высота которых не превы-
шает 1,3 м). Отдельно фиксируется количество всходов. Дополнительно на учет-
ных площадках рекомендуется регистрировать особи древесного яруса с диамет-
ром на высоте груди менее 6 см.
У каждого экземпляра древесных растений, относящегося к пологу возобнов-
ления, определяются следующие характеристики: видовая принадлежность, воз-
раст, высота, диаметр стволика у корневой шейки, категория жизненного состо-
яния. У особей старше 5 лет дополнительно измеряются: приросты главной оси
за последние 3 года, число и длина боковых побегов 2-го порядка в каждой
мутовке за последние три года. Кроме того, измеряется величина проективного
покрытия подроста по видам.
Определение возраста растений подроста проводится путем подсчета чис-
ла полных годичных приростов главной оси по заметным на коре стволика в виде
пояска следам чешуй верхушечных почек; если рост в высоту в текущем году не
завершен, прирост текущего года не учитывается. У сильно угнетенных особей
эти следы хорошо различимы только в верхней части стволика и очень плохо —
в нижней. Для повышения точности определения возраста (особенно это отно-
сится к подросту лиственных древесных растений) следует провести отбор 20-
50 контрольных экземпляров основных видов, формирующих полог подроста.
Контрольные экземпляры отбираются на территории, непосредственно прилега-
ющей к пробной площади. У каждого контрольного экземпляра возраст опреде-
ляется двумя способами: описанным выше и путем подсчета числа годичных
колец на поперечном срезе стволика, выполненном у корневой шейки. После-
дний метод является более точным, хотя и в' этом случае возможно некоторое
занижение возраста из-за отсутствия радиального прироста в отдельные годы
(выпадение годичных колец). Сопоставление возраста особей, определенного в
полевых условиях путем подсчета следов от почечных чешуй, и возраста, зафик-
сированного на поперечном срезе стволика в лабораторных условиях, позволяет
определить среднюю величину систематической ошибки полевого определения
и внести соответствующую поправку в полученные при работе на пробной пло-
щади значения возраста. При определении возраста растений подроста хвойных
в полевых условиях следует иметь в виду, что нижняя часть стволика в боль-
шинстве случаев скрыта в толще мохово-лишайникового яруса, слое опада, а
иногда — в верхней части лесной подстилки. Поэтому перед определением воз-
раста следует освободить главную ось и определить положение верхней грани-
58
цы гипокотиля, от которой и следует проводить отсчет возраста. Указанная гра-
ница (граница между гипокотилем и эпикотилем) хорошо заметна по смене глад-
кой коры гипокотиля на покрытую рубчиками (следы от первичной хвои и бра-
хибластов) кору надсемядольной части главного побега.
Высота растений подроста определяется как расстояние от корневой шейки
до конца последнего завершенного прироста главной оси или побега замещения
(если верхушечная почка погибла). В случае усыхания верхней части главной
оси и отсутствия побега замещения высота определяется как расстояние от кор-
невой шейки до последней живой боковой ветви.
У особей старше 5 лет отдельно регистрируется величина каждого прироста
главной оси за последние три года, а также число и длина боковых побегов 2-го
порядка в каждой мутовке за последние три года (приросты текущего года не
учитываются, если рост в момент проведения измерений не завершен). Высота
особей и величина приростов измеряется при помощи рулетки или линейки с
точностью до 0,1 см.
Диаметр стволика растений подроста определяется на уровне корневой
шейки при помощи штангенциркуля с точностью до 0,01 см.
Категория жизненного состояния подроста в полевых условиях определя-
ется на основе следующих критериев:
1) жизнеспособные особи — прогрессирующий или стабильный прирост глав-
ной оси за последние 3 года, нормальное развитие ассимиляционных органов
(отсутствие хлорозов, у хвойных — характерная для региона продолжительность
жизни хвои), отсутствие повреждений насекомыми и болезнями;
2) особи низкой жизнеспособности — регрессирующий прирост главной оси
за последние 3 года или отсутствие прироста (гибель верхушечной почки и усы-
хание верхней части главной оси), нарушение развития ассимиляционных орга-
нов (хлороз, у хвойных — снижение продолжительности жизни хвои), наличие
повреждений насекомыми и болезнями и механических повреждений;
3) нежизнеспособные особи — полностью усохшие или сохранившие не бо-
лее одной живой боковой ветви.
Проективное покрытие подроста древесных растений регистрируется на
учетных площадках отдельно для каждого вида с использованием рамки разме-
ром 1 х 1 м, разделенной на 100 квадратов по 100 см2, наименьшая оцениваемая
площадь принимается равной 1 см2 (0,01 %). Важно отметить, что в величине
проективного покрытия всегда присутствует размер анализируемой площади.
С, = а, / ад 100 %,
где а,- — площадь проекции z-того вида, ао — размер анализируемой площади
(площадь рамки). При участии в составе полога подроста нескольких видов и
взаимном перекрытии их крон, проективное покрытие каждого вида оценивает-
ся по доле его участия в формировании общего покрытия полога на площадке.
59
1.6.2. Характеристики травяно-кустарничкового
и мохово-лишайникового ярусов
Основными измеряемыми характеристиками травяно-кустарничкового и мо-
хово-лишайникового ярусов являются общее проективное покрытие, проектив-
ное покрытие отдельных видов, проективное покрытие отмерших частей рас-
тений, высота яруса, высота отдельных видов.
Общее проективное покрытие — доля (в процентах) видимой при взгляде
сверху площади проекции надземных частей всех растений яруса от всей учет-
ной площадки. Измеряется с помощью рамки, разделенной на 100 квадратов по
100 см2, наименьшая оцениваемая площадь при этом составляет 1 см2 (0,01 %).
Проективное покрытие вида — доля (в процентах) площади проекции над-
земных частей растений данного вида, не перекрытых выше расположенными
частями растений других видов, от площади учетной площадки. Сумма покры-
тий всех видов яруса при указанном методе их измерения всегда равна общему
проективному покрытию яруса.
Относительное покрытие вида — доля участия данного вида (в процентах)
в сложении общего проективного покрытия яруса.
В геоботанике наряду с понятием «общее покрытие яруса» используется по-
нятие «суммарное покрытие яруса». Оно не может быть измерено непосредствен-
но и рассчитывается как сумма проективных покрытий всех видов, независимо
от того, перекрываются они или нет вышерасположенными растениями других
видов. Величина суммарного покрытия яруса может быть больше величины об-
щего покрытия (и даже больше 100 %). Использование этой характеристики не
рекомендуется из-за низкой точности ее определения.
Для травяно-кустарничкового яруса проективное покрытие фотосинтезирую-
щих и нефотосинтезирующих частей растений оценивается отдельно.
Высота яруса и отдельных видов. Высота регистрируется для видов травя-
но-кустарничкового и мохово-лишайникового ярусов, доля участия которых в
формировании общего покрытия яруса на площадке составляет более 5 %. Она
определяется как средняя величина из 3-5 измерений для каждого вида. Измере-
ние высоты проводится от поверхности подстилки. Высота яруса в целом опреде-
ляется как средневзвешенная из высот основных видов яруса (доля участия кото-
рых в формировании общего покрытия яруса на площадке составляет более 5 %).
Проективное покрытие отмерших частей растений. Представляет собой
видимую при вертикальном взгляде площадь поверхности ветоши трав, мхов и
лишайников.
Кроме характеристик полога подроста, травяно-кустарничкового и мохово-
лишайникового ярусов на учетных площадках регистрируется относительная
площадь оснований стволов деревьев и пней, а также относительная площадь
нарушенной поверхности, занятая валежом, различными фракциями древесного
60
опада (хвои, листьев, мелких и крупных ветвей), экскрементами животных, от-
крытой лесной подстилкой, выходящими на поверхность валунами и минераль-
ными горизонтами почвы. Эти параметры имеют большое значение для харак-
теристики условий, в которых развивается растительность нижних ярусов лес-
ных сообществ на конкретных участках.
Следует отметить, что покрытие мохово-лишайникового яруса и покрытие рас-
полагающихся в пределах этого яруса нефотосинтезирующих частей древесных
растений и кустарничков, а также покрытие опада, открытой подстилки и валу-
нов в сумме составляет 100 %. Эта величина во избежание неточностей или
ошибок контролируется при измерении проективного покрытия напочвенного
покрова на каждой площадке. Измерением проективного покрытия опада и не-
фотосинтезирующих частей растений часто пренебрегают, что приводит к не-
верным оценкам проективного покрытия мохово-лишайникового яруса: его по-
крытие исследователи приравнивают к 100 %. На самом деле покрытие мохово-
лишайникового яруса равное или приближающееся к 100 % встречается крайне
редко — только в лишайниковых рединах. Измерение площади проекций отмер-
ших частей растений и нарушенной поверхности позволяет оценить роль раз-
ных факторов в происхождении нарушений, что является важным аспектом изу-
чения биологии лесных сообществ.
1.6.3. Характеристика условий местообитания растений нижних ярусов
и полога подроста лесных сообществ
При исследовании характеристик нижних ярусов и полога подроста в лесных
сообществах методом учетных площадок рекомендуется одновременно выполнить
измерение ряда характеристик, влияющих на их формирование В качестве пара-
метров, характеризующих условия светового и теплового режима, а также условия
почвенного питания и увлажнения рекомендуются следующие: проективное покры-
тие крон деревьев, относительная сумма площадей сечений и сквозистость древес-
ного яруса, степень каменистости почвы (глубина залегания валунного материала),
толщина лесной подстилки и верхних минеральных горизонтов почвы. Эти пара-
метры определяются для каждой учетной площадки или блока площадок.
Проективное покрытие крон деревьев — площадь проекции на площадку
расположенных над ней крон деревьев. Измерение этой характеристики выпол-
няется методом вертикальных секущих плоскостей, схема которого представле-
на на рис. 1.12 (см. стр. 62).
Относительная сумма площадей сечений древесного яруса измеряется в
одном из углов площадки или блока площадок (в месте расположения фиксиру-
ющего ее колышка) по методу Биттерлиха (описание метода представлено в
разд. 1.5).
Характеристики верхних горизонтов почв регистрируются в почвенных
прикопках размером 20 х 20 см и глубиной 30 см, которые располагают на сты-
61
Рис. 1.12. Измерение площади проек-
ции крон деревьев методом верти-
кальных секущих плоскостей
Линии а|-аг , bi—Ьг соотвествуют верти-
кальным плоскостям, которые проводят-
ся по рейкам, ограничивающим рамку.
Линии d|-d2 и е|—ез соответствуют верти-
кальным плоскостям, отграничивающим
площадь кроны дерева в пределах рамки.
Эти линии фиксируются на поверхности
рамки при помощи рейки и рулетки (или
двух реек)
ке площадок, если площадки расположе-
ны блоками по 4 или с одной из стород
площадки, если площадки расположены
по одной. В каждой прикопке проводит-
ся измерение толщины лесной подстил-
ки и подзолистого горизонта, а также ви-
димой толщины иллювиального горизон-
та. Если в верхней части подзолистого го-
ризонта выделяется темноокрашенный
гумусированный слой (переходный гори-
зонт А] Аг), его толщина также регистри-
руется. Толщина подстилки определяет-
ся от ее поверхности (после удаления
мохово-лишайникового покрова и слоя
свежего опада) до верхней границы вер-
хнего минерального горизонта. При на-
личии в толще подстилки или под под-
стилкой прослойки углей фиксируется
глубина ее расположения и толщина. Все
измерения проводятся на каждой из
4 сторон прикопки. Толщина каждого вы-
деленного генетического горизонта опре-
деляется как среднее из 4 измерений.
Глубина залегания валунного мате-
риала (рис. 1.13) измеряется на каждой
площадке при помощи металлического
щупа с заостренным концом длиной 50-
80 см и диаметром 1 см. По длине щупа
через 1 см делаются нарезки-деления. Измерения проводятся в 5 точках, четыре
из которых расположены по углам площадки и одна в центре. Острый конец щупа
с усилием погружается в почву до первого препятствия (ближайшего к поверх-
ности валуна или гальки). Глубина погружения отсчитывается от поверхности
подстилки и определяется по имеющимся на щупе делениям с точностью до 0,5
см. Глубина залегания валунного материала для каждой площадки определяется
как среднее из 5 выполненных измерений.
Выполненные на учетных площадках измерения основных характеристик ниж-
них ярусов лесных сообществ и параметров, влияющих на их формирование,
позволяют осуществить на их основе расчет производных характеристик. При
обработке данных необходимо определить средние для пробной площади значе-
ния всех измеренных параметров и рассчитать средние значения производных
параметров (среднего относительного проективного покрытия видов, среднего
62
прироста главной и боковых осей
растений подроста, средней высо-
ты яруса и т. д.). Можно рассчи-
тать для пробной площади значе-
ния различных индексов: видово-
го и пространственного разнооб-
разия разных ярусов, жизненно-
го состояния подроста и др. В
дальнейшем соответствующие
измерения и расчеты выполняют-
ся для всех постоянных пробных
площадей, которые в соответ-
ствии с целью исследований зак-
1 м
Рис. 1.13. Схема размещения точек измерения
глубины залегания валунного материала
ладываются в сообществах разной типологической принадлежности или в сооб-
ществах одного типа с разным уровнем какого-либо фактора (промышленного
загрязнения, давности последнего пожара или рубки). Это позволяет проанали-
зировать характер изменения различных характеристик подчиненных ярусов
лесных сообществ в зависимости от изменения величины характеристик других
ярусов или значений факторов местообитания, например степени каменистости
почв, толщины подстилки, сквозистости древесного яруса, концентрации загряз-
нителей в почве, давности последнего пожара и т. д. Принципы и методы мате-
матического анализа выполненных измерений и способы представления резуль-
татов рассмотрены в соответствующих разделах.
1.7. МЕТОДИКА ИЗМЕРЕНИЯ ХАРАКТЕРИСТИК ЭПИФИТНОГО
ЛИШАЙНИКОВОГО ПОКРОВА СТВОЛОВ ДЕРЕВЬЕВ 1
Эпифитные лишайники являются одним из компонентов бореальных лесных
сообществ. Они часто образуют большую биомассу и играют существенную роль
в круговороте веществ в биогеоценозах. Практическое значение этой группы
организмов определяется прежде всего высокой чувствительностью к измене-
нию химического состава атмосферного воздуха, что позволяет использовать их
в качестве биоиндикаторов.
Характеристики эпифитного лишайникового покрова тесно связаны с опре-
деленными экологическими факторами и закономерно изменяются в соответствии
с изменением условий среды. Такие факторы, как общее количество осадков,
относительная влажность воздуха, условия освещённости, скорость ветра, игра-
ют важную роль в фотосинтезе лишайников и во многом определяют их состо-
1 Автор В. В. Горшков
63
яиие (Горшков, 1990, 1991; Толпышева, 1998; Тарасова, 1998, 2000). Условия
окружающей среды могут оказывать как прямое, так и косвенное влияние. На-
пример, прямое воздействие на состояние эпипокрова могут оказывать густота
и протяженность кроны дерева, на котором проводятся описания. Низкая густая
крона дерева препятствует поступлению осадков на ствол и создает дополни-
тельное затенение. Примером показателей косвенного воздействия может слу-
жить соотношение мхов и лишайников в напочвенном покрове или проективное
покрытие травяно-кустарничкового яруса, характеризующие тип леса, а следо-
вательно, и условия освещенности и влажности под его пологом. Травяно-кус-
тарничковый ярус может оказывать и прямое влияние на эпифитный покров у
основания дерева, создавая дополнительное затенение.
Рекомендуемая методика количественного учета эпифитных лишайников и по-
казателей условий их местообитания основана на работах К. А. Рассадиной (1930);
О. Almbom (1948); J. J. Barkman (1958); И. Кишки (1984); описаниях традицион-
ных методов изучения растительности (Handbook off..., 1974, 1982, 1984), а также
методах, принятых в лесоведении и лесной таксации (Анучин, 1982).
Эпифитный лишайниковый покров так же, как и другие компоненты лесного
растительного сообщества, целесообразно изучать на постоянных (или времен-
ных) пробных площадях (см. разделы 1.2, 1.3). В пределах пробной площади
закладывают одну (или несколько) трансект, преимущественно с севера на юг. На
трансекте обследуются подряд все деревья диаметром более 5 см. Число деревь-
ев, обследованных на разных пробных площадях, должно составлять не менее 10.
На каждом дереве диаметром более 10 см измерение характеристик эпифит-
ного лишайникового покрова и условий его местообитания выполняется у осно-
вания дерева (на высоте 0-20 см) и на высоте 130-150 см на 4 сторонах, ориен-
тированных по основным румбам (север, восток, юг и запад) (рис. 1.14Г). На
деревьях диаметром менее 10 см измерения проводятся только с северной и
южной сторон ствола.
Измерение общего проективного покрытия и проективных покрытий всех
встреченных видов проводится при помощи рамок, размер которых пропорцио-
нален диаметру дерева. Для деревьев диаметром менее 10 см удобно использо-
вать рамку 5x5 см, для деревьев большего диаметра - 10x10 см, 10x20 см и 20x20
см. При помощи лески рамка делится на 100 или 200 квадратов, служащих шка-
лой для измерения проективного покрытия видов. В полевых условиях не все-
гда представляется возможным точно определить вид лишайника. В таком слу-
чае можно присвоить виду условное название (например, вид А, Б и т. д.), а за-
тем, взяв образец, установить точное название вида в лаборатории. Если же вы-
зывает затруднение даже само выделение отдельных видов, то можно ограни-
читься определением покрытия групп видов (например, накипных и кустистых
лишайников) и покрытием видов, которые наиболее широко распространены в
районе исследований и идентификация которых не вызывает затруднений.
64
б
Рис. 1.14. Методика измерений характеристик местообитания и эпифитного лишай-
никового покрова: а) измерение таксационных показателей деревьев и сквозис-
тости древесного яруса для конкретного участка эпифитного лишайникового по-
крова, б) расположение пробных площадок на стволах деревьев, в) расположение
площадок по окружности ствола, г) измерение угла наклона поверхности ствола
Обозначения: ha — высота дерева, hcr — высота нижней границы прикрепления кроны,
^bark — высота верхней границы грубой корки, тсг — радиус кроны, 7'нижн, Гсре4, Tsep]i —
области углов, где измерялась сквозистость древесного яруса, а — угол наклона поверх-
ности ствола, A-D — местоположение пробных площадок.
65
Кроме проективного покрытия эпифитных лишайников в каждом описании
регистрируются характеристики анализируемого участка на стволе (рис. 1.14):
параметры его расположения (высота от поверхности земли, сторона дерева),
угол наклона поверхности ствола, характеристики кроны дерева (высота при-
крепления, протяженность), сквозистость древесного яруса (для описаний на
высоте груди), проективное покрытие и высота травяно-кустарничкового яруса
(для описаний у основания дерева).
Для каждого дерева, на котором проводятся описания эпипокрова, измеря-
ются высота, возраст, диаметр, высота поднятия грубой корки.
Для всей пробной площади регистрируется давность последнего нарушения
(рубки, пожара), сомкнутость крон и относительная сумма площадей сечений
древесного яруса, доля лишайников в мохово-лишайниковом ярусе, покрытие
травяно-кустарничкового яруса.
1. Характеристики анализируемого участка ствола.
Высота от поверхности земли. Обычно описания проводят на высоте груди,
что соответствует высоте 130-150 см. На эту высоту при помощи шила крепит-
ся рамка.
Сторона дерева (румб) определяется по компасу.
Угол наклона поверхности ствола в месте описания измеряется горным ком-
пасом с точностью до 1 градуса (см. рис. 1.14В). Боковая сторона компаса при-
кладывается к стволу, затем по стрелке, показывающей отклонение от вертика-
ли, берется отсчет. Для эпифитного покрова это очень важная характеристика.
Положительно наклоненные поверхности получают больше влаги, чем поверх-
ности, имеющие отрицательный наклон. Кроме того, при прочих равных усло-
виях, поверхность, наклоненная положительно, получает больше света, чем на-
клоненная отрицательно.
Высота прикрепления, протяженность и ширина (радиус) кроны определя-
ют условия освещенности и увлажнения на стволе. Эти характеристики измеря-
ются с каждой стороны дерева, на которой проводятся описания эпипокрова.
Высота прикрепления кроны определяется по высоте прикрепления самой ниж-
ней ветви; протяженность кроны вычисляется как разность между высотой
дерева и высотой прикрепления кроны; ширина кроны соответствует длине са-
мой длинной ветви.
2. Характеристики дерева (высота, возраст, диаметр, высота границы
грубой корки), сквозистость древостоя (для данного конкретного участка ли-
шайникового покрова).
Высота дерева* 1. Определяется при помощи эклиметра (ЭВ1).
Возраст дерева*. Определяется подсчетом годичных колец по кернам, взятым
возрастным буром у основания ствола и на таксационной высоте (130 см от земли).
1 Подробная методика определения характеристик, помеченных звездочкой, приведе-
на в разделах 15, 1 6; 3 2
66
Диаметр дерева*. Измеряется при помощи мерной вилки у основания ствола
и на высоте 130 см от земли.
Сквозистость древесного яруса* определяется на высоте груди у каждого
ствола дерева с четырех сторон света в пределах углов от 0 до 90° к горизонту.
Высота границы грубой корки (высота границы между многослойной темной
и тонкой светлой коркой) — при помощи эклиметра или рулеткой при значени-
ях <1,5 м.
3. Характеристики лесного сообщества (пробной площади).
Давность последнего пожара*. Датировку пожаров выполняют на основе
анализа кернов и спилов, взятых с 5-8 поврежденных пожаром живых деревьев
в радиусе 50-100 м от территории пробной площади. Давность пожара опреде-
ляется как разность между текущим возрастом дерева и его возрастом в момент
повреждения камбия.
Относительная сумма площадей сечений древесного яруса* определяется при
помощи полнотомера Биттерлиха в 5-6 точках пробной площади.
Сомкнутость крон определяется глазомерно в процентах или долях едини-
цы. За 0 принимается полностью свободное от крон пространство между ство-
лами деревьев; 100%-ная сомкнутость (или 1) отмечается в тех случаях, когда
кроны деревьев полностью смыкаются. Плотность (густота) крон при этом не
учитывается.
Описания напочвенного покрова* проводят на временных площадках разме-
ром 1x1 м, расположенных по трансектам с юга на север через каждые 5-6 м.
Для анализа данных по напочвенному покрову достаточно иметь описания 10
площадок на каждой пробной площади.
Для выявления полного видового состава эпифитных лишайников деревьев
исследуются: 1) видовой состав в пределах учетных площадок; 2) поверхность
стволов и ветвей деревьев от 0 до 2 м от земли; 3) деревья, находящиеся в ради-
усе 10-100 м от пробной площади; 5) другие типы леса (маршрутный метод).
Все вышеперечисленные характеристики заносятся в журнал или бланк опи-
сания. В табл. 1.2 приведен пример бланка описания.
1.8. МЕТОДИКА ОПИСАНИЯ ПОЧВ.
КРАТКАЯ КЛАССИФИКАЦИЯ ЛЕСНЫХ ПОЧВ 1
Закладка почвенного разреза. Место для разреза выбирается с учетом осо-
бенностей микро- и мезорельефа в непосредственной близости от постоянной
пробной площади, но не в пределах ППП. На поверхности почвы лопатой отме-
чают границы в виде прямоугольника длиной 1,5-2 м и шириной 80 см. Разрез
копается до вскрытия материнской породы, что для почв подзолистого типа со-
ответствует 160-180 см. При сомнении в однородности почвообразующей поро-
67
Таблица 1.2. Примерный бланк описаний эпифитного лишайникового покрова ство-
лов сосен (список видов приведен в сокращении)
№ пр площади 1 1 1 1 1 1 1 1 1
№ описания 1 2 3 4 5 6 7 8 9
№ дерева 1 1 1 1 1 1 1 1 2
Румб* 1 2 3 4 1 2 3 4 1
Высота от земли, см 20 20 20 20 130 130 130 130 20
Размер рамки, дм2 4 4 4 4 3 3 3 3 4
Угол наклона, ° 7 1 10 6 2 1 1 0 -1
Общее покрытие, % 15 30 8 1 2 10 1 1 7
Пр покр видов, %
Imshaugia aleurites 1 2 1 1 1 1,5 1 1 1,5
Parmelopsis ambigua 4 10 3 1 3 — — 3
P hyperopia 8 9 — — — 3 — — 2
Hypogymnia physodes 1 5 4 — 0,5 — — 0,5
A 0,5 2 — — — 1 — — —
В 0,5 4 — — — 1 — — —
Гербарий + + + +
Примечание. * — румб. 1 — север; 2 — восток, 3 — юг; 4 — запад
ды, если глубиной разреза не вскрыты грунтовые воды, можно воспользоваться
приемом добуривания. Добуривание производится в открытом разрезе почвен-
ным буром после описания и взятия образцов.
Для лучшего описания профиля переднюю стенку разреза желательно распо-
лагать по направлению к солнцу. Место перед ней должно быть непритоптан-
ным Извлекаемую почву складывают по горизонтам с правой и левой стороны;
при этом желательно подстелить брезент, что помогает сохранить напочвенный
покров. Дерновый горизонт снимается небольшими пластами, которые склады-
ваются по порядку. При закрытии разреза каждый горизонт, по возможности, воз-
вращается на свое место.
Перед описанием разреза желательно сфотографировать профиль на цветную
фотопленку (рис. 1.15).
Описание почвенного разреза. Описание почвенного разреза проводят по
установленной схеме, начиная с лесной подстилки и заканчивая материнской по-
родой. Обязательно отмечают мощность каждого горизонта, его цвет, механи-
ческий состав, влажность, морфологические особенности, включения и пр. Же-
1 Авторы И В Лянгузова, В Т Ярмишко
68
лательно зарисовать схему располо-
жения горизонтов почв в почвенных
разрезах (рис. 1.16).
Строение почвенного профиля.
Горизонт Ао — лесная подстилка или
травяной войлок, представляющие
собой в разной степени разложив-
шийся опад растений. Этот горизонт
может быть расчленен на три подго-
ризонта (Ao,, Аог, Аоз) в зависимос-
ти от степени разложения органичес-
кого вещества. При этом Aqi образу-
ют остатки растений, сохраняющих
свое строение, Аоз — однородная
органическая масса, в которой опад
растений потерял признаки своего
строения.
Горизонт Aj — гумусовый, наибо-
лее темноокрашенный, в котором
происходит накопление органическо-
го вещества в форме гумуса, тесно
связанного с минеральной частью
почвы.
Если горизонт содержит большое _ , ,, _
р Рис. 1.15. Почвенная прикопка на постоян-
количество (от 30 до /0 /о) разложен- ной пробной площади
ных органических остатков, то гори-
зонт называется перегнойным.
Нередко горизонты Ао и А, невозможно разделить, в этом случае они описы-
ваются как один горизонт AqA|. Вместо горизонтов Aq и А, может иметь место
пахотный горизонт — Ап.
Горизонт А? — подзолистый, элювиальный, из которого вымываются раство-
римые вещества. Это сильно осветленный, бесструктурный или слоеватый рых-
лый горизонт. Он обогащен остаточным кремнеземом.
Горизонт В имеет иллювиальный характер, в него вмываются вещества из вер-
хних горизонтов. Обычно это бурый, охристо-бурый, красновато-бурый, уплот-
ненный оструктуренный горизонт. Могут выделяться три подгоризонта:
В] — подгоризонт с преобладанием гумусовой окраски;
В 2 — подгоризонт более слабой и неравномерной гумусовой окраски (он мо-
жет быть не выражен);
В} — подгоризонт окончания гумусовых затеков.
Горизонт Вк — в карбонатных почвах горизонт максимальной аккумуляции
69
Рис. 1.16. Лесные почвы на песках разного гене-
зиса дренированных равнин и пологих скло-
нов (Чертов, 1981)
А — ельник чернично-зеленомошный на грубогу-
мусных подзолистых гумусово-железо-иллювиаль-
ных почвах; Б — ельник чернично-кисличный на мо-
дергумусных подзолистых гумусово-железо-иллю-
виальных почвах
карбонатов (в виде налетов,
прожилок, конкреций).
Горизонт G — глеевый (в
почвах с избыточным увлажне-
нием). Он имеет характерную
окраску — сизую, серовато-го-
лубую, грязно-зеленую, а также
наличие ржавых и охристых пя-
тен.
Горизонт С — материнская
(почвообразующая) порода.
Выделяются также переход-
ные горизонты.
Горизонт А1А2 — прокра-
шен гумусом, но имеет призна-
ки оглеения.
Горизонт А2В — имеет чер-
ты А2 и В.
Для горизонтов с признака-
ми оглеения используется
строчная буква g: A2g, Bg. Тор-
фяной горизонт обозначается
буквой Г, торфянистый — Ат, дерновый — A<i, иллювиально-гумусовый — Вь,
иллювиально-железистый — В/.
Цвет почвы. Для определения окраски почвенного горизонта необходимо
установить преобладающий цвет, насыщенность этого цвета (темно-, светло-),
оттенки основного цвета, например буровато-светло-серый, белесовато-сизый и
т. п. Следует иметь в виду, что на интенсивность окраски влияет влажность по-
чвы Влажная почва обычно имеет более темную окраску, чем в сухом состоя-
нии Для достижения единообразия оценки цвета можно воспользоваться треу-
гольником Захарова (рис. 1.17).
Механический состав. Почва классифицируется по механическому составу.
В основе классификации лежит содержание физической глины в почве. Меха-
нический состав является очень важным свойством почвы, так как играет суще-
ственную роль в почвообразовательном процессе. С ним в значительной мере
связан режим влажности почвы. Механический состав почв можно определить
непосредственно в поле, различая шесть классов: песок, супесь, легкий суглинок,
средний суглинок, тяжелый суглинок, глина. Перед собственно определением
механического состава небольшой образец почвы смачивается водой и размеши-
вается до консистенции густого теста — вода из почвы не отжимается, но почва
блестит и мажется. Кусок «теста» раскатывается на ладони в шнур и сворачива-
70
ется в колечко. Толщина
шнура около 3 мм, а ди-
аметр кольца около 3 см.
По признакам, приведен-
ным на рис. 1.18, опреде-
ляется механический со-
став.
Влажность почвы.
Влажность почвы — ве-
личина крайне перемен-
ная. Она зависит от ме-
теорологических усло-
вий (количества осадков
и времени их выпадения
до описания почвенного
разреза), от уровня грун-
товых вод на момент
описания, от механичес-
кого состава и т. п. По
черный
гумус
FeOnHO SiOAIO
‘ ’ СаСО
Рис. 1.17. Треугольник Захарова, отображающий возник-
новение наиболее часто встречающихся окрасок почв
ла в определении влажности почвы. Однако интерес представляет наличие ка-
пельножидкой воды. Если из стенки ямы просачивается вода или даже накапли-
вается на дне ямы, то следует отметить глубину, на которой это происходит.
Структура почвы. Под структурой почвы понимается отсутствие или нали-
чие частиц более или менее четкой геометрической формы, на которые распада-
ется почва без значительного механического воздействия на нее. Выделяют сле-
дующие типы почвенной структуры.
Зернистая структура. Частицы почвы ясно ограниченной формы с шерохо-
ватой поверхностью диаметром 0,5-5,0 мм.
Комковатая структура. По сравнению с зернистой частицы большего диа-
метра и менее прочны, имеют неправильную округлую форму с несколько ше-
роховатой поверхностью.
Мелкокомковатая структура — диаметр частиц 0,5-3,0 см, крупнокомкова-
тая структура — диаметр частиц 3-5 см.
Глыбистая структура. Отдельности имеют неправильную форму, размер их
в поперечнике от 5 до 10 см и более.
Ореховатая структура Отдельности пирамидальной либо кубовидной фор-
мы, с острыми ребрами и плоскими гранями, в поперечнике 5-15 мм.
Призматическая структура. Отдельности вытянуты, с острыми ребрами,
плоскими гранями, часто покрыты темной блестящей пленкой. Размер по боль-
шей оси — 2-10 см, по меньшей — 1-5 см.
71
Механический состав
Шнур не образуется — песок
Зачатки шнура — супесь
Шнур дробится при раскатывании —
легкий суглинок
Шнур сплошной, кольцо при свертываний
распадается — средний суглинок
Шнур сплошной, кольцо с трещинами —
тяжелый суглинок
Шнур сплошной, кольцо цельное — глина
Вид образца в плане после раскатывания
Рис 1.18. Мокрый способ определения состава почв в поле
Столбчатая структура. Размер отдельностей по большей оси — 10-30 см,
по меньшей — 2-10 см. Такая структура характерна для солонцов.
Пластинчатая структура Она отличается правильными горизонтальными
слоями толщиной 1-5 мм. При разрушении почва превращается в мучнистую массу.
Неструктурные почвы. К ним относятся песчаные почвы, а также глинистые,
расплывающиеся во влажном состоянии, а после высыхания превращающиеся в
монолитный слой. При раскалывании такие почвы (не песчаные) образуют уг-
ловатые обломки.
Плотность почвы. По плотности почвы различаются на рассыпчатые (по-
чва обладает сыпучестью), рыхлые (лопата легко входит в почву, которая при
выбрасывании рассыпается, стенки ямы в сухом состоянии осыпаются), плот-
ные (лопата с трудом входит в почву), очень плотные (при рытье ямы требуется
лом или кирка).
Новообразования. К новообразованиям относятся локальные скопления ве-
ществ, отличающиеся по своей морфологии и составу от основной почвенной
массы Они могут быть в виде выцветов, налетов, корочек, примазок, потеков,
72
конкреций, прослоек. Новообразования — результат почвенных процессов, для
опытного почвоведа они могут быть важными диагностическими признаками
почв. Однако геоботаники, как правило, не владеют информацией для их рас-
шифровки и обычно не используют для анализа почвенного профиля.
Включения. К включениям относятся тела, находящиеся в почве и не свя-
занные с процессом почвообразования. Следует отмечать следы пожаров: золу,
угли, погребенную древесину.
Вскипание. На карбонатных почвах необходимо указывать горизонт (глуби-
ну), на которой обнаружено вскипание. Для этого на небольшой образец почвы
или на стенку ямы наносится несколько капель соляной кислоты (5-10%-ной).
Если образец содержит карбонаты, выделяются пузыри углекислоты — это и есть
вскипание.
Характер границ горизонтов. Отмечается переход от одного горизонта к
другому: четкий, плавный, язычками и т. п.
Почвенные прикопки При детальных исследованиях нижних ярусов раститель-
ности, полога подроста, а также для более подробного изучения неоднороднос-
ти почвенного покрова и отбора проб почв, кроме почвенного разреза, необхо-
димо проводить описание верхних горизонтов почв. Для этих целей с наружной
стороны ППП через каждые 10 м по периметру или в одном из углов учетных
площадок закладываются прикопки, в которых определяются толщина и морфо-
логические особенности подстилки и верхних горизонтов. Рекомендуется для
каждой прикопки показать на схеме расположение ближайших деревьев с указа-
нием породы и расстояния до ствола, что позволяет учесть влияние древесного
яруса на верхние горизонты почвы. Для получения репрезентативных данных о
толщине и физико-химических свойств лесной подстилки необходимо заклады-
вать не менее 10-20 прикопок на каждой ППП.
На основании описаний почвенного разреза и прикопок и результатов хими-
ческого анализа образцов определяется тип и подтип почвы на ППП, причем все
описания, схемы и результаты анализов включаются в паспорт ППП.
Для определения типов, подтипов почв, почвенных разностей следует восполь-
зоваться классификацией почв (Почвы СССР, 1979). Ниже приведен краткий оп-
ределитель типов и подтипов лесных почв (по почвенно-географическому райо-
нированию) (табл. 1.3, стр. 74-75)).
73
Таблица 1.3. Классификация лесных почв (Почвы СССР, 1979)
Типы почв
1 Почвы распространены на внепойменных территориях........................2
2 На поверхности почвы залегает органогенный горизонт, который сменяется
сизовато-серым или сизовато-темно-серым глеевым горизонтом................ 3
+ На поверхности почвы залегает органогенный горизонт, сменяющийся
подзолистым горизонтом А, ...............................................4
3 Мощность органогенного (торфяного) горизонта более 50 см, окрашен
в светлые буроватые тона. Тип торфяных болотных верховых почв.
+ Мощность органогенного (торфяного) горизонта более 50 см, он окрашен
в земные тона Тип торфяных болотных низинных почв.
4 На поверхности почв залегает органогенный горизонт, сменяющийся гумусовым
горизонтом А,, грубогумусной прослойкой А0А оподзоленным гумусовым
горизонтом A'A2, сменяющимся подзолистым горизонтом А2.....................5
5 . Иллювиальный горизонт В самый яркоокрашенный в профиле, уплотненный,
при суглинистом составе структура ореховатая, ореховато-комковатая или
призматическая. По трещинам и на поверхности структурных отдельностей
имеются белесая присыпка и коричневые натеки. Тип подзолистых почв.
+ Иллювиальный горизонт В глееватый, окрашен в грязные тона и имеет ясные
признаки оглеения в виде сизоватых и охристых пятен. В почвах на песках
горизонт В окрашен в темно-коричневые тона. Тип болотно-подзолистых почв.
Подтипы почв
Тип подзолистых почв
1 Под оторфованной лесной подстилкой из мхов, лесного опада и разнотравья
залегает грубогумусная прослойка А0Ар оподзоленный гумусовый горизонт А,А2,
либо подзолистый горизонт А2..............................................2
+ Под оторфованной лесной подстилкой из мхов, лесного опада и разнотравья
залегает гумусовый горизонт Ар сменяющийся подзолистым горизонтом А2.
Подтип дерново-подзолистых почв.
2 . Подзолистый горизонт А2 оглеен, имеет сизовато-светло-серый цвет
Подтип глееподзолистых почв.
+ Подзолистый горизонт А2 белесый, светло-серый, иногда светлый
палево-бурых тонов, не оглеен Подтип подзолистых почв.
74
Тип болотно-подзолистых почв
1 . Под органогенным горизонтом Ао либо с поверхности залегает гумусовый
или перегнойный горизонт А,................................. ... 2
+ Под органогенным горизонтом Ао залегает подзолистый горизонт А, . ... 3
2 Почвенный профиль оглеен лишь в верхней части, горизонт С не оглеен
Подтип торфянисто-подзолистых поверхностно-оглеенных почв.
+ Оглеение появляется в нижней части горизонта В или в горизонте С.
Подтип торфянисто-подзолистых грунтово-оглеенных почв.
3 Гумусовый горизонт почвы представлен дерновым горизонтом, содержащим
много корней растений.................................................. .4
+ Гумусовый горизонт почвы представлен перегнойным горизонтом, содержащим
значительное количество хорошо разложившихся органических остатков .. 5
4 Почвенный профиль оглеен лишь в верхней части, горизонт С не оглеен
Подтип дерново-подзолистых поверхностно-оглеенных почв.
+ Оглеение появляется лишь в нижней части горизонта В или в горизонте С
Подтип дерново-подзолистых грунтово-оглеенных почв.
5 . Почвенный профиль оглеен лишь в верхней части, горизонт С не оглеен
Подтип перегнойно-подзолистых поверхностно-оглеенных почв.
+ Оглеение появляется в нижней части горизонта В или в горизонте С
Подтип перегиойно-подзолистых грунтово-оглеенных почв.
Тип торфяных болотных верховых почв
1 Почва имеет органогенный торфяной горизонт мощностью 20-50 см, профиль
хорошо дифференцирован. Подтип болотных верховых торфяно-глеевых почв.
+ Почва имеет органогенный торфяной горизонт мощностью свыше 50 см, профиль
слабо дифференцирован. Подтип болотных верховых торфяных почв.
Тип торфяных болотных низинных почв
1. Мощность торфяного органогенного горизонта почвы не превышает 50 см.
Подтип болотных низинных торфяно-глеевых почв.
+ Мощность торфяного органогенного горизонта почвы свыше 50 см
Подтип болотных низинных торфяных почв.
75
РАЗДЕЛ 2. СПЕЦИАЛЬНЫЕ МЕТОДЫ ИЗУЧЕНИЯ
ЛЕСНЫХ СООБЩЕСТВ
2.1. МЕТОДЫ ИЗУЧЕНИЯ БИОЛОГИЧЕСКОЙ ПРОДУКТИВНОСТИ
ДРЕВЕСНОГО ЯРУСА, ПОЛОГА ПОДРОСТА И ПОДЛЕСКА 1
Создание и отмирание органической массы в растительных сообществах про-
текает с различной интенсивностью во времени. Скорость продукционного про-
цесса зависит от фазы вегетационного периода, погодных условий, давности пос-
леднего значительного нарушения (рубки, пожара, ветровала и др.), а также от
наличия хронических антропогенных воздействий (атмосферного загрязнения,
рекреации и др.). Это определяет необходимость динамического подхода к изу-
чению биологической продуктивности растительных сообществ.
Ниже представлен комплекс методов учета органической массы древесного
яруса, полога подроста и подлеска, который применялся автором при исследо-
вании состояния и динамики Европейских бореальных сосновых лесов.
Выбор пробных площадей
В соответствии с изложенными принципами для учета фитомассы и динамики
органического вещества древесного яруса лесных биоценозов в каждом изуча-
емом типе леса следует закладывать серию постоянных пробных площадей, ох-
ватывающих, по возможности, сообщества с древостоями разных классов воз-
раста (т. е. сообщества с разной давностью последнего катастрофического на-
рушения). В каждом классе возраста1 2 желательно иметь не менее двух пробных
площадей. Одна предназначается для проведения статистических учетов, дру-
гая — для дальнейших наблюдений (мониторинга). Особенно важно охарактери-
зовать первые три-четыре класса возраста древостоя (20-80 лет), т. к. в тече-
ние этого периода происходит наиболее существенное изменение величины фи-
томассы и годичной продукции эдификаторного яруса. В соответствии со вре-
менем восстановления основных параметров древесного яруса после полного
разрушения контрольные значения запаса фитомассы древостоя для соответству-
ющего типа леса могут быть определены в сообществах с давностью последнего
нарушения ~200 лет.
Для получения точных данных первостепенное значение имеет правильный
подбор пробных площадей (разд. 1.3).
1 Автор В Т. Ярмишко
2 Классы возраста для хвойных древесных пород выделяются через каждые 20 лет, для
лиственных — через 10 лет
76
Закладываемые пробные площади служат для выбора модельных деревьев,
для учета опада и отпада растений разных ярусов лесных сообществ. Пробная
площадь должна быть окружена защитной полосой шириной не менее 50 м, ко-
торая используется как рабочая зона для отбора модельных деревьев и учета фи-
томассы подчиненных ярусов сообществ.
Определение фитомассы древесного яруса
Выбор модельных деревьев. Выбор модельных деревьев для учета фито-
массы необходимо производить, строго следуя требованиям, изложенным в це-
лом ряде фундаментальных работ (Родин и др., 1968; Уткин, 1975; Вертикаль-
но-фракционное..., 1986; Бобкова, 1987 и др.).
На каждой пробной площади производят сплошной перечет и нумерацию (мас-
ляной краской или при помощи специальных металлических пластинок) деревь-
ев, а также определяют основные морфометрические параметры (диаметр, вы-
соту, высоту прикрепления кроны) всех растущих деревьев каждой породы и от-
дельно — параметры сухостоя. При помощи мерной вилки измеряют диаметр с
точностью до 1 см на высоте 1,3 м от корневой шейки в двух взаимно перпен-
дикулярных направлениях. В древостоях первых двух классов возраста измере-
ния проводятся, начиная ср ступени толщины 2 см с интервалами через 1 см,
третьего класса возраста и выше, — начиная со ступени толщины 6 см с интер-
валом 2 см, а при менее детальных исследованиях — 4 см. Учет деревьев гос-
подствующего и подчиненного компонентов1 производится раздельно. Затем,
пользуясь соответствующими таксационными справочниками (Третьяков и др.,
1965 и др.), вычисляют общую площадь сечения и средний диаметр отдельно
для деревьев господствующего и подчиненного компонентов каждой породы.
Данные о суммарной площади сечения используют для вычисления относитель-
ной полноты древостоя1 2. На основе этих данных определяется формула видо-
вого состава древостоя в процентах от общего числа деревьев и от общей пло-
щади сечения. Деревья, имеющие в формуле <5 %, в дальнейшем в расчет не
принимаются.
Одновременно с перечетом производят измерение высоты деревьев и высо-
ты прикрепления кроны (высоты прикрепления первой живой ветви) с помощью
высотомера (точность замеров — 0,1 м); для каждой ступени толщины выпол-
няется по три замера. На основе этих данных строится график зависимости вы-
сот деревьев от их диаметров (кривая высот). Далее по вычисленному средне-
му диаметру и кривой высот находят среднюю высоту деревьев и среднюю
1 К господствующему компоненту древостоя относят деревья I, II, III классов Крафта,
к подчиненному — IV и V классов.
2 Найденную сумму площадей сечения делят на сумму площадей сечения в данном
возрасте и бонитете при полноте 1,0, взятую из опытных таблиц по справочнику
77
высоту прикрепления кроны для господствующей части древостоя и для под-
чиненного компонента (для каждой породы отдельно).
Выбранные по этим показателям (среднему диаметру, средней высоте и сред-
ней высоте прикрепления кроны) модельные деревья служат основой для учета
фитомассы древостоя на пробной площади. Для господствующего и подчинен-
ного компонентов каждой породы берется по 2-3 модельных дерева. В целях
получения более точных данных можно рассчитать размеры и брать по 1 сред-
нему по высоте модельному дереву каждой породы для каждой ступени толщи-
ны (начиная с 6 см) или для каждого класса толщины, объединяющего 3 ступе-
ни толщины1. Чтобы не нарушать пробной площади, модельные деревья следу-
ет брать на территории окружающей ее рабочей зоны. Не рекомендуется брать
модельные деревья с края выделов. Согласно данным специальных исследова-
ний (Тюрин, 1945), погрешность при определении запасов фитомассы древосто-
ев методом средней модели по ступеням толщины составляет ±3 %.
Сроки взятия модельных деревьев определяются временем завершения фор-
мирования прироста текущего года. Обычно это середина августа и даже начало
сентября, однако в различных зонах и районах эти сроки могут изменяться.
Модельные деревья спиливают у самого основания, измеряют их длину, воз-
раст деревьев определяют по числу годичных колец. Со ствола обрубают все
ветви, разделяют ид на живые и отмершие и складывают на брезент. Крону при
этом желательно разделить на три равные части и ветви каждой трети сложить
отдельно.
Определение массы ствола. Из ствола выпиливают по два кружка на высо-
те 1, 1,3, 3, 5 м и т. д. через каждые два метра вплоть до вершины. В дальней-
шем одна серия кружков служит для изучения хода роста, вторая — для опреде-
ления химического состава и влажности1 2. Не следует брать пробы на анализ во
время дождя, а также в первые 3-4 дня после дождя.
Толщину кружков подбирают в зависимости от толщины ствола дерева с та-
ким расчетом, чтобы самые верхние отрезки ствола имели массу не менее 50 г.
Обе серии кружков взвешивают.
Содержание влаги в лесных растениях испытывает значительные колебания в
зависимости от сезона года, погодных условий, влажности почвы, влажности и
температуры воздуха и т. п. (Смирнов, 1961а, 1964). Кроме того, содержание вла-
ги в древесных растениях зависит от возраста, особенно это касается стволовой
древесины, влажность которой снижается с возрастом (Молчанов, 1971). Дан-
ные В. В. Смирнова (19616) показывают, что влажность хвои ели уменьшалась
1 В некоторых случаях рекомендуется кроме модельных деревьев, взятых для классов
толщины, срубить еще самое толстое и самое тонкое дерево.
2 Данные о содержании воды в отдельных частях деревьев, а также кустарников, тра-
вяно-моховом покрове имеют значение не только для вычисления массы сухого органичес-
кого вещества, создаваемого лесом, но и представляют самостоятельный интерес.
78
с увеличением возраста. Поэтому данные о содержании воды надо приводить в
составляемых ведомостях. Они могут быть использованы для расчетов элемен-
тного состава, энергетических процессов и т. д.
Анализ хода роста следует проводить на невысушенных спилах во избежание
их деформации. Распиленный на указанные выше отрезки ствол взвешивают и к
полученной массе прибавляют массу кружков. Таким образом получают сырую
массу ствола. Если деревья имеют очень большой диаметр (свыше 50 см) и взве-
шивание отрезков затруднительно, на основании измерений высоты и радиусов
вычисляют объем ствола. По справочникам (Ванин, 1949 и др.) находят плот-
ность древесины с учетом зональных особенностей (или определяют ее в лабо-
ратории из каждого отрезка ствола) и вычисляют массу ствола. В зависимости
от целей исследования можно отделить кору и определить отдельно массу коры
и массу ствола.
Для определения влажности древесины и химического анализа составляется
средняя проба древесины ствола массой около 0,5-1 кг. Для этого из каждого
кружка второй серии берется часть в форме сектора. Масса сектора определя-
ется на основе отношения квадратов радиусов данного кружка и самого малень-
кого кружка (или выпиленного из него сектора). Например, масса сектора 2-го
от вершины кружка определяется по формуле:
И J
где Р] — масса самого маленького кружка, Г] — его радиус; х — искомая масса
сектора 2-го от вершины кружка, гг — его радиус. Масса остальных секторов
определяется аналогично.
Для определения влажности древесины рекомендуется высушивать до аб-
солютно сухого состояния полностью всю среднюю пробу, далее используя ее
для химического анализа. На основе данных о сырой массе ствола и его влаж-
ности определяют абсолютно сухую массу ствола.
Определение массы листьев (хвои) и ветвей деревьев. Из каждой трети
кроны молодых деревьев (до 60 лет) отбирается по 1 средней по размеру ветви
из каждой мутовки для последующего отделения листьев или хвои и других уче-
тов. У деревьев старше 60 лет отбирают по 1-2 ветви из каждой трети кроны.
Отобранные средние ветви немедленно взвешивают1. После этого с них снима-
ют листья (хвою) и взвешивают снова. Таким образом определяют сырую мас-
су каждой из модельных ветвей. Иногда средние ветви, очищенные от листьев
(хвои), сортируют на две группы: мелкие ветви (<1 см в диаметре) и крупные
1 Предварительно с ветвей снимают эпифитные мхи, лишайники и грибы и, если это
предусмотрено задачами исследования, раздельно определяют их массу; берут средние
пробы на влажность и для химического анализа.
79
ветви. Затем ветви взвешивают отдельно по указанным группам, а после этого
измельчают. Крупные ветви распиливают на отрезки по 10-15 см. Из каждой
группы берут средние пробы (0,5-1,0 кг) для химического анализа и для опре-
деления влажности.
Сухие ветви учитываются отдельно, из них также берется средняя проба для
определения влажности и для химического анализа.
По разности массы облиственных (охвоенных) и безлистных ветвей опреде-
ляют общую массу листьев (хвои) на модельной ветви. Этот способ предпочти-
тельнее, чем непосредственное взвешивание листьев (хвои), которые за время
работы по обрыванию теряют много влаги. Зная общее число ветвей в соответ-
ствующей мутовке или в соответствующей трети кроны, определяют общую сы-
рую массу ветвей (или сырую массу крупных и мелких ветвей) и хвои для всего
модельного дерева. Масса побегов и хвои текущего года учитывается отдельно.
С этой целью с модельных ветвей отделяются приросты текущего года и взвеши-
ваются с хвоей и без хвои. Зная общее количество ветвей на дереве, путем расче-
та получают сырую массу побегов и хвои текущего года для всего дерева.
Листья (хвою) рассыпают на брезенте и берут из них средние пробы (0,5 кг)
для химического анализа и для определения влажности (для хвои текущего года
отдельно). Для определения влажности рекомендуется сушить среднюю пробу
листьев (хвои) целиком.
На основе данных о сырой массе и влажности ветвей и листьев (хвои) опре-
деляют их абсолютно сухую массу.
Надо помнить, что у большинства видов хвойных хвоя держится на ветвях
много лет (например, у ели сибирской в северной тайге до 23 лет, а у сосны
обыкновенной до 12 лет). С возрастом хвои изменяется ее влажность (Данилов,
1948), содержание химических веществ (Паршевников, 1959 и др.), энергия фо-
тосинтеза (Bourdeau, 1961) и другие характеристики. Поэтому во многих случа-
ях при проведении экологических исследований представляет интерес учет рас-
пределения массы хвои по возрастам. В. В. Смирной (1961а) предлагает следу-
ющий способ. Выбирают одну среднюю по показателям охвоения ветвь из каж-
дой трети кроны модельного дерева. Центральный и боковые побеги охвоенной
части ветви разрезают на участки годичных приростов (рис. 2.1). Затем взвеши-
вают охвоенные побеги каждого года и побеги с удаленной хвоей и определяют
сырую массу хвои каждого года. Берутся образцы для определения влажности
хвои разных лет. Зная сырую массу и влажность хвои каждого года, можно в
дальнейшем рассчитать ее абсолютно сухую массу.
Определение площади поверхности листьев (хвои). Для определения пло-
щади поверхности листьев совокупности деревьев отдельных пород в составе
древесного яруса листья из разных частей кроны каждого модельного дерева со-
ответствующей породы собирают в пачки по 50-100 штук черешками в разные
стороны (повторность 10-20-кратная). С помощью остро заточенной цилиндри-
80
ческой высечки (удобны спе-
циальные высечки для пыжей)
вырубается столбик на всю
толщину пачки (из крупных ли-
стовых пластинок можно вы-
сечь 2-3 образца). Высечки из
каждой пачки помещаются в
бюксы и взвешиваются в сы-
ром виде и после высушивания
до абсолютно сухого состоя-
ния. Зная число высеченных
кружков, площадь одного
кружка, среднюю массу, мож-
но вычислить площадь листь-
ев, приходящуюся в среднем на
единицу их массы. Далее, имея
данные об общей массе листь-
ев данной породы на пробной
Рис. 2.1. Схема описания размещения хвои со-
сны обыкновенной на побегах с учетом года
ее формирования в кроне
Условные обозначения: ЦП — центральный побег (ось
первого порядка ветвления); ЦП 1990 — хвоя 1990 г
на центральном побеге; БП2-1990 — хвоя 1990 г. на
боковом побеге второго порядка ветвления; БЗ— 1990
(БПЗ-1990) — хвоя 1990 г. на боковом побеге третье-
го порядка ветвления и т д.; Ml, М2, М3 — мутовки
соотвественно 1990,1989,1988 гг. формирования
площади, можно рассчитать площадь их поверхности на единицу площади рас-
тительного сообщества (листовой индекс).
Определение площади поверхности хвои можно проводить различными мето-
дами, например по методу, описанному И. В. Гулидовой (1959). Из разных час-
тей кроны модельных деревьев отбирается не менее 100 хвоинок, у каждой из
которых определяется длина. Далее у 30-50 хвоинок определяется величина пе-
риметра на поперечном срезе (рис. 2.2) под микроскопом с малым увеличени-
ем. Периметр рассчитывается
как средний из трех измерений,
выполненных в середине и с
разных концов хвоинки. После
соответствующих измерений
вычисляется величина площади
поверхности хвоинки путем пе-
ремножения средней длины и
периметра. Далее следует уста-
новить корреляцию между пло-
щадью поверхности и массой
хвоинок. Для этого берут на-
вески хвои от 0,2 до 1-2 г и
подсчитывают число хвоинок в
навеске. Зная площадь поверх-
ности хвоинки, рассчитывают
1
Рис. 2.2. Схематическое изображение хвои со-
сны обыкновенной
7 — общий вид хвои; 2 — поперечный разрез хвои по
линии к-л для измерения ширины и толщины хвои, а —
длина; б— ширина; в — толщина хвои
81
величину площади поверхности, приходящуюся на единицу массы. Затем полу-
ченную величину пересчитывают на массу хвои модельного дерева и массу хвои
древостоя на 1 га.
Определение массы генеративных органов. Для учета генеративных ор-
ганов могут быть использованы те же ветви, которые были взяты для определе-
ния количества листьев или хвои (различные методы учета числа генеративных
органов у хвойных охарактеризованы в разд. 2.4.1). При стационарных исследо-
ваниях учет должен производиться дважды: во время цветения (или формирова-
ния мужских и женских стробилов у хвойных) и в период созревания плодов и
семян (у хвойных учитываются двухлетние женские шишки).
На модельных ветвях определяется число генеративных органов (у хвойных
раздельно учитываются женские шишки текущего года и предшествующих лет).
При взвешивании собранных с ветвей шишек (или плодов у лиственных пород)
берутся образцы на влажность.
Для учета семенной продуктивности определяется среднее число семян в пло-
де (или шишке) и умножается на количество их на единицу площади. Общая мас-
са семян вычисляется по массе 100 штук: если семена взвешиваются в поле, то
берутся образцы на влажность для последующего пересчета на абсолютно су-
хую массу. При наличии у семян летучек, волосков и других приспособлений же-
лательно особо определить их массу.
После оценки основных характеристик древесного яруса при наличии доста-
точного объема данных может быть определен вид зависимости между диамет-
ром стволов и массой листьев (хвои), массой стволов и массой листьев, массой
листьев и массой ветвей, массой ветвей и массой стволов, массой стволов и мас-
сой корней. Установление таких зависимостей позволяет существенно облегчить
работу по определению общей фитомассы деревьев: достаточно установить мас-
су одной из частей дерева, чтобы по имеющимся уравнениям вычислить массу
других его частей. Естественно, что эти зависимости применимы в пределах ог-
раниченных регионов.
Определение фитомассы подроста и подлеска
Для определения фитомассы производят перечет растений полога подроста и
подлеска (кустарникового яруса) с определением средней высоты особей. Учет
можно производить на площадках размером 1 м2 (1 х 1 м) или 4 м2 (2 х 2 м) или
на профилях шириной 2 м, регулярно расположенных на территории пробной пло-
щади (подробнее см. разд. 1.4 и 1.6).
Затем для каждого из основных видов, формирующих полог подроста и под-
лесок, выбирают средние по высоте и развитию экземпляры (по 10 штук), кото-
рые срезают под корень. В срезанных экземплярах определяют массу ствола, вет-
вей (с выделением побегов текущего года), листвы (хвои с разделением на хвою
текущего года и старших возрастов), берут образцы для химического анализа и
82
определения содержания воды в целях последующего пересчета на высушенную
при 105 °C сухую массу. Найденные величины пересчитывают на число экземп-
ляров данной породы на площади 1 га. Затем определяют общую фитомассу всех
видов подроста и подлеска для данного типа леса при данной давности после-
днего нарушения.
Определение фитомассы подземных органов
Учет корней растений древесного яруса, полога подроста и подлеска.
У каждого спиленного модельного дерева господствующего яруса выделяют
площадку для сбора и учета корней. Размер площадки надлежит устанавливать,
исходя из средней площади питания одного дерева, которую определяют путем
деления величины пробной площади (в м2) на число растущих на ней деревьев
господствующего компонента древесного яруса. Затем площадки найденного раз-
мера закладывают вокруг модельных деревьев каждой породы, относящихся к
господствующему компоненту древостоя, таким образом, чтобы модельное де-
рево находилось в центре. Предпочтительнее закладывать площадки в форме
круга, однако допустимо использование квадратных площадок.
Расчет фитомассы корней на 1 га производится на основании определения их
массы у модельных деревьев каждой породы с учетом участия в составе гос-
подствующего компонента древостоя.1
Некоторая часть корней модельного дерева может выходить за пределы ус-
тановленной площадки, однако это компенсируется проникновением в пределы
площадки корней соседних деревьев. На площадке могут оказаться также дере-
вья подчиненного яруса, подрост и растения подлеска, корневые системы кото-
рых также подлежат учету.
Если площадь питания одного дерева меньше 3 м2, что имеет место в на-
саждениях I класса возраста, то следует для учета корней брать соответственно
больше деревьев с таким расчетом, чтобы учитываемая площадь была не ме-
нее 3 м2. Если средняя площадь питания значительно превышает указанную ве-
личину, что имеет место в древостоях старших классов возраста, то для учета
следует брать половину или соответственно четвертую часть площади питания
Для сбора корней по наружной стороне площадки копают траншею шириной
около 0,5 м и глубиной в зависимости от строения корневой системы (до 1,0-
1,5 м или больше). Почву из траншеи откидывают на наружную сторону на под-
стеленный брезент. Затем поверхность площадки очищают от растений травяно-
кустарничкового и мохово-лишайникового ярусов. Далее снимают на разостлан-
ный брезент слой лесной подстилки и последовательно один за другим минераль-
ные горизонты почвы и производят из них выбор корней древесных растений, ра-
1 При наличии достаточного количества весовых данных следует вычертить график
зависимости массы корневых систем от диаметра дерева По этому графику можно вы-
числять массу корней для деревьев промежуточных диаметров (Смирнов, 1962)
83
стений подлеска и кустарничков. В месте прохождения стержневого корня или
глубоко идущих якорных корней производят дополнительное углубление, чтобы
полностью извлечь все глубоко проникающие корни. Обычно корни учитывают
до глубины 1,5-2 м. Учет корней в более глубоких горизонтах почвы представ-
ляется очень сложной задачей, которую, однако, следует решать в тех случаях,
когда количество корней, проникающих на большую глубину значительно. Жи-
вые и мертвые корни учитываются раздельно. Полуразложившиеся остатки кор-
ней учитывают вместе с мертвыми корнями.
В зависимости от целей работы выборку корней можно производить из каж-
дого генетического горизонта почвы отдельно. Разделение корней по их принад-
лежности к разным видам древесных растений представляет собой достаточно
сложную задачу. Однако при большой практике исследовательской работы уда-
ется по цвету, внешнему виду, запаху и другим показателям отличить, например,
корни сосны от корней ели, корни черники от корней брусники и т. д. Заслужива-
ет внимания работа Л. П. Рысина и Г. П. Рысиной (1987), в которой приводятся
данные по морфоструктуре корневых систем более 200 видов лесных и лугово-
лесных травянистых растений.
Корни, собранные на брезент, отряхивают от частиц почвы и промывают во-
дой. Корни диаметром <10 мм промывают на ситах, верхнее — с ячейками 3 мм,
нижнее — 0,25 мм. Извлеченную почву после выбора из нее корней тщательно
перемешивают и определяют ее объем, используя определенную емкость, на-
пример ведро. Затем берут одно ведро почвы и отмывают оставшиеся мелкие
корни на сите с ячейками 0,25 мм. Полученную массу корней пересчитывают на
объем всей почвы и прибавляют к массе корней, непосредственно выбранных из
почвы. Запасы корней можно определять также путем отмывания всей почвы на
ситах, верхнее из которых имеет диаметр ячеек 3 мм, нижнее — 0,25 мм. Во всех
случаях сначала извлечение корней производят из половины площадки. Это де-
лается для зарисовки схемы распределения корней цо генетическим горизонтам.
Для этого на стенку траншеи, проходящей через середину площадки, в центре
которой находится модельное дерево, накладывают металлическую сетку с квад-
ратными делениями размером 10x10 см, после чего на миллиметровой бумаге
фиксируют распределение корней. Затем приступают к выбору корней из второй
половины площадки.
После сбора всех корней извлекают из ямы комель. Комель тщательно очи-
щают от приставшей к нему почвы (в необходимых случаях обмывают), взве-
шивают и выпиливают или вырубают из него образцы для химических анализов
и определения влажности.
Собранные корни после отмывки1 сушат в месте, защищенном от пыли. За-
тем их сортируют на крупные и мелкие, уточняя разделение по размерам, и взве-
шивают. Одновременно берут пробы на влажность. Границей для выделения круп-
ных и мелких корней считают условно диаметр 7 мм.
84
Для химического анализа корни отбирают отдельно по видам растений и нс
отмывают их водой во избежание выщелачивания химических элементов Для
очистки их ополаскивают спиртом на ситах (0,25 мм) Каждый образец для ана-
лиза должен быть массой около 0,5 кг.
Учет физиологически активных (деятельных) корней. Метод учета мас-
сы деятельных (сосущих) корней предложен А. Я. Орловым (1948), небольшие
изменения были внесены Е. М. Самойловой (1961). Метод заключается в сле-
дующем. С помощью металлического каркаса площадью 100 см2 и высотой 10
см берут послойно монолиты почвы объемом 1000 см3 (количество монолитов
определяется в каждом случае в зависимости от глубины распределения деятель-
ных корней, но не менее 150 шт. на пробной площади). Из каждого слоя препа-
ровальной иглой и пинцетом выбирают все корни исследуемого вида древесно-
го растения диаметром меньше 0,3 мм (при исследовании корневой системы
сосны приходится учитывать корни размером менее 0,5 мм); крупные корни при
этом отбрасывают. Выбранные корни в воде очищают от микориз и частиц по-
чвы и, снабдив этикеткой и завернув в кальку, помещают в 70%-ный раствор эти-
лового спирта. Почву монолита взмучивают в воде и переносят на сито с диа-
метром отверстий 0,25 мм. Остаток на сите содержит некоторое количество со-
сущих корней, ранее не выбранных. Из него берут две средние пробы массой до
0,5 г и также помещают в спирт. Оставшуюся часть доводят до воздушно-сухо-
го состояния и взвешивают. Пробы просматривают под бинокулярной лупой при
30-кратном увеличении и выбирают живые корни исследуемой древесной поро-
ды диаметром менее 0,3 мм. Массу корней, выбранных из пробы, пересчитыва-
ют на массу всего остатка. Корни, выбранные из монолита и помещенные в спирт,
также просматривают под лупой, отделяют корни большего диаметра, частицы
почвы и другие примеси, доводят до воздушно-сухого состояния и взвешива-
ют. Суммируя массу корней, отобранных непосредственно из монолита и из ос-
татка на сите, получают общую массу корней в 1000 см3. Суммарную массу
корней из всех монолитов затем пересчитывают на гектар. Следует отметить, что
в данную категорию попадают не только сосущие, но и мелкие проводящие корни;
поэтому правильнее говорить о живых физиологически активных мелких корнях.
Учет массы корней травянистых растений производится методом моно-
литов (размером 25 х 25 см) с отмывкой на ситах с ячейками 0,25 мм; повтор-
ность трехкратная. При этом древесные корни и корни кустарничков отбрасыва-
ются. Луковицы, клубни и корневища следует учитывать особо (не соединять их
с корнями). Также отдельно надо проводить их химический анализ.
1 Для окончательной отмывки мелких корней от песка, не прошедшего через сито 0,25 мм,
корни помещают в высокие стаканы или цилиндры Песчинки, более тяжелые, при этом
оседают на дно. Эту операцию можно делать после предварительной просушки корней, так
как сухие корни всплывают.
85
Годичная продукция древостоя
Годичная продукция надземных органов. Учет текущего прироста расте-
ний древесного яруса можно производить различными способами в зависимос-
ти от особенностей древостоев. Так, при исследовании одновозрастных древо-
стоев, имея серию пробных площадей разных классов возраста, получают вели-
чину массы древесины стволов, ветвей, корней, а также листьев или хвои модель-
ных деревьев определенного возраста. На основании этих данных строят кривую
(по горизонтальной оси откладывают возраст, по вертикальной — массу отдель-
ных фракций), по которой определяют изменение массы многолетних органов для
определенных отрезков времени. Затем вычисляют средний ежегодный прирост
древесины стволов, ветвей, корней. Годичный прирост хвои соответствует вели-
чине фитомассы хвои текущего года, у лиственных масса листьев соответству-
ет годичному приросту ассимиляционных органов. Умножением величин приро-
ста на число деревьев на одном гектаре находят средний годичный прирост раз-
личных фракций древостоя на площади 1 га.
В разновозрастных лесах (а при наличии всего одной пробной площади так-
же и в одновозрастных) годичный прирост древостоя по высоте, диаметру и
объему исчисляется на основании анализа хода роста принятыми в лесной такса-
ции методами по пяти- или десятилетним периодам (Тюрин, 1945; Ловелиус,
1979). Для этого используются выпиленные из модельных деревьев кружки. При
более точных учетах можно пользоваться сложной формулой срединного сече-
ния Губера, при менее точных — простой формулой Губера. Для перевода полу-
ченных показателей текущего прироста из объемных единиц в единицы массы
необходимо определить объемные массы той части древесных кружков, кото-
рая соответствует приросту за тот или иной период. К полученному значению
массы прироста древесины прибавляется масса коры (Пк), приходящаяся на этот
прирост, вычисляемая по формуле:
8 п
n‘=V’
где а — масса кружка без коры, 5 — масса коры кружка, пг — масса текущего
годичного прироста без коры (все выражается в граммах).
Годичный прирост ветвей по массе определяется на основании обмеров 2-3
средних ветвей, взятых из каждой трети кроны (удобно использовать те средние
ветви, которые брались для учета листьев или хвои). Ветвь (без листьев и коры)
взвешивается, измеряется ее длина (исключается прирост текущего года); из се-
редины ветви выпиливается кружок для упрощенного анализа прироста по диа-
метру (по простой формуле Губера), который условно принимается пропорцио-
нальным приросту по массе. Можно определить массу ветви с корой и затем, ус-
тановив для выпиленного кружка соотношение масс коры и древесины, вычис-
лить прирост всей ветви без коры по формуле:
86
пв=^
<5 ’
где Пв — прирост ветви по массе без коры, г; а — масса ветви без коры, г; 5 —
объем ветви без коры, см3; по — прирост ветви по объему, см3. Затем к полу-
ченной величине прироста ветви без коры прибавляется соответствующая ей ве-
личина прироста коры (по предыдущей формуле). Далее на основании определе-
ния прироста у нескольких ветвей вычисляется средняя арифметическая вели-
чина прироста (%) по следующей формуле:
Пср=”‘- 100.
о
где Пср — средняя арифметическая величина прироста, %; b — средняя масса
модельных ветвей с корой, г; Пк — средняя масса прироста модельных ветвей с
корой, г.
После этого средняя арифметическая величина прироста ветвей (%) умножа-
ется на общую массу ветвей из данной части кроны. К ней прибавляется масса
побегов текущего года у всех ветвей данной части кроны. Затем все данные сум-
мируются для всей кроны и пересчитываются на 1 га. Расчет текущего прирос-
та корней производится на основании процентного соотношения массы ствола,
ветвей и корней модельного дерева по формуле:
П = п‘к
где Пкр — прирост корней (единицы массы); пс — прирост стволов и ветвей (еди-
ницы массы); к — доля корней от суммарной массы ствола, ветвей и корней, %,
с — доля ствола и ветвей от суммарной массы ствола, ветвей и корней, %.
Прирост хвои рассчитывается, как было указано выше. Прирост листьев ли-
ственных пород принимается равным фитомассе листьев.
Для приближенного определения годичной продукции можно использовать
таблицы хода роста древостоев соответствующих регионов, где приведен ряд
показателей, характеризующих изменение числа стволов, текущего прироста
деловой древесины1 и т. д. Переведя объемные показатели прироста древесины
в единицы массы и зная соотношения между массой стволовой древесины, вет-
вей и корней, на основании массы структурных частей модельного дерева (вве-
дя поправку на полноту древостоя), можно рассчитать показатели массы приро-
ста стволов, ветвей и корней. Для получения величины полного прироста к этой
величине прибавляется масса прироста хвои (листвы), как указано выше.
Годичная продукция корней. Определение количественных показателей го-
дичного прироста корней древесных растений весьма трудный вопрос. В настоя-
щее время могут быть рекомендованы лишь косвенные методы для оценки при-
роста корней отдельных видов деревьев. Так, при наличии чистых древостоев
1 Последняя составляет около 75 % стволовой древесины
87
разных классов возраста в каждом из них определяется количество корней у
особей конкретного возраста. При этом выемку корней следует производить с
площади питания особи (см. подраздел «Определение фитомассы подземных
органов»). Повторность должна быть трех-пятикратная. Далее строят кривую (по
горизонтальной оси откладывают возраст, по вертикальной — массу корней) и
вычисляют средний прирост корней за 1 год. Одновременно с учетом живых
корней рекомендуется определять массу отмерших корней, что позволит уста-
новить их соотношение.
Второй способ (модификация метода, предложенного А. А. Молчановым и В.
А. Губаревой, 1965) состоит в следующем. Вычислив площадь питания дерева
определенного возраста, нужно очертить вокруг его основания круг, соответству-
ющий площади питания. Далее необходимо обозначить сектор равный */5 площади
питания и в пределах сектора до глубины распространения основной массы кор-
ней (1-1,5 м) извлечь послойно (по генетическим горизонтам) почву с корнями.
Затем из вынутой почвы отобрать все корни с разделением на живые и мертвые
и почву вновь засыпать в шурф, соблюдая последовательность генетических
горизонтов. С поверхности сектор обозначить системой колышков или ограни-
чить проволокой. На одной из таких площадок через два года (на другой — че-
рез три ит. д.) вновь произвести раскопку и определить массу живых и отмер-
ших корней. Получив данные по массе прироста корней (а также их отмирания)
за 2, 3, 4, 5 и т. д. лет для деревьев разных классов возраста, можно выявить
закономерности роста и отмирания корневых систем деревьев.
Годичная продукция растений полога подроста и подлеска
Довольно сложным является определение ежегодного прироста фитомассы
подлесочных видов и подроста древесных растений. В этих целях по годичным
кольцам на модельных экземплярах определяют средний возраст каждого вида
кустарников и растений в пологе подроста. Затем путем деления сухой массы
среднего модельного экземпляра (без листьев) на средний возраст определяют
средний прирост древесины (ствола, ветвей) этого экземпляра, а умножением на
число особей данного вида на 1 га вычисляют средний прирост древесины на
гектар. Суммируя прирост всех видов кустарников или видов, входящих в полог
подроста, находят общий прирост подроста и подлеска. Этот прием недостаточ-
но точен, так как годичный прирост в разном возрасте неодинаков. Однако из-за
незначительности массы подлеска и подроста по сравнению с массой древостоя
ошибка будет невелика.
При расчетах прироста подроста и подлеска можно пользоваться тем же ме-
тодом, что и при определении прироста ветвей.
Для определения общей массы органического вещества, синтезируемого под-
леском и подростом за год, к величине прироста их древесины следует приба-
вить массу листвы (хвои), произведенную этими породами за год.
88
2.2. МЕТОДЫ ИЗУЧЕНИЯ БИОЛОГИЧЕСКОЙ ПРОДУКТИВНОСТИ
НИЖНИХ ЯРУСОВ ЛЕСНЫХ СООБЩЕСТВ 1
Лесные сообщества являются многокомпонентными системами, которые ха-
рактеризуются сложной вертикальной и горизонтальной структурой. В составе
травяно-кустарничкового и мохово-лишайникового ярусов лесных сообществ
наблюдается большое разнообразие жизненных форм: кустарнички, полукустар-
нички, многолетние и однолетние травы, мхи и лишайники. В связи с этим изуче-
ние биологической продуктивности различных компонентов нижних ярусов тре-
бует применения разных подходов и вариантов методик.
2.2.1. Определение фитомассы
В пределах рабочей зоны пробной площади в регулярном порядке закладыва-
ют учетные площадки, размер которых зависит от характеристик анализируемых
ярусов (в частности, от размеров особей слагающих их видов). Необходимое
число площадок определяется планируемой точностью учета (см. разд. 4.1). В
некоторых случаях при изучении парцеллярной структуры сообществ использу-
ется типический принцип расположения учетных площадок (разд. 1.4). В этом
случае учетные площадки закладывают в пределах каждого из выделяемых ти-
пов парцелл в количестве, необходимом для получения достоверных данных о
величине фитомассы нижних ярусов данной парцеллы, независимо от занимае-
мой ею площади. Определение общей величины фитомассы нижних ярусов на
пробной площади производят с учетом площади, занимаемой каждой парцеллой.
С этой целью целесообразно провести предварительное картирование нижних
ярусов лесного сообщества на пробной площади, в процессе которого на мил-
лиметровую бумагу наносят контуры отдельных парцелл для последующего
вычисления занимаемой ими площади и определения общей фитомассы. В бо-
реальных лесных сообществах обычно выделяется не более 5 типов парцелл Для
получения репрезентативных результатов в пределах каждой парцеллы достаточно
заложить 5 учетных площадок. В этом случае точность учетов фитомассы со-
ставляет ±8-10 %.
На каждой учетной площадке проводят геоботаническое описание с опреде-
лением общего проективного покрытия травяно-кустарничкового и мохово-ли-
шайникового ярусов, проективного покрытия отдельных видов растений и изме-
ряют высоту ярусов (см. разд. 1.6).
Травяно-кустарничковый ярус. Для определения надземной фитомассы тра-
вяно-кустарничкового яруса наиболее часто используют площадки размером
1 Авторы В Т. Ярмишко, Е Н Андреева, Е А Мазная, М А Ярмишко
89
1 х 1 м или 0,5 х 0,5 м, границы которых отмечают при помощи рамки соответ-
ствующего размера. На каждой площадке срезают все особи растений (на уров-
не поверхности подстилки) и их части, попадающие в отграниченную рамкой пло-
щадь. Срезанные растения разбирают по видам и взвешивают. Для кустарничков
раздельно определяют массу листьев и стеблей. Листья и побеги текущего года
учитывают отдельно (см. подраздел 2.2.2). Определение массы отмерших рас-
тений каждого вида проводят отдельно.
Учет массы подземных органов растений травяно-кустарничкового яруса про-
изводится методом монолитов. Для этого закладывают площадки размером
25 х 25 см, на которых проводят отмывку корней из разных почвенных горизон-
тов на ситах с ячейками 0,25 мм. Площадки закладывают в 3-кратной повторно-
сти. Корни древесных растений при отмывке отбрасывают. Луковицы, клубни и
корневища следует учитывать отдельно, не соединяя их с корнями. Если необ-
ходимо проводить химические анализы корней, то их следует собирать вручную,
механически очищая от почвы, а затем ополаскивать на ситах спиртом.
Мохово-лишайниковый ярус. При определении фитомассы мохово-лишай-
никового яруса размер учетных площадок составляет от 0,25 х 0,25 м до 1 х 1 м
и зависит от состава и структуры мохово-лишайникового покрова. Необходимое
число площадок определяется исходя из характера мохового покрова и задан-
ной точности определения, оно может быть рассчитано на основе формулы, пред-
ставленной в разд. 4.
Мхи и лишайники срезают на уровне подстилки, разбирают по видам и взве-
шивают. Как правило, определяют массу каждого из основных доминантных
видов и суммарную массу остальных видов отдельно для мхов и лишайников. В
некоторых случаях можно ограничиться определением суммарных масс мхов и
лишайников. Стебли мхов и талломы лишайников рекомендуется разделять на
верхнюю (живую и растущую) и нижнюю (отмирающую и отмершую) части,
которые следует взвешивать раздельно.
Отмершую часть талломов лишайников от живой можно отличить по более
темной окраске и сплющенности подециев, связанной с расширением внутрен-
ней полости и утончением стенок.
Разделение стебля мхов на живую и отмершую части достаточно сложная
задача, т. к. на каждом стебле можно выделить 3 зоны: живую, переходную и
мертвую (Кильдюшевский, 1971). Переходная зона характеризуется чередова-
нием живых и мертвых листочков и постепенным или прерывистым изменением
их цвета. Существует способ разделения живой и мертвой частей, основанный
на изменении активности дыхательного фермента пероксидазы в анаэробных ус-
ловиях (Малышева, 1970). Качественная реакция с бензидином дает ярко-синее
окрашивание живых тканей. Отмершая часть стебля не окрашивается. Переход-
ная зона между сплошь окрашенными и неокрашенными частями делится попо-
лам (Грабовик, Антипин, 1982).
90
Величину надземной фитомассы определяют как в сыром, так и в сухом со-
стоянии. Для этого отбирают пробы отдельных фракций (побеги и листья кус-
тарничков, зеленые и отмершие части мхов, живые и отмершие части лишайни-
ков), которые взвешивают в сыром состоянии, затем их высушивают до посто-
янной массы при температуре 105 °C и снова взвешивают. Производят пересчет
сырой фитомассы в абсолютно сухую.
При проведении исследований на охраняемых территориях или учете надзем-
ной фитомассы и продуктивности редких и исчезающих видов рекомендуется
взятие навесок строго определенного объема (Кошкарева, 1989).
2.2.2. Определение годичной продукции
Травяно-кустарничковый ярус. Определение годичной продукции проводят
после завершения роста побегов в текущем году одновременно с учетом об-
щей фитомассы растений травяно-кустарничкового яруса на учетных площадках
(см. разд. 1.4). С этой целью надземную фитомассу кустарничков, срезанную на
учетной площадке, разбирают по видам, у каждого растения разделяют однолет-
нюю (текущего года) и многолетнюю части. Годичную продукцию составляют
побеги текущего года с листьями. У кустарничков граница побега текущего года
определяется по морфологическому признаку: круговому рубцу в месте прикреп-
ления чешуй верхней боковой почки, дающей начало побегу текущего года.
Дополнительным признаком может служить наличие у основания побега текущего
года концевой части прироста предшествующего года (например, у черники).
Величина годичной продукции может быть выражена в единицах массы в сы-
ром и (или) абсолютно сухом состоянии на единицу площади, обычно 1 м2. По-
беги текущего года с листьями каждого вида, собранные с каждой учетной пло-
щадки, взвешивают, определяя таким образом сырую массу. Для определения
абсолютно сухой массы пробы высушивают в сушильном шкафу при темпера-
туре 105 °C и снова взвешивают. Зная сырую и абсолютно сухую массу проб,
рассчитывают среднюю величину годичной продукции каждого вида кустарнич-
ков на единицу площади в изучаемом фитоценозе.
Годичная продукция травянистых растений принимается равной запасу фито-
массы надземных органов.
Мохово-лишайниковый ярус. Для определения величины годичной продук-
ции мхов необходимо знать среднее число побегов на единицу площади, массу
отрезка побега длиной 1 см и величину годичного линейного прироста побегов
каждого вида. При учете общей фитомассы мхов отбирают средние пробы по-
бегов каждого вида и определяют их сырую массу. В лабораторных условиях в
пробе подсчитывают количество стеблей определенного вида и от каждого по-
бега отделяют верхнюю часть длиной 1 см. Пробы высушивают до абсолютно
сухой массы при температуре 105 °C. Таким образом, для каждого вида опреде-
ляют массу отрезков побегов длиной 1 см и общую массу пробы в абсолютно
91
сухом состоянии, а также рассчитывают число стеблей на единицу массы. Соот-
ношение данных сырой и абсолютно сухой массы пробы позволяет оценить со-
держание влаги в ней.
Для определения линейного годичного прироста мхов разработаны различные
методы. Выбор метода обусловлен сроками исследования и анатомо-морфоло-
гическими особенностями мхов. Следует отметить, что величина годичного
линейного прироста мхов существенно варьирует в пределах синузий, поэтому
для более точной оценки роста особей необходимо брать образцы как из цент-
ральной, так и из периферической зон синузии (Тархова, 1971).
Метод перевязок. Метод перевязок заключается в перевязывании стеблей мхов
белой ниткой на конце побега или на 10-20 мм ниже верхушек стеблей (Корчагин,
1960), или на нижней границе ассимилирующей (зеленой) части побега (Тархова,
1969). Последнее позволяет учитывать не только прирост, но и отмершую за се-
зон часть стебля. Обычно перевязывают от 20-40 до 50-100 шт. стеблей (Грабо-
вик, Антипин, 1982; Максимов, 1982; Кошкарева, 1988; Грабовик, 1994). Для пе-
ревязки стеблей можно использовать капроновую или другую синтетическую нитку,
концы которой закрепляют на леске, натянутой между двумя колышками.
Метод имеет ряд недостатков, т. к. во время перевязки стеблей может быть
повреждена верхушка побега или нарушена естественная сомкнутость моховой
дерновинки (особенно у видов, имеющих жизненную форму с высокой плотно-
стью дерновинки), что приводит к остановке роста главного побега и формиро-
ванию латеральных побегов (Солоневич, 1966). Небольшая синузия при изуче-
нии динамики роста с использованием этого метода может быть полностью унич-
тожена или необратимо повреждена. При проведении многолетних наблюдений
на одних и тех же площадках неизбежно происходит небольшое смещение пере-
вязок с одних стеблей на другие, поэтому метод перевязок можно использовать
только при изучении достаточно больших по площади однородных участков
синузий мхов с ортотропным ростом. Кроме того, этот метод нельзя использо-
вать при изучении ритма прироста мхов в течение вегетационного периода.
Метод окрашивания. Метод окрашивания сходен с методом перевязок и
заключается в окрашивании стебля на определенном расстоянии от верхушки мха
(обычно 10 или 20 мм) (Корчагин, 1960). При помощи тонкой стальной прово-
локи наносится точка красной нитрокраской, которая сохраняется на стебле не
менее двух лет. Для определения прироста стебля через определенный период
времени измеряется расстояние между верхушкой и намеченной точкой. Метод
окрашивания менее трудоемок, более точен и при его использовании меньше
повреждаются верхушки стеблей по сравнению с методом перевязок. Метод
окрашивания чаще используют для измерений прироста водных и полуводных
мхов (Илометс, 1981).
Метод отметки положения кончика стебля на субстрате. При использо-
вании этого метода фиксируется положение кончика растущего стебля: у эпилит-
92
ных видов — несмывающейся краской, у эпифитных и эпиксильных видов — за-
рубками на древесине или вбитыми небольшими гвоздиками или булавками. Из-
менение положения конца стебелька за год относительно сделанной отметки по-
кажет величину годичного прироста (Корчагин, 1960). Этот метод наиболее при-
годен для измерения прироста у мхов и печеночников с плагиотропным ростом.
Метод реперов. Измерения линейного прироста проводят на трансектах (ли-
ниях), проходящих через участки болота с различными растительными сообще-
ствами. На каждом выбранном участке на расстоянии 10 см один от другого ус-
танавливают 20 реперов (Корчагин, 1960; Clymo, 1970). Репер представляет
собой коленчатую проволоку высотой 20 см с находящимся посередине коле-
ном длиной 2 см. Колено (горизонтальная часть колышка) устанавливают вро-
вень с верхушкой мха.
Замеры прироста (в мм) проводят ежемесячно в течение всего вегетацион-
ного периода. При невозможности проводить постоянные наблюдения число
измерений прироста можно сократить до трех (весной, летом и в конце осени).
Следует помнить, что длительность периода вегетации мхов зависит от средне-
суточной температуры воздуха в конкретные годы, например, сфагновые мхи
начинают развитие уже при температуре >+4 °C.
При исследованиях возможна утрата части колышков (например, в результате
посещения площадок лосями). Для того чтобы быстро восстановить утраченную
часть колышков, желательно составить схему расположения трансект на иссле-
дуемом участке. Во время наблюдений за состоянием мохового покрова следу-
ет производить замеры уровня грунтовых вод. Для этого выкапывают ямку и
через 20 мин измеряют расстояние до зеркала воды.
Метод реперов применяют только при определении прироста у мхов с ортот-
ропным ростом. Этот метод нельзя использовать при изучении мхов, у которых
происходит интенсивное разложение нижних частей стеблей, т. к. годичное нара-
стание дерновинки вверх у этих видов всегда меньше истинной величины годич-
ного прироста. Тем не менее этот метод наиболее популярен у болотоведов (Ило-
метс, 1981; Максимов, 1982; Бамбе, 1991; Вос, Kuzmina, 1990).
Морфологический метод. Для использования этого метода необходимо
иметь практический навык анатомо-морфологического анализа побегов у пред-
ставителей различных таксономических групп мхов. Существуют виды с четкой
морфологической границей годичных побегов. Например, для Hylocomium
splendens характерно симподиальное ветвление, когда рост верхушки главной оси
сильно отстает, а ее место занимает боковая ветвь, растущая в направлении глав-
ной оси (Словарь ботанических терминов, 1984); ежегодное образование ново-
го этажа происходит в верхней части прошлогоднего побега. У некоторых ви-
дов листостебельных мхов и печеночников годичные побеги можно различать
по величине и форме листьев, развивающихся в различное время вегетационно-
го периода. Границу между двумя соседними годичными побегами определяет
93
смена летних, более крупных листьев на стебле на весенние, более мелкие (Кор-
чагин, 1960). Наиболее четко граница заметна весной, когда молодой гаметофит
имеет окраску, контрастирующую с цветом перезимовавшего гаметофита. У ряда
видов (особенно у крупных представителей верхоплодных мхов) можно наблю-
дать изменение направления роста побегов. Зимой под тяжестью снегового по-
крова рыхлые высокие дерновинки мхов принимают наклонное положение, по-
этому побеги текущего года, растущие прямо вверх, расположены под углом к
наклоненному побегу предыдущего года.
У видов рода Polytrichum побеги текущего года начинают рост из верхушки
стебля, образуя его непосредственное продолжение. Такой моноподиальный ха-
рактер роста побегов свойственен мужским особям и только в очень редких
случаях женским, когда из гинецея (женского побега) не вырастает спорофит. У
верхоплодных мхов не каждый год на всех стеблях в дерновинке образуются
гаметангии, заканчивающие рост стебля. Многие стебли остаются стерильными,
поэтому весной продолжается рост осеннего побега, и граница незаметна. Для
установления линейного прироста необходимо сопоставить прирост у 15-20 экз.
стеблей одного и того же вида (равных по размерам и взятых обязательно из
одной и той же дерновинки), на которых заметны границы между приростами
(Корчагин, 1960).
Методика определения линейного годичного прироста лишайников слабо
разработана, и оценка этой величины может быть выполнена лишь с определен-
ной степенью приближения. Для определения годичного линейного прироста
кустистых лишайников, у которых в верхней части каждого подеция ежегодно
образуется только одно новое ветвление, используется морфологический метод
(Салазкин, 1937, Игошина, 1939; Андреев, 1954; Корчагин, 1960). Возраст жи-
вой части подеция и всей его сохранившейся части, включая отмершую, но еще
не разложившуюся, может быть определен по числу основных ветвлений (или
колен, заключенных между ними). Величина среднего ежегодного линейного
прироста каждого подеция кустистых лишайников рода Cladina может быть
оценена путем деления длины живой части главной оси подеция на число состав-
ляющих ее колен.
Линейный годичный прирост как кустистых, так и других форм лишайников
оценивают методом повторных измерений. Для определения прироста кустис-
тых или бокальчатых форм лишайников проводят перевязку подециев ниткой на
некотором расстоянии от верхушки (0,5-1 см). Нитка должна закрепляться на
подеции, находящемся во влажном состоянии, и располагаться в месте разветв-
ления подеция для того, чтобы она не смещалась. Учитывая наличие интерка-
лярного роста, следует закреплять не одну, а 2-3 нити на каждом подеции. У од-
ной из ниток оставляют длинные концы, к которым прикрепляют заметную бир-
ку с номером экземпляра. После закрепления нитки линейкой или циркулем точ-
но измеряют расстояние между верхушкой подеция и первой ниткой, а также меж-
94
ду первой и второй и т. д. Возможно использование вместо ниток отметок, на-
несенных на подеции черной тушью. Отметки делают на сухом лишайнике в не-
скольких местах, лучше через равные промежутки. Измерение расстояний меж-
ду сделанными отметками, как и в первом случае, проводят на влажном лишай-
нике. Второй способ более универсален (может использоваться для определе-
ния линейного прироста листоватых и трубчатых лишайников) и точен (можно
отмечать несколько десятков подециев каждого исследуемого вида). В журнал
заносят номера отдельных экземпляров, измеренные расстояния и дату. Повтор-
ное измерение расстояний между сделанными отметками проводят через год (2-
3 года) и определяют суммарное увеличение длины отрезков подеция за время
с момента начала эксперимента. Рассчитывают средний годичный линейный при-
рост. Для оценки возраста живой части лишайника общую длину живой части
каждого подеция делят на величину среднего годичного прироста.
Годичную продукцию каждого вида лишайников и общую годичную продук-
цию лишайникового покрова на единицу площади рассчитывают, исходя из дан-
ных по массе живой части лишайников и среднего возраста живой части подеци-
ев данного вида.
2.3. ОЦЕНКА ЗАПАСОВ РАСТЕНИЙ,
ИМЕЮЩИХ ХОЗЯЙСТВЕННОЕ ЗНАЧЕНИЕ 1
Основной задачей ресурсоведческих исследований является количественная
оценка запасов растений, имеющих хозяйственное значение (лекарственные, кор-
мовые, пищевые и другие растения). Биологический запас — величина сырье-
вой фитомассы всех экземпляров вида на исследуемой территории, выражается
в единицах массы (кг, ц, т). Эксплуатационный (промысловый, хозяйственный)
запас — условно принятая часть биологического запаса, которая реально может
быть заготовлена на данной территории с учетом биологических и экономичес-
ких факторов (Буданцев, Харитонова, 1999; Основные понятия и термины...,
2001). Таким образом, величины биологического и эксплуатационного запаса
значительно различаются.
До начала полевых работ необходимо составить наиболее полную эколого-
ценотическую характеристику заготавливаемого растения, т. е. установить, в ка-
ких растительных сообществах (типах леса) встречается данный вид, какие мес-
тообитания наиболее благоприятны для его произрастания. Такие сведения яв-
ляются исходными для планирования рациональных маршрутов обследования
района исследований и выбора метода оценки запасов вида.
1 Автор Е. А. Мазная
95
В научной литературе имеется ряд методических руководств и пособий по
определению запасов растительных ресурсов (Черкасов, 1979; Методы иссле-
дования ресурсов..., 1983; Борисова, Лошаков, 1985; Методика определения
запасов..., 1986; Принципы и методы рационального..., 1989; Буданцев, Хари-
тонова, 1999 и др.). Наиболее часто в ресурсоведческих исследованиях исполь-
зуют два метода оценки запасов сырья: 1) на конкретных зарослях', 2) на клю-
чевых участках. Первый метод более точен, дает репрезентативные сведения о
запасах, но более трудоемкий. Его целесообразно использовать при детальных
исследованиях растительных ресурсов. Для планирования возможных объемов
промышленных заготовок по районам и областям целесообразнее использовать
оценку запасов методом ключевых участков, который дает менее точные, но
более полные и стабильные данные.
2.3.1. Метод оценки запасов на конкретных зарослях
Заросль — совокупность особей одного вида, произрастающих в раститель-
ном сообществе на участке, пригодном для проведения промысловой заготов-
ки (Буданцев, Харитонова, 1999).
Изучение запасов на конкретных зарослях включает в себя выявление мес-
тонахождения зарослей, определение их площадей, оценку урожайности (плот-
ности запаса) в каждой заросли и расчет величины запасов сырья. Для установ-
ления местонахождения зарослей полезных растений используют картографичес-
кий материал (топографические средне- и крупномасштабные карты; геоботани-
ческие карты, а также лесоустроительные и землеустроительные материалы и
карты), который отражает размещение на обследуемой территории лесных, лу-
говых, болотных и других растительных сообществ.
Способы определения площади заросли зависят от характера произрастания вида.
При достаточно равномерном распределении вида по площади заросли и если за-
росль плотная, границы ее хорошо выражены, то очертания заросли приравнивают
к правильной геометрической фигуре (например, прямоугольнику, квадрату или
кругу) и измеряют параметры, необходимые для вычисления площади вобранной
фигуры (рис. 2.3а). В том случае, когда площадь заросли соответствует выделу на
плане лесонасаждений или землеустроительных планах (например, залежь или ле-
сопосадки), ее площадь устанавливают по указанным материалам.
Если вид представлен неравномерно, отдельными пятнами (например, пятна лан-
дыша в травяном покрове сосняка сложного), которые составляют менее 50 %
от всего участка или растительного сообщества, то сначала определяют площадь
всего участка. Затем оценивают площадь, занятую пятнами вида. Для этого уча-
сток пересекают параллельными и перпендикулярными маршрутными ходами, про-
ложенными через определенное число метров (10, 15, 20 м), на которых изме-
ряется расстояние, пройденное по пятнам (рис. 2.36). Далее суммируют длину
всех пятен по всем параллельным и перпендикулярным маршрутным ходам и вы-
96
числяют площадь пятен. Общую
площадь всех пятен рассматривают
как одну заросль и рассчитывают
долю площади, которую она занима-
ет от площади всего участка.
Плотность запаса сырья (уро-
жайность) — величина сырьевой
фитомассы, полученная с единицы
площади заросли, выражается в еди-
ницах массы (г, кг) на единицу пло-
щади (м2, га). Его можно опреде-
лить несколькими способами, выбор
которых зависит от жизненной фор-
мы изучаемого вида и от той части
растения, которая используется в
качестве растительного сырья. Для
определения плотности запаса над-
земных частей некрупных травянис-
тых и кустарничковых растений (ли-
стья ландыша, листья и побеги брус-
ники и т. д.) применяют метод учет-
ных площадок. При оценке урожай-
ности подземных органов или при
изучении крупных растений (деревья
и кустарники) используют метод
модельных экземпляров. Для низко-
граница заросли
граница однородного участка (сообщества)
маршрутные ходы
пятна черники
Рис. 2.3. Определение площади заросли (Бу-
данцев, Харитонова, 1999)
рослых травянистых растений в некоторых случаях возможно оценить урожай-
ность по их проективному покрытию.
Метод учетных площадок заключается в закладке серии учетных площадок
внутри заросли. Если изучаемый вид распределен по площади достаточно рав-
номерно, учетные площадки закладывают в регулярном порядке на определен-
ном расстоянии одна от другой так, чтобы по возможности охватить всю пло-
щадь заросли. Нельзя располагать учетные площадки субъективно, выбирая для
них «наиболее типичные места». В том случае, когда вид на участке представлен
отдельными пятнами или куртинами, учетные площадки закладывают только в их
пределах, определяют площадь, занимаемую пятнами, принимая ее при расчете
запаса за площадь заросли.
Размер учетных площадок определяется величиной взрослых экземпляров
изучаемого вида. Оптимальным считается такой размер площадки, при котором
на ней размещается не менее 5 взрослых экземпляров растений. Для травянис-
тых видов и кустарничков площадь учетных площадок обычно составляет от 0,25
97
до 4 м2. По форме площадки могут быть квадратными, круглыми или прямоу-
гольными.
Точность определения запаса сырья зависит от нескольких факторов. Она тем
выше, чем больше закладывается учетных площадок. Для получения более до-
стоверных сведений о плотности запаса сырья, лучше закладывать большее чис-
ло мелких площадок. Количество заложенных площадок считается достаточным,
если относительная ошибка среднего арифметического составляет не более 15 %.
В большинстве случаев для определения урожайности растений травяно-кустар-
ничкового яруса лесных сообществ достаточно заложить 25 площадок площа-
дью 1 м2. Ориентировочные данные о необходимом числе учетных площадок
можно получить на основании разницы между минимальной и максимальной мас-
сой сырья, собранного с одной площадки. Если разница между этими показате-
лями составляет свыше 15 раз, то необходимо заложить дополнительное число
учетных площадок. В зависимости от распределения вида по площади и его оби-
лия число учетных площадок для достижения заданной точности определения
варьирует. Чем равномернее вид распределен и больше его обилие, тем мень-
шее число учетных площадок следует закладывать.
Перед сбором сырья на каждой заложенной площадке определяют процент
проективного покрытия вида и/или подсчитывают число экземпляров. В зависи-
мости от целей исследования, т. е. определения эксплуатационного или биологи-
ческого запаса, подсчитывают либо только взрослые особи (товарные экземп-
ляры), либо учитывают все экземпляры, включая всходы, ювенильные и повреж-
денные растения. Собранное с каждой площадки сырье взвешивают с точнос-
тью до 5 %, получая таким образом данные о сырой массе. Затем сырье высу-
шивают до воздушно-сухого состояния и взвешивают. Результаты определения
плотности запаса (урожайности) сырья могут быть представлены в расчете на
сырую или сухую массу и выражены в виде среднего арифметического со сред-
неквадратичной ошибкой. Формулы для расчетов приведены в разд. 4.
При определении урожайности ягод проводят подсчет числа ягод на каждой
заложенной учетной площадке. Затем со всей заросли собирают средний обра-
зец ягод (не менее 1000 шт.), взвешивают их по 100 шт. в десятикратной по-
вторности и определяют среднюю массу 1 ягоды. Урожайность ягод (например,
в г/м2) вычисляют умножением среднего числа ягод на единице площади (м2)
на среднюю массу (г) 1 ягоды.
Метод модельных экземпляров используют: 1) когда у изучаемого расте-
ния можно легко определить границы экземпляра или побега (например, у видов
пижмы, зверобоя, шиповника и пр.); 2) для оценки урожайности подземных ор-
ганов (например, алтей, безвременник, кровохлебка и др.); 3) когда объектом
исследования являются крупные деревья и кустарники (рябина, малина и др.).
При оценке плотности запаса сырья методом модельных экземпляров необ-
ходимо установить численность товарных экземпляров или побегов на единицу
98
площади и среднюю массу сырья одного экземпляра (побега). Оценку этих по-
казателей проводят с точностью до 10%. Побег в качестве счетной единицы
используют в случаях, когда границы отдельных экземпляров невозможно чет-
ко определить или когда сбор сырья с одного экземпляра затруднен (например,
шиповник, рябина, липа и пр.).
Определение численности товарных экземпляров проводят на учетных площад-
ках, принципы закладки которых описаны выше. При обильной встречаемости и
относительно равномерном распределении вида обычно достаточно заложить 15-
20 площадок, при меньшем обилии и неравномерном распределении число пло-
щадок возрастает до 30-50 шт. Если численность растений невелика, т. е. на 1 м2
площади приходится менее одного экземпляра, то подсчет лучше проводить на
трансектах (см. разд. 1.4), заложенных вдоль маршрутных ходов в полосе ши-
риной 1-2 м. Возможно использование тех же маршрутных ходов, что и для оп-
ределения площади заросли (см. выше). Трансекты разбиваются на отрезки, на
которых подсчитывается численность товарных экземпляров. Отрезки должны
иметь тем большие размеры, чем реже встречается изучаемый вид. Обычно
достаточно 25^40 отрезков. Исходя из данных о численности товарных экземп-
ляров на каждой учетной площадке проводят расчет средней численности экзем-
пляров на единицу площади заросли.
Для определения сырьевой массы изучаемого растения на учетных площад-
ках или трансектах отбирают каждый второй, третий или пятый модельный экзем-
пляр (побег). Такой способ отбора является наиболее объективным, т. к. в этом
случае производят отбор товарных экземпляров без выбора «типичных». Число
модельных экземпляров зависит от степени их варьирования. При определении
массы подземных органов или соцветий бывает достаточно 40-60 модельных
экземпляров; для вегетативных органов — до 100 и более. У каждого модель-
ного экземпляра взвешивают сырьевую фитомассу, затем рассчитывают сред-
нее значение этого показателя. Проводить взвешивание всех модельных экзем-
пляров вместе и вычислять среднюю фитомассу модельного экземпляра путем
деления общей массы на число экземпляров недопустимо, т. к. этот метод ис-
ключает возможность статистической обработки полученных данных.
Плотность запаса сырья (урожайность) рассчитывают умножением средней
численности товарных экземпляров на единице площади на среднюю массу сы-
рья модельного экземпляра.
Метод проективного покрытия используется для определения урожайно-
сти низкорослых травянистых растений (например, чабрец, мать-и-мачеха и др.),
а также стелющихся форм (толокнянка, плаун и др.). С этой целью оценивают
среднее проективное покрытие вида в пределах заросли и выход массы сырья с
1 % проективного покрытия, так называемую «цену» 1 % проективного покры-
тия. Проективное покрытие определяют квадрат-сеткой или глазомерно. Первый
способ более трудоемкий, но дает более точные результаты. Квадрат-сетка пред-
99
ставляет собой рамку площадью 1 м2, разделенную проволокой или леской на
100 квадратов, т. е. каждый квадрат составляет 1 % площади. Рамку накладыва-
ют на растения и подсчитывают, сколько квадратов полностью или более чем
наполовину закрыто надземными частями изучаемого вида. Для определения
«цены» 1 % проективного покрытия на каждой площадке срезают и взвешивают
сырье с 1 дм2. Плотность запаса (урожайность) сырья равна произведению сред-
ней величины проективного покрытия заросли на среднюю «цену» 1 % проектив-
ного покрытия.
Глазомерный метод определения проективного покрытия является более про-
стым, но менее точным и доступен только опытным, натренированным исследо-
вателям. Оценивают проективное покрытие на каждой площадке, глядя на нее
сверху и подсчитывая, какую часть площадки займут надземные части исследуе-
мого растения, если они будут плотно примыкать друг к другу. Тренировку глазо-
мера можно проводить при работе с квадрат-сеткой или сеточкой Раменского.
После сбора фактического материала по соотношению проективного покры-
тия и урожайности сырья в разных растительных сообществах и разных экологи-
ческих условиях возможно его обобщение с использованием регрессионного ана-
лиза для составления расчетных таблиц с целью прогнозирования запасов расти-
тельного сырья в дальнейшем.
2.3.2. Метод изучения запасов растительного сырья на ключевых
участках
Ключевой участок — площадь, служащая эталоном данного типа угодий по
сырьевым запасам конкретного вида (Буданцев, Харитонова, 1999). Метод клю-
чевых участков следует применять лишь для определения ресурсов видов, име-
ющих четкую приуроченность к каким-либо элементам рельефа, определенным
типам угодий, типам леса и/или болот, границы которых показаны на картах, а
также господствующих или встречающихся со значительным обилием. Методом
ключевых участков можно определять запасы таких видов, как аир болотный,
толокнянка обыкновенная, черника, брусника и др.
Ключевые участки закладывают, используя лесоустроительные или землеус-
троительные материалы, либо по картам (геоботаническим, топографическим и
др-)-
Число таких участков должно быть достаточным для получения репрезента-
тивных данных о характере размещения и урожайности, расположенных на клю-
чевых участках зарослей. Обычно площадь ключевого участка составляет от
одного до нескольких км2 и зависит от неоднородности растительного покрова.
Чем выше неоднородность растительного покрова, тем большую площадь дол-
жен занимать ключевой участок. Ключевыми участками должно быть охвачено
не менее 10 % площади потенциально продуктивных угодий, на которых вид мо-
жет образовывать промысловые массивы.
100
На ключевых участках однородных по растительному покрову и с равномер-
ным распределением экземпляров по площади определение численности (про-
ективного покрытия) и урожайности проводят на трансектах (методика закладки
трансект описана выше). Затем рассчитывают среднюю урожайность сырья на
каждом ключевом участке с указанием типа угодья, для которого она характер-
на. После этого по типам угодий рассчитывают среднюю урожайность сырья.
Если растения размещены по площади участка неравномерно, то по маршрут-
ным ходам определяют средний процент площади, занятой растениями (или их
группами) в пределах ключевого участка, затем рассчитывают средний процент
площади, занятой зарослями, и определяют урожайность изучаемого вида в за-
рослях в пределах каждого ключевого участка. После этого вычисляют сред-
нюю урожайность для всех зарослей на всех ключевых участках.
Полученные первым или вторым способом данные о плотности запаса сырья
экстраполируют на площадь потенциально продуктивных угодий, которую опре-
деляют по картографическим материалам с помощью палетки. Когда в качестве
ключевых участков используют выделы типов леса с определенным составом,
возрастом, полнотой и бонитетом древостоя, то сведения об общих площадях
этих выделов можно взять из материалов лесоустройства.
2.3.3. Расчет объемов возможных ежегодных заготовок
При определении запасов сырья следует различать биологический и эксплуа-
тационный (промысловый) запасы. Биологический запас рассчитывается по верх-
нему пределу урожайности:
B = S- (Y + 2т),
где В — биологический запас, кг; S' — площадь заросли, га; Y — средняя уро-
жайность сырья, кг/га; т — ошибка среднего арифметического.
Эксплуатационный запас рассчитывается умножением площади заросли на
нижний предел величины урожайности (Y-2m). Для дикорастущих ягод эксплуа-
тационный запас составляет, как правило, 50 % от биологического (Черкасов,
1979). Эксплуатационный запас показывает количество экономически доступной
сырьевой продукции на площади участка заготовки, которое можно заготовить
при однократной эксплуатации заросли. Однако ежегодная заготовка на одной и
той же заросли возможна только в тех случаях, когда используются плоды. Во
всех остальных случаях с целью рационального использования и охраны природ-
ных ресурсов необходимо рассчитывать объемы возможных ежегодных загото-
вок. Для этого необходимо знать, за какой период времени происходит восста-
новление первоначальных параметров заросли. К настоящему времени прибли-
зительно для 30 видов лекарственных растений имеются экспериментальные
данные о сроках восстановления их запасов, для остальных видов можно лишь
указать приблизительную периодичность заготовок. Рекомендуемая ориентиро-
101
вочная периодичность заготовок для соцветий и надземных органов однолетни-
ков составляет один раз в 2 года, многолетников — один раз в 4—6 лет, а для
подземных органов — один раз в 15-20 лет (Буданцев, Харитонова, 1999).
Объем возможной ежегодной заготовки сырья (Квез) рассчитывается как
частное от деления эксплуатационного запаса сырья на оборот заготовки, вклю-
чающий год заготовки и продолжительность периода восстановления заросли:
v Е
г вез=---------
Л а гот + ^восст
где Е — эксплуатационный запас, кг; /iai0T — год заготовки; /В0Сст — продолжи-
тельность периода восстановления, лет.
Для объективного планирования объемов заготовок ягод часто рассчитыва-
ют доступный для освоения запас ягод. Доступный для сбора запас (урожай)
ягод вычисляют в процентах от эксплуатационного, при этом доступность тер-
ритории (%) определяется исходя из местных условий (населенность района ис-
следования, заболоченность территории, наличие транспортных путей и т. д.).
2.4. МЕТОДЫ ИЗУЧЕНИЯ СЕМЕННОЙ
ПРОДУКТИВНОСТИ РАСТЕНИЙ 1
Исследование характеристик генеративной деятельности растений является одним
из важных вопросов при изучении состояния их популяций, прогноза динамики и ус-
тойчивости лесных экосистем в целом в разных экологических условиях.
Семенное размножение растений в лесных сообществах контролируется боль-
шим числом взаимосвязанных факторов, которые условно можно подразделить
на две группы: абиотические и биологические. К числу наиболее важных абио-
тических факторов относятся метеорологические, эдафические, давность нару-
шения растительного сообщества, тип и интенсивность современных антропоген-
ных воздействий. Из биологических факторов следует прежде всего выделить
морфологические, анатомические, физиологические особенности семян, их ге-
терогенность, а также приспособления к диссеминации. Большое значение име-
ют параметры популяций видов, прежде всего возрастная и виталитетная струк-
тура, а также генеративность популяции — доля генеративных особей от их об-
щего числа.
Для изучения семенного возобновления в фитоценозе обычно ограничиваются
учетом производимых видом семян (Левина, 1981). При этом используется два
основных показателя: средняя семенная продуктивность — среднее число се-
1 Авторы И. В. Лянгузова, Н И. Ставрова
102
мян в расчете на одну счетную единицу (особь, парциальный куст, репродуктив-
ный побег) и общая семенная продуктивность — число семян, продуцируемых
растениями определенного вида, на единицу площади. Последняя характеристи-
ка зависит от доли генеративных особей в популяции и их средней семенной про-
дуктивности. Оба показателя существенно варьируют по годам в соответствии
с погодными условиями и естественными генеративными циклами. Поэтому до-
статочно точные данные о семенной продуктивности вида можно получить лишь
при многолетних наблюдениях на постоянных пробных площадях (для древес-
ных растений и кустарников) и постоянных учетных площадках (для растений
нижних ярусов сообществ) или на основе использования специальных методик,
позволяющих оценить средние многолетние величины продукции семян.
Для более углубленного изучения вопросов семенной продуктивности обя-
зательными показателями являются потенциальная семенная продуктивность,
под которой понимают число семязачатков на особь или генеративный побег, и
реальная семенная продуктивность — число спелых неповрежденных семян
на особь или генеративный побег. Следует помнить, что семенам дикорастущих
видов присуща высокая степень гетерогенности (неоднородности) семян, под
которой понимают любые различия семян одного и того же вида (Левина, 1981).
Более подробные сведенця о репродуктивной биологии растений можно полу-
чить из специальных работ, посвященных различным аспектам семенного размно-
жения растений (Левина, 1981; Злобин, 1968; 1989; 2000; Терехин, 1996; Нико-
лаева и др., 1999; Ходачек, 2000 и др.). Методические подходы к изучению се-
менного размножения отражены в публикациях Т. А. Работнова (1960), Р. Е. Ле-
виной (1960), И. Г. Строна (1962), И. В. Вайнагия (1973; 1974), Е. А. Ходачек
(1974) и др.
2.4.1. Изучение семеношения основных лесообразующих видов
хвойных древесных растений
Существует целый ряд методик, позволяющих оценить величину продукции
семян хвойных древесных растений в изучаемых лесных сообществах. Подроб-
ная методическая сводка по изучению семенной продуктивности древесных ра-
стений представлена в работе А. А. Корчагина (1960). Различные методы изуче-
ния семеношения хвойных описаны в работах Р. Сарваса (Sarvas, 1945, 1962),
П. Л. Горчаковского (1947, 1958), А. А. Молчанова (1950), В. Г. Нестерова
(1954), Т. П. Некрасовой (1955, 1957, 1960), В. П. Тимофеева (1959),
И. В. Семечкина (1963), С. А. Мамаева, В. М. Яценко (1965), М. Г. Романовско-
го (1976), Г. В. Стадницкого, Г. М. Сметанина (1985), В. И. Мосина (1987),
С. Н. Санникова (1992), В. В. Горшкова, Н. И. Ставровой (1999). Чаще всего ис-
пользуются различные модификации четырех основных методов.
Метод модельных деревьев (глазомерно-статистический). Различные вари-
анты этого метода были предложены и изучались еще в конце XIX — начале XX
103
века. Один из наиболее известных вариантов состоит в следующем. В пределах
каждой пробной площади предварительно производится глазомерная оценка оби-
лия семеношения (числа женских шишек) каждого дерева по 4-6-балльной шка-
ле. Затем для каждого выделенного класса обилия берут модельные деревья, на
которых пересчитывают все имеющиеся шишки и устанавливают среднее число
шишек на дереве каждого класса обилия. Эти величины умножают на число де-
ревьев, относящихся к соответствующему классу, и складывают. Полученное
значение общего числа шишек на всех деревьях в пределах пробной площади
позволяет рассчитать продукцию шишек на единицу площади сообщества. Отбор
модельных деревьев можно производить не только по классам обилия семеноше-
ния, но и по классам Крафта, по ступеням толщины или на основе определения
параметров «среднего дерева» в древостое. Как показали выполненные методи-
ческие исследования, удовлетворительная точность при использовании этого ме-
тода достигается только при выделении большого числа классов (до 10) и отбора
3-5 моделей для каждого класса. Таким образом, применение этого метода явля-
ется весьма трудоемким и требует рубки значительного числа деревьев.
Метод модельных ветвей (морфологический). Теория и первые варианты
применения этого метода были разработаны еще в начале XX века Н. С. Несте-
ровым (1914). При использовании этого метода могут решаться три задачи:
1) оценка относительных размеров семеношения текущего года, 2) прогноз се-
меношения в будущем году, 3) определение относительной величины семено-
шения за несколько прошедших лет по следам опавших плодов или шишек. Для
определения величины семеношения за прошлые годы методом модельных вет-
вей на основе более поздних исследований были сформулированы следующие
рекомендации для получения достаточно точных данных об относительной ве-
личине продукции шишек. Отбор ветвей производят с 15-20 модельных дере-
вьев (выбор которых проводится на основе одного из описанных выше принци-
пов) из двух соседних мутовок (у взрослых деревьев — чаще из 8-10-ой) жен-
ского (верхнего) генеративного яруса. Из каждой мутовки берут 4-5 ветвей, т. е.
всего 8-10 ветвей с одного дерева. Затем производят измерение длины модель-
ных ветвей и пересчет всех имеющихся двухлетних шишек и (или) шишек про-
шлых лет и следов опавших шишек, а также числа учтенных побегов разных лет
(1-, 2-, 3-летних и т. д.). Полученные данные позволяют рассчитать некоторые
относительные показатели: среднее число шишек на 1 м длины ветви или на
1 м длины побегов данного года; среднее число шишек за каждый учтенный год,
долю побегов данного года, несущих шишки. Примерную абсолютную величи-
ну продукции шишек сосны (в молодых древостоях) Т. П. Некрасова рекомен-
дует определять следующим образом: из каждой мутовки каждого модельного
дерева берут одну скелетную ветвь, с нее, в свою очередь, берут одну ветвь вто-
рого порядка, на которой и производят учет всех шишек и следов от опавших
шишек по годам. Зная число скелетных ветвей в каждой мутовке и число вет-
104
вей второго порядка на скелетной ветви, можно вычислить общее количество
шишек каждого года на каждой скелетной ветви, на каждом модельном дереве
и на всей пробной площади. Такое определение продукции шишек и семян, бе-
зусловно, очень трудоемко.
Метод семеномеров. Используется только при стационарных исследовани-
ях. Впервые был предложен в конце XIX века М. М. Орловым. Семеномеры
могут быть сделаны в виде ящиков закрытых сверху сеткой или воронок прямо-
угольной или округлой формы. Площадь семеномера может составлять от 0,25
до 4 м2. Чаще используют семеномеры площадью 0,25-1 м2. Семеномеры рас-
полагают на земле или на небольших столбиках. Как показали специально выпол-
ненные исследования, на пробной площади размером 0,25 га следует использо-
вать от 25 до 50 семеномеров размером 0,25 м2 или 15-25 семеномеров раз-
мером 0,5 м2, которые располагают в регулярном порядке по одному или груп-
пами по 4-5 штук. По оценке А. П. Тольского (1950), ошибка определения про-
дукции семян при использовании 50 семеномеров составляет ±15%. Семеноме-
ры устанавливают до начала опадения семян; в последующий период учет семян
производят каждые 3-7 дней до окончания их выпадения. Величина продукции
семян определяется по формуле
P = Spp-Ns /Ssm-Nsm;
где Р — величина продукции семян на единицу площади сообщества; Spp — раз-
мер пробной площади; Ng — общее число учтенных семян; Ssm — площадь по-
верхности одного семеномера (в тех же единицах, что и Spp)', Nsm — число се-
меномеров.
Метод учета опавших шишек. Предложен известным финским ученым
Р. Сарвасом (Sarvas, 1945, 1962). Впоследствии применялся рядом исследова-
телей в хвойных лесах разных географических регионов (Lehto, 1957; Семечкин,
1963; Санников, Санникова, 1985; Горшков, Ставрова, 1999). Именно этот ме-
тод рекомендуется использовать при проведении комплексных исследований
структуры и динамики лесных сообществ на сериях постоянных пробных
площадей. Метод обладает относительно невысокой трудоемкостью и позволяет
одновременно с измерением характеристик полога подроста на учетных площад-
ках получить данные о среднем многолетнем количестве семян, поступающем
ежегодно на единицу площади в пределах изучаемого сообщества. Учет числа
опавших шишек проводят на каждой площадке размером 1 х 1 м. Подсчитыва-
ют все неразложившиеся шишки (с чешуями, не отделяющимися от стержня),
находящиеся на поверхности и внутри толщи мохово-лишайникового покрова, а
также на поверхности и в верхней части горизонта лесной подстилки. Выявление
шишек, скрытых в толще мохово-лишайникового покрова и подстилки, прово-
дят прощупыванием вручную. Установленная величина средней плотности опав-
ших шишек позволяет оценить продукцию семян хвойных в пределах сообще-
105
ства. Она определяется из уравнения
А = А • (М-С / Т^,
где А — среднегодовая продукция семян на единицу площади; Dc — среднее
число опавших шишек на единицу площади; Nsc — среднее число семян в шиш-
ке; TL!j— время разложения шишки после опадения.
Для определения среднегодовой продукции семян на единицу площади лес-
ного сообщества необходимо дополнительно определить две величины: сред-
нее число семян в шишке и среднее время ее разложения в условиях района
исследований. На основе многолетних экспериментальных исследований Р. Сар-
васом (Sarvas, 1962) было установлено, что для сосны обыкновенной величина
переходного коэффициента (Nsc / Tcj) в западном секторе европейской тайги из-
меняется от 1,0-1,1 в северной подзоне до 1,6-1,7 — в южной. Среднее число
семян в шишке сосны в условиях европейской тайги составляет ~20 (17-26),
следовательно, среднее время разложения шишки изменяется от 20 лет в усло-
виях северной тайги до 12 лет — в южной.
2.4.2. Изучение семенной продуктивности растений нижних ярусов
лесных сообществ
Как уже отмечалось выше, достаточно точные данные о семенной продук-
тивности растений можно получить лишь при многолетних наблюдениях на точ-
но фиксированных площадках. При таком методическом подходе: а) достигает-
ся полная сравнимость между наблюдениями отдельных лет, поскольку они от-
носятся к одним и тем же площадкам; б) устраняется субъективизм в выборе
особей для определения средней семенной продуктивности, т. к. она определя-
ется у всех генеративных особей на определенной площади, в том числе и у уг-
нетенных, малозаметных экземпляров. При разовых оценках семенной продук-
тивности следует также закладывать учетные площадки, т. к. только в этом слу-
чае можно получить достоверные данные о продукции семян разных видов в
исследуемом фитоценозе. Методы подбора и закладки учетных площадок опи-
саны в разд. 1.4. Величина учетной площадки определяется размерами растений
и численностью генеративных особей, поэтому она может быть неодинаковой для
различных видов. Количество закладываемых площадок определяется точнос-
тью, с которой необходимо оценить семенную продуктивность, и пространствен-
ным размещением растений. При достаточно равномерном распределении осо-
бей по площади в лесных сообществах достаточно заложить 30-50 учетных пло-
щадок. В тех случаях, когда колебания в семенной продуктивности отдельных
особей особенно велики и (или) вид распределен по площади контагиозно, чис-
ло учетных площадок следует увеличить.
В качестве учетной единицы при оценке семенной продуктивности растений
обычно используют особь. Когда невозможно точно выделить отдельные осо-
106
би растений (часто у вегетативно-подвижных кустарничков), за счетную едини-
цу принимают парциальный куст (например, Vaccinium myrtillus, V vitis-idaea и
др.) или репродуктивный побег (например, Empetrum nigrum, Solidago lappomca
и др.).
На каждой учетной площадке подсчитывают раздельно генеративные и веге-
тативные особи изучаемых видов. Для каждой особи определяют ее семенную
продуктивность, полученные данные объединяют и рассчитывают среднюю се-
менную продуктивность растения. Общую семенную продуктивность рассчиты-
вают с учетом размера учетной площадки и выражают в числе семян на 1 м2.
Генеративность популяции вычисляют путем деления числа генеративных осо-
бей на общее число особей определенного вида.
Более универсальным методом определения средней семенной продуктивно-
сти является метод раздельного определения среднего числа: а) генеративных
побегов на растении; б) плодов на генеративных побегах; в) семян в плоде или
соплодии. Первые две величины большей частью легко определить путем под-
счета генеративных побегов и плодов у всех особей, встречающихся на учет-
ной площадке. Для определения среднего числа семян в одном плоде или со-
плодии отбирается средняя проба плодов (или соплодий) из числа собранных с
учетной площадки. Количество плодов, которые необходимо взять для средней
пробы, чтобы получить достаточно достоверные данные о среднем содержании
семян на один плод, целесообразно выяснить эмпирически.
Сбор плодов для определения в них числа семян следует проводить незадол-
го до их созревания, чтобы устранить возможность осыпания семян. Однако, если
в дальнейшем предполагается определение жизнеспособности семян, то сбор
семян следует проводить лишь после полного созревания семян.
Для более углубленного изучения вопросов семенной продуктивности наблю-
дения следует вести из года в год не только на одних и тех же площадках, но и
на тех же самых особях. В этом случае выбранные модельные особи маркиру-
ют этикетками. Для всех заэтикетированных экземпляров ежегодно фиксируют
их состояние и семенную продуктивность. Плоды с каждой особи собирают в
отдельные пакетики, снабженные номерами; подсчет семян обычно производят
позже. У видов с неравномерно созревающими плодами сбор плодов приходит-
ся проводить в 2-3 приема.
Работы по изучению семенной продуктивности растений важно сопровождать
выяснением качества и жизнеспособности семян (см. разд. 2.5).
107
2.5. МЕТОДЫ ОЦЕНКИ КАЧЕСТВА И ЖИЗНЕСПОСОБНОСТИ СЕМЯН 1
В термин качество семян специалисты различных областей знания вкладыва-
ют различные понятия, относящиеся к многочисленным свойствам семян, — это
может быть и анатомическое строение семян, их полноценность, посевные ка-
чества, способность к прорастанию, физиологическая спелость, сила семян и пр.
Разработка унифицированных методов определения качества семян как куль-
турных растений, так и дикорастущих видов остается насущной проблемой. В
различных странах исследования качества семян проводятся по программе Меж-
дународной ассоциации по испытанию семян (International Seed Test Association,
ISTA). Приведение принятых ISTA стандартных показателей качества семян обя-
зательно при международном обмене. Наряду с этим в разных странах создают-
ся государственные стандарты качества семян отдельных культур. В России также
разработаны «Правила отбора образцов и методы определения посевных качеств
семян» (1988) и ГОСТы для различных сельско- и лесохозяйственных культур.
Разработать какие-либо стандартные способы оценки качества семян дикорас-
тущих растений практически невозможно в силу разных причин (растянутость
периода созревания, а следовательно, и прорастания, наличие органического по-
коя, свойственного семенам многих видов, экотипическое разнообразие видов и
т. д.). Качество и жизнеспособность семян дикорастущих видов приходится ус-
танавливать опытным путем для каждого вида и даже для отдельных партий се-
мян.
Для характеристики качества семян используют различные показатели: про-
цент полнозернистости (выполненности), процент пустых семян в партии, зара-
женность семян патогенными организмами и т. д. Наиболее важным и общеупот-
ребительным показателем является всхожесть. Различают лабораторную и
полевую (грунтовую) всхожесть. Первая дает представление о потенциальных
возможностях семени, вторая характеризует способность семян прорастать и
давать нормальные растения в условиях производственного посева.
В общем смысле под лабораторной всхожестью понимают процент семян,
проросших при оптимальных условиях. Проросшими принято считать семена с
длиной корешка равной половине длины семени, у мелких семян — двойной дли-
не. Набухшие семена (когда они только увеличиваются в размерах, а иногда при
этом у них лопается кожура) или случаи ненормального прорастания семян (на-
пример, семядолями) учитываются отдельно и не включаются в подсчет про-
росших семян при определении всхожести. Следует упомянуть, что согласно фор-
мулировке, принятой ИСТА в 1976 г. (Seed Science & Technology, 1976), при оп-
1 Автор И. В Лянгузова
108
ределении лабораторной всхожести рекомендуется подсчитывать не просто про-
росшие семена, но образовавшие нормально развитые проростки. Тем не менее
под лабораторной всхожестью большей частью понимают просто способность
семян прорастать.
Грунтовая всхожесть — это количество всходов, появившихся на поверх-
ности почвы, выраженное в процентах от общего числа посеянных семян. Грун-
товая всхожесть практически всегда оказывается меньше лабораторной, однако
часто между этими показателями имеется корреляция.
Кроме вышеназванных существует еще ряд показателей, характеризующих
качество семян. Под энергией прорастания, согласно ГОСТ 13056.6-75, пони-
мается количество проросших семян за определенный (характерный для каждой
культуры) срок, выраженное в %. Скорость прорастания трактуется разными
авторами по-разному: как отношение числа проросших семян за определенный
промежуток времени к этому промежутку, или как среднее время прорастания
одного семени. Наряду с перечисленными показателями исключительное значе-
ние придается такому показателю, как сила семян (Лихачев, 1977, 1980, 1984 и
др.). Он необходим, чтобы оценить не только способность прорастать, но и по-
тенциальные возможности семян, во-первых, давать при посеве сильные расте-
ния, которые наиболее успешно противостоят стрессовым воздействиям и обес-
печивают высокий урожай, а во-вторых, возможность и целесообразность зак-
ладки их на хранение.
Оценка жизнеспособности семян производится различными методами (Жиз-
неспособность семян, 1978; Илли, 1982; Николаева и др., 1999 и др.). Безусловно,
наиболее надежным и достоверным способом определения жизнеспособности
является проращивание семян при благоприятных условиях. Однако это требует
определенного и подчас довольно длительного времени даже для семян, не на-
ходящихся в органическом покое, и практически малопригодно для покоящихся
семян, прорастающих только после специальной и, как правило, довольно дли-
тельной предпосевной подготовки. Трудности, связанные с проращиванием се-
мян, привели к необходимости разработки экспресс-методов оценки жизнеспо-
собности: метод витального окрашивания, тетразольный метод, рентгенография
и некоторые другие (Нелюбов, 1925; Фирсова, 1959; Мак-Кэй, 1978; Смирнова,
1978 и др.).
2.5.1. Лабораторное испытание всхожести
Прямым способом оценки жизнеспособности семян является определение их
всхожести, энергии и скорости прорастания, а также силы семян путем проращи-
вания в лабораторных условиях. Надежность и достоверность лабораторного
определения всхожести семян зависит от многих факторов: достаточного водо-
снабжения, подходящей температуры, газового состава атмосферы, а часто и
света. Кроме того, предварительно необходимо собрать доступные сведения о
109
биологии семян данного вида, глубине и типе покоя, степени развития зародыша
и т. п.
К настоящему времени накоплен обширный материал по способам и методам
лабораторных испытаний семян. Они отличаются многообразием используемых
субстратов, приспособлений и аппаратов для проращивания семян, а также спо-
собов предпосевной обработки семян. Кратко остановимся на наиболее распро-
страненных методиках лабораторных испытаний.
Проращивание семян обычно проводят на искусственных средах. В качестве
субстрата широко используется специальная бумага — достаточно толстая и рых-
лая, реже — просто фильтровальная. Семена раскладывают на поверхности бу-
маги (особенно если для прорастания необходим свет) или между сложенными
листами бумаги. Для не очень мелких семян рекомендуется следующий способ:
листы бумаги с разложенными на них семенами свертывают в рулоны, которые
устанавливают вертикально или слегка под наклоном в сосуды с водой. Преиму-
щество последнего способа состоит в том, что появляющиеся проростки рас-
тут нормально в соответствии с геотропической реакцией, что облегчает их оцен-
ку для определения силы роста семян. Кроме того, для проращивания семян
используют песок, предварительно просеянный для выравнивания размера час-
тиц и прокаленный для удаления органических примесей. Он широко применяет-
ся для достаточно крупных, но не пригоден для мелких семян. Семена высева-
ют, вдавливая в поверхность песка или засыпая слоем 1-2 см. Проращивание в
песке большинства видов семян проводят при влажности до 50-60 % его пол-
ной влагоемкости. В специальных исследованиях иногда используют разные пи-
тательные смеси.
При проращивании семян на фильтровальной бумаге часто используют чашки
Петри, в которых важно поддерживать оптимальное количество воды (для ча-
шек диаметром 9 см — в среднем 5 мл), а очень мелкие семена располагать на
«мостике» из предметных стекол на фильтровальной бумаге, конец которой опу-
щен в воду в чашке. Иногда для испытания всхожести семян используют специ-
альные термостаты или даже комнаты с регулируемыми условиями.
Длительность проращивания определяется необходимым для каждого вида
сроком, в течение которого в оптимальных условиях прорастает максимальное
количество жизнеспособных семян. Ход прорастания и появление нормально
развитых проростков регулярно регистрируют и такие проростки удаляют. Отста-
ющие в развитии экземпляры рекомендуется оставлять для оценки и выяснения
их дальнейшей судьбы. Они могут оказаться доброкачественными, хотя разви-
вающимися более медленно, или дефектными и ненормальными.
Для определения всхожести и силы роста семян в условиях, приближенных к
полевым, обычно в качестве субстрата используют песок. В этом случае семе-
на высевают в сосуды высотой примерно 12 см, заполненные на 8 см влажным
песком и засыпают сверху песком на глубину, соответствующую глубине задел-
110
ки в поле. Появление всходов регистрируют и оценивают в баллах степень раз-
вития проростков.
2.5.2. Определение жизнеспособности семян с помощью витальных
красителей
Впервые использование красителей для определения жизнеспособности семян
было предложено Д. Н. Нелюбовым (1925). Позже этот метод получил широ-
кое распространение. Предварительно замоченные семена освобождают от по-
кровов, разрезают вдоль или из них извлекают зародыш. Окрашивание можно
производить различными красителями, которые не проникают в живые ткани, но
быстро окрашивают мертвые ткани. Среди таких красителей наиболее часто ис-
пользуют водные растворы индигокармина (~0,2 %), а для зеленых зародышей —
кислый фуксин (—0,1—0,2 %). Время окрашивания семян или зародышей различ-
но для разных видов. К нежизнеспособным семенам относят семена с полнос-
тью окрашенным зародышем и с зародышем, у которого интенсивно окрашен
кончик корешка. Неокрашенные зародыши и те, у которых наблюдается окраши-
вание лишь поврежденных при изоляции участков или слабое окрашивание кон-
чика зародыша, относят к жизнеспособным.
2.5.3. Тетразольный метод
Впервые тетразольный ме-
тод для определения жизне-
способности семян был при-
менен G. Lakon (1949), впос-
ледствии получил широкое рас-
пространение для проверки се-
мян древесных пород (Лоба-
нов, Козелкова, 1958; Линник,
1959; Лобанов, 1964; Санкина,
Днепровский, 1974 и др.).
Этот метод основан на исполь-
зовании бесцветных солей тет-
разола, которые под влиянием
ферментов живых семян (де-
гидрогеназ) восстанавливают-
ся до трифенилформазана —
вещества, окрашенного в крас-
ный цвет. В мертвых тканях,
где отсутствуют активные фер-
менты, окрашивание не появ-
ляется. Обычно применяется
Рис. 2.4. Реакция зародышец красного клевера
на обработку тетразолом (Робертс, 1978)
1-4,10,11— жизнеспособные (окрашенные) зароды-
ши, остальные — нежизнеспособные
111
1%-ный водный раствор 2,3,5-трифенилтетразолхлорида или бромида при pH в
пределах от 6 до 7. Оценка жизнеспособности семян основана на интенсивности
окраски частей зародыша, жизненно необходимых для его роста и развития (ко-
рень, гипокотиль и т. п.). Поскольку строение зародыша у семян разных расте-
ний неодинаково, требуется разработка системы оценки результатов, подходящей
для конкретного вида. В качестве примера приведены результаты испытания ме-
тодом тетразола зародышей красного клевера (рис. 2.4). Однако необходимо
иметь в виду, что при использовании этого метода могут возникать артефакты в
результате повреждений, вызванных разными причинами, а также под влиянием
присутствующих в семенах микроорганизмов.
2.5.4. Люминесцентный метод
Наряду с упомянутыми биохимическими методами имеются различные дру-
гие способы быстрого определения качества и, в частности, жизнеспособности
семян: люминесцентный, окрашивание йодистым калием, солями селена и неко-
торыми другими веществами, а также по изменению окраски специальной бума-
ги, вызываемой эксудатами семян некоторых видов (Мак-Кэй, 1978).
Для определения доброкачественности сельскохозяйственных культур и дре-
весных растений довольно широкое распространение получил люминесцентный
метод, основанный на различном свечении семян разных видов растений при
ультрафиолетовом облучении (Борисевич, 1958; Маргайлик, 1959; Некрасов,
1973 и др.). Кроме того, жизнеспособные и нежизнеспособные семена одного
вида в ультрафиолетовых лучах светятся по-разному.
Увеличению контрастности свечения способствует прокрашивание срезов за-
родышей флуорохромами: акридин оранжевый, иодэозин, флуоресцеин, индиго-
кармин и др. Семена замачивают в течение 1-2 ч в теплой воде, освобождают
от покровов и обрабатывают в течение 20 мин в растворах люминесцирующих
красителей. Затем семена просматривают в ультрафиолетовом свете.
Люминесцентный метод определения жизнеспособности семян обладает удов-
летворительной точностью, отличается значительной быстротой, но имеет ряд
недостатков. Во-первых, семена даже сходных по морфологическим признакам
видов при люминесцентном анализе могут давать различное свечение. Поэтому
связь цвета люминесценции с жизнеспособностью семян устанавливают предва-
рительно для каждого вида сравнением с результатами определения жизнеспо-
собности другими методами (проращиванием или биохимическими). Во-вторых,
при люминесцентном анализе, как и при биохимических методах, требуется уда-
ление покровов семян, извлечение зародышей или приготовление срезов, что
ведет к уничтожению исследуемых семян.
112
2.5.5. Рентгенографический метод
Рентгеновский анализ позволяет, не вскрывая семян, быстро и точно опреде-
лить степень развития внутренних структур и наличие повреждений, а также оце-
нить жизнеспособность семян. Рентгеновское облучение семян в настоящее
время широко используется при анализе семян культурных и дикорастущих ви-
дов растений. Облучение обычно проводят мягкими лучами с длиной волны от
1 до 2,5 Е.
Использование рентгеновских лучей для определения качества семян у хвой-
ных растений впервые было предложено М. Simak, A. Gustavson (1953). В даль-
нейшем этот метод использовали для оценки качества семян различных видов
многие исследователи. Н. Г. Смирнова (1978) применила облучение семян рен-
тгеновскими лучами для оценки качества семян интродуцентов более чем 400 ви-
дов древесных растений. При этом она справедливо указала, что при дешифров-
ке рентгеновских снимков необходимо учитывать особенности строения семян
В частности, в зависимости от степени развития зародыша, характерной для се-
мян разных растений, ею было предложено делить их на 3 группы: 1) семена с
хорошо развитым эндоспермом и маленьким малодифференцированным или не-
доразвитым зародышем; 2) семена с развитым дифференцированным зародышем
и достаточно развитым эндоспермом; 3) се-
мена без эндосперма или почти без эндос-
перма, по с крупным зародышем, который
еще в период развития семени поглощает
эндосперм, и в его семядолях откладывают-
ся необходимые запасные вещества.
Для рентгенографии сухие семена (по
100-150 семян в 2-3 повторностях) реко-
мендуется раскладывать на липкую ленту,
что позволяет фиксировать их положение.
При облучении более плотные ткани семе-
ни поглощают больше рентгеновских лучей,
и на негативе они получаются светлыми. Пу-
стые семена или участки с внутренними де-
фектами, поглощая меньше рентгеновских
лучей, дают более темное изображение. В
зависимости от степени развития зародыша
и эндосперма семена каждой группы видов
на рентгенограммах (рис. 2.5) подразделяют
на классы (обычно 4—5 классов). Подсчиты-
вая количество семян каждого класса, опре-
деляют качество семян в партии.
Рис. 2.5. Фрагмент рентгенограм-
мы семян (негатив) Arctosta-
phylos uva-ursi
Классы развития эндосперма 1 — пу-
стое семя, 2 — заполнено менее поло-
вины полости семени, 3 — заполнено
от половины до 0,75 полости семени,
4 — заполнено более 0,75 полости, по
периферии остается незаполненное про-
странство, 5 — полость заполнена це-
ликом, эндосперм плотно прилегает к
семенной кожуре
ИЗ
Ценность данного метода состоит в том, что с его помощью удается полу-
чить представление о качестве семян большого числа образцов, которые затем
могут использоваться для получения растений, так как, во-первых, существует
корреляция между характеристикой семян, полученной с помощью рентгена и
данными прямого определения всхожести семян, а во-вторых, облучение не ока-
зывает на семена отрицательного действия.
Наряду с оценкой качества семян рентгенографию используют с целью опре-
деления жизнеспособности семян (Мак-Кэй, 1978). Для этого семена после за-
мачивания в воде (в течение примерно 16-24 ч) и удаления избытка воды пере-
носят на 1 ч в 20-30%-ный раствор хлористого бария. Подготовленные таким
образом семена подвергают мягкому рентгеновскому облучению. Рентгенограм-
мы исследуют с оценкой степени импрегнации тканей. Необходимо предваритель-
но подсчитывать число семян с плохо развитыми внутренними структурами, так
как они неспособны к нормальному прорастанию.
Для получения более четких рентгенограмм применяют контрастные агенты,
например урографин и умбрадил, а также используют различные приспособления
и методики обработки семян и рентгеновских пленок.
Таким образом, наиболее перспективным методом определения качества и
жизнеспособности семян является метод рентгенографии. Этот метод сохраня-
ет семена для посева, дает быстрые и точные результаты, позволяет судить об
особенностях развития и внутреннего строения семян, а также их жизнеспособ-
ности. Однако этот метод неприменим для мелких и очень мелких семян.
2.6. МЕТОДИКА ИЗУЧЕНИЯ БАНКА СЕМЯН В ПОЧВЕ 1
В почве различных фитоценозов содержится то или иное количество покоя-
щихся жизнеспособных семян разнообразных растений. Совокупность этих се-
мян называют почвенным банком семян или почвенным запасом семян. При этом
имеют в виду не только собственно семена, но и некоторые другие диаспоры
(односеменные плоды, отдельные части сложных и дробных плодов, содержа-
щие одно семя и т. д.).
В состав почвенного банка входят семена (диаспоры) различного происхож-
дения: 1) автохтонные, которые попали в почву с плодоносящих растений со-
временного фитоценоза; 2) реликтовые, которые принадлежат растениям прежне-
го фитоценоза, существовавшего ранее на данной территории; 3) заносные (ин-
вазионные), которые принесены различными способами извне (ветром, водой, жи-
вотными и т. д.).
1 Автор И. В Лянгузова
114
Исследование почвенного запаса семян дает материал для более разносторон-
него и глубокого понимания растительного сообщества и прежде всего истории
его развития. При помощи этих данных можно более отчетливо представить
факторы, обусловившие видовой состав и структуру современного растительного
сообщества. Кроме того, можно более обоснованно ответить на вопросы о воз-
можных изменениях в сообществе под влиянием тех или иных причин. Например,
как изменится видовой состав напочвенного покрова в результате антропоген-
ного воздействия (рубок ухода, пожаров, загрязнения окружающей среды и пр.).
Существуют разнообразные методы изучения банка семян в почве (Работнов,
1964, 1982; Петров, Груздева, 1974; Петров, 1989; Baskm, Baskin, 1998 и др.).
Каждый из них имеет как определенные преимущества, так и недостатки.
Основные методы, используемые при изучении почвенного банка семян, мож-
но подразделить на косвенные и прямые. Первые основаны на том, что о запасе
семян в почве судят по некоторым косвенным данным, но не берут почвенные
образцы. Такие данные были получены при изучении семенного возобновления
и зарастания свежих гарей (Комарова, 1986), а также при наблюдениях над появ-
лением всходов на опытных площадках в лесу, с которых удалена подстилка и
самый верхний тонкий слой почвы (Петров, 1977). В этом случае эксперимен-
тальное воздействие стимулирует прорастание семян, содержащихся в почвен-
ном слое. По числу появившихся на площадках всходов можно составить неко-
торое представление о численности и видовом составе семян, образующих поч-
венный банк.
О банке семян в почве можно в известной мере судить также по результатам
эксперимента другого рода, когда на опытных площадках в лесу перекапывают
почвенный слой и затем регистрируют появляющиеся всходы. Этот вид экспе-
риментального воздействия также вызывает прорастание семян, содержащихся
в почве.
Косвенные методы могут дать только самое общее, весьма приблизитель-
ное представление о банке семян. Прямые методы исследования дают более
точные результаты. В этом случае берут образцы почвы из разных слоев (от по-
верхности до известной глубины) и в них определяют содержание семян. Спо-
собы определения запаса семян в образцах различны. В одних случаях семена
специально извлекают из образцов, в других этого не делают.
Для извлечения семян иногда используют какую-нибудь тяжелую жидкость
(например, насыщенный раствор поваренной соли). Образец помещают в такую
жидкость, после чего семена всплывают на поверхность (обычно вместе с раз-
нообразными органическими остатками). Часто выбирают семена из почвенных
образцов вручную. Это очень трудоемко, требует больших затрат времени. Кроме
того, в процессе работы исследователь быстро утомляется и может не заметить
некоторые, даже достаточно крупные, семена. Наконец, очень мелкие, почти не-
различимые для глаза семена выбрать вообще практически невозможно.
115
В дальнейшем с извлеченными из почвы семенами поступают по-разному. Их
либо определяют до вида по внешним признакам и устанавливают жизнеспособ-
ность либо ставят на проращивание. В последнем случае видовую принадлеж-
ность семян определяют по проросткам.
В некоторых случаях почвенные образцы обрабатывают путем промывания их
водой на ситах с очень мелкими ячейками. Часть почвенной массы проходит
сквозь сито, часть вместе с семенами остается. Промывание приводит к значи-
тельному уменьшению объема образца, что облегчает последующую работу с
ним. Затем из остатка, накопившегося на сите, можно тем или иным способом
извлечь семена или поставить весь остаток на проращивание.
К числу методов, не связанных с извлечением семян из почвенных образцов,
относится метод проращивания семян в почве (Рысин, Рысина, 1965; Работ -
нов, 1982; Комарова, 1986; Петров, 1989 и др.). Взятые образцы почвы поме-
щают в условия, благоприятные для прорастания содержащихся в них семян (до-
статочно высокая температура, необходимая влажность и т. д.), а затем прово-
дят наблюдения за появлением всходов. По числу появившихся всходов судят о
запасе семян в почве. Достоинство этого метода состоит в том, что он выявля-
ет действительно живые семена, способные дать всходы. Метод хорош также
тем, что позволяет обнаружить не только более крупные семена, но и самые
мелкие, которые другими способами обнаружить почти невозможно. Недостат-
ком этого метода является очень малая суммарная площадь образцов (обычно
лишь доли квадратного метра). Это в значительной мере снижает репрезентатив-
ность получаемых данных. К числу недостатков относится также явная непол-
нота выявления видового состава и численности семян отдельных видов, т. к. в
почвенных образцах даже в самых благоприятных условиях температуры и влаж-
ности смогут прорасти семена не всех видов растений. Останутся непроросши-
ми, например, твердые семена некоторых бобовых, которым нужна специальная
термическая или химическая обработка. Не прорастут также семена, имеющие
глубокий покой и нуждающиеся в стратификации или специальной предпосевной
обработке. Таким образом, данные, полученные описанным методом, дают пред-
ставление не о всех живых семенах, а только о тех, которые могут прорасти при
определенных условиях проведения эксперимента.
Видовую принадлежность проросших семян устанавливают по появившимся
всходам. Для точного определения вида необходимо, чтобы всход достиг опре-
деленной стадии развития, при которой проявятся видовые признаки растения.
Такой стадии достигают далеко не все всходы. Та или иная часть их всегда поги-
бает в самом начале развития и их не удается определить до вида. Это также
следует отнести к недостаткам метода.
Недостатком метода является и то, что наблюдения за прорастанием семян в
почвенных образцах должны быть очень длительными (не менее двух вегетаци-
онных сезонов). Для работы требуется лабораторное помещение большой пло-
116
щади, чтобы разместить там многочисленные плоские ящики или кюветы с на-
ходящимися в них почвенными образцами.
Остановимся несколько более подробно на методике изучения почвенного
банка семян с помощью проращивания семян в почве.
Почвенный банк семян в каком-либо лесном фитоценозе, как правило, иссле-
дуют на достаточно большом участке площадью 0,1-0,25 га чаще всего на спе-
циально заложенной пробной площади. Подробно описывают растительное со-
общество с указанием обилия и жизненного состояния (образует генеративные
органы или вегетирует) растений всех ярусов, проводят описание почвенного
разреза с определением типа и механического состава почвы. В пределах этой
территории намечают 20 точек исследования, в которых отбирают почвенные
образцы. Точки исследования располагают по возможности равномерно по всей
пробной площади на расстоянии 3-4 м друг от друга в наиболее типичных мес-
тах, т. е. не у самого ствола дерева (на приствольном повышении), вне скопле-
ний побегов кустарников и т. д.
Для взятия почвенных образцов обычно используют специальный металли-
ческий бур определенного сечения. Образцы отбирают по отдельным, довольно
тонким слоям почвы (толщиной 2-5 см), либо по генетическим горизонтам по-
чвы. Чаще всего отбирают образцы до глубины 10 см, т. к. именно здесь во
многих типах леса содержится преобладающая часть семян, а в некоторых ти-
пах — весь почвенный банк семян полностью. Иногда образцы почвы отбира-
ют из более глубоких слоев (13-15, 18-20, 23-25, 28-30 см), чтобы устано-
вить, есть ли семена на соответствующей глубине и в каком количестве они
встречаются.
Дробное исследование почвы по слоям толщиной 2 см дает возможность
детально выявить особенности вертикального распределения семян в почве. А
это важно для суждения о происхождении семян и их возрасте.
Взятые образцы чаще всего сразу же помещают в чашки Петри или деревян-
ные ящики в зависимости от объема почвенного образца таким образом, чтобы
слой почвы не превышал 3-4 см. Сверху ящики прикрывают прозрачной поли-
этиленовой пленкой, хорошо пропускающей свет и вместе с тем препятствую-
щей случайному залету семян. Чашки или ящики с образцами выставляют на рас-
сеянный свет в теплом помещении, т. к. свет стимулирует прорастание покоящих-
ся семян в почвенных образцах. Периодически образцы почвы увлажняют. По-
являющиеся всходы периодически подсчитывают и определяют видовую при-
надлежность.
Наблюдения за появлением всходов в образцах продолжают до тех пор, пока
не перестают появляться новые всходы. Однако чаще всего эти наблюдения
проводят на протяжении двух вегетационных сезонов. Именно такая продолжи-
тельность наблюдений необходима для наиболее полного выявления видового
состава семян в почве и их численности.
117
При исследовании банка семян очень большое значение имеет время взятия
почвенных образцов. Необходимо брать образцы тогда, когда еще не успели
обсемениться растения в текущем году, т. е. в конце весны (конец мая — начало
июня). В этом случае в почвенные образцы попадут семена только прошлых лет,
а не текущего года.
Брать образцы как можно раньше важно и потому, что многие всходы в них
появятся в начале вегетационного периода. Они будут иметь возможность дос-
таточно хорошо развиться за теплый летний сезон, а это очень важно для опре-
деления видовой принадлежности всходов. «Поздние» всходы, появившиеся в
конце лета, развиваются медленно из-за более низких температур и недостатка
света. Многие из них погибают, прежде чем удастся определить их до вида.
Одна из самых больших трудностей при работе по изучению почвенного бан-
ка семян — определение видовой принадлежности всходов. Часто приходится
иметь дело с очень маленькими растениями, имеющими только 2 семядоли и 1-
2 настоящих листа. Определить такое растение до вида нередко довольно труд-
но. Специальных сводок, посвященных морфологии и биологии всходов и юве-
нильных растений, а также определителей растений на ранних этапах их жизнен-
ного цикла крайне мало. Существующими пособиями для определения всходов
(Csapody, 1968; Muller, 1978; Васильченко, 1960, 1979; и др.) по ряду причин не
всегда возможно воспользоваться. Прежде всего, нет никакой гарантии того, что
в данном пособии содержится материал по исследуемому растению. К тому же
пособия обычно довольно узкоспециализированы (определители всходов дере-
вьев и кустарников, определители всходов сорных растений и т. д.). Это тоже зат-
рудняет определение, т. к. зачастую неизвестно точно, с каким растением имеет
дело исследователь. Известную помощь в определении могут оказать рисунки
всходов, которые содержатся в пособиях (Рысина, 1973; Комарова, 1986 и др.).
Но и этот способ не очень надежен, поскольку всходы некоторых растений очень
сходны между собой. Еще один способ определения всходов — использование
гербарных образцов «эталонных» всходов, которые были предварительно выра-
щены из семян точно известного вида растений. Однако и такой способ опреде-
ления не всегда дает нужные результаты. Трудно, например, точно определить до
вида всходы некоторых злаков и осок, когда они находятся на ранних стадиях
развития, в возрасте нескольких недель или месяцев.
В целом же метод проращивания семян в почве, по мнению В. В. Петрова
(1989), дает вполне удовлетворительные результаты. Он довольно полно выяв-
ляет видовой состав и численность семян в почвенном банке (хотя и не с исчер-
пывающей полнотой). Результаты, полученные этим методом для разных лесных
фитоценозов, вполне сравнимы.
118
2.7. МЕТОДЫ ИЗУЧЕНИЯ ЦЕНОПОПУЛЯЦИЙ РАСТЕНИЙ 1
В последние годы при проведении экологического мониторинга стали широ-
ко использовать популяционный подход, который заключается в изучении цено-
популяций отдельных видов растений, слагающих растительное сообщество.
Ценопопуляционный анализ позволяет наиболее полно выявить специфичность
реагирования растений на различные экологические факторы как на уровне от-
дельных особей, так и на уровне популяций.
Ценопопуляция (ЦП) — это совокупность особей одного вида в пределах
однородного экотопа (Петровский, 1961). В связи с тем, что однородный эко-
топ занят одним фитоценозом, границы ценопопуляции совпадают с границами
фитоценоза.
В ценопопуляционных исследованиях используется много признаков, характе-
ризующих ценопопуляцию. Наиболее важными и реактивными из них являются
плотность — количество особей на единице площади, и гетерогенность, т. е.
наличие в составе ценопопуляции особей, различающихся по возрасту и жизнен-
ности (виталитету) (Ценопопуляции..., 1976; Куркин, Матвеев, 1981; Злобин,
1984а, б). Именно эти признаки наиболее полно отражают связь ценопопуляций
растений с экологическими факторами. Для оценки их варьирования в пределах
контуров ценопопуляции используют комплексно-статистический подход (Рысин,
Казанцева, 1975).
Для определения общего проективного покрытия вида, числа особей на еди-
нице площади, продуктивности, возрастного и виталитетного состава особей
ценопопуляции используют метод учетных площадок. Принципы закладки учет-
ных площадок описаны в разд. 1.4. Размер площадок зависит от размеров взрос-
лых особей исследуемого вида и должен быть не меньше суммарной площади
трех минимальных фитогенных полей взрослой особи. Размер фитогенного поля
особи можно установить по наибольшей проекции ее надземных или подземных
частей. Обычно при изучении ценопопуляций закладывают не менее 30-50 учет-
ных площадок.
Выбор счетной единицы. Изучение ценопопуляций растений требует анали-
за образующих их особей, и для растений ряда биоморф очень важен вопрос вы-
бора счетной единицы. Этот вопрос относительно легко решается, если особи
растений генеративного происхождения, т. е. формируются из семян, и четко от-
граничены друг от друга. У растений, возникающих в процессе вегетативного
размножения, за особь принимается система парциальных кустов, связанная меж-
ду собой подземными органами. В таких случаях за счетную единицу принято
1 Автор Е. А. Мазная
119
Таблица 2.1. Некоторые наиболее распространенные морфометрические парамет-
ры особей (Злобин, 1989а)
Статические параметры Условные обозначения Динамические параметры Условные обозначения
Общая фитомасса растения W Абсолютная скорость роста AGR
Фитомасса листьев (репродуктивных органов, корней, стеблей и т. д.) WL(WC, WRd Ws) Абсолютная скорость формирования поверхности листьев AGRa
Фитомасса отдельного листа (плода, семени) WL(WFr, WSm) Относительная скорость роста RGR
Площадь листьев (одного листа) 4 (az.) Относительная скорость формирования поверхности листа RGRa
Число листьев (цветков, плодов, соцветий, боковых ветвей.) N(Nfi, NFr, B) Продолжительность формирования листьев LAD
Высота растения h Продолжительность существования фитомассы BMD
Диаметр стебля d — —
считать отдельный парциальный куст (Злобин, 1961; 19846; Смирнова, 1976;
Куркин, Матвеев, 1981). Существует немалое число видов, у которых первич-
ная особь со временем распадается на вторичные особи. Учет численности этих
видов А. А. Уранов с соавторами (1970) предлагает проводить двумя способа-
ми. 1) учитывать только первичные особи; 2) считать каждую особь, в том чис-
ле вторичную.
Количественные признаки особей. Любая особь растений характеризует-
ся определенным набором качественных и количественных признаков, которые
выступают в качестве параметров ее морфологического статуса. Количествен-
ные признаки используют при исследовании онтогенетических адаптаций, при
оценке жизненного состояния и построении виталитетных спектров.
Количественные признаки, называемые морфометрическими параметрами (по-
казателями), разделяются на 2 группы: статические и динамические (табл. 2.1).
Первые характеризуют особи растений в определенный момент времени, тогда
как вторые оценивают темпы роста и формирования особей и их отдельных ча-
стей за определенный промежуток времени. Статические показатели несут инфор-
мацию о состоянии растений в данный момент времени, они характеризуют про-
шлые уровни активности растений, а следовательно, и условия их существова-
ния. Ценность динамических показателей состоит в том, что они в большей сте-
пени, чем статические параметры, отражают жизненность растений.
120
Таблица 2.2. Периодизация онтогенеза цветковых растений
Период Возрастное состояние растений' Индексы
Латентный Семена sm
Прегенеративный (виргинильный) Проростки (всходы) pl
Ювенильные J
Имматурные im
Виргинильные V
Генеративный Молодые генеративные gl
Зрелые (средние) генеративные g2
Старые генеративные g3
Постгенеративный (сенильный) Субсенильные ss
Сенильные s
Отмирающие sc
Примечание: 1 — При выделении возрастных состояний растений использована систе-
ма А. А. Уранова (1973).
Варьирование размеров особей и их отдельных структурных частей оценива-
ется с помощью коэффициента вариации (см. разд. 4).
Критерии выделения возрастных состояний особей. Возрастное состо-
яние — это совокупность возрастных признаков, характеризующих этап онтоге-
нетического развития особи. Онтогенез особи — это генетически обусловлен-
ная последовательность всех этапов ее развития от возникновения до старения
и отмирания. Полный онтогенез растений подразделяется на 4 периода и 11 воз-
растных состояний (табл. 2.2).
Отнесение растений к тому или иному возрастному состоянию производят на
основании комплекса признаков, таких как способ питания, наличие зародыше-
вых, ювенильных или взрослых структур и их количественные соотношения у
особи, способность к семенному или вегетативному размножению, соотноше-
ние и интенсивность этих процессов, соотношение процессов новообразования
и отмирания у особи, степень сформированности у особи основных признаков
биоморфы (табл. 2.3).
В зависимости от характера жизненной формы качественные признаки возра-
стных состояний имеют специфическое морфологическое выражение, поэтому
для характеристики возрастных состояний могут быть использованы также мор-
фометрические показатели (табл. 2.1), дающие количественную характеристику
ряда морфологических, анатомических, экологических и физиологических при-
знаков. В отличие от качественных признаков морфометрические показатели на
протяжении онтогенеза изменяются непрерывно. Максимальные значения боль-
121
Таблица 2.3. Наиболее общие качественные признаки возрастных состояний осо-
бей (Ценопопуляции..., 1976)
Индекс возрастного состояния Признак
Р1 Смешанное питание (за счет веществ семени и собственной ассимиляции первых листьев), наличие морфологической связи с семенем (или семядолей), наличие зародышевых структур, семядолей; первичного (зародышевого) корня и побега
J Простота организации, несформированность признаков и свойств, присущих взрослым растениям Наличие листьев иной формы и расположения, чем у взрослых особей, иной тип нарастания и ветвления (или отсутствие ветвления побегов), возможно изменение типа корневой системы Сохранение некоторых зародышевых структур (корня, побега), потеря связи с семенами; как правило, отсутствие семядолей
Im Наличие свойств и признаков, переходных от ювенильных растений к взрослым: развитие листьев и корневой системы переходного (полувзрослого) типа, появление отдельных взрослых черт в структуре побегов (например, смена типов нарастания, начало ветвления, появление плагиотропных побегов и т д.) Сохранение отдельных элементов первичного побега
V Появление основных черт типичной для вида жизненной формы Растения имеют характерные для вида взрослые листья, побеги и корневую систему. Генеративные органы отсутствуют
gi Появление генеративных органов Преобладание процессов новообразования над отмиранием, проявляющееся в разных формах В некоторых случаях (полурозеточные формы и др ) окончательное формирование взрослых структур.
g2 Уравновешивание процессов новообразования и отмирания Максимальный ежегодный прирост фитомассы, максимальная семенная продуктивность
gl Преобладание процессов отмирания над процессами новообразования; резкое снижение генеративной функции, ослабление процессов корне- и побегообразования. В некоторых случаях упрощение жизненной формы, выражающееся, в частности, в потере способности к образованию побегов разрастания.
Ss Полное отсутствие плодоношения. Резкое преобладание процессов отмирания над процессами новообразования Возможно упрощение жизненной формы, проявляющееся в смене способа разрастания или потере способности к ветвлению, вторичное появление листьев переходного (имматурного) типа.
s Накопление отмерших (прекративших рост) частей растения. Предельное упрощение жизненной формы, вторичное появление некоторых ювенильных черт организации (форма листьев, характер побегов и др ). В некоторых случаях полное отсутствие почек возобновления и других новообразований.
122
шинства морфометрических показателей, как правило, приходятся на особи, на-
ходящиеся в средневозрастном генеративном состоянии (g2).
Кроме возрастного состояния каждая особь ценопопуляции также может быть
охарактеризована календарным или абсолютным возрастом, т. е. отрезком вре-
мени с момента возникновения растения до момента наблюдения. Однако опре-
деление календарного возраста связано с определенными трудностями и не все-
гда возможно, например, для большинства травянистых растений. Для опреде-
ления календарного возраста деревьев, кустарников и кустарничков разработаны
специальные анатомические и морфологические методы.
В каждом возрастном состоянии растение пребывает в течение определенно-
го отрезка времени, при этом в пределах ценопопуляции возможно существова-
ние особей разного календарного возраста и одинакового возрастного состоя-
ния и наоборот. Сопоставление календарного возраста особей и их возрастного
состояния дает возможность установить темпы развития особей, т. е. период пре-
бывания особи в одном возрастном состоянии и время прохождения жизненно-
го цикла в целом, что представляет особый интерес при экологических исследо-
ваниях. Однако для ценопопуляционных исследований определение возрастного
состояния растений имеет большее значение, чем установление календарного
возраста, т. к. позволяет провести сравнительную оценку роли в сообществе осо-
бей как одного, так и разных видов.
Возрастной состав ценопопуляций. Распределение особей в ценопопуля-
ции по возрастным состояниям называется возрастным спектром или спект-
ром возрастных состояний. Спектр может быть выражен в абсолютных числах
или процентах от общего числа особей и представлен в виде таблицы, гистог-
раммы или графика.
По соотношению разных возрастных групп растений различают 3 типа ценопо-
пуляций: инвазионную, нормальную и регрессивную, — которые характеризуют
позицию и жизненность вида в среде исследуемого фитоценоза (Работнов, 1950).
Инвазионная ценопопуляция — это ценопопуляция, не способная к самопод-
держанию и зависимая от заноса зачатков извне. Такая ценопопуляция состоит
преимущественно из молодых (виргинильных) особей: 1) только из семян, зане-
сенных извне; 2) из семян и проростков; 3) из семян, проростков, имматурных и
виргинильных растений. Гистограмма возрастного спектра таких популяций обыч-
но имеет левостороннюю асимметрию.
Нормальная ЦП способна к самоподдержанию семенным или вегетативным
путем, либо тем и другим одновременно. В возрастном спектре таких ЦП содер-
жатся особи всех возрастных состояний (ЦП нормальная, полночленная') или
особи какого-либо возрастного состояния отсутствуют (ЦП нормальная, непол-
ночленная). Неполночленность ЦП может быть следствием влияния экзогенных
(внешних по отношению к ЦП) и эндогенных (биологических свойств вида) фак-
торов.
123
Регрессивная ЦП имеет повышенную долю субсенильных и сенильных рас-
тений при почти полном отсутствии виргинильных особей и не способна к само-
поддержанию. Такие ЦП недолговечны и со временем полностью выпадают из
сообщества. В гистограммах возрастных спектров таких популяций отмечается
правосторонняя асимметрия.
Различные стрессовые воздействия (например, атмосферное промышленное
загрязнение) на ценопоцуляции вполне четко проявляются в характере возраст-
ных спектров. Они становятся неполночленными и/или правосторонними.
Анализ численности особей различного возрастного состояния позволяет
определить индекс возрастности (коэффициент возрастности) ценопопуляции,
который рассчитывается по формуле:
где М— численность всей популяции, т, — численность конкретной возрастной
группы, к, — коэффициент возрастности конкретной возрастной группы:
кр = 0,0067, к, = 0,018, kim = 0,0474, kv = 0,1192, kg\ = 0,27, £gl_2 = 0,38, kg2 = 0,5,
kg2-3 = 0,62, kg3 = 0,731, kss = 0,8808, ks = 0,9819.
Показатель индекса возрастности изменяется от 0 до 1, при этом чем выше
его значение, тем старее ценопопуляция (Жукова и др., 1994).
Распределение особей ценопопуляции по календарному возрасту называется
возрастной структурой и так же, как возрастной спектр, может быть выраже-
но в абсолютных числах или процентах от общего числа особей и представлено
в виде таблицы, гистограммы или графика.
Оценка жизненности особей. Виталитетная структура ценопопуляций.
В результате изучения онтогенеза различных видов установлено, что особи од-
ного и того же возрастного состояния могут находиться на разных уровнях жиз-
ненности. Существуют различные способы оценки жизненного состояния осо-
бей, основанные на количественных признаках, реже используются качественные
признаки. Для наиболее информативных признаков состояния особей характерно
наличие высоких значений коэффициентов корреляции между функционально
значимыми параметрами. Для выявления таких признаков выборка должна быть
репрезентативной и включать в себя особи, находящиеся в одном и том же воз-
растном состоянии. Чаще всего для ценопопуляций нормального типа проводят
отбор особей средневозрастного генеративного состояния (g2). При первичном
изучении вида следует использовать не менее 10 статических и динамических
параметров. Значения морфометрических, ростовых и продукционных парамет-
ров каждой отдельной особи заносят в матрицы исходных данных. С использо-
ванием пакета статистических программ вычисляют матрицу коэффициентов кор-
реляции, в которой отмечаются значимые коэффициенты корреляции. Согласно
величине коэффициентов корреляции, ранжируют признаки по их информативно-
124
сти и выделяют наиболее значимые (ключевые), которые используют для оцен-
ки виталитета ценопопуляции в целом. На основе полученных данных строят кор-
реляционные плеяды. Набор ключевых признаков, используемых для оценки ви-
талитета особи, различен для разных видов и разных экологических ситуаций, но
наиболее часто используются фитомасса и площадь листовой поверхности.
Оценку виталитета ценопопуляций можно проводить одномерным и многомер-
ным способами (Злобин, 1989 а, б), когда ранжирование особей осуществляют
по одному или нескольким признакам.
Для определения виталитета ценопопуляции используют выборки из 25-50 ре-
зультатов измерений растений, ранжированных от минимального к максимально-
му значению. Простое расчленение ранжированного ряда на три равных интерва-
ла недопустимо, т. к. ряды распределения часто бывают асимметричными. Осо-
би группируют по трем классам виталитета: А — высший, В — промежуточный
и С— низший (Злобин, 1989 б). При отнесении особей к тому или иному классу
виталитета используют формулу х ± Sx-t, где х — среднее арифметическое,
5) — стандартная ошибка и t — стандартное значение критерия Стьюдента при
р = 0,05. Находят а — число особей, которые попадают в интервал более х + Sx-t
(высший класс виталитета), b — в интервал х ± Sxt (промежуточный), с — в
интервал х - Sx t (низший). Следует отметить, что более корректно вычисление
среднего арифметического по общей совокупности всех выборок всех изучае-
мых ценопопуляций. Исходя из значений классов виталитета, рассчитывают ин-
декс жизненности Q = (а + Ь) / 2.
По соотношению встречаемости в ценопопуляции особей разных классов ви-
талитета оценивают уровень ее жизнеспособности в конкретных условиях оби-
тания. Обычно результаты представляют в виде гистограммы: по оси абсцисс от-
кладывают классы виталитета, а по оси ординат — относительные частоты встре-
чаемости каждого класса. Затем оценивают симметричность виталитетных (раз-
мерных) спектров: спектр с правосторонней (положительной) асимметрией
((а + Ь)/2 < с) является признаком деградирующей, симметричный спектр
((а + Ь)/2 = с) соответствует равновесной, а спектр с левосторонней (отрица-
тельной) асимметрией ((а + Ь)/2 > с) — процветающей ценопопуляции, соот-
ветственно (рис. 2.6).
Пространственная структура ценопопуляций. Типы размещения особей по
площади обычно подразделяют на три группы: случайное, регулярное и контаги-
озное (пятнистое, групповое) (рис. 2.7) Существует два методических подхода
к анализу распределения особей в пределах ценопопуляции: 1) без учета поло-
жения площадки в пространстве; 2) с учетом положения площадки в простран-
стве (Василевич, 1972). Первый подход является наиболее распространенным ва-
риантом анализа пространственной структуры ценопопуляции. В этом случае по-
лученное эмпирическое распределение какого-либо показателя сравнивают с оп-
ределенным теоретическим распределением (Пуассона, биноминальным (бино-
125
0,8
Рис. 2.6. Виталитетный спектр ценопопуляций Vaccinium myrtillus L. в сосняках
лишайниково-кустарничковых, расположенных на различном расстоянии от
комбината «Североникель» (Кольский п-ов)
Ключевой признак — высота парциального куста. По горизонтали — классы виталитета:
А — высший; В— промежуточный; С— низший. По вертикали — частота встречаемости
класса
Рис. 2.7. Типы размещения особей по
площади (Бигон и др., 1989)
I — случайное; II — регулярное; III — кон-
тагиозное
миальным), нормальным и др.). Более
подробно эти распределения рассмот-
рены в разд. 4. Для анализа размещения
особей по площади в ценопопуляциях с
высокой плотностью особей наиболее
часто проводят сравнение с нормаль-
ным йли биномиальным распределени-
ем. Определение меры диссонанса
(Миркин, Розенберг, 1978) является
более общим тестом при анализе про-
странственной структуры ценопопуля-
ций. Мера диссонанса изменяется от 0
до 1. Чем меньше его величина, тем
ближе размещение вида к регулярному.
126
2.8. КОЛИЧЕСТВЕННАЯ ОЦЕНКА БИОРАЗНООБРАЗИЯ 1
Разнообразие — это одна из фундаментальных характеристик биологических
систем. Различают альфа-, бета- и гамма-разнообразие (Whittaker, 1977). Аль-
фа-разнообразие — это показатель сложности видового состава сообщества. Оно
может быть измерено числом видов на единицу площади или оценено по соот-
ношению долей участия видов. Бета-разнообразие — показатель, измеряющий
степень дифференцированности распределения видов по градиентам местооби-
таний, т. е. скорость изменения видового состава фитоценозов вдоль простран-
ственного или экологического градиента. Мерой оценки бета-разнообразия мо-
жет служить число установленных синтаксонов одного ранга (ассоциаций, субас-
социаций и т. д.). Гамма-разнообразие — показатель разнообразия растительно-
сти ландшафта, объединяющий альфа- и бета-разнообразие. Простейшим по-
казателем гамма-разнообразия является конкретная флора (набор видов в пре-
делах одного ландшафта) (Миркин и др., 1989). Для количественной оценки раз-
нообразия используют большое количество различных показателей, коэффици-
ентов и индексов, отражающих те или иные свойства сообществ (Миркин и др.,
1989; Мэгарран, 1992). Наиболее распространенными являются число видов,
индекс Шеннона — показатель общего разнообразия, индекс Симпсона — по-
казатель доминирования, индекс Пилу — показатель выровненное™. Все назван-
ные характеристики существенно зависят от площади, для которой они опреде-
лены (Грейг-Смит, 1967; Василевич, 1969; Мау, 1975; Одум, 1975; Баркман, 1989;
Мэгарран, 1992), поэтому необходимо всегда указывать, на какой площади оце-
нивали число видов или производили анализ индексов.
Одной из главных характеристик разнообразия является число видов (плот-
ность видов, видовое богатство) (N) — это наиболее понятная, непосредствен-
но измеряемая характеристика, не требующая сложных расчетов. Число видов
наиболее часто и широко используется при оценке разнообразия как отдельных
сообществ, так и для сравнения различных сообществ.
Большинство используемых для оценки разнообразия индексов рассчитыва-
ется на основе анализа долей участия видов в растительных сообществах. Ме-
тодика измерения характеристик компонентов лесных сообществ подробно рас-
смотрена в разд. 1.6.
2.8.1. Индексы разнообразия
Индекс Шеннона (Н) — энтропия (мера хаотичности) распределения отно-
сительных покрытий видов. Этот индекс используется для оценки общего раз-
1 Авторы И. Ю. Баккал, В. В Горшков
127
нообразия, т. е. для сравнения сообществ по числу входящих в них видов и по
равномерности распределения долей участия между видами. При этом виды,
имеющие незначительную долю участия оказывают большее влияние на вели-
чину индекса, чем доминанты. При одинаковом числе видов максимальная энт-
ропия (и, следовательно, максимальное значение индекса) наблюдается при ра-
венстве долей участия всех видов. С увеличением числа видов значения индек-
са увеличиваются.
W=-^5, Ins,, (1)
где 5, — доля участия /-го вида в формировании общего покрытия яруса, s, = c/ctol,
с, — проективное покрытие /-го вида: с;=а,/ао • 100 %, а, — площадь проекции
/-го вида, до — размер единичной анализируемой площади, ctot — общее проек-
тивное покрытие яруса: с,„, = , N— общее число видов на анализируемой пло-
щади.
Индекс Пилу (£) — показатель выровненности. Так же, как и индекс Шен-
нона, он характеризует распределение долей участия видов, но позволяет срав-
нивать видовое разнообразие сообществ, включающих в себя разное количество
видов. Например, величина индекса будет одинакова для сообществ, в которых
доли участия распределены поровну между двумя и двадцатью видами. Индекс
изменяется от 0 до 1. Его максимальное значение наблюдается при равенстве
долей участия всех видов.
Е = = 1п " ’ (2)
где Н— индекс Шеннона, рассчитываемый по формуле (1); N— общее число
видов на анализируемой площади.
Индекс Симпсона (5) — показатель доминирования. Его применение целе-
сообразно для выявления степени доминирования одного или нескольких видов
в сообществе. Наибольшее влияние на величину индекса оказывают виды, име-
ющие наибольшее относительное покрытие. При заданном числе видов значе-
ния индекса будут максимальны в случае доминирования только одного вида и
близких к нулю долях участия всех остальных видов.
S = t(s,y (3)
См. обозначения к формуле (1).
2.8.2. Коэффициенты сходства участков растительного покрова
Существует ряд коэффициентов, позволяющих оценить сходство различных
участков растительного покрова в пределах конкретного сообщества (или проб-
ной площади). В их число входят коэффициент сходства видового состава Жак-
кара (J1), количественный коэффициент Жаккара (УЕ 2 * * S), расстояние Евклида (£>е).
Расчет этих коэффициентов основан на попарном сравнении всех описаний (пло-
128
щадок). Перечисленные коэффициенты используют для оценки пространствен-
ного разнообразия, т. е. для выявления горизонтальной неоднородности яруса в
пределах конкретного сообщества (или пробной площади).
Коэффициент сходства видового состава Жаккара (J1) сравнивает между
собой участки растительного покрова по наличию одних и тех же видов без учета
их покрытий:
N
J' =------—------- (4)
na + nb-nmb’
где Naub — число видов общее для двух сравниваемых описаний (площадок) А
и В, Na, Nb — число видов соответственно в описаниях А и В.
Количественный коэффициент Жаккара (J2) и расстояние Евклида (£>с)
позволяют сравнить между собой участки растительного покрова по видовому
составу и проективному покрытию видов.
/ 2 = -°'____________ (->)
J N
У,(с,а +с,„ - min[c„,c4])
=Jix - с,,/ - . (б)
V i=J
где cla, Cib — проективные покрытия г-го вида в сравниваемых описаниях а и Ь.
Поскольку расстояние Евклида De, как следует из формулы (6), зависит от чис-
ла видов, участвующих в формировании покрытия компонента растительного по-
крова, то используется также нормированное расстояние Евклида .О"' "", величи-
на которого от числа видов не зависит:
2^ (с,« — )
цюгт _ л | =|____ ( / )
V
Несмотря на то, что количественный коэффициент Жаккара и расстояние Ев-
клида основаны на анализе одних и тех же характеристик растительного покрова
(водового состава и покрытия), результаты, полученные при их использовании в
качестве меры сходства, могут быть различными.
Рассмотрим это на примере сравнения двух пар описаний участков раститель-
ного покрова («1^1 — й2^2 и аз^з — ацЬц) (рис. 2.8). Количество осей на графи-
ке соответствует максимальному количеству видов в сравниваемых описаниях.
В нашем случае их две — ось покрытий вида «а» и ось покрытий вида «Ь». Ви-
довой состав на сравниваемых участках одинаков, разница в покрытиях видов а
иЬв обоих случаях составляет 6 %. Коэффициент Жаккара, вычисляемый по фор-
муле (5), для первой пары описаний равен J2 = 0,2; для второй пары опи-
саний ^2=10 ~^~ 0,45. Исходя из полученных значений коэффициента можно
сделать вывод, что описания а (6, и <72^2 имеют меньшее сходство в видовом
129
Проективное покрытие, %
Рис. 2.8. Схематичное сравнение 2 пар
участков растительного покрова с
использованием количественного
коэффициента сходства Жаккара и
расстояния Евклида (на примере
2-хмерного пространства)
Количественный коэффициент Жаккара
для пары а\Ь\ — a^b-i равен ,Р- = 0,2; для
пары ауЬу —a^b^.J2 = 0,45. Расстояния
Евклида De для обеих сравниваемых пар
составляют 8,4 %. Различия обусловле-
ны положением сообществ по отношению
к началу осей координат
составе и покрытии видов, чем описания
аз^з и ацЬц. Теперь проанализируем сход-
ство пар описаний, используя величину
расстояния Евклида. Эта величина равна
длине отрезков а\Ь\ — и <2363 —
Я4^4 и одинакова для обеих пар сравнива-
емых описаний (De ~8,4 %). Различия в
полученных результатах не несут биоло-
гического смысла и объясняются лишь
положением точек описаний по отноше-
нию к началу осей координат.
Эти примеры говорят об определенной
условности и некоторой степени необъек-
тивности результатов, полученных при
использовании различных коэффициентов.
Желание найти критерий, наилучшим обра-
зом отражающий реально наблюдающие-
ся условия, объясняет большое количе-
ство предлагаемых мер для оценки степе-
ни сходства объектов растительного по-
крова.
2.9. МЕТОДЫ ИЗУЧЕНИЯ МОХОВОГО ПОКРОВА 1
Моховой покров является одним из характерных компонентов бореальных
лесных сообществ. Ценотическая роль мохового покрова в лесных экосистемах
определяется его средообразующей способностью. Низкая теплопроводность
и высокая влагоемкость мохового покрова оказывают непосредственное влия-
ние на режим влажности и температуры, а также на многие химические процес-
сы в верхних горизонтах почвы. Отмирающие части мхов играют существенную
роль в формировании биогенного горизонта лесных почв — лесной подстилки.
Высота и плотность мохового покрова влияют на процессы семенного возоб-
новления растений в бореальных лесах.
Отличительной особенностью мохообразных является способность к фото-
синтезу при крайне низкой интенсивности фотосинтетически активной радиации
(ФАР), что определяет особую ценотическую и экологическую роль мохового
покрова в лесных сообществах с высокой сомкнутостью древесного яруса (Кан-
нукене, 1971).
1 Автор Е. Н. Андреева
130
Значительная роль мохового покрова в организации бореальных лесных со-
обществ определяет актуальность детальных исследований его состава, струк-
туры и динамики, а также экологических и индикационных характеристик.
2.9.1. Жизненные формы мхов и печеночников
Долгое время мхи рассматривались как единая жизненная форма, причем по-
нятия «жизненная форма» и «форма роста» первоначально не разделялись. Впер-
вые эти понятия были разделены в работах К. Magdefrau (1969, 1982), который
рассматривает жизненную форму мхов как форму роста группы особей, изменен-
ных внешними условиями в процессе онтогенеза. Однако, по нашему мнению,
следует учитывать и обратный процесс, поэтому жизненную форму мхов следу-
ет рассматривать как целостное множество взаимосвязанных особей, характери-
зующееся устойчивым развитием, сформированное под влиянием внешних ус-
ловий и обладающее, в свою очередь, высокой средообразующей способнос-
тью (Андреева, 1989, 1990а, б).
В большинстве случаев мохообразные имеют клоповую организацию, и только
незначительное число видов произрастает в виде совокупности отдельных осо-
бей. Прорастая, спора образует протонему, на которой формируется несколько
гаметофитов. Исследования показали, что в результате одновременно действу-
ющих процессов роста и отмирания лесных мхов в ненарушенных сообществах
толщина мохового покрова остается на протяжении длительного времени прак-
тически постоянной. Благодаря обильному ветвлению и последующему обособ-
лению побегов в самостоятельные особи (из-за перегнивания нижней части мха),
в моховом покрове не наблюдается изреживания при отмирании угнетенных осо-
бей. Кроме того, стебли мхов, полегающие под действием снега и талых вод,
через некоторое время дают начало многочисленным новым побегам. Таким
образом поддерживается целостность моховой дернины.
При определении жизненной формы внимание традиционно акцентировалось
на морфологических особенностях. Однако подробные исследования показали,
что не меньшее значение имеют анатомо-морфологические признаки (мощность
ризоидного войлока, длина гиалинового волоска, толщина клеточных стенок, на-
личие проводящих элементов и т. и.), находящиеся в тесной связи с условиями
местообитания, а также физиологические (способность к анабиозу, обезвожива-
нию и т. д.), биологические (тип жизненной стратегии) и ценотические (плотность
и высота дерновинок).
В табл. 2.4 представлена оригинальная система жизненных форм печеночни-
ков и мхов, разработанная и применяемая автором в течение десяти лет (Андре-
ева, 1989, 1990а). Всего выделено 11 типов жизненных форм, которые сгруппи-
рованы в шесть подгрупп, часто конвергентных по внешнему облику, но образо-
ванных разными классами мохообразных. Исключение составляют шнуровидная
и подушковидная жизненные формы: первая характерна только для печеночных
131
Таблица 2.4. Система жизненных форм лесных мхов
Название жизненной формы, представители Краткая характеристика Условия существования
1. Перинообразная (ПМ): Ptilidium ciliare, Plagiochila asplenioides, Trichocolea tomentella 2 Перинная (ЛМ): Hylocomium splendens, Pleurozium schreberi, Dicranum polysetwn рыхлые, высокие дерновинки, ФР — высокодерновинная (tall turjs), деревцевидная (dendroids); ТЖС—V ненарушенная лесная подстилка
3. Шнуровидная (ПМ). Cephalozia sp., Cephaloziella sp, Lophozia sp система тонких пучков между стебельками мхов; ФР — высокодерновинная (tall turfs); ТЖС—Р в толще дерновинок листостебельных мхов
4. Подушковидная (ЛМ). Polytrichum sp., Sphagnum sp., Dicranum sp. куполообразные уплотненные дерновинки; ФР — высокодерновинная (tall turfs), ТЖС —V лесная подстилка, торф
5 Войлочная (ПМ): Jamesoniella autumnalis, Jungermannia leiantha 6 Плетевидная (ЛМ). Brachythecium sp 7 . Плюшевая (ЛМ). Fissidens sp , Orthodicranum sp уплотненные невысокие дерновинки; ФР — сплетения (wefts); короткодерновинная (short turjs), ТЖС — VP трещины и щели между камнями, гнилая древесина, кора деревьев
8 Губчатовидная (ПМ): Barbilophozia barbata, Plagiochila porelloides, 9 Губчатая_(ЛМ): Orthotrichum sp., Racomitrium sp , Schistidium sp. полусферические рыхлые дерновинки; ФР — подушечная (cushions), ТЖС — Уэк. валуны, деревья, валеж
10 Корковая (ПМ): Cephaloziella sp , Scapania sp 11. Щеткообразная (ЛМ): Bryum argenteum, Funaria hygrometrica, Ceratodon purpureus уплощенные плотные или рыхлые дерновинки; ФР — короткодерновинная (short turfs)' ТЖС — Е пятна минерального грунта различного происхождения
Примечания: ПМ — печеночные мхи; ЛМ — листостебельные мхи; ФР — форма ро-
ста; ТЖС — тип жизненной стратегии: V — бриовиоленты, Р — бриопатиенты, Е — бри-
оэксплеренты, Уэк. — бриовиоленты, зависимые от экотопа. Английские термины форм
роста даны согласно работе К. Magdefrau (1982).
132
мхов, а вторая встречается исключительно у листостебельных мхов. Что же касает-
ся плетевидной и плюшевой жизненных форм, то они образованы двумя разными
группами листостебельных мхов и Обе конвергентны войлочной жизненной форме.
1. Перинообразная жизненная форма характерна для листостебельных мхов.
Отличается слабой средообразующей способностью и локальным распростра-
нением. Синузия этой жизненной формы (Plagiochila asplenioides, Trichocolea
tomentella) характерна для неморальных ельников, а перинообразная синузия
Ptilidium ciliare — характерный элемент северотаежных лесов (особенно ли-
ственничников), а также распространена в лесотундре.
2. Перинная жизненная форма листостебельных мхов распространена в боре-
альных лесах. Наличие хорошо развитой синузии перинной жизненной формы,
формируемой Pleurozium schreberi, Hylocomium splendens, Dicranum polysetum,
отражает устойчивое (ненарушенное) состояние лесного сообщества. Площадь
синузии может достигать нескольких десятков и даже сотен квадратных метров,
высота обычно колеблется от 20 до 40 (70) см и зависит как от сезона, так и от
климатических особенностей года. При внешних воздействиях на лесные сооб-
щества состав этой синузии меняется от олиго- или монодоминантного до поли-
доминантного. В случае разрушения лесного сообщества (вырубка, пожар, вет-
ровал и пр.) синузия этой жизненной формы деградирует: сначала резко сокра-
щаются ее размеры, а затем виды, слагающие синузию, либо интегрируются в
состав синузии другой жизненной формы, либо исчезают.
3. Шнуровидная жизненная форма печеночных мхов существует в виде сис-
тем тонких пучков, развивающихся между побегами листостебельных мхов, чаще
всего подушковидной жизненной формы. Между этими двумя жизненными фор-
мами существуют своего рода симбиотические отношения: стебельки печеноч-
ных мхов соединяют стебельки листостебельных мхов, увеличивая плотность дер-
новинки подушковидной жизненной формы, которая, в свою очередь, формиру-
ет среду для существования печеночных мхов, обеспечивая им сохранение вла-
ги и механическую опору.
4. Подушковидная жизненная форма листостебельных мхов характеризуется
плотным расположением стебельков и способностью к формированию наноре-
льефа в виде крупных кочек или гряд на болотах. Виды, образующие синузию
подушковидной жизненной формы (Polytrichum strictum, Sphagnum fuscum), ха-
рактеризуются высокой чувствительностью к антропогенным воздействиям.
5. Войлочная жизненная форма характерна для эпиксильных печеночников
(Jamesoniella autumnalis, Jungermannia leiantha), развивающихся на валеже и
опаде в условиях повышенной влажности, преимущественно в сфагновых ель-
никах и сырых мелколиственных лесах. Синузия этой жизненной формы может
встречаться и в других типах лесных сообществ, но только в микроместообита-
ниях с повышенным уровнем увлажнения (ямы, микропонижения, берега водо-
емов и пр.).
133
6. Плетевидная жизненная форма образуется преимущественно эпифитными
и эпиксильными видами листостебельных мхов, имеющими плагиотропный рост
(Brachythecium sp.). В некоторых случаях ее могут формировать виды, которые
обычно образуют перинную жизненную форму (Pleurozium schreberi,
Hylocomium splendens).
7. Плюшевая жизненная форма так же, как плетевидная, формируется листо-
стебельными мхами, но с ортотропной формой роста. Среди мхов, образующих
плюшевую жизненную форму, встречаются эпифитные (Orthodicranum
montanum), эпиксильные (Orthodicranum flagellare), эпигейные (Fissidens
osmundoides), эпилитные (Paraleucobryum longifolium) и копрофильные
(Splachnum rubrum) виды. Особенностью синузии этой жизненной формы явля-
ется то, что она всегда распространена локально.
8. Губчатовидная жизненная форма образуется видами печеночных мхов, спо-
собных к криптобиозу. Это могут быть эпигейные виды (Barbilophozia barbata),
но чаще эпилитные виды (Plagiochila porelloides), произрастающие в условиях
эпизодического увлажнения (например, на местах временных водотоков).
9. Губчатая жизненная форма листостебельных мхов характеризуется рых-
лым расположением стебельков, что способствует как быстрому поглощению
воды, так и быстрому обезвоживанию. Виды мхов этой жизненной формы
(Orthotrichum-sp., Racomitrium sp.) относятся к пойкилоксерофитам и способ-
ны во время полного обезвоживания находиться в состоянии криптобиоза, при
котором практически прекращается метаболизм. Эта жизненная форма обладает
значительной толерантностью к изменениям условий обитания. Следует заметить,
что морфологическое сходство дерновинок может привести к смешению двух
различных жизненных форм. Так, К. Мэгдефрау (Magdefrau, 1982) по сути объе-
динил в жизненной форме "cushions" две жизненные формы: подушковидную и
губчатую.
10. Корковая жизненная форма характерна только для печеночников. Части-
цы субстрата настолько тесно включены в архитектуру этой жизненной формы,
что практически невозможно отделить побеги печеночников (Cephaloziella sp.,
Frullania dilatata) от песчинок или коры деревьев. Эту жизненную форму мож-
но назвать стресс-толерантной, т. к. она способна выдерживать высокие концен-
трации химических элементов и сильную инсоляцию.
11. Щеткообразная жизненная форма листостебельных мхов аналогична кор-
ковой, т. к. ризоиды на стебельках верхоплодных мхов тесно оплетают элемен-
ты субстрата. Уникальная структура, образованная минеральными частицами по-
чвы и побегами мхов, создает возможность капиллярного тока, что позволяет
синузиям данной жизненной формы (Polytrichum piliferum, Bryum argenteum)
быть пионерами при зарастании открытых нарушенных субстратов.
Часто можно- наблюдать, что один и тот же вид в зависимости от условий
местообитания образует различные жизненные формы. Так, например, виды пе-
134
ченочных мхов шнуровидной жизненной формы (Mylia, Cladopodiella, Lophozia
и др.), обитающие между стебельками листостебельных мхов из родов Sphagnum
или Polytrichum, при уменьшении влажности образуют корковую жизненную
форму на сильно угнетенных дерновинках листостебельных мхов. В случае бла-
гоприятного изменения погодных условий происходит изменение их состояния:
листостебельный мох начинает интенсивно расти, образуя подушковидную жиз-
ненную форму, а печеночные мхи, соответственно, формируют шнуровидную
жизненную форму.
2.9.2. Типы жизненных стратегий мхов и печеночников
Типы жизненных стратегий по системе Раменского-Трайма отражают адапта-
цию растений к условиям местообитания (Миркин, Наумова и др., 2000). К пер-
вичным стратегиям (ценобиотические типы по Раменскому) относятся виолен-
ты (И), патиенты (Р) и эксплеренты (Е). Бриовиоленты являются эдификаторами
моховых сообществ, обладают сильно выраженным вегетативным ростом и
умеренным спороношением. Бриопатиенты способны существовать при слабой
обеспеченности ресурсами в условиях, создаваемых бриовиолентами и выдер-
живают сильную конкуренцию. Эти мхи характеризуются слабым вегетативным
ростом и спорадическим спороношением. Бриоэксплеренты способны быстро
захватывать территорию в нарушенных сообществах. Для них характерно обиль-
ное спороношение и наличие специальных органов вегетативного размножения.
В природе один и тот же вид может проявлять свойства разных типов стратегий;
в этом случае говорят о переходном типе стратегии (VP, VE, РЕ, ЕРЕ). Особен-
ностью мхов является их экологическая зависимость от определенных субстра-
тов, что привело к необходимости выделения помимо типичных стратегий их
экотопных вариантов (Иэк, Рэк, Еэк), например, эпифиты, эпилиты, эпиксилы и т. д.
Подробная характеристика южнотаежной бриофлоры по типам жизненных стра-
тегий приведена в работе Е. Н. Андреевой (2001).
2.9.3. Измерение характеристик мохового покрова
Изучение характеристик мохового покрова начинается с определения его об-
щего проективного покрытия и покрытия отдельных видов. Для определения
проективного покрытия используется метод учетных площадок, которые закла-
дывают в регулярном или случайном порядке вдоль параллельных трансект (см
также разд. 1.4, 1.6). Работа проводится с помощью рамки размером 20 х 50 см
(или другого подходящего размера) с масштабной сеткой. На каждой площадке
отмечают также высоту мохового покрова в сантиметрах (желательно отдельно
измерить высоту живого и мертвого слоев).
Второй этап — это выявление типов бриосинузий и определение характерис-
тик формирующих их видов. Синузия, согласно представлениям Т. М. Липпмаа
(Lippmaa, 1938; Липпмаа, 1946) и X. X. Трасса (1964, 1970), — это структурно
135
Таблица 2.5. Форма интегрального описания мохового покрова
Название синузии Параметры сииузии Тип местообитания Жизненная форма Тип жизиеииой стратегии Состав видов
площадь высота
и экологически обособленный элемент фитоценоза, более или менее гомоген-
ный по составу жизненных форм. Бриосинузия представляет собой участок мо-
хового покрова однородный по видовому составу и структуре. Эдификаторная
значимость отдельных видов в бриосинузии определяется величиной проектив-
ного покрытия и типом жизненной стратегии.
Рекомендуемая форма описания бриосинузий представлена в табл. 2.5.
Важной характеристикой при описании бриосинузий является тип местообита-
ния. И. Д. Кильдюшевский (1973) предложил вариант системы обозначения ти-
пов местообитаний бриосинузий, который может быть использован при компью-
терной обработке данных и обеспечивает сопоставимость результатов разных
исследователей. Каждый тип местообитания в этой системе обозначен числовым
индексом — двузначным (в более простых случаях) или трехзначным числом.
При детальном исследовании состава и структуры мохового покрова измеря-
ют следующие показатели для отдельных видов мхов: плотность побегов, вы-
сота (желательно отдельно измерить толщину живого и мертвого слоев), жиз-
ненное состояние, интенсивность спороношения.
Жизненность и интенсивность спороношения — важные критерии для оп-
ределения оптимума местообитания вида. Жизненность моховой дерновинки оце-
нивают по четырехбалльной шкале: крайне слабая (0), когда некроз и хлороз по-
бегов в пределах дерновинки достигают 70-90 %, но при благоприятных усло-
виях часть побегов все еще способна к регенерации; ослабленная (L), когда не-
кроз и хлороз побегов не превышают 50 %; нормальная (N), когда возможно про-
явление некротического или хлоротического повреждения отдельных побегов,
и высокая (Н), для которой характерно отсутствие повреждений.
В жизненном цикле мохообразных доминирует гаплоидная фаза, то есть раз-
меры и продолжительность развития гаметофита намного превышают аналогич-
ные характеристики спорофита, который, в сущности, паразитирует на гаметофи-
те. Изменение нормального жизненного цикла мохообразных в сторону увели-
чения значимости спорофита и ослабления гаметофита есть механизм приспособ-
ления к изменяющимся условиям и выживанию в экстремальных ситуациях
(Andrejeva, 1990; Ярмишко и др., 1998).
Интенсивность спороношения также можно оценивать по четырехбалльной
шкале: отсутствие спорогонов (0s), слабое спороношение (1s) — менее 10-20 %
спороносящих особей, нормальное спороношение (Ns) — от 20 до 40 % споро-
136
носящих особей, обильное спороношение (Ms) — более 40-50 % споронося-
щих особей. Интегральным показателем, характеризующим развитие мохового
покрова в целом, служит индекс спороношения (Андреева, 1989, 1990а)
Is = S / п, где S — число видов со спорогонами, п — общее число видов.
2.9.4. Особенности учета видового состава мхов и печеночников
на пробных площадях
До начала исследования видового состава мхов и печеночников в лесных
сообществах конкретного района необходимо составить предварительный спи-
сок видов мхов, используя имеющиеся литературные и гербарные материалы по
изучаемой территории. В такой предварительный список следует включать лишь
те виды, которые исследователь хорошо себе представляет. Кроме того, для
работы в полевых условиях рекомендуется составить справочный гербарий в виде
небольших образцов мхов, наклеенных на страницы полевого дневника.
При выполнении геоботанических описаний необходимо, по возможности,
максимально полно выявить видовой состав мхов, включая эпифиты, а также
виды, произрастающие на гниющей древесине (эпиксилы) и прочие. Для этого
рекомендуется изучить территорию всей пробной площади, а не ограничиваться
только обследованием заложенных учетных площадок. Это значительно облег-
чает определение видов при дальнейшей камеральной обработке и особенно
важно для мхов, имеющих незначительное обилие на учетных площадках, т. к.
исследование всей территории пробной площади увеличивает вероятность встре-
тить этот вид со всеми признаками, необходимыми для достоверного определе-
ния. Кроме того, это будет способствовать обнаружению спороносящих особей,
более объективной характеристике жизненных форм мхов в разнообразных эко-
топах, а также выявлению амплитуды экологической и морфологической вариа-
бельности видов. В некоторых случаях полезно обследовать и пограничную с
пробной площадью территорию.
В связи с тем, что разные виды мхов имеют различные сроки вегетационного
периода, для установления полного видового состава желательно проводить
инвентаризацию два или три раза в течение сезона с тем, чтобы учесть не только
многолетние виды мхов, но также весенние и осенние эфемеры. В случае одно-
разового посещения пробных площадей рекомендуется плохо развитые или не-
зрелые гаметофиты мелких мхов и печеночников сохранять живыми с последу-
ющим доращиванием в культуре. Это особенно важно для тех видов, определе-
ние которых требует наличие спорофита или спор (Ладыженская, 1948, 1950).
Основу видового состава мхов лесного фитоценоза формируют виды, эколо-
гические особенности которых максимально соответствуют данному типу мес-
тообитания. Доминанты мохового покрова — это прежде всего бриовиоленты,
для которых условия данного типа местообитания являются оптимальными. По-
этому нахождение бриовиолентов на пробной площади не составляет труда даже
137
на небольшом участке мохового покрова. Бриопатиенты, как правило, составля-
ют примесь в дерновинках доминантных видов. Для выявления бриопатиентов
требуется более широкий охват площади и обследование всех типов микромес-
тообитаний данного фитоценоза. Следует отметить, что бриопатиенты находят-
ся в динамическом равновесии с бриовиолентами и при изменении экологичес-
ких параметров (степень освещенности, увлажнения, трофности) под влиянием
внешних воздействий или внутренних процессов в лесном сообществе могут за-
нять позицию бриовиолентов.
Массовое присутствие в составе мохового покрова бриоэксплерентов харак-
теризует определенную степень нарушенности мохового покрова данного лес-
ного сообщества. В ненарушенных сообществах бриоэксплеренты характеризу-
ются небольшим обилием. Для их выявления необходимо вести целенаправлен-
ный поиск, а именно осмотреть максимальное число микроместообитаний в пре-
делах сообщества с естественным нарушением мохового покрова (ветроваль-
ные обнажения грунта, участки нарушенной поверхности, возникшие в результа-
те деятельности животных, пни, поваленные деревья и т. п.). Чем больше пло-
щадь, на которой учитывается флористическое богатство сообщества, тем боль-
шее количество видов (разумеется до известного предела) будет на ней зареги-
стрировано.
Известно, что зависимость числа выявленных видов от площади имеет лога-
рифмический характер, т. е. количество видов по мере возрастания суммарного
размера учетной площади сначала увеличивается довольно быстро, а в дальней-
шем изменяется незначительно. Площадь выявления видового состава сообще-
ства или его компонента — это та наименьшая площадь, на которой встречают-
ся все или почти все виды (не менее 90 %), произрастающие в сообществе (Ра-
менский, 1925). Константными называются виды, которые встречаются на 90-
100 % площади данной ассоциации. Площадь выявления видового состава мхов
зависит от размеров отдельных видов, характера жизненной формы, от разнооб-
разия микроместообитаний в пределах изучаемого сообщества и т. д. Площадь
выявления видового состава для разных типов сообществ различна и может быть
установлена только эмпирическим путем. Для этого закладывают серии увели-
чивающихся по размеру учетных площадок (1, 4, 16, 25, 36... 100, 400 м2 и т. д.).
Было установлено, что равные по размеру, но имеющие разную форму площад-
ки различаются по числу видов. Для выявления видового состава предпочтитель-
нее закладывать не квадратные, а ленточные учетные площадки; в этом случае
их общая площадь может быть значительно уменьшена (в 2 раза и более). Если
все константные виды ассоциации можно выявить на сравнительно небольшой
площади (1,4, 10 м2), то для выявления всего видового состава приходится об-
следовать площадь значительно большего размера.
138
2.10. МЕТОДЫ ИЗУЧЕНИЯ ПОДЗЕМНЫХ ЧАСТЕЙ РАСТЕНИЙ 1
При изучении лесных сообществ исследование подземных частей растений
столь же необходимо, как и надземных, так как сведения об их строении и фун-
кциях необходимы для оценки состояния и устойчивости растений в условиях
изменяющейся окружающей среды. Корневые системы закрепляют растения в
почве и обеспечивают им устойчивое положение. Они снабжают растение водой
и питательными веществами. Доказано, что в корнях происходит образование
сложных органических веществ из минеральных соединений (Сабинин, 1955;
Курсанов, 1976; Кулаева и др., 1957; Курсанов, Выскребенцева, 1960 и др.). Ус-
тановлено также, что корни могут поглощать из почвы углекислоту и углекис-
лые соли (Курсанов, 1976).
Строение подземных органов обусловлено видовыми особенностями расте-
ний и существенно зависит от условий произрастания. Однако изученность под-
земных частей растений значительно уступает изученности надземных органов,
что, по-видимому, связано с трудоемкостью и сложностью исследования и зна-
чительно меньшей хозяйственной ценностью.
2.10.1. Общие методические подходы к изучению корневых систем
растений
Задачи изучения подземных частей растений при проведении фундаменталь-
ных исследований могут быть очень разнообразны. Основными аспектами при
изучении подземных органов являются следующие:
1. Выявление особенностей строения, развития и взаимного влияния подзем-
ных органов растений разных видов, а также особей в популяциях одного вида;
2. Исследование строения и функционирования подземных частей растений в
лесных сообществах по фазам вегетационного сезона и по отдельным периодам
в онтогенезе в зависимости от экологических факторов;
3. Оценка изменения роста и развития корневых систем в зависимости от ус-
ловий произрастания и внешних воздействий.
В данном разделе приводится описание основных методов полевых исследо-
ваний подземных частей растений с указанием, какие из них наиболее пригодны
при проведении экологических исследований.
Согласно М. С. Шалыту (1960), в зависимости от целей и задач исследований
эти методы можно объединить в следующие 4 группы.
I. Качественные (морфологические) методы:
1) метод горизонтальной раскопки;
1 Автор В. Т. Ярмишко
139
2) метод траншей (траншейный метод);
3) метод полной раскопки или отмывки.
II. Методы количественного учета:
1) метод монолитов;
2) метод вольного монолита или проб;
3) методы дробного весового учета корней;
4) методы учетных рядов и площадок;
5) метод «бура» (метод взятия монолитов посредством бура);
6) метод среза (метод профиля);
7) метод модельных растений;
8) корреляционный метод.
III. Методы стационарных исследований возрастных изменений подзем-
ных частей растений:
1) метод наблюдения за ростом корней через щели или ниши в траншеях;
2) метод наблюдения за ростом корней: а) через стекла и остекленные рамы,
погруженные в почву; б) через застекленные стенки корневых домиков — под-
земных лабораторий; в) через стекла, вставленные в стенки наблюдательных
ящиков;
3) метод периодического обнажения подземных частей растений по фазам и
периодам их развития или одновременной раскопки разновозрастных растений.
4) метод «врастания».
IV. Методы определения поглощающей поверхности или физиологичес-
кой активности корневых систем:
1) метод адсорбции (метод определения поглощающей поверхности корней
по адсорбции из растворов метиленовой сини);
2) метод определения зоны поглощения корневой системы;
3) метод определения физиологической активности корневых систем по ско-
рости «плача» растений (скорости или интенсивности выделения пасоки);
4) метод потометра (метод определения активной поверхности корневых окон-
чаний по количеству всасываемой ими влаги).
Каждый из перечисленных методов разрабатывался для изучения определен-
ного круга вопросов, поэтому только комплексное использование ряда методов,
относящихся к различным вышеперечисленным группам, гарантирует достаточ-
но полное изучение подземных частей исследуемых растений.
2.10.2. Основные характеристики, определяемые при изучЛии
подземных органов растений
1. Строение и развитие корневых систем:
а) типы корневых систем, особенности строения, возрастные изменения, рит-
мика роста, степень пластичности, классификация типов корневых систем;
б) особенности строения и развития главного корня, направление роста, глу-
140
бина проникновения в почву, форма, сбежистость, интенсивность ветвления, ха-
рактер расположения боковых корней, явление остановки роста: формирование
стержневого корня из боковых корней, явления контракции и партикуляции;
в) строение и развитие боковых корней первого порядка, происхождение (бо-
ковые корни системы главного корня, придаточные боковые корни), типы боко-
вых корней и особенности их строения, расположение (на материнском корне или
подземных органах стеблевого происхождения), порядок и характер ветвления,
сбежистость, направление роста (угол расположения боковых корней первого
порядка по отношению к главному корню);
г) строение и развитие проводящих корней последующих порядков;
д) сосущая часть корневой системы и корневые мочки1, форма и типы сосу-
щих корней (корневых окончаний), возникающих на корнях первичного и вторич-
ного строения, рост и продолжительность жизни сосущих корней, наличие и типы
микоризы (эндотрофная, эктотрофная, экто-эндотрофная, перитрофная) и клубень-
ковых бактерий;
д) метаморфозы корня.
2. Строение подземных частей растений стеблевого происхождения:
а) типы подземных частей растений стеблевого происхождения: подземные
побеги, корневища, столоны и др.;
б) строение и развитие подземных частей растений стеблевого происхождения;
в) формирование придаточных корней на органах вегетативного размножения;
г) формирование и строение почек, развивающихся на подземных побеговых
органах.
3. Распределение корней и других подземных органов растений в почве:
а) распределение подземных органов в вертикальной и горизонтальной про-
екциях; наличие и характер ярусности; влияние экологических и ценотических
условий;
б) распределение массы подземных частей растений по почвенным горизонтам1 2.
4. Особенности роста подземных частей растений:
а) сезонный рост подземных органов;
б) многолетний рост подземных органов.
5. Соотношение между надземными и подземными органами растений:
а) площадь проекции кроны и площадь проекции корневой системы;
б) объем пространства, занимаемый кроной, и объем почвы, занимаемый кор-
невой системой;
в) масса надземной и подземной частей;
г) высота надземной части и глубина корневой системы.
1 Корневые мочки — проводящие корни последних порядков с находящимися на них вса-
сывающими и ростовыми корнями.
2 Методы изучения фитомассы подземных органов рассмотрены в разд. 2 1
141
6. Факторы внешнего воздействия на подземные органы растений:
а) антропогенная деятельность;
б) деятельность животных;
в) энтомовредители и патогенные грибы;
г) лесные пожары.
7. Наиболее важные показатели строения и развития подземных органов:
а) рабочая глубина проникновения корней (Казакевич, 1925) — глубина про-
никновения основной массы корней или глубина наиболее интенсивного ветвле-
ния корней;
б) коэффициент длины корней (Колесников, 1924) — суммарная величина
длины всех корней одного растения, деленная на общее число корней;
в) насыщенность почвы корнями (Погребняк, 1927) — отношение величины
объема корней по фракциям к величине объема почвы, из которой они извлека-
лись.
2.10.3. Описание основных полевых методов изучения подземных
органов растений
Изучение подземных частей растений следует начинать с подробного описа-
ния объектов исследования: лесных сообществ и характеристик условий место-
обитания (см. разд. 1.3-1.8).
Метод горизонтальной раскопки. Суть настоящего метода заключается в
послойном вскрытии подземных частей растений, начиная с поверхности и до глу-
бины залегания основной массы горизонтальных корней и подземных органов
стеблевого происхождения с последующей их зарисовкой и описанием (рис. 2.9).
Рис. 2.9. Поверхностное вскрытие корневых сис-
тем лиственницы сибирской на полуострове
Таймыр
Наиболее часто этот ме-
тод применяется совместно
с траншейным методом. В
сочетании оба метода во
многих случаях дают доста-
точно полное представление
о строении подземных час-
тей исследуемых растений
при значительно меньшей
затрате времени и труда по
сравнению с методом пол-
ной раскопки. После прове-
дения работ методом гори-
зонтальной раскопки и тран-
шей используют методы ко-
личественного учета (метод
монолитов, «бура» и т. д.).
142
Полевые исследования подземных частей большинства растений начинают, как
правило, с раскапывания их с поверхности (горизонтальной раскопки), потому что
только в результате такого подхода можно правильно установить необходимые
направления и длину траншей для последующей работы, а также наиболее раци-
онально наметить участки для взятия монолитов.
При помощи метода горизонтальной раскопки могут быть получены данные
по следующим вопросам:
1) строение, горизонтальное распределение подземных частей растений, зале-
гающих в верхних горизонтах почвы;
2) длина и диаметр поверхностных корней;
3) характер и энергия вегетативного размножения;
4) специфика развития сосущей части поверхностных корней и подземных ор-
ганов стеблевого происхождения.
Кроме того, можно получить данные о радиусе распространения корневых
систем, о площади питания исследуемого растения, о соотношении горизонталь-
ных проекций надземных и подземных частей, об особенностях взаимного рас-
положения корневых систем соседних особей (рис. 2.9).
Вскрытие подземных частей исследуемых растений производится послойно,
как правило, сухим способом (сухой раскопкой). Раскопку проводят при помо-
щи специальных инструментов. Значительно реже при вскрытии подземных час-
тей растений применяется их отмывка водяной струей.
Площадь раскопки определяется радиусом корневой системы растения. При
изучении корневых систем древесных пород или кустарников вскрывается толь-
ко их часть (половина или четверть). В естественных условиях П. К. Красильни-
ков (1983) рекомендует раскапывать корневые системы деревьев и кустарников
на половине их распространения со стороны основания ствола или куста, проти-
воположной месту, назначенному для рытья траншеи.
При изучении корневых систем травянистых растений удаление почвы необ-
ходимо проводить значительно более осторожно, ввиду значительно меньшей их
прочности. Вначале с поверхности почвы удаляют опад и лесную подстилку. В
том случае, если корни и корневища частично располагаются в подстилке, пос-
леднюю снимают до уровня расположения корней или корневищ, затем, продол-
жая постепенно удалять мертвый покров, очищают корневища и корни от при-
ставших к ним частиц. Минеральную часть почвы следует удалять на глубину 5-
15 (25) см в зависимости от глубины расположения основной массы поверхно-
стно залегающих корней. При более глубоком залегании корней рекомендуется
проведение серии последовательных (послойных) раскопок.
Одновременно с раскопкой проводят-описание и зарисовку подземных орга-
нов растений, залегающих в подстилке и почве. В каждом слое почвы раскопку
обычно проводят от базальных частей корневых систем к их периферии. Зари-
совка подземных частей растений производится на клетчатой или миллиметро-
143
вой бумаге в масштабе 1 : 5 или 1 : 10 в зависимости от густоты корневых сис-
тем. При зарисовках используются накладываемые на поверхность обнаженных
корневых систем масштабные сетки, представляющие собой легкую деревянную
(или металлическую) квадратную или прямоугольную раму с внутренними раз-
мерами 50 X 50, 50 х 100 или 100 х 100 см, на которой через каждые 5 или 10 см
натянута тонкая проволока, образующая ячейки 5x5 или 10x10 см. При вскры-
тии больших участков (порядка нескольких квадратных метров и более) их для
удобства зарисовок разделяют, используя колышки и шнур, на квадраты площа-
дью не более 1 м2 (рис. 2.9). Для того чтобы более точно представить загру-
женность почвы наиболее мелкими частями подземных органов растений, в верх-
нем почвенном горизонте на вскрытой площади зарисовывают в натуральную ве-
личину несколько площадок размером от 25 х 25 см до 100 х 100 см в зависи-
мости от объекта исследования. На рисунках по возможности фиксируют все кор-
ни, включая наиболее тонкие, а так-
Рис. 2.10. Метод траншей, использованный
при изучении корневых систем сосны
обыкновенной на Кольском полуострове
же подземные органы стеблевого
происхождения.
Метод траншей. Метод тран-
шей применяется для изучения рас-
пределения подземных частей от-
дельных растений или их совокуп-
ности в вертикальной плоскости.
Метод траншей заключается во
вскрытии подземных частей расте-
ний со стороны траншеи раскопкой
или отмывкой на заданную глубину
и последующей их зарисовкой и
описанием (рис. 2.10).
Метод траншей позволяет выя-
вить:
1) морфологические особеннос-
ти и возрастные изменения подзем-
ных частей растений и их зависи-
мость от эдафических факторов;
2) распределение подземных ор-
ганов растений по генетическим
горизонтам почвенного профиля;
3) наличие ярусности в распрос-
транении корневых систем расте-
ний, глубину проникновения в почву
основной массы и отдельных кор-
ней;
144
4) боковое простирание корней и других подземных органов;
5) площадь питания и объем питания (объем используемой почвы) изучаемых
растений;
6) взаимное распределение подземных частей совместно произрастающих ра-
стений, относящихся к одному или разным видам.
Траншеи закладывают вблизи оснований стволов (стеблей) изучаемых расте-
ний: у травянистых растений — на расстоянии 10-20 (30) см, у деревьев и кус-
тарников — на расстоянии 20-30 см. Для наиболее крупных деревьев расстоя-
ние может быть увеличено до 50 см. В плане траншеи должны быть прямоуголь-
ной или трапециевидной формы и иметь строго вертикальные стенки. Глубина
траншеи должна соответствовать глубине проникновения в почву корневых сис-
тем изучаемых растений или превышать ее на 10-20 см; длина траншеи при изу-
чении отдельных растений должна соответствовать диаметру их корневых сис-
тем. При изучении корневых систем деревьев и кустарников около каждого из
них закладывается отдельная траншея. При изучении травянистых растений одна
траншея может быть использована для исследования нескольких особей. При
изучении строения корневых систем совокупности растений в пределах сообще-
ства длина траншеи удлиняется до размеров, обеспечивающих получение необ-
ходимых данных о подземных органах всех растений, а ширина — до 70-100 см.
Цри рекогносцировочных ризологических исследованиях, когда требуются
ориентировочные данные о строении корневых систем, можно ограничиться толь-
ко траншейным методом исследования, без использования метода горизонталь-
ной раскопки и метода полной раскопки или отмывки.
Метод полной раскопки или отмывки. Метод полной раскопки или отмыв-
ки заключается в послойном обнажении всей подземной части растения раскоп-
кой (рис. 2.11) или
отмывкой.
Параллельно с
раскопкой ведут
зарисовки и под-
робное описание
обнажаемой под-
земной части ис-
следуемых расте-
ний. Зарисовку и
описание при ме-
тоде отмывки про-
водят обычно пос-
ле извлечения под-
земной части рас- рис 2ц. Полная раскопка корней лиственницы сибирской
тений из почвы. на полуострове Таймыр
145
Метод полной раскопки и отмывки дает наиболее точные данные о строении
подземных органов растений, их горизонтальном распространении, а метод тран-
шей — о вертикальном распределении корневых систем в различных генетичес-
ких горизонтах почвы.
После описания и зарисовки корневой системы можно получить количествен-
ные данные о массе, длине и площади поверхности корневых систем путем не-
посредственного измерения всех извлеченных из почвы корней. Такой способ
учета подземной массы применим только в отношении подземных органов от-
дельных особей.
Главными недостатками этого метода являются его трудоемкость, невозмож-
ность полного извлечения корневой системы большинства растений, сложная
увязка горизонтального и вертикального распределения корневой системы с поч-
венными горизонтами и ряд других моментов, что и вызвало во многих случаях
переход от метода полной раскопки или отмывки к методу горизонтальной рас-
копки и методу траншей.
Метод монолитов — основной метод учета подземной массы как отдель-
ных растений, так и растительных сообществ. При применении этого метода обыч-
но предварительно проводится горизонтальная или полная раскопка корневых си-
стем изучаемых растений, что позволяет наиболее рационально разместить мес-
та для взятия монолитов.
Метод монолитов заключается в учете массы или объема подземных частей
растений в определенном объеме почвы с последующим пересчетом получен-
ных данных на единицу площади.
Работа состоит из следующих этапов.
1. Установление необходимой повторности взятия почвенных столбов соот-
ветствующих размеров для получения достоверных данных с определенной сте-
пенью точности.
2. Выбор участков для взятия почвенных столбов с подземными органами
растений.
3. Описание растений, произрастающих на данных участках, и определение их
надземной массы.
4. Извлечение почвенных столбов (строго определенных размеров) из грунта
с разделением их на монолиты по генетическим горизонтам или по слоям опре-
деленной высоты.
5. Выделение подземных частей растений из монолитов (отмывкой или отбор-
кой и просеиванием с последующей очисткой или отмывкой).
6. Разборка и фракционирование подземных частей растений по видам, орга-
нам и толщине.
7. Определение массы, объема, длины, площади поверхности и других пока-
зателей у отдельных подземных органов растений и их фракций.
8. Увязка полученных данных с генетическими горизонтами.
146
9. Математическая обработка полученных материалов и пересчет полученных
данных на выбранную единицу площади (1 м2, га и т. д.).
10. Отбор образцов подземных частей растений для химических, анатомичес-
ких, физико-механических и других исследований.
Метод монолитов не дает четкого представления о структуре подземных ор-
ганов растений, но его использование позволяет получить количественные дан-
ные о распространении подземных частей растений (их массе, объеме, количе-
стве, длине и др.) в горизонтальной и вертикальной проекциях в почве по гене-
тическим горизонтам. Учет подземных частей растений ведется с разделением
или без разделения их по видам, органам и фракциям в зависимости от задач
исследования. В естественных сообществах значительную сложность представ-
ляет разделение наиболее тонких корней по видам. Этот метод отличается боль-
шой трудоемкостью.
Наиболее полное описание метода монолитов приведено в работе Н. А. Ка-
чинского (1925) при изучении корневых систем древесных пород и травянистых
растений, произрастающих на почвах подзолистого типа. Он предложил прово-
дить разделение почвенного столба по генетическим горизонтам, достаточно
точно определять объем почвенных столбов, тщательно выделять корни, разде-
ляя их на деятельные и недеятельные, и более детально учитывать надземную
часть.
В дальнейшем при применении метода монолитов Н. А. Качинского варьирова-
ли: 1) размеры почвенных столбов (Шайн, Чекмарева, 1940; Шайн, 1948; Рахте-
енко, 1949 и др.); 2) способы взятия почвенных образцов с подземными частями
растения (Калмыкова, 1938; Хайруллин, 1948; Шалыт, 1949, 1960; Лавриненко,
1952; Ильин, 1961; Колесников, 1962, 1972; Рустамов, 1963, 1968а, б, 1972; Ор-
лов, 1967 и др.); 3) способы выделения подземных частей растений из моноли-
тов; 4) размеры ячеек сит, применяемых для отмывки корней и других подземных
органов растений; 5) дробность фракций при разделении подземных органов рас-
тения; 6) способы получения тех или иных количественных показателей.
В настоящее время метод монолитов имеет большое число модификаций; это
в значительной степени обусловлено тем, что до сих пор еще не разработаны
стандартные приемы для учета подземных частей у отдельных жизненных форм
в различных условиях местообитания. Большая пестрота получаемых данных, по
мнению С. С. Шайна (1948), объясняется, помимо экологических условий и ви-
довых различий, погрешностями методики взятия почвенных монолитов и вы-
деления из них подземных частей растений. На практике каждый исследователь
в большинстве случаев по своему усмотрению решает вопрос количественного
учета подземных частей растений (размеры монолитов, число повторностей,
способы их взятия и т. п.), что затрудняет сравнение полученных данных.
Площадь монолитов чаще всего составляет от 0,01 до 0,25 м2 (с соответ-
ствующими объемами от 500 до 25000 см3 при высоте монолита 5 или 10 см).
147
Монолиты размером 25 х 25 см были использованы автором при изучении на-
сыщенности почвы корнями растений в лесных сообществах, значительно менее
точные данные получаются при использовании монолитов размером 0,1 х 0,1 м.
Высота монолитов обычно соответствует толщине генетических горизонтов
почвы. В тех случаях, когда в полевых условиях трудно установить границы меж-
ду генетическими горизонтами, почвенные столбы разделяют на слои опреде-
ленной высоты (5-10-15-20 см). При использовании буров или специальных
форм размеры почвенных образцов редко превышают 500-1000 см3.
До сих пор слабо изученным остается вопрос о необходимом количестве
почвенных образцов и оптимальном размещении монолитов (почвенных столбов),
обеспечивающих получение данных с определенной точностью. Поэтому перед
началом основных работ необходимо провести специальную методическую ра-
боту по определению варьирования количественных показателей. Расчет точно-
сти измерений и необходимого количества образцов приведен в разд. 4.
Как правило, подземные части растений выделяют из почвенных монолитов
путем отмывки через сита. Отмывка проводится разными способами:
1) струей воды с помощью шланга на специальных станках (Шалыт, 1949,
1960);
2) помещением сит в тазы или ванны с водой и последующим передвижени-
ем сит по дну этих сосудов (Чижов, 1931; Тарановская, 1957);
3) при помощи декантации (Саввинов, Панкова, 1942; Панкова, 1954);
4) установлением сит в проточной воде (Лапин, Польский, 1963);
5) помещением сит на определенную глубину в стоячем водоеме с периоди-
ческим покачиванием сит (Лапин, Польский, 1963);
6) на специальных отмывочных машинах.
Большинство исследователей к деятельным корням относит корни с диамет-
ром меньше 0,5-1-2 мм, хотя все подчеркивают, что это деление условно и что
к активным корням правильнее относить корневые окончания, на которых име-
ются корневые волоски, или использовать другие морфологические признаки. Но
так как выделение деятельных корней по морфологическим признакам сложнее
и требует больше времени, то этот способ используют значительно реже — пре-
имущественно при наиболее детальных работах. Живые корневые волоски воз-
можно обнаружить только при сильном увеличении или, с большей достоверно-
стью, по изменению fix окраски в результате поглощения органических красите-
лей (Колесников, 1962; Станков, 1964). По внешнему виду живые корневые во-
лоски молочно-белые, полупрозрачные, трубчатой формы, мертвые — темные,
коричневые или бурые, сдавленные, иногда скрученные.
Метод «вольного монолита», или метод «проб». Для учета деятельных
корней, а также для изучения морфологических и биологических особенностей
«корневых мочек» (совокупность конечных разветвлений мелких проводящих
корней с расположенными на них сосущими окончаниями) В. А. Колесников
148
(1947) предложил метод «вольного монолита», или метод «проб». Этот метод
отнесен нами к модификации метода монолитов, но он может быть также отне-
сен к группе методов стационарных исследований для изучения ритмики роста в
годичном цикле наиболее тонкой части корневой системы (всасывающих, пере-
ходных и проводящих корней последних порядков). Метод вольного монолита
В. А. Колесникова (1947) имеет много общего с методом А. Я. Орлова (1955а,
19556), используемым для определения количества сосущих корней древесных
пород в почве.
В основу метода вольного монолита положены следующие закономерности:
1) корневой коэффициент (Колесников, 1947), т. е. средняя длина корня древес-
ных пород определенного вида при одинаковом возрасте — величина постоян-
ная, и средняя длина корня в любой части корневой системы равна средней дли-
не корней в других частях корневой системы; 2) даже самые малые части корне-
вой системы — разветвления последних порядков (корневые мочки), достаточ-
но точно отражают закономерности роста и развития всей корневой системы, а
именно рост активных корней, особенности их морфологии, а также их реакцию
на воздействие окружающей среды; 3) соотношение между количеством актив-
ных и неактивных корней одинаково в пределах всей корневой системы.
Предварительно методом горизонтальной раскопки определяют расположе-
ние горизонтальных корней и места залегания основной массы корневых мочек.
На исследуемом участке выбирают 3 типичных средних дерева и намечают ме-
ста для взятия монолитов. В определенные сроки по периферии кроны деревь-
ев, где сосредоточена основная масса корневых мочек, отбирают пробы почвы.
Для этого выкапывают небольшую траншею таким образом, чтобы можно было
вырезать монолиты с площадью 20-25 см2 на трех глубинах: 0-35, 35-50, SO-
75 см. В зависимости от условий произрастания и вида растений поперечное се-
чение и высота монолитов, а также глубина их расположения могут меняться.
Обычно пробы почвы в 2—4 повторностях берут один раз в месяц от модельных
деревьев в течение всего вегетационного периода. Монолиты переносят в лабо-
раторию (лучше в банках с водой), где корни отмывают от почвы и выбирают из
монолитов неповрежденные системы корневых разветвлений последних поряд-
ков (корневые мочки) с 100-300 корнями (рис. 2.12). Такое количество корней
последнего порядка обычно имеется на 3-5 корневых разветвлениях предпос-
ледних порядков (обрастающих корней, корневых мочек, корневых сеток). За-
тем учитывают количество и измеряют длину проводящих (потемневших, корич-
невых), переходных (серого цвета, часто с оранжевым оттенком) и всасываю-
щих (утолщения белого цвета с корневыми волосками) корней. Корни измеряют
на стекле, покрытом водой, под которое подкладывают миллиметровую бумагу,
или же корни раскладывают на влажной материи или фильтровальной бумаге. В
последних двух случаях измерения проводят линейкой с точностью до 1 мм.
Затем проводят математическую обработку данных и вычисляют соотношения
149
Рис. 2.12. Плотное ветвистое обра-
зование микориз на корнях сосны
обыкновенной, произрастающей
при высоком уровне атмосферно-
го загрязнения (Кольский полуос-
тров)
по длине и количеству всасывающих, пе-
реходных и проводящих корней от их об-
щего количества.
Определение количества точек роста —
белых корневых окончаний, должно про-
водиться очень быстро, так как на возду-
хе белые корневые окончания быстро тем-
неют, что затрудняет их подсчеты. По дли-
не тонких проводящих корней и количе-
ству белых активных корней высчитывают
количество активных корней (новых точек
роста) на погонный метр корней корневой
мочки. Для каждой пробы корней при их
учете делают зарисовки отдельной корне-
вой мочки с нанесением на рисунок раз-
личной краской (коричневой и светло-жел-
той) проводящих и активных корней. Дан-
ный способ изображения дает образное
представление о характере роста корней
разных пород в определенные фазы разви-
тия и их реакцию на воздействия среды.
В настоящее время имеется много ва-
риантов метода вольного монолита, при-
меняемых при изучении древесных пород
в естественных условиях (Згуровская,
1958, 1962а, 19626; Колесников, 1962;
Волчков, 1964, и др.). Они различаются в основном размерами монолитов, спо-
собами измерения активных и неактивных корней, оформлением полученных
данных. Наиболее разработанной модификацией метода вольного монолита при
изучении сосущей части корневой системы древесных пород в естественных
условиях является модификация А. Я. Орлова (1955а, б, 1967).
Метод модельных растений заключается в полном или частичном извле-
чении из почвы подземных частей модельных растений, определении их массы
и на основании данных сплошного перечета растений на единицу площади вычис-
лении их биологической и хозяйственной продуктивности. Детально метод мо-
дельных растений был разработан при проведении учета массы корней береск-
лета (Юркевич, 1941; Моисеенко, 1947, 1949, 1951 и др.).
В настоящее врАы наиболее часто данный метод используется при определении
массы подземных частей стержнекорневых травянистых растений, кустарников и
древесных пород. Выделение подземных частей растений из почвы проводят двумя
способами: 1) методом полной раскопки, 2) методом крупных монолитов.
150
В наиболее типичных местах исследуемого растительного сообщества наме-
чают учетные площадки, размеры которых зависят от размеров и густоты раз-
мещения изучаемых растений и требуемой точности исследования. Наиболее
часто используемые размеры площадок: для травянистых растений — 1-25
(100) м2, для кустарников — 4-100 (625), для древесных пород — от 400-625
до 2500-5000 (10000) м2. На учетных площадках производят полный перечет
всех растений изучаемого вида при необходимости с подразделением на возрас-
тные группы. На основе данных перечета выбирают модельные растения: не
менее трех для каждой возрастной группы, средних по высоте и общему разви-
тию. Затем проводят выделение подземных частей модельных растений из по-
чвы. Тщательность выделения корневых систем обусловлена задачами, постав-
ленными перед исследователем. Для определения биологического запаса или био-
логической продуктивности подземных частей растения выкапывают по возмож-
ности все подземные органы данного растения. Для определения хозяйственно-
го запаса или хозяйственной продуктивности из почвы извлекают только часть
подземных органов, имеющих хозяйственное значение (обычно соответствую-
щих диаметров, согласно существующим стандартам).
Извлеченные подземные части растений очищают, отмывают, разделяют на
органы, а корни — на фракции в зависимости от их диаметра. Затем определяют
их массу в сыром и воздушно-сухом состояниях, а при более детальных иссле-
дованиях — в абсолютно сухом состоянии. На основании данных по модельным
растениям и перечета вычисляют биологическую и хозяйственную продуктив-
ность на единицу площади. Если модельные растения отбирали по возрастным
группам, то и все вычисления проводят по этим группам.
Метод «врастания» корней — сравнительно новый метод в ризологичес-
ких исследованиях. Обычно он используется при оценке фитомассы тонких фи-
зиологически активных корней растений, при определении интенсивности освое-
ния почвы корнями, при оценке воздействия рекреационных нагрузок на сооб-
щества, особенно в подземной сфере, и т. п. Мы применяли метод «врастания»
корней в процессе детальных исследований лесных экосистем, подверженных воз-
действию промышленного загрязнения. Суть его заключается в следующем. На
выбранном участке лесного сообщества пробуривали 60 шурфов (4 блока по 15
шурфов в каждом) диаметром 4 см и глубиной 40-45 см (рис. 2.13). В шурфы
аккуратно вставляли нейлоновые сетки с диаметром ячеек 7x7 мм, в которые
плотно набивали почву, предварительно просеянную через сито с диаметром от-
верстий 2 мМ. На уровне поверхности почвы сетки завязывали и слегка мульчи-
ровали. Каждая сетка была снабжена биркой, на которой была соответствующая
запись (№ участка, № блока, № шурфа и дата закладки сетки). Поскольку экспе-
римент был рассчитан на 2 года, то первую половину сеток-кернов (30 шт.) на
каждом участке извлекали через год, а остальные извлекали после 2-летней эк-
спозиции. Перед извлечением каждый керн обрезался длинным ножом по окруж-
151
с
Рис. 2.13. Схема размещения блоков сеток-кериов при иссле-
довании тонких физиологически активных корней растений
методом «врастания». Черными квадратами показаны мес-
та взятия почвенных монолитов. На нижней части рисунка
показаны стадии подготовки и проведения эксперимента
ности для предотвращения потери проникших сквозь сетку корней. Каждый из-
влеченный из почвы керн запаковывали в полиэтиленовый пакет и 'образцы дос-
тавляли в лабораторию. Извлеченные керны делили на 10-сантиметровые отрез-
ки и из соответствующих отрезков отмывали корни. Определяли массу корней,
интенсивность ветвления, морфометрические характеристики, жизнеспособность,
наличие микориз и т. д. Корни с микоризами разной формы и цвета фиксировали
в спирт-глицериновой смеси для дальнейших лабораторных исследований.
152
2.10.4. Методика отбора образцов активных корней для исследования
микотрофности растений
При комплексных ризологических исследованиях в ряде случаев возникает не-
обходимость сбора материала для последующего изучения микотрофности рас-
тений. Сбор образцов активных корней растений проводят согласно методике, раз-
работанной Н. В. Лобановым (1953). При помощи специальных инструментов об-
нажают корневые мочки изучаемого растения, при этом очень важно точно ус-
тановить их принадлежность к исследуемому растению. В большинстве случаев
корневые мочки берут из верхних почвенных горизонтов в пределах 20 см, наи-
более часто — с глубины 5-10 см. При детальных исследованиях — по генети-
ческим горизонтам.
При взятии образца отмечают следующие данные: место и дата, растение, его
описание, характеристика растительного сообщества, тип почвы, описание почвен-
ного профиля, глубина взятия образца с указанием генетического горизонта и т. п.
Для точного установления наличия или отсутствия микориз, а также их типа
обязателен просмотр поперечных и продольных срезов корневых окончаний под
микроскопом. Если просмотр образцов предполагается провести в день сбора,
то перевязанные цветными нитками образцы помещают в банку с водой. В слу-
чае более позднего микроскопического анализа материал фиксируют в 70-96%-
ном растворе спирта или в других фиксаторах. До фиксации необходимо указать
цвет корневых окончаний. Наиболее детальное изучение особенностей микориз
требует более тонкой фиксации и микротомных срезов. Подробное описание
методики полевых и лабораторных исследований микотрофности растений дано
в работах Н. В. Лобанова (1953), Н. М. Шемахановой (1962) и других авторов.
153
РАЗДЕЛ 3. МЕТОДЫ ИЗУЧЕНИЯ ЛЕСНЫХ СООБЩЕСТВ
В УСЛОВИЯХ НАРУШЕННЫХ МЕСТООБИТАНИЙ
3.1. ДИАГНОСТИКА ПОВРЕЖДЕНИЙ И ОЦЕНКА ЖИЗНЕННОГО
СОСТОЯНИЯ ДЕРЕВЬЕВ И ДРЕВОСТОЕВ В УСЛОВИЯХ
ПРОМЫШЛЕННОГО АТМОСФЕРНОГО ЗАГРЯЗНЕНИЯ 1
При изучении воздействия разнообразных антропогенных факторов на леса
оценка состояния последних производится в настоящее время по шкале катего-
рий состояния деревьев, согласно «Санитарным правилам в лесах Российской
Федерации» (1992). Эта шкала основана на описании повреждений деревьев от
насекомых и пожаров и не учитывает специфики воздействия промышленного
загрязнения атмосферы. Учитывая, что в последние десятилетия все большие
площади лесов подвергаются влиянию промышленного загрязнения атмосферы,
целесообразно составление специального методического руководства. Предла-
гаемые рекомендации выработаны в результате многолетней работы в лесах раз-
личных районов Российской Федерации, испытывающих влияние промышленно-
го атмосферного загрязнения и свободных от него.
3.1.1. Диагностические признаки жизненного состояния деревьев в
условиях атмосферного загрязнения
Воздействие атмосферного загрязнения на растение — сложное биохимичес-
кое явление, затрагивающее в первую очередь метаболические и физиологичес-
кие процессы и разрушающее ультраструктуру клеток листа. По мере разруше-
ния внутриклеточных структур начинают проявляться внешние, визуально наблю-
даемые повреждения и отклонения от нормы у ассимиляционных органов и дру-
гих частей растений. Степень воздействия загрязнителя на растение зависит не
только от его концентрации и продолжительности действия, но и от видовой при-
надлежности и толерантности растений к загрязнителю, от стадии онтогенеза, се-
зона года и состояния окружающей среды (температуры, влажности воздуха и
почвы, условий освещенности, ветра, условий минерального питания и пр.).
В процессе многолетних исследований жизненного состояния лесных фито-
ценозов были выявлены следующие диагностические признаки повреждения
деревьев промышленным загрязнением:
1) хлорозы и некрозы ассимиляционных органов (рис. 3.1);
2) снижение продолжительности жизни хвои (рис. 3.2);
3) снижение охвоенности крон с нарушением распределения фитомассы хвои
по высоте крон (рис. 3.2);
4) ускоренное отмирание ветвей основной кроны; снижение линейного приро-
ста оси ствола и ветвей (рис. 3.3);
1 Автор В Т Ярмишко
154
1 2 3 4 5 6
Рис. 3.1. Схематическое изображение ти-
пичных симптомов повреждений
хвои сосны обыкновенной под влия-
нием атмосферного загрязнения
Состояние хвои: 1 — здоровая, без измене-
ния цвета; 2 — здоровая, с отдельными ко-
ричневыми точками; 3 — поврежденная, кон-
чики хвои длиной 1,5-2 мм имеют некрозы, а
хлорозы занимают до 1/2 поверхности хвои;
4 — поврежденная, некрозы занимают >25 %
поверхности хвои, 5 — сильно поврежден-
ная, площадь поврежденной поверхности хвои
превышает 50 %; 6 — отмирающая, по-
врежденная поверхность хвои превышает
75 %
Рис. 3.3. Снижение интенсивности
линейного прироста ствола и бо-
ковых побегов у сосны обыкно-
венной на Кольском полуострове
Рис. 3.2. Сокращение продолжительно-
сти жизни хвои и изреживание крон
сосны обыкновенной под воздей-
ствием атмосферного загрязнения
Рис. 3.4. Короткоживущие побеги ели
из спящих почек придают вид
«ведьминых метел»
155
Рис. 3.5. Динамика радиального прироста по диамет-
ру сосны обыкновенной в 50-летних древостоях
в районах с разным уровнем атмосферного заг-
рязнения
По оси абсцисс — годы, по оси ординат — прирост по
диаметру, мм. Уровень атмосферного загрязнения: 1 —
фоновый, 2 — высокий
Рис. 3.6. Сухие 300-350-летние сосны вблизи ком-
бината «Североникель» на Кольском п-ве
5) ослабление побегооб-
разования вследствие от-
мирания ветвей основной
кроны с одновременным
усилением образования ко-
роткоживущих побегов из
спящих почек (рис. 3.4);
6) снижение радиально-
го прироста древесины
ствола (рис. 3.5) и скелет-
ных ветвей;
7) изменение габитуса
молодых деревьев, превра-
щение их в кустообразную
или псевдостланиковую
форму;
8) гибель деревьев
(рис. 3.6) с полной дегра-
дацией древостоя.
Для целей полевой инди-
кации загрязнения и оценки
жизненного состояния де-
ревьев наиболее часто ис-
пользуют первые пять при-
знаков.
Как отмечалось выше,
различия в химическом со-
ставе загрязнителей, их кон-
центрации, длительности ра-
зовых, эпизодических, сис-
тематических или постоян-
ных воздействий вызывают
неодинаковые ответные ре-
акции растений, при этом их
внешнее отображение час-
то неспецифично (или недостаточно специфично), поэтому без специальных хи-
мических анализов не всегда возможно отличить влияние одного загрязнителя
от другого. Кроме того, атмосферные выбросы промышленных предприятий
представляют собой, как правило, сложные смеси различных по химическому со-
ставу газов и твердых частиц. Их совместное действие на растение бывает си-
нергическим (взаимно усиливающим), просто суммарным или антагонистичес-
156
ким, а симптомы повреждения листьев несколько отличаются от симптомов вли-
яния главного загрязняющего агента, действующего в одиночку. Все это предоп-
ределило специфику принятого в данной работе подхода: описание повреждений
листьев дается не для одиночных загрязнителей, а для их смесей, типичных для
тех или иных промышленных предприятий.
3.1.2. Диагностические признаки повреждения ассимиляционных
органов деревьев при воздействии различных типов загрязнения
Основным диагностическим признаком повреждения ассимиляционных орга-
нов растений атмосферным загрязнением является наличие хлорозов и некрозов
на хвое или листьях. Однако этот признак недостаточно специфичен, поскольку
хлорозы и некрозы могут возникать в результате действия других факторов: не-
достатка или избытка питательных веществ почвы, высоких и низких темпера-
тур, засухи, подтопления корневых систем, а также в результате деятельности эн-
томовредителей и различных патогенов. Для выявления воздействия фактора ат-
мосферного загрязнения необходимо проводить обследование участков леса на
разном удалении от источника загрязнения.
В данном разделе диагностические признаки повреждений ассимиляционных
органов растений рассматриваются для наиболее распространенных источников
загрязнения: металлургических комбинатов, предприятий по производству мине-
ральных удобрений и тепловых электростанций, работающих на угле.
Медно-никелевые комбинаты
Основные загрязнители. Диоксид серы и тяжелые металлы (N1, Си, Со).
Возрастная чувствительность листьев. Наименьшей чувствительностью
обладают растущие хвоя и листья, наибольшей — полностью закончившие рост.
Старая хвоя обладает промежуточной чувствительностью, однако из-за усили-
вающихся с возрастом нарушений в структуре кутикулы и покрывающих ее вос-
ков происходит зимнее и ранне-весеннее иссушение хвоинок, и они преждевре-
менно опадают. Более сильные повреждения хвои прежних лет связаны также с
большей длительностью воздействия на нее загрязнителей.
Характер повреждений хвои и листьев. У хвои сосен и елей хлорозы жел-
товатого, желтого и оранжевого цвета в виде точек, поясков, пятен и сплошных
участков, охватывающих обычно верхнюю часть хвоинок. Некрозы белесовато-
го, коричневого или бурого цвета появляются сначала на кончиках хвоинок и рас-
пространяются к основанию. У сосен может наблюдаться деформация и неравно-
мерный рост хвоинок. Лиственницы более устойчивы к хроническому загрязнению,
но весьма чувствительны к высоким концентрациям выбросов. Хлорозы, перехо-
дящие в некрозы, возникают менее чем через сутки после аварийных выбросов.
У лиственных пород хлорозы желтого, кофейного или красноватого цвета,
межжилковые и краевые. При длительном загрязнении переходят в некрозы крас-
157
но-бурого, бурого или черного цвета. Крупные жилки листьев и часть прилегаю-
щего к ним мезофилла повреждаются позже других тканей.
Подрост хвойных и лиственных пород и невысокий подлесок, защищенные
пологом взрослого древостоя, повреждаются меньше, что особенно справед-
ливо для экземпляров, находящихся зимой под снегом.
Заводы по получению алюминия
Основные загрязнители. Соединения фтора, преимущественно фтористый
водород, и диоксид серы.
Возрастная чувствительность листьев. Наибольшей чувствительностью
обладают растущие хвоя и листья, наименьшей — закончившие рост.
Характер повреждений листьев. Типичные формы некрозов при воздействии
фтора — верхушечные и краевые, т. е. места наиболее активных процессов функ-
ционирования ассимиляционных органов. У хвойных пород некроз приурочен к
верхней части хвоинки и имеет буровато-красный цвет, за что получил название
«ожога верхушки хвои». Повреждение может охватывать треть, половину и даже
всю хвоинку. При нескольких сублетальных фумигациях некроз распространяет-
ся порциями к основанию хвоинок, причем число эпизодов загрязнения можно
определить по более темным пояскам, отделяющим некротические участки друг
от друга. Пораженная фтором ткань отделяется от зеленых, неповрежденных
частей хвои четкой, резкой границей, иногда с грязно-зеленой каймой.
Низкие концентрации при их повторном или продолжительном действии при-
водят к накоплению фтора в многолетней хвое, вызывают ее повреждение и
опадание. В первую очередь страдают старые и ослабленные деревья. Сосны,
ели и лиственницы относятся к чувствительным видам к данному типу загряз-
нения.
У лиственных пород возникновению некрозов обычно предшествуют верху-
шечные и краевые светло-зеленые хлорозы, меняющие затем окраску на крас-
но-коричневую, коричневую или бурую. У листьев сложной конфигурации некрозы
захватывают выемки и доли листа. Расширение повреждения идет вглубь и к
основанию листьев, так что впоследствии остаются нетронутыми несколько глав-
ных жилок с незначительной частью зеленого по цвету мезофилла листа. Сухие
некротированные части листьев выкрашиваются, оставляя на дереве живые ос-
татки листьев.
Предприятия по производству минеральных удобрений
Основные загрязнители. Диоксид серы, оксиды азота (в основном диоксид
азота), аммиак, фтор.
Возрастная чувствительность листьев. Фтор повреждает растущие листья,
диоксид серы — закончившие рост, таким образом, повреждаются хвоя и лис-
тья различного возраста и фаз роста.
158
Характер повреждений листьев довольно пестрый из-за участия несколь-
ких загрязнителей. Диоксид азота при одиночном действии — слабый токсикант,
вызывающий неспецифический, порой временный хлороз. В смеси с диоксидом
серы дает синергический, усиливающий повреждение эффект. При умеренных
воздействиях загрязнителей хвоя старых возрастов приобретает тусклый грязно-
зеленый цвет, при повторных эпизодах желтеет и преждевременно опадает. При
выбросах высокой концентрации на старой хвое и листьях возникают хлорозы,
тогда как на молодых наблюдаются некрозы оранжевого, красно-коричневого
или бурого цвета. Хлорозы и некрозы краевые, верхушечные, межжилковые. На
хвоинках наблюдаются точечные повреждения, пятна, темно-окрашенные пояс-
ки. Характерна частичная или полная дефолиация деревьев. У лиственных пород
отмечается более раннее пожелтение и опадание листьев.
Многолетние выбросы оксидов азота и аммиака вызывают изменения в поч-
венном содержании азота и приводят к бурьянистому разрастанию нитрофильных
растений.
Работающие на угле электростанции
Основные загрязнители. Диоксид серы, оксиды азота, кальций, калий, маг-
ний и различные микроэлементы.
Возрастная чувствительность листьев. Более чувствительны закончившие
рост хвоя и листья.
Характер повреждения листьев. Точечные или пятнистые хлорозы желто-
зеленого или желтого цвета, порой имеющие временный характер и исчезающие
после прекращения воздействия (например, перемене направления ветра) или
благоприятного изменения погоды (например, дожди после засушливого перио-
да).
Следует отметить, что в отличие от синергического или аддитивного (сум-
марного) действия диоксида серы в совокупности с тяжелыми металлами или
фтором, приводящего к полному распаду растительных сообществ и эрозии по-
чвенного слоя, примесь частиц щелочных металлов, необходимых для питания
растений (кальция, магния, калия) имеет антагонистический характер и снижает
неблагоприятное для деревьев действие избыточных количеств диоксида серы.
Леса вокруг небольших по мощности, а также крупных, но работающих на
низко сернистом топливе электростанций, в течение многих лет не обнаружива-
ют признаков внешних повреждений хвойных и лиственных пород. Угнетению и
гибели подвергаются чувствительные к SO2 лишайники и мхи. При хроническом
загрязнении несколько сокращается продолжительность жизни хвои. Полное улав-
ливание твердых частиц из труб электростанций приводит к созданию более жест-
кого воздействия оксидов серы и азота на растения. Неблагоприятные метеоро-
логические условия усиливают действие этого, довольно мягкого для древес-
ных растений типа загрязнения.
159
Таблица 3.1. Классификация хвои сосны обыкновенной по степени повреждения
атмосферным загрязнением
Площадь повреждения хвои, % Класс повреждения хвои Жизненное состояние хвои Краткое описание признаков повреждения хвои (изменения цвета)
0-5 0 здоровая Зеленая, без видимых признаков изменения цвета или повреждения, хвоя
6-10 1 здоровая Основной цвет хвои зеленый, однако на хвоинках могут встречаться светло-зеленые и коричневые точки микроскопических размеров. Приурочены они, как правило, к верхним частям и кончикам хвои.
11-25 2 слабо поврежденная На хвоинках встречаются некрозы кончиков хвои и хлорозы в виде пятен. Хвоинки могут частично приобретать светло-зеленый цвет с рассеянными по ним точечными некрозами («веснушки»).
26-50 3 умеренно поврежденная Цвет кончиков хвои (8-10 мм) с зеленого постепенно меняется на красно-коричневый или бурый Распространены верхушечные, краевые, пятнистые, в виде поясков, и точечные некрозы.
51-75 4 сильно поврежденная Начинает преобладать коричневый, красно- коричневый и серый цвет хвои. Здесь встречаются все типы некрозов.
>76 5 отмирающая или сухая Зеленого цвета практически нет, хвоинки деформированы, засыхают на побегах.
Сопоставление рассмотренных выше сведений о специфике повреждений дает
возможность убедиться, что, несмотря на некоторые отличия, атмосферные выб-
росы разных промышленных предприятий вызывают сходные симптомы. В ряде
случаев различи я в повреждениях нечетки и без дополнительных химических
анализов невозможно точно установить, какой именно фитотоксикант вызвал
повреждение.
Распространение хлорозов и некрозов у хвойных пород всегда происходит
однотипно: от кончика хвоинки к ее основанию. На основе многолетних иссле-
дований нами ра зработаны рекомендации по количественной оценке степени по-
вреждения хвои деревьев атмосферным загрязнением (табл. 3.1).
3.1.3. Диагностические признаки состояния деревьев в условиях
атмосферного загрязнения
Для рационалтьного ведения лесного хозяйства и применения санкций к пред-
приятиям, загрязняющим окружающую среду, необходимо количественно оце-
160
нить изменение жизненного состояния деревьев и древостоев. С этой целью, в
соответствии с принципами «Санитарных правил в лесах Российской Федерации»
(1992), разработана шкала оценки жизненного состояния деревьев в условиях
атмосферного промышленного загрязнения. В связи с существованием есте-
ственной дифференциации деревьев в древостоях необходимо проводить коли-
чественный учет состояния деревьев на постоянных пробных площадях, распо-
ложенных по профилю от фоновых (незагрязненных) условий до источника про-
мышленного загрязнения.
Шкала категорий жизненного состояния деревьев
1 — здоровое дерево (рис. 3.7). Деревья не имеют внешних признаков по-
вреждений кроны и ствола. Лишайниковый покров на стволах деревьев хорошо
развит, не имеет следов повреждения. Густота кроны обычная для деревьев I—II
классов Крафта. Мертвые и отмирающие ветви сосредоточены в нижней части
кроны; в верхней ее половине крупных отмерших ветвей нет или они малочис-
ленны и по периферии кроны не видны. Закончившие рост листья зеленого или
темно-зеленого цвета. Продолжительность жизни хвои типична для региона.
Любые повреждения листьев (хвои) незначительны (до 5 % их общей площади)
и не сказываются на состоянии дерева. При оценке состояния хвои на деревьях
мы используем классификации степени изменения цвета (табл. 3.2) и степени ее
жизненности (табл. 3.3).
Таблица 3.2. Классификация хвои сосны обыкновенной по степени изменения цвета
Площадь хвоинки, занятая хлорозами или некрозами, % Класс изменения цвета хвои Интенсивность изменения цвета хвои
0-5 0 нет
6-10 1 начальная
11-25 2 слабая
26-50 3 умеренная
51-75 4 сильная
>76 5 сплошная
Таблица 3.3. Классификация хвои сосны обыкновенной по степени жизненности
Доля хвоинок, изменивших цвет, % Класс изменения цвета Состояние хвои
0-10 0 здоровая
11-25 1 слабо поврежденная
26-50 2 умеренно поврежденная
>51 3 и4 сильно поврежденная
161
Рис. 3.7. Интенсивность охвоения и состояние крон ели европейской
1 — здоровое дерево, без видимых признаков повреждения хвои; 2 — дерево в условиях
слабых уровней атмосферного загрязнения, хвоя частично повреждена и начинает опадать;
3 —дерево в условиях средних уровней хронического загрязнения воздуха, хвоя интенсивно
повреждается и опадает (около 50%); 4 — дерево в условиях высоких уровней загрязнения
окружающей среды — сильно поврежденное (более 70 % хвои уже опало); 5 — отмираю-
щее или мертвое дерево.
2 — поврежденное дерево1 (рис. 3.7). В фоновых условиях (при отсутствии
загрязнения) обязателен хотя бы один из следующих признаков: снижение гус-
тоты кроны на 30-40 %; наличие 30-40 % мертвых и (или) усыхающих ветвей в
верхней половине кроны; повреждение 30-40 % хвои насекомыми, болезнями или
пожарами1 2. При загрязнении к числу обязательных диагностических признаков до-
бавляется наличие хлорозов и некрозов, занимающих не менее 6-10 % площади
всех листьев (хвои) дерева. Характерно также снижение продолжительности
жизни хвои3.
3 — сильно поврежденное дерево (рис. 3.7). При отсутствии загрязнения
(фоновые условия) обязателен хотя бы один из следующих признаков: сниже-
ние густоты кроны на 50-70 %; появление 50-70 % сухих и (или) отмирающих
1 Учитывая, что ухудшение жизненного состояния дерева при атмосферном загрязне-
нии, а часто и в силу других причин, вызывается повреждением ассимиляционного аппара-
та и других органов дерева, здесь и далее в шкале такие деревья называются поврежден-
ными. Термин «поврежденное дерево» используется здесь как синоним термина «ослаб-
ленное дерево».
2 Согласно «Санитарным правилам в лесах Российской Федерации» (1992), для листвен-
ных пород при этих видах повреждений «градации ослабленности устанавливаются в ос-
новном по степени усыхания кроны».
3 Этот показатель достаточно эффективен и легко определяем в северотаежных лесах,
где хвоя на деревьях.сосны сохраняется до 10-12 лет, а у ели — до 21-23 лет. По мере
снижения к южным широтам продолжительность жизни хвои у вечнозеленых деревьев сни-
жается и уже, например, в Ленинградской и Новгородской областях рассматриваемый по-
казатель практически «не работает».
162
ветвей в верхней половине кроны; отмирание вершины дерева; повреждение 50
70 % площади хвои насекомыми, болезнями или пожарами. У лиственных пород
возможно появление водяных побегов по стволу и в кроне. При загрязнении к
числу обязательных диагностических признаков добавляется наличие хлорозов
и некрозов, занимающих более 10 % площади всех листьев (хвои). Характерно
также существенное снижение продолжительности жизни хвои, резкое снижение
прироста дерева в высоту.
4 — отмирающее дерево (рис. 3.7). Основные признаки отмирания деревь-
ев одинаковы для контроля (фоновые условия) и при загрязнении: крона дерева
разрушена, ее густота составляет менее 15-20% по сравнению со здоровой;
более 70 % ветвей ее верхней половины сухие или усыхающие. Оставшиеся на
дереве листья хлоротичны — они бледно-зеленого, желтоватого или оранжево-
красного цвета. Некрозы имеют беловатый, коричневый или черный цвет. Мас-
совое распространение некрозов, возникающих при загрязнении, наблюдается не
всегда, поскольку большая часть полностью некротированных листьев (хвои)
быстро облетает.
5а — свежий сухостой. К нему относятся деревья, погибшие в течение пос-
леднего года. На дереве возможно наличие остатков сухой хвои или неопавших
сухих листьев.
56 — старый сухостой. К нему относятся деревья, погибшие в прошлые
годы. Постепенно утрачиваются ветви и кора.
Оптимальное время обследования состояния деревьев
Обследование древостоев хвойных пород с целью определения их жизненно-
го состояния возможно в любое время года, если деревья не покрыты снегом.
Наименьшее количество хвои отмечается весной, незадолго перед распускани-
ем почек, наибольшее — после завершения роста побегов и хвои. В таежной зоне
в естественных условиях (фон) наблюдаются два максимума некрозов: ранней
весной, при переходе, от отрицательных к положительным температурам и в кон-
це вегетационного периода. Оптимальное время для наблюдений и проведения
исследований — от второй половины июля до первой половины августа, когда
закончен рост побегов и вновь образовавшаяся хвоя становится восприимчивой
к воздействию большинства загрязнителей. Оценку жизненного состояния ли-
ственных древостоев в условиях загрязнения целесообразно проводить во вто-
рую половину лета, примерно за 2-3 недели до появления первых признаков по-
желтения фоновых (контрольных) древостоев. В первую половину вегетации,
когда листья еще не закончили рост, они редко обнаруживают некротические по-
вреждения (исключение составляют случаи очень высоких концентраций загряз-
нителей и воздействие фтористых соединений). Обследование деревьев в этот
период может привести к неправильной, заниженной оценке воздействия промыш-
ленных выбросов на лесные сообщества.
163
3.1.4. Оценка жизненного состояния древостоев
На основании данных, полученных в результате перечетов и оценки жизнен-
ного состояния деревьев на пробных площадях, рассчитываются индексы жиз-
ненного состояния древостоев. С этой целью деревьям той или иной категории
жизненности присваивают определенный балл: здоровым деревьям — 1,0; по-
врежденным — 0,7; сильно поврежденным — 0,4; отмирающим — 0,1; свеже-
му и старому сухостою — 0.
Расчет индекса состояния древостоя производят по формуле (Алексеев, 1990)
_ v, +0,7v2 +0,4v3 +0,1v4
v
где Iv — индекс жизненного состояния древостоя по объему деревьев; vj —
объем древесины здоровых деревьев лесообразователя (или лесообразователей)
на пробной площади, м3; V2, гз, v4 — то же для поврежденных (ослабленных),
сильно поврежденных и отмирающих деревьев соответственно; 0,7; 0,4 и 0,1 —
баллы поврежденных, сильно поврежденных и отмирающих деревьев; v — об-
щий запас древесины деревьев древостоя на пробной площади, м3 (включая объем
сухостоя).
При индексе от 1,7 до 0,8 жизненное состояние древостоя оценивается как
«здоровое», при индексе от 0,79 до 0,5 древостой считается «поврежденным»,
при индексе от 0,49 до 0,2 — «сильно поврежденным», при индексе от 0,19 и
ниже — «разрушенным» или «полностью деградированным».
При необходимости получения экспресс-информации допускается в качестве
ориентировочного показателя расчет индекса состояния древостоя по числу
деревьев. В этом случае
_ и, + 0,7/1, + 0,4/1, +0,1л4
п
где 1„ — индекс жизненного состояния древостоя по числу деревьев, п i — чис-
ло здоровых, «2 — поврежденных, пз — сильно поврежденных, /ц — отмираю-
щих деревьев лесообразователя или лесообразователей на пробной площади; п —
общее число деревьев (включая сухостой) на пробной площади.
Расчеты индекса состояния древостоя по числу деревьев более просты, чем
по запасу, но менее точны, поскольку не учитывают роль дерева тех или иных
размеров в древостое. Особенно искаженным оказывается состояние не подвер-
гавшихся уходу молодняков. Происходит это из-за того, что в обычных услови-
ях отпад формируется из отстающих в росте угнетенных деревьев, не играющих
существенной роли в сложении древостоя. При воздействии загрязнителей, на-
против, более сильное повреждение получают господствующие деревья, и ис-
пользование формулы 2 может привести к заниженной оценке вредоносности
промышленных выбросов.
164
3.2. ИСПОЛЬЗОВАНИЕ ЭПИФИТНОГО ЛИШАЙНИКОВОГО
ПОКРОВА ЛЕСНЫХ СООБЩЕСТВ ДЛЯ ИНДИКАЦИИ
АТМОСФЕРНОГО ЗАГРЯЗНЕНИЯ 1
Одной из важных экологических особенностей эпифитных лишайников явля-
ется их высокая чувствительность к атмосферному загрязнению (Barkrnan, 1958;
Somermaa, 1972; Wirth, 1980; Нильсон, Мартин, 1982; Нильсон, 1989). Это по-
зволяет использовать характеристики эпифитного лишайникового покрова для
биоиндикации промышленного загрязнения и долговременного мониторинга со-
стояния природной среды на территориях, занятых лесной растительностью
(Hawksworth, Rose, 1970; Somermaa, 1972; Gilbert, 1973; LeBlanc, Rao, 1975;
Wirth, 1980; Мартин, 1982, 1984; Крючков, Сыроид, 1984; Трасс, 1985; Горш-
ков, 1986, 1990, 1991; Pirmtsos et al., 1993).
Предлагаемая методика разработана в результате многолетнего изучения со-
стояния эпифитного лишайникового покрова в европейских северо- и южнота-
ежных сосновых лесах, подвергающихся воздействию промышленных выбросов
разной интенсивности и состава. В конкретных исследованиях методика может
быть реализована в полном или сокращенном объеме в зависимости от специ-
фики действующего фактора нарушения, характера изменения среды обитания
эпифитных лишайников, поставленных задач, квалификации исследователя.
Особенности выбора объектов исследования. Лихеноиндикационные иссле-
дования проводятся на постоянных пробных площадях, которые закладываются
вдоль одного или нескольких профилей на разном удалении от источника атмос-
ферного загрязнения. Система профилей создается таким образом, чтобы они рас-
полагались в разных направлениях от источника выбросов. Если закладывается один
профиль, то целесообразно, чтобы его направление совпадало с направлением вет-
ров, преобладающих в данном районе в течение вегетационного сезона.
Пробные площади в пределах каждого профиля закладываются в нескольких
точках, расположенных на разном расстоянии от источника выбросов. Число
точек, в которых проводятся лихеноиндикационные исследования, зависит от
поставленных задач (размеров территории, требуемой точности оценок и т. д.),
минимальное число точек — 3-4: две в пределах зоны загрязнения на разном
удалении от источника выбросов (опыт) и одна-две — в фоновых для региона
условиях (контроль). Если задачей исследования является зонирование террито-
рии по уровню промышленного загрязнения, на каждом из профилей необходи-
мо выбрать несколько (4-5) точек. Расположение точек определяется требова-
1 Автор В В Горшков
165
нием возможно более равномерного обследования территории и наличием лес-
ных массивов с определенными характеристиками сообществ (видовой состав,
возраст древесного яруса, типологическая принадлежность). В каждой точке ре-
комендуется закладывать не менее 2-3 пробных площадей. Подробнее о закладке
системы пробных площадей см. разд. 1.3. В качестве контрольных рассматрива-
ются пробные площади, расположенные на расстоянии не менее 40-50 км от всех
имеющихся на территории точечных источников промышленного загрязнения. При
индикации загрязнения среды, вызванного влиянием транспортных магистралей,
контрольные пробные площади могут располагаться от них на расстоянии >500 м.
Для обеспечения сопоставимости данных и получения корректных выводов
необходимо, чтобы лесные сообщества в точках исследования на территориях с
разным уровнем загрязнения (на разном расстоянии от источника выбросов в
пределах профиля) относились к одному и тому же типу леса и имели примерно
одинаковую давность последнего нарушения (см. разд. 1.1, 1.6), не связанного
с влиянием загрязнения — рубки, пожара, массового ветровала. В зависимости
от состояния лесной растительности на исследуемой территории могут рассмат-
риваться два основных возможных варианта системы пробных площадей: 1) в со-
обществах с разной давностью последнего пожара или рубки более 60-80 лет, в
которых в основном завершился процесс восстановления эпифитного лишайни-
кового покрова на деревьях, сохранившихся после пожара, или закончилось его
формирование на деревьях, выросших после рубки или пожара; 2) в восстанав-
ливающихся на месте вырубок и гарей молодых лесах с возрастом древостоя
менее 60 лет. В каждом из этих случаев изучение эпифитного лишайникового
покрова имеет определенную специфику.
Методика сбора данных при лихеноиндикационных исследованиях.
Измерение характеристик эпифитного лишайникового покрова на пробных пло-
щадях, расположенных по градиенту загрязнения, может выполняться на основе
общей методики изучения эпифитного лишайникового покрова, которая представ-
лена в разд. 1.7. Эта методика позволяет получить наиболее полную информа-
цию о составе и структуре эпифитного лишайникового покрова и экологических
особенностях отдельных видов лишайников в изучаемых типах лесных сооб-
ществ в фоновых условиях и на территории, подвергающейся воздействию ат-
мосферных выбросов, и в дальнейшем на основе сравнительного анализа этих
данных дать оценку изменения характеристик эпифитного лишайникового покрова
под воздействием фактора загрязнения.
Оценка состояния эпифитного лишайникового покрова. По характеру
изменения встречаемости и проективного покрытия под влиянием конкретных
источников загрязнения среди эпифитных лишайников может быть выделено
несколько (обычно 3-4) групп видов. Так, в составе эпифитной лишайниковой
биоты европейских бореальных сосновых лесов были выделены следующие
категории (группы) (Горшков, 1986, 1991; Ярмишко и др., 1998):
166
I — устойчивые, встречаемость и проективное покрытие которых не изменя-
ются или возрастают под влиянием загрязнения; эта группа выделяется вблизи
источников загрязнения с подщелачивающим эффектом выбросов на деревьях
с кислой (pH = 3,0-4,5) реакцией коры (сосна, ель, береза) или вблизи источни-
ков с подкисляющим эффектом выбросов на деревьях с субнейтральной
(pH = 5,0-5,5) реакцией коры (осина, тополь);
II — умеренно устойчивые, встречаемость и проективное покрытие которых
снижаются на расстоянии менее 3-5 (7-8) км от источника выбросов;
III — мало устойчивые, распространение которых снижается на расстоянии ме-
нее 20-30 км от источника выбросов;
IV — неустойчивые, встречающиеся только в фоновых районах.
В европейских северотаежных сосновых лесах, подвергающихся выбросам
крупного медно-никелевого комбината, ко II группе устойчивости были отнесе-
ны Cladonia spp., Lecidea flexuosa, Parmeliopsis hyperopta, P. ambigua; к III
группе — Hypocenomyce scalaris, Hypogymnia physodea, Ochrolechia
androgyna; к IV группе — Bryoria spp., Vulpicida pinastri, Imshaugia aleurites
(Горшков, 1991). Одни и те же виды в разных условиях загрязнения (при разной
интенсивности и разном составе выбросов) могут проявлять разную степень ус-
тойчивости и относиться к разным группам. В частности, в южнотаежных сосно-
вых лесах в окрестностях комбината по производству азотных удобрений лишай-
ник Hypogymnia physodes был отнесен к категории устойчивых, Imshaugia
aleurites и Parmeliopsis hyperopta — к категории умеренно устойчивых (Ярмиш-
ко и др., 1998).
Изменение состояния эпифитного лишайникового покрова по мере приближе-
ния к источникам промышленного загрязнения прослеживается по таким пара-
метрам, как:
1) среднее число видов лишайников в описании;
2) среднее суммарное проективное покрытие лишайников;
3) среднее покрытие диагностических (достоверно изменяющих покрытие под
действием загрязнения) таксонов;
4) встречаемость диагностических таксонов;
5) встречаемость описаний без лишайников;
6) среднее число видов лишайников на дереве;
7) общее число выявленных видов лишайников;
8) максимальная высота поднятия лишайников по стволу.
Указанные характеристики являются достаточно надежными диагностичес-
кими признаками. Последняя характеристика указывает на возрастание степени
повреждения эпифитного покрова по мере увеличения высоты над поверхностью
земли.
Точность определения таких показателей, как число видов лишайников в опи-
сании и общее число выявленных видов, существенно зависит от профессиональ-
167
ной подготовки исследователя, поэтому в качестве основных показателей состо-
яния эпифитного покрова в лесах с давностью последнего пожара или рубки бо-
лее 80 лет рекомендуется использовать характеристики 2, 3, 5. В некоторых слу-
чаях для оперативного получения общей оценки состояния эпифитного лишай-
никового покрова и лихеноиндикационного зонирования территории могут быть
использованы только две характеристики — 2 и 5.
В молодых восстанавливающихся лесных сообществах оценку состояния
эпифитного лишайникового покрова следует выполнять на основе использова-
ния характеристик 4, 6, 8. Это связано с тем, что параметры эпифитного лишай-
никового покрова (общее проективное покрытие, видовой состав и покрытие
отдельных видов) в значительной степени зависят от возраста деревьев: наибо-
лее существенные изменения происходят в течение первых 30 лет жизни дере-
ва, а стабилизация всех основных параметров отмечается в возрасте 60-80 лет
(Горшков, 1987; Тарасова, 2000). Кроме того, если система пробных площадей
заложена в восстанавливающихся молодых лесах, на каждой пробной площади
необходимо обследовать большее число деревьев, чем в коренных сообществах
(не 10-20, а не менее 30).
В европейских северотаежных сосновых лесах в качестве диагностических
таксонов предлагается использовать Imshaugia aleurites, Parmeliopsis ambigua,
P hyperopta, Lecidea flexuosa, Hypocenomyce scalaris, Cladonia spp. — у ос-
нования дерева, Hypogymnia physodes, Parmeliopsis ambigua, Imshaugia
aleurites, Bryoria spp — на высоте 1,3 м, а в молодых восстанавливающихся
лесах — Vulpicida pinastri. В южнотаежных сосновых лесах в качестве диагно-
стических таксонов могут быть использованы: Hypogymnia physodes,
Parmeliopsis ambigua, Parmelia sulcata, Cladonia spp , Imshaugia aleurites,
Hypocenomyce scalaris, Platismatia glauca, Xantoria spp , Physcia spp., si,
Pseudoevernia fuffuracea, Vulpicida pinastri и др.
Ограничения применимости диагностических признаков. Как в фоно-
вых условиях, так и в условиях загрязнения характеристики эпифитного лишай-
никового покрова существенно зависят от расположения участка на стволе де-
рева — высоты над землей и экспозиции, а также от угла наклона поверхности
ствола. Сопоставимость данных обеспечивается при использовании во всех точ-
ках исследования стандартной методики описания эпифитного лишайникового по-
крова (разд. 1.7): на двух фиксированных высотах (0-20 и 130-150 см) с четы-
рех сторон ствола, а также исключением из рассмотрения сильно наклоненных
деревьев (с углом наклона 10° и более).
Следует также принимать во внимание влияние низовых пожаров, во время
которых лишайниковый покров на стволах уничтожается. Полностью указанный
выше набор рекомендуемых диагностических признаков применим только к со-
обществам с давностью последнего пожара не менее 60-80 лет.
168
Таблица 3.4. Значения основных диагностических признаков эпифитного лишай-
никового покрова стволов сосен в окрестностях медно-никелевого комбината
(Кольский полуостров)
Признак Состояние лишайникового покрова
ненарушенный поврежденный разрушенный
Леса с давностью пожара или рубки более 80 лет
Среднее суммарное проективное покрытие (130-150 см), % 1,4-23 о, 1-0,5 . 0
Среднее суммарное проективное покрытие (0-20 см), % 9-32 3-10 <0,1-0,3
Встречаемость описаний без лишайников (130-150 см), % 0-38 66-95 100
Встречаемость описаний без лишайников (0-20 см), % 0-7 15-29 72-100
Среднее суммарное проективное покрытие видов II и Ш групп устойчивости (130-150 см), % 0,7-15 0-0,1 0
Восстанавливающиеся молодые леса
Среднее число видов лишайников на дереве 4,1-8,6 2,4-3,8 0-1,8
Встречаемость Vulpicida pinastri, % 30-100 0 0
Высота поднятия лишайников по стволу, см 24-200 4,6-21 0-4,6
Зоны состояния эпифитного лишайникового покрова. Дискретность
изменения отдельных характеристик лишайникового покрова при увеличении
уровня загрязнения позволяет выявить на исследуемой территории зоны с раз-
личным уровнем нарушения эпифитного лишайникового покрова (табл. 3.4) и оце-
нить относительный уровень загрязнения территории. Результаты специальных
исследований показывают, что наблюдается тесная взаимосвязь между разруше-
нием эпифитного лишайникового покрова и разрушением других компонентов
сообществ, поэтому состояние эпифитного лишайникового покрова позволяет
судить о состоянии лесных экосистем в зонах промышленного загрязнения (Мар-
тин, 1982; Крючков, Сыроид, 1984; Кишка, 1984; Горшков, Ярмишко, 1990;
Горшков, 1991).
169
3.3. ИСПОЛЬЗОВАНИЕ МХОВ ДЛЯ ИНДИКАЦИИ
ХАРАКТЕРИСТИК МЕСТООБИТАНИЯ1
Большинство мохообразных имеет широкую экологическую амплитуду, од-
нако существует немало видов мхов и печеночников, для благоприятного разви-
тия которых требуется наличие определенных эколого-ценотических условий.
Известно, что устойчивость болотных и лесных растительных сообществ
связана со стабильным состоянием доминантных видов мохового покрова. С
другой стороны, можно сказать, что устойчивое существование моховых сооб-
ществ — показатель нормального, ненарушенного состояния окружающей сре-
ды, например стабильное существование редких видов мхов или печеночников в
каком-либо конкретном местообитании. Появление же в моховом покрове сор-
ных видов, связанных с деятельностью человека, может быть показателем сте-
пени нарушения природной среды.
Для целей экспресс-бриоиндикации удобно использовать экологическую ха-
рактеристику видов мхов и печеночников. Подобная информация может содер-
жаться в имеющихся определителях и в специальных работах, но, к сожалению,
обычно она не охватывает всего разнообразия мхов и печеночников, встречаю-
щихся в лесных и болотных сообществах. В приведенном ниже списке представ-
лены широко распространенные и легко узнаваемые в поле виды мхов и пече-
ночников (необходимые диагностические признаки приводятся в описании видов),
обладающие видоспецифичной экологической приуроченностью. Данный список
видов является тем необходимым минимумом, который необходимо знать лю-
бому экологу, геоботанику или ботанику.
Водно-болотные мхи
Риччиа плавающая — Riccia fluitans L.
Класс — печеночники (Hepaticae). Семейство
— риччиевые (Ricciaceae).
Описание. Дихотомически ветвящийся слое-
вищный печеночник с линейными лопастями, от
1 до 5 см длины и 1 мм ширины, зеленый или жел-
товато-зеленый. Спороносит редко.
Экология. Встречается в небольших водо-
емах, прудах и канавах, на отмелях и старицах рек
и ручьев.
1 Автор Е. Н Андреева
170
Индикационная характеристика. Водный печеночник, встречающийся в
условиях повышенной эвтрофикации.
Сфагнум магелланский — Sphagnum magellanicum Brid.
Класс — мхи (Musci). Семейство — сфагновые (Sphagnaceae).
Описание. Дерновинки рыхлые или плотные, красноватые до пур-
пурных, в тени зеленые, но с красными вкраплениями. Стебель об-
лиственный, розовый, реже светло-зеленый; ветви собраны по 3-5 в
пучок, причем, две ветви всегда отстоящие, а остальные — свисаю-
щие; ветви вздуто облиственные (почкообразные). Ветви на верхуш-
ке стебля скучены в рыхлую головку. Спороносит редко.
Экология. Произрастает в сфагновых лесах и на олиготрофных и
мезотрофных болотах.
Индикационная характеристика. Содоминант мохового покрова
в сфагновых сосняках и на олиготрофных болотах.
Кукушкин лен обыкновенный — Polytrichum commune Hedw.
Класс — мхи (Musci). Семейство — политриховые
(Polytrichaceae).
Описание. Дерновинки мощные. Стебель прямостоячий, до 20-
40 см высоты, без ризоидного войлока. Листья шиловидно заост-
ренные (влажные — отогнуто отстоящие; сухие — прямо отсто-
ящие с отогнутой верхушкой) с высоким желтоватым или бело-
ватым стеблеобъемлющим основанием, плоскокрайные, до основа-
ния остропильчатые-, жилка выступает из верхушки листа в виде
красной зубчатой ости. Ножка спорогона до 12 см длиной, краснова-
тая. Коробочка призматическая с 4—6 острыми ребрами, вначале пря-
мостоячая, позднее горизонтальная. Колпачок густо-волосистый, зо-
лотисто-бурый.
Экология. Произрастает на влажных почвах в лесах, на лугах и на
болотах.
Индикационная характеристика. Показатель за-
болачивания.
Фонтиналис противопожарный — Fontinalis
antipyretica Hedw.
Класс — мхи (Musci). Семейство — фонтиналие-
вые (Fontinaliaceae).
Описание. Дерновинки плавающие, мощные, тем-
но-зеленые, коричневые до черных. Стебель до 50 см
длины, неправильно или кустисто разветвленный (во
171
влажном состоянии остротрехгранно облиственный), ветви острые или тупые.
Листья без жилки, острокилеватые, старые листья по линии киля обычно разор-
ваны. Спороносит только после спада воды или высыхания водоема.
Экология. Произрастает в стоячей и проточной воде при pH = 6,5-7,5.
Индикационная характеристика. Показатель чистой или умеренно чистой
воды.
Климациум древовидный — Climacium dendroides
(Hedw.) Web. et Mohr
Класс — мхи (Musci). Семейство — климациевые
(Climaciaceae).
Описание. Дерновинки рыхлые, зеленые или желто-зеле-
ные. Первичный стебель подземный, ползучий, с ризоидами;
вторичный стебель — прямостоячий до 15 см высоты, вни-
зу без ветвей; с нежными, пленчатыми низовыми листьями,
вверху древовидно разветвленный. Спороносит редко.
Экология. Произрастает на сырых лугах, окраинах болот,
черноольшаниках, ивняках, по берегам рек, ручьев и озер.
Индикационная характеристика. Показатель богатства
почр.
Хелодиум Бландова — Helodium blandowii (Web. et
Mohr) Warnst.
Класс — мхи (Musci). Семейство — хелодиевые
(Helodiaceae).
Описание. Дерновинки бледно- или желто-зеленые до бу-
рых. Стебель ползучий, восходящий или прямостоячий, 5-
15 см высоты, крупные, рыхлые, темно-зеленые или бурова-
тые, блестящие. Стебель перистый, густовойлочный (из-за
многочисленных нитевидных зеленых выростов стебля и
ризоидов)f с извилистыми, двурядно отстоящими, утонча-
ющимися к концам ветвями. Верхние ветви скученны и об-
разуют небольшую головку. Спороносит редко.
Экология. Произрастает на колоднике или среди сфагнов
в олиготрофных и мезотрофных болотах.
Индикационная характеристика. Вид переувлажненных
местообитаний; не выносит загрязнения.
172
Каллиергон гигантский — Calliergon giganteum
(Schimp.) Kindb.
Класс — мхи (Musci). Семейство — амблистегиевые
(Amblystegiaceae).
Описание. Дерновинки крупные, рыхлые, темно-зеленые
или буроватые, блестящие. Стебель прямостоячий, почти
перистый или неправильно ветвистый, реже простой, с прямы-
ми колючими, острыми верхушками стебля и ветвей. Лис-
тья рыхло сидящие, прямо отстоящие, вогнутые, с тупой, кол-
пачковидной, вверху часто разрывающейся верхушкой. Спо-
роносит редко.
Экология. Произрастает в сильно обводненных топяных
болотах, сплавинообразователь.
Индикационная характеристика. Содоминант эвтрофных и мезотрофных
болот.
Лесные мхи
Птилидиум красивейший — Ptilidium
pulcherrimum (G.Web.) Vain.
Класс — печеночники (Hepaticae). Семейство —
птилидиевые (Ptilidiaceae).
Описание. Дерновинки плотные, плоские, при-
жатые к субстрату, темно-оливковые или желто-
вато-бурые. Стебельки тонкие, лежачие, до 2 см дли-
ны, одиножды или дважды перисто-ветвистые, с ко-
роткими веточками. Листья глубоко разделенные на
доли, с длинными многочисленными ресничками по
краям. Двудомные. Половой диморфизм хорошо вы-
ражен: мужские дерновинки образованы более мел-
кими шнуровидными побегами с обильным ветвлени-
ем; женские дерновинки — с многочисленными выступающими булавовидными
перианциями, защищающими молодой спорогон. Спороносит часто.
Экология. Повсеместно произрастает на колоднике, валунах и на деревьях.
Индикационная характеристика. Эпифитный печеночник; чувствителен к
загрязнению воздуха.
173
bjt&ghp. Сфагнум оттопыренный — Sphagnum squarrosum Crome
Класс — мхи (Musci). Семейство — сфагновые (Sphagnaceae).
<r|gV Описание. Дерновинки рыхлые, мощные, зеленые, от сизо- до
Ijgr желтовато-зеленых. Стебель толстый, облиственный; ветви курчавые
Шкк (из-за отогнутых и загнутых назад верхушек веточных листьев),
собраны по 4—5 в пучок. На верхушке стебля ветви короткие, с огра-
ниченным ростом, скученны в рыхлую головку; наружные ветви го-
ловки узко заостренные. Спороносит часто.
Экология. Произрастает в сфагновых ельниках, в низинных и клю-
JmSL чевых болотах, на окрайках олиготрофных болот и на заболоченных
ягам лугах.
Индикационная характеристика. Показатель богатства почв.
V Дикранум многоножковый — Dicranum
polysetum Sw. Г
Класс — мхи (Musci). Семейство — дикрановые (Dicranaceae). /
Описание. Дерновинки крупные, мягкие, рыхлые, желтовато-зе- 1 /
леные, блестящие. Стебель прямостоячий, до 15 см высоты, с гу- Ufflk/
стым беловатым или бурым ризоидным войлоком. Листья до 1 см
длиной, прямо отстоящие до оттопыренных (сухие — сильно по-
перечно-волнистые), широколанцетные, остропильчатые, с узко-ту-
поватой верхушкой. Покровный лист обертывает от I до 5 ос-
нований ножек спорогонов.
Экология. Произрастает на влажных и сухих почвах в хвойных
лесах, на болотах и гарях.
Индикационная характеристика. Доминант мохового покро-
ва в зеленомошных сосняках.
Дикранум метловидный — Dicranum scoparium
Hedw.
Класс — мхи (Musci). Семейство — дикрановые
(Dicranaceae).
Описание. Дерновинки мягкие, рыхлые, темно-зеле-
ные, блестящие. Стебель слабоветвистый, 3-15 см вы-
соты, с умеренным беловатым или бурым ризоидным
войлоком; верхушка стебля оканчивается хохолком из
листьев. Листья серповидные, остропилъчатые, до
8 мм длиной, с широко-ланцетным основанием, посте-
пенно заостренные в длинную желобчато-шиловидную
верхушку; жилка на спинке вверху листа с 2-5 продоль-
ными зубчатыми пластиночками. Спорогоны одиночные.
174
Экология. Произрастает на деревьях, валунах, колоднике и почве в сухих и
влажных лесах.
Индикационная характеристика. Содоминант мохового покрова в зелено-
мошных сосняках.
Птилиум военное перо — Ptilium crista
castrensis (Hedw.) De Not.
Класс — мхи (Musci). Семейство — гипновые
(Нурпасеаё).
Описание. Дерновинки рыхлые, желтовато- или
светло-зеленые, блестящие. Стебель восходящий, до
20 см длины, густо и правильно перисто-ветвистый с
горизонтально отстоящими, плоско распростертыми
(на конце серповидно согнутыми) ветвями. Внешним
обликом напоминает вайю папоротника «страусо-
вое перо». Спороносит редко.
Экология. Произрастает в хвойных лесах, на по-
чве и на колоднике.
Индикационная характеристика. Гиперацидо-
фил; не выдерживает эвтрофикации среды.
Гилокомиум блестящий — Hylocomium
splendens (Hedw.) B.S.G.
Класс — мхи (Musci). Семейство — гилокомиевые
(Hylocomiaceae).
Описание. Дерновинки рыхлые, оливково-зеленые,
шелковисто-блестящие. Стебель восходящий до 20 см
длины с 5-8-этажным расположением дуговидно
изогнутых годичных побегов (новый побег закладыва-
ется перед верхушкой прошлогоднего побега). Годич-
ный побег в верхней части дважды или трижды перис-
то-ветвистый с двурядно отстоящими, плоско распрос-
тертыми ветвями. Спороносит редко.
Экология. Произрастает в хвойных и смешанных ле-
сах.
Индикационная характеристика. Сокращение чис-
ла годичных побегов до 1-3 — показатель нарушения
лесных экосистем.
175
Плевроциум Шребера — Pleurozium schreberi (Brid.)
Mitt.
Класс — мхи {Musci). Семейство — гилокомиевые
{Hylocomiaceae).
Описание. Дерновинки рыхлые, зеленые или желто-зе-
леные, блестящие. Стебель 10-15 см длины, восходящий,
вздуто облиственный, правильно перисто-разветвленный с
коричневато-красной корой, просвечивающей сквозь ли-
стья; на концах ветвей иногда могут образовываться ризо-
иды. Стеблевые листья черепитчатые, ложковидно-вогну-
тые. Спороносит редко.
Экология. Произрастает повсеместно на почве и на ко-
лоднике.
Индикационная характеристика. Доминант зелено-
мошных лесов.
Редкие виды
Трихоколеа войлочная — Trichocolea tomentella
(Ehrh.) Dum.
Класс — печеночники {Hepaticae). Семейство —
трихоколиевые {Trichocoleaceae).
Описание. Дерновинки рыхлые, бледно- или жел-
то-зеленые. Стебель от 3 до 12 см высоты, прямос-
тоячий дважды-трижды перисто-ветвистый, при
основании с редкими ризоидами. Листья рассечены
на 4 ланцетные лопасти; каждая лопасть несет
многочисленные длинные волоски из одного ряда кле-
ток, которые придают всему растению характерный
пушистый облик. Спороносит редко.
Экология. Произрастает во влажных ельниках.
Индикационная характеристика- Редкий вид
смешанных ельников.
Риччиокарпос плавающий — Ricciocarpos
natans (L.) Corda
Класс — печеночники {Hepaticae). Семейство —
риччиевые {Ricciaceae).
Описание. Дважды-трижды дихотомически ветвя-
щийся оливковый (до желто-зеленого) слоевищный пе-
ченочник, образующий неполные розетки с сердце-
видными лопастями. Диморфный вид. Плавающая
176
форма слабоветвящаяся, с крупными лентовидными темно-фиолетовыми зуб-
чатыми свисающими, брюшными чешуйками. Наземная форма чаще ветвяща-
яся с большим числом ризоидов. Лопасти достигают 14 мм длины и 3-7 мм
ширины, имеют центральный желобок и выемку на верхушке. На северо-западе
России не спороносит.
Экология. Плавающая форма приурочена к стоячим и слабопроточным во-
доемам.
Индикационная характеристика. Редкий вид. Устойчив к водному загряз-
нению.
Скорпидиум скорпионовидный — Scorpidiutn
scorpioides (Hedw.) Limpr.
Класс — мхи (Musci). Семейство — амблистегие-
вые (Amblystegiaceae).
Описание. Дерновинки рыхлые, желто- или темно-
зеленые до черных, блестящие. Стебель прямостоячий
или лежачий до 30 см длины, простой или вильчатый,
вместе с ветвями вздутосережчато облиственный,
с крючковидно согнутыми, верхушками стебля и вет-
вей. Листья черепитчатые, сильно выпуклые, серповид-
но изогнутые или прямые. Спороносит редко.
Экология. Произрастает в сильно обводненных эв-
трофных болотах, сплавино-образователь.
Индикационная характеристика. Редкий вид пе-
реувлажненных местообитаний.
Некера перистая — Neckera pennata
Hedw.
Класс — мхи (Musci). Семейство — не-
керовые (Neckeraceae).
Описание. Дерновинки крупные, светло-
или желто-зеленые, блестящие. Главный сте-
бель ползучий, нитевидный; вторичные
стебли перистые, восходящие, висячие, 3-
6 см длины. Ветви густо плоско облиствен-
ные, короткие или удлиненно утончающиеся.
Сухие листья сильно поперечно-волнистые,
несимметричные. На нижней (брюшной)
стороне ветвей развиваются многочислен-
ные, погруженные в обертку сидячие коро-
бочки.
177
Экология. Произрастает на стволах старых осин и широколиственных пород.
Индикационная характеристика. Отмечен на участках старых неморальных
ельников.
Виды нарушенных местообитаний
Маршанция многообразная —
Marchantia polymorpha L.
Класс — печеночники (Hepaticae). Семей-
ство — маршанциевые (Marchantiaceae).
Описание. Слоевище крупное, 2-20 см
длины и 1-2 см ширины, вильчато ветвяще-
еся, на верхней стороне имеется срединная
полоска темного цвета. Молодые слоевища
с выводковыми колокольчатыми корзиночка-
ми. Двудомное. Мужские подставки 0,5-2 см
длины, с восьмилопастной дисковидной голов-
кой. Женские подставки 2-10 см длины, с 9-
тилучевой головкой (у молодой подставки
лучи опущены, у зрелой — расположены го-
ризонтально). Спороносит часто.
Экология. Произрастает на ключевых и низинных болотах; на удобренной
земле, гарях, по стенкам канав, по берегам заболачиваемых речек, ручьев и озер.
Индикационная характеристика. Нитрофил; сорняк на огородах.
Кукушкин лен можжевельниковый — Polytrichum
juniperinum Hedw.
Класс — мхи (Musci). Семейство — политриховые
(Polytrichaceae).
Описание. Дерновинки рыхлые, сизо-зеленые. Стебель пря-
мостоячий до 10 см высотой, в основании с рыхлым бурым вой-
локом из ризоидов. Листья линейно-ланцетные с желтоватым
стебле-объемлющим основанием и цельными загнутыми
внутрь беловатыми пленчатыми краями; жилка выступает из
верхушки листа в виде короткой красновато-бурой ости. Ножка
спорогона до 6 см длиной, красновато-бурая. Коробочка удли-
ненно-призматическая с 4 острыми ребрами, вначале прямосто-
ячая, позднее — горизонтальная. Колпачок густо-волосистый,
светло-бурый или беловатый.
Экология. Произрастает на обнаженной почве, вырубках,
гарях и суходольных лугах.
Индикационная характеристика. Пионерный вид после пожаров.
178
Фунария влагомерная — Funaria hygrometrica Hedw.
Класс — мхи (Musci). Семейство — фунариевые
(Funariaceae).
Описание. Дерновинки рыхлые до 3 см высотой, от
бледно-зеленых до темно-зеленых. Верхние листья до
4 мм длины, почковидно скученные, коротко заострен-
ные. Ножка спорогона 3-5 (8) см длины, первоначально со-
гнутая, гигроскопичная; спирально скручивается в сухих ус-
ловиях и раскручивается во влажных. Коробочка горизон-
тальная или повислая, вздуто-косогрушевидная, горба-
тая, глубоко-бороздчатая, коричневая: крышечка широ-
кая, плоско-выпуклая, с оранжевым краем.
Экология. Произрастает повсеместно на обнаженной
почве. Нитрофил.
Индикационная характеристика. Массовое развитие обильно споронося-
щих дерновинок — показатель антропогенного воздействия.
Цератодон пурпурный — Ceratodon purpureus
(Hedw.) Brid.
Класс — мхи (Musci). Семейство — дитриховые
(Ditrichaceae).
Описание. Густые зеленые, желто-зеленые, корич-
невые или красновато-коричневые дерновинки, 2-3 см
высотой. Стебель прямостоячий. Сухие листья — при-
легающие, извилистые; увлажненные — прямо отстоя-
щие. Коробочка наклоненная до горизонтальной, сла-
бо согнутая, продолговатая, красно-бурая, блестя-
щая, полосатая (сухая — глубоко бороздчатая), на
пурпурно-красной или желтоватой блестящей нож-
ке. Крышечка тупоконическая.
Экология. Произрастает на природных (почва, де-
ревья, валуны) и искусственных (бетон, заборы, крыши,
кострища и т. п.)_ субстратах.
Индикационная характеристика. Массовое разви-
тие обильно спороносящих дерновинок — показатель
антропогенного воздействия.
Наибольшей индикационной значимостью обладают синузии мохообразных,
которые более точно, чем отдельный вид, входящий в данную синузию, отража-
ют условия местообитания. Действительно, синузия, по сравнению с отдельным
видом, обладает гораздо более узкой экологической амплитудой, что, собственно,
179
Таблица 3.5. Взаимосвязь синузий мхов с трофностью и увлажнением местообита-
ний (Буш, Аболинь, 1968)
Градиент трофности местообитания Градиент увлажнения местообитания
Кссро-мсзофитнос Мсзо-гигрофитнос Г игрофнтнос Г игро-гидрофитнос
Олиготрофное Ccratodon (Pohlia nutans, Leptobryum pyriforme) Sphagnum capillifolium (S russowii) Sphagnum fuscum (S rubellum, Dicranum bergeri, Mylia anomala) Sphagnum cuspidatum (S Jlexuosum)
Олиго- мсзотрофнос Dicranum scoparium Polytrichum commune Sphagnum mazellanicum Sphagnum fallax
(Hypnum cupressiforme, Orthodicranum monlanum, Plagiolhecium laetum) Plcurozium (Dicranum polyselum, Plilium crisla-castrensis, Polytrichum Juniperinum) - S anzustifolium (Wamstorjia fluitans)
(S cenlrale, Polytrichum slriclum, Aulacomnium paluslre)
Мезотрофнос Hvlocomium splendens Polytrichum longisetum Sphagnum zirzensohnu (S wulfianum, S paluslre) Sphagnum warnslorfii (S teres, Calliergon stramineum, Dicranum bonjeanii, Helodium blandowii) Sphagnum subsecundum (S contortum, S platyphyllum)
Мезо- эвтрофное Rhvtidiadelphus triauelrus - Thuidium erectum Climacium-Callicreonclla Limprihtia cossonii
Brachvlhecium oedipodium (Sanionia uncinata, Sphagnum squarrosum) (Bryum pseudolri- quetrum, Plagiomnium elatum, Sphagnum fimbrialum) (Campylium stellatum, Hamatocaulis vernicosus)
(Eurhynchium angustirele, Plagiolhecium denliculalum, Plagiomnium ajj'ine, Rhodobryum roseum, Cirriphyllum piliferum)
Эвтрофное Atrichum Plagiomnium cuspidalum Plagiomnium undulalum Callierzon cordifolium (Rhizomnium punctatum, Pseudobryum cinclidioides, Brachythecium rivulare, Marchantia polymorpha) Cratoncuron (Cratoneuron jllicinum, Palustriella commutata, Conocephalum conicum Philonotis jontana)
и создает уникальное своеобразие каждой синузии. Индикационное значение си-
нузий мхов или фитоценологических групп (Буш, Аболинь, 1968) для оценки влаж-
ности и трофности местообитаний показано в табл. 3.5. Следует отметить, что
моховые синузии, выделенные по результатам исследований в лесах Латвии, со-
храняют свое индикационное значение и для территорий северо-запада России.
Значение мохообразных для индикации антропогенного воздействия на раститель-
ные сообщества рассмотрено в работах Е. Н. Андреевой (1989, 19906).
180
3.4. ХИМИЧЕСКИЙ АНАЛИЗ ПОЧВ И РАСТЕНИЙ 1
Одна из основных задач при изучении состояния лесных экосистем состоит в
получении достоверной и всесторонней информации об уровнях загрязнения
окружающей среды. Для получения таких данных важен контроль качества ра-
боты на всех этапах определения загрязнения в растительных сообществах, на-
чиная с отбора проб. Неправильный выбор пунктов отбора, горизонтов почв,
растений и их частей, ошибки в организации и технике отбора проб приводят к
серьезным просчетам при получении окончательных результатов содержания
контролируемых ингредиентов в экосистеме. Погрешность результатов анализа,
обусловленная исключительно пробоотбором, может достигать сотен процентов,
что делает дальнейшее проведение анализа бессмысленным. Получение досто-
верной аналитической информации возможно только при тесном взаимодействии
специалистов-экологов и аналитиков.
Рассеивание промышленных выбросов в атмосфере зависит от метеорологи-
ческих факторов, в основном от направления и скорости ветра, наличия и высо-
ты температурных инверсий, количества осадков и т. д. Кроме того, имеют зна-
чение характеристики предприятий: их мощность и технология, система очист-
ных сооружений, высота труб, физико-химические свойства выбрасываемых ча-
стиц, их дисперсность и химический состав. В рассеивании и перераспределении
загрязнителей промышленного происхождения на земной поверхности важную
роль играют физико-географические особенности обследуемого района: рель-
еф местности, растительность, состояние поверхности почв и др.
3.4.1. Загрязнение почвы
В зависимости от состава почвообразующих пород, климата, рельефа мест-
ности, растительного покрова и других факторов естественное содержание ме-
таллов в верхних горизонтах почвы может существенно различаться даже для
одного типа почв. В настоящее время фоновый уровень содержания тяжелых
металлов является суммой естественного содержания тяжелых металлов с тем
добавлением металлов антропогенного происхождения, которое является след-
ствием глобального переноса загрязнителей от источников их выброса в окру-
жающую среду. Именно над этим уровнем выделяются очаги локальных высо-
ких концентраций в непосредственной близости от источника.
Загрязнение почвы в первую очередь определяется «розой ветров» данного
района и скоростью ветра. Наибольшие уровни загрязнения отмечаются в направ-
лении преобладающих ветров. При слабых ветрах обычно наблюдается сильная
1 Автор И. В. Лянгузова
181
вертикальная устойчивость атмосферы. При таких условиях примесь, поступая
из трубы, плохо рассеивается и переносится на большие расстояния. Усиление
ветра сопровождается усилением вертикального обмена в атмосфере, что при-
водит к переносу примеси от оси распространения к поверхности земли. При
дальнейшем увеличении скорости ветра до очень больших величин происходит
интенсивное рассеивание примеси в атмосфере, проникновение ее в верхние слои
тропосферы и дальнейший перенос. Особенно далеко переносятся высокодис-
персные аэрозоли. Так, например, в снежном покрове Антарктиды были обнару-
жены в значительных количествах тяжелые металлы и пестициды (Микроэлемен-
ты в природных водах..., 1974). Наибольшая плотность выпадения тяжелых
металлов от промышленного источника отмечается в его окрестностях обычно
в радиусе до 3-5 км от него. По мере удаления до 10-20 км концентрации тяже-
лых металлов уменьшаются примерно на порядок величины по сравнению с мак-
симальными, а иногда достигают фоновых значений. В случаях обследования почв
вокруг особо крупных металлургических комплексов фоновые содержания мо-
гут наблюдаться в направлении преобладающих ветров на расстоянии до 50, а
иногда и 100 км.
Особенностью изучения загрязнения почв металлами является то, что срав-
нительно большое количество металлов в почвах имеет естественное происхож-
дение, поэтому отделить металлы естественного происхождения от техногенно-
го очень сложно и даже не всегда возможно. Это затрудняет изучение их мигра-
ции в почвенном профиле. Распределение металлов в профиле загрязненной по-
чвы отличается от распределения в незагрязненной почве. В основном тяжелые
металлы техногенного происхождения сосредоточиваются в поверхностном 5-
10-сантиметровом слое. Часть металлов, попавших в почву, образует труднора-
створимые формы соединений с гумусовыми веществами, какая-то часть может
войти в состав поглощенных оснований, замещая кальций и магний, а часть — в
состав глинистых минералов с расширяющейся кристаллической решеткой. Все
они закрепляются в верхнем гумусовом слое, а оставшаяся часть металлов миг-
рирует в профиле почвы в растворенном виде с почвенным раствором, при ме-
ханическом перемещении в адсорбционном состоянии до большой глубины, при-
чем второй максимум содержания металлов чаще находится на границе с аллю-
виальным горизонтом.
При горизонтальной миграции наблюдается перераспределение металлов по
элементам ландшафтов. Накопление тяжелых металлов в трансаккумулятивном
и аккумулятивном ландшафтах в два-три раза больше, чем в почве элювиально-
го ландшафта (Вертинская и др., 1977).
Обследование почв для установления их промышленного загрязнения обыч-
но проводят принятым в почвоведении методом почвенно-геоморфологических
профилей (Бобовникова и др., 1980; Вертинская и др., 1983; Методические ука-
зания по контролю загрязнения почв, 1977), который позволяет систематизиро-
182
ванно, объективно и наиболее полно изучать почвы обширных территорий на
содержание в них тяжелых металлов.
Почвенно-геоморфологический профиль — это вытянутый в одном направ-
лении, узкий (несколько сотен метров) и длинный (несколько десятков километ-
ров) участок суши, пересекающий все элементы рельефа. Исходной точкой по-
чвенно-геоморфологического профиля служит источник загрязнения. По оцен-
кам Г. К. Вертинской и др. (1983), удвоение содержания металлов в верхнем
сантиметровом слое почвы в районе промышленных комплексов происходит
примерно в течение года, а в пахотном слое — за 10-20 лет, поэтому отбор проб
почвы вокруг промышленных комплексов проводится по полной и сокращенной
программам. По полной программе пробы отбираются один раз в пять лет по
восьми почвенно-геоморфологическим профилям (восьми румбам), один из
которых совпадает с преобладающим направлением ветра. По сокращенной про-
грамме отбор проб проводится ежегодно по четырем почвенно-геоморфологи-
ческим профилям. Если «роза ветров» близка к круговой, то расположение то-
чек отбора проб по всем направлениям при полной программе соответствует рас-
стояниям 0,5; 1; 2; 3; 4; 6; 8; 10; 20 и 30 км от источника выбросов, а при сокра-
щенной — расстояниям 1; 2; 3; 5 и 10 км. В случае вытянутой «розы ветров»
точки отбора по румбам целесообразно распределять пропорционально повто-
ряемости соответствующих направлений ветров. При этом для направлений с
малой повторяемостью ветров целесообразно не принимать во внимание преж-
де всего точки отбора на больших расстояниях.
Такой методический подход позволяет достаточно репрезентативно предста-
вить уровень загрязнения почв тяжелыми металлами и на основе данных хими-
ческого анализа провести экологическое картирование обширных территорий.
Однако для оценки состояния и устойчивости наземных экосистем данный ме-
тод имеет ограничения, т. к. при описанном способе отбора проб не учитывает-
ся тип растительности, расположенной по почвенно-геоморфологическому про-
филю.
При исследованиях состояния, продуктивности и устойчивости лесных экоси-
стем крайне важно, чтобы во всех точках отбора проб вокруг одного источника
все условия местообитания были одинаковыми (рельеф, тип почвы, тип расти-
тельности и пр.). Более подробно выбор ключевых участков и закладка посто-
янных пробных площадей для мониторинга состояния лесных экосистем описан
в разд. 1.3. Правильный и стандартный выбор места отбора проб определяет
сопоставимость и репрезентативность полученных результатов.
Отбор проб почв
Перед отбором проб почв проводят их описание (см. разд. 1.8).
Отбор проб почв проводят с учетом локальной неоднородности почвенного
покрова, приводящей к варьированию содержания микроэлементов.
183
Следует проявлять особое внимание при отборе проб по почвенному профи-
лю. Шри исследовании горизонтов почв, имеющих различного рода новообразо-
ваниж (карбонаты, гипс, оолиты), отбирают представительную пробу, характери-
зуюпвую горизонт в целом.
При полевых исследованиях уровней загрязнения почвы тяжелыми металла-
ми вазжно знать необходимое количество почвенных проб для составления пред-
ставительного смешанного почвенного образца. Этот вопрос все еще остается
сложным и слабоизученным. Для представительности смешанного образца не-
обходимо при усреднении брать достаточное число индивидуальных проб. При
изучении элементного состава почв Н. Г. Зырин и др. (1971) для получения до-
стове:рной информации о средней концентрации микроэлементов с вероятной
относительной погрешностью не выше 20 % рекомендуют брать не менее 4-5
индивидуальных проб в пределах почвенного контура в плакорных условиях; при
этом каждая из них смешивается из пяти проб, взятых с площади около 2 м2.
Известно, что вариабельность содержания металлов в почвах увеличивается
при гприближении к источнику загрязнения и является наименьшей в фоновых
районах. Проведя большую методическую работу по определению отбора не-
обходимого количества индивидуальных проб для получения представительно-
го среднего образца и различных способов отбора почвенных проб, Г. К. Вер-
тинская и др. (1983) пришли к выводу, что при массовом отборе образцов с це-
лью определения уровней загрязнения почв металлами можно отбирать пробы
почвенным, буром или лопатой и усреднять 10 проб.
В полевых условиях на большой кусок полиэтиленовой пленки помещают
почву одного из почвенных горизонтов из 10 индивидуальных прикопок, распо-
ложенных равномерно по территории пробной площади. Каждый образец тщатель-
но пе ремешивают лопатой или деревянной палочкой.
Для уменьшения количества отобранного материала до количества, необхо-
димого для пробы, применяют различные приемы и приспособления. Простей-
ший шз них — метод квартования, сущность которого видна на рис. 3.8. Почву
насыщают в виде конуса (рис. 3.8а) затем конус превращают в усеченную пира-
миду (рис. 3.86), которую делят на 4 части (рис. 3.8в). Заштрихованные части
отбра»сывают, а остальные 2 части соединяют и используют для следующего
квартования. Так продолжают до
тех пор, пока количество отобран-
ного материала не будет соответ-
ствовать необходимой величине
пробы. Обычно масса проб почвы
не превышает 100 г.
Отобранные для анализа пробы
почв помещают в чистые бумаж-
ные пакеты (восковая бумага или
184
пергамент) или мешочки из хлопчатобумажной ткани. В последнее время для
упаковки проб все шире используют полиэтиленовые мешочки, вызывающие
наименьшее загрязнение микроэлементами. В каждый пакет вкладывают этикет-
ку, на которой необходимо отметить:
1) название почвенного горизонта (Aq; AqA^; А2; Ви; Сит. д.);
2) мощность горизонта (0-2 см; 2-5 см и т. д.);
3) № пробной площади или ключевого участка;
4) дата взятия образца (число, месяц, год).
В зависимости от поставленной задачи на этикетке следует отметить допол-
нительную информацию. При исследовании загрязнения окружающей среды не-
обходимо указать расстояние и направление от источника загрязнения, при изу-
чении восстановительных процессов — давность последнего нарушения (пожа-
ра, рубки, мелиорации и пр.). Часто бывает целесообразно указать тип раститель-
ного сообщества или парцеллы.
В случаях более детального обследования (например, при изучении неодно-
родности напочвенного покрова) следует отбирать индивидуальные пробы в за-
висимости от поставленных задач.
Подготовка к химическому анализу и хранение проб почв
В лаборатории влажные образцы почвы необходимо высушить до воздушно-
сухого состояния в хорошо проветриваемом помещении. Образцы толщиной 1-
1,5 см раскладывают на бумагу (восковая, пергамент) на столах или полу. Комки
раздавливают фарфоровым пестиком или деревянной палочкой, отбирают кам-
ни, корни, шишки, ветки и другие неразложившиеся части растений. Периодически
почву перемешивают для более равномерного просушивания. Высушенные об-
разцы размельчают в ступке (лучше агатовой или яшмовой) и просеивают через
алюминиевое сито с диаметром отверстий 1 мм. Вместо металлических сит при
просеивании можно использовать шелковые, волосяные или из капроновой сет-
ки с пластмассовым или деревянным ободком.
Подготовленные к анализу пробы почв хранят в лабораторных условиях в
бумажных или полиэтиленовых пакетах с этикеткой.
Химический анализ почв
Для аналитической обработки собранных образцов почв обычно применяют
стандартные методы анализа почв. Классическими руководствами для физико-
химической характеристики почв являются «Руководство по химическому ана-
лизу почв» (Аринушкина, 1970) и «Агрохимические методы исследования почв»
(1975), а также «Практикум по почвоведению» (Гречин и др., 1964). Универсаль-
ными показателями физико-химических свойств почв являются:
1) pH водной вытяжки из почв;
2) pH солевой вытяжки;
185
3) гидролитическая кислотность почв;
4) сумма обменных оснований;
5) содержание обменных форм Са;
6) содержание обменных форм Mg;
7) содержание общего азота;
8) содержание общего углерода;
9) соотношение С : N;
10) содержание подвижных форм К2О;
11) содержание подвижных форм Р2О5;
12) общее содержание гумуса.
Важной характеристикой почвы является также ее механический состав.
В зависимости от целей и задач исследования проводят дополнительные хи-
мические анализы почв. С целью оценки уровня загрязнения окружающей сре-
ды и составления экологических карт обычно определяют валовые содержания
загрязнителей в почве. Для изучения миграции и трансформации загрязнителей
внутри почвенной толщи проводят определение их содержания по генетическим
горизонтам почвы. При оптимизации минерального питания растений анализиру-
ют количество NPK, а также содержание микроэлементов в пахотном слое по-
чвы для расчета необходимых доз удобрений. В случае оценки состояния, про-
дуктивности и устойчивости растительных сообществ в условиях промышлен-
ного загрязнения наиболее информативными будут данные о содержании подвиж-
ных форм тяжелых металлов в почве, т. к. именно эти соединения будут в пер-
вую очередь доступны для растений.
Почвы вместе с микроорганизмами играют роль универсального биологичес-
кого адсорбента, очистителя и нейтрализатора загрязнителей, минерализуют все
органические отходы и остатки. Благодаря этой функции почв в пределах био-
сферы в течение длительного времени (в историческом смысле) происходило
«самоочищение природы» отходов и остатков, производимых в постоянно воз-
растающих количествах населением и развивающейся промышленностью и выб-
расываемых в окружающую среду.
Как известно, основная масса корней растений в лесных биоценозах распола-
гается в верхних горизонтах почвы, а в лесных почвах (подзолах и подбурах)
только верхний органогенный горизонт является единственным аккумулятивным
горизонтом почв и, следовательно, основным источником питательных веществ
для растений. В связи с вышесказанным наиболее информативным горизонтом
почвы является верхний органогенный горизонт почвы, т. е. лесная подстилка.
Определение валовых количеств металлов в почвах
Разложение почвенных проб для анализа валовых количеств химических эле-
ментов обычно проводят двумя способами: 1) сплавление почвы со смесью буры
и соды; 2) кислотное разложение пробы почвы. Подробное описание методик
186
разложения проб почв и определения отдельных элементов приводится в «Ме-
тодических указаниях...» (1975; 1977; 1985; 1989).
Метод сплавления
Почвенную пробу переводят в раствор после сплавления со смесью буры и
соды в отношении 1:2. Этот способ позволяет полностью разложить пробу и
получить устойчивый раствор всех элементов.
Сущность метода. Навеску воздушно-сухой почвы помещают в платиновый
тигель и ставят в холодный муфель. Нагревают муфель и прокаливают тигли с
почвой до постоянного веса. Затем в тигель добавляют смесь буры и соды (1 : 2),
перемешивают содержимое стеклянной палочкой и сплавляют в муфеле при 950-
1000 °C. Горячий тигель вынимают из муфеля и осторожными вращательными
движениями распределяют плав тонким слоем по стенкам тигля. Охлажденный
тигель с плавом помещают в стакан, заливают кипящей дистиллированной водой
и нагревают на водяной бане до полного выщелачивания плава. Тигель вынима-
ют и тщательно обмывают горячей дистиллированной водой. В стакан при энер-
гичном перемешивании приливают НО (1 : 3) и продолжают перемешивать до
полного растворения гидроокисей. Раствор переносят в мерную колбу, после
охлаждения доводят до метки дистиллированной водой и перемешивают. В по-
лученном растворе определяют содержание металлов методом атомно-абсорб-
ционной спектрофотометрии.
Метод кислотного разложения
Выбор кислот при кислотном разложении почвы для определения общего со-
держания элементов обуславливается аналитической задачей. Наиболее часто
используют смесь плавиковой и соляной кислот.
Сущность метода Подготовленную к анализу воздушно-сухую почву допол-
нительно растирают до состояния пудры в агатовой или яшмовой ступке. Навес-
ку тонко растертой почвы помещают в платиновую чашку и прокаливают в му-
феле при температуре 500 °C. К охлажденной прокаленной почве, смоченной
дистиллированной водой, приливают последовательно НС1 (6 н) и концентриро-
ванную плавиковую кислоту и выпаривают почти досуха на этернитовой плитке.
Обработку смесью плавиковой и соляной кислот повторяют 2 раза. Прибавляют
концентрированную соляную кислоту для удаления фтора и выпаривают досуха.
К остатку приливают НС1 (6 н), закрывают часовым стеклом и нагревают на этер-
нитовой плитке до растворения осадка. Добавляют горячую дистиллированную
воду и фильтруют в мерную колбу. Фильтр промывают горячей водой, подкис-
ленной НС1 до удаления следов железа. После охлаждения раствор доводят до
метки дистиллированной водой и перемешивают. В полученном растворе опре-
деляют содержание металлов методом атомно-абсорбционной спектроскопии.
187
Определение подвижных форм элементов в почвах
С целью определения степени доступности различных элементов из почвы
растениям используют различные вытяжки из почв.
Вопрос о выборе универсального химического растворителя, позволяющего
учесть все влияния, крайне сложен и до сих пор не решен. Несмотря на долго-
летнюю работу, нет ни одной общепринятой вытяжки из почвы даже для одного
элемента питания в ограниченной зоне с одинаковыми почвенно-климатически-
ми условиями. Это связано с целым рядом обстоятельств:
1) растения используют не только подвижные, но и связанные формы элемен-
тов, но только из тех частиц почвы, которые непосредственно соприкасаются с
корневой системой;
2) растворители действуют на все частицы почвы в отличие от корней расте-
ний, поэтому возможно завышение результатов анализа;
3) несопоставимо время взаимодействия корневой системы и растворителей
с почвой (весь вегетационный сезон в первом случае и 1-24 ч в последнем);
4) имеется взаимовлияние различных химических элементов.
В связи с вышеизложенным к экстрагентам предъявляются следующие тре-
бования.
1. Учет взаимовлияния химических элементов. Его можно осуществить при
условии определения максимального числа элементов в одной вытяжке. Этому
условию удовлетворяют кислые буферные растворы и кислотные растворители.
2. Способность вытяжек отражать разнообразное влияние изменчивых свойств
почвы на поглощение различных элементов растениями. Этому требованию не
удовлетворяет практически ни одна из вытяжек.
Всесоюзным координационным совещанием по диагностике обеспеченности
почв микроэлементами и методам их определения в 1968 г. для извлечения под-
вижных форм Си, Мп, Zn, Со зональным агрохимическим лабораториям были
рекомендованы следующие вытяжки (табл. 3.6):
1) для некарбонатных и малокарбонатных (менее 4 % СО2) почв — вытяжки
по Пейве и Ринькису;
2) для карбонатных почв — вытяжка по Крупскому и Александровой;
3) для сероземов Средней Азии — вытяжка по Кругловой.
При разработке метода оптимизации минерального питания растений для оп-
ределения подвижных форм элементов в почве Г. Я. Ринькис и В. Ф. Нолендорф
(1977) выбрали вытяжку 1 н НС1, которая хотя и недостаточно полно отражает
влияние почвенных факторов, однако позволяет учесть взаимовлияние элемен-
тов и выгодна в методическом отношении, т. к. в ней можно определять все 13
элементов питания.
Экспериментально было установлено, что с увеличением кислотности экст-
р агента содержание подвижных форм тяжелых металлов в почвенной вытяжке
1 88
Таблица 3.6. Методика определения подвижных форм микроэлементов в почвах
Элемент Экстрагент Отноше- ние ПОЧВЫ к раствору Время взаимодей- ствия почвы с раствором Почвы, для которых применим метод Автор метода
Мп 0,1 н H2SO4 1 : 10 1 ч при взбалтыва- нии на ротаторе Некарбонатные и малокарбонатные почвы нечернозем- ной полосы, красноземы 1*
Си 1 н НС1 1 : 10 и н 1
Zn 1 н КС1 1 . 10 _ и _ 1
Со 1 н HNO3 1 : 10 и 1
Мп, Си, Zn, Со Ацетатный бу- ферный раствор с pH 4,8 1 : 10 _ п Карбонатные почвы нечерноземной по- лосы, почвы степ- ной, полустепной и пустынной зон 2
Мп, Си, Zn, Со Ацетатный бу- ферный раствор с pH 3,5 1 . 5 10 мин периодичес- кого взбалтывания от руки и 30 мин на ротаторе Почвы Средней Азии 3
В Н2О 1 . 5 5 мин при кипяче- нии Все почвы 4
Мо Оксалатный буферный раствор с pH 3,3 (реактив Тамма) 1 : 10 1 ч при взбалтыва- нии на ротаторе Все почвы 4
Примечание: 1 Пейве Я В, Ринькис Г Я. Методы быстрого определения доступных ра-
стениям микроэлементов (Си, Zn, Мп, Со, Мо, В) в почвах // Почвоведение — 1958. — № 9
2. Крупский Н. К., Александрова А М. К вопросу об определении подвижных форм микро-
элементов // Микроэлементы в жизни растений, животных и человека. Киев, 1964 3. Кругло-
ва Е. К. Прописи определения условно усвояемых растениями форм микроэлементов в кар-
бонатных почвах. Ташкент, 1970. 4. Веригина К. В, Добрицкая Ю. И., Целикова Н А. Про-
писи определения подвижных форм микроэлементов в некарбонатных почвах. М , 1969.
увеличивается (Чертов и др., 1990), что облегчает проведение химического ана-
лиза по обнаружению малых концентраций элемента. Кроме того, корреляцион-
ный анализ данных о содержании загрязнителей в различных вытяжках из почв и
растений показал, что наиболее высокие значения коэффициентов корреляции
наблюдаются между концентрациями тяжелых металлов в растениях и в соляно-
кислой вытяжке из верхнего органогенного горизонта почвы. Таким образом, для
целей экологического мониторинга состояния лесных экосистем в условиях про-
мышленного загрязнения окружающей среды целесообразно использовать в
189
качестве экстрагента 1 н НС1 для определения содержания подвижных форм
тяжелых металлов в почве.
3.4.2. Загрязнение растений
Повреждение растений токсикантами и уровень их накопления в растениях
определяются многочисленными факторами как биологическими, так и окружа-
ющей среды. Поступление тяжелых металлов в растения в значительной мере
зависит от химической природы металла (в больших количествах в растениях
могут накапливаться Zn, Cd, Со, Ni, Mo, Fe; слабо аккумулируются Pb, Cr, Hg),
концентрации и форм присутствия в почве (свободные ионы, комплексные со-
единения, нерастворимые минеральные и др. соединения), типа почвы, ее меха-
нического состава и физико-химических свойств, особенно pH и окислительно-
восстановительного потенциала. Здесь имеют значение и систематическое поло-
жение растений, и фаза развития растения, и его возраст, и даже, при всех прочих
одинаковых условиях, — индивидуальная природа самого растения и его отдель-
ных органов.
Отбор проб растений
Как отмечалось выше, на минеральный состав растений оказывают влияние
многочисленные факторы, поэтому крайне важно проводить отбор проб расте-
ний в сопоставимых однотипных условиях местообитания: положение в релье-
фе, тип почвы, тип растительного сообщества, таксационные характеристики
древостоя, видовой состав и строение напочвенного покрова и др. В связи с этим
в зависимости от поставленных задач проводят отбор:
1) ср едних проб растений по отдельным видам (чаще всего в конце вегетаци-
онного сезона);
2) смешанных проб всех растений с площадок 1 м2 (укосы) в нескольких ти-
пичных для исследуемой территории участках;
3) индивидуальных проб растений по одному экземпляру одного и того же
вида в пределах одной пробной площади.
При проведении детальных исследований состояния лесных сообществ в усло-
виях промышленного загрязнения закладывают пробные площади в сопоставимых
растительных сообществах, расположенных по градиенту загрязнения окружающей
среды. Подробное описание выбора и закладки пробных площадей приведено в
разд. 1.3. Для индикации загрязнения окружающей среды используют различные
виды высших и низших растений, а также отдельные их части и органы. Класси-
ческими аккумуляторами загрязнителей являются мхи и лишайники.
Наиболее часто в такого рода исследованиях применяют отбор средних проб
растений по отдельным видам. На территории пробной площади равномерно от-
бирают не менее 30 особей каждого вида растений травяно-кустарничкового и
мохово-лишайникового ярусов. При отборе проб растений прежде всего исклю-
190
чают загрязнение их почвой. Растения срезают, у них отделяют не только корни,
но и прикорневые листья, загрязненные почвой. Для очищения корней от почвы
их промывают водой или тщательно очищают щеточкой. В полевом журнале
отмечают фенофазу каждого вида и особенности его произрастания. В некоторых
случаях надземные части растений также промывают дистиллированной водой,
либо органическими растворителями. Растения расчленяют по органам (цветы,
листья, стебли, корни), сушат на воздухе в хорошо проветриваемом помещении
или в специальных сушильных шкафах. Затем пробы растений упаковывают в чис-
тую плотную бумагу или марлевый мешочек вместе с этикеткой, где указывают:
а) орган (цветы, семена, листья, стебли, корни и пр.);
б) название растения;
в) № пробной площади;
г) тип растительного сообщества;
д) расстояние и направление от источника загрязнения;
е) дату сбора (число, месяц, год);
ж) место сбора (область, район, село).
У деревьев пробы листьев (хвои) отбирают из средней части кроны, распо-
ложенной к источнику загрязнения. Многолетние побеги разделяют по годичным
приростам и разбирают хвою по возрастам. На этикетке дополнительно указы-
вают возраст хвои.
Одновременно отбирают пробы почвы и подстилки, описанным выше спосо-
бом.
Для определения относительной массы каждого вида в общей биомассе и для
устранения возможного пропуска видов-концентраторов используют методику
укосов. На наиболее представительных участках закладывают 5-10 площадок по
1 м2, при помощи секатора выстригают все растения на уровне подстилки. Взве-
шенную массу разбирают на группы (злаки, осоки, бобовые, разнотравье, мхи,
лишайники); взвешенные группы растений разбирают до видов; все экземпляры
выделенных видов взвешивают (все вместе) и отбирают на анализ среднюю про-
бу массой не менее 50 г. Пробы растений сушат на воздухе в хорошо проветри-
ваемом помещении и упаковывают в плотную чистую бумагу или марлевые ме-
шочки, вкладывая в каждый пакет этикетку с указанием вида растения, номера
пробной площади, расстояния и направления от источника загрязнения, даты и
места сбора. Одновременно с укосных площадок отбирают подстилку и почву.
Подготовка к химическому анализу и хранение проб растений
Высушивание, измельчение и хранение. При подготовке проб растений к
анализу обычными являются операции высушивания и последующего озоления.
Однако в результате этих процессов, особенно озоления, существенно изменя-
ется содержание химических элементов в анализируемых объектах. Высушива-
ние проб растений до воздушно-сухого (при 20-30 °C) или абсолютно-сухого
191
(при 100-120 °C) состоянии Чиводит к значительному концентрированию боль-
шинства химических элемйПВ, обычно в 2—5 раз. Кроме свободной и связан-
ной воды (в живом веществ[астений содержится от 30 до 98 % воды) при вы-
сушивании растений теряюИ’астворенные в воде и тканях растений газы, а так-
же некоторые химическиеэтменты, находящиеся в виде легколетучих соеди-
нений: I, Hg, Br, Se и др. Варивание при 105 °C и длительное хранение (в те-
чение 2 лет) проб растителИ«'О материала приводят к большим потерям ртути.
Не исключена возможное"!- 'о при высушивании и хранении растений могут
быть потери кадмия, таллил галура, полония и некоторых других элементов. По-
этому при проведении масс.зк анализов растений необходимо выбрать несколь-
ко типичных проб, изучить на их влияние условий хранения и высушивания (а в
дальнейшем и озоления) ыаэвультаты определения и внести соответствующие
поправки.
Для размельчения растирных образцов многие исследователи пользуются
кофейными мельницами. ОЬшая лабораторная мельница, в которой в процессе
измельчения происходит грене металла рабочих частей, для определения сле-
дов металлов непригодна. Цзельчение вручную пестиком в фарфоровых ступ-
ках не приводило к замети оуагрязнению пробы Fe, Си, Zn, В, Со, Мп, Мо, Са,
Na, Mg, Р, S, К, но этот пр1И°чень трудоемок (Карякин, Грибовская, 1979).
Хорошо известно, что тиИ измельченный растительный материал озоляет-
ся значительно хуже, чем, пример, целые листья, ветви, крупные части расте-
ний. Это объясняется тем, «растительный порошок обычно ложится доволь-
но плотным слоем и, такимм’азом, доступ кислорода в его нижние части зат-
руднен. Чтобы проба была наставительной, рекомендуется озолять несколько
порций слабо измельченны ртений, а золу объединять.
Минерализация расттлных проб. Химический анализ растений чаще
всего проводят после минерлзации органического вещества. Многочисленные
варианты способов озоленй ’Огут быть объединены в две основные группы:
сухое озоление и мокрое орние. При сухом озолении окисление органичес-
кой части пробы проходит >прсутствии кислорода воздуха под действием вы-
сокой температуры (на газегтгорелке, на электрической плитке, в электричес-
ких печах). Чаще всего проб покаливают в электрических печах — муфелях при
температуре 400-500 °C. И®гд озоление проводят при дополнительном введе-
нии кислорода. В некоторьиигодиках сухого озоления для предотвращения по-
терь определяемых элемента «ускорения процесса используют различные добав-
ки NH4NO3, Mg(NO3)2, KCLfr,hO2, HNO3, H2SO4 и др. Озоление мокрым спосо-
бом, т. е. нагревание пробы цеиущественно с концентрированными неорганичес-
кими кислотами, иногда в цристствии окислителя и катализатора, проводят при
более низкой температуре, («алетода имеют и недостатки, и преимущества.
Проведенный анализ обырой литературы, посвященной методикам минера-
лизации проб растительно!!! мтериала, приводит к мысли, что универсального
192
метода озоления пока не существует. Среди исследователей до сих пор нет еди-
ного мнения, какой же метод лучше. Выбор метода минерализации растительно-
го материала зависит прежде всего от определяемого элемента и его соедине-
ний в пробе, а также от выбора метода химического анализа. К тому же следует
учесть, что различные пробы требуют разной обработки. Как правило, разложе-
ние органических веществ кислотами используют для определения легколетучих
элементов или при работе с материалами, трудно озоляющимися сухим спосо-
бом или дающими мало золы.
Способы сухого озоления. Преимущества метода сухого озоления заклю-
чаются в том, что он удобен, прост и более доступен. Он не требует особого
внимания и не включает никаких реактивов, вносящих загрязнение. В процессе
сухого озоления происходит сначала обугливание озоляемого материала, а затем
его полная минерализация. При бурном протекании обугливания вместе с выде-
лением дыма и газов могут быть механические потери определяемых элемен-
тов. Поэтому рекомендуется помещать пробы в холодную муфельную печь и
затем постепенно доводить температуру до 450 °C. В начале озоления необхо-
дим небольшой приток воздуха, чтобы предотвратить осаждение углерода на золу.
При сухом сжигании имеется некоторая опасность загрязнения проб вещества-
ми, возгоняющимися с внутренней поверхности печи. В связи с этим рекомен-
дуется проводить озоление в муфельной печи, выложенной изнутри кварцем, а
платиновые тигли или чашки с озоляемым материалом, во избежание потерь
микроэлементов, прикрывать кварцевыми часовыми стеклами.
Но у метода сухого озоления имеются и недостатки. Некоторые органичес-
кие вещества крайне медленно озоляются при 400-450 °C, и пробы содержат уг-
леродный остаток даже после 72-часовой обработки. Кроме того, при сухом
озолении возможны потери определяемых элементов в результате уноса конвек-
ционными потоками не связанных между собой механически частиц золы, а так-
же в результате взаимодействия определяемых элементов со стенками чашек или
тиглей. Е. Сендел (1964) указывает, что при озолении проб с низким содержанием
минеральных веществ в кварцевых или фарфоровых чашках некоторое количество
определяемого элемента удерживается на поверхности чашки вследствие образо-
вания силиката. Особенно заметно это при пользовании старыми чашками с шеро-
ховатой поверхностью. При озолении в платиновой посуде легко окисляемые ме-
таллы, такие как свинец или железо, могут сплавляться с материалом посуды.
А. В. Карякиным и И. Ф. Грибовской (1979) не обнаружено потерь Мп, Mg,
Fe в процессе минерализации растительных проб, однако авторами установлено,
что при сухом озолении растений с предварительным обугливанием на электри-
ческой плитке могут быть потери Ni, Cr, V, РЬ и некоторых других химических
элементов.
Г. Я. Ринькис (1963) сравнил содержание Си, Zn, Со и Мо в навесках расте-
ний при различных способах обугливания: при медленном и быстром с пламенем.
193
Он пришел к выводу, что обугливание вполне можно проводить и ускоренными
темпами. В предложенном им ускоренном способе озоления парами азотной
кислоты (Ринькис, 1963) для быстрого обугливания пробы чашку с навеской
нагревают на электрической плитке и при появлении дыма содержимое чашки
поджигают. Обугливание должно быть полным. Не полностью обугленные образ-
цы при обработке парами азотной кислоты дают пламя высокой температуры, что
обуславливает термические и механические потери микроэлементов. После пре-
кращения выделения дыма чашку с обугленным материалом подвергают воздей-
ствию паров концентрированной азотной кислоты. Метод позволяет провести
быстрое озоление органического материала и сводит к минимуму потери мик-
роэлементов, обусловленные длительностью нагревания. Возможность загряз-
нения материала устраняется тем, что на оголяемый материал поступают только
пары HNO3. Несмотря на многие достоинства, этот метод все же не получил
широкого распространения, поскольку исключена возможность следить за ходом
озоления, и проверка полноты озоления материала занимает довольно много
времени.
Многие специалисты предпочитают смешивать высушенные образцы с неорга-
ническими окислителями. При этом озоление идет быстрее и более полно при низ-
ких температурах. Иногда пробу смешивают с насыщенным раствором Mg(NO3)2
для обеспечения более полного контакта озоляемого вещества с окислителем. В
некоторых случаях во время озоления с целью предупреждения потерь летучих
хлоридов (особенно при определении мышьяка и сурьмы) используют подщела-
чивание. Некоторые авторы для уменьшения потерь определяемых элементов и
ускорения процесса озоления добавляют к пробе раствор ацетата магния.
Большинство исследователей рекомендуют проводить озоление почв и рас-
тений при 450 -500 °C до полного окисления всего углерода озоляемого мате-
риала до СОг. Однако известны случаи, когда озоление проводят при указанной
температуре строго ограниченное время, например в течение 2 ч, 1 ч и т. д.
Следует отметить, что при совершенно одинаковых условиях для озоления
растений из разных семейств или из разных районов требуется неодинаковое вре-
мя. Так, для озоления злаков, собранных в пределах средней полосы Европейс-
кой части СССР, требуется иногда в два раза больше времени, чем для озоления
злаков из районов Средней Азии с повышенным содержанием кальция и щело-
чей. Озоление листьев ландыша протекает обычно на 3^1 ч медленнее, чем озо-
ление корней заманихи (Карякин, Грибовская, 1979). Наибольшие трудности и
затраты времени встречаются при озолении стеблей растений. При правильном
озолении полученная зола имеет белый, кремоватый, розовый или охристый цвет.
Количество времени, необходимое для полного озоления пробы, в каждом кон-
кретном случае разное.
Важным условием озоления является поддержание в муфельной печи строго
определенной температуры. Как уже отмечалось выше, большинство исследо-
194
вателей рекомендует озолять пробы при температуре не выше 500 °C, в то же
время в ряде случаев озоление проводят при более высокой температуре —1
600 °C и даже 800-900 °C.
Однако даже при строгом соблюдении температурного режима возможны
потери некоторых химических элементов, связанные с массой навески озоляе-
мого материала. При сжигании растительных проб массой 20 г потери Си дости-
гали 71 %, a Zn — 50 % по сравнению с потерями этих элементов при сжигании
пробы массой 2,5 г (Карякин, Грибовская, 1979). На потери меди и цинка влияет
содержание в растениях кремния и фосфора, причем наличие кремния увеличи-
вает, а фосфора уменьшает потери. Для устранения потерь предлагается озолять
небольшие навески растений массой до 2 г, распределяя озоляемый материал
тонким слоем. Для определения Со вполне допустимо сухое озоление от 10 до
20 г вещества, так как потери его при этом невелики. Совершенно недопустимо
озоление навесок биологического материала массой 50-100 г, так как при таком
приеме возможны большие потери некоторых микроэлементов.
Озоление в атмосфере кислорода является одним из вариантов сухого озоле-
ния. Его обычно проводят в герметически закрытом металлическом сосуде, а
именно в калориметрической бомбе в атмосфере кислорода под давлением. Для
предохранения попадания частичек бомбы в продукты сгорания внутренность
калориметрической бомбы и находящиеся в ней части должны быть покрыты
тонким слоем платины Хотя этот метод обеспечивает быстрое озоление и пол-
ное сохранение продуктов озоления, он пока не нашел широкого применения, так
как требует сложного оборудования. В методах с применением кислородной
бомбы высушенный образец сжигают электричеством в кислороде под давле-
нием 30-1000 бар. Озоление протекает в течение нескольких секунд, и золу лег-
ко извлекают промыванием бомбы кислотой. Для работы этим методом требу-
ется опытный исполнитель.
Другим вариантом метода сухого озоления является низкотемпературное
сжигание в кислородно-плазменном атомизаторе. Суть метода заключается в том,
что кислород, прошедший через высокочастотное поле, возбуждается в этом
поле и приобретает высокоокислительные свойства. Процесс озоления проходит
при температуре не выше 100°С, вследствие чего потери микроэлементов силь-
но уменьшаются. Недостатком метода является большая затрата времени на
озоление. Часто для полного озоления одной пробы требуется более 24 ч.
Метод мокрого озоления. Минерализацию проб мокрым способом, т. е. пу-
тем нагревания с концентрированными неорганическими кислотами, иногда в
присутствии катализатора и окислителя, достаточно широко используют при под-
готовке проб растений для определения в них микроэлементов. В настоящее
время известно много различных разлагающих смесей, однако до сих пор не
разработана единая смесь, которая бы хорошо разрушала все биологические
пробы с полным сохранением всех металлов. Предложенную еще в начале XX
195
века А. Нойманом смесь серной и азотной кислот для разрушения органическо-
го вещества применяют и по сей день. При использовании этой смеси обеспечи-
вается сохранение большинства металлов, но процесс разложения протекает
очень медленно. Если проба при этом содержит жиры, то их полное разложение
может занять 24 ч.
В типичном методе мокрого озоления пробы массой 2-5 г разлагают в 30 мл
смеси HNO3, HCIO4 и H2SO4 (3:2: 1) в колбе Кьельдаля. Смесь кислот добав-
ляют холодной и затем колбу медленно нагревают, чтобы обеспечить плавное
разложение без обугливания. После окончания разложения колбу нагревают силь-
нее, чтобы удалить избыток HNO3 и HCIO4 и уменьшить объем раствора. Про-
порции кислот в смеси могут быть самыми различными, например 10:4: 1.
В настоящее время рекомендуется применение хлорной кислоты для разру-
шения органического вещества. Горячая концентрированная HCIO4 представля-
ет собой сильный окислитель, при ее контакте с органическими веществами или
легко окисляющимися неорганическими веществами она взрывоопасна, поэто-
му применение ее связано с определенными условиями. Во многих лаборатори-
ях HCIO4 не применяют, поскольку она может быть опасна при работе в дере-
вянных вытяжных шкафах. Опасность уменьшается при избытке в смеси азот-
ной кислоты.
Для ускорения процесса озоления и снижения потерь предложены молибде-
новые, метаванадиевые катализаторы, смесь КМПО4 и H2SO4 и 50%-ная перекись
водорода.
Метод мокрого озоления более всего известен как метод разрушения орга-
нического вещества без потерь. В настоящее время он является обязательным
для определения мышьяка, сурьмы, ртути, кадмия и в ряде случаев — свинца и
цинка. Однако детальные исследования показали, что при выпаривании получен-
ных растворов досуха могут быть значительные потери сурьмы, мышьяка, бора,
хрома, германия, ртути, селена и цинка в результате испарения их летучих соеди-
нений, содержащихся в пробе или образовавшихся при взаимодействии с кисло-
тами. На основании детального изучения методов мокрого озоления с применени-
ем различных комбинаций HNO3, H2SO4 и HCIO4 и сравнения их с методами сухого
озоления был сделан вывод, что смеси HNO3 и HCIO4 (3 : 2) дают хорошие резуль-
таты для-определения Se, Zn, Ag, As, Cu, Co, Cd, Pb, Cr, Mo, Sr, Fe во многих образ-
цах, однако при определении ртути наблюдаются потери (Gorsuch, 1970).
К недостаткам метода мокрого озоления относится также загрязнение проб
определяемыми элементами за счет относительно большого количества приме-
няемых кислот и других агентов минерализации. Очистка этих агрессивных
средств неприятна и трудна. Кроме того, эти реагенты оказывают выщелачиваю-
щее действие на сосуды, в которых озоляется проба. Одним из недостатков
метода мокрого озоления является необходимость работы с небольшими навес-
ками гомогенных проб.
196
В целом рабочие затраты при использовании этого метода больше, чем при
сухом озолении. Из вышеизложенного становится понятно, почему для опреде-
ления микроэлементов при массовых анализах метод мокрого озоления менее
распространен.
Таким образом, в зависимости от поставленных задач необходим выбор оп-
тимального способа минерализации растительного материала. Выявление опти-
мальных условий озоления позволит избежать больших потерь определяемых
микроэлементов при наименьшем усложнении методики озоления, т. к. последнее
влияет на экспрессность анализа.
3.3.3. Методы количественного химического анализа
В распоряжении исследователя имеется целый ряд методов для получения
аналитической информации. Выбор метода будет основываться на таких показа-
телях, как скорость, удобство, правильность, наличие подходящего оборудова-
ния, число анализов, размер анализируемой пробы, содержание определяемого
компонента. Успех или неудача анализа часто зависят от выбора метода. К сожа-
лению, здесь не существует общего подхода; для правильного выбора метода
нужны здравый смысл и твердое знание преимуществ и ограничений разных до-
ступных методов анализа, последнее можно получить, изучив научную литера-
туру по данному вопросу.
Основная характеристика метода количественного определения — физичес-
кая величина, используемая в качестве меры интенсивности аналитического сиг-
нала. Поэтому приведенная в табл. 3.7 классификация количественных методов
химического анализа основывается именно на этих величинах.
Таблица 3.7. Классификация методов количественного анализа (Янсон, Путнинь, 1980)
Измеряемая физическая величина Второстепенные факторы Название метода
Масса а. Вещество получают путем электролиза б. Измерения осуществляют с помощью масс-спектрометров Гравиметрия: электрогравиметрия масс-спектрометрия
Объем а. Измеряют объем раствора реагента, израсходованного при титровании б. Измеряют объем газообразных продуктов реакции Волюмометрия: титриметрия газоволюмометрия
Равновесный потенциал электродов — Потенциометрия
197
продолжение таблицы 3 7
Поляризационное сопротивление электрода Измеряют поляризационное сопротивление капельного ртутного электрода Вольтамперометрия: полярография
Количество электричества — Кулонометрия
Электропроводность раствора — Кондуктометрия
Поглощение фотонов а. Измерения проводят с помощью колориметров б. Измерения проводят с помощью спектрофотометров в. Измеряют светопоглощение мутных растворов г. Измеряют светопоглощение атомами при высокой температуре Фотометрия: колориметрия спектрофотометрия турбидиметрия, нефелометрия атомно-абсорбцион- ный анализ
Испускание фотонов Испускание фотонов при нагревании в пламени Спектральный анализ: фотометрия пламени
Поглощение фотонов с последующим испус- канием других фотонов — Люминесцентный и флуоресцентный анализ
Скорость реакции — Кинетические методы анализа
Радиоактивность а. Применение радиоактивных атомов для определения соответствующих элементов б. Использование радиоактивности атомов, получаемых при радиоактивном облучении Радиометрические методы анализа: метод радиоактив- ных индикаторов радиоактивационный анализ
Переходы магнитных диполей между энергетическими уровнями, возникающими в магнитном поле а. Используются спины неспаренных электронов б. Используются спины ядер атомов Спектроскопия маг- нитного резонанса: спектроскопия электронного парамагнитного резонанса спектроскопия ядер- ного магнитного резонанса
198
В табл. 3.7 не включены отдельные варианты титриметрического анализа, на-
звания которых образуются по способу нахождения точки стехиометричности (эк-
вивалентности): потенциометрическое, амперометрическое, фотометрическое,
радиометрическое и т. д. титрования.
Для количественного определения содержания компонента измеряют интен-
сивность аналитического сигнала, вызванного данным компонентом (величина у,
рис. 3.9). Между интенсивностью сигнала у и содержанием (концентрацией) оп-
ределяемого компонента существует однозначная функциональная зависимость.
Она может быть выражена в виде соответствующей математической функции, на-
зываемой градуировочной
функцией, либо в виде соот-
ветствующего графика, назы-
ваемого градуировочным
графиком (градуировочной
кривой или прямой,
рис. 3.9).
В простейшем случае гра-
дуировочная функция имеет
вид у = Sc, где у интенсив-
ность аналитического сигнала;
Рис. 3.9. Градуировочный график
А — линейный, В—нелинейный
с — концентрация определяе-
мого элемента; S — коэффициент чувствительности. Если зависимость линейная
(рис. 3.9А), коэффициент чувствительности представляет собой постоянную ве-
личину, равную наклону градуировочной прямой. Чем больше коэффициент чув-
ствительности, тем меньшему содержанию соответствует частное значение ин-
тенсивности сигнала.
Если взаимосвязь между у и с нелинейная (рис. 3.9Б), восстановление значе-
ния концентрации по величине измеряемого сигнала осуществляется графичес-
кими или численными методами.
Среди многочисленных методов, используемых для химического анализа
объектов окружающей среды, пожалуй, наибольшее распространение для оцен-
ки уровня загрязнения почв и растений получил метод атомно-абсорбционной
спектрофотометрии
Сущность метода. Атомно-абсорбционная спектрофотометрия основана на
измерении уменьшения интенсивности излучения, прошедшего через анализиру-
емое вещество. Ослабление интенсивности излучения происходит за счет погло-
щения атомами рентгеновского, видимого или ультрафиолетового излучения и
перехода их в возбужденное состояние. Рентгеновское поглощение обусловле-
но возбуждением внутренних, не участвующих в образовании связей электронов,
вследствие чего характер рентгеновского атомного спектра не зависит от хими-
ческого состояния элемента в образце.
199
Рис. 3.10. Принципиальная схема атомно-адсорбционного спектрофотометра
I — стабильный источник излучения энергии — лампа с полым катодом; 2 — горелка-
атомизатор; 3 — монохроматор — устройство, позволяющее выделить ограниченную об-
ласть длин волн, 4 — детектор, превращающий энергию излучения в измеряемый электри-
ческий сигнал; 5 — индикатор сигнала— цифровой вольтметр, но можно подключить ком-
пьютер.
Напротив, для получения атомного спектра в видимой или ультрафиолетовой
области необходим переход элементов пробы в моноатомные частицы в газооб-
разном состоянии, поскольку поглощение в этой области обусловлено возбуж-
дением внешних валентных электронов. Таким образом, атомные спектры в ви-
димой или ультрафиолетовой'области снимают после атомизации образца. Спек-
тры испускания и поглощения атомизированного элемента состоят из относитель-
но небольшого числа дискретных линий, длины волн которых характерны для
данного элемента.
Принципиальная схема устройства атомно-абсорбционного спектрофотомет-
ра приведена на рис. 3.10.
Методы, основанные на атомной эмиссии или абсорбции, отличаются высо-
кой избирательностью, исключительной чувствительностью, скоростью и удоб-
ством; они относятся к наиболее селективным аналитическим методам. Этими
методами можно определять около 70 элементов. Чувствительность определе-
ния химических элементов лежит в интервале 1 CH-10-10 %.
3.3.5. Оценка достоверности аналитических данных
Обычно химический анализ одного образца проводят от 2 до 5 раз. Отдельные
результаты в такой серии параллельных измерений (выборке) редко совпадают,
поэтому возникает необходимость выбора центрального «лучшего» значения се-
рии. Центральной точкой серии может служить либо среднее, либо медиана.
Среднее, среднее арифметическое и выборочное среднее являются синони-
мами и представляют собой частное от деления суммы результатов отдельных
измерений на число измерений в выборке.
Медианой выборки называется результат, относительно которого число ре-
зультатов с меньшим и большим значением одинаково.
В идеальном случае среднее и медиана должны совпадать; однако чаще все-
го этого не происходит, особенно если число измерений в серии невелико.
Термин воспроизводимость используется для количественной оценки разбро-
са результатов. Эта величина характеризует близость по абсолютному значению
двух или более измерений, полученных в одинаковых условиях. Воспроизводи-
мость выражается несколькими способами.
200
Абсолютная воспроизводимость — это абсолютная величина отклонения от
среднего, т. е. разность между экспериментально найденным значением и сред-
ним из серии значений, включающей данное значение, без учета знака. Воспро-
изводимость можно также выразить отклонением от медианы. Мерой воспроиз-
водимости является также размах варьирования, или диапазон выборки, т. е.
разность между наибольшим и наименьшим результатами.
Двумя другими критериями вопроизводимости служат стандартное откло-
нение и дисперсия (см. разд. 4).
Относительная воспроизводимость — отношение абсолютного отклонения
к среднему арифметическому, выраженное в процентах.
Правильность или точность означает близость полученного значения к зна-
чению, принятому за действительное (истинное), и выражается ошибкой. Основ-
ное различие между правильностью и воспроизводимостью состоит в том, что
для оценки правильности сравнивают полученный результат с действительным
(истинным) или принятым за действительный, а для оценки воспроизводимости
проводят сравнение результата с другими результатами, полученными тем же
путем.
Правильность выражают абсолютной ошибкой, которая представляет собой
разность между наблюдаемыми значениями и значением, принятым за действи-
тельное (истинное). Часто более полезной величиной является не абсолютная, а
относительная ошибка, выражающая отклонение от действительного значения
в процентах.
Предел обнаружения — массовая концентрация определяемого элемента в
растворе, которая вызывает аналитический сигнал, равный его удвоенному сред-
нему квадратичному отклонению.
Чувствительность определения — минимальная концентрация элемента, вы-
зывающая 1%-ный аналитический сигнал.
Результаты химических анализов наиболее часто представляют в виде табли-
цы с указанием среднего значения содержания (концентрации) каждого элемен-
та, стандартной ошибки, размаха варьирования и числа проанализированных проб.
3.5. ПРИНЦИПЫ И МЕТОДЫ АНАЛИЗА ДАВНОСТИ
И ПЕРИОДИЧНОСТИ ПОЖАРОВ 1
Во время пожаров в результате воздействия высоких температур часто наблю-
дается частичное или полное повреждение и отмирание камбиального слоя ство-
лов деревьев. По краям поврежденного участка происходит образование каллус-
ной ткани, которая в процессе последующего периода жизни дерева постепенно
наплывает на поврежденный участок, закрывая его полностью или частично.
1 Автор В В Горшков
201
Рис. 3.11. Нижняя часггь ствола со-
сны, поврежденной» в результате
пожара
Обозначения, а — современная площадь
шрама, б — места отбор «а серии кернов
для определения давносдти последнего
пожара, в —-на заднем пхлане сильно уг-
нетенная особь сосны во^зрастом 117 лет
В случае полного зарастания поврежден-
ного участка его наличие может быть вы-
явлено только на спиле, при неполном за-
растании след повреждения (пожарный
шрам) регистрируется при визуальном ос-
мотре ствола (рис. 3.11).
Метод определения давности повреж-
дений на стволах деревьев основан на том,
что на участках с отмершим в результате
термического воздействия или механичес-
ки уничтоженным камбиальным слоем
прекращается образование клеток древе-
сины. Давность повреждения определяет-
ся как разность числа годичных слоев дре-
весины, сформировавшихся на неповреж-
денном и поврежденном участках ствола
(рис. 3.12). Этот метод широко использу-
ется для датировки пожаров (Мелехов,
1936, 1948; Молчанов, 1954; Zacknsson,
1977; Arno, Sneck, 1977; Madany et al.,
1982; Dieterich, Swetnam, 1984; Горшков,
1993; 1994, 2001; Kaennel, Schweingruber,
1995), а также для определения давности
механических повреждений стволов дере-
вьев с целью датировки различных при-
родных явлений, например при анализе пе-
риодичности схода лавин (Kaennel,
Schweingruber, 1995).
3.5.1. Анализ при знаков пожарного происхождения повреждений
стволов деревь-ев
Существующая неоопределенность при определении причины возникновения
повреждений стволов деревьев: температурное повреждение во время пожара
(рис. 3.12) или механическое повреждение непожарного происхождения
(рис. 3.13) вызывает нееобходимость анализа дополнительных признаков (Молча-
нов, 1954; Amo, Snec=k, 1977; Zackrisson, 1977; Madany et al., 1982; Dieterich,
Swetnam, 1984; Payette et al., 1989). Рассмотрим применимость основных из них
в условиях Европейского Севера.
Совпадение давно«сти повреждения у большого числа особей. Одним из
важных признаков пояасарного происхождения повреждений является точное со-
впадение давности их возникновения у нескольких деревьев.
202
Рис. 3.13. Участок поперечного сече-
ния ствола сосны со следами не по-
жарного повреждения
Обозначения' а/, а,—-участки ствола с по-
гибшим в результате неизвестных наруше-
ний камбием (по-видимому, падение погиб-
шего во время последнего пожара сосед-
него дерева); б — возраст дерева в год
отбора модели (131 год)
Рис. 3.12. Поперечное сечение ствола со-
сны со следами повреждения во вре-
мя последнего пожара. Давность пос-
леднего пожара 41 год
1 — общий вид; 2 — крупный план выделен-
ной области. Обозначения: а — возраст (65
лет) дерева во время повреждения пожаром,
б — возраст (106 лет) дерева в год отбора
модели; в — участок ствола, сформировав-
шийся в период после пожара (за 41 год); г —
светлым пунктиром и стрелками отграниче-
на область ствола, где камбий не был повреж-
ден во время пожара; д — пунктиром обозна-
чен слой поздней древесины, сформировав-
шейся в год повреждения дерева
Анализ частоты повреждений, выполненный на 260 поврежденных деревьях в
условиях Кольского полуострова, показал, что на каждые 100 лет жизни дерева
приходится в среднем ~1 повреждение, т. е. вероятность повреждения дерева в
течение конкретного года составляет ~10~2. Вероятность случайного совпаде-
ния давности повреждения нескольких деревьев равна произведению вероятно-
стей и для 5 деревьев составляет 1О~10. В континентальных районах Евразии с
высокой периодичностью пожаров при частоте повреждений деревьев -1/40 (Фу-
ряев, 1996) вероятность случайного совпадения давности повреждения 5 дере-
вьев составляет ~10-8. Таким образом, случайное совпадение повреждений у 5
203
деревьев является практически невероятным за время существования биосфе-
ры, которое составляет 109 лет.
Приведенные данные показывают, что точное совпадение давности повреж-
дения даже у 5 деревьев всегда означает неслучайность их повреждения и по-
зволяет с высокой вероятностью предполагать, что причиной этих повреждений
является пожар.
Наличие обугленных участков на: а) прилегающей к месту повреждения
части корки (при первом повреждении) или б) открытой древесине шрама (при
повторном и следующих повреждениях). Этот признак однозначно указывает на
наличие пожара. По мнению многих исследователей, достаточно даже одного из-
мерения (модельного дерева) для установления давности пожара при наличии
обугленных участков корки или древесины вблизи поврежденной области (Fritts,
1972; Zackrisson, 1977; Dieterich, Swetnam, 1984; Brown, Swetnam, 1994).
В лесах Кольского полуострова участки обугленной корки сохраняются до
90 лет после пожара, при этом их относительная площадь не превышает долей
процента. Уже через 60 лет после пожара у основания деревьев относительная
площадь обугленной корки составляет ~2 %, а в областях ствола, близких к по-
жарному шраму, обугленная кора практически не встречается1. Поэтому наличие
участков обугленной корки позволяет установить причину повреждения ствола
(при первом повреждении дерева пожаром) только при небольшой давности по-
жара (до 40-60 лет). При значительной давности пожара обугленные участки со-
храняются только на древесине шрамов от предшествующих пожаров.
Овальная (яйцевидная) или треугольная форма повреждения, при этом
широкая часть расположена в основании ствола. Этот признак многие исследо-
ватели лесов на территории Северной Америки рассматривают как характерный
для повреждения стволов деревьев пожарами (Amo, Sneck, 1977; Barret, Amo,
1988; Kaennel, Schweingruber, 1995). В европейских сосновых лесах из сосны
обыкновенной форма пожарного шрама с расширением к основанию дерева ре-
гистрируется, по данным И. С. Мелехова (1948), приблизительно в 50-60 % слу-
чаев, а в условиях Кольского полуострова — менее чем в 50 %. Таким образом,
форма повреждения не всегда является признаком, однозначно свидетельству-
ющим о причине возникновения повреждения. Следует отметить, что треуголь-
ная форма пожарных шрамов наблюдается обычно при большой сбежистости
ствола, когда диаметр ствола у основания дерева на 30-50 % больше, чем диа-
метр на высоте 1,3 м.
Наличие резкого изменения радиального прироста деревьев после по-
жара. На примере сосновых лесов (Pinus sylvestris, Р. resinosa Ait., Р. banksiana
Lamb.) европейского и американского Севера было показано, что может наблю-
1 В районах южной тайги уменьшение относительной площади обугленной корки до ве-
личин ~2-4 % наблюдается при давности пожаров -30-40 лет.
204
даться любой из вариантов изменения прироста после пожара: увеличение, умень-
шение или отсутствие изменений (Мелехов, 1948; Молчанов, 1954; Курбатский,
1970; Dieterich, Swetnam, 1984; Bergeron, Brisson, 1990; Арбатская, Ваганов,
1996; Горшков, 2001).
Анализ факторов, вызывающих повреждения стволов деревьев в бореальных
сосновых лесах, позволяет заключить, что главными признаками, которые дол-
жны использоваться при идентификации происхождения шрамов на стволах де-
ревьев, являются: 1) наличие обугленных участков корки (при первом повреж-
дении) и обугленных участков древесины (при втором повреждении) и 2) совпа-
дение давности повреждения у нескольких деревьев Отсутствие обугленных
участков на открытой поверхности мертвой древесины шрамов поврежденных
деревьев — признак отсутствия пожаров в течение периода с момента воз-
никновения повреждения до момента регистрации. Это утверждение обоснова-
но тем, что открытые участки древесины всегда обугливаются во время пожа-
ров, т. к. интенсивность и продолжительность горения в непосредственной бли-
зости от стволов деревьев в 1,5 -2 раза выше, чем на остальной территории из-
за повышенной сухости и более значительного запаса опада и подстилки.
3.5.2. Причины непожарных повреждений стволов деревьев
Повреждения камбия на стволах деревьев могут быть вызваны не только по-
жарами, но и другими причинами: деятельностью крупных животных, единичны-
ми и групповыми ветровалами (Формозов, Исаков, 1963; Zacknsson, 1977,
Kaennel, Schweingruber, 1995). Однако анализ плотности крупных животных
(лосей, медведей) в лесах Севера показывает, что она составляет не более
510-5 особей/га (Владышевский, 1980), поэтому животные, как правило, явля-
ются причиной повреждения единичных деревьев.
Массовые ветровалы приводят к полной гибели древесного яруса на значи-
тельной территории, но вызванные падением деревьев повреждения других осо-
бей регистрируются в пределах относительно небольшой полосы, окаймляющей
территорию ветровала (Турков, 1979; Quine, 1995). Падение деревьев при еди-
ничных и группо.вых ветровалах (ветровалы, захватывающие менее 3 % площа-
ди) вызывает повреждение стволов у единичных соседних деревьев.
Повреждение камбия на значительной части окружности ствола приводит к
гибели дерева, хотя в некоторых случаях в условиях Европейского Севера мо-
гут выживать деревья с повреждениями камбия, достигающими 75-90 % от ок-
ружности ствола (рис. 3.12). Погибшие в результате значительного термическо-
го повреждения деревья в течение 20 лет находятся в вертикальном положении.
Массовое падение погибших в результате пожара деревьев в сосновых лесах
отмечается через 15-20 лет (Молчанов, 1947). Доля вываленных деревьев (от
общего числа погибших) в лесах с этой давностью пожара составляет -50-60 %.
Большая часть оставшихся погибших деревьев падает в течение последующих
205
15-20 лет. Падение погибших во время пожара деревьев может вызывать меха-
нические повреждения стволов соседних деревьев в молодых восстанавливаю-
щихся после пожара лесных сообществах. Давность таких повреждений на 20-
40 лет меньше давности последнего пожара.
Повреждения стволов, вызванные падением соседних, погибших во время
пожара деревьев, в отличие от термических повреждений возникают не одновре-
менно, поскольку процесс вывала погибших деревьев растянут во времени. Время
падения каждого дерева определяется скоростью разложения древесины его
ствола и корней, которая, в свою очередь, зависит от условий микроместооби-
тания, степени засмоленности древесины. Одновременное падение всех погиб-
ших деревьев — событие невероятное1. Следовательно, вывал погибших во вре-
мя пожара деревьев может быть источником повреждения некоторых живых
деревьев в пределах сообщества, однако эти повреждения будут различаться по
времени возникновения. Напротив, повреждения, обусловленные гибелью кам-
бия при высоких температурах, совпадают по времени. Следует также отметить,
что данный тип повреждения деревьев не может рассматриваться как отдельное
природное явление, это последствие пожара, поскольку причиной массовой ги-
бели и падения деревьев являются огневые повреждения камбия.
Проведенный анализ показывает, что основным фактором, вызывающим до-
статочно массовое одновременное повреждение стволов деревьев, является
пожар Этот вывод подтверждают результаты многих исследователей (Ткачен-
ко, 1931; Мелехов, 1948, 1971; Корчагин, 1954; Молчанов, 1954; Siren, 1955;
Zacknsson, 1977; Kaennel, Schweingruber, 1995). Наиболее вероятным является
прямое повреждение в результате перегрева и гибели камбия во время пожара,
менее вероятным является механическое повреждение стволов в результате
массового падения погибших во время пожара деревьев.
Во время пожаров часть деревьев (от 30 до 100 %) обычно не повреждает-
ся. Поврежденные пожаром деревья в пределах одного и того же древостоя
могут нести на себе следы не всех пожаров, а только части из них. Таким обра-
зом, возможна ситуация, когда низовой пожар, уничтожив мохово-лишайниковый
и травяно-кустарничковый ярусы, не оставляет следов повреждения на деревь-
ях. Оценить вероятность таких случаев и разработать способы контроля продол-
жительности беспожарных периодов позволяет анализ предохраняющей функции
корки стволов деревьев.
1 Частичное совпадение давности повреждения возможно, потому что падение деревь-
ев — процесс дискретный, массовое падение будет наблюдаться в конкретные дни и годы
во время сильных ветров.
206
3.5.3. Анализ предохраняющей функции корки и возможности
повреждения камбия стволов деревьев во время пожаров
Для оценки предохраняющей функции корки использовано решение уравне-
ния теплопроводности Фурье (Смирнов, 1953; Ландау и др., 1965) в приближе-
нии относительно кратковременного воздействия пламени на ствол дерева, ко-
торое описывает градиент температуры, наблюдающийся на внешней и внутрен-
ней поверхности грубой корки (комплекса тканей на поверхности ствола, состо-
ящих из отмерших слоев клеток перидермы и вторичной флоэмы):
где Т(>, т) — градиент температуры [°C] внутреннего слоя грубой корки, дости-
гаемый за время воздействия огня t при толщине корки х и при исходной темпе-
ратуре внутренней части ствола и окружающего ствол воздуха Tq Tt„, . , —
температура на внешней поверхности грубой корки в период горения (темпера-
тура пламени), [°C]; t — время воздействия пламени (продолжительность горе-
ния), [с]; х — толщина корки, [м]; а — коэффициент, характеризующий темпера-
туропроводность, [м2 г1].
« = -, (2)
где X — теплопроводность, [Вт/(мтрад) • Дж/(м с град)]; с — теплоемкость (при
постоянном давлении), [Дж/(кг-град]; р — плотность, [кг м-3].
Уравнение (1) можно представить в виде:
^-K=(Tf~T0)e^, (3)
где — температура [°C] внутреннего слоя грубой корки, достигаемая за время
t воздействия огня при толщине корки х; То — температура воздуха, [°C]; Гу—
температура пламени, [°C]; t — время воздействия пламени, [с]; х — толщина
корки, [м]; а — коэффициент температуропроводности, [м2 с-1].
В отличие от использованного здесь приближенного решения существует
точное решение уравнения теплопроводности Фурье, где вместо экспоненциаль-
ной зависимости используется Гауссова функция ошибок (Ландау, Лившиц, 1954;
Johnson, 1992; Gutsel, Johnson, 1996):
(4)
где erf— Гауссова функция ошибок, остальные обозначения те же, что и для
уравнения (3). Анализ уравнений (3) и (4) показывает, что при оценке предохра-
1 Рассмотренный случай предполагает равновесное состояние, когда температура воз-
духа и температура ствола одинаковы.
207
няющей функции корки получаемые при использовании приближенного (3) и точ-
ного (4) решений уравнения Фурье величины различаются примерно на 10 %.
Поэтому предлагается использовать уравнение (3), более удобное для расчетов.
Температура пламени в кромке пожара в среднем составляет 500-600 °C (Гу-
лисашвили, 1931; Нестеров, 1939; Мелехов, 1947; Бачинский и др., 1951; Tunstall
et al., 1976; Gutsel, Johnson, 1996). Наименьшая зарегистрированная температу-
ра горения при распространяющемся пожаре составляет 210 °C — температура,
близкая к температуре воспламенения большинства лесных горючих материалов
(Гулисашвили, 1931; Курбатский, 1970); наивысшая, зарегистрированная при го-
рении локальных скоплений валежника и пней, — 880 °C (Нестеров, 1949; Меле-
хов, 1948).
Теплопроводность корки сосны обыкновенной и ели сибирской1 X = 0,065 Вт/
(мтрад); теплоемкость корки с = 2,7103 Дж/(гтрад); плотность корки р =
0,6-103 кг/м3; (Бачинский и др., 1951; Чайдлс, 1961) и, таким образом, темпера-
туропроводность а = 4-10-8 м2-с-1.
Продолжительность горения кромки пожара t в зависимости от интенсивнос-
ти пожара (т. е. главным образом от запаса сгораемого во время пожара органи-
ческого вещества) варьирует в пределах от 12 с при пожаре слабой интенсивно-
сти до 90 с при пожарах средней и сильной интенсивности (Фуряев, 1996).
Нетрудно убедиться, что при температуре Tf = 210 °C (наименьшей зарегист-
рированной во время пожаров температуре), времени горения t = 10 с (в 1,2 раза
меньшем, чем наименьшее зарегистрированное в исследовательских эксперимен-
тах (Фуряев, 1996)), толщине корки х = 1 мм (наблюдаемой у особей подроста
и сильно угнетенных деревьев) и максимально низкой температуре воздуха (0 °C),
при которой могут возникать лесные пожары (Мелехов, 1936; Нестеров, 1939;
Курбатский, 1964; Фуряев, 1996), температура на внутренней поверхности гру-
бой корки, согласно уравнению (3), составит:
=210°С е‘44ю-"м!с-'1ос = 210°С е-0,63 =210°С-0,55 = 115,5’С • (5)
Эта величина вдвое превышает температуру (~65 °C), при которой отмечает-
ся гибель камбия (Мелехов, 1948; Шешуков и др., 1979). Таким образом, все
особи с толщиной корки ~1 мм гибнут даже во время самого слабого (беглого)
пожара2.
Уравнение (3) позволяет оценить толщину грубой корки, необходимую для
предохранения камбия от повреждения и гибели при различных условиях пожа-
ра. Для этого необходимо, чтобы выполнялось неравенство:
Дрит - Тд > - Тд, (6)
1 Теплопроводность, теплоемкость и плотность корки сосны и ели совпадают с точнос-
тью до второго знака
208
Таблица 3.8. Расчетные величины толщины грубой корки, предохраняющей кам-
бий от гибели, при различной температуре воздуха, пламени и разной продол-
жительности горения
Продолжи- тельность горения t, с Температура воздуха
0°С 25 °C
Температура пламени, °C
167 210 400 600 880 167 210 400 600 880
Толщина корки, 10 3 м (мм)
10 1,7 1,9 2,3 2,6 2,8 2,1 2,3 2,6 2,8 3,0
18 2,3 2,6 3,1 3,4 3,7 2,9 3,1 3,6 3,8 4,1
50 3,9 4,3 5,2 5,7 6,2 4,8 5,1 5,9 6,4 6,8
90 (1,5 мин) 5,2 5,7 7,0 7,6 8,3 6,4 6,8 8,0 8,6 9,1
300 ( 5 мин) 9,4 10 13,0 14 15 11,5 12,5 14,5 155 17
600 (10 мин) 13 15 18 20 21 16,5 17,5 20,5 22 24
1800 (30 мин) 23 26 31 34 37 29 31 36 38 41
3600 (1 ч) 33 36 44 47 52 41 43 50 54 58
14400 (4 ч) 66 72 88 97 100 81 87 101 108 115
т,~тй
------4aZ
г-т;
где Гцрит — является температурой, при которой регистрируется гибель камбия
(65 °C), или, согласно уравнению (3), должно выполняться условие
(7)
В табл. 3.8 представлены результаты расчетов с использованием уравнения (3)
для крайних вариантов температур воздуха, при которых регистрируются пожа-
ры в условия севера таежной зоны в период с конца апреля по конец сентября —
О °C и 25 °C. Выбранные значения температур представляют минимальную
2 М. А. Шешуков с соавт. (1979) в процессе экспериментального исследования опреде-
лил температуру, при которой наблюдается гибель камбия. При помощи двух помещенных
под кору электродов измерялась величина электропроводности Температура определялась
по величине электропроводности на основе предварительно построенной калибровочной
Выполненное исследование показало: 1) при температуре 65 °C гибель камбия наблюдает-
ся мгновенно; 2) при температуре 60 °C гибель камбия наблюдается через 30 с; 3) при
температуре 55 °C гибель камбия наблюдается через 5 мин.
R. С. Hare (1965), А. М. Gill, D. N. Ashton (1968) и R. G. Vines (1968) на основании ис-
следований толщины корки поврежденных и не поврежденных пожаром деревьев решили
противоположную задачу, оценив температуру гибели камбия, составившую по их резуль-
татам ~60 °C.
209
(210 °C), переходную (400 °C), типичную (600 °C) и максимальную (880 °C) тем-
пературы горения (Гулисашвили, 1931; Нестеров, 1939; Мелехов, 1948; Martin,
1963; Tunstall et al., 1976; Gutsel, Johnson, 1996). Использованные значения про-
должительности горения представляют минимальное (18 с), среднее (50 с) и мак-
симальное (90 с) время горения кромки (Фуряев, 1996), а также возможные ва-
рианты локальных отклонений (5 мин — 4 ч) при сгорании скоплений сучьев,
крупного валежника и пней.
Таким образом, при прогнозе повреждения деревьев в результате низового
пожара необходимо учитывать следующее:
1. Толщина грубой корки стволов деревьев, предохраняющая камбий от по-
вреждений в большинстве складывающихся во время пожаров ситуаций, состав-
ляет 1 см. При толщине грубой корки >3 см для повреждения дерева необходи-
мо воздействие огня в течение более 30 минут — такая ситуация возможна в
ненарушенном человеческой деятельностью лесном сообществе лишь на очень
незначительной (менее 1 %) части площади.
2. Температура воздуха оказывает незначительное влияние на повреждаемость
деревьев во время пожара: различия в критических значениях толщины слоя гру-
бой корки, предохраняющей камбий от повреждения при температуре воздуха
0 °C и 25 °C, не превышает 12 %.
3. Изменение температуры горения оказывает незначительное влияние на по-
вреждаемость деревьев во время пожара. 50%-ое увеличение толщины слоя гру-
бой корки позволяет противостоять 4-кратному увеличению температуры пожа-
ра от 240 °C (минимальная) до 880 °C (максимальная).
4. Основными факторами, определяющими повреждение деревьев во время
пожара, являются толщина слоя корки и продолжительность горения, которая
зависит от общего запаса, структуры и свойств отдельных компонентов горю-
чих материалов в лесном сообществе.
3.5.4. Метод последовательного анализа возраста угнетенных деревьев
Основой метода последовательного анализа возраста угнетенных деревьев яв-
ляется то, что корка деревьев толщиной ~1 мм (или меньше) не предохраняет
камбий от воздействия высоких температур во время пожара. В лесных сооб-
ществах в большинстве случаев присутствуют особи древесных растений (вхо-
дящие в состав подроста или древостоя), минимальная толщина корки которых
не превышает 1 мм (рис. 3.11, 3.14).
Первым этапом анализа (рис. 3.15) является отбор на территории пробной пло-
щади и ближайшего ее окружения первой серии моделей угнетенных деревьев,
имеющих по окружности ствола участки с толщиной корки менее 1 мм. Затем
производится регистрация их возраста, диаметра у основания ствола и наличия
или отсутствия повреждений (нарушений радиального роста). Отсутствие нару-
шений радиального роста у таких особей и само их существование является сви-
210
детельством отсутствия пожа-
ров в течение периода их жизни
— I отрезка беспожарного пе-
риода.
Определение продолжитель-
ности II отрезка беспожарно-
го периода проводится на осно-
ве выявления в составе древес-
ного яруса второй серии мо-
дельных угнетенных деревьев,
параметры которых в начале
I беспожарного периода не пре-
вышали современных парамет-
ров особей первой серии угне-
тенных деревьев. Например, для
дренированных сосновых лесов
Европейского Севера эти пара-
метры составляют: возраст —
15-20 лет (при анализе отобран-
ных у основания кернов или спи-
лов) и диаметр у основания —
3 см. Деревья с такими парамет-
рами погибают в результате по-
жара любой интенсивности (Гор-
шков, 2001).
Продолжительность II от-
резка беспожарного периода
равна возрасту деревьев второй
серии в начале I беспожарного
периода.
Например, возраст деревьев
первой серии в момент прове-
дения исследований составляет
20 лет, следовательно, продол-
жительность I отрезка беспо-
жарного периода равна 20 го-
дам. Возраст деревьев второй
серии в начале первого отрезка беспожарного периода (20 лет назад) составлял
15 лет, следовательно, продолжительность II отрезка беспожарного периода
равна 15 годам. Последующие отрезки беспожарного периода устанавливаются
аналогично второму. Общая продолжительность беспожарного периода равна
Рис. 3.14. Диагональное и поперечное сечении
ствола угнетенной особи сосны без следов
повреждения камбия
Обозначения: а, б, в, г, д — радиусы, по которым
проводился подсчет числа слоев поздней древесины,
различия обусловлены наличием ложных, выпадаю-
щих или выклинивающихся колец
I период Ь.
2.
a.
II период
время, годы
Рис. 3.15. Метод последовательного (пошаго-
вого) анализа возраста угнетенных деревь-
ев для определения продолжительности бес-
пожарных периодов
211
Рис. 3.16. Поперечное сечение ствола сосны со
следами 2 пожаров. Давность последнего по-
жара 219±3 лет
Обозначения а — возраст (20 лет) дерева во время
повреждения первым пожаром; б — возраст (61 год)
дерева во время повреждения вторым (последним) по-
жаром; в — возраст (282± 1 года) дерева в год отбора
модели, г — участок ствола, сформировавшийся в пе-
риод между первым и вторым повреждением (за 41
год), di-ds — участок ствола, сформировавшийся пос-
ле последнего пожара (за 221±1 год), различия обус-
ловлены наличием ложных, выпадающих или выклини-
вающихся колец; е—обугленная древесина на поверх-
ности ствола в месте первого повреждения; 1 — свет-
лым пунктиром и стрелками отграничена область ство-
ла, где камбий не был поврежден во время первого по-
жара, 2 — области ствола с камбием, не поврежден-
ным во время второго (последнего) пожара
суммарной продолжительности
всех выявленных отрезков бес-
пожарного периода. Предельная
продолжительность определяе-
мого таким образом беспожар-
ного периода ограничена уста-
новленной (при наличии шрамов)
давностью пожара или макси-
мальным возрастом деревьев в
составе древесного яруса.
Следует отметить, что при
большой давности пожара (свы-
ше 150 лет) и стабилизировав-
шейся продуктивности характер-
ным является очень незначи-
тельный прирост деревьев по
диаметру, особенно это касается
угнетенных особей. В этих усло-
виях наблюдается явление фор-
мирования ложных годичных ко-
лец, выпадение или выклинивание
колец (Kaennel, Schweingruber,
1995).
В тех случаях, когда наблюда-
ется существенное различие
прироста по разным направлени-
ям окружности ствола (рис. 3.12,
3.16), подсчет числа годичных
колец проводится в области мак-
симального прироста. Например,
давность последнего пожара
(рис. 3.16) следует регистриро-
вать в области О 4 и dj. Такой ана-
лиз часто позволяет избежать
проблемы ложных и выклинива-
ющихся колец.
При анализе числа годичных колец в пределах одного спила по разным направ-
лениям могут быть получены разные результаты. Например, на спиле (см.
рис. 3.14) по 5 направлениям были полущены 5 разных результатов. В данном
случае полученные величины следует рассматривать как случайные и использо-
вать среднее значение с указанием его ошибки.
212
При отсутствии в составе древесного яруса сильно угнетенных деревьев, ми-
нимальная толщина корки которых не превышает 1 мм, продолжительность бес-
пожарного периода может быть оценена аналогичным образом на основе анали-
за расположенных на поверхности почвы корней деревьев. Использование кор-
ней при применении данного метода может быть рекомендовано для изучения
пожаров в лиственничных лесах, в которых отсутствие повреждений столов де-
ревьев и наличие повреждений корней наблюдается очень часто (Цуканов, Се-
менов, 1990).
При проведении любых геоботанических, экологических, фитоиндикационных
исследований на постоянных или временных пробных площадях обязательным
требованием является определение типа и давности последнего нарушения
В современный период пожары являются одним из наиболее распространенных
типов катастрофических внешних нарушений бореальных лесных сообществ. На
основе изложенных выше данных может быть предложена следующая методи-
ка определения давности пожара и контроля продолжительности беспожарных
периодов.
Давность пожара определяется на основе анализа спилов или возрастных кер-
нов, которые отбираются с 5 живых деревьев, имеющих пожарные поврежде-
ния (пожарные шрамы) стволов и расположенных в радиусе до 50-100 м от тер-
ритории пробной площади. При выборе модельных деревьев необходимо фик-
сировать наличие дополнительных признаков пожарного происхождения повреж-
дений, о которых говорилось выше, — в первую очередь наличие обугленных
участков корки ствола или древесины шрама. Для каждого модельного дерева
производится подсчет числа годичных слоев древесины в неповрежденном и
поврежденном пожаром секторах на спиле или на двух кернах, отобранных в
поврежденной и неповрежденной пожаром частях ствола. Давность пожара оп-
ределяется как разность числа годичных слоев, сформировавшихся на неповреж-
денном и поврежденном участках. В случае совпадения давности нарушения
камбиального слоя на 5 или более деревьях, зарегистрированную давность на-
рушения можно считать давностью пожара, поскольку вероятность случайного
совпадения непирогенного повреждения стволов у 5 деревьев имеет очень низ-
кую (-10-10) вероятность.
При отсутствии на территории пробной площади или в непосредственной бли-
зости от нее деревьев, имеющих пожарные повреждения стволов, определение
продолжительности беспожарного периода осуществляется на основе изложен-
ного выше метода последовательного анализа возраста угнетенных деревь-
ев. Предельная продолжительность беспожарного периода, которая может быть
точно зафиксирована при использовании данного метода, соответствует макси-
мальному возрасту деревьев в составе древесного яруса.
213
РАЗДЕЛ 4. ОСНОВНЫЕ ПРИНЦИПЫ
СТАТИСТИЧЕСКОЙ ОБРАБОТКИ ДАННЫХ1
В настоящем разделе изложены начальные сведения, необходимые для осво-
ения принципов корректной статистической обработки данных, собранных в про-
цессе исследования состояния и динамики растительных сообществ. Представ-
ленный раздел может рассматриваться только как дополнение к учебникам и
справочникам по статистике (Плохинский, 1961; 1978; Доэрфель, 1969; Закс,
1976; Лакин, 1980; Афифи, Эйзен, 1982; Зайцев, 1984; Бронштейн, Семендяев,
1986 и др.).
Одним из основных принципов в современной биологии, в том числе в гео-
ботанике и экологии, является принцип количественного подхода, т е. измере-
ние различных параметров исследуемого объекта — в данном случае расти-
тельного сообщества и характеристик его местообитания. Основы количествен-
ного подхода к исследованию растительных сообществ были заложены еще в
XIX — начале XX века при формировании лесной таксации, представляющей со-
бой комплекс методов количественной оценки характеристик древесного яруса
лесных сообществ. В последующий период этот подход получил широкое тео-
ретическое и практические развитие (Грейг-Смит, 1967; Василевич, 1969, 1972;
Раменский, 1971; Трасс, 1976; Дылис, 1978; Дыренков, 1984; Ипатов, 2001).
Количественный эмпирический материал, собранный в процессе полевых и
лабораторных исследований, представляет собой массивы числовых данных,
характеризующих различные параметры изучаемых объектов: растительных со-
обществ, их структурных компонентов (ярусов, синузий, популяций видов), от-
дельных особей и частей растений и т. д. Основная цель статистики и стати-
стических сравнений — унификация оценок разных исследователей по отноше-
нию к характеризуемым или сравниваемым объектам. В этом понимании статис-
тика аналогична системе измерения физических величин, позволяющей неопре-
деленные качественные понятия «большой» и «маленький» заменять числовыми
значениями, характеризующими изучаемый объект и позволяющими выполнить
сравнение с одинаковым для различных исследователей результатом. Равным
образом вместо экспертной оценки исследователя «средние значения характери-
стики различаются» на основе применения статистических сравнений будет по-
лучен числовой результат. Например, при применении однофакторного диспер-
сионного анализа результат статистического сравнения выглядит следующим
образом: средние значения-характеристики достоверно различаются при уровне
значимости а = 0,001. Или в математической терминологии: «нулевая гипотеза
о равенстве средних двух сравниваемых выборок отвергается по критерию Фи-
шера с уровнем значимости а = 0,001».
1 Авторы В. В Горшков, Н И Ставрова
214
Таким образом, анализ эмпирического материала с использованием аппарата
математической статистики позволяет решить следующие основные задачи:
1) получить обобщающую количественную оценку отдельных характеристик
объектов (средних значений, варьирования и т. д.); 2) установить достоверность
сходства или различия между параметрами различных объектов и данными, по-
лученными в разных исследованиях; 3) выявить законы временной динамики ха-
рактеристик объектов; 4) установить достоверность и тип взаимосвязи различ-
ных характеристик и степень их зависимости от факторов местообитания; 5) про-
гнозировать изменение параметров объекта за пределами изученной области.
Основные понятия
Генеральная совокупность и выборка. Генеральная совокупность — это
понятие статистического выборочного метода. В математической статистике
генеральной совокупностью называется множество каких-либо однородных эле-
ментов, из которого по определенному правилу выделяется некоторое подмно-
жество, называемое выборкой, которая является основным объектом статисти-
ческого анализа. В большинстве случаев выборка извлекается из генеральной
совокупности случайно. Для решения статистических задач нужна не сама гене-
ральная совокупность, а лишь те или иные характеристики соответствующей
функции распределения F(x). С точки зрения теории вероятностей, выборка из
бесконечной генеральной совокупности представляет собой наблюдаемые зна-
чения случайных величин, имеющих заданный закон распределения. При таком
истолковании термина «выборка» понятие генеральной совокупности оказывается
излишним, о чем свидетельствует резкое сокращение использования этого тер-
мина в научной литературе.
Многообразие объектов и задач при исследовании растительного покрова,
большие затраты времени на измерение отдельных характеристик, с одной сто-
роны, и существование в математической статистике определенных требований
к объему анализируемых числовых массивов (выборок), с другой стороны, при-
вели к разработке рекомендаций по объему выборок изучаемых объектов (на-
пример, количества модельных деревьев, числа учетных площадок и пр.) и жес-
ткому регламенту минимального числа измерений (например, числа повторнос-
тей в экспериментальных работах). В соответствии с законом больших чисел
ошибка полученных статистических оценок пропорциональна 1/4п , где п — чис-
ло измерений. Исходя из этого, а также из особенностей изучаемого объекта и
задач исследования определяется рекомендуемое или необходимое число изме-
рений. Наименьшее число измерений, позволяющее сделать корректное статис-
тическое сравнение по любому из критериев (включая не зависящие от типа рас-
пределения непараметрические критерии), равно 4. И именно это число измере-
ний (за исключением крайне трудоемких работ по измерению характеристик мо-
дельных деревьев) должно рассматриваться как минимальное.
215
В научных исследованиях в большинстве случаев рассматривается лишь не-
большая часть показателей, характеризующих выборку. В первую очередь, это
арифметическое среднее, стандартное отклонение, коэффициент вариации, а так-
же частотное распределение.
Среднее арифметическое (х) есть сумма всех наблюдений, поделенная на
число этих наблюдений:
(1)
где х, — результаты конкретных г-тых измерений, п — общее число измерений
характеристики.
Стандартное отклонение (s) (Закс, 1976), или среднее квадратическое от-
клонение (Плохинский, 1961; Зайцев, 1978), равно корню квадратному из сум-
мы квадратов отклонений, деленной на и - 1:
(2)
Квадрат стандартного отклонения называется дисперсией (s2).
Ошибка среднего (среднеквадратичная ошибка, стандартная ошибка) (т)
представляет собой частное от деления стандартного отклонения на квадратный
корень из числа наблюдений значения квадратов отклонений:
т -
(3)
Коэффициент вариации (V) Наряду со стандартным отклонением, одним из
очень часто используемых параметров распределения величины является коэф-
фициент вариации, представляющий собой нормированное стандартное отклоне-
ние. В исследовательских работах коэффициент вариации приводится либо в до-
лях единицы, либо в % от среднего значения:
Г= 1-100%, х^О (4)
х
Коэффициент вариации равен стандартному отклонению в том случае, когда
среднее значение равно единице. То есть коэффициент вариации есть относитель-
ная мера рассеяния при принятом за единицу среднем значении.
Показатель точности (относительная ошибка) (Р). Показатель точности рас-
считывается как процентное отношение ошибки средней и самой средней и, сле-
довательно, позволяет оценить точность определения последней. Показатель
точности определяется по формуле
Р = т/х 100 %, х*0.
(5)
216
Типы распределений
Одним из важных аспектов первичного представления и анализа данных явля-
ется анализ распределения значений характеристики в пределах выборки. Распре-
деления разделяются на два основных типа: дискретные и непрерывные в зави-
симости от характера изменения составляющих их случайных величин. Для по-
строения распределения выборка ранжируется от наименьшего значения х к наи-
большему. После этого область значений х разбивается на ряд классов. Число
классов выбирается в зависимости от объема выборки и точности измерения ве-
личин. Рекомендации по определению величины классовых интервалов и числа
классов можно найти в статистических руководствах (Плохинский, 1961; Закс,
1976; Зайцев, 1984). Для каждого класса подсчитывается число попадающих в
данный класс значений характеристики. При распределении величин по классам
принято следующее правило (Закс, 1976): к классу в интервале от а до b отно-
сятся все значения характеристики от а до Ь, исключая Ь, т. е. все значения х,,
подчиняющиеся выражению а<х,<Ь. Число значений изучаемой характеристи-
ки, относящихся к данному классу, называется его частотой. Относительная
частота (fj) класса определяется из следующего соотношения:
д=^.юо%, • (6)
где Ду — число значений характеристики, относящихся к данному классу. Полу-
ченные данные представляются в виде гистограммы распределения относитель-
ных частот (рис. 4.1), полигона распределения частот или в табличном виде.
Наряду с распределением относительных частот может быть построено распре-
Рис. 4.1. Гистограмма распределения относи-
тельных частот (распределение по высо-
те деревьев сосны обыкновенной в сосно-
вых лесах заповедника Кивач, Карелия, п
= 540)
Рис. 4.2. Гистограмма распределения накоп-
ленных частот (распределение накоплен-
ных частот высоты деревьев сосны
обыкновенной в сосновых лесах заповед-
ника Кивач, Карелия, п = 540)
217
Распределение значений накопленных частот представляется также в виде гра-
фиков (рис. 4. .2) или таблиц накопленных частот.
Выявление: типа распределения значений исследуемой характеристики (эмпи-
рического распределения) выполняется на основе его сравнения с известными
типами теоретических распределений. Среди распределений непрерывного типа
наиболее расппространенными в практике биологических исследований являются
нормальное, гпоказательное, логнормальное, среди распределений дискретного
типа — биномиальное распределение и распределение Пуассона (Закс, 1976;
Плохинский, 1978; Лакин, 1980; Зайцев, 1984). В первую очередь полученное
эмпирическое: распределение сравнивается с нормальным, которое является ос-
новным, т. к нв соответствии с теорией вероятностей к нему в пределе стремят-
ся все другие iизвестные типы распределений. Критерями соответствия выступа-
ют непарамегдшческие критерии %2 и/или Колмогорова-Смирнова.
Нормальнее распределение широко распространено и имеет важное значение
для теории итрактики биологических исследований (Зайцев, 1984). Нормальное
распределение характеризуется следующими свойствами: 1) симметричной ко-
локолообразиоой двускатной кривой распределения частот; 2) соотношением
средней арифметической и среднего квадратичного отклонения равным 3, вели-
чина коэффициента вариации при этом составляет примерно 33 %.
Показат&и&ное распределение отличается резко выраженной правосторонней
асимметриейроаспределения частот (крутое в левой части и плоское — в правой),
примерным раавенством среднего арифметического и среднего квадратического
отклонения и, i в соответствии с этим, коэффициентом вариации примерно равным
1 или 100 %.М\4ода (точка на оси абсцисс, соответствующая максимальной час-
тоте кривой [аеспределения) показательного распределения всегда равна 0. Сле-
дует отметить,,, что если изучаемая характеристика распределена по показатель-
ному закону, гто дисперсию и среднее квадратическое отклонение обычно рас-
считывают по । формулам (2) и (3) для нормального распределения. Соответствие
эмпирическое о распределения показательному при близких значениях среднего и
среднего квадроатического отклонения может быть проверено на основе величи-
ны соотношения s2/х2, которое должно быть меньше критического значения 1,56
(Зайцев, 1984|. Если соотношение превышает указанное критическое значение, то
эмпирическое ^распределение может быть аппроксимировано логнормальным.
Логнормальное (логарифмически нормальное) распределение характеризует-
ся двускатной I кривой распределения частот с умеренной положительной асим-
метрией (6ontes крутой в левой части и более пологой — в правой). В этом слу-
чае величинассреднего квадратического отклонения в отличие от нормального
распределен^ достаточно велика по сравнению со средним, и коэффициент ва-
риации существенно превышает 33 %. Дисперсия и среднее квадратическое от-
клонение таккхе, как и для показательного распределения, могут быть рассчита-
ны по формулам (2) и (3) для нормального распределения.
218
Биномиальное распределение (распределение Бернулли) требует выполнения
следующих условий: 1) вероятность определения характеристики в процессе
исследования остается постоянной; 2) величина дисперсии выборки меньше сред-
него арифметического, при этом характеристика может принимать только целые
положительные значения. Биномиальное распределение характерно для случаев,
когда исследуется Nn проб одинакового объема, равного п; для каждой пробы
учитывается доля вариант (р), обладающих данным признаком. Теоретические
частоты биномиального распределения показывают, какова вероятность появле-
ния проб с заданным числом или долей вариант с данным признаком. Параметра-
ми биномиального распределения являются пир. Для абсолютных частот при-
знака х = пр, s2=npq, q= 1 -р, для относительных частот — х=р, s2 = pqln.
При р = 0,5 биномиальное распределение характеризуется симметричной двускатной
кривой, при р > 0,5 кривая является более пологой слева, при р < 0,5 — справа.
Распределение Пуассона является результатом учета относительно редких,
случайных, взаимно независимых событий в единицу времени, длины, площади,
объема. Это распределение близко к биномиальному и образуется в случае, если
р — очень мало, a q близко к 1. Основным признаком распределения Пуассона
является то, что среднее арифметическое примерно равно дисперсии (х ~ s2).
Если справедливо s2 < х, то можно применять биномиальное распределение; в
противоположном случае можно использовать отрицательное биномиальное рас-
пределение. Следует отметить, что если для дискретного распределения х ~ s2,
то его можно аппроксимировать распределением Пуассона вплоть до больших
значений переменной х (Закс, 1976). При величине среднего (и соответственно
дисперсии) больше 1 вероятности по мере увеличения х сначала возрастают,
затем уменьшаются (т. е. имеет место двускатная кривая распределения частот).
С увеличением х асимметрия распределения стремится к 0, и оно становится
более симметричным. Максимум распределения приходится на ближайшее це-
лое число меньшее х . При х < 1 вероятности с ростом значений х уменьшают-
ся, т. е. имеет место предельная положительная асимметрия (односкатная кри-
вая с максимальной вероятностью для х = 0).
Отрицательное биномиальное распределение характерно для дискретных ве-
личин, вероятность появления которых в данной генеральной совокупности не
является постоянной. Распределение характеризуется превышением величины
дисперсии над величиной среднего (х2>х).
Если сравнение эмпирических и теоретических частот обнаруживает их соот-
ветствие, на полученную выборку распространяются закономерности, присущие
данному теоретическому типу распределения.
Статистические сравнения
Как отмечалось выше, при анализе полученных данных прежде всего прово-
дится сравнение эмпирического распределения с нормальным. Сравнение эмпи-
219
рического распределения с конкретным теоретическим распределением осуще-
ствляется на основе непараметрических критериев сравнения: критерия у2 и/или
критерия Колмогорова-Смирнова. Если на основе такого сравнения установле-
но, что распределение значений исследуемых характеристик в выборках соответ-
ствует нормальному, то сравнение статистических параметров двух или несколь-
ких выборок между собой может быть выполнено на основе параметрических
критериев', критерия Стьюдента или дисперсионного анализа. Если же эм-
пирические распределения не соответствуют нормальному, то для сравнения вы-
борок должны использоваться непараметрические критерии'. %2, Колмогоро-
ва-Смирнова или Тьюки, которые не зависят от типа распределения. Об этих кри-
териях будет сказано ниже.
Критерий Стьюдента (t) является одним из широко используемых в эмпи-
рических работах критериев. Он позволяет сравнить два средних значения, харак-
теризующих две выборки измерений, выполненных в разных условиях. Прове-
ряемая нулевая гипотеза — гипотеза о равенстве средних значений т\ = m2 гене-
ральных совокупностей, сравниваемых на основе двух выборок.
+ п2
(«! ~ 1)^ + («2 ~ 0^
+л, — 2
(8)
где х, , х2 — средние значения для первой и второй выборок, п\, П2 — число
наблюдений (измерений) в первой и второй выборках, si и S2 — оцененные стан-
дартные отклонения двух выборок. Данный вариант расчета критерия Стьюдента
используется при п\ Ф П2 и предположении о равенстве стандартных отклонений
двух генеральных совокупностей, которые оцениваются по сравниваемым выбор-
кам 5] = $2, при этом число степеней свободы v = п \ + П2~ 2. Нулевая гипотеза о
равенстве средних двух генеральных совокупностей отвергается, если оценен-
ное значение t превышает табличное для заданного числа степеней свободы и
выбранного уровня значимости, т. е. при условии t > tva. При использовании при-
кладных статистических пакетов программ в протоколе приводятся значения кри-
терия Стьюдента и уровень значимости, при котором отвергается нулевая гипо-
теза. Различия средних значений двух сравниваемых выборок считаются досто-
верными при уровне значимости р < 0,05.
Однофакторный дисперсионный анализ. Сравнение двух средних значе-
ний из нормально распределенных генеральных совокупностей, рассмотренное
выше, может быть распространено на любое число средних значений.
Пусть даны к выборок с объемом п, каждая и с общим числом элементов п.
Каждая выборка принадлежит к нормально распределенной генеральной со-
вокупности. Генеральные совокупности имеют равные, но неизвестные диспер-
сии. Среднее значение группы х,, а общее среднее — х .
220
Основой однофакторного дисперсионного анализа является то, что сумма
квадратов отклонений выборочных значений относительно общего среднего
(2общ) может быть разложена на 2 части:
1) сумму квадратов отклонений отдельных значений относительно групповых
средних, называемую суммой квадратов отклонений внутри групп (^внутри);
2) сумму квадратов отклонений групповых средних значений относительно об-
щего среднего, называемую суммой квадратов отклонений между группами (Омежлу)-
(?общ — ^внутри (2между (9)
£(*/, ~*)2 = ££(*,7 -г,)2 +'£п,(х, -х)2 .
i.J М 7 = 1 ..1
Число степеней свободы
Гобщ — л — 1 — гвнутри + умежДу — (и — £) + (£— 1). (Ю)
Отношение Q/v (где v — соответствующее Q число степеней свободы) назы-
вается в дисперсионном анализе средним квадратом (MQ). Если все выборки
принадлежат к одной и той же генеральной совокупности, то 0Внутри = бмежду Если
отношение F = MQMexay/MQBHyTpa больше критического значения F-распределе-
ния для V[ = к - 1, V2~ п - к и заданного уровня значимости a (F > Ftk..\. п-к, а)),
то среди групп имеются такие, которые имеют различные средние значения ц,. В
этом случае нулевая гипотеза Ц1 = Ц2 = ••• = Мл= = Мт- = И отклоняется.
Следует отметить, что такой результат не обязательно означает, что средние
значения всех выборок отличаются друг от друга, он свидетельствует о том, что
по меньшей мере два (или более) средних Ц, отличаются друг от друга, т. е. для
некоторого i подтверждается альтернативная гипотеза ц, Ф ц.
Однофакторный дисперсионный анализ, как следует из его названия, часто
используется для оценки достоверности изменения величины исследуемой ха-
рактеристики по градациям какого-либо фактора. В этом случае каждая группа
данных представляет выборку значений исследуемой характеристики при опре-
деленном уровне фактора (например, при определенной давности пожара, опре-
деленной концентрации загрязнителей в почве и т. д.). Указанный выше результат
интерпретируется таким образом: данный фактор достоверно влияет на величи-
ну исследуемой характеристики на уровне значимости а (при котором вычислен-
ное значение критерия Фишера F больше табличного F^-\,п-к, а)}- При обработ-
ке результатов измерений при помощи одного из стандартных пакетов статисти-
ческих программ для персональных компьютеров в итоговой таблице кроме ве-
личины критерия Фишера обычно указывается величина уровня значимости а, на
котором отвергается нулевая гипотеза. Для того чтобы влияние фактора было
признано достоверным, величина а должна быть менее 0,05, что соответствует
уровню значимости (вероятности ошибки) равному 5 % или доверительному
уровню равному 95 %.
221
Критерий Колмогорова-Смирнова. Если установлено, что эмпирическое
распределение исследуемой характеристики не соответствует нормальному, то
необходимо установить соответствие эмпирического распределения другому
теоретическому распределению. При анализе характеристик объектов исследо-
вания часто необходимо сравнить две независимые выборки измерений (два не-
зависимых распределения) и ответить на вопрос, относятся ли они к одной и той
же генеральной совокупности. В перечисленных случаях наиболее строгим кри-
терием однородности считается критерий Колмогорова-Смирнова. Принцип кри-
терия Колмогорова-Смирнова состоит в сравнении накопленных частот сравни-
ваемых распределений.
В качестве статистики служит максимальная разность (D) накопленных отно-
сительных частот двух распределений:
D = max/']' ‘ , = max^ — + -- j, (11)
где Т7/ ‘, С2' — относительные накопленные частоты, F\, F2 — абсолютные на-
копленные частоты, п\, П2 — объемы сравниваемых выборок.
Критические значения статистики критерия Колмогорова-Смирнова для дву-
стороннего критерия (т. е. для случая, когда сравниваются два эмпирических рас-
пределения) при п\ + П2 > 35 могут быть приближенно рассчитаны по формуле:
где К(а) — константа, зависящая от принимаемой вероятности ошибки а. Для
наиболее распространенных в биологических исследованиях уровнях значимос-
ти а=0,05, 0,01 и 0,001 значения К(а) составляют: А?(а)(0,05)=1,36;
К(а)(0,01)=1,63; К(а)(0,001)= 1,95. Величина К(а) рассчитывается по выражению:
К(а)=Л/-0,5-1п(а/2) . (13)
При Н] + П2— 35 рассчитанные по формуле (12) значения статистики D завы-
шены по сравнению с истинными.
Если определенное на основании двух выборок значение превосходит крити-
ческое, это свидетельствует о наличии значимой разницы между распределения-
ми двух сравниваемых выборок (или функциями накопленных вероятностей двух
сравниваемых выборок). Иными словами нулевая гипотеза о принадлежности двух
сравниваемых выборок к одной генеральной совокупности должна быть откло-
нена при заданном уровне значимости и принята альтернативная гипотеза о
том, что сравниваемые выборки принадлежат к разным генеральным совокупно-
стям. Двусторонность критерия означает, что результат применения критерия спра-
ведлив как по отношению к одной, так и по отношению к другой выборке.
В пакетах программ для персональных компьютеров при сравнении двух
выборок по критерию Колмогорова-Смирнова обычно рассчитывается вели-
222
чина максимальной разности накопленных частот и величина уровня значи-
мости, на котором гипотеза принадлежности двух распределений к одной
генеральной совокупности может быть отвергнута.
Следует отметить, что изначально критерий Колмогорова-Смирнова
был разработан для непрерывных величин, хотя в настоящее время руковод-
ства по статистике (Закс, 1976) рекомендуют использовать его как для срав-
нения непрерывных (например, измерения объема, массы и др.), так и для дис-
кретных (возраст, число лет) величин. Однако исходное предположение о
непрерывности распределения нашло отражение в алгоритме нахождения
максимальной разности, в связи с чем в пакете программ Statgraphics при
сравнении распределений дискретных величин статистика D — максималь-
ная разность накопленных относительных частот двух распределений, оце-
нивается неверно.
Помимо сравнения двух выборок критерий Колмогорова-Смирнова, как уже
говорилось, используется в качестве одностороннего критерия согласия при
сравнении эмпирического и теоретического распределений. Проверяемая нуле-
вая гипотеза — согласуется ли данное эмпирическое распределение с выбран-
ным теоретическим. Критические значения статистики критерия Колмогорова-
Смирнова для одностороннего критерия (при «[+ П2 > 35) могут быть прибли-
женно рассчитаны по формуле
В (а) = К(а) -71 / Л . (14)
Упрощенный (быстрый) критерий Тьюки (Tukey, 1959; Закс, 1976). Дан-
ный критерий базируется на положении, что две выборки измерений различают-
ся тем больше, чем меньше они пересекаются. Пусть первая выборка содержит
наименьшее значение, вторая — наибольшее. Тогда необходимо подсчитать:
Т\ — число значений в первой выборке (содержащей наименьшее для двух
сравниваемых выборок значение), меньших по величине наименьшего значения
второй выборки.
?2 — число значений во второй выборке (содержащей наибольшее для двух
сравниваемых выборок значение), превышающих по величине наибольшее зна-
чение в первой выборке.
Обе величины — Т\ и ?2 (каждая должна быть отлична от нуля) складывают-
ся и, таким образом, получается расчетное значение статистики Т = Т\+Т^. Если
одно из значений в сравниваемой выборке совпадает по величине с наименьшим
или наибольшим числом, то оно учитывается не как 1, а как 0,5.
Если объемы выборок примерно одинаковы,
Л] < «2 3 + 4 «1 /3, (15)
то критические значения Та статистики Тьюки составляют 7, 10 и 13 (двусторон-
ний критерий для уровней значимости соответственно а = 0,05, 0,01 и 0,001). То
223
есть при выполнении условия (25) и Т> 7 нулевая гипотеза о принадлежности
двух сравниваемых выборок одной генеральной совокупности отвергается на
уровне значимости а = 0,05 и т. д.
При значительном неравенстве выборок расчетное значение уменьшается на
определенную корректирующую величину, и суждение о нулевой гипотезе вы-
носится на основе сравнения Т - Т^оп- и Та:
при л2<2 «1; 7’СОГг = 1; (16)
(18)
при л2> 2 л,; 7’согг= (л2-«| + 1) / и, (17)
Критерий %2. Критерий х2 используется как для сравнения эмпирического и
теоретического распределений (т. е. в качестве критерия согласия), так и для срав-
нения двух эмпирических распределений. Основой метода сравнения распреде-
лений по критерию X2 является сравнение относительных частот по классам. Для
этого выборка подразделяется на к классов. Статистика критерия х2 определя-
ется как сумма по всем к классам квадратов разности наблюдаемых и ожидае-
мых частот, деленных на ожидаемые частоты,
Е, ’
где В, — наблюдаемая частота, Е, — ожидаемая частота,
Ei = NPi (19)
Число степеней свободы V = к - 1 - п, где к — число классов, п — число оце-
ниваемых при анализе выбранного типа распределения параметров распределе-
ния. Так, при сравнении эмпирического распределения с нормальным распреде-
лением ожидаемые частоты рассчитываются на основе 2 оцененных по выбор-
ке параметров — среднего и дисперсии (п = 2), при проверке согласия эмпири-
ческого распределения с распределением Пуассона п = 1. При сравнении двух эм-
пирических распределений число степеней свободы v = к - 1.
При использовании критерия х2 выборки должны быть не слишком малыми и
ожидаемые гипотетические частоты должны быть не ниже 4 (Е, > 4) (Закс, 1976).
Если они меньше, то их следует увеличивать до требуемого уровня объедине-
нием 2 и более соседних классов. При v > 8 и объеме выборки п > 40 значение
ожидаемой частоты может быть уменьшено до 11.
Критерий X2 служит для проверки гипотезы о том, что F%(x) = Fq(x), где
Fx(x) — функция распределения X, a F$(x) — заданное (теоретическое) распре-
деление. Согласно альтернативной гипотезе выборка принадлежит к неизвестно-
1 В некоторых руководствах по статистике на процедуру разбиения на классы наклады-
ваются более жесткие ограничения: необходимо, чтобы для граничных классов выполня-
лось неравенство пр,> 1, для остальных классов — пр,> 5 (Бронштейн, Семендяев, 1987).
224
му распределению Fx(x). Рассчитанное значение %2 сравнивается с табличным.
Нулевая гипотеза отклоняется, если рассчитанное значение равно табличному
X2 (v, а) или превышает его.
Корреляционный и регрессионный анализы
Рассмотренные ранее критерии позволяют оценить различия выборок сред-
них значений или других параметров. В то же время, перед исследователями ча-
сто стоят задачи по выявлению взаимосвязи между двумя характеристиками X и
Y. Прежде всего необходимо пары точек (*,-, и у,) нанести на координатную сетку
для того, чтобы получить общее предварительное представление о форме об-
лака точек (рис. 4.3). Корреляционный анализ экспериментальных данных позво-
ляет ответить на вопросы, существует ли между двумя характеристиками взаи-
мосвязь и насколько она сильна. Для ответа на вопрос, какова функциональная
зависимость между этими характеристиками, необходимо провести регрессион-
ный анализ.
Коэффициент корреляции Пирсона. Наиболее распространенным коэффи-
циентом, позволяющим судить о взаимосвязи X и Y, т. е. наличии стохастичес-
кой зависимости между X и Y, на основании анализа пар измерений этих величин
(*,, у,), является коэффициент
корреляции Пирсона (рис. 4.3).
Коэффициент корреляции (г) рас-
считывается на основании следу-
ющего выражения:
^(т,-х)(у,-у)
г = (20)
Проверяемая при корреляцион-
ном анализе нулевая гипотеза: две
случайные переменные стохасти-
чески независимы, т. е. оцененный
по выборке коэффициент корреля-
ции г не отличается от 0. Провер-
ка на значимость (относительно
нуля) коэффициентов корреляции
выполняется по таблицам, а при
использовании пакетов статисти-
ческих программ для компьюте-
ров уровень значимости приводит-
ся в протоколе результатов расче-
тов, выдаваемых программой. О
Рис. 4.3. Коэффициент корреляции при раз-
личном положении облака точек
1 — случайные переменные* иу не взаимосвяза-
ны; 2 — случайные переменные* и у взаимосвя-
заны, коэффициент г отличен от 0 при уровне зна-
чимости а > 0,05; 3 — между случайными пере-
менными * и у наблюдается строгая функциональ-
ная зависимость; 4 — между случайными пере-
менными* иу наблюдается криволинейная зави-
симость; при делении выборки на две области *|-
*2 и*2~*з в пределах каждой из них регистрирует-
ся достоверная взаимосвязь между случайными
переменными * и у г (*1*2) > 0 и г (*2*3) < 0
225
существовании статистической взаимосвязи говорят, если нулевая гипотеза от-
клоняется. Следует подчеркнуть, что существо и причины найденной статисти-
ческой взаимосвязи лежат вне статистических методов. Корреляция может быть
обусловлена: 1) существующей причинной зависимостью между % и Y, 2) общей
зависимостью от третьей величины, 3) неоднородностью данных или 4) быть чи-
сто формальной.
Иногда важной является задача попарного сравнения коэффициентов корреля-
ции. Такое сравнение проводится при помощи z преобразования Фишера (Закс,
1976). Предварительное преобразование коэффициентов корреляции (г) выпол-
няется по формуле:
Сравнение коэффициентов корреляции проводится по формуле:
z =11 ’1 ’ (22)
I-----------
у л, - 3 п2 - 3
где z — расчетное значение статистики, z, и z2— преобразованные значения ко-
эффициентов корреляции, л 1 и «2 — объемы выборок, для которых получены ко-
эффициенты корреляции. Критические значения статистики z, соответствующие
уровням значимости а = 0,05, 0,01 и 0,001, составляют соответственно 1,96;
2,58 и 3,29.
Важно отметить, что оценка значимости коэффициента корреляции и сравне-
ние двух коэффициентов корреляции аналогичны одностороннему и двусторон-
нему критериям. Коэффициент корреляции может значимо отличаться от нуля, но
при сравнении этого коэффициента корреляции с другим выборочным коэффи-
циентом, равным нулю, их различие может быть недостоверным.
Коэффициент ранговой корреляции Спирмена. Оценивание наличия вза-
имосвязи между парами характеристик на основе расчета коэффициента корре-
ляции предполагает нормальное распределение величин ХтлУ (бинормальная ге-
неральная совокупность). Ранговый коэффициент корреляции Спирмена позволяет
определить взаимосвязь, когда исследуемая выборка (по обоим параметрам)
относится к произвольному непрерывному распределению.
Для вычисления коэффициента ранговой корреляции Спирмена оба ряда ран-
жируются: п выборочных значений упорядочиваются по величине и обознача-
ются через x(i), Х(2), ...., х(„) и у(1), у(2)..., у(л) так, что
Х(1) < Х(2) — ... — Х(;) ... Х(п~),
У(1) <У(2) < ... <У(,) < ... <у(„),
при этом любое значение хц) (у(,)) называют ранговым значением или ранговой
статистикой. Номер выборочного значения называют рангом или ранговым чис-
лом. Значению х(,) (у^) соответствует ранг i. Равным ранговым значениям соот-
ветствует среднее ранговое число (если три числа 3,2; 3,2; 3,2 имеют ранги 1, 2,
226
3, среднее ранговое число составляет 2). Вычисление рангового коэффициента
корреляции Спирмена производится по формуле:
~Ч)2
rs= 1----------
»(»-!)
(23)
Если два ряда рангов равны, то разности равны нулю и rs~ 1. Если ряды ран-
гов обратны, то rs= -1. Данный критерий тем самым позволяет ответить на воп-
рос и о знаке имеющейся корреляции.
Проверяемая при этом нулевая гипотеза та же, что и при использовании коэф-
фициента корреляции Пирсона — две случайные переменные стохастически не-
зависимы. Проверка на значимость (относительно нуля) коэффициентов корре-
ляции Спирмена выполняется по таблицам или приводится в протоколе соответ-
ствующих статистических программ.
При л>30 значимость можно с достаточной точностью проверить на осно-
вании стандартного нормального распределения:
z = |rs| V« - 1, (24)
Критические значения статистики z, соответствующие уровням значимости
а = 0,05, 0,01 и 0,001, составляют соответственно 1,96; 2,58 и 3,29.
Регрессионный анализ. Любая форма связи между величинами может быть
выражена уравнением общего вида у =/(х), где признаку — зависимая перемен-
ная или функция от независимой переменной х, называемой аргументом. Изме-
нение функции в зависимости от изменений одного или нескольких аргументов
называется регрессией. Соответствие между аргументом и функцией может быть
задано таблицей, формулой, графиком и т. д. Для выражения регрессии служат эм-
пирические и теоретические ряды, их графики — линии регрессии, а также корре-
ляционные уравнения (уравнения регрессии) и коэффициент линейной регрессии.
Параметры регрессии выражают связь между X и Y двусторонне, учитывая
изменение средней величины ух признака Y при изменении значений х, признака
X, и наоборот, показывают изменение средней величины ху признака X по изме-
ненным значениям у, признака Y. Исключение составляют временные ряды, или
ряды динамики, показывающие изменение признаков по времени. Регрессия та-
ких рядов является односторонней.
Ряды регрессии, особенно их графики, дают наглядное представление о фор-
ме и тесноте корреляции между признаками, в чем и заключается их ценность.
Форма связи между признаками может быть разнообразной. Задача состоит в том,
чтобы любую форму связи выразить уравнением определенной функции (линей-
ная, параболическая), что позволяет получать нужную информацию о зависимо-
сти между переменными величинами Y и X, предвидеть возможные изменения
признака Y на основе известных изменений X.
227
Некоторые важные типы регрессионных уравнений в биологических
исследованиях
Уравнение линейной регрессии (рис. 4.4). Линейная регрессия характери-
зует наиболее простой и широко распространенный тип зависимости двух вели-
чин. Рассмотрим случай, когда зависимая переменная у — случайная перемен-
ная, значения входной (независимой) переменной заданы точно.
у = ах + Ь,
(25)
где а — размерный коэффициент [у, х"1], характеризующий скорость изменения
характеристики у по мере изменения характеристики х; b — коэффициент, имею-
щий размерность [у], равный значению функции при х = 0.
Рис. 4.4. Линейная регрессия (уравнение прямой)
1) а — угловой коэффициент, характеризует скорость изменения у по мере изменениях, то
есть а = (У2-у|)/(хг - х,); b — свободный член уравнения равный значению функции при
х = 0; 2) знак углового коэффициента определяет, возрастают ли значенияу с возрастанием х
(а и коэффициент корреляции положительны) или убывают (а < 0, г < 0)
В пакетах статистических программ оценка достоверности коэффициентов рег-
рессии выполняется на основе уровня значимости, приводимого в протоколе.
Линейная аппроксимация (приближенное описание) используется, когда изме-
нение (увеличение или уменьшение) зависимой переменной у во всем диапазоне
изменения независимой переменной х происходит с одинаковой скоростью. В
принципе, любая нелинейная зависимость может быть представлена как после-
довательность нескольких линейных приближений, различающихся по величине
коэффициента а в соответствии с разной скоростью изменения функции на раз-
ных участках диапазона изменения аргумента х.
Экспоненциальное уравнение (рис. 4.5):
у=се“х + ₽, (26)
где ос — размерный коэффициент [х“'], характеризующий скорость изменения ха-
рактеристики по мере увеличения значений х; Р — безразмерный коэффициент,
характеризующий значение характеристики в начальной точке (при х = 0); с —
228
единичный коэффициент, необходимый
для сохранения размерности [у].
Эспоненциальное уравнение может
быть записано в другом виде, например,
уравнение демографии А. Дж. Лотки
(Lotka, 1925), описывающее стационар-
ное состояние возрастной структуры по-
пуляции:
D(x) = Dotr^, (27)
где D(x) — плотность особей возраста х
[ед. га-1 год-1], Dq — начальная плот-
ность или плотность рожденных особей,
т — коэффициент, характеризующий
смертность особей [год-1].
Логистическое уравнение (рис. 4.6):
У max
l + e7+fk ’
Рис. 4.5. Кривая экспоненциального
уравнения
(28)
Рис. 4.6. Кривая уравнения логистической функции
где ymax — величина, соответствующая максимальному значению характеристи-
ки, достигаемому на стадии стабилизации; у и 0 — коэффициенты уравнения. Ве-
личина коэффициента 0 определяет размер области, где происходит основное из-
менение величины у (чем больше значение коэффициента 0, тем меньше размер
переходной области). Сам коэффициент 0 является фундаментальной константой
и имеет размерность х-1. Безразмерный коэффициент g характеризует значение
функции при х = 0. Чем больше коэффициент у, тем ближе значение функции при
х = 0 к максимальному значению при отрицательных величинах у или к нулевому
значению при положительных величинах коэффициента (коэффициенты у и 0 урав-
нения логистической кри-
вой всегда имеют проти-
воположный знак, при от-
рицательном коэффициен-
те у и положительном 0
значение функции по мере
увеличения аргумента
убывает, при положитель-
ном у и отрицательном
0 — возрастает).
Важной характеристи-
кой данной функции явля-
ется наличие областей, ог-
раниченных значениями х„
229
при которых абсолютная величина показателя степени |у+ Рх| составляет 0, 1, 2
и 3. При у+ Рх = 0 значение функции равно 1 /2утах; эта точка является характер-
ной точкой — точкой перегиба, значение х,- является характерным временем из-
менения характеристики. При изменении значения показателя степени от -1 до
+ 1 у,- изменяется соответственно от 73,5 до 26,4 % от утах, т. е. в этой области
происходит изменение величины у на 47 %. При изменении показателя степени
от -2 до +2 — соответственно от 88,5 до 11 % (77,5 % изменения величины у),
при изменении показателя степени от -3 до +3 — от 95,6 до 4 % (91,6 % изме-
нения величины у). Положение характерных точек позволяет оценить время вос-
становления изучаемой характеристики до заданных величин (в % от их стацио-
нарного значения), т. е. на 50, 77,5, 88,5 и 95,6 %. Значения х,-, определяющие
данные области изменения величины у, легко могут быть вычислены. Например,
х*и х~:
’_7 _-0 + У)
х+- р , X, - р . (29)
Уравнение затухающей экспоненты (рис. 4.7)
?=утах[1-^]> (30)
и затухающей экспоненты со сдвигом
У= утах[1- (31)
где утах — величина, соответствующая максимальному значению характеристи-
ки, достигаемому на стадии стабилизации, X — параметр, характеризующий ско-
Рис. 4.7. Кривая уравнения затухающей экспоненты
рость восстановления
характеристики [год-1];
к — сдвиг по оси х, ха-
рактеризующий запаз-
дывание начала восста-
новления характеристи-
ки.
При использовании
уравнения затухающей
экспоненты характер-
ный период восстанов-
ления — t\ = Т~ 1/Х
обозначает время вос-
становления характеристики на 63 % от утах; ?2= 27’ = 2/Х соответствует време-
ни восстановления характеристики на 86,5 % (относительное восстановление);
= 37’= 3/Х — на 95 % (время стабилизации).
В соответствии с уравнением затухающей экспоненты со сдвигом по оси х ха-
рактерные периоды времени восстановления — tt, и ц, за которые характери-
230
стика достигает значений, соответственно 63, 86,5 и 95 % от стационарной ве-
личины составляют:
1 2 3
t\~ Т~ --+ к, tz~ 2Т~ - + к, t^—3T~ — + к.
Л Л Л
В приведенных уравнениях данные значения степени восстановления характе-
ристики выбраны потому, что соответствуют целым значениям показателей сте-
пени и позволяют легко производить расчеты.
Заключение
Представленные в настоящем разделе некоторые базовые сведения, касаю-
щиеся анализа результатов измерений, выполняемых в процессе полевых и экс-
периментальных исследований, не исчерпывают всех возможных методов ста-
тистической обработки данных. Использование различных вариантов анализа
определяется спецификой изучаемых характеристик лесных сообществ, особен-
ностями их динамики и поставленными задачами. Следует обратить внимание на
способы представления результатов статистической обработки. Полученные оцен-
ки должны быть приведены с достаточной степенью подробности. Необходимо
иметь в виду рекомендацию о графическом представлении данных. Результаты
исследования временной динамики характеристик сообществ, их взаимной свя-
зи и изменения по градиентам факторов среды рекомендуется представлять в
системе координат в виде облака точек. Оценка распределения частот различ-
ных количественных градаций исследуемой характеристики может быть представ-
лена в виде гистограмм. Кривые на графиках проводятся не произвольно по точ-
кам, а в соответствии с уравнениями регрессии, описывающими характер изме-
нения изучаемого параметра. При невозможности аппроксимации данных в пре-
делах всего изучаемого диапазона изменения на основе единого типа кривой ре-
комендуется деление выборки на отдельные части на основе анализа облака то-
чек и выявления типа регрессионной зависимости для каждой части раздельно.
При отсутствии достоверного изменения характеристики по градиенту фактора
или оси времени (отсутствии различий между группами точек) на графике про-
водится прямая линия, параллельная оси х. Заключения об имеющихся достовер-
ных различиях между средними значениями, выборками, коэффициентами корре-
ляции и коэффициентами уравнений должны сопровождаться представлением
величин использованных критериев и уровней значимости, для которых сделана
оценка. Коэффициенты регрессионных уравнений использованных для аппрокси-
мации изучаемых зависимостей должны быть представлены с величиной их ошиб-
ки и оценкой степени достоверности (уровня значимости). Соответствующие
данные рекомендуется приводить в табличном виде. Выполнение указанных ре-
комендаций позволяет представить результаты исследования и обосновать его вы-
воды.
231
ОСНОВНАЯ ЛИТЕРАТУРА
Агрохимические методы исследования почв.— М., 1975. — 656 с.
Александрова В Д. Классификация растительности. Обзор принципов классификации и клас-
сификационных систем в разных геоботанических школах. — Л., 1969. — 275 с.
Алексеев В. А. (отв. ред.) Лесные экосистемы и атмосферное загрязнение. — Л.: Наука,
1990 а, —200 с.
Алексеев В. А. Некоторые вопросы диагностики и классификации поврежденных загрязнени-
ем лесных экосистем // Лесные экосистемы и атмосферное загрязнение. — Л., 1990а. —
С 38-54.
Андрееве. Н. Прирост кормовых лишайников и приемы его регулирования//Тр. БИН РАН.
Сер. III. Геоботаника. — 1954. — Вып. 9. — С. 11-74.
Андреева Е. И. Влияние атмосферного загрязнения на моховой покров северотаежных ле-
сов // Лесные экосистемы и атмосферное загрязнение. —Л., 1990а. — С. 159-172.
Андреева Е И. Динамика видового состава мхов // Влияние промышленного атмосферного
загрязнения на сосновые леса Кольского полуострова. — Л.: БИНРАН, 19906. — С. 133—
136.
Андреева Е Н. Структурные изменения мохового покрова // Влияние промышленного ат-
мосферного загрязнения на сосновые леса Кольского полуострова.—Л., 1990в. — С. 137-
140
Антанайтис В. В., Загреев В. В. Прирост леса. — М.: Лесн. пром-ть, 1969. — 240 с.
Анучин Н П Лесная таксация Изд 3-е. — М., 1971. — 509 с.
Аринушкина Е. В. Руководство по химическому анализу почв — М., 1970. —488 с.
Афифи А. А., Эйзен Ф. П. Статистический анализ. Подход с использованием ЭВМ. — М.,
1982,—488 с.
Баккал И Ю., Бобров А. Е., Горшков В В., Китсинг Л. И., Нешатаева В. Ю., Ярмишко
В Т. Объекты и методы исследований // Влияние промышленного атмосферного загряз-
нения на сосновые леса Кольского полуострова. — Л.: БИН РАН, 1990. — С. 15-32.
Баркман Я. Современные представления о непрерывности и дискретности растительного по-
крова и природе растительных сообществ в фитосоциологической школе Браун-Бланке //
Бот. журн. — 1989. — Т. 74, № 11, —С. 1545-1551.
Бигон М., Харпер Дж., Таунсенд К Особи, популяции и сообщества. — М., 1989. — 447 с.
Бобовникова Ц. И., Малахов С Г, Махонько Э. П. Система наблюдения и контроля за
загрязнением почвы // Миграция загрязняющих веществ в почвах и сопредельных сре-
дах,—Л, 1980, —С. 5-19.
Борисова Н А., Лошаков Л. А. Методические рекомендации по сбору и заготовке лекар-
ственного растительного сырья в Ленинградской области. — Л., 1985.
Буданцев А Л., Харитонова Н. П. Ресурсоведение лекарственных растений. Методичес-
кое пособие для студентов фармацевтического факультета. — СПб., 1999. — 87 с.
Булыгин Н Е., Ярмишко В Т. Дендрология. — СПб.: Наука, 2000. — 528 с.
Вайнагий И. В. О методике изучения семенной продуктивности растений // Бот. журн. —
1974. — Т. 59, № 6. — С. 826-831.
Варгас де Бедемар А. Исследования запаса и прироста лесонасаждений С.-Петербургской
губернии//Лесной журн. — СПб., 1848.
232
Василевич В И. К типологии лишайниковых и брусничных боров // Вести. ЛГУ. Сер Биоло-
гия. — 1961.—С. 5-15.
Василевич В. И. Статистические методы в геоботанике. — Л., 1969. — 232 с.
Василевич В И. Количественные методы изучения структуры растительности // Итоги на-
уки и техники. Ботаника. — М.: ВИНИТИ, 1972. — Т. 1. — С. 7-83.
Василевич В. И. О методах классификации растительности // Бот. жури. — 1985 — Т 70,
№ 12, —С. 1596-1604.
Васильченко И. Т. Всходы деревьев и кустарников: Определитель. — Л., 1960.
Вертикально-фракционное распределение фитомассы в лесах. / Под ред. С. Э. Вомперского,
А. И. Уткина. — М.: Наука, 1986. — 261 с.
Вертинская Г. К., Малахов С. Г., Махонько Э. П. Методика отбора проб почвы при конт-
роле загрязнения окружающей среды металлами //Труды ИЭМ. — 1983. — Вып. 11 (97)
— С. 94-100.
Воронин В И. Дендроиндикация в системе мониторинга лесов, подверженных воздействию
промышленных эмиссий // Лесопатологические исследования в Прибайкалье.—Иркутск,
1989,—С. 24-33.
Горчаковский П. Л. Новое в методике исследований динамики семеношения хвойных по-
род//Бот. журн. — 1958. — Т. 43, № 10.
Горшков В. В. Влияние атмосферного загрязнения окислами серы на эпифитный лишайнико-
вый покров северотаежных сосновых лесов // Лесные экосистемы и атмосферное загрязне-
ние.— Л.: Наука, 1990а. — С. 144—159.
Горшков В. В. Методика изучения эпифитного лишайникового покрова стволов сосен И Вли-
яние промышленного атмосферного загрязнения на сосновые леса Кольского полуостро-
ва,—Л.: БИН РАН, 19906, —С. 147-149.
ГоршковВ В. Напочвенный лишайниковый покров//Там же. — 1990в —С. 141-144.
Горшков В. В. Использование эпифитных лишайников для индикации атмосферного загряз-
нения: Методические рекомендации — Апатиты, 1991. — 38 с.
Горшков В В. Характеристики восстановления лесных экосистем после пожаров И ДАН
России. — 1993. — Т. 333, № 6. — С. 111-114.
Горшков В. В Изменение видового разнообразия напочвенных лишайников под действием
загрязнения в зависимости от давности пожара//ДАН России —1994. — Т. 334, № 5. —
С. 665-668.
ГоршковВ. В., СтавроваН. И. Влияние лесных пожаров на семенную продуктивность Pinus
sylvestris L. в сосновых лесах Кольского полуострова // Растительные ресурсы — 1999 —
Т. 35, вып. 1, —С. 38—46.
Грейг-Смит П. Количественная экология растений. — М., 1967. — 538 с.
Гречин И. П., Кауричев И. С., Никольский Н. Н, Панов Н. П., Поддубный Н. Н. Практи-
кум по почвоведению. — М., 1964. — 423 с.
Гудериан Р. Загрязнение воздушной среды. — М., 1979. — 200 с.
Деева Н М., Мазная Е. А., Ярмишко В. Т. Влияние атмосферного загрязнения на состояние
ассимиляционного аппарата растений сосновых лесов Кольского полуострова// Лесное
хоз.-во.—1992.—№ 10.—С.8.
Демьянов В. А. Метод изучения фитогенного поля древесных пород И Бот. журн — 1978. —
Т. 63,№9. — С. 1302-1308.
ДылисН. В. Основыбиогеоценологии. — М.: Изд. МГУ, 1978. — 151 с.
233
Дыренков С А. Методика выделения лесотипологических единиц и установления их потен-
циальной продуктивности // Методические указания к определению потенциальной про-
изводительности лесов. — М., 1973. — С. 51-59.
Дыренков С А. Роль признаков растительности и местообитаний в лесной типологии // Ле-
соведение.— 1982. — №4. — С. 31-37.
Дыренков С Л. Структура и динамика таежных ельников. — Л., 1984. — 182 с.
Жизнеспособность семян / Под ред. Е. X. Робертса. — М., 1978. — 415 с.
Жукова Л А , Ведерникова О И., Смирнова О. В , Торопова Н. А., Евстигнеев О. И.
Популяционная экология растений. — Йошкар-Ола, 1994. — 88 с.
Загреев В В., Сухих В И., Швиденко А 3, Гусев Н. И., Мошкалев А. Г. Общесоюзные
нормативы для таксации лесов. - М.: Колос, 1992. — 495 с.
Зайцев Г Н. Математическая статистика в экспериментальной ботанике. — М., 1984. —
424 с.
Закс Л. Статистическое оценивание. — М., 1976. — 598 с.
Заугольнова Л Б., Жукова Л. А., Бекмансуров М В. Методика проведения геоботаничес-
ких описаний // Полевой экологический практикум. — Йошкар-Ола, 2000. — Ч. 1. —
С. 39-47.
ЗлобинЮ. А К познанию строения клонов Vaccinium myrtillusL. // Бот. журн. — 1961. — Т.
46, вып 3, —С. 414-419.
Злобин Ю. А. Принципы и методы изучения ценотических популяций растений. — Казань,
1989а, — 146 с.
Злобин Ю А. Теория и практика оценки виталитетного состава ценопопуляций растений //
Бот. журн. — 19896. — Т. 74, № 6. — С. 769-780.
Злобин Ю А. Структура фитопопуляций // Успехи совр. биологии. — 1996. — Т. 116, № 2. —
С. 133-146.
Злобин Ю А Репродуктивное усилие и репродуктивный успех. Потенциальная и реальная
семенная продуктивность. Популяционное и ценотическое регулирование репродукции //
Эмбриология цветковых растений. Терминология и концепции. Т. 3. Системы репродук-
ции. — СПб., 2000. — С. 247-262, С. 428^132.
Зябченко С С. Сосновые леса Европейского Севера. — Л., 1984. — 247 с.
Илли И Э. Жизнеспособность семян // Физиология семян (под ред. А. А. Прокофьева). —
М, 1982, — С, 102-124.
Ильин С С. К методике изучения корневой системы растений // Бот. журн. — 1961, т. 46,
№ 10, —С. 1533-1537.
Ипатов В С. Отражение динамики растительного покрова в синтаксономических единицах //
Бот. журн. — 1990. — Т. 75, № 10. — С. 1380-1388.
Ипатов В С. Методы описания фитоценоза. — СПб., 2000. — 55 с.
Ипатов В С., Герасименко Г. Г. Основные теоретические подходы к динамической типо-
логии леса // Лесоведение. — 1992. — № 4.
Ипатов В С., Кирикова Л. А. Фитоценология. — СПб., 1999. — 316 с.
Ипатов В. С., Кирикова Л А., Бибиков В. И. Сквозистость древостоев (измерение и воз-
можность использования в качестве показателя микроклиматических условий под поло-
гом леса) // Бот. журн. — 1979. — Т. 64, № 11. — С. 1615-1624.
234
Исаченко Г. А. Методы полевых ландшафтных исследований и ландшафтно-экологическое
картографирование. — СПб., 1999. — 111 с.
Казимиров Н И., Волков А. Д., Зябченко С. С. и др. Обмен веществ и энергии в сосновых
лесах Европейского Севера. — Л., 1977 — 304 с.
Кишка И. Методика лихеноиндикационных исследований в Южной Польше // Международ-
ная школа по лихеноиндикации. — Таллин, 1984. — С. 75-77.
Колесников Б. П. Генетическая классификация типов леса и ее задачи на Урале // Тр. Ин-та
биологии УФАН СССР. — 1961. — Вып. 27. — С. 47-59.
Колесников В. А. Методика изучения корневой системы древесных растений. Изд. 2-е —
М., 1972. —152 с.
Комарова Т. А. Семенное возобновление растений на свежих гарях (леса южного Сихотэ-
Алиня). — Владивосток, 1986. — 222 с.
Корчагин А. А. Растительность северной половины Печоро-Ылычского заповедника // Тр.
Печоро-Ылычского заповедника. — 1940. — Вып. 2. — С. 5-412.
Корчагин А. А. Влияние пожаров на лесную растительность и восстановление ее после по-
жаров на Европейском Севере // Тр. БИН АН СССР. — 1954. — Сер. III, вып. 9.
Корчагин А. А. Еловые леса Западного Притиманья в бассейне реки Мезенской Пижмы (их
строение и возобновление)//Уч. Зап. ЛГУ. Сер. Геогр. — 1956. — Вып. 11 — С ill-
239.
Корчагин А. А. Методы учета семеношения древесных пород и лесных сообществ / Под
ред. Е.М. Лавренко, А. А. Корчагина И Полевая геоботаника. — М.-Л., 1960. — Т. II. —
С. 41-132.
Корчагин А А. Определение возраста и длительности жизни мхов и печеночников//Там же.
-М.-Л., I960, —TIL —С. 279-314
Корчагин А. А. Определение возраста и длительности жизни лишайников // Там же — М -
Л., I960, —Т.П,—С. 315-332.
Корчагин А. А. Видовой (флористический) состав растительных сообществ и методы его
изучения // Полевая геоботаника. — М.-Л , 1964. — Т. III. — С. 39-62.
Красильников И. К. Методика полевого изучения подземных частей растений (с учетом
специфики ресурсоведческих исследований). — Л.: Наука, 1983. — 208 с.
Курбатский Н П. Проблема лесных пожаров. Возникновение лесных пожаров. — М.: На-
ука, 1964. — С. 5-60.
Лакин Г. Ф. Биометрия. — М.: Высш, школа, 1980. — 293 с.
Левина Р. Е. Репродуктивная биология. — М., 1981. — 96 с.
Линник Е. Ф. Быстрые методы определения всхожести семян // Лесное хоз-во. — 1959 —
№ 1, —С. 85-89.
Липпмаа Т М.О синузиях // Сов. ботаника. — 1946. — № 3. — С. 139-145.
Лобанов Н. В. Микотрофность древесных растений. — М., 1953. — 232 с.
Лянгузова И. В. Содержание химических элементов в разных фракциях фитомассы сосны //
Влияние промышленного атмосферного загрязнения на сосновые леса Кольского полуост-
рова.—Л.: БИН РАН, 1990, —С. 48-55.
Лянгузова И. В, Меньшикова Г. П., Ярмишко М А. Загрязнение почв // Влияние промышлен-
ного атмосферного загрязнения на сосновые леса Кольского полуострова. — Л.: БИН
РАН 1990, —С. 38^48.
235
Мазная Е А Влияние промышленных выбросов на состояние и структуру ценопопуляций
liiccinium myrtillus L. и V vtis-idaea L. (Кольский полуостров) // Растительные ресурсы.
— 2001, —Т. 37, вып. 3, —С. 1-12.
Мак-КэйД Б Определение жизнеспособности//Жизнеспособность семян. — М., 1978.—
С.167-201.
Максимов А И. К вопросу о приросте сфагновых мхов // Комплексные исследования расти-
тельности болот Карелии. — Петрозаводск, 1982. — С. 170-179.
Малышева Т В. К методике разграничения живых и отмерших частей у мхов при учете их
фитомассы // Бот. журн. — 1970. — Т. 55, № 5. — С. 704—709.
Маргайлик Г И. Определение качества лесных семян люминесцентным методом // Лесное
хоз-во, — 1959, —№ 10, —С. 36-37.
Мартин Ю Л. Лихеноиндикация состояния окружающей среды // Взаимодействие лесных
экосистем и атмосферных загрязнителей. — Таллин, 1982.—Ч. 1. — С. 27-47.
Мелехов И С. Лесная типология. — М.: МЛТИ, 1976. — 73 с.
Мелехов И. С. Влияние пожаров на лес. — М.-Л.: Гос. лесотехн, изд-во, 1948. — 126 с.
Методика определения запасов лекарственных растений. — М., 1986.
Методические указания по контролю загрязнения почв / Под ред. С. Г. Малахова. — М.,
1977 —64 с.
Методические указания по определению Si, Al, Fe, Са, Mg, Ti, Мп в почвах атомно-абсорбци-
онным методом. — М., 1975. — 44 с.
Методические указания по определению микроэлементов в почвах, кормах и растениях ме-
тодом атомно-абсорбционной спектроскопии. — М., 1985. — 90 с.
Методические указания по определению тяжелых металлов в почвах сельского хозяйства и
продукции растениеводства. — М., 1989. — 87 с.
Методы исследования ресурсов полезных растений / Мат-лы 1 -ой школы по ботаническому
ресурсоведению. — Вильнюс, 1983. — 143 с.
Миркин Б. М Методы Браун-Бланке и современная отечественная фитоценология // Бюл.
МОИП. Отд. биол. — 1978. — № 3 — С. 76-87.
Миркин Б М, Розенберг Г. С Фитоценология. Принципы и методы. — М., 1979. — 212с.
Миркин Б М, Наумова Л. Г, Соломещ А. И. Современная наука о растительности. — М.,
2000 — 263 с.
Миркин Б М., Розенберг Л Г., Наумова Л Г. Словарь понятий и терминов современной
фитоценологии. — М.: Наука, 1989. — 223 с.
Моисеенко Ф П. Методы учета коры корней бересклета // Тр. ин-та леса АН СССР. —
1947 — Т. 1, —С. 95-98.
Молчанов А А. Быстрота разложения соснового и елового отпада // ДАН СССР. — 1947.
— Т. 56, №8. —С. 869-872.
МолчановА А. Влияние пожаров на древостой //Тр ин-талеса АН СССР. — 1954. — Т. 16.
— С 314-335.
Молчанов А А , Смирнов В. В. Методика изучения прироста древесных растений — М.:
Наука, 1967 —100 с.
Морозов Г. Ф. Учение о типах насаждений. Избранные труды. — М., 1971. — Т. 2. — 536 с.
Мэгарран А. Е. Экологическое разнообразие и его измерение. — М.: Мир, 1992. — 181 с.
Нешатаев В Ю. Проект Всероссийского Кодекса фитоценологической номенклатуры //
Растительность России — 2001. — № 1. — С. 62-70.
236
Нешатаев Ю. Н Методы анализа геоботанических материалов. — Л., 1987. — 192 с
Нешатаев Ю. Н, Нешатаева В Ю., Нешатаев В Ю. Методы сбора и обработки мате-
риала// Растительность Кроноцкого государственного заповедника (Восточная Камчат-
ка) / Тр. БИН РАН. — СПб., 1994. — Вып. 16. — С. 7-8.
Николаева М Г., Лянгузова И В., Поздова Л. М. Биология семян. — СПб., 1999. — 232 с
Нильсон Э. М., МартинЛ. Н. Эпифитные лишайники в условиях кислого и щелочного заг-
рязнения // Взаимодействие лесных экосистем и атмосферных загрязнителей. — Таллин,
1982, —Ч. 2, —С. 88-100.
НиценкоА А. Растительная ассоциация и растительное сообществ как первичные объекты
геоботанического исследования: сущность, свойства, методы выделения. — Л., 1971. —
184 с.
Одум Ю. Основы экологии. — М., 1975. — 740 с.
Орлов А. Я. К методике количественного определения сосущих корней древесных пород в
почве // Бюл. МОИП. Отд. биол. — 1955. — Т. 60, № 3. — С 93-102.
Орлов А. Я. Метод определения массы корней деревьев в лесу и возможность учета годич-
ного прироста органической массы в толще лесной почвы // Лесоведение. — 1967. —
№ 1, —С. 64-70.
Основные понятия и термины ботанического ресурсоведения.—Петрозаводск, 2001. — 104 с
Пегов Л. А. Исследование точности выборочного учета подроста под пологом древостоев //
Лесоведение.— 1992. — №4. — С. 51-59.
Петров В В. Банк семян в почвах лесных фитоценозов Европейской части СССР — М ,
1989, —175 с.
Петровский В. В. Синузии как формы совместного существования растений // Бот. журн.
— 1961,—Т. 46, №4.
Побединский А. В. Изучение лесовосстановительных процессов. — М., 1966. — 240 с
Поляков А. Н., Набатов Н. М. Основы лесоводства и лесной таксации. — М.: Лесн пром-
ть, 1983, —224 с.
Почвы СССР. 1979.
Правила отбора образцов и методы определения посевных качеств семян. — М., 1988. —
53 с.
Принципы и методы рационального использования дикорастущих полезных растений. —
Петрозаводск, 1989. — 168 с.
Программы для геоботанических исследований / Под ред. Б. А. Келлера, В. Н. Сукаче-
ва,—Л., 1932, —248 с.
Пушкина Н. М. Растительность сосновых гарей Лапландского заповедника и характер ее
восстановления//Тр. Лапландского гос. заповедника. — 1960. — Вып. 4. — С. 5-125.
Работное Т. А. Вопросы изучения состава популяций для целей фитоценологии // Проблемы
ботаники. — М., 1950. — Вып. 1.
Работное Т. А. Методы изучения семенного размножения травянистых растений в сообще-
ствах И Полевая геоботаника. — М.-Л., 1960. — Т. II. — С. 20—40.
Раменский Л. Г. Проблемы и методы изучения растительного покрова. Избр. работы. —
Л., 1971, —334 с.
Резолюция 3-го Всесоюзного совещания по классификации растительности (Ленинград, 19-
22 октября 1971). — Л., 1972. — 8 с.
Резолюция III Всесоюзного совещания по лесной типологии. — Красноярск, 1973. — 8 с.
237
Родин Л Е, Ремезов Н П., Базилевич Н И. Методические указания к изучению динами-
ки и биологического круговорота в фитоценозах. — Л.: Наука, 1968. — 143 с.
Романовский М Г Учет семеношения сосны обыкновенной по «внутренним» следам опав-
ших шишек//Научн тр. МЛТИ. — 1976. — Вып. 88. — С. 134—136.
Рысин Л П Сосновые леса Европейской части СССР. — М., 1975. — 212 с.
Рысин Л П Лесная типология в СССР. — М., 1982. — 216 с.
Рысин Л П, Казанцева Т Н Метод ценопопуляционного анализа в геоботанических ис-
следованиях//Бот журн.— 1975. — Т. 60, №2. — С. 199-209.
Рысин Л П, Комиссаров Е С, Маслов А А , Петерсон Ю В., Савельева Л И. Методи-
ческие предложения по созданию системы постоянных пробных площадей на особо ох-
раняемых территориях. — М : Наука. 1988 — 28 с.
Санитарные правила в лесах Российской Федерации. Министерство экологии и природных
ресурсов Российской Федерации. Комитет по лесу: Справ, пос. — М.: Экология, 1992. —
16с
Семенов БАК методике определения истинной ширины годичных слоев и содержания
поздней древесины в стволах сосны Крайнего Севера // Вопросы предтундрового лесо-
водства Европейской части СССР. — Архангельск, 1983. — С. 83-92.
Смирнова О В. Объем счетной единицы при изучении ценопопуляций растений различных
биоморф // Ценопопуляции растений (Основные понятия и структура). — М., 1976. —
С 72-80
Сочава В Б Классификация растительности как иерархия динамических систем // Геобо-
таническое картографирование.— Л., 1972 —С. 3-17.
Ставрова Н И. Влияние атмосферного загрязнения на семеношение хвойных пород // Лес-
ные экосистемы и атмосферное загрязнение. — Л.: Наука, 1990. — С. 115-121.
Сукачев В Н. Основы теории биогеоценологии // Юбил. сб., поев. 30-летию Вел. Окт. соц.
рев — М.-Л., 1947. — Ч. 2. — С. 283-304.
Сукачев В Н, Зонн С В. Методические указания к изучению типов леса. Изд. 2-е. — М .
Изд-во АН СССР, 1961. — 144 с.
Тарановская М П. Методы изучения корневых систем. — М., 1957. — 216 с.
Трасс X X. Вопросы теоретического обоснования метода синузий в фитоценологии // Изу-
чение растительности острова Сааремаа.— Тарту, 1964 —С. 82-111.
ТрассX X Классы полеотолерантности и экологический мониторинг// Проблемы экологи-
ческого мониторинга и моделирования экосистем. — Л., 1985. — Т. 7. — С. 122-137.
Третьяковы В. Лесная таксация: Конспект лекций. — Л.. ЛТА, 1957. — 299 с.
Третьяков Н В., Горский П В., Самойлович Г. Г. Справочник таксатора. — М.: Лесн.
пром-ть. 1965. — 459 с.
Тюрин А В Таксация леса. — М.: Гослесбумиздат, 1945. — 376 с.
УрановА А. Большой жизненный цикл и возрастной спектр ценопопуляций цветковых рас-
тений // V съезд ВБО: Тез. докл. — Киев, 1973. — С. 217-219.
Успенская И М. Определение массы мохового покрова в биогеоценозах широколиственно-
еловых лесов в связи с их парцелярной структурой // Бот. журн. — 1966. — Т. 51, № 12. —
С 1706-1709.
Уткин А И. Биологическая продуктивность лесов: методы изучения и результаты // Лесо-
ведение и лесоводство. — М.: ВИНИТИ, 1975. — Т. 1. — С. 9-189.
238
Федорчук В Н. О распознавании лесотипологических единиц // Лесоведение. — 1979. —
№ 4. — С. 76-82.
Федорчук В Н, Дыренков С А. Выделение и распознавание типов леса. Методические
указания. —Л., 1975. — 55 с.
Ценопопуляции растений (Основные понятия и структура) — М , 1976. — 216с
Черкасов А. Ф. Методика выявления и оценки ресурсов дикорастущих ягодников при лесо-
устроительных работах. —М., 1979. — 17 с.
ЧертовО Г. Экология лесных земель. — Л., 1981.— 191 с
Чертов О. Г., Лянгузова И. В, Друзина В Д, Меньшикова Г П Влияние на лесные
почвы загрязнения серой в комплексе с тяжелыми металлами // Лесные экосистемы и
атмосферное загрязнение. — Л , 1990. — С. 65-72
Шайн С. С .Чекмарева П. Г. О методе количественного определения корней трав при по-
мощи бура//Докл. ВАСХНИЛ.— 1940. — Вып 8. — С 22-23.
Шалыт М. С. Методика изучения морфологии и экологии подземной части отдельных рас-
тений и растительных сообществ // Полевая геоботаника. — М.-Л , 1960. — Т. II —
С. 369-447.
'Денников А. П. Введение в геоботанику. — Л , 1964 — 447 с
Чрмишко В Т. Прибор для измерения ширины годичных колец деревьев // Бот журн —
1983.’ — Т. 68, № 10. — С. 1428-1430.
Чрмишко В Т. Состояние сосновых древостоев // Влияние промышленного атмосферного
загрязнения на сосновые леса Кольского полуострова —Л • БИН РАН, 1990. — С. 104—
109.
Чрмишко В. Т. Сосна обыкновенная и атмосферное загрязнение на Европейском Севере —
СПб.: СПбГУ, 1997. — 210 с.
Чрмишко В. Т, Деева ЧЧ М., Мазная Е А , Леина Г Д. Влияние промышленных выбросов
на ассимиляционный аппарат Pinus sylvestris L и Vaccinium myrtillus L на Европейском
Севере И Растительные ресурсы. — 1995. — Вып. 3
Чстребов А Б, Познанская А. А. Анализ влияния древостоя на подрост в сосновых борах
Карелии//Бот. журн —1993 —Т. 78,№4. — С. 123-133.
\lmborn О. Distribution and ecology of some Scandinavian lichens // Botaniska Notiser. — 1948
— Vol. l,No2. — P. 1-252.
taisheva E Z. Bryophyte vegetation of Bashkiria (South Urals). II. Epiphytic and epixylic
communities of North-Eastern Bashkiria // Arctoa —1995. — Vol 4. — P. 55-63.
kiisheva E Z. Bryophyte vegetation of Bashkiria (South Urals). III. Epiphytic and epixyhc
communities of Western Bashkiria// Arctoa. — 2000. — Vol. 9. — P. 101-104
kiisheva E. Z, Solometch A I, Ignatova E A. Bryophyte vegetation of Bashkina, South Urals
1 Epiphytic and epixylic communities// Arctoa. — 1994. — Vol. 3. — P. 139-152.
•askin С C, Baskin J. M. Seeds. Ecology, Biogeography and Evolution of Dormancy and
Germination. — San Diego, London, Boston, NY, Sydney, Tokyo, Toronto, 1998. — 666 p.
'laun-BlanquetJ Pflanzensociologie. — Wien; New York, 1964. — 865 s.
a/anderA K. The theory offorest types//Acta Forest. Fenn. — 1926. — Vol. 29, No 3. — P. 1-
108
'IvmoR S The growth of Sphagnum: methods of measurements//J. Ecol.— 1970. — Vol 58,
No 1 — P 13-49
239
Dix R L , Svan J. M A. The role of disturbance and succession in upland forest at Candle Lake,
Saskatchewan // Can. J. Bot. — 1971. — Vol. 49, No 5. — P. 657-676.
Forest Condition in a Changing Environment. - The Finnish Case. — Klumer Academic Publishers,
2000, —Vol. 65, —378 p.
Fritts H C Tree rings and climate. — London etc.: Acad, press. — 1976. — 567 p.
Gilbert О L. Lichens and air pollution // The Lichens. / Eds. V. Ahmadjian, M. E. Hale. — N.Y.,
London, 1973, —P 443^172.
Gorsuch T T The destruction of organic matter. — Oxford, N.Y., Toronto, Sydney, 1970.—459p.
Handbook of vegetation science. Pt. 12. Application of vegetation science to forestry / Ed. G. Jahn.
— Boston, London, 1982 —405 p
Handbook of vegetation science. Pt. 4. Sampling methods and taxon analysis in vegetation science:
Releve surveys, “Vegetation saufnahmen”, floristic analysis of plant communities I Ed. R. Knapp.
— Boston, Lancaster, 1984. — 370 p.
Handbook of vegetation science. Pt. 6. Vegetation and environment / Eds B. R. Stain, W. D
Billings. — Growingen, 1974. — 193 p.
Hustich I The growth of Scots pine in northern Lapland, 1928-1977 // Ann Bot Fenn. — 1978. —
Vol. 15 —P 241-252.
Huttunen S. Interactions of disease and other stress factors with atmospheric pollution // Air pollution
and plant life. —1984 — P 321-356.
Johnson E A Fire and Vegetation Dynamics: Studies from the North American Boreal Forests.
Cambridge Studies in Ecology. — New York: Cambridge University Press, 1992. —129 p.
Kaennel M, Schweingruber F H Multilingual Glossary of Dendrochronology. — Haupt, Bern,
1995 — 467 p.
Lakon G The topographical tetrazolium method for determining the germinating capacity of seeds
// Plant Physiol. — 1949 — Vol. 24. — P. 389-394.
Leikola M The influence of environmental factors on the diameter growth of forest trees.
Auxanometnc study // Acta Forest. Fenn. — 1969. — Vol. 92. — 144 p.
Magdejrau K. Life forms of bryophytes // Bryophyte Ecology. — London, N.Y, 1982. — P. 45-
58
Madany M H, Swetnam T. W, West N. E. Comparison of two approaches for determining fire
dates from tree scars // For Sci. — 1982. — Vol. 28. — P. 856-861.
Pirintsos S A., Vokou D., Diamantopoulos J., Galloway D J An assessment of the sampling
procedures for estimating air pollution using epiphytic lichens as indicators // Lichenologist. —
1993, —Vol. 25, No 2, —P. 165-173.
Sarvas R Investigation on the flowering and seed crop of Pinus silvestris II Comm. Inst, forest,
fenn. — 1962. — Vol 53, —P. 1-198.
Siren G. The development of spruce forest on raw humus sites m northern Finland and its ecology
// Acta Forest. Fenmca. — 1955. — Vol. 62. — P. 1-363.
Whittaker R H. Evolution of species diversity in land communities // Evolution Biology. —1977. —
No 10. — P 1-67.
Wirth V Flechtenflora. — Stuttgart, 1980. — 552 p.
Zackrisson О Influence of forest fires on the North Swedish boreal forests // Oikos. — 1977. —
Vol. 29. — P 22-32.
240