


































































































































































































































































































































































































































































































































































































































































































































































































































Автор: Долгов В.В. Меншиков В.В.
Теги: семиология симптоматология признаки и симптомы заболеваний исследование диагностика пропедевтика общая диагностика медицина практическая медицина лабораторная диагностика издательство гэотар медиа
ISBN: 978-5-9704-2131-4
Год: 2012




Научное общество специалистов
лабораторной медицины
дсмок
КЛИНИЧЕСКАЯ ЛАБОРАТОРНАЯ
ДИАГНОСТИКА
НАЦИОНАЛЬНОЕ РУКОВОДСТВО
ИЗДАТЕЛЬСКАЯ ГРУППА
«ГЭОТАР-Медиа»
АССОЦИАЦИЯ
МЕДИЦИНСКИХ
ОБЩЕСТВ
ПО КАЧЕСТВУ
АСМОК
АССОЦИАЦИЯ
МЕДИЦИНСКИХ
ОБЩЕСТВ
ПО КАЧЕСТВУ
КЛИНИЧЕСКАЯ
ЛАБОРАТОРНАЯ ДИАГНОСТИКА
НАЦИОНАЛЬНОЕ РУКОВОДСТВО
В двух томах
Том II
Главные редакторы
д-р мед. наук В.В. Долгов,
чл.-корр. РАЕН, д-р мед. наук В.В. Меньшиков
Подготовлено под эгидой Научно-практического
общества специалистов лабораторной медицины
и Ассоциации медицинских обществ по качеству
Москва
ИЗДАТЕЛЬСКАЯ ГРУППА
«ГЭОТАР- Медиа»
2012
УДК 616-07(035)
ББК 53.4я81
К49
Национальное руководство рекомендовано
Наулно-практигеским обществом специалистов лабораторной медицины
и Российской медицинской академией последипломного образования
Минздравсоцразвития РФ в качестве улейного пособия
для последипломной подготовки вралей
К49 Клиническая лабораторная диагностика ; национальное руководство : в 2 т. - Т. II /
под ред. В.В. Долгова, В.В Меньшикова. — М.: ГЭОТАР-Медиа, 2012. - 808 с.
ISBN 978-5-9704-2131-4 (т. II)
ISBN 978-5-°704-2127-7 (общ.)
Национальное руководство по клинической лабораторной диагностике разработано и
рекомендовано Научно-практическим обществом специалистов лабораторной медицины.
В издании отражены все разделы клинических лабораторных исследований, представлен-
ные ведущими специалистами научных, образовательных и лечебно-профилактических
учрежпений Центрального. Севрро-Западного, Уральского, Сибирского. Северо-Кав-
казского федеральных административных округов.
В том II включены научные и практические материалы по актуальным проблемам
клинической иммунологии, бактериологии, вирусологии, микологии, паразитологии.
Представлены сведения как о повседневно применяемых аналитических технологиях,
так и о новых эффективных способах идентификации микроорганизмов и определения
их чувствительности к антимикробным агентам.
В отдельных главах представлены современные данные по иммуногематологии и
методам, применяемым в службе крови, по диагностике аутоиммунных заболеваний, а
также сведения о новом классе агентов межклеточной регуляции — цитокинах, и о роли
их определения при различных формах патологии.
Представленные сведения основаны на данных современной научной литературы,
рекомендациях профессиональных обществ специалистов, стандартах медицинской
помощи, многолетнем научно-практическом опыте авторов глав.
Руководство предназначено для сотрудников клинико-диагнжт ических лабораторий,
представителей различных клинических дисциплин, студентов медицинских образова-
тельных учреждений. Материалы руководства могут быть использованы как для базово-
го медицинского образования, так и для последипломной подготовки.
УДК 616-07(035)
ББК 53.4я81
Авторы, редакторы и издатели руководства предприняли максимум усилий, чтобы
обеспечить точность представленной информации, в том числе дозировок лекарствен-
ных средств. Учитывая постоянные изменения, происходящие в медицинской науке, мы
рекомендуем уточнять дозы лекарственных средств по соответствующим инструкциям.
Пациенты не могут использовать эту информацию для диагностики и самолечения.
Права на данное издание, принадлежат ООО Издательская группа «ГЭОТАР-Медиа».
Воспроизведение и распространение в каком бы то ни было виде ласти или целого издания не
могут быть осуществлены без письменного разрешения ООО Издательская группа «ГЭОТАР-
Медиа».
ISBN 978-5-9704-2131-4 (т, II)
ISBN 978-5-9704-2127-7 (общ.)
© Коллектив авторов, 2012
© ООО Издательская группа «ГЭОТАР-Медиа», 2012
© ООО Издательская группа «ГЭОТАР-Медиа»,
оформление, 2012
ОГЛАВЛЕНИЕ
Участники издания...................................................................6
Список сокращений..................................................................12
Глава 16. Лабораторная иммунология К.П. Кашкин. Ф.Ю. Гариб, Л.М. Скуинъ... 17
Врожденные факторы иммунной защиты..............................................17
Приобретенный иммунитет.........................................................25
Патология иммунной системы......................................................35
Иммунная система и воспаление...................................................48
Алгоритм лабораторного исследования иммунной системы............................61
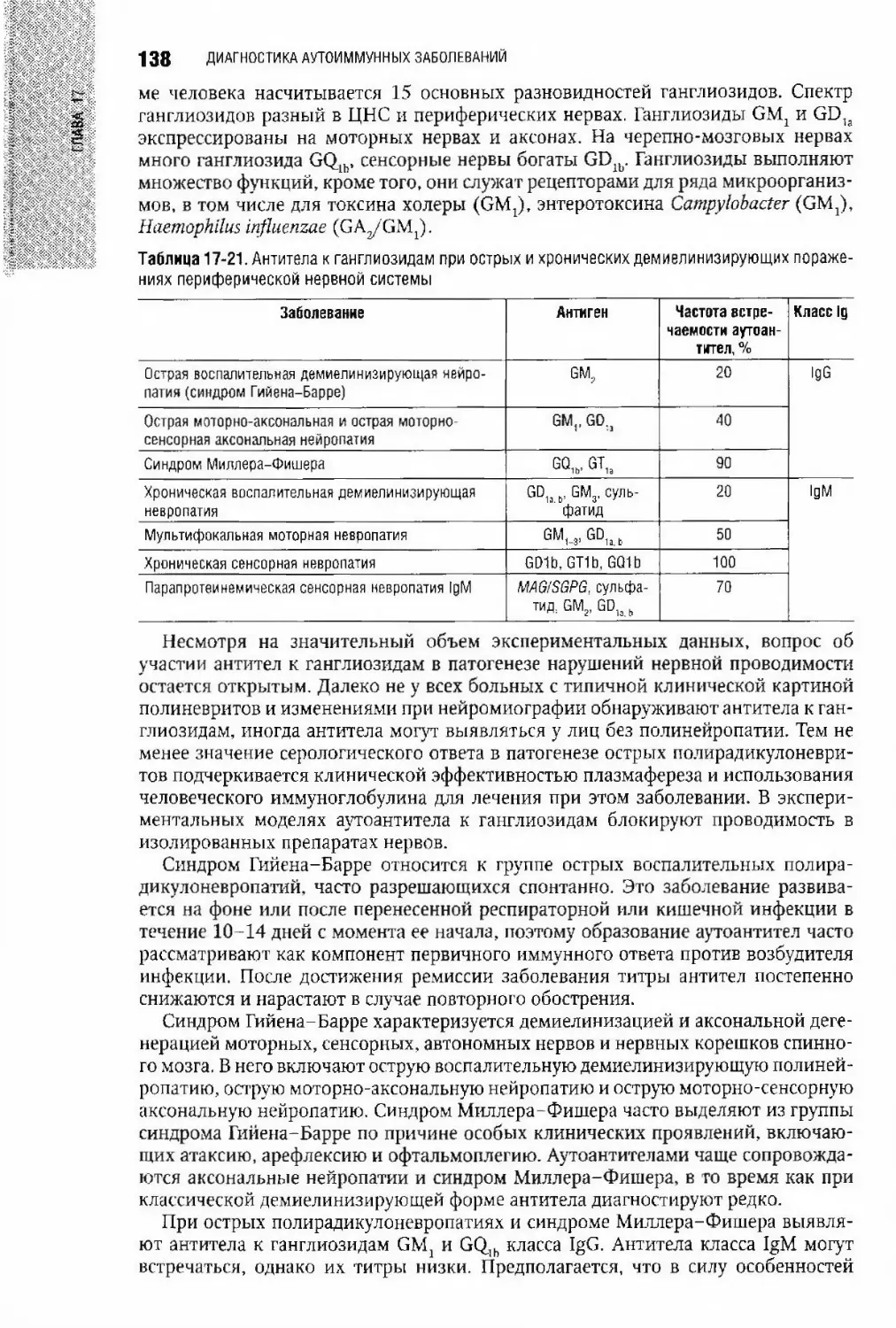
Глава 17. Диагностика аутоиммунных заболеваний С В. Лапин,
Арег А. Тотолян....................................................................6ь
Критерии, классификация и эпидемиология аутоиммунных заболеваний...............66
Антитела и аутоиммунные заболевания.............................................70
Лабораторные показатели при аутоиммунных заболеваниях...........................74
Диагностика ревматических заболеваний....................................... 146
Заключение.....................................................................174
Глава 18. Иммуногематология О.А. Тарасенко, О.Я. Волкова..........................176
Антигены эритроцитов...........................................................177
Иммуногематологическая безопасность трансфузионной терапии.....................179
Иммуногематологические патологические состояния................................185
Иммуногематологические лабораторные исследования...............................189
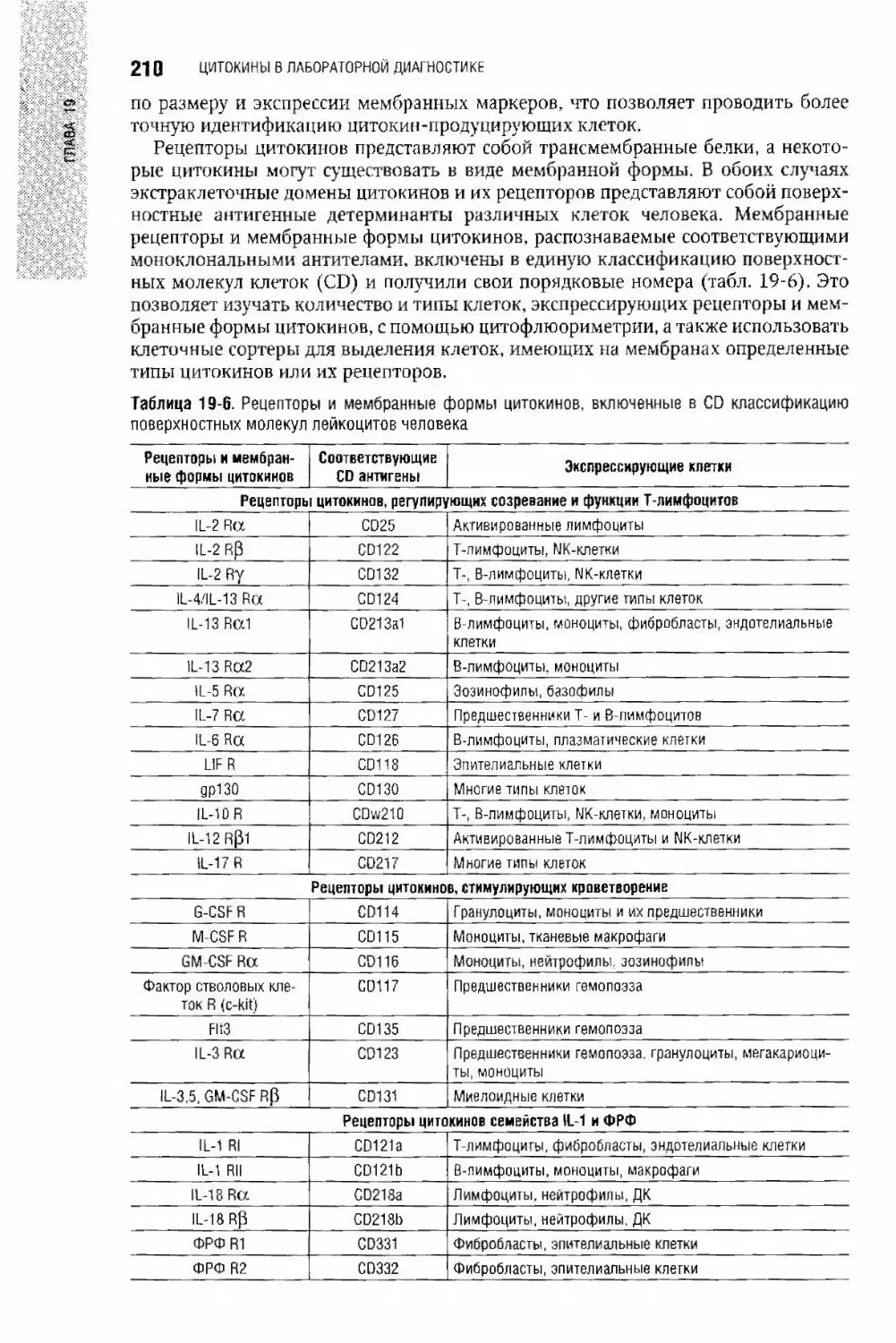
Глава 19. Цитокины в лабораторной диагностике А. С. Симбирцев,
Арег А. Тотполян..............................,......................................................... 193
Общие представления о цитокинах................................................194
Методы оценки функционирования системы цитокинов...............................198
Роль цитокинов в патогенезе заболеваний человека...............................217
Диагностическое значение отдельных цитокинов...................................220
Заключение.....................................................................228
Глава 20. Бактериологические исследования. Под редакцией А.Б. Жебруна.....230
Значение лабораторной диагностики в инфектологии А.Б. Жебрун...................230
Задачи бактериологической диагностики А.Б. Жебрун......................................232
Этиологическая диагностика бактериальных инфекций А.Г. Бойцов..............234
Принципы бактериологического исследования отдельных видов
биологическою материала и интерпретации их результатов
А.Г. Бойцов, А.В. Елисеев, Л.А. Кафтырева, Е.А. Оришак, Л.Ю. Нилова............239
Принципы идентификации бактерий А.А, Норин, А.Г. Бойцов........................283
Индикация антигенов возбудителей А.А. Норин, А.Г Бойцов........................287
Молекулярно-генетическая индикация бактерий О.В. Нарвская.
С.А. Егорова, М.А. Макарова, Л.А. Кафтырева....................................291
Индикация специфических антител Л.А. Кафтырева. М.А. Макарова.
С.А. Егорова................................................................. 295
Определение чувствительности бактерий к антимикробным препаратам
В.Д. Бабиков.............................................................................................................300
Глава 21. Частная микробиология. Под редакцией А.Б. Жебруна.......................312
I. ГРАМОТРИЦАТЕЛЬНЫЕ БАКТЕРИИ..................................................312
Хламидии Л.Б. Куляшова, Л.А. Березина, А.В. Закревская, А.Б. Жебрун,
В.А. Исаков...............................................................................................................312
Бордетеллы Н.Н. Курова, Г.Я. Цснева....................................................................323
4
ОГЛАВЛЕНИЕ
Легионеллы И.С. Тартаковский.....................................................329
Коксиеллы Н.К. Токарева?, О.А. Фрейлихман........................................335
Энтеробактерии Л А. Кафтырева, А.Г. Бойцов. МА. Макарова.........................342
Иерсинии Г.Я. Ценева, Е.А. Воскресенская, Г.И. Кокорина,
Е.А. Богумильгик, О.А. Бургасова.................................................365
Гемофильные бактерии А.Б. Жебрун ....................................................................375
Неферментирующие бактерии А.Г. Бойцов, О.Д. Васильев.............................380
Кампилобактеры А.А. Порин., З.Н. Матвеева........................................388
Хеликобактеры А.Б. Жебрун, А.В. Сварваль, Л.Б. Гонгарова, Р.С. Ферман............394
П. ГРАМПОЛОЖИТЕЛЬНЫЕ БАКТЕРИИ....................................................406
Стафилококки К.Г. Косякова.......................................................406
Стрептококки Артем А. Тотолян, ЛА. Бурова. А.В. Дмитриев, А.Н. Суворов.... 417
Пневмококковый стрептококк А.Б. Жебрун........................................................435
Менингококк А.Б. Жебрун..........................................................438
Листерии И.С. Тартаковский.......................................................445
Клостридии А.Г. Бойцов, Л.А. Кафтырева...........................................451
Лептоспиры НА. Стоянова, А.Н. Ваганова...........................................460
Коринебактерии Г.Я. Ценена, Л.А. Краева, Г.И. Беспалова, Е.А. Алексеева..........468
Глава 22. Некоторые инфекционные заболевания.......................................484
Микобактерии М.В. Шульгина. Б.И. Вишневский. Т.Ф. Оттен..........................484
Трепонемы А.М. Савигева, Е.В. Соколовский, Т.В. Красносельских,
Е.В. Шипицина....................................................................496
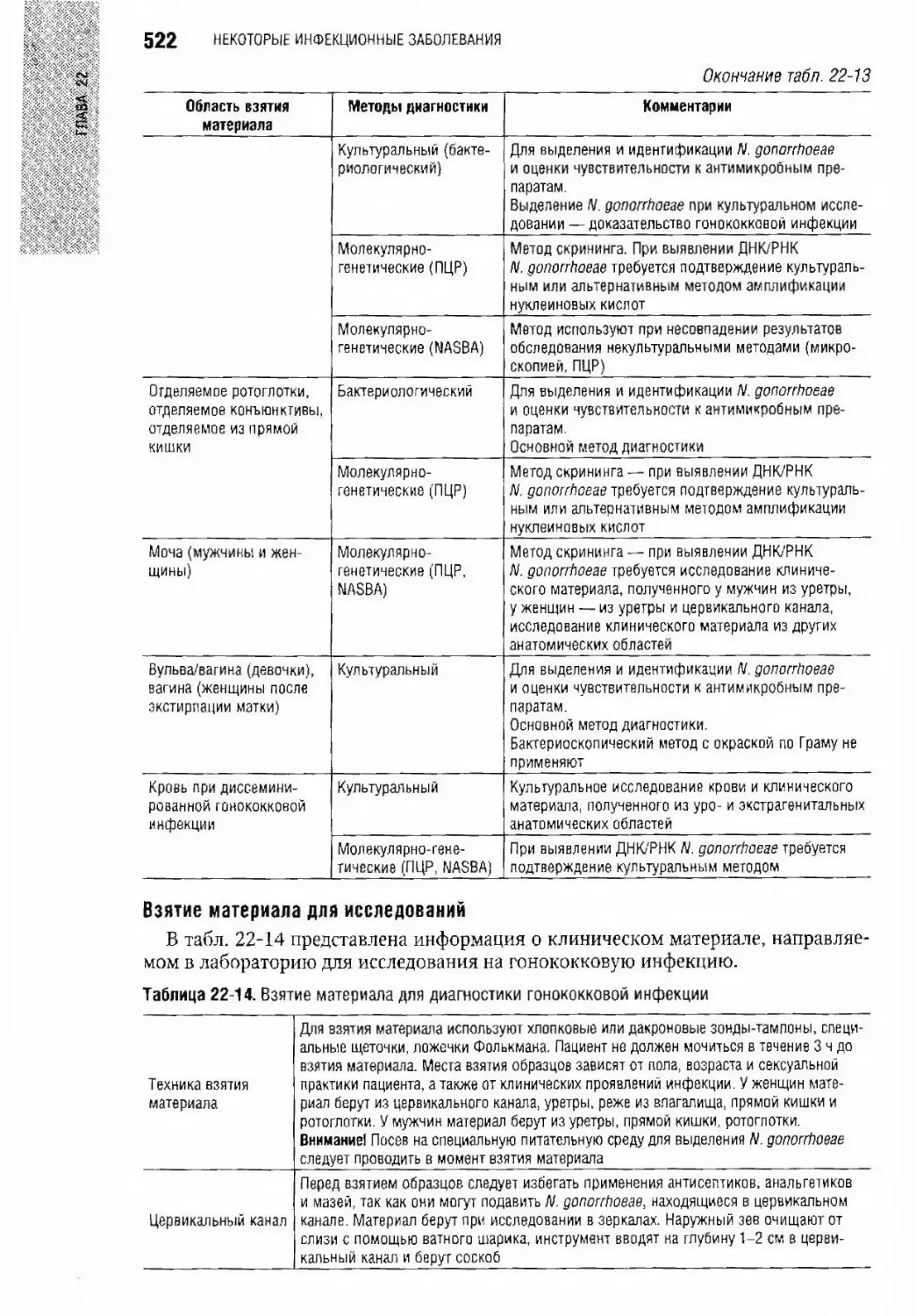
Гонококк А.М. Савигева, Е.В. Соколовский, А.В. Игнатовский, Е.В. Шипицина.... 519
Франсиселлы И.С. Мещерякова......................................................530
Глава 23. Вирусологические исследования Под редакцией А.Б. Жебруна.......... 542
Задачи диагностики вирусных инфекций А.Б. Жебрун.................................542
Организация работы лаборатории по диагностике вирусных
заболеваний С.С. Вашукова .......................................................545
Общие сведения о вирусах И.Н. Лаврентьева, С.П. Сухобаевская,
Л.Ф. Литвингук, А.Ю. Антипова....................................................548
Классические методы выделения и идентификации вирусов
С.П. Сухобаевская. Л.Ф. Литвингук, А.К. Сироткин. А.В. Семенов,
В.Н. Вербов, И.Н. Лаврентьева.................................................. 561
Молекулярно-биологические методы А.В. Семенов, Л.Б. Гонгарова....................613
Преаналитический этап при вирусологических исследованиях
И.Н. Лаврентьева....................................................................................................627
Обеспечение качества при вирусологических исследованиях А.В. Семенов.............633
Глава 24. Частная вирусология. Под редакцией А.Б. Жебруна..........................641
Аденовирусы И.Н. Лаврентьева.....................................................641
Астровирусы В.М. Малышев, ДА. Макаров............................................644
Вирус бешенства И.И. Яровая, П.В. Колотвина, МА. Кохновиг, С.В Грибенга..........645
Вирус гриппа С.С. Вашукова, Л.Б. Гонгарова.......................................649
Вирус иммунодефицита Г.И. Коровина...............................................653
Вирус клещевого энцефалита С. С. Вашукова, И.Н. Лаврентьева,
А.Ю. Аптипова....................................................................656
Вирус кори М.А. Бигурина......................................................................................659
Вирус краснухи И.Н. Лаврентьева. А.Ю. Антипова...................................663
Вирус парагриппа Л.П. Сухобаевская...............................................665
Вирус эпидемического паротита Е.О. Контарова, Н.В. Юминова.....................667
Вирусы — возбудители геморрагических лихорадок И.И. Яровая,
Л.А. Автушенко, Ф.С. Носков, Б.В. Вершинский, А.Б. Жебрун........................669
ОГЛАВЛЕНИЕ
5
Вирусы-возбудители гепатитов С.Л. Мукомолов...............................................673
Герпес-вирусы В А. Исаков, А.В. Закревская................................................680
Калицивирусы А.К. Сироткин...............................................................................691
Коронавирусы Л.Б. Гончарова................................................................................ 694
Респираторно-синцитиальный вирус И.Н. Лаврентьева, Л.П. Сухобаевская.....................699
Риновирусы Л.П Сухобаевская, А.Ю. Антипова700
Ротавирусы Л.В. Лялина, А.Н. Афанасьева...........................................................702
Парвовирус В19 А.Ю. Антипова..............................................................706
Папилломавирусы О.В. Нарвская .........................................................................708
Полиовирусы Н.И. Романенкова, М.А. Батурина.................................................714
Энтеровирусы Н.И. Романенкова, М.А. Батурина..............................................720
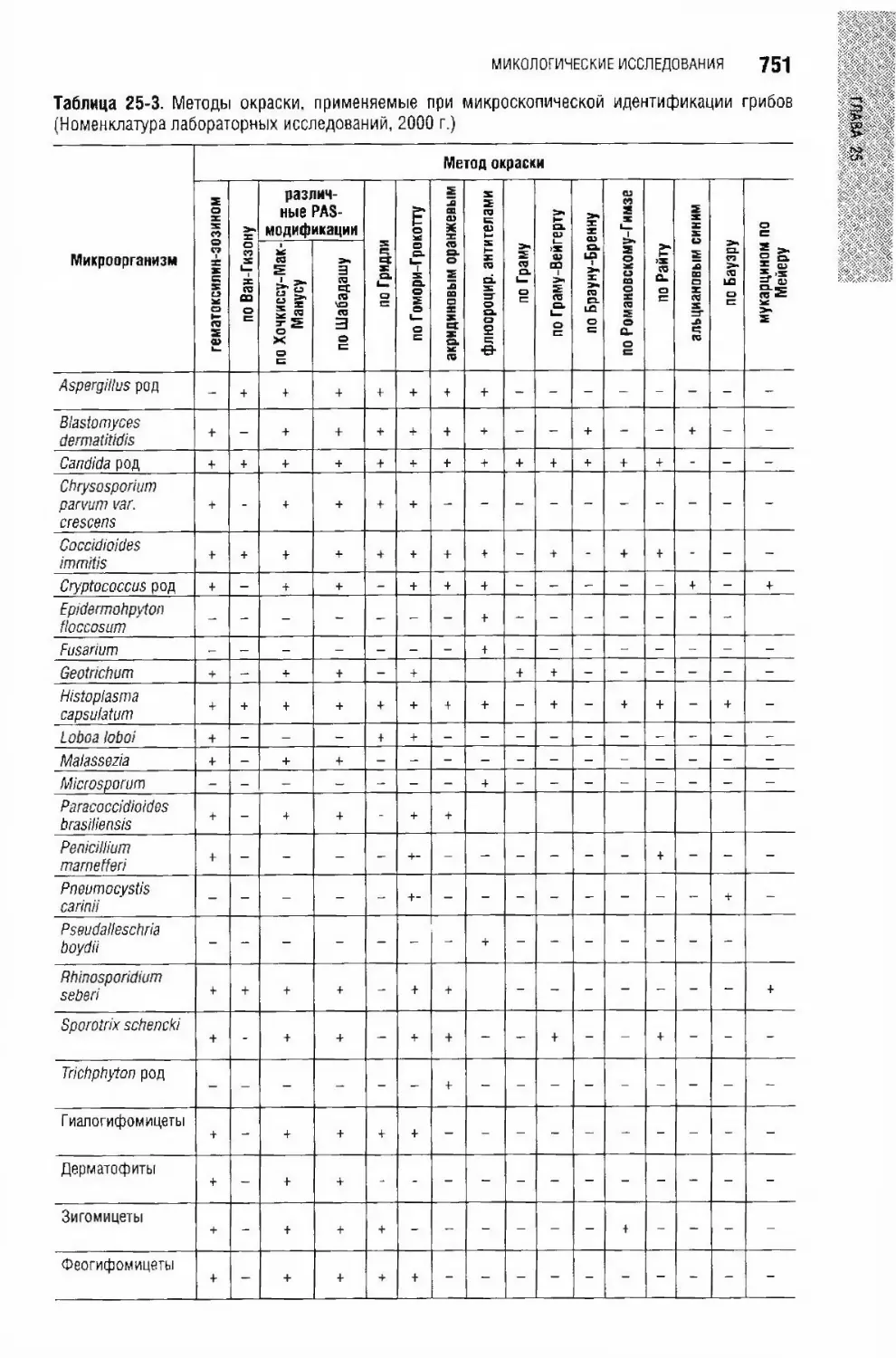
Глава 25. Микологические исследования Е.В. Липова, И.И. Глазка,
М.А. Мозжерова, В.Е. Колупаев ............................................................................-726
Систематика и классификация грибов........................................................726
Преаналитический этап лабораторной диагностики микозов....................................729
Макроскопические исследования.............................................................735
Микроскопические исследования.............................................................736
Культуральное исследование................................................................754
Определение антифунгеальной чувствительности..............................................760
Лабораторная диагностика поверхностных микозов............................................767
Критерии диагностики системных микозов....................................................767
Глава 26. Лабораторная диагностика паразитарных болезней А.С. Довгалев,
А.Е. Беляев, Т.Н. Константинова. Ю.П. Горбунова, Т.И. Авдюхина,
Т.М. Гузеева...............................................................................................................771
Кровепаразиты.............................................................................773
Паразиты в костном мозге..................................................................778
Паразиты в ликворе........................................................................778
Паразиты в лимфатических узлах............................................................780
Исследования паразитов в кале.............................................................781
Паразиты в дуоденальном содержимом........................................................791
Паразиты в моче...........................................................................792
Паразиты мокроты..........................................................................793
Паразиты отделяемого мочеполовых путей....................................................794
Паразиты в биопта гах тканей..............................................................795
Методы иммунодиагностики паразитарных заболеваний.........................................797
Предметный указатель.........................................................................801
УЧАСТНИКИ ИЗДАНИЯ
ГЛАВНЫЕ РЕДАКТОРЫ
Долгов Владимир Владимирович — д-р мед. наук, профессор, зав. кафедрой
клинической лабораторной диагностики ГОУ ДПО «Российская медицинская ака-
демия последипломного образования», Москва
Меньшиков Вадим Владимирович — д-р мед. наук, профессор, заслужен-
ный деятель науки РФ, член-корреспондент РАЕН, зав. лабораторией проблем
клинико-лабораторной диагностики НИЦ ГОУ ВПО «Первый Московский госу-
дарственный медицинский университет им. И.М. Сеченова» и экстремальной
хирургии, вице-президент ассоциации ортопедов и травматологов РФ.
АВТОРЫ И СОСТАВИТЕЛИ
Авдюхина Татьяна Николаевна - канд. мед. наук, доцент кафедры тропи-
ческих и паразитарных болезней ГОУ ДПО «Российская медицинская академия
последипломного образования*>, Москва
Автушенко Лариса Александровна — канд. мед. наук, врач-вирусолог ФГУЗ
«Северо-Западная противочумная станция» Роспотребнадзора, Санкт-Петербург
Алексеева Елена Андреевна — зам. зав. бактериологической лабораторией
Центра Госсанэпиднадзора Вологодской области, Вологда
Антипова Анастасия Юрьевна — мл. научн. сотр. ФГУН «Санкт-Петербург-
ский НИИ эпидемиологии и микробиологии имени Пастера» Роспотребнадзора
Афанасьева Анна Николаевна — вед. специалист-эксперт Управления
Федеральной службы в сфере защиты прав потребителя и благополучия человека
по Санкт-Петер бургу7
Вадиков Владимир Дмитриевич — д-р мед. наук, профессор, зав. кафедрой
микробиологии и микологии ГОУ ВПО «Санкт-Петербургская государственная
медицинская академия им. И.И. Мечникова»
Беляев Андрей Евгеньевич — канд. мед. наук, доцент кафедры тропических
и паразитарных болезней ГОУ ДПО «Российская медицинская академия последи-
пломного образования», Москва
Березина Людмила Александровна — канд. биол. наук. ст. научн. сотр.
лаборатории иммунологии ФГУН «Санкт-Петербургский НИИ эпидемиологии и
микробиологии имени Пастера» Роспотребнадзора
Беспалова Галина Ивановна — канд. мед. наук, доцент кафедры микробио-
логии и микологии ГОУ ВПО «Санкт-Петербургская государственная медицинская
академия им. И.И. Мечникова»
Бичурина Майна Александровна, д-р мед. наук, заслуженный деятель
науки РФ, зав. лабораторией этиологии и контроля вирусных инфекций ФГУН
«Санкт-Петербургски^ НИИ эпидемиологии и микробиологии имени Пастера»
Роспотребнадзора
Богумильчик Елена Александровна — мл. научн. сотр. лаборатории бактери-
альных капельных инфекций ФГУН «Санкт-Петербургский НИИ эпидемиологии
и микробиологии имени Пастера» Роспотребнадзора
Бойцов Алексей Геннадьевич — д-р мед. наук, профессор, зав. кафедрой
микробиологии, вирусологии и иммунологии ГОУ ВПО «Санкт-Петербургская
государственная медицинская академия им. И.И. Мечникова»
Бургасова Ольга Александровна — канд. мед. наук, доцент кафедры инфек-
ционных болезней и эпидемиологии Московского государственного медико-
стоматологического университета
Бурова Лариса Александровна — д-р мед. наук, вед. научн. сотр. отдела
молекулярной микробиологии НИИ экспериментальной медицины РАМН, Санкт-
Петербург
УЧАСТНИКИ ИЗДАНИЯ
7
Ваганова Анастасия Николаевна — биолог, сотр. лаборатории зооантропо-
нозных инфекций ФГУН «Санкт-Петербургский НИИ эпидемиологии и микро-
биологии имени Пастера» Роспотребнадзора
Васильев Олег Дмитриевич - канд. мед. наук, доцент кафедры микробиоло-
гии, вирусологии и иммунологии ГОУ ВПО «Санкт-Петербургская государствен-
ная медицинская академия им. И.И. Мечникова»
Вашукова Светлана Степановна — канд. биол. наук, врач клинической лабо-
раторной диагностики СПб ГУЗ «Городской консультативно-диагностический
центр (вирусологический)»
Вербов Вячеслав Николаевич — канд. хим. наук, руководитель отдела новых
технологий ФГУН «Санкт-Петербургский НИИ эпидемиологии и микробиологии
имени Пастера» Роспотребнадзора
Вершинский Борис Васильевич — канд. мед. наук, лауреат Государственной
премии СССР. Последняя должность — ст. науч. сотр. лаборатории зооантропоноз-
ных инфекций ФГУН «Санкт-Петербургский НИИ эпидемиологии и микробио-
логии имени Пастера» Роспотребнадзора
Вишневский Борис Израилевич — д-р мед. наук, профессор, зав. лаборатори-
ей микробиологии туберкулеза ФГУ «Санкт-11етебургский научно-исследователь-
ский институт фтизиопульмонологии»
Воскресенская Екатерина Александровна — канд. биол. наук, ст. научн, сотр.
лаборатории бактериальных капельных инфекций ФГУН «Санкт-Петербургский
НИИ эпидемиологии и микробиологии имени Пастера» Роспотребнадзора
Глазко Ирина Ивановна — канд. мед. наук, доцент курса лабораторной диа-
гностики и лабораторной микологии кафедры дерматовенерологии и клинической
микологии ГОУ ДПО «Российская медицинская академия последипломного обра-
зования». Москва
Гончарова Лариса Борисовна — канд. биол. наук, зав. вирусологическим
отделением СПб ГУЗ «Городской консультативно-диагностический центр (виру-
сологический)»
Горбунова Юлия Петровна — ст. лаб. кафедры тропических и паразитарных
болезней ГОУ ДПО «Российская медицинская академия последипломного образо-
вания». Москва
Грибенча Сергей Васильевич — д-р мед. наук, профессор, зав. лабораторией
иммунологии ГУ НИИ вирусологии им. Д.И. Ивановского, Москва
Гузеева Татьяна Михайловна — канд. мед. наук, доцент кафедры тропических
и паразитарных болезней ГОУ ДПО «Российская медицинская академия последи-
пломного образования», Москва
Дмитриев Александр Валентинович — д-р биол. наук, зав. лабораторией
функциональной геномики и протеомики микроорганизмов отдела молекулярной
микробиологии НИИ экспериментальной медицины РАМН. Санкт-Петербург
Довгалев Анатолий Семенович — д-р мед. наук, профессор, зав. кафедрой
тропических и паразитарных болезней ГОУ ДПО «Российская медицинская акаде-
мия последипломного образования», Москва
Егорова Светлана Александровна — канд. мед. наук, научн. сотр. лаборато-
рии кишечных инфекций ФГУН «Санкт-Петербургский НИИ эпидемиологии и
микробиологии имени Пастера» Роспотребнадзора
Елисеев Алексей Викторович — врач-бактериолиг бактериологической лабо-
ратории клинической больницы имени Петра Великого, Санкт-Петербург
Жебрун Анатолий Борисович — член-корреспондент РАМН, профессор, д-р
мед. наук, директор ФГУН «Санкт-Петербургский НИИ эпидемиологии и микро-
биологии имени Пастера» Роспотребнадзора
8 УЧАСТНИКИ ИЗДАНИЯ
Закревская Анна Васильевна — канд. мед. наук, вед. научн. сотр. лаборатории
иммунологии ФГУН «Санкт-Петербургский НИИ эпидемиологии и микробиоло-
гии имени Пастера» Роспотребнадзора
Игнатовский Андрей Викторович - канд. мед. наук, ассистент кафедры дер-
матовенерологии с клиникой ГОУ ВПО «Санкт-Петербургский государственный
медицинский университет им. акад И.П. Павлова»
Исаков Валерий Александрович — д-р мед. наук, академик РАЕН, профес-
сор кафедры инфекционных болезней и эпидемиологии < курсом ВИЧ-медицины
ГОУ ВПО «Санкт-Петербургский государственный медицинский университет
им. акад. И.П. Павлова», вед. науч. corp, лаборатории иммунологии ФГУН
«Санкт-Петербургский НИИ эпидемиологии и микробиологии имени Пастера»
Роспотребнадзора
Кафтырева Лидия Алексеевна — д-р мед. наук, зав. лабораторией кишеч-
ных инфекций ФГУН «Санкт-Петербургский НИИ эпидемиологии и микробио-
логии имени Пастера» Роспотребнадзора, главный специалист по бактериологии
Департамента здравоохранения Санкт-Петербурга, профессор кафедры ГОУ ДПО
«Санкт-Петербургская медицинская акад< мия последипломного образования».
Кашкин Кирилл Павлович — д-р мед. наук, профессор, академик РАМН, зав.
кафедрой иммунологии ГОУ ДПО «Российская медицинская академия последи-
пломного образования», Москва
Кокорина Галина Ивановна — врач-бактериолог отделения диагностики и
профилактики гепатитов ФГУН «Санкт-Петербургский НИИ эпидемиологии и
микробиологии имени Пастера» Роспотребнадзора
Константинова Татьяна Николаевна — канд. мед. наук, доцент кафедры тро-
пических и паразитарных болезней ГОУ ДПО «Российская медицинская академия
последипломного образования», Москва
Контарова Елена Олеговна — мл. научн. сотр. УРАМН «НИИВС имени
И.И. Мечникова» РАМН
Коровина Галина Ивановна — канд. биол. наук, врач клинической лаборатор-
ной диагностики Республиканской клинической инфекционной больницы, Санкт-
Петербург, Усть-Ижора
Косякова Карина Георгиевна - канд. мед. наук, ст. преподава гель кафедры
микробиологии, вирусологии и иммунологии ГОУ ВПО «Санкт-Петербургская
государственная медицинская академия им. И.И. Мечникова»
Кохнович Милана Анатольевна — мл. научн. сотр. ГУ НИИ вирусологии
им. Д.И. Ивановского, Москва
Краева Людмила Александровна — канд. мед. наук, ст. научн. сотр. лабора-
тории бактериальных капельных инфекций ФГУН «Санкт-Петербургский НИИ
эпидемиологии и микробиологии имени Пастера» Роспотребнадзора
Красносельских Татьяна Валерьевна — канд. мед наук, доцент кафедры дер-
матовенерологии с клиникой ГОУ ВПО «Санкт-Петербургский государственный
медицинский университет им. акад. И.П, Павлова»
Куляшова Лидия Борисовна — канд. мед. наук, вед. научн. сотр. лаборатории
иммунологии ФГУН «Санкт-Петербургский НИИ эпидемиоло] ии и микробиоло-
гии имени Пастера» Роспотребнадзора
Курова Наталья Николаевна — канд. мед. наук, ст. научн. сотр. лаборатории
бактериальных капельных инфекций ФГУН «Санкт-Петербургский НИИ эпиде-
миологии и микробиологии имени Пастера» Роспотребнадзора
Лаврент ьева Ирина Николаевна — д-р мед. наук, зав. лабораторией детских
вирусных инфекций ФГУН «Санкт-Петербургский НИИ эпидемиологии и микро-
биологии имени Пастера» Роспотребнадзора
Лапин Сергей Владимирович — канд. мед. наук, зав. лабораторией диа-
гностики аутоиммунных заболеваний НМЦ по молекулярной медицине ГОУ
УЧАСТНИКИ издания g
ВПО «Санкт-Петербургский государственный медицинский университет им. акад.
И.П. Павлова»
Лилова Елена Валерьевна — д-р мед. наук, профессор, зав. курсом лабора-
торной диагностики и лабораторной микологии кафедры дерматовенерологии и
клинической микологии ГОУ ДПО «Российская медицинская академия последи-
пломного образования», Москва
Литвинчук Людмила Филипповна — канд. биол. наук, вед. научн. сотр. ГУН
НИИ гриппа СЗО РАМН, Санкт-Петербург
Лялина Людмила Владимировна — д-р мед. наук, зав. отделом эпидемиоло-
гии ФГУН «Санкт-Петербургский НИИ эпидемиологии и микробиологии имени
Пастера» Роспотребнадзора
Макаров Дмитрий Александрович — мл. научн. сотр. ФГУН «Санкт-
Петербургский НИИ эпидемиологии и микробиологии имени Пастера»
Роспотребнадзора
Макарова Мария Александровна — канд. мед. наук, научн. сотр. лаборат ории
кишечных инфекций ФГУН «Санкт-Петербургский НИИ эпидемиологии и микро-
биологии имени Пастера» Роспотребнадзора
Малышев Владимир Васильевич — д-р мед. наук, руководитель группы
охраны окружающей среды ФГУН «Санкт-Петербургский НИИ эпидемиологии и
микробиологии имени Пастера» Роспотребнадзора
Матвеева Зоя Николаевна — канд. мед. наук, ст. научн. сотр. лаборатории
кишечных инфекций ФГУН «Санкт-Петербургский НИИ эпидемиологии и микро-
биологии имени Пастера» Роспотребнадзора
Мещерякова Ирина Сергеевна — д-р биол. наук, руководитель Центра
Минздравсоцразвития России по туляремии, НИИ эпидемиологии и микробио-
логии им. Н.Ф. Гамалеи РАМН. г. Москва
Мукомолов Сергей Леонидович — д-р мед. наук, профессор, зав. лаборато-
рией вирусных гепатитов ФГУН «Санкт-Петербургский НИИ эпидемиологии и
микробиологии имени Пастера» Роспотребнадзора
Мозжерова Марина Анатольевна — канд. мед. наук, ассистент курса лабора-
торной диагностики и лабораторной микологии кафедры дерматовенерологии и
клинической микологии ГОУ ДПО «Российская медицинская академия последи-
пломного образования», Москва
Нарвская Ольга Викторовна — д-р мед. наук, зав. лабораторией молеку-
лярной микробиологии ФГУН «Санкт-Петербургский НИИ эпидемиологии и
микробиологии имени Пастера» Роспотребнадзора, профессор кафедры медицин-
ской экологии и эпидемиологии человека имени Г.В. Хлопина ГОУ ДПО «Санкт-
Петербургская медицинская академия последипломного образования»
Нилова Людмила Юрьевна — канд. мед. наук, ст. преподаватель кафедры
микробиологии, вирусологии и иммунологии ГОУ ВПО «Санкт-Петербургская
государственная медицинская академия им. И.И. Мечникова»
Носков Фридрих Савельевич — д-р мед. наук, профессор, заслуженный
деятель науки РФ. Директор НИИЭМ имени Пастера (1992-1994 гг.). Последняя
должность — главный научный сотрудник ФГУН «Санкт-Петербургский НИИ
эпидемиологии и микробиологии имени Пастера» Роспотррбнадзора
Оришак Елена Александровна — канд. мед. наук, доцент кафедры микро-
биологии, вирусологии и иммунологии ГОУ ВПО «Санкт-Петербургская государ-
ственная медицинская академия им. И.И. Мечникова»
Оттен Татьяна Фердинандовна — д-р мед. наук, вед. научн. corp, лабо-
ратории микробиологии туберкулеза ФГУ «Санкт-Петербургский научно-
исследовательский институт фтизиопульмонологии»
10
УЧАСТНИКИ ИЗДАНИЯ
Пооин Александр Арнольдович - канд. мед. наук, доцент кафедры микро-
биологии, вирусологии и иммунологии ГОУ ВПО «Санкт-Петербургская государ-
ственная медицинская академия им. И.И. Мечникова»
Романенкова Наталия Ивановна — канд. мед. наук, вед. научн. сотр. ФГУН
«Санкт-Петербургский НИИ эпидемиологии и микробиологии имени Пастера»
Роспотребнадзора
Савичеоа Алевтина Михайловна — д-р мед. наук, профессор, руководитель
лаборатории микробиологии НИИ акушерства и гинекологии им. Д.О. Отта СЗО
РАМН, Санкт-Петербург
Сварваль Алена Владимировна — научн. сотр. лаборатории иммунологии
ФГУН «Санкт-Петербургский НИИ эпидемиологии и микробиологии имени
Пастера» Роспотребнадзора
Семенов Александр Владимирович — канд. биол. наук, зав. центральной
клинико-диагностической лабораторией ФГУН «Санкт'Петербургский НИИ эпи-
демиологии и микробиологии имени Пастера» Роспотребнадзора
Симбирцев Андрей Семенович — д-р мед. наук, профессор, зав. лабораторией
фармиммунологии Государственного НИИ особо чистых биопрепаратов ФМБА
России, Санкт- Петербург
Сироткин Алексей Константинович — канд. биол. наук, вед. научн. сотр.
ГУН НИИ гриппа СЗО РАМН, доцент Санкт-Петербургского государственного
университета
Скуинь Людмила Михайловна — канд. мед. наук, доцент кафедры иммуно-
логии ГОУ ДПО «Российская медицинская академия последипломного образова-
ния», Москва
Соколовский Евгений Владиславович — д-р мед. наук, профессор, зав. кафе-
дрой дерматовенерологии с клиникой ГОУ ВПО «Санкт-Петербургский государ-
ственный медицинский университет им. акад. И.П. Павлова»
Стоянова Наталья Александровна — канд. мед. наук, научн. сотр. лаборато-
рии зооантропонозных инфекций ФГУН «Санкт-Петербургский НИИ эпидемио-
логии и микробиологии имени Пастера» Роспотребнадзора
Суворов Александр Николаевич — д-р мед. наук, профессор, зав. лаборато-
рией молекулярной генетики патогенных микроорганизмов отдела молекулярной
микробиологии НИИ экспериментальной медицины РАМН, Санкт-Петербург
Сухобаевская Лариса Петровна — канд. биол. наук, ст. науч. сотр. ФГУН
«Санкт-Петербургский НИИ эпидемиологии и микробиологии имени Пастера»
Роспотребнадзора
Тартаковский Игорь Семёнович — д-р биол. наук, профессор, зав. лаборатори-
ей легионеллеза, УРАМН НИИ эпидемиологии и микробиологии им. Н.Ф. Гамалеи
РАМН, Москва
Токаревич Николай Константинович - д-р мед. наук, зав. лабораторией
зооантропонозных инфекций ФГУН «Санкт-Петербургский НИИ эпидемиоло-
гии и микробиологии имени Пастера» Роспотребнадзора, профессор кафедры
медицинской экологии и эпидемиологии человека имени Г.В. Хлопина ГОУ ДПО
«Санкт-Петербургская медицинская академия последипломного образования
Минздравсоцразвития РФ»
Тотолян Арег Артемович — д-р мед. наук, профессор, зам, директора ФГУН
«Санкт-Петербургский НИИ эпидемиологии и микробиологии имени Пастера»
Роспотребнадзора
Тотолян Артем Акопович — д-р мед. наук, профессор, академик РАМН, руко-
водитель отдела молекулярной микробиологии НИИ экспериментальной медици-
ны РАМН, Санкт-Петербург
УЧАСТНИКИ ИЗДАНИЯ
11
Ферман Раиса Семеновна — мл. научн. сотр. лаборатории иммунологии
ФГУН «Санкт-Петербургский НИИ эпидемиологии и микробиологии имени
Пастера» Роспотребнадзора
Ценева Галина Яковлевна — д-р мед. наук профессор, заслуженный дея-
тель науки РФ, зав. лабораторией бактериальных капельных инфекций ФГУН
♦Санкт-Петербургский НИИ эпидемиологии и микробиологии имени Пастера»
Роспотребнадзора
Шипицина Елена Васильевна — канд. биол. наук, ст. научн. сотр. лаборато-
рии микробиологии НИИ акушерства и гинекологии им. Д.0. Отта СЗО РАМН,
Санкт-Петербург
Шульгина Марина Владимировна — д-р биол. наук, вед. научн. сотр. отдела
стандартизации и контроля качества клинической лабораторной диагностики ФГУ
ГНИЦ профилактической медицины, зам. генерального директора по развитию
НП «Центр внешнего контроля качества», Москва
Юминова Надежда Васильевна — д-р биол. наук, зам. директора по научной
работе УРАМН «НИИ ВС имени И.И. Мечникова» РАМН
Яровая Ирина Ильинична — зав. вирусологической лабораторией ФГУЗ
«Северо-Западная противочумная станция» Роспотребнадзора, Санкт-Петербург
СПИСОК СОКРАЩЕНИИ
АБК — антитела к бокаловидным клеткам
Аг — антиген
АГМ А — антигладкомышечные антитела
АЕ — агглютинирующие единицы
АЗП — аутоиммунные заболевания печени
ЛИГ — аутоиммунный гепатит
АИЗ — аутоиммунные заболевания
АКА — антикератиновые антитела
АКЛА — антикардиолипиновые антитела
АМА — анти митохондриальные антитела
АМЦВ — антитела к модифицированному цитруллинированному виментину
АНА — антинуклеарные антитела
АНФ — атинуклеарный фактор
АнФА — антифилаггриновые антитела
АНЦА — антинейтрофильные цитоплазматические антитела
АОК — антитела к островковым клеткам
АПКЖ — антитела к париетальным (обкладочным) клеткам желудка
АПФ — антиперинуклеарный фактор
АСИТ — антиген-специфическая иммунотерапия
Ат — антитело
АФА — антифосфолипидные антитела
АФП — а-фетопротеин
АФС — антифосфолипидный синдром
АхР — ацетилхолиновый рецептор
АЦА — антицитруллиновые антитела
АЦЩ1 — антитела к циклическому цитруллин-содержащему пептиду
АЧТВ — активированное частичное тромбопластиновое время
АЭА — антиэндомизиальные антитела
АЭТА — ангиэндотелиальные антитела
БАЛ — бронхоальвеолярный лаваж
БНМ — белки наружной мембраны
БТШ — белок теплового шока
ВА — волчаночный антикоагулянт
ВГЧ-6 (HHV-6) — вирус герпеса человека 6-го типа
ВИЧ — вирус иммунодефицита человека
ВКК — внутрилабораторный контроль качества
ВКЭ — вирус клещевого энцефалита
ВПГ — вирус простого герпеса
ВПЧ (HPV) — вирус папилломы человека
ВРПВ — вакцинно-родственные полиовирусы
ВТД — внутритиповая дифференция
ВЭБ (EBV) — вирус Эпштейна-Барр
ГБ — синдром Гийена-Барре
ГВ — гранулематозный васкулит
ГВИ — герпес-вирусные инфекции
ГДК — глутаматдекарбоксилаза
ГИ — герпетическая инфекция
ГЛПС — геморрагическая лихорадка с почечным синдромом
ДБСТ — диффузные болезни соединительной ткани
ДВС — диссеминированное внутрисосудистое свертывание
ДК — дендритные клетки
СПИСОК СОКРАЩЕНИЙ 13
ДКВ — дискоидная красная волчанка
ДМ/ПМ — дерматомиозит/полимиозит
ИА — индекс авидности
ИБ — иммуноблот
ИППП — инфекции, передающиеся половым путем
ИФА — иммуноферментный анализ
ИФМ — флюоресцентная (люминесцентная) микроскопия
ИХТ — иммунохроматографический тест
ИЭМ — иммуноэлектронная микроскопия
КГЛ - крымская геморрагическая лихорадка
ККГЛ — крымская-Конго геморрагическая лихорадка
КОЕ — колониеобразующая единица
КУМ — кислотоустойчивые микобактерии
ЛПС — липополисахарид
М — молярность
МАК — мембраноатакующий комплекс
МАт — моноклональные антитела
МБК — минимальная бактерицидная концентрация
МБТ — микобактерии туберкулеза
ME — международные единицы
МИА — мультиплексный иммунный анализ
МИК — минимальная ингибирующая концентрация
МКТВ — Международный комитет по таксономии вирусов
МПК — минимальная подавляющая концентрация
МПО — миелопероксидаза
МР — методические рекомендации
МФА — микроскопический флюоресцентный анализ
НГОБ — неферментирующие грамотрицательные бактерии
НМФА — непрямой метод флюоресцирующих антител
НРИФ — непрямая реакция иммунофлюоресценции
НЯК — неспецифический язвенный колит
ОКИ — кишечные инфекции
ос/дсДНК — одно/двуспиральная ДНК
ОТ — обратная транскрипция
ПБА — патогенные биологические агенты
ПБЦ — первичный билиарный цирроз
ПИР — пирролидонил аминопептидаза
ПИФ — реакция прямой иммунофлюоресценции
ПМФА — метод флюоресцирующих антител
ПР-3 — протеиназа 3
ПТИ — пищевая токсикоинфекция
ПТО — посттрансфузионное осложнение
ПЦР — полимеразная цепная реакция
ПЭП — полиэндокринопатия
РА — ревматоидный артрит
РАИЛ — рецепторный антагонист интерлейкина-1
РВ — реакция Вассермана
РГА - реакция гемагглютинации
РИА — радиоиммунный анализ
РИФ — реакция иммунофлюоресценции
РМА — реакция микооагглютинации
PH — реакция нейтрализации
РИГА - реакция непрямой гемагглютинации
14 СПИСОК СОКРАЩЕНИЙ
РПГА — реакция пассивной гемагглютинации
PC — рассеянный склероз
РСВ — респираторно-синцитиальный вирус
РСК — реакция связывания комплемента
РТГА — реакция торможения гемагглютинации
рТТГ — рецептор тиреотропного гормона
РФ — ревматоидный фактор
СГВ - стрептококки группы В
СД — сахарный диабет
СЗСТ — смешанное заболевание соединительной ткани (синдром Шарпа)
СКВ — системная красная волчанка
СНЖ — спинномозговая жидкость
СП — санитарные правила
СС — сисэ емный склероз
СШ — синдром Шегрена
ТВП — тест волчаночной полоски
ТГ — тиреоглобулин
ТКИД — тяжелый комбинированный иммунодефицит
ТкТГ — тканевая трансглутаминаза
ТОРС — тяжелый острый респираторный синдром
ТП — тромбопоэтин
ТПО — тиреоидная пероксидаза
ТСЛП — тимический стромальный лимфопоэтин
ТТГ — тиреотропный гормон
ТЦД — тканевая цитопатогенная доза
УВЧ — электрическое поле ультравысокой частоты
ФВК — фосфовольфрамовая кислота
ФГА — фитогемагглютинин
ФГУП — федеральное государственное унитарное предприятие
ФСБ ОК — федеральная система внешней оценки качества
ЦИК — циркулирующие иммунные комплексы
ЦМВ — цитомегаловирус
ЦНС — центральная нервная система
ЦПД — цитопатическое действие
ЦПЭ — цитопатогенный эффект
ЭГДС — эзофагогастродуоденоскопия
ЭДТА — этилендиаминтетраацетат
ЭНА — экстрагируемые нуктеарные (ядерные) антигены
ЭП — эпидемический паротит
ЭПО — эритропоэтин
ЮРА — ювенильный ревматоидный артрит
ASCA - антитела к Saccharomyces cerevisiae
BCG — Bacille Calmette’-Guerin; БЦЖ
BMP — hone morphogenetic proteins; костные морфогенетические белки
CARS — compensatory) anti-inflammatory response syndrome; синдром компенсаторного
антивоспалительного ответа
CD — cluster of differentiation' кластер дифференцировки
CDC — Центр контроля заболеваемости
CREST-синдром — Calcinosis, Baynauld’s phenomenon. Esophagitis Sclerodactyly,
Telangiectasia — разновидность системного склероза, объединяющая подкожные
кальцинаты, синдром Рейно, нарушения моторики пищевода, склеродактилию и
тел еа н гиоэкта зии
CSF — colony-stimulationgfactor, колониестимулирующий фактор
СПИСОК СОКРАЩЕНИЙ 15
EGF — epidermal growth factor, эпидермальный фактор роста
ELISA — enzyme-linked immunosorbent assay; анализ твердофазный иммунофермент-
ный
ELISPOT — enzyme linked immunospot, твердофазный иммуноферментный анализ
синтеза цитокинов единичными клетками
FGF — fibroblast growth factor, фибробла( тный фактор роста
G-CSF — колониестимулирующий фактор для гранулоцитов
GM-CSF — колониестимулирующий фактор для гранулоцитов и моноцитов
HLA - human leucocyte antigen, человеческий лейкоцитарный антиген
1FN — интерферон
IL — интерлейкин
LC-1 — антитела к цитозольному антигену печени
LKM — антитела к микросомам печени-почек
M-CSF— колониестимулирующий фактор для моноцитов/макрофагов
МНС — главный комплекс гистосовместимости
MLST — мультилокусное генотипирование
ЫК(-клетки) — natural killer, естественный убийца
PDGF — platelet-derived growth factor, фактор роста, продуцируемый тромбоцитами
РТХЗ — пентраксин 3
RT-PCR — полимеразная цепная реакция с обратной транскрипцией
slCAM-1 — растворимая форма межклеточной адгезионной молекулы 1
SIL-2R — растворимая форма рецептора IL-2
SNP — single nucleotide polymorphism', полиморфизм вследствие замены единичных
нуклеотидов
TCR — антигенрасгознаюшие рецепторы
TGFp — transforming growth factor beta', трансформирующий фактор роста
Th — Т-лимфоциты-хелперы
TNF — tumor necrosis factor, фактор некроза опухоли
Глава 16
Лабораторная иммунология
Иммунная система защищает внутреннюю среду организма от
чужеродных и собственных антигенов, приобретающих призна-
ки чужеродности в силу’ различных обстоятельств. Способность
поддерживать собственные антигены жидкостей, клеток и тканей
позволяет человеку сохранить свою антигенную (иммунохимиче-
скую) индивидуальность. В защите от «чужого» участвуют клетки
органов и систем, а также гуморальные продукты жизнедеятельно-
сти этих клеток. Факторы иммунной защиты подразделяют на две
категории: врожденные (антиген-неспецифические) и приобретен-
ные (антиген-специфические).
ВРОЖДЕННЫЕ ФАКТОРЫ ИММУННОЙ ЗАЩИТЫ
Клетки врожденного иммунитета
Врожденные механизмы иммунитета препятствуют проникнове-
нию чужеродных антигенов в организм, нейтрализуют, разрушают и
выводят из организма чужеродные для него субстанции и клетки, не
«запоминая» их строения и не «узнавая» при повторных контактах
с ними. Врожденный иммунитет обеспечивается клетками фагоци-
тарной системы, соединительной и пограничных тканей, тромбо-
цитами, тучными, NK-клетками и т.д. В его поддержании участвуют
гуморальные факторы защиты: белки (острофазные, катионные и
системы комп лемента), медиаторы воспаления, цитокины, рецепто-
ры клеток и др. Врожденные факторы иммунной защиты вызывают
ранние реакции на чужеродные агенты при первичных и даже вто-
ричных контактах с ними. По изменению их содержания и активно-
сти судят о ранней реакции организма на чужеродные агенты
Увольнение в крови содержания полиморфноядерных лейкоцитов и
накопление юных форм этих клеток — ранняя реакция фагоцитов
на инфекционные агенты. Возрастает функциональная активность,
которую оценивают по фагоцитарной способности, а также по
изменению активности некоторых ферментов (миелопероксидаза,
глюкозо-6-фосфатдегидрогеназа) и увеличению концентрации в
крови белков, продуцируемых полиморфноядерными лейкоцитами
(лизоцим, прокальцитонин и др.). Полиморфноядерные лейкоциты
особенно значимы в качестве антимикробных агентов при инфек-
циях, вызываемых бактериями и грибами, а также при защите от
ГГАВ.« Ь
18 ЛАБОРАТОРНАЯ ИММУНОЛОГИЯ
капсульных микроорганизмов и возбудителей пиогенных инфекций. При дефектах
развития нейтрофильных лейкоцитов меняется их содержание в крови, возника-
ют морфологические и функциональные нарушения (табл. 16-1 и 16-2). В основе
отклонений — ферментопатии. нарушения созревания клеток, синтеза адгезивных
молекул, разнообразных рецепторов и др.
Таблица 16-1. Основные заболевания и состояния, сопровождающиеся нейтрофилией (количе-
ство нейтрофилов превышает 8x109/л крови)
Основные причины Интерпретация изменений
Влияние различных раздражителей (спортивные упражнения; адреналин; анестезия; пароксизмальная тахи- кардия. гипертермия) Быстрый переход гранулоцитов из краевого прилежания в циркули- рующую кровь за счет нейтрофилов, расположенных маргинально вдоль стенок мелких кровеносных сосудов или секвестрированных в селезенке
Введение различных ядов Связь со степенью тканевого распада
Лекарства и токсины (экстракты наперстянки, свинец, ртуть, бензол) Связь со степенью тканевого распада
Введение глюкокортикоидов или их избыточная продукция в организме (болезнь И цен ко-Кушинга) Нейтрофилия — следствие перехода нейтрофилов из краевого прилежания в сосудах в циркулирующую кровь в ответ на выбросы адреналина и адренокортикоидов
Острая бактериальная инфекция Стимуляция выброса нейтрофилов из костного мозга и их усилен- ная продукция
Неинфекционное воспаление (уре- мия. подагра) Гранулоцитоз вторичен по отношению к воспалительному процессу, связанному в одном случае с азотемией, в другом — с отложением кристаллов солей
Злокачественные опухоли Некроз быстрорастущих опухолей при недостаточности их кро- воснабжения. Некоторые опухоли (карциномы молочной железы, легких, почек, фибро- и липосаркомы) вырабатывают субстанцию, стимулирующую развитие нейтрофилии. Ларанеопластический синдром при солидных опухолях может вызвать нейтрофилию за счет секцеции нейтрофил-стимулирующего фактора роста
Восстановление костного мозга после агранулоцитоза Феномен превышения, характеризующийся гранулоцитозом (напри- мер, при лечении мегал области ой анемии)
Первичные гематологические забо левания (например, хронический миелолейкоз) Автономная продукция нейтрофилов
Спленэктомия Нейтрофилия вследствие нарушения секвестрирования гранулоци- тов в селезенке
Лейкемоидная реакция Диагноз исключения. Может сопровождать острые и хронические инфекции, интоксикации, злокачественные новообразования, включая метастазы в костный мозг (карцинома молочной и пред- стательной железы). Синдром Дауна может сопровождаться особой лейкемоидной реакцией, при которой картина крови и костного мозга неотличимы от характерных для острого миелолейкоза
Таблица 16-2. Основные заболевания и состояния, сопровождающиеся нейтропенией (количе-
ство нейтрофилов в крови менее 1,5x109/л)
Основные причины Интерпретация изменений
Лекарственные пое- параты Дозозависимая супрессия костного мозга (бензол, антиметаболиты, антраци- клины) или идиосинкразическая реакция (антитиреоидные, противосудорожные, антигистаминные препараты, фенотиазиды, сульфаниламиды и транквилизаторы)
Ионизирующее излу- чение Дозозависимая супрессия костного мозга
Вирусные инфекции Грипп, корь, ветряная оспа, краснуха, инфекционный гепатит, ВИЧ-инфекция
Бактериальные инфек- ции Тифоидная и паратифоидная лихорадка, туляремия, иногда бруцеллез, септице- мия
Коллагенозы Системная коасная волчанка: синдром Фелти, характеризующийся поражением суставов (ревматоидный артрит), спленомегалией и нейтропенией
ЛАБОРАТОРНАЯ ИММУНОЛОГИЯ 19
Окончание табл 16-2
Основные причины Интерпретация изменений
Аутоиммунная нейтро- пения Следствие образования антинейтрофильных антител
Нарушения гемопоэза Тяжелая недостаточность витамина В12 и фолиевой кислоты приводит к нару- шению созревания нейтрофилов в костном мозге. В периферической крови при этом встречаются большие гипеосегментированные нейтрофилы
Заболевания крови Апластическая анемия, острый лейкоз, миелодиспластический синдром
Воожденная патология Семейная нейтропения с аутосомно-доминантным типом наследования, цикличе- ская нейтропения с неясной этиологией; хроническая идиопатическая нейтропе- ния и синдром Костманна, при котором задерживается созревание нейтрофилов в костном мозге
Увеличение в крови количества эозинофилов и накопление продуктов их мета-
болизма (главный основной и эозинофильный катионный белок, эозинофильные
нейротоксин и пероксидаза) характерно для аллергических заболеваний, пара-
зитарных инфекций и опухолей (табл. 16-3). Эозинофилы при этой патологии
активно осуществляют свои эффекторные функции.
Таблица 16-3. Основные заболевания и состояния, при которых возможна эозинофилия (количс
ство эозинофилов в крови превышает 0,45x109/л)
Заболевания и состояния Комментарии
Аллергические заболе- вания Бронхиальная астма, крапивница, ринит, атопический дерматит, лекарственная аллергия, поллиноз и др.
Инфекционные заболе- вания Инфекции, сопровождающиеся стимуляцией Th2-зависимого иммунного ответа: гельминтозы (аскаридоз, стронгилоидоз, трихинеллез, токсокароз, филяриоз, шистоматоз и др.), протозоонозы (вызванные только Dientamoeba fragilis и Isospora belli), микозы (аспергиллез, кокциоидоз), вирусные инфекции (ВИЧ- инфекция, HTLV-1-инфекция)
Последствия медикамен- тозной терапии: -
генерализованные реак- ции Лекарственные препараты, стимулирующие продукцию IL-3, IL-5 и GM-CSF, усиливая эозинофилопоэз в костном мозге; нестероидные противовоспали- тельные препараты; антимикробные агенты; цитокины 1L-2 и GM-CSF
поражение легких Лекарственно индуцируемый легочный эозинофильный инфильтрат
поражение почек Острый интерстициальный нефрит, сопровождается эозинофилией и эозино- филурией (вызывают препараты бензатина бензилпенициллина и его произво- дные, сульфаниламиды, рифампицин, каптоприл, аллопуринол, ципрофлокса- цин и др.)
поражение сердца Острый некротизирующий эозинофильный миокардит может развиться как реакция на препараты ранитидин или клозапин
Миелоидная эозино- фильная лейкемия Симптомы заболевания сходны с миелопролиферативной патологией (высокая концентрация витамина В12, спленомегалия, анемия, миелоидная дисплазия, цитогенетические нарушения)
Гиперэозинофильный синдром Заболевание связано с дефектом генов а-рецептора к тромбоцит-зависимому ростовому фактору (хромосома 4) и сцепленным с Х-хромосомой дефектом развития эозинофилов. Синдром диагностирован у больных с генетически опо- средуемой высокой экспансией CD4+CD3~ Тбг-подобных лимфоцитов, выраба- тывающих IL-5; сопровождается гематологическими нарушениями, поражением кожи, сердечно-сосудистой и нервной системы, легких и др. органов
Неопластические заболе- вания (Т- и В-лимфомы, лимфома Ходжкина, миелоидные лейкемии, карциномы, аденокарци- номы и др.) При лимфоме Ходжкина и Т-клеточных лимфомах эозинофилия коррелирует с гиперпродукцией IL-5
20
ЛАБОРАТОРНАЯ ИММУНОЛОГИЯ
Окончание табл. 16-3
Заболевания и состояния К1 дментарии
Разнообразные пораже- ния легких Патология легких на вредных производствах сопровождается эозинофильной инфильтрацией легких и эозинофилией
Острая и хроническая эозинофильная пнев- мония Неясного генеза эозинофильная инфильтрация альвеол и интерстиция легких, эозинофилия. Вызывают лекарственные препараты, аллергены, табакокурение, высокая концентрация в воздухе химических вредностей и металлов
Моноциты крови и тканевые макрофаги участвуют в противоинфекционной
защите при хронических инфекционнв!х процессах, а также при инфекциях, воз-
будители которых облигатные или факультативные внутриклеточные паразиты
(вирусы, хламидии, риккетсии, микоплазмы и др.). Для этих заболеваний харак-
терно увеличение концентрации в крови моноиитов, возрастание их фагоцитарной
активности, способности в больших количествах продуцировать цитокины, оксид
азота, неоптерин и др. медиаторы воспаления (табл. 16-4).
Таблица 16-4, Основные заболевания и состояния, сопровождающиеся моноцитозом (количество
моноцитов в крови более 0,75x109/л)
Основные причины Интерпретация изменений
Инфекции Характерно для инфекций с гранулематозом (туберкулез, подострый бактери- альный эндокардит, сифилис, бруцеллез, висцеральные микозы, саркоидоз, инфекционный мононуклеоз) и на фоне нейтропении
Гематологические забо- левания Миелоидная метаплазия, лейкоз, множественная миелома, лимфома
Коллагенозы Ревматоидный артрит, системная красная волчанка, узелковый периартериит, полимиозит
Заболевания желудочно- кишечного тракта Язвенный колит, региональный энтерит, тропическая энтеропатия, паразитозы
Лекарственные средства Высокие дозы глюкокортикоидов
Дендритные клетки (ДК) миелоидного происхождения находятся в погранич-
ных тканях (клетки Лангерганса, интерстициальные-дермальные ДК) — важный
фактор врожденного иммунитета. Наряду с фагоцитарными функциями, они
способны обрабатывать захваченные антигены и представлять их фрагменты Т- и
В-лимфоцитам, NKT-клеткам, запуская антиген-специфический иммунный ответ.
Антиген-представляющие ДК могут стимулировать лимфоциты, направлять диф-
ференцировку антиген-активированных лимфоцитов в хелперные и регуляторные
клетки разных типов. Таким образом, ДК определяют характер иммунного ответа
на антиген (клеточный и/или гуморальный иммунный ответ).
Рецепторы фагоцитов реагируют на макромолекулы чужеродных клеток.
Лиганды для этих рецепторов — консервативные макромолекулы, синтезиоуемые
вирусами и клетками микроорганизмов, грибов и простейших. Они широко рас-
пространены среди представителей мира микроорганизмов, но отсутствуют на
клетках высших животных и человека. Антитела, комплемент, пентраксины и дру-
гие лиганды, к которым у «профессиональных» фагоцитов имеются соответствую-
щие рецепторы, обеспечивают избирательную активность клеток фагоцитарной
системы. Различные дефекты фагоцитов приводят к нарушению функции этого
звена врожденного иммунитета.
NK-клетки (англ, natural killer — естественный убийца) обладают цитотокси-
ческой активностью в отношении антиген но измененных собственных клеток.
Морфологически они представлены большими гранулярными лимфоцитами.
Количество NK-клеток в крови у взрослых достигает 15% лимфоцитов; они при-
сутствуют в костном мозге, лимфоидных opi анах, печени, легких и др.
Среди NK-клеток различают две субпопуляции: классические NK-клетки и
NKT-клеаки. NKT-клетки имеют антигенраспознаюший рецепторный комплекс
ЛАБОРАТОРНАЯ ИММУНОЛОГИЯ
21
(TCR CD3), сходный с таковым у Т-лимфоцитов. Набор антигенов, с которыми
могут взаимодействовать NKT-клетки, ограничен липидными и липопротеидны-
ми антигенами микроорганизмов. NK- и NKT-клетки способны продуцировать
TNF-a, IFN-y, GM-CSF, IL-12, они же индуцируют апоптотическую гибель клеток-
мишеней.
NK-клетки с высокой цитотоксической активностью отличаются большим
содержанием мембранных CD16 белков, а продуцирующие цитокины — экспрес-
сией CD56 поверхностных молекул. Цитотоксическая активность NK-клеток в
норме подавлена, активируются они при утрате ингибирующих их активность сиг-
налов. Пролиферацию, дифференцировку и функцию NK-клеток поддерживают
IL-15, IL-12, IL-2, IL-18, TNF-a, IFN-y и др. цитокины, а подавляют IL-10 и TGF-[1
Содержание В- и NK-клеток с учетом возраста представлено в табл. 16-5.
Таблица 16-5. Содержание В-лимфоцитов и NK-клеток в крови лиц разного возраста
Возраст СО19+В-клетки СОЗ О16~56* классические NK-клетки
% х108/л % х10в/л
0-3 мес 6-32 0,30-2.00 4-18 0,17-1,10
3-6 мес 11^1 0,43-3,00 3-14 0,17-0,83
6-12 мес 14-37 0,61-2,60 3-15 0,16-0 95
1-2 года 16-35 0,72-2,60 3-15 0,18-0,92
2-6 лет 14-33 0,39-1,40 4-17 0,13-0,72
6-12 лет 13-27 0,70-0,86 4-17 0,10-0,48
12-18 лет 6-23 0,11-0,57 3-22 0,07-0,48
Дефицит содержания или функциональная несостоятельность NK-клеток харак-
теризуется высокой чувствительностью человека к вирусам и тяжестью течения
таких вирусных инфекций, как цитомегаловирусная, опоясывающий лишай, про-
стой герпес, инфекционный мононуклеоз и др.
Гуморальные факторы врожденного иммунитета
Белки системы комплемента — важный гуморальный фактор врожденного
иммунитета. Эти белки присутствуют в сыворотке крови в неактивной форме и
приобретают иммунобиологическую активность в результате последовательной
активации компонентов системы комплемента. Образующиеся при этом комплек-
сы из нескольких белков системы комплемента и фрагменты белковых молекул
способны не только лизировать клетки микроорганизмов, паразитов, вирусные
частицы, соматические и др., но и в качестве эффекторных и регуляторных фак-
торов участвуют в воспалительных реакциях и антиген-специфическом иммунном
ответе.
Отдельные белки системы комплемента обозначают латинской буквой С и циф-
рами (Cl. С2), субъединицы белков и продукты их расщепления или активации —
дополнительно малыми латинскими буквами (например. Clq, Clr, СЗа).
Известны три главных пути активации системы комплемента: классический,
альтернативный и лектиновый (маннан-опосредованный).
При классическом пути активация системы инициируется Clq субкомпонен-
том комплемента в результате взаимодействия Clq с Fc-фрагментом конформа-
ционно измененных igM и IgG или с пентраксинами. В естественных условиях
конформационные изменения Fc-фрагментов IgM и IgG возникают при взаимо-
действии иммуноглобулинов с антигенами. Фиксированный на Ag-IgM и Ag-IgG
комплексах Clq взаимодействует с С1г и Cis субкомпонентами комплемента.
Последний активируется и, обладая активностью эстеразы, расщепляет С4 и С2.
Образующиеся С4Ь и С2а фрагменты в виде димерных молекул фиксируются на
22
ЛАБОРАТОРНАЯ ИММУНОЛОГИЯ
иммунных комплексах и как протеазы способны взаимодействовать с СЗ плаз-
мы,. а затем и расщеплять его на СЗа и СЗЬ субкомпоненты. Трехкомпонентный
комплекс С4Ь2аЗЬ способен взаимодействовать с С5 плазмы и, благодаря пеп-
тидазной активности, расщеплять С5 на субкомпоненты С5а и C5b. С образова-
ния С5Ь запускается активация компонентов, атакующих мембраны клеток. Эти
белки комплемента, последовательно взаимодействуя друг с другом, образуют
мультимолекулярный мембраноатакующий комплекс (МАК), способный атако-
вать липидные мембраны разнообразных клеток и, встроившись в них, нарушать
целостность клеточных мембран и соответственно вызывать лизис клеток. Сборка
МАК начинается с С5Ь. Фиксированная на мембране клетки молекула С5Ъ имеет
участок, взаимодействующий с белком С6. В результате такого взаимодействия
образуется эквимолекулярный комплекс С5ЬС6, который взаимодействует с С7,
образуя трехмолекулярный комплекс C5bC6C7. С7 плазмы крови характеризуется
амфофильными свойствами, и при взаимодействии с С5ЬС6 гидрофобный участок
С7 обеспечивает прочную фиксацию всего трехмолекулярного комплекса на фос-
фолипидах наружной клеточной мембраны.
Фиксированный на клетке С5ЬС6С7 трехмолекулярный комплекс служит рецеп-
тором для циркулирующего в крови С8. В результате фиксации на мембране клетки
С8 приобретает способность связывать циркулирующие в крови С9 и неэнзима-
тически катализировать их полимеризацию, после чего изменяется конформация
молекул этого белка, увеличивается число участков взаимодействия комплекса с
фосфолипидами мембраны и образуется полимерная белковая структура, по форме
напоминающая воронку или цилиндр. Такой цилиндр погружается в липидный
бислой наружной мембраны клетки-мишени. Снаружи стенки цилиндра образова-
ны гидрофобными, а внутренняя его сторона — преимущественно гидрофильными
аминокислотами полипептидов комплекса. Благодаря этому, вода смачивает вну-
треннюю поверхность стенок цилиндра и через отверстие в нем может легко посту-
пать из окружающей среды внутрь клеток, вызывая их гибель.
Мишенями для МАК комплемента могут быть клетки бактерий, грибов и про-
стейших, вирусные частицы, липосомы и другие структурированные образования,
а также собственные соматические клетки организма. Для активации комплемента
классическим способом против поверхностных антигенов клеток и частиц необхо-
димы иммунные или естественные антитела в виде иммуноглобулинов класса IgG
и IgM.
Лектиновый путь активации комплемента отличается от классического лишь
начальным инициирующим этапом. Б крови у людей и животных циркулирует
протеин, способный взаимодействовать с маннозой гликопротеинов и полиса-
харидов (англ. MBL — mannose binding lectin). Манноза в больших количествах
содержится в полисахаридах и гликопротеинах микроорганизмов и растений. При
внедрении микроорганизмов в макроорганизм MBL фиксируется на маннозных
остатках поверхностных полисахаридов и гликопротеинов. С иммобилизирован-
ным MBL взаимодействует сериновая протеаза крови, получившая название MBL-
ассоциированная протеаза (MASP). Протеаза в комплексе с MBL активируется и
приобретает способность расщеплять С4 и С2. В дальнейшем активация системы
комплемента осуществляется в той же последовательности и с образованием тех
же продуктов, что и при классическом пути.
Альтернативный путь активации системы комплемента также отличается
начальными этапами. Активация альтернативным способом осуществляется без
участия антител. Инициаторными молекулами выступают компоненты поверх-
ностных структур грамположительных и грамотрицательных микроорганизмов,
липополисахариды, тейхоевые кислоты клеточных стенок, полисахариды и кле-
точные стенки дрожжей и грибов, некоторые вирусы и вирус-инфицированные
клетки, клетки простейших.
ЛАБОРАТОРНАЯ ИММУНОЛОГИЯ 23
СЗ — компонент комплемента, который инициирует альтернативную актива-
цию всей системы и может подвергаться медленному спонтанному гидролизу тио-
эфирной связи молекулы. СЗ с измененной тиоэфирной связью не расщепляется на
СЗа и СЗЬ субкомпоненты, но приобретает свойства, делающие его тождественным
СЗЬ. Гидролизованный СЗ (СЗ-гидро), как и СЗЬ, взаимодействует с поверхност-
ными струк гурами чужеродных клеток.
Взаимодействие СЗЬ с клетками отличается определенной избирательностью и
легко осуществляется благодаря высокому сродству СЗЬ к поверхностным структу-
рам микроорганизмов, липополисахаридам клеточных стенок бактерий, зимозану
клеточных стенок дрожжей, полисахаридам чужеродных клеток. Высокое содер-
жание сиаловых кислот на мембране клеток человека и других млекопитающих
защищает соматические клетки от фиксации на их поверхности СЗЬ и альтерна-
тивной активации комплемента. Избирательность в фиксации СЗЬ на мембранах
чужеродных клеток, сопровождающаяся активацией системы комплемента аль-
тернативным способом, обеспечивает в организме примитивное распознавание
чужеродных (микробных) антигенов.
Каждый из путей активации системы комплемента регулируется в организме
относительно самостоятельно. Регуляцию активности системы осуществляют
молекулы, способные связать или инактивировать отдельные белки системы ком-
племента или, напротив, пролонгировать их пребывание в организме в активном
состоянии (пропердин).
В процессе активации системы комплемента образуется множество субкомпо-
нентов — продуктов оасщепления отдельных белков. Эти субкомпоненты, оста-
ваясь на мембране клетки или поступая в циркуляцию, обладают высокой имму-
нобиологической активностью. Рецепторы к субкомпонентам белков системы
комплемента есть в основном у клеток, участвующих в воспалительных процессах
и в иммунном ответе на антигены. Таким образом, образующиеся при активации
комплемента белковые субмолекулы — важные эндогенные регуляторы иммунно-
го ответа.
У новорожденных гемолитическая активность комплемента НС50 и содержание
СЗ и С4 составляет 50-75% взрослых людей, однако содержание в крови С8 и С9
не превышает 10% взрослых. К 3-месячному возрасту гемолитическая активность
комплемента в крови сопоставима с таковой у взрослых.
Врожденные и пи приобретенные дефекты синтеза отдельных компонентов ком-
племента характеризуются высокой чувствительностью к пиогенным и капсуль-
ным инфекциям и склонностью к возникновению аутоиммунных заболеваний.
Аутоиммунные заболевания при дефиците С4, С2 и СЗ компонентов развиваются
из-за нарушения выведения из циркуляции иммунных комплексов, в том числе
комплексов аутоантиген-аутоантитело (табл. 16-6). Повышенная активность ком-
племента, обусловленная дефектом ингибитора первого компонента комплемента
(Ci п11), проявляется развитием ангионевротического отека, решающим фактором
в патогенезе которого служат накапливающиеся в результате гиперактивации ком-
племента СЗа и С5а анафила токсины.
Таблица 16-6. Заболевания, ассоциирующиеся с недостаточностью белков системы комплемента
и их рецепторов
Белки Клинические проявления
Clq Системная красная волчанка и сходные синдромы, уртикарные васкулиты
Clr-CIs Системная красная волчанка и сходные синдромы, васкулиты
С2 Системная красная волчанка и сходные синдромы, гломерулонефриты. дерматиты, васку- литы
24
ЛАБОРАТОРНАЯ ИММУНОЛОГИЯ
Окончание табл. 16-6
Белки Клинические проявления
СЗ Аутоиммунные гломерулонефриты, коллагенозы, рецидивирующие пиогенные инфекции
С4 Системная красная волчанка и сходные синдромы
С5 Частые нейссериальные инсрекции, системная красная волчанка и сходные синдромы
С6; 07; С8; С9 Рецидивирующие нейссериальные инфекции
Пропердин Рецидивирующие пиогенные инфекции, молниеносное течение менингококкового сепсиса
Фактор D Рецидивирующие пиогенные инфекции
С1 ь inh Псевдоаллергический ангионевротический отек, склонность к аутоиммунным заболева- ниям
Фактор Н Системная красная волчанка и сходные синдромы, гломерулонефриты
Фактор I Рецидивирующие пиогенные инфекции и синдромы, подобные системной красной вол- чанке
CR1 Системная красная волчанка и сходные синдромы
CR3 Позднее отпадение пупочного канатика, рецидивирующие пиогенные инфекции, лейко- цитоз
DAF, HRF; CD59 Гемолиз
Нарушения синтеза таких ингибиторов системы комплемента, как DAF (англ.
decay acceleratingfactor — фактор, ускоряющий разрушение), HRF18 и HRF65 (англ.
homology restriction faster — фактор гомологической рестрикции), приводят к повы-
шенной активности комплемента в отношении аутологичных эритроцитов, что
клинически фиксируют у больных ночной пароксизмальной гемоглобинурией.
Именно поэтому лабораторное исследование белков системы комплемента реко-
мендуют проводить у лиц с системными аутоиммунными заболеваниями, при
хронических инфекциях, отеках и васкулитах неясного генеза.
С-реактивный белок и декамерный. протеин РТХЗ — пентраксины, являются
представителями гуморального врожденного иммунитета. С-реактивный белок —
представитель «коротких» белков семейства пектраксинов (М.м. 115 kDa), его
синтезируют гепатоциты под влиянием провоспалительных цитокинов IL-6,
IL-1, TNF-a. С-реактивный белок накапливается в крови и тканевых экссудатах
больных при острых и хронических инфекциях, травмах и других заболевани-
ях. У практически здоровых людей содержание С-реактивного белка в крови
не превышает 4 мг/л; при остром воспалении его уровень быстро возрастает
в течение первых часов. При бактериальном сепсисе, легионеллезе и тяжелых
васкулитах концентрация С-реактивного белка в крови превышает 300 мг/л; при
бактериальных инфекциях, острых васкулитах, некоторых лимфомах уровень
С-реактивного белка в крови варьирует в пределах 100-300 мг/л. При инфекцион-
ном мононуклеозе, цитомегаловирусной, герпетической и аденовирусной инфек-
циях, артритах, ревматоидной полимиалгии и большинстве лимфом содержание
С-реактивного белка в крови не превышает 100 мг/л. При большинстве вирусных
инфекций уровень С-реактивного белка в крови больных существенно не меня-
ется. Концентрация С-реактивного белка в крови больных отражает изменения в
развитии болезни за последние 6-8 ч.
С-реактивный белок способен взаимодействовать с остатками фосфорилхоли-
на в молекулах полисахаридов, лецитина, сфингомиелина, а также со свободной
ДНК разного происхождения, образуя мульти молекулярные комплексы. В них
С-реактивный белок взаимодействует с Clq компонентом комплемента и активи-
ЛАБОРАТОРНАЯ ИММУНОЛОГИЯ 25
рует комплемент классическим способом, а также опсонизирует об ьекты фагоци-
тоза и стимулирует активность NK-клеток.
РТХЗ — представитель длинных пентраксинов, декамерный протеин с М.м.
40 kDa, продуцируется активированными моноцитами и дендритными клетками,
способен взаимодействовать с маннозой гликопротеинов микробного и иного про-
исхождения. РТХЗ обладает опсонизирующей активностью, фиксируясь на объ-
ектах фагоцитоза, а также в комплексе с полисахаридами взаимодействует с Clq
белком комплемента и активирует комплемент классическим путем.
Таким образом, клеточные и гуморальные факторы врожденного иммуни-
тета обладают разнообразными средствами, позволяющими им противостоять
чужеродным агентам и участвовать в защите от внедрения, размножения и рас-
пространения в организме «чужого-»'-. Клетки врожденного иммунитета активно
взаимодействуют друг с другом и с клетками приобретенного иммунитета, что
делает процесс защиты высокоэффективным и надежным. Гуморальные факторы
врожденного иммунитета нейтрализуют и разрушают чужеродные агенты, способ-
ствуя их быстрому выведению из крови и организма.
ПРИОБРЕТЕННЫЙ ИММУНИТЕТ
Главные исполнители антиген-специфического иммунного ответа — лимфоци-
ты (гетерогенная популяция клеток). Они. не различаясь морфологически, суще-
ственно отличаются набором мембранных белков и функциональной активностью.
Количество их в крови может существенно меняться в зависимости от патологии
(табл. 16-7). Лимфоциты подразделяют на три большие популяции: Т-лимфоциты
(тимусзависимые), В-лимфоциты (костномозговые или бурсальные) и NK (нату-
ральные киллерные клетки).
Таблица 16-7. Основные заболевания и состояния, сопровождающиеся лимфоцитозом
Основные причины Интерпретация изменений
Вирусные инфекции Инфекционный мононуклеоз (в мазках крови обнаруживают атипичные монону- клеары) Инфекционный гепатит. Лимфоцитоз часто определяют при инфекциях, вызываемых вирусом Эпштейна-Барр, простого герпеса 2 типа, краснухи, адено- и цитомегаловирусом
Бактериальные инфекции Лимфоцитоз наблюдают при коклюше, хронических инфекциях (туберкулезе, бру- целлезе)
Заболевания крови Лимфаденопатия и спленомегалия сопровождают лимфому, хронический лимфо- пейкоз, волосатоклеточный лейкоз
Поспрансфузионный синдром Сопровождает лихорадка и спленомегалия. Причиной этого синдрома считают цитомегаловирус, пеоедающийся через лейкоциты донорской крови
Примечание. Количество лимфоцитов в норме имеет возрастные особенности. После второго
«возрастного» перекреста в лимфоцитарной формуле за лимфоцитоз принимают увеличение
абсолютного числа лимфоцитов в крови от 4,0х10ч/л и более. Абсолютный лимфоцитоз бывает
значительно реже относительного, встречающегося при гранулоцитопении.
СИСТЕМА Т-ЛИМФОЦИТОВ
Среди Т-лимфоцитов различают клетки Т-хелперные (Th), Т-регуляторные
(Тг) и Т-цитотоксические (Тс). Они фенотипически могут быстро меняться и
трансформироваться в Т-клетки с разными функциями. Тг обеспечивают регуля-
цию антиген-специфического иммунного ответа и функциональной активности
иммунной системы в целом. Эти клетки, продуцируя множество цитокинов, отве-
чают за взаимодействие иммунной системы с другими системами организма.
Современные методы идентификации Т-лимфоцитов и определения их принад-
лежности к тем или иным субпопуляциям основываются на исследовании мем-
бранных белков. Мембранные белки, иммунохимически охарактеризованные в
26 ЛАБОРАТОРНАЯ ИММУНОЛОГИЯ
качестве маркеров тех или иных клеток, получили обозначение CD (от англ, cluster
of differentiation - скопление дифференцировочных детерминант). Различные суб-
популяции Т-лимфоцитов отличаются друг от друга и от В-лимфоцитов набором
CD-протеинов, многие из которых — рецепторы клеток к различным лигандам.
Отдельные CD-протеины обозначают цифрами, например CD1, CD2, CD3 и т.д.
Распознавание антигенов Т-клетками происходит с помощью рецепторов,
способных специфически взаимодействовать с антигенными детерминантами и
одновременно с молекулами HLA, в комплексе с которыми находятся антигенные
детерминанты. В зависимости от строения антигенраспознающих рецепторов
(TCR), все Т-лимфоциты подразделяют на две субпопуляции: сфТ- (90-95%) и
уЗТ-клетки (5%). По расчетным данным, набор антигенных детерминант, с кото-
рыми могут взаимодействовать а|ЗТ-клетки, достигает 1018 вариантов молекул;
уЗТ-лимфоциты могут реагировать на антигенные детерминанты относительно
ограниченного репертуара (белки теплового шока, некоторые антигены микро-
организмов).
Т-клетки не реагиру ют на свободно циркулирующие антигены и могут взаимо-
действовать с ними только в том случае, если антигены подверглись обработке в
других клетках, а отдельные олигопептиды (детерминанты) обработанного анти-
гена представляются Т-лимфоцитам на поверхности вспомогательных клеток
(дендритных, макрофагов, В-лимфоцитов и др.) Фрагменты антигена на мембра-
не вспомогательных клеток представляются Т-клеткам посредством белков глав-
ного комплекса гистосовместимости (МНС) I класса для цитотоксических лим-
фоцитов или II класса для хелперных-регуляторных лимфоцитов (МНС-белки
от англ, major histocompatibility complex). У человека белки МНС обозначены как
HLA-белки (англ, human leukocyte antigens). Таким образом, Т-лимфоциты челове-
ка реагируют на чужеродные антигены, только если эти антигены ассоциированы
с HLA-бслками на мембранах любых клеток организма.
Кроме ар или уЗ полипептидных цепей, специфически взаимодействующих
с антигенной детерминантой, в построении антигенраспознающих рецепторов
Т-клеток участвует также несколько дополнительных полипептидых цепей. Их
принято обозначать как CD3 рецепторный комплекс. CD3 белки в антигенраспоз-
наюшем рецепторе Т-лимфоцитов выполняют несколько вспомогательных функ-
ций, в том числе проводят сигнал от взаимодействующего с антигеном рецептора
клетки в ядро Т-лимфоцита. Принадлежащие к разным клонам Т-лимфоциты
отличаются строением наружных концевых участков у оф и уЗ гетеродимеров, что
позволяет каждой Т-клетке с помощью TCR взаимодействовать лишь с определен-
ной, чаще какой-либо одной, антигенной детерминантой. Принципиально важно,
что в доантигенной дифференцировке Т-лимфоцитов в тимусе формируется TCR с
широким спектром специфичностей к антигенам, с которыми, вероятно, встретят-
ся Т-клетки на периферии.
В процесс контакта Т-клеток с антигеном с помощью TCR-CD3 рецепторного
комплекса вовлекаются также другие мембранные белки клетки, обозначаемые
как корецепторы CD4 и CDS. У зрелых Т-клеток экспрессия этих белков альтер-
нативная, клетки либо СО4чСП8*или CD4’CD8+. CD4+ белки обнаруживаются у
хелперных и регуляторных Т-лимфоцитов, CDS* молекулы — у цитотоксических
Т-клеток. CD4 белок на мембране Th-лимфоцитов способствует более прочному
связыванию антигенраспознающего рецептора лимфоцита с нагруженными анти-
геном белками II класса HLA (HLA-D, -DR, -DP, -DQ) вспомогательных клеток.
Белки II класса HLA в достаточных количествах, а также В-лимфоциты, экспресси-
руются преимущественно на мембранах антигенпредставляющих клеток иммунной
системы (дендритные клетки, макрофаги). Они фагоцитируют чужеродные анти-
гены. в эндосомах клеток расщепляют их до олигопептидов из 13-18 аминокислот
и образуют комплексы из олигопептидов антигена и белков HLA II класса. Такие
ЛАБОРАТОРНАЯ ИММУНОЛОГИЯ 27
комплексы затем встраиваются в мембрану вспомогательных клеток и, обладая
высокой иммуногенностью, представляются CD4 Т-лимфоцитам.
Полипептид CD8* обеспечивает более прочное взаимодействие антиген-
специфического рецептора Тс-лимфоцитов с антигеном, расположенным на
поверхности какой-либо соматической клегки в ассоциации с белками HLA
I класса (HLA-A, -В, -С). Поскольку белки HLA-A, -В, -С как трансплантаци-
онные антигены находятся на мембранах практически всех клеток организма,
любая клетка может представить антиген CD8+ Т-лимфоцитам в виде комплекса
из олигопептида антигена и белков HLA-A, -В, -С. В этом случае расщепление
молекулы (частицы) антигена до олигопептидов происходит в протеосомах
цитоплазмы любой инфицированной антигеном соматической клетки. Там же, в
цитоплазме, образуется высокоиммуногенный комплекс из коротких пептидов
антигена (8-10 аминокислотных остатков) и белков HLA I класса. На поверхности
ядросодержащих клеток содержание белков HLA I класса достигает 105 молекул,
так что каждая соматическая клетка может представлять множество антигенных
олигопептидов и вызывать против этих детерминант и себя ответ цитотоксических
CD8 Т-лимфоцитов.
Таким образом, в антигенной активации CD4+ и CD8+ субпопуляций
Т-лимфоцитов участвуют разнообразные клетки организма. Показано, что взаи-
модействие CD8* или CD4+ на поверхности Т-лимфоцитов соответственно с HLA
белками I или II класса вспомогательных клеток способствует образованию на
мембранах Т-лимфоцитов полимерных комплексов из TCR-CD3 белкпв и про-
ведению сигнала для активации Т-клеток. Несмотря на большое принципиальное
сходство в строении TCR-CD3 белков, у офТ- и убТ-клетки эти субпопуляции
существенно различаются функционально. У большинства уЗТ-клеток редко или
вовсе не экспрессированы белки CD4 и CD8, Эти клетки могут распознавать анти-
ген в комплексе с белком CD1 на поверхности антигенпредставляющей клетки. У
человека описано пять вариантов белков CD1, которые способны образовывать
комплексы и представлять у5Т-лимфоцитам ограниченные наборы антигенов.
yST-клетки цитотоксически активны, в качестве интраэпидермальных и интраэпи-
телиальных лимфоидных клеток в наибольших количествах представлены в коже,
слизистой оболочке тонкого кишечника и урогенитального тракта.
У Т-лимфоцитов имеются разнообразные адгезивные молекулы и рецепторы к
хемокинам. Посредством этих молекул клетка получает сигналы для направленной
миграции и взаимодействует с белками внеклеточного матрикса, с эндотелвалъ ны-
ми клетками сосудов лимфатических узлов и мукозных лимфоидных органов, с
клетками крови и белками комплемента. Адгезивные молекулы и рецепторы к
хемокинам обеспечивают накопление Т-лимфоцитов в очагах воспаления (грану-
лемы), лимфоклеточную инфильтрацию тканей, взаимодействие лимфоцитов друг
с другом и с клетками и молекулами врожденного иммунитета.
Большинство циркулирующих в крови Т-лимфоцитов находится в состоя-
нии покоя, но при контакте с соответствующим антигеном или под влиянием
других стимулов они активируются. Активированные лимфоциты отличаются
от покоящихся клеток возросшим содержанием в цитоплазме свободных ионоь
Са”, возросшим кислородным метаболизмом, повышенным синтезом РНК и бел-
ков, появлением на мембранах клеток новых белко и изменением содержания
ранее представленных. Все это приводит к превоащению покоящегося малого
лимфоцита в бластную клетку, которая затем делится 4-5 раз, воспроизводя все
фенотипические особенности исходной материнской клетки. При этом дочерние
клетки приобретают высокую иммунобиологическую активность, проявляя себя
в качест ве регуляторных или эффекторных Т-клеток. Мембранные маркеры акти-
вированных Т-клеток — CD25, CD54 (ТСАМ-1), CD69, CD70, CD71, белки 1ILA
II класса и некоторые другие молекулы. При активации на поверхности Т клеток
28 ЛАБОРАТОРНАЯ ИММУНОЛОГИЯ
увеличивается содержание CD2 и CD3, Из характерных для активированных
Т-лимфоцитов молекул выделяют: CD25 — а-иепь рецептора к IL-2, CD54 — инте-
грин, обес печивающий взаимодействие активированного Т-лимфоцита с другими
клетками иммунной системы, CD69 — ранний маркер активации разнообразных
лейкоцитов, CD71 — рецептор к сывороточному трансферрину.
Накопившиеся после антигенной активации иммунные лимфоциты по набо-
ру мембранных белков отличаются от так называемых наивных покоящихся
Т-клеток, которые, поступив из тимуса в кровь и лимфоидные органы, еще не
имели контакта с соответствующими антигенами. Маркеры наивных CD4’ или
CD8 Т-лимфоцитов — экспрессированные на мембране клеток CD45RA и CD62-L
белки. Белок CD45RA — изоформа тирозин-фосфатазы с М.м. 205-220 kDa. У
иммунных Т-лимфоцитов другая изоформа фермента — CD45RO с М.м. 180 kDa.
Важную роль в антигенной активации Т-клеток играет CD45, поскольку при его
отсутствии на поверхности клеток Т-лимфоциты не способны отвечать на анти-
генный стимул.
CD62L — L-селектин — обеспечивает узнавание наивными Т-лимфоцитами
вторичных лимфоидных органов и накопление их там. При антигенной активации
наивных Т-лимфоцитов важны костимулирующие сигналы, подаваемые клетке
через CD28 мембранный белок, посредством имеющихся белков CD80 и CD86 у
антигенпредставляющих клеток. Наивные Т-клетки без антигенной активации
редко делятся, а продолжительность их жизни составляет от нескольких недель до
10 лет. Содержание себпопуляций Т-лимфоцитов в крови с учетом возраста пред-
ставлено в табл. 16-8.
Таблица 16-8. Содержание в крови Т-лимфоцитов основных фенотипов у лиц разного возраста
Возраст СОЗ* (общий пул) CD4' (хеллеры/регулятори) CD8* 1киллеры/сулрессоры)
% хЮ’/л % хЮ’/Л % хЮ’/л
0-3 мес 53-94 2,5-5,5 35-64 1,640 12-28 0,56-1,7
3-6 мес 51-77 2 5-5,6 35-56 1,840 12-23 0,59-1.6
6-12 мес 49-76 1 9-5,9 31-56 1,443 12-24 0,5-1,7
1-2 года 53-75 2,1-6,2 32-51 1,3—1,3 14-30 0,62-2,0
2-6 лет 56-75 1 4-3,7 28-47 0,7-2,2 16-30 0,49-1,3
6-12 лет 60-76 1,2-2,6 31-47 0,65-1,5 18-35 0,37-1,1
12-18 лет 56-84 1.0-2,2 31-52 0.53-1,3 18-35 0.33-0.92
При антигенной активации наивных и иммунных Т-лимфоцитов связывание
антигена с TCR-CD3 белками лимфоцита должно быть дополнено сигналами,
подаваемыми Т-клетке через дополнительные костимулирующие рецепторы. У
наивных Т-лимфоцитов такой рецептор представлен CD28, а также рецепторами
к цитокинам и некоторым другим лигандам. Эти дополнительные команды могут
усиливать или угнетать пролиферацию Т-лимфоцитов или регулировать направ-
ление их дифференцировки.
Популяция CD4* Т-лимфоцитов человека неоднородна, ее подразделяют на
несколько субпопуляций регуляторных и хелперных Т-клеток. Среди последних,
в зависимости от их способности продуцировать те или иные наборы цитокинов,
различают ThO-, Thl- и Th2-, Th-17 и Tfh-клетки.
В крови у здоровых и неиммунизировапных людей среди Th-лимфоцитов
антиген-специфического клона преобладают ThO-клетки. При вакцинации наи-
вные ThO-клетки быстро превращаются в иммунные эффекторные ТЫ- или Th2-
лимфоциты. Маркерными цитокинами ТЫ-клеток служат продуцируемые ими
IFN-y и IL-2, а ТЬ2-лимфоцитов — IL-4, IL-5, IL-10 и IL-13. Дифференцировку
ThO-клеток в Thl- или ТЬ2-клетки определяют многие факторы и условия взаи-
модействия наивных ThO-лимфоцитов с антигеном. Важнейшее значение этой
ЛАБОРАТОРНАЯ ИММУНОЛОГИЯ
29
дифференцировки придают цитокинам, в особенности IL-12. Он определяет
дифференцировку ThO-клеток в ТЫ-лимфоциты, лишь в его отсутствие возмож-
на дифференцировка клеток в ТЬ2-лимфоциты. Продуцирование ТЫ- и Th2-
лимфоцитами тех или иных цитокинов определяет роль этих клеток в регуляции
анти ген-специфического ответа.
ТЫ-лимфоциты ответственны за клеточный антиген-специфический иммун-
ный ответ и функцию клеток фагоцитарной системы и NK-клеток, участвуют в
регуляции экспрессии генов IgM и IgG2 в В-клетках. Цитокины, продуцируемые
ТЬ2-лимфоцитами, поддерживают в организме биосинтез IgE и некоторых дру-
гих иммуноглобулинов, участвуют в аллергическом воспалении, активируя туч-
ные клетки (IL-9) и эозинофилы (IL-5). Синтезируемый ТЫ-лимфоцитами IFN-y
подавляет функцию и продукцию цитокинов ТЙ2-клетками, а продуцируемые
ТЬ2-лимфоцитами IL-4 и TL-5 угнетают синтез цитокинов ТЫ-лимфоцитами и
макрофагами. Таким образом, эти две популяции Th-лимфоцитов находятся в
реципрокных отношениях, каждая из них, поддерживая один оптимальный ари-
ант антиген-специфического ответа, угнетает другой.
В противовес ТЫ- и ТЬ2-лимфоцитам в крови обнаруживают несколько попу-
ляций Т-лимфоцигов, негативно регулирующих функцию врожденных и приоб-
ретенных клеточных факторов иммунной системы. Тг-клетки угнетают пролифе-
рацию СП4+ Т-лимфоцитов. Супрессия достигается как при прямом контактном
взаимодействии Тг-лимфоцитов с клетками, так и благодаря способности этих
клеток продуцировать иммуносупрессорные цитокины (TGF-p, IL-10). Тг-клетки,
наряду с лимфоцитами, угнетают функцию дендритных клеток и других клеток
вр< >жденного иммунитета. Среди Тг-клеток различают две субпопуляции. Первая
представлена натуральными клетками тимического происхождения (пТг), вто-
рая — адаптивными Т-клетка ми (аТг).
Tr-лимфоциты подавляют иммунный ответ на аутоантигены, опухолевые,
трансплантационные и некоторые инфекционные антигены. При их недостаточ-
ности в организме развиваются аутоиммунные заболевания (рассеянный склероз,
аутоиммунный полигландулярный синдром, диабет I типа и др.), аллергии и
лимфо-пролиферативная патология.
К эффекторным Т-клеткам крови относят Тс. Они вызывают лизис любых кле-
ток организма, несущих на поверхности антигенные детерминанты, к которым у
Тс-клеток имеется рецепторный комплекс TCR CD3+. Среди Тс-лимфоцитов пре-
обладают CD4 , CD8+ и офТ-лимфоциты. Количество их в крови у взрослых людей
может достигать 25% общего числа Т-лимфоцитов крови.
Около 4% CD4 CD8- Т-лимфоцитов крови имеют TCR-молекулы, построенные
с участием у- и 5-пептидных цепей. Эти клетки также обладают цитотоксической
активностью, но представление антигена клетками-мишенями в этом случае осу-
ществляется посредством CD1 белков. у5Т-клетки способны быстро синтезировать
цип окины, вызывающие воспалительную реакцию.
При антигенной активации цитотоксических Т-лимфоцитов вторые подтверж-
дающие активационные сигналы — сигналы, подаваемые Тс-лимфоцитам через
рецептор к IL-2. Источником IL-2 в этом случае служат участвующие в иммун-
ном ответе на этот же антиген ГЬ-лимфоциты (того же антиген-специфического
клона). Имении поэтому Тс-лимфоцитам для своей активации необходимо взаи-
модействие с Th-клеткам и. При получении активирующих и когтимулирующих
сигналов антиген-специфический клон Тс-лимфоцитов вступает в пролиферацию
и приобретает способность в качестве эффекторных клеток убивать клетки-
мишени, несущие антиген. Такой механизм иммунного цитолиза важен для проти-
вовирусного иммунитета, поскольку обеспечивает гибель любых клеток, несущих
на мембране вирусы или вирусные антигены в ассоциации с белками МНС I класса
или CD1 мембранными белками клеток.
30 ЛАБОРАТОРНАЯ ИММУНОЛОГИЯ
Цитотоксические лимфоциты активно участвуют в противоопухолевом и транс-
плантационном иммунитете, а также в защите от собственных клеток, несущих
белки теплового шока или гликолипидные микробные антигены. Тс-лимфоциты
осуществляют гибель клеток-мишеней путем выделения в их сторону цитоток-
сических белков (лимфотоксинов). К таким лимфотоксинам относят перфо-
рин (цитолизин) и некоторые сериновые протеазы. Перфорины напоминают
белки мембрано-атакующего комплекса комплемента: встраиваясь в мембрану
клетки-мишени, они образуют в ней поры, через которые в клетку-мишень из
Тс-лимфоцитов поступают гранзимы, активирующие каспазы. Последние раз-
рушают внутриклеточные мембраны, активируют эндонуклеазы и индуцируют
гибель клетки-мишени путем апоптоза.
Таким образом, для системы Т-лимфоцитов характерно исключительное раз-
нообразие образующих ее клеток, которые отличаются не только структурой
антигенраспознающих рецепторов, но и наборами ассоциированных с мембрана-
ми белков и рецепторов, спектром продуцируемых цитокинов и многими другими
признаками, а следовательно, разнятся функционально.
Показано, что набор экспрессируемых Th-лимфоцитами фенотипических мар-
керов может быстро меняться (при культивировании in vitro через 3-4 сут), клетки
могут нести фенотипические маркеры оппозитной популяции Th-клеток. Поэтому
при лабораторном анализе лимфоидной системы больных возрастает необхо-
димость проведения повторных анализов и осуществления иммунологического
мониторирования пациентов.
СИСТЕМА В-ЛИМФОЦИТОВ
Дифференцировка В-клеток происходит в костном мозге и предназначена для
формирования антигенсвязывающего рецептора В-лимфоцитов (BCR), BCR состо-
ит из двух тяжелых и двух легких полипептидных цепей, образующих молекулу
мембранного иммуноглобулина (mlg). На мембране клетки с молекулами inIg
связаны по две молекулы Iga- и IgP-полипептидных цепей, предназначенных для
проведения сигнала от BCR в ядро клетки после контакта с антигеном. BCR рас-
познает только одну антигенную детерминанту. Весь пул В-клеток обладает широ-
ким диапазоном специфичностей к аптигенам (до 109 антигенных детерминант).
Главный механизм формирования разнообразия специфичностей BCR — реаран-
жиривка (перестановка) вариабельных и константных участков генов тяжелых и
легких цепей иммуноглобулинов, постоянно происходящих у незрелых В-клеток
в костном мозге. Случайная реорганизация генов иммуноглобулинов приводит
к появлению рецепторов, специфичность которых направлена против не только
чужеродных антигенов, но и собственных молекул. Удаление таких аутореак-
тивных В-клеток происходит посредством апоптоза в костном мозге в процессе
негативной селекции, когда незрелые В-лимфоциты взаимодействуют с аутоанти-
генами организма.
Каждый этап дифференцировки клеток гемопоэтического ряда, в том числе
В-лимфоцитов, сопровождается экспрессией или исчезновением функциональных
молекул, которые могут служить маркерами стадии дифференцировки клеток
и их состояния. Важный маркер всех клеток-предшественников и перифериче-
ских В-лимфоцитов — CD 19 (на плазматических клетках он отсутствует) (см.
табл. 16-5). Он участвует в активации и пролиферации В-клеток. CD 19 экспресси-
руется на всех неопластических клетках при острых лейкозах В-клеточного про-
исхождения.
В-лимфоциты способны непосредственно взаимодействовать с антигенами.
Индуцируемая таким взаимодействием пролиферация и дифференцировка зрелых
В-клеток происходит в фолликулах и герминативных центрах вторичных лимфо-
идных органов, а также в лимфоидных тканях слизистых оболочек. После контак-
ЛАБОРАТОРНАЯ ИММУНОЛОГИЯ
31
та с антигеном в процессе размножения антиген-специфических В-лимфоцитов
повышается специфичность антигенсвязывающих рецепторов, а затем и переклю-
чение клеток с биосинтеза антигенраспознающих рецепторов в виде молекул IgM и
IgD на иммуноглобулиновые рецепторы иных классов (IgG. IgA, IgE). Антигенная
специфичность рецепторов возрастает. Переключение В-клеток на синтез рецеп-
торов в виде иммуноглобулинов иных классов осуществляется под влияни-
ем различных цитокинов, продуцируемых Th-лимфоцитами при кооперации с
В-клетками. Так. IFNy обеспечивает синтез В-клетками IgG2 или IgG3, TGFp — IgA,
IL-4 — IgE. В дальнейшем при дифференцировке лимфоцитов в плазматические
клетки сохраняется способность продуцировать те же иммуноглобулины, что и их
клетки- предшественники. В большинстве случаев плазматические клетки живут в
течение 2-3 нед, хотя описана популяция долгоживущих плазматических клеток,
сохраняющих жизнеспособность в течение года.
Среди зрелых В-клеток различают две субпопуляции. В^клетки маркированы
CD5T белками, а на В2-клетках CD5 отсутствует. В^клетки по своим характери-
стикам близки к клеткам врожденного иммунитета. Они локализуются в брюшной
и плевральной полостях и слизистых оболочках. В2-клетки составляют основной
пул циркулирующих в кровотоке и пребывающих в периферических лимфоидных
органах В-лимфоцитов.
Активированные антигеном В-лимфоциты способны дифференцироваться не
только в плазматические клетки, но и в В-клетки — носители иммунологической
памяти. Они могут сохраняться в организме в течение 8-10 лет.
С возрастом внутри пула периферических В-клеток происходит снижение числа
наивных В-лимфоцитов с пропорциональным увеличением В-клеток памяти
(CD27+). В связи с этим ответ антител на новый антиген у лиц пожилого и стар-
ческого возраста оказывается слабым и непродолжительным. По мере старения
в крови у людей возрастает спектр разнообразных аутоантител, представленных,
однако, в невысоких титрах.
Иммуноглобулины и Fc-рецепторы
Иммуноглобулины — белки, об падающие активностью антител, сходные по хими-
ческой структуре и по иммунохимической специфичности. Главными клетками-
продуцентами иммуноглобулинов считают плазматические клетки. Активность
антител выражена в способности иммуноглобулинов к высокоспецифичному
взаимодействию с антигеном с образованием мультимолекулярных комплексов
«антиген-антитело». Антитела обладают вторичными иммунобиологическими
свойствами и способны фиксироваться на клетках, взаимодействовать с белками
системы комплемента, усиливать активность макрофагов и цитотоксическое дей-
ствие NK-клеток, регулировать функцию лимфоцитов и др. Эндотелиальные клет-
ки, циркулирующие и резидентные макрофаги (в особенности печени и селезенки)
активно выводят из циркуляции, захватывают и разрушают комплексы «антиген-
антитело». Взаимодействуя с антигенами, токсинами и ферментами, антитела
блокируют их активные цен гры, нейтрализуют токсичность и угнетают энзимати-
ческую активность этих молекул. Микроорганизмы и паразиты с фиксированными
на них антителами утрачивают подвижность, если антитела взаимодействуют с их
жгутиками. Антитела против возбудителей препятствуют их прикреплению и вне-
дрению в различные клетки макроорганизма, в том числе в клетки пограничных
тканей, обеспечивая устойчивость к инфекционному заражению. Антительная
агрегация возбудителей на поверхности пограничных тканей препятствует перено-
су инфекта через слизистые покровы. F большинстве случаев антитела оказывают
на возбудителей микробостатическое действие. Иммобилизованные на антигене
антитела изменяют свою конформацию, что позволяет им фиксировать и активи-
32
ЛАБОРАТОРНАЯ ИММУНОЛОГИЯ
ровать белки системы комплемента, стимулировать активность фагоцитов и цито-
токсичность NK-клеток. Фиксированные на тканевых клетках антитела при взаи-
модействии с антигеном обеспечивают подачу клеткам активационных сигналов,
вовлекая соответствующие клетки в реакцию, направленную против чужеродного
агента. Фактически антитела в организме человека обеспечивают избирательное
разрушение и нейтрализацию различными факторами иммунитета чужеродных
антигенов — молекул и клеток.
Иммуноглобулины человека подразделяют на пять i рупп или классов, обозна-
чаемых IgG, JgM, IgA, IgD и IgE. Иммуноглобулины разных классов различаются
содержанием углеводов, молекулярной массой, первичной структурой, электрофо-
ретической подвижностью, продолжительностью жизни и скоростью обновления
в организме, способностью к трансплацентарному переносу, иммунобиологиче-
ской активностью (табл. 16-9).
Таблица 16-9. Некоторые свойства иммуноглобулинов человека
Иммуноглобулины IgG IgM IgA igD igE
Молекулярное строение:
Н-полипептидные цепи у1, у 2, уЗ. у4 ц а 5 £
L-полипептидные цепи кА кА кА кА кА
Молекулярная масса, kDa 150 900 160 180 190
Содержание в крови у взрослых людей, г/л 6,0-13 5 6,0-1,5 0,7-3,1 0,03-0,15 1,53-114 IU/мл
Т|2 — полупериод обновления, сут 23 5 6 2,8 2,5
Трансплацентарный перенос + - - - -
Антибактериальная активность + +++ + - -
Антивирусная активность + + +++ - -
Антипаразитарная активность iqg4 - - ++ +++
Иммуноглобулины классов G, D и Е представлены в крови м< аномерными моле-
кулами. IgM — молекула, состоящая из пя^и мономерных молекул, удерживаемых
вместе посредством специальной полипептидной J-цепочки. IgA могут быть в виде
двух молекулярных форм — мономерной и димерной. В последнем случае две
молекулы IgA удерживаются вместе также посредством J-полипептидной цепочки.
Несмотря на различия в свойствах, все мономерные иммуноглобулины и субье-
диницы в полимерных молекулах характеризуются единообразием организации.
Все они симметричны и состоят из двух тяжелых — «Н» (heavy — тяжелый) и
двух легких — «L» (light — легкий) полипептидных цепочек, удерживаемых друг
с другом посредством дисульфидных связей. Тяжелые полипептидные цепочки у
иммуноглобулинов разных классов обозначены соответствующими греческими
буквами: у-ценочки у IgG, ц-цепочки у IgM.
Отличающиеся строением вариабельные домены (VH- и VL-домены) у различ-
ных молекул иммуноглобулинов не только взаимодействуют с разными антигена-
ми, но и сами несут признаки аутоантигенной специфичности. Именно поэтому
против иммуноглобулинов могут накапливаться аутоантитела, реагирующие с
VH- и VL-доменами других антител как с новыми для организма антигенными
детерминантами. Под влиянием протеаз мономерные молекулы иммуноглобули-
нов расщепляются на три фрагмента. Два из них идентичны и образованы легкими
цепями и частью тяжелой цепи молекулы (Fab-фрагменты), а третий образован
ЛАБОРАТОРНАЯ ИММУНОЛОГИЯ
33
дисульфидными остатками тяжелых цепей иммуноглобулина (Fc-фрагмент).
Исследование активности фрагментов иммуноглобулинов, полученных при их
ферментативном расщеплении, позволило выявить роль отдельных фрагментов и
доменов молекулы в обеспечении различных проявлений иммунобиологической
активности антител. Гак, за способность антител взаимодействовать с антигеном
ответственен Fab-фрагмент молекулы. Строение и доступность для взаимодей-
ствий Fc-фрагмента молекулы определяет цитофильную активность антител, их
способность опсонизировать и фиксировать белки системы комплемента, активи-
ровать NK-клетки и В-лимфоциты.
Способность нативных иммуноглобулинов к вторичной иммунобиологи-
ческой активности возрастает при конформационных изменениях структуры
Fc-фрагмента, когда скрытые константные участки доменов тяжелой цепи оказы-
ваются на поверхности молекулы и доступны для взаимодействия. Такие конфор-
мационные изменения иммуноглобулинов в естественных условиях возникают в
ре зультате взаимодействия их Fab-фрагментов с соответствующими антигенами. В
комплексах с антигеном иммуноглобулины приобретают способность к побочной
иммунобиологической активности, стимулируя различные клетки организма и
взаимодействуя с разнообразными гуморальными факторами иммунитета.
Клетки, взаимодействующие с Fc-фрагментами иммуноглобулинов, имеют
соответствующие Fc-рецепторы (Fc-R). Описано три варианта клеточных рецеп-
торов к Fc-фрагментам IgG, которые представлены на разных клетках человека
и отличаются молекулярной массой и регуляторными свойствами. У моноцитов-
макрофагов обнаружен Fcy-RI-гликопротеин (CD64). Этот рецептор с высокой
аффинностью может взаимодействовать с субклассами IgG1 и IgG3 и менее активно
с IgG4, не взаимодействует с IgG2.
Fcy-RII (CD32) обнаруживают у макрофагов, нейтрофилов, эозинофилов,
В-лимфоцитов и тромбоцитов. Этот рецептор эффективно взаимодействует только
с конфирмационно измененными Fey-фрагментами преимушес гвенно IgGt и IgG3.
Сигналы через этот рецепторный белок усиливают фагоцитарную активность,
агрегацию и дегрануляцию тромбоцитов и подавляют пролиферацию и дифферен-
цировку В-лимфоцитов. Fc-RII нейтрофилов взаимодействуют не только с IgG, но
и с IgA.
Fcy^RIII (CD16) — гликопротеин, который определяют у NK-клеток, нейтро-
фильных лейкоцитов, макрофагов и Т-лимфоцитов. У этого рецептора низкая
аффинность, он взаимодействует преимущественно с IgGT и IgG3. Показано участие
Fcy-RIII в усилении фагоцитарной активности клеток и в реакциях антителозави-
симого цип олиза. Генетические нарушения экспрессии этого рецептора на клетках
описаны у больных с пароксизмальной ночной гемоглобинурией, что, как полага-
ют, приводит к сниженной активности соответствующих клеток и к повышенной
чувствительности больных к инфекции и к замедленному удалению из крови
иммунных комплексов.
У взрослых при вторичном иммунном ответе основные антитела — IgG, состав-
ляющие 75% сывороточных иммуноглобулинов Они наиболее медленно обнов-
ляемые: период полураспада их в крови составляет 23 сут. IgG человека подраз-
деляют на четыре субкласса (обозначают IgGr IgG2, lgG3 и IgG4), отличающиеся
первичной структурой Н-цепей в шарнирной области. Способность IgG разных
субкласеов фиксировать комплемент классическим путем убывает в следмющей
последовательности: IgG3 > IgG] > IgG,. IgG4 могут активировать комплемент аль-
тернативным путем, а не классическим.
В ответ на белковые антигены разного происхождения вырабатываются анти-
тела субкласса IgG, и в меньшей степени IgG3 и IgG4. Полисахаридные антигены
(например, капсульные полисахариды микроорганизмов) вызывают образование
34 ЛАБОРАТОРНАЯ ИММУНОЛОГИЯ
антител преимущественно субкласса IgG2. Вирусы вызывают образование антител
преимущественно субкласса IgG, и IgG3, а антигены паразитов — ^О4-антитела.
Врожденные дефекты биосинтеза иммуноглобулинов у людей могут распро-
страняться как на весь класс IgG, так и на отдельные субклассы. Дефект биосин-
теза IgG2 у детей выражается отсутствием или низким уровнем антител против
полисахаридов капсульных микроорганизмов, в результате чего для них харак-
терны хронические рецидивирующие инфекции дыхательных путей, возбудители
которых — микроорганизмы, образующие полисахаридные капсулы (в частности,
пневмококки и Haemophilus influenzae).
Содержание IgM в крови взрослых достигает 10% общего количества иммуно-
глобулинов. Антитела класса IgM накапливаются преимущественно при первичном
иммунном ответе, в начале продукции антител. Комплекс «IgM-антиген» наибо-
лее активно взаимодействует с Clq и эффективно активирует систему комплемента
классическим путем. IgM шрают важную роль в антибактериальном иммунитете.
Синтез IgM зависит от кооперации В-клеток с Th-лимфоцитами в меньшей сте-
пени, чем при продукции иммуноглобулинов других классов. Мономерные моле-
кулы IgM, будучи встроенными в мембраны В-лимфоцитов, служат, наряду с IgD,
антигенраспознающими рецепторами наивных В-клсток.
Содержание IgD в крови составляет 0,1-0.2% сывороточных иммуноглобули-
нов. Показано, что антитела против некоторых аутоантигенов (нуклеопротеины,
антигены щитовидной железы) — IgD, которые являются протективными анти-
телами при некоторых паразитарных инфекциях.
Содержание IgA в крови составляет 2,0 г/л (0,3 г/кг массы тела). В различ-
ных секретах и в смывах с неповрежденной кожи доминируют секреторные IgA
(slgA). Основную массу IgA, циркулирующих в крови, продуцируют плазмати-
ческие клетки в костном мозге, а с секретами выделяются синтезируемые плаз-
моцитами в лимфоидных органах подслизистых желудочно-кишечного тракта
и дыхательных путей. Все внутриклеточные, мембранные и большинство цир-
кулирующих в крови молекул IgA представлены в мономерной форме, а секре-
тируемые слизистыми оболочками IgA представлены димерными молекулами
(150-600 kDa). Секреторный фрагмент в slgA не только обеспечивает трансэпите-
лиальный перенос полимерных IgA из подслизистых в полости, но в дальнейшем
защищает их от действия протеаз хозяина и микробных протеаз на поверхности
слизистых оболочек.
У IgA описаны два субкласса — IgA, и IgAr Первый составляют около 80%
циркулирующих IgA, они преобладают среди slgA, секретируемых эпителиальны-
ми клетками слизистых оболочек дыхательных путей легких и верхних отделов
желудочно-кишечного тракта. Большую их концентрацию наблюдают в секрете
слизистых оболочек генитального тракта женщин и дистального отдела ЖКТ.
Содержание IgE в крови у здоровых индивидуумов составляет около 0,004%
общего количества сывороточных иммуноглобулинов. Базофильные лейкоциты
и тучные клетки обладают высокоаффинными рецепторами к Fc-фрагменту IgE,
поэтому даже при низком содержании IgE в крови эти клетки несут на поверхности
IgE, удерживаемый Fce-R. При контакте фиксированных на клетках IgE с соответ-
ствующим антигеном конформационно изменяется структура IgE, агрегируются
рецепторы к ним, что обеспечивает раздражение и дегрануляцию несущих IgE базо-
филов и тучных клеп ок. При этом освобождаются гистамины и другие компоненты
содержимого гранул — медиаторы аллергических реакций. IgE участвуют в антипа-
разитарном иммунитете, накапливаются при гельмиптозах и других паразитозах.
Поликлональные иммунные сыворотки и различные препараты нормальных
иммуноглобулинов давно и успешно используют в клинической практике. Так,
гомо- и гетерологичные иммунные сыворотки (антитела) применяют для лечения
и профилактики интоксикаций и инфекций, вызываемых различными микробами
ЛАБОРАТОРНАЯ ИММУНОЛОГИЯ
35
и вирусами. Внутривенно вводимые препараты иммуноглобулинов ока за пись неза-
менимыми при терапии врожденных или приобретенных дефектов их синтеза, при
острых и хронических бактериальных и вирусных инфекциях, развивающихся на
фоне нарушений функций иммунной и кроветворной систем. Иммуноглобулины,
назначаемые внутривенно в особенно высоких дозах, проявляют иммунорегуля-
торную активность. Они оказались эффективным средством лечения некоторых
аутоиммунных заболеваний (идиопатическая тромбоцитопеническая пурпура,
аутоиммунная гемолитическая анемия, полиомиозит, апластическая анемия),
гиперактивации макрофагальной системы (эндотоксический шок, токсическое
воспаление).
Таким образом, приобретенные факторы иммунитета накапливаются и активи-
руются при поступлении в организм соответствующих антигенов. Приобретенные
факторы иммунной защиты отличаются исключительно высокой специфично-
стью и обеспечивают реаю <ии иммунной системы строго на определенные анти-
генные детерминанты. Приобретенный иммунитет может сохраняться в течение
8-10 лет, для ею пожизненного поддержания необходима персистенция антигена
в организме или реиммунизация. Формирование приобретенного иммунитета осу-
ществляется кооперативно при взаимодействии вспомогательных клеток (кпетки
врожденного иммунитета) с Т- и В-лимфоцитами. Поэтому недостаточность
приобретенного иммунного ответа может быть связана не только с дефицитами
антиген-специфических клонов Т- и/или В-лимфоцитов, но и с нарушениями
кооперативного взаимодействия между клетками иммунной системы. Иммунная
система саморегулируется, для достижения ее нормального функционирования
крайне важны соотношение и активность не только эффекторных, но и регулятор-
ных антиген-специфических клеток.
ПАТОЛОГИЯ ИММУННОЙ СИСТЕМЫ
Первичные иммунодефициты
Первичными иммунодефицитами называют дефекты иммунной системы, воз-
никающие вследствие генетических поломок. В Международной классификации
болезней выделено в качестве нозологических единиц 36 первичных иммуноде-
фицитов. Вместе с тем, по данным ВОЗ, существует более 70 первичных иммуно-
дефицитных состояний. Для многих из них определен молекулярно-генетический
дефект. Членами Консультативного совета и Европейского общества по иммуноде-
фицитным заболеваниям ESID были разработаны и одобрены к распространению
рекомендации по первичным иммунодефицитным состояниям.
Настораживающие признаки:
• частые заболевания отитом (не менее 6-8 раз в течение 1 года):
• несколько подтвержденных серьезных синуситов (не менее 4-6 раз в течение
1 года);
• более двух случаев подтвержденных пневмоний;
• повторные глубокие абсцессы кожи или внутренних органов;
• потребность в длительной терапии антибактериальными препаратами для
купирования инфекции (2 мес или дольше);
• потребность во внутривенном вливании антибактериальных препаратов для
купирования инфекции;
• не менее двух инфекций (таких, как менингит, остеомиелйг, абсцессы
подкожно-жировой клетчатки, сепсис);
• отставание ребенка грудного возраста в росте и развитии;
• персистирующая молочница или грибковое поражение кожи у детей в воз-
расте старше 1 года:
36
ЛАБОРАТОРНАЯ ИММУНОЛОГИЯ
• наличие у родственников первичных иммунодефицитов, ранних смертей от
тяжелых инфекций или одного из вышеперечисленных симптомов.
Первичные иммунодефициты — обычно очень серьезные заболевания, но
далеко не всегда фатальны, в большинстве случаев их лечение возможно. Диагноз
первичного иммунодефицита можно поставить только при специальном обследо-
вании с использованием иммунологических, генетических и молекулярных мето-
дов анализа. Это очень важно сделать как можно раньше, чтобы предотвратить
тяжелые последствия заболевания.
Основные синдромы иммунодефицитов:
• инфекционный;
• аллергический;
• аутоиммунный;
• пролиферативный.
Инфекционный синдром характеризуется тяжелыми рецидивирующими муль-
тифокальными инфекциями, не отвечающими адекватно на проводимое лечение.
Это мот быть инфекции, вызванные бактериями, вирусами, грибами и парази-
тами. Входными воротами инфекции служат кожа, слизистые оболочки верхних
дыхательных путей, желудочно-кишечного и урогенитального трактов.
Аллергический синдром характеризуется кожными поражениями по типу экс-
судативного диатеза, атопического дерматита, нейродермита, повторными брон-
хитами с астматическим компонентом. При аутоиммунном синдроме характерны
симптомы ревматоидного артрита, васкулита, склеродермии и других системных
заболеваний соединительной ткани, аутоиммунной тромбоцитопении, аутоим-
мунного тиреоидита, сахарного диабета.
Пролиферативный синдром встречается в виде лимфо- и миелопролифератив-
ных заболеваний. Он связан с опухолевой трансформацией клеток иммунной
системы (лимфомы, лимфосаркомы, лимфогранулематоз, острый и хронический
лимфолейкоз).
НАРУШЕНИЯ ВРОЖДЕННОГО КЛЕТОЧНОГО ИММУНИТЕТА
Недостаточность фагоцитов составляет 10-15% всех первичных иммунодефи-
цитов. Она обусловлена нарушением пролиферации, дифференцировки, хемотак-
сиса нейтрофилов и макрофагов и собственно нарушением процесса фагоцитоза
(табл. 16-10). Клинически недостаточность фагоцитов отмечают со 2-го года
жизни, но может впервые диагностироваться существенно позже. Выраженная
недостаточность полиморфноядерных лейкоцитов приводит к развитию генера-
лизованной бактериальной инфекции. Больные с дефектом системы фагоцитов
нуждаются в длительном антибактериальном, противомикотическом и симптома-
тическом лечении.
Первичные дефекты Т-лимфоцитов
Изолированный дефект Т-системы иммуни тета составляет 5-10% всех первич-
ных иммунодефицитов. Характерны вирусные и микотические инфекции, которые
отмечают с первых дней жизни. Лечебные мероприятия при Т-клеточном иммуно-
дефиците определяются ведущим синдромом. Трансплантацией фетального тимуса
и введением тимических факторов корригируют Т-клеточные нарушения, связан-
ные с гипотрофией тимуса. В обобщенном виде патогенез, клиническая картина,
иммуно- и дифференциальная диагностика представлены в табл. 16-11 и 16-12.
Таблица 16-10. Некоторые врожденные нарушения клеток фагоцитарной системы
Наименование иммунодефицитного состояния Основные патогенетические факторы Клиническая картина Результаты иммунологического исследо- вания
Хроническая грануле- матозная болезнь. Тип наследования аутосомно- рецессивный {40%) И Х-сцепленный (60%) Нарушения метаболизма кислорода в гра- нулоцитах и моноцитах, а также образова- ния продуктов оксидативного взрыва и, как следствие, неспособность осуществлять киллинг захваченных микроорганизмов Рецидивирующие инфекционные заболевания, обуслов- ленные грибами и микроорганизмами, продуцирующими каталазу. При заражении обычно возникает чрезмерная воспалительная реакция. Характерны гнойные лимфаде- ниты, абсцессы печени, остеомиелит, поражение кожи и подкожно-жировой клетчатки, гранулемы Анемия, лейкоцитоз, высокая СОЭ, гипо- гаммаглобулинемия, нарушение функцио- нальной активности фагоцитов (НОТ-тест снижен)
Синдром Чедиака-Хигаси. Тип наследования аутосомно-рецессивный Нарушение хемотаксиса нейтрофилов, внутриклеточного транспорта протеинов (нарушение слияния фагосом с лизосома- ми), приводящее к неспособности клеток лизировать бактерии и длительной перси- стенции возбудителя внутри клетки Инфекции, вызываемые возбудителями Staphylococcus aureus, Escherichia, Pseudomonas, а также грибами рода Aspergillus и Candida. Часто частичный или полный альби- низм, светобоязнь, гипергидроз, тромбоцитопения, приво- дящая к кровотечениям Лейкопения или панцитопения нарушение хемотаксиса, фагоцитоза и внутрикле- точного киллинга бактерий, снижение уровня лизосомально-катионных белков нейтрофилов. Количество и функция Т- и В-лимфоцитов, уровень С1-компонента комплемента в норме
Гипер-1дЕ-синдром. Тип наследования неизвестен Нарушение хемотаксиса нейтрофилов из-за повышенной продукции IgE и осво- бождения гистамина при снижении продук- ции Th-1-зависимых цитокинов Повторные холодные абсцессы кожи, мягких тканей, легких, почек, печени, экзема, поражение костей (остео- миелит) на фоне нарушения обмена кальция и высокого уровня IgE в крови Высокое содержание в крови IgE и IgD, эозинофилия, нейтрофилия, повышенная продукция IL-4, нарушение хемотаксиса гранулоцитов
Синдром «ленивых» лейкоцитов. Тип насле- дования аутосомно- рецессивный Дефект рецепторов адгезии фагоцитов CD11 b, CD11с и CD11 а вследствие наруше- ния биосинтеза CD18; отсутствие у нейтро- филов рецептора для Е-селектина (CD15) Поражения пародонта, рецидивирующие пиогенные инфекции кожи, пазух носа, дыхательного тракта, некро- тические инфекции мягких тканей, септицемия Лейкоцитоз, нарушение хемотаксиса ней- трофилов и макрофагов
Таблица 16-11. Некоторые первичные иммунодефициты Т-лимфоцитов
Иммунодефицитное состояние Основные патогенетические факторы Клиническая картина
Врожденная аплазия тимуса (синдром Ди Джорджи, имму- нодефицит с гипопаратиреои- дизмом). Тип наследования неизвестен Нарушение эмбрионального развития (неправильное формирование органов, происходящих из III и IV фарингиальных карманов) до 12 недели гестации. Тимус редуцирован или полностью отсутству- ет, аномальное его расположение Хроническая инфекция вирусной, бактериальной, микотической и паразитарной этиологии; гипопара- тиреоз (гипокальциемия); пороки строения лица (низкопосаженные уши, рыбообразный рот, гиперте- лоризм, выемка ушного бугорка, микрогнотия); пороки сердца, дуги аорты
Окончание табл. 16-11
Иммунодефицитное состояние Основные патогенетические факторы Клиническая каршна
Хронический кандидоз кожи и слизистых оболочек. Тил наследования неизвестен Селективный дефицит ответа Т-клеток на антигены Candida Поражение грибами рода Candida слизистых оболочек рта, гениталий, кожи лица, волосистой части головы, туловища, конечностей, ногтей, ногтевых валиков. Молочница становится похожей на плоскую или эрозивную лейкоплакию. Язык утолщается, становится скротальным. формируется макрохеилит. В углах рта появляются трещины, покрытые налетами с инфильтрацией. I вариант — заболевание начинается с хронической кандидозной инфекции. II вариант — заболевание начинается с идиопатической эндокринопатии (гипопаратиреоз, гипокальциемия, тетания, Аддисонова болезнь, диабет сахарный, перницитозная анемия), а инфекция присоединяется позже
Таблица 16-12, Дифференциальная диагностика первичных иммунодефицитов Т-лимфоцитов
Иммун' дефицитное состояние Результаты иммунологического исследования Дифференциальная диагностика
заболевания отличительные признаки
Врожденная аплазия тимуса (синдром Ди Джорджи, имму- нодефицит с гипопаратиреои- дизмом) Количество лимфоцитов значительно снижено, Т-лимфоцитов резко снижено, или Т-клетки отсутствуют, реакция бласттранс- формации лимфоцитов на фитогемагглютинин снижена, реакция на аллогенные клетки снижена, количество В-лимфоцитов и обра- зование антител в норме или снижено, кальций в сыворотке крови резко снижен, фосфор в сыворотке крови повышен, активность естественных киллеров в норме Тяжелые врожденные пороки сердца Нет Т-клеточного иммунодефицита, гипокальциемия транзиторная
Тяжелая комбинированная иммунологическая недостаточ- ность При одинаковых изменениях Т-клеточного иммуни- тета не характерны гипокальциемия, врожденные пороки лица и сердца
Фетальный алкогольный синдром Анамнестические данные
Хронический кандидоз кожи и слизистых оболочек Количество Т-лимфоцитов в норме, реакция бласттрансформации лимфоцитов на фитогемагглютинин в норме, реакция на аллоген- ные лимфоциты и антигены в норме (кроме антигенов Candida'). гиперчувствительность замедленного типа на Candida снижена или отсутствует, образование МИФ чд антигены Candida снижено, количество Тс-лимфоцитов снижено, В-лимфоцитов — в норме, иногда IgA снижены или отсутствуют, нейтрофильный хемотаксис снижен, дефект макрофагов, концентрация кальция в сыворотке крови резко снижена, фосфора повышена, паратиреоидных гор- монов снижена Синдром Ди Джорджи Развивается е раннем возрасте, в отличие от хрониче- ского кандидоза кожи и слизистых оболочек, имею- щего постепенно прогрессирующий характер течения
Другие микотические пораже- ния кожи (хроническая трихо- фития, микоз, обусловленный Trichophyton rubrum) Определяются другие возбудители
Системная красная волчанка Определение специфических показателей (LE-клетки, антинуклеарный фактор, антитела к нативной ДНК и др.)
Энтеропатический акродер- матит Сопровождается блефароспазмом и фотофобией, нарушениями стула (стеаторея). В крови — дефицит цинка. Быстрое исчезновение симптоматики при получении препаратов цинка
Синдром Ядассона- Левандовского {многоформ- ный кератоз) Наследственный характер заболевания, семейный анамнез
ЛАБОРАТОРНАЯ ИММУНОЛОГИЯ 39
Нарушения воожденного гуморального иммунитета
Недостаточность системы комплемента выражена в дефектах эффекторных
контролирующих белковых систем, характерно нарушение опсонизации фагоци-
тоза и разрушение микроорганизмов. Клиническая картина дефектов комплемен-
та — иммунокомплексные заболевания, повышенная чувствительность организма
к гнойным инфекциям, а также особая предрасположенность к инфекциям, вызы-
ваемым двумя видами Neisseria (N. gonorrhoeae и N. meningitidis'). Наиболее тяжелые
последствия при нарушении функции комплемента (связано с недостаточностью
ингибитора С1 компонента комплемента), развивается наследственный ангионев-
ротический отек (табл. 16-13).
Таблица 16-13. Некоторые наследственные дефекты белков системы комплемента
Иммунодефицитное состояние Основные пато- генетические факторы Клиническая картина Результаты иммуно- логического иссле- дования
Недостаточность компонента комплемента Clq, г, s, компо- нентов комплемента С2 и С4. Тип наследования аутосомно- рецессивный Нарушение способности элиминировать иммунные ком- плексы Поражение кожи как при системной красной волчанке, васкулиты, дерматомиозит, полимиозиг, гломерулонефрит, рецидивирующие гнойные инфекции Количество компле- мента по 50% гемо- лизу нормальное или снижено, количество отдельных компо- нентов комплемента снижено или отсут- ствует, количество С1 ингибитора в норме
Недостаточность СЗ компонен- та комплемента. Тип наследо- вания аутосомно-рецессивный Нарушение аль- тернативного и классического пути активации комплемента, опсонизации и фагоцитоза Рецидивирующие гнойные инфекции
Недостаточность С5-С9 компонентов комплемента. Тип наследования аутосомно- рецессивный Генетический дефект мембрано- атакующего комплекса компонентов комплемента Рецидивирующая инфекция, вызываемая патогенами семей- ства Neisseria
Недостаточность компонента комплемента фактора Н (тип наследования аутосомно- рецессивный) фактора Д (тип наследования аутосомно- доминантный и аутосомно- рецессивный), недостаточность пропердина (Х-сцепленный тип наследования) Нарушение аль- тернативного пути активации комплемента Рецидивирующие гнойные инфекции, менингококковая и гонококковая инфекции, сеп- тицемия
Недостаточность С1 ингибитора Активация С2 и С4 компонентов комплемента, приводящая к образованию кининов и повышению сосудистой про- ницаемости Ангионееротический отек — ограниченный по площади, плотный, без ассоциации с крапивницей, особенно опасен при локализации на слизистой оболочке верхних дыхательных путей I вариант — количе- ство С1 ингибитора снижено, его функци- ональная активность в норме; II вари- ант — количество С1 ингибитора в норме, его функциональная активность снижена
Нарушения адаптивного гуморального иммунитета
Недостаточность гуморального иммунитета составляет около 60% всех пер-
вичных иммунодефицитов и характеризуется нарушением продукции антител.
Клинические проявления В-клеточных иммунодефицитов отмечают со второй
40
ЛАБОРАТОРНАЯ ИММУНОЛОГИЯ
половины первого года жизни, когда утрачиваются трансплацентарные IgG мате-
ри. Больные с обшим дефектом В-клеточной функции подвержены рецидивирую-
щим пиогенным инфекциям (табл. 16-14 и 16-15) и нуждаются в пожизненной
заместительной терапии антителосодержащими препаратами иммуноглобулинов
для внутривенного введения, которую проводят как в дозе насыщения, так и в
режиме поддерживающей иммунотерапии.
Комбинированные (В- и Т-клеточные) иммунодефицитные состояния
Комбинированная недостаточность гуморального и клеточного иммунитета
составляет 20-25% всех первичных иммунодефицитов. Дефекты функциониро-
вания Т-клеток могут быть причиной развития группы заболеваний, получившей
общее название «Тяжелые комбинированные иммунодефициты». Многогранное
участие Т-лимфоцитов в специфическом реагировании делает их ведущим компо-
нентом всей иммунной системы. Именно поэтому поражение Т-клеток приводит
к особо тяжелым формам иммунодефицита, при котором иммунная система не
способна сформировать адекватную специфическую защиту ко многим инфекци-
онным агентам. Больные с комбинированным иммунодефицитом обычно погиба-
ют в первые два года жизни. В табл. 16-16 и 16-17 систематизированы данные по
патогенезу, клинике, диагностике и дифференциальной диагностике комбиниро-
ванных иммунодефицитов.
Таблица 16-14. Первичные нарушения созревания и функций В-лимфоцит< в
Иммунодефицитное состояние Патогенетические факторы Клинические проявления
Сцепленная с Х-хромосомой а(гипо) гаммаглобулинемия (болезнь Брутона). Тип наследования Х-сцепленный Блокада дифференциоовки npeB-лимфоцитов из-за мутации гена, кодирующего тирозинкиназу Рецидивирующие бактериальные инфекции (отиты, бронхиты, пневмонии, менингиты, хронические конъюнктивиты, дерматиты), поражение десен, синдром мальабсорбции развитие ГЗТ. До возраста 9--12 мес болезнь не проявляется
Общий вариабельный иммунодефицит. Тип наследования аутосомно- рецессивный, аутосомно- доминантныи Блокада дифференцировки В-лимфоцитов в плазматиче- ские клетки из-за генетиче- ских дефектов Рецидивирующие инфекции носовых пазух, легких, бактериальные конъюнктивиты, синдром мальабсорбции, лямблиоз, холелитиаз, аутоим- мунные заболевания
Иммунодефицит с гипериммуноглобули- немией М. Тип наследо- вания Х-сцепленный и аутосомно-рецессивный Дефицит гена, коди- рующего молекулу CD40L на Т-лимфоцитах Вызываемые капсульными микроорганизмами рецидивирующие бактериальные инфекции. Характерны гиперплазия небных миндалин и периферических лимфатических узлов, гепато- спленомегалия, отставание в физическом раз- витии, артриты, агранулоцитоз
Селективный дефицит IgA. Тип наследования разный Блокада дифференцировки зрелых В-лимфоцитов в IgA- продуцирующие плазматиче- ские клетки из-за снижения костимуляции со стороны Т-лимфоцитов и клеточного микроокружения, а также дефицита TG-F[3 и IL 15, спо- собствующих выработке IgA Рецидивирующие инфекции слизистых обо- лочек, хронические отиты, бронхо-пневмонии, заболевания желудочно-кишечного тракта (энтериты, язвенный копит), аутоиммунные заболевания (ревматоидный артрит, системная красная волчанка, тиреоидит, пернициозная анемия, целиакия), онкологические заболевания (тимома, ретикуло-клеточная саркома, плоско- клеточный рак пищевода и легких)
Селективный дефицит субклассов IgG. Тип наследования неизвестен Депеция генов, отвечающих за константную часть тяже- лых цепей lg Рецидивирующие инфекции респираторного тракта, повторные пиогенные инфекции носо- вых пазух и легких, аутоиммунные заболевания
Транзиторная rwnoi ам- маглобулинемия у детей раннего возраста. Тип наследования неизвестен Задержка дифференцировки В-лимфоцитсп в плазматиче- ские клетки при нарушении Рецидивирующие бактериальные инфекции
Таблица 18-15. Дифференциальная диагностика врожденных нарушений адаптивного гуморального иммунитета
Иммунодефицитное сое, янье Результаты иммуноло) ического исследования Дифференциальная диагностика
заболевание отличительные признаки
Сцепленная с Х-хрБиосомой а(гипо) гаммаглобулинемия (болезнь Брутона) Снижение общего уровня иммуноглобулинов в сыворотке крови (до 2 г/л) и всех классов Ig, снижен первичный и вторичный ответ на антигенную стимуляцию и уровень нормальных анти- тел, В-лимфоциты отсутствуют Пролонгированная физиологическая гипо- гаммаглобулинемия При исследьвании в динамике уровень иммуноглобулинов повышается
Энтеропатия с потерей белка При интестинальной биопсии в кишечнике и других лим- фоидных тканях определяют нормальное количество В-лимфоцигов и внутриклеточного иммуноглобулина
Ювенильный ревматоид- ный артрит Уровень иммуноглобулинов повышен
Общий вариабельный иммунодефицит Общий уровень иммуноглобулинов ниже 3 г/л, уровень IgG ниже 2,5 г/л, IgA и IgM снижен до 0 г/л или определяют е следовых количествах, нормальных антител снижен, изогемагглютиьины отсутствуют или резко снижены, снижена способность выраба- тывать антитела на специфическую иммунизацию, количество В-лимфоцитов снижено или в норме, гиперчувстеительность замедленного типа снижена или отсутствует, реакция бласт- трансформации лимфоцитов на фитогемагглютинин снижена, количество Т-лимфоцитов снижено, снижена выработка in vitro IL-2, IL-4, IL-5, соотношение CD4/CD8 снижено, активность есте- ственных киллеров в норме Сцепленная с Х-хромосомой а(гипо) гаммаглобулинемия В интестинальном тракте и в периферических лимфоузлах отсутствуют плазматичские клетки, содержащие иммуно- глобулины, количество В-лимфоцитов резко снижено
Энтеропатия с потерей белка Отсутствие дефицита альбуминов, количество В-лимфоцитов нормальное. Диагностика затруднена если энтеропатия сопровождается снижением интестинальных лимфоидных клеток (обычно они в нормальном количе- стве)
Аутоиммунные заболе- вания Уровень иммуноглобулинов в норме или повышен
Хроническое неспецифи- ческое заболевание легких Проводить исследования на кистозный фиброз, хрониче- скую аллергию, дефицит al-антитрипсина
ВИЧ-инфекция Провести лабораторную диагностику Вь4Ч-инфекции
Иммунодефицит с гипериммуноглобули- немией М Резко повышен уровень IgM до 10,0 г/л, резко снижен уровень IgG и IgA, есть ответ на специфическую иммунизацию, титры изогемагглютининов снижены, Т-клеточный иммунитет в нозме - -
ЛАБОРАТОРНАЯ ИММУНОЛОГИЯ
ГЛАВАте
Окончание табл. 16-15 й
Иммунодефицитное состояние Результаты иммунологического исследования Дифференциальная диагностика
заболевание отличительные признаки
Селективный дефи- цит IgA Снижено количество IgA (<0,05 г/л), уровень IgG. IgM. IgD в норме или снижен, иногда снижен уровень !gG2, есть ответ на специфическую иммунизацию, в секрете уровень IgA снижен или равен 0, IgM секреторный повышен, образуются антитела против IgA, количество Т и В-лимфоцитов в норме (иногда количество Т-лимфоцитов снижено), гиперчувствительность замедленного типа, реакция бласпрансформации лимфоцитов на фитогемагглютинин и аллогенные лимфоциты в норме Атаксия-телеангиэктазия Нарушен клеточный иммунитет
Хронический кандидоз кожи и слизистых обо- лочек Преобладает специфический дефект клеточного иммуни- тета (на антиген Candida нет ответа Т-лимфоцитов и пода- влена выработка МИФ, выработка антител против Candida повышена)
Клеточный иммунодефи- цит с аномальным синте- зом иммуноглобулинов (синдром Незелофа) Дефект Т-клеточного иммунитета, нет выработки антител на специфическую иммунизацию
Селективный дефицит igG, Нет способности отвечать на полисахаридный антиген при нормальном ответе на белковые антигены
Лекарственный IgA- дефицит (прием противо- судорожных препаратов) Анамнестические данные о приеме лекарств
Селективный дефи- цит субклассов IgG Общий уровень иммуноглобулинов в норме или повышен, общий уровень IgG в норме или снижен, снижен уровень отдельных субклассов IgG (lgG2, lgG3, IgGJ, ответ на иммуниза- цию — от нормы до неспособности отвечать на полисахаридный антиген (IgG,), Т-клеточный иммунитет в норме Атаксия-телеангиэктазия Нарушен клеточный иммунитет
Селективный дефицит IgA Уровень IgA резко снижен, иногда определяют анти-lgA- антитела
Транзиторная гипо- гаммаглобулинемия у детей раннего воз- раста Снижен уровень IgA и IgM существенно снижен уровень IgG, низкий титр изогемагглютининов, снижена способность отве- чать на иммунизацию Сцепленная с Х-хромосомой а(гипо) гаммаглобулинемия Количество В-лимфоцитов резко снижено, в интестиналь- ном тракте и периферических лимфоузлах отсутствуют плазматические клетки, содержащие иммуноглобулины
ЛАБОРАТОРНАЯ ИММУНОЛОГИЯ
Таблица 1Б-16. Основные комбинированные (В- и Т-клеточные) иммунодефицитные состояния
Иммуш дефицитное состояние Основные патогенетические факторы Клинические проявления
Тяжелые комбинированные иммуно- дефицитные нарушения. Тип насле- дования аутосомно-рецессивный или Х-сцепленный Дефект на уровне стволовых клеток. Возможен врожденный дефект тимуса, отсутствие цитоки- нов или факторов дифференцировки Тяжелые хронические инфекции вирусной, бактериальной, микотической и прото- зойной природы (хроническая диарея, пневмонии, персистирующий кандидоз, отиты, сепсис). Гибель на первом году жизни
Дефекты проведения Т-клеточных мембранных сигналов и синтеза цито- кинов Врожденный дефект на уровне Т-кпеточных рецепторов, дефицит продукции IL-2 Рецидивирующие инфекции различной степени тяжести
Клеточный иммунитете нарушением синтеза иммуноглобулинов (синдром Незелофа, алимфоцитоз). Тип насле- дования аутосомно-рецессивный Нарушение IL-2 рецепторов с вторичным нару- шением развития тимуса Предрасположенность к рецидивирующим микотическим, паразитарным, бактериаль- ным, вирусным инфекциям. Признаки лимфаденопатии и гепатоспленомегалии
Иммунодефицит с атаксией- телеаы иэктазией (синдром Луи-Бар). Тип наследования аутосомно- рецессивный Дефект в хромосоме в локусе llq2Z^3 (atm), дефекты репарации ДНК Телеангиэктазии, мозжечковые нарушения и нистагм, неврологическая симпто- матика поражения экстрапирамидных путей задних столбов спинного мозга. Рецидивирующие инфекции дыхательного тракта. В пубертатном периоде развитие эндокринопатии (сахарный диабет). Позже — злокачественные новообразования (карцинома желудка, печени, яичников, лейкемия, неходжкинская лимфома)
Иммунодефицит с тромбоцитопенией, экземой, рецидивирующей инфекцией (синдром Вискотта-Олдрича). Тип наследования Х-сцепленный Нарушения в гене WAS в локусе Хр,1г2на корот- ком плече. Х-хромосомы, приводящие к ограни- чению экспрессии CD23 на поверхности клеток и аберрантному протеолизу CD23; дефицит гли- копротеина сиалофорина (CD43), приводящий к быстрому старению клеток Обнаруживают в основном у мальчиков. Склонность к рецидивирующим бактериаль- ным инфекциям (инфекции ЛОР-органов. дыхательного тракта, мочеполовой систе- мы, кожи, ЖКТ), экзема, геморрагический синдром, связанный с тромбоцитопенией
Иммунодефицит с тимомой (синдром Гуда). Тип наследования неизвестен Обнаруживают в основном у взрослых. Иммунодефицитное состояние может предше ствовать возникновению тимомы или возникает после ее обнаружения. Характерны рецидивирующие инфекции (носовых пазух, легких, мочеполовой системы, хрониче- ская диарея, дерматит, стоматит, септицемия), тимома, апластическая анемия, тром- боцитопения, сахарный диабет, амилоидоз, хронический гепатит, миастения
Иммунодефиците карликовостью из-за коротких конечностей. Тип наследования неизвестен - Предрасположенность к бактериальной, вирусной, микотической и протозойной инфекции. Прогрессирующая вакцинальная инфекция, повышенная чувствительность к инфекциям носовых пазух и легких, (батальной ветряной оспе. Короткие конечно- сти, гипоплазия хрящей и волос (тонкие светлые волосы)
Окончание табл. 16-16 *
Иммунодефицитное состояние Основные патогенетические факторы Клинические проявления
Иммунодефицит вследствие дефи- цита ферментов. Тип наследования аутосомно-рецессивный Дефицит аденозиндезаминазы и пуриннуклео- зидфосфорилазы приводит к нарушению ката- болизма пуринов с накоплением промежуточ- ных продуктов обмена. Повышенная активность этих продуктов обмена ингибирует функцию лимфоцитов Коррелируют с уровнем нарушения активности ферментов. В 85% случаев клини- ческая картина сходна с симптомами при тяжелых комбинированных иммунодефи- цитных нарушениях. Сочетается с патологией ребер, поперечных отростков и тел позвонков, изменениями в костях таза. При дефиците пуриннуклеозидфосфорила- зы — фатальные вирусные и вакцинальные инфекции
Дефицит 5'-нуклеотидазы как дифференци- ровочного маркера В-лимфоцитов. Как след- ствие — нарушение созревания В-лимфоцитов в периферической крови или снижение их количества Дефицит 5'-нуклеотидазы описан в связи с приобретенной гипогаммаглобулинемией. Х-сцепленной гипогаммаглобулинемией, синдромом Вискотта-Олдрича, селективным дефицитом IgA. Клинические проявления соответствуют данным заболеваниям
Дефицит транскобаламина II и витамин В-несущего протеина приводит к нарушению транспорта витамина В в клетки, в результате чего развивается гипогаммаглобулинемия При дефиците транскобаламина II и витамин В-несущего протеина отмечают сим- птомы макроцитарной анемии, гранулоцитопении, тромбоцитопении, лимфопении; синдром мальабсорбции
Дефицит биотин-зависимой карбоксилазы Кандидоз кожи и слизистых оболочек, атаксия, эпизоды сепсиса
Синдром «голых» лимфоцитов. Тип наследования аутосомно-рецессивный Отсутствие или значительное снижение экспрес- сии антигенов I и/или II класса HLA. Считают, что дефект связан с нарушением трансакти- вации регуляторного гена II класса HLA или с дефектом в ДНК-связывающем протеине, регу- лирующим транскрипцию генов II класса Н1А Оппортунистические инфекции, хроническая диарея, рецидивирующие вирусные инфекции, вирусная инфекция ЦНС, апластическая анемия
ЛАБОРАТОРНАЯ ИММУНОЛОГИЯ
Таблица 18-17. Дифференциальная диагностика комбинированных (В- и Т-клеточные) иммунодефицитных состояний
Иммунодефицитные состояния Результаты иммунологического исследования Дифференциальная диагностика
заболевание отличительный признак
Тяжелые комбиниро- ванные иммунодефи- цитные нарушения Лимфопения, количество Т-лимфоцитов резко снижено, реакция бласттранс- формации лимфоцитов на фитогемагглютинин отсутствует, число субпопуляций Т-лимфоцитов нарушено, нет ответа на Т-зависимые антигены, количество В-лимфоцигов снижено или отсутствует, активность NK в норме или снижена. При биопсии лимфатических узлов нет корково-медуллярной дифференцирое- ки, фолликулов, лимфоциты угнетены. Пои биопсии кишечника нет плазмати- ческих клеток Дефект преимущественно Т- или В-лимфоцитов Страдает и Т-, и В-клеточный иммунитет
Комбинированный иммунодефицит при дефиците аденозиндезаминазы Исследование активности аденозиндезами- назы позволяет провести дифференциаль- ную диагностику
Синдром Лет герера—Зиве Иммунологические показатели в норме
Синдром Оменна Иммунологические показатели в норме
Дефекты проведения Т-клеточных мем- бранных сигналов, дефицит продукции цитокинов Экспрессия Т-клеточных рецепторов снижена, реакция бласттрансформации лимфоцитов на фитогемагглютинин и антигены снижена, продукция IL-2, IL-3, IL-4, IL-5 снижена Тяжелые комбинированные имму- нодефицитные нарушения При стимуляции лимфоцитов in vitro нет экспрессии Т-клеточных рецепторов или не вырабатываются интерлейкины
Клеточный иммунитет с нарушением синтеза иммуноглобулинов (синдром Незелофа) Лимфопения или нормальное количество лимфоцитов, количество Т-пимфоцитов снижено, реакция бласттрансформации лимфоцитов на Фито- гемагглютинин и антигены снижена или отсутствует, ответ т-пимфоцитов на аллогенные лимфоциты снижен или отсутствует (иногда в норме), количество В-лимфоцитов в норме, нет выработки антител на специфическую иммуни- зацию, изогемагглютинины — отсутствуют или в норме. В лимфоузлах при- сутствуют плазматические клетки (при биопсии). В увеличенных лимфоузлах гранулемы с гистиоцитами и макрофагами Атаксия-телеангиэктазия а-Фетопротеин (АФП) повышен с первого года жизни
Синдром Вискотта-Олдрича Тромбоцитопения с рождения
Тяжелые комбинированные имму- нодефицитные нарушения Полное отсутствие Т- и В-лимфоцитов
Иммунодефицит при дефиците аденозиндезаминазы Отсутствие или резкое снижение уровня аденозиндезаминазы
Иммунодефицит с карликовостью Характерные клинические и рентгенологи- ческие данные
Синдром Ди Джорджи Эндокринопатия
Кандидоз кожи и слизистых обо- лочек Синтез антител в норме
СПИД Обнаружение вируса или антител к нему
Иммунодефицит с атаксией- телеангиэктазией (синдром Луи Барр) Реакция бласттрансформации лимфоцитов на фитогемагглютинин, смешанная культура лимфоцитов, Т-лимфоциты — в норме или снижены; лимфопения, ГЗТ отсутствует, количество В-лимфоцитов в норме, lgG2, lgG4, IgA снижено, IpE — снижено или отсутствует \у некоторых пациентов), выработка антител на специфические антигены снижена активность NK в норме, АФП повышен, уровень 17-глюкокортикоида снижен, а ФС гормона повышен, опоеделяются цитотоксические антитела к мозговой ткани и тимусу Синдром Незелофа Нет высокой концентрации АФП
Частичная церебральная атаксия Нет иммунологических нарушений
Селективный дефицит IgA Концентрация АФП в норме
Окончание табл. 16-17 *!
Иммунодефицитные состояния Результаты иммунологического исследования Дифференциальная диагностика
заболевание отличительный признак
Иммунодефицит с тромбоцитопенией, экземой, рецидиви- рующей инфекцией (синдром Вискотта- Олдрича) Тромбоцитопения, количество В-лимфоцитов и IgG в норме, IgM снижено, IgA и IgE повышено, изогемагглютинины отсутствуют или снижены, нет выработки антител на полисахаридный антиген, Т-клеточный иммунитет сначала в норме, затем снижается Идиопатическая тромбоцитопения Уровень всех иммуноглобулинов и изоге- > магглютининов, выработка антител на поли- р сахаридные антигены — в норме, крупные 5 размеры тромбоцитов р
Экзема с рецидивирующей инфек- цией у мальчиков Иммунологические показатели в норме при повышенных IgE, IgA, количество тромбо- £ цитов в норме =|
Иммунодефицит с тимомой (синдром Гуда) Количество иммуноглобулинов снижено, выработка антител на специфическую иммунизацию снижена или отсутствует, реакция бласттрансформации лимфо- цитов на фитогемагглютинин снижена, гиперчувстаительность замедленного типа снижена, супрессорная активность лимфоцитов повышена, тромбоцитопе- ния, гранулоцитопения. анемия Эритроцитарная аплазия, миасте- ния гравис Восстановление иммунитета после удаления § тимуса §
Иммунодефицит с карликовостью из-за коротких конечностей I вариант: снижено количество Т- и В-лимфоцитов, нет их функциональной активности. II вариант: снижены количество и функциональная активность Т-лимфоцитов, В-клеточный иммунитет в норме. Ill вариант: иммунитет Т-клеточный в норме, В-клеточный — отсутствует — —
Иммунодефицит вследствие дефицита ферментов Уровень аденозиндеэаминазы снижен, или фермент отсутствует, Т- и В-клеточный иммунитет (количественный и функциональный) снижен или отсутствует Тяжелые комбинированные имму- нодефицитные нарушения Уровень аденозиндезаминазы в норме
Уровень пуриннуклеозидфосфорилазы снижен, Т-клеточный иммунитет снижен, В клеточный иммунитет в норме Тяжелые комбинированные имму- нодефицитные нарушения Уровень пуриннуклеозидфосфорилазы в норме
Уровень 5'-нуклеотидазы снижен, количество и функциональная активность В-лимфоцитов снижены Х-сцепленная гипогаммаглобули- немия Нет снижения уровня 5'-нуклеотидазы
Синдром Вископа-Олдрича Нет снижения уровня 5'-нуклеотидазы
Селективный дефицит IgA Нет снижения уровня 5'-нукпеотидазы
Уровень транскобаламина II снижен, количество B-лимфоцитов и lg снижено, тромбоцитопения, анемия, гранулоцитопения СПИД Выявляют специфический вирус или анти- тела к нему
В)?-дефицитная анемия Микроцитарная анемия
Уровень биотин-зависимой карбоксилазы снижен, Т- и В-клеточный иммунитет (количественный и функциональный) снижен Хронический кандидоз кожи и сли- зистых оболочек Нет дефицита биотин-зависимой карбок- силазы
Атаксия Нет эффекта от лечения биотином, нет дефицита биотин-зависимой карбоксилазы
Синдром «голых» лимфоцитов Снижение экспрессии HLA (II или обоих классов), лимфопения, количество и функциональная активность Т- лимфоцитов снижены, ответ на антигены сни- жен, на митогены — в норме, количество В-лимфоцитов в норме или повышено Тяжелые комбинированные имму- нодефицитные нарушения При типировании HLA экспрессия антигенов 1 и/или II классов не снижена
ЛАБОРАТОРНАЯ ИММУНОЛОГИЯ
47
Вторичные иммунодефициты
Вторичные иммунодефициты обусловлены экзогенными факторами, харак-
теризуются стойкими клиническими и лабораторными признаками нарушения
функций иммунной системы, количественными и функциональными нарушения -
ми гуморального и клеточного иммунитета.
По клиническим признакам и лабораторным показателям вторичные и пер-
вичные иммунодефициты сходны. Принципиальное различие в том. что дефект
иммунной системы при вторичных иммунодефицитах — приобретенный.
Причины вторичных иммунодефицитов.
• Физиологический иммунодефицит:
< > новорожденных;
❖ пубертатного периода;
< при беременности:
❖ пожилых людей.
• Патологические:
❖ инфекционные:
- вирусные инфекции (острые и хронические персистирующие, ВИЧ);
- бактериальные инфекции:
- микотические инфекции;
- паразитарные инфекции;
> неинфекционные:
- нарушение питания (дефицит белка, витаминов и микроэлементов);
- хронические неинфекционные заболевания;
- злокачественные новообразования;
- аутоиммунные заболевания;
- интоксикации (эндогенные и экзогенные);
- нарушения метаболизма, дефицит энергопластического обеспечения
и нарушение регуляторных процессов (ожирение, кахексия, сахарный
диабет, тиреотоксикоз);
- стрессорные воздействия (травмы, операции, психические травмы,
спортивные перегрузки и др.);
- воздействие лекарственных препаратов (иммуносупрессоры, глюкокор-
тикоиды. антидепрессанты, химиотерапия);
- ожоговая болезнь:
- кровотечения;
- экологическое неблагополучие (воздействие неблагоприятных факто-
ров внешней среды — радиация, ксенобиотики).
Вторичные иммунодефициты разделяют по степени тяжести:
♦ компенсированные (с повышенной восприимчивостью к возбудителям
инфекций);
• субкомпенсированные (со склонностью к хронизации инфекционных про-
цессов);
• декомпенсированные (происходит развитие генерализованных инфекций,
индуцированных условно-патогенной флорой и злокачественными новооб-
разованиями).
Инфекционный синдром в «чистом виде» встречают в 50% случаев вторич-
ного иммунодефицита. Инфекционный анамнез ориентирован на определение у
пациента:
• упорно рецидивиру ющих вирусных и бактериальных инфекций ЛОР-органов,
респираторного и урогенитального тракта, ЖКТ;
• персистирующих вирусных инфекций;
• внутриклеточных бактериальных инфекций;
48 ЛАБОРАТОРНАЯ ИММУНОЛОГИЯ
• микотических инфекций;
• упорно рецидивирующих пиогенных инфекций кожи, подкожно-жировой
клетчатки и др. локализаций;
• генерализованных или тяжелых форм вирусных или бактериальных инфек-
ций;
• нарушений микробиоценоза открытых и закрытых слизистых оболочек.
Клинико-лабораторное обследование пациентов помогает оценить количе-
ственные и качественные характеристики, отражающие состояние иммунной
системы, выявить иммунодефицит и определить степень тяжести нарушений в
иммунной системе. Однако окончательный диагноз базируется, прежде всего, на
клинических проявлениях. Выбор лечения и прогноз его эффективности зависит
от комплексного подхода к такой патологии, как иммунодефицит.
ИММУННАЯ СИСТЕМА И ВОСПАЛЕНИЕ
В инфекционной клинике иммунологические обследования больных использу-
ют в основном с целью:
• диагностики инфекционного заболевания;
• прогнозирования характера течения и исхода болезни;
• определения у пациентов высокой степени риска возникновения инфекцион-
ных заболеваний;
• оценки эффективности проводимой терапии.
При благоприятном течении заболевания у больных формируется защитный
иммунитет. Динамика его выработки и роль факторов иммунной защиты при
разных инфекциях (и даже на разных этапах болезни) существенно различаются.
Именно поэтому схемы и объем иммунологического обследования больных раз-
ными инфекциями должны отличаться.
АНТИГЕН-НЕСПЕЦИФИЧЕСКИЕ ФАКТОРЫ ИММУННОЙ ЗАЩИТЫ
Антиген-неспецифические факторы иммунной защиты представлены клетками
пограничных тканей (кожа, слизистые оболочки дыхательных путей, пищевари-
тельного и урогенитального тракта), резидентными клетками разных органов и
тканей, эндотелиоцитами, а также клетками крови, различными циркулирующи-
ми и выделяемыми с секретами водорастворимыми молекулами. Характеристика
основных врожденных антиген-неспецифических факторов иммунной защиты
представлена в табл. 16-18.
Таблица 16-18. Антиген-неспецифические факторы иммунитета в противоинфекционной защите
Компоненты иммун- ной системы Иммунобиологическая активность
Клетки пограничных тканей (эпителий слизистых оболочек, кератоциты и др.) Препятствие на пути проникновения инфекционных агентов в организм; про- дукция слизи, микробиотических и микробицидных веществ; перенос антигенов в подслизистые оболочки кишечника и дыхательных путей; транспорт IgA и IgM из подслизистых оболочек в просвет дыхательных путей и кишечника, в секрет слизистых, слюнных, молочных желез; представление лимфоцитам антигенов в высокоиммунной форме
Дендритные клетки тканей и лимфоид- ных органов Захват и умерщвление инфекта; ферментативная обработка, транспортировка и представление антигенов лимфоцитам; депонирование антигена; инициация пер- вичного иммунного ответа; регуляция клеточного и/или гуморального иммунного ответа
Тучные клетки Инициация, поддержание и регуляция воспаления (в особенности аллергического) и репаоационных процессов; зкзоцитоз; продукция цитокинов и др. медиаторов воспаления
ЛАБОРАТОРНАЯ ИММУНОЛОГИЯ 49
Окончзние табл. 16-18
Компоненты иммун- ной системы Иммунобиологическая активность
Клетки разных тканей и органов Продукция цитокинов и гуморальных антиген-неспецифических факторов иммун- ной защиты (белки системы комплемента и острофазные, лизоцим и др.); удале- ние антигенов из циркуляции в организме; репарация повреждений; представление антигена Тс-лимфоцитам
Белки системы ком- племента Цитолиз, опсонизация; анафилатоксический и хемотаксический эффекты; регуля- ция воспаления и иммунного ответа; выведение из циркуляции в крови иммунных комплексов
С-реактивный белок Активация комплемента, опсонизация; взаимодействие с фосфорил-холиновыми производными полисахаридов и удаление их из крови
Нейтрофильные лей- коциты Фагоцитоз внеклеточных инфектов; экзоцитоз; участие в остром воспалении
Базофильные лей- коциты Экзоцитоз; поддержание и регуляция воспаления (особенно аллергического)
Эозинофильные лей- коциты Фагоцитарная и экзоцитарная антипаразитарная и противоопухолевая активность; участие в аллергическом воспалении; разрешение воспаления
Моноциты Фагоцитоз инфицированных соматических клеток; захват, обработка и представ- ление антигенов лимфоцитам; инициация антиген-специфического иммунного ответа; продукция цитокинов и других медиаторов воспаления и иммунного ответа; образование гранулем; удаление циркулирующих иммунных комплексов: репара ция повреждений
NK-клетки (есте- ственные киллеры) Антителозависимая клеточная цитотоксичность; продукция цитокинов
Эритроциты Сорбция циркулирующих антигенов и удаление их и циркулирующих иммунных комплексов из крови
Тромбоциты Уменьшение кровотока и ускорение свертывания крови в очагах воспаления, регу- ляция воспаления и репарации; удаление циркулирующих иммунных комплексов
Неповрежденные пограничные ткани — надежная преграда на пути про-
никновения инфектов внутрь организма. В слое эпителия слизистых оболочек
дыхательных путей и Ж КТ обнаружены М-клетки (от англ, microfold - микро-
загонщики), способные захватывать из слизи кишечника или дыхательных путей
водорастворимые антигены, вирусы и бактерии и в неизменном виде переносить
их в подслизистые. М-клетки со стороны подслизисз ых оболочек экспрессируют
на мембране переносимые антигены и обеспечивают взаимодействие с ними ден-
дритных клеток и макрофагов подслизистых. В свою очередь, последние не только
захватывают чужеродные антигены и микроорганизмы, но и убивают их, под-
вергают ферментативному расщеплению до олигопептидов, образуют комплексы
пептидов с внутриклеточными белками и представляют комплексы лимфоцитам
в высокоиммуногенной форме. При первичном попадании антигенов в организм
дендритные клетки захватывают и убиваю г инфекционных агентов. Захват анти-
гена стимулирует дифференцировку дендритных клеток и их миграцию из погра-
ничных тканей с током лимфы к ближайшим лимфатическим узлам.
Таким образом, дендпитные клетки осуществляют не только внутриклеточную
обработку антигенов, но и доставку их из пограничной ткани в лимфоидные орга-
ны. При первичном иммунном ответе в лимфоидных органах дендритные клетки
представляют антиген-специфическим наивным Т- и В-лимфоцитам низкомоле-
кулярные фрагменты процессированного антигена в ассоциации с белками HLA
II класса, инициируя антиген-специфический иммунный ответ.
Проникновение микроорганизмов через пограничные ткани активирует рези-
дентные клетки, которые включаются в локальный воспалительный процесс и
продуцируют хемотаксические вещества, цитокины, эйкозаноиды и другие медиа-
торы воспаления, запуская каскад воспалительных реакций.
50
ЛАБОРАТОРНАЯ ИММУНОЛОГИЯ
На ранних этапах развития воспаления с инфекционными агентами и продукта-
ми их жизнедеятельности взаимодействуют антиген-неспецифические гумораль-
ные факторы. Образующиеся при активации системы комплемента субкомпоненты
служат важными эндогенными эффекторами и регуляторами иммунной системы.
Фрагменты молекул белков системы комплемента — эндогенные опсонины, уси-
ливающие захват и завершенность фагоцитоза. Субкомпоненты комплемента Cis,
СЗа, С4а и С5а способны активировать тучные клетки и базофилы, вызывая их
дегрануляцию, активацию синтеза и секрецию медиаторов аллергического вос-
паления. Субкомпоненты СЗа и С5а вызывают сокращение гладкой мускулатуры
и повышают проницаемость сосудистой стенки, что позволяет называть их анафи-
лотоксинами.
Все клетки крови в той или иной степени участвуют в защите от чужого в каче-
стве эффекторных факторов иммунных реакций. Фагоцитарная система крови
представлена гранулоцитами и моноцитами. Среди гранулоцитов фагоцитарная
активность особенно выражена у нейтрофильных и эозинофильных лейкоцитов.
Нейтрофилы способны захватывать, убивать и переваривать разнообразные вне-
клеточно размножающиеся инфекционные агенты. Умерщвление захваченных
гранулоцитами и моноцитами микроорганизмов осуществляется за счет прямого
действия на них радикалов кислорода и множества дезинфектантов, образующих-
ся в фагоцитах при кислом pH и в присутствии галогенов хлора и йода с участи-
ем миелопероксидазы (хлорноватистая кислота, надйодная кислота, хлорамин
и др.). Для гранулоцитов характерна также экзоцитарная активность, в результате
которой лейкоциты выделяют в сторону микробной клетки и других объектов
содержимое внутриклеточных гранул. В их гранулах содержатся разнообразные
антимикробные продукты и активные гидролитические ферменты, а также гиста-
мин (базофилы), цитокины и другие медиаторы воспаления. Способность базофи-
лов продуцировать и при дегрануляции выделять гистамин, а также наличие у них
рецепторов к IgE делаю г базофилы, как и тучные клетки, важнейшими участника-
ми аллергического воспаления.
Моноциты определяют исход хронического воспаления и освобождают орга-
низм от внутриклеточно паразитирующих микроорганизмов. В отличие от поли-
морфноядерных лейкоцитов моноциты фагоцитируют инфицированные микро-
бами соматические клетки, прерывая размножение внутриклеточных инфектов.
Например, при хламидийной инфекции первая реакция иммунной системы в месте
внедрения возбудителя обязана нейтрофильным лейкоцитам, а в случае хрониза-
ции инфекции уничтожение возбудителя осуществляют моноциты, фагоцитирую-
щие инфицированные клетки. Недостаточность фагоцитарной активности моно-
цитов — главная причина хронического течения хламидийных инфекций. При
вирусных инфекциях решающая роль в уничтожении инфицированных вирусами
клеток также принадлежит моноцитам.
При хронических гранулематозных инфекциях моноциты крови в инфициро-
ванных органах дифференцируются в многоядерные эпителиоидные клетки, кото-
рые образуют вокруг возбудителя гранулемы. Гранулема ограничивает размно-
жение возбудителя, удерживает его в одном локусе и препятствует диссеминации
инфекта в организме. Нарушение способности моноцитов образовывать гранулему
(в частности, при ВИЧ-инфекции) делает больного беззащитным в отношении
микобактериальных инфекций.
Повышенное общее количество нейтрофильных лейкоцитов, появление в
крови юных форм этих клеток свидетельствует об остром воспалительном про-
цессе. Выявление нейтрофилов с токсической зернистостью или патологически
измененных клеток в крови больных указывает на серьезные изменения и даже
на несостоятельность этого важного механизма иммунной защиты. Увеличение в
крови содержания моноцитов может быть связано с вирусной или какой-либо дру-
ЛАБОРАТОРНАЯ ИММУНОЛОГИЯ 51
гой инфекцией, вызываемой внутриклеточными паразитами, и может свидетель-
ствовать о хроническом патологическом процессе. Для диагностики врожденных
заболеваний фагоцитарной системы (например, хроническая гранулематозная
болезнь) или для прогнозирования течения гнойных инфекций информативным
считают исследование миелопероксидазной активности фагоцитов — тест с гра-
нулами нитросинего тетразолия (НСТ-тест). Активность фагоцитарной системы
крови зависит от опсонизирующей или, наоборот, угнетающей фагоцитоз способ-
ности сыворотки крови больного. Накопление в крови эозинофилов может быть
следствием аллергической патологии, гельминтозов, онкологического процесса,
аутоиммунных заболеваний, врожденных иммунодефицитов и др.
К антиген-неспецифическим факторам иммунной защиты относят NK-клетки
крови. Основные мишени для них — клетки, инфицированные вирусами, опухо-
левые клетки, клетки с размножающимися внутриклеточно микробами. У таких
клеток-мишеней подавлена экспрессия и изменена пространственная конфигура-
ция ассоциированных с мембранами антигенов HLA (трансплантационные анти-
гены), что и активирует против них NK-клетки. Участие NK-клеток в защите от
инфекции особенно значимо в первые дни заболевания, когда у больного еще не
сформировались антиген-специфические иммунные реакции. Для оценки функ-
ции NK-клеток разработаны специальные тесты, получившие название реакций
антителозависимого клеточного цитолиза. В этой реакции цитолитическую гибель
клеток-мишеней осуществляют NK-клетки, а также соучаствующие в реакции
моноциты, эозинофилы и нейтрофилы, несущие Fc-R к IgG.
АНТИГЕН-СПЕЦИФИЧЕСКИЕ ФАКТОРЫ ИММУНИТЕТА
Антиген-специфические факторы иммунитета распознают и запоминают осо-
бенности молекулярной структура чужеродных антигенов и, благодаря этому,
узнают антиген при повторных контактах с ним. Антиген-специфические ком-
поненты иммунной системы — Т- и В-лимФоциты и антитела. Именно они обе-
спечивают формирование приобретенного иммунитета к инфекционным агентам.
Из всех клеток иммунной системы только лимфоциты способны распознавать
антиген, взаимодействовать с ним и обеспечивать формирование иммунологи-
ческой памяти. Антигенная активация лимфоцитов предполагает обязательное
взаимодействие нескольких клеток разного типа — антиген-представляющих
(А-клетки), Т- и В-лимфоцитов. При первичном иммунном ответе в качестве
А-клеток выступают дендритные клетки, эффективное взаимодействие имму-
нокомпетентных клеток происходит только во вторичных лимфоидных органах
(лимфатические узлы, селезенка, миндалины, пейеровы бляшки и др.). В первич-
ном ответе участвуют наивные Т- и В-лимфоциты, ранее не имевшие контакта с
антигеном и потому весьма требовательные к условиям, в которых осуществляется
представление антигена и ответ на него. В результате взаимодействия с антигенами
и пролиферации наивных лимфоцитов соответствующего клона накапливаются
иммунные лимфоциты, которые затем распределяются по всему организму. При
повторной иммунизации они могут взаимодействовать и отвечать на антиген не
только во вторичных лимфоидных органах, но и в любых других opiawax и тка-
нях. Всякий раз большая доля клеток размножившегося клона лимфоцитов спустя
некоторое время погибает апоптозом, но часть из них может дифференцироваться
в малые лимфоциты (клетки — носители иммунологической памяти), способные
без деления сохраняться в организме в течение нескольких лет. Вся последующая
иммунизация стимулирует быструю дифференцировку и деление соответствую-
щих иммунных антиген-специфических лимфоцитов и поддерживает сохранение
в организме иммунных лимфоцитов антигенреактивного клона (табл. 16-19).
Приобретенный иммунитет сохраняется до тех пор, пока имеется повышенное
содержание малых Т- и В-лимфоцитов антигенреактивного клона.
52
ЛАБОРАТОРНАЯ ИММУНОЛОГИЯ
Таблица 16-19. Иммунобиологическая активность антиген-специфических компонентов иммун-
ной системы (приобретенные факторы иммунной защиты)
Компоненты иммунной системы защиты Иммунобиологическая активность
Лимфоциты:
CD8* Т-клетки Цитотоксическая активность в отношении антиген-специфических клеток- мишеней (вирус-инфицированные, опухолевые, клетки трансплантатов, парази- тов, с поверхностными чужеродными антигенами)
CD4+ Т-клетки Контактная и опосредованная цитокинами регуляция функций клеток иммунной системы, участвующих в воспалении и антиген-специфическом клеточном и/или гуморальном иммунном ответе
CD19\ CD20+, СО2Г В-клетки Предшественники секретирующих антитела плазматических клеток: продукция цитокинов: эндоцитоз, процессинг и представление антигена Th-лимфоцитам, в особенности при вторичном иммунном ответе
Ig разных классов и суб классов Нейтрализация и удаление антигена, агрегация водорастворимых и корпускуляр- ных антигенов, опсонизация, инициация С- и NK-зависимого цитолиза, инициация аллергического воспаления (IgE), регуляция иммунного ответа
Популяция Т-лимфоцитов исключительно гетерогенна. Различают CD4-Th- и
СП8-Тс-лимфоциты. Тс-лимфоциты имеют рецепторы к антигенам. Взаимодействие
с ними активирует Т-лимфоциты и стимулирует их пролиферацию и дифференци-
ровку. Активированные Тс-лимфоциты, как и NK-клетки, выделяют в сторону
клетки с антигеном перфорины и гранзимы, в результате чего в клетке-мишени
образуются отверстия или включается программа их самоубийства (апоптоза).
Для накопления в организме Тс-лимфоцитов необходимо определенное время,
поэтому они включаются в процесс уничтожения инфицированных клеток позд-
нее, чем NK-клетки или другие антиген-неспецифические факторы иммунной
защиты. Именно эти клетки обеспечивают высокоспецифическую иммунную реак-
цию Т-лимфоцитов при вторичном иммунном ответе.
Т-лимфоциты прямым взаимодействием со вспомогательными клетками и
с другими лимфоцитами, а также посредством продуцируемых ими цитокинов
управляют иммунным ответом. Популяция CD4+ Т-лимфоцитов также весьма
неоднородна. В зависимости от того, какие наборы цитокинов они продуцируют,
среди Th-лимфоцитов различают субпопуляции клеток ThO, Thl, Th2 и Thl7 (Tri)
и Tr-лимфоциты. Дифференцировка ThO в иной тип Th-клеток зависит от многих
условий: свойств клеток, представляющих антиген Th-лимфоцитам; соотношения
антигенпрезентирующих А-клеток и Th-клеток; количества антигена на поверх-
ности А-клеток; набора цитокинов, воздействующих на Th-клетку; характера
костимулирующих сигналов, передаваемых Th-клеткам; клеток микроокружения
при взаимодействии А-клеток и Th-лимфоцитов.
Антигенная активация В-лимфоцитов приводит к их пролиферации и диф-
ференцировке в плазматические клетки или малые В-лимфоциты памяти.
Плазматические клетки живут в течение 2-3 нед. Плазматическая клетка может
продуцировать иммуноглобулины любого, но только какого-либо одного клас-
са. Исключение — начальный этап дифференцировки наивных В-лимфоцитов в
плазмоциты, когда клетки вначале синтезируют IgM, а затем переключаются на
синтез IgG. Некоторое время такие плазматические клетки могут одновременно
синтезировать антитела против того же антигена, но классов IgM и IgG. Выбор
класса иммуноглобулинов, которые будет продуцировать плазматическая клетка,
определяют Th-лимфоциты, секретирующие разные иммунорегуляторные цито-
кины. Эти цитокины воздействуют на В-лимфоциты, включая в них гены, контро-
лирующие синтез какого-либо определенного класса иммуноглобулинов.
ЛАБОРАТОРНАЯ ИММУНОЛОГИЯ
53
При инфекционных заболеваниях накапливающиеся в крови антитела облада-
ют множественной иммунобиологической активностью (табл. 16-20). Антитела
при взаимодействии с микробными или иными антигенами-токсинами способны
нейтрализовать их токсичность, а также обезвредить другие факторы патоген-
ности бактерий и вирусов. Взаимодействуя с чужеродными антигенами, антитела
способны образовывать мультимолекулярные иммунные комплексы из антигена и
антител, которые быстро выводятся из циркуляции.
Таблица 16-20. Участие антител в противоинфекционном иммунитете
Класс lg Механизмы защиты Возбудители инфекций
IgG, IgM Опсонизация Вирусы, микроорганизмы грибы
IgG, IgM, lgA*C Нейтрализация Вирусы
IgA Подавление адгезии Микроорганизмы, грибы
IgG, IgM'C Цитолиз Вирусы,микроорганизмы, паразиты
IgG Нейтрализация токсинов Микроорганизмы, грибы
IgG Подавление активности ферментов Вирусы, микроорганизмы
IgG, IgA Антителозависимый клеточный цитолиз Вирусы, грибы, паразиты
igE Активация тучных клеток Паразиты
IgA IgG Микробостатическое действие Микроорганизмы, микоплазмы
ОСОБЕНН СТИ ПРОТЕКТИВНОГО ИММУНИТЕТА ПРИ НЕКОТОРЫХ ИНФЕКЦИЯХ
В зависимости от свойств возбудителя, количества микроорганизмов, их способ-
ности размножаться и противостоять механизмам противоинфекционного иммуни-
тета исход заболевания определяет прежде всего активность эффекторных факторов
иммунной защиты, играющих при данной инфекции решающую роль. Так, уничто-
жение возбудителей гнойных инфекций, патогенов, размножающихся внеклеточно,
осуществляют, главным образом, нейтрофильные полиморфноядерные лейкоциты
(табл. 16-21). Нейтрофильные лейкоциты играют решающую роль в уничтожении
большинства капсульных микроорганизмов. Полисахариды капсул, в свою очередь,
угнетают фагоцитарную активность микро- и макрофагов и, таким образом, позво-
ляют микробам-возбудигелям избегать фагоцитоза и гибели в организме хозяина.
Нейтрализовать ингибирующую фагоцитоз активность капсульных полисахаридов
могут антитела к ним. Однако исключительное разнообразие строения капсульных
полисахаридов, даже у микроорганизмов одного и того же вида, затрудняет способ-
ность антител защищать макроорганизм от различных серовариантов возбудителя.
Именно поэтому активность нейтрофильных лейкоцитов и опсонизирующих фак-
торов сыворотки (антитела, комплемент, С-реактивный белок и др.) определяет
эффективную защиту от размножающихся внеклеточно инфектов.
Таблица 16-21. Факторы иммунитета, играющие решающую роль в исходе инфекционных забо-
леваний
Факторы Примеры инфекций
Гранулоциты Стафилококковая, стрептококковая, пневмококковая, менингококко- вая инфекции, гонорея
Моноциты,антитела и опсонины Хламидиозы, риккетсиозы, микоплазменная инфекция
Антитела Токсикоинфекции (анаэробные, дифтерия, холера)
Моноциты, антитела, Т-лимфоциты, NK-клетки Кандидоз, сисрилис, сальмонеллез, листериоз, вирусные инфекции
Моноциты, Т-лимфоциты Туберкулез, лепра, кокцидиоидоз
54 ЛАБОРАТОРНАЯ ИММУНОЛОГИЯ
Возбудители группы внеклеточных патогенов с помощью разнообразных меха-
низмов угнетают фагоцитоз, даже при захвате микроорганизмов фагоцитами
противостоят действию микробицидных факторов лейкоцитов. Так, поверх-
ностный М-белок и продуцируемые стрептококками А-стрептолизины О и S
угнетают фагоцитарную активность лейкоцитов, а лишенные М-белка штаммы
стрептококков отличаются низкой вирулентностью. У некоторых экзотокси-
нов стафилококков проявляются свойства суперантигенов, они могут активиро-
вать 5-25% Th-лимфоцитов крови больного. Такая олигоклональная активация
Th-лимфоцитов приводит к гиперпродукции цитокинов и последующей гиперак-
тивации моноцитов, что клинически выражено симптомокомплексами токсиче-
ского шока.
Антитела и другие опсонины играют важную роль в защите от внеклеточных
возбудителей инфекций. При нейссериальных инфекциях антитела к поверхност-
ным белкам и капсульным полисахаридам обеспечивают опосредованную компле-
ментом гибель микроорганизмов. За благоприятный исход пневмоний, бронхитов
и диссеминированных инфекций, вызываемых Haemophilus; influenzae, во многом
ответ ственны антитела против полисахаридных капсул. Чувс гвительность к инфек-
ции обратно коррелирует с уровнем бактерицидных антител. Наследственные
дефекты синтеза lgG2 создают условия для возникновения хронических инфекций
респираторного тракта.
Многие патогенные микроорганизмы (нейссерии, гемофильная палочка),
чтобы уменьшить антибактериальную активность антител, продуцируют про-
теазы, расщепляющие IgA-молекулы. Псевдомонады вырабатывают эласгазу,
которая инактивирует СЗа и С5а, снижая локальное воспаление. Важную роль в
регуляции функциональной активности клеток пограничных тканей и нейтро-
фильных лейкоцитов крови играют лимфоциты Thl7, продуцирующие IL-17.
Несостоятельш >сть CD4+ Th 17 может способствовать высокой чувствительности
больных к возбудителям инфекций и хроническому течению заболевания.
Таким образом, при пиогенных и других инфекциях, вызываемых внеклеточно
паразитирующими возбудителями, прогноз развития заболевания и его исход
определены активностью нейтрофильных лейкоцитов, уровнем антимикробных
антител и комплемента и других сывороточных опсонинов. В этих случаях целе-
сообразно исследовать приведенные факторы иммунной защиты.
Вторую группу инфекций составляют риккетсиозы, хламидиозы, легионелле-
зы, микоплазмозы. Возбудители этих инфекций размножаются внутриклеточно
и, поступая в разных формах из инфицированных клеток в окружающую среду,
инфицируют новые клетки. При заражении защиту от инфекции в местах внедре-
ния возбудителя осуществляют нейтрофильные лейкоциты. Однако при ее недо-
статочности возбудитель распространяется и накапливается в соматических клет-
ках. Уничтожение инфицированных клеток и находящихся в них возбудителей
осуществляют в основном циркулирующие и тканевые макрофаги. Оказавшиеся
в макрофагах возбудители стремятся выжить, подавляя у фагоцитов образование
фаголизосом (риккетсии, хламидии) и продукцию супероксида (хламидии). При
несостоятельности макрофагальной системы возбудители этой группы инфекций
размножаются в фагоцитах. Все это приводит к развитию хронического инфек-
ционного процесса. Микробные Т-суперантигены способны фиксироваться на
поверхности вспомогательных клеток с помощью HLA-белков II класса и одно-
временно взаимодействовать с р-полипептидной цепью антиген-распознающих
рецепторов Th-лимфоцитов. Взаимодействие Th-лимфоцитов с Т-суперантигенами
ангиген-неспецифично, с каждым Т-суперантигеном могут реагировать 5-
25% клонов Th-клеток. Следствие такой активации — повышенная продукция в
организме Th-цитокинов, которые, в свою очередь, вызывают гиперактиваиию с
накоплением у больных провоспалительных цитокинов. Клинически это выраже-
ЛАБОРАТОРНАЯ ИММУНОЛОГИЯ
55
но синдромом токсического шока. Олигоклональная активация Th-лимфоцитов
может приводить к развитию аутоиммунных процессов (синдром Рейтера) или к
апоптотической гибели активированных лимфоцитов. Свойствами Т-клеточных
суперантигенов обладают не только белки микоплазм, хламидий, иерсиний. но и
пирогенные экзотоксины стрептококков и стафилококков, а также вируса бешен-
ства и Эпштейна-Барр, цитомегаловируса.
Некоторые микробные белки способны взаимодействовать с иммуноглобули-
нами человека (но не с активным центром антител). Такие белки были названы
В-суперантигенами. Взаимодействуя с фиксированными на соматических клетках
иммуноглобулинами (тучные клетки, базофилы). В-суперантигены активируют
их и стимулируют синтез и высвобождение из этих клеток IL-4, IL-13, которые
обеспечивают дальнейшее развитие аллергического воспаления (например, кра-
пивницы). Свойствами В-суперантигенов обладают белок gp12U ВИЧ, L-протеин
клеточных стенок Pepto>treptococcus magnus, А-белок стафилококков. Вирусы гепа-
тита А, В, С и Е индуцируют в гепатоцитах синтез и секрецию сиалопротеина FV,
который может взаимодействовать с Н- и L-цепями иммуноглобулинов разных
классов. Связывание FV-белка с фиксированными на тучных клетках и базофилах
IgE приводит к освобождению из клеток гистамина и других медиаторов аллерги-
ческого воспаления и имитирует развитие у больных аллерген-опосредовапных
реакций. Способность возбудителей инфекций продуцировать В-суперантигены
привносит в клиническую картину заболевания симптомы, характерные для
аллергического воспаления.
При инфекциях, возбудители которых активно продуцируют экзотоксины
(анаэробные инфекции, дифтерия), течение и исход заболевания зависят от спо-
собности лимфоидной системы продуцировать антитоксические антитела. Такие
антитела, главным образом класса IgG, нейтрализуют токсины и обеспечивают
благоприятный исход заболевания. Однако формирование антитоксического
иммунитета не гарантирует освобождение макроорганизма от носительства,
поскольку для устранения бактерионосительства необходимо формирование
специфического иммунитета к поверхностным структурам бактерий, ответствен-
ным за их колонизацию.
Спирохеты, некоторые патогенные грибы и бактерии способны паразитировать
как внутриклеточно, так и внеклеточно. Вирусы размножаются только внутри-
клеточно, но могут длительное время пребывать и снаружи, и внутри клеток, не
оказывая цитопатогенного действия. У некоторых вирусов обнаружены гены,
продукты которых обладают антиапоптотическим свойством и способствуют дли-
тельной переживаемое™ инфицированных клеток и продукции вирусных частиц.
В уничтожении возбудителей инфекций этой группы участвует весь арсенал как
врожденных, так и приобретенных факторов иммунной защиты. Значимость
антиген-специфических механизмов противоинфекционного иммунитета возрас-
тает по мере прогрессирования заболевания, без участия эффекторных антиген-
специфических Th- и Тс-лимфоцитов выздоровление не наступает.
Особую группу заболеваний представляют инфекции, хроническое течение кото-
рых сопровождается образованием гранулем вокруг жизнеспособных возбудителей.
К таким гранулематозным инфекциям относят туберкулез, лепру, кокцидиоидный
микоз и др. Эти инфекции отличает длительная, часто пожизненная персистенция
возбудителей внутри образующихся гранулем. Стенки гранулем образованы эпи-
телиоидными или многоядерными клетками и препятствуют диссеминации воз-
будителя. Предшественники эпителиоидных клеток гранулем — активированные
макрофаги. Возбудители инфекций, будучи захваченными макрофагами, не поги-
бают, поскольку способны подавлять образование фаголизосом и противостоять
действию микробицидных агентов в макрофагах. В стенке гранулемы в больших
количествах представлены активированные ТЫ- и Тс-лимфоциты. Цитокины,
56
ЛАБОРАТОРНАЯ ИММУНОЛОГИЯ
продуцируемые Th] - лимфоцитами (INF-y, IL-12) и макрофагами (IL-12), под-
держивают высокую активность антиген-специфического Т-клеточного имму-
нитета и обеспечивают ремиссию заболевания. Несостоятельность макрофагов и
Т- лимфоцитов сопровождается обострением инфекции, диссеминацией возбуди-
телей и стимуляцией антиген-специфического гуморального иммунного ответа.
Антитела при этом выступают в основном как свидетели контакта иммунной
системы хозяина с возбудителем, не оказывают протективного действия и мало
влияют на течение и исход заболевания. При кокцидиоидном микозе снижение
активности Т-клеточного иммунитета (положительные пробы в реакции повы-
шенной чувствительности замедленного типа с кокцидоидином) и накопление
циркулирующих антител пролив гриба служат предвестниками скорой смерти
больного. При обследовании больных гранулематозными инфекциями прогно-
стическое значение имеют показал ели клеточного имму иного ответа: количество и
активность антиген-специфических Th]- и Тс-лимфоцитов, содержание локальных
и циркулирующих 1 h1 -цитокинов, активность NK-клеток и макрофагов.
При острой инфекции первую линию защиты образуют антиген-неспецифические
механизмы иммунитета. Так, при вирусной инфекции активное размножение
вируса обеспечивает быстрое накопление вирусных частиц в крови (виремия),
предшествующее противовирусной реакции иммунной системы. При ранней реак-
ции иммунной системы характерна активация в инфицированных клетках синтеза
INF-a/p, которые поступают в кровь и обеспечивают защиту еще не инфицирован-
ных клеток. В эти же сроки в крови накапливаются активированные NK-клетки,
способные к умерщвлению соматических клеток, несущих на своей поверхности
измененные вследствие нападения вирусов HLA-антигепы I класса. Накопление в
крови антиген-специфических Тс-лимфоцитов и противовирусных ан гител наблю-
дают позднее, их максимальное вовлечение в противовирусную защиту по времени
совпадает с уменьшением виремии или даже с полным исчезновением вируса. При
благоприятном исходе заболевания через 2,5-3 мес после заражения количество
Тс-лимфоцитов в крови снижается до исходного уровня, тогда как противовирус-
ные антитела могут определяться еще в течение нескольких месяцев.
При многих острых бактериальных и вирусных инфекциях антиген-
специфические резидентные и циркулирующие клеточные и гуморальные факторы
обеспечивают устойчивость к заражению, тогда как исход возникшего заболевания
зависит от активности формируемого приобретенного антиген-специфического
иммунного ответа.
При вирусном гепатите С и ВИЧ-инфекции хороший прогностический пока-
затель — накопление в остром периоде заболевания Т-лимфоцитов, специфич-
ных в отношении многих эпитопов поверхностных и внутренних белков вируса.
Накопление Т-клеток, даже в очень больших количествах, но реагирующих с
ограниченным числом эпитопов белков возбудителя, наоборот, свидетельствует
о недостаточности противовирусной защиты и предвещает хроническое течение
болезни.
Высокая иммунная реакция при вакцинации делает организм устойчивым к
заражению. Эффективность вакцинации возрастает, если применяют вакцины,
вызывающие формирование необходимого для данного заболевания клеточного
•/или гуморального иммунного ответа. Современные молекулярно-генетические
вакцины, в которых используются носители, доставляющие протективные ан гиге-
ны и активирующие определенные популяции регулятооных лимфоцитов, откры-
вают новые возможности для специфической профилактики и лечения инфекци-
онных заболеваний.
Таким образом, при различных заболеваниях и на разных стадиях инфекцион-
ного процесса прогностическое значение имеют исследования состояния опреде-
ленных антиген-неспецифических и антиген-специфических факторов иммунной
ЛАБОРАТОРНАЯ иммунология 57
защиты, а их оценка в динамике позволяет контролировать эффективность прово-
димой терапии.
Активность эффекторных механизмов иммунной защиты при инфекциях может
нарушаться не только вследствие их собственной несостоятельности, но и в
результате цисфункции регуляторных механизмов. Иммунная система располагает
множеством ауторегуляторных механизмов Существенную роль среди них имеют
регуляторные Th-лимфоциты и дендритные клетки, представляющие им анти-
гены. Посредством цитокинов и прямыми межклеточными взаимодействиями
Th-лимфоциты обеспечивают развитие антиген-специфического клеточного и/
или гуморального иммунного ответа, а также регулируют активность и участие
в противоинфекционном иммунитете разнообразных антиген-специфических
факторов. Особенности представляющих антиген дендритных клеток и условия
их взаимодействия с Th-лимфоиитами определяют тип первичного иммунно-
го ответа на соответствующие антигены (клеточный или гуморальный). При
повторном контакте с антигеном в ответе участвуют ранее накопившиеся Thl- или
ТЬ2-лимфоциты памяти. Представление антигена Th-клеткам при вторичном
иммунном ответе может осуществляться не только дендритными клетками, но и
любыми клетками, экспрессирующими белки HLA П класса.
Таким образом, при лабиоаторном обследовании больных инфекционны-
ми заболеваниями прежде всего необходимо оценить состояние эффекторных
антиген-неспецифических и антиген-специфических механизмов иммунитета,
играющих решающую роль в уничтожении возбудителей данной инфекции. Лишь
после этого исследуют содержание и активность регуляторных клеток и набор
продуцируемых ими цитокинов. Для прогнозирования течения инфекционно-
го процесса важно не однократное, а мониторинговое обследование больного с
оценкой состояния тех компонентов антиген-неспецифической и/или антиген-
специфической иммунной защиты, которые при данном заболевании на данном
этапе болезни играют решающую роль в уничтожении и выведении из организма
возбудителей инфекции.
ГИПЕРИММУННЫЕ ВОСПАЛИТЕЛЬНЫЕ ПРОЦЕССЫ
В воспалительных процессах разной этиологии участвуют как антиген-
неспецифические, так и антиген-специфические факторы иммунной защиты.
Обычно иммунный ответ при воспалениях протекает субклинически и обеспе-
чивает нейтрализацию антигена и удаление его из организма без формирования
клинически выраженных повреждений. В некоторых случаях характер и сила
иммунной реакции на антиген могут не соответствовать патогенетической актив-
ности антигена, при поступлении в организм абсолютно безвредных антигенов
формирующаяся антиген-специфическая иммунная реакция может приводить к
развитию тяжелых воспалительных процессов, сопровождающихся локальными
и генерализованными повреждениями тканей. Такие варианты гиперергических
воспалительных процессов принято называть иммунными воспалениями. За пато-
генез таких воспалений ответственны как антитела разных классов, так и эффек-
торные Т-лимфоциты. Различают четыре типа иммунопатологических процессов.
I тип гиперергических иммунопатологических реакций обязан накапливающимся
у пациентов IgE антителам к антигенам-аллергенам. От иммуноглобулинов дру-
гих классов IgE отличает высокая цитофильность. IgE фиксируются на клетках,
взаимодействуя с IgE-специфическими Fce-R. Описано два типа таких рецепто-
ров, отличающихся аффинностью взаимодействия с IgE. Если молекулы антигена
взаимодействуют с фиксированными на клетках несколькими молекулами IgE, то
происходит агрегация рецепторов к IgE, их конформация, активация связанных с
рецепторами протеинтирозинкиназ, которые, в свою очередь, фосфорилируют и
этим активируют внутриклеточные сигнальные молекулы.
58
ЛАБОРАТОРНАЯ ИММУНОЛОГИЯ
В развитии IgE- опосредованных реакций гиперчувст вительности I типа разли-
чают две фазы — раннюю и позднюю. Ранняя реакция развивается сразу после вве-
дения антигена-аллергена и обязана высвобождению из активированных тучных
клеток гистамина, триптазы и других медиаторов. При локальной аллергической
реакции через 4-6 ч после контакта с аллергеном развивается поздняя фаза реак-
ции (происходит инфильтрация очага воспаления эозинофилами, нейтрофилами
и, в меньшей степени, моноцитами, лимфоцитами и базофилами). Симптоматика
поздней реакции обязана медиаторам, высвобождаемым из активированных
эозинофилов и других клеток крови. При бронхиальной астме поздняя фаза про-
должается в течение 1-3 сут. Активированные эозинофилы играют ключевую
роль в симптоматике поздней фазы аллергического воспаления, В это время
количество эозинофилов в инфильтрате достигает 30% всех локально аккумули-
рованных клеток крови Под влиянием хемотактического фактора тучных клеток,
цитокинов и других медиаторов эозинофилы активируются, экспрессируют Fc-R к
IgE и IgG, рецепторы к цитокинам, компонентам комплемента и другим медиато-
рам. Активированные эозинофилы синтезируют и секретируют свойственные им
основные белки, ферменты, цитокины и другие молекулы. Активное вовлечение
эозинофилов в поздней фазе аллергического воспаления I типа придает реакции
некоторые специфические особенности, обязанные повреждению клеток тех
или иных тканей продуктами жизнедеятельности активированных эозинофилов.
Наряду с эозинофилами в патогенезе поздней фазы реакции участвуют нейтрофи-
лы, количество которых может достигать 30% кле гок инфильтрата. Накопление
нейтрофилов обязано хемотактическому действию на них нейтрофильного хемо-
тактического фактора и IL-8, продуцируемых активированными тучными клетка-
ми. В свою очередь нейтрофилы становятся источником разнообразных гидролаз,
лейкотриенов, цитокинов и других биологически активных молекул.
При бронхиальной астме наблюдаемые на ранней фазе вазодилатация и отек
обязаны гистамину, FGD2 и LTD4; бронхоконстрикция и секреция слизи — ЬТС4
и PGD2, высвобождаемые, главным образом, при дегрануляции тучных клеток.
Инфильтрация слизистых оболочек бронхов клетками крови в поздней фазе реак-
ции происходит в результате активации эндотелиальных клеток интерлейкинами
IL4, TNF-a, LTC4. Миграцию лейкоцитов из сосудов в слизистые оболочки вызыва-
ют PAR IL-5, IL-8, ЬТВ4, а активацию клеток в инфильтрате — IL-4, IL-5 и TNF-a.
Источником всех этих медиаторов поздней фазы служат как тучные клез ки, так и
активированные эпителиоциты, эндотелиоциты и лейкоциты.
Накапливаемые в очагах гиперактивности лимфоциты представлены преиму-
щественно Т-клетками, причем главным образом Тй2-фенотипа. Взаимодействие
активированных В-лимфоцитов с антигеном, опосредованное IgE, фиксирован-
ными на Fce-RII (CD23), усиливает эндоцитоз антигена и способность В-клеток
представлять антиген Т-лимфоцитам, но угнетает пролиферацию В-клеток и
синтез IgE. Фагоциты активно синтезируют оксид азота при взаимодействии CD23
молекул лимфоцитов с CDllb c моноцитов.
При протеолизе CD,3 внеклеточная часть рецептора может циркулировать в
крови в виде солюбилизированного sFce-RII, который сохраняет способность вза-
имодействовать с CD21 и mlgE на мембране В-лимфоцитов. Такое взаимодействие
стимулирует пролиферацию В-клеток. в особенности клонов клеток, продуци-
рующих IgE. Таким образом, сигналы, подаваемые клеткам через низкоаффинные
рецепторы к IgE, оказывают регуляторное воздействие на лимфоциты и клетки
фагоцитарной системы.
Опосредованная IgE гиперреактивность клинически может проявляться в
виде локальных и генерализованных патологических процессов. Пример генера-
лизованной реакции I типа — анафилактические реакции. IgE-опосредованные
анафилактические реакции возникают в результате взаимодействия с антигеном-
ЛАБОРАТОРНАЯ ИММУНОЛОГИЯ
59
аллергеном имеющихся у пациентов IgE-антител против некоторых медикаментов,
чужеродной сыворотки крови, морских продуктов, орехов, яда пчел, продуктов
ужаливания ос, шершней, москитов, и т.д. Генерализованные анафилактические
реакции характеризуются сосудистым коллапсом, изменением тонуса гладкой
мускулатуры бронхов, сосудов, сфинктеров, отеками и в случае запоздалого или
неправильного лечения могут привести к летальному исходу.
Примерами локальных IgE-опосредованных гиперергических реакций I типа
могут служить атопические риниты, бронхиальная астма, экзема, пищевая аллер-
гия. Антигенами (аллергенами), вызываюшими при этих заболеваниях продукцию
антител класса IgE, выступают’ белки и ферменты пыльцы растений, некоторые
экзотические пищевые продукты, метаболиты малопатогенных грибов и микро-
организмов, белки насекомых и др, Накапливаемые IgE фиксируются на Fce-RI
тучных клеток, особенно в пограничных тканях. При повторном поступлении
антигена через кожу и слизистые оболочки дыхательных путей и пищеварительно-
го тракта аллергены взаимодействуют с фиксированными на тучных клетках IgE,
что приводит к активации и дегрануляции тучных клеток.
Иммунное воспаление II типа обязано накоплению в организме антител, спо-
собных запускать процессы цитотоксического повреждения собственных клеток
организма. Примерами такого типа иммунных повреждений могут служить лекар-
ственная гемолитическая анемия и посттрансфузионные реакции при переливании
несовместимых эритроцитов. Некоторые антибиотики (бензилпенициллин, стреп-
томицин. цефалоспорины) в организме адсорбируются на эритроцитах, вызывая
против себя гуморальный иммунный ответ с продукцией антител класса IgM и
IgG. Антитела взаимодействуют с лекарственным препаратом на поверхности эри-
троцитов, образовавшиеся иммунные комплексы фиксируют и активируют белки
системы комплемента, В результате происходит опосредованный комплементом
лизис эритроцитов (гемолитическая анемия). Повторный контакт пациента с
теми же или антигенно сходными веществами поддерживает сформировавшийся
иммунно-патологический процесс.
Антигены, аналогичные группоспецифическим антигенам эритроцитов, часто
присутствуют у микроорганизмов кишечника человека. Против них у человека
вырабатываются антитела, чаще класса IgM, взаимодействующие с группоспе-
цифическитии антигенами эритроцитов. Против антигенов собственных эри-
троцитов, даже если микроорганизмы кишечника несут аналогичные антигены,
иммунный ответ не развивается, сохраняется иммунологическая толерантность.
При переливании эритроцитов несовместимой группы крови циркулирующие изо-
гемагглютинины пациента взаимодействуют с ними и обеспечивают комплементо-
посредованный гемолиз. Повторные трансфузии могут усиливать иммунный ответ
и стимулировать накопление изогемагглютининов класса IgG.
Клинически посттрансфузионные реакции могут развиваться немедленно или
отсроченно — через 2-6 сут. Немедленные реакции обычно обязаны анти-АВО
группе-специфическим IgM антителам. Отсроченные реакции обычно связаны с
иммунным ответом против других эритроцитарных антигенов и обеспечиваются
изогемагглютининами класса IgG. При немедленных реакциях лизис эритроцитов
происходит в просвете сосудов, сопровождаясь гемоглобинурией и накоплением в
крови свободного гемоглобина. Отсроченные реакции, в которых участвуют анти-
эритроцитарные антитела класса IgG, менее активно связывают комплемент, чем
антитела IgM, поэтому лизис эритроцитов не бывает столь катастрофическим и
может сопровождаться фагоцитозом частично поврежденных эритроцитов во вне-
сосудистых пространствах. Гемолитическая болезнь новорожденных, развивающа-
яся в результате вынашивания резус-негативной матерью резус-полижительного
плода, также обязана антиэритроцитарным антителам класса IgG.
60 ЛАБОРАТОРНАЯ ИММУНОЛОГИЯ
Третий тип иммунопатологических процессов формируется в результате образо-
вания в организме иммунных комплексов «антиген-антитело». В зависимости от
массы, размера агрегируемых в комплексы молекул, а также от места депонирова-
ния комплексов в организме могут развиваться локальные или генерализованные
патологические пооцессы. Антитела в иммунных комплексах способны фиксиро-
вать и активировать белки системы комплемента, приводить к гиперпродукции
СЗа, С4а, С5а, С5Ь и других субкомпонентов комплемента. Некоторые из этих
продуктов обладают активностью анафилотоксинов и способны повышать про-
ницаемость сосудов, другие активируют эндотелиоциты и оказывают хемотакси-
ческое и активирующее действие на нейтрофильные лейкоциты. В результате зоны
фиксации иммунных комплексов оказываются инфильтрированными нейтрофи-
лами, которые, в свою очередь, фагоцитируют иммунные комплексы, выделяют в
окружающую среду гидролитические ферменты и цитотоксические продукты. При
поступлении антигена через пограничные гкани иммунные комплексы образуются
в субмукозном и дермальном слое, так что активация там комплемента приводит к
вовлечению в патологический процесс тучных клеток.
Циркулирующие иммунные комплексы мигут оседать на базальных мембранах
кровеносных сосудов и гломерул почек. Более мелкие по размерам иммунные
комплексы проникают через базальные мембраны и локализуются в суббазальных
слоях. При фиксации иммунных комплексов базальные мембраны подвергаются
атаке активированными нейтрофилами и комплементом Это приводит к повреж-
дению как клеток базальных мембран, так и близлежащих тканевых клеток.
Активация комплемента можрт индуцировать агрегацию тромбоцитов, высвобож-
дение факторов свертывания крови и образование микротромбов.
Подкожное или внутрикожное введение антигена, против которого присутству-
ют циркулирующие антитела в высоких титрах, сопровождается образованием в
месте введения иммунных комплексов с развитием в течение 4-6 ч местного вос-
паления (феномен Артюса). Тяжесть воспаления может варьировать от локальной
гиперемии и отека до некроза тканей. Гиперергическая реакция типа феномена
Артюса может развиться в легких при поступлении антигенов (споры грибов и
бактерий, протеины животных и птиц) с вдыхаемым воздухом. В этом случае
воспаление затрагивает альвеолы и приводит к развитию аллергического альвео-
лита.
Поступление в кровь антигена вызывает образование в больших количествах
циркулирующих иммунных комплексов. Циркулирующие комплексы антиген-
антитело накапливаются на синовиальных мембранах суставов, в стенках сосудов
и гломерулах почек. Клинические проявления — васкулиты, артриты и гломеру-
лонефриты. В качестве антигенов, вызывающих продукцию антител в высоких
титрах и образование иммунных комплексов, выступают микроорганизмы про-
стейшие. аутологичные и чужеродные белки и нуклеиновые кислоты. Именно
поэтому генерализованные иммунопатологические процессы III типа характерны
для аутоиммунных заболеваний (системная красная волчанка, ревматоидный
артрит), лекарственной аллергии (например, к бензатина бензилпенициллину и
сульфаниламиду), инфекционных заболеваний (гепатиты, инфекционный моно-
нуклеоз, менингит, малярия) и сывороточной болезни. Сывороточная болезнь
развивается у пациентов, которым с лечебной целью вводили лошадиные имму-
ноглобулины против микробных токсинов. Внутримышечно введенные белки
чужеродной сыворотки поступают в кровь, где взаимодействуют с имеющимися
или вновь синтезируемыми антителами. В последнем случае заболевание может
развиться через 6-14 сут после пассивной иммунизации пациента. Для сыворо-
точной болезни характерны повышение температуры, лимфоаденопатия эритема,
артриты, генерализованные васкулиты и отеки, иногда гломерулонефриты.
ЛАБОРАТОРНАЯ ИММУНОЛОГИЯ
61
В отличие от предыдущих вариантов иммунопатологических процессов, IV тип
гиперреактивности обязан формированию в организме клеточного иммунного
ответа против антигена, в частности накоплению антиген-специфических Thl- и
CD8' Тс-лимфоцитов. В местах повторного поступления в организм антигена
накапливаются и активируются иммунные антиген-специфические Т-лимфоциты.
Активированные Thl клетки продуцируют и секретируют IL-2, IFN-y, факторы,
ингибирующие миграцию макрофагов, и другие цитокины. Под влиянием цито-
кинов в очаге накапливаются и активируются макрофаги, в свою очередь секрети-
рующие ферменты, NO, цитотоксические продукты и др. медиаторы воспаления.
Высвобождение этих продуктов приводит к локальному повреждению и некрозу
клеток тканей. Эта иммуновоспалительная реакция развивается через 48-72 ч,
поскольку требуется время для инфильтрации места введения антигена иммун-
ными лимфоцитами и моноцитами и для их активации. В отличие от гиперерги-
ческих реакций III типа, в патогенезе которых участвуют нейтрофилы, реакции
IV типа обязаны в основном мононуклеарным клеткам. Реакции IV типа имеют
особое значение в защите организма от внутриклеточно размножающихся микро-
организмов, вирусов и паразитов. В случае если макрофаги и Тс-лимфоциты не в
состоянии уничтожить инфицированные клетки и самих возбудителей, моноциты
в очаге могут трансформироваться в эпителиоидные клетки (основной клеточный
элемент гранулем).
Гиперергические реакции IV типа лежат в основе патогенеза контактных
дерматитов, развивающихся в результате повторных контактов пациентов с
различными химическими веществами и растительными белками (формаль-
дегид, тринитрофенол, никель, лаки для волос, компоненты косметических
средств и др.). Будучи малыми молекулами, эти вещества поступают транс -
кутанно, образуют комплексы с белками кожи, захватываются дендритными
и другими иммуно-вспомогательными клетками и вызывают Т-клеточную
антиген-специфическую иммунную реакцию. Повторное поступление этих
молекул в организм вызывает инфильтрацию места их введения иммунными
Т-лимфоцитами и макрофагами.
При внутрикожном введении антигена развитие замедленной (через 24-96 ч)
гиперергической реакции свидетельствует о наличии у пациента антиген-
специфических Т-лимфоцитов, т.е. пациент ранее уже имел контакт с соот-
ветствующим антигеном и ответил на него клеточной иммунной реакцией.
Внутрикожные тесты с микробными белками используют в клинике для выяв-
ления инфицированных ими ранее иммунизированных пациентов и для оценки
антиген-специфического клеточного иммунитета у больных.
АЛГОРИТМ ЛАБОРАТОРНОГО ИССЛЕДОВАНИЯ ИММУННОЙ
СИСТЕМЫ
Объем и качество иммунологического лабораторного обследования больных
должны быть достаточными для подтверждения диагноза, прогнозирования раз-
вития заболевания и контроля эффективности проводимой терапии.
Наиболее широко диагностические иммунолабораторные тесты используют для
выявления в биологических материалах от больных патогенетически значимых
антигенов или для оценки специфической реакции иммунной системы пациента
(накопление антиген-специфических лимфоцитов и антител на такие антигены,
как опухолевые, трансплантационные, аллергены, рецепторы, гормоны и др.).
При первичном контакте с инфекционным антигеном в крови пациентов нака-
пливаются антитела — IgM. При повторном контакте с инфекционным агентом
накапливаемые в крови антитела — IgG.
62 ЛАБОРАТОРНАЯ ИММУНОЛОГИЯ
Методы определения антигенов и антител к ним отличаются исключительно
высокой специфичностью и чувствительностью, что позволяет в биологических
материалах выявлять исследуемые молекулы, представленные в пико- и нанограм-
мовых количествах. Простота и относительная дешевизна серологического обсле-
дования больных обеспечили в современной клинике широкое распространение
лабораторных методик. Динамика накопления в крови антител к различным анти-
генам возбудителей инфекций позволяет прогнозировать течение заболевания и
его исход.
Наиболее трудоемко и объемно исследование состояния и диагностика заболе-
ваний самой иммунной системы. Алгоритм проведения таких исследований опре-
деляют клиническая картина заболевания и симптомокомплексы, характерные
для тех или иных нарушений функций иммунной системы. На первом этапе таких
обследований целесообразны скрининговые исследования, позволяющие выявить
количественные и качественные отклонения в состоянии основных компонентов
врожденного и приобретенного иммунитета больного (табл. 16-22).
Таблица 16-22. Скрининговые иммуно-лабораторные тесты в диагностике нарушений иммунной
системы
Компоненты иммун- ной системы Тесты
Система комплемента Определение активности комплемента в свежей сыворотке крови (CH Г|)
Фагоцитарная система Определение в крови содержания и морфологическая характеристика моноцитов и гранулоцитов разного типа (нейтрофильные, эозинофильные, базофильные)
Естественные килле- ры (МК-клетки) Определение содержания в крови NK-клеток
Система Т-лимфоцитов Определение в периферической крови содержания Т-лимфоцитов и их субпопуля- ций с маркерами: CD3*, С04+ и CD8'
Система В-лимфоцитов Определение в периферической крови содержания В-лимфоцитов. Определение з крови концентрации IgM, IgG, IgA и IgE. Определение в периферической крови содержания изогемагглютининов, антител, специфичных к антигенам вакцин, ранее использованных дня иммунизации пациента, а также циркулирующих иммунных комплексов
По данным клиник США, эффективность скрининговых исследований заболева-
ний иммунной системы достигает 75%, при этом стоимость их проведения и трудо-
вые затраты намного ниже, чем при осуществлении дополняющих исследований.
Иммунная система рано реагирует на самые разнообразные, особенно неблаго-
приятные воздействия на организм. Именно поэтому у больных степень участия
в реакции отдельных врожденных или приобретенных компонентов иммунной
защиты способствует постановке диагноза и прогнозированию развития патоло-
гического процесса (табл. 16-23).
Таблица 16-23. Факторы иммунитета, исследуемые при различных клинических проявлениях
нарушений иммунной системы
Заболевания Факторы врожденного иммунитета Факторы антиген-специфического (адаптивного) иммунитета
клеточные гуморальные клеточные гуморальные
Вирусные инфекции Содержание в крови NK-клеток и моно- цитов Содержание в крови интерферонов и неоптеринов Содержание в крови лимфоцитов, субпопу- ляций Т-лимфоцитов; исследование мар- керов активации Т-клеток и проли- феративного ответа Т-лимфоцитов на митогены и антигены Содержание IgM, IgG, IgA и антивирусных антител
ЛАБОРАТОРНАЯ ИММУНОЛОГИЯ 63
Поодолжение табл. 16-23
Заболевания Факторы врожденного иммунитета Факторы антиген-специфического {адаптивного) иммунитета
клеточные гуморальные клеточные гуморальные
Бактериальные инфекции Содержание и морфология ней- трофилов, моно- цитов, фагоцитарная активность нейтро- филов (хемотаксис, бактерицидность), признаки активации нейтрофилов (хеми- люминесценция, НСТ- тест); исследование адгезин.тозитивных клеток (CD11/CD18) Гемолитическая актив- ность комплемента, содержание СЗ и С4, опсонизирующая активность сыворотки крови; содержание в крови и биоматериа- лах провоспалитель- ных и антивоспали- тельных цитокинов, прокальцитонина, дефензинов Содержание в крови лимфоцитов, Т-лимфоцитов разных субпопуляций; иссле- дование маркеров активации Т-клеток и пролиферативного ответа Т-лимфоцитов на митогены и анти- гены; содержание В-лимфоцитов Содержание в крови IgM, IgG, IgA. субклассов IgG. изогемаг- глютининов и антибактериаль- ных антител
Микотические инфекции Содержание в крови нейтрофилов, моно- цитов и N КТ-клеток; фагоцитарная актив- ность нейтрофилов (хемотаксис, бактери- цидность), признаки активации нейтрофи- лов (хемилюминес- ценция, НСТ-тест) Содержание в крови дефензинов, гемо- литическая актив- ность комплемента, содержание СЗ и С4, опсонизирующая активность сыворотки крови; содержание провоспалительных цитокинов в биома- териалр Содержание в крови лимфоцитов. Т-лимфоцитов разных субпопуляций; иссле- дование маркеров активации Т-клеток и пролиферативного ответа Т-лимфоцитов на митогены и анти- гены Содержание антител и имму- ноглобулинов разных классов
Паразитарные инфекции Содержание в крови эозинофилов Содержание в крови эозинофильных про- теинов Содержание в крови лимфоцитов, субпопу- ляций Т-лимфоцитов, маркеры активации Т-клеток Содержание IgM IgG, IgA; содержание в крови общего и антиген- специфического IgE
Аллергические заболевания* Содержание в крови и бииматериалах эозинофилов, базо- филов аллерген- индуцированная активация- дегрануляция базо- филов Содержание в крови и биоматериалах и освобождение из кле- ток медиаторов аллер- гического воспаления (гистамин, триптаза, эйкозаноиды, основ- ные белки эозинофи- лов, цитокины — IL-4, IL-5, IL-13) Содержание в крови лимфоцитов, Т-лимфоцитов разных субпопуляций; иссле- дование маркеров активации Т-клеток и пролиферативного ответа Т-лимфоцитов на митогены и аллер- гены Содержание в крови общего и антиген- специфического igE
Аутоиммунная патология Содержание ней- трофилов (нейтро- филез — признак острой фазы вос- паления, нейтро- пения — признак аутоиммунного пора- жения нейтрофилов) Гемолитическая актив- ность комплемента, содержание СЗ и С4 компонентов компле- мента; содержание цитокинов Содержание в крови лимфоцитов, Т-лимфоцитов разных субпопуляций; иссле- дование маркеров активации у Т-клегок и пролиферативного ответа Т-лимфоцитов на митогены и специ- фические антигены Содержание антител к раз- нообразным аутоантигенам; исследование циркулирующих иммунных ком- плексов
64
ЛАБОРАТОРНАЯ ИММУНОЛОГИЯ
Окончание табл. 16-23
Заболевания Факторы врожденного иммунитета Факторы антиген-специфического (адаптивного) иммунитета
клеточные гуморальные клеточные гуморальные
Пролифера- тивные заболе- вания Содержание эозино- филов, нейтрофилов, моноцитов: имму- нофенотипирование лейкоцитов Содержание опухоле- вых маркеров в био- логическом материале Содержание в крови лимфоцитов, Т- и В-лимфоцитов и их субпопуляций, иссле- дование маркеров активации, проли- феративного ответа лимфоцитов на мито- гены и специфические антигены; иммунофе- нотипирование Содержание иммуноглобули- нов, М-протеина и парапротеинов в крови;иссле- дование белка Бенс-Джонса в моче
* Для диагностики аллергических заболеваний также оценивают реакции больного на различные
аллергены.
О вовлечении иммунной системы в острый процесс можно судить уже при рас-
смотрении результатов общего клинического анализа крови. На это указывает
увеличение в крови количества полинуклеарных лейкоцитов, повышение доли
юных клеток и ускоренная реакция оседания эритроцитов. Повышение в крови
содержания С-реактивного белка и других белков острой фазы, лизоцима, про-
кальцитонина также указывает на острое воспаление, причем накопление в крови
прокальцитонина особенно характерно для системных бактериальных инфекций
и бактериального сепсиса.
На вирусную природу острого воспаления указывает увеличение в крови содер-
жания NK-клеток и моноцитов, при котором моноциты отвечают на вирусы еще и
накоплением в крови неоптерина. Хронические инфекции, в особенности вызыва-
емые внутриклеточно размножающимися микроорганизмами или возбудителями
гранулематозных инфекций, также сопровождаются активацией и накоплением
моноцитов.
При наличии у больных симптомокомплексов, характерных для нарушений
приобретенного иммунитета, лабораторное обследование больных начинают со
скрининговых исследований, включающих оценку’ у больного общего количества
лимфоцитов, а также исследование содержания Т- и В-клеток. На скрининговом
уровне достаточно исследовать в крови у больных содержание Т-клеток с фено-
типическими маркерами CD3+, CD4~ (хелперные и регуляторые клетки), CD8+
(цитотоксические клетки).
О состоянии В-клеточного звена иммунитета можно судить косвенно по содер-
жанию в сыворотке крови обследованных иммуноглобулинов, изогемагглютини-
нов и антител к антигенам вакцин, ранее вводимых обследуемому.
Для уточнения причип и механизмов нарушения функций иммунной систе-
мы больного необходимо проведение аналитических диагностических тестов.
Анализы этого уровня предполагают оснащение лаборатории оборудованием и
обеспечение реагентами, позволяющими в автоматическом режиме исследовать
клетки крови больного по многим показателям и осуществить функциональные
тесты, оценивающие активность соответствующих клеток.
Аналитические исследования позволяют оценить у больного не только содер-
жание клеток определенного фенотипа, охарактеризовать их функциональную
активность, но и определить состояние центральных органов иммунной системы
и ассоциированной со слизистыми оболочками лимфоидной ткани (выход в кровь
наивных Т- и В-лимфоцитов) (табл. 16-24).
ЛАБОРАТОРНАЯ ИММУНОЛОГИЯ
65
Таблица 16-24. Аналитические тесты в диагностике нарушений иммунной системы
Показатели Тесты
Система ком- племента Определение в сыворотке крови содержания и активности компонентов комплемента (в особенности СЗ и С4) и ингибиторов комплемента (С1|п|1)
Фагоцитарная система Исследование микробицидной активности нейтрофилов и мононоцитов в крови; качественное и количественное исследование восстановления гранулоцитами и моно- ноцитами (НСТ-тест); количественное исследование отдельных этапов фагоцитарной активности лейкоцитов (подвижность и хемотаксис, адгезия, захват, образование фаго- лизосом, внутриклеточный киллинг, дегрануляция); исследование в очагах воспаления и в крови цитокинов — эффектооов и регуляторов воспаления; исследование аллерген- индуцированно! о высвобождения гистамина или лейкотриенов базофилами крови; исследование в крови эозинофильных протеинов (основного эозинофильного белка, катионного нейротоксина, эозинофильной пероксидазы, эозинофильного катионного белка)
Естественные киллеры (NK-клетки) Исследование общего количества NK-клеток и отличающихся высокой цитотоксичностью по отношению к клеткам миелоидной линии К-562 (CD16+M0ftf CD56'(//m), или активно продуцирующих цитокины (CD16'o7m СЕ 5frbrigh)
Система Т-лимфоцитов Определение в периферической крови содержания субпопуляций CD4+ и CD84 Т-клеток, различающихся функционально (активированные, наивные, иммунные, зрелые, незре- лые, памяти, апоптотические, с антигенраспознающими рецепторами типов у8 и ос(3 и др.); исследование содержания регуляторных Т-клеток (CD4+, CD25\ foxpj, Тг, клеток, Th1, Th2, Th17 и др. субпопуляций); исследование функциональной активности Т-клеток крови и их субпопуляций (пролиферативная активность, продукция цитокинов, цитоток- сическая активность и др.); исследование содеожания и функциональной активности антигенореакгивных (в том числе аутореактивных) Т-лимфоцитов периферической крови; исследование в крови определенных цитокинов и растворимых цитокиновых рецепторов
Система В-лимфоцитов Определение в периферической крови содержания функционально отличающихся субпопуляоий В-лимфоцигов (наивные, иммунные, активированные, зрелые, незрелые, памяти, апоптотические, несущие поверхностные иммуноглобулины разных классов); исследование функциональной активности В-лимфоцитов, парапротеинов и криоглобули- нов, определение в сыворотке крови содержания IgG и IgA разных субклассов, аллерген- специфических IgE; определение в секретах и смывах содержания секреторного IgA и иммуноглобулинов иных классов
При углубленном (аналитическом) исследовании лимфоидной системы оцени-
вают содержание в крови наивных и иммунных, покоящихся и активированных
клеток, хелперных, регуляторных и цитотоксических лимфоцитов, долгоживущих
клеток памяти и клеток, реализующих программу апоптоза.
При лабораторном исследовании лимфоидной системы возможно не только
определить содержание соответствующих клеток в крови и других биоматериалах,
но и оценить выраженность экспрессии на клетках соответствующих маркеров.
Оба этих показателя исследуют при анализе пула клеток с помощью набора моно-
клональных антител против мембранных молекул клеток с использованием про-
точного цитофлюориметра.
Уточнение причин и механизмов врожденной и приобретенной несостоятельно-
сти иммунной системы больного может потребовать постановки in vitro функцио-
нальных клеточных тестов, в которых оценивают способность различных клеток
иммунной системы реагировать на митогены, антигены и др. агенты, активируясь,
пролиферируя и дифференцируясь, продуцируя цитокины, хемокины, адгезивные
молекулы и экспрессируя разнообразные рецепторы к ним.
Глава 17
Диагностика аутоиммунных
заболеваний
КРИТЕРИИ, КЛАССИФИКАЦИЯ И ЭПИДЕМИОЛОГИЯ
АУТОИММУННЫХ ЗАБОЛЕВАНИЙ
Ведущий патогенетический механизм развития аутоиммунных
заболеваний — иммунный ответ по отношению к компонентам нор-
мальной человеческой ткани и органов. Для классификации ауто-
иммунных процессов применяют медико-биологические критерии
(табл. 17-1), но их сложно использовать в клинической практике,
поскольку для их понимания требуется большой объем научных
Таблица 17-1. Основы критериев аутоиммунных заболеваний (Rose—Вопа,1993)
________________________________________________________________________Прямые свидетельства____________________________________________________
Антитело-опосредованные__________________________________________________
♦ Аутоантитело в крови, влияющее на функции органа_____________________
• Аутоантитело, фиксированное в ткани__________________________________
• Иммунный комплекс, локализованный в ткани____________________________
• Воспроизведение заболевания при пассивном переносе lg________________
Клеточно-опосредованные ___ ________________ ___________________________
• Перенос Т-клеток в SCID-мышь с । каневым имплантатом_________________
• Цитотоксичность аутоагрессивных Т клеток in vitro ___ _________
___________________________Непрямые свидетельства______________________
Экспериментальная иммунизация _ _
Иммунизация антиидиотипическим антителом________________________________
Экспериментальмая дизрегуляция иммунной системы у животных______________
________________________Дополнительные свидетельства_____________________
Аутоантитела в крови______________________________________________________
Ассоциация с другими аутоиммунными заболеваниями________________________
Ассоциация с HLA-генотипом______________________________________________
Лимфоцитарная инфильтрация органа_______________________________________
Ответ на иммуносупрессивную терапию_____________________________________
Примечание. SCID — severe combinedimmunodefit iency (англ. - «тяжелый комби-
нированный иммунодефицит»).
данных. В то же время аутоиммунные заболевания (АИЗ) обладают
рядом четких клинических характеристик, к которым относятся
следующие:
ДИАГНОСТИКА АУТОИММУННЫХ ЗАБОЛЕВАНИЙ 67
❖ процессы неизвестной этиологии с хроническим системным/локальным вос-
палением;
о возникают у лиц с набором генов иммунного генеза (генов системы HLA);
заболевания поддаются терапии иммуносупрессивными препаратами;
❖ в крови и биологических жидкостях обнаруживают аутоантитела.
Аутоантитела, направленные против собственных молекул организма, зачастую
не связаны с непосредственным повреждением ткани. Чаще всего они являются
«свидетелями» аутоиммунного процесса. Поэтому определение аутоантител при
аутоиммунных заболеваниях служит важным инструментом в установлении и
последующем ведении потенциальных пациентов. Особое значение приобретает
обнаружение аутоантител на ранних этапах заболевания, когда морфологические
признаки разрушения ткани еще отсутствуют.
Наиболее известная классификация подразделяет АИЗ на органонеспецифи-
ческие и органоспецифические (табл. 17-2). Специфические АИЗ поражают кон-
кретные органы или ткани, это происходит при тиреоидите Хашимото или сахар-
ном диабете I типа. Органонеспецифические иммунные реакции могут поражать
несколько систем органов, как это происходит при ревматических заболеваниях.
В последнем случае нередко клиницисты определяют аутоиммунный ответ как
«системный», примером которого служит системная красная волчанка (СКВ).
Таблица 17-2. Классификация аутоиммунных заболеваний
Органоспецифические заболевания Рассеянный склероз
Зоб Хашимото
Первичная микседема
Тиреотоксикоз (болезнь Грейвса)
Пернициозная анемия
Первичный билиарный цирпоз
Аутоиммунный гепатит
Сахарный диабет I типа
Болезнь Крона
Язвенный колит
Аутоиммунная гемолитическая анемия
__________________________________Идиопатическая тромбоцитопеническая пурпура___________
Органонеспецифические заболевания Системная красная волчанка
Гранулематоз Вегенера
Склеродермия
Смешанное заболевание соединительной ткани
Дерматомиозит
Ревматоидный артрит___________________________________
По мере понимания механизмов развития аутоиммунных заболеваний эта клас-
сификация постепенно утрачивает свое значение, так как не отражает современно-
го представления об АИЗ.
АИЗ относят к редким патологиям, хотя их встречаемость достигает 10-20%
всех общетерапевтических заболеваний. К частым АИЗ, которые поражают
0,1-1% популяции, относят диффузный токсический зоб, тиреоидит, ревмато-
идный артрит, аутоиммунный гастрит, псориаз. Более редкие АИЗ встречаются
у 0.001-0,01% популяции, системные ревматические заболевания, первичные
гломерулонефриты, рассеянный склероз, витилиго, миастения, аутоиммунные
заболевания печени, аутоиммунная тромбоцитопения, пернициозная и гемоли-
тическая анемия, инсулин-зависимый диабет, саркоидоз, идиопатические увеиты.
Ряд клинических синдромов с классическим аутоиммунным патогенезом отме-
чают менее чем у 0,001% населения. Последняя группа представлена болезнью
Аддисона, синдромом Гудпасчера, синдромом Гийена-Барре.
АИЗ могут возникать в любом возрасте, хотя для каждой нозологической формы
имеются свои пики заболеваемости. Отдельно выделяют синдромы, которые воз-
68 ДИАГНОСТИКА АУТОИММУННЫХ ЗАБОЛЕВАНИЙ
никают у новорожденных от матерей с А ИЗ, обусловленные трансплацентарным
переносом патогенных аутоантител из организма матери. К таким транзиторным
состояниям относят врожденный гипертиреоз, неонатальную миастению и неона-
тальную красную волчанку. Примечательно, что женщины чаще болеют АЙЗ,
составляя, по разным оценкам, более 75% всех больных. Несмотря на множество
существующих теорий, причина выраженной половой предрасположенности оста-
ется малопонятной.
Определение термина «аутоантитело» и роль аутоантител
при аутоиммунных заболеваниях
Дать строгое определение термину «аутоантитело» далеко не так легко, как
кажется. Это связано с тем, что как направленность аутоиммунной реакции,
вызывающей синтез аутоантител, так и методы обнаружения аутоантител имеют
ряд условностей. С иммунологической точки зрения, «аутоантителами» являются
иммуноглобулины человека, способные специфически связывать антигенные эпи-
топы молекул собственного организма. Иммунный ответ, который лежит в основе
этой реакции, направлен на антигены вирусов, бактерий, экзогенных веществ, а
эпитопы собственных молекул становятся мишенями лишь в силу антигенного
сходства. Таким образом, сложно точно отделить аутоиммунный ответ от есте-
ственных реакций иммунной системы.
Данный парадокс еще больше усложняется тем фактом, что для обнаружения
аутоантител в лаборатории в большинстве случаев используют антигены, отли-
чающиеся от антигенов человеческого происхождения. Так, в качестве антиген-
содержащего субстрата применяют ткани или очищенные антигены животных,
перевиваемые клеточные линии и рекомбинантные белки, полученные на основе
человеческих генов в различных системах экспрессии белка. По своей антигенной
структуре рекомбинантные антигены значительно отличаются от нативных белков
тканей человека. Это обусловлено их третичной структурой гликозилированием,
а также присутствием аминокислотных последовательностей для эффективного
синтеза белка и его очистки. Именно поэтому одновременно с «аутоантителами»
при применении иммунологических методов часто определяют и иммуноглобу-
лины. реагирующие с чужеродными эпитопами. Более совершенным подходом
при выявлении аутоантител считают использование модельных «дизайнерских»
антигенов, структура которых основана на изучении иммунодоминантных моле-
кул аутоантигенов человека. В этом случае они могут сильно отличаться от
исходных мишеней. Наиболее революционным примером служит создание цикли-
ческого цитруллинированного пептида для диагностики ревматоидного артрита.
Используемый в качестве антигена цитруллиновый пептид был выбран в ходе
скрининга случайных пептидных библиотек, которые че имели ничего общего с
исходной последовательностью белка филлагрина, изучение которой позволило
установить характеристики иммунодоминантного эпитопа антицитруллиновых
антител. Для создания тест-системы все шире используют рекомбинантные
модельные белки, аминокислотная последовательность которых состоит из чере-
дования иммунодоминантных эпитопов антигена. В этом случае серологическая
реакция иммуноглобулина человека с таким «модельным» исскуственным анти-
геном уже не является аутоантителом в полном смысле этого слова. Поэтому
термин «аутоантитело» подразумевает определенную «серологическую» реакцию,
присущую тому или другому' аутоиммунному заболеванию. Результат этой серо-
логической реакции зависит от используемых лабораторных методов, каждый из
которых имеет свои особенности и ограничения.
ДИАГНОСТИКА АУТОИММУННЫХ ЗАБОЛЕВАНИЙ 69
Благодаря тому, что аутоантитела связываются с молекулами сходной структу-
ры или молекулами в составе одной ткани, клетки, органеллы или молекулярного
комплекса, их подразделяют на семейства. В большинстве случаев АИЗ, сопро-
вождающиеся образованием аутоантител одного семейства, имеют сходную кли-
ническую симптоматику и патогенез. Например, семейство антител к антигенам
цитоплазмы нейтрофилов направлено на разнородные по структуре и свойствам
белки гранул цитоплазмы лейкоцитов. В то же время большинство конкретных
разновидностей аутоантител этого семейства встречаются при системных васку-
литах. Наличие аутоантител на фоне АИЗ указывает на участие аутоантител в
механизмах аутоиммунной реакции. Однако прямое участие аутоантител в раз-
витии патофизиологических процессов, характерных для данного АИЗ, не всегда
представляется доказанным. В таком случае принято говорить об аутоантителах
как «свидетелях» иммунологических реакций.
Большинство разновидностей аутоантител сравнительно часто может быть
обнаружено у клинически здоровых лиц. К аутоантителам, которые могут стать
случайной находкой у клинически здоровых лиц, относят антинуклеарный фактор
(3%). ревматоидный фактор (3%), антитела к тиреопероксидазе (4%), антитела
к кардиолипину (1%). антитела к миокарду (5%), антитела к скелетной муску-
латуре (3%). Частота встречаемости многих аутоантител нарастает с возрастом.
Аутоантитела образуются на фоне инфекций, травмы и при новообразованиях,
которые не сопровождаются аутоиммунными проявлениями. В этом случае ауто-
антитела могут нести самостоятельную иммунную функцию, например участво-
вать в клиренсе тканевых антигенов. Антинуклеарный фактор часто диагностиру-
ют при болезни «трансплантат против хозяина» и солидных опухолях, что можно
объяснить аллоиммунным или противоопухолевым иммунным ответом. Это
доказывает, что в определенном смысле аутоантитела — компонент естественного
иммунного ответа, участвующий в удалении из системы циркуляции «состарен-
ных» белков
Высокое содержание аутоантител не всегда сопровождается высокой клиниче-
ской активностью АИЗ. Это указывает на то, что ряд характеристик аутоантител,
в том числе аффинность, количество, присутствие тканевого воспаления и других
реакций иммунной системы, крайне важны для проявления патогенетического
действия аутоантител. С другой стороны, при АИЗ может наблюдаться ситуация,
при которой антитела постепенно исчезают из сыворотки с увеличением длитель-
ности болезни. Так, антитела к островковым клеткам поджелудочной железы у
большинства больных сахарным диабетом I типа исчезают через 1 год после кли-
нического дебюта заболевания.
Для доказательства принципиальной роли аутоантител в развитии АИЗ необ-
ходимо соблюдение ряда формальных условий. Предполагают, что в пораженной
ткани можно обнаружить иммунные комплексы, состоящие из аутоантитела и
соответствующего аутоантигена. При моделировании заболевания на экспери-
ментальных животных сыворотка из организма больного животного или челове-
ка, перенесенная здоровому животному, или аутоантитело, полученное иммуни-
зацией аутоантигеном, приводит к изменению структуры и функции, схожему с
таковыми при оригинальном АИЗ. Аутоантитело должно оказывать действие на
клетки клеточных культур или изолированной ткани ex vivo. И, наконец, должна
существовать корреляция между титрами аутоантитела и активностью заболева-
ния. Далеко не для всех аутоантител могут быть соблюдены все эти требования.
Изученные механизмы участия аутоантител в иммунопатогенезе АИЗ представ-
лены в табл. 17-3.
70
ДИАГНОСТИКА АУТОИММУННЫХ ЗАБОЛЕВАНИЙ
Таблица 17-3. Патогенетические механизмы действия аутоантител при аутоиммунных заболева-
ниях
Механизм Основные аутоантитела
Взаимодействие с белковыми системами крови и биологических жидкостей Волчаночный антикоагулянт Антитела к C1q и C1INH Антитела к внутреннему фактору Касла
Взаимодействие с рецептором (слущивание, блокада или стимуляция) Антитела к ацетилхолиновому рецептору Антитела к рецептору тиреотропного гормона Антитела к ионным каналам
Комплемент-опосредованное разрушение ком- понентов клеток и ткани Антитела к эритроцитам Антитела к антигенам десмосом Антитела к антигенам базальной мембраны кожи Антитела к базальной мембране клубочка Антитела к ганглиозидам
Перекрестное связывание молекул и актива- ция клетки Антинейтрофильные цитоплазматические антитела
Отложение иммунных комплексов с реактив- ным воспалением Антитела к дсДНК и нуклеосомам Ревматоидный фактор Отложения полимерного Ig и комплемента в мезангии клубочка и стенках сосудов почки при IgA-нефропатии
Трансплацентарный перенос с возникнове- нием транзиторной врожденной клинической картины заболевания Антитела к SSA при СКВ Антитела к рецептору тиреотропного гормона Антитела к десмосомам Антитела к ацетилхолиновому рецептору
Опсонизация, фагоцитоз и изменение клирен- са молекул и клеток Антитела к гликопротеинам Hb/llla Антитела к окисленным липопротеинам
АНТИТЕЛА И АУТОИММУННЫЕ ЗАБОЛЕВАНИЯ
Ряд особенностей отличают методы обнаружения аутоантител от оцен-
ки содержания других биологических молекул в биологических жидкостях.
Аутоантитела представляют собой иммуноглобулины классов IgG, IgA, IgM,
специфически связывающиеся с антигенными эпитопами молекул человеческого
организма. Образование аутоантител представляет собой нормальный биологи-
ческий феномен, а связывание иммуноглобулинов с собственными антигенами
можно зафиксировать в сыворотке любого человека. В норме существует оха-
рактеризованная популяция СО5+ В-клеток, синтезирующая полиреактивные
низкоаффинные иммуноглобулины, способные связываться со многими антиге-
нами. Роль аутоантител в норме и причина их появления изучена недостаточно.
Вероятнее всего причиной образования аутоантител является перекрестная реак-
тивность иммуногенных эпитопов между антигенами инфекционных агентов и
собственными молекулами организма. Спектр антигенных стимулов, окружаю-
щих человека, постоянно меняется, что ведет к образованию низкоаффинных
непатогенных аутоантител.
Аутоантитела обнаруживают при инфекционных и онкологических заболева-
ниях, воспалительных процессах различного генеза, хроничес ких интоксикациях
(алкоголизме) либо на фоне приема лекарственных препаратов. Наличие ревма-
тоидного фактора при вирусном гепатите С, антимиокардиальных антител при
вирусных миокардитах указывает на большое значение инфекции в генезе аутоим-
мунных процессов. Встречаемость АНА несколько возрастает у пациентов с онко-
логическими заболеваниями, причем индукция АНА на фоне опухоли указывает
на лучший прогноз у онкологических больных. Встречаемость ряда аутоантител,
в частности ревматоидного и антинуклеарного фактора, нарастает в пожилом воз-
расте. Однако титры аутоантител, присутствующих в норме, низки и сравнительно
нестойки.
ДИАГНОСТИКА АУТОИММУННЫХ ЗАБОЛЕВАНИЙ 71
Возможность использования конкретной разновидности аутоантител в качестве
диагностического показателя определяется их встречаемостью при аутоиммунном
заболевании. Встречаемость тех аутоантител, которые используются для клини-
ческой диагностики, обычно составляет более 60-70% при патологии, менее 1%
в норме и при других заболеваниях. Хорошие клинике-лабораторные параметры
многих разновидностей аутоантител позволяют рассматривать их в качестве
лабораторных маркеров этих заболеваний, т.е. высокоспецифичных лаборатор-
ных тестов, обладающих исключительной диагностической информативностью.
Высокоспенифичными серологическими маркерами называют те разновидности
аутоантител, которые встречаются исключительно при этом заболевании. Так, к
высокоспецифичным серологическим маркерам относят анти-дсДНК и анти-Sm
при СКВ, антитела к Scl-70-антигену при склеродермии. Некоторые аутоантитела
могут быть высокоспецифичными маркерами одновременно нескольких групп
заболеваний. Например, антитела к цитоплазме нейтрофилов образуются как при
гранулематозных васкулитах, так и при воспалительных заболеваниях кишечни-
ка. Ряд аутоантител, характерных для аутоиммунных заболеваний, сравнительно
часто регистрируют у здоровых лиц без признаков аутоиммунного заболевания,
что значительно ограничивает их диагностическое использование. К примеру,
клиническое использование антител к скелетным мышцам при миастении или
антител к миокарду ограничено, поскольку они могут часто встречаться в норме
либо при неиммунных заболеваниях.
Обычно появление высоких титров специфических аутоантител у клинически
здоровых лиц рассматривается в качестве фактора риска развития аутоиммунного
заболевания. Аутоантитела в сыворотке крови могут обнаруживаться за несколько
месяцев до первых клинических проявлений заболевания. Так, при ревматоид-
ном артрите антицигруллиновые антитела MOiyr быть обнаружены за 1,5 года до
дебюта артрита, обнаружение антинейтрофильных антител предваряет развитие
гломерулонефрита на несколько месяцев.
Клиническое значение обнаружения аутоантител отчасти зависит от титров
содержания аутоантител в сыворотке обследуемого. Обычно высокое содержа-
ние антител указывает на высокую аффинность последних. Это, в свою очередь,
характеризует специфичность и выраженность иммунного ответа. Обнаружение
высоких титров антител, как правило, указывает на аутоиммунное заболевание, в
то время как низкие титры часто отмечаются в норме и не специфичны. В отсут-
ствие иммуносупрессивного лечения при аутоиммунных заболеваниях содержа-
ние аутоантител обычно сохраняется на высоком уровне в течение длительного
времени. При наличии антител на фоне инфекции или неиммунного воспаления
титры антител обычно нестойки. Аутоантитела могут быть индуцированы вос-
палительным, инфекционным, онкологическим процессом или спровоцированы
назначением определенных препаратов. Для того, чтобы отличить транзиторную
индукцию аутоантител от хронического присутствия аутоантител при аутоиммун-
ном заболевании, может потребоваться проведение повторного исследования.
Учитывая, что время полужизни сывороточных иммуноглобулинов составляет
около 3 мес, минимальное время между повторными исследованиями должно пре-
вышать полгода.
В то же время продукция аутоантител при аутоиммунных заболеваниях не
является постоянной величиной. Встречаемость аутоантител может нарастать с
увеличением длительности аутоиммунного заболевания, как это происходит при
ревматоидном артрите, или снижаться ниже порога детекции в течение первого
года заболевания при сахарном диабете. Аутоантитела синтезируются в составе
нормального пула иммуноглобулинов сыворотки крови плазмоцитами костно-
го мозга, лимфатических узлов и селезенки, однако в ходе иммунного ответа
при аутоиммунном процессе продукция антител в очаге воспаления может при-
72
ДИАГНОСТИКА АУТОИММУННЫХ ЗАБОЛЕВАНИЙ
обретать особые характеристики, например может измениться клональность
иммуноглобулинов. Например, при рассеянном склерозе отмечается выраженное
увеличение местного синтеза антител и иммунный ответ в ликворе становится
олигоклональным, а при смешанной криоглобулинемии в состав кпиоглобулинов
сыворотки входит моноклональный ревматоидный фактор.
Хотя большинство аутоантител служат свидетелями аутоиммунного воспале-
ния и их содержание не коррелируют с активностью заболевания, обнаружение
определенных типов аутоантител при ревматических заболеваниях указывает на
характерные особенности клинического течения. Так, у пациентов, имеющих в
сыворотке специфический набор антител, наблюдается особенное течение заболе-
вания, отличное от симптоматики больных, не имеющих этих антител. При СКВ.
сопровождающейся образованием антител к Ro/SS-A, гломерулонефрит встреча-
ется реже, чем у больных, имеющих высокий титр антител к дсДНК, но имеется
определенный риск развития поражений кожи и склонность к повышенной фото-
чувствительности. В связи с этим определение антинуклеарных антител (АНА)
позволяет врачу предсказать, а иногда и предотвратить развитие осложнений.
Динамика титра определенных видов антител в ходе заболевания имеет прогно-
стическое значение. Увеличение концентрации антител к дсДНК предшествует
вспышке СКВ, и ранняя патогенетическая терапия может уберечь больного от
развития клинических симптомов или уменьшить их остроту;
Выделение больных с тем или иным типом аутоантител в отдельную, часто
клинически гомогенную, группу подтверждается обнаружением определенно-
го HLA-генотипа больных с конкретным типом АНА. К примеру, пациенты с
дерматомиозитом, у которых в сыворотке крови обнаруживают антитела к Jo-1,
характеризуются исключительно HLA-DR3 (DRwl7) и HLA-DRw52-генотипом.
Таким образом, результаты исследования содержания аутоантител и их спектра
указывают на особенности функционирования иммунной системы и отражают ее
особую предрасположенность к формированию аутоиммунных ответов.
Благодаря единому патогенетическому механизму индукции аутоантител, кон-
кретные их разновидности могут быть сгруппированы в семейства, реагирующие
с мишенями одного рода. Например, все представители семейства АНА реагируют
с рибонуклеопротеиновыми антигенами ядер; аутоантитела, относящиеся к семей-
ству антинейтрофильных цитоплазматических антител, реагируют с набором
антигенов, локализованных в азурофильных гранулах цитоплазмы нейтрофилов.
Каждая разновидность аутоантител, даже направленная к одному’ антигену, пред-
ставляет собой поликлональную популяцию антител, представленную молекулами
иммуноглобулинов с разными физико-химическими свойствами. Это обусловлено
тем. что поликлональные антитела реагируют с разными эпитопами на поверх-
ности молекулы-антигена, причем сила связи каждой молекулы антитела с анти-
геном (аффинность) различна. Естественно, что в идеале иммунологический лабо-
раторный метод должен позволять установить не одну разновидность молекул
(как это происходит в большинстве традиционных лабораторных тестов), а спектр
аутоантител, направленных на разные антигены. Причем каждое антитело значи-
тельно отличается от других по заряду, молекулярной массе и силе связывания
с антигеном, которая зависит от условий реакции. Такого идеального метода не
существует. На практике для точного описания спектра аутоантител используют
несколько методов, каждый из которых имеет свои достоинства и недостатки. В
результате интерпретация результатов исследования одного лабораторного пока-
зателя зависит от метода, с помощью которого он определен.
Особенности обнаружения аутоантител приводят к тому, что один тест может
иметь до 10 вариантов ответов, включающих наличие аутоантитела, его титр, а
также несколько разнообразных типов связывания антител с субстратом. Обычно
на первом этапе тестирования применяют метод, обладающий максимальной
ДИАГНОСТИКА АУТОИММУННЫХ ЗАБОЛЕВАНИЙ 73
чувствительностью, в котором присутст вует максимально широкий спектр анти-
генов. Уточнение специфичности обнаруженных аутоантител возможно при
использовании подтверждающих тестов, назначенных по результатам первичного
тестирования.
Еще одной особенностью практического обнаружения аутоантител, вытекаю-
щей из биологической природы этого феномена, является то, что не существует
абсолютно корректного подхода к измерению содержания аутоантител. Дело
в том, что большинство иммунологических тестов построено на связывании
аутоантитела и антигена-мишени. Результат теста зависит как от концентрации
аутоантител, так и от аффинности аутоантитела и антигена. При аутоиммунных
заболеваниях специфические антитела обычно обладают большой аффинностью.
Однако большая концентрация низкоаффинных аутоантител может дать сигнал
более сильный, чем малая концентрации высокоаффинных специфических ауто-
антител. У больных с СКВ. у которых в результате поликлональной активации
В-клеток синтезируется большое количество полиспецифичных антител, наблюда-
ют ложноположительные реакции в серологических тестах для обнаружения анти-
тел к вирусу гепатита С. В то же время при волчаночном нефрите с нефротическим
синдромом в результате неселективной протеинурии утрачивается сывороточный
IgG. В результате при низком содержании IgG тесты по обнаружению аутоантител
могут быть ложноотрицательны. Обнаружение неспецифического и перекрестного
связывания аутоантител в некоторых иммунологических тестах, наличие низких
титров аутоантител в норме нередко требует выделения «серой зоны», или «зоны
сомнительных результатов».
В случае невысоких титров аутоантител, которые могут быть обусловлены
неспецифическим связыванием аутоантител или присутствием специфических
серологических маркеров, но в низкой концентрации, интерпретация результата
зависит от постановки клинической задачи. Если речь идет об исключении ревма-
тической природы заболевания в дифференциальной диагностике неревматиче-
ской патологии, то результат может быть интерпретирован как отрицательный. В
случае, когда тест используют для подтверждения ревматического заболевания у
больного с другими клиническими признаками ревматической патологии, резуль-
тат теста должен рассматрива гься как положительный.
Несмотря на определенные трудности в интерпретации результатов имму-
нологического обследования, а также большой объем информации, который
необходимо поинимать во внимание при анализе результатов обследования,
информативность иммунологических тестов очень высока. Особенности поста-
новки диагностической задачи и планирования иммунологического обследования
приведены в табл. 17-4.
Таблица 17-4. Постановка и решение задач лабораторной диагностики аутоиммунных заболева-
ний с помощью иммунологических методов
Клинический вопрос Диагностическая задача Назначение обследования
Как исключить диагноз? Ранняя диагностика и оценка риска Скрининговый тест, обладающий высокой чувствительностью
Как подтвердить диагноз? Дифференциальная диа- гностика Подтверждающий тест, обладающий высокой специфичностью
Границы нормы и патологии? Классификация заболе- вания Устанавливаются индивидуально на основа- нии опыта лаборатории и результатов контро- ля качества
Что значит для больного результат обследования? Прогноз исхода заболе- вания Данные литературы при анализе больших когорт больных
Как часто повторять тест? Контроль терапии Учитывать время клиренса исследуемых молекул
74
ДИАГНОСТИКА АУТОИММУННЫХ ЗАБОЛЕВАНИЙ
Планирование серологического обследования зависит от клинической задачи,
которую требуется решить клиницисту. В сьою очередь, назначение конкретных
тестов зависит от их клинико-лабораторных параметров.
Для скрининга используют тесты, отрицательный результат которых с большой
вероятностью позволяет исключить заболевание. Обычно такими параметрами
обладают методы, позволяющие одновременно определять антитела к антигенам
одного семейства. Так, назначение антинуклеарного фактора (АНФ) целесообраз-
но на этапе скрининга ревматических заболеваний, поскольку обычно диффуз-
ные заболевания соединительной ткани сопровождаются обнаружением АНА.
Отрицательный результат выявления АНФ делает диагноз СКВ или системной
склеродермии маловероятным.
Часто уже на этапе скрининга необходимо выполнять комбинации тестов,
параллельное назначение которых значительно снижает вероятность ошибочно-
го исключения диагноза. К примеру, в диагностике ревматоидного артрита уже
в дебюте этого заболевания рационально назначать определение ревматоидного
фактора (РФ) и антицитруллиновых антител.
Дифференциальная диагностика требует проведения тестов, обладающих боль-
шей специфичностью. Например, при обнаружении низких титров АНФ выявление
антител к дсДНК указывает на диагноз СКВ. Не менее важен вопрос о результатах
тестов, которые находятся в «серой зона». В случае скринингового теста результат,
находящийся в «серой зоне», следует интерпретировать как неспецифический, в
то время как у пациента с уже определенным диагнозом тот же результат следует
отнести к низкой, но значимой концентрации аутоантител. При решении задач
дифференциальной диагнос гики комбинации тестов делают результат обследова-
ния более убедительным. Так, диагностика анаифосфолипидного синдрома (АФС)
с помощью антифосфолипидных антител должна расширяться за счет параллель-
ного обследования для диагностики СКВ.
Комбинированное тестирование несомненно улучшает информативность обсле-
дования пациентов. Комбинированное тестирование, включающее параллельный
скрининг разных аутоиммунных заболеваний, имеющих общие черты, крайне
информативно при дифференциальной диагностике системных ревматических
заболеваний. Такой подход позволяет рекомендовать наборы тестов и комплекс-
ные алгоритмы обследования, что в итоге приводит к росту числа диагностических
находок.
ЛАБОРАТОРНЫЕ ПОКАЗАТЕЛИ
ПРИ АУТОИММУННЫХ ЗАБОЛЕВАНИЯХ
Выявление LE-клеток
В диагност ике аутоиммунных заболеваний к функциональным тестам относят
те, при которых измерение аутоантител основано на влиянии аутоантител на био-
логическую реакцию. Присутствие биологической активности антител представ-
ляет собой объективный показатель, в меньшей степени подверженный артефак-
там иммунометрических тестов. Недостатком функциональных методов является
плохая стандартизация и низкая воспроизводимость, трудоемкость и короткий
срок годности компонентов тест-системы. В результате практически всегда функ-
циональные тесты с течением времени заменяются на более простые и надежные
иммунометрические и серологические методы.
Классическим функциональным тестом детекции аутоантител считают обнару-
жение АНА с помощью метода выявления LE-клеток (клеток красной волчанки).
LE-клетки представляют собой лейкоциты крови, фагоцитировавшие ядра лимфо-
ДИАГНОСТИКА АУТОИММУННЫХ ЗАБОЛЕВАНИЙ 75
цитов. В основе этого феномена при СКВ лежит несколько механизмов. Прежде
всего при СКВ лимфоциты периферической крови претерпевают ускоренный
апоптоз, который рассматривается как попытка восстановления утраченной имму-
нотолерантности при этом заболевании. Фагоцитоз ядер лимфоцитов лейкоцитам
обусловлен наличием АЛА в сыворотке крови больных, которые опсонизируют
фрагменты ядер лимфоцитов для их поглощения лейкоцитами. Присутствие в
тесте сыворотки больного, содержащей АНА, является принципиальным для
обнаружения LE-клеток, что позволяет рассматривать этот тест среди других мето-
дов диагностики АНА.
Еще в начале XX в. в пунктатах костного мозга больных с СКВ были описаны
гиалиновые i ельца в цитоплазме фагоци тирующих клеток и лейкоциты перифе-
рической крови, нагруженные клеточным материалом. Этот феномен был смо-
делирован in vitro что привело к созданию лабораторного теста по определению
LE-клеток. Учет LE-клеток осуществлялся в мазке крови с помощью светового
микроскопа и гематологических красителей. За счет простоты и дешевизны со
временем этот метод получил широкое распространение. Однако не существует
единого рекомендуемого протокола обнаружения LE-клеток; из-за низкой чув-
ствительности и сложности стандартизации не рекомендуется его выполнение в
клинических целях. Учитывая низкие аналитические параметры метода, отсут-
ствие LE-клеюк при повторных анализах не может рассматриваться в качестве
повода для исключения возможного диагноза диффузной болезни соединительной
ткани. Выявление АНА с помощью АНФ полностью исключает необходимость
использования это! о устаревшего теста.
В середине прошлого века был описан сывороточный фактор, ответственный
за образование LE-клеток и стимулирующий фагоцитоз ядер клеток монону-
клеарами крови. Активность фактора наблюдалась во фракции гаммаглобули-
нов сыворотки крови. Это позволило для выявления АНА применить реакцию
непрямой иммунофлюоресценции или метод Кунса с использованием тканей
человека в качестве субстрата. Этот модифицированный тест на сегодняшний
день считают основным методом обнаружения АНФ в клинической практике.
Несмотря на то, что в аутоиммунной диагностике используют единичные функ-
циональные тесты, их роль остается значительной в исследовании патогенетиче-
ского действия аутоантител.
Антинуклеарный фактор
Определение АНФ — самое часто выполняемое лабораторное исследование
в практических иммунологических лабораториях, что отражает его значение
для диагностики аутоиммунных заболеваний. АНФ — семейство аутоантител,
связывающихся с рибонуклеиновыми кислотами и ассоциированными с ними
белками, Они встречаются более чем у 90% больных с диффузными болезнями
соединительной ткани, такими как СКВ, системный склероз (СС), смешанное
заболевание соединительной ткани (СЗСТ), синдром Шегрена (СШ) (табл. 17-5).
Представители этого семейства аутоантител могут быть обнаружены при инфек-
ционных, воспалительных и онкологических заболеваниях. Разнообразие пато-
логий, сопровождающихся появлением АНФ, заставляет заподозрить единство
иммунологических реакций в их патогенезе. В то же время встречаемость АНФ
достигает 1-5% у клинически здоровых лиц и несколько возрастает в пожилом
возрасте.
76 ДИАГНОСТИКА АУТОИММУННЫХ ЗАБОЛЕВАНИЙ
Таблица 17-5. Краткий перечень заболеваний, сопровождающихся наличием анти нуклеарного
фактора
Заболевания Встречаемость АНФ, %
Системная красная волчанка 95
Системный склероз 95
Синдром Шегрена 95
Дерматомиозиг, полимиозит 30-50
Смешанное заболевание соединительной ткани 100
Дискоидная красная волчанка 50-60
Локализованная склеродермия 50
Ревматоидный артрит 3(М0
Юношеский ревматоидный артрит (олиго- и полиартритная форма) 50-90
Аутоиммунный гепатит 95-100
Первичный билиарный цирроз 95-100
Узелковый полиартериит 10-20
Злокачественные заболевания 10-20
Хроническое невынашивание беременности 10-15
Пожилые (>65 лет) 10
Здоровые женщины 5
Общее содержание АНА в сыворотке крови больных обозначается как «анти-
нуклеарный фактор». Учитывая высокую чувствительность метода обнаружения
АНФ, нередко проводят равенство между присутствием АНА в крови больных
определенными заболеваниями и положительным результатом обнаружения
АНФ. Однако традиционно под названием «АНФ» подразумевают положитель-
ный результат иммунофлюоресцентного теста для обнаружения АНА. Другие
методы обнаружения АНА, например ИФА, значительно уступают непрямой
иммунофлюоресценции по чувствительности. Не рекомендуется называть терми-
ном «антинуклеарный фактор» показатели, определяемые другими методами.
Основным механизмом индукции АНА, по-видимому, является патологиче-
ский апоптоз кератиноцитов, лимфоцитов и других клеток, который возникает
при диффузных заболеваниях соединительной ткани. Апоптотические тельца,
содержащие в своем составе антигены АНА, напоминают по структуре вирусные
частицы и способны индуцировать специфические иммунные ответы. Антитела
могут иметь патогенетическое значение, что показано по отношению к антителам
к дсДНК при СКВ с поражением почек, но большинство разновидностей АНА
является, скорее, вторичным феноменом по отношению к деструкции ткани при
аутоиммунных заболеваниях.
К настоящему времени описано более 100 разновидностей АНА, направленных
против нуклеиновых кислот, гистонов, белков ядерной мембраны, компонентов
сплайсосомы, рибонуклеопротеинов, белков ядрышек и центромер Антитела к
цитоплазматическим антигенам, диагностируемые при поли- и дерматомиозите
взрослых, также причисляют к АНА. так как они направлены против рибопро-
теиновых структур, и в их возникновении предположительно участвуют сходные
патогенетические механизмы. Разнообразие типов АНА и уточнение клинико-
лабораторных корреляций позволило использовать АНА как показатель, позво-
ляющий клиницисту уточнить прогноз, течение и, зачастую, предупредить обо-
стрение заболевания.
Для обнаружения АНА применяют ряд показателей, которые могут измеряться
при помощи разных методов (табл. 17-6).
ДИАГНОСТИКА АУТОИММУННЫХ ЗАБОЛЕВАНИЙ 77
Таблица 17-6. Показатели и методы определения антинуклеарных антител
Показатель Определение Метод Цель определения
LE-клетки Опсонизирующие АНА к ядрам клеток Функциональный тест Скрининг АНА
АНФ АНА против нераство- римых и конформаци- онных антигенов ядра Непрямая иммунофлюорес- ценция Скрининг АНА
Антитела к экстрагируе- мому ядерному антигену АНА против раствори- мых антигенов, выде- ленных из клетки Двойная иммунодиффузия, контрэлектрофорез, ИФА Скрининг АНА
Антитела к дсДНК АНА против нуклеино- вых кислот ИФА, непрямая иммуно- флюоресценция, радиоиммун- ный анализ Определение кон- кретных разновид- ностей АНА
Антитела к рибонуклео- протеинам Ядерные мишени АНА: Sm, RNP, SS-A, SS-B. Jo-У, Sc/-70 и др. Двойная иммунодиффузия, контрэлектрофорез, ИФА, имму- ноблоттинг, радиоиммунный анализ Определение кон- кретных разновид- ностей АНА
Присутствие в сыворотке крови АНФ обусловливает обнаружение в крови кле-
ток красной волчанки (LE-клеток). Таким образом, LE-клетки можно рассматри-
вать в качестве вторичного показателя, указывающего на наличие АНА. Низкая
чувствительность и плохая воспроизводимость тестов, используемых для обнару-
жения LE-клеток, не позволяет им конкурировать с АНФ, постепенно эти тесты
полностью вытесняются серологическими методами обнаружения АНА.
АНТИНУКЛЕАРНЫЙ ФАКТОР И ТИПЫ СВЕЧЕНИЯ ЯДРА
Определение АНФ — основной метод обнаружения АНА. Результатом опреде-
ления АНФ является факт присутствия аутоантител в диагностическом титре,
конечный титр разведения сыворотки, отражающий аффинность и концентрацию
аутоантител, а также тип свечения ядра клетки. Характеристики конкретной мето-
дики зависят от субстрата, который может варьировать от криосцезов (крысиных
печени или почки) до культивируемых пролиферирующих клеточных линий (КЬ.
HeLa), а также метода фиксации ткани, который может влиять на сохранность
ряда ядерных антигенов АНА.
Криосрезы тканей крысы в качестве субстрата непрямой иммунофлюоресцен-
ции для обнаружения АНФ позволяют получить полуколичественный результат,
определяющий максимальный титр разведения сыворотки пациента, при котором
сохраняется свечение ядер. Это основной тест, который использовался для обнару-
жения АНА в клинической практике в течение многих лет, наряду с обнаружением
LE клеток. Основным недостатком использования данного метода считают отсут-
ствие в тканях грызунов ряда антигенов человека, что снижает чувствительность
теста. В последнее время его вытеснил ИФА.
Последующая модификация метода обнаружения АНФ связана с использова-
нием в качестве субстрата человеческой перевиваемой эпителиоидной клеточной
линии НЕр-2. Эта клеточная линия, полученная из аденокарциномы гортани
человека, представляет собой крупные полиплоидные неороговевающие плоские
эпителиоциты, образующие монослой на пласгике и стекле. Крупные ядра и при-
сутствие человеческих антигенов сделали метод непрямой иммунофлюоресценции
с использованием перевиваемой клеточной линии НЕр-2 основным методом обна-
ружения АНФ. В англоязычной литературе отсутствует единое название для этого
метода, иногда его для краткости обозначают как FANA (Fluorescent AnliNuclear
Antibody detection), в нашей стране сохраняется название этого показателя —
АНФ.
В качестве субстратов могут применяться и другие человеческие клеточные
линии, однако благодаря хорошей морфологии и удобству культивирования
78 ДИАГНОСТИКА АУТОИММУННЫХ ЗАБОЛЕВАНИЙ
именно линия НЕр-2 ст ала общепризнанным субстратом непрямой иммунофлюо-
ресценции. Ее применение улучшает чувствительность теста за счет яркой флюо-
ресценции даже при значительных разведениях сыворотки больного, а большое,
богатое эухроматином ядро позволяет точно описать тип свечения. Кроме того,
применение клеточной линии НЕр-2 способствует обнаружению антител Ro/
SS-A, а также антител к антигенам ядрышка, которые не выявляются при исполь-
зовании ткани лабораторных животных. Среди преимуществ НЕр-2 — высокая
частота деления клеток, позволяющая определять антитела к антигенам, экс-
прессирующимся только при делении клетки, и отсутствие клеточного матрикса
ткани, затрудняющего визуализацию специфического свечения, что характерно
для гистологических срезов.
При использовании тканей лабораторных животных диагностический титр
АНФ составляет 1:8-1:10, в случае применения клеточной линии НЕр-2 реко-
мендуется использовать исходный титр 1:40-1:80. При работе с такими титрами
частота встречаемости слабоположительных резульгатов у клинически здоровых
лиц составит не более 5% и в то же время не позволит пропустить значимые п итры
АНА у больных диффузными болезнями соединительной ткани. На фоне обостре-
ния ревматических заболеваний отмечают титры АНФ более 1'640, при ремиссии
титры снижаются до 1:160-1:320.
Существуют рекомендации по балльной оценке результатов выявления АНФ,
указывающей содержание «в крестах». Это позволяет лабораториям экономить
реактивы и уменьшить трудозатраты на выполнение исследований. Однако
конечный титр имеет более важное значение по сравнению с количеством связав-
шихся с субстратом аутоантител, так как непосредственно связан с аффинностью
их взаимодействия с антигеном. Желательно установить конечный титр у всех
«положительных больных», что позволяет уточнять присутствие в сыворотке
высокоаффинных аутоантител, которые более тегно связаны с активностью про-
цесса. Необходимо определять АНФ класса IgG. поскольку выявление АНФ, пред-
ставленных другими классами иммуноглобулинов, не имеет самостоятельного
диагностического значения.
Тип свечения ядра клетки значительно увеличивает информативность выявле-
ния АНФ. Использование клеток линии НЕр-2 позволяет охарактеризовать более
20 различных вариантов окрашивания ядра, которые зависят от спектра АНА,
присутствующих в исследуемой сыворотке. Однако для практической лаборатории
достаточно различать 6 основных вариантов свызывания аутоантител с антигенсо-
держащими структурами клетки.
Выделяют гомогенный, периферический, гранулярный (мелко-/кр\шно-),
ядрышковый, центромерный и цитоплазматический тип свечения ядра. Каждому
типу свечения присущи характерные признаки, которые позволяют отличить один
вариант от другого, а также набор антигенов, с которыми реагируют аутоантитела
в сыворотках больных. Описание типов свечения представляет ценную клини-
ческую информацию само по себе, кроме того, тип свечения может указывать на
необходимость проведения определенных лабораторных тестов в дальнейшем
(табл, 17-7).
Гомогенный тип свечения — аутоантитела реагируют с теми антигенами, кото-
рые распределены в ядре диффузно, т.е. входят в состав хроматина. Характерно,
что при обнаружении гомогенного типа свечения в делящихся клетках ярко окра-
шиваются конденсированные хромосомы. Основными структурными единицами
хроматина являются нуклеосомы — комплексы ДНК и гистонов. Таким образом,
гомогенный тип свечения предполагает наличие антител против нуклеосом, дсДНК
и антител к гистонам. Он встречается у больных с СКВ и лекарственной волчанкой,
ДИАГНОСТИКА АУТОИММУННЫХ ЗАБОЛЕВАНИЙ 79
а также у больных со склеродермией. Обычно обнаружение высокого титра АНФ
с гомогенным типом свечения свидетельствует о диагнозе СКВ.
Таблица 17-7. Тип свечения при непрямой иммунофлюоресценции и наличие антинуклеарных
антител
Тил свечения Специфичность антитела Характеристика антигена Заболевание
Гомогенный/перифе- рический дсднк ДНК хроматина СКВ
Гистоны НгН2Д,Н2в,Н3иН4 । Лекарственная СКВ
Атипический гомо- генный Ламины ядра Ламины А, В, С Аутоиммунный гепатит
Гранулярный Sm Белки В', В, D. Е. F, связанные с U, U8 RNP Маркер СКВ
RNP Белки А и С. связанные с U1 RNP СКВ.СЗСТ
PCNA'Ga 36 кДа-белок, связанный с ДНК- полимеразой 5 СКВ
Атипический грану- лярный Ro/SS-A 52, 60 кДа-белок. связанный с Y1-Y5 РНК СШ, СКВ
La/SS-B 48 кДа, связан с РНК-полимеразой 3 сш
Scl-70 70 кДа-антиген склеродермии сс
PCNA Белок, ассоциированный с ДНК полимеразой 6 СКВ
Центромерный Антицентромерный CENP А, В, С CREST
Ядрышковый Крупногранулярный Фибрилляции СС
Мелкогранулярный РНК-полимераза 1 сс
Мелкогранулярный РМ-1 (PM/Scl) пм/дм, сс
Ядрышковый, в зави- симости от клеточного цикла Ku Белки ядерного матрикса 70 и 80 кДа СКВ
Цитоплазматический ASM F-актин Аутоиммунный гепатит
AMA Пируват-декарбоксилазный ком- плекс Первичный билиар- ный цирроз
Анти Jo-1, PL-7. PL-12 и др. тРНК-сингетаэы ПМ/ДМ
Периферический тип свечения часто выделяют отдельно, хотя он являет-
ся разновидностью гомогенного типа свечения. Его обнаружение — артефакт
фиксации клеток, который приводит к перераспределению хроматина в ядре
на периферию Важно отличать периферический тип свечения от окрашивания
ядерной мембраны, которое встречается при аутоиммунных заболеваниях печени.
Периферический тин свечения характерен дпя больных с антителами к двуспи-
ральной ДНК и отмечается преимущественно у больных с СКВ.
Гранулярный тип встречается наиболее часто, но он нсспецифический. Иногда
этот тип свечения называют «крапчатым» или «сетчатым». Название «грануляр-
ный» более точно отражает этот феномен, поскольку в этом случае аутоантитела
реагируют с гранулами в ядре, представляющими надмолекулярные нуклеопроте-
иновые комплексы. Такие комплексы белков и нуклеиновых кислот осуществляют
в ядре ряд функций, необходимых для нормальной жизнедеятельности клетки.
К таким комплексам, в частности, относят сплайсосомы, которые осуществляют
пост транскрипционную перестройку мРНК, необходимую для синтеза белка на
рибосомах. В составе присутствует множество различных нуклеопротеинов, что
обусловливает многообразие антигенных мишеней при обнаружении грануляр-
ного типа свечения. К основным аутоантигенам, антитела к которым приводят к
80 ДИАГНОСТИКА АУТОИММУННЫХ ЗАБОЛЬВАНИЙ
визуализации гранулярного типа свечения, относят Sm, Ur-RNP, SS-A, SS-B анти-
гены и PCNA. Клетки в процессе деления утрачивают большинство сформирован-
ных нуклеопротеиновых комплексов, поэтому митотические фигуры в клеточной
линии при гранулярном типе свечения не окрашиваются.
Гранулярный тип свечения ядра отмечается у больных СКВ, СШ, СС, ДМ/ПМ,
РА и при ряде других аутоиммунных заболеваний. Низкие титры АНФ с грануляр-
ным типом преобладают в сыворотках крови у клинически здоровых лиц с АНФ
без признаков системного заболевания.
Ядрышковый тип флюоресценции бывает в тех случаях, когда антигены
ядрышка выступают в качестве мишеней АНА. Ядрышковый тип свечения харак-
терен для склеродермии и ее разновидностей. Ядрышковый тип флюоресценции
определяется у больных при наличии антител к компонентам ядрышка, таким как
РНК-полимераза 1, NOR, U3RNP, PM/Scl.
Центромерный тип флюоресценции наблюдают при образовании анти-
тел к центромерам хромосом, он обнаруживается только в делящихся клетках.
Присутствие данного типа флюоресценции характерно при CREST-варианте скле-
родермии.
Цитоплазматический тип свечения указывает на антитела к тРНК-синтетазам,
в частности к Jo-1, которые образуются при полимиозите. Кроме того, он выявля-
ется у больных с АНА, направленными против других компонентов цитоплазмы
клетки: антитела к актину при аутоиммунном гепатите, антитела к митохондриям
при первичном билиарном циррозе.
Нередко может встречаться сочетание нескольких типов свечения, к примеру
мелкогранулярного и ядрышкового, что характерно для антител к Scl-70. Более
того, часто в низких разведениях преобладает один тип свечения, например грану-
лярный, а при дальнейших разведениях проявляется гомогенный или центромер-
ный тип флюоресценции, что свидетельствует о наличии в сыворотке больного
различных видов АНА. Хотя тип свечения дает врачу определенные данные в
пользу того или иного диагноза, необходимо принимать во внимание сравнитель-
но низкую его специфичность и разнообразие встречающихся феноменов, что
требует проведения дальнейшего лабораторного обследования.
Обнаружение АНФ — основа для использования других лабораторных тестов,
уточняющих спектр АНА в крови больных. Учитывая крайнюю информативность
этого теста, дальнейшее тестирование и анализ результатов поел едущего обсле-
дования должны интерпретироваться в зависимости от результатов оценки типа
свечения и титра АНФ.
При использовании тканей лабораторных животных, таких как почка или
печень крысы, у 10% больных СКВ АНА не определяются или титры их очень
низки. Этих больных условно относят к группе «АНФ-негативной СКВ», для
уточнения диагноза у таких больных требуется дальнейшее обследование, прежде
всего определение антител к SS-A антигенам. Эти антигены обладают хорошей
растворимостью и могут утрачиваться из ядер клеток. Применение в качестве
субстрата клеточной линии НЕр-2 сокращает численность данной категории паци-
ентов, но не более чем на 5%. Необходимо учитывать, что при использовании тра-
диционной метанол-ацетоновой фиксации клеточной линии SS-A-антиген может
диффундировать в цитоплазму клетки, в результате чего титры АНФ значительно
снижаются.
Для устранения этих недостатков была создана трансгенная клеточная линия
НЕр-2000, в генотип которой был искусственно внесен ген SS-A антигена. За счет
этого в ее ядре отмечается гиперэкспрессия SS-A-антигена. что несколько уве-
личивает чувствительность выявления АНА. Однако даже использование транс-
генных клеточных линий не позволяет окончательно избавиться от проблемы
«АНФ-негативной СКВ». Даже при максимально скрупулезном обследовании до
ДИАГНОСТИКА АУТОИММУННЫХ ЗАБОЛЕВАНИЙ 81
2-3% пациентов с клинически очевидной СКВ не имеют диагностических концен-
траций АНФ.
Антинуклеарные антитела
Номенклатура конкретных разновидностей АНА зависит как от ядерных мише-
ней аутоантител, так и от методов, с помощью которых их определяют. Отдельно
выделяют АНА, реагирующие с нуклеиновыми кислотами, прежде всего ДНК:
антитела к водорастворимым антигенам, которые могут быть экстрагированы
водно-солевыми растворами, — антитела к экстрагируемому ядерному антигену
(ЭНА); антитела к нерастворимым в воде антигенам, а также конформационные
антигены, которые разрушаются при препаративном выделении из ядра клетки.
Уточнение антигенных мишеней АНА зависит от антигенного субстрата, который
применяется в иммунологических тесгах. Антигенсодержащий субстрат может
быть нативным, т.е. полученным при минимальной очистке, либо высокоочищен-
ным. Белки-антигены могут быть получены методами генной инженерии либо
получены химическими методами пептидного синтеза. Использование того или
другого метода очистки или синтеза аутоантигена зависит от его биохимической
природы и особенностей связывания с ним аутоантител. Рекомбинантные белки и
синтетические пептиды более удобны в работе, так как методы их получения легко
стандартизируются. Однако чем менее очищен антиген, тем больше он сохраняет
третичную структуру, поэтому взаимодействие аутоантитела более приближено к
условиям взаимодействия w vivo.
Большинство видов АНА направлено к сложным комплексам рибонуклеиновых
кислот и белков. Изучение АНА зачастую опережало открытие функциональных
белков и рибопротеиновых комплексов, что привело к возникновению двойной
номенклатуры при описании некоторых видов АНА, включающей как общепри-
нятое название аутоантитела, так и указание конкретного ядерного белка. Обычно
тип АНА получал название, в основе которого лежит имя пациента, у которого
данная разновидность АНА была впервые обнаружена. В основе более совре-
менной номенклатуры лежат клеточные мишени аутоантител, с которыми они
взаимодействуют. Так, антитела к антигену Ro/SS-A (Ro — по фамилии больного,
Robair, SS-A — Sjogren’s Syndrome A antigen) направлены против белков массой 52
и 60 кДа, связанных с РНК в составе сплайсосомы, а антитела к антигену
La/SS-B (La — по фамилии больного, Lapiere, Sjogren’s Syndrome В antigen) направ-
лены против белка, связанного с РНК-полимеразой 3. Номенклатура аутоантител,
основанная на фамилиях пациентов, постепенно уходит в прошлое, в современной
литературе доминирует название антигенов с указанием молекулярной массы,
например SS-A антиген молекулярной массы 52 и 60 кДа.
Может сложиться ситуация, что в одной сыворотке одновременно обнаружи-
вают антитела разных специфичностей, направленные к нескольким антинукле-
арным антигенам. Некоторые антитела часто определяются совместно, например
антитела к SS-A и SS-B либо IJ-RNP и Sm-антигенам. Эти ассоциации обуслов-
лены тем, что антигены входят в состав одного белкового комплекса, с которым
реагируют аутоантитела. Некоторые разновидности АНА могут часто встречаться
совместно за счет того, что они имеются при одном заболевании или при пере-
крестных синдромах.
Антитела к экстрагируемому нуклеарному антигену
В состав ЭНА входит фракция растворимых ядерных белков, которые могут
быть получены из ядра клетки. В его состав входит 6 основных рибонуклеопро-
теиновых антигенов АНА, таких как SS-A, SS-B, Sm, LJ-RNP, 5(7-70 и Jo-1, а также
ряд других минорных антигенов.
82 ДИАГНОСТИКА АУТОИММУННЫХ ЗАБОЛЕВАНИЙ
Использование ЭНА в методах «двойная иммунодиффузия» и «контрэлектро-
форез» может быть применено для скрининга этих специфичностей аутоантител.
Скрининговые методы с использованием ЭНА низкочувствительны и при сопо-
ставлении с АНФ позволяют диагностировать только 50-60% положительных
сывороток. Тем не менее высокая специфичность методов позволяет использовать
их в качестве методов сравнения по отношению к другим, более чувствительным
тестам.
Обычно обнаружение антител к ЭН А проводят совместно с определением АНФ
для того, чтобы не пропустить случаев «АНФ-негативной СКВ». Необходимо
отметить, что ИФА тест-системы позволяют проводить скрининговое обследо-
вание для обнаружения антител к ЭНА у ревматологических больных. Отличие
тест-систем для скрининга состоит в том, что в инкубационную лунку помещается
смесь антигенов, и положительный результат теста указывает на наличие у боль-
ного одной или одновременно нескольких разновидностей АНА.
С учетом большой чувствительности метода ИФА в целом его применение более
оправдано для скрининга АНА, однако необходимо помнить о большой частоте
развития неспецифических реакций при его применении. Существует 2 основные
разновидности ИФА для скринингового обнаружения антител к ЭНА. В одном
случае в лунку вносят тотальный ядерный экстракт, а в другом — смесь реком-
бинантных очищенных белков, представляющих собой основные разновидности
рибонуклеопротеиновых антигенов. Использование смеси рекомбинантных бел-
ков делает обследование более специфичным и улучшает стандартизацию выявле-
ния аутоантител. В то же время далеко не все антигены АНА могут быть нанесены
на ИФА-планшет, в результате чего комбинированные тесты пропускают до 40%
положительных результатов.
Антитела к двуспиральной ДНК
При СКВ антитела к двуспиральной ДНК (дсДНК) могут быть обнаружены у
40 -50% больных, они характеризуются гомогенным типом свечения ядра НЕр-2-
клеток при выявлении АНФ. Причиной их образования служит сенсибилизация
к компонентам ядерного материала, высвобождающегося из ядер клеток в ходе
апоптоза. Обнаружение антител к дсДНК способно «предсказать» развитие СКВ
еще до проявления развернутой картины заболевания. Так, при проспективном
наблюдении СКВ развивается в течение одного года после обнаружения антител
к дсДНК в радиоиммунном гесте у 70% пациентов, и только 15% таких пациентов
остаются здоровыми через 5 лет.
Антитела к дсДНК направлены на фосфодиэфирный скелет молекулы, связыва-
ние антител не зависит от ее конформации и нуклеотидной последовательности.
Взаимодействие основано на электростатических связях, которые чрезвычайно
чувствительны к pH и концентрации солей в растворе. В то же время антитела
к дсДНК (нативной) перекрестно реагируют с осДНК (денатурированной), что
затрудняет интерпретацию результатов теста, если в качестве антигена выступает
дсДНК, загрязненная осДНК. Низкоспецифичные антитела к осДНК, встречающи-
еся при различных аутоиммунных и инфекционных заболеваниях, включая СКВ.
лекарственную волчанку, ревматоидный артрит, хронический активный гепатит
и инфекционный мононуклеоз, распознают денатурированные одноцепочечные
фрагменты дсДНК. Их способность связываться со свободными пуриновыми и
пиримидиновыми основаниями в составе дсДНК приводит к тому, что антитела
к осДНК нередко определяют в тестах, направленных на определение антител к
дсДНК. Поэтому для обнаружения антител к дсДНК, которые встречаются только
при СКВ и входят в число наиболее специфичных маркеров этого заболевания,
требуется использовать тесты, которые позволили бы выявлять связывание анти-
тел только с дсДНК.
ДИАГНОСТИКА АУТОИММУННЫХ ЗАБОЛЕВАНИЙ 83
Для определения антител к дсДНК методом ИФА сложно сохранить нативную
структуру молекулы. Молекула дсДНК несет отрицательный заряд и не может быть
непосредственно нанесена на отрицательно зацяженный пластик иммукофермент-
ного планшета. Для того, чтобы нанести дсДНК на дно лунок, необходимо исполь-
зовать положительно заряженный белок протамин. С другой стороны, облучение
дсДНК позволяет сделать ее более реактоспособной и сенсибилизировать ИФА-
планшеты. Оба этих метода сенсибилизации приводят к деспирализации дсДНК
с образованием значительного числа односпиральных молекул. Это приводит к
тому, что большинство твердофазных иммуноферментных систем для обнаруже-
ния антител к дсДНК оказываются относительно неспецифичными. Практическим
ревматологам необходимо знать об этой особенности выявления антител к дсДНК
и с осторожностью интерпретировать низкие титры данного показателя.
Для решения проблемы специфичности обследования были разработаны мето-
ды НРИФ и радиоиммунный метод, которые обладают значительно более высо-
кой специфичностью.
Метод НРИФ основан на использовании клеток простейшего жгутикового
микроорганизма Crithidia tectUae* которые содержат неизмеренную молекулу
дсДНК. Этот микроорганизм относится к роду трипаносом, он неприхотлив и
хорошо поддается культивированию в лабораторных условиях. В основании жгу-
тика имеется кинетопласт, состоящий из гигантской митохондрии, содержащей
плотно упакованную кольцевую ДНК. Эта молекула митохондриальной дсДНК без
ассоциированной РНК и нуклеопротеинов — идеальный субстрат для непрямой
иммунофлюоресценции. При наличии у больного антител к дсДНК определяется
яркая флюоресценция в основании жгутика. Наличие гистонов в составе ДНК
кинетопласта может привести к ложноположителыюму результату у больного,
имеющего антитела к гистонам. Как и другие иммунофлюоресцентные методы,
тест с использованием С. lucilia позволяет получить только полуколичественный
результат, поэтому его целесообразно комбинировать с методом ИФА.
Наилучшим методом обнаружения антител к дсДНК считают метод радиоим-
мунного анализа, известного как тест Фарра (Farr test). Метод Фарра относится к
классическим методам, с помощью которых исследуют аффинность связывания
антитела с антигеном. Реакция связывания антител с дсДНК, меченной радиоак-
тивным изотопом, протекает в растворе, что позволяет предотвратить деспирали-
зацию ДНК. Образовавшиеся иммунные комплексы дсДНК и анти-дсДНК-антител
осаждаются с помощью насыщенного раствора сульфата аммония. Преципитат
многократно отмывается, я содержание радиоактивной метки считывают в счет-
чике радиоактивности. Результат теста выражают либо в процентном отношении
связывания дсДНК, либо в единицах радиоактивности. Благодаря высокой точ-
ности, тест Фарра применяют в качестве «золотого стандарта» количественного
определения антител к дсДНК. Большая часть исследований, указывающих на
роль концентраций антител к дсДНК в мониторинге активности заболевания,
выполнена с помощью радиоиммунного теста Фарра, но не всегда эти результаты
могут быть полностью подтверждены при использовании других тестов.
Новое поколение ИФА тест-систем для выявления антител к дсДНК позволяет
отчасти преодолеть проблему деспирализации молекул. К примеру, д чя сенсибили-
зации планшетов используют биотинилирование кольцевых плазмидных дсДНК,
в то время как дно планшетов покрывают авидином. Другой подход заключается
в обработке планшетов З^экзонуклеазой. разрушающей односпиральныс участки
Д1IK. Эти подходы позволяют добиться специфичности, близкой к 90-95%. Методы
ИФА имеют чувствительность около 45%, в то время как положительный тест
Фарра отмечается у 30% обследуемых, а НРИФ на С. luciha положителен менее чем
у 20% пациентов. Ряд практических лабораторий использует ИФА для скрининга
аутоантител, а специфичность связывания подтверждается посредством НРИФ.
84
ДИАГНОС ГИКА АУТОИММУННЫХ ЗАБОЛЕВАНИЙ
Антитела к дсДНК участвуют в патогенезе развития волчаночных васкулитов
и люпус-нефрита. Титры антител к дсДНК тесно коррелируют с концентрацией
IgG-содержащих циркулирующих иммунных комплексов (ЦИК) в сыворотке
крови больных СКВ. Исследования элюатов из почечных клубочков больных,
умерших от СКВ указывают на большую концентрацию антител к дсДНК в
элюате по сравнению с их концентрацией в сыворотке. Это позволило заподо-
зрить значение иммунных комплексов, содержащих антитела к дсДНК, в патоге-
незе поражения почек при СКВ. Обнаружение способности дсДНК связываться
с базальной мембраной клубочка способствовало укреплению теории об имму-
нокомплексном поражении почки при СКВ. приводящем к образованию иммун-
ных комплексов в клубочке in citu с активацией комплемента, потреблением его
сывороточных резервов и формированием воспалительных инфильтратов. У
большинства больных с активными почечными формами СКВ имеются антитела
к дсДНК. Для определения риска поражения почек рекомендуется обследование
всех больных СКВ на предмет выявления антител к дсДНК и гипокомплементе-
мии.
Аффинность антител к дсДНК в сыворотке представляет собой постоянно
меняющуюся величину. Так, при обострении гломерулонефрита содержание
высокоаффинных антител в сыворотке больного весьма незначительно, однако
в элюатах из почечных клубочков содержание высокоаффинных антител очень
высоко. Во время ремиссии титры высокоаффинных антител к дсДНК в сыворотке
постепенно возрастают, их уровни максимальны непосредственно перед вспышкой
гломерулонефрита. Поэтому динамике концентрации антител к дсДНК придают
большее прогностическое значение даже по сравнению с их абсолютным содер-
жанием. Нарастание титра антител к дсДНК (удвоение титра в течение 3 мес) и
гипокомплементемия предшествуют развитию гематурии и протеинурии в 90%
наблюдений. При выполнении биопсии почек у таких больных почти всегда обна-
руживают наличие значительных иммуноопосредованных изменений даже при
отсутствии клинических признаков обострения. Увеличение титра анти-дсДНК-
антител в течение нескольких недель служит предвестником вспышки СКВ. Раннее
распознавание таких состояний и активная стероидная/иммуносупрессивная тера-
пия играют существенную роль в снижении смертности и инвалидизации среди
больных с поражением почек при СКВ.
У пациентов с СКВ с высоким содержанием низкоаффинных антител в методе
ИФА чаще отмечается поражение центральной нервной системы.
Антитела к односпиральной ДНК
Антитела к осДНК встречаются при СКВ даже чаще, чем антитела к дсДНК,
и MOiyr быть обнаружены приблизительно у 70% больных с СКВ. Хотя они
неспецифичны для СКВ, их обнаружение в элюатах больных, умерших от гломе-
рулонефрита, может свидетельствовать о роли антител к осДНК в патогенезе пора-
жения почек при волчаночном нефрите. При других заболеваниях соединительной
ткани их наличие диагностируют с похожей частотой. Антитела к осДНК могут
быть обнаружены при множестве других патологических состояний, массивной
травме, инфекциях и у клинически здоровых лиц. Высока встречаемость антител
к осДНК у онкологических пациентов, получающих цитотоксическую терапию.
Неспепифичность обнаружения этих аутоантител в значительной степени ограни-
чивает их применение в клинической практике.
Определение в сыворотке больного антител к осДНК класса IgM имеет зна-
чение для диагностики дискоидной красной волчанки. При кожной форме вол-
чанки, сопровождающейся образованием антител к осДНК, отмечается высокая
частота развития системного заболевания в виде генерализованной формы СКВ.
Приблизительно 20% больных с кожной формой волчанки, негативных по АНФ при
ДИАГНОСТИКА АУТОИММУННЫХ ЗАБОЛЕВАНИЙ 85
проведении непрямой иммунофлюоресценции, имеют значительный титр антител к
осДНК класса IgM, Таких пациентов относят к группе «АНА-негативной СКВ».
Антитела к гистонам
Эти антитела реагируют с протеиновыми составляющими нуклеосом ДНК-
протеиновых комплексов. Антитела к гистонам реагируют с хроматином, диф-
фузно распределенным в ядре клетки, что приводит к гомогенному типу свече-
ния ядра при использовании клеток линии НЕр-2. Каждая нуклеосома состоит
приблизительно из 140 пар оснований! ДНК, обернутой вокруг ядра, из белков-
гистонов Н, , Н2В, Н3 и Н4; Нх покрывает ДНК в промежутках между нуклеосомами.
У 70% больных СКВ аутоантитела направлены на гисгоны Ц и Н2В, в то время как
А Н \ к другим типам гистонов встречаются значительно реже.
Высокие титры антител к гистонам характерны для лекарственной волчанки,
хотя эта зависимость не абсолютная. Они могут быть обнаружены совместно с
антителами к осДНК и использованы в целях ее диагностики, однако опреде-
ление антител к гистонам не входит в число распространенных лабораторных
тестов. Лекарственно-индуцированная волчанка является результатом побочного
действия фармацевтических препаратов. Список препаратов, при которых была
описана лекарственная волчанка, достаточно широк. Наиболее часто синдром
лекарственной волчанки отмечается на фоне приема фенитоина, хинидина,
гидралазина, метилдопы, прокаинамида, хлорпромазина и изониазида. Синдром
характеризуется развитием люпоидноподобною заболевания с лихорадкой и
полисерозитом. Лекарственно-индуцированная волчанка протекает, как правило,
без поражения почек. Лишь при приеме D-пенициллинамина^ описано развитие
гломерулонефрита, поражение кожи и появление антител к дсДНК. Для больных
лекарственной СКВ характерны очень высокие титры АНА, и почти у всех паци-
ентов имеются антитела к гистонам. Отмена лекарственного препарата приводит
к постепенному снижению титров аутоантител и их исчезновению из сыворотки
крови в течение 6 мес. Это позволяет рекомендовать при дифференциальной диа-
гностике лекарственной СКВ исследование парных титров АНФ в течение полу-
года после отмены препарата. Необходимо учитывать, что проявления васкулита
при лекарственных реакциях также могут быть обусловлены индукцией антиней-
трофил ьных цитоплазматических антител (АНЦА).
Антитела к гистонам в высоком титре диагностируют у больных со склеродер-
мией, а также у пациентов с аутоиммунным гепатитом, В низких титрах возможно
их обнаружение при ревматоидном и юношеском артрите, первичном билиарном
циррозе, инфекции вирусом Эпштейна-Барр, шизофрении, сенсорных нефропа-
тиях, моноклональных гаммапатиях и злокачественных новообразованиях. Они
часто встречаются у пожилых людей. Клинических особенностей у больных с этой
разновидностью АНА не обнаружено.
Антитела к нуклеосомам
Нуклеосомы представляют собой универсальные структуры хроматина и явля-
ются основной формой хранения эукариотической ДНК. Значительное содержа-
ние циркулирующих нуклеосом обнаруживается у больных с онкологическими
заболеваниями при цитостатической терапии, при септических состояниях, а
также у больных СКВ. Предполагается, что в основе этого явления лежит индук-
ция клеточного апоптоза. При СКВ установлен факт избыточного апоптоза ряда
клеток, в частности лимфоцитов периферической крови, эн цотелиальных клеток и
кератиноцигов. При апоптозе эндонуклеазы разрывают ДНК в промежутках между
нуклеосомами, последние выделяются в кровоток. Свободная дсДНК обнаружи-
вается в периферической крови только в составе нуклеосом. В настоящее время
нуклеосомы рассматривают в качестве основного иммуногена, стимулирующего
86 ДИАГНОСТИКА АУТОИММУННЫХ ЗАБОЛЕВАНИЙ
иммунный ответ Т- и В-клеток при СКВ. Из пула лимфоцитов периферической
крови больных можно выделить CD4+ клоны Т-клеток, специфичные к нуклео-
сомальным антигенам. Установлен ряд иммунодоминантных пептидов в составе
гистонов, индуцирующих пролиферативные ответы и секрецию цитокинов в куль-
турах лимфоцитов из крови больных СКВ. При контакте антиген-специфичных
CD4+ лимфоцитов с В-клетками в культурах in vitro наблюдается продукция
специфических ан гинуклеосомальных антител и антител к дсДНК. Антигены
некоторых вирусов способны ассоциироваться с нуклеосомами с образованием
иммуногенных комплексов, которые могут индуцировать бласттрансформацию
в культурах лимфоцитов периферической крови больных СКВ. Иммунизация
нуклеосомами лабораторных животных приводит к образованию АНА, антител к
ДНК, антифосфолипидных антител и появлению ряда морфологических призна-
ков волчаночного нефрита.
Антитела к нуклеосомам встречаются у 70% больных СКВ. Сравнительно редко
их обнаруживают у больных СШ и при СЗСТ (5-8%). Этот тип АНА не диагно-
стируют у больных другими заболеваниями соединительной ткани и здоровых
доноров, что позволяет отнести антинуклеосомные антитела в разряд высокоспе-
цифических маркеров СКВ. Высокие титры антинуклеосомных антител характер-
ны исключительно для больных активной СКВ, сопровождающейся нефритом, их
уровень положительно коррелирует с показателями активности заболевания. При
мониторинге их содержания было установлено, что титры антител к нуклеосомам
увеличиваются непосредственно перед вспышкой заболевания параллельно с раз-
витием гломерулонефрита. Нуклеосомы, находящиеся в крови, обладают высокой
аффинностью к гепарансульфату базальной мембраны клубочков и способны
откладываться на ней. приводя к связыванию антинуклеосомальных антител и
антител к дсДНК с последующим развитием гломерулонефрита.
При использовании ИФА тест-систем к нуклеосомам чувствительность состав-
ляет 50-60% при специфичности более 95%. Для диагностики СКВ у больных с
высокими титрами АНФ можно рекомендовать сочетанное обнаружение антител
к дсДНК и антител к нуклеосомам.
Антитела к рибонуклеопротеинам
Антитела к рибонуклеопротеинам (реагируют с комплексами полипептидов и
PH К) представляют собой подсемейство АНА, включающие анти-Sm, анти-от^АР
(U^-RNP), антн-Ro/SS-A, анти-Lc/SS-B. Все эти белки входят в состав сплайсо-
сом — надмолекулярных комплексов, осуществляющих перестройку мРНК. Эти
разновидности АНА суммарно встречаются при СКВ чаще, чем антитела к дсДНК.
Для всех АНА к рибонуклеопротеинам характерен гранулярный тип свечения
АНФ на клетках линии НЕр-2.
Концентрация антител к рибонуклеопротеинам в сыворотке больных диф-
фузными болезнями соединительной ткани (ДБСТ) необычайно высока. Если
антитела к дсДНК выявляются высокочувствительными методами, такими как
иммунофлюоресценция, радиииммунопреципитация и иммуноферментный метод,
то антитела к рибонуклеопротеинам определяются специфичными, но нечувстви-
тельными мет одами иммунодиффузии и контрэлектрофореза, В сыворотке боль-
ных антитела к рибонуклеопротеинам могут присутствовать как изолированно,
так и наряду с антителами к дсДНК. Клиническое значение антител к рибопротеи-
нам сложно переоценить в связи с их прогностическим значением. Они определя-
ют группу больных СКВ, у которых ведущим клиническим симптомом является
поражение кожи. Хотя поражения почек у этих больных не исключены, но частота
волчаночного гломерулонефрита в этой группе больных гораздо ниже, чем при
анти-дсДНК позитивной волчанке. Обнаружение рибонуклеопротеиновых АНА
при СШ указывает на вероят ность экстрагландулярных симптомов заболевания,
ДИАГНОСТИКА АУТОИММУННЫХ ЗАБОЛЕВАНИЙ 87
а наличие U^RNP позволяет отнести пациентов в группу смешанного заболевания
соединительной ткани.
Антитела к Sm-антигену
В 1966 г. Tan и Kunkel описали систему Sm-антигенов, реагирующих с антитела-
ми из сыворотки больной с фамилией Smith. Sm-антиген образует около 9 белков,
составляющих ядро snRNP (small nuclear ribonudeoprotdn) — рибопротеиновых
комплексов, включающих молекулы малой ядерной РНК (snRNA), и 10 ассоции-
рованных белков. Ядро комплекса, в которое входит Sm-антиген, покрыто белками
Ц-RNP-антигена. Антитела к белкам Sm-антигена (В/В’, D) встречаются изолиро-
ванно, в то время как антитела к белкам UX-RNP (70 кДа, А, С) могут встречаться
совместно с антителами к Sm. Как для антител к Sm, так и антител к U^RNP харак-
терен гранулярный тип свечения АНФ, который требует определения конкретных
мишеней АНА с помощью уточняющих тестов.
Антитела к Sm-антигену представляют высокоспецифичный, но сравнитель-
но редко встречающийся маркер СКВ (20-30% больных СКВ). Антитела к Sm
обнаруживают у 30-40% пациентов — представителей африканской и азиатской
расы, в европейской популяции встречаемость среди больных СКВ не выше 20%.
Важность обнаружения антител к Sm-антигену при СКВ обусловлена их абсолют-
ной специфичностью, что явилось причиной включения этой разновидности АНА,
наряду с антителами к дсДНК и LE-клетками, в классификационные критерии СКВ
Американской ассоциации ревматологов. Клиническими признаками, связанными
с наличием Sm-антител, являются прежде всего более агрессг Е ное течение забо-
левания, поражение ЦНС, волчаночные психозы и относительная сохранность
функции почек. Вопрос о необходимости мониторинга для прогнозирования обо-
стрения заболевания окончательно не решен, но в связи с низкой встречаемостью
антител к Sm практически не выполняется.
Антитела к snRNP (U}-RNP)
Антитела к snRNP направлены против белков А, С и 70 кДа, входящих в состав
рибонуклеопротеина, содержащего Ц-малую ядерную РНК. Антитела к snRNP
встречаются приблизительно у 30% больных СКВ (табл. 17-8), причем они обычно
встречаются совместно с антителами к Sm-антигену. Изолированное обнаружение
высоких титров этих антител характерно для гетерогенной группы больных с при-
знаками прогрессивного СС, СКВ и дерматомиозита. Это заболевание носит назва-
ние СЗСТ (mixed, connective tissue disease) или синдром Шарпа. Клиническая картина
при СЗСТ представлена разнообразными признаками склеродермии, включая
синдром Рейно, склеродактилию, нарушением моторики пищевода и фиброзирую-
щим поражением легких, миозитом, напоминающим таковой при полимиозите,
недеформирующим артритом и полисерозитом. СЗСТ характеризуется редкостью
поражения ЦНС и сохранностью функции почек, хорошим ответом на низкие
дозы глюкокортикоидов и. в целом, благоприятным прогнозом. СЗСТ представля-
ет собой частный случай так называемых перекрестных синдромов, сочетающих
клиническую картину различных заболеваний соединительной ткани.
Таблица 17-8. Диагностическое значение антител к sn-RNP (U^-RNP)
Л -луляция больных Комментарий
СЗСТ Сочетание миозита, склеродермии и волчанки
СКВ Часто развивается синдром Рейно, реже — поражение почек
Синдром врожденной красной волчанки (Жблокадй) Ребенок от $лЯЛ/Р-позитивной матери
Склеродермия Около 5% больных snRNP-позитивны
Полимиозит Около 10% больных S/JR/VP-ПОЗИТИВНЫ
88
ДИАГНОСТИКА АУТОИММУННЫХ ЗАБОЛЕВАНИЙ
Существует несколько диагностических критериев СЗСТ, основным из которых
является обнаружение антител к snRNP в сыворотке крови. Однако большинство
исследователей склоняются к тому, что клинические признаки большей части
больных с антителами к snRNP удовлетворяют классификационным критериям
СКВ, однако у них наблюдаются своеобразные клинические формы ее течения.
Характерно, что у 30-50% больных СЗСТ отмечается позитивный тест волчаноч-
ной полоски (ТВП) при иммунофлюоресцентном обследовании биопсий кожи.
Этот тест высокоспецифичен для СКВ, что лишний раз подчеркивает родство СЗСТ
и классической СКВ. У больных СКВ с антителами к snRNP обычно присутствует
синдром Рейно, признаки миозита и склеродактилия. Приблизительно у 5—10%
больных с классическими признаками прогрессивного СС и некоторых больных
с полимиозитом также присутствуют антитела к snRNP. У s/zWP-позитивных
матерей наблюдается высокий риск рождения ребенка с кожными признаками
врожденного люпоидного синдрома и врожденной AV-блокады. Поражение почек
также может встречаться у больных СКВ с антителами к snRNP, хотя ее встречае-
мое! ь значительно ниже, чем частота поражения почек, у больных с антителами
к дсДНК.
По всей видимости, СЗСТ — некий этап в развитии заболевания, после чего про-
исходит трансформация в определенную клиническую форму. Антитела к snRNP
могут встречаться у больных с клинической картиной диффузной склеродермии
или полимиозита. Так, при воспалительных миопатиях антитела к snRNP обна-
руживают приблизительно у 10% больных. У этих пациентов заболевание может
начаться с субфебрилитета, артрита и синдрома Рейно, в дальнейшем наблюдается
постепенная его прогрессия с развитием полимиозита. Схожая картина наблюда-
ется у больных ПМ при наличии антител к Ro/SS-A, что сопровождается клиниче-
ской картиной полимиозига, несущего черты СКВ и СШ.
В свою очередь, у больных с диагнозом СЗСТ имеются антитела к осДНК, гисто-
нам и антикардиолипиновые антитела с частотой, соответствующей таковой при
СКВ. РФ встречается приблизительно у 30% этих пациентов.
Тесты, направленные на обнаружение антител к snRNP, следует выполнять всем
больным, позитивным в иммунофлюоресцентном тесте, особенно с крупнограну-
лярным типом свечения. Поскольку нет данных, свидетельствующи к об изменении
их концентрации при обострении заболевания, определение их титра в динамике
не имеет важного клинического значения. Совместно с антителами к snRNP в 50?/»
случаев выявляются РФ и реже — антитела к другим рибонуклеопротеинам, таким
как Sm, SS-A и SS-B, а также антитела к дсДНК и к центромерам хромосом.
Антитела к snRNP способны проникать внутрь живых клеток и связываться
с внутриклеточными мишенями in vivo. При использовании метода проточной
цитометрии было показано, что антитела к snRNP способны проникнуть в ядра
половины живых клеток периферической крови, причем этот процесс не является
обычным захватом сывороточного IgG и зависит от сохранности Fc-фрагмента
молекулы иммуноглобулина. Аутоантитела могут проникать внутрь клетки путем
нарушения проницаемости клеточной мембраны либо в результате микропи-
ноцитоза иммуноглобулинов, связавшихся с антигенами, экспонированными
на поверхности клетки. В любом случае этот еще не изученный механизм про-
никновения антител в живые клетки, доказанный также в отношении антител к
топоизомеразе-1 при склеродермии, по-видимому, играет определенную роль в
иммунопатогенезе ДБСТ.
Антиуела к Ro/SS-A
Сывороточный фактор у больных с СШ, связывающийся с экстрактами слюн-
ных и слезных желез, был идентифицирован под названиями Ro и La, в соответ-
ствии с именами больных, у которых антитела были впервые выделены. Согласно
ДИАГНОСТИКА АУТОИММУННЫХ ЗАБОЛЕВАНИЙ g9
современной номенклатуре, эти разновидности антител получили названия SS-A
и SS-B (Sjogren's syndrome A and В antigens). Антитела Ro/SS-A направлены против
ядерных рибонуклеопротеинов 60 и 52 кДа, с которым связаны цитоплаз-
матические РНК, транскрибируемые РНК-полимеразой III. Антиген SS-А при-
сутствует в большинстве клеток человеческого организма, однако в наибольших
количествах в лимфоидных тканях, таких как тимус и селезенка. Антитела к Ro/
SS-A могут быть диагностированы всеми стандартными тестами, применяемыми
для обнаружения АНА. Контрэлектрофорез превосходит другие методы по чув-
ствительности (в том числе ИФА) и специфичности, которая достигает 100%, и
представляет собой оптимальный метод для выявления этой разновидности АНА в
клинической практике. Если субстратом для определения АНФ служат криосрезы
печени или почек крыс, то большинство больных с антителами к Ro/SS-A будут
негативны при проведении непрямой иммунофлюоресценции, что входит в поня-
тие «АНФ-негативная СКВ».
Антитела к Ro/SS-A имеются приблизительно у 50% больных СКВ, 70% боль-
ных дискоидной красной волчанкой (ДКВ), 60% больных с СШ и примерно
30-40% больных РА. Диагностически значимые титры этих антител могут встре-
чаться у 1% здоровых женщин в возрасте 20-45 лет. Антитела к антигену массой
60 кДа сравнительно чаще выявляют у больных СКВ, тогда как при СШ определя-
ют антитела против обоих антигенов массами 60 и 52 кДа.
Антитела к Ro/SS-A обычно встречаются в популяции больных СКВ с выра-
женной симптоматикой фотосенситивных кожных проявлений, близких ДКВ, что
обусловливает целесообразность активного поиска антител к Ro/SS-A при обсле-
довании больных с предполагаемой СКВ с выраженными проявлениями фото-
чувствительности (табл. 17-9). Часто такие больные описывают ожоги «сквозь
оконное стекло», что свидетельствует о фоточувствительности к низкоэнергетиче-
ским длинноволновым ультрафиолетовым волнам. Клинические проявления при
классической СКВ, сопровождающейся присутствием антител к Ro/SS-A, включа-
ют симптомы фоточувствительности, вторичный СШ у 10% больных, поражение
легких, лимфопению, реже — нефрит, тромбоцитопению и наличие РФ. Отмечено,
что антитела к Ro/SS-A встречаются при сравнительно позднем начале развития
СКВ после 50 лет, протекающей с вторичным СШ. Эта разновидность заболевания
характеризуется поражением легких, нейропсихическими проявлениями и реак-
циями фоточувствительности на фоне нормальной функции почек.
Таблица 17-9. Клинические формы системной красной волчанки, сопровождающиеся образова-
нием антител к Ro/SS-A массой 60 кДа
Клиническая форма Комментарий
АНА-негативная волчанка При прицельном обследовании 75% этих больных имеют антитела к Ro/SS-A
Подострая кожная волчанка Характерна выраженная фоточувствительность До 70% больных /to/SS-4-позитивны, нередко отмечается симптома- - тика СШ
Волчаноподобный синдром, свя- занный с недостаточностью фак- торов комплемента С, или С4 50-75% больных /to/SS-A-позитивны, часто обнаруживают фоточув- ствительные поражения кожи, низкая частота встречаемости антител к дсДНК
СКВ с поздним началом Начало волчанки в возрасте 55 лет, почти 90% больных Ro/SS-A- позитивны. Характерны кожные, нейропсихические и легочные про- явления СКВ
СКВсСШ Клинические признаки СКВ и СШ, низкая частота поражения почек, часто — выраженная фоточувствительность и кожные высыпания
Врожденная красная волчанка Антитела к Ro/SS-A обнаруживают у 100% матерей
Миозит У 5% больных с поли миозитом имеются антитела к Ro/SS-A
90 ДИАГНОСТИКА АУТОИММУННЫХ ЗАБОЛЕВАНИЙ
При классической ДКВ антитела к Ro/SS-A обнаруживают в 75% наблюдений.
У таких пациентов поражения кожи располагаются преимущественно в местах
подвергшихся инсоляции. Они представляют собой небольшие эритематозные
папуллы или бляшки, либо замкнутые циркулярные эритематозные высыпания. У
больных ДКВ отмечают наличие мышечно-суставного синдрома,, проявляющегося
артралгиями либо артритом.
Антитела к Ro/SS-A определяют у 98% матерей, дети которых страдают син-
дромом врожденной волчанки. В основе этого заболевания лежит проникновение
в кровь новорожденного через плаценту антител к Ro/SS-A. Основным признаком
врожденной волчанки является дерматоз, напоминающий ДЛЕ, и множество
системных и гематологических синдромов, включающих врожденную поперечную
АУ-блокаду, гепатит, гемолитическую анемию и тромбоцитопению. Среди клини-
ческих манифестаций врожденная атриовентрикулярная блокада — самое тяжелое
и необратимое осложнение, требует установки искусственного водителя ритма
непосредственно после рождения ребенка. Другие внесердечные проявления носят
транзиторный характер и разрешаются после Ь мес, что соответствует времени
истощения пула материнского IgG в крови ребенка. В эксперименте человеческий
иммуноглобулин, содержащий антитела Ro/SS-A, обладает прямым кардиотоксич-
ным действием в животных моделях ex vivo.
Около половины матерей детей с врожденной волчанкой обычно асимптомны
во время рождения ребенка или лишь изредка имеют проявления СШ, СКВ или
СЗСТ. После рождения детей у большинства асимптомных матерей через некото-
рое время разворачивается картина того или иного заболевания соединительной
ткани. Таким образом, все беременные с подозрением на системное заболевание
соединительной гкани должны быть лабораторно обследованы для выявления
группы риска врожденной красной волчанки.
Установлено, что у больных СШ имеющих антитела к Ro/SS-A, больных с позд-
ним началом СКВ, сопровождающимся наличием Ro/SS-А-антител и признаками
вторичного СШ, а также у больных с подострой кожной волчанкой отмечается
повышенная частота встречаемости определенных аллельных форм генов главно-
го комплекса гистосовместимости, в частности HLA-B8, DR3, DQ1/2, что может
указывать на общность генеза этих заболеваний.
Антитела к La/SS-B
Антитела к SS-B-антигену встречаются приблизительно у 10-15% больных
с СКВ и примерно у 50% больных с СШ. Антитела к La/SS-B направлены на
белки массой 48 и 43 кДа, связанные с транскриптами РНК-полимеразы 3. La/
SS-B-макромолекула связывает Ro/58-А-ассоциированные Y,_5 РНК, т.е. в ряде
случаев La/SS-B и Ro/SS-A представляют собой один антиген. Для антитела к La/
SS-B характерен крупногранулярный тип флюоресценции АНФ на линии HFp-2.
Антитела к La/SS-B в подавляющем большинстве случаев наблюдаются совместно
с антителами к Ro/SS-A, в то время как последние могут встречаться изолирован-
но. Совместное обнаружение этих видов АНА является основой лабораторной
диагностики СШ и обладает высокой специфичностью при данном заболевании.
СШ — ревматическое заболевание, поражающее слюнные и слезные железы,
приводящее к ксеростомии и ксерофтальмии. Его встречаемость у лиц 50-70-
летнего возраста достигает 5% всей популяции, в подавляющем большинстве СШ
поражает женщин. Часто СШ сопровождается экстрагландулярными проявле-
ниями, такими как васкулиты, а также на его фоне нередко отмечается развитие
лимфом. Первичный СШ встречается изолированно, но признаки СШ обнару-
живаются при РА, СКВ и других аутоиммунных заболеваниях в виде сочетания
ксеростомии и ксерофтальмии.
ДИАГНОСТИКА АУТОИММУННЫХ ЗАБОЛЕВАНИЙ 91
Предполагается, что антигены Ro/SS-A и La/SS-B концентрируются на поверх-
ности клеток эпителия конъюнктивы и желез у больных СШ, Сходное распреде-
ление этих антигенов отмечается в клетках перевиваемых линий, пораженных
ретровирусами, что позволяв т с определенной уверенностью предполагать вирус-
ную этиологию данного заболевания. У больных с антителами к La/SS-B отмечает-
ся выраженная инфильтрация слюнных желез иммуноглобулин-секретирующими
В-клегками, синтезирующими аутоантитела. Хотя данных за непосредственное
цитопатическое действие АНА при СШ не получено, слюнные железы, без сомне-
ния, являются основным местом продукции аутоантител, участвующих в образо-
вании ЦИК и развитии иммунокомплексной патологии.
Если антитела к Ro/SS-A встречаются при разных аутоиммунных заболеваниях,
то La/SS-B обычно указывают на наличие у больного СШ. Антитела к La/SS-B
крайне редко встречаются при СКВ, протекающей с выраженными проявлениями
вторичного СШ и сравнительной сохранностью функции почек. Комбинация Ro/
SS-A и La/SS-B нередко отмечается в сочетании с высокими титрами РФ, гипер-
гаммаглобулинемией и криоглобулинемией. Для больных с антителами к Ro/
SS-A и La/SS-B характерно раннее начало и большая длительность заболевания,
выраженное увеличение околоушных слюнных желез и значительная степень
лимфоцитарной инфильтрации слюнных желез. Титр антител к Ro/SS-A и La/SS-B
коррелирует со степенью лимфоцитарной инфильтрации малых желез. Их обнару-
жение у больного СШ может прогнозировать развитие таких экстрагландулярных
проявлений заболевания, как васкулит, лимфаденопатия, спленомегалия, анемия
и лейкопения. Таким образом, антитела к Ro/SS-A и La/SS-B отражают выражен-
ность патологического процесса при СШ и риск развития экстрагландулярных
проявлений. Однако обнаружение Ro/SS-A и La/SS-B при СШ характеризуется
сравнительно низкой чувствительностью.
Существенное значение в диагностике СШ играет определение антител к про-
токам слюнных желез, которые встречаются у 50-90% больных СШ Эти антитела
реагируют с цитоплазмой эпителиальной клетки Для их обнаружения применяют
метод НРИФ с использованием в качестве субстрата аутопсийных человеческих
тканей. Обычно у больного СШ, наряду с антителами к протокам, имеются анти-
тела к Ro/SS-A, La/SS-B, РФ и другие разновидности АНА.
Среди признаков аутоиммунного процесса при СШ характерно наличие антител
к париетальным клеткам слизистой оболочки желудка, нейтрофильным цитоплаз-
матическим антигенам и антител к митохондриям, что связано с поликлональной
активацией В-клеточного звена иммунитета, индуцирующей образование аутоа-
грессивных клонов плазмоцитов. Гиперстимуляция В-клеток при СШ является
причиной частого возникновения лимфопролиферативной патологии. Признаки
моноклональной В-клеточной пролиферации в виде образования моноклональ-
ного иммуноглобулина отмена ют у 60% больных с экстрагландулярными прояв-
лениями СШ и практически у всех больных обнаруживают свободные легкие цепи
иммуноглобулинов в моче.
Антитела к ядерному антигену пролиферирующих клеток (PCNA)
Антитела к PCNA-1 (proliferating cell nuclear antigen 1) взаимодействуют с белком
циклином массой 36 кДа, участвующим в контроле клеточного цикла. Этот белок
появляется в клетке перед митозом и подвергается деградации после деления.
Антитела при СКВ дают характерное гранулярное окрашивание митотических
клеток и характерную линию иммунопреципитации в геле. В экспериментах in vitro
антитела к PCNA способны ингибировать синтез ДНК в митотических клетках, что
указывает на их возможное патогенетическое значение при СКВ.
Данную группу антител обнаруживают у 3-5% больных СКВ. Они могут встре-
чаться при РА, при других ревматических заболеваниях их не диагностируют.
92
ДИАГНОСТИКА АУТОИММУННЫХ ЗАБОЛЕВАНИЙ
У больных СКВ с антителами к PCNA, наряду с типичными признаками волчан-
ки, выявляется диффузно-пролиферативный гломерулонефрит, поражение ЦНС
и тромбоцитопения. Характерно, что у больных с нейропатией и антителами
к PCNA-1 нет антител к дсДНК. Мониторинг содержания антител показал, что
увеличение титра антител к PCNA-1 сопутствует обострению гломерулонефрита,
стероидная терапия снижает их титр, У больных волча нкой отмечают увеличение
экспрессии PCNA-1 на мононуклеарах крови, преимущественно на CD4J и CD8+-
клетках. Процентное соотношение PCNA- 1-позитивных клеток коррелирует с
активностью заболевания.
Антитела к рибосомам и рибосомальному белку Р (RiboP)
Рибосомы явл яютс я цитоплазматическими органеллами, однако антитела к ним
причисляют к семейству АНА. Рибосомы — органеллы, состоящие из комплексов
белков и нуклеиновых кислот, их основная функция заключается в синтезе белка
на основании последовательности матричной РНК. При СКВ основными антиге-
нами служат фосфопротеиновые компоненты рибосом, прежде всего рибосомаль-
ные белки Ри, Рт и Р,. Антитела к рибосомам могут выявляться при использовании
флюоресцентного теста и характеризуются типичным свечением цитоплазмы
клетки. Иногда цитоплазматическое свечение может быть пропущено, больные
могут попадать в группу «АНФ-негативной СКВ», что требует более детального
обследования для обнаружения этой разновидности АНА.
Антитела к рибосомальному белку Р высокоспецифичны для СКВ, имеются у
10-20% больных. Антитела к рибосомам часто встречаются совместно с анти тела-
ми к 5?л-антигену. Антирибосомальные антитела обнаруживают преимущественно
у больных с обострением СКВ, с поражением ЦНС, особенно при волчаночном
психозе и депрессии, встречаемость антирибосомальных антител при этих состоя-
ниях составляет 50-80%. В диагностике поражения ЦНС при СКВ выявление
антител к рибосомальному белку Р целесообразно дополнять исследованием
ликвора для обнаружения олигоклонального IgG, который отражает иммуноо-
посредованное воспаление в ЦНС. Определение этих антител позволяет отличить
поражение ЦНС на фоне СКВ от психоза, вызванного приемом высоких доз кор-
тикостероидных гормонов.
Другие клинико-лабораторные взаимосвязи между обнаружением антител к
рибосомам и клинической картиной СКВ включают развитие дискоидных высы-
паний, фоточувствительность и афтозный стоматит, поражение печени у пациен-
тов с высокими титрами антирибосомальных антител.
Антитела к Ku/Ki и антитела к МАт-антигену
Ант и гела к Ku (Ki) взаимодействуют с белками ядерного матрикса массой 70 и
80 к Да. Они были впервые обнаружены у больных с перекрестным синдромом с
проявлениями полимиозита и склеродермии. Эти антитела встречаются у 10-40%
населения европейской популяции больных СКВ, причем в высоких титрах, пре-
вышающих 1:10 000. Их наличие диагностируют при СЗСТ, склеродермии и зна-
чительно реже - при полимиозите, РА, СШ, болезни Грейвса (30%) и первичной
легочной гипертензии (20%). Антитела к МАт направлены на нуклеарный кислый
антиген, чувствительный к протеолизу под воздействием трипсина, но устойчивый
к РНКазе и ДНКазе. Эта разновидность АНА встречается у 3-18% больных СКВ,
возможно, у больных с более активным течением заболевания.
Антитела к антигену Scl-70 (топоизомеразе-1)
При СС основной разновидностью АНА являются аутоантитела против анти-
гена ScZ-70. Эти антитела имеют основной мишенью топоизомеразу-1 массой
70 кДа. Топоизомераза представляет собой один из ферментов, участвующих в
механизмах деспирализации суперскрученной ДНК, необходимой для транс-
ДИАГНОСТИКА АУТОИММУННЫХ ЗАБОЛЕВАНИЙ 93
кринции генов. Антит енпая структура топоизомеразы хорошо изучена, основным
антигеном, который используют в методах ИФА и иммщюблота, является реком-
бинантный белок. Любой метод, применяемый для обнаружения антител к Scl-70,
может быть подтвержден непрямой иммунофлюоресценцией на клетках НЕр-2,
благодаря характерному двойному типу свечения, включающему мелкогрануляр-
ный и ядрышковый тип флюоресценции. Это обусловливает целесообразность
определения АНФ в качестве скринингового теста перед определением антител к
Scl-70.
Антитела к Sc/-70 представлены иммуноглобулинами классов IgG и IgA, однако
рутин ные лабораторные тесты позволяют обнаружить преимущественно антитела
класса IgG. Антитела к топоизомеразе встречаются при диффузной склеродермии
с поражением внутренних органов (СС). однако их наличие не исключает присут-
ствия перекрестных синдромов склеродермии с СКВ или СШ. Антитела к Scl-70
присутствуют у 70% больных СС. у 30% больных диффузной склеродермией и у
10% больных с локализованными формами этого заболевания. В единичных слу-
чаях могут встречаться у онкологических больных без признаков СС или склеро-
дермии У больных с диффузной склеродермией с антителами к Scl-70 чаще встре-
чаются рубцовые изменения на коже пальцев, развитие центрального склероза и
поражение почек и сердца.
Антитела к центромерам
Основным антигеном является центромерный белок CENP-B, который входит
в состав центромер — частей хромосомы эукариот, необходимых для образования
веретена деления при митозе. Ткани лабораторных животных слабо экспресси-
руют антигены центромерных антител, поэтому АНФ на клеточной линии НЕр-2
является основным методом обнаружения этих антител в клинической практике.
Антитела к центромерам хорошо визуализируются на НЕр-2-клетках благодаря
свечению в делящихся клетках, в которых хромосомы компактны. Если хромосомы
находятся в стадии спирализации, в ядре клетки флюоресцируют мелкие гранулы
либо наблюдается картина веретена деления, подчеркнутого свечением центро-
мер. Антицентромерные антитела обнаруживают в высоких титрах при использо-
вании линии НЕр-2, для которых типична большая частота клеточных делений.
Иммуноблоттинг имеет лишь незначительные преимущества в связи с большей объ-
ективизацией результатов теста. Антитела к центромерам и антитела к Scl-70 редко
встречаются совместно. Иммуноблоттнг показывает, что в 10% случаев у больных
с Scl-70 также имеются антицентромерные антитела. Часто у этих больных диагно-
стируют Ro/SS-A и La/SS-B, PM-Scl, антимитохондриальные антитела.
Антитела к центромерам являются серологическим маркером CREST-синдрома,
представляющего собой сравнительно доброкачественную форму диффузной
склеродермии, которая характеризуется подкожными кальцинатами, синдромом
Рейно, эзофагитом, склеродактилией и телеангиэктазиями на коже туловища.
Титры антител к центромерам не отражают активности заболевания Антитела
к центромерам выявляют у 50% больных с CREST, у 20% больных с СС и у
10% больных с изолированным синдромом Рейно. В этой группе пациентов
присутствуют типичные клинические признаки CREST-синдрома: отсутствие
рубцовых контрактур пальцев, более выраженный отек пальцев (по сравнению
со склерозом), кальциноз фасций и сухожилий. Больные с антителами к центро-
мерам старше, длительность заболевания у них обычно больше, чем у больных со
склеродермией без этих антител. Это связывают с тем. что у таких больных реже
отмечается поражение внутренних органов, реже встречаются атрофические про-
цессы на коже, но часто присутствует кальциноз и ишемическая гангрена пальцев
кисти. Практически у всех больных с антицентромерными антителами имеется
выраженный синдром Рейно.
94 ДИАГНОСТИКА АУТОИММУННЫХ ЗАБОЛЕВАНИЙ
Антицентромерные антитела присутствуют в сыворотке 20% больных с идио-
патическим синдромом Рейно, сопровождающимся артралгиями. У половины
из них через несколько лет развивается полная клиническая картина склеродер-
мии. Таким образом, антицентромерные антитела могут служить предвестником
склеродермии и могут быть использованы при дифференциальной диагностике
синдрома Рейно. Антицентромерные антитела не выявляют у здоровых лиц даже в
низких титрах, что позволяет рассматривать их обнаружение как предиктор разви-
тия заболевания. Патогенетическая роль аутоантител при склеродермии остается
под вопросом, изменения их титра не отражаются на активности аутоиммунного
процесса, однако указывают на определенные клинические проявления этого
заболевания.
Антитела к центромерам изредка встречаются при ряде других заболеваний,
сопровождающихся синдромом Рейно: СКВ, РА. тиреоидите Хашимото. первич-
ном билиарном циррозе.
Антитела к PM-Scl и другим антигенам ядрышка
АНА, направленные против компонентов ядрышка, таких как PM-Scl, U^-RNP
(фибрилларин), РНК-полимераза I, NOR90, часто обнаруживают при СС. Основным
методом обнаружения этих разновидностей АНА является прицельное определе-
ние АНФ с ядрышковым типом свечения.
Антитела к PM-Scl-антигену обнаруживают при сочетании клинических
признаков СС и НМ. Эта разновидность АНА направлена к антигенной системе
ядрышка, включающей более 16 белков с молекулярным весом от 20 до 110 кДа.
В случае наличия антител к PM/Scl приблизительно у 50% больных имеет место
сочетание миозита и СС, реже антитела встречаются на фоне классической кар-
тины ПМ или СС. Это сочетание получило название «склеромиозит», характе-
ризующийся миозитом и миалгиями в сочетании с кожными признаками дерма-
томиозита (периорбитальным отеком и папулами Готрона), наряду с синдромом
Рейно, склеродактилией и интерстициальным пневмонитом. У больных с анти-
телами к PM-Scl высок риск идиопатической легочной гипертензии, что приводит
к быстрой декомпенсации пациентов, поэтому такие больные требуют особого
внимания при оценке функции легких.
Антитела к РНК-полимеразам I, II и III встречаются у 20% больных с СС,
CREST и другими перекрестными синдромами. В иммунофлюоресцентном тесте
отмечают своеобразный точечный ядрышковый тип свечения. Их обнаруживают
изолированно, без антител к центромерам и Sd-70. Антитела к полимеразам свя-
заны с диффузным поражением кожи и внутренних органов, в том числе сердца и
почек.
Антитела к фибриллярину (U^noRNP) — одному из основных ядрышко-
вых рибонуклеопротеинов — диагностируют у 6-8% больных склеродермией.
Иммунофлюоресценция фиксирует анти ядрышковый тип свечения со значитель-
ной конденсацией и интенсивным свечением хроматина в клетках, находящихся в
процессе митоза. Антитела к фибриллярину встречаются у больных с агрессивным
прогрессирующим системным склерозом с вовлечением внутренних органов, в
том числе с поражением тонкого кишечника, легочной гипертензией, миозитом, и
указывают на неблагоприятный прогноз.
Антитела к РНКазе (RNAse MPR, Th snoRNP) направлены на один из ядрыш-
ковых рибонуклеопротеинов, связанных с процессингом митохондриальной РНК.
Они встречаются у 4—11*>о больных склеродермией. Их присутствие в сыворотке
типично для поражения тонкого кишечника, гипотиреоза, артрита и локализован-
ной кожной формы склеродермии.
Антитела к NOR90 взаимодействуют с белком массой 90 кДа, компонентом
ядрышкового организатора, участвующим в сборке ядрышка после митоза. При
ДИАГНОСТИКА АУТОИММУННЫХ ЗАБОЛЕВАНИЙ 95
выявлении АНФ в ядре обнаруживают 10-20 мелких дискретных точек, причем
интенсивность окраски меняется в зависимости от фазы жизненного цикла клетки.
Этот тип АНА неспецифичен при склеродермии и встречается при СКВ. РА, неко-
торых формах онкологических заболс ваний.
Миозмт-специфические антинуклеарные антитела: антитела к тРНК-синтетазам
(анти-Jo-l, PL-7, PL-12, EJ и 0J), антитела к Mi-2 антигену, SRP и р56
Заболевания, характеризующиеся воспалительным миозитом, представляют
полиморфную группу, характеризующуюся цитотоксическим ответом к компонен-
там поперечнополосатой мускулатуры. Их часто объединяют под названием «вос-
палительные миопатии». Среди наиболее часто выявляемых заболеваний, отно-
сящихся к воспалительным миопатиям, выделяют классический дерматомиозит,
миозит в рамках перекрестных синдромов, полимиозит и паранеопластический
полимиозит, а также миозит с внутриклеточными включениями.
Конкретные разновидности АНА при ПМ/ДМ условно подразделяют на миозит-
спсцифичные и миозит-ассоциированные, т.е. АНА, которые встречаются также
при других формах ДБСТ в рамках перекрестных синдромов. Классификация
аутоантител при миозите приведена в табл. 17-10. При воспалительных миопа-
тиях миозит-специфические антитела могут быть обнаружены в 40% случаев, в
то время как миозит-ассоциированные АНА обнаруживают у 20-30% больных.
Общая частота встречаемости АНА даже при развернутых фирмах с классической
картиной полимиозита не превышает 60%.
Таблица 17-10. Классификация антинуклеарных антител при воспалительных миопатиях
Разновидности АНА % обнару- жения Комментарий
Миозит- специфические антитела Антитела к тРНК- синтетазам (Jo-1, PL-7, PL-V2,EJ\aOS) 25 Маркеры «антисинтетазного синдрома», сочетающего характерные кожные и систем- ные проявления
Антитела к белку Mi-2 10 Встречаются при классическом дерматомио- зите с выраженными кожными проявлениями
Антитела к компонентам сигналраспознающей частицы SRP 3 Характерно тяжелое течение полимиозита с поражением дыхательных мышц и миокар- дитом
Миозит- ассоциированные антитела Антитела к PM-Scl 10 Встречаются у больных с перекрестным син- дромом, объединяющим признаки диффуз- ной склеродермии и полимиозита
Антитела к J-RNP 10 Маркер СЗСТ
Антитела к SS-A 3 0
К семейству миозит-спефицифических АНА относят 3 основные разновидности
аутоантител: антитела к аминоацил-тРНК-синтетазам, антитела к белку Mi-2 и
антитела к компонентам сигнал-распознающей частицы SRP.
Антисинтетазные антитела — наиболее обширное семейство миозит-
специфических аутоантител, насчитывает 5 основных представителей: Jo-1, PL-7,
PL-1'2, EJ и OJ, которые совместно встречаются у 20-30% больных ДМ/ПМ.
Антитела, взаимодействующие с Jo-1-антигеном, обнаруживают в несколько раз
чаще, чем другие антисинтетазные антитела. Эти антитела реагируют с гистидил-
тРНК-синтетазой; остальные антисинтетазные антитела направлены против
4-х различных типов тРНК-синтетаз: треониновой, аланиновой, глициновой, изо-
лейциновой. Антитела Jo-1 выявляют при использовании контрэлектрофореза и
ИФА, что позволяет широко применять определение этого типа АНА в лабора-
торной практике. Контрэлектрофорез обладает достаточной чувствительностью и
специфичностью для рутинной работы. Хотя антисинтетазные антитела не вклю-
чены в классификационные критерии ДМ. их обнаружение при соответствующей
96
ДИАГНОСТИКА АУТОИММУННЫХ ЗАБОЛЕВАНИЙ
клинической картине с высокой достоверностью подтверждает диагноз и, в ряде
случаев, делает ненужной биопсию ткани. Поскольку антитела к тРНК-синтетазам
обладают низкой чувствительностью при ДМ/ПМ, их отсутствие не может опро-
вергнуть диагноз ДМ или ПМ. Антитела к Jo-1 имеются у 15-20% больных ДМ/
ПМ. Предполагают, что более широкое внедрение тестирования в отношении дру-
гих антисинтетазных антител может удвоить эту цифру.
У всех больных с антисинтетазными антителами наблюдают очень сходную
клиническую картину, что привело к выделению особой клинико-лабораторной
рубрики — так называемого антисинтетазного синдрома, одной из разновидностей
перекрестных синдромов. Он характеризуется клинической картиной дермато-
миозита, но протекает более активно и требует назначения активной иммуносу-
прессивной терапии. Около 90% больных с антисинтетазным синдромом страдают
симметричным полиартритом, часто отмечается синдром Рейно. Для этих боль-
ных характерна особая форма поражения кожи, так называемая рука механика, с
гиперпигментацирй, гиперкератозом и трещинами по латеральным и пальмарной
поверхностям пальцев, напоминающая забитые грязью складки на коже рабочего.
Интерстициальное поражение легких или фиброзирующий альвеолиз присут-
ствуют у 50-75% больных с антителами к тРНК-синтетазам. Приблизительно у
1/5 больных с антисинтетазными антителами отмечают признаки другого забо-
левания соединительной ткани. Больные с антисинтетазными антителами хуже
поддаются терапии, и у них чаще возникают обостпения при снижении доз имму-
носупрессивных препаратов,
Антитела к Mi-2-антигену реагируют с ядерным антигеном, входящим в
состав ЭНА. Антитела к Mi-2 являются наиболее специфичным серологическим
маркером ДМ, а также встречаются при ювенильном ДМ. Эта разновидность АНА
может быть обнаружена благодаря гомогенному типу свечения АНФ. Данный тип
АНА обнаруживают у 10-20% больных ДМ с выраженными признаками пораже-
ния кожи. У таких больных наблюдают в целом благоприя гное течение заболева-
ния и хороший ответ на иммуносупрессивную терапию.
Антифосфолипидные антитела
ЛОЖНОПОЛОЖИТЕЛЬНАЯ РЕАКЦИЯ ВАССЕРМАНА ПРИ АНТИФОСФОЛИПИДНОМ
СИНДРОМЕ
Первым методом обнаружения антифосфолипидных антител стал серологи-
ческий тест для диагностики сифилиса, который был разработан Вассерманом в
Ш06 г. Основным антигеном экстракта бычьих сердец, требуемым для проведе-
ния реакции Вассермана (РВ), является фосфолипид, который выделен и назван
кардиолипином. Активное внедрение РВ для серодиагностики сифилиса приве-
ло к тому, чти была выделена категория пациентов с положительной реакцией,
однако признаков сифилиса у них не отмечалось. В то же время у этой категории
лиц с ложноположительным серологическим тестом была отмечена повышенная
частота встречаемости аутоиммунных заболеваний, прежде всего ДБСТ и СШ.
Этот феномен получил название «биологически ложноположительный тест на
сифилис».
Одновременно с ложноположительной РВ при СКВ в крови этих пациентов был
обнаружен сывороточный фактор., ко горый препятствовал свертыванию крови и
получил название «волчаночный антикоагулянт». Волчаночный антикоагулянт
(ВА) отсутствовал в плазме больных сифилисом и часто отмечался у пациентов с
СКВ. У больных с ВА был установлен характерный набор клинических признаков,
среди которых ведущими являются тромбозы артерий и вен, повторные выкиды-
ши и тромбоцитопения, составляющие симптомокомплекс антифосфолипидного
ДИАГНОСТИКА АУТОИММУННЫХ ЗАБОЛЕВАНИЙ 97
синдрома (АФС). Интерес к диагностике этого аутоиммунного заболевания зна-
чительно возрос в связи с разработкой тестов для выявления антифосфолипидных
антител (АФА).
АНТИТЕЛА К ФОСФОЛИПИДАМ И АНТИТЕЛА К КАРДИОЛИПИНУ
АФС представляет собой аутоиммунное нарушение коагуляции, составляющее
основную причину тромбозов у молодых людей и одну из ведущих причин при-
вычного невынашивания беременности. В основе АФС лежит образование АФА,
представляющих собой гетерогенную популяцию иммуноглобулинов, взаимодей-
ствующих с отрицательно заряженными, реже — с нейтральными фосфолипидами.
Эти антитела in vitro нарушают образование протромбинового комплекса (про-
тромбчназы), который состоит из фактора X, фактора V, фосфолипидов и кальция,
что приводит к гипокоагуляции и удлинению времени основных коагуляционных
тестов. При этом in vivo у больных АФС имеет место усиленное тромбообразование.
Нарушение функции протромбинового комплекса ведет к развитию тромбо-
тического состояния с изменением сосудисто-тромбоцитарного гемостаза и хро-
ническим потреблением факторов коагуляции, напоминающего ДВС-синдром.
При АФС тромбоз происходит на фоне обезвоживания, инфекции или венозного
застоя. Помимо коагулопатии, АФС сопровождается нарушениями микроциркуля-
ции. проявляющимися язвами кожи, микроангиопатией почек, кожным ливедо.
Название «антифосфолипидные антитела» не отражает самой сущности взаи-
модействия антифосфолипидных антител со своими антигенами, в котором наряду
с фосфолипидами важную роль играют белковые кофакторы этих аутоантител,
одним из которых является |3.,-гликопротеин-I ф2-ГП). Другими белковыми
кофакторами фосфолипидов могут быть протромбин, тромбомодулин, протеи-
ны С и S, фактор XI, аннексии V, кининоген, мембранные белки тромбоцитов
и эндотелиоцитов. Кроме того, антикардиолипиновые антитела (АКЛА) пере-
крестно реагируют с фосфатидилсерином и другими отрицательно заряженными
фосфолипидами, антителами к тромбоцитам, антителами к окисленным липопро-
теидам низкой плотности и с протромбином. Таким образом, АФа реагируют с
гетерогенной группой фосфолипидов и белковых антигенов сосудистого русла.
Предпочтительным тестом для обнаружения АКЛА служит метод ИФА. При
детекции АКЛА обязательным является наличие в тестируемом фосфолипидном
субстрате р2-ГП, который представляет собой белковый кофактор. Обычно он при-
сутствует в составе буферов для разведения сыворотки больных. С методической
точки зрения, метод определения АКЛА для диагностики АФС более корректно
было бы называть <ф2-ГП-зависимые антикардиолипиновые антитела», поскольку
действительной мишенью аутоантител является неоантиген, образующийся при
взаимодействии кардиолипина и р2-ГП.
АКЛА имеют несомненное патогенетическое значение в развитии коагулопатии.
В культурах АКЛА стимулируют выработку внутреннего фактора макрофагами.
АКЛА реагируют с тромбоцитами с их активацией, являются причиной тромбоци-
топений у большинства больных АФС. АКЛА активируют эндотелиальные клетки
с экспрессией факторов адгезии. Под действием АКЛА снижается антикоагулянт-
ная активность протеина С. Наконец, АКЛА увеличивают частичное тромбопла-
стиновое время in vitro — феномен, известный под названием «волчаночный анти-
коагулянт». В качестве антигена могут быть использованы другие отрицательно
заряженные фосфолипиды, в частности фосфатидилсерин или фосфатидилино-
зитол. Кардиолипин (дифосфатидил-глицерол) имеет значительную структурную
гомологию с другими фосфолипидами, поэтому в случае, если отсутствуют АКЛА,
обычно отсутствуют другие АФА, Целесообразность определения антител к другим
представителям фосфолипидов (фосфа гидилэтаноламину и протромбину) в прак-
тической диат ностике пока не доказана.
98 ДИАГНОСТИКА АУТОИММУННЫХ ЗАБОЛЕВАНИИ
Определение АКЛА более стандартизовано по сравнению с другими АФА: соз-
даны стандарты сывороток, содержащих АКЛА. Иногда эти стандарты называют
стандартами Harris, по имени ученого, который разработал единицы GPL для
АКЛА класса IgG и единицы MPL для АКЛА IgM. Одна единица представляет
связывающую активность 1 мкг аффинно очищенных поликлональных АКЛА.
Вторичный стандарт калибруется против первичных стандартов. Аутоантитела
каждого класса можно определять независимо друг от друга. Результаты обследо-
вания интерпретируют с помощью градации на низкие (менее 40 единиц), средние
(от 40 до 80 единиц) и высокие (более 80 единиц) титры антител. АКЛА могут
быть обнаружены как в сыворотке, так и в плазме крови. Положительный резуль-
тат определяется при среднем или высоком их титре, ее. при титре более 40 GPL
и MPL единиц. Наличие гетерогенных поликлональных АКЛА от различных
пациентов в стандартах приводит к различиям между коммерческими наборами.
Инактивация комплемента нагреванием может приводить к получению ложно-
положительных результатов, а повторное замораживание приводит к снижению
титров аутоантител. Согласно лабораторным классификационным критериям
АФС, пациент должен быть «положительным» как минимум по одному из тестов
(кардиолипину, [3,-ГП-1 или В А) при повторном определении по меньшей мере
через 12 нед. Наличие постоянно повышенных АКЛА в титре более 60 GPL может
предсказать присутствие у пациента ВА и положительных результатов при иссле-
довании на наличие антител к Р2-ГП-1.
Подавляющее число тест-систем для обнаружения АКЛА. на отечественном
рынке по-прежнему калибровано в единицах Harris. Очевидны недостатки единиц
Harris для стандартизации тест-систем. Фактически реактивы разных производи-
телей представляют собой разные методы, результаты которых сложно сопоста-
вить друг с другом. Для преодоления этих расхождений были предложены новые
стандарты АКЛА, основанные на моноклональных очеловеченных антителах
HCAL и EY2C9.
АКЛА в низких и средних титрах определяют у 30-40% пациентов с СКВ, что
позволяет использовать их в качестве одного из серологических маркеров этого
заболевания. Их выявление не коррелирует с возрастом больных, длительностью
заболевания или особыми чертами его течения, включая наличие полиартрита,
серозита или васкулита. В то же время наличие высоких титров АКЛА указывает
на риск развития вторичного АФС у больного СКВ. Однако СКВ не единственное
ревматол отческое заболевание, сопровождающееся АКЛА. Сравнительно часто
АКЛА диагностируют у больных СС, васкулитами и васкулитоподобными состоя-
ниями, например при синдроме Бехчета и болезни Крона. При отсутствии ДБСТ
заболевание рассматривают как первичный АФС.
АКЛА, относящиеся к изотипу IgG2, как правило, имеют максимальный титр
непосредственно перед развитием тромбоза и несколько снижаются сразу после
его возникновения, что свидетельствует об их потреблении в процессе коагуляции.
Увеличение титра АКЛА при развитии клинической картины тромбоза указывает
на АФС. В то же время АКЛА являются одними из наиболее распространенных
аутоантител, присутствующих у клинически здоровых лиц. Частота их обнару-
жения значительно выше частоты обнаружения АФС. Значимые концентрации
АКЛА возникают на фоне широкого ряда инфекционных заболеваний. Среди
наиболее частых индукторов АКЛА известны инфекции вирусами гепатита С,
ВИЧ. Эпштейна-Барр, парвовирусом В19, аденовирусом, стрептококком, спирохе-
тами, хеликобактером, сальмонеллами, лептоспирами, токсоплазмой. Обычно при
инфекциях появляются АКЛА класса IgM в низком титре, который плавно сни-
жается после выздоровления. Это требует повторных определений АКЛА с пере-
рывом в 1,5 мес, что соответствует времени обновления пула иммуноглобулинов
сыворотки. Онкологические заболевания, хронические интоксикации, в том числе
ДИАГНОСТИКА АУТОИММУННЫХ ЗАБОЛЕВАНИЙ Q9
алкоголизм, индуцируют образование АКЛА, что обусловливает сложность интер-
претации этого показателя. При этом не находят антител к р2-ГП-1 и не бывает
зафиксировано тромбозов.
АНТИТЕЛА К Р2-ГЛИК0ПР0ТЕИНУ-1
Р2-ГП-1 представляет собой гликозилированный белок плазмы крови, содержа-
ние которого в норме составляет 150-300 мкг/мл. Он обладает антикоагулянтной
активностью, способен связываться с апоптотическими тельцами и участвует в их
клиренсе из кровотока. Определение антител к |3?-ГП-1 было в 2006 г. включено
в лабораторные классификационные критерии АФС. р2-ГП-1 обладает непрямой
антикоагулянтной активностью за счет ингибирования протромбинового ком-
плекса и подавления активации тромбоцитов. Большую роль отводят р2-ГП-1 в
фагоцитозе апоптотических клеток и окисленных липопротеинов. При иммуни-
зации лабораторных животных р,-ГП происходит образование АФА, в результате
часто возникает резорбция плода, тромбоцитопения и увеличение активированно-
го частичного тромбопластинового времени (АЧТВ).
Антитела к р,-ГП-1 распознают исключительно человеческий белок и встреча-
ются у 70-80% больных с АФС. Антитела к Р2-ГП-1 низкоаффинны. их содержа-
ние обычно невелико. Связывание Р2ТП-1 на пластике ИФА-планшета приводит к
экспозиции скрытых эпитопов. Одной из основных проблем использования этого
маркера являются недостаточно сопоставимые результаты обследования, связан-
ные с отсутствием стандартизованных калибраторов. Коммерческие наборы кали-
бруются собственными калибраторами производителей, и результаты выражаются
в произвольных единицах. Несмотря на отсутствие стандартизации, в выявлении
антител к Р2-ГП наблюдается лучший межлабораторный консенсус относительно
точности этой методики по сравнению с определением АКЛА. На сегодняшний
день определение антител к р2-ГП-1 — полноправный лабораторный метод диа-
гностики АФС.
ВОЛЧАНОЧНЫЙ АНТИКОАГУЛЯНТ
АФА индуцируют тромбозы in vivo, но in vitro они приводят к замедлению
фосфолипид-зависимых коагуляционных тестов, прежде всего АЧТВ, и, в меньшей
степени, протромбинового времени. Выявление ВА — функциональный тест, кото-
рый позволяет оценить действие антител на каскад коагуляции, которые, в свою
очередь, предупреждая коагуляцию in vitro, сами ассоциируются с тромбозом.
Коагулогические тесты для обнаружения ВА подразделяют на скрининговые,
коррекционные и подтверждающие. Крайне важно соблюдение точности на преа-
налитическом этапе методики, поскольку присутствие тромбоцитов в исследуемой
плазме (они являются эндогенным источником фосфолипидов) может привести к
получению ложноотрицательных результатов теста.
В качестве скрининговых тестов используют АЧТВ с фосфолипид-зависимым
субстратом, коалиновое время свертывания и тест с ядом гадюки Рассела.
Положительным результатом считают удлинение фосфолипид-зависимого вре-
мени свертывания крови свыше нормы. Необходимо использование, по крайней
мере, двух коагуляционных тестов. В случае, если оба или хотя бы один из них ука-
зывает на наличие ВА, необходимо выполнить коррекционный (смешанный) тест,
т.е. повторить тест с добавлением плазмы здорового донора в соотношении 1:1.
Если удлинение времени свертывания есть результат недостаточности факторов
коагуляции, оно будет скорректировано до нормальных значений, в то время как
при наличии ВА коррекции не произойдет. Отсутствие коррекционного действия
плазмы здорового донора на время свертывания позволяет отличить действие ВА
от дефицита факторов свертывания.
100 ДИАГНОСТИКА АУТОИММУННЫХ ЗАБОЛЕВАНИЙ
Присутствие у больного специфического В А, зависимого от фосфолипидов,
и отсутствие в сыворотке других ингибиторов коагуляции проводят подтверж-
дающим тестом с добавлением избытка фосфолипидов. В качестве источника фос-
фолипидов применяют разрушенные повторным замораживанием-оттаиванием
отмытые тромбоциты, либо модифицированный аРТТ-реагент, в котором содер-
жится нейтральный фосфолипид в гексагональной форме, специфически связы-
вающий ВА.
Имеются значительные межлабораторные вариации в выполнении данного
типа исследований, поэтому референсные значения должны быть установлены
для каждого метода определения ВА и для каждого коагулометра. Независимо
от выбранных методов тестирования при определении ВА важно проведение
внутреннего лабораторного контроля качества с положительным и отрицатель-
ным BA-контролем для каждой партии реагентов. Стандартизация определения
ВА — насущная потребность, которую предполагается решить с разработкой моно-
клональных антител либо приготовлением референтных стандартов из лиофи-
лизированной плазмы больных. Как и в случае обнаружения АКЛА, необходимо
подтвердить стойкое присутствие ВА в сыворотке больного АФС во времени, так
как ВА может появляться на фоне вирусных и бактериальных инфекций либо под
действием ряда лекарственных препаратов.
Проводимая терапия антикоагулянтами, прежде всего гепарином*, ведет к
удлинению АЧТВ и других скрининговых тестов. Это затрудняет выявление ВА у
пациентов, у которых антикоагулянтная терапия была начата по жизненным пока-
заниям, В данном случае большую специфичность при диагностике АФС имеют
серологические тесты, основанные на определении анти-р2ТП-1-(зависимый ВА).
Этот тест признают в качестве наиболее перспективного подхода для оценки риска
тромбоза по сравнению с классическими методами определения ВА.
Ревматоидный фактор
При РА может быть обнаружено до 50 различных видов аутоантител, кото-
рые встречаются с различной частотой (табл. 17-11). Первой разновидностью
аутоантител, обнаруженных при РА, стал РФ. Основной мишенью РФ является
эпитоп Ga, расположенный в С^-С^ районе тяжелой цепи молекулы IgG под-
классов IgGp IgG2, IgG4, недалеко от сайта связывания со стафилококковым про-
теином А. К РФ причисляют антитела против IgG, представленные основными
классами иммуноглобулинов — IgG, IgM и IgA. Хотя РФ может быть представлен
любым классом иммуноглобулинов, однако турбидиметрические и агглютина-
ционные тесты помогают обнаружить в основном IgM-РФ. В латексном тесте
агрегированный человеческий IgG прикреплен к латексным частицам, которые
агглютинируют в присутствии РФ, Это быстрый легковыполнимый тест, однако
он дает большое количество ложнопозитивных результатов. Среди агглюти-
национных тестов классический тест Ваалера-Розе, основанный на пассивной
гемагглютинации с использованием эритроцитов барана, покрытых антиэритро-
цитарной сывороткой крови кролика, до сих пор не утратил своего значения. По
сравнению с латексным и другими агглютинационными тестами положительный
тест Ваалера-Розе является более специфичным для РА, поскольку аллотипиче-
ские антитела, образующиеся при беременности или после гемотрансфузий, не
взаимодействуют с кроличьим IgG. В ряде случаев, тем не менее, гетерофильные
антитела к эритроцитам барана, встречающиеся при инфекционном мононуклео-
зе и ряде других острых инфекций, способны привести к получению ложнополо-
жительного результата этого теста.
ДИАГНОСТИКА АУТОИММУННЫХ ЗАБОЛЕВАНИЙ 1 Q1
Таблица 17-11. Аутоантитела в лабораторной диагностике ревматоидного артрита
Название Источник антигена Мишень антитела
Ревматоидный фактор Иммуноглобулин IgG Ga-эпитоп в Рс-фрагменте
АНА Клетки линии НЕр-2 Анти-SS-A (52 кДа)
Антикератиновые антитела Ороговевающий слой эпите- лия пищевода Профилаггрин крысы
Антиперинуклеарный фактор Кератогиалиновые гранулы Филаггрин человека
Антифилаггриновые антитела Рекомбинантный Филлагрин человека
Антитела к циклическому цитруллин- содержащему пептиду (АЦЦП) Пептидный синтез Цитруллинированный кольцевой пептид
Цитруллинированные пептиды Пептидный синтез Несколько линейных пептидов
Модифицированный цитруллиниро- ванный виментин (MCV) Рекомбинантный Цитруллинированный виментин
Антитела к RA33 Рекомбинантный Рибонуклеопротеины ядра
Нефелометрическое и турбидиметрическое определение РФ — наиболее точ-
ный аналитический метод. Рассеивание света на формирующихся комплексах
«антиген-антитело» позволяет установить их концентрацию в сыворотке крови.
Этот тест является наиболее стандартизированным среди всех иммунологических
тестов, его применение позволяет быстро и точно получить результат в Ш/мл.
Результаты нефелометрического определения РФ хорошо согласуются с традици-
онным латексным тестом. Точное определение класса иммуноглобулинов, которым
представлен РФ, может быть выполнено с помощью метода ИФА. Убедительных
данных о целесообразности определения классов РФ в клинической практике нет.
Тем не менее отмечают, что IgG-РФ встречается при васкулитах, сопутствующих
РА и синдрому повышенной вязкости крови, поскольку он может участвовать в
образовании самоассоциирующихся иммунных комплексов. Для больных с IgA-
РФ характерно быстро прогрессирующее течение заболевания.
Для стандартизации тестов по обнаружению РФ ВОЗ разработан междуна-
родный стандарт сыворотки WHO1066, содержащий 100 IU/мл (international
units) РФ. Существующие стандарты сыворотки учитывают РФ класса IgM., таким
образом, в международных единицах (Ш/мл) может быть получен ответ для РФ,
представленного этим классом антител. В качестве популяционной нормы реко-
мендовано рассматривать концентрации РФ ниже 20 IU/мл. Частота встречаемо-
сти концентраций РФ, превышающих 20 IU/мл, у здоровых людей среднего воз-
раста составляет около 3% и увеличивается до 10-15% у пожилых старше 65 лет.
Концентрация РФ в сыворотке крови больного, превышающая 40 IU/мл, должна
рассматриваться как высокая и является высокоспецифичной для диагностики
РА. Обнаружение титров РФ более 40-50 Ш/мл указывает на высокий риск раз-
вития эрозивного артрита. У тех лиц, у которых был обнаружен высокий титр РФ,
отмечают значительный риск развития РА, так как обнаружение диагностических
титров РФ может предшествовать на несколько лет клиническим проявлениям
заболевания.
Определение РФ до сих пор является основным лабораторным методом диа-
гностики РА и служит основанием для выделения двух его основных клинике -
иммунологических разновидностей: серонегативного и серопозитивного РА. Более
того, отсутствие РФ при ряде воспалительных артритов позволяет выделить кли-
ническую группу серонегативных спондилоартропатий.
РФ обладает достаточно высокой чувствительностью и имеется у 60-80% боль-
ных РА. Однако в дебюте заболевания, на раннем этапе, РФ обнаруживают менее
чем у 25% больных, что существенно снижает его значение для ранней диагности-
ки этого заболевания. Однократного определения РФ на ранней стадии РА с отри-
цательным результатом недостаточно для того, чтобы исключить серопозитивную
102 ДИАГНОСТИКА АУТОИММУННЫХ ЗАБОЛЕВАНИЙ
клинико-иммунологическую форму РА.. Если диагноз РА подозревают или даже
он клинически подтвержден, при отрицательном результате теста по определению
РФ требуются повторные определения его титра каждые 6 или 12 мес. Этот срок
приблизительно соответствует времени обновления пула плазматических клеток,
способных синтезировать аутоантитела. Если же будет получен положительный
результат, то нет реальных предпосылок повторять определение РФ с течением
времени, поскольку титры аутоантител плохо отражают активность заболевания.
Значение повторных определений содержания РФ для контроля над течением
заболевания имеет несравненно меньшее значение, чем мониторинг острофазо-
вого ответа. Снижение титра РФ в плазме крови при успешной терапии может
отчасти объясняться цитостатическим эффектом применяемых препаратов. Тем не
менее у больных РА возможна сероконверсия как в одну, так и в другую сторону в
зависимости от активности и длительности заболевания, а также от проводимого
лечения. Сероконверсия из серопозитивной в серонегативную группу встречается
редко и обычно сопутствует полной клинической ремиссии заболевания.
Кроме низкой встречаемости РФ в дебюте заболевания, очевидным недостатком
РФ в качестве маркера РА является его сравнительно низкая специфичность, кото-
рая не превышает 60% (табл. 17-12). Фактически это означает, что каждый 3-4-й
положительный тест обнаруживается у больного без РА. Так, частота выявления
концентраций РФ, превышающих 20 МЕ/мл, у здоровых людей среднего возраста
составляет около 3% и увеличивается до 10-15% у пожилых лиц старше 65 лет.
Диагностические уровни РФ часто в< тречаются при СШ, гранулематозе Вегенера,
аутоиммунных поражениях печени. СКВ, криоглобулинемии, реже — при других
аутоиммунных заболеваниях, а также при хронических инфекциях (подостром
септическом эндокардите, туберкулезе, вирусном гепатите С). Хотя РФ входит в
классификационные критерии РА, его обнаружение не позволяет доказать нали-
чия РА при атипичной клинической картине, а отсутствие РФ в сыворотке боль-
ного не позволяет исключить диагноз РА.
Таблица 17 12. Частота встречаемости ревматоидного фактора при различных заболеваниях
Заболевание Приблизительная частота', %
Ревматоидный артрит 80
Синдром Шегрена 70
Смешанная криоглобулинемия 70
Системная красная волчанка 30
Полимиозит 20
Системный склероз 20
Подострый септический эндокардит 40
Цирроз печени 25
Инфекционный гепатит 25
Туберкулез 15
Саркоидоз 10
Сифилис 10
Здоровый контроль <5
Старость (>70 пет) 15
' — более 25 МЕ/мл.
Антицитруллиновые антитела
Антицитруллиновые антитела (АЦА), или антитела к циклическому цитруллин-
содержащему пептиду (АЦЦП), направлены против антигенных эпитопов, содержа-
щих аминокислоту цитруллин, которые возникают в белках синовиальной оболочки
ДИАГНОСТИКА АУТОИММУННЫХ ЗАБОЛЕВАНИЙ 1 (J3
под действием воспалительных ферментов нейтрофилов. Это семейство активно
расширяется, что обусловлено эффективностью АЦЦП для диагностики РА.
Антифилаггриновые антитела
Антифилаггриновые антитела (АнФА) исторически выделяют среди АЦА, к
которым относят антиперинуклеарный фактор (АПФ) и антикератиновые анти-
тела (АКА), — показатели, определение которых основано на НРИФ. Основной
антигенной мишенью АнФА является белок профилаггрин, который синтезирует-
ся в виде фосфорилированного полипептида, содержащего около 10 филаггрино-
вых повторов. Белок хранится в гранулах и подвергается ограниченному протео-
лизу, после чего положительно заряженные полипептиды филаггрина участвуют
в сборке цитокератинового скелета эпителиоцита. АнФА связываются с разными
антигенными мишенями, содержащими аминокис лоту' цитруллин, образующуюся
при дезаминировании аргинина с помощью фермента пептидиларгининдезамина-
зы (ПАД). К таким мишеням относят филаггрин, виментин, а также Sa-антиген,
выделенный из ткани плаценты, антитела к которому также причисляют к семей-
ству АЦА. Высокая активность ПАД наблюдается в воспаленной синовиальной
оболочке, что было показано при исследовании ее биоптатов у больных РА и на
моделях этого заболевания у лабораторных животных. Основным источником
ПАД в синовиальной оболочке могут быть полиморфноядерные лейкоциты,
содержащие этот фермент в цитоплазме, которые в значительном количестве
появляются в синовиальной жидкости при РА. Их разрушение приводит к высво-
бождению свободного фермента и его активации в присутствии ионов кальция.
Наличие цитруллинированных белков особенно характерно для воспалительно-
измененной синовиальной оболочки при РА, где они выслушают и в качестве
основной мишени АЦА. В качестве основного индуктора образования АЦА рас-
сматривают о,- и [1-цепи дезиминированного фибрина, которые в большом коли-
честве накапливаются в воспаленной синовиальной оболочке и локализуются с
отложениями цитруллинированных белков.
Антиперинуклеарный фактор (АПФ) был первым исследованным аутоан-
тителом относящимся к семейству АФА. Эта разновидность аутоантител получила
название из-за способности к связыванию с кератогиалиновыми гранулами в
цитоплазме клеток эпителия щеки человека, расположенными перинуклеарно.
Основным методом выявления АПФ служит НРИФ. Оптимальным тест-субстратом
для обнаружения АПФ считают эпителиальные клетки слизистой оболочки щеки
человека, в которых присутствует специфическое свечение крупных округлых
гранул (3- 8 на клетку), расположенных вокруг ядра. Выявление АПФ сопряжено
с рядом технических сложностей, что ограничивает широкое практическое при-
менение этого метода. Прежде всего, лишь около 5-10% здоровых лиц обладают
достаточным содержанием кератогиалиновых гранул в клетках эпителия щеки, и
для стандартизации метода необходим подбор подходящего донора. Кроме того,
кератогиалиновые гранулы быстро утрачивают свою антигенность при длитель-
ном высушивании клеток, большинстве типов химической фиксации, обработке
ферментами и детергентами.
Антикератиновые антитела (АКА) — другой представитель семейства анти-
филаггриновых антител. Их также определяют при помощи НРИФ, они реагиру-
ют с роговым слоем многослойного плоского ороговевающего эпителия средней
трети пищевода крысы. При использовании пищевода крысы в НРИФ выявляется
несколько типов окрашивания эпителия. Может визуализироваться окрашивание
клеток базального слоя эпителия, наблюдаться сетчатое окрашивание неорого-
вевающих слоев эпи гелия, а также гранулярное окрашивание stratum сотеит.
Для АКА характерен особый, линейный, тип окрашивания ороговевающих слоев
эпителия пищевода крысы, в то время как остальные типы окрашивания специфи-
104 ДИАГНОСТИКА АУТОИММУННЫХ ЗАБОЛЕВАНИЙ
ческого диагностического значения не имеют. Необходимо учитывать, что на
пищеводе могут1 быть также обнаружены аутоантитела к десмосомам и антитела
к базальной мембране, которые являются специфическими маркерами буллезных
дерматозов. Эти антитела также способны интерферировать с выявлением АКА и
должны быть дифференцированы от АКА.
Со времени первого описания АКА при РА, их диагностическое значение неод-
нократно подтверждалось. Встречаемость АКА в сыворотке крови больных РА
составляет около 45% при серопозитивном и 30% при серонегативном результате,
при специфичности, близкой к 100%. Несколько чаще обнаруживается АПФ, его
встречаемость составляет 45-61%. По сравнению с РФ для АКА и АПФ характер-
на более высокая специфичность, которая составляет 88-99%, при более низкой
чувствительности, не превышающей 60%. Полагают, что АПФ менее специфичен
по сравнению с АКА и чаще встречается при других ревматологических заболева-
ниях. Содержание АКА и АПФ у больных РА не коррелирует с титрами РФ, что
позволяет рассматривать эти антитела в качестве независимых серологических
маркеров заболевания. АФА встречаются у пациентов с серонегативным РА, что
значительно сокращает группу больных РА, у которых не выявляются специфиче-
ские серологические маркеры заболевания.
При РА описаны еще 2 представителя АФА — антитела к очищенному филаггри-
ну и антитела к Sa-антигену.
Антитела к филаггрину могул быть определены при помощи иммуноблот-
тинга с использованием нативных экстрактов человеческого или животного
филаггрина. Однако содержание цитруллина в антигенных экстрактах филаггрина
значительно различается, что обусловливает их антигенную гетерогенность и не
позволяет стандартизировать тесты для выявления АФА.
Антитела к Sa-антигену встречаются при РА приблизительно у 40% больных.
Их специфичность составляет 98%, что сближает их с АКА. Sa-аптиген пред-
ставляет собой белок с молекулярной массой 48-50 кДа, встречается в экстракте
человеческой плаценты, а также в тканях селезенки и ревматоидного паннуса.
Установлено, что Sa-антигену соответствует цитруллин-содержащая форма вимен-
тина — компонента цитоскелета клетки. В связи с этим антитела к Sa-антигену
стали относить к группе антицитруллиновых антител.
Антитела к циклическому цитруллиновому пептиду
Значительным достижением в изучении роли цитруллин-содержащих антиге-
нов при РА стало описание линейных иммунодоминантных пептидов филаггрина,
содержащих цитруллин. Всего было проанализировано 40 цитруллинированных
пептидов, результат исследований позволил выделить основной иммунодоминант -
ный пептид cfcl, в состав которого входит 19 аминокислот (306-324, Arg.?0—>Cit).
За счет замены серина на цистеин был создан cfcl-cyc — пептид, содержащий
цистеиновый мостик, или циклический цитруллинированный пептид (cyclic citrul-
linated peptide — ССР), более точно отражающий конформационную структуру
нативного белка, что увеличило сродство АЦЦП к этому антигену. Это позволило
использовать данный «неоантиген» для обнаружения АЦЦП. Этот случай стал
одним из первых примеров разработки диагностических антигенов для сероди-
агностики. В действительности циклический цитруллиновый пептид в организме
больного РА отсутствует, однако аутоантитела из сыворотки больного связыва-
ются с ним л vitro. Второе поколение тест-систем было создано с использованием
случайных пептидных библиотек, которые облегчили отбор цитруллинированно-
го антигена с улучшенными свойствами. Эти исследования позволили разработать
иммуноферментную тест-систему для выявления АЦЦП,
Иммуноферментпые тест-системы второго поколения стали золотым стандартом
определения АЦА в клинических лабораториях. Особенностью всех тест-систем для
ДИАГНОСТИКА АУТОИММУННЫХ ЗАБОЛЕВАНИЙ 105
обнаружения АЦЦП служит то. что антиген доступен коммерчески. Это стандарти-
зирует выявление АЦЦП, результаты международных программ внешнего контроля
качества, а также исследования, посвященные сопоставлению тест-систем АЦЦП
разных производителей, указывают на высокую сходимость параметров.
Метод второй генерации для выявления АЦЦП практически не уступает по
чувствительности РФ и позволяет обнаружить антитела у 70-75% пациентов с РА.
Таким образом, по чувствительности этот метод сопоставим с определением РФ,
однако он значительно превосходит последний благодаря специфичности, состав-
ляющей около 95%. Специфичность АЦЦП крайне высока, так как при других
воспалительных артропатиях и ревматических заболеваниях АЦЦП встречается
не чаще чем в 2-3% случаев. Среднее содержание у здоровых доноров крови в
10 раз меньше, чем рекомендуемая производителем граница нормы в 5 U/'мл.
Болыпим преимуществом АЦА является их образование до клинических симпто-
мов заболевания. Популяционные исследования, посвященные времени появления
аутоантител АЦЦП в ходе РА, показали, что АЦЦП могут быть обнаружены задолго
до развития клинических симптомов заболевания. Этот феномен был впервые
отмечен для АКА и позже подтвержден для АЦЦП. До появления клинических сим-
птомов заболевания в 34% случаев были обнаружены АЦЦП. У 19% здоровых на
момент обследования людей был обнаружен РФ класса IgM, причем в нескольких
случаях наличие антител предсказывало РА за 20 лет до появления первых симпто-
мов заболевания. Частота обнаружения антител значительно нарастала к момен-
ту; соответствующему 1,5 годам до постановки диагноза РА. После определения
минимального диагностического титра содержание антител обычно увеличивалось
вплоть до срока постановки диагноза. Ко времени постановки диагноза раннего РА
частота обнаружения АЦЦП и IgM-РФ сравнялась и достигла 70%. АЦЦП не только
обнаруживаются на ранних стадиях РА, но и отмечаются у больных РА с быстрым
рентгенологическим прогрессированием. АЦЦП позволяют предсказать развитие
деструкций приблизительно у 80% пациентов с РА и в сочетании с РФ представляют
важный инструмент для раннего распознавания больных РА с неблагоприятным
прогнозом заболевания уже на этапе постановки диагноза.
Антитела к модифицированному виментину
Основным конкурентом АЦЦП в лабораторной диагностике РА является дру-
гой представитель семейства АЦА — антитела к модифицированному цитрулли-
нированному виментину (АМЦВ), или anti-MCV. Тесты основаны на рекомби-
нантном белке виментине, одном из промежуточных цитофиламентов клетки.
Цитруллинирование виментина in vitro приводит к образованию большого коли-
чества иммунодоминантных эпитопов, число которых теоретически обеспечивает
лучшее связывание аутоантител и большую чувствительность тест-системы.
Тест-системы для обнаружения АМЦВ имеют чувствительность для выявления
РА 85%, что несколько превосходит чувствительность выявления РА с использо-
ванием АЦЦП. Последующие исследования различных групп ревматологических
больных с использованием АМЦВ обнаружили, что данные антитела встречаются
у 15-25% больных с системными заболеваниями, отмечаются у 5-10% больных
псориатическим артритом, могут встречаться у 20% пожилых лиц без признаков
РА. Таким образом, реальная специфичность при исследовании смешанных попу-
ляций для АМЦВ составляет не более 85-90%. Проблему низкой специфичности
можно отчасти решить при увеличении границы нормы тест-системы до 25-30 U/мл
при заявленной производителем норме в 20 U/мл. При таком изменении специ-
фичность обследования возрастает до 90-92%, однако чувствительность снижает-
ся до 60-70%. Таким образом, по основным клинике-лабораторным параметрам
anti-MCV близки АЦЦП, однако несколько менее специфичны, чаще встречаются
при других формах артритов.
106 ДИАГНОСТИКА АУТОИММУННЫХ ЗАБОЛЕВАНИИ
АНТИТЕЛА К ЯЛ-33
Для антител к RA-33 антигеном являются белки гетерогенных ядерных рибону-
клеопротеинов (hnRNP). Эти белки входят в состав сплайсосом наряду с антигена-
ми других рибонуклеопротеиновых АНА, в частности Sm и RNP. Антитела к RA-33
встречаются при РА в 30% случаев, а также отмечаются при СКВ и СЗСТ с частотой
10-15%. При СКВ и СЗСТ практически всегда они присутствуют совместно с анти-
телами к U^RNP. При других ревматических заболеваниях антитела к RA-33 прак-
тически не обнаруживают. В НРИФ при использовании клеток НЕр-2 они обычно
характеризуются мелко! ранулярным окрашиванием ядра, неотличимым от других
разновидностей АНФ. Определение АНФ не является надежным методом обнару-
жения данных аутоантител, так как антитела к RA-33 часто имеются в сыворотках,
в которых АНФ отсутствует. Оптимальным методом диагностики антител к RA-33
считают иммуноблоттинг, причем этот тип АНА содержится в сыворотке крови
больных в невысоких титрах (1:100), однако данные антитела могут быть выявле-
ны с помощью ИФА с использованием очищенного антигена.
Они присутствуют в сыворотке крови приблизительно у 20-30% больных с
серонегативным РА независимо от других серологических маркеров. Антитела к
RA-33 отмечаются на этапе раннего РА, могут встречаться до появления клини-
ческих признаков РА. При РА наличие этой разновидности АНА не зависит от
длительности заболевания, возраста и пола больных, а также от эффективности
проводимой терапии. Имеются данные, что у больных РА с антителами к RA-33
заболевание течет менее агрессивно и характеризуется лучшим прогнозом, что
позволяет использовать присутствие этих антител в прогнозировании исхода РА.
Аутоиммунологические показатели при васкулитах
Под аутоиммунными васкулитами понимают заболевания, связанные с вос-
палительным процессом в сосудистой стенке, без инфекционной или токсической
причины. Васкулиты подразделяют на первичные и вторичные, которые воз-
никают на фоне другого аутоиммунного заболевания. Общепринятой является
международная классификация первичных васкулитов, которая дала определение
10 формам первичных васкулитов и подразделила их в соответствии с калибром
поражаемых сосудов (табл. 17-13).
Таблица 17-13. Классификация васкулитов Chapel-Hill Consensus Conference (1994)
Тип васкулита Определение
Васкулиты крупных сосудов
Гигантоклеточный (височ- ный) васкулит Гранулематозный васкулит аорты и ее основных ветвей с поражением вет- вей наружной a. carotis. Больные старше 50 лет. Характерен выраженный острофазовый огвет
Болезнь Такаясу Гранулематозное воспаление аорты и ее основных ветвей. Больные младше 50 лет
Васкулиты сосудов среднего калибра
Узелковый полиартериит Некротизирующее воспаление артерий среднего и мелкого калибра, не сопровождающееся гломерулонефритом
Болезнь Кавасаки Артериит, поражающий артерии всех калибров. Характерно поражение коронарных сосудов. Чаще всего болеют дети
Васкулиты мелких сосуда
Гранулематоз Вегенера Гранулематозное воспаление, поражающее респираторный тракт и сопрово- ждающееся некротическим васкулитом мелких сосудов Характерен некро- тизирующий гломерулонефрит
Синдром Черджа-Сгоосс Богатое эозинофилами гранулематозное воспаление, поражающее респи- раторный тракт. Картина легочного некротизирующего васкулита, астмы и эозинофилии
ДИАГНОСТИКА АУТОИММУННЫХ ЗАБОЛЕВАНИЙ f 07
Окончание табл. 17-13
Тип васкулита Определение
Микроскопический поли- ангиит Некротизирующий васкулит с поражением мелких сосудов. Реже — признаки некротизирующего артериита с поражением мелких артерий. Некротизирующий гломерулонефрит и пульмонарный капиллярит
Пурпура Шенлейна-Геноха Васкулит с отложениями IgA, поражающий мелкие сосуды. Поражает кожу, суставы, ЖКТ и почки
Эссенциальный криоглобу- линемический васкулит Васкулит, сопровождающийся появлением криоглобулинов в сыворотке. Поражения кожи и почек
Кожный лейкоцитокластиче- ский васкулит Изолированный кожный ЦИК-васкулит без системных и почечных проявле- ний
Патогенез васкулитов крупных и средних сосудов опосредован преимуще-
ственно Т-клетками. абсолютно специфических серологических маркеров при
этих заболеваниях не описано. Антиэндотелиальные антитела с различной часто-
той встречаются при аортоартериите Такаясу и болезни Кавасаки, а также при
ряде васкулопатий, таких как гемолитико-уремический синдром, тромботическая
тромбоцитопеническая пурпура, болезнь Бехчета. Васкулиты крупных и средних
сосудов сопровождаются выраженным воспалением, выявление которого пред-
ставляет основную задачу лабораторных методов диагностики.
В соответствии с иммунопатогенезом васкулиты мелких сосудов могут быть
подразделены на гранулематозные и иммунокомплексные.
Гранулематозные васкулиты мелких сосудов, включающие синдром
Вегенера, микроскопический полиангиит, синдром Черджа-Стросс, имеют ряд
общих черт, к которым относят поражение почек по типу олигоиммунного экстра-
капиллярного гломерулонефрита с полулуниями, полиневрит и легочный капилля-
рит. Основным методом серологической диагностики гранулематозных васкулитов
является обнаружение АНЦА. Антитела этого семейства реагируют с белками азу-
рсфильных гранул нейтрофильных гранулоцитов. Значение АНПА в патогенезе и
диагностике гранулематозных васкулитов позволило выделить отдельную группу,
обобщенную под названием «АНЦА-ассоциированные васкулитьь. К ним относят
гранулематоз Вегенера, синдром Черджа-Стросс, микроскопический полиартери-
ит, быстропрогрессирующий гломерулонефрит с полулуниями. К этой группе забо-
леваний иногда причисляют синдром Гудпасчера, так как АНЦА могут отмечаться
у 10-15% пациентов с этим заболеванием, совместно с антителами к базальной
мембране клубочка, выявление которых — основной лабораторный метод его
диагностики. Хотя синдром Гудпасчера не относится к системным васкулитам, он
характеризуется геморрагическим альвеолитом и быстропрогрессирующим гломе-
рулонефритом, который сложно отличить от АНЦА-ассоциированного поражения
почек. В связи с этим определение АНЦА и антител к базальной мембране обычно
используют совместно.
Иммунокомплексные васкулиты обусловлены образованием иммунных ком-
плексов между аутоантителами и их антигенами в крови и сосудистой стенке.
Иммунные комплексы, которые можно обнаружить в сыворотке крови, называют
циркулирующими иммунными комплексами (ЦИК). Предполагается, что сниже-
ние растворимости ЦИК приводит к их отложению в сосудистой стенке и разви-
тию воспаления. Снижение растворимости при замедлении скорости кровотока и
температуры приводит к тому, что иммунными комплексами поражаются преиму-
щественно капилляры, прежде всего капилляры кожи. Криоглобулинемический
васкулит сопровождается появлением в крови особой разновидности ЦИК, кото-
рая требует особых методических подходов для их обнаружения.
Отдельно выделяют методы, позволяющие обнаружить отложения иммунных
комплексов и комплемента непосредственно в пораженной ткани. Обнаружение
отложений иммунных комплексов позволяет объективизировать иммуноком-
1 08 ДИАГНОСТИКА АУТОИММУННЫХ ЗАБОЛЕВАНИЙ
плексный процесс и обладает большей достоверностью по сравнению с методами
определения ЦИК.
АНТИЭНДОТЕЛИАЛЬНЫЕ АНТИТЕЛА
Антиэндотелиальные антитела (АЭТА) составляют гетерогенную группу анти-
тел, направленных к антигенным детерминантам эндотелиальных клеток. АЭТА
были впервые обнаружены у больных СКВ при использовании метода непрямой
иммунофлюоресценции на тканях лабораторных животных. При СКВ частота
встречаемости АЭТА составляет от 30 до 80%, в зависимости от используемого
метода и категорий больных. Методология обнаружения АЭТА далека от стан-
дартизации. Основные методы: непрямая иммунофлюоресценция на кратковре-
менных культ}-рах клеток эндотелия пуповины (HUVEC), а также методы ИФА и
иммуноблот.
АЭТА были обнаружены при многих первичных васкулитах и васкулопатиях.
Сравнительно часто их выявление связывают с болезнью Кавасаки, гранулематоз-
ными васкулитами, пурпурой Шенлейна-Геноха и вторичными васкулопатиями, в
частности с облитерирующим тромбангиитом (болезнь Бюргера), гемолитической
тромбоцитопенической пурпурой, антифосфолипидным синдромом. Наличие
АЭТА сравнительно неспецифично для определенной формы васкулита и рассма-
тривается в качестве общего диагностического маркера васкулитов и васкулопа-
тий. Целесообразность детекции АЭТА в клинической практике определяется их
выявлением только на фоне выраженной клинической активности заболевания,
что позволяет ориентироваться на их содержание для определения фазы заболе-
вания. Они могут играть определенную роль в патогенезе васкулитов в связи с тем,
что способны модулировать экспрессию эндотелиальных молекул адгезии и фак-
торов воспаления. АЭТА, обнаруженные в сыворотке крови больных СКВ, способ-
ны усиливать высвобождение IL-6 из эндотелиоцитов, таким способом участвуя в
патогенезе системной воспалительной реакции.
АЭТА имеются у большинства больных с активной формой АНЦА-
ассоциированных васкулитов. У детей с пурпурой Шенлейна-Геноха наличие
АЭТА класса IgA коррелирует с частотой поражения почек. Показано, что АЭТА
служат надежным маркером активности процесса при облитерирующем тромбан-
гиите. При синдроме Бехчета они встречаются у 50% больных, особенно у боль-
ных с выраженным кожным васкулитом.
Васкулиты и васкулопатии на фоне системных заболеваний соединительной
ткани также сопровождаются высокими титрами АЭТА. У больных СКВ при
выявлении высоких титров АЭТА чаще отмечают кожные признаки васкулита
и тяжелый синдром Рейно. У большинства больных с волчаночным нефритом и
гипокомплементемией имеются высокие титры АЭТА, что, по всей видимости,
обусловлено способностью аутоантител непосредственно взаимодействовать с
эпителиальными клетками почечного клубочка. Содержание АЭТА снижается при
успешной иммуносупрессивной терапии.
Склеродермия обычно сопровождается признаками генерализованной васку-
лопатии. Предполагают, что одной из причин развития микроангиопатии
при склеродермии могут являться АЭТА, отмечающиеся у 40-60% больных.
Антиэндотелиальные антитела при диффузных формах склеродермии ассоции-
рованы с высоким риском поражения легких, характеризующимся развитием
интерстициального фиброза и легочной гипертензии. У больных с АЭТА часто
отмечается выраженная ишемия пальцев и сосудисто-некротические изменения
кожи кисти.
Существует определенная связь между антифосфолипидными и антиэндоте-
лиальными антителами. АЭТА часто диагностируют у больных с АФС, невына-
шиванием беременности и идиопатическим ливедо. Антигены АКА, в частности
ДИАГНОСТИКА АУТОИММУННЫХ ЗАБОЛЕВАНИЙ *| Q9
комплексы фосфолипидов и Р2-ГП-1, способны связываться с поверхностью эндо-
телиальных клеток, что ведет к повреждению эндотелия, У женщин с повторными
выкидышами в анамнезе и предполагаемым АФС также обнаруживают АЭТА.
Наличие АЭТА характерно для вторичных ангиопатий: диабетической микроан-
гиопатии, сосудистого отторжения трасплантата.
Таким образом, АЭТА могут служить неспецифическим маркером ангиопатий
различного генеза, мониторинг их содержания в сывооотке крови больного с тече-
нием времени позволяет установить активность патологического процесса.
АНТИНЕЙТРОФИЛЬНЫЕ ЦИТОПЛАЗМАТИЧЕСКИЕ АНТИТЕЛА
Впервые в цитоплазме нейтрофилов антитела были обнаружены у больного с
быстропрогрессирующим гломерулонефритом. Затем АНЦА были идентифици-
рованы в качестве наиболее специфичного и чувствительного лабораторного мар-
кера гранулематоза Вегенера, что привело к активному изучению этого семейства
аутоантител и определению клинического значения их выявления при системных
васкулитах. Было обнаружено, что в качестве антигенов АНЦА выступают фер-
менты азурофнльных гранул нейтрофилов. К известным антигенным мишеням
АНЦА относят протеиназу-3, миелопероксидазу, лактоферрин, эластазу, белок BPI
и катепсин G.
Хотя механизм индукции АНЦА остается неизвестным, доказано, что связыва-
ние антинейтрофильных антител с соответствующими мишенями на активирован-
ных гранулоцитах приводит к преждевременной дегра нулянии клеток. Это нару-
шает трансэндотелиальную миграцию лейкоцитов из сосудистого русла и ведет к
образованию воспалительной гранулемы, составляющей основу морфологической
картины гранулематозных васкулитов.
Основным методом определения АНЦА является метод непрямой иммуноф-
люоресценции с использованием нейтрофилов донора, фиксированных этанолом,
что позволяет дифференцировать основные типы свечения аутоантител. По ана-
логии с диагностикой АНФ, при флюоресцентной микроскопии для обнаружения
АНЦА можно отметить несколько типов свечения цитоплазмы клеток. Каждому
из типов свечения соответствуют свои антигенные мишени. Диагностическим
титром АНЦА считают обнаружение антител при разведении сыворотки 1:40 и
более. Учитывая, что содержание АНЦА коррелирует с активностью заболевания,
в методе непрямой иммунофлюоресценции должен быть определен конечный
титр. При обострении гранулематозных васкулитов обычно отмечаются титры
АНЦА, превышающие 1:320. Одновременно с титром должен быть установлен тип
свечения цитоплазмы клетки.
Выделяют два основных типа свечения АНЦА — цитоплазматический и перину-
клеарный. Цитоплазматический тип АНЦА (цАНЦА) получил название благодаря
флюоресценции гранул, локализующихся в цитоплазме клетки между долями ядра
лейкоцита. При выявлении перинуклеарного типа свечения (пАНЦА) свечение
как бы очерчивает доли ядра лейкоцита, оставляя неокрашенным ядро клетки.
Обнаружение пАНЦА представляет собой последствие фиксации нейтрофилов
этанолом, которая ведет к преоаспределению гранул в цитоплазме, клетки, в
результате чего положительно заряженные антигены гранул притягивают отрица-
тельно заряженную ДНК ядра. Похожее свечение обнаруживается в присутствии
АНА, прежде всего антител к дсДНК и гистоновых белков. Нейтрофилы обладают
большинством антигенов АНА и являются субстратом для обнаружения АНФ,
Для того, чтобы отличить АНФ от специфического свечения АНЦА, необходимо
параллельно тестировать все сыворотки, положительные по антинейтрофильным
антигенам в отношении АНФ, что обеспечивает большую специфичность выявле-
ния антинейтрофильных аутоантител. Кроме двух основных типов свечения могут
быть отмечены атипичные варианты окрашивания клетки, которые обозначают
110 ДИАГНОСТИКА АУТОИММУННЫХ ЗАБОЛЕВАНИЙ
как Х-АНЦА. Обнаружение атипичных вариантов свечения АНЦА используют
в диагностике воспалительных заболеваний кишечника. Атипичные варианты
АНЦА описаны при язвенном колите (70%). болезни Крона (10%), аутоиммунном
гепатите (50%). первичном склерозирующем холангите (40%), первичном били-
арном циррозе (5%) и васкулитах при РА (5-10%).
Антигенная специфичность АНЦА может быть установлена не во всех сыво-
ротках, содержащих антинейтрофильные антитела, обнаруженные при помо-
щи непрямой иммунофлюоресценции. В соответствии с рекомендациями
Интернационального консенсуса (Savige, 1999), целесообразно определение спец-
ифичности АНЦА только после определения аутоантител с помощью непрямой
иммунофлюоресценции, что обеспечивает максимальную чувствительность опре-
деления антинейтр! (фильных антител (табл. 17-14).
Таблица 17-14. Антигенная специфичность основных типов антинейтрофильных цитоплазматиче-
ских антител и приблизительная частота их обнаружения
цАНЦА Протеиназа-3 90%
Миелопероксидаза 5%
Белок BPI 4%
лАНЦА Миелопероксидаза 70%
Лактоферрин 10%
Эластаза 8%
Катепсин G 5%
Протеиназа-3 2%
хАНЦА Лактоферрин 10%
Эластаза 10%
Неуточненные ангигены
Антитела к протеиназе 3 (ПР-3) преимущественно обнаруживаются при
цАНЦА-типе свечения. ПР-3 представляет собой сериновую протеиназу массой
29 кДа, которая локализуется в азурофильных гранулах нейтрофила наряду с
другими сериновыми протеиназами. Редко при выявлении цАНЦА-типа све-
чения отмечаются антитела к другим ферментам гранул, прежде всего к белку
BPI. цАНЦА/анти-ПР-3 антитела являются основной разновидностью антиней-
трофильных антител в сыворотке больных гранулематозом Вегенера. При этом
заболевании титр цАНЦА связан с его клинической активностью, и успешная
иммуносупрессивная терапия гранулематоза приводит к снижению содержания
антител ниже порога детекции.
Антитела к миелопероксидазе (МПО) определяются при перинуклеар-
ном типе иммунофлюоресценции. МПО — основной микробицидный фермент
азурофильных гранул, генерирующий кислородные радикалы. МПО обладает
выраженным положительным зарядом, что обусловливает ее перераспределение к
ядру при фиксации нейтрофилов этанолом. Кроме того, при перинуклеарном типе
флюоресценции часто отмечают образование АНЦА, направленных против других
ферментов нейтрофильных гранул, таких как лактоферрин, эластаза и катепсин G.
Антитела к МПО были впервые описаны у больных с идиопатическим быстро-
прогрессирующим олигоиммунным гломерулонефритом с полулуниями, который
представляет собой клиническую разновидность микроскопического полиангиита.
В отличие от антител к ПР-3, пАНЦ А/анти-МПО не обладают высокой специ-
фичностью для диагностики какого-либо конкретного васкулита и выявляются
при всех АНЦА-ассоциированных васкулитах и родственных им заболеваниях.
Антитела к МПО встречаются у большинства больных идиопатическим некро-
тизирующим васкулитом с полулуниями, при микроскопическом полиангиите, а
также при герпесвирусах без антител против ПР-3/цАНЦА.
ДИАГНОСТИКА АУТОИММУННЫХ ЗАБОЛЕВАНИЙ 111
При СКВ, благодаря высокой аффинности ДНК к МПО, циркулирующие в
криви рибонуклеиновые кислоты из разрушенных клеток способны связываться с
МПО-субстратом в иммуноферментных тестах для определения пАНЦА. что при-
водит к высокой частоте ложноположительных результатов.
Антинейтрофильные антитела к BPI (bactericidal permeability increasing protein}
диагностируют у 60% пациентов с муковисцидозом, при иммунофлюоресценции
они обладают цАНЦА-типом свечения. В качестве основного индуктора антител к
BPI рассматривают псевдомонадную инфекцию, которая часто сопутствует муко-
висцидозу.
Антитела к лактоферрину и катепсину G встречаются при васкулитах на фоне
РА, СКВ и при васкулитах при хроническом гранулематозном воспалении в случае
заболеваний кишечника. Высокая встречаемость антител к лактоферрину наблю-
дается у больных с аутоиммунными заболеваниями печени, такими как аутоим-
мунный гепатит и первичный билиарный цирроз. АНЦА, которые встречаются у
30-40% больных с первичным склерозирующим холангитом, обусловлены при-
сутствием антител к лактоферрину.
Антитела к нейтрофильной эластазе обнаруживают при СКВ, особенно при
неврологических проявлениях этого заболевания, а также они имеются у больных
РА и у больных с неспецифическим язвенным колитом.
АНТИТЕЛА К БАЗАЛЬНОЙ МЕМБРАНЕ КЛУБОЧКА
Антитела к базальной мембране клубочка направлены против неколлагеново-
го участка а3-цепи коллагена IV типа, антиген которого может быть обнаружен
в почках, легких, хрусталике, улитке, мозге и яичке. Коллаген IV типа является
основным компонентом базальных мембран альвеол и клубочка почки, что обу-
словливает специфическую клиническую картину синдрома Гудпасчера, сочетаю-
щую гломерулонефрит и геморрагический альвеолит. Поражение почек обычно
преобладает в клинической картине, что позволяет выделять лимитированную
форму синдрома Гудпасчера, сопровождающуюся развитием быстропрогресси-
рующего гломерулонефрита.
Антитела к базальной мембране непосредственно участвуют в патогенезе
поражения почек при этом заболевании, их введение лабораторному животному
приводит к развитию экстракапиллярного гломерулонефрита. Их высокие титры
могут быть обнаружены в элюатах из почек, а содержание антител в сыворотке
коррелирует с клинической активностью заболевания. При иммунофлюоресцент-
ном исследовании биопсий почек при синдроме Гудпасчера аутоантитела могут
быть обнаружены в виде линейных отложений иммуноглобулинов на базальной
мембране клубочков.
Основным методом определения антител к базальной мембране является ИФА
с использованием очищенного антигена. Обнаружение антител является диагно-
стическим для этого заболевания и позволяет дифференцировать от других при-
чин легочных кровотечений и гломерулонефрита. Раннее обследование больных с
синдромом Гудпасчера крайне важно для прогноза больных с этим заболеванием,
поскольку только назначение специфической терапии позволяет сохранить функ-
цию почек, а зачастую предотвратить гибель больного. Высокие титры антител к
базальной мембране могут сохраняться на этапе клинической ремиссии и посте-
пенно снижаются в течение нескольких месяцев. Пересадка почки может быть
рекомендована только серонегативным пациентам, так как при сохранении зна-
чимых титров антител к базальной мембране сохраняется высокий риск рецидива
заболевания в трансплантате.
Антитела к базатьной мембране почки могут сочетаться у пациентов с анти-
телами к МПО; клинически — с быстропрогпессирующим гломерулонефритом и
системными васкулитами.
112 ДИАГНОСТИКА АУТОИММУННЫХ ЗАБОЛЕВАНИЙ
Циркулирующие иммунные комплексы
Кожа, благодаря ее обильной васкуляризации и мелкому калибру сосудов, пора-
жается иммунокомплексными, лейкоцитокластическими васкулитами, названны-
ми васкулитами гиперчувствительности. Морфологическими признаками лей-
коцитокластического васкулита являются фибриноидный некроз стенок сосуда
с инфильтрацией полиморфноядерными нейтрофилами, которые разрушаются с
образованием характерного ядерного дебриса. Разновидность лейкоцитокласти-
ческих васкулитов — лимфоцитокластический вариант, при котором преобладает
лимфоцитарная инфильтрация.
Обычно лейкоцитокластический васкулит развивается на фоне другого ауто-
иммунного заболевания или представляет собой реакцию гиперчувствитель-
ности в отношении экзогенного агента. Все эти состояния характеризуются
появлением в сыворотке крови больного ЦИК, оседающих на стенках мелких
сосудов. Образование иммунных комплексов — часть нормальных процессов
взаимодействия антитела с антигеном. Появление экзогенного антигена в крови
и гуморальный иммунный ответ служат основной причиной формирования ЦИК.
В то же время иммунные комплексы могут состоять исключительно из иммуно-
глобулинов, например, при криоглобулинемиях и болезни Шенлейна-Геноха, что
обусловлено нарушением синтеза антител при этих заболеваниях.
Связывание экзогенного антигена со специфическими антителами приводит к
образованию иммунных комплексов, которые могут захватываться рецепторами
к Fc-фрагментам молекул иммуноглобулинов и к комплементу на клетках рети-
кулоэндотелиальной системы, обеспечивающих клиренс иммунных комплексов.
Иммунокомплексный васкулит может быть следствием образования избыточного
количества ЦИК, нарушения нормального клиренса ЦИК, а также сниженной
растворимости образующихся иммунных комплексов. В результате этого ЦИК
откладываются в стенке мелких сосудов, что приводит к активации системы ком-
племента, синтезу анафилотоксинов и цитокинов и к последующему образованию
лейкоцитарного инфильтрата.
Существует много лабораторных тест ов, направленных на обнаружение ЦИК
(табл. 17-15), но оптимального пока не существует. Это связано с полиморфизмом
иммунных комплексов, состав, масса и заряд которых непостоянны. Кроме того,
стандартизация ряда биологических тестов затруднена, в связи с чем чаще исполь-
зуют менее информативные физико-химические и биохимические методы. Хотя
выявление ЦИК признано полезным дополнительным лабораторным парам* тром
при СКВ и РА, использование конкретных тестов затруднено из-за их плохой
воспроизводимости, трудоемкости и низкой специфичности. В нашей стране в
подавляющем числе лабораторий применяют метод осаждения ЦИК при помо-
щи полиэтиленгликоля. Это грубый тест, позволяющий установить присутствие
в сыворотке комплексов большой молекулярной массы, Наилучшим тестом для
обнаружения ЦИК считают реакцию связывания Clq-компонента комплемента.
Таблица 17-15. Методы определения циркулирующих иммунных комплексов
Основа метода Тест
Физико-химические свойства Криопреципитация
Преципитация полиэтиленгликолем
Ультрацентрифугирование
Морфология и размер Непрямой фагоцитоз гранулоцитами
Связывание комплемента Связывание C1q
Связывание конглютинина
Антикомплементарная активность
ДИАГНОСТИКА АУТОИММУННЫХ ЗАБОЛЕВАНИЙ 11 3
Окончание табл. 17-15
Основа метода Тест
Связывание с антиглобулином Ингибирование активности РФ
Связывание рецепторами клеток крови {через Ес) Активация тромбоцитов
Связывание с рецепторами культур клеток (через Ес и рецептор комплемента) Тест на клетках Raji
Недостаточная надежность методов определения ЦИК в сыворотке приводит к
определенному скепсису использования этих методов. Кроме того, высокое содер-
жание ЦИК может обнаруживаться у клинически здоровых людей.
Альтернативным вариантом определения ЦИК является морфологическое
исследование биоптата ткани с помощью прямой иммунофлюоресценции для
обнаружения отложений иммуноглобулинов и компонентов комплемента. Если
в развитии заболевания подозревается иммунокомплексный процесс, то следует
рекомендовать проведение биопсии ткани с прямым иммунофлюоресцентным
исследованием.
КРИОГЛОБУЛИНЫ
Определение криоглобул инов стоит особняком среди других методов обнару-
жения ЦИК, поскольку является надежным методом диагностики криоглобулине-
мий с развитием иммунокомплесного васкулита.
Криоглобулины представляют собой ЦИК, состоящие исключительно из имму-
ноглобулинов и способные образовывать нерастворимые конъюгаты при темпе-
ратурах ниже температуры ядра человеческого тела. Их преципитация в мелких
сосудах кожи приводит к клинической картине криоглобулинемии, характеризую-
щейся геморрагическим васкулитом на конечностях, слабостью и артралгиями.
Реже при криоглобулинемиях наблюдается гломерулонефрит, лимфаденопатия
и спленомегалия, периферическая нейропатия и синдром Рейно. Наиболее часто
этот вид патологии встречается у 40 -50-летних людей, причем женщины болеют
в 3 раза чаще мужчин.
Определение криоглобулинов следует проводить в сыворотке, а не в плазме
крови, чтобы избежать возможной ошибки при обнаружении криофибриногена,
особого типа фибриногена, коагулирующего при низких температурах. Кровь
должна быть получена от больного непосредственно в лаборатории для того, чтобы
не допустить ее охлаждения; образование кровяного сгустка и центрифугирование
осуществляют при температуре 37 °C. Для обнаружения выпавшего криопреци-
питата в сыворотке крови требуется более 48 ч. Преципитация криоглобулинов
3 типа может занять до 5 дней. Поскольку даже нормальная сыворотка способна
за такое время образовывать нерастворимые преципитаты, то после обнаруже-
ния криопреципитата необходимо достичь его растворения путем повторного
нагревания до 37 °C. Результат преципитации криоглобулинов не всегда очевиден.
Агрегация криоглобулинов может происходить с образованием кристаллов белка,
хлопьев, геля. Для унификации содержания криоглобулинов после образования
осадка, преципитат необходимо отцентрифугировать в стандартных условиях, что
позволяет полуколичественно оценить объем криоглобулинов в виде криокрита.
Оценка криокрита может быть использована при контроле эффективности тера-
пии кри о гл о бул и нем и и.
Выделяют 3 типа криоглобулинемий. Криоглобулины первого типа характеризу-
ются присутствием в плазме моноклонального иммуноглобулина, обычно IgM или
IgG. Основными причинами его возникновения являются лимфопролифератив-
ные заболевания, прежде всего миелома, макроглобулинемия Вальденстрема. хро-
нический лимфоцитарный лейкоз и различные формы лимфом. Криоглобулины
имеются у 40% больных с миеломой, однако только у половины больных с
114 ДИАГНОСТИКА АУТОИММУННЫХ ЗАБОЛЕВАНИЙ
криоглобулинемией первого типа отмечают клинические проявления, связанные
прежде всего с увеличением вязкости крови, такие как головные боли, нарушения
зрения, геморрагии сосудов сетчатки и синдром Рейно.
При вторим типе описывают криоглобулины смешанного состава, включающие
моноклональный, компонент, обычно РФ класса IgM, и поликлональный IgG. Этот
тип криоглобулинемий часто встречается изолированно, представляя собой эссен-
циальную (смешанную) криоглобулинемию. Реже смешанная криоглобулинемия
является вторичной находкой у больных с вирусными инфекциями, заболеваниями
соединительной ткани, лимфопролиферативными процессами и скрытыми инфек-
циями. Основным инфекционным агентом, приводящим к развитию смешанной
криоглобулинемии, является гепатит С. Среди больных вирусным гепатитом С с
экстрапеченочными проявлениями значимые титры криоглобулинов обнаружива-
ются у 42% больных, у подавляющего большинства из них в сыворотке отмечают-
ся диагностические титры РФ. Криоглобулины диагностировали преимуществен-
но у больных с цирротической стадией вирусного гепатита. У 50% обследуемых с
криоглобулинемией отмечали клинические признаки геморрагического васкулита,
у 10% больных — мочевой синдром. Таким образом, смешанная криоглобулине-
мия характеризуется тяжелыми клиническими проявлениями, напоминающими
системный васкулит с триадой, сочетающей разнообразные поражения кожи — от
пурпуры до геморрагического некроза кожи, боли в суставах и гломерулонефрит,
встречающийся у 50% больных с криокритом. превышающим 3%.
Криоглобулинемия третьего типа является наиболее часто встречающейся раз-
новидностью данной патологии и представлена смешанными иммунными ком-
плексами поликлональных криоглобулинов, в составе которых обычно отмечается
поликлональный РФ класса IgM. Третий тип встречается у больных с ДБСТ, в
частности с СКВ, РА, СШ, аутоиммунными заболеваниями ЖКТ, постстрептокок-
ковым гломерулонефритом, подострым бактериальным эндокардитом, цитоме-
галовирусными инфекциями, инфекционным мононуклеозом. Особенно часто ее
обнаруживают у больных с хроническими вирусными гепатитами. Картина васку-
лита, сопровождающаяся обнаружением криоглобулинов третьего типа, может
стать первым симптомом этого заболевания.
ОПРЕДЕЛЕНИЕ ОТЛОЖЕНИЙ ИММУННЫХ КОМПЛЕКСОВ В БИОПТАТАХ КОЖИ И ТЕСТ
ВОЛЧАНОЧНОЙ ПОЛОСКИ
Прямая иммунофлюоресценция позволяет обнаруживать отложения иммунных
комплексов в ткани органов-мишеней. В диагностических целях наиболее часто
используют биопсию почек и/или биопсию кожи. Исследование биоптатов ткани
имеет четкое диагностическое значение и зачастую оказывается единственным
методом подтверждения диагноза при неоднозначной клинической и серологи-
ческой картине. При прямом иммунофлюоресцентном исследовании биоптатов
кожи при диффузной болезни соединительной ткани (ДБСТ) наблюдают 3 основ-
ных типа отложены*' иммуноглобулинов и комплемента: AHA in vivo, волчаноч-
ную полоску и отложения в стенках сосудов дермы. Феномен обнаружения АНА,
фиксированных непосредственно в ткани, обозначают как «антинуклеарные анти-
тела ш v/vo>>. AHA in vivo обычно представлены иммуноглобулинами класса IgG,
окрашивают ядра клеток эпидермиса кожи. Постоянно этот феномен встречается
при СКВ и СС.
В основе AHA in vivo лежит способность проникновения аутоантител в живую
клетку, что характерно именно для рибонуклеопротеиновых аутоантител. По ана-
логии с выявлением АНФ может быть исследован тип флюоресценции. Тип флюо-
ресценции дает дополнительные сведения: гранулярный тип свечения отмечается
при всех аутоиммунных заболеваниях, нуклеолярный тип свечения отмечается
только при склеродермии, а гомогенный тип свечения наблюдают преимуществен-
ДИАГНОСТИКА АУТОИММУННЫХ ЗАБОЛЕВАНИИ 11 5
но при СКВ. Обнаружение АНА в эпидермисе обычно связано с присутствием
антител к рибонуклеопротеинам, но не антител к дсДНК. При СШ характерный
тип нуклеарных/цитоплазматических депозитов IgG сопровождается наличием
аутоантител к Ro/SS-A и La/SS-B. Несмотря на то, что определение АНА в ткани
позволяет заподозрить СКВ, встречаемость этого феномена очень низка и не пре-
вышает 10% при исследовании биоптатов кожи.
Другим крайне характерным и постоянным признаком, который может быто
обнаружен при иммунофлюоресцентном исследовании биоптатов кожи, являются
крупно-гранулярные отложения иммуноглобулинов и комплемента под базаль-
ной мембраной эпидермиса. Обнаружение этого феномена в материале биопсий
клинически здоровой кожи больных СКВ носит название «тест волчаночной
полоски» (от англ, lupus band test} и отмечается только при СКВ. Благодаря частой
встречаемости и высокой специфичности ТВП прямое иммунофлюоресцентное
исследование биоптатов кожи при СКВ рекомендуется широко использовать для
диагностики этого заболевания.
ТВП представляет собой накопление иммунных комплексов in situ. Апоптоз
кератиноцитов при СКВ рассматривают в качестве источников иммуногенно-
го апоптотического материала, вызывающего гуморальный иммунный ответ.
Апоптотические тельца, которые образуются при разрушении базальных слоев
кератиноцитов, через базальную мембрану эпителия поступают в дерму, где они
связываются с аутоантителами, циркулирующими в крови. При окрашивании
ткани античеловеческой антисывороткой к иммуноглобулинам и факторам ком-
племента. меченной флюоресцентной меткой, при микроскопии выявляется яркая
прослойка крупных гранул иммунных комплексов.
Набор иммуноглобулинов в составе ТВП варьирует, более часто фиксируют
отложения IgM, реже — IgG и IgA. Факторы комплемента, прежде всего С3 ком-
понент, отмечаются чаще с отложениями IgM. Выраженность результатов ТВП
непостоянна и неоднородна на разных участках кожи. Частота выявления положи-
тельного ТВП выше при биопсии кожи открытых участков тела, подвергающихся
действию солнечных лучей, таких как предплечье, плечо и грудь. Рекомендуется
биопсировать неизмененную кожу, поскольку воспаление и рубцевание затрудня-
ют описание ТВП.
Положительный ТВП чаще отмечается у больных СКВ, и его чувствитель-
ность достигает 98% в диагностике тяжелых системных форм этого заболевания.
Выраженность депозитов IgG коррелирует с общей тяжестью заболевания, титром
антител к дсДНК и поражением почек. При дискоидной волчанке и формах СКВ с
преимущественным поражением кожи частота встречаемости ТВП ниже и состав-
ляет 50-60%, что превышает частоту обнаружения АНФ при дискоидной волчан-
ке. Морфологическая достоверность, легкость получения материала и высокая
специфичность этого метода делают его очень привлекательным для клинического
использования. При диагностике ДКВ ТВП оказался более чувствительным по
сравнению с обнаружением АНФ В 70% случаев основным иммуноглобулином,
который составляет 1ВП, оказывается IgM, реже выявляют IgG и IgA.
Наличие ТВП в редких случаях следует отличать от специфических отложений
IgA при герпетиформном дерматите Дюринга, который представляет собой раз-
новидность буллезных дерматозов и отмечается преимущественно у пациентов
с целиакией. При этом заболевании имеют место гранулярные отложения IgA
под базальной мембраной эпидермиса, располагающиеся на вершине сосочков
дермы.
При исследовании биоптатов кожи при ревматических заболеваниях могут
быть обнаружены гранулярные отложения иммуноглобулинов в стенках сосудов,
которые могут располагаться как в поверхностных, так и в глубоких сосудах.
Отложения иммуноглобулинов и факторов комплемента в сосудах дермы возмож-
116 ДИАГНОСТИКА АУТОИММУННЫХ ЗАБОЛЕВАНИЙ
ны при пурпуре Шенлейна -Геноха. герпетиформном дерматите Дюринга, лейко-
цитокластическом и криоглобулинемическом васкулите, СКВ и дерматомиозите
При дерматомиозите метод прямой иммунофлюоресценции позволяет обна-
ружить сложения мембраноатакующего комплекса (МАК) комплемента, пред-
ставленного отложениями терминальных компонентов системы комплемента
(С5Ь 9) в микроциркуляторном русле кожи. По диагностической значимости МАК
в капиллярах кожи под базальной мембраной может сравнит ься с ТВП при СКВ,
обладая специфичностью, достигающей 90% при сопоставимой чувствительности.
Сходные отложения МАК при дерматомиозите определяют в сосудах скелетных
мышц. Гистологическая картина поражения мышечной ткани при дерматомиозите
характеризуется выраженным васкулитом с периваскулярными инфильтратами,
состоящими из CD^-лимфоцитов. Некроз миоцитов происходит по периферии
мышечных волокон и возникает на фоне поражения сосудов. В мышечной ткани
отложения МАК характерны для зон с сохраненной гистологической структурой
мышечной ткани, в которой протекает активный воспалительный процесс. В
районах тотального фиброза мышцы отложения МАК не выявляются. По всей
видимости, разрушение капилляров под действием компонентов комплемента
приводит к склерозированию и разрушению мышечной ткани. Причиной образо-
вания МАК при дерматомиозите в сосудах кожи и мышц могут быть комплемент-
связывающие антиэндотелиальные антитела, которые диагностируют у 50%
больных дерматомиозитом. Образование МАК и антиэндотелиальных антител с
антигенами на поверхности эндотелиоцитов ведет к развитию васкулита в сосудах
кожи и мышц.
ДРУГИЕ ИММУНОЛОГИЧЕСКИЕ ПОКАЗАТЕЛИ ПРИ ВАСКУЛИТАХ
Помимо АЭТА, в качестве неспецифического маркера васкулитов разной этио-
логии применяют исследования факторов, продуцируемых поврежденной клеточ-
ной стенкой в ответ на иммунное поражение. С этой целью используют монитори-
рование содержания фактора VIII (фактора Виллебранда), тромбомодулина и
фибронектина — сывороточных белков, синтезируемых эядотелиоцитами в ответ
на повреждение. Практически у всех больных с активными формами васкулитов
отмечают повышенные уровни этих маркеров, что позволяет использовать их с
диагностической целью, нарастание их содержания в плазме крови свидетельству-
ет об активности васкулита.
Содержание фактора Виллебранда отражает непосредственное повреждение
эндотелиальных клеток. Однако результаты его определения у больных с васкули-
тами оказались неопределенными. Показано, что высокие уровни отмечаются при
активном заболевании и могут сохраняться в последующем, несмотря на клиниче-
ское улучшение. Последующие эксперименты in vitro показали, что фактор высво-
бождается как поврежденными, так и интенсивно делящимися эндотелиоцитами.
Это в какой-то степени объясняет неоднозначные клинические результаты.
Фибронектин представляет собой высокомолекулярный гликопротеин
матрикса соединительной ткани, включа ющийся в состав иммунных комплексов
и участвующий в их клиренсе. Его высокое содержание фиксируют при васкули-
тах, связанных с РА, СКВ, синдроме Кавасаки, васкулите при синдроме Бехчета,
оно отражает разрушение сосудистой стенки. Содержание фибронектина плазмы
снижено при эссенциальной смешанной криоглобулинемии, что говорит о его свя-
зывании в составе криоглобулинов.
Тромбомодулин — мембранный гликопротеин эндотелиоцита, и его выделе-
ние в кровоток сопутствует повреждению сосудистой стенки В качестве маркера
активности СКВ тромбомодулин превосходит растворимые молекулы адгезии,
SIL-2R, сывороточные IL-6, IL-10. антитела к дсДНК, острофазовые белки, СОЭ,
комплемент и иммуноглобулины, креатинин и АНА. Его высокое содержание в
ДИАГНОСТИКА АУТОИММУННЫХ ЗАБОЛЕВАНИЙ 117
плазме крови отмечается при пурпуре Шенлейна -Геноха, СЧШ и других АНЦА-
ассоциированных васкулитах.
Растворимые формы адгезионных молекул являются также продуктами
эндотелиальных клеток. Они выделяются при активации эндотелиальных клеток
при воспалении и потенциально могут использоваться в качестве показателей
активности васкулитов и близких васкулитам состояний. ICAM-1 — одна из
основных межклеточных молекул адгезии, осуществляющих взаимодействие лей-
коцитов и эндотелиоцитов. Ее экспрессия на мембране эндотелиоцита индуциру-
ется при воспалении. Кроме того, при активации клетки происходит слущивание
этих молекул с мембран клеток, приводящее к образованию растворимой формы
sICAM-1. Повышенные уровни sICAM-1 в сыворотке наблюдают при височном
васкулите, пурпуре Шенлейна-Геноха, АНЦА-ассоциированных и лейкоцито-
кластических васкулитах, активной СКВ, синдроме Бехчета, септическом шоке,
злокачественных новообразованиях и инфаркте миокарда. Васкулит, сопрово-
ждающий системные заболевания соединительной ткани, также приводит к уве-
личению содержания sICAM-1 У больных РА диагностируют значительно увели-
ченное содержание sICAM-1 в сыворотке крови, особенно высокие концентрации
sICAM-1 наблюдались у больных с клинической картиной иммунокомплексного
васкулита.
Серологические маркеры при аутоиммунных
поражениях печени
АНТИТЕЛА К ГЛАДКИМ МЫШЦАМ (F-АКТИНУ)
Антитела к гладким мышцам (АГМА) реагируют с миофиламентами в цито-
плазме гладкомышечных клеток. Существует несколько видов антител к гладким
мышцам, в том числе к фибриллярному (F) актину, тубулину и десмину. Среди
них только антитела к F-актину являются маркером аутоиммунного гепатита, в
то время как антитела к тубулину, десмину и виментину встречаются при ревма-
тических заболеваниях и вирусных инфекциях и самостоятельного клинического
значения не имеют.
Актин — глобулярный белок массой 46 кДа, который относят к микрифила-
ментам. Актин может существовать как в мономерной (М), так и в полимеризо-
ванной фибриллярной (F) форме. Связывание аутоантител зависит от конформа-
ции эпитопов в составе F-формы, поэтому выделение F-актина в нативном виде
затруднено. Это объясняет необходимость использования в НРИФ минимально
фиксированных криосрезов и сложность в разработке иммунохимических методов
для выявления аутоантител.
Стандартом выявления АГМА служит НРИФ на тройном субстрате, вклю-
чающем криосрезы желудка, почки и печени крысы. Антитела к актину окра-
шивают цитоплазму мышечных волокон, включая наружные мышцы желудка,
lamina muscular® mucosae, межгландулярные мышечные пучки слизистой оболочки,
стенки сосудов в подслизистом слое и почке, мезангиальные клетки клубочков.
Характерно, что антитела к актину обычно представлены классом IgG и встреча-
ются в титре, равном или более 1:80. Актиновые филаменты плохо экспрессиро-
ваны в НЕр-2 клетках, поэтому цитоплазматический тип свечения при наличии
антител к актину определяют редко. В отличие от него, филаменты виментина и
десмина обычно видны на НЕр-2 клетках и встречаются в титрах 1:20-1:40.
Для подтверждения антигенной специфичности антител к актину могут быть
использованы дополнительные субстраты, в том числе культуры фибробластов
или фибробластоподобных клеточных линий, обработанных винбластином, кото-
рый разрушает «неактиновые» филаменты.
118 ДИАГНОСТИКА АУТОИММУННЫХ ЗАБОЛЕВАНИЙ
Антитела к гладким мышцам с помощью НРИФ были обнаружены у пациентов
с хроническим активным гепатитом; у этих больных часто выявляют и другие
маркеры системных заболеваний — LE-клетки, АНФ, гипергаммаглобулинемию.
Определение АГМА позволило отделить эту форму гепатита от системных ревма-
тических заболеваний и дать название «аутоиммунный гепатит». На сегодняшний
день АГМА. направленные против F-актина, рассматривают в качестве наиболее
чувствительного маркера аутоиммунного гепатита I типа, хотя их встречаемость
при этом заболевании составляет только 55-70%.
АНТИТЕЛА К МИКРОСОМАМ ПЕЧЕНИ-ПОЧЕК
Антитела к микросомам печени/почек (liver-kidney microsomal autoantibodies —
LKM) образуются на фоне аутоиммунного гепатита II типа. Эта разновидность
аутоантител реагирует с проксимальными канальцами почки и цитоплазмой
гепатоцитов. В зависимости от антигенной мишени и связи с заболеванием LKM
разделяют на 3 типа, хотя в подавляющем большинстве случаев необходимым и
достаточным является определение основных аутоантител 1 типа. Антигенной
мишенью LKM-1 служит микросомальный цитохром системы р450 2D6. Антитела
2 и 3 типов крайне редко встречаются при лекарственно-индуцированных и вирус-
ных гепатитах и самостоятельного клинического значения не имеют. Их антиген-
ными мишенями также являются цитохромы и микросомальные ферменты.
Рекомендуемый метод обнаружения LKM-1 — НРИФ на тройном субстрате,
включающем криосрезы почки, печени и желудка крысы. На тканевом субстра-
те крысы LKM-1 антитела связываются с проксимальными канальцами почки и
клетками печени, однако, в отличие от митохондриальных антител, не реагируют
с эпителиальными клетками слизистой оболочки желудка и не окрашивают дис-
тальные канальцы почки. Кроме того, при сравнении с антителами к митохондри-
ям LKM-1 более выраженно связываются с тканью печени. Существуют методы
определения LKM-аутоантител с помощью ИФА и иммуноблоттинга.
Антитела к LKM-1 — основной серологический маркер аутоиммунного гепа-
тита II типа, которым болеют преимущественно дети старше 1 года и подростки.
АИГ-2, в отличие от АИГ I, чаще отмечается у девочек, заболевание протекает
более активно, чаще ведет к циррозу и нередко сопровождается другими ауто-
иммунными заболеваниями, в том числе диабетом, витилиго и аутоиммунными
тиреопатиями. Нередко фиксируют фульминантное начало, течение с периодами
обострения и затухания, в течение 2 лет развивается цирроз. В анализах крови
характерно умеренное повышение трансаминаз, снижение IgA сыворотки.
По сравнению с другими аутоантителами, которые используют для диагностики
аутоиммунных заболеваний печени, антитела к LKM-1 встречаются сравнительно
редко. Крайне редко антитела к LKM встречаются при вирусном гепатите С (1-4%)
и гепатите D. Совместно с антителами к LKM-1 при аутоиммунном гепатите И типа
обнаруживают антитела к цитозольному антигену печени (LC-1). Эти антитела
направлены к одному из белков цитоплазмы гепатоцита массой 230 кДа. Для их
определения может использоваться НРИФ на тройном субстрате тканей крысы,
однако антитела против LC-1 очень сложно обнаружить на фоне высоких титров
анти-LKM-l антител. Для диагностики этой разновидности аутоантител в практи-
ческой работе целесообразно использовать метод ИФА и иммуноблоттинг.
Для диагностики АИГ используют еще один редко встречающийся, но очень
специфичный серологический маркер, который представляет собой антитела к
растворимому печеночному антигену (soluble liver antigen — SLA). Аутоантитела к
SLA отмечаются в 10-20% всех случаев АИГ, при их наличии АИГ хуже поддается
иммуносупрессивному лечению.
ДИАГНОСТИКА АУТОИММУННЫХ ЗАБОЛЕВАНИЙ 119
АНТИТЕЛА К МИТОХОНДРИЯМ
Аутоантитела к митохондриям — семейство ан гимитохондриальных антител,
направленных против ферментов окислительного фосфорилирования вну гренней
мембраны митохондрий, впервые описаны при первичном билиарном циррозе
(ПБЦ). Основным антигеном является Е -субъединица 2-оксодегидрогеназного
комплекса, который участвует в цикле трикарбоновых кислот и синтезе жир-
ных кислот. Большинство митохондриальных аутоантигенов расположено на
внутренней митохондриальной мембране эукариотических и прокариотических
клеток. В процессе эволюции ферменты окислительного каскада не изменялись,
что позволяет использовать антигены крысы и скота в тестах для определения
антимитохондриальных антител (АМА). Субъединица Е2 — общая для нескольких
гликолитических ферментов (PDC, BCOADC, OGDC), в ее аминокислотной после-
довательности выделено 3 линейных иммунодоминантных эпитопа АМА, харак-
терных для ПБЦ. Пируват-декарбоксилазный комплекс осуществляет расщепление
липидов и подвержен действию многих гепатотоксичных веществ. Его антигенная
модификация играет ведущую роль в развитии аутоиммунного ответа при ПБЦ. В
то же время роль АМА в патогенезе заболевания не установлена, так как иммуни-
зация антигеном лабораторных животных не приводит к развитию ПБЦ.
Основным методом определения АМА является НРИФ на тройном субстрате,
включающем почку, желудок и печень крысы. При использовании тройного тка-
невого комплекса митохондриальные антитела окрашивают все богатые митохон-
дриями структуры — клетки канальцев почки и слизистой оболочки желудка, а
также цитоплазму гепатоцитов. Обработка мочевиной срезов слизистой оболочки
желудка уничтожает антигены митохондрий, что позволяет дифференцировать
АМА от антител к обкладочным клеткам желудка, которые также реагируют со
слизистой оболочкой.
Цитоплазма Нер-2-клеток богата митохондриями, поэтому уже на этапе выяв-
ления АНФ можно заподозрить присутствие АМА. Гранулярный тип свечения
АНФ отмечается у 30% больных с ПБЦ и может затруднять обнаружение цитоплаз-
матического свечения. Для определения антигенной специфичности АМА исполь-
зуют ИФА и иммуноблот.
Исторически выделяют 9 разновидностей аутоантител к антигенам митохон-
дрий, причем каждая разновидность обладает особенностями связывания с гисто-
логическими структурами на тройном тканевом субстрате (табл. 17-16). Антигены
митохондриальных антител разнообразны по природе. Так, AMA-Mt представля-
ют собой АФА, которые реагируют с внутренней мембраной митохондрий, богатой
кардиолипином.
Для первичного билиарного цирроза наиболее характерны антитела к М2, М4,
М,. и М9. Среди них АМА-М2 отмечаются у большинства больных билиарным
циррозом, поэтому обнаружение других антител представляет только научный
интерес. АМА-М2 обычно определяется в высоких титрах. Высокая диагностиче-
ская специфичность АМА обеспечивается их редким обнаружением при других
аутоиммунных заболеваниях, причем при ревматических процессах титры АМА
не превышают 1:80. АМА-М7 встречаются при миокардитах.
Существует ряд минорных серологических маркеров ПБЦ, которые относят к
семейству АНА, в том числе sp 100 и PML, а также gp210. Антитела к splOO и PML
являются антигенами АНФ со свечением точек в ядре, которые часто отмечаются
при ПБЦ. Антитела к gp2J0 реагируют с порами ядерной мембраны, в методе
НРИФ на НЕр-2 клетках обусловливают периферический тип свечения ядра. Эти
аутоантитела имеются у 10- 30% больных ПБЦ.
120 ДИАГНОСТИКА АУТОИММУННЫХ ЗАБОЛЕВАНИЙ
Таблица 17-16. Антитела к митохондриям
Разновидность АМА Основной антиген Особенности при проведении НРИФ на тройном субстрате Нозологическая форма
М, Кардиолипин Только дистальные канальцы почки АФС, СКВ и другие ревма- тические заболевания
Е,-субъединица пируват- декарбоксилазного комплекса (PDC-EJ Проксимальные и дистальные канальцы и слизистая оболочка желудка Первичный билиарный цирроз
м3 Не уточнен Клубочки, канальцы почки и слизи- стая оболочка желудка Лекарственная СКВ
Сульфит-оксидаза Неотличим от М2 Первичный билиарный цирроз
м5 Не уточнен Только проксимальные канальцы АФС и др. ревматические заболевания
Ч. Монамин-оксидаза В Связывается с клетками петли Генле Лекарственно- индуцированный гепатит
Саркозин- дегидрогеназа Окрашивает канальцы почки, но не связывается со слизистой оболоч- кой желудка Миокардит и кардиомио- патии
м. Не уточнен Преимущественно реакция со слизи- стой оболочкой желудка Первичный билиарный цирроз
м3 Гликоген- фосфорилаза Реакция только со слизистой обо- лочкой желудка Первичный билиарный цирроз
Серологические маркеры при аутоиммунных
поражениях желудочно-кишечного тракта
АНТИТЕЛА К ОБКЛАДОЧНЫМ (ПАРИЕТАЛЬНЫМ) КЛЕТКАМ ЖЕЛУДКА И ФАКТОРУ
КАСТЛА
Аутоиммунный гастрит (тип А) относится к частым аутоиммунным заболе-
ваниям ЖКТ. В 30% случаев он сопутствует аутоиммунным процессам в щито-
видной железе, алопеции и витилиго, хотя может встречаться самостоятельно.
Аутоиммунный гастрит протекает субклинически и редко является поводом
для обращения пациентов к врачу. В его основе лежит аутоиммунная реакция,
направленная против обкладочных (париетальных) клеток желудка. Обкладочных
клеток больше в теле желудка, поэтому обычно аутоиммунный гастрит характери-
зуется атрофией фундальных отделов по сравнению с антральным гастритом при
хеликобактерной инфекции. Антитела к обкладочным клеткам желудка (АПКЖ)
отмечаются у 90% больных с атрофическим гастритом и пернициозной анемией.
Обкладочные клетки желудка вырабатывают соляную кислоту и внутрен-
ний фактор Кастла, необходимый для всасывания витамина В12 в кишечнике.
Аутоантитела являются комплемент-фиксирующими и непосредственно уча-
ствуют в разрушении популяции обкладочных клеток и развитии атрофического
гастрита. Уничтожение популяции обкладочных клеток ведет к гипоацидному
гастриту, мальабсорбции, железодефицитной и пернициозной анемиям. Не отме-
чается взаимосвязи между обнаружением аутоантител к париетальным клеткам
желудка и антральным гастритом. Титры аутоантител не коррелируют с тяжестью
атрофии слизистой оболочки желудка, поэтому их мониторинг нецелесообразен.
Для выявления АПКЖ используют метод НРИФ на криосрезах фундальной
части желудка крысы или мыши. Антитела реагируют только с обкладочными
клетками желудка и проявляются характерным цитоплазматическим окрашивани
ем, которое соответствует расположению внутриклеточных канальцев. Основной
ДИАГНОСТИКА АУТОИММУННЫХ ЗАБОЛЕВАНИЙ f 21
антиген АПКЖ — [3-субъединица Н-, К-АТФазы, которая участвует в закислении
содержимого желудка. Рекомендуется использовать начальный титр сыворотки
1:40. Слизистая оболочка желудка богата митохондриями, поэтому она окраши-
вается антимитохондриальными антителами. Для дифференцировки АПКЖ и
АМА целесообразно параллельно использовать в качестве субстрата почку крысы,
с которой связываются только АМА. Существует метод с использованием 2М рас-
твора мочевины, которая, разрешая антигены АМА, позволяет дифференцировать
их от АПКЖ на слизистой оболочке желудка крысы.
Внутренний фактор (фактор Кастла) представляет собой гликопротеин массой
72 кДа, который секретируется в просвет желудка и служит для связывания и
транспорта в стенку кишки витамина В12. Аутоантитела к внутреннему фактору
способны вызывать нарушение его функции за счет блокады его связывания с
витамином, таким образом препятствуя адсорбции комплекса в тонком кишеч-
нике. Антитела к внутреннему фактору являются более специфичными по срав-
нению с АПКЖ при диагностике пернициозной анемии и дефицита витамина Вр.
Основным методом определения антител к внутреннему фактору является ИФА. ~
АНТИТЕЛА К ЭНДОМИЗИЮ И ТКАНЕВОЙ ТРАНСГЛУТАМИНАЗЕ
Антитела к эндомизию (АЭА) используются при диагностике целиакии; это
основной метод серологической диагностики целиакии, более надежный, чем
определение антител к глиадину. Основным антигеном АЭА служит тканевая
трансглутаминаза (ТкТГ), которая представляет собой кальций-связывающий
фермент. Основной функцией ГкТГ является образование перекрестных связей
между белками за счет реакции между глутамином в одном белке и лизином — в
другом. Существует 8 изоформ этого фермента, которые различаются по тканевой
локализации. В желудке преимущественно экспрессируется 2 тип Тк ГГ. Мишенью
антител при герпетиформном дерматите, который тесно связан с целиакией, явля-
ется 3 тип ТкТГ, который экспрессируется в коже. В норме активность этого фер-
мента максимальна в клетках базального слоя кишечного эпителия, а также в под-
слизистом слое. У больных целиакией отмечается повышенная экспрессия ТкТГ во
всех слоях слизистой оболочки кишечника. а-Глиадин на 60% состоит из амино-
кислоты глутамина, который подвергается дезаминированию под действием ТкТГ.
Дезаминирование глиадина приводит к образованию неоэпитопов, содержащих
отрицательно заряженную аминокислоту глутамат и имеющих высокое сродство к
HLA-DQ и OQg, носительство которых предрасполагает к развитию целиакии. Тем
не менее генотип HLA-DQ^ присутствует у 25-30% европейцев, однако далеко не у
всех развивается целиакия.
ТкТГ дезаминирует ряд белков соединительной ткани в составе эндомизиаль-
ных и ретикулиновых волокон. Таким образом, иммунный ответ на экзогенный
антиген ведет к образованию антител к а-глиадину, а также аутоантител против
аутоантигенов эндомизия, ретикулина и ТкТГ.
АЭА могут быть определены с помощью НРИФ на множестве антигенсодер-
жащих субстратов. Могут использоваться криосрезы пищевода, печени, желудка,
стенки мочевого пузыря обезьяны, крысиные почки и тонкая кишка. Еще одним
субстратом для обнаружения АЭА являются криосрезы человеческой пуповины.
Стандартным субстратом для НРИФ считают криостатные срезы пищевода обе-
зьяны. Обычно используют разведение сыворотки 1:5 и антисыворотку класса
IgA. Сыворотки с высокими титрами АЭА дают типичное окрашивание, которое
напоминает пчелиные соты. Первоначально в ИФА-системах для определения
антител к ТкТГ использовали антигены животных. Однако низкий уровень очист -
ки белка, а также наличие видоспецифичных эпитопов снижали специфичность
теста. Второе поколение ИФА было основано на рекомбинантной ТкТГ человека.
За счет высоких аналитических параметров эти ИФА-тесты в значительной сте-
122 ДИАГНОСТИКА АУТОИММУННЫХ ЗАБОЛЕВАНИЙ
пени вытеснили метод НРИФ для диагностики АЭА. Высокая чувствительность
ИФА тест-систем улучшает диагностику целиакии, однако тест-системы многих
производителей имеют низкую специфичность, особенно при дифференциальной
диагностике целиакии и других заболеваний ЖКТ. Изучение антигенной структу-
ры молекулы ТкТГ показало, что основной эпитоп антител при целиакии является
конформационным и образован ядром молекулы фермента. Аутоантитела к ТкТГ.
характерные для целиакии, подавляют активность фермента. В то же время анти-
тела к ТкТГ, встречающиеся у больных с аутоиммунным гепатитом, циррозом,
вирусными инфекциями, направлены к внешним доменам молекулы ТкТГ. Этим
может объясняться меньшая специфичность антител к ТкТГ по сравнению с
определением АЭА.
Для лабораторной диагностики целиакии применяют определение антител к
ТкТГ классов IgA и IgG. Наиболее специфичен тест для выявления антител к ТкТГ
класса IgA, которые имеются у 95% больных с дебютом целиакии. Аутоантитела
класса IgA могут быть обнаружены в крови, а также в других биологических жид-
костях, в том числе в слюне и желчи. Определение аутоантител класса IgG менее
специфично, так как они встречаются при других воспалительных заболеваниях
ЖКТ, однако они могут быть использованы для диагностики целиакии у лиц с
селективных дефицитом IgA.
АНТИТЕЛА К А-ГЛИАДИНУ КЛАССОВ IgG И IgA
Антитела к глиадину были основным методом иммунологической диагностики
целиакии. Строго говоря, глиадин не является аутоантигеном, однако иммунная
реакция против глиадина запускает каскад иммунных реакций, в результате кото-
рых образуются аутоантитела и развивается аутоиммунное заболевание.
Глиадин представляет собой алкогольрастворимую фракцию глютена, кото-
рый, в свою очередь, является компонентом глютенопектина злаковых. Глютен
обеспечивает эластичность мучного теста и. благодаря этому свойству, широко
используется для приготовления многих пищевых продуктов. В состав глютена
злаковых входит более 50 различных глиадиновых белков, которые подразделя-
ются на ос-, [%, у- и co-фракции. В связи с высоким содержанием пролина глиадины
сравнительно устойчивы к энзиматическому разрушению в ЖКТ. Изучение моле-
кулярной структуры белков в составе глютена привело к идентификации общей
для них линейной иммунодоминантной аминокислотной последовательности
QPFQPFP. Последний остаток глутамина в этой структуре служит мишенью для
фермента тканевой трансглутаминазы, под действием которой он превращается в
глутамат. Изменение заряда молекулы приводит к лучшему связыванию ее с HLA-
DQ^ и стимуляции иммунного ответа.
Для определения антител к глиадину используют метод ИФА. Антиген получа-
ется из алкогольрастворимой фракции глютена зерновых культур. В состав этой
фракции входит множество различных белков и последовательностей глиадина.
Значительные вариации в белковом составе, а также разнообразие антигенов объ-
ясняют отличия тест-систем различных производителей и низкую специфичность
этих тестов. Это ведет к тому, что антитела к глиадину значительно уступают по
клинико-лабораторной информативности АЭА и антителам к тканевой транс-
глутаминазе. Для диагностики целиакии применяют определение антител к глиа-
дину классов IgA и IgG. Антиглиадиновые антитела класса IgG чувствительны, но
очень неспецифичны, и их можно рассматривать как показатель, отражающий
воспаление в ЖКТ Антитела класса IgA имеются у 30-40% больных с целиакией.
Концентрация антител к сх-глиадину прогрессивно снижается после назначения и
соблюдения безглютеновой диеты.
Сравнительно недавно появились коммерческие тест-системы, основанные на
«модельном» антигене, содержащем многократно повторенные линейные имму-
ЦИАГНОСТИКА АУТОИММУННЫХ ЗАБОЛЕВАНИЙ 123
нодоминантные последовательности глиадина. Предполагается, что такой подход
значительно повысит диагностическую специфичность антител к глиадину и улуч-
шит стандартизацию определения этого показателя.
Антитела к Saccharomyces cerevisiae
В основе патогенеза болезни Крона лежит утрата толерантности к антигенам
пищи, в результате чего развивается иммунный ответ на антигены пищи и антиге-
ны содержимого кишки. Для диагностики болезни Крона используют определение
антител к пекарским дрожжам Saccharomyces cerevisiae (ASCA), которые являют-
ся естественным антигеном. Помимо антител к антигенам пекарских дрожжей,
при болезни Крона образуются антитела к мембранному белку ОгпрС кишечной
палочки и Pseudomonas fluorescens, При болезни Крона обнаруживают антитела
к полисахаридным последовательностям, входящим в состав клеточной стенки
многих организмов. Полисахаридные антитела называют ALCA, АМСА и АССА.
Их объединяют термином «антигликановые антитела». Встречаемость каждого
из этих маркеров не превышает 20%, поэтому их использование в диагностике
болезни Крона малоэффективно.
Основными иммуногенными эпитопами ASCA являются также компоненты
полисахаридов, в том числе маннотетроза и маннотриоза. Наличие ASCA может
объясняться перекрестной реакцией с распространенным дрожжевым грибком
С. albicans. Характерно, что С. albicans чаще высевают у больных воспалительными
заболеваниями кишечника и членов их семей. Основным методом определения
ASCA служит НРИФ на клетках дрожжей, хотя также могут быть использованы
тест-системы, основанные на методе ИФА.
Независимо от географического района и расовой принадлежности обследуе-
мых антитела к дрожжевым грибкам выявляют при болезни Крона с одинаковой
частотой, составляющей около 50% (40-70%). Характерно, что антитела к ASCA
чаще встречаются у родственников больных воспалительными заболеваниями
кишечника (20-25%), по сравнению с частотой показателя в здоровой популя-
ции (0-5%), и больных с неспсцифическим язвенным колитом (НЯК) (10-15%).
Антитела могут быть представлены IgG или IgA, обладая общей специфичностью
93%. При болезни Крона у пациентов с ASCA заболевание протекает тяжелее, что
связано с частыми эпизодами кишечной непроходимости, а также лучше отвечает
на терапию блокаторами TNF-a.
Антитела к бокаловидным клеткам
Антитела к бокаловидным клеткам (АБК) являются серологическим маркером
НЯК. Бокаловидные клетки кишечника производят муцин, который состоит из
осевого белка, покрытого полисахаридами. Его функция заключается в увлажне-
нии слизистой оболочки и обеспечении водно-солевого баланса. Муцин — основ-
ной антиген АБК, причем иммунодоминантные эпитопы обнаружены как в поли-
сахаридных остатках, так и в осевом белке муцина.
Антитела могут играть определенную роль в патогенезе НЯК, так как распро-
странение воспалительного поражения кишечника при НЯК отражает анатомиче-
ское распределение бокаловидных клеток в кишечнике. Их число минимально в
двенадцатиперстной кишке и увеличивается по направлению к прямой кишке, где
их число максимально. НЯК поражает прямую кишку и распространяется вверх
при увеличении активности заболевания. В норме бокаловидные клетки преоб-
ладают в криптах кишечника, а именно — в абсцессах крипт, очень характерных
для этого заболевания В то же время АБК отмечаются не у всех больных НЯК,
поэтому их рассматривают как вторичный феномен.
Стандартным методом диагностики АБК считают НРИФ на тонком кишечнике
приматов или новорожденных обезьян. Применение тонкого кишечника крысы
124 ДИАГНОСТИКА АУТОИММУННЫХ ЗАБОЛЕВАНИЙ
более чувствительно, однако приводит к увеличению числа ложноположительных
результатов. Характерное окрашивание проявляется дымчатыми «облачками» на
фоне слизистой оболочки кишечника. В качестве антигена может использоваться
перевиваемая клеточная линия муцин-продуцирующих бокаловидных клеток
HT29-18-N,
Встречаемость АБК при НЯК составляет 30-60% в зависимости от типа суб-
страта. При использовании совместно с АНЦА диагностическая чувствительность
достигает 60-70%.
Серологические маркеры при аутоиммунных эндокринопатиях
АНТИТЕЛА К ПЕРОКСИДАЗЕ ЩИТОВИДНОЙ ЖЕЛЕЗЫ
Тиреоидная пероксидаза (ТПО) — основной фермент гормоногенеза щитовид-
ной железы, изначально описана под названием «микросомальный антшен». До
сих пор существует название «антитела к микросомальной фракции щитовидной
железы», которое интерпретируется как антитела к слабо очищенной ТПО, хотя
микросомальная фракция содержит также белки митохондрий и ряд цитоплазма-
тических разновидностей АНА. В современных тест-системах используют высо-
коочищенный белок или рекомбинантную ТПО.
Пероксидаза находится на апикальной поверхности тиреоидных фолликуляр-
ных клеток. Она представляет собой основную антигенную мишень при тиреои-
дите Хашимото, против которой направлено большинство клеточных ответов при
этом заболевании.
До развития клинической картины заболевания в щитовидной железе можно
обнаружить выраженную клеточную инфильтрацию. Мононуклеарные инфильтра-
ты содержат большое количество В-клеток, Т-клеток и макрофагов. Формирование
инфильтратов сопровождается постепенной деструкцией этих фолликулов и
«попытками» железы регенерировать, что приводит к ее диффузному или узлово-
му увеличению и появлению зоба. Показана связь между антителами к ТПО и кли-
ническими особенностями заболевания: антитела чаще обнаруживают у субъектов
с узлами по сравнению с пациентами без узлов щитовидной железы.
Характерно, что аутоантитела к ТПО продуцируются главным образом лим-
фоцитарными инфильтратами щитовидной железы и только в малой степени —
клетками регионарных лимфатических узлов и костного мозга. Именно поэтому
аутоантитела к ТПО коррелируют с гистологической тяжестью тиреоидита и
увеличенными размерами железы на ранних стадиях заболевания. У больных с
высокой активностью аутоантител отмечается склонность к гипотиреозу. Титр
аутоантител к ТПО соотносится с продукцией в воспалительном инфильтрате
классических провоспалительных цитокинов, таких как IFN-y и TNF-oc, которые
могут быть связаны с повреждением щитовидной железы. Большинство антител к
ТПО относится к IgGj-субклассу, который считают хорошим активатором компле-
мента: продемонстрирована комплементопосредованная цитотоксичность на изо-
лированных тиреоидных клетках. Найдены отложения компонентов комплемента
в щитовидной железе больных, обусловленные его активацией под действием
антител к ТПО.
В подавляющем числе лабораторий для определения аутоантител к ТПО
используют ИФА и иммунохимические методы. Основным показанием к обследо-
ванию считают диагностику аутоиммунных заболеваний щитовидной железы. По
сравнению с антителами к тиреоглобулину антитела к ТПО более чувствительны и
специфичны, поэтому существует мнение, что дополнительное выявление антител
к тиреоглобулину только увеличивает цену обследования, не добавляя диагности-
ческой информативности. Антитела присутствуют у 75-90% пациентов с тиреои-
ДИАГНОСТИКА АУТОИММУННЫХ ЗАБОЛЕВАНИЙ 125
дитом Хашимото, у 60% пациентов с послеродовым тиреоидитом и примерно у
75% пациентов с болезнью Грейвса. Поскольку ТПО-антитела обнаруживают при
всех формах аутоиммунных заболеваний щитовидной железы, то они не могут
быть использованы для дифференциальной диагностики аутоиммунного тиреои-
дита и болезни Грейвса.
Другим показанием к определению этих антител является исключение ауто-
иммунного гипотиреоза во время беременности. Примерно 2% беременностей
протекает при пониженной тиреоидной функции. Беременность может «предъ-
являть» повышенные требования к тироксину, поэтому субклинический гипоти-
реоз обычно проявляется на ее фоне. Повышение уровня тиреостимулирующего
гормона или понижение свободного Т4 во время I триместра беременности может
сочетаться с нарушением нейропсихологического развития плода. Присутствие
антител к ТПО во время беременности указывает на увеличение риска развития
клинически значимого гипотиреоза и может рассматриваться как показание к
выполнению повторных измерений тиреостимулирующего гормона. Наличие
аутоантител к ТПО также увеличивает риск послеродовой тиреоидной дисфунк-
ции, Послеродовой гипотиреоз наблюдается примерно у 5% рожениц и характери-
зуется наличием аутоантител к ТПО. Это нарушение обычно наблюдают в первые
месяцы после родов, часто оно начинается с тиреотоксической фазы, сменяемой
пролонгированным гипотиреозом. Он может персистировать в течение многих
месяцев и примерно в 30% случаев может стать перманентным. Женщины с анти-
телами к ТПО во время беременности имеют 50% вероятность развития послеро-
довой тиреоидной дисфункции в сравнении только с 2% вероятностью у женщин
с негативными результатами исследования. Если антитела к ТПО выявляют
в Щ триместре беременности, то риск послеродовой тиреоидной дисфункции уве-
личивается и достигает 80%.
Достаточно противоречиво определение аутоантител для оценки субклиниче-
ского тиреоидита и гипотиреоза. Так же, как другие часто встречающиеся аутоан-
титела, например АНФ или РФ, антитела к ТПО часто обнаруживают у клинически
здоровых лиц. преимущественно у женщин среднего и пожилого возраста. Частота
таких находок варьирует в зависимости от возраста, географического и демогра-
фического состава популяции. Ряд исследователей указывают на встречаемость
аутоантител у 20% женск< >го населения. Этот феномен предлагается использовать
для выявления лиц с предрасположенностью к тиреоидиту и гипотиреозу. Наличие
антител к ТПО у клинически здоровых лиц может быть использовано как показа-
ние для обследования с целью диагностирования субклинического гипотиреоза,
характеризуемого повышенным уровнем тиреостимулирующего гормона при
нормальной концентрации свободного Т4. Имеются результаты популяционных
исследований, указывающих на то, что ежегодный риск развития гипотиреоза у здо-
ровых субъектов значительно увеличивается в присутствии аутоантител к ТПО. Эти
индивидуумы могут иметь повышенный риск развития явного гипотиреоза и/или
кардиоваскулярных болезней. Б связи с этим нужно проводить регулярное исследо-
вание функции щитовидной железы у лиц с высоким содержанием антител к ТПО.
Методов первичной профилактики тиреоидита не существует, а целесообраз-
ность регулярных гормональных обследований у лиц без клинических признаков
гипотиреоза представляется сомнительной с финансовой точки зрения, учитывая
естественную динамику функции щитовидной железы с течением времени.
Антитела к ТПО часто находят у больных другими аутоиммунными забо-
леваниями, такими как СКВ, РА, сахарный диабет I типа и болезнь Аддисона.
Гипофункция щитовидной железы присутствует примерно у 15% больных, полу-
чающих кордарон, и наличие антител к ТПО увеличивает риск развития гипо-
тиреоза. Кроме того, наличие антител к ТПО является фактором риска развития
гипотиреоза на фоне терапии литием.
&
126 ДИАГНОСТИКА АУТОИММУННЫХ ЗАБОЛЕВАНИЙ
АНТИТЕЛА К ТИРЕОГЛОБУЛИНУ
Исследования аутоиммунной реакции против тиреоглобулина (ТГ) в значитель-
ной степени определили понимание патогенеза всех аутоиммунных эндокринных
заболеваний. Доказано, что иммунизация кроликов, морских свинок или собак
очищенным ТГ того же вида или даже того же животного способна вызывать лим-
фоцитарный тиреоидит и образование аутоантител к ТГ. Аутоантитела к тирео-
глобулину были найдены у больных с болезнью Грейвса, тиреоидитом Хашимото,
раком щитовидной железы, некоторыми другими аутоиммуными заболеваниями,
а также они были обнаружены у значительного числа здоровых людей.
ТГ — главный компонент коллоида фолликул щитовидной железы. Он играет
основную роль в фиксации железой атомарного йода и синтезе йодированных
тиреоидных гормонов тироксина (Т4) и трийодтиронина (Тч). В норме йодируется
не более 25% тирозиновых остатков в составе тиреоглобулина.
Доказано, что йод пищи играет роль в модулировании аутоиммунного тире-
оидита, что базируется на клинических и эпидемиологических доказательствах
и экспериментальных исследованиях на животных, склонных к тиреоидиту.
Содержание йода в тиреоглобулине зависит от его поступлен ия с пищей. У экспери-
ментальных животных повышенное содержание йода в диете увеличивает тяжесть
тиреоидита. Роль йода в качестве одного из факторов развития аутоиммунного
процесса связана с его способностью менять конформационную структуру белка.
Высокойодированный ТГ представляется более антигенным, чем тиреоглобулин с
меньшим количеством йода.
Аутоантитела к ТГ рассматривают как непатогенные, главным образом, пото-
му. что введение сыворотки больных лабораторным животным не приводит к
тиреоидиту. Титр антител к тиреоглобулину обычно не коррелирует со степенью
тиреоидной дисфункции. В то же время сыворотка, содержащая антитела к ТГ,
может переносить заболевание при специальных условиях. Инъекция иммунной
сыворотки в щитовидную железу здоровых кроликов способна переносить ауто-
иммунный тиреоидит. По всей видимости, разрушение части клеток под действием
антителозависимой цитотоксичности способно привести к аутоиммунизации орга-
низма собственным ТГ.
Аутоантитела к ТГ представлены иммуноглобулином IgG и распределены
среди всех его 4 субклассов. Антитела класса IgG2 доминируют при тиреоидите
Хашимото, в то время как антитела класса IgG4 в большем объеме представлены
при болезни Грейвса, нетоксическом зобе и дифференцированной карциноме
щитовидной железы. Исторически антитела к ТГ измеряли многими иммунологи-
ческими методами, однако в настоящее время иммунохимические тесты и прежде
всего ИФА вытеснили все другие подходы.
Аутоантитела к ТГ чаще всего используют для подтверждения клинического
подозрения на тиреоидит Хашимото. их обычно определяют вместе с антитела-
ми к ТПО. Антитела к ТГ менее постоянны и ниже титром, чем антитела к ТПО.
Поскольку у всех больных тиреоидитом Хашимото в присутствии антител к ТГ
отмечаются и антитела к ТПО, последние являются наилучшим тестом для диа-
гностики тиреоидита. Чувствительность метода составляет 95-98%, поэтому
негативные результаты имеют высокое отрицательное предсказательное значение
и позволяют исключить диагноз тиреоидита.
Антитела к ТГ обладают низкой специфичностью, Они присутствуют в 40-50%
случаев болезни Грейвса, 20% случаев нетоксического зоба и при раке щитовидной
железы, а также у здоровых индивидуумов, особенно у пожилых женщин, у кото-
рых встречаемость аутоантител доходит до 10% случаев. Антитела были найдены
при нетиреоидных аутоиммунных болезнях таких как СШ, миастения, целиакия,
сахарный диабет I типа. Косвенно эти данные подтверждают, что тиреоидная
аутоиммунность — более распространенное явление, чем тиреоидит Хашимото.
ДИАГНОСТИКА АУТОИММУННЫХ ЗАБОЛЕВАНИЙ 127
Действительно, антитела к ТГ могут позволить оценить риск развития аутоим-
мунного тиреоидита. Когортное 20-летнее исследование, проведенное в Англии,
показало, что наличие тиреоидных антител является ведущим фактором риска при
идентификации лиц, которые имеют риск тиреоидита. Риск заболевания тиреои-
дитом в случае наличия аутоантител составляет около 3-5%. Кроме того, антитела
к ТГ полезны для определения риска развития послеродового тиреоидита.
Узкой областью клинического применения исследования на наличие антител к
ТГ является постоперационный мониторинг пациентов с дифференцированным
раком щитовидной железы. После тиреоидэктомии и абляции радиоактивным
йодом больных рецидив заболевания сопровождается увеличением свободного
тиреоглобулина в сыворотке. Поскольку тиреоглобулин — тканеспецифичный
белок, экспрессируемый исключительно тиреоидными клетками, повышенная
концентрация тиреоглобулина после операции указывает на рецидив рака или
метастазы. В этом случае антитела к ТГ приводят к заниженным концентрациям
свободного тиреоглобулина.
АНТИТЕЛА К РЕЦЕПТОРУ ТИРЕОТРОПНОГО ГОРМОНА
Рецептор тиреотропного гормона (рТТГ) экспрессируется преимущественно
на поверхности тиреоидных фолликулярных клеток, но также на лимфоцитах,
адипоцитах и фибробластах. Тиреотропный гормон (ТТГ) является регулятором
активности щитовидной железы. Сигнал от рТТГ запускает продукцию тиреоид-
ных гормонов и рост тиреоидных фолликулярных клеток. Избыточная активация
рТТГ при болезни Грейвса объясняет наличие тиреотоксикоза и зоба при этом
заболевании. Молекула рТТГ состоит из двух (А и Б) субъединиц гликопротеина.
Эффект связывания аутоантител с рТТГ может быть разнонаправленным. Так,
экстрацеллюлярная А-субъединица рецептора распознается тиреоидными стиму-
лирующими антителами, в то время как антитела, распознающие субъединицу Б,
локализованную вблизи клеточной поверхности, действуют как функционально
блокирующие антитела.
Стимулирующие антитела составляют основу болезни Грейвса — диффузно-
го токсического зоба. Блокирующие антитела связываются с рецептором, но не
вызывают его конформационных изменений, требуемых для сигнальной транс-
дукции. Они препятствуют связыванию ТТГ с рецептором, в результате чего воз-
никает снижение тиреоидной функции. Блокирующие антитела были обнаружены
у пациентов с атрофическим тиреоидитом Хашимото и у больных с болезнью
Грейвса, протекающей с офтальмопатией. Блокирующая активность аутоанти-
тел может отчасти объяснить колебания тиреоидной функции, наблюдаемой у
пациентов с болезнью Грейвса, и отсутствие корреляции между титром антител к
рецептору тиреотропина и тиреоидной функцией. Блокирующие антитела могут
проходить сквозь плаценту и вызвать транзиторный врожденный гипотиреоз.
Наконец, установлены нейтральные аутоантитела к рТТГ, которые связываются
с рецептором, но блокируют связывание тиреотропина или других рецепторных
антител. Нейтральные антитела определяют у пациентов с болезнью Грейвса. Их
патогенетическое значение остается неустановленным.
Нет сомнения в том, что антитела к рецептору являются патогенетически зна-
чимыми. Действительно, болезнь Грейвса — одно из немногих аутоиммунных
заболеваний, при котором имеются прямые доказательства его аутоиммунной
природы. Эти доказательства происходят от способности аутоантител переносить
болезнь. D.D. Adams, который обнаружил эти антитела, продемонстрировал повы-
шение функции собственной щитовидной железы после инъекции себе плазмы от
пациентов с болезнью Грейвса. Опосредованный перенос болезни Грейвса анти-
телами доказан также в периназ альном периоде, когда матери, страдающие болез-
128 ДИАГНОСТИКА АУТОИММУННЫХ ЗАБОЛЕВАНИЙ
нью Грейвса, передавали ее новорожденным, у которых транзиторно развивались
зоб и тиреотоксикоз.
Для определения аутоантител к рТТГ применяют радиоиммунный, ИФА и
иммунохимический методы. Ни один из этих методов не позволяет уточнить
присутствие в сыворотке крови стимулирующих или блокирующих антител.
Для этой цели был разработан функциональный тест на культурах тиреоцитов
животных, или LATS-тест (от англ, long acting thyroid stimulator). В его основе
лежит увеличение концентрации цАМФ в тиреоцитах под действием сыворот-
ки больного, добавляемой в культуральную среду. В связи с трудоемкостью и
сложностью стандартизации метод LAIS используют только в исследовательских
лабораториях.
Существует 4 возможных клинических показания для определения антител к
рецептору тиреотропина:
❖ диагностика транзиторной тиреоидной дисфункции у новорожденных
от матерей с тиреотоксикозом и прогнозирование неонатальной болезни
Грейвса;
• о определение антител аутоиммунного генеза у больных с изолированной
офтальмопатией без классических тиреоидных признаков (эутиреоидная
болезнь 1рейвса);
❖ предсказание рецидива тиреотоксикоза после прекращения 12-месячного
цикла антитиреоидной терапии;
о определение причины послеродового тиреотоксикоза.
Антитела к рТТГ высокоспецифичны, они крайне редко встречаются у здоровых
людей. Клиническая чувствительность составляет около 80%, то есть негативный
результат определения аутоантител не позволяет исключить заболевание. Низкая
чувствительность методов связана с тем, что аутоантитела к рецептору присутству-
ют в сыворотке в очень малой концентрации, на несколько порядков ниже, чем
уровни аутоантител к ТГ и ТПО.
АНТИТЕЛА К ОСТРОВКОВЫМ КЛЕТКАМ
Антитела к островковым клеткам (АОК) были впервые обнаружены у боль-
ных сахарным диабетом I типа (СД I) методом НРИФ на криостатных срезах
аутопсийной поджелудочной железы человека с 0 группой крови. Этот метод для
определения антиостровковых антител (от англ, islet cell antibodies — ICA) имеет
хорошую чувствительность и специфичность для СД I, но значительную межла-
бораторную вариацию. Поэтому был разработан стандарт сыворотки JDF (Juvenile
Diabetes Foundation), который стали использовать для стандартизации всех методов
выявления АОК.
АОК представляют собой гетерогенную группу антител, реагирующих с раз-
личными аутоантигенами в островковых р-клетках. Антигенами АОК является
обширная группа мембранно-ассоциированных и цитоплазматических белков
островковых клеток, причем ряд антигенов — общие для эндокринной и нерв-
ной ткани. В семействе АОК изучено несколько основных антигенов, таких как
глутамат-декарбоксилаза (ГДК), эндогенный инсулин, тирозиназа (IA-2), и около
20 минорных антигенов.
Непосредственная роль АОК в разрушении островковых клеток при сахарном
диабете остается спорной, однако была подтверждена способность антител фик-
сировать комплемент на островковых клетках, а также нарушать высвобождение
инсулина. В то же время АОК могут быть обнаружены у 0,5-2% контрольной
популяции. Терапевтический плазмаферез не оказывает действия на выработку
эндогенного инсулина в дебюте диабета. У новорожденных от матерей, больных
СД I, в крови которых присутствуют АОК, диабет не развивается.
ДИАГНОСТИКА АУТОИММУННЫХ ЗАБОЛЕВАНИЙ 129
Определение АО К — наиболее чувствительный серологический тест в диагно-
стике впервые выявленного СД I. Островковые антитела находят в 75-85% слу-
чаев СД I во время установления диагноза. Встречаемость значительно снижается
с увеличением возраста дебюта заболевания. Примерно 5-10% пациентов с сим-
птоматикой СД II могут иметь АОК на фоне инсулинозависимой формы диабета
взрослых, которая является разновидностью аутоиммунного диабета.
АОК могу г служить важным предиктором заболевания для родственнников
первой степени больных СД I. Среди них 2-5% оказываются положительными по
АОК, в то время как в контроле всего 0,4% испытуемых. Риск заболевания брата
или сестры максимален в течение 5 лет от момента заболевания пробанда.
АУТОАНТИТЕЛА К ИНСУЛИНУ И ТИРОЗИНАЗЕ IA-2
Антитела к инсулину у пациентов с СД I, не получавших экзогенный инсулин,
долгое время казались случайной находкой. Для диагностики СД I они были
впервые использованы у детей с впервые выявленным СД I до начала инсулиноте-
рапии. Антитела к инсулину у больных, уже получающих препараты инсулина, не
являются специфичными для СД I, поскольку в этом случае антигенные эпитопы
эндогенного и экзогенного инсулина несколько отличаются. Экзогенный челове-
ческий инсулин — плохой иммуноген, однако рекомбинантный инсулин, который
вводят в смеси с протамином или цинком, выражение иммуногенен и быстро
вызывает образование антител.
Аутоантитела, определяемые до назначения препаратов инсулина, т.е. к эндо-
генному инсулину, присутствуют у 20-40% больных с клиническим дебютом СД I.
Титр антител обратно пропорционален возрасту и уменьшается по мере разруше-
ния |3-клеток. В отличие от .других аутоантител к островковым антигенам, значе-
ние антител к инсулину максимально в возрасте до 10 лет, после чего аутоантитела
обнаруживают крайне редко. Антитела к инсулину присутствуют у большинства
детей до 5 лет, но только у 4% при начале СД I после 19-летнего возраста. Высокий
титр у малолетних детей связывают с тем обстоятельстом, что у них заболевание
протекает более агрессивно, с быстрым развитием зависимости от экзогенного
инсулина. При прогнозировании заболевания у близких родственников больных
отмечается слабая корреляция между определением антител к инсулину и риском
развития СД I, но она увеличивается, если они выявляются в комбинации с други-
ми антителами к островковым антигенам. Антитела к инсулину находят при ауто-
иммунных полиэндокринопатиях, при других аутоиммунных заболеваниях и при
назначении препаратов, содержащих тиоловое кольцо, таких как пеницилламин.
Долгое время основным методом обнаружения антител к инсулину был радио-
иммунный метод. Широкое внедрение ИФА привело к разработке соответ-
ствующих тест-систем, однако результаты сравнения методов показали, что
радиоиммунный метод имеет значительные преимущества, благодаря сохранению
нативной конформации белков.
Нейроэндокринный антиген IA-2 (ICA512) стал последним из обнаруженных
островковых антигенов, аутоантитела к которому используют в диагностике и про-
гнозировании сахарного диабета. Распространенный в нейроэндокринной ткани
белок IA-2 и его гомолог IA-2J3 (белок фо^рин) принадлежат к энзиматически
неактивным представителям семейства тирозинфосфатазы, участвующим в регу-
ляции секреции инсулина. Обе молекулы локализуются в секреторных гранулах
нейроэндокринных клеток и регулируют секрецию инсулина. Определение анти-
тел к IA-2 используют в прогнозировании развития СД I у родственников больных.
Основным методом выявления аутоантител к IA-2 служит иммунопреципитация с
358-метионин-меченого рекомбинантного человеческого IA-2 или конкурентный
радиоиммунный анализ. Метод ИФА малоспецифичен и не рекомендован для
клинического применения.
130 ДИАГНОСТИКА АУТОИММУННЫХ ЗАБОЛЕВАНИЙ
АНТИТЕЛА К ГЛУТАМАТ-ДЕКАРБОКСИЛАЗЕ
Аутоантитела к глутамат-декарбоксилазе (ГДК) были впервые обнаружены у
пациента с редким неврологическим заболеванием и СД I. Это неврологическое
заболевание носит название «синдром скованного человека» (англ, stiff-person
syndrome). Оно характеризуется прогрессирующей ригидностью нижних конеч-
ностей. Болезненные спазмы мышц при этом синдроме вызываются внезапными
стимулами, 10% больных страдают эпилепсией- Методом иммунопреципитации
антитела к ГДК были обнаружены в сыворотке у больных СД I, а также у экспе-
риментальных животных с диабетом. Антитела к ГДК были найдены при других
неврологических заболеваниях, таких как церебральная атаксия, рефрактерная к
лекарствам эпилепсия, небный миоклонус и болезнь Баттона. Кроме того, антите-
ла к ГДК обнаружили при тиреоидите Хашимото и диффузном токсическом зобе,
целиакии, болезни Аддисона и аутоиммунных полиэндокринопатиях.
Глутамат-декаобоксилаза — фермент, участвующий в синтезе у-оксибутировой
кислоты, которая контролирует высвобождение инсулина из секреторных гранул
р-клеток поджелудочной железы. ГДК не является специфичной для р-клеток
поджелудочной железы, поскольку присутствует в нервной системе и семенных
железах. Фермент присутствует в организме в двух изоформах: растворимая
форма — ГДК 67 (67 кДа) — распространена главным образом в нейронах, а нерас-
творимая — ГДК 65 (65 кДа) — присутствует в р-клетках поджелудочной железы.
ГДК65 — основная мишень аутоантител у пациентов с СД I, латентным аутоим-
мунным диабетом у взрослых, диабетом беременных и при синдроме скованного
человека. Помимо того, что она вызывает развитие гуморального иммунного
ответа, ГДК65 является мишенью для Т-клеточных ответов, обнаруживаемых как
у заболевших людей, так и на экспериментальных моделях диабета. Деструкция
р-клеток при диабете обусловлена активностью цитотоксических Т-клеток, чьи
антигенспецифичные рецепторы распознают р-клеточные пептиды, происходя-
щие из ГДК65.
В начале заболевания (при ювенильном начале) СД I 70-90% пациентов поло-
жительны по антителам к ГДК65. Распространенность антител к ГДК при диабете
беременных составляет 0-50% в разных исследованиях. Встречаемость аутоан-
тител в общей популяции составляет 1-2%, при этом у детей без клинических
признаков СД встречаемость аутоантител составляет до 4%, Определение антител
к ГДК в сыворотке может быть применено для идентификации лиц с риском раз-
вития СД I и для дифференциальной диагностики СД I и СД II.
Остается неясным, действительно ли наличие антител к ГДК у пациентов с СД II
влияет на клиническое течение заболевания. В некоторых исследованиях не было
найдено связи между антителами к ГДК и диабетическими осложнениями, в том
числе со стадией или тяжестью ретинопатии. Несколько исследований пациентов
с СД I и латентным аутоиммунным диабетом взрослых показали, что антитела к
ГДК после начала заболевания сохраняются в крови заболевших в течение 12 лет
и более. Снижение титров антител с течением времени отражает прогрессивную
потерю функции островковых клеток и снижение аутоиммунной реакции. Низки.!
уровень аутоантител указывает на большую давность заболевания с меньшей
агрессивностью аутоиммунного процесса. Высокая концентрация антител к ГДК65
у пациентов с латентным аутоиммунным диабетом указывает на высокий риск раз-
вития инсулиновой зависимости. Предсказательное значение антител для диагно-
за инсулиновой зависимости в пределах 6 лет наблюдения от начала заболевания
варьировало от 34% у лиц старше 55 лет до 84% у лиц моложе 34 лет. У родствен-
ников первой степени больных СД I частота обнаружения антител к ГДК65 варьи-
рует от 4 до 13%, В то же время предсказательное значение выявления аутоантител
при определении риска развития СД I не очень надежно: у родственников первой
степени родства оно варьирует от 20 до 60%. В крайнем варианте за 12-летнее
ДИАГНОСТИКА АУТОИММУННЫХ ЗАБОЛЕВАНИЙ 131
наблюдение за неидентичными близнецами в случае обнаружения антител к
ГДК65 риск развития СД I составил 88%.
Идентификация и молекулярное клонирование ГДК65 привели к разработке
метода определения антител к ГДК, который был стандартизован под руковод-
ством Immunology of Diabetes Society (IDS) и Centers for Disease Control and Prevention
(CDC). Разработан стандарт сыворотки ВОЗ для сравнения результатов в разных
лечебных учреждениях. Наилучшие параметры имеет конкурентный радиоиммун-
ный метод, использующий 3-’S- или 1251-меченую человеческую рекомбинантную
ГДК65: появились конкурентные иммунометрические системы для определения
антител к ГДК.
АУТОАНТИТЕЛА К КОРЕ НАДПОЧЕЧНИКОВ И К СТЕРОИДНОЙ 21 -ГИДРОКСИЛАЗЕ
Аутоиммунная болезнь надпочечников может развиваться как изолированная
форма болезни Аддисона или быть частью аутоиммунного полиэндокринного син-
дрома. Серологическим маркером поражения надпочечников являются аутоанти-
тела, реагирующие с ферментами стероидогенеза коры надпочечников. Ферменты
стероидогенеза относят к микросомальной фракции. Они обеспечивают окисление
гормональных предшественников и локализуются в цитоплазме клеток всех трех
слоев коры надпочечников. Основным методом определения антител к коре над-
почечников является НРИФ на нефиксированных криостатных срезах надпочеч-
ников человека и животных. Эти антитела представлены иммуноглобулинами IgG
и способны фиксировать комплемент. Мишенью антител к коре надпочечников
является 21-гидроксилаза. Инкубация сыворотки пациентов с болезнью Аддисона
с очищенной рекомбинантной 21-гидроксилазой человека приводит к потере
реактивности аутоантител к коре надпочечников. Разработаны методы радио-
иммунопреципитации и иммуноблоттинга, однако их использование ограничено
исследовательскими лабораториями, а для клинической диагностики их результа-
ты сопоставимы с результатами НРИФ. Чувствительность серологического иссле-
дования при болезни Аддисона в зависимости от длительности заболевания оце-
нивают примерно в 60-80%, а специфичность — в 98-100%. Определение антител
к стероидной 21-гидроксилазе у больных с дебютом болезни Аддисона чаще дает
положительный результат, чем у больных с длительным течением заболевания.
Кроме того, антитела к коре надпочечников и антитела к 21-гидроксилазе выявля-
ют при аутоиммунных полиэндокринопатиях 1 и 2 типа.
При применении НРИФ аутоантитела к коре надпочечников встречаются в 61%
наблюдений среди пациентов с аутоиммунной болезнью Аддисона разной дли-
тельности заболевания, у 6,7% пациентов с надпочечниковой недостаточностью в
результате туберкулеза и у 0,6% здоровых людей.
Особое значение имеет определение аутоантител у пациентов с полиэндокри-
нопатиями для прогнозирования развития надпочечниковой недостаточности.
Может быть рекомендовано выявление аутоантител к коре надпочечников у боль-
ных СД I моложе 10 лет, у родственников первой степени родства лиц, страдаю-
щих болезнью Аддисона, и у больных с первичной недостаточностью яичников.
Повышенный титр аутоантител к коре надпочечников, наличие хронического кан-
дидоза кожи и слизистых оболочек и/или гипопаратиреоза указывают на высокий
риск развития надпочечниковой недостаточности как компонента полиэндо-
кринопатии I или 2 типа. У этих пациентов необходимо регулярно обследовать
функцию коры надпочечников с интервалами каждые 6-12 мес для того, чтобы
предупредить жизнеугрожающие адреналовые кризы и вовремя начать замести-
тельную терапию.
132 ДИАГНОСТИКА АУТОИММУННЫХ ЗАБОЛЕВАНИЙ
АУТОАНТИТЕЛА К СТЕРОИДПРОДУЦИРУЮЩИМ КЛЕТКАМ ЯИЧКА, ЯИЧНИКА И
ПЛАЦЕНТЫ
Аутоантитела к стероидпродуцирующим клеткам — это иммуноглобулины
класса IgG, реагирующие с цитоплазматическими микросомальными антигенами
стероидпродуцирующих клеток. Окислительные ферменты, которые участвуют
в метаболизме стероидных гормонов, состоят из различных субъединиц цитох-
рома р450. Всего выделяют 3 фермента, которые являются мишенями аутоан-
тител: р450с21 (21-гидроксилаза), а также р450с17 (17-гидроксилаза) и p450scc
(табл. 17-17).
Таблица 17-17. Основные аутоантитела к ферментам стероидогенеза в эндокринных тканях
Фермент Кора надпочечник. Клетки Лейдига в яичке Клетки фолликулов яичника Плацента Заболевание
гломеруляоный слой ретикулярный слой
р450с17 - + + + Гипогонадизм у жен- щин, ПЭП 1
р450с21 + + - - + Болезнь Аддисона, ПЭП 2
p4t>0scc 4- + + + - Гипогонадизм у мужчин
Аутоантитела к стероидпродуцирующим клеткам могут играть определенную
роль в патогенезе аутоиммунных эндокринопатий. Так, сыворотки пациентов с
аутоиммунной боленью Аддисона, содержащие аутоантитела к стероидпродуци-
рующим клеткам, способны вызвать комплемент-зависимую цитотоксичность при
воздействии на текальные клетки яичников in vitro. Гистологическое исследование
яичников пациентов с аутоантителами к стероидпродуцирующим клеткам позво-
ляет обнаружить оофорит с лимфоцитарными инфильтратами.
Аутоантитела к стероидпродуцирующим клеткам реагируют с цитоплазмой
Лейдиговских клеток яичек, текальных клеток яичников, синцитиотрофобластами
плаценты, а также с желтым телом. Все позитивные сыворотки реагируют также с
корой надпочечников, которая содержит все разновидности стероидных фермен-
тов.
Антитела к стероидпродуцирующим клеткам находят в 100% случаев у боль-
ных с нормальным хромосомным набором и первичной аменореей (гипергонадо-
тропньш гипогонадизм) или вторичной аменореей в сочетании с аутоиммунной
болезнью Аддисона. Аутоантитела можно изредка обнаружить у лиц с вторичной
аменореей без признаков болезни Аддисона. Однако у этих больных обычно
выявляют аутоантитела к коре надпочечников, которые указывают на высокий
риск развития аутоиммунной болезни Аддисона. Аутоантитела к стероидпродуци-
рующим клеткам у мужчин, направленные к p450scc. применяют для диагностики
аутоиммунной тестикулярной недостаточности и задержки полового развития.
Основным подходом к определению аутоантител к стероидпродуцирующим
клеткам в практических лабораториях является НРИФ. Методом точной иденти-
фикации молекулярной мишени аутоантител служит радиоиммунопреципитация и
вестерн-блот. Оба этих метода используют преимущественно в научных лаборато-
риях. Помимо р450с17 и p450scc был описан ряд других аутоантител к антигенам
половых желез, но ни одно из них не имеет клинического или диагностического
значения.
При отсутствии болезни Аддисона у больных с идиопатической первичной недо-
статочностью яичников аутоантитела к стероидпродуцирующим клеткам опреде-
ляются крайне редко. Если они определяются, то это может служить предиктором
развития болезни Аддисона в дальнейшем. Кроме того, антитела к р450с17 связаны
с типом 1 полиэндокринопатий, протекающих с болезнью Аддисона, гипопарати-
ДИАГНОСТИКА АУТОИММУННЫХ ЗАБОЛЕВАНИЙ 133
роидизмом и слизистокожным кандидозом. Антитела к р450с21 присутствуют при
2 типе полиэндокринопатии, клиническая картина которого включает поражение
щитовидной железы, надпочечниковую недостаточность и аутоиммунный сахар-
ный диабет. Антитела к p450scc являются общим маркером задержки полового
развития и гипергонадотропного гипогонадизма. Обнаружение аутоантител у лиц
в группах риска позволяет использовать аутоантитела к стероидпродуцирующим
клеткам для оценки риска развития эндокрипопатии. У больных с различными
эндокринными заболеваниями, у которых были обнаружены аутоантитела к
стероидпродуцирующим клеткам надпочечника, имеется высокий риск развития
недостаточности половых желез и болезни Аддисона. У 35-40% женщин при обна-
ружении аутоантител к р450с17 и p4.50scc развивается недостаточность функции
яичников, что требует определения гормонсинтетической функции половых желез
в динамике.
СЕРОЛОГИЧЕСКИЕ МАРКЕРЫ ПРИ НЕВРОЛОГИЧЕСКИХ ЗАБОЛЕВАНИЯХ
Рассеянный склероз (PC) — наиболее частое аутоиммунное заболевание ЦНС.
В основе его патогенеза лежит выраженная активация забарьерной иммунной
системы, которая приводит к появлению фокусов инфильтрации в белом веществе
головного и спинного мозга с демиелинизацией нервных проводников и глио-
зом (склерозом). Очаги глиоза могут быть обнаружены магнитно-резонансной
томографией. Заболевание имеет разнообразную клиническую симптоматику,
типичным признаком которой является быстрая динамика неврологических про-
явлений. по мере прогрессирования процесса нарастает необратимый неврологи-
ческий дефицит. Системная воспалительная реакция при PC отсутствует, поэтому
для оценки иммунного воспаления необходим анализ цереброспинальной жид-
кости (СМЖ). Особенностью воспаления при демиелинизирующих заболеваниях
является то, что оно протекает за гематоэнцефалическим барьером (интратекаль-
но). Интратекальный иммунный ответ сопровождается рядом иммунологических
феноменов, в том числе изолированным синтезом иммуноглобулинов, появлением
неспецифических серологических реакций против распространенных вирусов, а
также синтезом олигоклонального иммуноглобулина.
Анализ СМЖ явлется основой для диагностики аутоиммунных и воспалитель-
ных заболеваний ЦНС (табл 17-18). Анализ СМЖ должен быть проведен в срок до
3 ч с момента взятия биоматериала. Для полноценной иммунологической оценки
необходимо получение парной пробы СМЖ и периферической крови.
Таблица 17-18. Лабораторною показатели при иммунологическом анализе цереброспинальной
жидкости
Показатель (единицы) Общий цитоз, кл/мкл Дифф, подсчет (Л/М/Н/П') Альбумин, Местный синтез lg, % обнаружения Олиго IgG, % обнару- жения
Норма <5 л >м <8 0 0
Рассеянный склероз 5-30 л >м >п <10 70 98
Бактериальный менингит 1000-30000 г »м, л 50-100 0 0
Туберкулезный менингит 50-2000 л = г = м 20-100 25 25
Нейроборрелиоз 50-1000 л >м >п 5-50 50 50
Вирусный энцефалит (HSV. VZV1 10-300 л >м >п <20 10 10
Ишемия 2-10 л >м <10 0 0
СКВ. болезнь Бехчета, нейросаркоидоз 50-200 л >м>п 5-50 50 50
'Л/М/Н/П — тип преобладающих клеток в ликворе (лимфоциты, моноциты, ней грофилы, плаз-
матические клетки).
134 ДИАГНОСТИКА АУТОИММУННЫХ ЗАБОЛЕВАНИЙ
ОЦЕНКА ИНТРАТЕКАЛЬНОГО ВОСПАЛЕНИЯ ПРИ ДЕМИЕЛИНИЗИРУЮЩИХ
ЗАБОЛЕВАНИЯХ
Первым признаком наличия интратекального воспаления считают умеренное
увеличение общего белка СМЖ. обусловленное увеличением местной продукции
иммуноглобулинов. Увеличение количества общего белка нередко не превышает
верхней границы нормы (>900 мг/л), поэтому требует использования более чув-
ствительных диагностических методов.
Поликлональная и олигоклональная активация синтеза иммуноглобулинов
является одной из форм подострого иммунного ответа, однако при большинстве
аутоиммунных заболеваний (СКВ, СШ и т.д.) она имеет системный характер.
При PC продукция иммуноглобулинов обусловлена неспецифической стимуля-
цией В-клеток, значительное содержание которых отмечают при цитологической
оценке состава СМЖ. Важным признаком, отражающим локальную индукцию
гуморального ответа в СМЖ при PC, является одновременное образование IgG-
антител к антигенам распространенных вирусов. Для обнаружения этой реакции
используют определение антител к антигенам вируса кори, герпеса зостер и вирусу
ветряной оспы. Этот феномен получил название «MZ\ реакция» (от первых букв
английских названий вирусов). Эти агенты могут рассматриваться в качестве
этиологической причины PC. однако одновременное появление антител против
них при отсутствии вирусов в организме указывает на анамнестическую реакцию
иммунной системы при неспецифической активации. Обнаружение MZV-реакции
многими исследователями рассматривается в качестве наиболее специфичного
лабораторного теста при PC, который положителен у 94% больных. Для оценки
MZV-реакции рекомендовано одновременное определение инфекционной серо-
логии в парных пробах СМЖ и сыворотки периферической крови, что позволяет
рассчитать индекс активности антител. Положительный результат оценивают как
индекс, указывающий на преобладание антивирусных антител в СМЖ по сравне-
нию с сывороткой (>1,5). Сложности интерпретации реакции как положительной
могут возникать, если обнаруживаются антитела лишь к одному возбудителю.
Несмотря на ценность данного теста, сложности в его выполнении и интерпрета-
ции затрудняют его широкое распространение для практической диагностики, он
остается вспомогательным методом даже в высокоспециализированных лабора-
ториях.
Более простой и надежный метод оценки неспецифической активации интрате-
кального гуморального ответа — определение относительных параметров секреции
иммуноглобулинов СМЖ. Метод изучения интратекальной продукции иммуно-
глобулинов СМЖ основан на общих иммунологических принципах исследования
проницаемости гематотканевых барьеров, определяющих местный иммунитет.
Для оценки состояния гематоэнцефалического барьера применяют изме-
рение альбумина ликвора и сывороточной его концентрации. Коэффициент про-
ницаемостх1 0,м1б определяют как отношение содержания альбумина в СМЖ к его
концентрации в сыворотке крови. Использование расчетного коэффициента обу-
словлено необходимостью нормализации концентраций альбумина СМЖ в связи
со значительной биологической вариацией уровней альбумина сыворотки. Этот
коэффициент позволяет оценить проницаемость тканевых барьеров и стандар-
тизировать определение локального синтеза других молекул воспаления, прежде
всего иммуноглобулинов. Нормальное значение этого показателя зависит от воз-
раста, составляет менее 5x10 3 в возрасте 20 лет и увеличивается до 8х10’3 в 60 лет.
Для PC изменения проницаемости гематоэнцефалического барьера не характерны,
и 0^пьб не превышает 10x10 3. Увеличение показателя 0яль6 отмечают при ряде вос-
палительных заболеваний, таких как гнойный бактериальный менингит, острый
нейроборрелиоз, синдром Гийена-Барре. Умеренное увеличение проницаемости
характерно для вирусных менингитов и полинейцопатий.
ДИАГНОСТИКА АУТОИММУННЫХ ЗАБОЛЕВАНИЙ 135
В отличие от альбумина, иммуноглобулины могут синтезироваться как внутри
ЦНС, так и вне ее. Для определения интратекального синтеза иммуноглобулинов
используют расчет коэффициентов для основных классов иммуноглобулинов
(Q{gG’ OjgA’ QigKf) на основе их концентраций в сыворотке крови и СМЖ. На прак-
тике достаточным является определение QgG, в то время как определение других
индексов самостоятельного значения не имеет. Для оценки локального синтеза
IgG применяют номограммы зависимости и Q основанные на математиче-
ской модели проницаемости гематоэнцефалического барьера. Номограмма позво-
ляет от нести результаты определения и Qj ( к одному из 4 вариантов, которые
соответствуют норме, локальному синтезу Igfe, нарушению функции барьера и
комбинации нарушений. PC обычно сопровождается выраженным увеличением
локального синтеза IgG при сохранности функций гематоэнцефалического барье-
ра. Выраженный локальный синтез IgG можно обнаружить у 70% больных с PC.
Очевидным преимуществом этого подхода к оценке местного иммунитета ЦНС
является его доступность для практических биохимических лабораторий, осна-
щенных анализаторами с возможностью турбидиметрии и нефелометрии.
Олигоклональный IgG и другие маркеры иммунного
воспаления центральной нервной системы
По мере хронизации любой специфический иммунный ответ из поликлональ-
ного становится олигоклональным, что о сражает клональную природу иммунной
системы с увеличением аффинности иммуноглобулинов по мере персистенции
антигена. Большая концентрация иммуноглобулинов плазмы крови маскирует
олигоклональные иммуноглобулины в периферической крови, хотя очевидно, что
олигоклональный синтез возникает при всех вариантах хронического или подо-
строго воспаления. В редких случаях олигоклональный IgG можно обнаружить
в у-фракции при электрофорезе белков крови при появлении трех и более моно-
клональных пиков.
Значительно чаще олигоклональный иммунный ответ обнаруживают при ана-
лизе биологических жидкостей с низкой концентрацией иммуноглобулинов. Так,
он может быть идентифицирован в слезной жидкости и в слюне больных с СШ, в
синовиальной жидкости при реактивных артритах. Сужение спектра иммунного
ответа возникает под действием хронической аутоантигенной стимуляции на фоне
аутоиммунного воспаления. Антигенная направленность олигоклональных имму-
ноглобулинов остается плохо изученной.
Метод изоэлектрофокусирования с последующим иммуноблоттингом — золо-
той стандарт иммунологической диагностики PC, признанный большинством
международных экспертов. Хорошие аналитические параметры этого теста делают
его наиболее удобным и надежным методом диагностики демиелинизирующих
заболеваний. В его основе определения клональности иммуноглобулинов СМЖ
методом электрофоретического изофокусирования лежит разделение белков в
соответствии с их изоэлектрической точкой в градиенте амфолитов — веществ,
которые под действием электрического тока формируют непрерывный градиент
pH. Молекулы фокусируются в формируемом градиенте, что позволяет отличить
молекулы одной молекулярной массы с небольшой разницей в заряде. Молекулы
человеческого IgG обладают зарядом в пределах от 6 до 9 единиц pH, который
определяется количеством остатков сиаловых кислот на Fc-домене молекулы.
Поскольку молекулы IgG, синтезируемые одним клоном плазмоцитов, идеи гичпы,
они накапливаются в одной тонкой полосе. При олигоклональном или монокло-
нальном синтезе количество идентичных молекул настолько велико, что полоску
можно различить на поликлональном фоне. Для детекции фокусированного
136 ДИАГНОСТИКА АУТОИММУННЫХ ЗАБОЛЕВАНИЙ
IgG применяют метод иммуноблота с окраской антисывороткой к IgG человека.
Иногда для лучшей визуализации типа синтеза может быть использована окраска
на к- и Х-легкие цепи.
Результат выявления олигоклонального IgG при рассеянном склерозе с помо-
щью изоэлектрофокусирования: в сыворотке крови определяется нормальный
поликлональный IgG, в ликворе отмечается выраженный олигоклональный
иммунный ответ, сопровождающийся синтезом олигоклонального IgG.
Тест является качественным, так как подсчитать число полос при олигоклональ-
ном типе крайне затруднительно, но это число не имеет клинического значения
Важной дополнительной информацией может служить изучение парной пробы
сыворотки крови. Результат оценивают на основании описания типа синтеза в
каждой из парных проб. Всего выделяют 5 вариантов синтеза иммуноглобулинов.
Для PC наиболее характерен 2-й тип синтеза, в то время как 3-й тип может встре-
чаться на фоне других аутоиммунных заболеваний, затрагивающих ЦНС, с мини-
мальной системной продукцией олигоклонального IgG (табл. 17-19).
Таблица 17-19. Типы синтеза IgG в цереброспинальной жидкости и сыворотке в дифференциаль-
ной диагностике демиелинизирующих заболеваний центральной нервной системы
Тип синтеза Синтез в ликворе Синтез f сыворотке Нозологическая форма
1 Поли- Поли- Норма или острое воспалительное заболевание
2 Олиго- Поли- PC в 85-95% случаев в дебюте заболевания
3 Олиго- Олиго- (меньше полос) PC, реже — СКВ, постинфекционные энцефалиты, саркоидоз
4 Олиго Олиго- (идентично ликвору) Генерализованные процессы с вовлечением ГЭБ: боррелиоз, нейросифилис, синдром Гийена—Барре, ВИЧ, грибковый менин- гоэнцефалит
5 Моно- Моно- Моноклональные гаммапатии
Примечание. PC — рассеянный склероз, СКВ — системная красная волчанка, ГЭБ — гематоэн-
цефалический барьер.
Олигоклональный IgG имеется у 93-98% больных с PC, его отсутствие с высо-
кой вероятностью исключает диагноз. Особенностью феномена олигоклонального
IgG является его стабильность Ни характер, ни число олигоклональных полос не
меняются с течением времени, поэтому мониторинг не требуется Тест может быть
отрицательным у единичных больных с глубоким залеганием очагов демиелини-
зации. Олигоклональный IgG обнаруживают у органиченного числа больных с
другими заболеваниями ЦНС.
АНТИТЕЛА К НЕЙРОНАМ ПРИ ПАРАНЕОПЛАСТИЧЕСКИХ ЭНЦЕФАЛИТАХ
Паранеопластические энцефалиты — группа редких синдромов, при которых
онкологические процессы (рак яичника, легкого) приводят к иммуноопосредо-
ванному энцефалиту, не связанному с объемным процессом и протекающему как
мозжечковая атаксия, энцефалит, рацикулоневропатия. Хотя эти заболевания
редки, эффективная их диагностика позволяет обнаружить опухоль на ранних
стадиях развития. У 70% больных неврологическая симптоматика опережает
диагноз онкологического заболевания. В основе образования аутоантител при
опухоли лежит перекрестная экспрессия опухолью антигенов нервной ткани.
Аутоантитела синтезируются местно, за гематоэнцефалическим барьером, поэто-
му в дебюте заболевания титры аутоантител в СМЖ выше, чем в сыворотке крови.
Антитела обладают цитотоксическим действем, с чем может быть связано выявле-
ние инфильтратов нервной ткани, а также дистрофия мозжечка и других отделов
мозга. В то же время экспрессия невральных антигенов в опухоли далеко не всегда
сопровождается паранеопластическим синдромом, что указывает на наличие дру-
ДИАГНОСТИКА АУТОИММУННЫХ ЗАБОЛЕВАНИЙ 137
гих факторов, определяющих аутоиммунную реакцию. Так, антитела к клеткам
Пуркинье мозжечка (Yo-1) присутствуют только у женщин.
Среди аутоантител к нейронам наиболее часто встречаются Yo, Hu и
Ri-аутоантитела (табл. 17-20). К другим часто встречающимся «паранеопла-
стическим» антителам относят антитела к кальциевым каналам при синдроме
Ламберта-Итона и антитела к скелетным мышцам при миастении на фоне опухоли
тимуса.
Таблица 17-20. Аутоантитела к нейронам при ларанеопластических нейропатиях
Аутоантитело Неврологическая симптоматика Опухоли
Анти Yo-1 (62 кДа) Мозжечковая дегенерация (атаксия), болеют исключительно женщины Яичник, молочная железа
Антитела к клеткам Пуркинье (не Yo-1) Мозжечковая дегенерация, мягко выраженная, мужчины поражаются чаще Лимфогранулематоз Ходжкина
Анти-Hu Сенсорная невропатия, кишечная непроходимость, энцефаломиелит, чаще болеют женщины Мелкоклеточный рак легкого
Анти-Ri Энцефалит ствола мозга Молочная железа, мелкоклеточ- ный рак легкого
Анти-CV, Хорея, атаксия, невропатия Рак бронха, тимома
Анти-Ма? Энцефалит ствола мозга, атаксия Семинома
Амфифизин Синдром «ригидного человека» Молочная железа
Для определения нейрональных антител используют НРИФ либо иммуно-
гистохимическую технику на криосрезах мозжечка обезьяны. Антитела диффе-
ренцированно связываются с нейронами разных отделов нервной системы. Так,
антитела к ядрам нейронов 1 типа (анти-Hu) направлены против ядер нейронов
центральной и периферической нервной системы, однако не окрашивают ядрыш-
ко и не реагируют с ядрами астродендроглии. Антитела к ядрам нейронов 2 типа
(анти-Ri) очень похожи на анти-Hu, однако реагируют только с ядрами нейронов
центральной нервной системы. Антитела к Yo-1-антигену окрашивают цитоплазму
гигантских грушевидных нейронов мозжечка (клеток Пуркинье).
Для комплексной оценки ларанеопластических аутоантител рекомендуется
использовать тканевой комплекс, включающий мозжечок обезьяны, желудок и
печень крысы. При оценке окрашивания желудка необходимо обращать внимание
на реакцию аутоантител с клетками мезентериального нервного сплетения, кото-
рая очень характерна для Hu-антител. Печень крысы используется для выявления
АНА, которые затрудняют исследование антител к ядрам нейронов.
После получения положительного результата НРИФ-теста можно рекомендо-
вать выполнение иммуноблота, который позволяет подтвердить наличие пара-
неопластических аутоантител. Анти-Ни-антитела реагируют с РНК-связывающим
белком HuD. Мишенью анти-КЬантител является белок массой 55 кДа — Nova-
антиген. Антигеном Yo-1 ан гител служит гидрофобный цитоплазматический белок
массой 62 кДа, однако при лимфогранулематозе образуются антитела к цитоплаз-
ме клеток Пуркинье, направленные против других белков цитоплазмы (не Yo-1).
АНТИТЕЛА К ГАНГЛИОЗИДАМ ПРИ ПОЛИНЕЙРОЛАТИЯХ
Гликолипидная оболочка миелинизированных нервных волокон служит мише-
нью антител к ганглиозидам, которые являются серологическими маркерами вос-
палительных полиневритов (табл. 17-21). Антитела к ганглиозидам направлены
против гликофосфолипидов, состоящих из липида церамида и полисахаридной
части 1-5 остатков гексоз, хотя бы одна из которых несет остаток сиаловой кис-
лоты. Именно остаток сиаловой кислоты позволяет отнести данный гликолипид к
ганглиозидам. Церамид заякорен в мембране клетки, в то время как гидрофильная
полисахаридная часть направлена кнаружи от клеточной мембраны. В организ-
138 ДИАГНОСТИКА АУТОИММУННЫХ ЗАБОЛЕВАНИЙ
ме человека насчитывается 15 основных разновидностей ганглиозидов. Спектр
ганглиозидов разный в ЦНС и периферических нервах, Ганглиозиды GMT и GDla
экспрессированы на моторных нервах и аксонах. На черепно-мозговых нервах
много ганглиозида GQlb, сенсорные нервы богаты GDlb. Ганглиозиды выполняют
множество функций, кроме того, они служат рецепторами для ряда микроорганиз-
мов, в том числе для токсина холеры (GMJ, энтеротоксина Campylobacter (GMT),
Haemophilus influenzae (G A^GMJ.
Таблица 17-21. Антитела к ганглиозидам при острых и хронических демиелинизирующих пораже-
ниях периферической нервной системы
Заболевание Антиген Частота встре- чаемости аутоан- тител, % Класс lg
Острая воспалительная демиелинизирующая нейро- патия (синдром Гийена-Барре) GM? 20 IgG
Острая моторно-аксональная и острая моторно сенсорная аксональная нейропатия GMr G0.a 40
Синдром Миллера-Фишера ет1а 90
Хроническая воспалительная демиелинизирующая невропатия GD1a.b’ GM3’ СУЛЬ’ фатид 20 IgM
Мультифокальная моторная невропатия ем,.,, GD1ab 50
Хроническая сенсорная невропатия GD1b, GT1b, GQ1b 100
Парапротеинемическая сенсорная невропатия IgM MAG/SGPG сульфа- тид. GM2, GDla b 70
Несмотря на значительный объем экспериментальных данных, вопрос об
участии антител к ганглиозидам в патогенезе нарушений нервной проводимости
остается открытым. Далеко не у всех больных с типичной клинической картиной
полиневритов и изменениями при нейромиографии обнаруживают антитела к ган-
глиозидам, иногда антитела могут выявляться у лиц без полинейропатии. Тем не
менее значение серологического ответа в патогенезе острых полирадикулоневри-
тов подчеркивается клинической эффективностью плазмафереза и использования
человеческого иммуноглобулина для лечения при этом заболевании. В экспери-
ментальных моделях аутоантитела к ганглиозидам блокируют проводимость в
изолированных препаратах нервов.
Синдром Гийена-Барре относится к группе острых воспалительных полира-
дикулоневропатий, часто разрешающихся спонтанно. Это заболевание развива-
ется на фоне или после перенесенной респираторной или кишечной инфекции в
течение 10-14 дней с момента ее начала, поэтому образование аутоантител часто
рассматривают как компонент первичного иммунного ответа против возбудителя
инфекции. После достижения ремиссии заболевания титры антител постепенно
снижаются и нарастают в случае повторного обострения.
Синдром Гийена- Барре характеризуется демиелинизацией и аксональной деге-
нерацией моторных, сенсорных, автономных нервов и нервных корешков спинно-
го мозга. В него включаю г острую воспалительную демиелинизирующую полиней-
ропатию, острую моторно-аксональную нейропатию и острую моторно-сенсорную
аксональную нейропатию. Синдром Миллера-Фишера часто выделяют из группы
синдрома Пгйена- Барре по причине особых клинических проя влений, включаю-
щих атаксию, арефлексию и офтальмоплегию. Аутоантителами чаще сопровожда-
ются аксональные нейропатии и синдром Миллера-Фишера, в то время как при
классической демиелинизирующей форме антитела диагностируют редко.
При острых полирадикулоневропатиях и синдроме Миллера-Фишера выявля-
ют антитела к ганглиозидам GM} и GQlb класса IgG. Антитела класса IgM могут
встречаться, однако их титры низки. Предполагается, что в силу особенностей
ДИАГНОСТИКА АУТОИММУННЫХ ЗАБОЛЕВАНИЙ 139
иммунного ответа возникает переключение классов тяжелой цепи иммуноглобу-
линов с синтезом патогенного IgG. Исследование генов тяжелых цепей иммуно-
глобулина G, направленных против GM ^ганглиозида, показало высокую частоту
соматических мутаций, что указывает на антигенспецифическую дифференциров-
ку иммунного ответа.
В качестве основной причины сенсибилизации иммунной системы к антигенам
ганглиозидов при острых полинейропатиях предполагается перекрестная иммун-
ная реакция с липополисахаридами бактерий ЖКТ. Синдрому Гийена-Барре часто
предшествует энтерит, вызванный Campylobacter jejuni Липополисахаиид С. jejuni
содержит ганглиозидподобную структуру, напоминающую по структуре моле-
кулу ганглиозида GMp которая способна стимулировать аутоиммунные ответы.
Другими известными индукторами синдрома Гийена-Барре являются цитомегало-
вирус, вирус Эпштейна-Барр, Mycoplasma pneumoniae и Haemophilus influenzae.
При хронических периферических нейропатиях антиганглиозидн ые антитела
представлены преимущественно IgM. Часто аутоантитела входят в состав моно-
клонального IgM-компонента на фоне «моноклональной гаммапатии неясного
значения». Моноклональный компонент может не обнаруживаться с помощью
электрофореза белков сыворотки, однако может быть определен при использова-
нии более чувствительного изоэлектрофокусирования.
Так называемый парапротеинемический нейропатический синдром, возни-
кающий на фоне парапротеинемии IgM, характеризуется клинической картиной
сенсорной полиневропатии с атаксией и офтальмоплегией. Невропатия является
основным клинически значимым признаком «моноклональной гаммапатии неяс-
ного значения» и может отмечаться у 5-20% больных. Хроническая сенсорная
невропатия при этом заболевании схожа с симптоматикой синдрома Миллера-
Фишера. При парапротеинемический невропатии отмечается присутствие в сыво-
ротке моноклонального IgM, направленною против ряда ганглиозидов, включая
GD]b, GQlb и GTlb, других гликолипидов, а также против миелин-ассоциированного
гликопротеина. Парапротеинемическая невропатия может сочетаться с парок-
сизмальной ночной гемоглобинурией, обусловленной образованием антител к
сиаловым остаткам гликолипидных антигенов эритроцитов (холодовыми агглю-
тининами).
Мультифокальная моторная невропатия с блоком проведения также относится
к редким хроническим полинейропатиям и сопровождается антителами к ган-
глиозиду GM, класса IgM в 50% случаев. Характеризуется асимметричным поли-
невритом с характерной нейромиографической картиной фокального нарушения
проводимости по моторным нервам.
Хроническая воспалительная демиелинизирующая полинеиропатия (CIPD)
представляет собой аналог хронического синдрома Гийена-Барре и только
в 20% случаев сопровождается антителами к СМ^ганглиозиду класса IgM.
Основным методом детекции антител к ганглиозидам является ИФАг. В каче-
стве более точного исследовательского метода может применяться тонкослойная
хроматография. Определение антиганглиозидных антител характеризуется низкой
чувствительностью, причина кроется в составе антигена. Примеси жирных кислот
в препаратах ганглиозидов повышают чувствительность тестов. Общепризнанных
стандартов антигена и стандартов аутоантител не существует, поэтому тесты раз-
личаются между собой.
Антитела к ганглиозидам можно рассматривать как одну из разновидностей
«антител к гликолипидам». Гликолипиды (точнее, гликосфинголипиды) представ-
ляют собой обширное семейство мембранных молекул. Гликолипиды имеют мно-
жество функций, в том числе ими представлены антигены групп крови, факторы
адгезии клеток, упаковка структуры миелина. Среди этой большой группы антиге-
нов при полиневритах обнаруживают антитела к сульфатированному глюкуронид-
140 ДИАГНОСТИКА АУТОИММУННЫХ ЗАБОЛЕВАНИЙ
пара глобозиду (SGPG), сульфатиду и галактоцереброзиду, которые не являются
ганглиозидами по химическому строению. Антитела к сульфатиду встречаются
при диабетической нейропатии, хронической воспалительной демиелинизирую-
щей полинейропатии и парапротеинемической сенсорной нейропатии. Антитела к
SGPG класса IgM встречаются при парапротеинемической сенсорной нейропатии.
По структуре полисахаридного остатка SGPG близок к миелин-ассоциированному
гликопротеину (MAG), поэтому антитела к двум молекулам часто диагностируют
вместе при парапротеинемической нейропатии.
Системные аутоиммунные заболевания — немаловажная причина развития
воспалительных полиневритов. Среди них выделяют системные ревматические
заболевания, васкулиты, АФС, аутоиммунные эндокринопатии, аутоиммунный
гастрит, парапротеинемии. Лабораторная и серологическая диагностика основно-
го заболевания — один из методов постановки диагноза.
АНТИТЕЛА К АЦЕТИЛХОЛИНОВОМУ РЕЦЕПТОРУ
Нарушение нервно-мышечной передачи при миастении обусловлено аутоанти-
телами, которые связываются с рецептором ацетилхолина на постсинаптической
мембране нервно-мышечного окончания (моторной пластинки). Никотиновый
(N-) ацетилхолиновый рецептор (АхР) является олигомерным трансмембранным
ионным каналом, который состоит из 6 субъединиц. При связывании ацетилхо-
лина на двух а-субъединицах рецептора открывается ионный канал, который
позволяет катионам (прежде всего натрию) поступать внутрь мышечной клетки.
Ток ионов обеспечивает возбуждение мембраны клетки, снижение трансмембран-
ного потенциала, открытие кальциевых каналов и, в конечном итоге, сокращение
мышечного волокна.
При миастении антитела непосредственно участвуют в патогенезе заболевания.
Введение сыворотки, содержащей антитела к АхР, лабораторному животному
воспроизводит клиническую картину миастении. У ряда лабораторных живот-
ных можно воспроизвести заболевание путем их иммунизации очищенным АхР.
Новорожденные от матерей, больных миастенией, в большинстве случаев перено-
сят транзиторную неонатальную форму заболевания, которая проходит в течение
месяца после истощения пула материнских антител к АхР.
Антитела к АхР снижают количество АхР нервно-мышечного окончания за счет
нескольких механизмов. Отмечается комплемент-опосредованный лизис пост-
синаптической мембраны со снижением числа моторных пластинок на мышце.
Под действием антител происходит димеризация и интернализация рецептора,
что сокращает его мембранный пул. В небольшой степени аутоантитела способ-
ны связываться с а-субъединицей АхР, приводя к его функциональной блокаде.
Эпитоп антител к АхР является конформационным, и хотя он локализован на
а-субъединице, только небольшое число аутоантител направлено на сайт связы-
вания ацет илхолина.
Основным методом определения антител к АхР служит радиоиммунопреципи-
тация, основанная на способности меченного 1251 а-бунгаротоксипа связываться
с ацетилхолиновым рецептором скелетной мышцы. Источником человеческого
АхР является оперативный материал или клетки перевиваемых клеточных линий
рабдомиосаркомы ТЕ671, которая содержит фетальный АхР. Несмотря на значи-
тельную гомологию гена АхР, достаточную чувствительность тестов обеспечивает
использование АхР человека или высших приматов. Тесты, основанные на методе
ИФА, не обеспечивают нативную укладку АхР, а потому недостаточно чувстви-
тельны.
У 15% больных миастенией антитела к АхР не обнаруживают. В то же время у
большинства серонегативных больных плазмаферез и иммуносупрессия оказы-
ваются высокоэффективными. Несмотря на отсутствие антител к АхР, введение
ДИАГНОСТИКА АУТОИММУННЫХ ЗАБОЛЕВАНИЙ 1 41
сыворотки крови этих больных лабораторным животным приводит к проявле-
ниям миастении. Это позволяет предположить присутствие в этих сыворотках
еще не изученных аутоантител, также влияющих на нервно-мышечную передачу.
Сравнительно недавно была обнаружена новая мишень аутоантител при сероне-
гативной миастении — MUSK-рецептор. Аутоантитела к ЛШЗК-рецептору явля-
ются показателем выбора при подозрении на серонегативную миастению. Для их
определения применяют метод радиоиммунного анализа.
Миастения — аутоиммунное заболевание, имеет два возрастных пика, один
из которых приходится на 15 лет, а другой начинается после 40. В 15% случаев
отмечается окулярная форма миастении, которая редко прогрессирует в генера-
лизованную форму. В остальных 85% случаев течение заболевания варьирует от
окулярной до генерализованной формы. Нередким симптомом миастении явля-
ются миастенические кризы, возникающие в результате инфекции или приема
препарата, с развитием острой мышечной слабости.
Антитела к АхР диагностируют у 80-85% больных миастенией, они обладают
высокой специфичностью. Содержание аутоантител к АхР снижается параллельно
эффективной терапии, уменьшение на 50% обычно соответствует выраженному
клиническому улучшению. У больных в стадии ремиссии могут сохраняться титры
антител выше диагностических. У 15% пациентов можно обнаружить опухоль
тимуса, а гиперплазия тимуса отмечается в 30-40% случаев. Тимэктомия является
операцией выбора при миастении в случае плохого ответа на иммуносупрессивную
терапию и в большинстве случаев улучшает клиническое течение заболевания.
Более 95% больных с тимомой имеют антитела к АхР, кроме того, в 30-50% из
них можно обнаружить антитела к скелетным мышцам. При рецидиве тимомы
отмечается нарастание титров антител к АхР и скелетным мышцам.
Еще одним нервно-мышечным заболеванием, имеющим паранеопластиче-
ский генез, является миастенический синдром Ламберта-Итона. Как и миасте-
ния, это заболевание характеризуется мышечной слабостью, сухостью во рту,
однако, в отличие от миастении, при нагрузке наблюдается «врабатывание» с
уменьшением мышечной слабости. Основной причиной развития этого миасте-
нического синдрома являются опухоли легких, молочной железы и яичников.
Нарушение нервно-мышечной проводимости обусловлено образованием анти-
тел к вольтажзависимым кальциевым каналам нервной ткани на пресинапти-
ческой стороне нервно-мышечного окончания. Вольтажзависимые кальциевые
каналы относят к трансмембранным белкам, состоящим из нескольких субъе-
диниц. Выделяют несколько субтипов и классов кальциевых каналов. При син-
дроме Ламберта-Итона имеют место антитела к P/Q-типам каналов, которые
контролируют высвобождение ацетилхолина в нервно-мышечном окончании,
а также каналам N-типа, которые участвуют в нейротрансмиссии в пределах
ЦНС. Последние встречаются почти исключительно на фоне мелкоклеточного
рака легкого.
Антитела к P/Q-каналам — маркер синдрома Ламберта-Итона, который обла-
дает чувствительностью около 95%. Они замедляют выделение ацетилхолина в
нервно-мышечном окончании, что нарушает работу моторной пластинки. Метод
определения антител к кальциевым каналам — радиоиммунный анализ экстрактов
нервной ткани с лигандом, меченным радиоактивной меткой. Техническая слож-
ность метода ограничивает его клиническое применение.
Антитела к ионным каналам могут встречаться совместно с другими паранео-
пластическими антителами. В 20-30% случаев антитела к кальциевым каналам
P/Q-типа обнаруживают при боковом амиотрофическом склерозе, однако их роль
при этом заболевании не установлена.
142 ДИАГНОСТИКА АУТОИММУННЫХ ЗАБОЛЕВАНИЙ
АНТИТЕЛА К СКЕЛЕТНЫМ МЫШЦАМ И МИОКАРДУ
Хотя антитела к скелетным мышцам и миокарду обнаруживают при разных
патологических процессах, основными антигенами выступают сократительные
белки и поверхностные рецепторы поперечнополосатой мускулатуры. Обе раз-
новидности аутоантител обладают выраженной перекрестной реактивностью и
требуют сочетаного определения.
Антитела к скелетным мышцам при миастении были впервые описаны как
комплемент-фиксирующие антитела, направленные против сократительных эле-
ментов скелетной мышцы. Исследование молекулярных мишеней аутоантител
показало их значительную гетерогенность. Аутоантитела связывают множество
сократительных белков, включая актин, актинин. миозин, титин (коннектин), а
также рианодиновый рецептор, представляющий собой кальциевый канал сарко-
плазматического ретикулума.
Появление аутоантител к скелетной мышце связано с аутоиммунной реакцией
против клеток тимуса. Поликлональные сыворотки и моноклональные антитела,
синтезированные на основе антительного спектра при миастении, связываются
как с антигенами скелетных мышц, так и с миоэпителиальными клетками тимомы.
Эта разновидност ь низкодифференцированных клеток тимуса сохраняет миоид-
ные элементы в цитоплазме, а также экспрессирует фетальный АхР. Экспрессия
а-субъединицы АхР возрастает при тимоме, причем ее синтез отмечается в кор-
тикальных эпителиальных клетках, окруженных незрелыми тимоцитами. Именно
нарушение цитоархитектоники тимуса с дефектом негативной селекции незрелых
CD1' Т-клеток и их постоянный контакт с миоэпителиальными клетками могут
лежать в основе возникновения аутоиммунных ответов при тимоме.
Основным методом определения антител к скелетным мышцам является НРИФ
на криосрезах мышц грызунов или человека. Другие методы обнаружения аутоан-
тител не позволяют охватить широкий спектр антигенов, поэтому используются
лишь для подтверждения. Определение конкретных мишеней аутоантител в кли-
нической диагностике в настоящее время не применяют. Пока гано, что антитела
к титину являются наиболее специфичным маркером пара неопластической миа-
стении, а антитела к рианодиновому рецептору указывают на тяжелое течение и
плохой прогноз заболевания.
Антитела к скелетным мышцам диагностируют методом НРИФ у 30-40% боль-
ных с классической миастенией, при паран еопл логической миастении и на фоне
тимомы они встречаются у 70-80% пациентов. Результат теста является положи-
тельным при обнаружении характерного окрашивания сократительных элементов
скелетной мышиы в титре более 1:20.
В пожилом возрасте встречаемость аутоантител постепенно нарастает, в то время
как у молодых больных миастенией антитела к скелетным мышцам регистрируют
редко (<10%). Поскольку при миастении существует надежный и чувствительный
серологический маркер, такой как антитела к АхР, то антитела к скелетным мыш-
цам имеют вспомогательное значение и используются только в серодиагностике
тимомы. Высокая чувствительность этих ауимнтител позволяет использовать
их для диагностирования тимомы на фоне миастении. Отрицательный результат
серологического теста позволяет с большой вероятностью исключить тимому у
лиц в возрасте 20-60 лет. Антитела к скелетным мышцам обнаруживают у части
пациентов с хронической болезнью «трансплантат против хозяина» при пересадке
костного мозга.
Антитела к миокарду отмечаются при поствирусных миокардитах, дилата-
ционной кардиомиопатии, синдроме Дресслера и ревмокардите. Спектр антиге-
нов антимиокардиальных антител по своей широте сопоставим с разнообразием
антигенов и антител к скелетным мышцам. Основными антигенами являются
а- и [3-изоформы тяжелой цепи сердечного миозина, тропомиозин, ряд митохон-
ДИАГНОСТИКА АУТОИММУННЫХ ЗАБОЛЕВАНИЙ 1 43
дриальных антигенов, в том числе белок-переносчик аденина (ANT) и ферменты
синтеза жирных кислот (BCKD). Рецепторный аппарат миокарда также становится
мишенью антител. Выявляют антитела к р, - адренорецепторам и М2-мускариновым
рецепторам.
Ведущая роль вирусной инфекции в патогенезе аутоиммунного поражения мио-
карда многократно подтверждена. Помимо результатов множества клинических
исследований, посвященных определению нуклеиновых кислот и антигенов в мио-
карде, разработано несколько животных моделей вирусиндуцированного миокар-
дита и дилатационной кардиомиопатии. Классическая модель миокардита, вызы-
ваемого вирусом Коксаки Вг сопровождается индукцией аутоантител к миозину.
При ряде бактериальных инфекционных заболеваний, таких как болезнь Чагаса
и стрептококковая инфекция, наблюдают образование лерекрестно-реагирующих
антител к соединительной ткани и миозину сердца.
Участие аутоантител в патогенезе миокардитов и миокардиопатии остается
спорным. Большая часть антигенов аутоантител является внутриклеточными,
поэтому они недосягаемы для циркулирующих аутоантител. Вероятным пато-
генетическим механизмом представляется стимуляция кардиомиоцитов через
^-адренорецептор, что приводит к нарушению их сократительной функции и раз-
витию тахиаритмий. Частота встречаемости антител к рх-адренорецептору выше у
носителей HLA-DR4, носительство которого ассоциировано с развитием идиопа-
тической дилатационной кардиомиопатии.
Основным клиническим методом обнаружения антител к миокарду счита-
ют непрямую иммунофлюоресценцию на миокарде грызунов или человека.
Выявление титров более 1:20 указывает на наличие клинически значимых ауто-
антител. Аутоантитела по-разному реагируют с миокардиальной тканью, поэтому
выделяют 2 типа окрашивания: сарколеммный и миофибриллярный. Антитела к
скелетной мышце, часто встречающиеся у пожилых лиц, при НРИФ также реагиру-
ют с сократительными белками миокарда и неотличимы от антимиокардиальных
антител. Целесообразно параллельное использование двух субстратов (мышцы и
миокарда) для дифференцировки этих специфичностей.
Аутоантитела к миокарду встречаются часто, чувствительность при дилата-
ционной кардиомиопатии достигает 90%. Однако встречаемость аутоантител у
здоровых доноров крови может достигать 10-20%, что затрудняет использование
аутоантител к миокарду в качестве подхода к дифференциальной диагностике
миокардиопатий.
Отсутствие коммерчески доступных тест-систем для определения антител к
миокардиальному миозину, ANT и p-адренорецептору не позволяет использовать
эти потенциально более специфичные маркеры в клинической практике.
Серологические маркеры при аутоиммунных поражениях кожи
АУТОАНТИТЕЛА ПРИ ПУЗЫРЧАТКЕ
Различают простую (вульгарную), листовидную, эритематозную и вегети-
рующую форму пузырчатки. В основе заболеваний этой группы пузырчатки
лежит образование внутриэпидермальных пузырьков, причиной которых явля-
ется антитело-опосредованное разрушение десмосом — межклеточных структур,
которые объединяют между собой клетки кожи. Десмосомы представляют собой
плотные межклеточные контакты, разрушение которых приводит к образованию
интраэпителиальных полостей и отделению клеток друг от друга (акантолитиче-
ские клетки). Особенностью аутоантител при пузырчатке является способность
активировать систему комплемента, в результате чего антитела имеют прямое
цитотоксическое действие.
144 ДИАГНОСТИКА АУТОИММУННЫХ ЗАБОЛЕВАНИЙ
При выполнении прямой РИФ в биоптатах кожи отмечаются инт раэпидермаль-
ные отложения IgG и Сч-компонента комплемента по межклеточным контактам
(десомомам) эпителиоцитов базальных слоев. Эти отложения напоминают ячейки
рыболовной сети, они обычно линейные, на большом увеличении можно отметить
мелкую гранулярность, которая соответствует морфологическому распределению
«плотных контактов» эпидермиса. Прямая РИФ является золотым стандартом,
поскольку позволяет обнаружить аутоантитела у всех пациентов с пузырчаткой,
в то время как НРИФ имеет чувствительность, не превышающую 90%. Биопсию
целесообразно проводить в области непораженной кожи на границе или рядом с
высыпным элементом. В биоптатах клинически не пораженной кожи часто отме-
чают интраэпидермальные полости с акантолитическими клетками, цитоплазма
которых особенно ярко окрашена иммуноглобулином.
Среди антигенов антидесмосомальных антител наиболее хорошо изучены гли-
копротеины десмосом: десмоглеин 1 и 3. Пациенты с листовидной пузырчаткой
имеют антитела против десмоглеина 1. При вульгарной пузырчатке и при ее гипер-
пролиферативном варианте (вегетирующая пузырчатка) образуются аутоантитела
к десмоглеину 3. Отличия в морфологии и клинической картине листовидной и
вульгарной пузырчатки обусловлены различиями в экспрессии двух основных
антигенов. Так, локализация пузырьков между верхними слоями эпидермиса при
листовидной и нижними слоями при вульгарной пузырчатке обусловлена экспрес-
сией десмоглеина 3 в преимущественно верхних слоях эпидермиса. В то же время
десмоглеин 1 отсутствует в эпидермисе слизистых оболочек, что обусловливает
отсутствие высыпаний на них при листовидной пузырчатке.
При пузырчатке выявляют широкий спектр аутоантител других специфично-
стей, в том числе десмоколлины 1-3, десмоплакины I и II, плакоглобин. аннек-
сины, Е-кадхерин и коллаген XVII. Перспективными аутоантигенами при пузыр-
чатке могут быть ацетилхолиновые рецепторы кератинопитов. Ряд исследований
показывает принципиальную роль этих антител в воспроизведении эксперимен-
тальных моделей пузырчатки.
Два редких пузырных заболевания кожи связаны с интраэпителиальными отло-
жениями IgA. К ним относят субкорнеальный пустулярный дерматоз и интраэ-
пидермальный нейтрофильный дерматоз. При этих заболеваниях основными
антидесмосомальными антигенами являются десмоглеины 1, 3 и десмоколлин 1.
Антитела класса IgA могут быть выявлены с помощью НРИФ на пищеводе обе-
зьяны в 50% случаев.
АУТОАНТИТЕЛА ПРИ БУЛЛЕЗНОМ ПЕМФИГОИДЕ
Пемфигоид представляет собой группу буллезных заболеваний кожи, включаю-
щую собственно буллезный пемфигоид, а также пемфигоид слизистых оболочек,
пемфигоид беременных, IgA-пемфигоид. В отличие от пузырчатки, при пемфигои-
де образуются антитела к гемидесмосомам базального слоя. Эти структуры связы-
вают между собой эпидермис и дерму. Как и при пузырчатке, антитела обладают
способностью фиксировать комплемент. В результате при разрушении гемидес-
мосом полости образуются на границе эпидермиса и дермы. Поскольку структуры
эпидермиса не нарушены, акантолитические клетки отсутствуют.
При прямой РИФ в биоптатах кожи отмечаются линейные отложения IgG по
базальной мембране кожи. Основными антигенами с тужат 2 белка гемидесмосом
массой 230 и 180 кДа. которые получили название BPAG1 и BPAG2 (от англ, bullous
pemphigus antigen). Вопрос о патогенетическом значении аутоантител не решен,
хотя показана возможность пассивного переноса пемфигоида животным с помо-
щью введения аутоантител человека. Тем не менее для образования субэпидер-
мальных полостей необходимо нейтрофильное воспаление с синтезом протеазных
ферментов. Вероятно, основной ролью аутоантител к антигенам гемидесмосом
ДИАГНОСТИКА АУТОИММУННЫХ ЗАБОЛЕВАНИЙ 145
является привлечение воспалительных клеток. Антитела к базальной мембране
кожи не коррелируют с активностью заболевания, поэтому их мониторинг не осу-
ществляется.
Для определения антител в сыворотке крови применяют метод НРИФ на
криосрезах пищевода обезьяны. Положительное окрашивание базальной мем-
браны кожи проявляется в виде яркой линии под клетками базального слоя.
Серологически обнаруживаются антитела у 75% больных с пузырными дерматоза-
ми, в то время как у большинства больных с начальными формами и поражением
слизистых оболочек антитела в крови отсутствуют.
Основной класс иммуноглобулинов при пемфигоиде — IgG (IgGT и IgG4). При
более редкой разновидности — линейном IgA буллезном пемфигоиде — основным
классом является IgA. При этой форме редко можно обнаружить аутоантитела в
крови, поэтому основой диагностики стало исследование биоптатов кожи.
АУТОАНТИТЕЛА ПРИ ДРУГИХ ЗАБОЛЕВАНИЯХ КОЖИ
Отложения IgG отмечаются при буллезной форме многоформной эритемы.
Антигенными мишенями являются белки гемидесмосомы десмоплакины. Они
служат основными конститутивными элементами десмосом и образуют внутри-
клеточную плотную десмосомальную пластину. Больные с антителами к десмопла-
кинам страдают от повторных высыпаний на слизистой оболочке рта и гениталий,
а также от обострений конъюнктивита. Антитела реагируют с эпитопом белка,
ответственным за связь с цитокератинами клетки.
При многоформной эритеме циркулирующие антитела обнаруживают не более
чем в 50% случаев; биопсия кожи является основой диагностики. Для уточнения
характера отложений используют «расщепление» кожи под действием высокой
концентрации NaCl (1М). При многоформной эритеме отложения IgG остаются на
эпидермальной части эпидермиса.
Буллезный эпидермолиз является заболеванием с преимущественным пора-
жением слизистых оболочек. Основной антиген — коллаген VII типа, который
связывает плотную пластинку гемидесмосом базального слоя эпидермиса с глу-
бокими слоями дермы. В сыворотке аутоантитела встречаются нечасто, поэтому
для диагностики применяют биопсию кожи. При использовании «солевого рас-
щепления» отложения иммуноглобулинов при буллезном эпидермолизе остаются
на дермальной части эпидермиса.
Герпетиформный дерматит (болезнь Дюринга) обычно характеризуется везику-
лопустулезной сыпью на фоне скрытых форм целиакии. При выполнении прямой
РИФ отмечают крупногранулярные отложения IgA в вершинах сосочков кожи.
В отличие от других буллезных дерматозов, при которых характер окрашенных
структур идентичен при прямой и непрямой РИФ, при герпетиформном дерматозе
при проведении НРИФ не выявляют антитела к антигенам кожи, но определяют
антитела к эндомизию и ретикулину. Еще одной специфической мишенью аутоан-
тител при герпетиформном дерматите служит эпидермальная трансглутаминаза,
антитела к которой могут иметь перекрестную специфичность с другими транс-
глутаминазами.
Витилиго — еще одно распространенное аутоиммунное заболевание кожи,
сопровождающееся появлениями аутоантител против антигенов кожи. Поскольку
витилиго часто сочетается с рядом аутоиммунных эндокринопатий, при этом забо-
левании можно обнаружить также аутоантитела к антигенам щитовидной железы,
надпочечникам, обкладочным клеткам желудка.
Аутоантитела к меланоцитам, циркулирующие в крови больных витилиго,
обладают цитотоксическим действием на культуры меланоцитов и способны
активировать комплемент. Установлен ряд мишеней аутоантител, среди них наи-
большее значение имеет 40 кДа-антиген витилиго (VIT40) и тирозиназа (69 кДа),
146 ДИАГНОСТИКА АУТОИММУННЫХ ЗАБОЛЕВАНИЙ
участвующая в синтезе меланина. Учитывая очевидную клиническую картину
этого заболевания и простоту постановки диагноза, серологическая диагностика
витилиго не разработана.
ДИАГНОСТИКА РЕВМАТИЧЕСКИХ ЗАБОЛЕВАНИЙ
Диффузные болезни соединительной ткани
Основной серологический тест в иммунологическом обследовании больных с
подозрением на заболевание соединительной ткани — определение АНФ с помо-
щью метода НРИФ. При подозрении на ДБСТ на первом этапе лабораторного
обследования должен быть определен титр АНФ на перевиваемой клеточной
линии НЕр-2. Более чем у 95% пациентов с обострением ДБСТ отмечают диагно-
стические титры этого показателя. Хотя встречаемость АНФ варьирует при диа-
гностике конкретных системных заболеваний, она превышает 90% при СКВ, СШ
и диффузных формах склеродермии, включая CREST-синдром. Несколько реже
АНФ обнаруживают при дискоидной красной волчанке, кожных формах склеро-
дермии. Частота встречаемости АНФ при воспалительных миопатиях, таких как
дерматомиозит и полимиозит, не превышает 50%.
При диагностике СКВ, СЗСТ. СШ и склеродермии отсутствие диагностических
титров АНФ или низкие титры, составляющие 1’40-1:80, делают диагноз ДБСТ
маловероятным. Принимая во внимание особенности метода обнаружения АНФ,
необходимо учитывать, что ряд распространенных антигенов АНА, прежде всего
антигены SS-A-антител, могут утрачиваться из ядра клетки в ходе фиксации.
Поэтому рекомендуется дополнительное обследование антител к экстрагируемому
ядерному антигену методом ИФА или диффузии в геле. В качестве альтернативно-
го варианта можно использовать селективные тесты, направленные на выявление
антител к SS-A антигену.
За счет высокой чувствительности ИФА-методов обнаружения антител к ЭН А
при совместном использовании с АНФ отрицательный результат такого тестиро-
вания позволяет исключить диагноз СКВ, СШ и диффузных форм склеродермии с
вероятностью около 98%.
Определение АНФ на клеточных линиях позволяет уточнить тип свечения
ядра клетки. Выявление высоких титров АНФ с гомогенным типом свечения ядра
клетки в большинстве случаев указывает на диагноз СКВ. Основным антигеном
при гомогенном типе свечения является дсДНК. Гомогенный тип свечения может
отмечаться при диффузной склеродермии и часто обусловлен наличием антител
к Scl-7Q ангигену. Титры гомогенного свечения АНФ при склеродермии обычно
ниже титров, отмечающихся при обострении СКВ,
Среди всех типов свечения при выявлении АНФ у лиц, обследуемых по поводу
ДБСТ, преобладает гранулярный тип. Он отмечается при СКВ, СШ. СЗСТ, склеро-
дермии, РА и других артропатиях. При СКВ и СЗСТ преимущественно устанавли-
вают крупногранулярный тип свечения ядра, связанный с обнаружением антител
к рибонуклеопротеиновым антигенам, таким как Sm и RNP. Поскольку СКВ и СШ
характеризуются выраженной гипергаммаглобулинемией, титры АНФ с грануляр-
ным типом свечения обычно выше, чем при других заболеваниях. Обнаружение
умеренных титров АНФ с гранулярным типом свечения может отмечаться у лиц
без признаков ДБСТ.
Наличие ядрышкового и центромерного типа окрашивания ядра характерно
для различных форм склеродермии. Антитела к центромерам, которые являются
маркером CREST-синдрома, отмечают в очень высоких титрах, обычно превыша-
ющих 1:2560. Диффузные формы склеродермии, протекающие с поражением вну-
тренних органов, практически в 100% случаев сопровождаются высокими титрами
ДИАГНОСТИКА АУТОИММУННЫХ ЗАБОЛЕВАНИЙ 147
АНФ. При локализованной склеродермии АНФ встречается у 30% больных, чаще
отмечается на фоне линейной склеродермии (40%), преимущественно у детей. При
бляшечной склеродерме встречаемость диагностических титров АНФ составляет
только 15%, причем антитела обнаруживают преимущественно у взрослых.
Цитоплазматический тип свечения описывают реже других, обычно он при-
сутствует в невысоких титрах. Нередко ложноотрицательные результаты опреде-
ления антител к антигенам цитоплазмы связаны с тем, что цитоплазматический
тип свечения имеет место наряду с более выраженным окрашиванием ядра клетки.
Большинство антигенов, определяющих цитоплазматический тип окрашивания
ядра, в частности рибосомы, диффузно распределены в ядре клетки, что затруд-
няет их обнаружение. Большинство положительных результатов обнаружения
цитоплазматического типа свечения обусловлено антителами к митохондриям,
которые представляют собой крупные органеллы, хорошо различимые среди дру-
гих структур цитоплазмы.
Тестирование на определение конкретных разновидностей АНА целесообразно
проводить только при получении положительного результата выявления АНФ
или антител к экстрагируемому ядерному антигену. Вероятность обнаружения
диагностических титров конкретных разновидностей АНА при отсутствии или
низких титрах АНФ очень низка. Основными разновидностями АНА, которые
отмечаются при низких титрах АНФ являются антигены SS-A и SS-B. Таким обра-
зом, использование уточняющих тестов целесообразно только при обнаружении
высоких титров АНФ (более 1:160) и/или антител к ЭНА, которые указывают на
присутствие антител к SS-A и SS-B.
Среди всего многообразия АНА у больных ДБСТ рекомендуется определение
антител к наиболее часто встречающимся разновидностям АНА. выявление кото-
рых имеет четкое диагностическое значение. Минимальный набор включает SS- А,
SS-B, Sm, U^-RNP, Scl-70 и Jo-1.
Наиболее объективные и специфичные результаты обследования могут быть
получены при использовании двойной иммунодиффузии и контрэлектрофореза.
Однако эти методы крайне трудоемки и требучот хорошо обученного персонала.
Метод ИФА для обнаружения этих разновидностей АНА является очень чувстви-
тельным, что может приводить к получению ложноположительных результатов. За
счет своей дешевизны, простоты и надежности ь большинстве иммунологических
лабораторий метод лайн-блота постепенно вытеснил другие методы обнаружения
АНА. Определение антител к дсДНК может быть оекомендовано только пациен-
там с СКВ, причем вероятность обнаружения этих антител выше при гомогенном
типе свечения АНФ. При других системных заболеваниях обнаружение антител к
дсДНК не показано. Учитывая высокую встречаемость АКА у больных СКВ, обна-
ружение низких титров АФА может быть использовано в качестве дополнительно-
го диагностического маркера при этом заболевании.
Все пациенты с ДБСТ должны хотя бы однажды быть обследованы для уточне-
ния спектра АНА. Титры всех аутоантител не являются постоянной величиной и
могут значительно меняться с течением времени, что справедливо по отношению
к АНА. присутствующих в сыворотке больных ДБСТ. Повторное обследование
для обнаружения АНФ и других АНА рекомендовано только при СКВ. При дру-
гих ревматических заболеваниях титр АНФ не коррелирует с активностью их
течения; данных, свидетельствующих о целесообразности повторных измерений
АНФ у больных с другими ДБСТ, не существует. Даже если у больнот о с типичной
клинической картиной ДБСТ был обнаружен высокий титр АНФ, то совсем не
обязательно, что дальнейшее иммунологическое обследование позволит устано-
вить конкретную разновидность АНА. Это связано с разнообразием АНА и несо-
вершенством используемых методов их определения.
148 ДИАГНОСТИКА АУТОИММУННЫХ ЗАБОЛЕВАНИЙ
Среди всех форм ДБСТ реже всего АНА встречаются при воспалительных мио-
патиях. Отсутствие АНФ не исключает наличия антител к синтетазам и другим
цитоплазматическим мишеням АНА. Результаты выявления АНФ при подозре-
нии на воспалительные миопатии следует интерпретировать с осторожностью.
Независимо от результатов обнаружения АНФ всем пациентам с ПМ/ДМ необхо-
димо провести специфические тесты, направленные на выявление антисинтетаз-
ных и других характерных разновидностей АНА.
Практически у 25% больных с диагнозом ДБСТ клиническая картина не укла-
дывается в классическое описание конкретного заболевания, и у них одновре-
менно отмечают признаки нескольких заболевании соединительной ткани. Такие
заболевания рассматривают как перекрестные синдромы, которые не могут быть
классидицированы на основании клинических критериев. Их прототипом явля-
ется СЗСТ или синдром Шарпа, речь о котором шла при обнаружении антител к
U^-RNPii который сочетает признаки всех основных заболеваний соединительной
ткани. Обнаружение у больного совокупности разнородных клинических при-
знаков в сочетании с выявлением одновременно иммунологических маркеров
нескольких ревматологических заболеваний заставляет предположить диагноз
перекрестного синдрома.
Тот факт, что при наличии определенных разновидностей АНА с большой
частотой отмечаются определенные клинические признаки и формы системных
заболеваний, послужил толчком к пересмотру определений и классификации
ряда нозологических форм. Существующие исследовательские данные однознач-
но указывают на то, что аутоантитела связаны с иммунопатогенезом системных
заболеваний и отражают особенности иммунного ответа. Это позволяет более
точно очерчивать клинические синдромы, характеризующиеся единым патогене-
зом. Так, на серологических показателях основано выделение ряда перекрестных
синдромов, в частности антисинтетазного синдрома. склеромиозита (табл. 17-22).
В основе современной классификации воспалительных миопатий лежит определе-
ние миозит-специфичных аутоантител.
Таблица 17-22. Серологические маркеры перекрестных синдромов в ревматологии
Признаки ДБСТ в клинической картине Основной серологический маркер
СШ/СКВ (СС/РА) Ro/SS-A, La/SS-B
СЗСТ U,-RNP
сс/пм PM-Scl, Ku
РА/СШ РФ, Ro/SS-A
Ангиосинтетазный синдром (ПМ/ДМ/интерстициальный пнев- монит/полиартрит) Jo-1, другие антисинтетазные антитела
Системные заболевания соединительной ткани, сопровождающиеся образо-
ванием АНА, представляют собой единое поле, объединяющее специфическую
клиническую симптоматику, иммунопатогенез и серологические маркеры. Это
характерно и для других аутоиммунных заболеваний, в частности аутоиммунных
эндокринопатий, при которых часто обнаруживается общность клинической
картины и совпадают иммунологические маркеры. В то же время АНА могут
обнаруживаться при заболеваниях как имеющих ревматическую природу, так
и не имеющих таковой. К последним относятся заболевания печени, вирусные
гепатиты, шизофрения, васкулиты, аутоиммунные тиреоидиты и болезнь Грейвса,
поликистоз, PC, пурпура Шенлейна-Геноха, юношеский РА, атопический дерма-
тит. миастения, паранеопластические синдромы, гиперэозинофильный синдром,
беременность, вирусные и бактериальные инфекции.
Таким образом, системные заболевания соединительной ткани объединяются
иммунологическими и, нередко, клиническими проявлениями с другими ауто-
ДИАГНОСТИКА АУТОИММУННЫХ ЗАБОЛЕВАНИЙ 149
иммунными заболеваниями, что позволяет проследить единые патогенетические
закономерности аутоиммунной патологии.
Иммунологическая диагностика системной красной волчанки
Встречаемость СКВ составляет 1:5000 женщин от 15 до 45 лет, что делает ее
одним из наиболее частых ДБСТ. Полиморфная клиническая картина, отсутствие
абсолютно патогномоничных морфологических и инструментальных критериев
этого заболевания позволяют рассматривать иммунологическое обследование в
качестве основного метода подтверждения диагноза СКВ. При использовании кле-
точной линии НЕр-2 АНФ выявляет АНА более чем у 98% больных с этим заболе-
ванием, что позволило использовать обнаружение АНФ в качестве самостоятель-
ного классификационного критерия СКВ. Серологические показатели, в том числе
антитела к дсДНК и антитела к Sm-антигену, благодаря высокой специфичности
также являются критерием этого заболевания
При применении человеческих клеточных линий в качестве субстрата для обна-
ружения АНА почти у всех больных СКВ фиксируют значительные титры АНА
(>1:160). При обострении отмечают крайне высокие титры АНФ на клеточной
линии НЕр-2, составляющие в среднем 1:1280-1:2540. Зачастую у больных СКВ
титры могут превышать 1:100 000. При успешной терапии титры АНФ снижаются,
составляя в среднем 1:320-1:640.
Отсутствие значимых титров АНФ у отдельных больных, имеющих типичные
клинические симптомы СКВ, является «яблоком раздора» между ревматологами
и сотрудниками лаборатории, что заставляет выделять условную группу пациентов
с «АНФ-негативной СКВ», При использовании тканей лабораторных животных,
таких как почка или печень крысы, до 10-15% больных, удовлетворяющих кри-
териям для диагноза СКВ, не имеют диагностических титров АНФ в сыворотке
крови. Одной из основных разновидностей АНА, которые обнаруживают у таких
пациентов, являются антитела к 55-А-антигенам. Эти антигены обладают хорошей
растворимостью и могут утрачиваться из ядер клеток.
Хотя применение в качестве субстрата клеточной линии НЕр-2 значительно
сокращает численность этой категории пациентов, серонегативными остаются до
5% больных. Необходимо учитывать, что при использовании меганол-ацетоновой
фиксации НЕр-2-клеток SS-A-антиген может диффундировать в цитоплазму клет-
ки, в результате чего титры АНФ значительно занижаются.
Для устранения этих недостатков была создана трансгенная клеточная линия
НЕр-2000, в генотип которой был искусственно внесен ген 55-А-антигена. За счет
этого в ее ядре отмечается гиперэкспрессия SS-A-антигена, что увеличивает чув-
ствительность выявления АНА. Однако даже использование трансгенных клеточ-
ных линий не позволяет окончательно избавиться от проблемы «АНФ-негативной
СКВ». Ложноотрицательные результаты определения АНФ могут быть получены
в присутствии АНА к антигенам цитоплазмы — рибосомальным антигенам и
антисинтетазным антигенам (Jo-1), а также в случае присутствия антител к фос-
фолипидам. Существует еще одна группа больных СКВ, у которых не отмечается
АНФ. Люди с врожденной недостаточностью компонентов комплемента (С2 и С4)
могут страдать кожным волчаночноподобным заболеванием с выраженной фото-
чувствительностью. При лабораторном обследовании у них отсутствуют сколько-
нибудь значимые титры АНФ и антитела к дсДНК, хотя в ряде случаев у них могут
быть обнаружены антитела к Ro/SS-A.
Определение типа свечения АНФ играет большую роль в определении прогно-
за при СКВ. Встречаемость конкретных типов свечения зависит от обследуемых
контингентов пациентов. При обследовании смешанных популяций, включающих
1 50 ДИАГНОСТИКА АУТОИММУННЫХ ЗАБОЛЕВАНИЙ
как кожные, так и системные формы заболевания, гомогенный и гранулярный тип
свечения отмечаются с одинаковой частотой.
Гомогенный тип свечения обусловлен присутствием антител к дсДНК и характе-
ризуется более агрессивным течением и большей вероятностью поражения почек.
Наряду с антителами к дсДНК, при гомогенном типе свечения обнаруживают
антитела к нуклеосомам и антиэндотелиальные антитела, которые также участву-
ют в патогенезе волчаночного нефрита.
Выделяют 2 основные разновидности гранулярного типа свечения у больных
СКВ. Крупногранулярный тип свечения присутствует у пациентов с антителами к
компонентам сплайсосом Sm и RNP. Наличие этих вариантов АНА сопровожда-
ется тяжелым течением заболевания с большой вероятностью поражения ЦНС,
цитопениями, выраженным синдромом Рейно. Мелкогранулярный тип свечения
обусловлен присутствием антител к SS-A и SS-B-антигенам, характеризуется
преимущественно кожными формами заболевания с выраженной фоточувстви-
тельностью.
Тест волчаночной полоски высокоспецифичен для системных форм СКВ и
преимущественно отмечается у пациентов с поражением почек и вторичным АФС.
В то же время обнаружение антител к фосфолипидам при СКВ не всегда сопут-
ствует клинической картине АФС. Низкие титры АКА зафиксированы у 40-50%
больных СКВ и представляют собой дополнительный диагностический критерий
этого заболевания.
Для оценки активности заболевания могт1 быть использованы титры АНФ,
антител к дсДНК, антител к нуклеосомам, антиэндотелиальных антител, содер-
жание сывороточных иммуноглобулинов и гаммаглобулинемия, содержание ком-
понентов системы комплемента, изменение уровней ЦИК и СОЭ. В отличие от
других заболеваний, концентрации С-реактивного белка при СКВ обычно лежат
в пределах нормы. Увеличение концентраций С-реактивного белка при СКВ ука-
зывает на присоединение инфекционного процесса и требует активного поиска
очагов скрытой инфекции.
Диагностика антифосфолипидного синдрома
АФС — аутоиммунное заболевание, поражающее преимущественно женщин
молодого возраста, проявляется рецидивирующими тромбозами, привычным
невынашиванием беременности, тромбоцитопенией и стойким наличием анти-
фосфолипидных антител. Диагноз АФС устанавливают на основании соответ-
ствия клинических и лабораторных находок диагностическим критериям этого
заболевания. В Сиднее в 2006 г. произошел последний пересмотр критериев этого
заболевания. На основе австралийских критериев в настоящее время строится
практическая диагностика АФС (табл. 17-23).
Таблица 17-23. Критерии антифосфолипидного синдрома, 2006 г.
Антифосфолипидный синдром может быть диагностирован, если у больного подтвержден один из клини-
___________________________ческих и один из лабораторных критериев заболевания’_
____________________________________________________________________________Клинические критерии________________________________________________________
» Сосудистый тромбоз"_________________________________________________________
Один или несколько эпизодов'" артериального, венозного тромбоза или тромбоза мелких сосудов"" в
любой ткани или органе. Тромбоз должен быть подтвержден КТ/МРТ, допплеровским исследованием или
морфологически. При морфологическом исследовании тромбоз не должен сочетаться с воспалительными
изменениями стенки сосуда________________________________________
• Невынашивание беременности
❖ Одна или более смерть плода от 10 и более нед беременности, подтвержденная нормальной
морфологией плода при ультразвуковом исследовании или морфологически
ДИАГНОСТИКА АУТОИММУННЫХ ЗАБОЛЕВАНИЙ 151
Окончание табл. 17-23
❖ Одни или более преждевременные роды морфологически нормального новорожденного на сроке от
34 нед беременности в результате тяжелой преэклампсии, эклампсии или плацентарной недостаточно-
сти.................................................................................
< ► Три или более необъяснимых спонтанных аборта на сроке до 10 нед беременности при исключении ана-
томических, гормональных и генетических причин невынашивания________________________
Лабораторные критерии*****__________________________________________________________
• Волчаночный антикоагулянт, обнаруженный два или более раз, с промежутком между исследованиями
не менее 12 нед с помощью комплекса из скринингового, подтверждающего и корректирующего коа-
гулогических тестов в соответствии с требованиями Международного общества изучения тромбозов и
гемостаза___________________________________________________________________________
• Антикардиолипиновые антитела классов IgG и/или IgM в сыворотке или плазме в среднем или высоком
титре (т.е. >40 GPL или MPL, или более 99 перцентиля здоровой популяции), повторно обнаруженные не
менее чем через 12 нед, выявленные при помощи стандартизованной ИФА тест-системы____
• Антитела к ^-гликопротеину классов <gG и/или IgM в сыворотке или плазме в титре более 99 перценти-
ля здоровой популяции, обнаруженные при помощи стандартизованной ИФА тест-системы___
Примечания экспертов к тексту критериев
* Необходимо избегать установки диагноза АФС, если положительные лабораторные результаты
и клинические проявления разделяют менее 12 нед или более 5 лет.
* * Присутствующие врожденные или приобретенные факторы риска тромбоза не являются при-
чиной исключения АФС у больного. Однако рекомендуется выделять отдельные группы паци-
ентов в зависимости от (а) отсутствия и (Ь) наличия дополнительных факторов риска тромбоза.
Такими факторами риска являются возраст (>55 у мужчин. >65 у женщин), наличие любых уста-
новленных факторов риска сердечно-сосудистых заболеваний (гипертензии, диабета, повышен-
ного холестерина ЛНП или низкого холестерина ЛВП, курения, наследственности, указывающей
на раннее начало сердечно-сосудистых заболеваний в семье, индекса массы тела более 30 кг/м2,
микроальбуминурии, снижения СКФ, врожденных тромбофилий, приема пероральных контра-
цептивов. нефритического синдрома, опухоли, иммобилизации и хирургической патологии).
* ** Эпизод тромбоза в прошлом может рассматриваться как положительный критерий, если он
был объективно подтвержден клинически.
* *** Тромбоз поверхностных ьен не включен в клинические критерии.
* **** Общепринятые признаки плацентарной недостаточности включают (1) отрицательные при-
знаки жизнедеятельности плода, (2) плохие формы кривой допплерограммы сосудов, указываю-
щие на признаки гипоксемии плода, (3) олигогидрамнион с индексом амниотической жидкости
менее 5 см, (4) постнатальный вес плода менее 10 перцентиля срока гестации.
* ***** Эксперты рекомендуют отдельно классифицировать больных АФС на следующие катего-
рии: (1) присутствует более 1 лабораторного критерия в любой комбинации, (2а) изолирован-
ный волчаночный антикоагулянт, (2b) только антитела к кардиолипину, (2с) только антитела к
р2-гп.
Выделяют несколько классификационных форм АФС. Прежде всего это первич-
ный или идиопатический АФС. возникающий у лиц моложе 45 лет. После этого
возраста большинство случаев АФС рассматривают как вторичный процесс по
отношению к основному заболеванию. Вторичный АФС развивается на фоне СКВ.
СШ, склеродермии, опухолей, ВИЧ и ВГС, приема лекарственных препаратов.
Еще одной разновидностью заболевания является катастрофический АФС, кото-
рый протекает как первая фаза ДВС с развитием полиорганной недостаточности.
В случае катастрофического ДВС диагноз часто ставят посмертно, в результате
обнаружения крайне высоких титров АКЛА. В то же время серонегативный АФС
характеризуется отсу гствием АКЛА при характерной клинической картине. В слу-
чае перенесенного эпизода тромбоза целесообразно проводить измерение АКЛА и
антител к р2-ГП через 3 нед, поскольку тромботический эпизод может быть связан
с потреблением больших количеств аутоантител.
Основу клинических признаков АФС составляют повторные венозные тромбо-
зы, локализуемые наиболее часто в глубоких венах голеней, что может ослож-
няться тромбоэмболией легочной артерии. Однако тромбозы могут развиваться
также в поверхностных венах и в любом другом венозном бассейне. Артериальные
тромбозы проявляются гангреной конечностей, инсультами, окклюзией аорты с
152 ДИАГНОСТИКА АУТОИММУННЫХ ЗАБОЛЕВАНИЙ
асептическим некрозом головок бедренной кости, инфарктом миокарда, почек и
других внутренних органов. Тромбы могут образоваться на клапанах и в полостях
сердца. Кожные проявления заключаются в язвах на голенях, кровоизлияниях
в подногтевом ложе и сетчатом ливедо. Сетчатое ливедо в виде поверхностной
синеватой венозной сеточки на голенях и бедрах особенно хорошо видно после
охлаждения.
Необъяснимые спонтанные аборты на ранних сроках или внутриутробная
гибель плода во II—III триместрах могут возникнуть при АФС без или на фоне
тяжелой преэклампсии. Тромбоцитопения при АФС умеренная, редко приводит
к осложнениям и не требует назначения лечения. Однако идиопатическая тром-
боцитопения, как и гемолитическая анемия, служат показанием к назначению
лабораторных тестов, направ пенных на выявление антифосфолипидных антител.
Клиническии спектр симптомов, которые связывают с АФС, достаточно широк
(табл. 17-24), большинство их них не были включены в итоговые критерии диа-
гностики АФС (2006), но активно обсуждаются в литературе.
Таблица 17-24. Клинические признаки антифосфолилидного синдрома
Проявления АФС Симптоматика Встречаемость, %
Периферический тромбоз Тромбоз глубоких вен, тромбоз артерий/вен 64
Невынашивание беременности Ранний/поэдний выкидыш, ранние роды 63
Ревматические жалобы Артралгия артрит 68
Неврологические проявления Мигрень, инсульт 66
Кожные проявления Livedo reticularis, язвы ног 40
Гематология Тромбоцитопения Гемолитическая анемия 30
Сердечные проявления Утолщение клапанов Инфаркт миокарда 27
Легочные пооявления ТЭЛА. легочная гипертензия 20
Отдельную клиническую форму АФС представляет собой катастрофический
АФС, или синдром Асерсона (Asherson’s syndrome), характеризующийся острой
полиорганной недостаточностью из-за внезапного развития множественных
тромбозов в микроциркуляторном русле различных органов. Опасность развития
катастрофического синдрома выше у женщин с па гологией беременности во время
инфекционных заболеваний, травм, в том числе после даже нетяжелых хирур-
гических вмешательств. Летальность при катастрофическом синдроме достигает
50% и существенно выше при таких осложнениях, как синдром диссеминирован-
ной коагуляции и/или системный воспалительный ответный синдром, которые
наблюдают в 15-20% случаев при данной форме заболевания. В терапии катастро-
фического синдрома используют прямые антикоагулянты, обменные переливания
замороженной плазмы, введение иммуноглобулина, высокие дозы стероидов,
антибиотиков, анти-СЭ20 моноклональные антитела. Для профилактики ослож-
нений при хроническом АФС назначают непрямые антикоагулянты, главным
образом варфарин, для поддержания МНО в пределах 2,0-3,0, что уменьшает риск
развития рецидивирующего венозного тромбоза на 80-90% независимо от нали-
чия антифосфолипидных антител. У лиц, перенесших инсульт, с единственным
положительным тестом ацетилсалициловая кислота и варфарин оказались одина-
ково эффективны в предупреждении повторных инсультов. Риск развития новых
тромбозов умеренно увеличен до 10% в год у женщин с повторными потерями
плода и оказывается более 10% у пациентов с венозными тромбозами, прекратив-
ших принимать антикоагулянты.
ДИАГНОСТИКА АУТОИММУННЫХ ЗАБОЛЕВАНИЙ 1 53
Серодиагностика АФС остается сложной проблемой. Положительные резуль-
таты определения АКЛА нередко отмечаются у клинически здоровых лиц, в том
числе пожилого возраста, при нормально протекающей беременности и при воспа-
лительных заболеваниях. В подавляющем большинстве подобных случаев присут-
ствие антител в невысоком титре — преходящий феномен, не ведущий к формиро-
ванию АФС, в этих случаях эпитоп отличен от эпитопа, типичного для АФС. Тесты
АКЛА, антитела к р,-ГП 1 и ВА предпочтительно выполнять совместно, так как
в случае положительного результата всех тестов увеличивается прогностическая
значимость обследования в отношении повторных тромбозов. Строгая значимая
связь обнаружена в нескольких ретроспективных исследованиях между тромбо-
зом и положительными результатами всех тестов (АКЛА антитела к р2-ГП 1 и
ВА) по сравнению с больными, которые оказались положительными в отношении
одного или двух тестов. Кроме того, имеются наблюдения, что пациенты с поло-
жительными результатами нескольких тестов сохраняют повышенные значения
при повторных исследованиях, что повышает вероятность у них диагноза АФС.
Проблема современной лабораторной диагностики заключается в том, что не все
тесты в равной степени стандартизованы и чувствительны, недостаточно и каче-
ство межлабораторных и внутрилабораторных стандартов. Назначение антикоа-
гулянтов препятствует обследованию пациентов. Так, АЧТВ нельзя использовать
для скрининга В А, если больной получает антикоагулянты по поводу имеющегося
тромбоза.
Для иммунологического подтверждения АФС требуется в каждом случае про-
ведение более одного теста, уровень антител должен был высоким, эти значения
должны сохраняться при повторном исследовании через 12 нед. Однако имеются
исключения. Так, при привычном невынашивании беременности с прогностиче-
ской целью достаточно определения антител к кардиолипину. Важно, что, согласно
существующим критериям диагностики АФС, для подтверждения этого аутоим-
мунного заболевания требуется обязательное сочетание хотя бы одного иммуно-
логического и одного клинического признака.
Обнаружение антител к фосфолипидам и развитие вторичного АФС — срав-
нительно частое явление у больных СКВ. До 50% больных с волчанкой в стадии
обострения имеют повышенное содержание АФА в крови. Высокие титры АФА
обнаруживают у 20% больных СКВ, их присутствие сопровождается клиниче-
скими проявлениями АФС: повышенной частотой тромботических осложнений,
значительным риском развития неврологических проявлений заболевания, син-
дрома Рейно и сетчатого ливедо. У больных СКВ, имеющих положительный ВА
и АКЛА, наблюдают высокую частоту выкидышей, венозных и артериальных
тромбозов, тромбоцитопении и анемии, легочной гипертензии и эндокардита
Либмана- Сакса, сетчатого ливедо с или без кожных изъязвлений. Высокие титры
АФА служат основой включения больного СКВ в группу риска по развитию тром-
ботических осложнений.
В подавляющем числе случаев серологические маркеры АФС обнаруживают
при СКВ, они относятся к случаям вторичного АФС при системных заболеваниях.
Распространенность первичного АФС в популяции до настоящего времени не уточ-
нена и должна быть скорректирована в связи с неспецифичностью обнаружения
АФА. У некоторой части пациентов формирование АФС может предшествовать
появлению признаков первичного соединительнотканного заболевания, как СКВ,
так и склеродермии, смешанного соединительнотканного заболевания, РА. Нужно
включать определение АНФ в качестве обязательного серологического теста при
обследовании пациентов с предполагаемым АФС. Частота поражения органа зре-
ния при соединительнотканных заболеваниях требует включить в обследование на
АФА для исключения данного синдрома больных с внезапными тромботическими
поражениями сосудов глаз. Однако даже позитивные результаты выявления АФА
154 ДИАГНОСТИКА АУТОИММУННЫХ ЗАБОЛЕВАНИЙ
требуют исключения других причин тромбоза, особенно в случае наличия других
факторов риска, в том числе курения, гиперлипидемии, атеросклероза, варикозной
болезни, приема пероральные контрацептивов.
Диагностика ревматоидного артрита
РА представляет собой наиболее часто встречающееся воспалительное заболе-
вание суставов. В североевропейской популяции его встречаемость достигает более
1%. РА представляет системное аутоиммунное заболевание, основным признаком
которого является воспаление синовиальных оболочек суставов, характеризуется
неумолимым прогрессирующим течением, хотя периоды ярких клинических про-
явлений чередуются с периодами относительного улучшения. Последствиями РА
является деградация суставного хряща с последующей эрозией косги, приводящие
к значительному разрушению сустава с потерей его функции. Помимо клинической
картины деформирующего артрита, у 20% больных отмечают внесуставные прояв-
ления заболевания. Диагноз РА устанавливают на основании клинических данных,
иммунологические тесты включены в качестве одного из 7 пунктов классификаци-
онных критериев РА Американского колледжа ревматологов (табл. 17-25)
Таблица 17-25. Классификационные критерии ревматоидного артрита 1988 г.
Утренняя скованность
Активный артрит более 3 суставов на момент обследования_________________________
Артрит_ суставов кисти__________________________________________________________
Симметричное поражение суставов_________________________________________________
Ревматоидные узелки_____________________________________________________________
Наличие ревматоидного фактора в сыворотке
Рентгенологические признаки периартикулярных поражений кости____________________
Классификация основывается на 7 критериях. Для постановки диагноза РА требуется 4 из 7 пере-
численных критериев. Критерии 1-4 должны персистировать не менее 6 мес.
Иммунологическое обследование особенно актуально на ранних стадиях болез-
ни, когда оно является единственным доступным методом объективизации кли-
нических предположений. Часто нетипичная клиническая картина в начале
заболевания, например олигоартрит и отсутствие костных эрозий, несмотря на
длительность жалоб пациента, а также тот факт, что применение активной терапии
в дебюте заболевания может замедлить быстрое развитие деформаций суставов,
делает лабораторные тесты чрезвычайно ценным инструментом дифференциаль-
ной диагностики. Корректный диагноз РА, поставленный на раннем этапе, служит
основой эффективной иммуносупрессивной терапии.
Дешевизна и доступность определения РФ по-прежиему делают его первым
серологическим тестом в обследовании пациентов с заболеваниями суставов.
Определение РФ в клинических лабораториях основано на использовании методов
латексагглютинации и турбидиметрии. Хотя турбидиметрия представляет собой
более чувствительный метод, однако ее сравнительно высокая чувствительность
снижает специфичность обследования. Целесообразно использовать в качестве
скрининга латексный метод определения РФ, что значительно удешевляет обсле-
дование больных. При этом чувствительность латексного теста должна составлять
не более 20-25 МЕ/мл. Положительные результаты латексных тестов необходимо
подтвердить количественными турбидиметрическими методами.
Отсутствие РФ при первом обследовании не исключает диагноза серонегатив-
ного РА. Однократного определения РФ на ранней стадии РА, принесшего отри-
цательный результат, недостаточно для того, чтобы исключить ранний РА. Если
диагноз РА подозревают или он даже клинически подтвержден при отрицательном
результате теста по определению РФ, требуется повторное определение ею титра
ДИАГНОСТИКА АУТОИММУННЫХ ЗАБОЛЕВАНИЙ 155
через 12 мес для определения серопозитивной формы этого заболевания. Этот
срок приблизительно соответствует времени обновления пула плазматических
клеток, способных синтезировать аутоантитела. Если же будет получен положи-
тельный результат, то нет реальных предпосылок повторять определение РФ с
течением времени.
Благодаря высокой специфичности наилучшими параметрами для ранней диа-
гностики обладают АЦЦП, так как их выявление имеет высокий фактор риска,
т.е. обнаружение повышает вероятность заболевания в 13 раз. В то же время мак-
симальной чувствительностью обладают АМЦВ, поэтому они могут быть реко-
мендованы для скрининга РА и исключения диагноза при неясной клинической
картине.
Хотя данные о применении RA33 в прогнозировании мягкого течения РА тре-
буют дальнейшего подтверждения, антитела к RA33 мот быть использованы
для диагностики серонегативного Ра. Совместное использование антител RA33 и
АМЦВ при серонегативном РА позволяет добиться чувствительности 81%.
Для оценки прогноза используют определение АК А, которые «предсказывают»
развитие деструктивного процесса в суставах. Пациенты с высокими титрами АКА
требуют более активной иммуносупрессивной терапии. Среди других методов
оценки прогноза РА — исследование РФ класса IgA методом ИФА. Большое время
полужизни сывороточного иммуноглобулина, а также постоянство иммунных
ответов не позволяет использовать серологические маркеры для мониторинга
течения заболевания.
ПРОГНОЗИРОВАНИЕ ИСХОДОВ РЕВМАТОИДНОГО АРТРИТА
Исходом РА является прогрессирующее снижение функциональной способ-
ности суставов, инвалидизация и сопутствующее снижение продолжительности
жизни пациентов. Средняя продолжительность жизни больных РА, не получавших
эффективного лечения, значительно меньше средней продолжительности жизни в
сопоставимой популяции. В то же время агрессивность и скорость прогрессирова-
ния РА широко варьируют. Общепризнано, что это заболевание весьма гетероген-
но — оно может протекать щадяще, в течение десятилетий не приводя к значимым
деструкциям, у одних больных, в то время как у других анкилоз суставов и инвали-
дизирующие деструкции развиваются за несколько первых лет заболевания.
На утрату функции сустава влияет степень местного воспаления и возникно-
вение признаков деструкции суставов. Персистирующее воспаление в суставной
полости является основной причиной развития костных изменений. Важным
прогностическим фактором считают раннее назначение базисной терапии, пода-
вляющей специфические иммунные ответы и местное воспаление в суставах, а
также препятствующее формированию органических изменений. Характерно, что
с течением времени лабораторная и клиническая активность заболевания снижа-
ются у большинства больных, в то время как выраженность костных изменений в
суставах нарастает.
Причина такой гетерогенности определяется генетическими факторами и
связана с определенным набором генов иммунной системы. Среди больных с РА
отмечается высокая частота встречаемости гомозигот по HLA-DR4 и DR1-аллелям
МНС, особенно В1*0401. Было обнаружено, что в генах HLA, ассоциированных с
развитием РА, присутствует определенная последовательность аминокислот, так
называмый общий эпитоп (shared epitope), которая входит в вариабельную петлю
3 (VR^) р-цепи HI А II, Определение этого эпитопа может быть использовано в
диагностике РА. Появление специфических аутоантител при РА определяется осо-
бенностями иммунной системы пациента и тесно связано с HLA-генотипом боль-
ного. Имеется тесная взаимосвязь между встречаемостью аллелей HLA-DR (DR4)
и выявлением РФ. АПЦП и антифилаггриновых антител. Аллель HLA-DRB104*01
156 ДИАГНОСТИКА АУТОИММУННЫХ ЗАБОЛЕВАНИЙ
обладает высокой аффинностью к цитруллинированным эпитопам филаггрина,
что указывает на тесную связь между генотипом МНС и образованием АФА.
В то же время патогенетическая роль аутоантител при РА остается спорной.
Существование серонегативных форм заболевания, значительные вариации в
содержании аутоантител у больных не позволяют сделать однозначного заключе-
ния об их значении в поражении суставов. Тем не менее гуморальный иммунитет
тесно связан с патогенезом РА, аутоантитела отражают определенные особенности
патогенеза заболевания. Одним из наиболее «элегантных» доказательств важ-
ности гуморального иммунитета в патогенезе РА стал успешный опыт лечения
больных РА с помощью ритуксимаба (МАБТЕРА) — терапевтического гумани-
зованного антитела, направленного против СО20-маркера В-клеток. Элиминация
В-клеток вызывает клинико-лабораторную ремиссию у 60% больных РА. Кроме
того, наблюдалось снижение уровней ау гоантител (РФ и АФА) в сыворотке крови,
что тесно коррелировало с динамикой и активностью заболевания, и рецидив
заболевания сопровождался увеличением титров сывороточных аутоанти тел.
Этот факт указывает на важность определения сывороточных концентраций
аутоантител для оценки клинических и иммунопатогенетических особенностей
заболевания. Наличие аутоантител при РА может отражать специфическую
реактивность иммунной системы, серологический статус больного может опреде-
лять особенности заболевания и указывать на его исход. Это делает возможным
использование аутоантител для выделения группы больных с неблагоприятным
прогнозом.
Общепризнано, что серопозитивный по РФ РА харакз еризуется более тяжелым
поражением суставов и чаще сопровождается экстрасуставными проявлениями по
сравнению с cepoHei ативным РА. Обнаружение РФ при РА в концентрации более
40 Ш/мл в дебюте заболевания — один из факторов неблагоприятного прогноза
РА, связанный с более ранним развитием суставных деструкций и инвалидизацией
пациента в то время как у серонегативных больных заболевание протекает более
легко. Ревматоидные узелки и другие проявления васкулита отмечают исключи-
тельно у серопозитивных пациентов Более того, инвалидизация и смертность от
РА выше именно в группе серопозитивных больных. Все же частая встречаемость
деструктивного полиартрита у пациентов без РФ указывает на недостатки исполь-
зования РФ в качестве единственного маркера тяжести течения РА.
АЦЦП, по-видимому, присущи основные характеристики антифилагтриновых
антител: они обнаруживаются на ранних стадиях РА и в целом более типичны для
подгруппы больных с быстрым рентгенологическим прогрессированием. АЦЦП
позволяют предсказать развитие деструкций приблизительно у 2/3 пациентов с
деструктивным процессом и в сочетании с РФ представляют важный инструмент
для распознавания больных РА с неблагоприятным прогнозом заболевания уже на
этапе постановки диагноза.
Другим важным фактором является раннее назначение базисной терапии,
получившее название «перевернутой терапевтической пирамиды», при которой
более активные препараты назначают раньше. Время начала базисной терапии
от момента возникновения симптомов заболевания являе гея одним из основных
факторов, влияющих на дальнейшее формирование деструкций суставов. Ранняя
«агрессивная» терапия невозможна без достоверной диагностики РА. Базисные
препараты небезопасны, их назначение связано с большим числом побочных
эффектов. В то же время традиционно используемые критерии РА Американского
колледжа ревматологов недостаточно чувствительны для диагностики раннего РА.
Высокая встречаемость и прогностическое значение АЦЦП позволили вкаючить
их в разрабатываемые критерии раннего РА. Применение этих критериев позволя-
ет как поставить диагноз РА, так и установить прогноз развития деструкций суста-
вов, причем антитела к циклическому цитруллин-содержащему пептиду обладают
ДИАГНОСТИКА АУТОИММУННЫХ ЗАБОЛЕВАНИЙ f57
наибольшим удельным весом для диагностики эрозивного артрита. Таким обра-
зом, серологические тесты — одна из основных возможностей объективизации
диагноза, они могут служить основой для раннего назначения высокоэффектив-
ной базисной терапии.
Другие формы воспалительных артритов
Ювенильный ревматоидный артрит (ЮРА) представляет отдельную группу
ревматических заболеваний. Основными маркерами ЮРА, особенно поли- и оли-
гоартикулярной формы, служат РФ и АНФ. Серопозитивная по РФ полиартрити-
ческая форма ЮРА наиболее близко напоминает РА взрослых. При полиартрити-
ческой форме ЮРА диагностические титры АНФ отмечают у 25% больных.
При олигоартикулярной форме ЮРА, особенно при раннем начале заболевания,
АНФ встречается гораздо чаще, и его встречаемость достигает 70-80%. Эта форма
заболевания возникает преимущественно у младенцев женского пола и сопро-
вождается артритом и высоким риском раз1 ития аутоиммунного иридоциклита.
Определение АНФ может быть использовано при дифференциальной диагностике
гематогенного остеомиелита и олигоартикулярной формы ЮРА. Если увеличение
сывороточных уровней острофазовых маркеров С-реактивного белка в сочетании
с выраженной лихорадкой свидетельствует, скорее, о септическом поражении
сустава у детей, то затяжное течение артрита с низким острофазовым ответом
является показанием к определению АНФ. Наряду с АНФ и РФ при подозрении на
ЮРА необходимо определять иммуноглобулины сыворотки, поскольку гипергам-
маглобулинемия является независимым маркером ЮРА.
Спондилоартропатии представляют группу заболеваний, характеризующуюся
поражением суставов осевого скелета. Совокупная встречаемость этих заболева-
ний в популяции составляет около 2%, что позволяет отнести их к одним из наи-
более часто встречающихся ревматических заболеваний. Спондилоартропатии у
детей и взрослых требуют иммуногенетического обследования для обнаружения
антигена HLA-B27. Наиболее тесно ассоциированы с HLA-B27 анкилозирующий
спондилит (95%), синдром Рейтера (80%) и недифференцированные спондилоар-
тропатии (70%). При артритах на фоне воспалительных заболеваний кишечника,
псориатическом арз рите и реактивном артрите встречаемость HLA-B27 не превы-
шает 30%.
Большинство спондилоартропатий возникает на фоне воспалительного пора-
жения слизистой оболочки и кожных покровов, ч го ведет к увеличению объемов
антигенного материала, поступающего из внешней среды. В основе иммунопато-
генеза всех состояний лежит нарушение иммунологической толерантности при
презентации антигенов, опосредованной HLA-B27. Присутствие генетической
предрасположенности к заболеванию приводит к возникновению аутоиммунного
артрита даже тогда, когда индуцирующий фактор уже перестал действовать.
Аутоиммунные васкулиты
Под аутоиммунными васкулитами понимают заболевания, связанные с некро-
тизирующим воспалительным поражением сосудистой стенки без инфекционной
или токсической причины. Васкулиты подразделяют на первичные и вторичные,
которые возникают на фоне другого аутоиммунного заболевания. Хотя существует
ряд удобных клинических классификаций васкулитов, общепринятой является
международная классификация первичных васкулитов, разработанная в Чапелл-
Хилл в J994 г. Эта классификация дала определение 10 формам первичных
васкулитов и подразделила их в соответствии с калибром поражаемых сосудов
(см. табл. 17-13).
158 ДИАГНОСТИКА АУТОИММУННЫХ ЗАБОЛЕВАНИЙ
Ч1А-А *’
Однако, помимо морфологического значения, данная классификация предо-
ставляет ключ к диагностике этих заболеваний с помощью иммунологических
тестов (рис. 17-1). До недавнего времени отсутствие других диагностических
подходов, кроме клинических и морфологических, в значительной мере затруд-
няло их выявление. Это нередко приводило к диагностическим ошибкам в связи
с полиморфной клинической картиной и сходством симптоматики с системными
заболеваниями соединительной ткани и другими неревматическими заболева-
ниями. Проведение иммунологического обследования на раннем этапе позволяет
сократить время постановки окончательного диагноза и избавить от анализа зача-
стую неоднозначных клинических данных.
Рис. 17-1. Алгоритм использования иммунологических тестов в диагностике системных васку-
литов.
Абсолютно специфических серологических маркеров при этих заболевани-
ях не описано. Патогенез васкулитов крупных и средних сосудов опосредован
преимущественно Т-клетками. Антиэндотелиальные антитела с различной часто-
той встречаются при аортоартериите Такаясу и болезни Кавасаки, а также при
ряде васкулопатий, таких как гемолитико-уремический синдром, тромботическая
тромбоцитопеническая пурпура, болезнь Бехчета. Васкулиты крупных сосудов
сопровождаются выраженным воспалительным ответом, установление которого
представляет основу лабораторных методов их диагностики, наряду с морфоло-
гическим и инструментальным обследованием. Васкулиты сосудов среднего кали-
бра, прежде всего узелковый полиангиит, являются инфекционно-зависимыми и
не сопровождаются образованием специфических аутоантител. В соответствии с
иммунопатогенезом васкулиты мелких сосудов могут быть подразделены на грану-
лематозные и иммунокомплексные.
Гранулематозные васкулиты мелких сосудов, включающие синдром Вегенера,
микроскопический иолиангиит, синдром Черджа-Стросс, имеют ряд общих черт, к
которым относят поражение почек по типу олиго иммунного экстракапиллярного
гломерулонефрита с полулуниями, полиневрит и легочный капиллярит. Основным
методом серологической диагностики гранулематозных васкулитов служит обна-
ружение АНЦА. Антитела этого семейства реагируют с белками азурофильных
ДИАГНОСТИКА АУТОИММУННЫХ ЗАБОЛЕВАНИЙ 159
гранул нейтрофильных гранулоцитов. Значение АНЦА в патогенезе и диагностике
гранулематозных васкулитов позволило выделить отдельную группу, обобщенную
под названием «АНЦА-ассоциированные васкулиты». К ним относят гранулема-
тоз Вегенера, синдром Черджа-Стросс, микроскопический полиартериит, быстро-
прогрессирующий гломерулонефрит с полулуниями. К этой группе заболеваний
иногда причисляют синдром Гудпасчера, поскольку АНЦА могут отмечаться у
10-15% пациентов с этим заболеванием совместно с антителами к базальной
мембране клубочка, определение которых — основной лабораторный метод его
диагностики. Хотя синдром Гудпасчера не является системным васкулитом, он
характеризуется геморрагическим альвеолитом и быстропрогрессирующим гломе-
рулонефритом, который сложно отличить от АНЦА.-ассоциированного поражения
почек. В связи с этим обнаружение АНЦА и антител к базальной мембране почек
обычно используют совместно.
Иммунокомплексные васкулиты обусловлены образованием иммунных ком-
плексов между аутоантителами и их антигенами в крови и сосудистой стенке.
Иммунные комплексы, которые можно обнаружить в сыворотке крови, называют
ЦИК. Предполагается, что снижение растворимости ЦИК приводит к их отложе-
нию в сосудистой стенке и развитию воспаления. Снижение растворимости при
замедлении скорости кровотока и температуры приводит к тому, что иммунными
комплексами поражаются преимущественно капилляры, прежде всего капилляры
кожи. Криоглобулинемический васкулит сопровождается выходом в кровь особой
разновидности ЦИК, которая требует специальных методических подходов для их
обнаружения.
Отдельно выделяют методы, позволяющие обнаружить отложения иммунных
комплексов и комплемента непосредственно в пораженной ткани. Определение
отложений иммунных комплексов позволяет объективизировать иммуноком-
плексный процесс и обладает большей достоверностью по сравнению с методами
выявления ЦИК.
Васкулиты крупных сосудов
В основе патогенеза васкулитов крупных сосудов, к которым относят аортоар-
териит Такаясу и височный артериит, а также ревматическую полимиалгию, лежит
поражение сосудистой стенки, что обусловливает большую площадь вовлеченно-
го эндотелия. Синтез эндотелием провосналительных цитокинов, прежде всего
TNF-ct и IL-6, приводи^ к формированию выраженного острофазового ответа,
исследование которого используют в диагностике васкулитов крупных сосудов.
Так, при аортоартериите Такаясу, височном артериите и ревматической полими-
алгии отмечают чрезвычайно высокие уровни С-реактивного белка и значительно
повышенные значения СОЭ.
Диагноз ревматической полимиалгии и гигантоклеточного артериита
ставят на основе обнаружения значительно повышенной СОЭ (до 100 мм/ч)
при использовании метода Вестергрена и увеличения С-реактивного белка более
40 мг/мл. При височном артериите и ревматической полимиалгии только у 1%
больных, не получавших специфической терапии, СОЭ и С-реактивный белок
находятся в пределах нормы. СОЭ больше 30 мм/ч имеет место у 95% больных в
активную фазу этих заболеваний и у 20% больных вне активности заболевания.
На фоне иммуносупрессивной терапии уменьшение значений острофазовых реак-
тантов происходит с различной скоростью. К примеру, при лечении височного
артериита кортикостероидными гормонами уровни фибриногена и С-реактивного
белка снижаются быстрее всего, у 70% больных они достигают нормальных зна-
чений в течение 2 нед от начала терапии. Изменение СОЭ происходит значительно
медленнее, у половины больных этот показатель нормализуется через 2 нед после
160 ДИАГНОСТИКА АУТОИММУННЫХ ЗАБОЛЕВАНИЙ
начала лечения. Нормальные значения СОЭ наблюдают через 5 нед у 75% боль-
ных. Скорость снижения СОЭ наиболее точно соответствует снижению активности
височного артериита и ревматической полимиалгии, что позволяет использовать
значения СОЭ для мониторинга активности заболевания и контроля терапии.
Узелковый полиартериит — редкая форма системных васкулитов. Он мижет
развиться в течение I года после перенесенного вирусного гепатита В. Обычно
происходит самоизлечивание, рецидивирует заболевание крайне редко. Несмотря
на то что морфологически узелковый полиартериит проявляется гранулематозным
воспалением, сходным с гранулемами при микроскопическом полиангиите. АНЦА
и другие специфические серологические маркеры при классическом узелковом
полиартериите отсутствуют. Обычно его обострение сопровождается гипокомпле-
ментемией. а также увеличением концентраций острофазовых белков. В сыворот-
ке крови пациентов обычно обнаруживают признаки текущей ВГВ-инфекции в
виде HBsAg и HBeAg.
Бо гсезнь Кавасаки поражает преимущественно детей до 10 лет. Клинические
симптомы включают лихорадку, лимфаденопатию, кожную сыпь, характерное
покраснение языка и слизистой оболочки рта, а также васкулит, поражающий
коронарные артерии. Большая часть случаев болезни Кавасаки возникает спора-
дически, однако описаны вспышки этого заболевания, которые указывают на ее
инфекционный характер. Антиэндотелиальные антитела при этом заболевании
отмечаются сравнительно часто и коррелируют с его активностью.
Гранулематозные васкулиты и быстропрогрессирующий
гломерулонефрит
Гранулематозные васкулиты - группа заболеваний, морфологическая карти-
на которых представлена образованием тканевых гранулем, ассоциированных
с мелкими сосудами, а также поражением клубочка в виде некротизирующего
олигоиммунного гломерулонефрита с полулуниями. Тесная взаимосвязь между
обнаружением АНЦА и основными формами гранулематозных васкулитов и
значение АНЦА в их патогенезе позволили обобщить их в виде группы АНЦА-
ассоциированных васкулитов.
Цитоплазматический тип свечения АННА, обусловленный образованием
антител к протеиназе-3, является чувствительным и специфичным маркером
гранулематозного васкулита (ГВ), тяжелого заболевания, проявляющегося триа-
дой с гранулематозным поражением носоглотки, дыхательных путей, легких и
почек. Высокие титры антител к ПР-З/цАНЦА встречаются у 95% больных с
развернутой картиной активного ГВ с поражением почек, в то время как лока-
лизованные формы ГБ, поражающие верхние дыхательные пути, реже сопрово-
ждаются аутоантителами. Встречаемость АНЦА при локализованных формах ГВ
не превышает 50-60%. Несмотря на высокую специфичность выявления АНЦА
при ГВ, цАНЦА могут встречаться при паранеопластическом и лекарственном
васкулите.
Титры цАНЦА при ГВ являются клинически значимыми и коррелируют с актив-
ностью заболевания. Высокий титр служит основанием для патогенетической
иммуносупрессивной терапии. Титр антител увеличивается в среднем за месяц до
начала очередного обострения заболевания. У больных в фазе ремиссии титры
антител очень низки или вообще не определяются. Ежемесячный мониторинг
содержания цАНЦА может быть использован для предсказания рецидива забо-
левания, и патогенетическая терапия может предупредить развитие заболевания.
Показано, что терапия, проводимая на основании мониторинга цАНП А, способна
эффективно предо гвращать возникновение рецидивов ГВ.
ДИАГНОСТИКА АУТОИММУННЫХ ЗАБОЛЕВАНИЙ 161
Снижение титра аутоантител может занять до полугода, С-реактивный белок
лучше отражает уменьшение активности аутоиммунного воспаления на фоне
эффективной терапии. У пациентов с ГВ, у которых сохраняются высокие титры
АНЦА, несмотря на проводимую иммуносупрессивхзую терапию, высок риск
повторного обострения заболевания.
Преобладающей формой васкулитов у больных с пАНЦА/анти-МПО считают
микроскопический полиангиит. По гистологической картине микроскопический
полиангиит сходен с узелковым полиартериитом, но не сопровождается образова-
нием гранулем и поражает сосуды мелкого калибра. Клинически он характеризу-
ется сочетанным поражением легких и почек, представленным легочным капил-
ляритом и экстракапиллярным гломерулонефритом с полулуниями. Кроме того,
приблизительно у 1/3 больных с антителами к МПО отмечается изолированный
олигоиммунный гломерулонефрит с полулуниями, который не сопровождается
внепочечными проявлениями.
Как и в случае ГВ, титр пАНЦА/анти-МПО максимален во время активно про-
текающего гломерулонефрита. Затихание процесса или подавление активности
васкулита при эффективной цитостатической терапии приводит к элиминации
антител у 95% больных, причем у 75% больных сероконверсия происходит в тече-
ние ближайших 6 мес плслр достижения клинической ремиссии процесса.
Другая клиническая форма гранулематозных васкулитов, которая сопровожда-
ется пАНЦА/анти-МПО, - это синдром Черджа-Стросс. Клиническая симптома-
тика этого заболевания представлена легочным васкулитом с эозинофильными
инфильтратами, астмой и гиперэозинофилией. Антитела к МПО выявляют у 75%
больных. Помимо АННА, при этом заболевании часто обнаруживают антиэндо-
телиальные антитела и повышенные уровни эозинофильных катионных белков
(ЕСР) в сыворотке крови.
Изолированный быстропрогрессирующий васкулит без признаков внепочеч-
ной патологии представляет отдельное заболевание в структуре гранулематозных
васкулитов. Его следует дифференцировать от гломерулонефрита при синдроме
Гудпасчера, при котором отсутствует образование гранулем.
Морфологически эти две формы полулунного гломерулонефрита очень схожи,
однако при синдроме Гудпасчрра присутствуют линейные отложения IgG по
базальной мембране почек, которые соответствуют отложениям антител к базаль-
ной мембране клубочка. При АНЦА-ассоциированном гломерулонефрите, напро-
тив. отложений иммуноглобулинов в клубочке не наблюдается, поэтому его назы-
вают олигоиммунным. АНЦА обнаруживают приблизительно у 75% пациентов с
изолированным быстропрогрессирующим гломерулонефритом и у 15% больных
также отмечают антитела к базальной мембране клубочков.
Как пАНЦА/МПО, так и цАНЦА/ПР-3 встречаются при быстропрогресси-
рующем гломерулонефрите с приблизительно одинаковой частотой. У небольшой
категории больных с олигоиммунным i ломерулонефритом с полул} ниями антител
к МПО и ПР-3 не обнаруживают, однако могут встречаться антите на к нейтро-
фильной эластазе.
Основным показанием для определения АНЦА в ревматологической и нефро-
логической практике принято считать подозрение на системный гранулематозный
васкулит. Совокупная частота обнаружения основных разновидностей АНЦ А при
васкулитах представлена в табл. 17-26.
162 ДИАГНОСТИКА АУТОИММУННЫХ ЗАБОЛЕВАНИЙ
Таблица 17-26. Частота обнаружения антинейтрофильных цитоплазматических антител при гра-
нулематозных васкулитах
Заболевание Встречаемость цАНЦА/ анти-ПРЗ, % Встречаемость пАНЦА/ анти-МПО, %
Идиопатический гломерулонефрите полулуниями 50 50
Микроскопический полиангиит 10 70
Гранулематоз Вегенера 90 10
Синдром Черджа-Стросс 10 75
Классический узелковый полиартериит 5 15
Синдром Гудпасчера н.д. 10
Очевидно, что не существует абсолютной специфичности того или иного типа
АНЦА при конкретных формах васкулитов. Замечено, что изолированные формы
быстропрогрессирующего гломерулонефрита, сопровождающегося антителами
к ПР-3, в целом протекают более агрессивно. В частности, наблюдается более
быстрое снижение функции почек, чаще диагностируют экстраренальные прояв-
ления васкулита, в том числе образование тканевых гранулем. Также чаще отмеча-
ют обострения заболевания.
Показания к обследованию на АНЦА представлены в табл. 17-27.
Таблиц? 17-27. Диагностический алгоритм по применению определения антинейтрофильных
цитоплазматических антител в клинической практике
Иммунофллоресцентный тест для определения АНЦА необходимо выполнять при следующих состояниях
• Наличие видимых признаков васкулита — пурпуры, некрозы кончиков пальцев и т.д.___
• Наличие более или менее типичных признаков синдрома Вегенера, Черджа-Стросс и других АИЗ, сопро-
вождающихся образованием АНЦА _____________________________________________________
• Быстропрогрессирующий гломерулонефрит (совместно с антителами к базальной мембране клубочка)
• Признаки воспалительной реакции (лихорадка, миалгии, увеличение СОЭ или появление С-реактивного
белка) без видимой причины или локализации воспалительного процесса________________
После получения положительного ответа в методе непрямой иммунофлюоресценции должен
быть выполнен иммуноферментный тест для установления конкретной специфичности АНЦА.
Целесообразно одновременное исследование на АНФ и АНЦА. в слу®е высоких титров АНФ
нужно проводить дифференциальную диагностику ДБСТ.
Развернутое серологическое обследование пациентов должно включать опреде-
ление АНЦА. АНФ. а также антител к базальной мембране клубочка. При обнару-
жении АНЦА во флюоресцентном тесте необходимо выполнить подтверждающие
тесты, включающие обнаружение антител к МПО и ПР-3. Обследование на АНФ
позволяет избежать получения ложноположительных результатов пАНЦА, а
также исключить наличие системного ревматического заболевания. Определение
антител к ЕМК у больного с быстропрогрессирующим гломерулонефритом с изме-
нениями на рентгенограмме легких или гемофтизе позволяет пос гавить диагноз в
пользу синдрома Гудпасчера и назначить патогенетическую терапию.
Существует два возрастных пика развития быстропрогрессирующего гломе-
рулонефрита, на 3-й и 7-й декаде. Синдром Гудпасчера с одинаковой частотой
поражает женщин и мужчин, однако поражения легких чаще отмечают у мужчин.
Среди больных с синдромом Гудпасчера 10-15% пациентов имеют пАНЦА.
Выявление аутоантител во многом определяет как терапевтическую тактику
ведения пациентов с быстропрогрессирующим гломерулонефритом, так и общий
прогноз заболевания. Худший прогноз относительно сохранности функции почек
имеет место при обнаружении АНЦА к ПР-3, сравнительно лучший — при обна-
ружении антител к МПО.
ДИАГНОСТИКА АУТОИММУННЫХ ЗАБОЛЕВАНИЙ 163
Поскольку все гранулематозные васкулиты протекают с продромальным перио-
дом. который длится от нескольких месяцев до одного года, диагностика АНЦА,
АНФ и антител к БМК должна входить в число лабораторных тестов для диффе-
ренциальной диагностики лихорадок неясного генеза. На продромальном этапе в
клинической картине гранулематозных васкулитов лидирует полиневрит, который
обнаруживают у 70-80% пациентов. Быстрое развитие почечной недостаточности
и кровохарканье составляют основу клинической картины при обострении систем-
ных васкулитов и синдроме Гудпасчера. Часто клиническая симптоматика может
развиваться более смазанно, включая появление легочных инфильтратов, гемату-
рии, анемии и одышки. У пожилых пациентов отмечают плавное нарастание уре-
мии. У больных с почечно-легочными синдромами необходимо проводить диф-
ференциальную диагностику с другими формами вторичных гломерулонефритов,
таких как постстрептококковый гломерулонефрит, пурпура Шенлейна-Геноха,
СКВ и смешанная криоглобулинемия.
Возможно использование быстрых тестов для обнаружения антител к БМК,
МПО и ПР-3 методом лайн-блота. Ряд хроматографических тестов позволяет
получить результат в течение L5 мин непосредственно у постели больного Эти
тесты предназначены только для исключения заболевания в сомнительных
случаях. Недостаточные показатели их специфичности и чувствительности тре-
буют использования независимого развернутого лабораторного тестирования.
Целесообразность их использования определяется важностью раннего нача-
ла терапии, так как в отсутствие лечения системных васкулитов и синдрома
Гудпасчера смертность превышает 75%. В случае развития олигурии почечная
функция восстанавливается редко, даже несмотря на эффективную иммуносу-
прессивную терапию. Быстрая диагностика синдрома Гудпасчера позволяет рано
назначить плазмаферез, который является основой лечения этого заболевания.
Если удалось добиться ремиссии синдрома Гудпасчера и исчезновения антител к
базальной мембране клубочка, то заболевание редко рецидивирует. Среди паци-
ентов с легочно-почечным синдромом менее чем в 20% случаев обнаруживают
антитела к базальной мембране клубочка, у большинства оппеделяют антитела к
МПО и ПР-3.
ИММУНОКОМПЛЕКСНЫЕ ВАСКУЛИТЫ
Иммунокомплексные васкулиты поражают ткани с наибольшим капиллярным
руслом, прежде всего капилляры кожи. При патоморфологическом исследовании
при иммунокомплексных васкулитах имеет место выраженная лейкоцитарная
инфильтрация дермы, что обычно описывается морфологами как лейкоцитокла-
стический васкулит. Наиболее часто лейкоцитокластический васкулит характери-
зуется геморрагической пурпурой, разрешающейся с образованием характерной
пигментации кожи, которая позволяет отличить ее от других разновидностей кож-
ных высыпаний. Другой формой иммунокомплексных кожных высыпаний явля-
ется панникулит, или васкулит, протекающий в подкожной жировой клетчатке.
Эта форма проявляется узловатой эритемой, ревматическими узелками узелками
Рандю- -Ослера, Габердена.
Первичные иммунокомплексные васкулиты представлены тремя основными
нозологическими формами, в том числе пурпурой Шенлейна-Геноха, криогло-
булинемическим васкулитом и лейкоцитокластическими васкулитами. Большая
часть лейкоцитокластических васкулитов является вторичной, возникает на фоне
заболеваний, сопровождающихся поступлением в кровяное русло больших коли-
честв экзогенного антигена. Этиологические причины лейкоцитокластических
васкулитов чрезвычайно разнообразны (табл. 17-28).
164 ДИАГНОСТИКА АУТОИММУННЫХ ЗАБОЛЕВАНИЙ
Таблица 17-28. Основные причины вторичных иммунокомплексных васкулитов
> Лекарственные васкулиты (пенициллины, сульфаниламиды, аллопуринол, тиазиды, ретиноиды)_
* Сывороточная болезнь (сыворотки, моноклональные антитела, цитокины, стрептокиназа)_____
• Хронические бактериальные инфекции (подострый септический эндокардит, бронхоэктатическая
болезнь)_________________________________________________________________________________
* Вирусные инфекции (ВГС, ВГВ, ВИЧ, ЦМВ)_________________________________________________
« Паранеопластические процессы при солидных опухолях_____________________________________
• Гематоонкопогия (миелома, лимфомы)_____________________________________________________
• Системные заболевания соединительной ткани_____________________________________________
• Аутоиммунные заболевания (воспалительные заболевания кишечника)_ ______ _______
• Эссенциальная криоглобулинемия_________________________________________________________
» Пурпура Шенлейна-Геноха и lgA-неФоопатия (болезнь Берже)
Поражение микроциркуляторного русла кожи иммунокомплексными васку-
литами обусловлено медленным кровотоком в капиллярах кожи, механической
нагрузкой и снижением температуры. Все эти факторы способны нарушать рас-
творимость иммунных комплексов и приводят к их преципитации на стенке сосу-
дов. Иммунокомплексные васкулиты часто поражают сосуды нервов, что ведет к
моно- или полиневриту. В тяжелых случаях может развиться поражение почек,
причиной котсрого является избыточное содержание ЦИК в крови. Классические
формы иммунокомплексного поражения почек — IgA-нефропатия при пурпуре
Шенлейна-Геноха и мезангиокапиллярный гломерулонефрит при эссенциальной
криоглобулинемии. При обследовании больших контингею ов больных с острым
иммунокомплексным васкулитом было установлено, что у 40% из них отмечаются
изменения в мочевом осадке, у 30% — артралгии, у 5% — абдоминальный син-
дром. Воспалительная симптоматика, включая лихорадку и миалгии, присутствует
только у 10% больных.
Пурпура Шенлейна-Геноха поражает преимущественно детей и подростков,
однако ее признаки могут появиться в любом возрасте. Основным патогенетиче-
ским механизмом в развитии этого заболевания служит синтез аномального IgA,
обладающего способностью к самоагрегации. В результате в мезангии клубочка и
стенках мелких сосудов почки наблюдают выраженные отложения полимерного
иммуноглобулина и комплемента. Обнаружение ЦИК при данном заболевании
неинформативно. Основной метод диагностики пурпуры Шенлейна-Геноха —
анализ биоптатов ткани с иммунофлюоресцентным исследованием, которое
позволяет обнаружить отложения иммунных комплексов. В биоптате почки
отмечается мембранозный гломерулонефрит с мезангиальными отложениями IgA
и СЗ-компонента комплемента. Исследования биоптата кожи позволяют обнару-
жить гранулярные отложения IgA в стенках сосудов дермы.
Чаще всего выраженный лейкоцитокластический васкулит отмечается при
эссенциальной смешанной криоглобулинемии, связанной с появлением в сыворот-
ке крови криоглобулинов 2-го типа с активностью РФ. Основная причина эссенци-
альной криоглобулинемии — гепатит С, который часто протекает субклинически.
Частота развития криоглобулинемии у больных гепатитом С составляет 10-15%.
Персистенция вируса гепатита С в клетках ретикулоэндотелиальной системы ведет
к поликлональной стимуляции иммунной системы и синтезу аутоантител. При
эссенциальной смешанной криоглобулинемии РФ практически всегда присут-
ствует в составе криопреципитата, как правило, в высоких титрах. Это позволяет
использовать выявление РФ в качестве метода скрининга криоглобулинемии.
При эссенциальной криоглобулинемии имеется поражение кожи в виде пурпу-
ры, преимущественно локализованной на коже нижних конечностей. Классическим
клиническим признаком криоглобулинемии является триада Мельтцера, которая
ДИАГНОСТИКА АУТОИММУННЫХ ЗАБОЛЕВАНИЙ 165
включает пурпуру, артралгию и слабость, к ним нередко присоединяется гломеру-
лонефрит и периферическая невропат ия. Выраженность клинических признаков не
зависит от абсолютного содержания криоглобулинов, так как даже незначительное
количество криоглобул ина может приводить к характерной клинической симпто-
матике. Динамика содержания криоглобулинов позволяет оценить эффективность
проводимой патогенетической терапии, которая включает иммуносупрессивные и
противовирусные препараты. Плазмаферез снижает количество криоглобулинов,
однако не может быть использован в качестве монотерапии.
В диагностике иммунокомплексных васкулитов применяют и другие иммуноло-
гические тесты, в том числе определение содержания иммуноглобулинов сыворот-
ки, компонентов комплемента и острофазовых реактантов. Нормальные показате-
ли этих тестов не позволяют исключить диагноз иммунокомплексного васкулита.
Наблюдение за динамикой содержания ЦИК может быть использовано при
оценке клинической активности иммунокомплексных заболеваний, поэтому долж-
но выполняться периодически, наряду с определением СОЭ. С-реактивного белка.
При СКВ и РА содержание ЦИК достоверно коррелирует с активностью процесса
и может применяться с целью мониторинга состояния пациента. Одновременно с
содержанием ЦИК необходимо выполнять тесты, оценивающие систему компле-
мента, такие как СН50, СЗ и С4. Потребление комплемента, наряду с увеличением
содержания ЦИК, служит признаком текущей активности иммунокомплексного
васкулита и требует принятия активных мер.
Диагностика аутоиммунных поражений печени
Хотя аутоиммунные заболевания печени (АЗП) встречаются реже, чем вирус-
ные гепатиты, вероятность наличия АЗП необходимо предполагать у каждого
пациента, у которого отсузствуют серологические маркеры вирусных гепатитов
или отмечается тяжесть клинической симптоматики, которая не характерна для
обычной клинической картины вирусного гепатита. Другой причиной обследова-
ния для обнаружения серологических маркеров АЗП является дифференциальная
диагностика с другими невирусными заболеваниями печени, в том числе с мета-
болическими поражениями и онкологическими заболеваниями желчевыводящих
путей.
Основным серологическим методом диагностики АЗП принято считать НРИФ
с использованием «тройного субстрата», который включает криосрезы печени-
почки-желудка крысы. В иностранной литературе его часто назвают LKS-субстрат,
по первым буквам английских названий органов «liver-kidney-stomach». Нередко
поставщики предлагают все три субстрата для НРИФ в одном криосрезе, что
облегчает визуализацию положительных реакций.
Комбинированный метод LXS-субстрата позволяет провести в одной реак-
ции скрининг наиболее часто встречающихся аутоантител, поэтому может быть
и( пользован для первичного обследования больных с разными АИЗ. В случае при-
менения этого метода в одной реакции можно выявить, включая АНФ, антитела к
гладким мышцам, антитела к митохондриям, антитела к LKM и антитела к обкла-
дочным клеткам желудка. Другой причиной использования комбинированного
субстрата является удобство идентификации аутоантител. Действительно, анти-
тела к митохондриям и антитела к обкладочным клеткам желудка практически
идентично реагируют со слизистой оболочкой желудка. В то же время антитела к
митохондриям реагируют с богатыми митохондриями клетками канальцев почки
и гепатоцитами, а антитела к обкладочным клеткам эти клетки не окрашивают
(табл. 17-29). Тестирование на LKS-субстраге целесообразно дополнять другими
серологическими тестами для определения аутоантител (рис. 17-2).
166 ДИАГНОСТИКА АУТОИММУННЫХ ЗАБОЛЕВАНИЙ
Подозрение на аутоиммунное заболевание печени
нРИФ на LKS-субстрате
+
АНФ на НЕр2 клетках
Дополнительное тестирование
на редкие антигены
SlA/LP, LC1,
gp210, PML, sp100
Обследование на АНЦА
АИГ 1-го типа АИГ2-гатипа ПБЦ ПСХ Гастрит типа А
АГМА, АНФ, SLA/LP, I IgGAM LKM, LC1. АНФ АМА М2, АНФдр210, PML sp100 АНЦА АНФ АПКЖ, антитела к фактору Касла
Рис. 17-2. Алгоритм обследования больных с подозрением на аутоиммунное заболевание печени.
Таблица 17-29. Использование криосрезов ^S-субстрата тканей крысы в методе непрямой имму-
нофлюоресценции для обнаружения аутоантител при аутоиммунных заболеваниях печени
Заболевание Тип аутоантител Тканевой субстрат
желудок почка печень
АЗП, СКВ или другое АИЗ АНФ Свечение ядер клеток Свечение ядер клеток Свечение ядер клеток
Аутоиммунный гепатит 1-го типа Антитела к гладким мышцам (F-актин) Свечение собствен- ной пластинки сли- зистой оболочки Свечение стенки сосудов и мезанги- альных клеток Свечение стенок сосудов и протоков печени
Аутоиммунный гепатит 2-го типа (детский) Антитела к микро- сомам печени- почки (LKM) Нет окрашивания Свечение цитоплаз- мы клеток канальцев почки Свечение центро- лобулярных гепато- цитов
Первичный билиарный цирроз Антитела к мито- хондриям АМА-М2 Свечение цитоплаз- мы клеток слизистой оболочки желудка Свечение цитоплаз- мы клеток канальцев почки Свечение в цитоплаз- ме гепатоцитов
Аутоиммунный гастрит (тип А) Антитела к обкла- дочным клеткам желудка Свечение цитоплаз- мы клеток слизистой оболочки желудка Нет окрашивания Нет окрашивания
Аутоиммунный гепатит (АИГ) 1-го типа — редкое заболевание с популя-
ционной частотой около 0,01%, встречается у лиц возраста 15-40 лет, носителей
HLA-S8 и DR3, несколько чаще — у женщин. Иногда АИГ сочетается с другими
аутоиммунными заболеваниями, в том числе с НЯК, РА, вторичным «сухим син-
дромом* и аутоиммунными полиэндокринопатиями. АИГ 1-го типа протекает с
выраженной поликлональной активацией иммунной системы, которая встречает-
ся при ограниченном числе заболеваний, включая СКВ и СШ. Иммунологическая
диагностика основана на определении АНФ, АГМА и повышенных концентрациях
IgG сыворотки. Поскольку эти маркеры встречаются чаще всего, их выявление
называют «диагностической триадой* АИГ (табл. 17-30).
ДИАГНОСТИКА АУТОИММУННЫХ ЗАБОЛЕВАНИЙ 167
Таблица 17-30. Диагностические критерии аутоиммунного гепатита
Критерий Диагностическая граница Баллы
АНФ пли АГМА >1:40 1
АНФ или АГМА или LKM или SLA' >1:80 2
>1:40
Выявлены в ИФА или лайн-блоге
IgG (турбидиметрия) >Верхняя граница нормы для используемой тест-системы 1
IgG (турбидиметрия) >Превышение на 1,1 верхней границы нормы для используе- мой тест-системы 2
Гистология печени Гепатит, сопоставимый с морфологией АИГ 1
Типичный АИГ 2
Отсутствие маркеров вирусных гепатитов Маркеры отсутствуют 2
Сумма баллов >6 — диагноз АИГ вероятен
Сумма баллов >7 — диагноз АИГ установлен_____________________________________
— при выявлении хотя бы одного аутоантитела в титре выше диагностической границы при-
сваивается 2 балла.
Клиническая картина этого заболевания развивается постепенно, с появлением
дурноты, слабости, потери аппетита, миалгиями, желтухе и и кожными высы-
паниями. Клинические симптомы сопровождаются значительным увеличением
активности трансаминаз и высокими иммуноглобулинами сыворотки крови. В
гистологической картине АИГ 1-го типа присутствует лимфоцитарная инфильтра-
ция портальных трактов, некрозы в центре печеночных долек. Характерно хро-
ническое течение с частыми обострениями, которые без лечения приводят к раз-
витию цирроза. Обычно АИГ быстро и эффективно контролируется стероидными
гормонами, поэтому при ранней диагностике имеет хороший прогноз. На фоне
адекватной иммуносупрессивной терапии титры АГМА постепенно снижаются,
однако мониторинг этого показателя не рекомендован.
ПБЦ поражает женщин в 10 раз чаще, чем мужчин, причем частота забо-
левания растет с увеличением возраста. В связи с отсутствием эффективной
терапии и быстрой прогрессией заболевания ПБЦ остается основной причиной
пересадки печени в европейских странах. У подавляющего числа больных ПБЦ
(95%) в сыворотке крови можно обнаружить АМА-М2, направленные к пируват-
декарбоксилазному комплексу. Очень небольшое количество больных ПБЦ явля-
ются серонегативными, что делает АМА одним из наиболее надежных методов
диагностики этого заболевания. Урсодезоксихолиевая кислота (Урсофальк), кото-
рую используют для лечения ПБЦ, несколько снижает титры АМА. АНФ опреде-
ляют у 60-80% больных ПБЦ, причем в 1/3 случаев он обладает характерным
типом свечения «точек в ядре». Антигенами этой разновидности АНФ являются
белковые антигены splOO и PML.
Первичный склерозирующий холангит (ПСХ), в отличие от ПБЦ, пора-
жает преимущественно мужчин среднего возраста. Нередко холангит развивается
на фоне НЯК, что роднит оба заболевания. Как и при язвенном колите, при ПСХ
обнаруживают АНЦА, которые можно зафиксировать у 60-70% пациентов. Для
диагностики ПСХ может быть использовано выявление АНФ и АНЦА, которые
отмечаются у 50-60% больных.
Использование метода НРИФ на тройном LKS-субстрате позволяет также выяв-
лять случаи аутоиммунного гастрита, который сопровождается аутоиммунными
эндокринопатиями. Обнаружение антител к фактору Кастла у больного с высоким
титром антител к париетальным клеткам желудка может указывать на высокий
риск развития Вр-дефицита с характерными гематологическими и неврологиче-
скими признаками.
168 ДИАГНОСТИКА АУТОИММУННЫХ ЗАБОЛЕВАНИЙ
Серологическая диагностика целиакии
Целиакия — иммуноопосредованная непереносимость cz-глиадина — белка,
который является основным белком клейковины злаковых культур. Наряду с
дефицитом лактазы и пищевыми аллергиями целиакия является наиболее частой
причиной пищевой непереносимости. Целиакия сопровождается рядом кожных
проявлений, в том числе герпетиформным дерматитом.
Высокая чувствительность ИФА тест-систем для определения антител к ТТГ
позволила значительно улучшить диагностику целиакии, особенно ее асимпто-
матичных форм. Популяционные исследования показали, что в Северной Европе
0,5-1% населения имеют стертые признаки этого заболевания. Действительно, у
многих больных целиакией отсутствует типичная клиническая картина, включаю-
щая диарею, повышенное газообразование, запор, однако большинство проблем
связано с мальабсорбцией, в том числе с железодефицитом, остеопенией, плохой
эмалью зубов и рядом других расстройств метаболизма. С повышенной частотой
целиакия отмечается у больных с эндокринными заболеваниями, такими как СД I,
аутоиммунный тиреоидит, а также при селективном IgA-дефиците и синдроме
Дауна.
Для первичной диагностики целиакии рекомендуется сочетанное выявление
анти-ТТГ класса IgA, а также определение сывороточной концентрации IgA для
установления лиц с IgA-дефицитом. Б связи с большей специфичностью АЭА
после положительного результата анти-ТТГ IgA теста целесообразно определение
АЭА. Положительный результат обоих тестов почти со 100% вероятностью ука-
зывает на целиакию. В случае отрицательного АЭА-теста результат рассматривают
как сомнительный, и может быть рекомендовано определение генотипа HLA-
DQ2/8. Определение антител к ТТГ класса IgG целесообразно только у больных с
IgA-дефицитом, так как у лиц с нормальным уровнем IgA в крови результаты этого
теста малоспецифичны. У большинства лиц с предрасположенностью к целиакии,
не ограниченных в потреблении глиадин-содержащих продуктов, появление анти-
тел к ТТГ происходит в возрасте 4-5 лет.
Антитела к глиадину IgA/G остаются тестом резерва, поскольку обладают боль-
шим числом ложноположительных результатов. Одним из вариантов их клиниче-
ского использования может быть диагностика целиакии в возрасте моложе 5 лет,
до образования антител к ТТГ и АЭА.
После серологической оценки у серопозитивных пациентов необходимо выпол-
нение биопсии тонкого кишечника. У небольшого числа лиц с анти-ТТГ и АЭА
признаки хронического воспаления могут отсутствовать, что служит основанием
для диагностики у них «скрытой целиакии» и долгосрочного мониторинга. У
большинства серопозитивных людей аутоантитела могут быть обнаружены за
несколько лет до развития клинической симптоматики.
Безглютеновая диета остается основным методом лечения целиакии. Ее соблю-
дение в течение года приводит как к исчезновению симптоматики, так и к выра-
женному снижению титров аутоантител.
Контроль диетотерапии несколько удобнее проводить с помощью аТТГ,
поскольку тесты имеют количественный результат. Целесообразность определе-
ния антител класса IgG диктуется высокой частотой встречаемости целиакии у
лиц с селективным дефицитом продукции IgA. Б этой группе (1:400-1:1000 попу-
ляции) снижена или отсутствует продукция IgA, поэтому необходимо определение
антител других классов.
Герпетиформный дерматит представляет собой заболевание, часто возникаю-
щее на фоне целиакии. Его ведущим симптомом являются зудящие везикулярные
высыпания на разгибательных поверхностях. При выполнении пРИФ на биопта-
тах кожи больных герпетиформным дерматитом отмечаю^ отложения IgA на вер-
ДИАГНОСТИКА АУТОИММУННЫХ ЗАБОЛЕВАНИЙ 169
шинах сосочков кожи. В сыворотке крови больных герпетиформным дерматитом
можно обнаружить все характерные для целиакии антитела, включая глиадино-
вые, ретикулиновые, эндомизиальные и антю ела к тканевой трансглутаминазе.
Аутоиммунный сахарный диабет
Распространение диабета в развитых странах быстро растет. Согласно данным
Международной диабетической федерации (/Г»Е). в настоящее время 194 млн
(5,1%) взрослых лиц 20-79 лет страдают диабетом, и к 2025 г. это число возрас-
тет до 280 млн человек. Наибольшая распространенность зафиксирована в США
(7,9%) и Европе (7,8%). СД II типа (СД II) составляет 85 -95% всех случаев диа-
бета. Примерно 49 млн больных имеют СД I, в том числе дети и подростки. Эти
данные определяют огромную важность лабораторной диагностики и определения
поогноза этого тяжелого заболевания. Хотя основное внимание фокусируется на
росте заболеваемости СД II, параллельно растет и число случаев СД I. Зачастую
дифференциальная диагностика между СД I и СД II не представляет проблемы.
Однако, наряду с типичными проявлениями СД I в виде острого начала и похуда-
ния, чаще отмечается стертая картина у полных людей и пациентов с медленным
развитием симптоматики. Распространение ожирения в развитых странах приво-
дит к росту заболеваемости СД II как у подростков, так и у детей. Именно поэтому
клиническое значение диабетических аутоантител с течением времени только
нарастает
Клинические признаки СД I являются исходом аутоиммуно-обусловленного
разрушения р-клеток без видимых повреждений соседних эндокринных островко-
вых клеток. К началу заболевания уже разрушено 80-90% р-клеток. При диабете
концентрации всех разновидностей аутоантител максимальны к моменту клиниче-
ского дебюта. Встречаемость аутоантител значительно снижается в течение 1 года
заболевания, что соответствует прогрессивной аутоиммунной деструкции остров-
ков. Феномен исчезновения аутоантител с прогрессией заболевания при сахарном
диабете уникален для всех АИЗ, при которых титры аутоантител обычно повыша-
ются с увеличением активности и длительности заболевания. Этот феномен плохо
исследован, и исчезновение аутоантител сложно объяснить. Несомненно, что сни-
жение титров АОК характеризуется снижением остаточной функции р-клеток. В то
же время на фоне заболевания показана способность островков к саморегенерации
и синтезу небольшого количества эндогенного инсулина в течение заболевания у
большинства больных.
Аутоиммунный диабет имеет выраженный наследственный компонент. Так,
риск развития диабета у однояйцевого близнеца составляет 30%, риск развития
диабета при заболевании близкого родственника — 10%. Использование аутоан-
тител позволяет уточнить риски развития заболевания и является основой иссле-
дований для предупреждения развития СД I.
Для прогнозирования развития диабета применяют комбинированное опреде-
ление АОК, ГДК65, IA-2 и инсулина. Аутоантитела против островковых аутоан-
тигенов имеют специфичность около 100% с чувствительностью 60-80%, в зави-
симости от исследуемой популяции и возраста. В отличие от других аутоантител,
встречаемость островковых аутоантител выше у детей и подростков и значительно
снижается к 20-летнему возрасту. Более чем у 95% детей находят, по меньшей
мере, одну разновидность аутоантител в дебюте диабета, и более 90% из них
имеют несколько типов аутоантител. Обнаружение у ребенка нескольких разно-
видностей островковых аутоантител прогнозирует приблизительно на 50% риск
развития СД I в пределах 5 лет и 80% в пределах 10 лет.
Наиболее широко используемой скринирующей стратегией для уточнения
риска у родственников больного СД I — сестер, братьев и детей — является
170 ДИ Al НОСТИКА АУТОИММУННЫХ ЗАБОЛЕВАНИЙ
определение у последних аутоантител к ГДК65 и IA-2. Если один из этих скри-
нирующих тестов оказывается позитивным, определяют антитела к инсулину
и островковые аутоантитела, а также толерантность к глюкозе. Такой подход
позволяет предсказать развитие СД I с точностью до 90% в пределах 5-30 лет.
Было показано, что родственники первой линии, позитивные только по одно-
му аутоантителу, имели риск развития СД I примерно 10-20% в течение 5 лет.
В исследовании DPT-1 98% родственников первой степени родства, у которых
развился СД I, имели одно антитело или более, и 80% имели два и более типа
аутоантител. Таким образом, 2 обнаруженных аутоантитела свидетельствуют о
5-летнем риске около 70%, а 3 аутоантитела указывают на 100% 5-летний риск
развития СД I.
Островковые аутоантитела, определяемые при родах у женщин, уточняют риск
развития СД после родов. Риск СД I после родов прогрессивно увеличивается с
17% для одного типа аутоантител до 61% для двух и, наконец, до 84% для трех
типов аутоантител. На ранних стадиях заболевания обнаруживают антитела к
ГКД65 и инсулину.
Успешный метод исследования для предупреждения развития СД I пока не обна-
ружен. На сегодняшний день нет успешных результатов превентивного лечения
СД I у человека, несмотря на разработку ряда эффективных методов в экспери-
ментах на животных. Продолжаются исследования, направленные на первичную
профилактику диабета у ближайших родственников. Работа в этом направлении
также должна помочь установить факторы внешней среды и предложить превен-
тивные мероприяз ия для всех АИЗ.
Аутоиммунные полиэндокринопатии
Полиэндокринопатии (ПЭП) представляют собой сочетание одной аутоим-
мунной эндокринопатии с другой или с ассоциированным аутоиммунным забо-
леванием. Выделяют несколько типов ПЭП, которые иногда делят на ПЭП детей
(1 тип) и ПЭП взрослых (2 и 3 типы). В основе НЭП 1-го и 2-го типов лежит ком-
бинация недостаточности коры надпочечников с тиреопатией или СД I. Однако
I ЕЭП 1 и ПЭП 2 отличаются между собой по возрасту дебюта (до 20 и после 20 лет
соответственно), комбинации синдромов и типов наследования. Эти заболевания
также носят название синдромов Шмидта и Карпентера. Наиболее частый, 3 тип,
ПЭП включает другие сочетания эндокринопатий между собой и эндокринопатий
с другими аутоиммунными заболеваниям, такими как целиакия, витилиго, алопе-
ция, аутоиммунный гастрит. АИГ. Характерно, что их реальная частота в десятки
раз превосходит расчетную, основанную на случайном сочетании эндокринопа-
тий. Так, встречаемость антител к тиреопероксидазе у больных СД I составляет
около 10%, с ежегодным риском развития тиреоидита — 10—20%.
Помимо поражения надпочечников, СД I и тиреопатии, ПЭП 1 характеризу-
ется клинической картиной кандидоза, гипопаратиреоидизма и гипогонадизма.
Иммунодефицит, который приводит к рецидивирующему кандидозу слизистых
оболочек, обычно дебютирует первым, после чего присоединяется гипопарати-
реоидизм.
До 1% людей в европейской популяции имеют ПЭП, преимущественно 3 тип
заболевания. Пик заболеваемости ПЭП 2 и 3 приходится на 3-4-ю декаду жизни,
и преобладают они у женщин. При ПЭП взрослых дебюты отдельных эндокрино-
патий могут быть разделены несколькими годами, что затрудняет клиническую
диагностику.
Существует выраженная наследственная предрасположенность к развитию всех
типов ПЭП, которая связана с генами иммунного ответа. Хотя ПЭП взрослых
имеет полигенный характер, гены HLA играют основную роль в наследовании
ДИАГНОСТИКА АУТОИММУННЫХ ЗАБОЛЕВАНИЙ 171
этого заболевания. В отличие от взрослой формы. ПЭП 1-го типа является моно-
генным заболеванием с аутосомно-рецессивным вариантом наследования.
Наиболее часто синдромы ПЭП диагностируют на фоне СД I. Среди больных
СД I до 1/4 пациентов имеют клинические признаки другого эндокринного или
аутоиммунного заболевания. Наиболее частые сочетания эндокринопатии пред-
ставлены СД I и тиреопатиями (15%), тиреопатиями, витилиго и аутоиммунным
гастритом (10%). Реже отмечаются сочетания СД I и витилиго, тиреопатий и над-
почечниковой недостаточности, гипогонадизма и алопеции и, наконец, СД и над-
почечниковой недостаточности.
Диагностика синдромов ПЭП основана на серологическом исследовании и
оценке функции соответствующих эндокринных желез. Обнаружение аутоан-
тител у пациентов, а также их родственников с любой формой эндокринопатии
позволяет предсказать развитие заболевания и уточнить группы риска. Повторное
тестирование каждые 3-5 лет позволяет обеспечить достаточную уверенность.
Перечень серологических маркеров для клинического использования приведен в
табл. 17-31.
Таблица 17-31. Лабораторное обследование при аутоиммунных полиэндокринопатиях
Заболевание Аутоантиген Тканевой/клетг <ный субстрат Другие тесты
Сахарный диабет 1 типа ГДК, IA-2, инсулин, АОК р-Клетки Глюкоза натощак
Диффузный токсиче- ский зоб рПГ Тиреоциты ПГ, свободный Т4
Тиреоидит ТПО, ТГ Тиреоциты ТТГ, свободный Тд
Гипопаратиреоидизм Са-зависимый рецептор Паратиреоидная железа Паратгормон
Надпочечниковая недо- статочность Р450С21, Р450с17, P450SCC Кора надпочечников Утренний кортизол, АКТГ
Гипогонадизм Р450С17, P450SCC Клетки Лейдига/теки ФСГ, ЛГ, тестостерон
Аутоиммунный гастрит Н*. К+-АТФаза Обкладочные клетки желудка Витамин В12, железо, ожсс
Пернициозная анемия Фактор Кастла Белок Витамин В1?, железо
Целиакия Тканевая трансглутами- наза, а-глиадин Тонкий кишечник, пищевод Эндоскопия с биопсией слизистой оболочки
Аутоиммунный гепатит F-акгин. LKM-1 Печень/почка/желудок АЛТ, ACT, ГГТП,билирубин
Алопеция Тирозингидроксилаза Волосяные фолликулы -
Витилиго Тирозиназа Меланоциты кожи -
Примечание. ТТГ — тиреотропный гормон, АКТГ — адренокортикотропный гор-
мон, ФСГ — фолликулостимулирующий гормон, ЛГ - лютеинизирующий гормон, Г1ТП —
у-глутаматтранспептидаза.
Диабет дебютирует до надпочечниковой недостаточности. Детей с СД I целе-
сообразно обследовать на предмет наличия антител к тканевой трансглутаминазе
и эндомизию, чтобы уточнить наличие субклинической формы целиакии. При
надпочечниковой недостаточности целесообразно повторное тестирование на
антитела к ТПО, ГДКА65, АОК. В случае обнаружения аутоанти гел к островковым
антигенам может быть рекомендовано проведение глюкозотолерантного теста
даже при нормальных показателях глюкозы натощак. Встречаемость антител к
обкладочным клеткам желудка отмечается у больных с тиреоидитом и составляет
20-40%, что позволяет использовать этот тест для скрининга ПЭП у больных с
аутоиммунными эндокринопатиями.
172 ДИАГНОСТИКА АУТОИММУННЫХ ЗАБОЛЕВАНИЙ
Неврологические заболевания
Для иммунологической диагностики заболеваний, поражающих разные отде-
лы нервной системы, используют широкий спектр тестов (табл. 17-32). PC и
другие демиелинизирующие поражения ЦНС обусловлены иммунной реакцией,
протекающей за гистогематическим барьером. В этом случае иммунный ответ
протекает местно, что делает невозможным его обнаружение в периферической
крови. При этом заболевании наблюдается повышенный синтез иммуногло-
булинов за гистогематическим барьером, кроме того, вновь синтезируемые
иммуноглобулины приобретают новые свойства, прежде всего они становятся
о лигоклональными, что отличает их от поликлональных иммуноглобулинов,
синтезирующихся в норме или при остром воспалительном ответе. Выявление
олигоклонального IgG в СМЖ позволяет проводить дифференциальную диа-
гностику демиелинизирующих заболеваний центральной нервной системы.
Иммунологический анализ СМЖ — основа диагностики воспалительных про-
цессов за гематоэнцефалическим барьером.
Таблица 17-32. Диагностика аутоиммунных неврологических заболеваний
Паранеопластический энцефалит Антитела к Hu, Yo-1, Ri, Ма2, CV2, амфифизину
Демиелинизирующие поражения ЦНС Олигоклональный IgG в ликворе и сыворотке
Демиелинизирующие поражения нервов Антитела к ганглиозидам CM.. GQ h GD1h
Полиневриты АН® дсДНК, АНЦА, АКЛА, криоглобулины
Полимиозит АНФ, лайн-блотс определением анти-Jol, RNP, PM-Scl, Mi 2, Ku
Миастения Антитела к скелетным мышцам
Антитела к нейронам встречаются в системной циркуляции при редких пара-
неопластических поражениях ЦНС. В этом случае за счет перекрестной реакции
системный противоопухолевый иммунный ответ поражает нервную ткань. Другим
вариантом аутоантител, направленных против компонентов нервной гкани, явля-
ются антитела к ганглиозидам. Разрушая миелиновую оболочку нервов, они
приводят к развитию демиелинизирующих полирадикулоневропатий и перифе-
рических полиневритов. При обследовании пациентов с полиневритами следует
использовать иммунологические тесты для диагностики ДБСТ и васкулитов, так
как ревматические заболевания нередко дебютируют с картины периферического
моно- или полиневрита.
Миастения — нервно-мышечное заболевание, при котором аутоантитела к АхР
и другим антигенам скелетной мышцы приводят к нарушениям нервно-мышечной
передачи. В отличие от миастении, при воспалительных миопатиях в циркуляции
можно обнаружить ряд АНА.
Заболевания кожи
Среди аутоиммунных состояний с поря женим кожи можно выделить 4 основные
группы (табл. 17-33): системные заболевания с преимущественным поражением
кожи, реакции против интраэпидермальных антигенов, буллезные дерматозы с
реакциями против антигенов базальной мембраны кожи, заболевания с реакциями
против антигенов дермы.
ДИАГНОСТИКА АУТОИММУННЫХ ЗАБОЛЕВАНИЙ 173
Таблица 17-33. Классификация аутоантител при поражениях кожи на фоне аутоиммунных забо-
леваний
Форма Нозологическая форма Основные аутоантигены
Системные АИЗ с преимущественным поражением кожи СКВ, кожные формы КВ дсДНК, нуклеосомы, Sm. SS-A (52 и 60 кДа), SS-B, U,-RNP. C1q, фосфолипиды
Дерматомиозит тРНК-синтетазы (Jo-1 и др.), Mi-2, Ku
Системный склероз CENT-B. Sc/-70, РМ-Sc/
Васкулиты Криоглобулины, PR3, МРО, РФ, зндотелио- циты и т.д.
АИЗ с ответом против внутриэпидермальных антигенов Вульгарная пузырчатка Десмоглеины 3
Листовидная пузырчатка Десмоглеины 1
IgA-пузырчатка Десмоглеины 1 и 2, десмоколин 1
Витилиго VIT40, тирозиназа
АИЗ с ответом против антигенов базальной мембраны кожи Буллезный пемфигоид, пемфигоид слизистых оболочек, герпес бере- менных 230 кДа-белок (BPAG 1) и 180 кДа-белок (BPAG 2)
Многоформная эритема Десмоплакины I и II
АИЗ с ответом против дермальных антигенов Буллезный эпидермолиз Коллаген VII и ламин-а,
Герпетиформный дерматит Эпидермальная трансгулатминаза, глиадин, зндомизий
Примечание. BPAG • bullous pemphigoid antigen (англ.) — антиген буллезного пемфигоида.
Из-за большой поверхности кожи и ее богатого сосудистого русла поражения
кожи возникают при многих АИЗ и васкулитах. Кожа часто поражается при вол-
чанке. склеродермии, дерматомиозите и васкулитах. Апоптоз клеток кожи являет-
ся одним из индукторов аутоиммунных реакций, связанных с образованием АНА.
Именно поэтому большинство разновидностей АНА при системных заболеваниях
имеют взаимосвязь с различными кожными проявлениями.
Большую группу заболеваний кожи составляют пузырные дерматозы. Деление
на пузырчатку (pemphigus) и пемфигоид основано на локализации пузырьков в
толще эпидермиса или по базальной мембране, что обусловливает ряд морфо-
логических и клинических феноменов, характерных для этих заболеваний. К
буллезным дерматозам с субэпидермальной везикуляцией причисляют также
многоформную эритему, буллезный эпидермолиз и herpes gestationis, который
представляет собой кожное проявление целиакии. Еще одно аутоиммунное забо-
левание, сопровождающееся изолированной иммунной реакцией против эпители-
альных клеток, — это витилиго.
Аутоантитела встречаются при всех АИЗ с поражением кожи, что определяет
их значение в диагностике. В состав лабораторно-клинического обследования
больных с кожными заболеваниями должна входить биопсия кожи с прямой РИФ
(пРИФ). Благодаря легкости получения материала и четкому морфологическому’
строению, изучение отложений иммуноглобулинов и факторов комплемента в
биоптатах кожи очень информативно и должно проводиться в комплексе с серо-
логическим исследованием.
пРИФ биоптатов непораженной кожи в диагностике пузырных дерматозов
обладает почти 100% чувствительностью и является золотым стандартом обсле-
дования. Оно весьма информативно, так как позволяет обнаружить изменения,
характерные для основных АИЗ кожи. Исследуют уже существующие отложения
иммуноглобулинов и комплемента в структурах кожи. В отличие от морфологиче-
ской оценки высыпного элемента с помощью световой микроскопии, для пРИФ
необходим участок с полностью сохраненной морфологией кожи. Рекомендуется
взятие биопсии при помощи одноразовых дерматологических «пробойников»
диаметром 3-4 мм, при использовании которых косметический дефект мини-
174 ДИАГНОСТИКА АУТОИММУННЫХ ЗАБОЛЕВАНИЙ
мален. Биопсию доставляют в лабораторию в специальной транспортной среде
Михаэлиса, содержащей стабилизаторы иммуноглобулинов.
При иммунофлюоресцентном исследовании биопсии кожи могут быть обна-
ружены антитела к межклеточным контактам (пузырчатка/, базальной мембране
(пемфигоид, пемфигоид беременных, линейный IgA-дерматоз, пемфигоид сли-
зистых оболочек, буллезный эпидермолиз). IgA в сосочках при герпетиформном
дерматите Дюринга, «волчаночная полоска» IgM/IgG по базальной мембране,
отложения IgA в сосудах дермы при пурпуре Шенлейна-Геноха. Основные диа-
гностические находки при биопсии кожи перечислены в табл. 17-34.
Таблица 17-34. Основные диагностические феномены при проведении реакции прямой иммуно-
флюоресценции биоптата кожи
Структура кожи Форма отложений 1д Заболевание
Эпидермис Межклеточные отложения 1д6 и разрывы в верхних слоях Листовидная пузырчатка
Межклеточные отложения IgG и разрывы в глубоких слоях Вульгарная пузырчатка
Межклеточные отложения IgA Интраэпидермальныи пустулярный дер- матоз
Флюоресценция ядер клеток AHA in vivo
Дермо- эпидермальный стык Линейные отложения IgG Пемфигоид
Линейные отложения IgA Линейный IgA-пемфигоид
Отложения на эпидермальной стороне при солевом расщеплении Пемфигоид, многоформная эритема
Отложения на дермальной стороне при соле- вом расщеплении Буллезный эпидермолиз
Гранулярные отложения всех классов, чаще IgM «Волчаночная полоска» при СКВ
Гранулярные отложения IgA в сосочках кожи Герпетиформный дерматит
Сосуды дермы IgA-отложения в стенках сосудов Пурпура Шенлейна-Геноха
IgG-отложения в стенках сосудов Кожный васкулит, криоглобулинемия, системные заболевания
ЗАКЛЮЧЕНИЕ
Лабораторные иммунологические тесны охватывают весь спектр клинических
задач, касающихся вопросов диагностики, назначения терапии и прогноза аутоим-
мунных заболеваний (табл. 17-35). Однако ни один тест, как клинический, так и
лабораторный, не может рассматриваться изолированно, в отрыве от совокупной
клинике-лабораторной картины заболевания.
Информативность и надежность данных иммунологического теста во многом
зависят от взаимодействия клинициста и сотруд] гика лаборатории. Врач должен
знать основы лабораторных методов, их недостатки и преимущества. Тесная связь
между клиницистом и врачом клинической лабораторной диагностики позволяет
улучшить точность диагностики установить «слабые места» лабораторной служ-
бы и способствовать их устранению. Только активное сотрудничество с иммуноло-
гической лабораторией способно обеспечить клиницисту надежные, осознанные
результаты анализов и повысить эффективность их использования в клинической
практике.
ДИАГНОСТИКА АУТОИММУННЫХ ЗАБОЛЕВАНИЙ 175
Таблица 17-35. Клиническое применение иммунологических лабораторных тестов
Клиническая задача Пример теста
Скрининг аутоиммунного заболевания АНФ
Подтверждение диагноза Определение АЦЦП при артрите
Определение прогноза заболевания Исследование спектра АНА при системных забо- леваниях
Риск развития заболевания Определение антител к антигенам островковых клеток у родственников больных сахарным диа- бетом I типа
Контроль активности заболевания АНЦА при гранулематозных васкулитах
Основание для терапии с целью профилактики обо- стрения Мониторинг титров антител к дсДНК у больных СКВ
Контроль эффективности иммуносупрессивной тера- пии Изменение С-реактивного белка, СОЭ, биомаркеры
Дифференциальная диагностика обострения основно- го заболевания и сопутствующих осложнений Компоненты комплемента, С-реактивный белок, прокальцитонин
Глава 18
Иммуногематология
Иммуногематология — учение об антигенах клеток крови и
антителах к ним. Клетки крови и плазма характеризуются огром-
ным разнообразием антигенов: антигены лейкоцитов, тромбоцитов,
эритроцитов, нейтрофилов, плазменных белков; набор у каждого
человека строго индивидуален. Поэтому в клинических ситуациях,
подразумевающих возможность контакта человека с чужеродны-
ми для него антигенами других людей, иммуногематоло! ические
знания необходимы. Антигенную совместимость надо учитывать
в трансфузиологии при переливании крови или гемокомпонент ов.
в трансплантологии — при пересадке органов и тканей, в акушер-
стве — при беременности.
Абсолютно идентичных индивидуумов не бывает (исключение —
однояйцовые близнецы), поэтому любая трансфузия, трансплан-
тация, беременность сопровождаются сенсибилизацией теми или
иными антигенами. Конечный результат сенсибилизации — выра-
ботка антител к чужеродным антигенам. Взаимодействие антител и
антигенов приводит к активации, инактивации или гибели клеток,
несущих соответствующие антигены. Клиническими проявления-
ми подобных реакций являются посттрансфузионные реакции и
осложнения, отторжение трансплантага, гемолитическая болезнь
плода и новорожденного и другие патологические состояния.
В связи с этим выполнение любого вида гемокомпонентной терапии
или аллогенной трансплантации органов и тканей должно сопро-
вождаться соответствующими иммуногематологическими иссле-
дованиями крови доноров и реципиентов. Такие же исследования
необходимо выполнять при каждой беременности и родах для
крови матери и ребенка.
Полный спектр антигенов клеток крови определить невозможно,
что вносит ограничения для иммуногематологической апробации
крови донора и реципиента, включающей только те антигены и
антитела, клиническим значением которых пренебречь невозмож-
но. Прежде всего это антигены эритроцитов, несовместимость по
которым может стать причиной серьезных гемолитических кон-
фликтов — мать-плод или реципиент-донор.
ИММУНОГЕМАТОЛОГИЯ 177
АНТИГЕНЫ ЭРИТРОЦИТОВ
Изучение антигенов эритроцитов и анализ иммуногенных свойств показали их
различную способность к сенсибилизации в процессе трансфузий или при бере-
менности. Расположив частоту’ встречаемости антител к антигенам эритроцитов
в убывающем порядке, получили шкалу иммуногенности антигенов эритроци-
тов, которую также можно назвать шкалой приоритета трансфузионно опасных
эритроцитарных антигенов. В настоящее время шкала с учетом только основных
клинически значимых антигенов систем АВО, Резус и Келл выглядит следующим
образом: А, В > D > К > с > С.М Е > е.
СИСТЕМА АВО
Система АВО состоит из двух основных компонентов — антигенов А и В, пред-
ставленных на эритроцитах как по одному, так и вместе — группы А (II), В (III),
АВ (IV); отсутствие антигенов на мембране эритроцитов обозначают символом
«О» — группа 0 (1). Уникальное свойство системы АВО — наличие 8 норме анти-
тел к отсутствующим антигенам. Эти антитела называют изогемагглютининами
анти-А, анти-В (или устаревшее обозначение аи|3). Сочетание антигенов А и В на
эритроцитах и антител к ним в сыворотке крови человека определяет его принад-
лежность к четырем основным группам крови (табл. 18-1).
Таблица 18-1. Основные группы крови человека системы АВО
Группа крови по системе АВО Антигены системы АВО Антитела системы АВО
А В Антн-А Анти-В
А (П) + - - +
В (III) - + + -
АВ (IV) + + - -
0 (I) - - + +
Антигены А и В представляют собой неоднородные структуры, определяющие
наличие большого количества вариантов антигенов системы АВО. Так, структуру
антигена А описывают более чем 30 вариантами: Ar А2, А,, А4 А27, Ах, Aend и др.
На эритроцитах могут быть представлены только некоторые из перечисленных
вариантов антигена; при этом возможна выработка антител к отсутствующим
частям антигенной мозаики. Наиболее иммуногенна часть А ; при ее обнаружении
на эритроцитах человека говорят о второй А(П) или четвертой АВ (IV) группе
крови. Диагностика подгруппы А возможна только при использовании специаль-
ного реагента — анти-Аг В случаях наличия антигена А без выявления варианта
Aj — обобщенный вариант А2, обозначается подгруппой А2(П) или A2B(IV) соот-
ветственно. Выявляемые антитела к отсутствующим частям антигена А(АЦ назы-
вают экстрагглютининами и обозначают как анти-Aj или о ,
Возможное присутствие у реципиентов с подгруппой А, экстрагглютинина
анти-А1 обусловливает назначение им гемокомпонентов с обязательным исклю-
чением антигена А. Реципиентам с А (II) назначают эритроцитсодержашие гемо-
компоненты группы 0(1), а реципиентам с A?B(IV) — группы ВЦП) или 0(1). При
невозможности выбора указанных гемокомпонентив трансфузии эритроцитсо-
держащих сред группы крови А(П) или АВЦУ) проводят, только используя метод
индивидуального подбора.
СИСТЕМА РЕЗУС
Система состоит из 75 антигенов, каждый из которых имеет различное кли-
ническое значение. Выделяют 5 наиболее важных антигенов системы Резус (Rh):
D, С, с, Е, е. Самый сильный иммуноген системы — антиген D. Термины «резус-
положительный» и «резус-отрицательный» относят к наличию или отсутствию
178
ИММУНОГЕМАТОЛОГИЯ
8Л ¥8Ш
антигена D на мембране эритроцита индивидуума. Распространенность антигена
Rh (D) в европейской популяции составляет 85%. Rh(D)-отрицательное население
России составляет 15%. В связи с тем что антиген D системы Резус обладает силь-
ными антигенными свойствами, его определение обязательно как для доноров,
так и для реципиентов гемокомпонентов наряду с определением группы крови
системы АВО. В отличие от системы АВО, в норме к отсутствующим антигенам
системы Резус (в том числе и антигену D) антитела не вырабатываются. Для выра-
ботки антител обязательно должен произойти контакт клеток иммунной системы
с чужеродным антигеном (сенсибилизация), что возможно при трансфузии несо-
вместимой по этому антигену крови или антиген-конфликтной беременности.
При повторном контакте с антигеном, вызвавшим сенсибилизацию, вырабатыва-
ются антитела, приводящие к серьезным гемолитическим посттрансфузионным
осложнениям и гемолитической болезни плода и новорожденного. Реципиентам
с антигеном D обычно переливают Rh(D)-положительные гемокомпоненты,
хотя с учетом всех клинически значимых антигенов системы Резус (С, с, Е, е)
возможно использование и Rh(D)-отрицательных трансфузионных сред. Rh(D)-
отрицательным реципиентам для предотвращения иммунизации и последующих
тяжелых осложнений назначают трансфузии только Rh(D)-отрицательных гемо-
компонентов.
Несмотря на то, что определение антигена D в клинической практике исполь-
зуют уже давно, в некоторых случаях возможны затруднения. Обычно такие
отклонения в определении антигена D обозначают специальным термином D'J.
Основные причины появления антигена Du: несовершенство реагентов и методов
определения, генетически обусловленная неоднородность антигена.
Эпитопы нормального антигена D:
< • все эпитопы присутствуют у Rh(D)-положительных индивидуумов;
❖ все эпитопы отсутствуют у Rh(D)-отрицательных индивидуумов;
❖ в некоторых случаях у индивидуумов отсутствует один или несколько эпито-
пов — частичный D (partial D).
В некоторых случаях анти-П-антитела находят у небольшого числа Rh(D)-
положительных субъектов; показано, что в таких ситуациях эритроциты несут
только часть эпитопов антигена D. При этом к утраченным эпитопам могут
вырабатываться антитела; анти-В-аллоантитела реагируют практически со всеми
эритроцитами ИЬ(В)-положительных индивидуумов, имеющих нормальный
(полный) антиген D. Продукция анти-П-антител индивидуумами с частичным
антигеном О — достаточно редкие случаи, тем не менее это является причиной
серьезных клинических осложнений, включая гемолиз после гемотрансфузии и
тяжелую гемолитическую болезнь новорожденных. Соответственно все варианты
антигена Du должны быть обнаружены в случаях, когда его присутствие может
вызвать иммунологическую угрозу.
Доноры гемокомпонентов. Гемокомпоненты доноров с антигеном Du мар-
кируют как Rh(D)-положительные в связи с тем, что при их переливании Rh(D)-
отрицательным индивидуумам может произойти стимуляция выработки анти-
тел и развиться посттрансфузионные осложнения. Б то же время при внесении
данных о групповой (АВО) и резус-принадлежности в документ донора с анти-
геном Du рассматривают как потенциального реципиента, в связи с чем его резус-
принадлежность в этом случае маркируют как Rh(D)-отрицательную.
Беременные. Определение антигена D11 следует проводить для беременных,
одновременно выполняя скрининг антител. Женщины, имеющие слабый антиген D,
не нуждаются в дородовых и послеродовых инъекциях иммуноглобулина анти-
резус (D).
Н оворожденные Rh(D)-отрицательных матерей. Определение антигена D'1
должно быть выполнено у новорожденных Rh(D)-отрицательных матерей. Если
ИММУНОГЕМАТОЛОГИЯ 179
новорожденный имеет антиген Du, то Rh(D)-отрицательной матери назначают
иммунопрофилактику иммуноглобулином анти-резус. Определение антигена Du
для беременных и новорожденных Rh(D)-отрицательных матерей связано исклю-
чительно с назначением иммуноглобулина анти-резус (D). Как реципиентов гемо-
компонентов их считают Rh(D)-отрицательными. Для реципиентов определение
антигена Du необязательно. Если Rh(D)-принадлежность больного определена как
Rh(D)-отрицательная стандартными методами, то тест для выявления антигена Du
не проводят, так как переливать будут Rh(D)-отрицательные гемокомпоненты.
Это предотвращает сенсибилизацию по антигену D, но не является гарантией
безопасности по другим эрит роцитарным ан гигенам.
ИММУНОГЕМАТОЛОГИЧЕСКАЯ БЕЗОПАСНОСТЬ
ТРАНСФУЗИОННОЙ ТЕРАПИИ
В стандартных случаях нет необходимости в широком типировании антигенов
эритроцитов для доноров и реципиентов гемокомпонентов. В норме к антигенам
всех эритроцитарных систем, за исключением системы АВО, антител быть не
должно. По этой причине принцип безопасности трансфузий эритроцитсодержащих
гемокомпонентов возможно осуществлять за счет выявления предшествующей
сенсибилизации, что достигается обязательным для всех доноров и реципиентов
скринингом антиэритроцитарных антител. Антитела к антигенам эритроцитов
принято классифицировать с учетом их клинического значения. Под клини-
чески значимыми антителами подразумевают антитела, способные вызывать
in vivo разрушение эритроцитов, имеющих на мембране соответствующий анти-
ген. С существованием этих антител связаны механизмы развития гемолити-
ческой болезни новорожденных, посттрансфузионных гемолитических реак-
ций, осложнений или укорочение времени выживания перелитых эритроцитов.
При отсутствии антител у реципиента (отрицательный результат скрининга)
эритроцитсодержащие среды выбирают только с учетом группы крови по системе
АВО и Rh(D)-принадлежности. Положительный результат скрининга говорит об
«опасном» реципиенте; трансфузии гемокомпонентов осуществляют только по
индивидуальному подбору, с обязательным исключение.м антигена, к которому
выявлена сенсибилизация.
Реципиентам с запланированными многократными трансфузиями гемоком-
понентов (пациенты гематологических и онкологических стационаров, на гемо-
диализе, реципиенты органов и тканей) целесообразно выполнять типирование
эритроцитов по антигенам С, с, Е, е системы Резус, антигену К системы Келл и
другим для точных рекомендаций по выбору гемокомпонентов для трансфузий.
Индивидуальный подбор таким реципиентам проводят не только с учетом выяв-
ления предшествующей сенсибилизации, но и с обязательным учетом фенотипа
эритроцитов для исключения при выборе гемокомпонентов антигенов, способных
стать причиной появления клинически значимых антител.
РЕАГЕНТЫ И МЕТОДЫ ДЛЯ ОПРЕДЕЛЕНИЯ ГРУППЫ КРОВИ ПО СИСТЕМЕ АВй,
РЕЗУС(О)-ПРИНАДЛЕЖНОСТИ
Для определения антигенов системы АВО изначально использовали реагенты,
полученные из сыворотки крови доноров с установленной групповой принадлеж-
ностью. Это были проверенные и специальными способами окрашенные поли-
клональные реагенты, содержащие антитела к отсутствующим у данного донора
антигенам системы АВО. Такие диагностикумы называют стандартными изогемаг-
глютинирующими сыворотками. Для получения реагента, позволяющего опреде-
лять резус(В)-принадлежность, долгое время использовали поликлональные
реагенты, приготовленные из сыворотки крови резус(Э)-отрицательных доноров-
180
ИММУНОГЕМАТОЛОГИЯ
добровольцев, иммунизированных резус(D)-положительными эритроцитами дру-
гих доноров. В конце 80-х гг. XX в. альтернативой реагентам, полученным из крови
человека, стали моноклональные антитела — продукт биотехно готического произ-
водства, позволивший получить стандартные реагенты к антигенам эритроцитов.
Моноклональные антитела выпускают под различными торговыми названиями
(наиболее распространены в РФ «Цоликлоны». «Медиклоны», «Трансклоны»).
Стандартные изогемагглютинирующие сыворотки и моноклональные антитела
работают в реакции прямой агглютинации, образуя с эритроцитами, несущими
соответствующий антиген, агтлютинаты — комплексы, видимые невооруженным
глазом. Для поликлональных антител при определении резус(Э)-принадлежности,
как правило, необходимы дополнительные реагенты для проявления результатов
неспецифического (33% раствор полиглюкина, 10% раствор желатина) или специ-
фического (антиглобулиновый реагент) взаимодействия антигенов и антител.
Для полноценного определения группы крови по системе АВО необходимо,
наряду с определением антигенов, выявлять существующие в норме антитела к
отсутствующим антигенам. Для определения антител используют консервиро-
ванные стандартные эритроциты доноров группы А (II) и В (III). Определение
антигенов эритроцитов системы АВО стандартными изогемагглютинируюшими
сыворотками или моноклональными антителами называют прямой реакцией
для определения труппы крови. При применении совместно с ними стандартных
эритроцитов (обратная реакция) говорят о перекрестном методе определения.
Диагностика подгруппы А, возможна только с использованием специального реа-
гента анти- А .
Определение резус(D)-принадлежности с поликлональными и моноклональны-
ми реагентами проводят различными методами. При этом выявление вариантов
антигена Du требует дополнительного тес гирования.
Наиболее точный метод диагностики антител к антигенам эритроцитов —
непрямой антиглобулиновый тест (НАГТ), Важная составляющая адекватного
скрининга антител — использование образцов стандартных эритроцитов группы
0 (I), типированных по всем клинически значимым системам эритроцитов.
В настоящее время, наряду с серологическими методами диагностики антигенов
эритроцитов и антиэритроцитарных антител (самый точный из которых — метод
агглютинации в геле), используют и молекулярно-биологическую диагностику.
Применение этих методов ограничено ввиду трудоемкости и высокой стоимости.
ПОРЯДОК ВЫПОЛНЕНИЯ иммуногематологической апробации
КРОВИ ДОНОРОВ
Для всех доноров крови или ее компонентов, необходимы иммуногематологиче-
ские исследования; при их проведении всех исследуемых рассматриваю! как доно-
ров гемокомпонентов. При внесении данных о групповой и резус-принадлежности
в удостоверение личности или иной документ донора рассматривают как реципи-
ента гемокомпонентов.
1. Первично перед донацией группу крови по системе АВО определяют простой
реакцией (используя стандартные изогемагглютинируюшие сыворотки или моно-
клональные антитела). Результат записывают в карту донора с обязательным ука-
занием серий используемых реагентов и даты определения. Для кадровых доноров
при повторных донациях данную процедуру можно не проводить.
2. Во время донации с применением закрытых систем взятия крови необходим
контрольный образец крови донора для выполнения иммуногематологических
исследований. Для апробации используют пробирки с активатором свертывания
крови или антикоагулянтом на основе ЭДТА или цитрата Na объемом не менее
5 мл (требования к образцу крови определяет лаборатория, в которой проводят
исследования). Пробирку маркируют одновременно с заготовленной гемопродук-
ИММУНОГЕМАТОЛОГИЯ 181
цией и передают с направлением на исследование, имеющим аналогичную марки-
ровку, в КДЛ.
3. В КДЛ врач, прошедший специальную подготовку по иммуногематологии, у
переданного образца крови донора выполняет определение группы крови ( АВО)
перекрестным методом, резус-принадлежности (D), типирование по антигенам С,
с, Е, е системы Резус и антигена К системы Келл, исследование на наличие алло-
иммунных антиэритроцитарных антител. Результаты определения фиксируют в
рабочем журнале, электронной картотеке и выносят на лицевую сторону карты
донора с датой и подписью врача.
4. Наличие слабой агглютинации при выявлении антигена А и/или экстраг-
глютинина анти-А] требует дополнительной верификации подгруппы антигена А
(Aj) с реагентами анти-А1 и анти-Н. При выявлении экстрагглютинина анти-Aj
учреждение службы крови не выдает для переливания консервированну ю кровь
или полученные из этой дозы крови плазму, криопреципитат и концентрат тром-
боцитов.
5. Обязательно определение слабых вариантов антигена D (Du) у доноров. При
наличии слабых вариантов антигена D (Du) резус-принадлежность донора считают
положительной.
6. Типирование антигенов эритроцитов С, с, Е, е, К систем Резус и Келл у доно-
ров обязательно: проводят исследование 3 раза (однократно при каждой донации)
различными сериями типирующих реагентов или различными реагентами. При
совпадении результатов Резус/Келл (Rh/K) фенотип считают установленным и
при последующих донациях не определяют. При использовании гелевой техноло-
гии для типирования указанных антигенов допускают двукратное исследование.
Аналогично осуществляют типирование антигенов эритроцитов других систем,
выполняемое по мере необходимости.
7. Полученные данные о групповой (АВО) принадлежности с учетом подгруп-
пы А . резус-принадлежности (D), Rh/K-фенотипе эритроцитов донора выносят
на этикетку эоитроцитсодержашего гемокомпонента. Резус-принадлеж] юсть гемо-
компонентов маркируют как положительную при наличии антигенов D (или D"), С
или Е системы Резус или как отрицательную при их отсутствии. Данные о полном
типировании антигенов С, с, Е, е системы Резус и антигена К системы Келл должны
быть представлены в Rh/K-фенотипе.
8. Скрининг антиэритроцитарных аллоантител обязателен для всех доноров
гемокомпонентов, независимо от групповой и резус-принадлежности крови.
Исследование проводят при каждой донации. При обнаружении антиэритроци-
тарных аллоантител в образце донорской крови учреждение службы крови не
выдает для переливания консервированную кровь или полученные из этой дозы
крови плазму, криопреципитат и концентрат тромбоцитов. Допускается донорство
эритроцитов. Плазма доноров с аллоантителами может быть использована для
изготовления препаратов и иммуногематологических диагностикумов.
9. Для выяснения специфичности антител проводят их идентификацию.
Идентификацию антител считают завершенной при соответствии результатов
реакций с панелью эритроцитов результатам типирования антигенов.
10. Справку о наличии антиэритроцитарных антител с указанием их специфич-
ности и рекомендаций по выбору гемокомпонентов для переливания выдают на
руки донору для предъявления при госпитализации. В рекомендациях указывают,
что данному человеку в дальнейшем как реципиенту для переливания необходимо
выбирать эритроцитсодержащие гемокомпоненты только с отсутствием антигена,
к которому выявлены антитела, даже если антитела повторно не выявляют. Такая
справка должна всегда быть при доноре для обеспечения безопасности трансфу-
зий, в том числе при экстренных госпитализациях.
182
ИММУНОГЕМАТОЛОГИЯ
ПОРЯДОК ВЫПОЛНЕНИЯ ИММУНОГЕМАТОЛОГИЧЕСКОЙ АПРОБАЦИИ КРОВИ
РЕЦИПИЕНТОВ
Всем пациентам, которым может потребоваться переливание крови или ее ком-
понентов, должны быть выполнены иммуногематологические исследования.
• Первично группу крови по системе АВО определяет лечащий (или дежурный)
врач простой реакцией (стандартными изогемагглютинирующими сыворот-
ками или моноклональными антителами). Результат записывают в медицин-
ской карте больного и направлении в КДЛ с обязательным указанием серий
используемых реагентов, даты и фамилии врача, проводившего определение.
В направлении указывают также фамилию, имя и отчество больного, возраст,
отделение, номер палаты и номер медицинской карты.
• При использовании закрытых систем взятия крови должен быть взят кон-
трольный образец венозной крови для выполнения иммуногематологиче-
ских исследований. Для апробации могут быть использованы пробирки с
активатором свертывания крови или антикоагулянтом объемом не менее
5 мл (требования к образцу крови определяет лаборатория, в которой про-
водят исследования). Пробирку маркируют и передают с направлением на
исследование в КДЛ.
• В КДЛ врач, прошедший специальную подготовку по иммуногематоло-
гии, определяет группу крови (АВО) перекрестным методом и резус-(П)-
принадлежность, исследует на наличие аллоиммуиных антител к антигенам
эритроцитов в переданном образце крови больного. Результаты определения
фиксируют в рабочем журнале и направлении на исследование (при нали-
чии информационных систем результат анализа выдают на специальном
бланке лаборатории). Выдача ответа КДЛ возможна лишь в случае совпа-
дения результатов первичного и повторного определения. Если результаты
не совпадают, проводят повторные исследования из вновь заготовленного
образца крови больного.
• Наличие экстрагглютинина анти-А1 и/или слабой агглютинации при выявле-
нии антигена А требует дополнительной верификации подгруппы антигена А
(А^ с реагентами анти-А и анти-Н.
• При наличии слабых вариантов антигена D (Du) резус-принадлежность реци-
пиента считают отрицательной.
• Скрининг аллоиммуиных антител к антигенам эритроцитов обязате-
лен для всех реципиентов гемокомпонентов при плановых трансфузи-
ях (независимо от групповой (АВО) и резус-(D)-принадлежности крови).
Исследование выполняют в КДЛ перед каждой вновь назначаемой транс-
фузией. Обнаружение антиэритроцитарных аллоантител — основание для
назначения реципиенту индивидуального подбора по антигенам эритроцитов
при трансфузиях эритроцитсодержащих гемокомпонентов.
• Для выяснения специфичности антител образец крови реципиента передают
в референсную (экспертную) лабораторию, где проводят идентификацию
антител.
• Справку о наличии антител с указанием их специфичности, фенотипа эритро-
цитов и рекомендаций по выбору гемокомпонентов для переливания выдают
на руки реципиенту для предъявления при последующих госпитализациях. Б
рекомендациях указывают, что данному человеку в дальнейшем как реципи-
енту для переливания необходимо выбирать эритроцитсодержащие гемоком-
поненты только с отсутствием антигена, к которому выявлены антитела, даже
если антитела повторно не выявляют. Такая справка находится у пациента и
предъявляется для обеспечения безопасности трансфузий, в том числе при
экстренных госпитализациях.
ИММУНОГЕМАТОЛОГИЯ 183
• После получения ответа из КДЛ лечащий врач выносит данные о группе
(АВО) и резус-принадлежности крови на лицевую сторону медицинской
карты больного, проставив дату вынесения и свою подпись. Бланк ответа
вклеивают в медицинскую карту? пациента.
• Если больной не относится к категории «опасных» реципиентов или реци-
пиентов группы риска по данным анамнеза и отсутствуют аллоантитела к
антигенам эритроцитов (отрицательный результат скрининга антител), то
эритроцитсодержащие гемокомпоненты переливают с учетом совместимо-
сти по группе крови (АВО) и резус-принадлежности (D). Перед трансфузией
обязательно определение врачом группы крови (АВО) донора и реципиента и
постановка проб на совместимость.
• При экстренных показаниях к трансфузии гемокомпонентов и отсутствии
возможности выполнения иммуногематологическиго исследования в КДЛ
(ночное время, выходные и праздничные дни) исследование групповой
(АВО) и резус-принадлежности (D) проводит дежурный врач; при этом в обя-
зательном порядке заготавливают образец крови больного до трансфузии,
который передают в КДЛ в часы ее работы.
ИН ДИ ИДУАЛЬНЫЙ ПОДБОР ПО АНТИГЕНАМ ЭРИТРОЦИТОВ
Индивидуальный подбор эритроцитсодержащих гемокомпонентов — это
предтрансфузионные пробы на совместимость сыворотки (плазмы) крови реци-
пиента с эритроцитами донора (или донороь) гемокомпонентов. выполненные
в условиях специализированной лаборатории адекватными методами. Такой
подбор может быть рекомендован всем реципиентам гемокомпонентов. Группы
пациентов, для которых индивидуальный подбор эритроцитсодержащих гемо-
компонентов обязателен при каждой плановой трансфузии эритрицитсодержа-
щих гемокомпонентов:
реципиенты с отягощенным акушерским и трансфузионным анамнезом:
❖ реципиенты с положительным результатом скрининга антител;
❖ сенсибилизированные реципиенты (по данным анамнеза), независимо от
выявления антител в настоящий момент;
< > новорожденные;
< > пациенты педиатрических стационаров;
❖ беременные, роженицы, родильницы;
❖ реципиенты с многократными трансфузиями в анамнезе:
< > пациенты при неэффективности трансфузий эритроцитсодержащих гемоком-
понентов;
о пациенты гематологических и онкологических стационаров;
❖ пациенты отделений гемодиализа;
❖ реци пиенты органов и тканей.
Индивидуальный подбор осуществляют из совместимых с реципиентом по
группе крови (АВО), резус-принадлежности (D) образцов крови доноров, отри-
цательных по антигену, к которому в данный момент или ранее были выявлены
антитела у пациента.
Исследования образцов крови доноров и реципиента при выполнении индиви-
дуального подбора в специализированной лабора гории:
повторное определение групповой (АВО) и резус-принадлежности (D);
❖ скрининг и идентификация антиэритроцитарных аллоиммунных антител у
реципиента;
проба на совместимость сыворотки (плазмы) крови реципиента и эритро-
цитов каждого образца крови доноров гемокомпонентов в непрямом анти-
глобул иновом тесте (НАГТ) для выявления несовместимости, вызванной
клинически значимыми антителами.
184 ИММУНОГЕМАТОЛОГИЯ
Результаты исследований фиксируют в журнале индивидуального подбора
крови. Ответ выдают на бланке индивидуального подбора с обязательным ука-
занием фамилии, имени и отчества, группы крови (АВО), резус-принадлежности
(D), специфичности выявленных и ранее обнаруженных антител реципиента.
Также в нем отражают сведения об идентификационных номерах, группах крови
(АВО), резус-принадлежности (D) и фенотипе по соответствующему антигену всех
подобранных образцов крови доноров гемокомпонентов с обязательным указа-
нием методов индивидуального подбора, даты и подписи врача, осуществлявшего
исследование.
Для трансфузий используют только гемокомпоненты, совместимые по антиге-
нам эритроцитов. Несовместимость в ходе индивидуального подбора — противо-
показание к трансфузии гемокомпонента от данного донора. Бланк индивидуаль-
ного подбора передают в ЛПУ вместе с гемокомпонентом и вклеивают в историю
болезни реципиента при осуществлении трансфузии данного гемокомпонента.
Перед трансфузией гемокомпонентов по индивидуальному подбору врач, осу-
ществляющий трансфузию, обязательно определяет групповую принадлежность
крови доноров и реципиента. Убедившись в том, что они совпадают с данными
истории болезни и бланка индивидуального подбора, врач проводит биологиче-
скую пробу на совместимость и осуществляет трансфузию гемокомпонентов.
ЛАБОРАТОРНЫЕ ИССЛЕДОВАНИЯ ПРИ ПОСТТРАНСФУЗИОННЫХ РЕАКЦИЯХ
ГЕМОЛИТИЧЕСКОГО ТИПА
Лабораторные исследования при подозрении на посттрансфузионное осложне-
ние (ПТО) должны быть направлены, с одной стороны, на подтверждение диагноза
гемолитической реакции, с другой — на выявление причин и подбор совместимых
гемокомпонентов для последующих трансфузий.
К первой группе относят тесты, подтверждающие гемолиз: определение кон-
центрации гемоглобина и гематокрита. Показатели острого внутрисосудистого
гемолиза — гемоглобинурия, гемоглобинемия и гипербилирубинемия за счет
неконъюгированного билирубина.
Вторая группа тестов направлена на выяснение причины иммуногематологиче-
ской несовместимости, приведшей к ПТО. Здесь необходимо тщательное исследо-
вание образцов крови донора и реципиента, причем для реципиента дополнитель-
но изучают образец крови, взятый до переливания. Во всех трех образцах крови
проводят повторное определение групповой (АВО) и резус(О)-принадлежности,
фенотипирование эритроцитов, скрининг и идентификацию аллоантител. Для
этих исследований используют НАГТ, непрямую реакцию Кумбса. Кроме этого, из
образца крови больного после трансфузии проводят прямой антиглобулиновый
тест (ПАГТ, прямую пробу Кумбса,); исследование используют для выявления
антител или компонентов комплемента, фиксированных на поверхности эритро-
цитов.
Проведение прямой пробы Кумбса показано при следующих состояниях:
аутоиммунный гемолиз;
4 гемолитическая болезнь новорожденных;
лекарственная иммунная гемолитическая анемия;
❖ гемолитические посттрансфузионные реакции и осложнения.
Для определения специфичности антител используют элюат с поверхности сен-
сибилизированных эритроцитов. Если в образце крови больного, взятом до пере-
ливания, обнаружены антитела, их специфичность совпала с полученной в элюате,
при этом у больного не выявлен антиген (или антигены), к которому определены
антитела, то диагноз ПТО подтвержден и причина установлена (специфичность
антигена). В случае подозрения на посттрансфузионное осложнение гемолитиче-
ского типа для последующих трансфузий такому реципиенту необходимо выби-
ИММУНОГЕМАТОЛОГИЯ 185
рать эритроцитсодержащие гемокомпэненты только по индивидуальному подбору
в непрямом антиглобулиновом тесте (непрямой реакции Кумбса) с обязательным
исключением в донорской крови антигена — причины ПТО,
ИММУНОПАТОЛОГИЧЕСКИЕ ПАТОЛОГИЧЕСКИЕ СОСТОЯНИЯ
Лабораторная диагностика гемолитической болезни плода
и новорожденного
Гемолитическая болезнь плода или новорожденного — патологическое состоя-
ние, обусловленное несовместимостью матери и плода (новорожденного) по
антигенам эритроцитов. Подобная несовместимость может быть вызвана практи-
чески любым антигеном, унаследованным ребенком от биологического отца, при
условии отсутствия этого антигена у матери. Выработка антител (за исключением
антител к антигенам системы АВО) происходит при контакте чужеродного антиге-
на с клетками иммунной системы матери, что возможно только при беременности
или трансфузии. При этом сенсибилизация происходит при первой беременности
(трансфузии), а антитела вырабатываются при повторном контакте с данным
антигеном. По этой причине первая беременность обычно протекает без клини-
ческих признаков конфликта. Механизм иммунизации чужеродным для матери
антигеном запускается в результате трансплапентарных геморрагий, характерных
для любой беременности и обычно составляюших не более 1 мл. Количество крови
плода, достаточное для иммунизации, может попасть в кровь матери в момент
родов или при плодоразрушающих процедурах. Риск иммунизации высок при
амниоцентезе или кордоцентезе. В процессе внутриутробного развития антиге-
ны эритроцитов формируются довольно рано; так, антигены системы АВО плода
полностью сформированы уже к 8-9-й неделе беременности. При несовместимо-
сти по антигенам системы АВО гемолитическая болезнь новорожденных может
возникнуть даже во время первой беременности в связи с тем. что антитела к этим
антигенам постоянно присутствуют в крови матери.
Гемолитическая болезнь плода наиболее вероятна для конфликта мать-плод по
антигену D системы Резус. Для несовместимости по другим антигенам эпитроцитов
характерно развитие гемолитической болезни новорожденного. Клиническая кар-
тина болезни определяется интенсивностью гемолиза эритроцитов. У плода разви-
вается гемолитическая анемия, в тяжелых случаях приводящая к сердечной недо-
статочности, водянке и внутриутробной гибели. Снижение гематокрита в образце
крови плода до 18% и менее, а также совокупность других критериев оценки его
состояния — показание для внутриутробного переливания эритроцитсодержащих
гемокомпонентов Для переливания в этом случае выбирают отмытые, лейко-
фильтрованные и облученные эритроциты группы 0(1). совместимые по антигенам
эритроцитов с сывороткой крови матери в непрямом антиглобулиновом тесте. При
необходимости такие трансфузии повторяют каждые 2-3 нед.
Гипербилирубинемия у плода, как правило, не возникает, так как билирубин
свободно проникает через плаценту и выводится организмом матери. У новорож-
денного, напротив, наибольшую опасность представляет гипербилирубинемия,
поскольку она может привести к билирубиновой энцефалопатии.
При несовместимости по антигенам эритроцитов сразу после рождения опреде-
ляют группу крови (АВО), резус(Б)-принадлежность, уровень гемоглобина и
билирубина в пуповинной крови. После внутриутробного заменного переливания
резус-принадлежность крови новорожденного может быть определена как Rh(D)-
отрицательная из-за присутствия в кровеносном русле большого количества
перелитых донорских эритроцитов. С эритроцитами новорожденного проводят
186
ИММУНОГЕМАТОЛОГИЯ
прямую пробу Кумбса; при положительной пробе определяют, к каким эритро-
цитарным антигенам направлены антитела. При гипербилирубинемии (уровень
билирубина более 20 мкмоль/л у доношенных и 12-16 мкмоль/л у недоношен-
ных) применяют светолечение и заменное переливание крови. Для светолечения
используют ультрафиолетовый спектр, способствующий превращению билируби-
на в водорастворимые продукты. Эритроциты, покрытые антителами, в некоторых
случаях разрушаются медленно, что может привести к анемии при отсутствии
гипербилирубинемии даже через 3-6 нед после рождения.
Гемолитическая болезнь новорожденных, вызванная несовместимостью по
антигенам системы АВО, может возникнуть даже у ребенка, рожденного от первой
беременности, Иногда в первые сутки жизни появляется желтуха, причем гемолиз
бывает выражен интенсивнее, чем при несовместимости по антигенам системы
Резус. Прямая проба Кумбса с эритроцитами новорожденного отрицательна или
слабо положительна. Антитела, элюированные с эритроцитов новорожденного,
агглютинируют эритроциты доноров с группами крови А (II) или В (III) и АВ (IV).
В мазке крови новорожденного выявляют сфероциты, иногда фрагментированные
эритроциты. Гемолитическую болезнь новорожденных, вызванную несовмести-
мостью по антигенам системы АВО, бывает трудно отличить от наследственного
микросфероцитоза. Если необходимо заменное переливание, то используют эри-
троцитсодержащие гемокомпоненты группы О (I). индивидуально подобранные по
сыворотке крови матери, и плазму группы АВ (IV). Интенсивное раннее лечение
позволяет снизить риск осложнений.
Риск иммунизации антигеном D системы Резус в процессе беременности
существенно снижается, если резус-отрицательной матери с отсутствием анти-
тел вводят иммуноглобулин анти-резус(Э). Первично его назначают в конце
II триместра беременности, второй раз — в первые 72 ч после рождения резусо-
положительного ребенка. Антирезусный иммуноглобулин также необходимо вво-
дить после аборта и амниоцентеза.
Иммуногематологические исследования в акушерстве и неонатологии играют
большую роль как для обеспечения безопасности гемокомпонентного лечения,
так и для диагностики гемолитических процессов. Их правильное и своевременное
выполнение дополнит и подтвердит клинический диагноз, а также позволит про-
контролировать адекватность лечебных меооприятий. Все беременные должны
проходить плановые иммуногематологические исследования с целью выявления
конфликта по эритроцитарным антигенам между матерью и плодом, определе-
ния риска развития гемолитической болезни плода и новорожденного и воз-
можной профилактики резус-сенсибилизации. Определение группы крови по
системе АВО перекрестным методом и резус(О)-принадлежности выполняют
два раза в процессе беременности (рис. 18-1). Обязательно выявление слабых
вариантов антигена О (Du). При обнаружении слабых вариантов антигена Du
резус-принадлежность беременных считают отрицательной, но назначение имму-
ноглобулина анти-резус (D) противопоказано. Скрининг аллоиммунных анти-
тел к антигенам эритроцитов производят независимо от резус-принадлежности
беременной не реже 1 раза в триместр. Скрининг антиэритроцитарных аллоан-
тител проводят в непрямом антиглобулиновом тесте (непрямой реакции Кумбса).
Обязательно проведение скрининга антител перед назначением иммуноглобули-
на анти-резус Rh(D)-отрицательным женщинам, как в процессе беременности,
так и после родов и абортов. При обнаружении антител специфичности анти-D
назначение иммуноглобулина анти-резус (D) противопоказано. Обнаружение
антиэритроцитарных аллоантител — показание к определению их титра и назна-
чению повторных исследований (не реже 1 раза в месяц) в течение беременности
для определения динамики титров антител и прогноза развития гемолитической
болезни плода и новорожденного. Если обнаружена несовместимость крови бере-
ИММУНОГЕМАТОЛОГИЯ 187
менной и биологического отца ребенка по антигенам системы АВО и есть указания
на наличие гемолитической болезни новорожденного по системе АВО в анамнезе,
то целесообразно определение титров иммунных антител (IgG) системы АВО в
динамике по аналогичной схеме (см. рис. 18-1). Важно учитывать, что гемолити-
ческая болезнь новорожденного при несовместимости по системе АВО характерна
не только для новорожденных от матерей группы О (I), но и от матерей группы
А (И) и В (III). При несовместимости, когда мать и огец имеют группу А (II) и В
(III), отмечают преобладание тяжелых степеней гемолитической болезни ново-
рожденного, требующих срочного заменного переливания для лечения.
Наличие антизритроцитарных аллоантител — признак возможного конфликта
матери и плода по антигенам эритроцитов, а увеличение титра антител определен-
ной специфичности в динамике с большой вероятностью указывает на иммуноло-
гический конфликт при данной беременности. При этом необходимо учитывать
не абсолютную величину титра антител, а только динамику. Титром антител
называют последнее разведение сыворотки, в котором обнаруживают антитела.
Соответственно такие разведения, полученные в различных лабораториях и раз-
личными методами, могут существенно различаться. Это зависит прежде всего
от чувствительности и специфичности используемых лабораторных методик.
Сравнивать можно только значения титра, полученные в одном и том же мето-
де определения, но с учетом того, что определенная ошибка может находиться
и в пределах погрешности метода. По этой причине достоверным изменением
величины титра принято считать его снижение или увеличение более чем на две
ступени.
В случае выявления аллоиммунных антител, при выполнении заменных транс-
фузий индивидуальный подбор по антигенам эритроцитов необходим беременной
и плоду (новорожденному). При необходимости внутриутробных переливаний
плоду выбирают эритроцитсодержащие гемокомпоненты группы О (I), совмести-
мые в индивидуальном подборе по антигенам эритроцитов с сывороткой матери,
Рис. 18-1. Антенатальный скрининг антител.
188 ИММУНОГЕМАТОЛОГИЯ
даже если предварительному исследованию на группу крови (АВО) подвергался
образец крови плода, полученный методом кордоцентеза. Антитела системы АВО
у новорожденных могут частично или полностью отсутствовать. В этих случаях
заключение о групповой принадлежности делают по антигенам эритроцитов.
Верификацию подгруппы антигена А у детей до 18 мес не производят. При обнару-
жении слабых вариантов антигена А у новорожденных для трансфузий выбирают
эритроцитсодержащие гемокомпоненты без антигена А.
Во избежание ошибок при определении групповой (АВО) и резус-принадле-
жности новорожденных из образца пуповинной крови необходимо исключить
химеризм по антигенам эритроцитов (наличие двух популяций эритроцитов, одна
из которых принадлежит матери, а другая — ребенку). Если выявляют химеризм,
то определение необходимо повторить из образца крови, взятого непосредственно
у новорожденного. Определение антител, фиксированных на эритроцитах (прямая
реакция Кумбса), выполняют новорожденным при подозрении на гемолитиче-
скую болезнь. Положительный результат прямой реакции Кумбса подтвержцает
гемолитический конфликт, отрицательный же его не исключает. Скрининг алло-
иммунных антител к антигенам эритроцитов проводят для установления причин
гемолитической болезни и разработки точных рекомендаций по выбору гемо-
компонентов для возможных трансфузий. Одновременно необходимо определять
антитела в сыворотке крови матери. Обнаружение антиэритроцитарных аллоан-
тител — показание к опреде пению их специфичности (идентификации). В этом
случае новорожденный нуждается в индивидуальном подборе эритроцитов для
заменных переливаний по сыворотке матери и собственной сыворотке крови.
Иммунные гемолитические анемии
В норме время жизни эритроцитов составляет 90-120 дней. При гемолизе,
связанном с повреждением эритроцитов, оно укорачивается. Причинами гемо-
лиза могут быть как дефекты эритроцитов, так и внешние воздействия. Диагноз
гемолитической анемии основан на совокупности клинических данных и лабо-
раторных показателей: содержание гемоглобина; гематокрит: количество рети-
кулоцитов; морфология эритроцитов; концентрация билирубина, гаптоглоби-
на; активность лактатдегидрогеназы; определение времени жизни эритроцитов.
Пммуногематологические исследования помогают определить, имеет ли гемолиз
иммунную природу, а при ее подтверждении отнести иммунную анемию к опреде-
ленному типу. Иммунный гемолиз обусловлен выработкой антител к эритроци-
тарным антигенам с последующим разрушением эритроцитов фагоцитозом или за
счет активации комплемента. Иммунный гемолиз может быть вызван антителами
как аллогенной, так и аутологичной природы, а в некоторых случаях — антитела-
ми к лекарственным препаратам. Другие причины гемолиза, которые важны при
дифференциальной диагностике иммунных гемолитических анемий: врожденные
дефекты мембраны эритроцитов; механическое повреждение эритроцитов (напри-
мер, при микроангиопатии); инфекции; врожденная недостаточность ферментов
эритроцитов (гемоглобинопатии). Типичные примеры аллоиммунных гемолити-
ческих анемий — посттрансфузионные реакции и осложнения гемолитического
типа, гемолитическая болезнь плода и новорожденного.
Аутоиммунные гемолитические анемии — заболевания, при которых аутоан-
титела к антигенам эритроцитарной мембраны укорачивают время жизни как
собственных, так и перелитых эритроцитов. Причины выработки аутоантител к
эритроцитам:
о фиксация гаптена (например, лекарственного средства) или высокомолеку-
лярных антигенов (например, бактериальных) на эритроцитах;
❖ нарушение функции Т-супрессоров:
ИММУНОГЕМАТОЛОГИЯ 1&Q
❖ изменение структуры эритроцитарных антигенов;
< > перекрестные реакции между бактериальными и эритроцитарными антиге-
нами;
❖ нарушение функции В-лимфоцитов. характерное для гемобластозов и колла-
генозов.
В зависимости от антиэритроцитарных аутоантител выделяют три группы этого
заболевания: тепловую аутоиммунную гемолитическую анемию, болезнь холодо-
вых агглютининов и пароксизмальную ночную гемоглобинурию.
В лабораторной диагностике аутоиммунных процессов большую роль играют
иммуногематологические исследования, связанные с выявлением и определением
характеристик аутоантител. Эти исследования, конечно, не могут объяснить при-
роду аллоиммунизации, но их с успехом в течение многих лет применяют совмест-
но с клиническими данными для определения типа аутоиммунной гемолитической
анемии и выбора оптимальной тактики лечения. В табл. 18-2 обобщены основные
виды иммуногематологических исследований, применяемых для диагностики
аутоиммунного гемолиза.
Таблица 18-2. Иммуногематологические особенности аутоиммунных гемолитических анемий
Cl.'Wr-
Особенности Тип аутоиммунной гемолитической анемии
тепловая болезнь холодовых агглютининов пароксизмальная ночная гемоглобинурия
Оптимальная температу- ра выявления in vitro 37 °C 0 4 СС 0-4 °C — фиксация антител; 37 °C — гемолиз
Класс иммуноглобу- линов Преимущественно IgG, редко IgA, IgM igM IgG
Результат ПАГТ IgG, реже 03 сз СЗ
Специфичность антиге- нов, к которым выявля- ются антитела Системы Резус, Келл, LW, U, Епа, Wrb I р
Тип антител Неполные. Чаще поликло- нальные, реже монокло- нальные Агглютинины, иногда гемо- лизины. Моноклональные при первичной форме болезни, поликлональные — при вторичной Двухфазные гемолизины. Поликлональные
Наиболее характерные иммуногематологиче- ские особенности Наличие ауто- и алло- антител. Трудности индивидуального подбора эритроцитов Спонтанная холодовая агглютинация эритроцитов в образце крови Антитела Доната- Ландштейнера
ИММУНОГЕМАТОЛОГИЧЕСКИЕ ЛАБОРАТОРНЫЕ
ИССЛЕДОВАНИЯ
Методы исследования антигенов эритроцитов
и антиэритроцитарных антител
При выполнении иммуногематологических исследований обязателен ежеднев-
ный внутрилабораторный контроль качества, а также участие во внешнем контро-
ле качества выполняемых исследований.
Определение группы крови по системе АВО
Прямая реакция с моноклональными антителами (изогемагглютинирующими
сыворотками).
190 ИММУНОГЕМАТОЛОГИЯ
Перекрестный метод с моноклональными антителами (изогемагглютинирую-
щими сыворотками) и стандартными эритроцитами Ар В, 0.
Прямая и/или перекрестная реакция в гелевом тесте.
Прямая и/или перекрестная реакция в микропланшетной технологии.
Определение резус-принадлежности (антиген D)
Реакция прямой агглютинации с моноклональными антителами анти-D.
Реакция с универсальным реагентом анти-резус (D).
Реакция конглютинации с 10% раствором желатина.
Непрямой антиглобулиновый тест (непрямая реакция Кумбса).
Реакция с моноклональными антителами в гелевом тесте.
Реакция с моноклональными антителами в микропланшетной технологии.
Типирование антигенов эритроцитов
Реакция прямой агглютинации с моноклональными антителами соответствую-
щей специфичности.
Реакция конглютинации с 10% раствором желатина и антителами соответ-
ствующей специфичности.
Непрямой антиглобулиновый тест (непрямая реакция Кумбса).
Реакция с моноклональными антителами в гелевом тесте.
Реакция с моноклональными антителами в микропланшетной технологии.
Скрининг аллоиммунных антиэритроцитарных антител
Непрямой антиглобулиновый тест (пробирочная или гелевая методика) с
панелью стандартных эритроцитов, состоящей не менее чем из трех видов кле-
ток, типиров|нных по всем клинически значимым антигенам. Применение пула
(смеси) случайных образцов эритроцитов для скрининга антиэритроцитарных
антител не допускается.
Идентификация аллеиммунных антиэритроцитарных антител
Непрямой антиглобулиновый тест (пробирочная или гелевая методика) с пане-
лью стандартных эритроцитов, состоящей не менее чем из десяти видов к ictok.
чипированных по всем клинически значимым антигенам.
Типирование антигена (или антигенов) эритроцитов реципиента, к которому
предположительно обнаружены антитела
Идентификацию антител считают завершенной при соответствии результатов
реакций с панелью эритроцитов результатам фенотипирования эритроцитов
исследуемого образца крови.
Определение иммунных антител системы АВО
Непрямой антиглобулиновый тест (пробирочная или гелевая методика) со
стандартными типированными эритроцитами Ах и/или В после разрушения есте-
ственных антител 5% рас гвором унитиола или прогреванием при 70 °C.
Определение аутоантител
Прямой антиглобулиновый тест (пробирочная или гелевая методика).
ИММУНОГЕМАТОЛОГИЧЕСКИЕ ОСОБЕННОСТИ ТЕРАПИИ КОНЦЕНТРАТОМ
ТРОМБОЦИТОВ
Тромбоциты — носители антигенов системы АВО, системы HLA (Human Leukocyte
Antigens) I класса, а также специфических тромбоцитарных антигенов системы
НРА (Human Platelet Antigens).
HLA-система человека — комплекс гликопротеиновых молекул, которые экс-
прессируются на поверхности большинства ядросодержащих клеток; это наиболее
полиморфная из всех известных антигенных систем. HLA-антигены локализованы
ИММУНОГЕМАТОЛОГИЯ 191
на гликопротеинах клеточной мембраны, кодированных тесно сцепленными гена-
ми, расположенными в сегменте р21.3 короткого плеча хромосомы 6. Комплекс
состоит из нескольких классов, разделенных более чем на 250 локусов (генов и
псевдогенов), 21 из которых прямо связаны с трансплантацией. Локусами кодиру-
ются около 900 аллелей I и II классов. Антигены HLA играют важнейшую роль в
регуляции иммунного ответа на чужеродные антигены и сами являются сильными
антигенами.
В настоящее время широко используют две номенклатуры системы HLA:
старая основана на результатах иммунологических реакций, таких как серологи-
ческое типирование или смешанная культура лейкоцитов in vitro; новая исполь-
зует молекулярно-биологические данные о специфических последовательностях
нуклеотидов в аллелях.
Иммунологическая номенклатура включает название системы (HLA); локус, к
которому принадлежит данный антиген, обозначают большими буквами латин-
ского алфавита (А, В, С, DR, DP, DQ); цифра обозначает непосредственно антиген
(HLA-A,, HLA-EL).
Подобрать донора, полностью совместимого с реципиентом по антигенам
системы HLA, очень сложно, поскольку число комбинаций, составленных более
чем из 100 антигенов этого семейства, чрезвычайно велико. Вероятность найти
полностью совместимого донора составляет от 1:1000 до 1:1 000 000 в зависимо-
сти от распространенности того или иного антигена HLA. Вероятность подбора
полностью совместимого донора среди родных братьев и сестер составляет 1:4; это
связано с тем, что гены HLA наследуются по законам Менделя.
Система НРА диаллельна; имеет аллели «а» и «Ь» и состоит из 16 локусов —
HPA-l-HPA-14w, 15,16w. В настоящее время изучено молекулярное строение всех
32 генов НРА, входящих в новую номенклатуру, в то время как антигенов системы
НРА открыто только 22. Это связано с тем, что в локусах HPA-6w-HPA-14w, 16w
серологически можно определить только редко встречающиеся антигены аллеля
«Ъ»; к часто встречающимся антигенам аллеля «а» антисыворотки до настоящего
времени не получены. Расшифровка молекулярной структуры генов НРА стала
возможной благодаря использованию метода полимеразной цепной реакции
(ПЦР).
Популяционные различия людей по антигенам систем HLA и НРА способствуют
аллоиммунизации пациентов при проведении трансфузионного лечения компо-
нентами крови, прежде всего тромбоцитами. Переливание концентрата тромбоци-
тов — эффективный метод лечения и профилактики геморрагических осложнений,
обусловленных снижением числа тромбоцитов в периферической крови вслед-
ствие аплазии кроветворения или их дисфункцией. Отсутствие скорректирован-
ного посттрансфузиинного прироста тромбоцитов через 18 ч после их трансфузии
называют рефрактерностью. Рефрактерность может быть обусловлена как имму-
нологическими, так и неиммунологическими причинами. Неиммунологическая
рефрактерность связана с повышенным потреблением тромбоцитов, неадек-
ватно пополняемым при их переливании, что происходит при кровотечениях
различной локализации, лихорадке, тромбофлебите, сепсисе, спленомегалии.
Иммунологическая рефрактерность к трансфузиям концентрата тромбоцитов обу-
словлена антителами, выработанными в результате несовмес гимости по антигенам
систем АВО, HLA и НРА реципиентами в ответ на переливание соответствующих
антигенов доноров. Аллоиммунизация пациентов являясь основной причиной
развития иммунологических реакций негемолитического типа, в том числе и
иммунологической рефрактерности, может привести к полному отсутствию кли-
нического эффекта от переливания тромбоцитов. К счастью, состояние иммуноло-
гической рефрактерности возникает редко.
192 ИММУНОГЕМАТОЛОГИЯ
Основной метод профилактики — соблюдение принципа совместимости кон-
центрата тромбоцитов с реципиентом по системе АВО. При этом желательно
учитывать также резус(П)-принадлежность крови, из которой были получены
тромбоциты. Наличие в тромбоконцентоате примеси эритроцитов может привести
к аллоиммунизации и выработке антител к антигену D. Наибольшую роль в разви-
тии рефрактерности отводят антигенам системы HLA. Полагают, что возникнове-
ние признаков рефрактерности при переливании АБО-совместимых тромбоци гов
обусловлено в первую очередь аллоантителами специфичности анти-HIA (А) или
(В). При этом обеспечение реципиента концентратом тромбоцитов от доноров,
совместимых по этим антигенам, позволяет добиться желаемого эффекта от транс-
фузий. Достаточно часто встречают и сочетанную аллоиммунизацию антигенами
систем HLA и НРА. Лишь малая толика в развитии иммунологической рефрактер-
ности к трансфузиям концентрата тромбоцитов принадлежит несовместимости
собственно по антигенам тромбоцитов системы НРА.
Ввиду трудности подбора совместимых доноров при развитии рефрактерности
к трансфузиям концентрата тромбоцитов целесообразно осуществлять плановое
HLA- и НРА-типирование доноров тромбоцитов для создания банков данных.
Подобные данные являются подчас единственной возможностью для больного с
рефрактерностыо найти совместимого донора концентрата тромбоцитов.
Глава 19
Цитокины в лабораторной
диагностике
ОБЩИЕ ПРЕДСТАВЛЕНИЯ О ЦИТОКИНАХ
Цитокины г- эндогенные полипептидные и белковые медиато-
ры межклеточного взаимодействия, регулирующие эмбриональное
развитие, некоторые нормальные физиологические функции орга-
низма, защитные реакции при внедрении патогенов и при развитии
опухолей, (Ьормирование аллергических, аутоиммунных и иных
иммунопатологических процессов и восстановление поврежденных
тканей при травмах. Именно поэтому совершенно очевидно, что
цитокиновая регуляция имеет огромное значение и в норме, и при
патологии. Известно более 200 индивидуальных веществ, принад-
лежащих к семейству цитокинов: интерфероны, интерлейкины с
исторически сложившимися порядковыми номерами, ростовые и
колониестимулирующие факторы, хемокины, медиаторы из группы
фактора некроза опухолей, трансформирующие ростовые факторы
и некоторые другие молекулы.
Учение о цитокинах
Современное учение о цитокинах имеет четыре независимых
источника.
Первый и наиболее важный источник — учение о лимфокинах.
Это направление активно развивалось в середине 1960-х гг., когда
было показано, что белки, продуцируемые лимфоцитами, регули-
руют рост и функцию различных лейкоцитов. Начало учению о
лимфокинах было положено исследованиями, демонстрирующими
способность супернатантов, полученных от сенсибилизированных
лимфоцитов после их стимуляции специфическим антигеном, угне-
тать миграцию нормальных макрофагов. Фактор, ответственный за
это, был назван фактором угнетения миграции макрофагов (тес-
rophage migration inhibitory factor). В дальнейшем был введен термин
«лимфокин», означающий растворимые клеточные факторы, кото-
рые продуцируются сенсибилизированными лимфоцитами во время
их взаимодействия со специфическим антигеном и ответственны за
развитие клеточно-опосредованных иммунологических реакций.
Примером классического лимфокина может быть интерлейкин-2
(IL-2), играющий центральную роль в регуляции роста и функции
Т-клеток. Затем показано, что лимфоциты мот продуцировать
1 94 ЦИТОКИНЫ В ЛАБОРАТОРНОЙ ДИАГНОСТИКЕ
один или несколько митогенных факторов для других лимфоцитов. Так, митоген-
активированные мононуклеары человека продуцируют фактор, поддерживающий
рост Т-лимфоцитов. Этот фактор в настоящее время известен как IL-2. Однако
вскоре выяснилось, что моноциты также являются источником белков (моно-
кинов), которые могут модулировать функции лейкоцитов. Первый цитокин
моноцитарно-макрофагального происхождения — фактор некроза опухоли {tumor
necrosis factor — TNF). Кроме прямого цитотоксического эффекта по отношению к
некоторым опухолевым клеткам т vitro, TNF обладает множеством других биоло-
гических эффектов в отношении различных лейкоцитов и тканевых клеток. Среди
первых цитокинов моноцитарного происхождения был описан фактор актива-
ции лимфоцитов (lymphocyte activating factor), известный в настоящее время как
интерлейкин-1 (IL-1). Другие исследователи описали этот же фактор под такими
названиями, как митогенный белок, лейкоцитарный пироген, эндогенный пиро-
ген, фактор, активирующий В-клетки, эндогенный медиатор лейкоцитов и др.
Второй источник — учение об интерферонах. Интерферон впервые был описан
как фактор, который продуцируется вирусиндуцированными клетками, способ-
ными вызывать клеточную резистентность к вирусным инфекциям. Затем был
открыт вирус-ингибирующий белок (interferon gamma — IFNy), продуцируемый
митоген-активированными Т-лимфоцитами. IFNy, продуцируемый Т-клетками
и естественными киллерами, по структуре полностью отличается от большого
семейства IFNa/P, продуцируемого множеством различных клеток: моноцитами,
естественными киллерами, В-лимфоцитами, различными негемопоэтическими
клетками. Интерфероны были определены как факторы роста и дифференцировки
клеток иммунной системы и клеток других систем организма человека. В резуль-
тате граница между лимфокинами/монокинами и интерферонами стала размытой,
сегодня понятно, что интерфероны - это цитокины.
Третьим источником являются факторы роста гемопоэтических клеток (коло-
ниестимулирующие факторы; colony-stimulationgfa ctors — CSF). Их основной функ-
цией является обеспечение пролиферации и дифференцировки гемопоэтических
клеток, а название отражает способность формировать гранулоцитарные или
моноцитарные колонии. Эти факторы также регулируют некоторые функции зре-
лых клеток, что размывает границы между ними и лимфокинами/монокинами.
Наконец, четвертым источником являются факторы роста негемопоэтических
клеток: фактор роста, продуцируемый тромбоцитами (platelet-derived growth fac-
tor — PDGF), эпидермальный (epidermal growth factor — EGF) и фибробластный
(fibroblast growth factor — FGF) факторы роста, трансформирующий фактор роста
(tran forming growth factor p — TGFp).
Как упоминалось выше, лимфокинами называют продукты сенсибилизиро-
ванных лимфоцитов при воздействии на них специфического антигена. Однако
в связи с тем. что со временем способность продуцировать аналогичные факторы
была обнаружена и у других типов клеток, термин «лимфокины > было предложе-
но заменить на «цитокины». Кроме того, в 1979 г. на Втором международном сове-
щании по лимфокинам был утвержден термин «интерлейкины» для растворимых
белков, обеспечивающих взаимодействие между различными популяциями лейко-
цитов. В качестве первого шага на Совещании были приняты IL-1 и IL-2. которые
ранее имели множество различных названий. Несмотря на то, что множество
цитокинов относятся к семейству интерлейкинов, часть факторов сохраняют свои
старые названия, отражающие лишь одну из функций этих плейотропных агентов:
CSF, IFN, TNF и т.д. Таким образом, цитокины — регуляторные белки, секретируе-
мые лейкоцитами крови и другими клетками организма человека, плейотропные
эффекты которых включают регуляцию клеток иммунной системы и модуляцию
воспалительной реакции.
ЦИТОКИНЫ В ЛАБОРАТОРНОЙ ДИАГНОСТИКЕ 195
Цитокины имеют ряд общих биохимических и функциональных характери-
стик, отличающих их от других классов регуляторных молекул, среди которых
важнейшими считаются следующие: характерные биохимические свойства (цито-
кины представляют собой белки или полипептиды с молекулярной массой от 5 до
50 кДа), проявление биологической активности в пикограммовых концентрациях,
отсутствие тканевой и антигенной специфичности, плейотропность и взаимозаме-
няемость биологического действия, проведение сигнала путем взаимодействия со
специфическими клеточными рецепторами, формирование цитокиновой сети, В
связи с этим цитокины могут быть выделены в новую самостоятельную систему
регуляции, существующую наряду с нервной и эндокринной системами поддержа-
ния гомеостаза, причем все три системы тесно взаимосвязаны и взаимозависимы.
Характерные признаки цитокинов
• В основном цитокины — простые белки или гликопротеины с молекулярной
массой до 30 кДА; лишь некоторые цитокины являются олигомерами с высо-
кой молекулярной мае сой.
• Продукция цитокинов регулируется различными индукторами на уровне
транскрипции или трансляции.
• Продукция цитокинов кратковременна, а радиус действия обычно короткий
(типичное действие аутокринное или паракринное, но не эндокринное).
• Цитокины реализуют свои эффекты через специфические высокоаффинные
рецепторы с Kd=10?-1012 М.
• Фенотипически действие цитокинов приводит к повышению (или к сниже-
нию) скорости клеточной пролиферации, к изменению клеточной дифферен-
цировки и проявлению различных функций соматических клеток.
• Хотя набор биологических эффектов различных цитокинов может разли-
чаться, мишенями для большинства цитокинов являются гемопоэтические
клетки.
В табл. 19-1 представлены отличительные признаки гормонов и цитокинов.
Таблица 19-1. Отличительные признаки гормонов и цитокинов
Характеристика ГорМОНв! Цитокины
Клетки-продуценты Специализированные Различные типы клеток
Эффекты Характерные для каждого гормона Структурно различные цитокины обладают сходными биологическими эффектами
Клетки-мишени Строго определенные Для одного цитокина различные типы клеток
Радиус действия Большой (эндокринный тип действия) Короткий (аутокринный или паракринный тип дей- ствия)
Как следует из вышесказанного, цитокины и гормоны имеют много общего,
однако между ними есть существенные отличия. Классические гормоны про-
дуцируются специализированными клетками: например, инсулин продуцируется
0-клетками поджелудочной железы, гормон роста — гипофизом, паратгормон —
паращитовидными железами и т.д. Цитокины, наоборот, выделяются менее специ-
ализированными клетками Кроме того, различные типы клеток могут синтезиро-
вать один и тот же цитокин. Например, IL-1 продуцируют моноциты, макрофаги,
мезангиальные клетки, естественные киллеры (NK-клетки), Т- и В-лимфоциты,
нейтрофилы, эндотелиальные клетки, мышечные клетки, фибробласты, астроци-
ты, клетки микроглии. Однако существуют исключения: IL-2, IL-5 и лимфотоксин
продуцируются исключительно Т-лимфоцитами. Одно из основных отличий цито-
кинов от гормонов — это наличие у цитокинов, имеющих структурное сходство,
принципиально различных биологических эффектов (например, TNF-a и TNF-0;
1 96 ЦИТОКИНЫ В ЛАБОРАТОРНОЙ ДИАГНОСТИКЕ
IL-1P и IL-lp). Кроме того, один и тот же цитокин проявляет многообразные био-
логические эффекты по отношению к различным клеткам-мишеням (табл. 19-2).
Таблица 19-2. Основные характеристики действия цитокинов
Характеристика Комментарии
Плейотропность Большинство цитокинов имеют много клеток-мишеней
Избыточность эффектов Один цитокин имеет множество биологических эффектов
Сичергизм/антагонизм Одновременное влияние на клетки двух или нескольких цитокинов может при- вести к качественно различным эффектам
Цитокиновый каскад Цитокины могут повысить (или снизить) продукцию других цитокинов
Модуляция рецепторов Цитокины могут повысить (или снизить) экспрессию рецепторов для других цитокинов
Несмотря на некоторые различия, цитокины, факторы роста и гормоны форми-
руют большое семейство внеклеточных сигнальных молекул со сходным механиз-
мом действия. Так, рецепторы для некоторых цитокинов и гормонов [IL-2, IL-3,
IL-4, IL-5, IL-6, IL-7, гранулоцитарно-макрофагальный колониестимулирующий
фактор (GM-CSF), гранулоцитарный колониестимулирующий фактор (G-CSF),
эритропоэтин, пролактин, гормон роста] имеют сходную с груктуру. Также струк-
турная гомология была описана для рецепторов к PDGF и макрофагальному коло-
ниестимулирующему фактору (M-CSF). Более того, сходные механизмы обеспечи-
вают передачу сигнала от рецепторов (к гормонам, факторам роста, цитокинам) к
клеточному ядру и приводят к сходным биологическим эффектам.
Роль цитокинов в регуляции функций организма может быть разделена на
4 основные составляющие:
• регуляция эмбриогенеза, закладки и развития органов, в том числе органов
иммунной системы;
• регуляция отдельных нормальных физиологических функций;
• регуляция защитных реакций организма на местном и системном уровне;
• регуляция процессов регенерации тканей.
Экспрессия генов отдельных цитокинов происходит стадиоспецифически на
определенных этапах эмбрионального развития. Факторы стволовых клеток,
трансформирующие ростовые факторы, цитокины семейства TNF и хемокины
регулируют дифференцировку и миграцию различных клеток и закладку органов
иммунной системы. После этого синтез некоторых цитокинов может не возобнов-
ляться, тогда как другие продолжают регулировать нормальные физиологические
процессы или участвуют в развитии защитных реакций.
Несмотря на то, что большинство цитокинов являются типичными индуци-
бельными медиаторами и в постнатальном периоде не синтезируются клетками
вне воспалительной реакции и иммунного ответа, некоторые из этих веществ не
подпадают под это правило. В результате конститутивной экспрессии генов часть
из них синтезируется постоянно и в достаточно больших количествах находится
в циркуляции, регулируя пролиферацию и дифференцировку отдельных типов
клеток в течение всей жизни. Примером такого типа физиологической регуляции
функций цитокинами может быть постоянно высокий уровень эритропоэтина и
некоторых CSF для обеспечения гемопоэза.
Цитокины, в первую очередь, регулируют развитие местных защитных реак-
ций в тканях с участием различных типов клеток крови, эндотелия, соедини-
тельной ткани и эпителиев. На тканевом уровне цитокины ответственны за
развитие воспаления, а затем регенерации тканей. При развитии системной
воспалительной реакции (острофазового ответа) цитокины оказывают влия-
ние практически на все органы и системы организма, участвующие в регуляции
гомеостаза. Попадание цитокинов в циркуляцию безусловно, означает, что мест-
ЦИТОКИНЫ В ЛАБОРАТОРНОЙ ДИАГНОСТИКЕ 197
ная защита не справилась с патогеном, требуется включение системной воспали-
тельной реакции для предотвращения его распространения и развития сепсиса.
Регуляция защитных реакций организма цитокинами происходит не только в
рамках иммунной системы, но и на уровне целостного организма, где цитокины
осуществляют связь между иммунной, нервной, эндокринной, кроветворной
и другими системами и служат для их вовлечения в организацию и регуляцию
единой защитной реакции.
Цитокины играют ключевую роль в регуляции врожденного иммунитета. Не
менее важное значение цитокины имеют в формировании и развитии реакций
специфического иммунитета, прежде всего связанных с регуляцией пролиферации
и дифференцировки Т- и В-лимфоцитов. Сейчас не вызывает сомнения разделе-
ние популяции Т-лимфоцитов на два принципиально разных класса: Т-хелперы
и цитотоксические Т-лимфоциты. Благодаря дальнейшим исследованиям функ-
ций активированных антигеном Т-лимфоцитов хелперов выяснилось, что эта
популяция клеток также неоднородна и может быть разделена на подклассы по
функциональной активности. Эта активность связана, главным образом, с син-
тезируемыми цитокинами. Оказалось, что Т-хелперы различаются между собой
именно набором продуцируемых цитокинов. Активация ТЫ, секретирующих IL-2
и IFNy, ведет к стимуляции, главным образом, функций Т-лимфоцитов и макро-
фагов и к развитию клеточного типа ответа, тогда как синтез Th2 IL-4, IL-5, IL-10
и IL-13 стимулирует преимущественно гуморальное звено иммунитета. Защита
организма от внутриклеточных патогенов происходит с помощью формирования
клеточного иммунитета, зависимого от ТЫ, тогда как внеклеточные патогены
нейтрализуются и выводятся из организма благодаря гуморальному иммунитету,
поддерживаемому Th2.
Направление антигензависимой дифференцировки Т-лимфоцита хелпера
зависит от присутствия в межклеточном пространстве цитокинов, синтезируе-
мых, главным образом, антигенпредст являющими клетками и определяющими
путь дифференцировки активированных антигеном наивных Т-лимфоцитов в
разные типы Th клонов. У человека описаны, по крайней мере, 3 типа Th-клонов,
из них клоны ТЫ и Th2 почти полностью соответствуют аналогичным клонам
Т-лимфоцитов мышей. Дифференцировка Th! индуцируется IL-12, который син-
тезируется макрофагами и дендритными клетками после взаимодействия моле-
кулярных структур патогенов с толл-подобными рецепторами (toll-like receptors}
данных клеток. IL-12 взаимодействует со своими специфическими рецепторами
на ThO и запускает активацию сигнального пути с участием транскрипционного
фактора T-bet, являющегося основной внутриклеточной молекулой для диффе-
ренцировки ТЫ.
Th2 дифференцируются, главным образом, под действием IL-4, который может
синтезироваться базофилами и тучными клетками в ответ на проникновение в
ткани аллергенов, а также под действием цитокина ТСЛП (тимический стромаль-
ный лимфопоэтин), синтезируемого эпителиальными клетками в коже и легких и
активирующего функции миелоидных дендритных клеток (ДК), в свою очередь
активирующих Th2. Интересно, что сам IL-4, синтезируемый Th2, является основ-
ным фактором в определении дифференцировки стимулированных антигеном
наивных CD4* (CD — cluster of differentiation} T-клеток в Th2.
Изучение активации Т-лимфоцитов хелперов человека привело к появлению
более сложной схемы развития Th-клонов. Выяснилось, что IL-23, еще один цито-
кин семейства IL-12, ответственен за дифференцировку особого типа Th-клонов,
продуцирующих IL-17 и получивших название ТЫ7, Вместе с IL-23 в дифферен-
цировке ТЫ7 принимают участие IL-6, IL-la и IL-lp. Цитокины семейства IL-17
стимулируют синтез ряда провоспалительных цитокинов макрофагами и за счет
этого усиливают воспалительную реакцию, изначально вызванную патогеном.
ГЛАВА Н
198 ЦИТОКИНЫ В ЛАБОРАТОРНОЙ ДИАГНОСТИКЕ
В последнее время описаны также клоны Th человека, дифференцирующиеся под
действием третьего члена семейства IL-12 — IL-27 и синтезирующие преимуще-
ственно IL-10. Значение дифференцировки этих клонов при встрече с антигеном
может заключаться в негативной регуляции активации клонов Thl, Th2 и Thl7 и в
подавлении воспалительной реакции любого типа. Кроме того, существуют клоны
П1, относящиеся к естественно встречающимся регуляторным Т-лимфоцитам, экс-
прессирующим поверхностные маркеры CD4, CD25 и транскрипционный фактор
FuxP3. Дифференцировка этого типа Th, по-видимому, происходит в тимусе под
влиянием тимического стромального лимфопоэтина, секретируемого клетками
эпителия медуллярной зоны тимуса, а также IL-2, синтезируемого как в тимусе, так
и на периферии, хотя окончательная ясность в схеме дифференцировки регулятор-
ных Т-лимфоцитов отсутствует.
МЕТОДЫ ОЦЕНКИ ФУНКЦИОНИРОВАНИЯ
СИСТЕМЫ ЦИТОКИНОВ
Изучение системы цитокинов как т vivo, так и in vitro проводится в зависимости
от конкретных задач, стоящих перед лабораторией, и включает следующие основ-
ные подходы.
• Генетический анализ на предмет отсутствия мутаций в генах цитокинов, их
рецепторов и белков внутриклеточных сигнальных систем запуска синтеза и
передачи сигнала.
• Анализ полиморфизма генов цитокинов.
• Изучение экспрессии генов цитокинов.
• Изучение уровня продукции цитокинов клетками в культуре.
• Определение концентраций цитокинов в биологических жидкостях.
• Изучение синтеза цитокинов на уровне отдельных клеток.
• Изучение синтеза цитокинов в тканях.
Все перечисленные методы могут быть разделены на три основные группы,
появившиеся в следующем порядке по мерс развития исследований системы цито-
кинов и развития методологических подходов в целом:
1) методы оценки биологической активности цитокинов;
2) иммунохимические методы;
3) молекулярно-биологические методы.
Биологические методы
Первые эксперименты, приведшие к открытию и начальной характеристике
цитокинов, были связаны с изучением их биологической активности. Так, на
основании изучения противовирусной активности в культурах зараженных виру-
сами фибробластов, был открыт интерферон. Несомненно, биологические мето-
ды измерения уровня цитокинов являются очень ценными для исследователей.
Эти методы отличаются достаточно высокой чувствительностью и, с биологиче-
ской точки зрения, являются более правильными, поскольку измеряют только
биологически активные формы цитокинов, а следовательно, наиболее адекватно
отражают работу системы цитокинов in vivo. В связи с этим уже в первые годы
изучения цитокинов были разработаны несколько тестов с использованием
первичных культур клеток для оценки разнообразных биологических функций.
Однако определение биологической активности цитокинов с использованием
первичных культур клеток не соответствовало одному из главных требований —
стандартизации методик анализа. Именно поэтому вскоре для биологического
тестирования многих цитокинов были получены длительно культивируемые
ЦИТОКИНЫ В ЛАБОРАТОРНОЙ ДИАГНОСТИКЕ 199
трансформированные линии клеток, отвечавших на действие цитокинов изме-
нением пролиферации, дифференцировки или функциональной активности
(табл. 19-3). Тем не менее во всех случаях стандартизация обеспечивается
сравнением биологического действия изучаемого цитокина с международными
стандартными образцами, выпускаемыми Международной лабораторией биоло-
гических стандартов ВОЗ в Национальном институте биологической стандарти-
зации и контроля (Великобритания), которая выпускает и рассылает по запросу
охарактеризованные по биологической активности стандартные образцы цито-
кинов человека.
Таблица 19-3. Клеточные линии и первичные культуры клеток для оценки биологической актив-
ности цитокинов (Meager, 2006, с дополнениями)
Цитокины Клеточные линии Тканевое проис- хождение Способ ОЦЕНКИ актив- ности Другие цитокины, актив- ные в данном тесте
IL-1 Первичные куль- туры Тимоциты мыши Пролиферация IL-2, IL-6; IL-4, TNF. видо- специфично
D10S Лимфома Пролиферация IL-2
IL-2 CTLL-2 Лимфома Пролиферация IL-15, TGFp
IL-3 TF-1 Костный мозг Пролиферация IL-4, 5,9.13, GM-CSF
IL-4 CTLL-2 Лимфома Пролиферация, видо- специфично IL-15, TGF₽
IL-6 В9 Миелома Пролиферация IL-11, IL-13
IL-7 2Е8 Лимфома Пролиферация -
IL-8 Первичные куль- туры Нейтрофилы Миграция Некоторые CXC хемокины
IL-12 KIT-225 Т-лимФома Пролиферация IL-2.4,7,15, 21
IL-18 KG-1 Миелолейкемия Синтез IFNy IL-12, IL-21
TNF L929 Фибробласты Цитотоксичность Лимфотоксин
GM-CSF TF-1 Эритролейкемия Пролиферация IL-8, 4,5.9,13, ЭПО
IFNa Первичные куль- туры Фибробласты пупо- вины человека Антивирусная актив- ность Другие типы IFN, видо- специфично
А-549, 2D9. Daudi Карцинома легких, глиобластома, лим- фома Антивирусная актив- ность, продукция белка МхА
IFNy А-549 Карцинома легких Антивирусная актив- ность Другие типы IFN,видо- специфично
WiDr Карцинома толстой кишки Подавление пролифе- рации -
Как следует из табл. 19-3, большинство используемых в биологических тестах
клеточных линий не обладают строгой специфичностью в отношении одного
цитокина, а могут1 отвечать на действие еще некоторых медиаторов, что существен-
но снижает специфичность определения биологической активности цитокинов
в культуральных средах и биологических жидкостях, где обычно одновременно
содержится несколько десятков цитокинов. В то же время эти клеточные линии
могут быть идеальным инструментом для оценки биологической активности
рекомбинантных цитокинов. Во многих лабораториях производят искусственные
клеточные линии, отвечающие на отдельные цитокины, путем трансфекиии кле-
ток генами рецепторов данных цитокинов. Трансфецированные клетки приобрета-
ют способность специфически отвечать на действие низких концентраций нужных
цитокинов. Однако многие клетки экспрессируют рецепторы большого числа
цитокинов, поэтому метод не в состоянии обеспечить абсолютную специфичность
тестирования.
200 ЦИТОКИНЫ В ЛАБОРАТОРНОЙ ДИАГНОСТИКЕ
Клеточные линии, используемые для оценки уровня цитокинов человека,
имеют различное происхождение. Большинство таких линий получены от паци-
ентов с лейкемией либо при лабораторно индуцированной лейкемии. Некоторые
цитокины человека не являются видоспецифичными, и для их изучения можно
использовать клеточные линии мышиного происхождения. Уровень цитокинов
в образцах оценивается в основном по их способности стимулировать или инги-
бировать пролиферацию клеток. Существует несколько способов измерения про-
лиферации клеток:
1) подсчет клеток;
2) оценка синтеза ДНК по включению радиоактивно меченного тимидина.
Альтернативой радиоизотопным методам служат методы оценки клеточного
метаболизма как индикатора пролиферации, основанные на восстановлении раз-
личных солей тегразолия.
Кроме измерения пролиферативной активности для оценки уровня некоторых
цитокинов, таких как TNF. используют методы, основанные на цитотоксической
активности цитокинов. Здесь с помощью витальных красителей оценивают цито-
токсический эффект по отношению к некоторым клепочным линиям. Для оценки
инп ерферонового статуса используют методы, основанные на ан гивирусной актив-
ности интерферонов. При этом используются различные клетки-мишени, зара-
женные вирусом, на которых оценивается антивирусная активность интерферонов
путем визуальной оценки жизнеспособности клеток или с помощью витальных
красителей. Для оценки биологический активности хемокинов, в частности IL-8,
применяют методы хемотаксиса, где определяют их способность привлекать
клетки, измеряя при этом количество клеток, мигрировавших чецез мембрану
Бойдена, либо длину пробега клеток в агаре.
В целом все методы оценки биологической активности цитокинов in vitro могут
быть объединены в 5 основных групп.
1. Оценка пролиферативной активности клеток (стимуляция или подавление).
2. Оценка цитотоксичности.
3. Определение экспрессии мембранных рецепторов, синтеза других цитоки-
нов.
4. Оценка влияния на функциональную активность клеток, что всегда зависит
от типа клеток и от изучаемого цитокина, например оценка хемотаксиса либо
оценка завершенности фагоцитоза.
5. Оценка противоинфекционного действия, как в случае интерферона.
Несмотря на очевидные достоинства, биологические методы имеют и ряд суще-
ственных недостатков:
❖ они являются трудоемкими и сложными в исполнении;
< требуют значительно больше времени до получения конечного результата;
требуют особых стерильных условий и оборудования для постановки;
❖ являются менее специфичными, чем иммунохимические методы.
Иммунохимические методы
Биологическая оценка активности цитокинов обладает целым рядом ограни-
чений в области специфичности, воспроизводимости и точности (табл. 19-4).
В связи с этим определение уровней цитокинов в различных биологических
средах чаще всего проводят с использованием иммунохимических методов,
основанных на применении поликлональных, а в настоящее время, главным
образом, моноклональных антител, специфически взаимодействующих с моле-
кулами цитокинов.
ЦИТОКИНЫ В ЛАБОРАТОРНОЙ ДИАГНОСТИКЕ 201
Таблица 19-4. Сравнительный анализ методов определения цитокинов
Характеристика Биологические методы Иммунохимические методы
Что выявляет Только биологически активные молекулы Число доступных эпитопов (биологиче- ски активные и неактивные молекулы)
Специфичность Недостаточная (биологические активности различных цитокинов интерферируют); необходимо подтверждать с помощью ней- трализующих антител Очень высокая
Интерференция Специфические и неспецифические натив- ные ингибиторы цитокинов,антицитокино- вые антитела Белки сыворотки крови, компоненты комплемента, антицитокиновые анти- тела ревматоидный фактор
Чувствительность Высокая Очень высокая
Точность Низкая (CV может достигать 100%) Высокая (CV в пределах 5-10%)
Длительность 14 дня Несколько часов
Иммунохимические методы чрезвычайно разнообразны и представлены сле-
дующими видами методик количественного определения концентрации раство-
римых цитокинов в различных биологических жидкостях.
• Иммуноферментный анализ в различных модификациях (ИФА; Enzyme-
linked. Immunosorbent Assay — ELISA).
• Радиоиммунный анализ (РИА). Чувствительность метода достаточно высо-
ка, но в последние годы рад иоиммун элегические методы используются все
реже и заменяются ИФА, чтобы избежать работы с изотопами и уйти от
использования дорогостоящих счетчиков радиоактивности.
• Мультиплексный иммунный анализ (МИА), позволяющий с помощью моно-
клональных антител одновременно определять до 100 белков с использо-
ванием флюоресцентных и других меток, в том числе с применением тех-
нологии микрочастиц или микрошариков, покрытых антителами, а также
микрочипов (Bioplex, Multiplex analysis, Biochip Array Technology). В этом
варианте несколько цитокинов анализируют с использованием одного био-
чипа одновременно, что существенно экономит время и реагенты, однако
стоимость оборудования для проведения такого анализа пока еще слишком
высока.
• Цитофлюориметрические методы для анализа экспрессии поверхностных
молекул клеток (мембранных форм цитокинов и рецепторов цитокинов) и
определения цитокинов в цитоплазме клеток.
• Метод оценки продукции цитокинов единичными клетками в культуре
(ELISPOT).
• Иммуноцитохимия и иммуногистохимия для оценки содержания цитокинов
в цитоплазме клеток на мазках и на срезах тканей.
ИММУНОФЕРМЕНТНЫЙ АНАЛИЗ
Наибольшее распространение получили иммуноферментные диагностические
тест-системы, позволяющие проводить количественный анализ содержания цито-
кинов в любых биологических жидкостях и обладающие достаточно высокой
чувствительностью для определения уровней цитокинов у человека и живот-
ных. Принцип работы большинства современных тест-систем заключается в
использовании «сэндвич»-варианта твердофазного иммуноферментного анализа
(рис. 19-1). Для реализации этого варианта используют два моноклональных
антитела с различной эпитопной специфичностью к определяемому цитокину.
Первое из этих антител иммобилизовано на твердой фазе (внутренняя поверх-
ность лунок) 96-луночного пластикового микропланшета стандартного формата
для постановки ИФА. На первом этапе анализа исследуемый образец вносится в
лунки, где происходит специфическое связывание определяемого антигена (цито-
202 цитокины В ЛАБОРАТОРНОЙ диагностике
А
Нормальная Ложноположительная Угнетение
реакция реакция реакции
Твердая Флуоресцентно-
фаза ИФА меченная
микрочастица
для МИА
Рис. 19-1. Базовые принципы «сэндвич»-анализа на твердой фазе: А — слева ИФА. справа
микрочастицы; В — слева нормальная реакция, посередине — ложноположительная реакция
вследствие интерференции с ревматоидным фактором IgM класса, справа — угнетение реакции
путем блокирования сайтов связывания иммуноглобулинами животных.
кина) моноклональными антителами, сорбированными на поверхности лунок.
После этого производится отмывка всего несвязавшегося с аптителами материала
буфером с детергентом. Этот важный этап позволяет удалить все неспецифически
связывающиеся молекулы, но не влияет на специфическое высокоаффинное взаи-
модействие антител с определяемым цитокином. На второй стадии анализа свя-
завшийся цитокин взаимодействует со вторыми антителами, конъюгированными с
биотином. При этом количество связавшегося конъюгата прямо пропорционально
количеству цитокина в исследуемом образце. На последней стадии анализа в лунки
вносят авидин-пероксидазу, взаимодействующую f биотином на вторых анти-
телах. После этого в лунки вносят особый субстрат для фермента пероксидазы,
меняющий цвет под действием фермента, например тетраметилбензидин, в резуль-
тате чего происходит ферментативная реакция и окрашивание раствора в лунках.
Возможно также использование наборов из трех антител, когда вторые антитела
(например, поликлональные кроличьи антитела к цитокину) на последнем этапе
взаимодействуют с антивидовыми антителами, конъюгированными с ферментом.
В результате образуется «сэндвич» из двух или трех антител и молекулы антигена
между ними.
Количественную оценку концентрации цитокина в пробах проводят, сравнивая
результаты с калибровочной кривой зависимости оптической плотности раствора
от концентрации стандартного образца цитокина. Иммуноферментные методы
высокоспецифичны, быстры (время постановки иммуноферментного анализа
составляет менее 5 ч) и относительно просты в исполнении. Порог чувствительно-
сти для таких тест-систем достигает 0,5 пкг/мл. В настоящее время в России широ-
ко доступны коммерческие варианты иммуноферментных систем для определения
широкого спектра цитокинов.
Большинство иммуноферментных тест-систем адаптированы для работы с
любыми биологическими жидкостями. Для измерения уровней цитокинов на
системном уровне используют сыворотку или плазму крови. Несмотря на короткий
(минуты) период жизни и индуцибельный тип синтеза большинства цитокинов, в
сыворотках крови здоровых доноров иногда определяются незначительные уров-
ни цитокинов. Обычно их концентрации находятся на пределе чувствительности
иммуноферментного метода, но при различных заболеваниях, связанных с острой
или хронической продукцией цитокинов, сывороточные уровни цитокинов ста-
новятся значительно выше. Однако, поскольку цитокины являются локальными
медиаторами, более целесообразно измерять их уровни в интересующих исследо-
ЦИТОКИНЫ В ЛАБОРАТОРНОЙ ДИАГНОСТИКЕ 203
вателя тканях после экстракции тканевых протеинов in vitro или в биологических
жидкостях. При развитии воспаления, аллергии или аутоиммунных процессов зна-
чимые уровни цитокинов могут быть определены в слезной жидкости, жидкости
десневых карманов, смывах из полостей, моче, спинномозговой жидкости и даже
в конденсате выдыхаемого воздуха.
Анализ данных определения цитокинов показывает что благодаря исполь-
зованию различных реагентов в ИФА и различиям в протоколах исследования,
в разных лабораториях могут получаться не вполне сопоставимые результа-
ты, даже если при этом используют одни и те же международные стандарты.
Подобные различия можно объяснить использованием в тест-системах разных
пар антител. Используемые в ИФА тест-системах антитела могут связываться
с одними и теми же цитокиновыми молекулами, однако антитела могут не рас-
познавать фрагменты цитокиновых молекул или их формы-предшественники.
Для создания антител обычно используют рекомбинантные формы цитокинов,
которые могут отличаться от естественных форм. Это может приводить к раз-
личиям в аффинитете связывания определяемых молекул с разными антителами.
И хотя, как считается, ИФА является «золотым стандартом» для определения
цитокинов, в настоящее время возможность определения таких сигнальных
молекул с помощью этого метода ограничена. Это, прежде всего, касается одно-
временного исследования уровней большого числа цитокинов для более полной
характеристики цитокиновой регуляции различных биологических процессов в
норме и при патологии.
Таким образом, основными достоинствами ИФА являются:
о высокая специфичность;
высокая чувствительность (низкий предел детекции);
о хорошая во< производимость;
❖ относительно низкая себестоимость анализа;
❖ достаточно большой срок годности наборов.
С другой стороны, к основным недостаткам ИФА следует отнести:
возможность исследования только одного парамет ра;
❖ достаточно большой объем исследуемого образца (100-200 мкл и в дублях —
200-400 мкл);
> относительно продолжительную процедуру выполнения анализа (протокол
опыта включает достаточно большое число этапов инкубации, отмывок, при
этом ручной труд способствует повышению вероятности погрешностей в
постановке ИФА тест-систем);
❖ сложности, связанные с динамическим рядом определения параметров
(калибровочные кривые) и с интерпре гацией данных, превышающих разре-
шающую способность калибровочного графика.
МУЛЬТИПЛЕКСНЫЙ ИММУННЫЙ АНАЛИЗ
Для оценки уровней цитокинов в различных биологических жидкостях наи-
более распространенными методами мулы индексного иммунного анализа (МИА)
являются мультиплексные методы, в основе которых лежит «сэндвич»-метод в
разных комбинациях — FAST Quant Search Light и xMAP. Каждый из этих методов
имеет некоторые различия по ряду параметров: определение доступных аналитов,
возможность использования новых аналитов, динамический ряд оценки, чувстви-
тельность метода, стоимость оборудования, цена расходных материалов.
Все эти критерии следует учитывать при выборе метода исследования уровней
цитокинов. Поскольку результаты определения биологически активных веществ
в каждом методе могут существенно различаться, считается, что проводить ста-
тистический анализ данных, полученных с помощью разных методов, нецеле-
сообразно. Для каждого отдельного исследования следует выбирать только одну
204 цитокины В ЛАБОРАТОРНОЙ диагностике
какую-нибудь технологию для того, чтобы полученные данные и выводы были
репрезентативными и соответствовали требованиям доказательной медицины.
Измерение уровня одного какого-либо цитокина для характеристики различ-
ных процессов, протекающих в организме, является абсолютно недостаточным,
так как не позволяет получить более или менее объективную информацию о
комплексных биологических взаимодействиях на клеточном уровне как в норме,
так и при патологии. В этой связи совершенно очевидно, что роль повышения или
снижения уровня одного какого-либо цитокина в межклеточных взаимодействиях
необходимо рассматривать с учетом изменения уровней других цитокинов, при-
сутствующих в этом же исследуемом биологическом образце.
В табл. 19-5 приведен сравнительный анализ характеристик МИА и ИФА мето-
дов определения цитокинов.
Таблица 19-5. Сравнительные характеристики ИФА и МИА методов определения цитокинов
Сравниваемые параметры ИФА МИА
Чувствительность Высокая-—<10 пг/мл Высокая — несколько пг/мл
Определение Количественное Количественное
Специфичность Высокая Высокая
Выявление Биологически активных и неактивных молекул Биологически активных и неактив- ных молекул
Время проведения анализа Относительно небольшое (день) Более быстрый анализ (часы)
Коэффициент вариации 5-10% 10-15%
Интерференция Отсутствует Отсутствует
Протокол исследования Относительно простой Простой
Объем образца Относительно большой (100-200 мкл на 1 определение) Небольшой (50 мкл на все определе- ния — до 100 параметров)
Стимуляция детекции Отсутствует Присутствует
Срок хранения тест-системы Небольшой Более продолжительный
Разрешающая способность Определение 1 цитокина в одной оиопробе Одновременное определение до 100 цитокинов в 1 биопробе
ОГРАНИЧЕНИЯ И КРИТИЧЕСКИЕ МОМЕНТЫ В ОПРЕДЕЛЕНИИ ЦИТОКИНОВ
С ПОМОЩЬЮ ОСНОВНЫХ ИММУНОХИМИЧЕСКИХ МЕТОДОВ
Сбор и хранение биопроб
В зависимости от задач клинической лабораторной диагностики цитокины
определяют в различных биологических жидкостях (сыворотке крови, церебро-
спинальной жидкости, амниотической жидкости, в экссудатах, жидкости бронхо-
альвеолярного лаважа, смывах, мокроте и др.). Особое внимание следует уделять
условиям сбора и хранения биологических образцов, поскольку на этом этапе
возможны грубые ошибки, которые в дальнейшем могут привести к неправиль-
ным результатам оценки и соответственно неверной интерпретации полученных
данных. При повторных циклах размораживания-замораживания биопроб в
результате действия протеиназ молекулы цитокинов подвержены разрушению.
При неправильном сборе биологического материала не исключена возможность в
условиях ex vivo освобождения цитокинов из клеток либо связывания их с клеточ-
ными рецепторами, что может повлиять на полученные результаты. Если центри-
фугирование крови для получения сыворотки не выполнить за короткий срок, на
результаты оценки цитокинов может существенно повлиять способность тромбо-
цитов, как во время процесса свертывания крови, так и при центрифугировании,
освобождать целый ряд цитокинов и хемокинов, включая IL-1, IL-6, CXCL8/IL8,
CCL5/RANTES.
ЦИТОКИНЫ В ЛАБОРАТОРНОЙ ДИАГНОСТИКЕ 205
При анализе цереброспинальной, амниотической, синовиальной, жидкости
бронхоальвеолярного лаважа, а также супернатантов клеток, перед заморажива-
нием образцов необходимо избавиться от клеток. Связывание цитокинов с их рас-
творимыми рецепторами также может повлиять на полученные результаты оценки
уровней цитокинов в биопробах и не отразить истинную концентрацию цитокинов
в анализируемых образцах. Этот эффект может происходить при специфическом
связывании IL-1 и IL-6 с их растворимыми рецепторами. Не исключена возмож-
ность связывания цитокинов, например IL-1, IL-6, TNFa аутоантителами, что
также может приводить к получению неверных, искаженных результатов.
Гетерофильные антитела
При анализе уровней цитокинов в сыворотке и плазме крови наибольшее число
ложноположительных результатов можно объяснить присутствием в этих биоло-
гических жидкостях гетерофильных антител. Эти антитела вырабатываются про-
тив неизвестных, недостаточно полно охарактеризованных антигенов и обычно
обладают низкой авидностью. Такие низкоавидные антитела могут взаимодей-
ствовать с различными иммуноглобулинами и присутствуют у 40% здоровых лиц.
а при различных аутоиммунных заболеваниях (например, у больных ревматоид-
ным артритом) их количество существенно возрастает. Гетерофильные антитела
могут включать антитела IgG, IgM и IgE и присутствуют в сыворотке и плазме
крови, а также в других биологических жидкостях, в которых обычно проводится
определение уровней цитокипов с помощью «сэндвич»-методов, используемых
как в ИФА, так и в МИА-технологнях. Поскольку эти гетерофильные антитела
могут связываться с компонентами иммуноаналитических систем, перекрестное
связывание может приводить к ложноположительным результатам, реже — к
ингибиции связывания (см. рис. 19-1).
Для того чтобы исключить интерференцию с гетерофильными антителами, в
настоящее время разработан ряд подходов. Поскольку гетерофильные антитела
могут связываться с антителами разных видов животных, можно проводить пре-
инкубацию образца со смесью сывороток разных видов животных, лишь после
этого ставить методы оценки уровней цитокинов. Выбор сывороток животных
зависит от используемых в тест-системах антител (антитела мыши, кролика, овцы
и других животных). Однако при таком подходе возможны и другие осложнения:
в смеси сывороток крови животных разных видов антитела могут взаимодей-
ствовать друг с другом, что может приводить к неэффективному связыванию с
гетерофильными антителами человека в исследуемых образцах. Таким образом,
этот подход имеет ограничения. У больных ревматоидным артритом или у детей
с ювенильным идиопатическим артритом в сыворотке крови в больших количе-
ствах могуч присутствовать IgM- и IgG-аутоантитела, в частности ревматоидный
фактор. Существуют методы, позволяющие снизить уровни ревматоидного факто-
ра в исследуемых образцах крови таких больных. Для этого используют протеин L.
В противоположность протеину А или протеину G, этот протеин обладает способ-
ностью к высокоаффинному связыванию с IgM и IgG и меньшей аффинностью
связывания с IgA.
Гетерофильные антитела также могут взаимодействовать с компонентами
супернатантов клеток, а именно с компонентами сыворотки крупного рогатого
скота или аутологичной сыворотки человека, так как эти сыворотки обычно
добавляют в культуральную среду, что способствует росту7 и дифференцировке
культивируемых клеток.
Общее содержание белка и pH
Время достижения равновесия на разных этапах анализа зависит от концентра-
ции белков, например более низкое содержание белка может приводить к более
206 ЦИТОКИНЫ В ЛАБОРАТОРНОЙ ДИАГНОСТИКЕ
высокой интенсивности люминесценции. Если определение цитокинов проводит-
ся в биологических жидкостях с высоким содержанием белка (например, в цере-
броспинальной жидкости, амниотической, синовиальной и других), стандартные
кривые необходимо строить с использованием буфера с сопоставимым содержа-
нием белка. Кроме концентрации белка, pH исследуемых образцов также может
влиять на результаты анализа уровней цитокинов. Например, низкие уровни pH
мочи могут приводить к конформации молекул цитокинов, причем некоторые
молекулы менее устойчивы к таким изменениям среды (например, структура моле-
кулы TNFa подвергается конформации при низких pH), в то время как молекулы
других цитокинов (например, IFNy) остаются достаточно устойчивыми в широком
диапазоне pH — от 4,8 до 9,5.
Перекрестное связывание
Существует определенная степень гомологии между молекулами антител и
цитокиновыми молекулами, поэтому можно предположить, что при проведении
анализа антитела могут распознавать не только молекулы цитокинов, но и другие
белковые молекулы, присутствующие в биопробе. Поичем при мультиплексном
анализе, в котором используют очень большое число всевозможных антител,
вероятность такого перекрестного связывания значительно выше, чем при ИФА.
Стандартные кривые и вычисления результатов
Среди прочих факторов, влияющих на точность количественной оценки уров-
ней цитокинов, важную роль играют используемые стандартные образцы и
соответственно стандартные кривые. Обычно такие кривые состоят из серии
определений с известными количествами рекомбинантных цитокинов (от 3 до
12 определений). В ИФА часто используют модель линейной регрессии. Если
уровни исследуемых цитокинов находятся в пределах линейного ряда стандар гной
кривой, такая модель линейной регрессии является приемлемой для вычисления
результатов. Однако, если подушенные данные выходят' за пределы линейной части
стандартной кривой, образцы следует разбавлять и анализировать повторно, учи-
тывая их разведение.
Предел детекции в МИА ниже, чем в ИФА, даже если в обеих технологиях
используются одни и те же пары антител, а динамический ряд измерения —
шире. Более широкий динамический ряд определений (3-4-логарифмическая
шкала) в МИА позволяет минимизировать число биологических проб, для кото-
рых необходимо проводить повторное определение уровней цитокинов (в ИФА
обычно используют 1-2-логарифмическую шкалу). Поскольку ряд определений
в стандартной кривой достаточно широк, линейная модель не всегда приемлема,
и поэтому для вычисления уровней цитокинов в биопробах чаще используется
нелинейная регрессионная модель.
Определение цитокинов в клинической практике
Определение цитокинов в различных биологических жидкостях обычно
используют для оценки активности воспаления, определения степени поляриза-
ции иммунного ответа, оценки эффективности проводимой терапии и прогноза
заболевания. В ряде случаев уровни отдельных цитокинов могут коррелировать
с тяжестью течения заболевания и прогнозом. Классическим примером может
служить уровень интерлейкина-6 в плазме крови больных с тяжелыми травмами:
высокие уровни IL-6 являются предикторами развития полиорганной недоста-
точности. Другим примером может служить определение высоких уровней IL-8
в цереброспинальной жидкости больных с лептоменингиальными метастазами
солидных опухолей — как предиктора скорой гибели больного.
Однако при большинстве заболеваний определение одного какого-либо цитокина
обычно является совершенно недостаточным для диагностики или оценки прогноза
ЦИТОКИНЫ В ЛАБОРАТОРНОЙ ДИАГНОСТИКЕ 207
заболевания. Для оценки состояния иммунной системы у таких больных необходи-
мо определять достаточно большой спектр цитокинов, позволяющий оценить осо-
бенности цитокиновой регуляции в каждом конкретном компартмечте.
Благодаря сложным механизмам регуляции синтеза цитокинов, изменение экс-
прессии одного какого-либо цитокина обычно приводит к изменениям уровней
других цитокинов. Причем скорость синтеза цитокинов может быть различной
и зависит от целого ряда физиологических характеристик организма благодаря
тесной кооперации трех основных адаптационных систем: нервной, эндокринной
и иммунной. В этой связи определение различных цитокинов в биологических
образцах лучше проводить одновременно, в мультиплексном режиме оценки,
чтобы избежать возможных ошибок в интерпретации полученных данных.
Определение продукции цитокинов клетками
Определение продукции цитокинов мононуклеарами периферической крови in
vitro как при стимуляции, так и без нее является альтернативой определения плаз-
матического уровня цитокинов. Мононуклеары периферической крови служат в
качестве удобной модели для оценки продукции цитокинов в основном из-за их
доступности. При активации in vitro такими агентами, как фитогемагглютинин
(ФГА) или бактериальный липополисахарид (ЛПС), мононуклеары перифериче-
ской крови секретируют в культуральную среду целый спектр провоспалительных
цитокинов: IL-1, IL-6, IL-8, TNF и другие. Измерение уровня цитокинов в био-
логических жидкостях дает возможность оценить уровень цитокинов, синтезиро-
ванных клетками организма, в норме или при развитии патологии. Определение
уровня цитокинов в супернатантах мононуклеаров периферической крови обеспе-
чивает надежную оценку способности клеток продуцировать цитокины. Этот тест
можно считать одним из важнейших методов оценки функционального состояния
клеток иммунной системы. Он стоит в одном ряду с оценкой бластной трансфор-
мации лимфоцитов или определением фагоцитарной активности клеток.
Определение синтеза цитокинов изолированными клетками в культуре может
быть легко стандартизовано, так как в тесте используют стандартные препара-
ты индукторов синтеза цитокинов, культуральные среды и другие реагенты, а
сама оценка уровня цитокинов в культуральной среде может быть проведена как
биологическими методами, так и стандартизованными иммуноферментными
тест-системами, а также и их комбинациями. Более того, в супернатантах клеток,
стимулированных одним либо двумя стандартными индукторами, может быть
измерен одновременно целый ряд разнообразных цитокинов, позволяющих не
только оценить общий уровень синтеза цитокинов индивидуума, но и определить
цитокиновые профили, например поляризацию Т-хелперных клонов по харак-
терному профилю синтезируемых цитокинов, соотношение синтеза агонистов и
антагонистов и т.д. Кроме того, в культуре клеток можно оценить синтез цитоки-
нов в ответ на специфический антиген, что может иметь неоценимое значение в
диагностике силы и развития иммунного ответа при ряде инфекционных заболе-
ваний, при вакцинации, аллергической сенсибилизации и других патологических
состояниях. В культуре имеется возможность определения продукции цитокинов
у предварительно криоконсервированных клеток, полученных либо от разных
доноров, либо в разное время и собранных вместе для более удобного, экономич-
ного и надежного серийного мониторинга при клинических испытаниях.
Методика определения синтеза цитокинов изолированными клетками доста-
точно проста и заключается в следующем. Свежую венозную кровь, взягую в
емкость с гепарином (20 МЕ/мл), тщательно перемешивают и разводят в 5 раз
средой RPM1 1640 с добавлением 2 мМ глутамина и 80 мкг/мл гентамицина,
обычно к 0,6 мл крови добавляют 2,4 мл среды RPMI 1640. В качестве индуктора
208 ЦИТОКИНЫ В ЛАБОРАТОРНОЙ ДИАГНОСТИКЕ
синтеза группы провоспалительных цитокинов (IL-1. IL-6, TNF), IL-10, IFNa и
хемокинов могут быть использованы препараты ЛПС в концентрации 1-10 мкг/
мл, например отечественный ЛПС-содержащий препарат продигиозан*. В каче-
стве индукторов синтеза IL-2, IL-4, IFNy и других Т-клеточных цитокинов может
быть использован ФГА-М (Sigma*) в конечной концентрации 50 мкг/мл. ФГА
является универсальным индуктором и может быть использован также и для
индукции синтеза провоспалительных цитокинов. В 96-луночный планшет для
культивирования клеток («COSTAR» или аналогичный) вносят индуктор по
100 мкл на лунку, в контрольные лунки (для оценки спонтанной продукции)
вносят по 100 мкл среды RPMI 1640, затем во все лунки добавляли по 100 мкл
разведенной, как описано выше, крови. Культивирование проводят при 37 °C в
5% СО2в течение 24 ч, после чего осторожно отбирают супернатанты и проводят
исследование содержания цитокинов в биологическом или иммуноферментном
тесте. У больных подсчитывают количество лейкоцитов и формулу крови по
стандартным методикам. Полученные данные о концентрациях цитокинов в
супернатантах пересчитывают на 1 млн лейкоцитов либо мононуклеаров, лим-
фоцитов или других интересующих клеток в 1 мл с учетом произведенных раз-
ведений крови. При необходимости супернатанты могут храниться при -20 °C до
6 мес при однократном замораживании.
Многие провоспалительные цитокины в норме не должны циркулировать в
криви. Тем не менее описаны случаи увеличения уровней провоспалительных
цитокинов, в частности IL-1, в плазме крови при стрессовых ситуациях и тяжелой
физической нагрузке. По-видимому, присутствие невысоких уровней этих цитоки-
нов в плазме крови здоровых доноров можно объяснить следующими причинами.
Во-первых, они могут появляться в циркуляции в результате некоторых нормаль-
ных физиологических процессов жизнедеятельности организма, либо под влияни-
ем сгрессорных факторов неинфекционной природы. Во-вторых, в ряде случаев
данные уровни цитокинов могут являться проявлением вяло текущих скрытых
воспалительных процессов, а также иммунопатологических состояний, включая
ранние стадии аутоиммунных и аллергических заболеваний еще до появления
клинических проявлений. Наконец, ряд цитокинов в плазме крови могут быть свя-
заны с белками переносчиками и различными ингибиторами. При этом цитокины
могут не обладать биологической активностью, но выявляться иммунохимически
с помощью специфических антител.
Определение уровней цитокинов в сыворотке крови имеет ряд сложностей из-за
наличия неспецифических факторов связывания, которыми являются различные
сывороточные белки. Цитокины IL-1, IL-2 и другие могут взаимодействовать с
а,-макроглобулином, образуя комплексы, которые могут маскировать иммуно-
реактивность цитокинов. Наряду с неспецифическим связыванием существует и
специфическое. Уровни цитокинов, измеренные иммуноферментных методом,
могут быть неинформативными из-за присутствия растворимых рецепторов для
цитокинов. В этих случаях специфическое взаимодействие цитокина с рецептором
ведет к кажущемуся исчезновению его из крови. Циркулирующие аутоантитела к
цитокинам являются другими специфическими ингибиторами, присутствующими
в сыворотке некоторых индивидуумов, например у лиц, получавших цитокиновую
терапию.
Уровни цитокинов в плазме периферической крови отражают текущее состоя-
ние работы иммунной системы и развития защитных реакций, то есть синтез
цитокинов клетками организма in vivo. Определение уровней продукции цитоки-
нов изолированными клетками in vitro показывает их функциональное состояние.
Спонтанная продукция цитокинов в культуре свидетельствует о том, что клетки
уже активированы in vivo в результате развития воспаления или иммунопатологи-
ческих процессов. В то же время активация клеток может произойти вследствие
ЦИТОКИНЫ В ЛАБОРАТОРНОЙ ДИАГНОСТИКЕ 209
манипуляций в процессе выделения. Иногда это происходит из-за несоблюдения
стерильности при заборе крови или при использовании культуральных сред и
растворов реаге нтов. загрязненных ЛПС. Индуцированная продукция цитокинов
позволяет оценить потенциальные возможности активации клеток, что очень
важно для оценки иммунологической реактивности. Сниженная индуцированная
продукция цитокинов т vitro может служить одним из признаков иммунодефи-
цитного состояния. Именно поэтому оба варианта изучения уровней цитокинов в
циркуляции либо продукции лейкоцитами важны с точки зрения характеристики
работы иммунной системы.
Преимущество метода индукции синтеза цитокинов в культурах цельной крови
имеет ряд преимуществ по сравнению с индукцией цитокинов в культурах клеток,
выделенных на градиенте плотности. В первую очередь это простота и связанная
с этим более высокая степень стандартизации процедуры. Во-вторых, стимуляция
клеток происходит в присутствии собственной плазмы и лучше отвечает задаче
наиболее полного воспроизведения условий синтеза цитокинов в организме. Тем
не менее в ряде случаев разделение клеток на фракции позволяет более точно
определить продукцию цитокинов определенными популяциями лимфоцитов,
моноцитов или гранулоцитов.
В этой связи одним из наиболее перспективных современных методов анализа
продукции цитокинов изолированными клетками является метод ELISPOT, пред-
ставляющий собой своеобразный гибрид метода индукции синтеза цитокинов
в культуре и обычного иммунофсрментного анализа.. Суть метода заключается
в том, что выделенные клетки культивируют в присутствии индуктора синтеза
цитокинов в микропланшетах, дно которых покрыто антителами к интересующе-
му цитокину, как это делается на первом этапе ИФА. В процессе культивирования
клетки, находящиеся на дне планшета, синтезируют цитокины, которые тут же
связываются с сорбированными на пластике антителами. Таким образом, оказы-
вается, что цитокин соединяется с сорбированными антителами в тех местах, где
его синтезировали клетки. Дальнейшие этапы анализа проводятся, как в стандарт-
ном варианте ИФА, с той лишь разницей, что при постановке ELISPOT исполь-
зуется нерастворимый цветной субстрат ферментативной реакции, выпадающий
в осадок там, где цитокин связался с сорбированными антителами В результате
на дне планшета образуются окрашенные пятна, соответствующие местам, где
происходил синтез цитокинов клетками. Отсюда и произошло название метода —
ELISPOT как комбинация метода ELISA с конечным формированием окрашенного
пятна (spot). Пятна легко видны под малым увеличением микроскопа, их количе-
ство относительно исходного числа клеток и разный размер позволяют оценить
интенсивность синтеза цитокинов на уровне отдельных клеток. В настоящее время
разработаны варианты метода с использованием субстратов разных цветов для
оценки синтеза 2-3 цитокинов одновременно и созданы приборы для автомати-
ческого подсчета к анализа размеров пятен, что очень важно для стандартизации
метода ELISPOT.
Наиболее точную характеристику отдельных клеток, продуцирующих цитокины,
дает метод цитоплазматического окрашивания цитокинов в варианте цитофлюо-
риметрического анализа. Мембранные формы цитокинов и рецепторы цитокинов
легко определяют на поверхности клеток с помощью меченных флюорохромами
антител в обычном варианте цитофлюориметрии аналогично другим поверхност-
ным молекулам клеток. Для выявления цитоплазматических цитокинов требуется
достижение состояния проницаемости или пермебиализация мембран для обе-
спечения проникновения антител в клетки. Для этой цели используется сапонин в
сочетании с блокаторами обратного транспорта (брефелдин А или монензин) для
предотвращения экскреции антител из цитоплазмы клеток. Метод выгодно отли-
чается от метода ELISPOT возможностью дополнительной характеристики клеток
21 о ЦИТОКИНЫ В ЛАБОРАТОРНОЙ ДИАГНОСТИКЕ
гПАЧг IP
по размеру и экспрессии мембранных маркеров, что позволяет проводить более
точную идентификацию цитокин-продуцирующих клеток.
Рецепторы цитокинов представляют собой трансмембранные белки, а некото-
рые цитокины могут существовать в виде мембранной формы. В обоих случаях
экстраклеточные домены цитокинов и их рецепторов представляют собой поверх-
ностные антигенные детерминанты различных клеток человека. Мембранные
рецепторы и мембранные формы цитокинов, распознаваемые соответствующими
моноклональными антителами, включены в единую классификацию поверхност-
ных молекул клеток (CD) и получили свои порядковые номера (табл. 19-6). Это
позволяет изучать количество и типы клеток, экспрессирующих рецепторы и мем-
бранные формы цитокинов, с помощью цитофлюориметрии, а также использовать
клеточные сортеры для выделения клеток, имеющих на мембранах определенные
типы цитокинов или их рецепторов.
Таблица 19-6. Рецепторы и мембранные формы цитокинов, включенные в CD классификацию
поверхностных молекул лейкоцитов человека
Рецепторы и мембран- ные формы цитокинов Соответствующие CD антигены Экспрессирующие клетки
Рецепторы цитокинов, регулирующих созреззние и функции Т-лимфоцитов
IL-2 Ra CD25 Активированные лимфоцит ы
IL-2 Rp CD122 Т-лимфоциты, NK-клетки
IL-2 Ry CD132 Т-, В-лимфоциты, NK-клетки
IL-4/IL-13 Rtt CDT24 Т В-лимфоциты, другие типы клеток
IL-13 R0t1 CD213a1 В-лимфоциты, моноциты, фибробласты, эндотелиальные клетки
IL-13 Ra2 CD213a2 В-лимфоциты. моноциты
IL-5 Ra CD125 Эозинофилы, базофилы
IL-7 Ra CD127 Предшественники T- и В-лимфоцитов
IL-6 Ra CD126 В-лимфоциты, плазматические клетки
LIF R CD118 Эпителиальные клетки
gp13O CD13O Многие типы клеток
IL-10 R C0w21Q Т-, В-лимфоциты, NK-клетки, моноциты
IL-12 R(31 CD212 Активированные Т-лимфоциты и NK-клетки
IL-17 R CD217 Многие типы клеток
Рецепторы цитокинов, стимулирующих кроветворение
G-CSFR CD114 Гранулоциты, моноциты и их предшественники
M-CSF R CD115 Моноциты, тканевые макрофаги
GM CSF Ra CD116 Моноциты, нейтрофилы, эозинофилы
Фактор стволовых кле- ток R (c-kit) CD117 Предшественники гемопоэза
Flt3 CD135 Предшественники гемопоэза
IL-3 Ra CD123 Предшественники гемопоэза, гранулоциты, мегакариоци- ты, моноциты
IL-3,5, GM-CSF Rp CD131 Миелоидные клетки
Рецепторы цитокинов семейства IL-1 и ФРФ
IL-1 Rl CD121a Т-лимфоциты, фибробласты, эндотелиальные клетки
IL-1 RII CD121b В-лимФоциты, моноциты, макрофаги
IL-18 Ra GD218a Лимфоциты, нейтрофилы, ДК
IL-18 Rp CD218b Лимфоциты, нейтрофилы, ДК
ФРФ R1 CD331 Фибробласты, эпителиальные клетки
ФРФ R2 CD332 Фибробласты, эпителиальные клетки
ЦИТОКИНЫ В ЛАБОРАТОРНОЙ ДИАГНОСТИКЕ 211
Окончание табл. 19-6
Рецепторы и мембран- ные фирмы цитокинов Соответствующие CD антигены Экспрессирующие клетки
ФРФ нз CD333 Фибробласты, эпителиальные клетки
ФРФ R4 CD334 Фибробласты, эпителиальные клетки
Цитокины и рецепт! семейства TNF
TNF RI CD120a Моноциты, гранулоциты
TNF RII CD120b Моноциты, гранулоциты, лимфоциты
Fas CD95 Апоптотические клетки
Fas лиганд CD178 Активированные Т-лимфоциты
0X40 CD134 Активированные Т-лимфоциты
0X40 лиганд CD252 Активированные В-лимфоциты ДК эндотелиальные клетки
4-1ВВ C0137 Т-лимфоциты
CD30 лиганд CD153 Активированные Т-лимфоциты
CD40 лиганд CD154 Активированные Т-лимфоциты, базофилы, тучные клетки
TRAIL CD253 Активированные Т-, В-лимфоциты, моноциты
TRAIL R1 CD261 Лейкоциты, опухолевые клетки
TRAIL R2 CD262 Многие типы клеток
TRAIL R3 CU263 Многие типы клеток
FRAIL R4 00264 Многие типы клеток
TRANCE, RANKL CD254 Активированные Т-лимфоциты, остеобласты, стромальные клетки
RANK, TRANCE-R CD265 Многие типы клеток
APRIL CD256 Миелоидные клетки
BlyS, BAFF CD257 Миелоидные клетки
LIGHT CD258 Активированные Т-лимфоциты, незрелые ДК
1WEAK-R CD266 HUVEC
BAFF-R CD268 Т-, В-лимфоциты
Рецепторы хемокинов
CCR1 00191 Т-лимфоциты, моноциты, стволовые клетки
CCR2 CD192 Моноциты. Т-. В-лимфоциты, базофилы, эндотелиальные клетки
CCR3 CD193 Эозинофилы, базофилы, Т-лимфоциты, ДК
CCR4 CD194 Т-лимфоциты, базофилы, моноциты, тромбоциты
CCR5 CD195 Моноциты, Т-лимфоциты
CCR6 CD196 Т-, В-лимфоциты, ДК
CCR7 CD197 Т-лимфоциты
CCR8 CDW198 Т-лимфоциты, моноциты
CCR9 CDw199 Т-лимфоциты
CXCR1 00181 Нейтрофилы, Т-лимфоциты
CXCR2 CD182 Моноциты, гранулоциты, Т-лимфоциты
CXCR3 CD183 Активированные Т-лимфоциты и NK-клетки
CXCR4 CD184 Моноциты. Т-, В-лимфоциты, эндотелиальные клетки, ДК
CXCR5 CD185 Т-, В-лимфоциты
CXCR6 GD186 Т-лимфоциты
212 ЦИТОКИНЫ В ЛАБОРАТОРНОЙ ДИАГНОСТИКЕ
Определение уровней цитокинов в тканях
Несмотря на то что определение уровня цитокинов в сыворотке или плазме
является более простым в исполнении, определение уровня цитокинов в тканях
может дать дополнительную информацию о состоянии местного иммунитета.
Так как цитокины — это локальные медиаторы, то методы определения уровня
цитокинов в месте воспаления или органного повреждения и репарации могут
быть более биологически значимыми, чем определение их уровня в перифериче-
ской крови. Однако все эти методы требуют получения биопсии ткани и являются
трудоемкими. Простейший подход — это диссоциация ткани и приготовление
тканевого экстракта, который затем тестируется методом ИФА. Однако этот метод
не дает информации о том, какие клетки продуцируют цитокины и где они локали-
зованы в ткани. Кроме того, для корректной интерпретации результатов требуется
нормальная ткань в качестве контроля.
ИММУНОГИСТОХИМИЧЕСКИЙ МЕТОД
Широко распространенными методами детекции цитокинов в тканях являются
иммуногистохимические методы с использованием специфических поликлональ-
ных или моноклональных антител к цитокинам. С помощью этих методов цитоки-
ны могут быть обнаружены в цитоплазме клеток — в мазках клеточных суспензий,
приготовленных разными способами на предметных стеклах в виде обычного
гематологического мазка, с помощью цитоцентрифуги, способом тонкой капли и
другими методами. Кусочки тканей, полученные с помощью биопсии или во время
операции, обычно глубоко замораживают в жидком азоте, а затем готовят срезы
толщиной 4-6 мкм в криостате при температуре -20 °C. Приготовленные и высу-
шенные мазки или срезы тканей фиксируют этанолом, метанолом или пара-
формальдегидом (4% раствор в натрий-фосфатном буфере. pH 7,2-7,4) 20 мин
при +4 СС, затем отмывают в PBS (pH 7,2-7,4), в течение 10 мин, высушивают и
хранят при +4 °C до проведения реакции.
Чаще всего изучение локализации цитокинов в клетках проводят методом
непрямой иммуногистохимии с использованием специфических антител к цито-
кинам, а на втором этапе — антивидовых антител, меченных флюорохромами
для люминесцентной микроскопии или ферментами для световой микроскопии.
Важно, что в случае флюоресцентных меток антитела к цитокинам могут быть
использованы для их выявления с помощью цитофлюориметра, причем в дан-
ном варианте возможно выявление как цитоплазматических цитокинов, так и их
мембранных форм. В случае ферментов используют биотинилированные анти-
видовые антитела, а на последнем этапе комплексы стрептавидин-пероксидаза
либо стрептавидин-щелочная фосфатаза. При проведении цветной реакции при-
меняют нерастворимые субстраты, выпадающие в осадок в месте взаимодействия
антител с цитокинами, то есть в месте локализации цитокинов в клетках. В случае
световой микроскопии препараты могут быть докрашены гематоксилином и неко-
торыми другими красителями, что позволяет достаточно хорошо различать струк-
туру ткани. Кроме того, существуют методы иммуногистохимического выявления
отдельных цитокинов на парафиновых срезах, дающих наиболее полное сохране-
ние морфологической структуры исследуемых тканей.
Молекулярно-биологические методы изучения цитокинов
Определение экспрессии генов цитокинов в клетках-продуцентах по накопле-
нию мРНК служит еще одним подходом к оценке их синтеза. Появление мРНК в
клетках или тканях обычно детектируется путем гибридизации со специфически-
ми олигонуклеотидными зондами, длина которых варьирует от нескольких пукле-
ЦИТОКИНЫ В ЛАБОРАТОРНОЙ ДИАГНОСТИКЕ 21 3
отидов до целой кДНК, соответствующей гену искомого цитокина. Эти зонды
содержат радиоактивные, флюоресцентные или ферментные метки, в зависимости
от метки применяют соответствующий метод для выявления тканевой локализа-
ции мРНК. При использовании метода гибридизации in situ на свежих криосрезах
тканей или мазках клеток для выявления локализации мРНК цитокинов могут
использовать комплементарные РНК зонды для РНК-РНК гибридизации. Метод
позволяет определять количество и тип клеток, экспрессирующих цитокины.
Недостатком метода является тот факт, что в ряде случаев наличие мРНК совсем
необязательно отражает присутствие соответствующего полипептида цитокина в
клетке: процессы транскрипции и трансляции могут регулироваться независимо.
Накопление большого количества мРНК может не сопровождаться последова-
тельным синтезом белка. Если мРНК не транслируется, что часто бывает при ряде
патологических состояний, то искомый цитокин будет отсутствовать. Кроме того,
возможно появление различных изоформ мРНК в результате альтернативного
сплайсинга. Таким образом, для получения более полной информации о содержа-
нии цитокинов в тканях необходимо, видимо, использовать комбинацию методов,
позволяющих оценить как экспрессию мРНК, так и синтез данного цитокина в
клетке.
Новейшим количественным методом для определения цитокиновой мРНК явля-
ется метод полимеразной цепной реакции с обратной транскрипцией (RT-PCR).
Этот метод отличается очень высокой чувствительностью и специфичностью, как
и все тесты, основанные на методике ПЦР. В настоящее время разработаны многие
модификации метода, такие как ПЦР в реальном времени { real-time PCR), микро-
чипы (microarray), позволяющие анализировать одновременно несколько сотен
генов, и другие. Гибридизация с олигонуклеотидными зондами, комплементарны-
ми участкам единичных нуклеотидных замен в генах цитокинов, с использованием
амплификации методом ПЦР применяется и для анализа аллельного полимор-
физма. Одной из последних разработок является использование аптамеров — оли-
гонуклеотидов, полученных по технологии SELEX, для количественной оценки
уровней цитокинов. Существует модификация ИФА, где вместо антител в качестве
распознающих цитокины молекул используются аптамеры.
РОЛЬ ЦИТОКИНОВ В ПАТОГЕНЕЗЕ ЗАБОЛЕВАНИЙ ЧЕЛОВЕКА
Врожденные нарушения цитокиновой регуляции
Мутации генов цитокинов и регуляторных молекул в большинстве случаев при-
водят к снижению функциональной активности цитокинов и нарушению развития
защитных реакций организма. Наследственные нарушения в системе цитокиновой
регуляции затрагивают многие ключевые медиаторы и их рецепторы, приводя к
тяжелым клиническим проявлениям разнообразных видов иммунодефицитных
состояний. В то же время отдельные мутации могут приводить к повышенному
синтезу цитокинов либо нарушению регуляторных взаимодействий в цитокиновой
сети и к возникновению связанных с этим аутоиммунных, аутовоспалительных и
лимфопролиферативных состояний.
Одним из ярких примеров цитокин-зависимого первичного иммунодефицита
служит наследственный дефект гена у-цепи рецептора IL-2. Отдельные субъе-
диницы рецепторного комплекса IL-2 являются общими для IL-2 и некоторых
других цитокинов. Так, у-цепь служит общей субъединицей рецепторов для IL-2,
IL-4, IL-7, IL-9, IL-15 и IL-21. У человека мутации в гене у-цепи рецептора IL-2
(делеции аминокислотных остатков 62 и 81) связаны с развитием сцепленного с
Х-хромосомой синдрома тяжелого комбинированного иммунодефицита (ТКИД).
Это заболевание характеризуется отсутствием или значительным падением содер-
214 ЦИТОКИНЫ В ЛАБОРАТОРНОЙ ДИАГНОСТИКЕ
жания Т-лимфоцитов и резко сниженными показателями клеточного и гумораль-
ного иммунитета.
Клинически ТКИД является гетерогенной группой заболеваний, связанных с
нарушением функционирования Т-лимфоцитов, сочетающегося с различными
вариантами дефицита В-лимфоцитов, NK-клеток и других типов лейкоцитов.
Кроме ТКИД, развивающегося как следствие дефекта у-цепи рецептора IL-2,
известны как минимум еще два варианта этого заболевания, вызванных дефекта-
ми генов JAK3 и а-субъединицы рецептора IL-7, Клинические симптомы ТКИД,
связанного с дефицитом киназы JAK3, неотличимы от проявлений иммунодефи-
цита, связанного с дефектом у-цепи. Это объясняется участием киназы JAK3 в
проведении внутриклеточного активационного сигнала от у-цепи рецепторного
комплекса IL-2. По сути два описанных генетических дефекта нарушают один и
тот же путь активации лимфоцитов на разных уровнях, что, естественно, приводит
к одинаковым функциональным нарушениям и клиническим проявлениям имму-
нодефицитных состояний. Дефицит а-субъединицы рецептора IL-7 приводит к
несколько иной картине ТКИД. У больных с этим генетическим дефектом наблю-
далось снижение количества Т-лимфоцитов, но отмечены нормальные уровни
В-лимфоцитов и NK-клеток. В данном случае нарушения касаются проявлений
функциональной активности только одного цитокина, IL-7, стимулирующего
созревание предшественников лимфоцитов. Однако отсутствие зрелых Т-клеток
оказывается достаточной причиной для развития тяжелых проявлений ТКИД.
У детей с клиническими проявлениями ТКИД дифференциальная диагностика
дефектов в системе цитокинов молекулярно-биологическими методами может
иметь важное значение для правильной постановки диагноза.
Нарушения в системе IL-12, II,-23, IFNy и их рецепторов составляют другое
врожденное нарушение цитокиновой регуляции, связанное с дефектом врожденно-
го иммунитета. В результате взаимодействия с патогенами макрофаги и дендрит-
ные клетки начинают синтезировать несколько разных групп цитокинов, включая
цитокины семейства IL-12 (IL-12, IL-23, IL-27), нужные для запуска дифференци-
ровки незрелых Т-лимфоцитов, Синтез 1L-12 параллельно с представлением анти-
гена стимулирует дифференцировку Т-клеток в направлении Т-лимфоцитов хел-
перов 1 типа, приводящую к началу экспрессии генов цитокинов, среди которых
одним из главных регуляторов клеточного иммунитета является IFNy. Нетрудно
представить, что любые сбои в функционировании этой последовательной систе-
мы развития защитных реакций могут привести к серьезным нарушениям имму-
нитета. в первую очередь связанным с защитой от микобактерий. В приведенном
пути регуляции начальных этапов развития защитных реакций известны пять раз-
личных генетических дефектов, затрагивающих цитокины, их рецепторы и моле-
кулы сигнальных путей активации клеток. Это ген субъединицы р40, являющейся
одной из двух субъединиц гетеродимерных цитокинов IL-12 и IL-23, ген р-1 цепи
рецептора IL-12, служащей обшей субъединицей рецепторов для IL-12 и IL-23,
гены субъединиц 1 и 2 рецептора IFNy и ген внутриклеточной сигнальной молеку-
лы STAT1 (signal transducer and activator of transcription T), участвующей в передаче
сигнала от рецепторов IFNy.
Описано более 50 пациентов с полным рецессивным вариантом дефицита
рецептора IL-12, приводящего к отсутствию нормальной экспрессии этой рецеп-
торной субъединицы, резкому снижению ответа на IL-12 и IL-23 и продукции
IFNy. Клинически это проявлялось повышенной восприимчивостью к инфекциям,
вызываемым микобактериями и сальмонеллами, и частично могло быть компен-
сировано назначением препаратов IFNy. Нарушения продукции IFNy со сходными
клиническими проявлениями наблюдались у детей с дефектами гена субъединицы
р40 IL-12/IL-23, этот иммунодефицит дозозависимо корректировался назначени-
ем рекомбинантного IL-12. При правильной оценке цитокиновой недостаточности
ЦИТОКИНЫ В ЛАБОРАТОРНОЙ ДИАГНОСТИКЕ 215
и адекватной терапии клинический прогноз у детей с такими дефектами достаточ-
но благоприятный, особенно в случая к вторичных инфекций, протекающих менее
тяжело. Наследственные иммунодефицитные состояния, связанные с дефектами
генов рецепторов IFNy, подразделяются на полные дефициты рецепторов, которые
приводят к полному отсутствию ответа на этот цитокин, частичные рецессивные
и частичные доминантные дефициты рецепторов, возникающие в результате раз-
личных мутаций и делений генов рецепторов 1 и 2 к IFNy. Тяжесть клинических
проявлений иммунодефицита во всех этих случаях различна, однако всегда связа-
на с развитием инфекций, вызванных микобактериями, реже сальмонеллами или
Listeria monocytogenes.
ФУНКЦИОНАЛЬНЫЙ ПОЛИМОРФИЗМ ТЕНОР ЦИТОКИНОВ КАК ВАРИАНТ РАЗВИТИЯ
НАСЛЕДСТВЕННЫХ НАРУШЕНИЙ ЦИТОКИНОВОЙ РЕГУЛЯЦИИ
Мутации в генах цитокинов приводят к серьезным нарушениям иммунитета.
У человека подобные мутации встречаются достаточно редко, что может свиде-
тельствовать о важнейшей роли цитокинов в осуществлении защитных реакций.
Тем не менее накапливается все больше данных о выявлении функционального
полиморфизма генов цитокинов, связанного с заменами единичных нуклеотидов
(полиморфизм вследствие различия единичных нуклеотидов; single nucleotide poly-
morphism — SNP), вызывающих количественные изменения функционирования
соответствующих генов, в отличие от мутаций, полностью выключающих функции
кодируемых ими белков. SNP являются наследственными генетическими измене-
ниями. Накапливается все больше данных о том, чго эти изменения могут оказы-
вать достаточно серьезное влияние на протекание защитных реакций. В отличие
от мутаций в генах цитокинов, SNP не удаляются естественным отбором, а боль-
шей частью сохраняются в популяции, что делает их важными диагностическими
маркерами, которые могут быть определены с использованием молекулярно-
биологических методов. В последнее десятилетие опубликовано огромное коли-
чество данных о выявлении полиморфизма генов цитокинов человека, их рецеп-
торов и регуляторных молекул. Часть выявленных аллельных вариантов генов
обусловливает снижение либо увеличение проявлений биологической активности
кодируемых белков, что позволяет говорить именно о функциональном поли-
морфизме. Связь обнаруженных изменений с конкретной патологией у человека
можно проиллюстрировать следующим примером.
Для гена TNF описано несколько различных SNP, два из которых связаны с раз-
нонаправленными изменениями уровня продукции биологически активного TNF.
Это нуклеотидные замены в положениях —308 (G->A) и —238 (G-^A) в нетранс-
лируемой области гена в зоне промоутерных участков. Наличие полиморфного
аллеля (-308А) приводит к увеличению эффективности транскрипции гена и уве-
личению продукции TNF в 2-5 раз, по сравнению с нормальным вариантом Связь
функционального полиморфизма гена TNF с развитием патологии прослеживает-
ся при изучении распределения аллелей этого гена среди африканцев, болеющих
малярией. При обследовании более 1000 человек оказалось, что полиморфный
аллель (-308А), связанный с повышенной продукцией TNF, в 4 раза чаще встре-
чается у больных с наиболее тяжело протекающей церебральной формой маля-
рии с более сильным развитием системного воспалительного ответа и в 7-8 раз
чаще у больных с развившимися впоследствии тяжелыми нарушениями нервной
системы. Более широкие изучение распределения аллелей гена TN F среди жителей
эндемичных по малярии регионов Африки привело к поразите тьным результатам,
показавшим, что среди африканцев частота носительства данного полиморфного
аллеля составляет лишь 5%, тогда как среди белых европейского происхождения
она достигает 30%. Данные говорят в пользу того, что высокопродуцирующий
аллель гена TNF в случае заболевания малярией приводит к повышенному синтезу
216 ЦИТОКИНЫ В ЛАБОРАТОРНОЙ ДИАГНОСТИКЕ
TNF и более выраженному развитию системной воспалительной реакции, чаще
приводящей к гибели заболевших или к тяжелым осложнениям. По-видимому,
в данном случае имеет место результат давления определенного патогена на
популяцию за счет механизма естественного отбора, происходит это за счет суще-
ствования наследственных вариантов гена одного из главных провоспалительных
цитокинов. Естественно, что диагностическое определение наличия указанного
полиморфного аллеля гена TNF может дать очень важную информацию для про-
гноза и своевременного назначения адекватной терапии больных с инфекционны-
ми заболеваниями и гнойно-воспалительной патологией.
Функциональный полиморфизм генов цитокинов обусловливает индивиду-
альные наследственные колебания уровня их продукции, которые влияют на раз-
витие и исход инфекционных заболеваний и иммунопатологических процессов.
Именно поэтому полиморфизм генов цитокинов может считаться вариантом
наследственных иммунодефицитных состояний. Безусловно, обнаружение SNP
свидетельствует лишь о предрасположенности индивидуумов к тому или иному
заболеванию, однако их определение позволяет прогнозировать риск развития
патологии и подбирать адекватную терапию. Подобные изменения в перспективе
должны заноситься в генетический паспорт индивидуума.
Изменения цитокинов при сепсисе
Клинические проявления сепсиса связаны с развитием системного воспали-
тельного ответа организма на инфекцию. Это послужило основой для введения
термина «синдром системного воспалительного ответа» (systemic inflammatory
response syndrome), применимого к обозначению воспалительного ответа на уровне
организма при различных состояниях, включая сепсис с бактериемией, а также
состояниях без определяемой бактериемии, например травма и острые воспали-
тельные заболевания отдельных органов (острый панкреатит).
Син гез большинства цитокинов, участвующих в регуляции воспаления, возрас-
тает 1 гри развитии сепсиса и коррелирует с уровнями других лабораторных тестов
(С-реактивный белок и другие), тяжестью клинических проявлений и исходом
заболевания. Лабораторные данные определения уровней цитокинов свидетель-
ствуют о том, что чем выше были уровни цитокинов при поступлении больных с
диагнозом «сепсис» в отделение интенсивной терапии, тем выше оказался уровень
смертности, причем это относилось как к группе провоспалительных цитокинов
(TNF, IL-1, IL-6, IL-8), так и к антивоспалительным цитокинам, таким как IL-10
и рецепторный антагонист интерлейкина-1 (РАИЛ). По-видимому, в начале
развития сепсиса происходит активация различных типов лейкоцитов и других
синтезирующих цитокины клеток и индукция или увеличение уровня экспрессии
генов нескольких цитокинов без избирательной активации синтеза только про-
воспалительных медиаторов или только цитокинов, ограничивающих развитие
воспаления.
Тем не менее именно провоспалительные цитокины обусловливают развитие
патологических изменений при сепсисе. В высоких концентрациях они способны
вызывать нейроэндокринные изменения, нарушения терморегуляции, активацию
эндотелия, приводящую к увеличению выхода жидкости в ткани, расширению
сосудов и падению артериального давления (коллапс), диссеминированному
внутрисосудистому свертыванию крови, полиорганной недостаточности и гибели
больного. Среди провоспалительных цитокинов более всего перечисленные пато-
логические изменения способен вызывать TNF, часто называемый «медиатором
смерти». Уровень IL-ip также повышен в плазме крови больных сепсисом, однако
это обнаружено не во всех исследованиях. Самые высокие уровни IL-lp выявлены
у больных с менингококковым сепсисом. У них концентрации IL-ip коррелируют
ЦИТОКИНЫ В ЛАБОРАТОРНОЙ ДИАГНОСТИКЕ 217
с выраженностью бактериемии, тяжестью клинических проявлений и развитием
септического шока. Другой представитель семейства IL-1 — IL-18 также повы-
шается у больных с сепсисом, его уровень коррелирует с клиническими проявле-
ниями сепсиса. Наиболее высокая степень корреляции с клинической картиной
сепсиса получена при исследованиях уровней IL-6 и IL-10. Именно концентрация в
плазме крови IL-6 соответствовала клинической картине сепсиса по шкале тяжести
состояния критических больных (APACHE II) и могла быть критерием для про-
гнозирования исхода заболевания. Уровень этого цитокина также коррелирует с
другими маркерами септического процесса: С-реактивным белком, компонентом
комплемента СЗа, лактатом и TNF.
Если в кровотоке количество цитокинов повышается, то их продукция изо-
лированными лейкоцитами больных сепсисом снижена по сравнению с уровнем
продукции тех же медиаторов лейкоцитами здоровых доноров. Это касается про-
воспалительных цитокинов и хемокинов, а также цитокинов Т-лимфоцитов хелпе-
ров и первого, и второго типов: IL-2, IFNy, IL-5. IL-10, т.е. сниженной оказывается
продукция большинства цитокинов. Вероятно, это связано с общими явлениями
анергии после интенсивного синтеза, так как такое падение продукции цитокинов
описано не только при бактериальном сепсисе, но и после травм и массивных ожо-
гов, когда в первые дни после ожога происходит очень интенсивный синтез этих
веществ в ответ на травму («цитокиновый шторм»), а затем наступает резкое паде-
ние уровней их продукции. Эти данные ставят вопрос об источнике повышенного
уровня цитокинов в плазме крови больных сепсисом, так как период существова-
ния большинства цитокинов в циркуляции обычно очень короткий (например,
период полужизни IL-1 в плазме крови составляет всего 7,5 мин), а повышенные
уровни цитокинов у больных сохраняются несколько суток.
Важно, что антивоспалительные цитокины (IL-10, РАИЛ и TGF0) и раствори-
мые рецепторы TNF и IL-6 тоже синтезируются в повышенных количествах при
сепсисе. Средние уровни IL-10 и РАИЛ выше при септическом шоке по сравнению
с сепсисом и выше у умерших больных по сравнению с выжившими. Наиболее
высокие уровни IL-10 выявляют у больных с неблагоприятным течением сепсиса и
плохим прогнозом, причем чем выше соотношение уровней IL-10 и TNF, тем хуже
прогноз. Данные о гиперпродукции антивоспалительных цитокинов у больных
сепсисом привели к появлению понятия «синдром компенсаторного антивос-
палительного ответа» (compensatory1 anti-inflammatory response syndrome — CARS).
Считают, что CARS развивается вслед за системной воспалительной реакцией,
выполняя функции ограничения гиперпродукции воспалительных цитокинов и
системного воспаления, но у некоторых больных это может приводить к i гммуно-
супрессии. Экспериментальные данные не полностью подтверждают эту гипотезу,
так как провоспалительные и антивоспалительные цитокины у большинства боль-
ных синтезируются одновременно, а их функции трудно оценивать однозначно.
Цитокины в низких концентрациях необходимы для правильного формирования
местного воспаления, более высокие дозы вызывают развитие системной воспали-
тельной реакции, но патологически высокие концентрации приводят к состоянию
септического шока и гибели организма.
РОЛЬ ЦИТОКИНОВ В РАЗВИТИИ АУТОИММУННОЙ ПАТОЛОГИИ
При развитии иммунопатологических процессов нарушения в работе иммун-
ной системы и клинические проявления во многом зависят от баланса клонов
Thl и Th2, а также Thl7, основанном на соотношении продукции регуляторных
цитокинов. Хроническая несбалансированность активации Т-хелперных клонов
приводит к развитию иммунопатологических состояний, в частности связанных
с проявлениями аутоиммунитета. Главным образом, аутоиммунные нарушения
21 8 ЦИТОКИНЫ В ЛАБОРАТОРНОЙ ДИАГНОСТИКЕ
развиваются при избыточной активации воспалительной реакции и клеточного
иммунитета. Однако роль различных Т-хелперных клонов в развитии аутоиммун-
ных заболеваний гораздо сложнее. При ряде аутоиммунных процессов в области
поражения тканей доминирует продукция цитокинов ТЫ, а симптомы аутоиммун-
ного воспаления могут быть подавлены с помощью введения животным цитоки-
нов, продуцируемых Th2. В то же время при других аутоиммунных расстройствах
в развитии патологического процесса важную роль играет гиперактивация не
только ТЫ, но и Th2.
Ревматоидный артрит (РА) — аутоиммунное заболевание, поражающее,
главным образом, синовиальную оболочку, хрящевую и костную ткань суставов.
Цитокины являются одними из главных медиаторов развития патологических
изменений в ткани суставов при РА, выступая в роли посредников между актива-
цией иммунной системы в результате индукции аутоиммунитета, развитием хро-
нического воспалительного процесса и деструкцией суставов.
У больных РА в синовиальной оболочке обнаружены плазмоцитоидные и мие-
лоидные дендритные клетки, которые вместе с синовиальными макрофагами син-
тезируют широкий спектр цитокинов, способных индуцировать дифференцировку
наивных Т-лимфоцитов в любом направлении. В синовиальной жидкости при ана-
лизе экспрессии генов цитокинов по уровню мРНК и при иммуноцитохимическом
анализе показано, что клетки синовиальной оболочки синтезируют IL-12 и IL-1S,
направляющие дифференцировку Т-лимфоцитов в сторону ТЫ, IL-4 — в сторону
Th2, a IL-1, IL-6, IL-23 — в сторону Thl7. В отличие от экспериментов на живот-
ных, где доказана патогенетическая роль ТЫ 7 в развитии артрита, у больных РА
пока отсутствуют убедительные доказательства преимущественной активации
одного из типов Т-хелперов.
Ключевыми медиаторами развития воспаления в области суставов у больных РА
служат провоспалительные цитокины, в первую очередь TNF и IL-1. Повышенная
экспрессия TNF практически всегда обнаруживается в синовиальной оболочке
воспаленных суставов у этих больных. TNF вызывает функциональную активацию
синовиальных фибробластов, эндотелиальных клеток и привлеченных в ткань
сустава лейкоцитов, синтез цитокинов и других медиаторов, развитие воспаления
и ремоделирование ткани сустава с развитием патологических изменений и нару-
шением функции. Не менее важна также и роль цитокинов семейства IL-1 (IL-lct и
IL-1[3), экспрессия которых тоже обычно повышена у больных РА.
Увеличение экспрессии провоспалительных цитокинов при РА сопровождается
снижением продукции антивоспалительных и регуляторных цитокинов, еще боль-
ше усугубляя нарушение нормального баланса медиаторов с разной направленно-
стью действия. У больных РА отмечено снижение экспрессии РАИЛ, IL-10, IL-4,
IL-2, помимо прочего участвующего в генерации регуляторных Т-лимфоцитов, а
также растворимых рецепторов для TNF 1 типа и для IL-11 типа, способных свя-
зывать избыток этих цитокинов. Среди других цитокинов в развитии воспаления
активное участие принимают IL-6, IL-15, IL-18 и IL-32. Их роль в патогенезе РА
доказана в экспериментальных исследованиях, и эти цитокины считаются одними
из потенциальных мишеней для проведения антицитокиновой терапии у больных
этим заболеванием.
Повышенный синтез цитокинов в области воспаленного сустава может приво-
дить к увеличению их концентрации в крови. Уровни провоспалительных цитоки-
нов, например IL-1 р, в сыворотке крови больных РА коррелируют с клиническими
проявлениями заболевания и могут служить достаточно информативными диа-
гностическими маркерами, появление которых иногда опережает развитие сим-
птомов поражения суставов. Органные проявления РА также сопровождаются зна-
чительными изменениями синтеза провоспалительных цитокинов в затронутых
аутоиммунным процессом тканях. При многих аутоиммунных состояниях возрас-
ЦИТОКИНЫ В ЛАБОРАТОРНОЙ ДИАГНОСТИКЕ 219
тание концентрации провоспалительных цитокинов в кровотоке ведет к развитию
системных проявлений их биологического действия. В этом случае значение оцен-
ки уровня цитокинов связано с тем, что хроническое увеличение уровня IL-1, TNF
и IL-6 в циркуляции приводит к развитию нарушения метаболизма костной ткани
на уровне организма, ведущего к появлению клинических признаков остеопороза
трубчатых костей. Так, IL-6 стимулирует синтез острофазовых белков в печени, а
TNF вмешивается в метаболизм липидов, вызывая кахексию. Хронически высокий
уровень провоспалительных цитокинов также вызывает центральные нарушения,
связанные с развитием депрессии и снижением познавательной активности.
Цитокины и аллергия
Аллергические состояния (аллергический конъюнктивит, пишевая аллергия,
IgE-ассоциированный атопический дерматит, аллергический ринит и астма) явля-
ются генетически детерминированной группой заболеваний, характеризующихся
повышенной способностью В-лимфоцитов синтезировать антитела класса Е (IgE),
направленные против особой группы антигенов, называемых аллергенами. IgE вза-
имодействуют с высокоаффинными мембранными FceRl-рецепторами базофилов
и тучных клеток, что приводит к дегрануляции и выбросу вазоактивных аминов, в
первую очередь гистамина, а также хемокинов и провоспалительных цитокинов,
индуцирующих развитие аллергического воспаления. Попадающие в организм
аллергены приводят к преимущественной активации клонов Т-лимфоцитов,
синтезирующих набор цитокинов, характерных для Т-хелперов 2 типа. Развитие
иммунного ответа с преимущественным синтезом антител класса IgE регулируется
Th2 путем синтеза и секреции IL-4, IL-5 и IL-13.
Однако первый источник цитокинов при развитии аллергического воспаления
в тканях — тучные клетки, которые содержат цитокины IL-4, IL-13, TNF в грану-
лах. Эти цитокины попадают в межклеточное пространство при дегрануляции в
первые минуты после контакта связанного тучными клетками IgE с аллергеном.
Они обладают провоспалительной активностью, вызывают усиление продукции
хемокинов и усиление экспрессии молекул адгезии на эндотелии, что ведет к
активному привлечению в ткани эозинофилов, базофилов и Т-лимфоцитов. В
дальнейшем источником циз окинов в тканях могут быть базофилы, тучные клет-
ки и другие лейкоциты, активация которых сопровождается экспрессией генов,
синтезом и секрецией IL-3, IL-4, IL-5, IL-6, TNF и GM-CSF. В периферической
крови больных различными формами аллергии существенно выше число клонов
Т-лимфоцитов, синтезирующих цитокины. CD41-лимфоциты больных аллергией
продуцируют IL-4 и IL-5 в ответ на бактериальные антигены, которые у здоровых
людей обычно вызывают развитие типичного ответа по типу активации Thl. По
мере развития заболевания аллергенспецифические Т-лимфоциты все более нака-
пливаются в органах, где происходит развитие аллергического воспаления.
Лимфоциты периферической крови и лимфоциты бронхоальвеолярных лава-
жей, полученные от больных бронхиальной астмой, секретировали повышенные
количества цитокинов Th2 (IL-4, IL-5, IL-13), концентрации которых коррели-
ровали с выраженностью клинических симптомов, сывороточными уровнями
IgE и содержанием эозинофилов в крови и бронхоальвеолярном лаваже. Таким
образом, у больных с бронхиальной астмой и в периферической крови, и в ткани
легких продемонстрирована активация Th2 с увеличением уровней секретируемых
ими цитокинов.
У детей, склонных к аллергии, сразу после рождения не наблюдается усиления
продукции цитокинов Th2, но впоследствии происходит активация их синтеза. У
детей с генетическим анамнезом аллергии (наличие аллергических заболеваний
у ближайших родственников) наблюдают повышение уровней продукции IL-4,
220 ЦИТОКИНЫ В ЛАБОРАТОРНОЙ ДИАГНОСТИКЕ
IL-5 и IL-13, постепенно нарастающих от момента рождения к 1-2 годам, такое
повышение служит ранним прогностическим фактором развития клинических
симптомов аллергии в возрасте 2-6 лет. С другой стороны, показано, что снижен-
ные уровни IFNy у детей трехмесячного возраста ассоциированы с более высокой
частотой клинических проявлений астмы в годовалом возрасте, а аллергенспе-
цифическая продукция IFNy лимфоцитами в культуре снижена у детей с атопией.
Обнаружение IFNy в пуповинной крови во время родов связано с низким риском
развития астмы и других вариантов аллергии при наблюдении за этими детьми
вплоть до шестилетнего возраста. Приведенные данные подтверждают роль поля-
ризации Т-хелперных клонов в сторону гиперактивации Th2 в патогенезе аллергии
и свидетельствуют о том, что повышенная продукция цитокинов ТЫ может иметь
защитное значение от развития аллергических заболеваний. Определение уровней
и продукции цитокинов у детей раннего возраста может быть важным элементом
диагностики предрасположенности к развитию аллергических заболеваний.
ДИАГНОСТИЧЕСКОЕ ЗНАЧЕНИЕ ОТДЕЛЬНЫХ ЦИТОКИНОВ
Уникальная роль системы цитокинов в регуляции функций организма и их уча-
стие в патогенезе широкого круга заболеваний делают перспективным использо-
вание определения их синтеза как маркера развития инфекционных, иммунодефи-
цитных заболеваний, иммунопатологических процессов и рака. Диагностическая
ценность цитокинов связана с несколькими факторами. Прежде всего большин-
ство цитокинов являются растворимыми циркулирующими медиаторами, их
концентрацию легко оценить в плазме крови и других биологических жидкостях.
Многие цитокины появляются при активации иммунитета, который, в свою оче-
редь, можно оценить по уровню этих цитокинов. Синтез цитокинов сопровождает
развитие защитных реакций. Во врожденном иммунитете это ответ клеток на
патоген-ассоциированные молекулы, в приобретенном иммунитете — это ответ на
антиген, показатель активации лимфоцитов. Изменение синтеза различных групп
цитокинов на местном уровне позволяет оценить развитие инфекционного, аллер-
гического и аутоиммунного типов воспаления. Изменение уровней цитокинов в
плазме крови позволяет оценить развитие системной воспалительной реакции и
работу иммунной системы в целом. Определение синтеза цитокинов отдельных
семейств позволяет оценить практически все ключевые процессы работы иммун-
ной системы, а также диагностировать нарушения кроветворения, состояние
противоопухолевой резистентности, отклонения нормального эмбрионального
развития, процессы обмена веществ и другие параметры. Ниже приводятся дан-
ные по диагностическому значению определения синтеза и уровней отдельных
цитокинов, играющих наиболее важную роль в регуляции нормальных физиоло-
гических функций организма, защитных реакций и в развитии иммунопатологии.
Нормальные значения концентраций цитокинов приведены в табл. 19-7.
Таблица 19-7. Нормы содержания цитокинов в сыворотке и уровни их продукции клетками пери-
ферической крови здоровых людей
Цитокины Концентрация в сыворотке крови, пг/мл Продукция клетками крови в культуре, пг/мл
спонтанная индуцированная
IL-1oc 0-50 0-50 100-1000
IL-10 0-50 0-50 1000-5000
РАИЛ 300-700 0-500 1000-5000
IL-2 0 0 2-10 Ед/мл
IL-4 0-50 30-50 100-400
IL-6 0-50 30-50 1000-3000
IL-8 0-50 30-100 1000-5000
ЦИТОКИНЫ В ЛАБОРАТОРНОЙ ДИАГНОСТИКЕ 221
Окончание табл. 19-7
Цитокины Концентрация в сыворотке крови, пг/мл Продукция клетками кровг в культуре, пг/мл
спонтанная индуцированная
IL-10 0-50 30-50 1000-3000
TNF-a 0-5 30-50 500-3000
IFN-a 0-50 30-50 100-500
IFN-y 0-10 10-100 1000-5000
G-CSF 10-100 30-50 1000-5000
M-CSF 1800-7000 н.д. н.Д.
GM-CSF 10-100 Н.д. н.Д.
ЭПО 10-50 МЕ/л Н.Д. н.Д.
ТПО 50 Н.Д. н.Д.
Интерлейкин-1 бета
В плазме крови большинства здоровых людей не удается обнаружить IL-1.
Однако примерно у трети здоровых людей в плазме могут появляться незначи-
тельные количества этого цитокина (С-50 пг/мл), так же как и некоторых других
провоспалительных цитокинов. Причины появления малых количеств провпс-
палительных цитокинов в плазме крови здоровых людей не совсем понятны.
Возможно, это связано с присутствием биологически неактивных либо связанных
с белками-переносчиками молекул цитокинов, которые определяются иммуно-
химическими методами. Определить биологическую активность цитокинов в
плазме или сыворотке крови чрезвычайно сложно из-за присутствия большого
количества различных биологически активных веществ, как стимуляторов, так и
ингибиторов, мешающих проведению анализа. Увеличение уровня IL-113 отмечают
при стрессовых ситуациях и тяжелой физической нагрузке.
IL-lp является одним из основных провоспалительных цитокинов, поэтому
повышение уровня его спонтанной или индуцированной продукции клетками в
культуре либо повышение уровня IL-lp в сыворотке крови может свидетельство-
вать об активации клеток или развитии воспаления. В связи с этим возрастание
уровня является признаком заболеваний, имеющих в своей основе вос-
паление, причем независимо от типа воспаления. Так, рост концентрации IL-1[3
наблюдается при инфекционных заболеваниях, где острота процесса коррели-
рует с содержанием IL-lp и на местном, и на системном уровнях. Повышенная
концентрация IL-1 обнаружена в эпителиальных смывах при локальных острых
вирусных инфекциях, в содержимом полости абсцессов и флегмон. На примере
изучения продукции IL-1 у больных с пневмонией показано, что уровень этого
вещества в сыворотке и его продукция мононуклеарами крови возрастают в острой
фазе болезни, но оба показателя были ниже у пациентов пожилого возраста.
По-видимому, с возрастом продукция IL-1 может снижаться, что может являться
одной из причин более низких показателей иммунитета в старости. При выра-
женном инфекционном процессе, сопровождающемся изменениями показателей
острофазового ответа, концентрация IL-1 коррелирует с подъемом температуры
тела, повышением уровня других цитокинов и достигает наивысшего значения
у лихорадящих септических больных. Таким образом, развитие инфекционных
заболеваний у человека может сопровождаться повышением уровня IL-1, синтез
которого, очевидно, связан с развитием защитных механизмов.
Концентрация IL-1 может также повышаться при развитии иммунопатологи-
ческих состояний. Увеличение уровней IL-10 отмечено при аллергии, в том числе
при обострении бронхиальной астмы, и при аутоиммунной патологии, например
при диабете 1-го типа, при болезни Крона, ревматоидном артрите. Высокие титры
222 цитокины В ЛАБОРАТОРНОЙ диагностике
IL-1 обнаружены в суставной полости при активных формах ревматоидного артри-
та. а содержание IL-10 в плазме коррелирует с выраженностью поражения суставов
и возрастает в случаях осложнений заболевания. Следовательно, развитие ряда
хронических иммунопатологических процессов у человека может сопровождаться
повышением уровня IL-1 вследствие активации иммунной системы.
Рецепторный антагонист интерлейкина-1
Рецепторный антагонист интерлейкина-1 (РАИЛ) уникален среди цитокинов
семейства IL-1 тем. что он в норме присутствует в плазме периферической крови в
достаточно высоких концентрациях, иногда до 1 нг/мл. Это связано с конститутив-
ной экспрессией гена РАИЛ и его постоянным синтезом тканевыми макрофагами
и гепатоцитами. По-видимому, присутствие РАИЛ в циркуляции выполняет роль
своеобразного буфера, блокирующего действие эндогенного IL-1 и защищающего
организм от резкого увеличения уровня IL-1 в плазме крови в результате развития
воспалительных процессов в тканях, которые в той или иной степени эпизодиче-
ски возникают при попадании в организм патогенов. Несмотря на поддержание
постоянного уровня синтеза, при развитии воспаления происходит усиление экс-
прессии гена РАИЛ и существенное повышение его продукции. Индукция синтеза
РАИЛ происходит под действием тех же стимулов, что и IL-1, однако регуляция
продукции IL-1 и РАИЛ существенно различается. Уровень РАИЛ характеризует
антивоспалительный ответ, что может иметь значение при развитии сепсиса. В то
же время показано, что соотношение продукции IL-1|3 и РАИЛ может быть изме-
нено в стороны увеличения уровней РАИЛ при вторичных иммунодефицитных
состояниях, связанных с развитием гнойно-воспалительной патологии.
Интерлейкин-2
В норме в плазме крови существуют ингибиторы биологической активности
IL-2, что практически исключает его определение методом биологического тести-
рования. Использование иммунохимических методов анализа более предпочти-
тельно, но также имеет свои особенности. Дело в том, что природный и рекомби-
нантный IL-2 отличаются по расположению эпитопов, узнаваемых антителами к
данному цитокину. В природном IL-2 вследствие гликозилирования происходит
экранирование иммунодоминантного эпитопа, к которому в основном вырабаты-
ваются антитела при иммунизации рекомбинантным негликозилированным IL-2.
В результате эти антитела очень плохо распознают природный IL-2 при его иссле-
довании иммунохимическими методами. В современных иммуноферментных
тест-системах это обязательно учитывается, и в них используют только монокло-
нальные антитела, распознающие другие общие для обоих вариантов молекулы
эпитопы. При развитии патологии уровень IL-2 в плазме периферической крови
повышается незначительно. Гораздо более информативные данные получают при
изучении изменений его спонтанной и индуцированной продукции изолирован-
ными лимфоцитами в культуре, в том числе в культурах клеток цельной крови.
Повышение спонтанной продукции 1L-2 может быть при лимфопролифератив-
ных и аутоиммунных заболеваниях. Снижение индуцированной продукции IL-2
клетками крови может быть следствием иммунодефицитных состояний разной
этиологии, в первую очередь Т-клеточного иммунодефицита. Развитие СПИДа
характеризуется неуклонным снижением количества Т-хелперных лимфоцитов с
постепенным развитием тяжелой иммунологической недостаточное] и. Одним из
важных проявлений и ммунодефицитного состояния является падение продукции
IL-2, которое на фоне снижения синтеза и других цитокинов Т-лимфоцитов ведет
к серьезным нарушениям функций всех звеньев иммунной системы.
ЦИТОКИНЫ В ЛАБОРАТОРНОЙ ДИАГНОСТИКЕ 223
Интерлейкин-4
В крови здоровых людей IL-4 практически не определяется. Появление IL-4
в сыворотке крови сопровождает развитие аллергических реакций и активацию
противоинфекционного иммунитета по типу преимущественного развития Th2.
Так, у пациентов, страдающих гельминтозами, обнаружено повышение уровня
IgE, который коррелирует с повышенной секрецией IL-4. Дисре, уляция секреции
IL-4 является ключевой в развитии аллергической патологии. Стимулированная
продукция IL-4 мононуклеарами периферической крови у пациентов с агопией и
лимфоцитами бронхоальвеолярных лаважей, полученных от больных бронхиаль-
ной астмой, значительно выше, чем у здоровых субъектов. Уровень IL-4 (так же
как уровни IL-5 и IL-13) коррелировал с выраженностью клинических симптомов
бронхиальной астмы, сывороточными уровнями IgE и содержанием эозинофилов
в крови и лаважах. Однако при длительно текущих хронических аллергических
процессах может наблюдаться снижение продукции IL-4 лимфоцитами, причем
иногда эти изменения сочетаются со снижением общего IgE в сыворотке крови.
Интерлейкин-13
IL-13 во многом повторяет биологическую активность IL-4. Оба цитокина
являются продуктами Th2 и участвуют в развитии аллергии, но в настоящее время
считается, что IL-13 в большей степени, чем IL-4, связан с развитием тяжелых
симптомов аллергии у человека. Повышение концентрации 1L-13 обнаружено в
бронхоальвеолярных лаважах, а экспрессия гена IL-13 — в биопсийном материале
из легких больных бронхиальной астмой. Уровни IL-13 падали при проведении
аллергенспецифической иммунотерапии (АСИТ) и при терапии глюкокортикои-
дами и коррелировали с клинической картиной заболевания.
Интерлейкин-5
При аллергических заболеваниях у человека количество эозинофилов повыше-
но в системном кровотоке, и в органах-мишенях развивается аллергическое вос-
паление, например в коже при атопическом дерматите или в легких при бронхи-
альной астме. Это коррелирует с усилением продукции IL-5 Th2 и увеличением его
концентрации в тканях. При аллергии биологическое действие IL-5 заключается
в усилении дифференцировки костномозговых предшественников эозинофилов и
увеличении числа зрелых эозинофилов, стимуляции хемотаксиса эозинофилов, их
функциональной активации и замедлении апоптоза. В связи с этим IL-5 считается
одним из основных цитокинов, участвующих в иммунопатогенезе аллергии.
У человека описан идиопатический гиперэозинофильный синдром, характери-
зующийся увеличением количества циркулирующих эозинофилов, эозинофиль-
ной инфильтрацией органов, развитием асептического воспаления и связанной с
этим мультиорганной недостаточностью, в тяжелых случаях приводящей к гибели
больного. Это заболевание свя зано, главным образом, с гиперпродукцией IL-5 —
основным фактором, стимулирующим созревание и активацию эозинофилов, а
также других лигандов общей бета-цепи рецептора: IL-3 и GM-CSF. принимающих
участие в развитии эозинофильного ростка кроветворения.
Интерлейкин-6
В норме отмечаюд лишь минимальный уровень экспрессии гена и накопления
мРНК IL-6 в органах, и его практически не удается обнаружить в плазме крови.
Экспрессия гена IL-6 происходит под влиянием попадающих в организм вирусов,
224 цитокины В ЛАБОРАТОРНОЙ диагностике
бактерий и их продуктов, а также провоспалительных цитокинов. IL-6 является
типичным ранним индуцибельным цитокином, быстро накапливающимся в цир-
куляции при встрече с патогенами. В ряде исследование показано, что гиперпро-
дукция IL-6 у человека ведет к патологии, связанной с развитием аутоиммунных
и воспалительных процессов. Повышенные уровни IL-6 обнаружены в синовиаль-
ной жидкости больных ревматоидным артритом, увеличение его синтеза отмечено
при миксоме сердца и у больных с миеломной болезнью. Видимо, главное диа-
гностическое значение IL-6 связано с тем, что именно его уровень наиболее точно
коррелирует с тяжестью клинических проявлений при сепсисе,
Интерлейкин-7
Биологическая активность IL-7, главным образом, связана с его действием в
качестве ростового фактора для предшественников В- и Т-лимфоцитов. IL-7 в
небольших количествах продуцируется стромальными клетками костного мозга,
эпителием тимуса, кератиноцитами и клетками фетальной печени. II-7 синте-
зируется стромальными клетками костного мозга и тимуса конститутивно, обе-
спечивая постоянное созревание и обновление Т-лимфоцитов. Возможно, в орга-
низме существует гомеостатический механизм регуляции числа Т-лимфоцитов
на периферии, построенный по механизму отрицательной обратной связи. У
больных с ВИЧ-инфекцией уровни IL-7 в плазме отрицательно коррелировали с
числом Т-лимфоцитов периферической крови, возрастая при падении количества
Т-клеток. Отмечено также увеличение концентрации IL-7 в циркуляции при дру-
гих заболеваниях, сопровождающихся лимфопенией.
Интерлеикин-8
У здоровых людей в сыворотке крови IL-8 выявляется крайне редко. Спонтанная
продукция IL-8 мононуклеарами крови наблюдается у 61,5%, индуцированная —
у 100% здоровых доноров. Увеличение уровня 1L-8 в биологических жидкостях
описано при различных заболеваниях, сопровождающихся нейтропенией и/
или нейтрофильной инфильтрацией тканей очага воспаления. В случае введения
в организм ЛПС в плазме крови увеличивается уровень циркулирующего 1L-8.
То же наблюдается после введения добровольцам IL-I или TNF, а при сепсисе
титры сывороточного IL-8 положительно коррелируют с тяжестью заболевания
и летальным исходом. IL-8 может быть важен в модуляции некоторых органных
дисфункций. При многих заболеваниях легких, сопровождающихся воспалением
в легочной ткани, хемокины активно синтезируются альвеолярными макрофагами
и находятся в повышенной концентрации в лаважной жидкости. При респира-
торном дистресс-синдроме взрослых IL-8 обнаруживается в бронхоальвеолярной
лаважной жидкости, его концентрация прямо пропорциональна количеству ней-
трофилов и смертности у пациентов. Высокие уровни IL-8 и других хемокинов в
легочной ткани и лаважах описаны при бронхиальной астме и фиброзе легких.
IL-8 определяется в больших количествах в области псориатических бляшек.
Кроме IL-8, в области бляшек обнаружены и другие СХС-хемокины: GROa и
1Р-10. Основным источником IL-8 и указанных хемокинов служат кератиноци-
ты. Кроме того, хемокины синтезируются эндотелиальными клетками и самими
привлеченными в очаг воспаления нейтрофилами. Повышенная продукция хемо-
кинов приводит к еще большему привлечению лейкоцитов и формированию хро-
нического очага асептического воспаления. При ревматоидном артрите высокие
уровни IL-8 определяются в суставной жидкости и коррелируют с активностью
процесса и с количеством нейтрофилов и других клеток в жидкости. Источником
TL-8 в полости сустава служат, главным образом, синовиальные клетки. По мере
ЦИТОКИНЫ В ЛАБОРАТОРНОЙ ДИА1НОСТИКЕ 225
прогрессии заболевания в сыворотках пациентов появляются аутоантитела к IL-8,
относящиеся к IgG классу.
Интерлейкин-10
В клинических исследованиях продемонстрировано, что у больных с аллергией
снижена частота встречаемости клонов Т-хелперных лимфоцитов, синтезирую-
щих IL-10, и снижены уровни IL-I0 в сыворотке крови. Более того, обнаружены
инверсные взаимоотношения между уровнем IL-10 и тяжестью клинических про-
явлений астмы и других типов аллергии. В настоящее время есть данные о том, что
при АСИТ успех терапии коррелировал со снижением аллергенспецифического
Th2 ответа и появлением аллергенспецифических Т-лимфоцитов, секретирующих
IL-10.
Интерлейкин-15
IL-15 синтезируется различными типами клеток во многих органах. У чело-
века IL-15 синтезируется моноцитами/макрофагами, эпителиальными клетками
стромы костного мозга и другими эпителиальными клетками, но мРНК IL-15 не
обнаружена в активированных Т- и В-лимфоцитах. Синтез IL-15 активируется под
действием IFN 1 типа, IFNy, двухцепочечной РНК, ЛПС и других производных
патогенов. Экспрессия мРНК IL-15 выявлена во многих типах клеток, но IL-15
крайне редко обнаруживается в супернатантах клеточных культур или в плазме
крови. Для IL-15 человека описано несколько вариантов мРНК, образующихся в
результате альтернативного сплайсинга. Все эти варианты транслируются в одина-
ковые формы зрелого IL-15, но скорость трансляции и варианты продукции белка
отличаются.
Фактор некроза опухоли
Для определения уровня TNF в биологических жидкостях следует учитывать,
что биологически активная молекула этого цитокина состоит из трех одинаковых
полипептидных субъединиц, объединенных в гомотример. Только такая трехмер-
ная структура биологически активна, и только она должна определяться специфи-
ческими моноклональными антителами, используемыми в современных диагно-
стических системах. Часть антител к TNF могут связывать и мономерные формы,
которые могут находиться в плазме и биологических жидкостях в достаточно
высоких концентрациях, не проявляя биологической активности. Определение
общей концентрации мономеров вместе с биологически активными тримерами
TNF приводит к ложным заключениям о значительном повышении уровня TNF
в исследуемых образцах, хотя иногда очевидно, что неправильно определенные
высокие уровни TNF просто несовместимы с жизнью пациента. В связи с этим
для оценки уровня TNF следует использовать только те диагностические системы,
которые позволяют определять именно биологически активный цитокин.
В сыворотке крови здоровых доноров биологически активный TNF определяет-
ся в очень низких концентрациях (0-5 пг/мл), однако выделенные мононуклеары
способны продуцировать TNF в больших количествах. У пациентов с сепсисом
TNF определяется в плазме крови, и его концентрация коррелирует с тяжестью и
исходом заболевания. У пациентов с бактериальными менингитами тяжесть забо-
левания коррелирует с концентрацией TNF в цереброспинальной жидкости, хотя
высокие концентрации TNF не коррелируют с его высокими системными концен-
трациями. В ряде случаев повышение уровня TNF и других провоспалительных
цитокинов в плазме крови позволяет заподозрить заболевания, прямо не свя-
226 ЦИТОКИНЫ В ЛАБОРАТОРНОЙ ДИАГНОСТИКЕ
занные с нарушениями иммунитета или с инфекционной патологией. Например,
повышение уровня TNF наблюдается при инфаркте миокарда, длительное уве-
личение уровней TNF, IL-lp и IL-6 в плазме периферической крови может быть
предвестником остеопороза.
Некоторые цитокины, члены семейства TNF, используются для диагностики
рака. Определение концентрации растворимого Fas вместе с анализом других
антигенов применяется в диагностике рака мочевого пузыря. В целом в диагности-
ке рака все больше начинают использоваться методы, позволяющие одновременно
определять сразу несколько цитокинов. Это связано с тем, что в диагностике все
большее значение приобретает анализ не единичных факторов, включая цитоки-
ны, а изучение паттернов изменений десятков и более медиаторов. В случае цито-
кинов это особенно важно с учетом формирования в организме цитокиновой сети.
Сейчас такие методы широко применяют для исследования наиболее значимых
среди цитокинов диагностических маркеров рака, для изучения молекулярных
механизмов патогенеза рака, эффективности действия противораковых препара-
тов, в том числе во время клинических испытаний.
Интерферон-альфа
Участие IFNa в патогенезе множества вирусных заболеваний не вызывает
сомнений. С развитием терапии рекомбинантным IFNct приобрели большую диа-
гностическую ценность исследования интерферонового статуса, который отра-
жает характер течения вирусной инфекции, позволяет прогнозировать исход
заболевания и эффективность лечения. При изучении роли IFNa в патогенезе
гриппа показана зависимость между сывороточным уровнем IFNa и тяжестью и
продолжительностью гриппозной инфекции. У больных гриппом, осложненным
пневмонией, и при затяжном течении гриппа наблюдалось значительное сниже-
ние уровня IFNa к периоду ранней реконвалесценции, что связывают с развитием
астенического синдрома и что указывает на целесообразность применения средств
с иммуномодулирующей активностью в этом периоде. Снижение продукции IFNa
отражает наличие нарушений противовирусной защиты организма.
Особую значимость приобретают исследования уровня IFNa для изучения
фармакокинетики препаратов рекомбинантного IFNa. После типичной внутри-
мышечной или подкожной инъекции от 3 млн до 50 млн единиц рекомбинант-
ного pIFNa пиковая концентрация от 100 до 1000 ед/мл достигается через 4-6 ч.
Однако индивидуальные различия в уровне эндогенного IFNa, времени пиковой
концентрации и времени полу жизни препаратов этого цитокина делают необходи-
мыми исследования фармакокинетики препарата у каждого отдельного пациента
для обеспечения оптимальной схемы терапии.
Интерферон-гамма
Оценка уровня IFNy в культуре клеток крови имеет значение для определения
поляризации иммунного ответа по типу активации клонов Т 1 Этот тип иммунно-
го реагирования характерен для клеточного ответа, вызванного так называемыми
внут риклеточными патогенами. В последнее время разработаны два теста, позво-
ляющие на основании уровня продукции IFNy в культуре клеток периферической
крови оценить наличие туберкулезной инфекции. Принцип метода состоит в акти-
вации синтеза интерферона двумя специфическими антигенами из Mycobacterium
tuberculosis, которые отсутствуют в вакцинном штамме БЦЖ (Bacille Calmette-
Gu rin — BCG) и других микобактериях. Одна модификация метода основана на
выявлении синтезирующих IFNy клеток методом ELISPOT, в другом варианте
уровень IFNy в супренатанте клеток оценивается методом ИФА Клинические дан-
ЦИТОКИНЫ В ЛАБОРАТОРНОЙ ДИАГНОСТ ИКЕ 227
ные указывают, что оба теста более специфичны, чем стандартный кожный тест,
гак как исключают перекрестную реактивность, обусловленную предшествующей
вакцинацией. По чувствительности метод ИФА сравним с кожным тестом, а метод
ELISPOT его значительно превосходит.
Гранулоцитарный колониестимулирующий фактор
У здорового человека уровень G-CSF в плазме крови очень низок, но при разви-
тии инфекционных заболеваний его концентрация резко возрастает. Повышенные
уровни G-CSF также наблюдают при наследственной нейтропении и лекарствен-
ной аплазии костного мозга, что указывает на существование механизма обрат-
ной связи между количеством нейтрофилов на периферии и уровнем продукции
G-CSF. Исключение составляет аутоиммунная нейтропения у детей, где уровень
G-CSF не отличался от уровня у здоровых доноров, хотя количество лейкоцитов в
периферической крови было значительно снижено.
Моноцитарный колониестимулирующий фактор
М-CSF находится в плазме периферической крови в очень высокой для цитоки-
нов концентрации — от 1,8 до 7,1 нг/мл. Кроме того, он постоянно присутствует в
моче, спинномозговой и амниотической жидкости. Повышение уровня эндогенно-
го М-CSF наблюдается при травмах, инфекционных заболеваниях, беременности
и некоторых формах рака.
Г ранулоцитарно-макрофагальный колониестимулирующий
фактор
В норме GM-CSF синтезируется в незначительных количествах и практически
от сутствует в плазме периферической крови. При развитии воспаления он начина-
ет активно синтезироваться Т-лимфоцитами, макрофагами, эндотелиальными и
эпителиальными клетками в ответ на компоненты клеточной стенки патогенов и
провоспалительные цитокины. Уровень GM-CSF может быть повышен в области
очага воспаления, хотя при сепсисе с развитием синдрома системной воспалитель-
ной реакции концентрация GM-CSF редко оказывалась повышенной в отличие от
G-CSF, уровень которого значительно возрастал у септических больных. Клетки
некоторых типов злокачественных опухолей кроветворной системы секретируют
все разновидности CSF, IL-3 и фактор роста стволовых клетик. Концентрации этих
цитокинов могут быть повышены у больных с гематологическими формами рака
и служить диагностическими маркерами стадии развития опухолей и ответа на
терапию.
Эритропоэтин
Регуляция дифференцировки и поддержания функциональной активности
красного ростка кроветворения является важнейшей функцией, связанной с
обеспечением жизнедеятельности организма. В нормальных физиологических
условиях экспрессия гена эритропоэтина (ЭПО) происходит конститутивно.
Цитокин постоянно находится в плазме крови в невысоких концентрациях в
пределах 10-15 МЕ/л. Это обеспечивает поддержание нормального гемопоэза, в
ходе которого из костного мозга в периферическую кровь за одну минуту выходят
1,8x10’ ретикулоцитов, что требуется для поддержания концентрации 25х1012 цир-
кулирующих эритроцитов. Период су шествования эритроцитов составляет около
228 цитокины В ЛАБОРАТОРНОЙ диагностике
120 сут, после чего происходит их утилизация в о< новном с помощью макрофагов
селезенки. Такая высокая скорость образования эритроцитов может увеличивать-
ся еще в 10-12 раз при серьезных кровопотерях; это сочетается с возрастанием
уровня экспрессии гена ЭПО в клетках продуцентах и с ростом его концентрации
в плазме до 10 000 МЕ/л. Период полужизни ЭПО в плазме человека составляет
от 6 до 10 ч. В норме ЭПО выводится через почки и обнаруживается в неизменен-
ном виде в моче, попадая туда из плазмы, а не путем прямой почечной экскреции
после синтеза в почечной ткани. Определение уровня ЭПО в плазме крови либо в
моче используется для мониторинга его концентрации при анемиях различного
генеза: при почечной недостаточности, травмах, при анемиях у беременных и у
недоношенных детей. Уровень ЭПО может быть диагностическим критерием для
полицитемии (polycythemia vera), у больных с этим диагнозом концентрация ЭПО
(метод ИФА) ниже границы нормального значения в 3.3 МЕ/л.
Тромбопозтин
Основная биологическая функция тромбопоэтина (ТП) заключается в уско-
рении созревания предшественников тромбоцитопоэза, тогда как для усиления
пролиферации этих клеток требуется его совместное действие с другими гемо-
поэтическими факторами. Основным местом продукции ТП является печень,
высокий уровень экспрессии мРНК ТП обнаружен также в почках и селезенке.
ТП может синтезироваться и мегакариоцитами; тромбоциты имеют депо ТП в
своих гранулах. При активации тромбоцитов в процессе свертывания крови ТП
высвобождается из гранул, его концентрация возрастает у больных с синдромом
внутрисосудистого свертывания крови. Это может служить механизмом активации
тромбоцитарного ростка кроветворения для образования новых тромбоцитов.
Тромбоцитопении, часто сопутствующие циррозу печени и почечной недостаточ-
ности, не всегда связаны с недостатком выработки ТП, у этих больных уровень
ТП в плазме периферической крови практически не отличается от его концентра-
ции в плазме крови здоровых доноров, составляющей около 50 пг/мл. В других
исследованиях показано, что уровень ТП повышен в случае тромбоцитопении при
циррозе печени, осложненном миелодиспллогическим синдромом или возникшем
как следствие вирусного гепатита.
Трансформирующий ростовой фактор [3
TGF[3 может быть выделен из нормальных тканей, например из тромбоцитов
или плаценты человека и из опухолевых клеток. TGFp, а также другие члены этого
обширного семейства, костные морфогенетические белки (bone morphogenetic pro-
teins — BMP) и активины экспрессируются в эндометрии, секретируются в полость
матки и принимают участие в регуляции имплантации и постимплантационном
развитии плода, а также регулируют синтез плацентарных гормонов. Во время
беременности ингибины и активины синтезируются клетками плаценты и могут
служить маркерами плацентарной патологии, включая преэклампсию и угрозу
выкидыша.
ЗАКЛЮЧЕНИЕ
Таким образом, в настоящее время не вызывает сомнения, ч го цитокины слу-
жат важнейшими мишенями иммунодиагностики широкого круга заболеваний
человека. Цитокины являются антиген-неспецифическими факторами. Именно
поэтому специфическая диагностика инфекционных, аутоиммунных и аллергиче-
ских заболеваний с помощью определения уровня тех или иных цитокинов невоз-
ЦИТОКИНЫ В ЛАБОРАТОРНОЙ ДИАГНОСТИКЕ 229
можна. Тем не менее оценка содержания цитокинов в биологических жидкостях и
изучение их продукции клетками дает неоценимую помощь для характеристики
состояния иммунной системы организма. Изучение уровней цитокинов позво-
ляет получить информацию о функциональной активности различных типов
иммунокомпетентных клеток, о тяжести воспалительного процесса, его перехо-
де на системный уровень и прогнозе (например, уровень некоторых цитокинов
коррелирует с тяжестью клинических проявлений и летальностью при сепсисе);
о соотношении процессов активации типов Т-хелперов, что очень важно при
дифференциальной диагностике ряда инфекционных и иммунопатологических
процессов; о стадии развития ряда аллергических и аутоиммунных заболеваний,
когда, например, повышение уровня IL-1 в плазме крови опережает клинические
проявления при ревматоидном артрите и системной красной волчанке. В диагно-
стическую практику все более входят методы молекулярно-биологического ана-
лиза, позволяющие выявлять первичные генетические дефекты и полиморфизм
по единичным нуклеотидам в системе цитокинов, связанные с патогенезом целого
ряда заболеваний, в том числе таких широко распространенных, как туберкулез.
Такие же примеры можно привести по использованию любого из перечисленных
выше методов анализа состояния системы цитокинов. Каждый метод по-своему
хорош, и его использование определяется конкретной задачей диагностического
исследования.
В настоящее время очевидно, что цитокины являются важнейшими фактора-
ми патогенеза инфекционных заболеваний и иммунопатологических процессов.
Поэтому определение уровней синтеза эндогенных цитокинов и их концентраций
в биологических жидкостях можно использовать для целей иммунодиагностики,
изучения патогенеза заболеваний, адекватного назначения иммунотерапии. Все
это говорит о том. что оценка функционирования системы цитокинов позволяет
по-новому подойти к изучению состояния иммунной системы организма в клини-
ческой практике.
СПИСОК РЕКОМЕНДУЕМОЙ ЛИТЕРАТУРЫ
Кетлинский С.А., Симбирцев А.С. Цитокины. — СПб.: Фолиант. 2008. — 552 с.
Румянцев А.Г., Морщакова Е.Ф., Павлов А.Д. Эритропоэтин. — М.; ГЭОТАР-мед, 2002. —
399 с.
Сесь Т П., Тотолян А.А. Возможности мультиплексных технологий, основанных на при-
менении микрочастиц, в клинической лабораторной диагностике. Лабораторная диагности-
ка России 2007/2008: Справочник. — СПб.: Человек, 2007. — С. 51-56.
Тотолян А.А., Фрейдлин И.С. Клетки иммунной системы. — СПб.: Наука, 1999, — 231 с.
Фрейдлин И.С., Тотолян А.А. Клетки иммунной системы. — СПб.: Наука, 2001. — 309 с.
Глава 20
Бактериологические
исследования
ЗНАЧЕНИЕ ЛАБОРАТОРНОЙ ДИАГНОСТИКИ
В ИНФЕКТОЛОГИИ
Основной принцип диагностики инфекционных болезней —
единство трех составляющих: клинической диагностики, эпидемио-
логической диагностики и инструментально-лабораторной диагно-
стики.
Методы инструментальной и лабораторной диагностики, в свою
очередь, могут быть подразделены на общие (анализы крови и
мочи, рентгенография, ультразвуковое исследование) и специфи-
ческие (бактериологические, вирусологические, серологические,
молекулярно-биологические), позволяющие раскрыть природу
заболевания, характер его течения и прогноз, обосновать стратегию
и тактику лечебных и профилактических мер.
Значение лабораторной диагностики в инфектологии неуклонно
возрастает в силу трех объективных причин:
О эволюции инфекционных агентов;
о прогресса молекулярной биологии и молекулярной медици-
ны;
< быстрого развития средств и методов лабораторного анализа.
Эволюция инфекционных болезней носит глобальный характер,
ее отличают контрастные тенденции в динамике различных групп
заболеваний.
• Инфекции, контролируемые средствами вакцинопрофилак-
тики и санитарно-гигиеническими мерами, теряют массо-
вый характер. В стадии ликвидации находятся полиомиелит,
корь, минимизирована заболеваемость дифтерией, коклюшем,
брюшным тифом, малярией, устойчиво снижается частота
шигеллезов, сальмонеллезов и эшерихиозов.
• В структуре заболеваемости классическими инфекциями уве-
личивается относительное число стертых, атипичных и суб-
клинических форм.
• На фоне успехов в борьбе с инфекциями сохраняется риск
возврата массовых инфекционных болезней вследствие недо-
работок в прививочной работе, генетического дрейфа пато-
генов, распространения антибиотике- и химиорезистентных
штаммов микроорганизмов.
БАКТЕРИОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ 231
• Ускоряется процесс выявления новых патогенов (около 50 видов за послед-
ние 40 лет), вызывающих болезни с малоизученной клинической картиной.
• Угроза возникновения пандемий «в любом месте и в любое время» по заклю-
чению экспертов ВОЗ, становится постоянной вследствие глобализации
миграционных потоков населения и появления «новых» патогенов (напри-
мер, пандемия ВИЧ-инфекции).
• Продолжается или нарастает неблагополучие по социальнозначимым инфек-
циям (ВИЧ-инфекции, ИППП, туберкулезу, вирусным гепатитам), которым
свойственны длительный латентный период и, часто, скрытое течение, кото-
рое эпидемиологически и прогностически столь же опасно, как и манифест-
ное.
• По мере вовлечения в эпидемический процесс новых для России движущих
факторов (наркомании, рискованного социального поведения, коммерче-
ского секса) обостряется проблема множественной инфекции — смешан-
ной, сочетанной, суперинфекции (ВИЧ-инфекция и ВИЧ-ассоциированные
инфекции, вирусные гепатиты и ИППП), которые могут маскировать друг
друга и требуют полной этиологической расшифровки.
Перечисленные особенности придают новый смысл специфической лаборатор-
ной диагностике. Из средства подтверждения она все более превращается в инстру-
мент первичного выявления инфекционных заболеваний и состояний (латентной
инфекции, бессимптомной инфекции, носительства). Наряду со специфической
диагностикой по клиническим показаниям все большее значение придается доно-
зологической диагностике инфекций, а также сходным направлениям — прена-
тальной, перинатальной и постнатальной диагностике.
Прогресс молекулярной биологии и молекулярной медицины обогащает спец-
ифическую лабораторную диагностику новыми подходами и критериями для
идентификации патогенов и оценки состояния больных. Сиквенс генома возбу-
дителей болезней и широкий фронт молекулярно-биологических исследований
позволяют:
❖ получить новые данные о генетическом разнообразии бактерий и вирусов;
< использовать в клинической и лабораторной практике генотипическую
систему классификации микроорганизмов;
❖ пополнить знания о географическом распределении различных генотипов
микроорганизмов:
❖ выделить генетические маркеры пато1енности, химио- и антибиотикорези-
стентно сти;
о - определить молекулярные маркеры фаз инфекционного процесса, в том
числе более ранние признаки инфекции, чем IgM;
> выявить важную для диагностики динамику созревания иммунного ответа.
Объектом определения все чаще становятся не только сами бактерии, вирусы,
их антигены или токсины, но и специфические элементы генома патогенов (ДНК,
РНК). В связи с этим впервые возникла возможность быстрой детекции некуль-
тивируемых и труднокультивируемых микроорганизмов прямо в патологическом
материале, а также возможность быстрого определения антибиотикорезистентно-
сти бактерий с помощью выявления характерных мутаций в их геноме. Развитие
средств и методов лабораторного анализа в инфектологии носит комплексный и
системный характер:
< быстро расширяется реагентная база в результате создания и внедрения
новых рекомбинантных продуктов, моноклональных антител и специфиче-
ских праймеров;
❖ непрерывно совершенствуются методики, основанные на принципах ИФА,
иммуноблоттинга, ПЦР и технологии биологических микрочипов;
232 БАКТЕРИОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ
❖ создаются и обновляются высокопроизводительные автоматизированные и
компьютеризированные приборы и оборудование.
Общий результат новых разработок:
❖ повышение чувствительности и специфичности тестов;
< стандартизация и улучшение воспроизводимости методов;
❖ минимизация требуемого объема исследуемого материала;
возможность поточной обработки большого числа образцов.
По быстроте получения результата современные методы, основанные на молеку-
лярных подходах, превосходят классические методы бактериологического и виру-
сологического исследований. Используя некоторые модификации ПЦР, можно
обнаружить от 1 до 10 нуклеиновых кислот бактерий или вирусов в исследуемом
образце, что приближается к величине заражающей дозы патогенов. Достижение
и преодоление подобного порога чувствительности — давняя стратегическая цель
прогресса в лабораторной диагностике.
ЗАДАЧИ БАКТЕРИОЛОГИЧЕСКОЙ ДИАГНОСТИКИ
Отмеченное выше многообразие проблем современной инфектологии и возрас-
тающие методические возможности формируют разнообразие и многоплановость
задач современной диагностики бактериальных инфекций. Задачи лабораторной
диагностики бактериальных инфекций состоят в:
❖ этиологической расшифровке инфекционного заболевания;
❖ идентификации биологического вида бактерии-возбудителя;
о идентификации и дифференциации вирулентных (патогенных) вариантов
возбудителя (С. diphtheriae, Cl. diftcile, Е. coll, У. enterocolitica);
❖ генотипировании, субтипировании, риботипировании, VNTR-типировании.
сполиготипировании и другой детальной характеристике бактерий в целях
идентификации эпидемических штаммов и клонов, госпитальных штаммов
(М. tuberculosis, S. typhi, С. diphthenae. Е. cod, Salmonella spp.);
установлении клинических фаз инфекции, выявлении острой (ранней) инфек-
ции, дифференциации инфекции и состояния поствакцинального иммуните-
та (при коклюше, иерсиниозе, бруцеллезе, туляремии, боррелиозе);
о контроле эффективности лечения и эрадикации инфекции (при туберкулезе,
сифилисе, дифтерии, острых кишечных инфекциях, сальмонеллезах, брюш-
ном тифе, инфекции, вызванной Н. pylori');
❖ выявлении и дифференциации суперинфекции, рецидива, реинфекции (при
туберкулезе, инфекции, вызванной II. pylori, других хронических инфекци-
ях);
< > выявлении ВИЧ-индикаторных инфекций (ВИЧ-ассоциированных инфек-
ций у ВИЧ-инфицированных пациентов);
о- выявлении оппортунистических инфекций у больных на фоне иммуносу-
прессивной терапии после трансплантации органов и тканей и у пациентов с
хроническими вирусными гепатитами;
❖ контроле антибиотикорезистентности (первичной, вторичной) штаммов воз-
будителей бактериальных инфекций
Специальные задачи исследований по клиническим показаниям включают:
❖ исследование фекалий на дисбактериоз;
< = исследование биоценоза влагалища.
Основными задачами донозологической диагностики бактериальных инфекций
являются:
❖ скрининговые и профилактические исследования по выявлению сифилиса:
- обследование лиц, направляемых на стационарное лечение, на работу в
эпидемиологически значимые коллективы и производства;
БАКТЕРИОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ 233
- обследование лиц, относящихся к группам риска (согласно действующим
федеральным и местным нормативным документам);
❖ скрининговые и профилактические исследования по выявлению больных
дифтерией и носителей С. diphtheriae:
- обследование лиц, направленных в детские дома, дома ребенка, интернаты
психоневрологического профиля, противотуберкулезные детские санато-
рии, а также лиц, поступающих на работу в такие учреждения;
- обследование больных с подозрением на дифтерийную инфекцию при
ларингитах, ларинготрахеитах, крупе, ангинах, заглоточном абсцессе,
инфекционном мононуклеозе;
❖ профилактические исследования на острые кишечные инфекции:
- обследование работников предприятий молочной, мясной, пищевых
отраслей промышленности, общественного питания, торговли, водоснаб-
жения, водоотведения и очистки населенных мест, детских и подростко-
вых учреждений всех профилей;
- обследование лиц, направляемых в учреждения и отделения психоневро-
логического (психосоматического) профиля, дома престарелых, интерна-
ты и другие типы закрытых учреждений с круглосуточным пребыванием;
- обследование эпидемически значимых контингентов на брюшной тиф и
носительство S. typhv,
* обследование на носительство патогенного стафилококка лиц, поступающих
на работу в детские молочные кухни, предприятия пищевой промышленно-
сти.
Задачи пренатальной, перинатальной и постнатальной диагностики инфекци-
онных заболеваний сводятся к:
< выявлению или исключению TORCH-комплекса (токсоплазмоза, краснухи,
цитомегаловирусной инфекции, герпеса) и других инфекций (вирусных гепа-
титов В и С, ВИЧ-инфекции, сифилиса, хламидиоза, гонококковой инфек-
ции, листериоза) у беременных;
❖ определению иммунной защищенности беременных против инфекций
TORCH-комплекса;
о пределению группы риска перинатальной инфекции TORCH-комплекса;
о выявлению внутриутробного инфицирования плода;
< ► обоснованию медицинских показаний для искусственного прерывания бере-
менности в связи с выявлением инфекционных заболеваний беременных и
перинатальной инфекции;
< > выявлению рисков интранатального инфицирования плода;
о дифференциации перинатальной, интранатальной и постнатальной инфек-
ции новорожденных.
Лабораторное заключение по результатам бактериологического, серологиче-
ского или молекулярно-биологического исследования все шире используется в
качестве основания для постановки окончательного диагноза инфекционного
заболевания или состояния (латентной инфекции, бактерионосительства). Это
становится возможным при интеграции лабораторной диагностики с другими
медицинскими дисциплинами, правильной постановке задачи и выборе эффектив-
ного алгоритма обследования.
К обязательным условиям успешной работы также относятся:
❖ использование адекватных методов исследования;
< > наличие системы внутрилабораторного и внешнего контроля качества;
❖ взаимодействие лабораторных служб учреждений лечебно-профилакти-
ческого и санитарно-эпидемиологического профиля;
❖ готовность лабораторий к постоянной модернизации методической базы и
приборного обеспечения.
234 БАКТЕРИОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ
ЭТИОЛОГИЧЕСКАЯ ДИАГНОСТИКА
БАКТЕРИАЛЬНЫХ ИНФЕКЦИЙ
Основные задачи клинической микробиологии:
о выявление этиологии инфекционного процесса;
❖ выбор рациональных средств этиотропной терапии.
Эти задачи тесно связаны между собой, и без успешного решения первой невоз-
можно решение второй.
Диагностический процесс в клинической микробиологии складывается из четы-
рех основных этапов:
❖ формулировки задачи и выбора метода исследования;
❖ выбора, взятия исследуемого материала, его хранения и транспортировки;
< > проведения исследований;
❖ анализа полученных результатов.
Важно, что из четырех указанных этапов только один (и то не во всех слу-
чаях) полностью выполняется в лаборатории. Формулировка задачи исследо-
вания — прерогатива лечащего врача. В выборе метода исследования, вида иссле-
дуемого материала функции врача-микробиолога сугубо консультативные. Взятие
и транспортировку материала чаще осуществляет персонал лечебного учреждения
и только в особых случаях персонал лаборатории. Анализ полученных результа-
тов лечащий врач проводит совместно с врачом-микробиологом, однако право и
обязанность постановки этиологического диагноза принадлежат первому из них.
11менно поэтому ответственность за успех микробиологического исследования
не может быть возложена исключительно на лабораторию. Достоверность и кли-
ническая значимость получаемой информации определяется согласованностью
действий лечащего врача и врача-микробиолога. Эффективное взаимодействие
возможно только на основе взаимопонимания, когда обе стороны учитывают
потребности друг друга и трудности, которые возникают при решении поставлен-
ных задач перед каждым из них.
Формулировка задачи исследования подразумевает создание рабочей гипотезы
о возможной этиологии патологического процесса у данного больного. Задача
исследования определяет как выбор метода исследований, так и вид исследуемого
материала и отражается в бланке направления, который прикладывается к матери-
алу, доставляемому в лабораторию. Многообразие патогенных микроорганизмов
определяет многообразие методов, используемых для диагностики инфекционных
заболеваний. Каждый из них имеет свои достоинства и недостатки, но идеаль-
ного метода, одинаково эффективного при диагностике любого патологического
процесса, не существует. Также ни один из существующих методов не позволяет
определять в исследуемом материале все микроорганизмы одновременно. Именно
поэтому врач-микробиолог, который, как правило, не видит больного, должен
получить представление о предполагаемой этиологии патологического процесса,
определенной на основании клинических и эпидемиологических данных.
Для лабораторной диагностики заболеваний инфекционной природы применя-
ют следующие методы:
❖ методы, основанные на выявлении инфекционных агентов (бактерий, виру-
сов, грибов, простейших):
- микроскопические методы (в том числе бактериоскопический), базирую-
щиеся на прямой визуализации возбудителя в патологическом материале
с помощью различных приемов микроскопии;
- культуральные методы (в том числе бактериологический), главная состав-
ляющая которых — культивирование возбудителя на питательных средах,
в организме лабораторных животных или в культурах тканей с целью
выделения его чистой культуры и последующей идентификации;
БАКТЕРИОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ 235
- методы, позволяющие обнаружить в исследуемом материале продукты,
синтезированные микроорганизмами (например, летучие жирные кис-
лоты при диагностике инфекций, обусловленных неспорообразующими
анаэробами, или токсин при диагностике ботулизма):
- иммунологические методы индикации антигенов возбудителей в исследуе-
мом материале;
- генетические методы, основанные на обнаружении нуклеиновых кислот
возбудителя в исследуемом материале;
о - методы выявления активного иммунного ответа — чаще всего по нарастанию
титра антител к антигенам возбудителя (серодиагностика) или сенсибилиза-
ции (аллергодиагностика);
❖ неспецифические лабораторные тесты, по характеру отклонения которых
можно заподозрить патологические изменения, характерные для инфекци-
онных процессов определенной этиологии (например, изменение активности
трансаминаз при вирусных гепатитах).
Успех выделения чистой культуры микроорганизма определяется правильно-
стью выбора питательной среды и условий культивирования. Универсальной пита-
тельной среды, использование которой гарантирует выделение любого микро-
организма, не существует. Обнаружение возбудителей в исследуемом материале,
содержащем значительное количество сопутствующей микрофлоры (например,
фекалиях), требует применения элективных питательных сред, предназначенных
для выделения конкретных видов микроорганизмов. Для некоторых бактерий
необходимы особые условия культивирования (микроаэрофильные — для кампи-
лобактеров, анаэробные — для клостридий и бактероидов, атмосфера, обогащен-
ная углекислым газом, — для нейссерий). Все это следует учитывать при направле-
нии материала в лабораторию, так как успех исследования во многом определяется
правильностью предварительного диагноза лечащею врача и, следовательно,
правильностью поставленной перед лабораторией задачи. Например, если лабо-
раторию не просили специально выполнить дополнительные исследования фека-
лий на Campylobacter spp. или С/, difficile, ответ «патогенные микроорганизмы не
обнаружены» может означать, что необходимые исследования не проводили и
указанные виды бактерий не искали.
Метод исследования необходимо выбирать с учетом всего комплекса диагности-
ческих и лечебных процедур, проводимых данному’ больному. Например, исполь-
зование бактериологического метода на фоне антибиотикотерапии будет заведомо
малоэффективным. Методы, не позволяющие дифференцировать живые и «убитые»
микроорганизмы (ПЦР, РИФ), следует с осторожностью использовать при контроле
результатов лечения. Подобные исследования необходимо проводить не ранее, чем
через несколько недель после окончания этиотропной терапии, так как погибшие
микробные клетки или их антигены могут длительное время сохраняться в организ-
ме и выявляться с помощью указанных методов. Постановка кожной аллергической
пробы в интервале между взятием двух парных сывороток при проведении серо-
диагностики может привести к увеличению титра антител, которое обусловлено не
развитием заболевания, а экзогенным введением аллергена.
Выбор вида исследуемого материала зависит от конкретною заболевания и
преимущественной локализации возбудителя на данним этапе развития патологи-
ческого процесса. Классический пример, подтверждающий значение обоснованно-
го выбора материала в зависимости от этапа патогенеза болезни, — брюшной тиф.
При этой инфекции на различных этапах бактериологического анализа сначала
исследуют кровь, а затем фекалии.
Важно осущест ви гь взятие проб в оптимальные сроки. Например, при сальмонел-
лезе вероятность выделения возбудителя из фекалий на 2-й неделе заболевания в
2 раза, а на 3-й в 6,8 раза меньше, чем при обследовании на 1-й неделе. При коклюше
236 БАКТЕРИОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ
вероятность обнаружения возбудителя в носоглоточной слизи на 1-й неделе заболе-
вания составляет 95%, на 4-й — 50%, а на 5-й приближается к нулю.
Процедуры отбора проб для бактериологического исследования зачастую
достаточно сложны технически, а правильность их выполнения имеет решаю-
щее значение. Например, нарушение правил взятия крови ведет к контаминации
пробы микроорганизмами с кожи или из окружающей среды и может стать при-
чиной ошибочного этиологического диагноза.
Сложность процедуры должна оправдывать ценность получаемой информации.
Наиболее эффективный способ получения мочи для бактериологического иссле-
дования, максимально гарантирующий отсутствие контаминации посторонней
микрофлорой, — надлобковая пункция мочевого пузыря. Однако на практике
такой способ используют редко из-за травматичности для пациента, ограничива-
ясь исследованием средней порции свободно выпущенной мочи.
В большинстве случаев время от момента взятия материала до начала иссле-
дования ограничено (часто оно не должно превышать 2 ч). Если существует
необходимость увеличения времени от момента взятия материала до начала
исследования в лаборатории, при использовании некоторых методов диагностики
материал можно консервировать, В случаях, когда условия его доставки не могут
быть выполнены, следует отказаться от использования данного исследования и
попытаться найти метод, в меньшей степени зависящий от особенностей транс-
портировки.
Проведение исследований в лаборатории регламентировано национальными
стандартами и инструкциями фирм-изготовителей реагентов и диагностикумов.
Строгое соблюдение правил, определяющих технологию диагностических про-
цессов, обеспечивает унификацию исследований и возможность сопоставления
данных, полученных в различных учреждениях.
Анализ результатов исследований включает оценку:
<> достоверности результатов;
<> полноты полученной информации:
4- этиологической значимости обнаруженных микроорганизмов.
Достоверность результатов анализов обеспечивает осуществление в лабо-
раториях работ по внешнему и внутреннему контролю качества исследований.
Внешний контроль осуществляют с помощью внешних инспекций, включающих
решение диагностических задач. По результатам таких проверок лаборатории про-
ходят лицензирование на право проведения диагностических исследований.
Внутренний контроль качества должен непрерывно проводиться в лаборатории
и призван оградить пациента и лечащего врача от получения ложноположитель-
ных или ложноотрицательных результатов, возникающих из-за допущенных в
ходе работы ошибок. Его осуществляют с помощью входного контроля реактивов
и питательных сред, использования эталонных штаммов микроорганизмов, иссле-
дования заведомо положительных и отрицательных проб. В некоторых случаях
контрольные образцы исследуют одновременно с каждой пробой. При этом учет
результатов теста осуществляют только при получении ожидаемого результата
контролей.
Достоверность результатов определяется не только качеством работы лабо-
ратории, но и соблюдением правил взятия материала и его доставки. К сожа-
лению, далеко не всегда с помощью лабораторных методов удается выявить
ошибки, допущенные на данном этапе исследований, но иногда это возможно.
Например, обнаружение в пробе мокроты при микроскопии буккального эпителия
и отсутствия лейкоцитов указывает на примесь значительного количества слюны.
Бактериологическое исследование такого образца нецелесообразно.
Анализ полноты полущенной информации подразумевает решение вопроса о
достаточности результатов выполненных тестов для постановки этиологического
БАКТЕРИОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ 237
диагноза и необходимости проведения дополнительных исследований. В ряде олу-
чаев дополнительные исследования могут быть проведены без повторного взятия
материала. Например, могут быть определены факторы вирулентности выделен-
ного возбудителя или поставлены дополнительные серологические реакции при
исследовании плазмы крови. В других случаях план дальнейших исследований
должен быть согласован с лечащим врачом.
Точная этиологическая диагностика многих форм инфекционной патологии
(Ку-лихорадка, гастроэнтероколиты) крайне затруднена или даже невозможна без
комплекса лабораторных исследований. В то же время результат лабораторного
анализа не является «истиной в последней инстанции» и окончательный диа-
гноз больному ставит лечащий врач на основании всего комплекса клинических
данных, включающего результаты объективных методов исследования, жалобы
больного, анамнестические данные и т.д.
При оценке диагностической значимости бактериологического исследования
необходимо помнить о неравнозначности положительного и отрицательного резуль-
татов. Если обнаружение микроорганизма в исследуемом материале однозначно
свидетельствует о его присутствии в организме больного в момент исследования, то
отрицательный результат не всегда свидетельствует об отсутствии микроорганиз-
ма. В ходе специального эксперимента на волонтерах, зараженных вирулентными
штаммами шигелл и заболевших дизентерией, возбудитель удалось выделить толь-
ко в 70% случаев. Причины, по которым возбудитель не удается выделить от боль-
ного определенным инфекционным заболеванием, многообразны. К ним относятся
неравномерность распределения микроорганизмов в общей массе исследугмого
материала и изменение интенсивности выделения возбудителей из организма боль-
ного по времени. Поэтому вероятность обнаружения патогенных микроорганизмов
резко возрастает при увеличении кратности обследования больного и увеличении
числа исследованных видов материала. Так, для выявления возбудителя у больных
острым трихомониазом необходимо трехкратное исследование нативных и окра-
шенных препаратов, а при хронических формах — пятикратное.
Таким образом, отрицательный результат бактериологического исследования,
особенно однократного, не является достаточным основанием для исключения
инфекционного заболевания. Факт обнаружения патогенного микроорганизма без
связи с конкрет ными обстоятельствами не всегда является достаточным основани-
ем для постановки конкретного диагноза. Это обусловлено широким распростра-
нением бактерионосительства при ряде нозологических форм. Значимость факта
обнаружения патогенного микроорганизма во многом определяется видом иссле-
дуемого материала. Например, выявление возбудителей брюшного тифа в крови,
соскобе из розеол или моче имеет большую диагностическую ценность, чем изо-
ляция S. typhi из фекалий или желчи. В последних двух случаях вероятно наличие
другого инфекционного заболевания на фоне носительства S. typhi.
Наибольшую сложность представляет трактовка результатов бактериологиче-
ского исследования в случае обнаружения условно-патогенных микроорганизмов,
многие из которых являются представителями нормальной микрофлоры. При
этом следует говорить о доказательстве этиологической роли выделенного микро-
организма как об особом этапе диагностического исследования. Универсальные
критерии для оценки диагностической значимости факта обнаружения тех или
иных условно-патогенных микроорганизмов отсутствуют. Наиболее часто прини-
мают во внимание следующие обстоятельства:
> количество микроорганизмов данного вида в исследуемом материале;
< отсутствие в исследуемом материале патогенных микроорганизмов.
❖ выделение данного микроорганизма в монокультуре или в ассоциации;
< > частота находок данного микроорганизма в том же исследуемом материале у
здоровых лиц;
238 БАКТЕРИОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ
повторное выделение одного вида микроорганизмов на протяжении всего
заболевания и его исчезновение по мере выздоровления;
❖ выявленное с помощью серологических исследований нарастание титра
антител к данному виду микроорганизма;
❖ одновременное обнаружение определенного вида микроорганизмов у ряда
пациентов со сходной клинической картиной и аналогичным источником
(фактором) заражения.
Не менее сложна оценка результатов серологических исследований. Только в
немногих случаях (сифилис, вирусные гепатиты) диагностическое значение имеет
сам факт обнаружения антител определенной специфичности. Чаще диагноз ста-
вят с помощью одного из двух диагностических критериев — диагностического
титра или нарастания титра антител. Диагностический титр — условная величи-
на титра антител в сыворотке крови к конкретному возбудителю, превышение
которой может быть расценено как признак заболевания. Величину диагности-
ческого титра устанавливают эмпирически для каждого заболевания и каждой
серологической реакции При этом в случае применения для диагностики одного
заболевания нескольких серологических реакций, величины диагностических
титров для каждой из них могу г не совпадать, так как тесты отличаются по своей
чувствительности. Также различия в величинах диагностических титров возмож-
ны при применении одной и той же реакции с использованием разных диагности-
ческих препаратов, поскольку в этом случае могут выявляться антитела к разным
антигенам. Величина диагностического титра приведена в инструкции-вкладыше,
которая имеется в каждой коробке с диагностическим препаратом. Для определе-
ния нарастания титра антител необходимо исследовать две сыворотки больного,
взятые с интервалом в 7-10 дней. Важно помнить, что обе сыворотки необходимо
исследовать в одной и той же серологической реакции. Таким образом, для опенки
результата врачу недостаточно знать абсолютную величину титра антител, также
необходима информация о виде поставленной серологической реакции. Данная
информация еще более важна, если учесть, что динамика изменения титра анти-
тел, определяемая в различных тестах, при некоторых заболеваниях не совпадает.
Например, при сыпном тифе с помощью РИГА антитела удается выявить в высоких
титрах на ранних сроках заболевания и в период ею разгара, но не после выздо-
ровления. В РСК, напротив, титр антител нарастает медленнее, но сохраняется
высоким длительное время после выздоровления. Таким образом, сопоставляя
данные, полученные с помощью различных серологических реакций, врач в ряде
случаев может получить ценную дополнительную информацию.
Современные методы серодиагностики позволяют определить концентрацию
в плазме крови иммуноглобулинов отдельных классов определенной специфич-
ности. После первого контакта с микроорганизмом индуцируется образование
специфических IgM. Титры IgM быстро нарастают и достигают максимума, а затем
относительно быстро снижаются. IgG появляются позже, но дольше сохраняют-
ся в высоких титрах. При повторном контакте с тем же антигеном (вторичный
иммунный ответ) IgM вырабатываются в незначительном количестве, реакция
гуморального иммунитета проявляется увеличением титров IgG. В связи с этим
по соотношению IgM и IgG можно отличить первичный иммунный ответ (острое
заболевание) от вторичного (рецидив или повторное заражение). О различиях в
характере иммуноглобулинов, образуемых при первичном и вторичном иммунных
ответах, следует помнить при оценке результатов, полученных с помощью неко-
торых современных иммунологических методов (например, ИФА). Большинство
выпускаемых тест-систем ИФА для серодиагностики позволяют определить титр
IgM или IgG. Соответственно и лаборатория предоставляет врачу информацию не
о титре антител вообще, а о титре иммуноглобулинов соответствующего класса.
В зависимости от конкретной клинической ситуации интерпретация факта обна-
БАКТЕРИОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ 239
ружения высоких или низких титров IgM и IgG будет отличаться. В частности,
низкие титры или отсутствие IgM не всегда можно расценивать как факт, свиде-
тельствующий об отсутствии заболевания.
ПРИНЦИПЫ БАКТЕРИОЛОГИЧЕСКОГО ИССЛЕДОВАНИЯ
ОТДЕЛЬНЫХ ВИДОВ БИОЛОГИЧЕСКОГО МАТЕРИАЛА
И ИНТЕРПРЕТАЦИИ ИХ РЕЗУЛЬТАТОВ
Кровь
Показаниями к проведению бактериологического исследования крови служат:
о клиническая картина сепсиса;
о лихорадочные состояния неустановленной этиологии;
О пневмонии;
> подозрение на инфекционные заболевания — брюшной тиф и паратифы,
сальмонеллезы, бруцеллез, возвратный тиф, лептоспирозы, малярию, эпи-
демический менингит, пневмококковые инфекции, ПТИ (при наличии лихо-
радки), стафилококковые и стрептококковые инфекции, сибирскую язву,
туляремию, чуму.
Количество независимо отбираемых проб крови и время их взятия определя-
ется лечащим врачом в зависимости от особенностей клинического состояния
больного (табл. 20-1). Как правило, от пациентов берут не менее 2-3 проб крови
из разных сосудов или разных участков одного сосуда. Это необходимо для диффе-
ренциации истинной бактериемии от случайной контаминации крови.
У больных, получающих антибактериальные препараты, пробы необходимо
собирать непосредственно перед введением (приемом) следующей дозы препара-
та. При наличии лихорадки оптимально взятие крови на фоне повышения темпе-
ратуры тела (не на пике лихорадки!).
Таблица 20-1. Рекомендуемое число отбираемь1х проб крови в зависимости от клинической
ситуации
Клиническое состояние Количество проб Примечания
Острый сепсис., менингит, остеомиелит, артрит, острые пневмонии и пиелонеф- рит 2 Из 2 сосудов или 2 участков кровеносного сосуда перед началом антибактериальной терапии
Лихорадка неясного генеза 4 2 пробы из разных кровеносных сосудов (разных участков сосуда), через 24-36 ч — еще 2 пробы
Инфекционный эндокардит в период обострения 2 Из 2 участков сосудов (2 различных сосудов) в течение первых 1 -2 ч лихирадки (не на пике лихооадки!) и до начала антибактериальной терапии
Подострый или вялотекущий инфекци- онный эндокардит 3-6 Сначала 3 пробы с интервалом 15 мин и более, если все пробы отрицательные, на 2-е сутки — еще 3 пробы
Инфекционный эндокардит на фоне антибактериальной терапии 6 По 2 отдельные пробы в каждый из 3 дней с положительной клинической динамикой антибак- териальной терапии
Вялотекущий сепсис на фоне антибакте- риальной терапии 6 6 проб в течение 48 ч; пробы необходимо собирать непосредственно перед введением (приемом) следующей дозы антибактериального препарата
240 БАКТЕРИОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ
Взятие проб крови
Пробы для определения наличия в крови биологических агентов получают
пункцией периферических вен, артерий или из пятки (у новорожденных детей).
С этой целью можно пользоваться стерильными одноразовыми или стеклянны-
ми шприцами объемом 20 мл (для взрослых пациентов) или 10 мл (для детей).
Применение вместо них вакуумных пробирок, представляющих собой стерильные
полностью закрытые системы, гарантирует от посторонней контаминации проб
во время их взятия, транспортировки и хранения. Сбор проб из постоянного
внутривенного или внутриартериального катетеров допускается только в случа-
ях подозрения на возникший катетерный сепсис или отсутствия возможности ее
получения венопункцией.
Выбранный для пункции кровеносного сосуда участок кожи дезинфицируют
тампоном, смоченным 70% этиловым спиртом, а затем другим тампоном, смочен-
ным 1 -2% раствором йода или другим дезинфектантом, круговыми движениями,
начиная от центра, в течение 30 с. Нельзя пальпировать сосуд после обработки
кожи перед введением иглы. Пункцию сосуда проводят после полного высыхания
обработанного участка кожи. После взятия пробы и извлечения иглы место ее
вкола обрабатывают 70% этиловым спиртом.
Пробы крови в вакуумные пробирки берут в следующем порядке:
❖ короткий конец двойной иглы для вакуумных пробирок освобождают от пла-
стиковой защиты и укрепляют иглу в держателе;
< > снимают пластиковую защиту с длинного конца иглы и пунктируют сосуд;
о вакуумную пробирку вводят в держатель до прокола крышки пробирки «про-
бирочным» концом двойной иглы;
о по окончании вакуумной аспирации крови снимают пробирку с иглы и поме-
щают в штатив для немедленной транспортировки в лабораторию для посева;
❖ извлекают иглу из вены, демонтируют из держателя при помощи демон-
тажного профиля специальной емкости для отходов (в отсутствие тако-
вой — вручную, предварительно осторожно закрыв иглу пластиковой
защитой).
После пункции кровеносного сосуда (независимо от того, какими средствами
ее проводили), для предотвращения возможного раздражения (ожога) с участка
кожи пациента стирают остатки йода с помощью тампона, смоченного 70% эти-
ловым спиртом.
При использовании флаконов промышленного изготовления со средой и реа-
гентами, нейтрализующими антибиотики и разрушающими форменные элементы
крови, или без них, отечественного и импортного производства, у взрослых полу-
чают с помощью шприца 10-20 мл крови, у детей — в зависимости от возраста
(табл. 20-2).
Таблица 20-2. Рекомендуемый для проведения бактериологического исследования объем проб
крови детей
Возраст, лет Объем «р -Ви, вносимой в 2 флакона, мл
<2 2
2-5 8
6-10 12
>10 20
Исследование материала
Сразу после взятия кровь засевают на питательные среды в соотношении 1:10—
1:60 для устранения бактерицидного действия крови посредством ее разведения.
Техника посева зависит от вида используемых питательных сред.
БАКТЕРИОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ 241
На среды лабораторного приготовления, расфасованные во флаконы с ватными
пробками, посев проводят следующим образом:
- у- снимают иглу со шприца;
открывают флакон над пламенем спиртовки;
> вносят половину пробы крови из шприца во флакон;
< обжигают горлышко и пробку флакона в пламени спиртовки;
о закрывают флакон пробкой;
< > осторожно, чтобы не замочить пробку, перемешивают содержимое флакона
круговыми движениями;
< повторяют указанные операции со вторым флаконом.
Флаконы со средами промышленного изготовления с резиновыми либо пласти-
ковыми пробками засевают кровью согласно инструкции производителя:
> при наличии на флаконах защитных колпачков их удаляют (отгибают), не
открывая при этом пробки;
< > пробки флаконов обрабатывают 70% этиловым спиртом (использование
йодной настойки определяется рекомендациями производителя питательных
сред);
❖ кровь в равных объемах вносят в аэробную и анаэробную питательные
среды, прокалывая пробки флаконов.
Посевы крови, сделанные вне лаборатории, немедленно доставляют в послед-
нюю, оберегая от охлаждения.
Бактериоскопию крови широко применяют для диагностики инфекционных
заболеваний, сопровождающихся бактериемией (например, возвратных тифов).
Кровь для бактериоскопического исследования при правильном типе лихорад-
ки берут за 2 ч до ее предполагаемого наступ тения, при неправильном типе — в
вечернее время.
Для бактериоскопического исследования готовят мазок и препарат толстой
капли.
• Для приготовления мазка каплю крови диаметром 2-3 мм наносят на обе-
зжиренное предметное стекло вблизи от тооца. Перед каплей крови ставят
под углом 45° стекло со шлифованным краем и плавным движением равно-
мерно распределяют материал по поверхности. Применение специального
пластикового шпателя с микрофибрами позволяет получать одинаковые по
толщине мазки и избегать случайного травмирования кожи медицинских
работников при проведении данной процедуры.
• Для приготовления препарата толстой капли на стекло наносят каплю крови
диаметром около 5 мм и распределяют ее с помощью иглы или пипетки с
образованием диска диаметром 10-15 мм. Толщина капчи должна позволять
видеть через нее газе гный шрифт.
• Иногда мазок и толстую каплю готовят на одном стекле. Для этого на поверх-
ность приготовленного, как описано выше, мазка до его высыхания наносят
еще одну каплю крови, которая, как правило, сама растекается в правильный
диск необходимой толщины.
• Препараты высушивают (толстая капля сохнет в течение 2-3 ч) и доставляют
в лабораторию с соблюдением необходимой осторожности.
Бактериологическая лаборатория при исследовании посевов крови выдает
ответ с отрицательным результатом, если рост микроорганизмов отсутствовал в
течение 10 дней инкубации. При подозрении на наличие у больного внутрисосуди-
стой инфекции или бруцеллеза, что должно отмечаться в направлении на исследо-
вание, возникает необходимость пролонгированной до 1 мес инкубации посевов.
Рутинный посев крови не обеспечивает выявления ряда микроорпнизмов. Их
перечень, а также способы выявления приведены в табл. 20-3.
242 БАКТЕРИОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ
Таблица 20-3. Микроорганизмы, обнаружение которых в крови требует применения специальных
диагностических приемов
Микроорганизм Специальные приемы выявления
Borrelia spp. В. recurrenfis, В. persica. В. latyschewwii (возбудители возвратных тифов) — бактериоскопия мазкое крови, окрашенной по Романовскому-Гимзе В. burgdorferi — посев на специальные среды (на практике не применяется)
Brucella spp. Применение специальных питательных сред инкубация в атмосфере с 5-10% углекислого газа
Cardiobaclerium hominis Инкубация в атмосфере, обогащенной углекислым газом, пересев с жидких питательных сред на плотные, содержащие в своем составе дрожжевой экстракт
Legionella spp. Пересев с обычных аэробной и анаэробной сред для первичного посева крови на угольно-доожжевой агар для легионелл
Leptospira spp. Микроскопия препаратов цитратной крови, посев крови (по 5 капель) в 3-5 пробирок с дистиллированной водой или специальной питательной средой, постановка биопробы
Mycobacterium spp* Посев осадка центрифугированной лизированной крови, использование автоматических бактериологических анализаторов
Bartonella (Rochalimaea) spp. Применение специальных методов предпосевной обработки крови, пита- тельных сред и приемов инкубации
Streptobacillus moniliformis Микроскопия препаратов цитратной крови, применение специальных мето- дов предпосевной обработки крови, инкубация в атмосфере с повышенным содержанием углекислого газа
' Исследование крови целесообразно проводить у больных с иммунодефицитом при подозрении
на генерализованную инфекцию, обусловленную атипичными микобактериями.
Существуют автоматизированные системы для индикации микробного роста в
посевах крови. Их применение позволяет ускорить получение результата и спо-
собствует более раннему назначению адекватного лечения. Детекцию микробного
роста осуществляют по продукции углекислого газа, изменению pH, электропро-
водности, окислительно-восстановительного потенциала или мутности питатель-
ной среды (одновременно могул учитываться несколько параметров).
Спинномозговая жидкость
Показанием к проведению исследования СМЖ служит подозрение на нейроин-
фекцию.
При подозрении на гнойный менингит в лабораторию направляют:
< СМЖ для первичного посева, бактериоскопии и иммунологических исследо-
ваний в количестве не менее 1,0 мл:
< СМЖ в 0,1% полужидком сывороточном агаре (среда «обогащения»);
❖ препарат «толстой капли» крови для бактериоскопии;
❖ посев крови (см, выше);
❖ пробу крови в объеме не менее 2,0 мл для серологических исследований
(P1IIA, ВИЭФ, латекс-агглютинация и др.).
Взятие проб СМЖ
Пробы СМЖ берут посредством люмбальной пункции, которую целесообразно
проводить при поступлении больного в стационар или до начала лечения.
Для выполнения этой врачебной манипуляции необходимы специальная под-
готовка и допуск. Пункцию и взятие материала проводят с соблюдением правил
асептики, медперсонал работает в масках и стерильных перчатках.
Следует придерживаться следующего порядка получения проб СМЖ.
БАКТЕРИОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ 243
< > Больного укладывают на бок на кровать, у которой максимально опущена
передняя сторона. Голову пациента прижимают к его груди, а ноги — к живо-
ту, спина должна быть максимально согнута.
< > Определяют анатомические ориентиры, необходимые для выбора места
пункции. Пункцию проводят между поясничными (Гщ-Ц.) и пояснично-
крестцовыми (L^-Sj) позвонками.
о Обрабатывают область пункции раствором антисептика, а затем 70% этило-
вым спиртом.
❖ Пальпируют точку пункции и вводят в эту область раствор анестетика:
< Проводят пункцию иглой со вставленным мандреном до возникновения ощу-
щения «провала в пустоту».
< • Извлекают мандрен,
о Первую порцию СМ.Ж (1 мл) собирают в обычную пробирку для проведения
общего ликворологического исследования.
❖ Вторую порцию СМЖ (1,0 мл), предназначенную для бактериологического
исследования (бактериоскопии, посева на питательные среды и иммуноло-
гических тестов), собирают в стерильную одноразовую пробирку с плотно
закрывающейся крышкой.
Третью порцию СМЖ (0,5 мл) засевают в 5,0 мл подогретого в термостате 0,1%
полужидкого сывороточного агара непосредственно у постели больного
о При необходимости проведения ПЦР берут четвертую пробу СМЖ (0,2 мл) в
специальную одноразовую пробирку.
❖ При проведении упомянутых выше манипуляций нельзя касаться руками
краев канюли, иглы, пробирки, класть пробку.
❖ Материал доставляют в бактериологическую лабораторию немедленно после
забора в теплом виде (при температуре 37 °C в термосумках или термокон-
тейнерах).
f При невозможности немедленной доставки допускается хранение проб СМЖ
в термостате (при 37 °C) в течение не более 18 ч.
< При необходимости проведения дополнительных вирусологических иссле-
дований пробы СМЖ помещают в холодильник при температуре 2-8 °C или
замораживают.
Исследование материала
Микроскопия — один из важнейших методов исследования СМЖ, позво-
ляющий максимально быстро получить необходимую информацию. Прозрачную
СМЖ перед исследованием центрифугируют при 2500-3000 об./мин (1500xg)
в течение 15-20 мин и готовят мазки из осадка. Из мутной СМЖ мазки можно
готовить без центрифугирования. Высушенные мазки фиксируют в пламени спир-
товки и окрашивают метиленовым синим и/или по Граму, а при подозрении на
туберкулез — по Циль-Нильсену. Препараты микроскопируют с помощью иммер-
сионного объектива. При обнаружении М. tuberculosis на основании микроскопии
выдается окончательный ответ, во всех остальных случаях — предварительный,
предполагающий уточнение по результатам применения культурального метода
диагностики туберкулеза.
Для выявления Criptococcus neojormans на предметном стекле смешивают одну
бактериологическую петлю осадка СМЖ с индийскими чернилами, накрывают
препарат предметным стеклом и микроскопируют с помощью объектива х40.
Криптококки под микроскопом выглядят как инкапсулированные, сферические,
почкующиеся дрожжевые формы. Результаты микроскопического исследования
немедленно сообщают лечащему врачу.
В лаборатории посевы инкубируют при температуре 37 °C в течение 24-48 ч в
атмосфере, содержащей 5-10% углекислого газа. При наличии роста на плотных
питательных средах проводят оценку культуральных свойств колоний, готовят
244 БАКТЕРИОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ
мазок и окрашивают его по Граму, определяют оксидазную и каталазную актив-
ность и, в зависимости от полученного результата, проводят дальнейшую иден-
тификацию возбудителя и определение чувствительности к антимикробным пре-
паратам.
При наличии признаков роста в 0,1% полуЖ идком сывороточном агаре проводят
высев на чашки с шоколадным агаром. Дальнейшая работа по выделению чистой
культуры, ее идентификации и определению чувствительности к антибиотикам
строится в зависимости от характеристик обнаруженных микроорганизмов.
При подозрении на туберкулезный менингит по капле осадка засевают как
минимум в 3 пробирки со средой Левенштейна-Йенсена. Пробирки затыкают
резиновыми пробками или парафинируют ватно-марлевые пробки. Посевы инку-
бируют в течение 6 нед с ежедневным просмотром.
Оценка результатов
СМЖ в норме стерильна, поэтому диагностически значимо обнаружение
любого микроорганизма. Необходимо учитывать возможность случайной кон-
таминации, которую можно заподозрить при отрицательных результатах бакте-
риоскопического исследования СМЖ и отсутствии роста на плотных питательных
средах в сочетании с ростом на жидких питательных средах. При подозрении на
контаминацию исследование следует повторить. Диагностически значимо повтор-
ное выделение того же вида микроорганизмов.
Превалирующие этиологические агенты при менингитах меняются в зави-
симости от возраста больных и сопутствующих обстоятельств. У новорожден-
ных бактериальный менингит чаше всего вызывают стрептококки групп В и D,
L. monocytogenes, энтеробактерии, а у детей первых 3 мес жизни также N. meningiti-
dis, Str. pneumoniae и Н. influenzae. У детей старше 3 мес наиболее важными этио-
логическими агентами являются N. Meningitidis, Str. pneumoniae и Н. influenzae. Для
взрослых N. meningitidis и Str. pneumoniae сохраняют этиологическое значение, а
Н. influenzae в старших возрастных группах вызывает менингиты только при нали-
чии предрасполагающих факторов — алкоголизма, наркомании, иммунодефицита.
Важно, что листерии также могут вызывать менингиты в старших возрастных
группах. В начальном периоде листериоза СМЖ серозная, а на поздних сроках
заболевания — гнойная. Ппи серозных менингитах следует в первую очередь
исключить туберкулез. Менингит, возникший после травм и нейрохирургических
операций, могут вызывать различные условно-патогенные микроорганизмы —
S. aureus, S. epidermidis, Str. Pyogenes, P. aeruginosa. E. coli. K. pneumoniae, Serratia
marcescens.
Раневое отделяемое, аспират и биоптаты кожи
и подлежащих тканей
Показанием к проведению исследования служит выявление у пациента при-
знаков гнойно-воспалительного процесса в ране, на коже и в прилегающих к ней
тканях.
Взятие исследуемого материала
Взятие материала осуществляет врач, как правило, при проведении перевязки
или операции. Технология взятия образца для микробиологического исследова-
ния во многом определяется клинической ситуацией и не может быть унифициро-
вана. Исследуемым материалом может служить экссудат, аспират из раны, мазки
(тканевая жидкость, пропитавшая тампон), биоптаты. Направление на исследова-
ние гноя или струпов нецелесообразно.
БАКТЕРИОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ 245
Пробы берут с соблюдением правил асептики. Кожу вокруг раны или над оча-
гом воспаления обрабатывают 70% этиловым спиртом и 1-2% настойкой йода
(обязательно необходимо смыть 70% этиловым спиртом во избежание ожогов)
или другим разрешенным антисептиком. При необходимости удаляют с помощью
стерильной салфетки некротические массы, детрит, гной. Использовать растворы
антисептиков для снятия повязки или обработки раневой поверхности до взятия
материала нельзя.
Взятие материала из ран
Отбор проб проводят после обработки раны вышеописанным способом. Для
взятия материала, предназначенного для бактериологического и микроскопи-
ческого исследований, пользуются стерильными зондами-тампонами (предпо-
чтительнее промышленного изготовления, а не самостотельно изготовленными,
вискозными или дакроновыми, а не ватными).
Прокатывают каждым зондом-тампоном по раневой поверхности от центра к
периферии в течение 5-10 с; гампон должен равномерно пропитаться тканевой
жидкостью; манипуляцию необходимо проводить максимально осторожно, не
травмируя ткани, появление даже следов крови недопустимо, так как кровь обла-
дает бактерицидными свойствами.
Взятие материала для микроскопии обязательно, так как в ряде случаев она
более информативна по сравнению с бактериологическим исследованием; зонд-
тампон. предназначенный для микроскопического исследования, помещают в
пустую пробирку.
Зонд-тампон с материалом для бактериологического исследования помещают в
пробирку с транспортными средами Эймса или Стюарта.
Взятие раневого отделяемого из дренажей
При наличии в ране дренажей отделяемое из них собирают стерильными шпри-
цем или одноразовой пастеровской пипеткой в объеме 1-2 мл и помещают в сте-
рильную пробирку или пробирку с транспортными средами Эймса или Стюарта
Можно пропитать отделяемым из дренажа стерильный зонд-тампон и погру-
зить его в пробирку с транспортной средой.
Взятие аспирата;
❖ материал забирают после обработки кожи;
> после высыхания дезинфектанта врач с помощью одноразового шприца
объемом 3-5 мл и иглы №22 или №23 берет аспират из глубины раны;
❖ если имеется везикула, берется жидкость и клетки у основания дефекта;
❖ если попытка взятия аспирата не удалась, подкожно вводят стерильный
0,9% раствор натрия хлорида и повторно пытаются взять аспират;
❖ если и эта попытка оказывается неудачной, через иглу в шприц набирают
жидкую питательную среду или 0,9% раствор натрия хлорида в небольшом
количестве, ополаскивают иглу и используют полученную жидкость в каче-
стве исследуемого материала;
❖ при наличии в ране дренажей из них забирают шприцем 1-2 мл отделяемого;
❖ аспират из шприца, сняв иглу, переливают в пробирку с транспортной средой
Эймса, Стюарта или другой питательной средой, обеспечивающей сохране-
ние неспорообразующих анаэробов;
❖ можно пропитать аспиратом стерильный тампон и побузить его в транс-
портную среду;
❖ если гарантирована немедленная доставка материала в лабораторию, допу-
скается транспортировать материал непосредственно в шприце с иглой, надев
на нее защитный колпачок.
Взятие биоптатов:
п ри сборе материала в процессе операции кусочки ткани (3-5 мл) помещают
в стерильные контейнер или стеклянную емкость;
246 БАКТЕРИОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ
> необходимо добавить 3-5 мл 0,9% раствора натрия хлорида для предотвра-
щения высыхания материала.
Подозрение на анаэробную инфекцию возникает при наличии неприятного
гнилостного запаха раневого отделяемого, некроза тканей, глубоких абсцессов,
газа в тканях или клинической картины газовой гангрены, черного окрашивания
экссудата. В таких случаях материал для бактериологического исследования соби-
рают:
< > во флаконы или в пробирки с агаровой основой и анаэробной атмосферой с
индикатором для ее контроля, в которых доставляют аспирационный мате-
риал;
о в специальный мешок или систему пробирок, из которых удален кислород;
< транспортную систему для анаэробов со стерильным зондом-тампоном.
Для доставки материала, собранного тампоном, можно использовать пробирку
с ядровой основой и анаэробной атмосферой с индикатором для ее контроля, про-
бирку с катализатором, генерирующим анаэробную атмосферу после введения там-
пона в пробирку, пробирку с анаэробной атмосферой и редуцированной средой.
При отсутствии специальных систем и сред допустимо использование пробирок
и других контейнеров с редуцированной тиогликолевой средой, тиолевым бульо-
ном, а также со средами, содержащими гемин и менадиол. Контейнеры с материа-
лом должны быть плотно закрыты пробками или крышками.
Исследование материала
Материал, взятый с помощью одного из зондов-тампонов, используют для
приготовления мазка, который окрашивают по Граму При микроскопии мазка
оценивают количество микр<)бных клеток и характеризуют их морфологические и
тинкториальные свойства. Микроскопия мазков из раны — ценный диагностиче-
ский прием, позволяющий быстро получить представление о наиболее вероятной
этиологии процесса, а также осуществить выбор оптимальных питательных сред
для дальнейшего бактериологического исследования. Выполнение данной про-
цедуры должно стать неотъемлемой частью ведения инфекционного больного.
Микроскопическое исследование позволяет обнаружить труднокультивируемые
микроорганизмы, выявление которых при бактериологическом исследовании
маловероятно или требует длительной инкубации.
Материал, взятый другим зондом-тампоном, используют для посева на 5%
кровяной агар, среду для контроля стерильности и сахарный бульон, а в случае
исследования на анаэробные инфекции — на специальные среды (среду Китт-
Тароцци, агар и бульон Шеллера, тиогликолевую среду, анаэробный кровяной
агар с амикацином или гентамицином и др.). Посев на чашку Петри проводят
методом «тампон-петля». Тампоном проводят по поверхности питательной среды
дорожку от одного края чашки Петри до другого (через ее центр), а затем вторую
параллельно первой. После этого материал рассевают бактериологической петлей
штрихами, перпендикулярными дорожкам. Посевы инкубируют при температуре
37 °C в течение 18-24 ч в аэробных и/или анаэробных условиях в зависимости
от предполагаемого типа инфекции При отсутствии роста в 1-е сутки инкубацию
продолжают. При отсутствии роста в посевах на 5-е сутки дают отрицательный
ответ.
Оценка результатов
Оценка количества микроорганизмов в раневом отделяемом важна для прогно-
зирования течения процесса. Установлено, что при уровне микробной обсеменен-
ности раны выше 105 КОЕ на 1 г ткани или 1 мл раневого отделяемого нагноение
развивается даже в жизнеспособных тканях. При наличии в ране некротизиро-
ванных тканей, гематом и инородных тел развитие гнойного процесса возможно
БАКТЕРИОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ 247
и при более низких уровнях обсемененности. Кроме того, при уровне обсеменен-
ности раны, превышающем 105 КОЕ на 1 г ткани, резко возрастает угроза развития
сепсиса. Однако большинство существующих методов количественной оценки
трудоемки и недоступны практическим лабораториям.
При одновременном обнаружении в материале нескольких видов микроорга-
низмов ведущее значение имеют виды, обнаруженные в наибольшем количестве.
Отделяемое верхних дыхательных путей
Показания к взятию и исследованию проб: воспалительные заболевания верх-
них дыхательных путей.
Взятие материала
Отделяемое верхних дыхательных путей берут с помощью стерильных зондов-
тампонов.
Исследование материала
Посев на кровяной или шоколадный агар без элективных добавок проводят
зондом-тампоном, которым была взята проба. Сначала материал втирают в пита-
тельную среду, поворачивая тампон вокруг своей оси на участке размером 2x1 см,
Затем частыми непрерывающимися штрихами засевают оставшуюся поверхность.
Стандартная схема исследования отделяемого верхних дыхательных путей
предусматривает посев материала из глотки, носа и ротовой полости только на
5% кровяной агар. Основной возбудитель бактериальных инфекций верхних
дыхательных путей, включая скарлатину, тонзиллиты, фарингиты, — Str. pyogenes.
К сожалению, большинство практических лабораторий используют 5% кровяной
агар, приготовленный с применением не овечьей, а человеческой крови. На данной
питательной среде стрептококки растут крайне плохо, что делает исследование
малоэффективным. Кроме того, Str. pyogenes сохраняют чувствительность к пени-
циллинам и цефалоспоринам, что делает в большинстве случаев излишним опре-
деление чувствительности выделенных штаммов к антимикробным препаратам. В
настоящее время доступны готовые питательные среды в чашках Петри и пробир-
ках стандартно высокого качества, в том числе кровяной агар, приготовленный с
применением овечьей крови.
О необходимости выявления таких видов микроорганизмов, как N. meningitidis,
С. diphtheriae Н. influenzae, врач должен уведомить лабораторию заранее. В этом
случае для посева будут использованы специальные питательные среды.
Диагностику некоторых специфических форм ангин осуществляют прежде всего
с помощью бактериоскопического исследования. Микроскопия мазков необходи-
ма при диагностике ангины Людвига (тяжелая гнилостно-некротическая флегмона
полости рта полимикробной этиологии) и ангины Симановского Пласта-Венсана
(некротическая ангина, вызываемая симбиозом Fusobacterium spp. и Treponema
vincentii).
Оценка результатов
Оценка этиологической значимости обнаруженных в дыхательных путях микро-
организмов осложняется обилием нормальной микрофлоры и широким рас-
пространением носительства патогенных микроорганизмов. Нормальная микро-
флора полости рта и глотки характеризуется высокой степенью разнообразия и
включает Streptococcus spp. (включая Str. viridans и Str. pneumoniae), Neisseria spp..
Staphylococcus spp., Corynebacterium spp., Haemophilus spp... Lactobacillus spp., Candida
spp., Mycobacterium spp., Treponema spp. У лиц преклонного возраста и пациентов с
иммунодефицитом в составе микрофлоры ротоглотки возрастает количество гра-
мотрицательных микроорганизмов, включая Е. coli, Klebsiella spp., Acinetobacter spp.,
Pseudomonas spp.
248 БАКТЕРИОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ
Среди болезней верхних дыхательных путей бактериальной этиологии лиди-
рующее положение занимают ангины. В этиологии бактериальных ангин ведущая
роль принадлежит стрептококкам группы А. Почти в 80% случаев ангины бакте-
риальной этиологии вызывает Str. pyogenes, реже — стафилококки (самостоя гельно
или в ассоциации со стрептококками).
В этиологии эпиглоттитов у детей ведущая роль принадлежит Н. influenzae.
а у взрослых — Str. pyogenes и Str. pneumoniae. Ларингиты в 90% случаев имеют
вирусную этиологию, но могут быть обусловлены Str. pyogenes, а у взрослых также
Moraxella catarrhalis.
Возможная причина острых ларингобронхитов — Mycoplasma pneumoniae.
С особой осторожностью следует подходить к диагнозу «гонококковый фарин-
гит*. Данный диагноз может быть поставлен только с помощью бактериоло-
гического метода при условии точной видовой идентификации N. gonorrhoeas.
Это обусловлено присутствием значительного числа сапрофитных видов нейс-
серий в составе нормальной микрофлоры полости рта. Их дифференциация от
N. gonorrhoeas по морфологии невозможна, а с помощью ПЦР и других современ-
ных методов — не всегда достоверна.
Рецидивирующие заболевания верхних дыхательных путей могут вызывать
штаммы S. aureus, продуцирующие ^-лактамазы, Н influenzae и B.fragilis.
При оценке результатов бактериологического исследования необходимо учиты-
вать, что часто первичные поражения респираторного тракта вызывают вирусы,
а секундарные бактериальные инфекции осложняют их течение.
Мазок со слизистых ооолочек полости носа для диагностики носительства
S. Aureus
Показания к взятию и исследованию проб: диагностика носительства золоти-
стого стафилококка. S. aureus распространен повсеместно, колонизирует у чело-
века слизистую оболочку носа, брюшную полость и кожу подмышечной области
(уровень носительства среди клинически здоровых лиц составляет 15-30%);
обычно длится несколько недель или месяцев, принимая хронический характер
у персонала медицинских учреждений и людей, которым часто делают инъекции.
При обострении инфекции регистрируют появление на коже гнойников, осложне-
ние заживления ран, бактериемию, эндокардит, пневмонию, артриты, остеомие-
лит, перитонит, поражения мочевыводящего тракта, синдромы ошпаренной кожи
и токсического шока, пищевые токсикоинфекции.
Взятие материала
Материал берут у пациента одним зондом-тампоном из двух носовых ходов,
доставляя в лабораторию в кратчайшие сроки (он должен быть засеян на пита-
тельные среды в течение 2 ч).
Исследование материала
Для посева материала используют желточно-солевой агар, агар Байрда-Паркера,
колумбийский CNA агар и другие элективные и селективные среды для стафило-
кокков. Посев проводят зондом-тампоном, которым забирали материал. При
посеве тампон многократно поворачивают вокруг своей оси.
Оценивают биохимическую активность изолятов. При необходимости дополни-
тельно определяют ферментацию маннита в анаэробных условиях или ДНКазную
активность. Для дифференциации транзиторного и злостного (резидентного)
носительства оценивают чувствительность выделенной культуры к фузидовой
кислоте.
Оценка результата
S. aureus в течение 18-24 ч образует на плотных средах крупные (диаметром
2-5 мм), выпуклые, гладкие колонии с ровными и более светлыми по сравнению с
центральной частью краями: они чаще окрашены в желт ый цвет (он может варьи-
БАКТЕРИОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ 249
ровать от белого до оранжевого). Если выделенная культура обладает лецитовети-
лазой и плазмокоагулазой, ее идентифицируют как S. aureus. Резидентные штаммы
бактерии идентифицируют по устойчивости к фузидовой кислоте.
Сосксб со слизистых оболочек передних отделов полости носа для выявления
клебсиелл
Показания к взятию и исследованию проб: диагностика озены и риносклеромы.
Взятие материала
В случае инфильтрационной формы риносклеромы материал берут с помо-
щью бактериологической петли и носового или носоглоточного зеркала прямо с
инфильтрированной слизистой оболочки, слегка повреждая ее.
У больных озеной или атрофической формой риносклеромы взятие материала
проводят петлей или стерильным зондом-тампоном, причем предпочтительнее
последний. При всех видах и формах кпебсиеллезной инфекции одновременно
берут материал из обеих половин носа и глотки, а при локализации процесса в
гортани, трахее и бронхах — также методом кашлевых пластинок.
Исследование материала
Озену и риносклерому диагностируют по результатам обнаружения их возбуди-
телей в мазках взятого материала. В сомнительных случаях прибегают к изоляции
бактерий на питательных средах (Эндо, Левина, Плоскирева, Мак-Конки и др.)
с последующей их идентификацией по культуральным, морфологическим и био-
химическим признакам.
Оценка результата
Обнаружением в мазках и посевах К. pneumoniae spp. ozaenae (палочка Абеля)
и К. pneumoniae spp. rhinoscleromatis (палочка Фриша-Волковича) с последующей
идентификацией изолятов по биохимической активности, а палочки Фриша-
Волковича в реакции связывания комплемента подтверждают диагноз озены или
риносклеромы соответственно, однако следует учитывать возможность носитель-
ства.
Слизь из носоглотки и носа при подозрении на дифтерию
Показания к взятию и исследованию проб: подозрение на дифтерию.
Взятие материала
• Материал берут натощак или через 3-4 ч после еды. При антибиотикотера-
пии исследование проводят не ранее чем через 3 дня после отмены антибак-
териального препарата.
• Материал с миндалин и из носа берут разными стерильными зондами-
тампонами: один предназначен для приготовления мазков, а другой — для
посева на питательные среды. Если имеются налеты, материал забирают с
границы пораженных и здоровых тканей, слегка нажимая на них тампоном.
• Материал должен быть доставлен в лабораторию в течение 3 ч (в транспорт-
ной среде) или сразу после взятия. В холодное время года при транспорти-
ровке материал следует оберегал ь от замораживания. При транспортиривке
на дальние расстояния допускается использование тампонов, пропитанных
5% раствором глицерина в дистиллированной воде и простерилизованных
автоклавированием.
Исследование материала
Мазки окрашивают по Граму и Нейссеру. Первый из названных методов
неспецифичен, но позволяет косвенно идентифицировать непатогенные Корине-
бактерии, располагающиеся в виде палисада или китайских иероглифов. Окраска
по Нейссеру выявляет типичные зерна Бабеша-Эрнстэ При проведении рутинных
исследований ограничиваются бактериоскопией.
250 БАКТЕРИОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ
В сомнительных случаях прибегают к выделению чистой культуры (это делают
только в лабораториях, имеющих разрешение на работу с данным возбудителем).
Для первичного посева используют среду Клауберга II, кровяной теллуритовый
агар (или другие среды с теллуритом), хинозольную среду Бучина. Рекомендуется
одновременно высевать материал на обычный кровяной агар, так как не все штам-
мы бактерии способны расти на упомянутых выше средах.
Чашки со средой предварительно согревают при комнатной температуре или в
термостате (в течение 15-20 мин). Чашки с посевом исследуемого материала или
пробирки с транспортной средой можно поместить для подращивания в термостат
при температуре 37 °C на 15-18 ч, после чего доставить в лабораторию.
Посев проб, взятых от одного пациента, проводят на одну чашку - на половину
чашки засевают материал из ротоглотки, на другую половину — материал из носа.
Для посева мазка с кожи или проб, взятых из других мест, используют еще одну
чашку. Материал сначала втирают в питательную среду со всех сторон тампона
на участке площадью 2x1 см2, а затем этим же тампоном засевают оставшуюся
поверхность половины чашки частыми неперекрывающимися штрихами, не отры-
вая тампон от поверхности питательной среды и не изменяя его положения.
На средах с теллуритом возбудитель дифтерии формирует серовато-черные
колонии с ровными или изрезанными краями, которые постепенно приобретают
металлический блеск. Диаметр колоний различных типов бактерии различен:
1,5-2,5 мм (тип grans), 1,5-2.0 мм (тип ntitis) и 0,5-0,75 мм (тип Intermedius).
Токсигенность возбудителя раньше определяли биопробой на морских свинках
и в реакции преципитации (тесте Илека-Оухтерлони); в настоящее время с этой
целью пользуются ПЦР, позволяющую выявлять ген tox.
Оценка результатов исследований
Инфекцию диагностируют в большинстве случаев на основании результатов
бактериологического исследования, в сомнительных случаях прибегают к куль-
туральному методу. Этиологически значимо обнаружение только токсигенных
штаммов С. diphtheriae.
Слизь с задней стенки носоглотки при подозрении на менингококковую инфекцию
Показания к взятию и исследованию проб: подозрение на менингококковую
инфекцию. Носительство N. meningitties широко распространено (до 30% здоро-
вых людей, а во время вспышек этот показатель возраст ает в 2-3 раза). Входными
воротами инфекции служит носоглотка.
Взятие материала
Для диагностики менингококковой инфекции берут слизь с задней стенки носо-
глотки пациента натощак или через 3-4 ч после еды. При взятии проб предпочти-
тельнее пользоваться не ватными, а вискозными зондами-тампонами.
• Шпателем надавливают на корень языка, что ведет к подниманию небной
занавески.
• Зонд-тампон заводят за мягкое небо и 2-3 раза проводят по задней стенке
носоглотки
• При извлечении зонд-тампон не должен касаться зубов, слизистой оболочки
щек и языка.
• Помещают зонд-тампон в пустой или содержащий транспортную среду
Стюарта контейнер.
Исследование материала
При наличии такой возможности материал сразу же засевают на предваритель-
но подогретые питательные среды (сывороточный агар, сывороточный агар с
линкомицином, кровяной агар, шоколадный агар с ростовыми добавками, среду
Мюллера-Хинтона с овечьей кровью, селективную среду Тайера-Мартина). При
посеве на чашки Петри материал втирают на поверхности небольшого участка
БАКТЕРИОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ 251
(1x2 см) питательном среды всеми сторонами тампона, затем этим же тампоном
засевают штрихами (с отрывом) всю поверхность среды.
При транспортировке посевов или взятого материала в лабораторию пользуются
термосумками. Если время транспортировки материала в лабораторию превышает
3 ч, то зонд-тампон с материалом помещают в транспортную среду Стюарта. При
отсутствии готовой транспортной среды можно использовать стерильную пептон-
ную воду с линкомицином (0.5 мг/100 мл).
Оценка результата
Исследование носоглоточной слизи позволяет выявлять потенциальные источ-
ники инфекции. Для диагностики гнойного менингококкового менингита следу-
ет исследовать СМЖ и кровь. О каждом случае заболевания менингококковой
инфекцией, подозрении на заболевание или носительство менингококка врачи
всех специальностей, медицинские работники лечебно-профилактических, дет-
ских подростковых и оздоровительных организаций, независимо от ведомствен-
ной принадлежности и организационно-правовой формы, а также медицинские
работники, занимающиеся частной медицинской деятельностью, в течение 2 ч
сообщают по телефону и затем в течение 12 ч посылают экстренное извещение
по установленной форме в территориальный центр государственного санитарно-
эпидемиологического надзора по месту регистраци] i заболевания (независимо от
места проживания больного)
Слизь с задней стенки носоглотки при диагностике коклюша
Показания к взятию и исследованию проб: диагностика коклюша.
Обследованию подлежат дети и сотрудники детских учреждений, кашляющие
более 7 дней без установленной причины или контактировавшие с источником
инфекции.
Взятие материала
Исследуемым материалом могут служить мазки с задней стенки глотки или
слизь, полученная методом кашлевых пластинок,
• Мазки с задней стенки глотки берут с помощью изогнутых под прямым углом
зондов-тампонов с металлическим стержнем. Одновременно рекомендуется
использовать два зонда-тампона - сухой и увлажненный. Сначала материал
забирают сухим тампоном, а затем смоченным забуференным 0,9% раство-
ром хлорида натрия. Материал, взятый сухим тампоном, нужно сразу засеять
на питательные среды (казеиново-угольный агар, молочно-кроьяной агар,
среду Борде-Жангу и др.). Посев материала, взятого влажным тампоном, на
перечисленные среды можно провести в лаборатории.
• Для диагностики коклюша, помимо исследования мазков из носоглотки,
пользуются методом кашлевых пластинок. С этой целью открытую чашку
Петри с питательной средой удерживают в вертикальном положении на рас-
стоянии 5-10 см ото рта в течение 5-6 кашлевых толчков.
Исследование материала
Возбудителя коклюша можно обнаружить в мазках клинического материала в
реакции иммунофлюоресценции. В рутинной практике лабораторной диагностики
коклюша большее значение придают изоляции В. pertussis па питательных средах с
последующей идентификацией по морфологическим и культуральным свойствам,
а также в реакции агглютинации.
Оценка результатов исследований
Бактериологическая диагностика коклюша эффективна только в первые 1-2
недели заболевания при условии отсутствия антибиотике терапии. При получении
отрицательного результата для подтверждения диагноза можно использовать
ПЦР. а начиная с 3-4-й недели — серологические методы.
252 БАКТЕРИОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ
Отделяемое нижних отделов дыхательных путей
Мокрота
Показания к взятию и исследованию проб: воспалительные заболевания ниж-
них отделов дыхательных путей, сопровождающиеся отделением мокроты.
Взятие материала
Предпочтительно исследование утренней порции мокроты. Перед сбором мокро-
ты больному предлагают почистить зубы и прополоскать рот кипяченой водой, а
также предупреждают о недопустимости попадания в мокроту слюны и носовой
слизи. Мокроту собирают в стерильный широкогорлый контейнер или «карман-
ную плевательницу».
Особенности взятия мокроты для бактериоскопической диагностики
туберкулеза
Мокроту собирают трижды: в 1-й день — в присутствии медработника, на 2-й
день — проинструктированным пациентом самостоятельно, на 3-й день пациент
приносит собранный во 2-й день материал и мокроту забирают в третий раз в при-
сутствии медработника.
< > Пациента заранее предупреждают о важности получения именно мокроты, а
не слюны или носовой слизи, а также о необходимости перед сбором мате-
риала почистить зубы и прополоскать рот кипяченой водой.
о Медработник в маске, резиновых перчатках и резиновом фартуке распола-
гается за спиной пациента, таким образом, чтобы направление движения
воздуха было от него к пациенту, снимает крышку с контейнера для сбора
мокроты и передает его пациенту.
< Пациенту рекомендуют держать контейнер как можно ближе к губам и сразу
же сплевывать в него мокроту по мере ее откашливания; кашель может быть
индуцирован с помощью нескольких глубоких вдохов.
♦ По завершению сбора мокроты медработник оценивает ее качество, плотно
закрывает контейнер и заполняет направление на исследование.
❖ Срок хранения материала в холодильнике без добавления консервирующих
средств не должен превышать 48-72 ч.
< > При отсутствии мокроты накануне вечером или рано утром в день, намечен-
ный для сбора материала, пациенту назначают отхаркивающее средство или
раздражающие ингаляции.
Усилить отделение мокроты можно с помощью ингаляции, для проведения
которой используют раствор следующего состава: в 1 л стерильной дистиллиро-
ванной воды растворяют 150 г хлорида натрия и 10 г натрия бикарбоната. Больной
должен вдохнуть 30-60 мл подогретого до 43-45 °C раствора в течение не менее
10-15 мин.
Собранный таким образом материал не подлежит консервации и должен быть
исследован в тот же день. В направлении обязательно отмечают, что материал был
получен после ингаляции.
Исследование материала
Для приготовления мазка из мокроты, распределенной тонким слоем по
дну чашки Петри, с помощью двух игл или пинцета выбирают и переносят на
середину предметного стекла слизистые или гнойные комочки. Затем, согласно
классическому методу «двух стекол», берут второе стекло и накладывают поверх
первого. Комочки мокроты растирают между двумя стеклами до тех пор, пока не
получится достаточно тонкий мазок. При этом во избежание инфицирования ру к
работающего и на первом, и на втором стеклах мазки не должны занимать более
1/2—2/3 поверхности. Препараты высушивают и фиксируют над пламенем. Один
БАКТЕРИОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ 253
из мазков окрашивают по Граму, второй микроскопируют неокрашенным для
выявления друз актиномицетов или мицелия грибов.
Данная техника сопряжена с высоким риском для медперсонала, особенно в
случаях туберкулеза. При подозрении на туберкулез рекомендуется пользоваться
для приготовления мазков мокроты деревянной палочкой, которую перед рабо-
той разламывают пополам. С помощью сломанных конпов палочки из разных
участков образца мокроты выбирают 2-3 наиболее плотных небольших гнойных
комочка, переносят их на стекло, при необходимости разминают и равномерно
распределяют тонким слоем в центре стекла на поверхности приблизительно раз-
мером 1x2 см в виде овала. Использованные палочки сбрасывают в контейнер с
дезинфицирующим раствором.
При использовании для приготовления мазка бактериологической петли или
игл существует риск формирования аэрозоля мокроты при их обжигании в пла-
мени после использования. Для предупреждения формирования аэрозоля инстоу-
мент предварительно очищают от остатков мокроты 2-3-кратным погружением
в банку с песком, залитым этиловым спиртом, и только после этого прожигают в
пламени горелки. Песок длительно остается пригодным для этой цели при условии
периодического обновления раствора этилового спирта с таким расчетом, чтобы
его уровень превышал уровень песка не менее чем на 3 см.
Приготовленные мазки помещают на 15-30 мин на лотки (подносы) и высу-
шивают при комнатной температуре. Фиксация влажных мазков недопустима.
Для фиксации мазок трижды медленно проводят через верхнюю треть пламени
горелки. Продолжительность пребывания мазка в пламени не должна превышать
3-5 с,
В зависимости от задач исследования готовят несколько мазков, которые окра-
шивают по Граму. Цилю-Нильсену или микроскопируют в нативном виде для
выявления мицелия грибов и друз актиномицетов. При исследовании большого
количества образцов (более 30 в день) вместо окраски по Цилю-Нильсену для
выявления кислотоустойчивых микроорганизмов (КУМ) целесообразно исполь-
зовать окраску флюоресцентными красителями с последующей люминесцентной
микроскопией.
Первоначально препарат мокроты микроскопируют под малым увеличением
микроскопа (объектив х10) для оценки правильности взятия материала и степени
его загрязнения слюной, а затем проводят микроскопию мазка под большим уве-
личением (иммерсионный объектив х90 или хЮО) для обнаружения бактерий.
Бактериологическое исследование мокроты проводят качественным или коли-
чественным методом.
При использовании качественного метода мокроту выливают в чашку Петри и с
помощью игл и пинцета выбирают 2-3 гнойных комочка. Комочки мокроты трех-
кратно в 3 чашках Петри отмывают стерильным 0,9% раствором хлорида натрия
для удаления сопутствующей микрофлоры, наслоившейся в верхних дыхательных
путях и полости рта. Затем их переносят на питательную среду и тщательно равно-
мерно растирают шпателем по ее поверхности. Посевы инкубируют при темпе-
ратуре 37 °C. Промывные воды бронхов засевают аналогичным образом, но без
предварительной отмывки гнойных комочков.
В основе количественного метода исследования лежит высев на плотные пита-
тель ные среды разведений, приготовленных из гомогената мокроты. Из много-
численных методов гомогенизации мокроты наиболее удобный и безопасный
для медперсонала — использование муколитика ацетилцистеина. В этом случае
в неметаллический сосуд вносят равные объемы мокроты и 0,5% раствора аце-
тилцистеина и тщательно перемешивают пипеткой с резиновой грушей в течение
1-2 мин. Ацетилцистеин не только не угнетает рост Str. pneumoniae и Н. influenzae,
но и стимулирует развитие последней из-за устранения ингибирующего действия
254 БАКТЕРИОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ
муцина. Посев гомогената мокроты может быть проведен по методу Gold, анало-
гично посеву мочи.
Оценка результатов исследований
Обнаружение при микроскопировании под малым увеличением микроскопа в
препарате мокроты большого количества эпителиальных клеток, особенно при
отсутствии гранулоцитов, свидетельствует о том, что материал был взят непра-
вильно (с примесью слюны).
Для повышения обьективности оценки может быть использована одна из при-
веденных ниже шкал. Использование каждой из них предполагает микроскопию
окрашенного по Граму мазка из гнс. ной части мокроты под малым увеличением
(объектив х10). При использовании шкалы Barlett просматривают 20 -30 полей
зрения, давая каждому оценку и суммируя баллы (табл. 20-4). Затем рассчитывают
средний балл — складывают все начисленные баллы и делят сумму на количество
просмотренных полей. Если полученный результат равен 0 или меньше, вероят-
ность воспаления мала или мокрота, вероятно, контаминирована в ротовой поло-
сти. Такие образцы не подлежат бактериологическому исследованию, и взятие
мокроты необходимо повторить.
Таблица 20-4. Схема Barlett оценки правильности взятия мокроты
Тип клеток Количество клеток в поле зрения при микроскопии с объ- ективом х10 Балл
Нейтрофилы <10 0
10-25 +1
>25 +2
Присутствие слизи +1
Эпителиальные клетки 10-25 -1
>25 -2
При оценке мокроты по Murray Washington принадлежность образца к опреде-
ленной группе устанавливается по табл. 20-5. При этом бактериологическому
исследованию подлежат только образцы, принадлежащие к 4-5-й группе.
Таблица ?0-5. Схема Murray Washington оценки правильности взятия мокроты
Группа Количество клеток в поле зрения при микроскопии с объективом хЮ
эпителиальные клетки лейкоциты
1 25 10
2 25 10-25
з 25 25
4 10-25 25
5 <10 25
Если образец мокроты значительно контаминирован слюной, то взятие мокро-
ты необходимо повторить.
При бактериоскопии мазков мокроты в первую очередь оценивают общую кар-
тину микрофлоры: наличие скоплений грамположительных кокков (Staphylococcus,
Micrococcus), цепочек грамположительных кокков (Streptococcus), мелких ланце-
товидных, окруженных зоной неокрасившейся капсулы (S. pneumoniae), грамо-
трицательных кокков (Neisseria), грамотрицательных палочек с закругленными
концами, окруженных капсулой в виде светлого ореола (Klebsiella и др.), грамотри-
цательных палочек (Е. coli, Р. aeruginosa и др.); мелких грамотрицательных палочек
в виде скоплений (Haemophylus), микобактерий.
БАКТЕРИОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ 255
По морфологическим и тинкториальным признакам при микроскопии мазка,
окрашенного по Граму, можно ориентировочно идентифицировать следующие
микроорганизмы, присутствующие в мокроте;
скопления грамположительных кокков — Staphylococcus spp., Micrococcus spp:
Ъ цепочки грамположительных кокков — Streptococcus spp;
О ланцетовидные грамположительные диплококки, окруженные капсулой, —
Str. pneumoniae-,
❖ грамотрицательные диплококки — Neisseria spp.:
грамотрицательные палочки — Enterobacteriaceae, Pseudomonasspp., Haemophilus
spp., неспорообразующие анаэробы;
грамотрицательные палочки с закругленными концами, окруженные капсу-
лой, — Klebsiella spp.:
мелкие полиморфные грамотрицательные палочки в виде скоплений —
Нает орк i lus spp.;
обрывки тонкого несептированного ветвящегося мицелия фиолетового цвета,
споры темно-фиолетового цвета и друзы розового цвета — Actinomyces spp.
Для острой инфекции характерно наличие в мокроте 1-2 видов бактерий, рас-
положенных вблизи гранулоцитов. Микроорганизмы-контаминанты, попавшие в
мокроту из ротовой полости, чаще расположены вблизи эпителиальных клеток.
Диагностически значимо обнаружение трех и более пневмотропных микроор-
ганизмов в большинстве полей зрения. Анализ результатов микроскопии мазка
мокроты в сочетании с клинико-рентгенологическими особенностями течения
заболевания позволяет поставить ранний клинике-бактериологический диагноз
более чем у 80% больных острой пневмонией или, по крайне:i мере, облегчает
эмпирический выбор антибактериального препарата на основании преобладания
грамположительной или грамотрицательной микрофлоры.
Необходимо подчеркнуть важность бактериоскопического исследования
мокроты при деструктивных пневмонитах (абсцесс легкого, гангрена легкого).
Опытный микробиолог на основании морфологии микроорганизмов, выявляемых
при бактериоскопии, может высказать более точное суждение об этиологии про-
цесса, чем это возможно при анализе результатов рутинного бактериологического
метода. Это связано с тем, что посев на питательные среды, обычно применяемые
в настоящее время в практических лабораториях, не всегда позволяет обнаружить
облигатные анаэробы, играющие ведущую роль при данной патологии. Кроме
того, результаты целенаправленного бактериологического исследования на неспо-
рообразующие облигатно анаэробные микроорганизмы можно получить только
на 7-8-й день.
На основании первичного осмотра посевов ориентировочно оценивают числен-
ность микроорганизмов в мокроте, пользуясь следующими критериями:
❖ I степень— очень скудный рост — рост единичных колоний (до 10);
❖ II степень — скудный рост — рост 10-25 колоний;
о- III степень — умеренный рост — рост множества сосчитываемых колоний (не
менее 50);
❖ IV степень — обильный рост — сплошной рост несосчитываемых колоний.
I и II степени роста свидетельствуют о загрязнении исследуемого материала или
бактерионосительстве, а III и IV — об этиологической значимости данного микро-
организма.
Изоляты бактерий, выделенные из мокроты в посевах на питательных средах,
идентифицируют по культуральным, морфологическим, тинкториальным, биохи-
мическим свойствам и в ряде случаев по антигенной структуре.
256 БАКТЕРИОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ
Материал из брони
Показания к взятию и исследованию проб: воспалительные заболевания ниж-
них отделов дыхательных путей при отсутствии мокроты.
Взятие материала
Из бронхов для исследований берут смывы, соскобы и биоптаты.
Концентрация микроорганизмов в смывах в 10-1000 раз ниже, чем в мокроте,
поэтому их бактериологическое исследование имеет меньшую диагностическую
ценность
Наиболее просто получить трахеобронхиальный смыв следующим методом:
❖ гортанным шприцем с помощью аппарата Боброва вводят в трахею около
10 мл стерильного 0,9% раствора хлорида натрия и при кашле собирают тра-
хеобронхиальный смыв;
❖ маленьким детям в трахею вводят через катетер 5-10 мл 0,9% раствора хло-
рида натрия и затем шприцем аспирируют трахеобронхиальный смыв.
С помощью бронхоскопии удается получить:
< > бронхоальвеолярный лаваж (предпочтительно);
смыв с бронхов (низкая чувствительность при диагностике пневмоний);
❖ соскоб с бронхов (более значим, чем смыв);
❖ биоптаты.
Смыв с бронхов или бронхоальвеолярный лаваж выполняют, вводя шприцем
через биопсийный канал бронхоскопа стерильный 0,9% раствор хлорида натрия
отдельными порциями объемом 5-20 мл. Перед введением каждой следующей
порции аспирируют шприцем жидкость и переносят в стерильный контейнер.
При этом для каждой новой порции используют новый контейнер. По решению
лечащего врача можно объединить отдельные смывы, взятые в различных участ-
ках легких. В направлении указывают общий объем введенного 0,9% раствора
хлорида натрия.
Для получения соскоба с бронхов через биопсийный канал бронхоскопа вво-
дят телескопический двойной катетер с обработанным полиэтиленгликолем или
другим соответствующим реактивом дистальным концом для предотвращения
контаминации пробы. Материал помещают в пустой стерильный контейнер и в
пробирку с тиогликилевой средой (средой СКС) или специальный консервант для
облигатных анаэробов.
Биоптаты доставляют в лабораторию в пробирках типа «Эппендорф», запол-
ненных стерильным 0,9% раствором хлорида натрия или тиогликолевой средой.
Исследование материала
Делают посев взятого материала на кровяной агар, шоколадный агар, желчно-
солевой агар, среды Эндо, Сабуро и другие, выбор которых определяется предпо-
лагаемой этиологией болезни.
Опенка результатов
Оценку результатов бактериологического исследования материала, взятого из
бронхов, проводят на основании анализа информации о видовом сост аве микро-
флоры (по данным бактериоскопического и бактериологического исследований),
численности отдельных видов микроорганизмов, стабильности их выделения во
времени, клинических дачных и результатах лечения.
При оценке результатов бактериологического исследования необходимо при-
нимать во внимание определяющее значение некоторых видов микроорганизмов
в типичных клинических ситуациях:
> крупозная пневмония — Str. pneumoniae',
ъ лобарная пневмония — Str. pneumoniae, Klebsiella spp.. Staphylococcus spp.',
❖ очаговые пневмонии — Str. pneumoniae, H. influenzae, энтеробактерии, P. aerug-
inosa-,
БАКТЕРИОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ 257
❖ первичная атипичная пневмония — микоплазмы, легионеллы или хлами-
дии;
< пневмония на фоне хронического бронхита — Н. influenzae, Streptococcus spp.',
-о- пневмония на фоне гриппа — Staphylococcus spp., Str. pneumoniae, H. influenzae-,
> аспирационная пневмония — энтеробактерии, Р. aeruginosa, анаэробы;
< > пневмония на фоне иммунодефицита — энтеробактерии, Staphylococcus spp.
Госпитальная пневмония у новорожденных (в первые 7 дней жизни) чаще
обусловлена Streptococcus spp , Е. coli, реже — 8. aureus, Chlamydia trachomatis,
Pseudomonas spp.
У больных муковисцидозом в бронхах и легких наиболее часто обнаруживают
Р. aeruginosa. Основную роль в этиологии острых инфекционных заболеваний лег-
ких играют неспорообразующие анаэробы и Staphylococcus spp.
Острые бронхиты и бронхиолиты вызываются, как правило, вирусами или
микоплазмами, но со 2-3-го дня заболевания осложняются присоединением бакте-
риальной инфекции, чаще всего обусловленной Str. pneumoniae или Н. influenzae.
Главную роль в этиологии внебольничных пневмоний играет Str. pneumoniae.
С помощью количественного посева проб можно уточнить этиологию острой
пневмонии, пользуясь следующими критериями:
< обнаружение монокультуры Str. pneumoniae в крови или в плевральном экс-
судате;
❖ выделение Str pneumoniae или Н. influenzae в трахеобронхиальном смыве в
титре 106 КОЕ/мл;
❖ обнаружение в трахеобронхиальных смывах небольшого количества (менее
106или 104 КОЕ/мл) Str. pneumoniae при обследовании больных острой пнев-
монией на поздних сроках болезни и на фоне этиотропной антибактериаль-
ной терапии с последующим снижением содержания Str. pneumoniae не менее
чем в 100 раз;
❖ выделение в 2-3 последовательных исследованиях трахеобронхиальных
смывов, взятых с интервалом 2-5 дней, одного и того же вида условно-
патогенных бактерий в титре 104 КОЕ/мл (при возникновении вторичного
инфекционного процесса в сочетании с выявлением местной или системно-
иммунологической реакции организма на данный микроорганизм).
Пневмококкозную этиологию пневмонии подтверждает обнаружение пневмокок-
кового антигена в крови, плевральном экссудате или моче, обнаружение в реак-
ции непрямой иммунофлюоресценции специфических сывороточных антител в
титре 1:320 (дети в возрасте 0-3 лет) или 1:640-1:1280 (взрослые), установление
4-кратного или более значительного нарастания первоначального титра антипнев-
мококковых антител.
Этиологию внебольничных пневмоний с помощью стандартных методов лабо-
раторного исследования не удается установить в 30-50% случаев. Это может быть
обусловлено следующими причинами:
о невозможностью обнаружения ряда инфекционных агентов (Mycuplasma
pneumoniae, Chlamydia pneumoniae, I egionella pneumoniae) рутинными методами;
❖ трудностью дифференциации возбудителей болезни от резидентной микро-
флоры;
о- проведением этиотропного лечения ряда больных до начала бактериологи-
ческого исследования.
Желчь
Показания к проведению исследования:
❖ диагностика брюшного тифа и брюшнотифозного хронического бактерионо-
сительства;
258 БАКТЕРИОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ
❖ воспалительные заболевания желчною пузыря и желчных протоков (холе-
циститы, холангиты, желчнокаменная болезнь).
Взятие материала
Желчь получаю г с помощью зондирования или во время операции при пункции
желчного пузыря.
При зондировании над пламенем спиртовки собирают в стерильные пробирки
3 порции желчи (по 10-12 мл) — для бактериологического исследования пред-
почтительна средняя порция. Пробы должны быть доставлены в лабораторию в
течение 1-2 ч.
При наличии у больного дренажа собирают из него пробу шприцем, предвари-
тельно тщательно обработав участок пункции. Нельзя собирать пробу желчи из
дренажного мешка.
Пробы желчи, а также гноя и аспирата из печеночных абсцессов также можно
собирать во время операции. Материал из очага воспаления направляют в лабора-
торию в контейнере с транспортной средой для анаэробов или в полностью запол-
ненном шприце, надев на него стерильную иглу с защитным чехлом.
Исследование материала
При бактериологическом исследовании по 0,1 мл каждой порции желчи засева-
ют на кровяной агар, по 0,5 мл — на среду Эндо.
Для выделения сальмонелл желчь засевают в соотношении 1:9 в селенитовый
бульон, а также инкубируют в термостате нативную желчь. Затем в течение 3 дней
ежедневно производят высев как с селенитового бульона, так и с нативной желчи
на висмут-сульфитный агар и другие среды, предназначенные для культивирова-
ния S. enterittdis.
Для выделения анаэробов желчь высевают в 1 пробирку со средой для контро-
ля стерильности и пи в 2 пробирки со средой Китта-Тароцци. Одну из засеянных
пробирок со средой Китта-Тароцци прогревают на водяной бане для уничтожения
неспорообразующих микроорганизмов. Наблюдение за посевами ведут в течение
5 днеч.
Оценка результатов
В норме желчь стерильна. Полученная в результате дуоденального зондиро-
вания. она практически всегда содержит микроорганизмы, попавшие в нее из
ротовой полости и верхних отделов пищеварительного тракта. На контаминацию
желчи микрофлорой полости рта указывает обнаружение в ней нейссерий и дрож-
жеподобных грибов. Стафилококки и стрептококки значительно чаще обнаружи-
вают в желчи, полученной при зондировании, чем в образцах, взятых при пункции
желчного пузыря. Поэтому наиболее достоверны результаты бактериологического
исследования проб желчи, полученных во время операции.
При инфекциях из желчи в 70-80% случаев высевают Е. coli, Enterococcus spp.,
несколько реже К. pneumoniae, Enterobucter spp., а также сальмонеллы (при
временном и хроническом бактерионосительстве). Из анаэробных микроорга-
низмов выделяют Cl. perfringens, в 10-20% случаев желчнокаменной болезни —
Peptococcaceae spp. В отдельных случаях встречается смешанная аэробная и анаэ-
робная инфекция.
При оценке результатов необходимо учитывать количество микроорганизмов
в материале. При планируемом оперативном вмешательстве прогностически
неблагоприятным считается обнаружение в желчи бактерий (энтеробактерий,
псевдомонад, бактероидов) в титре выше 103 КОЕ/мл. Выделение значительных
количеств S. aureus может свидетельствовать о печеночном или поддиафрагмаль-
ном абсцессе.
БАКТЕРИОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ 259
Моча
Показания к проведению исследования:
❖ установление этиологии болезней мочевой системы;
< выявление бактерионосителей.
Взятие материала
Мочу для бактериологического исследования берут при естественном мочеиспу-
скании. катетером, из участка подвздошной кишки, использованной для создания
искусственного пузыря, надлобковой пункцией мочевого пузыря, билатеральной
катетеризацией мочеточника с использованием цистоскопа и другими методами,
при которых сведена к минимуму возможность контаминации проб посторонней
микрофлорой.
Независимо от способа получения мочи ее следует высевать на питательные
среды не позднее чем через 2 ч после сбора. При отсутствии такой возможности
пробы хранят не более 24 ч в холодильнике (4 °C) или консервируют борной
кислотой. Следует помнить, что моча — прекрасная питательная среда для многих
бактерий, поэтому чем позднее после ее сбора проводят посев на питательные
среды тем менее достоверным будет результат бактериологического исследова-
ния.
Взятие средней порции мочи при естественном мочеиспускании
Показания к проведению:
4 подозрение на воспалительные заболевания почек и мочевого пузыря;
< > диагностика брюшного тифа (с конца 2-й недели заболевания), лептоспиро-
за.
Взятие материала
В стерильную посуду собирают среднюю порцию свободно выпущенной мочи
в объеме 3-5 мл. Женщинам рекомендуется сдавать 2-3 порции мочи в течение
2-3 дней. Перед взятием проб мочи больной дотжен совершить тщательный туа-
лет наружных половых органов. Это редкая ситуация, когда медработник не может
контролировать соблюдение правил сбора материала. Поэтому важно подробно
объяснить пациенту правила сбора мочи. Целесообразно напечатать инструкции
на обратной стороне бланка направления.
Инструкция для мужчин:
❖ тщательно вымыть руки;
ъ тщательно вымыть половой член теплой водой с мылом и высушить стериль-
ной салфеткой;
❖ обнажить головку полового члена и выпустить небольшую порцию мочи;
< > прервать мочеиспускание и выпустить порцию мочи в стерильный контей-
нер;
< > закрыть контейнер и передать в лабораторию.
Инструкция для женшин:
❖ тщательно вымыть руки;
вымыть половые органы, используя стерильные марлевые салфетки и теплую
мыльную воду, в направлении спереди назад;
о промыть половые органы еще раз теплой водой и вытереть стерильной сал-
феткой; отверстие влагалища желательно закрыть стерильным ватным там-
поном;
на протяжении всей процедуры держать половые губы раздвинутыми;
0- помочиться, выпустив первую порцию мочи;
❖ выпустить порцию мочи в стерильный контейнер;
❖ закрыть контейнер и передать в лабораторию.
260 БАКТЕРИОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ
Инструкция для диц, собирающих мочу маленьких детей:
< > дать ребенку выпить воды или другой жидкости;
< вымыть руки с мылом, сполоснуть водой, высушить;
- у мальчиков — тщательно промыть половой член и оттянуть крайнюю
плоть, область заднего прохода вымыть теплой водой с мылом и сполос-
нуть теплой водой, высушить стерильной марлевой салфеткой;
- у девочек — тщательно промыть отверстие мочеиспускательного канала, а
также промежность и область заднего прохода теплой мыльной водой или
жидким мы пом. сполоснуть теплой водой, высушить стерильной марлевой
салфеткой;
❖ усадить ребенка на колени помощника
- у мальчиков — при мочеиспускании держать крайнюю плоть оттянутой
для предотвращения контаминации пробы мочи;
- у девочек — держать наружные половые губы на расстоянии друг от друга
в процессе мочеиспускания;
< спустить небольшое количество мочи в специальную посуду для утилиза-
ции;
< собрать среднюю порцию мочи (10-15 мл) в специальный одноразовый кон-
тейнер с завинчивающейся крышкой.
Взятие мочи катетером
Показания к проведению:
> отсутствие возможности получения мочи естественным путем;
<► большая вариабельность получаемы х результатов;
4 необходимость дифференциации очага инфекции (мочевой пузырь или
почки).
Взятие материала
❖ Если мочевой пузырь заполнен, перед катетеризацией пациент должен
частично его опорожнить
4 Проводят туалет наружных половых органов теплой водой с мылом и высу-
шивают их стерильными марлевыми салфетками, затем вводят катетер в
мочевой пузырь.
о- Собирают из катетера первые 15-30 мл мочи в специальную посуду для ути-
лизации, после чего заполняют на 1/3-1/2 стерильную емкость, в которой
мочу доставляют в лабораторию.
4 Для уточнения локализации инфекции мочевой пузырь опорожняют кате-
тером, промывают раствором антисептика и через 10 мин забирают пробы
мочи (при инфекциях мочевого пузыря моча остается стерильной).
Билатеральная катетеризация мочеючника под контролем цистоскопии
Показание к проведению — определение очага инфекции в мочевыводящих
путях.
Взятие материала
4 Перед проведением цистоскопии пациент должен освободить заполненный
мочевой пузырь.
4 Следует обработать наружные половые органы, области промежности и
заднего прохода, как указано выше.
❖ Ввести цистоскоп в мочевой пузырь и собрать 5-10 мл мочи в стерильный
одноразовый кон гейнер.
❖ Маркировать стерильный одноразовый контейнер символом
КМП (Катетерная Моча Пузыря), затем оросить пузырь стерильным физио-
логическим раствором.
БАКТЕРИОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ 261
❖ После орошения пузыря и введения катетера собрать истекающую жидкость,
держа концы обоих катетеров над открытым контейнером.
❖ Тщательно промаркировать контейнер символом ПЖП (Промывная
Жидкость Пузыря).
❖ Провести мочеточниковые катетеры к среднему отделу каждого мочеточника
или почечных лоханок без дополнительного введения жидкости; открыть
цистоскоп, чтобы освободить мочевой пузырь.
❖ Не использовать для анализа первые 5-10 мл мочи из каждого катетера в
мочеточнике.
❖ Собрать 4 последующих пары проб (5-10 мл каждая) прямо в стерильный
одноразовый контейнер или соответствующие стерильные специальные
емкости.
❖ Тщательно промаркировать емкости с материалом символами ПП-1, ЛП-1,
ПП-2, ЛП-2, где ПП — Правая Почка. ЛП — Левая Почка. Все пробы передать
в лабораторию на посев.
Сбор мочи из участка подвздошной кишки, использованной для создания
искусственного пузыря
Показания к проведению:
< = жалобы больного на недомогание;
о- лихорадка.
Взятие материала
< Отсоединяют мочеприемное устройство и опорожняют его.
❖ Осторожно очищают отверстие тампоном, смоченным 70% этиловым спир-
том. а затем 1-2% раствором йода или другого дезинфектанта, разрешенного
к применению для данных целей.
❖ Удаляют избыток йода 70% этиловым спиртом для предотвращения ожога
тканей.
Вводят катетер и собирают мочу в стерильную емкость.
Надлобковая пункция мочевого пузыря
Показания к проведению:
о острая задержка мочи;
❖ получение проб мочи у новорожденных и маленьких детей.
Взятие материала
Этим способом могут быть получены наиболее достоверные результаты. Однако
процедура сопряжена с опасностью для здоровья больного, и ее проводят только
по клиническим показаниям.
Техника надлобковой пункции:
❖ перед пункцией пациент, если это возможно, должен освободить пузырь, так
как имеющаяся в нем жидкость снижает титр микроорганизмов в моче;
❖ дезинфицируют кожу над лобком (лучше после бритья);
❖ через небольшой разрез, сделанный в эпидермисе, шприцем с иглой аспири-
руют мочу (10-15 мл) из пузыря;
❖ переносят мочу из шприца в стерильный одноразовый контейнер, отправляе-
мый в лабораторию.
Исследование материала
Решение о необходимости выполнения бактериологического исследования
мочи может быть принято на основании скрининговых методов, позволяющих
относительно быстро определить количество микроорганизмов в моче. К их числу
относят микроскопию мазков мочи и химические тесты.
Микроскопическое исследование мазков мочи, окрашенных по Граму, позволяет
выявить бактериурию при концентрациях микроорганизмов 105 КОЕ/мл и выше.
262 БАКТЕРИОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ
Наличие одного или более лейкоцитов в поле зрения — характерный признак
инфекции мочевыводящих путей. Присутствие в мазках мочи, взятых у женщин,
большого количества спущенного эпителия может быть признаком контаминации
мочи влагалищной микрофлорой и указывает на необходимость повторного сбора
материала.
Препарат для микроскопии готовят из нативной мочи. На предметное стекло
наносят с помощью пипетки каплю мочи (50 мкл). Для ускорения высыхания пре-
парата, без существенной потери чувствительности, объем капли можно уменьшить
до 10 мкл и использовать для ее нанесения калиброванную петлю. Высушенные
препараты фиксируют в пламени и микроскопируют с масляной иммерсией, про-
сматривая не менее 50 полей зрения по диаметру капли. Положительным считает-
ся результат, при котором в среднем в одном поле зрения присутствуют 2 и более
КОЕ.
Химические тесты основаны на выявлении ферментов микроорганизмов или
лейкоцитов в моче. В нашей стране рекомендованы к применению два ускоренных
метода исследования мочи, основанных на регистрации активности микробных
ферментов, — нитритный тест и ТТХ-тест.
Нитритный тест основан на выявлении с помощью реактива Грисса нитритов,
которые образуют из нитратов микроорганизмы семейства Enterobactei iacea.
Появление красного окрашивания при добавлении к 1 мл мочи 1 мл реактива
свидетельствует о присутствии бактерий в количестве не менее 105 КОЕ/мл. Для
повышения эффективности теста до его постановки в мочу добавляют нитраты и
инкубируют в термостате при температуре 37 СС в течение 2-4 ч.
ТТХ-тест основан на преобразовании хлористого тринитрозолия под действием
микробных ферментов в красный нерастворимый в воде трифенилформазан. При
постановке теста к 2 мл мочи добавляют 0,5 мл рабочего раствора ТТХ. Смесь
инкубируют в термостате при температуре 37 °C в течение 4 ч, Появление красно-
го осадка на дне пробирки свидетельствует о присутствии бактерий в количестве
более 105 КОЕ/мл.
Применение упомянутых скрининговых методов позволяет снизить затраты на
расходные материалы, однако значительно увеличивает трудоемкость исследова-
ния.
Для оценки степени бактериурии пользуются методом секторных посевов.
Калиброванной бактериологической петлей диаметром 2 мм, емкостью 0.005 мл
берут каплю мочи и делают 30-40 штрихов на сектор А чашки Петри. Прожигают
петлю и производят 4 штриха через сектор А в сектор I. Аналогичным образом
из сектора I засевают сектор II. а из сектора II — сектор III (рис. 20-1). Чашки
Рис. 20-1. Секторный посев мочи.
инкубируют при температуре 37 °C в течение
24 ч. Посев проводят на питательный агар,
5% кровяной агар, среды Эндо, CLED или
хромогенный агар (Uriselect и др.). При отсут-
ствии признаков бактериального роста инку-
бацию посевов увеличивают до 3 сут. Учет
результатов проводят по специальной таблице
(табл. 20 6).
Альтернатива методу штрихового посева —
посев с помощью пипетки. В этом случае на
центр чашки с питательной средой наносят
каплю мочи объемом 10 мкл и распределя-
ют 4 штрихами бактериологической петли
от центра к периферии. Затем всю поверх-
ность питательной среды равномерно засе-
вают частыми штрихами. После инкубации
БАКТЕРИОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ 263
проводят оценку результатов — при росте на чашке 10-100 колоний концентрация
бактерий в моче 103 КОЕ/мл, 101-1000 колоний — 104 КОЕ/мл, более 1000 коло-
ний — 105 КОЕ/мл,
Таблица 20-6. Учет результатов посева мочи секторным методом
Количество колоний Количество микроорганизмов в исследуемом материале, КОЕ/мл
А I II 111
1-6 - - - <1000
8-20 - - - 3000
20-30 - - - 5000
30-60 - - - 10 000
70-80 - - - 50 000
100-150 5-10 - - 100 000
Невозможно под- считать 20-30 - - 500 000
Невозможно под- считать 40-60 - - 1 000 000
Невозможно под- считать 100-150 10-20 - 5 000 000
Невозможно под- считать Невозможно под- считать 30-40 - 10 000 000
Невозможно под- считать Невозможно под- считать 60-80 Единичные колонии 100 000 000
За рубежом широкое распространение получили дипслайды — пластинки с
питательными средами, погружаемые в мочу. На две стороны пластинки могут
наноситься разные питательные среды: обычно одна служит для оценки титра
бактерий в моче, а другая для их идентификации (по ферментации глюкозы, экс-
прессии ферментов, образованию индола, морфологии колоний и другим призна-
кам). Это позволяет идентифицировать наиболее важные уропатогенные бактерии
(в сомнительных случаях проводят дополнительные биохимические анализы
изолятов, выросших на средах пластинки). Основным недостатком дипслайдов
служит невозможность оценки степени бактсриурии, когда титр бактерий в моче
превосходит 105 КОЕ/мл — в этом случае часто отмечают газонный рост бактерий.
Этот недостаток устранен в дипстрике — аналогичном приспособлении, в котором
посев осуществляется не погружением пластинки со средами в мочу, а штриховым
дозированным методом с помощью пластикового кольца с зубцами. Внедрение
подобных систем позволяет перенести значительную часть работы из лаборатории
в кабинет врача, где проводится посев, инкубация и первичная оценка результатов.
Посевы, в которых количество бактерий меньше диагностически значимой вели-
чины, уничтожают, а оставшиеся направляют в лабораторию для точной иденти-
фикации микроорганизмов и определения их чувствительности к антимикробным
препаратам.
Оценка результатов
В норме в мочевом пузыре моча стерильна, однако во время прохождения через
нижнюю треть уретры происходит ее контаминация нормальной микрофлорой.
Поэтому при оценке результатов исследования на первый план выходят количе-
ственные критерии, позволяющие дифференцировать бактериурию, связанную с
264 БАКТЕРИОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ
патологическим процессом, от естественной контаминации. У мужчин достаточно
исследовать одну пробу мочи. У женщин вследствие высокого риска случайной
контаминации целесообразно исследовать серию из 2-3 проб - достоверность
положительного результата 1 пробы составляет 80%, 2 проб — 90%. 3 проб —
100%, при условии, что результаты всех проб одинаковы. При оценке диагности-
ческой значимости результата в России используются критерии, предложенные
Е.Х. Кассом в 1960 г.:
> титр микроорганизмов, не превышающий 103 КОЕ/мл, — естественная кон-
таминация мочи:
❖ титр бактерий от 103 до 105 КОЕ/мл — сомнительный результат, необходимо
повторение исследования:
о титр бактерий, равный или превышающий 105 КОЕ/мл, — диагностический
уровень.
В настоящее время эти нормы пересматриваются, так как они занижены и
служат причиной недовыявления большого числа инфекций мочевого тракта. В
соответствии с «Европейским руководством по анализу мочи» (2000 г.) диагности-
ческими уровнями бактериурии следует считать:
при изоляции монокультуры Е. coli и S. saprophyticus —> 103 КОЕ/мл;
❖ при изоляции монокультуры (Enterobacter spp., Enterococcus spp., Klebsiella spp.,
P. mirabilis, P. aeruginosa, Citrobacter spp., M. morganii. P. vulgaris, Serratia spp.,
S. aureus, C. urealyricum, Haemophilus spp., Pneumococci spp.) — > 104 КОЕ/мл;
о - при изоляции смешанной культуры двух бактерий — > 10s КОЕ/мл.
Изоляцию 3 и большего числа микроорганизмов считают результатом случай-
ной контаминации тестируемых проб мочи. При оценке результатов этого этапа
бактериологического анализа делают поправку на способ, которым была получена
моча, и наличие или отсутствие у пациента симптоматики (табл. 20-7)
Таблица 20-7. Оценка диагностической значимости бактериурии (по Norrby S.R.)
Инфекции мочевыводящих путей Диагностическое значение (количеств! бактерий, КОЕ/мл)
Острый неосложненный цистит у женщин
Грамотрицательные бактерии >103
Staphylococcus spp. >102
Острый неосложненный пиелонефрит
Грамотрицательные бактерии >104
Staphylococcus spp >10э
Осложненные инфекции и инфекции у мужчин >10<
Пациенты с бессимптомной бактериурией >105 в двух пробах
В отдельных случаях воспалительные процессы в мочевыводящей системе могут
протекать и при низких уровнях бактериурии, например на фоне форсированно-
го диуреза (низкий удельный вес мочи), антибиотикотерапии или при низких
(менее 5) значениях pH мочи. В этом случае постановке диагноза помогает анализ
видового состава микрофлоры. Обнаружение Е. coli, S. saprophyticus, Enterobacter
spp., Enterococcus spp., Klebsiella spp., P. mirabilis, P. aeruginosa расценивают как небла-
гоприятный признак. Выявление инфекции менее патогенных и непатогенных
бактерий не служит основанием для постановки этиологического диагноза.
БАКТЕРИОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ 265
Пробы из мочеполовых органов
Твердый шанкр, папулы, розеолы
Показания к взятию и исследованию проб: диагностика сифилиса.
Взятие материала
• Если до обследования пациент применял дезинфицирующие или прижигаю-
щие среде гва, перед взятием материала для бактериологического анализа ему
рекомендуют в течение 12-24 ч прикладывать примочки с 0,9% раствором
хлорида натрия.
• За 2-3 ч до исследования на поверхность эрозии или язвы накладывают на
15-20 мин примочку с гипертоническим (10%) раствором хлорида натрия, а
затем — с 0,9% раствором хлорида натрия.
• Поверхность эрозии, язвы, эрозированной папулы или бляшки дважды осто-
рожно протирают ватным тампоном, смоченным стерильным 0,9% раствором
хлорида натрия.
• Осторожно массируют поверхность очага прокаленной и остуженной бактерио-
логической петлей, шпателем или ложкой Фолькмана до появления прозрачной
или слегка опалесцирующей тканевой жидкости. Можно осторожно сдавить края
эрозии или язвы руками в перчатках для увеличения объема жидкости.
• При возникновении кровотечения его останавливают с помощью марлевого
тампона, увлажненного 0,9% раствором хлорида натрия, и операцию повторя-
ют. В пробе не должно быть даже следов крови!
• Из папул и розеол при скарификации с помощью скальпеля получают тканевый
сок.
• Небольшую каплю тканевой жидкости бактериологической петлей переносят
на чистое обезжиренное предметное стекло без царапин, накрывают покровным
стеклом и сразу же микроскопируют с помощью темнопольного микроскопа.
Исследование материала
Взятый материал исследуют посредством темнопольной микроскопии, реакции
иммунофлюоресценции, а также методами молекулярной диагностики на наличие
Т. pallidum (детальное описание этих методов приведено в главе 26).
Оценка результатов
Обнаружение трепонем с характерной морфологией и типом движения свиде-
тельствует в пользу диагноза сифилиса. Дифференцировать Т. pallidum от сапро-
фитных трепонем, входящих в состав нормальной микрофлоры, можно в реакции
иммунофлюоресценции или ПЦР.
Материал из уретры
Показания к проведению исследования:
❖ острые и хронические заболевания уретры;
❖ острые и хронические заболевания половых органов;
❖ бесплодие.
Взятие материала
• Перед взятием материала необходимо вымыть половые органы теплой водой
или 0,9% раствором хлорида натрия, удалить с помощью ватного тампона
свободно стекающие выделения.
• Материал для микроскопического исследования берут с помощью бакте-
риологической петли, желобоватого зонда, стеклянной глазной палочки или
маленькой ложки Фолькмана. В последние годы широкое распространение
получили специальные одноразовые зонды с ворсистой головкой, позво-
ляющие отбирать материал качественно и с минимальной травма гизацией
слизистой оболочки.
ГДАЙ 20
266 БАКТЕРИОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ
• При обильном отделяемом материал собирают с помощью стеклянной
палочки или края шлифованного стекла и сразу готовят мазки на предмет-
ных стеклах. В лабораторию нужно направлять не менее 2 мазков от одного
больного.
• При скудном отделяемом больному предлагают до исследования не мочиться
в течение 4-8 ч, а затем массируют уретру, стараясь выдавить собравшийся в
глубине секрет.
• Материал для бактериологического исследования забирают с помощью
бактериоло! ической петли Для предупреждения его контаминации сапро-
фитной микрофлорой, в большом количестве ирисутст вующей в дистальном
отделе уретры, в мочеиспускательный канал вводят стерильную ушную
воронку, через которую бактериологической петлей собирают отделяемое из
глубоких отделов уретры.
• Если планируется посев с целью выделения N. gonorrhoea^ необходимо
использовать вискозные, дакроновые и альгинатно-кальциевые зонды-
тампоны, поскольку хлопковая вата может быть токсичной для этой бак-
терии. Зонд-тампон осторожно вводят в уретру и оставляют на 10 с. После
извлечения его помещают в транспортную среду (например, среду Эймса с
углем). Материал должен быть доставлен в лабораторию в течение 12 ч. Не
допускается его охлаждение ниже 30 °C.
При подозрении на трихомоноз наиболее эффективно исследование смывов из
уретры, получаемых с помощью пипетки или стеклянной трубки, снабженной
резиновой грушей.
• На стерильную стеклянную трубку (пипетку) надевают резиновый баллон-
чик и набирают в нее 0,5-1,0 мл стерильного теплого раствора Рингера.
Трубку вставляют в наружное отверстие уретры, раствор несколько раз вво-
дят в канал и аспирируют в трубку. Затем материал переносят в стерильную
пробирку типа «Эппендорф» и используют для приготовления препаратов
или посева на питательные среды.
Исследование материала
Методы, применяемые для исследования взятого материала, зависят от предпо-
лагаемого возбудителя заболевания и могут включать бактериоскопию, посевы
на питательные среды, иммунологические и молекулярные лабораторные тесты,
предназначенные для обнаружения микроорганизмов, их антигенов или нуклеи-
новых кислот.
Оценка результатов
Оценка этиологической значимости выделенных микроорганизмов при уретри-
тах сопряжена со значительными сложностями, особенно при уретритах негоно-
рейной этиологии.
Полиэтиологичность уретритов диктует необходимость проведения целого
комплекса лабораторных исследований, направленных на выявление возможно
более широкого круга микроорганизмов — потенциальных этиологических аген-
тов. Этиологическую диагностику в большинстве случаев может обеспечить при-
менение следующих методов:
микроскопии мазков, окпашенн ых метиленовым синим и по Граму; для обна-
ружения N. gonorrhoeas, Gardnerella vaginalis. Trichomonas vaginalis и грибов
рода Candida spp.;
посева отделяемого из уретры и секрета половых желез для выявления
N. gonorrhoeae и условно-патогенных бактерий, трихомонад и грибов:
• о цитологического исследования соскобов из уретры, окрашенных по
Романовскому-Гимзе, для выявления внутриэпителиальных включений хла-
мидий.
БАКТЕРИОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ 267
В настоящее время для выявления хламидий применяют также реакцию имму-
нофлюоресценции и ПЦР, а для обнаружения уреаплазм — культуральный метод
или количественный вариант ПЦР.
• Существует возможность развития смешанной инфекции, когда одновремен-
но этиологически значимы несколько возбудителей. Поэтому обследование
больного нельзя прекращать после обнаружения одного вида патогенов, не
исключив одновременного присутствия других.
• Чувствительность используемых в лабораториях методов выявления пато-
генных микроорганизмов из уретры недостаточно высока. Например, при
трихомонозе у 3-20% больных возбудитель удается обнаружить только
после 5-кратного обследования. В связи с этим для уверенности в отрица-
тельном результате исследования необходимо выполнять неоднократно и, по
возможности, с параллельным использованием нескольких методов.
• Обнаружение уреаплазм, Candida spp. и условно-патогенных бактерий не
всегда свидетельствует об их этиологической роли, так как данные микро-
организмы часто входят в состав нормальной микрофлоры уретры.
Для оценки этиологической роли уреаплазм были предложены следующие кри-
терии:
< > обнаружение возбудителя в титре выше 104 КОЕ/мл в секрете предстатель-
ной железы или более 103 КОЕ/мл в моче;
❖ обнаружение в посевах уреаплазм при одновременном отсутствии других
патогенных микроорганизмов и нарастании титра соответствующих антител
при исследовании парных сывороток.
При оценке этиологической роли Candida spp. учитывают их титр в первой пор-
ции мочи — его диагностический уровень составляет 1(Р КОЕ/мл.
Уретриты, вызванные условно-патогенными бактериями, встречаются крайне
редко. Диагностика этих заболеваний основана на сведениях о титре бактерий в
исследованном материале, постоянстве видового состава микрофлоры и ее моно-
морфизме, наличии и выраженности фагоцитарной реакции и отсутствии других
инфекционных агентов. Эти критерии можно использовать только в комплексе,
при отсутствии хотя бы одного из них диагноз сомнителен.
Уретрит у мужчин и женщин — полиэтиологичное заболевание. Его причиной
могут быть как патогенные, так и условно-патогенные микроорганизмы, входящие
в состав нормальной микрофлоры.
Гонококковый уретрит может быть р уверенностью диагностирован с помощью
бактериоскопического метода при обнаружении в мазках грамотрицательных
диплококков с характерной морфологией, расположенных внутри лейкоцитов
(незавершенный фагоцитоз). При отсутствии внутриклеточно расположенных
N. gonorrhoeae точность диагностики снижается. В этих случаях для подтверждения
диагноза необходимо использовать бактериологический метод и ПЦР. У девочек
(до наступления менархе) и женщин (в менопаузе) диагноз гонореи устанавливают
только на основании бактериологического исследования.
Эффективность диагностики можно повысить с помощью «провокаций» перед
взятием исследуемого материала.
Бактериоскопический метод малоэффективен для диагностики трихомонадных
уретритов. В данном случае оптимально проведение культурального исследова-
ния смывов из уретры, которое по числу находок более результативно, чем ПЦР.
Уретриты, вызванные С. trachomatis, широко распространены, проявляются, как
правило, скудной клинической симптоматикой, однако могул сопровождаться и
признаками острого воспаления, сходными с симптомами гонореи.
Если хламидии обнаружены с помощью флюоресцирующих антител, в том числе
моноклональных (ПИФ, РИФ), результат следует подтвердить с помощью ПЦР.
Целесообразность выполнения культурального исследования дискутабельна.
268 БАКТЕРИОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ
Язвы половых органов
Показания к взятию и исследованию проб: изъязвления на коже и слизистой
оболочке полового члена, за исключением подозрения на твердый шанкр.
Язвенные поражения гениталий мот быть обусловлены Haemop} ilus ducreyeyi
(шакроид), С. irachomatis серологических типов L1-L3 (венерическая лимфогра-
нулема, четвертая венерическая болезнь), Calymmatobacteiium grajuilomatis (доно-
ваноз, паховая гранулема).
Взятие ма гериала
• Очищают поверхность язвы с помощью стерильного тампона, смоченного
0,9% раствором хлорида натрия. Стерильной марлевой салфеткой обтирают
поверхность язвы до появления тканевой жидкости. Следует избегать появ-
ления крови!
• Первую порцию жидкости удаляют салфеткой, затем руками в перчатках
сдавливают язву у основания до появления прозрачной или слегка опалесци-
рующей тканевой жидкости.
• Тканевую жидкость аспирируют одноразовым шприцем с иглой, закрывают
иглу защитным колпачком и как можно быстрее направляют в лаборато-
рию.
Исследование материала
Для выявления Н. ducreyeyi тканевую жидкость засевают на шоколадный агар
или кровяной агар с овечьей кровью, а также готовят мазки, окрашенные по
Граму.
С. trachomatis серологических типов L1-L3 выявляют с помощью культурально-
го метода, ПЦР, РИФ, ИФА.
Для выявления Cdlymmatobacterium granulomatis необходимо исследовать окра-
шенные по Райту и Гимзе мазки грануляционной ткани или биоптаты.
Оценка результатов
В пользу шанкроида свидетельствует обнаружение в мазке грамотрицательных
палочек овоидной формы, расположенных параллельными цепочками («железно-
дорожные пути»), группами («стайки рыб») или парами, однако бактериоскопиче-
ский метод недостаточно чувствителен и, если имеется такая возможность, следует
пользоваться для постановки окончательного диагноза культуральным методом и
ПЦР.
При диагностике венерической лимфогранулемы культуральный метод считают
«золотым стандартом». Однако он трудоемок и требует специальной подготовки
исследователей, а также особых условий проведения работы. Поэтому в настоящее
время им практически не пользуются для рутинной диагностики инфекции С. tra-
chomatis, отдавая предпочтение другим диагностическим методам, особенно ПЦР.
Для выявления Са1уттатоЬастегшт granulomatis необходимо исследовать мазки
грануляционной ткани или биоптатов. В препаратах, окрашенных по Райту и
Гимзе, удается обнаружить биполярно окрашенные цитоплазматические включе-
ния — тельца Донована, что служит основанием для диагностики паховой грану-
лемы.
Материал из простаты
Показание к проведению исследования — установление этиологии хронических
простатитов.
Взятие материала
Непосредственное взятие проб из предстательной железы возможно лишь при
хирургической операции, что во многих случаях неприемлемо. Поэтому для диа-
гностики бактериальных простатитов и установления их этиологии, как правило,
собирают 1-ю и 2-ю порции мочи, с помощью массажа получают секрет предста-
тельной железы, а затем собирают 3-ю порцию мочи.
БАКТЕРИОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ 269
• Перед взятием материала для исследований пациент должен провести тща-
тельный туалет наружных половых органов с помощью теплой воды, мыла и
стерильных марлевых салфеток.
• Больному предлагают помочиться в стерильный контейнер. Собрав 10-20 мл
мочи, он должен продолжить мочеиспускание в емкость для сброса. Контейнер
помечают как пробу №1.
• Не опорожняя полностью мочевой пузырь, паииент собирает 10-20 мл мочи
в другой стерильный контейнер, который помечают как пробу №2.
• Врач проводит массаж простаты через задний проход и, если удается полу-
чить сок простаты, готовит из него мазок на стекле.
• Больной собирает первые 10 мл мочи в третий стерильный контейнер, кото-
рый помечают как пробу №3.
Исследование материала
Из полученного материала готовят мазки, которые исследуют под световым
микроскопом в нативном виде и после окраски. При обнаружении в пробах бак-
терий проводят посев материала на соответствующие питательные среды с после-
дующей идентификацией выделенных изолятов.
Оценка результатов
Первая проба мочи характеризует состояние микрофлоры уретры, вторая —
мочевого пузыря, третья — простаты.
На наличие у пациента хронического бактериального простатита указывают:
ъ обнаружение лейкоцитов в моче после массажа простаты в количестве более
10 в 1 мл и в соке простаты более 10 в поле зрения;
♦ увеличение количества бактерий в 3-й пробе мочи более чем в 10 раз по срав-
нению со 2-й пробой;
о обнаружение в 3-й пробе микроорганизмов, отсутствовавших в 1 -й и 2-й про-
бах.
Наиболее вероятная причина хронических бактериальных простатитов — гра-
мотрицательпые уропатогенные бактерии (Е. coll, Bsbsiella spp.. Pseudomonas spp.),
мгнее вероятная — грамположительные бактерии (E. faeailis, Staphylococcus spp.).
Широко дискутируется роль хламидий в этиологии простатитов. Также в каче-
стве возможных кандидатов на роль этиологических агентов в различных публи-
кациях упоминаются облигатно-анаэробные бактерии, уреаплазмы, пропионобак-
терии, хеликобактеры и даже лактобактерии.
Отделяемое женских половых органов
Показания к проведению исследования:
< • диагностика инфекций женских половых органов:
обследование женщин, у половых партнеров которых диа! ностирован уре-
трит.
Взятие материала
Минимальная схема обследования женщины должна включать бактериоскопи-
ческое исследование мазков проб, полученных из:
❖ уретры (диагностика ИППП);
❖ заднего свода влагалища (оценка состояния влагалищного биоценоза, диа-
гностика вагинозов и вагинитов);
< > цервикального канала (диагностика ИППП).
Если все три мазка направляют в лабораторию на одном стекле, они должны
быть четко промаркированы.
При необходимости, дополнительно отбирают пробы для культурального
исследования и ПЦР.
При воспалении бартолиниевых желез проводят их пункцию.
270 БАКТЕРИОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ
Из вульвы и влагалища материал забирают зондами-тампонами. В последнем
случае придерживаются следующих правил.
• Материал для анализа получают до проведения мануального влагалищного
исследования.
• Зеркало и подъемник вводят во влагалище и с помощью стерильной салфетки
убирают избыток выделений и слизи.
• Материал собирают с заднего свода или с патологически измененных участ-
ков влагалища двумя стерильными зондами-тампонами.
• Первый зонд-тампон возвращают в пробирку-тубсер или помещают в сте-
рильную пробирку и как можно быстрее доставляют в лабораторию для про-
ведения бактериологического исследования.
• Второй зонд-тампон используют для приготовления мазка. Мазок наносят
на 2 предметных стекла. Если одновременно планируется исследование мате-
риала из цервикального канала или уретры все 2-3 мазка можно сделать на
одном стекле (в лабораторию в этом случае доставляют 2 стекла, на каждом
из которых находятся мазки из всех обследуемых биотопов). Мазки марки-
руют, высушивают на воздухе и, поместив в чашки Петри или специальные
транспортные контейнеры, доставляют в лабораторию.
Из цервикального канала материал забирают следующим образом:
• Обнажают шейку матки с помощью зеркала (лучше стерильного одноразо-
вого), убирают избыток выделений и слизи стерильной марлевой салфеткой
или ватным шариком, смоченными стерильным 0,9% раствором хлорида
натрия или дистиллированной водой, и высушивают стерильной сухой мар-
левой салфеткой.
• Если предполагается бактериологическое исследование (на гонорею, хла-
мидиоз, микоплазмоз, уреаплазмоз и т.д.), пользуются тонким стерильным
зондом-тампоном (цервикальной щеточкой). Его аккуратно вводят в цер-
викальный канал на глубину 1,0-1,5 см, вращают в течение 10 с, не касаясь
стенок влагалища, и сразу же погружают в транспортную среду (среду Эймса
с углем).
При подозрении на трихомоноз материал должен быть доставлен в лабораторию в
кратчайшие сроки, а на гонорею — в течение 12 ч; при подозрении на эти инфекции
не допускается охлаждение проб. В остальных случаях при отсутствии возможно-
сти быстрой доставки проб в лабораторию допускается их хранение в холодильни-
ке (4-8 °C) 24-48 ч. Для проведения ПЦР материал помещают в микропробирку с
лизирующим буфером или 0,9% раствором хлорида натрия.
Материал из матки получают с помощью специального инструмента — шприца-
аспиратора. После прохождения зондом цервикального канала в полости матки рас-
крывают наружную оболочку зонда и аспирируют шприцем содержимое матки.
Взятие для исследования материала из придатков матки проводят во время опе-
ративного вмешательства.
Исследование материала
Схема исследования отделяемого женских половых органов должна соответ-
ствовать предполагаемой этиологии болезни.
Состояние влагалищной микрофлоры оценивают 3 методами:
❖ бактериоскопическим;
ф полуколичественным бактериологическим;
< > количественным бактериологическим.
В рутинной лабораторной практике обычно пользуются первыми двумя мето-
дами.
Оценка результатов
Основные критерии оценки состояния бактериальной микрофлоры получен-
ных результатов приведены в табл. 20-8.
БАКТЕРИОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ 271
Таблица ;*й-8. Микроскопическая характеристика биоценоза влагалища
Состояние биоценоза Описание признаков Нозоло! ические формы
Нормоценоз Преобладание лактобацилл, отсутствие грам отрицательных бактерий, спор, мицелия, псевдогифов, лейкоцитов, единичные «чистые» эпителиальные клетки Типичное состояние нормального биотопа влагалища
Промежу- точный тип Умеренное или незначительное количество лактобацилл, наличие грамположительных кокков, грамотрицательных палочек, обна- ружение лейкоцитов, макрофагов, эпителиальных клеток Часто наблюдается у здоровых женщин, редко сопровождается жалобами и клинически- ми проявлениями
Дисбиоз влагалища Незначительное количество или полное отсутствие лактобацилл, обильная, полиморфная грамотрицательная и грамположитель- ная палочковая и кокковая микрофлора, обнаружение ключевых клеток, количество лейкоцитов вариабельно, отсутствие или незавершенность фагоцитоза, полимикробная картина мазка Бактериальный вагиноз
Вагинит Большое количество лейкоцитов, макрофагов, эпителиальных клеток, выраженный фагоцитоз Неспецифический вагинит
Обнаружение N. gonorrhoeae Гонорея
Обнаружение Trichomonas vaginalis Трихомониаз
Обнаружение мицелия, псевдогифов, спор Микотический вагинит
Бактериальный вагиноз, или дисбиоз влагалища, — нарушение состояния вла-
галищной микрофлоры, характеризующееся избыточным развитием ряда микро-
организмов {Gardnerella vaginalis, Mobiluncus spp., Prevotella spp., Mycoplasma hominis)
и замещением ими лактобактерий.
Развитию бактериального вагиноза могут способствовать различные эндоген-
ные и экзогенные факторы:
< изменение гормонального статуса;
о- снижение иммунологической реактивности;
о предшествующая антибактериальная терапия;
❖ воспалительные заболевания мочеполового тракта.
Диагноз бактериального вагиноза ставят на основании совокупности клиниче-
ских и лабораторных данных (критерии Амсела):
о- обильные гомогенные выделения, прилипающие к стенкам влагалища:
❖ pH влагалищного секрета более 4,5;
❖ рыбный запах вагинальных выделений непосредственно или после добавле-
ния нескольких капель 10% гидроксида калия (аминный тест);
❖ обнаружение диагностически значимых клеток.
Для постановки диагноза «бактериальный вагиноз» достаточно наличия 3 из 4
указанных признаков.
Факт обнаружения в материале из влагалища Gardnerella vaginalis с помощью
бактериологического метода или ПЦР не является основанием для постановки
диагноза «гарднереллез», так как данный микроорганизм входит в состав нор-
мальной микрофлоры влагалища. Использование бактериологического метода
или ПЦР целесообразно только на этапе планирования беременности при наличии
выкидышей в анамнезе, так как некоторые метаболиты гарднерелл могут прово-
цировать бесплодие.
Истинный гонорейный вагинит развивается у беременных, женщин в период
менопаузы и до полового созревания (при щелочной реакции влагалищного секре-
та и при истончении эпителия влагалища). При этом обнаружение N. gonorrhoeae
при бактериоскопическом исследовании влагалищных мазков осложняется отсут-
272 БАКТЕРИОЛ01 ИЧЕСКИЕ ИССЛЕДОВАНИЯ
ствием ключевого признака — внутриклеточного расположения грамотрица-
тельных диплококков. Поэтому для подтверждения диагноза предпочтительно
использование ПЦР и бактериологического метода. У девочек (до наступления
менархе) и женщин (в менопаузе) диагноз гонореи устанавливают только на осно-
вании бактериологического исследования.
Трихомониазный вагинит может быть диагностирован с помощью микро-
скопического метода (предпочтительна микроскопия нативного неокрашенного
препарата), культурального метода и ПЦР. Наиболее высокой чувствительностью
обладает культуральный метод, однако его исследование затруднено при одновре-
менном присутствии во влагалищной слизи трихомонад и Candida spp.
Наиболее частая причина кандидозного вагинита — С. albicans, реже — С. glabrata,
еще реже — С. tropicalis. Прочие виды Candida spp. (С. crusei, С. parapsilosis) вызы-
вают вагинит очень редко. В подавляющем большинстве случаев для диагностики
кандидозного вагинита достаточно бактериоскопического исследования (микро-
скопия нативного неокрашенного препарата или мазка, окрашенного по Граму).
При этом следует помнить, что Candida spp. входит в состав нормальной микро-
флоры влагалища и обнаружение отдельных клеток при отсутствии клинической
симптоматики не является основанием для назначения этиотропного лечения.
К культуральному исследованию прибегают для подтверждения диагноза при
отрицательном результате микроскопии и наличии симптомов заболевания, а
также для видовой идентификации возбудителя при подозрении на нетипичную
этиологию кандидоза. Диагностически значимо обнаружение Candida spp. в титре
не менее 10 000 КОЕ/мл. В связи с этим информативность качественных модифи-
каций ПЦР сомнительна.
Пробы из органов слуха
Отделяемо? наружного слухового прохода, среднего и внутреннего уха
Показания к взятию и исследованию проб: воспалительные заболевания уха.
Взятие материала
Из наружного слухового прохода
• Обрабатывают кожу 70% этиловым спиртом и промывают стерильным 0,9%
раствором хлорида натрия.
• Тампоном, смоченным с серильным 0,9% раствором хлорида натрия, удаляют
соринки и корки из ушного канала.
• Материал для исследования берут зондом-тампоном, интенсивно (но осто-
рожно, чтобы не повредить барабанную перепонку!) вращая его в наружном
слуховом проходе.
• Зонд-тампон помещают в пробирку с транспортной средой (например, сре-
дой Эймса с углем), а при ее отсутствии — в пустую стерильную пробирку. В
последнем случае материал доставляют в лабораторию немедленно.
• При подозрении на отомикозы предпочтительнее взятие соскоба стерильным
аттиковым зондом или костной ложкой. Материал помещают между двумя
обезжиренными предметными стеклами.
• Если гнойное отделяемое имеет густую сливкообразную консистенцию, то к
нему добавляют несколько капель 30% гидроксида калия и оставляют под
покровным стеклом на 1-2 ч.
Из среднего и внутреннего уха
• Берут пробы (пунктаты и другой материал) во время операций. Пунктаты
доставляют в лабораторию в закрытом шприце с предварительно удаленным воз-
духом, а образцы ткани — в транспортной среде Кэри-Блейр или в стерильном
одноразовом контейнере с завинчивающейся крышкой.
БАКТЕРИОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ 273
Парацентез барабанной перепонки провидят для микробиологической диагно-
стики инфекций среднего уха только в случаях, когда лечение отита неэффективно
или катаральное воспаление среднего уха имеет торпидный характер.
• Если целостность барабанной перепонки не нарушена:
< очищают наружный канал с помощью тампона, смоченного 70% этиловым
спиртом, с последующей обработкой стерильным 0,9% раствором хлорида
натрия;
аспирируют шприцем жидкость из барабанной полости;
< переносят материал в пробирку с консервантом или доставляют материал
в шприце, удалив из него воздух и надев на иглу защитный колпачок.
• Если целостность барабанной перепонки нарушена, материал собирают с
помощью стеоильного ушного зеркала и зонда-тампона. Зонд-тампон со взя-
тым материалом помещают в транспортную среду или в пустую стерильную
пробирку. В последнем случае материал должен быть доставлен в лаборато-
рию немедленно.
Исследование материала
В зависимости от задач исследования выполняют микроскопию препаратов (при
увеличении в 100, 200 и 400 раз), окрашенных по Романовскому-Гимзе, Граму и
другими методами, посев на питательные среды (кровяной агар с овечьей кровью,
шоколадный агар, среду Сабуро, тиспликолевую среду и др.), идентификацию
выделенных изолятов бактерий и оценку их чувствительности к антимикробным
препаратам.
Оценка результатов
Воспаление наружного уха могут вызывать бактерии S. aureus, Str pyogenes,
Р. aeruginosa, энтеробактерии, Т. pallidum. М. tuberculosi, а также грибы родов
Aspergillus, Penicillium. Mucor, Candida spp.
Возбудители острого среднего отита чаще всего — Str. pneumoniae и Н. influenzae,
значительно реже — Moraxellacatarrhaiis, S. aureus. Str. pyogenes, иногда — Mycoplasma
pneumoniae, Chlamydia trachomatis и Chlamydophila pneumoniae.
При проведении дифференциально-диагностических исследований следует
учитывать, что воспаление наружного и среднего уха нередко вызывают вирусы.
При хроническом гнойном среднем отите выделяют Р. aeruginosa, грамотрица-
телъные палочки семейства Enterobacteriaceae, S. aureus.
У пациентов с хроническим гнойным средним отитом, протекающим с разви-
тием внутричерепных осложнений, важный этиологический фактор — анаэробная
микрофлора, преимущественно Bacteroides spp.
Пробы из глаз
Показания к взятию и исследованию проб: воспалительные заболевания глаз.
Взятие материала
За 6-8 ч до сбора материала отменяют проводимое лечение.
Отделяемое собирают стерильными зондом-тампоном или стеклянной палоч-
кой.
• Несколько раз проводят этими приспособлениями по слизистой оболочке
нижней переходной складки, края век.
• При язье собирают материал с роговицы после обезболивания,
• При «уголковом конъюнктивите» собирают материал с уголков век.
• При блефарите гнойные корочки снимают пинцетом и собирают отде-
ляемое из язвочек у основания ресниц, с внутренней поверхности нижнего
века (движением в направлении от наружного к внутреннему углу!" глазной
щели). Пациент должен придерживать веки руками, чтобы при моргании
ресницы не касались тампона. При скудном отделяемом тампон предва-
274 БАКТЕРИОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ
рительно смачивают стерильным физиологическим раствором. Избыток
влаги отжимают о внутреннюю поверхность емкости, после чего собирают
материал.
• Отделяемое из слезного мешка получают выдавливанием после осторожного
массажа.
При пресептальном целлюлите (флегмоне век) обрабатываю'1 кожу 70% этило-
вым спиртом и 1-2% раствором йода, йодоформом или другим дезинфектантом,
разрешенным к применению в установленном порядке. Затем проводят сбор мате-
риала:
О - при отсутствии открытой раны — после прокола или надреза скальпелем
верхнего или нижнего век;
❖ при наличии открытой раны — шприцем с иглой.
При эндофтальмите проводят пункцию глаза, объем отбираемого внутриглаз-
ного содержимого - 0,3-0,5 мл.
У пациентов, у которых воспаление роговицы развилось на фоне применения
контактных линз, целесообразно взять смывы с последних.
• Больной снимает контактные линзы, не прикасаясь руками к их внутренней
поверхности, и укладывает их на дно стерильной чашки Петри внутренней
поверхностью вверх.
* Смывы забирают с помощью стерильного тампона или зонда-тампона,
увлажненного 0,9% раствором хлорида натрия.
• Тампон/зонд-тампон помещают в пробирку с транспортной средой или
немедленно доставляют в лабораторию в стерильной пробирке.
Более корректную информацию о возбудителе воспалительного процесса полу-
чают при микробиологическом анализе соскобов, а не смывов.
• Перед получением соскоба закапывают в глаз 1-2 капли пропаракаина
гидрохлорида или другого анестетика.
• Несколькими резкими движениями в одном направлении с помощью специ-
ального стерильного шпателя делают соскоб с конъюнктивы или роговицы.
При проведении манипуляции глаз должен быть открыт. Надо внимательно
следить за тем, чтобы при сборе пробы не касаться ресниц.
При бактериальном эндофтальмите (гнойном воспалении сосудистой и сетча-
той оболочки глаз с формированием абсцесса в стекловидном теле) пробу сте-
кловидного тела (1-2 мл) собирают с помощью тонкоигольной аспирации или
получают при проведении витректомии (удалении патологически измененного
стекловидного тела). Предварительно берут соскоб с конъюнктивы для контроля
возможной контаминации образца сопутствующей микрофлорой. Пробу можно
транспортировать, поместив в транспортную среду или непосредственно в шприце,
удалив воздух и иглу, а затем закрыв шприц стерильной резиновой пробкой или
надев на иглу защитный колпачок.
При каналикулите (воспалении малых протоков, через которые происходит
отток слез с конъюнктивы в слезный мешок) прижимают внутреннюю часть века,
чтобы получить гной.
В месте взятия упомянутых выше материалов из них готовят мазки (не менее 2
из каждой пробы).
• Наносят материал на чистое обезжиренное предметное стекло круговыми
движениями на площадь диаметром 1 см.
• После подсыхания на воздухе мазки фиксируют в течение 5 мин в 95%
метиловом/эгиловом спирте или посредством нанесения 0,1 мл ацетона до
полного испарения. Стекло с фиксированным мазком можно хранить при
температуре -10 °C не более 7 сут.
• Окрашивают препараты по Романовскому-Гимзе.
БАКТЕРИОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ 275
Полученные любым из описанных выше методов пробы предпочтительнее
доставлять в лабораторию в пробирках с транспортной средой или, в крайнем слу-
чае, в пустых стерильных пробирках. Пробирки с материалом маркируют инфор-
мацией о том, из какого глаза он был взят. Параллельно из полученного материала
готовят мазки. Также материал может быть непосредственно после взятия засеян
на кровяной и шоколадный агар, тиогликолевую среду, агар Сабуро — для этого
лучше пользоваться средами в пробирках или чашках Петри промышленного про-
изводства. Засеянный материал вместе с приготовленными мазками передают для
дальнейших исследований в лабораторию.
Исследование материала
В лаборатории проводят бактериоскопию мазков (если они не были сделаны в
месте обследования пациента, то их готовят из доставленных проб), посев на пита-
тельные среды, идентификацию изолятов бактерий по морфологическим, тинкто-
риальным, культуральным, биохимическим свойствам (а в ряде случаев по анти-
генной структуре), оценку их чувствительности к антимикробным препаратам.
Оценка результатов
Результаты лабораторных анализов оценивают с учетом данных анамнеза, кли-
нических проявлений заболевания и видов выделенных микроорганизмов.
Определенное представление о вероятной этиологии конъюнктивита можно
составить на основании цитологического исследования соскобов с конъюнктивы,
окрашенных по Романовскому-Гимзе (табл. 20-9).
Таблица 20-9. Зависимость результатов цитологического исследования от этиологии конъюн-
ктивита
Клетки или включения Природа конъюнктивита
аллергический бактериальный вирусный хламидийный
Нейтрофилы - + + +
Эозинофилы + - - -/+
Лимфоциты + - + +
Плазматические клетки - - - +
Включения в цитоплазме - - - 4-
Включения в ядре - - + (герпес) -
Поражение конъюнктивы может быть обусловлено бактериями, вирусами, гри-
бами или простейшими.
Наиболее частая причина бактериальных конъюнктивитов — Staphylococcus spp.
(при остром конъюнктивите чаще всего обнаруживается S. aureus, при хрониче-
ском — S. epidermidis). S. aureus поражает и другие отделы глаза. У новорожденных
один штамм стафилококков часто вызывает одновременно заболевания глаз,
пупка, полости рта и носа.
N. gonorrhoeae — причина конъюнктивита у детей всех возрастных групп, а нс
только новорожденных. Среди коринебактерий наиболее тяжелые формы конъ-
юнктивитов, при которых в патологический процесс может вовлекаться роговица,
вызывают С. diphtheriae.
Энтеробактерии (Klebsiella spp., Proteus spp., E. coif), а также P. aeruginosa — воз-
можная причина конъюнктивитов как у взрослых, так и у новорожденных.
У взрослых конъюнктивиты данной этиологии развиваются, как правило, после
химического или механического повреждения конъюнктивы, а у новорожден-
ных — при инфицировании во время родов.
Moraxella lacunata вызывает острый эпидемический конъюнктивит Коха-Уикса.
При диагностике этого заболевания вследствие сложности культивирования
моракселл можно ограничиться обнаружением грамотрицательных диплобакте-
рий в мазках.
276 БАКТЕРИОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ
На фоне длительной антибиотикотерапии может наблюдаться колонизация
конъюнктивы Candida spp. и Aspergillus. Необходимо учитывать возможность
профессионального носительства Candida spp., например у пекарей и молочни-
ков. Причиной заболеваний глаз также мот быть и другие грибы — Nocardia,
Penicillium, Mucor.
Распространенность хламидийных конъюнктивитов среди отдельных групп
населения может достигать 30%. Чувствительность микроскопического метода
диагностики хламидийного конъюнктивита невелика. Это заболевание вызывают
серовары, Е, F, G, Н. 1, Y, К Chlamydia trachomatis, т.е. серовары, которые вызыва-
ют урогенитальный хламидиоз. В связи с этим для диагностики хламидийного
конъюнктивита могут быть использованы те же методы, что и для диагностики
урогенитального хламидиоза, в первую очередь ПЦР. Помимо соскобов с конъюн-
ктивы, целесообразно также исследовать материал из половых органов.
Чаще всего развитие бактериальных кератитов обусловлено Staphylococcus spp..
Streptococcus spp.. Pseudomonas spp. При этом наиболее тяжелые поражения с образо-
ванием язв вызывает Р. aeruginosa. Реже причиной кератитов становятся грамотрица-
тельные микроорганизмы родов Escherichia, Neisseria, Proteus, Moraxclla. Вялотекущие
хронические эндофтальмиты могут быть обусловлены Propionobacterium acnes, а
эндофтальмиты у наркоманов, внутривенно вводящих героин, — Bacillus ccreus.
В спорадических случаях кератиты вызывают Nocardia spp.
Возбудители эндофтальмита — Staphylococcus spp.. Streptococcus spp., Pseudo-
monas spp. и другие грамотрицательные палочки. Причиной посттравматических и
эндогенных эндофтальмитов также могут быть Clostridium spp. и Bacillus spp.
Иридоциклиты в настоящее время чаще всего связаны с герпетической инфек-
цией (вирус простого и опоясывающего герпеса), но также могут быть обусловле-
ны Т. pallidum.
Развитие дакриоцититов чаще всего обусловлено Str. pneumoniae, S. aureus и
EL influenzae.
Воспаление протоков слезных желез может быть обусловлено анаэробными
микроорганизмами - Bacteroides spp., Fusobacterium spp., Peptostreptococcus spp. или
Actinomyces spp.
Заболевания глаз, связанные с ношением контактных линз, могут быть вызваны
Bacillus spp., Serratia spp., Acanthamoeba spp.
При пресептальном целлюлите этиологическое значение имеют S. aureus,
S. pyogenes, Н. influenzae, у детей в возрасте от 6 мес до 1,5 лет — Н. influenzae, при
травмах — Clostridium spp.
При орбитальном целлюлите (системная инфекция глубоких тканей глазницы)
аспират из зоны поражения получают с помощью шприца и исследуют, как описа-
но выше. Параллельно целесообразно провести исследование крови на гемокуль-
туру.
При послехирургических инфекциях чаще выделяют 5. aureus, при инфекциях
вследствие панофтальмита — S. aureus, Str. pneumoniae, Р. aeruginosa. при проник-
новении инфекции из околоносовых пазух — Н. influenzae, S. aureus. Str. pyogenes,
Str. pneumoniae.
При дакриоадените (воспалении слезной железы) берут мазок с конъюнктивы
без пунктирования слезной железы. Основные возбудители дакриоаденита —
S. aureus, Str. pyogenes, Str. pneumoniae.
При дакриоцистите (воспалении слезного мешка) материал собирают тампоном
с конъюнктивы, после чего для получения экссудата сдавливают слезный мешок и
собирают материал тампоном или шприцем. Материал помещают в транспортную
среду или сразу- засевают на кровяной и шоколадный агар и в тиогликолевую среду,
а также используют для приготовления мазков. Наиболее частая причина дакрио-
цистита — S. aureus, Str. pyogenes. Str. pneumoniae, H. influenzae.
БАКТЕРИОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ 277
Основные возбудители каналикулита — Actinomyces israelii, Propionobacterium
propionicus, Moraxella spp,, дифтероиды и зеленящие стрептококки.
Фекалии
Показания к взятию и исследованию проб:
❖ острые и хронические кишечные инфекции любой этиологии;
❖ дисбактериоз.
Взятие исследуемого материала
Сбор фекалий после дефекации
• Судно тщательно моют для удаления следов дезинфектантов,
• На его дно помещают лист чистой плотной бумаги.
• Больного просят совершить акт дефекации, предварительно предупредив о
нежелательности попадания в фекалии мочи.
• Пробу фекалий отбирают сразу после акта дефекации. При наличии патоло-
гических примесей необходимо выбрать участки, содержащие слизь, гной,
хлопья, но без следов крови. Образцы жидких фекалий отбирают с помощью
стеклянной трубки с резиновой грушей.
• Полущенный материал помещают:
❖ в пустой стерильный контейнер и доставляют в лабораторию в течение 2 ч:
- объем забираемого материала должен составлять нс менее 2 г;
- при оформленном стуле оптимально взятие материала в объеме грец-
кого ореха, при жидком стуле его слой в посуде должен быть не менее
2 см;
> в консервант (пробы для диагностики острых кишечных инфекций, вызы-
ваемых условно-патогенными бактериями, и дисбактериоза забирают и
транспортируют без консерванта в транспортной среде) (табл. 20-10):
- объем забираемого материала не должен превышать 1/3 объема консер-
ванта;
- фекалии должны быть тщательно гомогенизированы в консерванте с
помощью проволочной петли или стеклянной палочки;
- образцы могут храниться в холодильнике в течение 1 сут.
Наиболее подходящей емкостью для сбора фекалий является стерильный одно-
разовый контейнер с широким горлом и завинчивающейся крышкой, содержащий
пластиковую ложечку-шпатель, вмонтированную в крышку контейнера.
Если фекалии жидкие (например, профузная диарея), их собирают:
4- стерильной одноразовой серологической или пастеровской пипеткой с широ-
ким носиком и пипетора:
< стерильным катетером со стерильным стеклянным наконечником с одной
стороны и грушей, предварительно простерилизованной или обработанной
70% этиловым спиртом и тщательно ополоснутой стерильным физиологиче-
ским раствором, - с другой.
Жидкими фекалиями контейнер заполняют не более чем на 1/3 объема для
предохранения от разбрызгивания материала при вскрытии емкости в лаборато-
рии. Бели фекалии оформленные, плотные, то в контейнер помещают 3-4 ложечки
(1,5-2 г). Материал, собранный с несвежего белья, анализу не подлежит.
В случае отсутствия в лаборатории одноразовой посуды порцию фекалий поме-
щают в специально подготовленную стерильную емкость.
278 БАКТЕРИОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ
Таблица 20-10. Консерванты (транспортные среды), наиболее часто используемые для транс-
портировки фекалий
Название консерванта Для каких бактерий применяют
Глицериновая смесь Энтеробактерии, кроме иерсиний; предпочтительна для транспор- тировки шигелл
Боратно-буферный раствор Энтеробактерии
Фосфатная буферная смесь Энтеробактерии
0,9% раствор хлорида натрия Энтеробактерии, кампилобактерии
0,1% пептонная вода Кампилобактерии
Тиогликолиевая среда (среда СКС) Кампилобактерии
Среда Кэри-Блеер Энтеробактерии
При вынужденном удлинении сроков доставки материала в лабораторию
жидкими фекалиями пропитывают полоску промокательной бумаги или другого
гигроскопичного материала и герметично упаковывают в пластиковый пакет для
предотвращения высыхания. На таких полосках холерные вибрионы способны
сохраняться в течение 4-5 нед или более.
Взятие ректальных мазков
• Ректальные мазки берут одноразовыми зондами-тампонами на металличе-
ском или пластиковом стержне, вмонтированными в сухие стерильные про-
бирки или пробирки с транспортной средой. Использование многоразовых
петель из алюминия, стали, титана или других нержавеющих материалов,
выдерживающих стерилизацию, допускается только в экстремальных усло-
виях (реанимационные больные, маленькие дети).
• Больного просят лечь на бок с приведенными к животу ногами и ладонями
развести ягодицы.
• Тампон вводят в задний проход на глубину 4-5 см и, аккуратно вращая его,
собирают материал с крипт ануса.
• Осторожно извлекают тампон и погружают его в консервант или транспорт-
ную среду. Транспортировка тампона без этих сред не допускается.
Для забора материала у больных с обильным водянистым стулом пользуются
резиновым катетером, один конец которого вводят в прямую кишку, а другой опу-
скают в банку. Жидкие фекалии стекают в сосуд свободно или при легком надавли-
вании на переднюю брюшную стенку.
При обследовании беременных женщин на носительство стрептококка группы В
(Str. agalactiae) материал берут из влагалища и прямей кишки двумя разными
зондами-тампонами. Затем оба тампона могут быть погружены в 1 пробирку с
транспортной средой типа Эймса или Стюарта.
Взятый материал переносят во флакон или пробирку с 1% пептонной водой.
В сопроводительном документе указывают, для каких исследований направлен
материал в лабораторию. Если цель исследований не указана или указана не кон-
кретно, в лаборатории будет выполнена совокупность исследований, позволяющая
выделить и идентифицировать только шигеллы, сальмонеллы, энтеропатогенные,
энтеротоксигенные и энтероинвазивные эшерихии.
Для исследования на дисбактериоз материал доставляют в лабораторию в тече-
ние 4 ч при условии забора его в большом объеме и транспортировки при низкой
температуре.
Исследование материала
В лаборатории проводят бактериоскопию мазков (если они не были сделаны в
месте обследования пациента, то их готовят из доставленных проб), посев на пита-
тельные среды, идентификацию изолятов бактерий по морфологическим, тинкто-
риальным, культуральным, биохимическим свойствам (а в ряде случаев по анти-
генной структуре), оценку их чувствительности к антимикробным препаратам.
БАКТЕРИОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ 279
Оценка полученных результатов
Обнаружение в фекалиях даже патогенных микроорганизмов не всегда свиде-
тельствует о наличии заболевания и может быть обусловлено бактерионоситель-
ством. Поэтому оценка результатов должна проводиться с учетом клинической
картины. Для установления наличия острой кишечной инфекции, обусловленной
условно-патогенными микроорганизмами, необходимо учитывать следующие
критерии:
< > отсутствие специфических признаков заболевания, обусловленного патоген-
ными микроорганизмами;
о - обнаружение условно-патогенного микроорганизма в фекалиях в титре выше
Iflff КОЕ/г;
❖ повторное обнаружение того же микроорганизма в возрастающей концентра-
ции;
< исчезновение условно-патогенного микроорганизма из фекалий или сниже-
ние его титра на фоне проводимого лечения;
о нарастание титра антител к данному условно-патогенному микроорганизму
более чем в 4 раза при одновременном присутствии его в фекалиях в концен-
трации выше 105 КОЕ/г.
Вопрос о диагностической ценности исследования микрофлоры толстой кишки
в настоящее время широко дискутируется, а существующие руководства по диагно-
стике дисбактериоза кишечника нередко содержат противоречивую информацию,
особенно в вопросах сбора и транспортировки материала, техники исследования
и трактовки результатов. Очевидно, что и результаты исследований, полученные
разными лабораториями, могут различаться. Существует множество причин
расхождения результатов, одна из которых — невозможность соблюдения регла-
ментированных сроков доставки в условиях практического здравоохранения.
Считается, что исследование должно начинаться не позднее 2 ч с момента забора
материала. Нормирование времени доставки материала обусловлено тем, что
вскоре после акта дефекации состав микрофлоры в силу нестандартных условий
может изменяться. При интерпретации результатов, полущенных при анализе дли-
тельно сохранявшихся проб, необходимо учитывать возможность размножения
условно-патогенных микроорганизмов в образце.
Для адекватной трактовки цифр, приводимых в ответе, необходимо иметь
представление о точности данного метода. Ответ, выдаваемый лечащему врачу,
не содержит информации о погрешности измерений, что может послужить при-
чиной как гипердиагностики дисбиотических состояний, так и неправильной
оценки динамики изменений состояния биоценоза кишечника в ходе лечения.
Титрационный посев в полужидкие среды, используемый большинством лабора-
торий для определения количества бифидобактерий (а часто и лактобактерий) в
1 г фекалий, характеризуется невысокой точностью.
При использовании традиционного однорядного метода для высева каждого раз-
ведения доверительный интервал рассчитывается с помощью умножения (верхняя
граница) и деления (нижняя граница) наиболее вероятного числа на коэффициент
14,45. Например, если наиболее вероятное число составляет 10ч КОЕ/г, истинное
количество бифидобактерий находится в интервале от 6.9х107до 1,4х1010 КОЕ/г.
Повысить точность метода можно с помощью увеличения числа рядов титрова-
ния, что применяется в санитарной микробиологии. При этом число рядов титро-
вания достигает 10, а расчет наиболее вероятного числа осуществляют по таблице
Мак-Креди, разработанной ня основании методов вариационной статистики. При
использовании трехрядного метода значение упомянутого коэффициента снижа-
ется с 14,45 до 4,68. На практике это означает, что исследование, проведенное в
разных лабораториях, или повторное исследование в одной и той же лаборатории
может привести к расхождениям в количестве бифидобактерий или лактобакте-
280 БАКТЕРИОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ
рий, которое могут трактовать как признак динамики патологического процесса.
Для предотвращения подобных ситуаций необходимо иметь представление о точ-
ности используемого метода, и при наличии расхождений в 1 или даже 2 порядка
рассматривать их как несущественные. При необходимости максимально точного
определения количества бифидо- и лактобактерий можно рекомендовать исполь-
зование трехрядного метода, что особенно актуально при контроле эффективно-
сти проводимого лечения. Однако трехрядный метод не может рассматриваться
как рутинный вследствие значительного повышения стоимости исследования и
существенного увеличения трудозатрат. Именно поэтому только согласованная
работа лечащих врачей и врачей-микробиологов позволяет скорректировать так-
тику исследования и выбор методик в каждом конкретном случае. Сотрудничество
специалистов на различных уровнях лечебного процесса — один из залогов адек-
ватной трактовки получаемых результатов.
Если лаборатория работает только с плотными питательными средами и
анаэробной техникой, необходимо принимать во внимание возможное снижение
титров анаэробных микроорганизмов в силу особенностей использования плотных
питательных сред. Однако ошибка метода в данном случае также уменьшается.
В фекалиях здоровых людей идентифицируют, по меньшей мере, 15 групп
микроорганизмов — бифидобактерий, лактобактерий, бактероидов, энтерококков,
фузобактерий, эубактерий, пептострептококков, клостридий, типичных Е. coli.
лактозонегативных Е. coli, гемолитических Е. coli, других условно-патогенных
энтеробактерий (Klebsiella spp., Enterobacter spp., Hafnia spp., Serratia spp., Proteus spp.,
Morgan ella spp., Providencia spp., Citrobacter spp.), S. aureus, сапрофитного и эпидер-
мального стафилококков, Candida ьрр., неферментирующих бактерий (Pseudomonas
spp., Acinetobacter spp.). Приходится констатировать слабость позиций традицион-
ных представлений о микрофлоре кишечника вследствие неполной информации
о микробиоценозе. Очевидно, что 15 групп микроорганизмов недостаточно для
описания всего многообразия микрофлоры кишечника. Например, количество
метанобразующих архебактерий из рода Methanobrcvibacter в фекалиях достига-
ет 109 КОЕ/г, что сопоставимо с численностью бифидобактерий и бактероидов.
Нельзя исключить присутствия в составе кишечной микрофлоры и некультиви-
руемых in vitro видов бактерий. Дать исчерпывающую характеристику состава
нормальной микрофлоры кишечника невозможно даже в научных, а тем более
в практических лабораториях. Можно надеяться, что по мере расширения круга
наушных знаний станет возможно установить конкретные клинические показа-
ния для определения отдельных групп бактерий. Например, при обследовании
больных с дисбиозами на фоне антибиотикотерапип обязательно выявление ток-
синов Cl. difficile. Однако вряд ли имеется необходимость бесконечно расширять
круг определяемых микроорганизмов, поскольку исследование на дисбактериоз
носит прикладной, а не описательный характер. По-видимому, следует уделять
внимание только тем группам бактерий, количество которых в той или иной сте-
пени влияет на тактику лечения. Такими показательными микроорганизмами для
взрослых являются гемолитические бактерии, условно-патогенные энтеробакте-
рии, Staphylococcus spp. и Candida spp. У детей в возрасте до 1 года на первый план
выходят Staphylococcus spp. и условно-патогенные энтеробактерии.
В существующих руководствах часто избыточной детализации подвергает-
ся описание Е. coli Рекомендуется определять общее количество эшерихий,
количество эшерихий с выраженной и со сниженной ферментативной активно-
стью, количество лактозонегативных, гемолитических, неподвижных эшерихий.
Однако, учитывая тот факт, что получаемые результаты статистически недосто-
верны. а изучение данных свойств весьма трудоемко, целесообразно объединение
всех критериев, описывающих атипичные свойства Е. coli, в один — «кишечные
палочки со сниженной ферментативной активностью» или «кишечные палочки с
БАКТЕРИОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ 281
измененными свойствами». Гемолитические Е. coli при этом выносят в о гдельную
графу - «гемолитические микроорганизмы»
Учитывая сомнительность разделения бактерий родов Lactococcus и Lactobacillus
только на основании морфологии и трудности идентификации Lactobacillus spp.
из-за слабой выраженности многих тестовых реакпий и зависимости их от соста-
ва питательной среды и условий культивирования, целесообразно объединение
критериев «молочнокислые палочки» и «молочнокислый стрептококк» в один —
«лактобактерии».
Подобные разночтения весьма несущественны, однако их следует принимать во
внимание, трактуя результаты, полученные при исследовании на дисбактериоз, в
разных лабораториях.
Рвотные массы и промывные воды
Показания к взятию и исследованию проб; подозрение на пищевые отравления,
холеру, сальмонеллез.
Взятие исследуемого материала
• Рвотные массы отбирают в количестве 50-100 мл (если это невозможно, то в
меньшем количестве), промывные воды — в объеме 100 -200 мл.
• Материал должен быть доставлен в лабораторию в кратчайшие сроки.
• После рвоты больному дают прополоскать рот теплой водой, тяжелым или
ослабленным больным очишают полость рта ватным тампоном, смоченным
водой или слабым раствором гидрокарбоната натрия или перманганата
калия.
Исследование материала
Проводят теми же методами, что и фекалий.
Оценка диагностической значимости полученных результатов
При расследовании пищевых отравлений параллельно с исследованием рвот-
ных масс исследуют остатки пищевых продуктов. Окончательное заключение
делают на основании сопоставления полученных результатов.
Аутопсированный материал
Показания к взятию и исследованию проб; исключение бактериальных инфек-
ций как возможных причин смерти.
Взятие исследуемого материала
Материал для бактериологического исследования необходимо забирать в тече-
ние 12 ч после смеоти больного, достоверность исследований материала, полу-
ченного позже 24 ч. резко снижается. Для взятия проб целесообразно привлекать
врачей-бактериологов. Выбор исследуемого материала зависит от предполагаемо-
го диагноза (табл. 20-11).
Таблица Z0-11. Объекты для исследования при подозрении на смерть от бактериальных заболе-
ваний
Заболевание Объект исследования
Ангина Плаун-Венсана Пленки и отделяемое с пораженных участков слизистых оболочек (зева носа, половых органов, глаз) и кожи, кровь
Бруцеллез Кровь, моча, внутренние органы (печень, селезенка), мокрота, гной, экссудат из пораженных тканей и органов, молочные железы (у женщин)
Брюшной тиф Паратифы Кровь из сердца, желчь, внутренние органы (печень, селезенка, легкие, почки, головной мозг), мезентериальные лимфатические узлы, содержимое толстой и тонкой кишки
Возвратный тиф Кровь, внутренние органы (селезенка, печень, головной мозг), СМЖ, моча
282 БАКТЕРИОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ
Продолжение табл. 20-11
Заболевание Объект исследования
Газовая гангрена Кусочки ткани из области ран и пораженных тканей и органов, перитонеальная жидкость, секрет шейки матки, кровь, внутренние органы, а также инородные тела в случае их обнаружения
Гемофильная инфекция Слизь носоглотки, мокрота, кровь, гной, моча, СМЖ, внутренние органы (лег- кие), плевральный экссудат
Гнойно-септические инфекции (инфек- ции, вызванные Staphylococcus spp., Streptococcus spp., P. aeruginosa) Гной, экссудат, кровь, участки пораженных тканей и органов, соскобы со слизи- стой оболочки зева, региональные лимфатические узлы
Гонококковая инфекция Отделяемое уретры, внутренние органы (прямая кишка), экссудат суставов и брюшины Внутренние органы (предстательная железа) (у мужчин), внутренние органы (влагалище, шейка матки, придатки матки) (у женщин)
Дизентерия Содержимое нижних отделов толстой кишки, кровь, моча, внутренние, мезенте- риальные лимфатические узлы
Дифтерия Пленки и отделяемое с пораженных участков слизистых оболочек (зева, носа, половых органов, глаз), кожи, кровь
Клебсиеллезная инфек- ция Слизь носоглотки, мокрота, кровь, гной, моча СМЖ, внутренние органы (лег- кие), плевральный экссудат
Коклюш Слизь носоглотки, внутренние органы (легкие)
Лептоспироз Кровь, внутренние органы (селезенка, печень, головной мозг), СМЖ, моча
Листериоз Кровь, внутренние органы (селезенка, печень мозг)
Менингококковая инфекция СМЖ. мозговые оболочки, кровь, слизь из зева, тканевая жидкость, гной
Орнитоз Внутренние органы (кусочки легких из пневмонических очагов, селезенка), плев- ральный экссудат
Пневмококковая инфекция Слизь носоглотки, мокрота, кровь, гной, моча, СМЖ, внутренние органы (лег- кие), плевральный экссудат
Сап Кусочки кожи и слизистой оболочки в изъязвленных участках, лимфатические узлы, кусочки мышечной ткани в области абсцессов, внутренние ор< аны. кровь
Сибирская язва Пораженные участки кожи и других тканей, лимфатические узлы кровь, СМЖ, костный мозг, экссудат, внутренние органы
Сифилис Отделяемое слизистых оболочек, кусочки тканей в области поражения, кровь, СМЖ, внутренние органы
Столбняк Гной, кусочки ткани из пораженных областей, старых рубцов, кровь, внутренние органы (печень, селезенка), инородные тела в виде кусочков орудия травмы
Сыпной тиф Кровь, кусочки кожи с элементами сыпи, внутренние органы
Пищевые токсикоин- фекции, вызванные Е. coll Содержимое тонкой кишки, кровь из сердца, внутренние органы {печень, селе- зенка, легкие, почки), мезентериальные лимфатические узлы
Пищевые токсикоин- фекции, вызванные Salmonella spp. Кровь из сердца, внутренние органы (печень, селезенка, легкие, почки), содер- жимое желудка и тонкой кишки, мезентериальные лимфатические узлы
Пищевые токсикоин фекции, вызванные Proteus spp. Содержимое тонкой и толстой кишки
Пищевые токсикоин- фекции, вызванные Staphylococcus spp. Содержимое тонкой и толстой кишки
Пищевые токсикоин- фекции, вызванные Cl. perfringens Содержимое тонкой кишки (200-300 мл) перитонеальная жидкость, внутренние органы (печень), мезентериальные лимфатические узлы, кровь из сердца
Пишевые токсикоин- фекции, вызванные Cl. botulinum Кровь, внутренние органы (печень, отрезки тонкой кишки, желудок с содержи- мым, головной мозг)
БАКТЕРИОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ 283
Окончание табл. 20-11
Заболевание Объект исследования
Туберкулез Мокрота, мазки из гортани, кусочки легких и других внутренних органов (в зави- симости от характера поражения), моча, СМЖ
Туляремия Слизь зева, мокрота, кровь, кусочки пораженной кожи, СМЖ, лимфатические узлы (бубоны), костный мозг, внутренние органы (селезенка, печень, легкие)
Холера Внутренние органы (3 отрезка тонкой кишки, желчный пузырь, печень, селезен- ка), лимфатические узлы
Трупную кровь берут из сердца до извлечения головного мозга. Пробы крови
получают из желудочка сердца. Выбранный участок дезинфицируют, протирая
стерильной салфеткой, смоченной 3% раствором перекиси водорода. Антисептик
удаляют стерильной салфеткой, смоченной 0,9% раствором хлорида натрия, или
с помощью прижигания ткани шпателем. Затем стерильным шприцем аспирируют
не менее 10 мл крови, которую немедленно засевают на питательные среды, как
было описано выше. Если в полости сердца кровь свернулась или отсутствует, ее
берут с соблюдением тех же условий из полой, бедренной или яремной вены.
Изъятие проб внутренних органов проводят после прижигания их поверхности
раскаленным шпателем. Кусочки легких, печени, головного мозга и других органов
вырезают размером 1x1x2 см (3-5 см3). Кусочки головного мозга следует вырезать
до извлечения его из полости черепа. Кусочки легких вырезают из прикорневой
области и из середины каждой доли (не следует брать кусочки из гипостатических
участков). Кусочки печени вырезают из каждой доли. Желчный пузырь изымают
целиком с содержимым после наложения лигатур. Для исследования содержимого
кишечника берут 2-3 отрезка тонкой кишки длиной 15-20 см между предвари-
тельно наложенными двойными лигатурами. Гной из вскрытых полостей, СМЖ
и другие жидкости отбирают и транспортируют, как описано в соответствующих
разделах.
Исследование материала
Из взятых проб в первую очередь изготавливают мазки и мазки-отпечатки.
Их высушивают на воздухе. Мазки фиксируют в пламени спиртовки, если пред-
полагаемый метод исследования не предполагает другого. При необходимости
бактериоскопический анализ дополняют посевом проб на питательные среды с
последующей изоляцией и идентификацией бактерий.
ПРИНЦИПЫ ИДЕНТИФИКАЦИИ БАКТЕРИЙ
Идентификация выделенной чистой культуры бактерий в лабораториях, как
правило, предполагает ее видовую идентификацию. Для решения эпидемио-
логических вопросов необходимо проведение внутривидового типирования. В
настоящее время для этого обычно используют серотипирование, фаготипирова-
ние, хемотипирование и генотипирование. Однако в некоторых случаях вполне
достаточно определения рода микроорганизмов или их принадлежности к опреде-
ленной группе. Для доказательства этиологической роли выделенного микроорга-
низма в отдельных случаях также требуется постановка дополнительных тестов.
Например, при обнаружении С. diphtheriae, необходимо дополнительно опреде-
лить токсигенность выделенного штамма.
Основное условие правильной идентификации бактерий, независимо от исполь-
зуемого метода. — наличие чистой культуры. Именно поэтому отколу колонии и
последующей макро- и микроскопической проверке чистоты культуры необходи-
мо уделять особое внимание. Следует избегать откола сразу нескольких колоний
даже при их кажущейся однотипности. Рекомендуется использовать одноразовую
стерильную бактериологическую петлю с маленькой головкой. Если пользуются
многоразовыми металлическими бактериологическими петлями или иглами, то их
284 БАКТЕРИОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ
рекомендуется охлаждать после прожигания в пламени прикосновением к пита-
тельной среде в зоне, где нет признаков роста.
В отдельных случаях даже откол отдельно лежащей колонии не гарантирует
получения чистой культуры. Например, колонии слизеобразующих бактерий
могут содержать клетки других микроорганизмов, захваченные вместе со сли-
зью. Культуры актиномицетов и представителей Bacillus spp. могут содержать
контаминанты в пространствах между цепочками клеток. При раннем отборе
колоний с селективной питательной среды загрязнение может быть обусловле-
но захватом контаминантов, находящихся в подавленном, но жизнеспособном
состоянии. Даже на несслективных питательных средах откол колонии не сле-
дует проводить слишком рано из-за возможного наличия медленно растущих
микроорганизмов.
Работа по идентификации бактерий начинается до выделения чистой культу-
ры с изучения культуральных свойств — характера роста микроорганизмов на
питательной среде. Культуральные свойства бактерий, выращенных на плотной
питательной среде, включают форму и размер колоний, наличие пигмента, спо-
собность к росту при определенной температуре и концентрации углекислого газа
и кислорода на среде определенного состава. Помимо культуральных свойств, до
выделения чистой культуры иногда определяют морфологические и тинкториаль-
ные свойства (например, с помощью микроскопии мазка из колонии, окрашенного
по Граму), некоторые биохимические свойства (например, наличие оксидазы и
каталазы), антигенные свойства (например, с помощью РА или РИФ) и некото-
рые другие признаки. В отдельных случаях этого бывает достаточно для выдачи
предварительного ответа о виде обнаруженного микроорганизма, но чаще всего
изучение указанных признаков позволяет только наметить путь дальнейших
исследований, которые выполняют с чистой культурой.
Решающее значение при идентификации большинства видов бактерий имеет
изучение их ферментативной активности. В классическом варианте с этой целью
пользуются набором дифференциально-диагностических сред. Однако в послед-
ние годы все чаще для идентификации бактерий применяют коммерческие тест-
системы, обеспечивающие высокую точность исследований и сокращение времени
на получение результата.
Существует несколько подходов к идентификации бактерий. Традиционный,
сложившийся еще в начале века способ предусматривал постепенное приближение
к ответу? по ветвям дихотомического информационного дерева.
При таком подходе применяется ограниченное количество пробирочных
(чашечных) тестов. Одними и теми же тестами можно пользоваз ься неоднократно
на разных ветвях информационного дерева. Очень важным при таком подходе
оказывается ранжирование тестов по их значимости и выбор ключевых тестов.
Это связано со следующими обстоятельствами:
о изучаемый признак должен встречаться (или отсутствовать) у всех штаммов
данного вида, т.е. в 100% случаев, в противном случае атипичные по данному
признаку штаммы никогда не будут правильно идентифицированы;
воспроизводимость всех тестов должна приближаться к 100%.
Вторая система идентификации, построенная на принципах нумерической
(числовой) таксономии, получила широкое распространение в связи с развити-
ем вычислительной техники, а также разработки коммерческих тест-систем, что
позволило реализовать эту систему на уровне диагностических лабораторий.
Нумерическая таксономия базируется на следующих принципах:
❖ необходимо изучить максимально возможное количество признаков (в совре-
менных коммерческих тест-системах их число может достигать 25-30);
< все изучаемые признаки имеют равное диагностическое значение;
БАКТЕРИОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ 285
результат идентификации выглядит как коэффициент соответствия свойств
выделенной культуры свойствам эталонных штаммов.
Характеристика некоторых тест-систем для биохимической идентификации
бактерий представлена в табл. 20-12.
Таблица 20-12. Системы биохимической идентификации бактерий
Идентифицируемые микроорганизмы Тест-система Количество тестов Время инкуба- ции, ч Фирма-производитель
1 2 з 4 5
Энтеробактерии ПБДЭ 20 18-24 НПО «Диагностические систе- мы», Нижний Новгород
ММТ Е-1, ММТ Е-2 24 (12+12) 24 НПО «Аллерген», Ставрополь
Enterotube II 15 18-24 Becton Dickinson, США
Micro-ID 15 4 Organon Teknika, США
API Rapid 20E 21 4 BioMerieux. Франция
ЭНТЕРО-тест 1 и 2 24 18-24 Lachema, Чехия
Энтеробактерии и НГОБ’ API 20Е 21 24—48 BioMerieux, Франция
GN-IDENT 29 4-13 BioMerieux, Франция
Crysta1 E/NF 30 18-20 Becton Dickinson, США
НГОБ' НЕФЕРМ-тесг 12 48 Lachema, Чехия
API NFT 20 24-48 BioMerieux, Франция
RapID NF Plus 17 4 Innovative Diagnostic Systems, США
Грамотрицательные бактерии Crystal Rapid Stool/ Enteric 30 18-20 Becton Dickinson, США
Oxi/Ferm 9 24-48 Becton Dickinson, США
NEG ID Type 2 34 15-42 Baxter Diagnostics, США
Rapid NEG ID 2 36 2 Baxter Diagnostics, США
Анаэробы АНАЭРО-тест 12 48 Lachema, Чехия
AN-IDENT 28 4“ BioMerieux, Франция
API 20A 21 24 BioMerieux, Франция
RapID ANA II 18 4-6'’ Innovative Diagnostic Systems, США
Rapid Anaerobe 24 4" Baxter Diagnostics. США
Streptococcus spp. и Entero coccus spp. СТРЕПТО-тест 12 24 Lachema, Чехия
API 20 Strep 20 4-24 BioMerieux, Франция
Staphylococcus spp. и Micrococcus spp. ПБДС 20 24 НПО «Диагностические систе- мы», Нижний Новгород
СТАФИ-тест 8 24 Lachema, Чехия
API Staph-IDENT 10 5 BioMerieux, Франция
API Staph 20 24 BioMerieux, Франция
Neisseria spp. и Moraxella spp. НЕЙССЕРИЯ-тест 8 24 Lachema, Чехия
Neisseria Enzyme Test 3 0,5 Carr-Scarborough, США
QuadFERM + 4 2 BioMerieux, Франция
Neisseria spp. и Haemophilus spp. NH-IDENT 15 4 BioMerieux, Франция
RapID NH 13 4(1)’“ Innovative Diagnostic Systems, США
Грамположительные кокки и листерии POS ID 27 18—48 Baxter Diagnostics, США
Коринебактерии API Coryne 20 24 BioMerieux, Франция
’ НГОБ — неферментирующие грамотрицательные бактерии.
" Инкубация в аэробных условиях.
Для N. gonorrhoeas.
WO
286 БАКТЕРИОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ
Существует два основных типа таких систем.
Наиболее широкое распространение получили индивидуальные системы, рас-
считанные на идентификацию одной культуры. К ним относятся отечествен-
ные тест-системы ПБДЭ, ПБДС производства НПО «Диагностические системы»
(Нижний Новгород), семейство систем API производства фирмы BioMerieux
(Франция) и многие другие.
Второй тип систем идентификации основан на использовании многорядных
планшетов, в которых каждый ряд лунок позволяет идентифицировать одну
культуру. Как правило, используют стандартные 96-луночные полистироловые
планшеты, аналогичные тем. какие применяются в ИФА. К системам данного
типа относятся наиболее широко распространенные в нашей стране тест-системы
фирмы Lachema (Чехия), а также идентификационные наборы ММТ Е производ-
ства НПО «Аллерген» (Ставрополь).
Идентификация выделенной культуры состоит из следующих этапов;
приготовления суспензии и инокуляции лунок;
- о инкубации:
❖ учета результатов тестов;
❖ оценки результатов.
Для приготовления суспензии, как правило, используют стерильный 0,9%
раствор хлорида натрия, однако в некоторых случаях необходимо использовать
специальные буферы или инокуляционные среды, входящие в состав набора.
Концентрацию микроорганизмов в суспензии определяют по оптическому стан-
дарту мутности. Неправильное приготовление суспензии (слишком высокая или
низкая концентрация микроорганизмов, использование старой культуры) может
привести к некорректному срабатыванию системы.
В зависимости от конструктивных особенностей диагностического набора для
инокуляции могут быть использованы градуированные пипетки, микродозаторы,
пастеровские пипетки. В некоторых тест-системах инокуляция осуществляется
методом погружения или при помощи специального инокулирующего стержня.
Инкубацию тест-систем проводят в обычных термостатах или в специальных тер-
мостатируемых устройствах, входящих в комплект некоторых автоматизирован-
ных систем идентификации. При идентификации анаэробных микроорганизмов
обычно требуется создание бескислородной атмосферы, однако некоторые тест-
системы, основанные на выявлении предсинтезированных ферментов, инкубиру-
ют в аэробных условиях. Температура и время инкубации, указанные в инструкции
фирмы-изготовителя, должны выдерживаться максимально точно. Несоблюдение
этих параметров — одна из наиболее частых причин неправильной идентифика-
ции кулЕлуры.
Учет результатов осуществляют визуально или с помощью специальных считы-
вающих устройств — «ридеров». Их использование позволяет избежать субъек-
тивизма при оценке реакции и в итоге повышает достоверность идентификации.
В некоторых случаях (реакция Фогеса-Проскауэра, тесты на нитратредуктазу.
фосфатазу, индол) перед учетом результата теста в лунку необходимо добавить
специальные проявляющие реактивы. При этом необходимо использовать только
реактивы, входящие в состав диагностического набора.
После учета всех тестов производится кодирование результатов. В простейшем
случае каждый положительный признак обозначают знаком «+», а отрицатель-
ный — знаком «-». Такое кодирование без использования компьютера делает
заключительный этап идентификации весьма продолжительным и трудоемким.
В современных системах идентификации для определения «профиля» (кода)
культуры используемые тесты разбивают на триады. Положительный результат
каждого из тестов, входящих в триаду, оценивается как 2й, где п = 2, 1 или 0, т.е.
положительный результат первого теста в триаде кодируется цифрой 4 (22), вто-
БАКТЕРИОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ 287
рого — цифрой 2 (21), третьего — цифрой 1 (2°). Отрицательный результат теста
всегда обозначается цифрой 0. При сложении результатов тестов в триаде получа-
ют значение от 0 (все тесты в триаде отрицательны) до 7 (все тесты в триаде поло-
жительны). Каждая триада дает одну цифру «профиля» (табл. 20-13). Полученный
«профиль» может быть использован для последующей компьютерной обработки
или для ручной идентификации с помощью кодовой книги.
Таблица 20-13. Профиль культуры
» 20
СТДФИ-тест ГЛИ САХ ТРЕ МАН У АЦ Ф Н К
Результаты теста + + - - + + + + +
Цифровые значения 4 2 0 0 2 1 4 2 1
Профиль 6 3 7
637 - S. epidermidis
ГЛИ - глицерин. САХ — сахароза. ТРЕ - трегалоза, МАН - манит, У — уреаза. АЦ — ацетоин,
Ф — фосфатаза, Н — нитраты, К - каталаза.
Производители тест-систем для биохимической идентификации микроорга-
низмов, как правило, разрабатывают и специальное программное обеспечение
для компьютерной обработки результатов. Основу таких программ составляет
база данных, содержащая частотную матрицу признаков для идентифицируе-
мых микроорганизмов. Фирменные программы можно применять только с той
тест-системой, для которой они разработаны. Однако существуют различные
программы-оболочки, позволяющие пользователю самостоятельно вносить
частотные характеристики признаков. Примером может служить бесплатно рас-
пространяемая программа Id Вас: 1.1. которая доступна на сервере Каролинского
университета (Швеция) по адресу http://www.ki.se/labmed/clim.
ИНДИКАЦИЯ АНТИГЕНОВ ВОЗБУДИТЕЛЕЙ
Выявление в клиническом материале антигенов бактерий являются одним из
основных способов диагностики инфекционных болезней, а обнаружение специ-
фических антигенов у изолятов микроорганизмов позволяет идентифицировать
их на родовом, видовом и серотиповом уровнях. Дтя решения этих задач применя-
ют разнообразные тесты. В современной лабораторной практике наиболее широ-
кое применение из них получили иммунофлюоресцентный, иммуноферментный
и иммунохроматографический анализы, а также различные варианты реакции
агглютинации.
иммунофлюоресцентный анализ
Принцип метода основан на использовании а нтител, коньки ированных с флюо-
ресцентным красителем (в отечественной практике чаще всего — флюоресцеин
изотиоцианатом). Учет результатов реакции осуществляют визуально, с помощью
люминесцентного микроскопа или специальных приборов — флюориметров.
Варианты метода при учете результатов с помощью люминесцентного микроскопа
в различных модификациях под разными названиями — реакция иммунофлюорес-
ценции (РИФ), прямой иммунофлюоресцентный анализ (ПИФ), метод флюорес-
центного анализа (МФА) — широко применяются в клинической микробиологии.
Популярность РИФ объясняется экономичностью, производством широкого
спектра диагностических наборов и быстротой получения ответа. В данном тесте
применяют как поликлональные сыворотки, так и моноклональные антитела.
Чаще всего РИФ используют для быстрого обнаружения возбудителя в патоло-
гическом материале. В этом случае из тестируемого материала готовят мазок на
предметном стекле, как для обычной микроскопии. Препарат фиксируют мети-
288 БАКТЕРИОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ
левым спиртом, ацетоном или другим химическим реагентом, иногда входящим в
состав диагностического набора. На поверхность фиксированного мазка наносят
меченные флюоресцеин изотиоцианатом сыворотки или моноклональные анти-
тела. При непрямой РИФ препарат сначала обрабатывают сывороткой против
искомого антигена, а затем мечеными антителами к иммуноглобулинам, исполь-
зованным на первом этапе теста. Поскольку непрямая РИФ — разновидность гете-
рогенного анализа, один этап реакции отделяют от другого промыванием. Если
прямая РИФ применима для обнаружения антигенов, то непрямая РИФ — как для
обнаружения антигенов, так и для серологической диагностики инфекций (напри-
мер, сифилиса).
Основной недостаток РИФ — субъективность.
Классические критерии специфичности РИФ:
о характерная морфология, размеры и расположение возбудителя в мазке;
<> периферический характер свечения объекта;
о цвет флюоресценции;
❖ интенсивность флюоресценции.
При исследовании крупных объектов (например, трихомонад и гарднерелл)
данные критерии позволяют получить достоверный результат. Однако элемен-
тарные тельца хламидий и микоплазмы имеют размеры, находящиеся на пределе
разрешающей способности люминесцентного микроскопа. При этом оценка мор-
фологии микроорганизмов затруднена, а свечение теряет периферический харак-
тер. Остающихся критериев явно недостаточно для уверенной идентификации
наблюдаемого микроорганизма. Именно поэтому субъективный характер учета
реакции предъявляет особые требования к квалификации проводящего исследова-
ния медицинского персонала.
Альтернатива субъективному визуальному методу учета результатов — аппарат-
ный метод учета.
ИММУНОФЕРМЕНТНЫЙ АНАЛИЗ
Иммуноферментный анализ (ИФА) — одно из наиболее бурно развивающихся
направлений прикладной иммунологии. В основе данного метода лежит присоеди-
нение к антителам ферментной метки, что позволяет учитывать результат реакции
антиген-антитело по появлению ферментативной активности или изменению ее
уровня.
Предложены десятки вариантов реакций, основанных на данном принципе,
однако наиболее широкое распространение в диагностике инфекционных заболе-
ваний получил твердофазный гетерогенный ИФА. Чаще всего для его осуществле-
ния применяют 96-луночные полистироловые планшеты, так как на полистироле
можно легко адсорбировать антитела, а с помошью специальной обработки — и
различные антигены.
При использовании ИФА для поиска антигена на стенках лунок адсорбиру-
ют соответствующие антитела. Исследуемый материал вносят в лунку, при этом
антиген, взаимодействуя с антителами, фиксируется в ней. Лунки тщательно про-
мывают буферным раствором для удаления неадсорбировавшихся веществ. Затем
в лунки вносят антитела против искомого антигена, меченные ферментом. Чаще
всего в качестве фермента-метки используют пероксидазу хрена. После инкуба-
ции лунку снова промывают, чтобы удалить избыток меченых антител, если они
не удерживаются комплексом антиген-антитело, образовавшимся на предыдущем
этапе. На заключительном этапе в лунки вносят хромогенный субстрат (напри-
мер, о-фенилендиамин). Ферментативное расщепление субстрата приводит к
образованию окрашенных продуктов реакции. Таким образом, изменение окраски
содержимого лунки свидетельствует о положительном результате (произошло
взаимодействие антигена и антитела).
БАКТЕРИОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ 289
При серологической диагностике используют полистироловые планшеты, на
стенки лунок которых заранее с помощью тех или иных методов адсорбируют
антиген. Для предотвращения неспецифического связывания антител с полисти-
ролом оставшаяся свободной поверхность полистирола подвергается специальной
обработке. Исследуемую сыворотку вносят в лунку планшета. При этом гомоло-
гичные антигену антитела прикрепляются к нему. Неприкрепившиеся антитела
удаляют промыванием. Затем в лунки вносят антитела против иммуноглобулинов
человека, меченные ферментом. Если в исследуемой сыворотке присутствовали
искомые антитела, то на данном этапе они выступают в роли антигенов, с кото-
рыми реагируют меченые антитела. Добавление после промывки хромогенного
субстрата позволяет провести учет реакции по развивающемуся окрашиванию в
лунках.
Несомненное преимущество ИФА перед классическими иммунологически-
ми реакциями (РА. РСК, РИГА) — возможность количественного анализа без
использования метода серийных разведений. Интенсивность окраски в лунке
пропорциональна количеству фермента и, следовательно, количеству искомого
ингредиента (антигена или антител). Измеряя оптическую плотность жидкости
в лунке и сравнивая ее с контролем, можно определить концентрацию искомого
компонента. В этом случае для учета результатов применяют специальные прибо-
ры («ридеры»), представляющие собой спектрофотометры с вертикальным ходом
луча, специально приспособленные для работы со стандартными полистироловы-
ми планшетами.
Планшеты с «нагруженными» на их поверхность антигенами или анз ителами
служат основой коммерческих диагностических наборов для ИФА. 96-луночные
панели получили очень широкое распространение. Для работы в данном формате
разработаны специальные многоканальные автоматические пипетки, позволяю-
щие вносить реактивы одновременно в несколько лунок панели, а также автома-
тические и полуавтоматические устройства для промывания лунок. Существуют и
специальные автоматы, позволяющие осуществить все этапы анализа практически
без участия человека.
Использование %-луночцых планшетов экономически целесообразно при про-
ведении массовых исследований в крупных иммунологических лабораториях. При
единичных исследованиях такой формат невьподен, поскольку проведение тестов
в отдельных лунках планшета не позволяет использовать многоканальные пипет-
ки, промывающие устройства и другие приспособления, облегчающие проведение
анализа. Разведенные реагенты имеют ограниченный срок годности, поэтому все
более широкое распространение получили планшеты, состоящие из стрипов —
полосок по 8 или 12 лунок. собираемых в формат 96-луночного планшета в про-
извольном порядке. При этом можно задействовать только часть из 96 позиций и
параллельно выявлять различные антигены (антитела).
Указанных недостатков лишена разновидность ИФА, получившая название
dot-ELISA. При этом варианте антигены, необходимые для выявления антител
в плазме крови больного, адсорбированы не в лунках, а на зубцах пластиковой
гребенки. Для проведения исследования необходимо отломить необходимое
количество зубцов, а тестируемые сыворотки внести в первый ряд лунок специ-
альной панели, входящей в состав диагностического набора. Изначально лунки
панели, содержащие все необходимые компоненты в готовом к употреблению
виде, герметично закрыты фольгой. Прокалывая фольгу и последовательно пере-
нося гребенку из одного ряда лунок в другой, выполняют все этапы реакции. С
помощью описываемой системы удается провести ИФА даже вне лаборатории при
отсутствии высококвалифицированного персонала и дорогостоящего оборудова-
ния. Продолжительность исследования составляет от 45 мин до 1,5 ч. Учет резуль-
татов возможен без применения специальных приборов с помощью прилагаемой
290 БАКТЕРИОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ
к набору цветной шкалы-компаратора. Еще одно несомненное достоинство опи-
сываемого диагностического набора — наличие системы внутреннего контроля.
Тест-системы ImmunoComb — быстрые и простые тесты для диагностики многих
инфекционных болезней, в том числе хламидиоза и хеликобактериоза.
Наиболее полно возможности ИФА раскрываются при использовании для его
постановки моноклональных антител. Это позволяет достигать высокой чувстви-
тельности в сочетании с исключительной специфичностью по сравнению с други-
ми серологическими реакциями.
Еще одно достоинство ИФА при серологической диагностике — возможность
определения классов специфических антител (IgG, IgM, IgA), что имеет значение
при определении фазы инфекционного процесса и его остроты.
ИММУНОХРОМАТОГРАФИЧЕСКИЙ АНАЛИЗ
Иммунохроматографию также можно отнести к группе реакций с мечеными
антителами. При этом в качестве метки используют окрашенный латекс или части-
цы коллоидного золота. В реакции применяют антитела к выявляемому антигену,
адсорбированные на микрочастицах, и антитела к тому же антигену, иммобилизо-
ванные в виде полосы на хроматографической бумаге. В данной реакции существу-
ет внутренний контроль (антивидовые антитела, закрепленные в виде полосы на
хроматографической бумаге).
Иммунохроматография может использоваться как для экспресс-индикации
антигенов в пробе, так и для идентификации выделенных чистых культур.
На рынке имеется значительный ассортимент хроматографических систем как
отечественного, так и импортного производства, позволяющих диагностировать
большое количество бактериальных болезней, в том числе гонорею, хеликобакте-
риоз, стрептококкозы.
Тест-системы, основанные на иммунохроматографическом принципе, обладают
высокой специфичностью, позволяют получить результат в течение 5-10 мин и
могут быть использованы не только в лабораториях, но и при первичном обсле-
довании пациента. Поэтому их основное назначение — скрининг. Однако чувстви-
тельность ряда таких тестов недостаточно высока, чтобы обеспечить выявление
всех инфицированных определенным возбудителем пациентов без применения
дополнительных методов лабораторной диагностики.
РЕАКЦИИ АГГЛЮТИНАЦИИ
Различные варианты реакции агглютинации (РГА) широко применяются в
лабораторной практике благодаря простоте постановки и учета результатов реак-
ции, возможности получения последних в течение короткого ввемени (от 30 с
до нескольких минут), отсутствию необходимости в применении какой-либо аппа-
ратуры.
Паиболее часто РГА проводят на предметном стекле, смешивая небольшие
объемы тестируемого материала и содержащего специфические антитела реагента.
После их перемешивания смесь становится мутной и остается такой при отсутствии
в пробе антигенов, реагирующих со специфическими антителами. Положительная
реакция проявляется выпадением агглютинатов на фоне просветления смеси. Учет
результатов теста проводят невооруженным глазом или с помощью лупы.
РГА пользуются для выявления антигенов возбудителей чумы, бруцеллеза,
холеры, коклюша, клостридиозов, менингококкового менингита, иерсиниозов и
ряда других болезней. Данный тест остается «золотым стандартом» серотипирова-
ния энтеробактерий. Особенно сложна и важна пробема серотипирования сальмо-
нелл, поскольку в настоящее время число известных серотипов подвида Enteritidis
превысило 2,5 тыс. Определение антигенной формулы изолята сальмонелл в реак-
ции агглютинации предоставляет обширную информацию как для клиницистов,
БАКТЕРИОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ 291
так и для эпидемиологов, поскольку позволяет судить не только об опасности для
пациента выделенного возбудителя, но и установить потенциально возможные
источники инфекции и проследить всю эпидемиологическую цепочку. Благодаря
этому, появляется возможность прерывания последней, что предотвращает воз-
никновение новых вспышек сальмонеллеза. Для серотипирования S. enteritidis по
схеме Кауфмана-Уайта применяют диагностические наборы, в состав которых
входят сыворотки, полученные от иммунизированных кроликов. Для устранения
неспецифической реактивности сыворотки сорбируют гетерогенными культурами
сальмонелл. Такая технология эффективна, но не лишена недостатков (невозмож-
ность получения специфических антисывороток к ряду антигенов, достаточно
высокий уровень перекрестной реактивности с гетерогенными антигенами ряда
сорбированных сывороток и др.). Поэтому наряду с гипериммунными сорбиро-
ванными сыворотками в настоящее время применяют моноклональные антитела,
специфичность и авидность которых значительно выше.
Часто в лабораториях пользуются одним из вариантов реакции агглютинации—
реакцией латексагглютинации. Его впервые использовали в 1956 г. для диагности-
ки ревматоидного артрита, а сейчас отечественные и зарубежные фирмы выпу-
скают наборы реагентов для диагностики в реакции латексагглютинации свыше
100 инфекционных болезней. Принцип метода такой же, как и других вариантов
реакции агглютинации. При использовании данного теста для обнаружения анти-
генов микроорганизмов применяют латексные частицы, покрытые специфически-
ми антителами. Чтобы легче было учитывать результаты теста без применения
автоматической аппаратуоы, частицы латекса должны иметь достаточно большой
диаметр. В противном случае пользуются латексными частицами, окрашенными
обычными или флюоресцирующими красителями.
МОЛЕКУЛЯРНО-ГЕНЕТИЧЕСКАЯ ИНДИКАЦИЯ БАКТЕРИЙ
В клинической лабораторной диагностике инфекционных болезней все шире
применяют молекулярно-генетические методы, основанные на выявлении фраг-
ментов генома микроорганизмов.
Наибольшее распространение получили методы амплификации нуклеиновых
кислот, в число которых входит и ПЦР. Разработаны многочисленные варианты и
форматы постановки ПЦР, позволяющие добиться оптимального результата при
решении различных диагностических задач. В условиях клинической диагности-
ческой лаборатории для проведения ПЦР применяют диагностические наборы
коммерческих фирм, которые включают необходимые реагенты и растворы для
проведения всех этапов ПЦР в зависимости от ее разновидности и формата.
Существуют следующие разновидности ПЦР:
ПЦР с «горячим» стартом;
< мультиплексная (мультипраймерная) ПНР;
гнездовая ПЦР;
> ПЦР с обратной транскрипцией (ОТ-ПЦР);
< ПЦР в режиме реального времени.
ПЦР в режиме реального времени проводят в следующих форматах:
❖ с применением интеркалирующих флюоресцентных красителей;
> с использованием меченных флюоресцентными красителями олигонуклео-
тидных зондов, комплементарных участку ампликона (гибридизационно-
флюоресцентная детекция).
Формат исследования может быть качественным или количественным.
ПЦР в режиме реального времени позволяет использовать одновременно
несколько зондов, меченных разными флюоресцентными красителями. Данный
вариант ПЦР в режиме реального времени получил название мультиплексного.
292 БАКТЕРИОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ
При этом возможна одновременная детекция нескольких продуктов ПЦР в одной
пробе. Например, в пробе фекалий пациента с клиническими проявлениями остро-
го кишечного заболевания возможно выявление ДНК разнообразных возбудит е-
лей (набор АмплиСенс ОКИ обеспечивает идентификацию шителл, сальмонелл,
кампилобактерии, ротавирусов группы А, норовирусов генотипа 2, астровирусов и
аденовирусов группы F).
Внедрение в повседневную практику клинико-диагностических лаборато-
рий молекулярно-генетических методов исследования позволяет проводить
экспресс-диагностику инфекционных заболеваний и, в некоторых случаях, диф-
ференциальную диагностику и контроль эффективности проводимого лечения.
Высокая чувствительность и относительная простота исполнения позволяет
широко использовать методы амплификации нуклеиновых кислот микроорга-
низмов 1—IV групп патогенности в клинической микробиологии для решения
различных задач:
❖ как метод экспресс-диагностики при исследовании биолоз ического материа-
ла, взятого от человека, с целью выявления ДНК (РНК) микроорганизмов
I—IV групп патогенности и их количественной оценки;
как метод специфической индикации патогенных биологических агентов в
объектах окружающей среды и пищевых продуктах:
❖ как ускоренный предварительный тест при выполнении культурального и
биологического методов исследования и для идентификации чистой культу-
ры;
< для определения эпидемиологической значимости изолятов на основании
выявления генетических маркеров вирулентности и патогенности, а также
маркеров устойчивости к антимикробным препаратам;
о - для таксономической характеристики штаммов на основании выявления
специфических видовых, родовых и других маркеров;
❖ для генотипирования штаммов с целью определения их происхождения;
❖ для прогнозирования течения инфекционного заболевания и оценки эффек-
тивности проводимого лечения.
По результатам качественного или количественного анализа выдают ответ
о наличии в пробе специфических фрагментов ДНК или РНК, гомологичных
определенному участку генома возбудителя инфекционного заболевания, а также
о присутствии в пробах генетических маркеров или генетически модифицирован-
ных ДНК.
Для некоторых бактерий использование традиционных культуральных методов
диагностики проблематично в силу различных причин:
о чувствительности к условиям забора и транспортировки клинического мате-
риала;
❖ необходимости дополнительных специальных факторов роста;
< > медленного роста на питательных средах.
Идентификация микроорганизмов традиционными методами (как правило, с
использованием различных биохимических тестов) может приводить к получению
ошибочных данных вследствие недостаточной стандартизации условий тестирова-
ния и субъективности интерпретации результатов. Использование молекулярно-
генетических тестов позволяет избежать этих проблем.
Наиболее распространенный подход — применение специфических прайме-
ров, комплементарных определенной последовательности ДНК, характерной
только для данного вида микроорганизмов. Также возможно использование
универсальных праймеров для амплификации ДНК, присутствующей у всех
микроорганизмов определенной таксономической группы. Дальнейший анализ
амплифицированной ДНК с помощью различных методов (секвенирование,
метод полиморфизма длины рестрикционных фрагментов, метод одноцепочеч-
БАКТЕРИОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ ?93
ного конформационного полиморфизма) позволяет провести молекулярную
идентификацию бактерий.
Детекция генов, ответственных за продукцию Факторов вирулентности раз-
личных микроорганизмов, особенно актуальна при идентификации условно-
патогенных бактерий для подтверждения их этиологической роли в возникнове-
нии заболевания.
В настоящее время проблему отсутствия полного набора сывороток для клас-
сического типирования эшерихий по О- и Н-антигенам можно решить с помощью
ПЦР для детекции факторов вирулентности, наличие которых свидетельствует об
этиологической роли данного микроорганизма. Например, обнаружение у эшери-
хий генов, кодирующих инвазивность, продукцию энтеротоксинов или веротокси-
нов, позволяет отнести выделенный изолят к определенному патотипу.
Использование ПЦР для определения чувствительности или устойчивости
микроорганизмов к антимикробным препаратам, а также определение меха-
низмов и выявление генетических маркеров резистентности необходимо в тех
случаях, когда невозможно использовать традиционные фенотипические методы
(Af tuberculosis) или необходимо подтвердить механизм резистентности, выявлен-
ный фенотипическими методами (MRSА, БЛРС-продуцирующие энтеробактерии,
МБЛ-продуцируюшис Pseudomonas spp.). Как правило, для анализа используют
ДНК возбудителя, выделенного в чистой культуре, однако в некоторых случаях
можно исследовать образец клинического материала.
В настоящее время рекомендовано использовать ПЦР для подтверждения и
выявления генетических маокеров резистентности при идентификации следую-
щих клинически значимых микроорганизмов:
❖ стафилококков, устойчивых к оксациллину (детекция гена тесА, кодирующе-
го дополнительный пенициллин-связывающий белок 2а);
< > энтерококков, устойчивых к ванкомицину (детекция генов vanA, vanB, vanC,
vanD, обусловливающих синтез модифицированной полипептидной цепи
пептидогликана);
❖ энтеробактерий, устойчивых к цефалоспоринам III—IV поколения [детекция
Ыа генов, кодирующих продукцию р-лактамаз расширенного спектра дей-
ствия (например, СТХ-М), и выявление мутаций в генах р-лактамаз ТЕМ-,
SHV- и ОХА-типа, расширяющих спектр активности данных ферментов];
❖ энтеробактерий, включая возбудителя брюшного тифа, а также и грампо-
ложительные кокки, устойчивые к хинолонам [детекция точечных мутаций
в хромосомных генах субъединиц А и В ДНК-гиразы (гены gyrA и gjrB) и
субъединиц С и Е топоизомеразы IV (генырагС ирагЕ)\;
НГОБ, устойчивых к карбапенемам (детекция генов, кодирующих продук-
цию основных типов металле-р-лактамаз — VIM, IMP. SPM, GIM, SIM).
Ограничения использования ПЦР для определения механизма резистентно-
сти — отсутствие данных о конкретных механизмах резистентности или наличие
одновременно нескольких механизмов резистентности.
Данные о механизме резистентности, полученные с помощью ПЦР, позволя-
ют прогнозировать эффективность антимикробных препаратов без определения
чувствительности возбудителя фенотипическими методами. Например, штаммы
S. aureus, имеющие ген тес A (MRS А), устойчивы ко всем р-лактамным антибио-
тикам.
Примеры использования ПЦР для выявления генетических механизмов разви-
тия резистентности к антибактериальным препаратам представлены в
табл. 20-14.
294 БАКТЕРИОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ
Таблица 20-14. Генетические маркеры резистентности к антимикробным препаратам
Микроорганизм Антимикробный препарат Генетические маркеры резистентности
MRSA Оксациллин Наличие гена теса, кодирующего дополнительный пени- циплинсвязывающий белок 2а
Энтерококки (Е. faecalis, Е. faecium) Ванкомицин Наличие генов vana, vanb, vane или vand, обусловливаю- щих синтез модифицированной поли пептидной цепи пептидогликана
Энтеробактерии, проду цирующие |3-лактамазы расширенного спектра действия (Е. coli, Klebsiella pneumoniae) Цефалоспорины III—IV поколения Наличие генов Ыа, кодирующих продукцию Р-лактамаз расширенного спектра действия (например, СТХ-М), выявление мутаций в генах р-лактамаз ТЕМ-, SHV- и ОХА-типа расширяющих спектр активности данных ферментов
НГОБ (Р. aeruginosa, acinetobacter spp.) Карбаяенемы Наличие генов, кодирующих продукцию основных типов металло-р-лактамаз (vim, imp, spm, gim, sim)
Энтеробактерии (включая возбудителя брюшного тифа) и грамположительные кокки Хинолоны Наличие точечных мутаций в хромосомных генах субъе- диниц А и В ДНК-гиразы (гены дуга и gyrb) и субъединиц С и Е топоизомеразы IV (гены pare и pare)
М. tuberculosis Изониазид Мутации в гене катапазы-пероксидазы (Каф), реже в генах inha и ahpc
Рифампицин Мутации в гене р-субъединицы РНК-полимеразы (rpob)
Зтамбугол Мутации в гене embb. кодирующем синтез арабинога- лактана
Стрептомицин Мутации в генах 16S ррнк (гга) и рибосомного белка S12 (rpsTj
Пиразинамид Мутации в гене пиразинамидазы (рпса)
Фторхинолоны Мутации в гене субъединицы А ДНК-гиразы (дуга)
Выпускаются коммерческие тест-системы для анализа образцов различных
биологических материалов на наличие ДНК возбудителей различных инфекци-
онных заболеваний. Исследумые микроорганизмы разделяют в зависимости от их
принадлежности на разные группы патогенности:
• I группа патогенности — У. pestis;
• II группа патогенности — В. anthracis, Brucella spp., Р pseiidumallei, Francisella
tularensis, Vibrio cholerac,
• III группа патогенности — Bordetella pertussis, C. diphtheria, возбудители
острых кишечных инфекций (Cl. difficile, E. coll 0157:H7 и другие группы диа-
реегенных эшерихий, Salmonella spp.. Shigella spp., Campylobacter spp., H. pylori
астровирусы, ротавирусы, калицивирусы), L. pneumophila, Leptosptia inter-
rogans, L. monocytogenes, M. tuberculosis complex, N. gonorrhoeas, N. meningitidis,
T. pallidum, У. pseudotuberculosis',
• IV группа патогенности — Bacteroides spp. (возбудители сепсиса и гной-
ных инфекций), Haemophilus spp. (возбудители менингита, пневмоний и
ларингитов), возбудители ИППП (Ureaplawa spp., Chlamydia trachomatis,
N. gonorrhoeas, T. pallidum, Trichomonas vaginalis, Mycoplasma genitalium,
Mycoplasma hominis, Gardnerella vaginalis).
Перечень инфекционных заболеваний и микроорганизмов, для выявления
которых используют молекулярно-генетические методы диагностики, представ-
лен в табл. 20-15.
БАКТЕРИОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ 295
Таблица 20-15. Бактериальные инфекции и возбудители, для выявления которых используют
молекулярно-генетические методы диагностики
Инфекционное заболевание Возбудители
Урогенитальные инфекции Chlamydia trachomatis N. gonorrhoeae T. pallidum Trichomonas vaginalis Mycoplasma genitalium Mycoplasma hominis Ureaplasma spp. Gardnerella vaginalis Candida albicans
Инфекции респираторного тракта C. diphtheria Streptococcus spp. L. pneumophila
Туберкулез M. tuberculosis complex
Нейроинфекции N. meningitidis Neisseria spp. Haemophilus spp. Listeria monocyiugenes
Кишечные инфекции Campylobacter spp. (термофильная группа) H. pylori Shigella spp. и энтероинвазивные штаммы E. coli Salmonella spp. Y. enterocolitica Y. pseudotuberculosis Cl. difficile
Особо опасные и природно- очаговые инфекции Viorio cholerae B. anthracis Brucella spp Leptospira spp. Borrelia burgdorferi sense lato [B. burgdorferi seosu stricto, B. afzelii, B. garinii)
ИНДИКАЦИЯ СПЕЦИФИЧЕСКИХ АНТИТЕЛ
Выявление антител к антигенам микроорганизмов в сыворотке крови и ликво-
рах (спинномозговом и др.) пациентов является одним из основных способов диа-
гностики инфекционных заболеваний. При заболеваниях бактериальной природы
определяют наличие у пациентов антител к антигенам и токсинам возбудителей. В
ряде случаев с помощью серологической диагностики можно оценивать эффектив-
ность проводимого лечения или вакцинации. С помощью серологических тестов
возможно выявление переболевших лиц и бактерионосителей, пациентов со
стертыми и субклиническими формами заболевания, что обеспечивает получение
более полной и обоснованной ин формации для оценки активности инфекционно-
го и эпидемического процессов.
ВЗЯТИЕ МАТЕРИАЛА ДЛЯ ИССЛЕДОВАНИЯ
Для серологических исследований чаще всего используют сыворотку, которая
должна быть прозрачной, без примеси эритроцитов и следов гемолиза. Кровь заби-
рают натощак или не ранее чем через 6 ч после еды, при условии, что больной не
употреблял жирной пищи, включая яйцепродукты. В против ном случае сыворотка
может быть мутной (хилезной) и непригодной для исследования. Кровь собирают
с соблюдением правит асептики в стерильную посуду. После образования сгустка
его обводят и оставляют на несколько часов в холодильнике (2-4 °C) для более
полного отделения сыворотки. Взятие проб можно проводить в вакуумные одно-
разовые пробирки (пустые или с ускорителями свертывания крови). Полученную
296 бактериологические исследования
сыворотку можно хранить при температуре 2-8 °C в течение 2-3 дней. Для более
длительного хранения сыворотки замораживают при температуре -20 °C и ниже.
Следует избегать их многократного размораживания и замораживания.
При диагностике некоторых заболеваний (например, сифилиса) и необходи-
мости длительной транспортировки материала в лабораторию допускается высу-
шивание сыворотки. Для РСК сыворотки высушивают на поверхности сложенной
вдвое вощеной бумаги или целлофана. На полосу (8x10 см) такой подложки
в 4 места вносят по 0,5 мл сыворотки и по 3 капли 40% раствора сахарозы,
оставляя подсыхать при комнатной температуре. На краю подложки указывают
Ф.И.О. пациента, объем сыворотки и дату. Носитель складывают в виде аптеч-
ного пакетика, помещают в конверт и направляют в лабораторию. Такие высу-
шенные пробы необходимо исследовать в течение 5 дней в теплое время года и в
течение 10 дней — в холодное.
Известно большое количество серологических тестов, которыми диагностируют
инфекционные болезни. Техника их постановки описана в инструкциях по приме-
нению соответствующих тест-систем и диагностикумов.
При выборе серологической реакции принимают во внимание:
❖ доступность необходимых реагентов и оборудования для ее постановки;
сроки получения ответа;
ее чувствительность и специфичность.
В ряде случаев применение определенной реакции в конкретной ситуации —
следствие своеобразного компромисса. Отсутствие идеальной серологической
реакции диктует необходимость в некоторых случаях использовать для диагно-
стики не один, а несколько тестов, в том числе скрининговые, диагностические и
экспертные. Например, реакция микропреципитации, используемая для сероло-
гической диагностики сифилиса, обладает невысокой специфичностью, однако
она дешева и удобна в постановке. Наиболее специфичная и чувствительная при
сифилисе серологическая реакция (РИФ-абс), напротив, сложна в постановке и
требует применения малодоступных реагентов. В связи с этим реакцию микро-
преципитации используют как скрининговую, и выявленные в ней положительные
результаты подлежат подтверждению в более специфических реакциях, таких как
ИФА и РИГА. Если результаты используемых реакций неоднозначны или не соот-
ветствуют клиническим данным, используют РИФ-абс как экспертную реакцию.
К постановке дополнительных реакций, подтверждающих достоверность получен-
ного положительного результата, также приходится прибегать при диагностике
ВИЧ-инфекции и вирусных гепатитов.
Диагностические критерии
Сам факт обнаружения антител в сыворотке больного не является достаточным
основанием для постановки диагноза. Только в немногих случаях (например, при
сифилисе) диагностическое значение имеет сам факт обнаружения антител опреде-
ленной специфичности. Постоянно контактируя с различными антигенами, в том
числе и с антигенами возбудителей инфекционных заболеваний, организм чело-
века насыщен огромным разнообразием антител. Поэтому клиническая интер-
претация результатов серологических тестов во многом зависит от результатов
повторных исследований. Данное, обстоятельство подчеркивает необходимость
серологического исследования парных сывороток, полученных в острой стадии
заболевания и в период выздоровления. У любого пациента с неясным диагнозом
взятую в начале болезни сыворотку замораживают, чтобы при необходимости
провести параллельное исследование с сывороткой, полученной в более поздние
сроки, для оценки динамики антительного ответа.
Для постановки этиологического диагноза на основании показаний серологиче-
ских тестов, как правило, пользуются одним из двух диагностических критериев:
БАКТЕРИОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ 297
о выявление так называемого диагностического титра при однократном иссле-
довании сыворотки — количество антител должно быть не ниже определен-
ного (дискриминантного) значения;
достоверное нарастание (в 4 раза и более) титра антител в парных пробах
сыворотки крови, взятых у пациента с интервалом в 7-14 дней.
Первый из указанных критериев — статический показатель, превышение
которого расценивают как признак инфекционного заболевания. Величина диа-
гностического титра устанавливается эмпирически для каждого заболевания и
для каждой серологической реакции. При использовании для диагностики одно-
го заболевания нескольких серологических тестов, величины диагностических
титров для каждого из них могут не совпадать, так как разные серологические
реакции различаются своей чувствительностью. Различия в величинах диагности-
ческих титров также возможны при применении одной серологической реакции,
в результате использования разных диагностических препаратов, так как в этом
случае могут выявляться антитела к разным антигенам. Величину диагностиче-
ского титра обычно указывают в инструкции вкладыше, которая имеется в каждой
упаковке с диагностическими тест-системами или диагностическим препаратом.
Часто величину диагностического титра устанавливают эмпирически в процессе
создания каждой новой серологической тест-системы. Его значение зависит от
вида заболевания и типа серологической реакции. Кроме того, он может разли-
чаться в тест-системах, изготовленных разными фирмами.
При ряде заболеваний (шигеллезы, сальмонеллезы) значение диагностического
титра в определенной степени условно, так как зависит не только от перечисленных
выше объективных причин, но и от конкретной эпидемиологической ситуации,
когда риск заражения возбудителями и широта их распространения в популяции
варьируют, что приводит к уменьшению или повышению титра соответствующих
антител у здоровых лиц (так называемые фоновые или анамнестические антитела)
и, следовательно, к разной оценке диагностического титра.
Диагностический титр антител может быть выявлен в сыворотке здоровых
людей и обусловлен профилактической вакцинацией, перенесенной в прошлом
инфекцией в любой форме или иммунизацией антигенами, которые могут попа-
дать в организм с загрязненными пищей или водой. В зависимости от эпидемиче-
ской ситуации величина диагностического титра различна на разных территориях.
Ее нужно устанавливать на каждой территории по результатам титрования сыво-
роток 100 здоровых лиц. Большое значение имеет определение диагностического
титра IgG, поскольку их наличие свидетельствует о значительном специфическом
антигенном стимуле и более позднем сроке заболевания.
Более достоверен второй критерий (нарастание титра антител), позволяющий
проследить развитие иммунного ответа у конкретного пациента. В этом случае
диагноз заболевания можно установить даже при аномально низком уровне
иммунного ответа, что, в свою очередь, связано с индивидуальными особенностя-
ми некоторых больных. У таких людей при постановке серологической реакции
уровень антител не достигает диагностического титра, однако будет зафиксирова-
но нарастание титра антител при исследовании парных сывороток.
При некоторых заболеваниях (микоплазмоз) достоверное нарастание титра
антител регистрируют редко, что делает проблематичным возможность их сероло-
гической диагностики. Диагностически достоверным считается нарастание титра
антител в 4 раза и более. Нарастание титра в 2 раза, как правило, в расчет не при-
нимается, поскольку может быть связано с ошибкой постановки реакции (ошиб-
ки, допущенные при разведении сыворотки, несоблюдение объемов реагентов,
нарушение температурного режима, инкубация реакционной смеси и др.). В таких
случаях тест необходимо провести повторно.
298 БАКТЕРИОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ
Недостаток этого диагностического критерия — относительно большой срок,
необходимый для получения ответа. Как правило, при его использовании диагноз
подтверждают (или отвергают) к концу 2-3-й недели заболевания. В некоторых
случаях' такая диагностика носит ретроспективный характер.
ОПРЕДЕЛЕНИЕ ИММУНОГЛОБУЛИНОВ РАЗНЫХ КЛАССОВ
Большое диагностическое значение имеет определение соотношения спец-
ифических иммуноглобулинов разных классов. Это позволяет определить
стадию заболевания и, в некоторых случаях, прогнозировать его течение и воз-
можный исход. Современные методы серологической диагностики позволяют
определить концентрацию иммуноглобулинов отдельных классов определен-
ной специфичности. Когда первый контакт с микроорганизмом индуцирует
иммунный ответ, первыми появляются IgM. Их титры быстро нарастают и
достигают максимума, а затем относительно быстро снижаются. IgG появля-
ются позже, но дольше сохраняются в высоких тиграх. При повторном кон-
такте с тем же антигеном (вторичный иммунный ответ) IgM вырабатываются
в незначительном количестве, реакция гуморального иммунитета проявляется
прежде всего увеличением титров IgG. Именно поэтому по соотношению IgM
и IgG можно отличить первичный иммунный ответ (острое заболевание) от
вторичного (рецидив или повторное заражение). О различиях в характере
вырабатываемых иммуноглобулинов при первичном и вторичном иммунном
ответе следует помнить при оценке результатов, полученных с помощью неко-
торых современных иммунологических методов, например ИФА. Большинство
выпускаемых тест-систем для серодиагностики с помощью ИФА позволяют
определить титр IgM или IgG. Соответственно и лаборатория сообщает врачу
не титр антител вообще, а титр иммуноглобулинов соответствующего класса.
В зависимости от конкретной клинической ситуации интерпретация высоких
или низких титров IgM и IgG будет различаться. В частности, низкие титры
или отсутствие IgM не всегда можно расценивать как факт, свидетельствующий
в пользу отсутствия заболевания.
Наличие специфических IgM в сыворотке — ранний маркер инфекции, свиде-
тельствующий о развитии острой фазы заболевания. IgM можно выявить на 3-5-й
день болезни, пик их нарастания приходится на 2-ю неделю от начала болезни,
затем происходит снижение титра антител. Специфические IgG можно определять
через 7-10 дней после начала заболевания и позже. Их наличие отражает общую
картину положительного иммунного ответа при острой, хронической или перене-
сенной инфекции, причем часто после выздоровления на протяжении многих лет
IgG могут сохраняться в крови в низких титрах. Увеличение и снижение их титров
происходит медленнее по сравнению с IgM. Обнаруживаемые у новорожденных
детей IgG. как правило, имеют колостральное происхождение.
В настоящее время раздельное определение иммуноглобулинов различных
классов (IgM, IgG, IgA) используют достаточно часто. Достоверно можно выявить
титр IgG. используя редуцирующие вещества, например меркаптаны (меркаптоэ-
танол, меркаптопурин), цистеин и димеркаптопропансульфонат натрия, которые
избирательно инактивируют IgM. Различия в величинах титров антител в натив-
ной и обработанной таким реактивом сыворотке позволяют судить о наличии или
отсутствии IgM. Полное исчезновение антител в пробах сывороток, обработан-
ных редуцентом, свидетельствует о присутствии IgM и острой фазе заболевания.
Отсутствие IgM в отдельных сыворотках не является основанием для исключения
инфекционного процесса. В этом случае необходимо дополнительно исследовать
парные пробы сывороток в динамике заболевания В сыворотках, после обработки
которых не снижается титр антител, доминируют IgG, что свидетельствует о более
поздних сроках заболевания. Уменьшение титра антител в несколько раз после
БАКТЕРИОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ 299
обработки редуцентом свидетельствует о том, что антитела представлены смесью
IgM и IgG.
Поэтому для подтверждения специфичности результатов, получаемых при
исследовании одной сыворотки, необходима постановка данной реакции в дина-
мике заболевания с последующим определением выраженных серологических
сдвигов в титрах специфических IgG. Необходимо одновременно исследовать
парные сыворотки, из которых первая взята в остром периоде заболевания (не
позднее 3—5-го дня болезни), а вторая — через 7-10 дней. Диагноз ставят на осно-
вании увеличения титра антител во второй сыворотке по сравнению с первой в 4
раза и выше.
Современные диагностические наборы, основанные на «сандвич-методе», зна-
чительно упростили данную процедуру. Используемые в таких наборах монокло-
нальные антитела взаимодействуют только с иммуноглобулинами определенного
класса (IgM, IgG, IgA).
Определение IgG
Наиболее простой и доступный способ дифференциации IgM и IgG — метод раз-
рушения IgM редуцирующими веществами. Однако он не позволяет количественно
определить IgM, если их содержание в сыворотке меньше или равно концентрации
IgG. Метод определения IgG основан на избирательном разрушении редуцирую-
щими веществами макроглобулиновых антител, к которым относятся молекулы
IgM. IgM под действием редуцентов расщепляются с разрывом дисульфидных
связей молекул белка, диссоциируя при этом на 6-8 мономеров, не обладающих
иммунной активностью. IgG под действием данных веществ не разрушаются.
Обработка сыворотки цистеином
Для такой обработки сывороток готовят ex tempore раствор солянокислого
или «свободного» цистеина из расчета 15.0 мг цистеина на 1,0 мл растворителя.
Раствор цистеина можно хранить не более 30-60 мин, поэтому сначала готовят
необходимые разведения сывороток. Исследуемую сыворотку в разведении 1:5
(в 0,9% растворе хлорида натрия) прогревают на водяной бане при температуре
56 °C в течение 30 мин и делят на 2 одинаковые по объему порции. К первой из
них добавляют равный объем раствора цистеина, ко второй — равный объем 0,9%
раствора хлорида натрия. Вещества смешивают в стерильных пробирках, которые
плотно закрывают воздухонепроницаемыми пробками. Объем пробирок должен
быть таким, чтобы между содержимым и пробкой оставалось минимальное сво-
бодное пространство для предотвращения инактивации цистеина кислородом воз-
духа. Пробирки с обработанными и необработанными сыворотками выдерживают
в термостате при температуре 37 °C в течение 18-20 ч. Затем обе пробы сыворотки
исследуют в серологических реакциях с диагностикумами по общепринятой мето-
дике, начиная с разведения 1:10.
Обработка сыворотки димеркаптопропансульфонатом натрия
Для обработки сывороток 5% раствор димеркаптопропансульфоната натрия
разводят в 4 раза 0,9% раствором хлорида натрия до получения концентрации
1,25. Полученный 1,25% раствор димеркаптопропансульфоната натрия можно
хранить в стерильной стеклянной посуде с хорошо притертой воздухонепрони-
цаемой пробкой при температуре 4 СС в течение недели. Исследуемую сыворотку в
разведении 1:5 (в 0,9% растворе хлорида натрия) прогревают на водяной бане при
температуре 56 °C в течение 30 мин и делят на 2 равные порции. К первой порции
добавляют равный объем 1,25% раствора димеркаптопропансульфоната натрия,
ко второй — равный объем 0,9% раствора хлорида натрия. Вещества смешивают
в стерильных пробирках, котооые плотно закрывают воздухонепроницаемыми
пробками. Обе порции сыворотки оставляют на 18 ч при комнатной температуре.
300 БАКТЕРИОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ
а затем исследуют с диагностикумами по общепринятой методике, начиная с раз-
ведения 1:10. Возможно сокращение срока инактивации IgM до 2 ч, однако в этом
случае необходимо инкубировать сыворотки в термостате при температуре 37 °C.
Оценка результатов
Результаты серологических тестов необходимо интерпретировать с учетом
дополнительной информации о пациенте, включая данные о предшествующей
иммунизации, перенесенных заболеваниях, вероятности контакта с определенны-
ми возбудителями или их антигенами. Ни один метод лабораторной диагностики
не может заменить клинической оценки заболевания лечащим врачом. Например,
при тифо-паратифозных заболеваниях даже при самом тщательном серологиче-
ском обследовании в 20-25% случаев получают отрицательные результаты (отсут-
ствие диагностически значимых титров антител и их нарастания) при наличии
бактериологического подтверждения диагноза. Отрицательные серологические
реакции (серореверсия) могут наблюдаться на фоне этиотропной терапии, напри-
мер при использовании антибактериальных препаратов. Поэтому отрицательные
результаты серологического исследования при наличии специфических клини-
ческих симптомов не являются основополагающим критерием для отмены диа-
гноза.
ОПРЕДЕЛЕНИЕ ЧУВСТВИТЕЛЬНОСТИ БАКТЕРИЙ
К АНТИМИКРОБНЫМ ПРЕПАРАТАМ
ПОНЯТИЕ 0 ТИПАХ И СТЕПЕНИ ЛЕКАРСТВЕННОЙ УСТОЙЧИВОСТИ БАКТЕРИЙ
К АНТИБИОТИКАМ
Природная устойчивость — генетически обусловленное отсутствие чувствитель-
ности микроорганизма к антимикробным препаратам. При этом точки приложе-
ния действия препарата («мишени») в микробной клетке отсутствуют или недо-
ступны для него. Пример природной устойчивости — резистентность микоплазм
и псевдомонад к бензилпенициллину или резистентность бактерий к антигрибко-
вым препаратам. Природная устойчивость не требует рутинного лабораторного
подтверждения.
Приобретенная устойчивость обусловлена изменением генома микробной
клетки в результате мутаций или связана с получением новой генетической
информации при конъюгации (передача генетического материала в результате
своеобразного «полового» процесса у микроорганизмов), трансдукции (пере-
дача генетического материала с помощью фагов) или трансформации (непосред-
ственная передача фрагмента ДНК донора реципиентной клетке) с последующей
селекцией устойчивых штаммов. Данный термин применяют в том случае, когда в
чувствительной к определенному антимикробному препарату популяции микро-
организмов обнаруживают устойчивые штаммы.
Уровень природной устойчивости к антимикробным препаратам у «диких»
штаммов микроорганизмов может быть достаточно точно предсказан на основа-
нии видовой идентификации, а наличие и уровень приобретенной устойчивости
у «госпитальных» штаммов могут быть установлены только после выявления
определенными методами чувствительности возбудителей к антибиотикам.
По степени чувствительности к антимикробным препаратам микроорганизмы
принято разделять на три группы:
<- чувствительные микроорганизмы:
о промежуточные микроорганизмы;
❖ устойчивые (резистентные) микроорганизмы.
БАКТЕРИОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ 301
Существует два подхода к интерпретации этих понятий — клинический и
микробиологический.
В соответствии с клиническим подходом, основанным на оценке эффективности
антибактериальной терапии, к чувствительным относятся возбудители, рост кото-
рых в организме больного прекращается при введении обычных терапевтических
доз антибиотика. В группу промежуточных входят штаммы, для угнетения роста
которых требуется назначение максимально допустимых доз антимикробного пре-
парата. Хороший лечебный эффект в отношении возбудителей с промежуточной
чувствительностью можно получить при применении обычных терапевтических
доз препарата в том случае, если очаг инфекции локализуется в тканях, в которых
антибиотик накапливается в высоких концентрациях. К устойчивым относятся
микроорганизмы, для подавления роста которых требуются чрезвычайно высо-
кие концентрации химиопрепаратов, недостижимые в организме из-за опасно-
сти проявления их токсических свойств. Таким образом, с точки зрения врачей,
чувствительными к антибиотику считаются такие микроорганизмы, на которые
он оказывает бактериостатическое или бактерицидное действие в концентрации,
достигаемой в очаге инфекции.
Микробиологический подход к разделению чувствительных, промежуточных
и устойчивых микроорганизмов основап на анализе эффективности подавления
антибиотиком роста исследуемых штаммов в питательной среде (выявление
«пограничных» концентраций антимикробных препаратов, изучение диаметров
зон задержки роста бактерий вокруг дисков с химиопрепаратами).
При оценке чувствительности возбудителей следует иметь в виду, что клиниче-
ская и микробиологическая эффективность антимикробных препаратов совпада-
ют далеко не всегда.
КЛАССИФИКАЦИЯ МЕТОДОВ ОПРЕДЕЛЕНИЯ ЧУВСТВИТЕЛЬНОСТИ
МИКРООРГАНИЗМОВ К АНТИМИКРОБНЫМ ПРЕПАРАТАМ
Методы, применяемые в микробиологической лаборатории для определения
чувствительности (устойчивости) возбудителей к антимикробным препаратам,
принято разделять на несколько групп.
• Методы серийных разведений антимикробных препаратов в питательной
среде:
метод серийных разведений антибиотика в жидкой питательной среде;
метод серийных разведений препарата в плотной питательной среде;
❖ макрометод (в пробирках);
❖ микрометод (в планшетах).
• Метод «пороговых» («пограничных») концентраций.
• Диффузионные методы;
< диско-диффузионный метод;
о Е-тест (эпсилометрический).
• Метод выявления толерантности микроорганизмов к антимикробным пре-
паратам.
• Специальные методы обнаружения устойчивости микроорганизмов к анти-
микробным препаратам:
> метод выявления р-лактамаз микроорганизмов с использованием нитро-
цефинового диска;
< специальные методы обнаружения устойчивости Staphylococcus spp. к окса-
циллину и ванкомицину;
специальные методы обнаружения устойчивости Enterococcus spp. к амино-
гликозидам и ванкомицину;
о специальные методы обнаружения устойчивости Str, pneumoniae к бензил-
пенициллину;
302 БАКТЕРИОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ
о- методы выявления р-лактамаз расширенного спектра действия у энтеро-
бактерий;
❖ выявление продукции металло-р-лактамаз у грамотрицательных бактерий
методом двойных дисков с ЭДТА.
Методы определения чувствительности микроорганизмов к антимикробным
препаратам позволяют проводить выбор оптимального антибиотика для лечения
конкретного больного, а также разраба гыват ь тактику эмпирической антимикроб-
ной терапии для отдельных нозологических форм инфекционное патологии на
основании микробиологического мониторинга за уровнем антибиотикорезистент-
ности в конкретных регионах или медицинских учреждениях.
Основные параметры, влияющие на выбор метода оценки ангибиотикоре-
зистентности микроорганизмов, — точность и воспроизводимость результатов
исследования, а также трудоемкость и стоимость метода. Результаты исследования
во многом зависят от состава и pH питательной среды, величины посевной дозы,
возраста культуры, условий культивирования и некоторых других факторов.
Необходимое условие получения точных и воспроизводимых результатов —
максимальная стандартизация выбранного метода исследования. В некоторых
странах (США, Германия, Франция) для определения чувствительности микро-
организмов к антибиотикам существуют собственные (национальные) стандарты,
однако в большинстве стран мира в настоящее время используют методы, раз-
работанные Институтом клинических и лабораторных стандартов США (Clinical
and Laboratory Standards Institute, CLST), который ранее назывался Национальным
комитетом по клинико-лабораторным стандартам (National Committee for Clinical
Laboratory Standards, NCCLS). В странах Евросоюза существует Европейский
комитет по определению чувствительности к антибиотикам (European Committee
for Antimicrobial Susceptibility Testing, EUCAST), созданный Европейским обще-
ством по клинической микробиологии и инфекционным болезням (European
Society) for Clinical Microbiology and Infectious Diseases, ESCMID). В 2004 г. главным
государственным санитарным врачом РФ утверждены и введены в действие
«Методические указания по определению чувствительности микроорганизмов к
антибактериальным препаратам» (МУК 4.12 1890-04), в основе которых лежат
стандарты CLSI (NCCLS).
Отсутствие единых международных критериев при определении чувствитель-
ности микроорганизмов к антимикробным препаратам приводит к тому, что
штаммы, оцениваемые по одним стандартам как чувствительные, по другим стан-
дартам оказываются резистентными. Для целого ряда патогенов методы изучения
чувствительности к антимикробным препаратам не стандартизированы, а обосно-
ванные критерии оценки полученных результатов отсутствуют. Проводить изуче-
ние чувствительности таких микроорганизмов к антибиотикам не рекомендуется.
Штаммы возбудителей с необычным типом резистентности, не описанным в науч-
ной литературе (например, резистентность Str. pyogenes к бензилпенициллину),
требуют проведения повторной тщательной идентификации и отправляются для
проверки в специализированные лаборатории.
ХАРАКТЕРИСТИКА МЕТОДОВ ОПРЕДЕЛЕНИЯ ЧУВСТВИТЕЛЬНОСТИ
МИКРООРГАНИЗМОВ К АНТИМИКРОБНЫМ ПРЕПАРАТАМ
Методы серийных разведений антимикробного препарата в питательной среде
Методы серийных разведений антимикробных препаратов в питательной
среде — количественные, они позволяют определить минимальную ингибирую-
щую концентрацию (МИК) и минимальную бактерицидную концентрацию (МБК)
антибиотика, которые измеряются в ЕД/мл или в мкг/мл. В качестве синонима
МИК нередко используют термин «минимальная подавляющая концентрация»
БАКТЕРИОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ 303
(МПК). В зависимости от объема питательной среды выделяют методы серийных
макро- и микроразведений (в бактериологических пробирках и в пластиковых
микропланшетах соответственно), а в зависимости от ее состава — методы серий-
ных разведений в жидкой и плотной питательных средах.
Для выявления МИК определенную концентрацию изучаемой культуры засе-
вают на питательные среды, содержащие 2-кратные разведения исследуемого
антимикробного препарата, после инкубации посевов в термостате учитывают
результат. Величину МИК определяют по максимальному разведению антибиоти-
ка, задерживающему рост возбудителя в питательной среде. Для выявления МБК
из пробирок с отсутствием видимого роста микроорганизмов производят высев
на плотные или жидкие питательные среды. За величину МБК принимают наи-
меньшую концентрацию антибиотика в пробирке, высев из которой не дал роста
культуры вследствие гибели исследуемых микроорганизмов (рис. 20-2).
Рис. 20-2. Схема определения чувствительности возбудителей к антимикробным препаратам
методом серийных разведений антибиотика в жидкой питательной среде.
Особенность методов серийных разведений — недостаточно точное выявление
значения МИК. На самом деле истинная величина МИК находится в промежутке
между ее величиной, определенной во время опыта, и величиной, уступающей ей в
2 раза и не вызвавшей задержки роста исследуемого микроорганизма. Например,
если замедление роста возбудителя произошло при концентрации антибиотика
64 мкг/мл, то истинное значение МИК будет находиться в промежутке между 32
и 64 мкг/мл. Для более точного определения величины МИК следует провести
дополнительные исследования в указанном диапазоне концентраций антибио-
тика.
Для эффективной антимикробной терапии бактериальных инфекций требуется
создать в очаге воспаления концентрацию антибиотика, превышающую МИК (для
получения бактериостатического эффекта) или превышающую МБК (для полу-
чения бактерицидного эффекта). Если при назначении максимально допустимых
(терапевтических) доз препарата его концентрация в очаге воспаления окажется
меньше выявленной in vitro МИК, то возбудитель инфекции следует отнести к
категории устойчивых, а для лечения больного подобрать другой антибиотик.
Таким образом, сведения о величине МИК антибиотика позволяют лечащему
врачу грамотно рассчитать дозы и режимы введения антимикробного препарата.
Как показывает практика, для получения хорошего терапевтического эффекта
концентрация антимикробного препарата в крови должна в 5-10 раз превышать
его МИК в отношении возбудителя. Методы серийных разведений антибиотиков в
жидких и плотных питательных средах — одни из самых информативных, однако
они весьма трудоемки и дороги.
304 БАКТЕРИОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ
Метод «пороговых» концентраций
Достаточно простой и экономичный способ определения чувствительности
микроорганизмов к антимикробным препаратам — метод «пороговых» концен-
траций. В его основе лежит представление о двух «пороговых» («пограничных»)
концентрация к антибиотика, разделяющих штаммы микроорганизмов на три
категории — чувствительные, промежуточные и устойчивые (табл. 20-16).
Таблица 20-16. Разделение штаммов микроорганизмов на чувствительные, промежуточные и
резистентные в соответствии со значениями «пороговых» концентраций антимикробных пре-
паратов (4 и 64 мкг/мл)
1 мкг/мл 2 мкг/мл 4 мкг/мл 8 мкг/мл 16 мкг/мл 32 мкг/мл 64 мкг/мл 128 мкг/мл 256 мкг/мл
S чувствительные I промежуточные R резистентные
При этом меньшая концентрация (в данном примере — 4 мкг/мл) разграничи-
вает чувствительные и промежуточные штаммы, а большая (в данном примере —
64 мкг/мл) — промежуточные и устойчивые штаммы возбудителей. При выборе
значений пограничных концентраций для антимикробных препаратов учитывают
микробиологические, клинические и фармакологические факторы. Для каждого
антибиотика установлены свои «пороговые» концентрации в отношении опреде-
ленного микроорганизма. Эти концентрации не являются постоянными величина-
ми и периодически пересматриваются в специальных комитетах (CLSI, EUCAST)
по мере изменения чувствительности популяций возбудителей, а результаты
публикуются в соответствующих нормативных документах.
Метод «пороговых» концентраций в большинстве случаев предполагает исполь-
зование готовых коммерческих тест-систем, в основу которых заложен принцип
микрообъемных технологий. Тест-система представляет собой планшет с двумя
рядами лунок, в каждой из которых содержатся лиофилизированные растворы
антибиотиков в питательной среде в объеме 0,1 мл (рис. 20-3). При этом каждый
антибиотик представлен двумя лунками (верхней и нижней), в которых содержат-
ся соответствующие «пороговые» концентрации.
Результаты определения чувствительности микроорганизмов к антимикробным
препаратам методом «пороговых» концентраций можно учитывать невооружен-
ным глазом или использовать специальные автоматизированные системы. В зави-
симости от типа применяемых панелей учет результатов и анализ проводят через
18 24 ч или через 4-6 ч инкубации.
Диффузионные методы
Диско-диффузионный метод
Наиболее простой и доступный для большинства практических бактериологи-
ческих лабораторий способ оценки чувствительности возбудителей к антимикроб-
ным препаратам — диско-диффузионный метод.
Принцип метода основан на способности антимикробных препаратов диффун-
дировать из пропитанных ими бумажных дисков в питательную среду и угнетать
Рис. 20-3. Микрообъемная панель для определения чувствительности стафилококков к анти-
микробным препаратам методом «пороговых» концентраций.
БАКТЕРИОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ 3Q5
рост колоний бактерий. Сопоставление полученных в результате исследования
величин диаметров зон задержки роста возбудителей вокруг дисков с антибио-
тиками с табличными данными, изложенными в соответствующих нормативных
документах, позволяет отнести исследуемые штаммы микроорганизмов к катего-
риям чувствительных (S), промежуточных (I) или устойчивых (R) к соответствую-
щим антимикробным препаратам. При этом величина диаметра зоны задержки
роста исследуемой культуры тесно связана со значениями МИК тестируемого
антибиотика для S, I и R штаммов. Таким образом, диско-диффузионный метод
позволяет не только определить чувствительность возбудителя к антимикробным
препаратам, но и помочь врачу в выборе режима дозирования антибиотика с уче-
том его МИК.
Чаще всего диско-диффузионный метод используют для определения чув-
ствительности к антибиотикам микроорганизмов семейства Enterobacteriaceae,
Р. aeruginosa, Acinetobacter spp., Staphylococcus spp., Enterococcus spp., Haemophilus spp.,
N. gonorrhoeae, Streptococcus spp. Применяют диски только с теми антимикробными
препаратами, которые обладают клинически подтвержденной эффективное гью в
отношении соответс гвующих возбудителей инфекции.
Определение чувствительности микроорганизмов к антибиотикам с помощью
диско-диффузионного метода включает следующие этапы:
о выделение чистой культуры возбудителя;
❖ приготовление рабочей суспензии исследуемого микроорганизма (иноку-
лята);
❖ посев стандартно: « суспензии возбудителя на плотную питательную среду;
❖ наложение дисков с антимикробными препаратами на поверхность плотной
питательной среды с помощью стерильного пинцета или диспенсера;
инкубация посевов в термостате в течение 18-48 ч:
измерение диаметра зон задержки роста исследуемого микроорганизма
вокруг дисков с антибиотиками с последующей интерпретацией полученных
данных.
Важно, что для выявления устойчивости возбудителей к антимикробным пре-
паратам стандарт CLSI предусматривает определенный набор питательных сред —
агар Мюллера-Хинтона, бульон Мюллера-Хинтона, гемофилезную среду (НТМ).
Выбор этих питательных сред обусловлен целым рядом обстоятельств — хоро-
шими ростовыми качествами, высокой воспроизводимостью и точностью резуль-
татов исследования, оптимальной концентрацией различных ионов (кальция,
магния), низким содержанием ингибиторов антимикробных препаратов, а также
большим опытом тестирования на них различных микроорганизмов во многих
странах мира. Эти среды можно готовить самостоятельно из сухих, однако лучше
применять готовые среды, качество и стандартность которых гарантированы про-
изводителем.
Следует учитывать, что диско-диффузионный метод не дает надежных результа-
тов при определении чувствительности микроорганизмов к плохо диффундирую-
щим в питательную сре^ полипептидным антибиотикам (полимиксины, гликопеп-
тиды). Именно поэтому при использовании этих препаратов для лечения больных
с генерализованной инфекцией для определения чувствительности необходимо
применять методы серийных разведений или «пороговых» концентраций.
Опыт работы с врачами-бактериологами лечебно-профилактических учрежде-
ний и центров гигиены и эпидемиологии свидетельствует о наличии типовых оши-
бок при постановке и учете результатов диско-диффузионного метода, в том числе:
❖ несоблюдение режимов хранения картриджей и флаконов, содержащих
диски с антимикробным препаратом;
❖ использование чашек с неадекватным (большим или меньшим) объемом
питательных сред;
306 БАКТЕРИОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ
отсутствие во многих бактериологических лабораториях адекватного стан-
дарта мутности, эквивалентного 0,5 по стандарту МакФарланда и соответ-
ствующего плотности посевной дозы 1-2x10* КОЕ/мл;
❖ неадекватный подбор дисков с антимикробными препаратами, неравномер-
ное распределение их на поверхности питательной среды и увеличение их
оптимального количества на одной чашке (не более 6 дисков на чашке диа-
метром 90-100 мм);
4- несоблюдение трех «правил 15 минут*:
- посев стандартной взвеси микроорганизмов на чашку с плотной питатель-
ной средой не позднее 15 мин с момента приготовления инокулята;
- подсушивание засеянной чашки с питательной средой не более 15 мин при
комнатной температуре;
- помещение засеянной чашки в термостат не позднее 15 мин после нанесе-
ния дисков с антибиотиками;
о- отсутствие или нарушение адекватного контроля воспроизводимости и точ-
ности результатов определения чувствительности к антибиотикам с исполь-
зованием референтных штаммов микроорганизмов;
❖ слабое знание нормативных документов и литературы по методам определе-
ния чувствительности микроорганизмов к антимикробным препаратам.
Эпсилометрический метод (Е-тест)
Разновидность диффузионного метода определения чувствительности микро-
организмов к антибиотикам — эпсилометрический метод (Е-тест), позволяющий
точно и быстро определять МИК антимикробных препаратов.
Вместо бумажного диска в Е-тесте предусмотрено использование полимер-
ной полоски размером 5x50 мм, откалиброванной в соответствии с градиентом
концентрации антибиотика. Концентрация антимикробного препарата в полоске
может варьировать в зависимости от исследуемого антибиотика в следующих диа-
пазонах — от 0,02 до 32 мкг, от 0,016 до 256 мкг и от 0,064 до 1024 мкг.
Полоски с антибиотиками укладывают на засеянную поверхность плотной
питательной среды, после инкубации посевов в термостате учитывают резуль-
тат. Значение МИК антибиотика определяют в том месте, где эллипсовидная
зона задержки роста вплотную подходит к полимерной полоске (рис, 20-4).
Достоинства метода — возможность быстрого выявления МИК антибиотиков и
простота выполнения, недостатки — высокая стоимость полимерных полосок.
Метод выявления толерантности бактерий к антимикробным препаратам
Впервые понятие «толерантность» появилось в медицинской литературе после
выделения мутантного штамма Str. pneumoniae, который, в отличие от типового
штамма, не терял жизнеспособности при воздействии концентрации бензилпени-
циллина, в несколько раз превышающей МИК, По существу, бензилпенициллин
действовал на данный штамм Str. pneumoniae как бактериостатический препарат.
Рис. 20-4. Полимерная полоска для определения чувствительности микроорганизмов к анти-
микробным препаратам с помощью эпсилометрического метода (Е-теста).
БАКТЕРИОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ 307
Толерантность — утрата чувствительности микроорганизмов к бактерицидному
действию антибиотика при сохранении чувствительности к бактериостатическому
действию. В настоящее время феномен толерантности к антибиотикам описан у
нескольких десятков видов бактерий, большинство из которых трамположительны.
Для выявления феномена толерантности микроорганизма к антимикробным
препаратам определяют МИК и МБК антибиотика ме годом серийных разведении
в жидкой питательной среде. Если МБК превышает МИК в 32 раза и более, микро-
организм считают толерантным к исследуемому антимикробному препарату.
Поскольку данный препарат не обладает клинической эффективностью, для лече-
ния больного подбирают другой антибиотик.
Специальные методы обнаружения устойчивости бактерий к антимикробным
препаратам
Необходимость разработки и внедрения в практику микробиологических лабо-
раторий специальных методов обнаружения устойчивости микроорганизмов к
антимикробным препаратам связана с тремя основными причинами:
❖ диско-диффузионный метод не обладает 100% чувствительностью и не
позволяет выявить все антибиотикорезистентные штаммы возбудителей;
❖ сроки определения антибиотикорезистентности микроорганизмов с помо-
щью диско-диффузионного метода часто не удовлетворяют врачей;
< из-за трудоемкости методы серийных разведений антимикробных пре-
паратов не получили широкого распространения в лабораториях лечебно-
профилактических учреждений.
Метод выявления |3-лактамаз микроорганизмов с использованием
нитроцефинового диска
Тест с нитроцефином*1 используют для быстрого выявления р-лактамаз у
Staphylococcus spp., Haemophilus spp., Enterococcus spp., N. gonorrhoeae, MoraxeUa
catarrhalis и некоторых других бактерий. Возбудители, продуцирующие
р-лактамазы, гидролизуют содержащийся в бумажном диске хромогенный цефа-
лоспориновый антибиотик нитроцефин^, в результате происходит изменение
цвета диска. Положительный результат теста указывает на наличие у исследуемых
микроорганизмов р-лактамазы и свидетельствует об устойчивости возбудителей к
пенициллинам. Отрицательный результат указывает на отсутствие у исследуемых
штаммов бактерий р-лактамазы, однако не всегда свидетельствует о чувстви-
тельности к пенициллинам. Это объясняется тем, что устойчивость возбудите-
лей к р-лактамным антибиотикам может быть связана с другими механизмами
резистентности (например, с изменением структуры пенициллин-связывающих
белков). Достоинства метода — быстрота получения результатов, высокая чув-
ствительность и специфичность. Основной недостаток метода, препятствующий
широкому внедрению его в лабораторную практику, — высокая стоимость ком-
мерческих дисков.
Специальные методы обнаружения устойчивости Staphylococcus spp. к
оксациллину и ванкомицину
Скрининговые тесты определения чувствительности S. aureus к оксациллину
и ванкомицину основаны на использовании плотных питательных сред (среда
MRSA, представляющая собой агар Мюллер-Хинтона с 4% хлорида натрия и
6 мкг/мл оксациллина, агар на сердечно-мозговой вытяжке с 6 мкг/мл ванкоми-
цина соответственно). Появление роста исследуемых штаммов стафилококков на
этих селективных средах свидетельствует об устойчивости к соответствующему
антимикробному препарату.
Диско -диффузионный метод не позволяет выявлять штаммы стафилококков с
пониженной чувствительностью к ванкомицину, поэтому применение его с этой
целью не предусмотрено стандартами CLSI (2009).
308 БАКТЕРИОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ
Для быстрого выявления оксациллин-резистентных стафилококков (ORS)
разработаны тесты, позволяющие обнаруживать ген тес А в ПЦР, а также
пенициллин-связывающий белок 2а в реакции латексагглютинации (находящий-
ся в клеточной стенке ORS дополнительный пенициллин-связывающий белок 2а
кодируется геном тес А и обладает низким аффинитетом к р-лактамам).
Специальные методы обнаружения устойчивости Enterococcus spp. к
аминогликозидам и ванкомицину
Скрининговые тесты для выявления уровней резистентности Enterococcus spp.
к аминогликозидам включают диско-диффузионный метод (с использованием
дисков, содержащих 120 мкг гентамицина или 300 мкг стрептомицина), а также
чашечный и планшетный методы (на чашках Петри с сердечно-мозговым агаром
и в лунках полистиролового планшета с сердечно-мозговым бульоном соответ-
ственно), Концентрация антибиотиков в плотных и жидких питательных средах
составляет 500 мкг/мл для гентамицина и 1000-2000 мкг/мл для стрептомицина.
Наличие роста исследуемых штаммов энтерококков вокруг дисков с гентамици-
ном (стрептомицином) или на указанных питательных средах свидетельствует
о наличии у возбудителей высокого уровня резистентности к аминогликозидам.
Энтерококки с низким уровнем устойчивости считают потенциально чувстви-
тельными к синергическим комбинациям пенициллинов и аминогликозидов (при
отсутствии высокого уровня резистентности к пенициллинам), а штаммы с высо-
ким уровнем резистентности к аминогликозидам устойчивы к данным комбина-
циям.
Достаточно простым и чувствительным способом выявления ванкомицин-
резистентных энтерококков (VRE) является скрининговый тест на сердечно-
мозговом агаре с 6 мкг/мл ванкомицина. Рост 2 и более колоний исследуемых
штаммов энтерококков в месте посева расценивают как предварительный положи-
тельный результат на наличие VRE, однако необходимо подтверждение (диффе-
ренциация с обладающими низким уровнем резистентности к этому антибиотику
Е. gallinarum, Е. casseliflavus, E.flavescens).
Специальные методы обнаружения устойчивости Str. pneumoniae к бен-
зилпенициллину
Несмотря на то, тгго практически повсеместно отмечают рост устойчивости
Str. pneumoniae к р-лактамным антибиотикам, бензилпенициллин остается одним
из наиболее важных антимикробных препаратов для лечения пневмококковой
инфекции. Основной механизм устойчивости пневмококков к бензилпенициллину
и другим р-лактамам заключается в появлении мутантной ДНК в генах, кодирую-
щих пенициллин-связывающие белки.
Для выявления чувствительности Str. pneumoniae к бензилпенициллину исполь-
зуют диски с оксациллином (1 мкг), поскольку диски с бензилпенициллином не
всегда позволяют определить штаммы с промежуточным уровнем резистентности.
Str. pneumoniae с зоной задержки роста вокруг диска с оксациллином >20 мм счита-
ют чувствительными к бензилпенициллину и другим р-лактамным антибиотикам
(ампициллину, амоксиклаву, цефотаксиму, имипенему) и не тестируют с данными
дисками. При выявлении колоний Str. pneumoniae с диаметрами зон задержки
роста вокруг дисков с оксациллином <19 мм необходимо определить чувстви-
тельность к бензилпенициллину, меропенему и цефалоспоринам III поколения
(цефотаксиму или цефтриаксону) методом серийных разведений, поскольку такая
зона задержки роста характерна как для устойчивых, так и для промежуточных и
чувствительных штаммов
Методы выявления р-лактамаз расширенного спектра действия у энте-
робактерий
р-Лактамазы — группа бактериальных ферментов, способных расщеплять
р-лактамные антибиотики, содержащие в структуре циклическую амидную связь.
БАКТЕРИОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ 309
Внедрение в клиническую практику цефалоспоринов III поколения привело к
появлению и широкому распространению в лечебных учреждениях нового класса
[3-лактамаз расширенного спектра действия {Extended Spectrum Beta-Lactamases,
ESBL). Характерные особенности этих ферментов:
о способность расщеплять пенициллины и цефалоспорины (за исключением
цефамицинов);
❖ чувствительность к ингибиторам (клавулановая кислота, сульбактам, тазо-
байтам);
> эпидемический характер распространения (локализация генов резистентно-
сти в плазмидах).
В настоящее время описано свыше 200 различных [3-лактамаз расширенного
спектра действия. Продукция ESBL у госпитальных штаммов микроорганизмов
ведет к формированию резистентности ко всем пенициллинам, цефалоспоринам
и монобактамам, что является причиной клинической неэффективности этих пре-
паратов. Бактерии, продуцирующие [З-лактамазы расширенного спектра действия,
нередко проявляют ассоциированную устойчивость к антибиотикам других групп
(мультирезистентность). Например, резистентность данных микроорганизмов к
гентамицину в некоторых лечебных учреждениях достигает 80%, а к ципрофлок-
сацину — 40-60%. В данной ситуации карбапенемы — практически единственные
антибиотики, сохраняющие клиническую эффективность в отношении этих муль-
тирезистентных возбудителей.
Общепринятых рекомендаций в отношении подлежащего тестированию на
ESBL перечня микроорганизмов в настоящее время не существует. Чаще всего
исследования на наличие этих ферментов проводят у госпитальных штаммов
Klebsiella spp., Е. coli и некоторых других энтеробактерий, у которых выявлена
пониженная чувствительность (МИК >2 мкг/мл или соответствующее умень-
шение диаметра зоны задержки роста) хотя бы к одному из цефалоспоринов
III поколения. Эффективность выявления ESBL с помощью традиционного
диско-диффузионного метода остается крайне низкой. В связи с этим разра-
ботаны специальные фенотипические методы их обнаружения, основанные
на эффекте подавления активности [3-лактамаз в присутствии клавулановой
кислоты (метод двойных дисков, обнаружение ESBL с помощью Е-теста, тест
Oxoid, метод CLSI). Однако следует подчеркнуть, что ни один из данных мето-
дов не обеспечивает 100% детекцию ESBL у грамотрицательных аэробных
бактерий.
Выявление продукции металл о-[3-лактамаз у грамотрицательных бак-
терий
Одна из частых причин устойчивости неферментирующих грамотрицательных
бактерий (Acinetobacter spp.. Pseudomonas spp,) к [3-лактамным антибиотикам —
образование металле-[3-лактамаз.
Металле-[З-лактамазы принадлежат к молекулярному классу В и содержат в
активном центре фермента ионы двухвалентного цинка (Zn2+). В настоящее время
описано 5 генетических типов приобретенных металло-£-лактамаз — GIM, IMP,
SIM. SPM и VIM. У госпитальных штаммов грамотрицательных бактерий чаще
всего обнаруживают типы IMP и VIM. Металле-13-лактамазы обладают высокой
каталитической активностью и разрушают практически все 0-лактамные анти-
биотики, включая цефалоспорины III- IV поколения и карбапенемы. Металло-|3-
лактамазы способны быстро распространяться в условиях лечебного учреждения,
поскольку кодирующие их гены локализованы внутри подвижных генетических
элементов. Сцепление в микробной клетке генов металло-[3-лактамаз с другими
детерминантами резистентности приводит к появлению мультирезистентных
штаммов грамотрицательных бактерий.
310 БАКТЕРИОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ
Приобретенные металло-р-лактамазы чаще всего обнаруживают у НГОБ и
значительно реже — у энтеробактерий. Stenotrnphomonas maltophilia и Burkholderia
cepacia продуцируют видоспецифические метал ло-р-лактамазы, обусловливающие
природную резистентность данных микроорганизмов к карбапенемам.
Активность металло-р-лактамаз не подавляется классическими ингибитора-
ми сериновых р-лактамаз (клавулановая кислота, сульбактам, тазобактам), для
их инактивации требуется применение этилендиаминтетраацетата (ЭДТА) или
2-меркаптопропионовой кислоты. Для выявления металле-р-лактамаз исполь-
зуют метод двойных дисков с ЭДТА или ПЦР. Наиболее значимый маркер про-
дукции металло-Р-лактамаз — пониженная чувствительность или резистентность
грамотрицательных бактерий к карбапенемам.
Метод двойных дисков с ЭДТА
Метод двойных дисков с ЭДТА основан на выявлении синергизма между
р-лактамными антибиотиками (цефтазидимом, имипенемом и меропенемом) и
ЭДТА, который связывает ионы цинка в активном центре металло-р-лактамаз и
подавляет действие этих ферментов в отношении р-лактамов. При определении
чувствительности грамотрицательных бактерий диско-диффузионным методом
в присутствии ЭДТА отмечают расширение зон задержки роста микроорганизмов
вокруг дисков с цефтазидимом, имипенемом и меропенемом. Использование дис-
ков с несколькими р-лактамными антибиотиками повышает чувствительность
метода, поскольку некоторые штаммы грамотрицательных бактерий могут про-
являть синергизм только с одним из перечисленных выше антимикробных пре-
паратов.
Чистую культуру исследуемого микроорганизма доводят до мутности 0,5 по
стандарту МакФарланда и с помощью тампона засевают на среду Мюллера-
Хинтона в чашке Петри. После подсыхания инокулума в центр чашки помещают
пустой диск, а на расстоянии 15 мм между центрами дисков укладывают диски с
цефтазидимом (30 мкг), имипенемом (10 мкг) и меропенемом (10 мкг). На пустой
диск наносят 5 мкл 0,5 М раствора ЭДТА (pH 8,0). Посевы инкубируют в термо-
стате при температуре 35 °C в течение 16-18 ч. Параллельно с исследованием
испытуемых культур проводят анализ контрольных штаммов микроорганизмов.
Расширение зоны подавления роста бактерий между хотя бы одним из дисков с
р-лактамными антибиотиками и диском с ЭДТА указывает на продукцию металло-
р-лактамаз у исследуемого штамма (рис. 20-5).
Чувствительность и специфичность обнаружения металло-р-лактамаз методом
двойных дисков с ЭДТА составляет 96,4 и 99,1% соответственно.
Рис. 20-5. Выявление продукции металло-р-лактамаз у грамотрицательных бактерий методом
двойных дисков с ЭДТА. А и Б — отсутствие металло-Р-лактамаз у исследуемых штаммов бак-
терий.
БАКТЕРИОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ 311
Рис. 20-5. Окончание. В и Г — наличие металл о-^-лактамаз у исследуемых штаммов бактерий.
Выявление металло-р-лактамаз с помощью мультиплексной ПЦР
Обнаружение металло-р-лактамаз с помощью ПЦР основано на выявлении наи-
более распространенных типов генов (IMP и УШ), кодирующих эти ферменты. В
реакции используют 2 пары праймеров к внутренним участкам генов Ыащ 4 и Ыа 1Н.
Продукты амплификации детектируют с помощью электрофоретического анализа,
а также в режиме реального времени. Метод мультиплексной ПЦР для выявления
мет алли-р-лактамаз грамотрицательных бактерий обладает 100% чувствительно-
стью и специфичностью.
Глава 21
Частная микробиология
I. ГРАМОТРИЦАТЕЛЬНЫЕ БАКТЕРИИ
ХЛАМИДИИ
ТАКСОНОМИЯ
Хламидии относятся к семейству Chlamydiaceae а-протеобакте-
рий. Ранее хламидии подразделяли на 3 вида: Chi. trachomatis (воз-
будитель трахомы-лимфогранулемы, урогенитального хламидиоза
и ряда других болезней человека), Chi. psittaci (возбудитель орни-
тоза и хламидиозного аборта овец), Chi. pneumoniae (возбуди-
тель пневмонии человека). Сравнение 16S и 23S рибосомальной
РНК, биохимических, серологических и экологических свойств
этих видов привело к изменению классификации. Стали выделять
9 видов, разделенных на 2 рода: Chlamydia (Chi. trachomatis, Chi. mun-
darum и Chi, suis) и Chlamydophila (Chi abortus. Chi. cavtae, Chl.felus,
Chl.pecurum, Chi. pneumonia и Chi. psittaci).
В этом разделе приведена детальная информация о Chi. trachoma-
tis, а также о принципах и методах ее диагностики.
ОБЩАЯ ХАРАКТЕРИСТИКА CHL TRACHOMATIS
Chi. trachomatis — грамотрицательная бактерия, являющаяся
облигатным внутриклеточным паразитом человека. Биологическим
своеобразием хламидий, которое заключается в энергозависимом
паразитизме и уникальном цикле развития, определяется самостоя-
тельное положение данных микроорганизмов в системе прокариот.
Хламидии имеют сходную структуру с липополисахаридным
антигеном и распознаются моноклональными антителами, специ-
фичными к трисахаридному фрагменту alphaKdo-(2-8)-alphaKdo-
(2-4)-alphaKdo липополисахарид (ЛПС). Именно поэтому многие
из них в ПИФ и серологических тестах идентифицируются как
Chi. trachomatis. Chi. trachomatis имеет 2 биовара и 15 сероваров,
вызывает трахому, урогенитальные заболевания, некоторые формы
артрита, конъюнктивит и пневмонию новорожденных. Возбудитель
урогенитального хламидиоза — Chi. trachomatis сероваров D, Da. Е,
F, G, Н, I, la,J, К.
Хламидии отличаются от других бактерий уникальным жиз-
ненным циклом, который включает последовательную смену двух
ЧАСТНАЯ МИКРОБИОЛОГИЯ 313
высокоспециализированных форм, адаптированных для внутри- и внеклеточного
существования.
Элементарные тельца — внеклеточная инфекционная форма хламидий.
Элементарные тельца имеют размер 250-500 нм, овальную форму, окружены
ригидной малипроницаемой клеточной стенкой и содержат плотный нуклеотид
и обезвоженный протопласт, что обеспечивает резистентность к факторам внеш-
ней среды при переносе от клетки к клетке и от одного хозяина к другому. Цикл
развития хламидии происходит только внутри вакуолей, связанных с мембраной
клетки-хозяина, где элементарные тельца последовательно преобразовываются в
более крупные — ретикулярные.
Ретикулярные тельца обладают полиморфизмом и могут иметь округлую,
овальную, полулунную форму, не имеют постоянного размера (величина варьиру-
ет от 300 до 1000 нм), не обладают инфекционными свойствами, не имеют нуклео-
тида, проницаемы для трипсина и разрушаются ультразвуком. Ретикулярные
тельца — так называемая вегетативная внутриклеточная форма, способная к росгу
и делению.
В клинических исследованиях показано, что одни антибактериальные средства
могут действовать на стадии превращения элементарных телец в ретикулярные,
другие — во время бинарного деления ретикулярных телец. Элементарные тельца
обычно локализуются в крупных внутриклеточных включениях, ретикулярные —
в образованиях, ограниченных мембраной.
Инку бационный период хламидиоза длится 5-30 дней с момента заражения (в
среднем 10-14 дней). Первый этап инфекционного процесса — адсорбция элемен-
тарных телец хламидий на плазмалемме чувствительной клетки организма хозяи-
на под действием электростатических сил. Внедрение хламидии происходит путем
эндоцитоза в течение 7-10 ч. При этом элементарные гельца, внедряясь в клетку,
используют для роста и размножения клеточную АТФ и через 6-8 ч превращаются
в ретикулярные тельца. Хламидии способны синтезировать в небольших количе-
ствах собственную АТФ, отдельные аминокислоты, накапливать гликоген, однако
в основном они существуют за счет макроергических соединений и биологиче-
ских субстратов клетки хозяина. В процессе роста и деления ретикулярных телец
образуются микроколонии — хламидийные включения (тельца Гальберштадтера-
Провачека). В течение 18-24 ч развития они локализуются в цитоплазматическом
пузырьке, образованном из мембраны клетки-хозяина.
Вступая во взаимодействие с чувствительными клетками, хламидии являются
активными паразитами, используя различные факторы патогенности. Проникнув
в клетку, жизнеспособные хламидии проявляют строго специфическую актив-
ность, направленную против лизосом с окружающей их фагоцитарной вакуолью,
ограниченной участком цитоплазматической мембраны клетки-хозяина, отде-
лившимся при поглощении фагоцитом возбудителя инфекции. Таким образом,
они выключают важнейший защитный механизм клетки хозяина — фагоцитоз,
обеспечивая себе возможность дальнейшего размножения в цитоплазматическом
включении клеток макроорганизма. В патогенезе урогенитальных хламидиозов,
помимо непосредственного действия возбудителя на инфицированные клетки
эпителия, выстилающего мочеполовые пути, повреждения окружающих тканей
гидролитическими лизосомальными ферментами, выбрасываемыми из инфици-
рованных клеток в период размножения микроорганизма, и токсического влияния
продуктов аутолиза разрушенных клеток, существенное значение имеет токсиче-
ская активность, свойственная всем хламидиям. В микроколониях может содер-
жаться от 100 до 500 элементарных телец.. Процесс созревания и трансформации
ретикулярных телец путем превращения в элементарные тельца занимает 36-42 ч.
Полный цикл репродукции происходит за 48-72 ч и завершается разрушением
пораженной клетки. Хламидии могут высвобождаться из инфицированной клетки
314
ЧАСТНАЯ МИКРОБИОЛОГИЯ
путем экзоцитоза, сохраняя ее жизнеспособность, что приводит к бессимптомному
течению хламидийной инфекции.
При неблагоприятных биохимических условиях продолжительность жизненно-
го цикла хламидий может значительно увеличиваться В результате размножения
Chi. trachomatis в инфицированных эпителиальных клетках развивает ся воспали-
тельный процесс, выраженность которого зависит от состояния местного и обще-
го специфического и неспецифического иммунитета. После инвазии хламидий
образуется комплемент, который приводит к стимуляции гранулоцитов. Эта вос-
палительная реакция сопровождается локальной гибелью клеток и разрушением
ткани, что дополняется высвобождением фосфолипазы А2 и простагландинов, что
приводит к отеку и гиперемии слизистой оболочки, нарушению целостности эпи-
телиального слоя с частичной десквамацией эпителия. Превращение ретикуляр-
ных телец в элементарные не только требует энергии, но и зависит от присутствия
биологических субстратов, среди которых важнейшее место занимает триптофан.
В процессе воспаления лимфоциты выделяют цитокины, в том числе у-ИФН,
который индуцирует внутриклеточную индоламиндиоксигеназу, разрушающую
триптофан. Снижение содержания триптофана приводит к гибели хламидий или
замедлению процессов репликации, при этом в эпителиальной клетке начинают
накапливаться неинфекционные, неделящиеся ретикулярные тельца. Цикл раз-
вития хламидий приостанавливается, однако, несмотря на неблагоприятные усло-
вия, ретикулярные тельца сохраняют свою жизнеспособность (персистируют).
Дальнейшее развитие и исход данного состояния зависят от уровня содержания
триптофана. Уменьшение содержания у-ИФН и увеличение внутриклеточного
пула триптофана приводит к редифференцировке персистирующих форм хлами-
дий в инфекционные элементарные т ельца, их выходу из клетки и продолжению
инфекционного процесса.
Клеточная стенка хламидий состоит из двух двойных мембран — внутренней
(цитоплазматической) и наружной, что обеспечивает прочность клеточной стенки.
Антигенные свойства хламидий определяются внутренней мембраной, которая
представлена липополисахаридами. ЛПС имеют две антигенные детерминанты.
Первая — родоспецифическая детерминанта, реактивная половина которой пред-
ставлена 2-кето-З-дезоксиоктановой кислотой, используется при диагностике
заболевания иммунофлюоресцентными методами с применением специфических
антител. Вторая — антигенная детерминанта — дает перекрестные реакции с неко-
торыми грамотрицательными бактериями.
Во внутреннюю мембрану интегрированы так называемые белки наружной
мембраны (ОМР). На основной белок наружной мембраны (МОМР) приходится
60% общего количества белка. Остальная антигеннная структура представлена
белками наружной мембраны второго типа — ОМР-2 (насыщенным цистеином
белком с молекулярной массой 60 кДа). МОМР и ОМР-2 определяют видо- и
типоспецифичность хламидий, хотя из-за наличия участков с высоким сход-
ством внутри рода возможны перекрестные реакции между различными видами
(табл. 21-1).
Таблица 21-1. Антигены Chi. trachomatis (по Mardh Р.А., 1990)
Антиген Химический состав Примечания
Общий для разных хламидий ЛПС Три различных антигенных домена
Видоспецифический Белок Более 16 различных компонентов 155 кДа у Ch!, trachomatis, эпитопы в белке 40 кДа, белок теплового шока lisp 60
Типоспецифический Белок Эпитопы в 40 кДа протеине (МОМР), протеине 30 кДа у серотипов А и В
ЧАСТНАЯ МИКРОБИОЛОГИЯ 315
МОМР и богатые цистеином белки соединены дисульфидными связями.
Обнаружено 5 генов дисульфидсвязанных изомераз, играющих роль при диф-
ференциации элементарных телец в ретикулярные. У Chi. trachomatis выявлено
9 генов, кодирующих ОМР. МОМР считается первичным местом взаимодействия
с Т-лимфоцитами у Chi trachomatis. Соответственно гуморальный иммунный ответ
направлен против вариабельных поверхностно-экспонированных доменов дан-
ного белка. Присутствие МОМР Chi. trachomatis в ткани маточных труб женщин
связывают с бесплодием и хроническим сальпингитом, а также с билатеральной
окклюзией маточных труб. Средний титр антихламидийных IgG значительно
выше у пациентов с МОМР по сравнению с больными, у которых данный анти-
ген не выявлялся. ОМР-2 Chi. trachomatis также является участком связывания с
иммунной системой. Инвазивность хламидий связывают со строением углеводной
части главного ЛПС их внешней мембраны. Выявлены токсичность и спермицид-
ные свойства ЛПС клеточной стенки хламидий для сперматозоидов.
В составе хламидий имеется белок теплового шока (БТШ), который по анти-
генному составу гомологичен БТШ человека. Персистирующие формы хламидий
способствуют образованию высокоиммуногенного БТШ с молекулярной массой
60 кДА (БТШ-60). При развитии инфекционного процесса значительно повыша-
ется синтез БТШ хламидий для защиты от внешних иммунологических механиз-
мов. Иммунный ответ организма хозяина на БТШ может быть как защитным, так
и патологическим.
Существует мнение, что образуемые в организме хозяина противохламидий-
ные антитела одновременно являются аутоантителами к собственному БТШ-60,
который представляет собой один из первых белков, синтезируемых в организме
женщины эпителиальными клетками decidua bazdlis после оплодотворения. На
ранних стадиях беременности у женщины с хронической хламидийной инфекцией
экспрессия БТШ-60 может реактивировать лимфоциты, сенсибилизированные
хламидийным БТШ-60, что приводит к оттоожению эмбриона. Данный механизм
может иметь существенное значение в патогенезе женского бесплодия. Доказано
наличие клинических и гистологических корреляций иммунореактивности к
Chi. trachomatis и эпитопам БТШ-60 у женщин с эктопической беременностью и
женщин с самопроизвольными выкидышами. У них регистрировались специфи-
ческие антитела к 13 синтетическим пептидам, соответствующим главным эпи-
топам БТШ-60. Сальпингит был обнаружен примерно у половины пациенток с
внематочной беременностью, его наличие коррелировало с выявлением антител
к двум эпитопам БТШ-60. Пациентки с эктопической беременностью имели боль-
ший уровень антихламидийных IgG по сравнению с больными с внутриматочной
локализацией беременности. В группе инфицированных Chi. trachomatis женщин,
у которых выявлялись специфические антитела к эпитопам БТШ-60, отмечалась
большая частота возникновения сальпингита и спаечного процесса в области при-
датков матки. Пациентки, у которых выявлялись специфические антитела только
к Chi. trachomatis, не отличались от серонегативных женщин по данным катего-
риям, поэтому полагают, что индукция спаечного процесса в области придатков
матки в присутствии Chi. tracnomatis определяется наличием специфических анти-
тел к БТШ-60.
КЛИНИЧЕСКИЕ ПРОЯВЛЕНИЯ ИНФЕКЦИИ
Chi. trachomatis вызывает у человека ряд заболеваний: трахому, паратрахому,
венерическую лимфогранулему, урогенитальный хламидиоз, конъюнктивит и
пневмонию новорожденных.
Резервуаром инфекции служит человек (животные в естественных условиях к
этой хламидии невосприимчивы). Около 5% мужчин и 10% женщин являются
бессимптомными носителями.
316 ЧАСТНАЯ МИКРОБИОЛОГИЯ
Основные пути передачи — половой, перинатальный и бытовой (при близких
контактах с источником инфекции).
Трахома и паратрахома. Заболевание распространено повсеместно: ежегодно
в мире заболевает до 400-500 млн человек, из которых у 10-20 млн трахома ста-
новится причиной потери зрения. В нашей стране заболеваемость резко снизилась
после массовых профилактических мероприятий в 20-30-х гг.
Инкубационный период колеблется от 8 до 16 дней. Чаще поражаются оба
глаза. Болезнь носит хронический характер и протекает постадийно:
• на первой стадии резко выражено воспаление, на фоне гиперемии и инфиль-
трации конъюнктивы в области переходных складок и век появляются круп-
ные фолликулы и сосочки;
• на второй стадии часть фолликулов распадается, часть сливается друг с дру-
гом, что придает конъюнктиве студенистый вид, появляются рубцы. На этой
стадии больные наиболее контагиозны;
• на третьей стадии превалирующим становится процесс рубцевания конъ-
юнктивы, хотя сохраняются признаки воспалительного процесса, роговица
мутнеет, может формироваться заворот век;
• на четвертой стадии конъюнктива имеет белесый вид, с большим количе-
ством звездчатых по форме рубцов, признаки воспаления отсутствуют.
Интенсивность симптоматики может варьировать в широких пределах, а
описанные выше клинические проявления в ряде случаев бывают стертыми и
труднораспознаваемыми. Одну из форм болезни с наиболее доброкачественным
течением принято называть паратрахомой. Этот синдром, вызываемый уроге-
нитальными сероварами возбудителя, начинается с острого конъюнктивита, для
которого характерны резкая гиперемия конъюнктивы век и переходных складок,
ее отек и инфильтрация. Типично появление крупных рыхлых фолликулов, рас-
положенных рядами в нижнем своде; в дальнейшем фолликулы могут сливаться,
образуя горизонтально расположенные валики. Однако, в отличие от трахомы,
фолликулы конъюнктивы полностью исчезают без образования рубцов. Болезнь
протекает остро, изредка принимая подострое или хроническое течение.
Урогенитальный хламидиоз. Урогенитальный хламидиоз проявляется двумя
основными формами:
• уретритом, который начинается незначительным зудом, дизурией, умерен-
ным появлением серозных, реже серозно-гнойных выделений из уретры.
Первоначально поражается передний отдел уретры, в последующем инфек-
ция может распространяться восходящим путем, поражая предстательную
железу. Симптоматика этой формы инфекции может значительно меняться
при смешанном течении хламидиоза с другими венерическими болезнями
(в первую очередь трихомонозом), поскольку трихомонады фагоцитируют
Chi. trachomatis, перенося в непораженные отделы уретры;
• цервицитом, проявляющимся серозно-гнойным воспалением задней части
влагалища и шейки матки.
Урогенитальный хламидиоз нередко вызывает осложнения со стороны мочепо-
ловой и других систем органов. Из таких осложнений наиболее часто регистрируют
эпидидимит, орхоэпидидимит (способен вызывать бесплодие), реже стриктуру уре-
тры, геморрагический цистит и системные поражения, в том числе синдром Райтера.
Последний характеризуется последовательным развитием у больного уретрита,
поражений глаз (конъюнктивита, иридоциклита, увеита) и реактивного артрита.
Клиническая карт ина разных форм урогенитального хламидиоза и их ослож-
нений неспецифична, напоминает симптоматику ИППП другой этиологии. Кроме
того, инфекция нередко протекает бессимптомно. Это диктуют необходимость
использования лабораторных методов диагностики для постановки окончатель-
ного диагноза.
ЧАСТНАЯ микробиология зп
Перинатанальный хламидиоз новорожденных. Возникает в результате
заражения детей во время родов. Может протекать бессимптомно (чаще всего
возбудитель локализуется в носоглотке). При остром течении инфекции у детей
в первые несколько суток после рождения развивается конъюнктивит (данный
синдром часто называют бленнореей с включениями). Прогрессирующая коло-
низация носоглотки может вызывать у младенцев в течение первых 3 мес жизни
пневмонию.
ЛАБОРАТОРНАЯ ДИАГНОСТИКА
Лабораторная диагностика инфекции С/?/, trachomatis включает выявление
антигенов хламидий в реакции иммунофлюоресценции (РИФ) и одноэтап-
ном иммунохроматографическом тесте (ИХТ), выделение их на лабораторных
моделях перевиваемых клеточных культур, обнаружение генома хламидий
молекулярно-биологическими методами и выявление специфических антител в
крови.
Диагноз хламидиоза считается подтвержденным при положительном резуль-
тате метода культуры клеток или двух других методов. Важно выбрать из них
надежные, которые позволят правильно поставить диагноз и своевременно прове-
сти и проконтролировать специфическое лечение. В спорных случаях предлагают
использовать еще один дополнительный метод — арбитражный.
Для того чтобы правильно выбрать способ диагностики инфекций, ассоцииро-
ванных с Chi. trachomatis, необходимо сравнить методы, которыми пользуется каж-
дая конкретная лаборатория, оценить их чувствительность, специфичность, про-
гностическую значимость положительного и отрицательного результата. Пррвое
требование к эффективному исследованию при подозрении на хламидиоз — выбор
надежных диагностических тестов. В табл. 21-2 представлены различные методы
лабораторной диагностики хламидийной инфекции, рекомендуемые ECDC, указа-
ны их достоинства и недостатки.
Таблица 21-2. Методы диагностики хламидиоза
Диагностические методы Паьимущества Недостатки
Тесты амплификации нуклеи- новых кислот (ПЦР, SDA, TDA) Высокая чувствительность (90-95%), возможность использования для исследования образцов мочи и вуль- вовагинальных смывов, в том числе взятых пациентками самостоятельно, при экстра^енитальной локализации хламидиоза, в частности ректальной Высокая стоимость, возможны ложноположигельные результаты, не лицензированы для эксграге- нитальной локализации
ИФА Может быть адаптирован для единич- ных исследований, низкая стоимость Низкая чувствительность (40-70%), не подходит для иссле- дования образцов мочи и само- стоятельно взятых образцов
Культуральный метод Возможность использования для исследования всех типов образцов, высокая специфичность Низкая чувствительность (60-80%), высокая стоимость, трудоемкость, требует высокой квалификации исследователя, не подходит для массовых иссле- дований
РИФ Возможность использования для исследования всех типов образцов, быстрое получение ответа Низкая чувствительность при исследовании образцов мочи, трудоемкость, необходимость подтверждения
Описаны новые варианты Chi. trachomatis, которые могут не определяться стан-
дартными методами. В этих случаях необходимо повторное исследование образца
с использованием нескольких методов.
318
ЧАСТНАЯ МИКРОБИОЛОГИЯ
Взятие материала для исследований
Эффективность диагностики хламидийной инфекции зависит от правильности
взятия клинического материала, условий его доставки в лабораторию и исполь-
зования высококачественных диагностических тестов. Для обнаружения хлами-
дий исследуют различный клинический материал, чаще всего мазки-соскобы со
слизистой оболочки уретры у мужчин и из цервикального канала и слизистой
оболочки уретры у женщин. С внедрением методов молекулярной биологии стало
возможным получение неинвазивных клинических образцов, таких как свободно
выпущенная моча у мужчин и отделяемое влагалища у женщин. При необходимо-
сти материал для исследования берут из прямой кишки, носоглотки, конъюнктивы
нижнего века. У детей исследуют отделяемое конъюнктивы нижнего века, задней
стенки глотки, вульвы у девочек По клиническим показаниям возможно иссле-
дование биопсийных и операционных материалов. Материал можно получать с
помощью ложечки Фолькманна, специальной щеточки или дакронового зонда-
тампона.
Качество полученного клинического материала зависит от общего состояния
пациента на момент взятия мазка. Наиболее информативен клинический матери-
ал, полученный при условиях, когда:
• исследование проводилось при клинических признаках заболевания;
• пациент не использовал лекарственные средства местного применения в
последние 48-72 ч;
• больной не принимал антибактериальные препараты в последние 3-4 нед;
• у женщин при исследовании отделяемого или мазков-соскобов из урогени-
тального тракта взятие образцов проводилось приблизительно в середине
менструального цикла (если заболевание не имеет явных проявлений) или в
дни, когда отсутствовали кровянистые выделения (при обострении инфекци-
онного процесса);
• у мужчин взятие образцов из уретры проводилось при задержке мочеиспу-
скания в течение 1,5-2 ч.
Несоблюдение этих условий может влиять на качество исследования и искажать
его результаты.
Для исследования материалов, полученных из цервикального канала, уретры,
прямой кишки, носоглотки, конъюнктивы, а также биопсийных и операционных
материалов, чаще пользуются культуральным методом., ПИФ, ПЦР и ИФА. Для
исследования первой порции мочи и отделяемого влагалища применяют только
ПЦР.
При взятии материала для этих исследовании рекомендуют придерживаться
приводимых ниже рекомендаций.
Взятие материала для РИФ и ИХТ. У мужчин при наличии выделений из уре-
тры поверхность головки полового члена и область наружного отверстия уретры
должны быть очищены с помощью марлевого тампона, а крайняя плоть отведена
назад для предупреждения контаминации материала посторонней микрофлорой.
Специальную цитощеточку вводят на 1,5-2 см в уретру, вращают е течение 15 с и
выводят.
У женщин образец из уретры берут так же. как и у мужчин. При большом количе-
стве выделений наоужное отверстие уретры должно быть предварительно очищено
с помощью ватного тампона. Образец из шейки матки берут с помощью влагалищ-
ного зеркала и дакронового зонда-тампона, одноразовой цервикальной щеточки
или ложечки Фолькманна. Необходимо тщательно очистить наружное отверстие
цервикального канала от влагалищных выделений с помощью большого марлевого
тампона, что предотвратит контаминацию проб посторонней микрофлорой. После
введения в цервикальный канал на 1-2 см зонд-тампон вращают несколько раз.
Исследуемый материал переносят с зонда-тампона на предметное стекло.
ЧАСТНАЯ МИКРОБИОЛОГИЯ 319
Для взятия мазка с конъюнктивы необходимо провести зондом-тампоном по
нижнему веку.
Для взятия мазка из носоглотки специальным тампоном проводят по задней
стенке глотки.
Взятый материал помещают на обезжиренное предметное стекло вращатель-
ным движением, при этом цитощеточкой прокатывают по стеклу.
Препараты высушивают на воздухе при комнатной температуре.
Взятие материала для культурального исследования. Клинический мате-
риал, предназначенный для изоляции хламидий в культуре клеток, помещают
в стерильные контейнеры со специальной транспортной средой для хламидий.
Правила взятия материала аналогичны приведенным выше. Для забора материала
используют цитощеточки и зонды-тампоны, которые помещают в специальные
пробирки с транспортной средой. При отсутствии транспортной среды промыш-
ленного производства ее готовят из среды RPMI-1O40. в которую добавляют 5%
фетальной сыворотки ити сыворотки крови крупного рогатого скота, антибио-
тики (бензилпенициллин, стрептомицин, гентамицин, канамицин) и преднизолон
(3 мкг/мл). Среды, не предназначенные для этой цели, могут быть токсичными
для клеток, что приводит к ложноотрицательным результатам исследования.
Взятие материала для молекулярно-биологических исследований.
Клинический материал для молекулярно-биологических исследований собирают
в пробирки типа «Эппендорф» с транспортной средой, рекомендуемой произво-
дителем тест-системы.
Первую порцию свободно выпущенной мочи в количестве 5-10 мл собирают
после тщательного туалета гениталий в стерильную сухую пробирку.
Взятие крови для серологического исследования. Для серологического
исследования в сухую пробирку без консерванта берут капиллярную или венозную
кровь объемом 1,0-1,5 мл. Предпочтительнее нользоваться специальными ваку-
умными пробирками, предотвращающими заражение проводящего венепункцию
персонала и гемолиз отбираемых проб. С этой целью можно применять вакуумные
пробирки без наполнителей или с ускорителями свертывания крови.
Транспортировка материала. Материалы, предназначенные для культураль-
ной диагностики, необходимо хранить в холодильнике при температуре 4-6 СС и
в течение 4 ч доставить в лабораторию. Пробы доставляют в лабораторию по воз-
можности в течение 1,5-2 ч после забора и пи хранят при 2-6 °C в течение 1 сут.
При необходимости их можно заморозить при температуре -20 °C и доставить в
лабораторию в течение 48 ч, обложив льдом. Исследование требует сохранения
живых хламидий, поэтому колебания температуры при транспортировке могут
сыграть решающую роль в получении достоверного результата. Во избежание
этого транспортировку проб осуществляют в термосумках; с помошью специаль-
ных индикаторов можно судить об изменениях в них температуры.
Препараты, предназначенные для исследования иммунолюминесцентными
методами, необходимо в тот же день доставить в лабораторию. В случае необходи-
мости препараты, фиксированные в течение 1- 2 мин холодным ацетоном, можно
хранить при температуре 6 °C в течение 3 дней, а при температуре -20 °C — 1 мес.
Образцы с материалом для ПЦР можно хранить не дольше 3 дней до отправки
в лабораторию при температуре 4 °C.
Пробы мочи должны быть доставлены в лабораторию не позднее 24 ч после
сбора. При необходимости данные образцы можно хранить в течение 1 мес при
температуре -20 °C.
Кровь для серологического исследования доставляют в лабораторию в течение
4 ч. После отделения плазмы крови материал хранят в течение 3-4 мес при темпе-
ратуре -20 °C.
320 ЧАСТНАЯ МИКРОБИОЛОГИЯ
Обнаружение возбудителя в клиническом материале
Люминесцентной микроскопией выявляют хламидии в мазках клинического
материала и зараженных культур клеток, приготовленных на предметных стеклах.
Высушенные на воздухе препараты фиксируют охлажденным до 4 °C ацетоном
в течение 10 мин. Затем их обрабатывают специфическими моно- или поликло-
нальными антителами, меченными флюоресцеином. Дальнейшую обработку
препаратов, учет и опенку результатов проводят в соответствии с инструкцией к
диагностической тест-системе.
Чувствительность и специфичность РИФ при анализе клинического материала
на наличие хламидий составляют 80-90 и 98-99% соответственно. Однако он не
предназначен для исследования материалов, полученных из прямой кишки, носо-
глотки, а также проб мочи. Для его проведения нужны дорогостоящие реактивы
(иммунофлюоресцируюшие антитела) и специальная аппаратура.
Одноэтапные хроматографические тесты находят все более широкое при-
менение в медицинской практике. Их преимущества состоят в простоте, низкой
стоимости, возможности получения результата в течение 5-10 мин (это дает воз-
можность использовать их непосредственно у постели больного или во время его
первичного осмотра в поликлинике). Разработаны такие тесты и для диагностики
инфекции Chi. trachomatis. На нитроцеллюлозных тест-полосках сорбирован конъ-
югат частиц коллоидного золота с моноклональными антителами к липополисаха-
ридному антигену бактерии. При наличии в тестируемом клиническом материале
мочи, смывов из половых органов и уретры конъюгат формирует цветную полосу.
Данный метод не позволяет дифференцировать инфекции разных хламидий,
поскольку их липополисахариды дают перекрестные реакции. Другой существен-
ный недостаток выпускаемых в настоящее время гест-систем ИХТ состоит в их
невысокой чувствительности, что позволяет использовать их преимущественно
для скрининга: отрицательный результат ИХТ не доказывает отсутствия у пациен-
та хламидиозной инфекции и нуждается в подтверждении результатами исследо-
вания проб другими методами.
Изоляция и идентификация возбудителя
Один из наиболее объективных методов лабораторной диагностики инфекции —
выделение Chi. tracnomatis из пораженных тканей в культуре клеток. Он трудое-
мок и длителен — для получения ответа требуется от 7 до 14 дней. В то же время
культуральный метод уступает другим методам в чувствительности, которая при
исследовании одного образца из цервикального канала составляет 75- 80% по
сравнению с РИФ и 40-60% по сравнению с ПЦР. Изоляты, выделенные в культу-
ре клеток, идентифицируют в РИФ или ИФА.
Клеточные культуры перевиваемых линий (McCoy, L929) первоначально выра-
щивают во флаконах из нейтрального стекла или пластика в питательной среде
RPMI-1640 с добавлением 10% фетальной сыворотки и антибиотиков. После фор-
мирования на стенке флакона клеточного монослоя питательную среду полностью
удаляют, клетки промывают раствором Хенкса, покрывают подогретым до 37 ’С
раствором Версена и помещают флакон в термостат при температуре 37 °C на
несколько минут до набухания клеток. Затем раствор Версена полностью уцаляют
и вносят свежую питательную среду. Количество вносимой питательной среды
должно быть в 4 раза меньше ростовой. Клетки тщательно суспендируют и под-
считывают концентрацию с помощью камеры Горяева. Посевная концентрация
клеток составляет 150 000-200 000 клеток в 1 мл. Суспензию клеток разливают по
2 мл в стеклянные флакончики с пластинками-подложками из покровных стекол.
Инкубируют клетки в термостате при 37 °C в наклонном положении под углом
7-10° до образования клеточного монослоя
Перед заражением все флаконы просматривают под малым увеличением инвер-
тированного микроскопа, отбраковывая те из них, которые не имеют сплошного
ЧАСТНАЯ МИКРОБИОЛОГИЯ 321
клеточного монослоя. Из флаконов с клеточными культурами, предназначен-
ных для выделения хламидий, полностью удаляют питательную среду. Каждый
исследуемый материал внося г не менее чем в 2 флакона в таком объеме, чтобы
пластинка с клетками была полностью покрыта (не менее 0,3 мл). Пробирки
на 1,5-2 ч помещают в термостат для адсорбции хламидий, затем исследуемый
материал удаляют, монослой клеток однократно промывают раствором Хенкса и
вносят во флаконы поддерживающую среду без антибиотиков с 2% сывороткой
и 3 мкг/мл преднизолона. Флаконы помещают в термостат при 36 °C на 72 ч.
После окончания срока инкубации пластинки извлекают из флаконов, промывают
0.9% раствором NaCl, высушивают, приклеивают к предметному стеклу клеем БФ-2
клеточным монослоем вверх и фиксируют в охлажденном ацетоне. Выявление
хламидий в препаратах клеточных культур проводят с помощью ПИФ.
Выделение ChL trachomatis в культуре клеток позволяет не только диагностиро-
вать инфекцию, но и определить чувствительность выделенного изолята к антими-
кробным препаратам, что необходимо для выбора наиболее эффективных средств
лечения пациента. С этой цель ю проводят 2 слепых пассажа клинических образцов
на клеточных культурах с увеличением числа заражаемых флаконов в первом
пассаже до 4, во втором - до 10. Перед заражением культуры клеток исследуемый
материал подвергают предварительному замораживанию и оттаиванию для более
полного выхода возбудителя в питательную среду. После третьего пассажа из
одного флакона вынимают пластинку-подложку и проводят оценку степени нако-
пления хламидий с помощью иммунофлюоресцентного анализа. Из оставшихся
9 флаконов готовят смешанную пробу. При накоплении хламидий в контрольной
пробирке более чем на 3+ проводят анализ на антибиотикорезистентность.
Для этого флаконы с культурой клеток засевают изолятом хламидий и поме-
щают в термостат при температуре 36 °C. Начиная с первых суток в них вносят
антибиотики в среде RPMI-1640 в трех концентрациях — среднетерапевтической,
в 2 раза большей и в 2 раза меньшей. Например, при среднетерапевтической суточ-
ной дозе доксициклина 0,1 г тестируют разведения 50,100 и 200 мкг/мл. На каж-
дое из них берут не менее 2 флаконов. Через 5 сут пластинки-подложки извлекают
из флаконов, промывают 0,9% раствором NaCl, высушивают, закрепляют на пред-
метном стекле, фиксируют, окрашивают люминесцирующей сывороткой и оце-
нивают антибиотикорезистентность с помощью люминесцентной микроскопии.
О чувствительности хламидий к антибиотикам судят по отсутствию в препарате
люминесцирующих морфологически типичных элементов.
Молекулярная диагностика
Наиболее перспективными и высокочувствительными методами обнаружения
хламидий являются молекулярно-биологические — ПЦР, реакция гибридиза-
ции, РНК-детекция, SDA, NASBA. ПЦР, проводимая в режиме реального вре-
мени (ПЦР-РВ), позволяет совместить амплификацию с детекцией накопления
ее продуктов в период проведения реакции. В реакционную смесь добавляют
флюоресцентно-меченые зонды (ФМ-зонды), при появлении специфического
продукта амплификации зонды гибридизуются с ним, вызывая увеличение уровня
флюоресцентного сигнала, который считывается флюориметрическим детекто-
ром; обработка сигнала ведется с помощью программного обеспечения прибора.
Применение ПЦР-РВ не предполагает постамплификационный анализ продуктов
реакции и извлечение контаминационно опасного содержимого пробирок. Общее
время исследования позволяет получить результат уже через 1,5-2 ч после посту-
пления клинического материала в лабораторию. Данный метод обеспечивает
количественную оценку содержания ДНК микроорганизма в клинической пробе,
так как установлена прямая зависимость исходного количества а модифицируемой
ДНК от начала регистрации флюоресцентного сигнала. На основании этого были
разработаны протоколы определения концентрации ДНК-возбудителей.
322 ЧАСТНАЯ МИКРОБИОЛОГИЯ
Существует корреляция между выраженностью клинических проявлений при
хламидийной инфекции и концентрацией Chi. trachomatis в отделяемом урогени-
тального тракта. При анализе причин неудачи лечения некоторых пациентов с
хламидиозом было выдвинуто предположение о возможной связи между исхо-
дной высокой плотностью колонизации хламидиями и формированием гетероти-
пической устойчивости хламидий к антибактериальным препаратам. Возможно,
определение исходной концентрации ДНК Chi. trachomatis позволит прогнозиро-
вать ответ на антибактериальную терапию и выбирать наиболее оптимальную
схему лечения. Выявление низкой концентрации ДНК Chi. trachomatis в отделяе-
мом урогенитального тракта позволит более объективно оценивать и объяснять
несовпадающие результаты амплификационных и неамплификационных методов
диагностики хламидийной инфекции.
В качестве альтернативы ПЦР, в основе которой лежит принцип удвоения
генетического материала при делении клеточных систем, предложен метод, моде-
лирующий in vitro репликацию ретровирусов, где мишенью для амплификации
служит одноцепочечная молекула РНК. Метод, названный Self-Sustained Sequence
Replication, или 3SR. базируется на конкурентном действии 3 ферментов, участвую-
щих в ретровирусной репликации, — обратной транскриптазы, рибонуклеазы Н
(РНКазы Н) и ДНК-зависимой РНК-полимеразы. Реакция протекает с участием
специфических праймеров при постоянной температуре. В результате реакции
амплификации за 60-90 мин образуется до 10у- Ю12 копий РНК. На основе мето-
дики 3SR была разработана технология ПЦР под названием Nucleic Acid Sequence-
Based Amplification (NASBA). В настоящее время NASBA — запатентованное назва-
ние технологии, на базе которой выпускаются коммерческие наборы реагентов с
торговой маркой «NucliSens» («bioMerieux*», Франция). Особенности технологии
NASBA имеют преимущество перед ПЦР при диагностике инфекционных агентов,
в том числе и хламидий, так как мишенью для ЗЗК-амплификации в отличие от
ПЦР служит молекула РНК, а использование при диагностике бактериальных
инфекций рибосомной РНК может давать некоторые преимущества.
• Количес гво копий рибосомной РНК может варьировать от нескольких сотен
до нескольких десятков тысяч в одной клетке, обеспечивая даже при мини-
мальной концентрации бактериальных клеток в пробе достаточно высокую
концентрацию мишеней для амплификации.
• РНК — гораздо менее стабильный по сравнению с ДНК материал, и результа-
ты диагностики на основе NASBA могут более адекватно отражать эффектив-
ность проводимой антибактериа пьнлй терапии.
Поскольку ДНК Chi. trachomatis благодаря своей стабильности может доста-
точно долго выделяться из урогенитального тракта после успешно проведенного
лечения, контроль излеченное™ рекомендуют проводить через 3-4 нед после его
окончания. В то же время РНК достаточно быстро деградирует при гибели и раз-
рушении клетки, именно поэтому на основании РНК-диагностики можно более
точно судить о наличии или отсутствии жизнеспособных возбудителей и текущей
хламидийной инфекции. Результаты при изучении возможности использования
NASBA как инструмента контроля эффективности лечения показали, что РНК
Chi. trachomatis не определялась ни у одного больного через неделю после окон-
чания лечения, в то время как ДНК обнаруживали у некоторых больных на про-
тяжении 2-3 нед. Таким образом, NASBA позволяет не только добиться высокой
чувствительности при обнаружении Chi. trachomatis, но и выявлять жизнеспособ-
ные микроорганизмы.
Серологическая диагностика
Серологические исследования относят к вспомогательным методам диагности-
ки хламидиоза. Они позволяют определить стадию и характер течения заболева-
ЧАСТНАЯ МИКРОБИОЛОГИЯ 323
ния. Это особенно важно при восходящей и персистирующей инфекции, которая
поддерживает хроническое течение болезни на протяжении многих месяцев и лет
и повреждает или разрушает ткани и органы. Методы ретроспективной диагности-
ки инфекции незаменимы при затруднениях взятия материала для обнаружения
антигена (например, у детей). В их основе лежит выявление специфических анти-
тел, которые накапливаются в крови и секретах в процессе иммунного ответа на
внедрение возбудителя инфекционного заболевания.
Специфическую реактивность в отношении к хламидиям проявляют 3 класса
антител — IgM, IgG и IgA. Накопление каждого из них происходит через раз-
личные промежутки времени от начала иммунного ответа и зависит от характера
инфицирования (первичного или вторичного). При первичном инфицировании
сначала появляются IgM. затем IgG и в последнюю очередь IgA. По мере угасания
иммунного ответа снижение концентрации антител каждого из классов проис-
ходит в той же последовательности.
Иммунный ответ при вторичном инфицировании характеризуется быстрым
нарастанием титра IgG и IgA и практически полным отсутствием IgM. Таким обра-
зом. при остром инфекционном процессе у больных можно обнаружить IgM или
быстронарастающие титры IgG и IgA. При хронической инфекции выявляются
специфические IgG и IgA, концентрации которых не изменяются на протяжении
длительного времени. В ряде случаев («3%) при хронически протекающих инфек-
циях образованию специфических IgG предшествует регистрация невысоких
титров 1g \ в течение нескольких недель. У больных с бессимптомным течением
заболевания определение IgA в постоянно низких титрах в течение длительного
времени свидетельствует о персистенции микроорганизма. Низкие постоянные
титры специфических IgG указывают на давно перенесенную хламидийную
инфекцию.
При острой инфекции диагностическое значение имеет обнаружение специфи-
ческих IgM, при хронических формах хламидиоза следует определять IgA и IgG.
При интерпретации полученных данных необходимо помнить, что обнаружение
специфических антител однозначно не свидетельствует об инфицированное™
хламидиями, так же как и отрицательные результаты серологических тестов не
исключают наличия текущей или перенесенной хламидийной инфекции. Для того
чтобы оценить динамику изменений титров антител различных классов, в клини-
ческой практике часто исследуют парные пробы сыворотки крови, полученные от
пациента с интервалом 2-3 нед.
В процессе лечения 2-3-кратное снижение титров IgG и IgA указывает на его
эффективность.
Существующие коммерческие тест-системы позволяют определять специфиче-
ские IgM-, IgA-, IgG-антитела к хламидиям в сыворотке крови. Серологические
методы могут быть рекомендованы для обследования детей, при осложненном
течении инфекционного процесса у взрослых, когда другие методы выявления
возбудителя или его антигена неприемлемы (например, при локализации возбуди-
теля в маточных трубах), а также при проведении массовых эпидемиологических
исследований.
БОРДЕТЕЛЛЫ
ТАКСОНОМИЯ
Возбудитель коклюша, В. pertussis, относится к роду Bordetella, который вклю-
чает еще 8 видов — В. ansorpii, В. avium, В. bronchiseptica, В. hinzii. В. holmesii,
В. parapertussis, В. petrii, В. trematum.
Род Bordetella входит в состав р-протеобактерий.
324 ЧАСТНАЯ МИКРОБИОЛОГИЯ
ОБЩАЯ ХАРАКТЕРИСТИКА БОРДЕТЕЛЛ
Бактерии рода Bordetellae — мелкие (0,2-0,5х0.5-2,0 мкм) грамотрицательные
коккобациллы. В мазках часто биполярно окрашенные, одиночные или в парах,
реже в цепочках.
Все бордетеллы, за исключением В. petrii, строгие аэробы, требовательные к
условиям роста.
Основные биохимические свойства бордетелл, которые используются в прак-
тической работе при изучении выделенных культур, представлены в табл. 21-3.
В. pertussis наименее активна ферментативно (стабильно положительный тест на
оксидазу). В. parapertussis вырабатывает ферменты тирозиназу и уреазу, не образу-
ет оксидазы. Наиболее активна В. bronchiseptica — вырабатывает уреазу, оксидазу,
утилизирует цитраты, восстанавливает нитраты до нитритов.
Таблица 21-3. Дифференцирующие признаки видов рода Bordeteila
S. pertussis В. parapertussis В. bronchiseptica В. avium В. hlnzii В. hoimesii В. trematum В. petrii В. ansorpii
Рост на кровяном агаре - + + + + + +
Рост на агаре Мак- Конки - + + + + + + + +
Оксидаза + - + + + - - + -
Восстановление нитратов - - + - - - + - -
Уреаза - + (24 ч) (4% - ± - - - -
Подвижность - - + + + - + - +
АНТИГЕННАЯ СТРУКТУРА В. PERTUSSIS
Коклюшный токсин является важнейшим фактором патогенности В. pertus-
sis. Это экзотоксин, белок с молекулярной массой 117 000 Да, состоящий из двух
функциональных частей (А и В) и пяти структурных субъединиц. Участок А соот-
ветствует субъединице S1 и обладает ферментативной активностью, ингибирует
клеточную аденилатциклазу. Участок В состоит из субъединиц S2-S5, отвечает за
присоединение токсина к рецепторам клеток-мишеней. Токсин обладает высокой
иммуногенностью, в инактивированной форме включен в состав всех бесклеточ-
ных коклюшных вакцин. Определение антител к коклюшному токсину методом
ИФА применяют для диагностики коклюша и контроля эффективности вакцина-
ции.
Филаментозный гемагглютинин — поверхностный белок, участвующий в
адгезии, обладает протективными свойствами. Включен в состав бесклеточных
коклюшных вакцин. В ряде коммерческих ИФА-тест-систем для диагностики
коклюша предлагают определять уровень антител различных классов к антиген-
ному комплексу, в состав которого входят гемагглютинин и коклюшный токсин.
В отличие от токсина, гемагглютинин не строго специфичен для В. pertussis, присут-
ствует также у В. parapertussis, может давать перекрестные реакции с Н. influenzae,
С. pneumoniae и рядом других бактерий.
Пертактин — белок наружной мембраны, относится к системе адгезинов, про-
дуцируемых бактериями при попадании в человеческий организм. Обладает про-
тективными свойствами, входит в состав ряда бесклеточных коклюшных вакцин.
ЧАСТНАЯ МИКРОБИОЛОГИЯ 325
Аденилатцик паза-гемолизин — комплекс экзофермента аденилатциклазы с
токсином гемолизином. Фермент при попадании в клетки катализирует образова-
ние цАМФ. Токсин — основной фактор патогенности, действующий на начальном
этапе инфекции, также с ним связывают протективные свойства комплекса.
Агглютиногены — поверхностные белки, ответственные за выработку агглю-
тинирующих антител. У бордетелл выделено 16 агглютиногенов. Седьмой агглю-
тиноген — общеродовой, а первый — основной видовой для В. pertussis. В зависи-
мости от наличия в бактериальной клетке 2-го и 3-го агглютиногена выделяют
четыре серотипа Б. pertussis — 1.2.0,1.0.3,1.2.3 и 1.0.0. Также у В. pertussis выделяют
4,5,6,13,15 и 16-й агглютиногены. Видовой агглютиноген для В. bronchiseptica —
12-й. для В. parapertussis — 14-й, а 8-й, 9-й и 10-й агглютиногены — общие для
В. bronchiseptica и В. parapertussis, 11-й — встречается у В. bronchiseptica.
Понятие об агглютиногенах тесно связано с фимбриями (Пт;. Выделяют Fim 2,
Fim 3, Firn X. Фимбрии включены в состав некоторых бесклеточных коклюшных вак-
цин. Серотипирование В. pertussis основано на агглютинации бактериальных клеток:
в отечественной практике — монофакторными сыворотками (антителами к агглю-
тиногенам) в РГА на стекле, в зарубежной — моноклональными антителами к фим-
бриальным антигенам в реакции агглютинации в микропланшете. Для диагностики
коклюша в России проводят РГА с цельноклеточным диагностикумом, в которой
определяются агглютинирующие антитела в первую очередь к агглютиногенам.
Липополисахарид — в его состав входят два липида — А и X. С X-фракцией
связана биологическая активность липополисахарида. Он обладает множествен-
ными функциями, в том числе выраженной иммуногенностью. Также с ним связы-
вают реакзогенность клеточной вакцины.
Трахеальный цитотоксин — фрагмент пептидогликана клеточной стенки.
Проявляет разнообразные биологические свойства — пирогенность, адъювант-
ность, артритогенность, стимуляцию продукции ИЛ-1. Токсин поражает тра-
хеальные эпителиальные клетки и вызывает цилиогтаз. При этом нарушается
мукоцилиарный клиренс (первая линия защиты) и создаются условия для перси-
стирования возбудителя инфекции.
Дермонекротизирующий токсин обладает сосудосуживающей активностью,
у экспериментальных животных вызывает снижение прироста массы тела, атро-
фию селезенки, ишемические поражения или некроз кожи. Его роль в развитии
заболевания остается неясной.
Все вышеперечисленные факторы присутствуют у свежевыделенных штаммов
В pertussis. Однако при хранении на искусственных питательных средах прояв-
ляется изменчивость возбудителя. Установлено, что в процессе сапрофитизации
В. pertussis проходит четыре фазы — свежевыделенный микроб (гладкий штамм),
обладающий высокими вирулентными и иммуногенными свойствами, относится к
1-й фазе. По мере перехода к 4-й фазе постепенно утрачиваются иммуногенность,
вирулентность, изменяются культурально-биологические свойст ва.
РАСПРОСТРАНЕНИЕ И ЭПИДЕМИОЛОГИЯ
В. pertussis патогенна для человека и является возбудителем коклюша,
В. parapertussis вызывает у людей паракоклюш — коклюшеподобное заболевание.
В. bronchiseptica — возбудитель заболеваний дыхательных путей у многих мле-
копитающих (в том числе собак и свиней), встречается также бессимптомное
носительство. У человека В. bronchiseptica редко вызывает заболевание, однако
описаны случаи, когда у пожилых людей, заразившихся от домашних животных,
В. bronchiseptica вызывала длительный кашель. В. avium — возбудитель ринотра-
хеита птиц. В. ansorpii, В. hinzii, В. habnesii, В. trcmatum редко вызывают заболевания
у людей, главным образом у пациентов с иммунодефицитом. Возможно также бес-
симптомное носительство В. hinzii у домашней птицы. Единственный представи-
тель рода, выделенный из окружающей среды (из речного осадка), — В. petr'o
326 ЧАСТНАЯ МИКРОБИОЛОГИЯ
КЛИНИЧЕСКИЕ ПРОЯВЛЕНИЯ ИНФЕКЦИИ
Коклюш — заболевание, длящееся нс менее 2 нед, без явлений интоксикации и
повышения температуры тела. Протекает циклично со сменой ряда периодов.
Инкубационный период продолжается от 3 до 14 дней (в среднем 7-8 дней).
Предсудирожный период протекает от 3 до 14 дней, проявляется сухим навязчи-
вым кашлем на фоне нормальной температуры тела.
Период судорожного кашля (ПСК) продолжается от 2-3 до 6-8 нед и более, харак-
теризуется типичными приступами судорожного кашля. Приступ кашля представ-
ляет следующие друг за другом дыхательные толчки на выдохе, прерываемые сви-
стящим судорожным вдохом (репризой), возникающим при прохождении воздуха
через суженную вследствие ларингоспазма голосовую щель. Заканчивается приступ
отхождением вязкой, стекловидной мокроты или рвотой. Приступу кашля может
предшествовать аура (чувство страха, беспокойство, чихание, першение в горле).
Приступы могут быть кратковременными или продолжаться 2-4 мин. Возможны
пароксизмы — концентрация приступов кашля в течение короткого периода вре-
мени. При типичном приступе кашля больной приобретает характерный вид: лицо
краснеет, затем синеет, становится напряженным, набухают вены шеи, лица и
головы, отмечается слезотечение. Язык высовывается до предела, кончик его под-
нимается кверху. В результате трения уздечки языка о зубы и ее чрезмерного растя-
жения происходит надрыв или образование язвочки — патогномоничный симптом
коклюша. При гладком течении заболевания температура тела остается нормаль-
ной. Характерно постепенное развитие симптомов заболевания с максимальным
учащением и утяжелением ПСК на 2-й неделе болезни. Возможно присоединение
специфических осложнений на 3-й неделе и неспецифических осложнений на фоне
развития вторичного иммунодефицита — на 4-й неделе ПСК.
Период обратного развития (ранней реконвалесценции) — от 2 до 8 нед. на фоне
улучшения самочувствия кашель ст ановится реже и постепенно теряет типичный
характер.
Период реконвалесценции (поздней) продолжается от 2 до 6 мес и характеризу-
ется состоянием гиперреактивности с возможным развитием приступообразного
кашля при интеркуррентных заболеваниях или эмоциональных нагрузках.
ЛАБОРАТОРНАЯ ДИАГНОСТИКА
Схема лабораторной диагностики коклюша представлена в табл. 21-4.
Таблица 21-4. Рекомендуемая схема лабораторной диагностики коклюша
Обследуемые 1-2 нед от начала заболевания 3-4 нед Более 4 нед
без антибмо- тикотерапии на фоне анти- биотикотерапии без антнбмо- тикотерапии на фоне анти- биотикотерапии без антибиоти- котерапии или на ее фоне
Непривитые дети до 1 года БМ', ПЦР ПЦР БМ,ПЦР ПЦР серология Серология
Непривигые дети старше 1 года БМ,ПЦР ПЦР ПЦР серология (БМ") Серология (ПЦР) Серология
Привитые дети ПЦР, БМ ПЦР ПЦР серология (БМ) Серология (ПЦР) Серология
Подростки, взрос- лые ПЦР, БМ ПЦР ПЦР серология (БМ) Серология (ПЦР) Серология
* БМ — бактериологический метод.
** Эффективность метода, указанного в скобках, у данной группы пациентов существенно сни-
жается.
ЧАСТНАЯ МИКРОБИОЛОГИЯ
327
Изоляция и идентификация возбудителя
Бактериологическое исследование следует проводить в ранние сроки заболева-
ния (не позднее начала 3-й недели) двукратно ежедневно или через день. Позднее
высеваемость возбудителя на питательные среды резко снижается. Она зависит от
следующих факторов:
• срока обследования;
• кратности исследования;
• соблюдения правил взятия, транспортировки и посева материала;
• качества используемых материалов и сред.
Взятие материала. Материал забирают со слизистой оболочки задней стен-
ки глотки зондом-тампоном (поскольку вата может действовать на возбудителя
коклюша бактерицидно, лучше пользоваться вискозным или дакроновым зондом-
тампоном). Назофарингеальные мазки получают с помощью мягкого тонкого
зонда, который вводят по ходу нижней носовой раковины и штриховым движени-
ем собирают материал с задней стенки глотки. Ретрофарингеальные мазки берут
с задней стенки глотки через рот, опуская тампон как можно ниже, не касаясь
миндалин, неба и языка. Материал можно забирать методом кашлевых пластинок.
Оптимально высевать пробы на питательные среды сразу после их взятия.
Питательные среды. Для выделения бордетелл пользуются плотными
питательными средами — картофельно-глицериновым (среда Борде-Жангу) и
казеиново-угольным (КУА) агаром. Оптимально добавление в среды лошадиной
или овечьей крови; в среду Борде-Жангу — 20%, в КУА — 10%. Добавление чело-
веческой крови или эритроцитарной массы дает худшие результаты, их можно
частично или полностью заменить плазмой крови крупного рогатого скота (5%).
Также можно добавить питательную среду № 199 (1%).
Для ингибирования посторонней флоры используют цефалексин (40 мг/л
среды). Б отличие от бензилпенициллина, цефалексин не подавляет рост В, pertus-
sis и более эффективен в отношении сопутствующей флоры.
Посевы инкубируют при температуре 35-37 °C без добавления углекислого газа
в условиях повышенной влажности.
Общая продолжительность бактериологического исследования составляет
5-7 дней;
• 1-й — прямой посев на плотную среду с добавлением антибиотика;
• 2-3-й — осмотр посевов;
• 4-6-й — идентификация бордетелл.
Бордетеллы формируют:
• на среде Борде-Жангу — выпуклые, гладкие, блестящие колонии серебряно-
го цвета, напоминающие капли ртути, окруженные зоной гемолиза;
• на КУА — выпуклые, гладкие колонии серого цвета с жемчужным, желтова-
тым или беловатым оттенком, маслянистые, легко снимаются петлей.
В. parapertussis за счет образования пигмента вызывает потемнение питательных
сред, содержащих кровь, образует бурую подложку. Температура выращивания
бордетелл 35-37 °C, при этом колонии разных видов бордетелл формируются в
различные сроки; более продолжительный рост наблюдается у В. pertussis.
При обнаружении на среде подозрительных колоний проводят выделение
чистой культуры путем отсева изолированных колоний на чашку Петри с одной
из упомянутых выше сред (без антибиотика), используя отдельные ее сектора
(вместо обычных чашек Петри можно использовать 2- или 4-секционные). При
обнаружении нескольких однотипных колоний делают мазки, окрашивают по
Граму и микроскопируют. Возможно проведение РГА на стекле со специфиче-
скими видовыми неадсорбированными сыворотками, по возможности — с адсор-
бированными монорецепторными сыворотками к агглютиногенам L и 14. При
значительном числе колоний можно поставить пробу Заксе на наличие уреазы. На
328
ЧАСТНАЯ МИКРОБИОЛОГИЯ
основании данных изучения колоний, морфологии бактерий в мазках, положи-
тельного результата РГА на 3-и сутки можно выдать предварительный ответ. При
отсутствии роста подозрительных колоний на плотной среде посевы помещают в
термостат еще на 24-48 ч.
Видовую принадлежность выделенных культур устанавливают на основании
комплекса типичных морфологических, биохимических, антигенных и других
свойств.
Для оценки биохимических свойств проводят посевы чистой культуры на среду
с тирозином (определение наличия тирозиназы), пробой Закса или посевом на
бульон Хеттингера определяют наличие уреазы. При выделении чистой культуры
В. bronchisepti^a определяют утилизацию цитратов на среде Симмонса и подвиж-
ность путем посева в полужидкий агар (0,5% агар-агар). В табл. 21-3 приведен
набор биохимических тестов, позволяющих дифференцировать различные виды
бордетелл.
Серовар коклюшного микроба определяют в РГА на стекле с монорецепторны-
ми сыворотками к агглютиногенам (факторам) 2 и 3.
На основании изучения морфологии колоний и клеток чистой культуры, а
также результатов биохимических тестов и РГА с монорецепторными сыворотка-
ми дают окончат ельный положительный ответ или спустя 6 сут исследования при
отсутствии подозрительных колоний — отрицательный.
Исследование атипичных штаммов. Бордетеллы способны существовать
в 4 фазовых состояниях, переходя из одной фазы в другую и изменяя свои
культурально-морфологические свойства. Такие штаммы образуют более круп-
ные колонии с плоской поверхностью, западающим центром, имеют беловатый,
бурый, желтоватый или слегка розоватый и зеленоватый цвет, маслянистую
консистенцию. В мазках обнаруживают более длинные или раздутые овоидные
палочки, палочки с ослизненным концом, формы в виде стрептобацилл. Чистая
культура, выделенная из таких колоний, обладает типичными биохимическими
и серологическими свойствами В ряде случаев могут быть выделены грамотри-
цательные бактерии, сходные по морфилогии и биохимическим свойствам, но не
агглютинируемые монорецепторными сыворотками и хорошо растущие на про-
стых питательных средах. Учитывая наличие атипичных штаммов коклюшного
микроба, рекомендуют при выделении чистой культуры проводить отсев с пита-
тельной среды не менее пяти колоний.
Молекулярная диагностика
В условиях высокого охвата детей прививками против коклюша снижается
эффективность бактериологического исследования и возрастает роль ПЦР как
быстрого (в течение 4-6 ч) метода, позволяющего обнаружить ДНК возбудителя
у привитых детей на более поздних стадиях заболевания и на фоне ангибиоти-
котерапии. ПЦР может давать положительный результат при наличии мертвых
и нежизнеспособных клеток возбудителя, поэтому подтверждение диагноза дан-
ным методом после антибиотикотерапии не является основанием для назначения
повторного курса антибактериальных препаратов. ДНК может обнаруживаться
приблизительно в течение 1 мес после успешного лечения антибактериальными
препаратами и на фоне клинического выздоровления, поэтому ПЦР не реко-
мендуют использовать для подтверждения эффективности лечения вскоре после
завершения курса антибиотикотерапии, как это принято при бактериологическом
исследовании, Существует методика обнаружения путем амплификации нуклеи-
новых кислот только живых микроорганизмов (NASBA), но в настоящее времч
тест-системы для диагностики коклюша в данном формате не разработаны.
Дня анализа используют мазки с задней стенки носо- или ротоглотки или
носоглоточный аспират. Мазки берут сухим стерильным одноразовым тампоном
ЧАСТНАЯ микроьиология 329
(оптимальный материал — дакрон, можно использовать хлопок или вискозу, осно-
вание — алюминий или пластик) в индивидуальной пластиковой пробирке. После
взятия материала тампон помещают обратно в пробирку. До анализа материал
можно хранить в течение 3 сут при температуре 2-8 °C или в течение месяца при
температуре -18-24 °C (без размораживания). Транспортируют при температуре
2-8 °C.
Для анализа тампон погружают в одноразовую пробирку типа «Эппендорф» с
1,5 мл транспортной среды, вращают его в течение 10-15 с, избегая разбрызгива-
ния раствора, затем отжимают избыток жидкости, прижимая к стенке пробирки,
тампон вынимают, пробирку закрывают и центрифугируют. Если тампон хранили
при температуре -18-24 °C, перед исследованием его необходимо разморозить
при комнатной температуре в течение 30 мин. ПЦР проводят в соответствии с
инструкциями к наборам реагентов.
Серологическая диагностика
В условиях массовой вакцинопрофилактики существенно снизилось количество
тяжелых форм заболевания, участились случаи позднего обращения за медицин-
ской помощью, нередко уже на фоне проводимой антибиотикотерапии. При этом
возросло значение серологических гестов как средств поздней (ретроспективной)
диагностики. Необходимо учитывать, что у привитых детей может быть повышен
уровень антител к В. pertussis или его компонентам (поствакцинальный имму-
нитет). Именно поэтому серологическое исследование не следует применять в
течение одного года после вакцинации против коклюша. Также не рекомендуют
использовать серологические методы диагностики у детей в возрасте до 6 мес, так
как у них могут сохраняться материнские антитела.
РГА — классический метод серологической диагностики. Реакцию проводят
в агглютинационных пробирках. Положительным результатом считают нарас-
тание титра специфических антител в 4 раза и более — первую пробу сыво-
ротки берут не менее чем через 2 нед после начала заболевания, вторую — не
менее чем через 2 нед после первой. РГА по чувствительности уступает ИФА.
Использование корпускулярных диагностикумов может приводить к перекрест-
ным реакциям с антителами к возбудителю паракоклюша, а также некоторым
другим бактериям.
ИФА. Метод более чувствителен, чем РГА. Позволяет определять антитела
различных классов (IgM, IgA, IgG). В различных тест-системах выявляют анти-
тела к разным антигенам либо к их комплексам — корпускулярному комплексу,
КТ и ФГА, только КТ. Наиболее специфично определение антител к коклюшному
токсину. За положительный результат принимают выраженную сероконверсию по
IgG или IgA (повышение или понижение уровня иммуноглобулинов при повтор-
ном исследовании через 2-4 нед не менее чем в 4 раза, если в инструкции к тест-
системе нет других указаний). Определять IgM к В. pertusw в плазме крови можно
у непривитых больных для быстрой серологической диагностики коклюша без
исследования парных сывороток.
ЛЕГИОНЕЛЛЫ
ТАКСОНОМИЯ
Legionella является единственным родом семейства Legionellaceae, относящимся к
у-притеобактериям. Общепринятая классификация рода имеет генетическую осно-
ву. Штаммы легионелл о сносят к одному виду, если уровень гибридизации ДНК
между ними при оптимальных условиях превышает 70%, В частности, гомология
штаммов L. pneumophila составляет 82-99%, а некоторых других видов легионелл —
40-70%. Для видовой идентификации легионелл используют рестрикционный
330
HACTl 1АЯ МИКРОБИОЛОГИЯ
анализ, амплификацию и секвенирование ДНК, анализ состава клеточной с генки
и серологические методы. Большинство данных методов доступно только для
специализированных референс-лабораторий. Биохимические тесты имеют крайне
ограниченное значение, так как легионеллы не Ферментируют углеводы.
В настоящее время известно около 50 видов легионелл, 22 из которых игра-
ют роль в инфекционной патологии человека. Более 95% случаев легионеллеза
ассоциировано с видом L. pneumophila. Другие виды легионелл (L. micdadei, L. long-
beachae, L. dumoffii, L. bozemanii), как правило, вызывают заболевание при наруше-
ниях клеточного иммунитета или на фоне сопутствующей патологии.
Вид L. pneumophila включает 16 серогрупп. Чаще всего легионеллез вызывают
серогруппы 1, 4 и 6, причем превалирует серогруппа 1, с которой связано свыше
80% регистрируемых случаев легионеллеза (серогруппы 4 и 6 выявляют в 5-10%
случаев болезни).
ОБЩАЯ ХАРАКТЕРИСТИКА ЛЕГИОНЕЛЛ
Легионеллы — грамотрицательные палочки диаметром 0,5-0,7 мкм, длиной
2-5 мкм. Иногда встречаются нитевидные формы длиной до 20-25 мкм. Не обра-
зуют эндоспор, микроцист и капсул: аэробы. Подвижны вследствие наличия одно-
го, двух или большего числа жгутиков. Не ферментируют углеводы, разжижают
желатин, не образуют уреазу, не восстанавливают нитраты, каталазоположитель-
ны, оксидазовариабельны.
Легионеллы не растут на обычных питательных средах (кровяном агаре, среде
Мак-Конки), что связано с потребностью возбудителя в L-цистеине и растворимом
пирофосфате железа. Для выделения легионелл используют модификации буфер-
ного угольно-дрожжевого агара, содержащего L-цистеин, растворимый пирофос-
фат железа и а-кетоглютаровую кислоту (среда BCYE-a).
Все виды легионелл рас гут во влажной атмосфере при температуре 35 °C. Рост
некоторых видов стимулируется наличием в атмосфере 2,5-3% углекислого газа.
Колонии легионелл вырастают при первичном выделении в течение 3-6 сут.
Молодые колонии обычно имеют вросший центр, гранулярную или блестящую
поверхность.
В лабораторных условиях чувствительность L. pneumophila к стандартным
дезинфицирующим и антисептическим препаратам не отличается от большинства
неспорообразующих бактерий. 3% раствор хлорамина вызывает гибель возбуди-
теля через 10 мин при его концентрации в суспензии 109 КОЕ/мл. L. pneumophila не
кислотоустойчива, не окрашивается по Цилю-F ильсену.
Несмотря на то что микроорганизм высокочувствителен к действию различных
химических и физических агентов и требователен к температуре и pH питательной
среды, в подходящих условиях водной среды L pneumophila способна сохраняться
достаточно долго. В природной водной среде значительно расширяется диапазон
физико-химических и биологических условий существования микроорганизма. Об
этом свидетельствует выделение возбудителя из водоемов с различными физиче-
скими, химическими и биологическими характеристиками.
ЭПИДЕМИОЛОГИЯ
Прошло 35 лет со времени первой зарегистрированной эпидемической вспышки
данного заболевания в Филадельфии (США). Тогда, летом 1976 г., из 4400 участ-
ников конгресса организации «Американский легион» у 221 (5%) развилась тяже-
лая пневмония. Из них умерли 34 (15,4%) человека. Первый штамм легионелл,
выделенный от умершего пациента, был принят за типовой штамм Legionella pneu-
mophila в разрабатываемой классификации легионелл как новый род бактерий.
В названии бактериального агента увековечены первые жертвы инфекции и обо-
значена пневмотропность возбудителя.
ЧАСТНАЯ МИКРОБИОЛОГИЯ 331
В целом уровень заболеваемости легионеллезом в мире невелик, однако спора-
дические случаи и десятки эпидемических вспышек болезни ежегодно выявляют в
различных странах мира.
Типичные примеры:
• аукцион цветов в Голландии (1999) — 188 человек заболели, зарегистрирова-
но 16 летальных исходов:
• дельфинарий в Австралии (1999) — 98 заболевших, 9 летальных исходов:
• гостиничный комплекс в Норвегии (2001) — 28 заболевших, 7 летальных
исходов;
• общественные здания в Испании (2002) — 470 заболевших, 9 летальных
исходов;
• промышленные предприятия вс Франции (2004) — 86 заболевших;
• промышленные предприятия в Норвегии (2005) — 55 заболевших, 10 леталь-
ных исходов
Крупные вспышки легионеллеза за последнее время отмечались в следующих
регионах:
• Верхней Пышме Свердловской области. Россия (2007) — более 100 заболев-
ших, 5 летальных исходов;
• Ульме, Германия (2010) — 65 заболевших, 5 летальных исходов.
Механизм передачи легион ел лезной инфекции — аспирационный, путь переда-
чи — воздушно-капельный, основной фактор передачи — мелкодисперсный аэро-
золь. Легионеллез — типичный пример техногенных инфекций, обусловленных
активным использованием в промышленности и быту циркулирующих замкнутых
водных систем, источников бактериального аэрозоля. Практически все крупные
эпидемические вспышки и многие спорадические случаи легионеллеза связаны
с распространением мелкодисперсного аэрозоля, содержащего легионеллы и
генерируемого бытовыми, медицинскими или промышленными водными систе-
мами. Сочетание высокой концентрации легионелл в водной среде с источниками
мелкодисперсного аэрозоля позволяет возбудителю попасть в респираторные
отделы легких человека, где происходит контакт с альвеолярными макрофагами, в
которых вирулентные штаммы возбудителя активно размножаются. При анализе
эпидемической заболеваемости установлено, что у людей, находившихся в зоне
действия мелкодисперсного аэрозоля, содержащего легионеллы, пневмония раз-
вивается в 5-10%, а лихорадка Понтиак — в 80-100% случаев.
При спорадическом легионеллезе и отдельных нозокомиальных вспышках
заболевания возможна аспирация воды, контаминированной легионеллами. без
образования аэрозоля. Спорадический легионеллез выявляют, как правило, у
пациентов среднего и пожилого возраста на фоне действия факторов риска (куре-
ния, сопутствующих заболеваний, иммуносупрессивной терапии, первичных и
вторичных иммунодефицитов).
Легионеллы вызывают 2-6% всех пневмоний и до 10-15% атипичных пнев-
моний. У детей легионеллез выявляют редко, обычно на фоне сопутствующей
патологии.
Легионеллез распространен повсеместно, его выявляют круглогодично, однако
пик заболеваемости приходится на летние месяцы.
В последние годы особое значение стали придавать проблеме легионеллеза,
возникающего во время путешествий и диагностируемого, как правило, по воз-
вращении. Более 30% случаев спорадического легионеллеза и многочисленные
эпидемические вспышки в гостиницах, нередко с летальным исходом, послужили
основой для создания единой международно, i системы эпидемиологического над-
зора за стучаями легионеллеза, связанного с путешествиями.
Риск возникновения внутрибольничного легионеллеза определяется не только
возможностью контаминации легионеллами систем водоснабжения, кондициони-
332
ЧАСТНАЯ МИКРОБИОЛОГИЯ
рования. но и наличием восприимчивых к инфекции пациентов с нарушениями
клеточного иммунитета. В онкологических отделениях или отделениях транс-
плантации органов при контаминации легионеллами водных систем доля легио-
неляеза в этиологической структуре внутрибольничных пневмоний составляет
15-20%, летальность при данной инфекции — 30-40%. Помимо L. pneumophila,
внутрибольничную инфекцию нижних дыхательных путей часто вызывает вид
L. micdadei.
КЛИНИЧЕСКИЕ ПРОЯВЛЕНИЯ ИНФЕКЦИИ
Известны две клинические формы легионеллеза — болезнь легионеров и лихо-
радка Понтиак. Ведущая клиническая форма легионеллеза — болезнь легионеров.
Эта форма характеризуется четко очерченной клинической картиной воспаления
легочной ткани. Инкубационный период обычно составляет 2-10 дней. Высокая
лихорадка сопровождается ознобом, интенсивными болями в груди. Одышка на
фоне массивной воспалительной инфильтрации легочной ткани и вовлечения в
патологический процесс плевры появляется в 1-й день болезни и прогрессирует
при неадекватном лечении. У 20-30% заболевших легионеллезом развивается
острая дыхательная недостаточность, требующая респираторной поддержки.
В ряде случаев возникают инфекционно-токсический шок и острая почечная недо-
статочность. Лихорадка продолжается 12-15 дней. Симптомы поражения верхних
дыхательных пулей отмечают редко. Кашель первоначально малопродуктивный,
но с течением времени может приобретать продуктивный характер с отделением
слизисто-гнойной мокроты. У 25% больных наблюдают симптомы поражения
желудочно-кишечного тракта.
При общем клиническом анализе крови выявляют относительную или абсо-
лютную лимфопению на фоне умеренного лейкоцитоза, увеличение СОЭ до
60-80 мм/ч. При биохимическом анализе крови отмечают цитолиз, диспротеине-
мию со снижением содержания альбуминов, гипонатриемию, гипофосфатемию.
При рентгенографии органов грудной клетки в 60-70% случаев уже на ранней
стадии болезни визуализируются oflHOvTopOHHne пневмонические инфильтра-
ты. В дальнейшем отмечается прогрессирование инфильтративных изменений с
вовлечением в патологический процесс новых сегментов легких. Летальность при
болезни легионеров составляет 8-25%. У пациентов с иммунологическими нару-
шениями летальность может достигать 60%.
Другая клиническая форма легионеллеза — респираторная лихорадка — впер-
вые описана у служащих и посетителей Департамента здравоохранения в Понтиаке
(США). Данный ОРЗ-подобный синдром, называемый также лихорадкой Понтиак,
характеризуется 1-2-дневной лихорадкой, сухим кашлем, катаральными явле-
ниями в носоглотке, головными и мышечными болями. При нахождении в зоне
распространения аэрозоля, содержащего легионеллы, заболевание развивается в
90-100% случаев. Инкубационный период лихорадки Понтиак обычно состав-
ляет 36-48 ч. Болезнь протекает, как правило, в нетяжелой форме и завершается
выздоровлением в короткие сроки (2-7 дней). Детальны? исход при лихорадке
Понтиак не регистрировался. Носительство и персистенция легионелл в организме
человека не описаны.
ЛАБОРАТОРНАЯ ДИАГНОСТИКА
Для микробиологов легионеллез — труднодиагностируемая инфекция.
Возможно, относительно невысокий уровень выявляемой заболеваемости связан
с несовершенством методов лабораторной диагностики и высокой стоимостью
реагентов для эффективной изоляции легионелл.
ВОЗ рекомендует для лабораторной диагностики болезни легионеров три груп-
пы методов. Диагноз легионеллеза при острой инфекции нижних дыхательных
частная микробиология ззз
путей (клинически и рентгенологически подтвержденной пневмонии) считается
установленным:
• при выделении легионелл из отделяемого респираторного тракта или легоч-
ной ткани;
• 4-кратном или большем нарастании титра антител к L. pneumophila, выявляе-
мых в реакции непрямой иммунофлюоресценции;
• определении специфического растворимого антигена легионелл в моче
иммунохроматографическим методом или с помощью ИФА.
При отсутствии первой пробы сыворотки, взятой в ранние сроки болезни,
выявление высокого уровня антител к L. pneumophila (1:128 и выше) в одиночной
пробе сыворотки методом непрямой иммунофлюоресценции позволяет поставить
предположительный диагноз легионеллеза. Остальные методы применяют как
вспомогательные.
Для диагностики легионеллеза исследуют клинический материал, взятый в
стерильных условиях до 7-го дня болезни (отделяемое, полученное при бронхо-
скопии, плевральный экссудат, биопсийный или аутопсийный материал). Около
1-2 мл плеврального экссудата или отделяемого, полученного при бронхоскопии,
помещают в стерильную пробирку, плотно закрытую резиновой пробкой, и хранят
при 4 °C не более суток. Биопсийный или аутопсийный материал помещают в сте-
рильные флаконы, плотно закрытые пробками, и хранят или транспортируют при
температуре 4 °C в течение суток. Длительное (до года) хранение проб возможно
только при температуре -70 °C.
Изоляция и идентификация возбудителя
Для выделения легионелл из клинического материала применяют буферный
угольно-дрожжевой агар с p-кетоглютаровой кислотой (среда BCYE-a). кото-
рая оптимальна для выделения и культивирования легионелл в соответствии со
стандартом ISO11731E. Эту среду также можно приготовить непосредственно в
лаборатории, используя следующие компоненты: дрожжевой экстракт — 10 г;
активированный уголь —2 г; L-цистеин — 0,4 г; пирофосфат железа растворимый —
0,25 г; ACES-буфер — 10 г; агар — 15 г; p-кетоглютаровую кислоту — 0,25 г; дис-
тиллированную воду — 980 мл.
Все компоненты среды, кроме L-цистеина и пирофосфата железа, добавляют
к 980 мл дистиллированной воды, растворяют и доводят pH до 6,95 с добавле-
нием 1 N раствора гидроксида калия. Среду автоклавируют в течение 15 мин при
температуре 121 °C. Затем ее охлаждают до 50 °C на водяной бане и добавляют
свежеприготовленные растворы L-цистеина (0,4 г в 10 мл дистиллированной
воды) и растворимого пирофосфата железа (0,25 г в 10 мл дистиллированной
воды), профильтрованные через мембранный фильтр (величина пор — 0,3 мкм).
Питательную среду быстро разливают в чашки Петри, слегка взбалтывая флакон
для равномерного распределения активированного угля. Разлитую питательную
среду хранят в пластиковых или металлических контейнерах в холодильнике не
более 2 нед.
Легионеллы на среде BCYE-a инкубируют при температуре 35 °C в атмос фере,
содержащей 2,5% углекислого газа, при влажности около 65% в течение 2 нед.
Рост колоний в посевах клинического материала проявляется не ранее чем через
4- 5 сут. Максимальное количество видимых колоний отмечается на 8-10-е сутки.
Подозрительные колонии пересевают на ту же питательную среду и на среду,
не поддерживающую рост возбудителя (контрольную). В качестве контрольной
среды обычно используют среду BCYE-a без добавления L-цистеина и пирофос-
фата железа или соответствующей ростовой добавки фирмы-производителя.
Наличия роста во втором пассаже на среде BCYE-a при отсутствии роста на кон-
трольной среде обычно достаточно для подтверждения выделения бактерий рода
334 ЧАСТНАЯ МИКРОБИОЛОГИЯ
Legionella. Если отмечается рост на контрольной среде, определяемый микроорга-
низм не относится к роду Legionella.
Легочная ткань часто бывает контаминирована микрофлорой, быстрый рост
которой при посеве на питательные среды не позволяет выделить чистую культуру
легионелл. Для ингибирования роста посторонней микрофлоры в питательную
среду добавляют цефамандол (4 мкг/мл), полимиксин В (80 ЕД/мл) и анизо-
мицин (8 мкг/мл). Цефамандол можно заменить ванкомицином (0.5 мкг/мл).
Цефамандол по сравнению с ванкомицином подавляет рост некоторых видов
легионелл, не продуцирующих «-лактамазы, включая L. micdadei. В отличие от
стандартной ростовой добавки, обязател ьно содержащей L-цистеин и пирофосфат
железа, антимикробная добавка, подавляющая рост посторонней микрофлоры,
может различаться по составу у различных фирм-производителей, а также в зави-
симости от степени контаминации исследуемого материала.
Также существуют другие методы подавления роста посторонней микрофлоры
в клиническом материале:
• прогревание на водяной бане в течение 30 мин при температуре 50 СС;
• 5-минутная экспозиция НС1/КС1 буфером (pH 2,2) (используют для выделе-
ния легионелл из сильно контаминированных образцов).
Поскольку морфоло! ия легионелл типична для грамотрицательных бактерий, а
методы окраски неспецифичны, окраска по Граму или Романовскому-Гимзе мало-
ипформативна.
Легионеллы обладают оксидазой и каталазой, разжижают желатин, вызыва-
ют гидролиз гиппурата натрия, не ферментируют углеводы, не восстанавливают
нитраты до нитритов, не обладают уреазной активностью. В лабораторной прак-
тике в последние годы для идентификации выросших колоний используют реак-
цию латекс-агглютинации, проводимую с помощью соответствующих наборов,
или реакцию иммунофлюоресценции. Выпускают несколько вариантов наборов,
позволяющих выявить колонии L. pneumophila серогруппы 1 и серогрупп 2-14,
а также родоспецифический антиген, характерный для колоний других видов
легионелл. Этого вполне достаточно для быстрой идентификации возбудителя в
клинической лаборатории. Постановка РГА на стекле с суспензией предполагае-
мой колонии летонелл занимает не более 15 мин.
Серологическая диагностика
Помимо бактериологических, для диагностики легионеллеза широко применя-
ют иммуносерологические методы. Антигенная структура легионелл достаточно
сложна. Специфичность серогрупп обусловлена липополисахаридным антигеном.
Разработанные в настоящее время методы иммунодиагностики основаны на обна-
ружении в плазме крови больных специфических антител, а моче или отделяемом
респираторного тракта — антигена легионелл
Наиболее распространенный и доступный практическим лабораториям метод —
выявление специфических антител в плазме крови методом непрямо! иммуноф-
люоресценции. Плазму крови пациентов с подозрением на легионеллез берут в
первые дни и на 14-21-й день болезни Кровь из вены в количестве 3-5 мл поме-
щают в стерильную пробирку, после свертывания крови сгусток отслаивают от
стекла пастеровской пипеткой и пробирку выдерживают при комнатной темпера-
туре в течение 5-6 ч. Отделившуюся плазму помещают в стерильную пробирку и
хранят при температуре 4 ' С.
Диагноз на легионеллез подтверждается, если наблюдается не менее чем 4-крат-
ное нарастание титров антител в реконвалесцентных сыворотках по сравнению с
первой сывороткой, взятой в острый период заболевания. Если проанализировать
динамику изменения титров антител не представляется возможным, предвари-
тельный диагноз ставят по уровню специфических антител (1:128) в одиночной
сыворотке. В качестве контроля используют положительную (титр антител —
ЧАСТНАЯ МИКРОБИОЛОГИЯ 335
не менее 1:256) и отрицательную (титр антител 1:16) плазму крови человека.
Качество антигена может быть проверено в реакции непрямой иммунофлюорес-
ценнии с разведениями плазмы крови кролика, иммунизированного антигеном
L. pneumophila.
Экспресс-диагностика
Для экспресс-диагностики легионеллеза применяют ИФА. позволяющий выяв-
лять растворимый антиген легионелл в моче в острый период заболевания (первые
10 дней). В международных испытаниях, проводимых в 1998-1999 гг. под эгидой
Европейской рабочей группы по легионеллезу, доказаны высокие чувствитель-
ность и специфичность данного метода при инфекции серогруппы 1 L. pneumophila.
В результате ИФА был включен в стандарты диагностики легионеллеза в качестве
достаточного для окончательного подтверждения диагноза инфекции, вызванной
L. pneumophila серогруппы 1. В настоящее время в 90% случаев первичный диагноз
легионеллеза ставят на основании обнаружения легионеллезного антигена в моче.
При определении антигенов L pneumophila других серогрупп чувствительность и
специфичность ИФА несколько ниже. Для других видов легионелл ИФА не при-
меняют.
Разработан иммунохроматографический метод для быстрого определения
антигена легионелл в моче. Метод практически не уступает по специфичности и
чувствительности ИФА. не требует специального оборудования и широко при-
меняется клиническими микробиологами при диагностике тяжелых пневмоний.
К достоинствам иммунохроматографического метода можно отнести и наличие
аналогичной тест-системы для быстрого выявления в моче растворимого антигена
Str. pneumoniae, что позволяет быстро провести дифференциальную диагностику’
тяжелой пневмонии.
КОКСИЕЛЛЫ
ТАКСОНОМИЯ
В течение длительного времени коксиелл считали родственными риккетсиям,
относящимся к а-протеобактериям. В восьмом издании «Определителя бактерий»
Берджи род Coxiella отнесен к порядку Rickettiales, семейству Rickettsiaceae, трибе
Rickettsiaea вместе с родами Rickettsia и Rochalimaea. Однако данная классификация
отражает только фенотипические характеристики. Согласно современной таксо-
номии прокариотических организмов, основанной на анализе строения генома,
филогенетическое положение Coxiella bumeh. определено иначе: род Coxiella отне-
сен к у-протеобактериям, в который входят и легионеллы [10J.
ОБЩАЯ ХАРАКТЕРИСТИКА С. BURNETII
С. burnetii — облигатный внутриклеточный паразит. Это маленькая грамо-
трицательная бактерия (размером 0,2-0Лх0.4-0,1 мкм). Может размножаться
в фаголизосомах эукариотических клеток в кислой среде, имеет сложный цикл
развития, включающий споровые и вегетативные формы. У вегетативных форм в
период роста и размножения бактерий при электронной микроскопии выявляют
трехслойную оболочку. В неблагоприятных для роста и размножения условиях
коксиеллы переходят в стадию покоящихся форм, которые характеризуются
уплотнением всех элементов бактериальных клеток и утолщением их оболочек.
Присутствие данного микроорганизма не оказывает выраженного повреждающего
действия на клетки хозяина, что способствует длительной персистенции коксиелл
и хроническому течению инфекции. Для коксиелл характерна высокая резистент-
ность к физическим и химическим воздействиям, например к облучению ультра-
фиолетовыми лучами или действию дезинфицирующих растворов. Они выдер-
336 ЧАСТНАЯ МИКРОБИОЛОГИЯ
живают нагревание до 90 °C более 1 ч и стандартные процедуры пастеризации
молока (гибель коксиелл достигается лишь при его кипячении не менее 10 мин). В
молочнокислых продуктах и свежем мясе зараженных животных, хранящихся при
температуре 4 °C, коксиеллы сохраняют жизнеспособность в течение месяца, а в
замороженном мясе — в течение более длительною срока.
С burnetii могут находиться в различных фазовых состояниях аналогично
энтеробактериям. В естественных условиях (в организме клещей и животных)
коксиеллы находятся в 1-й фазе, при длительном лабораторном культивирова-
нии в развивающихся куриных эмбрионах или в клеточных культурах переходят
во 2-ю фазу. Это сопровождается мутационными изменениями поверхностных
липополисахаридов. Биологические свойства коксиелл, находящихся в разных
фазовых состояниях, существенно различаются. Например, коксиеллы 2-й фазы
менее вирулентны и обладают меньшей протективной активностью по сравнению
с коксиеллами 1-й фазы.
Геном С. burnetii характеризуется рядом особенностей, которые обусловливают
как специфику выбора нуклеотидных последовательностей-мишеней для моле-
кулярной диагностики, так и методов типирования данного возбудителя. Длина
генома С. burnetii в зависимости от штамма колеблется от 1,6 до 2,4 млн п.н. Геном
включает хромосому, имеющую циркулярное строение, и плазмидную последова-
тельность. организованную в плазмиду или представленную плазмидоподобными
участками в хромосоме.
В целом геном С. burnetii характеризуется наличием многочисленных генов,
ассоциированных с адгезивностью, инвазивностью, способностью к модуляции
клеток хозяина, детоксикации бактериальной клетки С. burnetii. Данные гены
обладаю г высокой видовой специфичностью.
Важная особенность генома С. burnetii, которая определяет его пластичность. —
наличие большого количества мобильных элементов. Они представлены встра-
ивающимися последовательностями (IS), неравномерно распределенными по
хромосоме, и включают 21 копию изотипа IS110, 1 копию IS1111, 5 копий IS30
и 3 копии ISAsl, а также 3 дегенеративных транспозазных гена. Одна из их важ-
нейших особенностей IS в геноме С. burnetii — фланкирование так называемых
островков патогенности.
Организация части генома, кодирующей транспорт и метаболизм у коксиелл.
характеризуется высоким содержанием транспортеров, что определяется образом
жизни возбудителя. Более 25% транспортеров у коксиелл задействованы в систе-
ме лекарственного выброса, по сравнению с другими протеобактериями их доля
значительно увеличена.
Гены синтеза компонентов, обеспечивающих функции липополисахарида —
липида А и 2-кето-З-деоксиоктулосонических кислотных компонентов липопо-
лисахаридов, расположены в разных участках хромосомной ДНК, а гены синтеза
олигосахаридного ядра липополисахарида и наружных повторяющихся единиц
О-антигена, напротив, компактно локализованы в двух регионах. Гены, кодирую-
щие синтез белков данной группы обладают высокой видовой специфичностью.
У коксиелл отмечены многочисленные гены, б низкие по функции к эукариоти-
ческим: ген, кодирующий гомолог десатуразы жирных кислот, и две стеролредук-
тазы, вовлеченные в синтез холестерола у эукариот. Возможно, данные ферменты
способствуют ткане- и органотропизму, которыми обладает С. burnetii.
Важнейшую роль в обеспечении жизненного цикла коксиеллы играют гены,
кодирующие факторы патогенности возбудителя. Адгезивность и инвазивность —
определяющие факторы успешного размножения коксиелл. Хотя у коксиелл
отсутствуют гены, кодирующие типичные структуры адгезии, регистрируются
гены, кодирующие 13 анкирин-повторсодержащих белков, которые могут быть
вовлечены в процесс прикрепления к клетке организма-хозяина.
ЧАСТНАЯ МИКРОБИОЛОГИЯ 337
У коксиелл идентифицировано четыре типа п лазмид, которые могут быть пред-
ставлены в геноме, — QpHl, QpRS, QpDV и QpDG и выделена группа бесплазмид-
ных штаммов, у которых плазмидоподобные нуклеотидные последовательности
интегрированы в хромосому. В разделении плазмид на типы участвуют «ядро
плазмидного сиквенса» с неопределенной функциональной ролью и уникальные
регионы. Данные регионы содержат, в частности, ген cbbE, предположительно
кодирующий штаммоспецифичный фактор вирулентности С. burnetii.
КЛИНИЧЕСКОЕ ЗНАЧЕНИЕ
Коксиеллы характеризуются выраженной полиадаптивностью. Они обитают в
организме разных видов беспозвоночных и позвоночных животных, в том числе
далеко отстоящих друг от друга в систематическом плане. Наиболее существен-
ное значение для экологии коксиелл и эпидемиологического проявления очагов
данной инфекции имеет связь возбудителя с иксодовыми клещами, дикими и
сельскохозяйственными животными. Животные являются главными источни-
ками коксиеллезной инфекции. С. biimetv в большом количестве содержатся в
плаценте и выделяются при окоте или отеле с околоплодной жидкостью, молоком
и экскрементами. Крупные вспышки ку-лихорадки в большей степени свчзаны с
мелким, чем с крупным рогатым скотом. Домашняя птица, собаки, кошки и серые
крысы также сравнительно широко вовлечены в пепь циркуляции коксиелл. Эти
животные довольно часто являются источником спорадических заболевании или
небольших вспышек ку-лихорадки. Напротив, непосредственная роль природ-
ных очагов коксиеллеза в качестве источника инфекции для людей ограничена.
Заболевания ку-лихорадкой редко связаны с посещением естественных угодий
и нападением кровососущих клещей. Больной человек, как правило, не является
источником коксиелл. Однако заражение возбудителем ку-лихорадки возможно
при переливании донорской крови или внутривенном употреблении наркотиков,
что аргументирует необходимость большего внимания в отношении коксиеллез-
ной инфекции у данных групп населения.
Ведущее значение при заражении коксиеллами имеют аспирационный кон-
тактный и алиментарный пути. Весьма часто вспышки ку-лихорадки обусловлены
вдыханием полевых частиц, инфицированных С. burnetii, при этом заражение
людей может произойти на значительном расстоянии от источника инфекции.
С. burnetii способны проникать в организм человека через поврежденную кожу
или слизистые оболочки глаз (например, при уходе за животными или обработке
сырья животного происхождения), через желудочно-кишечный тракт при упо-
треблении молока и молочных продуктов, реже — загрязненной воды, овощей и
фруктов.
Высокая патогенность для людей и устойчивость коксиелл к воздействию внеш-
ней среды позволяют рассматривать этот микроорганизм в качестве потенциаль-
ного агента для биологического терроризма.
Ку-лихорадку регистрируют у людей в течение всего года. Подъем заболеваемо-
сти отмечают в весенне-летние месяцы, что связано с сезоном окота и отела сель-
скохозяйственных животных. Мужчины болеют несколько чаще, чем женщины.
К «угрожаемым» в отношении ку-лихорадки профессиям относятся сельскохо-
зяйственные рабочие, пастухи, чабаны, доярки, зоотехники, ветеринары, рабочие
шерстеобрабатывающих и меховых фабрик. В последнее время среди заболевших
ку-лихорадкой увеличивается доля лиц, не относящихся к этим профессиям,
поскольку все больше людей проводят свободное время за городом и содержат
домашних животных.
Заболевания ку-лихорадкой людей или природные и хозяйственные очаги
данной инфекции выявлены во всех регионах мира, ее диагностиоуют во всех
странах, где проводятся соответствующие клинические исследования. В Европе
338
ЧАСТНАЯ МИКРОБИОЛОГИЯ
многочисленные вспышки ку-лихорадки были зарегистрированы во Франции,
Испании, Швейцарии, Великобритании, Германии, Словакии, Болгарии. В России
ку-лихорадка выявлена более чем в 50 субъектах Федерации.
По официальным данным, с 1957 г. (начало регистрации) в России заболели
ку-лихорадкой около 13 000 чел. В разные годы количество больных существенно
различалось, при этом прослеживается тенденция к снижению регистрации случа-
ев ку-лихорадки. В настоящее время в России имеются реальные предпосылки для
расширения очагов ку-лихорадки и увеличения заболеваемости. К важнейшим из
них относятся:
• ликвидация специализированных животноводческих комплексов с налажен-
ной системой ветеринарного контроля:
• выпас домашних животных на неблагополучных в отношении ку-лихорадки
территориях;
• ослабление в последние годы контроля за ку-лихорадкой животных;
• бесконтрольная миграция скота из пораженных коксиеллами хозяйств;
• расширение круга лиц, занимающихся разведением сельскохозяйственных
или домашних животных непрофессионально или участвующих в переработ-
ке с ырья животного происхождения.
Неблагоприятная ситуация усугубляется отсутствием оценочно-прогности-
ческой информации о природных и антропогенных факторах, способствующих
формированию и эпидемическому проявлению очагов ку-лихорадки.
КЛИНИЧЕСКИЕ ПРОЯВЛЕНИЯ ИНФЕКЦИИ
Клиническая диагностика спорадических случаев ку-лихорадки весьма слож-
на из-за выраженного полиморфизма клинических проявлений и отсутствия
патогномоничных симптомов болезни. У большинства инфицированных кокси-
еллами людей болезнь протекает со слабовыраженными симптомами или в лег-
кой форме. Как правило, при спорадических заболеваниях коксиеллезная этио-
логия инфекции остается нераспознанной. Легкое течение болезни не исключает
возможность ее перехода в хроническую форму и возникновения осложнений.
Среднюю и тяжелую формы болезни регистрируют у 6-8% больных, как прави-
ло. именно этих больных госпитализируют. Средняя и тяжелая формы болезни
характеризуются лихорадкой, сопровождающейся сильной головной болью,
ознобом, общей слабостью, мышечными и суставными болями, нередко пораже-
нием легких и печени.
Инкубационный период варьирует от 3 до 39 дней, однако наиболее часто он
составляет 10-21 день. Начало болезни при тяжелом и среднем течении, как пра-
вило, острое, с внезапным ознобом и повышением температуры тела до 39-40 °C в
1-й день заболевания. В течение дня происходят значительные колебания темпе-
ратуры. У больных с отягощенным преморбидным фоном, например поражением
сердечных клапанов, а также у пациентов с неадекватным или поздно начатым
лечением ку-лихорадка может перейти в хроничесю ю форму. Эндокардит — самое
частое и самое тяжелое клиническое проявление хронической формы заболева-
ния.
Для лечения острой формы ку-лихорадки используют тетрациклины (доксици-
клин), фторхинолоны, а при противопоказаниях к этим препаратам — макролиды
и сульфаниламиды (ко-тримоксазол). Антикоксиеллезная активность многих
лекарственных препаратов значительно возрастает при использовании лизосо-
мальных агентов, повышающих pH в фаголизосомах, в которых персистируют
коксиеллы. Для этиотропного лечения коксиеллезного эндокардита целесообраз-
но длительное комбинированное применение доксициклина и гидроксихлорохи-
на. После выписки из стационара реконвалесцент должен находиться под диспан-
серным наблюдением не менее года.
ЧАСТНАЯ МИКРОБИОЛОГИЯ 339
ЛАБОРАТОРНАЯ ДИАГНОСТИКА
Лабораторная диагностика ку-лихорадки строится на изоляции и идентифика-
ции возбудителя, его антигенов и/или фрагментов генома, а также обнаружении
специфических антител.
Изоляция и идентификация ви збудителя
Прямое подтверждение инфицирования коксиеллами — изоляция возбудителя.
Клиническим материалом как для выделения, так и для обнаружения коксиелл
служат кровь, моча, СМЖ, мокрота, а при хроническом течении болезни — био-
птаты печени, сердечных клапанов. Взя гие биологического материала для выде-
ления возбудителя при остром течении болезни желательно проводить на высоте
лихорадки до назначения антибиотиков обладающих антикоксиеллезной актив-
ностью, или в первые дни нормализации температуоы.
В целях выделения коксиелл за рубежом проводят комплекс микробиологиче-
ских исследований:
• биопробу на морских свинках массой 250+25 г;
• исследование плазмы крови инокулированных биологическим материалом
морских свинок иммунологическими методами для обнаружения антител
к С. burnetii, а также органов этих животных на наличие коксиелл (методы
описаны ниже);
• заражение развивающихся куриных эмбрионов или культуры клеток;
• идентификацию выделенной культуры микроорганизмов.
Выделение С. burnetii для диагностики ку-лихорадки у людей в России практи-
чески не применяют. Это обусловлено тем, что, с одной стороны, все исследования
с материалом, подозрительным на заражение коксиеллами, могут осуществляться
лишь в специализированных лабораториях, имеющих разрешение на работу с
возбудителями II группы патогенности, с другой стороны, их трудоемкостью и
длительностью. В некоторых странах, например во Франции, выделение коксиелл
из организма больного применяют для определения чувствительности данного
микроорганизма к различным антибиотикам и оптимизации этиотропного лече-
ния хронических форм коксиеллезной инфекции (как правило, коксиеллезного
эндокардита). В России такие исследования пока не проводятся.
Выявление антигенов возбудителей
В соответствии с методическими рекомендациями «Серологические методы
диагностики риккетсиозов» (1988), коксиеллы в биологических объектах, в том
числе в клиническом материале, выявляют прямым методом флюоресцирующих
антител (ПМФА) или в реакции непрямой гемагглютинации (РИГА) с эритроци-
тарным иммуноглобулиновым диагностикумом. Однако их применение в наших
лабораториях затруднено из-за отсутствия отечественных диагностикумов.
В настоящее время для обнаружения в клиническом материале антигенов
С. burnetii может быть использована разработанная в НИИЭМ им. Л Пастера
иммуноферментная тест-система. В ней использованы поликлональные анти-
тела, сорбированные на поверхности лунок сгрипов. Образующийся комплекс
«ан гитело-антиген» выявляют с помощью пероксидазного конъюгата на основе
поликлональных антител по изменению окрашивания содержимого лунок на
этапе ферментного превращения субстратного раствора. Иммуноферментная тест-
система позволяет выявлять 10-50 нг/мл корпускулярного антигена С. burnetii
1-й фазы, что составляет 3,78х105-1,89х106 клеток возбудителя в 1 мл. Ее чув-
ствительность превосходит чувствительность ПМФА более чем в 6-12 раз, а чув-
ствительность РИГА — более чем в 16-32 раза. Это позволяет выявить С. burnetii в
организме экспериментально зараженных животных как в первые дни заражения,
так и в течение более длительно] о периода.
340
ЧАСТНАЯ МИКРОБИОЛОГИЯ
Молекулярная диагностика
Молекулярно-биологические методы для определения С. burnetii имеют ряд
преимуществ перед иммунобиологическими методами:
• большие чувствительность и специфичность;
• относительно короткое время получения результата (1-3 ч);
• отсутствие повышенных требований к лаборатории по показателю инфекци-
онной защиты после ci адии выделения ДНК;
• возможность диагностики ку-лихорадки на ранних сроках заболевания;
• возможность обнаружения целевой ДНК в материале даже в случае гибели
возбудителя;
• универсальность результатов.
Возможности молекулярно-биологических методов значительно расширились
вследствие распространения практики секвенирования генома микроорганизмов
с возможностью свободного доступа к этим данным в сети Интернет, благодаря
банку генотипов «GenBank» и другим базам данных нуклеотидных последователь-
ностей. Наиболее широко применяют ПЦР для мониторинга распространения
С. burnetii в организме и контроля эффективности проводимого лечения, а также
для санитарно-гигиенической проверки продуктов питания и источников воды,
идентификации возбудителя во внешней среде и выявления его переносчиков и
естественных резервуаров. К настоящему времени известен ряд праймеров, при-
меняемых для выявления С. burnetii в различных клинических образцах, в том
числе парафинированных, включая материал из сердечных клапанов, сосудистой
аневризмы, мокроты, СМЖ, биоптата печени, молока, плаценты, ткани плода и
плодной жидкости, крови. Выделение ДНК проводят стандартными методами
(фенол-хлороформной экстракцией, сорбцией), в том числе с использовани-
ем стандартных коммерческих наборов, предназначенных для выделения ДНК
из соответствующего типа биологических образцов. В качестве мишени для
амплификации используют многочисленные нуклеотидные последовательности.
Во многих лабораториях для диагностики ку-лихорадки применяют праймеры
IS1111, амплифицирующие ген транспозазы, который входит в состав инсерцион-
ного домена высокой копийности (19-31 повторов на хромосому). Применение
праймеров к данному гену позволяет увеличить выход продукта амплификации
и значительно повысить чувствительность ПЦР. Используют также праймеры к
генам 16S rRNA, 23S гРНК, внутреннему транскрибируемому спейсеру 16S-23S
гРНК. генам супероксиддисмутазы (sod), CbbE, rpoB, coml и гену изоцитратдеги-
дрогеназы (icd).
Наиболее эффективный молекулярно-биологический метод выявления кок-
сиелл Бернета — ПЦР в формате реального времени (ПЦР-РВ). Данный метод
обладает не только большей чувствительностью по сравнению со стандартной
ПЦР, но и позволяет установить соотношения между концентрацией целевой
ДНК и количеством ПЦР-продукта, вырабатываемого в процессе амплификации.
ПЦР-РВ проводят в двух модификациях, различающихся принципом генерации
репортерной флюоресценции. В соответствии с первым принципом в качестве
интеркалирующих красителей используют SYBR Green I, SYTO-13, РОРО-3 и др.
Второй принцип предполагает использование технологий (например, «TaqMan»,
«Molecular Beacons» и «LightCycler»), основанных на применении меченных
флюорофорами олигонуклеотидных последовательностей. В настоящее время
для выявления С. burnetii с помощью ПЦР-РВ в качестве мишени для праймеров
используют различные гены — sod, сот 1, icd, ген транспозазы IS1111.
Серологическая диагностика
Выявление антител к С. burnetii, равно как и непосредственное выделение дан-
ного возбудителя, возможно только в лабораториях, имеющих специальное разре-
ЧАСТНАЯ микробиология 341
шение, что существенно ограничивает круг практических лабораторий, выполняю-
щих эти исследования. Напротив, серологические исследования, предназначенные
для выявления антител к С. burnetii в плазме крови больных, не требуют особых
условий и могут осуществляться во многих лабораториях. Для лабораторной диа-
гностики случаев острого заболевания ку-лихорадкой исследование плазмы крови
целесообразно проводить на самых ранних сроках заболевания. Отрицательные
результаты серологических реакций или низкие титры антител к коксиеллам
2-й фазы диктуют необходимость повторного исследования плазмы крови, полу-
ченной от больного, через 7-10 дней после забора первой пробы и сравнения
результатов одновременного исследования двух проб. Появление антител к С. bur-
netii 2-й фазы или нарастание титров этих антител во второй пробе в 4 раза и более
свидетельствуют о коксиеллезной природе заболевания.
Для выявления антител к С. burnetii используют реакцию связывания ком-
племента (РСК), реакцию агглютинации (РГА) в различных модификациях и
непрямой метод флюоресцирующих антител (НМФА), при этом параллельное
применение антигенов из коксиелл 1-й и 2-й фаз существенно повышает эффек-
тивность серологического исследования. С помощью РСК, как правило, не удается
обнаружить антитела к С. burnetii в ранний период заболевания. Именно поэтому
однократное обследование больных в данной серологической реакции во время
лихорадки или в период ранней реконвалесценции в большинстве случаев оказы-
вается недостаточно эффективным. Для более ранней серологической диагности-
ки целесообразно применение РГА и НМФА.
Для повышения эффективности серологической диагностики ку-лихорадки в
НИИЭМ им. Л. Пастера была разработана тест-система для ИФА. Ее особенность
заключается в том, что за счет химической модификации коксиеллезного анти-
гена 1-й фазы удалось существенно повысить активность твердофазного сорбента.
При этом обеспечивается выявление антител к различным детерминантам, так
как происходит пространственная демаскировка антигена С. burnetii 2-й фазы при
сохранении или даже активации антигена ксдемелл 1-й фазы. Разработанная тест-
система ИФА обладает значительно большей чувствительностью по сравнению с
другими серологическими тестами. С ее помощью удается обнаружить антитела
к С. burnetii как на ранних стадиях болезни, так и на протяжении нескольких лет
после выздоровления.
Способность серологических тестов обнаруживать антитела к С. burnetii в
течение нескольких лет после перенесенной ку-лихорадки обосновывает необ-
ходимость дифференцировать следовые иммунологические реакции от недав-
него антигенного раздражения. С этой целью постановку РСК целесообразно
проводить одновременно с антигенами С. burnetii, находящимися в разных
фазовых состояниях. Антитела к С. burnetii 2-й фазы выявляются раньше, чем к
С. burnetii 1-й фазы, и определяются при остром течении болезни, как правило,
за более длительный период времени. Именно поэтому выявление в РСК анти-
тел к С. burnetii 1-й фазы на ранних сроках болезни (первые 2-3 нед) с высокой
степенью вероятности свидетельствует об инфицировании данным возбудителем
в прошлом.
Дифференциация антител по классам иммуноглобулинов также позволяет
ориентировочно судить о сроках, прошедших с момента заражения С. burnetii.
Обнаружение IgM к С. burnetii 2-й фазы с помощью НМФА на ранних стадиях
заболевания, как правило, позволяет серологически распознать коксиеллезную
этиологию инфекции без повторного исследования плазмы крови в динамике.
Обнаружение в данной реакции высоких титров IgG (1:800 и выше) к С. burnetii
1-й фазы — показатель хронической инфекции.
342 ЧАСТНАЯ МИКРОБИОЛОГИЯ
ЭНТЕРОБАКТЕРИИ
Более 100 лет представители данного семейства находятся в центре внимания
медицинской микробиологии. Первоначально интерес к энтеробактериям был
обусловлен принадлежностью к ним ведущих возбудителей острых кишечных
инфекций (дизентерии, эшерихиозов, брюшного тифа, паратифов и других саль-
монеллезов), а также чумы, лсевдотуберкулеза и кишечного иерсиниоза. На про-
тяжении последних десятилетий знания об этиологической роли энтеробактерий
в патологии человека претерпели существенные изменения. Установлено значение
широкого спектра представителей семейства в этиологии гнойно-септических
внутрибольничных и респираторных заболеваний, инфекций мочевыводящих
путей. Отмечается неуклонный рост доли грамотрицательных микроорганизмов,
в том числе энтеробактерий, в этиологической структуре бактериемий, гнойно-
септических и внутрибольничных инфекций.
ХАРАКТЕРИСТИКА СЕМЕЙСТВА ENTEROBACTEHIACEAE
Семейство Enterobacteriaceae включает 44 рода у-протеобактерий (жирным
шрифтом выделены роды, имеющие медицинское значение).
Arsenophonus Erwinia Parrtoea Samsonia
Brenneria Escherichia Pectobacterium Serratia
Buchnera Ewmgella Photorhabdus Shigella
Budvicia Hafnia Plesiomonas Sodalis
Buttiauxella Klebsiella Pragla Tatumella
Calymmatobacterium Kluyvera Proteus Thorsellia
Citrobacter Leclercia Provlaencla Trabulsiella
Cedecea Lemlnorella Rahnelia Wigglesworthia
Dickeya Moeilerella Raoultelia Xenorhabdus
Edwardsietla Morganella Saccharobacter Yersinia
Enterobacter Obesumbacterium Salmonella Yokenella
Представители семейства Enterobacteriaceae — грамотрицательные, не обра-
зующие спор и микроцист тонкие палочки среднего размера, не содержащие
включений, имеющих диагностическое значение, обладающие и не обладающие
капсулой, подвижные перитрихи (за исключением Tatumella) или неподвижные.
Некислотоустойчивы Факультативные анаэробы, хорошо растут на обычных
питательных средах. По способности расти на питательных средах минимально-
го состава принадлежат к прототрофам (используют глюкозу как единственный
источник углерода) и ауксотрофам, требующим для роста в качестве добавок
витамины или аминокислоты. Питание осуществляют за счет органических соеди-
нений. Энергию получают путем мембранного и дыхательного фосфорилирова-
ния. Каталазоположительны, за исключением Shigella dysenteriae и Xenofhabdus
nematophilus. Оксидазонегативны, за исключением Plesiomonas shigelloides.
Проявляют ферментативную активность в отношении углеводов (ферментируют
их до кислоты или до кислоты и газа), многоатомных спиртов, органических
кислот и их солей, аминокислот. Нитраты редуцируются в нитриты, за исключе-
нием некоторых штаммов Erwinia и Yersinia Ферментативные реакции положены в
основу дифференциации энтеробактерий на роды, виды и подвиды.
Энтеробактерии обладают следующими антигенами:
• соматическим О-антигенным комплексом:
• Н-антигенами (подвижными бактериями);
• поверхностными, капсульными антигенами (Klebsiella spp., Е. coli, S. typhfy
• общим антигеном энтеробактерий (СА), за исключением Erwinia chrysan-
themi.
Представители многих родов характеризуются различными факторами виру-
лентности (адгезинами, инвазинами. токсинами), которые кодируются генами,
ЧАСТНАЯ МИКРОБИОЛОГИЯ 343
локализованными в островках патогенности (расположены в хромосоме), или
генами, локализованными в плазмидах, бактериофагах и других мобильных гене-
тических элементах.
КЛИНИЧЕСКИЕ ПРОЯВЛЕНИЯ ИНФЕКЦИИ
Энтеробактериями вызываются разнообразные заболевания. Клиническое зна-
чение различных представителей семейства Enterobacteriaceae неравнозначно.
Шигеллами, сальмонеллами, диареегенными эшерихиями, иерсиниями вызыва-
ются инфекционные заболевания, способные к широкому эпидемическому рас-
пространению (острые кишечные инфекции, псевдотуберкулез, чума, брюшной
тиф). Практически все энтеробактерии могут вызывать пищевые токсикоин-
фекции (ПТИ) из-за способности размножаться в пищевых продуктах. Многие
представители семейства Enterobacteriaceae нередко являются причиной инфекций
мочевыводящих путей, респираторных заболеваний, послеоперационных ослож-
нений, сепсиса, менингитов. Они способны вызывать тяжелые внутрибольничные
инфекции, особенно у пациентов с ослабленным иммунитетом, детей раннего
возраста и недоношенных. Энтеробактерии также могут вызывать аппендицит,
поражение суставов. Данные заболевания часто обусловлены представителями
родов Citrobacter, Enterobacler. Esherichia, Klebsiella, Morganella, Proteus, Providencia,
Serratia.
Два представителя энтеробактерий — E. coll и К. pneumoniae — входят в число
бактерий, являющихся индикаторами развития резистентности к антимикробным
препаратам в Европе. Определение этиологической значимости условно патоген-
ных энтеробактерий основано на количественной характеристике выявляемого
микроорганизма.
ЛАБОРАТОРНАЯ ДИАГНОСТИКА
Преаналитический этап
Материалом для бактериологического исследования служат кровь, слизь из зева
и носа, гной, экссудат, раневое отделяемое, пунктат различных органов, отделяе-
мое цервикального канала, мокрота, желчь, моча, фекалии, СМЖ.
Пробы крови, как правило, засевают на питательные среды непосредственно
у постели больного. Для взятия и транспортировки проб, исследуемых на нали-
чие энтеробактерий, следует использовать стерильные тубсеры, зонды-тампоны,
одноразовые стерильные емкости (пробирки) с герметично завинчивающейся
крышкой.
Если материал нельзя доставить в лабораторию немедленно после взят ия пробы,
для сохранения жизнеспособности энтеробактерий целесообразно использовать
специальные транспортные системы с питательной средой Выбор транспортной
среды с учетом вида материала, цели исследования и технических возможностей
осуществляют сотрудники лаборатории. К числу наиболее универсальных транс-
портных сред относятся среды Стюарта и Эймса (с углем или без него). Для транс-
портировки фекалий также допустимо использовать среду Кэри-Блеер, глицери-
новую (за исключением бактерий рода Yersinia) и фосфатную буферную смесь,
боратно-буферный раствор или 0,9% раствор натрия хлорида. Транспортировка
материала должна осуществляться в максимально короткие сроки, как правило, в
течение 1,5-2 ч.
Каждая проба должна иметь четкую маркировку, а также направление, содер-
жащее информацию, необходимую для получения адекватного результата и его
правильной интерпретации (вид и цель исследования, дата и время получения
материала).
344 ЧАСТНАЯ МИКРОБИОЛОГИЯ
Аналитический этап
Бактериологическое исследование доставленного в лабораторию материала на
наличие в нем энтеробактерий обычно включает:
• посев проб на питательные среды и выделение чистой культуры;
• идентификацию выросших колоний по биохимическим (ферментативным)
свойствам (определение рода, вида, биовара);
• идентификацию патогенных энтеробактерий (шигелл, сальмонелл, эшери-
хий) по антигенной характеристике (определение серогруппы, серовара);
• определение чувствительности к антимикробным препаратам и выявление
клинически значимой резистентности к препаратам выбора для лечения.
Большинство видов энтеробактерий нетребовательны к питательным средам и
хорошо растут на простом питательном агаре, кровяном агаре и других питатель-
ных средах. Энтеробактерии образуют круглые, слегка выпуклые, средней вели-
чины, влажные, блестящие, прозрачные в проходящем свете колонии с ровными
краями (гладкие S-формы) или более плоские, сухие, с неровными волнистыми
краями (шероховатые R-формы). Диссоциация на S- и R-формы чаще проявляется
спонтанно на питательных средах, содержащих углеводы.
Выделение энтеробактерий из материалов, богатых собственной микрофлорой,
может быть затруднено и требует применения специальных дифференциально-
диагностических, обогащенных, элективно-дифференциатьных, хромогенных
и других питательных сред. Наиболее широкое распространение получили
дифференциально-диагностические питательные среды с лактозой в качестве
дифференциального фактора. Они универсальны, позволяют выделить подавляю-
щее большинство видов энтеробактерий, дифференцируя выросшие колонии по
отношению к лактозе.
В настоящее время разные авторы предложили многочисленные варианты про-
писей подобных питательных сред, отличающиеся друг от друга применяемым
индикатором pH, питательной основой и технологией приготовления. Широкое
распространение получили три среды данного класса — Эндо, Левина и Мак-
Конки.
• Среда Эндо — слабоселективная дифференциально-диагностическая пита-
тельная среда с лактозой, фуксином и сульфитом нагрия. Последние два
компонента, помимо функции индикатора, обладают слабым ингибирующим
действием по отношению к грамположительным микроорганизмам.
• Среда Левина (эозин-метиленблау-лактоза-агар) отличается от среды Эндо
индикатором, в качестве которого используются эозин и метиленовый синий.
Данная питательная среда также обладает слабым ингибирующим действием
по отнопк нию к грамположительным бактериям.
Существует разновидность — дифференциально-диагностическая питательная
среда, называемая агаром с эозин-метиленовым синим (ЭМС). В ее состав, кроме
лактозы, входит сахароза. Это необходимо учитывать, так как в литературе ее
часто некорректно идеи гифицируют со средой Левина. К числу выпускаемых за
рубежом дифференциально-диагностических сред с лактозой относится агар с
бромкрезол-фиолетовым, отличающийся от среды Левина типом индикатора.
• Агар Мак-Конки отличается от соеды Эндо индикатором, в качестве кото-
рого используется нейтральный красный, и наличием желчных солей. Данная
дифференциально-диагностическая питательная среда обладает более выра-
женными элективными свойствами по сравнению со средой Эндо, однако
спектр вырастающих на ней микроорганизмов достаточно широк. Кроме
энтеробактерий, на этой питательной среде растут энтерококки, стафилокок-
ки, псевдомонады, аэробактер и некоторые другие устойчивые к желчи бак-
терии. Во многих странах данная дифференциально-диагностическая среда
широко используется при исследовании на энтеробактерии, энтерококки, а
ЧАСТНАЯ МИКРОБИОЛОГИЯ 345
также для дифференциации некоторых видов микроорганизмов, например
пастерелл.
Разработаны и выпускаются различными фирмами многочисленные варианты
среды Мак-Конки с более узкой целевой направленностью, что необходимо учи-
тывать при проведении исследований.
Агар Мак-Конки с сорбитом применяют для целенаправленного поиска
Е. coli 0157:Н7 — основного представителя группы энтерогеморрагических
эшерихий. Штаммы Е. coli 0157:Н7 не ферментируют сорбит и на этой среде
образуют бесцветные или бледно-розовые колонии, в отличие от большин-
ства эшерихий. у которых вследствие ферментации сорбита колонии на
этой среде имеют розовый цвет, а на среде Эндо — лактозоположительны.
Использование данной среды повышает частоту обнаружения Е. coli 0157:Н7,
поскольку сорбитотрицательных штаммов эшерихий значительно меньше,
чем лактозоположительных.
Для выделения отдельных родов (видов) энтеробактерий разработаны узко-
специализированные элективные среды. Производится селективная среда для
выделения микроорганизмов родов Proteus, Providencia и Morganella, содержащая
хлоргидрат алкилэтилглицина (поверхностно-активное вещество), который пода-
вляет рост других энтеробактерий.
• Висмут-сульфит агар — высокоселективная дифференциально-диагно-
стическая среда для выделения сальмонелл. Входящие в ее состав брилли-
антовый зеленый и висмут ингибируют рост грамположительных бактерий,
а также многих энтеробактерий. Дифференцирующее действие основано на
том. что образуемый сальмонеллами сероводород из сульфата железа вызы-
вает почернение индикатора (сульфита висмута), именно поэтому бактерии,
продуцирующие сероводород, формируют черные колонии с металлическим
блеском, а среда вокруг колоний окрашивается в черный цвет. Бактерии, не
продуцирующие сероводород, могут вырастать в виде мелких бесцветных,
зеленоватых или коричневых колонии.
Висмут-сульфит агар выпускают в сухом виде. Предпочтительнее включать в
исследование питательную среду, выдержанную перед посевом при температуре
4 °C в течение не менее 24 ч. Посевы инкубируют в термостате при температуре
37 °C с ежедневным просмотром через 24 и 48 ч.
• Среда Плоскирева — селективная питательная среда для выделения шигелл
и сальмонелл. Входящие в состав среды желчные соли, бриллиантовый
зеленый и йод полностью подавляют рост грамположительных бакте-
рий и способность роения протея, а также задерживают рост эшерихий.
Дифференцирующий фактор — лактоза.
Жидкие среды обогащения применяют до посева на плотные питательные
среды для увеличения первоначального количества энтеробактерий в биоматериа-
ле. Их принцип действия состоит в подавлении химическими веществами (тетра-
тионатом натрия, кристаллвиолетом, малахитовым зеленым), входящими в состав
среды, роста Е. coli при отсутствии воздействия на патогенные энтеробактерии.
При подавлении конкурирующего действия сопутствующей микрофлоры патоген-
ные энтеробактерии развиваются быстрее.
Наиболее распространены среды обогащения для выделения сальмонелл.
Компоненты, определяющие избирательный рост сальмонелл, — соли селенистой
кислоты, желчь и соли желчных кислот (преимущественно дезоксихолат натрия),
соли магния, препараты йода, а также некоторые красители (бриллиантовый зеле-
ный).
• Среда Мюллера относится к числу признанных сред, способствующих пре-
имущественному накоплению сальмонелл. При добавлении к тетратионато-
вому бульону тиосульфата натрия и раствора Люголя образуется тетратионат
ГЛАВА 21
346 ЧАСТНАЯ МИКРОБИОЛОГИЯ
натрия, который является ингибитором для эшерихий и другой микрофлоры
кишечника, но не влияет на развитие большинства сероваров сальмонелл.
Питательная среда готовится ex tempore.
• Селенитовый бульон обеспечивает избирательный рост сальмонелл за счет
действия гидроселинита натрия.
• Магниевая среда содержит селективный фактор — соли магния. Питательная
среда имеет ряд модификаций, однако ингибирующий эффект основан на
дегидратирующем действии магния хлорида. Также селективность среды
усиливают введением в ее состав малахитового зеленого.
• Желчный бульон и среда Раппопорта являются средами обогащения, в
них используются свойства желчи, их, как правило, применяют для посева
крови при лабораторной диагностике брюшного тифа и паратифов А, В и С.
• Среда Кауфмана сочетает ингибирующие свойства желчи и бриллиантового
зеленого.
Приготовленные в лаборатории жидкие питательные среды обогащения долж-
ны подвергаться внутрилабораторному контролю для опенки ростовых и инги-
бирующих свойств. В настоящее время отечественным лабораториям доступны и
готовые питательные среды для энтеробактерий, обладающие стандартно высо-
ким качеством.
Холодовое обогащение используют для обнаружения иерсиний — посевы инкуби-
руют в забуференной пептонной воде или пептонно-калиевой среде в условиях холо-
дильника. Для выделения У. enterocolitica производится ряд элективных питательных
сред с селективными добавками, содержащими цефазолин, иргазан и новобиоцин.
Дифференциацию бактерий проводят по ферментации маннита. Выпускаются сла-
боэлективные среды, в которых содержится, в частности, один только иргазан; они
пригодны для выделения как У. enteroculitica, так и У. pseudotuberculosis.
В последние годы все более широкое распространение дтя идентификации энте-
робактерий получили хромогенные среды. Принцип действия данных сред осно-
ван на выявлении специфических или уникальных ферментов микроорганизмов.
Большинство хромогенных сред содержат селективные добавки, подавляющие
рост посторонней микрофлоры. Использование хромогенных сред позволяет уско-
ренно (в течение суток) выделять и одновременно идентифицировать наиболее
частые и значимые в клинической практике виды энтеробактерий, практически
без проведения дополнительных тестов (или используя 1-2 теста, выполняемых в
течение нескольких часов в пределах первых суток).
Разработаны и выпускаются хромогенные среды для колиформных бактерий и
видовой идентификации Е. coli (хромогенная среда для выявления (З-глюкуронидазы
и р-галактозидазы), Salmonella spp. (хромогенная среда с пропиленгликолем) и др.
НИИЭМ им. Л. Пастера производит две хромогенные питательные среды для
одноэтапного выделения и прямой идентификации бактерий — среду «Протеус
ППМ» для выявления родов Proteus, Providencia и Morganella (принцип действия
основан на выявлении специфичной для данного вида энтеробактерий триптофан-
дезаминазы), а также среду «Klebsiella 5-АСК» для идентификации рода Klebsiella
(принцип действия основан на обнаружении уникального фермента клебсиелл —
5-аминосалицилатдекарбоксилазы).
ИДЕНТИФИКАЦИЯ ЭНТЕРОБАКТЕРИЙ ПО БИОХИМИЧЕСКИМ СВОЙСТВАМ
Для доказательства принадлежности микроорганизма к семейству Entero-
bacteriaceae достаточно поставить три теста — на оксидазу, редукцию нитратов и
OF-тест (среда Хью-Лейфсона). Энтеробактерии — оксидазоотрицательны (кроме
Plesiomonas sfugelloides), редуцируют нитраты и ферментируют глюкозу.
Для ориентировочной межродовой дифференциации осуществляют дальней-
шую идентификацию выросших колоний по ферментативным свойствам с приме-
ЧАСТНАЯ МИКРОБИОЛОГИЯ 347
нением полиуглеводных питательных сред (среда Клиглера, Олькеницкого, трех-
сахарного железного агара, лизин-железного агара, среды Ресселя с мочевиной).
Данные многофункциональные питательные среды позволяют судить о микро-
организме сразу по нескольким признакам. Как правило, они предназначены
для дифференциации энтеробактерий по способности ферментировать глюкозу,
лактозу, а также образовывать газ и сероводород.
При расщеплении углеводов образуется кислота, что приводит к изменению
цвета индикатора. Соотношение углеводов в питательных средах подобрано таким
образом, что при ферментации глюкозы изменяет цвет на желтый только столбик
среды. Пожелтение скошенной части среды свидетельствует об утилизации лак-
тозы или сахарозы. Если при ферментации углеводов образуется газ, то в толще
среды появляются разрывы.
Также в состав питательных сред входит тиосульфат, который под действием
микробной тиосульфатредуктазы (ею обладают Salmonella, Proteus, Citrobacter)
восстанавливается в сульфит с образованием сероводорода. Входящие в состав
среды соли железа вступают с сероводородом в реакцию, приводя к образованию
сульфида железа в виде преципитата черного цвета. Именно поэтому при образо-
вании сероводорода среда чернеет.
Более сложный состав имеет среда Олькеницкого (вариант трехсахарного агара
с мочевиной). Присутствие в среде мочевины позволяет определить наличие у
бактерий уреазы.
С помощью полиуглеводных питательных сред ориентировочно определяют
родовую принадлежность и намечают ход дальнейшего анализа (табл. 21-5).
Удобнее использовать среды Клиглера и с мочевиной (по Кристенсену) или
среду Олькеницкого, однако на последней при ферментации мочевины изолятами
затруднен учет других биохимических свойств.
Тесты, используемые для дальнейшей межродовой дифференциации клиниче-
ски значимых и малоизученных родов семейства Enterobacteriaceae, представлены
в табл. 21-6 и 21-7.
Из числа сложных питательных сред, которые предназначены для применения
с этой целью, следует упомянуть двухфазную среду «Энтеро Плюс» (ее название и
состав запатентованы компанией «Новамед», Израиль). Она позволяет одновременно
определять подвижность изолятов энтеробактерий и наличие у них способности:
• расщеплять орто-нитрофенил галактозидазу на глюкозу и орто-нитрофенил;
• ферментировать глюкозу;
• образовывать сероводород;
• образовывать индол;
• образовывать газ;
• разлагать мочевину.
Для точной видовой идентификации энтеробактерий требуется выполнение
многочисленных биохимических тестов, нередко с использованием дорогостоящих
химических субстратов. С этой целью применяют специальные диагностические
тест-системы. Принцип действия большинства из них одинаков: об утилизации
или расщеплении возможно минимизированного объема субстрата в питательной
среде судят по изменению цвета специального индикатора.
Таблица 21-5. Первичная дифференциация родов семейства Enterobacteriaceae на полиуглевод-
ных средах
Энтеробактерии Глюкоза (газ) Лактоза Сахароза Продукция сероводорода
Enterobacter cloacae subsp. cloacae Citrobacter spp. Klebsiella spp. Raoultelia spp. + + + -
348 ЧАСТНАЯ МИКРОБИОЛОГИЯ
Окончание табл. 21-5
Энтеробактерии Глюкоза {газ) Лактоза Сахароза Продукция сероводорода
Citrobacter spp. Proteus myxofaciens + - + -
Morganella spp. Citrobacter spp. + - - -
Citrobacter spp. + + - -
Proteus vulgaris + - + +
Proteus mirabilis + - - +
Yersinia enterocolitica - - + -
Yersinia pseudotubercuiosis - - - -
Budvica aquatica - - - +
Buttiauxella spp. Citrobacter spp. Edwardsiella ictaluri Enterobacter taylorae Escherichia spp. Hafnia aivei Salmonella paratyphi A Yokenella regensburgei + - - -
Buttiauxella agrestis Citrobacter spp. Enterobacter mmipressuralis Escherichia coli + + - -
Enterobacter sakazaKii Escherichia coli Klebsiella mobilis Kluyvera spp. Rahneila aquatltis Serratia rubidaea + + + -
Edwardsiella hoshinae Enterobacter hormaechei Serratia spp. + - + -
Salmonella spp. Leminorella grimontii Citrobacter spp. + - - +
Budvica aquatica Hafnia alvei биовар 1 Edwardsiella ictaluri Escherichia coli неактивная Obesumbacterium proteus биогруппа 1 Plesiomonas shigelloides Shigella spp. Yersinia pestis Yersinia ruckeri Ewingeila americana - - - -
Ewingella americana — + - -
Moellerella wisconsensis - + -
Edwardsiella hoshinae Enterobacter hormaechei Pantoea disperse Serratia spp. Tatumella ptyseos - - + -
Leminorella richardii Pragia fontium Salmonella typhi Budvica aquatica - - - +
Примечание, «+» — положительная реакция; — отрицательная реакция.
Таблица 21-6. Биохимическая дифференциация клинически значимых родов семейства Enterobacteriaceae
Тест или субстрат Escherichia Shigella Edwardsieila Citrobacter Salmonella Klebsiella Enterobacter Hafnla Serratia Proteus Providencia Morganeila Yersinia Erwinia
Ферментация глюкозы кг, к к1 кг кг кг кг кг кг, к кг, к кг кг кг к к
Образование кислоты из лак- тозы + -/(+) - + -/.+ + + + + - - - - ±
Образование сероводорода - - + ± +! - - - - + - - - -
Образование индола + -/+ + —/+ - -/+ - - - + 4> + -/+ -/+
Цитрат (среда Симмонса) - - - + + + 4- + + + 4- - - +
Фенилаланиндезаминаза - - - - - - - - - + 4- 4- - -
Лизиндекарбоксилаза ± - + - + + -/+ + + - - - - -
Орнитиндекарбоксилаза -/+ - ± -/+ - - 4- 4- + - - + + -
Проба с метиловым красным + + + + + -А - -/+ + 4- 4- 4- + ±
Реакция Фогеса-Проскауэра - - - - - + + -/+ + - - - - -/+
Ферментация адонитола -/+ - - -/+ - 4- -/+ - -/+ - -/+ - - -
Ферментация инозита + *•* - -/+ - + -/+ - -!+ - - - - -
Ферментация сорбита + - - + + + + - + - -/+ - ± —
Ферментация рамнозы + -/+ - + + + + + -/+ - ±
Подвижность + - 4- ± ± - 4- + + + + + -А4 + S
Гидролиз мочевины - - - - - (+) (+) - -/+ + +6 -J+
Примечание: о «кг>> — кислота-газ; «к» — кислота; «+» или •}-/+* — разные биовары или виды рода; «(+)» — замедленная или слабовыраженная реакция. § 1 Кроме S.flexneri 6. о 2 Кроме S. typhi. § 3 Кроме некоторых редко встречающихся серовариантов. 1 При темпемйгуре 25 °C. 5 Кроме У. pestis.
со
Таблица 21-7. Биохимическая дифференциация малоизученных родов семейства Enterobacteriaceae
Тест или субстрат Cedecea Ewlngella Kluyvera Leclercia Moellerella Pantoea Pragia Rahnella Tatumella Xenorhabdus Yokenella
Образование индола - - + -г - - - - - - -
Проба с метиленовым красным + + + + + -/+ + + - - +
Реакция Фогеса-Проскаузра + + - - - + - + - - -
Цитрат (среда Симмонса) + + + - + + + + - - +
Образование сероводорода - - - - - -/+ + - - - -
Гидролиз мочевины - - - - - - - - - - -
Фенилаланиндезаминаза - - - - - - - + + - -
Лизиндекарбоксилаза - - - - - - - - - +
Орнитиндекарбоксилаза -/+ - + - - -/+ - - - - +
Подвижность + -/+ + + - + + - - + +
Ферментация глюкозы с выделением газа + + + - - - + - - +
Ферментация адонитола - - - + + - - - - - -
Ферментация инозита - - - - - -/+ - - - - -
Ферментация лактозы -/+ -/+ + + + - - + - - -
Ферментация маннита + + + + ± + - + - - +
Ферментация рамнозы - - + + - + - + - - +
Ферментация сахарозы + - + -/+ + + - + - - -
Ферментация сорбита -/+ - -/+ - - - - + - - -
Ферментация целлобиозы + - + + - + - + - - +
Ферментация мальтозы + - + + + 4- - + - +
Пигмент - - - ж - Ж - - - К, Ж. 0 -
Примечание. «Ж» — желтый; «К» — коричневый; «О» — оранжевый
ЧАСТНАЯ МИКРОБИОЛОГИЯ 351
С технической точки зрения, доступные тест-системы можно разделить на три
группы.
• Тест-системы, содержащие высушенную смесь «субстрат-индикатор-среда»
в специальном планшете. Таких тест-систем большинство. Суспензию тести-
руемого микроорганизма вносят в лунки планшета. Преимущество данных
тест-систем — высокая технологичность, недостаток — невозможность выбо-
ра отдельных тестов, планшет всегда расходуется полностью.
• Тест-системы, содержащие жидкую смесь «субстрат-индикатор-среда»,
которая раскапывается в лунки планшета непосредственно перед примене-
нием. Преимущество данных тест-систем — экономичность, так как набор
тестов для каждой культуры может варьировать по мере необходимости,
недостаток — увеличение трудозатрат.
• Тест-системы, в которых смесью «субстрат-индикатор-среда» пропитаны
бумажные диски. Перед применением диски в асептических условиях рас-
кладывают пинцетом в пробирки, содержащие взвесь исследуемого микро-
организма. Для данных тест-систем характерны максимальные трудозатраты
при возможности неограниченно варьировать набор тестов.
По времени учета результатов все тест-системы можно разделить на две
группы:
• позволяющие полечить результат через 4-5 ч инкубации;
• требующие инкубации в течение 18-24 ч.
Первая промышленная тест-система, сохранившая свое значение в качестве
«золотого стандарта» для идентификации энтеробактерий, — тест-система «API
32Е» («Bio Merieux»). Она позволяет в течение 18 ч определить с помощью одной
панели результаты 20 тестов. Дополнительно определяют цитохромоксидазу,
окисление и ферментацию глюкозы, подвижность. Объектом идентификации
может быть изолированная колония со среды первичного посева. Фирма про-
изводит тест-систему «Rapid 32Е» для идентификации энтеробактерий, которая
позволяет получить результат в течение 4 ч.
Тест-системы «Enterotest 16», «Энтеротест 24», «ЭнтероСкрин» и «Энтеро-
Рапид24» («Эрба-Лахема с.р.о.») включают тесты для родовой идентификации
энтеробактерий — опредетения уреазы, р- и D галактозидазы, липазы, фенила-
ланиндезаминазы, орнитиндекарбоксилазы, лизиндекарбоксилазы, образования
индола, сероводорода, ацетоина, ферментации маннитола, цитрата и инозитола,
адонитола, эскулина, малоната натрия, рамнозы, целлобиозы, сахарозы, сорбита,
глюкозы, дульцитола и трегалозы, а также определения аргининдегидролазы.
Система «Enterotest 16» («Эрба-Лахема с.р.о.») предназначена для идентификации
наиболее значимых в клинической практике энтеробактерий в течение 18-24 ч с
помощью проведения 16 тестов — определения уреазы, фенилаланиндезаминазы,
лизиндекарбоксилазы, орнитиндекарбоксилазы, образования индола, сероводо-
рода, ферментации цитрата, эскулина, малоната, инозитола, адонитола, целлобио-
зы, сахарозы, сорбита, маннитола и трегалозы.
В настоящее время наряду с импортными выпускаются и многочисленные оте-
чественные тест-системы для идентификации энтеробактерий, в том числе MMTF
1 и ММТЕ 2 (НПО «Аллерген, Ставрополь), СИБ-набор № 2 (НПО «Микроген»
предприятие «ИмБио», Нижний Новгород), ПБДЭ (НПО «Диагностические
системы», Нижний Новгород), «РАПИД-ЭНТЕРО-200» и «РАПИД-ЭНТЕРО-50»
(НИИЭМ им. Л. Пастера. Санкт-Петербург).
Способ идентификации культуры по биохимическим тестам зависит от тех-
нических возможностей лаборатории. Следует отметить, что существующие диа-
гностические таблицы позволяют идентифицировать только основные, хорошо
известные роды энтеробактерий. Использование диагностических ключей (осо-
бых графических схем) дает возможность сократить трудоемкость, но иногда за
лг vwu .
352
ЧАСТНАЯ МИКРОБИОЛОГИЯ
счет снижения точности. Наиболее современные способы идентификации — кодо-
вые книги и компьютерные программы.
Новые возможности быстрой идентификации микроорганизмов, включая
энтеробактерии, с одновременным определением чувствительности к антибак-
териал ьным препаратам открываются при использовании автоматизированных
микробиологических анализаторов. Высокая пропускная способность и авто-
матизация основных этапов работы снижают трудоемкость и значительно рас-
ширяют спектр идентифицируемых микроорганизмов. В табл. 21-8-21-13 при-
ведены тесты для меж- и внутриродовой идентификации клинически значимых
энтеробактерий.
Таблица 21-8. Внутриродовая дифференциация рода Citrobacter
Тест или субстрат С. freundii С. diversus С. amalnnaticus
Образование сероводорода Ы - -
Образование индола - + +
Малонат натрия - + -
Орнитиндекарооксилаза [-] + +
Образование кислоты из адонита - + -
Таблица 21-9. Внутриродовая дифференциация рода Escherichia
Тест или субстрат Е. СОИ F. hermand F. vitlneris E. tergusoni F. blattae
Образование индола + + - + -
Орнитиндекарбоксилаза d + - + +
Ферментация дульцита d [-] - d -
Ферментация адонита - - - + -
Ферментация сорбита + - - - -
Ферментация целлобиозы - + + + -
Использование малоната - - [+] d +
Образование желтого пигмента - + d - -
Таблица 21-10. Внутриродовая дифференциация рода Serratia
Тест S. entomophila S. ficaria S. fonticola S. grimesii S. Ilquefaciens S. marcescens s. odorifera биогруппы 1 S. odorifera би группы 2 s. plymuthica S prote- awacuians S. rubidaea
Реакция Фогеса-Проскауэра + d — d + + d + Ш f+1 +
Лизиндекарбоксилаза — — + + + + + + + d
Аргининдекарбоксилаза — — — + - - - - - - -
Орнитиндекарбоксилаза — + + + + + - - + -
Гидролиз желатина + — + + + + + d + 4-
Ферментация малоната — — [+1 _ - — — - - - - +
Образование газа из О-глюкозы — — f+1 + [+1 d - Ы d + d
Ферментация D-адонита — — + — d d d - +
Ферментация L-арабинозы - + + + + - + + + + +
Ферментация целлобиозы — + — — — — + + [+] - +
Ферментация дульцита — — — - — — - - -
Ферментация лактозы — r-1 + - — — d + [±]__ - +
Ферментация мелибиозы — d 4- + bl - + + 4- + 4-
Ферментация рафинозы — d + + 1+1 — 4- — + + +
Ферментация L-рамнозы — d [+1 — f-1 — + + - d -
Ферментация D-сорбита — + + + + + + + d [±J -
Ферментация сахарозы + + bl + + 4- + - + + +
Ферментация D-ксилозы d + r+1 + + — + + + + +
Ферментация метил-0-глюкозида — - + - — — - d - -
Дезоксирибонуклеаза + + - + [+1 + + + + + +
Липаза [-1 f+1 - + f+1 4- d d d + +
Пигмент (красный, розовый, оран- жевый) - - - d - - d - +
Примечание:
«-» — 0—10% штаммов положительны:
«[-]» — 11-25% штаммов положительны,
«d» — 26-75% штаммов положительны;
«[+]» — 76-89% штаммов положительны;
«+» - 90-100% штаммов положительны.
Таблица 21-11. Биохимическая дифференциация родов Proteus, Providencia и Morganella morganii
1 ест Proteus Morganella morganii Providencia
mirabllls myxofa- ciens penned vulgaris alkali- faoiens heim- bacbae rettgeri rustlgianii sluartii
Реакция Фогеса-Проскауэра d 4- — — — - - — - -
Цитрат (среда Симмонса) d + — [-1 - + — + -[-] 4
Образование сероводорода + - d 4 — - — - —
Образование индола - - - + + 4 - 4 4 4
Образование газа из D-глюкозы 4 + • d [+] 4 — - d -
Гидролиз мочевины + 4 + 4 + - 4 - d
Орнитиндекарбоксилаза 4 — - - + - - — — —
Гидролиз желатина 4 + d + — - — - — -
Ферментация D-адонита - — — - - 4 + 4 - -
Ферментация D-арабита - - - — — - 4 + - -
Ферментация глицерина d + d d — H — d - d
Ферментация миоинозита — — — - - d 4 - 4
Ферментация мальтозы — + + 4 - — d — - -
Ферментация D-маннозы - — — — + + + 4 4 4
Ферментация метил-О-глюкозида — + [+1 d - - - - - -
Ферментация 1-рамнозы - - - - - 4 d - -
Ферментация сахарозы [-1 + + + — [-] - d d
Ферментация трегалозы + 4 d d - - - 4
Ферментация D-ксилозы + - + + - — - - -
Дезоксирибонуклеаза d + d M - - — - - -
Липаза + + d [+1 - - - — - -
Просветление среды с тирозином + — + + + + + + 4 4
Роение + + d + - - - - - -
Примечание:
«-» - - 0-10% штаммов положительны;
354 ЧАСТНАЯ МИКРОБИОЛОГИЯ
«[-]»• — 11-25% штаммов положительны;
«Ф> — 26-75% штаммов положительны:
«[+]» — 76-89% штаммов положительны;
«т» — 90-100% штаммов положительны.
ЧАСТНАЯ микробиология 355
Таблица 21-12. Дифференциальная характеристика видов и подвидов Salmonella
Вид £ ent erica $. bongon
Подвид enterica salamae arizonae diarizonae houtenae hidica
Обозначение 1 II Illa lllb IV VI V
Характеристика
Дульцит + + - - - d 4-
p-Галактозидаза - - + + - d +
Малонат - + + + - - -
Желатин - + + + + + -
Сорбит + + + + + - +
Рост с KCN - - - - + - +
D (Ц-тартрат + - - - - - -
Галактуронат - + - + + + +
у-Глютамил- трансфераза + + - + + + +
р-Глюкоронидаза d d - + - d -
Му кат + + + - (70%) - + +
Салицин - - - - + - -
Лактоза - - - (75%) + (75%) - d -
Лизис фагом 01 + + - + - + d
Естественный резервуар Теплокровные животные Хладнокровные животные и окружающая среда
Примечание:
«+» — 90% или более положительных реакций;
— 90% или более отрицательных реакций:
— разные серовары дают различные реакции.
Таблица 21-13. Внутриродовая дифференциация рода Klebsiella
Тест KI oxytoca KI. pneumoniae
subsp. ozaenae subsp. pneu- moniae subsp. rhinosclero- matis
Образование индола 4- - - -
Проба с метиленовым красным [-] + L_ H +
Гидролиз мочевины + - + -
Лизиндекарбоксилаза + d +• -
Использование малоната + - + +
Образование газа из D-глюкозы + d + -
Образование кислоты из 5-кетоглюконата + - - -
Образование кислоты из О-сорбозы + - - -
ИДЕНТИФИКАЦИЯ ЭНТЕРОБАКТЕРИЙ ПО АНТИГЕННОЙ ХАРАКТЕРИСТИКЕ
Для энтеробактерий один из этапов идентификации — изучение антигенной
структуры выделенного микроорганизма (серологическая идентификация).
В настоящее время достаточно глубоко изучено антигенное строение мношх
энтеробактерий, однако диагностические препараты для серологической иденти-
фикации разработаны только для бактерий родов Salmonella, Shigella, Escherichia
и Yersinia. Штаммы трех видов шигелл (S. dysentrriae, S.flexneri, S. boydii) диффе-
ренцируются на серовары в зависимости от типовых антигенов. Штаммы S.flex-
neri дифференцируются на подсеровары в зависимости от групповых антигенов
(табл. 21-14).
356 ЧАСТНАЯ МИКРОБИОЛОГИЯ
Таблица 21-14. Классификационная схема бактерий рода Shigella
Группа Вид Серовар Подсеровар
А Shigella dysenteriae 1-15 -
В Shigella ffe.xneri 1 1a, 1b
2 2a. 2b
3 3a, 3b
4 4a, 4b
5 5a, 5b
6 -
Х-вариант -
Y-вариант -
С Shigella boydii 1-19 -
D Shigella sonnei 1 -
Серологическую классификацию сальмонелл проводят на основе междуна-
родной схемы Кауфмана-Уайта, включающей названия и антигенную струк-
туру известных к настоящему времени серологических вариантов сальмонелл.
Серогруппа штамма определяется по комплексу О-антигенов, серовар внутри
каждой серогруппы — по наличию Н-антигенов.
У эшерихий серогруппа устанавливается по сочетанию О- или OK-антигенов, а
по комплексу содержания ОН или ОКН-антигенов определяется серовар штамма.
Серологическая идентификация, как правило, начинается с изучения выделен-
ной культуры в РГА на стекле с поливалентными О-сыворотками. Для сальмонелл
промышленно выпускаются поливалентные адсорбированные О-сыворотки к
сальмонеллам групп А, В, С, Д, Е и редких групп. Стали доступны моноклональные
антитела, позволяющие идентифицировать сальмонелл на родовом, групповом и
серотиповом уровнях. Некоторые из этих моноклональных антител уникальны,
поскольку к выявляемым им антигенам сальмонелл (особенно жгутиковым) не
удается получить специфичные гипериммунные сорбированные сыворотки.
Для шигелл выпускаются сыворотки для определения S. dysenteriae* S. flex-
пеа + S. sonnet, 5. boydii.
При серотипировании эшерихий вначале используют поливалентные сыво-
ротки (ОКА, ОКБ, ОКС, ОКД и ОКЕ), а затем проводят РГА с моновалентыми
О-сыворотками (или иммуноглобулинами), входящими в состав поливалентной
смеси, давшей положительный результат. Для эшерихий выпускается дополни-
тельная панель моновалентных ОК-сывороток.
Разработаны сыворотки для дифференциации иерсиний — иерсиниозные (ОЗ;
04,32; 04,33; 05,27; 06,30; 06,31; 07,8; 09; 013,7), псевдотуберкулезные (серо-
типы 1 и 3), а также сыворотка к вирулентным иерсиниям, позволяющая диффе-
ренцировать патогенные и непатогенные У. enterocolitica в РГА. Серологическую
идентификацию культур проводят строго по инструкции производителя.
КРАТКАЯ ХАРАКТЕРИСТИКА ОТДЕЛЬНЫХ РОДОВ ЭНТЕРОБАКТЕРИЙ
В настоящее время клиническая значимость некоторых родов и видов, недавно
включенных в семейство Enterobacteriaceae, еще не определена и требует уточне-
ния.
• Род Budvicia (В. aquatica). Сероведородпродуцируюшие водные бактерии.
В медицинской практике встречаются крайне редко. Обитают в незагрязнен-
ной воде поверхностных водоемов. Имеются единичные сообщения о выде-
лении из крови и мочи пациентов с иммунодефицитом.
• Род ButtiauxeUa (В. agrestis, В. Ъгеппегае, В. ferragutiae, В. gaviniae, В. izardii.
В. noackiae, В. wannboldiae). Чрезвычайно редко встречаемые в медицинской
ЧАСТНАЯ МИКРОБИОЛОГИЯ 357
практике бактерии. Обнаружены у моллюсков и улиток, в том числе съедоб-
ных. Имеются сообщения об обнаружении в клиническом материале — моче
и раневом отделяемом.
Род Cedecea (С. davisae, С. lapagei, С. neteri, С. species 3, С. species 5). Выделяют
в основном из дыхательных путей человека. Оппортунистические инфекции
вызывают редко. Дифференцируют с представителями родов Ewingella и
Serratia.
Род Citrobacter (С. amalonaticus, С. freundii, С. koseri (diversus), С. farmed,
С. sedlakii. С. rodentium, С. youngae, С. braakii, С. werkmanii, С. gillenii, С. murl-
imae\. Обнаруживают в фекалиях и моче человека, воде водоемов и других
объектах окружающей среды. Могут быть отнесены к представителям нор-
мальной микрофлоры кишечника. Описаны многочисленные случаи острых
кишечных инфекций (спорадических и вспышек), обусловленных данными
микроорганизмами. Заболевания протекают по типу гастроэнтеритов, дис-
пепсий. Бактерии рода Citrobacter выделены в качестве этиологического
агента при инфекциях моче- и желчевыводящих путей, отитах, остеомие-
литах. Используя короткий ряд биохимических тестов, бактерии данного
рода сложно отличить от эшерихий и сальмонелл. На практике достаточ-
но часто приходится дифференцировать от сальмонелл. При этом важно
помнить, что цитробактеры имеют ряд общих антигенов с сальмонеллами.
В дифференциации сальмонелл и цитробактеров особое значение имеет
отсутствие у сальмонелл лизиндекарбоксилазы. Мультирезистентные штам-
мы S. typhimudum, вызывавшие внутрибольничные инфекции в 1977-1982 гг.
в ряде территорий РФ, нередко давали отрицательный результат на средах с
лизином. Цитробактеры подразделяются по О-антигену на 42 серогруппы,
внут ри которых возможна дифференциация по отдельным О - и Н-антигенам.
Однако серотипирование данных микроорганизмов в лабораторной практи-
ке в настоящее время не применяют.
Род Edwardsiella (Е. hoshinae, Е. ictalun, Е. tarda, Е. tarda биогруппы 1).
Наиболее широко распространены в тропических и экваториальных странах.
Часто выделяют от рептилий. Е. tarda патогенны для рыб. Вопрос о патоген-
ности данных микроорганизмов для людей остается спорным. Имеются дан-
ные о большей частоте обнаружения бактерий данного рода у больных с диа-
рейным синдромом, по сравнению со здоровыми лицами. Описаны случаи их
выделения при абсцессах печени, эмпиеме плевры, сепсисе новорожденных,
гнойно-септических осложнениях.
Род Enterobacter (Е. cloacae subsp. cloacae. Е. cloacae subsp. dissolvens, E. amni-
genus биогруппы 1, E. amnigenus биогруппы 2, E. asburiae, E. gergoviae, E. hor-
maechei, E. sakazakii, E. intermedins. E. nimipressuralis, E. taylorae, E. cancerogenus,
E. pyrinus, E. kohei, E. aerogenes). Первоначально были описаны как возбудите-
ли инфекций насекомых, в частности саранчи. Вопрос о патогенности энте-
робактеров для человека долгое время оставался открытым. Однако в послед-
ние годы накоплен значительный материал, свидетельствующий, что данные
микроорганизмы могут вызывать острые кишечные инфекции, инфекции
моче- и желчевыводящих путей, сепсис новорожденных; энгеробактеры
рассматриваются как важный возбудитель нозокомиальных инфекций. В
некоторых регионах по частоте выделения от госпитализированных боль-
ных данные микроорганизмы уступают только эшерихиям и клебсиеллам.
Наиболее часто у человека обнаруживают Е. aerogenes. Описаны госпиталь-
ные вспышки, обусловленные Е. cloacae. Е. sakazakii - возбудитель паренте-
ральных инфекций у детей раннего возраста. В последние годы нередко реги-
стрировались эпидемиологически документированные спорадические с лучаи
и вспышки септических инфекций и некротизирующего энтероколита у детей
358
ЧАСТНАЯ МИКРОБИОЛОГИЯ
первого года жизни, в первую очередь недоношенных, новорожденных с
низкой массой гела или иммунодефицитом. У детей регистрировали также
менингиты и бактериемию (летальность в ряде случаев достигала 80%). Все
случаи заболеваний были связаны с употреблением детских молочных сме-
сей, контаминированных Е. sakazakii. При этом установлено, что в первую
очередь Е. sakazakii может попадать в сухие молочные смеси с контаминиро-
ванными ингредиентами, добавляемыми после сушки, или из окружающей
среды в процессе упаковки готового продукта. Идентификацию проводят
по совокупности морфологических, биохимических и других признаков,
определяющих принадлежность к виду Е. sakazakii Прина цлежность изолята
к Е. sakazakii осуществляют на основании его развернутых биохимических
характеристик штаммов. Дифференциацию следует проводить с близкород-
ственными представителями энтеробактерий — Е. cloacae и Р. agglomerans.
Ведущие дифференциальные признаки Е. sakazakii — способность к образо-
ванию желтого пигмента при культивировании на неселективных средах при
температуре 25 °C и отсутствие ферментации D-сорбита. Для дифференциа-
ции Е. sakazakii от сорбитвариабельных и обладающих желтым пигментом
представителей рода Р. agglomerans используют тест разжижения желатина.
В качестве дополнительного теста при идентификации Е. sakazaki возможно
определение О',-глюкозидазной активности на хромогенных питательных сре-
дах, содержащих 4-нитрофенил-а-В-глюкопиранозид или 5-бромс-4-хлоро-
З-индолил-а-В-глюкопиранозид.
• Род Escher chia (Е. coli, Е. hermannii, Е vulneris, E.fergusom., Е. ЫаИаё). Данный
род объединяет обширную гетерогенную группу энтеробактерий, различаю-
щихся по биологическим, серологическим и морфологическим свойствам,
антагонистической активности, продукции энтеро- и иитотоксинов, патоген-
ности и другим признакам. Эшерихии — представители облигатной микро-
флоры кишечника человека.
Клиническое значение имеют конкретные серологические варианты и биовары
Е. coli, способные вызывать острые кишечные инфекции, внекишечные формы
эшерихиозов. протекающие в форме менингита, сепсиса, пиелонефрита, цистита,
пневмонии и других заболеваний. Полиморфизм клинической картины обуслов-
лен наличием разнообразных факторов вирулен гности. Е. coli относится к наи-
более часто встречаемым возбудителям внутрибольничных инфекций, вызывая
гнойно-септические осложнения в послеоперационном периоде, бактериемию,
инфекции ЦНС, диарею. В санитарной микробиологии Е. coli — санитарно-
показательный микроорганизм, свидетельствующий о фекальном загрязнении
окружающей среды. Результаты лабораторной диагностики, основанной на огра-
ниченном количестве фенотипических тестов, не позволяют с уверенностью оце-
нить этиологическую значимость штаммов Е. coli даже с учетом их принадлежно-
сти к определенной серологической группе.
Антигенная структура Е coli сложна. Для построения антигенно-диагностических
схем и серологической идентификации наиболее важное значение имеют О-, К- и
Н-антигены. Остальные антигенные комплексы представляют интерес в основном
для установления механизмов патогенеза, выявления факторов вирулентности эше-
рихий в генетических исследованиях. По сочетанию О- или ОК-анлггенов все эше-
рихии разделяют на серологические группы (серогруппы), а по спектру содержания
ОН- или ОКН-антигенов — на серологические варианты (серовары). К настоящему
времени известно 175 О-, 56 Н- и 100 К-антигенов, однако это еще не предел, так как
постоянно выделяют штаммы, которые не идентифицируются диагностическими
сыворотками, содержащими антитела к известным О-, Н- и К-антигенам.
Для эшерихий характерно сходство антигенных связей по О-антигену между
серогруппами, а также с другими патогенными и условно патогенными энтеробак-
ЧАСТНАЯ МИКРОБИОЛОГИЯ 359
териями (Shigella spp., Salmonella spp., Klebsiella spp.). Трудности при идентификации
эшерихий связаны с тем, что схемы их серотипирования неполны и несовершен-
ны. Типирование также затруднено необходимостью использования большого
набора диагностических сывороток, что, в свою очередь, связано с высокими эко-
номическими затратами.
Серологическая идентификация основана на выявлении О- или OK-антигенов в
РГА развернутой или на стекле.
Среди диареегенных эшерихий особое значение имеет Е. coli О157:Н7, проду-
цирующая энтеротоксины, которые вызывают гемоколиты с возможными ослож-
нениями в виде гемолитико-уремического синдрома, почечной недостаточности
и тромбоцитопенической пурпуры. Данные энтеротоксины представлены двумя
разновидностями (I и II) и по структуре и активности напоминают энтеротоксин,
продуцируемый S. dysenteriae 1 (именно поэтому данные энтеротоксины называют
шигаподобными). В настоящее время выпускается одноэтапный иммунохромато-
графический тест, предназначенный для скрининга на наличие в фекалиях Е. coli
О157:Н7. Он позволяет получить результат в течение 5-10 мин.
В медицинской микробиологии в зависимости от вызываемых заболеваний и
наличия определенных факторов вирулентности патогенные эшерихии разделяют
на две большие группы:
• эшерихии, обусловливающие развитие острых кишечных инфекций:
• эшерихии, вызывающие развитие инфекций внекишечной локализации.
Эшерихии, обусловливающие развитие острых кишечных инфекций (диарее-
генные эшерихии), подразделяют на шесть условных групп:
• ЕРЕС — энтеропатогенные эшерихии (Е. coli О18ас, 020, 025, 026, 044, 055,
075, 086, 091, 0111, 0114, 0119, 0125, 0126, 0127, 0128 0142, 0158);
перечисленные 0-группы включают 38 сероваров;
• EIEC — энтероинвазивные эшерихии (в основном неподвижные Е. coli 028ас,
029, 0112, 0121, 0124, 0136, 0143, 0144, 0152. 0159, 0164, 0167);
• ЕТЕС — энтеротоксигенные эшерихии (Е. coli О6:Н16, 08:Н9. 011:Н27,
015:Н11, О20:Н-, О25:Н42, О25:Н-, О27:Н7, О78:Н11, О78:Н12, О128ас:Н7,
О148:Н28, О149:НЮ, О159:Н20, О173:Н-);
• ЕНЕС — энтерогеморрагические эшерихии (Е. coli0157:H7,0157:Н- О26:Н11,
О103:Н2, 0111:Н-, О113:Н21, а также более 100 сероваров групп 01-0172);
• ЕАЕС — энтероагтрегативные эшерихии (Е. coli 07:Н-, 077;Н18, 086;Н-,
0126:Н27, О127:Н2):
• ПАЕС — диффузно-аггрегативные эшерихии.
Каждая группа диареегенных эшерихий принадлежит к определенным серо-
варам, характеризуется индивидуальными генетическими детерминантами виру-
лентности и специфическими клиническими проявлениями вызываемой острой
кишечной инфекции.
Эшерихии, вызывающие инфекции внекишечной локализации, так же как и
диареегенные эшерихии, подразделяют на 3 патогруппы:
• MENEC — менингиальные;
• SEPEC — септицемические;
• UPEC — уропатогенные.
Накопленные данные свидетельствуют о необходимости пересмотра принципов
лабораторного типирования эшерихий — включение на первом этапе определения
маркеров вирулентности (генетических детерминант) с последующим серотипиро-
ванием для обоснования этиологической роли.
Значительный прогресс в лабораторной диагностике энтеробактерий, в том
числе эшерихий, может быть связан с внедрением быстрого и надежного метода
(RFLP) на основе анализа полиморфизма длин фрагментов рестрикции амплифи-
цированных генов rfb и/ИС, контролирующих синтез ЛПС О-антигена и флагел-
360
ЧАСТНАЯ МИКРОБИОЛОГИЯ
лина жгутиков соответственно. Данный метод получил название «молекулярное
серотипирование». так как он поддерживает, уточняет и углубляет общепринятую
классификацию серотипирования микроорганизмов, а также унифицирует и уде-
шевляет определение серологической принадлежности штаммов с помощью сыво-
роток. Используя данный метод, можно провести серологическую идентификацию
неподвижных и находящихся в R-форме штаммов.
Острые кишечные инфекции диареегенных эшерихий характеризую гея преиму-
щественным поражением желудочно-кишечного тракта с развитием интоксика-
ции и диарейного синдрома, реже — генерализацией патологическою процесса.
Возбудители острых кишечных инфекций отличаются от штаммов непатогенных
Е. coli наличием определенных факторов патогенности, их генетической детерми-
нацией. принадлежностью к конкретному серологическому варианту и особен-
ностями патогенеза и клинико-эпидемиологических проявлений вызываемых
заболеваний.
• Род Ewingella (Е. americana). Выделяют из мокроты, отделяемого ран,
крови человека. Возможный возбудитель оппортунистических инфекций.
Дифференцируют от бактерий рода Cedecea.
• Род Hafnia (Н. alvei, Н. alvei биовар 1). Гафнии нередко обнаруживают у мле-
копитающих, включая человека, в пищевых продуктах, особенно в молочных,
а также в объектах окружающей среды. Вопрос об их значении в патологии
человека нуждается в дальнейшем изучении. Имеются данные о возможной
роли данных микроорганизмов при диарейных заболеваниях, сепсисе, в том
числе неонатальном.
• Род Klebsiella (К. oxytoca, К pneumoniae subsp. pneumoniae, К. pneumoniae
subsp. ozaenae, К. pneumoniae subsp. rhinoscleromatis, K. granulomatis). Главная
отличительная особенность клебсиелл — наличие капсулы. Классификация
этих микроорганизмов неоднократно пересматривалась, в настоящее время
в род Klebsiella входят 4 вида, причем 3 первоначально входивших в этот род
вида (К. pneumoniae, К. rhinoscleromatis и К. ozaenae) теперь считают 3 под-
видами К. pneumoniae. В последние годы представления о роли клебсиелл, в
первую очередь К. pneumoniae, в патологии человека значительно углубились.
Клебсиеллы — возбудители внутрибольничных инфекций, различных забо-
леваний новорожденных. В клиническую практику введен особый термин —
«клебсиеллез» для обозначения заболеваний, протекающих по типу острого
сепсиса или с локальным поражением отдельных органов. Клебсиеллы широ-
ко распространены в окружающей среде, что связано с высоким уровнем
устойчивости к антибиотикам. Для них характерна более высокая устойчи-
вость к действию дезинфектантов. Для родовой идентификации клебсиелл
НИИЭМ им. Л. Пастера выпускает хромогенную среду «Klebsiella 5-АСК»,
на которой только эти микроорганизмы образуют зоны черно-коричневой
окраски вокруг колонии.
* Род Klu yvera (К. ascorbata, К. cryocrescens, К. imermedius, К. cochleae, К. geor-
giand). К. cochleae выделяют из улиток и слизней. Несколько видов обна-
ружены в пищевых продуктах, почве, сточных водах, а также в соста-
ве нормальной микрофлоры желудочно-кишечного тракта человека. Эти
микроорганизмы выделяются из патологического материала, преимуще-
ственно из дыхательных, мочевыводящих путей, фекалий, эпизодически из
крови. Оппортунистические инфекции вызывают редко.
• Род Leclercia (L. adecarboxylata). Встречаются в пище, воде. Выделены из
мокроты, слюны, крови человека. Описана этиологическая роль данных
микроорганизмов при септицемиях, пиелонефритах, раневых инфекциях,
септических артритах. Могут продуцировать р-лактамазы широкого спектра
действия.
ЧАСТНАЯ МИКРОБИОЛОГИЯ 361
Род Lemmorella (L. grimontii, L. richardii). Выделяют из фекалий и мочи чело-
века. Могут вызывать инфекции моче выводящих путей, раневые инфекции,
первичную бактериемию, перитонит, инфекции дыхательных путей.
Род Moellerella (М. wisconsensis). Малоизученный род энтеробактерий.
Обнаруживают в воде, пищевых продуктах, ротовой полости животных.
У человека выделены из фекалий, крови, бронхиального аспирата, желчи.
Род Morganella (Л/. morgami subsp. morganii, М. morganii subsp. sibonii, M. mor-
ganii биовар 1). До 1984 г. M. morganii принадлежала к роду Proteus (Р. mor-
ganii). М. morganii — условно патогенный микроорганизм, входящий в состав
нормальной микрофлоры кишечника. Может вызывать различные инфекци-
онные процессы — сепсис, абсцесс головного мозга, эндоофтальмит, менин-
гит, раневые инфекции. Также может быть причиной внутрибольничных
инфекций.
Род Obesumbacterium (О. proteus биогруппы 2). Малоизученные энтеро-
бактерии. Возможный контаминант пивных дрожжей и сусла. Клиническая
значимость неизвестна.
Род Pantoea (Р. agglomerans; синонимы: Enterobacter agglomerans, Erwinia
herbicola. Erwinia milletiae, P. dispersd). P. agglomerans (ранее E. agglomerans) —
частая причина экзогенных инфекций. Данные микроорганизмы широко
распространены у растений и часто бывают причиной загрязнения ваты.
Интенсивно размножаются при температуре 4 °C, в том числе в инъекцион-
ных растворах и препаратах крови, хранящихся в холодильнике.
Род Plesiomonas (Р dugelloides). Плезиомонады — частая причина гастроэн-
теритов у пациентов, связанных с морской водой и упигреблением морепро-
дуктов, в частности устриц. Также могут вызывать бактериемию, септический
артрит, остеомиелит, менингит, эндокардит, целлюлит, перитонит, холеци-
стит, панкреатит. Описано возникновение инфекции у пациентов, находя-
щихся на перитониальном диализе.
Род Pragia (Р. ftntium). Малоизученные энтеробактерии. Выделяются из
воды, фекалий. Значение в патологии человека неизвестно.
Род Proteus (Р. mirabilis, Р. myxofaciens, Р.реппеп Р. vulgaris). Характерная осо-
бенность данного рода — высокая подвижность и способность к ползучему
росту на плотных средах. В наибольшей степени роение присуще двум видам —
Р vulgaris и Р. mirabilis, однако этот феномен может быть индуцирован и у
других видов. Наиболее надежный тест для определения принадлежности к
роду Proteus — определение фенилаланиндезаминазы.
Род Providencia (Р. alcalifaciens, Р. heimbachae, Р. rettgeri, Р. rustigianii, Р. stu-
artii). У людей выделены из мочи, фекалий, крови, а также из ротоглотки,
промежности, подмышечной впадины, отделяемого ран. Р. stuartu или реже
Р. rettgeri — частая причина инфекций мочевыводящих путей у пациентов
с длительно установленным катетером. Р. alcalifaciens, Р. rettgeri и Р. stuartii
могут вызвать диарею. Представители данного рода могут быть причиной
внутрибольничных инфекций, часто обладают множественной лекарствен-
ной устойчивостью.
Род Rahnella (R. aquatilis). Широко распространены во внешней среде (воде,
почве). R. aquatHis обнаружена в мясе, пресноводной рыбе, пиве, паренте-
ральном питании и молочных продуктах. Размножение в рыбе может вести к
накоплению в ней опасных концентраций гистамина. У человека выделены из
крови, отделяемого послеоперационных ран, мочи, мокроты, бронхиальных
смывов, фекалий, £. aquatilis необходимо дифференцировать от £. agglomerans
вследствие сходства их биохимических свойств.
Род RaoulteUa (R. planticola, R. terrigena, R. ornithinolytica) — данные бактерии,
размножаясь в рыбе, так же как и микроорганизмы рода Rahnella, способны
362
ЧАСТНАЯ МИКРОБИОЛОГИЯ
преобразовывать гистидин в гистамин, что может приводить к пищевым
отравлениям.
• Род Salmonella (S. enterica, S. bongori). S. enterica подразделяют на 6 подви-
дов. Клиническое значение для человека имеют представители вида S. enter-
ica subsp. enterica, который является самым многочисленным. Сальмонеллы
подразделяются на серогруппы по О-антигенам и на серовары — по О- и
Н-антигенам. Современная схема Кауфмана-Уайта насчитывает 2557 серова-
ров, объединенных в 67 серогрупп. Заболевание у людей вызывают не более
150 сероваров, а широко распространены не более 20 из них.
Сальмонеллезы — типичные зооантропонозы, за исключением брюшного тифа
и паратифов В последние 20 лет наиболее часто сальмонеллез у людей вызывается
S. enteritidis. Заболевания связаны с контаминированными продуктами промыш-
ленного птицеводства (мясом птицы, яйцами). Другие серовары (S. typhimurium,
S. derby, S. infantis, S. london, S. haifd) также могут вызывать спорадические заболе-
вания и парентеральные инфекции. Мультирезистентные штаммы многих серова-
ров способны вызывать внутрибольничные инфекции с контактно-бытовым путем
передачи.
Сальмонеллы — подвижные микроорганизмы, продуцирующие сероводород,
игтдолотрицательные. Для их выделения разработаны специальные среды обо-
гащения (селенитовая среда, среды Мюллера, Кауфмана), которые увеличивают
частоту^ положительных находок. Для посева используют висмут-сульфит агар,
SS-arap. Промышленно выпускается поливалентный А-, В-, С-, Д-, Е-бактериофаг,
который может быть использован как подтверждающий тест при диагностике
сальмонеллезов.
• Род Serratia (S. entomophila, S.ficaria, S. fonticola, S grimesii, S. liquefaciens,
S. marcescens, S. marcescens биовар 1, S. odorifera биогруппы 1, S. odorifera био-
группы 2, S. plymuthica, S, proteamaculans, S. rubidaed). Наиболее характерная
особенность бактерий рода Serratia — способность образовывать характер-
ный красный пигмент — продигозин. Однако наличие пигмента наблюдают
не у всех представителей этого рода. Продукция пигмента во многом опреде-
ляется условиями культивирования. Именно поэтому при исследовании
клинического материала и объектов внешней среды нередко встречаются
беспигментные серрапии, идентификация которых сопряжена со значитель-
ными трудностями. Наиболее значимые для медицинской микробиологии
виды — S, marcescens и S. liquefaciens. Описаны случаи сепсиса, обусловленного
S. odorifera биогруппы 1. S. marcescens может вызывать инфекции мочевыво-
дящих и дыхательных путей, бактериемию, конъюнктивит, эндокардиты,
раневую инфекцию. Вирулентность серраций обусловлена наличием плазмид,
кодирующих факторы патогенности (цитотоксины, гемолизины), а также
определяющих устойчивость к антимикробным препаратам. S. liquefaciens —
наиболее частая причина бактериемии и гнойно-cem ических осложнений в
центрах гемодиализа.
• Р д Shigella (S. dysenteriae, S.flexneri. S. boydii, S. sonnet). Бактерии данного
рода трудно отличить от неподвижных, лактозоотринательных анаэроген-
ных эшерихий. Для достоверной идентификации необходимо проведение
серологического типирования по О-антигенам в РГА на стекле. Для всех
представителен рода Shigella характерна высокая частота резистентности к
антибактериальным препаратам. Идентификация шигелл включает биохи-
мическое и серологическое типирование.
Шигеллы — возбудители острых кишечных инфекций человека, а также бак-
териальной дизентерии. Характерный признак дизентерии — диарея с кровью,
однако в 50% случаев шигеллы вызывают острую диарею без крови, которую кли-
нически невозможно отличить от диареи, вызванной другими энтеробактериями.
ЧАСТНАЯ МИКРОБИОЛОГИЯ 363
Тяжелое, длительное течение заболевания и летальные случаи отмечаются при
дизентерии, вызванной S. dysenteriae 1 (дизентерия Григорьева-Шига), и дизенте-
рии Флекснера.
• Род Tatumella (Т. ptyseos). Редко встречаемые энтеробактерии. У людей чаще
всего обнаруживают в мокроте. Клиническая значимость требует уточнения.
• Род Yersinia (У. enterocolitica, У. frederiksenii, У. intermedia, У. krtstensenn,
У. moUaretii, Y.pestis, Y. pseudotuberadosis, У. rohdei, У. ruckerl).
Медицинское значение имеют следующие виды:
• У. pestis — возбудитель чумы (относится к 1-й группе опасности, диагностику
в обычных лабораториях не проводят);
• У. pseudotuberculosis — возбудитель псевдотуберкулеза (дальневосточной
скарлатиноподобной лихорадки или экстраинтестинального иерсиниоза по
МКБ-10);
• У. enterocolitica — подразделяется по биохимическим свойствам на 6 биова-
ров, из которых биовар 1А считается непатогенным, биовары 1В-5 вызыва-
ют кишечный иерсиниоз (энтерит, вызванный У. enterocolitica, по МКБ-10).
Иерсинии биовара 1А расщепляют салицин за 24 ч, обладают пиразинамида-
зой (положительный ПИР-тест). Тест на гидролиз эскулина вариабелен, у пато-
генных штаммов всегда отрицателен. По О-антигену иерсинии подразделяют на
29 сероваров. Заболевания вызываются сероварами ОЗ и 09, редко 08. 05,27 и
некоторыми другими. Для выделения У. enterocolitica производят ряд элективных
сред, используют холодовое обогащение — посевы инкубируют в холодильнике в
забуференной пептонной воде или пептонно-калиевой среде. Высевы проводят на
2-3, 5 и 7-й день.
Нативный материал или среда обогащения могут быть подвергнуты дополни-
тельно щелочной обработке: 1 петлю диаметром 3-4 мм обогатительной среды
вносят в 0,2 мл свежеприготовленного раствора гидроксида калия (9,5 мл 0,5%
раствора натрия хлорида + 0,13 мл 40% раствора гидроксида калия) на 1-2 мин,
затем проводят высев на плотную питательную среду (среду Эндо или специаль-
ные элективные среды для иерсинии).
• Род Yokenella (У. regensburgei, синоним: Koserella trabulsii). Редко встречае-
мый род семейства bnterobactenaceae. У. regensburgei выделены из кишечника
насекомых, воды, крови, мочи, фекалий и суставной жидкости человека. По
биохимическим свойствам практически идентичны бактериям рода Hafnia.
ОПРЕДЕЛЕНИЕ ЧУВСТВИТЕЛЬНОСТИ ЭНТЕРОБАКТЕРИЙ К АНТИМИКРОБНЫМ
ПРЕПАРАТАМ
Заключительный этап лабораторного исследования — определение чувстви-
тельности выделенных и идентифицированных бактерий к антимикробным пре-
паратам. Хотя в целом энтеробактерии чувствительны ко многим антимикробным
препаратам, микроорганизмы некоторых родов (Enterobacter, Proteus, Klebsiella,
Е coli) характеризуются природной устойчивостью к ряду р-лактамных анти-
биотиков, которая связана с продукцией хромосомно кодируемых р-лактамаз.
Сочетанная резистентность к антимикробным препаратам разных групп нередко
встречается у патогенных и условно-патогенных энтеробактерий.
Внутрибольничные инфекции чаще обусловлены мультирезистентными к анти-
биотикам штаммами различных энтеробактерий. Появление и распространение
приобретенных механизмов устойчивости определяет необходимость тестирова-
ния выделенных возбудителей к набору антимикробных препаратов. Выбор кон-
кретных препаратов для лечения пациента зависит не только от вида выделенного
микроорганизма, но и от локализации инфекционного процесса.
В табл. 21-15 указан перечень препаратов для тестирования штаммов энтеро-
бактерий выделенных при инфекциях различной локализации.
364 ЧАСТНАЯ МИКРОБИОЛОГИЯ
Таблица 21-15. Рекомендуемый перечень антибактериальных препаратов для определения чув-
ствительности бактерий семейства Enterobacteriaceae
Препараты первого ряда Дополнительные препараты
Энтеробактерии, выделенные при внекишечных инфекциях
Ампициллин Ингибиторзащищенные пенициллины (ампицил- лин + сульбактам или амоксициллин + клавулановая кислота) Цефапоспоримы III поколения (цефотаксим или цефтриаксон) Цефтазидим Гентамицин Фторхинолоны (норфлоксацин, ципрофлоксацин или офлоксацин) Карбапенемы(имипенем или меропенем) Цефепим Цефоперазон + сульбактам Тикарциллин + клавулановая кислота Второй цефалоспорин III поколения (цефтриаксон или цефотаксим) Цефокситин Амикацин Цефуроксим Оральные цефалоспорины II-III поколения
Энтеробактерии выделенные при кишечных инфекциях
Ампициллин Ко-тримоксазол Хинолоны (налидиксовая кислота) Фторхинолоны (норфлоксацин, ципрофлоксацин или офлоксацин) Цефалоспорины III поколения (цефотаксим или цефтриаксон) Хлорамфеникол Тетрациклины (тетрациклин или доксициклин)
Энтеробактерии, выделенные при внебольничных инфекциях мочевыводящих путей
Ампициллин Ингибиторзащищенные пенициллины (амоксицил- лин + клавулановая кислота) Ко-тримоксазол Фторхинолоны (норфлоксацин, ципрофлоксацин или офлоксацин) Фосфомицин Нитрофурантоич Цефуроксим Цефалоспорины III поколения (цефотаксим или цефтриаксон) Гентамицин Амикацин
Необходимость такого разделения связана с особенностями фармакокинетики
отдельных антимикробных препаратов в желудочно-кишечном тракте и мочевы-
водящих путях, а также различиями в их клинической эффективности.
Два вида энтеробактерий (£. coli и К. pneumoniae) входят в число семи видов
микроорганизмов, принятых в качестве индикаторов развития антимикробной
резистентности в Европейской системе надзора и контроля за антимикробной
резистентностью (EARSS).
В настоящее время нередко штаммы Е. coli и К. pneumoniae продуцируют
^-лактамазы расширенного спектра, что обеспечивает данным штаммам высокую
резистентность к цефалоспоринам III—IV поколения. Сочетанная резистентность
таких штаммов к фторхинолонам создает серьезные трудности выбора эффектив-
ной этиотропной терапии.
СПИСОК РЕКОМЕНДУЕМОЙ ЛИТЕРАТУРЫ
Джонсон Д.Р., Каплан Э. Лабораторная диагностика инфекций, вызванных стрептокок-
ком 1 руппы А. — М.: Медицина, 1908. — 118 с.
Долгих Т.И. Краткая характеристика возбудителей хламидиоза // Яковлев В.М ,
Новиков А.И. Сосудистый эндотелий и хламидийная инфекция. — М.: Медицина, 2000. —
С. 127-153.
Европейские стандарты диагностики и лечения заболеваний передаваемых половым
путем. — М.: Медлит, 2004. — 272 с.
Же б рун А.Б. Инфекция Helicobacter pylori. — СПб.: Феникс, 2006. — 380 с.
Методики клинических лабораторных исследований: Справочное пособие: Т. 3 / Под ред.
В В. Меньшикова. — М.: Лабора, 2009. — 880 с
Методические указания МУК 4.2.1887-04 «Лабораторная диагностика менингококковой
инфекции и гнойных бактериальных менингитов». Утверждены главным государственным
санитарным врачом Российской Федерации 4 марта 2004 г.
ЧАСТНАЯ микробиология 365
МУК 4.2.22-17-07 < Методические указания по выявлению бактерий Legionella pneumophila
в объектах окружающей среды». — 2007.
Определитель бактерий Берджи: В 2 т.: Пер. с англ. / Под ред. Дж. Хоулта, Н. Крига,
П. Снята и др. - М., 1997. - Т. 1. - С. 195-196, 227-229, 255-258.
Тартаковский И.С., Малеев В.В., Ермолаева С.А. Листерии: роль в инфекционной патоло-
гии человека и лабораторная диагностика. — М.. 2002.
Тимченко В.Н., Бабаченко И.В., Ценева Г.Я. и др. Эволюция коклюшной инфекции у
детей. - СПб.: ЭЛБИ-СПб., 2005. - 192 с.
Тотолян А.А., Суворов А.Н., Дмитриев А.В. Стрептококки группы В в патологии челове-
ка. — СПб.: Человек, 2009. — 212 с.
Фтизиатрия: Национальное руководство / Под ред. М.И. Перельмана. — М.: ГЭОТАР-
Медиа, 2007. — 512 с.
Черкасский Б.Л. Частная эпидемиология. — М., 2002.
Шурыгина И.А., Чеснокова М.В., Климов В.Т. и др. Псевдотуберкулез. — Новосибирск:
Наука, 2003. — 319 с.
Этиологическая роль хламидий при заболеваниях репродуктивной сферы: Аналитический
обзор СПб. НИИЭМ им. Л. Пастера / Под ред. А.Б. Жебрупа. - СПб., 2007. —56 с.
Якубович А.И., Корепанов А.Р. Урогенитальный хламидиоз. — Иркутск, 2006. —108 с
Malfertheiner Р„ Megraud F., Moran С. Guidelines for the managemenc of the H. pylori infec-
tion. European Gastroenterology Review. — 2005.
ИЕРСИНИИ
ТАКСОНОМИЯ
Род Yersinia относится к семейству Enterobacteriaceae и включает 14 видов, из
которых 3 — У. pestis, У. pseudotuberculosis, У. enterocolitica — являются возбудителя-
ми инфекционных болезней. Род Yersinia назван в честь французского микробио-
лога А. Йерсена, открывшего в 1894 г. возбудителя чумы (Y.pestis).
Возбудитель псевдотуберкулеза — У. pseudotuberculosis — вызывает у лаборатор-
ных животных в условиях экспериментального заражения патологоанатомические
изменения внутренних органов, сходные с наблюдаемыми при туберкулезе, поэто-
му бактерию назвали псевдотуберкулезной иерсинией. У человека она вызывает
заболевание, сопровождаемое энтеритами, энтероколитами и тяжелой генера-
лизованной инфекцией с множественными поражениями отдельных органов и
систем.
ОБЩАЯ ХАРАКТЕРИСТИКА Y. ENTEROCOLITICA И Y. PSEUDOTUBERCULOSIS
Иерсинии — грамотрицательные, не образующие спор палочки, кокки или ово-
иды, обладающие перитрихиальными жгутиками. При культивировании У. entero-
colitica и У. pseudotubercuiosis в течение 48 ч при температуре 37±1 °C в культурах
обнаруживают бактерии без жгутиков, что коррелирует с потерей подвижности.
Инкубирование культур в течение 24-48 ч при температуре ниже 30 °C (26±2 СС)
ведет к восстановлению подвижности бактерий.
В мазках из бульонных культур У. pseudotuberculosis может располагаться цепоч-
ками по 2-5 клеток, У. enterocolitica цепочек не образует. Размеры отдельных
бактерий при культивировании на искусственных питательных средах составляют
0,8-1,2 мкм в длину и 0,5-0,8 мкм в ширину. Иерсинии хорошо окрашиваются
всеми анилиновыми красителями. В молодых культурах, выращенных при темпе-
ратуре 26±2 °C, преобладают кокковидные бактерии, в старых отмечается тенден-
ция к полиморфизму, особенно при температуре 37+1 °C на различных плотных
питательных средах (например, агаре Эндо).
К настоящему времени по О-антигену выделяют 21 серотип У. pseudotuberculosis.
Чаще всего заболевания вызываются всемирно распространенными штаммами,
принадлежащими к серотипу 0:3 (биотипу 4), 0:5,27 (биотипам 2 и 3), 0:9 (био-
типу 2) и «американскими» штаммами 0:8 (биотипом 1В). Антигенное родство
366 ЧАСТНАЯ МИКРОБИОЛОГИЯ
(по О-антигену) У. pseudotuberculosis и большинства серотипов У. enterocolitica
выражено довольно слабо. Однако выявлено значительное сходство в антиген-
ной структуре У. pseudotuberculosis серотипа 3 и У. enterocoliiica «американских»
штаммов серотипов 0:8, 0:18 и 0:21. Наиболее выпаженны антигенные связи
между представителями различных серотипов внутри вида У. enterocolitica, а
также между У. enterocolitica серотипа 0:9 и бруцеллами благодаря сильному
сходству в строении О-специфических полисахаридных цепей липополисахарида.
Описано антигенное сходство У enterocolitica и других представителей семейства
Enterobactenaceae {Salmonella, Citrobacter, Escherichia, Shigella, Klebsiella, Enterobacter.
Serratia). Перекрестные реакции между данными видами могут затруднять получе-
ние строго специфичных диагностических препаратов.
В составе наружной мембраны патогенных иерсинии определены БНМ, коди-
руемые как хромосомными генами, так и генами, расположенными на плазмиде
вирулентности pYV 42-48 МДа. БНМ — факторы защиты возбудителя от действия
иммунной системы макроорганизма.
Иерсинии продуцируют каталазу, не продуцируют оксидазу и ферментируют
глюкозу, как и другие представители семейства Enterobactenaceae. Их основные
биохимические свойства, учитываемые в практике рутинных лабораторных иссле-
дований, представлены в табл. 21-16.
У. pseudotuherculosis не ферментируют лактозу, сахарозу, раффинозу, целло-
биозу, дульцит, инозит, сорбит, адонит, инулин и D-арабинозу, не продуцируют
фенилаланиндезамидазу, орнитин- и лизиндекарбоксилазу, не образуют индол и
ацетилметилкарбинол (реакция Фогеса-Проскауэра). Ферментируют до кислоты
глюкозу, галактозу, а-арабинозу, левулезу, маннозу, ксилозу, рамнозу, мальтозу,
трегалозу, маннит; редуцируют нитраты, метиленовый синий; вступают в реакцию
с метиленовым красным, образуют каталазу, гидролизуют мочевину и эскулин.
Y. enterocolitica не ферментируют лактозу, рамнозу, раффинозу, не продуцируют
фенилаланиндезаминазу, лизиндекаобоксилазу и аргининдегидролазу. Реакция
Фогеса-Проскауэра положительная при 26±2 °C и отрицательная при 37±1 °C.
Гидролизуют мочевину, продуцируют орнитиндекарбоксилазу, ферментируют
сахарозу и мальтозу. По способности ферментации сорбита, ксилозы, салицина,
образованию индола и наличию липазы У. enterocolitica делится на 6 биотипов.
Другие виды рода Yersinia выделяют из фекалий, смывов с окружающей среды,
органов и тканей грызунов. Они, как У. enterocolitica и Y, pseudotuberculosis, хоро-
шо растут на питательных средах для выделения иерсиний. Их также необходи-
мо дифференцировать, используя основные биохимические тесты и субстраты
(табл. 21-16).
Иерсинии — гетеротрофные факультативно-анаэробные микроорганизмы. Они
неприхотливы, растут не только на обычных питательных средах, но и на средах
с обедненным составом (пептонной воде) или «голодных» средах (синтетических
средах, фосфатно-буферном растворе, 0,9% растворе NaCl). Температурный фак-
тор — один из важнейших, определяющих изменчивость микробов и устойчивость
их во внешней среде. Температура культивирования иерсиний влияет на размеры
клеток, темпы размножения, скорость метаболических процессов; оптимальная
температура роста составляет 26±2 °C. Способность иерсиний к более высоким
темпам размножения при низких температурах позволила на практике использо-
вать холодовое обогащение для их выделения из материала, контаминированного
сопутствующей микрофлорой, особенно колиформными бактериями. В жидкой
соеде (питательном бульоне, пептонной воде, синтетических средах) иерсинии
образуют равномерное помутнение среды, иногда нежную пленку (У. pseudotu-
berculosis) или нежное пристеночное кольцо (У. enterocolitica). При дальнейшем
культивировании, особенно при 37 ЭС, среда просветляется и образует слизистый
и крошковидный осадок. На плотных средах иерсинии образуют колонии типов
Таблица 21-16. Основные биохимические свойства бактерий рода Yersinia, дифференцирующие их внутри рода и от других представителей семейства
Enterobacteriaceae
Тест или субстрат Y. pestis Y. pseudotuber- culosis К enterocolltlca Биотипы Y. frederiksenii Y. intermedia К knstenseoil Y. ruckeri К atdovae С rohdei Y. mollaretti Y. bercovieri ЧАСТНАЯ МИКРОБИОЛОГИЯ
II III IV V
IA IB
Гидролиз мочевины — + + 4- + + 4- + l 1±1__ [+1 _И_ — В [-] ... в
Образование индола - — + + в - - - 4- 4- В - -
Подвижность (22-24 °C) - + + + + + 4- 4- + 4- 4- в 4- + 4- 4-
(37 °C) - - - - - - - - - - - - - - - -
Реакция Фогеса- Проскауэра (22-24 °C) — — 4- 4- 4- 4- 4- + 4- - - + - - —
(37 °C) — — - - — — — - - — - - - - - -
Фенилаланиндезаминаза — — - — - — - — - - - - - - - -
Орнитиндекарбоксилаза - — + + 4- + 4- в 4- + + 4- в _ы _[+]_ О+]_
Лизиндекарбоксилаза - — - - - - - - - - - В - - -
Липаза (твин-80) — - 4- + — - - - в _Б_ - В - - - -
Цитрат Симмонса (22-24 °C) — — - - — — - - в 4- - + + - - —
Гидролиз эскулина (24 ч) в + (+1 — — - — — [+]-._ + — - - - - [-1
Образование кислоты при ферментировании угле од ов
0-ксилоза + + + 4- 4- 4- — В 4- 4- [+1 - в в В 4-
Мальтоза 1+1 + 4- + + + 4- + + + + 4- - - в 4-
О-маннитол + + + 4- 4- 4- 4- 4- + + 4- 4- 1+1 4- + 4-
Мелибиоза [-1 в — - — - - - - _Ы_ - - В -
L-рамноза в - - — - — - 4- 4- - - - - - -
Рафиноза - 1-1 — — - — — — В В — — - в - -
Салицин (24 ч) в [-1 4- — — - - - + + _ы - - - [-]
Сахароза - + + + + + в 4- 4- - _L-J _ + 4- 4-
D-сорбит в - 4- + 4- 4- 4- - + 4- 4- в в 4- 4- 4-
Трегалоза + + + + - 4- + 4- 4- _[+!_ 4- + 4-
Примечание: со
«+«► -- положительная реакция у 90% штаммов и более; «[+]» — положительная реакция у 76-89% штаммов; «В> — положительная реакция у 26-75% штам- о»
мов; ч[-]» — положительная реакция у 11-25% штаммов; «-» - отрицательная реакция у 90% штаммов и более.
368
ЧАСТНАЯ МИКРОБИОЛОГИЯ
S (гладкий), R (шероховатый) и SR (переходный). Например, У. pseudotuberculosis
при пониженной температуре растут в S-форме, но накопление культуры происхо-
дит медленно, а при повышенной (более 28 °C) активно диссоциируют в R-форму;
в посевах наблюдается полиморфизм колоний по размеру, форме и цвету.
У. pseudotuberculosis и некоторые представители У. enterocolitica имеют большое
сходство, проявляющееся в их инвазивности, способности к внутриклеточному
размножению, что коррелирует с генерализацией инфекционного процесса. Для
большинства выделенных от больных штаммов У. enterocolitica характерны адге-
зия, колонизация на поверхности кишечного эпителия и энтеротоксигенность с
образованием больших количеств термостабильного энтеротоксина. Патогенные
У. pseudotuberculosis и У. enterocolitica обладают широким набором факторов пато-
генности, детерминируемых хромосомными и плазмидными генами. Способность
У. pseudotuberculosis к адгезии и инвазии в клетки эпителия кишечника зависит
от экспрессии хромосомного йт-гена, кодирующего белок наружной мембра-
ны инвазии с молекулярной массой 108 кДа. Системная диссеминация микро-
ба в организме хозяина связана с функционированием системы поглощения
ионов железа, названной иерсиниабактином. Хромосомный сегмент, содержащий
гены иерсиниабактина, называется островом высокой патогенности (the High
Pathogenicity Island — HPI). Установлено, что HPI содержат штаммы, циркулирую-
щие в Европе. Австралии, Северной Америке. Они. как правило, обусловливают
гастроинтестинальные проявления псевдотуберкулезной инфекции. Показано, что
У. pseudotuberculosis синтезируют суперантиген (Yersinia Pseudotuberculosis-derwed
Mitogen — YPM), ответственный за поликлональную активацию Т-лимфоцитов,
и гм пер продукцию провоспалительных цитокинов. Около 98% штаммов, выделен-
ных от больных на Дальнем Востоке, в Японии и Корее, содержат суперантиген,
вызывающий системное поражение органов и ткачей. Патогенные У. enterocolitica
имеют белки наружной мембраны (БНМ), обеспечивающие им адгезивные и инва-
зивные свойства. Один из них — инвазии — имеет молекулярную массу 92 кДа и
кодируется хромосомным mv-геном. Другой хромосомный ail-ген кодирует белок
адгезии и инвазии Ail с молекулярной массой 17 кДа. Гены HPI не выявлены у непа-
тогенных У. enterocolitica биотипа 1А и у низкопатогенных штаммов биотипов 2-5,
однако они определяются у всех высокопатогенных штаммов биотипа 1В.
Унтеротоксигенность У. enterocolitica биотипов 1В и 2-4 связана с экспрессией
хромосомного ysfA-гена. У непатогенных представителей биотипа 1А обнаружен
у$ГВ-ген.
Плазмидой вирулентности иерсиний pYV 42-48 МДа обладают штаммы
У. pseudotuberculosis и У. enterocolitica. Плазмида вирулентности кодирует белок
наружной мембраны Yad А, являющийся ведущим адгезином иерсиний, а также
белки наружной мембраны Yop и аппарат их секреции III типа Ysc (Yop secretion),
которые представляв it собой комплекс факторов, участвующих в нейтрализации
иммунного ответа макроорганизма. Аппарат секпеции III типа Ysc включает не
менее 28 белков и позволяет иерсиниям вводить синтезируемые белки в цитоплаз-
му клетки-мишени при тесном контакте без проникновения в нес,
Плазмида с молекулярной массой 82 МДа обнаружена только у У. pseudotubercul
шжеротипа 1. Показано, что штаммы, имеющие плазмиду pVM 82 МДа, вызывают
более тяжелое течение псевдотуберкулеза.
КЛИНИЧЕСКИЕ ПРОЯВЛЕНИЯ ИНФЕКЦИЙ Y. ENTEROCOLITICA
И Y. PSEUDOTUBERCULOSIS
У. enterocolitica вызывают гастроэнтерит, илеит, энтероколит, псевдоаппенди-
цит. Нарушения деятельности пищеварительного тракта сопровождаются диареей,
болями в животе и лихорадкой. Иногда при этой инфекции развиваются узловатая
эритема и острый артрит.
ЧАС ждя микробиология 369
Псевдотуберкулез регистрируют в спорадических случаях. Он проявляется
брызжеечным аденитом и аппендицитоподобным синдромом.
Полиморфизм и неспецифичность клинической картины обоих иерсиниозов
диктует необходимость их дифференциации от широкого круга инфекционных,
паразитарных и неинфекпионных заболеваний, в том числе скарлатины, кори,
краснухи, энтеровирусной инфекции, лептоспироза, вирусных гепатитов, три-
хинеллеза, брюшного тифа, острых кишечных инфекций различной этиологии
(сальмонеллеза, кампилобактериоза, шигеллеза), инфекционного мононуклеоза,
бруцеллеза, туляремии, геморрагических лихорадок, острого аппендицита. Иногда
иерсиниозы приходится дифференцировать от менингококковой инфекции, грип-
па, сыпного тифа, ревматизма, геморрагического васкулита. Нередко при подо-
зрении на иерсиниоз приходится исключать наличие у пациентов лимфосаркомы,
опухолей толстой кишки, внелегочного туберкулеза.
ЛАБОРАТОРНАЯ ДИАГНОСТИКА
Окончательный диагноз иерсиниозов ставят на основании результатов лабо-
раторных исследований — выделения на питательных средах и идентификации
возбудителя обнаружения в клиническом материале антигенов, а также генома
У. pseudotuberculosis и У. enterocolitica, антигенов, выявления у пациентов серокон-
версии.
Изоляция и идентификация У. pseudotuberculosis и Y. enterocolitica
Одно из важнейших условий качественного бактериологического исследова-
ния — правильный сбор материала и его подготовка к исследованию. Правила
взятия и подютовки исследуемого материала представлены в табл. 21-17.
Таблица 21-17. Материалы для исследования и их подготовка
Вид исследуемого материала Количество, сроки Правила взятия, подготовка к исследованию бактериологиче- ским методом и методом ИФА Правила взятия, подготовка к ПЦР
Материал от больных
Фекалии1 0,5-1.0 г на протяжении всего периода заболевания Стерильной палочкой из судна или зондпм-тампоном из прямой кишки. Помещают в 5 мл среды накопления (ПК) В одноразовый стерильный контейнер с 5 мл забуфе- ренного 0,9% физраство- ра (ЗФР) — pH 7,2-7.,4, ложечкой, прилагаемой к контейнеру
Моча’ 20-30 мл средней пор- ции утренней мочи е первые 7 дней болез- ни Засевают осадок после центрифуги- рования или отстаивания. Для ИФА осадок исследуют нативно, затем помещают в 5 мл среды накопления В одноразовый стерильный контейнер
Смыв из зева В первые 3 дня болезни Натощак с задней стенки глотки и корня языка зондом-тампоном, смоченным в 0,9% растворе натрия хлорида. Зонд-1 ампон помещают в 5 мл среды накопления Зонд-тампон помещают в одноразовую стерильную пробирку с 5 мл ЗФР
Кровь 5-10 мл в первые 3 дня болезни Стерильно из локтевой вены; сгу- сток измельчают, помещают 1 мл крови в 5 мл среды накопления В одноразовую стерильную пробирку
Операционный или секционный материал
Червеобразный отросток 1-2 Г Стерильно измельчают, 1 мп суспензии помещают в 5 мл среды накопления В одноразовую стерильную пробирку с 5 мл ЗФР
370
ЧАСТНАЯ МИКРОБИОЛОГИЯ
Окончание табл. 21-17
Вид исследуемого материала Количес-яо, сроки Правила взятия, подготовка к исследованию бактериологиче- ским методом и методом ИФА Правила взятия, подготовка к ПЦР
Мезентериальные лимфоузлы, сино- виальная оболочка и другие органы и ткани 1-2 г Стерильно измельчают, 1 мл суспензии помещают в 5 мл среды накопления В одноразовую стерильную пробирку с 5 мл ЗФР
Синовиальная жид- кость 1-2 мл Стерильно из суставной сумки, 1 мл помещают в 5 мл среды накопления В одноразовую стерильную пробирку с 0,5 мл ЗФР
Желчь 1-2 мл Собирают в стерильную пробирку 0,5-1.0 мл. помещают в 5 мл среды накопления
Кишечное содер- жимое 0,5-1.0 г Собирают в стерильную пробирку 0,5-1,0 мл, помещают в 5 мл среды накопления В одноразовую стерильную пробирку с 5 мл ЗФР
Сгусток крови 0 5-1,0 МЛ Стерильно измельчают. 1 мл поме- щают в 5 мп среды накопления В одноразовую стерильную пробирку с 5 мл ЗФР
' Высеваемость иерсиний наиболее высока в первые 3 -5 дней болезни и до назначения анти-
бактериальных препаратов.
Схема бактериологическою исследования включает обогащение исследуе-
мого материала при низкой температуре, использование дифференциально-
диагностических сред для выделения и идентификации культур, определение их
биотипа, серотипа, патогенности.
Несмотря на низкую эффективность, прямой посев нативного материала на плот-
ные питательные среды проводят, как правило, при подозрении на инфицирован-
ность иерсиниями продуктов питания или при групповых заболеваниях, вызванных
употреблением инфицированных продуктов. Для этой цели целесообразно исполь-
зовать дифференциально-диагностическую среду для выделения иерсиний с бром-
тимоловым синим (СБТС).
При холодовом обогащении исследуемый материал в пептонно-калиевой среде (ПК)
помещают в холодильник и выдерживают в нем до первого положительного посева, но
не более 10 дней. Высев на плотные питательные среды проводят со среды ПК на 2-3,
5-7 и 10-е сутки. Его проводят петлей из верхней трети слоя среды (но не с п< >верхно-
сти). Перед посевом содержимое пробирок не взбалтывают. При использовании среды
Эндо или исследовании сильно контаминированных посторонней микрофлорой проб
проводят щелочную обработку (в 0,2 мл исследуемого материала вносят 0,2 мл свеже-
приготовленного 0.72% раствора гидроксида калия в 0,5% растворе натрия хлорида,
перемешивают, выдерживают в течение 1-1,5 мин и высевают на плотную питатель-
ную среду). Методика щелочной обработки основана на относительной резистентно-
сти иерсиний к щелочам по сравнению с другими микроорганизмами.
Для первичной изоляции и культивирования иерсиний чаще всего используют спе-
циальную дифференциально-диагностическую среду' для иерсиний (СБТС), а также
среды Хотгингера, Эндо, Олькеницкого, цефсулоидин-иргазан-новобиоци! ювый агар.
Посевы на этих средах инкубируют при температуре 26+2 СС и осматривают еже-
дневно (со вторых суток), регистрируя появление подозрительных по внешнему виду
колоний (табл. 21-18).
Видовую принадлежность выделенных культур устанавливают на основании
комплекса типичных морфологических, культуральных, биохимических и анти-
генных свойств. Для предварительного отбора используют стабильный биохи-
мический признак — наличие уреазы. Положительный уреазный тест позволяет
отличить иерсинии ст других видов патогенных кишечных бактерий — шигелл,
сальмонелл, эшерихий. Отсев подозрительных колоний проводят на скошенный
столбик среды Олькеницкого штрихом и уколом в столбик.
Таблица 21-18. Характеристика колоний иерсиний на плотных питательных средах
Вид среда Хопингера Среда Эндо СБТС Следа Олькеницкого Примечание
24 ч 48 ч 24 ч 48 ч 24 ч 48 ч Столбик Скошенная часть
К pseudo- tuberculosis Колонии диаме- тром 0,1-0,2 мм, голубовато-белые, полупрозрачные, с ровными краями, слизистые, со слабоприпод- нятым центром (при осмотре под микроскопом имеют вид пира- мид или со сходя- щимися к центру гранями) Колонии диаметром 0,2-0,5 мм, прозрачные,с ровным краем. В полиморфной популяции — часть колоний бугристая, с неровными фестончатыми краями Колонии мел- кие (росинча- тые) круглые, выпуклые, блестящие, бесцветные Колонии диаметром 0,1-0,2 мм, выпуклые, блестящие, с ровными краями, бес- цветные Колонии мелкие кру- глые, могут иметь слегка желтоватую окраску Колонии диа- метром 1-2 мм, голубовато- зеленые, с фестон- чатыми краями, выпуклые, с при- поднятым центром и суховатой мато- вой поверхностью на синем фоне среды Малиновый Малиновый На СБТС через 48 ч колонии Е. coli — ярко-желтые, диаметром 2-3 мм, сочные, выпуклые; Sh. flexneri — желтые, диа- метром 1-2 мм сочные, плоские
Y. enterocolitica Колонии диа- метром от 0,1-0,2 до 0,5 мм, кру- глые, выпуклые, с ровным краем, полупрозрачные, с голубоватым опенком мягкой консистенции Колонии диа- метром 0,3- 0,5 мм, более выпуклые, с ровным краем, прозрачные Такие же Такие же, но с розоватым опенком Колонии диа- метром 2-4 мм, голубовато- зеленые, выпу- клые, сухие, с матовым налетом Малиновый, слег- ка желтоватый в самой нижней части столбика Малиновый
ЧАСТНАЯ МИКРОБИОЛОГИЯ
372
ЧАСТНАЯ МИКРОБИОЛОГИЯ
Выделение антигенов возбудителей
Методы, направленные на выявление антигенов иерсиний, наиболее пер-
спективны для ранней диагностики их инфекций. Основные их преимущества —
быстрота получения результатов, возможность обнаружения антигенов погибших
микробов, что особенно важно при обследовании больных, которым проводили
антибиотикотерапию.
Материал для исследования, сроки и правила его взятия представлены в
табл. 21-17. Исследование проб проводят в 1-е сутки их полугения и затем на 3-5-е
сутки после подращивания при температуре 4 °C в среде ПК. Непосредственно
перед исследованием материал инактивируют 30 мин при 56±1 °C, центрифугиру-
ют при 3000 об./мин 15 мин и берут для исследования надосадочную жидкость.
Для выявления антигенов иерсиний в различных субстратах применяют реак-
цию ко-агглютинации (РКоА) и иммуноферментный анализ (ИФА).
РКоА. Реакция предназначена для выявления микробных антигенов в материале,
взятых у пациентов в течение первой недели болезни, а также связанных анти-
генов в составе иммунных комплексов. Тест основан на способности белка А
золотистого стафилококка соединяться с Fc-фрагментом IgG. Именно поэтому в
таких антительных диагностикумах свободные, ориентированные наружу Fab-
фрагменты иммуноглобулинов соединяются со специфическими антигенами иер-
синий. Реакцию учитывают визуально.
ИФА. Принцип теста основан на образовании комплекса «антиген-антитело;»,
выявляемого с помощью второго антитела, меченного пероксидазой хрена (конъ-
югата). Имеются коммерческие иммуноферментные тест-системы для выявления
антигенов иерсиний псевдотуберкулеза на основе антител к синтетическим олиго-
пептидам — фрагментам инвазина (белка наружной мембраны) У.pseudotuberculosis,
ответственного за адгезию и инвазию возбудителя на начальных этапах инфекци-
онного процесса.
РГА. Реакцию проводят на стекле со свежевыделенными культурами бактерий,
выращенными на среде Хоттингера при температуре 37±1 и 26±2 °C (по 18-24 ч
при каждом температурном режиме). Для экспресс-индентификации pYV-i- штам-
мов используют сыворотку к вирулентным иерсиниям (СВИ), а для серотипиро-
вания — диагностические сыворотки к У. enterocolitica серотипов 0:3, 0:4, 0:4,32;
0:4,33; 0:5, 0:5,27; 0:6,30; 0:6,31; 0:7.8; 0:9, 0:13, 0:13,7 и серотипы I и III
У. pseudotuberculosis.
Молекулярно-генетическая диагностика
Позволяет выявлять иерсинии в тестируемых пробах на основании обнаруже-
ния определенных участков генома этих бактерий. Наибольшее применение нашла
ПЦР. При диагностике псевдотуберкулеза и кишечного иерсиниоза ПЦР позволя-
ет выявить специфические участки генов, локализованных на плазмиде вирулент-
ности иерсиний pYV или хромосоме, кодирующих ряд факторов патогенности.
Основная трудность ПЦР заключается в том, что при исследовании клинического
материала реакция может ингибироваться биологическими продуктами деграда-
ции тканей, ферментами, полисахаридами и другими компонентами исследуемого
образца. Именно поэтому перед постановкой реакции необходимо подготовить
пробы к исследованию описанными ниже способами.
Кровь. 50 мкл нативной крови смешивают со 100 мкл лизирующего буфера
(10 мМ Трис-НС1, pH = 8,0, 0,5% Твин-20,100 мкг/мл протеиназы К) и инкубиру-
ют в течение 1 ч при температуре 56 °C, затем протеиназу К инактивируют нагрева-
нием до 95 СС 10 мин, смесь центрифугируют 3 мин при 5000 об./мин. Для анализа
в ПЦР достаточно 2-3 мкл надосадочной жидкости.
Моча. 10-20 мл мочи центрифугируют 10 мин при 2000 об./мин. К осадку
добавляют 300 мкл стерильной дистиллированной воды и прогревают в течение
ЧАСТНАЯ МИКРОБИОЛОГИЯ 373
10 мин до 100 °C. Центрифугируют при 5000 об./мин. Для исследования в ПЦР
берут 1-3 мкл надосадочной жидкости.
Фекалии. Подготовка Фекалий основана на щелочной обработке, при которой
большая часть посторонней микрофлоры погибает и удаляется как супернатант
при центрифугировании, а относительно устойчивые к щелочи иерсинии выжива-
ют, что обеспечивает снижение концентрации ингибиторов в пробе:
• исследуемый материал суспендируют в ФБР в соотношении 1:10 и центри-
фугируют при 500-1000 об./мин в течение 1,5-2 мин для осаждения грубых
частиц или отстаивают в течение 2-3 ч при комнатной температуре;
• 0,2-0,5 мл верхней фазы переносят в лунку полистиролового планшета для
серологических реакций и смешивают с таким же объемом 0,72% раствора
гидроксида калия;
• после 25-30 с экспозиции проводят посев материала на СБТС и сразу же в
лунки планшета добавляют 0,2-0.5 мл бульона Хотгингера для прекращения
действия щелочи как селективного агента;
• материал из лунки переносят в пробирку типа «Эппендорф» и центрифугиру-
ют на микроцентрифуге при 10 000-12 000 об./мин в течение 3-5 мин;
• надосадочную жидкость удаляют, осадок дважды промывают дистиллиро-
ванной водой;
• к осадку добавляют 20 мкл деионизованной воды, прогревают в течение
10 мин до 100 °C, центрифугируют при 12 000 об./мин в течение 2 мин.
Для ПЦР используют 1-3 мкл надосадочной жидкости. Для выделения ДНК
из операционного или секционного материала применяют комплект реагентов
«ДНК-сорб-В» производства ФГУН ЦНИИЭ Роспотребнадзора.
ПЦР дает возможность получить сигнальный результат в течение первых суток
исследования, тогда как наиболее короткий срок бактериологического выделения
культуры и идентификации иерсиний составляет 5-8 сут.
ПЦР-анализ можно использовать при 3-этапном варианте бактериологического
исследования (рис 21-1).
Двукратно отрицательные результаты ПЦР позволяют прекратить дальнейшее
бактериологическое исследование.
Серологическая диагностика
Антитела к возбудителям псевдотуберкулеза и энтероколитного иерсиниоза
появляются в крови уже на 1-й неделе болезни, а существенное повышение их
концентрации до диагностических уровней происходит к началу 3-й недели забо-
левания. Этими сроками регламентировано обязательное исследование парных
сывороток крови. Достоверным считается 4-кратное и большее нарастание титра
антител при исследовании парных проб сыворотки. Через 2 мес начинается сни-
жение концентрации антител, а через 6 мес они присутствуют в крови пациентов
в минимальных количествах. Наиболее выраженная динамика антителообразова-
ния наблюдается у больных с заболеванием средней тяжести. При легком течении,
особенно при гастроэнтеритической форме, динамика ан гител выражена слабо, их
начинают регистрировать в диагностических титрах с конца 2-й недели заболева-
ния.
Кровь для серологических исследований берут натощак, с соблюдением правил
асептики. Плазму крови получают общепринятыми методами. Парные сыворотки
желательно исследовать одновременно, поэтому первую сыворотку до постановки
следует разделить на несколько объемов и заморозить. Замораживать и размора-
живать сыворотки можно однократно, так как при повторной разморозке антитела
разрушаются.
Серологическую диагностику иерсиниозов проводят в РИГА, ИФА и иммуноблот-
тинге.
374
ЧАСТНАЯ МИКРОБИОЛОГИЯ
Рис. 21-1. ПЦР при 3-этапном варианте бактериологического исследования.
РИГА. Наиболее часто применяемый в лабораториях метод серологической диа-
гностики иерсиниоззв. Для постановки теста пользуются лиофилизированными
эритроцитарными диагностикумами, в частности псевдотуберкулезным — для
выявления антител к Y, pseudotuberculosis серотипа 1 и кишечноиерсиниозным —
для выявления антител к У. enterocolitica серотипов 0:3 и 0:9 (производства ФГУП
«Санкт-Петербургский научно-исследовательский институт вакцин и сывороток
и предприятие по производству бактерийных препаратов»). Эти диагностикумы
представляют собой формалинизированные эритроциты барана, сенсибилизиро-
ванные специфическими полисахаридными антигенами иерсиний. Агглютинация
эритроцитов происходит в результате взаимодействия специфических антител
исследуемой сыворотки и антигенов, иммобилизованных на поверхности эри-
троцитов. Диагностикумы выпускают в комплекте с несенсибилизированными
эритроцитами барана для оценки специфичности реакций с исследуемыми сыво-
ротками и специфическими кроличьими сыворотками для контроля активности
диагностикумов. Постановку РИГА проводят макро- и микрометодами. Для про-
ведения реакции макрометодом используют полистироловые пластины с лунками
вместимостью 2 мл. Для постановки РИГА микрометодом удобны одноразовые
микропланшеты с U-образными лунками. Перед постановкой исследуемые сыво-
ротки прогревают при температуре 56 °C в течение 30 мин для устранения неспе-
цифических гемагглютининов.
ИФА. Для постановки ИФА используют тест-системы, предназначенные для
выявления IgA, IgM и IgG к БНМ иерсиний, кодируемым плазмидой вирулентно-
сти pYV и ответственным за проявление патогенных свойств иерсиний. Высокие
чувствительность и специфичность метода достигаются благодаря использова-
нию в качестве антигенов, сорбированных в лунках полистироловых планшетов.
ЧАСТНАЯ МИКРОБИОЛОГИЯ 375
комбинации рекомбинантных БНМ иерсиний (YopD, YopH, YopE, YopM). Тест-
сисгемы представляют собой многокомпонентные наборы реагентов для поста-
новки непрямого ИФА на твердофазном носителе.
Иммуноблоттинг. Позволяет выявлять антитела классов А и G к отдельным
антигенам У. enterocolitica путем постановки ИФА на нитроцеллюлозных мембра-
нах. В качестве антигенов используют БНМ иерсиний (YopM, YopH, YopD, YopN,
YopP, YopE, LcrV), специфичных для вирулентных штаммов. Очищенные БНМ раз-
деляют по молекулярной массе методом электрофореза в полиакриламидном геле
и переносят на нитроцеллюлозою мембрану тестовых стрипов в виде отдельных
полос. При постановке ИФА стрипы инкубируют с образцом разведенной сыво-
ротки или плазмы крови пациента. Если образец положительный, специфические
антитела будут связываться с антигенами Для визуализации комплекса «антиген-
антитело» проводят инкубацию с использованием ферментного конъюгата — при
наличии антител на мембране видны полосы различной интенсивности. Метод
имеет высокую специфичность и применяется для диагностики хронических форм
иерсиниозов.
ГЕМОФИЛЬНЫЕ БАКТЕРИИ
ТАКСОНОМИЯ
Род Haemophilus относится к семейству Pasteurellaceae у-протеобактерий. Он
включает не менее 15 видов, 8 из которых могут быть выделены из организма
человека.
Эпидемиологическое значение имеют следующие виды гемофильных бактерий:
— Н. Influenza — поражает детей, вызывая гнойный менин! ит, острые отит и
синусит, пневмонию, хронический бронхит, гнойный конъюнктивит, эндо-
кардит, сепсис;
— Н. ducreyi — возбудитель мягкого шанкра;
— Н. parainfluenzae — обостряет течение хронического бронхита, выделяется
при подостром инфекционном эндокардите;
— Н. aphrophilus — достаточно редко изолируют из абсцессов, крови, плевраль-
ной жидкости и СМЖ. Встречается в материале из ротовой полости, верхних
дыхательных путей;
— Н. segnis — встречаются при раневой инфекции, могут быть выделены из
крови и плевральной жидкости;
Данный раздел посвящен 2 видам 1емофильных бактерий — Н. ducreyi и Н. Influ-
enza, инфекции которых имеют наибольшее клиническое значение для человека.
Н. influenzae
ОБЩАЯ ХАРАКТЕРИСТИКА
Н. influenzae — грамотрицательная, факультативно-анаэробная, неподвижная,
не образующая спор коккобацилла, имеющая размеры 0,3-0,4х1-1,5 мкм. Она
склонна к полиморфизму и может трансформироваться в вытянутые (нитевидные)
формы. Некоторые штаммы (по оценке разных авторов — от 5 до 30%) образуют
капсулу. По капсульному антигену различают 6 серотипов (a, b, с, d, е, f). Их иден-
тифицируют в РГА. Довольно часто встречаются и нетипируемые в данном тесте
изоляты.
Н. influenzae способна расти в значительном диапазоне температуры — от 23
до 43 °C, оптимальная температура культивирования — 37 °C. Для размножения
требует присутствия в питательной среде гемина (фактора X) и никотинамидаде-
ниндинуоклеотида (фактора V).
376 ЧАСТНАЯ МИКРОБИОЛОГИЯ
Не ферментирует сахара, кроме глюкозы: ее он разлагает до янтарной, молоч-
ной и уксусной кислот.
ЭПИДЕМИОЛОГИЯ
Н, influenza распространена повсеместно, а в ряде стран эта инфекция эндемич-
на. В большинстве случаев воротами инфекции и местом персистенции бактерии
в организме человека служит носоглотка. Возбудитель передается воздушно -
капельным и контактным путями.
Наиболее опасен серотип b (Hib), способный вызывать у детей сепсис, менингит,
пневмонию, эпиглоттит. Этот серотип занимает третье место (после N, meningiv-
dis и Str. pneumoniae) в структуре этиологических факторов гнойных менингитов.
В связи с этим разработана и включена в национальный календарь прививок
вакцинация против Hib-инфекции, Наиболее подвержены Hib-инфекции дети в
возрасте от 2 мес до 5 лет, у которых генерализованная форма инфекции сопро-
вождается высокой летальностью (в довакцинальный период она составляла 30%,
по данным ВОЗ). Тяжелые формы Hib-инфекции встречаются и у взрослых людей
(в нашей стране заболеваемость взрослой части населения в 4-5 раз ниже, чем
детей), хотя чаше взрослые являются носителями Н. influenzae.
Н. influenzae биовара aegypticus вызывает гнойный конъюнктивит и синдром
бразильской пурпурной лихорадки.
КЛИНИЧЕСКИЕ ПРОЯВЛЕНИЯ ИНФЕКЦИИ
Характер клинических проявлений зависит от локализации инфекции.
Острый эпиглоттит начинается у детей остро и внезапно. Характерный при-
знак — увеличенный, гиперемированный и отечный надгортанник. Заболевание
протекает тяжело и часто завершается летальным исходом вследствие прогрес-
сирующей асфиксии (дети погибают, если не удается своевременно провести им
трахеостомию).
Менингит развивается вследствие диссеминации инфекции по лимфатической
системе при бессимптомном носительстве. У выздоровевших детей нередко оста-
ются стойкие неврологические нарушения (умственная отсталость, гидроцефалия,
слепота).
Распространение инфекции по респираторному тракту и ЛОР-органам ведет к
развитию острых отитов и синуситов, пневмонии и хронического бронхита.
Гнойный конъюнктивит притекает у детей тяжело и проявляется системными нару-
шениями (повышением температуры, лимфаденопатией, невралгией, бессонницей).
При легком конъюнктивите (синдроме розовых глаз) появляются незначительные
серозные истечения, которые со временем становятся серозно-гнойными. При тяже-
лом конъюнктивите отмечают сильное раздражение глаз, слезотечение, отек век со
слизисто-гнойным или фибринозно-гнойным отделяемым, светобоязнь.
Синдром бразильской пурпурной лихорадки — тяжелая септицемия, клиниче-
ски напоминающая менингококцемию. Характерны системные поражения, обу-
словленные интоксикацией, и сыпь (без петехий). Летальность достигает 70%.
ЛАБОРАТОРНАЯ ДИАГНОСТИКА
Для постановки окончательного диагноза в лабораторию отсылают материал,
соответствующий клинической форме инфекции, — СМЖ, кровь, плевральную
жидкость, СЖ, перикардиальную жидкость, отделяемое из гайморовых и других
пазух, мокроту, гной из абсцессов соскоб со дна язв при мягком шанкре.
Бактериологический анализ должен влючать бактериоскопию мазков проб, их
посев на питательные среды, определение чувствительности выделенных изолятов
Н. influenzae к антибактериальным препаратам. В последнее время все чаще для
диагностики инфекции стали применять экспресс-методы обнаружения антигенов
Н. influenza и ПЦР.
ЧАСТНАЯ МИКРОБИОЛОГИЯ 377
В лабораторию клинический материал доставляют в транспортных средах.
Для транспортировки носоглоточной слизи подходит 2% пептонная вода (pH ~
7,2+7,4) с линкомицином (1 мг/мл). Зонд-тампон со слизью погружают в среду,
закрывают пробирку пробкой и доставляют в лабораторию, где тампон отжимают
о стенки пробирки и проводят посев на чашку с сывороточным агаром.
Бактериоскопия
В препаратах толстой капли СМЖ или крови после окраски водно-спиртовым
раствором метиленового синего выявляют мелкие полиморфные бактерии (кок-
ковидной, овальной или нитевидной формы), расположенные в лейкоцитах или в
свободном виде и формирующие цепочки различной длины.
В мазках мокроты, окрашенных по Граму, выявляют полиморфные грамотрица-
тельные бактерии внутри полиморфноядерных нейтрофилов или вне клеток. Для
учета результатов необходимо уточнить, действительно ли исследуемый материал
является мокротой, а не слюной или отделяемым носоглотки (в мокроте в боль-
ших количествах должны присутствовать альвеолярные макрофаги).
Изоляция возбудителя
Посевы исследуемого материала инкубируют в течение 24 ч при температуре
37 °C и повышенной влажности в атмосфере, содержащей 5-10% углекислого газа.
На шоколадном агаре Н. influenzae образует серые, слизистые, блестящие
мелкие колонии диаметром 0,2-2,0 мм. Часто определяется характерный запах
индола (мышиный). Колонии R-форм Н, influenzae очень мелкие, серовато-белого
цвета, зернистые, с неровными краями.
На полужидком питательном (0,1% сывороточном) агаре признаки роста гемо-
фильных палочек отсутствуют, хотя они способны сохраняться на данной среде
до 48 ч.
На сывороточном агаре Н. influenzae не растет в отличие от N, meningitidis и
Str. pneumoniae.
При посевах крови на двухфазную питательную среду признаки роста гемо-
фильных бактерий в жидкой фазе среды отсутствуют, тогда как на плотной фазе
отмечается формирование колоний.
При бактериоскопии колоний Н. influenzae в мазках, окрашенных по Граму,
выявляют грамотрицательные полиморфные палочки (от овальных кокковидных
до типичных тонких палочек), различные по длине и образующие нитевидные
цепочки. Нередко отмечают наличие капсулы.
Подозрительные на Н. influenzae колонии тестируют на сахаролитическую и
ферментативную активность. Коммерческие наборы позволяют провести биохи-
мическую идентификацию в течение 2 ч.
Также из колоний, подозрительных на Н. influenzae, делают пересев на чашки
с шоколадным агаром для получения чистой культуры. Потребность бактерий в
факторах роста определяют посредством посева чистых культур на сывороточный
агар. После посева на поверхность агара накладывают диски с V и X факторами
роста (расстояние между дисками — 5 мм). Колонии Н. influenzae растут только в
пространстве между дисками с V и X факторами.
Экспресс-диагностика
Экспресс-метод определения Н. influenzae — реакция латекс-агглютинации.
Материал для исследования — нативная СМЖ или кровь. Идентификацию
Н. influenzae серотипа b осуществляют с помощью реакции латекс-агглютинации
с соответствующим латекс-диагностикумом. Для дифференциальной диагностики
Н1Ь-ин(Ьекции с инфекцией, вызванной другими гемофильными и морфологиче-
ски сходными бактериями, используют различия культурально-биохимических
свойств (табл. 21-19, 21-20).
Таблица 21-19. Биохимические свойства различных видов рода Haemophilus
Вид Гемолиз* Каталаза Оксидаза Порфирино- вый тест Потребность в факторах роста Ферментация
глюкозы фруктозы сахарозы лактозы маннозы
Н. influenzae - + + - V, X + - - - -
Н. parainfluenzae - + 4- + V + + + - +
Н. haemoliticus + + + - V. X + + - - -
Н. parahaemoliticus + + + + V + + - -
Н. aprophilus - - - + - + + + + +
Н, paraprophilus - - + + V + + + +
Н. segnis - 4- - + V + + + - -
Н. ducreyi - + + - X - - - - -
’ На кровяном агаре с эритроцитами лошади.
ЧАСТНАЯ МИКРОБИОЛОГИЯ
ш
ео
Таблица 21-20. Дифференциальная диагностика гемофильных бактерий с другими морфологически сходными микробами
Микроб Рост на простых пита- тельных средах Морфология Каталаза Оксидаза Редукция нитратов Уреаза ^-Галактозидаза (тест с ONPG) Гемолиз
Н. influenzae - Мелкие палочки + - + + - -
Н. haemoliticus - Мелкие палочки + - + - + +
Н. parainfluenzae - Мелкие палочки + - + - + -
N. meningitidis — Кокки + + - - - -
N. mucosa + Кокки + + + - - -
N. sicca + Кокки + + - - - -
N. subfiava 4- Кокки + 4- - - - -
N. perflava 4- Кокки + + + - - +
Moraxella catarrhalis + Кокки + + + - - -
Acinetohacter spp. + Палочки + - - - - -
Str. pneumoniae - Кокки - - - - 4" +
ЧАСТНАЯ МИКРОБИОЛОГИЯ 379
Существуют коммерческие наборы для определения специфических фрагмен-
тов генома Н. influenzae типа b в СМЖ и крови больного методом ПЦР.
ОПРЕДЕЛЕНИЕ ЧУВСТВИТЕЛЬНОСТИ К АНТИМИКРОБНЫМ ПРЕПАРАТАМ
Особенность анализа — особые требования к питательной среде (обязатель-
ное наличие V и X факторов роста). Хорошо зарекомендовала себя питательная
среда НТМ (Haemophilus Test Medium). Ее готовят в лаборатории из бульона и агара
Мюллера -Хинтона. После рагтворения основы в нее добавляют дрожжевой экстракт
(до конечной концентрации 5 мг/мл) и раствор гематина (до конечной концентра-
ции 15 мг/л). Сначала готовят основной раствор гематина: 50 мг порошка вносят в
100 мл 0.01 N раствора гидоиксида натрия (0.01 моль/л) и нагревают, тщательно
перемешивая до полного растворения. В подготовленную для автоклавирования
среду на 1 л вносят 30 мл основного раствора гематина. После автоклавирования в
охлажденную до 48-50 °C питательную среду асептически вносят раствор никоти-
намидадениндинуклеотида до конечной концентрации 15 мг/л. Раствор никотина-
мидадениндинуклеотида стерилизуют фильтрацией через мембранные фильтры с
размером пор 0,45 мкм. При определении чувствительности к сульфаниламидам и
триметоприму в охлажденную до 48-50 СС питательную среду асептически вносят
раствор тимидинфосфорилазы до конечной концентрации 0,2 ЕД/мл.
Для контроля антибиогикорезистентности используют диско-диффузионный
метод или Е-тест.
Н. ducreyi
ОБЩАЯ ХАРАКТЕРИСТИКА
Н. ducreyi — факультативный аэроб, требующий для своего роста фактор X, а не
V. Бактерия оксидазоположительна и каталазоотрицательна. Уреазной и орнитин-
декарбоксилазной активностью не обладает, глюкозу не ферментирует, индол не
образует.
ЭПИДЕМИОЛОГИЯ
Мягкий шанкр в течение последних десятилетий в России практически не
регистрируют (за исключением случаев заражения за рубежом). Инфекция рас-
пространена в Африке, странах Ближнего Востока и Юго-Восточной Азии, а также
Центральной и Южной Америки. Кроме того, мягкий шанкр регистрируют в пор-
товых городам Великобритании, Португалии, Италии и США.
КЛИНИЧЕСКИЕ ПРОЯВЛЕНИЯ ИНФЕКЦИИ
Инкубационный период составляет 3-10 дней, однако через 12-18 ч после зара-
жения на месте внедрения возбудителя образуется красное пятно Через 2-5 дней
оно превращается в изъязвленную папулу, а затем пустулу (язву). Язва мягка на
ощупь и болезненна, что отличает ее от сифилитического твердо! о шанкра. Она
имеет тонкие, неровные, подрытые края, а ее дно покрыто желтоватым сальным
налетом и обильно кровоточит. Первоначально язва по размерам соответствует
зерну чечевицы, но в течение 3 нед ее диаметр увеличивается до 2 см. Мягкие шан-
кры могут быть множественными, в этом случае они часто сливаются, формируя
гигантские ползучие язвы. Одновременно развивается регионарный лимфаденит,
сопровождаемый в 40% случаев нагноением пораженных лимфатических узлов.
В неосложненных случаях язвы спонтанно заживают в течение 1-2 мес.
ЛАБОРАТОРНАЯ ДИАГНОСТИКА
Бактериоскопия
При типичной клинической картине лабораторная диагностика заключается в
бактериоскопии отделяемого язвы. Мазки окрашивают по Граму, Цилю-Нильсену
380 ЧАСТНАЯ МИКРОБИОЛОГИЯ
или метиленовым синим. При обнаружении в мазках грамотрицательных палочек
с типичным расположением группами («косяки рыб») или параллельными цепоч-
ками («железнодорожные пути») диагностируют мягкий шанкр. В сомнительных
случаях (при характерной симптоматике и отрицательном результате бактерио-
скопии) проводят посев клинического материала на питательные среды в целях
изоляции возбудителя болезни. Также в сомнительных случаях больному прово-
дят провокационный тест: инокулируют подозреваемому в заражении Н. ducreyi
убитую вакцину, приготовленную из этой бактерии. Такая инъекция при мягком
шанкре индуцирует появление нового очага поражения. Ранее тест проводили,
используя вместо вакцины отделяемое язвы, взятое у того же больного.
Изоляция возбудителя
В качестве обогатительной среды используют полужидкий 0,1% сывороточный
агар. При его приготовлении сыворотку инактивируют прогреванием при темпе-
ратуре 56 сС.
Нативный (отделяемое язв) или обогащенный материал на среды с обяза-
тельным содержанием Х-фактора, посевы инкубируют в атмосфере, содержащей
5-10% углекислого газа, при повышенной влажности. Лучше всего применять
шоколадный агар с 5% овечьей сыворотки крови.
Н. ducreyi формирует колонии плотной консистенции, трудноотделяемые от
поверхности плотных питательных сред.
НЕФЕРМЕНТИРУЮЩИЕ БАКТЕРИИ
ТАКСОН! МИЯ И ОБЩАЯ ХАРАКТЕРИСТИКА
Неферментирующие грамотрицательные бактерии (НГОБ) — нетаксономи-
ческая группа микроорганизмов, неспособных к продукции кислоты на трехса-
харном агаре, не ферментирующих глюкозу в О-/F-тесте, не растущих или плохо
растущих в анаэробных условиях. Она включает разнообразные роды бактерий, не
связанных между собой филогенетически или генетически. Причины ее формиро-
вания носят скорее утилитарный, чем биологический характер. Скудность инфор-
мации о фенотипических признаках многих из этих бактерий создает проблемы
как при идентификации, так и при определении таксономического положения
НГОБ. Наиболее достоверные результаты могут быть получены при использова-
нии молекулярно-генетических методов или при хромотографическом исследова-
нии состава клеточных жирных кислот. На практике для первичной идентифика-
ции НГОБ целесообразны изучение морфологических и тинкториальных свойств
(мазок по Граму), проведение О-/Т-теста на среде Хью-Лейфсона, выявление
подвижности, анализ кулогуральных свойств на плотных средах (табл. 21-21).
Для углубленной идентификации в условиях практических лабораторий наиболее
рационально применение коммерческих идентификационных систем [«НЕФЕРМ-
тест 24» («Lachema»), API 20 NE («BioMerieux») или аналогичных.].
Таблица 21-21. Критерии первичной идентификации НГОБ
Род O'/F-тест Пиг- мент Окси- даза Нитрат- редук- таза Подвиж- ность Рост на МПА Рост на SS-arape
Achromobacter В, Е, F/Ochrobactrum anthropi 0 - + + + + +
Acidovorax delafieldii 0 + + + + -
Acinetobacter 0 или NO - - - - + -/+
Agrobacterium 0 + + + + - -
ЧАСТНАЯ МИКРОБИОЛОГИЯ 381
Окончание табл. 21-21
Род O/F-тест Пиг- мент Окси- даза Нитрат- редук- таза Подвиж- ность Рост на МПА Рост на SS-arape
Alcaligenes NO' - + + + + +
Ciuyseomor.as luteola 0 + - + + + +
Comamonas spp. NO + + + + + +
Flavimonas oryzibabitans 0 + - - + + ±
Flavobacterium 02 + + - + + -
Kingeila F - + + - - -
Moraxella NO - + -/+ - - -А
Oligella NO - + + - + -
Pseudomonas 0 + + - + + +
Xanthomonas maltophilia 0 + - + + + +
Примечание:
О - окисление глюкозы на среде Хью-Лейфсона.
NO — отсутствие реакции.
F — фермент ация.
«+» — наличие признака.
<?-» — отсутствие признака.
«-/+» и + — признак вариабелен у отдельных видов или штаммов
1 Alcaligenes xyiosoxidans subsp. xylosoxidans - О.
2 Flavobacterium odoratum — NO.
3 Строго говоря, ферментирующие глюкозу кингеллы не относятся к НГОБ. но, учитывая, что,
во-первых, К. kingae ранее входила в род MoraxeUa, а во-вторых, невозможность выявления фер-
ментации глюкозы на трехсахарном агаре, данный род включен в настоящую таблицу,
Род Achromobacter подразделяют на три биовара — В, Е, F. Биовары А, С, D сегод-
ня рассматривают как биовары нового вида Ochrobactrum anthropi. Представители
рода — грамотрицательные подвижные палочки. Ферментируют глюкозу, ксилозу,
сахарозу, мальтозу (реакции могут протекать медленно). Растут на среде Мак-
Конки и селективном агаре для сальмонелл и шигелл. На плотных средах образуют
колонии, напоминающие колонии энтеробактерий, но более мелкие. Оксидазо-,
каталазо- и уреазоположительны. Утилизируют цитрат на среде Симонса, гидроли-
зуют эскулин, восстанавливают нитрат. Индол не образуют. Ochrobactrum anthropi
обладают сходными свойствами и могут быть дифференцированы по морфо-
логии жгутиков и отсутствию способности ряда штаммов к гидролизу эскули-
на. Эскулинположительные штаммы Ochrobactrum anthropi дифференцируют от
Achromobacter только по составу клеточных жирных кислот с помощью хромото-
графии. В связи с этим, с практической точки зрения, уместно говорить об условной
группе — Achromobacter и Ochrobactrum anthropi. Данные микроорганизмы могут
быть обнаружены в различных видах клинического материала, включая кровь.
Чувствительны к хлорамфениколу, ципрофлоксацину, гентамицину и тобрамицину
Acidovorax spp. — грамотрицательные подвижные палочки, требовательные к
питательным средам. Плохо растут или вообще не растут на среде Мак-Конки.
Ферментируют глюкозу, оксид азо- и каталазоположи гельны. Встречаются в почве.
Описаны случаи выделения из центрального венозного катетера (A. delafieldif),
мочи, отделяемого раны (A. temperans).
Род Acinetobacter семейства Neisseriaceae объединяет неподвижные аспорогенные
короткие грамотрицательные палочки. В стационарную фазу роста и на неселек-
тивных агаровых средах доминируют коккобактерии. Оксидазоотрицательны,
каталазоположительны. Это единственный оксидазоотрицательный род семейства
Neisseriaceae. Хорошо растут на простых питательных средах, образуя гладкие оваль-
ные колонии, напоминающие колонии энтеробактерий, но меньшего размера.
382 ЧАСТНАЯ МИКРОБИОЛОГИЯ
До недавнего времени в составе данного рода выделяли один вид — A. calcoa-
ceticus с несколькими биоварами. В настоящее время с помощью геносистемати-
ки род подразделен на 17 геновидов, часть из которых получила наименования
прежних биоваров, а часть осталась безымянной (табл. 21-22). Дифференциация
отдельных геновидов недоступна для практических лабораторий. В нашей стране,
согласно приказу М3 СССР № 535 от 22.04.1985 «Об унификации микробиоло-
гических (бактериологических) методов исследования, применяемых в клинико-
диагностических лабораториях лечебно-профилактических учреждений», преду-
смотрена идентификация двух видов — A. calcoaceticus и A. Iwoffii (табл. 21-23).
Таблица 21-22, Геновиды рода Acinetobacter
Номер геновида Наименование Окисление глюкозы Гемолиз Желатин Температура роста, °C
37 41 44
1 A. calcoaceticus 100 0 - 100 0 0
2 A. baumannii 95 0 - 100 100 100
3 100 0 - 100 100 0
4 A. haemoliticus 52 96 100 100 0 0
5 А. junii 0 0 0 юо 90 0
6 66 100 100 100 0 0
7 A. johnsonИ 0 0 0 0 0 0
3/9 A. Iwofii 6 0 0 100 0 0
10 100 0 0 100 0 0
11 0 0 0 100 0 0
12 33 0 0 100 0 0
13 100 100 100 89 0 0
14 100 100 100 10 0 0
15 0 100 100 100 0 0
16 0 100 100 75 0 0
17 0 100 100 100 0 0
Таблица 21-23. Дифференциация бактерий рода Acinetobacter, согпасно приказу Ns 535 М3 СССР
Признак A. calcoaceticus A. Iwofii
Оксидаза - -
Каталаза + +
Глюкоза + -
Лактоза + -
10% лактоза + -
Достаточно подробно изучена антигенная структура A. calcoaceticus — выявле-
ны капсульные. К- и О-антигены. По К-антигену различают 33 серологические
группы, по О-антигену — 28 сероваров. Обнаружены сходные (перекрестнореаги-
рующие) антигены с некоторыми видами Moraxella, стрептококками групп В и С,
пневмококком типа 20. хламидиями. Описано 34 серовара A. baumannii и 26 серо-
варов генетического вида 3. Однако на практике серотипирование данных микро-
организмов не проводят.
Acinetobacter spp. широко распространены в водоемах, сточных водах и почве.
Устойчивы в окружающей среде, способны выживать на сух )й поверхности до
4 нед. Входят в состав микрофлоры тела человека, чаще колонизируя кожу между
пальцами ног и в паховой области (особенно у проживающих в жарком и влаж-
ном климате), желудочно-кишечный тракт, уретру. Acinetobacter spp. могут быть
ЧАСТНАЯ МИКРОБИОЛОГИЯ
383
причиной госпитальных инфекций. Часто вызывают пневмонии, особенно у
больных, находящихся на искусственной вентиляции легких, реже — первичные
бактериемии и септицемии, менингиты, эндокардиты, абсцессы головного мозга
и легких, эмпиему плевры, медиастиниты, урологические катетерные инфекции и
перитониты ’ диализных больных. В настоящее время особое внимание уделяют
A. baumannii. В основном это связано с раневыми инфекциями у пострадавших во
время боевых действий.
Чувствительность отдельных штаммов Acinetobacter к антибактериальным
препаратам варьирует, но большинство из них подавляются гентамицином,
тобрамицином, амикацином и уреидопенициллинами. такими как пиперациллин.
Чувствительность к тетрациклинам непредсказуема, большинство штаммов устой-
чивы к бензилпенициллину, ампициллину, цефалоспоринам, эритромицину и хло-
рамфениколу. При тяжелых системных инфекциях следует применяют антибио-
тики из группы аминогликозидов. При локализованных абсцессах, вызываемых
данными микроорганизмами, необходимо хирургическое лечение.
В зарубежной литературе появились многочисленные сообщения о широ-
ком распространении мультирезистентных штаммов A. baumannii. На 45-й
Междисплинарной конференции по антимикробным препаратам и химиотерапии
(ICAAC) эти штаммы получили название «грамотрицательные MRSA». Также
существует мнение, что инфицирование ран A. baumannii может способствовать
последующему развитию инфекции, обусловленной MRSA.
Agrobacterium spp. — прямые или слегка изогнутые палочки. Расположены один оч-
но или попарно. Нетребовательны к питательным средам. Оксидазоположительны,
синтезируют уреазу, не образуют индол. Некоторые виды патогенны для расте-
ний — вызывают опухолевые образования у растений. Вследствие способности
A. tumefaciens трансформировать растительные клетки данная бактерия активно
используется для привнесения генетического материала с целью генетической
модификации растений. A. radiobacter — единственный вид. который не вызывает
тканевых разрастаний на стеблях растений. В редких случаях эти бактерии способ-
ны вызывать инфекции у пациентов с иммунодефицитом (особенно при наличии
постоянного катетера).
Alcaligenes — короткие палочки или коккобактерии, перетрихий, спор и капсул
не образуют. Строгие аэробы, однако некоторые штаммы могут существовать в
анаэробных условиях в присутствии нитратов или нитритов. Продуцируют ката-
лазу и оксидазу. Индол не образуют, углеводы не ферментируют. К питательным
средам нетребовательны, хорошо растут на простых средах, образуя колонии,
сходные с колониями энтеробактерий. Широко распространены в природе.
Встречаются в воде, почве, пищевых продуктах. Подразделяются на две. группы —
несахаролитические и сахаролитические.
Из числа несахаролитических видов наибольшее медицинское значение имеет
A.faecalis (фекальный щелочеобразователь). Входит в состав микрофлоры кишеч-
ника. При посеве фекалий образует на дифференциальных питательных средах
лактозонегативные колонии, сходные с колониями патогенных энтеробактерий.
От последних A.faecalis легко отличить по росту на трехсахарном агаре, сопрово-
ждаемом защелачиванием столбика и скошенной части среды.
A. odorans черезвычайно схож с A. faecalis, что заставляет некоторых авторов рас-
ценивать его в качестве биовара, в связи с чем название микроорганизма часто заклю-
чают в кавычки — «А. odorans». Отличается от A. faecalis способностью к выделению
фруктового запаха при росте на плотных питательных средах и формированием зеле-
ной зоны вокруг колоний при росте на кровяном агаре. К данной группе также при-
надлежит A. denitrificans. Эти микроорганизмы нередко обнаруживают в различных
видах исследуемого материала при септицемиях, инфекциях желче- и мочевыводя-
щих истей. Однако чаще всего они являются случайными контаминантами.
384 ЧАСТНАЯ МИКРОБИОЛОГИЯ
К числу сахаролитических видов принадлежит Alcaligenes xylosoxidans subsp.
xylosoxidans. Они способны ферментировать глюкозу и ксилозу, но не окисляют
мальтозу. Через сутки культивирования образуют на плотных средах очень мелкие
колонии, которые резко увеличиваются в размере через 48 ч вследствие продукции
слизи. Alcaligenes xylosoxidans subsp. xylosox'dans рассматривают как относительно
частый этиологический агент при внутрибольничных септицемиях. Причина
больничных вспышек — инфузионные растворы, датчики давления и растворы
антисептиков. Эти микроорганизмы обычно чувствительны к пиперациллину и
цефтазидиму и резистентны к аминогликозидам, ципрофлоксацину и цефепиму.
Chryseomonas luteola — грамотрицательные, подвижные, оксидазоотрицатель-
ные палочки. Нетребовательны к питательным средам. Образуют гладкие или
шероховатые колонии, иногда колонии желтого цвета, прикрепленные к среде.
Трехсахарный агар при росте не изменяют. Широко распространены в окру-
жающей среде. Часто могут быть выделены из различных объектов в стацио-
нарах. Их связывают с инфицированием катетеров, сепсисом и перитонитом на
фоне перитониального диализа, а также возникновением смешанных инфекций.
Чувствительны к широкому кругу цефалоспоринов, аминогликозидов, тетраци-
клинов и хинолонов.
Comamonas spp. — грамотрицательные палочки, подвижные благодаря поляр-
но расположенным жгутикам. Обладают оксидазой активностью, редуцируют
нитраты, но без образования газообразных продуктов. Не продуцируют кислоту
из глюкозы и других углеводов (С. acidovorans ферментирует фруктозу и маннит на
среде Хью-Лейфсона, чем отличается от прочих видов). С. acidovorans и С. testos-
terone способны к росту на минеральной среде с ацетатом в качестве единственного
источника углерода. С. terrigena нуждаются в факторах роста — метионине и нико-
тинамиде. Сапрофиты, обитатели воды и почвы. Однако могут быть выделены из
пищевых продуктов, госпитальной среды, клинических образцов, но, как правило,
не имеют самостоятельного этиологического значения. Описаны отдельные слу-
чаи септицемии, эндокардитов, внутрисосудистой инфекции, связанные с данны-
ми микроорганизмами.
Flavimonas oryzihabitans — грамотрицательные подвижные палочки. Нетре-
бовательны к питательным средам. На плотных средах образуют колонии желтого
цвета. Трехсахарный агар при росте не изменяют. Оксидазоотрицательны. Эскулин
не гидролизуют. Нитрат, как правило, не восстанавливают. Возможно, имеют
этиологическое значение при ассоциированных с применением катетеров инфек-
циях, перитонитах на фоне перитониального диализа. Чувствительны к широкому
кругу цефалоспоринов, аминогликозицов, тетрациклинов и хинолонов. Широко
распространены во внешней среде. Могут быть выделены из различных видов кли-
нического материала, вв лючая кровь, мочу и отделяемое слизист ых оболочек глаз.
В условиях стационаров F. oryzihabitans связывают с инфицированием катетеров,
сепсисом и перитонитом на фоне перитониального диализа, а также возникнове-
нием смешанных инфекций. Чувствительны к широкому кругу цефалоспоринов,
аминогликозидов, тетрациклинов и хинолонов.
Flavobacterium/Chryseobacterium — таксономическое положение многих видов
микроорганизмов, входящих в состав семейства Flavobacteriaceae, в настоящее
время пересмотрено. Представители рода Flavobacterium, имеющие клиническое
значение, включены в состав нового рода, получившего название Chryseobacterium.
Наибольшее медицинское значение имеют виды С. meningo^epticum и С. indologenes,
меньшее — С. balustinum. С. gleum, С. indoltheticum и С. scopbihalmum.
Chryseobacterium spp. — грамотрицательные, неподвижные, длинные, тонкие,
немного изогнутые палочки. Ката л азо- и оксидазоположительны, гидролизуют
желатин и эскулин. Все штаммы Chryseobacterium spp. дают слабоположительную
реакцию на индол, в связи с чем для их выявления следует использовать метод
ЧАСТНАЯ МИКРОБИОЛОГИЯ 385
Эрлиха как наиболее чувствительный, Chryseobacterium хорошо растут на про-
стых питательных средах, кровяном и шоколадном агаре, образуя колонии уже
в течение 24 ч при температуре 35-37 °C, образуя желтый или оранжевый пиг-
мент различной интенсивности. Колонии С. meningosepthum крупные (диаметром
1-2 мм), с гладкой поверхностью, чаще имеют бледно-желтую окраску. Колонии
С. indologenes — насыщенного темно-желтого цвета. Способность к росту на средах
с желчью и желчными кислотами вариабельна. Гемолитическая активность раз-
лична. При росте на трехсахарном агаре могут вызывать закисление среды через
3-7 сут.
Микроорганизмы рода Chryseobacterium широко распространены в окружающей
среде (их обнаруживают в почве, воде), а также в различных пищевых продуктах
(сыром мясе, молоке). В стационарах они контаминируют различные объекты и
поверхности. Присутствие Chryseobacterium в клиническом материале, как прави-
ло, представляет собой колонизацию, а не инфекцию, однако они могут быть при-
чиной инфекционного процесса, в первую очередь у ослабленных больных.
Иск точение составляет С. meningosepticum, вызывающи менингит у новорож-
денных, преимущественно недоношенных детей, в первые 2 нед жизни. Описаны
нозокомиальные вспышки менингита у новорожденных, находящихся в отделе-
ниях реанимации и интенсивной терапии. С. meningosepticum может вызывать у
взрослых бактериемию, которая чаще всего имеет транзиторный характер, однако
крайне редко у госпитализированных взрослых пациентов С. meningosepticum
может вызывать эндокардит (особенно у больных с протезированными клапана-
ми), целлюлит, раневые и интраабдоминальные инфекции, эндофтальмит, синусит
и бронхит.
Chryseobacterium обладают природной устойчивостью к аминогликозидам,
пенициллинам, цефалоспоринам, карбазепамам, тетрациклину, хлорамфениколу,
чувствительны к ванкоми) щну, рифампицину, клиндамицину.
Kingetta — род, объединяющий короткие кокковидные палочки, расположенные
парами или в цепочках, неподвижные. Ферментируют глюкозу (к среде реко-
мендуют добавить 1-2 капли кроличьей сыворотки на 3 мл среды). К. kingae, в
отличие от К. denifrificans, также ферментируют мальтозу. Оксидазоположите тьны,
каталазоотрицательны. Требовательны к питательным средам. На кровяном агаре
К. kingae, как правило, дают гемолиз. К. denitrificans часто образуют ямки в агаре.
Кингеллы часто обнаруживают в мазках из зева. К. denitrificans клинического зна-
чения не имеют. К. kingae — условно патогенный микроорганизм, представляющий
опасность прежде всего для маленьких детей. Большинство штаммов чувствитель-
ны к различным антибактериальным препаратам, включая бензилпенициллин.
Moraxella, род семейства Neisseriaceae, — грамотрицательные палочко- или кок-
ковидные неподвижные бактерии, расположены в мазках парами или короткими
цепочками. Отдельные виды (М. catarrhalis) имеют фимбрии, что обеспечивает
«дергающую* подвижность. Все моракселлы образуют капсулу. На кровяном и
шоколадном агаре растут в виде очень мелких гладких или шероховатых колоний
с зоной гемолиза или без нее. Сахара не ферментируют, индол и сероводород не
образуют. Синтезируют каталазу и оксидазу. Для моракселл характерна высокая
чувствительность к fi-лактамам, за исключением М. catarrhalis, продуцирующих
р-лактамазу. Межвидовая дифференциация моракселл достаточно сложна, тру-
доемка и длительна (табл. 21-24). Бактерии рода Moraxella — представители нор-
мальной микрофлоры слизистых оболочек верхних дыхательных путей человека
и некоторых животных, однако в ряде случаев могут быть причиной различных
заболеваний.
М. catarrhalis вызывает гнойный отит, острые синуситы, хронические бронхиты,
внебольничные пневмонии. Микроорганизм часто выделяют из мокроты больных
с хронической обструктивной болезнью легких и рассматривают как одну из воз-
386
ЧАСТНАЯ МИКРОБИОЛОГИЯ
можных причин его обострения. Описаны случаи менингитов и эндокардитов,
вызываемых М. catarrhalis.
М. lacunata вызывает конъюнктивит наружных углов глаза (конъюнктивит
Коха-Уикса).
Таблица 21-24. Свойства видов рода МогахеНа, встречаемых у человека
ВИД Уреаза на среде Кристен- сена Утилизация цитрата на среде Симмонса Фенил- аланин дезами- наза Желати- наза Редукция Рост в присут- ствии 5% желчи Гемолиз
нитра- тов нитри- тов
М. lacunata — - -/+ ± + - - -/+
М. lincolnii - — - - - - - -/+
М. nonliquefaciens - - - - + - - -
М. phenylpyruvica + - + - + - + -
М. osloensis - - - + - - -
М. atlantae - - — - - - + -
М. catarrhalis - - + - + + - -/+
Oligella - мелкие грамотрицательные кокковидные палочки. О. ureolytica под-
вижна, О. urethralis неподвижна и плохо обесцвечивается при окраске по Граму.
Сахара не ферментируют. Оксидазо- и каталазоположительны. Гемолитическая
активность варьирует у разных штаммов. Oligella spp. выделяются из мочи и поло-
вых органов, при этом Oligella ureolytica наиболее часто обнаруживают у мужчин
(предполагается участие этой бактерии в патогенезе уретритов).
Род Pseudomonas объединяет грамотрицательные, подвижные, не образую-
щие спор и капсул, оксидазо- и каталазопозитивные палочки. Аэробы, однако в
некоторых случаях способны использовать нитрат в качестве акцептора электро-
нов и, следовательно, расти в анаэробных условиях. Большинство псевдомонад
нетребовательны к питательным средам и не нуждаются в органических факторах
роста. Некоторые виды — факультативные автотрофы, способные использовать в
качестве источника энергии водород и углекислый газ. Многие псевдомонады спо-
собны к синтезу пигментов, из которых наиболее известен пиоцианин — пигмент
синегнойной палочки. Микроорганизмы данного рода чрезвычайно широко рас-
пространены в природе. Многие из них — представители аутохтонной микрофло-
ры почвы. Среди них встречаются бактерии, патогенные для растений, животных
и человека. Род Pseudomonas объединяет свыше 20 видов, многие из них способны
вызывать заболевания у людей. Однако наибольшее медицинское значение имеет
Р. aeruginosa — возбудитель синегнойной инфекции. Остальные псевдомонады,
которые могугг быть выделены при различных формах патологических процессов,
принято называть необычными (см. ниже).
Р. aeruginosa — грамотрицательная палочка с одним, реже двумя жгутиками,
расположена в мазках из чистой культуры одиночно, парами или короткими
цепочками. Капсула отсутствует, однако при определенных условиях культивиро-
вания способна вырабатывать капсулоподобное вещество — внеклеточную слизь,
которая тонким слоем окружает микробную клетку. Из мокроты часто выделяют
мукоидные штаммы, образующие повышенное количество слизи независимо от
условий культивирования. Внеклеточная слизь — один из факторов патогенности
Р, aeruginosa. Как и многие псевдомонады, Р. aeruginosa по потребностям в пита-
тельных веществах приближается к олигокарбофилам. Растет при минимальных
количествах органических веществ. Р. aeruginosa способна размножаться в раз-
личных растворах медицинского назначения, а также длительно сохраняться
в некоторых видах антисептических растворов, например фурацилина. Однако
ЧАСТНАЯ МИКРОБИОЛОГИЯ 387
синегнойная палочка высокочувствительна к высушиванию- действию высокой
температуры и хлорсодержащих дезинфектантов.
Наиболее важная биологическая особенность Р aeruginosa — способность к
продукции водорастворимого сине-зеленого пигмента — пиоцианина. Также пода-
вляющее большинство штаммов Р. aeruginosa синтезируют флуоресцин — пигмент,
флюоресцирующий в ультрафиолетовых лучах. В отличие от пиоцианина способ-
ностью к синтезу флуоресцина обладают и другие псевдомонады, что снижает диа-
гностическую ценность данного признака. Отдельные штаммы Р. aeruginosa могут
вырабатывать и другие пигменты — красный (пиорубин), черный (пиомеланин),
желтый (а-оксифеназин) либо не синтезировать пигменты вообще. Отсутствие
пигмента, а также нетипичные пигменты резко затрудняют диагностику. Доля
беспигментных штаммов Р. aeruginosa составляет до 18%, атипичные пигменты
образуют 6-37% штаммов. Для усиления способности Р. aeruginosa к продукции
пигментов используют специальную питательную среду «Кинг А», в состав кото-
рой входят пептон, глицерин, агар и минеральные соли. Некоторые штаммы синег-
нойной палочки образуют на этой среде пигмент красного цвета. На поверхности
жидких питательных сред Р aeruginosa образует серовато-серебристую пленку.
На плотных средах синегнойная палочка образует колонии 5 типов:
• плоские колонии неправильной формы
• ишерихиоподобные колонии;
• складчатые колонии («цветок маргаритки»):
• слизистые, редко дающие пигментацию при первичном выделении, образую-
щие слизистый налет, со временем приобретающие зеленую окраску;
• карликовые, которые проявляются только через 18 ч.
На среде Эндо Р. aeruginosa образует колонии бледно-розового цвета, небольших
размеров. На кровяном агаре ее колонии окружены зоной гемолиза. Очень часто
можно наблюдать феномен «радужного лизиса» культур, который характеризу-
ется наличием нежного блестящего металлического налета и зон лизиса. Данный
феномен настолько специфичен для Р. aeruginosa, что его можно использовать
для ее идентификации. Отдельные штаммы отличаются друг от друга по виду зон
«радужного лизиса», что делает данный Феномен полезным для внутривидового
типирования. Посевы синегнойной палочки имеют специфический запах, который
сравнивают с запахом земляничного мыла, жасмина или карамели.
На селективном псевдомонадном агаре с иргазаном Р. aeruginosa образует в
первые 18 ч инкубации зеленоватые колонии, которые в последующем становятся
сине-зелеными.
На агаровых средах с цетримидом (бромидом цетилтриметиламмония) усили-
вается образование синегнойной палочкой пиоцианина, что облегчает идентифи-
кацию ее колоний по желто-зеленому или синему цвету.
Синегнойная палочка обладает низкой сахаролитической активностью, фер-
ментируя только глюкозу. Протеолитическая активность, напротив, сильно выра-
жена. Р aeruginosa имеет сложную антигенную структуру, обладает О-антигеном, а
также жгутиковым Н-антигеном. В целях стандартизации методов серотипирова-
ния была разработана международная схема, предусматривающая подразделение
Р. aeruginosa пи О-антигену на 17 серогрупп. Р. aeruginosa — один из основных воз-
будителей гнойно-воспалительных процессов, особенно в условиях стационара.
Инфекции, вызываемые Р aeruginosa, потенциально опаснее вызванных другими
условно патогенными бактериями. Они развиваются у пациентов с ожогами,
острым лейкозом, муковисцидозом, находящихся по разным причинам на искус-
ственной вентиляции легких. Инфекция обычно локализуется в местах скопления
и застоя жидкости: в трахеостомах, нижних отделах легких, постоянных катетерах
мочевого пузыря, мокнущих ранах. Актуальна проблема колонизации Р. aeruginosa
сосудистых катетеров. При любой локализации первичного очага инфекции, обу-
388 ЧАСТНАЯ МИКРОБИОЛОГИЯ
словленной Р. aeruginosa, возможно развитие бактериемии. Инфекции, вызванные
синегнойной палочкой, плохо поддаются лечению вследствие множественной ее
резистентности, передаваемой R-плазмидами.
Необычные псевдомонады. Отличаются от описанных выше тем. что могут вызы-
вать инфекцию у людей только при значительном снижении уровня естественной
резистентности. К их числу относятся P.fluorescens, Р. stutzeri и Р maltophilia.
Клиническое значение Р. fluorescens определяется двумя обстоятельствами —
способностью размножаться при температуре 4 °C и колонизировать кожу здо-
ровых людей. Возможна случайная контаминация Р. fluorescens донорской крови.
При этом количество бактерий в крови будет увеличиваться, несмотря на хранение
в холодильнике, что может привести к токсическому шоку при гемотрансфузии.
Однако в большинстве случаев обнаружение Р. fluorescens следует расценивать
как случайную контаминацию. Одновременное обнаружение в крови нескольких
пациентов Р. fluorescens или Р. stutzeri может свидетельствовать о контаминации
инъекционных растворов в стационаре.
Р. stutzeri — редкая причина заболеваний человека. Описаны случаи обнару-
жения этого микроорганизма при бактериемии у больных с иммунодефицитом,
пневмониях у алкоголиков и пациентов, находящихся на гемодиализе вследствие
контаминации им воды аппаратов.
Р. maltophilia — вторая по распространенности, после синегнойной палочки,
псевдомонада. Встречается в воде, почве, на овощах. Является частым контамина-
тим дистиллированной воды и растворов, в том числе некоторых дезинфектантов.
В госпитальных условиях часто выделяется от больных на фоне антибиотикоте-
рапии. Обычно ее обнаруживают при исследовании отделяемого из дыхательных
путей, влагалища, мочи, гноя, крови в ассоциации с другими микроор!анизмами.
Считают, что патогенетическое значение Р. maltophilia сомнительно.
К основным группам антибактериальных препаратов, обладающих клинически
значимой антипсевломонадной активностью, относятся [3-лактамы, аминоглико-
зиды и фторхинолоны.
Xantomonas (Stenotrophamonas) maltophilia. Систематическое положение данного
вида бактерий неоднократно подвергалось пересмотру. Первоначально он был
отнесен к роду Pseuaomonas (Pseudomonas maltophilia). Позже его переместили в рпд
Xantomonas на основании данных ДНК-гибридизации. Однако из-за существенных
отличий в биологических свойствах (лофотрих, не имеющий желтого пигмента
ксантомонанедина, непатогенен для растений, растет при 37 °C) предложено
выделить его в отдельный род Stenctrophomonas. X. maltophilia — короткие прямые
палочки с полярным пучком жгутиков. Формируют большие гладкие колонии с
неровными краями. На плотной среде с тирозином образуют коричневый водо-
растворимый пигмент, на кровяном агаре под зоной роста появляется зеленовато-
обесцвеченная зона. Рост большинства штаммов нуждается в метионине. Для изо-
ляции X. maltophilia применяют селективные среды с имепенемом.
КАМПИЛОБАКТЕРЫ
ТАКСОНОМИЯ
Семейство Campylobacteraceae объединяет е-протеобактерии двух родов:
Arcobacter и Campylobacter. Наиболее значимы в патологии человека термофиль-
ные кампилобактеры — С. jejuni, С. coli и С. lari, входящие в состав последнего из
названных родов.
ОБЩАЯ ХАРАКТЕРИСТИКА
Кампилобактеры представляют собой тонкие, изогнутые (от легкого загиба до
спиралеобразной формы) грамотрицательные бациллы. Концы бактериальных
ЧАСТНАЯ микробиология 389
клеток обычно утончены. В мазках культур они расположены одиночно, в цепоч-
ках зигзагообразной или спиральной формы разной длины, а также парами в
форме буквы S или крыльев летящей чайки. При контакте с воздухом, воздействии
повышенной температуры и старении культур бактерии округляются. Размер бак-
териальных клеток С. jejuni и С. coli составляет 0,5-5,0 х 0,2-0,5 мкм. Они имеют
на одном или обоих полюсах по жгутику, обеспечивающих им стремительное
перемещение в жидких средах по винтообразной траектории.
Кампилобактеры — микроаэрофилы и капнофилы. С. jejuni, С. coli и С. lari
относят к группе термофильных кампилобактеров, так как они лучше растут при
температуре 37-43 °C, а при 25 °C их рост прекращается.
ЭПИДЕМИОЛОГИЯ
Природный резервуар термофильных кампилобактеров — различные живот-
ные и птицы, в том числе сельскохозяйственные. Частота обнаружения С. jejuni у
кур колеблется от 70 до 100%, и в экономически развитых странах именно куры
рассматриваются как основной источник инфекции. В странах с низким уровнем
жизни на первом месте находится водный путь передачи инфекции. Отсутствие
достоверной информации не позволяет сделать какие-либо выводы о характере
эпидемического процесса в России, но отмечается выраженная гиподиагностика
кампилобактериоза.
КЛИНИЧЕСКИЕ ПРОЯВЛЕНИЯ ИНФЕКЦИИ
Инфекция термофильных кампилобактеров наиболее часто проявляется у
человека колиэнтеритом. Непатогномоничность его симптоматики не позволя-
ет поставить окончательный диагноз на основании результатов клинического
обследования пациента. В то же время некоторые симптомы являются безуслов-
ным показанием к проведению целенаправленных исследований для выявления
данной группы микроорганизмов. К ним относится обнаружение крови и слизи
в фекалиях, то есть симптомы гемоколита. Обычно кампилобактериозный энте-
роколит протекает благоприятно и заканчивается полным выздоровлением.
Генерализация процесса как одно из грозных осложнений встречается редко (как
правило, у детей и пожилых пациентов). У пациентов с ослабленным иммунитетом
инфекция может приобретать генерализованную форму и вызывать системные
клинические нарушения.
Другой тип осложнений, регистрируемых при инфекциях термофильных кам-
пилобактеров, — аутоиммунные процессы, в основном синдром Гийена-Барре
(прогрессирующий восходящий обратимый паралич, связанный с демиелиниза-
цией нервных волокон).
ЛАБОРАТОРНАЯ ДИАГНОСТИКА
Окончательный диагноз кампилобактериозов ставят на основании обнаруже-
ния в клиническом материале возбудителя и/или его антигенов, а также участков
генома методами, которые описаны ниже. Серологическая диагностика при этих
инфекциях низкоэффективна.
Сбор и транспортировка исследуемого материала
Для исследований в лабораторию направляют пробы фекалий, мазки и смывы
из прямой кишки пациента. Последние получают с помощью зонда-тампона.
Учитывая микроаэрофильность кампилобактеров и, следовательно, их низкую
выживаемость в присутствии кислорода, зонд-тампон с пробой необходимо
погружать в транспортную среду даже при непродолжительной транспорти-
ровке. Наилучшей транспортной средой для кампилобактеров является среда
Кэри-Блэйра. Практически аналогичными свойствами обладают среды Стюарта
и Эймса. Тиогликолевая среда и 0,1% пептонная вода не являются транспортам-
390 ЧАСТНАЯ МИКРОБИОЛОГИЯ
ми средами, поскольку содержат питательные вещества, которые могут привести
к нежелательному размножению контаминантов в процессе транспортировки и
сделать невозможным выделение кампилобактеров. Сторонники использования
тиогликолевой среды для транспортировки в качестве аргумента приводят данные
об эффективности Campylo thioglycollate medium, однако данная среда содержит
селективную добавку Блейзер-Вонг и предназначена для транспортировки и
последующего селективного обогащения кампилобактеров.
Если материал доставляют в лабораторию в течение 2-4 ч, нативные фекалии
можно транспортировать в контейнере без консерванта.
Бактериоскопия
Бактериоскопический метод диагностики кампилобактериозного колиэнтерита
упоминается в различных руководствах, хотя его вряд ли можно рекомендовать
диагностическим лабораториям. Препараты, в которых кампилобактеры в иссле-
дуемом материале находятся в виде чистой культуры, — скорее исключение, чем
правило.
Обнаружение антигена хламидий в клиническом материале
Для обнаружения кампилобактеров в фекалиях можно использовать одноэтап-
ные иммунохроматографические тесты (ИХТ). Хотя они не позволяют диффе-
ренцировать выявляемые кампилобактеры на видовом уровне, их применяют для
скрининговых целей. Материалом для исследования служит суспензия фекалий.
Отрицательный результат ИХТ не исключает наличия у пациента кампилобакте-
риозной инфекции, для подтверждения этого необходимы дополнительные иссле-
дования другими тестами.
Изоляция и идентификация возбудителя
Длительное время кампилобактеры относили к некультивируемым бактериям,
однако после определения оптимальных для них состава питательных сред и усло-
вий культивирования бактериологический метод диагностики стал основным.
Во многих зарубежных лабораториях на первом этапе бактериологического
исследования кампилобактериозов применяют обогатительные среды (Джефри,
Абейта-Ханта-Барка, обогатительный бульон с оксиразой, бульоны Болтона,
Весли, Пресли).
Выделение чистой культуры проводят с использованием одного из трех мето-
дических подходов:
• посева на селективные питательные среды;
• посева на неселективную питательную среду с помощью фильтров;
• подвижного роста на полужидкой питательной среде.
Последний способ выделения чистой культуры не нашел широкого применения
при культивации кампилобактеров.
Для ингибиции роста контаминантов в селективные среды добавляют цефо-
перазон, цефалотин, цефазолин, эритромицин, колистин, бацитрацин, поли-
миксин В, новобиоцин, колимицин, ванкомицин, рифампицин, амфотерицин В.
циклогекс емид. Поскольку некоторые виды С. jejuni и С. coli проявляют высокую
чувствительность к цефалотину, колистину и полимиксину В, в предназначенные
для работы с ними среды эти антибиотики не добавляют. Создано несколько удач-
ных селективных смесей, которые выпускаются микробиологическими фирмами
(табл. 21-25, 21-26).
Селективные среды удобны в использовании, но не в одинаковой мере при-
годны для разных кампилобактеров. Это делает актуальным, с одной стороны,
поиск новых селективных добавок, а с другой — развитие альтернативных методов
выделения кампилобактеров.
ЧАСТНАЯ микробиология 391
Альтернатива селективных сред — фильтрационная техника выделения. Описано
использование мембранных и ядерных фильтров с диаметром пор 0,45-0,55 мкм.
Мембранные фильтры хорошо работают при высоких концентрациях кампило-
бактеров и резко уступают селективным средам по чувствительности при иссле-
довании реконвалесцентов и пациентов с легкими и субклиническими формами
заболевания. Это связано с особенностями работы данных фильтров и их высокой
сорбционной способностью. Попытка повысить эффективность за счет увеличе-
ния размера пор (до 0,65 мкм), как правило, сопровождается снижением селек-
тивности и обильным ростом сопутствующей микрофлоры. Ядерныо мембраны
лишены недостатков, присущих мембранным фильтрам. Они имеют стандартные
по диаметру короткие прямые поры, не обладают сорбционной способностью и
более эффективно фракционируют бактерии по размеру. Однако они могут сильно
электризоваться, что затрудняет работу с ними. Ядерные мембраны изготавли-
ваются на заказ и поступают в виде полотна, которое в лаборатории необходимо
нарезать, упаковать и стерилизовать.
Независимо от типа используемых фильтров технология посева одинакова. На
неселективную среду накладывают стерильный фильтр. На поверхность филь-
тра стеклянной палочкой или пипеткой наносят капли суспензии исследуемого
материала в транспортной среде или стерильном 0,9% растворе натрия хлорида
(1:5-1:10). Через 30 мин фильтры аккуратно удаляют, не допуская попадания
исследуемого материала на питательную среду.
Таблица 21-25. Селективные добавки для кровяного агара для кампилобактеров
Добавка Sklrrow Blaser-Wang Butzler Preston
Полимиксин В, МЕ/л 2500 2500 5000
Колистин, МЕ/л 10 000
Цефалотин, мг/л 15
Цефазолин, мг/л 15
Ванкомицин, мг/л 10 10
Бацитрацин, МЕ/л 25 000
Рифампицин мг/л 10
Триметоприм, мг/л 5 5 10
Амфотерицин В, мг/л 2 10 10
Таблица 21-26. Селективные добавки для угольных сред для кампилобактеров
Доьаака CCDA CAT Karmali
Цефоперазон. мг/л 32 8 32
Тейкопланин, мг/л 4
Ванкомицин, мг/л 20
Амфотерицин В, мг/л 10 10 10
В качестве ростовых сред применяют содержащие (например, среду Скирроу) и
не содержащие (среды Кармаля, Болтона и др.) кровь.
Наиболее перспективны угольные среды, поскольку их применение позволяет
отказаться от использования дорогостоящей и дефицитной овечьей крови.
Другая альтернатива кровяных сред — хромогенные среды, разработка которых
активно ведется многими фирмами.
Оптимальная газовая среда для кампилобактеров содержит 5-7% кислорода,
10% углекислого газа и 83-85% азота. В некоторых руководствах рекомендуют
добавлять до 10% водорода, но для возбудителей кишечных форм кампилобак-
териоза в этом нет необходимости. Для создания указанной атмосферы широко
392 ЧАСТНАЯ МИКРОБИОЛОГИЯ
применяют газогенерирующие пакеты различных конструкций. В зарубежной
практике используются каталитические системы с гидроборидом натрия в каче-
стве источника атомарного водорода, который затем соединяется с кислоцодом
воздуха на катализаторе, или бескаталитические сухие системы на основе аскор-
биновой кислоты. Данные газогенераторы не нашли в нашей стране широкого
применения из-за их высокой стоимости. Отечественные газогенерирующие паке-
ты, работающие по принципу пирогаллолового поглотителя, позволяют добиться
хорошего результата при существенно меньших затратах.
Использование химических газогенераторов позволяет обойтись упрощенными
микроанаэростатами, не имеющими манометров и клапанов. Такие устройства
очень удобны в работе, они надежно и легко герметизируются, долговечны и зна-
чительно дешевле классических анаэростатов.
Температура инкубации зависит от поставленных задач и используемой среды.
Температура 37 °C теоретически позволяет выделить не только термофильные
кампилобактеры. С другой стороны, при темперач уре 42-43 °C достигается допол-
нительный селективный эффект и упрощается последующая идентификация выде-
ленных кампилобактеров. Инкубацию на среде Карма ли следует проводить при
температуре 42 °C, в то время как на CCDA-arape кампилобактеры лучше растут
при температуре 37 °C. Обычно время инкубации составляет 48 ч, однако иногда
рекомендуют просматривать посевы первый раз через 24 ч. Такая тактика повы-
шает стоимость исследования, поскольку требует использования дополнительного
газогенерирующего пакета.
Кампилобактеры могут образовывать на плотных средах два типа колоний.
Нежные, влажные, неправильной формы крупные колонии, имеющие тенденцию
к слиянию и напоминающие капли конденсата, легко различимы даже на фоне
обильного роста сопутствующей микрофлоры. Они образованы типичными по
морфологии клетками кампилобактеров. Их обнаружение на специальной среде,
инкубируемой в специальных условиях, позволяет с очень большой долей вероят-
ности уже на данном этапе говорить о выделении бактерий рода Campylobacter, а
если инкубация проводилась при 42 °C — о термофильных кампилобактерах.
Возможно образование и колоний другого типа. Мелкие, выпуклые правильной
округлой формы колонии образованы в основном кокковидной формой камлило-
бактеров. В мазке можно обнаружить небольшое количество типичных по мор-
фологии бактериальных клеток и сделать вывод о выделении кампилобактеров.
Как правило, закончить идентификацию в данном случае не удается, поскольку
невозможно провести субкультивирование таких ш гаммов.
Для изучения морфологических свойств хорошо подходит окраска фvкcинoм,
приготовленным из карболового фуксина Циля путем разведения его водой в
соотношении 1:3-1:5. Вместо окраски по Граму с успехом применяют КОН-тест.
Иногда при обильном росте колоний прямо в чашке Пе гри с первичным посевом
можно поставить оксидазный и каталазный тесты общепринятым методом. Для
завершения идентификации необходимо пересеять культуру на неселективный
агар. Здесь предпочтительнее использовать кровяной агар (пусть даже с донорской
кровью), поскольку угольная среда не всегда пригодна для субкультивирования.
Например, гемин и пируват, входящие в состав среды Кармали, находятся не в
основе среды, а в селек гивной добавке. Лучше, если кровяной агар разлит и ско-
шен в пробирках, однако приготовление скошенного кровяного агара не является
обычной лабораторной практикой и поэтому чаще пересев проводят на сектора
в чашке Петри (с этой целью удобно пользоваться полисекционными чашками).
Не рекомендуют делить чашку более чем на 6 секторов. Штрихи желательно рас-
полагать близко друг к другу, чтобы по крайней мере в начале сектора получить
обильный сливной рост. После получения культуры можно продолжить иденти-
фикацию. Поскольку в большинстве случаев первичные посевы инкубируются при
частная микробиология зэз
42 °C, в температурном тесте нет необходимости. Ключевой идентификационный
тест — тест на гидролиз гиппурата. На нем необходимо остановиться подробно,
поскольку в последнее время был выпущен ряд документов, в которых выполне-
ние данного теста описано неправильно.
Для выполнения теста необходимо приготовить 1% раствор натрия гиппурата
в 0,9% растворе натрия хлорида. Полученный раствор разливаю! в пробирки по
0,4 мл. Пробирки закрывают пробками и замораживают в морозильной каме-
ре бытового холодильника (-18 °C) Герметично закрытые пробирки можно
хранить до года. Большое значение имеет выбор пробирок. Это должны быть
узкие пробирки, очень удобно использовать полипропиленовые пробирки типа
«Эппендорф». По мере надобности пробирки размораживают и используют для
выполнения теста. Для этого в растворе гиппурата суспендируют испытуемую
культуру. Необходимо получить густую суспензию (как правило, достаточно
2 петель культуры). Пробирки помещают в термостат при температуре 37 °C на 2 ч.
Пока идет инкубация, готовят проявляющий раствор нингидрина. По окончании
инкубации на суспензию бактерий в водном растворе гиппурата надо очень осто-
рожно наслоить 0,2 мл нингидринового реактива. Верхний и нижний слой нельзя
перемешивать! Пробирки помещают в тепмостат и инкубируют в течение 10 мин.
В случае положительного результата наблюдается интенсивное фиолетовое окра-
шивание верхней (неводной) фазы. Если окрашивания нет, результат оценивают
как отрицательный. При слабом окрашивании рекомендуют повторить тест,
увеличив плотность бактериальной суспензии. Окрашивание всего содержимого
пробирки свидетельствует об очень высокой активности фермента или о ложно-
положительном результате реакции из-за перемешивания растворов. Для диффе-
ренцировки данных вариантов рекомендуют после наслаивания нингидринового
реактива просматривать пробирки через 3, 5 и 10 мин. При ложноположительной
реакции окраска развивается сразу во всем объеме раствора, а при резко положи-
тельном результате сначала появляется окраска в виде кольца и лишь затем это
кольцо, расширяясь, распространяется на весь объем раствора. Показания цветовой
реакции неспецифичны, поскольку нингидрин взаимодействует не только с глици-
ном, продуктом расщепления гиппурата, но и с любой другой аминокислотой.
Положительный тест с гиппуратом свидетельствует о выделении С. jejuni.
В случае отрицательного результата необходимо продолжить исследование для
дифференцировки гиппуратотрицательных термофильных кампилобактеров —
С. coh и С. lari. Для их идентификации используют тест гидролиза индоксилаце-
тата. Применяемое в прошлом определение устойчивости к налидиксовой кислоте
утратило свою актуальность из-за распространения резистентных к данному пре-
парату штаммов С. coli.
Таким образом, для идентификации термофильных кампилобактеров в боль-
шинстве случаев достаточно всего нескольких простых пестов, доступных любой
лаборатории (табл. 21-27).
Таблица 21-27. Дифференциация термофильных кампилобактеров
Тесты С. jejuni C. coli C. lari
ssp. jejuni ssp. doylei
Гидролиз гиппурата + 4- - -
Устойчивость к налидиксовой кислоте S [ s SR R
Устойчивость к цефалотину s R S R
Гидролиз индоксилацетата + + + -
Используемые для идентификации кампилобактеров тесты основаны на выяв-
лении конституциональных (пресинтезированных) ферментов. Это облегчает
идентификацию, поскольку не требует длительной инкубации.
394
ЧАСТНАЯ МИКРОБИОЛОГИЯ
Коммерческие тест-системы для идентификации кампилобактеров не нашли
широкого применения. Api Campy — практически единственный пример таких
систем. Он состоит из двадцати микропробирок, собранных в стрип обычного для
систем Api-формата. Стрип разделен на две части. Первая часть содержит обыч-
ные тесты для определения ферментативной активности (уреаза, нитратредуктаза,
индоксил эстераза, гиппураза, гамма глютамил трансфераза, ТТХ-редуктаза, пиро-
лидонил ариламидаза, аргинин ариламидаза. аспартат арила мидаза, щелочная
фосфатаза, продукция сероводорода), вторая — ассимиляционные тесты и тесты
на резистентность (ассимиляция глюкозы, сукцината, ацетата, малата, пропиона-
та, цитрата, чувствительность к налидиксовой кислоте, цефалотину и эритромици-
ну). Такой широкий спектр определяемых свойств позволяет надежно идентифи-
цировать практически любую культуру кампилобактеров. Однако в большинстве
случаев получаемая информация избыточна, учитывая высокую стоимость тест-
системы, ее вряд ли можно рекомендовать практическим лабораториям.
Экспресс-иммунодиапюстика
Существующие тест-системы, принцип действия которых основан на обнару-
жении антигена кампилобактеров в исследуемом материале, можно разделить на
2 группы:
• тест-системы для ИФА в классическом формате:
• одноэтапные иммунохроматографические тесты.
ИФА не нашел широкого применения из-за его невысокой чувствительности при
обнаружении антигена кампилобактеров в фекалиях. Иммунохроматографические
тест-системы также малоприменимы, когда необходимо прямое исследование
фекалий. Однако данные тест-системы очень хорошо зарекомендовали себя при
обнаружении антигена в среде обогащения и широко применяются в санитарно-
бактериологических исследованиях пищевых продуктов.
Молекулярно-генетическая диагностика
В последние годы в качестве ускоренного метода диагностики различных
инфекционных процессов все чаще рекомендуют использовать генетические мето-
ды (ПЦР, NASBA). При расследовании вспышек данные методы могут оказаться
незаменимыми, в перспективе не исключается их применение в повседневной диа-
гностической практике.
Серологическая диагностика
Методы серологической диагностики никогда широко не применялись при
диагностике кампилобактериозов. Основными препятствиями служат большое
антигенное разнообразие возбудителей, быстрое и доброкачественное течение
заболевания, отсутствие достоверного нарастания титров антител у значительной
части больных.
Используемые в серологической диагностике кампилобакгериоза тест-системы
для ИФА или непрямой РИФ позволяют определять как общее содержание анти-
тел, так и количество различных их классов (IgG, IgM. IgA). Поскольку основной
возбудитель компилобактериоза — С. jejuni, диагностические наборы рассчитаны
на обнаружение антител именно к данному возбудителю, реже — суммарно к
С. jejuni и С. coh.
ХЕЛИКОБАКТЕРЫ
ТАКСОНОМИЯ
Согласно последней редакции руководства «Bergey’s Manual of Systematic
Bacteriology» (2009), семейство Campylobacteraceae, так же как и семейство
ЧАСТНАЯ МИКРОБИОЛОГИЯ 395
Helicobacteraceae, относится к е-протеобактериям. Семейство Hellcohacteraceae
включает рода Helicobacter и WoltneUa. Из всех видов рода Helicobacter только
Н. pylori широко распространен среди населения (хотя имеются отдельные сообще-
ния о случаях инфекции у людей и других видов бактерий данного рода).
Род Helicobacter включает 23 зарегистрированных биологических вида бакте-
рий, в том числе И. pylor Наряду с официально признанными видами в различных
базах данных представлены еще десятки неклассифицированных штаммов, жду-
щих определения своего места в систематике бактерий.
ОБЩАЯ ХАРАКТЕРИСТИКА Н. PYLORI
Н. pylori — грамотрицательная бактерия, имеющая вид палочки слегка изо-
гнутой, спиралевидной или S-образной формы, длиной до 3 мкм и диаметром
0,5-1,0 мкм. При электронной микроскопии наружная стенка выглядит гладкой.
Группа жгутиков (как правило, не больше 5) расположена на одном из концов
бактерии. Жгутики покрыты чехлом, имеют концевое утолщение (луковицу).
Наличие жгутиков обеспечивает подвижность Н. pylori в жидких и гелеобразных
средах. Наряду с палочковидной формой существует кокковая, которую микро-
организм принимает при старении культуры, ее хранении и воздействии неблаго-
приятных факторов внешней среды.
ЭПИДЕМИОЛОГИЯ
Инфекция, вызванная Н. pylon, — одна из самых распространенных инфек-
ций человека. Она вызывает развитие хронического гастрита, является важ-
нейшим фактором патогенеза язвенной болезни желудка и двенадцатиперстной
кишки, лимфомы желудка низкой степени злокачественности (мальтомы). а также
некардиального рака желудка. Уничтожение (эрадикация) Н. pylori приводит к
исчезновению воспалительного инфильтрата в слизистой оболочке желудка, зна-
чительному снижению частоты рецидивов язвенной болезни желудка и двенадца-
типерстной кишки, гистологической ремиссии мальтомы желудка и, возможно, к
существенному уменьшению риска возникновения рака желудка.
КЛИНИЧЕСКИЕ ПРОЯ «ЛЕНИЯ ИНФЕКЦИИ
Н. pylori играет ключевую этиопатогенетическую роль при язвенной болезни
желудка и двенадцатиперстной кишки и хронических гастритах.
Два заболевания внежелудочной локализации этиопатогенетически связаны с
Н. pylori — идиопатическая тромбоцитопения и железодефицитная анемия неяс-
ной этиологии. При этих патологиях имеются показания к диагностике и эрадика-
ции Н. pylori (3-е Маастрихтское соглашение).
Широко обсуждалась связь инфекции, вызванной Н. pylori, с другими нозологиче-
скими синдромами, однако она не доказана. Воспалительные заболевания кишечни-
ка, аденома толстой кишки, вероятно, не связаны с инфицированием Н. pylori.
Пациент ы с синдромом раздраженного кишечника, язвенным колитом и болез-
нью Крона, как правило, реже инфицированы Н pylori по сравнению с общей
популяцией.
Заболевания сердечно-сосудистой системы, реактивный артрит, ряд кожных
заболеваний (узловатое пруриго, крапивница, некоторые формы экземы) слабо
коррелируют с инфекцией, вызванной Н. pylori.
ЛАБОРАТОРНАЯ ДИАГНОСТИКА
Ни один метод не является самостоятельным «золотым стандартом» для диа-
гностики инфекции Н. pylori. Предлагают использовать сочетание выделения
чистой культуры Н. pylori и гистологического исследования биоптата слизистой
оболочки желудка, что соответствует рекомендациям Европейской рабочей груп-
пы по исследованиям Н. pylori.
396 ЧАСТНАЯ МИКРОБИОЛОГИЯ
Методы диагностики инфекции принято подразделять на инвазивные [требую-
щие проведения эзофагогастродуоденоскопии (ЭГДС) с биопсией слизистой обо-
лочки желудка или двенадцатиперстной кишки] и неинвазивные (не требующие
бипсии). По характеру исследования тесты могут быть сгруппированы следующим
образом,
• Определение микроорганизма, его генома или продуктов жизнедеятельности
в биоптатах слизистой оболочки и других материалах от больных:
<>- бактериологическое исследование;
о гистологическое исследование;
определение антигенов Н. pylori в фекалиях;
о- быстрые уреазные тесты;
о- дыхательный уреазный тест;
о ПЦР-детекция элементов генома Н. pylori.
• Серологическая диагностика — определение специфического иммунного
ответа;
о определение антител в плазме крови;
❖ иммуноблоттинг (выявление антител к CagA- и Vac А-антигенам Н. pylori):
о- определение антител в выделениях и секретах (моче, слюне).
Из 1ляция и идентификация возбудителя
Самый достоверный метод лабораторной диагностики Н. pylori включает выде-
ление и идентификацию возбудителя. Бактериологический метод позволяет не
только идентифицировать штамм Н. pylori, но и уточнить его чувствительность
к антибиотикам, что позволяет выбрать наиболее эффективную схему лечения
пациента. Это особенно важно в тех странах, где высока частота выявления
штаммов, резистентных к метронидазолу. Широкое применение метронидазола
для лечения язвенной болезни в России в качестве «препарата с репаративной
активностью» привело к формированию крайне высокой резистентности Н. pylori
к данному препарату.
Бактериологическому исследованию подлежат биоптаты слизистой оболочки
желудка и двенадцатиперстной кишки, а также материал, полученный во время
оперативных вмешательств по поводу язвенной болезни желудка и двенадцати-
перстной кишки. Биоптаты берут во время эндоскопии в условиях строгой асепти-
ки. Пробы резецированных органов берут сразу после операции в области тела и
антрального отдела желудка, луковицы двенадцатиперстной кишки. Биопсийный
материал помещают в пробирку типа «Эппендорф» со стерильным 20% раствором
глюкозы и хранят до отправки в холодильнике при 4 °C. Необходимо следить за
тем, чтобы биоптат был полностью погружен в раствор глюкозы, так как прили-
пание его к стенке пробирки может привести к потере жизнеспособности микроба.
В растворе глюкозы Н. pylori сохраняет жизнеспособность в течение 3-4 ч, В тече-
ние этого времени биопсийный материал должен быть доставлен в лабораторию
для посева.
В качестве транспортных сред моя но использовать 0,9% раствор NaCl, тиогли-
колевую среду для контроля стерильности, транспортную сухую среду для кампи-
лобактерий. Важно помнить, что время транспортировки и обработки биопсийно-
го материала всегда ограничено и является серьезным фактором, лимитирующим
успешность выделения Н. pylori.
Биопсийный материал подлежит обработке и посеву сразу после доставки в
лабораторию. Наилучшие результаты получают при посеве не позднее 2-4 ч после
взятия биоптата. При увеличении интервала времени между взятием материала и
посевом до 5-6 ч успешность выделения Н. pylon уменьшается в несколько раз.
В стерильную фарфоровую ступку вносят 100-200 мкл стерильного 0,9% рас-
твора NaCl; стерильной микробиологической петле й или пастеровской пипеткой
ЧАСТНАЯ МИКРОБИОЛОГИЯ 397
переносят в ступку биоптат из пробирки с транспортной средой. Тщательно расти-
рают биоптат пестиком, распределяют полученный гомогенат по имеющимся чаш-
кам Петри и втирают в агар шпателем (тщательно, но не пересушивая). Остатки
биоптата переносят в пробирку с реактивом для определения уреазной активно-
сти. Каждый биоптат высевают параллельно на две чашки Петри с питательной
средой, не содержащей и содержащей антибиотики соответственно. Б первую из
упомянутых сред добавляют следующие антибиотики: ванкомицин (6 мкг/мл),
триметоприм (2 мкг/мл, в форме спиртового раствора) и амфотерицин В или
налидиксовую кислоту (2-10 мкг/мл).
В качестве основы питательных сред для выделения и культивирования Н. pylori
используют колумбийский и сердечно-мозговой агар. Перед использованием в них
добавляют 5-7%. сыворотки или дефнбринированную кровь лошади, а также 1%
раствор «IsoVitalex». Применение других питательных сред менее эффективно.
Инкубацию посевов осуществляют в микроаэрофильных условиях при концен-
трации кислорода около 5%. Для этих целей удобно использовать анаэростаты
системы «GasPaclOO* (вмещает до 12 пластмассовых чашек Петри однократного
применения) или «GasPakl50> (вмещает до 36 чашек Петри). После загрузки анаэ-
ростаты размещают в термостате при 37 °C, оптимальной для роста Н. pylori.
Срок формирования колоний при первичном высеве составляет 5-7 сут. На
кровяной питательной среде на 5-7-е сутки Н. pylori формирует мелкие, круглые,
гладкие, прозрачные, влажные колонии диаметром около 1 мм. При необходи-
мости дальнейшего изучения первичные колонии пересевают на кровяной агар,
содержащий ростовые добавки и антибиотики, и инкубируют в микроаэрофиль-
ных условиях в течение 4-6 сут при температуре 37 °C. Колонии Н. pylori, полу-
ченные в результате первичного посева биопсийного материала, используют для
приготовления мазков и окраски их по Тфаму, а также для выполнения уреазного,
оксидазного и каталазного тестов.
Решение вопроса о принадлежности выделенной культуры к роду Helicobacter
выносят на основании характерной морфологии выделенных колоний, а также
микроскопии культуры в мазке, окрашенном по Граму, и наличия характерных
биохимических свойств (продукции уреазы, каталазы и оксидазы).
Окрашенные по 1раму мазки микроскопируют под иммерсией обычным спосо-
бом. При микроскопии Н. pylori выглядят как тонкие, изогнутые, нежно-розовые
палочки. На 6-8-е сутки культивирования и при многократных пассажах могут
появиться признаки старения культуры — появляются менее изогнутые палочки,
кокковые формы.
Тест на способность к продукции каталазы выполняют следующим образом:
материал агаровой культуры суспендируют в капле 3% раствора перекиси водоро-
да на поверхности предметного стекла, через 3-5 с во взвеси каталазопозитивной
культ ры происходит образование пузырьков газа. Положительным контролем
может служить суточная агаровая культура Е. coli, отрицательным — любой стреп-
тококк.
Тест на способность к продукции цитохромоксидазы выполняют следующим
образом: каплю 1% водного раствора тетра метилпарафенилендиамина гидрох-
лорида наносят на исследуемые колонии непосредственно на поверхности агара в
чашке Петри. Через 20-30 с оксидазопозитивные колонии окрашиваются в тем-
ный цвет. Этот же тест можно выполнять на фильтровальной бумаге, на которую
петлей наносят агаровую культуру и сверху каплю реактива. Через 5-10 с пози-
тивная культура приобретает пурпупную или черную окраску. Контрилями служат
суточные агаровые культуры Е. coli (отрицательный контроль) и псевдомонад
(положительный контроль).
Тест на присутствие уреазы в биоптате или в агаровой культуре выполняют с
помощью реактивов, содержащих мочевину и индикатор. В присутствии уреазы
398
ЧАСТНАЯ МИКРОБИОЛОГИЯ
происходит гидролиз мочевины до аммиака, pH среды сдвигается в щелочную сто-
рону и при этом изменяется цвет индикатора. Стандартной средой для определе-
ния уреазной активности является бульон с 2% мочевины по Кристенсену. На этой
питательной среде реакция становится положительной через 6-24 ч. Для ускоре-
ния реакции предложен ряд модификаций: замена бульона фосфатным буфером,
увеличение содержания мочевины до 6%, использование в качестве основы гелео-
бразных субстратов. Это позволяет сократить время исследования до 15-20 мин
у 75% больных с бактериологически документированным хеликобактериозом.
В остальных случаях реакция становится положительной в течение 4-18 ч. Более
поздние реакции рассматривают как сомнительные.
Совпадение результатов бактериологического метода и уреазного теста
при исследовании биоптатов отмечают в подавляющем большинстве случаев.
Изменение цвета реактива пци внесении живой культуры Н. pylori наступает
быстро — от 5-30 с до 3 мин в зависимости от количества взятых на исследование
колоний. Более медленно, как правило, в данном тесте реагируют кокки, протеи
и другие уреазоположительные бактерии. Уреазный тест с изучаемой культу-
рой можно ставить в пробирках для микропроб типа «Эппендорф» с крышками
объемом 1.5 см или в ячейках микропланшетов для иммунологических реакций.
При использовании пробирок типа «Эппендорф» крышки необходимо закрывать
сразу же после выполнения реакции, а ячейки планшетов для иммунологических
реакций заклеивают прозрачной клейкой лентой. Нельзя оставлять пробирки
открытыми и закрывать ячейки планшетов общей крышкой, так как в этом случае
аммиак, выделяющийся в воздух из материала положительной пробы, будет рас-
творяться в жидкости соседних пробирок или ячеек, изменяя pH среды. В каждое
исследование включается контроль чистого реактива для оценки изменения его
цвета в течение срока учета уреазного теста.
При ясной клинической картине, в случаях, когда морфология выделенных
колоний и результаты окрашивания мазков по Граму не вызывают сомнении, для
решения вопроса о принадлежности выделенной культупы к Н. pylori можно огра-
ничиться только выполнением уреазного теста.
Определение чувствительности к антибиотикам
Вопрос эрадикационной терапии инфекции, вызванной Н. pylori, является
актуальным. Одна из основных причин неудач при эрадикации — резистентность
И. pylori к применяемым антибактериальным препаратам. Описана резистент-
ность И. pylori ко всем группам антибиотиков, которые используются в схемах
противохеликобактерной терапии, — макролидам, производным нитроимидазола,
р-лактамам, тетрациклинам и нитрофуранам. Основное внимание уделяется рези-
стентности к кларитромицину как основному антибиотику, используемому для
эрадикации инфекции, вызванной Я. pylori. Согласно Маастрихтскому соглаше-
нию, определение резистентности к кларитромицину рекомендуют, если устойчи-
вость Н. pylori к данному антибиотику в популяции достигает 15-20%. а также при
неэффективности двух курсов эрадикации. Механизм развития резистентности
Н. pylori к кларитромицину заключается в точечных хромосомных мутациях, при-
водящих к замене нуклеотидов в различных участках 23S рРНК, что приводит к
нарушению связывания антибиотика с клетками-мишенями. В настоящее время
описано более 20 мутаций, определяющих резистентность Н. pylori к кларитроми-
цину.
При определении чувствительности к антибиотикам применяют классические
методы исследования чистой культуры И. pylori и молекулярно-биологические
методы. Бактериологические методы определения чувствительности к антибиоти-
кам подразделяют на методы серийных разведений (в агаре или бульоне) и диффу-
зионные (диско-диффузионный и метод Е-тестов). Методы серийных разведений
ЧАСТНАЯ МИКРОБИОЛОГИЯ 399
и Е-тесты позволяют определить минимальную ингибирующую концентрацию
антибиотика. Наиболее прост и удобен в использовании диско-диффузионный
метод, который основан на регистрации диаметра зон подавления роста Н. pylori
вокруг бумажного диска с антибиотиком. Диско-диффузионный метод — полуко-
личественный и позволяет подразделить штаммы Н. pylori на три группы — hvb-
ствительные, слабочувствительные и устойчивые. Оценку результатов проводят
с использованием критериев, разработанных на основе корреляции значений
диаметров зон подавления роста и минимальной ингибирующей концентрации
антибиотика. Метод агаровых разведений и Е-тест имеют сходные результаты при
анализе чувствительности к различным антибиотикам, однако оба метода непри-
годны для контроля резистентности штаммов Н. pylor к метронидазолу. Только
метод определения чувствительности с помощью бумажных дисков оказался при-
емлемым для исследования чувствительности к макролидам и метронидазолу.
Молекулярные методы включают тесты, основанные на амплификации 23S
рРНК и гибридизации (FISH-анализ). Они позволяют быстро получить ответ,
однако имеют ряд недостатков. В ряде случаев были получены результаты, рас-
ходящиеся с диско-диффузионным методом. Результаты молекулярного иссле-
дования направлены на поиски именно той мутации, которая изучается в данной
работе. Полученные данные могут не отражать реальный уровень конкретных
мутаций, ответственных за возникновение резистентности. Оценивая чувствитель-
ность Н. pylori к кларитромицину молекулярными методами, следует учитывать,
что не всегда существует прямая зависимость между выявленной мутацией, ответ-
ственной за резистентность к кларитромицину, и фенотипическим проявлением
данной мутации.
Дифференциальная диагностика
Наиболее важна дифференциация Н. pylori от С. jejuni. Другие представители
рода Helicobacter крайне редко выделяются от человека. В отличие от кишечного
патогена С. jejuni который способен расти при высоких концентрациях желчи (до
10%). Н. pylori чувствителен к желчи. Его размножение задерживается на несколь-
ко логарифмов при наличии в составе питательной среды 0,5% желчи и полностью
прекращается при наличии 10% желчи. Н. pylori, как правило, не размножается
в присутствии 1% глицерина, его рост ингибируется при небольшом увеличении
концентрации NaCl (до 1,5%), большинство штаммов резистентны к хлориду три-
фенилтетрозолия в концентрациях до 1 мг/мл.
Дифференциальные признаки Н. pylori, Н. mustelea, С. jejuni представлены в
табл. 21-28.
Таблица 21-23. Биохимические и культуральные характеристики Н. pylori, Н. mustelea, С. jejuni
Свойства Н. pylori Н. mustelae С. jejuni
Оксидаза + + +
Каталаза + + +
Уреаза + + -
Гидролиз гиппурата - - +
Редукция нитратов - + +
Гам магл ютами лтраи спепти даза + + -
Щелочная фосфатаза + 4- +
Подвижность в бульоне + + +
Подвижность в агаре - - +
Рост в микроаэрофипьных условиях при температуре 25 °C - - -
Рост в микроаэрофипьных условиях при температуре 30-37 °C + + +
Рост в микроаэрофипьных условиях при температуре 42 °C - + +
400
ЧАСТНАЯ МИКРОБИОЛОГИЯ
Окончание табл. 21-28
Свойства Н. pylori Н. mustelae С. jefunl
Рост при содержании в атмосфере 10% углекислого газа + — +
Рост в анаэробных условиях - + +
Толерантность к 3;5% раствору натрия хлорида - - -
Толерантность к 0,5% глицина + +
Толерантность к 1% глицина - + +
Толерантность к 1% желчи - - +
Чувствительность к налидиксовой кислоте р ч ч
Чувствительность к цефалотину ч р р
Чувствительность к метронидазолу ч ч р
Гистологическое исследование
Метод гистологического исследования биоптатов широко применяют для диа-
гностики заболеваний желудочно-кишечного тракта. Открытие Н. pylori значи-
тельно увеличило количество ЭГДС с гистологическим исследованием слизистой
оболочки желудка во всем мире. При комплексной морфологической оценке
гистологической картины патолог может определить наличие Н. pylori на поверх-
ности эпителиального слоя, в подслизистом пространстве и слизи, основываясь
на типичной морфологии данного микроорганизма. Успех диагностики во многом
зависит от правильного выполнения биопсии. Одиночный образец, взятый на рас-
стоянии 2 -3 см от привратника в области малой кривизны, обеспечивает чувстви-
тельность определения Н. pylori не менее 90%, два образца (из слизистой оболочки
антрального отдела и тела желудка) значительно повышают точность результата,
4 биоптата делают ее практически 100%.
Гистологическое исследование — субъективный метод, что может приводить к
ошибкам при выявлении и идентификации Н. pylori. Однако опытные патологи,
используя шкалу оценок, предусмотренную усовершенствованной Сиднейской
классификацией гастритов, достигают специфичности определения Н. pylori на
уровне 95-98%.
Существующие методы окраски тканевых срезов по Вартину-Старри, Гимзе,
Генте обеспечивают примерно равные результаты по чувствительности и специ-
фичности определения Н. pylon. Достаточно широко применяют иммуногисто-
химический метод обработки срезов со специфическими мечеными антителами к
Н. pylori. Его предлагают считать «золотым стандартом» диагностики инфекции,
вызванной Н. pylori. Однако в сравнительных исследованиях показано, что более
простые методы окраски, в частности модифицированная окраска по Гимзе, мало
уступают иммуногистохимическому методу, более практичны и доступны.
Иммунологическая экспресс-диагностика
Методом ИФА с использованием поликлональных антител к Н. pylori можно
выявлять антигены Н. pylori в фекалиях. Его чувствительность в долечебный
период, по обобщенным данным клинических центров мира, составляет 92,4%,
специфичность — 91,9%, а в постэрадикационный период наблюдения за пациен-
тами — 88,8 и 94,5% соответственно. Этот метод применяют как для первичной
диагностики инфекции, вызванной И. pylon, так и для контроля эрадикации.
В настоящее время выпускается несколько тест-систем, основанных на прин-
ципах ИФА и иммунохроматографии. В этих тест-системах использованы моно-
клональные антитела к Н. pylon, сорбированные на твердой фазе, что позволило
повысить чувствительность и в еще большей степени специфичность.
При диагностике этими тест-системами инфекции, вызванной Н. pylori у паци-
ентов с атрофическим гастритом, получают результаты, сопоставимые с сероло-
ЧАСТНАЯ МИКРОБИОЛОГИЯ 401
гическим и гистологическим исследованиями, однако по чувствительности они
уступают уреазному дыхательному тесту.
К числу факторов, снижающих чувствительность экспресс-методов иммуно-
логической диагностики инфекции, относятся кровотечение из верхних отделов
желудочно-кишечного тракта, прием ингибиторов протонного насоса, антибио-
тикотерапия, атрофия и метаплазия слизистой оболочки желудка.
Ускоренные уреазные тесты
Экстраординарная активность уреазы Н. pylori послужила основанием для
разработки быстрого и селективного определения этой бактерии в биоптатах.
Принцип метода заключается в том, что выделяемая микробом уреаза расщепляет
мочевину, которая содержится в смеси реагентов, куда помещают биоптат, выде-
ляющийся при этом аммиак сдвигает pH среды в щелочную сторону и изменяет
цвет индикатора (фенолового красного). Существует много коммерческих наборов
для быстрого уреазного теста. Чувствительность и специфичность уреазного теста
ощутимо различны в до- и постэрадикационный период. До лечения чувстви-
тельность достигает 80-95% (ложноотрицательные ответы составляют 5-20%),
специфичность — 95-3 00% (ложноположительные ответы составляют 0-5%).
После курса эрадикации Н. pylori чувствительность уреазного теста явно недо-
статочна для обнаружения остаточной инфекции. Именно поэтому уреазный тест
непригоден для документирования факта эрадикации возбудителя.
Другое ограничение применения уреазного теста — состояние ахлоргидрии и
прием антисекреторных препаратов. В данных условиях тест ндостаточно чувстви-
телен вследствие подавления активности уреазы Я. pylon в среде с высоким pH.
Третье серьезное ограничение для использования уреазного теста — кровотече-
ние из верхних отделов желудочно-кишечного тракта, при котором чувствитель-
ность теста резко снижается. Кровотечение мало влияет на точность дыхательного
уреазного теста с мочевиной, меченной изотоном.
Чувствительность уреазного теста снижается (как и бактериологического и
гистологического методов и дыхательного уреазного теста) при распространенной
атрофии и метаплазии слизистой оболочки желудка из-за сокращения или потно-
го прекращения колонизации Н. pylori.
Несмотря на наличие нескольких коммерческих вариантов уреазного теста,
многие лаборатории готовят реактивы для его выполнения самостоятельно.
Наиболее прост в приготовлении и удобен для работы с Н. pylori реактив следую-
щего состава: мочевина — 2 г, 0,5% раствор феноловый красный — 10 мл, 0,01 М
фосфатный буферный раствор (pH = 6,0) — до конечного объема 100 мл. Реактив
хранят при температуре 4-8 °C в темноте (в посуде из темного стекла) не более
2 нед. Для ускорения реакции концентрация мочевины может быть увеличена до
6 г. Цвет реактива должен быть темно-желтым.
Перед исследованием реактив по 3-5 капель разливают в пробирки для микро-
проб или ячейки планшета для иммунологических реакций, затем туда помещают
биоптаты или бактериологической петлей вносят выделенные культуры Н. pylon.
Учет реакции проводят в течение 18 ч. Реакция идет при комнатной температуре,
причем каждая пробирка или ячейка должны быть закрыты во избежание выхода
образующегося аммиака в окружающую среду и растворения его в соседних про-
бах.
Тесты, основанные на определении уреазы, особенно полезны в тех случаях,
когда требуется быстрый метод диагностики инфекции, вызванной Н. pylori.
В настоящее время уреазный гест — один из самых дешевых и простых. С помо-
щью уреазных тестов обычно удается идентифицировать до 75-80% Н. pylori-
позитивных больных. Таким пациентам при необходимости можно назначить
лечение в течение 1 ч после проведения ЭГДС.
ГЛАВАМ'
402 ЧАСТНАЯ МИКРОБИОЛОГИЯ
Уреазный дыхательный тест
Уреазный дыхательный тест (Urea Breath Test — UBT) основан на регистра-
ции активности фермента уреазы Н. pylori. В норме ткани желудка не содержат
уреазы. Пациент принимает внутрь слабый раствор или таблетку уреазы (75-
100 мг), меченной изотопом углерода 14С или 13С. При наличии Н. pylon бакте-
риальная уреаза расщепляет мочевину до аммиака и углекислого газа, которые
всасываются в кровь, углерод выводится с выдыхаемым воздухом, в котором
регистрируют наличие изотопа углерода 14С или 13С. Радиоактивный изотоп
14С применяют все реже, так как существуют ограничения даже для малых доз
радиоактивного излуЭДния в послеоперационном периоде, во время беременно-
сти, при детском возрасте пациентов, а также во время приема антисекреторных
препаратов. Изотоп 13С лишен радиоактивности и определяется методом масс-
спектрографии, который широко используется в зарубежных исследованиях
в целях эпидемиологической и клинической диагностики благодаря высокой
точности. Чувствительность и специфичность уреазного дыхательного теста
достигают 95%.
На чувствительности уреазного дыхательного теста отрицательно сказывает-
ся любое лечение, подавляющее или ограничивающее рост Н. pylori. Результаты
существенно занижаются и могут быть ложноотрицательными при применении
ингибиторов протонного насоса. Именно поэтому необходимо прекратить прием
антисекреторных препаратов как минимум за 2 нед до теста. Распространенная
метаплазия и атрофия слизистой оболочки желудка при далеко зашедшей инфек-
ции. вызванной Н.pylori, снижают чувствительность уреазного дыхательного теста,
как и других методов, основанных на определении микроорганизма и продуктов
его жизнедеятельности. Кровотечение из верхних отделов желудочно-кишечного
тракта не влияет на результаты уреазного дыхательного теста.
Эмпирически было установлено, что прием лимонной кислоты перед тестом
повышает выделение меченого углекислого газа с выдыхаемым воздухом.
Показано, что тот же эффект дает прием яблочного сока и стандартного раство-
ра соляной кислоты с pH =3,0 (умеренно кислая среда оптимизирует активность
уреазы). Помимо ложноотрицательных, могут регистрироваться и ложноположи-
тельные результаты из-за наличия сопутствующей микрифлоры. Такие ситуации
редки, однако они возможны при уреазоположительной микрофлоре в полости
рта и ротоглотке. Именно поэтому рекомендуют тщательное полоскание рта
и горла перед проведением 1 С-дыхательного теста. Коммерческие препараты
13С-мочевины в последние годы стали изготовлять в дозированных упаковках —
капсулах или в виде инкапсулированных таблеток. Это способствует стандартиза-
ции теста и воспроизводимости его результатов.
Молекулярная диагностика
Методы молекулярной биологии все шире внедряются в клиническую практику,
успешно конкурируя с бактериологической диагностикой Н. pylori. Биологический
материал для проведения ПЦР — биоптаты слизистой оболочки желудка или
двенадцатиперстной кишки, фекалии, слюна, зубной налет и желудочный сок
(последние 4 вида патологического материала непригодны для бактериологиче-
ского исследования).
Тесты, основанные на ПЦР, высокочувствительны и способны определять от 10
до 100 клеток Н. pylori. Объект детекции — участки уникальных генов Н. pylori —
cagA. vacA b ген уреазы. ПЦР позволяет в относительно короткий срок установить
факт инфицировапности пациента и установить генотип штаммов Н. pylori —
CagA+ или CagA- для принятия решения о тактике лечения, а также сделать
выводы о прогнозе заболевания. Процесс канцерогенеза наиболее интенсивно
происходит при инфицировании Н. pylori с генотипом sl/ml VacA. Разработаны
ЧАСТНАЯ МИКРОБИОЛОГИЯ 403
модификации ГЩР для прямого определения cagA- и гасД-генов Н. pylori в образ-
цах биопсийного материала, а также в образцах, залитых парафином.
Генотипирование с помощью ПЦР дает возможность типировать и диффе-
ренцировать штаммы Н. pylori, что позволяет осуществить их эпидемиологи-
ческое изучение, отличать случаи реинфицирования от рецидива инфекции.
Разработанный недавно метод FRET-ПЦР (Fluorescence Resonance Energy Transfer)
позволяет обнаруживать Н. pylori в желудочных биоптатах и фекалиях с чувстви-
тельностью и специфичностью до 98%. Модификация данного теста использована
для прямого определения резистентности Н. pylori к кларитромицину в образцах
биопсийного материала.
Серологическая диагностика
Длительное персистирование Н. pylori в организме человека приводит к нако-
плению специфических антител, которые могут быть выявлены с помощью
многих серологических тестов, наиболее часто применяют ИФА. Долгое время
серологическим методам диагностики инфекции, вызванной Н. pylori, придавали
ограниченное значение, основная сфера их применения — эпидемиологические
исследования и массовый скрининг. Однако классический ИФА. для выявления
в плазме крови больных специфических антихеликобактерных иммуноглобули-
нов (IgG, IgM, IgA) совершенствуется и его чувствительность и специфичность
в настоящее время достигают 80 -90%. Результатом этого стало более широкое
использование ИФА для первичной диагностики инфекции, вызванной Н. pylori.
К недостаткам данного метода можно отнести 1-2-суточный интервал между
моментом взятия кцови у пациента для исследования и временем получения
ответа, а также отсутствие возможности контролировать результаты лечения
сразу же после его окончания, так как в случае эрадикацни Н. pylon необходим
определенный (достаточно длительный) промежуток времени, в течение которого
концентрация специфических антихеликобактерных иммуноглобулинов в плазме
крови достоверно снизится. Можно говорить о своеобразной «инерционности»
гуморального звена иммунной системы, когда возбудителя в организме уже нет,
а специфические антитела к нему в крови обнаруживают в диагностически зна-
чимом титре. Оптимальный вариант исследования в таком случае — сравнение
титров специфических антихеликобактерных антител до лечения и через 2-3 мес
после его окончания. Однако при первичной диагностике инфекции, вызванной
Н. pylori, при положительном результате серологического теста и ясной клини-
ческой ситуации (эндоскопически выявленная язвенная болезнь желудка или
двенадцатиперстной кишки, эрозивный гастрит) информативно и однократное
исследование.
Ложноотрицательные результаты серологических тестов могут быть обусловле-
ны отсутствием иммунологической компетентности (больные, в организме которых
не синтезируются антитела к антигенам Н. pylori), ранней стадией инфицирования
или недостаточной чувствительностью используемого серологического теста.
Твердофазный ИФА — наиболее распространенный метод серологической диа-
гностики инфекции, вызванной Ц. pylori, — неинвазивный, простой, чувствитель-
ный и специфичный тест.
В практических целях чаще всего при проведении ИФА в качестве антигенов
используют суммарную смесь антигенов Н. pylori, но есть и специализированные
варианты ИФА для определения антител к CagA-белку, VacA-цитотоксину, уреазе
Н. pylori. Наиболее широко применяется метод определения IgG антител к CagA-
белку.
Обнаружение антител к Н. pylori свидетельствует об инфицированное™ дан-
ным возбудителем и может указывать на хронический гастрит типа В. язвенную
болезнь желудка и двенадцатиперстной кишки. Серопозитивны до 95% взрослых
404 ЧАСТНАЯ МИКРОБИОЛОГИЯ
больных с хеликобактерной инфекцией (у детей данный показатель ниже, поэто-
му серологический метод ограниченно применим при обследовании пациентов
детского возраста). Напряженность иммунного ответа, как правило, коррелирует с
массивностью обсеменения слизистой оболочки желудка Н. pylori и патоморфоло-
гической картиной процесса. Имеется корреляция и с клиническими проявления-
ми патологии, однако она менее выражена. Одно из преимуществ серологических
тестов — возможность выявления факта инфицирования не только при манифест-
ных, но и при субклинических формах, а также в стадии ремиссии заболевания.
Целесообразность применения серологических тестов для первичного обследова-
ния больных или проведения скрининга (сероэпидемиологических исследований)
не вызывает сомнений. Однако лечение инфекции, вызванной Н. pylori, не следует
назначать только на основании данных серологической диагностики, рекоменду-
ют проведение ЭГДС для уточнения диагноза, в том числе и у детей.
Ограничения при оценке показателей серологических тестов:
• тесты положи гельны у новорожденных вследствие трансплацентарной пере-
дачи антител, хотя ребенок может быть и не инфицирован Н. pylori (серопо-
зитивность сохраняется приблизительно до 6-месячного возраста);
• з есты остаются положительными в течение 2-6 мес после санации (эрадика-
ции возбудителя и при достоверном отсутствии его в тканях по результатам
бактериологического исследования);
♦ тесты отрицательны на ранних стадиях инфекции (до 60 то дня после зара-
жения), пока гуморальный ответ еще не развился.
Иммуноблоттинг. Западный блоттинг использовали при создании наборов
реагентов для детекции антител к CagA- и VacA-белкам Н. pylori (Recombinant
ImmunoBlot Assay — RIBA, Western Blot, Helicoblot и др.). Методы сочетают электро-
форетическое разделение антигенов Н. pylori и последующую встречную преципита-
цию в геле с антителами исследуемой сыворотки. Коммерческие тест-системы для
иммуноблоттинга обеспечивают более высокую точность диагностики инфекции,
вызванной Н. pylori, по сравнению с обычным ИФА. Частота выявления Н. pylori у
больных достигает 97,5%. Особенно ощутимы преимущества иммуноблоттинга в
случаях ретроспективной диагностики инфекции, вызванной Н, pylori, у больных с
отрицательным результатом стандартного ИФА — при атрофическом гастрите, раке
желудка в ассоциации с гастритом и раке желудка без признаков гастрита.
Определение антител в слюне, моче и капле крови. В некоторых зарубеж-
ных странах разработаны и выпускаются ИФА-тест-системы для определения
антител к Н. pylori в слюне, моче и капле крови. Сообщения об информативности
данных тестов крайне противоречивы, их использование в клинической практике
не рекомендовано Маастрихтскими соглашениями по лечению инфекции, вызван-
ной Н. pylori.
КЛИНИЧЕСКОЕ ПРИМЕНЕНИЕ МЕТОДОВ ДИАГНОСТИКИ
Первичная диагностика (доэрадикационный период)
Международный (Маастрихтский) консенсус по лечению инфекции, вызванной
Н. pylori, определил «строго рекомендуемые» показания к антихеликобактерной
терапии:
♦ язвенная болезнь желудка и двенадцатиперстной кишки (как в фазе обостре-
ния, так и ремиссии), включая осложненную язвенную болезнь;
• MALT-лимфома;
• атрофический гастрит;
• состояние после резекции желудка по поводу рака;
• наличие родственников первой степени родст ва с диагностированным раком
желудка;
• желание пациентов (после подробной консультации лечащего врача).
ЧАСТНАЯ МИКРОБИОЛОГИЯ 4Q5
В 3-м Маастрихтском соглашении указывается на необходимость дифференци-
рованного подхода к эрадиканионной терапии при неисследованной и исследован-
ной диспепсии. Пациентам с неисследованной диспепсией рекомендуют стратегию
«test-and-treat» («выявить и лечить»). Пациентам с инфекцией, вызванной Н. pylori,
и исследованной неязвенной (функциональной) диспепсией также рекомендуют
эрадикацию Н. pylori. Подчеркивается, что «test-and-treat» — стратегия выбора у
всех взрослых пациентов с функциональной диспепсией в популяции с высокой
распространенностью Н. pylori, в то время как эффективность данной стратегии
при низкой распространенности Н. pylori (<20%) невысока. В таких случаях пред-
почтение отдают более дешевой эмпирической антисекреторной терапии.
Назначению эрадиканионной терапии должно предшествовать лабораторное
подтверждение инфицирования Н. pylori. У пациентов с эндоскопически выяв-
ленной патологией или с известным диагнозом заболеваний, ассоциированных с
Н. pylori (язвенной болезни, карциномой, лимфомой), берут биоптаты слизистой
оболочки с последующим лабораторным исследованием с применением гисто-
логического и бактериологического методов, а также быстрого уреазного теста,
Допускается, что положительный результат быстрого уреазного теста с биоптатом
может быть основанием для начальной терапии инфекции, вызванной Я. pylori,
даже при отсутствии предыдущего тестирования неинвазивными методами.
Неинвазивные тесты выполняют в тех случаях, когда биопсию не проводили,
но они могут быть использованы и в дополнение к гистологическому, бактериоло-
гическому исследованиям биоптата при возможностях лаборатории или при кли-
нических показаниях. Взгляды относительно приемлемых неинвазивных тестов
диагностики инфекции, вызванной Н.pylon, претерпели значительные изменения
с момента выхода 1-го Маастрихтского соглашения. В соглашении 1996 г. для
первичной диагностики инфекции, вызванной Н. pylori, были рекомендованы
дыхательный уреазный тест и определение IgG в плазме крови с помощью ИФА
(ELISA). Тесты с каплей цельной крови отклонены, как малочувствительные
Во 2-м Маастрихтском соглашении (2000) внесены существенные коррективы,
которые сохраняют свое значение и сегодня. Подтверждена строгая рекомендация
использовать дыхательный уреазный тест, а в качестве альтернативного метода
рекомендуют тест на выявление антигенов Н. pylori в фекалиях. ELISA-тест на
антитела Н. pylori в плазме крови, с поправками на его недостаточную чувстви-
тельность. рекомендован для стран с высокой распространенностью Н. pylon.
В 3-м Маастрихтском соглашении (2005) регламентируются следующие специаль-
ные случаи, когда необходимо серологическое исследование: кровоточащая язва,
атрофия слизистой оболочки желудка, лимфома MALT-типа и лечение ингибито-
рами протонного насоса и антибиотиками. В этих случаях частично ингибирована
жизнедеятельность Н.pylori или же инфекция самокупирована, и основные тесты —
дыхательный уреазный тест и тест на выявление антигенов Н. pylori в фекалиях —
часто дают ложноотрицательные результаты. Тесты на антитела в слюне и моче
неприемлемы для первичной диагностики инфекции, вызванной Н. pylori.
Постэрадикационный период
Рекомендации о проведении специфической диагностики инфекции, вызванной
Я. pylori, в постэрадикационном периоде были выработаны после серьезных дис-
куссий. Во 2-м Маастрихтском соглашении введено положение о целесообразно-
сти контроля эрадикации в постэрадикационном периоде. Это было обусловлено
накоплением данных о нередких случаях неэффективности лечебных схем с после-
дующими рецидивами основного заболевания.
В качестве контрольных тестов на наличие инфекции, вызванной Н. pylori, в
постэрадикационном периоде Маастрихтские соглашения рекомендуют два мето-
да — дыхательный уреазный тест или тест на антигены Н. pylori в фекалиях, осно-
406 ЧАСТНАЯ МИКРОБИОЛОГИЯ
ванный на определении моноклональных антител. Специально оговаривается, что
серологические тесты неприемлемы для контроля излеченности пациентов от инфек-
ции, вызванной Н. pylon. Это обусловлено длительной циркуляцией специфических
антител даже после полной ликвидации инфекционного процесса в организме.
Контрольные тесты — дыхательный тест или тест на антигены в фекалиях —
могут быть проведены не ранее чем через 4 нед после прекращения эрадикацион-
ного лечения. Как минимум за 1 нед до выполнения тестов должны быть отменены
антисекреторные препараты.
При выполнении ЭГДС по клиническим показаниям в постэрадикационном
периоде и в любом случае при осложненной язвенной болезни, раке желудка, лим-
фоме MALT-типа, локальной резекции раннего рака желудка рекомендуют взятие
биоптатов слизистой оболочки из антральной части и тела желудка и выполнение
гистологического и бактериологического исследований для выявления Н, pylori.
В 3-м Маастрихтском соглашении (2005) контроль антибиотикорезистентности
Н. pylori в постэрадикационном периоде рекомендуют при отсутствии эффекта
после двух курсов антибиотикотерапии. Рекомендации Маастрихтских соглаше-
ний по поводу специфической диагностики инфекции, вызванной Н. pylori, дают
хорошую основу для стандартизации подходов, но в то же время требуют клини-
ческого мышления и рациональных решений при различной патологии.
II. ГРАМПОЛОЖИТЕЛЬНЫЕ БАКТЕРИИ
СТАФИЛОКОККИ
ТАКСОНОМИЯ
Согласно второму изданию «Руководства по систематической бактериологии
(Берги) [«Bergey’s Manual of Systematic Bacteriology» (2009)], род Staphylococcus
исключен из семейства Micrococcaceat и вместе с родамиJeotgaltcoccus, Macrococcus
и Salinicoccus выделен в новое семейство — Staphylococcaceac, в котором к стафило-
коккам относятся 32 вида и 12 из них имеют подвиды.
В медицинской микробиологии стафилококки принято подразделять на две
большие группы — коагулазоположителъные и коагулазоотрицательныс. Первая
группа имеет наибольшее медицинское значение.
ОБЩАЯ ХАРАКТЕРИСТИКА
Стафилококки — неподвижные грамположительные кокки, которые вследствие
деления в нескольких плоскостях в мазках расположены в виде беспорядочных
скоплений, иногда напоминающих виноградные грозди. Спор не образуют, но
могут образовывать капсулы in vitro и in vivo. Подавляющее большинство стафило-
кокков — факультативные анаэробы, за исключением S. saccharolyticus и S. aureus
subsp, anaerobes.
Стафилококки — хемоорганотрофы. каталазоположительны, содержат цитох-
ромы, но обычно оксидазоотрицагельны. Обладают выраженной биохимической
активностью — восстанавливают нитраты до нитритов или газообразного азота,
гидролизуют белки, гиппурат, жиры, твины, расщепляют большое количество
углеводов в аэробных условиях с образованием уксусной кислоты и незначитель-
ных количеств СО2. Однако эскулин и крахмал, как правило, не гидролизуют,
индол не образуют. Стафилококки лизируются под действием лизостафина, но
устойчивы к лизопиму.
Температурный оптимум — 30-37 °C, но могут расти в широком температур-
ном диапазоне — от 6,5 до 45 °C при pH = 4,2-9,3 и в присутствии 40% желчи.
ЧАСТНАЯ МИКРОБИОЛОГИЯ 407
Стафилококки нетребовательны к питательным средам, обладают галотолерант-
ностью и хорошо растут в присутствии 5-10% раствора NaCl. что широко исполь-
зуется при целенаправленном их выявлении в различных видах материалов от
больных и из объектов внешней среды.
ЭПИДЕМИОЛОГИЯ
Стафилококки широко распространены в окружающей среде: их постоянно
обнаруживают на предметах обихода, в воздухе и воде, в составе нормальной
микрофлоры кожи и слизистых оболочек, в фекалиях людей и ряда животных.
На поверхности кожи и слизистых оболочек человека встречается до 15 видов
стафилококков (в том числе S. aureus), локализованных более чем в 20 биотопах,
основными из которых считают слизисту ю оболочку нижних носовых ходов, зева,
кожу подмышечных впадин и паховой области. Количество видов, имеющих меди-
цинское значение, несколько больше, так как причиной заболеваний в ряде слу-
чаев являются стафилококки, паразитирующие у животных. К числу таких видов
относят S. schleiferi и S. intermedius, которые обнаруживают у хищных животных и
могут вызывать инфекции у человека при укусах. Виды, встречаемые у травоядных
животных, — S. hyicus, S. chromogenes, S. sciuri, S. lentus, S. vitulus, S. caseolyticus —
выявляют в молочных и мясных продуктах
Особое значение имеют стафилококки, и особенно S. aureus, как возбудите-
ли внутрибольничных инфекций. Эпидемиологическая ситуация осложняется
широким распространением в стационарах и появлением во внебольничной среде
клинических изолятов S. aureus, которые способны вызывать разнообразные кли-
нические формы внутрибольничных инфекций, включая бактериемию, внутри-
больничную пневмонию, синдром септического шока, септический артрит, остео-
миелит, требующие длительного и дорогостоящего лечения. S. aureus используют
при санитарно-микробиологических исследованиях для оценки биологической
безопасности многих объектов окружающей среды — воздуха, воды, почвы, пище-
вых продуктов, фармацевтической и косметической продукции.
КЛИНИЧЕСКИЕ ПРОЯВЛЕНИЯ ИНФЕКЦИИ
Коагулазоположительные стафилококки. Золотистый стафилококк,
S. aureus, традиционно относят к числу патогенных микроорганизмов. Тем не
менее данный вид чрезвычайно широко распространен в популяции людей и
может быть обнаружен у 15-30% здоровых лиц Особенно часто выявляют у пер-
сонала медицинских учреждений.
S. aureus вызывает у человека различные виды гнойно-септических инфекций
(ГСП): кожные гнойничковые и раневые инфекции, сепсис эндокардиты, вне-
больничные пневмонии, артриты, остеомиелиты, перитониты, инфекции моче-
выводящих путей. Многообразие клинических форм обусловлено наличием у ста-
филококков различных факторов вирулентности, таких как токсины, в том числе
гемолизины, ферменты, факторы адгезии и защиты от фагоцитоза. Клинические
проявления ГСИ обычно неспецифичны, диагноз следует подтверждать бакте-
риологическим исследованием материала от больных и при необходимости из
объектов окружающей среды в целях выделения и идентификации чистой куль-
туры возбудителя. Кроме ГСИ, S. aureus может вызывать особые нозологические
формы, обусловленные главным образом наличием у данного вида экзотоксинов:
синдромы «ошпаренной кожи» и «ошпаренных младенцев», пищевые токсикоин-
фекции (ПТИ), синдром токсического шока (СТШ).
Синдром «ошпаренных младенцев» (болезнь Риттера Фон Риттерштайна,
эпидемическая пузырчатка новорожденных) возникает у младенцев, инфициро-
ванных штаммами, продуцирующими токсины — эксфолиатины А и В. Это осо-
бые экзотоксины белковой природы, обладающие узконаправленным действием
4Q8 ЧАСТНАЯ МИКРОБИОЛОГИЯ
на десмосомы клеточных мембран, которыми клетки скреплены друг с другом.
Заболевание высококонтагиозно и может приводить к летальному исходу.
Синдром «ошпаренной кожи», описанный у детей старшего возраста и взрос-
лых, также характеризуется эритемой, образованием пузырей и общей интокси-
кацией организма.
СТШ обусловлен штаммами S, aureus, продуцирующими особый экзотоксин
(TSST-1), механизм действия которого основан на стимуляции Т-лимфоцитов,
усилении продукции ФИО и ИЛ-1. СТШ встречается, в первую очередь, у молодых
женщин, использующих определенные типы влагалищных тампонов с высокой
сорбционной способностью. Однако описаны случаи данного заболевания у жен-
щин вне менструаций и у мужчин, например, при тампонаде раны.
ПТИ связаны с действием стафилококковых энтеротоксинов — экзотоксинов
белковой природы, которые по антигенным свойствам подразделяются на 6 серо-
варов. Одно из важнейших свойств энтеротоксинов стафилококков — термоста-
бильность (устойчивость к воздействию температуры 100 °C), особенно экзоток-
синов В и С. Несмотря на описанные в литературе многочисленные биологические
эффекты энтеротоксинов, механизм их действия остается неизвестным.
Особая форма инфекционного процесса — носительство S. aureus на слизистой
оболочке носа и зева, которое у персонала лечебных учреждений может явиться
причиной внутрибольничного инфицирования пациентов.
Коагулазоотрицательные стафилококки вызывают инфекционные заболева-
ния значительно реже. S epidermidis может быть причиной сепсиса, эндокардитов,
артритов, глазных инфекций и инфекций мочевыводящих путей. Данные микро-
организмы вызывают значительную часть кардиоваскулярных инфекций, связан-
ных с длительной установкой внутривенных катетеров и операциями на открытом
сердце, а также перитонитов в отделениях перитониального диализа.
S. saprophyticus, являясь представителем нормальной микрофлоры кожи и
слизистых оболочек человека, имеет этиологическое значение при заболеваниях
мочевыводящих путей, особенно у молодых, сексуально активных женщин, и
негонококковых уретритов и простатитов у мужчин. Описаны случаи раневых
инфекций и сепсиса, вызванных S. saprophyticus.
S. haemolyticus может вызывать эндокардиты, перитониты, септицемии, инфек-
ции мочевыводящих путей и раневые инфекции.
Существуют данные о возможной этиологической значимости при различных
патологических состояниях S. hommis, S. warned, S. capitis, S. simulans, S. cohnii,
S. xylosus, S. saccharolyticus, S. lugdunensis, S. schleiferi, как правило, на фоне иммуно-
дефицита различной природы и выраженности.
MRSA- и MRSE-ассоциированные инфекции могут начинаться на фоне лечения
антибактериальными препаратами, в том числе аминогликозидами и цефалоспо-
ринами, а неадекватная антибиотикотерапия при тяжелых внутрибольничных
инфекциях значительно ухудшает прогноз заболевания. Природа резистент-
ности к антибиотикам пенициллинового ряда выяснена неокончательно, но,
по-видимому, связана с изменениями, возникающими в специфических пени-
циллинсвязывающих белках клеточной стенки стафилококков. Резистентные
стафилококки, обладая всеми характерными морфологическими, культураль-
ными, физиологическими и биохимическими свойствами вида, имеют свои био-
логические особенности.
• Уникальный биохимический механизм резистентности обеспечивает устой-
чивость ко всем полусинтетическим пенициллинам и цефалоспоринам.
• Такие штаммы способны «аккумулировать» гены антибиотикорезигтентно-
сти и именно поэтому нередко обладают устойчивостью к нескольким клас-
сам антибактериальных препаратов одновременно, тем самым значительно
затрудняя лечение пациентов.
ЧАСТНАЯ МИКРОБИОЛОГИЯ 409
Чаще всего препаратом выбора в таких случаях является ванкомицин, однако в
последние годы появились сообщения о ванкомицинрезистентных штаммах сна-
чала среди S. haemolyticus, а затем среди S. epidermidis.
ЛАБОРАТОРНАЯ ДИАГНОСТИКА
Исследование на стафилококки включает:
• прямую микроскопию биоматериала;
• посев материала на питательные среды и выделение чистой культуры;
• идентификацию выделенных микроорганизмов по морфологическим, тин-
кториальным, культуральным и биохимическим признакам (определение
рода и вида);
• внутривидовое типирование по антигенным свойствам или лизабельности
типовыми стафилококковыми бактериофагами;
• выявление носительства S. aureus
• иммунологическую или биологическую индикацию стафилококковых экзо-
токсинов;
• определение чувствительности/резистентности к антимикробным препара-
там и выя вление метициллинрезистентных штаммов.
МИКРОСКОПИЧЕСКОЕ ИССЛЕДОВАНИЕ
Бактериоскопическим методом исследуют СМЖ, аспират из суставов, отделяе-
мое из ран, абсцессов. Следует помнить, что при титре стафилококков в материале
менее 1и5-106 КОЕ/мл данный метод неприемлем.
Изоляция стафилококков
Стафилококки нетребовательны к питательным средам и могут культивиро-
ваться на большинстве питательных сред общего назначения, используемых при
посеве клинического материала. Для выделения этих микроорганизмов разрабо-
тан ряд специальных элективных и элективно-дифференциальных питательных
сред. В качестве элективного фактора в составе питательных сред для стафилокок-
ков используют гипертонический раствор NaCl. Примеры таких сред — солевой
агар и солевой бульон.
Желточно-солевой агар используют для выделения стафилококков чаще других
элективно-дифференциальных сред. Для ее приготовления в расплавленный и
остуженный до 45-50 °C 10% солевой агар вносят желточную взвесь. Желточную
взвесь чаще всего готовят в лабораториях путем смешивания 1 желтка куриного
яйца, взятого в асептических условиях, с 200 мл стерильного 0,9% раствора NaCl
и взбалтывания содержимого с соблюдением правил асептики. Колонии микро-
организмов, обладающих лецитовителлазой активностью, на данной питательной
среде окружены зоной опалесценции — «радужного венчика».
Молочно-солевой и молочно-жечточно-солевой агары позволяют выявить про-
теолитическую активность стафилококков.
Солевой агар с маннитом, широко используемый за рубежом, содержит 7.5%
раствор NaCl, маннит и феноловый красный. Коагулазопозитивные стафилококки
образуют на данной среде колонии, окруженные желтой зоной, коагулазонегатив-
ные — красной или пурпурной.
Стафилококковая среда № ПО — элективно-дифференциальная среда, содер-
жащая 7,5% раствор NaCl, желатин, маннит, лактозу и индикатор pH. Колонии
коагулазопозитивных стафилококков на данной среде желтого цвета, окружены
зоной разжижения желатина.
Спеда Байрда-Паркера содержит хлорид лития и глицин в сочетании с теллу-
ритом натрия, а также эмульсию куриного желтка в качестве дифференциального
фактора. Данная питательная среда высокоспецифична для изоляции коагулазо-
410 ЧАСТНАЯ МИКРОБИОЛОГИЯ
позитивных стафилококков из пищевых продуктов и, в отличие от многих других
сред, обеспечивает выделение стафилококков в течение 24 ч.
Бульон Джилотти-Сантони предназначен для выделения S. aureus из пищевых
продуктов. Содержит факторы роста (пируват натрия и маннит) и элективные
добавки (хлорид лития и глицин в сочетании с теллуритом натрия). Для предупре-
ждения роста микрококков на поверхность среды после посева наливают масло,
Агар Кранера — высокоэлективная питательная среда для выделения стафи-
лококков из пищевых продуктов. Содержит элективные факторы — тиоционат
натрия, маннит, лития хлорид, актидион и азид натрия.
Солевой агар с маннитом обеспечивает селективную изоляцию 5. aureus за счет
высокого (7,5%) содержания в этой среде натрия хлорида.
На неселективных питательных средах стафилококки в течение 24 ч инку-
бации образуют колонии диаметром 1-3 мм, а при продолжении инкубации до
3 сут — диаметром до 3-8 мм. У пигментообразующих видов колонии могут быть
окрашены в золотистый, кремовый или желтый цвет. Наиболее интенсивно пиг-
ментообразование проявляется на питательных средах, обогащенных белками
(молочном, желточном, сывороточном агарах). Пигмент стафилококков нераство-
рим в воде, поэтому окрашивается только сама колония, а не среда вокруг нее.
Идентификация стафилококков
На практике идентификация выделенных стафилококков, как правило, сводит-
ся к дифференциации трех видов — S. aureus, S. epidermidis и 5. sapropkyticus, при
этом сначала первый вид дифференцируют от двух оставшихся, а затем прово-
дят дифференцировку коагулазоотрицательных стафилококков. На практике для
дифференциации стафилококков используют небольшое количество признаков —
пигмент, лецитовителлазу, плазмскоагулазу, ДНКазу, ферментацию или окисление
маннита, фосфатазу. Схема идентификации представлена на рис. 21-2
Указанная схема идентификации в подавляющем большинстве случаев вполне
удовлетворяет потребностям лечащих врачей, однако носит несколько условный
характер. При необходимости более точной видовой дифференциации следует
использовать дополнительные тесты и ключи, преимущественно в соответствии
с «Руководством по систематической бактериологии (Берги) [«Bergey’s Manual of
Systematic Bacteriology л (2009)].
Выделение из патологического материала коагулазоотрицательных стафило-
кокков не может являться безусловным основанием для признания их этиоло-
гической значимости. В пользу этиологической роли данных микроорганизмов
свидетельствуют:
S. aureus
Коагулаза (+),
лецитовителлаза и/или пигмент (+)
Маннит анаэробно
или
ДНКаза
Коагулаза (-),
лецитовителлаза и/или пигмент (-)
Тесты на:
устойчивость к новобиоцину,
фосфатазу и окисление маннита
Рис. 21-2. Схема идентификации S. aureus.
ЧАСТНАЯ МИКРОБИОЛОГИЯ 411
• выделение чистой культуры микроорганизма (для случайной контаминации
более характерны смешанные культуры);
• повторное выделение тех же штаммов или их комбинации в течение заболе-
вания.
Тесты, используемые для доказательства принадлежности к роду Staphylococcus,
представлены в табл. 21-29.
Таблица 21-29. Дифференциация грамположительных аэробных и факультативно-анаэробных
кокков
Признак Staphylococcus Streptococcus Micrococcus Stomatococcus
Расположение клеток Группы, пары Цепочки, пары Группы, тетрады Группы, иногда пары, обычно в капсулах
Подвижность - - - -
Каталаза + - +
Рост в присутствии 10% раствора натрия хлорида + - - -
Анаэробная фермента- ция маннита + ND - +
Аэробная ферментация глицерина + ND - ND
Примечание
«+» — 90% положительных по данному свойству штаммов.
«-» — 90% отрицательных по данному свойству штаммов.
«d» — 11-89% положительных по данному свойству штаммов.
«ND» — тест не имеет значения для дифференциации вида.
Для внутривидовой дифференциации стафилококков используют несколько
тестов, из которых наиболее актуальный — коагулазный. Данное исследование —
ключевое при идентификации стафилококков в практических лабораториях. Для
выявления плазмокоагулазы могут быть использованы два метода: пробирочный
тест, с помощью которого выявляют свободную плазмокоагулазу, и тест на стекле,
с помощью которого определяют клампинг-фактор (слайд-тест). Данные тесты
неодинаковы по своему значению. При идентификации S. aureus пробирочный тест
следует расценивать как окончательный, а слайд-тест — как быстрый ориентировоч-
ный. Слайд-тест рекомендуют использовать при идентификации новых видов ста-
филококков S. lugdunensis и 5. schleifcri, которые не обладают свободной коагулазой.
Для определения плазмокоагулазы используют пробирочную реакцию плаз-
мокоагуляции. Для ее выполнения необходима цитратная лиофилизированная
кроличья плазма, которую перед использованием разводят согласно указаниям
на этикетке. Для выполнения реакции одну петлю агаровой культуры суспенди-
руют в небольшом количестве плазмы и инкубируют пробирки в термостате при
температуре 37 °C. Учет реакции проводят через 1, 2,4 и 18 ч. Появление сгустка
в течение первых 4 ч расценивается как положительный результат, отсутствие в
течение 18 ч — как отрицательный. В качестве контролей используют тест с заве-
домо положительной культурой и пробирку с неинокулированной плазмой.
Выявление связанной коагулазы (клампинг-фактора) проводят в основном с
помощью нагрузочных реакций, наборы для которых выпускают многие фирмы-
производители. Преимущество данных реакций — быстрота получения ответа,
однако отрицательный результат не является полностью достоверным, так как
лишь 97% штаммов продуцируют одновременно свободную и связанную коагула-
зу. Для повышения достоверности отрицательного результата используют особые
комбинированные тесты. Также могут наблюдаться ложноположительные резуль-
таты при тестировании культур, выращенных на питательных средах, содержащих
гипертонический раствор NaCl.
412 ЧАСТНАЯ МИКРОБИОЛОГИЯ
Термостабильная нуклеаза (ДНКаза) — фермент, расщепляющий фосфодиэ-
фирные связи как РНК, так и ДНК. Учет ДНКазной активности проводят на плот-
ной питательной среде с добавлением высокополимерной ДНК. Вокруг колоний
штаммов, продуцирующих ДНКазу, наблюдается просветление среды.
Для оценки способности ферментировать маннит в анаэробных условиях при-
меняют предварительно регенерированную среду Хью-Лейфсона под слоем вазе-
линового масла; наблюдение за посевами ведут в течение 5 дней.
Ферментацию маннита в аэробных условиях (окисление) определяют на плот-
ной питательной среде с индикатором ВР или среде Хью-Лейфсона, содержащих
1% маннита, через 24 ч инкубации при 37 °C. На ферментацию маннита указывает
появление желтого окрашивания вокруг колонии.
Тест на фосфатазу основан на изменении окраски какого-либо фосфорорга-
нического соединения под действием данного фермента. Реагенты, чаще всего
динатриевую соль паранитрофенилфосфата или фенолфталеинфосфат, вводят в
состав питательного агара, на поверхность которого в виде бляшек или штрихов
засевают испытуемые культуры. При использовании соли паранитрофенилфос-
фата непосредственно после инкубации вокруг колонии появляется интенсив-
ное желтое окрашивание. При добавлении фенолфталеинфосфата дополнитель-
но используют раствор аммиака, который приводит к розовому окрашиванию
колоний.
В настоящее время широко представлены наборы для биохимической иден-
тификации стафилококков, позволяющие одновременно дифференцировать
выделенные культуры от других грамположительных кокков, например такие,
как СТАФИ-тесты 16 и 24 («Эрба-Лахема»). Набор содержит стрипованные
панели с высушенными питательными средами и субстратами для 16 или 24
биохимических тестов и позволяет провести идентификацию при визуальном
или фотометрическом считывании. Системы дополнены ускоренными тестами
для определения продукции ацетоина, пирролидонилариламидазы и выявле-
ния цитохромоксидазы в виде диагностических полосок. После инкубации при
температуре 37 “С в течение 24 ч инокулированных стрипов СТАФИ-теста 16 и
добавления реактивов проводят учет результатов по цветовой таблице интер-
претации. При использовании СТАФИ-теста 24 добавления реактивов после
инкубации не требуется. При окончательной идентификации следует учитывать
всю дополнительную информацию: данные микроскопии, характер колоний,
наличие пигмента, гемолиз.
Интерпретация результатов биохимической дифференцировки коагулазополо-
жительных и наиболее значимых коагулазоотрицательных стафилококков пред-
ставлена в табл. 21-30, 21-31.
Внутривидовое типирование стафилококков проводят по антигенным свой-
ствам или фаготипированию типовыми стафилококковыми бактериофагами.
Типирование по антигенным свойствам можно проводить с помощью реакции
агглютинации, однако ни одна из предложенных к настоящему времени схем не
получила широкого распространения, что связано главным образом с трудностью
приго говления типовых адсорбированных сывороток и отсутствием единого под-
хода к принципу серотипироьания и способам его осуществления.
Фаготипированию — наиболее доступный и широко используемый метод вну-
тривидового типирования S. aureus. Фаготипирование стафилококков имеет свои
особенности:
• штаммы стафилококков, как правило, лизируются несколькими фагами,
поэтому термин «фаготип» означает штамм, взаимодействующий с одним
или несколькими типовыми фагами;
• стафилококки, выделенные в различных географических широтах, значи-
тельно лучше типируются так называемыми местными фагами:
ЧАСТНАЯ МИКРОБИОЛОГИЯ 413
• стафилококки, выделенные от людей и животных, различаются по фаготи-
пам, поэтому существуют особые наборы фагов применительно к стафило-
коккам, полученным из разных источников.
В нашей стране для фаготипирования 5. aureus используют Международный
набор из 23 типовых стафилококковых фагов, выпускаемый Институтом эпиде-
миологии и микробиологии им. Н.Ф. Гамалеи, который следует дополнять бакте-
риофагами из местных коллекций.
Мазок или смыв из передних отделов носа и зева засевают на желточно-
сывороточный агар, после инкубации проводят количественный учет и идентифи-
кацию S. aureus. Выделенные штаммы фаготипируют для определения категории
носительства.
Для выделения чистой культуры стафилококка с персистентными свойствами
предлагают использовать элекз ивную питательную среду с фузидином и изучать
наиболее информативные признаки — антилизоцимную, антиинтерфероновую и
антикомплементарную активность. Описана методика балльной оценки получен-
ных результатов с помощью диагностической модели по формуле дискриминант-
ной функции. Представленный метод и способы диагностики стафилококкового
носительства позволяют провести однократное исследование и повысить эффек-
тивность диагностики на 15-20%.
Выявление носительства S. aureus у медицинского персонала не является обя-
зательным. однако данное исследование регламентируется как один из разделов
микробиологического мониторинга возбудителей внутрибольничных инфекций.
Обнаружение стафилококковых экзотоксинов
Диагностика СТШ основана на выявлении TSST-1 токсина с помощью РИА,
ИФА или реакции латекс-агглютинации. Стафилококковый энтеротоксин выявля-
ют одним из двух методов — биологическим или иммунологическим.
Биологический метод определения энтеротоксинов основан на заражении котят
4-8-недельного возраста. Взрослые кошки, а также щенки менее восприимчивы.
Биопробу на котятах проводят путем скармливания подозрительного продукта
или 5-суточной молочной культуры 2-3 котятам натощак или реже на взрослых
кошках путем внутривенного введения водного экстракта питательного агара, на
котором в течение 48 ч выращивалась испытуемая культура. При положительном
результате через 0.5-5 ч наблюдается возникновение рвоты.
Иммунологические методы выявления энтеротоксинов и предложенные для
этого реакции преципитации, РИГА используют для качественного определения
стафилококковых энтеротоксинов в продовольственном сырье и пищевых продук-
тах животного происхождения. Гвердофазовый ИФА имеет предел обнаружения
стафилококковых энтеротоксинов в диапазоне массовых концентраций от 0,2 до
2 мкг/кг (в зависимости от типа энтеротоксина и вида продукта), это ниже уровня
стафилококкового энтеротоксина типа А, чаще всего обусловливающего развитие
ПТИ у человека. Метод позволяет обнаруживать стафилококковые энтеротоксины
типа А или В при раздельном определении, а также энтеротоксины типов А, В, С. D
и Е при одновременном определении с помощью различных наборов реагентов.
Определение чувствительности к антимикробным препаратам
Проводят методом стандартных дисков. При оценке чувствительности выде-
ленных штаммов стафилококков в первую очередь необходимо тестировать пре-
параты, имеющие основное клиническое значение, — ^-лактамы, макролиды,
фторхинолоны, аминогликозиды и ванкомицин.
Препараты выбора для лечения стафилококковых инфекций — р-лактамные
антибиотики. Определение чувствительности к р-лактамам должно включать
выполнение двух тестов:
414
ЧАСТНАЯ МИКРОБИОЛОГИЯ
• определение чувствительности к бензилпенициллину или выявление продук-
ции р-лактамаз (пенициллиназ);
• определение чувствительности к оксациллину или выявление ПСБ2а или
кодирующего его гена тесА.
При интерпретации результатов следует принимать во внимание, что:
• штаммы стафилококков, лишенные механизмов резистентности, чувстви-
тельны ко всем р-лактамным антибиотикам:
• р-лактамазы с гафилококков способны гидролизовать природные и полусин-
тетические пенициллины, за исключением оксациллина;
• штаммы стафилококков, обладающие ПСБ2а, клинически устойчивы ко всем
р-лактамным антимикробным препаратам, маркером наличия ПСБ2а явля-
ется устойчивость к оксациллину.
Лабораторные методы идентификации и типирования MRSA можно разделить
на фено- и генотипические. Фенотипические методы позволяют изучить характе-
ристики, экспрессируемые микроорганизмами, генотипические методы направ-
лены на исследование структуры ДНК. Основанием к проведению исследований
служат как данные клинического мониторинга внутрибольничных инфекций,
эпидемиологического мониторинга за уровнем внутрибольничных инфекций,
вызванных S. aureus, так и данные микробиологического мониторинга, отражаю-
щие чувствительность клинических изолятов к антибактериальным препаратам.
Идентификацию MRSA проводят:
• диско-диффузионным методом с использованием дисков, содержащих 1 мкг
оксациллина;
• с помощью скрининг-теста на агаре Мюллера-Хинтона, содержащего 4%
раствор хлорида натрия и 6 мг/л оксациллина (MRSA-скрининговый агар).
Оба метода имеют свои недостатки. В первом случае к категории метициллин-
резистентных штаммов могут быть ошибочно отнесены клинические изоляты
с гиперпродукцией [3-лактамазы, то есть стафилококки с пограничным уровнем
резистентности, минимальная подавляющая концентрация (МПК) оксациллина
для которых колеблется в пределах 1,0-2,0 мкг/мл. Во втором случае возникают
трудности при идентификации MRSA относящихся к первому классу по фенотипу
гетерорезистентности. Скрининг на агаре — высокочувствительный и специфич-
ный метод, легко выполнимый в обычных бактериологических лабораториях.
Тест проводят с использованием особой селективной питательной среды, содержа-
щей 4% раствор NaCl, 6 мг/л оксациллина, 1% маннита и индикатор pH. На среду
проводят массивный посев, так как в материале может содержаться небольшое
количество MRSA. Резистентными считают штаммы, для которых МПК оксацил-
лина составляет 4 мг/л и более.
Популяции резистентных стафилококков гетерогенны по профилю лекар-
ственной устойчивости, и резистентные микроорганизмы, которые растут мед-
леннее, могут остаться незамеченными. Данный факт диктует необходимость
использования специальной элективной среды и особых технических приемов
при определении чувствительности к антибактериальным препаратам. Удобством
использования отличаются хромогенные среды, в которых используются особые
субстраты, специфически расщепляемые штаммами MRSA, что придает колониям
характерное окрашивание. Дополнительно в состав таких сред входят специаль-
ные ингибиторные добавки для подавления сопутствующей микрофлоры, а также
ростовые добавки для стафилококков, такие как пируват натрия.
Идентификация резистентности возможна путем определения продукции ПСБ2а
методом латекс-агглютинации с помощью готовых тест-систем. В случае получе-
ния сомнительных результатов при использовании данного метода необходимо
прямое определение гена тесА с использованием ПЦР, являющейся «золотым
стандартом» для идентификации MRSA.
Таблица 21-30 Дифференциация коагулазоположительных стафилококков
Виды стафило- кокков Пигмент Рост анаэ- робно Рост аэробно Коагулаза Хлопья Термо- нуклеаза Гемолиз Фосфа- таза Ацетоин (ФП) Продукция кислоты в анаэробных условиях
маннит мальтоза трегалоза манноза
S. aureus subsp. aureus + + + + + + + + + + + +
S. aureus subsp. afiaerobius - + - + - + + + - ND - -
S. intermedius - + + + d 4- d + - (d) ы + +
S. hyicus - + + + - + - + - - - + +
S. scfileiferi subsp coagualns - + + - + + + V - - +
S. delphini - + + + - - + - (+) + - +
Примечание:
«+>> — 90% положительных по данному свойству штаммов.
«—» — 90% отрицательных по данному свойству штаммов.
«dt — 11-89% положительных ио данному свойству штаммов.
«ND» — тест не имеет значения для дифференциации вида.
() — характеристика выполняется в большинстве случаев.
V — высокая вариабельность.
И увуш
ЧАСТНАЯ МИКРОБИОЛОГИЯ 415
Таблица 21-31. Дифференциация клинически значимых коагулазоотрицательных стафилококков
Ниды стафило- кокков Пигмент Рост анаэ- робно Рост аэробно К пагулаза Гемолиз Фойатаза Нпчобиоцин Продукция кислоты в анаэробных условиях
маннит мальтоза трегалоза манноза
S. epidermidis - + + - (d) + S - + - +
S. warned d + + - (d) - S + + + -
S. haemoliticus d 4- + - + - S d + + -
S. hominis subsp. hominis d + + - - - S - + d -
S. hominis subsp. hominis novobiosepticus - - + - - - R - + - -
S. saprophyticus subsp. saprophyticus d - + - - - R d + + -
416 ЧАСТНАЯ МИКРОБИОЛОГИЯ
Примечание:
«+» — 90% положительных по данному свойству штаммов.
«-» — 90% отрицательных по данному свойству штаммов.
«Ф> — 11-89% положительных по данному свойству штаммов.
«ND» — тест не имеет значения для дифференциации вида.
«S »— чувствительны.
«R» — резистентны.
ЧАСТНАЯ МИКРОБИОЛОГИЯ 417
Показания к проведению молекулярно-генетической идентификации MRSA:
• сомнительные результаты при определении чувствительности к оксациллину
с помощью фенотипических методов;
• тяжелые случаи внутрибольничных инфекций (бактериемия, сепсис), а
также случаи низкой клинической эффективности при использовании окса-
циллина или антибиотиков цефалоспоринового ряда при лечении осложне-
ний. вызванных 5. aureus:
• выделение клинических изолятов 5. aureus, устойчивых к 2-3 антибактери-
альным препаратам, помимо 0-лактамов, но чувствительных к оксациллину
при определении диско-диффузионным методом:
• увеличение частоты внутрибольничных инфекций, вызванных S. aureus.
Полученные при определении чувствительности к антимикробным препаратам
результаты следует трактовать следующим образом:
• штаммы стафилококков, чувствительные к бензилпенициллину, считаются
чувствительными ко всем р-лактамным антибиотикам, препараты выбора —
природные и аминопенициллины;
• при выявлении продукции р-лактамаз и чувствительности к оксациллину
микроорганизм считается резистентным к природным и полусинтетическим
пенициллинам, препараты выбора — оксациллин, ингибиторзащищенные
пенициллины и цефалоспорины I—II поколения.
Эпидемиологический надзор за MRSA — составная часть эпидемиологического
надзора за внутрибольничными инфекциями, центральное звено которого —
молекулярно-генетический мониторинг. Основанный на его данных эпидемиоло-
гический анализ позволяет не только правильно оценивать, но и прогнозировать
эпидемические ситуации, посредством заблаговременных противоэпидемических
мероприятий предотвращать вспышки внутрибольничных инфекций, вызванных
MRSA.
СТРЕПТОКОККИ
ТАКСОНОМИЯ
Стрептококки формируют род Streptococcus из семейства Streptococcaceae (поря-
док Lactobacillales типа Firmiuutes).
В молекулярном анализе показано, что видовое распределение стрептококков
по гемолитической активности, размеру колоний, наличию групповых и типовых
антигенов не всегда согласуется с данными анализа их геномов. По фенотипу
стрептококки подразделяют на 0-гемолитические и не-0-гемолитические. Первая
группа включает пиогенные виды — Str. pyogenes, Str. agalactiae, Str. dysgalactiae, Str.
equi и Str. cams. Видовое обозначение более точное, поскольку пиогенные стреп-
тококки включают и отдельные не-0-гемолитические виды (например. Str. dysga-
lactiae, подвид aysgaiacnae). близкие к Str. anginosus, который отнесен к зеленящим
стрептококкам.
Ангинозные стрептококки на плотных питательных средах формируют срав-
нительно мелкие колонии (<0,5 мм) в отличие от пиогенных стрептококков, фор-
мирующих более крупные колонии (>0.5 мм). Пиогенные, или 0-гемолитические,
стрептококки группируются по Лэнсфилду в соответствии со свойствами поли-
сахаридных антигенов. Однако, если группа В по Лэнсфилду ограничена лишь
видом Str. agalactiae, групповой полисахарид А по Лэнсфилду обнаруживают как у
Str. pyogenes, так и у Str. dysgalactiae подвида equisimilis и Str. anginosus. Связь между
видовой принадлежностью штаммов и другими антигенами по Лэнсфилду еще
более сложная (табл. 21-32),
Таблица 21-32. Фенотипическая характеристика Р-гемолитических стрептококков [Manual of Clinical Microbiology (Editor in chief: Patrick R. Murray). — 9th ed. —
Washington: ASM Press, 2007. — Vol. 1. — P. 412-429]
Виды Группа no ЛэнсФилду Размер колонии, мм Хозяин Тест на бацитрацин САМР-тест ПИР-тест Образование ацетона (VP-тест) Гидролиз гиппурата Образование
трегалозы сорбитола
Str. pyogenes A >0,5 Человек + - + - - + -
Str. agalactiae В >0,5 Человек, корова - + - - + В -
Str. dysgalactiae, подвид. dysgalactiae A, C, G. L >0,5 Животные - - - - - + В
Str. dysgalactiae, подвид equisimilis C >0,5 Человек - - - - - + -
S. equi, подвид equi G >0,5 Животные - - - - - - -
S. equi, подвид zooepidemicus A, C, G. F >0.5 Животные, человек - - - - - - +
S. canis E, P, U, V >0,5 Собака, чело- век - + - - - В -
S. anginosus Негруппируемые <0.5 Человек - - - + - + -
S. porcinus Негруппируемые >0.5 Свинья, чело- век - + + в + +
Примечание:
«-» — отрицательный результат:
«+>> — положительный результат;
«В» — вариабельный результат.
ЧАСТНАЯ МИКРОБИОЛОГИЯ
ЧАСТНАЯ МИКРОБИОЛОГИЯ 419
Вторая группа — непиогенные виды, в основном а-гемолитические и негемоли-
тические, но к ним также относят отдельные виды р-гемолитических стрептокок-
ков. Данное заключение сделано на основании анализа нуклеотидных последова-
тельностей, кодирующих гены 16S рРНК. К непиогенным стрептококкам относят
Str. mitis, Sir. anginosus, Str. mutans, Str. salivarius и Str. bovis.
Среди а-гемолитических видов отдельную группу составляет Str. pneumonias,
отличающийся от других зеленящих стрептококков по чувствия ельности к оптохи-
ну и растворимости в желчи. В группу Str. anginosus входят три вида — Str. anginosus,
Str. constellatus и Str. intermedins.
ОБЩАЯ ХАРАКТЕРИСТИКА
Бактерии рода Streptococcus грамположительны и каталазоотрицательны, имеют
диаметр около 2 мкм и способны к росту в виде цепочек различной длины в жидких
питательных средах. Строение клеточной стенки типично для грамположитель-
ных бактерий. В ее состав входят пептидогликан, содержащий глюкозаминовую и
мурамино зую кислоты, и галактозамин в качестве вариабельного компонента.
Стрептококки — факультативные анаэробы. Для роста видов, относящихся к
Str. viridians и Str. pneumoniae, необходима инкубация в атмосфере, содержащей
5% углекислого газа. Температурный оптимум роста — 37 °C. Для быстрого роста
стрептококков в питательную среду добавляют кровь или сыворотку. Добавление
глюкозы и других углеводов в жидкую питательную среду усиливает рост стрепто-
кокков, но снижает pH среды. Именно поэтому следует применять забуференные
питательные среды.
ЭПИДЕМИОЛОГИЯ
Стрептококки распространены повсеместно. Отдельные виды являются ком-
менсалами и нормальными компонентами микрофлоры кожи. При высокой виру-
лентности стрептококки могут колонизировать различные ткани без видимых
клинических проявлений. Во многих случаях возбудитель обнаруживали даже
при бессимптомном течении заболевания. В то же время их инфекции могут про-
являться как высоколетальные инвазивные болезни. Трудно найти человека, кото-
рый в течение жизни не перенес ту или иную форму стрептококковой инфекции.
Вызываемые стрептококками инфекции значимы с медицинской точки зрения,
по объему социально-экономического ущерба вызывают потери, которые пре-
восходят потери от других нозологических форм. Не существует органов и систем
организма человека, которые не поражаются стрептококками. Они политропны и
именно поэтому актуальны для многих разделов медицины. В XX в. человечество
дважды сталкивалось с крупными вспышками эпидемически значимых заболева-
ний: в 1920-е гг. в Юго-Восточной Азии произошла вспышка «стрептококковой
гангрены», унесшей миллионы жизней, а 1986-96 гг. ознаменовались вспышками
инфекций, вызванных стрептококками групп А и В (так называемыми flesh eating),
в Америке, Западной Европе и Скандинавии. Эти заболевания регистрируют в
настоящее время в виде спорадических случаев во многих странах мира, напоми-
ная об угрозе стрептококковой инфекции, а наблюдаемая смена циркулирующих
типов возбудителей указывает на возможность нового роста заболеваемости.
Стрептококковые инфекции не являются классическими зоонозами, хотя для
большинства видов характерен определенный хозяин. Описаны случаи передачи
инфекции от животного к человеку (Str. suis. Str. agalactiae). Однако гено- и фено-
типический анализ штаммов животного и человеческого происхождения пока-
зал, что штаммы, вызывающие инфекцию у человека, отличаются от штаммов,
вызывающих инфекцию у животных. Анализ штаммов группы В (Str. agalactiae)
выявил существенные генетические различия между изолятами различного про-
исхождения, Человеческие штаммы стрептококков групп С и G принадлежат к Str.
420
ЧАСТНАЯ МИКРОБИОЛОГИЯ
dysgalactiae подвиду equisimilis, в то время как штаммы от животных, относящиеся
к тем же серогруппа м, принадлежат к Str. dysgalactiae, подвиду dysgalactiae, Str. canis,
Str. equi, подвиду equi и Str equi, подвиду zooepidemicus. Резервуаром для штаммов
Str. dysgalactiae подвида equisimilis является человек, и передача инфекции осущест-
вляется только внутри человеческой популяции.
Заражение стрептококками происходит различными путями. Для патогенных
видов (Str. pyogenes и Str. pneumoniae) характерен воздушно-капельный и контакт-
ный пути передачи. Передача кариогенных Str. mutans осуществляется от матери к
ребенку в младенческом возрасте через слюну. Данный путь типичен и для неона-
тальной инвазивной инфекции, вызываемой Str. agalactiae. Часто заражение про-
исходит гематогенным путем или во время родов. Описан контактный путь пере-
дачи неонатальной инвазивной инфекции ребенку от медперсонала родильного
отделения. Причиной эндогенных инфекций могут быть зеленящие стрептококки,
являющиеся частью нормальной микрофлоры слизистой оболочки полости рта.
Проблема разнообразия факторов патогенности стрептококков остается акту-
альной, является предметом организованного регулярного и периодического
мониторинга заболеваемости, в том числе и с помощью методов молекулярной
диагностики. В странах Евросоюза в настоящее время действуют единые програм-
мы по мониторингу стрептококков и вызываемых данными микроорганизмами
заболеваний. Концепция надзора за инфекционной патологией и заболеваемо-
стью обычно строится на показателях распространенности возбудителя инфек-
ции, уровнях заболеваемости, инвалидизации и смертности и размере ежегодно
наносимого стране ущерба. Ошибки в данном подходе приводят к искажениям
и недооценке регистрируемых показателей. В России из большого количества
вызываемых стрептококками заболеваний официально регистрируется только
скарлатина. Б США ежегодно регистрируется до 25 млн больных стрептококкоза-
ми, затраты на лечение оцениваются в 1 млрд долларов в дополнение к еще боль-
шему ущербу от трудовых потерь. Если экстраполировать эти данные на Россию,
то ежегодно следует ожидать до 13-15 млн больных при развитии осложнений
в 0,45 млн случаев. На основании специфики и особенностей стрептококкозы
следует отнести к инфекциям, для которых социальный и экономический ущерб
достаточно выражен.
Заболеваемость инфекциями стрептококковой этиологии находится в поле
зрения ВОЗ: в мире постоянно повышаются уровни лабораторной диагностики,
технологии лечебных и профилактических вмешательств, изучаются вопросы
патогенеза и молекулярной эпидемиологии, значительный прогресс достигнут
в исследовании генетики стрептококков. В настоящее время несколько стран
приступили к созданию рекомбинантных вакцин на основе белковых факторов
патогенности стрептококков групп А и В. Заболеваемость стрептококкозами фор-
мипуется преимущественно за счет инфекций, вызываемых стрептококками групп
А и В, и в меньшей степени — стрептококками групп С, F и G.
КЛИНИЧЕСКИЕ ПРОЯВЛЕНИЯ ИНФЕКЦИИ
Стрептококки гпуппы A (5/г. pyogenes)
У человека Str. pyogenes колонизируют преимущественно слизистую оболочку
полости рта и кожу, что приводит к формированию первичных очагов, являю-
щихся резервуарами передачи инфекции. Str. pyogenes могут быть причиной как
поверхностных, так и глубоких поражений, обусловленных патогенными и токси-
ческими свойствами данного микроорганизма. Str. pyogenes вызывают тонзиллофа-
рингиты, импетиго, скарлатину, синдром токсического шока, бактериемию, сепсис
и поражение мягких тканей (рожистое воспаление, некротизирующий фасциит,
миозит). Помимо перечисленных заболеваний, Str. pyogenes могут вызывать остео-
миелит, септические артрит, пневмонию, менингит, эндокардит и перикардит.
ЧАСТНАЯ МИКРОБИОЛОГИЯ 421
Осложнениями стрептококковой инфекции являются острый гломерулонефрит,
ревматическая лихорадка и хорея. Доминирующие М-типы стрептококков, вызы-
вающих данные нозологические формы, — штаммы серотипов М1 и М3. Наиболее
часто поражаются дети и пожилые пациенты. В группу риска входят больные
сахарным диабетом, вирусными инфекциями, пациенты с хроническими сердечно-
сосудистыми и легочными заболеваниями, иммунодефицитом, наркоманы.
Стрептококки группы В (Str. agalactiae)
Str. agalactiae известен в качестве причины неонатальных инфекций, описанных
в двух клинических проявлениях, а именно как ранняя неонатальная инфекция
(сепсис, пневмония), развивающаяся в первые 7 дней жизни новорожденного, и
более поздняя отсроченная инфекция (менингит, сепсис), развивающаяся между
7-м днем и 3-м месяцем жизни ребенка. Фактор риска неонатального инвазивно-
го заболевания — колонизация урогенитального тракта и толстой кишки матери
Str. agalactiae, выявляемая у 10-35% беременных. У взрослых инвазивная форма
инфекции протекает в виде пневмонии, бактериемии, эндокардита, инфекций
мочеполового тракта, поражений кожи и мягких тканей, остеомиелита. Данные
заболевания развиваются в послеродовом периоде, а также у пациентов с сахар-
ным диабетом, злокачественными новообразованиями, иммунодефицитом.
СтрептококкигруппСиС (Str. dysgalactiae, подвид equisimilis)
Str dysgalactiae, подвид equisimilis, вызывает инфекции, схожие с инфекциями,
вызываемыми Str. pyogenes. Str. dysgalactiae, подвид equisimilis, ассоциирован с инфек-
циями дыхательных путей. кожи и мягких тканей и инвазивными инфекциями,
такими как некротизирующий фаспиит. синдром токсического шока. Описаны слу-
чаи постстрептококкового гломерулонефрита и острой ревматической лихорадки,
связанные с Str. dysgalactiae подвида equisimilis и Str. equi подвида zooepidemicus.
Группа Str. anginosus
Стрептококки видов Str. anginosus, Str. constellatus и Str. intermedins входят в состав
нормальной микрофлоры глотки, урогенитального и кишечного тракта. С этими
стрептококками связывают развитие абсцессов головного мозга, носоглотки и
брюшной полости. Str. constellatus подвида pharyngis — причина фарингитов. По
групповому антигену штаммы группы S. anginosus относят к серогруппам А. С, F, G
и нетипируемым.
Группа Str. mitis
Стрептококки данной группы (Str. mitis, Str. sanguinis. Str. parasanguinis, Str. gor-
donii, Str. cristatus, Str. oralis, Str infantis, Str. peroris и Str pneumoniae) входят в состав
нормальной микрофлоры слизистой оболочки полости рта, кишечного тракта и
наружных родовых путей женщин. Их обнаружение в крови при отсутствии кли-
нических проявлений заболевания не требует антибактериальной терапии вслед-
ствие транзиторного характера бактериемии. Данные микроорганизмы могут
также входить в состав нормальной микрофлоры кожи. Стрептококки данной
группы часто становятся причиной эндокардитов, сепсиса или пневмонии на фоне
иммуносупрессорной химиотерапии.
В эюй группе Str. pneumoniae занимают особое место среди зеленящих стрепто-
кокков по своему клиническому значению и отличию от стрептококков Str. tnitis.
Именно поэтому детальная харак герметика Str. pneumoniae должна быть описана
отдельно, с учетом их высокой эпидемиологической и клинической значимости.
Str. pneumoniae чаще всего выделяют из дыхательных путей. Данные микроор-
ганизмы — возбудители пневмоний, менингитов, отитов, синуситов и в редких
случаях перитонитов. Колонизация дыхательных путей нередко сопровождается
длительным носительством Str. pneumoniae у детей.
.«a. z
422 ЧАСТНАЯ МИКРОБИОЛОГИЯ
Группа Str. saiivarius
Стрептококки группы Str. saiivarius включают Str. saiivarius и Str. vestibularis. Str.
saiivarius могут вызывать бактериемию, эндокардит и менингит. Str. vestibularis не
связывают с конкретной патологией. Их выделение из крови в значительной сте-
пени коррелирует с интерстициальной неоплазией.
Группа Str. mutans
Str. mutans и Str. sobrinus обычно выделяют из полости рта. Str. mutans — этиоло-
гический агент зубного кариеса, Str. criceti и Str. ratti крайне редко обнаруживают
у человека, в то время как Str. downed Str. ferns, Str. macacae и Str. hyovagi nalis встре-
чаются только у животных.
Группа Str. bovis
Данная группа стрептококков включает Str. equinis, Str. gallolyticus, Str. infantanus
и Str. alactolyticus, которые обнаруживают в крови пациентов с бактериемией,
сепсисом и эндокардитом, а также при интерстициальных новообразованиях (Str.
gallolyticus, подвид gallolyticus) и менингите (Str. gallolylicus, подвид pastenanus).
Наибольшие таксономические изменения произошли именно в этой группе стреп-
тококков.
Другие стрептококки, обнаруживаемые в патологическом материале
Некоторые виды, исходно являющиеся патогенными для животных, время от
времени обнаруживают у людей, имеющих тесный контакт с животными. Str. suis,
Str. porcinus и Str. iniae относятся к данной категории микробов Str. suis — патоген
свиней, однако в некоторых случаях встречается у больных людей при менингите
и бактериемии. Str. suis может быть положительным по антигенам R, S или Т. Str.
porcinus (группы Е, Р. U или V по Лэнсфилду) также является патогеном свиней.
Str. iniae патогенен для рыб и не относится ни к одной из серогрупп по Лэнсфилду.
Данный вид выявляют при инфекции мягких тканей, эндокардите и менингите у
людей, занятых обработкой рыбы.
ЛАБОРАТОРНАЯ ДИАГНОСТИКА
Лабораторная диагностика стрептококковых инфекций человека строится на
традиционных принципах бактериологии, предусматривающих обнаружение воз-
будителя в клиническом материале (с этой целью применяют бактериоскопию,
изоляцию на питательных средах, выявление антигенов стрептококков иммуно-
логическими тестами и их генома методами молекулярной диагностики) и выяв-
ление у пациентов специфической сероконверсии.
Особенности лабораторной диагностики стрептококковых инфекций обуслов-
лены многообразием этих бактерий, что диктует необходимость применения
различных методов их идентификации и типирования. На заключительном этапе
лабораторных исследований встает задача определения чувствительности выде-
ленного стрептококка к антимикробным препаратам для выбора оптимальной
схемы лечения пациента.
Бактериоскопия
Поскольку стрептококки хорошо воспринимают анилиновые красители, для
окраски мазков клинического материала используют обычные методы (в первую
очередь, окрашивание по Граму).
Обнаружение стрептококков в клиническом материале
В качестве экспресс-методов диагностики инфекций стрептококков групп А и
В в настоящее время ряд фирм производит одноэтапные иммунохроматографи-
ческие тесты. Они позволяют быстро (в течение 5-10 мин) проводить исследова-
ЧАСТНАЯ МИКРОБИОЛОГИЯ 423
ние клинического материала, высокоспецифичны и чувствительны. Основное их
назначение — скрининг. Получение в этих тест-системах отрицательного резуль-
тата не исключает наличие у обследуемого пациента стрептококковых инфекций
(это необходимо подтвердить другими лабораторными методами диагностики).
Изоляция и идентификация азбудителя
Гемолитические стрептококки весьма требовательны к питательным средам.
В качестве стандартных сред обычно используют бульон Тодда-Хевитта и колум-
бийский кровяной агар. При выращивании стрептококков группы А для их
М-типирования к бульону Тодда-Хевитта добавляют неопептон, который пода-
вляет активность ферментов, гидролизующих М-белок. Добавление к бульону
Тодда-Хевитта 2-5% экстракта дрожжей стимулирует рост бактерий. Культуры
стрептококков из очагов с вероятным присутствием грамотринательной флоры
лучше растут в присутствии антибиотиков. Колумбийский кровяной агар, содержа-
щий колистин (10 мкг/мл) и налидиксовую кислоту (15 мкг/мл), не поддерживает
рост грамотрицательных бактерий, но на нем хорошо растут стафилококки или
коринебактерии. При замене налидиксовой кислоты на оксолиновую (5 мкг/мл)
подавляется рост грамотрицательных бактерий, стафилококков и коринебакте-
рий Считается, что комбинация триметоприма (1,25 мкг/мл) и сульфаметоксазо-
ла (23,76 мкг/мл) в кровяном агаре способствует улучшению роста стрептококков
группы А. Сочетание неомицина с полимиксином В не оказывает существенного
влияния на рост стрептококков группы А. но подавляет размножение других
бактерий, часто присутствующих в материале, собранном из верхних дыхательных
путей. Добавление в жидкую питательную среду гентамицина (8 мкг/мл) и нали-
диксовой кислоты (15 мкг/мл) улучшает рост стрептококков группы В.
При посеве исследуемого материала с тампона на чашку с кровяным агаром сле-
дует прокатать тампон по участку размером 3x3 см у края чашки. От этого места
материал следует распределить петлей в разных направлениях для получения
изолированных колоний. Проколы петлей внутрь агара позволяют получить рост
в глубине, что усиливает р-гемолиз штаммов стрептококков, которые в аэробных
условиях слабо экспрессируют гемолизин. Глубинный гемолиз характерен для
стрептолизина-О, чувствительного к кислороду. При попытке выделения бактерий
из «скудного» материала, кроме первичного посева, желательно выполнит ь парал-
лельный посев на жидкие селективные питательные среды или среды обогащени я
При обследовании здоровых носителей использование среды обогащения увели-
чивает частоту выявления стрептококков на 15-20% по сравнению с результатами
первичных посевов.
Гйой из абсцессов, жидкость из зоны рожистого воспаления или другой иссле-
дуемый материал с предположительно низким содержанием возбудителя можно
сразу посеять в обогащенную жидкую питательную среду. При инфекции глотки
посевы слюны могут содержать гемолитические стрептококки, однако более
достоверные результаты дают посевы материала из гло гки.
Если ставится задача выделения из исследуемого материала только гемолити-
ческого стрептококка, лучше использоват ь посев на кровяной агар с кристаллвио-
летом, который угнетает рост стафилококков. Селективные питательные среды,
содержащие антибиотики, подавляют рост всех грамотрицательных микроорга-
низмов, не оказывая нежелательного эффекта на формирование колоний стреп-
тококков. Благодаря угнетению роста конкурирующих микробов селективные
питательные среды повышают частоту выделения гемолитических стрептококков
в работе с исследуемым материалом, содержащим незначительное количество
гемолитических стрептококков.
Инкубация материала при 35-37 °C в течение 12 ч в аэробных условиях на
чашках с кровяным агаром (предпочтительнее использовать кровь овцы или
424
ЧАСТНАЯ МИКРОБИОЛОГИЯ
лошади) — общепринятый стандарт выделения гемолитических стрептококков
групп А, В, С, F и G. Скорость роста стрептококков групп А и В можно увеличить,
инкубируя засеянные чашки в атмосфере, содержащей 5% углекислого газа, или в
анаэробных условиях. Анаэробная среда стимулирует рост стрептококков и ока-
зывает угнетающее действие на стафилококки, гемофильную палочку и корине-
бактерии. При этом ускоряется формирование зон гемолиза вследствие продукции
стрептолизина-О. Чашки, в которых после 12 ч инкубации отсутствуют признаки
роста стрептококка, рекомендуют инкубировать еще 24-48 ч.
Большинство типичных колоний стрептококка группы А имеют округлую
форму с ровным краем и диаметром 0,5-2 мм и более.
Существуют три типа колоний стрептококков — слизистые, матовые и бле-
стящие. Слизистые колонии обычно более крупные по размеру, часто сливаются
и выглядят как капельки воды. Такие колонии характерны для штаммов, про-
дуцирующих большое количество гиалуроновой кислоты для построения кап-
сулы. Матовые колонии, иногда называемые подслизистыми, часто являются
результатом подсыхания слизистых колоний, плоские, с неровной поверхностью.
Блестящие колонии имеют меньший размер и куполообразны.
Размер колоний обычно варьирует у штаммов разных групп. р-Гемолитические
стрептококки пиогенной группы (Str. pyogenes. Str. agalactiae, Str. dysgalactiae под-
вида equisimilis) образуют при росте на плотной питательной среде колонии раз-
мером более 0,5 мм после 12-часовой инкубации. Для 0-гемолитических форм
Str. anginosus характерны колонии размером менее 0,5 мм, обладающие специфи-
ческим запахом ириски или карамели из-за продукции диацетила. Str. agalactiae
образуют более крупные колонии с относительно малой зоной гемолиза в отличие
от остальных р-гемолитических стрептококков пиогенной группы. Встречаются
не гемолитические формы Str. agalactiae, схожие с энтерококками. Размер зоны
гемолиза вокруг колоний Str. pyogenes обычно превышает диаметр колонии в
2-4 раза. У некоторых штаммов зона гемолиза шире, у других представлена узким
кольцом.
Большинство р-гемолитических стрептококков пиогенной группы вызывают
полный лизис эритроцитов. Однако вокруг колоний некоторых штаммов появ-
ляется зеленая окраска, напоминающая а-гемолиз. У таких штаммов полный
лизис эритроцитов происходит после длительной инкубации или инкубации в
анаэробных условиях. При изучении гемолитических и морфологических свойств
стрептококков следует учитывать влияние компонентов сред, толщину агара, кон-
центрацию и происхождение крови, атмосферу инкубации. Использование крови
человека не рекомендуют, так как она часто содержит антитела к стрептококкам и
отрицательно влияет на их инкубацию.
Негемолитические серые колонии типичны для Str. bovis и Str. salivarius.
Тщательное исследование морфологии и гемолитических свойств колоний
важно для подтверждения диагноза стрептококковой инфекции и выделения
штаммов. Для дифференциации стрептококков от других гемолитических бакте-
рий следует прибегать к микроскопии окрашенных по Граму мазков из выросших
колоний. С этой же целью применяют тест на каталазу (стрептококки каталазоо-
трицательны).
Идентификация и типирование
Групповая идентификация стрептококков по Лэнсфилду
Группоспецифический антиген большинства стрептококков — полисахарид,
локализованный в их клеточной стенке. Его можно опредепить посредством
группоспецифической сыворотки с помощью реакции преципитации, РГА, имму-
нофлюоресценции и ИФА. Существуют разные методы экстракции групповых
полисахаридов, в том числе соляной и азотистой кислотами, формамидом, авто-
ЧАСТНАЯ МИКРОЬИОЛОГИЯ 425
клавированием взвеси стрептококков в 0,9% растворе NaCl, а также с применени-
ем ферментов и фаг-ассоциированного лизина.
Реакция преципитации имеет ряд вариантов: кольцепреципитацию, преци-
питацию в капиллярах, реакцию двойной встречной диффузии или иммуноэ-
лектрофореза в геле. Реакция агглютинации может быть выполнена как прямая
агглютинация, коагглю гинация или латекс-агглютинация (с применением взвеси
нагруженных группоспецифическими антителами стафилококков, содержащих
белок А. либо калиброванных частиц латекса). Групповой антиген В по Лэнсфилду
встречается только в штаммах Str. agalacticae. Групповой антиген F позволяет иден-
тифицировать штаммы, принадлежащие к Str. anginosus. Групповые антигены А, С
и G указывают на необходимость дальнейшей видовой идентификации штаммов с
использованием молекулярно-генетического анализа.
В настоящее время существует много видов экспресс-анализа для дифференциа-
ции стрептококков в зависимости от обнаружения группового антигена непосред-
ственно в первичном материале. Наиболее применимы тесты, в которых исполь-
зуют группоспецифическе IgG, фиксированные на частицах калиброванного
латекса или на взвеси клеток S. aureus, несущих на поверхности белок А в качестве
рецептора для IgG. При этом время, необходимое для диагностики, сокращается до
10-2С мин, что играет важную роль при своевременном назначении эффективной
антибиотикотерапии.
Фенотипическая идентификация стрептококков
Описываемые ниже фенотипические тесты служат дополнением к групповой
идентификации стрептококков.
А. Бацитрациновый тест. Str. pyogenes, в отличие от других 0-гемолитических
стрептококков, обладает высокой чувствительностью к бацитрацину. Тест на
бацитрацин можно использовать для идентификации Str. pvogenes, поскольку
|3-гемолитические штаммы других видов стрептококков, которые могут содержать
групповой антиген А, устойчивы к бацитрацину. Считается, что если вокруг дисков
с бацитрацином (0,04 ME), помещенных на поверхность агара, отсутствует рост
[3-гемолитических стрептококков, испытуемый штамм относится к Str. pyogenes.
Размер зоны задержки роста вокруг диска не имеет значения. Оптимальна инкуба-
ция чашек в течение 18-20 ч при 36 °C в атмосфере, содержащей 5% углекислого
газа. Существуют коммерческие диски с бацитрацином. Однако необходимо иметь
в виду, что опубликованы данные по обнаружению в Европе бацитрацинрези-
стентных изолятов Str. pyogenes.
Б. ПИР-тест. Метод определения групповой принадлежности Str. pyogenes
основан на способности стрептококковой лирролидониламинопептидазы (ПИР)
гидролизовать 1-пирролидонил-р-нафтиламид до р-нафтиламина, который в при-
сутствии специального реагента окрашивается в красный цвет. Около 98% штам-
мов Str. pyogenes дают положительную ПИР-реакцию. Хотя Str. iniae и Sir. porcinus
также ПИР-позитивны, они крайне редко встречаются в клиническом материале,
поскольку являются видами, ассоциированными с патологией животных. Ряд
бактерий каталазоотрицательных родов (Enterococcus, Aerococcus, Abiotrophia.
Gemella, Lactococcus) также дают положительную ПИР-реакцию. Однако они не
могут помешать определению Str. pyogenes из-за своей характерной морфологии.
Широкий спектр ПИР-положительных несгрептококковых бактерий ограничи-
вает применение данного теста. Существует ряд коммерческих вариантов ПИР-
теста, имеющих одинаковые чувствительность и специфичность, которые можно
использовать в бактериологической диагностике.
В. Гиппурат-гидролизный тест. Способность к гидролизу гиппурата — допол-
нительный тест для идентификации Str. agalactiae. При его выполнении суспензию
бактериальных клеток в 1% растворе гиппурата натрия выдерживают в течение 2 ч
при температуре 35 °C. Продукт гидролиза гиппурата определяют по появлению
426 ЧАСТНАЯ МИКРОБИОЛОГИЯ
пурпурного цвета после добавления нингидринового реагента. Помимо Str. agalac-
tiae, гидролизовать гиппурат способны некоторые зеленящие стрептококки.
Г. Тест Фогеса-Проскауэра. Тест выявляет образование ацетона при фер-
ментации глюкозы штаммами Str. anginosus с групповыми антигенами А, С или
G по Лэнсфилду и позволяет дифференцировать их от штаммов с идентичными
групповыми антигенами, но принадлежащими к другим видам стрептококков.
Тестируемый штамм инокулируют в 2 мл VP-среды и инкубируют при 35 °C в
течение 6 ч. Затем в пробирку добавляют 5% раствор а-нафтола и 40% раствор
гидроксида калия и выдерживают при комнатной температуре в течение 30 мин.
Положительный результат определяют по появлению розово-красного цвета.
Д. [З-Глюкуронидазный тест. Выявление [3-глюкуронидазной активности у
штаммов Str. dysgalactiae подвида equisimilis, содержащих групповой антиген С или
G по Лэнсфилду, позволяет дифференцировать их от штаммов Str. anginosus с теми
же групповыми антигенами. Для выполнения экспресс-теста используют коммер-
ческие наборы.
Е. Оптохиновый тест. Тест используют в основном для идентификации опто-
хинчувствительных штаммов Str. pneumoniae. Наряду с чувствительными к опто-
хину штаммами регистрируются и резистентные штаммы Str. pneumoniae, а также
оптохинчу ветвит ельные изоляты Str. mitis. Диски с оптохином наносят на поверх-
ность кровяного aiapa и инкубируют в течение 18-20 ч в атмосфере, содержащей
5% углекислого газа. Оптохинчувствительные изоляты образуют зоны подавления
роста не менее 14 мм с 6-миллиметровым диском, содержащим 5 мкг оптохина,
или зоны подавления роста не менее 16 мм с 10-миллиметровым диском. При
размере зоны подавления роста 9-13 мм с 6-миллиметровым диском необходимо
повторное тестирование штамма с использованием чистой культуры. Для выпол-
нения теста используют коммерческие диски с оптохином.
Ж Колориметрический тест StrepSwift. Представляет собой карточку с 3
тестовыми окошками для выявления у тестируемой культуры пироглютаматами-
нопептидазы, лейцинаминопептидазы и эскулиназы. По наличию этих ферментов
можно определить принадлежность изолята к грамположительным каталазонега-
тивным коккам и провести их первичную дифференциацию. Результаты оценивают
через 10 мин после внесения суспензии исследуемой культуры в тестовые окошки.
Серотипирование стрептококков
Серотипирование стрептококков группы А. Типирование стрептококков
существенно при изучении нефрито- и ревматогенности штаммов группы А и
эпидемиологическом надзоре за вспышками инфекции. Разделение Str. pyogenes на
серотипы основано на идентификации белковых антигенов, известных как Т-, OF-
и М-белки. Наиболее важен М-белок — высокоспецифичный антигенный маркер
и ведущий фактор вирулентности Str. pyogenes. Постинфекционный иммунитет к
стрептококкам группы А связан с формированием типоспецифических антител
к М-белку. Некоторые М-типы стрептококков продуцируют и другое типоспеци-
фическое вещество, названное фактором опалесценции. Поскольку антигенная
специфичность М-белка и фактора опалесценции коррелируют у 30% типов стреп-
тококков. оба белка важны для диагностики.
А. Т-типирование. Т-типирование проводят в реакции РГА на стекле, исполь-
зуя набор анти-Т-сывороток, как пулированных (Т, U, W, X, Y, Z), так и моно-
валентных. Производятся коммерческие наборы обоих вариантов сывороток.
Однако область их применения достаточно узка. Для реакции применяют ста-
бильные суспензии стрептококков. Спонтанно агглютинирующие культуры сле-
дует предварительно обработать трипсином. Существует около 20 вариантов
Т-антигенного паттерна штаммов.
Б. Типирование с использованием реакции опалесценции сыворотки.
Наличие или отсутствие фактора опалесценции (OF-фактора) — постоянная
ЧАСТНАЯ микробиология 427
характеристика любого М-типа Str. pyogenes Все М-типы можно разделить на
ОБ-положительные и ОБ-отрицательные. Фактор опалесценции выявляют в надо-
садочном слое бульонной культуры и в экстрактах из культур стрептококков, полу-
ченных обработкой соляной кислотой или додеиилсульфатом натрия. В качестве
су6с грата для активности OF-фактора применяют лошадиную, бычью или свиную
сыворотку. OF-типирование основано на специфическом подавлении реакции
опалесценции сыворотки антисывороткой, содержащей типоспецифичные анти-
OF-антитела.
Существуют различные методы OF-типирования: агаровый, пробирочный и
микропланшетный. В клинической практике OF-типирование не используют, так
как для определения данного фактора необходимо поддерживать коллекцию из
30 анти-ОБ-сывороток, что под сил}- только высокоспециализированным центрам.
К недостаткам OF-типирования относится и невозможность применения метода
для идентификации наиболее распространенных OF-отрицательных штаммов Str.
pyogenes.
В. М-типирование. Серотипирование, основанное на антигенных различиях
М-белков, связанных с гетерогенностью аминокислотной последовательности их
N-терминальной области, стало основным (особенно в эпидемиологии) методом
внутривидовой классификации штаммов Str. pyogenes. В настоящее время известно
более 130 М-серотипов Str. pyogenes. Недостатки этого метода: необходимость
большого количества типовых сывороток высокого качества, невозможность
типирования штаммов, слабо экспрессирующих М-белок и экспрессирующих
М-белок неизвестного типа.
М-типирование осуществляют с помощью нескольких подходов.
• Реакцию преципитации в геле между М-антигеном и типоспецифической
антисывороткой используют для идентификации М-типа Str. pyogenes.
Реакция считается положительной, если солянокислый экстракт стрепто-
коккового штамма дает четкую полосу преципитации в геле с единственной
адсорбированной М-типоспецифической антисывороткой.
Этот метод позволяет использовать и неадсорбированные М-антисыворотки
(при наличии соответствующих контрольных проб), что является одним из преи-
муществ реакции преципитации в агаровом геле.
• Бактерицидный тест, основу которого составляют антифагоцитарный эффект
М-белка стрептококка и его нейтрализация типоспецифическими антитела-
ми.
Существует два типа бактерицидного теста.
Непрямой тест служит для идентификации или подтверждения М-типа иссле-
дуемого штамма. Для его постановки используют нормальную кровь человека,
максимально свободную от типовых антител, а добавляемая гипериммунная анти-
М-сыворотка должна опсонизировать исследуемый штамм, то есть способствовать
фагоцитозу клеток.
В прямом тесте не используются типовые сыворотки. По способности исследуе-
мого штамма размножаться в нормальной крови при отсутствии типоспецифиче-
ских анти-М-антител определяют содержание М-белка. С помощью данного теста
можно резко увеличить количество М-белка в штаммах Str. pyogenes с исходно
малым его содержанием.
Постановка бактерицидного теста довольно трудоемка и занимает много вре-
мени. Его используют при особых обстоятельствах, например для верификации
новых М-типов стрептококков. Важная сфера применения бактерицидного теста —
отбор штаммов для производства типовых анти-М-сывороток.
С ростом количества постоянно обнаруживаемых новых М-серотипов (к насто-
ящему моменту обнаружено более 130 иммунологических вариантов М-белка) в
мире не осталось практической лаборатории, способной выполнить полноценную
428 ЧАСТНАЯ МИКРОБИОЛОГИЯ
серодиагностику штаммов Str. pyogenes по М-белку. Более привлекательными ста-
новятся методы молекулярно-генетического типирования, поскольку их выпол-
нение не требует широкой панели иммунных сывороток, подготовки стандартных
инокулюмов, хранения при температуре -70 °C, подбора универсальных доноров,
не имеющих антител к большинству серотипов стрептококков, как для выполне-
ния бактерицидного теста.
Серотипирование стрептококков группы В. Если для отнесения штамма к
стрептококкам группы В (СГВ) достаточно обнаружение у него группового анти-
гена, то для внутривидовой идентификации штаммов СГВ используют особен-
ности наружной капсулы, которая рассматривается в качестве основного фактора
вирулентности. Среди СГВ долгое время выделяли три серотипа — I, II, III, затем
девять серотипов — la, lb, II, III, IV, V, VI, VII, VIII. Недавно был обнаружен новый
антиген, обозначенный номером IX. Капсула всех известных типов СГВ включает
остатки глюкозы, галактозы и N-ацетилнейраминовой (сиаловой) кислоты; в кап-
суле присутствует N-ацетилглюкозамин, за исключением типов VI и VIII. Рамноза
обнаружена в капсуле серотипа VIII. На антигенные свойства типового полисаха-
рида влияет не только состав, но и порядок соединения компонентов в полимер-
ную структуру, а также их соотношение. Например, в составе капсулы клинически
значимого типа V глюкоза, галактоза, N-ацетилглюкозамин и сиаловая кислота
присутствуют в соотношении 3:2:1:1, а в составе капсулы типа II — те же компо-
ненты, но в соотношении 2:3:1:1.
Рутинные методы для внутривидового типирования — серологическая иденти-
фикация штамма СГВ нейтрализацией в капиллярах или преципитацией в агаре.
Эти методы просты в выполнении, однако имеют недостатки, обусловленные
необходимостью получения очищенных моноспецифических сывороток ко всем
или как минимум к основным типам, — такая диагностика возможна только в про-
филированных научных учреждениях. Увеличение числа серотипов СГВ еще более
осложняет возможности серотипирования.
В диагностике СГВ можно использовать определение белковых антигенов.
Ранее у СГВ были обнаружены два поверхностных пептида, |3- и а-антигены.
Исходно ошибочное представление, что данные антигены представляют собой
антигенные варианты одного и того же белка (белка С), привело к обозначению
штаммов разных серотипов как С.( или Ср. Генотипированием доказано, что эти
антигены самостоятельные, генетически независимые белки. Белок С встречается
в виде набора как минимум пяти различных иммунологичес ких вариантов, к кото-
рым, помимо самого а-антигена, относятся а- и й/6-подобные белки, а-Антигены
семейства представлены у 70-80% штаммов СГВ. [3-Антиген — рецептор иммуно-
глобулинов А, иммунологически однороден и встречается лишь у 30-40% штам-
мов, преимущественно принадлежащих к серотипам I и II.
Молекулярно-генетические методы диагностики
Появление новых молекулярных методов внутривидовой диагностики стреп-
тококков, таких как е/л/и-типирование, мультиплексное типирование но генам
капсульных полисахаридов и MLST-типирование, вероятно, снизит в ближайшем
будущем диагностическое значение типирования стрептококков, основанного на
детекции поверхностных белков иммунологическими методами.
Emm-генотипирование Str. pyogenes. Антигенные различия М-белков
обусловлены гетерогенностью аминокислотной последовательности их
N-терминальной области. Это свойство определяется гетерогенностью нуклеотид-
ной последовательности б'-конца кодирующего М-белок гена етт. В связи с этим
для классификации штаммов Str. pyogenes разработан метод ewm-типирования,
основанный на анализе нуклеотидной последовательности 5’-конца гена етт. Для
его выполнения из изучаемого штамма экспресс-методом или традиционной фенол-
ЧАСТНАЯ МИКРОБИОЛОГИЯ 429
Праймер етт seq2 emm-тип emm-подтип
Секвенирование
Праймер №2
ПЦР
Праймер №1
I----------
Сигнальный пептид 90 пар нуклеотидов 60 пар
Рис. 21-3. Схема определения emm-тила и тл-подтипа.
хлороформенной экстракцией выделяют геномную ДНК. Затем с помощью прай-
меров 5’-TArT(C/G)GCTTAGAAAATTAA-3’ и 5’-GCAAGTTCTTCAGCTTGTTT-3’
(праймеры № 1 и № 2) амплифицируют фрагмент гена етт (рис. 21-3). С помо-
щью праймера 5’-TATTCGCTTAGAAAATTAAAAACAGG-3’ (праймера етт seq2)
осуществляют секвенирование гипервариабельного участка на 5’-конце гена етт.
Для ежда-типирования используют 150 п.н. амплификата, кодирующие фрагмент
М-белка, начиная с его первой аминокислоты. Подушенную последовательность
(150 п.н.) сравнивают с имеющимися в базе данных (ftp://ftp.cdc.gov/pub/inieciiuus_
diseases/biotech/tsemni) последовательностями етлтп-типов. Для вывода о принадлеж-
ности штамма определенному em/я-типу достаточна последовательность гена етт
длиной 90 п.н., кодирующая первые 30 аминокислот зрелого М-белка (см. рис. 21-3).
Если гомология данного участка с последовательностями известных ет/л-генотипов
мгнее 92%, Центр контроля заболеваемости присвоит штамму новый едал-генотип.
ЕТкот-подтип определяют по последовательности с 91-го по 150-й нуклеотид
амплификата. Любые мутации и единичные нуклеотидные замены в данной после-
довательности — основание для присваивания нового едам-подтипа. Например,
аббревиатура «етт4.1» означает, что штамм Str. pyogenes относится к типу етт4 и
подтипу етт4.1.
Стандартизованность метода ewm-типирования и воспроизводимость результа-
тов позволяют проводить надежный мониторинг генотиов Str. pyogenes, выделен-
ных в разные периоды времени в разных регионах. В Африке наиболее распро-
странены штаммы Str pyogenes emml2 и етт75, в Азии — emml и етт12. Наиболее
инвазивные штаммы Str pyogenes в Азии и Латинской Америке — emml и етт12,
в США на второе место выходят штаммы типа еттЗ. С кожными заболеваниями
в Азии чаще ассоциированы штаммы emml и етт2, в Африке — етт80, emmlOO,
етт'28 и етт74. в Латинской Америке — етт83 и етт53. В России проведены
обследования детских учреждений и воинских коллективов. В частности, обна-
ружено, что доминирующими ете/л-типами Str. pyogenes, выделенными от детей
в Санкт-Петербурге, являются еттЗ (31%), етт!2 (19%), етт4 (18%) и етт!
(17%). Подобная информация позволяет в определенной мере оценить эпидемио-
логическую ситуацию и циркуляцию возбудителей различных типов.
Технологию emm-типирования применяют в основном для штаммов Str. pyo-
genes. Однако в базу данных етлтл-типов внесен и ряд елг/л-типов стрептококков
ceporpvnn С и G, технология анализа которых аналогична. Данные о типировании
стрептококков групп С и G разрознены и не систематизированы, основной массив
данных доступен только для штаммов Str. pyogenes.
Гено типирование стрептококков группы В. Генетические структуры, детер-
минирующие синтез капсулы, компактно локализуются в одном участке генома
СГВ и представлены 16-18 однонаправленными генами. Большинство генов
характеризуются высокой консервативностью. Типичный капсульный локус имеет
следующую структуру:
• регуляторные гены;
430
ЧАСТНАЯ МИКРОБИОЛОГИЯ
• гены, кодирующие длину полимерной цепи и ее экспорт;
• гены, кодирующие ферменты, определяющие сборку компонентов капсулы
(гликозилтрансферазы);
• гены, кодирующие ферменты, определяющие полимеризацию (полимеразы);
• гены, кодирующие белки, отвечающие за транспорт и синтез моносахароь.
В средней части капсульного регулона локализуются наиболее гетерогенные
участки — гены cspG до cspK, кодирующие ферменты, в основном гликозилтранс-
феразы, отвечающие за межструктурные связи компонентов капсулы и опреде-
ляющие их антигенные различия.
Генетические различия между серотипами позволили разработать молекулярно-
диагностические подходы, основанные на мультиплексной ПЦР, которая пред-
ставляет собой стандартную ПЦР с одновременным использованием набора
праймеров к генам, кодирующим различные капсульные антигены. Праймеры
подбирают таким образом, чтобы по размеру продукта ПЦР было возможно сде-
лать заключение о принадлежности исследуемого штамма к определенному серо-
типу. Этот подход основан на применении двух наборов праймеров. Первая смесь
содержит праймеры на серотипы la, lb, II, III и IV. а вторая — на серотипы V. VI,
VII и VIII. Праймеры dltS-F и dltS-R на ген dltS, специфичные для СГВ, предлагают
использовать в качестве положительного контроля (табл. 21-33). Таким образом,
для определения типа штамма СГВ достаточно выполнения трех П1 IP. что доступ-
но любой современной диагностической лаборатории.
Мультилокусное типирование ст рептококков. Один из наиболее признан-
ных в настоящем приемов молекулярной диагностики — мультилокусное генотипи-
рование (MLST). Этот метод заключается в сравнительном анализе нуклеотидных
последовательностей наиболее гетерогенных фрагментов ДНК штаммов одного
вида. В настоящее время известны базы данных MLST (http://www.mlst.net. http://
pubmlst.org), позволяющие проводить эпидемиологический мониторинг штаммов
многих патогенных микроорганизмов, MLST обладает многими преимуществами
по сравнению с другими методами типирования. Для него характерны высокие
специфичность и достоверность, абсолютная воспроизводимость результатов,
практически полная стандартизовапность. Метод MLST информативен и надежен
для эпидемиологического анализа любых микроорганизмов. В базы данных для
анализа включена информация о нуклеотидных последовательностях для Str. pyo-
genes, Str. agalactiae. Str. pneumoniae, Str. oralis. Str. uberis, Str. suis и Str. zooepidemicus.
Для каждого вида обычно выбирают по 7 маркерных генов. В целях сравнения
штаммов конкретные фрагменты данных генов (размером 400-500 п.н.) амплифи-
цируют с помощью ПЦР и секвенируют. По установленной нуклеотидной последо-
вательности каждого гена определяют его аллель, а суммарным набором аллелей
7 генов для каждого из штаммов характеризуется его мультилокусный генотип
(MLST-тип, ST-тип или ST-sequence genotype). Например, если штамм Str. pyogenes
характеризуется набором аллелей 7 генов 1-1-1-1-1-1-1, то такой набор аллелей
соответствует ST-типу № 1, а если штамм имеет набор аллелей 15-14-7-7-19-3-1,
то ei о относят к ST-типу № 73. Обнаружение каждого нового аллеля любого из
маркерных генов автоматически приводит к регистрации нового ST-типа.
Таблица 21-33. ДНК-праймеры и предполагаемые продукты ПЦР для типирования штаммов СГВ
Праймер Последовательность праймеров Ген Размеры ампли- кона, TH. Номер GenBank
la-F la-R GGTCAGACTGGATTAATGGTATGC GTAGAAATAGCCTATATACGTTGAATGC cpslaH cpslaH 521 и 1826 AB028896
Ib-F Ib-R 1AAACGAGAATGGAATATCACAAACC GAATTAACTTCAATCCCTAAACAATATCG cpsIbJ cpsIbK 770 AB050723
ll-F ll-R GCTTCAGTAAGT AHGTAAGAC GATAG nCTCTAGGAAATCAAATAATTCTATAGGG cps2K cps2K 397 AY375362
ЧАСТНАЯ МИКРОБИОЛОГИЯ 431
Окончание табл. 21-33
Праймер Последовательность праймеров Ген Размеры ампли- кона, п.н. Номер GenBank
Ill-Fa TCCGTACTACAACAGAC ГСАТСС ср$1а/2/31 1,826 AF163833
lll-Ra AGTAAGCGTCCATACATTCTATAAGC cps1a/2/3J
IV-F IV R GGTGGTAATCCTAAGAGTGAACTGT CCTCCCCAATTTCGTCCATAATGGT cpsfN cps4N 578 AF355776
V-F GAGGCCAA1CAGTTGCACGTAA cps50 701 AF349539
V-R AACCTTCTCCTTCACACTAATCCT cps5O
Vl-F GGrtCTTGAGATGGCAGAAGGTGAA cpsbl 487 AF337958
Vl-R CTGTCGGACTATCCTGATGAATCTC cpsSI
Vll-F CCTGGAGAGAACAAT GTCCAGAT cps7M 371 AY376403
Vll-R GCTGGTCGTGATTTCTACACA cps7M
Vlll-F AGGTCAACCACTATATAGCGA cps3J 282 AY375363
Vlll-R TGTTCAAATTCCGCTGACTT cps8J
ditS-F AGGAaTACCAGGCGATGAACCGAT (fits 952 AL766853
dltS-R TGCTCTAATTCTCCCCTTATGGC dltS
В табл. 21-34 приведены маркерные гены и соответствующие праймеры,
используемые для мультилокусного типирования трех наиболее патогенных для
человека видов стрептококка — Str. pyogenes, Str. agalactiae и Str. pneumoniae. Для
каждого вида существует свой специфический набор генов и праймеров. В отличие
от Str. pyogenes и Str. pneumoniae, для маркерных генов Str. agalactiae используют по
две пары праймеров — внешнюю, позволяющую амплифицировать относительно
протяженный участок гена (от 589 до 859 п.н.), и внутреннюю, с помощью которой
происходит секвенирование искомых последовательностей (от 459 до 501 п.н ) на
матрицах ДНК, полученных с использованием внешней пары праймеров.
К началу 2010 г. в базах данных зарегистрировано 569 MLST-типов для Str. pyogenes,
482 MLST-типа для Str. agalactiae и 4586 MLST-типов для Str. pneumoniae. Например,
инвазивные штаммы Str. pyogenes генотипа emml часто характеризуются типом ST-28,
а инвазивные штаммы генотипа еттпЗ — типом ST-15. Среди штаммов Str. agalactiae
наиболее часто встречаются типы ST-1, ST-9, ST-17, ST-19 и ST-23, что указывает на
наличие неизвестных селективных преимуществ для адаптации к организму человека.
Высокая стандартизация метода MLST позволяет сравнивать друг с другом штаммы,
выделяемые в различных регионах и в разные годы. При этом становится возможным
оценивать уровень эпидемиологического контроля за циркуляцией штаммов.
Серологическая диагностика
Выявление стрептококковых антител — диагностический показатель, позво-
ляющий отличать активную инфекцию и бактерионосительство. Это важно для
постстрептококковых заболеваний, таких как ревматическая лихорадка и гломеру-
лонефрит, которые почти всегда сопровождаются увеличением содержания анти-
стрептококковых антител в плазме крови. Считается, что 4-кратное повышение
титров антистрептококковых антител свидетельствует о перенесенной стрептокок-
ковой инфекции. Наиболее показательно выявление антител к стрептолизину-0 и
ДНКазе Б.
Уровень антител к каждому из этих антигенов определяют с помощью реак-
ции нейтрализации. Концентрация антител к сгрептолизину-0 (ASO) достигает
максимума в течение 3-6 нед от начала заболевания. Обычно увеличение содер-
жания ASO наблюдают при инфекции верхних дыхательных путей, вызванной
Str. pyogenes, в то время как стрептодермия не приводит к их повышенной про-
дукции. Штаммы Str. dysgalactiae подвида equisimilis также способны к продукции
стрептолизина-О однако при вызванных ими инфекциях уровень ASO не повы-
шается и, следовательно, не является диагностическим критерием.
Таблица 21-34. Праймеры, используемые для мультилокусного типирования
Ген Кодируемый белок Прямой праймер (5-3 ) Обратный праймер (5-3’) Размер ампли- кона, П.Н.
Streptococcus pyogenes
дк> Глюкозокиназа GGCATTGGAATGGGATCACC TCTCCTGCTGCTGACAC 498
gtr Белок транспорта глутамина GAGGTTGTGCTGATTATTGG G3AAAGCCCATTTCATGAGTC 450
mur! Глутамзтрацемазз TGCTGACTCAAAATGTTAAAATGATTG GATGATAATTCACCGTTAATGTCAAAATAG 438
mutS Белок репарации неправильно спаренных нуклеотидов GAAGAGTCATCTAGTTTAGAATACGAT AGAGAGT TGTCACTTGCGCGTTTGATTGCT 405
recP Транскетолаза GCAAATTCTGGACACCCAGG CTTTCACAAGGATATGTTGCC 459
xpt Ксантинфосфорибо- зилтрансфераза TTACTTGAAGAACGCATCTTA ATGAGGTCACTTCAATGCCC 450
yiqL Ацетип-КоА-ацетилтрансфераза TGCAACAGTATGGACTGACCAGAGAACAAGATGC CAAGGTCTCGTGAAACCGCTAAAGCCTGAG 434
Streptococcus agalactiae
adhP Алкогольдегидрогеназа GTTGGTCATGGTGAAGCACT (GGTGTGTGCCATACTGAHT) ACTGTACCTCCAGCACGAAC (ACAGCAGTCACAACCACTCC) 672 (498)
pheS Фенилаланил-тРНК-синтетаза gattaaggagtagtggcacg (ATATCAACTCAAGAAAAGCT) TTGAGATCGCCCATTGAAAT (TGATGGAATTGATGGCTATG) 723 (501)
atr Белок транспорта глутамина CGATTCTCTCAGCTTTGTTA ^ATGGTTGAGCCAATTATTTC) AAGAAATCTCTTGTGCGGAT (CCTTGCTCAACAATAATGCC) 627 (501)
glnA Глутамин синтетаза CCGGCTACAGATGAACAATT (AATAAAGCAATGTTTGATGG) CTGATAATTGCCATTCCAGG (GCATTGTTCCCTTCATTATC) 589 (498)
sdhA Сериндегидратаза AGAGCAAGCTAATAGCCAAC (AACATAGCAGAGCTCATGAT) ATATCAGCAGCAACAAGTGC (GGGACTTCAACTAAACCTGC) 646 (519)
glcK Глюкозокиназа CTCGGAGGAACGACCATTAA (GGTATCTTGACGCTTGAGGG) CTTGTAACAGTATCACCGTT (ATCGCTGCTTTAATGGCAGA) 607 (459)
tki Транскетолаза CCAGGCTTTGATTTAGTTGA (ACACTTCATGGTGATGGTTG) AATAGCTTGTTGGCTTGAAA (TGACCTAGGTCATGAGGTTT) 859 (480)
ЧАСТНАЯ МИКРОБИОЛОГИЯ 433
Окончание табл. 21 -34
434 ЧАСТНАЯ МИКРОБИОЛОГИЯ
Среди четырех продуцируемых стрептококками ДНКаз наиболее значим иммун-
ный ответ организма на ДНКазу В. Концентрация этих антител достигает макси-
мума только через 6-8 нед, однако их уровень сохраняется дольше по сравнению
с уровнем ASO, что свидетельствует о перенесенной инфекции.
Повышение титров антител к продуктам стрептококков можно расценивать как
результат недавнего инфицирования. Однако их однократное определение может
ввести в заблуждение. Повышенная концентрация ASO определяется только у
80-85% больных ревматизмом. Одновременное определение антител к ДНКазе
В увеличивает чувствительность этого подхода до 97%. Прием антибиотиков в
острую фазу инфекции значительно снижает выраженность иммунного ответа,
увеличение концентрации антител может быть гораздо ниже. Нормальное значе-
ние титров антител зависит от возраста пациента. Здоровые лица после перенесен-
ной инфекции могут иметь титр антител выше нормальных величин. Длительное
наблюдение за концентрацией антистрептококковых антител после достижения
пикового значения малоинформативно в диагностическом и прогностическом
плане независимо от того, сохраняется титр на высоком уровне или снижается.
Для отслеживания тенденции титров к возрастанию более информативно опреде-
ление антител одновременно в нескольких образцах плазмы крови, взятых с
недельным интервалом. При каждом определении концентрации антител следует
использовать контрольные сыворотки с известным титром или стандарт антител.
Сравнение титров возможно между лабораториями, использующими одинаковые
стандарты или идентичные контрольные сыворотки.
Определение чу ствительности стрептококков к антимикробным препаратам
Бензилпенициллин остается препаратом выбора при лечении большинства стреп-
тококковых инфекций. Пенициллинрезистентных штаммов 0-гемолитических
стрептококков не зарегистрировано ни в одной из стрептококковых референс-
лабораторий ВОЗ. Однако с учетом довольно высокой аллергии на бензилпени-
циллин, обнаруживаемой более чем у 10% пациентов, существуют альтернативные
антибактериальные препараты для лечения стрептококковой инфекции —
макролиды. Необходимо отметить, что частота формирования резистентности к
макролидам среди клинических изолятов Str. pyogenes и Str. agalactiae значительно
возрастает как в США, так и в странах Европы, что коррелирует с их широким
использованием в клинической практике в последние годы. У 0-гемолитических
стрептококков и Str. pneumoniae неизвестна устойчивость к ванкомицину. Однако
имеются данные о ванкомицинрезистентности изолята, принадлежащего к Str.
bovis.
В связи с формированием резистентности к бензилпенициллину у Str. pneumo-
niae и других а-гемолитических стрептококков не рекомендуют испод ьзовать дан-
ный антибиотик при начале лечения. Бензилпенициллин применяют для лечения
инфекций, вызванных данными стрептококками, только при их установленной
чувствительности. Имеются сообщения о высокой частоте пенициллинрезистент-
ных штаммов среди изолятов, принадлежащих к видам Str. mitts и Str. saiivarius.
Применение фторхинолонов для лечения инфекций, вызванных Str. pneumoniae,
привело к формированию резистентности данного вида к фторхинолонам.
Для определения чувствительности стрептококков к антибактериальным пре-
паратам используют следующие методы:
• определение чувствительности с использованием коммерческих дисков с
определенной концентрацией антибиотика - диско-диффузионный метод;
• метод серийных разведений антибиотика в бульоне, позволяющий опреде-
лить его минимальную подавляющую концентрацию;
• метод Е-тестов с использованием пластиковой полоски размером 5x50 мм с
нанесенным градиентом концентрации антибиотика.
ЧАСТНАЯ МИКРОБИОЛОГИЯ 435
При тестировании чувствительности к антибактериальным препаратам предпо-
чтительнее использовать агар Мюллера-Хинтона с добавлением 5% дефибрини-
рованной бараньей крови. Толщина слоя кровяного агара в чашке Петри должна
быть 4 мм. Имеет значение качество дисков с антибиотиками. Зону задержки роста
после инкубации чашки в течение 18-20 ч при 36-37 °C измеряют с помощью ЖМЖ
линейки, причем необходимо измерять диаметр зоны подавления роста. По вели-
чине зоны задержки роста интерпретируют полученные результаты. Обязательно
использование контрольных штаммов. Этот подход применяют в бактериологиче-
ских лаборат ориях мира в качестве рутинного.
ПНЕВМОКОККОВЫЙ СТРЕПТОКОКК
ТАКСОНОМИЯ
Пневмококковый стреп гококк (Str. pneumoniae) относится к роду Streptococcus, так-
сономическое положение которого приведено выше, в разделе «Стрептококки».
ОБЩАЯ ХАРАКТЕРИСТИКА
Str. pneumoniae — грамположительная мелкая бактерия, клетки которой раз-
мером 0,5-2,0 мкм имеют овальную или ланцетовидную форму. Неподвижные,
расположены попарно или цепочками, спор не образуют, большинство штаммов
имеют капсулу, легко переходят в L-форму.
Биохимическая активность Str. pneumoniae сходна с активностью зеленя-
щих стрептококков других видов. Ферментируют глюкозу, лактозу, мальтозу и
сахарозу до кислоты. Также ферментируют инулин, не ферментируют маннит,
салицин и сорбит. Ферментация углеводов ведет к образованию молочной кис-
лоты, газ не выделяется. Каталазоотрицателен, способен лизировать эритроциты.
Факультативный анаэроб. На искусственных питательных средах способен расти
при температуре 22-45 °C (оптимум — 37 °C).
Продуцирует токсины — гемолизин, лейкоцидин. М-белок Str pneumoniae обла-
дает антинейтрофильной активностью. Полисахаридная капсула защищает бакте-
рию от фагоцитоза и опсонизирующего де11ствия антител. Продукция ферментов —
гиалуронидазы и пептидазы — способствует распространению возбудителя в ткани
пораженного органа
Известно свыше 90 серотипов Str. pneumoniae., различающихся по строению
антигенов полисахаридной капсулы (не менее 40 серогрупп). Повсеместно выяв-
ляются нетипируемые изоляты, поэтому серотипирование Str. pneumoniae не полу-
чило распространения в клинической диагностике.
Str. pneumoniae чувствительны к физическим и химическим факторам окружаю-
щей среды, в том числе к дезинфектантам. Способны длительно сохранять жизне-
способность при низких температурах.
КЛИНИЧЕСКИЕ ПРОЯВЛЕНИЯ ИНФЕКЦИИ
Str. pneumoniae вызывает ряд патологических процессов, которые объединены
общим понятием пневмококковой инфекции:
• пневмонию (до 70% внебольничных пневмоний);
• острый средний отит (до 25% отитов);
• гнойный менингит (5-15% бактериальных менингитов);
• бактериальный эндокардит (около 3% инфекционных эндокардитов);
• плеврит;
• артрит;
• конъюнктивит.
Нередко пневмококковая инфекция развивается как осложнение других,
как правило вирусных, заболеваний. Пневмококки могут быть выявлены в
436 ЧАСТНАЯ МИКРОБИОЛОГИЯ
материале из носоглотки у практически здоровых детей и взрослых — бакте-
рионосителей.
ЛАБОРАТОРНАЯ ДИАГНОСТИКА
Лабораторная диагностика пневмококковой инфекции включает следующие
этапы:
• бактериоскопию;
• посев материала на питательные среды:
• серологическую диагностику:
• экспресс-диагностику;
• молекулярную диагностику;
• биологическую пробу (введение патологического материала внутрибрюшин-
но белым мышам).
При гнойных менингитах необходима дифференциальная диагностика с менин-
гококковой, гемофильной инфекцией и другими бактериальными менингитами.
При пневмонии проводят дифференциальную диагностику с инфекционными про-
цессами, вызванными Н. influenzae, Mycoplasma pneumoniae, М. catarrhalis, S. aureus,
L. pneumophila, Klebsiella pneumoniae, P. aeruginosa, Chlamydia pneumoniae, а также энте-
робактериями, ацинетобактериями, анаэробами и патогенными грибами.
Материалы для лабораторных исследований
При гнойных менингитах исследуют СМЖ и кровь.
При пневмонии и других клинических формах пневмококковой инфекции
материалом для исследования служат мокрота, аспираты из бронхов, плевральная
жидкость, кровь, отделяемое из среднего уха, СЖ.
В некоторых странах при поступлении пациента в стационар рекомендуют
брать пробы крови для серологического исследования и хранить сыворотку в
замороженном виде. При тяжелом течении пневмонии и неэффективности анти-
биотикотерапии, а также по эпидемическим показаниям через 7-10 дней получа-
ют повторный образец крови и проводят серологическую диагностику атипичных
пневмоний — легионеллезной, микоплазменной, хламидийной, коронавируснои.
В тех же целях используют дополнительные методы — ПЦР, определение легио-
неллезного антигена в моче.
Для проведения ПЦР материал (СМЖ, кровь и мокроту) собирают в стерильные
емкости с плотно закручивающейся крышкой без транспортной среды. Хранение
и транспортировка этих проб пои комнатной температуре допускаются в течение
4 ч, при температуре 4 °C — в течение суток.
Бактериоскопия
При бактериоскопии мазка нативной СМЖ или препарата толстой капли, окра-
шенных водно-спиртовым раствором метиленового синего, выявляют диплококки
ланцетовидной или овальной формы, как правило, окруженные общей капсулой.
Бактерии могут располагаться в виде коротких цепочек.
Мазки мокроты окрашивают по Граму и микроскопируют с иммерсионным
объективом. При их просмотре обращают внимание на наличие альвеолярных
макрофагов, всегда обнаруживаемых в мокроте, и гранулоцитов, появляющихся
в значительных количествах при воспалительных процессах в нижних отделах
дыхательного тракта. Отсутствие гранулоцитов и альвеолярных макрофагов
и большое количество эпителиальных клеток указывают на примесь слюны в
исследуемом материале и необходимость повторения исследования. Str. pneu-
moniae быстро перевариваются фагоцитами, именно поэтому более вероятно их
расположение вблизи гранулоцитов, а не внутрилейкоцитарно. Str. pneumoniae в
мазках мокроты имеют вид грамположительных мелких ланцетовидных клеток,
окруженных зоной неокрасившейся капсулы.
ЧАСТНАЯ МИКРОБИОЛОГИЯ 437
Изоляция возбудителя
Str. pneumoniae растет на питательных средах, обогащенных углеводами, содер-
жащих кровь, сыворотку, асцитическую жидкость.
СМЖ высевают на среду обогащения (0,1% сывороточный агар) и шоколадный
агар, кровь — на двухфазную среду.
Учитывая возможность возникновения при пневмониях гранзиторной бактерие-
мии, исследуют кровь методом гемокультуры. Гнойные комки мокроты, аспират из
бронхов и тому подобный патологический материал высевают в чашки Петри на 5%
кровяной или шоколадный агар. Высеваемость пневмококков из мокроты возрас-
тает при использовании селективных питательных сред, содержащих ингибиторы
роста сопутствующей микрофлоры (среды с гентамицином в его конечной концен-
трации 5 мг/л или колумбийского агара с колистином и налидиксовой кислотой).
Посевы инкубируют при 37 °C и содержании в атмосфере 5-10% углекислого
газа. После 18-24 ч инкубации посевы просматривают под стереомикроскопом
или под лупой. Учитывают количество и морфологию выросших колоний, готовят
мазки и окрашивают их по Граму.
При наличии Str. pneumoniae в ис следуемом материале через сутки инкубации
при температуре 37 °C и содержании в атмосфере 5-10% углекислого газа:
'Z на 0.1% сывороточном агаре рост пневмококков начинается в верхней
части среды, постепенно распространяясь вниз;
J на двухфазной среде через 24 ч отмечается помутнение жидкой части
среды и образование мелких, не склонных к слиянию колоний на поверх-
ности плотной фазы;
J на сывороточном агаре определяются нежные, мелкие, не сливающиеся
колонии, часто уплошенные, с углублением в центре;
J на шоколадном агаре наблюдаются мелкие, не сливающиеся колонии,
окруженные гемолитической зоной желто-зеленого цвета;
S на кровяном агаре определяются мелкие, плоские, не сливающиеся коло-
нии с зоной зеленящего гемолиза.
Если в мазках из подозрительных на Str. pneumoniae колоний определяются
грамположительные диплококки, из таких колоний делают отсевы на сывороточ-
ные или кровяные среды для получения чистой культуры.
При достаточном количестве колоний, выросших на сывороточном или шоко-
ладном агаре, допускается их отсев на дифференциально-диагностические среды.
Колонии, подозрительные на Str. pneumoniae, отсевают на сектора чашек с кро-
вяным агаром. Выросшие культуры используют для контроля чувствительности
к желчи и антибиотикорезистентности. Если колоний мало, проводят отсев на
чашку с сывороточным или шоколадным агаром для нак опления чистой культуры,
с которой выполняют дальнейшую идентификацию бактерий.
Если на чашках с прямыми посевами патологического материала рост отсут-
ствует, проводят высевы из среды обогащения (0,1% сывороточного агара) на
шоколадный или сывороточный агар, повторяя высевы в течение 5 дней инкуба-
ции или до появления первых признаков роста.
На 3-й день исследования учитывают характер гемолиза на секторах с кровяным
агаром, готовят мазки и окрашивают их по Граму. При сплошном росте на секторах
уже в этот день может быть выполнена проба на чувствительность к желчи: на агар
с выросшими колониями помещают диски, пропитанные 20% раствором желчи,
через 1-2 ч инкубации при температуре 37 °C колонии пневмококков вокруг дис-
ков лизируются и образуется зона просветления шириной 1-2 мм.
Биохимическое исследование
Дифференциацию Str. pneumoniae от других видов стрептококков проводят с
помощью проб с опгохином и дезоксихолатом.
438 ЧАСТНАЯ МИКРОБИОЛОГИЯ
Проба с оптохином. Выполняют газонный посев исследуемой культуры
на чашку с кровяным ат аром. Таким же образом высевают контрольные тест-
культуры — положительный контроль (Str. pneumoniae) и отрицательный контроль
(Str. salivarius). Затем на поверхность агара помещают диски, содержащие 5 мкг
оптохина.
Через 18-24 ч инкубации посевов, содержащих Str. pneumoniae, при температуре
37 °C вокруг дисков с оптохином проявляется зона отсутствия роста диаметром
более 14 мм при диаметре диска 6 мм и более 16 мм при диаметре диска 10 мм.
Ингибирования Str. salivarius вокруг дисков не наблюдается.
Чувствительность подозрительных на Str. pneumoniae культур к желчным кисло-
там контролируют желчным тестом или дезоксихолатной пробой.
Проба с дезоксихолатом. В 0,85% растворе NaCl готовят суспензию иссле-
дуемой культуры с мутностью 1 по стандарту мутности и вносят по 0,5-1,0 мл в
две пробирки. В одну из пробирок (тест) добавляют 0.15 мл 10% раствора дезок-
сихолата натрия, в другую пробирь.у (контроль) — 0,15 мл 0,85% раствора NaCl.
Содержимое перемешивают, выдерживают при 35 °C в течение 1-2 ч и сравнивают
мутность суспензий в обеих пробирках.
Просветление суспензии в тест-пробирке указывает на принадлежность куль-
туры к Str. pneumoniae (дезоксихолат активизирует аутолиз бактериальных клеток
пневмококков, что не свойствено стрептококкам других видов). Важно помнить,
что около 15% штаммов Str. pneumoniae не лизируются в данных условиях или
лизируются не полностью.
Экспресс-диагностика
Реакцию латекс-агглютинации рекомендуют в качестве одного из методов обна-
ружения Str. pneumoniae при гнойных менингитах. Встречный иммуноэлектрофо-
рез, ИФА, иммунофлюоресцентный анализ применяют для определения пневмо-
кокков или их антигенов в патологическом материале в качестве дополнительных
методов диагностики пневмококковой инфекции.
Молекулярно-генетические методы диагностики
В качестве основного метода молекулярной диа!ностики используют ПЦР.
В ней исследуют СМЖ, кровь и мокроту.
ОПРЕДЕЛЕНИЕ ЧУВСТВИТЕЛЬНОСТИ К АНТИБАКТЕРИАЛЬНЫМ ПРЕПАРАТАМ
Исследование проводят методом серийных разведений (в бульоне или агаре)
или диско-диффузионным методом. В качестве жидкой питательной среды исполь-
зуют бульон Мюллера-Хинтона с добавлением лизированной крови лошади.
В качестве плотной питательной среды (для метода серийных разведений и диско-
диффузионного метода) используют агар Мюллера-Хинтона, содержащий 5%
дефибринированной крови барана.
МЕНИНГОКОКК
ТАКСОНОМИЯ
Менингококк (Neisseria meningitides) — бактерия рода Neisseria, семейства
Neisseriaceae. Помимо N. meningitidis, род включает N. gonorrhoeae1 и непатогенные
виды — N. sicca, N. mucosa, N. subfiava, N.flavescens, нередко выделяемые из носо-
глоточной слизи, а также N. lactamica, которая в редких случаях вызывает гнойные
отиты, синуситы. Различают серогруппы менингококка — А, В, С, D. Н. I, К, L, X,
Y. Z, 29Е и W-135.
1 Информация по этой инфекции приведена в следующей главе.
ЧАСТНАЯ микробиология 439
ОБЩАЯ ХАРАКТЕРИСТИКА
N. meningitidis — грамотрицательный диплококк, неподвижный, образующий
капсулу, аэроб. Проявляет слабую биохимическую активность — ферменти-
рует только глюкозу и мальтозу с образованием кислоты без газа, оксидазо-,
каталазоположителен, желатин не разжижает. Во внешней среде неустойчив.
В носоглоточной слизи сохраняется до 2 ч, погибает при температуре 50 °C в
течение 5 мин, при температуре 10 °C — ь течение 2 ч. Оптимальная температура
размножения менингококка — 37 °C, при температуре 35 °C развитие бактерий
резко замедляется.
КЛИНИЧЕСКИЕ ПРОЯВЛЕНИЯ ИНФЕКЦИИ
N. meningitidis — безусловный патоген, возбудитель менингококковой инфек-
ции, которая может протекать в следующих клинических формах:
• генерализованная менингококковая инфекция:
❖ гнойный бактериальный менингит;
о менингоэнцефалит;
менингококцемия;
• локализованная инфекция.
<> назофарингит;
<> носительство менингококка;
• редкие формы:
❖ артрит;
❖ эндокардит;
о иридоциклит;
< пневмония.
Наибольшую опасность представляет генерализованная менингококковая
инфекция, которая примерно в 30% случаев протекает в гипертоксической форме
с высокой частотой летальных исходов.
В этиологической структуре спорадических случаев гнойных бактериальных
менингитов (ГБМ) менингококковая инфекциязанимаетпервоеместо. Следующие
по частоте этиологические факторы ГБМ — Str. pneumoniae, Н. influenzae серо-
типа b. М. tuberculosis. У новорожденных и детей в возрасте до 1 мес ГБМ, как
правило, обусловлены стрептококками группы В, эшерихиями, листериями. При
черепно-мозговых травмах, нейрохирургических и челюстно-лицевых операциях
возможно возникновение ГБМ как осложнения основного заболевания. В этих
случаях ведущая роль принадлежит Str. pneumoniae, стафилококкам, колиформ-
ным бактериям и Р. aeruginosa, однако возможно развитие молниеносных форм
менингококковой инфекции при не диагностированном менингококковом назофа-
рингите или носительстве менингококка. Следовательно, незамедлительная иден-
тификация возбудителя необходима для выбора этиотропной терапии больных
ГБМ. В этнологической структуре генерализованной менингококковой инфекции
более 90% случаев обусловлены менингококками серогрупп А, В и С.
ЛАБОРАТОРНАЯ ДИАГНОСТИКА
Материалы для лабораторных исследований
Основной патологический материал для исследований при генерализованных
формах менингококковой инфекции и других ГБМ — СМЖ, кровь и моча. Для
бактериологического подтверждения менингококкового назофарингита и носи-
тельства менингококка необходимо исследование носоглоточной слизи.
Тяжелый симптомокомплекс клинических проявлений в первые дни болезни
указывает на необходимость экстренного незамедлительного проведения иссле-
дований по определению этиологии заболевания с обязательным использованием
методов экспресс-диагностики.
440 ЧАСТНАЯ МИКРОБИОЛОГИЯ
Данное исследование включает;
• микроскопию препарата толстой капли крови и СМЖ,
• первичный бактериологический посев СМЖ и крови с последующей иден-
тификацией нейссерий и определением чувствительности к антимикробным
препаратам;
♦ реакцию латекс-агглютинации для обнаоужения полисахаридного (группо-
специфического) антигена менингококка в СМЖ, крови и моче:
• встречный иммуноэлектрофорез для определения антигенов менингококка в
СМЖ или плазме крови;
• ИФА для определения антигенов менингококка в СМЖ или плазме крови;
• ПЦР для выявления генетического материала менингококка в СМЖ или
плазме крови;
• серологическое исследование для обнаружения группоспецифических анти-
тел к менинп жоккам в сыворотке крови больных с помощью РИГА (РПГА)
или ИФА.
СМЖ получают методом люмбальной пункции или пункции желудочков мозга
и распределяют следующим образом:
• 1 мл направляют на ликворо- и цитологическое исследование;
• 1 мл направляют на бактериологический посев (если он не выполнен у посте-
ли больного), бактериоскопию, реакцию латекс-агглютинации, встречный
иммуноэлектрофорез, ИФА для определения антигенов менингококков;
• 0,5 мл высевают на чашку с шоколадным агаром непосредственно у постели
больного;
• 0,5 мл вносят в пробирку со средой обогащения (5 мл 0,1% полужидкого
сывороточного питательного агара) в момент выполнения люмбальной
пункции;
• 0,2 мл в пробирке типа «Эппендорф» незамедлительно направляют на
молекулярно-генетическое исследование (допускается хранение образцов
материала при температуре 4 °C не более 3 сут).
Кровь отбирают методом венепункции. Взятую пробу распределяют следую-
щим образом:
• для препарата толстой капли;
• 5-10 мл крови у взрослых (2,0-2,5 мл — у детей, 1-2 мл — у новорожденных)
направляют на бактериологическое исследование для выделения и иденти-
фикации чистой культуры и определения чувствительности к антимикроб-
ным препаратам;
• 3-5 мл крови направляют в лабораторию для получения сыворотки и
выполнения встречного иммуноэлектрофореза, реакции латекс-агглю-
тинации, ИФА для определения антигенов менингококков и серологиче-
ских реакций (РИГА, ИФА) на наличие антител к менингококкам серо-
групп А. В, С.
Носоглоточную слизь берут натощак стерильным зондом-тампоном (предпо-
чтительнее вискозным или дакроновым, так как ватные тампоны часто бывают
токсичными для менингококка). Зонд-тампон заводят за мягкое небо в носоглотку
и проводят несколько раз по ее задней стенке. Взятые пробы доставляют в лабо-
раторию в транспортных средах с линкомицином, Стюарта, Кэри-Блэйр, Эймса
(если срок транспортировки превышает 3 ч).
Материал, предназначенный для бактериологического исследования на менин-
гококковую инфекцию, доставляют в лабораторию незамедлительно в контейне-
рах или гермосумках, поддерживая в них температуру 37 °C. До момента посева
материала допускается его хранение при 37 °C не более 12 ч.
Нативную СМЖ для реакции латекс-агглютинации, встречного иммуноэлек-
трофореза и ПЦР допускается хранить при температуре 4 °C в течение суток.
ЧАСТНАЯ МИКРОБИОЛОГИЯ 441
Оста гки СМЖ и плазмы крови хранят при температуре 4 °C в течение недели на
случай дополнительных исследований.
Материалы (СМЖ, кровь), хранившиеся при комнатной температуре или в
холодильнике, как правило, непригодны для культивирования менингококков.
Бактериологическое исследование
Спинномозговая жидкость Немедленно после доставки в лабораторию
СМЖ центрифугируют при 2500-3000 об./мин (1500 g) в течение 10-15 мин.
Над осадочную жидкость осторожно отсасывают стерильной пипеткой и затем
используют для проведения реакции латекс-агглютинации или встречного имму-
ноэлектрофореза, позволяющих выявить группоспецифические антигены менин-
гококка.
Толстую каплю СМЖ наносят на предметное стекло, дают свободно растечься
и высушивают при 37 °C или при комнатной температуре. Препарат окрашивают
без предварительной фиксации водно-спиртовым раствором метиленового синего
в течение 2-3 мин и осторожно промывают водой, затем просушивают и микро-
скопируют, просма гривая при необходимости не менее 20 полей зрения. Отмечают
наличие и характер расположения (внеклеточно, внутрилейкоцитарно), морфоло-
гию и тинкториальные свойства бактерий.
Затем проводят посев на питательные среды (если он не был выполнен у постели
больного): примерно по 2 капли осадочной фракции СМЖ стерильной пипеткой
высевают на чашку с предварительно прогретым в термостате шоколадным агаром
и в пробирку с предварительно подогретым 0,1% полужидким сызороточным ага-
ром (среда обогащения).
Если СМЖ получена в недостаточном количестве (1-2 капли) из нее готовят
толстую каплю, а к остатку материала в пробирке добавляют 1 мл подогретого
до 37 °C питательного бульона, перемешиваю! и сеют на шоколадный и в 0,1%
полужидкий агар. Чашки и пробирки с посевами немедленно помещают в термо-
стат и инкубируют при 37 СС и содержании в атмосфере 5-10% углекислого газа.
Через 24 и 48 ч посевы проверяют на наличие роста бактерий. При его наличии на
чашках с агаром готовят мазки и окрашивают по Граму. Определяют каталазную
и оксидазную активность, делают посев на сывороточный агар для получения
чистой культуры.
При наличии роста на 0,1% полужидком сывороточном агаре с него проводят
высев на шоколадный агар и продолжают работу по выделению чистой культуры
и идентификации возбудителя, как описано выше.
На 3-й день исследования контролируют чистоту’ культур, выросших на сыворо-
точном агаре, учитывают наличие или отсутствие пигментообразования. выполня-
ют посев на желточную среду (для более точного контроля пигментообразования),
делают контрольный посев на бессывороточную среду и посевы на сахаролитическ^по
активность. При достаточном количестве чистой культуры уже в этот день может быть
выполнена РГА группоспецифическими менингококковыми антисыворотками.
На 4-й дено учитывают результаты посевов предыдущего дня — чистоту куль-
туры, наличие или отсутствие пигментообразования на желточной среде, наличие
или отсутствие роста на бессывороточной среде, сахаролитическую активность,
выполняют окончательную идентификацию серогруппы менингококка в РГА.
По результатам контролей выдают окончательный ответ. Чистую культу-
ру менингококка используют для контроля антибиотикорезистентности.
Исследование первичных посевов на среду обогащения завершают на день позже
(на 5-й день).
Кровь. Толстую каплю крови готовят у постели больного нанесением 1 капли
крови на середину предметного стекла, распределяя ее так, чтобы диаметр
составлял примерно 1 см. Высушивают на воздухе и доставляют в лабораторию.
442 ЧАСТНАЯ МИКРОБИОЛОГИЯ
Окрашивают, не фиксируя, водно-спиртовым раствором метиленового синего
в течение 2-3 мин, осторожно промывают водой и подсушивают на воздухе.
Просматривают под иммерсией не менее 20 полей зрения, регистрируя внеклеточ-
но и внутриклеточно расположенные бактерии.
Посев крови проводят во флаконы с двухфазной питательной средой в соотно-
шении 1 часть крови на 10 частей жидкой фазы среды. В настоящее время доступ-
ны готовые двухфазные среды, содержащие факторы роста V. X и витамины.
Флаконы с посевами инкубируют при температуре 37 °C в течение 5 сут.
ежедневно контролируя микробный рост (наличие помутнения, хлопьевидного
осадка, образования газа, пленки, колоний на плотной фазе питательной среды).
Независимо от наличия или отсутствия признаков роста через 48 ч инкубирования
проводят посев жидкой фазы среды на шоколадный агар.
При признаках роста готовят мазки из бульона двухфазной среды и колоний,
выросших на агаре под жидким слоем, окрашивают по Граму в модификации
Калины, а также пповодят посев на шоколадный агар. Если в мазках, окрашенных
по Граму, обнаруживают бактерии, отличающиеся от менингококков, пневмокок-
ков и гемофильных палочек, помимо посева на шоколадный агар проводят посев
на элективные питательные среды.
Из колоний, выросших на шоколадном агаре, готовят мазки, окрашивают по
Граму, определяют оксидазную и каталазную активность, осуществляют отсев на
сывороточный агар для получения чистой культуры. Затем исследование ведут,
как описано выше (3-й, 4-й день исследования).
Носоглоточная слизь. Посев проб носоглоточной слизи выполняют неза-
медлительно после доставки в лабораторию. Материал, содержащийся на зонде-
тампоне, втирают на % площади чашки с сывороточным агаром и 14 чашки с селек-
тивным агаром (содержащим линкомицин). Оставшиеся площади агара на чашке
засевают штриховым методом с помощью бактериологической петли, начиная
штрихи с места посева тампоном.
Чашки инкубируют при температуре 37 °C, через 22-24 ч их просматривают.
При наличии роста оценивают характер колоний, предварительно дифференцируя
колонии N. meningitidis и непатогенных нейссерий. На сывороточном агаре и агаре
с линкомицином менингококки формируют через 24 ч роста плоские или слегка
выпуклые, нежные, голубовато-опалесцир^ющие колонии диаметром 1 мм с ров-
ными краями.
/V. subfiava и N.flavescens образуют более крупные (1-2 мм в диаметре) колонии
желтого цвета, окраска которых усиливается при выдержке на све гу.
N. mucosa образует мутно-белые, слизистые колонии диаметром 1-2 мм.
N. sicca формирует мелкие, сухие, плохо снимающиеся колонии желтоватого
цвета.
N. lactamica образует круглые колонии желтоватого цвета с ровными краями,
диаметром 1-1,5 мм.
М. catarrhalis формирует круглые, выпуклые колонии беловатого цвета, диа-
метром 1,5-2 мм.
Из колоний, подозрительных на менингококк, осуществляют отсевы на сектора
чашек или пробирки с сывороточным агаром для получения чистой культуры и
дальнейшей идентификации. Параллельно проводят посев на бессывороточный
питательный агар, на котором растут сапрофитные виды Neisseria и не растут
N. meningitidis и N. gonorrhoeae.
Идентификация N. meningitidis Идентификацию менингококка проводят
на основании исследования морфологических, тинкториальных. культуральных,
ферментативных и антигенных свойств чистой культуры изолята.
При бактериоскопии толстой капли крови или СМЖ, окрашенной метиленовым
синим, на голубом фоне выявляются кокки и диплококки бобовидной формы,
ЧАСТНАЯ МИКРОБИОЛОГИЯ 443
обращенные друг к другу вогнутыми сторонами. Весьма характерно внутрилейко-
цитарное расположение бактерий, но часто встречается и внеклеточное располо-
жение или их сочетание в различной степени.
На чашках с шоколадным агаром менингококки образуют нежные, полупро-
зрачные, сероватые колонии диаметром 1-2 мм, маслянистой консистенции
(легко снимаются петлей), не изменяют цвета среды.
На 0,1% полужидком сывороточном агаре вызывают интенсивное помутнение
верхней части питательной среды.
На двухфазной питательной среде вызывают помутнение ее жидкой фазы и
развитие полупрозрачных колоний на поверхности плотной фазы. На чашках с
сывороточным агаром менингококк формируют плоские или слегка выпуклые,
нежные, голубоватые колонии с ровными краями и гладкой поверхностью.
На питательных средах без добавления сыворотки или крови животных менин-
гококки в первых генерациях не растут.
Для окрашивания мазков культур менингококка рекомендуют использование
метода Грама в модификации Калины. Его проводят следующим образом: на обе-
зжиренное стекло наносят каплю 0,15 М раствора натрия хлорида в ней суспенди-
руют исследуемую бактериальную культуру, затем в каплю суспензии вносят каплю
спиртового раствора бриллиантовой зелени. Препарат подсушивают и фиксируют
над пламенем затем наливают основной краситель на 1,5-2 мин. Краску сливают,
препарат промывают водой, затем отмывают спиртом до видимого отхождения
краски и немедленно промывают водой. После этого докрашивают препарат
водным раствором фуксина в течение 2 мин, промывают водой, просушивают и
просматривают под иммерсией при большом увеличении. Грамотрицательные
бактерии окрашиваются в ярко-розовый цвет, грамположительные — в сине-
черный.
В мазках, окрашенных по Граму, менингококки, как и другие нейссерии, выде-
ленные на искусственных питательных средах, грамотрицательны, полиморфны
по размерам и форме (округлая или овальная форма, размеры 0,6-1 мкм). Парное
классическое расположение и форма в виде диплококков бобовидной формы,
обращенных вогнутыми сторонами друг к другу, характерны только для изолятов
менингококка, выделенных из СМЖ или крови.
Культуры менингококков, выращенные на сывороточном или шоколадном
агаре, оксидазо- и каталазоположительны. Сахаролитическая активность различ-
на у разных видов нейссерий (табл. 21-35), что позволяет провести внутривидовую
идентификацию менингококков.
Таблица 21-35. Сахаролитическая активность нейссерий
Вид Углеводы
глюкоза мальтоза лактозз сахароза
N. meningitidis + + - -
N. gonorrhoeas + - - -
N. ubffeva + + -
N. sicca + + - +
N. flavescens - - - -
N. lactamica + + + -
N. mucosa + + - +
N. catarrhalis - - - -
В настоящее время имеются коммерческие системы биохимической идентифи-
кации нейссерий, которые могут заметно ускорить и упростить исследование.
Внутривидовую идентификацию — определение серогруппы менингококка —
выполняют в РГА чистой культуры возбудителя с помощью агглютинирующих
444
ЧАСТНАЯ МИКРОБИОЛОГИЯ
сывороток серогрупп А, В, С, X. Y, Z, W-135, 29Е или с помощью группоспецифи-
ческих гест-систем для ИФА.
Экспресс-диагностика
Для ускоренного обнаружения антигенов (капсульных полисахаридов) менин-
гококка в биологическом материале (СМЖ и крови) применяют:
• реакцию латекс-агглютинации;
• ветвенный иммуноэлектрофорез;
• ИФА.
На фоне антибиотикотерапии экспресс-тесты могут быть более эффективны,
чем бактериологическое исследование, так как регистрируют в исследуемом пре-
парате не только жизнеспособные, но и разрушенные бактерии.
Для выполнения экспресс-тестов применяют коммерческие тест-системы, заре-
гистрированные в России в установленном порядке.
Реакцию латекс-агглютинации ставят на ст екле, смешивая капли надосадочного
слоя СМЖ. прогретой до 100 °C в течение 5 мин, со специфическими реагентами
(серогрупп А, В, С, Y, W-135). Учет реакции проводят через 2 мин после смеши-
вания компонентов. Тест позволяет получить ответ в течение 5-15 мин с момента
выполнения люмбальной пункции.
Встречный иммуноэлектрофорез выполняют по общепринятой методике в 1%
агаре на мединал-вероналовом буферном растворе (pH = 8,6). В качестве материа-
ла для исследования используют СМЖ (цельную, осадок или надосадочную жид-
кость), а также плазму крови. Коммерческие группоспецифические антименинго-
кокковые преципитирующие антисыворотки вносят в лунки со стороны анода, а
исследуемый материал — со стороны катода. Продолжительность исследования
составляет 20-30 мин, что позволяет (с учетом подготовительных процедур) полу-
чить ответ через 1-1,5 ч.
ИФА. для определения антигенов менингококков предпочтительнее выполнять
с осветленной СМЖ или плазмой крови больного в соответствии с инструкциями
к ИФА-тест-системам. Продолжительность ИФА составляет не менее 2 ч.
Молекулярно-генетические методы диагностики
Определение участков генома менингококка в ПЦР — более чувствительный
метод диагностики по сравнению с бактериологическим исследованием, не требует
присутствия жизнеспособных микробных клеток в исследуемом материале и, сле-
довательно, пригоден для диагностики на фоне антибиотикотерапии.
Материалом для исследования служат СМЖ, кровь, носоглоточная слизь,
аутопсийные материалы, наиболее целесообразно исследование СМЖ и крови.
Пригодны образцы сразу же после их получения, а также хранившиеся при тем-
пературе 4 °C в течение 3 сут, при температуре -20 °C — в течение нескольких
месяцев, температуре -70 °C - неограниченно долго.
Для выполнения теста необходимо стандартное оборудование для ПЦР или
ПЦР в режиме реального времени и коммерческие наборы реагентов. Выполнение
процедур и учет результатов должны строго соответствовать инструкциям по
применению соответствующих наборов для ПЦР-диагностики менингококковой
инфекции. Существуют коммерческие отечественные ПЦР-наборы для диагно-
стики менингококковой инфекции и дифференциальной диагностики гнойных
менингитов, вызванных менингококками, пневмококками (Str. pneumoniae) и
гемофильными палочками (Я influenzae).
Серологическая диагностика
Серологические тесты (РИГА, ИФА) выполняют функцию вспомогательных
методов диагностики менингококковой инфекции. При отрицательных показани-
ях других тестов (например, на фоне антибиотикотерапии) они, тем не менее, могут
дать полезную информацию при исследовании парных сывороток, полученных в
ЧАСТНАЯ МИКРОБИОЛОГИЯ 445
первые 3 сут и затем на 10-12-е сутки болезни. Антитела при менингококковой
инфекции группоспецифичны, появляются в крови на 5-7-е сутки заболевания, их
высокий уровень сохраняется в течение месяца и более.
При оценке результатов серологического исследования необходимо учитывать,
что в Российской Федерации и ряде других стран существуют вакцины против
менингококков серогрупп А и С, которые могут применяться по эпидемическим
показаниям.
Интерпретация результатов лабораторных исследований
Обнаружение менингококков, их антигенов или генома в СМЖ или плазме
крови больных указывает на генерализованную менингококковую инфекцию.
Обнаружение менингококков, их антигенов и генома только в материале из
носоглотки пациента не является достаточным доказательством менингококковой
этиологии менингита, менингоэнцефалита или сепсиса у больного и расценивается
как свидетельство локализованной формы менингококковой инфекции или носи-
тельства менингококка.
Присутствие антител к менингококкам в плазме крови может быть результа-
том перенесенного менингококкового назофарингита, бактерионосительства или
предшествовавшей вакцинации менингококковыми вакцинами (последнее отно-
сится к антителам против менингококков серогрупп А и С).
Четырехкратное и более нарастание антител в РИГА, а также сероконверсия по
данным ИФА при исследовании парных сывороток — ретроспективное доказа-
тельство менингококковой инфекции.
Определение чувствительности к антибактериальным препаратам
Менингококки высокочувствительны к бензилпенициллину, который остается
препаратом выбора для лечения менингококкового менингита и менингоэнцефа-
лита. В зарубежной литературе имеются сообщения о появлении пенициллинрези-
стентных штаммов менингококков. При менингококковом сепсисе (менингокок-
цемии) необходимы и другие схемы антибиотико терапии.
Контроль антибиотикорезистентности менингококков осуществляют мето-
дами серийных разведений в агаре или бульоне или методом Е-теста. Диско-
диффузионный метод не рекомендуют.
Для выполнения Е-теста требуется агар Мюллера-Хинтона с добавлением 5%
овечьей крови, взвесь чистой суточной культуры менингококка с мутностью 0.5 по
стандарту мутности, стрипы Е-тестов бензилпенициллина, цефотаксима, цефтри-
аксона. хлорамфеникола, доксициклина, рифампицина и триметоприма (возмож-
ны другие стрипы по показаниям). В суспензии менингококка смачивают ватный
тампон, слегка отжимают его о стенку пробирки и проводят посев тампоном на
чашку со средой, тщательно втирая бактериальную взвесь для равномерного рас-
пределения по всей плошади чашки. Затем на поверхность агара помещают стрипы
Е-тестов (на чашку диаметром 90 мл необходимо 2 стрипа. 150 мм — 5 стрипов).
Посевы инкубируют при температуре 37 °C и содержании в атмосфере 5%
углекислого газа в течение 24 ч. Результаты учитывают согласно инструкции по
применению Е-тестов.
ЛИСТЕРИИ
ТАКСОНОМИЯ
В 9-м издании определителя Берджи род Liberia относят к 19-й группе микро-
организмов — грамположительные, неспорообразующие палочки правильной
формы. Анализ гомологии нуклеотидных последовательностей ДНК листерий и
близких к ним микроорганизмов показал, что листерии филогенетически наибо-
446 ЧАСТНАЯ МИКРОБИОЛОГИЯ
лее близки к роду Brochotrix и вместе с данным родом занимают промежуточное
положение между бациллами и лактобациллами, несколько более удалены от
стрептококков, а также родов Ku/thica, GemeUa и Erysepelothrix. Из шести извест-
ных в настоящее время видов листерий (I. monocytogenes, L. seeligeri, L. wehhimeri.
L. innocua, L. tvanovti, L. gray) только L. monocytogenes патогенна для человека и
животных, a I. ivanovii — только для животных.
ОБШАЯ ХАРАКТЕРИСТИКА
Листерии — грамположительные короткие палочки с закругленными концами,
иногда почти кокки, одиночные или в коротких цепочках (0,4-0,5х0,5-2,0 мкм),
реже образуют длинные нити. Ферментируют глюкозу, кагалазоположительны.
оксидазоотрицательны, образуют цитохромы. Оптимальная температура роста
составляет 30 -37 °C, могут расти как при температуре 20-25 °C. так и при темпера-
туре 4 °C. При температуре 20-25 °C листерии подвижны за счет немногочислен-
ных перитрихиальных жгутиков. При температуре 37 °C жгутики, как правило, не
образуются и листерии неподвижны. Не кислотоустойчивы, не образуют споры и
капсулу, факультативные анаэробы, хемоорганотрофы.
Литстерии терморезис гентны в связи:
• с внутриклеточным паразитизмом, снижающим чувствительность к теплово-
му воздействию;
• феноменом теплового шока, когда короткая экспозиция при 44-48 ЭС повы-
шает устойчивость бактерий к термообработ ке в бульоне, молоке и мясе.
Для полного восстановления частично инактивированных повышенной темпе-
ратурой листерий важное значение имеет инкубация в течение 6-9 ч в неселектив-
ной питательной среде при температуре 20-40 °C. Более низкая температура (5 и
12 °C) снижает уровень восстановления. Такие компоненты питательной среды,
как углеводы, соли, глицерин или маннит, способствуют восстановлению клеток,
тогда как фруктоза и хлорид аммония ускоряют их деградацию.
Высокая устойчивость к изменениям pH, концентрации солей, а также способ-
ность к росту в аэробных условиях позволяют листериям размножаться в пищевых
продуктах с длительным сроком хранения.
ЭПИДЕМИОЛОГИЯ
L. monocytogenes распространена повсеместно и представляет собой зооноз с
природной очаговостью и многочисленными источниками инфекции. Основной
природный резервуар — грызуны, насекомоядные, птицы и жвачные.
Основные пути передачи инфекции — контактный (через кожу и слизистые
оболочки), алиментарный, аэрогенный, трансмиссивный (через кровососущих
насекомых).
В связи с низкой патогенностью возбудителя заболеваемость носит спорадиче-
ский характер.
Для листериоза характерна сезонность — пик заболеваемости приходится на
зимне-весенний период,
КЛИНИЧЕСКИЕ ПРОЯВЛЕНИЯ ИНФЕКЦИИ
L. monocytogenes прот екает у человека остро, подостро, хронически и абортивно,
вызывая ряд синдромов:
• ангинозно-септический (с мононуклеозом);
• окулогландулярный;
• септико-тифозный.
• нервный:
• листериоз беременных;
• аборт и рождение мертвых детей;
• септико-гранулематозный (сепсис новорожденных);
ЧАСТНАЯ МИКРОБИОЛОГИЯ 447
• менингит новорожденных;
• уретральный.
Прогноз при листериозе серьезный у взрослых он в 30% случаев заканчивается
летально (а у людей с иммунодефицитом этот показатель вдвое выше).
Особую опасность представляет внутриутробное заражение плодов, обычно
приводящее к аборту или рождению мертвых детей вследствии интенсивного
поражения (образивания абсцессов и гранулем) в легких, печени и ЦНС. У выжив-
ших внутриутробно инфицированных листериями новорожденных в течение 3 нед
после родов развивается менингит, приводящий в 50-90% случаев к летальному
исходу.
У взрослых людей с иммунодефицитом и алкоголиков инфекция наиболее часто
проявляется менингитом (55%), септицемией (25%) и эндокардитом (7%).
ЛАБОРАТОРНАЯ ДИАГНОСТИКА
Для диагностики листериоза применяют иммунологические, бактериологи-
ческие и серологические методы. ПЦР используют в качестве дополнительного
метода при исследовании клинического материала и для быстрой идентификации
изолятов листерий.
Материалы для лабматорных исследований
При подозрении на листериоз исследованию подвергают:
• кровь и СМЖ — при сепсисе, менингите и менингоэнцефалите;
• меконий — пои заболеваниях новорожденных;
• околоплодную жидкость, плаценту; отделяемое родовых путей.
При отягощенном акушерском анамнезе, патологическом течении беременно-
сти, воспалении яичников и шеики матки исследования на наличие листерий —
обязательный компонент расширенного обследования генитального тракта. При
аутопсии мертворожденного или погибшего в первые дни жизни ребенка бакте-
риологическому исследованию подвергают кусочки головного мозга, печени и
селезенки. При клинических формах листериоза исследуют содержимое абсцесса,
экссудат, СМЖ, перитонеальную и перикардиальную жидкости, отделяемое глаз и
носоглотки. Исследование фекалий, за исключением случаев развития гастроэнте-
ритов, имеет скрининговое, а не диагностическое значение.
Биологический материал для микробиологической диагностики листериоза сле-
дует забирать в максимально стерильных условиях, тщательно соблюдая правила
асептики. Пробы для посева на питательные среды необходимо брать до начала
антибиотикотерапии или в интервалах между приемом антибактериальных пре-
паратов (не менее 12 ч). Отобранный материал должен быть в кратчайшие сроки
доставлен в микробиологическую лабораторию.
Обнаружение листерий в клиническом материале
С помощью одноэтапного иммунохроматографического теста (ИХТ) можно
проводить скрининг клинического материала на наличие листерий. Он позволяет
получить результат в течение 5-10 мин, прост и недорог. Основным недостатком
теста является его недостаточно высокая чувствительность, поэтому получение
отрицательного результата не служит доказательством отсутствия инфекции и тре-
бует подтверждения результами исследования клинического материала, взятого от
того же пациента, другими лабораторными методами. Кроме того, в выпускаемой в
настояшее время тест-системе ИХТ можно исследовать только фекалии.
Изоляция и идентификация возбудителя
Важнейший элемент современных схем выявления листерий — применение
селективных питательных сред. Эти среды повысили эффективность выделения
листерий из клинического материала и продуктов питания по сравнению с ранее
448
ЧАСТНАЯ МИКРОБИОЛОГИЯ
применяемыми кровяным агаром, мясопептонным агаром или агаром Хоттингера
с теллуритом калия и полимиксином В.
Наибольшее распространение в качестве элективных сред для выделения листе-
рий получили среды «Оксфорд» и «Палкам».
На среде «Оксфорд» через 24 ч инкубации посевов появляются мелкие (диа-
метром 1 мм) сероватые колонии, окруженные черным ореолом. Через 48 ч инку-
бации колонии становятся более темными, около 2 мм в диаметре, черный ореол
сохраняется.
На среде «Палкам» через 24 ч инкубации регистрируются мелкие, серовато - или
оливково-зеленые колонии диаметром 1 мм, часто с черным центром. Колонии
также окружены харак герным черным ореолом на красном фоне агара. Через 48 ч
инкубации отмечаются колонии до 1,5-2 мм в диаметре с характерным вогнутым
центром, зеленоватая окраска и черный ореол сохраняются.
Последующий анализ характерных колоний, утилизирующих эскулины, позво-
ляет на основании ограниченного числа тестов идентифицировать культуру
L. monocytogenes.
Посев на селективные питательные среды выполняют как непосредственно
из клинического материала, так и после предварительного накопления микро-
организмов с помощью питательной среды обогащения. Посевы инкубируют в
течение 48 ч при температуре 30 °C. Рост листерий сопровождается утилизацией
эскулина, что вызывает почернение среды. Для последующего анализа подозри-
тельные на патогенные листерии полупрозрачные небольшие колонии (2-3 мм в
диаметре) с черным ореолом пересевают на питательные среды, рекомендуемые
для идентификации листерий. Для пересева выделенных колоний рекомендуют
использование триптон-соевого агара с дрожжевым экстрактом следующего соста-
ва: триптон — 17 г. пепсиновый гидролизат сои - 3 г, дрожжевой экстракт - 6 г,
декстраза — 2,5 г, NaCl — 5 г, гидрофосфат калия — 2,5 г, агар — 15 г.
После селекции выбор питательных сред для пересева выделенных колоний
уже не имеет принципиального значения, для культивирования листерий можно
использовать достаточно широкий спектр традиционных питательных сред.
Предполагаемые колонии листерий, выросшие на селективных дифференциально-
диагностических питательных средах, для дальнейшей идентификации высевают
на триптон-соевый агар с дрожжевым экстрактом, мясопептонный агар с 1%
глюкозой или ЛФГА. Посевы инкубируют при температуре 30 °C в течение 24 ч.
На триптон-соевом агаре с дрожжевым экстрактом колонии листерий выпуклые,
бесцветные, непрозрачные, росинчатые, диаметром 1-2 мм, легко снимаются с
поверхности агара и образуют гомогенную суспензию при переносе в солевой рас-
твор.
На мясопептонном агаре с 1% 1люкозы и ЛФГА колонии листерий образуют
мелкие, сероватые, полупрозрачные, росинчатые колонии.
На этапе идентификации используют совокупность методов, позволяющих:
• окончательно подтвердить принадлежность выделенной культуры к Listeria
spp.;
• установить принадлежность выделенной культуры Listeria spp. к патогенному
для человека виду L. monocytogenes, используя тесты для идентификации дан-
ного вида и его дифференциации от непатогенных листерий.
Первая I руппа методов включает:
• окраску по Граму и микроскопию колоний;
• выявление каталазной активности;
• определение подвижности культуры при температуре 22 и 37 °C.
При окраске по Грам\ 16-24-часовой культуры листерии выглядят как короткие
грамположительные палочки. В старых культурах морфология листерий часто
вариабельна (бактерии могут приобретать форму диплококков или дифтероидов).
ЧАСТНАЯ МИКРОБИОЛОГИЯ 449
Именно поэтому результаты окраски по Граму мазков-отпечатков из клинического
материала не отличаются высокой степенью достоверности.
Для оценки каталазной активности на поверхность колонии наносят кашпо
3-5% свежего водного раствора перекиси водорода. Появление пузырьков газа
указывает на наличие каталазы. Все виды Listeria spp. каталазоположительны.
Для идентификации Listeria spp. информативен тест на подвижность, в котором
используется высокая подвижность клеток при температуре 22-25 °C и неподвиж-
ность или слабая подвижность при 37 °C Для исследования берут колонии с чашек,
инкубированных при температуре не выше 30 °C. Для определения подвижности
используют висячую или раздавленную каплю, которую микроскопируют с помо-
щью масляного иммерсионного или фазово-контрастного объектива. Листерии,
выросшие при температуре 22 СС, выглядят как тонкие короткие палочки с враща-
тельной или подпрыгивающей активностью.
В последние годы для определения подвижности листерий чаще используют
посев уколом в полужидкий агар. При температуре 22 °C для подвижной культуры
листерий характерен специфический рост в форме зонтика. Помимо полужидких
модификаций мясопептонного агара или триптон-соевого агара существуют спе-
циальные коммерческие среды («Listeria Motility medium») для определения под-
вижности листерий.
Вторая группа методов, необходимых для видовой идентификации L. monocy-
togenes, включает определение восстановления нитратов до нитритов; фермента-
цию трех углеводов — маннита, ксилазы и рамназы: определение способности к
|3-гемолизу; выполнение КАМП-теста, выявление лецитиназной активности на
средах с активированным углем и без него.
В качестве дополнительных ме годов используют серотипирование, ПЦР с прай-
мерами на листериозин или лецитиназу, биопробу на мышах.
Серологическая диагностика
В лабораторной диагностике листериоза используют два серологических
метода — РСК с инактивированным цитоплазменным антигеном и РИГА с эритро-
ци гарным антигенным диагностикумом. Однако серологические реакции недоста-
точно специфичны для диагностики данного заболевания. Первичный диагноз с
помошью одних только серологических реакций ненадеже н. Их можно проводить
для выявления больных с бессимптомным течением заболевания и бактерионо-
сителей в неблагополучных по листериозу хозяйствах, где наличие заболевания
установлено с помощью клинико-эпизоотологических и бактериологических
исследований. Это связано с тем, что большинство случаев листериоза наблюда-
лось именно в неблагополучных животноводческих хозяйствах у людей, контак-
тировавших с больными животными. В последнее время е условиях длительного
хранения и транспортировки продукты питания часто контаминированы листе-
риями, основным путем передачи листериозной инфекции стал пищевой (сыры,
мясные и куриные полуфабрикаты, копчености, морепродукты), В организме
человека происходит нарастание титров антите л к L monocytogenes при отсутствии
клинических проявлений, причем антитела могут длительно сохраняться после
элиминации листерий из организма. Именно поэтому серологические методы
можно использовать для постановки предполагаемого диагноза. Окончательное
подтверждение диагноза листериоза у людей возможно только после выделения
чистой культуры возбудителя из клинического материала.
Наиболее перспективны для серологической диагностики листериоза имму-
ноферментные тест-системы, высокая специфичность которых основывается
на определении антител к рекомбинантным белкам — секретируемому фактору
патогенности листерий (листериозину-О) или другим белковым антигенам
(JrpA, JnlB). Для оценки эффективности диагностики листериоза с помощью
£
450 ЧАСТНАЯ МИКРОБИОЛОГИЯ
данного метода необходимо сравнение с результатами бактериологического
метода.
Критерии постановки диагноза листериоза
Диагностика листериоза представляет значительные трудности в связи с много-
образием клинических проявлений инфекции и их сходством с рядом других,
более распространенных заболеваний. Именно поэтому диагноз листериоза не
может быть установлен только на основании характерной клинической картины.
Немаловажное значение имеют эпидемиологические и анамнестические данные.
Следует также учитывать особенности производственной деятельности и быта
больных, которые могут оказывать влияние на частоту встречи с источником
листериоза. При обследовании женщин (особенно беременных) следует обращать
внимание на патологию беременности и предшествующих родов. Не меньшую роль
играют анамнестические указания, а также обнаружение клинико-лабораторных
признаков заболеваний и состояний, позволяющих отнести обследуемого к группе
повышенного риска развития листериоза. Однако характерные анамнестические
и эпидемиологические данные даже в сочетании с типичной клинической кар-
тиной позволяют поставить только предположительный диагноз. Вероятность
диагноза возрастает, если при типичном поражении ЦНС при микроскопии СМЖ,
полученной при диагностической люмбальной пункции, обнаруживают грампило-
жительные палочки или на 2-й неделе болезни в плазме крови пациента уровень
специфических антител превышает диагностический титр.
Окончательный диагноз листериоза ставят, если характерная эпидемиологи-
ческая и клиническая картина сочетается с обнаружением L. monocytogenes в био-
логических жидкостях организма, которые в норме ci ерильны (крови, СМЖ, СЖ,
плевральной жидкости). Выделение L. monocytogenes из фекалий и мочи имеет
меньшую диагностическую ценность, так как листерии нередко обнаруживают у
здоровых лиц и инфекция имеет условно-патогенный характер. При доказанной
эпидемиологической связи типичной клинической картины заболевания с уста-
новленным источником инфекции положительные результаты бактериологиче-
ского исследования также служат основанием для подтверждения окончательного
диагноза листериоза.
Большую диагностическую ценность при листериозе беременных и новорож-
денных приобретает выделение L. monocytogenes из секретов половых органов,
амниотической жидкости и плацентарной ткани. При легальных исходах и невоз-
можности прижизненного выделения возбудителя принимают во внимание типич-
ные патоморфологические изменения. Особенно характерно обнаружение в раз-
личных органах признаков листериозного ромбэнцефалита и другой специфичной
для листериоза патологии, Выявление листерий в трупных материалах успешно
далеко не всегда и имеет ретроспективное значение.
Спектр заболеваний, с которыми проводят дифференциа пьную диагностику
листериоза, широк, что объясняется многообразием клинических форм листери-
озной инфекции. Наиболее распростоанены у взрослых листериозные менингиты
и менингоэнцефалиты, их следует дифференцировать от инфекций, вызываемых
Str. pneumoniae, N. meningitidis, M. tuberculosis и другими бактериями. Листериозные
менингиты и менингоэнцефалиты чаще регистрируют у пожилых пациентов.
В отличие от менингитов нелистериозной этиологии, у больных гораздо чаще
наблюдаются судороги, нарушение координации, тремор и психические расстрой-
ства.
Листериоз новорожденных дифференцируют от врожденной цитомегалии, ток-
соплазмоза, сифилиса, стреги ококкового сепсиса, гемолитической болезни ново-
рожденных, внутричерепной родовой травмы. Для листериоза характерны диссе-
минированные узелковые экзантемы, фебрильная лихорадка и прогрессирующее
ЧАСТНАЯ МИКРОБИОЛОГИЯ 451
нарушение кровообращения. Учи гывают анамнестические данные о клинических
признаках заболевания матери в последнюю треть беременности, которая окончи-
лась преждевременными родами, а также обнаружение специфических гранулем
в плаценте. Листериоз беременных необходимо дифференцировать от острых
респираторных инфекций и вялотекущего пиелонефрита (пиелоцистита) нелисте-
риозной этиологии.
Гастроинтестинальная форма листериоза отличается от острых кишечных
инфекций другой этиологии относительно более тяжелым течением, выражен-
ностью лихорадки, интоксикацией, нередким исходом в специфический сепсис,
менин! ит или менингоэнцефалит.
Септическую форму листериоза дифференцируют от сепсиса, вызываемого
другими, чаще грамотрицательными бактериями. Вследствие быстрого развития
заболевания и отсутствия патогномоничных признаков решающее значение в
постановке правильного диагноза отводится результатам бактериологических
исследований. Определенную роль также играют анамнестические и эпидемиоло-
гические данные.
КЛОСТРИДИИ
ТАКСОНОМИЯ
Род Clostridium принадлежит к семейству Clostridiaceae и объединяет более
150 видов, точная идентификация которых требует применения специальных
методов и возможна только в специализированных лабораториях.
Клиническая классификация этих бактерий предполагает разделение их на
3 группы:
• нейротоксигенные;
• энтеротоксигенные;
• гистолитические.
Такая классификация имеет ряд недостатков (например, одни и те же виды бак-
терий могут в соответствии с ней относится к разным группам), однако ее удобно
использовать в практических целях.
ОБЩАЯ ХАРАКТЕРИСТИКА
Клостридии — крупные, грамположительные (в молодых культурах), обра-
зующие споры, облигатно анаэробные подвижные (перитрихии) или неподвижные
палочки с закругленными или заостренными концами. Некоторые из них в орга-
низме человека способны образовывать капсулу. Диаметр спор превышает диаметр
бактериальной клетки, поэтому при центральном расположении споры палочка
приобретает форму веретена, а при дистальном — теннисной ракетки или барабан-
ной палочки. Отдельные виды рода Clostridium под влиянием различных факторов
могут приобретать кокковидную, спиралевидную и даже нитевидную форму. В маз-
ках клостридии чаще расположены попарно или короткими цепочками. Способны
продуцировать сильные экзотоксины. Многие штаммы спорулируют слабо, дают
слабую положительную окраску по 1раму. Такие культуры могут быть ошибочно
отнесены к родам Eubacterium, Bifidobacterium, Bacteroides или Fusobacterium,
КЛИНИЧЕСКИЕ ПРОЯВЛЕНИЯ ИНФЕКЦИИ
Патогенные для человека клостридии способны вызывать газовую гангрену,
столбняк, ботулизм, псевдомембранозный энтероколит и другие заболевания, свя-
занные с поражением различных органов и систем. Некоторые виды клостридий
могут быть причиной пищевых токсикоинфекций. Патогенность представителей
рода Clostridium обусловлена продукцией экзотоксинов. Этиологический диагноз
устанавливают на основании клинических и микробиологических данных.
452 ЧАСТНАЯ МИКРОБИОЛОГИЯ
Газовая гангрена относится к группе раневых анаэробных инфекций, возни-
кает в результате травм и ранений. Этиология газовой гангрены разнообразна.
В зависимости от патогенных свойств клостридий заболевание может быть вызва-
но одним из возбудителей (наиболее вирулентные виды — CL perfringens, Cl. novyi,
Cl. septicum; менее вирулентные — Cl. histolyticum, Cl. bifermentans. Cl. sporogenes,
Cl.failax). Слабовирулентные виды, такие как Cl. tertium, Cl. sordellu, Cl. butiricum,
неспособны самостоятельно вызвать газовую гангрену, однако, присоединяясь к
более вирулентным видам, они значительно ухудшают течение болезни.
Cl. difficile вызывают два типа заболеваний, возникающих на фоне интенсив-
ной и поодолжительний антибиотикотерапии, — антибиотикоассоциированную
диарею и псевдомембранозный колит (ПМК). В течение первого года жизни
Cl. difficile обнаруживают у 50% новорожденных, затем их распространенность
снижается, у детей старше 2 лет их выявляют менее чем в 4% случаев, так как при
отсутствии селекционного давления антибиотиков они вытесняются нормальной
микрофлорой кишечника. На фоне антибиотикотерапии количество Cl. difficile
резко возрастает, особенно у пациентов, находящихся в стационаре. Фактор, пред-
располагающий к развитию заболевания, — противоопухолевые и антимикробные
препараты. Особенно часто развитие данных заболеваний связано с применением
пенициллинов широкого спектра действия (ампициллина, амоксициллина), клин-
дамицина, цефалоспоринов. Возможно носительство Cl. difficile среди взрослых,
в том числе среди медицинских работников. Патогенез заболеваний обусловлен
действием двух экзотоксинов возбудителя — А и В (токсин А — энтеротоксин, ток-
син В — цитотоксин). Большинством токсигенных штаммов продуцируются оба
токсина, однако описаны штаммы, вырабатывающие только один из токсинов.
Некротические энтероколиты клостридиальной этиологии регистрируются
у новорожденных и расцениваются как госпитальные инфекции. Чаще болеют
недоношенные или дети, родившиеся с малой массой тела. У взрослых заболе-
вание известно под названием «кишечная гангрена». Описаны вспышки данной
инфекции. Причина заболевания — Cl. perfringens типа С. Патогенез заболевания
обусловлен действием р-токсина Cl. perfringens типа С. Токсин обладает леталь-
ным действием и вызывает очаговый паралич отек, геморрагии и сегментарный
гангренозный некроз кишечной стенки. У новорожденных клиническая картина
некротического энтероколита проявляется вздутием живота, отказом от кормле-
ния, появлением крови в испражнениях. Заболевание прогрессирует до развития
перфораций, перитонита и сепсиса.
ПТИ вызывают Cl. perfringens типа А. Способность вызывать заболевание обу-
словлена наличием энтеротоксина. Как правило, заболевания связаны с употре-
блением мясных блюд и развиваются через 7-30 ч после употребления пищи. ПТИ
проявляется бо;1ями в животе и диареей (часто пенистой). Заболевание возникает
при попадании в организм человека около 10s живых Cl. perfringens типа А, кото-
рые в щелочной среде тонкого кишечника образуют споры. Процесс споруляции
сопровождается высвобождением энтеротоксина.
СД botulinum вызывает ботулизм, который подразделяют на четыре категории:
• классический (пищевой), протекающий как ПТИ, при котором накопление
токсина Cl. botulinum происходит в пищевом продукте;
• новорожденных, при котором ботулотоксин вырабатывается в кишечном
тракте младенца;
• раневой — редчайшая форма, обусловленная продукцией токсина возбуди-
телем, размножающимся на поверхности раны; описаны случаи развития
ботулизма после внутривенных инъекций:
• неопределенно классифицированный, когда развитие заболевания не удается
связать ни с одним из указанных Факторов (раневой инфекцией, пищевым
продуктом); возникает у взрослых и детей старше 1 года.
ЧАСТНАЯ МИКРОБИОЛОГИЯ 453
Во всех случаях ведущее звено патогенеза — ботулотоксин. По антигенным
свойствам различают 7 типов ботулотоксина, обозначаемых латинскими буквами
А, В, С, D, Е, F, G. Наиболее опасен ботулотоксин типа А, наименее — типа Е. Кроме
Cl. botulinum, ботулотоксин или подобные ему нейротоксины могут вырабаты-
вать и некоторые другие виды кл< ютридий: 07. buryricum — ботулотоксин типа Е,
Cl baratii — ботулотоксин типа F.
Возникновение столбняка обусловлено попаданием спор Cl. tetani в рану из
почвы или объектов внешней среды. Столбняк новорожденных возникает при
инфицировании во время родов вне ЛПУ или при перерезке и перевязке пупоч-
ного канатика нестерильным инструментом или материалом. Патогенез обуслов-
лен способностью возбудителя к продукции двух очень сильных экзотоксинов —
тетаноспазмина (разрушает эритроциты) и тетанолизина (вызывает спастиче-
ский паралич, действуя на синапсы мотонейронов спинного мозга, поражает
ЦНС).
Виды рода Clostridium, имеющие медицинское значение, представлены в
табл. 21-36.
Таблица 21-36. Клостридии, имеющие медицинское значение, и заболевания, вызываемые ими
Группа клостридий Заболевание Возбудители
Гмстолитические Газовая гангрена Наиболее патогенные: Cf. perfringens типа А, Cl. septicum, Cl. novyi
Менее патогенные: Cl. histolyticum, Cl. bifermentans. Cl. sporogenes. Cl fallax
Часто вызывают заболевание в ассоциации: Cl. tertium, Cl. but‘ricum, Cl. sordellii
Продуценты нейроток- СИНОВ Столбняк Cl. tetani
Ботулизм Cl. botulinum
Продуценты энтероток- СИНОВ Пищевые токсикоинфекции Cl. perfringens типа A
Некротические энтероко- литы Cl perfringens типа C
Псевдомембранозный энте- роколит Cl. difficile
ЛАБОРАТОРНАЯ ДИАГНОСТИКА
Лабораторная диагностика клостридиозов включает два параллельных направ-
ления:
• выделение возбудителя и его видовую идентификацию (в ряде случаев необ-
ходима количественная оценка содержания бактерий в пробе);
• выявление токсинов возбудителей.
Первый этап бактериологического исследования включает микроскопию натив-
ного материала, посев его на питательные среды и биологическую пробу на лабо-
раторных животных. При микроскопии оценивают морфологичес кие особенности
и тинкториальные свойства микроорганизмов, наличие спор и их расположение
в бактериальной клетке (окраска по Граму). Подвижность определяют при иссле-
довании висячей капли, наличие капсулы — при окраске мазков по Бури или по
Бури-Гинсу.
На втором этапе бактериологического исследования проводят оценку культу-
ральных свойств выросших на питательных средах микроорганизмов и повторную
микроскопию чистой культуры.
На третьем этапе бактериологического исследования продолжают изучение
биологических свойств (культуральных и ферментативных), а также выявляют
токсинообразование.
454 ЧАСТНАЯ МИКРОБИОЛОГИЯ
Материалы для наборам рных исследований
При подозрении на инфекции клостридий в лабораторию отсылают раневое
отделяемое, гной, кусочки пораженных тканей, кровь, фекалии, секционный мате-
риал (кусочки пораженных тканей, пунктаты лимфатических узлов, отделяемое
из раны, кровь из полостей сердца), хирургические инструменты, перевязочный
и шовный материал, пищевые продукты. При подозрении на газовую гангрену
исследуют кусочки патологически измененных тканей, отобранные на границе со
здоровой тканью, раневое отделяемое и экссудат, венозную кровь. При некротиче-
ских энтероколитах, ПТИ и ботулизме новорожденных исследуют фекалии, полу-
ченные с помощью клизмы. При пищевом ботулизме исследованию подлежат про-
мывные воды желудка, плазма крови, при раневом ботулизме — плазма крови.
При столбняке взятие материала после заживления раны сопряжено с трудно-
стями. Если место ранения известно, исследуют отделяемое раны, кусочки тканей.
Когда входные ворота инфекции неизвестны, материал берут из царапин, ссадин,
старых, длительно незаживающих ран. При столбняке, развившемся после родов
или аборта, исследуют отделяемое из влагалища и матки, при пупочном столбня-
ке — отделяемое из пупочного канатика. На секции отбирают кусочки тканей из
предполагаемого очага, а также кровь, кусочки печени и селезенки.
Материал для исследования собирают в стерильную посуду или контейнеры
с плотными пробками и не позднее 3 ч доставляют в лабораторию. В качестве
транспортных сред могут быть использованы питательные среды, содержащие
редуцирующие агенты (тиогликолят натрия, цистеин). Материал, помещенный в
транспортную систему, должен быть доставлен в лабораторию в течение 12-18 ч.
При необходимости материал можно сохранять в замороженном состоянии в
течение 1-2 дней.
Для микроскопического и бактериологического исследования материал заби-
рают отдельно.
Изоляция клостридий
В питательные среды для выделения клостридий для понижения окислительно-
восстановительного потенциала входят различные редуцирующие факторы —
кусочки паренхиматозны х органов (среда Китта-Тароцци), тиогликолевая кисло-
та или ее соли (тиогликолевая среда, СКС).
В плотные агаровые среды вносят различные дифференциально-диагности-
ческие добавки — кровь для выявления гемолиза (среда Цейсслера), яичный жел-
ток для выявления лецитиназы, молоко, сыворотки, углеводы, мочевину и другие
вещества для выявления различных ферментативных свойств. Посевы на плотных
средах (в чашках Петри) инкубируют в бескислородной среде, используя гермети-
чески закрывающиеся сосуды (аьаэростаты), в которые помещают специальные
химические поглотители кислорода, выпускаемые промышленностью, — газоге-
нерирующие пакеты (Gaz-Pack).
Полужидкие среды разливают в пробирки высоким столбиком, сверху на среды
наносят вазелиновое масло слоем 4-5 мм Перед посевом среды, разлитые высо-
ким столбиком, регенерируют прогреванием на кипящей водяной бане в течение
10—15 мин для удаления кислорода, зат ем остужают до 40-45 °C.
Для обеспечения роста и проявления токсигенных свойств большинства пато-
генных клостридий в основном применяют культивирование при 37 °C и значени-
ях pH = 7,0^-7,4.
Биохимическая идентификация
Патогенные клостридии характеризуются различными биохимическими свой-
ствами и отличаются по сахаролитической и протеолитической активности.
Некоторые виды отличаются высокой сахаролитической активностью, другие
частная микробиология 455
обладают большой протеолитической активностью. Некоторые патогенные кло-
стридии малоактивны в биохимическом отношении. Биохимические свойства
определяют на полужидких средах, разлитых в пробирки высоким столбиком и
содержащих различные тест-субстраты, или в коммерческих индикаторных тест-
системах. Дифференциальная характеристика клинически значимых видов кло-
стридий представлена в табл. 2J -37.
Диагностика газовой гангрены, вызванной Ci. perfringens
А. Экспресс-диагностика. Включает световую микроскопию (мазков, окра-
шенных по Граму и Бури, препарата висячей капли), иммунофлюоресцентный
метод (проводят с применением специфической люминесцирующей сыворотки
« Антиперфрингенс»).
Экспресс-анализ позволяет получить ориентировочный положительный ответ
через 1,5-2 ч после поступления материала в лабораторию. Основанием для
него служит выявление неподвижных крупных грамположительных палочек с
обрубленными краями, обладающих капсулой, а также регистрация флюорес-
цирующего желто-зеленого свечения после обработки мазка люминесцирующей
сывороткой «Антиперфрингенс».
Ускоренная диагностика. Ускоренный анализ предусматривает применение
подращивания и получение предварительного результата через 4-6 ч культи-
вирования при температуре 42 "С. Для этого пробу биологического материала
подращивают на мясном или печеночном бульоне с 0,08% цистеина, лакмусовом
молоке с цистеином или на тиогликолевой среде с последующей идентификацией
возбудителя с помощью микроскопии. На лакмусовом молоке (по Минкевичу)
через 2-4 ч культивирования проб, содержащих Cl. perfringens, при температуре
42 °C в результате бурной ферментации молока и разложения лакмуса в кислой
среде отмечаются характерные изменения — кирпично-красный, пронизанный
пузырьками газа творожистый (губкообразный) сгусток казеина и прозрачная
сыворотка. На среде Вильсона-Блера через 4-6 ч инкубации отмечается почерне-
ние среды и появление множественных разрывов агара вследст вие интенсивного
газообразования, что свидетельствует о росте CL perfringens.
Стандартная диагностика. Полная схема бактериоло! ического исследова-
ния осуществляется параллельно с экспресс- и/или ускоренной диагностикой.
Кровь в соотношении 1:10 засевают на специальные жидкие питательные среды,
предназначенные для анаэробов. Другие жидкие пробы засевают в количестве
5-10 капель на дно пробирок с питательными средами. Каждый образец засевают
в 3 пробирки. Одну пробирку перед инкубацией в термостате (при температуре
37 °C) прогревают до 75-80 ’С в течение 20 мин (данная процедура позволя-
ет избавиться от присутствующих в пробе вегетативных форм сопутствующей
микрофлоры). Вторую (непрогретую) пробирку инкубируют при температуре
37 °C, третью — при температуре 42 °C. Инкубация при 42 °C позволяет выявить
С?, perfringens через 1-2 ч. Исследование прогретого и непрогретого материала в
дальнейшем выполняют параллельно.
При посеве в пробирки с высоким столбиком агара для получения роста
отдельных колоний исследуемый материал запаянной пастеровской пипеткой
(тампоном) последовательно погружают в 5-8 пробирок с питательной средой,
тщательно перемешивая содержимое каждой. Плотные материалы гомогенизиру-
ют с небольшим количеством среды и засевают на плотные и жидкие питательные
среды. Дополнительно в жидкие среды можно внести кусочки исследуемого мате-
риала.
В качестве плотной питательной среды может быть использована среда
Цейсслера (кровяной агар с глюкозой). Посевы на плотные среды в чашках Петри
инкубируют в анаэробных условиях 24-48 ч при температуре 37 °C. Посевы на
ГГЖЙ
Таблица 21-37. Биологические свойства некоторых видов клостридий
Виды клостридий Положение спор Подвиж- ность Реакция с яичным желтком (на агаре) Ферментация Восстано- вление нитратов Гидролиз Индол
лецити- наза липаза протеолиз глюкоза маннит лактоза эскулин желатин казеин
Cl. botulinum типа А СТ + - + + + - - - + + + -
Cl. botulinum типа В СТ + - + - + - - - + + + -
С/, difficile ст/т V - - - + + - - + - -
Cl. histolyticum ст + - - - - - - - - + + -
Cl. novyi тип A ст + + + - + - - - V + - -
Cl. perfringens (СТ) - + - - + - + + V + ± -
Cl. septicum ст - - - + - + + + + -
Cl. sordellii ст + + - + + - - - - + + +
Cl. sporogenes ст + - + + + — - - + + + -
Cl. tetani т + - + - - - - - + - V
ЧАСТНАЯ МИКРОБИОЛОГИЯ
Примечание:
<СТ» — субтерминальное положение спор.
«(.СТ)» - субтерминальное положение спор наблюдается редко.
«Т» — терминальное положение спор.
«+» — тест положительный.
«-» — тест отрицательный
«V» — тест вариабельный.
«±» — реакция проявляется у 11 - 25% штаммов.
ЧАСТНАЯ МИКРОБИОЛОГИЯ 457
жидкие питательные среды начинают просматривать через 2 ч инкубации. При
признаках роста на жидких средах проводят высев на плотные среды с последую-
щим изучением колоний. Откол колоний со столбиков агара проводят прокалыва-
нием среды пастеровской пипеткой или предварительно извлекают столбик агара
из пробирки в чашку Петри. Для этого, удерживая пробирку в наклонном положе-
нии над чашкой Петри, подогревают ее конец, что приводит к расплавлению агара
и выталкиванию среды из пробирки его парами.
Клостридии идентифицируют по культуральным, морфологическим и био-
химическим свойствам, а при возможности — дополнительно путем постановки
реакции нейтрализации на лабораторных животных.
Характерные признаки Cl. perfringens:
• быстрый рост на питательных средах;
• черный цвет колоний с почернением среды вокруг них на железосульфитной
среде Вильсона-Блера через 4-6 ч инкубации;
• позеленение колоний на плотных питательных средах по мере пребывания на
воздухе;
• створаживание молока с просветлением сыворотки и образованием губкоо-
бразного сгустка оранжево-красного цвета, содержащего пузырьки газа, на
лакмусовом молоке по Минкевичу через 1-3 ч после посева.
На кровяном агаре колонии Cl. perfringens нередко окружены двумя зонами гемо-
лиза, что обусловлено гемолитическим действием гетерогенных а- и р-токсинов.
Биологические свойства основных возбудителей газовой гангрены и столбняка
представлены в табл. 21-38.
Таблица 21-38. Биологические свойства основных возбудителей газовой гангрены и столбняка
Виды клостридий Капсула Подвижность Лецитиназа ИНДОЛ Ферментация
лактозы сахарозы маннита
Cl. perfringens + - + - + + +
Cl. novyi типа А - + + - - - -
Cl. septicum - + - - + - -
Cl. tetani - + - 4- - - -
Принципы диагностики газовой гангрены и определения токсигенности кло-
стридий представлены на рис. 21-4, 21-5. Для определения чувствительности к
антибактериальным препаратам следует использовать метод серийных разведений
или Е-тест, поскольку критерии оценки для диско-диффузионного метода не раз-
работаны.
Диагностика некротического энтероколита, вызванного Cl. perfringens
Лабораторная диагностика основана на выделении Cl.perfringensтнпз С из фека-
лий, при летальном исходе — из секционного материала.
Диагностика пищевой токсикоинфвкции, вызванной Cl. perfringens
Лабораторная диагностика клостридиозной ПТИ включает следующие этапы:
У экспресс-анализ на наличие энтеротоксина A Cl. perfringens в фекалиях
больных в коммерческих тест-системах;
J микробиологический анализ, проводимый в целях обнаружения
Cl perfringens типа А в подозреваемом пищевом продукте и фекалиях
больных, оценка энтеротоксигенности выделенных Cl. perfringens.
Важно выявление спор Cl. perfringens в количестве более 106 клеток/г в пробах
фекалий, взятых в течение первых 24 ч с момента начала заболевания.
Готовят десятикратные разведения пробы с последующим высевом на питатель-
ные среды и подсчетом выросших колоний. Это необходимо, так как выделение
458
ЧАСТНАЯ МИКРОБИОЛОГИЯ
Исследуемый материал
маслом 1-3 ч, 42 °C
условиях 24 ч, 37°C
маслом 24 ч, 37 °C
Определение
чувствительности к АБП
Откол колоний на жидкие среды, инкубация под вазелиновым маслом
Инкубация 6—18 ч, 37 =С
Определение токсина биохимические свойства
PH на белых Реакция
мышах с лецитовителлином
Рис. 21-4. Принцип диагностики газовой гангрены.
токсина происходит при споруляции. Для данного исследования пробу перед
титрованием прогревают для уничтожения вегетативных форм.
Диагноз ставят на основании выделения из фекалий больных Cl. perfringens
типа А с энтеротоксигенными свойствами и обнаружения в них энтеротоксина А
Cl. perfringens.
Диагностика инфекции, вызванной Cl. difficile
Основные диагностические критерии ПМК:
• клиническая картина колита;
• присутствие в фекалиях токсина;
• наличие в слизистой оболочке толстой кишки псевдомембран слизево-
лейкоцитарной структуры, внешне похожих на фибринозные мембраны при
дифтерии.
Токсин С/, difficile можно выявить:
• по специфическому цитопатогенному эффект}- в культуре клеток челове-
ческих легочных фибробластов, который нейтрализуется антитоксином к
CZ. sordellii или Cl. difficile',
• методом ИФА или встречного иммуноэлектрофореза (данные методы менее
чувствительны и специфичны по сравнению с тестом на культуре тканей);
ЧАСТНАЯ МИКРОБИОЛОГИЯ 459
Рис. 21-5. Принцип выявления токсигенности возбудителей газовой гангрены.
• иммунохроматографическим методом (простая процедура анализа, доступ-
ная для воспроизведения в кабинете врача).
Принцип действия иммунохроматографического метода основан на взаимодей-
ствии токсина с соответствующими моноклональными антителами, находящимися
в тест-полоске, что проявляется изменением цвета в месте реакции. Не требуется
дополнительного оборудования, тесты могут храниться при температуре от 2 до
30 °C длительное время. С помощью выпускаемых в настоящее время иммуно-
хроматографических тест-систем можно выявить токсин А или одновременно
токсины А и В.
Бактериологический метод имеет вспомогательное значение и применяется
главным образом для эпидемиологических целей. В случае выделения Cl. difficile
из фекалий необходимо определить наличие продукции токсина одним из вышеу-
казанных методов.
Днагноит ботулизма
Диагностика ботулизма основана на выявлении и идентификации ботулоток-
сина. Основной метод диагностики — биоприба на белых мышах. Исследование
материала от больных регламентировано «Инструкцией о порядке расследова-
ния, учета и проведения лабораторных исследований в учреждениях санитарно-
эпидемиологической службы при пищевых отравлениях» №1135-73. Исследование
пищевых продуктов выполняют в соответствии с ГОСТ 10444.7-86 «Продукты
пищевые. Методы выявления ботулинических токсинов и Cl. botulinums.
460
ЧАСТНАЯ МИКРОБИОЛОГИЯ
Рис. 21-6. Принцип лабораторной диагностики столбняка
Диагностика столбняка
Лабораторные исследования при столбняке проводят в двух направлениях —
для обнаружения столбнячного токсина и возбудителя.
Исследуемый материал растирают в ступке со стерильным песком, добавляют
двойной объем 0,9% раствора натрия хлорида и полученную суспензию разделяют
на две части: первую оставляют при комнатной температуре для экстрагирования
токсина, вторую засевают на бульон Мартена или Вейнберга и кровяной агар.
Посевы инкубируют в анаэробных условиях в течение 10 дней, через каждые 2 дня
делают мазки, окрашивают по Граму и микроскопируют. При появлении роста
на бульоне проводят высев на кровяной агар. На поверхности кровяного агара
CZ. tetani дает нежные, прозрачные колонии в виде капель росы, которые быстро
распространяю гея и покрывают всю чашку слегка заметной сетчатой пленкой.
Перед посевом на половину чашки можно нанести антитоксическую сыворотку.
Это позволяет ориентировочно идентифицировать культуру по угнетению гемоли-
за. Решающий фактор при идентификации — обнаружение столбнячного токсина.
С этой целью может быть использован одноэтапный иммунохроматографический
тест. Внутривидовая дифференциация основана на определении антигенных
свойств вырабатываемого токсина в реакции нейтрализации на лабораторных
животных.
Принцип лабораторной диагностики столбняка представлен на рис. 21-6.
ЛЕПТОСПИРЫ
ТАКСОНОМИЯ
Leptospira interrogans sensu lato — возбудитель лептоспироза людей и животных.
Она относится к роду Leptospira, принадлежащему к самостоятельному семейству
Leptospiracea в порядке Spirochaetales. Род Leptospira включает также Leptospira bifl-
ЧАСТНАЯ МИКРОБИОЛОГИЯ 461
exasensu lato — сапрофитические лептоспиры, свободно живущие преимуществен-
но в водной среде и не способные вызва гь заболевания людей и животных.
Систематика и номенклатура лептоспир включает две различные классифика-
ционные системы — серологическую и генотипическую.
В серологической систематике таксономическим критерием служит антигенный
состав. Основным таксоном является серовар. В качестве стандартного метода
определения сероваров используют реакцию микроагглютинации (РМА) с пере-
крестной абсорбцией антител. Серологическая классификация имеет большое
практическое значение, так как иммунитет при лептоспирозе серовароспецифи-
чен. Идентифицировано более 268 серовариантов патогенных лептоспир, объеди-
ненных по серологическим связям в 25 серогрупп. Из этого обширного количе-
ства Leptospira interrogans наибольшее значение имеют возбудители серогрупп
Ictcrohaemorrhagiae, Camcola. Grippoiyphosa, Pomona, Tarassovi, Sejroe, Australis.
В генетической систематике лептоспир выделено 8 геномовидов лептоспир —
I. Interrogans, L. alexanderi, L. kirschneri, L. borgpetcrsenii, L. weiflii, L. noguchii, L. santa-
rosai. I. genomospecies 1, из которых наиболее распространены три — £. interrogans,
L. kirschneri. L. borgpetersenii.
ОБЩАЯ ХАРАКТЕРИСТИКА
Лептоспиры занимают особое положение в иерархии эубактерий, отличаясь
уникальной архитектоникой клетки и своеобразным способом движений, позво-
ляющим идентифицировать их на надродовом уровне исключительно на основа-
нии морфологических критериев.
Лептоспиры — спирохеты диаметром 0,1-0,16 мкм и длиной от 6 до 20 мкм,
имеют характерную спиралевидную конфигурацию и специфические локомотор-
ные органел лы. С помощью электронной микроскопии выделены три основных
структурных элемента:
• осевая нить (аксистиль);
• цитоплазматический цилиндр, винтообразно закрученный на осевой нити;
• наружная оболочка.
Оболочка лептоспир типична для грамотрицательных бактерий, в том числе
и других спирохет, имеет цитоплазматическую мембрану и слой пептидогли-
кана, ассоциированного с наружной мембраной. В сослав осевой нити входят
два периплазматических жгутика, закрепленных на разных полюсах клетки и
направленных к ее центру навстречу друг другу. Осевая нить расположена между
наружной оболочкой и цитоплазматическим цилиндром и выполняет локомотор-
ную функцию, напоминая этим жгутики других бактерий. В клетке присутствуют
ламеллярные мембранные органеллы, а также включения, различающиеся по виду
и функции (гликоген, полифосфаты).
Нуклеотид расположен эксцентрично. Геном лептоспир состоит из двух кольце-
вых хромосом, имеющих размер 4500 и 350 т.п.н. В i еноме лептоспир обнаружены
гены, кодирующие факторы патогенности такие как способность к быстрой инва-
зии и диссеминации, устойчивость к фагоцитозу, адгезивность, опосредованную
белками, входящими в состав наружной мембраны лептоспир, и последующую
колонизацию поверхности гепатоцитов и нефротелия, цитотоксичность. Роль
цитотоксического фактора, имеющего существенное значение в патогенезе леп-
тоспироза, играет липопротеин LipL32. Присутствие LipL32 усиливает процесс
гемолиза, осуществляемого выделяемой патогенными лептоспирами сфинго-
миелиназой Н (SphH). У всех патогенных лептоспир обнаружен ген, кодирую-
щий сфингомиелиназоподобный протеин LK73.5, обладающий цитотоксической
активностью по отношению к эндотелиальным клеткам. Выявлены гены, коти-
рующие ферменты, отвечающие за синтез липополисахарида, обладающего вос-
палительным и цитотоксическим эффектом. Эти гены формируют кластер rfb.
462
ЧАСТНАЯ МИКРОБИОЛОГИЯ
Изменчивость локуса rfb и различия в сос гаве геномов семейства degT, которые
также принимают участие в синтезе липополисахарида, обусловливают его вариа-
бельность, с чем, в свою очередь, связано существование большого количества
серогрупп и сероваров лептоспир.
Лептоспиры слабо окрашиваются анилиновыми красителями. Неокрашенные
клетки при светлопольной микроскопии не видны, но хорошо заметны в темном
поле или при фазово-контрастной микроскопии. При микроскопии в темном поле
лептоспиры имеют вид тонких серебристых нитей с крючками на одном или обоих
концах и обладают разнообразными формами подвижности. Активная подвижность
способствует проникновению возбудителя в организм животных или человека.
Лептоспиры — облигатные аэробы, хемоорганотрофы. Источник углерода и
энергии для них — жирные кислоты с длинными углеродными цепями. Они могут
утилизировать неорганические соли аммония как главный источник азота, а также
пуриновые азотистые основания.
Являясь типичными гидрофилами, лептоспиры способны длительное время
выживать во внешней среде при повышенной влажности. В воде открытых bi >до-
емов они сохраняют жизнеспособность от 7 до 30 дней, во влажной почве — до
270 дней, а на пищевых продуктах — от нескольких часов до нескольких дней.
Лептоспиры быстро погибают под воздействием прямых солнечных лучей, а
нагревание до 56-58 °C убивает их в течение 25-30 мин. К низким температурам
данные микроорганизмы устойчивы и сохраняют жизнеспособность после дли-
тельного замораживания. 0,1% раствор соляной кислоты и 0,5% раствор фенола
инактивируют лептоспиры в течение 20 мин, а активный хлор в дозе 0,3-0,8 мг/л —
после двухчасовой экспозиции. Для обеззараживания чистых культур лептоспир
рекомендуют 2% раствор соляной кислоты с экспозицией 24 ч, а патологического
материала — 5% раствор фенола с экспозицией 2 ч.
ЭПИДЕМИОЛОГИЯ
Лептоспироз относится к зооантропонозам, распространенным повсеместно
среди многих видов диких и домашних животных. Наиболее высокий уровень
заболеваемости людей отмечается в странах с влажным субтропическим и тропи-
ческим климатом. Основными источниками лептоспирозной инфекции для людей
являются дикие мелкие млекопитающие — грызуны (полевки, мыши, крысы) и
насекомоядные (ежи, землеройки), домашние животные — крупный и мелкий
рогатый скот, свиньи, лошади, собаки, а также пушные звери, содержащиеся в
клетках, — лисицы, песцы, нутрии. Инфекционный процесс у большинства живот-
ных протекает в хронической форме с длительным и интенсивным выделением
лептоспир с мочой (от нескольких месяцев до года).
Патогенным лептоспирам свойственна гостальная (хозяинная) специфичность.
Возбудители отдельных сероваров циркулируют преимущественно в популяциях
одного или огоаниченного количества видов животных, являющихся их основ-
ными резервуарами. Животные-лептоспироносители выделяют возбудителей во
внешнюю среду, инфицируя воду открытых водоемов, почву, корма, пищевые про-
дукты и другие объекты внешней среды. В организм человека лептоспиры прони-
кают через поврежденный кожный покров и неповрежденные слизистые оболочки
глаз, носа, полости рта, желудочно-кишечного и мочеполового трактов. Основной
путь передачи инфекции — водный, меньшее значение имеют контактный и пище-
вой. Больной человек - «биологический тупик» для возбудителя лептоспирозной
инфекции и не имеет эпидемиологического значения как ее источник. Территории,
на которых выявлены заболевания или носительство лептоспир среди диких или
домашних животных, носят название очагов лептоспироза и потенциально опасны
для человека. Их подразделяют на природные, хозяйственные (антропургические)
и смешанные.
ЧАСТНАЯ МИКРОБИОЛОГИЯ 463
КЛИНИЧЕСКИЕ ПРОЯВЛЕНИЯ ИНФЕКЦИИ
Лептоспироз — зооан гропонозная инфекция, протекающая у человека в виде
острой лихорадки с выраженной интоксикацией, поражением почек, печени, ЦНС,
развитием геморрагического синдрома. Лептоспироз относится к категории тяже-
лопротекающих инфекций с высокой частотой летальных исходов при некоторых
этиологических формах (Icterohaemorrha&ae, Canicula) и нередкими отдаленными
клиническими последствиями.
Клиническое течение лептоспироза имеет выраженную цикличность, опреде-
ляемую пятью фазами развития инфекционного процесса.
• Первая фаза - внедрение лептоспир в месте входных ворот (через повреж-
де иную кожу и слизистые оболочки), проникновение их в лимфатическую, а
затем в кровеносную систему, кратковременная бессимптомная лептоспире-
мия и дальнейшая паренхиматозная диффузия, преимущественно в печень,
почки, надпочечники, селезенку и легкие. В паренхиматозных органах
происходит интенсивное размножение лептоспир. Клинически данная фаза
соответствует инкубационному периоду, который продолжается от 3 дней до
3 нед.
• Вторая фаза — повторный выход лептоспир в кровь с последующим нако-
плением продуктов их метаболизма и распада. Из крови лептоспиры вновь
попадают в ткани, проявляя тропизм к печени, почкам, надпочечникам и
оболочкам головного мозга. Длительность второй фазы составляет 3-7 дней,
клинически она соответствует начальному периоду заболевания и проявляет-
ся выраженной интоксикацией.
• Третья фаза — максимальное развитие токсемии и поражение внутренних
органов. Развиваются генерализованный капилляротоксикоз, синдром дис-
семинированного внутрисосудистого свертывания (ДВС) с возникновением
кровотечений и кровоизлияний во внутренние органы. Клинически третья
фаза патогенеза соответствует периоду разгара заболевания (конец 1-й —
начало 3-й недели болезни).
Гемодинамические нарушения и действие токсинов лептоспир, образуемых
в результате их гибели под влиянием накапливающихся антител, ведут к пора-
жению печени и почек. В тяжелых случаях развивается острая почечная недо-
статочность (ОПН). Дегенеративные изменения гепатоцитов, холестаз, гемолиз
эритроцитов — причины появления желтухи. При глубоких органных поражени-
ях возникает сочетанная острая почечно-печеночная недостаточность (ОППН),
являющаяся наряду с инфекционно-токсическим шоком (ИТШ) основной при-
чиной смерти при лептоспирозе. Неблагоприятные исходы могут быть связаны
с развитием респираторного дистресс-синдрома взрослых (РДСВ), геморраги-
ческого синдрома, пневмонии, менингита, а также полиорганной недостаточ-
ности.
• Четвертая фаза — нестерильная стадия иммунитета. Количество специфи-
чески.,. антител в плазме крови нарастает, но лептоспиры еще сохраняются в
органах, чаще всего в почках. По времени данная фаза соответствует 3-4-й
неделе болезни.
• Пятая фаза — формирование серовароспецифического иммунитета, соот-
ветствует 5-6-й неделе болезни, клинически определяют период реконвалес-
ценции.
ЛАБОРАТОРНАЯ ДИАГНОСТИКА
Результаты лабораторного исследования играют решающую роль для под-
тверждения лептоспирозной этиологии заболевания, так как дифференциальная
диагностика лептоспироза представляет значительные затруднения вследствие
полиморфизма клинических проявлений.
464 ЧАСТНАЯ МИКРОБИОЛОГИЯ
Лабораторная диагност ика лептоспироза основана на комплексе микробиоло-
гических, иммунологических и молекулярно-генетических методов. Успех лабора-
торной диагностики лептоспироза в значительной степени зависит от правильного
и оптимального по срокам взятия материала от больного, для чего необходимо
ориентироваться на особенности патогенеза данной инфекции.
Материалы для исследования — кровь, моча и спинномозговая жидкость
(СМЖ) больного, а при летальных исходах — паренхиматозные органы, транс-
судат грудной и брюшной полостей.
На 1-й неделе болезни на высоте лихорадки (в период лептоспиремии) целесоо-
бразно использовать методы, направленные на выявление возбудителя; микроско-
пию цитратной крови, посев крови, определение ДНК лептоспир в крови методом
ПЦР и заражение лабораторных животных (метод биопроб).
С конца 1~й и в начале 2-й недели определяют специфические антитела, кото-
рые можно выявить с помощью иммунологических реакций, — РМА, макроагглю-
тинации на стекле (РГА), иммуноферментного анализа (ИФА). Начиная со 2-й
недели болезни при менингеальных симптомах в целях обнаружения лептоспир
исследуют СМЖ, с 3-й недели — мочу.
Бактериоскопия
Микроскопируют кровь, мочу, СМЖ, паренхиматозные органы, транссудат.
Поскольку лептоспиры плохо воспринимают окраску анилиновыми красителями,
проводят микроскопию живых возбудителей в темном поле. Из исследуемого
материала готовят раздавленную каплю, для чего наносят его на предметное стек-
ло и сверху накрывают покровным стеклом. На верхнюю линзу конденсора микро-
скопа наносят каплю дистиллированной воды. Микроскопию производят с приме-
нением темнопольного конденсора, объектива х40, окуляра х10 и осветителя.
Для бактериосколического исследования 2 мл венозной крови больного непо-
средственно после ее взятия смешивают с 4 мл 1,5% раствора натрия цитрата. Смесь
центрифугируют при 3000 об./мин в течение 15 мин для осаждения эритроцитов.
Надосадочную жидкость центрифугируют при 10 000 об./мин в течение 30 мин.
Полученный осадок микроскопируют, просматривая 15-20 полей. Микроскопия
крови — простой и доступный для любой лаборатории способ быстрой и ранней
диагностики лептоспироза. Однако отрицательные результаты не исключают леп-
тоспирозную этиологию заболевания.
Мочу и СМЖ исследуют либо в нативном виде, либо после центрифугирова-
ния в гом же режиме, чго и кровь. Ткань паренхиматозных органов массой 1-2 г
измельчают в ступке с 3-4 мл 0,9% раствора натрия хлорида и отстаивают 1-2 ч
при 4 °C или центрифугируют при 3000 об./мин в течение 15 мин. Надосадочную
жидкость микроскопируют.
Изоляция и идентификация возбудителя
Для выделения лептоспир от больного — кровь, мочу (среднюю порцию), СМЖ
в количестве 5-15 капель засевают последовательно в 3 пробирки с 5 мл питатель-
ной среды, желательно непосредственно у постели больного. При посеве тканей
трупа во избежание загрязнения посевов разогретым на спиртовке шпателем
осторожно прижигают поверхност ь органа. Затем в этом месте проводят прокол
пастеровской пипеткой на глубину до 0,5 см, забирают межтканевую жидкость с
элементами ткани и последовательно переносят в 3 пробирки с питательной сре-
дой. При посеве необходимо соблюдать правила асептики. Посевы помещают в
термостат и культивируют при температуре 28-30 °C.
Лептоспиры относятся к медленнорастущим микроорганизмам. Период инку-
бации для их максимального накопления составляет в среднем 6-14 дней, но
может достигать 4 нед и более. Размножаясь в питательного среде, лептоспиры не
изменяют ее внешнего вида. Среда остается прозрачной и слегка опалесцирующей.
ЧАСТНАЯ МИКРОБИОЛОГИЯ 465
Для выявления роста лептоспир через 4-5 дней из содержимого каждой про-
бирки готовят раздавленную каплю и микроскопируют. При обнаружении роста
лептоспир проводят их пересев (по 0,5 мл) на свежую питательную среду для
последующего накопления и идентификации выделенных культур. Пробирки, где
рост лептоспир не выявлен, вновь помещают в термостат и подвергают повторно-
му анализу через каждые 10 дней. При отсутствии роста в течение 3 мес посевы
считают отрицательными.
Идентификацию выделенных штаммов до серогруппы проводят с помощью
набора групповых агглютинирующих сывороток. Определение серовара выделен-
ных лептоспир осуществляют с помощью перекрестной РМА. Для этого необходи-
мо располагать гипериммунными сыворотками против испытуемого и эталонных
штаммов сероваров, входящих в серогруппу. В связи со сложностью и трудоемко-
стью данного исследования выделенные штаммы следует направлять для иденти-
фикации в Сотрудничающий центр ВОЗ и Минздрава России по лептоспирозам на
базе лаборатории лептоспирозов ГУ НИИЭМ им. Н.Ф. Гамалеи РАМН.
Молекулярно-генетическая диагностика
1Ш Р наиболее перспективна для обнаружения ДНК лептоспир в биологических
жидкостях и органах.
Кровь для исследования берут при венепункции в сухие стерильные пробирки
или вакуумные системы в количестве не менее 1 мл без добавления каких-либо
веществ. Для получения плазмы после взятия крови необходимо оставить закры-
тую пробирку в вертикальном положении в течение 15-20 мин. Отбор следует
проводить из фракции плазмы крови или из пограничного слоя между плазмой и
осевшими форменными элементами.
Для взятия мочи в стерильный контейнер собииают среднюю порцию свободно
выпущенной мочи в количестве 3-5 мл.
СМЖ получают при люмбальной пункции в стерильные сухие пробирки в коли-
честве не менее 1 мл.
Для исследования трупного материала берут кусочки паренхиматозных органов
(печени, почек) размер< >м 1x1x0,5 см. Каждый кусочек помещают в отдельную сте-
рильную банку с притертой пробкой или пластмассовой крышкой или стерильную
чашку Петри. Следует избегать загрязнения горлышка банок инфицированным
материалом и строго следить за тем, чтобы дезинфекционные растворы не попали
на кусочки органов или в банку.
Отбор проб из образцов любого вида желательно проводить не позднее чем
через сутки после взятия материала от больного. До этого момента образец сле-
дует хранить при температуре 4 °C. Отбирают пробы в сухие чистые одноразовые
пробирки типа «Эппендорф». Отобранную пробу можно хранить до момента
выделения ДНК не более 24 ч при температуре 4 °C, а при необходимости более
длительного хранения---20 °C.
ПЦР характеризуется высокими специфичностью, чувствительностью (от 10
до 1000 клеток в пробе) и диагностической эффективностью на первой неде-
ле заболевания (начиная с первых суток) даже на фоне антибиотикотерапии.
Именно поэтому данный тест можно рекомендовать как метод ранней экспресс -
диагностики лептоспироза. При тяжелом течении инфекции на фоне отсроченно-
го синтеза специфических антител ПЦР является методом выбора для быстрого
лабораторного подтверждения клинического диагноза.
Отрицательный результат ПЦР не должен быть основанием для прекраще-
ния диагностического поиска. Диагностическая чувствительность лабораторных
исследований при лептоспирозе может быть значительно повышена за счет парал-
лельного использования серологического теста (РМА) и ПЦР независимо от фазы
заболевания.
466 ЧАСТНАЯ МИКРОБИОЛОГИЯ
Серологическая диагностика
Основное место в лабораторной диагностике лептоспироза у людей занимает
серологический метод исследования, осуществляемый в основном с помощью
реакции микроагглютинации (РМА) с живыми культурами лептоспир. Данный
тест, несмотря на его трудоемкость, необходимость работать с живыми культурами
лептоспир нескольких серогрупп и проводить учет реакции под микроскопом, до
сих пор остается «золотым стандартом» в мировой практике благодаря высоким
чувствительности и специфичности. РМА позволяет не только определить уровень
антител к лептоспирам, но и установить серогруппу возбудителя, что важно для
эпидемиологического анализа.
Для серологического скрининга можно использовать более простые тесты
(иммуноферментный анализ, РИГА, реакцию слайд-агглютинации) с родоспеци-
фическими антигенами лептоспир. Антитела к лептоспирам появляются в крови
больных в конце первой — начале второй недели болезни и достигают максималь-
ного накопления на 14-17-е сутки. Иногда при тяжелом течении инфекционного
процесса, особенно при интенсивной антибиотикотерапии, у больных развивается
иммуносупрессия. В таких случаях при явной клинической картине лептоспироза
антитела могут не выявляться или обнаруживаться через 2-3 мес после начала
болезни.
Кровь больного необходимо исследовать на наличие антител к лептоспирам
не менее 2 раз — при поступлении больного в стационар (даже если это 1-й день
болезни) и через 7 дней. При необходимости исследование следует повторить на
3-4-й неделе заболевания. Если при типичной клинической картине лептоспироза
и характерном эпидемиологическом анамнезе результат серологического иссле-
дования отрицательный, необходимо повторить исследование перед выпиской и
через 2 мес после выписки из стационара в период диспансерного наблюдения за
реконвалесцентом.
Реакция микроагглютинации. Для постановки РМА используют живые куль-
туры лептоспир с хорошей подвижностью клеток, без спонтанной агглютинации
и посторонних примесей (осадка, контаминации посторонней микрофлорой), с
накоплением 5х107 клеток в 1 мл (50-100 лептоспир в поле зрения микроскопа
при увеличении в 400 раз). Каждую сыворотку исследуют со всеми референтными
штаммами, представляющими серогруппы патогенных лептоспир, циркулирующих
в данной местности. Для диагностики лептоспироза на территории Российской
Федерации рекомендуют стандартный набоц из 13 штаммов, принадлежащих к
10 серогруппам (табл. 21-39).
При реакции микроагглютинации сыворотку больного разводят в 10 и 50 раз
0,9% раствором натрия хлорида (1:10 и 1:50 соответственно) для исключения
феномена прозоны. В случае положительной реакции сыворотку титруют для
определения конечного разведения, в котором обнаруживают агглютинаты.
Таблица 21-39. Набор эталонных штаммов лептоспир для РМА, рекомендуемый дня диагности-
ческих исследований на лептоспироз в Российской Федерации (Методические указания. — М,:
Минздрав России, 2002)
Серогоуппа Серовар Эталонный штамм
Icterohaemorrhagiae Copenhagen! M20
Javanica Poi Poi
Hanka Hanka'
Canicola Canicola Hond Utrecht IV (Каширский)"
Aotumnalis Autumnalis Akiyami A
Australis Australis Ballico
ЧАСТНАЯ МИКРОБИОЛОГИЯ 467
Окончание табл. 21-39
Серогруппа Серовар Эталонный штамм
Pomona Pomona Pomona
Mozdok П.О. 5621
Grippotyphosa Grippotyphosa Moskva V
Sejroe Saxkoehmg Mus 24
Wolffi 3705
Bataviae Djatzi HS26
Tarassovi Tarassovi Перепелицин
' Циркуляция лептоспир серовара Hank а установлена только в природных очагах Приморского
края, поэтому соответствующий штамм рекомендуют для включения в диагностическую панель
только на данной территории.
" В скобках указан отече< твенный аналог.
Разведенную сыворотку и живые культуры лептоспир смешивают в равных
объемах по 50 мкл в лунках полистироловых планшетов. Контроль — культуры
лептоспир, смешанные с равным объемом физиологического раствора. После экс-
позиции в течение 30 мин при комнатной температуре проводят учет реакции. Для
этого из каждого разведения сыворотки готовят раздавленную каплю и микроско-
пируют в темном поле при увеличении в 200 или 400 раз. Вследствие добавления к
испытуемой сыворотке равного объема культуры лептоспир разведение сыворотки
удваивается, то есть сыворотка, разведенная 1J0 с культурой, становится разве-
денной 1:20. Феномен агглютинации проявляется образованием различных форм
агглютинатов — в виде паучков, клубочков, кос. Реакцию оценивают по условной
3-балльной шкале:
• «+» — Уз лептоспир агглютинированы (по сравнению с контролем);
• «++» — Уз лептоспир агглютинированы,
• «-Н-+» - агглютинированы почти все лептоспиры.
Для серологического исследования в РМА используют нативную сыворотку.
Доказательство лептоспирозной этиологии заболевания — нарастание титров
антител даже в невысоких разведениях (при первом исследовании антитела не
обнаружены, при втором — титр 1:20). Во время вспышки лептоспироза при
выявлении у больных антител в титрах 1:100 и выше достаточно однократного
исследования.
При исследовании сывороток крови больных в 60% испытуемых образцов,
особенно на 1-2-й неделе заболевания, наблюдаются межгрупповые реакции
(сыворотка реагирует с лептоспирами нескольких серогрупп), что затрудняет
установление этиологического агента. В таких случаях следует считать, что
заболевание обусловлено лептоспирами той серогруппы, к которой определяют-
ся антитела в наиболее высоком разведении сыворотки. Исследование парных
сывороток также помогает установить серогруппу возбудителя, так как на более
поздних сроках заболевания специфические антитела синтезируются только к
нему.
Реакция макроагглютинации (слайд-агглютинация). Для предварительно-
го тестирования сывороток больных с подозрением на ле птоспирозную инфекцию
и для массового скрининга можно использовать реакцию макроагглютинации на
стекле (слайд-агглютинацию). В данной реакции используют родоспецифический
антиген, позволяющий выявлять антитела к различным серогруппам лептоспир.
Сыворотку, дающую положитрльную реакцию агглютинации, в дальнейшем
исследуют в РМА. Для диагностики лептоспироза используют тесты, основанные
на реакции непрямой гемагглютинации. Система, включающая эритроциты, свя-
занные с антигеном, полученным путем разрушения клеток лептоспир, позволяет
выявлять антитела к различным серовариантам лептоспир. Использование вместо
46Я
ЧАСТНАЯ МИКРОБИОЛОГИЯ
эритроцитов синтетических микрокапсул и латексных частиц позволяет создавать
более доступные системы диагностики, в основе которых лежит тот же принцип,
что и в случае реакции непрямой гемагглютинации. Преимущество синтетических
микрокапсул перед эритроцитами заключается в их химической стабильности.
Латексные гранулы обладают теми же преимуществами, что и синтетические
микрокапсулы. В различных системах, основанных на агглютинации латекса, в
качестве антигена используют молекулы липопротеина LipL41, выделенные из
рекомбинантного штамма лептоспир, или клетки лептоспир, разрушенные термо-
лизом.
Нитроцеллюлозные тест-полоски для выявления антител к лептоспирам с нане-
сенным антигеном лептоспир применяют в том числе и для скрининговых иссле-
дований. Они не требуют особых условий хранения и высокого уровня подготовки
персонала. В качестве антигена можно использовать термически разрушенные
клетки лептоспир.
Иммуноферменгный анализ. В конструировании тест-систсм для ИФА при-
меняют антигены различного состава, в том числе разрушенные термолитически
или ультразвуком клетки патогенных лептоспир. Созданы специфичные системы
на основе рекомбинантных белков для определения содержания IgM, IgG и IgA.
В состав таких систем входят молекулы липопротеина LipL32, консервативный
домен иммуноглобулиноподобных белков LigA и LigB. белков поверхностной
мембраны MPL17 и MPL21, а также рекомбинантной белковой молекулы, вклю-
чающей эпитопы порина OmpLl и липопротеинов LipL21, LipL32, LipL41. LipL45.
Применение фрагмента молекулы LigA и порина OmpLl в качестве антигенов
для ИФА делает диагностику более точной, поскольку при вакцинации антитела
к данным антигенам не вырабатываются в отличие от антигенов, выявляемых в
РМА, титры антител к которым могут достигать уровня, сопоставимого с титром
антител при острой инфекции. Антитела к LigB также ассоциированы с острым
течением лептоспироза, их содержание в плазме крови в период реконвалеспенции
снижается быстрее, чем содержание антител, выявляемых в РМА. Недостаток диа-
гностических систем, основанных на выявлении антител к LipL32 и LipL41, — воз-
можность перекрестной реакции с плазмой крови больных сифилисом и болезнью
Лайма, а также вирусным гепатитом.
КОРИНЕБАКТЕРИИ
ТАКСОНОМИЯ
Род Corynebactenum входит в состав семейства Corynebactenaceae. относяще-
гося к порядку Actinomycetales. Наиболее значимые виды рода Corynebacterium —
С. diphtherias. С. ulcerans, С. pseudotuberculosis, С. tyiatum, С. minutisslmum, С, xerosis,
С. pseudodiphihenticum, С. glucuronolyticum, С. renale, С. cistitidis.
Основной признак систематики коринебактерий — биохимический состав
клеточной стенки (наличие коротких цепей миколовой кислоты с 32-36 атомами
углерода и мезодиаминопимелиновой кислоты в качестве диаминокислоты пепти-
догликана).
ОБЩАЯ ХАРАКТЕРИСТИКА
Коринебактерии — грамположительные, неравномерно окрашиваемые пря-
мые или слегка изогнутые палочки размером 1,5-8х0.3-0,8 мкм. Многие из них
способны формировать внутриклеточные гранулы волютина (полиметафосфаты).
Спор не образуют, но имеют микрокапсулу. Неподвижны. Кислотонеустойчивы.
Каталазоположительны. Обладают смешанным (бродильным и дыхательным)
метаболизмом. Факультативные анаэробы.
ЧАСТНАЯ МИКРОБИОЛОГИЯ 469
ЭПИДЕМИОЛОГИЯ И КЛИНИЧЕСКИЕ ПРОЯВЛЕНИЯ ИНФЕКЦИИ
Коринебактерии широко рас пространены. Большинство из них — облигатные
паразиты слизистых оболочек человека и животных.
С. diphtheriae. Вызывает у человека классическую дифтерию верхних дыха-
тельных путей, сопровождаемую тяжелой интоксикацией. В 1884 г. Леффлер не
только установил этиологическую роль данного возбудителя, но и доказал воз-
можность бактерионосительства без клинических симптомов.
Входные инфекции — чаще всего слизистые оболочки миндалин, реже — слизи-
стые носа и гортани, очень редко — конъюнктива глаз, наружные половые органы,
поврежденная кожа. Клинические симптомы дифтерии обусловлены воздействием
дифтерийного экзотоксина и проявляются в виде местных и общих нарушений.
Наибольшая выраженность действия токсина наблюдается при формах дифтерии,
протекающих с развитием тяжелых осложнений (миокардите, полирадикулонев-
ритах).
Инкубационный период при дифтерии составляет от 3 до 7 дней. Клиническая
картина болезни разнообразна. Классификация дифтерии основана на локали-
зации и распространенности местного поражения (налета) и оценке тяжести
заболевания по выраженности интоксикации или степени нарушения функции
пораженного органа (стеноза гортани). В зависимости от локализации различа-
ют дифтерию зева, носа, гортани, глаза, уха, половых органов, кожи; выделяют
комбинированные формы дифтерии с локализацией местного поражения в двух
и более органах (дифтерию зева и носа, зева и гортани). Диагностика атипичных
форм дифтерии основывается на данных лабораторного исследования (выделении
дифтерийных палочек из материала, взятого из места поражения) и обнаружении
частых параллельных симптомов поражения зева или носа.
С. ulcerans. Является одним из возбудителей мастита у коров. У людей ухажи-
вающих за инфицированным скотом, может вызывать дифтериеподобное заболе-
вание.
С. pseudotuberculosis. Бактерия известна как возбудитель ряда заболеваний
мышей, овец, лошадей и других теплокровных животных. У человека в редчайших
случаях может вызывать абсцесс глотки.
С. minutissimum. Возбудитель эритразмы, бактериемии, эндокардита чело-
века,
С. striatum. Представитель нормальной микрофлоры слизистой оболочки
глотки, но может выделяться и из молока коров, больных маститом. Двадцати-
четырехчасовая культура детальна для мышей при внутривенном введении.
У человека может вызывать плевральные и легочные абсцессы.
С. xerosis. Нормальный обитатель кожи и слизистых оболочек человека, впер-
вые был изолирован из материала, взятого из слезного канала. У человека в редких
случаях вызывает вялотекущий конъюнктивит, эндокардит, пневмонию и раневую
инфекцию. Последние регистрируют у больных с тяжелыми сопутствующими
заболеваниями или принимавших глюкокортикоиды.
С. pseudodiphtheriticum (ложнодифтерийная палочка). Нормальный обита-
тель слизистых оболочек человека, однако у пациентов с ослабленным иммуни-
тетом может вызывать эндокардит, лимфаденит, кожные поражения, инфекцию
мочевыводящих путей.
С. glucuronolyticum, Выделена от людей с заболеваниями мочеполовой систе-
мы (в том числе простатитом и уретритом).
С. renale. Часто обнаруживают на коже вымени здоровых коров, может вызы-
вать у жвачных цистит и пиелит. Могут вызывать цистит и пиелонефрит у челове-
ка, чаще у женщин. У людей обычно поражает почки.
С. cistitidis. Обнаруживают в моче здорового крупного рогатого скота.
Может вызывать геморрагические циститы и инфекции препуция. У человека
470 ЧАСТНАЯ МИКРОБИОЛОГИЯ
наиболее часто колонизирует кожу и слизистые оболочки, особенно в обла-
сти промежности. Чаще выделяют у женщин. Вызывает повышение pH мочи и
образование камней в мочевыводящих путях. Является одним из возбудителей
геморрагического цистита, пиелонефрита и бактериемии у пациентов старшего
возраста, имеющих в анамнезе урологическую патологию, перенесенное опе-
ративное вмешательство или длительный прием антибактериальных препара-
тов широкого спектра действия. Отмечены случаи госпитальных бактериемий
и пневмоний.
Сведения об отдельных видах недифтерийных коринебактерий, этиологическая
роль которых в возникновении патологических состояний является доказанной,
приведены в табл. 21-40.
Таблица 21-40. Пейзаж недифтерийных коринебактерий, выделенных при различных заболева-
ниях (Краева Л.А., 20)0)
Ns Диагноз Исследуемый материал Коринебактерий
1 Дифтерия зева Слизь из зева С. xerosis С. paurumetabolicum
2 Дифтерия наружных половых органов Соскоб из влагалища С. urealyticum
3 Назофарингит Слизь из зева и носа С. pseudodiphtheriticum С. auris С. xerosis С. paurumetabolicum С. ulcerans
4 Ринит Слизь из носа С. pseudoaiphtheriticum С. auris С. haemolyticum С. ulcerans
5 Тонзиллит Слизь из зева С. pseudodiphtheriticum С. xerosis С. paurumetabolicum С. ulcerans С. pseudotuberculosis
6 Бронхит Мокрота С. striatum С. auris С. xerosis
7 Пневмония Мокрота С. pseudotuberculosis С. xerosis
8 Аллергический дерматит Смыв с поверхности кожи С pseudodipbtheriticum С. minutissimum
9 Дерматоз Смыв с поверхности кожи С. minutissimum С naemolyticum
10 Офтальмит Мазок из глаза С. macginleyi
11 Аднексит Мазок из влагалища С. xerosis
12 Вульвовагинит Мазок из влагалища С glucuronolyticum С. urealyticum
13 Уретрит Моча С. glucuronolyticum С. urealyticum
14 Пиелонефрит Моча С. urealyticum
15 Незаживающая рана Отделяемое раны С. pilosum
16 Отит Отделяемое из уха С. minutissimum С. auris
Всего 16 14 видов рода Corynebacterium
ЧАСТНАЯ МИКРОБИОЛОГИЯ 471
ЛАБОРАТОРНАЯ ДИАГНОСТИКА
Материалы для лабораторных исследований
Для хорошей высеваемости С. diphtheriae большое значение имеют правиль-
ные взятие и доставка материала. Выполнять данные действия должен специ-
ально обученный медперсонал. Исследование проводят с диагностическими
и профилактическими целями, а также по эпидемиологическим показаниям.
От больных с подозрением на заболевание дифтерией материал забирают как
можно скорее, не позднее 12 ч с момента обращения в ЛПУ. Для исследования
берут пленки и слизь с миндалин, слизистой оболочки зева и носа. При подо-
зрении на дифтерию редкой локализации (глаз, ухо, кожа, рана, влагалище)
забирают отделяемое с пораженных участков и одновременно со слизистой обо-
лочки зева и носа. При видимых поражениях материал рекомендуют брать на
границе здоровой и пораженной ткани, слегка нажимая зондом-тампоном Если
видимые поражения отсутствуют, следует взять как можно больше материала с
миндалин. Взятие проб проводят натощак или через 2 ч после еды, перед этим не
следует пить и полоскать рот. Материал со слизистой оболочки зева и носа соби-
рают отдельными зондами-тампонами, при подозрении на дифтерию редкой
локализации дополнительно используют еще один зонд-тампон. Если доставка
материала в лабораторию будет осуществлена быстро (менее чем за 3 ч), исполь-
зуют сухие стерильные зонды-тампоны. Если время доставки больше, возможны
следующие варианты:
• забирают материал зондом-тампоном, смоченным в 5% растворе глицерина в
0,9% растворе натрия хлорида и простсрилизованным при 0,5 атм в течение
30 мин,
• взятие материала осуществляют сухим зондом-тампоном, который погружа-
ют в транспортную среду.
Использование в качестве транспортной среды питательного бульона с добавле-
нием сыворотки позволяет применять экспресс-методы выявления дифтерийного
токсина — ИФА или РИГА.
В случае исследования трупного материала целесообразно взять пробу с мин-
далин, слизистой оболочки гортани и полостей носа. Локализация возбудителя во
внутренних органах встречается крайне редко. В осенне-зимний период весь мате-
риал доставляют в лабораторию в термосумках или термоконтейнерах. Каждая
пробирка должна быть пронумерована с указанием источника исследуемого мате-
риала (слизистая оболочка зева, носа, отделяемое раны). В прилагаемом списке
соответственно номеру указывают Ф.И.О., возраст и домашний адрес больного,
название учреждения, отправившего материал, цель исследования (диагности-
ческое, по эпидпоказаниям, профилактическое), дату и время взятия материала,
Ф.И.О. медицинского работника, взявшего материал, и подпись врача.
Изоляция и идентификация возбудителя
Схема бактериологического исследования
Первый день. Материал от одного лица засевают на чашку с дифференциально-
диагностической средой (кровяно-теллуритовым агаром, Клауберта II и др.) и
чашку с кровяным агаром, используя при этом половину чашки для посева мате-
риала со слизистой оболочки носа, а вторую половину — со слизистой оболочки
зева. Если проба взята из очага редкой локализации, добавляют еще одну чашку с
кроеяно-теллуритовой средой. Необходимо следить за тем, чтобы чашки не были
холодными.
При посеве материал втирают в питательную среду, поворачивая и обжимая
тампон со всех сторон на площади 2x1 см, а затем частыми непрерывными движе-
ниями штрихуют всю поверхность, не меняя положения тампона (не поворачивая
его). Такая методика позволяет получать изолированные колонии, которые можно
472 ЧАСТНАЯ МИКРОБИОЛОГИЯ
использовать для дальнейшей идентификации, что сокращает длительность ана-
лиза.
Засеянные чашки и пробирки с транспортной средой помещают в термостат. Из
транспортной среды на следующие сутки проводят высев на среды для первичного
выделения (на практике такой высев делают через 6-8 ч),
На обезжиренных стеклах готовят несколько мазков, поворачивая тампон раз-
ными сторонами, высохшие мазки фиксируют в пламени и окрашивают щелочной
метиленовой синькой Леффлера. При микроскопии просматривают как можно
больше полей зрения (до 100). Если материал был взят с пораженных участков,
бактерии, подозрительные на принадлежность к роду Corynebacterium, удается
обнаружить в 40-50% случаев. Особенно целесообразно использовать бактерио-
скопию первичного материала при подозрении на дифтерию раны, так как в этом
случае первичный посев может дать отрицательный результат из-за местного при-
менения лекарственных средств.
Морфологические признаки, характер расположения микроорганизмов не
позволяют установить видовую принадлежность, но дают возможность предпо-
ложительно отнести выделенную культуру к роду коринебактерий.
Второй день. Начинают с просмотра чашек с первичным посевом. Проводить
его лучше с помощью стереомикроскопа, чтобы не пропустить подозрительные
колонии, которые следует отобрать для дальнейшей идентификации по всем
тестам. Подозрительные на принадлежность к С. diphtheriae колонии на кровяно-
теллуритовых средах выпуклые, круглые (1-2 мм), черно-серого цвета, на кровя-
ном агаре — круглые, непрозрачные, слабого кремового оттенка.
Микроскопию мазков из типичных колоний не проводят, но из сомнительных
колоний препараты готовят обязательно, окрашивая их щелочной метиленовой
синькой Леффлера. Коринебактерии, выросшие на средах с ингибитором, могут
быть полиморфными, однако сохраняют присущее им характерное расположение.
При обнаружении на чашках только стафилококков, дрожжей и споровых палочек
дальнейшее исследование прекращают.
При работе следует пользоваться бактериологической петлей небольшого диа-
метра (1-2 мм). Дальнейший ход исследования зависит от того, какое количество
подозрительных колоний выросло на средах первичного посева.
• Если выросла только одна подозрительная колония, ее засевают на среду
для определения токсигечности и, не обжигая петли, в столбик среды Пизу
для определения цистиназной активности. Дальнейшее изучение культуры
можно проводить из «бляшки», выросшей на среде для определения токси-
генности.
• Если выросло много подозрительных колоний, приступают к изучению ток-
сигенных свойств, используя при этом как можно больше колоний (не менее
10), изучая как чистые культуры из изолированных колоний, так и смешан-
ные по 5-6 культур в одной «бляшке». При этом важно, что в исследуемом
материале могут находиться одновременно токсигенные и нетоксигенные
штаммы С. diphtheriae.
При обнаружении на чашках первичного посева большого количества подо-
зрительных колоний в столбик среды Пизу вносят 5-6 однотипных колоний.
Положительный результат — почернение по ходу укола и образование коричне-
вого «облачка» сульфида свинца — учитывается через 3-6 ч при использовании
модифицированной среды Пизу.
Дополнительно можно выполнить тест на ферментацию мочевины (по методу
Заксе или с использованием индикаторного диска) с 5-6 однотипными коло-
ниями. Результат учитывают через 30 мин-2 ч инкубации при температуре 37 °C.
Подозрительные колонии отсевают на сывороточный бульон и после 5-24 ч под-
ращивания используют РИГА или ИФА для определения дифтерийного токсина.
ЧАСТНАЯ микробиология 473
Культуру из изолированной колонии засевают на среду накопления (косячок
сывороточного агара).
По характерной морфологии клеток, виду колоний, положительной пробе на
цистиназу, отрицательной на уреазу, положительному результату ИФА или РИГА
можно выдать предварительный ответ об обнаружении культуры, подозрительной
на С. diph.fhen.ae.
Чашки с первичным посевом вновь помещают в термостат еще на сутки.
Третий день. Просматривают чашки с посевами на токсигенность в проходя-
щем свете и на темном фоне, с лупой и без нее. Выделенная культура считается
токсигенной, если образовавшиеся линии преципитации контрольного и иссле-
дуемого штаммов, сливаясь концами, плавно переходят одна в другую. Если испы-
туемый штамм дал несколько преципитационных линий, специфической следует
считать наиболее четкую.
При отсутствии преципитационных линий чашки дополнительно инкубируют
еще 24 ч, но не больше, так как на более поздних сроках возрастает вероятность
появления линий неспецифической преципитации.
При наличии линий специфической преципитации, положительной пробе на
цистиназу и отрицательной на уреазу выдают документированный предваритель-
ный ответ, что выделена культура токсигенного штамма С. diphtheriae.
Культуру, выросшую на сывороточном косячке или на среде Пизу, после про-
верки чистоты засевают на среды с мочевиной, пиразинамидом и углеводами
(глюкозой, сахарозой и крахмалом), а также нитратную среду.
Просматривают повторно чашки с первичными посевами. Если отсутствует рост
колоний, подозрительных на С. diphtheriae, выдают окончательный отрицательный
ответ. Если в сопроводительном документе было указано, что материал был взят от
больного с клинической картиной дифтерии, исследование необходимо повторить.
К настоящему времени существуют данные о том, что рост С. diphtheriae из пато-
логического материала иногда можно наблюдать через 72 ч и более после посева.
Именно поэтому к срокам выдачи отрицательного ответа через 48 ч, определен-
ным в нормативных документах, следует относиться осторожно.
Четвертый день. Повторно просматривают чашки с посевами на токсиген-
ность. Учитывают результаты ферментации углеводов, характер изменения сред с
мочевиной, пиразинамидом и нитратами.
При отсутствии специфических преципитационных линий через 48 ч инкуба-
ции, но положительной пробе на цистиназу, ферментации глюкозы до кислоты,
отрицательными тестами на уреазу и пиразинамидазу, сахарозу, выделенную куль-
туру идентифицируют как нетоксигенный штамм С. diphtheriae.
Если предварительный ответ о выделении токсигенного штамма уже был выдан
на 3-й день, то на основании учета результатов всех биохимических тестов окон-
чательный ответ с указанием биовара выдается на 4-й или 5-й день исследования
в зависимости от использованного варианта теста на ферментацию крахмала.
Последний на среде Гисса занимает 48 ч, а с использованием индикаторного диска —
24 ч. Предварительный ответ может быть изменен после окончания бактериоло-
гического исследования.
В 1997 г. были опубликованы данные, которые могут привести к переоценке
значимости цист иназного теста для идентификации С. diphtheriae. Впервые было
выделено 4 штамма, которые не обладали цистиназной активност ью. При этом у
всех культур токсигенные свойства были выявлены только на 2-е сутки По всем
другим тес гам они имели типичные свойства, характерные для С. diphtheriae gravis.
При проведении ПЦР принадлежность данных культур к С. diphtheriae была под-
тверждена.
Дифференциацию других видов проводят на основании оценки результатов
биохимических тестов с использованием прилагаемых таблиц и схем.
474 ЧАСТНАЯ МИКРОБИОЛОГИЯ
Культурально-морфологические особенности коринебактерий
Ниже приведены культурально-морфологические особенности наиболее часто
встречаемых у человека коринебактерий.
С. diphtheriae. По морфологии колоний, выросших на крпвяно-теллуритовых
средах, морфологии палочек в мазках, отношению к ферментации крахмала и
редукции нитратов С. diphtheriae делят на четыре культурально-биохимических
варианта — gravis, mitis, intermedius, belfanti.
C, diphtheriae gravis на кровяно-теллуритовых средах через 24 ч вырастают в
виде черно-серых сухих колоний, крошащихся под бактериологической петлей.
Диаметр колоний 1,5-2,0 мм. В микроскопических препаратах, приготовленных
из этих колоний, видны короткие полиморфные палочки с небольшим количе-
ством волютина. Ферментируют крахмал.
С. diphtheriae mitis на кровяно-теллуритовых средах через 24 ч роста формируют
черные круглые выпуклые гладкие колонии диаметром 1,5-2,0 мм. В мазках —
длинные изогнутые палочки с большим количеством волютина. Крахмал не фер-
ментируют.
С. diphtheriae intermedins на кровяно-теллуритовых средах через 24 ч выраста-
ют в виде серо-черных нежных круглых плоских колоний размером 0,5-1,0 мм
В мазках — длинные, бочкообразной формы палочки с небольшим количеством
волютина. Крахмал не ферментируют.
С. diphtheriae belfanti на кровяно-теллуритовых средах через 24 ч формируют
черно-серые округлые гладкие колонии среднего размера — 1-2 мм. Крахмал не
ферментируют, в отличие от других вариантов С. diphtheriae не редуцируют нитра-
ты.
В настоящее время на кровяно-теллуритовых средах признаки дифференци-
рованного роста и различия по морфологии палочек в мазках не выявляются.
Остается только один четкий дифференцирующий тест — ферментация крахма-
ла. По данному признаку С. diphtheriae intermedius не отличается от С. diphtheriae
mitis, к тому же этот вариант практичес ки отсутствует в естественной циркуляции.
Именно поэтому описание С. diphtheriae intermedins из многих официальных доку-
ментов исключено. С. diphtheriae belfanti также относится к редко встречаемым
вариантам.
С. ulcerans. На кровяно-теллуритовых средах через 1 сут вырастает в виде коло-
ний, похожих на С. diphtheriae. В микроскопических препаратах — мелкие овоиды
и коккоподобные палочки.
С. pseudotuberculosis. Палочки размером 0,5-0,6x1-3 мкм с метахроматиче-
скими гранулами. На кровяном агаре через 24 ч формируют колонии кремового
цвета, с матовой поверхностью и небольшой зоной гемолиза. В микроскопических
препаратах — палочки размером 0,5-0,6x1-3 мкм с метахроматическими грану-
лами.
С. minutissimutn. На плотных пи гательных средах через 24 ч формируют глад-
кие блестящие колонии размером 1 мм. Если колонии выросли на сывороточном
агаре, то под лампой Вуда (длина волны 365 нм) они светятся розовым светом. В
мазках — палочки (0,3-0,6x1-2 мкм) с перешейком на одном из сужающихся кон-
цов с метахроматическими гранулами.
С. striatum. На кровяном агаре через 48 ч формируют мелкие, блестящие, кру-
глые колонии со слабой зоной гемолиза в глубине. Некоторые штаммы продуци-
руют зеленовато-желтый пигмент, диффундирующий в среду. В микроскопических
препаратах — палочки кокковидной формы (0,25-0,5x2-3 мкм) с небольшим
количеством волютина.
С. xerosis. На плотных питательных средах через 1 сут формируют мелкие
(1 мм) колонии с гладкой или шероховатой поверхностью. Колонии могут бы гь
окрашены в разные оттенки желтого цвета.
ЧАСТНАЯ МИКРОБИОЛОГИЯ 475
С. pseudodiphtheritu ит. Хорошо растут на питательных средах без добавления
крови и сыворотки. На плотных средах через 1 сут формируют серовато-белые или
кремовые колонии слизистой консистенции с блестящей поверхностью, В микро-
скопических препаратах — палочки правильной формы, размером 0,3-0,5x05-
2 мкм, с небольшим количеством волютина.
С. glucuronotyticum. На агаре с кровью через 24 ч вырастают в виде желтоватых
выпуклых колоний диаметром 1,0-15 мм без зоны гемолиза.
С. renale. Хорошо растут на средах с сывороткой, на агаре с кровью дают зону
гемолиза, окружающую желтовато-прозрачные колонии.
С. cistitidis. Даже на сывороточном агаре растут скудно, давая белые колонии.
В микроскопических препаратах — мелкие прямые или слегка изогнутые палочки,
с характерным расположением.
Идентификационные биохимические свойства коринебактерии
При идентификации выделенных культур коринебактерий основными призна-
ками служат тесты:
• на цистиназу;
• пирамидазу;
• уреазу;
• нитратредуктазу;
• ферментацию углеводов (глюкозы, сахарозы, крахмала).
С. diphtheriae не расщепляют мочевину, редуцируют нитраты (за исключением
С. diphtheriae belfantt), продуцируют цистиназу, не обладают пиразинамидазной
активностью, ферментируют глюкозу и мальтозу до кислоты. С. diphtheriae gravis,
ферментируют крахмал. Подавляющее большинство штаммов не ферментируют
сахарозу. Имеются сведения о выделении штаммов С. diphtheriae, не обладающих
цистиназой.
Выпускаются коммерческие диски с мочевиной и сахарами, тест-системы
для биохимической идентификации С. diphtherae. определения биохимических
свойств и дифференцирования коринебактерий внутри рода и другие изделия для
микробиологической идентификации С. diphtheriae и определения антимикробной
активности.
Информация о биохимических тестах представлена ниже и в табл. 21-41, 21-42.
Выявление цистиназы Культуры, принадлежащие к С. diphtheriae, продуциру-
ют цистиназу. Активность этого фермента оценивают в модифицированной среде
Пизу.
2,7 г АГВ-агара вносят в 90 мл дистиллированной воды и кипятят до полного
растворения. Готовят 10 мл 5% раствора натрия гидрокарбоната, в него вносят
0,1 г L-цистина и кипятят, пока навеска не растворится. 2 мл кипящего раствора
цистина вливают в 90 мл АГВ-агара, доводят pH до 7,7. стерилизуют при 0,5 атм в
течение 10 мин, охлаждают до 50 °C и последовательно добавляют 10 мл лошади-
ной сыворотки, 1 мл 10% раствора уксуснокислого свинца и 1,5 мл 10% раствора
гипосульфита натрия (готовят непосредственно перед использованием в стериль-
ных пробирках, растворяя вещества в стерильной дистиллированной воде и про-
гревая на водяной бане при температуре 90 °C в течение 30 мин).
Среду разливают в узкие пробирки столбиком высотой 3-4 см. Посев проводят
уколом. Результаты учитывают через 3-5 ч инкубации. В процессе роста культуры,
продуцирующей цистиназу, цистин расщепляется, а образующийся сероводород
вступает в реакцию с уксуснокислым свинцом, образуя сернистый свинец черно-
коричневого цвета, При этом наблюдается почернение среды по ходу укола и
появление коричневого «облачка» в радиусе 1 см от него. Выполнение положи-
тельного и отрицательного контроля обязательно,
Выяв пение пиразинамидазы. В нашей стране тест широко не используется (в
отличие от зарубежных лабораторий). Наряду с тестом на цистиназу он позволяет
476 ЧАСТНАЯ МИКРОБИОЛОГИЯ
дифференцировать три потенциально токсигенных вида (С. diphtheriae, С. ulcerans,
С. pseudotuberculosis) от других коринебактерий
2 г пиразинамида растворяют в 1 мл дистиллированной воды. Стерилизуют,
пропустив через мембранный фильтр с диаметром пор 0,45 мкм. Хранят при тем-
пературе -20 °C. В порошкообразном состоянии пиразинамид токсичен, поэтому
не следует допускать его контакта с кожей и попадания на слизистую оболочку
верхних дыхательных путей.
Раствор сульфата аммонийного железа в присутствии пиразинамида бесцветен.
В качестве реактива используют 20% раствор сульфата аммонийного железа в сте-
рильной дистиллированной воде, хранят при температуре -20 °C.
В 0,25 мл раствора пиразинамида вносят несколько капель густой взвеси суточной
бак сериальной культуры. Инкубируют при температуре 37 СС в течение 4-24 ч, после
чего в каждую пробирку прибавляют по одной капле 20% раствора сульфата аммоний-
ного железа. При наличии пиразинамидазы в пробирке появляется красно- оранжевое
окрашивание, при отсутствии фермента среда остается бесцветной или слегка желтеет.
Выполнение положительного и о грииательного контроля обязательно.
Выявление уреазы Пробу на наличие у изолята уреазы осуществляют
несколькими способами:
• методом Заксе;
• методом индикаторных дисков.
Метод Заксе
Раствор А: 2 г мочевины растворяют в 2 мл 96% этилового спирта, добавляют
4 мл дистиллированной воды.
Раствор Б: 0,1 г КН2РО, + 0,1 г К,НРО4 + 0,5 г натрия хлорида + 1 мл 0,25% рас-
твора фенолового красного + до 100 мл дистиллированной воды.
Раствор Б автоклавируют при 0,5 атм в течение 30 мин.
Перед исследованием берут 1 часть раствора А и 19 частей раствора Б, смеши-
вают и разливают по 0,1 мл в стерильные агглютинационные пробирки. Вносят
1-2 петли исследуемой культуры и помещают пробирки в термостат при темпе-
ратуре 37 °C на 30 мин. При наличии уреазы среда приобретает ярко-малиновое
окрашивание.
Метод индикаторных дисков
В 0,3 мл густой взвеси суточной культуры в 0,9% растворе натрия хлорида (pH =
7,3±0,1) вносят диск с мочевиной и индикатором, инкубируют при 37 °C, учитывают
результаты через 40 мин-2 ч. При наличии уреазы мочевина расщепляется, образу-
ющийся аммиак подщелачивает среду и белый цвет диска меняется на малиновый.
Выявление нитратредуктазы. Способность С. diphthei >ае восстанавливать
нитраты в нитриты — дополнительный тест, позволяющий дифференцировать ее
от С. ulcerans. Исследуемую культуру инкубируют в течение 24 ч в питательном
бульоне с 0,1% раствором нитрата натрия, после чего добавляют 2-3 капли реак-
тива Грисса. В случае образования нитритов среда окрашивается в красный цвет.
Реактив Грисса состоит из двух растворов: № 1 — 0,8 г сульфаниловой кисло-
ты растворяют в 100 мл 30% раствора уксусной кислоты при слабом нагревании;
№ 2 — 0,6 г а-нафталамина растворяют в 100 мл 30% раствора уксусной кислоты.
Растворы хранят в посуде из темного стекла с притертой пробкой, перед использо-
ванием смешивают в равных объемах.
Оценка ферментации углеводов. Густую взвесь суточной культуры
в 0,9% растворе натрия хлорида (pH = 7,3±0,1) разливают в пробирки по 0,3 мл.
В каждую пробирку вносят соответствующий диск (с глюкозой, сахарозой, крах-
малом). Инкубируют при температуре 37 °C. При ферментации глюкозы и сахаро-
зы диски меняют цвет через 5-6 ч. Для учета ферментации крахмала через 18-24 ч
инкубации в пробирку дополнительно вносят индикаторный диск с йодом. При
отсутствии амилазы он интенсивно синеет через 1-2 мин.
Таблица 21-41. Дифференциальные признаки видов рода Corynebacterium по биохимическим тестам
Вид микроорганизма Глюмза Мальтаза Манноза Сахароза Крахмал Уреаза Тирозин Фосфатаза Пиразин MK. Нитраты
С. accolens + - X X - X +
С. atermentans - - - - - - + -
С. amycolatum + (+) + d - d - +
С. argentoratense + - - - - + -
С. auris - - - - - + -
С. bovis + + - - - - - + + - -
С. саНипае + + + + - + + -
С. cystitidis + + - - + + - - + - -
С. diphtheriae belfanti + + - - - - - - - -
С. diphtheriae gravis + + - - + - - - - +
С. diphtheriae intermedins + + - - - - - - - +
C. diphtheriae mitis + + - - - - - - - +
C. glucuronolyticum + X - + X + X
C. glutanrcum + + + + - + + +
C. flavescens + - + - - - - - + -
C. jeikeium + d - - - - - - - -
C. kutscheri + + + + + + - - + - +
C. macginleyi + - X + - - +
C. matruchotii + + + + - d - + - +
C. minutissimum + + d +• - - + + - -
C. mycetoides + - - - + - -
C. paurometaboiicum - - - - - - - + - -
C. pilosum + + + - + + - - + - +
C. propinquum - - - - - X +
C. pseudodiphtherit/cum - - - - - + - - + - +
C. pseudotuberculosis + + + d - + - - + d
ЧАСТНАЯ МИКРОБИОЛОГИЯ
Окончание табл. 21-41
Вид микроорганизма Гпюкоза Мальтоза Манноза Сахароза Крахмал Уреаза Тирозин Фосфатаза Пиразин М.К. Нитраты
С. renale + - - - + + -
С. striatum + + + - + - + + + -
С. ulceraus + + - - + - -
С. urealyticum - - - - + + -
С. vitarumen + + + + - + - + + +
С. xerosis + - + + - - - — + - +
Примечание:
<<+» — позитивная реакция.
«-» — нега тивная реакция
«(+)» - 80-89% штаммов позитивны.
«б» — 21-79% штаммов позитивны.
«х» — данные отсутствуют.
МК — проба с метиленовым красным.
478 ЧАСТНАЯ МИКРОБИОЛОГИЯ
Таблица 21-42. Биохимические свойства возбудителя дифтерии, других патогенных и часто встречаемых коринебактерий
Тест С. diphtheriae gravis С. diphtheriae mills С. diphtheriae intermedins C. diphtheriae belfanti C uicerans C. pseudo- tuberculosis C. xerosis C. pseudodiph- thertticum
Ферментация
Глюкоза + + + + + + + -
Мальтоза + + + + + + -
Сахароза - - - - - - + -
Крахмал + - - - + - (5%) - -
Активность ферментов
Уреаза - - - + + + -
Цистиназа + + + + + + - -
Нитратредуктаза + + + - - + (1%) + +
Пиразинамидаза - - - - - + +
ЧАСТНАЯ МИКРОБИОЛОГИЯ 479
Выявление дифтерийного токсина
При использовании классических лабораторных методов выделения и иден-
тификации С. diphtheriae получение ответа возможно через 48-72 ч от начала
исследования Это обусловливает необходимость внедрения новых подходов к
диагностике дифтерии, особенно при определении основного фактора патоген-
ности микроорганизма — дифтерийного токсина, что позволяет сократить время
исследования, а также использовать более чувствительные и специфичные методы
диагностики. С этой целью предложены генотипические, фенотипические и био-
логические методы индикации дифтерийного токсина:
• тест Илека;
• иммуноферментный анализ;
• реакция пассивной гемагглютинации;
• иммунохроматограф ический тест;
• полимеразная цепная реакция.
Тест Илека
В основе теста лежит реакция встречной иммунодиффузии в агаровом геле: анти-
ген (дифтерийный токсин) и антитоксин, нанесенные на полоску фильтровальной
бумаги или бумажные диски, диффундируют навстречу друг другу. В месте, где они
встречаются, формируется полоса преципитации в виде белой линии, или «усов».
Для того чтобы не затруднять диффузию токсина и антитоксина, а образуемые
линии преципитата хорошо визуализировались, питательный агар должен быть
не очень плотным (не более 1 ,5%) и прозрачным. Поскольку длительная термиче-
ская обработка ухудшает качество питательной среды, стерилизовать агар следует
однократно текучим паром в течение 30 мин.
Обязательно выполнение параллельных контролей с тест-штаммами С. diph-
theriae tox+ (NCTC № J0648), C. diphtheriae tox± (NCTC № 3984), C. diphtheriae
tox- (NCTC № 10356). В качестве контрольных не следует брать производствен-
ные штаммы, так как они могут давать ярко выраженные линии неспецифической
преципитации, трудноотличимые от специфических. При выполнении теста лучше
применять диагностический антитоксин, очищенный от примесей ферментолизом
и специфической сорбцией. Диагностическая антитоксическая сыворотка менее
специфична. В ней. помимо антител к дифтерийному токсину, содержа гея антите-
ла к другим компонентам микробной клетки, что может привести к образованию
неспецифических линий преципитации. Последние обычно слабее специфических,
однако затрудняют учет реакции.
С помощью трафарета «бляшки» культур располагают на одинаковом рас-
стоянии не более 0,5-0,7 см от диска или антитоксин-стрипов (полосок филь-
тровальной бумаги, пропитанных антитоксином). Линии преципитации могут
отсутствовать и у токсигенных штаммов, если они засеяны далеко от диска или
антитоксин-ci рипов.
Приготовление чашек со средой. Основой среды могут служить бульон
Хоттингера, мартеновский бульон, специальная сухая среда «Коринетоксагар».
Среду однократно стерилизуют текучим паром в течение 30 мин, охлаждают до
50 °C и добавляют 20% лошадиной сыворотки или 30% сыворотки крупного рога-
того скота. Разливают в стерильные чашки Петри. Хранят в холодильнике не более
суток.
Приготовление бумажных дисков с дифтерийным антитоксином. Диски
из фильтровальной бумаги стерилизуют при 2 атм в течение 30 мин, подсушива-
ют и пропитывают антитоксином, разведенным по инструкции: 1 мл стерильной
дистиллированной воды на 500 ME сухого антитоксина, что достаточно для про-
питки 80-100 дисков. Затем диски повторно подсушивают в термостате в течение
18-24 ч и хранят в холодильнике в плотно закрытом флаконе с влагопоглотителем
до 1 года.
480
ЧАСТНАЯ МИКРОБИОЛОГИЯ
На поверхность среды в чашке Петри раскладывают стерильным пинцетом
4 диска с антитоксином, при необходимости исследования большого количества
культур в центр чашки можно поместить 5-й диск. Вокруг каждого диска на рас-
стоянии 6-7 мм засевают «бляшки* размером 6-7 мм из 24-часовых культур
(2 «бляшки* контрольные и 3 испытуемые из одного анализа, 2 «бляшки» испы-
туемых культур из отдельных колоний, 3-я - смесь 5-6 колоний).
Прит отселение антитоксин-стрипов. Полоски фильтровальной бумаги раз-
мером 0,7x8 см стерилизуют при 2 атм в течение 30 мин, подсушивают и пропиты-
вают антитоксином из расчета 0,25 мл (125 ME) на одну полоску. Подсушивают в
термостате в течение 18-24 ч и хранят в холодильнике в плотно закрытом флаконе
с влагопоглотителем до 1 года.
Антитоксин-стрип берут стерильным пинцетом и помещают на поверхность
питательной среды в центр чашки Петри. Культуру засевают в виде «бляшек» диа-
метром 7-8 мм по обе стороны от полоски на расстоянии 5 мм. Расстояние между
«бляшками» -7-8 мм. На одну чашку засевают не более 10 «бляшек» — 4 кон-
трольные и 6 испытуемых культур. Испытуемые культуры состоят из изолирован-
ных колоний и смеси 5-6 колоний.
Учет результатов. Результаты теста учитывают через 24-48 ч.
Испытуемая культура С. diphtheriae токсигенна, если линия преципитации,
образованная ею, сливается со специфической линией преципитации контрольно-
го штамма или плавно переходит в нее. Испытуемая культура не токсигенна, если:
• отсутствует линия преципитации, образованная испытуемой культурой, при
наличии специфической линии преципитации у контрольного штамма;
• линия преципитации, образованная испытуемой культурой, перекрещивает-
ся с линией преципитации контрольного штамма;
• линия преципитации, образованная испытуемой культурой, перекрещивает-
ся со специфической линией преципитации и сливается с неспецифической
линией преципитации контрольного штамма.
Последний вариант можно наблюдать при использовании противодифтерийной
сыворотки через 48 ч инкубации.
Хранение контрольных культур. Для постановки основных тестов при
идентификации С. diphtheriae обязательно применяют контрольные штаммы. Для
этой цели используют С. diphtheriae gravis (токсигенный NCTC № 10648+, слабо-
токсигенный NCTC № 3984± и нетоксигенный NCTC № 10356-) Штаммы могут
быть получены из лаборатории бактериальных инфекций НИИЭМ им. Л. Пастера
(Санкт-Петербург) или Национального центра, НИИЭМ им. Габричевского
(Москва). Культуру в течение 24 ч инкубируют на 0,1-0,2% питательном агаре
с 10% сыворотки. При росте заливают слоем стерильного вазелинового масла.
Хранят культуру в холодильнике. Пересевают 1 раз в 3 мес. При выполнении тестов
используют суточные культуры. При снижении токсигенных свойств контрольные
штаммы пассируют 3-4 раза через полужидкий питательный агар с добавлением
0,2% глюкозы и 10% крови. Хорошие результаты дает пассаж культур через 199-ю
среду с добавлением 20% сыворотки крупного рогатого скота.
Иммуноферментный анализ
На первом этапе реакции моноклональные антитела к концевому участку
В-фрагмента дифтерийного токсина (МкАт) сорбируют в лунках полистироловой
панели в течение 2 ч при температуре 37 “С и в течение 18-24 ч в холодильнике.
Затем трижды промывают 0,05% раствором «Твин-20» в 0.9% растворе натрия
хлорида. Промывную жидкость из лунок каждый раз тщательно удаляют. Данный
раствор используют и для приготовления разведений исследуемых проб, конъю-
гата и анатоксина.
В сенсибилизированные лунки вносят исследуемые и контрольные пробы.
В качестве контролей используют дифтерийный анатоксин, токсигенную и неток-
ЧАСТНАЯ МИКРОБИОЛОГИЯ 481
сигенную культуры С. diphtheriae. Исследуемый материал, положительный и отри-
цательный контроль готовят накануне или за 5-6 ч до проведения ИФА. Тампоны
с исходным материалом или 2-3 колонии с чашек первичного посева инкубируют
в сывороточном бульоне в течение 18 ч, при необходимости время инкубации
можно сократить до 5-6 ч. Параллельно на сывороточном бульоне инкубируют
и контрольные штаммы. Культивирование бактерий в полужидком питательном
агаре при постановке ИФА и РИГА не проводят. В лунки полистироловой панели
вносят по 0,1 мл верхнего слоя среды культивирования, инкубируют при 37 °C
в течение 1 ч, после чего лунки трижды тщательно промывают При наличии в
пробе дифтерийный токсин взаимодействует с МкАт, образуя иммунный ком-
плекс. Затем вносят антитоксический конъюгат, меченный пероксидазой хрена
(ПкАт), и инкубируют в течение 1 ч при 37 °C, при этом свободные антигенные
детерминанты дифтерийного токсина взаимодействуют с ним. Образуется ком-
плекс «МкАт-дифтерийный токсин-ПкАт», который проявляют индикаторной
субстратной смесью, состоящей из перикиси водорода и 5-аминосалициловой
кислоты (хромогенного субстрата, растворимого в воде). ПкАт восстанавливает
перекись водорода до воды, при этом выделяется кислород, который окисляет
хромогенный субстрат, приобретающий коричневый цвет. Окраска может варьи-
ровать от красновато-коричневой в положительных пробах до бесцветной или
желтовато-золотистой — в отрицательных. Реакцию можно ставить в качествен-
ном и количественном варианте.
Высокие чувствительность и специфичность метода позволяют выявить дифте-
рийный токсин в концентрации 4-8 нг/мл. Преимущество ИФА — обнаружение
токсина в жидкой питательной среде после первичного накопления материала,
минуя выделение чистой культуры.
Реакция непрямой гемагглютинации
РИГА ставят с антительным эритроцитарным дифтерийным диагностикумом
(эритроцитами, на поверхности которых сорбированы антитела к дифтерийному
токсину). Тампоны с исходным материалом или 2-3 колонии с чашек первичного
посева инкубируют в сывороточном бульоне в течение 18 ч, при необходимости
можно сократить время инкубации до 5-6 ч. Хорошие результаты дает пересев
колоний с чашек первичного посева на среду 199 с добавлением 20% СКРС.
В случае положительного результата, после того как антиген (дифтерийный
токсин) вступит в реакцию с антителами, сорбированными на эритроцитах, в
лунке реакционной панели образуется «зонтик» из склеенных эритроцитов. При
отсутствии специфического антигена эритроцитарный диагностикум осаждается
на дне лунки в виде плотной капли («пуговки»).
С учетом чувствительности эритроцитарного диагностикума, входящего в
комплект для постановки реакции (равного, как правило, 0,03 Lf/мл токсина),
и стандартности препарата токсигенность у исследуемых штаммов С. diphtheriae
можно определить количественно. Выполнение РИГА обязательно сопровождает-
ся контролями — с дифтерийным анатоксином и токсигенными и нетоксигенными
культурами С. diphtheriae, выращенными аналогично исследуемым культурам.
Чувствительность метода не уступает ИФА, но он значительно проще в выпил не-
нии.
Применение ИФА и РИГА для индикации дифтерийного токсина позволяет на
1-2 сут сократить время выдачи ответа, а также выявить штаммы с низким уров-
нем токсинообразования.
Иммунохроматографический тест (ИХТ)
ИХТ — тест с иммунохроматографичес кими полосками относится, как и ИФА,
к быстрым фенотипическим методам. Сущность ИХТ заключается в том, что
при наличии в образце дифтерийного токсина происходит его взаимодействие с
поликлональными специфическими антителами, фиксированными на нитроцел-
482 ЧАСТНАЯ МИКРОБИОЛОГИЯ
люлозной мембране. На следующем этапе образовавшийся комплекс вступает в
соединение с моноклональными антителами к A-фрагменту дифтерийного токси-
на, меченными коллоидным золотом. При положительной реакции на мембране
образуется полоска красного цвета, отсутствующая в образцах с отрицательным
контролем. Результаты оцениваются через 15 мин.
В исследованиях показано совпадение результатов, полученных с использова-
нием теста Илека и ИХТ. Однако, используя последний, можно получить ответ
через 3 ч после отбора колоний, то есть в течение рабочего дня. Для этого доста-
точно инкубировать подозрительные колонии или мазки из зева в оптимальной
питательной среде (ГРМ бульон + 20% сыворотки) в течение 3 ч и опустить туда
нитроцеллюлозную полоску.
Стандартизация посевной взвеси и время инкубации — необходимые условия,
особенно для слаботоксигенных штаммов. Если готовят взвесь с чашек первичного
посева, она должна содержать 1х108 КОЕ/мл. Одночасовая (для ИФА) и 3-часовая
(для ИХТ) инкубация в жидкой питательной среде позволяет избежать ложноо-
трицательных результатов.
Для прямого определения токсигенности при исследовании мазков время инку-
бации на оптимальной питательной среде необходимо увеличить до 6-18 ч.
Полимеразная цепная реакция
ПЦР относится к молекулярно-генетическим методам исследования. Она дает
возможность получить более быстрый и легко интерпретируемый ответ по срав-
нению с фенотипическими тестами Основной недостаток метода — получение
информации о наличии в хромосоме С. diphtheriae tax-гена, при отсутствии данных
о типе гена (экспрессируемый или «молчащий»).
Выполнение ПЦР со специфическими праймерами проводят в режиме ампли-
фикации, описанном в инструкции по применению тест-системы для выявле-
ния ДНК токсигенных и потенциально токсигенных штаммов С. diphtheriae.
Амплифицируется участок A-фрагмента гена дифтерийного токсина. Исходным
материалом служат бактерии, выращенные на плотной питательной среде, или
мазки из зева и носа. Методика включает предварительную подготовку образцов с
последующим включением в циклы ПЦР.
Отрицательный результат П ЦР используют для быстрого исключения неток-
сигенных штаммов, тогда как положительный результат требует дополнительных
фенотипических тестов (Илека, ИФА, РИГА), которые дадут окончательный ответ
о токсинопродукции, чго приводит к задержке получения окончательного ответа.
Сер^ югическая диагностика
Для определения антитоксических противодифтерийных антител (АТ-Ат) в
нашей стране используют реакцию непрямой (пассивной) гемагглютинации.
В большинстве стран мира титр АТ-Ат определяют в ИФА, который имеет ряд
преимуществ перед РИГА. В практическом применении этот метол удобен тем, что
позволяет оценивать титр АТ-Ат в международных единицах (МЕ/мл), что соот-
ветствует международному стандарту.
При выполнении ИФА с диагностической целью у больных определяют сум-
марные АТ-Ат. За положительный результат принимают 3-4-кратное их нараста-
ние. В случаях бактерионосительства уровни АТ-Ат существенно не изменяются.
Дополнительное преимущество ИФА — возможность одновременного определе-
ния на одном планшете как суммарных АТ-Ат, так и высокоавидных (истинных,
протективных) антител. Чем выше авидность антител, тем выше способность
антител нейтрализовывать определенные вирусы или бактерии. Принцип опреде-
ления авидности антител заключается в разрушении слабых связей между анти-
геном и антителом с помощью различных химических агентов и последующем
определении оставшихся после обработки комплексов АГ-АТ. При определении
ЧАСТНАЯ микробиология 483
высокоавидных противодифтерийных АТ-Ат в качестве агента для удаления низ-
коавидных антител используют раствор роданистого калия.
Высокоавидными считаются антитела, индекс авидности которых выше 30%.
Индекс авидности определяют по формуле:
ОП(Е)
индекс авидности = рщА) х Ю0%,
где ОП (Е) — оптическая плотность в лунке, обработанной роданистым калием;
ОП (А) — оптическая плотность в лунке, обработанной обычным промывочным
раствором, используемым в тест-системе.
СПИСОК РЕКОМЕНДУЕМОЙ ЛИТЕРАТУРЫ
Джонсон Д.Р., Каплан Э. Лабораторная диагностика инфекций, вызванных стрептокок-
ком группы А. — М.; Медицина, 1998. — 118 с
Долгих Т.И. Краткая характеристика возбудителей хламидиоза // Яковлев В.М.,
Новиков А.И. Сосудистый эндотелий и хламидийная инфекция. — М.; Медицина. 2000. —
С. 127-153.
Европейские стандарты диагностики и лечения заболеваний, передаваемых половым
путем. — М.: Медлит, 2004. — 272 с.
Жебрун А.Б. Инфекция Helicobacter pylori. — СПб.: Феникс, 2006. — 380 с.
Методики клинических лабораторных исследований: Справочное пособие: Т. 3 / Под ред.
В.В Меньшикова. — М.; Лабора, 2009. — 880 с.
МУК 4.2.1887-04 «Лабораторная диагностика менингококковой инфекции и гнойных
бактериальных менингитов». Утверждены Главным государственным санитарным врачом
Российской Федерации 4 марта 2004 г.
МУК 4.2.22-17-07 «Методические указания по выявлению бактерий Legionella pneumophila
в объектах окружающей среды». — 2007.
Определитель бактерий Берджи: Б 2 т.: Пер. с англ. / Под ред. Дж. Хоулта, Н. Крига,
П. Снита и др. - М., 1997. - Т. 1. - С. 195-196, 227-229. 255-258.
Тимченко В.Н., Бабаченко И.В., Ценева Г.Я. и др. Эволюция коклюшной инфекции у
детей. - СПб.: ЭЛБИ-СПб., 2005. - 192 с.
Тотолян А.А., Суворов А.Н., Дмитоиев А.В. Стрептококки группы В в патологии челове-
ка. — СПб.: Человек, 2009. — 212 с.
Фтизиатрия: Национальное руководство / Под ред. М.И. Перельмана. — М.; ГЭОТАР-
Медиа, 2007. — 512 с.
Шурыгина И.А., Чеснокова М.В., Климов В.Т. и др. Псевдотуберкулез, — Новосибирск.
Наука, 2003. — 319 с.
Энтеробактерии / Под. ред. В.И. Покровского. — М.: Медицина, 1985. — 320 с.
Этиологическая роль хламидий при заболеваниях репродуктивной сферы: Аналитический
обзор СПб. НИИЭМ им. Л. Пастера / Под ред. А.Б. Жебруна. — СПб., 2007. — 56 с.
Якубович А.И., Корепанов А Р. Урогенитальный хламидиоз. — Иркутск, 2006. —108 с.
European Guidelines for Control and Prevention of travel associated Legionnaires Disease. —
2002.
Garrity, G M., Bell J. A.. Lilburn T. Family II Helicobacteraceae fam. nov. — 2nd ed. — New York:
Springer, 2005.
Legionella and the prevention of Legionellosis. — WHO, 2007.
Malfertheiner P„ Megraud F„ Moran C. Guidelines for the management of the H. pylori infec-
tion. — European Gastroenterology Review, 2005.
Manual of Clinical Microbiology' (Editor in chief: Patrick R.Mmray). — 9th ed. — Washington:
ASM Press, 2007. - P. 412- 429.
Manual of clinical microbiology' / Editor in chief P.R.Murray. — 9th ed. — Washington: ASM
Press, 2007. - P. 688-697.
Глава 22
Некоторые инфекционные
заболевания
МИКОБАКТЕРИИ
Туберкулез — инфекционное заболевание. Это заболевание,
являвшееся одной из основных причин смертности в Средние века,
широко распространено в мире и в наши дни.
Обнаружение в 1882 г. Р. Кохом бациллы, признанной возбуди-
телем туберкулеза, легло в основу лабораторной диагностики этого
заболевания.
ТАКСОНОМИЯ
Род Mycobacterium входит в состав семейства Mycobacteriaceae,
которое совместно с семейством Cory neb acteriaceae относится к
порядку hctinomycetales.
Определяющие свойства микобактерий — кислою- и щело-
чеустойчивость, высокое содержание липидов в клеточной стенке
[воска, входящие в состав клеточной стенки, содержат растворимые
в хлороформе миколовые кислоты с длинными разветвленными
цепями углерода (от 60 до 90 атомов)], а также высокое (62-70%)
содержание нуклеотидных пар «гуанин + цитозин» в составе ДНК.
По своему биохимическому составу микобактерии приближают-
ся к другим микроорганизмам, способным синтезировать миколо-
вые кислоты, — Nocardia, Rodncoccus и Corynebacterium. В настоящее
время данный род насчитывает около 100 видов микобактерий.
Большинство из них — сапрофиты, широко распространенные в
окружающей среде.
В клинической микробиологии для классификации патогенных
микобактерий используют понятие «комплекс», объединяющее
виды микобактерий с одинаковой клинической значимостью, когда
более тонкая дифференциация несущественна. Виды микобакте-
рий, вызывающие туберкулез у человека и животных, объединены
в комплекс М. tuberculosis (М. tuberculosis complex), включающий
М. tuberculosis. М. bovis, М. microti. М. africanurn и М. canettii, а также
М. саргае и М. pmnipedii, филогенетически родственные М. bovis и
М. microti. С помощью молекулярно-генетических методов установ-
лена генетическая неоднородность глобальной популяции М. tuber-
culosis, которая представлена многочисленными генетическими
линиями (семействами) — Beijing, LAM, Haarlem, Т.
Другие микобактерии, вызывающие различные микобактериозы,
отнесены к группе нетуберкулезных микобактерий (НТМБ), вклю-
НЕКОТОРЫЕ ИНФЕКЦИОННЫЕ ЗАБОЛЕВАНИЯ 485
чающей следующие комплексы: М. avium (М. avium, М. intracellular? и М. scrofy-
laceum), M.fortuitum (M.fortuivum и M. chelonae), М. terrac {М. terrae, М. trivial? и
М. nonchromogenicum). К роду Mycobacterium относят также возбудителя проказы —
М. leprae — и возбудителя язвенных поражении Бурули — М. ulcerans. Для иден-
тификации видов микобактерий используют биологические, биохимические и
молекулярно-генетические методы.
На основе различий в культуральных свойствах выделяют четыре группы мико-
бактерий.
• Группа I — фотохромогенные микобактерии (потенциально патогенные —
М. asiaticum, М. kansasii, М. marinum. М. simiae). Колонии микобактерий дан-
ной группы, не пигментированные при инкубации в темноте, приобретают
ярко-желтую или желто-оранжевую окраску после выдерживания на свету.
Растут медленно (кроме М marinum).
• Группа II — скотохромогенные микобактерии (от греч. skotos - «тем-
нота»). Б темноте образуют пигмент, скорость роста — 30-60 дней. Данная
группа включает сапрофитные микобактерии М. aquae и М. gordonae, обнару-
живаемые в водоемах, и условно-патогенные микобактерии М. scrofulateum,
вызывающие поражение лимфатических узлов и легких у детей,
• Группа III — нефотохромогенные микобактерии. Не образуют пигмент
или имеют бледно-желтую окраску колоний, которая не усиливается на свету.
Растут от 2-3 до 5-6 нед. К данной группе относят условно-патогенные мико-
бактерии М. avium, М. uitracellulare, М. xenopi, сапрофитные микобактерии
М. terrae, а также М. gasm. М. hattey и Af. bruiiense. М. avium (микобактерии
птичьего типа), обнаруживаемые в воде и почве, могут быть патогенны для
человека, лабораторных и домашних животных (например, свиней).
• Группа IV — быстрорастущие микобактерии. Не образуют пигмента на
свету. Растут быстро (до 10 дней). К данной группе относят широко рас-
пространенные в природе условно-патогенные микобактерии М. fortuitum и
сапрофитные микобактерии М. phlei и М. smegmatis.
ОБЩАЯ ХАРАКТЕРИСТИКА
Микобактерии — тонкие, слегка изогнутые или прямые палочки размером
1-10x0,2-0,6 мкм, иногда ветвящиеся наподобие мицелия. Микобактерии —
аэробы, спор не образуют, неподвижны. Клеточная стенка микобактерий имеет
строение и химический состав, характерные длч грамположительных бактерий.
Пептидогликан клеточной стенки содержит мезодиаминопимелиновую кислоту;
аланин, глутаминовую кислоту, глюкозамин, мурамовую кислоту, арабинозу и
галактозу. Однако высокое содержание липидов не позволяет использовать для
окрашивания микобактерий обычные анилиновые красители, поэтому применя-
ют специальные методы, способствующие проникновению красителя в клетку.
Окрашенные микобактерии не обесцвечиваются при последующей обработке
кислотой и спиртом. При окраске карболовым фуксином микобактерии тубер-
кулеза имеют вид тонких, слегка изогнутых палочек малиново-красного цвета с
зернистой структурой.
Продолжительность роста микобактерий на плотных питательных средах
варьирует от 2 до 60 сут. Оптимальная температура инкубации — 30-45 °C.
Колонии могут быть шероховатыми, плотными (R-форма) или гладкими, влажны-
ми (S-форма). На жидких питательных средах микобактерии растут на поверхно-
сти в виде сухой пленки. Диффундирующий пигмент встречается редко. Колонии
некоторых видов имеют пигментацию от слабо-желтого до оранжевого цвета.
Фотохромогепным видам микобактерий требуется экспозиция на свету для обра-
зования пигмента, скотохромогенные виды образуют пигмент независимо от
действия света.
486 НЕКОТОРЫЕ ИНФЕКЦИОННЫЕ ЗАБОЛЕВАНИЯ
ЭПИДЕМИОЛОГИЯ
Микобактерии широко распространены в окружающей среде, многие из них
являются сапрофитами, а остальные способны вызывать заражение человека и/
или животных. Особенно опасны для человека микобактерии группы М. tuber-
culosis Резервуаром последних служит больной человек. Заражение происходит
чаще всего аэрогенным путем, реже через кожу и слизистые оболочки. В спо-
радических случаях регистрируют трансплацентарное инфицирование плода.
Заражение туберкулезными микобактериями не всегда влечет за собой развитие
патологического процесса. Большое значение при этом играют условия жизни и
трудовой деятельности, полноценность питания, состояние иммунной системы
людей.
По данным ВОЗ, в 2003 г. в мире выявлено 8,4 млн новых случаев туберкулеза;
ежегодно от туберкулеза умирают 2-3 млн человек, или 5000 человек ежедневно.
Около трети населения нашей планеты инфицированы микобактериями туберку-
леза. Сегодняшний этап характеризуется нарастанием распространенности тубер-
кулеза, вызванного лекарственно-устойчивыми штаммами микобактерий, а также
увеличением количе( тва иммунодефицитных состояний. По статистическим дан-
ным, риск возникновения туберкулеза у ВИЧ-инфицированных увеличивается
примерно в 170 раз. Большинство летальных случаев больных СПИДом проис-
ходит от сочетанной с ним туберкулезной инфекции.
Заболеваемость туберкулезом россиян превышает 80 на 100 тыс. человек, при-
чем в Дальневосточном, Сибирском и Уральском федеральных округах она, по
данным Минздравсоцразвития РФ, на 25-50% выше. Не менее 2,5 млн наших
соотечественников больны туберкулезом, а смертность от него составляет полови-
ну всех летальных исходов, вызываемых инфекционными болезнями.
КЛИНИЧЕСКИЕ ПРОЯВЛЕНИЯ ИНФЕКЦИИ
Туберкулез может проявля гься легочной и нелегочными формами, что обуслов-
лено тем, каким путем произошло заражение. Локализацией инфекции определя-
ется характер клинической картины болезни.
Вследствие медленных роста и размножения микобактерий, а также иммунного
ответа на них организма клинические проявления туберкулеза часто регистрируют
спустя длительное время после заражения.
Термином «латентный туберкулез» принято обозначать стадию или форму
туберкулезной инфекции, характеризуемую отсутствием клинических проявлений
инфекции и относительно свободной диссеминацией микобактерий по организму.
У подростков и взрослых с ослабленным иммунитетом такая диссеминация может
вести к развитию милиарного туберкулеза, характеризуемого образованием грану-
лем в разных органах. Наиболее часто генерализация туберкулезных поражений
происходит после прорыва содержимого гранулем в кровоток.
ЛАБОРАТОРНАЯ ДИАГНОСТИКА
Микробиологические исследования проводят в целях выявления больных
туберкулезом, для верификации диагноза, контроля эффективности и коррекции
химиотерапии, оценки эффективности лечения.
Лабораторные исследования для диагностики туберкулеза можно разделить на
две группы:
v' для постановки или подтверждения диагноза «туберкулез»;
J для характеристики выделенного возбудителя заболевания.
Классический микробиологический метод диагностики туберкулеза включает
микроскопию и культуральное исследование. К числу дополнительных современ-
ных ускоренных методов относят культивирование микобактерий туберкулеза в
автоматизированных системах и молекулярно-генетические исследования. Кроме
НЕКОТОРЫЕ ИНФЕКЦИОННЫЕ ЗАБОЛЕВАНИЯ 487
того, достигнуты определенные успехи в серологической диагностике туберкулеза,
так как стали доступны тест-системы, предназначенные для скрининга.
К первой группе относятся прямые методы, позволяющие выявить непо-
средственный этиологический агент заболевания. К прямым методам относятся
традиционные методы микробиологической диагностики (микроскопия и посев)
и ГШР-диагностика, позволяющая определять наличие ДНК-возбудителя в диа-
гностическом материале.
Косвенные методы, направленные на выявление последствий воздействия этио-
логического агента на организм больного, имеют относительно низкое распростра-
нение при лабораторной диагностике туберкулеза. К этим методам можно отнести
иммуноферментный метод определения уровня иммуноглобулина G, который все
шире применяют в странах с низким уровнем заболеваемости туберкулезом.
Исследования, относящиеся ко второй группе, направлены на характеристику
выделенного возбудителя: определение его видовой принадлежности (доказа-
тельство принадлежности к микобактериям туберкулезного комплекса) и спектра
лекарственной чувствительности. В настоящее время для характеристики клини-
ческих штаммов микобактерий туберкулеза применяют как традиционные микро-
биологические и биохимические, так и современные молекулярно-биологические
методы.
Для обеспечения сравнимости эпидемиологических показателей в различных
регионах страны и мира необходима стандартизация процедур исследований,
формы представления и регистрации результатов. В связи с этим в Российской
Федерации разработаны и утверждены унифицированные методы микробиологи-
ческих исследований для диагностики туберкулеза, обязательные для применения
в клинических лабораториях всех уровней и любого ведомственного подчинения.
Дополнительно регламентируются формы регистрации микроскопических иссле-
дований для диагностики и контроля химиотерапии туберкулеза.
Материалы для лабораторных исследований
Лаборатория противотуберкулезной службы должна быть готова получить
любой биологический материал на исследование в соответствии с приложением 1
к приказу Минздравсоцразвития № 690 от 02.10.2006, учетная форма № 05-ТБ/у.
Мокрота. Чаще всего для диагностики туберкулеза исследуют мокроту боль-
ного. У пациента с продуктивным кашлем получение мокроты не представляет
трудностей.
Диагностический материал при внелегочном туберкулезе
Асептитески собранные жидкости. Биологические жидкости (ликвор, пери-
кардиальную, синовиальную, асцитическую, кровь, костный мозг) следует соби-
рать в асептических условиях в стерильный контейнер. К жидкостям, имеющим
склонность к свертыванию, добавляют стерильные гепарин (0,2 мг/мл) или окса-
лат калия (0,01-0,02 мл 10% нейтрального оксалата на 1 мл материала). Материал
доставляют в лабораторию немедленно.
Асептитески собранные ткани помещают в стерильные контейнеры без
добавления консервантов. В случае длительной транспортировки ткани необходи-
мо поместить в стерильный физраствор, обложить сухим льдом или охладить при
температуре 4-15 “С. Следует немедленно доставлять материал в лабораторию.
Заведомо контаминированные материалы
Moza. Мочу (среднюю часть или всю утреннюю порцию) собирают в стериль-
ную посуду после тщательного туалета наружных половых органов. Анализ мочи
на микобактерии должен предусматривать обязательное троекратное исследова-
ние. В лаборатории мочу центрифугируют, используя метод накопления осадка.
Особенностью существования М. tuberculosis в жидкостях является способность их
долгое время находиться во взвешенном состоянии. В связи с этим рекомендуют
488 НЕКОТОРЫЕ ИНФЕКЦИОННЫЕ ЗАБОЛЕВАНИЯ
центрифугирование при 3000 g всего материала, а не его донной фракции, полу-
чаемой после отстаивания в естественных условиях.
Сбор суточной мочи для бактериологического исследования не практикуется.
Установлено, что при хранении мочи более 1 ч после ее сбора количество микроб-
ных клеток неа щцифической гноеродной и гнилостной микрофлоры увеличива-
ется в несколько раз. Ферменты жизнедеятельности этой флоры могут угнетать
способность микобактерий к росту; Не следует проводить микроскопические иссле-
дования моги из-за частой встречаемости в ней кислотоустойчивых сапрофитов.
Менструальная кровь. Исследование менструальной крови требует особого
методического подхода. Наличие в этом материале большого количества про-
теолитических, фибринолитических и других ферментов обусловливает необ-
ходимость незамедлительной доставки материала в лабораторию и тщательной
его обработки, так как менструальная кровь, с одной стороны является весьма
подходящим материалом для развития гноеродной и гнилостной флоры, а с дру-
гой — благодаря обилию ферментов неблагоприятно влияет на жизнеспособность
микобактерий. Менструальную кровь следует собирать не тампоном, а вакуумным
отсосом или колпачком Кафки. Исследуют ее так же, как кровь или другие мате-
риалы с примесью крови.
Исследованию подлежат следующие виды диагностического материала; мокро-
та, отделяемое верхних дыхательных путей после аэрозольных ингаляций, про-
мывные воды бронхов, бронхоальвеолярные смывы, материал, получаемый при
бронхоскопии и биопсии, экссудат, моча, кровь, СМЖ, резецированные ткани,
мазки.
Материал следует собирать с соблюдением правил инфекционной безопасности
в стерильные флаконы с плотно завинчивающимися крышками. Флаконы объе-
мом 40-50 мл (диаметр отверстия - не менее 30 мм) должны быть изютовлены
из ударостойкого материала, легко расплавляться при автоклавировании, быть
удобными в обращении, прозрачными или полупрозрачными.
Чаше всего материалом для выделения туберкулезных микобактерий служит
мокрота. Для повышения эффективности исследования необходимо проводить
сбор материала до начала химиотерапии, утреннего приема пищи и лекарствен-
ных препаратов. Желательно собрать не менее 3 проб утренней мокроты в течение
3 дней подряд. Сбор мокроты следует проводить в специально выделенном для
этих целей отдельном, хорошо вентилируемом помещении (пункте сбора мокро-
ты), оснащенном бактерицидными лампами и средствами дезинфекции, или на
открытом воздухе. Правильно собранная мокрота (3-5 мл) имеет слизистый или
слизисто-гнойный характер. Контейнер с порцией мокроты достаточного объема
(не менее 3-5 мл), содержащей уплотненные или гнойные комочки без слюны,
тщательно закрывают завинчивающейся крышкой, затем контейнер маркируют и
помещают в специальный бикс для транспортировки в лабораторию.
Наилучшим вариантом является использование для сбора мокроты одноразо-
вых пластиковых прозрачных контейнеров объемом 30-50 мл. Материал, из кото-
рого изготовлен такой контейнер, должен расплавляться (деформироваться) при
автоклавировании (121 °C).
При проведении исследования методом посева для повышения его эффектив-
ности время между сбором любого материала и его обработкой в лаборатории не
должно превышать 24 ч. При отсутствии такой возможности материал хранят в
холодильнике (4-8 °C не более 72 ч). Добавление к пробам консервантов (10%
раствора трехзамещенного фосфата натрия, 1 % раствора цетилпиридина хлорида,
2-3% раствора борной кислоты) позволяет увеличить срок их хранения в холо-
дильнике до 5-7 дней. Допускается замораживание пробы не более 1 раза.
Ни один из указанных выше методов не является оптимальным, своевременная
доставка является единственным залогом сохранности материала.
НЕКОТОРЫЕ ИНФЕКЦИОННЫЕ ЗАБОЛЕВАНИЯ 489
В лаборатории должны быть разработаны и документально оформлены прави-
ла выбраковки материала, включающие:
❖ отказ от приема первичных проб без необходимой идентификационной доку-
ментации и маркировки (недопустимо проведение исследований на основа-
нии устного запроса, без соответствующего письменного направления);
❖ поведение в отношении проб неудовлетворительного качества;
❖ меры при обнаружении поврежденных или негерметично закрытых контей-
неров.
Бактериоскопия
Целями бактериоскопии клинического материала являются обнаружение мико-
бактерий при подозрении на туберкулез и оценка эффективности химиотерапии
больных им людей.
В клинико-диагностических лабораториях обшей лечебной сети, где проводят
только микроскопические исследования, используют метод прямой микроскопии
мазка непосредственно из клинического материала (мокроты, мочи, гноя). При
этом обнаружение микобактерий туберкулеза возможно с вероятностью 50%, если
в 1 мл мокроты содержится более 5000 микробных клеток. Если в тестируемой
пробе содержится менее 104 микобактерий/мл, обнаружить их в мазке трудно.
Результативность микроскопии повышается при исследовании нескольких проб
мокроты, взятых от одного пациента, и при концентрировании диагностического
материала, например, центрифугированием. В последнем случае мазок готовят из
обогащенного осадка, подготовленного из деконтаминированного материала для
культурального исследования.
Для того чтобы обеспечить выживание достаточной части микобактериальной
популяции, необходимо использовать щадящие методы обработки, позволяющие,
с одной стороны, подавить быстрорастущие гноеродные и гнилостные микроорга-
низмы, а с другой - максимально сохранить жизнеспособность присутствующих
в материале микобактерий.
Для унификации результатов исследования необходимо использовать для гомо-
генизации и деконтаминации диагностического материала стандартные методы,
а для посевов на готовые жидкие среды — методы, рекомендуемые производите-
лем.
Следующие биологические жидкости и ткани не нуждаются в деконтами-
нации, если они были взяты в стерильные флаконы с соблюдением правил
асептики:
❖ спинномозговая, синовиальная и друтие жидкости из закрытых полостей;
костный мозг:
гной из холодных абсцессов;
< резецированные ткани (за исключением материала аутопсии);
о пунктаты печени и лимфатических узлов, а также материалы биопсий (при
отсутствии свища)
Отрицательный результат микроскопического исследования не исключает диа-
гноз туберкулеза, поскольку в мокроте может содержаться мало туберкулезных
микобактерий (это бывает связано с объективными факторами, например фор-
мой, стадией и локализацией инфекции, а также с субъективными факторами —
неправильным взятием материала и плохой подготовкой мазка).
Для обнаружения в мазке кислотоустойчивых микобактерий (КУМ) использу-
ют различные методы окрашивания, из которых в целях унификации процедуры
рекомендуют два: метод Циля-Нильсена и флюорохромные красители.
При окраске методом Циля-Нильсена приготовленный на предметном стекле
мазок фиксируют над пламенем и окрашивают карболовым фуксином, который
490 НЕКОТОРЫЕ ИНФЕКЦИОННЫЕ ЗАБОЛЕВАНИЯ
проникает в микробную клетку через клеточную стенку, богатую липидами и
восками, при одновременном воздействии нагревания и протравливающего
действия фенола. Последующая обработка мазка 25% раствором серной кисло-
ты или 3% солянокислым спиртом приводит к обесцвечиванию всех некисло-
тоустойчивых элементов мазка, которые докрашивают 0,3% раствором мети-
ленового синего. КУМ, не воспринимающие обычные аналиновые красители,
окрашиваются в малиново-красный цвет, другие микроорганизмы, клетки и
прочие элементы — в голубой. Мазок высушивают при комнатной температуре
и микроскопируют.
Для исследования мазков, окрашенных по Цилю-Нильсену, используют свето-
вой микроскоп с иммерсионным объективом (х90 или хЮО) и окуляром с 7- или
10-кратным увеличением. Исследуют 100 полей зрения (достаточно для выявле-
ния единичных микобактерий), отмечая количество обнаруженных КУМ. При
отрицательном результате просматривают еще 200 полей зрения. Разрешающая
способность метода — 50 000 микробных клеток в 1 мл материала.
Результаты микроскопии оценивают полуколичественно (табл. 22-1).
Таблица 22-1. Оценка результатов микроскопического исследования при окраске по Цилю-
Нильсену
Количество КУМ Количества полей зрения для просмотра Форма записи результата Интерпретация результата
КУМ не обнаружены в 300 полях зрения 300 Отрицательный Отрицательный
1-2 КУМ в 300 полях зрения 300 Следует повторить исследование Результат не оценивается
1-9 КУМ в 100 полях зрения 100 — КУМ в 100 полях зрения’ Положительный
10-99 КУМ в 100 полях зрения 100 1+‘” Положительный
1-10 КУМ в 1 поле зрения 50 2+" Положительный
10 КУМ в 1 поле зрения 20 3+” Положительный
1 Указать точное количество КУМ.
” Соответствие градаций:
❖ точное количество - единичные КУМ в препарате;
❖ «1 + » - единичные КУМ в поле зрения;
v «2+» — умеренное количество КУМ;
❖ <<3->> — значительное количество КУМ.
Окраска флюорохромными красителями (аурамином или родамином) повы-
шает эффективность микроскопии на 15%. Мазки для люминесцентной микро-
скопии готовят из осадка, полученного путем обработки материала детергентом
(для освобождения микобактерий от окружающей их слизи), с последующими
отмыванием или нейтрализацией и центрифугированием при 3000 g. При ультра-
фиолетовом облучении окрашенные клетки начинают светиться на темном фоне —
КУМ имеют вид золотистых палочек, сапрофитные микобактерии окрашены в
зеленый цвет. Яркость и контрастность изображения позволяют уменьшить сум-
марное увеличение микроскопа в 4-10 раз, что способствует расширению поля
зрения и сокращению времени просмотра препарата. При этом люминесцентная
микроскопия позволяет обнаружить измененные клетки микобактерий, утра-
тившие кислотоустойчивость под влиянием химиотерапии и не выявляемые при
окраске по Цилю-Нильсену.
Качественная и эффективная окраска флюорохромными красителями требу-
ет обязательного соблюдения кислотности (pH) мазка, а также освобождения
микобактерий от окружающей их слизи, которая препятствует проникновению
НЕКОТОРЫЕ ИНФЕКЦИОННЫЕ ЗАБОЛЕВАНИЯ 491
красителя в микробную клетку. Несоблюдение этих условий приводит к снижению
эффективности люминесцентной микроскопии и получению ложноотрицатель-
ных результатов исследования.
Микроскопические методы обнаружения микобактерий широко применяют
для раннего выявления наиболее эпидемически опасных больных — бактериовы-
делителей. Преимущества данного метода — высокая специфичность (8^-100%),
быстрота получения результата (1-2 дня) и экономичность. Микроскопическое
исследование не позволяет провести идентификацию (выявление видовой принад-
лежности) и определить лекарственную чувствительность выделенных КУМ.
При отрицательном результате анализа мокроты ответ по форме 05-ТБ/у не
выдается, пока не проведено исследование всех трех (в крайнем случае двух) проб
мокроты.
О положительном результате анализа лаборатория сообщает направившему на
исследование материал врачу немедленно.
Изоляция микобактерий
Культуральный метод, в отличие от микроскопического, имеет высокую чув-
ствительность (с его помощью удается обнаруживать в пробах микобактерии,
когда их титр >10 живых бактерий/мл). Это способствует увеличению (более
чем на 15-20%) количества впервые выявленных больных на ранней стадии
заболевания, когда туберкулез еще относительно хорошо поддается лечению.
Получение чистой культуры позволяет осуществить идентификацию, определе-
ние лекарственной чувствительности и изучение вирулентности туберкулезных
микобактерий.
Существенные недостатки культурального метода — длительность (срок ожи-
дания результатов посева на плотных питательных средах может достигать 3 мес),
более высокая стоимость и сложность обработки диагностического материала.
Диагностический материал после гомогенизации и деконтаминации центрифу-
гируют (обогащение осадка), подвергают нейтрализаций и высевают на поверх-
ность плотных питательных сред или вносят в пробирки с жидкими (полужид-
кими) питательными средами. Из оставшейся части осадка готовят мазки для
микроскопии. Для достоверной клинической интерпретации результатов микро-
биологического исследования необходимо параллельно проводить культуральное
и микроскопическое исследования из одной и той же пробы диагностического
материала.
Инокулированные пробирки инкубируют в термостате при температуре 37 °C
в течение 2 сут в горизонтальном положении (для равномерного всасывания
материала в питательную среду), затем переводят в вертикальное положение, гер-
ме! ично закрывают резиновыми или силиконовыми пробками и выдерживают в
термостате при температуре 37 °C в течение 10-12 нед, осуществляя регулярный
еженедельный просмотр. При каждом просмотре регистрируют срок визуально
наблюдаемого роста со дня посева, интенсивность роста (количество КОЕ), загряз-
нение посева посторонней микрофлорой (такие пробирки удаляют), отсутствие
видимого роста <пробирки оставляют в термостате). Отрицательный результат
регистрируют по окончании периода инкубации.
Для культивирования микобактерий используют плотные, полужидкие и жид-
кие питательные среды. Для повышения результативности посева рекомендуют
применять одновременно 2-3 питательные среды разного состава.
В качестве стандартной питательной среды для первичного выделения воз-
будителя туберкулеза и определения его лекарственной чувствительности ВОЗ
рекомендует среду Левенштейна-Йенсена. В нашей стране, помимо нее, при
бактериологическом исследовании на туберкулез пользуются средой «Финн-2».
Это плотная яичная среда сложного состава, на которой рост микобактерий
492 НЕКОТОРЫЕ ИНФЕКЦИОННЫЕ ЗАБОЛЕВАНИЯ
проявляется на 20-96-е сутки. Появившиеся в последние годы агаровые среды
Миддлбрука (агары 7Н10,7Н11) позволяют быстрее обнаружить рост туберкулез-
ных микобактерий (в течение 2-4 нед) и обеспечивают лучшие возможности для
изучения морфологии колоний, чем среды на яичной основе.
Туберкулезные микобактерии формируют на плотных питательных средах
колонии с характерной морфологией. Например, М. tuberculosis на плотных пита-
тельных средах обычно образуют шероховатые колонии (R-форма) различной
величины и формы — сухие, морщинистые, цвета слоновой кости или слегка
пигментированные. Под влиянием противотуберкулезного лечения изменяется
характер роста — появляются гладкие, влажные колонии (S-форма).
Комплекс специальных исследований помогает отличить микобактерии тубер-
кулеза от нетуберкулезных микобактерий и кислотоустойчивых сапрофитов.
Положительный ответ дают после обязательного микроскопического исследо-
вания мазка из колоний, окрашенного по Цилю-Нильсену: микобактерии в пре-
парате выглядят как ярко-красные палочки, расположенные одиночно или груп-
пами, образующими скопления в виде жгутов и кос (рис. 22-1, см. цв. вклейку).
В молодых культурах, особенно выделенных от больных, которым проводили
длительные курсы химиотерапии, микобактерии полиморфны (кокковидные,
удлиненные, напоминающие мицелий грибов).
При выделении микобактерий количественно оценивают интенсивность их
роста:
о (+) — 1-20 КОЕ в пробирке — скудное бактериовыделение;
(++) - 21-100 КОЕ в пробирке — умеренное бактериовыделение;
(+++) — более 100 КОЕ в пробирке — обильное бактериовыделение.
Ценное прогностическое значение при туберкулезе имеет характеристика жиз-
неспособности возбудителя, которую оценивают по двум показателям — скорости
роста и времени, которое прошло от момента посева до выделения культуры
М. tuberculosis. Низкую степень жизнеспособности микробной популяции отме-
чают при росте менее 20 КОЕ в пробирке при продолжительности роста более
30 дней, высокую — при росте 100 КОЕ и более в пробирке при продолжитель-
ности роста менее 30 дней.
Преимущество культурального метода — выделение чистой культуры
М. tuberculosis, что позволяет провести идентификацию, определение лекарствен-
ной чувствительности, вирулентности и жизнеспособности возбудителя.
Культуральная диагностика в настояшее время претерпевает изменения, свя-
занные с внедрением в практику автоматизированных систем культивирования
М. tuberculosis — «MGIT-BACTEC-960» и «МВ/Bact». Преимущества систем на
основе жидких питательных сред — полная или частичная автоматизация, уско-
ренное выявление роста микобактерий, возможность определения лекарственной
чувствительности к противотуберкулезным препаратам 1-го ряда и некоторым
противотуберкулезным препаратам 2-го ряда.
В системе MGIT {Micobacteria Growth Indicating Tube), предназначенной для
использования в приборе «ВАСТЕС-960». микобактерии культивируются в специ-
альных пробирках с жидкой питательной средой на основе модифицированной
среды «Middlebrook-7Н9», включающей пищевые добавки для стимуляции роста
микобактерий и смесь антибактериальных препаратов для подавления сторонней
микрофлоры. Рост М. tuberculosis на данной среде удается обнаружить уже через
1-2 нед в зависимости от исходного количества бактериальных клеток в исследуе-
мом материале. Снижение концентрации кислорода в пробирке при размножении
М. tuberculosis вызывает усиление флюоресценции, которая становыгся видимой
при облучении пробирок ультрафиолетовым светом и регистрируется автомати-
чески. Р1нтенсивность свечения выражена в единицах роста (ЕР). Компьютерный
анализ кривых роста помогает оценить ростовые свойства различных видов мико-
НЕКОТОРАЯ ИНФЕКЦИОННЫЕ ЗАБОЛЕВАНИЯ ДЭЗ
бактерий, в том числе нетуберкулезных. Посев материала на жидкие питательные
среды обязательно должен сопровождаться посевом на среду Левенштейна-
Йенсена, которая играет роль дублера в тех редких случаях, koi да на других средах
М. tuberculosis не дают роста.
В результате внедрения автоматизированных систем культивирования время
появления росла микобактерий значительно сократилось и составляет 11 и
19 дней для «ВАСТЕС-960» и <-MB/Bact-> соответственно против 33 дней на стан-
дартной плотной питательной среде.
Наряду с очевидными преимуществами такие системы культивирования мико-
бактерий имеют и ряд существенных недостатков, в том числе высокую стоимость
аппаратуры и повышенный риск заражения лабораторного персонала.
Идентификация микобактерий
Питательные среды, используемые для культивирования микобактерий, не
являются строго селективными, поэтому выделенная культура подлежит иденти-
фикации (определению вида). Необходимость дифференциации микобактерий
обусловлена различиями в патогенезе, течении и исходе туберкулеза и микобакте-
риозов, а также наличием у различных видов микобактерий природной устойчи-
вости к некоторым противотуберкулезным препаратам. Актуальность дифферен-
циации микобактерий увеличивается с ростом частоты регистрации случаев ВИЧ,
ассоциированных с микобактериозами.
Для бактерий М. tuberculosis характерна совокупность культуральных призна-
ков, отличающих их от нелуберкулезных микобактерии
❖ медленный рост (более 3 нед);
❖ рост при температуре 35-37 °C;
о отсутствие пигментообразования (колонии имеют цвет слоновой кости);
❖ кислотоустойчивость при окраске по Цилю- Нильсену;
❖ положительный ниациновый тест.
< положит ельный нитратредуктазный тест;
< отсутствие термостабильной каталазы (68 °C);
❖ отсутствие роста на среде Левенштейна-Йенсена, содержащей 1000 мкг/мл
салициловокислого натрия, 500 мкг/мл паранитробензойной кислоты, 5%
раствор натрия хлорида;
❖ рост в присутствии 1-5 мкг/мл тиофен-2-карбоксиловой кислоты.
В большинстве случаев для дифференциации Як tuberculosis и М. bovis достаточ-
но использовать ниациновый тест на наличие нитратредуктазы и пиразинамида-
зы, а также тест на способность роста на среде, содержащей 2 мкг/мл гидразида
тиофен-2-карбоксиловой кислоты (2 мкг/мл).
Оценка лекарственной чувствительности туберкулезных микобактерий
Лекарственную чувствительность туберкулезных микобактерий определяют в
следующем порядке:
❖ до начала лечения однократно для определения стратегии и тактики лечения
больного;
❖ при выделении от больного культур микобактерий из различных видов
материала (мокроты, бронхоальвеолярного лаважа, мочи, экссудат ов, СМЖ)
исследуют все выделенные культуры;
❖ в конце интенсивной фазы лечения при отсутствии клинико-рентгеноло-
гической динамики;
при необходимости изменения схемы лечения в случае отсутствия негатива-
ции мокроты, повторного выделения культуры микобактерий после негати-
вации мокроты, резкого увеличения количества КУМ в мазке после первона-
чального снижения
494 НЕКОТОРЫЕ ИНФЕКЦИОННЫЕ ЗАБОЛЕВАНИЯ
Определение лекарственной чувствительности микобактерий проводят с помо-
щью микробиологических методов абсолютных концентраций (разведений на
плотной или жидкой питательной среде), пропорций, коэффициента резистент-
ности. Из числа рекомендуемых Комитетом по химиотерапии ВОЗ в России в
качестве унифицированного метода определения лекарегвенной чувствительности
микобактерий наибольшее распространение получил непрямой метод абсолют-
ных концентраций. При его использовании осуществляют дозированный посев
суспензии (10 млн микробных клеток) чистой культуры микобактерий в про-
бирки с питательной средой Левенштейна-Йенсена, содержащие определенные
концентрации химически чистых субстанций противотуберкулезных препаратов,
контрольную пробирку с питательной средой без противотуберкулезных препа-
ратов и пробирку с 1000 мкг/мл салициловокислого натрия или 500 мкг/мл пара-
нитробензойной кислоты для выявления роста нетуберкулезных микобактерий.
Засеянные пробирки инкубируют в термостате при температуре 37 °C. Результат
регистрируют через 3-4 нед.
Поскольку выделение возбудитель из диагностического материала на пита-
тельных средах требует не менее 1-1,5 мес, результаты определения лекар-
ственной чувствительности получают не ранее чем через 2-2,5 мес после посева
материала.
Культура микобактерий считается чувствительной к определенной концентра-
ции противотуберкулезного препарата в питательной цэеде. если число выросших
колоний микобактерий не превышает 20 при обильном росте в пробирке с пита-
тельной средой без препаратов. Для различных лекарственных средств установле-
на критическая концентрация, при которой наблюдается размножение критиче-
ской доли популяции микобактерий (табл. 22-2).
Таблица 22-2. Концентрации препаратов используемые при определении чувствительности
микобактерий туберкулеза методом абсолютных концентраций на среде Левенштейна-
Йенсена
Препарат Концентрация, мкг/мл
критическая предельная
Препараты 1-го ряда
Де1 идрострептомицин 10 25
Изониазид 1 10
Рифампицин 40 80
Эгамбутол 2 5
Препараты 2-го ряда
Канамицин 30 50
Протионамид (этионзмид) 30 50
Циклпсерин 30 50
Капреомицин 30 50
Офлоксацин 2 10
Аминосалициповая кислота 1 1
Кри герий устойчивости культуры микобактерий — рост 20 колоний и более на
питательной среде с препаратом в критической концентрации. Если число коло-
ний приближается к 20. необходимо отметить пограничный характер результата
определения чувствительности исследуемой культуры микобактерий.
Рекомендуемая критическая концентрация пиразинамида, который широко
используется в современной химиотерапии туберкулеза, составляет 200 мкг/мл.
Однако до сих пор не существует универсального метода определения лекарствен-
ной чувствительности к данному препарату; поскольку его антибактериальная
НЕКОТОРЫЕ ИНФЕКЦИОННЫЕ ЗАБОЛЕВАНИЯ 495
активность проявляемся только в кислой среде (pH <6), препятствующей росту
большинства штаммов микобактерий.
Критические концентрации препаратов существенно различаются при исполь-
зовании различных методов определения лекарственной чувствительности.
Для определения порога устойчивости противотуберкулезные препараты целе-
сообразно тестировать не только в критических, но и в дополнительных концен-
трациях. Это дает возможность врачу варьировать дозы препаратов и режимы
химиотерапии за счет повышения дозы, изменения метода введения и исполь-
зования аддитивных и синергидных свойств различных комбинаций противо-
туберкулезных средств.
Альтернативные методы определения лекарственной устойчивости микобакте-
рий (методы пропорций и коэффициента резистентности) позволяют установить,
какая часть популяции микобактерий в процентном отношении устойчива к дан-
ному препарату при визуальной оценке роста. Данные методы, хотя и являются
более достоверными, особенно при оценке чувствительности к противотуберку-
лезным препаратам 2-го ряда, значительно более трудоемки, поэтому в практиче-
ской деятельности используются редко.
Для ускорения исследований рекомендуют использование прямого метода
абсолютных концентраций, который заключается в одновременном посеве диа-
гностического материала [бактериоскопически положительного с массивностью
бактериовыделения не менее (++)] на контрольные питательные среды и среды с
соответствующими противотуберкулезными препаратами.
Перспективный метод определения лекарственной чувствительности микобак-
терий — модифицированный метод пропорций с использованием автоматизиро-
ванной системы «MGIT-BACTEC-960», которая позволяет проводить исследова-
ние популяций микобактерий, выращенных на плотных питательных средах и в
пробирках MGIT. В последнем случае существенно сокращается время исследова-
ния: данные о лекарственной чувствительности культуры могут быть получены уже
через 3 нед с момента сбора диагностического материала по сравнению с 3 мес при
использовании традиционного метода абсолютных концентраций. Относительно
высокая стоимость исследования компенсируется своевременностью получения
результатов при интенсивной фазе химиотерапии.
Молекулярно-генетические методы идентификации микобактерий
и оценки их лекарственной устойчивости
ПЦР — быстрый и эффективный метод выявления микобактерий в биоло-
гическом материале. Чувствительность ПЦР достаточно высока, теоретически
возможна детекция даже одной бактериальной клетки в образце. Отечественные
диагностические тест-системы для выявления ДНК губеркулезных микобактерий
в ПЦР позволяют исследовать образцы диагностического материала из мокроты,
бронхоальвеолярных смывов, плевральной и синовиальной жидкости, СМЖ
мочи, аспирата из матки, биоптатов ткани легких, лимфатических узлов. В подоб-
ных тест-системах используют праймеры, фланкирующие участок инсерционного
элемента IS6110, который присутствует в геноме всех видов микобактерий, входя-
щих в состав комплекса М. tuberculosis.
ПЦР чаще применяют для ускоренного (в течение суток) выявления микобакте-
рий туберкулеза в образцах диагностического материала. Чувствительность ПЦР
при исследовании мокроты и поомывных вод бронхов больных с подтвержденным
диагнозом туберкулеза легких, по данным литературы, составляет 92.6% против
88,9 и 52,4% при использовании культуральной и микроскопического методов
соответственно. Специфичность всех методов составляет 99%. ДНК микобак-
терий. выделенную при ПЦР, обнаруживают при эффективной химиотерапии в
496 НЕКОТОРЫЕ ИНФЕКЦИОННЫЕ ЗАБОЛЕВАНИЯ
течение более длительного времени по сравнению с выявлением микобактерий
при люминесцентной микроскопии (в среднем на 1,7 мес) и бактериологическом
исследовании (на 2,5 мес), Высокие чувствительность и специфичность — большое
преимущество ПЦР особенно при диагностике и дифференциации внелегочных
форм туберкулеза, при которых клинико-рентгенологические и бактериологиче-
ский методы малоэффективны.
Серологическая диагностика
Антигенная структура туберкулезных микобактерий претерпевает значитель-
ные изменения на разных стадиях инфекции и при разных формах болезни, что
влечет соответствующие изменения гуморального иммунного ответа у заражен-
ных ими людей. В связи с этим длительно не удавалось разработать достаточно
чувствительные и специфичные методы серологической диагностики туберкулеза.
В настоящее время стали доступны одноэтапные экспресс-методы серологической
диагностики туберкулеза на основе иммунохроматографического и иммунофер-
ментного анализов. Большинство современных тест-систем основаны на выяв-
лении в сыворотке крови пациентов антител к лммунодоминантному антигену
микобактерий А60. Чувствительность теста близка к бактериоскопическому мето-
ду диагностики туберкулеза, однако возможности серологического метода диа-
гностики туберкулеза шире, так как он позволяет выявлять и нелегочные формы
туберкулезной инфекции, а также их доклиническую стадию.
Принципы безопасности лабораторных исследований на микобактериозы
Заболеваемость туберкулезом медицинских работников значительно превы-
шает показатели заболеваемости туберкулезом населения России. Опасность
заражения существует как среди медицинского персонала противотуберкулезных
стационаров, так и среди сотрудников бактериологических лабораторий, рабо-
тающих непосредственно с возбудителем. Распространение и заражение тубер-
кулезом в противотуберкулезных учреждениях происходит преимущественно
ингаляционным путем. Заражение восприимчивых лиц может происходить
при ингаляции всего 10 микробных клеток. Персонал противотуберкулезных
учреждений должен быть обучен работе с инфекционными больными и инфици-
рованным материалом. Для предупреждения инфицирования существуют инди-
видуальные средства защиты и технические приспособления, обеспечивающие
защиту работников лаборатории от контакта с микобактериями туберкулеза.
К индивидуальным средствам защиты относятся закрытый халат или моющийся
костюм с фартуком, маска или противонылевой и противодымовой респиратор,
защитные перчатки.
В лаборатории любую манипуляцию с жидкой суспензией или культурой
микобактерий туберкулеза следует проводить в шкафу биологической защиты.
Необходимо избегать возникновения аэрозолей при работе с инфицированным
материалом. Для работы с микобактериями туберкулеза следует использовать
ламинарные боксы биологической защиты 1-го и 2-го класса, защищающие
лаборанта от заражения при условии правильного обращения с оборудованием
(аэрозоли обеззараживаются и устраняются с помощью вытяжной вентиля-
ции).
ТРЕПОНЕМЫ
ТАКСОНОМИЯ
Трепонемы относятся к порядку Spirochaetales, семейству Treponemataceae, роду
Treponema. У человека вызывают инфекцию Т. pallidum, Т. carateum и Т. vincentii.
НЕКОТОРЫЕ ИНФЕКЦИОННЫЕ ЗАБОЛЕВАНИЯ А97
Ранее к опасным для человека видам трепонем относили Т.pertenue и Т. bejel, одна-
ко на основании изучения генома эти бактерии отнесли к подвидам Т. pallidum.
В этом разделе приведена детальная информация о Т. pallidum подвида pal-
lidum — возбудителе сифилиса.
ОБЩАЯ ХАРАКТЕРИСТИКА
Т. pallidum подвида pallidum — тонкий спиралевидный микроорганизм, имею-
щий длину от 6 до 20 мкм и диаметр в поперечном сечении 0,2 мкм. Спирохета
имеет обычно 8-14 равномерных завитков с закругленными вершинами и острым
углом между соседними завитками, амплитуда завитков уменьшается по направ-
лению к концам. Это грамотрицательный микроорганизм, плохо окрашиваемый
анилиновыми красителями (за что трепонема и получила название «бледная»).
Неокрашенные, нефиксированные трепонемы из-за малого диаметра не видны в
обычном световом микроскопе.
Основным способом размножения трепонемы является поперечное деление.
Делится она сравнительно медленно: каждые 30-32 ч. При усиленном размноже-
нии трепонема может делиться не только пополам, но и на большее количество
сегментов вплоть до одного завитка.
При неблагоприятных для существования условиях, например под воздействи-
ем антибиотиков, антител, фагоцитов и т.д., бледная трепонема из спириллярной
формы патогенного существования переходит в устойчивые формы выживания.
Их образование — не только способ сохранения возбудителя в организме в тече-
ние дли тельного времени, но и метод размножения инфекции. В случае благо-
приятного изменения условий существования трепонема получает возможность
вновь реверсировать в патогенную форму. К устойчивым формам выживания
бледной трепонемы относя г цисты, L-формы, зерна (гранулы) и полимембранные
фагосомы.
Бледная трепонема является тканевым, внутриклеточным паразитом, которого
активно фагоцитируют клетки организма-хозяина. Особенностью фагоцитоза при
сифилисе является то, что он осуществляется не только макрофагами и нейтро-
фильными лейкоцитами, но и «непрофессиональными» фагоцитами — лимфо-
цитами, плазматическими клетками, фибробластами, шванновскими клетками,
перицитами нервных волокон, эндотелиоцитами капилляров. Однако фагоцитоз
бледной трепонемы обычно остается незавершенным: фагоцитированные бакте-
рии не только сохраняют жизнеспособность, но и даже способны к внутриклеточ-
ному и внутриядерному паразитированию (эндоцитобиозу).
Бледная трепонема является факультативным анаэробом, поэтому она преиму-
щественно тропна и активно размножается в нервной, лимфоидной и соедини-
тельной тканях. Кровь не является благоприятной средой для жизнедеятельности
трепонем, но их можно обнаружить в крови, особенно в определенные периоды
заболевания, характеризуемые массовым размножением и диссеминацией воз-
будителей (периоды «трепонемного сепсиса»). Трепонема имеет сложный анти-
генный состав. Описано более 20 разнообразных антигенов, сконцентрированных
главным образом в клеточной стенке и цитоплазматической мембране. К каждому
из этих антигенов на разных этапах течения сифилиса вырабатываются антитела
различных классов. Наибольшее значение для практики имеют белковые и липид-
ные антигены, поскольку серологическая диагностика сифилиса основана преиму-
щественно на выявлении антител именно к этим антигенам.
При посеве материала, полученного от больного, на искусственные питательные
среды роста бледной трепонемы получить не удается: при культивировании она
утрачивает патогенность. Кроме того, культуральные трепонемы имеют морфоло-
гические и антигенные отличия от Т. pallidum подвида pallidum.
498 НЕКОТОРЫЕ ИНФЕКЦИОННЫЕ ЗАБОЛЕВАНИЯ
Вне организма бледная трепонема очень чувствительна к внешним воздействи-
ям. Оптимальной для ее жизнедеятельности является температура тела человека
(37 °C). Она плохо переносит нагревание, воздействие ультрафиолетового облуче-
ния, быстро погибает при высушивании. Т. pallidum подвида pallidum устойчива к
низкой температуре: даже после глубокого и длительного замораживания содер-
жащие ее биологические субстраты сохраняют заразность для экспериментальных
животных. Бледная трепонема устойчива также во влажной среде вне организма,
например в нативном препарате она может сохранять подвижность в течение
нескольких суток. Бактерия очень чувствительна к традиционным антисептикам,
кислотам и щелочам, мылам, солям тяжелых металлов.
ЭПИДЕМИОЛОГИЯ
Заболевание регистрируют повсеместно, особенно высока его инцидентность в
развивающихся странах, В прошлом веке рост заболеваемости сифилисом отме-
тили в 30 -40-х гг., а затем он снижался до конца 80-х гг. В последующий период
повсеместно (в том числе и в России) происходил рост заболеваемости сифили-
сом.
Резервуаром инфекции служит больной человек. Возбудитель болезни переда-
ется в большинстве случаев половым, реже контактным путем. Наибольшую опас-
ность в плане распространения инфекции представляют люди, находящиеся на
первых 2 стадиях болезни, а в последующем они практически теряют инфекцион-
ность. Возможно внутриутробное заражение плодов (с 5-го месяца беременности),
а также заражение детей во время родов.
КЛИНИЧЕСКИЕ ПРОЯВЛЕНИЯ ИНФЕКЦИИ
Бледная трепонема вызывает у человека сифилис. Это заболевание в большин-
стве случаев передается половым путем и характеризуется хроническим волно-
образным течением с периодами обострений, чередующихся с длительными
периодами бессимптомного течения инфекции.
Первичный сифилис. Инкубационный период сифилиса длится от 9 до
90 дней. Первичная стадия заболевания характеризуется следующим комплексом
клинических симптомов: первичной сифиломой, регионарным склераденитом,
специфическим лимфангитом, специфическим полиаденитом, продромальными
явлениями.
Первичная сифилома — первое клиническое проявление заболевания, воз-
никающее на месте инокуляции бледных трепонем (в области входных ворот
инфекции). Первичная сифил* ма (первичный аффект, твердый шанкр) — чаще
единичная эрозия или язва, безболезненная, без признаков воспаления вокруг,
сравнительно плохо васкуляризирована, имеет в основании четко ограниченный
инфильтрат. Возникновению эрозивного или язвенного дефекта предшествует
появление небольшого гиперемического воспалительного пятна, которое вскоре
превращается в папулу. Эти изменения протекают бессимптомно и, как прави-
ло, остаются незамеченными ни пациентом, ни врачом. Вскоре после появления
папулы покрывающий ее эпидермис (эпителий) подвергается распаду и формиру-
ется эрозия или язва (первичная сифилома). Глубина дефекта при этом зависит от
выраженности и характера тканевой реакции на внедрение возбудителя.
Наиболее частые локализации первичной сифиломы у мужчин: в венечной
борозде, на внутреннем и наружном листках крайней плоти, на головке полового
члена, в области уздечки, наружного отверстия уретры и на слизистой оболочке
уретры, реже — на стволе и у основания полового члена, на мошонке. На половых
органах женщин первичные сифиломы чаще всего локализуются на малых и боль-
ших половых губах, в области задней спайки, клитора, на шейке матки.
НЕКОТОРЫЕ ИНФЕКЦИОННЫЕ ЗАБОЛЕВАНИЯ 409
Первичный аффект может остаться незамеченным при локализации в прямой
кишке, внутри уретры. Он может иметь нетипичные клинические проявления
вследствие осложнения вторичной инфекцией. Первичный аффект спонтанно
регрессирует в течение 3-8 нед, обычно не образуя рубца. Первичная сифилома
часто сопровождается умеренным увеличением регионарных лимфатических
узлов. Ь случае локализации первичного аффекта в области наружных половых
органов отмечают билатеральное ограниченное увеличение паховых лимфатиче-
ских узлов умеренной плотности (паховую лимфоаденопатию). Регионарный лим-
фаденит возникает обычно спустя 5-8 дней после появления первичного аффекта
и является почти постоянным спутником последнего, У некоторых пациентов в
конце первичного периода развиваются полиаденит и продромальный синдром,
обусловленный интоксикацией в результате массовой гибели трепонем в крово-
токе.
Дифференциальную диагностику первичной сифиломы проводят с пузырько-
вым лишаем, неспецифическими эрозиями и язвами различного генеза, изъяз-
вивши мися злокачественными новообразованиями кожи и слизистых оболочек,
шанкриформной пиодермией, мягким шанкром или шанкроидом
Вторичный сифилис. Вторичный сифилис является наиболее контагиозной
стадией заболевания и характеризуется следующим комплексом клинических
проявлений: пятнистым сифилидом (сифилитической розеолой), папулезным
сифилидом, папуло-пустулезным сифилидом. сифилитической алопецией (облы-
сением), сифилитической лейкодермой.
Высыпания появляются на коже и слизистых оболочках — розеолезный, папу-
лезный, папул о-пустулезный, везикулезный варианты сифилидов. Сыпь обычно
равномерно распространена по всему телу, включая ладони рук и подошвы ступ-
ней ног. На участках тела с повышенной влажностью (например, в паховой и
перианальной областях, в складках) папулезные высыпания могут образовывать
плоские вегетирующие бляшки, называемые широкими кондиломами. В некото-
рых случаях на слизистых оболочках могут появиться папулезные высыпания с
мацерированной молочно-белой, опалесцирующей поверхностью. Эти слизистые
бляшки имеют тенденцию к слиянию. Элементы сыпи высокоинфекционны,
любой физический контакт (сексуальный или несексуальный) с высыпаниями на
коже и слизистых оболочках может привести к заражению. Сыпь обычно исчезает
без лечения через несколько недель или месяцев. Кроме высыпаний отмечают
выпадение волос (очаговое или диффузное), нарушение пигментации (лейкодер-
му). При вторичном сифилисе в ряде случаев появляются признаки поражения
нервной системы, внутренних органов и опорно-двигательного аппарата.
Скрытый сифилис. Если пациент во вторичной стадии сифилиса не получает
лечения, все видимые проявления заболевания постепенно регрессируют, сифилис
переходит в раннюю скрытую стадию. На этой стадии он остается потенциально
заразным, так как трепонемы в его организме сохоаняются. Примерно через год
после начала заболевания количество грепонем в организме снижается, пациент
вступает в следующую стадию сифилиса, известную как поздний скрытый сифи-
лис. Выделение этих стадий имеет большое значение для диагностики и лечения
сифилиса. Часто невозможно определить точную продолжительность заболева-
ния, и если есть сомнения относительно времени заражения, пациента следует
рассматривать как больного с поздним скрытым сифилисом.
Третичный сифилис. После многих лет скрытого течения, при отсутствии
лечения заболевание переходит в наиболее деструктивную стадию — третичный
сифилис. При этом может наблюдаться поражение кожи и слизистых оболочек,
морфологической основой которого является гранулематозное воспаление, а
клинические проявления характеризуются бугорковыми и гуммозными высыпа-
ниями. Помимо кожи и слизистых оболочек, могут поражаться сердце, печень,
500 НЕКОТОРЫЕ ИНФЕКЦИОННЫЕ ЗАБОЛЕВАНИЯ
нервная система, опорно-двигательный аппарат. Нейросифилис может протекать
как бессимптомно, так и проявляться в форме менинговаскулярчого сифилиса,
спинной сухотки или прогрессирующего паралича. Кардиоваскулярный сифилис
характеризуется мезаортитом. который вначале протекает бессимптомно, а затем
осложняется недостаточностью аортального клапана, аневризмой аорты, главным
образом восходящей, стенозом устьев коронарных артерий. Третичный сифилис
протекает недоброкачественно и может приводить к летальному исходу.
Сифилис беременных. При нелеченом сифилисе у беременных инфекция
может передаваться плоду: в 25% случаев беременность заканчивается мертво-
рождением или ранней гибелью новорожденного: в 40-70% случаев рождаются
дети, больные врожденным сифилисом, который может протекать с симптомами
или бессимптомно.
При постановке беременной на учет должно быть проведено скрининговое
обследование на сифилис, которое повторяют во II и в III триместрах беремен-
ности. Если женщина не состояла на учете по беременности, обследование на
сифилис проводят во время родов. Новорожденных не выписывают из родильного
дома, пока не получен результат анализа на сифилис, особенно если во время анте-
натального наблюдения серологический статус матери был сомнительным.
Врожденный сифилис. Врожденный сифилис возникает в результате инфи-
цирования плода во время беременности трансплацентарным путем от больной
сифилисом матери. Обычно внутриутробное заражение плода происходит на
четвертом-пятом месяце беременности. Бледные трепонемы проникают в орга-
низм плода через пупочную вену, лимфатические щели пуповины или через
поврежденную плаценту. Беременность может закончиться мертворождением
либо рождением живого ребенка с проявлениями заболевания, возникающими
сразу после родов или несколько позднее.
Как и приобретенная инфекция, врожденный сифилис подразделяется на
ранний и поздний. Некоторые дети с ранним врожденным сифилисом могут
иметь клинические проявления заболевания при рождении, но у большинства
они развиваются в сроки от 2 нед до 3 мес после рождения. У некоторых ново-
рожденных возможно скрытое течение инфекции. Клинические проявления при
раннем врожденном сифилисе: кожа морщинистая, серо-желтой окраски, со спец-
ифическими высыпаниями, в том числе характерными только для врожденного
сифилиса (сифилитическая пузырчатка, диффузные папулезные инфильтрации).
К другим проявлениям раннего врожденного сифилиса относятся сифилитический
остеохондрит Вегнера, сифилитический ринит Гохзингера, лихорадка, увеличение
печени и селезенки, анемия и различные пороки развития. В дальнейшем (через
год и в более поздние сроки) могут развиться симптомы позднего врожденного
сифилиса, включающие поражение костей, зубов, органов зрения и слуха, мозга.
К признакам, патогномоничным для позднего врожденного сифилиса, относят
триаду Гетчинсона: паренхиматозный кератит, лабиринтную глухоту (специфиче-
ский лабиринтит) и зубы Гетчинсона.
ЛАБОРАТОРНАЯ ДИАГНОСТИКА
Общепринятые критерии лабораторной диагностики сифилиса представлены в
табл. 22-3.
Материалы для лабораторных исследований
Пробы для бактериоскопии. Для обнаружения Т. pallidum подвида pallidum
методами темнопольной микроскопии и прямой иммунофлюоресценции (ПИФ) у
пациентов берут серозное отделяемое с поверхности эрозии, язвы, мацерирован-
ной или экскориированной, эрозированной папулы или бляшки.
НЕКОТОРЫЕ ИНФЕКЦИОННЫЕ ЗАБОЛЕВАНИЯ 501
Таблица 22-3. Лабораторные критерии диагностики сифилиса
Диагноз Методы
Окончательный Выявление Т. pallidum в клиническом образце с использованием темнопольной микроскопии, ПИФ-ТР*, а также валидированных МАНК* или эквивалентных им методов
Предварительный Возможно использование двух видов серологических тестов: положительного нетрепонемного (например, RPR, РМП, VDRL*). подтвержденного трепонемного (TPHA, ТРРА, ИФА, РИФ-абс.*)
’ ПИФ-ТР — прямая иммунофлюоресиенция с антителами к Т. pallidum-, МАНК — методы ампли-
фикации нуклеиновых кислот; RPR — Rapid Plasma Reagin test (тест на быстрые реагины плазмы):
РМП — реакция микропреципитации; VDRL — Venerea! Disease Research Laboratory test TPHA —
T. Pallidum Haemagglutination As^y (реакция пассивной гемагглютинации); TPPA' — T. Pallidum
Passive Particle Agglutination assay (реакция пассивной агглютинации частиц); ИФА — иммунофер-
ментный анализ; РИФ-абс. — реакция иммунофлюоресценции.
Отбор проб осуществляют после тщательной и осторожной очистки поверх-
ности поражений марлевым тампоном, смоченным физиологическим раствором.
Очистку проводят с целью механического удаления трепонем-комменсалов и дру-
гих компонентов микрофлоры, которые могут затруднить исследование. Важно
при данной процедуре не травмировать ткани и не вызвать кровотечения, так как
присутствие в мазках лейкоцитов, эритроцитов и клеточного детрита значительно
усложняет обнаружение бледной трепонемы
Для получения тканевой жидкости используют несколько методов.
• Метод раздражения. Очаг поражения (твердый шанкр, мокнущая папу-
ла, широкая кондилома и т.д.) тщательно очищают ватой или тампоном,
смоченным стерильным физиологическим раствором. Стерильной ложкой
Фолькмана или петлей делают осторожные, чтобы не вызвать кровотечения,
поглаживающие движения по поверхности очага поражения. Через некото-
рое время его поверхность становится блестящей в результате просачива-
ния тканевой жидкости. Она прозрачная или слегка опалесцирует. В случае
появления кровотечения его следует остановить, прижав ватный тампон к
поверхности элемента, после чего продолжать поглаживающие движения.
Небольшая примесь крови к тканевой жидкости допустима.
• Метод сдавливания После очистки поверхности эрозию или язву сдавли-
вают с боков пальцами в перчатках или пинцетом, при этом стимулируется
выделение серозного экссудата, который собирают с поверхности бакте-
риологической петлей или путем аккуратного прикладывания к поверхности
эрозии/язвы покровного стекла. Полуденное серозное отделяемое смешива-
ют на предметном стекле с равным количеством физиологического раствора,
накрывают покровным стеклом
• Метод скарификации. К этому методу прибегают при исследовании сухих
элементов (розеолы, лентикулярных папул, энителизированных эрозий).
Поверхность исследуемого элемента скарифицируют, при этом неизбежно
появляется капиллярное кровотечение, которое останавливают марлевым
тампоном со стерильным физиологическим раствором. Посредством акку-
ратного раздражения скарифицированной поверхности получают серозную
жидкость. Этот метод по эффективности значительно уступает предыду-
щим. Исследование может дать ложноотрицательный результат вследствие
применения пациентом местных антисептиков, антибиотиков или при
присоединении вторичной инфекции. В этом случае назначают примочки с
физиологическим раствором, после чего (на следующие сутки) исследова-
ние повторяют. В некоторых случаях материал для микроскопической диа-
гностики берут из увеличенного регионарного лимфатического узла, чаще
пахового.
502 НЕКОТОРЫЕ ИНФЕКЦИОННЫЕ ЗАБОЛЕВАНИЯ
• Пункция лимфатического узла. Пунктируют плотный подвижный лим-
фатический узел без признаков воспаления. Место прокола обрабатывают
йодом, узел фиксируют двумя пальцами левой руки, а правой вкалывают в
узел иглу, соединенную со шприцем, в котором находится небольшое коли-
чество стерильного физиологического раствора. Прокол проводят у одного
из полюсов железы и проталкивают иглу по направлению к длинной оси
до противоположного полюса, при этом выпуская содержащийся в шприце
физиологический раствор. Затем иглу медленно выводят из узла, одновре-
менно оттягивая поршень из шприца. Правильность проведения пункции
подтверждается наличием в пунктате лимфоцитов.
Кровь. Пробы крови предпочтительнее брать с помощью вакуумных систем в
пустые пробирки или пробирки с ускорителем свертывания. Пробирки с кровью
осторожно транспортируют в лабораторию в специальном закрытом контейнере
при комнатной темпера гуре. При невозможности немедленного проведения ана-
лиза пробирки с кровью хранят в холодильнике при 4-8 °C.
В идеале образцы крови, полученные без ускорителя свертывания, инкубируют
в термостате при 37 °C или при комнатной температуре для образования сгустка
фибрина, затем помещают в холодильник при 4 °C на 30 мин. Пробы, взятые в
вакуумные пробирки с ускорителем свертывания, центрифугируют 10 мин при
скорости вращения ротора 3000 об./мин. Наличие в таких пробирках геля или
гранул создает барьер между сывороткой и остальными фракциями крови.
При необходимости хранения в течение длительного срока сыворотку разлива-
ют в несколько пробирок типа «Эппендорф» в объеме 0,5-1,0 мл и замораживают:
при -18-20 °C их можно хранить 1-1,5 мес, при -65-80 °C срок хранения проб
не ограничен. Не допускают оттаивания и повторного замораживания сыворотки
крови; при необходимости повторного исследования используют дублирующую
пробирку с образцом. Оттаивание сыворотки крови для проведения анализов про-
водят при комнатной температуре или при температуре 8+2 °C,
Перед исследованием сыворотки оттаивают (если пробы хранили в заморожен-
ном виде), тщательно перемешивают и инактивируют прогреванием при 56 °C в
течение 30 мин.
Спинномозговая жидкость В норме СМЖ бесцветна и поозрачна. Изменения
цвета и прозрачности СМЖ обусловлены примесями в ликворе (патологическими —
при заболеваниях, кровью — при травме кровеносных сосудов во время взятия
проб). СМЖ получают посредством спинномозговой пункции, которую выполня-
ет врач-невролог. Показаниями к проведению исследования СМЖ при сифилисе
служат:
о - наличие неврологических симптомов на любой стадии болезни;
о сифилитическое поражение органов зрения, сердечно-сосудистой системы;
< третичный или врожденный сифилис;
< скрытые формы сифилиса.
Спинномозговую пункцию обязательно следует проводить всем пациентам:
о с ВИЧ-инфекцией и положительными серологическими реакциями на сифи-
лис;
<> неврологическими симптомами при подозрении на возможную этиологиче-
скую роль сифилитическом инфекции в развитии этих симптомов.
Спинномозговую жидкость подвергают следующим анализам:
< > цитологическому;
определению содержания белка;
о- исследованию в реакциях микропреципитации (РМП), пассивной гемаг-
глютинации (РПГА), иммунофлюоресценции (РИФ-абс) и Venereal Disease
Research Laboratory test (VDRL).
НЕКОТОРЫЕ ИНФЕКЦИОННЫЕ ЗАБОЛЕВАНИЯ 503
Обнаружение возбудителя в клиническом материале
Микроскопия в темном поле. Исследование на бледную трепонему в темном
пиле проводят с помощью светового микроскопа с конденсором темного поля.
Метод темнопольной микроскопии основан на известном феномене Тиндаля,
проявляющемся визуализацией светящихся пылинок в луче света. Необходимые
условия создают путем замены обычного конденсора микроскопа особым конден-
сором, в котором центральная часть затемнена, проникновение света про-
исходит через узкую щель. При боковом освещении на темном поле достигается
отраженное свечение всех твердых частиц, в том числе бледной трепонемы. Для
получения темного поля необходим сильный источник света.
Темнопольная микроскопия может быть использована для диагностики пер-
вичного, вторичного и раннего врожденного сифилиса. Кроме отделяемого мок-
нущих сифилидов, приготовить препарат для микроскопии можно из пунктата
регионарных лимфатических узлов, спинномозговой, амниотической жидкости.
Присутствие пепатогенных трепонем-комменсалов Г. reftingers, Т. phagedenis (reit-
еп) в урогенитальном тракте. Т. denticola в полости рта делает затруднительным
и малодостоверным исследование с материалом, полученным с проявлений на
слизистой оболочке полости рта или прямой кишки, так как морфологически
эти трепонемы сходны с Т. pallidum. При необходимости исследования мате-
риала, полученного из этих локализаций, лучше отдать предпочтение ПИФ или
молекулярно-биологическим метода м.
На темном фоне можно также увидеть нейтрофильные лейкоциты (имеют
вид округлых, ярко светящихся зернистых образований), лимфоциты (серовато-
темные, тускло светящиеся клетки округлой формы), клетки эпителия (круглые
или неправильной формы, значительно крупнее лейкоцитов и ярко светящиеся),
эритроциты (темные элементы круглой формы, окаймленные светящейся поло-
ской размером чуть меньше лейкоцитов). Если эритроцитов много, поле становит-
ся слишком светлым, трепонемы видны хуже, могут скрываться под эритроцита-
ми. В этом случае материал лучше взять повторно.
Требуется определенный опыт для распознавания в препарате Т. pallidum,
которая выглядит как очень тонкая спираль длиной 6-20 мкм и толщиной
0,13-0,15 мкм, с 8-12 равномерными завитками глубиной до 0,5-0,7 мкм и может
быть легко перепутана с микроорганизмами-комменсалами. Т. pallidum выполняет
различные виды движений, среди которых можно выделить характерные для
патогенной трепонемы: сгибательные, чаще всего посередине длины ее тела, и
маятникообразные (рис. 22-2). Морфологические характеристики и наличие
Виды движения бледной трепонемы
в нативном поапаоате
Непатогномоничные для Т palladium
Рис. 22-2. Виды движения бледной трепонемы в нативном препарате.
504 НЕКОТОРЫЕ ИНФЕКЦИОННЫЕ ЗАБОЛЕВАНИЯ
движений, характерных для бледной трепонемы, являются диагностическими
признаками Т. pallidum.
Если после однократного исследования бледную трепонему обнаружить не уда-
лось, поражение следует промыть физиологическим раствором и затем провести
повторное исследование. Однако даже многократный отрицательный результат
такого исследования не позволяет исключить сифилис. В подобных случаях необ-
ходимо провести серологическое исследование.
Метод прямой иммунофлюоресценции. Метод ПИФ предусматривает
прямое выявление Т. pallidum в образце при обработке клинического материала
конъюгатами специфических моноклональных антител с флюоресцирующими
красителями. Преимуществами метода являются возможность дифференциации
патогенных трепонем от непатогенных при исследовании проб, взятых со
слизистых оболочек полости рта и прямой кишки, фиксация препаратов перед
транспортировкой в лабораторию. Этим методом можно также исследовать
материал, полученный при биопсии или аутопсии.
Препараты исследуют с использованием люминесцентного микроскопа при
увеличении х200, х400 и хЮОО. При этом клетки окрашиваются в красно-
оранжевый или желтый цвет, бледная трепонема флюоресцирует ярко-зеленым
цветом, сохраняя все морфологические признаки спирохеты.
Молекулярно-генетические методы. Б настоящее время не существует
коммерческих тестов для выявления ДНК Т. pallidum. Все самостоятельно
изготовленные тест-системы необходимо валидировать с помощью не менее
10 положительных образцов, полученных от серопозитивных пациентов с
поражениями, в которых наличие трепонем подтверждено методом темнопольной
микроскопии, а также 10 отрицательных образцов, полученных от серонегативных
пациентов.
В ПЦР можно исследовать соскобы шанкров и других высыпаний (папул,
широких кондилом и т.д.), спинномозговую, амниотическую жидкость,
сыворотку крови. При проведении реакции необходим как отрицательный,
так и положительный контроль. В качестве положительного контроля можно
использовать суспензию трепонем, полученную из яичек инфицированного
кролика и разведенную до концентрации приблизительно 100 трепонем/мл,
тогда как препарат из яичек неинфицированного кролика можно использовать в
качестве отрицательного контроля. ПЦР служит хорошим методом для диагности-
ки сифилиса при небольшом количестве трепонем в исследуемом материале. Этот
тест пока не нашел применения в практике рутинных лабораторных исследований
на сифилис, его применяют только в исследовательских целях. Он высокоспеци-
фичен, чувствителен и дает воспроизводимые результаты при правильных прове-
дении и подготовке образцов.
Разработаны и другие неамплификационные молекулярно-генетические мето-
ды диагностики сифилиса (в том числе гибридизация нуклеиновых кислот и дот-
гибридизация), однако они менее чувствительны, чем ПЦР.
Методы, используемые для установления предварительного диагноза
А. Нетрепонемные тесты
В качестве антигена в нетрепонемных тестах применяют смесь кардиолипина,
лецитина и холестерина. Они позволяют проводить скрининг большого количе-
ства людей, оценку эффективности терапии. В основном они основаны на фло-
куляции (выпадении хлопьев в реакционной смеси). Позволяют обнаруживать
IgG- и IgM-антитела к липидам клеточной стенки бледной трепонемы, которые
появляются в крови примерно через 1 нед после формирования твердого шанкра.
Достоинствами нетрепонемных тестов являются низкая стоимость, техническая
простота и скорость получения результата исследования. Возможность постанов-
НЕКОТОРЫЕ ИНФЕКЦИОННЫЕ ЗАБОЛЕВАНИЯ 505
ки реакции в количественном варианте с определением титра антител позволяет
использовать их в случае реинфекции/рецидива и для оценки эффективности
терапии. Ограничением к применению являются довольно низкая чувствитель-
ность и наличие ложноположительных результатов при скрининге популяций с
низкой инцидентностью заболевания.
Информация, характеризующая наиболее часто применяемые нетрепонемные
реакции (РМП RPR, тест VDRL), представлена в табл. 22-4.
Таблица 22 4. Чувствительность и специфичность нетрепонемных методов серологической диа-
гностики сифилиса
Тип реакции Чувствительность при разных стадиях сифилиса, % Специфичность, %
первичный вторичный скрытый ранний скрытый поздний
РМП 81 (70-90) 91 94(88-100) 70 (57-80) 98 (93-99)
VDRL 78 (59-87) 100 95 (88-100) 71 (37-94) 98 (96-99)
RPR 86 (77-100) 100 98 (95-100) 73 (57-85) 98 (93-99)
Примечание. РМП — реакция микропреципитации; VDRL — Venereal Disease Research Laboratory test;
RPR - реакция быстрых реагинов плазмы (Rapid Plasma Reagin Test).
РМП является модификацией VDRL и применяется в большинстве стран
Содружества Независимых Государств (СНГ).
Реакция микропреиипитации
Качественный вариант. Для исследования используют плазму, инактивиро-
ванную сыворотку или СМЖ. При добавлении к плазме, сыворотке крови или
ликвору больного сифилисом эмульсии кардиолипинового антигена (холестерин-
кардиолипин-лецитинового комплекса) происходит агглютинация и образуется пре-
ципитат (комплекс «антиген-ан гителл»), выпадающий в виде хлопьев белого цвета
РМП выполняют в лунках иммунологического планшета с округлым дном для
макротитрования или в лунках с плоским полированным дном планшетов, изго-
товленных из органического стекла (плексигласа) толщиной 0,8-1,0 см. Реакция
протекает при постоянном перемешивании реакционной среды: при покачивании
планшета вручную или на платформе шейкера (орбитального ротатора). При
отсутствии антител в исследуемом образце внешний вид реакционной среды не
изменяется — она остается опалесцирующей.
О наличии в образце биологического материала антител-преципитинов свиде-
тельствует образование хлопьев преципитата, одновременно происходит просвет-
ление реакционной среды. Величина частиц преципитата, оцениваемая визуально,
является мерой полуколичественного учета содержания антител и может быть
выражена в условных единицах — от одного до четырех плюсов (табл. 22-5).
При максимальной выраженное ги ответа (4+) дальнейшую полуколичественную
оценку содержания антител проводят путем тестирования последовательных раз-
ведений исследуемого образца патологического материала (титрование). Учет
проводят при боковом освешении планшета. Планшет держат в руках и слегка
покачивают; для более четкой регистрации под планшетом размещают лист чер-
ной бумаги и применяют лупу двукратного увеличения.
Полуколичественный вариант. Все образцы биологического материала,
давшие при скрининге положительный результат, необходимо исследовать в
полуколичественном варианте РМП. В лунках планшета проводят серию последо-
вательных двукратных разведений каждого образца: от цельного биологического
материала к 1:2, 1:4 — до 1:128 (восемь разведений) или 1.512 (десязь разведе-
ний). Учет проводят по стандартной методике. При этом титром антител считают
последнее разведение, где сформировался преципитат.
506 НЕКОТОРЫЕ ИНФЕКЦИОННЫЕ ЗАБОЛЕВАНИЯ
Таблица 22-5. Оценка результатов РМП
Результат Описание
Резко положительный (4+) Крупные хлопья преципитата белого цвета распределяются равномерно по всему объему лунки либо имеют тенденцию к расположению по перифе- рической части лунки, при этом реакционная среда практически полностью прозрачная
Положительный (3+) Хлопья средней величины распределены по всему объему лунки, реакцион- ная среда имеет белесоватый опенок
Слабоположительный (?+ и 1+) Мелкие хлопья преципитата распределены по объему лунки, реакционная среда имеет белесоватый опенок
Сомнительный (±) Наличие очень мелких хлопьев, сомнение в наличии преципитата; при пока- чивании планшета частицы антигена демонстрируют перламутровые перели- вы реакционной среды. При получении подобного результата рекомендуют повторное исследование данного образца биологического материала в цельном виде и в разведениях (для исключения феномена эоны) и нового образца (парные сыворотки), полученного через 7-10 дней, для контроля динамики процесса
Отрицательный Преципитата нет, реакционная среда непрозрачная, при покачивании план- шета частицы кардиолипинового антигена перемещаются, формируя перла- мутровые переливы белого цвета в центре лунки
Источники ОШИБОК ПРИ ВЫПОЛНЕНИИ РМП
• Иск лючение из реакции контрольных сывороток крови, в частности слабопо-
ложительных.
• Неравномерная концентрация антигена в эмульсии из-за недостаточного
перемешивания ее перед использованием.
• Бактериальное загрязнение эмульсии.
• Нарушение сроков и условий хранения плазмы и сыворотки крови, антигена
и его эмульсии, растворов.
• Использование при постановке реакций загрязненных пробирок, пипеток,
планшет, растворов.
• Нарушение техники взятия крови из пальца (изменение соотношения цитра-
та Na и крови).
• Замена трехзамещенного цитрата натрия двухзамещенным.
Эти погрешности могут приводить к появлению как ложноотрицательных, так
и ложноположительных результатов реакции.
Тест на определение быстрых плазменных реагинов (RPR)
При добавлении к плазме или сыворотке крови больного сифилисом суспензии
стабилизированного кардиолипинового антигена, адсорбированного на мелко-
дисперсных угольных частицах, происходит образование комплексов «антиген-
антитело», собирающих частицы угля в более крупные агрегаты; при этом резуль-
тат реакции можно учитывать невооруженным глазом.
Тест RPR выполняют на специальных картонных или пластиковых карточках
одноразового использования, на которых выдавлены неглубокие лунки диа-
метром 18 мм. Перед началом исследования проводят разметку карточек: руч-
кой или карандашом надписывают номер исследуемого образца сыворотки или
плазмы крови в соответствии с записями в журнале регистрации результатов
RPR -теста. В лунки карточек вносят образцы исследуемой сыворотки или плазмы
крови и суспензию антигена. Для более эффективного образования комплексов
«антиген-антитело» содержимое лунок перемешивают в течение 8 мин при ком-
натной температуре на платформе орбитального шейкера. При отсутствии анти-
тел в исследуемом образце внешний вид реакционной среды не изменяется, она
остается гомогенной.
НЕКОТОРЫЕ ИНФЕКЦИОННЫЕ ЗАБОЛЕВАНИЯ 5Q7
Качественный вариант определения быстрых плазменных реагинов.
Учет результатов проводят при хорошем освещении, карточку располагают на
столе или держат в руках и слегка покачивают; для более четкой регистрации при-
меняют лупу двукратного увеличения. О наличии в образце биологического мате-
риала реагиновых антител свидетельствует образование частиц флокулята раз-
личной величины, одновременно происходит просветление реакционной среды.
Принципы оценки результатов теста изложены в табл. 22-6.
Таблица 22-6. Оценка результатов теста на определение быстрых плазменных реагинов
Результат Описание
Положительный Крупные и средней величины агрегаты угольных частиц черного цвета располо- жены по всему объему лунки либо имеют тенденцию к расположению ближе к периферической части лунки, при этом реакционная среда практически полно- стью прозрачная
Слабоположительный Редкие и мелкие агрегаты угольных частиц, хлопья преципитата распределены по периферии реакционной лунки, реакционная среда имеет гомогенную струк- туру
Отрицательный Видимые агрегаты угольных частиц в пробе отсутствуют, реакционная среда гомогенной структуры либо частицы угля собираются в центральной части лунки, формируя пятно черного цвета
Образцы сыворотки или плазмы крови, показавшие при скрининге положи-
тельные и слабоположительные результаты, исследуют полуколичественным
вариантом (титрованием).
У некоторых пациентов с очень высоким содержанием антител возможно полу-
чение слабоположительного ответа в RPR-тесте с неразведенным биологическим
материалом (проявление феномена прозоны); причина этого явления в том, что
переизбыток антител препятствует образованию крупных хлопьев преципитата.
В этих случаях рекомендуют исследование разведенных образцов биологического
материала.
Полуколичественное определение быстрых плазменных реагинов Полу-
количественную RPR проводят посредством титрования проб и применяют в сле-
дующих случаях:
о- для исследования образцов сыворотки или плазмы крови, показавших при
скрининге положительные и слабоположительные результаты;
❖ если данные RPR расходятся с результатами др}тих тестов на сифилис;
в случае прямого указания лечащего врача.
Исследуют серию разведений цельного биологического материала с 1:2 до 1:16
или до 1:512. Титром антител считают последнее разведение, показавшее положи-
тельный результат при RPR-тестировании.
Оценка диагностической значимости нетрепонемных тестов. Нетре-
понемные тесты не являются строго специфическими реакциями на сифилис,
так как предназначены для выявления антикардиолипиновых антител. В сыво-
ротке крови людей реагиновые антитела могут транзиторно обнаруживаться
при системных поражениях паренхиматозных органов (печени, почек, легких),
при миокардите, атеросклерозе, острых вирусных инфекциях, включая гепатит,
ветряную оспу и корь, при малярии, после вакцинации и во время беременности.
Хроническую персистенцию наблюдают при заболеваниях соединительной ткани,
лепре, онкологии, применении внутривенных наркотиков и в пожилом возрасте.
Количество ложноположительных результатов увеличивается при исследовании
образцов крови пожилых людей.
Нетрепонемные тесты используют для первоначального скрининга, они стано-
вятся положительными примерно на шестой неделе после инфицирования, поэто-
му часть пациентов с положительными результатами темнопольной микроскопии
508 НЕКОТОРЫЕ ИНФЕКЦИОННЫЕ ЗАБОЛЕВАНИЯ
Первичный
Ранний скрытый
°енний сифилис
Третичный
Вюричный
Испе-
ченный
Правильно
проле-
ченный
Поздний сифилис
Поздний
скрытый
Положительный
нетрепонемный
антительный тест
Рис. 22-3. Реактивность нетрепонемных серологических тестов в зависимости от стадии сифили-
са и влияния проведенного лечения (схема).
Отрицательный
нетрепонемный
антительный тест
(до 40%) при первичном сифилисе могут быть серонегативными. Количественные
варианты тестов используют для определения эффективности терапии. После
адекватного лечения раннего сифилиса титры антител в нетрепонемных тестах
должны значительно снижаться или результаты тестов становиться отрицатель-
ными. Тем не менее даже при адекватном лечении поздних стадий заболевания в
последующем титры антител могут стойко сохраняться, снижаться или персисти-
ровать на прежнем уровне, но никогда не увеличиваться (рис, 22-3).
Б. Трепонемные тесты
К трепонемным реакциям относятся ИФА, РПГА или ее разновидность — метод
микрогемагглютинации Т. pallidum (МГА-ТР), реакция иммобилизации бледных
трепонем (РИБ Г), РИФ, а также метод ИБ и тест с использованием хроматографи-
ческих полосок [«анализ по месту лечения» (point-of-care)}. В этих тестах применя-
ют антиген бледной трепонемы (в реакциях РИФ, РИБТ), рекомбинантные белки,
полученные генно-инженерным способом, или искусственно синтезированные
пептиды (ИФА, РПГА, модифицированный иммуноблоттинг и иммунохромато-
графический тест).
Трепонемные тесты используют для подтверждения результатов нетрепонем-
ных тестов. В популяции с низкой инцидентностью заболевания их можно исполь-
зовать для скрининга, так как в настоящее время возможна автоматизации ИФА.
Несомненные достоинства этих диагностичеких тестов — высокие чувстви-
тельность и специфичность (табл. 22-7). Экспресс-тесты можно использовать в
местах оказания медицинской помощи. Однако трепонемные тесты непригодны
для контроля эффективности терапии, так как даже после выздоровления многих
Таблица 22-7. Чувствительность и специфичность трепонемных методов серологической диа-
гностики сифилиса
Тил реакции Чувствительность при разных стадиях сифилиса, % Специфичность, %
первичный вторичный скрытый ранний скрытый поздний
РИФ-абс. 84(70-100) 100 100 96 (93-100) 97(94-100)
МГА-ТР 76 (64-90) 100 97 (94-100) 97 (94-100) 99 (98-100)
Применение. РИФ-абс. - реакция иммунофлюоресценции; МГА-ТР — реакция микрогемагглю-
тинации к Т. pallidum.
НЕКОТОРЫЕ ИНФЕКЦИОННЫЕ ЗАБОЛЕВАНИЯ 5Q9
пациентов их сыворотка крови продолжает давать положительную реакцию на
протяжении длительного времени (иногда в течение всей жизни). И трепонемные,
и нетрепонемные тесты могут давать положительный результат при невенериче-
ских трепонематозах.
Иммунофлюоресцент ные тесты (РИФ)
Для серологической диагностики сифилиса применяют несколько вариантов
РИФ. Наиболее чувствительным из них («золотым стандартом») является РИФ с
абсорбцией (РИФ-абс.). По данным ВОЗ. чувствительность РИФ-абс.-теста равна
70-100% при первичном и 96-100% — при вторичном и позднем сифилисе, тогда
как его специфичность составляет 94-100%. РИФ-абс.-тест часто применяют для
подтверждения диагноза. Поскольку он относительно дорогой, трудоемкий и тре-
бует наличия в лаборатории люминесцентного микроскопа, его не рекомендуют
применять в качестве скринингового теста.
При его проведении исследуемую сыворотку разводят 1:5 и обрабатывают сор-
бентом для удаления неспецифических антител.
РИФ-абс.-DS-TecT отличается от РИФ-абс. тем, что к разбавителю сыворотки
добавляют краситель (обычно родамин) для создания контрастного фона ярко-
зеленым спиральным трепонемам.
Для ранней диагностики врожденного сифилиса используют тесты на основе
флюоресцирующих антител, разработанные для детекции специфических IgM.
Большие молекулы IgM неспособны проходить через плаценту. Их обнаруживают
в крови плода только при нарушении плацентарного барьера или при активной
продукции инфицированным плодом. По этой причине выявление специфических
IgM-антител в сыворотке крови новорожденных детей считают свидетельством
врожденного сифилиса. К сожалению, специфичность данного теста невысока,
так как может происходить неспецифическое связывание коньки ата. Разработано
несколько вариантов РИФ для выявления при сифилисе специфических IgM-
анти гел:
РИФ-абс.-IgM — выявляет антитела к специфическим IgM-антителам чело-
века;
198-^М-РИФ-абс. — содержащиеся в исследуемой сыворотке антитела
разделяют на мини-колонке и исследуют в данном тесте фракцию 19S-lgM.
Для теста РИФ-абс. необходимы:
❖ исследуемая сыворотка;
о антиген. Суспензию Т. pallidum (штамм Nichol) получают из яичек инфици-
рованного кролика через 7 дней поете инокуляции. Кроме того, существуют
коммерческие препараты — предметные стекла с нанесенными на них трепо-
немами;
❖ сорбент — готовят из культур непатогенных трепонем Рейтера без добавле-
ния консерванта;
❖ конъюгат, меченный флюоресцеином. Сыворотку получают от животных,
иммунизированных фракцией иммуноглобулинов человека, и метят флюо-
ресцеином;
о- фосфатный буфер.
Тест проводят следующим образом. На тонких обезжиренных предметных сте-
клах обводят oкpvжнocти (по 10 на каждом стекле) диаметром 0,7 см. Суспензию
Т. pallidum наносят в цен гр каждого круга и равномерно распределяют по нему
вращательными движениями. Препараты антигенов сушат на воздухе и фиксиру-
ют. Исследуемые инактивированные сыворотки разводят 1:5 сорбентом, который
разбавтяют до заранее определенного титра. Разведенные сыворотки наносят на
антиген так, чтобы мазок был покрыт равномерно. Выдерживают препараты во
влажной камере инкубатора. После инкубации стекла дважды тщательно про-
мывают в фосфатном буфере, после чего высушивают на воздухе. Затем на мазки
51 о НЕКОТОРЫЕ ИНФЕКЦИОННЫЕ ЗАБОЛЕВАНИЯ
наносят конъюгат, меченный флюоресцеином, и повторно инкубируют во влаж-
ной камере. После инкубации образцы промывают в фосфатном буфере, высу-
шивают на воздухе, наносят на мазок каплю монтирующей жидкости, накрывают
его покровным стеклом и исследуют с помощью люминесцентного микроскопа с
кварцево-ртутной лампой при увеличении х400. При учете реакции оценивают
степень флюоресценции трепонем:
❖ 4+ (зелено-желтая флюоресценция) — резко положительный образец;
о 3+ (яркая флюоресценция) — положительный;
❖ 2+ (слабая флюоресценция) — слабоположительный;
❖ 1+ (видимое окрашивание) — минимально-реактивный;
❖ отсутствие флюоресценции — отрицательный.
Учет результатов РИФ-абс. носит субъективный характер, следствием чего в
ряде случаев (особенно при отсутствии у специалиста, проводящего тест, доста-
точного опыта) получают ложноположительные результаты, Неспецифическую
положительную реакцию могут давать также сыворотки крови пациентов с ауто-
иммунными заболеваниями и лепрой.
Для выявления антител в СМЖ при подозрении на нейросифилис предложена
модификация — РИФ-ц-тест. Используют те же антиген и конъюгат. Исследуемый
образец СМЖ не инактиьируют и тестируют в неразведенном виде.
Реакция пассивной гемагглютинации (РПГА)
Для проведения теста применяют эритроцитарный диагностикум — формали-
низированные и таннизирэианные эритроциты барана или птиц (чаще куриные),
которые сенсибилизировали обработанным ультразвуком антигеном Т. pallidum
или рекомбинантными белками бледной трепонемы (TpN15, TpN17, TpN4?).
Коммерческие тест-наборы для РПГА обычно включают эритроцитарный диа-
гностикум, контрольные эритроциты, буфер для разведения, положительный и
отрицательный контроль и 96-луночные планшеты с U-образным дном.
В лунки планшета вносят образцы исследуемой сыворотки крови и эритроци-
тарный диагностикум. При наличии специфических антител в сыворотке крови
пациента происходит их взаимодействие с антигенами бледной трепонемы и обра-
зование пространственных структур иммунных комплексов «антитело-антиген -
эритроцит», которые под действием силы тяжести постепенно опускаются вниз,
распределяются по всей поверхности дна лунки и формируют характерную кар-
тину «перевернутою зонтика». Реакцию учитывают визуально через 60-120 мин.
При использовании более крупных эритроцитов птиц получают более четкую кар-
тину, учет результатов проводят в более ранние сроки — через 45- 60 мин.
В зависимости от количества иммунных антител, содержащихся в исследуемом
образце, изображение «перевернутого зонтика» варьирует от максимального,
занимающего всю поверхность дна лунки, до небольшого участка в центральной,
самой низко расположенной его части, с просветлением в центре и формировани-
ем более интенсивного кольца из осевших эритроцитов по периферии.
При оценке результатов РПГА опираются на форму и характер осадка эритроци-
тов на дне лунок иммунологического планшета (табл. 22-8).
При отсутствии в образце специфических ан гител и при проведении реакции
с интактными эритроцитами (контроль) образования иммунных комплексов не
происходит: эритроциты постепенно оседают на дно лунки, формируя фигуру в
виде компактного пятна или «пуговки», иногда с незнании ельным просветлением
в центре.
РПГА проста в выполнении, не требует специального оборудования и не занима-
ет длительного времени; высокочувствительна и специфична. Реакция становится
положительной спустя 3-4 нед после появления твердого шанкра. В зависимости
от стадии заболевания чувствительность РПГА колеблется от 76% при первичном
до 100% при вторичном сифилисе и 94-97% при скрытых формах.
НЕКОТОРЫЕ ИНФЕКЦИОННЫЕ ЗАБОЛЕВАНИЯ 511
Таблица 22-8. Оценка результатов РПГА для выявления Т. pallidum
Результат Описание
Резко положительный (4+) Эритроциты расположены ровным слоем по всему дну лунки — высокое содержание специфических антител в пробе
Положительный (3+) Эритроциты расположены на оольшей части дна лунки, по периферии осадка формируется заметное кольцо из эритроцитов
Слабоположительный (2+) Эритроциты расположены на небольшой части дна лунки, в центральной части формируется плотное кольцо из осадка эритроцитов с заметным просветлением в центральной части
Неопределенный (1+ и +) Наличие небольшого рыхлого осадка эритроцитов с нечеткими краями в центральной части лунки, с незначительным просветом в центре на сво- бодном от эритроцитов окружающем фоне — такие образцы рекомендуют исследовать повторно, иногда со свежей порцией сыворотки, полученной через 1-2 нед
Отрицательный Компактный осадок в центральной части дна лунки на чистом окружаю- щем фоне
Антитела-аплютинины выявляют в крови переболевших сифилисом людей в
течение длительного времени, поэтому РПГА нельзя рекомендовать для диффе-
ренциальной диагностики реинфекции или определения остроты инфекционного
процесса (рис. 22-4).
Преимущества РПГА перед РИТ, РИФ состоят в доступности готовых тест-
систем, отсутствии надобности в живой патогенной бледной трепонеме, возмож-
ности автоматизации реакции.
Разновидностью РПГА является ТРРА, в которой используют несенсибилизи-
рованные и сенсибилизированные частицы латекса вместо эритроцитов (реакция
пассивной агглютинации с частицами латекса или ТРРА). Тест проводят так же,
как РПГА, но результаты оценивать легче. Реагенты для постановки ТРРА более
стабильны.
Иммуноферментный анализ (ИФА)
Принцип теста состоит в фиксации на поверхности лунок полистиролового
планшета иммунных комплексов, образующихся при взаимодействии антител
больного сифилисом с антигенами бледной трепонемы, с последующим выявлени-
ем их в цветовой реакции с помощью специфических конъюгатов и соответствую-
щих субстратно-хромогенных добавок,
Первичный
Ранний сифилис
Вторичный
Ранний скрытый
------Поздний сифилис
Поздний
скрытый
Третичный
Рис. 22-4. Реактивность трепонемных серологических тестов в зависимости от стадии сифилиса
и влияния проведенного лечения (схема).
512 НЕКОТОРЫЕ ИНФЕКЦИОННЫЕ ЗАБОЛЕВАНИЯ
Для выполнения теста применяют различные антигены:
о обработанную ультразвуком суспензию Т. pallidum’,
о рекомбинантные — получают генно-инженерными технологиями путем вне-
дрения в геном бактериальной клетки (например, Е. coli) гена, ответственного
за синтез определенного антигена Г. pallidum, с последующим наращиванием
бактериальной массы продуцирующего микроорганизма, разрушением этих
клеток, выделением и очисткой антигена;
❖ пептидные — получают в результате последовательного химического синтеза
антигенных эпитопов белков Т. pallidum. Наиболее пригодными для теста
оказались иммунодоминантные участки белков бледной трепонемы с моле-
кулярной массой 15,17 и 47 кДа.
Классический (непрямой) вариант ИФА
• Внутреннюю поверхность лунки иммунологического планшета сенсибилизи-
руют антигенами бледной трепонемы.
• При внесении в лунки и инкубировании в них исследуемых сывороток
специфические антитела образуют иммунные комплексы, которые остаются
связанными после отмывания буферным раствором (1 фаза).
• В реакцию добавляют конъюгат, представляющий собой антитела к иммуно-
глобулинам человека, меченные ферментом (пероксидазой хрена) (II фаза).
• После повторного отмывания буфером наличие сложных иммунных
«сэндвич»-комплексов выявляют внесением в лунки раствора перекиси водо-
рода (субстрата), расщепляемой ферментом конъюгата, и хромогена, кото-
рый меняет цвет при взаимодействии с атомарным кислородом (III фаза).
• Насыщенность цвета реакционной среды пропорциональна количеез ву реа-
гирующих антител к Т. pallidum, что можно измерить фотометрически.
Ловушечный вариант ИФА
Его применяют в основном для выявления антител различных классов:
поверхность лунки иммунологического планшета сенсибилизируют аффин-
но очищенными антителами к иммуноглобулинам человека определенного
класса (например, IgM, IgG или IgA);
содержащиеся в исследуемой сыворотке иммуноглобулины соответствую-
щего класса связываются с антителами, сенсибилизированными на стенках
лунки иммунологического планшета (I фаза);
<> присутствие среди иммобилизованных иммуноглобулинов специфических
антител определяют внесением антигенов бледной трепонемы, конъюгиро-
ванных с ферментом (II фаза):
❖ результаты визуализируют добавлением в реакцию хромоген-субстратной
смеси (III фаза).
Модифицированный двухфазный вариант ИФА
Этот вариант ИФхА применяют для выявления суммарных антител, специфиче-
ских к определенному антигену, независимо от их класса:
< специфические сывороточные антитела взаимодействуют одновременно с
антигеном, фиксированным на поверхности лунок планшета для ИФА и тем
же антигеном, меченным ферментом, при совместном инкубировании испы-
туемого образца и конъюгата (I фаза);
< образование иммунных комплексов определяют с помощью субстрата
(П фаза).
Оценка результатов ИФА. Результаты ИФА-тестов носят качественный
характер. Их учитывают автоматически по величине цифровых показателей
оптической плотности (ОП) в лунках с испытуемыми сыворотками. При каж-
дом выполнении теста рассчитывают критическое значение ОП и серую зону
(интервал, лежащий в пределах критической ОП, ±10%). Результат теста счита-
ют позитивным, если ОП исследуемой пробы располагается выше серой зоны;
НЕКОТОРЫЕ ИНФЕКЦИОННЫЕ ЗАБОЛЕВАНИЯ 513
сомнительным, если ОП находится в пределах серой зоны, отрицательным, если
ОП ниже серой зоны,
К сожалению, все тест-системы ИФА невозможно привести к единому стандар-
ту. Один и тот же образец сыворотки может давать в разных тест-системах разный
уровень окрашивания раствора субстрата. Это зависит от конструкции самой тест-
системы, используемого антигена, концентрации конъюгата, а также величины
критической ОП.
Иммуноблоттинг
Для диагностики сифилиса применяют несколько вариантов иммуноблоттин-
га — западный блоггинг-тест «Innolia».
Эти тесты дают легковоспроизводимые результаты. Они относительно просты
в выполнении и интерпретации результатов. Тем не менее они достаточно доро-
ги, занимают много времени и требуют высокой квалификации лабораторных
работников. Позволяют определять наличие в тестируемых пробах антител сразу
к нескольким антигенам Т. pallidum. Применение высокоочищенных рекомбинант-
ных и пептидных антигенов снижает до минимума неспецифическую реактив-
ность сывороток. Применение иммуноблоттинга дает возможность отказаться от
трудоемких и субъективных методов, связанных с опасными для исследователя
манипуляциями (перевивка штаммов Т. pallidum, работа с патогенной бледной тре-
понемой при выполнении реакции). Всем этим определяется предпочтение имму-
ноблоттинга другим трепонемным тестам для диагностики сифилиса, в частности
РИФ и в особенности — РИБТ (см. ниже). Существуют технические возможности
для инструментальной оценки интенсивности окрашивания антигенных полос на
Стрипах (сканирование) с автоматизацией аналитического процесса.
Западный блоттинг. Данный тест проводят с применением коммерческих и
самостоятельно приготовленных тест-систем. Они представляют собой стрипы,
содержащие специфические компоненты экстракта Т. pallidum, которые разделя-
ют электрофорезом. Стрип инкубируют с сывороткой пациента. При наличии в
ней специфических антител они связываются участками стрипа, содержащими
гомологичные антигены возбудителя. Для выявления участка связывания стрипы
инкубируют с антителами к иммуноглобулинам человека, меченными пероксида-
зой хрена. Наличие конъюгата (а следовательно, и антител к Т. pallidum) определя-
ют по действию фермента на субстрат, дающий цветовую реакцию. В иммуноблот-
тинге можно выявлять специфические IgG- и IgM-антитела.
Модифицированный иммуноблоттинг (тест «Innolia»). Основу теста для
проведения линейного иммуноблоттинга in vitro в целях обнаружения специфи-
ческих антител против Т. pallidum в образцах сыворотки или плазмы крови
человека представляют нейлоновые тест-полоски (стрипы), фиксированные на
подложке из нейтрального пластика. На каждый стрип в виде дискретных линий
нанесены антигенные детерминанты Т. pallidum: три рекомбинантных белка
(TpN47, TpN17 и TpN15) и один синтетический пептид (ТшрА). Кроме того, на
каждом стрипе имеются четыре контрольные линии; одна антистрептавидиновая
и три калибровочные линии, по интенсивности окрашивания соответствующие
Зт, 1+, ± {cut-off). Калибровочная линия, соответствующая 3+, содержит иммо-
билизованные антитела против IgG человека и одновременно служит контро-
лем.
Оценка результатов иммуноблоттинга. Для суждения о достоверности
(валидности) проведенного исследования учет реакции начинают с контролей:
на стрипе, инкубированном с положительным контролем, должно происходить
окрашивание всех 4 полос с трепонемными антигенами не менее чем на 1+, а на
стрипе, тестированном с отрицательным контролем, не допускается окрашивания
антигенных полос; возможно фоновое окрашивание слабее критического уровня.
Затем приступают к оценке испытуемых проб (табл. 22-9).
514 НЕКОТОРЫЕ ИНФЕКЦИОННЫЕ ЗАБОЛЕВАНИЯ
Таблица 22-9. Оценка результатов модифицированного иммуноблоттинга (теста «Innolia»)
Результат Описание
Отрицательный Все антигенные полосы на стрипе не окрашены или одна из полос окра- шена с интенсивностью «±»
Положительный Две или более антигенные полосы на стрипе окрашены с интенсивно- стью «±» или выше
Сомнительный Одна антигенная полоса на стрипе окрашена с интенсивностью «1+» или выше (при получении такого ответа рекомендуют повторное исследова- ние новой порции крови с интервалом в несколько недель)
Одноэтапные иммунохроматографические тесты (ИХТ)
Эти тест-системы предназначены для диагностики сифилиса на основании
исследования цельной капиллярной крови. Они недороги, выпускаются в виде
нитроцеллюлозных тест-полосок и позволяют получить результат в течение 5 мин
(его устанавливают по появлению на полоске цветовой полосы) и пригодны для
применения в месте первичного обследования пациента. Их рассматривают как
прорыв в диагностике, потому что они позволяют установить этиологию болезни
и назначить лечение во время первого визита пациента к врачу. К сожалению,
производимые в настоящее время одноэтапные ИХТ предназначены только для
серологической диагностики сифилиса, а это значит, что их положительный
результат свидетельствует о том. что человек в течение жизни болел сифилисом и
необязательно болен в момент исследования.
Реакция иммобилизации бледных трепонем
РИБТ является одним из классических тестов для определения специфических
трепонемных антител. Суспензию живых трепонем смешивают с тестируемой
сывороткой. При наличии в последней специфических антител трепонемы утра-
чивают подвижность. Однако РИБТ — достаточно сложный, трудоемкий и
дорогостоящий анализ, требующий высокой квалификации персонала и наличия
вивария. Тест обладает низкой чувствительностью, особенно на ранних стадиях
сифилиса. К тому же РИБТ нельзя проводить при наличии в крови больного тре-
понемоцидных антибиотиков.
В связи с этим не рекомендуют применение данного метода при рутинной диа-
гностике сифилиса.
Основные причины ошибочных показаний серологических тестов
А. Ложноположительные результаты
Основные причины ложноположительных результатов трепонемных тестов.
• Технические погрешности, допущенные при сборе, доставке, хранении проб
сыворотки крови, а также ыри выпо мнении тестов.
• Эндемичные трепонематозы, такие как эндемичный сифилис (вызываемый
Г. palladium var. endemicum), фрамбезия (Г. реПепие) и пинта (Г. carateum).
Пациентов с положительными серологическими реакциями на сифилис из
стран с эндемичными трепонематозами необходимо обследовать на сифилис
и получить противосифилитическое лечение в качестве профилактической
меры, если их раньше не подвергали такому лечению.
В трепонемных тестах биологические ложноположительные реакции могут про-
являться при аутоиммунных заболеваниях. ВИЧ-инфекции и во время беременно-
сти (РИФ-абс. чаще, чем РПГА/микроРПГА). Ложноположительные результаты
ИФА и РПГА могут проявляться у пациентов с инфекционным мононуклеозом
(особенно на фоне высокого уровня гетерофильных антител). Существует опреде-
ленная взаимосвязь между ложноположительными реакциями в РИФ-абс и под-
НЕКОТОРЫЕ ИНФЕКЦИОННЫЕ ЗАБОЛЕВАНИЯ 515
твержденным диагнозом системной, дискоидной и лекарственной красной волчан-
ки, В этих случаях иногда наблюдают атипичную («бисерную») флюоресценцию.
Для устранения этих типов ложноположительных реакций проводят абсорбцию
сыворотки на тимусной ДНК теленка для удаления анти-ДНК-антител.
Ложноположительные результаты РИФ-абс. могут возникать из-за ошибки в
выборе сорбента, используемого в данном тесте для удаления перекрестно реаги-
рующих антител к групповым, родовым или семейственным антигенам: абсорбция
сыворотки рейтеровской трепонемой необходима для снижения количества лож-
ноположительных результатов.
Б. Ложноотрицательные результаты
Основные причины ложноотрицательных серологических реакций при диагно-
стике сифилиса следующие.
• Эффект «прозоны» — нетрепонемные агглютинационные тесты при вто-
ричном сифилисе могут давать слабоположительную или отрицательную
реакцию, обусловленную избытком антител, когда нормальная реакция
«антиген-антитело» не завершается либо блокируется.
• Отрицательный результат серологических реакций на сифилис регистрируют
у ВИЧ-инфицированных пациентов или пациентов с признаками иммуноде-
фицита другого происхождения.
• Эффект диагностического «окна» обусловлен отрицательными результатами
серологических реакций на сифилис, наблюдаемыми в период первичного
сифилиса, и запоздалой выработкой антител.
Исследование СМЖ на наличие антител к Т. pallidum
Для обнаружения специфических антител в СМЖ пациентов, подозреваемых в
заражении Т. pallidum, используют РМП, РИФ-ц и ИФА. Тесты проводят с цельным
пиквором. РМП проявляет высокую специфичность, но при некоторых вариантах
нейросифилиса она недостаточно чувствительна.
Показания к применению серологических реакций
Скрининговое обследование населения на сифилис рекомендуют проводить с
помощью РМП, ИФА или РПГА в зависимости от местной эпидемической ситуа-
ции и возможностей местной лабораторной службы. При этом в популяциях с
низким уровнем инфекции (соматических стационарах, поликлиниках, кабинетах
медицинских осмотров) обследование предпочтительнее проводить с помощью
трепонемного теста, так как при этом можно сразу выделить группу больных или
пациентов, перенесших в прошлом сифилис. Результаты скрининга подтвержда-
ются данными нетрепонемных тестов.
Скрининговое обследование на сифилис группы лиц с высоким уровнем пре-
валентности инфекции в популяции (работников коммерческого секса, заключен-
ных) целесообразно начинать с нетрепонемного теста. В каждом случае положи-
тельного ответа для подтверждения необходимо проводить трепонемные тесты.
Ввиду возможности выявления поздних форм сифилиса среди больных в
офтальмологических, психоневрологических, кардиологических стационарах
необходимо использовать РМП в сочетании с ИФА, РПГА, РИФ (любая из реак-
ций на выбор) в зависимости от возможностей клинико-диагностических лабора-
торий лечебно-профилактических учреждений.
В связи с высоким уровнем заболеваемости сифилисом в Российской Федерации
беременных следует подвергать профилактическому обследованию на сифилис
трижды: при постановке на учет, на 18-20-й и 32-й неделе беременности.
Лиц с клиническими проявлениями первичного сифилиса надо обследовать в РМП
(качественный и количественный вариант), РИФ или, при наличии такой возможно-
сти, в ИФА-IgM (если тест-система зарегистрирована) либо 19S-IgM-FTA-abs.
516 НЕКОТОРЫЕ ИНФЕКЦИОННЫЕ ЗАБОЛЕВАНИЯ
Лица с клиническими проявлениями, подозрительными на наличие вторичного
сифилиса, должны быть обследованы с помощью РМП (качественный и количе-
ственный вариант) в сочетании с РПГА, ИФА или РИФ (любым из трепонемных
тестов на выбор).
Лиц с клиническими проявлениями, подозрительными на наличие позднего
сифилиса (третичного, висцерального, в особенности с поражением сердечно-
сосудистой системы, опорно-двигательного аппарата и т д.), обследуют с помощью
нетрепонемного теста, а также двух трепонемных тестов: ИФА или РИФ, а также
РПГА или РИБТ.
Лиц без клинических проявлений заболевания, бывших в половом или тесном
бытовом контакте с больными заразными фирмами сифилиса, надо обследовать
как больных с подозрением на первичный сифилис.
Лиц без клинических проявлений заболевания с подозрением на скрытый сифи-
лис необходимо обследовать с помощью нетрепонемного, а также двух трепонем-
ных тестов. При этом возможны следующие сочетания: ИФА или РИФ + РИБТ:
ИФА или РИФ + РПГА.
Новорожденных от матерей, перенесших сифилис и не получивших адекват-
ную терапию либо проходивших лечение от сифилиса во время беременности,
обследуют с помощью нетрепонемного теста, а также тестов на выявление ранней
инфекции — РИФ, 19S-IgM-FTA-abs или ИФА-IgM.
Новорожденных от матерей, перенесших сифилис и прошедших полный курс
лечения, обследуют с помощью РМП (или RPR). При положительном результате
дополнительно применяют ИФА, IgG-ИФА, РИФ, 19S-IgM-FTA-abs или ИФА-
IgM.
Детей при подозрении на поздний врожденный сифилис обследуют с помощью
РМП и двух трепонемных тестов: ИФА, РИФ + РПГА или РИБТ.
Алгоритмы и критерии диагностики сифилиса. Достоверным диагностиче-
ским лабораторным критерием сифилиса является обнаружение патогенной блед-
ной трепонемы с помощью прямой микроскопии, ПИФ или ПЦР при клинических
симптомах ранней стадии сифилиса. Серологические методы при клинических
симптомах сифилиса также достаточно достоверно подтверждают диагноз.
С осторожностью следует относиться к результатам серологического обследова-
ния пациента, если клинические симптомы заболевания отсутствуют. Предлагают
следующий алгоритм обследования пациента, у которого диагноз сифилиса может
быть установлен впервые (на основании жалоб и объективных симптомов, сви-
детельствующих о возможном сифилисе, либо у лица, обследуемого по контакту
с больным сифилисом). Если у пациента есть клинические проявления ранних
стадий заболевания, проводят исследования на бледную трепонему в темном поле,
ПИФ или ПЦР. При положительном результате диагностируют сифилис, при
отрицательном — исследования проводят повторно. Пациента обследуют серо-
логически по одной из предложенных выше схем. Если при этом скрининговый
метод (РМП или трепонемный тест) дает положительный результат, проводят под-
тверждающий трепонемный или нетрепонемный серологический тест и пациента
направляют к дерматовенерологу.
Если у пациента скрытая форма сифилиса, диагноз будет, скорее всего, установ-
лен на основании результатов серологического исследования. В таком случае на
первом этапе необходимо провести количественный нетрепонемный тест, после
чего пациента направляют к дерматовенерологу для обследования, лечения и
наблюдения.
Если у пациента выявлен скрытый сифилис при серологическом скрининге с
использованием трепонемного или нстрепонемного теста, применяют количе-
ственный нетрепонемный тест (или соотвтетственно трепонемный). Результаты
анализа (титр) фиксируют в карте больного и назначают ему лечение. Повторные
НЕКОТОРЫЕ ИНФЕКЦИОННЫЕ ЗАБОЛЕВАНИЯ 51
количественные серологические тесты проводят на 3, 6, 9 и 12-м месяце после
лечения.
У пациентов, получивших лечение по поводу ранних стадий сифилиса (первич-
ного, вторичного или раннего скрытого), регистрируют положительные реакции
при обследовании в трепонемных тестах и отрицательные — в нетрепонемн ых (в
случае успешного результата лечения).
Рецидив заболевания может проявляться появлением сыпи на коже или слизи-
стых оболочках на фоне возрастания титров нетрепонемных тестов.
В случае реинфекции отмечают значительное (четырехкратное и большее)
повышение титров нетрепонемных тестов. Повторное обследование с использова-
нием трепонемных тестов не имеет диагностического значения, так как они будут
положительными после первой инфекции.
При подозрении на ранний врожденный сифилис у новорожденного обследу-
ют мать и ребенка. Критерием подтвержденного раннего врожденного сифилиса
является обнаружение бледной трепонемы с помощью темнопольной микроско-
пии, ПИФ или ПЦР в пробах, взятых у ребенка (кожные поражения, слизистая
оболочка носа, пупок), а также из плаценты.
При отрицательных результатах темнопольной микроскопии о предполагаемой
врожденной инфекции свидетельствуют положительные результаты трепонемных
тестов на сифилис в крови ребенка (19S-IgM-FTA-abs, IgM-ИФА, IgM-РИФ) и зна-
чительное (в четыре и более раз) превышение количества антител, выявляемых в
нетрепонемных тестах у ребенка в сравнении с матерью.
После окончания лечения больного по поводу сифилиса проводят повторные
серологические исследования (обычно через 3, 6, 9 и 12 мес) в количественном
варианте РМП. Критерием успешно проведенного лечения является наступаю-
щая в течение этого времени «негативация» нетрепонемного теста или снижение
титров антител не менее чем в 4 раза в сравнении с исходными. Необходимо пом-
нить, что у пациентов с поздними стадиями сифилиса после проведенного лечения
высокие титры антител могут сохраняться и после 12 мес. Дополнительное лече-
ние назначают только в случае выявления нетрепонемными тестами сероконвер-
сии в этот период.
По окончании лечения сифилиса пациент должен быть обследован с помощью
количественного нетрепонемного теста. При этом показателем эффективности
терапии считают снижение титра в 4 и более раз в течение 1 года наблюдения. Для
оценки эффективности лечения применяют тест-систему одного и того же произ-
водителя.
Интерпретация результатов серологических тестов на сифилис приведена в
табл. 22-10.
СПИСОК РЕКОМЕНДУЕМОЙ ЛИТЕРАТУРЫ
Дмитриев Г.А. Сифилис. Дифференциальный клинико-лабораторный диагноз. — М.:
Медицинская книга. 2004. — 363 с.
Department of Health and Human Sendees. Centers for Disease Control and Prevention.
Sexually transmitted diseases treatment guidelines // MMWR. - 2006. - Vol. 55. - 94 p.
Domeika M. Laboratory7 Diagnostics for Non-Viral Sexually Transmitted Infections in St.
Petersburg, Russia: Current Situation and Hallmarks for Improvements //J. Eur. Acad. Dermatol.
Venereol. (EADV). - 2008. - Vol. 22. - P. 1094-1100.
Goh B.T., van Voorst Vader P.C. European guideline for the management of syphilis // Int. J.
STD AIDS. - 2001. - Vol. 12. - Suppl. 3. - P. 14-22.
Herring A., Ballard R., Mabey D., Peeling R.W. WHO/TDR Sexually Transmitted Diseases
Diagnostics Initiative. Evaluation of rapid diagnostic tests: syphilis // Nat. Rev. Microbiol. -
2006. - Vol. 4. - N 2. - Suppl. - P. 33-40.
Larsen S.A.. Pope V.. Johnson R.E., Kennedy EJ. Jr. A Manual of Tests for Syphilis. -
Washington: American Health Association, 1998.
' ГЛАВА 22
Таблица 22-10. Интерпретация результатов серологических тестов на сифилис
Клинические проявления сифилиса Документы о проведе- нии лечения ИФА (суммарные анти- тела - IgM + IgG) IqM-ИФА iqG- ИФА РПГА РМП (RPR) Интерпретация результатов
1 + - + + - - - Первичный сифилис
2 - - + + - - - Инкубационный период или ЛПР
3 + - + +/- + + + Сифилис (стадия в соответствии с клинической картиной)
4 - - + +/- + + + Скрытый сифилис
5 - + + +/- + + + Состояние после приведенного лечения. Серорезистентность?
6 - + + - + + - Состояние после проведенного лечения Дальнейшему лечению и наблюдению не подлежит
7 - - + - + + - Скрытый сифилис, возможно, поздний или состояние после пеленесенной в прошлом инфекции
8 - - + + + + - Скрытый сифилис
9 - + + + + + - Реинфекция или состояние вскоре после окончания лечения, или ЛПР на IgM
10 + + + +/- + + +/- Реинфекция или клинический рецидив
11 - + + - +/- +/- - Состояние после проведенного лечения Дальнейшему лечению и наблюдению не подлежит
12 - - + - +/- +/- - Состояние после проведенного лечения или ЛПР
13 - - - - - - + ЛПР
14 - - + + - - + Инкубационный период или ЛПР
НЕКОТОРЫЕ ИНФЕКЦИОННЫЕ ЗАБОЛЕВАНИЯ
НЕКОТОРЫЕ ИНФЕКЦИОННЫЕ ЗАБОЛЕВАНИЯ 519
Lewis D.A., Young Н. Syphilis // Sex. Transin. Infect. — 2006. — Vol. 82. — Suppl. IV. —
P. 13-15.
Palmer H.M., Higgins S.P, Herring A.J., Kingston M.A. Use of PCR in the diagnosis of early
syphilis in the United Kingdom // Sex. Transm. Infect. — 2003. — Vol. 79. — P. 479-483.
Mabey D., Peeling R.W., Ballard R. et al. Prospective, multi-centre clinic-based evaluation of
four rapid diagnostic tests for syphilis // Sex. Transm. Infect. — 2006. — Vol. 82. - Suppl. 5. —
P. 13-16.
Ratnam S. The laboratory diagnosis of syphilis // Can. J. Infect. Dis. Med. Microbiol. — 2005. —
Vol. 16. - N 1 - P. 45-51.
ГОНОКОКК
ТАКСОНОМИЯ
Гонококк (Neisseria gonorrhoeae) — бактерия из рода Neisseria, относящегося к
семейству Neisseriaceae.
ОБЩАЯ ХАРАКТЕРИСТИКА
N. gonorrhoeae — грамотрицательный аэробный диплококк, имеет форму кофей-
ных зерен размером 1,25-1.6x0,7-0,8 мкм, обращенных вогнутой поверхностью
друг к другу. Образует капсулу, проявляющую иммуногенные и антифагоцитарные
свойства.
N. gonorrhoeae продуцирует оксидазу и каталазу, в отличие от других представи-
телей рода Neisseria ферментирует глюкозу и не ферментирует мальтозу, сахарозу
и лактозу. Ферментация глюкозы происходит с образованием кислоты без газа.
Измененные бактерии, в том числе L-сЬормы, не ферментируют углеводы, не про-
являют протеолитическую активность, не продуцируют аммиак, сероводород и
индол, не вызывают гемолиз на средах, содержащих кровь.
N. gononhoeae малоустойчива к факторам внешней среды: гибнет при высушива-
нии, в мыльной воде, под действием антисептиков и антибактериальных препара-
тов, при нагревании выше 56 °C, под влиянием прямых солнечных лучей.
ЭПИДЕМИОЛОГИЯ
Гонорея — антропонозная инфекция. Вне организма человека возбудитель
быстро погибает. Основной путь заражения — половой. Различают генитальную
гонорею (поражение мочеполовых органов), экстрагенитальную (поражение пря-
мой кишки, глотки, конъюнктивы) и метастатическую (диссеминированную) —
осложнение первых двух вариантов. Гонококковая инфекция может протекать без
осложнений или с осложнениями (табл. 22-11).
Таблица 22-11. Основные клинические проявления неосложненной гонококковой инфекции
Пациенты Клинические проявления
Женщины Цервицит, уретрит, бартолинит
Мужчины Уретрит, баланопостит
Мужчины и женщины Фарингит, конъюнктивит, проктит
Дети Уретрит, вагинит, конъюнктивит, арингит, проктит
Новорожденные и младенцы Офтальмия новорожденных, вульвовагинит
Заражение от полового партнера с экстрагенитальной локализацией инфекции
при генитально-оральных контактах приводит к гонорейному тонзиллиту (тон-
зиллофарингиту), при генитально-анальных контактах — к гонорейному прокти-
ту. Внеполовое заражение возможно при прямом контакте:
❖ при попадании или затекании отделяемого в глаза, на слизистую оболочку
полости рта или прямой кишки;
при прохождении новорожденного через родовые пути матери.
520 НЕКОТОРЫЕ ИНФЕКЦИОННЫЕ ЗАБОЛЕВАНИЯ
У детей описаны случаи гонококкового менингита, которому7 нередко предшеству-
ет конъюнктивит, а также сепсис и артрит новорожденных. В данных случаях зараже-
ние могло произойти гематогенным путем или через инфицированные околоплодные
воды. Возможен интранатальный путь заражения. Непрямое внеполовое заражение
встречается редко при очень тесном бытовом контакте маленького ребенка с больной
матерью (девочек — через общие с матерью предметы личной гигиены).
После выздоровления при новом контакте с источником инфекции возможно
повторное заражение — реинфекция. Помимо реинфекции, может возникать супе-
ринфекция (обычно при изолированном хроническом очаге).
КЛИНИЧЕСКИЕ ПРОЯВЛЕНИЯ ИНФЕКЦИИ
Инкубационный период составляет 1-14 дней. Затем во многих случаях гоно-
кокковой инфекции возникает развернутая клиническая картина гонорейной
инфекции, основное проявление которой — гнойные выделения. N. gonorrhoeae,
распространяясь по слизистой оболочке (восходящий путь), лимфо- или гемато-
генно, может вызывать поражение органов малого таза, брюшной полости вплоть
до развития гонококкового сепсиса с септицемией и септикопиемией.
В соответствии с МКБ-10 гонококковая инфекция встречается в следующих
формах:
- > А54.0 — гонококковая инфекция нижних отделов мочеполового тракта без
абсцедирования парауретральных и придаточных желез (например, уретрит,
цистит, вульвовагинит, цервицит);
❖ А54.1 — гонококковая инфекция нижних отделов мочеполового тракта с
абсцедированием парауретральных и придаточных желез (например, гоно-
кокковый абсцесс больших вестибулярных желез);
о А54.2 — гонококковый пельвиоперитонит и другая гонококковая инфекция
мочеполовых органов (например, эпидидимит, орхит, простатит, воспали-
тельные заболевания органов малого таза у женщин);
❖ А54.3 — гонококковая инфекция глаз (например, конъюнктивит, иридоци-
клит, гонококковая офтальмия новорожденных);
А54.4 — гонококковая инфекция костно-мышечной системы (например,
артрит, бурсит, остеомиелит, синовит, теносиновит);
❖ А54.5 — гонококковый фарингит;
о А54.6 — гонококковая инфекция аноректальной области;
А54.8 — другие клинические проявления гонококковой инфекции (абсцесс
мозга, эндокардит, менингит, миокардит, перикардит, перитонит, пневмония,
сепсис, поражение кожи).
Осложнения гонококковой инфекции многообразны — эпидидимит, аднексит,
артрит и др.
В ряде слуачев инфекция N. gonorrhoeae протекает бессимптомно, при этом воз-
будитель размножается только на поверхности слизистых оболочек, не внедряясь
в ткани и не вызывая воспаления.
В табл. 22-12 представлены показания к обследованию на гонококковую инфек-
цию.
Таблица 22-12. Лица, подлежащие обследованию на гонококковую инфекцию
Пациенты Показания к обследованию
Мужчины Жалобы на гнойные или слизисто-гнойные выделения из уретры, зуд уретры, сим- птомы дизурии. Боли в области придатка яи^ка или яичка. Боли и выделения из прямой кишки; признаки проктита. Воспалительные изменения в области наружного отверстия уоетоы, парауретраль- ных ходов. Признаки воспаления предстательной железы Признаки конъюнктивита
НЕКОТОРЫЕ ИНФЕКЦИОННЫЕ ЗАБОЛЕВАНИЯ 521
Окончание табл. 22-12
Пациенты Показания к обследованию
Женщины Воспалительные заболевания мочеполовой сферы, гнойные или слизисто-гнойные выделения из влагалища, проявления аднексита, проктита, вульвовагинита, церви- цита. Жалобы на появившиеся субъективные расстройства в области половых органов (зуд, жжение при мочеиспускании, боли в низу живота, усиление белей, кровяни- стые выделения). Признаки конъюнктивита. Эрозии шейки матки. Бесплодие, привычные выкидыши, преждевоеменные роды в анамнезе. Направляемые на прерывание беременности. Беременных обследуют трижды: ❖ 1-е обследование — при постановке на учет О 2-е обследование — при сроке 27-30 нед. 3-е обследование — при сроке 36-40 нед. Вне указанных сроков обследование беременных пооводят по показаниям (появле- ние выделений из половых путей, субъективные жалобы). В гинекологических стационарах обследуют всех женщин, не обследованных до госпитализации, перед назначением антибактериального лечения. В родильных домах обследуют всех рожениц без обменных карт, родильниц с осложненным течением послеродового периода, лучше на 5-6-й день после родов
Новорожденные С гнойным конъюнктивитом или вульвовагинитом. При подтверждении гонококковой этиопогии конъюнктивита или вульвовагинита необходимо обследование родителей
Дети(девочки) С симптомами вупьвовагини'1а, вагинита
Пациенты Вступавшие в половой контакт с больным гонореей. Проходящие обследование на другие ИППП. С диагностированным трихомонозом до и после лечения. Декретированных профессий при проведении обязательных предварительных (при поступлении на работу) и периодических медицинских осмотров в соответствии с утвержденными регламентирующими документами. Подвергшиеся сексуальному насилию
Группы риска, в которых целесообразно применение скрининга с использовани-
ем ДНК-/РНК-методов, включают:
< > лиц, занимающихся коммерческим сексом;
❖ мужчин-гомосексуалистов;
❖ беременных;
ъ лиц моложе 25 лет, имеющих многочисленных половых партнеров.
ЛАБОРАТОРНАЯ ДИАГНОСТИКА
Для лабораторной диагностики гонококковой инфекции применяют следующие
методы (табл. 22-13):
❖ микроскопический (бактериоскопия);
❖ культуральный (бактериологический);
о молекулярно-генетический.
Таблица 22-13. Рекомендуемые методы диагностики гонококковой инфекции
Область взятия материала Методы диагностики Комментарии
Отделяемое уретры (муж- чины), отделяемое эндоцервикса или уретры (женщины) Бактериоскопический с окраской по Граму Выявление грамотрицательных диплококков, рас- положенных вне и внутри лейкоцитов, оценка тяже- сти воспаления. Не может быть использован в качестве единственно- го метода диагностики
522 НЕКОТОРЫЕ ИНФЕКЦИОННЫЕ ЗАБОЛЕВАНИЯ
Окончание табл. 22-13
Область взятия материала Методы диагностики Комментарии
Культуральный (бакте- риологический) Для выделения и идентификации /V. gonorrhoeae и оценки чувствительности к антимикробным пре- паратам. Выделение N. gonorrhoeae при культуральном иссле- довании — доказательство гонококковой инфекции
Молекулярно- генетические (ПЦР) Метод скрининга. При выявлении ДНК/РНК N. gonorrhoeae требуется подтверждение культураль- ным или альтернативным методом амплификации нуклеиновых кислот
Молекулярно- генетические (NASBA) Метод используют при несовпадении результатов обследования некультуральными методами (микро- скопией, ПЦР)
Отделяемое ротоглотки, отделяемое конъюнктивы, отделяемое из прямой кишки Бактериологический Для выделения и идентификации /V. gonorrhoeae и оценки чувствительности к антимикробным пре- паратам. Основной метод диагностики
Молекулярно- генетические (ПЦР) Метод скрининга — при выявлении ДНК/РНК А/, gonorrhoeae требуется подтверждение культураль- ным или альтернативным методом амплификации нуклеиновых кислот
Моча (мужчины и жен- щины) Молекулярно- генетические (ПЦР, NASBA) Метод скрининга — при выявлении ДНК/РНК А/, gonorrhoeae требуется исследование клиниче- ского материала, полученного у мужчин из уретоы, у женщин — из уретры и цервикального канала, исследование клинического материала из других анатомических областей
Вульва/вагина (девочки), вагина (женщины после экстирпации матки) Культуральный Для выделения и идентификации /V. gonorrhoeae и оценки чувствительности к антимикробным пре- паратам. Основной метод диагностики. Бакгериоскопический метод с окраской по Граму не применяют
Кровь при диссемини- рованной гонококковой инфекции Культуральный Культуральное исследование крови и клинического материала, полученного из уро- и экстрагенитальных анатомических областей
Молекулярно-гене- тические (ПЦР, NASBA) При выявлении ДНК/РНК N gonorrhoeae требуется подтверждение культуральным методом
Взятие материала для исследований
В табл. 22-14 представлена информация о клиническом материале, направляе-
мом в лабораторию для исследования на гонококковую инфекцию.
Таблица 22-14. Взятие материала для диагностики гонококковой инфекции
Техника взятия материала Для взятия материала используют хлопковые или дакроновые зонды-тампоны, специ- альные щеточки, ложечки Фолькмана. Пациент не должен мочиться в течение 3 ч до взятия материала Места взятия образцов зависят от пола, возраста и сексуальной практики пациента, а также от клинических пооявлений инфекции. У женщин мате- риал берут из цервикального канала, уретры, реже из влагалища, прямой кишки и ротоглотки У мужчин материал берут из уретры, прямой кишки, ротоглотки. Внимание! Посев на специальную питательную среду для выделения N. gonorrhoeae следует проводить в момент взятия материала
Цервикальный канал Перед взятием образцов следует избегать применения антисептиков, анальгетиков и мазей, так как они могут подавить N. gonorrhoeae, находящиеся в цервикальном канале. Материал берут при исследовании в зеркалах. Наружный зев очищают от слизи с помощью ватного шарика, инструмент вводят на глубину 1-2 см в церви- кальный канал и берут соскоб
НЕКОТОРЫЕ ИНФЕКЦИОННЫЕ ЗАБОЛЕВАНИЯ 523
Окончание табл. 22 14
Уретра Область наружного отверстия мочеиспускательного канала очищают с помощью марлевого тампона или шарика. У мужчин при отсутствии явных выделений слегка массируют уретру в направлении к наружному отверстию, чтобы получить экссудат. Если экссудат не получен, вводят уретральный зонд-тампон на глубину 1-2 см. осторожно вращая, и оставляют зонд-тампон в уретре в течение 5-10 с. У женщин при отсутствии явных выделений массируют уретру, прижимая ее к лонному сочле- нению и используя ту же технику, что и для мужчин
Прямая кишка Вводят ватный тампон на глубину 3 см в прямую кишку и вращают его в течение 10 с, чтобы получить экссудат из крипт прямо внутри анального кольца. Если про- исходит фекальное загрязнение, тампон следует выбросить и взять другой для получения пробы
Конъюнктива Проводят тампоном по внутренней поверхности конъюнктивы нижнего века
Влагалище Вагинальные пробы следует брать у женщин, перенесших экстирпацию матки, и у девочек (w'rgo). У девочек материал берут тампоном из преддверия влагалища. У женщин материал берут при осмотре в зеркалах с передней или боковой стенки
Ротоглотка Материал берут в утреннее время, пациенту не разрешают чистить зубы и прини- мать пищу до взятия материала. Материал берут зондом-тампоном из тонзилляр- ных крипт и задней стенки глотки
Предстательная железа Пациент должен помочиться перед взятием материала. Проводят ректальный мас- саж предстательной железы от периферии к центру. Собирают на тампон экссудат из отверстия уретры
Транспортировка Образцы следует направлять в лабораторию как можно быстрее (непосредственно после посева). Материал необходимо транспортировать в лабораторию при темпе- ратуре не ниже 18 °C. При нарушении правил транспортировки вероятность выделе- ния N. gonorrhoeae резко снижается
Применение любрикантов, спермицидных препаратов, спринцеваний и гор-
мональных контрацептивов может отрицательно влиять на жизнеспособность
N. gonorrhoeae, а следовательно, и на результаты бактериологического исследова-
ния проб, которые берут у пациентов, подозреваемых в заражении этой бактерией.
Тампоны для взятия материала могут обладать антибактериальными свойствами.
Вата, альгинат кальция, ненасыщенные жирные кислоты, находящиеся в ватных
волокнах, хлорсодержащие отбеливатели, смола, входящая в состав деревянных
палочек, клей, используемый для соединения тампонов с крышками, также ток-
сичны для культуры N. gonorrhoeae, поэтому материал предпочтительнее брать
вискозными зондами-тампонами.
Поскольку N. gonorrhoeae чувствительна к воздействию факторов внешней среды
(высушиванию, высокой температуре, кислороду), рекомендуют сразу проводить
посев материала, полученного от пациентов. Если это невозможно, материал
доставляют в лабораторию в транспортной среде (Стюарта, Эймса, Кэри-Блеера).
Взятые пробы в кратчайшие сроки доставляют в лабораторию и инокулируют в
питательные среды; посев должен быть проведен не позднее 24 ч (в исключитель-
ных случаях — 48 ч) после сбора проб. Транспортировку материала осуществляют
при температуре не ниже 18 °C.
Бактериоскопия
При использовании микроскопического метода у женщин и детей устанавли-
вают предварительный диагноз, у мужчин при исследовании проб из уретры —
окончательный.
Преимущества бактериоскопического метода:
быстрота получения рез льтата, простые условия транспортировки;
❖ несложность выполнения, низкая стоимость:
о высокие чувствительность и специфичность для мазков из уретры мужчин
(референс-метод).
524 НЕКОТОРЫЕ ИНФЕКЦИОННЫЕ ЗАБОЛЕВАНИЯ
Недостат ки бактериоскопического метода:
субъективность оценки результата (зависит от опыта специалиста, проводя-
щего микроскопическое исследование, качества взятия материала, окраски,
используемого микроскопа):
н изкая чувствительность для цервикальных проб (фарингеальные и ректаль-
ные пробы не подходят для микроскопического исследования);
< невозможность использования для решения юридических вопросов;
низкая чувствительность при ранней диагностике, бессимптомном тече-
нии инфекции, а также при выявлении некоторых аукситипов и вариантов
N. gonorrhoeae’,
Если препарат доставляют в лабораторию нефиксированным, перед окрашива-
нием его фиксируют 96% этиловым спиртом или троекратным проведением стекла
через пламя горелки.
Фиксированные препараты можно хранить при комнатной температуре в тече-
ние нескольких дней.
Препараты окрашивают метиленовым синим и по Граму. Нельзя выносить
заключение по результатам просмотра мазков, окрашенных только одним из этих
методов. Окраска метиленовым синим позволяет сделать заключение о воспале-
нии и морфологии бактерий.
При окраске по Граму возможно выявление грамотрицательных диплококков.
При микроскопии препаратов оценивают наличие эпителия, количество лейко-
цитов, эритроцитов, морфологию бактерий (лактобацилл, кокков, коккэбацилл),
наличие вне- и внутриклеточно расположенных диплококков.
Мазки, окрашенные метиленовым синим или по Граму, оценивают при увели-
чениях в 100 и 1000 раз. При малом увеличении определяют адекватность взя-
тия из соответствующего анатомического участка, наличие в тестируемой пробе
примесей, а также выбирают участок препарата для дальнейшего исследования.
Микроскопия при большом увеличении позволяет выявить и оценить воспали-
тельную реакцию и наличие микроорганизмов. При микроскопировании с иммер-
сией при увеличении в 1000 раз подсчет лейкоцитов следует проводить в 5 полях
зрения. При этом особое внимание уделяют поиску внутриклеточных диплокок-
ков в лейкоцитах.
Диагноз уретрита у мужчин ставят на основании обнаружения 4 лейкоцитов
и более в поле зрения микроскопа при увеличении в 1000 раз. При цервиците
количество полиморфноядерных лейкоцитов повышено (более 10 при увеличе-
нии в 1000 раз). При клиническом исследовании следует принимать во внимание
наличие или отсутствие слизисто-гнойных цервикальных выделений (зеленовато-
желтого гноя со слизью на белом тампоне).
При исследовании вагинального мазка у женщин необходимо помнить, что
количество лейкоцитов зависит от индивидуальных особенностей организма,
срока менструального цикла, наличия внутриматочной спирали. Именно поэтому
для диагностики слизисто-гнойного цервицита рекомендуют использовать оба
критерия, то есть наличие клинических проявлений и воспалительный характер
цервикального мазка.
Диагноз уретрита у женщин подтверждается обнаружением более 10 лейкоци-
тов в поле зрения при увеличении в 1000 раз. Если во влагалище или шейке суще-
ствует патологический воспалительный процесс, при этом наблюдают обильные
выделения из влагалища, уретральный мазок всегда загрязнен материалом этих
выделений (клетками плоского эпителия, лейкоцитами, вагинальной микрофло-
рой) и не годится для дальнейшей оценки, поскольку не имеет ничего общего с
содержимым уретры.
Диагноз гонореи на основании бактериоскопии ставят по 3 признакам возбуди-
теля болезни:
НЕКОТОРЫЕ ИНФЕКЦИОННЫЕ ЗАБОЛЕВАНИЯ 525
< форме бактериальных клеток;
о их расположению;
❖ окраске.
Только наличие всех трех признаков позволяет поставить диагноз (табл. 22-15).
Если же отсутствует хотя бы один из них. переходят к культуральному исследова-
нию.
Я. gonorrhoeae — диплококк, имеющий форму кофейного зерна и расположенный
попарно вогнутыми сторонами друг к другу. Решающее значение при микроскопи-
ческой диагностике гонореи имеет расположение диплококков. N. gonorrhoeae в
основном находятся вну гри лейкоцитов и эпителиальных клеток.
На основании обнаружения диплококков, расположенных вне клеток, микро-
скопический диагноз гонореи не может быть поставлен.
Я. gonorrhoeae окрашивается по Граму в красно-розовый цвет (грамотрицатель-
ный диплококк), при этом ядра лейкоцитов и эпителиальных клеток окрашивают-
ся в фиолетовый цвет.
Таблица 22-15. Клиническая интерпретация результатов микроскопического исследования
Пол Источник исследуемого материала Микробы и «ключевые» клетки Лейкоциты в поле зрения микроскопа при увели те- нии в 1000 раз Диагностическая гипотеза
Мужской Уретра Не обнаружены 5 и более Здоров
Внутриклеточные грам- отрицательные дипло- кокки 5 и более Гонококковый уретрит
Другие микроорганизмы 5 и более Негонококковый уретрит
Женский Влагалище Лактобациллы Количество зависит от фазы менструального цикла Л:Э = 1:1 Здорова
«Ключевые» клетки Л;Э <1:1 Бактериальный вагиноз
Candida spp. Л:Э >1:1 Кандидозный вагинит
Т. vaginalis Л:Э >1:1 Тоихомонадный вагинит
Другие микроорганизмы Л:Э>1:1 Неспецифический вагинит
Женский Цервикальный канал Внутриклеточные грамо- трицательные диплокок- ки не обнаружены <10 Здорова
Внутриклеточные грамо- трицательные диплокок- ки обнаружены ID и более Гонококковый цер- вицит? Необходимо дальней- шее обследование
Другие микроорганизмы >10 Церлицит? Необходимо уточнение этиологии
Женский Уретра Внутриклеточные грамо- трицательные диплокок- ки не обнаружены <5 Здорова
Внутриклеточные грамо- трицательные диплокок- ки обнаружены >5 Гонококковый уретрит? Необходимо дальней- шее обследование
Другие микроорганизмы >5 Уретрит? Необходимо уточнение этиологии
526 НЕКОТОРЫЕ ИНФЕКЦИОННЫЕ ЗАБОЛЕВАНИЯ
Изоляция и идентификация возбудителя
Бактериологическое исследование на гонорею проводят по упомянутым выше
показаниям бактериоскопии и в следующих случаях:
- о- у детей, так как у них встречается большое количество непатогенных нейссе-
рий, особенно на слизистой оболочке полости рта, глотки и гениталий;
❖ у женщин, так как диагностическая чувствительность микроскопии гениталь-
ных мазков у данной категории пациентов низкая:
у больных с бессимптомной и экстрагенитальной гонореей (глотки, прямой
кишки, гонорейным конъюнктивитом);
❖ у половых партнеров пациентов с доказанной гонореей (если при микроско-
пии не удалось выявить возбудителя);
п ациентов с гонореей после окончания лечения (не ранее 10 дней) и при сня-
тии с учета;
❖ с целью окончательной идентификации нейссерий;
❖ для определения чувствительности нейссерий к антибиотикам;
- о- при фенотипической или генотипической идентификации N. gonorrhoeae;
< > в случае сексуального насилия при запросе следственных органов или
судебно-медицинских экспертов.
Ответ по результатам посева при выделении чистой культуры и ее идентифика-
ции выдается через 5 дней после взятия пробы.
Принцип метода заключается в выделении чистой культуры N. gonorrhoeae с
использованием специальных питательных сред и последующей идентификацией
возбудителя по биохимическим и морфологическим признакам.
Преимущества бактериологического метода:
о высокие чувствительность, специфичность;
❖ подходит большинство типов образцов;
❖ позволяет определять чувствительность к антимикробным препаратам.
Недостатки бактериологического метода:
❖ при нарушении технологии исследования снижаются чувствительность и
специфичность;
т ребует много времени по сравнению с методами, основанными на амплифи-
кации ДНК/РНК;
о более высокая стоимость по сравнению с микроскопией, однако последняя не
может применяться для идентификации N. gonorrhoeae и постановки оконча-
тельного диагноза гонореи (см. ниже).
Быстрый посев на питательные среды рекомендуют для сокращения экспози-
ции исследуемого материала с токсическими веществами и предотвращения его
высыхания.
TV. gonorrhoeae весьма требовательна к составу питательных сред. Она может
расти только на средах, содержащих определенные концентрации аминокислот,
пуринов и пиримидинов, а также усваиваемых бактерией источников энергии
(глюкозы, пирувата или лактата).
В качестве селективных компонентов в питательные среды (агар Тайера-
Мартина и др.)-, предназначенные для изоляции N. gonorrhoeae, добавляют антибак-
териальные препараты, подавляющие рост посторонней микрофлоры (линкоми-
цин, ванкомицин, колистин, нистатин, амфотерицин В, триметоприм). Некоторые
ингредиенты питательных сред (крахмал, уголь, кровь, дрожжи) действуют как
адсорбирующие вещества и могут ингибировать рост N. gonorrhoeae.
Полученный биологический материал необходимо посеять одновременно на
селективную и неселективную питательные среды, так как некоторые штаммы
N. gonorrhoeae чувствительны к ванкомицину и триметоприму, входящим в состав
селективной среды. Необходимо использовать эффективные селективные куль-
НЕКОТОРЫЕ ИНФЕКЦИОННЫЕ ЗАБОЛЕВАНИЯ 527
туральные среды, которые должны быть валидированы и сертифицированы для
культивирования N. gonorrhoeae.
Культивирование N. gonorrhoeae проводят на чашках Петри диаметром 90 мм с
количеством среды не менее 20 мл на чашку или диаметром 100 мм с количеством
среды 25 мл на чашку. Возможно также использование чашек диаметром 40-60 мм
с толщиной агара не менее 4.0-4,5 мм. Недопустимо использовать пробирки с
питательной средой для выделения и идентификации N. gonorrhoeae. После при-
готовления чаши со средой помешают в термостат при температуре 36±1 °C на 1 ч
для удаления конденсата
Клинический материал от каждого пациента следует засевать на отдельную
чашку Петри. При использовании больших чашек (диаметром 90-100 мм) мате-
риал из уретры и цервикального канала у женщин можно засевать на одну чашку в
разные ее секторы с соответствующей маркировкой (в этой ситуации удобно при-
менять многосекционные чашки Петри). Материал из уретры мужчин засевают на
отдельную чашку.
Исследуемый материал зондом-тампоном наносят на поверхность, равную при-
мерно % поверхности среды чашки или сектора. Затем стерильной бактериологи-
ческими петлей или шпателем материал распределяют по оставшейся поверхности
питательной среды штриховыми движениями в 3-4 разных направлениях в целях
создания условий для роста отдельно расположенных колоний N, gonorrhoeae.
Чашки немедленно помещают в анаэростат в атмосферу, содержащую 5±2% угле-
кислого газа и с влажностью 70%, который ставят в термостат при температуре
36±1°C.
Посевы просматривают после 18-24 ч и повторно при отсутствии роста после 48
и 72 ч инкубации. При отсутствии признаков роста через 72 ч инкубации наблю-
дение прекращают. При выявлении характерных колоний проводят первичную и
видовую идентификацию.
Алгоритм культивирования N. gonorrhoeae с использованием коммерческой
культуральной системы:
> дают агару нагреться до комнатной температуры:
❖ осторожно, стараясь не повредить слой ат ара. вынимают его из контейнера;
4- наносят клинический материал на агар, впащая щеточку или тампон по
поверхности агара;
< с помощью стерильного пинцета вынимают таблетку, генерирующую угле-
кислый газ, и помещают ее в контейнер;
❖ влажный воздух внутри контейнера инициирует процесс выделения углекис-
лого газа, необходимого для успешного роста N. gonorrhoeae;
• у при внесении агара в контейнер нельзя прикасаться к таблетке;
о контейнер маркируют;
❖ инкубируют при температуре 36±1 °C в течение 48 ч.
При обнаружении колоний с типичной для гонококков морфологией проводят
предварительную идентификацию. При необходимости проводят подтверждаю-
щую идентификацию для постановки окончательного диагноза гонореи.
Первичную идентификацию нейссерий осуществляют посредством:
- о визуальной оценки вида колоний:
❖ микроскопии подозрительных колоний, окрашенных по Граму;
< оксидазного теста.
В течение 18-24 ч инкубации N. gonorrhoeae образует выпуклые прозрачные
колонии серо-белого цвета, диаметром 0,5-1,0 мм. При дальнейшей инкубации
колонии увеличиваются в размерах (до 3,0 мм) и уплощаются. Нередко на одной
чашке встречаются колонии различной морфологии.
Большие трудности возникают при идентификации N gonorrhoeae в посе-
вах из ротоглотки, так как при этом часто наблюдается рост N. meningitidis и
528 НЕКОТОРЫЕ ИНФЕКЦИОННЫЕ ЗАБОЛЕВАНИЯ
непатогенных нейссерий, колонии которых схожи с колониями N. gonorrhoeae.
Колонии N. meningitidis — голубоватые, непатогенных нейссерий — беловатые, а
N. gonorrhoeae — бесцветные, слизистые. Размер, цвет, морфология и консистенция
колоний MOiyr варьировать в зависимости от применяемой питательной среды.
Идентифицировать возбудителя можно только при определении сахаролитиче-
ских свойств или с помощью других тестов, подтверждающих видовую принад-
лежность микроорганизма.
При рутинной диагностике обнаружение оксидазоположиз ельных грамотрица-
тельных диплококков считают достаточным для подтверждения их принадлеж-
ности к виду N. gonorrhoeae. Для определения цитохромоксидазы используют один
из рекомендуемых ниже методов.
• На подозрительную колонию наносят каплю оксидазного реагента (1%
водный раствор тетраметил-р-фенилендиамина). Быстрое изменение цвета
реагента (обычно в течение 5-10 с) на сине-фиолетовый и его сохранение
дольше 30 с позволяет считать тест положительным. Реагент, используемый
в оксидазном тесте, убивает N. gonorrhoeae, поэтому дальнейшая работа с
исследуемыми колониями невозможна.
• Полоску фильтровальной бумаги смачивают несколькими каплями реагента,
затем на нее бактериологической петлей помещают материал из подозри-
тельной колонии. Данный способ позволяет сохранить колонию для прове-
дения дальнейших исследований.
• Существуют коммерческие диски и полоски, содержащие диметил-р-
фенилендиамина гидрохлорид. Платиновой петлей или стерильной стеклян-
ной палочкой испытуемую колонию наносят на диск, предварительно слегка
смоченный дистиллированной водой. Темно-лиловое или синее окрашива-
ние, появляющееся через 10-30 с, позволяет считать тест положительным,
отсутствие изменения цвета — отрицательным.
Для учета результатов необходимо соблюдение временного интервала (10-
30 с). Высокая чувствитетьность оксидазного теста сочетается с низкой специфич-
ностью. Оксидазоположительными могут быть и другие бактерии, выделяющие
цитохромоксидазу, поэтому необходимо проводить микроскопию оксидазополо-
жительных колоний, окрашенных по Граму.
Для микроскопии берут материал из подозрительной колонии, смешивают с
небольшой каплей 0,9% раствора NaCl на предметном стекле, высушивают на воз-
духе, фиксируют в пламени и окрашивают по Граму.
При исследовании мазков из культур учитывают морфологию, расположение и
окраску N. gonorrhoeae. После 18-24 ч культивирования N. gonorrhoeae представ-
ляют собой скопления грамотрицательных кокков и диплококков, имеющих более
компактное расположение в центре и разреженное неравномерное распределение
по периферии.
Наряду с интенсивно окрашенными в розово-красный цвет кокками встречаются
и бледно окрашенные формы. Более старые культуры (48 ч и более) часто тяжело
интерпретировать, так как отмечается большое количество полностью лизирован-
ных клеток. По мере старения культуры увеличивается полиморфизм бактерий.
Для контроля качества окрашивания препарата необходимо оценить изменение
окраски в зависимости от толщины мазка. При правильной окраске наблюдает-
ся изменение цвета от фиолетового к розовому. В первичной культуре наряду с
Я, gonorrhoeae могут встречаться штаммы стафилококков и стрептококков, кото-
рые легко теряют свою первоначальную фиолетовую окраску при обесцвечивании
спиртом и в процессе окраски по Граму приобретают розово-красный цвет. В этом
случае они ошибочно могут быть приняты за N, gononhoeae.
Препараты, в которых обнаруживают i рамотрицательные кокки и диплококки
из культуры, необходимо хранить в лаборатории в течение 3 мес.
НЕКОТОРЫЕ ИНФЕКЦИОННЫЕ ЗАБОЛЕВАНИЯ 529
При выделении оксидазоположительных грамотрицательных диплокок-
ков проводят видовую идентификацию иммунологическими и молекулярно -
генетическими методами на основании оценки их ферментативной активности.
По ферментативной активности изоляты N. gonorrhot ае можно отличить от дру-
гих нейссерий и схожих видов бактерий. Ферментативную активность N. gonorrhoeae
изучают с помощью ростозависимых и ростонезависимых тестов. При проведе-
нии ростозависимых тестов углеводы и другие субстраты (например, феноловый
красный) вводят непосредственно в питательную среду, на которой осуществля-
ется рост исследуемой культуры В настоящее время рекомендуют использование
ростонезависимых тестов, которые дают возможность получить более быстрый (в
течение нескольких часов) и специфический результат, позволяя идентифициро-
вать исключительно N. gonorrhoeae и исключить другие непатогенные нейссерий.
Ростонезависимые тесты предназначены для определения ферментов N. gonorrhoeae
в концентрированном инокуляте, полученном из чистых колоний.
Иммунологические меи ды диагностики г- порей и идентификации N. gonorrhoeae
В настоящее время доступны одноэтапные хроматографические тест-системы
(ИХТ), которые в течение 5-10 мин позволяют идентифицировать N. gonorrhoeae
в клиническом материале и суспензии чистой культуры. В первом случае их при-
меняют для скрининга инфекции; получение положительного результата со 100%
вероятностью свидетельствует о наличии в тестируемой пробе гонококка. Однако
^тувствительность этих тестов недостаточно высока, чтобы исключить инфек-
цию при получении отрицательного результата. При идентификации изолятов
N. gonorrhoeae такой недостаток ПХТ практического значения не имеет.
РИФ и коагглютинации, а также ИФА проводят только в референс-лабораториях
для внутривидовой идентификации N. gonorrhoeae. Эти иммунологические тесты
высокочувствительны и специфичны, особенно при использовании реагентов,
приготовленных из моноклональных антител (к белку РгоВ наружной мембраны
и другим антигенам N. gonorrhoeae).
Молекулярно-генетическая диагностика гонореи и идентификация N. gonorrhoeae
Для обнаружения N. gonorrhoeae в клиническом материале и идентификации ее
изолятов могут быть использованы молекулярно-биологические методы — ПЦР,
Strand Displacement Amplification (SDA), Transcription Mediated Amplification (TMA), a
также методы, основанные на гибридизации ДНК и РНК.
Существуют коммерческие тест-системы, позволяющие выявлять N. gonorrhoeae
с высокими чувствительностью и, как правило, специфичностью. Однако резуль-
таты, полученные при их использовании, следует оценивать с учетом данных
анамнеза, так как в неко торых случаях ДНК N. gonorrhoeae сохраняется в клиниче-
ском материале в течение 2-3 нед после окончания лечения гонореи.
Молекулярно-генетические методы оптимальны для исследования образцов,
полученных неинвазивным способом, однако их чувствительность и специфич-
ность вариабельны. Чувствительность молекулярно-генетических методов обыч-
но выше при исследовании мочи мужчин по сравнению с мочой женщин. Наличие
ингибиторов в некоторых пробах, а также отсутствие у ряда штаммов N. gonorrhoeae
нуклеотидной последовательности, являющейся мишенью для молекулярно-
генетических методов, могут приводить к снижению их чувствительности.
Известна недостаточная специфичность некоторых методов гибридизации
ДНК/РНК: они дают большое количество ложноположительных результатов
вследствие перекрестных реакций с условно-патогенными видами нейссерий
(N. lactamica. N. cinerea, N. subflava).
Высокую специфичность проявляет ПЦР, проводимая с праймерами, РогА-
псевдогена Я, gonorrhoeae. Она позволяет дифференцировать N. gonorrhoeae и
N. meningitidis.
530 НЕКОТОРЫЕ ИНФЕКЦИОННЫЕ ЗАБОЛЕВАНИЯ
Для всех экстрагенитальных образцов или для определения чувствительности к
антимикробным препаратам следует проводить культивирование с последующей
идентификацией нейссерий до вида. В настоящее время не существует лицензиро-
ванных молекулярно-биологических методов для исследования ректальных или
фарингеальных образцов. Методы гибридизации ДНК/РНК могут быть использо-
ваны для видовой идентификации N. gonorrhoeae из выделенных культур.
Оценка чувствительности изолятов N. gonorrhoeae к антимикробным препаратам
Во всем мире увеличивается резистентность N. gonorrhoeae к большинству анти-
бактериальных препаратов, что обусловлено мутациями в хромосомах и плазми-
дах бактерии, определяющих резистентность к пенициллинам и тетрациклинам.
Однако существуют географические различия в резистентности N. gonorrhoeae.
Именно поэтому в алгоритме лечения следует учитывать данные национального
и местного мониторинга чувствительности N, gonorrhoeae к антимикробным пре-
паратам. Необходимо постоянное проведение эпидемиологических исследований
референс-лабораториями для получения точной региональной и национальной
информации по антибиотикорезистентности гонококков.
Показания к определению чувствительности к антимикробным препаратам:
❖ эпидемиологическое тестирование, регулярно проводимое референс-
лабораториями, для оценки устойчивости N. gonorrhoeae к антимикробным
препаратам на местном, региональном и национальном уровнях;
❖ отсутствие эффекта стандартной антибактериальной терапии при лечении
гонококковой инфекции;
❖ мониторинг клинической эффективности стандартных рекомендуемых алго-
ритмов лечения гонококковой инфекции;
изучение антигонококковой активности новых антимикробных препаратов.
Однако определение чувствительности 2V. gonorrhoeae к антибиотикам не реко-
мендуют использовать в качестве рутинного исследования при лечении гонореи у
всех пациентов
Определение чувствительности N. gonorrhoeae к антимикробным препаратам
проводят с помощью:
❖ диско-диффузионного метода;
❖ метода серийных разведений в агаре;
❖ Е-теста.
Качественные тесты — диско-диффузионный и молекулярно-генетические,
позволяющие определить генетические маркеры, кодирующие резистентность
N. gonorrhoeae к определенным классам антибиотиков.
Количественные тесты определения чувствительности — метод серийных раз-
ведений в агаре и Е-тест позволяют выявить МПК антибактериального препарата
(наименьшую концентрацию антибиотика, при которой подавляется видимый
рост исследуемого микроорганизма).
Минимальный набор антибактериальных препаратов для определения анти-
биотикочувствительности N. gonorrhoeae должен состоять из наиболее характер-
ных представителей различных групп антибиотиков, применяемых для лечения
гонореи (бензилпенициллин, цефтриаксон, ципрофлоксацин, спектиномицин,
азитромицин, тетрациклин).
ФРАНСИСЕЛЛЫ
ТАКСОНОМИЯ
Возбудитель туляремии — Francisella tularensis — относится к роду Francisella,
семейству Francisellacae, подклассу у-протеобактерий. В настоящее время род
Francisella представлен двумя видами' Francisella tularensis и Francisella philomiragia.
НЕКОТОРЫЕ ИНФЕКЦИОННЫЕ ЗАБОЛЕВАНИЯ 531
В пределах вида Е tularensis четко выделяются четыре подвида: F. tularensis novi-
cida, F. tularensis tularensis (неарктический, американский или тип A), F. tularensis
mediasiatica (среднеазиатский), F. tularensis holarctica (голарктический или тип В).
Последний подвид включает три биологических варианта: японский биовар, био-
вар I Erys (эритромицинчувствительный) и биовар И Eryk (эритромицинрезистент-
ный).
Внутривидовая дифференциация возбудителя туляремии основана на разли-
чиях подвидов и биоваров по ряду фенотипических и генотипических признаков:
биохимической активности, составу высших жирных кислот, степени патоген-
ности для человека и животных, чувствительности к некоторым антибиотикам,
а также на особенностях экологии и ареале возбудителя. На территории России
распространен голарктический подвид F. tularensis holarctica, циркуляция которого
осуществляется главным образом грызунами и зайцеобразными, а также иксодо-
выми клещами и через воду.
Для подвидов туляремийной бактерии характерны различные ареалы распро-
странения. F. tularensis tularensis известен только на территории Северной Америки,
F. tularensis holarctica — на территории стран Северного полушария, F. tularensis
niediasiatk а — только в районе поймы рек Чу и Или (Казахстан) и в дельте реки
Амударьи (Узбекистан), F. tularensis novicida — только на территории Северной
Америки.
ОБЩАЯ ХАРАКТЕРИСТИКА ВОЗБУДИТЕЛЯ
F. tularensis представляет собой мелкие, грамотрицательные, кокковидныс (или
эллипсоидные) плеоморфные палочки размером 0,2-0,7x0,2 мкм (для подвидов
tularensis, holarctica, mediasiatica) и 0,7x1,7 мкм (для подвидов novicida, philomiragia),
окруженные слизистым (капсульным) веществом, неподвижные, не образующие
спор. Морфология клеток типична в логарифмической фазе роста на жидких
средах, тогда как в инфицированных эукариотических клетках и в стационарной
фазе роста клетки часто вытянуты, могут иметь нитевидную форму, а их размер
не превышает 600 нм в диаметре. Клеточная стенка имеет гладкие контуры. Для
клеточной стенки и капсулоподобного вещества туляремийного микроба харак-
терно повышенное, по сравнению с другими грамотрицательными бактериями,
содержание липидов.
F. tularensis относят к факультативным анаэробам. Оптимальный рост наблю-
дается только в аэробных условиях. Температурный оптимум 36-37 °C. Бактерии
кульгивируют на сложных агаровых или желточных средах с добавлением ткане-
вых экстрактов, цистеина, кроличьей дефибринированной крови и других пита-
тельных веществ. Включение в питательную среду полимиксина В или пеницилли-
на подавляет рост других микроорганизмов. Возбудитель туляремии не растет на
обычных питательных средах (мясопептонном агаре и бульоне), что служит одним
из признаков при идентификации культуры.
Возбудитель туляремии обладает слабой биохимической активностью, фер-
ментирует до кислоты глюкозу, мальтозу, левулезу и маннозу. Бактерия имеет
следующие ферменты: каталазу, глутаминазу, аспарагиназу, дез- и трансаминазу,
цитруллинуреидазу; индол не образует. Бактерия образует сероводород. Различия
в биохимической активности — один из признаков дифференциации подвидов
туляремийного микроба.
Патогенность возбудителя туляремии зависит от его подвидовой принадлеж-
ности. Наибольшей вирулентностью для человека и кроликов обладает F. tularensis
subsp. tularensis'. LD5!. для кроликов — 101 м.к.; F. tularensis subsp. holarctica и F. tula-
rensis subsp. mediasiatica характеризуются меньшей вирулентностью для человека и
кроликов: LD кроликов — более 106м,к. Возбудитель F. tularensis novicida обладает
пониженной вирулентностью: LD50 для кроликов — более 106 м.к. Вирулентность
ж
532 НЕКОТОРЫЕ ИНФЕКЦИОННЫЕ ЗАБОЛЕВАНИЯ
для человека недостаточно известна. Различия в патогенности — один из призна-
ков дифференциации подвидов туляремийной бактерии.
Причина высокой патогенности Е tularensis j\q сих пор неясна. Из числа воз-
можных факторов вирулентности рассматривают ЛПС, белок размером 23 кДа,
ферменты циклов биосинтеза пуринов, белки теплового шока, белки патогенно-
сти, регуляторные белки и другие факторы.
ЛПС гуляремийной бактерии — основной имм)кодоминантный антиген и
мишень специфического антительного ответа макроорганизмов на возбудитель.
На выявлении специфических туляремийных антител к эпитопам ЛПС основана
серологическая диагностика туляремии у человека и животных. Структура и анти-
генная специфичность ЛПС F. tularensis трех основных подвидов (кроме Е tularensis
novicida) идентичны.
Возбудитель туляремии проявляет значительную устойчивость во внешней
среде, особенно при низких температурах. Так, в речной воде при температуре
1 °C он сохраняется до 9 мес, в замороженной воде (при температуре -5 °C) — до
10,5 мес, В зимний период сохраняет жизнеспособность в воде мелких проточных
водоемов не менее 5 мес. Во влажной почве при температуре 4 ‘С он сохраняется
свыше 4 мес, а при температуре 23-25 °C — до 2,5 мес. При подсыхании почвы
сроки сохранения бактерий сокращаются до 10 сут. В естественно инфицирован-
ном иле при температуре 7 °C туляремийные бактерии остаются жизнеспособными
более 3 мес. Длительность инфицирования продуктов питания зависит от темпера-
туры хранения: в молоке, сливках при температуре 8-15 °C сохраняется до 8 сут,
в замороженном молоке — ботее 3 мес. В молочнокислых продуктах бактерии
быстро отмирают. В условиях эксперимента туляремийные бактерии сохранялись
на зерне и соломе при температуре -5 °C — до 192 сут, при температуре 8-12 °C —
до 56 сут, а при температуре 20-30 СС — 19 сут. В естественных условиях в зара-
женной ржаной соломе (при эпизоотии на грызунах) наличие жизнеспособных
бактерий отмечалось с января по май. В замороженных трупах грызунов бактерии
сохраняются 4-6 мес, при комнатной температуре отмирание бактерий в них про-
исходит в течение 5—10 сут. В высушенных шкурках инфицированных водяных
крыс туляремийные бактерии при темпера гуре 15-20 СС могут сохраняться до
20 сут.
При выраженной устойчивости в условиях низких темпера iyp возбудитель
туляремии весьма чувствителен к физическим и химическим факторам (солнеч-
ным, ультрафиолетовым лучам, ионизирующей радиации, высокой температуре).
Он чувствителен к аминогликозидам, стрептомицину, гентамицину, канамицину,
тетрациклинам, хлорамфениколу и хинонам, но резистентен к пенициллинам,
цефалоспоринам и полимиксину.
ЭПИДЕМИОЛОГИЯ
Туляремия — зоонозная природно-очаговая инфекция, по уровню регистрируе-
мой заболеваемости занимает относительно скромное место в структуре инфекци-
онной патологии человека в России. Однако природные очаги туляремии широко
распространены на большей части территории нашей страны и постоянно прояв-
ляют эпидемиологическую активность.
Актуальность проблемы обусловлена различными факторами и особенностями
эпидемиологического проявления инфекции, а также свойствами возбудителя.
Возбудитель туляремии — один из самых инфекционных патогенных микро-
организмов; его относят к так называемым критическим биологическим агентам
категории А — потенциальным агентам биологического оружия и биотерроризма.
Возбудитель туляремии обладает высокой патогенностью для человека: 10-50
бактерий при инокуляции или ингаляции приводят к развитию инфекционного
процесса при почти 100% восприимчивости организма человека к инфекции.
НЕКОТОРЫЕ ИНФЕКЦИОННЫЕ ЗАБОЛЕВАНИЯ 533
Заражение человека происходит в результате укусов инфицированными кро-
вососущими членистоногими (комарами, клещами, слепнями); при употреблении
продуктов питания и воды, зараженной возбудителем; через кожные покровы и
слизистые оболочки при соприкосновении с больными или павшими грызунами и
зайцами, а также при раздавливании инфицированного насекомого на коже; при
вдыхании аэрозоля, образующегося в результате переработки зерна и корнеплодов
(свекла), перекладки сена и соломы, контаминированных выделениями больных
грызунов.
КЛИНИЧЕСКИЕ ПРОЯВЛЕНИЯ ИНФЕКЦИИ
Туляремия — острое лихорадочное заболевание токсико-аллергического, реже
септического характера. Широкий круг животных — источников инфекции и раз-
нообразие путей заражения людей обусловливают полиморфизм клинических
проявлений этого заболевания.
По Международной статистической классификации болезней (10-му пере-
смотру ВОЗ, 1995) выделяются следующие клинические формы туляремии:
ульцерогландулярная (язвенно-бубонная, кожно-бубонная), гландулярная (бубон-
ная), окулогландулярная (глазно-бубонная, офтальмическая), легочная (тора-
кальная), ангинозно-гландулярная (ангинозно-бубонная), желудочно-кишечная
(абдоминальная), генерализованная и неуточненная. В последующем использова-
на номенклатура клинических форм, указанная в скобках и применяемая в клини-
ческой практике РФ.
По тяжести инфекционного процесса различают легкие, средней тяжести и
тяжелые формы туляремии. По длительности течения различают острую, затяж-
ную и рецидивирующую формы туляремии.
От пути внедрения возбудителя в организм в значительной мере зависит после-
дующее развитие той или иной клинической картины. Инкубационный период
при туляремии в среднем составляет 3-7 сут с отклонениями в большую или
меньшую сторону (от нескольких часов до 10-15 дней) в зависимости от дозы
заражения и входных ворот инфекции. Для туляремийной инфекции нехарактерно
наличие продромального периода. Независимо от клинической формы туляремия
характеризуется острым (редко постепенным) началом с познабливанием или
потрясающим ознобом и резким повышением температуры тела до 38-40 °C и
выше. Для начального периода заболевания характерны слабость, головная боль,
головокружение, снижение аппетита; язык обложен серовато-белым налетом,
наблюдаются мышечные боли разной локализации, нарушение сна, повышенная
потливость, отставание пульса от темпера гуры (относительная брадикардия), тен-
денция к снижению артериального давления.
Продолжительность начального периода общих клинических проявлений
составляет 2-3 сут, затем обнаруживают признаки той или иной клиниче-
ской формы. Длительность лихорадочного периода составляет 2-3 нед и более.
Продолжительность этого периода зависит от клинической формы, тяжести забо-
левания и времени начала специфического лечения.
Температура быстро нарастает спустя 2-3 сут после начала болезни.
Продолжительность лихорадки колеблется от 5-7 до 30 сут, в среднем составляя
2-3 нед. Фаза выздоровления сопровождается непостоянным субфебрилитетом.
Продолжительность боиезни составляет в среднем от 16 до 30 сут. При преобла-
дающем поражении лимфатической системы, то есть развитии бубонов, болезнь
затя1ивается до 2-3 мес. Лимфадениты занимают центральное место в клиниче-
ской картине туляремии.
Поскольку начальный период заболевания не имеет патогномоничных сим-
птомов, для диагностики туляремии большое значение имеет тщательный эпиде-
миологический анамнез. Для туляремии характерно отсутствие контагиозное™, то
534 НЕКОТОРЫЕ ИНФЕКЦИОННЫЕ ЗАБОЛЕВАНИЯ
есть опасности заражения здоровых людей от больного человека. Это позволяет
госпитализировать больного в терапевтическое или другие специализированные
клинические отделения.
Язвенно-бубонная форма туляремии. Эта форма туляремии развивается
в результате трансмиссивного заражения — укуса слепня, комара, клеща или
других кровососущих членистоногих. Для язвенно-бубонной формы характер-
но наличие язвочки, или первичного аффекта, в месте внедрения возбудителя.
Трансмиссивный путь заражения типичен для летнего времени года. Образование
язвочки всегда сопровождается образованием лимфаденита — увеличением и
умеренной болезненностью лимфатических узлов. Локализация увеличенных
лимфатических узлов связана с местом внедрения возбудителя. Эта форма туляре-
мии протекает обычно легко и значительно реже имеет среднюю степень тяжести.
Длительность заболевания обычно не превышает 2-3 нед, особенно при своевре-
менном специфическом лечении.
Бубонная форма туляремии Лимфаденит при туляремии, как правило, раз-
вивается в результате контактного заражения, то есть при обработке и разделке
тушек инфицированных животных. Чаще заражение происходит через кожу рук
с последующим вовлечением в воспалительный процесс группы локтевых и под-
мышечных лимфатических узлов. Лимфаденит развивается через 2-3 сут после
начала болезни, лимфатические узлы постепенно увеличиваются и достигают мак-
симальных размеров через 5-8 сут. Исход лимфаденита, как и при других клини-
ческих формах, может быть различным — от полного рассасывания до нагноения
с последующим самопроизвольным или хирургическим вскрытием, заканчиваю-
щимся рубцеванием.
Глазно-бубонная форма туляремии. Развивается в результате заражения
через слизистую оболочку глаза. Характерен острый специфический конъюнкти-
вит с сильным слезотечением, выраженной отечностью век, слизисто-гнойным
отделяемым. Специфическое поражение глаз при туляремии, как правило, одно-
стороннее и сопровождается воспалительной реакцией со стороны предушных и
подчелюстных лимфатических узлов и формированием лимфаденита. Заражение
происходит при умывании водой из инфицированных источников, а также при
случайном внесении инфекции руками в глаз. Эта форма туляремии протекает
довольно тяжело. Длительность заболевания, возможные осложнения и исход
конъюнктивита зависят от своевременно начатого специфического лечения.
Легочная форма туляремии (торакальная). Для легочной формы туляре-
мии характерно развитие первичного воспалительного процесса в легких. Частота
развития пневмонии колеблется от 11 до 30%. Заражение происходит аспираци-
онным путем при обмолотах и других сельскохозяйственных работах (перекладке
ометов, скирд, стогов, подвозке кормов, обслуживании животноводческих ферм),
связанных с вдыханием инфицированной пыли. Такой путь заражения может
наблюдаться на предприятиях по переработке сельскохозяйственных продуктов
при использовании сырья, инфицированного выделениями больных грызунов или
загрязненного их трупами (мельницах, элеваторах, крахмально-паточных заводах
и т.п.). В редких случаях аспирационное заражение туляремией может произойти в
условиях специализированных лабораторий. В результате аспирационного зараже-
ния патологический процесс развивается непосредственно в легких. Клиническая
картина при этом характеризуется пневмонией (очаговой, сегментарной, лобар-
ной, диссеминированной), бронхопневмонией, бронхитом. В отдельны:; случаях
процесс может ограничиться трахеитом. Наиболее тяжело протекает пневмониче-
ский вариант, который характеризуется резким повышением температуры тела до
39-40 °C. Лихорадочная реакция может протекать в виде двух волн, хапактерна
одышка. Наблюдается сухой кашель, позже выделяется небольшое количество
слизистой или слизисто-гнойной вязкой мокроты, а в последующем возможно
НЕКОТОРЫЕ ИНФЕКЦИОННЫЕ ЗАБОЛЕВАНИЯ 535
появление примеси крови. Выражены общая интоксикация, состояние адинамии,
профузные поты, особенно в ночные часы. Нередко больных беспокоят загрудин-
ные боли, связанные с вовлечением в воспалительный процесс трахеобронхиаль-
ных лимфатических узлов. Наблюдается увеличение печени, селезенки, ослабле-
ние сердечной деятельности. В периферической крови отмечается лейкоцитоз, а в
тяжелых случаях — лейкопения и ускорение оседания эритроцитов. Туляремийная
пневмония не характеризуется какими- либо патогномоничными симптомами,
поэтому в начальный период болезни постановка диагноза бывает затруднена.
Заболевание продолжается 4-6 нед и более. Продолжительность болезни зависит
от степени тяжести и сроков начала специфического лечения.
Желудочно-кишечная форма туляремии (абдоминальная). Абдоминаль-
ная форма возникает в результате алиментарного (орального) заражения при
употреблении продуктов, инфицированных выделениями больных грызунов,
недостаточно термически обработанного мяса зайца и при употреблении воды
из инфицированных источников. Развитие абдоминальной туляремии проис-
ходит при массивной дозе заражения, так как желудочный сок с нормальным или
повышенным содержанием соляной кислоты — мощное препятствие на пути воз-
будителя туляремии. Симптомы этой клинической фоомы — высокая температура
тела и боли в животе разной интенсивности и продолжительности. Язык обложен
суховатым серо-белым налетом, характерны тошнота, рвота, проявления общей
интоксикации, вовлечение в процесс мезентериальных лимфатических узлов в
стенке тонкого кишечника. Может наблюдаться увеличение печени и селезенки.
По сравнению с другими клиническими формами туляремии абдоминальная
форма протекает наиболее тяжело. Симптомы этой формы сходны с симптомами
брюшного тифа. Для дифференциальной диагностики решающее значение имеют
данные эпидемиологического анализа.
Ангинозно-бубонная форма туляремии. Заражение происходит через рот
при употреблении продуктов, инфицированных больными грызунами, их выде-
лениями в различных хранилищах, при употреблении воды из инфицированных
источников при попадании в них больных грызунов или их трупов. Ангинозно-
бубонная форма развивается также при употреблении недостаточно термически
обработанного мяса зайца. Входными воротами инфекции служит слизистая
оболочка миндалин. Для этой формы характерна специфическая ангина, чаще
односторонняя. Одновременно с развитием ангины наблюдается увеличение под-
челюстных. переднешейных лимфатических узлов. Продолжительность туляре-
мийной ангины колеблется от 8 до 24 сут.
Генерализованная форма туляремии (тифоидная, септическая).
Характеризуется тяжелым течением, развитием общих симптомов болезни.
Выражен токсикоз, иногда отмечаются потеря сознания, бред, адинамия, сильные
головные и мышечные боли. Лихорадка волнообразная, продолжительностью до
3 нед и более На коже отмечаются высыпания, поражаются сосуды. Сыпь наблю-
дается в течение 8-12 сут. Высыпание нередко сопровождается припухлостью
суставов (кистей, стоп), болезненностью, отечностью пальцев. Печень и селезенка
увеличены. Выздоровление наступает медленно, возможны рецидивы болезни.
Эта форма заболевания также возникает при внутрилабораторном заражении.
ДИАГНОСТИКА
Туляремию диагностируют на основании данных эпидемиологического анали-
за клинического обследования пациента, аллергической пробы и лабораторных
исследований взятых у него материалов. Важную роль играет эпидемиологи-
ческий анамнез, который должен быть детальным и исчерпывающим. Особое
внимание необходимо уделять информации о вакцинации против туляремии в
прошлом и ее сроках, данным о заражении этой инфекцией ранее, выяснению про-
536 НЕКОТОРЫЕ ИНФЕКЦИОННЫЕ ЗАБОЛЕВАНИЯ
фессиональной занятости и ряду других факторов, которые необходимы для того,
чтобы в каждом отдельном случае подтвердить или исключить диагноз туляремии.
Последнее необходимо для определения тактики лечения, а также проведения
профилактических мероприятий. Б случаях с наружно выраженными клиниче-
скими проявлениями (язвенно-бубонная, бубонная, глазно-бубонная, ангинозно-
бубонная формы) и наличием эпидемиологических данных диагноз туляремии не
должен вызывать затруднений. Однако при легочной и абдоминальной формах
туляремии большое значение имеют анамнестические данные и результаты лабо-
раторного анализа.
В ряде случаев при позднем начале лечения заболевание принимает затяжной
характер с возможными рецидивами. Рецидив может наступить как вскоре после
перенесенного заболевания (4-6 мес), так и через 2-6 лет и позже. Чаще всего
рецидив проявляется в виде воспаления лимфатических узлов на месте перво-
начального регионарного лимфаденита. Воспаление лимфатических узлов при
рецидивах развивается постепенно, без повышения температуры тела и ухудше-
ния общего состояния больного. Несколько позже развиваются субфебрилитет,
слабость, возможны потливость, нарушение сна. Лимфатические узлы обычно не
достигают таких размеров, как при первичном заболевании, болезненность их не
выражена. Исход лимфаденита при рецидивах может быть таким же. как и при
первичном заболевании, однако нагноение происходит значительно реже.
Патогенез туляремийных рецидивов малоизучен, но предполагают, что их раз-
витие обусловлено длительной персистенцией возбудителя в организме, В каждом
конкретном случае постановки диагноза «рецидив туляремии» необходимо устано-
вить связь с ранее перенесенным заболеванием и исключить возможность повтор-
ного заражения, а также проследить за наличием антител в крови и нарастанием их
титра. Имеются также немногочисленные данные, указывающие на возможность
хронического течения туляремии, но этот вопрос нуждается в уточнении.
Спорадическая заболеваемость туляремией и полиморфизм клинических
проявлений создают затруднения при постановке диагноза Особенно сложно
поставить диагноз в начальном периоде болезни, когда ее часто путают с грип-
пом, брюшным или сыпным тифом, пневмонией и другими заболеваниями. На
более поздних сроках туляремию следует дифференцировать от сибирской язвы,
язвенно-некротической ангины, листериоза, гнойных лимфаденитов и лимфан-
гоитов, паротита, бубонной формы чумы, бруцеллеза, доброкачественного лим-
(Ьоретикулеза и содоку.
Бубонную форму туляремии следует отличать от бубонной формы чумы.
При чуме более выражен токсикоз, состояние больного значительно тяжелее.
Туляремийный лимфаденит отличается выраженными контурами, меньшей болез-
ненностью по сравнению с чумными бубонами, относительно умеренным периа-
денитом, отсутствием сильных осложнений и благоприятным исходом. Язвы при
туляремии также менее болезненны, чем при чуме. Для туляремийной пневмонии
характерно отсутст те кровавой мокроты (за редким исключением). Больные при
туляремии неконтагиозны.
Гнойные банальные лимфадениты (стафилококковые и стрептококковые)
характеризуются резкой болезненностью, ранним нагноением по сравнению с
туляремией, развитием периаденита и лимфангоита. При туберкулезных лимфа-
денитах начало болезни постепенное, с субфебрильной температурой, лимфати-
ческие узлы плотные, болезненность их отсутствует. Как правило, лимфатические
узлы не достигают таких размеров, как при туляремии. Туляремийные лимфаде-
ниты в отличие от лимфаденопатии, наблюдаемой при бруцеллезе, отличаются
строгой привязанностью к входным воротам инфекции.
В отличие от дифтерии, ангина при туляремии характеризуется более острым
началом, обычно односторонней локализацией и нечастым распространением
НЕКОТОРЫЕ ИНФЕКЦИОННЫЕ ЗАБОЛЕВАНИЯ 537
налетов за пределы миндалин. Туляремийные высыпания чаще симметричны и
более полиморфны по сравнению с брюшным или сыпным тифом.
При паротите шейные лимфатические узлы увеличены незначительно.
Отмечаются резкая болезненность при надавливании на козелок уха и выражен-
ная отечность одной половины лица.
Поскольку туляремию относят к острым лихорадочным заболеваниям, может
возникнуть необходимость дифференциальной диагностики с такими болезнями,
как малярия, ку-лихорадка, сепсис, сифилис, туберкулез и паротит.
Нужно помнить, что трудность постановки диагноза туляремии на основании
клинических признаков обусловливает необходимость комплексной диагностики
с учетом данных эпидемиолот ического анализа, клинических наблюдений и лабо-
раторного исследования.
Аллергическая проба
У больных туляремией кожно-аллергическая реакция становится положитель-
ной обычно спустя 3-5 сут после начала болезни, реже — в более поздние сроки и
сохраняется в течение многих лет, поэтому аллергический метод служит основой
для ранней или ретроспективной диагностики. Этот метод строго специфичен.
У привитых живой вакциной пациентов также возникает аллергическая реак-
тивность, но она развивается медленнее (через 10-15 сут после вакцинации).
Аллергический ответ у вакцинированных пациентов сохраняется 5-6 лет, иногда
дольше и может служить методом определения сохранности поствакцинального
иммунитета. Кожную аллергическую реакцию выполняют как накожно, так и вну-
трикожно с соответствующим тулярином.
Накожная туляриновая проба проста в исполнении и до сих пор используется
на практике. Однако введение даже убитого антигенного препарата небезопасно
для организма обследуемого, поэтому вместо нее в настоящее время все чаще при-
меняют лабораторный метод оценки аллергической реактивности.
ЛАБОРАТОРНАЯ ДИАГНОСТИКА
Лабораторную диагностику туляремии проводят в соответствии с методиче-
скими указаниями «Эпидемиологический надзор за туляремией» МУ 3.1.2007-05.
Она основана на комплексном подходе, направленном на выявление как живых
бактерий, так и специфических антигенов или специфических антител против
туляремийного микроба. Основную роль играют серологические и аллергические
тесты, а также биопроба на лабораторных животных. Бактериологические методы
не всегда эффективны, что обусловлено особенностями течения инфекции у чело-
века, малой обсемененностью органов и тканей возбудителем. Выделение возбуди-
теля наиболее вероятно в течение первых 2-3 нед от начала заболевания и реже в
более поздние сроки. Его эффективность во многом зависит от правильности сбора
и доставки исследуемо! о материала.
Для лабораторной диагностики от подозреваемых в заболевании туляремией
людей берут содержимое бубона, материал из зева, конъюнктивы глаза, отделяе-
мое язвы, мокроту; кровь и сыворотку крови: постсмертно — увеличенные лимфа-
тические узлы, измененные участки легких и селезенки.
При эпизоотологических исследованиях материалом для лабораторных иссле-
дований служат:
о пробы, полученные от диких млекопитающих, или их трупы, остатки трупов,
подснежные гнезда грызунов, продукты жизнедеятельности млекопитаю-
щих, погадки птиц, помет хищных млеке питающих, а также солому, мякину,
талую воду и другие объекты, загрязненные выделениями грызунов, воду? из
естественных водоемов и колодцев, гидробионтов, членистоногих (преиму-
щественно иксодовых клещей), мелких эктопаразитов, собранных с млеко-
питающих (вшей, гамазовых и краснотелковых клещей, блох);
538 НЕКОТОРЫЕ ИНФЕКЦИОННЫЕ ЗАБОЛЕВАНИЯ
о кровососущие двукрылые (комары, слепни и др.), отловленные в районах
вспышек болезни;
❖ сыворотка крови домашних животных (при соответствующих эпизоотологи-
ческих и эпидемиологических показаниях).
Серологическая диагностика
РГА. Самый распространенный и наиболее точный метод серологической диа-
гностики туляремии Реакцию применяют для постановки диагноза, а также для
изучения иммунологического состояния пациентов, привитых против туляремии.
При серологической диагностике туляремии у человека диагностическим считают
титр 1:100 и выше. При этом обязательно нужно прослеживать его нарастание.
При отрицательной или сомнительной реакции, а также при положительной
реакции только в низких разведениях сыворотки крови с подозрением в заболе-
вании туляремией людей рекомендуют повторить исследование сыворотки через
7-10 сут. Нарастание титра сыворотки в 2-4 раза и выше служит основанием для
постановки окончательного диагноза туляремии.
РПГА. Чувствительный метод серологической диагностики туляремии Его
используют как для ранней, так и для ретроспективной диагностики, а также для
определения иммунологического состояния привитых пациентов. У больных
туляремией антитела обычно обнаруживают в конце первой или на второй неделе
заболевания. Через 1-1,5 мес титры РПГА достигают максимальных показате-
лей — 1:10 000-1:20 000 и выше, после чего снижаются и на уровне 1:100-1:200
сохраняются длительное время.
У привитых пациентов антитела также обнаруживают постоянно, однако в более
низких титрах, не превышающих 1,2000-1:5000 через 1-1,5 мес после вакцинации.
Титры сохраняются в течение нескольких лет на низком уровне — 1:2-1:80.
Антигеном для постановки РПГА служит туляремийный эритроцитарный
диагностикум, который представляет собой формалинизированные эритроциты
барана, сенсибилизированные туляремийным антигеном; выпускается в жидком
и сухом виде. Специфичность положительного результата, полученного в РПГА,
может быть проверена с помощью трехкомпонентной реакции торможения пас-
сивной гемагглютинации.
ИФА на твердофазном носителе. Метод используют для диагностики туля-
ремии у больных и переболевших людей, а также для определения иммунитета
у вакцинированных против туляремии. У больных туляремией специфические
антитела обнаруживают в ИФА через 6-10 суп1 после начала болезни и достигают
максимальных показателей через 4-7 нед. Позже уровень антител снижается,
однако они сохраняются в крови в течение долгого времени после болезни (более
10 лет). ИФА обеспечивает более раннюю и эффективную по сравнению с РГА и
РПГА иммунологическую диагностику туляремии. Титры анти гел у больных и вак-
цинированных колеблются в ппеделах от 1:400 до 1:40 000 и выше и, как правило,
в 10 20 раз превышают таковые в РГА и РПГА.
ИФА проводят в диагностической иммуноферментной тест-системе для опреде-
ления специфических антител. Выявление последних в сыворотках людей проис-
ходит за счет специфического взаимодействия антигена туляремийного микроба,
адсорбированного на планшете, с туляремийными антителами в исследуемой
сыворотке, Образовавшийся комплекс «антиген-антитело» определяют с помо-
щью антител против иммуноглобулинов человека, меченных пероксидазой.
В соответствии с назначением тест-система представляет собой набор ингреди-
ентов для иммуноферментного анализа на твердофазном носителе. Она включает
туляремийный липополисахарид, антитела против иммуноглобулинов человека,
меченных пероксидазой, контрольную положительную, контрольную отрица-
тельную сыворотку, набор солей, необходимых для приготовления буферных рас-
НЕКОТОРЫЕ ИНФЕКЦИОННЫЕ ЗАБОЛЕВАНИЯ 539
творов, субстрат О-фенилендиа мин; твин-20; бычий сывороточный альбумин и
планшеты для ИФА однократного применения.
Непрямой метод флюоресцирующих антител (НМФА). При приме-
нении НМФА для обнаружения антител к туляремийному микробу готовят
мазки 1 2-суточной агаровой культуры возбудителя из взвеси с концентрацией
1х10ч м.к./мл. При исследовании сыворотки больного с подозрением на туляре-
мию диагностическим считают титр не менее 1:40.
Тест оценки аллергической реактивности in vitro. Этот тест заслуживает
внимания как эффективный и безопасный для пациента метод выявления и оцен-
ки гиперчувствительности. Он основан на феномене разрушения лейкоцитов сен-
сибилизированного организма в присутствии специфического антигена. Реа кция
лизиса лейкоцитов становится положительной в первые дни после вакцинации
или заболевания и удерживается в течение многих лет после выздоровления
или вакцинации (у переболевших — до 40 лет), а после вакцинации — не менее
5-7 лет.
Изоляция и идентификация возбудителя
Бактериологические методы диагностики туляремии имеют второстепенное
значение и не всегда эффективны, что определяется особенностями течения
инфекции у человека с малой обсемененностью органов и тканей возбудителем.
Следует учитывать, что не в любом материале, взятом от больного, содержатся
живые туляремийные бактерии. Кроме того, выделение возбудителя наиболее
вероятно в течение первых 2-3 нед от начала заболевания и реже в более поздние
сроки. Именно поэтому лишь в редких случаях удается изолировать возбудителя
туляремии из патологического материала от больных людей (содержимого язвы
на коже, содержимого бубона и т.д.).
Кроме того, изоляцию и идентификацию возбудителя гшяремии можно про-
водить только в специально оборудованных режимных лабораториях. Забор и
доставку патологического материала в лабораторию осуществляют с соблюдением
предосторожностей и правил работы с особо опасными инфекциями II группы
патогенности.
Биопроба. Биопробу проводят на белых мышах и морских свинках. Эю
самый чувствительный метод обнаружения F. tularensis в любом исследуемом
материале, поскольку зараженные им лабораторные животные заболеваю г туля-
ремией со смертельным исходом при подкожном введении единичных бактерий.
Однако при обследовании больных даже биологический метод диагностики не
всегда надежен, так как не в любом образце материала, взятом от больного, име-
ются жизнеспособные туляремийные бактерии. В отделяемом кожной язвы и в
пунктате из пораженного лимфатического узла возбудитель туляремии может
быть обнаружен в течение 3 нед от начала заболевания, реже позднее. Известны
случаи выделения возбудителя из мокроты, материала, взятого из зева (с мин-
далин) или конъюнктивы глаза, В редких случаях удается выделить F. tularensis
из крови, но обычно не позднее первой декады болезни. Культуру туляремийных
бактерий от биопробных животных выделяют посевом на специальные пита-
тельные среды.
Изоляция в питательных средах. Для изоляции F. tularensis из патологи-
ческого материала применяют специальные среды, в том числе свернутую жел-
точную среду, желточно-агаровую среду, различные варианты агаровых сред
содержащих цистеин, глюкозу, кровь и другие факторы роста. Скорость роста
бактерии в посевах зависит от ее титра в исследуемом материале: при посеве сла-
боинфицированного материала рост отдельных колоний возможен в отдаленные
сроки, поэтому посевы следует выдерживать в термостате при 37 °C в течение не
менее 10 сут.
540 НЕКОТОРЫЕ ИНФЕКЦИОННЫЕ ЗАБОЛЕВАНИЯ
Идентификация. Идентификацию свежевыделенных туляремийных культур
осуществляют на основании совокупности следующих признаков:
< > морфологии клеток и грамотрицательной окраски в мазках:
< > характера роста на желточной среде или специальных агаровых средах;
- о отсутствия роста на простых мясопептонных средах;
о агглютинации туляремийной сывороткой;
❖ специфического свечения в реакции иммунофлюоресценции;
о способности вызывать гибель белых мышей и морских свинок при зараже-
нии испытуемой культурой с характерными для туляремии патологоана-
томическими изменениями в органах и выделением культуры возбудителя
туляремии — F. tularensis.
В качестве дополнительного метода диагностики туляремии у человека реко-
мендуют обнаружение специфической ДНК в патологическом материале методом
молекулярной биологии с использованием ПЦР. Перечень тест-систем для диа-
гностики туляремии представлен в таблице 22-16.
Оценка результатов лабораторных методов диагностики туляремии.
Диагноз туляремии устанавливают на основании сопоставления результатов
серологических исследований, представленных обычно реакцией РГА или РПГА.
Другие серологические методы, например ИФА, применяют в сомнительных
случаях, а также для подтверждения достоверности результатов, полученных
другими методами (РГА или РПГА). Выделение возбудителя от больного, как
было указано выше, не всегда эффективно и доступно только в специально осна-
щенных лабораториях, имеющих санитарно-эпидемиологическое разрешение на
работу с возбудителем туляремии (II группа патогенности микроорганизмов).
Кожная проба может служить методом ранней диагностики, так как становится
положительной уже через 3-5 сут после начала болезни. Однако в случаях, когда
имеются противопоказания к применению накожного тулярина, прибегают к
методу аллергодиагностики in vitro — реакции лейкоцитолиза. Однако следует
иметь в виду, что методы аллергодиагностики не позволяют дифференцировать
свежие случаи заболевания от анамнестических реакций, поскольку специфиче-
ская клеточная реактивность у переболевших и привитых пациентов сохраняется
многие годы.
В связи с вышесказанным нецелесообразно основывать диагностику туляремии
на результатах алллергических и серологических методов диагностики.
Выявление антител к туляремийной бактерии при первом обследовании боль-
ного в титрах 1:50—1:100 при отсутствии нарастания титра при последующем
исследовании не дает основания для постановки окончательного диагноза туляре-
мии. Именно поэтому кровь больного исследуют по крайней мере дважды: первый
раз немедленно после обращения больного за медицинской помощью и второй —
спустя неделю после первого исследования. Если при повторном исследовании
антитела не обнаружены или титр сыворотки остался без изменений, кровь боль-
ного нужно исследовать в третий раз спустя неделю после второго исследования.
Нарастание титра антител в РГА и РПГА подтверждает диагноз туляремии, а его
отсутствие указывает на анамнестический характер реакции.
Следует также учитывать, что раннее начало лечения с применением антибио-
тиков (на первой неделе болезни) приводит к снижению титра антител у пациен-
тов, вакцинированных против туляремии или ранее перенесших это заболевание.
Основанием для диагностики свежего случая туляремии может служить лишь
выраженное (в 2 раза и более) нарастание титров антител в сыворотке, а также
наличие характерных клинических проявлений болезни.
Окончательный диагноз туляремии ставят на основании сопоставления результа-
тов серо-аллергического обследования больного с клинико-эпидемиологическими
данными.
НЕКОТОРЫЕ ИНФЕКЦИОННЫЕ ЗАБОЛЕВАНИЯ 541
Таблица 22-16. Перечень коммерческих противотуляремийных препаратов и тест-систем, произ-
водим ыхв Российской Федерации
№ Наименование Назначение Производитель
1 2 3 4
1 Вакцина туляремийная живая сухая Профилактика туляремии ФГУП НПО «Микроген» М3 РФ, г. Омск
2 Аллерген туляремийный жидкий для накожного применения (тулярин) Определение иммунологической структуры населения, про- живающего на энзоотичных по туляремии территориях Тот же
3 Диагностикум туляремийный для объемной и кровяно-капельной реакций агглютинации, жидкий Идентификация возбудителя туляремии; серологическая диа- гностика туляремии Тот же
4 Сыворотка диагностическая туля- ремийная сухая Идентификация возбудителя туляремии; компонент диагно- стических тест-систем ФГУЗ <Иркутский госу- дарственный научно исследовательский противо- чумный институт Сибири и Дальнего Востока»
5 Диагностикум эритроцитарный туляремийный иммуноглобули- новый сухой Выявление туляремийных анти генов и антител в различных объектах окружающей среды Научно-исследовательский институт микробиологии МО РФ, г. Киров
6 Диагностикум эритроцитарный туляремийный антигенный жидкий Серологическая диагностика туляремии; выявление туляре- мийных антигенов и антител в различных объектах окружаю- щей среды фнуз «Ставропольский научно-исследовательский противочумный институт»
7 Иммуноглобулины диагностиче- ские флюоресцирующие сухие Идентификация (индикация) возбудителя туляремии НИИЭМ им. Н.Ф. Гамалеи РАМН (филиал «Медгамал»)
8 Набор реагентов для амплифика- ции ДНК F. tularensis holarctica Идентификация возбудителя туляремии Лаборатория «Изоген», РФ, г. Москва
9 FT-агар Для культивирования и выделе- ния туляремийного микроба ФГУН ГНЦ ПМБ, г. Оболенск
10 Селективная добавка к ЕТ-агару Для культивирования и выделе- ния туляремийного микооба ФГУН ГНЦ ПМБ, г. Оболенск
СПИСОК РЕКОМЕНДУЕМОЙ ЛИТЕРАТУРЫ
Лабораторная диагностика опасных инфекционных болезней: Практическое руковод-
ство / Под общ ред. Г.Г. Онищенко, В.В. Кутырева. — М.: Медицина, 2009. — 472 с.
Методические указания. Профилактика инфекционных болезней. Эпидемиологический
надзор за туляремией. МУ 3.1.2007-05. — М., 2005. — 59 с.
Мещерякова И.С. Таксономия, идентификация и иммунологическая диагностика возбу-
дителя туляремии: Автореф дис.... д-ра биол. наук. — М., 1990. — 47 с.
Мещерякова И.С. Туляремия // Эволюция инфекционных болезней в России в XX веке /
Под общ. ред. В.И Покровского, Г.Г. Онищенко, Б.Л. Черкасского. — М., 2003. — С. 432-450.
Никифоров В.В., КареткинаГ.Н. Туляремия: от открытия до наших дней //Инфекционные
болезни. - 2007. - Т. 5. - № 1. - С. 67-76.
Тчляремия / Под общ. ред. Н.Г. Олсуфьева, Г.П. Руднева. — М., 1960. — 439 с.
Dennis D.T., Ingiesby T.V., Henderson D.A. et al. Tularemia as a Biological Weapon //JAMA. —
2001. - Vol. 285. - N 21. - P. 2763-2773.
Ellis J., Oyston C.F., Green M., Titball R.W. Tularemia // Clin. Microb. Rev. — 2002 — Vol. 1. —
P. 631-646.
Olsufjev N.G., Meshcheryakova I.S. Subspecific Taxonomy of Francisella tularensis McCoy and
Chapin 1912 // International Journal of Systematic Bacteriology. — 1983. — Vol. 1. — P. 872-
874.
Sjostedt F. Family XYII. Francisellaceae, genus I. francisella. Bergey s Manual of Systematic
Bacteriology / Brenner D.J., ed. — New York, 2003. — P. 201-210.
Глава 23
Вирусологические исследования
ЗАДАЧИ ДИАГНОСТИКИ ВИРУСНЫХ ИНФЕКЦИЙ
Лабораторная диагностика вирусных инфекций в последние годы
востребована практически при всех видах медицинской помощи.
Это обусловлено возрастанием удельного веса вирусных инфекций в
общей структуре инфекционной заболеваемости, прогрессом меди-
цинской вирусологии и быстрым развитием технологий лаборатор-
ной диагностики и лабораторной техники.
Развитие глобального эпидемического процесса показывает, что
заболевания вирусной этиологии эволюциониру ют быстрее бакте-
риальных. В структуре 50 инфекций, возникших на земном шаре в
течение последних 40 лет, ведущее место занимают именно вирус-
ные инфекции — 60%, тогда как новые заболевания бактериальной,
паразитарной и прионовой этиологии в совокупности составляют
лишь 40%,
Риски новых пандемий, которые держат в постоянном напря-
жении международные и национальные органы здравоохранения,
также обусловлены патогенами вирусной природы (ВИЧ, СПИД,
грипп, SARS и др.). Из социально значимых инфекционных болез-
ней наименее управляемы и трудноизлечимы заболевания вирусной
этиологии: ВИЧ-инфекция, вирусные гепатиты. Основная доля
острых кишечных инфекций неустановленной этиологии обуслов-
лена вирусными агентами — энтеровирусами, норавирусами, астро-
вирусами и др. Среди онкогенных инфекционных агентов также
преобладают вирусы — вирусы папилломы человека, гепатитов В и
С, Эпштейна-Барр, HTLV, ВИЧ и др.
Большая распространенность и увеличивающееся разнообразие
вирусных заболеваний приводят к тому, что перечень нозологи-
ческих форм, подлежащих обязательной этиологической расшиф-
ровке, постоянно расширяется. Поскольку многие из вирусных
инфекций протекают в латентных и атипичных формах (которые
прогностически не менее опасны, чем манифестные), методы спец-
ифической диагностики вирусных инфекций все шире проникают
в клиническую практику, становятся эффективным инструментом
скрининговых программ и массовых профилактических исследова-
ний, иными словами, инструментом донозологической диагности-
ки, а также пре-, пери- и постнатальной диагностики.
ВИРУСОЛОГИЧЕСКИЕ исследования 543
Прогресс медицинской вирусологии приводит к быстрому накоплению и углу-
блению знаний о генетике вирусов и патогенезе вирусных инфекций. Работы по
молекулярной генетике вирусов позволили охарактеризовать их внутривидовую
гетерогенность, определить генотипы, субтипы, клоны, квазивиды, рекомбинанты
и более точные модификации вирусного генома. На базе этих знаний формируется
современная классификация вирусов, котоцая часто более эффективна, чем клас-
сификация на основе серотипирования.
Накапливаются знания о географическом распределении вирусных геноти-
пов. 1еном вирусов, обладающих потенциалом пандемического распространения,
детально исследуется лабораториями, аккредитованными ВОЗ. Информация и
сами штаммы вирусов доводятся до национального здравоохранения всех стран.
Это открывает путь для идентификации вирусов, их географического происхожде-
ния, путей и факторов передачи.
Благодаря тому что геном наиболее актуальных вирусных агентов полностью
расшифрован, появилась возможность индикации и идентификации специфиче-
ских элементов генома непосредственно в материале от больных. В дополнение к
трудоемким методам выделения вирусов в культурах клеток вирусологи получили
методы прямого определения вирусной ДНК или РНК в первичном патологиче-
ском материале. Это, в свою очередь, стимулировало разработку целого семейства
диагностических методов на основе молекулярно-биологических подходов.
Новые знания о молекулярных маркерах вирусных инфекций, их динамике в тече-
ние инфекционного процесса дали возможность дифференцировать клинические
фазы заболевания — от латентной стадии до хронической инфекции или состояния
реконвалесценции. Характерные маркеры и их комбинации, а также период их при-
сутствия в крови пациента могут быть использованы для контроля эффективности
лечения и прогнозирования клинического исхода инфекции. В дополнение к тради-
ционному критерию острой стадии инфекции ио IgM-иммунному ответ}' найдены и
апробированы подходы, основанные на оценке авидности антител.
Присутствие низкоавидных антител в сыворотке крови пациента используют в
качестве показателя острой краснушной, герпетической, ВЭБ-инфекции и др.
Технологии специфической диагностики вирусных инфекций развивались
опережающими темпами по сравнению с другими видами лабораторной диагно-
стики Массовый характер к цинических и скрининговых исследований потребовал
создания бысп рых, точных, специфичных тестов высокопроизводительных авто-
матизированных технических средств для их осуществления. Массовые исследова-
ния по выявлению ВИЧ-инфекции и вирусных гепатитов стали двигателем общего
прогресса в области иммуноферментного анализа (ИФА) и методов, основанных
на полимеразной цепной реакции (ПЦР). В настоящее время повсеместно идет
оснащение лабораторий современным оборудованием для ИФА и ППР, В перспек-
тиве — появление технических средств для компьютеризированной биочиповой
диагностики вирусных инфекций.
Возрастающее многообразие вирусных инфекций и их клинических форм, с
одной стороны, и широкий круг доступных аналитических тестов — с другой,
усложняют алгоритм диагностики и в то же время позволяют решать постоянно
растущий перечень диагностических задач. Задачи лабораторной диагностики
вирусных инфекций.
• Этиологическая расшифровка инфекционного заболевания.
• Идентификация биологического вида вируса-возбудителя.
• Идентификация генотипа, субтипа, госпитального штамма.
• Идентификация онкогенных генотипов вируса.
• Идентификация вакцинных и «диких» вариантов вируса.
• Дифференциация острой (первичной), ранее перенесенной инфекции или
состояния постпрививочного иммунитета.
544 ВИРУСОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ
• Дифференциация клинических фаз инфекции.
• Дифференциация обострений и состояния ремиссии.
• Кош роль эффективности лечения вирусных инфекций.
• Количественное определение вирусной нагрузки.
• Выявление С'ПИД-индикаторных и СПИД-ассоциированных инфекций.
• Выявление микст-инфекции.
• Выявление оппортунистических инфекций у пациентов после транспланта-
ции органов и тканей.
• Прогноз клинического исхода инфекции.
Задачи донозологической диагностики вирусных инфекций.
• Скрининговые и профилактические исследования по выявлению ВИЧ-
инфекции:
❖ у доноров:
Ь беременных;
пациентов с заболеваниями, передающимися половым путем;
• о лиц. употребляющих наркотические вещества:
❖ пациентов, обследуемых по клиническим показаниям;
❖ лиц, обследуемых при эпидемиологических расследованиях;
❖ лиц, находящихся в местах лишения свободы;
❖ медицинского персонала;
> иностранных граждан.
« Скрининговые и профилактические исследования по выявлению вирусных
гепатитов В и С в группах риска.
Задачи лабораторной пре-, пери- и постнатальной диагностики инфекционных
заболеваний.
• Выявление или исключение инфекции TORCH-комплекса, вирусных гепати-
тов В и С, ВИЧ-инфекции, сифилиса, хламидиоза, гонококковой инфекции,
листериоза у беременных.
• Определение иммунной защищенности беременных против инфекций
TORCH-комплекса (токсоплазмоза, краснухи, цитомегаловирусной инфек-
ции, герпеса).
• Определение группы риска перинатальной инфекции TORCH-комплекса.
• Выявление внутриутробной инфекции.
• Обоснование медицинских показаний к искусственному прерыванию бере-
менности в связи с выявлением инфекционных заболеваний беременных и
перинатальной инфекции.
• Выявление рисков интранатального инфицирования плода.
• Дифференциация пери-, интра- и постнатальной инфекции новорожденных.
Многообразие и сложность задач, которые решает современная лабораторная
диагностика вирусных инфекций, свидетельствуют, что она давно перестала быть
средством подтверждения диагноза. В возрастающем числе случаев она становится
инструментом первичного выявления инфекции (в манифестной ее форме и тем
более в атипичной или субклинической). Более того, лабораторное заключение все
чаще становится основанием для окончательного медицинского диагноза. Следует
подчеркнуть, что в полной мере этого достигают при соблюдении ряда обязатель-
ных условий, к числу которых относятся:
• интеграция лабораторной диагностики с другими клиническими дисциплинами;
• соблюдение правил организации диагностических лабораторий:
• наличие системы внутрилабораторного и внешнего контроля качества;
• взаимодействие лабораторных служб лечебно-поофилактических и санитар-
но-противоэпидемических учреждений;
• рациональное сочетание классических и новых лабораторных методик;
• готовность лабораторий к модернизации технологий и лабораторной техники.
Рис. 22-1. Характерное расположение М. tuberculosis в препарате в виде жгутов и кос (окраска по
Цилю-Нильсену).
Рис. 23-12. Специфическая флюоресценция в клетках, инфицированных вирусами: а — герпеса
II типа; б — гриппа A (H.N2).
Рис. 23-14. Схема гибридизационного анализа в растворе с захватом гибридов «зонд-мишень»
моноклональными антителами: а — после тепловой денатурации образуются гибриды синте-
тического РНК-зонда (красный) с молекулой-мишенью ДНК (синий); б — РНК-ДНК-гибриды
захватываются сорбированными на твердой Фазе анти-РНК-ДНК-Ат; в — добавляют вторые
анти-РНК-ДНК-Ат, меченные щелочной фосфатазой, в реакционный объем приливают субстрат;
образующийся продукт выявляют хемилюминесцентной реакцией (технология Digene Corp.,
Qiagen Coro).
Рис. 23-18. Схема исследования по методу разветвленной ДНК (пояснения см. в тексте).
ВИРУСОЛОГИЧЕСКИЕ исследования 545
ОРГАНИЗАЦИЯ РАБОТЫ ЛАБОРАТОРИИ ПО ДИАГНОСТИКЕ
ВИРУСНЫХ ЗАБОЛЕВАНИЙ
Работа по лабораторной диагностике инфекционных вирусных заболеваний
подразумевает исследование клинического материала для диагностики, предупре-
ждения и лечения болезни или оценки состояния здоровья человека. Организация
работы лаборатории, осущесч вляющей лабораторную диагностику инфекционных
вирусных заболеваний, должна обеспечивать биологическую безопасность для
персонала, окружающей среды и пациентов, а также качество исследований и ком-
петентность персонала.
Вопросы биологической безопасности лаборатории регламентируются
Санитарно-эпидемиологическими правилами, устанавливающими требования к
организационным, санитарно-противоэпидемическим мероприятиям при работе
С ПБА. Основные документы, регламентирующие деятельность лабораторий по
показателям безопасности для здоровья, принципам и методикам исследований:
• санитарные нормы (СН), санитарные правила (СП);
• государственные и отраслевые стандарты (ГОСТ, ОСТ);
• методические указания (МУ);
• методические рекомендации;
• методическое письмо;
• инструкции по применению;
• технические условия.
Деятельность каждой лаборатории, каждого структурного подразделения
выстраивается в соответствии с используемой группой ПБА: I—II или III-IV.
Помещения лаборатории подразделяют на «заразную» зону, в которой осущест-
вляют манипуляции с ПБА и их хранение, и «чистую», где не проводят работы с
ПБА. На границе «чистой» и «заразной» зон обязательно располагают санитарный
пропускник. Должны быть предприняты меры для предупреждения перекрестного
заражения. Соответствующим образом разделяют секции лаборатории, в которых
осуществляется деятельность несовместимых видов. Вход в лабораторию устраи-
вают таким способом, чтобы несанкционированный доступ в помещения лабора-
тории был невозможен. Система связи внутри лаборатории должна соответство-
вать размеру и сложности устройства помещений и обеспечивать -*ффективную
передачу сообщений.
Отводят специальные помещения и создают условия для хранения проб,
образцов, микроорганизмов (штаммов), реагентов и расходных материалов (цен-
трализованно и на рабочих местах), регистрационных журналов и результатов
исследований (на бумажных и электронных носителях), необходимых для работы
документов, руководств, методических рекомендаций и др.
Таким образом, устройство отдельных помещений и всей лаборатории в целом,
а также условия работы в лаборатории должны соответствовать выполняемым
задачам с учетом уровня ПБА.
В лаборатории должное внимание следует уделять биологической безопасно-
сти и удалению отходов. В каждой вирусологической лаборатории должна быть
программа производственного контроля, регламентирующая объем и перечень
лабораторных исследований на стерильность воздуха, рабочих поверхностей
и др., характер используемой воды и процедуры для проверки отсутствия отри-
цательного влияния окружающей среды (температура в помещениях, влажность,
запыленность, электромагнитные помехи, радиация, уровень звука, вибрации
и т.п.) на проведение исследований и работу персонала, При отсутствии возмож-
ности выполнения указанных исследований собственными силами следует при-
влекать центры государственного санитарно-эпидемиологического надзора для
осуществления исследований и испытаний. В отношении хранения диагностиче-
546 ВИРУСОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ
ских препаратов и реагентов в лаборатории необходимо разработать программу
«холодовой цепи».
Лабораторное оборудование должно соответствовать целям и задачам, включая
подготовку и обработку проб, спецификацию исследований различными методами и
хранение При установке и повседневном применении оборудования строго выпол-
няют эксплуатационные требования в соответствии с техническими паспортами.
Руководство лаборатории разрабатывает программы, обеспечивающие регулярную
поверку поиборов, реагентов и аналитических систем, в том числе программы профи-
лактического обслуживания приборов. Действующие инструкции по использованию
и эксплуатации приборов должны быть легкодоступны персоналу лаборатории.
В лаборатории необходимо принимать адекватные меры для обеззараживания
оборудования перед использованием, восстановлением или списанием и после
них. При использовании компьютеров или автоматических анализаторов (для
регистрации, обработки, передачи, хранения или поиска данных исследований)
лаборатория должна обеспечить следующие условия:
J компьютерные программы, включая встроенные в оборудование, должны
быть документированы и оценены как пригодные к использованию;
необходимо разработать процедуры для защиты сохранности данных в
любое время;
J компьютерные программы следует защищать от несанкционированного
доступа, повреждений и разрушений случайными (необученными) лицами.
Оборудование, включая компьютеры и компьютерные программы, стандартные
образцы, реагенты и аналитические системы необходимо защищать от поправок и
подделок, которые могут исказить результаты исследований,
В лаборатории должна быть разработана и внедрена действенная система
управления качеством лабораторных исследований, включающая внутрилабора-
торный и внешний конгроль качества. Все мероприятия по контролю качества
излагают в документе «Руководство по качеству», которое создают в каждой
отдельной лаборатории. Возможно назначение лица, ответственного за качество.
«Руководство по качеству» должно определять политику, процессы, процедуры,
инструкции, корректирующие действия при выявлении исследований, не соответ-
ствующих требованиям.
Принципы применения лабораторных методов
диагностики вирусных инфекций
Цель специфической лабораторной диагностики состоит в определении в кли-
ническом материале возбудителя заболевания (его антигена, генома) и/или анти-
тел к возбудителю инфекции в образцах периферической крови. Тестирование воз-
будителя возможно на ранних сроках заболевания, сразу после инкубационного
периода, в то время как обнаружение антител (в плане диагностики) несколько
отсроченно, так как для их формирования должно пройти определенное время.
Основная задача — ранняя диагностика с целью этиологической расшифровки
заболевания для определения тактики лечения, проведения эпидемиологических
мероприятий в очаге и др. Для достижения указанной цели необходимо знать пато-
генез заболевания, а также пути передачи инфекции и механизмы заражения.
Для обнаружения возбудителя в клиническом материале используют сле-
дующие методы диагностики:
• электронную микроскопию, позволяющую обнаруживать вирионы возбуди-
теля в клиническом материале;
• выделение вируса, позволяющее выявить и накопить возбудитель; реализу-
ется путем заражения клиническим материалом от больного чувствительных
культур клеток, куриных эмбрионов или лабораторных животных;
ВИРУСОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ 547
• метод флюоресцицующих антител (МФА), позволяющий напрямую опреде-
лять возбудителя заболевания в нативных соскобах. секционном материале;
• метод иммуноферментного анализа (ИФ^), позволяющий идентифициро-
вать антигены возбудителя в клиническом материале;
• метод полимеразной цепной реакции (ПЦР), позволяющий обнаруживать
нуклеиновые кислоты (РНК, ДНК) генома возбудителя в клиническом мате-
риале.
Выбор метода выявления возбудителя зависит от целей, задач и оснащенности
лаборатории.
Электронная микроскопия имеет бесспорные преимущества в случае ? новых»
заболеваний, для лабораторной диагностики которых не разработано диагности-
ческих средств. Трудоемкий процесс выделения вируса требует больших мате-
риальных затрат (заражение культуры ткани, куриных эмбрионов, содержание
вивария для заражения лабораторных животных) и времени. Оба указанных
метода, как правило, редко используют в диагностических лабораториях, чаще — в
крупных научно-исследовательских институтах.
Такие методы, как МФА, ИФА и ПЦР. широко практикуются в различных диа-
гностических лабораториях. Применение МФА, достаточно простого и быстрого
диагностического метода, ограничивается лишь методикой забора материала. От
правильного забора материала, который осуществляют, как правило, клиницисты,
зависит достоверность результата, так как для исследования необходимо доста-
точное количество зараженного эпителия (носоглоточных соскобов при ОРВИ,
урогенитальных соскоблв при ИППП). Метод ИФА, позволяющий обнаружить
антиген, используют для диагностики тех заболеваний, которые характеризу-
ются накоплением огромного количества возбудителя в клиническом материале
(например, HBsAg в сыворотке крови или ротавирусов в фекалиях).
Наиболее перспективным для определения возбудителя в биологических суб-
стратах путем обнаружения его генома (РНК, ДНК) считают метод ПЦР, который
в настоящее время широко применяют в диагностике СПИДа, гепатитов, герпети-
ческих. клещевых инфекций, инфекций, передаваемых половым путем (ИППП),
гриппа A/В, включая высокопатогенный грипп А птиц (H5/II7), грипп А свиней
(Hl-swine).
Для определения специфических антител, формирующихся в организме
человека в течение болезни в целях элиминации возбудителя, используют раз-
личные методы:
• эритроцитарные — РТГА, РПГА. РИГА, РСК (определение титров антител);
• иммуноферментного анализа (ИФА) (определение классов антител — IgM,
IgG, IgA).
Классические серологические эритроцитарные методы тестирования антител
базируются на титровании испытуемых сывороток и определении титров антител к
возбудителю в образцах крови пациентов. Лабораторный диагноз ставят на основа-
нии диагностического титра антител (РСК) или нарастания (сероконверсии) титров
антител (четырехкратного и более) в парных образцах крови (РТГА, РСК и др.).
В настоящее время серологические эритроцитарные методы используются все
реже из-за нестандартности основного компонента реакции — эритроцитов крови
различных животных: куриных — для диагностики гриппозной и парагриппозной
инфекций методом РТГА, морской свинки — для диагностики эпидемического
паротита, голубиных — для диагностики краснухи и т.д.
По этой причине для получения достоверных результатов следует выполнять
два основных правила:
исследовать парные сыворотки антител (забранные в остром периоде
болезни, при первичном обращении пациента и через 10-14 дней от взя-
тия первого образца крови);
548 ВИРУСОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ
обе сыворотки (первую и вторую, взятые с интервалом в две недели)
исследовать одновременно, после поступления в лабораторию второго
образца крови.
Несмотря на некоторые недостатки (нестандартность эритроцитов, необхо-
димость предварительной обработки сывороток для разрушения неспецифиче-
ских ингибиторов, огсроченность лабораторного диагноза из-за взятия второй
сыворотки, невозможность определения классов выявляемых антител), при ряде
инфекций, например при диагностике гриппа и ОРВИ, РТГА до сих пор остает-
ся основным и единственным методом серологической диагностики указанных
заболеваний. Современный ИФА позволяет тестировать антитела лишь к вирусам
гриппа А, не различая их по штаммам, тогда как РТГА раздельно выявляет анти-
тела к вирусам гриппа А — A (H^N , HN , Н Nj-swine), A., (H3N2) и др., обнару-
живает и более тонкие штаммовые различия циркулирующих вирусов гриппа А,
претерпевающих постоянные дрейфовые изменения.
Метод ИФА. в отличие от эритроцитарных методов, позволяет определять
классы специфических антител (IgM- и IgG-антитела), что составляет его главное
преимущество и обеспечивает более быструю лабораторную диагностику по одно-
му образцу крови. Кроме того, все компоненты системы ИФА стандартизованы.
Тем не менее для получения достоверных результатов серологической лаборатор-
ной диагностики методом ИФА и его разновидностей (ИХЛ, ИХЭЛ) необходимо
придерживаться следующих правил.
• Для выявления IgM-антител предпочтительнее использовать технологии
ц-capture (ц-захвата) в целях снижения риска получения ложноположитель-
ных результатов,
• Одновременно определять в сыворотке IgG- и IgM-антитела (для достовер-
ности и снижения количества ложноположительных результатов тестирова-
ния IgM-антител).
• Применять количественные методы оценки уровня IgG-антител (для выявле-
ния стадии реактивации инфекционного процесса, особенно при отсутствии
IgM-антител, эффективности поствакцинального иммунитета).
• Использовать тест на авидность IgG-антител (в целях установления давности
заболевания и достоверности определения IgM-антител).
ОБЩИЕ СВЕДЕНИЯ О ВИРУСАХ
Структурные компоненты и химический состав вирусов
Характеристика структуры и химического состава вирусов дана применительно
к вириону как наименьшей составляющей (элементарной частицы) вируса.
Простые вирионы представлены двумя составляющими: нуклеиновой кислотой
и белком — капсидом. Термин «капсид» произошел от латинского capsa — «фут-
ляр». Нуклеиновая кислота простых вирионов включена в капсид и непосред-
ственно связана с ним, поэтому используют термин «нуклеокапсид». В состав
сложных вирионов, помимо капсида, входит двухслойная оболочка — фрагмент
мембраны клетки-хозяина,
В составе вириона присутствует только один тип нуклеиновой кислоты. Вирусный
геном может быть представлен одно- или двухцепочечными (одно- или двухнитевы-
ми) молекулами РНК или ДНК. Нуклеиновые кислоты могут быть как линейными
так и кольцевыми. Если геном вируса состоит не из одной, а из нескольких молекул
нуклеиновой кислоты, речь идет о сегментированном геноме.
В структуре геномной ДНК присутствует несколько участков, функции которых
заключаются в обеспечении репликации, транскрипции и транспорта вирусной
ВИРУСОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ 549
ДНК к месту ее упаковки в капсид. Основной репликативной формой ДНК служат
двухцепочечные кольцевые молекулы, что далеко не всегда совпадает с организа-
цией вирионной ДНК. Вирионная форма ДНК содержит определенные последо-
вательности, которые в результате рекомбинации переводят ее в репликативную
форму. Генетическая информация может быть сосредоточена последовательно
на одной кодирующей цепочке ДНК или мозаично — на двух цепях молекул.
Протяженность ДНК различных вирусов широко варьирует — от 2-3x103 до
2,5x10s нуклеотидных оснований.
Размер вирусной РНК редко превышает 2-Зх104 нуклеотидных основа-
ний. РНК как носитель генетической информации — уникальная особенность
вирусов. РНК могут обладать положительной (позитивной) полярностью, что
обеспечивает синтез вирусспецифических белков на рибосомах клеток, либо
отрицательной (негативной) полярностью. В первом случае функцию инфор-
мационной РНК выполняет сам геном вируса. Во втором случае для синтеза
вирусных белков необходимы комплементарные РНК-транскрипты, которые
являются информационными и синтезируются в процессе транскрипции (то
есть синтеза цепочек РНК, комплементарных геномной) при участии вирусной
транскриптазы.
Особую группу составляют двухнитевые РНК-содержащие вирусы (семей-
ство Reoviridae), в частности ротавирусы. Вторичная структура РНК этой группы
вирусов представляет собой двойную спираль, основанную на комплементарных
взаимодействиях между цепями. Двухспиральные РНК обладают более высокой
термостабильностью, чем ДНК, что обусловливает устойчивость этой группы
вирусов к температурному фактору. Вторая их особенность — фрагментарность
генома. Так, геном ротавируса содержит 11 фрагментов.
Особое положение занимают и представители семейства Retroviruiae. имеющие в
составе генома две идентичные молекулы РНК (диплоидный набор).
Генетическая информация, заложенная в ДНК или РНК вириона, может обеспе-
чить формирование как единичных структурных белков (например, трех у вируса
краснухи), так и большого их количества (более 30 у вируса осповакцины).
Укладка нуклеиновых кислот в вирионе строго упорядочена и осуществляется
путем образования комплексов с белком капсида.
Капсид построен из белковых субъединиц, которые собраны строго определен-
ным образом в соответствии с относительно простыми геометрическими принци-
пами. Существуют два типа организации, при которой идентичные асимметричные
субъединицы, такие как молекулы белка, могут соединиться друг с другом таким
образом, чтобы находиться на равном расстоянии от некоего центра: спираль-
ная сборка или формирование замкнутых белковых оболочек. Соответственно
существуют два вида капсидов: спиральные и замкнутые, то есть изометрические.
Капсиды всех вирусов относятся к одной из этих категорий.
Спиральные капсиды без внешней оболочки содержатся в вирионах многих рас-
тений и некоторых фагов. У представителей ряда семейств РНК-содержащих виру-
сов позвоночных (Paramixoviridae, Rabdoviridae, Coronavlridae и др.) спиральный
нуклеокапсид находится в полости, образованной в процессе почкования через
плазматическую мембрану клетки. Эта внешняя оболочка состоит из белковой и
липидной оболочек.
Изометрические (квазисферические) капсиды по форме почти идентичны сфере.
Однако при электронной микроскопии показано, что капсиды подобной организа-
ции представлены не сферой, а правильными многогранниками. Икосаэдрические
капсиды РНК-содержащих вирусов позвоночных семейств Astroviridae, Caticiviridae,
Nodaviridae, Picomoviridae не имеют липидной оболочки. У представителей семей-
ства Togaviridae капсид окружен липидной оболочкой.
550 ВИРУСОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ
Особая структура ну клеокапсида отмечается у вириона РНК-содержащего
вируса иммунодефицита человека ЗИЧ-1 (семейство Retroviridae). Он обладает
кубической симметрией ц. согласно современным представлениям, имеет форму
усеченного конуса в отличие от других вирусов с икосаэдрической структурой
нуклеокапсида. Для ДНК-содержащих вирусов позвоночных характерен икосаэ-
дрический нуле леока пси д.
Вирусспецифические белки разделяют на структурные, входящие в состав
вириона, и неструктурные, принимающие участие в репродукции вируса в инфи-
цированной клетке, но не входящие в состав зрелого вириона.
Структурные белки можно разделить на капсидные и сунеркапсидные. Капси-
дные белки простых вирусов имеют низкую молекулярную массу (5-15 кДа) и обра-
зуют в комплексе с нуклеиновой кислотой нуклеокапсид. Следующая группа капсид-
ных белков — среднего размера полипептиды (молекулярная масса — 15-40 кДа),
которые выполняют защитную для нуклеокапа иного комплекса функцию.
Функции поверхностных гликопротеинов различны. Они могут обладать гемаг-
глютинирующей, нейраминидазной активностью, содержат эпитопы, ответствен-
ные за выработку различных видов антител. Поверхностные белки обеспечивают
адсорбцию вируса на чувствительных клетках, а также участвуют в сборке зрелых
вирионов.
Внешние оболочки сложных вирусов позвоночных формируются в составе
плазматической или ядерной мембраны клетки. Для обозначения липопротеино-
вой оболочки вириона применяют термин «пеплос» (от греч. — «мантия»), а для
поверхностных белков (гликопротеинов), выступающих над поверхностью пепло-
са, — термин «пепле меры». Толщина двухслойной липидной мембраны составляет
в среднем 5-6 нм.
Химический состав вириона определяется, помимо нуклеиновых кислот и бел-
ков, наличием в его структуре липидов и полисахаридов. Липиды входят в состав
сложных вирусов, локализуются, как правило, в оболочке. Состав липидов отра-
жает тип клеток, использованных вирусом для репродукции. В среднем липиды
различных вирионов обеспечивают 15-35% сухого веса вирусной частицы и пред-
ставлены в основном фосфолипидами (до 60-70%). Основные функции липидной
оболочки — стабилизация структуры вириона и защита от воздействия факторов
внешней среды. Липиды также определяют конформацию поверхностных белков.
Обработка липидсодержащих вирусных суспензий органическими растворителя-
ми приводит, за некоторым исключением (семейства Poxviridae, Hepadnoviridac), к
потере инфекционности вирусных частиц.
Углеводный состав вириона определяется поверхностными гликопротеинами и
представлен преимущественно глюкозой, галактозой, маннозой, N-ацетилнечра-
миновой кислотой, N-ацетилглюкозамином, N-ацетилгалактозамином, фукозой.
Суммарная доля углеводов в вирионах различных вирусов может достигать
16-13%.
Принципы классификации, таксономия вирусов
Первые попытки классифицировать вирусы предпринимались в 1940-х гг. по
принципам, принятым в микробиологии в отношении бактерий. Эта система ока-
залась несостоятельной. В 1966 г. на Международном микробиологическом кон-
грессе в Москве был учрежден Международный комитет по номенклатуре вирусов
(МКНВ). В 1973 г. он был переименован в Международный комитет по таксоно-
мии вирусов (МКТВ), или International Committee on Taxonomy of Viruses (ICTV).
МКТВ кодифицирует и поддерживает систематику вирусов и таксономическую
базу данных — The Universal Virus Database (ICTVdB).
ВИРУСОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ 551
Была выбрана следующая схема подразделения царства вирусов: отдел, класс,
порядок, семейство, подсемейство, род и вид.
Порядок в настоящее время является высшим таксоном для вирусов. В поря-
док объединяются семейства, обладающие определенными общими свойства-
ми. Семейства выделяются на основании общности происхождения вирусов.
Таксономическую группу подсемейства можно использовать только при необхо-
димости решения иерархических задач. Семейство (или подсемейство) состоит
из родов. Род — группа родственных видов вирусов, обладающих сходными свой-
ствами, часто различающихся только по кругу восприимчивых хозяев или уровню
патогенности.
Порядок, семейство, подсемейство, род обозначаются одним словом, которое
пишется с заглавной буквы и заканчивается на -virales, -viddae, -wrinae и-virus соот-
ветственно. Наименование вида может содержать несколько слов, но не должно
состоять только из слова virus или названия хозяина. Первое слово названия вида
пишут с заглавной буквы.
В основу классификации вирусов положены их морфологические, физико-
химические, молекулярные и экологические особенности. Главными критериями
служат тип и структура нуклеиновой кислоты (РНК или ДНК), наличие липопро-
теиновой оболочки, стратегия вирусного генома, размер и морфология вириона,
круг восприимчивых хозяев, географическое распространение, антигенные свой-
ства.
На основании перечисленных признаков до 2007 г. выделяли три порядка —
Caudovirales, Mononegavirales и Nidovirales. В 2008 г. были добавлены порядки
Herpesvirales и Picornavirales, а в 2009 г. — порядок Tymovirates.
Порядок Caudovirales образован тремя семействами: Му overdue, Podovmdae и
Siphovindae. Их представители являются патогенами архей и бактерий и содержат
одну молекулу линейной двухнитевой ДНК.
Порядок Mononegavirales включает семейства Bo/naviridae, Filoviridae,
Paramyxoviridae и Rhabdoviridae. Вирусы этого порядка имеют оболочку и линей-
ную несегментированную однонитевую РНК негативной полярности.Три семей-
ства — Coronaviridae, Arteriviridae и Ronivitidae — объединены в порядок Nidovtrales
и характеризуются линейной несегментированной однонитевой РНК позитивной
полярности. Поражают позвоночных животных.
К порядку Herpesvirales относятся семейства Alloherpesviridae, Herpesvtridae,
Malacoherpesviridae. Представители порядка содержат единственную несегмен-
тированную линейную двухнитевую молекулу ДНК и имеют сложное строение
вирусной частицы.
Порядок Picornavirales, по данным 2009 г,, объединяет пять семейств: Dicistro-
viridae (два рода), Iflaviridae (один род), Marnaviridae (один род), Picornavtridae
(12 родов) и Secoviridae (7 родов). Общими для всех представителей этого порядка
являются отсутствие оболочки и однонитевая РНК позитивной полярности.К
порядку Tymovirates относятся четыре семейства: Alphaflexiviridae (6 родов),
Betaflexiviridae (7 родов), Gammaflexiviridae (один род), Tymovvidae (три рода).
Вирусы этого порядка поражают преимущественно растения.
Ни в один из известных порядков не входит 66 семейств вирусов.
База данных основных семейств и родов вирусов, представители которых
вызывают заболевания у позвоночных животных и человека. ICTVdB (2009)
(табл. 23-1), обновляется по мере накопления новых сведений, поэтому таксоно-
мическое положение некоторых вирусов пересматривается. В частности. в 2008 г.
Human coronavirus ОС43 и Human enteric coronavirus относились к роду Coronavirus,
который в настоящее время получил статус подсемейства.
Таблица 23-1. Основные семейства и роды, представители которых вызывают заболевания у позвоночных животных и человека (ICTVdB, 2009)
Геном Порядок Семейство, подсемейство Род Вирусы, патогенные для человека
Двухцепочечная ДНК Herpesvirales Alloherpesviridae ictalurivirus -
Batrachovirus -
Cyprinivirus -
Ictalurivirus -
Salmonivirus -
Herpesviridae Alphaherpesvirinae 1 Ito virus -
Mardivirus -
Simplexvirus Вирус герпеса человека 1, 2 (ВПГ 1, ВПГ 2)
Vari cellovirus Human herpesvirus 3 (герпес-зостер, вирус герпеса челове- ка 3)
Unassigned -
Betaherpesvirinae Cytomegalovirus Human herpesvirus 5 (цитомегаловирус, вирус герпеса чело- века 5-го типа)
Muromegalovirus -
Proboscivirus -
Roseolovirus Human herpesvirus 6 (вирус герпеса человека 6-го типа)
Human herpesvirus 7
Unassigned -
Ga mmaherpes virina e Lymph ocrypto virus Human herpesvirus 4 (вирус Эпштейна-Барр, или вирус гер- песа 4-го типа)
Macavirus -
Perea virus -
Rhadinovirus
Unassigned -
Unassigned Unassigned -
Malacoherpesviridae Ostreavirus -
552 ВИРУСОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ
Продолжение табл. 23-1
Геном Порядок Семейство, подсемейство Род Вирусы, патогенные для человека
Семейства, не классифи- цированные по порядку Adenoviridae Atadenovirus -
Aviadenovkus -
Ichtadenovkus -
Maslaaenovirus Human adenovirus А, В, C, D, E. F (аденовирус человека С, аденовирусы человека 1-3, 7, 8,12,18, 31-го типов)
Siadenovkus -
Asfarviridae Asfivkus -
Indoviridae Chloriridovkus -
Iridovkus -
Lymphocystivirus -
Megalocytivkus -
Ranavirus -
Mimivkidae Mimivirus АРМУ
Papillomaviridae AlphapapHlomavirus Human papillomavirus 2,6, 7,10,16,18,26, 32, 34 53,54.61,71 ™ Human papillomavirus-cand 90 5
Betapapillomavkus Human papillomavirus 5. 9, 49 g Human papillomavirus-cand 92, cand 96 g
Deltapapillomavirus “ m
Epsilonpapilloma virus — §
Etapapillomavirus
Gammapapilloma virus Human papillomavirus 4, 48, 50. 60 88 ё
lotapaplllomavirus g
Kappapapdlomavkus — 5
Lambdapapitloma virus
Mupapiilomavirus Human papillomavirus 1,63
Mupapiilomavirus Human papillomavirus 41 сл
Продолжение табл. 23-1
ш
Геном Порядок Семейство, Род Вирусы, патогенные для человека
подсемейство
Omicronpapillomavirus -
Pipapillomavirus -
Thetapapillomavirus -
Xipapillomavirus -
Zetapapilllomavirus -
Poiyomaviridae Polyomavirus Human polyomavirus
Poxviridae Avipoxvirus -
Chordopoxvirinae Capripoxvirus -
Cervidpoxvirus -
Leporipoxvirus -
Molluscipoxvirus -
Orthopoxvirus Вирус осповакцины
Parapoxvirus Вирус паравакцины
Suipoxvirus —
Yatapoxvirus -
Unassigned -
Entomopoxvirinae Alphaentomopoxvlrus -
Betaentomopoxvirus -
Gammaentomopoxvirus -
Unassigned -
ВИРУСОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ
сл
Продолжение табл. 23-1
Геном Порядок Семейство, Род Вирусы, патогенные для человека
подсемейство
Одноцепочечная ДНК Семейства, не классифи- цированные по порядку Circoviridae Circovirus -
Cyrovirus -
Parvoviridae Bwvidensovirus -
Densovirinae Densovirus -
itera virus -
Pefudensovirus -
Parvovirinae Amdowrus -
Boca virus Бокавирусы
Dependovirus -
Erythrovirus Human parvovirus В19 (парвовирус В19)
Parvovirus -
Unassigned Anellovirus -
Двухцепочечная ДНК с обратной транскриптазой Семейства, не классифи- цированные по порядку Hepadnaviridae Avihepadnavirus -
Orthohepadnavirus Hepatitis В virus (вирус гепатита В человека)
Двухцепочечная РНК Семейства, не классифи- цированные по порядку Birnaviridae Aquabirnavirus S
Avibimavirus 2=3 - о
Blosnavirus - _с
Entomobirnavirus
Reovindae Cardoreovirus S
Sedoreovirinae Mimoreovirus 3
OrSvirus g
Phytoreov/rus от
Rotavirus
Seadornavirus -
BIBS
Продолжение табл. 23-1
Геном Порядок Семейство, Род Вирусы, патогенные для человека
подсемейство
Spinareovirinae Aquareovirus -
Coltivirus -
Cypovirus -
Dinovernavirus -
Fijivirus -
Idnoreovirus -
Mycoreovirus -
Orthoreovirus -
Oryza virus -
Одноцепочечная РНК негативной полярности Mononegav/rales Bornavindae Bornavirus -
Filovirirfae Ebolavirus Вирус тамды Вирус Заира (Эбола)
Marburgvirus Вирус озера Виктория
Paramyxoviridae Avulavirus -
Paramyxovirinae Henipavirus Вирус Нипах
Вирус Хендра
Morbillivirus Measles virus (вирус кори)
Respirovirus Human parainfluenza virus 1 (вирус парагриппа 1-го типа) Human parainfluenza virus 3 (вирус парагриппа 3-го типа)
Rubulavin/s Human parainfluenza virus 2 Human parainfluenza virus 4 (вирусы парагриппа 2-го и 4-го типа) Mumps virus (вирус эпидемического паротита)
Pneumovirinae Metapneumovirus Human metapneumovirus (метапневмовирусы)
Pneumovirus Human respiratory syncytial virus (респираторно- синцитиальный вирус человека)
ВИРУСОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ
Геном Порядок Семейство,
подсемейства
Rhabdoviridae
Семейства, не классифи- цированные по порядку Arenaviridae
Bunyaviridae
Orthomyxoviridae
Продолжение табл. 23-1
— Род Вирусы, патогенные для человека
Cytorhabdovirus -
Epnemerovirus -
Lyssavirus Вирус бешенства
Novirhabdovirus -
Nucleorhabduvirus
Vesiculovirus -
Unassigned -
Arenavirus Вирус Мачупо (Боливийская лихорадка) Вирус Хунин (Аргентинская лихорадка) Вирус Гуанарито (Венесуэльская лихорадка) Вирус Сабиа (Бразильская геморрагическая лихорадка) Вирус лимфоцитарного хориоменингита
Hantavirus Вирус Хантаан, Пуумала, Добрава, Сеул, Син-Номбре
Nairovirus Вирус крымской геморрагической лихорадки
Orthobunyavirus Вирусы Инко, Тягиня, зайца-беляка
Phlebovirus Вирус лихорадки долины Рифт
Tospovirus О
Infiuenzavirus A influenza A virus (грипп А субтипов Н1N1, H2N2, H3N2)
Influenzavirus В Influenza В virus (грипп В)
Influenzavirus C Influenza С virus (грипп С) §
Isavirus - S
Thogotovirus - §
§
ЖМШ® «Ж
Продолжение табл. 23-1 J1
Геном Порядок Семейство, подсемейства Род Вирусы, патогенные для человека
Одноцепочечная РНК с обратной транскриптазой Семейства, не классифи- цированные по порядку Retroviridae Alpharetrovirus -
Orthoretrovirinae Betaretrovirus 8
Deltaretrovirus _ о
Epsilonretrovirus Вирус саркомы кожи
Gammaretrovirus
Lentivirus Human immunodeficiency virus 1 == Human immunodeficiency virus 2 (вирусы иммунодефицита g человека 1,2) g
Spumaretrovirinae Spumavirus Пенящийся вирус человека g
Одноцепочечная РНК позитивной полярности Nidovirales Arterivirldae Arterivirus X - s: из
Coronaviridae Coronavirinae Alphacoronavirus Human coronavirus 229E Human coronavirus NL63
Betacoronavirus Human coronavirus HKU1
Gammacoronavirus -
Torovirinae Bafinivirus -
Torovirus Human Torovirus
Roniviridae Okavirus -
Dicistroviridae -
Picornavirales Iflaviridae -
Marnaviridae -
Picornaviridae Aphthovirus -
Avihepatovirus -
Cardiovirus B«.pyc Сырдарьи
Энцефагомиокардит
Геном
Порядок
Семейство,
подсемейства
Secoviridae
Astroviridae
Семейства, не классифи-
цированные по порядку
Caliciviridae
Продолжение табл. 23-1
Род Вирусы, патогенные для человека
Enterovirus Human enterovirus А, В, C, D Human rhinovirus A, В ЕСНО-вирусы (31-й тип, наиболее актуальные — 11-й и 31-й) Е70, Е71 Коксаки А (23 типа, наиболее актуальные — 9,16, 24-й типы) Коксаки В (6 типов) Полиовирусы Полиомиелит 1-3-го типа
Erbovirus -
Hepatovirus Hepatitis A virus
Kobuvirus -
Parechovirus Human parechovirus (парэховирус человека)
Sapelovirus -
Senecavirus —
Teschovirus -
Tremovirus "" ОТ
Avastrovirus ~ §
Mamastrovirus Human astrovirus (5 типов астровируса человека) m
Lagovirus
Nebovirus
Norovirus Норфолк (4 типа) §
Sapovirus Т=з О
Vesivirus - 5
сл
Окончание табл. 23-1
Генам Порядок Семейство, подсемейство Род Вирусы, патогенные для человека
Flaviviridae Flavivirus Вирус геморрагической лихорадки Алхурма Вирус желтой лихорадки Вирус киассанурской лесной болезни Вирус Денге 4-го типа Вирус омской геморрагической лихорадки
Hepacivirus Hepatitis С virus (гепатит С)
Pestivirus -
Nodaviridae Alphanodavirus -
Betanodavirus -
Togaviridae Aiphavirus Вирус Карельской лихорадки Вирус Чикунгунья Semliki Forest virus
Rubivirus Rubella virus (вирус краснухи)
Hepeviridae Hepe virus Hepatitis E virus (гепатит E)
Unassigned Deltavirus Hepatitis delta virus (дельтавирус)
ВИРУСОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ
ВИРУСОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ 561
Кроме крупных таксономических категорий, используют понятия «штамм»,
«изолят», «вариант», «кластер» для разделения «особей» внутри вида. Для удоб-
ства систематизации каждому вирусу присваивают цифровой код, состоящий из
8 групп по одной, две или по три цифры в каждой. Первая группа обозначает поря-
док, вторая — семейство, третья — подсемейство, четвертая — род, пятая — вид.
шестая — подвид, седьмая — серотип, восьмая группа цифр соответствует номеру
штамма или изолята (табл. 23-2).
Таблица 23-2. Пример присвоения цифрового кода, по данным ICTVdB (2006)
Influenza A virus, A/Puerto Rico/g/jM (Hfl,) isolate
Порядок 00.
Семейство 00.046. Orthomyxovirldae
Подсемейство 00.046.0.
Род 00 046 0.01. Influenzavirus А
Вид 00.046.0.01.001. Influenza A virus
Подвид 00.046.0.01.001.00.
Серотип 00.046.0.01.001.00.001 ИД
Штамм или изолят 00.046.0.01.001.00.001.001. A/Puerto Rico/8/34
КЛАССИЧЕСКИЕ МЕТОДЫ ВЫДЕЛЕНИЯ
И ИДЕНТИФИКАЦИИ ВИРУСОВ
ЛАБОРАТОРНЫЕ ЖИВОТНЫЕ
В современной вирусологии животные продолжают играть важную роль при
изучении вопросов патогенеза и патоморфологии экспериментальных вирусных
инфекций, становления и развития иммунитета, а также при выделении и инди-
кации вирусов Для экспериментальных целей используют, как правило, чисто-
породные линии мышей и других животных, свободных от многих латентных
инфекций и высокочувствительных к экспериментальному заражению различны-
ми вирусами.
Латентные инфекции, котовые могут быть распространены среди лаборатор-
ных животных, характеризуются тем, что возбудитель не проявляет себя кли-
нически в привычных для животного условиях содержания, В случае снижения
резистентности организма возбудитель начинает активно размножаться, вызывая
манифестную форму заболевания. Заражение животных при вирусологических
исследованиях — тот стрессовый фактор, который способен вызвать переход
латентной инфекции в острую.
В практике работы КДЛ манипуляции с лабораторными животными в настоя-
щее время применяют крайне редко, если речь не идет о специализированных
лабораториях по диагностике высокопатогенных зооантропонозных инфекций,
например противочумных станциях или антирабических лабораториях.
РАЗВИВАЮЩИЕСЯ КУРИНЫЕ ЭМБРИОНЫ
Куриные эмбрионы как система для культивирования вирусов известны
с 30-х гг. XX в. и до настоящего времени используются для первичного выделения
вирусов от больных и из объектов внешней среды для последующего культивиро-
вания вирусов в лаборатории, идентификации вирусов и антител, приготовления
вакцин и диагностикумов.
Метод культивирования вирусов в куриных эмбрионах имеет ряд преимуществ
по сравнению с культивированием на лабораторных животных. Данный метод
позволяет избежать трудностей, связанных с возможным спонтанным инфици-
562 ВИРУСОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ
рованием животных рядом вирусных инфекций. Кроме того, при использовании
куриных эмбрионов получают значительно большее количество вируса, чем от
лабораторных животных; потери эмбрионов вследствие гибели при заражении
незначительны и сводятся к минимуму при соблюдении правил асептики, опреде-
ленном навыке работы и соблюдении условий инкубирования. Скорлупа и под-
скорлупная оболочка надежно защищают эмбрион от микробного заражения
извне. Развивающиеся эмбрионы высокочувствительны к большому количеству
вирусов. Кроме того, куриные эмбрионы — легкодоступный материал в связи с
широким развитием промышленного птицеводства, не требуют ухода и кормления
в лабораторных условиях.
Заражение эмбрионов проводят в асептических условиях. Скорлупу над воздуш-
ным пространством обрабатывают йодированным спиртом или фламбируют, то
есть обрабатывают смоченным в спирте и горящим ватным тампоном. Пользуются
стерилизованными инструментами.
Существуют различные способы заражения куриных эмбрионов. Чаще всего
материал вводят в аллантоисную и амниотическую полости, хорионаллантоисную
оболочку, реже — в желточный мешок. Выбор способа заражения, температура
и сроки инкубирования зависят от биологических особенностей вируса и цели
исследования. Вводят 0,1-0,2 мл инфекционного материала Независимо от спо-
соба заражения яйца помещают в специальные подставки тупым концом вверх и
ставят в термостат для дальнейшего культивирования.
О наличии вируса в аллантоисной и амниотической жидкостях, полученных из
зараженных эмбрионов, судят по реакции гемагглютинации, которую ставят либо
капельным методом на стекле, либо титрованием путем двукратных разведений
вируссодержащей жидкости. Индикатором наличия вирусов в хорионаллантоис-
ной и амниотической оболочках служат фокусные специфические поражения или
выявление гемагглютинирующего антигена.
В случае положительных результатов проводят серологическое типирование
выделенного вируса с помощью РТГА, нейтрализации в куриных эмбрионах или
РСК. При получении небольшого количества вируса выполняют дополнительн ые
его пассажи в тех же условиях с последующей индикацией. Если после трех после-
дующих пассажей в эмбрионах вирус в исследуемых материалах не обнаруживает-
ся, результат считают отрицательным.
КЛЕТОЧНЫЕ КУЛЬТУРЫ
Открытие в конце 1940-х гг. Дж. Эндерсом, Т. Уэллером, Ф. Роббинсом воз-
можности культивировать живые клетки вне организма и поддерживать в них
репродукцию вируса полиомиелита стало подлинно революционным событием
для науки в целом и для вирусологии в частности. Культура клеток до сих пор счи-
тается наиболее совершенной лабораторной системой, предназначенной для куль-
тивирования вирусов. Чаще всего в вирусологической практике клеточные куль-
туры используют для обнаружения вирусов и их выделения из патологического
материала, а также для изготовления вакцин и диагностикумов, культивирования
эталонных и вновь выделенных вирусных штаммов в лабораториях и музейных
коллекциях, титрования вирусного материала и в качестве тест-объекта в реакции
нейтрализации.
Для нормального существования и развития клеток вне организма требуются
сложный комплекс физико-химических факторов и выполнение ряда условий.
Первое условие — строгое соблюдение стерильности при работе, поскольку пита-
тельные среды, используемые для культивирования клеток, служат таким же
питательным субстратом для различных микроорганизмов. Вследствие высокой
чувствительности клеток к солям тяжелых металлов и остаткам моющих средств
особое внимание следует уделять качеству воды и всех ингредиентов, составляю-
ВИРУСОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ 563 И
ших основу солевых растворов и питательных сред, а также качеству и способам
обработки посуды, употребляемой для выращивания клеток.
Растворы. Наиболее широко известные и чаше всего применяемые в работе с Щ
клеточными культурами растворы Хенкса и Эрла готовят на бидистиллированной
воде с добавлением различных солей и глюкозы. Ряд государственных и частных
предприятий производит готовые стерильные растворы для работы с культурами
клеток.
Сбалансированные солевые растворы обеспечивают сохранение pH, осмотиче-
ского давления, концентрацию необходимых минеральных веществ. Кроме того,
их используют при приготовлении диспергирующих растворов для снятия клеток
с поверхности, на которой они культивируются:
• 0.25% раствор трипсина на фосфатном буфере;
• раствор Версена — 0,02% раствор натриевой соли этилендиаминтетрауксус -
ной кислоты на растворе Хенкса.
Питательные среды Естественные питательные среды, состоящие из смеси
солевых растворов Хенкса или Эрла, сыворотки, гидролизатов лактальбумина и
крови животных и т.д., в настоящее время используются редко из-за невозмож-
ности точно контролировать количество и качество их компонентов.
В период 1950-1960-х гг. основные усилия исследователей были сосредото-
чены на создании питательных сред и методик перевода клеток из организма в
культуру. Основная проблема заключалась в поиске факторов культивирования,
обеспечивающих жизнеспособность клеток и поддержание их пролиферативной
активности in vitro. Этот трудный и принципиально важный этап завершился в
1955 г. созданием первых синтетических питательных сред Игла и № 199. Эти
среды имели химически точную композицию, включающую более 60 компонен-
тов (20 аминокислот, 17 витаминов, компоненты нуклеиновых кислот, источники
липидов, гормоны, минеральные соли и другие вещества). С добавлением 5-10%
сыворотки крови эти среды обеспечивали эффективный рост многих клеточных
культур. Состав этих сред оказался настолько удачно и точно подобранным, что
среды Игла и № 199 широко используют и в настоящее время.
Наиболее часто применяемые питательные среды для культур клеток: основ-
ная среда Игла — ВМЕ, минимальная среда Игла — МЕМ и MEMS, среда Игла
в модификации Далбекко — DMEM, среда № 199, среда RPMI 1640 и др. Во все
питательные среды и некоторые растворы добавляют индикатор pH феноловый
красный в концентрации, не оказывающей токсичного действия на клетки и виру-
сы, но позволяющей определить факт закисления среды продуктами метаболизма
клеток и необходимость замены ее на свежую. Для предотвращения появления и
развития посторонней микрофлоры в среды добавляют антибиотики: пенициллин
и стрептомицин по 100 ЕД/мл или канамицин по 200 мкг/мл. Для предотвращения
роста плесени рекомендуют натриевую соль нистатина — 100 мкг/мл среды.
Для культивирования клеток, особенно в первые дни роста, используют
ростовые питательные среды, содержащие 2-10% сыворотки крови животных
(чаще всего крупного рогатого скота). Это обязательный компонент ростовых
пита гельных сред, так как в состав сыворотки входит целый набор биологически
активных веществ, необходимых для роста клеток in vitro. Так, фракция альбу-
минов и фетуин способству ют прикреплению клеток к поверхности стекла или
пластика. Кроме того, в сыворотке содержатся разнообразные ростовые факто-
ры, способствующие активному распластыванию, росту и размножению клеток.
Особенно пенной добавкой к питательным средам служит сыворотка эмбрионов
коров и других животных, как правило, не содержащая противовирусных анти-
тел в отличие от сывороток крови взрослых животных. Необходим строгий
контроль сывороток на стерильность и токсические свойства по отношению к
клеточным культурам.
564 ВИРУСОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ
В вирусологической практике применяют следующие виды культуры клеток:
J первично-трипсинизированные;
J диплоидные:
J перевиваемые.
Первично-трипсинизированные культуры клеток
Их можно получить практически из любого органа и ткани животных и птиц
(кур, кроликов, собак, коров, овец, свиней, хомяков, мышей, морских свинок, обе-
зьян), а также из извлеченных у человека при операции. Чаще всего используют
почки, легкое, кожу, тестикулы, тимус и др. Поскольку способность к размно-
жению клеток т vitro зависит от степени дифференциации тканей, лучше брать
эмбриональные орыны. Клеточные культуры могут быть получены из нормаль-
ных и злокачественно перерожденных тканей, состоящих из эпителиальных и
фибробластных, а также смешанных типов клеток. Выбор ткани для конкретно-
го вирусологического исследования определяется чувствительностью ее клеток
in vitro к тестируемому вирусу.
Для получения первичных клеток органы не позднее 3-4 ч после извлечения в
асептических условиях трипсинизируют.
Куриные эмбрионы должны быть 7-10-дневного возраста, при использо-
вании эмбрионов от лабораторных животных асептически извлекают матку.
Органы крупных взрослых животных берут при их забое на мясокомбинатах.
Обработанные дезинфицирующими растворами или спиртом, промытые физио-
логическим раствором с большим количеством антибиотиков органы помещают в
чашки Петри для последующего диспергирования. Наиболее часто применяемый
метод диспергирования тканей — их обработка протеолитическими ферментами
(трипсином, панкреатином, коллагеназой и др.). Обычно используют метод трип-
синизации тканей.
Ткань, подготовленную для трипсинизации, тщательно измельчают ножница-
ми, переносят в стерильную колбу, отмывают физиологическим раствором или
буферным раствором для трипсинизации и заливают диспергирующим раствором
(например, подо1ретой до 37 С'С смесью 0,15% раствора трипсина и равных объе-
мов фосфатного буфера с 0,02% раствором Версена: pH = 7,2^-7,4). Трипсинизацию
проводят на магнитной мешалке. Через каждые 10-15 мин клеточную взвесь сли-
вают и помещают в холодильник, а оставшуюся ткань заливают новой порцией
диспергирующего раствора. Процедуру повторяют до полного истощения измель-
ченной ткани. Полученные сливы объединяют, фильтруют и центрифугируют при
600 об./мин в течение 20 мин.
Надосадочную жидкость сливают, осадок тщательно пипетируют в небольшой
порции ростовой среды, подсчитывают количество клеток в полученной взвеси в
камере Горяева. Для этого к 1 мл взвеси клеток в питательной среде добавляют
равный объем 0,1% раствора кристаллического фиолетового, приготовленного
на 0,1 М растворе лимонной кислоты, перемешивают. Подсчитывают количество
живых клеток (имеющих ядро и неповрежденную цитоплазму, вследствие чего они
не окрашиваются красителем) в двух камерах. Определяют среднее количество
клеток. Количество клеток в 1 мл (X) рассчитывают по формуле:
Х = (Ах 2x1000)/0,9,
где А — среднее количество клеток в камере
После подсчета количества клеток взвесь разводят ростовой питательной средой
до концентрации 200 000-300 000 клеток в 1 мл и разливают в пробирки, матрасы
или роллерные флаконы. Значительная часть жизнеспособных клеток, получен-
ных в результате ферментативного диспергирования, прикрепляется к субстрату
и начинает размножаться. На этом этапе культивируемые клетки определяются
ВИРУСОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ 565
как первичные. В своем разви гии клеточные культуры проходят фазы адаптации,
логарифмического роста, стационарную и старения (постепенной гибели клеток).
В процессе роста и размножения клетки полностью покрывают ростовую
поверхность сосуда и формируют монослой. Он может быть однослойным, если
клетки при тесном соприкосновении друг с другом прекращают делиться (эффект
контактной ингибиции), или не строго многослойным, состоящим из нескольких
слоев, если клетки полущены из злокачественно трансформированных тканей, для
которых не имеет критического значения феномен контактной ингибиции. Как
правило, монослой формируется через 3-5 дней, клетки сохраняют жизнеспособ-
ность в течение 7-21 дня в зависимости от вида ткани и ряда других факторов.
Питательную среду меняют по мере ее загрязнения продуктами обмена веществ,
которые вызывают изменение pH и отрицательно влияют на нормальный метабо-
лизм клеток, что может привести к их гибели. Для культивирования вируса исполь-
зуют молодые клеточные культуры, только что сформировавшие монослой.
Диплоидные культуры клеток
Основные сведения о специфике цитопатического действия многих вирусов
были получены на первично-трипсинизированных культурах. По мере расши-
рения их ассортимента и внедрения в практику стали очевидными недостатки
первичных культур: трудоемкость изготовления, нестандартность различных
партий первичных клеток к тому или иному вирусу, относительно высокая частота
контаминации исходного материала. Первично-трипсинизированные культуры
не пересеваются и не масштабируются, это модели одноразового использования с
проблемами сохранения и транспортировки.
При субкультивировании и пассировании клеток оказалось возможным в соот-
ветствующих условиях формировать популяции фибробластоподобных клеток,
размножающихся с большой скоростью и сохраняющих более чем в 75% попу-
ляции исходный набор хромосом. Эти клеточные культуры получили название
диплоидных клеточных культур. Как правило, это культуры фибробластов из сое-
динительной ткани. Стали возможными их масштабирование, криоконсервирова-
ние и контроль на отсутствие экзогенных контаминантов до начала вирусологи-
ческих экспериментов. Существенный недостаток этих моделей — ограниченный
срок жизни. Количество генераций зависит от вида животного, из которого была
получена диплоидная клеточная линия. Например, для фибробластов эмбриона
человека срок жизни составляет 50+10 пассажей. Это значение может существенно
варьировать как в одну, так и в другую сторону у диплоидных линий от разных
эмбрионов.
Широкое распространение в медико-биологических исследованиях и практиче-
ской вирусологии получили уникальные по своим свойствам диплоидные линии
фибробластов легкого эмбриона человека Wi-38 (1964) и MRC-5. Эти культуры
характеризуются широким спектром чувствительности к различным вирусам и
высокой пролиферативной активностью (табл. 23-3,23-4). Благодаря своевремен-
ному созданию и сохранению посевных криобанков эти линии все еще остаются
актуальными и эффективными клеточными моделями, несмотря на то, что в соот-
ветствии со своим происхождением они имеют ограниченный срок жизни.
Таблица 23-3. Перечень клеточных линий согласно видовому и тканевому происхождению
Ns п/п Линия Видовое и тканевое происхождение
1 А-204 Человек, рабдомиосаркома
2 А 549 Человек, карцинома легких
3 Alexander Человек, карцинома печени
4 AUBEK Бык, почка
566 ВИРУСОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ
Продолжение табл. 23-3
N" п/п Линия Видовое и тканевое происхождение
5 В 95-8 Мармозетка, трансформированные вирусом Эпштейна-Барр лейкоциты, продукция вируса Эпштейна-Барр
6 BALB/3T3 Clone А31 Мышь, эмбрион, фибробласты
7 BGM Африканская зеленая мартышка, почка
8 ВНК-21 Сирийский хомячок, почка
9 ВНК-21 clone 13 Клон линии ВНК-21
10 ВНК-21 (Clone 13) V Сублиния клона ВНК-21 clone 13
11 BSC-1 Африканская зеленая мартышка, почка
12 Chang conjunctive Человек, конъюнктива
13 Chang liver Человек, печень
14 CV-1 Африканская зеленая мартышка, почка
15 CV-3 Африканская зеленая мартышка, почка
16 FL V Человек, амнион,сублиния FL
17 FECH (ФЭЧ) Человек, эмбрион, фибробласты
18 FLECH 385/13 (ФЛЭЧ 385/13) Человек, эмбрион, фибробласты легкого
19 FHHK-4R Макака-резус, эмбрион, почка
20 GMK-AH-1 Африканская зеленая мартышка, почка
21 HAK Сирийский хомячок, почка
22 HeLa Человек, карцинома шейки матки
23 HeLa KD, HeLa S3, HeLa 229, HeLaV Сублинии HeLa
24 HEp-2 Человек, эпидермоидная карцинома гортани
25 HL-60 Человек, Т-клеточная лейкемия
26 НН (ГН) Человек, гипернефрома
27 IMR-32 Человек, нейробластома
28 KB Человек, карцинома ротовой полости
29 KBV Сублиния КВ
30 LLC-MK2 Макака-резус, почка
31 L-41 Человек, моноцитарная лейкемия
32 L-132 Человек, эмбрион, легкое
33 L-929 Мышь, соединительная ткань, клон линии L
34 MA-104 Макака-резус, почка
35 Me Coy Мышь, гепатома
36 MDBK Бык, почка
37 MDCK Собака, почка
38 МЕС (M3K) Кролик, эмбрион, мышцы
39 Molt-3 Человек, Т-лимсЬобластная лейкемия
40 Molt-4 Человек, Т-лимфобластная лейкемия
41 MRC-5 Человек, эмбрион, фибробласты легкого
42 MT-4 Человек, совместное культивирование лимфобластов крови сердца и Т-клеток больного лейкемией
43 MV 1LU (NBL-7) Норка, легкое
44 Namalva Человек, лимфома Беркитта
45 NGUK-1 (НГУК-1) Крыса, невринома
46 NIH/3T3 Мышь, эмбрион, фибробласты
47 PEO (ПЭО) Овца, эмбрион, почка
ВИРУСОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ 567
Окончание табл. 23-3
№ п/п Линия Видов e и тканевое происхождение
48 PES (ПЭС) Свинья, эмбрион,почка
49 РК (15) Свинья, почка
50 РО-2 (ПО-2) Овца, ночка
51 Р0-ТК-50 (ПО-ТК-50) Гибридные культуры: овца, клетки почки и лимфоциты
52 PO-TK-xLO (ПО-ТК-хЛО) Гибридные культуры: овца, клетки почки и лимфоциты
53 PO-TK-xSO (ПО-ТК-хСО) Гибридные культуры: овца клетки почки и спленоциты
54 PPES (ППЭС) Свинья, почка
55 PS Свинья, почка
56 Raji Человек, лимфома Беркипа
57 RAC х KMFCH (RAC x КМФЧ) Гибридная культура клеток: мышь RAC, опухоль и человек, кожно-мышечные фибробласты
58 RD Человек, рабдомиосаркома
59 RH K-13/3 Человек, почка, клон линии RH
60 RK-13 Кролик, почка
61 RPMI 2650 Человек, карцинома носовой перегородки
62 SEK (СЭК) Кролик, эмбрион, сердце
63 SPEV(СПЭВ) Свинья, эмбрион,почка
64 Sirk Кролик, роговица глаза
65 Taurus-1 (Taypyc-1) Теленок, почка
66 ТЕК(ТЭК) Крупный рогатый скот, эмбрион, тимус
67 T24 Человек, карцинома мочевого пузыря
68 T98G Человек, глиобластома
69 T1387 Человек, лимфобластная лейкемия
70 Vero Африканская зеленая мартышка, почка
71 Vero K19 Клон линии Vero
72 Vero (V), Vero 76. Vero C1008 Сублинии Vero
73 U-937 Человек, гистиоцитарная лимфома
74 WI-38 Человек, эмбрион, фибробласты легкого
75 293 Человек, эмбрион, клетки почки трансформированы ДНК адено- вируса типа 5 (Ad 5)
76 4647 Африканская зеленая мартышка, почка
Таблица 23-4. Перечень клеточных линий, рекомендуемых для культивирования вирусов
Вирусы Культура клеток
Вирусы гриппа: A (H1N1), А (НД), A (НД), А (НД), В и С МДСК, Vero MDBK, ВНК-21, SPEV (СПЭВ), WI-38. MRC-5, FLECH 385/13 (ФЛЗЧ 385/13), NIH-3T3, Chang Conjunk, НАК
Аденовирусы (1-31-го типа) HeLa, НЕр-2, Vero, МДСК, GMK-AH-1, РК(15), HN (ГН), L-41, А549, МАт-104, IMR-32 (12-й тип), FLV, HelaKD (7-й тип), HeLaS3 (5-й тип), КВ (3-й, 7-й типы), 293, ВНК-21 clone13, Chang liver. L 132, FLECH 385/13 (ФЛЭЧ 385/13)
Пневмовирусы (респираторно- синцитиальные) WI-38, MRC-5, FLECH 385/13 (ФЛЭЧ 385/13). МАт-104, HeLa, HEp-2, Vero, L-41, FLV, BSC-1, HN (ГН), L 132, KBV
Вирусы парагриппа (1-4-го типа) Vero, L-41, МАт-104, HEp-2, HN (ГН), RD. L-929, A549, BSC-1, Taurus-1 (Taypyc-1), FLECH 385/13 (ФЛЗЧ 385/13), FLV
Коронавирусы WI-38, MRC-5, FLECH 385/13 (ФЛЭЧ 385/13), Vero, T9qG, BALB/3T3 Clone A31, SPEV (СПЭВ)
Риновирусы HeLaV, BGM, WI-38, MRC-5, FLV, KBV, AUBEK, LLC-MK2
568 ВИРУСОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ
Окончание табл. 23-4
Вирусы Культура клеток
Морбилливирусы (вирус кори) HeLa, BSC-1, Vero, L-41, Molt-4, BGM, KBV, FLECH 385/13 (ФЛЭЧ 385/13)
Вирусы эпидемического паротита HeLa. KB, HEo-2, BGM, BSC-1, Molt-4
Вирус простого герпеса человека (1-го и 2-го типа) A549, PH K-13/3, Hfiua, HEp-2, HN (ГН). MAt-104, L-41, Vero, KB, WI-38, MRC-5, FLECH 385/13 (ФЛЭЧ 385/13). BHK-21, NIH/3T3, RD, Vero (V), IMR-32, Chang liver, BSC-1, BALB/3T3 clone A31
Вирус ветряной оспы RK-13, Vero, NIH/3T3. BHK-21. HEp-2, BSC-1, CV-1, BAlB/ЗТЗ clone A31, RD, MV 1LU (NBL-7)
цитомегаловирус WI-38, MRC-5. MV 1 LU (NBL-7) Vero. HN (ГН), A549. RD MAt- 104, Vero(V), МЕК (МЭК), SEK (СЭК)
Вирус Эпштейна-Барр BHK-21 Clone13, Vero
Полиовирусы (1, 2, 3-го типа) HeLa, HEp-2, BSC-1, Vero, RD. Chang Conjn, GMK-AH-1, CV-1, Chanq liver, BGM, HN (ГН). L41, МАт-104, A549, KB, RPMI 2650, WI-38, MRC-5, LLC-MK2, L132, FLECH 385/13 (ФЛЭЧ 385/13)
Коксаки А {1-24-го типа) HeLa, HEp-2, RD, BGM, RK-13, FLV
Коксаки В (145-го типа) HeLa, HEp-2, PK(15), МДСК, IMR-32, BGM, BHK-21 clone13
Вирусы ECHO HeLa, HEp-2, RD, Vero, BGM. RK-13, FLV, FLECH 385/13 (ФЛЭЧ 385/13)
Гепатит А Alexander, Vero (V), CV-1, BSC-1,4647, T 24, LLC-MK2, FRHK-4R
Гепатит В Vero, CV-1, А549
Вирус краснухи WI-38. FLECH 385/13 (ФЛЭЧ 385/13), Vero, GMK-AH-1, Sirk, BHK-21, RK-13
Ротавирусы МАт-104, МДВК, 4647, RD, BGM, SPEV (СПЗВ)
Реовирус BGM, PK(15), Vero, PS. MDCK, HeLa, SV-1, MV 1LU (NBL-7), BHK-21clone13
Вирус везикулярного стоматита WI-38, FLECH 385/13 (ФЛЭЧ 385/13), MRC-5. BSC-1, Vero, IMR- 32, LLC-MK2, Namalva. BHK-21 clone 13, RPMI 2650, BALB/ЗТЗ clone A31, Chang Fver, HeLaS3, HN (ГН), Alexander, MDCK, L 929, RD, Me Coy, В 95-8
Бешенство BHK-21 clone 13. BHK-21 (clone 13) V, MV 1LU (NBL-7), NGUK-1 (НГУК-1), ТЕК (ТЭК)
Псевдобешенство BHK-21 clone 13, BHK-21 (clone 13) V, MV 1 LU (NBL-7), NGUK-1 (НГУК-1), ТЕК (ТЭК)
Вирус осповакцины RK-13, МАт-104, Hei_a, МДСК, RD, BGM, PK(15), MV 1LU (NBL-7)
Вирус натуральной оспы WI-38, MRC-5, HAK, FECH (ФЭЧ)
Аренавирусы: лимфоцитарного хориоме- нингита, вирусы Пичинде, Манула BHK-21, МДСК, Vero, Vero K-19
Вирусы геморрагических лихора- док: Ласса, Эбола, Марбург, Хунин, Карельской и Омской лихорадки Vero 76, Vero C1008
Арбовирусы (L-вирусы): западный, вос- точный и венесуэльский энцефалиты, лихорадки Чикуньгунья и леса Семлики PES (ПЭС), FLV. HeLaV, Vero, CV-3, SPEV (СПЭВ), L-929, Raji, MAt-104, KBV. T-1387, Namalva, Molt-3, Molt-4, В 95-8, RHK- 13/3, BSC-1, LLC-MK2, MDCK, PS
Флавивирусы: клещевой и японский энцефалиты, лихорадка Денге, желтая лихорадка SPEV (СПЭВ), PPES (ППЭС), MAt-104, BSC-1, RHK-13/3, PEO (ПЭО), Vero K-19, NGUK-1 (НГУК-1), PES (ПЭС), FLECH 385/13 (ФЛЭЧ 385/13)
Буньявирусы: калифорнийская лихорадка BHK-21, A-549, LLC-MK2. Vero
Вирусы иммунодефицита человека 1 и 2 HL-60, U-937 (1-й и 2-й типы), Molt-3, МТ-4
Лейковирусы, вирус Т-клеточчой лейке- мии человека, вирус SV-40 MV 1LU (NBL-7), CV-1, BSC-1, Balb/ЗТЗ clone 31, Vero. NIH/3T3, HL-60, А-204, HN (ГН)
Прионы NGUK-1 (НГУК-1), PO-2 (ПО-2), PO-TK-50 (ПО-ТК-50), PO-TK- xLO (ПО-ТК-хЛО), PO-TK-xSO (ПС TK-xCO), RACxKMF4
ВИРУСОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ 569
Большинство диплоидных линий получено из тканей 5-6-месячных челове-
ческих плодов после искусственного аборта, а также из амниотической оболочки
от рожениц при нормальных родах. В последнее время применение диплоидных
линий клеток человека обсуждается в специальной литературе с точки зрения меди-
цинской этики. Это связано с рекомендациями, сформулированными Комитетом
по биоэтике ЮНЕСКО в отношении исследований на человеческих эмбрионах,
полученных позднее 14 сут эмбрионального развития. Некоторые положения
этого документа применимы к исследованиям по выделению и использованию
диплоидных линий клеток человека Это в первую очередь наличие письменного
информированного согласия женщины, подлежащей операции, отсутствие ком-
мерческой заинтересованности со стороны медицинского персонала, проводящего
прерывание беременности, строго медицинские цели исследования.
Перевиваемые культуры клеток
Ограниченный срок жизни диплоидных линий побудил исследователей к поис-
ку новых клеточных культур. Были найдены постоянные клеточные линии с нео-
граниченным сроком жизни. Фактическое «бессмертие» этих моделей обусловле-
но трансформацией генетического материала клеток. Линии, как правило, имеют
онкогенное происхождение или созданы вследствие спонтанной трансформации
нормальных клеток при культивировании in vitro. Получить перевиваемые куль-
туры клеток можно как из здоровых, так и из опухолевых тканей животных путем
длительных пересевов первично-трипсинизированных клеток (характеризуемых
высокой активностью роста) в заданном режиме культивирования. Клетки пере-
виваемых культур имеют гетероплоидный набор хромосом, одинаковую форму и
стабильность в условиях культивирования in vitro.
В лабораториях перевиваемые культуры клеток поддерживают путем пересе-
вов из одного сосуда в другой с обязательной заменой питательной среды. После
удаления из сосуда для культивирования ростовой питательной среды клетки
монослоя снимают с поверхности стекла или пластика с помощью диспергирова-
ния 0,02% раствором Версена, 0,25% раствором трипсина или смесью (1:1) этих
растворов. Диспергирующее действие раствора Версена обусловлено тем, что
он связывает двухвалентные катионы (Mg~+, Са++), которые обеспечивают при-
крепление клеток к стеклу и формирование монослоя. Под действием раствора
Версена клетки округляются и отслаиваются от стекла. Через 3-5 мин выдержива-
ния монослоя под подогретым до 37 Ю диспергирующим растворим его удаляют,
оставляя небольшую порцию для последующего периодического омывания клеток
до момента их сползания. Далее во флакон добавляют небольшое количество
ростовой питательной с реды, тщательно пипетируют сползший монослой и берут
пробу для подсчета клеток в камере Горяева. Полученную клеточную взвесь раз-
водят ростовой питательной средой до необходимой концентрации (100-200 тыс.
клеток в 1 мл) и разливают в пробирки, матрасы или флаконы для последующего
культивирования при 37 °C в течение 2-4 дней до образования сплошного моно-
слоя. Клетки можно не подсчитывать в камере Горяева, а расгевать их привычным
для данной культуры клеток коэффициентом от 1:2 до 1:8 и более.
Клеточные культуры, как правило, культивируют в статических условиях,
используя для этого пробирки, матрасы и флаконы различной емкости, а также
специальные культуральные 6- и 96-луночные пластиковые планшеты. Для полу-
чения больших количеств клеточной массы используют технику роллерного или
суспензионного культивирования.
Перевиваемые клеточные культуры имеют ряд преимуществ по сравнению с
первичными. Они служат современной эффективной альтернативой биологиче-
ским моделям (куриным эмбрионам, лабораторным животным), включая эти-
ческий компонент исследований. Постоянные культуры и их клональные линии
570 ВИРУСОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ
обеспечивают стандартные условия размножения вирусов в отличие от первич-
ных культур, представляющих смешанные популяции клеток. Это уникальная по
степени контролипуемости система. Разработанные методы контроля клеточных
культур обеспечивают идентификацию практически любых экзогенных контами-
нантов. Благодаря относительной простоте приготовления перевиваемых культур
существенно экономятся труд и материальные средства.
Перевиваемые клетки, как правило, обладают широким спектром чувствитель-
ности к вирусам (см. табл. 23-4).
Практика медицинских и биологических центров мира и многолетний опыт
работы с постоянными клеточными линиями показали, что для этих моделей
характерна межвидовая и внутривидовая контаминация, а также контаминация
микоплазмами и вирусами. В ряде работ описана подмена одних линий другими.
Появились публикации о проблемах селекции и возникновении множества субли-
ний, оазличающихся между собой, но имеющих одно наименование.
Стало очевидным, что надежное использование постоянных клеточных линий
невозможно без создания специализированных центров, основной задачей кото-
рых являлось бы хранение линий, имеющих определенную характеристику
(паспорт). Эта проблема решена путем создания Национальных коллекций кле-
точных культур. Создание Российской коллекции клеточных культур (РККК)
обусловило значительный прогресс в решении проблем стандартизации вирусоло-
гических исследований.
В настоящее время РККК объединяет 9 специализированных коллекций,
сосредоточенных в научно-исследовательских институтах. Сохраняется около
2000 клеточных линий, из которых 700 являются референтными, депонированны-
ми в связи с патентованием, 170 — имеют промышленное значение.
Основная сторона деятельности РККК — обеспечение эталонным клеточным
материалом научных, производственных и практических учреждений России
и зарубежья. Каждая линия в РККК охарактеризована согласно принятому
Международному стандарту; которого придерживаются все Национальные кол-
лекции мира. Ниже приведен для примера образец паспорта коллекционной линии
MDCK — одной из важных и распространенных вирусологических моделей.
Паспорт культуры клеток MDCK
Происхождение: собака, почка.
Proc.Soc.Exp.Biol.Med. 1958. 98: 574.
Морфология: эпителиоподобная.
Способ культивирования: монослойный.
Условия культивирования:
среда - EMEM, ВМЕ или DMEM;
сыворотка — эмбриональная бычья 10%;
другие компоненты — NEAA 1% (ЕМЕМ);
процедура пересева — снятие клеток с использованием трипсина 0,25%, раство-
ра Версена 0,02% (1:3);
кратность рассева — 1:3-1:10, оптимальная плотность — 1-Зх104клеток/см2;
криоконсервация — ростовая среда (можно добавить 30% эмбриональной
бычьей сыворотки), 10% DMSO, 1х106клеток/мл в ампуле.
Жизнеспособность после криоконсервации: 90% (окраска трипановым
синим на нулевом пассаже).
Контроль контаминации: бактерии, грибы и микоплазма не обнаружены.
Контроль видовой идентичности: кариологический анализ.
Кариология: 2п = 78, пределы изменчивости по количеству хромосом — 75-83,
модальное количество хромосом — 78-80, количество маркеров — 1-2, крупные
субметацентрические хромосомы, в некоторых клетках имеются 1-2 средние
ВИРУСОЛОГИЧЕСКИЕ исследования 571
мета- или субметацентрические хромосомы (рутинная окраска), количество поли-
плоидов — 0,6%.
Эффективность клонирования: 35% (АТСС).
Другие характеристики: чувствительность к вирусам — везикулярный стома-
тит; коровья оспа; Коксаки В-5; реовирусы 2, 3; аденовирусы 4, 5; грипп А, В, С;
чума собак; арбовирусы; аренавирусы; везикулярная экзантема свиней; инфекци-
онный гепатит собак.
Область поименения; вирусология, биотехнология, клеточная биология.
Коллекции: АТСС CCL 34; ЕСАСС 84121903; 85011435; НИИ вирусологии
РАМН; ЕСКК; ИНЦ РАН.
Микроскопические и культуральные методы
ПРЯМАЯ МИКРОСКОПИЯ ПРИ ИДЕНТИФИКАЦИИ ВОЗБУДИТЕЛЯ В КУЛЬТУРЕ КЛЕТОК
Микроскопическое исследование инфицированных тканей проводят в целях
выявления в них патологических изменений или обнаружения вирусного материа-
ла. Группа микроскопических методов включает обычную световую микроскопию
(вирусоскопию и обнаружение внутриклеточных включений с помощью цитоло-
гических методов) и микроскопию в ультрафиолетовом свете с использованием
феномена флюоресценции.
Методы обычной световой микроскопии, как правило, достаточно просты,
однако в настоящее время они имеют ограниченное диагностическое значе-
ние, поскольку могут быть применены при немногих вирусных заболеваниях.
Основная задача этих методов — обнаружение в фиксированных и окрашенных
тканях патологических и патогномоничных изменений или вирусного антигена,
а в мазках или препаратах-отпечатках, изготовленных из материала тканей, жид-
костей и экссудатов, — вирусного агента или его антигенных компонентов, телец-
включений и измененных клеток.
ВИРУСОСКОПИЯ
Только крупные вирусы, размеры которых превышают 150 нм, могут быть
обнаружены с помощью прямого микроскопического исследования. Для этого
используют окрашенные мазки и отпечатки. Одним из лучших способов окраски
препаратов считают серебрение по Морозову. Окраска позволяет увеличить раз-
мер вируса с помощью осаждения на него частиц серебра. С помощью световой
микроскопии возможно обнаружение вирионов оспенных вирусов. Поскольку
они являются гигантами (достигая 300-390 нм) по сравнению с вирионами
других вирусов, их удается увидеть в световом микроскопе, хотя и на пределе
видимости.
Вирусоскопия как быстрый ориентировочный метод лабораторной диагностики
вирусных заболеваний все же не является точным методом для установления при-
роды возбудителя. Для этого требуются другие методы индикации и идентифика-
ции вирусов.
ЦИТОЛОГИЧЕСКИЕ МЕТОДЫ
Эти методы редко применяют в диагностике вирусных инфекций, но при ряде
инфекций они по-прежнему востребованы. При репродукции некоторых вирусов
в клетках формируются вирусные тельца-включения, которые могут быть вну-
триядерными (как правило, у ДНК-содержащих вирусов) или цитоплазматиче-
скими (у РНК-содержащих). Некоторые вирусы способны вызывать образование
телец-включений обоих типов. Появляются они в результате скопления в клетке
большого количества вирионов, изменения или дегенерации клеточного содержи-
мого, избытка вирусных белков, не встроившихся по разным причинам в состав
572 ВИРУСОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ
вирионов. Размер телец-включений колеблется от едва различимого до величины
клеточного ядра, а количество — от одиночных до 10-12 на клетку.
Для обнаружения телец-включений материалы аутопсии, биопсии, мазки и
отпечатки (посмертные или прижизненные), а также препараты культур клеток
подвергают специальной окраске с последующей микроскопией. Для включений,
образуемых разными типами вирусов, методы окраски различны. Среди множе-
ства рецептов окраски можно выделить универсальные (обзорные), к которым
относится окраска гематоксилин-эозином.
Цитоплазматические тельца-вклюгения Бабеша-Негри, формируемые вирусом
бешенства, при окраске любым, методом визуализируются в гистологических срезах
головного мозга человека или животных; они настолько характерны, что позволяют
установить диагноз. Типичные изменения в ткани печени (полученной путем висцеро-
томии или на секции) указывают на желтую лихорадку. При венерической лимфогра-
нулеме элементарные тельца можно обнаружить в гнойном материале из бубонов.
В цитоплазме эпителиальных клеток в результате репродукции вируса оспы
птиц образуются тельца Баллингера.
Внутриядерные включения обнаруживают в перевиваемых эпителиоидных
культурах клеток L или EL, выращенных на покровных стеклах и зараженных
вирусом болезни Ауески, окрашенных гематоксилин-эозином.
Несколько сложнее искать такие включения в препаратах из культур эпителио-
идных клеток, выращенных на покровных стеклах и зараженных оспой соответ-
ствующего вида, после аналогичной окраски.
В большинстве случаев обнаружение вирусных телец-включений служит вспо-
могательным методом диагностики. При энцефалите, например, патогистологиче-
скими методами нельзя точно установить этиологию, хотя возможно использовать
их для дифференцирования инфекционного и неинфекционного процессов или
некротического процесса и демиелинизации, наблюдающейся при посгвакциналь-
ных энцефалитах. Именно поэтому патологоанатомам, берущим материал для
гистологического исследования, рекомендуют параллельно осуществлять сбор
материала для выделения вируса.
МЕТОДЫ МИКРОСКОПИЧЕСКИЙ ИНДИКАЦИИ ВИРУСОВ В КЛЕТОЧНЫХ КУЛЬТУРАХ
Способность вирусов вызывать специфическую деструкцию клеток делает воз-
можным использование культуры клеток в вирусологии для первичного обнару-
жения и выделения вирусов из патологического материала (рис. 23-1 и 23-2). При
правильном подборе культур клеток, соблюдении условий заражения клеточного
монослоя и культивирования вирусов можно выявить их уже на первых пассажах.
Индикаторы наличия вируса в инфицированных культурах клеток:
• развитие специфической для исследуемого вируса дегенерации клеток и кле-
точного монослоя;
• образование бляшек;
• выраженная интерференция вирусов;
• появление внутриклеточных включений;
• подавление метаболизма клеток (цветовая проба);
• положительная гемадсорбция;
• положительная гемагглютинация;
• иммунофлюоресцентные признаки наличия вируса или вирусных антигенов;
• электронно-микроскопические критерии наличия вируса.
Индикация вирусов по цитопатическ му действию
О размножении вируса в культуре клеток обычно судят по цитопатическому
действию (ЦПД), выражаемому морфологическими изменениями в зараженных
культурах клеток, которые наблюдают при ежедневном просматривании под
малым увеличением микроскопа (объектив — х8-19, окуляр — х7-10).
ВИРУСОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ 573
Эти изменения (по сравнению с состоянием клеток в незаряженных культурах)
могут охватывать весь монослой или отмечаться как разного размера очаги изме-
ненных клеток на фоне нормального монослоя. Фирмы ЦПД зависят от биологи-
ческих свойств вируса, вида клеток, дозы заражения, условий культивирования
и т.д. 1ак, некоторые из аденовирусов проявляют ЦПД через 2-3 сут после зараже-
ния, а респираторно-синцитиальный вирус — через 1-2 нед.
Формы ЦПД можно сгруппировать в три наиболее различающихся между собой
варианта: фрагментация клеток, округление клеток и симпластообразование.
Фрагментация — разрушение клеток на отдельные фрагменты, которые, теряя
связь с подложкой (стеклом или пластиком), попадают в культура тьную жидкость
в виде клеточного детрита (вируса везикулярного стоматита).
Округление — потеря клетками способности удерживаться на подложке,
вследствие чего они принимают округлую (шаровидную) форму, отделяются от
стекла или пластика и свободно плавают в культуральной среде, постепенно поги-
бая (энтеровирусы, аденовирусы и др.).
Симпластообразование — образование многоядерных гигантских клеток
в результате слияния цитоплазмы и амитотического деления клеток (от 3-5 до
нескольких десятков). При этом клеточные оболочки соседних клеток раство-
ряются. их цитоплазмы сливаются, образуя большую клетку, в которой ядра либо
сосредоточиваются в центре, либо располагаются по периферии. Такие гигантские
многоядерные клетки называются симпластами. Механизм слияния клеток объ-
ясняется нарушением процесса деления клеток под влиянием вируса или рас-
творением клеточных оболочек под действием фермента лецит иназы некоторых
вирусов.
Иногда в клеточных культурах может наблюдаться развитие очагов клеточной
пролиферации, состоящих из слоев интенсивно делящихся клеток. При неопла-
Рис. 23-1. Нормальный монослой клеток перевиваемых клеточных культур MDCK, Таурус, HeLa,
ВНК-21.
574 ВИРУСОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ
Рис. 23-2. Клетки фибробластов легкого эмбриона человека (ФЛЭЧ) и почки эмбриона человека
(ПЭЧ), зараженные виоусами гриппа и геопеса или парагриппа типов 2 и 3.
стической трансформации пораженных клеток в монослое возникают плотные
фокусы трансформации белого цвета, различных величины и формы (вирус сар-
комы Рауса).
Некоторые вирусы, размножаясь в клеточных культурах, не вызывают ЦПД,
что может быть обусловлено природой вируса или типом клеток, которые исполь-
зовали для его культивирования (вирус бешенства, вирус краснухи в диплоидных
клетках человека и др.). Клетки остаются жизнеспособными, однако со временем
в них понижается интенсивность клеточного деления и несколько меняется мор-
фология.
Характерное для каждого типа исследуемых вирусов ЦПД может развиваться в
культурах клеток при заражении средними дозами вирусов (0,1-1,0 ТЦД5,,/клет-
ка), когда инфицированные клетки не погибают сразу, а продолжают функцио-
нировать еще некоторое время, достаточное для формирования специфических
образований. При больших дозах вирусов (>1.0 ТЦД50/клетка) клетки гибнут
достаточно быстро с возникновением генерализованной инфекции и полной
деструкции монослоя. При малых дозах заражения (<0,1 ТЦД50/клетка) инфи-
цирование клеток происходит медленно и вызывает медленно прогрессирующую
нетипичную дегенерацию клеток.
Эпителиальные клетки, как правило, более чувствительны к цитопатическому
действию вируса, чем фибробластные, что и проявляется более оанним развитием
морфологических изменений и наступлением тотальной дегенерации.
Цитопатическое действие вируса в культурах клеток необходимо дифферен-
цировать от неспецифической дегенерации клеточного монослоя, которая может
быть обусловлена естественным старением клеток, токсическим действием ино-
кулируемого материала, контаминацией клеток посторонними вирусными и бак-
териальными объектами.
вирусологические исследования 575
Поскольку большинство вирусов способно оказывать ЦПД, такой метод инди-
кации вируса в клеточных культурах применяют очень широко. Иногда в первом
пассаже ЦПД может отсутствовать вследствие репродукции незначительного
количества вируса, которое неспособно вызвать ярко выраженный цитопатиче-
ский эффект. Требуется несколько «слепых» пассажей для накопления достаточ-
ного количества вируса или утверждения об его отсутствии в материале.
Гемадсорбция
Реакцию гемадсорбции впервые обнаружили Фогель и Щелоков (1957) в куль-
туре ткани, инфицированной вирусом гриппа. Суть явления заключается в том. что
вирусы, в вирионе которых присутствует гемагглютинин, обладают рецепторами,
комплементарными рецепторам эритроцитов некоторых животных (морской
свинки, обезьян) и человека группы крови 0. При репродукции вирусов в клеточ-
ных культурах еще до проявления ЦПД находящиеся у поверхности клеток вирусы
и их фрагменты соединяются с рецепторами эритроцитов настолько прочно, что
последние не смываются питательной средой или буферным раствором и образуют
на монослое клеток скопления.
Методика гемадсорбции: на 2-4-й день культивирования вируса в клеточной
культуре берут одну пробирку с зараженными клетками и одну контрольную, не
содержащую вируса. Культуральную жидкость из них удаляют, вносят 0,2 мл 0,5%
суспензии отмытых эритроцитов и оставляют на 15-30 мин при комнатной тем-
пературе. 37 или 4 °C. Затем эритроциты отмывают физиологическим раствором
и просматривают под малым увеличением микроскопа. В кон грольной пробирке
эритроциты полностью отмываются физиологическим раствором или проплыва-
ют свободно в поле зрения. В зараженной пробирке эритроциты остаются прикре-
пленными к поверхности клеток. В этом случае реакция считаете я положительной.
В зависимости от типа вируса и вида клеток локализация эритроцитов может быть
различной. Так, при заражении клеток вирусом гриппа эритроциты, как правило,
расположены очагами и скоплениями, вирусом парагриппа — диффузно на слое
клеток, вирусом африканской чумы свиней — в виде ожерелья только по перифе-
рии клеток.
Гемагглютинация
Некоторые вирусы могут агглютинировать эритроциты. Это явление впервые
обнаружил Херст в 1941 г.
В основе агглютинации лежит способность гемагглютинина, одного из
поверхностных вирусных белков, взаимодействовать с рецепторами на поверх-
ности эритроцитов. Каждая вирусная частица содержит множество молекул
гемагглютинина, образующего шиповидные выступы, которые при доста-
точно высокой концентрации вируса могут формировать крупные агрегаты с
эритроцитами, по структуре напоминающие сеть. Появление таких агрегатов
хорошо наблюдается в лунке с U-образным дном. Нормальные эритроциты при
оседании из взвеси образуют компактный осадок в виде диска на дне лунки, а
агглютинированные — «зонтик» из тонкой пленки равномерно склеившихся
эритроцитов.
Для постановки реакции гемагглютинации (РГА) могут быть использованы
аллантоисная и амниотическая жидкости, суспензия хорионаллантоисных обо-
лочек куриных эмбрионов, взвеси и экстракты из культур клеток или органов
животных, зараженных вирусами. Последовательность постановки РГА:
J готовят 1% суспензию отмытых эритроцитов того вида животных или
человека, который способен агглютинировать исследуемый вирус;
J готовят двукратно возрастающие разведения вируса в физиологическом
растворе;
576 ВИРУСОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ
J смешивают разведения вируса с эритроцитами в равных объемах (напри-
мер, по 0,4 мл), одновременно ставят контроль эритроцитов (смесь рав-
ных объемов физиологического раствора и суспензии эритроцитов);
J смеси выдерживают необходимое время при определенной температуре в
зависимости от свойств изучаемого вируса;
J оценивают интенсивность гемагглютинации в крестах.
Результаты РГА учитывают после полного оседания эритроцитов в контроле.
Время их оседания может варьировать в зависимости от видовой принадлежности
эритроцитов, процентного содержания их во взвеси и температуры, при которой
проводится реакция.
С помощью РГА нельзя определить в препарате точное количество вируса или
инфекционных единиц, однако ГАЕ служит удобной единицей для сравнения
вирусных препаратов. Кроме того, для любого конкретного штамма вируса можно
оценить соотношение между ГАЕ и другими единицами измерения количества
вируса (например, инфекционной активности).
Более экономный метод постановки РГА — микровариант этой реакции, кото-
рый проводят в микропланшетах для титрования. В этом случае необходимо учи-
тывать, что 1 ГАЕ определяется как количество вируса в микрообъеме (0,05 мл),
агглютинирующее 50% эритроцитов в 0.05 мл 1% суспензии.
Образование бляшек
Из локальных повреждений, вызываемых вирусами, хорошо известны так
называемые бляшки в клеточных культурах или оспины на хорионаллантоисных
оболочках куриных эмбрионов, зараженных покс-вирусами и некоторыми дру-
гими. Метод обнаружения вируса путем получения бляшек технически сложнее
других и применяется в основном для титрования вируса. Бляшки способны
образовывать вирусы везикулярного стоматита, полиомиелита, Коксаки, гриппа
и др.
Принцип метода состоит в следующем. Однослойные культуры клеток, выра-
щенные в культуральных планшетах, заражают цельной вирусной суспензией
(выделение, типирование вируса) или в последовательных десятикратных раз-
ведениях (титрование вируса), обеспечивают контакт вируса с клетками в течение
необходимого для разных типов вирусов времени. Затем удаляют неадсорбиро-
вавшийся вирус, промывая несколько раз монослой клеток раствором Хенкса,
и покрывают его слоем специального питательного агара. Благодаря агаровому
покрытию размножение и распространение вируса ограничиваются только перво-
начально инфицированными и соседними с ними клетками, в сплошном слое
живых клеток появляются ограниченные островки дегенерации клеток, погибших
в результате воздействия вируса. Эти островки визуализируются, так как содер-
жащийся в агаровом покрытии или добавленный непосредственно перед учетом
результатов титрования нейтральный красный окрашиз^ет только живые клетки,
поэтому в матрасе на ровном красновато-розовом фоне появляются бесцветные
пятна — бляшки. Каждая бляшка соответствует одной инфекционной частице
вируса. При небольшой плотности частиц, когда бляшки не могут сливаться или
перекрывать друг друга, эффект бляшкообразования можно использовать для
достаточно точного количественного определения инфекционности вирусов или
нейтрализующей активности антител.
Цветовая проба
Цветовая проба основана на способности вирусов изменять метаболическую
активность зараженных клеток. Под влиянием метаболитов клеточных культур
pH питательной среды в незаряженных культурах сдвигается в кислую сторону,
а в случае заражения вирусом сохраняется примерно на первоначальном уровне
вирусологические исследования 577
из-за гибели клеток под воздействием вируса. Колебания pH улавливаются по
изменению цвета фенолового красного, добавляемого в среду в качестве инди-
катора. Для реакции используют клетки перевиваемых линий: HeLa, НЕр-2, FL,
СОЦ и др. Метод не отличается высокой достоверностью, и его редко применяют
на практике.
Интерференция вирусов
Метод, основанный на интерференции вирусов, предполагает, что одни из
вирусов, размножаемые в культуре клеток, способны снижать или блокировать
размножение других вирусов. Чаще всего этот метод используют для обнаружения
вирусов, не оказывающих ЦПД в клеточных культурах, или когда задержка раз-
вития ЦПД обусловлена другими причинами. Применяют метод и при титровании
вирусов, не вызывающих отчетливого ЦПД.
Клеточную культуру заражают испытуемым вирусом; по истечении нескольких
дней, достаточных для его репродукции, вводят стандартную дозу (100 ТЦД50)
«индикаторного» вируса, обладающего выраженной цитопатогенностью или спо-
собного вызывать гемадсорбцию. После инкубации зараженных культур в течение
времени, достаточного для проявления ЦПД «индикаторного» вируса, оценивают
наличие обусловленных им цитопатических изменений. Если в культуое клеток
не обнаруживают ЦПД, это означает, что имела место репродукция испытуемого
вируса. Если наблюдается выраженное ЦПД «индикаторного» вируса, то испытуе-
мого вируса в клетках нет.
Наиболее широко данный метод используют при работе с вирусом краснухи. На
роль «индикаторного» вируса в этом случае больше всего подходит вирус везику-
лярного стоматита.
Внутриклеточные включения
При размножении многих вирусов в эпителиоподобных или фибробластопо-
добных клеточных культурах появляются особые внутриклеточные включения
в цитоплазме или ядре. Для выявления этих включений клетки выращивают на
покровных стеклах, в пробирках или специальных флаконах, заражают испытуе-
мым материалом. Через необходимое для данного вируса время инкубации при
оптимальной температуре культивирования готовят препараты, окрашивая их
красителями или флюорохромами.
Хорошо обнаруживаются тельца-включения в препаратах, окрашенных гемато-
ксилин-эозином. Готовят препараты, предварительно фиксируя клеточный слой на
покровных стеклах в смеси Дюбока-Бразиля-Буэна, фиксаторе Карнуа, Ценкера
или в других фиксаторах, а затем окрашивая, чаще всего по Романовскому-Гимзе.
Эта окраска универсальна и в разных вариантах позволяет визуализировать все
виды внутриклеточных включений, которые обычно оксифильны и красятся в
розовый или сиреневый цвет. При окраске по Манну, также хорошо выявляющей
цитоплазматические и внутриядерные включения в клетках, зараженных различ-
ными вирусами, цвет включений ярко-красный. Выбор окраски определяется в
значительной степени типом вируса, поражающего клетки.
Б качестве флюорохрома для обнаружения внутриклеточных включений
используют акридиновый оранжевый. При этом можно работать как с нативными
препаратами, так и с фиксированными в течение 10-15 мин в ацетоне. Включения,
содержащие ДНК, дают голубовато-зеленое свечение на сероватом фоне цитоплаз-
мы, РНК-содержащие включения — красновато-оранжевые.
Как и при вирусоскопии, выявление внутриклеточных включений в культиви-
руемых клетках служит только ориентировочным методом быстрой лабораторной
диагностики вирусных инфекций.
578 ВИРУСОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ
Электронная микроскопия
Особенности электронной микроскопии в сравнении с другими методами
диагностики вирусных инфекций
В 1960-1970-е гт. с изобретением метода негативного контрастирования с
помощью электронной микроскопии (ЭМ) были открыты сотни вирусов. Однако
с развитием молекулярных методов диагностики, таких как ИФА и ПЦР, сфера
применения ЭМ значительно сузилась. Снижение роли ЭМ в вирусологической
диагностике обусловлено рядом недостатков этого метода: относительно низкой
чувствительностью, невысокой производительностью и необходимостью исполь-
зования дорогостоящего оборудования.
Вместе с тем в крупных диагностических центрах развитых стран ЭМ практи-
куется наравне с другими методами. В некоторых случаях ЭМ служит методом
выбора при определении этиологии новых, неизвестных ранее заболеваний. Так,
в середине 1990-х гг. с помощью ЭМ были открыты высоколетальные парамиксо-
вирусы Hendra и Nipah, в 2003 г. удалось выявить этиологический агент атипичной
пневмонии, впоследствии получивший название «коронавирус SARS».
Основные преимущества ЭМ для диагностики вирусных инфекций следующие.
• Широкий спектр выявляемых вирусных агентов. Электронно-микроскопи-
ческое исследование позволяет по морфологическим особенностям вирусных
частиц идентифицировать инфекционный агент до уровня семейства, иногда
до рода. В большинстве случаев этого достаточно для постановки диагноза.
Кроме того, можно определить серотип вируса по эффекту агрегации вирус-
ных частиц в присутствии специфической сыворотки (иммуноэлектронная
микроскопия). С помощью ЭМ выявляют не только известные, но и новые
вирусные агенты, а также случаи сочетанных инфекций. Природная измен-
чивость вирусов не влияет на результат электронно-микроскопического
исследования, в отличие от таких методов, как ИФА и ПЦР.
• Быстрота диагнск тики. Если материалом для исследования служит вируссо-
держащая жидкость (экстракт фекалий, содержимое кожных везикул, культу-
ральная среда), то для определения возбудителя обычно требуется не более
10-15 мин с момента получения материала.
• Достоверность положительных результатов. Обнаружение вирусных частиц
и возможность количеогвенИРй оценки их содержания в препарате делают
ЭМ-диагностику особенно достоверной. Вирусологи сталкиваются как с лож-
ноположительными, так и с ложноотрицательными результатами ИФА и ПЦР,
но положительные результаты ЭМ всегда воспринимаются с доверием. В силу
вышеуказанного ЭМ считается «золотым стандартом» диагностики. В настоящее
время в вирусологической диагностике ЭМ применяют в следующих случаях.
А. Идентификация новых и неизвестных вирусных агентов. К ним следует
отнести в первую очередь особо опасные вирусные инфекции, такие как гемор-
рагические лихорадки, вызываемые вирусами семейства Filovtridae. Материалом
для исследования в данном случае служат инфицированные клеточные культуры
или секционный материал. Занос возбудителей подобного рода в северные страны
возможен с импортом экзотических животных или культур клеток животного про-
исхождения. В последнее время участились сообщения о случаях инфицирования
людей поксвирусами животных. Описано несколько случаев заражения людей пок-
свирусами от экзотических грызунов, содержащихся в неволе, а также от кошек и
других домашних животных. Материалом для исследования в данном случае может
быть содержимое кожных везикул, поскольку концентрация вирусных частиц в них
достаточно высока. Если была предприняла попытка выделения инфекционного
агента на клеточной культуре, материалом могут служить вируссодержащая культу-
ральная среда или сами клетки.
ВИРУСОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ 579
Б. Диагностика вирусных кишечных инфекций человека. В данном слу-
чае применение ЭМ обусловлено тем, что при сходных клинических признаках
заболевание может быть вызвано несколькими (более 10) различными вирусами,
которые легко определяются в фекалиях на основании морфологических призна-
ков. Далеко не все агенты вирусных гастроэнтеритов можно идентифицировать с
помощью ИФА или ПЦР. Например, генетическая вариабельность норавирусов
настолько велика, что даже новейшие тест-системы ПЦР не в состоянии выявить
весь спектр циркулирующих генотипов. Некоторые агенты, например торовирусы
и кишечные коронавирусы человека, обнаруживают только с помощью ЭМ, так
как тест-системы к ним не разработаны.
В. Технологический контроль качества стандартизированных клеточ-
ных линий и вакцинных препаратов. Клеточные линии могут быть контами-
нированы самыми различными вирусными агентами, внесенными с поддерживаю-
щей средой, сывороткой животных, загрязненными реактивами и пр. В данном
случае ЭМ оказывается незаменимым методом, позволяющим идентифицировать
контаминацию и выявить источник заражения.
Основные методы ЭМ-диагностикм, применяемые на практике
• Позитивное контрастирование улътратонких срезов применяют при иссле-
довании биопсий, секционного материала либо инфицированной культуры
клеток. Изготовление ультратонких срезов требует времени (от нескольких
часов до нескольких сут ок) и навыка, поэтому в диагностической практике
этот метод используют редко.
• Негативное контрастирование — наиболее широко практикуемый метод.
Используют при исследовании вируссодержащих жидкостей, таких как
культуральная среда, моча, сыворотка крови, экстракт фекалий, содержимое
везикул, бронхолегочный лаваж, ликвор и т.д. (рис. 23-3-23-9). В настоящее
Рис. 23-3. Интактные трехслойные частицы
ротавируса в фекалиях ребенка с острым
гастроэнтеритом. Вверху справа - частица с
проникшим внутрь контрастирующим веще-
ством (негативное контрастирование, мас-
штабный отрезок —100 нм).
Рис. 23-4. Двухслойные вирионы ротавируса в
фекалиях ребенка с острым гастроэнтеритом
(негативное контрастирование, масштабный
отрезок — 100 нм).
580 ВИРУСОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ
Рис. 23-5. Скопление вирионов норовируса
в фекалиях ребенка с острым гастроэнтери-
том. Отдельные частицы с проникшим внутрь
контрастирующим веществом имеют форму
ромба (вверху справа) (негативное контрасти-
рование, масштабный отрезок — 100 нм).
Рис. 23 Б. Скопление частиц аденовируса в
фекалиях ребенка с острым гастроэнтеритом
(негативное контрастирование. масШ|Эбный
отрезок —100 нм).
* *
.#
# ♦ * * «
Л'♦'***.'*>
я #. X “
* ' *
• * * •** -S*
ж* * > *
*♦*
S ыЬ. * .4ft VC.
*
♦ ф'
• «
-----5%
100 тв
Рис 23-7. Скопление частив астоовиру-
са в фекалиях ребенка с гастроэнтеритом.
Некоторые частицы имеют звездообразную
структуру (негативное контрастирование, мас-
штабный отрезок —100 нм).
Рис 23-8. Вирион коронавируса в фекалиях
ребенка 3 лет с острой диареей. Видны харак-
терные для коронавируса крупные булавовид-
ные пепломеры длиной около 20 нм (нега-
тивное контрастирование ФВК, масштабный
отрезок —100 нм).
ВИРУСОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ 581
время метод не1ативного контрастиро-
вания в различных модификациях при-
меняют в ведущих лабораториях мира.
Для приготовления одного препарата
в целях просмотра требуется в среднем
5-7 мин.
Каплю вируссодержащей суспензии поме-
щают на поверхность парафильма в пла-
стиковой чашке Петри. На каплю опускают
медную сетку с углеродной либо формвар-
углеродной подложкой. После адсорбции в
течение 15-60 с сетку отмывают в дистил-
лированной воде. Препарат контрастируют
1,5% раствором калиевой или натриевой соли
фосфорно-вольфрамовой кислоты (pH = 6,7).
Для выявления поксвирусов в качестве кон-
трастирующего вещества следует применять
1% водный раствор уранилацетата, так как он
лучше сохраняет морфологические особен-
ности их поверхности. Затем сетку с препара-
том высушивают при комнатной температуре.
Таким же способом готовят препараты на
подложках, модифицированных синтетиче-
скими полимерами. Готовые препараты про-
сматривают под электронным микроскопом
при инструментальном увеличении хЮ ООО-
Рис. 23-9. Вирионы энтеровируса в
фекалиях ребенка 2 лет с гастроэн-
теритом (негативное контрастирование
ФВК, pH = 6,7, масштабный отрезок —
100 нм).
50 ООО, при этом обращают особое внимание на скопления вирусных частиц.
Этиологические агенты заболевания идентифицируются по морфологическим
признакам. Помимо формы частиц, большое значение имеет точное определение их
размера. Современные микроскопы снабжены цифровыми камерами и программа-
ми, позволяющими измерять частицы непосредственно во время просмотра.
Методы повышения чувствительности ЭМ
Чувствительность ЭМ ниже в 103-104раз, чем ПЦР. но уступает ИФА незна-
чительно. На практике в большинстве случаев чувствительность ЭМ составляет
около 106-107 частиц на 1 мл, однако она может быть повышена путем концентра-
ции вирусных частиц в образце либо непосредственно на подложке.
Ключевой момент приготовления препаратов негативного контрастирования —
адсорбция вирусных частиц на сетках-подложках. Количество частиц в поле зре-
ния и соответственно чувствительность диагностики можно увеличить двумя спо-
собами: I) повышением содержания вирусных частиц в препарате; 2) улучшением
сорбционных свойств подложки.
Широко используемым методом повышения содержания частиц в едини-
це объема препарата является ультрацентрифугирование. Плавучая плотность
вирионов лежит в пределах 1,2-1,4 г/мл, поэтому для концентрации вирус-
ных частиц применяют их осаждение высокоскоростным центрифугированием.
Предварительно осветленную вирусную суспензию в количестве 2-10 мл осажда-
ют при 80 000-120 000 g в течение 1-1,5 ч, затем осадок ресуспендируют в неболь-
шом объеме буферного раствора (50-100 мкл) и готовят препараты негативного
контрастирования. Таким образом достигается увеличение чувствительности ЭМ
в 50-100 раз.
Содержание вирусных частиц в препарате можно повысить путем диффузии
жидкости в агар. В простейшей модификации метод диффузии применяют еле-
582 ВИРУСОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ
дующим образом. Каплю осветленной вирусной суспензии наносят на поверхность
агарового блока (1-2% агарозы) толщиной 2-3 мм и сверху на каплю помещают
сетку с подложкой. За 20-30 мин экспозиции жидкость впитывается в агар, в
результате чего концентрация вирусных частиц в образце возрастает в 10-15 раз.
Непосредственно перед тем как вся жидкость впитается, сетку снимают с препара-
та, отмывают водой и контрастируют.
В другой модификации метода используют гибкие микропланшеты с лунками,
залитыми агаром на три четверти. В этом случае сетку кладут в лунку поверх агара
лицевой стороной вверх, затем на сетку наносят препарат в количестве около
100 мкл так, чтобы заполнить лунку доверху, и выжидают 20-40 мин, пока жид-
кость не впитается в агар. Сетку снимают, очищают нижнюю сторону от остатков
агара фильтровальной бумагой, отмывают в капле воды и контрастируют.
В сравнительных исследованиях было показано, что ультрацентрифугирование
эффективнее метода диффузии в агар приблизительно в 1.3 раза, но диффузия
быстрее, экономичнее и безопаснее с точки зрения возможности заражения персо-
нала. Кроме того, для метода диффузии требуется меньшее количество исходного
материала.
Другое направление повышения чувствительности электронно-микроскопи-
ческой диагностики состоит в увеличении сорбционной способности подложек.
Классическими методами в этом плане служат иммтноэлектронная микроскопия
(ИЭМ) и обработка углеродных подложек в камере тлеющего разряда.
Для концентрации вирусных частиц на подложке методом ИЭМ требуется сыво-
ротка реконвалесцентов. В классическом варианте каплю сыворотки в разведении
около 1:1000 смешивают с каплей вируссодержащей жидкости и инкубируют во
влажной камере (в закрытой чашке Петри) в течение получаса. Вирусные частицы
образуют комплексы с антителами и в виде агломератов оседают на подложку.
Такие агломераты гораздо легче обнаружить под электронным микроскопом, чем
отдельные вирусные частицы.
Метод особенно эдфек гивен при обнаружении мелких вирусов и был с успехом
применен для идентификации вируса гепатита А.
Повышения адсорбционных свойств подложек по отношению к широкому спек-
тру вирусных агентов достигают с помощью гидрофилизации подложек различ-
ными методами. Свежеприготовленные углеродные подложки гидрофобны, из-за
чего вирусные частииы (обладающие, как правило, гидрофильными рецепторны-
ми группами) плохо адсорбируются и распределяются на подложке крайне нерав-
номерно. Описан способ повышения сорбционных свойств подложек с помощью
обработки слабым раствором полилизина, однако намного эффективнее вместо
полилизина использовать линейные поликатионы, в частности полиаллиламин.
Обработка подложек полиаллиламином позволяет увеличить чувствительность
ЭМ в среднем в 5-10 раз.
Таким образом, электронная микроскопия остается одним из наиболее эффек-
тивных методов в диагностике вирусных инфекций, так как позволяет одновре-
менно выявлять все известные, а также новые вирусы.
Метод иммунофлюоресценции
Флюоресцентная (люминесцентная) микроскопия (ИФМ) — широко приме-
няемый метод лабораторной диагностики вирусных инфекционных заболеваний.
ИФМ позволяет выявить присутствие возбудителя в биологическом материале
пациента, а также определить антитела к нему. ИФМ осуществляют с помощью
специального микроскопического оборудования — флюоресцентного микроскопа
(рис. 23-10). Наиболее часто используют в ИФМ флюорофор флюоресцеинизо-
тиоцианат (ФИТЦ, или FITC).
ВИРУСОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ 583
I
Фильтр
возбуждения
Объектив
Фильтр эмиссии
4 (пропускает свет от флюорофора)
Образец
Рис. 23-10. Принципиальное устройство люминесцентного (флюоресцентного; микроскопа.
К сожалению, применяемые в настоящее время флюорофоры подвержены
эффекту светового выцветания: при многократном облучении возбуждающим
светом светимость препарата необратимо снижается вплоть до полного исчез-
новения, что связано с взаимодействием возбужденных молекул флюорофора с
атомами кислорода. Консервация препарата в специальных бескислородных сре-
дах помогает замедлить процесс выцветания, но не останавливает его. Вследствие
этого создание музея препаратов флюоресцентной микроскопии, подобно гисто-
логическому музею, представляется весьма проблематичным. Один из вариантов
решения проблемы — качественная фотофиксация образцов с формированием
цифрового архива.
Современные флюоресцентные микроскопы оборудованы набором светофиль-
тров, позволяющим использовать все основные флюорофиры, применяемые в
лабораторной диагностике методом ИФМ. Кроме того, флюоресцентный микроскоп
может работать как световой, что полезно при оценке морфологии препарата.
Наиболее распространенный вариант ИФМ в клинической лабораторной диа-
гностике вирусных инфекций — иммунофлюоресцентная микроскопия, когда для
визуализации используют не флюорохромы как красители, а антитела, конъюги-
рованные с флюорохромом. Метод применяют r двух видах.
• Препарат окрашивают раствором антител, конъюгированных с флюорофо-
ром (рис. 23-11), после чего подвергают микроскопическому анализу (пря-
мой ИФМ).
• Препарат обрабатывают раствором антител против какого-либо антигена
возбудителя, отмывают в целях удаления несвязавшихся антител и повтор-
но окрашивают раствором, содержащим конъюгированные с флюорофором
антитела (конъюгат) против первых антител. В большинстве про гоколов реко-
мендуют дополнительно обработать материал синькой Эванса для снижения
неспецифической флюоресценции. В некоторых коммерческих наборах синька
Эванса уже входит в состав лиофилизированного конъюгата. После этого пре-
парат подвергают микроскопическому анализу (непрямой ИФМ).
К достоинствам прямого варианта ИФМ следует отнести простоту выполнения
и относительную быстроту получения результатов. Однако прямой ИФМ имеет
ряд недостатков, в том числе меньшую чувствительность.
Непрямой ИФМ более сложен (требуется несколько инкубаций и отмывок),
что повышает риск лабораторной ошибки и требует более высокой квалификации
584 ВИРУСОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ
Первичные Ат,
меченные ФИТИ
Образовавшиеся комплексы
Аг-Ат-конъюгат излучают
специфическое свечение
Схема прямого М<ЭД
Схема непрямого МФА
Рис. 23-11. Схема постановки прямого и непрямого микроскопического флюоресцентного ана-
лиза.
Образовавшиеся комплексы
Аг-Ат-конъюгат излучают
специфическое свечение
персонала. Вследствие увеличения количества этапов возрастает и общее время
исследования. Однако непрямой ИФМ обладает неоспоримым достоинством:
его чувствительность значительно выше, чем у прямого ИФМ, благодаря воз-
можности одновременного связывания сразу нескольких антител конъюгата с
Fc-фрагментом первых антител. Вследствие такого мультисвязывания увеличива-
ется сила флюоресцентного сигнала и. следовательно, чувствительность диагно-
стического метода в целом.
Методы прямого и непрямого ИФМ используют для непосредственного выяв-
ления антигенов вируса в препаратах. Основу диагностикумов составляют полн-
или моноклональные антитела, получаемые от иммунизированных животных
либо методом гибридомных технологий. В качестве примера типа диагностиче-
ских наборов, зарегистрированных в РФ, можно привести наборы для диагности-
ки герпеса I—II типа и цитомегаловируса в урогенитальных соскобах, наборы для
диагностики гриппа и наиболее значим ых респираторных инфекций в мазках или
аспиратах из носоглотки, а также в секционном материале, наборы для выявления
антигенов вируса гепатита В в биоптате печени и т.п.
С помощью непрямого ИФМ можно определять в качественном и количествен-
ном вариантах специфические антитела к возбудителям инфекционных вирусных
заболеваний. Для установления присутствия в сыворотке крови антител к анти-
генам вируса (например, возбудителя геморрагической лихорадки с почечным
синдромом) используются предварительно сорбированные на предметном стекле
клетки из клеточной культуры, зараженной вирусом. При инкубации с сывороткой
пациента, за раженного вирусом ГЛПС, содержащиеся в ней антитела связываются
с антигенами вируса. Не связавшиеся за время инкубации антитела и другие ком-
поненты сыворотки удаляются при отмывке. Образовавшиеся комплексы «Аг-Ат»
ВИРУСОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ 585
выявляются при инкубации с конъюгатом. В случае положительного результата
при микроскопическом анализе видны специфически светящиеся зернистые вну-
триклеточные включения.
При последовательном титровании сыворотки можно установить титр иссле-
дуемых противовирусных антител по максимальному разведению, при котором
наблюдается специфическое свечение. Определение титра противовирусных анти-
тел методом непрямого ИФМ в динамике (обычно через 2-3 нед после иссле-
дования первой сыворотки) позволяет выявить нарастание титра при развитии
заболевания, снижение — завершении инфекционного процесса или постоянный
титр при наличии анамнестических антител.
Таким образом, с помощью ИФМ можно обнаружить не только антигены виру-
сов в микроскопируемом биологическом материале, но и антитела к исследуемым
вирусам, а также провести количественную оценку их содержания в динамике.
Несмотря на очевидные достоинства метода (простоту исполнения, относи-
тельную быстроту окрашивания, отсутствие повышенных тпебований к оснащен-
ности лаборатории и т.п.), существует ряд факторов, ограничивающих широкое
использование ИФМ в лабораторной диагностике. Как все микроскопические
методы, ИФМ характеризуется субъективизмом интерпретации результата и тре-
бует высокой квалификации персонала. При микроскопии разными операторами
даже одного и того же препарата можно получить противоречивые результаты.
Кроме того, повышенные требования предъявляются к взятию материала для
исследования. Например, неправильный забор клеток из носоглотки (недостаточ-
ное количество или полное отсутствие эпителиальных клеток) ведет к получению
ложноотрицательного результата при ИФМ. Неверная оценка локализации све-
чения (внеклеточное, а не внутриклеточное) может вызвать появление ложнопо-
ложительных результатов, например при определении антител к вирусу ГЛПС в
сыворотке крови
Система внутрилабораторного контроля качества микроскопических иссле-
дований методом ИФМ должна включать постановку положительных и отрица-
тельных контролей, контролей чистоты антител и конъюгата, проверочные тесты
для оператора на панели аттестованных препаратов. Подобные меры помогают
значительно снизить частоту неверных результатов и уменьшить субъективность
оценки препарата специалистом, проводящим микроскопический анализ методом
ИФМ.
Методические аспекты иммунофлюоресцентного анализа
Для метода ИФМ характерны простота постановки анализа и возможность
быстрого (1-2 ч) обнаружения широкого круга вирусных антигенов или опреде-
ления титца специфических антител к какому-либо вирусу.
ИФМ, как и многие другие методы твердофазного анализа, заключается в
последовательном проведении нескольких стадий. От методически правильного
выполнения каждой стадии анализа во многом зависит достоверность результатов,
а также чувствительность и специфичность метода.
Для различных вариантов ИФМ (прямого, непрямого методов) можно выде-
лить следующие общие этапы:
забор и подготовку клинических материалов;
J приготовление фиксированных препаратов;
Vх формирование на фиксированных препаратах соответствующих иммуно-
химических комплексов, содержащих флюоресцентную метку;
удаление неспецифически связавшихся антител;
J микроскопическую визуальную детекцию вирусных антигенов;
J интерпретацию результатов анализа.
586 ВИРУСОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ
Взятие и подготовка клинических материалов к исследованию
Объектом исследования в ИФМ могут быть клинические материалы из очагов
поражения: соскобы со слизистых оболочек и кожных покровов, носоглоточные
смывы, экссудаты, моча, мокрота, содержимое везикул, органы и ткани умерших
людей и животных, экспериментально зараженные культуры клеток. В практиче-
ском здравоохранении ИФМ используют в основном для обнаружения вирусных
возбудителей, вызывающих острые респираторные инфекции (ОРВИ), заболева-
ния урогенитальной сферы и офтальмологические инфекции.
Сроки взятия материала. Оптимальные сроки взятия материала при ОРВИ —
первые три дня от начала заболевания. На более поздних сроках происходит
элиминация вирусов в секреты дыхательных путей, что снижает их концентра-
цию в пробах и тем самым значительно уменьшает результативность анализа.
Лабораторные исследования следует проводить до начала терапии или исключить
обработку зон поражения антисептиками за 1-2 сут до взятия ма гериала.
Особенности взятия материала. Важным условием получения достоверных
результатов ИФМ служит наличие в пробе достаточного количества интактных
клеток из очага поражения. Посторонние компоненты (слизь, кровь, гной, жир),
дающие интенсивную неспецифическую флюоресценцию, должны содержаться в
образцах в минимальных количествах.
При диагностике ОРВИ соскобы берут со слизистых оболочек из задних, глу-
боколежащих отделов носоглотки, где находится основная диагностическая ткань
для детекции вирусов — цилиндрический мерцательный эпителий. Материал
может быть дополнен образцами из переднего отдела носа, выстланного плоским
эпителием, на котором возможна колонизация клеток бактериальной флорой.
При конъюнктивитах соскобы берут из каждого глаза отдельно. Как правило,
количество эпителиальных клеток в этих образцах незначительно, поэтому образ-
цы соскобов с конъюнктив сразу наносят на предметные стекла.
При заболеваниях мочеполовой сферы делают соскобы со слизистой оболочки
уретры, заднего свода влагалища, канала шейки матки. При заболеваниях пред-
стательной железы исследуют ее сок. При налижи везикул, эрозий на слизистых
и кожных покровах получают соскобы из оснований измененных участков тканей.
Для повышения достоверности диагностики следует брать образцы из нескольких
очагов поражения.
Транспортировка и хранение клинических материалов. Возбудители вирусных
инфекций находятся внутри эпителиальных клеток. До фиксации клеток суспен-
зию держат в пробирках с 0,01 М фосфатно-солевым буферным раствором (pH =
7,0-^7,2 (ФСБ) не более 8-12 ч при температуре 4-6 °C). Замораживание образцов
не допускается.
Приготовление клеточной суспензии. Пробирки с клеточной суспензией в ФСБ
встряхивают и концентрируют с помощью центрифугирования при 1000 об./мин
в течение 10 мин. Надосадэчную жидкость удаляют, а осадок клеток ресуспенди-
ру ют в минимальном объеме ФСБ.
При необходимости следует предварительно оценить концентрацию клеток в
материале. Для этого готовят пробный мазок. 0,1-0,2 мл суспензии наносят на обе-
зжиренное предметное стекло, высушивают, фиксируют спиртом в течение 5-10 мин
при комнатной температуре, окрашивают по Романовскому-Гимзе и микроско-
пируют. При низком содержании клеток, что часто бывает в образцах соскобов из
уретры, сока i [ростаты, слизистой оболочки носа у детей, материал концентрируют.
При высокой концентрации клеток (в гинекологическом материале, соскобах со
слизистой оболочки ротовой полости, языка) материал разбавляют ФСБ.
Приготовление фиксированных препаратов
Для приготовления препаратов в ИФМ используют нефлюоресцирующие
стекла или полимерные слайды, покрытые поли-Ь-лизином. Если стекла имеют
ВИРУСОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ 587
ободки лунок, полученные методом травления, стекла обезжиривают в 50% сер-
ной кислоте в течение 1 ч, затем промывают 30 мин проточной водой и хранят в
дистиллированной воде. В тех случаях, когда ободки тунок на стеклах нанесены
нерастворимыми цветовыми полимерами, стекла обезжиривают замачиванием в
течение 1 ч в смеси Никифорова (50% спирта и 50% эфира по объему).
В зависимости от конкретных задач клеточную суспензию наносят на стекло в
виде четко ограниченных капель объемом 15-20 мкл в количестве, соответствую-
щем числу тестируемых антигенов. Если необходима более углубленная, уточняю-
щая диагностика при хронических и осложненных вирусных инфекциях, готовят
дублирующие мазки. Мазки-от печатки из плотных проб получают, прикладывая
свежий срез исследуемого органа (ткани) к поверхности стекла. Для гистологиче-
ских образцов используют нефиксированные криостатные срезы.
При низкой концентрации клеток, наличии гноя, жировых включений и детри-
та в клинических образцах для увеличения количества адсорбированных клеток
стекла предварительно обрабатывают раствором поли-Ь-лизина (молекулярная
масса 70-90 кДа). внося в каждую лунку по 10 мкл 1% раствора. Инкубируют не
менее 5 мин, промывают дистиллированной водой и высушивают.
Мазки после высушивания фиксируют охлажденными органическими раство-
рителями или обработкой сшивающими реагентами. Выбор способа фиксации
зависит от локализации определяемых аитиг енов. При фиксации органическими
растворителями (метанолом, метанолом/ацетоном 1:1, ацетоном) стекла с высу-
шенными мазками погружают в заранее охлажденный до 10-20 °C органический
растворитель на 10-20 мин. Органические растворители должны быть высокой
степени очистки (хч. осч). Необходимо иметь в виду, что при этом частично уда-
ляются растворимые и мембраносвязанные антигены.
Для предотвращения вымывания антигенов вместо органических растворителей
используют сшивающие реагенты. Для этого на мазок наносят 15-20 мкл 1-4% рас-
твора параформальдегида или раствор, содержащий 3% параформальдегида и
0,02% глутарового альдегида, с последующей 15-минутной инкубацией. Обработка
сшивающими реагентами позволяет увеличить чувствительность метода, что важно
в случае низкого содержания вирусных антигенов в клинических образцах.
Зафиксированные мазки хранят при температуре 4-6 °C в течение 2-3 мес или
при -20 °C — до года. Хранение препаратов в некоторых случаях важно для ретро-
спективного анализа, контроля эффективности терапии и сохранения диагности-
чески ценных, уникальных препаратов.
Формирование на фиксированных препаратах иммунохимических ком-
плексов, содержащих флюоресцентную метку
При фиксации на каждый из мазков наносят по 25-50 мкл рабочих растворов
соответствующих специфических антител. В случае прямого варианта ИФМ мазки
обрабатывают антителами, содержащими флюоресцентную метку. При непрямом
варианте ИФМ обработку мазков проводят в две стадии. Сначала наносят анти-
вирусные антитела или иммунную сыворотку, затем сформировавшийся комплекс
сантиген-антитело» выявляют с помощью специфического антивидового ФИТЦ-
конъюгата. Определяющим моментом на этих стадиях анализа служит специфич-
ность реакций, характеризуемая величиной констант связывания комплекса
«антиген-антитело» и кинетикой процессов.
Основной параметр, детерминирующий чувствительность и специфичность
ИФМ, — аффинность применяемых реагентов. Время этой стадии анализа во
многом зависит от скорости достижения равновесия в специфических реакциях
и, как правило, составляет 30-60 мин при комнатной температуре и 20-40 мин —
при температуре 36 ° С.
В качестве флюоресцентной метки наиболее часто используют флюоресцеин.
Его активная форма — флюоресцеинизотиоционат (ФИТЦ) — в физиологических
588 ВИРУСОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ
условиях реагирует с аминогруппами специфических иммуноглобулинов, образуя
флюоресцентный конъюгат. Флюоресцеин имеет широкий спектр поглощения в
ультрафиолетовой и синей областях спектра и зеленый свет испускания. Это обу-
словило его преимущество перед другими флюорэхромами, так как зеленый свет
излучения редко наблюдается при аутофлюоресценции клеток тканей
Важная характеристика качества конъюгата — молярное соотношение ФИТЦ/
белок. В экспериментальных данных показано, что отношение значений опти-
ческой плотности поглощения ФИТЦ (ОП — 490 нм) и оптической плотности
поглощения белка (ОП — 280 нм) должно быть 5-6 для конъюгатов, используе-
мых в прямом варианте ИФМ, и 3-4 — для конъюгатов, применяемых в непрямом
варианте ИФМ.
Удаление неспецифически связавшихся антител
После каждого этапа инкубации раствора специфических антител на фиксиро-
ванных мазках следует этап промывки с помощью раствора ФСБ. Ее цель заклю-
чается в устранении неспецифических взаимодействий.
Более результативная промывка обеспечивается добавлением в промывной
раствор неионных детергентов типа «Твин-20», что позволяет осуществлять
эффективную конкуренцию между белок-белковыми взаимодействиями и взаи-
модействиями «белок-детергент». Поскольку все эти процессы равновесные, для
более полного устранения неспецифической сорбции необходима многократная
промывка мазков. Это способствует снижению уровня неспецифической и фоно-
вой флюоресценции.
Для уменьшения неспсцифической сорбции из-за перекрестно реагирующих
антигенов в состав конъюгата вводят неспецифические иммуноглобулины, род-
ственные антителам в конъюгате. Например, если конъюгат получен на основе
кроличьих антител, в состав конъюгата вводят до 1% нормальной кроличьей
сыворотки.
Микроскопическая визуальная детекция вирусных антигенов
Учет результатов ИФМ осуществляют с помощью люминесцентных микро-
скопов. укомплектованных иммерсионным объективом (х90), сине-фиолетовыми
светофильтрами возбуждения и запирающим светофильтром. В качестве иммер-
сионных жидкостей применяют 90% водный раствор глицерина с показателем
преломления 1,434, нефлюоресцирующее иммерсионное масло и диметилфталат.
Использование иммерсионных жидкостей позволяет повысить разрешающую спо-
собность микроскопа, так как они имеют более высокий показатель преломления,
чем воздух. В результате регистрируемые лучи света, выходя из препарата, не рас-
сеиваются и попадают в объектив микроскопа.
При учете результатов ИФМ необходимо иметь в виду, что при длительном
освещении препарата имеет место снижение интенсивности специфической флюо-
ресценции (выцветание) вследствие фотохимических реакций метки — ФИТЦ,
Этот процесс можно существенно замедлить, добавив на пробу 90% глицерина и
накрыв покровным стекле м. Рекомендуют проводить все стадии анализа после
добавления конъюгата при минимально возможном освещении.
Для длительного хранения окрашенных эталонных или демонстрационных
препаратов следует каждый мазок накрыть покровным стеклом, края которого
загерметизировать лаком.
Интерпретация результатов анализа
При интерпретации результатов ИФМ необходимо оценивать специфичность
флюоресценции и знать морфологические свойства изучаемого объекта.
При триппе и ОРВИ анализируют мазки из слизистой оболочки носоглотки.
Несмотря на клеточный полиморфизм, клетки легко типируются. Диагностически
значимы клетки цилиндрического эпителия: удлиненные, с мерцательным аппа-
ратом и базально расположенным ядром, а также круглые и овоидные клетки с
ВИРУСОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ 589
относительно большим ядром. Диагностически достоверно обнаружение в мазке
не менее трех клеток цилиндрического эпителия с характерной зеленой флюо-
ресценцией, Не следует учитывать свечение в крупных полигональных клетках
плоского эпителия с небольшим, центрально расположенным ядром и в ярко све-
тящихся лейкоцитах.
Свечение антигенов вируса гриппа, адено- и герпес-вирусов наблюдается в ядре
и цитоплазме (рис. 23-12, см. цв. вклейку). Характерно многообразие свечений:
глыбчатое, гранулярное и диффузное; возможно тотальное свечение. Антигены
парагриппа, корона- и PC-вирусов локализуются в цитоплазме, при этом ядра
клеток остаются темными. При инфекциях, вызванных вирусами герпеса, пара-
гриппа и PC-вирусом, возможно обнаружение многоядерных клеток (симпластов).
Р1нтактные клетки не окрашиваются и имеют слабое свечение.
При хронических инфекциях вирусные антигены определяются в стадии обо-
стрения. При смешанных вирусно-бактериальных инфекциях в хронической
стадии вирусные антигены выявляю гея не всегда. Дополнительная информация о
возможной вирусной инфекции может быть получена при изучении рутинной мор-
фологии мазков, окрашенных по Романовскому-Гимзе. Обнаруживают гигантские
одно- и многоядерные клетки, внутриядерные и цитоплазматические включения,
воспалительную инфильтрацию тканей при отсутствии или небольшом количестве
бактериальной флоры, деструктивные изменения, апоптотические клетки и клетки
со светлым ободком вокруг ядра — так называемые совиные глаза. Перечисленные
признаки могут свидетельствовать о иитопатогенном действии вирусных агентов
в исследуемых образцах. На основании цитолог ической патологии можно опреде-
лить направление дальнейшего поиска вирусных антигенов: провести окрашива-
ние дублирующих препаратов с помощью непрямого варианта ИФМ, имеющего
на порядок более высокую чувствительность, или осуществить исследование с
помощью других методов.
1аким образом, высокая диагностическая ценность ИФМ может быть достигну-
та при соблюдении следующих правил:
• образцы клинических материалов, полученные из очагов поражения, долж-
ны содержать достаточное количество характерных для диагностируемых
тканей интактных клеток и минимум воспалительных и деструктивных ком-
понентов ткани;
• техника взятия патологического материала должна быть безопасна для паци-
ента и клинициста;
• доставку клинических образцов следует осуществлять в рекомендуемые
сроки (8-12 ч) с использованием специальных контейнеров и транспортной
среды при температурном режиме 4-8 °C;
• диагностические наборы и реагенты, применяемые в ИФМ, должны иметь
сертификаты;
• врач клинической лабораторной диагностики должен иметь соответствую-
щую специализацию и опыт цитологическс>й работы.
Серологические методы
ТРАДИЦИОННЫЕ МЕТОДЫ СЕРОДИАГНОСТИКИ
В основе всех серологических реакций лежит взаимодействие антигена и анти-
тела, Существуют две модификации серологических реакций.
Обнаружение антител в сыворотке крови больного
В этом случае из двух компонентов реакции неизвестным является сыворот-
ка крови, содержащая антитела, поэтому для исследования требуется заведомо
известный антиген.
590 ВИРУСОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ
Для диагностики вирусных инфекций в качестве антигенов традиционно исполь-
зуют аллантоисную, амниотическою жидкости, суспензии хорионаллантоисных
оболочек и желточных мешков куриных эмбрионов, жидкую и клеточную (экс-
тракты) фракции культур клеток, а также гомогенаты органов животных, содержа-
щие определенные вирусы. Существуют коммерческие антигены для постановки
реакций торможения гемагглютинации (РТГА) и связывания комплемента (РСК).
Положительный результат реакции свидетельствует о наличии в сыворотке
крови антител, гомологичных используемому антигену. Отрицательный результат
указывает на отсутствие антител.
Для получения достоверных результатов серологической диагностики необхо-
димо изучать так называемые парные сыворотки крови больного, взятой в пер-
вые дни и через определенный период времени (2-3 нед) от начала заболевания.
Четырехкратное и более увеличение титра антител во второй сыворотке по срав-
нению с первой имеет диагностическое значение.
Установление родовой, видо ой и типовой принадлежности вируса
Неизвестный компонент реакции в этом случае — антиген. Исследование требу-
ет постановки реакции с заведомо известными иммунными сыворотками, которые
получают в результате длительной иммунизации животных (кроликов, морских
свинок, крыс, лошадей, ослов, баранов и др.) соответствующими, уже известными
антигенами. Иммунные сыворотки должны обладать специфичностью и высоким
содержанием антител. Уровень антител определяют в любой серологической реак-
ции с разводе ниями антител или антигенов. Последнее из разведений, вызывающее
отчетливый положительный результат, называется титром сыворотки. Сыворотка
может быть специфичной в пределах рода, вида или типа и соответственно реаги-
ровать с гомологичным антигеном и не реагировать с гетерологичным.
Для серологической идентификации вирусов выпускают нативные сыворот-
ки и адсорбированные. Неадсорбированные сыворотки отличаются высокими
титрами антител (до 1:12 500-1:25 000), но не свободны от антител к микробам
и неспецифических ингибиторов. Адсорбированные сыворотки обладают строгой
специфичностью, но низкими титрами и не подлежат разведению.
РЕАКЦИЯ ГЕМАГГЛЮТИНАЦИИ
Механизм реакции прямой гемагглютинации (РГА) основан на природной
комплементарности участвующих в ней компонентов, а именно на способности
эритроцитов склеиваться при адсорбции на них определенных антигенов. Реакция
не является серологической, так как протекает без участия иммунной сыворотки.
РГА используют для индикации вирусных агентов, титрования вируссодержащих
материалов и ориентировочной диагностики некоторых вирусных инфекций.
В серологических исследованиях РГА применяют при выборе рабочей дозы анти-
гена, используемого в реакции торможения гемагглютинации.
Для постановки реакции требуются: 1) растворимый антиген; 2) испытуемая
или стандартная иммунная сыворотка; 3) эритроциты; 4) изотонический раствор
натрия хлорида (физиологический раствор) (pH = 7,24-7,4); 5) пипетки градуиро-
ванные и пастеровские, 6) пластинки с лунками из органического стекла.
Используют эритроциты животных (барана, обезьяны, морской свинки), птиц
(петуха, гуся, голубя), человека — I (0) группы. Кровь собирают в стерильный
флакон со стеклянными бусами и дефибринируют, энергично встряхивая флакон в
течение 10-15 мин, или немедленно смешивают с равным объемом антикоагулянта
(2,5% раствором цитрата натрия или модифицированным раствором Алсевера —
2,5% глюкозы, 0,42% натрия хлорида, 0,83% цитрата натрия; pH = 6.4). Взвесь
эритроцитов в растворе антикоагулянта можно хранить 2-3 нед при 4 °C.
Непосредственно перед применением осадок эритроцитов трижды отмывают,
каждый раз добавляя 10-кратные объемы физиологического раствора, перемети-
ВИР\СОЛОГИЧЕСКИЕ исследования 591
вая и осаждая эритроциты путем центрифугирования при 1000 об./мин в течение
10 мин и затем сливая или отсасывая надосадочную жидкость и заменяя ее све-
жим физиологическим раствором. Отмытые неразведенные эритроциты можно
использовать в течение 48 ч при условии хранения их на холоде, в виде 0,25- 5,0%
взвеси в физиологическом растворе — в течение 1-7 дней. Нельзя допускать гемо-
лиза или спонтанной агглютинации эритроцитов.
Постановку реакции гемагглютинации осуществляю! путем последовательного
двукратного разведения 0.2 мл антигена (вируса) физиологическим раствором в
лунках с округлым гладким дном панелей из плексигласа, начиная с 1:2 до 1:1024
или более вплоть до предполагаемого титра данного антигена. Затем в этот же ряд
лунок с разведенным антигеном добавляют по 0.2 мл физиологического раствора
и по 0,4 мл 0,25-1,0% взвеси эритроцитов. Одновременно осуществляют контроль
качества взвеси эритроцитов. Для этого к 0,4 мл физиологического раствора
добавляют 0,4 мл той же, что и в основном опыте, взвеси эритроцитов. Панели
энергично встряхивают и оставляют при комнатной температуре или при 4 °C на
30-60 мин. Эритроциты кур оседают за 30 мин, эритроциты животных — медлен-
нее, обычно в течение часа. В эти сроки и учитывают РГА.
Результат оценивают в плюсах:
«++++» — реакция положительная, образуется характерный осадок эритроци-
тов в виде тонкой пленки с неправильными, слегка волнистыми краями, рас-
пределяющийся равномерным слоем по всему дну, похожий на перевернутый
зонтик;
«+++s> — реакция положительная, осадок эритроцитов несколько менее широ-
кий, но отчетливый, на дне лунки едва намечается кольцевая зона из осевших
эритроцитов;
«++» — реакция неотчетливо положительная, «зонтик» эритроцитов заметно
меньше, на дне лунки формируется более плотная кольцевая зона;
«+» — реакция слабоположительная, отмечается значительное оседание эри-
троцитов на дне лунки в виде плотного диска с зернистой каймой;
«-» — реакция отрицательная, на дне лунки образуется плотный осадок эритро-
цитов в виде диска с ровными краями.
Титром вируса считается то наибольшее его разведение, при котором еше
наблюдается полная агглютинация эритроцитов на 4 или 3 плюса. В контрольных
лунках эритроциты должны формировать плотный диск с ровными краями.
РЕАКЦИЯ НЕЙТРАЛИЗАЦИИ
Одним из основных серологических методов, применяемых в вирусологиче-
ской практике, считаемся реакция нейтрализации. Используют ее для диагностики
заболеваний вирусной этиологии, оценки противовирусных, профилактических и
диагностических препаратов.
Реакция нейтрализации основана на способности антител, присутствующих в
иммунных сыворотках, предотвращать размножение вируса в различных чувстви-
тельных системах: в организме животных, клеточных культурах, куриных эмбрио-
нах. Применяют для типирования выделенных вирусов и титрования антител в
сыворотках переболевших.
Для постановки реакции требуются:
S лабораторные животные, кульгуры клеток или развивающиеся куриные
эмбрионы;
J инфекционный вирус известного типа (при серологической диагностике
инфекции) или исследуемый (при идентификации выделенного вируса):
S сыворотка исследуемая (при серологической диагностике инфекции) или
типоспецифическая (при идентификации выделенного вируса);
J шприц объемом 1 мл;
592
ВИРУСОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ
J пробирки и штативы к ним или 96~луночные полистироловые планшеты с
плоским дном для культивирования клеточных культур;
J градуированные пипетки по 1 и 5 мл;
J изотонический раствор натрия хлорида или забуференный физиологиче-
ский раствор для работы с клеточными культурами;
S термостат с температурой 37 °C или СО,-инкубатор.
Источником вируса для. постановки реакции могут служить вируссо держащие
культуральные жидкости, а также экстракты из куриных эмбрионов и орга-
нов зараженных животных. Вируссодержащий материал должен быть гомоген-
ным, освобожденным от крупных частиц путем центрифугирования при 1500-
3000 об./мин в течение 15 мин. Для постановки реакции нейтрализации необ-
ходимо использовать вируссодержащий материал с известной активностью, чего
достигают предварительным его титрованием Сроки хранения оттитрованной
вирусной суспензии зависят от типа вируса и указаны при описании лабораторной
диагностики каждой инфекции.
Сыворотки для реакции нейтрализации в зависимости от цели постановки
реакции могут быть полущены от специально вакцинированных животных либо от
людей, в чьих сыворотках необходимо определить содержание нейтрализующих
антител для верификации или постановки диагноза. Поскольку' окончательным
подтверждением диагноза служит нарастание титра антител к вирусу, следует брать
сыворотки от больных в начале острого периода заболевания и спустя 1-2 нед (так
называемые парные сыворотки).
Для постановки реакции нейтрализации с большинством вирусов сыворотки
не требуют специальной обработки. К некоторым же вирусам в сыворотках крови
человека и животных содержатся неспецифические термолабильные и термоста-
бильные ингибиторы, нейтрализующие инфекционную активность вирусов. Для
инактивации неспецифических термолабильных ингибиторов сыворотки крови
человека, обезьян, лошадей, морских свинок прогревают 30 мин при температуре
56 °C, белых крыс и кроликов — при 60 °C, петухов — при 58 °C. (Методы высво-
бождения сывороток от неспецифических термостабильных ингибиторов приве-
дены при описании лабораторной диагностики каждой инфекции.) Обработанные
таким образом сыворотки используют сразу либо сохраняют в холодильнике при
4-6 °C в течение 1-3 дней или в замороженном виде в течение более длительного
времени, необходимого для сбора парных сывороток.
Возможны две модификации реакции, проводимой в асептических условиях:
J возрастающие разведения суспензии, содержащей вирус, соединяют в рав-
ном объеме с постоянной дозой сыворотки;
J возрастающие разведения сыворотки соединяют с постоянной дозой
вируса.
Постановка реакции нейтрализации на лабораторных животных
Пробирки расставляют в штативы параллельными рядами. Количество проби-
рок в каждом параллельном ряду должно соответствовать числу разведений вируса
и превышать его биологический титр. Количество рядов должно соответствовать
числу взятых в опыт сывороток и контролей. Необходимыми контролями служат:
1) нормальная сыворотка; 2) активность вируса, взятого в опыт. Для первого
контроля в ряд пробирок вносят нормальную сыворотку, для второго — изотони-
ческий раствор натрия хлорида до 0,5 мл каждого ингредиента в пробирку, сыво-
ротки при этом не разводят или разводят 1:5.
Во вспомогательном ряду готовят 10-кратные разведения вирусной суспензии,
наливая в каждую пробирку по 4,5 мл изотопического раствора натрия хлорида.
В первую пробирку добавляют 0,5 мл вирусной суспензии, осторожно спуская ее
по стенке пробирки и не касаясь пипеткой жидкости в пробирке (это необходимо,
ВИРУСОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ 593
чтобы не внести излишки вируса, остающиеся на стенках пипетки, и не исказить
истинное разведение вируса); пипетку сбрасывают в емкость с дезинфицирую-
щим раствором. Другой пипегкой тщательно перемешивают содержимое первой
пробирки, набирают 0,5 мл суспензии и переносят ее во вторую пробирку и так
далее до последней пробирки ряда. Начиная с наибольшего разведения, вирусную
суспензию переносят в объеме 0,5 мл в соответствующие пробирки параллельных
рядов, содержащие в таком же объеме специфические сыворотки, нормальную
сыворотку или изотонический раствор хлорида натрия.
Пробирки со смесями встряхивают и оставляют на 1-2 ч при комнатной тем-
пературе или в термостате либо выдерживают 18-20 ч при 4 °C. По истечении
времени контакта смеси вводят лабораторным животным для индикации инфек-
ционного вируса: на каждое разведение берется не менее четырех животных.
Способ введения, доза и время учета результатов варьируют в зависимости от
используемого животного и свойств вируса (приведены в описании лабораторной
диагностики каждой инфекции).
О наличии инфекционного вируса в смеси судят по гибели лабораторных
животных. Проводят учет результатов и рассчитывают по методу Рида и Менча
титры вируса в смеси с нормальной и в смеси с испытуемой сывороткой, за кото-
рые принимают предельное разведение вирусной суспензии, вызвавшее гибель
50% зараженных животных. Это разведение рассматривают как одну дозу LD.O.
Для каждой исследованной иммунной сыворотки высчитывают индекс нейтра-
лизации (максимальное количество летальных доз вируса, нейтрализованное дан-
ной иммунной сывороткой по сравнению с контрольной, — IN). Индекс нейтрали-
зации вычисляют по разности логарифма титра вируса с нормальной и иммунной
сывороткой. Например: 5,22 - 1,77 = 3,45. По таблице антилогарифмов находят
соответствующее число, равное 28 18, которое выражает в нашем случае индекс
нейтрализации иммунной сыворотки, взятот в опыт. IN от 1 до 10 принято считать
отрицательным, от 10 до 50 — сомнительным, выше 50 — положительным.
При серологической идентификации выделенного вируса к различным, обычно
двукратным, разведениям известной типоспецифической сыворотки иммунизиро-
ванных животных добавляют постоянное количество предварительно тщательно
оттитрованного вируса (обычно 100 LD50). В качестве контроля используют не
содержащую антител нормальную сыворотку, полученную от соответствующего
животного до иммунизации. Продолжительность контакта вируса с сывороткой и
температура инкубации варьируют в зависимости от исследуемого вируса. После
инкубации смесь вируса с сывороткой вводят животным. Нейтрализующую актив-
ность сыворотки выражают ее титром, то есть тем наибольшим разведением,
которое предотвращает гибель половины инокулированных животных. Тип вируса
устанавливают по типоспецифической сыворотке, которая показала наивысший
индекс нейтрализации.
При исследовании парных сывороток больного с целью ретроспективной диа-
гностики инфекции каждую из сывороток исследуют с несколькими типами виру-
сов. Контролем в реакции может служить сыворотка больного, взятая в острой
стадии заболевания. Возбудителем заболевания считают вирус того серологиче-
ского типа, по отношению к которому вторая из парных сывороток показала наи-
больший индекс нейтрализации.
Постановка реакции нейтрализации в куриных эмбрионах
Реакцию нейтрализации в куриных эмбрионах проводят двумя способами:
J с учетом результатов реакции по гемагтлютиницующей активности;
J с учетом результатов по обнаружению морфологических изменений в
виде оспин на хорионаллантоисных оболочках.
Соблюдают общие правила предварительного определения биологической
активности вируса, разведения вируса или сыворотки, соединения ингредиентов,
594 ВИРУСОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ
времени контакта и температуры, при которой его осуществляют. Способ введе-
ния полученных смесей зависит от тропизма вируса к тканям куриного эмбриона.
Так, вирус гриппа и Ньюкасла вводят в аллантоисную полость, вирусы вакцины и
натуральной оспы — на хорионаллантоисную оболочку, ряд вирусов, вызываю-
щих энцефалиты, — в желточный мешок. На каждый вариант разведения берут по
4-6 эмбрионов, вносят по 0,1-0,2 мл смеси вируса и сыворотки.
При постановке реакции первым способом используют 10-11-дневные эмбрио-
ны. После 2-3 дней инкубации пци 35-37 ПС эмбрионы вскрывают и определяют
наличие вируса в аллантоисной жидкости каждого эмбриона с помощью реакции
гемагглютинации.
Если в реакции используют постоянную дозу сыворотки и возрастающие раз-
ведения вируса, то оценивают титры вируса в опыте и контроле, а затем индекс
нейтрализации испытуемых сывороток. За титр вируса принимают предельное
ею разведение в смеси с сывороткой, вызвавшее у 50% зараженных эмбрионов
образование гемагглютининов при наличии их в аллантоисной жидкости эмбрио-
нов, зараженных вирусом в смеси с физиологическим раствором или нормальной
сывороткой.
Если же в реакции применяют постоянную дозу вируса (обычно 100 или
1000 ИДЧ|) и возрастающие разведения сыворотки, то нейтрализующую актив-
ность последней выражают ее титром, то есть наибольшим разведением, предот-
вратившим развитие гемаюлютинации в половине инфицированных эмбрионов.
Постановку реакции вторым способом осуществляют путем внесения по 0,1 мл
подготовленных суспензий на хорионаллантоисную оболочку 4-6 эмбрионов
12-дневного возраста. Через 48 ч инкубации при 37- 39 °C эмбрионы вскрывают,
подсчитывают количество оспин (бляшек) на каждой хорионаллантоисной оболоч-
ке и находят среднеарифметическое для каждого разведения сыворотки или вируса.
За титр вируса или антител сыворотки принимают предельное их разведение, в
котором наблюдается погашение 50% и более оспин, образуемых в контроле с рабо-
чим разведением вирусной суспензии в смеси с физиологическим раствором.
Постановка реакции нейтрализации в культурах клеток
При реакции нейтрализации в культурах клеток ход реакции такой же, как и при
постановке ее на лабораторных животных. Опыты проводят в (трого асептических
условиях.
В эксперименте могут быть использованы культуры различных линий переви-
ваемых клеток, а также культуры первично-трипсинизированных тканей, но такие,
в которых вирус проявляет отчетливое цитопатическое действие. Источником
вируса могут служить культуральная жидкость или экстракт инфицированных и
дезинтегрированных культур клеток, подвергнутых повторному 3-5-кратному
замораживанию и оттаиванию.
Инфекционный материал освобождают от клеточного детрита путем центрифу-
гирования при 2000-3000 об./мин в течение 10-15 мин. Затем в поддерживающей
среде или физиологическом растворе готовят десятикратные разведения вирусной
суспензии в количестве, превышающем предельную биологическую активность
вируса на 1-2 разведения. По 0,1 мл каждого разведения вируса вносят в четыре
пробирки с однослойной культурой клеток, в которых предварительно заменяют
ростовую среду на поддерживающую.
Зараженные культуры клеток инкубируют при 37 СС в течение 5-14 сут в
зависимости от типа вируса. Далее оценивают результаты по цитопатическому
действию вируса и рассчитывают одну ТЦД50, то есть то предельное разведение
вируса, которое вызывает развитие ЦПД в 50% зараженных культур клеток.
В реакции нейтрализации используют 10 или 100 ТЦД50, содержащиеся в 0,1 мл
вирусной суспензии.
ВИРУСОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ 595
Для серологической идентификации выделенного вируса постоянное его разве-
дение в объеме 0,5 мл добавляют к равному объему двукратных разведений испы-
туемых типослецифических и нормальной сывороток. Параллельно осуществляют
контроль активности вируса и токсичности сыворотки.
Контроль активности вируса. То же рабочее разведение вируса соединяют
анало1ичным образом с физиологическим раствором. Во второй модифика-
ции постановки реакции нейтрализации в целях типирования выделенного
вируса и одновременной серологической диагностики инфекции для контро-
ля правильности выбранного рабочего разведения вируса его разводят в 10,
100 и 1000 раз и каждое разведение вносят в объеме 0,1 мл в контрольные
пробирки с культурой клеток, добавляя по 0,9 мл поддерживающей питатель-
ной среды или физиологического раствора.
Контроль токсичности сыворотки. Сыворотку в исходном разведении соеди-
няют с физиологическим раствором. Пробирки со смесями инкубируют при
37 °C или при комнатной температуре в течение 1-2 ч. В пробирках с культе-
рами клеток того же типа, который использовался для титрования инфекцион-
ной активности вируса, предварительно меняют ростовую среду на поддержи-
вающую, добавляя по 0,8 мл в ь аждую пробирку. Остав. гяют четыре пробирки
с незараженными культурами клеток, внося в них по 1,0 мл поддерживающей
питательной среды (контроль состояния клеток в течение опыта).
После инкубации каждую из опытных и контрольных смесей последовательно
вносят по 0,2 мл в четыре пробирки с культурами клеток, содержащими поддержи-
вающую питательную среду. Инокулированные и контрольные культуры клеток
помещают в гермосзат при 35-37 °C и периодически проверяют состояние клеток
под микроскопом. В процессе относительно короткого (недельного) инкубирова-
ния питательную среду можно не менять, при более длительном инкубировании
питательную среду заменяют на свежую. При развитии специфической дегенера-
ции клеток в контрольных культурах, зараженных вирусом без добавления сыво-
ротки, проводят учет результатов. Определяют предельное разведение сыворотки,
нейтрализовавшее цитопатическое действие вируса в 50% зараженных культур, и
принимают его за титр антител для каждой из испытуемых сывороток. Тип выде-
ленного вируса соответствует типу той иммунной сыворотки, которая оказывается
специфической для выделенного вируса, то есть предупреждает развитие специфи-
ческой дегенерации клеток в наиболее высоком титре.
При постановке реакции нейтрализации с целью ретроспективной диагностики
заболевания используют вторую модификацию, когда предварительно определя-
ют цитопатогенный титр всех серологических типов вирусов, которые необходимо
взять в реакцию для исследования парных сывороток. Приготовленных во вспо-
могательном ряду двукратных разведений каждой сыворотки больного должно
быть достаточно для соединения со всеми типами вирусов. В пробирки каждого
ряда добавляют равные объемы вирусов в рабочей дозе, равной 100 ЦПД50 вируса
в 0,1 мл Одновременно ставят необходимые контроли активности примененных
доз всех типов вирусов, цитотоксичности сывороток и их разведений. Условия
проведения реакции аналогичны предыдущим. Об этиологическом значении одно-
го из примененных типов вируса судят по наличию четырехкратного и большего
прироста титра антител во второй сыворотке больного по сравнению с первой.
Реакция нейтрализации с учетом результатов по гемадсорбции
Реакцию нейтрализации гемадсорбции выполняют в культуре клеток, исполь-
зуют при работе с гемадсорбирующими вирусами для типирования выделенных
вирусов и титрования антител в парных сыворотках больных.
Вначале определяют гемадсорбирующий титр вируса, для чего 10-кратные
разведения вируса в поддерживающей питательной среде или физиологическом
596 ВИРУСОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ
растворе вносят по 0,1 мл в пробирки с чувствительными к вирусу клеточными
культурами, после предварительной замены ростовой питательной среды на под-
держивающую. Каждым разведением вируса заражают не менее двух лунок. Срок
и температура инкубации в термостате зависят от оптимальных условий культиви-
рования того или иного вируса.
Результаты титрования учитывают с помощью реакции гемадсорбции. Для этого
во все пробирки добавляют по 0,2 мл 0.4% вз1 еси отмытых эритроцитов, чувстви-
тельных к гемагглютинирующему действию данного вируса in vitro. Пробирки
выдерживают 20-30 мин при температуре, оптимальной для проявления гемаг-
глютинирующей активности вируса. Затем монослой клеток 1-2 раза промывают
физиологическим раствором и, слегка покачивая или вращая, просматривают под
малым увеличением микроскопа.
При вирусной гемадсорбции эритроциты прочно фиксируются на клетках, пора-
женных вирусом, и образуют характерные скопления островкового типа или рас-
пределяются диффузно, но не уплывают при покачивании пробирки. На поверх-
ности клеток, не содержащих вируса, эритроциты не адсорбируются и свободно
проплывают в поле зрения. Предельное разведение вирусной суспензии, давшее
положительный результат гемадсорбции в половине зараженных культур, прини-
мают за одну гемадсорбирующую единицу. В опыт берут 10-100 ТЦД50 вируса.
Сыворотки инактивируют 30 мин при 56 °C и готовят двукратные разведения
(от 1:10 до 1:1280 и далее по необходимости). К двукратным разведениям сыво-
ротки в объеме 0,2 мл добавляют равный объем вирусной суспензии, содержащей
10 или 100 гемадсорбирующих единиц вируса. Смеси выдерживают 1-2 ч при
комнатной температуре или 18 ч — при 4 °C. Затем по 0,2 мл каждой смеси вносят
в две пробирки с культурами клеток того же вида, что и при определении гемад-
сорбирующего титра вируса. Контролируют также гемадсорбирующую активность
взятой в опыт дозы вируса, соединенной с поддерживающей питательной средой
или физиологическим раствором.
Пробирки помещают в термостат и при появлении в отдельных контрольных
культурах, зараженных вирусом, четкого цитопатического эффекта в них вносят
по 0,2 мл 0,4% взвеси чувствительных к гемагглютинирующему действию виру-
са эритроцитов, выдерживают 20-30 мин при той же температуре, при которой
определяли гемадсорбцию в контрольных культурах, и учитывают результаты
путем микроскопирования.
При нейтрализации вируса специфическими антителами эритроциты не адсор-
бируются на зараженных клетках. Положительная гемадсорбции наблюдается в
культурах клеток с контролем вирусной активности, а также в культурах, заражен-
ных смесями, в которых вирус нейт рализован антителами частично или совсем не
нейтрализован. За титр антител в сыворотке принимают то ее предельное разведе-
ние, которое нейтрализовало гемадсорбирующую активность взятой в опыт дозы
вируса в 50% зараженных клеточных культур.
Типирование выделенных вирусов и исследование парных сывороток больных
для ретроспективной диагностики инфекций проводят так же, как во второй моди-
фикации реакции нейтрализации по цитопатическому эффекту
РЕАКЦИЯ ТОРМОЖЕНИЯ ГЕМАГГЛЮТИНАЦИИ
Реакцию торможения гемагглютинации (РТГА) применяют для диагностики
вирусов, обладающих способностью агглютинировать эрш ооциты. Соединение
вирусных антигенов с антителами иммунной сыворотки приводит к подавлению
этой способности, выражаемому в задержке (торможении) феномена агглюти-
нации эритроцитов. С помощью РТГА проводят титрование вируссодержащих
материалов, идентификацию и типизацию вирусных антигенов, серологическую
диагностику вирусных инфекций.
ВИРУСОЛОГИЧЕСКИЕ исследования 597
Для постановки реакции используют:
s эритроциты человека (0 группы), млекопитающих (чаще всего морской
свинки) или птиц (кур, гусей, голубей), трижды отмытые физиологиче-
ским раствором и приготовленные в виде 0,5-1,0% суспензии;
J антиген — вируссодержащий материал, полученный из инфицированных
куриных эмбрионов, культуры клеток или животных и освобожденный от
крупных частиц путем центрифугирования, отстаивания или фильтрова-
ния;
антитела — специфические иммунные сыворотки, освобожденные от
неспецифических ингибиторов вирусной гемагглютинации (способы уда-
ления ингибиторов — см. «Ингибиторы гемагглютинации»);
J обычный физиологический раствор или забуференный физиологический
раствор pH - 7,0ч-7,4. Последний готовят путем растворения 8,5 г NaCl,
0,56 г NaHPO4 и 0,14 г КН2РО4 в 1 л дистиллированной воды и используют
для разведения антигенов (кроме арбовирусных), иммунных сывороток и
приготовления суспензии эритроцитов необходимой концентрации.
Для РТГА используют пластины из органического стекла с лунками. Вначале
титруют антиген в реакции гемагглютинации в целях выбора необходимого рабо-
чего оазведения, которое, как правило, должно содержать 4 агглютинирующие
единицы (АЕ) в 0,2 мл физиологического раствора. Для некоторых вирусных
антигенов используют 2 или 1 АЕ (вирус оспы, кори соответственно) или 8 АЕ
(арбовирусы). Антиген разводят физиологическим раствором так, чтобы полу-
чить 4 АЕ в 0,2 мл. Число, соответствующее титру вируса 1:640, определенному в
объеме 0,2 мл, делят на 4 и получают: 640:4=160, то есть антиген нужно развести
в 160 раз.
Подготовленное для РТГА рабочее разведение антигена контролируют допол-
нительно, чтобы убедиться, что в 0,2 мл его содержится 4 АЕ. Для этого исполь-
зуют два ряда по четыре лунки. Во вторую, третью и четвертую лунки первого и
второго ряда наливают по 0,2 мл физиологического раствора, а в первую лунку
второго ряда — 0,4 мл. В первую и вторую лунку первого ряда и в первую лунку
второго ряда вносят по 0,2 мл рабочего разведения антигена. Титрую г его в первом
ряду, начиная со второй лунки ряда, путем переноса 0,2 мл перемешанного рас-
твора в следующую лунку; из последней лунки 0.2 мл удаляют. В первой лунке
второго ряда смесь антигена и физиологического раствопа перемешивают, 0,2 мл
смеси удаляют и титруют антиген, как в первом ряду. Затем во все .гунки вносят по
0,2 мл физиологического раствора и по 0,4 мл 1% взвеси эритроцитов. В допол-
нительные четыре лунки третьего ряда для контроля эритроцитов добавляют
по 0,4 мл физиологического раствора и по 0,4 мл взвеси эритроцитов. Пластин ы
встряхивают и оставляют при комнатной температуре до оседания эритроцитов в
контрольных лунках.
Пои правильно приготовленном рабочем разведении антигена в первой, второй
и третьей лунках первого ряда, содержащих 4, 2 и 1 АЕ соответственно, а также
в первой и второй лунках второго ряда, содержащих 3 и 1.5 АЕ соответственно,
должна наблюдаться агглютинация эритроцитов. В остальных лунках агглютина-
ция может быть незначительной или совсем отсутствовать. При ином характере
агглютинации в лунках необходимо внести коррективы в рабочее разведение
антигена, добавив необходимое количество антигена или физиологического рас-
твора и заново перетитровав полученное разведение.
Ингибиторы гемагглютинации
Неспецифические термолабильные и термостабильные ингибиторы агглютина-
ции обладают способностью соединяться с вирусами и тормозить агглютинацию
эритроцитов. Ингибиторы агглютинации содержатся в сыворотках человека и
598 ВИРУСОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ
животных или появляются при длительном хранении сывороток в жидком состоя-
нии на холоде.
От термолабильных ингибиторов освобождаются путем прогревания сыворо-
ток в течение 30 мин при 56 °C или 20 мин при 60 ’С (при этом, чтобы избежать
возможного свертывания, цельную сыворотку перед инактивацией разводят
физиологическим раствором 1:10).
Для устранения термостабильных ингибиторов используют методы обработки
сывороток углекислотой, перйодатом калия, каолином и эритроцитами, фильтра-
том холерного вибриона.
Обработка сывороток углекислотой. Обычно такой обработке подвергают
противогриппозные сыворотки. Цельную сыворотку разводят дистиллирован-
ной водой 1:10 и пропускают через нее углекислый газ 5-7 мин до интенсивного
помутнения. Вместо газообразной кислоты можно использовать сухой лед, кото-
рый небольшими кусочками опускают в дистиллированную воду в течение 10 мин
и затем этой водой, насыщенной углекислотой, разводят сыворотки 1:10.
Обработанную и помутневшую сыворотку освобождают от образовавшихся
хлопьев белка, содержащих неспецифические ингибиторы, путем центрифугиро-
вания при 1500 об./мин в течение 10 мин. Надосадочную жидкость отсасывают
и добавляют 1/10 объема 8,5% раствора NaCl для восстановления в сыворотке
изотонической концентрации хлорида натрия. Сыворотку, обработанную таким
образом, не нужно прогревать, так как она уже освобождена и от термостабиль-
ных, и от гермолабильных ингибиторов, при этом титр специфических антител
практически не снижается.
Обработка сывороток перйодатом калия (КЮ4). Обычно этот способ обра-
ботки применяют для противогриппозных сывороток. Готовят 0.05 М раствор
К1О4 на дистиллированной воде. 1.15 г К1О4 добавляют в 100 мл дистиллирован-
ной воды и подогревают для лучшего растворения на водяной бане при 70-80 °C
в течение 2-5 мин. Хранить приготовленный раствор можно 6 мес при 4 °C. Для
удаления осадка, образовавшегося при стоянии на холоде, раствор перйодата
калия повторно подогревают на водяной бане. Неразведенные сыворотки сначала
прогревают при 56 °C 30 мин. а затем смешивают с раствором перйодата капия в
равных объемах и оставляют для контакта на 2 ч при комнатной температуре. Для
нейтрализации активности перйодата к смеси добавляют 5% раствор глюкозы в
соотношении 1:1. При РТГА следует учитывать, что разведение сыворотки после
обработки соответствует 1:4.
Обработка сывороток каолином и эритроцитами. Способ применяют для
обработки противокоревых, краснушных и парагриппозных сывороток. К 0,2 мл
сыворотки добавляют 0,8 мл физиологического раствора и 1 мл 25% взвеси као-
лина в физиологическом же растворе. Смесь выдерживают 20 мин при комнатной
температуре, периодически встряхивая, и центрифугируют при 1000 об./мин
10 мин. Сыворотки отсасывают и на каждый миллилитр добавляют 0,1 мл 50%
взвеси эритроцитов. Смесь выдерживают 1 ч при 4 °C, эритроциты осаждают
центрифугированием при 1000 об./мин в течение 10 мин, сыворотку считают раз-
веденной 1:10.
РТГА
Реакцию применяют для определения титра антител в сыворотках от больных
(по известному вирусу) или для установления вида или типа вируса (по известной
сыворотке). Готовят двукратные разведения испытуемых сывороток от больных
или типоспецифических иммунных сывороток в физиологическом растворе от
1:10 до 1:640 и выше (при ожидании более высокого титра антител) и разливают
по 0,2 мл в лунки плексигласовой панели. В каждую лунку добавляют по 0,2 мл
рабочего разведения антигена, содержащего 4 АЕ.
ВИРУСОЛОГИЧЕСКИЕ исследования 599
Одновременно ставят три контроля:
S контроль эритроцитов, для чего к 0,4 мл физиологического раствора
добавляют 0,4 мл используемой взвеси эритроцитов;
J контроль используемой сыворотки на способность агглютинировать эри-
троциты — к 0,2 мл исходного разведения сыворотки добавляют 0,2 мл
физиологического раствора и 0,4 мл взвеси эритроцитов;
контроль специфичности антигена (при ретроспективной диагностике
инфекций) со стандартной иммунной сывороткой — к 0,2 мл иммунной
сыворотки в разведении 1:10 добавляют 0,2 мл рабочего разведения анти-
гена и 0,4 мл взвеси эритроцитов.
Кроме того, ставят контроль титра рабочего разведения антигена, как описано
выше.
Панели встряхивают, выдерживают в течение 30 мин-2 ч при комнатной тем-
пературе и вносят по 0,4 мл 0,25-1,0% взвеси эритроцитов, использованной для
определения гемагглютинирующего титра антигена. Панели снова встряхивают и
оставляют до оседания эритроцитов в контрольных лунках при той температуре,
при которой оценивали гемагглютинирующий титр антигена. Если используемая
сыворотка вызывает агглютинацию эритроцитов, ее обрабатывают одним из ука-
занных выше способов в целях истощения гемагглютининов.
Результаты РТГА учитывают по характеру оседания эритроцитов. В разведе-
ниях сыворотки, содержащей высокую концентрацию специфических антител,
гемагглютинирующая активность антигена полностью подавляется, и эритроциты
оседакг на дно лунки, образуя четкую точку, как и в контроле. По мере разведения
сыворотки количество антител в ней уменьшается и больше проявляется гемаг-
глютинирующая активность антигена: появляется «зонтик» из склеивающихся
эритроцитов. За титр антител в сыворотке принимают предельное ее разведение, в
котором наблюдается полное подавление гемагглютинирующей активности анти-
гена.
При использовании РТГА для серологической диагностики вирусных инфек-
ций парные сыворотки больных титруют одновременно с несколькими типовыми
антигенами, специфичность которых заранее проверена эталонными типоспе-
цифическими сыворотками. Для каждого антигена предварительно определяют
гемагглютинирующий титр и готовят рабочее разведение, содержащее 4 АЕ в
0,2 мл. Ход реакции аналогичен. Четырехкратное и более увеличение титра анти-
тел к одному из антигенов во второй сыворотке по сравнению с первой имеет
диагностическое значение и свидетельствует о том, что заболевание было вызвано
вирусом этого серологического типа.
При использовании РТГА для определения вида или типа вируса по известной
иммунной сыворотке готовят двукратные разведения типоспецифических сыворо-
ток, которые в объеме 0,2 мл соединяют с равным объемом рабочего разведения
антигена, содержащего 4 АЕ. Дальнейший ход реакции обычный. Типовую при-
надлежность выделенного вируса устанавливают по его соответствию специфиче-
ской иммунной сыворотке, показавшей наивысший титр анти гел к этому вирусу.
РЕАКЦИЯ НЕПРЯМОЙ (ПАССИВНОЙ) ГЕМАГГЛЮТИНАЦИИ
Реакция непрямой гемагглютинации (РИГА) основана на том, что эритроциты
животных и человека после обработки дубильной (таниновой) кислотой при-
обретают способность адсорбировать на своей поверхности бетковые молекулы
антигенов, которые затем агглютинируются специфическими иммунными сыво-
ротками либо молекулами сывороточных белков, которые могут агглютиниро-
ваться при взаимодействии с гомологичными антителами. По чувствительности
и специфичности РИГА часто превосходит друг ие серологические реакции, такие
как реакция связывания комплемента.
600 ВИРУСОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ
В лабораторной диагностике РИГА используют как для определения и иден-
тификации неизвестного возбудителя заболевания, так и для ретроспективной
серологической диагностики путем обнаружения антител в парных сыворотках
больного.
Методика постановки РИГА включает следующие основные этапы:
S приготовление взвеси эритроцитов, обработанных формалином и тани-
ном;
приготовление антигена и определение его рабочей дозы;
J сенсибилизацию эритроцитов антигеном и постановку основного опыта.
Для реакции чаще всего используют эритроциты барана, обезьян или человека
группы крови 0. Предпочтительны формалинизированные эритроциты, поскольку
они не лизируются в присутствии танина и при адсорбции антигена.
Необходимо приготовить следующие растворы:
J физиологический раствор (0,85% раствор NaCl):
S фосфатно-буферные растворы (ФБР) с pH = 7,2 и 6,4. Вначале готовят два
раствора: 26,85 г двузамещенного фосфата натрия (Na2HPO4) в 1 л воды;
20,4 г однозамещенного фосфата калия (КН2РО4) в 1 л воды.
Эти растворы можно использовать в течение 7-10 дней при условии хранения
в холодильнике. Непосредственно перед постановкой реакции готовят забуфе-
ренные физиологические растворы. Для получения раствора с pH = 7,2 к 100 мл
физиологического раствора добавляют 7,61 мл раствора двузамещенного фосфата
натрия и 23,9 мл раствора однозамещенного фосфата калия. Для приготовления
физиологического раствора с pH = 6,4 к 100 мл физиологического раствора при-
бавляют 32,3 мл раствора двузамещенного фосфата натрия и 67,7 мл раствора
однозамещенного фосфата калия.
Формалинизирование эритроцитов. Кровь барана собирают асептически и
дефибринируют во флаконе со стеклянными бусами. Эритроциты трижды отмы-
вают физиологическим раствором с pH = 7.2 и готовят из отмытого осадка 8%
суспензию. К одному объему 8% взвеси эритроцитов добавляют один объем 3%
формалина (pH = 7.2, который устанавливается с помощью 0,1н раствора NaOH в
физиологическом растворе). Колбу со смесью помещают в термостат при 37 °C на
18- 20 ч, при этом первые 4-6 ч эритроциты перемешивают на магнитной мешал-
ке. Затем их четыре раза отмывают и готовят 10% суспензию эритроцитов на
забуференном физиологическом растворе с pH = 7,2. К этой суспензии добавляют
мертиолят натрия до 1:10 000. Формалинизированные эритроциты разливают по
небольшим флаконам и сохраняют при 2-4 °C в течение 6 мес.
Обработка эритроцитов танином. Раствор танина в физиологическом раство-
ре с pH = 7,2 (1:20 000) готовят непосредственно перед обработкой эритроцитов;
танин должен обладать хорошей растворимостью, хранить его следует в темной
посуще в прохладном и сухом месте. Из осадка трижды отмытых физиологическим
раствором эритроцитов готовят 2,5% взвесь в ФБР с pH = 7,2. Смешивают равные
объемы взвеси эритроцитов и свежеприготовленного раствора танина и оставляют
на 15 мин при 37 °C в термостате или водяной бане или на 30 мин при комнатной
температуре. Затем смесь центрифугируют 10-20 мин при 3500-2000 об./мин,
надосадочную жидкость сливают, а осадок эритроцитов промывают тем же ФБР и
доводят до исходной концентрации 2,5%.
Сенсибилизация эритроцитов. I мл обработанной танином 2,5% суспен-
зии эритроцитов смешивают с 1 мл антигена и 4 мл буферной смеси (pH = 6.4).
Сенсибилизацию проводят в течение 2 ч при 37 °C. Затем эритроциты отмывают
два-три раза физиологическим раствором путем центрифугирования 1-2 мин при
1000-1500 об./мин. Разведение сенсибилизированных танин изированных эри-
троцитов для основного опыта с испытуемой сывороткой осуществляют нормаль-
ной кроличьей сывороткой 1:100 (для стабилизации взвеси).
ВИРУСОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ 601
Реакция непрямой гемагглютинации для детекции уровня антител.
Перед титрованием испытуемых сывороток необходимо определить рабочую дозу
антигена, способную при адсорбции на эритроцитах обеспечить максимальный
титр соответствующей иммунной сыворотки. Вначале сенсибилизируют антиген,
для чего его разводят ФБР (pH М7.2) и соединяют с эритроцитами, разведенными
тем же ФБР таким образом, чтобы получить четыре различных разведения анти-
гена.
Реакцию учитывают по четырехплюсовой системе через 2 ч инкубирования пла-
стин при температуре, оптимальной для взаимодействия ингредиентов реакции:
«++++» — реакция положительная, эритроциты равномерно распределены по
всей лунке пластины в виде зонтика с неровными краями;
«+++» — реакция положительная, на фоне склеившихся в виде пленки эритро-
цитов в центре имеется небольшое кольцо из осевших эритроцитов;
«++» — реакция положительная, меньшая степень агглютинации с большим
количеством несклеившихся эритроцитов;
«+» — реакция слабоположительная, наблюдается очень незначительная сте-
пень агглютинации, почти все эритроциты оседают в виде небольшого
диска;
«-» — реакция отрицательная, как и в контроле, скопление эритроцитов имеет
вид плотного диска или пуговки.
Рабочей дозой антигена считают дозу, при которой иммунная сыворотка реа-
гирует в реакции гемагглютинации на «++» в самом большом разведении и при
полном отсутствии гемагглютинации в контролях.
В каждом опыте следует контролировать все ингредиенты реакции. В связи с
этим в РИГА для исключения неспецифических реакций предусмотрены следую-
щие контроли.
Контроль специфичности реакции. Для этого реакцию ставят с заведомо
специфичными иммунными сыворотками, которые разводят нормальной
кроличьей сывороткой (1:100) от 1:10 до 1:1280 и разливают по 0,4 мл в
лунки пластин. К разведениям сыворотки добавляют по 0,2 мл эритроци-
тов, сенсибилизированных рабочим разведением используемого антигена.
При учете результатов в этом контроле реакция должна быть положитель-
ной.
Контроль стабильности и отсутствия спонтанной агглютинации танини-
зированных эритроцитов. Ставят в две лунки со взвесью сенсибилизиро-
ванных и несенсибилизированных эритроцитов в разведенной нормальной
сыворотке. В этих лунках реакция должна быть отрицательной.
Титром антител считают последнее разведение сыворотки, которое вызывает
склеивание эритроцитов при почти полном просветлении надосадочной жидко-
сти (++).
РЕАКЦИЯ СВЯЗЫВАНИЯ КОМПЛЕМЕНТА
Реакция связывания комплемента (РСК) представляет собой достаточно высо-
кочувствительный метод лабораторного исследования in vitro и применяется для
обнаружения соответствующих антител при бактериальных и вирусных инфекци-
ях, а также для типирования вирусов.
РСК — непрямая двухсистемная гетерологичная пятикомпонентная реакция.
В ней участвуют две системы «Аг-Ат» различной специфичности и система ком-
племента. Первая система — «Аг-Ат» — опытная, вторая — «АГ1-АТ1» — инди-
каторная.
РСК основана на том, что при взаимодействии в опытной системе Аг и Ат (в
исследуемой сыворотке содержатся антитела, гомологичные антигену) образуется
специфический комплекс «Ar t Ат», который адсорбирует (связывает) комплемент.
602 ВИРУСОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ
При отсутствии специфического сродства между Аг и Ат (в сыворотке отсутствуют
антитела к соответствующему антигену) комплекс «Аг+Ат» нс формируется и ком-
племент остается свободным.
Однако процесс взаимодействия комплемента с опытной системой является
невидимым, поэтому^ на втором этапе в реакцию вводят в горую, заведомо специ-
фическую индикаторную систему «АГ1+АТ1» (так называемую гемолитическую
систему, представляющую собой смесь эритроцитов барана и гемолитической
сыворотки, содержащей ан гит ела — гемолизины — против эритроцитов барана),
которая свидетельствует о результатах РСК.
Если в первой системе связывание комплемента происходит (положительный
результат), в индикаторной системе не возникает задержки гемолиза эритроци-
тов.
Если адсорбции комплемента в опытной системе не наступает (отрицатель-
ный результат РСК), комплемент вовлекается в реакцию между гемолитической
сывороткой и чувствительными эритроцитами. Вследствие этого в индикаторной
системе появляется гемолиз различной интенсивности вплоть до полного лизиса
эритроцитов, оцениваемый плюсами (от одного до четырех).
РСК может быть использована для обнаружения в сыворотке больного спец-
ифических антител с помощью известных антигенов (диагностика вирусных
инфекций и аутоиммунных состояний); для выявления и идентификации в мате-
риале специфического антигена при тех же заболеваниях посредством иммунной
сыворотки, содержащей специфические антитела. Следует, однако, учитывать, что
комплементсвязывающие антитела образуются при некоторых инфекциях раньше
и исчезают быстрее, чем нейтрализующие антитела. В некоторых случаях при
заболеваниях легкой и средней тяжести комплементсвязывающие антитела могул
не обнаруживаться вовсе.
Для постановки РСК требуются следующие материалы и ингредиенты:
S Аг, не обладающий антикомплементарностью и гемотоксичностью. Его
строго оттитровывают и берут в рабочей дозе;
S Ат — иммунная сыворотка, освобожденная от комплемента и лишенная
гемотоксичности;
✓ комплемент, взятый в рабочей дозе;
v' АП — эритроциты барана, отмытые в физиологическом растворе и взя-
тые в опыт в виде 0,5-3,0% взвеси в том же или в специальном буферном
растворе, употребляемом при работе с данным типом вируса;
S ATI — гемолитическая (ан гиэритроцитарная) сыворотка, освобожденная
от комплемента;
J физиологический раствор с pH = 7,2+7,4;
s лабораторная посуда: градуированные пипетки, пробирки, цилиндры, ста-
каны или колбы для разведений (употребляют только в РСК; не следует
использовать для иных целей); оборудование и аппаратура: центрифуга,
водяная баня, термостат, штативы.
Постановка РСК требует большой предварительной работы. Накануне основно-
го опыта готовят необходимые ингредиенты реакции, лабораторную посуду.
Ант иген. В качестве Аг в РСК может быть использован любой вируссодержа-
щий материал, полученный из органов лабораторных животных, куриных эмбри-
онов и клеточных культур, инфицированных различными вирусами. Антигенами
в РСК служат убитые вирусные агенты, очищенные различными способами и не
обладающие антикомплементарными свойствами, то есть неспособные фикси-
ровать комплемент при отсутствии специфической сыворотки. Для выявления
антикомплементарности антигена в предварительном опыте титруют комплемент
в присутствии этого антигена. Допускается употребление антигена, подавляющего
активность комплемента не более чем на 30%.
ВИРУСОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ 603
Аг не должен обладать гемотоксическими свойствами. Двойная доза антигена
не должна давать даже следов гемолиза рабочего объема взвеси бараньих эритро-
цитов.
Антигены из аллантоисной и амниотической жидкостей инфицированных
эмбрионов являются высокоактивными и высокоспецифическими, не требуют
трудоемкой предварительной обработки. Перед постановкой РСК вируссодержа-
щую жидкость осветляют путем центрифугирования и разводят 1:10 раствором,
который используют для постановки опыта с данным типом вируса, или физио-
логическим раствором.
Вируссодержащий материал, полученный при культивировании вируса в кле-
точной культуре, после развития максимально выраженного цитопатического
эффекта подвергают троекратному замораживанию и оттаиванию, осветляют
путем центрифугирования в течение 15-30 мин при 1500-3000 об./мин, надоса-
дочную жидкость используют в качестве антигена.
Антигены хранят при 4 °C, в замороженном состоянии или лиофильно высу-
шенными. Активность антигенов устанавливают в предварительных опытах
титрования со специфической сывороткой, избранной в качестве стандарта.
Антитело. В РСК используют сыворотки животных и людей (больных и
реконвалесцентов). Проследить за динамикой титра антител позволяют парные
сыворотки.
Для получения иммунной сыворотки у больного берут кровь из вены в объеме
5-10 мл, обычно натощак или спустя 6 ч после приема пищи. Сыворотку инак-
тивируют прогреванием при 56 °C в течение 30 мин. В результате разрушают
комплемент и добиваются необходимой стабилизации коллоидов сыворотки.
Инактивированную сыворотку хранят при 4 °C в течение 5-6 дней. При необхо-
димости более длительного хранения (2-3 нед) к сыворотке после инактивации
добавляют 2% сухой борной кислоты. Длительно хранящиеся сыворотки перед
постановкой РСК снова инактивируют.
Пшериммунные сыворотки, содержащие комилементсвязывающие антитела, полу-
чают путем иммунизации лабораторных животных (морских свинок, белых крыс,
кроликов, коз, обезьян и др.) по определенным схемам. Не существует общепринятых
схем иммунизации, для каждого типа вируса предлагаются специальные схемы.
Существует ряд общих моментов, которые следует учитывать при использова-
нии различных сывороток в РСК. Одно из затруднений, с которыми приходится
сталкиваться при постановке РСК, — антикомплементарность сывороток, то есть
способность связываться с комплементом при отсутствии антигена и тем самым
задерживать гемолиз. Антикомплементарность может быть обусловлена длительно-
стью хранения сывороток или свойством, присущим сывороткам некоторых видов
животных, например кроликов. Перед постановкой реакции сыворотки инактиви-
руют прогреванием на водяной бане в течение 30 мин: сыворотки морской свинки и
кролика — при 56 °C, человека и обезьян — при 58 °C, крыс — при 60 °C, собак — при
65 °C. В двух последних случаях перед прогреванием сыворотки разводят 1:2 или 1:3
дистиллированной водой или физраствором, чтобы избежать коагуляции белков.
Если после прогревания сыворотки все же сохраняют антикомплементарные
свойства, их необходимо обработать одним из следующих методов:
S однократным замораживанием при -15...-20 °C с последующим удалени-
ем образовавшегося флоккулята путем центрифугирования;
S разведением комплементом в соотношении: 1 часть комплемента +
9 частей сыворотки; смесь выдерживают 18-20 ч при 4-6 °C и инактиви-
руют прогреванием;
S обработкой углекислотой методом, используемым для удаления неспец-
ифических ингибиторов гемагглютинации, после чего сыворотки прогре-
вают, как обычно.
604 ВИРУСОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ
В сыворотках иммунизированных или переболевших животных наряду со спец-
ифическими антителами образуются и антитела к тем тканям, из которых была
приготовлена вируссодержащая суспензия для иммунизации, особенно при много-
кратных инъекциях антигена. При постановке РСК эти антитела могут приводить
к появлению ложноположительных результатов. Этого можно избежать, если для
иммунизации использовать антиген, полученный в одной инфицированной ткани,
а антиген для последующей РСК готовить в гетерологичной ткани. Иногда при-
ходится прибегать к истощению сыворотки нормальной тканью, для чего к раз-
веденной физиологическим раствором сыворотке (в соотношении 1:1) добавляют
20% гомогенизированной гомологичной ткани, выдерживают 2 ч при 37 °C, 18 ч
при 4-6 °C и центрифугируют при 3000 об./мин в течение 30 мин. Удаляют остаток
тканевого антигена путем обработки углекислотой по описанной выше методике и
полученную сыворотку прогревают при температуре 56 °C в течение 30 мин.
Эритроциты. В РСК в качестве первого компонента гемолитической системы
могут быть использованы эритроциты любых животных и человека. Однако преи-
мущественно применяются эритроциты барана. Их получают путем пункции ярем-
ной вены здорового животного в возрасте от 2 до 5 лет. Кровопускание у баранов
можно проводить не чаще одного раза в 10 дней, до 200 мл, в течение 1-1,5 лет,
после чего нужно брать других животных.
В реакции применяют 0,5-3,0% взвесь эритроцитов. При 4 °C эритроциты
могут сохраняться 5-7 дней. Для хранения эритроцитов в течение более длитель-
ного времени прибегают к их консервированию.
Гемолитическая сыворотка. Второй компонент гемолитической системы
получают путем 3-4-кратной внутривенной иммунизации кроликов 50% взвесью
эритроцитов барана в количестве 2-5 мл с промежутками 2-3 дня. На 6-7-й день
после окончания иммунизации берут небольшое количество крови и проверяют
активность сыворотки (титр ее должен быть не ниже 1:1200).
Титр гемолитической сыворотки определяют следующим образом. Готовят
основное разведение сыворотки 1:100 (0.1 мл сыворотки + 0,9 мл физиологиче-
ского раствора) и используют его для получения дальнейших разведений. В ряд
лунок пластины последовательно отмеривают по 0,2 мл разведений сыворотки,
начиная с 1:1000; добавляют в лунки 0,2 мл комплемента 1:10, затем по 0,2 мл
3% взвеси эритроцитов и доливают в каждую лунку по 0,4 мл физиологического
раствора. Смесь ингредиентов помещают в термостат на 1 ч при 37 СС. За титр
гемолитической сыворотки принимают ее разведение, которое вызывает полный
гемолиз эритроцитов.
В реакции связывания комплемента следует использовать гемолитическую
сыворотку в трое- или четырехкратном титре; например, если титр получен-
ной сыворотки 1:1200. то ее троекратный титр соответствует разведению 1:400.
Употребление гемолитической сыворотки в таком титре обеспечивает надежное
функционирование гемолитической системы, если учитывать, что реакция пяти-
компонентная. При титре 1:1200 и выше проводят тотальное кровопускание.
Сыворотку прогревают при 56 СС в течение 30 мин, для консервации добавляют
до 0,5% сухой борной кислоты, титруют, разливают по ампулам и хранят в холо-
дильнике при 4 °C. В настоящее время выпускают сухие стандартные препараты
гемолитической антибараньей сыворотки с титром 1:1200, 1:1500 и т.д. Сухие
препараты гемолитической сыворотки разводят стерильным физиологическим
раствором coi ласно инструкции и используют в работе.
Для получения гемолитической системы смешивают равные объемы 3% эри-
троцитов барана и разведенной до троекратного титра гемолитической сыворотки.
Их смешивают непосредственно перед добавлением гемолитической системы в
реакцию, делают это быстро, причем гемолитическую сыворотку приливают к
взвеси эритроцитов (а не наоборот) и некоторое время энергично перемешивают
ВИРУСОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ 605
полученную взвесь. Используют свежеприготовленную гемолитическую систему в
течение двух дней.
Источник комплемента. В качестве комплемента — неспецифического термо-
лабильного фактора, присутствующего в сыворотке свежей крови человека, боль-
шинства млекопитающих и птиц, — в ряде иммунологических реакций используют
сыворотку морских свинок, поскольку в их крови комплемент обнаруживают чаще
и в наибольшем количестве, чем у других животных. Отечественные биофабрики
выпускают сухой препарат комплемента.
В реакцию связывания комплемента берут свежую, полученную не более 48 ч
назад сыворотку крови 4-5 животных, самцов или небеременных самок. Сыворотку
можно хранить в замороженном состоянии. Для длительного (до 6 мес) сохране-
ния комплемент в сыворотке морских свинок консервируют путем добавления на
каждые 10 мл сыворотки 0,4 г борной кислоты и 0,5 г сернокислого натрия. Сухой
(лиофилизированный) комплемент активен в течение нескольких лет.
Титрование комплемента. Поскольку комплемент — очень лабильная суб-
станция. чувствительная к солнечному свету; теплу и т.д., за день перед каждой
постановкой основного опыта РСК следует проводить его титрование, его осу-
ществляют в том же объеме, в котором будут выполнять основной опыт РСК. Для
постановки реакции комплемент разводят основным раствором (физраствором
или специальным буферным раствором) 1:10 и соединяют последовательно коли-
чества комплемента от 0,05 до 0,5 мл с интервалом 0,05 мл в каждой пробирке,
доводя общий объем в каждой из них до 1 мл.
Одновременно осуществляют контроли всех ингредиентов, входящих в реак-
цию:
• контроль комплемента (к 0,2 мл комплемента в соотношении 1:10 добавляют
0,2 мл 3% взвеси эритроцитов барана и 0,6 мл основного раствора):
• контроль эритроцитов (к 0,2 мл 3% взвеси эритроцитов добавляют 0,8 мл
основного раствора);
• контроль гемолитической сыворотки (к 0,2 мл сыворотки, разведенной до
троекратного титра, добавляют 0,2 мл 3% взвеси эритроцитов и 0,6 мл основ-
ного раствора).
В контрольных лунках не должен наступать гемолиз эритроцитов.
За единицу комплемента принимают наименьшее его количество, которое спо-
собно гемолизировать эритроциты в используемой гемолитической смеси. Полной
рабочей единицей комплемента для постановки основного опыта считают коли-
чество его во второй пробирке с полным гемолизом (0,2 мл), то есть на 20-30%
больше по отношению к титру комплемента с учетом возможных антикомплемен-
тарных свойств антигенов. Для постановки РСК на холоде в качестве рабочей дозы
используют 1,5-2 полные дозы комплемента, то есть 3,5-4,0 мл комплемента,
разведенного 1:10, или 0,35-0,4 мл неразведенного.
Далее рассчитывают количество комплемента, требуемое для всего опыта. Если
одна рабочая доза составляет, например, 0,4 мл неразведенного комплемента, а
использовать в опыте предполагается 100 лунок, необходимое количество соста-
вит 4 мл. Поскольку ингредиенты берутся в реакцию в объеме 0,2 мл, на 100 лунок
потребуется 8 мл разведенного комплемента, то есть к объему самого комплемента
(4 мл) следует добавить еще 4 мл основного раствора, в котором ставится вся реак-
ция. Для более корректной постановки РСК с учетом возможных антикомплемен-
тарных свойств антигенов и сывороток комплемент титруют в присутствии всех
используемых антигенов и сывороток.
Затем устанавливают рабочую дозу антигена, для чего в три ряда лунок разли-
вают по 0,2 мл возрастающих двукратно разведений антигена. К лункам первого
ряда прибавляют по 0,2 мл специфической сыворотки в разведении, соответствую-
щем ее четырехкратному титру, к лункам второго ряда — по 0,2 мл нормальной
606 ВИРУСОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ
сыворотки в том же разведении (контроль антигена на специфичность) и в лунки
третьего ряда вносят по 0,2 мл основного раствора (контроль антигена на анти-
комплементарность). Дополнительно ставят контроль сыворотки (без антигена).
Во все лунки добавляют по 0,2 мл комплемента в двукратном титре, смесь поме-
щают на 1 ч в термостат при 37 СС или на ночь при 4 °C.
Гемолитическую систему, приготовленную непосредственно перед использова-
нием, предварительно выдерживают 30 мин при 37 СС, затем добавляют по 0,4 мл
во все лунки, пластины ставят в термостат на 30 мин и учитывают результаты
после наступления гемолиза эритроцитов в контролях.
За единицу антигена принимают его наибольшее разведение, давшее с типо-
специфической сывороткой задержку гемолиза на два плюса. В качестве рабочего
разведения антигена используют его наименьшее разведение (содержащее не
менее двух антигенных единиц), при котором он реагирует со специфической
сывороткой и не задерживает гемолиз в присутствии нормальной сыворотки и
основного раствора. Контрольный антиген применяют в РСК в том же разведении,
что и типовой.
Методика проведения основного опыта. В соответствии с целями в реакции
связывания комплемента при выполнении основного опыта используют либо
известный стандартный антиген, либо известную стандартную сыворотку. В зави-
симости от этого осуществляют контроли на антикомплементарность и гемоток-
сичность соответственно испытуемых сывороток или минимальных разведений
неизвестного антигена.
Схема выполнения основного опыта с испытуемой сывороткой:
• сыворотка в разведениях от 1:10 до 1:1280 и более по необходимости —
0,2 мл;
• антиген в рабочей дозе — 0,2 мл;
• комплемент в рабочей дозе — 0,2 мл;
• встряхивание, инкубирование при 37 °C в течение 1 ч или при 4 ,;‘С 18-24 ч;
• гемолитическая система — 0,4 мл:
• встряхивание, термостат при 37 °C в течение 0,5-1 ч до наступления гемолиза
в контроле.
Результаты учитывают по системе плюсов:
« ++++» — гемолиз эритроцитов отсутствует — надосадочная жидкость абсо-
лютно прозрачная, эритроциты в осадке;
« +++» — гемолиз эритроцитов составляет примерно 25% — надосадочная жид-
кость слегка окрашена, осадок;
« ++» — гемолиз эритроцитов составляет 50% — надосадочная жидкость окра-
шена интенсивно, но осадок ясно виден;
« +» — гемолиз эритроцитов составляет 75% — над осад очная жидкость окраше-
на интенсивно, осадок заметен, но слабо:
« -» — полный гемолиз эритроцитов — отрицательная реакция.
Титром испытуемой сыворотки считают наибольшее ее разведение, давшее
задержку гемолиза.
Контролями к основному опыту служат:
• контроль сыворотки без антигена (0,2 мл сыворотки в наименьшем разведе-
нии + 0,2 мл комплемента + 0,2 мл основного раствора + 0,4 мл гемолитиче-
ской системы);
• контроль антигена без сыворотки (0,2 мл антигена + 0,2 мл комплемента +
0,2 мл основного раствора + 0.4 мл гемолитической системы).
Взаимодействие антигена с антителом осуществляют либо при 37 °C в течение
1 ч (горячий способ), либо при 4 -6 °C в течение 18-24 ч. При холодовом способе
проведения реакции отмечена ее большая чувствительность.
Схема выполнения основного опыта с испытуемым антигеном:
ВИРУСОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ 607
• типоспецифические и нормальная сыворотки в двукратных разведениях — по
0,2 мл каждого разведения;
• антиген в исходном разведении — по 0,2 мл:
• комплемент в рабочей дозе — 0,2 мл;
• встряхивание, инкубирование при 37 °C в течение 1 ч или при 4 °C в течение
18-24 ч;
• гемолитическая система — 0,4 мл:
• встряхивание, термостат при 37 °C в течение 0,5-1 ч до наступления гемолиза
в контроле.
Контролируют все ингредиенты реакции аналогично предыдущему опыту,
условия проведения реакции те же. Результаты оценивают обычным способом.
Типовую принадлежность выделенного вируса устанавливают по специфической
сыворотке, показавшей наивысший титр антител к этому вирусу.
При ретроспективной диагностике в РСК исследуют парные сыворотки от боль-
ных с рядом типовых антигенов и контрольным антигеном. Каждый из исполь-
зуемых антигенов должен быть проконтролирован предварительно на специфич-
ность и оттитрован для выбора рабочего разведения. Дальнейший ход реакции
аналогичен ранее описанному, за исключением того, что для каждого из типовых
антигенов выбирают необходимое рабочее разведение комплемента.
Иммуноферментный анализ: модификации метода
Иммуноферментный анализ (ИФА), или твердофазный иммуноферментный
анализ (тИФА), — метод клинической лабораторной диагностики, выявляющий
антигены возбудителей вирусных инфекционных заболеваний или антитела к ним.
Метод основан на специфическом взаимодействии «антиген-антитело» и после-
дующей визуализации образовавшихся комплексов «Аг-Ат» с помощью фермен-
тативной реакции, при которой конъюгированный с антителом или антигеном
фермент превращает добавленный в реакционную смесь субстрат в окрашенный
продукт. Скорость накопления окрашенного продукта ферментативной реакции
пропорциональна концентрации специфических комплексов «Аг-Ат» и учитыва-
ется с помощью фотоме гра.
Главными компонентами тест-систем для ИФА служат твердая фаза (композиция
антигенов или антител), закрепленная на подложке, конъюгат (один из распростра-
ненных типов — антитело с ковалентно «пришитым» ферментом типа пероксидазы
хрена или иной пероксидазы), субстрат (хромоген) и промывочный буфер.
Реакцию проводят в лунках полистироловых планшетов (наиболее распростра-
нен формат 96-луночного планшета размерности 8x12, хотя существуют планшеты
и 384-луночного формата), где на дне каждой лунки (твердая фаза ИФА) сорби-
рованы антитела к определяемым антигенам вируса или антигены вируса, если
устанавливают содержание антител к этим антигенам в биологическом образце.
Кроме планшетного формата твердой фазы, в автоматических ИФА-анализаторах
также применяют пробирочный формат, где Ат/Аг сорбируются на дно одиночной
пробирки, в которой проводится реакция, либо на поверхность полистиролового
шарика, вносимого в реакционную пробирку. Подобный подход удобен, посколь-
ку позволяет выполнить исследование одиночного образца. Кроме того, площадь
поверхности сферы в четыре раза больше площади круга одинакового диаметра,
поэтому при одинаковых линейных размерах можно сорбировать на твердую фазу
в 4 раза больше Ат/Аг, что увеличивает чувствительность анализа.
ИФА состоит из нескольких этапов. На первом этапе инкубируют исследуемые
сыворотки с предалсорбированчыми на твердой фазе (дно и стенки реакционной
емкости) антигенами (если определяют присутствие антител к вирусным белкам
в сыворотке). Несвязавшиеся Ат и другие компоненты сыворотки удаляют про-
608 ВИРУСОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ
мывкой буферным раствором, а образовавшиеся комплексы «Аг-Ат» остаются в
реакционном объеме.
На втором этапе добавляют конъюгированные с ферментом анти-Ат, специфич-
ные против IgG человека (если определяют IgG-Ат, соответственно возможно
определение IgM-, IgA-, IgE-Ат). Несвязавшийся конъюгат также удаляют про-
мывкой. Для выявления сформировавшихся комплексов добавляют субстрат.
В результате пероксидазной активности субстрат разлагается с образованием
окрашенного продукта. Колориметрическую реакцию останавливают добавлени-
ем раствора кислоты. Окрашенный раствор фотометрируют.
Последующее развитие метода ИФА привело к появлению ряда его модифи-
каций. Разновидности тест-систем, поименяемые в лабораторной диагностике
инфекционных вирусных заболеваний, можно разделить следующим образом:
• по схеме выполнения исследования:
❖ непрямой ИФА,
«сэндвич»-ИФА;
❖ конкурентный ИФА;
• по сорбированной твердой фазе:
❖ антительные (например, для выявления HBS Аг);
❖ антигенные (например, для выявления HBS Ат);
к омбинированные (например, для одновременною выявления антиге-
на р24 и антител к ВИЧ);
• по типу сорбированного антигена:
❖ лизатные (на твердую фазу сорбируется вирусный лизат из клеточной
культуры);
❖ рекомбинантные (используются генно-инженерные рекомбинантные
белки или фрагменты белков вирусов);
< > пептидные (сорбируются синтетические эпитопы вирусных антигенов);
• по специфичности конъюгата:
о- поликлональный (реагирует с антителами всех типов IgM, IgG, IgA);
❖ моноклональный (реагирует только с одним типом антител);
• по формату твердой фазы:
> планшетные:
пробирочные (для автоматических ИФА-анализаторов).
Кроме того, отдельно следует упомянуть такие разновидности иммунологиче-
ского качественного и количественного определения антигенов и антител вирусов
как иммунохемилюминесцентный анализ (ИХЛА) и иммуноэлектрохемилюми-
несцентный анализ (ИЭХЛА). Оба эти метода также близки к ИФА, поскольку в
их основе лежит тот же принцип взаимодействия антигена с антителом с исполь-
зованием твердой фазы. Основное отличие заключается в замене ферментативной
реакции (для визуализации комплексов «Аг-Ат») люминесцентной меткой что
значительно повышает чувствительность метода и снижает вероятность получения
ложноположительных результатов вследствие уменьшения количества конъюгата
в реакционном объеме.
Метод прямого ИФА (рис. 23-13) используют в иммуногистохимических иссле-
дованиях (например, при выявлении антигенов вируса гепатита В в биоптате пече-
ни). При прямом ИФА требуется специальный субстрат, дающий нерастворимый
окрашенный продукт.
Непрямой ИФА и «сэндвич»-ИФА — основные варианты метода в диагностике
инфекционных вирусных заболеваний. Возможности метода позволяют опреде-
лять как антигены вирусных частиц (например, HBS Аг), так и антитела к ним
(см. рис. 23-13). Основное отличие заключается в композиции твердой фазы: для
выявления антител к вирусным белкам на подложку сорбируется Аг вируса, для
обнаружения вирусных антигенов — моноклональные Ат.
ВИРУСОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ 609
Субстрат
Прямой ИФА
Непрямой ИФА
(выявление Ат)
Рис. 23-13. Основные варианты иммуноферментного анализа.
Непрямой ИФА Сэндвич-ИФА
(выявление Аг)
Состав твердой фазы ИФА претерпел значительные изменения по мере разра-
ботки новых вариантов анализа. Эволюция тИФА связана с методами получения
вирусных Аг: в тест-системах I поколения использовали лизатные Аг. II — реком-
бинантные, III — пептидные.
Лизатные антигены получали из клеточной культуры, зараженной вирусом.
Вирусные частицы накапливали, выделяли, очищали, инактивировали. Суспензию
инактивировали и наносили на подложку лунки. Такие тест-системы облада-
ют невысокими чувствительностью и специфичностью, на сегодняшнй день их
используют в основном в научно-исследовательских лабораториях.
С развитием генно-инженерных технологий молекулярного клонирования полу-
чили распространение рекомбинантные тест-системы, в которых ген определяемого
белка вируса встраивается в геном бактериальной клетки (чаще всего Е. coli), что
позволяет синтезировать рекомбинантные белки практически в любых количе-
ствах. Подобный подход сразу сделал метод тИФА доступным как с точки зрения
производства тест-систем, так и с точки зрения экономичности. Рекомбинантные
системы обладают более высокой чувствительностью по сравнению с лизатными,
поскольку возможно получение значительных количеств Аг для нанесения на под-
ложку. Специфичность тоже повышается, так как на твердую фазу наносят не смесь
лизированных белков вируса, а единичный белок или его часть.
Основная технологическая трудность связана с необходимостью очистки реком-
бинантного белка от бактериальных белков, особенно если клонирование ведется
в Е. coll. Даже малейшая примесь бактериальных белков может привести к появ-
лению ложноположительных результатов, поскольку антитела к белкам эшерихий
содержатся в сыворотке крови большинства людей.
Получение более специфических антигенов возможно при применении баку-
ловирусных векторов, культивируемых в клеточных культурах насекомых, что
значительно снижает вероятность перекрестных реакций.
Наиболее специфичны синтетические антигены, полученные путем химическо-
го синтеза полипептидных цепочек. Преимущество тест-систем на синтетических
пептидах состоит в возможности создания композиции из нескольких пептидов
к разным иммунодоминантным эпитопам одного антигена или нескольких анти-
генов разных геновидов вируса. Подобный подход позволяет выявлять антитела
к вирусным белкам различных вариантов вируса (например, к разным генотипам
вируса гепатита С). Кроме того, синтетические пептиды не содержат примесей,
способных привести к ложноположительным результатам ИФА.
61 о ВИРУСОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ
Для сокращения периода серологического «окна», что особенно важно для
максимально раннего установления факта инфицирования такими социально
значимыми инфекциями, как ВИЧ или гепатит С, были разработаны комбиниро-
ванные тест-системы IV поколения. Твердая фаза комбинированных тест-систем
состоит не только из синтетических Аг поверхностных вирусных белков, но и из
сорбированных на подложке моноклональных антител к ранним вирусным бел-
кам. циркулирующим в сыворотке до образования диагностических Ат в период
серологического «окна» (Аг р24 — у ВИЧ, core Аг у ВГС и т.п.), что дополнитель-
но сокращает сроки выявления инфицированных лиц. Например, для лизатных
ИФА-тест-систем период серологического «окна» для ВГС-инфекции составлял
около 150 дней, для рекомбинантных — 80, для синтетических — 70 дней, а ком-
бинированные иммунохемилюминесцентные тест-системы способны достоверно
устанавливать факт инфицирования уже через 60 дней.
Отдельно стоит упомянуть тест-системы для определения антител класса IgM к
Аг вирусных частиц. Одна из существенных проблем ИФА-диагностики вирусных
инфекционных заболеваний состоит в специфическом и достоверном определении
антител класса IgM как маркеров ранней инфекции. К сожалению, при опреде-
лении IgM-Ат высока частота ложноположительных результатов. Особенно эта
проблема актуальна при диагностике ВУИ (краснухи, цитомегалии, герпеса, пар-
вовируса В19) у беременных, онкологических больных.
Причина низкой достоверности и специфичности выявления IgM-Ат заключа-
ется в следующем: IgM-Ат имеют более низкую авидность по сравнению с IgG-Ат
(конкуренция за сайты связывания на твердой фазе, приводящая к получению
ложноотрицательных результатов), перекрестные реакции в присутствии ревма-
тоидного фактора, гетерофильных аутоантител и т.п. Для специфического и досто-
верного определения IgM-Ат, согласно рекомендациям ВОЗ и CDC. следует при-
менять тест-системы с так называемым IgM-захватом (IgM-capfure, или и-capture).
Основное отличие этих тест-систем — состав твердой фазы, на которую сорбиру-
ются не Аг, против которых определяются IgM-антитела, а анти-IgM-Ar (анти-ц-
Ат). Именно поэтому при более специфичном установлении IgM-Ат сначала про-
исходит связывание всей фракции IgM-Ат на подложке твердой фазы, а уже потом
специфически выявляются IgM-антитела против определяемого антигена.
Следующий важный компонент тИФА — конъюгат. Наиболее распространен-
ным видом конъюгата, широко применяемым в тест-системах для выявления
антител к антигенам вирусных частиц, служат анти-Ат, то есть антитела против
выявляемых Ат. Анти-Ат обычно видоспецифичны (например, мышиные про-
тив IgG человека) и направлены на Fc-фрагмент определяемых Ат. В свою оче-
редь, к анти-Ат ковалентно «пришит» фермент из семейства пероксидаз (чаще
пероксидаза хрена). Можно использовать иные высокоавидные белки, например
авидин-биотиновые или стрептавидин-биотиновые конъюгаты. Последний вари-
ант предпочтительнее, поскольку позволяет значительно повысить чувствитель-
ность тест-системы. В отличие от конъюгата Ат-пероксидазы, у стрептавидин-
биотинового комплекса на одно связанное меченое антитело приходится не одна,
а три молекулы фермента, что существенно усиливает специфический сигнал и,
как следствие, повышает чувствительность тест-системы для ИФА. Современные
конъюгаты обладают высокой стойкостью в окружающей среде и очень большой
активностью. Необходимо тщательно избегать загрязнения рабочей зоны конъю-
гатом при выполнении ИФА, поскольку даже следовые его количества, попавшие в
реакцию, способны привести к появлению ложноположительных результатов.
Следующий компонент ИФА-тест-систем — субстрат, необходимый для раз-
вития окрашивания реакционного раствора. Первоначально для этих целей при-
меняли ОФД (ортофенилендиамин), превращавшийся под действием пероксидазы
конъюгата в окрашенный продукт. Интенсивность окрашивания измеряли при
ВИРУСОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ 611
длине волны фотометра 492 нм. Поскольку ОФД — нестойкий и высокотоксичный
субстрат с канцерогенным потенциалом, в современных вариантах ИФА в качестве
субстрата используют ТМБ (3,3’,5,5’-теграметилбензидин). Под действием перок-
сидазы ТМБ образует в растворе продукт голубого цвета. При остановке реакции
с помощью кислоты продукт реакции гидролизуется с развитием желтого окра-
шивания с максимумом поглощения 450 нм. Ин генсивность окрашивания прямо
пропорциональна количеству комплексов «Аг-Ат».
Хемилюминесцентный (ХЛИА) и электрохемилюминесцентный (ИЭХЛА) ана-
лизы, в отличие от иммуноферментного, не нуждаются в субстрате, поскольку
для выявления образовавшихся комплексов «Аг-Ат» используют не колориме-
трическую, а люминесцентную реакцию. Вместо комплекса «анги-Ат-фермент»
конъюгатом служит комплекс «анти-Ат-люминофор». Этот комплекс может
быть достаточно сложным и включать антитела, конъюгированные, например, с
рутениевым комплексом «рутений-трис(2.2’бипиридил)-дихлорид», способным
испускать световые волны строго определенной длины (620 м) при пропускании
электрического импульса через раствор или при облучении раствора УФ -светом
Поскольку на один фотон света, поглощенного рутениевым комплексом, прихо-
дится один фотон испускаемого света, количество световых импульсов, детекти-
руемых в ИЭХЛА, прямо пропорционально количеству связанных антител, что
позволяет вести значительно более точный учет связанных анти -Ат.
Кроме того, люминесцентная детекция по чувствительности существенно пре-
восходит колориметрическую, что дополнительно увеличивает чувствительность
метода. Измерение испускаемого иммунным комплексом света дает возможность
легко проводить количественный учет интенсивности свечения. К существенным
преимуществам указанных методов относится и более низкое по сравнению с
классическим вариантом анализа количество конъюгата, что снижает вероятность
ложноположительного результата.
Учитывая, что интенсивность окрашивания раствора в лунке планшета для
ИФА прямо пропорциональна количеству образовавшихся комплексов «Аг-Ат»,
возможно исследование биологического материала методом ИФА как в качествен-
ном, так и в количественном варианте. Качественный вариант ИФА позволяет
установить факт присутствия или отсутствия определяемого аналита (например,
HBsAg) в материале. С помощью количественного варианта ИФА не только
определяют факт наличия или отсутствия аналита в материале, но и оценивают
его количество.
Для некоторых маркеров вирусных инфекций достаточно качественного
определения, например, при установлении факта инфицирования (антитела к
ВИЧ или анти-ВГС-антитела), в других случаях целесообразно проводить коли-
чественный ИФА (например, определение напряженности поствакцинального
иммунитета против гепатита В — анти-НВз-Ат или краснухи — анти-Rubella
IgG-Ат). Количественный ИФА требует измерения заранее охарактеризованных
калибраторов (контролей с аттестованными значениями аналита) в одном и
том же опыте, что и измерение исследуемого материала. Полученные аттесто-
ванные значения аналита используют для построения калибровочной кривой,
с помощью которой можно вычислить содержание исследуемого аналита в
образце. При проведении ИФА в ручном режиме калибровочную кривую строят
на специальной графической бумаге либо с использованием компьютерных про-
грамм. При работе на ИФА-анализаторе эту функцию выполняет программное
обеспечение, управляющее анализатором. Достоверно определить количество
аналита методом ИФА можно только в тех линейных пределах, которые указаны
в инструкции к тест-системе и установлены производителем. Экстраполирование
данных за линейные пределы измерения, указанные в инструкции к тест-системе,
не допускается.
61 2 ВИРУСОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ
Выполнение ИФА при вирусологических исследованиях требует тщательного
соблюдения условий качественной работы, соответствующих санитарных норм
и правил, действующих нормативных документов, а также инструкции фирмы-
производителя к тест-системе. Этими нормативными документами и инструк-
циями не только определяются основные требования к выполнению исследований
методом ИФА. но и регламентируются алгоритмы исследований и мероприятия по
внутрилабораторному контролю качества.
Согласно основным методическим рекомендациям по проведению ИФА. изло-
женным в отечественных и зарубежных руководствах, специализированных изда-
ниях, к критическим параметрам, влияющим на конечный результат, следует отне-
сги процесс пробоподготовки, лабораторное оборудование и приборы, качество
расходных материалов, культуру лабораторной работы, условия внешней среды в
помещениях, где выполняется анализ, и т.п.
Например, при отборе проб следует избегать гемолизированных или гиперли-
пидемических сывороток, поскольку их исследование может дать ложноположи-
тельные результаты. Нельзя использовать наконечники повторно при отборе или
разведении сывороток во время пробоподготовки, пользоваться стеклянными
пипетками и т.п. Если требуется предварительное разведение сывороток, дополни-
тельный планшет следует применять только однократно.
Пристальное внимание следует уделять точности автоматических пипеток
(погрешность не должна превышать 5%, в том числе по каналам многоканальной
пипетки), чистоте и стабильной работе автоматических промывателей (объем
дозируемых жидкостей по разным каналам промывания также не должен превы-
шать 5%). Необходимо периодически проверять истинную температуру в термо-
статах для инкубации и холодильниках, где хранятся гест-сиг гемы. Для этих целей
внутрь оборудования помещают отдельный термометр и ведут журнал контроля
температуры в термостате (холодильнике).
Для точности измерения оптической плотности следует периодически прово-
дить проверку планшетных спектрофотометров, своевременно заменять источник
света и следи гь за тем, чтобы погрешность измерения по разным каналам не пре-
вышала 10% (оптимальная погрешность — 5%). Оптическую плотность измеряют
на спектрофотометре с прогретой не менее 30 мин лампой.
Из расходных ма сериалов и вспомогательного оборудования для приготовле-
ния растворов конъюгата и субстрата следует использовать специально выделен-
ные и помеченные стеклянные емкости. Употреблять их для других целей нельзя.
Исключается повторное применение пластиковых наконечников для автоматиче-
ских пипеток.
Необходимо тщательно контролировать качество дистиллированной воды в
лаборатории. Нормальной считается дистиллированная вода с pH - 5,5-5-6,5 и
удельной проводимостью не более 5 мкСим/см. Особенно важно качество воды
при работе с автоматическими ИФА-анализаторами Для некоторых моделей ана-
лизаторов качество получаемой в стандартных перегонных дистилляторах воды
может оказаться недостаточным, в таком случае рекомендуют установить систему
водоподготовки с применением дополнительных систем очистки воды.
При постановке ИФА обязательно соблюдают инструкцию к тест-системе.
Всегда следует использовать реагенты одно! серии или партии. Все реагенты долж-
ны быть комнатной (18-25 °C) температуры, поэтому при хранении диагностиче-
ских наборов в холодильнике необходимые компоненты следует предварительно
довести до рекомендуемой температуры. При выполнении анализа нельзя касаться
дна планшета руками, а дна и стенки лунок — наконечником пипетки Следует
следить за тщательностью и равномерностью промывки всех лунок планшета, осо-
бенно после инкубации с конъюгатом. При работе с растворами субстрата нужно
помнить, что применяемые в ИФА хромогены светочувствительны и длительная
ВИРУСОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ 613
экспозиция на свету может привести к неконтролируемому разложению субстрата.
Добавление стоп-реагента (чаще всего, это раствор серной или соляной кислоты)
не останавливает полностью процесс окисления, поэтому длительная инкубация
даже «остановленного» ИФА может исказить результаты анализа.
При учете результатов спектрофотометрии всегда контролируют визуаль-
но степень окрашивания и состояние фо гометрируемых лунок и сверяют ее с
результатами измерения. Это поможет избежать искажения результатов при
появлении таких артефактов анализа, как пузыри воздуха в лунке или нали-
чие посторонних предметов в лунке. Нельзя дотрагиваться до дна планшета,
поскольку даже малейшие следы от рук могут повлиять на результаты измерения
и увеличить значения ОП.
При анализе результатов руководствуются критериями, изложенными в инструк-
ции к тест-системе, и трактуют полученные результаты согласно ей. Учитывая
погрешности, вносимые средствами измерения (пипетками и спектрофотоме-
тром), границами серой зоны считают 15% значения ОП критической (cut-off}.
Все результаты, попадающие в серую зону следует признавать сомнительными и
подвергать повторному исследованию.
Соблюдение вышеперечисленных правил позволит получать достоверные и
надежные результаты при исследовании биологического материала на маркеры
вирусных инфекций.
МОЛЕКУЛЯРНО-БИОЛОГИЧЕСКИЕ МЕТОДЫ
Внедрение методов молекулярной биологии в повседневную диагностическую
практику в 90-е гг. XX в. — одно из наиболее революционных событий в клиниче-
ской лабораторной диагностике, сопоставимое по значимости разве что с широким
внедрением тИФА в серологическую диагностику десятилетием раньше. Бурное
развитие молекулярной биологии на рубеже XX и XXI вв позволило не только сде-
лать эти методы доступными для большинства разделов клинической лаборатор-
ной диагностики, но и включить их в большинство диагностических алгоритмов
вирусных инфекционных заболеваний.
С помощью молекулярно-биологических методов возможны идентификация
инфекционных агентов, определение их количества в биологических жидкостях
(клетках организма), дифференциальная диагностика генетических вариантов
возбудителей, принятие решения о начале противовирусной терапии, подбор
оптимальной схемы терапии, контроль эффективности терапии, выявление мута-
ций резистентности к противовирусным препаратам и т.п.
К молекулярно-биологическим методам можно отнести все методы клиниче-
ской лабораторной диагностики, используемые для идентификации возбудителей
вирусных инфекционных заболеваний определение специфических нуклеотид-
ных последовательностей нуклеиновых кислот (ДНК/РНК) возбудителей. Как
следует из определения, все молекулярно-биологические методы — это методы
прямого выявления возбудителей, то есть положительный результат молекулярно-
биологических исследований свидетельствует о выявлении самого возбудителя, а
не следов его контакта с иммунной системой организма.
В основе всех молекулярно-биологических методов лежит способность двух-
цепочечных молекул нуклеиновых кислот к взаимному комплементарному свя-
зыванию. Вторая особенность нуклеиновых кислот — возможность синтеза ДНК/
РНК в условиях in vitro практически неограниченное количество раз. Первая
особенность позволяет специфически идентифицировать генетический материал,
присущий только определяемому организму и никакому другому; вторая особен-
ность — амплифипировать участок нуклеиновой кислоты, специфичный для дан-
ного типа возбудителя, в количестве, достаточном для аппаратной (например, для
614 ВИРУСОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ
ПЦР в режиме реального времени) или визуальной (для электрофоретического
варианта ПЦР) детекции.
Молекулярно-биологические методы, как правило, состоят из трех основных
этапов:
S выделения препарата нуклеиновой кислоты из биологического образца;
J проведения исследования;
J детекпии результатов и их интерпретации.
Современные технологии позволяют значительно упростить и автоматизиро-
вать наиболее трудоемкий первый этап исследования, связанный с выделением
тотального препарата нуклеиновой кислоты. К сожалению, этот ответственный
этап анализа автоматизируется значительно медленнее, чем этап проведения
исследования (максимально автоматизированный на данный момент) или этап
детекции и интерпретации результатов. Эволюция систем выделения ДНК/РНК
идет двумя путями:
J упрощением методов выделения нуклеиновых кислот путем снижения коли-
чества операций, появлением систем для быстрого («экспресс») выделения
нуклеиновых кислот, повышением эффективности выделения и чистоты
нуклеиновых кислот;
J появлением автоматических станций для экстракции нуклеиновых кислот с
возможностью настраиваемых протоколов выделения.
Эти процессы протекают параллельно, но следует отметить, что более пер-
спективным считают внедрение автоматизированных систем экстракции нуклеи-
новых кислот, значительно повышающее производительность лаборатории и
качество исследований Особенно важно применять автоматизированные систе-
мы экстракции нуклеиновых кислот с настраиваемыми протоколами выделения,
поскольку различные варианты молекулярно-биологических методов предъ-
являют разные требования к материалу. Например, для ПЦР с амплификацией
длинных фрагментов нуклеиновых кислот вируса предпочтительнее исполь-
зовать более мягкий вариант выделения тотального препарата нуклеиновых
кислот, поскольку при таких условиях получают больше высокомолекулярных
нуклеиновых кислот.
Для большинства молекулярно-биологических методов достаточно относитель-
но простых методов экстракции нуклеиновых кислот. Наибольшее распростране-
ние имеют следующие методы выделения: гуанидинизотиоциана гный с сорбцией
на силикагеле и применение детергентов с высаливанием ДНК/РНК из водного
раствора в присутствии этанола или изопропанола. Обычно не применяют допол-
нительные методы очистки или фракционирования препарата и для исследования
берут тотальный препарат РНК/ДНК.
Наиболее часто в лабораторной диагностике инфекционных вирусных забо-
леваний практикуют два подхода; методы молекулярной гибридизации и методы
амплификации нуклеиновых кислот.
К методам гибридизации следует относить:
v' гибридизацию в растворе или на твердом носителе;
S гибридизацию in situ (ISH, FISH);
S метод разветвленной ДНК (branch DNA).
Методы амплификации нуклеиновых кислот:
J полимеразная цепная реакция (PCR);
лигазная цепная реакция (LCR);
самопроизвольная репликация последовательностей (3SR);
J транскрипциинно-опосредованная амплификация (ТМА);
Vх амплификация с замещением цепи (SDA);
реакция транскрипционной амплификации (NASBA).
ВИРУСОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ 615
Гибридизационные молекулярно-биологические методы
Гибридизационная детекция нуклеиновых кислот вирусных частиц основана на
способности синтетических зондов связываться с гомологичными участками нукле-
иновых кислот вируса, после чего образовавшиеся гибриды «зонд-нуклеиновые
кислоты» определяются. За время, прошедшее с внедрения в молекулярную био-
логию гибридизационных методов исследования (в 70-х гг. XX в.), они получили
существенное развитие как в техническом, так и в методологическом аспектах.
Многие из гибридизационных методов утратили свое значение либо стали частью
иных, методически более совершенных способов клинической лабораторной диа-
гностики. Например, гибриды «зонд-мишень» в современной диагностической
лаборатории выявляют либо с помощью гибридизационно-иммуноферментного
анализа, либо более чувствительным и перспективным иммунохемилюминесцент-
ным методом.
Гибридизационным методом можно исследовать как клеточный материал
(мазок, соскоб), так и биологические жидкости (плазму крови, сперму, СМЖ
и т.п.). Молекулярные комплексы «зонд-мишень» формируются при реакции
гибридизации, при которой сначала молекулы нуклеиновых кислот подвергают
тепловой денатурации с последующим охлаждением до температуры образова-
ния комплементарных связей (гибридов) между цепями нуклеиновых кислот. На
первом этапе из пробы выделяют тотальный препарат нуклеиновых кислот.
В реакционный объем вносят тотальный препарат нуклеиновых кислот, выде-
ленный из образца, и синтетические зонды, комплементарные к специфическо-
му участку генома возбудителя (рис. 23-14, см. цв. вклейку). Смесь подвергают
денатурации нагреванием до 95 °C и охлаждению для образования гибридов
«зонд-мишень». Гйбриды захватываются сорбированными на твердой фазе Ат.
Несвязавшиеся с зондами молекулы удаляют промывкой, а в реакционный объем
добавляют Ат к двухспиральным нуклеиновым кислотам (зонды могут быть как
РНК, так и ДНК), выполняющим роль конъюгата. Подобную технологию захва-
та гибридов «зонд-NK» (hybrid-capture) применяют, например, для выявления
нуклеиновых кислот таких инфекционных агентов, как ВИЧ или ВПЧ высокого
канцерогенного риска.
Практикуя подобный подход, можно достаточно быстро исследовать значи-
тельное количество образцов, поскольку исследование в жидкой фазе позволяет
автоматизировать процесс и привести его в планшетный или иной высокопроиз-
водительный формат.
В случае гибридизационного исследования на твердой фазе один из ком-
понентов реакции (зонд) сорбируется на поверхности реакционного объема
(рис. 23-15). В качестве твердой фазы наиболее часто выступают нитроцеллю-
лозная или нейлоновая мембрана, либо поверхность лунки планшета. В одном
из вариантов метода сначала проводят реакцию гибридизации с образованием
гибрида «зонд 1-мишень», затем в реакционную смесь добавляют зонд 2, ком-
плементарный к соседнему участку мишени и имеющий определенную метку
(ферментную или флюоресцентную). После этого детектируют образовавшие-
ся комплексы «зонд 1-мишень-зонд 2». Эта схема анализа носит название
«сэндвич»-гибридизации по аналогии с «сэндвич»-ИФА.
Данный метод анализа с некоторыми модификациями применяют преиму-
щественно на этапе детекции продуктов ПЦР, при исследовании методом ПЦР
с гибридизационно-иммуноферментной детекцией. Отличие этого метода от
«сэндвич»-гибридизации заключается в использовании на этапе выявления ком-
плекса «зонд 1-мишень» не меченого зонда 2, а антител к двухцепочечным гибри-
дам, конъюгированных со стрептавидин-биотиновым комплексом, меченным
ферментом (пероксидазой или щелочной фосфатазой).
616 ВИРУСОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ
Рис. 23-15. Схема «сэндвича-гибридизации с гибридизационно-ферментной детекцией.
Еще одна интересная модификация гибридизационного анализа — технология
анализа линейных проб (LiPA — Line Probe Assay). LiPA особенно эффективна при
анализе генетических мутаций, генотипировании вирусов (гепатита В и С, напри-
мер), выявлении генетически обусловленной лекарственной резистентности (ВИЧ,
ВГВ и т.п.). В общих чертах метод анализа линейных проб заключается в гибри-
дизационном анализе предварительно амплифицированного продукта. Однако в
отличие от ПЦР, где амплифицируется преимущественно консервативная область
генома, в LiPA для анализа мутационных замен или генетической гетерогенности
популяции (выявление генотипов) амплифицируется генетически гетерогенная
часть вирусного генома. Полученные в результате ПЦР биотинилированные
ампликоны гибридизируют с иммобилизованными на нитроцеллюлозном стрипе
олигонуклеотидными зондами (рис. 23-16), комплементарными различным гено-
типам вируса. Несвязавшиеся ампликоны удаляют отмывкой. После связывания
конъюгата («щелочная фосфатаза-стрептавидин») и добавления субстрата в месте
нанесения специфического зонда появляется полоса, как и в иммунном блоте.
По сравнению с иными технологиями обнаружения мутационных замен
(рестрикционным анализом полиморфных фрагментов, частичным секвениро-
ванием. пиросеквенированием), LiPA позволяет относительно быстро и дешево
получить результаты. Ограничение метода заключается в возможности выявить
только уже охарактеризованные мутационные замены, в отличие, например,
от секвенирования.
Следующим этапом развития гибридизационного анализа на твердой фазе стала
гибридизация in situ. Метод основан на проведении процессов гибридизации непо-
средственно на срезе или в мазке с использованием меченых ДНКу'ГНК-зондов.
В зависимости от вида метки (ферментной или люминесцентной) различают
несколько видов гибридизации in situ, в том числе гибридизацию in situ с флюо-
ресцентной детекцией (FISH — Fluorescent In Situ Hybridization). Метод применяют
преимущественно для выявления ДНК-содержащих вирусов, поскольку вирусная
РНК менее устойчива к разрушению, особенно на стадии пробоподготовки.
ВИРУСОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ
617
А
Б
retttlttttteee
ееееее
ееееее
Генотип
Рис. 23-16. Метод анализа линейных проб (LiPA): А — схема исследования; Б — образец гибри-
дизационных стрипов для генотипирования ВГВ (tittp://www.innogenetics.be).
Метод позволяет установить наличие частиц вируса в фиксированных и заклю-
ченных в формалин препаратах, пунктатах (биоптатах) ткани, что особенно важно
при патоморфологическом анализе либо просто в соскобе. Гибридизация in situ
базируется на принципах твердофазной гибридизации и гибридизации в растворе.
Чувствительность метода гибридизации in situ ограничена возможностью проникно-
вения зондов внутрь клеток для связывания с искомой мишенью. По этой причине
после фиксации клеток соскоба (мазка, биоптата и т.п.) осуществляют разрушение
фиксированных клеток и высвобождение внутриклеточной ДНК (рис. 23-17). Процесс
гибридизации проходит в тканях, при этом зонд снабжен флюоресцентной меткой.
После промывки материала и удаления несвязавшихся молекул зондов приступают к
детекции, для чего мазок или срез анализируют в флюоресцентном микроскопе.
С помощью гибридизации in situ наиболее удобно определять вирусные частицы
(которые представлены большим количеством копий) в инфицированных клет-
ках, например, ВПЧ высокого канцерогенного риска в цервикальных соскобах.
либо выявлять оккультные формы гепатита В.
618 ВИРУСОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ
Рис. 23-17. Схема гибридизационного анализа in situ с флюоресцентной детекцией (FISH). На
первом этапе образец фиксируют на твердой фазе/предметном стекле (А); второй этап — раз-
рушение клеточных мембран и высвобождение молекул ДНК (Б), после чего образец подвергают
денатурации (В) и гибридизации с меченными флюорохромом ДНК/Р НК-зондам и (Г, Д); специфи-
чески связавшиеся зонды детектируют с помощью люминесценции (Е).
К достоинствам методов гибридизационного анализа относят высокую спец-
ифичность, крайне малый риск контаминации, универсальность методов для
любых типов микроорганизмов.
Недостатками всех гибридизационных методов следует считать относительно
невысокую чувствительность (по сравнению с амштификационными методами
молекулярно-биологических исследований), сравнительно высокую стоимость
гибридизационных зондов (для повышения специфичности необходимо синтези-
ровать длинные зонды), ограниченность применения для различных биологиче-
ских субстратов или объектов внешней среды. Кроме того, большинство современ-
ных методов исследования с использованием гибридизационного анализа требует
длительных инкубаций гибридизационной смеси, что значительно удлиняет время
исследования, а в некоторых случаях делает невозможным получение результата
в течение одного рабочего дня. Некоторые гибридизационные методы, такие как
гибридизация in situ, методически сложны, требуют высокой квалификации пер-
сонала и не могут применяться для скрининговых исследований большого коли-
чества образцов.
Многие из перечисленных выше недостатков устранены в методе разветвленной
ДНК с использованием принципа умножения сигнала (в отличие от амплификаци-
онных методов, основанных на принципе умножения мишени).
Схема исследования по методу разветвленной ДНК (рДНК) следующая
(рис. 23-18, см. цв. вклейку). На твердой фазе сорбируется зонд 1 (улавливающий),
комплементарный к ДНК/РНК-мишени (а). После реакции гибридизации (б) все
несвязавшиеся молекулы нуклеиновых кислот удаляются отмывкой, а в реакци-
онную смесь добавляют зонд 2 (детектирующий), специфический к иному участку
мишени, чем зонд 1, и усиливающие зонды (в). Особенность усиливающих зон-
дов — самокомплементарность и комплементарность к зонду 2, в результате чего
происходит многократная гибридизация усиливающих зондов самих на себя с
возникновением своеобразной структуры типа «дерево» (г). Усиливающие зонды
ВИРУСОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ 619
связываются с молекулами люминофора (д). Благодаря явлению самогибридиза-
ции усиливающих зондов наступает эффект амплификации (умножения) сигнала.
На одну молекулу мишени приходится один детектирующий, один улавливающий
и многие тысячи и десятки тысяч усиливающих зондов, каждый из которых связан
с несколькими молекулами люминофора, что позволяет усилить детектируемый
сигнал от единственной молекулы-мишени, В ходе реакции детекции (люми-
несцентной, хемилюминесцентной или иной) в реакционной смеси выявляют
специфическое свечение (е).
Подобным методом можно идентифицировать ДНК/РНК различных возбуди-
телей инфекционных заболеваний (ВИЧ, ВГВ, ВГС и др.). Поскольку величина
детектируемого сигнала прямо пропорциональна исходному количеству молекул-
мишеней, связавшихся с улавливающим зондом, при наличии охарактеризован-
ных количественных стандартов, возможно не только качественное выявление
вирусных агентов инфекционных заболеваний методом рДНК, но и оценка их
количественного содержания в биологических образцах.
Амплификационные молекулярно-биологические методы
В амплификационных методах детекции используется принцип накопления
(амплификации) исходной матрицы (фрагмента мишени) до достижения опреде-
ляемых значений положительного сигнала. Наибольшее применение в практике
лабораторной диагностики получила полимеразная цепная реакция (ПЦР).
Полимеразная цепная реакция
За последние несколько лет метод ПНР и его модификации стали занимать одно
из ведущих мест в современной лабораторной диагностике. Для ПЦР характерны
такие уникальные свойства, как высокие специфичность, чувствительность, уни-
версальность и короткое время исследования.
В медицине ПЦР применяют для диагностики инфекционных заболеваний,
в том числе вызванных агентами, трудноподдающимися культивированию, для
определения устойчивости микрофлоры к антибиотикам, пренатальной диагно-
стики и диагностики наследственных заболеваний тестирования донорской крови
на вирусные патогены, при различных видах генотипирования, определении
отцовства, для выявления мутаций и т.д.
Высокие чувствительность и специфичность, непосредственное обнаружение
инфекционного агента и возможность проведения генотипирования определя-
ют широту области применения метода ПЦР в клинической диагностике: для
ранней диагностики инфекционных заболеваний у серонегативных пациентов,
когда лечение наиболее эффективно; выявления персистирующих, латентных и
рецидивирующих форм инфекций; контроля эффективности лечения; разрешения
сомнительных результатов серологических исследований; эпидемиологических
исследований; диагностики оппортунистических инфекций, часто протекающих
на фоне иммунодефицита, вследствие чего постановка диагноза только по резуль-
татам серологических исследований может быть затруднена.
Санитарно-эпидемиологические службы применяют ПЦР для контроля микро-
биологического загрязнения окружающей среды и продуктов питания, а также для
выявления генетически модифицированных источников пищи.
Принцип метода основан на обнаружении в материале специфичных фрагмен-
тов ДНК (РНК) различных биологических объектов, их избирательном синтезе до
концентрации, при которой их легко детектировать, и дальнейшем определении
продуктов реакции амплификации — ампликонов.
Дезоксирибонуклеиновая кислота (ДНК) — универсальный носитель генети-
ческой информации у всех существующих на Земле организмов, за исключением
620 ВИРУСОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ
РНК-содержащих вирусов. Уникальное свойство ДНК заключается в ее способно-
сти удваиваться после расплетения спирали и расхождения нитей ДНК. Удвоение
ДНК (репликация) осуществляется по принципу комплементарности ферментом
ДНК-полимеразой. Для того чтобы фермент начал свою работу, требуется нали-
чие небольшого начального двухцепочечного фрагмента ДНК. Такой фрагмент
образуется при взаимодействии короткого одноцепочечного фрагмента ДНК,
называемого праймером, с комплементарным участком соответствующей цепи
родительской ДНК, Репликация протекает одновременно на двух нитях ДНК, но
наращиваются они в противоположных направлениях. В результате репликации
из одной двухцепочечной молекулы ДНК получаются две двухцепочечные, каждая
из которых содержит одну нить от материнской молекулы ДНК и вторую, дочер-
нюю, — вновь синтезированную.
Таким образом, цикл репликации ДНК включает три основные стадии:
J расплетение спирали ДНК и расхождение нитей (денатурацию);
J присоединение праймеров;
S достраивание дочерней цепи ДНК.
В ПЦР эти процессы осуществляются в пробирке в циклическом режиме.
Переход от одной стадии реакции к друтой достигается изменением температуры
инкубационной смеси. Для ПЦР необходимы:
✓ два синтетических олигонуклеотидных праймера (длиной примерно по
20 нуклеотидов), комплементарные участкам ДНК из противоположных
цепей, фланкирующим последовательность-мишень (праймеры ограни-
чивают фрагмент ДНК, который будет миллионы раз скопирован фермен-
том Taq-ДНК-полимеразой. присоединяющейся к 3' -концам праймеров и
достраивающей их до заданной длины в несколько сотен пар оснований);
J ДНК-мишс нь;
S термостабильная ДНК-полимераза, не теряющая активности при темпе-
ратуре 95 иС;
J четыре дезоксирибонуклеотида;
J буферная система для эффективной работы ДНК-полимеразы, обязатель-
но содержащая ионы магния.
Основные этапы ПЦР:
S подготовка пробы биологического материала, которая заключается в
выделении ДНК или РНК;
S собственно полимеразная цепная реакция (ПЦР-амплификация);
J детекция продукта ПЦР (амплифицированной нуклеиновой кислоты).
Пробоподготовка (выделение ДНК или РНК из клинического матеоиа-
ла). На данном этапе образцы клинического материала подвергают специальной
обработке, во время которой происходят лизис клеток, устранение белковых,
полисахаридных, липидных компонентов. Для этого используют различные
методы [например, сорбентный метод, заключающийся в сорбции ДНК (РНК)
на сорбенте после лизиса клеток, многократной огмывке нуклеиновых кислот и
последующей элюции ДНК (РНК) буферным раствором и др.]. В результате такой
обработки получают раствор, содержащий всю ДНК и/или РНК из клинического
материала. Набор для выделения ДНК (РНК) обычно выбирают в зависимости от
вида клинического образца.
Детально методики выделения ДНК (РНК) из клинических образцов описаны
в инструкциях, прилагаемых производителем к набору реагентов для выделения
нуклеиновых кислот.
Полученный раствор ДНК можно хранить в течение недели при температуре
2-8 °C и до года при температуре -60 °C. Раствор очищенной РНК хранению не
подлежит, его следует сразу направлять на исследование (пробы можно сохранять
только в виде полученных с помощью обратной транскрипции растворов кДНК).
ВИРУСОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ 621
Типичная ПЦР-амплификация состоит в многократном повторении следую-
щих трех реакций: денатурации, ренатурации, синтеза.
Денатурация. Первый этап ПЦР заключается в тепловой денатурации образца
ДНК выдерживанием его при температуре 94-95 °C. Кроме ДНК, в реакционной
смеси содержатся в избытке два праймера, термостабильная ДНК-полимераза Taq
(выделяют из бактерий Thermus aquaticus) и четыре дезоксирибонуклеотида.
Ренатурация. Температуру реакционной смеси понижают до 50-60 °C, при этом
праймеры гибридизуются с комплементарными последовательностями ДНК (исхо-
дной расплавленной ДНК-матрицы) с образованием коротких двухцепочечных
ДНК-фрагментов, необходимых для начала работы фермента полимеразы (отжиг
праймеров). Каждый из праймеров гибридизуется на одной из двух цепей ДНК-
матрицы таким образом, что концы праймеров, которые способны удлиняться,
направлены навстречу друг другу. Присоединившись к противоположным цепям
молекулы ДНК, праймеры ограничивают собой тот ее участок, который будет в
дальнейшем многократно удвоен или амплифицирован. Длина такого фрагмента,
который называется ампликоном, обычно составляет несколько сот нуклеотидов.
Синтез. Температуру реакционной смеси повышают до 70-72 °C — величины,
оптимальной для работы ДНК-полимеразы Taq. Происходит синтез фрагмента
комплементарной (дочерней) цепи ДНК (в качестве строительного материала
используют находящиеся в смеси четыре дезоксинуклеотидтрифосфата). Синтез
фрагментов дочерних цепей ДНК протекает одновременно на обеих цепях мате-
ринской ДНК.
Далее цикл повторяется снова. Образовавшиеся в первом цикле амплифика-
ции фрагменты цепей ДНК служат матрицами для второго цикла амплификации,
фрагменты, образовавшиеся в первых двух циклах, — для третьего и т.д. Таким
образом, вновь синтезированные фрагменты служат матрицами для синтеза новых
цепей ДНК в следующем цикле амплификации — цепная реакция. В результате
количество копий фрагмента увеличивается в геометрической прогрессии и про-
исходит накопление ампликонов в растворе по формуле: 2п, где п — количество
циклов амплификации.
Все реакции проводят в пробирках, размещенных в термостате. Разработка
программируемых термостатов (термоциклеров, или амплификаторов), которые
по заданной программе осуществляют циклическую смену температур, создала
предпосылки для широкого внедрения метода ПЦР в практику лабораторной
клинической диагностики. Смену температурных режимов и их поддержание эти
приборы выполняют автоматически. Продолжительность каждого цикла обычно
составляет несколько минут.
Для синтеза праймеров, специфичных в отношении исключительно ДНК-
мишени. нужно знать нуклеотидную последовательность ДНК предполагаемого
патогенного микроорганизма. Основным критерием подбора праймеров считают
комплементарность матрице. В этом случае в ходе ПЦР будет амплифицироваться
только специфический фрагмент ДНК, длина которого равна суммарной длине
двух праймеров и фрагмента ДНК между ними. Подбор оптимальной температуры
отжига праймеров на матрице — важный фактор, влияющий на эффективность и
специфичность амплификации.
Модификация метода ПЦР — проведение обратной транскрипции на матрице
РНК. Обратная транскрипция (ОТ) — метод, позволяющий получать на матрице
РНК соответствующую цепь комплементарной ДНК (кДНК). ОТ-ПЦР используют
для выявления генетического материала РНК-содержащих возбудителей (напри-
мер, вируса гепатита С). В реакции участвует фермент, синтезирующий кДНК на
матрице РНК, — обратная транскриптаза (ревертаза). В результате получается
раствор, содержащий ДНК-копию РНК-вируса. Затем кДНК включается в ампли-
фикацию, то есть в этом случае анализ протекает в два этапа. ОТ-ПЦР можно
В22 ВИРУСОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ
выполнять как в различных пробирках (в две стадии), так и в одной пробирке
(совмещенная ОТ-ПЦР) без существенной потери чувствительности.
Детекция продуктов ПЦР -амплификации. Существуют различные способы
детекции результатов ПЦР.
Присутствие специфических ПЦР-пподуктов (ампликонов) определяют элек-
трофоретическим разделением амплификационной смеси в окрашенном бро-
мистым этидием агарозном геле Бромистый этидий связывается с фрагмента-
ми двухцепочечных ДНК, которые проявляются в виде светящихся полос при
УФ-облучении геля (с длиной волны 290-330 нм). Для визуализации и ультрафио-
летового подсвечивания полос ДНК используют специальный прибор — трансил-
люминатор, а полученные результаты документируют фотографированием (реко-
мендуют хранить в компьютерной базе данных не менее трех лет).
Фрагменты ДНК в агарозном геле разделяют в зависимости от их молекулярной
массы (размера). Специфичность полосы амплифицированной ДНК под1вержда-
ется ее положением по отношению к маркерам молекулярной массы и положению
фрагмента положительного контроля амплификации. Дополнительные доказа-
тельства специфичности ампликона можно получить методами рестрикционного
анализа, гибридизации и прямого секвенирования.
К технологиям, альтерна гивным электрофоретическому способу детекции про-
дуктов амплификации, относят модифицированные системы анализа с использо-
ванием флюоресцентных красителей, при этом продукты реакции анализируют с
помощью флюориметрии. Пробирки из амплификатора поступают в детектор, где,
не открывая их, регистрируют флюоресценцию. Полученные результаты сохраня-
ют в компьютерной базе данных. Уровни флюоресценции оказываются пропор-
циональными количеству образовавшихся специфических продуктов ПЦР. В таких
вариантах анализа флюоресценцию регистрируют после окончания реакции, и поэ-
тому метод не является количественным. Важно то, что результаты ПЦР фиксируют
по наличию флюоресценции в закрытых пробирках. Таким образом, решается одна
из основных проблем ПЦР — проблема контаминации ампликонами.
Real-time PCR. Один из самых современных вариантов метода — ПЦР в режи-
ме реального времени. В основе этого варианта лежит количественная детекция
флюоресцентного сигнала, который увеличивается пропорционально количеству
ПЦР-продукта. Результат реакции можно регистрировать и наблюдать на экране
монитора компьютера непосредственно в процессе ПЦР-амплификации.
Для количественных вариантов исследований в режиме ПЦР в реальном време-
ни требуются соответствующие стандарты, которые используют для построения
калибровочных кривых после проведения ПЦР. Используя эти кривые, можно
вычислить неизвестное исходное количество копий ДНК (РНК) в образцах.
Преимущества ПЦР в реальном времени:
• регистрация интенсивности флюоресценции позволяет судить о количестве
инфекционного агента в пробе:
• исключается главная причина ложноположительных результатов — рассеи-
вание продуктов амплификации с аэрозолями и персоналом в процессе гель-
электрофоретической детекции:
• требуется всего несколько часов дня получения результата (быстрота анализа);
• наличие дополнительного специфического зонда, комплементарного вну-
треннему участку амплифицируемого фрагмента, снижает риск получения
ложноположительных результатов и увеличивает чувствительность анализа;
• появляется возможность проводить множественную ПЦР — регистрировать
в одной пробирке наличие нескольких инфекционных агентов, используя
различные флюоресцентные красители;
• появляется возможность идентифицировать в продукте амплификации оди-
ночные нуклеотидные замены.
ВИРУСОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ 623
Преимущества ПЦР как метода диагностики инфекционных заболеваний сле-
дующие.
Прямое определение наличия возбудителей. Выявление специфического участка
ДНК-возбудителя методом ПЦР дает прямое указание на присутствие возбудителя
инфекции. Многие традиционные методы диагностики, например иммунофер-
ментный анализ, детектируют белки-маркеры, являющиеся продуктами жизне-
деятельности инфекционных агентов, что лишь опосредованно свидетельствует о
наличии инфекции.
Высокая специфичность. В исследуемом материале идентифицируют уникаль-
ный, характерный только для данного возбудителя фрагмент ДНК. Специфичность
задается нуклеотидной последовательностью праймеров, что сводит к минимуму
риск получения ложных результатов в отличие от иммунологических методов ана-
лиза, где нередки ошибки в связи с перекрестно-реагирующими антигенами.
Высокая чувствительность. ПЦР-анализ позволяет обнаруживать возбудителей
инфекционных заболеваний в тех случаях, когда другими методами (иммуноло-
гическими. бактериологическими, микроскопическими) это сделать невозможно.
Чувствительность ПЦР-анализа приближается к 10 -100 клеткам в пробе.
Универсальность процедуры выявления различных возбудителей. Материалом для
исследования методом ПЦР служит ДНК возбудителя, что позволяет диагности-
ровать несколько возбудителей в одном образце (сходством химического состава
всех нуклеиновых кислот определяются унифицированные методы выделения
ДНК/РНК). Для ПЦР-диагностики практически всех инфекционных заболеваний
могут быть использованы один набор оборудования, универсальные процедуры
пробоподготовки и постановки анализа.
Высокая скорость получения результата. Для ПЦР-анализа не требуется выде-
ления возбудителя в культуре, что значительно сокращает время исследова-
ния. Унифицированные методы выделения нуклеиновых кислот, автоматизация
амплификации и детекции продуктов реакции дают возможность проведения
полного анализа в течение одного рабочего дня.
Возможность диагностики не только острых, но и латентных инфекций.
Особенное значение метод ПЦР имеет для диагностики труднокультивируемых и
персистирующих микроорганизмов, с которыми часто приходится сталкиваться
при латентных и хронических инфекциях. ПЦР-диагностика также очень эффек-
тивна в отношении возбудителей с высокой антигенной изменчивостью (напри-
мер, вирусов гриппа).
Ограничения метода ПЦР:
У амплифицируется ДНК как живых, так и неживых микроорганизмов, что
обусловливает определенные требования к интерпретации и срокам про-
ведения исследований при контроле эффективности лечения (контроль
следует осуществлять спустя промежуток времени, в течение которого
происходит полная элиминация возбудителя);
теоретически существует вероятность перекрестных реакций (например,
в результате неадекватного подбора праймеров), что может приводить к
появлению ложноположительных результатов.
Взятие клинического материала. Взятие материала, его предварительная
обработка, хранение и перевсзка, передача исследуемого материала в другие
организации осуществляются согласно инструктивно-методическим документам,
регламентирующим выполнение исследований для каждого вида возбудителя
инфекций, инструкциям к наборам реагентов и в соответствии с действующими
Санитаоными правилами (СП).
В целях предотвращения разрушения нуклеиновых кислот рекомендуют
использовать специальные транспортные среды, разработанные и рекомендо-
ванные производителями наборов реагентов в зависимости от вида материала.
624 ВИРУСОЛОГ ИЧЕСКИЕ ИССЛЕДОВАНИЯ
Перевозку и хранение материала следует осуществлять в условиях холодовой цепи
с обеспечением контроля установленного температурного режима с помощью
термоиндикаторов. В зависимости от транспортной среды, вида материала и тем-
пературного режима сроки перевозки и хранения материала могут варьировать.
Ввиду вероятности разрушения ДНК или РНК в клиническом материале сроки
хранения и транспортировки материала должны быть минимальными. При невоз-
можности доставить образцы в ПЦР-лабораторию в течение требуемого времени
их замораживают (-20 °C). Допускается только однократное кратковременное
замораживание-оттаивание материала.
Материалом для ПЦР может служить практически любой биологический обра-
зец. Пробы крови (плазмы) используют в качественных и количественных иссле-
дованиях, пробы сыворотки крови — только при качественном ПЦР-анализе. Для
исключения контаминации клинический материал необходимо забирать с помо-
щью стерильных одноразовых инструментов (шприцев, соответствующих зондов,
пробоотборников и пр.) в одноразовые стерильные пластиковые контейнеры
(например, пробирки типа «Эппендорф»).
Клинические образцы следует хранить отдельно от реагентов.
Организация работы ПЦР-лаборатории
Требования к организации работ в ПЦР-лабораториях обобщены и сформули-
рованы в нормативных документах и методических указаниях. Методическими
указаниями определяются принципы организации лаборатории и этапы выполне-
ния анализа с использованием ПЦР: взятие проб, первичная обработка, хранение,
условия перевозки, обеззараживание материала, выделение нуклеиновых кислот,
проведение обратной транскрипции и/или амплификации, учет и регистрация
результатов исследования биологического материала, пищевых продуктов, материа-
ла из обьектов окружающей среды. Методическими указаниями регламентируется
деятельность персонала лабораторий при выполнении исследований с помощью
методов амплификации нуклеиновых кислот, проводимых с использованием заре-
гистрированных в установленном порядке наборов реагентов и оборудования.
ПЦР-циагностика инфекций связана с проблемой, обусловленной высокой чув-
ствительностью метода, — возможностью контаминации. Правильная организация
ПЦР-лаборатории имеет принципиальное значение для получения достоверных
результатов и существенно зависит от метода детекции продуктов амплификации.
При использовании метода горизонтального электрофореза этапу детекции следу-
ет уделять большое внимание, поскольку он является основным источником кон-
таминации. Помещение для электрофореза должно располагаться в месте, исклю-
чающем попадание ампликонов в другие технологические зоны лаборатории.
Кроме того, необходимы отдельный сотрудник, тщательно продуманная система
вентиляции, отсутствие возможности перемещения пробирок, штативов и прочего
оборудования из комнаты для электрофореза в другие помещения.
Сотрудник, занимающийся ПЦР-диагностикой, в своей работе сталкивается с
двумя видами контаминации:
J перекрестной контаминацией от пробы к пробе (например, в процессе
обработки клинических образцов), приводящей к появлению спорадиче-
ских ложноположительных результатов;
J контаминацией продуктами амплификации (ампликонами), имеющей
наибольшее значение, поскольку в процессе ПЦР ампликоны накапли-
ваются в огромных количествах и служат идеальными продуктами для
реамплификации.
Контаминация следовыми количествами ампликонов посуды, автоматических
пипеток и лабораторного оборудования, поверхностей лабораторных столов или
даже поверхности кожи сотрудников лаборатории приводит к появлению система-
тических ложнопиложительных результатов.
ВИРУСОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ 625
Как правило, определить источник контаминации бывает очень трудно, ликви-
дация контаминации требует значительных затрат времени и средств. Соблюдение
требований, предъявляемых к планировке помещений ПЦР-лабораторий и про-
ведению самих анализов, исключает возможность контаминации и получения
ложноположительных результатов.
В каждом помещении ПЦР-лаборатории должен быть набор реагентов, автома-
тических пипеток, наконечников, пластиковой и стеклянной посуды, лабораторно-
го оборудования, халатов и перчаток, используемых только в данном помещении и
не выносящихся в другие ПЦР-помещения. Оборудование, материалы и инвентарь
в каждой комнате соответствующим образом маркируют.
При исследованиях с использованием методов амплификации нуклеино-
вых кислот обязательно обеспечение поточности движения материала, проб
нуклеиновых кислот, продуктов амплификации. Работа в лаборатории должна
быть организована в одном направлении: от пре-ПЦР-помещений к пост-ПЦР-
помещению.
В лаборатории, использующей ПНР в диагностических целях, осуществляют
внутрилабораторный контроль качества исследований с периодичностью, зави-
сящей от объема работы и определяемой руководителем лаборатории, но не реже
одного раза в квартал. Лаборатория должна участвовать в установленном порядке
в мероприятиях (программах) по внешней оценке качества лабораторных иссле-
дований по конкретным нозологическим формам не реже одного раза в год.
Внутрилабораторный и внешний контроль качества лабораторных исследо-
ваний осуществляют путем анализа шифрованных аттестованных контрольных
панелей, содержащих положительные и отрицательные пробы. В ходе внутрила-
бораторного контроля качества используют аттестованные на наличие аналита
(его количества) панели производителей коммерческих наборов или внутрила-
бораторные аттестованные образцы, содержащие и не содержащие нуклеиновые
кислоты конкретных возбудителей в различной концентрации, стабильные в усло-
виях хранения.
Следует отметить, что широкое использование ДНК-диагностики ни в коей мере
не отменяет методы иммунохимических исследований (ИФА, ПИФ, РИФ и др.), а
дополняет их. Комплексное обследование пациента различными методами дает
врачу-клиницисту более полную информацию как о наличии инфекционного аген-
та. так и об иммунном статусе, что позволяет более точно ставить диагноз, назна-
чать соответствующее этиотропное лечение и вести его последующий контроль.
Биочипы
Биологические микрочипы — одно из наиболее быстроразвивающихся экспери-
ментальных направлений современной молекулярной биологии и биотехнологии.
Биочипы, подобно электронным микрочипам, обрабатывающим огромные масси-
вы цифровой информации, предназначены для получения и обработки больших
объемов молекулярно-биологической информации при проведении многопараме-
трического анализа микрообразца биологического материала.
Основа биочипов — матрипа микроячеек на плоскости, каждая из которых
содержит молекулярные зонды, специфичные к одной из множества биологи-
ческих молекул или их фрагментов (например, последовательностям ДНК или
РНК, белкам). Молекулярными зондами могут быть олигонуклеотиды, фрагмен-
ты геномной ДНК, РНК, белки, полипептиды, антитела, лиганды, олигосахариды
и т.д.
Применение биочипов позволяет оперативно обнаруживать микроорганизмы
и вирусы, выявлять мутации, лекарственно-устойчивые формы возбудителей
инфекционных заболеваний, идентифицировать индивидуальные генетические
626 ВИРУСОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ
особенности пациента, определять предрасположенность к наследственным и
онкологическим заболеваниям, переносимость некоторых курсов терапии и т.д.
Существенный момент при этом — возможность анализировать со^ни и тысячи
образцов в кратчайшие сроки.
Размеры ячеек современных микрочипов составляют примерно 10-100 микрон,
а значения концентраций анализируемых макромолекул находятся в пределах
около 1 пмоль-10 мкмоль. Общее число ячеек на чипе может колебаться от
нескольких сотен до 103-105, линейные размеры чипа — несколько квадратных
сантиметров.
В поверхностных матричных биочипах молекулярные зонды иммобилизуют-
ся на поверхности мембран или пластинок из стекла, пластика, полупроводника
или металла. В гелевых биочипах зонды иммобилизуются или в слое полиакри-
ламидного геля толщиной около 20 микрон, нанесенного на специально обрабо-
танную поверхность стекла (такие чипы производит фирма «PerkinElmer»), или
в полусферических гидрогелевых ячейках диаметром примерно 100 микрон (как
в чипах, разработанных в НИИ молекулярной биологии им. В.А. Энгельгардта
РАН).
В основе принципа работы всех молекулярных биочипов с иммобилизованны-
ми зондами лежит способность биологических макромолекул к молекулярному
узнаванию (высокоспецифичному избирательному связыванию с другими моле-
кулами). Так, например, образование двухцепочечных ДНК подчиняется прави-
лу комплементарное™, а молекулы антител способны различать определенные
структуры на поверхности белка — антигенные эпитопы. Биологические микро-
чипы прошли стадию лабораторных испытаний и применяются в диагностических
лабораториях.
Время анализа при использовании биочипов сокращается с двух месяцев до
одного дня, что позволяет оперативно назначать резервные терапевтические
средства тем больным, у которых обнаруживают формы туберкулеза, устойчивые
к обычно применяемым антибиотикам. Впервые в мире метод быстрой идентифи-
кации лекарственно-устойчивых форм туберкулеза, основанный на оригинальной
технологии биочипов, был сертифицирован для медицинского применения в
России. Как показали результаты анализа более 3000 пациентов, надежность мето-
да превышает 90%. Созданная на основе биологических микрочипов тест-система
«ТБ-Биочип» с 2005 г. поставляется в противотуберкулезные центры России и
передана для проведения медицинских исследований и диагностики в Центр по
контролю над заболеваниями (США).
Для выполнения анализа образец крови пациента или другой исследуемой
жидкости подвергают предварительной обработке, в процессе которой нахо-
дящиеся в нем молекулы метят флюоресцирующим красителем. Затем образец
наносят на биочип, помещенный в специальную микрокамеру. Каждый тип
молекул образца взаимодействует со специфичными к ним зондами, локализо-
ванными в индивидуальных ячейках микрочипа, что может быть зарегистриро-
вано по интенсивности свечения соответствующей ячейки биочипа. По картине
свечения множества ячеек биочипа специальный прибор-анализатор определяет
количественно наличие характеристических последовательностей ДНК, РНК
или набора белков в образце.
Миниатюрность, быстрота выполнения, экономичность расходования реаген-
тов и биологических образцов, возможность выполнения сотен и тысяч исследо-
ваний одновременно, характерные для новой технологии, определяют ее преиму-
щества и перспективы дальнейшего развития.
ВИРУСОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ 627
ПРЕАНАЛИТИЧЕСКИЙ ЭТАП
ПРИ ВИРУСОЛОГИЧЕСКИХ ИССЛЕДОВАНИЯХ
Правила сбора биологического материала для вирусологических исследований
и его транспортировка в микробиологические лаборатории регламентирована
действующими нормативными доку ментами.
Сбор материала следует проводить как можно раньше, при появлении первых
симптомов болезни, так как период вирусовыдепения при многих острых инфекци-
ях ограничен несколькими днями (3-5 сут). Сбор материала при аутопсии необхо-
димо осуществлять по возможности в самые ранние сроки после смерти пациента.
Все пробы должны быть получены с соблюдением правил асептики. Взятие
клинического материала выполняют стерильными одноразовыми инструментами.
Работу следует проводить в стерильных одноразовых перчатках. Каждый обра-
зец помещают в отдельный стерильный одноразовый флакон или в контейнер с
завинчивающейся крышкой. Могут быть использованы стерильные конические
центрифужные пробирки с плотно закрывающимися пробками. Объем пробирки
должен быть достаточным для внесения 1-5 мл транспортной среды.
Клинический материал для ПЦР-диагностики необходимо забирать в пробирки
с транспортной средой (если таковая необходима), предоставляемой фирмой-
производителем тест-систем или реагентов. Для сбора пробы зондом-тампоном
используют стерильные тампоны (вискозные или дакроновые) на пластиковой или
алюминиевой основе. Не допускается применение тампонов с алгинатом кальция и
деревянной осью. Жидкость везикул аспирируют туберкулиновыми шприцами.
Кровь собирают в стерильные одноразовые или стеклянные пробирки с анти-
коагулянтом (гепарином, ЭДТА, цитратом натрия). Сразу после взятия необходи-
мо плотно закрыть флаконы, пробирки, контейнеры с биоматериалом, не касаясь
их внутренней поверхности и внутренней поверхности крышек, не допуская зазора
и смятия внутренней части крышки.
Пробы биологического материала следует доставить в лабораторию неза-
медлительно; если такая возможность отсутствует, должно быть обеспечено хра-
нение биоматериала в холодильнике. Температура хранения определяется видом
биоматериала. Транспортировку проб, хранившихся при температуре менее О °C,
осуществляют в термоконтейнерах, исключающих оттаивание биологического
материала; охлаждающие элементы перед транспортировкой клинического мате-
риала следует замораживать до нужной температуры.
При переносе клинического материала из пробирок, флаконов, контейнеров в
новые необходимо использовать отдельные одноразовые стерильные наконечни-
ки с аэрозольными барьерами. Открывая флаконы, пробирки, контейнеры в про-
цессе работы, нельзя выполнять резких движений во избежание разбрызгивания
и расплескивания клинического материала, что может привести к контаминации
проб и рабочих поверхностей.
Для детекции нуклеиновых кислот вирусов следует использовать следующие
образцы биологического материала; кровь, мочу, отделяемое уретры и урогрни-
тального тракта, фекалии, цереброспинальную, слезную жидкость, отделяемое
конъюнктивы и т.д. (табл. 23-5). Эти же разновидности образцов могут быть
использованы для выделения вирусов.
Кровь
Взятие крови следует проводить натощак или через 3 ч после приема пищи из
локтевой вены одноразовой иглой (диаметром 0,8-1,1 мм) в специальную вакуум-
ную систему типа «Vaculetie®» с 6% ЭДТА. После взятия крови пробирку следует
несколько раз плавно перевернуть для перемешивания пробы с антикоагулянтом
и промаркировать.
628 ВИРУСОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ
Таблица 23-5. Образцы клинического материала для выделения и идентификации вирусов
Проба Вирусы-возбудители
Кровь цельная ВИЧ, герпес-вирусы 1,2, 3, 4, 5, 6-го типов, парвовирус В19, энтеро- вирусы
Кровь периферическая (клетки) ВИЧ, герпес-вирусы 1.2 3 4, 5, 6-го типов, парвовирус В19, энтеро- вирусы
Кровь периферическая (плазма) Герпес-вирусы 3, 4, 5-го типов, вирусы гепатитов А, В, С, D, Е, G, ККГЛ, краснухи, парвовирус 819, энтеровирусы
Кровь пуповинная Герпес-вирусы 1, 2, 3, 5, 6-го типов, вирус краснухи, парвовирус В19
Соскоб из цервикального канала Герпес-вирусы 1, 2. 5-го типов, вирус папилломы человека
Соскоб из уретры Герпес-вирусы 1,2, 5 го типов
Соскоб из эрозивно-язвенных элементов Герпес-вирусы 1,2, 3-го типов
Моча Аденовирус, герпес-вирусы 1, 2, 5-го типов, вирус краснухи, полиови- рус, вирус эпидемического паротита
Амниотическая жидкость Герпес-вирусы 1, 2,3,4, 5, 6-го типов, вирус краснухи, парвови- рус В19
Бронхоальвеолярный лаваж Герпес-вирус 6-го типа
Мазок из носоглотки Аденовирус, вирусы гриппа А, В, краснухи, парагриппа 1, 2, 3-го типов, респираторно-синцитиальный вирус
Мазок из ротоглотки Аденовирус, герпес-вирусы 1. 2,4, 6-го типов, вирусы гриппа А, В, краснухи, парвовирус В19, вирусы парагриппа 1-4-го типа, респираторно-синцитиальный вирус, SARS
Мокрота Герпес-вирус 5 го типа, вирусы гриппа А. В, парагриппа 1, 2, 3-го типов, респираторно-синцитиальный вирус
Отделяемое конъюнктивы Аденовиоус
Плацента Герпес вирусы 3, 4, 5, 6-го типов, краснухи, парвовирус В19
Слюна Аденовирус, вирус бешенства, герпес-вирусы 1, 2. 3,4, 5, 6-го типов, вирус краснухи, парвовирус В19, вирус эпидемического паротита, SARS
Спинномозговая жидкость Герпес-вирусы 1,2, 3. 4, 5, 6-го типов, вирусы краснухи, клещевого энцефалита, парвовирус В19, энтеровирусы, вирус эпидемического паротита
Фекалии Аденовирус группы F, вирус гриппа H5N1, астровирус, норавирус, ротавирус, флавивирус, энтеровирусы, SARS
Аутопсийный материал Астровирус, вирусы гриппа А, В, H1N1sw, H5N1, герпес-вирусы 1, 2, 3, 4, 5, 6-го типов, вирус коаснухи, ККГЛ, парвовирус В19, норавирус, ротавирус флавивирус, энтеровирусы, вирус клещевого энцефалита, SARS
Предобработка. Плазму крови следует получать центрифугированием про-
бирок с цельной кровью при 3000 об./мин (800-1600 g) в течение 20 мин при
комнатной температуре. После центрифугирования, используя наконечники с
аэрозольным барьером, необходимо отобрать не менее 1 мл плазмы в стерильные
пробирки типа «Эппендорф» на 2 мл.
Клетки крови (лейкоцитарную фракцию цельной крови) следует отбирать после
центрифугирования цельной крови и удаления плазмы. С помощью наконечников
с аэрозольным барьером собирают лейкоцитарную массу с поверхности осадка
клеток в объеме 0,2 мл и переносят в стерильную пробирку типа «Эппендорф».
Условия хранения. Образцы цельной крови хранят при температуре:
• 20-25 °C — не более 6 ч с момента получения материала для количественно-
го, в течение 12 ч — для качественного определения нуклеиновых кислот;
• 2-8 °C — не более одних суток как для качественного, так и для количествен-
ного определения нуклеиновых кислот.
ВИРУСОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ 629
Замораживание образцов цельной крови недопустимо.
Образцы плазмы крови можно сохранять при температуре 2-8 °C в течение
5 сут, -16...-20 °C — в течение года.
Допускается однократное замораживание-оттаивание материала. Образцы,
предназначенные для длительного хранения, необходимо квотировать в отдель-
ные стерильные пробирки.
Транспортировка. Осуществляют в специальном термоконтейнере с охлаж-
дающими элементами или в термосе со льдом.
Образцы крови транспортируют при температуре 2-8 °C в течение 6 ч с момента
взятия материала для количественного; 12 ч — для качественного определения
нуклеиновых кислот.
Образцы плазмы транспортируют при температуре 2-8 °C в течение 3 сут, в
замороженном виде — в течение 1 сут.
Материал из урогенитального тракта женщин
Материал следует получать из нескольких точек: эпителия цервикального кана-
ла (эндоцервикса), вагинального отделяемого; при необходимости — из уретры и
эрозивно-язвенных поражений.
Перед забором материала необходимо удалить слизь с поверхности шейки
матки стерильным тампоном. Соскоб эпителия проводят отдельными для каж-
дой локализации одноразовыми стерильными цервикальными цитощетками или
универсальными зондами. Рабочую часть цитощетки с собранным материалом
следует обломить в области насечки и оставить в пробирке.
Отделяемое уретры получают с помощью универсального зонда. Рабочую часть
зонда с собранным материалом следует обломить в области насечки и оставить в
пробирке.
Материал из разных локализаций помещают в отдельные пробирки с транс-
портной средой. Для скрининга и диагностики папилломавирусной инфекции
материал собирают с помощью комбинированного цервикального зонда для одно-
временного взятия материала из эндо- и экзоцервикса. Во всех случаях допус гимо
присутствие примесей в виде слизи и крови.
Пробирки необходимо промаркировать.
Предобработка. Образцы, взятые в транспортную среду, тщательно пере-
мешивают на вортексе до полного растворения слизи и образования гомогенной
суспензии клеток.
Образцы соскоба эпителия из эндо- и экзоцервикса, взятые в пробирку с транс-
портной средой, следует тщательно перемешать на воргексе для растворения слизи
и получения однородной суспензии; затем, используя наконечник с аэрозольным
барьером, отобрать 0,5 мл образца, перенести в пробирку типа «Эппендорф»,
центрифугировать при 10-12 тыс. об./мин в течение 5 мин, удалить 0,4 мл надо-
садочной жидкости, осадок перемешать с 0,1 мл оставшейся жидкости.
Условия хранения. Клинический материал из урогенитального тракта хранят
при температуре 18-25 °C в течение 48 ч, 2-8 °C — в течение 14 сут, -70 °C — дли-
тельно. Допускается однократное замораживание-оттаивание материала.
Транспортировка. Осуществляют в специальном контейнере с охлаждающи-
ми элементами или в термосе со льдом при температуре 2-8 °C в течение 5 сут, в
замороженном виде — в течение 1 сут.
Материал из урогенитального тракта мужчин
Перед взятием материала следует обработать головку полового члена в области
наружного отверстия уретры тампоном, смоченным стерильным физиологическим
раствором. Отделяемое уретоы получают с помощью специального зонда, который
вводят на глубину 1-2 см и несколькими вращательными движениями проводят
630 ВИРУСОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ
соскоб эпителиальных клеток. Зонд переносят в пробирку со специальной транс-
портной средой, обламывают рабочую часть в области насечки и оставляют в про-
бирке. Допустимо присутствие примесей в виде слизи, крови и гноя.
Предобработка. Образцы, взятые в транспортную среду, необходимо тщатель-
но перемешать на вортексе до полного растворения слизи и получения гомогенной
суспензии клеток.
Условия хранения. Клинический материал из урогенитального тракта хранят
при температуре 18-25 °C в течение 48 ч, 2-8 °C — в течение 14 сут.
Транспортировка. Ос> ществляют в специальном контейнере с охлаждающи-
ми элементами или в термосе со льдом при температуре 2-8 °C в течение 5 сут, в
замороженном виде — в течение 1 сут.
Моча
Забирают первую порцию утренней мочи в количестве 20-30 мл в специальный
сухой стерильный флакон объемом 50 мл с крышкой, маркируют.
Условия хранения. Нативные пробы мочи хранят при температуре 18 -25 °C
в течение 6 ч, 2-8 °C — в течение 1 сут, предварительно обработанные образцы
мочи — при температуре 2-8 °C в течение 1 сут. -20 °C — в теиение 7 сут, -70 СС —
длительно. Допускается однократное замораживание-оттаивание проб.
Транспортировка. Осуществляют в специальном контейнере с охлаждающи-
ми элементами при температуре 2-8 °C в течение 1 сут, в замороженном виде — в
течение 1 сут.
Фекалии
Пробы отбирают из предварительно продезинфицированной емкости, из
нескольких мест, одноразовой лопаткой, прикрепленной к крышке флакона для
забора материала, маркируют.
Предобработка. Для получения осветленного экстракта фекалий используют
нагивные образцы водянистой консистенции или свежеприготовленную суспен-
зию, или суспензию, подвергнутую замораживанию с глицерином.
Приготовление фекальной суспензии осуществляют следующим образом. В цен-
трифужные пробирки (объемом 1,5 мл) типа «Эппендорф», соответствующие
количеству образцов фекалий, вносят по 0,8 мл стерильного физиологического
раствора, после чего отдельными одноразовыми наконечниками (или одноразо-
выми лопатками) в каждую пробирку добавляют по 0,1 мл образца и тщательно
перемешивают на вортексе до образования гомогенной суспензии. Ее осветляют
центрифугированием при 12 000 об./мин (10 000 g) в течение 5 мин. Супернатант
отбирают в отдельную емкость, смешивают с равным объемом отрицательного
контрольного образца и используют для выделения нуклеиновых кислот.
Суспензию фекалий с глицерином готовят при необходимости длительного хра-
нения материала. Для этого к 10-20% суспензии фекалий добавляют глицерин до
конечной концентрации 10-15%, тщательно гомогенизируют на вортексе и после
экспозиции с глицерином в течение 40 мин пробы замораживают.
Условия хранения Пробы фекалий хранят при температуре 18-25 °C в тече-
ние 6 ч, 2-8 °C — в течение 3 сут.
Фекальную суспензию с глицерином и осветленный фекальный экстракт хранят
при температуре -20 °C в течение 7 сут, при температуре -70 °C — длительно.
Транспортировка Осуществляют в специальном контейнере с охлаждающи-
ми элементами или в термосе со льдом.
Образцы нативных фекалий транспортируют при температуре 2-8 °C в течение
6 ч.
Фекальную суспензию с глицерином и осветленный фекальный экстракт транс-
портируют в замороженном состоянии в течение 1 сут,
ВИРУСОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ 631
Спинномозговая жидкость
Спинномозговую жидкость забирают с помощью одноразовых игл в одноразовые
пластиковые сухие пробирки в количестве не менее 1 мл. Пробирки маркируют.
Предобработка. Спинномозговую жидкость центрифугируют для концентри-
рования содержащих вирусы клеток. Осадок и 100 мкл супернатанта используют
для выделения нуклеиновых кислот
Условия хранения Пробы спинномозговой жидкости хранят при темпера-
туре 18-25 “С в течение 6 ч, 2-8 СС — в течение 1 сут. -20 °C — в течение 30 сут,
-70 °C — длительно. Допускается однократное замораживание-оттаивание проб.
Транспортировка. Осуществляют в специальном контейнере с охлаждающи-
ми элементами при температуре 2-8 °C в течение 6 ч, в замороженном виде — в
течение 1 сут.
Слезная жидкость
Материал объемом не менее 0,5 мл забирают с помощью одноразовых пласти-
ковых пипеток в одноразовые сухие стерильные пластиковые пробирки (напри-
мер, «Эппендорф*). Для усиления слезотечения проводят провокацию слезото-
чивым веществом (например, нашатырным спиртом). Пробирки с материалом
маркируют.
Условия хранения. Пробы слезной жидкости хранят при температуре 18-25 °C
в течение 6 ч, 2-8 °C — в течение 1 сут, -20 °C — в течение 7 сут, -70 °C - длитель-
но. Допускается однократное замораживание-оттаивание проб.
Транспортировка Осуществляют в специальном контейнере с охлаждающи-
ми элементами при температуре 2-8 °C в течение 6 ч, в замороженном виде — в
течение 1 сут.
Отделяемое (инъюнктивы
Материал получают под местной анестезией (2 капли раствора дикаина) с помо-
щью зонда с сухим стерильным ватным тампоном. Оттягивая нижнее веко, прово-
дят несколько раз по конъюнктиве, захватывая внешний и внутренний углы глаза.
Материал помещают в специальную пробирку с транспортной средой; нижнюю,
рабочую часть зонда обламывают, оставляя ее в пробирке. Пробы маркируют.
Условия хранения. Пробы отделяемого конъюнктивы хранят при температуре
18-25 °C в течение 6 ч, 2-8 °C — в течение 3 сут.
Транспортировка. Осуществляют в специальном контейнере с охлаждающи-
ми элементами при температуре 2-8 °C в течение 6 ч.
Замороженный клинический материал транспортируют в специальном контей-
нере с сухим льдом в течение 1 сут.
Мазки со слизистпй оболочки полости носа
Отделяемое со слизистой оболочки полости носа следует забирать сухим сте-
рильным ватным тампоном на пластиковой основе. Тампон вводят по наружной
стенке носа на глубину 2-3 см до нижней носовой раковины. После этого тампон
слегка опускают, вводят в нижний носовой ход, делают вращательное движение
и удаляют вдоль наружной стенки носа. Далее, погрузив рабочую часть зонда
в транспортную среду, помещенную в стерильную одноразовую пробирку типа
«Эппендорф», обламывают стержень, оставляя рабочую часть зонда в пробирке.
Пробы маркируют.
Условия хранения. Мазки со слизистой оболочки полости носа хранят при
температуре 18-25 °C в течение 6 ч, 2-8 °C — в течение 3 сут. При необходимости
более длительного хранения температура не должна превышать -16 °C.
Транспортировка. Осуществляют в специальном контейнере с охлаждающими
элементами при температуре 2-8 °C в течение 1 сут. Замороженный клинический
материал транспортируют в специальном контейнере с сухим льдом в течение 1 сут.
632 ВИРУСОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ
Смывы из полости носа
Взятие материала осуществляют в положении больного сидя, с отклоненной
назад головой. В оба носовых хода с помощью одноразового шприца вводят по
3-5 мл стерильного физиологического раствора. Промывную жидкость из обоих
носовых ходов собирают через стерильную воронку в одну стерильную емкость.
Пробы маркируют.
Условия хранения. Смывы из полости носа хранят при температуре 2-8 °C в
течение 6 ч, -20 °C — в течение 7 сут, -70 °C — длительно.
Транспортировка. Осуществляют в специальном контейнере с охлаждающи-
ми элементами при температуре 2-8 °C в течение 6 ч, в замороженном виде — в
течение 1 сут.
Мазки из ротоглотки
Отделяемое забирают сухим стерильным ватным тампоном на пластиковой
основе, делая вращательные движения по поверхности миндалин, небных дужек,
задней стенки ротоглотки. Далее, погрузив рабочую часть зонда в транспортную
среду, помещенную в стерильную одноразовую пробирку типа «Эппендорф», обла-
мывают стержень, оставляя рабочую часть зонда в пробирке. Пробы маркируют.
Условия хранения Мазки из ротоглотки хранят при температуре 18-25 °C в
течение 6 ч, 2-8 °C — в течение 3 цуг. При необходимости более длительного хра-
нения температура не должна превышать -16 °C.
Смывы из ротоглотки
Предварительно больной прополаскивает полость рта водой. После этого про-
водит тщательное повторное полоскание с помощью 10 мл физиологического
раствора в течение 10-15 с. Жидкость собирают в стерильный флакон. Пробу
маркируют.
Условия хранения. Смывы из ротоглотки хранят при температуре 18-25 °C в
течение 6 ч, 2-8 °C — в течение 3 сут. При необходимости более длительного хра-
нения температура не должна превышать -16 °C.
Транспортировка. Осуществляют в специальном контейнере с охлаждающи-
ми элементами или в термосе со льдом при температуре 2-8 °C в течение 6 ч, в
замороженном виде — в течение 1 сут.
Мокрота
Забирают в количестве не менее 1 мл в одноразовые стерильные флаконы с
широким горлом и завинчивающейся крышкой. Пробы маркируют
Условия хранения. Пробы мокроты хранят при температуре 18-25 °C в тече-
ние 6 ч, 2-8 °C — в течение 3 сут. При необходимости более длительного хранения
температура не должна превышать -16 °C.
Слюна
Предварительно больной троекратно прополаскивает полость рта физиологи-
ческим раствором. Слюну в количестве не менее 1 мл собирают в сухие стерильные
одноразовые пробирки с завинчивающейся крышкой. Пробы маркируют.
Условия хранения. Пробы слюны хранят при температуре 18-25 °C в течение
6 ч, 2-8 °C — в течение 1 сут.
Транспортировка. Осуществляют в специальном контейнере с охлаждающи-
ми элементами при температуре 2-8 °C в течение 6 ч.
Бронхоальвеолярный л ан аж, или промывные воды бронхов
Взятие материала проводят в одноразовые, плотно завинчивающиеся пробирки
объемом 50 мл. Пробы маркируют.
ВИРУСОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ 633
Условия хранения. Предварительно обработанные пробы хранят при темпера-
тур< 2-8 °C в течение 1 сут, -20 °C — в течение 7 сут, -70 °C — длительно.
Транспортировка. Осуществляют в специальном контейнере с охлаждающи-
ми элементами или в термосе со льдом при температуре 2-8 °C в течение 6 ч, в
замороженном виде — в течение 1 сут.
Биопсийный и аутопсийный материал
Материал следует забирать из зоны предполагаемого местонахождения воз-
будителя инфекции, поврежденной ткани или из пограничной с поврежденным
участком зоны.
Кусочки ткани диаметром не более 5 мм помещают в одноразовые стерильные
пробирки типа «Эппендорф», содержащие 0,5 мл специальной транспортной
среды. Пробирки маркируют.
Условия хранения. Предварительно обработанные пробы хранят при темпе-
ратуре 18-25 °C в течение 6 ч, 2 -8 °C — в течение 3 сут. При необходимости более
длительного хранения температура не должна превышать -16 °C,
Транспортировка. Осуществляют в специальном контейнере с охлаждающи-
ми элементами при температуре 2-8 °C в течение 6 ч.
Замороженный клинический материал транспортируют в специальном контей-
нере с сухим льдом в течение 1 сут (только для материала, подозрительного на
наличие ДНК-содержащих вирусов).
ОБЕСПЕЧЕНИЕ КАЧЕСТВА
ПРИ ВИРУСОЛОГИЧЕСКИХ ИССЛЕДОВАНИЯХ
Высокое качество лабораторных исследований может быть обеспечено только
постоянным поддержанием работы системы контроля качества.
Проблемы контроля качества изучаются лабораторной квалиметрией — систе-
мой научных и практических методов оценки качества лабораторных иссле-
дований. Современная лабораторная квалиметрия предполагает применение в
клинико-диагностической лаборатории системы тотального контроля качества —
обеспечения качества. Суть подхода состоит в том, что в обеспечение качества
включен не только процесс измерения того или иного параметра (например,
определение антител к ВИЧ в сыворотке), но и все остальные компоненты и струк-
туры лабораторного процесса, прямо или косвенно влияющие на достоверность
результатов.
Система обеспечения качества — многокомпонентная структура, характеризую-
щаяся следующими параметрами:
организационной структурой учреждения;
J персоналом лаборатории (штатом, квалификацией специалистов, профес-
сиональной подготовкой и переподготовкой и т.п.);
оборудованием (в том числе метрологическим контролем и техническим
обслуживанием);
4 материально-техническим и кадровым обеспечением процесса контроля
качества;
/ мероприятиями по контролю качества;
J информационной сист емой лаборатории и управлением данными;
системой документооборота;
J системой надзора за внештатными ситуациями и мероприятиями по
устранению возможных последствий подобных ситуаций;
J системой обеспечения внедрения мер по контролю качества;
S системой работы с ЛПУ и пациентами (потребителями лабораторной
услуги);
634 ВИРУСОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ
J системой оценки качества работы лаборатории (внешней и внутренней):
S мероприятиями по биобезопасности и организации труда специалистов на
рабочем месте.
Только по результатам комплексной оценки всех компонентов системы обе-
спечения качества можно объективно оценить качество клинико-диагностической
лаборатории. Все остальные подходы к оценке качества являются либо неполны-
ми, либо субъективными.
Описывая систему обеспечения качества клинической лаборатории, выпол-
няющей исследования по диагностике заболеваний, вызванных вирусными инфек-
циями, следует начать с организационной структуры лаборатории. В современном
крупном лабораторном учреждении при составлении организационно-штатной
структуры следует предусмотреть выделение ответственного за систему контроля
качества. В средних и малых клинико-диагностических лабораториях эту функ-
цию может выполнять заместитель заведующего лабораторией. Функциональные
обязанности лица, ответственного за систему контроля качества, должны отражать
все аспекты дея гельности, задачи, полномочия, механизм отчетности и контроля
деятельности, связанной с внедрением «тотального» контроля качества, а также
механизмы интегрирования системы контроля качества клинико-диагностической
лаборатории в систему контроля качества лечебно-профилактического учрежде-
ния и/или местной системы здравоохранения.
Несмотря на значительную автоматизацию клинико-диагностических лабора-
торий, выполняющих исследования маркеров вирусных инфекций, важнейшая
роль в организации контроля качества принадлежит персоналу, непосредственно
воплощающему идеи организации системы контроля качества на своем рабочем
месте. Именно поэтому необходимо постоянно проводить занятия, семинары, пре-
зентации, посвященные повышению качества лабораторных исследований. В этом
должны принимать участие все сотрудники лаборатории, включая врачебный,
средний, младший медперсонал, и работники вспомогательных подразделений
(регистратура, инженерно-технический и снабженческий персонал).
Материально-техническое оснащение КДЛ, выполняющей вирусологические,
серологические и молекулярно-биологические исследования маркеров вирусных
инфекций, подразумевает наличие сложного аппаратно-приборного комплекса
с высокоточными системами измерения. Даже незначительные погрешности в
работе комплекса могут вызвать крайне серьезные лабораторные ошибки. Только
при поддержании лабораторного оборудования в исправном, полностью соот-
ветствующем техническим параметрам анализа состоянии можно обеспечить
высококачественное выполнение лабораторных исследований. Система контроля
качества подразумевает постоянный мониторинг состояния средств измерения
в лаборатории, что должно включать не только быстрое устранение поломок
оборудования, но и их предотвращение. Для этого организуют регулярное про-
филактическое обслуживание аппаратного комплекса внешними техническими
специалистами. Годовой (полугодовой, квартальный и т.п.) график профилактиче-
ского обслуживания оборудования разрабатывают в каждой КДЛ и согласовывают
его с соответствующими техническими службами. Назначают ответственного за
соблюдение графика.
Внедрение системы обеспечения качества в работу КДЛ требует финансовых
и материально-технических затрат. Сюда следует включать не только необходи-
мость учитывать реактивы и расходные материалы, затрачиваемые на ежеднев-
ный контроль качества, но и затраты на профилактику оборудования, обучение
персонала, организацию и совершенствование лабораторной информационной
системы, организацию документооборота, лабораторного процесса на каждом
рабочем месте и т.п. Эти необходимые расходы следует учитывать при планиро-
вании бюджета КДЛ, составлении заявок на реактивы и расходные материалы,
ВИРУСОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ 635
планировании рабочего времени и т.п. В современных зарубежных лабораторных
учреждениях референсного уровня затраты на создание и поддержание системы
тотального контроля качества достигают 12-15% общелабораторного бюджета.
Собственно мероприятия по контролю качества — центральная составляющая
системы. Мероприятия выполняют на двух уровнях — внешнем и внутреннем.
Участие во внешней системе контроля качества обязательно, поскольку значи-
тельная доля выполняемых в КДЛ исследований маркеров вирусных инфекций
связана с социально значимыми инфекциями (СПИДом, гепатитами, ИППП, ВУИ
и т.п.). Внешний уровень оценки необходим для определения степени сопостави-
мости результатов разных лабораторий и соответствия их установленным нормам
точности. Количество программ Федеральной системы внешней оценки качества
(ФСВОК) постоянно возрастает и включает разделы, посвященные не только
идентификации серологических маркеров инфекционных заболеваний, обуслов-
ленных вирусами, но и молекулярно-биологическим методам (например, детекции
ДНК и РНК вирусов гепатитов В, С и ВИЧ). Любая КДЛ должна (при оптимальном
варианте развития системы контроля качества) стремиться участвовать в макси-
мально возможном для учреждения количестве программ ФСВОК.
Внутренний уровень контроля качества (уровень учреждения) - система меро-
приятий, принятая в лаборатории для постоянного слежения за всеми этапами
лабораторной работы и позволяющая оценивать качество измерений, выполняе-
мых в КДЛ. Внутренние мероприятия обязательны для всех исследований, прово-
димых в лаборатории, и необходимы для установления соответствия получаемых
результатов критериям их приемлемости. Внутрилабораторный контроль качества
(ВЛКК) — обязанность персонала КДЛ.
Регулярная внешняя оценка качества и постоянный внутренний контроль каче-
ства дополняют, но не заменяют друг Друга. Задачи внешней оценки качества —
выявление систематических ошибок лабораторных методов и обеспечение един-
ства и сопоставимости результатов измерений в системе здравоохранения в целом.
Задачи ВЛКК — обеспечение стабильности аналитической системы, обнаружение
и устранение случайных и систематических погрешностей.
Для ряда выявляемых в КДЛ маркеров вирусных инфекций доступны охарактери-
зованные контрольные материалы с заранее определенным количественным содер-
жанием исследуемого аналита (HD^Ag, HBsAb, CMV IgG и т.п.). Производством таких
материалов занимаются как референтные научно-исследовательские центры, так и
производители тест-сис гем. Использование таких контрольных материалов позволя-
ет в полной мере осуществлять все мероприятия по ВЛКК при проведении исследова-
ний для диагностики инфек пи энных заболеваний, обусловленных вирусами.
Подобный контрольный материал должен проходить валидацию в конкретных
условиях КДЛ, несмотря на то что определение исследуемого аналита в охаракте-
ризованных контрольных материалах проводилось либо в референс-лаборатории,
либо производителем тест-систем. Таким образом, контрольный материал с уста-
новленным (аттестованным) значением должен быть исследован в нескольких
повторах с применением тех тест-систем и анализаторов, которые используются в
повседневной практике КДЛ.
При валидации контрольного материала в конкретной лаборатории возможно
получение результата, отличающегося от аттестованного. Причин такого отклоне-
ния может быть несколько: дизайн тест-системы отличается от такового референс-
тест-системы (например, при измерении НВ A.g в лунке планшета сорбированы
антитела к иным, чем в референсной тест-системе, эпитопам вирусного белка),
используется иной принцип измерения (например, не хемилюминесценция, при-
меняемая во многих тест-системах референсного уровня, а тИФА) и т.п. Валидация
позволяет учесть систематическую ошибку измерения и внести соответствующий
поправочный коэффициент в результаты.
636 ВИРУСОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ
^охарактеризованные контрольные материалы также проводятся вне КДЛ и
отличаются от охарактеризованных тем, что концентрация аналита в них неиз-
вестна (например, высоко- или слабоположительная сыворотка) или установ-
лена нереференсным методом и не подвергалась валидации по общепринятому
международному стандарту. Неохарактеризованные материалы в обязательном
порядке подлежат процедуре валидации в лаборатории с определением соб-
ственной внутрилабораторной концентрации аналита. При этом установленная
концентрация аналита в таком контрольном материале может отличаться от
истинной, что обусловливается систематической ошибкой, вносимой исполь-
зуемыми тест-системами и анализаторами. Тем не менее даже такой контрольный
материал позволяет проводить ВЛКК и эффективно выявлять случайные ошиб-
ки, возникающие в процессе количественного измерения исследуемого аналита.
Однако применение неохарактеризованных контрольных материалов не дает воз-
можности устранить систематическую погрешность количественных измерений,
выполняемых в лаборатории.
Третий тип контрольных материалов, применяемых при проведении мероприя-
тий по ВЛКК, — так называемые домашние контроли, то есть пулированная сыво-
ротка, собранная в лаборатории, охарактеризованная и аликвотированная для
ежедневного использования. Это наименее предпочтительный тип контрольных
материалов, поскольку при их приготовлении не исключается не только система-
тическая ошибка измерения, но и вероятность случайной ошибки (использование
при пулировании ложноположительных или ложноотрицательных образцов).
Прибегать к домашним контролям следует только в том случае, когда недоступны
контрольные материалы первых двух типов.
Основные требования, предъявляемые к домашним контрольным материалам:
• контрольный материал должен быть слабоположительным, то есть рабочее
разведение Аудированной сыворотки проводят до получения слабоположи-
тельного результата;
• количественное значение аналита должно быть меньше, чем у положитель-
ного контроля, но больше ОПк ит (cut-off)\ обычно применяют значение
2 х ОИ ;
• значение измеряемого аналита должно находиться на линейном участке
калибровочной кривой.
Для приготовления контрольного материала сначала определяют необходимое
количество контрольного материала исходя из прогнозируемых потребностей в
лабораторном анализе на длительный срок (от полугода до года). Затем вычис-
ляют обшее количество партий контрольного материала, которые будут извле-
каться из хранилища (обычно длительное хранение контрольных материалов
для серологических исследований осуществляется при -20 °C, для молекулярно-
биологических — при -70 °C и ниже). Для непосредственного использования в
течение нескольких дней допускается кратковременное хранение контрольных
материалов для детекции антигенов вирусных частиц или антител к ним при тем-
пературе 4 °C в рабочей зоне (но не более 1 нед).
На следующем этапе определяют рабочее разведение высокоположительной
аудированной сыворотки и приготавливают необходимое количество контроль-
ного материала (разводить лучше негативной человеческой сывороткой/плаз-
мой). Для предотвращения бактериального пророста в контрольный материал
следует вносить консервант (например, азид натрия).
Следующий этап — валидация контрольного материала. Для этого прово-
дят тесты на стабильность (измеряют содержание аналита после 1, 2, 3 и 4 нед
хранения в замороженном состоянии) и межсерийную вариацию (определяют
содержание аналита в разных партиях хранения). Удовлетворительными считают
недостоверное снижение содержания аналита при хранении не менее 4 нед и меж-
ВИРУСОЛОГИЧЕСКИЕ исследования 637
серийную вариацию не более 20% (предпочтительно — не более 20%) в разных
партиях хранения. Валидированный домашний контрольный материал аликво-
тируют (размер аликвоты соответствует используемому в исследовании объему
анализируемой пробы), маркируют и хранят. Применяют домашний контрольный
материал, так же как и другие типы контрольных материалов, постоянно, при
каждой постановке анализа.
Существенной проблемой ВЛКК остается отсутствие регламентирующих доку-
ментов для контроля качества неколичественных методов исследования. Проблема
особенно остро стоит для клинических лабораторных исследований заболеваний,
обусловленных вирусными инфекциями. В отличие от иных разделов клинической
лабооаторной диагностики (биохимии, гематологии и пр.), лабораторная диа-
гностика инфекционных заболеваний, в том числе вирусных, основана (особенно
на этапе скрининга) на качественном методе измерения, поскольку достоверного
качественного определения аналита (например, антител к ВИЧ или ДНК папил-
ломавируса 16/18-го типа) достаточно для постановки лабораторного диагноза.
Однако даже получение положительного или отрицательного результата исследо-
ваний позволяет применить математический аппарат лабораторной квалиметрии
для качественных методов исследования. Современные методы лабораторной
диагностики, такие как ИФА и ПЦР в режиме реального времени, используют
для учета результатов аппаратуру, выдающую их в количественном выражении.
При стабильной концентрации аналита и корректном выполнении качественного
варианта ИФА эти параметры будут одинаковыми (или близкими), что позво-
ляет использовать их для построения контрольных карт и оценки по правилам
Вестгарта.
Однако следует учитывать, что для статистического анализа следует брать не
абсолютные значения полученных параметров, а относительные, например при
построении контрольной карты результатов качественного определения HBsAg в
качестве численного значения берут коэффициент серопозитивности [КС — соот-
ношение оптической плотности (ОП) контрольного материала и ОП ]. В даль-
нейшем полученные данные анализируют так же, как и данные количественных
методов исследования, с применением аналогичных критериев интерпретации.
Что позволяет оценить применение подобного подхода к ВЛКК? Так, если при
выполнении качественного исследования по данным контрольной карты получено
значение КС ниже среднего более чем на 2S, можно предполагать, что в результате
случайной ошибки произойдет недовыявление слабоположительных образцов.
Соответственно следует внимательно проанализировать образцы со значениями
близкими, но ниже ОПкрит, можно проанализировать их повторно. Если же при
качественном исследовании получено значение КС выше среднего более чем на 2S
по данным контрольной карты, следует предполагать, что в результате случайной
ошибки произойдет выявление ложноположительных образцов.
Таким образом, применение системы статистического анализа результатов
ВЛКК для количественных методов исследования позволяет повысить достовер-
ность результатов и для качественных методов.
Следующий компонент системы тотального контроля качества — система
управления данными в лаборатории. Правильно построенная работа с данными
позволяет существенно снизить количество ошибок на пре- и посганалитическом
этапах лабораторного процесса. Ключевым звеном этого компонента системы
обеспечения качества современного ЛПУ служит лабораторная информационная
система, ориентированная на работу с данными пациентов. Наиболее сложный
участок работы КДЛ с точки зрения управления данными — регистрация проб и
ввод данных пациентов. Правильно разработанная форма заявки на лабораторные
исследования, соблюдение требований к ее заполнению, переход к электронной
документации, штрих-кодирование образцов позволяют избежать множества кан-
638 ВИРУСОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ
целярских ошибок при выполнении лабораторных исследований на преаналити-
ческом этапе. Доставка результатов исследования конечному потребителю (ЛПУ
или пациенту) в электронном виде снижает количество канцелярских ошибок
лабораторного процесса на постаналитическом этапе, сокращает полное время на
выполнение исследования и повышает качество лабораторных исследований.
Правильно сконфигурированная лабораторная информационная система необ-
ходима как для рутинной лабораторной работы, так и для развития системы
контроля качества. Рационально организованная база данных пациентов позво-
ляет проводить адекватный статистический анализ выполненных исследований
и планировать мероприятия по ВЛКК на перспективу, в том числе с точки зрения
материально-технического обеспечения процесса. Разумеется, компьютеризация
лабораторного процесса предъявляет и повышенные требования к квалификации
персонала лаборатории. Таким образом, система обеспечения качества — много-
компонентный процесс сетевого типа со сложными комплексными связями между
отдельными частями системы.
Для своевременного информирования сотрудников о мероприятиях по обеспече-
нию качества, об изменениях в работе лаборатории, для предотвращения дублирова-
ния документов, их утраты или неверной адрес ации в КДЛ должна быть разработана
внутренняя система документооборота. Она должна регламентировать порядок изве-
щения сотрудников и способ их ознакомления с тем или иным документом опреде-
лять порядок и сроки хранения лабораторной документации и лид. за это i лъетствен-
ных. Это также один из компонентов системы тотального контроля качества.
Следующий компонент системы тотального контроля качества — система над-
зора за внештатными ситуациями и мероприятиями по устранению возможных
последствий подобных ситуаций. К сожалению, невозможно избежать внештат-
ных ситуаций в работе КДЛ (например, аварийное отключение электропитания,
поломка анализатора, сбой в работе сервера или рабочей станции и т.п.), но можно
подготовиться к ним и минимизировать их последствия. Для этого разрабатывают
систему надзора за возможными внештатными си гуациями. В целях обеспечения
качественной работы КДЛ в подобной ситуации необходим набор внутренних
регламентирующих документов, указывающих порядок действий во внештатных
ситуациях, лиц, ответственных за их устранение, предпочтительные варианты обе-
спечения бесперебойной работы лаборатории и т.п.
Следует учитывать, что даже правильно организованная система ВЛКК, обучен-
ный персонал, разработанная система документации по системе обеспечения каче-
ства будут чисто формальными признаками, если нет функционирующей системы
обеспечения внедрения мер по контролю качества. Необходимо четко определить
и неукоснительно вводить в повседневную работу КДЛ все компоненты системы
тотального контроля качества лабораторных исследований. Система обеспечения
внедрения мер по обеспечению качества регламентирует порядок внедрения меро-
приятий по контролю качества, определяет ответственных за это лиц и устанавли-
вает сроки исполнения мероприятий по внедрению мер.
Любая диагностическая лаборатория — учреждение, оказывающее диагности-
ческие услуга конечному потребителю. Как правило, качество работы КДЛ на
пре- и постаналитическом этапах определяется еще одним компонентом системы
обеспечения качества — системой работы с потребителями лабораторной услуги
(ЛПУ и пациентами). Все усилия персонала лаборатории на аналитическом этапе
легко могут быть сведены к нулю, если ответ не доходит до конечного потребителя
или доходит, но не к тому потребителю. Особенно это важно с точки зрения норм
законности и медицинской этики, поскольку часто результаты диагностических
исследований инфекционных заболеваний, обусловленных вирусами, имеют край-
не важное социальное и психологическое значение (например, результат исследо-
вания на ВИЧ, гепатиты В и С и т.п.).
ВИРУСОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ 639
При этом недостаточно только четко и достоверно выполнять лабораторные
исследования, необходимо делать их понятными врачу-клиницисту. Например,
при количественных исследованиях на определение IgG-антител к вирусным
инфекциям с нестерильным иммунитетом (герпесу I—II типа, цитомегаловирусу
и т.п.) полезно указывать граничные значения искомой величины, их интерпрета-
цию или лабораторный диагноз. Врач клиники может не знать граничные значе-
ния, и тогда результат исследования потеряет для него ценность. Кроме того. КДЛ
должна активно взаимодействовать с Л ПУ: не только получать образцы и отсылать
ответы, но и проводить постоянную работу по повышению уровня осведомленно-
сти сотрудников ЛПУ об особенностях клинической лабораторной диагностики
вирусных инфекционных заболеваний, появлении новых видов исследований,
особенностях интерпретации результатов исследований и т.п. В лаборатории
должны быть разработаны и утверждены нормативы сроков выполнения всех
видов исследований, порядок выдачи ответов, организации консультирования
сотрудников ЛПУ и пациентов по результатам исследования биологического
материала, присланного в лабораторию, и т.п. Этот компонент системы контроля
качества тесно взаимосвязан с сис темой управления данными и организационной
структурой учреждения.
Мероприятием по оценке качества может быть периодическое тестирование
зашифрованных панелей образцов сотрудниками лаборатории. Внешнюю оценку
контроля качества могул проводить представители профессионального сообще-
ства, имеющие высокую квалификацию, или сотрудники референс-лаборатории,
занимающиеся диагностикой инфекционных: вирусных заболеваний. Подобные
проверки помогают выявить недочеты в работе системы качестьа КДЛ и своевре-
менно их устранить.
Мероприятия по биобезопасности и организации труда специалистов на рабочем
месте — еще один компонент системы обеспечения качества лабораторной работы.
Большинство исследований, проводимых при рутинной диагностике инфекцион-
ных вирусных заболеваний, связаны с анализом потенциально инфекционного
материала, относящегося ко II—IV группе патогенности. Разумеется, коллектив
КДЛ обязан неукоснительно соблюдать все меры биобезопасности, стремясь при
этом к максимальной достоверности и точности исследований. Эффективная рабо-
та системы обеспечения качества лабораторных исследований должна учитывать
требования санитарных норм и правил.
Таким образом, современный подход к организации системы обеспечения каче-
ства клинических лабораторных исследований для диагностики инфекционных
заболеваний должен быть тотальным, то есть повсеместным. В него включают
все стороны деятельности лабораторного учреждения. Контроль качества должен
быть непрерывным (ежедневным) и касаться всех видов исследований, проводи-
мых в лаборатории. В работу по обеспечению качества должен быть вовлечен весь
персонал лаборатории.
СПИСОК РЕКОМЕНДУЕМОЙ ЛИТЕРАТУРЫ
Вирусология. Методы. — М.: Мир, 1988. — 344 с.
Вопросы общей вирусологии / Под ред. И.Н. Жилинской. — СПб.; Псков: Дом Печати,
2007. - 376 с.
Методы культивирования клеток / Под ред. Г.П. Пинаева и Н.С. Богдановой. — СПб.:
Изд-во политехнического ун-та 2008. — 278 с.
Мирзабеков А.Д., Прокопенко Д.В., Чечеткин В.Р. Применение матричных биочи-
пов с иммобилизованной ДНК в биологии и медицине // Информационные медико-
биологические технологии / Под ред. В.А. Княжева и К.В. Судакова. — М.: ГЭОТАР-МЕД,
2002. - С. 166-198.
Организация работы лабораторий, использующих методы амплификации нуклеиновых
кислот при работе с материалом, содержащим микроорганизмы I—1V группы патогенности:
Методические указания МУ 1.3.2569-09. — М.: Госсанэпиднадзор. 2009.
640 ВИРУСОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ
Поздеев О.К. Медицинская микробиология / Под ред. В.И. Покровского. — М.: ГЭОТАР-
МЕД, 2002. - 765 с.
Приказ М3 РФ № 45 от 07.02.2000 г. «О системе мер по повышению качества клинических
лабораторных исследований в учреждениях здравоохранения Российской Федерации?-.
Приказ М3 РФ от 21.10.2002 г. № 322 «О применении в практике здравоохранения
иммуиоферментных тест-систем для выявления поверхностного антигена вируса гепатита В
(HBsAg) и антител к вирусу гепатита С (анти-ВГС) в сыворотке крови человека».
Санитарно-эпидемиологические правила «Безопасность работы с микроорганизмами
III—IV группы патогенности (опасности) и возбудителями паразитарных болезней» СП
1.3.2322-08. — Федеральная служба по надзору в сфере защиты прав потребителей и благо-
получия человека, 2008.
Санитарно-эпидемиологические правила «Безопасность работы с микроорганизмами
I—II группы патогенности (опасности)» СП 1.3.1285-03. — Минздрав России, 2003.
Санитарно-эпидемиологические правила «Порядок выдачи санитарно-эпидемиологи-
ческого заключения о возможности проведения работ с возбудителями инфекционных
заболеваний человека I—IV группы патогенности (опасности), генно-инженерно-модифи-
цированными микроорганизмами, ядами биологического происхождения и гельминтами»
СП 1.2.1318-03. — Минздрав России, 2003.
Чандлер Д., Роберсон Р. Оптическая и электронная микроскопия в медицине и в биоло-
гии: Пер. с англ. — М.: Интеллект, 2009. — 705 с.
Чечеткин В.Р., Прокопенко Д.В., Макаров А.А., Заседателев А.С. Биочипы для медицин-
ской диагностики // www.NANORF.RU Российские нанотехнологии. Исследования и раз-
работки. - 2006. - Т. 1. - № 1-2. - С. 13-28.
Edwards К„ Logan J., Saunders N. Real-time PCR: An Essential Guide. — Wymondham:
Horizon Bioscience, 2004. — 346 p.
Глава 24
Частная вирусология
АДЕНОВИРУСЫ
Таксономия
Семейство Adenoviridae (аденовирусов) включает четыре рода:
Atadenovirus, Aviadenovirus. Mastadenovirus, Siadenovirus.
Внутри родов представители семейства распределяются по груп-
пам в зависимости от природного хозяина. Круг хозяев включает
пресмыкающихся, земноводных, птиц, млекопитающих, человека.
В одну группу могут быть объединены несколько подгрупп и серо-
типов вируса.
Аденовирусы, патогенные для человека, относятся к роду
Mastadenovirus и обозначаются аббревиатурой HAdV (Human
AdenoVirus). Распределение аденовирусов человека по тканевому
тропизму в зависимости от подгруппы и серотипа вируса представ-
лено в табл. 24-1.
Таблица 24-1. Аденовирусы человека
Подгруппа Серотипы Тропизм
А 12, 18, 31-Й Кишечный и респираторный тракты
В 3. 7, 11, 14, 16,21,34, 35, 50-й Респираторный тракт
С 1. 2, 5, 6-Й Респираторный и кишечный тракты
D 8-10,13,15,17,19, 20, 22-30. 32. 33, 36-39, 42-49, 51-Й Ткани глаза
Е 4-й Респираторный тракт
F 40, 41-й Кишечный тракт
Роль аденовирусов в инфекционной патологии человека различ-
на. Вирусы подгрупп А (31-й серотип), В (7, 11, 21, 34, 35-й сероти-
пы), С (1, 2, 5-й серотипы) могут быть причиной тяжелых пневмо-
ний, бронхитов и бронхиолитов у больных с иммунодефицитами
разной этиологии. Вирусы подгрупп В (3, 7,14, 21, 34, 35-й сероти-
пы) и С (1, 2-й серотипы) могут вызывать острую респираторную
инфекцию. В развитии эпидемического кератоконъюнктивита веду-
щая роль принадлежит аденовирусу 8-го серотипа (подгруппа D).
Представители подгрупп А, С и F — возбудители острых кишечных
вирусных инфекций. Распространение аденовирусной инфекции
может осуществляться воздушно-капельным, алиментарным и кон-
тактным путями.
642 частная вирусология
Характеристика вируса
Аденовирусы являются причиной около 8% ’ гех клинически выраженных
вирусных инфекций человека. Строение вириона представлено на рис. 24-1.
Вирион аденовируса не имеет липидной оболочки. Масса его 150-180 МДа.
Зрелый вирион состоит из капсида, фибрилл, сердцевины (core) и связанных с
ними белков. Капсид аденовирусов имеет диаметр 70-90 нм и форму икосаэдра, от
12 вершин которого отходят нити — фибриллы длиной 9-78 нм. Капсид построен
из 252 капсомеров, 240 из них — гексоны, образующие 20 риангулярных граней;
еще 12 — пептоны. Гексон соседствует с шестью такими же субъединицами и состо-
ит из трех одинаковых белковых молекул. Каждый пентон состоит из пяти белко-
вых молекул и окружен перипентонными гексонами. Фибриллы состоят из трех
идентичных гликозилированных полипептидов, выполняющих функцию рецеп-
торов, с помощью которых осуществляется адсорбция вируса на чувствительных
клет ках. Тип симметрии капсида — кубический.
Геном аденовируса представлен двухнитевой линейной ДНК. В процессе репро-
дукции вируса синтезируются ранние (неструктурные) белки, необходимые для
репликации вирусной ДНК, и поздние — структурные белки вириона. Белок II
является с груктурной единицей гексона; белок III — структурная единица пентона;
белок Ilia участвует в ассоциации перипентонных гексонов с пептоном; IV — гли-
козилированный полипептид, структурная субъединица фибриллы; IX — ассо-
циирован с центральной частью триангулярной грани икосаэдра; белки VI и VIII
расположены с внутренней стороны капсида и являются структурами, повышаю-
щими его устойчивость; белки сердцевины — V, VII, X — способствуют правильной
укладке вирионной ДНК внутри капсида. Молекулярная масса структурных бел-
ков различна: от 10 (белок X) до 120 (белок II) кДа.
Вирион обладает антигенной активностью, ассоциированной с белками гексона,
пентона и фибриллы. Антиген А (гексон) — род о- и группоспецифический анти-
Рис. 24-1. Строение аденовируса. Нехоп {antigen 4) — гексон (антиген A). Penton base — основа-
ние пентона. Penton {antigen В) — пентон (антиген В). Fibre {antigen С) — фибрилла (антиген С).
ЧАСТНАЯ ВИРУСОЛОГИЯ 643
ген, общий для всех представителей семейства Adenovlfidae. Антиген В (пентон)
обладает эндонуклеазной активностью и обусловливает токсические действие аде-
новирусов. Антиген С (фибриллы) — типоспецифический антиген, обладающий
гемагглютинирующей активностью. Вирусней грализующие антитела вырабатыва-
ются против антигенных детерминант антигенов А и С.
Аденовирусы относительно устойчивы к факторам внешней среды: время инак-
тивации при температуре 56 °C составляет 30 мин. При температуре 36 °C вирус-
ные частицы сохраняют активность в течение 7 сут; при 23 °C — в течение 14 сут.
Аденовирусы хорошо переносят низкие температуры и лиофилизацию, устойчивы
к изменению pH и органическим растворителям (эфиру, хлороформу и др.).
Лабораторные методы диагностики
Лабораторная диагностика аденовирусной инфекции основана:
• на детекции вирусных частиц, антигенов или вирусспецифической ДНК;
• определении вирусспецифических антител или регистрации их прироста.
Материалами для исследований являются мазок из носоглотки, мазок из рото-
глотки, отделяемое конъюнктивы, слюна, амниотическая жидкость, ау гопсийный
материал. При подозрении на кишечную инфекцию аденовирусной этиологии
исследуют фекалии больного. Забор клинического материала следует проводить
как можно раньше, желательно при появлении первых клинических симптомов
заболевания, не позднее 3-4-го дня болезни. Аутопсийный материал следует полу-
чать как можно раньше после смерти пациента. Для серологических исследований
используют следующие материалы.
• Одну сыворотку крови больного, полученную на 3-4-й день заболевания
(для определения IgM-фракции антител). Наличие IgM-антител, независимо
от их уровня, является показателем острой инфекции.
• Парные сыворотки крови больного, полученные в ранний период и на стадии
реконвалесценции. Четырехкратный прирост Ат свидетельствует о положи-
тельном диагнозе.
Аденовирус, его антигены и ДНК могут быть обнаружены в клинических образ-
цах следующими методами.
• Просвечивающей ЭМ вирусных частиц.
• Выделением вируса на культуре клеток с последующей идентификацией в
иммунологических реакциях.
• Методом иммунофлюоресценции (МИФ).
• Иммуноферментным анализом для выявления вирусных антигенов (ИФА).
• Иммунохроматографией.
• ПЦР.
Антитела к аденовирусу определяются в следующих серологических реакциях.
• Реакции связывания комплемента (РСК).
• Реакции нейтрализации.
• ИФА для выявления вирусспецифических антител.
Алгоритм лабораторного исследования определяется тяжестью клинических
проявлений болезни. При развитии острой диареи у детей младшего возраста или
тя желой бронхопневмонии у больных с иммунодефицитами, когда чрезвычайно
важно своевременное установление диагноза, предпочтение отдается методам
ранней диагностики. Как правило, это МИФ, ИФА для выявления антигенов виру-
са, ИФА для выявления IgM-антител, ПЦР. В сомнительных случаях используют
подтверждающие тесты.
Серологическую диагностику аденовирусной инфекции проводят, как правило,
в РСК с использованием парных сывороток больного, полученных с интервалом
2-3 нед. В качестве, по сути, метода ретроспективной диагностики реакцию свя-
644 ЧАСТНАЯ ВИРУССЛО! ИЯ
зывания комплемента используют для диагностики клинически легких случаев
заболевания, проведения дифференциальной диагностики как подтверждающий
тест.
Лабораторная диагностика аденовирусной инфекции может быть основана на
выделении вируса в культуре клеток HeLa или Нер-2 с последующим типирова-
нием в РСК или PH. Нужно иметь в виду, что выделение и последующая иденти-
фикация вируса - процедура трудоемкая и длительная. Для накопления вируса
в количестве, достаточном для иммунологических реакций, зачастую требуется
несколько (2-3) последовательных (слепых) пассажей исходного клинического
материала в культуре клеток. Как прави ио, выделение аденовируса в культуре кле-
ток используют как подтверждающий тест.
Дифференциальная диагностика
Дифференциальную диагностику следует проводить с другими острыми заболе-
ваниями, обусловленными вирусами. Это респираторно-синцитиальная инфекция,
парагрипп 1,2, 3-го типов, грипп А и В, а также острые гастроэнтериты, возбудите-
лями которых являются астровирусы, калицивирусы, коронавирусы, ротавирусы,
торавирусы, энтеровирусы.
Как и при других инфекциях, диагноз устанавливают по совокупности клиниче-
ских. эпидемиологических данных и лабораторных тестов.
АСТРОВИРУСЫ
Астровирусы — безоболочечные, икосаэдрической формы вирусы с одноните-
вой (информационной) РНК. Семейство Astroviridae подразделяется на два рода
в соответствии с решением Международного комитета по таксономии вирусов;
Mamastrovirus, который поражает млекопитающих, и Avastrovirus, поражающий
птип.
Размеры вириона составляют от 27 до 38 нм в диаметре и обладают уникаль-
ными пяти- или шестиконечными звездами, различимыми под электронным
микроскопом на своей поверхности, которые и дали название семейству (от греч.
astron — «звезда»), Они содержат одну положительную нить РНК размером 7,2 kb,
которая содержит три открытые рамки считывания (ОРС); ОРС 1а и 1b, кодирую-
щую неструктурные белки, и ОРС 2, кодирующую структурный белок. Описано
несколько вирусных белков, включая белок с молекулярной массой 87 кДа, кото-
рый взаимодействует с моноклональными антителами, специфичными для вирус-
ного капсида. Известны еще три структурных белка (около 30 кДа). Анализ вирус-
ного генома позволил выделить астровирусы в новое семейство РНК-содержащих
вирусов — Astroviridae.
На основе серологических данных было выделено восемь серотипов астровиру-
сов человека (HAastV-l-HAastV-8), из которых доминирующим оказался первый.
Все восемь антигенных разновидностей были адаптированы к росту на клеточных
культурах, в которых используется трипсин как дополнение к питательной среде.
Была клонирована ДНК (cDNA) человеческого астровируса первого серотипа
(оксфордский шгамм). Когда закрытый (capped) посредник, синтезированный
на основе этой ДНК, использовался для заражения культуры клеток СаСо-2, раз-
вивалась инфекция. Небольшой размер астровирусного генома наряду с хорошим
ростом на тканевых культурах в экспериментах и пригодностью ДНК-клона для
исследований сделали человеческий астровирус первого серотипа подходящей
системой для изучения сборки вируса.
ЧАСТНАЯ ВИРУСОЛОГИЯ 645
Устойчивость во внешней среде
Астровирусы относительно устойчивы к критическим изменениям pH и тем-
пературы, к воздействию ультрафиолетового излучения и уровню хлора, обычно
определяемому в плавательных бассейнах. После двухчасовой экспозиции астро-
вирусов в воде с концентрацией свободного хлора 1 мг/л наблюдается остаточная
инфекционность. Астровирусы характеризуются длительной жизнеспособно-
стью в объектах окружающей среды (пористых и непористых поверхностях) в
температурном диапазоне от 4 до 20 °C и высокой влажности. По показателям
жизнеспособности в окружающей среде астровирусы превосходят полиовирусы и
аденовирусы, немного уступают ротавирусам и вирусу гепатита А. Таким образом,
контактно-бытовой путь распространения астровирусов может играть существен-
ную роль во вторичной передаче возбудителя.
Методы диагностики заболевания
Клинический материал, используемый для диагностики, — фекации больного.
Для вирусологического исследования применяют ИФА или ПЦР, для чего
пробы фекалий собирают в стерильную стеклянную посуду. Если образцы фекалий
не берут сразу в работу, пробы фекалий для исследования на астровирусы необхо-
димо хранить в морозильной камере.
Методы исследования:
• ОТ-ПЦР — позволяет определять РНК всех 8 серотипов вируса.
• РНК-гибридизация.
• ИФА - для выявления антигенов вируса.
Дифференциальная диагностика
Ротавирус (группа А), кишечные аденовирусы, калицивирусы, энтеровирусы.
Алгоритм исследования: ИФА РНК-гибридизация ОТ-ПЦР электронная микро-
скопия.
ВИРУС БЕШЕНСТВА
Таксономия
Вирус бешенства принадлежит к отряду Monanzgavirales, семейству Rhabdoviridae.
Семейство Rhabdoviridae представлено наиболее распространенными в природе
вирусами. Оно охватывает шесть родов и многочисленные неклассифицирован-
ные вирусы, которые поражают позвоночных, беспозвоночных и растения. Среди
рабдовирусов наибольшее значение для патологии человека и животных имеют
лиссавирусы. Глобальный характер распространения лиссавирусов, высокие пато-
генность и летальность, отсутствие средств лечения развившихся заболеваний
выдвигают лиссавирусы на одно из первых мест по значимости для здоровья
человека и животных. На основании решения МКТВ от 2009 г. род Lyssavirus пред-
ставлен 11 видами (табл. 24-2).
Среди лиссавирусов существует перекрестная антигенная связь на уровне
нуклеопротеина. Уровень гомологии последовательности аминокислот достигает
78-93%. Это позволяет создавать общий для всех лиссавирусов диагностический
препарат (метод флюоресцирующих антител).
64Б ЧАСТНАЯ ВИРУСОЛОГИЯ
Таблица 24-2. Род Lissavirus
Виды Генотип Вирусы Источники выделения Ареал распространения
1 Вирус бешенства: дикие и вакцинные ВБ Млекопитающие Европа, Азия, Америка (Северная, Южная), Африка
2 Лагос-Бат Плотоядные, летучие мыши, собаки, кошки Центральная и Южная Африка
3 Мокола Землеройки, человек, собаки, кошки Центральная и Южная Африка
4 Дувенхаге Насекомоядные, летучие мыши, человек Южная Африка, Зимбабве
5 Лиссавирус европейских летучих мышей 1-го типа Насекомоядные, летучие мыши, человек Европа, Россия
6 Лиссавирус европейских летучих мышей 2-го типа Насекомоядные, летучие мыши, человек Европа
7 Лиссавирус австралийских летучих мышей Плотоядные и насе- комоядные, летучие мыши, человек Австралия. Филиппины
8 Аоаван Насекомоядные, летучие мыши Кыргызстан
9 Западнокавказский вирус летучих мышей То же Кавказский регион
10 Иркут То же Восточная Сибирь
11 Худжанд То же Таджикистан
Характеристика вируса
Вирионы имеют пулевидную форму, их длина составляет около 180 нм, а диа-
метр — 75 нм. Вирион состоит из несегментированного генома, представленного
одной молекулой спиралеобразно скрученной негативной РНК и пяти структур-
ных белков: нуклеопротеина (N), фосфопротеина (Р), матричного белка (М),
гликопротеина (G) и FHK-зависимой РНК полимеразы или большого белка (L —
Large protein). Геномная РНК неинфекционна.
Инфекционностью обладает нуклеокапсид, или рибонуклеопротеид, состоя-
щий из геномной РНК, тесно скрученной с тремя внутренними белками (L, N, Р).
Нуклеокапсид «одет» мембранными белками (М. G) и бислойной липидной обо-
лочкой, через которую проступают гликопротеиновые шипы длиной 8.3 нм.
Низкие температуры консервируют вирус. Температура 23 °C инактивирует
его через 28-53 дня, 50 °C — через 1 ч, 60 СС — за 5-10 мин, 70 °C — мгно-
венно. Высушивание без вакуума инактивирует вирус в течение 10-14 дней.
В гниющем материале он гибнет через 15 дней. УФ-лучи убивают за 5-10 мин.
Дезинфицирующие вещества (растворы формальдегида, гидроксида натрия, хлор-
ной извести) действуют на ВБ губительно. Вирус быстро инактивируется липидо-
растворителями (2% мыльным раствором) и 0,1% раствором трипсина.
Все лиссавирусы. кроме Лагос-Бат, вызывают у людей летальные энцефалиты,
клинически неотличимые (за исключением вируса Мокола).
Бешенство — острое нейровирусное заболевание, передаваемое человеку через
укус собаки, лисы, волка и других плотоядных животных, зараженных в природе, в
свою очередь, также через укус. Вирус бешенства — единственный из царства виру-
сов, который поражает всех теплокровных млекопитающих, в том числе человека,
в глобальном масштабе с наивысшей летальностью (100%).
ЧАСТНАЯ ВИРУСОЛОГИЯ 647
Лабораторные методы диагностики
Быстрая и своевременная диагностика, основанная на надежных и своевремен-
ных экспресс-методах, является критической:
• для немедленного оказания укушенному антирабической помощи и преду-
преждения развития у человека абсолютно смертельного бешенства;
• оценки эпизоотической ситуации по бешенству среди домашних и диких
животных, а также эффективности мер борьбы с бешенством животных.
Диагностика бешенства у человека бывает прижизненной и постмортальной.
Прижизненная диагностика базируется:
• на определении антигена N-белка в криостатных срезах кожи (волосяного
мешка) методом иммунофлюоресценции;
• обнаружении РНК в криостатных срезах кожи (волосяного мешка), моче,
образцах слюны, слезной и цереброспинальной жидкостях методом
ОТ-ПЦР;
• выделении вируса in vivo и in vitro, а также определении антител в крови, цере-
броспинальной жидкости невакцинированных пациентов.
Эффективность прижизненной диагностики низка и варьирует от нуля до 45%,
поэтому перечисленные выше методы повторяют до установления диагноза.
ПРЯМОЙ МЕТОД ФЛЮОРЕСЦИРУЮЩИХ АНТИТЕЛ
Прямой метод флюоресцирующих антител (ПМФА) — детекция антигена
(нуклеопротеина) вируса бешенства с помощью флюоресцирующих моноклональ-
ных антител, рассматривается как «золотой стандарт» диагностики бешенства.
Феномен иммунофлюоресценции дает возможность выявлять специфический
вирусный ангиген-нуклеопротеин в инфицированных клетках при обработке их
мечеными антителами. Учитывая высокую степень гомологии нуклеотидов нукле-
опротеина представителей рода лиссавируса, флюоресцирующий моноклональный
иммуноглобулин к нуклеопротеину вируса бешенства является высокоспецифиче-
ским для всех 11 видов лиссавирусов. Методом флюоресцирующих антител (МФА)
также выявляют внутриклеточные цитоплазматические включения нуклеопро-
теина в отпечатках или мазках головного мозга, микротомных срезах головного и
спинного мозга, а также в культуре клеток.
Из каждого предназначенного для исследования образца готовят два стекла,
а из образцов, взятых от животных, нанесших укусы человеку, готовят двойные
стекла; их просматривают два опытных микроскописта-исследователя. В поле
зрения микроскопа наблюдаются включения, светящиеся интенсивным ярко-
зеленым цветом на фоне темного «неба». В каждом отпечатке-мазке просматри-
вают 25-50 полей. Форма зеленых включений варьирует от мелких точечных
до крупных овальных, круглых или удлиненных образований (от 2 до 10 мк).
Антигенспецифичные зеленые включения легко отличаются от желтой, белой и
золотой аутофлюоресценции. Время, необходимое для постановки диагноза МФА,
варьирует от 60 до 80 мин.
МОЛЕКУЛЯРНАЯ ДИАГНОСТИКА И ИССЛЕДОВАНИЯ
ПЦР и его модификации, в том числе ПЦР в реальном времени, высокочувстви-
тельные методы, однако ПЦР не рекомендуется экспертами ВОЗ для рутинной
диагностики бешенства из-за возможности получе ния ложноположительных или
ложноотрицательных результатов.
Секвенирование
Секвенирование генов N- и G-белков, а также полного генома используют для
идентификации вирусов группы бешенства и вариантов вирусов, циркулирующих
в определенной географической зоне, классификации вирусов по родам, видам
648
ЧАСТНАЯ ВИРУСОЛОГИЯ
(генотипам) и филогенетическим группам. Секвенирование не имеет отношения к
рутинной диагностике.
Выделение вируса
Выделение вируса с последующей его идентификацией в МФА используют как
для прижизненной, так и для постмортальной диагностики бешенства.
Выделение вируса in vivo проводят путем его культивирования в мозгу молодых
мышей или в культуре клеток. Для заражения используют мозг умершего челове-
ка , павших или гуманным способом забитых бешенных или подозрительных на
бешенство животных. Из кусочков мозга (аммонова рога, ствола мозга, мозжечка
и больших полушарий) готовят 10% взвесь, которую используют для заражения в
мозг 5-10 мышей или культуру клеток. За мышами наблюдают 30 дней на предмет
заболевания или гибели. Мозг заболевших или павших мышей немедленно иссле-
дуют методом МФА для подтверждения диагноза бешенства. При наличии вируса
в инфекционном материале заболевание мышей наступает на 7-20-й день. Для
сокращения срока наблюдения используют семью новорожденных мышей и после
5-го дня после заражения ежедневно забивают по одной-две мыши и исследуют
методом МФА.
Выделение вируса in vitro связано с труднопрогнозируемой продолжительно-
стью инкубационного периода зараженных животных. Этот недостаток устраня-
ется при инокуляции вируса бешенства в культуру чувствительных перевиваемых
клеток нейробластомы мыши или ВНК-21 (96- или 24-луночных планшетах) с
последующей детекцией антигена. Спустя 18 ч-7 дней в цитоплазме зараженных
клеток формируются включения нуклеопротеина, которые выявляются методом
МФА. Таким образом, метод выделения вируса бешенства в культуре клеток имеет
значительное (во времени) преимущество по сравнению с методом in vivo.
Иммуноферментный анализ
Иммуноферментный анализ (ИФА) может быть использован для диагностики
бешенства, однако риск получения неспецифических ложноположительных и
ложноотрицательных результатов делает его применение ограниченным, в осо-
бенности когда касается укусов человека подозрительными на бешенство живот-
ными.
Постмортальная диагностика
Посмертная диагностика бешенства у человека высокоэффективна и базируется
на выделении вируса (m vivo или in vitro), детекции нуклеопротеина (антигена),
вирусной РНК или ее фрагментов. Для транспортировки аутопсийного материала
могут быть использованы низкие температуры (ниже нуля) и консервирование
головного мозга в 50% глицерине. К «золотым стандартам» относятся два метода:
выделение вируса (in vivo и in vitro) и детекция антигена (нуклеопротеина) вируса
бешенства прямым методом флюоресцирующих моноклональных антител (МФА).
Прямой быстрый иммуногистохимический тест
Другим, равным по качеству МФА, методом детекции антигена вируса бешенства
является прямой быстрый иммуногистохимический тест, основанный на взаимо-
действии биотинконъюгированных моноклональных антител с нуклеопротеином
вируса бешенства. Для исследования используют кратковременно фиксированные
формалином (10 мин) отпечатки свежего, замороженного или сохранившегося в
глицерине головного мозга животных, больных бешенством. Для прямого быстро-
го иммуногистохимического теста не требуется дорогостоящего оборудования,
достаточно светового микроскопа. В поле зрения выявляются вирусспецифиче-
ские включения таких же размеров и формы, как и в МФА. Метод предназначен
для использования в вирусологических цент рах, полевых условиях.
ЧАСТНАЯ ВИРУСОЛОГИЯ 649
ВИРУС ГРИППА
Таксономия
Вирусы гриппа (Mixovirus influenzae} относятся к семейству Orthomyxovu idae,
имеют сегментированный геном и подразделяются на типы/роды. Известны три
типа вируса гриппа: А, В и С (роды Influenzavirus A, Influenzavirus В, Influenzavirus С
соответственно). Тип вируса определяется антигенными свойствами двух отно-
сительно стабильных внутренних структурных белков, нуклеопротеина (NP) и
матричного (М) белка. Подтипы/субтипы вируса гриппа А классифицируются по
антигенным свойствам поверхностных гликопротеинов, гемагглютинина (Н) и
нейраминидазы (N).
Вирус гриппа — возбудитель острого заболевания с воздушно-капельным меха-
низмом передачи, протекающего с явлениями общей интоксикации и поражением
респираторного тракта. У вирусов гриппа, выделенных от человека, установлено
наличие трех подтипов антигена Н (Hl, Н2, НЗ) и двух подтипов антигена N (N1
и N2). Вирусы гриппа, циркулирующие среди населения до 1957 г., имеют общую
антигенную формулу: A (H1N1), с 1957 по 1968 г. — A (H2N2), а с 1968 г. — А
(H3N2'. Вирусы гриппа человека типа А трех данных субтипов (H1N1, H2N2 и
H3N2) ассоциированы с масштабными эпидемиями и пандемиями.
Вирусы гриппа А инфицируют не только человека, но и многие виды млеко-
питающих (лошадей, свиней и др.) и птиц, главным образом водоплавающих.
Вирусы гриппа, циркулирующие среди людей, домашних и диких животных и
птиц, способны к обмену генетическим материалом в процессе реассортации.
Однако межвидовая передача вируса гриппа А является сложным многофактор-
ным процессом и происходит крайне редко (например, инфицирование людей
вирусами гриппа А птиц субтипов Н Np H7N7 в странах Юго-Восточной Азии).
Возникновение абсолютно новых субтипов вируса (антигенный «шифт») про-
исходит через нерегулярные интервалы времени и только с вирусами гриппа
типа А; они вызывают пандемии гриппа (в XX в. зарегистрированы три панде-
мии: в 1918.1957 и 1968 гг.).
Вирусы гриппа В значительно реже вызывают масштабные эпидемии, а вирусы
гриппа С ассоциированы со спорадическими случаями заболевания или неболь-
шими локальными вспышками.
Частые мутации генов, кодирующих поверхностные гликопротеины вирусов
гриппа А и В, приводят к появлению новых вариантов, в связи с чем была раз-
работана международная система кодировки, по которой каждый вариант вируса
получает свой код:
1 — обозначение типа вируса (А, В или С);
2 — географическое место выделения;
3 — порядковый номер выделенного в данном году в лаборатории вируса/номер
культуры;
4 — год выделения;
5 — обозначение антигенного подтипа.
Примеры; штамм А/Пекин/262/95 (H1N1), штамм А/Япония/305/57 (Н.,Гф),
штамм А/Сидней/5/97 (H,N,); штамм В/Яманаши/1о6/98).
Если вирус был выделен у животного (а не у человека), после указания типа
вируса указывают сокращенное название животного.
Характеристика вирусов
Вирионы вирусов гриппа А и В имеют сферическую структуру и размер около
100 нм, хотя встречаются и нитевидные формы. В состав частиц вируса гриппа А
650 ЧАСТНАЯ ВИРУСОЛОГИЯ
входят порядка 1% РНК, 70% белков, 20% липидов и 5- 8% углеводов. Вирионы
с молскУяяРН()й массой 250 МДа имеют липопротеидную оболочку. Три вирусных
белка (у вируса гриппа В — 4) пронизывают липидный слой и образуют внешнюю
поверхность вириона. Два из них - вирусные гликопротеиды: гемагглютинин
(ДА) и нейраминидаза (NA). Они формируют поверхность вирусной частицы.
Именно против НА и NA направлен антивирусный иммунитет.
У вируса гриппа С в оболочке присутствует только один гликопротеид, НЕ,
совмещающий функции гемагглютинина и ацетилэстеразы.
НА, NA и ИЕ являются трансмембранными белками. Их гидрофобные транс-
мембранные участки пронизывают липидный слой оболочки вируса. Третий
трансмембранный белок вирусной оболочки обозначен у вируса гриппа А как М2,
у вируса гриппа В — ВМ2, у вируса гриппа С — СМ2. Внутренняя оболочка вируса
образована белком Ml, наиболее обильно представленным в вирусной частице (до
3000 молекул на вирион у вируса гриппа А). Нуклеокапсид у вирусов гриппа пред-
ставлен рибонуклеопротеидными тяжами, содержащими вирусную РНК и белки.
Генетический материал вируса представлен одноцепочечной минус-цепью РНК,
состоящей из восьми фрагментов, кодирующих вирусные белки. Каждый сегмент
РНК кодирует один или два вирусных белка.
Вирус гриппа обладает слабой устойчивостью к действию физических и хими-
ческих факторов, разрушается при комнатной температуре в течение нескольких
часов (в воздухе погибает примерно через 4 ч, на поверхности предметов — через
1-4 сут), в то время как при низких температурах (от -25 °C до -70 °C) сохраня-
ется несколько лет. Благодаря устойчивости к низким температурам может сохра-
няться во внешней среде до 3 нед. Вирус гриппа быстро погибает при нагревании
(прогревание при температуре 50-60 СС вызывает инактивацию вируса в течение
нескольких минут), высушивании, при воздействии небольших концентраций
хлора, озона, ультрафиолетовой радиации (во внешней среде вирус гриппа доволь-
но быстро погибает под действием солнечного света). Вирус чувствителен к дей-
ствию обычных дезинфектантов.
Лабораторные методы диагностики
КЛИНИЧЕСКИЙ МАТЕРИАЛ ДЛЯ ЛАБОРАТОРНОГО ИССЛЕДОВАНИЯ
В слизи, выделяемой больным из носоглотки при чихании и кашле, вирус
гриппа может сохраняться довольно длительное время. Для выделения виру-
сов гриппа исследуемым клиническим материалом являются мазки из носа, зева
и задней стенки глотки, смывы из носовой части глотки, секционный материал
(фрагменты трахеи, бронхов, легких). Иногда для выделения вирусов используют
кровь и цереброспинальную жидкость. Оптимальным сроком забора клинического
материала являются первые 2-3 сут от начала заболевания. Для транспортировки
забранного материала в лабораторию используют среду с белковым стабилизато-
ром, способствующим более длительному сохранению инфекционной активности
вируса гриппа. Это могут быть раствор Хенкса или среда № 199, среда Игла с 0,5%
бычьего сывороточного альбумина или с 0,5% желатина или даже с 0,5% снятого
(обезжиренного) молока. Можно применять забуференный физиологический рас-
твор — 0,01 М фосфатно-буферный раствор, pH = 7,2+7,4 + 0,15 М натрия хлорида
с 0,5% бычьего сывороточного альбумина. Перед выездом за исследуемым матери-
алом среду разливают в пробирки или флаконы по 0,5 мл, добавляют по 400 ЕД/мл
пенициллина, 200 мкг/мл стрептомицина, 100 мкг/мл неомицина, 100 мкг/мл
микостатина.
Материалом для серологической диагностики гриппа служат одна или две (пар-
ные) сыворотки крови.
ЧАСТНАЯ ВИРУСОЛОГИЯ
651
МЕТОД ФЛЮОРЕСЦИРУЮЩИХ АНТИТЕЛ (МФА)
Метод флюоресцирующих антител (МФА) является достаточно простым мето-
дом быстрой диагностики (2-3 ч от момента доставки клинического материала в
лабораторию), при котором вирусные включения обнаруживают в мазках эпите-
лия слизистой оболочки носа в течение первых дней заболевания.
Используют его и для выявления антигенов вируса гриппа в пробах секцион-
ного материала путем исследования отпечатков кусочков ткани легкого, а также
трахеи и бронхов со специфическими гипериммунными люминесцентными сыво-
ротками к вирусам гриппа A (HjNJ, A (H,N2) и В.
МЕТОД ИММУНОФЕРМЕНТНОГО АНАЛИЗА
Метод иммуноферментного анализа (ИФА), направленный на обнаружение
антигенов вирусов гриппа в клиническом материале носоглотки и верхних дыха-
тельный путей, не получил широкого применения из-за недостаточной чувстви-
тельности и ограниченной возможности тестирования подтиповых/штаммовых
различий вирусов гриппа A (H1N1, H3N1, H3N2, H5N1 и т.д.). Именно поэтому
в целях поиска возбудителя при гриппозной инфекции метод ИФА был быстро
вытеснен методом ПЦР, преимуществом которого являются чувствительность,
специфичность, возможность быстрой диагностики гриппа и дифференциальной
диагностики штаммов.
ПОЛИМЕРАЗНАЯ ЦЕПНАЯ РЕАКЦИЯ
ПЦР является идеальным методом определения малых и сверхмалых количеств
РНК вирусов гриппа в различном клиническом материале (нативном — мазки из
носоглотки, ротоглотки, мокрота, бронхоальвеолярный лаваж, секционном —
ткань легкого, трахеи).
В случае выявления РНК-содержащих вирусов используют модификацию
метода — ОТ-ПЦР, когда на первой стадии исследования происходит фермента-
тивный синтез копий комплементарной ДНК с матрицы РНК и полученные к ДНК
фрагменты используют для получения специфических копий. Продукты реакции
анализируют электрофоретически или с использованием флюориметрии.
СЕРОЛОГИЧЕСКИЕ ТЕСТЫ
Серологические тесты основаны на способности вируса гриппа агглютинировать
человеческие или куриные эритроциты, а специфические антитела ингибируют
этот процесс Именно поэтому основным серологическим методом для определе-
ния противогриппозных антител является реакция торможения гемагглютинации
(РТГА), позволяющая различать подтипы и штаммы вирусов гриппа.
Материалом для исследования служат парные образцы сывороток крови, взя-
тые с интервалом 10-14 дней, которые исследуют одновременно. Лабораторный
диагноз ставят на основании сероконверсии — четырехкратного и более прироста
титров антител в парных образцах крови.
Таким образом, оптимальный алгоритм лабораторного подтверждения кли-
нического случая гриппа представляется следующим. В ранней стадии заболе-
вания проводят прямую идентификацию вирусных антигенов в носоглоточных
мазках (соскобах) с использованием иммунофлюоресцирующих антител (МФА)
и/или с помощью метода ПЦР (путем амплификации выделенной вирусной РНК).
Инфицирование также может быть подтверждено демонстрацией специфическо-
го серологического ответа при исследовании парных сывороток крови больного
методом РТГА.
Что касается выделения вирусов гриппа из носоглоточных секретов и другого
клинического материала путем заражения культуры клеток или куриных эмбрио-
нов, эти процедуры все реже применяют в диагностических лабораториях, но они
652
ЧАСТНАЯ ВИРУСОЛОГИЯ
необходимы для накопления и изучения циркулирующих штаммов вируса гриппа
и используются, как правило, в научно-исследовательских институтах.
Дифференциальная диагностика
Дифференциальную диагностику гриппа необходимо проводить с острыми
респираторными заболеваниями, рядом других инфекций, которые по симптомам
интоксикации и катаральных явлений напоминают грипп. Это парагрипп, аде-
новирусная инфекция, риновирусная инфекция, корь в продромальном периоде,
начальный (преджелтушный) период вирусного гепатита, паратиф А.
Вирус гриппа H1N1 (swine 2009)
ТАКСОНОМИЯ
Вирус гриппа свиней типа A (H1N1), выделенный от больных в Мексике и
США весной 2009 г. (впервые в Калифорнии), получил официальное название
Калифорния/Н1К1/04/2009, он является новым вирусом гриппа А, который
ранее не выявлялся в Северной Америке. Этот вид вируса изначально называли
свиным гриппом, поскольку в лабораторном тестировании было показано, что
многие гены этого нового вируса схожи с вирусом гриппа, который обычно встре-
чается у свиней Северной Америки. Анализ нуклеотидных последовательностей в
базе «GenBank» показал, что наибольшее сходство (95%) по гену гемагглютинина
он показал со штаммом вируса гриппа A/swine/Indiana/P12439/00(HlN2) — типич-
ным представителем североамериканской линии свиного гриппа, циркулирующей
в США с 1999 г.
Новый вирус включает гены вируса гриппа, свойственного свиньям в Европе и
Азии, а также гены птиц и человека, что позволяет назвать его трое-четырехкратным
реассортантом. Пандемия гриппа A/H1N1 (комбинация антигенов которого ранее
не встречалась у вирусов гриппа, вызывающих заболевания людей или свиней)
была объявлена ВОЗ 11 июня 2009 г. В соответствии с рекомендациями ВОЗ,
предпочтительным методом лабораторной диагностики нового пандемического
гриппа A/HlNl(sw2009) является ПЦР.
КЛИНИЧЕСКИЙ МАТЕРИАЛ ДЛЯ ЛАБОРАТОРНОГО ИССЛЕДОВАНИЯ
Для лабораторного исследования у людей лучше всего подходят образцы, взя-
тые из верхних дыхательных путей. Следует брать следующие образцы: из глуби-
ны носовых отверстий (назальный мазок), носоглотки (носоглоточный мазок),
носоглоточный аспират, аспират из зева или бронхиальный аспират, секционный
материал. При взятии мазков из полости носа и ротоглотки для исследования
методом ПЦР на пандемический свиной грипп нельзя использовать зонды на
деревянной основе и с хлопковыми тампонами. Для данного вида исследования
рекомендуют совмещать мазки из полости носа и ротоглотки в первой пробирке
с транспортной средой (фосфатно-солевым буферным раствором, 0,1 моль/л) и
исследовать как один образец.
Обязательному вирусологическому исследованию подлежит секционный мате-
риал от умерших с диагнозом «пневмония» или «острая респираторная инфек-
ция». При подозрении на пандемический грипп A/HlNl(sw2009) исследования
рекомендуют проводить в два э гапа.
• Этап 1. Исследование биологического материала на присутствие вирусов А и
В методом ПЦР. При получении положительного результата по обнаружению
вируса гриппа А на первом этапе образцы к ДНК, в которых была обнаружена
РНК Influenza virus А, исследуют далее.
ЧАСТНАЯ ВИРУСОЛОГИЯ 553
• Этап 2. Для лабораторного подтверждения заболевания пандемиче-
ским гриппом нового варианта A/HlNl(sw2009) (подобного штамму А/
Калифорния/04/2009) следует использовать набор реагентов для иденти-
фикации вируса гриппа типа A/H1N1 (sw) методом ПЦР с гибридизационно-
флюоресцентной детекцией.
Подтвержденным случаем инфекции, вызванной вирусом гриппа типа А/
HlNl($w), считается тот, при котором у человека имеет место острое респиратор-
ное заболевание с лабораторно подтвержденной инфекцией, вызванной вирусом
гриппа свиней типа A(H1N1), с помощью проведения одного или более из выше-
перечисленных исследований. При отрицательных результатах после второго
этапа исследования для дальнейшего типирования следует использовать набор
реагентов для идентификации вирусов гриппа А, вызывающих сезонный грипп
человека H1N1 и H3N2. Если образец не типируется по одной из мишеней (НА
или NA), материал считается подозрительным на содержание нового антигенного
варианта вируса гриппа А и его следует направлять в референс-лабораторию для
дальнейшего исследования.
Биобезопасность
Диагностическую лабораторную работу с клиническими образцами от пациентов,
подозреваемых на инфекцию, вызванную вирусом гриппа A (H1N1) свиного проис-
хождения, следует проводить в условиях BSL-2 с использованием соответствующих
средств индивидуальной защиты. Все манипуляции с клиническими образцами
необходимо проводить внутри сертифицированного бокса биобезопасности.
ВИРУС ИММУНОДЕФИЦИТА
Таксономия
Согласно данным Международного комитета по таксономии вирусов 2002 г.,
ВИЧ относится к семейству Retroviridae, подсемейству Orthoretrovirinae, роду
Lent! virus. Уникальной особенностью представителей семейства Retroviridae явля-
ется наличие фермента — обратной транскриптазы, которая осуществляет синтез
ДНК на матрице геномной РНК. ДНК затем встраивается в геном клетки и назы-
вается провирусом. Известно о двух типах ВИЧ: ВИЧ-1, получивший наибольшее
распространение в мире, и ВИЧ-2, встречающийся реже. Штаммы ВИЧ-1 в связи
со значительным генетическим разнообразием отнесены к трем группам: М — наи-
более многочисленной и включающей девять субтипов от А до Н, а также N и О.
Характеристика вируса
Вирионы ВИЧ имеют округлую форму и диаметр 100-120 нм. Наружная мем-
брана вириона, позаимствованная от клетки-хозяина, пронизана собственными
белками вируса. Их два — трансмембранный гликопротеин gp 41 и поверхностный
гликопротеин gp 120. Они связаны между собой нековалентной связью и форми-
руют отростки на поверхности вирионов. Нуклеоид вируса имеет характерную
вытянутую форму. Оболочка нуклеоида построена из молекул белка, называемого
р24. Между наружной оболочкой вириона и нуклеоидом существует каркас тол-
щиной 5-7 чм, состоящий из молекул белка — матриксного протеина р17. Внутри
нуклеоида содержатся две одноцепочные молекулы РНК. белок р7, связанный с
РНК. и комплекс ферментов (обратная транскриптаза, РНКаза Н, интеграза и про-
теаза). Геномная РНК ВИЧ содержит структурные (gag, pol, env) и регуляторные
гены.
654
ЧАСТНАЯ ВИРУСОЛОГИЯ
Все белки вириона ВИЧ способны вызывать иммунный ответ как в организме
человека, так и при иммунизации лабораторных животных. Наиболее иммуноген-
ными белками вирусной частицы являются поверхностные гликопротеины gp 120
и gp 41, а также их предшественник gp 160. Антитела к этим белкам появляются
относительно рано, выявляются у 98% инфицированных и более стабильны, чем
антитела к другим антигенам.
ВИЧ-1 инактивируется следующими средствами: изопропиловым спиртом,
этиловым спиртом, гипохлоридом натрия, перекисью водорода, лизолом и др.
Считается, что для инактивации инфекционности ВИЧ для тестов in vitro доста-
точно широко описанного прогревания вируссодержащего материала при 56 °C
в течение 30 мин. Однако имеются данные об инфицировании ВИЧ 6 пациентов
с гемофилией, получивших в качестве заместительной терапии фактор VIII, при-
готовленный из скринированной крови и прогретый при 60 °C в течение 30 ч.
Другими словами, в ситуации in vivo инфекционно опасными могут быть не только
полностью интактные вирионы ВИЧ, но и внутри капсидный интеграционный
комплекс (то есть геномная РНК вируса и ферменты, обеспечивающие ее обрат-
ную транскрипцию и интеграцию в геном).
Методы диагностики ВИЧ подразделяются на две группы: прямые и непрямые.
В первом случае обеспечивается обнаружение либо самого возбудителя, либо его
структурных компонентов, к которым относятся белки и нуклеиновые кислоты —
РНК или ДНК. Во втором случае диагностика строится на ответной реакции орга-
низма на вторжение возбудителя. К этой группе относятся серологические мето-
ды, направленные на обнаружение специфических к возбудителю антител (Ат).
Исторически первыми были разработаны и применены в практике серологические
методы диагностики ВИЧ.
Лабораторные методы диагностики
СЕРОЛОГИЧЕСКИЕ МЕТОДЫ
Серологические методы являются ведущими в диагностике ВИЧ. К ним отно-
сятся иммуноферментный анализ (твердофазный ИФА) и иммуноблот (ИБ). ИФА
характеризуется очень высокой чувствительностью (практически 100%) и служит
для скрининга образцов. Для подтверждения серопозитивности используют ИБ.
так как специфичность этого теста превосходит ИФА.
При использовании серологических методов (ИФА, ИБ) основной проблемой
является наличие серонегативного «окна» — периода от момента заражения до
появления Ат. С помощью ИФА-тестов 1, II поколения обнаруживали Ат через
6-12 нед. Применение современных тестов (III, IV поколения) позволило сокра-
тить серонегативный период до 3-4 нед. Более того, тесты IV поколения позволя-
ют одновременно выявлять суммарные антитела к ВИЧ-1 и антиген р24 ВИЧ-1 в
сыворотке (плазме) крови человека. Раннее появление р24 в крови (через 2 нед)
связано с активной репликацией вируса во время острой фазы инфекции. Когда
начинают обнаруживать Ат к ВИЧ, антиген р24 часто уже не выявляется в резуль-
тате образования комплекса между Аг и Ат.
Выявление Ат к ВИЧ-1 группы 0 и ВИЧ-2 осуществляют путем сорбции на
твердой фазе пептидов, воспроизводящих иммунодоминантные эпитопы белков
оболочки этих вирусов.
По рекомендации ВОЗ, положительные результаты при ИФА-диагностике
ВИЧ-инфекции следует подтверждать методом ИБ, то есть иммуноферментным
анализом с выявлением антител к индивидуальным белкам вируса. В классиче-
ском варианте ИБ иммуносорбентом служат полоски (стрипы) нитроцеллюлозной
мембраны с сорбированными на них методом электропереноса индивидуальными
ЧАСТНАЯ ВИРУСОЛОГИЯ
655
ИФА 1
(скрининг)
ИФА 2
(скрининг)
Иммуноблоттинг
(подтверждающий
тест)
Диагноз
подтвержден
Консультирование и оценка недавнего
риска инфицирования ВИЧ
Повторное
обследование
в ИФА/иммуноблоте
через 1-3-6 мес
или проведение
ДНК ПЦР
Диагноз исключен
Рис. 24-2. Алгоритм серодиагностики ВИЧ-инфекции.
белками вируса. Вся совокупность нативных антигенов вируса распределена по
поверхности стрипа, занимая на нем зоны в соответствии с молекулярной мас-
сой каждого антигена. При наличии в исследуемой сыворотке соответствующие
антитела связываются с антигенами в соответствующих зонах. Образовавшиеся
комплексы «Аг-Ат» выявляются с помощью стандартных приемов ИФА: связы-
вания комплекса «Аг-Ат» с конъюгатом и затем ферментативного расщепления
субстрата с появлением окраски. В идеале необходимо присутствие Ат против всех
белков, но в реальности наблюдаются разные профили присутствия Ат: к одним
белкам они присутствуют, к другим — нет. В связи с этим имеются критерии оцен-
ки результатов ИБ, которые приводятся в инструкции к тесту.
Учитывая возможность с помощью тестов IV поколения выявлять антиген р24
ВИЧ-1, дополнительно появились тесты для подтверждения наличия р24 ВИЧ-1.
Алгоритм исследования на ВИЧ-инфекцию включает определение Ат методом
ИФА с последующим подтверждением в ИБ. Обобщенная диагностическая схема
представлена на рис. 24-2.
МОЛЕКУЛЯРНЫЕ МЕТОДЫ
В настоящее время уже трудно представить чтобы лабораторное сопровождение
ВИЧ-инфицированных пациентов осуществлялось без применения молекулярных
методов. В первую очередь, это относится к полимеразной цепной реакции (ПЦР).
Их широкое применение в отечественных лабораториях началось относительно
недавно (с 2004 г.). Они необходимы:
• для диагностики ВИЧ-инфекции у детей до года, рожденных ВИЧ-инфи-
цированными женщинами;
• количественного определения РНК ВИЧ в 1 мл плазмы крови (вирусная
нагрузка);
• определения лекарственной устойчивости ВИЧ.
Диагностика ВИЧ-инфекции у детей до года
Практически все дети, рожденные ВИЧ-инфицированными женщинами, явля-
ются серо позитивными в тесте на Ат к ВИЧ, при этом лишь 15-30% из них в
действительности инфицированы. Длительность циркуляции материнских Ат
в крови детей составляет в среднем 9-12 мес, и в этот период методы сероди-
агностики неинформативны. Применение ПЦР-тестов, предназначенных для
качественного обнаружения провирусной ДНК ВИЧ в мононуклеарных клетках
периферической крови (МКПК), позволяет осуществить раннюю (<4 мес) диагно-
стику ВИЧ-инфекции у детей. Алгоритм исследования предусматривает первичное
656
ЧАСТНАЯ ВИРУСОЛОГИЯ
тестирование в возрасте 1 мес. При отрицательном результате повторное тести-
рование проводят в возрасте >4 мес. При положительном результате повторное
тестирование (подтверждение) осуществляют в максимально короткий срок после
первичного.
КОЛИЧЕСТВЕННОЕ ОПРЕДЕЛЕНИЕ РНК ВИЧ В 1 МЛ ПЛАЗМЫ
Появление новых препаратов для лечения ВИЧ-инфекции требует быстрых
методов оценки эффективности лечения. Для достоверных изменений клини-
ческих и иммунологических показателей требуется длительное время (годы).
Первые количественные методы были внедрены в практику в 1996-1997 гг.
Достаточно быстро было показано, что уровень вирусной нагрузки (ВН) является
прогностическим маркером прогрессирования заболевания и информативен при
оценке эффективности терапии. Это предопределило их быстрое и повсеместное
внедрение в практику для лабораторного мониторинга ВПЧ-инфицированных
пациентов. Был определен алгоритм обследования пациентов с целью своевремен-
ного начала терапии и оценки ее эффективности (табл. 24-3).
Таблица 24-3. Определение ВН у ВИЧ-инфицированных пациентов
Цель Сроки обследования Критерии начала терапии и оценки ее эффективности
Мониторинг естественного течения Каждые 3-6 мес >50 000 копий/мл
Контроль эффективности тера- пии Фон —
через 1 мес, Снижение ВН >1.5 lg
далее каждые 3-6 мес Снижение ВН <50 копий/мл
Обследование беременных 24 нед ВН определяет схему профилактики
36 нед Снижение ВН <50 копий/мл
Определение лекарственной устойчивости ВИЧ
Основной причиной неэффективности комбинированной антиретровирусной
терапии, включающей не менее трех препаратов, является формирование устойчи-
вости БИЧ к одному или более препаэатам. Установить, какой препарат (препараты)
в данной схеме неэффективен, позволяет тест на резистентность. Его применение
фактически обеспечивает индивидуальный подход к лечению каждого пациента.
Принцип: методом секвенирования определяют нуклеотидную последователь-
ность участка гена pol исследуемого образца ВИЧ, кодирующего два вирусных
фермента: обратную транскриптазу и протеазу. Именно против этих ферментов
направлено действие большинства лекарств. Прилагаемые программы позволяют
сравнить полученную последовательность ДНК с референс-последовательностью
для выявления генетических отличий (мутаций), оценить значение этих мутаций
для возникновения устойчивости к тому или иному препарату и сформировать
«отчет» о лекарственной устойчивости.
ВИРУС КЛЕЩЕВОГО ЭНЦЕФАЛИТА
Таксономия
Вирус клещевого энцефалита (ВКЭ) относится ко 2-й группе патогенности.
Основным переносчиком являются иксодовые клеши, возможно заражение людей
через употребление молока инфицированных животных.
Вирус клещевого энцефалита, названный в международной классификации
«Tick-borne encephalitis virus», относится к роду Flavivirus, семейству Flaviviridae.
ЧАСТНАЯ ВИРУСОЛОГИЯ 657
Является прототипным представителем одноименной серогруппы вирусов. Вирусы
этой группы характеризуются наличием типоспецифичных и серогруппспецифич-
ных антигенных детерминант. Типоспецифичные антигенные детерминанты вовле-
чены в нейтрализацию, опосредованную антителами. Выделяют Дальневосточный
(или Восточно-Сибирский), Европейский (или Западно-Европейский) и Сибирский
генотипы ВКЭ.
Вирион ВКЭ сферической формы. 50 нм в диаметре, состоит из липидсодержа-
щей оболочки и нуклеокапсида, который имеет икосаэдрический тип симметрии,
30 нм в диаметре. Геном вируса представлен единственной несегментирован-
ной одноцепочечной линейной РНК позитивной полярности, длиной 10 488-
ДО 976 н.о., кодирующий один полипротеид-предшественник десяти белков. Три
белка являются структурными. Полипротеиды Е и М образуют пепломеры —
выступы на поверхности липидной оболочки. Структурный полипептид С форми-
рует капсид. Структурный белок Е отвечает за связывание вируса с клеточными
рецепторами, определяя тропизм и вирулентность, является основным антигеном
ВКЭ. Семь неструктурных белков: NS1, NS 2А, NS 2В, NS 3, NS 4А, NS 4В и NS 5 —
обеспечивают репликацию вируса в клетке. Кроме того, в процессе репродукции
вируса образуются три коротких полипептида: CTHD. Рги, 2К, которые обеспечи-
вают транспортировку структурных белков и правильность сборки.
Внутри-, межочаговая и региональная генетическая гетерогенность ВКЭ обу-
словлена структурным разнообразием эпитопов каждого из антигенов вируса.
Вирионы КЭ обладают гемагглютинирующей и комплементсвязывающей актив-
ностью. Вызывает агглютинацию эритроцитов гусей.
Характеристика вируса
Вирион имеет молекулярную массу 60х106Да. Лиофилизированный, а также
замороженный вирусный субстрат сохраняет патогенные свойства в течение
нескольких лет, хотя при этом снижаются его вирулентные свойства. Вирус КЭ
хорошо переносит низкую температуру, при -60 °C сохраняет ся длительное время.
Малоустойчив к высоким температурам. При комнатной температуре (16-18 °C)
сохраняется в течение 10 дней, при 37 СС — сутки, при 50 СС инактивируется в
течение 20 мин. Вирус чувствителен к кислотам, лучше переносит слабощелочную
среду. Желудочный сок нормальной кислотности при 37 °C вызывает отчетливую
инактивацию вируса через 30 мин и полную — через 2 ч. В коровьем молоке при
температуре холодильника вирус не снижает своей активности в течение 2 нед,
длительное время (не менее 2 мес) сохраняется в сметане.
Инактивируется при воздействии дезинфицирующих веществ (5% хлорной
извести, 1% раствора квасцов, 3-5% раствора серной кислоты, 3-5% раствора
лизола) в течение нескольких минут, при воздействии прямых солнечных лучей,
ультрафиолетового излучения — в течение 10-15 мин.
Лабораторные методы диагностики
Вирусы выделяют из крови больных, взятой в первые дни болезни (идеаль-
но — в первые 3 сут болезни), цереброспинальной жидкости, реже носоглоточно-
го секрета и мочи, взятых в первые 5 дней. Пробы крови и ликвора целесообразно
заморозить для разрушения клеточных элементов. Выявление ВКЭ в суспензии
мозга возможно только при ранней гибели больного (не позднее 7 сут).
Лаборат орная диагностика клещевого энцефалита основана на обнаружении в
клиническом материале специфических антител, а также вируса КЭ, его антигенов
или нуклеиновой кислоты (РНК ВКЭ) в биологических субстратах.
658 ЧАСТНАЯ ВИРУСОЛОГИЯ
СЕРОЛОГИЧЕСКИЕ МЕТОДЫ
В практической диагностике преобладает использование серологических мето-
дов, то есть определение антител к ВКЭ в РТГА, РСК и иммунохимического —
ИФА.
До недавнего времени из-за простоты выполнения ведущими были эритроци-
тарные методы: РТГА и РСК. Известно, что гемагглютинирующие антитела при
КЭ появляются в первые дни заболевания, достигая максимальных значений на
2-3-й неделе от начала заболевания. В крови реконвалесцентов их уровень сни-
жается, но они сохраняются практически пожизненно. Комилементсвязывающие
(КС) антитела появляются в крови гораздо позже — лишь на 2-3 нед заболевания,
у части больных (до 60%) они отсутствуют до 4-5-й недели, наивысших пока-
зателей достигают к 6-9-й неделе заболевания. Комплементсвязывающие анти-
тела сохраняются в крови переболевших недолго — около года. По процентному
содержанию в популяции КС-антител с определенной долей вероятности можно
судить о популяционном иммунитете к инфекции (уровне «естественного проэпи-
демичивания» населения). Следует учитывать наличие среди циркулирующих в
природе штаммов вируса антигенно-дефектных вариантов, которые характеризу-
ются отсутствием гемагглютинирующего и преципитирующего антигенов и слабой
иммуногенностью на фоне высокой инфекционной активности вируса.
Эритроцитарные методы характеризуются рядом недостатков:
• нестандартностью методики из-за использования гусиных эритроцитов;
• необходимостью предобработки сывороток пациентов козлином и эритро-
цитами для удаления ингибиторов гемагглютинации;
• отсроченностью лабораторного диагноза из-за необходимости исследования
парных сывороток, взятых в первые дни заболевания (или госпитализации)
и через 2 нед.
Этих недостатков не имеет метод ИФА, главным преимуществом которого я вля-
ется возможность раздельного определения специфических IgM- и IgG-антител.
IgM-антитсла появляются в крови в первые дни заболевания, со 2-3-й недели к
ним присоединятся IgG-антитела, которые постепенно замещают IgM-антитела
и остаются в крови пожизненно. Однократное определение вирусспецифических
иммуноглобулинов класса IgM следует считать достоверным свидетельством в
пользу диагноза клещевого энцефалита. Анализ Е-антигена ВКЭ используют для
определения инфицированное™ клеща, снятого с укушенных. Период циркуляции
вируса КЭ в крови больного обычно соответствует продолжительности лихора-
дочного состояния. В этот период (особенно в первые 3-5 дней лихорадки) вирус
обнаруживают в крови, спинномозговой жидкости, в мозгу умерших.
ВИРУСОЛОГИЧЕСКИЕ МЕТОДЫ
Для выделения вируса применяют метод биопробы на беспородных белых
мышах и культивирование в культуре перевиваемых линий клеток СПЭВ, LLC-
МК, Vero. ВНК-21, Hela и др. Монослой клеток через 48-72 ч культивирования
изучают методом МФА. В цитоплазме инфицированных клеток обнаруживают
включения в виде зеленой массы или гранул.
Для проведения биопробы вируссодержащий материал (образцы крови, СМЖ.
разведенной ФБР мочи) вводят интрацеребрально 2-6-суточным мышам.
Вследствие чрезвычайной трудоемкости, длительности выделения и иденти-
фикации возбудителя КЭ на мышах или культуре ткани, жестких требований к
хранению и доставке материала в лабораторию, а также соблюдения особых усло-
вий противоэпидемического режима (ВКЭ относится к возбудителям II группы
патогенности) вирусологический метод выделения вируса КЭ из клинического
материала для диагностики используют все реже. Даже при соблюдении всех необ-
ходимых предосторожностей и исследовании материала в первые 4 дня заболева-
ЧАСТНАЯ ВИРУСОЛОГИЯ 659
ния частота выделения вируса из крови и спинномозговой жидкости не превышает
40%.
Существенным достижением последних лет стало использование метода ПЦР
для обнаружения РНК вируса КЭ в клиническом материале от пациентов: крови
и спинномозговой жидкости. Преимуществом метода является его высочайшая
чувствительность, а также возможность применения в клинико-диагностических
лабораториях.
При дифференциальной диагностике следует учитывать возможность заболе-
вания пациента японским энцефалитом, геморрагической лихорадкой с почечным
синдромом, энтеровирусным менингитом, клешевым риккетсиозом, болезнью
Лайма (боррелиозом) и др.
ВИРУС КОРИ
Характеристика вируса
Вирус кори относится к роду Morbillivirus, семейству Paramyxoviridae и является
близким родственником вируса чумы крупного рогатого скота и собак.
Вирионы кори полиморфны и имеют размеры от 100 до 300 нм. Геном вируса
представлен несегментированной одноцепочечной РНК отрицательной полярно-
сти, с молекулярной массой 4,5х106, состоит из шести генов, кодирующих восемь
белков. Порядок генов следующий: N-ген, кодирующий нуклеокапсидный белок;
Ртен, кодирующий фосфопротеин Р, два неструктурных белка — С и V; М-ген,
кодирующий мембранный белок; F-ген, с которого считывается белок слияния:
Н-ген, кодирующий гемагглютинин; L-ген, кодирующий главный полимеразный
компонент — L-белок. Молекулярная масса вирусспецифических белков колеблет-
ся в пределах от 21 (белок С) до 210 (белок L) кДа. Вирусная РНК. белки N, L и Р
формируют спиралевидный нуклеокапсид, окруженный липидной оболочкой, на
поверхности которой определяются два вида гликопротеинов: белки гемагглю-
тинина (димеры Н-белка) и белки слияния (тетрамеры F-белка), частично погру-
женные в липидную мембрану. Длина каждого из гликопротеинов 22-24 нм.
В слиянии с оболочкой вирионов принимает участие поверхностная мембрана
инфицированной клетки. Парамиксовирусы обладают повышенным сродством к
клеточным рецепторам, содержащим сиаловую кислоту.
В делом у вируса кори стабильный геном. Вакцинные штаммы имеют незна-
чительные отличия, а дикие вирусы более вариабельны. В настоящее время в
мире циркулирует несколько генетических вариантов этого вируса, которые были
определены методами молекулярной эпидемиологии в процессе изучения путей
передачи возбудителя.
Вирус кори термолабилен, чувствителен к действию детергентов, быстро инак-
тивируется в кислой среде (pH = 2^-4) При 56 °C инактивируется в течение 30 мин.
Сочетанная обработка твином-80 и эфиром позволяет получить более устойчивый
во внешней среде, лишенный инфекционное™ гемагглютинирующий антиген.
Типичное течение кори сопровождается высокой температурой и сыпью.
Инкубационный период заболевания длится в среднем около 10 дней (от 7 до
18) с момента инфицирования дыхательных путей до повышения температуры.
Первоначальная репликация вируса происходит в эпителиальных клетках трахеи
и бронхов, затем вирус проникает в локальные лимфатические узлы. В дальней-
шем инфекция через моноциты с кровотоком переносится в другие органы и ткани:
селезенку, тимус, легкие, печень, почки, соединительную ткань и кожу. Репликация
вируса продолжается в этих органах. В продромальной стадии заболевания вирус
обнаруживают в секреторных жидкостях из носа и глаз, в моче и крови.
6Ь0 ЧАСТНАЯ ВИРУСОЛОГИЯ
Иммунный ответ на натуральную инфекцию
Анти тела впервые можно выявить в момент появления сыпи. После натуральной
инфекции они выполняют свои защитные функции в течение всей жизни. Сначала
вырабатываются IgM-антитела, затем IgG и IgA в сыворотке крови и секреторных
жидкостях. Как IgM-, так и IgG-антитела начинают вырабатываться практически
одновременно в момент появления сыпи. Однако максимальный уповень содер-
жания IgM-антител приходится на 7-10-й день от момента появления сыпи, а
затем он начинает стремительно снижаться. Спустя 8 нед после появления сыпи
IgM-антитела выявляются только в исключительных случаях. В общем, наличие
IgM-антител свидетельствует о первичной инфекции диким или вакцинным виру-
сом. Однако отсутствие IgM-антител в образце, особенно в первые 3 дня с момента
появления сыпи, не исключает инфицирования, а может свидетельствовать о
низкой чувствительности использованного теста. Уровень IgG-антител достигает
максимального значения спустя 2 нед после появления сыпи, затем он постепенно
снижается. В отличие от IgM-, IgG-антитела выявляются в течение долгих лет
после инфекции.
Клинический материал для исследования
Вирус кори можно выделить на продромальной стадии заболевания и на стадии
высыпания из мочи, назофарингеальных секретов и лимфоцитов периферической
крови. Интенсивность экскреции вируса быстро падает. Обнаружение и иденти-
фикация вируса кори на культуре клеток могут занять несколько недель. Однако
выделение изолятов вируса кори позволяет в дальнейшем проводить их генети-
ческий анализ и сравнивать штаммы, полученные из различных мест в разные
годы. Таким образом, можно получить исчерпывающую информацию об их про-
исхождении и проследить пути передачи вируса. Именно поэтому для проведения
молекулярно-эпидемиологического анализа необходимо иметь коллекции мест-
ных штаммов вируса из эндемичных стран.
Для получения корректных результатов и облегчения их интерпретации боль-
шое значение имеет правильно выбранное по отношению к появлению клиниче-
ских симт омов кори время взятия образца. Поскольку IgM-ИФА-диагностические
тесты являются максимально чувствительными в промежу гке между 4 и 28 днем
от момента появления сыпи, один образец крови, полученный в эти сроки, счита-
ется адекватным для серологической диагностики кори. Забор повторного образца
сыворотки крови может понадобиться в случаях, если:
• первый образец крови, предназначенный для выявления IgM, был получен
в течение первых 4 дней от момента появления сыпи и показал при прове-
дении ИФА отрицательный результат. Лаборатория может запросить взятие
повторного образца крови для исследования, поскольку предыдущий резуль-
тат мог быть ложноотрицательным,
• клиницист нуждается в подтверждении окончательного диагноза у больного,
исследование сыворотки которого сначала показало отрицательный резуль-
тат.
Повторный образец крови для исследования можно взять в любой день в про-
межутке между 4 и 28 днем с момента появления сыпи. Взятие повторного образца
через 10-20 дней позволяет провести не только повторное тестирование на нали-
чие IgM-антител, но и количественный анализ увеличения титров IgG-антител.
Стерильную сыворотку можно хранить в холодильнике при 4-8 °C не более
7 дней, для более длительного хранения ее можно заморозить при -20 °C,
транспортировать в лабораторию с замороженными холодильными элементами.
Повторное замораживание и оттаивание сыворотки недопустимо.
ЧАСТНАЯ ВИРУСОЛОГИЯ 661
Устойчивость во внешней среде
Вирус кори чрезвычайно чувствителен к повышению температуры, поэтому
образцы материалов, предназначенные для выделения вируса, должны быть
транспортированы в лабораторию как можно быстрее и в условиях строгого
соблюдения холодовой цепи. Выделение вируса не рекомендуют использовать в
качестве метода первичной диагностики кори Можно ограничиться выделением
вируса только в специализированных лабораториях для генетического анализа
полученных штаммов.
Лабораторные Me i оды диагностики
Для диагностики кори рекомендуют использовать серологические методы,
основанные на выявлении вирусспецифических антител в одной или парных
сыворотках. Однако вирус кори может быть выделен из различных клинических
образцов путем культивирования его на культурах клеток или с применением
методов молекулярной биологии. Методы, основанные на выделении вируса,
непригодны в качестве диагностических тестов. В основном их используют для
генетической характеристики вирусов кори в молекулярно-эпидемиологических
исследованиях.
СЕРОЛОГИЧЕСКАЯ ДИАГНОСТИКА
Для диагностики кори используют следующие серологические методы:
• выявление вирусспецифических IgM-антител;
• количественное определение специфических иммуноглобулинов в крови для
установления их значимого прироста при исследовании парных сывороток,
полученных от больного в остром периоде и на стадии реконвалесценции
Выявление вирусспецифических IgM-антител
Вирусспецифические IgM-антитела достоверно свидетельствует об острой све-
жей инфекции. IgM-антитела также вырабатываются при первичной вакцинации,
и их уровень снижается быстрее, чем уровень IgM, вырабатываемых в ответ на
инфицирование диким вирусом. Вакцинные и дикие вирусспецифические IgM-
антитела не могут быть разделены посредством серологических исследований.
В этом случае для интерпретации полученных результатов следует обратиться к
истории вакцинации кэнкретного пациента. Для выявления вирусспецифических
IgM-антител к кори используют метод ИФА.
Количественное определение суммарных специфических иммуноглобулинов
Реакция нейтрализации вируса — реакция нейтрализации цитопатического
действия вируса кори (PH) — требует наличия двух парных образцов сывороток
крови, взятых от больного в остром периоде и на стадии реконвалесценции, пока-
зывает 100% чувствительность по отношению к клинически подтвержденным
случаям. Этот тест требует высококвалифицированного персонала, имеющего
опы г работы с культурами клеток. Результаты анализа могут быть получены через
10 дней после получения сыворотки реконвалесцента.
Реакция торможения гемагглютинации (РТГА) требует наличия двух
парных образцов сывороток крови от больного в остром периоде и на стадии
реконвалесценции и показывает 98% чувствительность у вакцинированных. Этот
тест также не является легким в применении, поэтому требует высококвалифи-
цированного персонала, имеющего опыт работы в области вирусной серологии,
наличие свежих эритроцитов обезьян. Результаты анализа могут быть получены
через 2 дня после получения сыворотки реконвалесцента.
662 ЧАСТНАЯ ВИРУСОЛОГИЯ
ПОЛИМЕРАЗНАЯ ЦЕПНАЯ РЕАКЦИЯ
Амплификацию РНК вируса кори после обратной транскрипции проводят толь-
ко в специализированных лабораториях для решения специальных задач. ПЦР не
применяют в целях рутинной лабораторной диагностики.
Алгоритм проведения лабораторной диагностики кори представлен в
табл. 24-4.
Таблица 24-4. Лабораторная диагностика кори в клинических материалах
Тип теста Цель исследования и источник материала Метод Примечания
Серологические методы иссле- дования Выявление вирус- специфических IgM- антител Непрямой IgM-ИФА IgM «двойной сэнд- вич» ИФА ИФА для выявления IgM в слюне Для IgM-ИФА требуется только один образец сыворотки; доступны коммерче- ские диагностические наборы Вариант «двойного сэндвича» является более прогрессивной модификацией непрямого ИФА Определение IgM методом ИФА в слюне ооычно требует дополнительной стадии концентрации в целях повышения чув- ствительности и специфичности реакции до необходимого уровня
Выявление вирус- специфических IgG- антител IgG-ИФА Требуется наличие парных образцов сыворотки от одного больного с интерва- лом 10—14 дней: доступны коммерческие диагностические наборы
Выявление сум- марных вирус- специфических иммуноглобулинов в сыворотке коови Реакция нейтрализа- ции (PH) Реакция торможения гемагглютинации (РТГА) PH полезна для определения иммунитета против кори; требуется техническое обеспечение для работы с клеточными культурами и парных образцов сыворот- ки; получение результата возможно через месяц Требуется постоянное наличие свежих эритроцитов обезьян и парных сыво- роток; получение результата возможно через 3 нед Оба последних метода требуют больших временных затрат
Выделение вируса Из мочи, назофа- рингеальных секре- тов и лимфоцитов периферической крови Культуры клеток и иммунофлюорес- ценция Требуются особая тщательность при сборе и транспортировке образцов, специальное оборудование для работы с культуоами клеток и иммунофлюорес- ценции
Обнаружение и генетическая характеристика вирусной РНК Из мочи, назофа- рингеальных секре- тов и лимфоцитов периферической крови Из изолятов дикого вируса ПЦР с обратной транскрипцией и сек- венирование (анализ последовательностей нуклеотидов) Требуется специальное оборудование; опасность перекрестных реакций Анализ последовательностей необходим для генотипирования диких вирусов
Для диагностики кори ВОЗ рекомендует использовать метод иммуноферментного
анализа (ИФА), предназначенный для обнаружения вирусспецифических IgM-
антител.
ЧАСТНАЯ ВИРУСОЛОГИЯ 663
ВИРУС КРАСНУХИ
Характеристика вируса
Вирус краснухи (Rubella virus) определен как единственный представитель рода
Rubivirus, семейства Togaviridae.
Вирион диаметром 40-70 нм состоит из липопротеидной оболочки и нуклео-
капсида с однонитевой, линейной, несегментированной, инфекционной (плюс-
нить) молекулой РНК. Геном длиной 9960 нуклеотидов кодирует пять белков, из
них NS1 и NS2 — неструктурные белки, С, Ы и Е2 —структурные белки вируса
краснухи. Капсид икосаэдрической формы собирается из молекул С-белка, свя-
занного с геномной РНК. Из липидной мембраны клетки-хозяина со встроенными
оболочечными гликопротеинами Е1 и Е2, которые образуют пепломеры, форми-
руется оболочка вириона. Белок Е1 является гемагглютинином и имеет эпитопы,
индуцирующие образование гемагглютинирующих и вируснейтрализующих анти-
тел. Гликопротеид Е2 содержит эпитоп, индуцирующий образование вирусней-
трализующих антител. Молекулярная масса зрелой вирусной частицы составляем
3500-5200 кДа.
Вирус краснухи термолабилен: при температуре 56 °C инактивируется через
30 мин, при 70 °C— через 4, а при 100 °C— через 2 мин. Наличие в среде белка
(альбумина, сыворотки) или сульфата магния повышает устойчивость вируса
к воздействию высоких температур. При -20 °C вирус быстро разрушается, но
сохраняется несколько лет при температуре -60 СС и ниже, а также при лиофиль-
ном высушивании в присутствии стабилизатора.
Во внешней среде вирус нестоек, быстро погибает при изменениях pH (<6,8 и
>8,0), под влиянием ультрафиолетовых лучей, эфира, формалина и других дезин-
фицирующих веществ и детергентов. Обработка твином-80 защищает гемагглюти-
нины от действия эфира. В присутствии белка вир\с можно повторно заморажи-
вать и размораживать без потери инфекционных свойств.
Вирус способен размножаться на многих клеточных культурах, но цитопа-
тическое действие оказывает лишь на немногих, в частности на перевиваемых
культурах клеток BHK-21, RK-13, Vero. Оптимальная температура культиви-
рования — 35 °C. Вирус краснухи агглютинирует эритроциты голубей, гусей,
однодневных цыплят, людей с группой крови 0 (1), обладает гемолитическими
свойствами.
Вирус является возбудителем острой антропонозной инфекции с воздушно-
капельным механизмом передачи, характеризуемой лихорадкой, легкими ката-
ральными явлениями, макулопапулезной сыпью, увеличением лимфатических
узлов. Размножается в слизистых оболочках носоглотки и заднешейных, затылоч-
ных, заушных лимфатических узлах. Вирус обладает выраженным тератогенным
действием. Острая форма краснухи рассматривается как показание к прерыванию
беременности в I триместре. Клинический диагноз краснухи требует лабораторно-
го подтверждения, так как инфекция характеризуется большим количеством бес-
симптомных форм (30-50%) Несмотря на наличие 13 генотипов вируса красну хи,
образующих две филогенетические группы (клайд 1 и клайд 2), серологических
различии между штаммами не выявлено, выделяют один серотип. После краснухи
формируется пожизненный иммунитет.
Клинический материал для лабораторного исследования
Клиническим материалом являются сыворотка крови, сухая капля крови, носо-
глоточные смывы, мазки из зева, слюна, ликвор, моча, образцы биопсии и аутоп-
сии. Вирус выделяется из носоглотки за 7-10 дней до появления сыпи, но вероят-
664 ЧАСТНАЯ ВИРУСОЛОГИЯ
ность его выделения наиболее высока в первые 3 сут с момента появления сыпи.
Дети с врожденной кпаснухой могут выделять вирус с экскретами на протяжении
первого года жизни, но частота выявления вируса со временем снижается — от
80% (1-2 мес) до 11% (12 мес).
Носо- и ротоглоточные смывы и соскобы должны быть доставлены в лаборато-
рию для исследования в течение 48 ч при температуре 2-8 СС. Допускается транс-
портировка при -20 °C. Если доставить образцы для исследования в указанные
сроки невозможно, их хранят в транспортной среде при —70 °C. При отсутствии
коммерческой транспортной среды можно использовать питательную среду для
культур клеток (DMEM, RPMI, Игла).
Мониторинг циркулирующих штаммов предполагает генотипирование изолята.
Для этого предпочтительнее использовать носоглоточные смывы или цельную
кровь. Носоглоточные образцы мигут быть взяты тремя способами, которые пред-
ставлены в порядке уменьшения вероятности выделения вируса:
• аспирацией носового секрета физиологическим раствором;
• смывами из горла;
• носоглоточными или ротоглоточными соскобами.
Рутинная диагностика инфекции основана преимущественно на определении
вирусспецифических антител.
Иммунный ответ на инфекцию
Как IgM-, так и IgG-антитела начинают вырабатываться практически одновре-
менно в момент появления сыпи. Однако максимальный уровень содержания IgM-
антител приходится на 2-ю неделю от момента появления сыпи, а затем он быстро
снижается. Спустя 8-9 нед после появления сыпи IgM-антитела выявляются
только в исключительных случаях. Наличие IgM-антител свидетельствует о пер-
вичной инфекции либо диким, либо вакцинным вирусом. Отсутствие IgM-антител
в образце, в особенности в первые 3 дня с момента появления сыпи, не исключает
инфицирования, а может свидетельствовать о низкой чувствительности исполь-
зованного теста. Уровень IgG-антител достигает максимального знаиения спустя
4-5 нед после появления сыпи, затем он постепенно снижается. IgG-антитела
выявляются в течение долгих лет после инфекции.
Постановка лабораторного диагноза у больного основана:
• на выявлении IgM-антител, если сыворотка получена от больного в период
4-28 сут от начала высыпания, за исключением случаев, когда больной полу-
чил вакцину против краснухи в период до 8 нед, предшествующих забору
образца;
• сероконверсии (четырехкратном приросте) IgG-антител при условии, что
второй образец сыворотки крови получен спустя не менее 10 сут от первого,
полученного в острой фазе болезни, за исключением случаев, когда больной
получал вакцину против краснухи в период до 8 нед, предшествующих забору
образца.
Если сыворотка крови получена позднее 28 сут от начала заболевания, диагноз
устанавливают по высокому уровню IgG-антител. Если сыворотка от больного с
клиническим диагнозом «краснуха» получена ранее 3 сут, в случае отрицатель-
ного результата определения IgM-антител исследование следует повторить через
несколько суток.
Для серодиагностики краснухи применяют и реакцию торможения гемагглю-
тинации (РТГА). Однако для постановки реакции необходимы свежие эритроци-
ты голубей или однодневных цыплят, реакция достаточно трудоемка и требует
использования парных сывороток, полученных с интервалом 2 нед.
ЧАСТНАЯ ВИРУСОЛОГИЯ 665
Дифференциальная диагностика
Рребуется дифференциальная диагностика с корью, эпидемическим паротитом,
гриппом, парагриппом, аденовирусной, респираторно-синцитиальной инфекция-
ми, скарлатиной, инфекционной эритемой, медикаментозными или аллергически-
ми высыпаниями, сывороточной болезнью, энтеровирусными и другими инфек-
циями, парвовирусом В19, вирусом герпеса человека 6-го типа, вирусом Экхо и
вирусом Коксаки, инфекционным мононуклеозом, розовым лишаем.
Лабораторные методы диагностики
Особую значимость имеет лабораторная диагностика краснухи у беременных.
Показаниями к лабораторному обследованию беременной являются:
• пренатальный скрининг беременных на ЕУИ, в том числе на краснуху;
• контакт беременной с больным краснухой или недиагностированным экзан-
тематозным заболеванием.
При скрининговом обследовании определяют уровень IgG-антител. Защитным
является титр 15 МЕ/мл. При высоком уровне IgG (более 100 МЕ/мл) женщина
считается защищенной от краснушной инфекции, при отсутствии IgG-антител —
относится к группе риска.
При обследовании контактных по краснухе беременных следует проводить
лабораторную диагностику как можно раньше, желательно в первые дни контакта
или первый день обращения к врачу. Следует одномоментно исследовать сыво-
ротку крови на наличие IgG- и IgM-фракции антител. Низкий или средний уро-
вень IgG (до 100 МЕ/мл) свидетельствует о перенесенной в прошлом инфекции.
Высокий уровень IgG может свидетельствовать как о наличии инфицирования,
так и о «бустер-эффекте» в отпет на повторную встречу с вирусом. При отсутствии
IgM и IgG в сыворотке, полученной в первые дни контакта, женщину относят к
группе риска и обследуют повторно в конце инкубационного периода, продол-
жительность которого не превышает 21 день. Наличие IgM-антител, высокий
уровень IgG свидетельствуют об инфекционном процессе. Необходимо учитывать,
что реактивные на IgM результаты обследования беременной могут являться
ложноположительными за счет ревматоидного фактора. Кроме того, гипердиаг-
ностика может иметь место в случае тестирования лиц, инфицированных вирусом
Эпштейна-Барр, парвовирусом и др. Целесообразно применять ИФА, основанный
на определении индекса авидности специфических IgG-антител. Малоавидные
IgG-антитела — показатель острой инфекции, высокоавидные характерны для
анамнестических антител. Низкоавидные IgG-антитела сохраняются в течение
более длительного срока по сравнению с вирусспецифическими антителами класса
IgM. Фактором, подтверждающим наличие инфекционного процесса, является и
прирост IgG-антител в динамике. Для диагностики краснухи и диагностики СВК в
целях воспроизводимости и стандартности результатов при динамическом наблю-
дении наиболее целесообразно использовать тест-системы одного производителя.
ВИРУС ПАРАГРИППА
Характеристика вируса
Вирусы парагриппа отнесены ко второму из пяти родов подсемейства
Paramyxovirinae, семейства Paramyxoviridae, порядка Mononegavirales, выделенному
на основании наличия у вирусов оболочки нуклеокапсида и единственной несег-
ментированной, линейной (минус-нить), однонитевой молекулы РНК. Human
parainfluenza virus I и Human parainfluenza virus III включены в род Resgirovrrus.
666 ЧАСТНАЯ ВИРУСОЛОГИЯ
Human parainfluenza virus II и Human parainfluenza virus IV (4a и 4b) входят в род
Rubulavirus.
Вирион подсемейства Paramyxovinnae сферической или плеоморфной формы,
диаметром 120-300 нм, состоит из нуклеокапсида 12-17 нм в диаметре и обо-
лочки. Нуклеокапсид спиральной симметрии, имеющий свернутую шарообразную
форму, состоит из РНК и трех белков; белка N — основного белка нуклеокапсида,
белка Р — фосфопротеида, белка L — РНК-зависимой РНК-полимеразы. Белки Р и
L составляют полимеразный комплекс, отвечающий за инициацию транскрипции.
Белок L, наиболее крупный из трех, осуществляет синтез РНК (инициацию, элон-
гацию и терминацию). Наружная липопротеидная оболочка содержит два транс-
мембранных белка: HN (или Н) — представлен тетрамерами, необходимыми для
прикрепления вирусной частицы к остаткам сиаловой кислоты в олигосахаридах
клетки-мишени; гликопротеин F — является тримером и обеспечивает слияние
вирусной оболочки с клеточной мембраной.
Для рода Rubulavirus характерно наличие гемагглютинина (Н), гликопротеина
слияния (F), негликозилированного трансмембранного белка (SH) и магриксных
протеинов.
Известным в настоящее время четырем типам вирусов парагриппа, выделенных
от человека, несвойственна вариабельность антигенной структуры. Нет общих
антигенов, хотя некоторый уровень перекрестной реактивности возможен с
разными серотипами. Вирусы обладают гемагглютинируюшей и комплементсвя-
зывающей активностью. Парагриппозные вирусы агглютинируют эритроциты
человека (группы 0), кур, морских свинок, обезьян. Специфические иммунные
сыворотки тормозят реакцию гемагглютинации. Гемагглютинирующие и компле-
ментсвязывающие антитела нс являются строго специфичными.
Вирусы парагриппа нестойки во внешней среде, при комнатной температуре
сохраняются не более 4 ч, полная их инактк вация происходит при нагревании до
50 °C в течение 30-60 мин, при 56 СС — через 15-30 мин. Кислая среда действует
на них губительно, щелочная — способствует сохранению активности. Вирусы
чувствительны к действию обычных дезинфектантов.
Парагриппозные вирусы репродуцируются в клетках эпителия дыхательных
путей, вызывая литическую инфекцию эпителиальных клеток. In vitro вирусы
парагриппа размножаются в первичных эмбриональных клетках почек обезьян,
диплоидных человеческих фибробластах, перевиваемых клетках LLC-MK2 с
добавлением трипсина (протеазы необходимы для репликации вируса пара-
гриппа), в клетках НЕК вирусы парагриппа продуцируются без цитопатогенного
эффекта. Продукция вирусов парагриппа в клетках МК, HeLa, HEp-2, Vero сопро-
вождается формированием синцитиев. Наиболее интенсивным цитопатическим
действием (ЦПД) обладает вирус Сендай, который вызывает быструю и полную
дегенерацию клеточного монослоя. Остальные типы парагриппозных вирусов
вызывают медленно развивающиеся цитопатические, не приводящие к деструкции
всего монослоя изменения: округление, зернистое перепождение клеток или обра-
зование вакуолизированных многоядерных симпластов.
Вирусы парагриппа являются возбудителями острых респираторных заболе-
ваний примерно у 20% взрослого населения и до 30% — у детей. Клинические
проявления варьируют от легких поражений верхних дыхательных путей до забо-
леваний нижних отделов легких: ларингита, ложного крупа, бронхита, пневмоний.
Наиболее тяжело протекают заболевания, вызванные ВПГЧ-3, который является
этиологическим фактором 60-70% заболеваний нижних отделов дыхательных
путей у детей первых полутора лет жизни. Передача осуществляется воздушно-
капельным путем. Вирусы типов I, II и III распространены повсеместно и вызыва-
ют заболеваемость в любое время года. Тип IV выделен только в США.
ЧАСТНАЯ ВИРУСОЛОГИЯ 667
Лабораторные методы диагностики
Материалом для исследования являются мазки со слизистой оболочки, носо-
глоточные мазки и смывы, мокрота, моча, кровь. Раннюю диагностику проводят
на основании обнаружения вирусспецифического антигена в эпителиальных клет-
ках слизистой оболочки носа больных методом иммунофлюоресценции.
СЕРОЛОГИЧЕСКАЯ ДИАГНОСТИКА
Серологическую диагностику парагриппозной инфекции проводят в РТГА, РСК
и PH, используют парные сыворотки, взятые с интервалом 10-14 дней. Могут
быть перекрестные реакции с вирусами паротита и кори. В сомнительных случаях
диагноз устанавливают на основе подтверждающих тестов.
Лабораторная диагностика парагриппозной инфекции может быть проведена
с помощью выделения вируса; в практических целях для этого использую! пере-
виваемые кульгуры клеток (преимущественно HeLa, НЕр-2). Для предварительной
индикации вируса может быть использован феномен гемадсорбции эритроцитов
морской свинки на зараженных клетках. При выраженной гемадсорбции в зара-
женных культурах проводят последующее его типирование в РТГА, РСК или PH.
Выделение вируса парагриппа в культуре клеток используют как подтверждающий
тест.
Парагриппозную инфекцию необходимо дифференцировать от аденовирусной,
респираторно-синцитиальной, гриппа А и Б, бактериальных инфекций респира-
торного гракта. Окончательный диагноз устанавливают по совокупности клини-
ческих, эпидемиологических данн ых и лабораторных тестов.
ВИРУС ЭПИДЕМИЧЕСКОГО ПАРОТИТА
Характеристика вируса
Этот вирус отнесен к семейству Paramyxoviridae, роду Rubulavirys. Геном вируса
эпидемического паротита (ЭП) — несегментированная одноцепочная PI IK отри-
цательной полярности, длиной в 15 384 нуклеотида. Геном вируса включает семь
генов. Гены вируса распределены в следующем порядке от 3’- к 5’-концу: NP, V\P, М,
F, SH, HN, L. Генами F и HN кодируются два поверхностных белка: гемагглютинин-
нейраминидаза (HN) и белок слияния (F), которые отвечают за прикрепление и
слияние вирусной оболочки с мембраной клетки. Кроме того, HN-белок обладает
и нейраминидазной активностью, которая позволяет образуемым вирионам отде-
ляться от клеточной поверхности, предотвращать агрегацию вирусных частиц.
HN- и F-антитела обладаю! вирусней грализующей активностью.
Другие пять белков вируса ЭП расположены внутри вириона, образующие-
ся к ним антитела не обладают протективной активностью. Нуклеокапсидный
NP-белок придает спиральную нуклеокапсидную структуру, контактируя с вновь
синтезированной геномной РНК. РНК-транскриптазная активность приписыва-
ется как фосфопротеину (Р), так и большому полимеразному белку (L). Показано,
что матриксный (М) белок участвует в сборке вирусных частиц и отпочковании
вирионов от клеточной поверхности. Два неструктурных белка (V и Г) кодируются
P-геном и являются продуктами синтеза транскрипции матричной РНК. V-белок
ответственен за уход от интерферонопосредованного антивирусного ответа, бло-
кируя сигналы интерферона и ограничивая его выработку Роль 1-белка в репли-
кации вируса пока неизвестна.
SH - трансмембранный белок, отвечает, как и V-белок, за уход от антивирус-
ного ответа хозяина, однако механизм ухода другой. SH-белок блокирует TNF-a-
опосредованный сигнал апоптоза.
668 ЧАСТНАЯ ВИРУСОЛОГИЯ
Вирионы вируса эпидемического паротита в подавляющем большинстве пред-
ставляют собой сферические и нитевидные структуры, их диаметр колеблется в
диапазоне от 100 до 300 нм. Липидная оболочка обволакивает спиралевидную
нуклеокапсидную структуру, состоящую из РНК и белка. На поверхности вирио-
на вируса ЭП присутствуют два поверхностных гликопротеина (HN- и F-белки),
частично включенных в липидную мембрану, и, как в случае с вирусом кори, оба
или один из белков контактир\ ет с белком М, выстилающим липидную оболочку
вируса изнутри. В свою очередь, М-белок контактирует с нуклеокапсидной спира-
левидной структурой.
Вирус ЭП обладает гемагглютинируюшей, нейраминидазной, гемолитиче-
ской и симпластообразуюшей активностью. Культивирование вируса ЭП про-
водят на развивающихся куриных эмбрионах и культурах клеток (первично-
трипсинизированных и клетках перевиваемых линий). Вирусом ЭП заражают
куриные эмбрионы в амниотическую или аллантоисную полость и инкубируют
5 дней при температуре 33-35 °C.
Вирус ЭП малоустойчив во внешней среде, полностью инактивируется при
55-60 °C (в течение 20 мин), а также под действием ультрафиолетового облучения,
0,1% раствора формалина, 1.0% раствора лизола, спирта и эфира. При хранении
при -4 °C инфекционность вируса ЭП снижается в течение нескольких дней,
при -20 °C — нескольких недель, а при -50-70 °C — в течение многих месяцев.
В мире циркулирует 12 генотипов вируса ЭП: A, Y/K, F, В, G, a. D, С, L, Н и
выделенный в особую группу Ленинград-3. Различия в нуклеотидных последо-
вательностях у разных штаммов составляют от 6 до 19%. Такая вариабельность
в нуклеотидной последовательности (определяется по SH-гену) диких штаммов
вируса ЭП может быть использована для дифференциации эндемичных и ввезен-
ных штаммов, а также в случае учета поствакцинальных осложнений, таких как
серозные менингиты и энцефалиты.
При заболевании ЭП противопаротитные антитела обнаруживают на 1-4-й день
после появления первых клинических признаков, концентрация их быстро нарас-
тает и достигает максимального уровня к 40-50-му дню болезни. Первыми появ-
ляются антитела класса М. с ними почти одновременно — низкоавидные антитела
G2-класса. К концу первого месяца антител класса М практически не обнаружива-
ют. Большинство продуцируемых антител у человека — антитела к нуклеокапсиду
вируса, отсутствие этих антител — точный индикатор того, что человек не болел,
не болеет и не был привит. При ЭП в защите от первичной инфекции и реинфек-
ции велика роль антител, которые образуются в ответ на поверхностные антигены
вируса, — антитела к поверхностным HN- и F-белкам, а также секреторных анти-
тел класса А, которые участвуют в защите от инфекции на уровне входных ворот.
Все эти антитела обладают вируснейтрализующей активностью.
Диагностика эпидемического паротита
Диагностика ЭП включает исследование мочи, слюны, носоглоточных смывов
и/или крови. Оптимальным клиническим материалом для идентификации ЭП
являются носоглоточные смывы или слюна. В случае развития серозного менин-
гита осуществляют взятие цереброспинальной жидкости (СМЖ) в объеме 2-3 мл.
Работа с СМЖ по выделению вируса ЭП должна начаться немедленно, в крайнем
случае допускается хранение при температуре -70 °C чуть более суток. Процедуру
получения клинических материалов при ЭП следует проводить либо в больнице,
либо в процедурном кабинете поликлиники, в особых случаях — на дому у паци-
ента.
В типичных случаях клинически поставить диагноз «эпидемический паротит»
нетрудно. Определенные трудности представляет диагностика легких (стертых
ЧАСТНАЯ вирусология 669
и атипичных) и особенно инаппарантных (субклинических) форм паротитной
инфекции. Инаппарантные формы (25-40% всех заболевших) невозможно выя-
вить без специфических лабораторных исследований.
Необходимо дифференцировать ЭП от заболеваний, клиническим проявлением
которых могут быть отеки. Так, отеки могут быть следствием заражения вирусами
Коксаки, парагриппа или вирусом Эпштейна-Барр. Кроме того, очень часто боль-
ных с подозрением на ЭП приходится проверять и на наличие вируса герпеса 6 го
типа, адено-, энтеро- и парвовирусов.
Лабораторные методы диагностики
Среди лабораторных методов диагностики ЭП используют очень редко гисто-
химический, чаще вирусологический и молекулярно-генетический, но предпо-
чтение отдается серологическим. Лабораторная диагностика стала обязательной
составляющей системы эпидемиологического надзора за ЭП.
Вирусологический метод основан на раннем выделении (при первых клиниче-
ских признаках ЭП) возбудителя из лейкоцитов больного на различных культурах
клеток, например Vero, Л-41, Hela. Вирус ЭП также в первые 5 дней заболевания
обнаруживают в клинических образцах, таких как смывы входного отверстия сте-
нового протока, слюна, моча, спинномозговая жидкость
Все чаще, особенно для верификации случая «эпидемический паротит», кроме
вирусологического используют молекулярно-генетические методы — ПЦР с обрат-
ной транскрипцией или ПЦР в реальном времени и секвенирование (анализ после-
довательностей нуклеотидов). Использование метода обратно-транскриптазной
ПЦР (ОТ-ПЦР) подтвердило его преимущество над чувствительностью имму-
ноферментного (ИФ) метода для выявления IgM. С помощью ОТ-ПЦР и ПЦР в
реальном времени можно не только дифференцировать ЭП от других инфекций,
но и провести анализ последовательностей нуклеотидов для генотипирования вак-
цинных штаммов (особенно при возникновении неврологических осложнений),
отслеживать циркуляцию, изменчивость вирусов ЭП, выявлять завозные шгаммы,
подтверждать прекращение трансмиссии вируса на территории региона.
Противопаротитные антит ела определяют с помощью серологических методов:
реакции связывания комплемента (РСК), реакции нейтрализации (PH), которая
до сих пор является «золотым стандартом», реакции торможения гемагглютина-
ции (РТГА) и др. Наиболее перспективным в последние годы считается иммуно-
ферментный анализ (ИФА) В нашей стране для диагностики ЭП используют в
основном непрямой ИФА. Он улавливает весь спектр антител, высокочувствите-
лен (чувствительность находится в пределах 15±5 нг), специфичен (106%), прост
и быстр в постановке Воспроизводимость метода 99,5%. Особенно он оказался
удобным для экспресс-диагностики ЭП. Однако возможно получение ложнополо-
жительных результатов, дальнейшая разработка новых ИФТС, новых подходов к
их конструированию очень актуальна.
ВИРУСЫ - ВОЗБУДИТЕЛИ ГЕМОРРАГИЧЕСКИХ ЛИХОРАДОК
Таксономия
Вирусы, вызывающие геморрагические лихорадки, принадлежат к разным
семействам и родам: ареновирусы — к семейству Arenoviridae (Хунин, Мачупо
Ласса), филовирусы — к семейству Filoviridae (Эбола, Марбург), флавовирусы
семейства Togaviridae (желтая лихорадка, ГЛ Денге), хантавирусы и найровирус —
к семейству bungaviridae (ГЛ с почечным синдромом, крымско-конголезская ГЛ).
Вирусы отдельных семейств различаются стпоением, величиной и формой вирио-
670 ЧАСТНАЯ ВИРУСОЛОГИЯ
нов, видом генома, скоростью репродукции, антигенными свойствами, патогенно-
стью для людей и животных.
Вирусные геморрагические лихорадки — зоонозные (антропонозные) природно-
очаговые, особо опасные инфекции, возбудители которых вызывают тяжелые
острые заболевания, протекающие с выраженным геморрагическим синдромом.
Эпидемиологические проявления этих инфекций, как правило, характеризу-
ются возникновением вспышек или эпидемий той или иной интенсивности.
Естественные механизмы распространения вирусных геморрагических лихо-
радок обусловливают постоянный риск инфицирования сотен тысяч людей,
создавая серьезную угрозу здоровью и жизни населения, тяжелые социально-
экономические последствия.
В соответствии с классификацией вирусов человека и животных, действующей
на территории Российской Федерации, возбудители вирусных геморрагических
лихорадок относятся к I—II группе патогенности.
Диагностические исследования материала, подозрительного на заражен-
ность микроорганизмами I—II группы патогенности, выполняют лаборатории,
имеющие санитарно-эпидемиологическое заключение о возможности проведе-
ния соответствующих работ и лицензию на деятельность, связанную с исполь-
зованием возбудителей особо опасных инфекционных заболеваний человека.
В Российской Федерации работа с патогенными для человека микроорганиз-
мами регламентирована пакетом нормативных правовых актов, утвержденных
Главным государственным санитарным врачом, обязательных для выполнения
всеми учреждениями и организациями страны независимо от их ведомственной
принадлежности.
Крымская геморрагическая лихорадка
ТАКСОНОМИЯ
Крымская геморрагическая лихорадка (КГЛ) — зоонозная природно-очаговая
арбовирусная инфекционная болезнь преимущественно с трансмиссивным и
контактным механизмами передачи возбудителя. Резервуаром и переносчиками
вируса являются более 3 видов иксодовых клещей.
Ютиническая картина КГЛ определяется поражением нервной и сосудистой
систем человека. Инкубационный период длится от 1 до 14 дней (чаще 2-7). В кли-
ническом течении, как и при других геморрагических лихорадках, выделяются три
периода, последовательно сменяющих друг друга: предгеморрагический, геморра-
гический, реконвалссцентный. Возможно инаппарантное течение инфекции.
Возбудителем КГЛ является РНК-содержащий вирус крымско-конголезской
геморрагической лихорадки (ККГЛ), относящийся к семейству Bunyavindae, роду
Nairovirui. В соответствии с принятой в России классификацией патогенных для
человека микроорганизмов вирус ККГЛ относится ко II группе патогенности.
Вирус ККГЛ содержит одноцепочечную РНК имеет сферическую форму и внеш-
нюю липопротеиновую оболочку, диаметр 90-100 нм. РНК вируса ККГЛ сег-
ментирована и состоит из малого, среднего и большого сегментов, кодирующих
нуклеокапсидный белок, оболочечные гликопротеины и белки полимеразного
комплекса соответственно. Малый и большой сегменты РНК высококонсерватив-
ны. Их нуклеотидные последовательности используют как для разработки тест-
систем, выявляющих РНК вируса ККГЛ в пробах различного происхождения, так
и для филогенез ического анализа. Средний сегмент РНК вариабелен, даже в одной
генетической группе наблюдаются значительные различия в нуклеотидной после-
довательности у различных штаммов вируса ККГЛ.
ЧАСТНАЯ ВИРУСОЛОГИЯ 671
УСТОЙЧИВОСТЬ ВО ВНЕШНЕЙ СРЕДЕ, К ФИЗИКО-ХИМИЧЕСКИМ ФАКТОРАМ
Вирус малоустойчив в окружающей среде. Инактивируется при кипячении момен-
тально, при температуре 60 °C разрушается в течение 30 мин, при 37 °C - через 20 ч,
при 45 °C — через 2 ч. Лиофилизированная культура сохраняется до 2 лет. Вирус
чувствителен к ультрафиолетовому облучению и дезинфицирующим средствам.
ЛАБОРАТОРНЫЕ МЕТОДЫ ДИАГНОСТИКИ
Лабораторная диагностика КГЛ основана на выявлении вируса или его структур-
ных компонентов, исследовании динамики титра антител и проводится с исполь-
зованием вирусологических, иммунологических и молекулярно-генетических
методов. Для выявления вируса в организме больных людей берут кровь (на 5-7-й
день болезни для вирусологического и серологического исследований и на 10-
14-й день для серологического исследования).
Отбор проб материала от людей (трупов), а также подозрительных больных
осуществляется медицинским персоналом в соответствии с требованиями санитар-
ных правил «Безопасность работы с микроорганизмами I—II групп патогенности*.
КЛИНИЧЕСКИЙ МАТЕРИАЛ
Материалом для выделения вируса ККГЛ являются цельная кровь, плазма,
сгусток крови, секционный материал (пе«ень, легкие, селезенка, почки, головной
мозг), полущенный в ранние сроки от начала заболевания (до 7-го дня), то есть в
период вирусемии. Эффективность вирусологических исследований в этот период
очень высока и приближается к 100%. При нормализации температуры тела воз-
можность выделения вируса резко снижается, а при появлении в крови вирусспе-
цифических 1g класса G (10-14-й день) — становится невозможной.
Вирус ККГЛ размножается в перевиваемых клетках линий LLC-MK2, ВНК-
21, Vero без цитопатического эффекта. При исследовании монослоя зараженных
клетик с помощью МФА через 24-48 (72) ч вирусный антиген выявляется в
цитоплазме клеток в виде светящихся мельчайших зернышек и гранул различной
величины, сливающихся в диффузну ю массу.
Вирус довольно трудно адаптируется к лабораторным животным, у которых
инфекция часто протекает бессимптомно. Исследуемый материал вводят интраце-
ребрально двенадцатидневным сосункам белых мышей. Инкубационный период
при первичном заражении составляет 5-12 дней. Через 24 и 48 ч часть мышей
забивают и из мозга готовят мазки-отпечатки для МФА и суспензии для РИГА.
При получении отрицательных результатов исследования этого материала за
оставшимися мышами наблюдают еще трое суток, после чего подвергают их мозг
такому же исследованию.
СЕРОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ
Исследования на обнаружение антигена или определение антител к вирусам
II группы патогенности в связи с отсутствием регламентированных методов инак-
тивации вирусов проводят только в боксированном помещении или в боксе био-
логической безопасности.
Выявление вирусного антигена и специфических антител класса М (IgM) к виру-
су ККГЛ в сыворотке (плазме) крови человека проводят с целью ранней дифферен-
циальной диагностики КГЛ в клинических и эпидемиологических исследованиях.
Подтверждением диагноза является нарастание уровня антител класса G (IgG) в
парных сыворотках крови (интервал — 10-14 сут). Серодиагностику заболевания
осуществляют с помощью ИФА, РАО, РСК, РДПА, РТНГА и непрямого МФА.
МОЛЕКУЛЯРНО-ГЕНЕТИЧЕСКИЕ ИССЛЕДОВАНИЯ
Для выявления в исследуемом материале специфичной РНК вируса ККГЛ
используют метод полимеразной цепной реакции с обратной транскрипцией
672
ЧАСТНАЯ ВИРУСОЛОГИЯ
(ОТ-ГШР). Учет результатов проводят методом электрофореза в агарозном геле
или в режиме реального времени.
ПЦР-диагностика в режиме реального времени позволяет не только определить
РНК вируса ККГЛ в крови, но и установить уровень и продолжительность вире-
мии, а также оценить эффекгивность противовирусной терапии.
ОЦЕНКА РЕЗУЛЬТАТОВ СПЕЦИФИЧЕСКОЙ ЛАБОРАТОРНОЙ ДИАГНОСТИКИ
Выявление РНК и/или антигена ККГЛ в исследуемом материале, взятом на
ранних сроках заболевания (до 5-7-го дня), свидетелоствует об инфицирован-
ности больных и в совокупности с данными эпидемиологического анамнеза, кли-
нической картины и результатами клинической лабораторной диагностики может
считаться основанием для постановки диагноза «КГЛ».
IgM-антитела появляют ся на 5-7-й день от начала заболевания, достигая мак-
симальных титров (1:20 000 и более) на 2-3-й неделе, и постепенно исчезают к
90-м суткам. Специфические IgG-антитела появляются на 7-1 Ой день, достигая
пика к концу 2-го месяца, держатся в высоких титрах 6-8 мес, постепенно снижа-
ясь к концу года, у некоторых больных сохраняются годами. Для постановки диа-
гноза необходимо четырехкратное и более нарастание титра IgG при исследовании
парных сывороток. При обнаружении IgM в титре 1:800 и более и IgG в любом
титре диагноз «КГЛ» считается подтвержденным.
Геморрагическая лихорадка с почечным синдромом
Геморрагическая лихорадка с почечным синдромом (ГЛПС) — зоонозная
природно-очаговая инфекционная болезнь с преимущественно аэрогенным
(воздушно-капельным и воздушно-полевым) механизмом заражения людей.
Возможно также алиментарное заражение при употреблении продуктов питания,
контаминированных экскрементами грызунов, и контактное — через поврежден-
ные участки кожи.
На территории России циркуляция хантавирусов отмечена в популяциях
47 видов животных, из них основные — рыжая полевка, полевая мышь, серая и
черная крысы, разные виды серых полевок.
ХАРАКТЕРИСТИКА ВОЗБУДИТЕЛЯ
Возбудитель ГЛПС относится к семейству Bunyaviridae, роду Hantavirus, сероти-
пы: Хантаан, Сеул, Пуумала, Добрава. Тула, Хабаровск, Топографов, возбудитель
геморрагической лихорадки с легочным синдромом. Вирионы хантавирусов сфе-
рической формы, диаметром 90-120 нм, морфологически однородны. Хантавирус
имеет липидную оболочку, три фрагмента РНК-отрицагельной полярности, два
поверхностных гликопротеина и нуклеокапсидный белок (NP). Малый сегмент (S)
генома хантавируса кодирует NP-белок, средний сегмент (М) кодирует два поверх-
ностных гликопротеина: G1 и G2, а самый большой (L) — сегмент белка полиме-
разы. NP-белок ответственен за продукцию антител и проявляет высокую кросс-
реактивность по отношению к другим штаммам хантавирусов. Поверхностные
белки гликопротеина индуцируют низкий иммунологический ответ, но проявляют
специфическую активность в отношении других штаммов.
Вирус чувствителен к эфиру, хлороформу, ацетону, бензолу, ультрафиолетовым
лучам, дезинфицирующим веществам, обычно применяемым для уничтожения
вирусов.
ЛАБОРАТОРНЫЕ МЕТОДЫ ДИАГНОСТИКИ
Вирус размножается в легких диких грызунов, не вызывая у них заболевания.
Методом выявления антигена вируса в суспензиях легких грызунов является
иммуноферментный анализ (ИФА).
ЧАСТНАЯ вирусология 673 I
Культивирование вируса осуществляют в перевиваемых культурах клеток кар-
циномы легких человека или VeroE6. |
Основой лабораторной диагностики ГЛПС является серологическая. Материалом |
для исследования является сыворотка крови больных ГЛПС или подозритель- |
ных на это заболевание, взятая в разные сроки: на начальном этапе заболевания
и через 2-3 нед. Основным методом серодиагностики ГЛПС является непрямой |
метод флюоресцирующих антител (МФА). Специфические иммуноглобулины в |
диагностических титрах выявляются уже через 3 дня после начала заболевания, |
достигая максимума на 8-25-й день болезни, регулярно обнаруживаются 3-4 мес,
сохраняются у реконвалесцентов от 6 мес до 2-3 лет.
Выявление РНК хантавирусов в клинических и полевых материалах осу-
ществляют методом полимеразной цепной реакции с обратной транскрипцией
(ОТ-ПЦР). Для подтверждения диагноза «ГЛПС» все результаты серологической
и молекулярно-генетической диагностики должны быть сопоставлены с клиниче-
скими и эпидемиологическими данными.
ВИРУСЫ - ВОЗБУДИТЕЛИ ГЕПАТИТОВ
Вирус гепатита А
ТАКСОНОМИЯ
Вирус гепатита A (HAV — Hepatitis A Virus) — возбудитель гепатита А челове-
ка — относится к роду Hepatovirus, семейству Picomaviridae. Изоляты HAV подраз-
деляются на шесть генотипов, которые обозначают римскими цифрами от I до VI.
Генотипы вируса I, II, III вызывают заболевание у человека, а генотипы IV, V VI — у
обезьян. Различия между изолятами HAV, выделенными в различных регионах
мира, по нуклеотидной последовательности геномной РНК составляют 15-25 и
7,5% на уровне генотипов и субтипов соответственно. При этом аминокислотные
последовательности изолятов HAV достаточно консервативны, чем объясняет-
ся наличие всего одного серотипа данного ВВГ. Штаммы вируса, выделенные в
России, принадлежат к субтипам IA, IB, IIIА.
ХАРАКТЕРИСТИКА ВОЗБУДИТЕЛЯ
Морфологически HAV представляет собой сферические частицы диаметром
27-30 нм, построечные по типу’ икосаэдрической симметрии и лишенные обо-
лочки. Вирусный геном представлен линейной одноцепочечной РНК, состоящей
из приблизительно 7500 нуклеотидных оснований, с общей молекулярной массой
порядка 2,2 Md. В нем определяются: 5’ — нетранслируемая область; единичная
рамка считывания, кодирующая структурные и неструктурные белки, и короткая
3’ — нетранслируемая область, заканчивающаяся поли-А-трактом. Геном HAV
имеет положительную полярность, то есть вирусная РНК непосредственно выпол-
няет матричные функции. Капсид HAV построен из множественных копий четы-
рех структурных протеинов, обозначаемых как VP1, VP2, VP3 и VP4.
HAV является одним из наиболее устойчивых вирусов человека к факторам
внешнего воздействия. При 60 СС он полностью сохраняется в течение 1 ч и лишь
частично инактивируется за 10-12 ч, однако кипячение приводит к разрушению
вируса за несколько минут. HAV остается инфекционным в течение, по крайней
мере, одного месяца после высушивания на твердой поверхности в условиях
обычного помещения (25 ° С и 42% влажности). Он устойчив к растворителям
жиров, таким как эфир, хлороформ и др. Эффективным методом стерилизации
материалов, содержащих HAV, является автоклавирование. Дезинфицирующие
средства также достаточно эффективны: хлорамин в концентрации 2,0-2,5 мг/л
полностью устраняет инфекционность ВГА за 15 мин, перманганат калия
674 ЧАСТНАЯ ВИРУСОЛОГИЯ
(30 мг/л) — за 5 мин, 3% формалин — за 5 мин (при 25 °C), йодистые соединения
(3 мг/л) — за 5 мин.
ЛАБОРАТОРНЫЕ МЕТОДЫ ДИАГНОСТИКИ
Специфическую лабораторную диагностику гепатита А осуществляют по выяв-
лению вируса, антигена вируса, антител классов IgM и IgG, а также РНК H.AV.
Клиническими образцами для лабораторной диагностики являются фекалии и
кровь больного. Вирус и его антиген могут быть выявлены в фекалиях с помищью
иммуноэлектронной микроскопии, твердофазного радиоиммунного, иммунофер-
ментного и хемилюминесцентного анализа. В связи с тем что массивная экскреция
вируса имеет место до развития симптомов заболевания и быстро прекращается
после появления желтухи, методы определения вируса и его антигена для диагно-
стики ГА используют сравнительно редко.
Значительно большее применение в диагностике ГА имеют методы определения
антител классов IgM и IgG к вирусу в сыворотке крови. Выявление анти-HAV-IgM
является «золотым стандартом» лабораторной диагностики гепатита А. Эти анти-
тела становятся определяемыми с конца инкубационного периода и продолжают
циркулировать до 3-6 мес. Диагноз ГА может основываться и на определении
антител класса IgG, но в этом случае необходимо исследовать сыворотку крови
больного дважды с интервалом не менее 2 нед. В случае увеличения титра анти-
HAV-IgG в четыре и более раз можно сделать заключение о наличии у пациента ГА.
Специфические антитела к вирусу ГА можно определить с помощью радиоиммун-
ного, иммуноферментного или хемилюминесцентного анализа.
В последние годы широкое распространение, для диагностики ГА получила
полимеразная цепная реакция, которая позволяет выявить РНК вируса в образцах
фекалий или крови больного. В силу большего удобства работы с сыворотками
крови именно эти клинические образцы исследуют наиболее часто РНК HAV в
сыворотке крови выявляется у 80% больных в течение первого месяца после появ-
ления желтухи, а у некоторых пациентов — даже 2 мес и более.
Вирус гепатита В
ХАРАКТЕРИСТИКА ВОЗБУДИТЕЛЯ
Вирус гепатита В (HBV- Hepatitis В virus) — возбудитель гепатита В. основной
представитель семейства гепаднавирусов.
HBV (частица Дейна) — сферическая частица диаметром 42 нм, состоит из
ядра — нуклеоида, имеющего форму икосаэдра диаметром 28 нм, вну три которого
находятся двухцепочечная ДНК, концевой белок и фермент ДНК-полимераза.
В состав серцевинного белка — HBcAg входит HBeAg. Внешняя оболочка образо-
вана поверхностным антигеном вируса гепатита В — HbsAg.
Геном HBV представлен двухцепочечной кольцевой молекулой ДНК — наимень-
шей из всех ныне идентифицированных ДНК. ДНК HBV состоит приблизительно
из 3 200 нуклеотидов. Наружная минус-цепь длиннее внутренней плюс-цепи на
15-45%. Минус-цепь в двухнитевой части имеет разрыв в 5’-конце, к которому
ковалентно присоединен белок. ДНК HBV содержит четыре гена (S, С, Р, X) и
регуляторные последовательности, ответственные за синтез белков и репликацию
вируса. Открытые рамки считывания определенных генов частично перс крывают
друг друга, что обеспечивает высокую информационную емкость генома.
Популяция HBV достаточно гетерогенна. Серологическими методами было
показано существование субтипов поверхностного антигена вируса — HBsAg. При
этом имеется одна общая детерминанта а, которая в комбинации с двумя дополни-
тельными детерминантами d/y и w/r образует четыре субтипа HBsAg: adw, ay w, adr,
ayr. В настоящее время описаны новые варианты детерминант, и общее количество
ЧАСТНАЯ ВИРУСОЛОГИЯ
675
субтипов достигло девяти. На основе анализа нуклеотидных последовательностей
S-гена изоляты HBV, выделенные в различных регионах мира, объединили в
восемь основных генотипов, которые обозначили заглавными буквами латинско-
го алфавита: А, В, С, D, Е, F, G и Н. Следует отметить, что полного соответствия
между генотипами HBV и серотипами HBsAg не установлено. Так, изоляты HBV,
относящиеся к четырем совершенно разным генотипам — А, В, С и F, могут иметь
один и тот же серотип adw2. В России доминирующим генетическим вариантом
HBV является генотип D, но в некоторых регионах страны от пациентов выделены
вирусы, относящиеся к генотипам А и С.
При электронной микроскопии определяются две морфологические формы:
частицы, имеющие оптически плотное ядро, содержащие ДНК HBV, и неполные
частицы — без ДНК. Концентрация вирусных частиц в сыворотках крови с нали-
чием HBsAg колеблется от 10 частиц в 1 мл до количеств, недоступных выявлению
с помощью электронной микроскопии. Некоторые сыворотки крови с наличием
HBV инфекционны даже в разведениях 10 7—10 8.
Инфекционность HBV в сыворотке крови сохраняется при 30-32 °C в течение
6 мес, при -20 ЭС — 15 лет, после прогревания до 60 СС — 4 ч, при 98 °C — в течение
1 мин сохраняется частично, а через 20 мин исчезает полностью, при обработке
сухим жаром (-Н60 °C) разрушается в течение 1 ч; обработка {3-пропиолактоном
в сочетании с ультрафиолетовым облучением снижает инфекционность HBV-
содержащей плазмы примерно в 10 млн раз. Частицы HBV чувствительны к эфиру
и неионным детергентам, которые разрушают внешнюю оболочку вириона, осво-
бождая при этом нуклеокапсид.
Многочисленные попытки культивирования HBV в различных клеточных куль-
турах были неудачны.
КЛИНИЧЕСКИЙ МАТЕРИАЛ
Поскольку вирус в огромных количествах содержится в крови больных остры-
ми и хроническими формами гепатита В, основным клиническим материалом для
лабораторной диагностики инфекции является сыворотка крови. Серологические
маркеры вируса (HBsAg, HBeAg) и его геномный материал (ДНК HBV) могут
быть обнаружены в конце инкубационного периода, однако такая возможность
диагностики крайне ограничена, поскольку в этот период отсутствуют специфи-
ческие симптомы поражения печени и больные не обращаются за медицинской
помощью.
ЛАБОРАТОРНЫЕ МЕТОДЫ ДИАГНОСТИКИ
Специфическая лабораторная диагностка острых форм гепатита В основана на
выявлении в сыворотке крови антигенов HBV (HBsAg, HBeAg) и антител к серце-
винному антигену вируса класса IgM (IgM анти-НВс) методами радиоиммунного,
иммуноферментного или хемилюминесцентного анализа. Эксперты ВОЗ считают,
что при лабораторной диагностике острых форм герпес-вирусов предпочтение
должно отдаваться определению IgM анти-НВс. Только при отсутствии возмож-
ности детекции этого маркера лабораторный диагноз может опираться на данные
о наличии в сыворотке крови HBsAg. Обнаружение HBeAg в сыворотке крови
коррелирует с присутствием HBV ДНК. В случае выздоровления маркеры вируса
HBsAg, HBeAg и HBV ДНК перестают выявляться в сыворотке крови, а на смену
приходят серологические маркеры сероконверсии к антигенам вируса — анти-
HBcIgG, анти-НВе, анти-HBs. Протективной защитой обладают анти-HBs.
В случае выявления HBsAg в течение 6 мес и более диагностируется хрони-
ческая HBV-инфекция. Маркерами активно текущей хронической инфекции
являются HBeAg, HBV ДНК и IgM анти-НВс. Со временем у большинства боль-
ных с хронической инфекцией происходит сероконверсия HBeAg в анти-НВе.
676
ЧАСТНАЯ ВИРУСОЛОГИЯ
Длительная персистенция HBeAg является неблагоприятным фактором развития
цирроза печени. ДНК HBV может быть выявлена в сыворотке крови с помощью
технологий амплификации нуклеиновых кислот (полимеразной цепной реакции,
технологии разветвленной ДНК — bDNA, NASBA и др.).
Вирус гепатита С
ХАРАКТЕРИСТИКА ИРУСА
Вирус гепатита С (HCV — Hepatitis С Vims) — возбудитель гепатита С. Ранее
обозначали как вирус, вызывающий гепатит ни А. ни В, передающийся паренте-
рально. Результаты клонирования и полного секвинирования РНК ВГС, а также
физико-химические характеристики вируса позволили отнести ВГС к семейству
Flavivindae, выделив в отдельный род Hepacivims.
Определены следующие характеристики вируса: диаметр вириона — от 30 до
60 нм; чувствителен к действию хлороформа, коэффициент седиментации — не
менее 150 S. ВГС инактивируется при температуре 60 °C — в течение 30 мин, а
при 100 °C — за 2 мин. Геном ВГС представлен однонитевой линейной молеку-
лой РНК положительной полярности протяженностью около 9400 нуклеотидов.
Организация генома HCV подобна таковой геномов других флавивирусов. В РНК
HCV выделяют зоны, кодирующие структурные и неструктурные (функциональ-
ные) белки. Структурные белки представлены нуклеокапсидным белком (core —
С), двумя оболочечными белками (Е1 и Е2), считающимися гликопротеидами
внешней оболочки HCV (Env). Они играют роль в прикреплении и проникновении
вируса в клетку. Область РНК ВГС, кодирующая неструктурные белки, состоит из
следующих участков: NS2, NS3, NS4a, NS4b, NS5a и NS5b. Все эти белки являются
необходимыми для процесса репродукции вируса. Они несут антигенные детерми-
нанты, на которые вырабатываются антитела.
HCV характеризуется самой высокой генетической вариабельностью генома
среди возбудителей вирусных гепатитов. Все изоляты HCV группируются в шесть
главных генотипов, обозначаемых арабскими цифрами от 1 до 6 с гомологией
нуклеотидных последовательностей в РНК между ними менее 72% Внутри каж-
дого генотипа выделяют субтипы, которые включают изоляты. состоящие из
квазивидов. На принадлежность изолята к определенному субтипу указывают
прописные латинские буквы, которые пишут вслед за арабской цифрой: HCVla,
HCVlb. HCVlc, HCV2a и т.д. В настоящее время известно более ста субтипов HCV.
Уровень гомологии между различными субтипами внутри генотипа 72-86%. При
генотипировании HCV, проведенном в различных регионах мира, установлено,
что наибольшее распространение имеют генотипы la, lb, 2а, 2Ь и За. В России
наибольшее распространение имеют два субтипа — 1b и За. Остальные субтипы
обнаруживают сравнительно редко.
С точки зрения молекулярной эпидемиологии важно знать, что изоляты гено-
типа TV доминируют в Северной Африке, генотипа V — в Южной Африке, гено-
тип VI — в Юго-Восточной Азии.
ЛАБОРАТОРНЫЕ МЕТОДЫ ДИАГНОСТИКИ
Лабораторная диагностика ГС основана на определении антител к различным
белкам вируса и выявлении РНК вируса ГС.
Серологическая диагностика
Определение антител (анти-HCV) осуществляют с помощью (ИФА) тест-систем,
которые позволяют определять антитела к широкому спектру белков вируса ГС
(core, NS3, NS4, NS5), и их специфичность достигает 99%. В связи с отсутствием
общепринятых стандартов четких данных о чувствительности различных тест-
ЧАСТНАЯ ВИРУСОЛОГИЯ 677
систем привести нельзя. Тем не менее считается, что этот показатель достигает
97-98% при обследовании иммунокомпетентных лиц.
В последние годы разработаны тесты для экспрессного определения анти HCV,
основанные на иммупох имических и других принципах. Указанные тесты облада-
ют значительно ме.чылими чувствительностью и специфичностью по сравнению
со стандартным ИФА, поэтому результаты, полученные при их применении, могут
считаться сугубо предварительными и подлежат обязательному подтверждению.
Несмотря на высокую специфичность современных ИФА-систем, они могут
показать ложноположительные результаты при тестировании образцов крови,
поэтому требуется использование подтверждающих тестов и методов. Одним из
признанных методов подтверждения присутствия антител является иммуноблот-
тинг. Иммуноблоттинг (англ, biot — «пятно») основан на дифференцированном
определении антител к различным антигенам вируса (С, NS3, NS 4 и NS5), которые
в виде дискретных фракций (пятен) фиксируются на нитроцеллюлозных полосках
и после инкубации с исследуемой сывороткой проявляются мечеными антителами
так же, как в обычном ИФА. Результат считаете я положительным при выявлении
антител к двум и более антигенам.
Молекулярно-биологическая диагностика
Молекулярно-биологическая диагностика (определение РНК HCV) включает
качественную, количественную характеристику РНК вируса и определение гено-
типической принадлежности исследуемых изолятов вируса.
Качественное определение РНК НСV
Качественное определение РНК HCV осуществляют с помощью классической
обратно транскриптазной полимеразной цепной реакции (ОТ-ПЦР), ПЦР в реаль-
ном времени или транскриптазоопосредованной амплификации. Диагностические
препараты, основанные на перечисленных технологиях, должны определять не
менее 50 МЕ/мл РНК HCV.
Количественная характеристика РНК
Количественная характеристика РНК может быть дана с помощью конкурент-
ной ПЦР, ПЦР в реальном времени или технологии рДНК (bDNA).
Стандартным (классическим) методом генотипирования вируса ГС является
прямое секвенирование NS5B- или El-регионов генома с последующим сравни-
тельным анализом полученных последовательностей нуклеотидов с известными
(прототипными) последовательностями. Этот подход позволяет определить не
только генотип изолята, но и его субтиповую принадлежность и отличие (сход-
ство) от других циркулирующих изолятов. Он является основой молекулярной
эпидемиологии ГС.
В клинической практике достаточно определить генотип вируса. Это может
быть осуществлено с помощью коммерческих тестов, определяющих различия в
последовательностях нуклеотидов 5‘-нетранслируемои области.
Специфическая лабораторная диагностика острого гепатита С
Использование серологических и молекулярно-биологических методов осу-
ществляют одновременно. На основании различных сочетаний анти-HCV и РНК
HCV выделяют четыре маркерных профиля.
• Положительный результат определения РНК HCV при отрицательном результа-
те определения анти-HCV указывает на начало острой инфекции, когда антитела
еще не успели выработаться; они появятся через несколько дней/недель.
• Положительный результат определения РНК HCV и анти-HCV. В этом случае
сложно разделить острый ГС и обострение хронического ГС. Кроме этого
возможен острый гепатит иной этиологии на фоне хронического ГС.
678
ЧАСТНАЯ ВИРУСОЛОГИЯ
• Отрицательный результат при определении РНК HCV и анти-HCV, скорее
всего, свидетельствует, что у обследуемого пациента с клинической картиной
острого гепатита не подтверждается диагноз гепати га С.
• Отрицательный результат определения РНК HCV при положительном резуль-
тате определения анти-HCV — такой маркерный профиль обычно нехаракте-
рен для острого ГС, Пациенты должны быть повторно обследованы на РНК
через несколько недель, поскольку отсутствие РНК может быть временным
при развитии хронического процесса. Выявление анти-HCV при постоянно
отрицательных результатах определения РНК чаще всего является свиде-
тельством перенесенного в прошлом ГС, завершившегося выздоровлением.
Кроме того, возможен постоянный ложноположительный результат исследо-
вания на анти-HCV, причины которого не совсем ясны.
Специфическую лабораторную диагностику хронического гепатита С также
проводят с помощью серологических и молекулярно-биологических методов. При
клинико-биохимических проявлениях хронического гепатита текущий хрониче-
ский ГС документируется наличием в крови как анти-HCV, так и РНК HCV.
У пациентов с хроническим ГС, который развился на фоне иммунодепрессив-
ных состояний (больные на хроническом гемодиализе, пациенты с агаммаглобу-
линемией и др.), результат определения анти-HCV может быть отрицательным.
Важно отметить, что оба маркера ГС (анти-HCV и РНК HCV) не имеют прогно-
стического значения, так как не коррелируют с выраженностью воспалительного
процесса в печени и фиброза печени.
Антивирусную терапию назначают только при положительном результате
исследования на РНК HCV. Перед началом терапии определяют генотип вируса,
так как от этого зависят продолжительность лечения и дозы назначаемых препа-
ратов. Для мониторирования эффективности лечения до начала лечения опреде-
ляют количественную характеристику РНК вируса (вирусную нагрузку), которую
повторно контролируют на 12-й неделе лечения в случае лечения пациента с
генотипом 1.
В случае лечения пациентов с гено типами 2 или 3, а также 4, 5 или 6 проводят
качественное определение РНК HCV перед началом и в конце лечения, а также
спустя 24 нед после завершения лечения для установления очищения от вируса.
Вирус гепатита D
ТАКСОНОМИЯ
Вирус гепатита D (HDV — Hepatitis Delta virus) — возбудитель гепатита D (дельта-
гепатита). HDV не принадлежит ни к одному из известных семейств вирусов
животных. По своим свойствам этот вирус наиболее близок к вироидам и сател-
литным вирусам растений. Он является частью трансмиссивного агента (5-агента,
или ВГЭ), вызывающего гепатит, отличный от других вирусных гепатитов.
ХАРАКТЕРИСТИКА ВОЗБУДИТЕЛЯ
HDV - сферическая частица со средним диаметром 36 нм (с колебаниями
от 28 до 39 нм), состоящая из ядра (HDAg) и внешней оболочки, образованной
поверхностным антигеном вируса В (HBsAg). Геном ВГЮ представлен одноните-
вой циклической молекулой РНК, состоящей приблизительно из 1700 нуклеоти-
дов. Анализ нуклеотидных последовательностей РНК ВГО нескольких изолятов
выявил их частичную гетерогенность (от 72,1 до 92,0% гомологии). На этом осно-
вании изоляты HDV были сгруппированы в три генотипа, обозначаемые римскими
цифрами I, II, III,
При нагревании ВГВ происходит укрепление связи HDAg с РНК, которая
становится устойчивой к действию мочевины, гуанидина или меркаптоэтанола.
ЧАСТНАЯ ВИРУСОЛОГИЯ В79
8-Антиген устойчив к нагреванию, действию кислот и нуклеаз, но разрушается в
присутствии щелочей и протеаз. Он обладает антигенной активностью и может
быть использован для конструирования диагностических препаратов.
Вирус гепатита D не может участвовать в развитии гепатитной инфекции без
одновременной репликации вируса гепатита В. Этим фактом определяются две
возможные формы их взаимодействия: одновременного инфицирования HBV и
HDV — коинфекция и инфицирования HDV носителя вируса гепатита В — супе-
ринфекция. Характер взаимосвязи этих вирусов определяется не только исполь-
зованием HBsAg для формирования внешней оболочки HDV, но и другими,
до конца не понятными взаимодействиями. Так, HDV ингибирует репликацию
HBV, приводя к уменьшению экспрессии HBcAg и HBsAg и угнетению актив-
ности ДНК-полимеразной активности в течение острой инфекции. Одним из
возможных объяснений этого факта являются данные о стимуляции 8-вирусом
внутриклеточного синтеза интерферона, который вызывает ингибицию репли-
кации HBV.
8-Вирус может быть обнаружен в гепатоцитах и в крови больных с острой и
хронической 8-инфекцией. Маркерами, свидетельствующими о наличии 8-вируса
в исследуемом материале, служат 8-антиген и/или РНК HDV.
Для обнаружения 8-антигена в биопсийном или аутопсийном материале при-
меняют метод флюоресцирующих антител и иммунопероксидазные методы.
В сыворотке крови 8-антиген определяют с помощью иммуноферментного, хеми-
люминесцентного или радиоиммунного анализа лишь только после тою, как
исследуемый материал обрабатывают неионными детергентами (твином, трито-
ном и др.), разрушающими наружную оболочку вируса, тем самым делая доступ-
ными антигенные детерминанты HbAg, Кроме того, диагностическое значение
имеет выявление антител классов IgM и IgG к 8-антигену.
Детекцию РНК HDV проводят с помощью обратно-транскриптазной полиме-
разной цепной реакции, позволяющей выявлять до 103 молекул РНК HDV.
Вирус гепатита Е
ТАКСОНОМИЯ
Вирус гепатита Е (HEV — Hepatitis Е Virus) — возбудитель гепатита Е, ранее
известного как гепатит ни А, ни В, с фекально-оральным механизмом передачи.
По основным фенотипическим признакам HEV мог бы быть причислен к семей-
ству калицивирусоь, однако состав и организация его генома полностью исключа-
ют эту таксономическую принадлежность. Можно лишь предполагать, что HEV в
единственном пока числе представляет отдельный род вирусов.
ХАРАКТЕРИСТИКА ВОЗБУДИТЕЛЯ
Частицы ВГЕ — округлые образования диаметром около 32 нм (вариации от 27
до 34 нм), построенные из идентичных структурных элементов и лишенные обо-
лочки. Вирусным геномом является одноцепочечная РНК позитивной полярности.
Клонирование генома вируса позволило характеризовать РНК HEV по химиче-
скому строению и генным функциям. Полноразмерная РНК насчитывает около
7500 нуклеотидных оснований и включает три независимые открытые рамки
считывания (ОРС), каждая из которых кодирует синтез определенного белка или
группы белков.
HFV хорошо сохраняется при температуре -20 °C и ниже, однако хранение при
температурах выше 0 °C и особенно последовательное замораживание-оттаивание
ведет к быстрому разрушению вируса. Он чувствителен к воздействию хлор- или
йодсодержащих дезинфектантов.
г чку >
680 ЧАСТНАЯ ВИРУСОЛОГИЯ
ЛАБОРАТОРНЫЕ МЕТОДЫ ДИАГНОСТИКИ
Лабораторная диагностика гепатита Е основана на определении антител к виру-
су классов IgM и IgG.
ГЕРПЕС-ВИРУСЫ
Таксономия
Семейство герпесвирусов (Herpesviridae — от греч. herpes — «ползучая») объеди-
няют пантропность к органам и тканям, пожизненная персистенция в организме
человека и способность вызывать многообразные манифестные формы заболева-
ния в условиях возникновения иммунодефицита. Вирусы семейства Hetpesviridae
широко распространены в природе. В настоящее время выделено и классифици-
ровано свыше 100 представителей этого семейства, вызывающих заболевания у
человека, диких и домашних животных, земноводных и пресмыкающихся. Все
герпес-вирусы ДНК-содержащие, сходные по морфологии, типу нуклеиновой
кислоты, способу репродукции в ядрах инфицированных клеток и размерам, а
также по способности индуцировать латентную, острую и хроническую инфекции
у человека.
По решению Комитета экспертов по таксономии в настоящее время разработана
и принята классификация герпес-вирусов, где семейство герпес-вирусов подразде-
ляют на три подсемейства — а, 0, у. В табл. 24-5 представлены и основные клини-
ческие формы инфекции, обусловленные герпес-вирусами.
Таблица 24-5. Герпес-Вирусы человека и основные клинические формы инфекций
Подсемейство Род Вирусы Клинические формы инфекции
Альфа-герпес- вирусы {Alphaherpes virinae) Вирусы простого герпеса (Simplexvirus) Вирус простого герпеса 1-го типа Н. Simplex Virus type 1 (HSV-1) Лабиальный герпес. Герпес кожи и слизистых оболочек. Офтальмогерпес. Пневмониты. Генитальный герпес. Герпетические энцефалиты
Вирусы простого герпеса 2-го типа Н. Simplex Virus type 2 (HSV-2) Генитальный герпес. Неонатальный герпес
Вирусы ветряной оспы — опоясы- вающего герпеса (Varicellovirus) Вирусы ветряной оспы Varicella Zoster Virus type 3 (VZV) Ветряная оспа. Опоясывающий герпес
Бега-герпес- вирусы {Betaherpes- virinae) Цитомегаловирус (Cytomegalovirus) Цитомегаловирус Cytomegalovirus type 5 (CMV) Врожденные поражения ЦНС. Ретинопатии, пневмониты. Гепатиты,сиаладениты
Вирусы, образую- щие розеолы (Roseolovirus) Вирус герпеса человека 6A типа Human HerpesVvirus type 6A (HHV-6A) Лимфотропные вирусы (предпола- гают этиологическую связь ВГЧ-6В с внезапной экзантемой)
Вирус герпеса человека 6В типа Human HerpesVirus type 6В (HHV-6B)
Вирус герпеса человека 7-го типа Human HerpesVirus type 7(HHV-7) Предполагают этиологическую связь с синдромом хронической усталости
ЧАСТНАЯ вирусология 681
Окончание табл. 24-5
Подсемейство Род Вирусы Клинические формы инфекции
Гамма-герпес- вирусы (Gammatierpes- virinae) Лимфогропные вирусы (Lymphocryptovirus) Вирус Эпштейна-Барр Epsteina-Barr Virus type 4 Инфекционный мононуклеоз. Лимфома Беркипа. Гемофагоцитарный синдром. Лимфопролиферативные заболевания: лимфогранулематоз; неходжкинские лимфомы Рак носоглотки, желудка, слюнных желез, злокачественная лейкоплакия полости рта. Лейомиосаркома
Вирус, ассоцииро- ванный с саркомой Капоши (Rhadinovirus) Вирус герпеса человека 8-го типа Human HerpesVirus type 8 (HHV-8) Саркома Капоши у ВИЧ- серонегативных людей. Саркома Капоши, ассоциированная с ВИЧ-инфекцией и СПИДом. Лимфопролиферативные заболе- вания: • лимфома первичного экссудата; • многоочаговое заболевание Кастле мана
Примечание: Н. — Herpes.
АЛЬФА-ГЕРПЕС-ВИРУСЫ
Альфа-герпес-вирусы (Alphagerpesvirinae) характеризуются коротким циклом
репродукции с цитопатическим эффектом в клетках инфицированных культур,
патогенны для большинства хозяев. Обычно персистируют в центральной нерв-
ной системе — в сенсорных ганглиях, поддерживая латентную инфекцию, которая
нередко проявляется как периодически обостряющееся заболевание.
БЕТА-ГЕРПЕС-ВИРУСЫ
Бета-герпес-вирусы (Bethaherpesvirinae) вызывают манифестную и латентную
инфекции в слюнных железах, почках и других органах. Могут быть причиной
генерализованных поражений у новорожденных и взрослых при иммунодефицит-
ных состояниях.
ГАММА ГЕРПЕС-ВИРУСЫ
Гамма-герпес-вирусы (Gammaherpesvirinae) характеризуются тропизмом к В-
или Т-лимфоцитам (но репродуцируются в основном в В-клетках), лимфоидным
клеткам, в которых они способны размножаться и длительно персистировать. Эти
вирусы нередко являются причинами тяжелых, смертельных лимфом и лейке-
мий. Патогенные для человека вирусы — вирус Эпштейна-Барр (ВГЧ-4) и ВГЧ-8
(см. табл. 24-5).
Характеристика вирусов
Размеры зрелого вириона — 120-200 нм в диаметре (вирион цитомегаловиру-
са — 150-300 нм), имеет сферическую форму, четыре структурных компонента:
• сердцевину — геном представлен вирусной ДНК — линейной двухнитевой
молекулой с коротким и длинным компонентами, различными по величине
и нуклеотидному составу, молекулярной! массой от 80-100х106 (ВПГ) до
145х106 (ЦМВ), в составе имеет до 80 генов, подгруппы — a, b, g:
• суперкапсид (внешнюю оболочку), пронизанный гликопротеиновыми шипа-
ми, — это типоспецифические антигены, по которым определяют отдельные
серотипы герпес-вирусов;
682 ЧАСТНАЯ ВИРУСОЛОГИЯ
• внутреннюю оболочку (тегумен — покрытие) — расположена между супер-
капсидом и капсидом, толщина различна у разных вирусов, непробиваемая
для лучей электронного микроскопа;
• капсид — построен из 162 капсомеров, диаметр составляет 100-110 нм, орга-
низован по типу кубической симметрии, представлен группоспецифическими
для каждого подсемейства антигенами, образованными белками нуклеокап-
сида.
В структуре вириона более 30 структурных белков (гликопротеинов — gp), семь
из которых (В, С, D, Е, F, G, X) находятся на поверхности и вызывают образова-
ние вируснейтрализующих антител. Капсид содержит 6 белков (гликопротеидов),
неструктурные белки (их десятки), в том числе вирусспецифический фермент
тимидинкиназу (ТК), ДНК-полимеразу и белок, связанный с ДНК.
Вирусная ДНК состоит из 80 генов, подгруппы — a, b, g
• а-гены транскрибируются РНК-полимеразой и не требуют для транскрипции
присутствия синтезированных de novo белков:
• продукты Ь-генов — вирусспецифическая ДНК-полимераза и ТКаза, необхо-
димые для биосинтеза ДНК вириона:
• g-гены — структурные белки вириона, представлены мембранными гликопро-
теидами А, В, С, D, Е, F. G, которые играют важную роль в иммунопатогенезе
герпеса (проникновение ВПГ в клетку, Феномен образования симпластов), а
также индуцируют формирование вируснейтрализуюших и комплементсвя-
зывающих антител.
ОСОБЕННОСТИ СТРУКТУРЫ НЕКОТОРЫХ ГЕРПЕС-ВИРУСОВ
Вирус Эпшгейна-Барр (ВЭБ) является представителем онкогенных ДНК-
содержащих вирусов, его капсид диаметром 120-150 нм окружен оболочкой,
содержащей липиды. В процессе репликации вируса экспрессируется свыше
70 различных вирусспецифических белков, однако к настоящему времени выделе-
ны группы иммуногенных белков, определение антител к которым дает возмож-
ность дифференцировать стадию инфекции, а именно;
• ЕА (Early Antigen) — ранний антшен, включает белки р54, р138;
• EBNA-1 (Epstein-Barr Nuclear Antigen) — ядерный антиген, белок р72;
• VCA (Vital Capsid Antigen) — капсидный антиген (поздний антиген), включает
комплекс белков р150, р!8, р23; к настоящему времени показано, что имму-
нодоминантными белками в этом комплексе являются р18 и р23;
• LMP (Latent Membrane Protein) — латентный мембранный белок (комплекс
продуктов ранних и поздних антигенов), gpl25.
Знание сроков появления антигенов ВЭБ и антител к нему позволяет докумен-
тировать латентную, острую и хроническую ВЭБ-инфекнии. Известны два штамма
ВЭБ, неразличимые серологически: тип 1 (А) и тип 2 (В).
Структура ВГЧ-6 расшифрована, молекулярно-биологическими и иммуноги-
стохимическими методами выделены два изолята (подтипы А и В) и обнаружена
гомологичность вируса с ЦМВ человека — более 50%. ВГЧ-6 имеет суперкап-
сидную липидсоцсржащую оболочку и электронно-плотный икосаэдрический
нуклеокапсид размером 95-105 нм, состоящий из 162 капсомеров, содержащих
геном вируса, представленный двухнитевой ДНК. Размер вируса — 160-200 нм, он
состоит из 25-29 полипептидов с молекулярной массой от 19 до 200 кДа, часть из
которых представлена клеточными белками. Специфические последовательности
групп А и В вируса используют для ПЦР-диагностики. ВГЧ-6 и ВГЧ-7 по своей
морфологической структуре схожи с другими герпес-вирусами, по составу ДНК
существенно отличаются от них. ВГЧ-7 и IJMB имеют общий фрагмент генома.
ВГЧ-8 содержит последовательности, гомологичные двум известным онкогенным
1 ерпес-вирусам — герпес-вирусу саймири и ВЭБ.
ЧАСТНАЯ ВИРУСОЛОГИЯ 683
Таким образом, герпес-вирусы имеют общие признаки; наличие ДНК, сходство по
морфологии, способу репродукции, размерам, способность поражать лимфоидные
клетки и ткани, пантропность и персистенцию в клетках различных органов и систем,
клинический полиморфизм инфекционного процесса и индую щи репродукции дру-
гих вирусов, а также способность вызывать мутационные i [роцессы в соматических
клетках человека. В то же время каждый вирус достаточно отличается по антигенным
и биологическим свойствам, способности к репродукции в определенных тканях и
клетках организма, вызывая при этом различные клинико-патогенетические формы
заболевания.
Устойчивость во внешней среде
ВПГ термолабильны, инактивируются при температуре 50-52 °C через 30 мин,
неустойчивы к действию физических и химических факторов, сравнительно
легко разрушаются под воздействием ультрафиолетовых и рентгеновских лучей,
разрушаются органическими растворителями, детергентами, протеолитическими
ферментами. Этиловый спирт, фосфатаза, желчь, эфир и другие детергенты быстро
инактивируют ВПГ. При низких температурах (от -20 до -70 °C) вирус сохраняет-
ся годами и десятилетиями. После лиофильного высушивания ВПГ не теряет своей
активности более 10 лет.
Клинический материал для лабораторного исследования
Исследование на ГВИ проводят с учетом анамнеза, клинико-эпидемиологи-
ческих данных. Материал для исследований может быть различным (табл. 24-6).
Помимо манифестных форм герпетической инфекции (ГИ), известны ее бессимптом-
ные и скрытые формы (нераспознанный герпес атипичного течения — 60%, бессим-
птомный герпес — 20%). Материалом для исследования при подозрении на нерас-
познанный герпес являются соскобы клеток, СМЖ, аспираты, биоптаты, кровь.
Таблица 24-6. Материалы, исследуемые при выделении герпес-вирусов в зависимости от лока-
лизации поражений
Локализация поражений Содержимое везикул Соскоб клеток СМЖ Аспират из бронхов Биоптат Кровь
1 2 3 4
Кожа + +
Глаза + +
Гениталии + + +
Анус + + +
Рот + + +
ЦНС + + + +
Легкие + + +
Печень + +
Врожденный герпес + + + + +
Соскоб клеток: 1 — со слизистой оболочки полости рта, зева: 2 — конъюнктивы; 3 — слизистой
оболочки влагалища, шейки матки; 4 — слизистой оболочки прямой кишки.
Лабораторная диагностика герпес-вирусной инфекции
Лабораторные методы диагностики герпес-вирусных инфекций в настоящее
время применяют только в тех случаях, когда необходимо уточнить этиоло-
684
ЧАСТНАЯ ВИРУСОЛОГИЯ
гию или оценить динамику заболевания. Наиболее часто уточнение этиологии
заболевания требуется при поражении органов зрения (увеите, конъюнктивите
и кератоконъюнктивите), ЦНС (серозном энцефалите и менингите) и половых
органов (наружных половых органов, цервиците, уретрите), так как не только
герпес-вирусы, но и довольно большое число других вирусов вызывают острые и
рецидивирующие заболевания этих органов. Важно лабораторно обследовать на
герпес-вирусы пациентов с неблагополучным акушерским анамнезом, подозрени-
ем на микст-инфекции, пациентов со вторичной иммунологической недостаточно-
стью, особенно при лимфаденопатии, длительной лихорадке. В подобных случаях
ошибка при выборе этиопатогенетических препаратов и их доз зачастую приводит
к серьезным негативным последствиям. Лабораторный контроль течения герпес-
вирусных заболеваний необходим при диссеминации процесса, у беременных.
В этих ситуациях малоинформативны методы определения титров противогер-
петических антител, которые являются часто косвенным свидетельством актив-
ности процесса и специфического иммунного ответа, в то время как для уточнения
этиологии, стадии и течения инфекционного процесса врачи должны располагать
сведениями о наличии или отсутствии самих герпес-вирусов или их антигенов.
Вирусологический метод
Вирусологический метод является «золотым стандартом» в диагностике ГВИ.
Отличительной особенностью данного метода является высокая степень досто-
верности полученных результатов, а также высокие чувствительность (85-100%)
и специфичность (100%), возможность получения чистой культуры возбудителя
для его дальнейшего изучения, в частности испытания чувствительности к антиви-
русным препаратам; недостаток — длительность проведения (от 2 до 5-14 дней).
Культивирование вируса осуществляют на чувствительной культуре клеток Vero.
Ускоренный вирусологический метод
В настоящее время используют и ускоренный вирусологический метод выяв-
ления вируса. Сущность метода заключается в том, что материал, полученный от
больных, культивируют одновременно с культурой клеток Vero в течение 24 ч,
после чего клетки культуры фиксируют в холодном ацетоне. Далее проводят выяв-
ление антигенов вируса в реакции непрямой иммунофлюоресценции (РНИФ).
На фиксированные препараты наносят моноклональные антитела (МКА) к ВПГ.
Затем препараты инкубируют во влажной камере в течение 1 ч при температуре
37 "С и промывают проточной водой. После промывки на препараты наслаивают
антивидовой конъюгат — меченные ФИТЦ (флюоресцеинизотиоцианатом) анти-
тела против иммуноглобулинов мыши — и помещают на 30 мин при температуре
37 СС. Окрашенные препараты исследуют с помощью люминесцентного микро-
скопа. При микроскопии определяют вирусспецифические включения в виде обра-
зований с зеленой или желто-зеленой флюоресценцией в ядре и/или цитоплазме
эпителиальных клеток. Включения могут иметь зернистую, гомогенную или
смешанную структуру. Ценность данного метода состоит в том, что он является
пока единственным методом, позволяющим выбрать антивирусный препарат для
лечения ГИ и оценить эффективность противовирусной терапии через 1,5-2 мес
после окончания лечения. Результаты лабораторного исследования готовы в тече-
ние 2 сут.
Методы прямой цитологической индикации
Самыми первыми методами диагностики ГВИ являлись методы прямой цито-
логической индикации герпес-вирусов в образцах первичного патологического
материала. При наличии вируса в пораженных тканях цитологический метод
позволяет обнаружить гигантские округлые многоядерные клетки со специфиче-
скими внутриядерными включениями; при ЦМВ — гигантские округлые клетки
ЧАСТНАЯ ВИРУСОЛОГИЯ 685
с ядром, занимающим почти всю цитоплазму, увеличенным за счет гомогенно-
го включения («совиный глаз»). Показанием к проведению цитологического
метода исследования ГИ является бессимптомное течение, а также острая фаза
заболевания. Простота метода заставляет использовать его и до настоящего вре-
мени. однако большинство авторов признают его несовершенным и ненадежным.
Недостатки метода — невозможность дифференцирования первичной инфекции и
рецидивирующей, а также невозможность уточнения типа ВПГ, являющегося при-
чиной заболевания. Кроме того, чувствительность и специфичность не превышают
75% по сравнению с методом выделения вируса в культурах тканей и зависят от
получения достаточного количества клеток для исследования, то есть правильного
забора материала.
Иммунофлюоресцентный mgi од
Иммунофлюоресцентный метод основан на обнаружении в образцах пер-
вичного материала от больных светящихся комплексов антигена возбудителя
(липополисахарида или основного белка наружной мембраны) со специфиче-
скими антителами с использованием люминесцентного микроскопа. При люми-
несцентной микроскопии вирусы герпеса имеют специфическое интенсивное
изумрудно-зеленое свечение. Антиген вируса может быть обнаружен в ядре, ядре и
цитоплазме неизмененных клеток. Многие авторы считают результаты внутрикле-
точной флюоресценции положительными при наличии не менее трех светящихся
антитенсо держащих клеток. Достоинствами РИФ являются невысокая стоимость,
высокие специфичность (90%) и чувствительность (55-75%), быстрота получения
результатов (в пределах 2 ч), а недостатком — зависимость результатов от качества
забора материала, стадии заболевания и квалификации врача-вирусолога.
Методы лабораторной диагностики
Следует отметить, что для повышения надежности диагностики генитального
герпеса, особенно в случаях субклинических и маломанифестных форм герпеса,
рекоменд) ют использовать в работе два-три метода лабораторной диагностики,
особенно при обследовании беременных, женщин с неблагополучным акушерским
анамнезом, лиц с неуточненным гинекологическим диагнозом.
В последние годы большое распространение приобрели диагностические тесты
для определения антигенов и антител к герпес-вирусам, основанные на принципе
ИФА, а также ПЦР-методики выявления в материале от больных ДНК герпес-
вирусов.
Определение противовирусных антител
Противовирусные антитела классов IgG, IgA, IgM в сыворотке крови опреде-
ляют с помощью ИФА. Простота, высокие специфичность и чувствительность, а
также невысокая стоимость этого метода и возможность определения стадии забо-
левания сделали его наиболее распространенным в диагностике ГИ. Недостатком
метода является возможность получения ложиоположительных результатов за
счет перекрестных реакций с нормальными антителами.
После внедрения вируса герпеса в организм человека начинается синтез спец-
ифических иммуноглобулинов класса М (IgM), которые определяются на 4-6-й
день после инфицирования и достигают максимального значения к 15-20-м сут-
кам. С 10-14-х суток начинается продукция специфических IgG, несколько позд-
нее IgA. IgA-антитела сохраняются в организме человека недолго (3 мес), IgG-
антитела — в течение всей жизни. Диагностическое значение при первичной ГИ
имеет выявление IgM и/или четырехкратное увеличение титров специфических
иммуноглобулинов G (IgG) в парных сыворотках крови, полученных с интервалом
14-21 дней. Рецидивирующий герпес обычно протекает на фоне высоких пока-
зателей IgG-антигел, свидетельствующих о постоянной антигенной стимуляции
686
ЧАСТНАЯ ВИРУСОЛОГИЯ
организма больного. Нередко диагностика активной фазы инфекции по четы-
рехкратному приросту титра IgG-антител вызывает затруднения, поскольку титр
IgG-антител может увеличиваться достаточно быстро (в течение 1-2 су г) после
проявления симптомов заболевания. В случае обнаружения IgG-антител яюбого
уровня рекомендуют тестировать сыворотки на IgM-антитела, особенно к ЦМВ.
Определение низкоавидных антител при герпес-вирусных инфекциях
Единственным способом, позволяющим сразу и достоверно диагностировать
первичную инфекцию, является определение индекса авидности (ИА) специфи-
ческих антител. Тест на определение авидности IgG-антител позволяет установить
точный момент инфицирования и разграничить первичщто, реинфекцию или
реактивацию инфекционного процесса, ллидность специфических IgG при серо-
диагностике возбудителей инфекционных заболеваний определяют различными
методами: агглютинацией, радиоиммунным методом, РСК, ИФА, электроблоттин-
гом и электрофорезом.
Чаще определяют показатель индекса авидности с помощью ИФА. Суть метода
заключается в следующем. В процессе инкубации образцов сыворотки крови с
адсорбированными на планшете антигенами образуются иммунные комплексы.
После промывания планшета в часть лунок добавляют раствор, который способ-
ствует удалению «ранних» IgG с низкой авидностью (денатурирующее вещество).
Затем вносят антиглобулиновый конъюгат и контролируют его связывание с ком-
плексом «антиген-антитело» с помощью раствора хромогена. После остановки
ферментативной реакции измеряют спектрофотометром оптическое поглощение
окрашенного раствора. Наличие в исследуемом образце вирусспецифических IgG-
антител с низкой авидностью определяется по снижению интенсивности окраши-
вания в сравнении с лунками, обработанными раствором сравнения В качестве
денатурирующего вещества используют диэтиламид, тиоционат калия, глюта-
ральдегид. мочевину. Показано, что обработка 8 М раствором мочевины является
самым простым методом измерения авидности IgG, однако этот метод менее чув-
ствителен, чем с применением диэтиламида, так как он не позволяет определять
низкоавидные антитела спустя 3 мес после первичного инфицирования.
Индекс авидности (ИА) антител сывороток рассчитывают по формуле:
ИА = ОПХ х 1С0 / ОП2,
где ОПХ — оптичы кая плотность в лунках с антигенами после обработки денатури-
рующим агентом: ОП2 — оптическая плотность в лунках с той же сывороткой после
обработки раствором сравнения.
Учет реакции: показатель ИА ниже 30-50% (в зависимости от особенностей
тест-системы) указывает на свежую первичную инфекцию. Показатель ПА,
равный или превышающий 50%, свидетельствует о присутствии в сыворот-
ке высокоавидных антител — маркеров перенесенной в прошлом инфекции.
Показатель авидности антител в пределах 31-49% может свидетельствовать
о поздней стадии первичной инфекции или недавно перенесенной инфекции
(ранней паст-инфекции) только при условии выявления высоких концентраций
антител. Интерпретировать результаты определения ИА необходимо в строгом
соответствии с рекомендациями фирмы-производителя, так как величина ИА
для одной и той же стадии заболевания может колебаться в широких пределах
в зависимости от способа разрушения иммунного комплекса и природы исполь-
зуемого денатуранта.
Таким образом, если в исследуемой сыворотке крови при наличии или отсут-
ствии IgM-антител, обнаруживаются IgG-антитела с низкой авидностью, то это
свидетельствует о первичной (недавней) инфекции. Присутствие высокоавидных
антител IgG указывает на вторичный иммунный ответ в случае попадания возбу-
ЧАСТНАЯ ВИРУСОЛОГИЯ 687
дителя в организм или обострения (реактивации) заболевания. Тест на авидность
прост в исполнении, доступен для большинства иммунологических лабораторий.
Метод иммуноблота
Метод иммуноблота (Westernblot, Line-blot) — новое поколение подтверж-
дающих тестов на инфекции. Представляет собой индивидуальный оценочный
шаблон, имеется возможность автоматизации исследования.
Лабораторная диагностика заболеваний, вызываемых ВПГ 1-го и 2-го типа,
включает комплекс исследований по выявлению специфического иммунного отве-
та (антител), определению вирусной ДНК и вирусных белков (антигенов), а также
подтверждению природы патогена культуральным методом (по цитопатогенному
эффект}' вируса в перевиваемой культуре клеток).
В серологических тестах на ВПГ, направленных на выявление антител против
ВПГ 1-го и 2-го типа, показана низкая диагностическая значимость из-за неодно-
значности клинической трактовки. Нарастание титров антител к ВПГ происходит
медленно, в течение нескольких недель, при этом могут одновременно определяться
специфические антитела классов IgM и IgG. Впоследствии антитела к ВПГ класса М
могут циркулировать в крови от нескольких месяцев до нескольких лет. Этот фено-
мен связывают с нарушением иммунитета у инфицированных ВПГ лиц. Кроме того,
в случае реактивации ВПГ в крови могут вновь появиться специфические антитела
класса М. С другой стороны, у иммунокомпрометированных пациентов при реци-
дивах инфекции ВПГ антитела к вирусу (как IgM, так и IgG) могут не выявляться.
Ложноотрицательный результат теста на анти-ВПГ класса М может быть у новорож-
денных. Тем не менее серологическое исследование необходимо проводить методом
парных сывороток (с интервалом 14-21 день). Отсутствие в первой и появление во
второй сыворотке противогерпетических IgM или нарастание титров специфиче-
ских IgG в четыре раза и более могут быть показателем острой инфекции ВПГ.
Схематически интерпретация результатов исследования сывороток на антитела
к ВПГ 1-го и 2-го типа может быть представлена следующим образом: низкий и
средний уровень IgG-антител — ремиссия, высокий уровень — обострение. Если
обнаружены высокие уровни IgG-антител к герпес-вирусам, рекомендуют допол-
нительно тестировать сыворотки методом ПЦР, который может достоверно ука-
зать на присутствие вируса в организме.
Метод прямой иммунофлюоресценции
Метод прямой иммунофлюоресценции (ПИФ) с поли- или моноклональными
антителами против ВПГ-1 и ВПГ-2 при исследовании первичного материала от
больных дает представление о морфологических свойствах клеток и изменении
локализации антигенов ВПГ Кроме прямых признаков поражения клеток вируса-
ми герпеса (специфического свечения), имеются косвенные признаки герпетиче-
ской инфекции по данным ПИФ.
• агрегация ядерного вещества, отслаивание кариолеммы;
• наличие так называемых ядер-дырок, когда от ядра клетки остается только
одна кариолемма;
• наличие внутриядерных включений — телец Каудри.
При постановке ПИФ врач получает не только качественную, но и количествен-
ную оценку состояния инфицированных клеток.
Метод иммуноферментного анализа
При ИФА определяют антигены с помощью антител, меченных ферментом.
Преимуществом ИФА по сравнению с РИФ являются большая степень чувстви-
тельности, стандартизация и автоматизация исследований, снижение частоты
ложных, субъективных ошибок, связанных с квалификацией врача-лаборанта.
Чувствительность метода составляет 70-80%.
688 ЧАСТНАЯ ВИРУСОЛОГИЯ
Метод полимеразной цепной реакции
В настоящее время является основным диагностическим способом в практи-
ческом здравоохранении для выявления ДНК вируса. При этом для выделения
ДНК ВПГ может быть использован любой биологический материал: клетки, био-
логические жидкости, ткани и т.п. В настоящее время разработаны методы коли-
чественного определения вирусной ДНК в тестируемом образце. На основании
полученных данных можно оценить форму инфекционного процесса. Так, если
количество ДНК превышает 1000 копий геном-зквивалента (г/э) на 10 лейкоци-
тов периферической крови, это может свидетельст вовать о развитии диссемини-
рованной инфекции.
Лабораторная диагностика ЦМВ-инфекции
Лабораторная диагностика ЦМВ-инфекции включает несколько под-
ходов:
• выявление инфекционных вирусных частиц или антигенов;
• определение вирусной ДНК;
• определение иммунного ответа организма;
• выявление прямого цитостатического действия вируса, выделенного из био-
логических субстратов пациента, на перевиваемую культуру клеток.
Серологические тесты на ЦМВ являются основными для установления инфи-
цирования данным вирусом. Однако серопозитивность отнюдь не является осно-
ванием к заключению, что болезнь действительно вызвана ЦМВ. Показано, что
высокие титры а нти-ЦМВ-антител обнаружены как у здоровых носителей, так и у
больных с острым течением инфекции. Селективное определение специфических
антител против ЦМВ классов М и G также не позволяет с полной уверенностью
дифференцировать острую инфекцию от реактивации хронической. Именно
поэтому определение вирусных антигенов и ДНК ЦМВ становится необходимым
тестом для диагностики заболевания этим вирусом.
Метод ПЦР широко используют д пя диагностики ЦМВ в любых биологиче-
ских тканях и жидкостях, в том числе и биопсийном материале, фиксированном
формалином. Этот метод превышает по своей чувствительности все имеющиеся
аналоги. Для увеличения диагностической и прогностической значимости приме-
няют количественные методы исследования для определения вирусной нагрузки
ЦМВ в крови методом ПЦР в формате реального времени, что позволяет оценить
эффективность противовирусной терапии.
Лабораторная диагностика ВЭБ-инфекции
Лабораторная диагностика ВЭБ-инфекции базируется на цитологическом иссле-
довании крови или костного мозга, серологических исследованиях (определении
гетерофильных и противовирусных антител) и ПЦР.
Серологическая диагностика ВЭБ-инфекции представляет определенные труд-
ности. Она основана на выявлении специфических антител к нескольким анти-
генам вируса (VCA, ЕА, EBNA), однако не существует единого надежного теста,
позволяющего получать достоверную информацию для постановки окончатель-
ного диагноза.
На ранних стадиях развития инфекции в образцах сыворотки крови пациента
обнаруживают IgM и IgG к нуклеарному антигену (IgM-VCA и IgG-VCA), Активная
фаза инфекционного мононуклеоза также характеризуется продукцией низкоа-
видных антител IgG к VCA и в большинстве случаев наличием IgG, специфичных к
комплексу ранних антигенов. IgM-антитела к нуклеарному антигену (EBNA) могут
служить индикатором острой инфекции. Однако ряд авторов отмечают частое воз-
ЧАСТНАЯ ВИРУСОЛОГИЯ 689
никновение ложноположительных реакций вследствие неспецифической реактив-
ности. Показано, что антитела к ядерному белку EBNA-1 р72 являются маркерами
только паст-инфекции.
Появление IgM анти-VGA-антител может происходить с опозданием или вооб-
ще не происходить. С другой стороны, IgM анти-VCA могут выявляться в течение
нескольких месяцев после первичного инфицирования, повторно появляться при
реактивации инфекции, а также при мононуклеозах иной этиологии (ЦМВ, токсо-
плазмозе. гепатите А, ВИЧ).
Возможны варианты интерпретации результатов серологического исследова-
ния больных с ВЭБ-инфекцией с учетом качественных характеристик тест-систем
фирм-изготовителей (табл. 24-7).
Таблица 24-7. Варианты интерпретации результатов серологического исследования больных ВЭБ"
Стадии ВЭБ-инфекции Низкоавидные антитела Маркеры ВЭБ-инфекции
VCA-JqG VCA-IgM EA-IgG FBNA-IgG
Инкубационный период или отсутствие инфекции - - - - -
Предранняя первичная инфекция + + - - -
Ранняя первичная инфекция + + + - -
Острая первичная инфекция (инфекци- онный мононуклеоз) + + + + -
Поздняя первичная инфекция - + +/- (<0,3) + + или -
Хроническая инфекция - + - + -
Атипичная первичная инфекция - + + - -
Ранняя паст-инфекция - + - + +
Поздняя паст-инфекция - + - - +
Реактивация - + + + +
Примечание. ЕА - ранний антиген: EBNA — ядерный антиген; VCA — капсидный антиген.
«-» — отсутствие антител.
«+/-» — вероятное присутствие антител
«+» — присутствие антител.
После положительного результата ИФА на антитела к ВЭБ ставят подтверж-
дающую реакцию иммуноблоттинга, в которой определяют наличие антител к
отдельным маркерным белкам ВЭБ (р — протеины): р23, р54, р72 (наличие этого
белка указывает на возможность размножения ВЭБ), р138 и др, Выявление белка
VCA 125 указывает на раннюю фазу инфекции. В разгар болезни и на этапе завер-
шения острого процесса появляются VCA 19. О поздней фазе свидетельствует
высокоспецифический маркер VCA 22, который выявляется самостоятельно или
одновременно с EBNA-1 (р79). Последний белок длительно присутствует у пере-
болевших и свидетельствует о перенесенной инфекции.
ПЦР-методы могут быть использованы для качественного и количественного
определения ДНК ВЭБ.
Лабораторная диагностика инфекции, вызванной вирусом
герпеса человека 6-го типа (ВГЧ-6, ВГЧ-6А, ВГЧ-6Б)
Серологические методы, используемые цля диагностики герпетической инфек-
ции этого типа, имеют целый ряд недостатков и редко помогают в диагностике
клинической манифестации. Определение титра специфических IgM используют
для диагностики острой инфекции или реактивации. Не у всех детей с первичной
690
ЧАСТНАЯ ВИРУСОЛОГИЯ
инфекцией отмечается продукция антител IgM, примерно 5% здоровых взрос-
лых имеют антитела IgM к ВГЧ-6. В связи с тем что практически у всех взрослых
выявляют IgG к ВГЧ-6, обнаружение специфических антител в одном образце не
значимо. К тому же, повышение их титра не указывает на новую инфекцию или
реактивацию. Возможно также выявление перекрестно реагирующих антител к
другим ДНК-вирусам, особенно ВГЧ-7. Доступные в настоящий момент сероло-
гические тест-системы не позволяют дифференцировать варианты А и В ВГЧ-6.
У детей диагноз первичной инфекции ВГЧ-6 требует обнаружения виремии (изо-
ляции ВГЧ-6 в моноядерных клетках периферической крови) и существенного
нарастания показателей серологических тестов. ВГЧ-6-виремия наблюдается
относительно редко у здоровых детей по гравнению с переносящими первичную
инфекцию. Изоляция ВГЧ-6 требует культивации со стимулированными клетка-
ми крови пуповины и последующей идентификации на оборудовании, которое
доступно только исследовательским лабораториям. Трудны для диагностики пер-
систирующая и латентная инфекции.
ДНК ВГЧ-6 можно обнаружить в лимфоцитах периферической крови или в
других тканях методом ПЦР. Однако обнаружение ДНК ВГЧ-6 в этих тканях не
всегда указывает на первичную инфекцию, наиболее часто это является проявле-
нием персистирующей инфекции, развившейся после первичной инфекции и не
сопровождающейся виремией. Обнаружение ДНК ВГЧ-6 в плазме и определение
высокого титра вируса методом ПЦР в реальном времени — более чувствительный
метод для диагностики первичной инфекции (около 90%), однако это может сви-
детельп вовать и о реактивации инфекции ВГЧ-6.
Лабораторная диагностика инфекции, вызванной вирусом
герпеса человека 7-го типа (ВГЧ-7)
Множество различных заболеваний ассоциируется с ВГЧ-7-инфекцией, но
очень мало доказательств их причинной связи. ВГЧ-7 может быть кофактором, но
зачастую его присутствие — результат высокой частоты персистенции и особен-
ностей выделения ВГЧ-7. Значительно чаще (до 85%) ВГЧ-7 может выделяться
из слюны здоровых индивидуумов, в активизированной пуповинной крови или
мопонуклеарах периферической крови.
Для серологической диагностики HHV-7 используют различные методы, наи-
более часто применяют ИФА, разработаны также иммуноблоттинг и реакция ней-
трализации. Для исключения перекрестной реактивности между ВГЧ-6 и ВГЧ-7
перекрестно реагирующие антитела ВГЧ-6 исключают адсорбцией с зараженными
ВГЧ-6 клеточными лизатами.
ДНК ВГЧ-7 обнаруживают с помощью ПЦР в слюне, моноядерных клетках
периферической крови и других биологических жидкостях. Разработаны каче-
ственные и количественные методы ПЦР. К сожалению, любое из этих исследова-
ний в отдельности не позволяет дифференцировать первичную инфекцию ВГЧ-7
от реактивации и взаимодействия с другими вирусами герпеса. У детей изоляция
ВГЧ-7 из мононуклеаров периферической крови плюс сероконверсия обычно ука-
зывают на первичную инфекцию. IgM к ВГЧ-7 не могут быть использованы для
диагностики первичной инфекции, поскольку их определяют у здоровых людей,
ранее перенесших инфекцию.
Таким образом, хотя ВГЧ-7 близок с ВГЧ-6 по морфологии и геному, он отли-
чается по биологической значимости как причинный фактор заболеваний у детей.
Возможно, наиболее интересными особенностями являются многообразное и
частое взаимодействие с другими герпес-вирусами и его потенциальная роль
как кофактора или модулятора ассоциированных болезней, например, у ВИЧ-
инфицированных и у пациентов с другими иммунодефицитами
ЧАСТНАЯ ВИРУСОЛОГИЯ 601
Лабораторная диагностика инфекции, вызванной вирусом
герпеса человека 8-го типа (ВГЧ-8)
Для диагностики ВГЧ-8-инфекции преимущественно используют серологи-
ческие исследования и ПЦР. ДНК В14 8 можно обнаружить в ткани саркомы
Капоши, мононуклеарах периферической крови. Для обнаружения антител про-
тив антигенов ВГЧ-8 разработаны различные серологические методы, прежде
всего ИФА, иммуноблоттинг. Некоторые сыворотки от здоровых людей содержат
антитела, которые перекрестно реагируют с антигенами ВГЧ-8 и могут привести к
ложноположительным результатам.
Диагностика множественной герпес-вирусной инфекции
Возможно одномоментное инфицирование несколькими вирусами семейства
Herpesviridae. Нередко этому подвержены пациенты с клинико-лабораторными
проявлениями вторичной иммунологической недостаточности (онкогематоло-
гические, онкологические, ВИЧ-инфицированные). При этом прогностически
наиболее значимым можно считать комплексное одномоментное выявление ДНК
ВПГ-1, ЦМВ и ВГЧ-6.
Для адекватной терапии ГВИ недостаточно только выявить возбудителя забо-
левания. Необходимо иметь представление о характере течения инфекционного
процесса. Таким образом, диагноз ГВИ следует устанавливать по совокупности
анамнестических, эпидемиологических, клинических данных и доступных лабо-
раторных методов исследования: цитоморфологического исследования мазков для
обнаружения многоядерных гигантских клеток и внутриклеточных включений;
выделения вируса на хорионаллантоисной оболочке куриных эмбрионов или в
культуре клеток; иммуноферментного анализа для определения гитра специфиче-
ских противовирусных антител (и антигенов вирусов); определения вирусной ДНК
с помощью ПЦР, подтверждающей реакции иммуноблота.
КАЛИЦИВИРУСЫ
Таксономия
Калицивирусы человека относятся к семейству Caliciviridae. Генетическим
материалом калицивирусов является однонитевая плюс-нить РНК размером
около 7,5x103 о н. В состав семейства Caliciviridae входят роды Norovfrus, Sapovirus,
Lagovirus и Vesivirus. В инфекционной патологии человека имеют значение только
представители родов Norovirus и Sapovirus. Ранее их называли Норуолк- и Саппоро-
подобными вирусами, в настоящее время они получили название норовирусов и
саповирусов соответственно. И те и другие вирусы вызывают острое кишечное
заболевание, причем норовирусы поражают все возрастные группы населения, а
саповирусы, как правило, только детей в возрасте до 5 лет. Норовирусная инфек-
ция индуцирует слабый и нестойкий иммунный ответ, чем и объясняется широкое
распространение норовирусных инфекций во всех возрастных группах.
Норовирусы и саповирусы отличаются высоким антигенным и генетическим
разнообразием. К настоящему времени идентифицированы норовирусы пяти раз-
личных геногрупп, каждая из которых подразделяется на несколько генетических
кластеров. Норовирусы I, II и IV геногруппы вызывают заболевание у человека.
Наибольшее значение в эпидемиологии имею г норовирусы геногруппы GII, в осо-
бенности кластера GII.4. Саповирусы также делятся на несколько геногрупп. Для
человека имеют значение саповирусы 4 групп: GI, GII, GIV и GV.
692
ЧАСТНАЯ ВИРУСОЛОГИЯ
До настоящего времени никому из исследователей не удалось культивировать
калицивирусы человека, поэтому их интенсивное изучение началось лишь в конце
XX — начале XXI в. с появлением методов обратной генетики Секвенирование и
клонирование геномов калицивирусов позволило разработать тест-системы ПЦР
и наборы ИФА, направленные на выявление консервативных этитопов наружных
белков калицивирусов. Широкое применение новых диагностических методов в
значительной мере заполнило существующий ранее пробел в расшифровке этио-
логии острых кишечных инфекций (ОКИ).
Характеристика вирусов
Вирионы норовирусов — безоболочечные округлые частицы диаметром
26-32 нм. обладающие икосаэдрической симметрией. Значительный разброс
в размерах указывает на генетическое разнообразие и недостаточную изучен-
ность норовирусов. В препаратах, полученных методом негативного контрасти-
рования. вирусные частицы имеют неровную поверхность (feathery — «пуши-
стый»). Норовирусы, принадлежащие к различным генетическим кластерам, могут
несколько различаться по морфологии. Например, вирус Торонто долгое время
был известен под названием мини-реовирус, что отражает особенности его мор-
фологии. Детализированное изображение частиц норовирусов удалось получить
с помощью криоэлектронной микроскопии. Было показано, что капсид норови-
русов состоит из 180 молекул белка VP1, кодируемого рамкой считывания ORF2.
Белок VP1 формирует миниатюрные арки на поверхности вириона. Внутри вирус-
ной частицы находи гея геномная РНК в комплексе с белком VPg и субгеномная
РНК, кодирующая белки капсида.
Саповирусы человека по морфологии сходны с вирусом везикулярной экзан-
темы свиней, послужившим прототипом для семейства Caliciviridae. Название
семейства происходит от латинского слова calix — «чаша, бокал». Подобно норо-
вирусам, частицы саповирусов представляют собой безоболочечные белковые
капсиды, содержащие РНК- Капсиды саповирусов также образованы белком
VP1 с молекулярной массой около 60 кДа. Частицы саповирусов под определен-
ным углом зрения имеют вид звезды Давида с чашеобразными углублениями
(calix) на поверхности. Диаметр частиц в среднем составляет 30 -34 нм. Пустые
вирусные капсиды, присутствующие в клинических образцах, не имеют столь же
характерной морфологии, как полные частицы, и могут быть ошибочно приняты
за капсиды норовирусов. Саповирусы содержат однинитсвую плюс-РНК |ss(+)
RNA], которая, так же как и у норовирусов, имеет три рамки считывания. Первая
рамка (ORF1) кодирует полипротеин, расщепляющийся на неструктурные белки,
в том числе РНК-зависимую РНК-полимеразу; ORF2 кодирует мажорный белок
капсида VPli ORF3 кодирует минорный структурный РНК-связывающий белок.
В отличие от норовирусов, рамка считывания капсидного белка ORF2 слита с рам-
кой считывания неструктурных белков ORF1.
Вирионы калицивирусов не содержат липидов, поэтому они устойчивы к обра-
ботке эфиром, органическими растворителями и слабыми детергентами. Вирусы
инактивируются прогреванием при 60 °C в течение 30 мин или кипячением при
100 °C в течение 3-5 мин. При замораживании калицивирусы сохраняют инфек-
ционную активность в течение длительного времени. Хлорирование и обработка
сильнодействующими окислителями и детергентами, четвертичными аммонийны-
ми соединениями подавляют инфекционную активность калицивирусов.
К настоящему времени установлено, что норовирусы отвгтственны за пода-
вляющее большинство вспышек острого небактериального гастроэнтерита. По
различным оценкам, доля норовирусов в этиологии небактериальных ОКИ в
Европе и США составляет 80-95%. В России массовую диагностику калицивирус-
ЧАСТНАЯ ВИРУСОЛОГИЯ 693
ных инфекций не проводят, однако в выборочных исследованиях показано, что
норовирусы широко распространены в разных регионах нашей страны, причем в
циркуляции доминируют те же генотипы норовирусов, что и во всем мире. В част-
ности, в 2002-2008 гг. в России, как и во всем мире, доминировали норовирусы
кластера GII.4.
В отличие от норовирусов, саповирусы поражают в основном детей в возрасте
до 5 лет. наиболее распространены в младшей возрастной группе от 0 до 6 мес.
Количество саповирусных гастроэнтеритов у детей в среднем в три-четыре раза
меньше норовирусных и варьирует от 1—10% в европейских странах до 30% в
Японии. Иногда отмечаются вспышки саповирусных гастроэнтеритов у взрос-
лых, по-видимому, связанные с появлением антигенно отличающихся штаммов.
Основным источником саповирусных инфекций могут быть контаминированная
вода и пищевые продукты.
Калицивирусная инфекция передается i основном фекально-оральным путем
и через предметы, загрязненные рвотными массами В фекалиях и рвотных мас-
сах больных содержится до 1010 частиц на мл. В то же время инфекционная доза
крайне мала и составляет около 10-20 вирусных частиц. Источником инфекции
могут служить также контаминированные фекалиями пищевые продукты и вода.
Одним из наиболее загрязненных калицивирусами продуктов питания являются
двустворчатые моллюски, употребляемые в сыром виде.
Клинический материал для лабораторного исследования
Клиническим материалом для диагностики служат фекалии пациентов, боль-
ных калицивирусной инфекцией. Содержание частиц калицивирусов в фекалиях
наиболее высоко в первые дни заболевания, затем оно быстро падает. Для диа-
гностики с помошью электронной микроскопии желательно обеспечить забор
материала в период острого заболевания, то есть в первые 2-3 дня. Поскольку
чувствительность тест-систем ПЦР много выше электронной микроскопии, для
анализа методом ПЦР достаточно провести забор материала в течение недели с
момента начала заболевания. Фекалии отбирают в стерильные пластиковые либо
стеклянные флаконы и хранят до исследования при 4 °C в течение 3-14 дней. При
необходимости более длительного храпения материал замораживают при темпе-
ратуре ниже -20 °C и размораживают перед исследованием. Желательно избегать
повторного замораживания-оттаивания, так как это снижает количество вирусной
РНК в материале. Для диагностическою исследования можно использовать также
рвотные массы.
Лабораторные методы диагностики
Калицивирусная инфекция в среднем протекает легче, чем ротавирусная,
однако по клиническим признакам ее невозможно дифференцировать от про-
чих вирусных ОКИ. Сходное по клинической картине заболевание у детей может
быть вызвано аденовирусами группы F. астровирусами, ротавирусами, корона-
вирусами и торовирусами. Примечательно, что для калицивирусной инфекции,
в особенности у взрослых, характерны внезапное начало и многократная рвота.
Продолжительность заболевания составляет 1-3 сут.
В диагностике калицивирусных гастроэнтеритов наиболее широкое распро-
странение во всем мире получили различные тест-ситемы ПЦР, в том числе ПЦР в
реальном времени (RT-PCR). Преимуществом тест-систем ПЦР являются высокие
чувствительность и специфичность. Однако в сил} высокой изменчивости не все
калицивирусы удастся выявить этим методом. По различным оценкам, от 5 до 20%
циркулирующих штаммов калицивирусов не выявляются методом ПЦР.
694 ЧАСТНАЯ ВИРУСОЛОГИЯ
Разработаны быстрые иммуноферментные тесты для выявления норовирусной
инфекции, в которых используют антитела к рекомбинатным белкам норовирусов.
И ФА-тест-системы последнего поколения отличаются высокой специфичностью,
но недостаточно высокой чувствительностью, поэтому они пригодны в основном
для выявления этиологии вспышек ОКИ при наличии достаточно большого коли-
чества образцов, взятых в период острого заболевания.
КОРОНАВИРУСЫ
Таксономия
Коронавирусы — семейство Coronavindae — РНК-содержащие вирусы, отли-
чительным признаком которых при исследовании под электронным микро-
скопом является наличие особых периферических короноподобных выростов.
Коронавирусы распространены повсеместно, инфицируют птиц и млекопитаю-
щих, включая людей Представители первых двух групп рода Coronavirus вызывают
заболевания у млекопитающих, третьей — только у птиц. Заражение происходит
воздушно-капельным и фекально-оральным путем. До 2003 г. эти вирусы не счита-
лись особо патогенными для человека, хотя и отмечались случаи коронавирусной
пневмонии у людей с различными иммунодефицитными состояниями. При этом у
животных коронавирусы могут вызывать серьезные заболевания. Представители
семейства Coronaviridae имеют узкий спектр хозяев, однако, по-видимому, некото-
рые из них способны преодолевать межвидовой барьер.
Характеристика вирусов
Вирионы коронавирусов имеют сфероидную форму (80-200 нм) и снабжены
липидной оболочкой. Выступы на поверхности вириона (пепломеры) образованы
тримсрами белка S, который расщепляется на две субъединицы: S1 и S2. У неко-
торых представителей рода Coronavirus имеется дополнительный поверхностный
гликопротеид НЕ, обладающий гемагглютинирующей и эстеразной активностью.
Трансмембранные белки М и Е играют важную роль в сборке и стабильности
вирионов. Нуклеокапсид образован белком N, который при взаимодействии с
геномной РНК образует спиральный нуклеоид в центральной части нуклеокап-
сида. Геном коронавирусов представлен одноцепочечной РНК размером 17,6-
31х103н.о. позитивной полярности, которая представ аяет собой инфекционную
мРНК.
Основными клетками-мишенями коронавирусов являются эпителиальные
клетки и макрофаги, с рецепторами на поверхности которых связывается S-белок.
Сборка вирионов происходит в цитоплазме инфицированных клеток на мембра-
нах эндоплазматического ретикулума. Основную роль в формировании зрелого
вириона играют трансмембранные белки М и Е.
Инфицирование коронавирусами вызывает формирование антител против
эпитопов, расположенных на S-, М-, N- и НЕ-антигенах. Антитела против
М-белка выявляются в реакции связывания комплемента. S- и НЕ-белки содержат
основные эпитопы для нейтрализующих и антигемагглютинирующих антител
Детерминанты клеточного иммунного ответа находятся в составе N-белка.
В отношении резистентности коронавирусов, патогенных для человека, имеют-
ся ограниченные сведения. Коронавирусы инактивируются средствами из группы
окислителей (хлор- и кислородсодержащими), спиртами, фенолами и их произ-
водными, альдегидами, четвертичными аммониевыми соединениями. Известно,
что 5% раствор формальдегида и 70% этиловый спирт через 5 мин инактивируют
коронавирус.
ЧАСТНАЯ ВИРУСОЛОГИЯ 695
Тяжелый острым респираторный синдром
Тяжелый острый респираторный синдром (ТОРС, атипичная пневмония) впер-
вые был выявлен на юге Китая (в ноябре 2002 г.) и в течение нескольких месяцев
получил глобальное распространение, поразив более 30 стран мира. Проведенный
комплекс противоэпидемических мероприятий позволил остановить распростра-
нение ТОРС.
В апреле 2003 г. ВОЗ объявила, что этиологическим агентом ТОРС является
новый патоген — вирус SARS (SARS-CoV). относящийся к семейству коронавиру-
сов (Coronaviridae, род Coronavirus), но не родственный ни одному из известных
штаммов этого вируса.
ХАРАКТЕРИСТИКА ВИРУСОВ
К настоящему времени геном трех штаммов коронавируса, с которым связыва-
ют развитие ТОРС, полностью расшифрован. Он представлен последовательно-
стью из 29 727 нуклеотидов и включает одиннадцать открытых рамок считывания
(ORF), расположение которых в целом типично для коронавирусов. Однако пока-
зано отличие нуклеотидных последовательностей генов, кодирующих структурные
белки вируса от остальных представителей семейства Coronaviridae, позволяющие
провести четкую границу между вирусом ТОРС и другими коронавирусами. Кроме
того, в геноме TOPC-CoV не обнаружен ген НЕ, присутствующий у коронавирусов
второй и некоторых представителей третьей группы. Вопрос о происхождении
вируса ТОРС до сих пор остается открытым, потенциальный предшественник дан-
ного вируса неизвестен.
УСТОЙЧИВОСТЬ ВО ВНЕШНЕЙ СРЕДЕ И К ФИЗИКО-ХИМИЧЕСКИМ ФАКТОРАМ
Вирус ТОРС сохраняет жизнеспособность на пластиковых поверхностях в
течение 48 ч после высушивания. Находящийся в фекалиях больных вирус ТОРС
сохраняет жизнеспособность на протяжении 1-2 дней (до 4 дней при диарее), а
в моче при комнатной температуре вирус стабилен, по крайней мере, в течение
1-2 дней. Вирус ТОРС более стабилен при комнатной температуре по сравнению
с другими коронавирусами, инфицирующими человека. Вирус чувствителен к дей-
ствию ультрафиолетовых лучей, высоким температурам. Прогревание при 56 °C
инактивирует вирус.
КЛИНИЧЕСКИЙ МАТЕРИАЛ ДЛЯ ЛАБОРАТОРНОГО ИССЛЕДОВАНИЯ
Вирус ТОРС присутствует в фекалиях, отделяемом верхних дыхательных путей
и моче больных. Надежность лабораторных диагностических тестов для детекции
SARS-CoV-инфекции зависит от типа получаемого клинического образца, сроков
и способов его получения. Важным фактором является взятие материала хорошо
обученным персоналом Полученные клинические образцы, согласно рекоменда-
циям ВОЗ, должны быть аликвотированы на три образца:
• один образец исследуют в диагностической лаборатории:
• второй (не вскрывают) — в национальной референс-лаборатории;
• третий — сохраняют для передачи в ВОЗ, референс-лабораторию по верифи-
кации SARS в случае, если возникнет такая необходимость.
Вирус SARS отнесен ко II группе патогенности, и все работы, с ним связанные,
требуют обеспечения уровня безопасности BSI3. Сбор, транспортировку и подго-
товку проб клинического и секционного материала от больных и подозрительных
на заражение вирусом SARS осущес гвляет медицинский персонал с соблюдением
правил противоэпидемического режима в строгом соответствии с требованиями
безопасности работ с микроорганизмами I-II группы патогенности.
Сбор клинического материала и все последующие манипуляции с ним осущест-
вляю г стерильными одноразовыми инструментами в стерильные одноразовые
696
ЧАСТНАЯ ВИРУСОЛОГИЯ
флаконы, пробирки, контейнеры. Все виды работ проводят в одноразовых меди-
цинских перчатках. Все материалы, доставляемые в лабораторию, должны быть
последовательно дважды упакованы.
• В транспортную емкость (плотно закрывающиеся пробирки или флаконы
с завинчивающимися крышками). Плотно закрытый верхний конец транс-
портной емкости вместе с крышкой для надежности заклеивают и помещают
в пластиковый пакет подходящего размера вместе с небольшим количеством
любого адсорбирующего материала, например ваты. Пластиковый пакет сле-
дует заклеить или запаять.
• Два или более образца от одного пациента могут быть упакованы в один пла-
стиковый пакет. Не допускается упаковывать образцы материалов от разных
людей в один и тот же пакет.
• Заклеенные пакеты с образцами помещают внутрь дополнительного пла-
стикового контейнера с завинчивающейся крышкой. Дважды упакованные
образцы материалов от разных пациентов могут быть транспортированы в
одном дополнительном контейнере.
• Описание особенностей конкретного образца, полную информацию и доба-
вочные сведения, касающиеся образцов, а также способ транспортировки и
название лаборатории назначения приклеивают к наружной стенке дополни-
тельного контейнера.
• Плотно закрытые пластиковые контейнеры с заклеенными крышками поме-
щают в термоизолирующий контейнер, укомплектованный охлаждающими
термоэлементами.
• Каждую пробу материала сопровождают бланком направления, прикреплен-
ным к наружной стенке контейнера.
• Транспортировку проб клинического материала в референс-лабораторию для
дальнейшего исследования в целях подтверждения результатов осуществляет
нарочный, информированный о правилах доставки материала.
Материал для исследования методом ПЦР забирают в стерильные пробирки с
герметично завинчивающимися крышками. Консервант не добавляют.
• Забор крови проводят из локтевой вены в количестве 4,5 мл стерильным шпри-
цем, в который предварительно вносят 0,5 мл стерильного 4% раствора натрия
цитрата и переносят в пробирку. Аккуратно перемешивают покачиванием.
• Мазки и смывы из носоглотки, бронхоальвеолярные смывы, плевральную
жидкость забирают в количестве 5 мл. Мазки забирают тампонами и поме-
щают в стерильные пробирки с 5 мл физиологического раствора. Не реко-
мендуют использовать тампоны на деревянной палочке, так как они могул
содержать субстраты, ингибирующие ПЦР.
• Секционный материал: исследуют легкие, сегментарные бронхи, кровь, печень,
селезенку. Кусочки органов массой 5-10 г помещают в стерильные, герметично
закрывающиеся емкости. Кровь, смывы с дыхательных путей хранят и транс-
портируют в лабораторию для исследований при температуре -20 °C.
ВОЗ рекомендует проводить взятие мазков и смывов из носоглотки в течение
первой недели заболевания, в эти же сроки проводят забор образцов плазмы или
сыворотки для серологических исследований в острой фазе заболевания. Образцы
смывов с дыхательных путей и образцы фекалий для ОТ-ПЦР можно исследовать
в течение двух недель от начала заболевания. Повторный забор образцов крови
для серологических исследований проводят через 2-3 нед, парные сыворотки
исследуют одновременно.
ЛАБОРАТОРНЫЕ МЕТОДЫ ДИАГНОСТИКИ
Клинические случаи ТОРС у детей и взрослых подтверждают, что данное забо-
левание не имеет специфических клинических признаков. Эксперты CD С (США,
ЧАСТНАЯ ВИРУСОЛОГИЯ ь97
Атланта) и ВОЗ сформулировали критерии, позволяющие при подозрении на
ТОРС сузить диагностический поиск.
Вероятный случай ТОРС — наличие тяжелого респираторною синдрома неиз-
вестной этиологии и эпидемиологических критериев (контакта с лицами с подо-
зрением на ТОРС, пребывание в регионе эпидемии); лабораторные критерии
могут быть подтверждающими, отрицательными, неопределенными. Случай,
подозрительный на ТОРС, — наличие умеренного респираторного синдрома
неизвестной этиологии и эпидемиологических критериев (контакта с лицами с
подозрением на ТОРС, пребывание в регионе эпидемии), лабораторные критерии
могут быть подтверждающими, отрицательными, неопределенными. В связи с
этим первоочередному обследованию при указанной клинической симптоматике
подвергаются липа, недавно посещавшие регионы, в которых регистрировались
случаи ТОРС. Лабораторные исследования в целях диагностики ТОРС следует
проводить также у лиц без клинических проявлений заболевания, при тесном кон-
такте с подозреваемым на ТОРС больным.
Вирусологические методы диагностики
Вирусологические методы диагностики ТОРС — молекулярно-биологические
методы (ОТ-ПЦР) и выделение вируса в культуре клеток. В связи с существенны-
ми трудностями культивирования и выделения коронавирусов наиболее эффек-
тивные диагностические тесты, доступные в настоящее время, — тесты, которые
обеспечивают информацией о вирусном геноме, в частности ОТ-ПЦР или ПЦР в
режиме реального времени. Значение ПЦР-тестов заключается также в том, что их
можно использовать для относительно ранней диагностики заболевания (в 1-10-е
сутки заболевания).
Культура клеток является очень требовательным методом, однако и единствен-
ным способом выявить наличие живого вируса.
Серологические методы
Серологические методы определения антител к вирусу ТОРС — непрямая реакпия
иммунофлюоресценции (НРИФ) или ИФА (ELISA). Несмотря на то что у некоторых
пациентов антитела уже могут определяться во время острой фазы заболевания (то
есть в течение 14 дней после появления симптомов), окончательная интерпретация
результатов серологического исследования возможна только для образцов, полу-
ченных после 21-го дня с момента появления симптомов. Следовательно, сероло-
гические тесты не могут быть использованы для выявления заболевания на ранней
стадии инфекции, что особенно важно для предотвращения ее paenpot гранения
Первичное исследование материала от пациентов (умерших) методом ОТ-ПЦР
и ИФА проводят на базе учреждений Госсанэпиднадзора. При получении положи-
тельных результатов исследований материал направляют в референс-лабораторию
для подтверждающего тестирования и депонирования. При этом получение отри-
цательных результатов в подтверждающих тестах служит основанием для исклю-
чения диагноза ТОРС.
При попущении положительных результатов подтверждающих тестов диагноз
ТОРС считают установленным. Результаты исследований в кратчайшие сроки
передают в соответствующие учреждения Госсанэпиднадзора, а материал для изо-
ляции вируса и сыворотку^ крови переболевших направляют в Центр специальной
лабораторной диагностики и лечения особо опасных и экзотических инфекци-
онных заболеваний. Полученную в ходе исследования комплементарную ДНК,
образуемую в результате реакции обратной транскрипции ДНК с участием фер-
мента ревертазы, направляют в Центр молекулярной диагностики инфекционных
заболеваний (Центральный НИИ эпидемиологии, Москва) для проведения секве-
нирования, которое обладает абсолютной информативностью при установлении
диагноза ТОРС.
698 ЧАСТНАЯ ВИРУСОЛОГИЯ
АЛГОРИТМ ИССЛЕДОВАНИЯ
При обследовании пациента до 10-го дня от начала заболевания используют
метод ОТ-ПЦР двукратно с интервалом 5 дней. Для повышения результатив-
ности анализа методом ОТ-ПЦР необходимо исследовать как минимум три вида
клинического материала: мазок из носоглотки (или смыв), мокроту и фекалии.
Дополнительно также может быть исследована плазма крови. Каждую пробу кли-
нического материала тестируют троекратно.
При получении хотя бы одного положительного результата пробу (читают
первично-положительной. Дополнительное тестирование проб методом ИФА в
этом случае не проводят. Пробы материала и полученную кДНК отправляют в
референс-лабораторию.
При отрицательных результатах ОТ-ПЦР проводят исследование методом ИФА
с тестированием сывороток взятых на 10, 20 и 30-й день заболевания Результат
считают1 положительным при выявлении сероконверсии или увеличении титра
антител в фазе реконвалесценции не менее чем в черыре раза по сравнению с
острой фазой. При получении положительного результата ИФА пробы клиниче-
ского материала отправляют в референс-лабораторию.
При начале обследования пациента после 10-го дня заболевания проводят
параллельное исследование материала методом ОТ-ПЦР двукратно с интервалом
5 дней и тестирование парных сывороток, взятых на 10, 20 и 30-й день заболева-
ния методом ИФА. При получении положительного результата при любом методе
исследования пробы клинического материала и полученную кДНК отправляют в
референс-лабораторию.
При двукратных отрицательных результатах ОТ-ПЦР с интервалом 5 дней и
отрицательных результатах ИФА на 10, 20 и 30-й день заболевания диагноз ТОРС
может быть исключен.
Секционный материал (ткани легкого, сегментарные бронхи, кровь, образцы
фекалий), собранный в кратчайшие сроки после наступления смерти, тестируют
методом ПЦР. Каждый вид секционного материала тестируют троекратно. В случае
получения хотя бы одного положительного результата пробу считают первично-
положительной и в кратчайшие сроки направляют в референс-лабораторию в
целях подтверждения (верификации).
Полный перечень и характеристика тест-систем для диагностики ТОРС и
рекомендации к их использованию представлены на сайте ВОЗ: www.who. Use of
Laboratory Methods for SARS Diagnosis; www.who.Severe Acute Respiratory Syndrome
(SARS), Laboratory diagnostic tests.
ДИФФЕРЕНЦИАЛЬНАЯ ДИАГНОСТИКА
Клинические проявления (слабость, лихорадка, озноб и одышка) при SARS
принципиально не отличаются от симптомов ОРВИ или гриппа. Однако, в случае
наблюдения за пациентом с подобными симптомами, обязательно необходимо
учитывать эпидемиологические данные, например недавнее посещение районов
мира, эндемичных по SARS, таких как Китай, Япония. Вьетнам, Сингапур и другие
страны Юго-Восточной Азии. Также обязательно следует выяснить, находился ли
пациент в контакте с заболевшим SARS. Таким образом, для установления диагно-
за SARS всегда необходимо принимать во внимание совокупность факторов, таких
как клинические симптомы заболевания, эпидемиологические данные и результа-
ты лабораторных тестов.
ЧАСТНАЯ ВИРУСОЛОГИЯ 699
РЕСПИРАТОРНО-СИНЦИТИАЛЬНЫЙ ВИРУС
Характеристика вируса
Респираторно-синцитиальный вирус (РОВ) человека был выделен от детей с
пневмонией и бронхиолитом. Два штамма вируса были изолированы на пере-
виваемых культурах клеток КВ, Chang. Заболеванию подвержены дети раннего
возраста, у которых инфекция, вызванная респираторно-синцитиальным вирусом,
может послужить причиной смерти.
PC-вирус относится к семейству Paramyxoviridae, подсемейству Pneumovirinac,
роду Pneumovirus. Помимо РСВ человека, к роду Pneumovirus относятся РС-вирус
крупного рогатого скота и вирус пневмонии мышей.
Вирион имеет, как правило, шарообразную форму, размеры его колеблются в
диапазоне от 65 до 800 нм, встречаются нитевидные формы. Имеется наружная
липопротеиновая оболочка с двумя трансмембранными гликопротеинами, обра-
зующими выступы длиной 12 нм. Геном вируса представлен линейной однонитевой
РНК отрицательной полярности, уложенной внутри нуклеокапсида спиральной
симметрии. Нуклеокапсид содержит геномную РНК и три белка: N, Р, L, РНК протя-
женностью 15—19x103 н.о. содержит десять генов, кодирующих одиннадцать белков,
при этом две открытые рамки считывания имеются в гене, кодирующем белок М2.
В процессе репродукции вируса, которая происходит в цитоплазме чувствительных
клеток, синтезируются ранние (неструктурные) и поздние (структурные) белки:
• неструктурные белки NS1 и NS2;
• белки нуклеокапсида: белок N и фосфопротеин Р, участвующие в транскрип-
ции вирусной РНК; РНК-зависимая РНК-полимераза — белок L;
• матриксные белки М и М2, ассоциированные как с липидным слоем, так и
с нуклеокапсидом; М2 необходим также для осуществления транскрипции
вирусного генома;
• три поверхностных белка: F — белок слияния, G — поверхностный глико-
протеин, белок SH. Поверхностные белки отвечают за слияние вирусной
оболочки и клеточной мембраны, проникновение вируса в клетку. Белок G
детерминирует иммунный ответ на PC-вирусную инфекцию.
В состав зрелого вириона входя г восемь белков.
В отличие от других представителей семейства Paramyxoviridac. PC-вирус не
содержит ни гемагглютинина, ни нейраминидазы. Антигенная активность вируса
определяется его способностью стимулировать образование вируснейтрализую-
щих и комплементсвязывающих антител. Существенных антигенных различий
между штаммами РСВ и группами А и В, различающимися по гликопротеиду G, не
выявлено; можно говорить о наличии одного серотипа вируса.
РСВ характеризуется низкой устойчивостью к факторам внешней среды и
физико-химическим факторам. Вирус чувствителен к действию эфира, кислот и
детергентов, переносит однократное замораживание при -70 °C. Вирионы инакти-
вируются при 55 °C в течение 5 мин, при 37 °C — в течение 24 ч. Вирус полностью
разрушается при pH = 3,0, лучше переносит щелочную среду. Оптимальное значе-
ние pH для сохранения инфекпионности - 7,5.
Лабораторная диагностика
Диагностика респираторно-синцитиальной инфекции основана:
• на детекции вирусных частиц, антигенов или вирусспецифической РНК;
• определении вирусспецифических антител или регистрации их прироста.
Материалом для исследований I группы являются мазик из носоглотки, мазок
из ротоглотки, мокрота и аутопсийный материал. Взятие клинического материала
700 ЧАСТНАЯ ВИРУСОЛОГИЯ
следует проводить как можно раньше,, желательно при появлении первых клини-
ческих симптомов заболевания, не позднее 3-го дня болезни. Аутопсийный мате-
риал следует получать как можно раньше после смерти пациента.
Для серологических исследований используют:
• одну (для определения IgM-фракции антител) сыворотку крови больного,
полученную на 3-4-й день заболевания:
• парные сыворотки крови больного, полученные в ранний период и на стадии
реконвалесненции.
РСВ, его антигены и РНК могут быть обнаружены в клинических образцах сле-
дующими методами:
• трансмиссионной электронной микроскопией вирусных частиц:
• выделением вируса в культуре клеток с последующей идентификацией в
иммунологических реакциях;
• методом флюоресцирующих антител;
• иммуноферментным анализом для выявления вирусных антигенов;
• полимеразной цепной реакцией.
Антитела к РСВ могут быть определены следующими серологическими мето-
дами:
• реакцией связывания комплемента;
• реакцией нейтрализации;
• иммуноферментным анализом.
Лабораторную диагностику проводят для подтверждения клинического диа-
гноза, дифференциации PC-инфекции от ОРВИ иной этиологии. Предпочтение
отдается методам быстрой диагностики: МФА, ПЦР и IgM-ИФА. Осуществляют
также определение нарастания титра специфических антител в процессе болезни в
РСК. а при необходимости — выделение вируса в клеточных культурах.
РИНОВИРУСЫ
Характеристика вирусов
Риновирусы (HRVs) (от греч. rhinos — «нос») относятся к роду семейства
Picornaviridae, порядку Picomavitales На основании сродства к определенным кле-
точным рецепторам, устойчивости к антивирусным компонентам и генетического
родства выделяют три вида:
• Human rhinovirus А (мажорная группа, сродство к I САМ-1-рецептору):
• Human rhinovirus В (минорная группа, сродство к LDL-репептору);
• Human rhinovirus С. По структуре единственного типоснецифического антиге-
на выделяют 113 иммуноразнородных групп. Группоспецифический антиген
отсутствует.
Вирион состоит из нуклеокапсида икосаэдрической формы, диаметром 30 нм,
собран из двенадцати капсомеров, образованных четырьмя полипептидами: VP1,
VP2, VP3 и VP4. Геном, размером 7,2-8,5х103 н.о. состоит из инфекционной
несегментированной одноцепочечной моноцистронной +РНК, кодирует один ген.
Транслируемый белок обладает протеолитической активностью и разрезается на
три полипро^еина: Pl, Р2 и РЗ. Белок Р1 — предшественник структурных полипро-
теидов VP1, VP2, VP3 и VP4. Потомки Р2 способствуют синтезу вирусного белка
после выключения синтеза клеточных белков, а вирусная репликаза и ферменты,
образуемые из РЗ, необходимы для контроля генома клетки-хозяина. Репликация
пикорнавирусов происходит в цитоплазме, цикл репродукции очень короткий,
составляет примерно 8 ч. Риновирусы имеют общий рецептор ICAM-1, отвечаю-
щий за тканевый тропизм.
ЧАСТНАЯ ВИРУСОЛОГИЯ 7Q1
Риновирусы являются наиболее частой причиной острых нетяжелых респира-
торных заболеваний, характеризуемых насморком, ангиной, реже кашле м и проте-
кающих, как правило, без повышения температуры. Имеют воздушно-капельный
и орально-фекальный пути передачи. В большом числе случаев вызывают бессим-
птомную инфекцию: доказаны случаи одновременной инфекции двумя, тремя и
даже четырьмя энгеровирусами.
Риновирусы устойчивы к эфиру, дезоксихолату, различным детергентам.
Чувствительны к кислым значениям pH, действию ультрафиолетового облучения,
высушиванию. Обработка 0,3% формальдегидом, 0,1 N соляной кислотой или
свободным остаточным хлором в концентрации 0,3- 0,5 мг/л ведет к быстрой
инактивации, однако присутствие органических веществ может оказывать защит-
ное действие. Вирусы относительно термолабильны. Добавление к вирусной
взвеси хлористого магния в одномолярной концентрации сохраняет титр вируса
при 50 °C в течение часа. В замороженном состоянии энтеровирусы хранятся в
течение многих лет, при 4-6 °C — несколько недель, при комнатной температу-
ре — несколько дней, выдерживают многократное замораживание и оттаивание
без потери активности.
По способности к репродукции в клетках приматов риновирусы разделяют на
две группы: Н и М. Риновирусы группы Н размножаются и вызывают цитопа-
тические изменения в ограниченной группе диплоидных клеток человеческого
эмбриона (ФЛЭЧ) и специальной линии (R) клеток HeLa. Риновирусы группы М
размножаются и вызывают цитопатические изменения в клетках почек обезьян,
почек эмбриона человека и различных перевиваемых клеточных линиях челове-
ческих клеток (HeLa, BGM). Оптимальная температура культивирования ринови-
русов составляет 33 °C.
Лабораторная диагностика
Основным клиническим материалом являются носоглоточные смывы, кровь,
содержимое синусов (носовых пазух). Возможны взятие различных тканей и жид-
ких сред организма и сбор фекалий, как и для других энтеровирусов. Носоглоточные
смывы забирают в первые 3-4 дня от начала заболевания. Образцы крови заби-
рают в остром периоде болезни и в стадии реконвалесценции. Парные сыворотки
исследуют в одном опыте.
Лабораторную диагностику риновирусов проводят редко ввиду легкости тече-
ния заболевания. Основными методами лабораторного подтверждения ринови-
русной инфекции являются:
• обнаружение РНК риновируса с помощью ПЦР:
• выделение вируса (в культуре клеток или на лабораторных животных).
Детекция РНК риновирусов в назальных смывах, крови, цереброспинальной
жидкости и материалах из верхних дыхательных путей методом ОТ-ПЦР обла-
дает большей чувствительностью, бьдтротой и позволяет выявлять вирусы, не
размножающиеся в культуре клеток. Имеется множество модификаций методи-
ки. Используют праймеры для 5’-нетранслируемой области вирусного генома.
Молекулярное типирование риновирусов основано на определении нуклеотид-
ной последовательности части генома, кодирующей белок капсида VP1. Серотип
исследуемого изолята определяют сравнением полученных последовательностей с
имеющимися в генном банке последовательностями прототипных энгеровирусов
человека. Этот метод несоизмеримо ускоряет типирование и может быть исполь-
зован для идентификации изолятов, которые трудно или невозможно типировать,
используя обычные иммунологические и вирусологические методы. Метод особо
полезен для оценки родства штаммов во время вспышек.
702 ЧАСТНАЯ ВИРУСОЛОГИЯ
Выделение вируса требует большего времени, но дает однозначный ответ на
вопрос об этиологии заболевания и позволяет использовать выделенный вирус
для последующих исследований. Для увеличения вероятности выделения вируса
из исследуемого материала целесообразно использовать не менее двух видов куль-
тур клеч ок. одной из которых должна быть культура клеток RD. Риновирус может
быть выделен в клеточной культуре, Wro, через 2-7 сут после заражения с после-
дующим определением специфических антител к выделенному штамму.
СЕРОЛОГИЧЕСКАЯ ДИАГНОСТИКА
Диагностика заболевания серологическими методами требует большого коли-
чества исследований со всеми известными серотипами энтеровирусов, поэтому
ее предпринимают только при данных, позволяющих предполагать тип этиоло-
гического агента (характерной клинической картине, выделении и идентифика-
ции вируса от больного, вспышках заболевания, обусловленного установленным
типом вируса). Диагноз инфекции подтверждается значительным подъемом титра
специфических антител, диагностически значимым считают четырехкратный и
более подъем титра во второй пробе.
При наличии диагностических типоспецифических иммунных сывороток воз-
можны серологические исследования с помощью реакции нейтрализации, РИФ,
реакции связывания комплемента, реакции преципитации в агаре, метода имму-
нофлюоресценции или подавления гемагглютинации. Наиболее чувствительным,
специфичным и распространенным способом идентификации является реакция
нейтрализации.
РОТАВИРУСЫ
Характеристика вирусов
Ротавирусы относятся к семейству Reoviridae, роду Rotavirus, который объединя-
ет большое количество сходных по морфологии и антигенной структуре вирусов,
вызывающих гастроэнтерит у человека, млекопитающих и птиц. Ротавирусная
инфекция имеет широкое распространение во всем мире. До начала программ вак-
цинации претив этой инфекции в развитых странах около 80% детей в возрасте до
5 лет болели гастроэнтеритом ротавирусной этиологии. Ежегодно от ротавирусной
инфекции в мире умирают 500-600 тыс. детей в возрасте до 5 лет.
В составе ротавирусов обнаружены четыре антигена, основной из них — груп-
повой антиген, обусловленный белком внутреннего капсида. С учетом группоспе-
цифических антигенов все ротавирусы делятся на семь серогрупп: А, В, С, D, Е, F,
G. Ротавирусы одной группы имеют общий групповой антиген, который выявляют
иммунологическими методами: иммуноферментным анализом, иммунофлюорес-
ценцией, иммунной электронной микроскопией и др. Большинство ротавирусов
человека и животных относятся к группе А. Внутри группы А существуют под-
группы и серотипы. Гетерогенностью ротавирусов объясняются повторные забо-
левания этой инфекцией. Вероятность повторного заражения на первом году
жизни составляет около 30%. К двум годам почти 70% детей болеют ротавирусной
инфекцией дважды, 40% — трижды, 20% детей — четыре раза. Известно, что при
последуюших заражениях заболевание протекает легче или в виде бессимптомной
инфекции.
Вирион имеет сферическую форму (от лат. rota — «колесо»), диаметр от
65 до 75 нм. В вирионе выявляется электронно-плотный центр диаметром
38-40 нм (сердцевина) и две белковые оболочки: наружный и внутренний капси-
ды. Сердцевина содержит внутренние белки и генетический материал, представ-
ленный двухнитевой фрагментированной РНК, которая составляет 14-22% массы
ЧАСТНАЯ ВИРУСОЛОГИЯ 703
вириона. Геном ротавирусов человека и животных состоит из одиннадцати сегмен-
тов, которые могут быть разделены при электрофорезе в полиакриламидном геле
или агарозе. Сегментарность генома позволяет ротавирусам легко обмениваться
гомологичными участками РНК. Ротавирусы не имеюг суперкапсидной липопро-
теиновой оболочки и не содержат липидов.
Наружный слой включает два структурных вирусных белка (VP): VP4 (расще-
пленный протеазой белок, или Р-протеин) и VP7 (гликопротеин, или G-протеин).
Эти два белка определяют серотип вируса и считаются важными для разработки
вакцин, поскольку они являются основными антигенами, вовлеченными в про-
цесс нейтрализации вируса для обеспечения защитного эффекта. Поскольку два
сегмента гена, кодирующих эти белки, могут расщепляться независимо, в практике
применяются методы исследования (ПЦР), позволяющие определить оба геноти-
па — Р и G. В различных странах и в разные годы в одной стране могул быть рас-
пространены вирусы, содержащие различные комбинации генотипов — Р и G; Р[8]
G1, Р[4]G2, Р[8]G3, Р[8]G4, P[8]G9, Р[6]G9, Р[1 l]G10 и др.
Ротавирусы относительно стабильны в окружающей среде. По устойчивости
к дезинфектантам и выживаемости во внешней среде ротавирусы приближаются
к энтеровирусам, они не погибают при обычном хлорировании воды в голов-
ных водопроводных сооружениях, выживают в водопроводной воде до 60 дней,
на различных объектах внешней среды — от 10-15 до 30 дней (в зависимости
от температуры и влажности воздуха). В фекалиях ротавирусы сохраняются от
нескольких недель до 7 мес, на фруктах — 5-30 дней, на тканях из хлопка и шер-
сти — 12-45 дней, на различных поверхностях — до 10 дней. Ротавирусы устой-
чивы к эфиру, хлороформу, детергентам, воздействию ультразвука, гипохлорита,
они не разрушаются при многократном замораживании-оттаивании. Ротавирусы
быстро инактивируются фенольными соединениями, крезолом, формалином.
Дая быстрой инактивации требуется 4-10% формальдегид. Самым эффективным
дезинфектантом для них является этиловый спирт. Вирус утрачивает инфекцион-
ность при кипячении, обработке сильными кислотами и щелочами (инфекционная
активность сохраняется в диапазоне pH = 3,0-9,0). Протеолитические ферменты
(панкреатин, трипсин, эластаза и др.) усиливают инфекционную активность виру-
са, что используют для культивирования вируса в культурах клеток.
Лабораторная диагностика
Наиболее информативным материалом для лабораторной диагностики
ротавирусной инфекции являются фекалии. В лабораторной диагностике при-
меняют три основных подхода:
• непосредственное исследование материала на наличие вирусного антигена
или нуклеиновых кислот;
• изоляцию и идентификацию вируса из клинического материала;
• серологические исследования, основанные на установлении значительного
прироста антител в течение болезни.
При любим выбранном подходе к диагностике инфекции одним из важнейших
факторов является качество исследуемого материала. Для прямого анализа образ-
ца или для изоляции вируса исследуемый материал должен быть получен в самом
начале заболевания, когда возбудитель еще экскретируется в относительно боль-
ших количествах и не связан антителами, а объем образца должен быть достаточен
для прямого исследования. Важную роль в успешной диагностике играет среда, в
которую забирают материал, а также условия транспортировки и хранения образ-
цов.
Методы лабораторной диагностики включают обнаружение ротавирусов с
помощью электронной микроскопии в первые 3 дня от начала болезни, а также
704 ЧАСТНАЯ ВИРУСОЛОГИЯ
их антигенов в фекалиях больного на 4-5-й день болезни с помощью реакции
ко-агглютинации (твердофазной), латекс-агглютинации, иммуноферментного
анализа. Серологические методы — реакция нейтрализации вируса и реакция тор-
можения гемагглютинации с ротавирусным антигеном — позволяют обнаружить
специфические антитела в крови со второй недели болезни. Определение антител к
ротавирусам класса IgM с использованием непрямого варианта иммуноферментно-
го анализа позволяет получить результат более оперативно (в день исследования).
Дифференциальную диагностику проводят с астровирусной, норовирусной,
аденовирусной инфекциями. Ротавирусы необходимо дифференцировать также
от частиц бактериофагов и бактериальных жп'тикон. встречаемых в фекальном
материале.
Прямые методы диагностики
Прямые методы диагностики — это методы, которые позволяют обнаружить
вирус, вирусный антиген или вирусную нуклеиновую кислоту непосредственно в
клиническом материале, они являются наиболее быстрыми (2-24 ч).
ЭЛЕКТРОННАЯ МИКРОСКОПИЯ
Для успешного определения вируса его концентрация в пробе должна быть
примерно 1х106 частиц в 1 мл. Забор проб фекалий проводят в остром перио-
де заболевания, желательно в течение первых трех суток от начала проявления
клинических признаков болезни, когда содержание вирусных частиц достигает
10—10° в 1 мл. Важнейшее значение имеет соблюдение условий предохранения
материала от контаминации. В последующие дни заболевания, по мере нормали-
зации стула, выделение вируса резко сокращается, поиск его затруднен, требуется
предварительное осаждение с помощью высокоскоростного центрифугирования с
последующим негативным контрастированием. Для диагностических исследова-
ний готовят 10-20% суспензию фекалий в стерильном изотоническом растворе,
тщательно гомогенизируют и осветляют центрифугированием в течение 30 мин
при 3000 об./мин, добавляют антибиотики и хранят при -20 °C до исследования.
В препаратах при электронной микроскопии ротавирусы встречаются в виде
дву- и однокапсидных частиц. Диаметр двукапсидных частиц — 70 нм. Наружный
и внутренний капсиды соединены между собой короткими выростами, из-за чего
частица по внешнему виду напоминает колесо. Диаметр однокапсидных частиц —
60 нм. Они лишены сердцевины и ограничены лишь капсидным слоем.
ИММУННАЯ ЭЛЕКТР -ИНАЯ МИКРОСКОПИЯ
Одним из вариантов ЭМ., используемым в диагностических целях, является
иммунная электронная микроскопия (ПЭМ). В результате взаимодействия анти-
тел с вирусами образуются komi глексы, которые после негативного контрастирова-
ния легче обнаружить. В присутствии иммунной сыворотки образуются компакт-
ные агрегаты из 3-4 полных и пустых частиц. Также наблюдаются одиночные
частицы, покрытые слоем иммунных глобулинов.
Иммуноэлектронная микроскопия более чувствительна, чем электронная.
Электронная микроскопия является стандартом для оценки специфичности и
чувствительности современных методов исследования и коммерческих тест-
систем.
МЕТОДЫ ВЫЯВЛЕНИЯ РНК РОТАВИРУСОВ
Электрофорез
При электрофорезе в полиакриламидном геле или агарозе происходит распре-
деление сегментов генома с образованием характерного профиля, состоящего из
ЧАСТНАЯ ВИРУСОЛОГИЯ 705
четырех классов генов. Картина распределения сегментов РНК на классы явля-
ется уникальной именно для данной группы кишечных вирусов. Обнаружение
специфического профиля является прямым указанием на наличие ротавирусов в
исследуемом материале. Чувствительность метода составляет 97,6% по сравне-
нию с методом электронной микроскопии, позволяет выявлять РНК в пробах с
разрушенными вирионами и дифференцировать электрофоретип РНК на уровне
штаммовых вариаций. По количеству cei ментов внутри классов генов метод
электрофореза геномной РНК позволяет идентифицировать ротавирусы анти-
генных групп А, В и С.
Полимеразная цепная реакция
Выявление РНК ротавируса осуществляют методом ПЦР. Для разработки прай-
меров была выбрана в достаточной степени консервативная область гена белка
NSP4. Для выявления РНК ротавирусов в клиническом материале и объектах
окружающей среды применяют, как правило, ПЦР с электрофоретической детек-
цией продуктов амплификации в агарозном геле.
Иммуноферментный анализ
ИФА может быть использован как для определения антигена (на твердую
фазу — дно лунки полистиролового планшета — наносят антитела), так и для
определения антител (в этом случае на твердую фазу наносят антигены). В усо-
вершенствованных иммуноферментных тест-системах для исключения неспеци-
фических ложноположительных результатов предусматривается возможность
подтверждающего теста.
ИММУНОХИМИЧЕСКИЕ МЕТОДЫ ИССЛЕДОВАНИЯ
Иммуноцитохимическая детекция вируса
Метод основан на иммунохимическом определении вирусного антигена, обра-
зуемого в клетках в результате их заражения вируссодержащим материалом.
Преимущество метода по сравнению с ИФА заключается в более высокой чувстви-
тельности, а также возможности определить и количественно оценить инфекцион-
ность исследуемой пробы.
Опыт проводят на перевиваемых клетках почки зеленой мартышки в лунках
плоскодонных панелей. Тестируемый материал разводят в среде Игла-МЕМ,
активируют в течение 1 ч при 37 °C в присутствии 10 мкг/мл трипсина и вносят
в четыре лунки. Положительным контролем служат лунки, в которые внесен
материал, заведомо содержащий ротавирус. Отрицательные контроля — лунки
с незараженными клетками и лунки, в которые внесен материал, аналогичный
исследуемому, но заведомо не содержащий ротавируса. В лунки вносят иммунную
антиротавирусную сыворотку кролика. После отмывания панели в лунки вносят
меченные пероксидазой антитела против IgG-кролика (коммерческий препарат),
затем субстрат пероксидазы 3-амино-этилкарбазол. Учет реакции проводят под
световым микроскопом с малым увеличением. Лунки с отрицательными кон-
тролями не должны содержать окрашенных клеток. Локальные скопления окра-
шенных клеток представляют собой фокусы вирусной инфекции.
Агглютинационные методы
Агглютинационные методы основаны на способности частиц с адсорбирован-
ными на них специфическими антителами образовывать комплексы (агглюти-
наты) в присутствии антигена ротавирусов группы А. Образуемый агглютинат
позволяет проводить визуальную оценку результата анализа. Простота выпол-
нения, скорость получения результата, низкая стоимость анализа привлекают к
этим методам большинство практических лабораторий.
706 ЧАСТНАЯ ВИРУСОЛОГИЯ
ПАРВОВИРУС В19
Характеристика вируса
Парвовирус В19 является возбудителем повсеместно распространенной антро-
понозной инфекции. Обладает тератогенным действием, в 20% случаев инфици-
рование беременных парвовирусом приводит к внутриутробной гибели плода или
сопровождается неиммунной водянкой плода.
Human parvovirus В19 относится к семейству Panoviridae (от лат, parvum —
«маленький»), подсемейству Parvovirinae (парвовирусы позвоночных), роду
Erythrovirus (от лат. crythros — «красный»). Это семеш тво не классифицировано по
порядку, характерными особенностями Parvoviridae являются организация генома
в виде однонитевой ДНК и небольшие размеры вирионов. Род Erythrovirus выделен
на основании генетической карты вируса и ввиду особого сродства к клеткам-
предшественникам эритроцитов.
Вирион В19, диаметром от 22 до 24 нм, состоит из капсида с заключенной вну-
три линейной однонитевой молекулой ДНК, не имеет оболочки. Для парвовируса
определен икосаэдрический тип симметрии. Зрелая вирусная частица имеет моле-
кулярную массу 5,6x106. Капсид состоит из 60 капсомеров, образованных двумя
белками: VP1 и VP2. Минорный белок VP1 имеет несколько консервативных
участков, обеспечивает транспорт и локализацию капсида в ядре клетки и пред-
положительно функцию фосфолипазы A. VP2 является основным структурным
белком капсомеров, обеспечивает сборку вирионов. На N-тепминальных концах
структурных белков VP1 и VP2 находятся антигенные сайты связывания с лим-
фоцитами, индуцирующие образование вируснейтрализуюших антител. Главный
неструктурный фосфорилированный белок NS1 участвует в сайтспецифическом
ДНК-связывании, обладает АТФазной, нуклеазной, транскрипционной и хели-
казной активностью, обеспечивает репликацию вирусного генома и контроль экс-
прессии генов клетки-хозяина. В процессе репродукции вируса в чувствительных
клетках синтезируются еще три неструктурных белка. Их функция неизвестна.
Геном парвовируса В19 представлен линейной одноцепочечной инфекционной
ДНК длиной 5596 н.о. Выделено три генотипа парвовируса человека В19. Генотип I
включает изоляты В19, А6, Au и Wi, ко II генотипу относятся Lain и К71, а штамм
V9 принят за прототип III генотипа. Внутри генотипа I различие нуклеотидных
последовательностей геномов межд} штаммами не превышает 6%. Генотипы II и
III отличаются от первого составом белков капсида и патогенетическими свойства-
ми. Несмотря на 3% дивергенцию аминокислотного уровня у разных штаммов В19
выделяют один серотип.
В19 устойчив к эфиру, спирту, желчи, трипсину, изменению pH среды, сохраняет
инфекционную активность пос ле прогревания при температуре 56 °C в течение
часа, термостабилен при 80 °C. Инактивируется кипячением, р-пропиолактоном,
глутаральдеждом, гипохлоридом. у-излучением. Воздействие 0,1% формалина не
оказывает инактивирующего действия, но в 0,5% растворе формалина или щелочи
вирионы разрушаются в течение суток.
Обладая высокой устойчивостью к физическим факторам среды, паровирус В19
представляет большую опасность в качестве контаминанта препаратов крови и транс-
плантатов. В острой фазе инфекции может присутствовать в крови в очень высокой
концентрации — до 1012 ГЭ/мл. Кроме того, парвовирус В19 может длительно (до
года) сохраняться в крови при отсутствии каких-либо клинических симптомов
Лабораторная диагностика
Исследовать на наличие парвовируса В19 можно различные клинические образ-
цы: сыворотку крови, носоглоточные смывы, амниотическую жидкость, кровь
ЧАСТНАЯ ВИРУСОЛОГИЯ 7Q7
плода, образцы тканей, костный мозг, лейкоциты, клетки амниотической, плев-
ральной жидкости и др.
Правила забора, хранения и транспортировки наиболее часто используемых
клинических образцов не отличаются от традиционных для работы с вирусинфи-
цированным материалом. Взятие амниотической жидкости осуществляют в сте-
рильную пробирку типа «Эппендорф» при проведении амниоцентеза по стандарт-
ной методике. Полученный образец амниотической жидкости ресуспендируют.
Автоматическим дозатором с наконечником с аэрозольным барьером забирают
1 мл материала в новую стерильную пробирку типа «Эппендорф» и центрифу-
гируют при 8000-9000 g (12-13 тыс. об./мин) в течение 10 мин. Надосадочную
жидкость забирают, используя наконечник с аэрозольным барьером, оставляя над
осадком 200 мкл жидкости, и ресуспендируют материал на вортексе. Допускается
хранение амниотической жидкости и предобработанного материала в течение
1 сут при температуре от 2 до 8 °C, 1 мес — не выше -16 °C. Амниотическую жид-
кость можно длительно хранить при -70 °C.
Лабораторная диагностика парвовирусной инфекции основана на использо-
вании цитологических, вирусологических (идентификация вируса, его антигенов
или нуклеиновых кислот) и серологических методов.
ЦИТОЛОГИЧЕСКОЕ ИССЛЕДОВАНИЕ
Цитологическое исследование мазков крови или костного мозга показано у
пациентов с клинически значимой анемией или транзиторным апластическим
кризом. Проводят подсчет количества ретикулоцитов и определение изменений в
клетках миелоидного и мегакариоцитного рядов. Характерно появление гигант-
ских пронормобластов (25-32 мкм) с эозинофильными внутриядерными включе-
ниями, вакуолизацией цитоплазмы, выступами по типу «собачье ухо».
ВИРУСОЛОГИЧЕСКИЕ МЕТОДЫ
К вирусологическим методам диагностики парвовируса В19 относятся элек-
тронная микроскопия, выделение вируса в культуре клеток, реакция гемагглюти-
нации.
Выделение вируса in vitro возможно на первичных культурах клеток человека и
обезьян: культурах клеток костного мозга, печени плода, пуповинной крови, пери-
ферической крови, линии мегакариобластных клеток человека МВ-02 и UT-7/
Еро-Sl, линии клеток эритроидной лейкемии человека JK-1 и KU812Ep6. Заражать
следует монослой на стадии 30-50% или в момент пересева. Для инициации
репликации требуется присутствие эритропоэтина. Инфекция ци голитическая.
Вирус может быть обнаружен в пробах с помощью электронной микроскопии,
реакции гемагглютинации. В Японии (Red Cross Blood Center) применяют метод
рецепторопосредованной гемагглютинации для скрининга вируса В19 в образцах
донорской крови. Тест занимает 3 ч, но чувствительность ниже чем 105копий/мл.
Изоляцию вирусной ДНК осуществляют методами гибридизации in situ, част-
ным случаем которой является прямая гибридизация, и ПЦР. Метод гибридизации
in situ успешно применяют для локализации специфических вирусных нуклеи-
новых кислот внутри морфологически целых клеток. Он заключается в том, что
меченные дигоксигенином пробы иммунохимически обнаруживают вирусную
ДНК из клинического образца с помощью антидигоксигенин поликлональных
Fab-фрагментов, меченных алкалин фосфатазой или пероксидазой хрена, при
добавлении хромогенного субстрата. В результате действия ферментов образу-
ется окрашенный продукт. Чувствительность метода составляет от 1,5x105 до
ЗхЮ4 копий генома/мл.
Таким образом, можно исследовать аспират костного мозга у пациентов с апла-
стическим кризом или гипопластической анемией, эмбриональные ткани, раз-
личные клеточные линии. Можно использовать длительно хранящиеся образцы,
708 ЧАСТНАЯ ВИРУСОЛОГИЯ
которые были зафиксированы в 4% растворе параформальдегида, дегидратирова-
ны в разведениях этанола, высушены и хранились при 4 °C. Применение люми-
несцентных субстратов в гибридизации in situ позволило повысить чувствитель-
ность метода, В случае прямой гибридизации, обычно в формате slot blot или dot
blot (точка-пятно), применяют пробу полноразмерной вирусной ДНК. меченной
32Р, дигоксигенином или биотином, и хемилюминесцентные субстраты. Анализ
занимает около 30 ч. Чувствительность этого метода составляет приблизительно
105 копий генома/мл. Чувствительность методов увеличивается в ряду: dot blot-
гибридизация — колориметрическая in ^-гибридизация — люминесцентная in
situ-гибридизация. Таким образом, можно выявить все известные изоляты парво-
вируса В19,
СЕРОЛОГИЧЕСКАЯ ДИАГНОСТИКА
Основные методы серологической диашостики направлены на выявление
специфических антител к парвовирусу. Это МФА, иммуноэлектрофорез, РИА,
ИФА. Стандартным методом является ИФА на основе рекомбинантных антигенов
В19, IgM определяют с помощью тест-систем с сорбированным на твердую фазу
рекомбинантным структурным белком капсида VP2, a IgG — белком VP1. При
беременности IgM-тест может дать ложноположительные результаты. В этом слу-
чае требуется подтверждение с помощью вестерн-блота.
При выборе метода диагностики важно помнить, что только у 50% беременных
выявляется IgM к В19 во время постановки клинического диагноза водянки плода.
у пациентов с преходящими апластическими кризами IgM-антитела обнаружива-
ют, а у больных с иммунодефицитом и хронической анемией специфических анти-
тел нет, но выявляют вирусную ДНК.
В случае низкого уровня виремии более чувствительным является метод ПЦР.
Рекомендуют одновременное использование нескольких пар праймеров, так
как не все праймеры могут выявить штаммы парвовируса трех генотипов. ПЦР-
исследование предполагает использование плазмы периферической и пуповинной
крови, амниотической жидкости, слюны, носоглоточных смывов и мазков из
зева.
Для решения специальных задач возможно использование усовершенствован-
ною метода ПЦР с защитой гибридизации (PCR-HPA). Суть метода заключается
в инкубировании продуктов ПЦР (праймеров к участку VP1-VP2) вместе с допол-
нительными пробами, меченными acridinium ester. Учитывают люминесцентный
сигнал от акридиниума в люминометре. Для детекции вируса этим методом требу-
ется сорок циклов амплификации и НРА. Таким способом может быть обнаружена
единственная копия ДНК парвовируса В19 в 10-pl образца. Время исследования
составляет 6 ч.
Дифференциальная диагностика
Требуется дифференциальная диагностика с корью, краснухой, скарлати-
ной, внезапной экзантемой, энтеровирусными инфекциями, псевдотуберкулезом,
системными заболеваниями, токсико-аллергическими состояниями.
ПАПИЛЛОМАВИРУСЫ
Характеристика вирусов
Papillomaviridae — семейство ДНК-содержащих вирусов простого строения,
обычно именуемых папилломавирусами. Папилломавирусы адаптированы к опре-
деленному виду хозяев. Несколько сотен видов (типов) папилломавирусов пора-
ЧАСТНАЯ ВИРУСОЛОГИЯ 799
жают млекопитающих, птиц, рептилий, вызывая у них образование доброкаче-
ственных опухолей — папиллом (бородавок).
Вирус папилломы человека (ВПЧ, Human Papilloma Virus — HPV) вызывает
широкий спектр повреждений эпителия кожи и слизистых оболочек аноурогени-
тальной области, верхних дыхательных путей, реже полости рта, пищевода, пря-
мой кишки, конъюнктивы глаза у человека.
Таксономически ВПЧ подразделяют на роды, обозначаемые греческими
буквами (а, 0, у и т.д.), виды (обозначают арабскими цифрами и буквой рода,
например, а9), типы (генотипы, обозначаемые арабскими цифрами, например
16, 18 и др.). В настоящее время идентифицировано более ста генотипов ВПЧ.
Большинство типов ВПЧ, вызывающих поражения слизистой оболочки аноуро-
генитальной области, относят к роду a (Alpha). Филогенетическая и эпидемио-
логическая классификации ВПЧ, ассоциированных с карциномой шейки матки,
представлены в табл. 24-8.
ВПЧ родов 0 (beta) и у (gamma) тропны к ороговевающему эпителию.
Инфицирование ВПЧ рода 0 чаще всего протекает бессимптомно, однако у
лиц, страдающих некоторыми наследственными болезнями (Epidermodysplasia
Verruciformis — EV) и иммунодефицитами, может привести к развитию добро-
качественных и злокачественных опухолей плоского эпителия. Цитопатическое
действие ВПЧ других родов — у (gamma), ц (ти) и v (пи) — может проявляться в
виде доброкачественных разрастаний эпителия (папиллом, кондилом).
Вирионы имеют относительно малые размеры, внешняя липидная оболочка
отсутствует. Геном ВПЧ представляет собой кольцевую двухцепочечную молекулу
ДНК протяженностью около 8000 пар нуклеотидов, покрытую белковым капси-
дом. Капсид имеет форму икосаэдра и сформирован 72 пентамерами протеина L1,
с которым ассоциирован протеин L2. В составе ДНК имеется три функциональные
области: Early (Е), Late (L) и Upstream Regulatory Region (URR). Область E содержит
гены, кодирующие шесть «ранних» белков: Е1, Е2, Е4, Е5, Е6, Е7, которые участву-
ют в репликации вирусной ДНК и формировании вирионов в инфицированных
клетках эпителия. Все гены расположены только на одной из цепей ДНК. Белки Е6
и Е7 определяют онкогенные свойства вируса, подавляя клеточные белки р53 и
pRb — супрессоры опухолей. Гены области L кодируют «поздние» структурные
белки вирусного капсида: L1 и L2. Кодирующие участки генома разделены неко-
дирующей областью URR, которая регулирует экспрессию генов, репликацию ДНК
и сборку вирионов.
Таблица 24-8. Филогенетическая и эпидемиологическая классификация ВПЧ, ассоциированных
с карциномой шейки матки
Род о, виды Тип Плоскоклеточный рак, % Аденокарцинома,% Уровень риска
А1 32, 42 Низкий
АЗ 61, с62, 72 Низкий
81 0,04 Низкий
83 0,04 Низкий
84 Не определен
А 5 26 0,22 Высокий
51 0,75 0,54 Высокий
69 0,26 Не определен
82 Высокий
аб 30 Не определен
53 0,04 Высокий
56 1,09 Высокий
66 0,19 Высокий
710
ЧАСТНАЯ ВИРУСОЛОГИЯ
Окончание табл. 24-8
Род а виды Тип Плоскоклеточный рак, % Аденокарцинома,% Уровень риска
а? 18 11.27 37,30 Высокий
39 0,82 0,54 Высокий
45 5,21 5,95 Высокий
59 1,05 2,16 Высокий
68 0,37 Высокий
70 Не определен
аЭ 16 54,38 41,62 Высокий
31 3,82 1.08 Высокий
33 2,06 0,54 Высокий
35 1.27 1,08 Высокий
52 2,25 0,54 Высокий
58 1.72 Высокий
67 Не определен
А10 6 0,07 Низкий
11 0.07 Низкий
13, 44 0.04 Низкий
55 Низкий
74 Не определен
А11 34 Высокий
73 0,49 Высокий
А13 54 Низкий
А14 90 Низкий
А15 71 Низкий
Клеточная пролиферация и дифференцировка, сопровождаемые репликаци-
ей вирусного генома, транскрипцией и трансляцией вирусных белков и сборкой
вирионов, морфологически проявляются в виде койлоцитоза и появления визу-
ально наблюдаемых папиллом и кондилом. Именно эти участки кожи опасны в
отношении контактного заражения. Инфекционные вирусные частицы попадают
в окружающую среду по мере слущивания клеток поверхностных слоев эпителия.
Это характерно для папиллом и для легкой степени повреждения (дисплазии) эпи-
телия шейки матки, вызванных ВПЧ низкого и высокого канцерогенного риска.
Персистенция ВПЧ высокого канцерогенного риска обусловливает поврежде-
ния плоского и железистого эпителия (особенно в зоне трансформации) шейки
матки. Встраивание вирусной ДНК в ДНК клетки сопровождается утратой способ-
ности вируса к репликации при обязательном сохранении транскрипции генов Е6
и Е7. Взаимодействуя с клеточными белками (опухолевым супрессором р53 и др.),
белки — продукты генов Е6 и Е7 — нарушают стабильность клеточного генома,
процессы дифференцировки клеток и стимулируют их пролиферацию.
Папилломавирусы термостабильны, устойчивы к эфиру, хлороформу, детерген-
там, низким значениям pH и ультрафиолетовому облучению, инактивируются при
рентгеновском облучении.
ВПЧ распространяется контактным путем (в том числе половым), поражая кож-
ные покровы и слизистые оболочки (табл. 24-9). Типы ВПЧ, ассоциированные с
поражениями кожи и слизистых оболочек, представлены в табл. 24-9.
Таблица 24-9. Типы ВПЧ, ассоциированные с поражениями кожи и слизистых оболочек (Villiers,
1989)
Клинические проявления Типы ВПЧ
Поражения кожи
Подошвенные бородавки 1,2,4
Обычные бородавки 2, 4, 26, 27, 29, 57
Плоские бородавки 3, 10, 28, 49
ЧАСТНАЯ ВИРУСОЛОГИЯ 711
Окончание табл. 24-9
Клинические проявления Типы ВПЧ
Бородавки Бютчера 7
Бородавчатая эпидермодисплазия 5,8, 9,10,12,15, 19, 36
Небородавчатые кожные поражения 37, 36
Поражения слизистых оболочек гениталий
Остроконечные кондиломы 6,11,42-44, 54
Некондиломатозные поражения 6, 11, 16. 18, 30, 31, 33, 34, 35, 39, 40, 42. 43. 51,52, 55, 56, 57-59, 61, 64. 67-70
Карцинома 16, 18, 31,33, 35, 39, 45, 51,52. 56, 58, 59, 66, 70
Экстрагенитальные поражения слизистых оболочек
Папиллома гортани 6, 11.30
Карцинома шеи, языка 2, 6,11,16,18, 30
Лабораторная диагностика
Лабораторная диагностика папилломавирусной инфекции основана на
молекулярно-генетических методах обнаружения фрагментов генома ВПЧ непо-
средственно в цервикальных соскобах или биоптатах ткани. Методы культивиро-
вания ВПЧ не разработаны.
Среди высокоразрешающих технологий детекции нуклеиновых кислот самое
широкое распространение получила полимеразная цепная реакция (ПЦР).
ПЦР-тест-системы, обладающие высокими аналитической и диагностической
чувст витальностью и специфичностью, используют не только для выявления, но и
для определения в исследуемом материале количественного содержания (вирусной
нагрузки) и дифференциации (генотипирования) ВПЧ ВКР основных клинически
значимых генотипов (16, 18, 31, 33, 35, 39, 45, 51, 52, 56. 58, 59), ответственных
более чем за 94% случаев тяжелых цервикальных дисплазий и рака шейки матки,
а также для обнаружения ВПЧ 6 и 1].
Количественное определение содержания широкого спектра генотипов ВПЧ
ВКР обеспечивает возможность дифференцированной интерпретации лабора-
торных данных с учетом показателя вирусной нагрузки (Ig-количества копий
ДНК вируса в 105 клеток). Это позволяет прогнозировать характер течения ВПЧ-
инфекции (наличие транзиторной инфекции, развитие дисплазии или высокий
риск ее прогрессирования), оценивать эффективность терапии. Количество вируса
менее 3 lg-копий ДНК ВПЧ ВКР на 105клеток считают клинически малозначимым,
ассоциированным с минимальным риском развития дисплазии и спонтанной эли-
минацией вируса: 3-5 lg-копий ДНК на 105 клеток — клинически значимое коли-
чество вируса: 5 и более lg-копий ДНК на 105 клеток — повышенное количество
вируса, отражающее наличие дисплазии или высокий риск ее развития. Снижение
вирусной нагрузки более чем на 1 1g за 6 мес рассматривают в качестве маркера
транзиторной инфекции.
Генотипирование, направленное на выявление и дифференциацию ДНК ВПЧ
ВКР, имеет большое значение по ряду причин. Во-первых, выявление нескольких
генотипов вируса ассоциировано с более высоким риском длительной перси-
стенции возбудителя и менее благоприятным прогнозом течения заболевания.
Во-вторых, сохранение генотипа (спектра генотипов) ВПЧ ВКР через год после
первого тестирования свидетельствует о хронической (персистирующей) инфек-
ции, поскольку повторное инфицирование тем же генотипом вируса практически
не встречается. Наконец, выявление ВПЧ генотипов 16 и 18, обладающих наибо-
лее высокой онкогенностью, требует особой тактики ведения пациентов, которым
рекомендуют сразу проводить кольпоскопическое исследование.
712 ЧАСТНАЯ ВИРУСОЛОГИЯ
Существуют также неамплификационные (гибридизационные) методы выявле-
ния ДНК ВПЧ, которые пока не нашли широкого применения в России,
Основной задачей диагностики папилломавирусной инфекции, обусловленной
ВПЧ высокого канцерогенного риска, является раннее выявление предраковых
изменений эпителия, для чего дополнительно используют цитологический и
гистологический методы, с помощью которых выявляют наличие характерных для
папилломавирусной инфекции изменений эпителия, дисплазий, рака. Выявление
ВПЧ в диагностическом материале особенно важно на стадии латентной инфек-
ции, когда цитологическое и кольпоскопическое исследования неэффективны.
Вместе с тем установлено, что присутствие вируса даже без признаков дисплазии
эпителия сопровождается 300-кратным возрастанием риска развития карциномы
шейки матки.
ЦИТОЛОГИЧЕСКОЕ ИССЛЕДОВАНИЕ
Информативность цитологического метода определяется качеством взятия и
обработки материала для исследования и уровнем подготовки врача-цитолога.
Материал для цитологического исследования следует брать с экзоцервикса, с
участка на границе многослойного плоского и цилиндрического эпителия церви-
кального канала и из нижней трети эндонервикса с помощью гцетки-эндобраша
до гинекологического осмотра или кольпоскопии при условии отсутствия воспа-
лительных заболеваний. Наносить материал следует тонким слоем на специально
обработанное обезжиренное стекло. Для получения адекватного результата мазок
должен быть немедленно обработан во избежание высыхания клеток, поскольку
их структура может необратимо деформироваться и трактовка такого материала
будет неправильной. Фиксированные препараты окрашивают разными способами-
но классической является окраска по Папаниколау. При микроскопии препара-
тов, окрашенных этим методом, учитывают специфический комплекс признаков,
характеризующих ядро и цитоплазму клеток, а также их расположение (разроз-
ненное, в виде пластов и др.).
При папилломавирусном поражении эпителия в цитологических мазках выяв-
ляют оксифильно и базофильно окрашенные клетки поверхностного и проме-
жуточного слоев эпителия, двухъядерные клетки и многоядерные симпласты,
койлопиты и дискератоциты. Койлоциты представляют собой клетки много-
слойного плоского эпителия промежуточного типа, имеющие одно или несколько
гиперхромных ядер неправильной формы, оптически светлую зону вокруг ядра,
цитоплазму в виде узкого ободка по периферии клетки. Среди типичных койлоци-
тов выявляют клетки, в которых наблюдают мелкие единичные вакуоли в околг»я-
дерной зоне. Подобные клетки ассоциированы с легкой дисплазией.
При тяжелой дисплазии обнаруживают дискератоциты — поверхностные клетки
многослойного плоского эпителия небольших размеров с гиперхромными ядрами
и эозинофильной цитоплазмой с различной степенью ороговения, которые чаще
расположены комплексами; двухъядерные клетки — клетки в состоянии амитоза,
многоядерные клетки.
Интерпретация результатов цитологического исследования мазков с шейки
матки представляет большие сложности для практикующего врача в связи с
одновременным использованием сразу нескольких классификаций повреждений
эпителия шейки матки. Для лучшего понимания соотношения терминов, при-
меняемых в различных классификационных схемах, предлагают использовать
нижеследующую таблицу (табл. 24-10).
Самой распространенной классификацией при оценке результатов цитологиче-
ского исследования является классификация Папаниколау:
• 1-й класс — атипические клетки отсутствуют, нормальная цитологическая
картина;
ЧАСТНАЯ ВИРУСОЛОГИЯ 713
Таблица 24-10. Морфологические и цитологические классификации эпителиальных поражений
шейки матки
Морфологические классификации Цитологические классификации
морфологическая оценка CIN system (1966) Папаниколау (1947) Bethesda system (2001)
Норма, изменения отсут- ствуют Норма 1 -й класс Нормальная цитологическая картина
Незначительные клеточные изменения Незначительные клеточ- ные изменения 2-й класс Доброкачественные! клеточ- ные изменения, воспаление. Признаки различных инфек- ций. Радиационные, преактив- ные, репаративные изменения
Плоскоклеточные поли- морфные изменения (воспа- ление, инфекция и т.п) Плоскоклеточные поли- морфные изменения (вос- паление, инфекция и т.п.) 2-й класс ASCUS (АПНЗ) плюс поли- морфные изменения
Папилломавирусные изме- нения (койлоцитоз) Папилломавирусные изме- нения (койлоцитоз) 2-й класс LSIL (Н-ПИП)
Слабая дисплазия CIN 1 3-й класс LSIL (Н- ПИП)
Дисплазия средней степени СИМ 11 3-й класс HSIL (В-ПИП)
Дисплазия высокой степени CIN III 3-й класс HSIL (В-ПИП)
Carcinoma in situ (CIS) CIN III 4-и класс HSIL (В-ПИП)
Инвазивный РШМ Инвазивный РШМ 5-й класс Инвазивный РШМ
Атипия клеток железистого (эндоцервикального) эпи- телия Атипия клеток железисто- го (эндоцервикального) эпителия Атипия клеток железистого эндоцервикального) эпителия (AGUS)
Adenocarcinoma Adenocarcinoma Adenocarcinoma
Примечание. CIN — Cervical Intraepithelial Neoplasia', ASCUS — Atypical Squamous Cells of
Undetermined Significance (АПНЗ — атипия плоскоклеточная неопределенной значимости); LSIL —
Low-Grade Squamous Intraepitelial Lesions (Н-ПИП — плоскоклеточное интраэпителиальное пора-
жение низкой степени выраженности); HSIL — High-Grade Squamous Intraepitelial Lesions (В-ПИП —
плоскоклеточное интраэпителиальное поражение высокой степени выраженности).
• 2-й класс — незначительное изменение клеточных элементов (небольшое
увеличение ядра, появление клеток метаплазированного эпителия), обуслов-
ленное воспалительным процессом во влагалище и/или шейке матки;
• 3-й класс — единичные клетки с изменениями соотношения ядра и цитоплаз-
мы, дискариоз (требуется повторение цитологического исследования или
гистологическое исследование биоптата):
• 4-й класс — отдельные клетки с явными признаками озлокачествления
(увеличение ядер, базофильная цитоплазма, неравномерное распределение
хроматина);
• 5-й класс — большое количество атипических (раковых) клеток. Диагноз
злокачественного новообразования не вызывает сомнений.
Верификация злокачественных эпителиальных опухолей шейки матки возмож-
на лишь при гистологическом исследовании, которое является «золотым стандар-
том», но вторичным методом диагностики в силу инвазивности и трудоемкости.
Цитологические критерии, позволяющие опытному врачу-цитологу прийти к
заключению о поражении цервикального эпителия ВПЧ, выявляют лишь в 30%
всех случаев инфицирования, исключая недоступную для морфологической диа-
гностики латентную форму инфекции. Вместе с тем установлено, что присутствие
вируса даже без признаков дисплазии эпителия сопровождается 300-кратным воз-
растанием риска развития карциномы шейки матки.
В этой связи ВОЗ и многие международные организации по борьбе с онколо-
гическими заболеваниями рекомендуют использовать молекулярно-генетические
714 ЧАСТНАЯ ВИРУСОЛОГИЯ
исследования — тест на ВПЧ при первичном скрининге у женщин старше 30 лет
в сочетании с цитологическим исследованием или в качестве самостоятельного
теста в странах., где плохо организованы программы цервикального цитологиче-
ского скрининга, а также для мониторинга лечения умеренной и тяжелой дис-
плазии (CIN II и CIN III), карциномы in situ и инвазивного рака. Данный метод
весьма полезен и для разрешения сомнительных результатов цитологического
исследования как при первичном выявлении неопластических изменений, так и
у пациенток, подущающих лечение по поводу дисплазии и рака. Контингенты и
кратность исследования на носительство ВПЧ высокого канцерогенного риска с
помощью ВПЧ-теста достаточно освещены в литературе.
Использование цитологического исследования совместно с ВПЧ -тестом позво-
ляет повысить выявляемость предрака и рака шейки матки до 96-99%. а также
увеличить интервалы между регулярными обследованиями до 5-7 лет у пациен-
ток с отрицательным результатом ВПЧ-теста (включая группы ASC-US и CINI).
Однако использование этого метода как диагностического критерия для неопла-
стических процессов шейки матки приводит к значительной гипердиагностике, так
как примерно в 80% случаев инфицирование имеет кратковременный характер и
заканчивается спонтанным выздоровлением и элиминацией вируса. Таким обра-
зом, обнаружение ДНК ВПЧ не позволяет в большинстве случаев прогнозировать
развитие цервикального рака. Однако ВПЧ-тест имеет большую прогностическую
значимость, особенно если на фоне ВПЧ-инфекции уже имеется картина диспла-
зии эпителия шейки матки.
Верификация диагноза злокачественных эпителиальных опухолей шейки матки
возможна лишь при патоморфологическом исследовании, которое позволяет
также судить о степени поражения эпителия при клинической и субклинической
формах ВПЧ-инфекции по наличию цитопатического действия вируса.
ПОЛИОВИРУСЫ
Характеристика вирусов
Полиовирусы относятся к семейству Picomaviridae (от греч. pico — «малый» и
та — «содержащий РНК»), к роду Enterovirus. Существует три серотипа полио-
вирусов; тип 1, тип II, тип III. Типовым представителем рода Enterovirus и семе. -
ства Picomaviridae является вирус полиомиелита — штамм Mahoney полиовируса
типа 1.
Вирусы полиомиелита принадлежат к числу самых мелких из известных
РНК-содержащих вирусов. Вирусные частицы круглой формы, диаметром
22-30 нм, с икосаэдрической симметрией, не имеют липидной оболочки.
Высокоструктурированная белковая оболочка полиовируса состоит из шестиде-
сяти субъединиц, каждая из которых формируется из четырех различных поли-
пептидных цепей, внутри нее расположен вирусный геном — одна молекула рибо-
нуклеиновой кислоты (РНК). Геном вируса полиомиелита представляет собой
позитивную одноцепочечную РНК, состоящую примерно из 7500 нуклеотидов.
Четыре капсидных полипептида (VP1-VP4) являются продуктами протеолити-
ческого расщепления полипротеина-предшественника. З’-конец РНК полиадени-
лирован, а к 5’-концу ковалентно присоединен низкомолекулярный белок VPg.
Согласно трехмерной структуре полиовируса, большинство аминокислот, распо-
ложенных на поверхности вирусной частицы, принадлежат белку VP1, белки VP2
и VP3 частично экспонированы, а белок VP4 является полностью внутренним
белком. Геномная РНК полиовируса инфекционна и служит матричной РНК для
синтеза вирусных белков. Большая часть РНК полиовируса, кроме двух нетранс-
лируемых областей на 3'- и 5?-конце, представляет собой одну протяженную
ЧАСТНАЯ ВИРУСОЛОГИЯ 715
открытую рамку считывания, кодирующую один большой белок — полипротеин,
который образуется в процессе трансляции в цитоплазме инфицированных кле-
ток. Затем в результате каскада протеолитических расщеплений, катализируемых
двумя различными протеазами, из полипротеина образуются все функционально-
активные структурные белки вирусного капсида (от VP1 до VP4) и неструктурные
белки, включая протеин 3D, функционирующий как РНК полимераза — фермент,
ответственный за репликацию геномной РНК,
Механизм проникновения вируса в цитоплазму клетки и освобождение в ней
вирусного генома остаются неясными. Полиовирус связывается со специфическим
белком цитоплазматической мембраны — рецептором полиовируса (PVR; CD155),
относящимся к белковому суперсемейству иммунвглобулинов. Прикрепление к
рецептору обусловливает изменения в структуре капсида, необходимые для попа-
дания генома в цитоплазму. Ни один дцугой пикорнавирус не способен использо-
вать этот белок как клеточный рецептор.
Высокая частота ошибок РНК-полимеразы при репликации РНК. составляющая
в среднем НУ4 на одно основание, способствует появлению нуклеотидных замен
в геноме полиовируса, а отсутствие механизмов коррекции этих ошибок может
привести к накоплению точечных мутаций в процессе репликации полиовирусов.
Эволюция полиовирусов помимо мутаций, связана также с феноменом молекуляр-
ной рекомбинации. Введение в организм оральной полиомиелитной вакцины в виде
смеси полиовирусов трех серотипов обеспечивает возможность одновременного
инфицирования одной клетки кишечного эпителия вирусами разных серотипов, ’гго
ведет к реализации молекулярной рекомбинации между геномами полиовирусов
разных серотипов и появлению штаммов, являющихся межтиповыми рекомбинан-
тами. Это является основой появления межтиповых рекомбинантных вакцинных
штаммов у больных с вакциноассоциированным паралитическим полиомиелитом.
Межтиповые рекомбинантные штаммы были обнаружены и у здоровых вакциниро-
ванных детей. Феномен рекомбинации регистрируется также между вакцинными и
дикими штаммами полиовирусов одного серотипа (внутритиповые рекомбинанты)
у больных вакциноассоциированным паралитическим полиомиелитом.
Полиовирусы сходны с энтеровирусами, но отличаются от других пикорнавиру-
сов по физическим и химическим свойствам — плавучей плотности и устойчивости
к слабым кислотам. Частины полиовируса имеют плавучую плотность 1,34 г/мл
и коэффициент седиментации 156 S. Они относительно устойчивы к нагреванию
(в присутствии катионов магния). В течение 1-3 ч полиовирусы устойчивы в
кислой среде (pH = 3,05,0). Жизнеспособны при pH = 3,0 и ниже; относительно
устойчивы к действию ряда детергентов и дезинфектантов, включая мыло, неион-
ные детергенты, эфир, хлороформ и другие растворители жиров. Вирусы сохраня-
ют жизнеспособность в течение нескольких недель при 4 °C и нескольких дней при
комнатной температуре, но быстро инактивируется под действием высушивания,
ультрафиолета, высокой температуры, формалина и свободного хлора.
Клинический материал для лабораторного исследования
Поскольку полиовирусы размножаются в кишечном тракте в течение несколь-
ких недель после инфицирования организма, пробы фекалий наиболее пригодны
для выделения вируса. Менее пригодны глоточные тампоны, так как полиовирусы
обнаруживают в глотке на протяжении 7-10 дней после начала болезни. Из цере-
броспинальной жидкости полиовирусы выделяются редко.
Эффективность вирусологической диагностики в большой степени зависит от
правильного и своевременного сбора клинических проб, а также от соблюдения
оптимальных условий их транспортировки в лабораторию. Пробы фекалий от
больных ОВП должны быть взяты в ранние сроки (до 14 дней) от начала паралича.
716 ЧАСТНАЯ ВИРУСОЛОГИЯ
От больного берут две пробы фекалий (около 8 г каждая) с интервалом 24-48 ч,
так как содержание вируса в фекачиях может колебаться. Пробы помещают в
пластиковую пробирку с завинчивающейся крышкой. После сбора пробы следует
хранить в холодильнике, если их транспортируют в вирусологическую лаборато-
рию — сразу же после сбора. Если это невыполнимо, пробы следует заморозить
при -20 °C и пересылать в замороженном виде Доставку проб осуществляют
любым видом транспорта, обязательно в термоконтейнере с хладоэлементами.
Срок доставки проб в лабораторию не должен превышать 72 ч от даты забора.
К пробам прилагается направление, в котором обязательно отражается приви-
вочный анамнез ребенка (количество полученных доз вакцины и дата последней
прививки). В вирусологической лаборатории проверяют и регистрируют состоя-
ние полученной пробы (достаточный объем, отсутствие протечки и нарушения
холодовой цепи). Каждая проба получает идентификационный номер, который
вносится в рабочий журнал, в направление и этикетку на флаконе с крышкой.
Лабораторная диагностика
Вирусологические исследования материала от больных полиомиелитом и
острыми вялыми параличами (ОВП) осуществляют только аккредитованные ВОЗ
вирусологические лаборатории, входящие в глобальную лабораторную сеть ВОЗ
по диагностике полиомиелита. Семь лабораторий из глобальной сети находят-
ся в Российской Федерации и образуют лабораторную сеть РФ по диагностике
полиомиелита. Это Национальная лаборатория по диагностике полиомиелита
в Институте полиомиелита и вирусных энцефалитов им. М.П. Чумакова РАМН
(НЛ). которая одновременно является Региональной референс-лабораторией ВОЗ
(РРЛ). В лабораторную сеть РФ также входят субнациональные вирусолотческие
лаборатории, работающие в составе региональных центров эпидемиологического
надзора за полиомиелитом и ОВП в г. Москве, Хабаровском, Ставропольском краях.
Свердловской, Омской областях, Санкт-Петербургском НИИЭМ им. Л. Пастера.
Все вирусологические исследования проводят в соответствии с Руководством ВОЗ.
СП 3.1.2343-08 и МУК 4.2.2410-08 с соблюдением требований контейнмента —
обеспечения безопасного лабораторного хранения полиовирусов.
Обработку поступивших для исследования проб фекалий проводят в ламинар-
ном шкафу 2-го класса защиты с использованием хлороформа, к действию которо-
го энтеровирусы устойчивы. Помимо бактерий и грибов хлороформ удаляет также
потенциально токсичные липиды и способствует диссоциации вирусных агрегатов.
Для приготовления 20% фекальной суспензии устойчивые к хлороформу полиэ-
тиленовые центрифужные пробирки емкостью 50 мл маркируют в соответствии с
номером пробы. В каждую пробирку вносят 10 мл фосфатно-солевого буферного
раствора (ФСБ) с антибиотиками, 1 г стеклянных бусин и 1 мл хлороформа. В про-
бирку вносят 2 г фекальной пробы. Плотно закрывают центрифужную пробирку и
тщательно встряхивают в течение 20 мин в механическом шейкере или вручную.
Центрифугируют 20 мин при 1500 g в центрифуге с охлаждением. Надосадочнук*
жидкость каждой пробы переносят в два маркированных флакона с винтовой
крышкой. Если жидкость непрозрачна, обработку хлороформом повторяют. После
обработки пробы надосадочную жидкость переносят в две пластиковые пробирки,
одну пробирку оставляют на хранение при -20 °C, другую используют для иссле-
дования в культуре клеток.
Для выделения и идентификации полиовирусов ВОЗ рекомендует 2 линии кле-
ток: L20B и RD. L20B - рекомбинантная линия мышиных клеток (й-клеток), соз-
данная для селективного выделения вирусов полиомиелита. Мышиным клеткам
этой линии генно-инженерным путем придана способность экспрессировать чело-
веческий рецептор полиовируса. RD — клетки, полученные из рабдомиосаркомы
ЧАСТНАЯ ВИРУСОЛОГИЯ 717
человека. Третья линия клеток (Нер-2, Cincinnati), полученная из эпидермоидной
карциномы человека, ранее была рекомендована ВОЗ для работы в лабораторной
сети ВОЗ и продолжает использоваться для проведения серологического мони-
торинга напряженности иммунитета к полиовирусам. Клетки L20B избирательно
чувствительны к полиовирусам, которые вызывают в них характерный цитопато-
генный эффект (ППЭ). Клетки RD высокочувствительны к полиовирусам, которые
также хорошо размножаются на клетках Нер-2
Выделение и идентификацию полиовирусов и других энтеровирусов проводят
в соответствии с алгоритмом, описанным в Руководстве ВОЗ по лабораторным
исследованиям полиомиелита. Обработанную пробу фекалий вносят на клетки
L20B и RD. которые просматривают в течение 7 дней, при отсутствии ЦПЭ про-
водят слепой пассаж и наблюдение продолжают еще 7 дней. Проба считается
отрицательной, если после двух пассажей ЦПЭ обнаружить не удалось. Общий
срок наблюдения за зараженными культурами должен быть не менее 14 дней. При
наличии ЦПЭ на линии клеток L20B в реакции микронейтрализации на этих же
клетках определяют серотип полиовируса, штамм направляют в РРЛ для внутри-
типовой дифференциации. Если вирус не типирован, его также направляют в РРЛ
для идентификации. При отсутствии ЦПЭ на клетках L20B, но наличии ЦПЭ на
клетках RD проводят пассаж на клетках L20B. Наличие ЦПЭ на этой клеточной
культуре свидетельствует о возможном выделении полиовируса, и все последую-
щие процедуры проводят, как описано выше. Если ЦПЭ наблюдается только на
клетках RD (RD+, a L20B-), следует определить тип энтеровируса в клетках RD.
Идентификацию полиовирусов проводят с помощью реакции микронейтрали-
зации с набором сывороток, специфичных к серотипам I, II и III полиовирусов.
В опыте по нейтрализации вирусную суспензию смешивают в равном объеме с
диагностическими сыворотками в лунках планшета для микронейтрализации,
инкубируют 1 ч яри 36 °C, добавляют суспензию клеток, помещают в термостат,
проводят ежедневное микроскопирование до получения окончательного результа-
та идентификации (как правило, в течение 5 сут).
Подготовку рабочих разведений сывороток проводят в соответствии с инструк-
циями производителя. Проведение теста микронейтрализации, интерпретацию
результатов выполняют в соответствии с Руководством ВОЗ по лабораторным
исследованиям полиомиелита.
Типирование выделенного вируса проводят на той культуре, на которой он
выделен. Если вирус выделен и на клетках RD, и на клетках L20B, проводят иден-
тификацию на обеих культурах, при этом в первую очередь исследуют штамм,
выделенный на L20B.
Иммунные сыворотки с высокими титрами антител смешивают с 100 ККИД50
выделенного штамма неизвестного вируса. Вирусная суспензия с ЦПЭ на «+++»
или «++++» содержит примерно 105-106 ККИД50 в 50 мкл. В данном тесте исполь-
зуют разведения 10’ ’ и 10 4 в целях экономии времени и материалов для предва-
рительного титрования каждого штамма. Титрование штамма включают в каждый
тест. Если вирус выделен и на клетках RD, и на клетках L20B, шп амм, выделенный
на L20B, исследуют первым, чтобы наиболее важный результат получить наиболее
быстро. Если полученный результат не убедителен, типируют штамм, выделенный
на клетках RD. Штамм, полученный на L20B, направляют в НЛ/РРЛ для дальней-
ших исследований, в ходе которых:
• пассируют 2 раза, общее впемя наблюдений — не менее 14 дней;
• если в клетках L20B из проб, собранных в неэндемичных или недавно
эндемичных странах, выделяется нетипируемый вирус, об этом информируют
регионального координатора лабораторной работы и руководителя
национальной программы ликвидации полиомиелита;
• неполиомиелитные энеторовирусы типируют по запросу РПИ.
718
ЧАСТНАЯ ВИРУСОЛОГИЯ
При интерпретации результатов реакции нейтрализации (табл. 24-11) важно
иметь в виду, что в лунках контроля клеток должен быть сплошной клеточный
монослой, а в лунках контроля вируса — полный ЦПЭ. Титрование должно под-
твердить, что использованное в опыте количество вируса в каждом из разведений
было в пределах 32-320 ККИД50 (то есть титр вируса составлял 101-5—102-5/50 мкл,
что соответствует 104,5 -1065в суспензии при выделении вируса). Результаты теста
следует оценивать по разведению, которое соответствует истинному титру вируса
в опьпе. Смесь иммунных сывороток, которая подавляет развитие ЦПЭ, указы-
вает тип вируса или типы вирусов в смеси. Неспособность вируса размножаться в
присутствии смеси иммунных сывороток является результатом его нейтрализации
одной из сывороток, входящих в смесь.
Таблица 24-11. Интерпретация результатов реакции нейтрализации при идентификации полио-
вируса
Полно 1+2+3 Полно 1+2 ПОЛНО 1+3 Полно 2+3 Идентификация вируса
0 0 0 + Полиовирус типа 1
0 0 + 0 Полиовирус типа 2
0 + 0 0 Полиовирус типа 3
0 0 + + Смесь полиовирусов типов 1 и 2
0 + 0 + Смесь полиовирусов типов 1 и 3
0 + + 0 Смесь полиовирусов типов 2 и 3
0 + + + Смвсь 3 типов полиовирусов
+ + + + Неполиомиелитный вирус или смесь полиовируса с другим энтеровирусом
Примечание. «+» — наличие ЦПЭ; «О» — отсутствие ЦПЭ.
Все штаммы полиовирусов, выделенные из любых источников, должны пройти
внутритиповую дифференциацию с помощью двух методов — антигенного и одно-
го молекулярного, поскольку важно определить, является ли он диким, вакцинным
или относится к вакцинно-родственным полиовирусам (ВРПВ). Установлена воз-
можность длительной репликации в организме и длительной экскреции штаммов
ВРПВ всех трех серотипов полиовирусов, а также такая важная характеристика
ВРПВ, как их способность к длительной циркуляции за счет передачи от человека
к человеку. Вспышки острых вялых параличей, вызванные штаммами вакцинного
происхождения, которые циркулировали среди населения в течение нескольких
лет, наблюдали в разных географических зонах.
Согласно определению ВОЗ, вакцинно-родственные полиовирусы, значительно
дивергицовавшие от вакцинных предков, представляют собой «полевые» изо-
ляты вакцинного происхождения, имеющие ке менее 1% нуклеотидных отличий
от родительских вакцинных штаммов, выявленных при полном секвенировании
участка генома VP1. Трудно проследить во времени наблюдаемые антигенные
изменения этих штаммов. Все изоляты, у которых была проанализирована полная
нуклеотидная последовательность участка VP1 генома, можно четко разделить на
три категории. Штамм считается вакцинным, если он имеет менее 1% различий с
эталонным вакцинным штаммом Сэбина. Штамм расценивается как ВРПВ, если
он имеет от 1 до 15% различий с вакцинным штаммом Сэбина; как дикий, если
он имеет более 15% различий с вакцинным штаммом. Единственное исключение
составляют дикие родители вакцинных штаммов, которые, согласно этим опреде-
лениям, являются вакцинными. Все «вакцинные» вирусы и большинство ВРПВ
расцениваются как вакцинные с помощью молекулярных методов ВТД. Некоторые
вакцинные вирусы, все ВРПВ и дикие вирусы расцениваются как невакцинные
с помощью антигенных методов внутритиповой дифференции (ВТД). Любой
ЧАСТНАЯ ВИРУСОЛОГИЯ 713
вирус, идентифицированный как вакцинный обоими методами, является вакцин-
ным. Любой вирус, идентифицированный обоими методами как дикий, является
диким. Любой вирус, для которого получены противоречивые результаты двумя
методами ВТД, указывающие на принадлежность штамма к дикому или вакцинно-
родственному полиовирусу, должен быть в течение 7 дней ст момента получения
результатов охарактеризован с помощью секвенирования участка генома VP1.
Такие вирусы обычно являются вакцинными, но могут также оказаться ВРПВ.
Проведение и интерпретацию результатов тестов выполняют в соответствии
с Руководством ВОЗ по лабораторным исследованиям полиомиелита. Методы
иммуноферментно! о анализа (ИФА) и диагностической ПЦР нашли наиболее
широкое применение.
Метод ИФА является антигенным методом ВТД, основанным на использовании
перекрестно адсорбированных антисывороток. Этот метод применяют только
по отношению к монотидоспецифическим штаммам. В случае выделения смеси
полиовирусов двух или трех типов их необходимо предварительно разделить.
Принцип метода: титровальные планшеты с IgG-антителами (бычьей сывороткой,
иммунной к полиовирусам типов I, II и III) в лунках инкубируют после внесения
идентифицированного штамма полиовируса определенного серотипа, подготов-
ленного для исследования. Затем инкубируют планшеты с внесенной типоспеци-
фической перекрестно адсорбированной иммунной сывороткой кролика. Удаляют
промыванием несвязавшуюся сыворотку и добавляют меченные пероксидазой
IgG-антитела к иммуноглобулинам G кролика, чтобы выявить связанную кроли-
чью сыворотку. Институт в Билтховене комплектует наборы реагентов, необходи-
мых для проведения ИФА, и рассылает их по разнарядке ВОЗ.
Метод диагностической ПЦР, разработанный в Центре контроля инфекци-
онных болезней в США, относится к молекулярным методам ВТД. Принцип
метода — определение присутствия в образце РНК полиовируса. ПЦР прово-
дят с идентифицированными штаммами полиовирусов определенного серотипа.
Интерпретация результатов
• Все праймеры (-) — не энтеровирус.
• Пан-энтеровирус (+), все другие (-) — энтеровирус (не полиовирус).
• Пан-энтеровирус (+), пан-полиовирус (+). 3 типа Сэбина (-). 1 или более
серотипов (+) — дикий полиовирус выявленного типа (типов).
• Пан-энтеровирус (+), пан-полиовирус (+), 1 или более типов Сэбина (+),
соответствующий серотип (серотипы) (+) — вакцинный полиовирус (полио-
вирусы) выявленного типа (типов).
• Пан-энтеровирус (+), пан-полиовирус (+), 1 или более типов Сэбина (+),
больше серотипов (+), чем типов Сэбина, — смесь дикого и вакцинного
полиовирусов различных типов.
При синтезе комплементарной ДНК и реакции ПЦР используют наборы олиго-
нуклеотидных праймеров с различной специфичностью. Используют наборы прай-
меров как специфические для энтеровирусной группы, так и типоспецифические
по отношению к каждому типу полиовирусов Сэбина. Дополнительно используют
праймеры, способные специфически амплифицировать все полиовирусы, а также
все известные штаммы каждого из трех типов по отдельности. Такие комбинации
праймеров, как специфические для всех энтеровирусов (PanEV), специфические
для всех полиовирусов (PanPV), специфические к каждому серотипу полиовируса
(Sero 1, Sero 2 и Sero 3), специфические к вакцинным штаммам каждого серотипа
(Sabin 1, Sabin 2, Sabin 3), дают полную характеристику штаммов полиовирусов и
подтверждают идентификацию типа каждого штамма. Центр контроля инфе кци-
онных болезней (Атланта, США) комплектует наборы реагентов (исключая фер-
менты), необходимых для проведения ПЦР на полиовирусы. ВОЗ распространяет
их в сети полиомиелитных лабораторий.
720
ЧАСТНАЯ ВИРУСОЛОГИЯ
Метод ВТД с моноклональными антителами является антигенным и основан
на реакции нейтрализации с использованием панели моноклональных анти-
тел, разработанной в Институте им. Л. Пастера в Париже. Панель состоит из
6 моноклональных антител (1о и 1о, 2о и По, Зо и Шо), специфичных к дикому
и вакцинному типам полиовирусов серотипов I, II и III соответственно. Рабочие
разведения моноклональных антител определяют в соответствии с результатами
нейтрализации эталонных штаммов полиовирусов. Принадлежность исследуемо-
го штамма полиовируса того или иного серотипа к дикому или вакцинному типу
определяют на основании индекса нейтрализации, рассчитанного как разница
между логарифмами титров вируса, полученных в отсутствии и присут ствии моно-
клональных антител. Положительным считается индекс нейтрализации не менее
3. Исследуемый штамм классифицируется как вакцинный или как невакцинный в
зависимости от индекса нейтрализации, полученного в реакции нейтрализации с
соответствующими моноклональными антителами, специфичными к вакцинному
или дикому типу полиовируса. Штаммы, которые не удается нейтрализовать моно-
клональными антителами ни к вакцинному, ни к дикому типу вируса, определяют
как не нейтрализуемые штаммы.
Для окончательной идентификации штаммов полиовирусов необходимо исполь-
зовать два метода внутритиповой дифференциации полиовирусов: антигенный и
молекулярный. Комбинация антигенного и молекулярного методов ВТД выбрана
для работы в сети полиомиелитных лабораторий ВОЗ, поскольку эти методы осно-
ваны на различных принципах. Иммунофермент ный анализ (антигенный метод,
основанный на использовании поликлональных перекрестно адсорбированных
иммунных сывороток) позволяет выявлять антигенные различия между дикими
и вакцинными полиовирусами. Диагностическая полимеразная цепная реакция
(молекулярный метод, основанный на использовании олигонуклеотидных прай-
меров с разной специфичностью) позволяет выявлять различия в РНК полиови-
русов.
ЭНТЕРОВИРУСЫ
Таксономия
Энтеровирусы относятся к семейству Picomaviriaae (от pico — «малый» и та —
«содержащий РНК»), роду Enterovirus. Согласно последней классификации вирусов
(Международного комитета по таксономии вирусов, 2003), основанной на геном-
ных характеристиках вирусов, неполиомиелитные энтеровирусы (НПЭВ) челове-
ка объединены в четыре группы (вида): А, В, С, D (табл. 24-12). Различные серо-
типы неполиомиелитных энтеровирусов отнесены к этим группам (табл. 24-13).
В филогенетическом плане энтеровирусы группы С и полиовирусы представляют
один вид энтеровирусов.
Таблица 24-12. Семейство Picomaviridae
Род и входящие в него виды Количество серотипов
Род Enterovirus
Вирус полиомиелита 3
Энтеровирус человека А 16
Энтеровирус человека В 52
Энтеровирус человека С 10
Энтеровирус человека D 3
Обезьяний энтеровирус 20
Бычий энтеровирус 2
ЧАСТНАЯ ВИРУСОЛОГИЯ 721
Окончание табл. 24-12
Род и входящие в него виды Количество серотипов
Свиной энтеровирус А 1
Свиной знтеровирус В 2
Род Hepatovirus
Вирус гепатита А человека (бывший энтеровирус 72) Вирус гепатита А обезьян 1
Род Rhinovirus
Риновирус человека А 18
Риновирус человека В 3
Неклассифицированные 82
Род Cardiovirus
Вирус знцефаломиокардита мышей 1
Вирус Тейлора мышей 2 или 3
Род Teschovirus
Вирус Тешенской болезни свиней 10
Род Aftovirus
Вирус ящура 7
Вирус ринита лошадей А 1
Род Parechovirus (бывшие вирусы ECHO 22 и ECHO 23) 3
Род Erbovirus
Вирус ринита лошадей 1
Род Kobuvirus
Вирус крупного рогатого скота 1
Вирус Aichi 1
Примечание. Приведенная в таблице классификация основана на геномных характеристиках
вирусов. По-види.мому7 будут считаться отдельными родами еще несколько энтеровирусов живот-
ных. Большое количество энтеровирусов (не менее 20, в том числе пронумерованные энтеровиру-
сы человека типов 92, 93, 96-101) еще не классифицировано.
Таблица 24-13. Таксономическая классификация неполиомиелитных энтеровирусов человека
Группа Количество серотипов Относятся серотипы
Знтеровирус человека А 16 Коксаки А 2-8,10.12,14.16 Знтеровирус 71,76, 89-91
Знтеровирус человека В 52 Коксаки А 9, Коксаки В 1-6. ECHO 1-7, 9,11-21, 24-27, 29-33 Энтеровирусы 69, 73-75, 77—88, 95
Энтеьовирус человека С 10 Коксаки А 1,11,13, 15.17-21.24
Знтеровирус человека D 3 Энтеровирусы 68,70, 94
В настоящее время определена полностью или частично нуклеотидная после-
довательность геномов многих энтеровирусов. Все энтеровирусы оказались сход-
ными по общей схеме организации генома, хотя и имеются различия у вирусов
разных видов и серотипов.
Характеристика вирусов
Вирусные частицы энтеровирусов круглой формы, диаметром 22-30 нм,
имеют икосаэдричесю ю симметрию и не имеют липидной оболочки.
Высокоструктг рированная белковая оболочка вируса состоит из 60 субъединиц,
каждая из которых формируется из четырех различных полипептидных цепей,
722 ЧАСТНАЯ ВИРУСОЛОГИЯ
внутри нее расположен вирусный геном — одна молекула рибонуклеиновой
кислоты (РНК). Геном вируса представляет собой позитивную одноцепочечную
РНК, состоящую примерно из 7500 нуклеотидов. Четыре капсидных полипептида
VP1-VP4 являются продуктами протеолитического расщепления полипротеина-
предшественника. З’-конец РНК полиаденилирован, а к 5’-концу ковалентно
присоединен низкомолекулярный белок VPg. Большая часть РНК энтеровиру-
сов, кроме двух нетранслируемых областей на 3’- и 5’-конце, представляет собой
одну протяженную открытую рамку считывания, кодирующую один большой
белок — полипротеин, который образуется в процессе трансляции в цитоплазме
инфицированных клеток. В результате каскада расщеплений, катализируемых
протеазами, полипротеин расщепляется на структурные белки капсида (от VP1 до
VP4) и неструктурные белки, включая протеин 3D. функционирующий как РНК
полимераза — фермент, ответственный за репликацию геномной РНК.
Устойчивость во внешней среде
Энтеровирусы быстро погибают при температуре выше 50 °C (при 60 °C они
погибают в течение 6-8 мин, при 65 °C — 2,5 мин, при 80 °C — 0,5 мин, при
100 °C — мгновенно). Добавление к вирусной взвеси хлористого магния сохраняет
титр вируса при 50 °C практически неизменным в течение часа. В заморожен-
ном состоянии активность энтеровирусов сохраняется в течение многих лет, при
хранении в обычном холодильнике (4-6 °C) — в течение нескольких нед, а при
комнатной температуре — на протяжении нескольких дней. Они выдерживают
многократное замораживание и оттаивание без потери активности.
Энтеровирусы быстро разрушаются под воздействием ультрафиолетового облу-
чения, при высушивании, кипячении. Быстро инактивирует вирусы раствор йода.
Обработка 0,3% формальдегидом, 0,1 N НС1 или свободным остаточным хлором
в концентрации 0,3-0,5 мг/л ведет к быстрой инактивации энтеровирусов, однако
присутствие органических веществ может оказывать защитное действие. Перекись
водорода в дозе 6,8 мг/л инактивирует энтеровирусы в воде за 30 мин. Этиловый
спирт (70% и более высокой концентрации) и формальдегид (0,3%) применя-
ют для дезинфекции в отношении энтеровирусов при экспозиции не менее 3 ч.
Энтеровирусы устойчивы в кислой среде (pH = 3,0^-5,0). Эфир, дезоксихолат и
другие детергенты, разрушающие ряд вирусов, не оказывают влияния на энтеро-
вирусы.
Лабораторная диагностика
Для лабораторной диагностики энтеровирусной инфекции в зависимости от
особенностей клинической картины заболевания используют СМЖ (при клини-
ческих показаниях для люмбальной пункции), смыв из ротоглотки/носоглотки,
отделяемое конъюнктивы, везикул, язв при герпангине, образцы фекалий. При
необходимости может быть исследован аутопсийный материал: ткани головного,
спинного, продолговатого мозга, печени, легких, миокарда, лимфоузлы, содержи-
мое кишечника и ткань кишечной стенки (в зависимости от особенностей клини-
ческой картины заболеваний).
При клинических показаниях для взятия стерильных типов клинического мате-
риала их следует включать в исследование в обязательном порядке.
Эпидемиологический и вирусологический надзор за энтеровирусными инфекци-
ями является важным направлением и видом дополнительного надзора в системе
мероприятий по профилактике полиомиелита. С 2006 г. в Российской Федерации
введена обязательная регистрация всех случаев энтеровирусной инфекции, в том
числе энтеровирусного менингита. Надзор за энтеровирусными инфекциями
ЧАСТНАЯ ВИРУСОЛОГИЯ 723
(ЭВИ) включает мониторинг заболеваемости ЭВИ, в том числе энтеровнрусного
менингита (формы № 1 и № 2 федерального государственного статистического
наблюдения), и слежение за циркуляцией неполиомиелитных энтеровирусов
путем исследования материала от больных ЭВИ и проб из объектов окружающей
среды.
Лабораторная диагностика базируется на классических вирусологических мето-
дах: выделении неполиомиелитных энтеровирусов на чувствительных культурах
клеток и идентификации выделенных энтеровирусов в реакции нейтрализации
на культурах клеток с помощью типоспецифических сывороток Используются
молекулярно-биологические методы диагностики: полимеразная цепная реакция
для выявления РНК энтеровирусов в материале от больных и идентификация
энтеровирусов с помошью молекулярного типирования путем секвенирования
области вирусного генома,, кодирующей белок капсида VP1.
Вирусологические исследования материала от больных энтеровипусными
инфекциями осуществляют вирусологические лаборатории Центров гигиены и
эпидемиологии в субъектах Российской Федерации. Вирусологические исследова-
ния материала от больных ЭВИ проводят так же, как при диагностике полиомие-
лита.
Для выделения и идентификации энтеровирусов ВОЗ рекомендует линию кле-
ток RD — клетки, полученные из рабдомиосаркомы человека. Многие серотипы
вирусов ECHO и некоторые вирусы Coxsackievirus А вызывают ЦПЭ в клетках RD.
Линию клеток Нер-2 (Cincinnati), полученную из эпидермоидной карциномы чело-
века, также используют для выделения энтеровирусов. На клетках Нер-2 хорошо
размножаются вирусы Coxsackievirus В, а также аденовирусы. Непилиомиелитные
энтеровирусы (Coxsackievirus А) могут размножаться в клетках L20B и вызывать
ЦПЭ. Ряд штаммов энтеровирусов могут быть выделены только in vivo с использо-
ванием чувствительных лабораторных животных.
Выделение и идентификацию неполиомиелитных энтеровирусов проводят в
соответствии с Руководством ВОЗ по лабораторным исследованиям полиомиели-
та, поскольку при исследовании в целях поиска энтеровирусив возможно выде-
лить полиовирусы (у детей, привитых против полиомиелита в течение последних
2 мес). Используют три линии клегок, на которых можно выделить большинство
неполиомиелитных энтеровирусов: L20B и RD и Нер-2. Культуры после зараже-
ния просматривают в течение семи дней, при отсутствии ЦПЭ проводят слепой
пассаж и наблюдение продолжают еще семь дней. Проба считается отрицательной
и культура подвергается уничтожению, если после двух пассажей ЦПЭ не обнару-
жен. Общий срок наблюдения за зараженными культурами должен быть не менее
L4 дней.
При наличии ЦПЭ на линии клеток L20B в реакции микронейтрализации на
этих же клетках определяют серотип полиовируса, после чего штамм направляют
в РРЛ для внутритиповой дифференциации. Если вирус не типирован, его также
направляют в РРЛ для идентификации.
При отсутствии ЦПЭ на клетках L20B, но наличии ЦПЭ на клетках RD, про-
водят пассаж на клетках L20B. Наличие ЦПЭ на этой клеточной культуре свиде-
тельствует о возможном выделении полиовируса, и все последующие процедуры
проводят, как описано выше. Если ЦПЭ наблюдается только на клетках RD (RD+,
a L20B-), следуеа определить серотип неполиомиелитного энтеровируса в реакции
нейтрализации.
Для идентификации штаммов энтеровирусов используют реакцию микроней-
трализации в культуре клеток с использованием иммунных сывороток к вирусам
ECHO и Coxsackievirus. Большое количество неполиомиелитных энтеровирусов
делает весьма трудоемкой постановку реакции нейтрализации с моноспецифиче-
скими сыворотками Моноспецифические сыворотки к неполиомиелитным энте-
724 ЧАСТНАЯ ВИРУСОЛОГИЯ
ровирусам производят в Институте полиомиелита и вирусных энцефалитов имени
М.П. Чумакова (Москва).
В глобальной лабораторной сети ВОЗ по диагностике полиомиелита для иден-
тификации неполиомиелитных Энтеровирусов используются смеси иммунных
сывороток приготовленных по перекрывающей схеме, которые позволяют иден-
тифицировать большинство энтеровирусов, используя всего девять смесей (пулы
ВОЗ А-G). Смеси иммунных к большинству наиболее часто встречаемых вирусов
ECHO и Coxsackievirus производят в Национальном институте охраны здоровья и
окружающей среды в Бильтховене (Нидерланды) и рассылают в полиомиелитные
лаборатории ВОЗ.
Каждый вирус исследуют со смесью сывороток к трем типам полиовирусов,
смесью антисывороток к вирусам Коксаки В1-В6 и с семью смесями (A- G) анти-
сывороток к 20 типам вирусов ECHO и вирусу Коксаки А 9. Вирусы, которые не
типировалисъ этим набором иммунных сывороток, возможно, присутствуют в
исследуемом материале в виде агрегатов, и полной нейтрализации не происходит.
Можно повторить типирование после обработки материала хлороформом (при-
мерно 10% объема). Замораживание может привести к агрегированию вируса,
поэтому типирование проводят после обработки материала до замораживания.
Идентифицируют вирус по результатам предотвращения ЦПЭ соответствующими
смесями сывороток. Интерпретации результатов помогает схема типичных для каж-
дого энтеровируса итогов нейтрализации, которая прилагается к каждому набору.
Наряду с классическими вирусологическими методами выделения энтеровиру-
сов на культурах клеток и их идентификации в реакции нейтрализации с помощью
диагностических сывороток используют молекулярно-биологические методы —
ПЦР и секвенирование генома вирусов.
Применение ПЦР для выявления РНК энтеровирусов в материале от больных
рекомендуют в следующих случаях.
• При необходимости проведения исследований большого количества образ-
цов в случае развития вспышки энтеровирусной инфекции.
• При осуществлении эпидемиологического надзора за энтеровирусами как
элемента скрининга в сочетании с методиками молекулярного типирования
(определения серотипа) энтеровирусов и/или вирусологическими исследова-
ниями.
• При оперативном надзоре за определенными серотипами энтеровирусов,
ассоциированными со вспышками заболеваний (EV71-HFMD). с примене-
нием генотипоспецифических ПЦР-тест-систем или с последующим опреде-
лением серотипа энтеровируса путем секвенирования генома вируса.
• В случае, когда энтеровирусы не вызывают ЦПЭ на культуре клеток. При
этом также необходимо определить серотип энтеровируса путем секвениро-
вания генома.
Молекулярно-диагностические работы осуществляют в соответствии с норма-
тивными документами. Проведение и интерпретацию результатов реакции выпол-
няют в соответствии с инструкцией производителя.
ПЦР — анализ для выявления РНК энтеровирусов в материале от больных —
проводят в несколько этапов: выделение РНК из исследуемого материала; получе-
ние комплементарной ДНК в реакции обратной транскрипции; проведение ПЦР-
амплификации; детекция продуктов ПЦР с помощью электрофореза в агарозном
геле. Если используемая ПЦР-тест-система не позволяет идентифицировать энте-
ровирусы до уровня серотипа, существует вероятность пропустить присутствую-
щие в пробе полиовирусы. Именно поэтому при обнаружении РНК энтеровируса
методом обратной транскрипции и ПЦР необходимо провести выделение вируса
на культуре клеток и его идентификацию в реакции нейтрализации или провести
идентификацию энтеровируса молекулярными методами.
ЧАСТНАЯ ВИРУСОЛОГИЯ 725
При выделении из исследуемой пробы полиовируса необходимо направить его
в аккредитованную ВОЗ вирусологическую лабораторию, входящую в лаборатор-
ную сеть Российской Федерации по диагностике полиомиелита для подтвержде-
ния результатов и проведения внучритиповой дифференциации.
Молекулярное типирование энтеровирусов основано на определении нуклео-
тидной последовательности в области генома, кодирующей белок капсида VP1.
Типирование энтеровирусов методом амплификации и прямого секвенирования
нуклеотидных последовательностей участка вирусного генома VP1 обладает
высокими показателями специфичности и может проводиться с применением
лабораторных методик, не имеющих государственной регистрации на территории
РФ. Серотип и генотип энтеровируса определяют сравнением полученной нуклео-
тидной последовательности с имеющимися в банке данных NCB1 (l<tp://www.
ncbi.nlm.njh.gov/) последовательностями прототипных штаммов энтеровирусов
человека.
Глава 25
Микологические исследования
СИСТЕМАТИКА И КЛАССИФИКАЦИЯ ГРИБОВ
Последние два десятилетия характеризуются неуклонным ростом
распространенности микологических инфекций, отмечается увели-
чение эпидемиологической роли видов с природной резистентно-
стью к антифунгеальным препаратам, накопление штаммов с при-
обретенной резистентностью,
С точки зрения клинической лабораторной диагностики целе-
сообразно учитывать уровень патогенности/степень биологической
угрозы, которую представляет тот или иной гриб. В России возбу-
дителей инфекционных заболеваний классифицируют по группам
патогенности (I—IV по убыванию) в зависимости от степени их
социально-эпидемиологической опасности, В странах Европы и
США классифицируют возбудителей по степеням биологического
риска, который они представляют для людей.
В клинической практике прогностическая значимость той или
иной микологической инфекции определяется риском развития
системных патологических процессов. Наибольшее значение в кли-
нической практике имеют грибы, относящиеся к трем родам:
Aspergillus;
о- Cryptococcus;
< Candida.
На фоне широкого распространения этих микроорганизмов про-
исходит развитие системных процессов. Смертность от системного
кандидоза, по данным многих исследователей, может достигать при
отсутствии своевременной диагностики и терапии от 55 до 70%,
Знание наиболее часто выявляемых в биоматериалах различных
локализаций возбудителей позволяет рационально планировать
этап лабораторной диагностики пациента с подозрением на микоз,
избежать дорогостоящих, но малоинформативных методов, сокра-
тить время, затрачиваемое на обследование.
Лабораторный мониторинг антифунгеальной чувствительности
наиболее распространенных возбудителей микологических инфек-
ций позволяет сформировать систему обоснованной эмпирической
терапии максимальной эффективности.
МИКОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ 727
Физиология грибов
Грибы — аэробные организмы, являющиеся гетеротрофами, неспособны к
фотосинтезу, неподвижны. Толстые клеточные стенки грибов препятствуют актив-
ному поглощению питательных веществ, поэтому питание грибы осуществляют с
помощью абсорбции из окружающей среды. Такой тип питания предполагает рост
гриба на источнике питательных веществ или внутри него. Большая площадь вса-
сывающей поверхности создается сетью гиф Нерастворимые вещества, например
кератин, грибы переваривают с помощью экзоферментов за пределами клетки.
Азот, сера, фосфор и ионы магния и других металлов, составляя основу питания
гриба, могут поглощаться им в неограниченной форме. Интенсивность роста
грибов зависит от концентрации питательных веществ, часто рост лимитируется
источником углерода. Высокие температуры (до 60 °C) и сильный холод не исклю-
чают присутствия грибов. В природе они преобладают над бактериями в средах
с более высокой концентрацией водорода (pH = 3,5+6,5). Наряду с этим многие
грибы развиваются в чистых культурах при pH = 6,5+8,5 так же хорошо, а иногда
даже лучше, чем в кислой среде. Нейтральная или слабощелочная реакция среды,
как правило, способствует развитию их конкурентов, прежде всего бактерий.
Жизненный цикл гриба можно охарактеризовать следующими этапами:
о прорастанием (заражением для грибов — абсолютных патогенов);
-о распространением или размножением.
Непосредственно после прорастания тело гриба состоит из нерасчлененного
протопласта (таллома, колонии) или сходных друг с другом, окруженных клеточ-
ной стенкой единиц (гифов), которые питаются и размножаются в значительной
степени независимо друг от друга — вегетативная фаза. Процессы дифференци-
ровки и формирования различных специализированных органов размножения
составляют репродуктивную фазу жизненного цикла гриба. Рост и развитие гриб-
ной колонии изменяют окружающую среду. Она может обедняться питательными
веществами, высыхать или разжижаться; наряду со сдвигами pH, окислительно-
восстановительного потенциала и осмолярности начинают влиять ингибирую-
щие (антибиотики) и стимулирующие (ростовые факторы, гормоны) факторы.
Отклоняются от исходных значений концентрации кислорода и диоксида углеро-
да. Выживанию грибов это обычно не\ грожает, так как вследствие эволюционного
отбора у них сформировались реакции на изменяющиеся условия существования.
Физиологические ограничения (постепенное высыхание, дефицит отдельных ком-
понентов питания и др.) вызывают переход от первичного метаболизма к вторич-
ному, обычно к образованию покоящихся и репродуктивных органов.
Если условия изменяются незначительно и постепенно, форма гриба (размер
клеток, толщина их стенки, степень агрегации гиф, образование хламидоспор
и т.д.) модифицируется также плавно, затрагивая лишь незначительные части
таллома. При глубоких или длительных изменениях среды происходят качествен-
ные перестройки метаболизма. Вегетативная фаза завершается, и начинается
репродуктивное развитие. Внешне для ра жых таксономических групп грибов это
выражается в образовании воздушного мицелия, конидий, плодовых гел. К концу
такого развития в экстремальном случае вес части вегетативной грибницы пре-
вращаю гея в покоя щиеся либо расселительные структуры или отмирают, то есть
ответной реакцией гриба на ухудшение условий окружающей среды становится
активное формирование «переживающих структур»?, а не перевод метаболизма в
«дремлющую» фазу. Вариабельность ответных реакций грибов на изменения раз-
личных параметров среды обитания является источником многочисленных диа-
гностических и прогностических ошибок, а также неправильной оценки эффек-
тивности терапии.
728 МИКОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ
Возбудители микозов, наиболее часто выявляемые
при исследовании биопс ических материалов
Для правильной интерпретации результатов микологических исследований
одним из определяющих моментов является знание типичных биотопов различ-
ных микромицетов в организме человека (табл. 25-1). Накопленный к настоящему
времени клинический опыт, знания о биологии и эпидемиологии грибов позво-
ляют разделить получаемую с помощью лабораторных методов информацию о
нахождении грибов, их различных биосубстратов на три группы.
• Виды грибов, нахождение которых в любом биологическом материале явля-
ется достоверным признаком микоза (дерматофиты, пневмоцисты, бласто-
мицеты).
• Грибы, нахождение которых в материале в норме стерильных биологических
жидкостей и тканей свидетельствует о заболевании с учетом данных клиники
и анамнеза пациента (криптококки, аспергиллы и другие плесневые грибы).
• Грибы-оппортунисты и комменсалы, имеющие представительство на несте-
рильных субстратах (кожных покровах, слизистых оболочках), об этио-
логической роли которых в возникновении заболевания свидетельствует
нахождение их в количестве, значительно превышающем обычные значения
(грибы рода Candida).
Перед взятием образца важно понимать, какой материал должен быть взят и
что предполагается обнаружить.
Таблица 25-1. Возбудители микозов, наиболее часто выявляемые при исследовании различных
биологических материалов
Возбудитель Исследуемый биологический материал
кровь ликвор материал из грудной, брюшной полостей стеклпзидное тело гной, отделяемое язв, абсцессов материал из ран, огогов мокрота, БАЛ моча волосы ногти соскобы чешуйки кожи Соскобы СО СЛИЗИСТЫХ оболочек
Aspergillus spp. - + + + + + - - 4- + -
Candida spp. + + + + + + + + - 4- + 4-
Coccidio'des immitis - - -
Cryptococcus spp. + + - - + - + + - - - -
Epidermohpyton spp. - 4- -
Fusarium 4- - + - - 4- - - - 4- + -
Malassezia spp. - 4- 4- -
Microsporum spp. - + - 4- -
Pneumocystis carinii - - - - - - + - - - - -
Scedosporium apiospermum - - - - + - + - - - - -
Sporotrix schencki - - - - 4- - + - - -
Trichphyton spp. + - - - - — - - + 4- -
Trichosporof! spp. - - - - - - 4- + + - -
Г иалогиФомицеты - -
Дерматофиты - -
Зигомицеты - - - + + - - - - -
Феогифомицеты - -
МИКОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ 729
ПРЕАНАЛИТИЧЕСКИЙ ЭТАП
ЛАБОРАТОРНОЙ ДИАГНОСТИКИ МИКОЗОВ
С учетом потенциальной опасности взятие любого исследуемого в лаборатории
биологического материала следует проводить с применением правил асептики.
Сбор материала
КОЖА
Сбор материала с кожных поражений проводят с помощью соскоба, мазка,
отпечатка липкой лентой. Предварительно следует обработать пораженное место
тампоном, смоченным 70% этиловым спиртом, особенно если на этот участок
кожи были нанесены крем, пудра или иные косметические препараты. Такая обра-
ботка позволяет уменьшить контаминацию исследуемого материала посторонней
бактериальной флорой.
• Соскоб — наиболее частый способ сбора материала с кожи. Для соскоба при-
меняют тупые скальпели, кюретки (во избежание травм кожи), предметное
стекло. Взятие материала оптимально проводить с наружного края пораже-
ний, так как здесь наиболее высоко содержание жизнеспособных, активно
размножающихся грибов. В случае пузырных высыпаний необходимо взять
для исследования покрышки свежих пузырьков, пузырей, обрывки мацери-
рованного эпидермиса.
• Мазок берут с участков кожи в области естественных складок, с мацериро-
ванных очагов, при выраженном воспалении. Взятие материала проводят
стерильным тампоном, смоченным водой, физиологичес ким раствором или
жидкой питательной средой.
• Отпетаток с помощью липкой лепты применяют при отсутствии видимого и
скрытого шелушения патологического кожного очага. Прозрачную липкую
ленту прижимают к очагу, затем удаляют и кладут липкой стороной на пред-
метное стекло, тщательно расправляя ее.
Собранные методом соскоба чешуйки кожи помещают в чашки Петри для транс-
портирования соскобов с маркировкой дна чашки и доставляют в лабораторию.
НОГТЕВЫЕ ПЛАСТИНКИ
Перед взятием материала поверхность ногтя обрабатывают тампоном, смочен-
ным 70% этиловым спиртом. Сбор материала проводят скальпелем, лезвием брит-
вы, маникюрными ножницами, кусачками, бором или фрезой бормашины.
При поверхностной форме онихомикоза делают соскобы с пораженной части
ногтевой пластинки до здоровых слоев ногтя, не пораженных грибом. Оптимально
использовать для анализа средние слои ногтевой пластины.
При локализации патологического процесса в дистально-латеральных зонах
ногтя для исследования берут срезы ногтевой пластинки. Желательно делать срез
во всю толщу ногтя и наиболее далеко от его свободного края. Кооме того, необ-
ходимо исследовать подногтевые роговые гиперкератотические массы, по воз-
можности захватывая наиболее проксимальные и глубокие слои. Поверхностные
слои соскобов ногтевых пластин и гиперкератотических масс не используются в
исследовании из-за высокой контаминации бактериальной микрофлорой.
В случае подногтевой локализации очага в проксимальной части ногтевой пла-
стинки взятие материала проводят бормашиной. Ногтевую пластинку просверли-
вают бором. Образующуюся пыль собирают с помощью отсоса.
730 МИКОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ
волосы
Перед взятием материала проводят выбор очага с помощью лампы Вуда. Волосы
эпилируют пинцетом. Коротко обломанные волосы из очагов можно извлечь
иглой. При фавусе взятие волоса проводят из центра скутулы, при микроспории,
инфильтративно-нагноительной трихофитии — с периферии очага.
Собранный материал (соскобы и срезы ногтевых пластинок, волосы) помещают
в стерильные чашки Петри с маркировкой дна и транспортируют в лабораторию.
ЯЗВЫ ВАРИКОЗНЫЕ, ИШЕМИЧЕСКИЕ И ДИАБЕТИЧЕСКИЕ. ПРОЛЕЖНИ
Мазок с поверхности раны существенно менее информативен, чем проба, аспи-
рированная шприцем.
Материал получают следующим образом:
❖ очищают поверхность раны стерильной салфеткой, смоченной стерильной
дистиллированной водой;
о используя лезвие скальпеля, получают соскоб с периферической границы
раны.
Помещают материал в стерильный контейнер с завинчивающейся крышкой или
специально смонтированную стерильную стеклянную емкость, причем для предо-
хранения пробы от высыхания добавляют в емкость 1-3 капли дистиллированной
воды и сразу же передают пробу в лабораторию.
СЛИЗИСТЫЕ ОБОЛОЧКИ
Полость рта
Сбор проб со слизистой оболочки ротовой полости проводят утром натощак до
приема пищи после полоскания рта теплой кипяченой водой. Взятие материала
проводят с помощью стерильных влажных тампонов соскобами с пораженной
поверхности. Пленки и налеты со слизистой оболочки снимают стерильной лож-
кой Фолькмана, стоматологическим шпателем или бактериологической петлей.
Слизистые оболочки урогенитального тракта
При воспалительной симптоматике урогенитального тракта образец из уретры
у мужчин берут бактериологической петлей объемом 1 мкл либо дакроновой щет-
кой/тампоном; при выделениях из уретры поверхность головки полового члена и
область наружного отверстия уретры должны быть очищены с помощью марле-
вого тампона, крайняя плоть отведена назад для предупреждения контаминации.
При отсутствии свободных выделений необходимо попросить пациента слегка
помассировать уретру скользящими движениями от основания пениса к его голов-
ке. После введения пластиковой петли в уретру на 1-2 см необходимо плоскость
«глазка» петли двигать к отверстию, слегка нажимая на стенки уретры. Не реко-
мендуют вращать петлю во время процедуры взятия образца в связи с болезнен-
ностью этой манипуляции для пациента. Образец из препуциального мешка берут
дакроновым тампоном.
У женщин на исследование берут1 соскобы из уретры, цервикального канала, с
заднего и боковых сводов влагалища, свободно стекающие выделения из заднего
свода влагалища. Дня получения проб со слизистых оболочек уретры, влагалища
(заднего свода), цервикального канала рекомендуют использовать одноразовый
стерильный инструментарий: уретральный пропиленовый зонд с синтетическим
ворсом — «сваб» (вискозой или дакроном), а также специальные щеточки-
браши — cervix brush и vola brush. Для получения полноценного материала из шейки
матки рекомендуют использовать одноразовый стерильный инструментарий,
например шпатель Айре: для беременных, женщин в менопаузе и после ионизации
шейки матки — специальные браши. представляющие собой щеточки с множе-
ственными щетинками, расположенными по спирали.
МИКОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ 731
После обнажения шейки матки в зеркалах тщательно очищают шейку от секре-
тов вагины и слизи с помощью ватного тампона, смоченного стерильным физио-
логическим раствором или стерильной водой. После этого щет< >чку (стерильный
зонд-тампон, или зонд-тампон транспортного коллектора со средой) осторожно
вводят в цервикальный канал на глубину 1.0-1,5 см, не касаясь стенок влагалища.
Вращая любой из перечисленных выше инструментов несколько раз вокруг оси,
захватывают материал — клсгки, экссудат по периметру цервикального канала.
Сбор пробы из уретры у женщин осуществляют через 1 или 2 ч после мочеи-
спускания. Стимулируют образование отделяемого легким массажем уретры через
влагалище, собирают образовавшееся отделяемое стерильным зондом-тампоном.
Соскобы слизистой оболочки пищевода проводят при фиброгастродуоденоскопии.
МОКРОТА
Сбор мокроты проводят утром сразу же после пробуждения, натощак. Пациент
должен предварительно почистить зубы и прополоскать полость рта кипяченой
водой, что позволяет механически удалить основную часть вегетирующей в рото-
вой полости микрофлоры и остатки пищи. Предупреждают больного, чтобы он не
собирал в контейнер слюну или носоглоточное отделяемое. Пациенту предлагают
глубоко прокашляться; если кашель сухой, непродуктивный, можно попытаться
стимулировать появление мокроты с помощью ингаляций.
Если же пациент не выделяет мокроту или выделяет ее только эпизодически и
в скудном количестве, накануне вечером и рано утром в день сбора мокроты сле-
дует дать ему отхаркивающее средство или применить раздражающие ингаляции.
Последние провоцируют усиление секреции бронхов, кашель и отделение мокро-
ты. Собранный таким образом материал не подлежит консервации, поэтому при-
готовление мазков из такого материала необходимо проводить в день его сбора.
Для аэрозольных ингаляций используют портативные или стационарные
аэрозольные ингаляторы. Для ингаляции рекомендуют использовать раствор,
в 1 л которого содержится 150 г натрия хлорида (NaCl) и 30 г натрия бикарбо-
ната (Na2CO3). Для предотвращения загрязнения раствора кислотоустойчивыми
сапрофитами, которые нередко содержатся в водопроводной воде и могут при-
вести при микроскопическом исследовании к получению ложноположительного
результата, используют дистиллированную воду. Для провокации выделения
мокроты необходимо вдохнуть от 30 до 60 мл подогретого до 42-45 °C физиоло-
гического раствора. Продолжительность ингаляции должна составлять не менее
10-15 мин.
Поскольку вдыхаемый во время ингаляции раствор вызывает усиленную сали-
вацию еще до появления кашля и отделения мокроты, в первые минуты после
завершения процедуры пациент должен сплюнуть слюну в специально приготов-
ленный лоток с 5% раствором хлорамина или другого дезинфицирующего сред-
ства и только после этого собрать мокроту для исследования. В связи с тем что
аэрозольная ингаляция вызывает выделение водянистого секрета, напоминающе-
го по консистенции слюну, во избежание выбраковки материала в направлении и
на флаконе с материалом должна быть обязательная маркировка, указывающая на
то, что материал получен после аэрозольной ингаляции.
Образцы мокроты должны быть исследованы не позднее 2 ч после сбора.
Допускается хранение образцов в холодильнике при 4 СС. Суточная мокрота не
подлежит микологическому исследованию из-за быстрой контаминации бактери-
альной микрофлорой,
БРОНХОАЛЬВЕОЛЯРНЫЙ ЛАВАЖ
Сбор лаважной жидкости проводят при промывании бронхов с помощью фибро-
бронхоскопа. Шприцем вводят через биопсийный канал бронхоскопа отдельными
Ш-
732 МИКОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ
порциями стерильный небактериостатический (официальный) физиологический
раствор (общий объем — от 5-20 до 100 мл).
Пеоед введением следующую порцию физиологического раствора осторожно
отсасывают введенной частью шприца в стерильный одноразовый контейнер с
завинчивающейся крышкой или в стерильную одноразовую или стеклянную про-
бирку с пробкой или оставляют в закрытом шприце, предварительно удалив из
него воздух (как правило, 50-70% введенного физиологического раствора нахо-
дится в лаваже).
Каждую отсасываемую порцию собирают в отдельную посуду.
По окончании процедуры соединяют пробы, полученные из одного и того же
участка. Пробы из разных участков (например, правой верхней доли легкого и
правой нижней доли) следует соединять вместе только после консультации с леча-
щим врачом.
Транстрахеальная аспирация малоинформагивна и чревата частыми осложне-
ниями.
МОЧА
Исследуют среднюю порцию мочи, собранную утром у пациентов без катери-
зации мочевого пузыря. Перед сбором пробы необходимо тщательно промыть
наружные половые органы и область заднего прохода теплой кипяченой водой;
особое внимание при этом следует уделять обработке отверстия мочеиспуска-
тельного канала у мужчин (преддверию влагалища — у женшин) для уверенности,
что при проведении процедуры проба не будет дополнительно контаминирована
микробами. Для обработки не допускается использование дезинфектантов, так как
при попадании в пробу они могут ингибировать рост микроорганизмов.
Пробу передают в лабораторию не позднее чем через 2 ч с момента сбора. Для
культурального исследования используют осадок центрифугата мочи. Не допуска-
ется использование пробы из суточной мочи.
Для исследования не принимают мочу из мочеприемников, подкладного судна,
пеленок. У детей сбор мочи проводят методом надлобковой аспирации.
КРОВЬ
При подозрении на фунгемию с помощью венепункции периферических вен
(чаще вены локтевого сгиба) или из пятки у новорожденных в асептических
условиях бе руг 10 мл венозной крови (у детей — 5 мл) либо, при отсутствии
положительного результата исследования и нарастании симптоматики, исследуют
артериальную кровь.
При подозрении на эндокардит и вялотекущий сепсис с маленькой (10-30 КОЕ/
мл) концентрацией возбудителя в циркуляции:
❖ при остром процессе собирают две пробы из двух участков различных сосу-
дов в течение первых 1-2 ч подъема температуры тела (не на пике темпера-
туры!) и до начала терапии;
❖ при подостром или вяло текущем течении собирают в первый день три пробы
с интервалом 15 мин и более. Если все пробы отрицательны, на вторые сутки
после посева собирают еще три пробы;
у пациентов с эндокардитами, получающими антибиотики, собирают по две
отдельные пробы в каждый из трех дней с положительной клинической дина-
микой терапии;
у больных, в комплекс терапии которых включены антибиотики, собирают
шесть проб в течение 48 ч; пробы необходимо собирать непосредственно
перед введением (приемом) следующей дозы препарата;
< при наличии у больного лихорадки неясного генеза первоначально собира-
ют две пробы из разных кровеносных сосудов (двух участков сосуда), затем
МИКОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ 733
через 24-36 ч еще две пробы на фоне повышения температуры тела (не на
пике температуры!).
Все гемфлаконы с образцами венозной и/или артериальной крови, взятые у
пациентов с подозрением на фунгемию или сепсис, сохраняют при температуре
37 ’С в течение 7-10 сут для выявления медленно растущих видов.
Исследованию подлежат также постоянные центральные катетеры.
ПРОБЫ ДЛЯ ОПРЕДЕЛЕНИЯ ОБСЕМЕНЕННОСТИ СОСУДИСТОГО КАТЕТЕРА
При подозрении на катетерассоциированную инфекцию материал собирают
следующим образом:
о очищают кожу вокруг катетера стерильной марлевой салфеткой, смоченной
70% этиловым спиртом;
❖ с соблюдением правил асептики удаляют катетер;
❖ стерильными ножницами отрезают около 5 см от дистального конца катете-
ра;
о помещают отрезанный кусок в сухую стерильную одноразовую пробирку с
пробкой, или стеклянн} ю пробирку, или стерильный одноразовый контейнер
с завинчивающейся крышкой (для сбора мокроты);
о немедленно передают материал в лабораторию, для предохранения от высы-
хания можно добавить несколько капель стерильного физраствора.
СПИННОМОЗГОВАЯ жидкость
При люмбальной пункции собирают 3-5 мл ликвора. Сбор проб проводят мед-
ленным заполнением трех пробирок тремя порциями материала для исследования
в лабораториях. Используют только стерильные пробирки с плотно закрывающи-
мися крышками (одноразовые с пробкой или стеклянные со стерильной резино-
вой пообкой).
Из трех пробирок с материалом, полученным при люмбальной пункции, всегда
отправляют пробирку с самым мутным содержимым, как правило, это вторая про-
бирка в процессе сбора пробы. Материал центрифугируют, полученный суперна-
тант исследуют серологическими методами, а осадок микроскопируют или иссле-
дуют культуру.
ЖИДКОСТИ СТЕРИЛЬНЫХ ПОЛОСТЕЙ ОРГАНИЗМА
Плевральную, внутрибрюшинную жидкости, собранные путем аспирации или
с помощью дренирования, собирают в стерильные пробирки с добавлением гепа-
рина в разведении 1:1000. Дренажную жидкость при перитонеальном диализе
собирают без добавки гепарина. Осадок центрифугата исследуют культуральными
методами
Минимальный объем жидкости, который необходимо направить в лаборато-
рию для выявления грибов (кроме крови, ликвора, мочи), составляет 10-12 мл.
При этом следует иметь в виду, что больший объем пробы увеличивает возмож-
ность обнаружения патогена.
КОСТНЫЙ мозг
При подозрении на глубокий микоз (например, гистоплазмоз, криптококкоз)
3-5 мл аспирата костного мозга помещают в стерильную пробирку с 0,5 мл сте-
рильного раствора гепарина 1:1000. Посев материала на питательную среду про-
водят сразу после взятия материала.
ГНОЙ
Взятие гноя из абсцессов кожи и свищевых ходов проводят стерильными шпри-
цем и иглой. Из вскрывшимся абсцессов и с поверхности язв взятие материала
проводят бактериологической петлей, ложкой Фолькмана, пастеровской пипет-
734 МИКОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ
кой. Если взятие материала осуществляют тампонами, что нежелательно, следует
погружать тампон как можно глубже. При наличии в гное зерен и сгустков их обя-
зательно берут на исследование.
Отделяемое дренажей, используемых для активной аспирации полостей, отса-
сывают стерильным шприцем с плотным поршнем в объеме 2-4 мл. Затем на
заполненный шприц надевают стерильную иглу, закрытую стерильным ватным
тампоном, удаляют из шприца избыток воздуха. Ватный тампон сбрасывают в
дезинфицирующий раствор. Конец иглы вкалывают в стерильную резиновую
пробку и в таком виде шприц с материалом доставляют в лабораторию.
ЯЗВЫ И УЗЕЛКОВЫЕ УТОЛЩЕНИЯ
Мазок с поверхности язвенного дефекта существенно менее информативен, чем
проба, выполненная методом кюретажа. Очищают поверхность язвы 70% этило-
вым спиртом, затем дезинфицируют 1-2% раствором йода или другим дезинфек-
тантом. Раствор иода удаляют после проведения процедуры стерильной салфет-
кой, смоченной 70% этиловым спиртом, чтобы избежать ожога пациента.
• Сухой стерильной салфеткой удаляют некротические массы, детрит, гной.
• Кюретажной ложкой выскабливают основание язвы или узелкового утолще-
ния.
• Помещают материал в стерильный контейнер.
При наличии экссудата в язве или узелке рекомендуют собирать пробу с помо-
щью шприца во избежание контаминации материала.
БИОПТАТ
Материал для патоморфологического исследования берут из середины и краев
очага поражения с соблюдением стандартных правил асептики. Пробы, получен-
ные во время операции, помещают в емкость с анаэробной средой, в стерильную
пробирку с тиогликолевой средой, закрытую стерильной резиновой пробкой, либо
в стерильную сухую одноразовую пробирку типа «Эппендорф».
Материал помещают в специальный штатив из материалов подлежащих стери-
лизации в автоклаве, и немедленно отправляют в лабораторию.
В некоторых странах Евросоюза (Италии, Франции) при заборе биоптата при-
нято использовать кроме флаконов со средами для аэробов и анаэробов также
отдельный флакон с жидкой средой Сабуро для выявления микологических
инфекций.
Транспортировка
Материал, предназначенный для культурального исследования микологических
инфекций, можно хранить в транспортной среде или физрастворе при комнатной
температуре не более 4 ч. предохраняя от воздействия прямых солнечных лучей и
тепла.
Материал для гистологического исследования можно хранить до 48-72 ч в
холодильнике при температуре 4-8 ’С.
Допускается пересылка материала в замороженном состоянии без консервации.
Недопустимо после размораживания повторное замораживание диагностического
материала, что ведет к нарушению структуры и типкториальных свойств грибных
клеток. Исследуемый материал транспортируют в специальной таре или металли-
ческих биксах.
Каждая проба материала должна быть промаркирована, иметь соответствую-
щее направление, на котором указываются фамилия, инициалы, возраст или год
рождения пациента, штрих-код или идентификационный номер истории болезни,
идентификационный номер образца, локализация и тип образца, дата и время
МИКОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ 735
взятия материала. На самом контейнере с образцом указывается идентификацион-
ный номер образца или штрих-код. При отсутствии вышеназванной информации
образец не подлежит исследованию и выбраковывается.
МАКРОСКОПИЧЕСКИЕ ИССЛЕДОВАНИЯ
Определение грибов в нативных препаратах визуально
Piedrata В случае поражения волос плесневыми грибами Trichosporon oxides и
Т inkin, являющимися возбудителем белой пьедры, диагностику проводят методом
осмотра пораженных участков волос под лупой.
Макроскопически обнаруживают:
< мягкие гранулы, легко отделяющиеся от волоса:
❖ «футляры» вдоль стенки волоса.
Гриб поражает только кутикулу волоса и растет перпендикулярно поверхности
дистальной части волос.
Определение грибов в нативных препаратах с лампой Вуда
(люминесцентное исследование)
Методика основана на феномене люминесцентного свечения некоторых грибов
при воздействии ультрафиолетового излучения с длиной волны 290-320 нм.
Исследование проводят в затемненном помещении с помощью лампы Вуда.
Метод эффективен для обнаружения в нативных образцах клинического материа-
ла следующих видов грибов:
• > Malassezia (Pityrospnrum) — чешуйки кожи, роговой слой эпидермиса;
> Microsporum — волосы;
❖ Trichphyton schontetnii — волосы;
❖ Trichphyton mentagiophytes - чешуйки кожи (свечение светло-зеленое или нет
флюоресценции).
Перед исследованием очаги необходимо очистить от остатков мазей, корок;
проводимая перед исследованием мазевая терапия может служить источником
ложноотрицательных результатов.
При поражении волос грибами рода Microsporum наблюдают ярко-зеленое све-
чение. причем светятся как остистые, так и пушковые волосы. При исследовании
свежих очагов свечение может отсутствовать, что связано с незначительным по
площади поражением волоса. В таких случаях свечение во тоса можно обнаружить
в прикорневой части после эпиляции. Погибшие волосы сохраняют способность к
свечению.
Исследование дерматофитий с помощью лампы Вуда является вспомогатель-
ным диагностическим методом и служит для определения распространенности
поражения волосяного покрова, оценки эффективности проводимой терапии,
эпидемиологического контроля контактных лиц, особенно при исследовании в
эпидемиологических очагах (детских коллективах), выявления инфекции или
миконосительства у животных.
При обследовании пациентов с отрубсвидным лишаем метод люминесцентного
свечения в лучах лампы Вуда используют для диагностики очагов поражения в
области волосистой части головы, которые могут не иметь клинической симпто-
матики в виде шелушащихся желтовато-бурых пятен. Очаги имеют зеленовато-
желтое, желтовато-коричневое или красновато-желтое свечение. В этом случае
своевременное назначение терапии в области волосистой части головы предотвра-
щает вероятность рецидива.
736 МИКОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ
МИКРОСКОПИЧЕСКИЕ ИССЛЕДОВАНИЯ
Метод используют в клинико-диагностических лабораториях при первичном
обследовании лиц с подозрением на заболевание микозом. Во многих случаях,
особенно при поверхностных микозах, микроскопическое исследование состав-
ляет диагностический минимум, дополняя клинико-анамнестические данные
пациента. Однако следует помнить, что результаты микроскопии не могут быть
единственным основанием для постановки клинического диагноза. Клинический
материал для нативного микроскопического анализа возбудителей микозов пред-
ставлен в табл. 25-2.
Таблица 25-2. Клинический материал для нативного микроскопического исследования различных
возбудителей микозов
Клинический материал Дрожжевые и дрожжепоцобные грибы
Candida Crypto- coccus Geotri chum Malassezia (Pityro- sporum) Rhodo tonila Saccharo- myces cervlsla Tricho- sporon
Соскобы С КОЖИ - + + + + -
Соскобы с высыпаний + - + - - - -
Соскобы со слизистой оболочки полости рта + - + - - - -
Ногтевые пластинки + - + - - - -
Мокрота + + + - + - -
Моча + + + - + - -
Желчь 4- - - - - - -
Сок простаты + - - - - - -
Спинномозговая жид- кость + + - - - - -
Пунктат из закрытых полостей + - - - - - -
Мицелиальные грибы
Клинический материал дермато- фиты феогафо- мидеты гиалогифомицеты аскомицеты зигомицеты
Aspergillus Penicillium Piedraia portal
Гной - + - - - +
Волосы - - - - - -
Отделяемое свищей + - - - - -
Соскобы с кожи + + - - - +
Ногтевые пластинки + + + + - +
Мокрота - + + + - +
Пунктат из закрытых очагов - + + - - -
Гной — + + - - +
Волосы + - - - + -
Отделяемое пазух носа - - - - - +
МИКОЛОГИЧЕСКИЕ исследования 737
Окончание таАл. 25-2
Клинический материал Blastomyces dermatitfdis Димор Coccidtoides immitis фные гр Laboa loboi ибы Paraco- ccidioide;: brasiliensls Sporotrix schencki Неклассифици- рованные патоген- ные грибы Rhinosporl- ditimseberi
Соскобы с высыпных элементов + - - - - -
Отделяемое язв + - + + + -
Отделяемое свищей + + - - - -
Отделяемое абсцессов - + - - + -
Отделяемое полипоо- бразных очагов - - - - - +
Мокрота + - - + + -
Экссудаты - - + - - -
Спинномозговая жид- кость + - - - -
Гной - - - + + -
Моча + - - - - -
Хотя микроскопическое исследование до настоящего времени является обяза-
тельным, его информативная ценность в зависимости от нозологической формы
микоза и характера исследуемого биологического материала значительно разли-
чается. В крови, спинномозговой жидкости, пунктатах костного мозга возбудитель
может присутствовать в очень малых количествах, недостаточных для визуализа-
ции. Кроме того, особенности жизненного цикла возбудителей многих системных
микозов делают их пребывание в центральном кровотоке очень кратковременным
(«пульсирующие» выбросы), что также затрудняет верификацию диагноза мето-
дом микроскопии.
Резко различается трактовка результатов в случае обнаружения грибов — абсо-
лютных патогенов и условно патогенной микофлоры, особенно в локализациях,
являющихся в норме нестерильными, (кожа, слизистые оболочки). Микроскопия
может считаться качественным методом исследования при обнаружении грибов
в стерильных в норме биологических средах организма и в материале органов и
тканей.
При исследовании нестерильных субстратов на условно патогенные грибы
трактовка результатов микроскопии более разноречива. Нахождение оппортуни-
стической флоры в больших количествах, свидетельствующих о массивной коло-
низации субстрата, можно считать основанием для проведения культурального
исследования. Микроскопия не позволяет осуществить родовую идентификацию
возбудителя, что негативно отражается на выборе тактики терапии, неадекватное
лечение, в свою очередь, может приводить к хронизации микоза, отбору и нако-
плению резистентных штаммов в популяции.
Преаналитические предосторожности
Образцы, предназначенные для микроскопического исследования, должны
сопровождаться письменным направлением лечащего врача. По устной просьбе,
не подтвержденной соответствующими документами, исследования не проводят.
Не следует проводить исследование безымянных проб или проб с неправильно
оформленными направлениями.
738 МИКОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ
Бланки направлений должны быть правильно оформлены, а каждая получен-
ная проба диагностического материала — правильно промаркирована. На бланке
направления на исследование необходимо отметить время доставки материала в
лабораторию и фиксировать любые задержки в поступлении проб, что имеет осо-
бое значение при отрицательных результатах.
Для предупреждения случаев внутрилабора горного заражения необходимо
свести к минимуму возможность образования аэрозолей. В лабораториях при
микроскопическом исследовании основным источником образования инфекцион-
ных аэрозолей являются манипуляции по приготовлению мазков. Аэрозоли могут
образовываться в процессе следующих манипуляций.
о приготовления мазков путем нанесения материала на предметное стекло и
распределения его по поверхности стекла:
< прожигания не очищенных от остатков материала бактериологических
петель, которые используют для переноса материала на стекло и приготовле-
ния мазка;
< попыток фиксировать над горелкой невысохший влажный мазок, что приво-
дит к вскипанию и разбрызгиванию частичек материала;
< > незамеченном в процессе работы попадания капель материала на рабо-
чую поверхность столов и отсутствия последующей дезинфекции рабочих
поверхностей.
Приготовление мазков из нативного материала
При приготовлении мазка для микроскопического исследования необходимо
добиваться правильной его толщины. Если мазок слишком тонкий и содержит
мало материала, при микроскопическом исследовании можно получить лож-
ноотрицательный результат, Если мазок слишком толстый, это затрудняет его
фиксацию, материал недостаточно плотно прикрепляется к стеклу и может быть
частично или полностью удален при окраске. Кроме того, толстый мазок плохо
просматривается при микроскопическом исследовании. Не рекомендуют готовить
мазки способом растяжки материала между двумя предметными стеклами. Такой
способ сопровождается образованием биологически опасного аэрозоля.
Через правильно приготовленный неокрашенный мазок можно читать газетный
шрифт, расположенный позади стекла на расстоянии 5-10 см, Ограниченная пло-
щадь мазка (около 1x2 см в центре стекла) значительно повышает безопасность
манипуляции и последующей микроскопии, так как периферические части и ребра
предметного стекла остаются не загрязненными инфекционным материалом.
Микроскопия нативных препаратов
Перед исследованием плотного биологического материала необходимо измель-
чить крупные фрагменты и просветлить препарат.
Жидкий биологический материал просматривают в неокрашенном состоянии в
воде, изотоническом растворе или с применением просветления.
Просветляющие жидкости
• Спирто-глицериновая смесь:
❖ этиловый спирт — 1 часть;
< > глицерин — 2 части;
❖ дистиллированная вода — 2 части.
• Раствор Люголя:
к ристаллический йод — 1 г;
❖ калия йодид — 2 г;
❖ вода — 150 мл.
МИКОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ 739
Методика приготовления жидкого нативного препарата
На предметное стекло пипеткой или петчей наносят каплю материала, затем
2 капли просветляющей жидкости, накрывают покровным стеклом.
Микроскопируют сначала при увеличении х400, дающем возможность увидеть
скопления клеток, псевдомицелий, мицелий и другие элементы гриба, а затем —
при большом (хЮОО с масляной иммерсией), когда возможно характеризовать
отдельные клетки.
Методики просветления плотного нативного препарата
При исследовании тканей с высоким содержанием кератина просветление
препарата осуществляют добавлением капли 10-20% раствора щелочи (КОН
или NaOH). Экспозиция чешуек кожи 2 ч, волос — 30-40 мин, ногтей — 24 ч.
Подогревание патологического материала перед микроскопированием не реко-
мендуют, так как это может привести к выпадению кристаллического осадка солей
кальция, затрудняющего исследование (микроскопически сходного со спорами), и
разрушению грибковых элементов.
Для просветления можно использовать также смесь спирта с глицерином или
раствора Люголя двойной крепости (кристаллического йода — 1 г, йодистого
калия — 2 г, воды — 150 мл). На исследуемый инокулят (чешуйки, соскобы с кожи
и высыпаний, волосы) наносят каплю 10% КОН. или смеси спирта с глицерином
(спирта и глицерина — по 2 части, воды — 4 части), или раствора Люголя двойной
крепости (кристаллического йода — 1 г, йодистого калия — 2 г, воды — 150 мл).
Препарат накрывают покровным стеклом. Экспозицию проводят по указанным
временнь'тм интервалам.
МЕТОДИКА ОБОГАЩЕНИЯ
Чешуйки кожи и срезы с ногтей, помещенные в центрифужную пробирку, запи-
вают 20-30% раствором КОН на 10-12 ч для чешуек кожи и 24 ч — для ногтей.
Полученную гомогенную желеобразную массу тщательно перемешивают и поме-
щают хорошо просветленный патологический материал на предметное стекло.
Критерием хорошей подготовки материала считается потеря им очертаний и рас-
плющивание при сдавливании между предметным и покровным стеклом.
При микроскопии нативных препаратов могут быть хорошо видны строение
мицелия, расположение спор и отдельные клеточные элементы грибов. Однако
при применении этого метода возможны серьезные ошибки, так как клеточные
элементы могут1 расцениваться как элементы грибов.
Так, большую клетку с вакуолями в цитоплазме можно принять за сферулу кок-
цидиоидоза с эндоспорами, а округлую клетку с кажущимся двухконтурным очер-
танием — за бластоспору дрожжевого гриба. Именно поэтому на основании только
нативного препарата ставить диагноз микоза нежелательно, его необходимо соче-
тать с исследованием окрашенных мазков и микробиологическими тестами.
Микроскопия окрашенного препарата
Окраски с одним красителем называются простыми, а с использованием
нескольких красителей и обесцвечивающих компонентов — сложными.
МЕТОДЫ ОКРАСКИ ДРОЖЖЕВЫХ ГРИБОВ
• Метиленового синего водный раствор (2,5 г метиленового синего, 3 мл ледя-
ной уксусной кислоты, 10 мл воды):
о смешивают 30 мл насыщенного спиртового раствора метиленового синего
со 100 мл 0,01% раствора гидроксида калия;
4 мазки окрашивают в течение 3-5 мин;
740 МИКОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ
Ф ополаскивают в проточной воде;
❖ дифференцируют Зев 0,5% растворе уксусной кислоты;
❖ ополаскивают в проточной воде.
• Метиленового синего 1% спиртовый раствор (2,5 г метиленового синего,
100 мл 95% этанола), окрашивают мазки в течение 1-3 мин.
• Метиленового синего с лактофенолом раствор (0,05 г метиленового синего,
100 мл лактофенола), окрашивают препарат в течение 30-60 мин.
• Фуксина 1% водный раствор, окрашивают мазки в течение 0,5-1,0 мин.
• Генциан иолета 1% спиртовой раствор окрашивают мазки в течение 2-3 мин:
смешивают 10 мл насыщенного спиртового раствора генцианвиолета со
100 мл 2,5% раствора фенола (смесь хранится долго);
❖ мазки окрашивают в течение 2-5 мин через фильтровальную бумагу;
❖ промывают в проточной воде, сушат;
❖ исследуют при масляной иммерсии.
МЕТОДЫ ОКРАСКИ ДЕРМАТОФИТОВ
• 1% раствор хлоразола черного в диметилсульфоксиде.
• 5% раствор метиленового синего в дистиллированной воде (1:9).
• 0,3% раствор конго красного в 5% растворе лаурилсульфата натрия.
ОКРАСКА С ИНДИЙСКОЙ ТУШЬЮ (ВЫЯВЛЕНИЕ CRYPTOCOCCUS NEOFORMANS В
СПИННОМОЗГОВОЙ ЖИДКОСТИ)
К капле патологического материала добавляют каплю туши, смешивают и
накрывают покровным стеклом. При микроскопии на темном фоне под большим
увеличением наблюдают почкующиеся клетки криптококка, окруженные широ-
ким светлым ободком.
Поскольку сходную микроскопическую картину демонстрируют лейкоциты
(сегментоядерные, полиморфноядерные), этот метод окраски в настоящее время
применяют редко, чаще используя реакцию латекс-агглютинации на выявление
ксиломананового антигена криптококков в ликворе.
ОКРАСКА ГЕМАТ0КСИЛИН-Э03ИН0М
Используемые реактивы:
• раствор Вейгерта I:
4- 1г гематоксилина:
❖ 100 мл 96% этилового спирта.
• раствор Вейгерта II:
о 4 мл раствора полуторахлористого железа (FeC13);
1 мл концентрированной соляной кислоты;
<> 95 мл дистиллированной воды.
ЖЕЛЕЗНЫЙ ГЕМАТОКСИЛИН ВЕЙГЕРТА
Составляют по мере надобности из двух основных растворов.
Перед применением смешивают оба раствора точно в равных объемах. Оба
основных раствора могут сохраняться длительное время. Смешанный раствор
можно применять лишь несколько дней.
Методика окраски гематоксилин-зозином
• Смешивают растворы Вейгерта I и II.
• Окрашивают препараты 1-2 мин.
• Промывают водопроводной водой.
• Проводят дифференцировку препаратов путем отмывания избыточной кра-
ски в 0,1% растворе солянокислого спирта.
• Тщательно промывают водопроводной водой.
МИКОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ 741
ОКРАСКА ПО ГРАМУ
Метод основан на способности микроорганизмов удерживать краситель кри-
сталлический фиолетовый или обесцвечиваться в спирте в зависимости от химиче-
ской структуры клеточной стенки. По результатам окрашивания микроорганизмы
подразделяют на две группы: грамположительные (после окраски генпиановым
фиолетовым и связывания с йодом не обесцвечиваются в органических раство-
рителях, оставаясь сине-фиолетовыми) и грамотрицательные (обесцвечиваются
за счет высокого содержания липидов и отсутствия тейхоевых кислот в клеточной
стенке; становя гея розово-красными после подкрашивания сафранином или фук-
сином).
Используемые реактивы
Раствор кристаллического фиолетового по Эрлиху: 1,2 г кристаллического
фиолетового растворяют в 12 мл 96% спирта и добавляют 100 мл свежеприготов-
ленной анилиновой воды, полученной при смешивании 2 мл анилина со 100 мл
воды. Эта красящая смесь может храниться 2 нед. Генциановый фиолетовый, кри-
сталлический фиолетовый и метиловый фиолетовый принадлежат к красителям
трифенилметанового ряда, что позволяет использовать их в равной степени при
окраске по методу Грама.
Красящие смеси для грамположительных бактерий:
❖ 10 г кристаллического фиолетового + 100 мл спирта;
❖ L0 г генцианового фиолетового + 100 мл спирта;
10 г метилового фиолетового + 100 мл спирта;
о 1 мл насыщенного спиртового раствора кристаллического фиолетового +
10 мл дистиллированной воды;
❖ 0,25 мл насыщенного спиртового раствора кристаллического фиолетового +
100 мл 5% раствора уксусной кислоты;
< > 8-9 г метиленового синего -г 100 мл спирта,
< 30 мл насыщенного раствора метиленового синего в спирте + 1 г гидроксида
калия + 99 мл дистиллированной воды.
Красящие смеси для грамотрицательных бактерий:
❖ 8-9 г основного фуксина + 100 мл 96% спирта;
❖ 1 мл насыщенного спиртового раствора основного фуксина + 10 мл дистилли-
рованной воды;
< 1 г основного фуксина + 1 г фенола + 0,5 мл глицерина + 10 мл 96% спирта +
100 мл дистиллированной воды;
о 0,2-0,5% водный раствор сафранина либо 0,25% раствор сафранина в 10%
спирте.
Методика окраски
• Наиболее распространенный в настоящее время метод фиксации мазков
над пламенем горелки потенциально опасен для персонала возможностью
образования аэрозолей инфицированного материала. Также возможно разру-
шение структуры грибов, что приводит к ложноотрицательным результатам
исследования, поэтому рекомендуют фиксацию в парах формалина (в сосуде
Коплина) в течение 15 мин.
• Наливают на фиксированный мазок карболовый или анилиновый ген-
циановый фиолетовый (или кристаллический фиолетовый) на 2-3 мин.
Во избежание осадков окрашивают через фильтровальную бумагу. Очень
удобно работать с полосками фильтровальной бумаги, предварительно про-
питанной 1 % спиртовым раствором одного из вышеуказанных красителей и
затем высушенной (метод Синева). Такую окрашенную и высушенную бумагу
742 МИКОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ
накладывают на мазок и смачивают 1-2 каплями дистиллированной воды.
Продолжительность окрашивания остается прежней (2-3 мин).
• Сливают краску, аккуратно удаляют фильтровальную бумагу и быстро опо-
ласкивают в водопроводной воде (15-30 с).
• Мазок заливают на 1-2 мин раствором Люголя или другим йодистым раство-
ром, например по Граму (2 г йодида калия растворяют в 2-3 мл дистиллиро-
ванной воды, добавляют 1 г кристаллического йода, доливают до 300 мл., а по
Вейгерту — до 100 мл дистиллированной воды).
• Раствор сливают, мазок промывают водой.
• Дифференцируют 96% спиртом, наливая и сливая его, пока отходит синяя
краска или наиболее тонкие участки препарата не станут бледно-серого цвета
(приблизительно 20-60 с). Во время дифференцировки препарат покачива-
ют. Если вместо спирта использовать ацетон, то промывание продолжается
30 с. Можно дифференцировать смесью спирта и ацетона (1:1) 30 с.
• Промывают стекла в проточной или дистиллированной воде 1-2 мин.
• Дополнительно окрашивают (для выявления грамотрицательной группы
бактерий) либо 0.5% водным раствором нейтрального красного, либо раз-
веденным (в 10 раз) карболфуксином Циля — 1-3 мин. Возможна доокраска
сафранином (0,2-0,5% водным раствором или 0,25% раствором в 10% спир-
те), основным коричневым (бисмаркбрауном) либо пиронином J или В (2-5
мин).
• Промывают в проточной воде 5 с и высушивают фильтровальной бумагой.
В классической прописи при окраске по Граму не промывают мазки в воде ни
после окраски генциановым фиолетовым (кристаллическим фиолетовым), ни
после обработки раствором Люголя. ограничиваются лишь сливанием растворов.
Предлагаемое промывание в воде имеет целью только чистоту в работе и позволя-
ет добиться лучшего контрастирования препарата.
Грамположительные микроорганизмы окрашиваются в иссиня-черный (темно-
синий. сине-фиолетовый) цвет, грамотрицательные — в розовый, красный или
коричневый в зависимости от красителя; ядра клеток — ярко-красные, цитоплаз-
ма — розовая, нейтрофилы, мононуклеарные клетки, эпителий и другие мате-
риалы окрашиваются как грамотрицательные. При правильной окраске препарат
оранжево-красного цвета на тонких участках, лилово-фиолетового — на толстых.
Дрожжеподобные грибы (псевдомицелий, дрожжевые клетки) грамотрицатель-
ны и окрашиваются в ярко-фиолетовый цвет.
Во всех модификациях окраски по Граму лучше всего окрашиваются моло-
дые (почкующиеся и ветвящиеся) формы гриба с рыхлой клеточной стенкой.
Нитчатые структуры видны менее отчетливо, а иногда и совсем не окрашиваются.
Кроме того, отрицательной чертой окраски по Граму является значительное коли-
чество артефактов, в результате чего некоторые тканевые структуры (например,
фибрин в материале язвенного дефекта) могул быть приняты за грибы. Избежать
этой неоднозначности в интерпретации результатов микроскопии позволяет окра-
ска препарата по Брауну-Бренну.
ОКРАСКА ПО БРАУНУ-БРЕННУ
Фиксация: 10% формалин.
Методика окраски
• Мазки, отпечатки или депарафинированные срезы заливают 1 мл 1% раство-
ра кристаллического фиолетового и добавляют 5 капель 5% раствора натрия
бикарбоната. Слегка перемешивают в течение 1 мин (растворы можно пред-
варительно перемешать).
• Промывают проточной водой.
• Обработывают раствором Люголя в течение 1 мин,
МИКОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ 743
• Прополоскивают в воде и просушивают фильтровальной бумагой.
• Обесцвечивают смесью равных частей эфира и ацетона до отхождения кра-
ски.
• Обесцвечивают рабочим раствором основного фуксина в течение 1 мин.
• Промывают водой и ополаскивают в ацетоне.
• Окрашивают 0,1% растворим пикриновой кислоты в ацетоне, пока препарат
не станет желто-розового цвета.
• Быстро ополаскивают в ацетоне, потом в смеси «ацетон-ксилол», затем в
ксилоле.
• Заключают препарат в бальзам.
Грамположительные микроорганизмы — лилово-синие, грамотрицательные —
красные. Ядра клеток красно-коричневого цвета. Другие тканевые структуры
желтого цвета.
Наиболее употребительными в рутинной лабораторной практике являются
методы окраски метиленовым синим и по Граму. Такой выбор оправдан простотой
методики, отсутствием многоэтапной длительной процедуры окрашивания, чет-
кости и контрастности получаемого микроскопического препарата, небольшими
трудозатратами на проведение исследования.
При просмотре препаратов, содержащих большое количество фибрина, богатый
клеточный материал, желательно применение окраски по Брауну Бренну и ШИК-
реакции.
В практике исследования биоматериала из бронхолегочного тракта (мокроты,
бронхоальвеолярного лаважа) рекомендуют дополнительную окраску PAS-методом
(ШИК-реакцией) для выявления истинных грибов-эумицетов и окраску по Цилю-
Нильсену или модификацию по методу Киньона для выявления кислотоустойчи-
вых микроорганизмов, в первую очередь микобактерий туберкулеза, возбудителей
лепры — микобактерий Хансена, нокардий и спорообразующих дрожжей.
При микроскопическом исследовании препаратов — отпечатков органов, цен-
трифуга тов, гистологических срезов рекомендуют осуществлять окраску биологи-
ческого материала тремя методами;
❖ окраской PAS-методом (ШИК-реакцией);
о- окраской по Цэаму или модификациями по Граму-Вейгерту, Боголепову,
методом Брауна-Бранна для выявления сопутствующей бактериальной
микробиоты, актиномицетов и нокардий;
< >• окраской по Цилю-Нильсену или модификацией по методу Киньона для
выявления кислотоустойчивых микроорганизмов, нокардий и спорообра-
зующих дрожжей.
ОКРАСКА ПО РОМАНОВСКОМУ-ГИМЗЕ
Готовый жидкий краситель перед окрашиванием мазков разводят из рас-
чета 1-2 капли красителя на 1 мл дистиллированной воды. Мазки окрашивают
20-25 мин при 37 °C во влажной камере. После окрашивания мазки промывают в
проточной воде, сушат на воздухе и исследуют при масляной иммерсии.
Красящую смесь Романовского-Гимзы в виде порошка (коммерческий краси-
тель) растворяют в смеси равных объемов метилового спирта и глицерина (800 мг
красителя на 100 мл растворителя). Краситель растворяется плохо, поэтому лучше
его растереть с растворителем в количестве 300 мг на 100 мл, а затем, помешивая,
добавлять краситель до получения нужной концентрации. Приготовление краси-
теля часто занимает несколько дней. Важно в качестве растворителей использо-
вать химически чистый метиловый спирт и глицерин, так как примеси ухудшают
свойства красителя. Вместо ме гилового спирта можно применять 100% этиловый
спирт. Приготовленную красящую смесь хранят в сухом прохладном месте в плот-
но закрытом сосуде.
744 МИКОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ
Методика окраски
• Мазки, фиксированные в метиловом спирте, окрашивают раствором (1 мл
готовой жидкой краски + 2 мл основного буферного раствора + 47 мл дис-
тиллированной воды) в лечение 40-120 мин. Используют фосфатный буфер,
но pH буфера зависит от вида мазка: для мазка костного мозга — 5,8-6,0, для
мазка крови — 0,4-65.
• Ополаскивают в дистиллированной воде.
• Высушивают и исследуют при иммерсии.
Бактерии окрашиваются в фиолетово-красный цвет, цитоплазма клеток — в
голубой, ядра — в красный.
ШИК-РЕАКЦИЯ И ОКРАСКА НА НЕЙТРАЛЬНЫЕ ПОЛИСАХАРИДЫ
Эти методы окраски основаны на выявлении в стенках грибов нейтральных
полисахаридов при окислении гликалиевых групп углеводов йодной или хромовой
кислотой. В стенках всех эумицетов (истинных грибов) в той или иной концен-
трации имеется глюкан-маннановый комплекс. При окраске материала, содер-
жащего грибы, фуксинсернистой кислотой (PAS-реакция, реакция шифф-йодная
кислота — ШИК) происходит окрашивание стенки гриба в пурпурно-красный
или фиолетовый цвет. Считают, что в основном окрашивается маннан, глюкан
PAS-отрицателен, а хитин дает слабую непостоянную окраску. Благодаря высокой
концентрации полисахаридов в клеточных стенках грибов их окраска значительно
ярче, чем окружающих их биологических сред человеческого организма.
Эумицеты PAS-положительны и хорошо окрашиваются, причем как вегети-
рующие, так и дегенеративные, в том числе и фагоцитированные элементы гриба.
Прокариоты являются PAS-отрицательными.
В лабораторной практике используют модификации PAS-метоца: окраску по
Хочкиссу-Мак-Манусу и окраску по Шабадашу.
Методика окраски по Хочкиссу-Мак-Манусу
• Стекла помещают в раствор йодной кислоты на 5-10 мин.
• Ополаскивают в дистиллированной воде 3 4 мин.
• Помещают препараты в фуксинсернистую кислоту на 10-30 мин, затем про-
мывают в дистиллированной воде.
• Тщательно промывают в сернистой воде.
• Окрашивают гематоксилином Гарриса, затем дифференцируют 1% водным
раствором соляной кислоты и промывают в щелочной воде (для восстанов-
ления синеватой окраски ядер).
Этапы окраски 1, 3 и 4 целесообразно проводить в затемненной оранжевой
посуде (реактивы подвержены быстрому разложению на свету).
Значительным недостатком модифицированной методики PAS-реакции по
Хочкиссу-Мак-Манусу является быстрое разложение иодной кислоты. Этого
недостатка лишена методика окраски на полисахариды по Шабадашу с примене-
нием перйодата калия.
Методика окраски по Шабадашу
• Стекла помещают в раствор перйодата калия на 5-10 мин.
• Ополаскивают в дистиллированной воде 3-4 мин.
• Помещают препараты в фуксинсернистую кислоту на 10-30 мин, затем про-
мывают в дистиллированной воде.
• Тщательно промывают в сернистой воде.
• Окрашивают гематоксилином Гарриса, затем дифференцируют 1% водным
раствором соляной кислоты и промывают в щелочной воде (для восстанов-
ления синеватой окраски ядер).
МИКОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ 745
При окраске этими методами грибы окрашиваются в красно-фиолетовый
или пурпурный цвет, ядра клеток синевато-серого цвета, общий фон бледно-
фиолетовый. Недостатком этой методики является интенсивное окрашивание
тканевых элементов, наиболее выраженное в очагах распада.
Методика окраски по Гридли
10% формалин.
• Предметные стекла с исследуемым материалом (мазками, отпечатками или
депарафинированными срезами) опускают на 1 ч в 4% хромовую кислоту,
затем промывают 5 мин в проточной воде.
• Погружают препараты в реактив Фельгена, приготовленный по Колеману
(растворяют 1 г основного фуксина в 200 мл кипяченой воды, отфильтро-
вывают и прибавляют 2 г K2S2O5 — метабисульфита калия и 10 мл нормаль-
ного раствора соляной кислоты). Дают обесцветиться в течение суток, затем
добавляют 0.5 г активированного угля, встряхивают и фильтруют через бума-
гу. Правильно приготовленный раствор должен быть бесцветным.
• Стекла ополаскивают в сернистой кислоте 3 раза по 2 мин (10% метабисуль-
фат натрия — 6 мл, нормальная серная кислота — 5 мл, дистиллированная
вода — 100 мл), после этого промывают в проточной воде.
• Погружают препараты на 15 мин в альдегидно-фуксиновый раствор (фук-
син основной -1г, этиловый спирт 70% — 200 г, паральдегид — 2 мл,
соляная кислота — 2 мл). Раствор оставляют на 3 дня для вызревания.
Приготовленный раствор должен быть синего цвета.
• Смывают избыток краски этиловым спиртом и промывают водой.
• Подкрашивают метаниловым желтым в течение 2-5 мин (метаниловый жел-
тый — 0.25 г, ледяная уксусная кислота — 0,25 г, дистиллированная вода —
100 мл).
• Промывают препараты в воде, обезвоживают 96% этиловым спиртом, про-
светляют в ксилоле и заключают в бальзам.
При окраске по Гридли удается получить яркое контрастное окрашивание
трибов, в том числе и грамотрицательных Нити окрашиваются в темно-голубой
цвет, споры приобретают оттенки от темно-розового до пурпурного, общий фон
желтый. Лучше всего окрашиваются псевдомицелий и стенки эндоспор. Плохо
окрашивается мицелий аспергилл, лучше всего — старые клетки.
Преимуществом окрасок по Шабадашу и Гридли является равномерное окраши-
вание грибов, четко выявляются фагоцитированные элементы грибов; окрашива-
ются также и их мертвые частицы.
Окраска по Шабадашу и Хочкиссу-Мак-Манусу хорошо выявляет молодые
формы грибов, особенно эндоспоры и бластоспоры, что позволяет успешно диф-
ференцировать вид возбудителя. Однако в дегенеративно измененных формах
некоторых грибов (например, криптококков) содержание полисахаридов снижа-
ется, количество PAS-реакций резко падает, и поэтому окраска на полисахариды
не всегда дает положительные результаты.
Почти не выявляются по этим методикам возбудители мукороза.
Поскольку при этих методиках окрашиваются как тканевые элементы, так и
бактерии, богатые полисахаридами, необходимо параллельно окрашивать пре-
параты гематоксилин-эозином, чтобы дифференцировать грибы от тканевых
элементов.
РЕАКЦИЯ СУЛЬФАТИРОВАНИЯ
Основой метода является предварительное сульфатирование препаратов с
последующим окрашиванием толуидиновым синим, который дает метахромати-
ческую окраску грибов в красный цвет.
746 МИКОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ
Методика окраски
• Препараты помещают в абсолютный спирт на 5 мин и высушивают на воз-
духе в течение 5 мин
• Помещают препараты в сульфатирующий реактив на 5 мин. Сульфатирующий
реактив получают при смешивании охлажденной концентрированной серной
кислоты с предварительно замороженным диэтиловым эфиром в соотноше-
нии 1:1.
• Затем препараты кладут на 3 мин в 3% уксусную кислоту.
• Окрашивают 0,01% раствором толуидиновот о синего в 3% уксусной кислоте
в течение 1 мин.
• По окончании окраски промывают в 3% уксусной кислоте 1 мин, после чего
помещают на 1 мин в 95% этиловый спирт.
Метод сульфатирования позволяет выявить очень молодые формы грибов,
которые далеко не всегда могут быть выявлены с помощью PAS-метода. Особенно
информативен этот метод при диагностике кокцидиоидоза.
МЕТОД ИМПРЕГНАЦИИ СЕРЕБРОМ
Сущность метода состоит в окислении нейтральных мукополисахаридов три-
оксидом хрома с последующей окраской продуктов окисления нитрата серебра
в щелочном растворе уротропина. Применение комбинации «уротропин-нитрат
серебра» вместо реактива Шиффа (PAS-реакции) вызывает более сильное окра-
шивание клеточной стенки грибов, окружающие же компоненты препарата
окрашиваются значительно слабее. Стенки грибковых клеток окрашиваются в
коричнево-черные цвета. Высокая контрастность окраски позволяет выявлять
грибы в некротических и обызвествленных очагах.
Методика окраски по Гомори-Грокотту
Фиксация препарата
10% формалин.
• Депарафинированные срезы доводят до воды.
• Препараты опускают на 1 ч в 5% хромовую кислоту.
• Промывают 10 мин в воде.
• Помещают препараты в 1% водный раствор бисульфата натрия на 1 мин для
освобождения от остатков хромовой кислоты, затем вновь промывают в про-
точной воде 5 мин.
• Трижды ополаскивают в дистиллированной воде.
• Погружают препараты в раствор уротропин-нитрата серебра на 1,5-2 ч при
температуре 45 °C или на 30-60 мин при 58-60 °C (по 25 мл уротропин-
нитрата серебра и дистиллированной воды, 1 мл 5% раствора буры).
Препараты должны стать слегка коричневыми.
• Хорошо промывают срезы в дистиллированной воде.
« Тонируют в 0,1% растворе хлорного золота в течение 2-5 мин.
• Промывают в дистиллированной воде.
• Закрепляют окраску погружением препаратов на 1-2 мин в 2% водный рас-
твор тиосульфата натрия.
• Докрашивают в рабочем растворе светлого зеленого 30-45 с.
• Дегидратируют и заключают препараты в бальзам.
Приготовление уротропин-нитрата серебра: 5 мл 5% раствора нитрата серебра
смешивают со 100 мл 3% раствора уротропина. Вначале образуется беловатый
осадок, исчезающий при помешивании, и раствор становится прозрачным и бесц-
ветным. При комнатной температуре раствор не теряет своих свойств до 14 дней, в
холодильнике может храниться свыше месяца.
микологические исследования 747
Грибы окрашиваются в черный цвет, контрастируя с общим фоном. Муцин
гемно-серого цвета. Внутренние части гиф грибов — темно-розового цвета.,
окружающие ткани — светло-зеленого. Хотя, по мнению многих исследователей,
метод Гомори -Грокотта является лучшим для выявления патогенных грибов, он
не лишен недостатков, основными из которых являются длительность процедуры
и потребность в дорогостоящих реактивах (хлорном золоте). Помимо этого из-за
бледности фона невозможно определить отношение гриба к окружающим структу-
рам. Эритроциты могул стать темно-коричневыми и симулировать клетки грибов.
Также возможны выпадение осадков серебра и слабое окрашивание фона. Важным
ограничением является высокая квалификация врача лабораторной диагностики
как в отношении техники выполнения окраски, так и просмотра препаратов из-за
опасности спутать элементы паразитирующего гриба с тканевыми структурами,
содержащими полисахариды.
К числу несомненных достоинств относится высокая выявляемость с его помо-
щью возбудителей мукороза, плохо окрашиваемых при других методиках. Хорошо
выявляются также омертвевшие элементы грибковых клеток, причем не только в
очагах воспаления, но и в старых обызвествленных очагах.
ОКРАСКА АЛЬЦИАНОВЫМ СИНИМ ПО МОУРИ
• Депарафинируют срезы и доводят до воды.
• Помещают на 1-2 ч в 0,5-2% раствор альцианового синего на 3% уксусной
кислоте или на 0,1% буфере (наЛия цитрат — НС1) с pH = 2,2+2,4.
• Промывают в 3% уксусной кислоте, затем в воде.
• Проводят через спирты и заключают в бальзам.
Капсулы Cryptococcus neoformans и Cryptococcus albldus окрашиваются в ярко-
голубой цвет.
ОКРАСКА МУЦИКАРМИНОМ ПО МЕЙЕРУ
Применяют для дифференциации криптококков. Иногда при этом методе слабо
окрашиваются клетки б тастомицетов, однако их без груда различают от интенсив-
но окрашивающихся криптококков. Исследованию подлежат парафиновые срезы
тканей.
Фиксация препарата
Любым способом.
Методика окраски
• Препарат депарафинируют ксилолом, затем абсолютным и 95% этанолом.
• Ополаскивают дистиллированной водой.
• Окрашивают в течение 7 мин гематоксилином Вейгерта.
• Промывают водопроводной водой 5-10 мин.
• Помещают в разбавленный раствор муцикармина на 30-60 мин. Интенсивность
окраски проверяют при микроскопировании контрольного препарата.
• Окрашивают метаниловым желтым в течение 1 мин.
• Ополаскивают препарат дистиллированной водой, затем 95% этанолом.
• Препарат обезвоживают, просветляют, заключают в бальзам.
Муцин капсулы криптококков окрашивается в темно-розовый или красный
цвет, ядра — в черный, а фон — в желтый цвет.
Mei оды окраски с флюоресцирующими красителями
Метод основан на наблюдении микроскопических объектов с использованием
их способности к свечению. По сравнению с методами обычной микроскопии
исследование в свете люминесценции обладает рядом преимуществ:
< > цветовым свечением;
748 МИКОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ
> высокой степенью контрастности светящихся объектов на темном фоне;
❖ значительно большей площадью поля зрения.
Суть люминесцентной микроскопии заключается в том, чго объекты (гриб-
ковые клетки), окрашенные флюорохромами, под действием облучения ультра-
фиолетом испускают излучение в видимом спектре света. В случае, когда объект
не окрашен специальными красителями, ультрафиолетовый свет, проходя через
объектив и попадая на препарат, поглощается молекулярными структурами объ-
екта и остается невидимым или почти невидимым для человеческого глаза. Если
же какие-либо объекты окрашены специальным красителем, молекулы красителя
под действием ультрафиолета возбуждаются и начинают испускать кванты света в
длинноволновой области, иначе говоря — светиться. В этом случае клетка стано-
вится источником света определенною спектра и хорошо видна на общем темном
контрастном фоне препарата.
Поскольку собственная флюоресценция грибов невелика, для ее усиления при-
меняют красители — акридиновый желтый или акридиновый оранжевый.
За счет свечения вокруг клетки образуется ореол, благодаря которому види-
мые размеры светящейся клетки превышают ее физические размеры. В связи с
этим окрашенные флюорохромными красителями препараты обычно исследуют
при увеличении х250-450, тогда как окрашенные обычными красителями — при
увеличении х800-1000. Разница в увеличении позволяет микроскописту видеть
одновременно в 4-10 раз большее поле зрения, что существенно сокращает время,
необходимое для просмотра мазка, Наряду с этим при люминесцентной микроско-
пии отмечается большая резкость и контрастность микроскопической картины,
что повышает комфортность микроскопического исследования. Это делает метод
люминесцентной микроскопии особенно ценным при исследовании материапа с
малым содержанием грибковых клеток.
При окраске флюорохромными красителями нельзя подогревать мазки и поль-
зоваться фильтровальной бумагой. Промывание мазков в процессе окраски следу-
ет проводить только дистиллированной водой, так как водопроводная вода содер-
жит соединения хлора, которые могут изменять флюоресценцию, Микроскопию
проводят по возможности сразу же после окончания процедуры окраски люминес-
центными красителями и высыхания мазков. В случае невозможности проведения
немедленной микроскопии окрашенные мазки рекомендуют сохранять, прикрыв
черной бумагой во избежание ослабления флюоресценции. При окраске не следует
делать толстых мазков, так как это затрудняет обесцвечивание и фиксацию мазка
на стекле.
При окраске мазков не следует помещать на штатив («рельсы») и окрашивать
одновременно более 12 стекол. Соприкосновение стекол боковыми краями может
привести к переносу краски и контаминации микроорганизмами с одного стекла
на соседние.
Окрашенные для исследования флюорохромными красителями мазки непри-
годны, если при окраске:
❖ в положительном контрольном мазке не обнаруживаются ярко окрашенные
бактериальные клетки или они имеют тусклую флюоресценцию;
❖ отрицательные контрольные мазки содержат флюоресцирующие объекты;
❖ фон недостаточно темный или имеет флюоресцентное свечение.
ПРОСТАЯ ЛЮМИНЕСЦЕНТНАЯ ОКРАСКА
Методика окраски
• Окпашивают препараты гематоксилином в течение 5 мин.
• Промывают в воде 3-4 мин.
• Окрашивают водным раствором акридинового оранжевого (1:1000).
МИКОЛОГИЧЕСКИЕ исследования 749
• Ополаскивают в дистиллированной воде в течение 30 мин.
• Обрабатывают в 95% этаноле и абсолютном спирте (2 раза по 2-3 мин).
Раствор акридинового оранжевого готовят по следующей прописи:
❖ безводный двузамещепный натрия фосфат (Na,HPO4) — 0.01 г;
дистиллированная вода — 100 мл;
4- акридиновый оранжевый — 0,01 г.
Растворяют натрия фосфат в дистиллированной воде. Добавляют акридиновый
оранжевый.
Препараты просматривают в обычном световом микроскопе с электроосве-
тителем. В качестве входного используют на конденсере голубой, а в качестве
окулярного — желтый фильтры, которые подбирают таким образом, чтобы они
полностью перекрещивались. Лучшие результаты дает просмотр в люминесцент-
ном микроскопе.
При этом одной из важных особенностей работы с мазками, окрашенными
флюорохромами, является то, что они не подлежат длительному наблюдению
в выбранном поле зрения, так как интенсивность люминесценции окрашенного
объекта быстро снижается. Другой особенностью работы является необходимость
строгого соблюдения режима работы люминесцентного микроскопа.
При нанесении иммерсионного масла капля должна свободно упасть на стек-
ло (при этом желательно немного смазать иммерсионным маслом фронтальную
линзу объектива). Пипетка капельницы масла ни в коем случае не должна сопри-
коснуться с мазком. Это может привести к переносу материала с одного мазка на
другой, к загрязнению иммерсионного масла и получению ложноположительного
результата микроскопического исследования.
При микроскопии хорошо выявляется свечение грибов на темном фоне, под-
крашенном эозином. Аспергиллы дают свечение зеленого цвета, дрожжевые
грибы рода Candida — желто-зеленого, криптококки — красного, бластомице-
ты — желто-зеленого, гистоплазмы — красно-желтого, возбудитель риносно-
ридиоза — красного цвета. Возбудители нокардиоза при использовании этого
метода не выявляются.
ОКРАСКА АКРИДИНОВЫМ ОРАНЖЕВЫМ ПО ДАРТУ-ТЕРНЕРУ
Методика окраски
• Мазки фиксируют в смеси равных частей диэтилового эфира и 95% этилово-
го спирта в течение 1 ч.
• Проводят через спирты нисходящей концентрации (80, 70,50%) и дистилли-
рованную воду, окуная мазок по 5 раз в каждую порцию.
• Окунают 4 раза в 1% уксусную кислоту.
• Переносят в дистиллированную воду на 2 мин.
• Помещают на 3 мин в ацетатный буфер (pH = 3,8).
• Переносят в 0,01% забуференный раствор акридинового оранжевого на 3 мин.
• Дифференцируют 4 мин в ацетатном буфере (pH = 3,8).
• Протирают концы предметных стекол, а затем осторожно удаляют избыток
буферного раствора фильтровальной бумагой.
• Заключают под покровное стекло в буферный раствор и оставляют на 2 мин.
Препарат исследуют с помощью флюоресцентного микроскопа сразу либо,
если мазок оставляют надолго, держат в темноте и парафинируют края
покровного стекла. Флюоресценция сохраняется в течение 3 мес.
Бактерии, дрожжи, грибы рода Candida окрашиваются в красный цвет, парази-
ты — в красный с желтыми ядрами, лейкоциты — в ярко-зеленый.
750 МИКОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ
МЕТОДИКА ОКРАСКИ С ФЛЮОРОХРОМОМ ДЛЯ ВЫЯВЛЕНИЯ ДЕРМАТОФИТОВ
Флюорохром в смеси с 2% раствором гидроксида натрия в диметилсульфоксиде
(1:9) с предварительным просветлением патологического материала.
ЭКСПРЕСС-ДИАГНОСТИКА ДЕРМАТ1ФИТ0В С ФЛЮОРОХРОМАМИ
БЕЗ ПРОСВЕТЛЕНИЯ
♦ Препарат окрашивают буферным раствором акридинового оранжевого с экс-
позицией 5 мин
• 0,1% бланкофор.
• Окрашивают с калькофлюором белым.
• 1% uvitex2b.
Методика окраски калькофлюором белым
Приготовление красителя:
< растворяют 0,1 г калькофлюора белого (М2Ю и Эванса голубого 0,05 г в
100 мл дистиллированной воды;
❖ тщательно перемешивают;
хранят в темной посуде при комнатной температуре.
Процедура окраски:
❖ добавляют 1 каплю раствора калькофлюора белого и 1 каплю раствора КОН
к исследуемому образцу на предметном стекле;
❖ накрывают образец покровным стеклом;
< просматривают в УФ-свете, используя К530 возбуждающий фильтр и В12
барьерный фильтр (или G365 возбуждающий фильтр и LP барьерный
фильтр).
Флюорохром поглощается структурами гриба, и при микроскопии под люминес-
центным микроскопом элементы гриба флюоресцируют зеленым или голубовато-
белым цветом (в зависимости от сочетания фильтров) на тусклом красноватом
фоне. Ошибки в диагностике могут’ быть связаны со свечением других полисаха-
ридов препарата, не входящих в хитиновые структуры гриба.
МЕТОД ФЛЮОРЕСЦИРУЮЩИХ АНТИТЕЛ (РЕАКЦИЯ
ИММУНОФЛЮОРЕСЦЕНЦИИ - РИФ)
Наибольшее распространение в практике этот метод имеет при исследовании
биологических жидкостей. С его помощью можно обнаружить корпускулярные,
грибные антигены, установить наличие и локализацию антигенов, высвободив-
шихся при разрушении грибов в организме.
К числу недостатков метода относится необходимость иметь для диагностики
каждого вида, а иногда и штамма гриба соответствующую меченую иммунную
сыворотку, сложную в получении. При диагностике микозов с неизвестным воз-
будителем в лаборатории необходимо иметь набор готовых иммунных высоко-
специфичных сывороток против различных наиболее распространенных пара-
зитирующих грибов, что весьма дорого и возможно только в условиях крупных
профильных лечебных учреждений.
В табл. 25-3 суммированы методы окраски, применяемые при микроскопиче-
ской идентификации грибов.
МИКОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ 751
Таблица 25-3. Методы окраски, применяемые при микроскопической идентификации грибов
(Номенклатуоа лабораторных исследований, 2000 г.)
Микроорганизм Метод окраски
гематоксилин-эозином по Ван-Гизону различ- ные PAS- модификации по Гридли по Гомори-Грах 1тту | акридиновым оранжевым флюороцир. антителами по Граму по Граму-Вейгерту по Брауну-Бренну по Романовскому-Гимзе по Райту альциановым синим по Баузру мукарцином по Мейеру
по Хочкиссу-Мак- Манусу по Шабадашу 1
Aspergillus род - + + + -1- + + + -
Blastomyces dermatitidis + - + + + + + + - - + - - + - -
Candida род + + + + + + + + + + + + + - - -
Chrysosporium parvum var. crescens + - + + + + -
Coccdioides immitis + + + + + + + + - + - + + - - -
Cryptococcus род + - + + - + + + - - - - - + - +
Epidermohpyton floccosum +
Fusarium +
Geotrichum + - + + - + + + - - - - - -
Histoplasma capsulatum + + + + + + + + - + - + + - + -
Loboa loboi + - - - + + - -
Malassezia + - + + -
Microsporum +
Paracoccidioides brasiliensis + - + + - + +
Penicillium marnefferi + - - - - +- - - - - - - + - - -
Pneumocystis carinii +- - + -
Pseudalleschiia boydii +
Rhinosporidium seberi + + + + - + + - - - - - - - +
Sporotrix schencki + - + + - + + - - + - + - - -
Trichphyten род - -
Гиалогифомицеты + - + + + + -
Дерматофиты + - + + - - -
Зигомицеты + - + + + - - - - - - + - - - -
Феогифомицеты + - + + + + -
752 МИКОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ
Причины ошибок при микроскопических исследованиях.
Структуры, имитирующие патогенные грибы
ИМИТАТОРЫ ГРИБОВ В ПРЕПАРАТАХ КОЖИ И ЕЕ ПРИДАТКОВ (ВОЛОС, НОГТЕЙ),
СЛИЗИСТЫХ ОБОЛОЧЕК
• Волокна текстиля лежат отдельно от клеток биологического материала, круп-
нее. чем гифы грибов, неравномерной толшины. не септированы.
• Кристаллы щелочи. Имеют вид тонких волокон с двойной стенкой, но, в
отличие от мицелия грибов, не имеют общей оболочки для всех элемен-
тов нити, не ветвятся, имеют стекловидную прозрачность, мономорфны.
Добавление под покровное стекло капли дистиллированной воды приводит к
растворению кристаллов.
• Жировые капли — имитируют дрожжевые клетки, часто встречаются в плохо
просветленном материале.
• Зерна кератогиалина могут быть приняты за споры грибов, однако они мель-
че по размерам и расположены внутри клеток кожи, что устанавливают при
вращении микровинта микроскопа.
• Мозаичный гриб — артефакт, образующийся при кристаллизации КОН при
чрезмерном нагревании препаратов. В нем нет четкого деления на клетки. Он
представлен в виде гомогенных светлых элементов, различных по форме и
размерам, имеющих сетчатое деление. При подогревании препарата или его
хранении в течение 1-3 дней «мозаичный гриб» исчезает.
• Пузырьки воздуха.
ИМИТАТОРЫ ПАТОГЕННЫХ ГРИБОВ В СРЕЗАХ ТКАНИ
Имитаторы нитчатой структуры
• Нити фибрина (при окраске по Граму-Вейгерту).
• Вытянутые и уплощенные волокнистые фиброциты.
• Нити полисахаридов стенок капилляров при PAS-методе окраски.
• Базальные мембраны, щеточная кайма эпителия (PAS-метод).
• Коллагеновые и эластические волокна при импрегнации серебром по Гомори-
Грокотту.
• Ретикулярные волокна.
• Вытянутые и уплощенные волокнистые фиброциты.
• Тонкие обызвествленные волокна интерстиция. имитирующие коккоподоб-
ный мелкозернистый мицепий нокардий.
Округлые и сферические внутри- и внеклеточные имитаторы грибов
• Русселевские тельца, амилоидные тельца.
• Обесцвеченные, вспученные и видоизмененные эритроциты.
• Зерна гемосидерина при импрегнации серебром.
• Микроглобулярные преципитаты рибонуклеиновой кислоты.
• Скопления муцинозных веществ в эпителиальных клетках, макрофагах, ино-
гда расположенные внеклеточно, выявляемые при окраске на мукополисаха-
риды.
• Выпадающие осадки красителей.
• Жировые капли
• Округлые образования, обнаруживаемые при гранулематозных образовани-
ях и в гигантских клетках.
МИКОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ 753
Имитаторы патогенных грибов, напоминающие сферулы или спорангии
Цитоплазматические вакуоли макрофагов, симулирующие сферулу с эндоспо-
рами.
Обеспечение качества микроскопических исследований
Важным элементом обеспечения качества выполняемых исследований является
регулярное осуществление контроля качества микроскопических исследований,
который достигается систематическим наблюдением за проводимой в лаборато-
рии работой и оценкой ее эффективности. Контроль качества осуществляется на
всех технологических’ этапах микроскопического исследования и включает:
• оценку качества поступающих биопроб;
• контроль за соблюдением рецептуры и методики приготовления реагентов и
красителей;
• наблюдение за соблюдением методических приемов:
❖ при приготовлении мазков (включая качество предметных стекол);
окраске мазков;
❖ проведении микроскопического исследования;
❖ учете и регистрации результатов.
Кроме того, элементами внутрилабораторного контроля качества являются:
< проверка правильности организации рабочих мест (приема и регистрации
материала, приготовления и окраски мазков, микроскопирования);
❖ соблюдение правил и сроков хранения реагентов и красителей;
❖ настройка оборудования;
❖ микроскопирование положительных и отрицательных контрольных образ-
цов;
своевременность и точность передачи результатов в учреждение, направив-
шее мат ериал для исследования.
Успех применения контроля качества обеспечивается:
< > регулярным его применением в лабораторном подразделении;
❖ правильно обученными, заинтересованными и ответственными работниками.
❖ рациональным применением регламентированных методов;
J регулярным анализом допущенных ошибок и немедленным их исправлением.
Одной из форм обеспечения качества микроскопических исследований является
использование в ряду клинических мазков двух дополнительных неокрашенных
контрольных мазков, один из которых заведомо является положительным, а вто-
рой — отрицательным. Просмотр начинают с контрольных, затем просматривают
клинические мазки.
Заключение по результатам микроскопического
исследования
Результат микроскопического исследования патологического материала фикси-
руется в специальном лабораторном журнале.
Заключение оформляется для выдачи врачу-клиницисту в виде бланка, в кото-
ром должны быть отражены патологический процесс (количество лейкоцитов при
воспалении, пролиферация, фиброз и др.), количественный и качественный состав
микрофлоры, а также наличие специфических возбудителей микоза. Заключение
формулируется не в виде диагноза, а как развернутое описание микроскопической
картины препарата. Формулировка диагноза в заключении по результатам микро-
скопического исследования недопустима, так как она является прерогативой леча-
щего врача, а не лабораторного работника.
754 МИКОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ
КУЛЬТУРАЛЬНОЕ ИССЛЕДОВАНИЕ
Культуральное исследование является значительно более чувствительной мето-
дикой, чем микроскопия. Выделение чистой культуры гриба применяют для иден-
тификации рода и вида гриба, подтверждения диагноза микотического процесса
при отрицательных данных микроскопии.
Оптимальные значения pH среды для культивирования большинства видов гри-
бов находятся в диапазоне 5,8-6,5.
Для получения чистой культуры и устранения контаминации сопутствующей
бактериальной флорой используют различные селективные добавки к средам.
С этой целью наиболее часто применяют различные антимикробные вещества и
антибактериальные антибиотики.
При культуральном исследовании мазок и посев проводят обязательно из одной
и той же порции материала. Это основное требование правильного микробиологи-
ческого исследования.
ТРЕБОВАНИЯ К ДОСТАВКЕ ПРОБ БИОМА1ЕРИАЛА В МИКРОБИОЛОГИЧЕСКУЮ
ЛАБОРАТОРИЮ
Все собранные пробы отправляют в микробиологическую лабораторию немед-
ленно после получения, за исключением случаев использования емкостей с транс-
портировочными средами, разрешенными к применению для этих целей в уста-
новленном порядке.
Это необходимо:
< для сохранения жизнеспособности возбудителей и возможности выделения
микроорганизмов, требующих особых условий культивирования;
❖ предотвращения избыточного роста быстрорастущих и активных микроорга-
низмов;
❖ поддержания соотношения исходных концентраций изолятов при наличии в
пробе микробных ассоциаций:
❖ сокращения времени контакта пробы с некоторыми антисептиками, исполь-
з\ емыми местно, которые могут обладать антифунгеальной активностью;
о объективизации клинического диагноза заболевания и оценки результатов
терапии.
Если отделяемого всего несколько капель, его переносят из шприца в небольшую
ем» ость или в пробирку с транспортной средой немедленно после получения.
Получение у пациента любой пробы, требующей использования инвазивных
методов, должен проводить врач (исключение — проба крови, которую может
собирать процедурная медицинская сестра).
Поступивший в лаборатопию материал перед обработкой и посевом следует
зарегистрировать в лабораторном регистрационном журнале. Каждой пробе
материала необходимо присвоить порядковый регистрационный лабораторный
номер, который должен использоваться при всех последующих лабораторных
исследованиях.
Этап пробоподготовки
Патологический материал делят на гри части: для микроскопии, культурал! -
ного исследования и повторного изучения. Засеваемый материал предварительно
измельчают.
ПРИНЦИПЫ ПРЕДПОСЕВНОЙ ОБРАБОТКИ ДИАГНОСТИЧЕСКОГО МАТЕРИАЛА
Все процедуры по переносу материала из одной посуды в другую при обработке
и посеве проводят только стерильными пипетками. Запрещается перенос мате-
микологические исследования 755
риала переливанием его через край флаконов, пробирок или иной лабораторной
посуды. Все реактивы, используемые при приготовлении растворов для обработки
диагностических материалов, должны иметь степень очистки не менее категории
«химически чистый».
Выбор алгоритма культурального исследования
Основной средой для получения чистой культуры грибов в медицинской мико-
логии является среда Сабуро. В нее вводят различные селективные добавки в зави-
симости от типа исследуемого образца.
Для исследования мазков со слизистых оболочек носо- и ротоглотки, урогени-
тального тракта, при исследовании кала на дисбактериоз используют среду Сабуро
с хлорамфениколом, гентамицином, реже используют среду, содержащую комби-
нацию этих антибиотиков. При культуральном исследовании материалов кожи и
ее придатков в качестве добавки в среду Сабуро вводят актидион (циклогексимид).
который обеспечивает селективный рост дерматофитов. Дерматофиты относятся
к медленно растущим грибам, и формирование колоний для некоторых из них
может наблюдаться лишь к концу 5-7-дневного периода инкубации. В течение
этого периода необходимо обеспечить в инкубаторе достаточную влажность во
избежание пересыхания среды. На практике хорошо зарекомендовал себя метод
культивирования дерматофитов с использованием двухфазной среды (твердая
фаза — агаризованная среда Сабуро с актидионом, жидкая фаза — бульон Сабуро с
цитратом или сердечно-мозговой бульон) во флаконе для гемкультуры. Актидион
из твердой фазы диффундирует в жидкую среду и препятствует росту бактериаль-
ной флоры, а жидкая фаза препятствует пересыханию агара и обогащает агаризо-
ванную среду в зонах роста.
При симптоматике иммунодефицитных состояний и подозрении на поражение
ЦНС образцы спинномозговой жидкости и сыворотки или плазмы крови высевают
в гемфлакон и одновременно проводят их исследование на наличие растворимого
ксиломаннанового антигена методом латекс-агглютинации или иммунофермент-
ного анализа. При получении положительного результата в качестве подтверж-
дающего метода осуществляют пересев обогащенной культуры из гемфлакона на
специализированные агаризованные среды для выявления криптококка (среда
L-DOPA). Оценивают оксидазную активность полученных колоний.
Оценка результатов посева
При оценке результатов культурального исследования необходимо соблюдать
следующие правила:
❖ посевы, выполненные в течение одного дня, помещают в отдельные ящики
или штативы с указанием даты посева и размещают их в термостате в порядке
номеров регистрации в хронологическом порядке, не нарушая его при еже-
дневных просмотрах;
О наблюдение за посевами и просмотр засеянных пробирок проводят ежедневно;
у при оценке результатов регистрируют следующие параметры:
- появление роста — срок появления, начиная со дня посева;
- интенсивность роста — количество колоний;
- загрязнение посева посторонней микрофлорой;
- отсутствие роста.
Соблюдение этих правил позволяет, во-первых, своевременно выявлять макро-
скопически видимый рост грибов или загрязняющей микрофлоры, а во-вторых, на
основании регистрации сроков появления роста и его особенностей осуществлять
первичную идентификацию грибов.
756 МИКОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ
Идентификация выделенных культур грибе в
В лабораторно-диагностической практике используют культурально-
морфологические критерии идентификации:
< вначале оценивают макроморфологические характеристики роста культуры
гриба на агаровых средах, а затем микроморфологию гриба;
❖ в затруднительных случаях проводят изучение ферментативной активности
культуры гриба.
В изучение макроморфологии входят:
❖ оценка скорости роста и созревания;
< структуры колонии (пушистая, шерстистая, войлочная, бархатистая, паути-
нистая. клочковатая, мучнистая);
поверхности (плоская, складчатая, бугристая, куполообразная, зональная);
пигментации;
❖ наличие экссудата на поверхности колонии.
Микроморфологию грибной культуры изучают по препаратам, приготовлен-
ным следующим способом:
о на предметное стекло наносят каплю жидкости для приготовления препара-
тов (спирта, глицерина, дистиллированной воды в равных частях);
< в нее помещают кусочек грибницы, вырезанный микологической лопаточкой
из колонии в виде треугольного сектора от центральной части к периферии;
❖ двумя препаровальными иглами расправляют вырезанный кусочек во избе-
жание образования воздушных пузырьков;
❖ накрывают покровным стеклом
Аспергиллез
ВОЗБУДИТЕЛИ
Aspergillus fumigatu, A.flavus, A. niger, A. terreus; A. nidulans и др.
ХАРАКТЕРИСТИКА КУЛЬТУРЫ
• Aspergillus fumigatus — колонии гладкие, бархатистые или шерстисто-
клочковатые, цвет от различных оттенков зеленого до темно-бурого.
Обратная сторона колоний бесцветная или желтая.
❖ Микроскопия:
- конидиеносцы гладкие, зеленоватые;
- септированные или несептированные;
- размеры 2-8 мкм в ширину и 300-500 мкм в длину.
- конечное вздутие (везикул) обратно-фляжковидное;
- в верхней половине находятся стеригмы, расположенные в один ряд,
размером 2-6x3-6 мкм, пригнуты кверху;
- каждая стеригма отчленяет цепочку конидий;
- гриб термофильный, хорошо растет при 37 °C, некоторые штаммы —
при 52 °C.
• A.flavus — колонии клочковатые, зеленовато-желтого цвета. Обратная сторо-
на колоний желтоватая.
❖ Микроскопия:
- конидиеносцы неокрашенные, шероховатые, везикул округлый;
- стеригмы однорядные, в малых головках, старые — двухрядные, с ко-
лонками конидиальных цепочек;
- конидии грушевидные или круглые, 3-6 мкм в диаметре, склероции
обильные, вначале белые, затем бурые;
- рост культуры - при 25- 37 °C.
микологические исследования 757
• A. niger — колонии отличаются быстрым ростом, цвет вначале белый, со
временем становятся желтыми, с мелкими черными головками; через 4-
5 дней вся поверхность колоний становится черного цвета. Обратная сторона
колоний светло-желтая. Стеригмы расположены в два ряда друг над другом
и заканчиваются цепочками конидий с гладкой или бородавчатой поверхно-
стью, 4-5 мкм в диаметре. Культура растет при 37 СС.
• A. terreus — культура растет при 25-37 °C, колонии имеют клочковатую (до
вельветовой) поверхность, иногда пушистые, с радиальными полосками от
желтовато-бурого до шоколадно-коричневого цвета.
❖ Микроскопия.
- конидиеносцы длинные и тонкие, гладкие, одинаковой ширины по всей
длине, размером 5-6x100-250 мкм;
- стеригмы расположены в два ряда на споровых головках;
- конидии гладкие, эллипсоидные;
- культура растет при 25-37 “С.
• A. nidulans — колонии темно-зеленого цвета с участками от оранжевого до
желтого в местах образования плодовых тел. Обратная сторона колоний пур-
пурная.
❖ Микроскопия:
- конидиеносцы гладкие, короткие, коричневого цвета, размером 60-
150 мкм:
- конидии размером 3-4 мкм, шаровидные, шероховатые;
- плодовые тела коричневые, сферические, окружены большим количе-
ством крупных стекловидных клеток;
- аскоспоры красновато-коричневые, чечевицеобразные, размером 4-
5 мкм.
Кандидоз
ВОЗБУДИТЕЛИ
Основными возбудителями являются С. albicans, С. tropicalis, С. parapsilosis,
С. glabrata и С. crusei. Значительно реже выявляются С. lusitaniae, С. guillermondii,
С. rugosae и др.
ХАРАКТЕРИСТИКА КУЛЬТУРЫ
< • С. albicans на среде Сабуро имеет вид блестящих кремово-белых колоний.
❖ При микроскопии колония состоит из округлых, овальных почкующихся
клеток 3-10 мкм в диаметре.
< По периферии колоний могут встречаться нити псевдомицелия.
❖ На обедненных средах формирует толстостенные хламидоспоры.
❖ Биохимические характеристики С. albicans представлены в табл. 25-4.
Mycotorula — шаровидный тип роста. Круглые бластоконидии образуют шаро-
видные верти циллы.
Mycocandida — ветви псевдомицелия образуют фигуру в виде елки, появление
бластоконидий запаздывает (рудиментарный тип роста).
Mycotoruloides — крыловидный тип роста. Крыловидные вертициллы. состоя-
шие из бластоконидий, расположены на перетяжках псевдомицелия с обеих сто-
рон поочередно.
Candida — цепочечный тип роста. Ветви псевдомицелия оканчиваются цепочка-
ми бластоконидий.
Blastodendrion — древовидный тип роста. Псевдомицелий, состоящий из поли-
морфных клеток, в виде деревца. Шаровидные вертициллы могут отсутствовать.
758 МИКОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ
Таблица 25-4. Биохимические характеристики наиболее часто встречаемых видов Candida
Криптококкоз
Возбудители
Cryptococcus neoformans [rar. neoformans ~ серотип D; var. grubu ~ серотип A; var
gattii = серотипы В, C; vangrubii/van neoformans (диплоид) = серотипы A. D].
Характеристика культуры
• C. neoformans — колонии на сусло-агаре или агаре Сабуро при 37 °C белые,
желтоватые; на среде Штайба или агаризованной среде с L-DOPA — темно-
коричневые, гладкие, блестящие, слизисто-тягучие. Чувствительность куль-
турального метода диагностики — 70%. Преобладают дрожжевые клетки с
одной почкой. Размеры капсулы различаются в зависимости от штамма и
состояния иммунной системы организма человека У ВИЧ-инфицированных
в стадии СПИД встречаются бескапсульиые формы или клетки с истончен-
ной капсулой, которые при пересеве вновь приобретают капсулу. У тканевых
форм всех видов криптококков капсула хорошо выявляется в препаратах,
окрашенных индийской тушью (чувствительность 40 -70%), альциановым
синим или основным коричневым.
• С. albidus — колонии блестящие, гладкие, тонкие, крсмовси о цвета.
❖ Микроскопия:
- почкующиеся сферические или овальные клерки со стенками различной
толщины, размером 3-5x8-10 мкм;
- гифы и псевдомицелий отсутствуют.
• С. laurentii — колонии влажные, слизистые, кремового цвета, по мере старе-
ния приобретают палевый охристый оттенок.
о Микроскопия:
- почкующиеся сферические или эллипсоидные клетки, размером
2-3x5,5-7,0 мкм;
- гифы и псевдомицелий отсутствуют.
Биохимическая характеристика культур возбудителей криптококкоза, используе-
мая при дифференциальной диагностике методом пестрого ряда, представлена в
табл. 25-5.
МИКОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ 75Q
Таблица 25-5. Дифференциальная диагностика различных видов криптококка по ферментатив-
ным свойствам
Возбудитель Ферментативные свойства
ассимиляция сахаров нитрат уреаза циклотексемнд рост при 37 °C
лактоза мелибиоза мелицтоза
С. neoformans - + - + - +
С. albidus + + + + - +
С. laurentii + + - + + ±
Зигомикозы
ВОЗБУДИТЕЛИ
Низшие грибы, относящиеся к классу Zygomycetes: Rhizopus spp. (R. arrhizus,
R. microsporus var. rhizopodiformis, R. microsporus var. oligosporus, R. stolonifer), Absrdia
spp. (A. corimbifera, A. coerulea), Mucor spp. (AL tircineUoides, AL hiemalis, AL indicus,
AL racemosus, AL ramosissibus), Rhyzomacor pttssibilis.
* Rhizopus spp. — колонии быстро растут при 37 °C. Воздушный мииелий обиль-
ный, серовато-белый, шерстистый, затем по мере старения меняет цвет от
желтовато-кооичневого до коричневого. Субстратный мицелий несептиро-
ванный, бесцветный, спориносцы неветвяшиеся, одиночные или группами
с хорошо развитыми ризоидами. Спорангии с коричневыми или черными
стенками, содержат округлые или угловатые спорангиоспоры.
• R. arrhizus — наиболее частый возбудитель зигомикоза. Колонии войлочные,
неокрашенные или (при спороношении) пестрые, темно-серого цвета, с хоро-
шо развитым мицелием, высотой 0.8 -1,5 см.
Дерматомикозы (дерматофитии)
Повсеместно распространены, встречаются во всех возрастных группах населения.
ВОЗБУДИТЕЛИ
Наибольшее эпидемиологическое значение имеют 10 видов дерматоминетов:
Trichophyton rubrum, Т. mentagrophytes, Т. ajelloi, Т. tonsurans, Т. violaceum, Microsporum
canis, M. audouinii, M.femigineum, AL gypseum. Epidermophytonfloccosum.
ХАРАКТЕРИСТИКА КУЛЬТУРЫ
• Trichophyton rubrum — культуры полиморфны, вначале белые, бархатистые,
позже могут быть пурпурно-красными, мучнистыми или кожистыми с кон-
центрическими зонами красно-коричневого или фиолетового оттенка, пуши-
стыми белыми с розовыми бороздками, белыми мучнистыми, окруженными
пурпурной полосой, кожистыми розовато-коричневого или коричневого
цвета. Обратная сторона культур красного или малинового цвета. Мицелий
ровный, септированный, диаметром 2-3 мкм. Микроконидии обильные,
каплевидные, удлиненные, грушевидные, размером 3-5 мкм, расположены
по бокам мицелия. Макроконидии состоят из 5-6 клеток, образуются на кон-
цевых нитях, тупоконечные, встречаются не у всех штаммов. Хламидоспоры
интеркалярные и концевые.
• Т. mentagrophytes', по морфологическим особенностям выделяют два варианта:
Т. mentagrophytes var. gypseum и Т. mentagrophytes var. interdigitate.
• T. mentagrophytes var. gypseum — колонии гипсовидно-мучнистые. Мицелий ров-
ный, ветвистый, сеп тированный. Микроконидии округлые или грушевидные,
расположены гроздьями по бокам мицелия. В зрелых культурах нередко обра-
760 МИКОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ
зуются спирали из 10-12 завитков. Макроконидии веретенообразные, состоят
из 3-5 клеток, свободный конец закруглен.
• Т. meal aerophytes var. inicrdigitole — зрелые колонии белые, бархатистые, цвет
от розового до коричневого, с ровной, реже радиально исчерченной поверхно-
стью, с углублением в центре. Мицелий длинный, ветвистый, септированный,
с тонкими завитками и спиралями. Микроконидии округлые, расположены по
бокам мицелия. Макроконидии встречаются редко Часто наблюдается плео-
морфизм культур.
Идентификация грибов чистой культуры
по ферментативной активности
Для этой цели разработаны коммерческие наборы. В частности, набор
«Ауксаколор 2» - система идентификации, основанная на принципе утилизации
сахаров, позволяющая идентифицировать 31 вид дрожжей и один вид одно-
клеточных водорослей Protothea wickerhamii. Рост грибов становится видимым в
результате изменения цвета pH-индикатора. Набор включает 16 биохимических
характеристик, распределенных в лунках: в 13 лунках находятся метаболиче-
ские тесты, содержащие сахара (глюкозу, мальтозу, сахарозу, галактозу, лактозу,
раффинозу, инозитол, целлобиозу, трегаллозу, адонитол, мелезитозу, ксилозу,
арабинозу), имеется 3 ферментных теста, включая тест на обнаружение фенолок-
сидазной активности Cryptococcus neoformans. Для исследования используют 24-
или 48-часовую чистую культуру, выращенную на среде Сабуро или хромогенной
среде. Большинство дрожжей могут быть идентифицированы в пределах 24 ч, но
иногда результаты необходимо считывать через 48-72 ч. Рост дрожжеподобных
микроорганизмов регистрируется по изменению цвета индикатора от голубого до
желтого и помутнению раствора в лунке.
ОПРЕДЕЛЕНИЕ АНТИФУНГЕАЛЬНОЙ ЧУВСТВИТЕЛЬНОСТИ
Методики определения чувствительности
к антифунгеальным препаратам
В настоящее время разработан ряд объективных методик для определения п
vitro чувствительности широкого спектра дрожжевых и плесневых грибов к анти-
фунгеальной терапии. В настоящее время сделан целый ряд шагов по увеличению
стандартизации и улучшению клинической интерпретации данных методов. Это
нашло отражение в стандартах NCCLS, описывающих референсные методы для
определения чувствительности дрожжевых и плесневых грибов. NCCLS постоянно
развивает референсную методологию определения антифунгеальной чувствитель-
ности, используя многочисленные воспроизводимые исследования. Разработка и
практическое внедрение новых методов определения антифунгеальной чувстви-
тельности осуществляются с использованием контрольных штаммов грибов при
сопоставлении с результатами референсных методов NCCLS.
СТАНДАРТ NCCLS М27 А2
Референсный метод для определения антифунгеальной чувствительности
дрожжевых грибов методом последовательных разведений в жидкой среде
Стандарт NCCLS М?7 А2 распространяется на исследования дрожжевых три-
бов, включая грибы рода Candida, Cryptococcus (С. neoformans и С. gattii) с
использованием макро- или микропоследовательных разведений в жидкой среде.
МИКОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ 761
В качестве среды рекомендуют использовать RPMI-1640 (сложную многокомпо-
нентную среду с добавками 0,2% глюкозы, pH = 7,0). Инокулят стандартизуется
по мутности. Инкубацию осуществляют при 35 °C в течение 24 ч для грибов рода
Candida или 48-72 ч — для требовательных грибов, таких как криптококк. Метод
микроразведений осуществляют в лунках стандартного 96-луночного микроплан-
шета, при этом сопоставляют рост грибов в лунках с различной концентрацией
препарата и контрольной лункой. Ингибирование оценивают в баллах от 0 до 4 по
следующей шкале:
> 0 — содержимое лунки оптически прозрачно;
< I — легкая «дымка»;
❖ 2 — существенное снижение мутности суспензии по сравнению с лункой, не
содержащей антифунгеального препарата;
❖ 3 — слабое снижение мутности по сравнению с лункой, не содержащей анти-
фунгеального препарата;
❖ 4 — мутность совпадает с мутностью в лунке, не содержащей антифунгсиль-
ного препарата.
Минимальной ингибирующей концентрацией (МИК) для амфотерицина В*
считается наименьшая концентрация, при которой результат оценивается как
0 (содержимое лунки оптически прозрачно). МИК для азольных препаратов и
5-флюороцитозинар является минимальная концентрация, при которой результат
оценивается как 2 (существенное снижение мутности).
Разработаны коммерческие тест-системы для определения чувствительности
дрожжевых грибов, основанные на модификации метода последовательных раз-
ведений, описанного в стандарте NCCLS М27 А2. Исследование выполняют, как
правило, в микропланшетном варианте.
Интерпретация результат»
Для большинства штаммов грибов рода Candida МИК амфотерицина В* коле-
блется от 0,25 до 1,0 мк^/мл, однако следует отметить, что метод NCCLS М27 не
позволяет определить разницу между резистентными штаммами и штаммами,
для которых минимальная ингибирующая концентрация больше 1 мкг/мл. И те
и другие предлагают считать резистентными по отношению к амфотерицину В*.
В настоящее время существуют жидкие среды, которые позволяют определить
МИК амфотерицина В*, в том числе и для устойчивых штаммов, но, к сожалению,
состав таких сред не поддается стандартизации, результаты с использованием раз-
ных партий этих сред могут различаться.
Имеются критерии интерпретации результатов определения чувствительно-
сти для трибов рода Candiaa по отношению к флюконазолу^, итраконазолу*,
5-флюороцитозину^. Эти критерии также могут быть использованы для крипто-
кокков.
Ошибки в определении МИК
Некоторые азольные препараты, в особенности флюконазол*’, демонстрируют
следующий феномен: мутность спеды снижается при увеличении концентрации
препарата, но суспензия не становится прозрачной (частичное ингибирование
роста в широком спектре концентраций антимикотика). Для большинства изоля-
тов различия между результатами, зафиксированными после 24 ч и 48 ч инкуба-
ции, минимальны, однако некоторые штаммы демонстрируют резкое увеличение
МИК в зависимости от времени инкубации. В общем случае результаты для грибов
рода Candida фиксируют через 24 ч инкубации. Попытка разрешить эту ситуацию
отражена в описании процедуры контроля качества NCCLS М27 А2, которая
в настоящее время является референсным по отношению ко всем остальным
методам, в том числе по отношению к нескольким коммерческим тест-системам,
762 МИКОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ
которые в настоящее время широко применяются в лабораторно-диагностической
практике для определения чувствительности к антифунгеальным препаратам.
NCLLS М44-Р
Определение антифунгеальной чувствительности дрожжевых грибов с помощью
диско-диффузионного метода
В стандарте М44-Р описана методология для исследования чувствительности
грибов рода Candida диско-диффузионным методом. Документ включает значения
диаметров зон ингибирования роста для флюконазола” и рекомендации по про-
цедуре контроля качества для флюконазола^ и вориконазола*’. В качестве твердой
среды для постановки диско-диффузионного метода в документе рекомендуется
использовать агаризованную среду Мюллера-Хинтона, обогащенную 0,2% глю-
козы и содержащую краситель метиленовый синий в концентрации 0,5 мкг/мл.
Среда Мюллера-Хинтона отличается высокой степенью стандартизации состава
и высокой воспроизводимостью результатов. Добавка глюкозы обеспечивает
условия роста большинства дрожжевых грибов, наличие метиленового синего в
составе среды контрастирует границу зоны ингибирования роста. Готовую среду
разливают в чашки Петри, pH готовой среды — 7,2-7,4. Посев осуществляют
методом сплошного газона, плотность инокулята стандартизована по мутности.
На поверхность среды апплицируются бумажные диски стандартного диаметра,
содержащие антифунгеальные препараты. Чашку с культурой инкубируют в тече-
ние 24 ч при 35 °C. Если по истечении 24 ч наблюдается скудный рост культуры,
необходимо продолжить инкубацию в течение дополнительных 24 ч. В течение
инкубации происходит диффузия антифунгеального препарата от диска в толщу
среды с образованием концентрационного градиента. Критерием определения
чувствительности служит диаметр зоны ингибирования роста.
Метод является простым и недорогим средством для скрининговых исследо-
ваний. Однако рекомендуют все культуры, зарегистрированные как устойчивые,
дополнительно исследовать методом последовательных разведений М27 А2 в
качестве подтверждающего.
Имеются коммерческие тест-системы, основанные на методах, схожих с диско-
диффузионным. В частности, вместо диска используют бумагу, содержащую гради-
ент концентрации антимикробного препарата. Этот градиент позволяет осущест-
влять непосредственное определение МИК. Выпущены наборы для исследования
антифунгеальных агентов, включая амфотерицин В*, кетоконазол*, итракозол*,
вориконазол*’, 5-флюороцитозинр, флюконазолр.
В перспективе диско-диффузионный метод может быть адаптирован для иссле-
дования других грибов, включая плесневые, но получаемые результаты нуждаются
в стандартизации с точки зрения интерпретации с использованием референсных
методов NCCLS.
NCCLS М38-А
Референсный Mei од последовательных разведений в жидкой среде
для определения чувствительности к антифунгеальным препаратам
мицелиеобразующих грибов
В стандарте описан метод исследования чувствительности к антимикотикам
плесневых грибов, которые способны вызывать системные инфекции, включая
Aspergillus spp., Fusarium spp., Pseudallescheria (Scedosporium) spp., зигомицеты и
другие пат огенные плесени. Рекомендуют использовать жидкую среду RPMI-1640
с той же формулой, что и в стандарте NCCLS М27 А2 для дрожжевых грибов.
Однако, в отличие от дрожжей, приготовление инокулята плесневых грибов сле-
дует осуществлять с помощью спектрофотометра. Необходимая концентрация,
МИКОЛОГИЧЕСКИЕ исследования 763
обеспечивающая воспроизводимость значений МИК, колеблется в пределах
0,4х104-0,5х104 КОЕ/мл, при этом оптическая плотность (ОП), измеренная на
530 нм, будет зависеть от морфологии исследуемого вида грибов. Необходимая
концентрация Aspergillus spp. и Sporothrix spp. при 0,09-0,11 ЕД ОП для Fusarium,
Pseudallescheria (Scedosporiim) и Rhizopus spp. ОП должна составлять 0,15-0,17 ЕД,
а для гистиоплазмы — 0,2 ЕД ОП. Допускается использование малого количества
твин-20 в качестве осушающего агента, облегчающего приготовление инокулята
Aspergillus. Культуру инкубируют при 35 °C. Чтение результатов проводят через 24
ч для Rhizopus spp., через 48 ч — для Aspergillus spp., Fusanum, Sporothrix spp. и боль-
шинства других, и через 72 ч — для медленно растущих плесневых грибов, таких
как Pseudallescheria (Scedosporium) spp.
При оценке результатов используют условную шкалу баллов от 0 до 4, при этом
мутность каждого образца оценивают относительно контроля, не содержащего
антифунгеальный препарат:
0 — образец оптически прозрачный, рост отсутствует;
Ф 1 - слабый рост, 25% мутности контрольного образца;
2 — существеное ингибирование роста, 50% мутности контроля;
3 - слабое ингибирование роста. 75% мутности контроля;
❖ 4 — нрт ингибирования
МИК амфотерицина Вт, итраконазола*, вориконазолар и позаконазола*3 явля-
ется минимальная концентрация, при которой результат оценивается как 0 (среда
оптически прозрачна). МИК флюконазола5’, кетоконазола и 5-флюоропитозина^
является минимальная концентрация, при которой наблюдается 50% снижение
мутности по сравнению с контрольным образцом (результат оценивается как 2).
Интерпиетация результатов
Минимальная ингибирующая концентрация амфотерицина В* обычно опреде-
ляется в пределах 0,5-2,0 мкг/мл, однако некоторые виды плесневых грибов,
такие как Aspergillus terreus, Acremonium stratum, Pseudallescheria hoydil и Scedosporium
prolificarts, демонстрируют более высокие значения МИК — от 2 до 16 мкг/мл.
МИК флюконазола*3 для большинства плесневых грибов превышает 64 мкг/мл.
Исключение составляют некоторые плесневые грибы, обладающие диморфизмом,
и дерматофиты. Значение МИК итраконазола*, вориконазола53 и позаконазола^
обычно колеблется в широких пределах — от 0,03 до 16 мкг/мл.
Основными проблемами при исследованиях плесеней являются затруднения
стандартизации инокулята, низкая скорость роста, отсутствие спорообразования,
характерное для некоторых видов, которое не лает возможности получения стан-
дартизованного инокулята.
Проблема интерпретации результатов: методика исследования антифунгеаль-
ной чувствительности плесневых грибов технологически более капризна, чем
дрожжевых Коммерческие тест-системы отсутствуют.
ПРОФИЛЬ АНТИФУНГЕАЛЬНОЙ ЧУВСТВИТЕЛЬНОСТИ
Результаты определения чувствительности грибов к антимикотическим препа-
ратам представлены в табл. 25-6-25-8.
Таблица 25-6. Профиль антифунгеальной чувствительности патогенных для человека грибов
Вид гриба Амф. В, мг/мл Флю, мг/мл Вори, мг/мл ^ра. мг/мл Кето, мг/мл Тербин, мг/мл 5-флю, мг/мл Каспо
Candida albicans Ч 0,5 Ч 0,5-1.0 Ч 0,06 4 0,25 Ч 0,25 В 0,03->128 4 1 ч
Candida famata 4 0,5 ЧДЗ 4-8 4 0,06 4 0,25 4 0,25 Нет данных 4 1 4
Candida glabrata 4 0,5 ЧДЗ 2-32 4 1-2 ЧДЗ 0.5-1,0 В 1-4 У >128 4 0,5 4
Candida guilliermondii 4 0,5 ЧДЗ 4-8 4 0,25-0,5 4 0,5 4 0.5-1 В 6.25-100 Ч 0,125-0,2 Ч
Candida kef yr Ч 0,5 Ч 0,5-1,0 4 0,03 4 0,25-0,5 4 0.25 У 0,5-50 6 0,03-16 4
Candida krusei Ч 0.5 1 У 32-64 4 0.5-1 4ДЗ 0,5 11,0 У 50->100 В 8-32 Ч
Candida iusitamae В 0,5-2 4 0,5-1,0 4 0.06 4 0,25 4 0,25 Нет данных 4 0,125 Ч
Candida parapsilosis Ч 0.5-1 Ч 1-2 4 0,06 4 0,125-0,5 4 0,5 4 0.25-2 41 Ч
Candida tropicalis Ч 0,5 ЧДЗ 1-8 4 0,25 Ч 0,25 В 0,5-1,0 В 10-128 41 4
Cryptococcus neoformans Ч 0,5-1 ЧДЗ 2-8 4 0,25 ЧДЗ 0,5 4 0.25 4 0 25 4 2 -8 У
Malassezia urfur Ч 4 4 Ч 4 4* Ч Нет данных
Saccharomyces cerevisiae 4 0,5 4 2-8 4 0,03 4 0,5-1,0 4 0,5 Нет данных 4 0,125 Нет данных
Trichosporon beigellii Ч 1,0 4 2-8 Ч 0,125 Ч 0,125 Ч 0,5 В 0,05-128 I 8-16 Нет данных
МИКОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ
Примечания: Ч — чувствительный; ЧДЗ — чувствительный или дозозависимый; У — устойчивый; В - - вариабельный; П — промежуточный.
Амф. В — амфотерицин В*; Флю — флюконазол1'; Вори — вориконазолИтра - - итраконазол*; Кето — кетоконазол*; Тербин — тербинафин* (только для топи-
ческой формы): 5-флю — 5-флюцитозин*'; Каспо — кааюфунгин.
Таблица 25-7. Профиль антифунгеальной чувствительности для дерматофитов
Вид।риба Тербинафин Флюконазол Итраконазол Кетоконазол Гризеофульвин Вармкоьазил
Trichophyton spp. 4 4 ч 4 4 4
Epidermophytia floccosum 4 4 ч 4 4 4
Microsporum spp. 4 Ч ч Ч Ч 4
МИКОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ 765
Таблица 25-8. Профиль антифунгеальной чувствительности для патогенных плесневых грибов
Вид гриба Амф. В, мг/мл Флю, мг/мл Вори, мг/ мл Итра, мг/ мл Кето, мг/ Мл Тербии, мг/ мл 5-флю, мг/мл Каспо
Aspergillus fumigatus 4 1,0 У >100 4 0,5 ЧДЗ 0,5 У8 4 0,5 У >64 4
Aspergillus flavas Ч 1-2 У >100 4 0,5-1 чдз 0.5-1,0 У8 4 0,5 У >64 Ч
Pseudalle- scheria boydii [=Scedospo- rium apiospermun] В 1-16 В 8-64 4 0,5 В 0,5-4,0 В М У 10-100 У >64 Нет данных
Scedosporium prolificans У >16 У >64 4 0,5-1 V >8 У >16 У >100 У >64 Нет данных
Fusarium sp. 11^1 У >64 Ч 1-4 У >8 У >16 У 0,5-128 У >64 Нет данных
Bipoiads sp. 4 0.25 У >64 4 0,25- 0,5 4 0,5 4 0,5 4 0,5-1,0 У >64 Нет данных
Cladophialo- phora sp. 41,0 В 4-64 41,0 4 0,125 В Ч 0,25-1 4 0,25 Нет данных
Exopfiiala spp. 4 0,25 У 64 4 0-25 4 0,5 В 0,5-2 4 0,06-2,0 В 4-64 Нет данных
Phialophora spp- 4 0,5-1 У 16-64 4 0,25 4 0,5 В 0.5 4 1 Нет данных Нет данных
Paecilomyces spp- 8 0,5-16 У64 4 0,5-1.0 В 0.5- 8,0 Нет данных В 1-64 В 0,5-64,0 Нет данных
Penicillium spp- 4 0,5-1,0 В 4-64 4 0,5 4 0,5 41 4 0,5-4.0 В 0,5 Нет данных
ВЫВОДЫ
• Основные практические затруднения при определении чувствительности гри-
бов к антифунгеальным препаратам связаны со стандартизацией приготовле-
ния инокулята и определением конечной концентрации. Проверку плотности
и стерильности инокулята следует осуществлять при каждом исследовании,
то есть параллельно с тестированием высевают стандартный объем готового
инокулята. Через 24 ч инкубации подсчитывают количество колоний. Если
их слишком мало или слишком много, рекомендуют повторить тест.
• В настоящее время существуют модификации метода определения МИК, в
которых на стадии регистрации результатов применяют цветовые индика-
торы или регистрация автоматизирована, с ис пользованием микропланшет-
ного ридера или автоматического анализатора зон ингибирования роста, что
позволяет избежать субъективных ошибок при получении количественных
данных. Для увеличения клинической эффективности исследования часто
полученный результат антифунгеальной чувствительности подтверждают
другим методом, особенно в тех случаях, когда имеются сомнения в досто-
верности полученного результата.
• Современная стадия развития лабораторных методов определения ан гифун-
геальной чувствительности позволяет лаборатории осуществлять эти иссле-
дования в рутинной практике. Однако для определения чувствительности
необходима культура, полущить которую не всегда возможно. Видовая иденти-
фикация часто дает возможность предсказать чувствительное гь вида in vitro.
• Вопрос, какие клинические образцы должны быть исследованы, остается
открытым. Надо ли исследовать только образцы, в которых грибы являются
клинически значимыми, или осуществлять лабораторный скрининг всех изо-
f.W4 25
766 МИКОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ
лятов? Ответ на этот вопрос может быть получен для каждого отдельного
ЛПУ с учетом анализа накопленных данных о превалирующих возбудителях,
их видовом составе и антифунгеальной чувствительности при участии спе-
циалистов по химиотерапии, клинических микробиологов и клиницистов.
Тест-системы для серологической диагностики
микотических инфекций
Серологическая диагностика важна тем, что позволяет определить высокие титры
антител или циркулирующие антигены у пациентов с иммунодепрессией, которые
относятся к одной из групп риска по развитию системных микозов. Классическая
микробиологическая диагностика системных микозов, вызванных Candida,
подтверждается повторным выделением грибов из биологических образцов,
которые в норме стерильны (крови, мочи и т.д.). Однако интерпретировать
результаты, получаемые при выращивании культур грибов, трудно, так как грибы
часто выделяются в виде единичных колоний, а при распространенной инфекции
гемокультура может оставаться отрицательной.
Обнаружение маннанового антигена, циркулирующего в сыворотке крови,
позволяет усовершенствовать диагностику кандидодной инфекции, особенно у
иммуносупрессивных пациентов.
Коммерческий тест «Pastorex® Candida* позволяет определить в сыворотке
полисахарид Candida (маннан), используя простой метод агглютинации латекс-
ных частиц. Маннан — основной компонент стенки грибов и основной циркули-
рующий антиген. В тесте используются латексные частицы, сенсибилизированные
моноклональными антителами к маннану. Порог чувствительности теста 2,5 нг/мл.
В случае если сыворотка положительная, можно провести полуколичественный
анализ путем серийных разведений супернатанта в глициновом буфере.
Тест «Pastorex® Aspergillus* предназначен для качественного и полуколиче-
ственного определения полисахарида Aspergillus (галактоманнана) в сыворотке,
основан на методе агглютинации латексных частиц. В тесте используются латекс-
ные частицы, сенсибилизированные моноклональными антителами к галактоман-
нану. Порог чувствительности — 15 нг/мл.
Качественный и полуколичественный тест «Pastorex® Cryptococcus» основан на
определении капсульного полисахарида Cryptococcus neoformans (гликуронокси-
ломаннан) в биологических жидкостях (сыворотке, спинномозговой жидкости,
бронхоальвеолярном лаваже, моче) с помощью реакции агглютинации. Б тесте
используются латексные частички, покрытые моноклональными антителами
к глюкуроноксиломаннану, который является главным компонентом капсулы
Cryptococcus neoformans.
Тест «Platelia Aspergillus ИФА» — одноэтапный ИФА-анализ в «сэндвич»-
формате для определения галактоманпанового антигена грибов рода Aspergillus в
сыворотке крови. Иммунологическая диагностика, основанная на определении
антител, не адаптирована для случаев иммунодефицита. Аспергиллы крайне редко
изолируются из гемокультуры, часто отсутствуют в образцах мокроты. Диагностика
аспергиллеза часто основывается на неспецифических критериях или на признаках,
соответствующих поздним клиническим проявлениям (клинических симптомах,
радиологических методах исследования и т.д.). Внедрение серологического
анализа растворимых антигенов в сыворотке пациента дает возможность улучшить
диагностическую ситуацию, добиваясь высокой специфичности на самых ранних
стадиях заболевания.
Одноэтапный «сэндвич»-иммуноферментный тест «Platelia Candida Аг» в
микропланшетном формате позволяет качественно или количественно определить
МИКОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ 767
уровень циркулирующего в сыворотке маннанового антигена. Чувствительность
теста сильно зависит от вида изолированного микроорганизма.
Двухэтапный непрямой иммуноферментный тест «Platelia Candida Ат» в микро-
планшетном формате позволяет обнаружить антитела к маннановому антигену
дрожжевых грибов рода Candida, главному компоненту стенки грибов этого рода.
Маннан является маркером кандидоза и представляет собой высокоиммуноген-
ный полисахарид. Тест позволяет количественно оценить уровень антиманна-
новых антител в сыворотке человека. Уровень чувствительности различается в
зависимости от вида изолированного гриба.
Диагностика инвазивного кандидоза должна быть основана на сочетании тестов
на определение как антител в маннану, так и самого циркулирующего в крови ман-
нанового антигена в тесте «Platelia Candida Ат». Сочетание этих двух тестов позво-
ляет поставить ранний лабораторный диагноз, повысить чувствительность мето-
дов лабораторной диагностики. В целом сочетание клинических и лабораторных
результатов позволяет снизить рис к ятрогенных факторов риска. Комбинация этих
двух тестов может стать важной составляющей системы клинико-лабораторного
мониторинга в ходе лечения.
ЛАБОРАТОРНАЯ ДИАГНОСТИКА ПОВЕРХНОСТНЫХ МИКОЗОВ
Микозы кожи
Микоз гладкой кожи:
❖ микроскопия чешуек, соскобов из очагов поражения;
ъ культуральное исследование материала из очага поражения;
> видовая идентификация и определение антифунгеальной чувствительности
(при рецидивирующем течении и/или резистентности к стандартной анти-
фунгеальной терапии).
Микоз кистей, стоп. Онихомикоз:
микроскопия чешуек, соскобов из очагов поражения, соскобов и срезов пора-
женной ногтевой пластинки;
культуральное исследование материала из очага поражения;
< > видовая идентификация и определение антифунгеальной чувствительности
(при рецидивирующем течении и/или резистентности к стандартной анти-
фунгеальной терапии).
Микоз волосистой части головы:
о люминесцентное исследование с помощью лампы Вуда;
❖ микроскопия чешуек, волос из очагов поражения;
❖ культуральное исследование материала из очага поражения;
ъ видовая идентификация и определение антифунгеальной чувствительности
(при рецидивирующем течении и/или резистентности к стандартной анти-
фунгеальной терапии).
Микозы слизистых оболочек:
• > микроскопия соскобов из очагов поражения;
о видовая идентификация и определение антифунгеальной чувствительности
(при рецидивирующем течении и/или резистентности к стандартной анти-
фунгеальной терапии).
КРИТЕРИИ ДИАГНОСТИКИ СИСТЕМНЫХ микозов
Микотический бронхит:
❖ рентгенография или компьютерная томография легких;
бронхоскопия, бронхоальвеолярный лаваж (БАЛ);
768 МИКОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ
о- микроскопия БАЛ, мокроты с окраской флюорохромами, материала биоп-
сии;
о культуральное исследование БАЛ, мокроты с окраской флюорохромами,
материала биопсии;
❖ патоморфологическое исследование материала биопсии.
Инвазивный микоз легких:
рентгенография или компьютерная томография легких;
< > бронхоскопия, бронхоальвеолярный лаваж (БАЛ);
❖ биопсия оиагов поражения (чрезбронхиальная, открытая, трансторакальная);
❖ микроскопия БАЛ, мокроты, биоптата с окраской флюорохромами;
❖ культуральное исследование БАЛ, мокроты, биоптата с окраской флюорох-
ромами;
❖ патоморфологическое исследование биоптата;
о видовая идентификация выявленного возбудителя;
о определение антигена Aspergillus в сыворотке крови («Pastorex» или «Platelia
Aspergillus BioRad »);
< определение специфических антитея к Aspergillus (при хроническом некроти-
зирующем аспергиллезе).
Синусит:
клинические и рентгенологические признаки острого синусита;
❖ микроскопия аспирата или биоптата с окраской люминесцентными красите-
лями;
4 культуральное исследование аспирата или биоптата;
❖ патоморфологическое исследование аспирата или биоптата;
< определение антигена Aspergillus в сыворотке крови (тест латекс-агглютина-
ции «Pastorex» или «Platella Aspergnlus BioRad»).
Эзофагит:
❖ нахождение бляшек на стенках пищевода при эзофагоскопии;
❖ нахождение грибов в окрашенных микроскопических препаратах;
< > рост грибов при культуральном исследовании биопсийного материала.
Микотическая инфекция мочевыводящих путей:
< микроскопическое и культуральное исследование мочи с ростом грибов,
выявление более 1х103 КОЕ/мл правильно собранной мочи;
❖ компьютерная томография и ультразвуковое исследование почек;
о цистоскопия (по показаниям);
❖ видовая идентификация и определение антифунгеальной чувствительности
(при рецидивирующем течении и/или резистентности к стандартной анти-
микотической терапии).
Фунгемия:
❖ однократное выявление грибов в культуре крови в период подъема темпера-
туры тела.
Острый диссеминированный кандидоз:
❖ фунгемия в сочетании с культуральными или патоморфологическими при-
знаками поражения глубоких тканей, включая подкожную клетчатку.
Эндофтальмит:
❖ микроскопическое, культуральное, патоморфологическое исследование биоп-
сийного материала (пробы жидкости стекловидного тела в объеме 1-2 мл),
собранной путем тонкоигольной аспирации);
❖ повторное культуральное исследование крови 2 раза в день в течение не
менее 2 дней;
❖ видовая идентификация и определение антифунгеальной чувствительности
❖ повторное определение антигена в сыворотке крови (тест латекс-
агглютинации).
микологические исследования 759
Абсцесс или остеомиелит:
- о- рентгенологические (компьютерно-томографические/магнитно-резонансные)
признаки остеомиелита;
< > выявление грибов при микроскопическом, культуральном, патоморфологи-
ческом исследовании аспирата или биопсийного материала;
< > повторное культуральное исследование крови 2 раза в день в течение не
менее 2 дней;
< видовая идс нтификация и определение антифунгеальной чувствительности;
❖ повторное определение антигена в сыворотке крови (тест латекс-
агглютинации).
Менингит:
люмбальная пункция с определением давления спинномозговой жидкости;
❖ микроскопия ликвора;
❖ культуральное исследование ликвора;
микроскопическое и культуральное исследование крови, материала из пред-
полагаемых очагов инфекции, мазков из носоглотки, среднего уха, собран-
ных в день проведения люмбальной пункции;
❖ видовая идентификация и определение антифунгеальной чувствительности
выявленного возбудителя;
❖ определение антигена Crypticoccus neoformans в спинномозговой жидкости или
крови с помощью теста латекс-агглютинации («Pastorex Crypticoccus BioRad»;
«Immuno-Mycologics Inc»; «Meridian Diagnostic Inc»; «Murex Diagnostic») или
теста ELISA («Meridian Diagnostic Inc»).
Хронический диссеминированный гепатолиенальный кандидоз.
• Возможный:
❖ персистирующая или интермиттирующая лихорадка после завершения
периода нейтропении в сочетании с клинической симптоматикой пораже-
ния печени, селезенки, почек и характерными ультразвуковыми
(компьютерно-томографическими или магнитно-резонансно-томографи-
ческими) признаками поражения этих органов.
• Доказанный:
❖ вышеуказанные симптомы;
о положительный результат культурального исследования крови (до появ-
ления признаков поражения печени, селезенки или почек);
-о патоморофологические признаки кандидоза в биопсийном материале из
очагов поражения.
Аспергиллез:
о микроскопическое и культуральное исследование мокроты, бронхоальвео-
лярного лаважа, биопсийного материала;
о определение антител методом количественного электрофореза;
❖ определение антигена (тестом латекс-агглютинации «Pastorex BioRad»):
❖ определение антигена методом ИФА («Platelia Aspergillus BioRad»);
определение £-1.3-D («Fungitec G-test»).
Бластомикоз:
микроскопия гноя, мокроты. БАЛ, мочи;
- о- культуральное исследование гноя, мокроты, БАЛ, мочи;
о определение антител методом иммунодиффузии.
Кокцидиоидомикоз:
❖ микроскопия и культуральное исследование мокроты, синовиальной жидко-
сти, гноя и центрифугата спинномозговой жидкости;
о кожная проба с кокцидиоидином или сферулином;
❖ определение IgM в сыворотке крови методом латекс-агглютинации, преципи-
тации или иммунодиффузии;
770 МИКОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ
< определение IgG в сыворотке крови методом фиксации комплемента или
иммунодиффузии;
=> определение антител в ликворе при подозрении на менингит.
Криптококкоз:
< > микроскопия и культуральное исследование црнтрифугата спинномозговой
жидкости, крови, мочи, секрета предстательной железы и биопсийного мате-
риала;
определение антигена Crypticoccus neoformans в спинномозговой жидкости,
крови, моче с помощью теста латекс-агглютинации («Pastorex Crypticoccus
BioRad»; «Immuno-Mycologics Inc.»; «Meridian Diagnostic Inc.»: «Murex
Diagnostic») или теста ELISA («Meridian Diagnostic Inc.»').
Гистоплазмоз;
о микроскопия окрашенных мазков крови, мокроты, БАЛ, гноя;
культуральное исследование крови, мокроты, костного мозга, гноя, биопсий-
ного материала;
> определение антигена в крови, моче, спинномозговой жидкости, БАЛ радио-
иммунным методом или тестом ELISA;
❖ определение антител методом фиксации комплемента или иммунодиффу-
зии.
Му кор микоз:
о микроскопия отделяемого из участков некроза, мокроты, БАЛ, биопсийного
материала:
❖ культуральное исследование материала с очагов на твердом небе, отделяемо-
го придаточных пазух носа, мокроты, БАЛ.
Паракокцидиоидомикоз:
❖ микроскопия и культуральное исследование гноя, мокроты, материала из
гранулематозных поражений;
❖ определение антител методом фиксации комплемента или тестом ELISA.
Пенициллиоз (возбудитель — Penicillium mamefferi):
< > микроскопия окрашенных по Райту7 мазков костного мозга, материала кож-
ных элементов, биоптатов лимфатических узлов.
Споротрихоз:
< микроскопия и культуральное исследование гнойного отделяемого, биопсий-
ного материала.
Гиалогифомикозы
• Фузариоз:
❖ культуральное исследование крови, биопсийного материала, материала
пораженных ногтей.
• Феогифомикозы,
❖ Синусит — поражение придаточных пазух носа {Altemaria, Bipolaris,
Curvularia, Exs^ohiluim):
- микроскопия отделяемого из носа, биопсийного материала, аспирата;
- культуральное исследование отделяемого из носа, биопсийного мате-
риала, аспирата.
о Поражение головного мозга (Cladovhialophora)'.
~ культуральное исследование биопсийного материала придаточных па-
зух носа, аспирата, мокроты, БАЛ.
< > Системный микоз, вызванный Malassezia furfur (Pityrosporum):
- микроскопия мазков крови;
- культуральное исследование крови на средах, обогащенных липидами;
- культуральное исследование дистального фрагмента венозного катетера
на средах, обогащенных’ липидами.
Глава 26
Лабораторная диагностика
паразитарных болезней
Паразитарные болезни развиваются в результате проникновения
в организм гельминтов, простейших, членистоногих, использующих
его как место обитания и источник питания. Паразитарные болезни
часто протекают без выраженных и специфических клинических
симптомов и признаков, однако при снижении иммунитета, в част-
ности при СПИДе, токсоплазмоз, пневмоцистоз, криптоспоридиоз,
акантамебиаз и другие паразитарные болезни являются причинами
летальных исходов. Пациенты жалуются на утомляемость, слабость,
повышенную температуру. При кишечных паразитозах часто разви-
ваются нарушения пищеварения: диарея, метеоризм, гиперсалива-
ция. тошнота, рвота, боли в животе. При пневмоцистозе поражения
легких протекают в виде пневмонии, бронхопневмонии вплоть до
развития пневмоторакса. При висцеральном лейшманиозе пора-
жаются клетки ретикулоэндотелиальной системы, наблюдается
гепатоспленомегалия. Для ряда паразитарных болезней характерны
поражения кожи и слизистых оболочек: кожный лейшманиоз, неко-
торые филяриидозы (онхоцеркоз, дирофиляриоз).
Некоторые паразитарные болезни при поздней диагностике и
отсутствии адекватного лечения представляют серьезную опасность
для здоровья и жизни пациентов: тропическая малярия, висце-
ральный лейшманиоз, трихинеллез, цистицеркоз, диссеминирован-
ные формы стронгилоидоза, эхинококкозы, диоктофимоз, амебные
абсцессы.
Основой диагностики паразитарных болезней являются результа-
ты лабораторных исследований, которые позволяют выявить непо-
средственно возбудителей, антигены возбудителей болезней или
антитела к ним. Возбудители паразитарных болезней могут быть
обнаружены при исследовании крови, фекалий, мочи, мокроты,
спинномозговой жидкости, пунктатов органов и тканей. Лечащий
врач, включающий в диагностический алгоритм паразитологиче-
ские исследования, должен учитывать следующие обстоятельства:
о заболеваемость паразитарными болезнями продолжает оста-
ваться высокой, вероятность обнаружения возбудителей также
высока;
❖ паразитарные болезни, такие как аскаридоз, трихоцефа-
лез, энтеробиоз, токсоплазмоз, лямблиоз, криптоспоридиоз,
пневмоцистоз, встречаются почти во всех регионах мира.
Лейшманиозы, филяриидозы, трипаносомозы, шистосомозы
772 ЛАБОРАТОРНАЯ ДИАГНОСТИКА ПАРАЗИТАРНЫХ БОЛЕЗНЕЙ
имеют зональное или региональное распространение. Это связано с осо-
бенностями жизненных циклов паразитов, в которых участвуют животные,
поэтому для установления предварительного диагноза следует учитывать,
не выезжал ли пациент в регионы, эндемичные в отношении того или иного
паразитоза;
некоторые паразитарные болезни намного чаще встречаются у детей, чем у
взрослых (аскаридоз, трихоцефалез, токсокароз, энтеробиоз);
❖ существуют паразитарные болезни, которые чаще встречаются у пациентов
со сниженным иммунитетом (диссеминированные формы стронгилоидоза,
криптоспоридиоз, пневмоцистоз, энцефалиты, вызванные сапрофитными
амебами). У пациентов со сниженным иммунитетом мот развиваться тяже-
лые формы тех инвазий, которые обычно протекают в виде легких и бессим-
птомных случаев (лямблиоз, гимен олепидоз);
о- следует учитывать, что период, когда при инвазии возбудитель не может быть
выявлен паразитологическими методами (паразитологической инкубацией),
может быть весьма долгим. При аскаридозе, описторхозе, дифиллоботриозе
этот период может продолжаться несколько недель;
< > ряд паразитарных болезней вообще не может быть выявлен прямыми
паразитологическими методами, в этих случаях применяют молекулярные
методы диагностики для обнаружения ан гигенов, антител, ДНК и РНК воз-
будителей.
Таким образом, подходы к диагностике паразитарных болезней весьма разно-
плановы.
Для обнаружения разных видов возбудителей требуется применение разных
методов паразитологических исследований, поэтому в направлении на исследо-
вание необходимо указать, какой паразитоз подозревается у пациента. Например,
в случае указания в направлении «кал на яйца гельминтов» лабораторией будет
проведено именно это исследование, в то время как возбудитель стронгилоидоза
может быть обнаружен только специальным методом исследования фекалий. Для
выявления ооцист криптоспоридий требуется специальная окраска фиксирован-
ных препаратов. Кровь для поиска возбудителей малярии в виде толстой капли
или тонкого мазка на стекле и мазки костного мозга для поиска лейшманий окра-
шивают по Романовскому-Гимзе. У пациентов возможно одновременное наличие
нескольких паразитарных инвазий, например гельминтов и простейших, малярии
и лейшмании. По этой причине иногда приходится проводить исследование раз-
ных материалов от одного пациента.
Диагноз паразитарной болезни ставят на основании обнаружения в исследуе-
мом материале особей паразитов, фрагментов особей, разнообразных стадий их
развития. В ряде случаев при получении сомнительных результатов требуется
повторное исследование, при подозрении на некоторые паразитарные болезни
(лямблиоз, криптоспоридиоз и др.) однократное исследование не дает уверен-
ности в отсутствии возбудителя и, следовательно, в предоставлении клиницисту
заключения об его отсутствии.
Следует иметь в виду что большинство гельминтозов не передаются непосред-
ственно от одного человека к другому, поэтому пациенты неопасны в качестве
источника возбудителя для окружающих. Из этого общего правила есть несколь-
ко исключений: энтеробиоз, гименолепидоз, вызываемый карликовым цепнем,
стронгилоидоз и свиной цепень, при котором серьезную опасность представляет
заражение онкосферами с последующим развитием цистицеркоза. Следует пом-
нить, что инвазированные кишечными простейшими являются источниками
возбудителей и могут передавать инфекцию окружающим. Малярия может пере-
даваться от человека к человеку не только через комаров, но и с кровью (при
ЛАБ0РА1 ОрНАЯ ДИАГНОСТИКА ПАРАЗИТАРНЫХ БОЛЕЗНЕЙ 773
переливании крови, эритроцитарной массы, при использовании инструментария,
контаминированного кровью),
Возбудители паразитарных болезней относятся к патогенным биологическим
агентам (ПБА) III и IV группы патогенности, чем определяется соответствующий
режим работы паразитологических лабораторий (подразделений), выполняющих
диагностические исследования.
КРОВЕПАРАЗИТЫ
Кровь является местом обитания на определенной стадии развития следующих
паразитов: простейших — малярийных плазмодий (Р. vivax, Р. falciparum, Р. ovale,
Р. malanae), бабезий (Babesia spp.); гельминтов — личинок филяриид (Wuchererta
bancrofti, Brupia malays). В некоторых случаях в крови могут быть обнаружены
висцеротропные лейшмании (Leishmania donovar.i подвид infantum, Lefhmania
donovanl), однако исследование крови не является основным методом их диагно-
стики.
Неспецифические изменения крови при паразитарных
болезнях
Эозинофилия. Многие паразитарные болезни сопровождаются повышением
количества эозинофилов в периферической крови (анизакидоз, анкилостомидозы,
описторхоз, стронгилоидоз, токсокароз, трихинеллез, фасциолез, филяриидозы
цистицеркоз, шистосомозы, эхинококкозы и др.).
Анемия встречается при многих паразитарных болезнях: анкилостомидо-
зах, дифиллоботриозе, трихоцефалезе, фасциолопсидозе, малярии, висцеральном
лейшманиозе, трипаносомозе, бабезиозе. При малярии в начальной стадии анемия
нормохромная и нормоцитарная: между приступами лихорадки и после лечения
отмечается ретикулоцитоз. В дальнейшем наблюдается повышенное количество
полихроматофилов, имеют место анизоцитоз, пойкилоцитоз. При висцеральном
лейшманиозе анемия резко выражена, содержание гемоглобина прогрессивно
снижается и может доходить до 30 г/л, увеличивается количество ретикулоци-
тов. При анкилостомидозах развивается железодефицитная анемия. Содержание
гемоглобина падает, развиваются анизоцитоз, пойкилоцитоз, отмечается низкое
среднее содержание гемоглобина в эритроците. У некоторых больных при тяже-
лой анемии уровень гемоглобина падает до 30-50 г/л, количество эритроцитов
ниже 1,5х1012/л.
При дифиллоботриозе анемия протекает по типу пернициозной, характерен
мегалобластический тип кроветвопения. В периферической крови появляют-
ся мегалобласты, изредка нормобласты, встречаются эритроциты с тельцами
Жолли и кольцами Кебота. Отмечаются анизопойкилоцитоз, полихроматофилия.
Количество эритроцитов может быть от 1,5х1012/л до 0,5х1012/л, а концентрация
гемоглобина — 70-110 г/л. Лейкоцитоз с последующей лейкопенией и относи-
тельным моноцитозом характерен для многих протозоозов и гельминтозов. На
ранних стадиях трихинеллеза, токсокароза и стронгилоидоза. при анизакидозе
лейкоцитоз может достигать 30х10*/л. При амебном гепатите и амебном абсцессе
описан длительный лейкоцитоз до 25-30x107л со сдвигом лейкоцитарной фор-
мулы (до 70-80% сегментоядерных лейкоцитов). У больных с тяжелым течением
тропической малярии встречается нейтрофильный лейкоцитоз, в периферической
крови отмечаются анэозинофилия и моноцитоз.
Лейкопения до 0,2-0,6х10ч/л вплоть до агранулоцитоза может развиться в
терминальной стадии висцерального лейшманиоза. При висцеральном лейшма-
?S 774 ЛАБОРАТОРНАЯ ДИАГНОСТИКА ПАРАЗИТАРНЫХ БОЛЕЗНЕЙ
ниозе лейкопения сочетается с анемией, 1ранулоцитопелией, тромбоцитопенией,
gt имеют место относительный моноцитоз, лимфоцитоз (70-90%), анэозинофилия.
Лейкопения также наблюдается при кишечных шистосомозах, анкилостомидозах.
Igj; При малярии может наблюдаться лейкопения с относительным моноцитозом, в
р\ х лихорадочный период характерен лейкоцитоз с палочкоядерным сдвигом
Биохимические изменения крови. При поражениях печени при описторхозе
отмечают повышение уровня прямого и непрямого билирубина. Диспротеинемию
обнаруживают при малярии, висцеральном лейшманиозе, токсокарозе, шисто-
сомозах, многокамерном эхинококкозе. Гипопротеинемию со снижением уровня
альбуминов выявля ют при анкилостомидозах.
Малярийные плазмодии
Малярию у человека вызывают 4 вида возбудителей, относящихся к роду
Plasmodium: Р. falciparum, Р. vivax, Р. malariae и Р. ovale. Первый вызывает тропиче-
скую малярию, которая может протекать злокачественно и без лечения закончить-
ся летально. Трех-, четырехдневная и овале-малярия отличаются относительно
доброкачественным течением. В последнее десятилетие в России около 99%
случаев малярии вызваны либо Р. falciparum (завозится из-за рубежа, в основном
из тропической Африки), либо Р. vivax (завозится в основном из Азии, временами
передастся в пределах России).
Биология возбудителей
Малярийные плазмодии паразитируют поочередно в двух хозяевах: в самке
комара рода Anopheles и в человеке. Во время укуса комар вводит в кровяное русло
человека спорозоиты, которые уже через несколько минут попадают в печень. В
гепатоцитах происходит экзоэритроцитарная шизогония, длящаяся не менее 7 сут.
При трехдневной и овале-малярии часто спорозоиты превращаются в гипнозоиты.
которые пребывают в неактивном состоянии; эта фаза обычно продолжается 6-
12 мес. Во время пребывания в печени малярийные паразиты никак себя не про-
являют. Из печени продукты экзоэритроцитарной шизогонии выходят в кровь и
внедряются в эритроциты — так начинается фаза эритроцитарной шизогонии. Все
патологические проявления малярии (лихорадочные приступы, анемия, спленоме-
галия, полиорганные поражения при тропической малярии) связаны только с эри-
троцитарной шизогонией. В этом процессе сменяются бесполые стадии мерозоита,
трофозоита и шизонта, из которых опять образуются мерозоиты. Единичный цикл
эритроцитарной шизогонии от мерозоита до мерозоита продолжается 3 сут при
черырехдневной малярии и 2 сут — при других видах. В результате одного выхода
паразитов из печени таких последовательных циклов обычно бывает от одного до
нескольких десятков, пока эритроцитарная шизогония не будет прервана иммун-
ным процессом. Однако в силу нестойкости иммунитета пациент может вновь
заболеть либо в результате нового заражения, либо за счет активации гипнозоитов
в печени. Из некоторых мерозоитов образуются и половые клетки — гаметоциты,
которые в организме человека не делятся и, следовательно, не оказывают патоген-
ного воздействия.
Дифференциальная диагностика
Диагностика малярии основывается на обнаружении в препарате крови боль-
ного любых стадий плазмодиев. При этом следует учитывать важные отличия
эритроцитарной шизогонии и гаметоцитогонии у различных видов малярийных
плазмодиев.
При Р. falciparum оболочка эритроцита вскоре после вторжения в него паразита
приобретает способность прилипать к эндотелию сосудов. Вследствие этого эри-
ЛАБОРАТОРНАЯ ДИАГНОСТИКА ПАРАЗИТАРНЫХ БОЛЕЗНЕЙ 775
троциты с паразитами, кроме самых молодых (на стадии кольца), задерживаются
в венулах внутренних органов (секвестрируются), поэтому в капиллярной крови
из бесполых форм обнаруживаются только самые молодые трофозоиты (кольца).
Гаметоциты созревают в венулах внутренних органов и впервые выплывают в
периферическое русло только с 10-12-го дня болезни. Гаметоциты Р. falciparum
отличаются большой продолжительностью индивидуальной жизни (до 6 нед),
отчего их продолжают обнаруживать и после прекращения эритроцитарной шизо-
гонии в результате лечения. Именно поэтому при тропической малярии крайне
важно указывать, обнаружены ли в препарате только кольца, кольца вместе с
гаметоцитами (указание, что болезнь имеет давность 12 дней и более) либо только
гаметоциты (указание, что образование новых бесполых паразитов прекратилось,
но пациент остается заразен для комаров). Обнаружение гаметоцитов ^falciparum
после излечения больного — обычное явление, оно не является показателем
неэффективности лечения. При других видах малярии с самого начала болезни
в периферической крови обнаруживают гаметоциты и бесполые паразиты всех
возрастов, Гаметоциты после созревания живут лишь в течение нескольких часов,
поэтому нет нужды специально отмечать их присутствие или отсутствие.
Методы исследования
Основным методом диаг ностики малярии является исследование толстой капли
крови, окрашенной по Романовскому-Гимзе. Исследование тонкого мазка явля-
ется вспомогательным методом и проводится, как правило, для уточнения вида
малярийных паразитов. Используют иммерсионный объектив х90 и окуляр х7.
Принцип метода. Объем исследуемой крови в шестой капле в 20-40 раз боль-
ше, чем в тонком мазке. По этой причине положительный ответ можно дать после
исследования мазка или толстой капли, а отрицательный — только после иссле-
дования толстой капли. При подозрении на малярию необходимо просмотреть
не менее 200 полей зрения толстой капли. Это позволяет выявить паразитемию
порядка 5-10 паразитов в 1 мкл крови. В препаратах крови обнаруживают внутри-
эритроцитарные стадии развития паразитов: трофозоиты, шизонты и гаметоциты.
Внеэритроцитарные стадии — мерозоиты, существуют короткое время, поэтому
они обнаруживают редко.
Трофозоиты (от греч. trovhe — «питание») — питающиеся гемоглобином, расту-
щие стадии с одним ядром. Шизонты (от греч. schizo — «делюсь») — делящиеся
стадии, имеюшие 2 ядра и более. Трофозоиты подразделяются на 5 возрастных
классов, границы между которыми достаточно условны: самый ранний класс
(1) — кольца, затем (2) юные, (3) полувзрослые, (4) взрослые и (5) трофозоиты,
готовящиеся к делению, или прешизонты. Шизонты подразделяются на незрелые
и зрелые, иногда называемые также морулами.
Обнаружение плазмодиев. Доказательством наличия малярийного плазмо-
дия служит обнаружение даже одного паразита на любой стадии развит ия. Однако
следует с осторожностью относиться к находке единственного образования,
похожего на кольцевидный трофозоит в толстой капле, так как эта находка может
быть результатом сочетания посторонних элементов, случайно попавших в иссле-
дуемый материал, например кокков, частиц крас ки и пр. В сомнительных случаях
кровь пациента должна быть взята и исследована повторно.
Видовая идентификация плазмодиев обязательна для формулирования клини-
ческого и эпидемиологического прогноза, выбора лечения. Морфология плазмоди-
ев различных видов в тонком мазке практически полностью сохранена, в толстой
капле форма изменяется в процессе дегемоглобинизации при окраске нефиксиро-
ванной пробы крови. В практической работе можно использовать определитель,
разработанный А.Е. Беляевым (табл. 26-1). Для постановки диагноза сначала
решают, является ли данное образование малярийным паразитом (пункт ]).
77В ЛАБОРАТОРНАЯ ДИАГНОСТИКА ПАРАЗИТАРНЫХ БОЛЕЗНЕЙ
При положительном решении переходят к пункту 2 и выбирают одно из двух
утверждений. Если в конце выбранного утверждения стоит название вида — это и
есть искомый диагноз, если цифра — это указание, к какому пункту определителя
следует перейти. Процесс повторяют, пока не дойдут до названия вида.
При обнаружении возбудителей малярии обязательно определение интенсив-
ности паразитемии. Это дает информацию для прогноза заболевания и оценки
его эффективности. Достаточно точный метод сводится к подсчету среднего числа
паразитов на одно поле зрения толстой капли.
Таблица 26-1. Определитель возбудителей маляоии (по А.Е. Беляеву)
В тонком мазке
1. Образование, лежащее на фоне эритроцита, с одним или несколькими ядрами розового, вишневого или рубинового цгета с цитоплазмой голубого или синего цвета Plasmodium spp.
2. Имеются только кольцевидные трофозоиты (кольца) и/или полулунные гаметоциты Р. falciparum
Имеются также более крупные трофозоиты и/или шизонты 3
3 Пораженные эритроциты такого же или меньшего диаметра, чем непораженные. Зернистость отсутствует. Трофозоиты компактные Р. malariae
Пораженные эритроциты увеличены, имеется зернистость 4
4. Зернистость мелкая и обильная. Трофозоиты обычно с ясно выраженными псевдопо диями. Ядра относительно мелкие Р. vivax
Зернистость относительно крупная и редкая. Трофозоиты компактные. Ядра крупные Р. ovale
В толстой капле
1. Образование мельче сегментоядерного лейкоцита, имеет одно или несколько ядер (обычно розового, вишневого или рубинового цвета) и цитоплазму (обычно голубого или синего цвета) Plasmodium spp.
2. Имеются только кольцевидные трофозоиты (кольца) и/или полулунные гаметоциты P. falciparum
Имеются также более крупные трофозоиты и/или шизонты 3
3. Трофозоиты неправильной формы, часто разорваны на мелкие фрагменты. Могут при- сутствовать остатки пораженных эритроцитов розового цвета P. vivax
Трофозоиты компактные и никогда не разрываются на мелкие фрагменты 4
4. Обычно имеются осгатки пораженных эритроцитов розового цвета. Ядра крупные P. ovate
Остатков пораженных эритроцитов не наблюдается. Ядра мелкие P. malariae
Выделяется 5 классов паразитемии, при этом численность паразитов в каждом
последующем классе в 10 раз выше, чем в предыдущем, образуя, таким образом,
логарифмическую шкалу:
J «+» — 1-10 в 100 полях зрения;
J «++» — 1-10 в 10 полях зрения;
/ «+++-» — 1-10 в 1 поле зрения;
J «++++» — 10-100 в 1 поле зрения;
Z «+++++» — более 100 в 1 поле зрения.
Если паразитов много, подсчет ведут в 10 полях зрения, если мало — в 100, а
при очень высокой численности достаточно и одного поля. В случае Р. falciparum
численность указывают отдельно для бесполых форм и гаметоцитов, например,
кольца «+++», гаметоциты «+».
Более точным является метод подсчета численности паразитов по отношению
к количеству лейкоцитов. Для этого необходимо параллельно определить числен-
ность лейкоцитов. Например, в толстой капле на 100 лейкоцитов найдено 20 парази-
тов, при этом известно, что у пациента 8000 лейкоцитов в 1 мкл крови. Составляют
пропорцию:
8000x20/100 = 1600 паразитов.
При очень высокой паразитемии ее интенсивность может быть охарактеризова-
на количеством паразитов на 100 эритроцитов, подсчитанных в тонком мазке.
ЛАБОРАТОРНАЯ ДИАГНОСТИКА ПАРАЗИТАРНЫХ БОЛЕЗНЕЙ 777
Оформление диагн за
В ответе лаборатории указывают:
< какой препарат исследован (толстая капля или мазок);
4- количество исследованных полей зрения;
❖ вид паразита;
< стадии паразита для Р. falciparum',
❖ уровень паразитемии.
Разработаны и применяются в некоторых случаях на практике экспресс-методы
диагностики малярии.
Микрофилярии
Большой группой тропических гельминтов филярий вызываются у человека
8 нозоформ филяриидозов. Клинические проявления весьма разнообразны и в
зависимости от вида возбудителя включают поражение лимфатических сосудов,
кожи, подкожной клетчатки, органов брюшной полости. Филяриидозы характе-
ризуются длительным инкубационным периодом, медленным развитием болезни
и продолжительным ее течением.
Взрослые паразиты (4-10 см в длину; обитают в разных органах и тканях, где
отрождают личинки — микрофилярии. Отрождаемые личинки концентрируются
в крови (или в коже), где находятся длительно. Переносчиками филярий явля-
ются кровососущие насекомые (мошки, слепни, комары, мокрецы). Численность
микрофилярий в крови связана с активностью переносчиков в разное время суток,
например, для микрофилярий Brugia malayi и Wuchereria hancrofti характерен ноч-
ной пик численности, для Loa loa — дневной.
Лабораторная диагностика основывается на обнаружении (в определенное
время суток) и идентификации микрофилярии в препаратах крови. Исследуют:
❖ толстую каплю периферической крови;
❖ мембранные фильтры после фильтрации крови;
< > осадок гемолизированной венозной крови.
Идентификация микрофилярий. В нативных препаратах устанавливают
наличие микрофилярий змеевидной формы с характерной подвижностью, раз-
меры в зависимости от вида возбудителя составляют ] 80-280 мкм в длину, 5-
10 мкм в ширину. Видовую идентификацию проводят при окраске препаратов по
Романовскому-Гимзе или, что предпочтительнее, по Гайденгайну. Основные мар-
керы: наличие чехлика, расположение ядер в теле, форма хвостовой части.
Бабезии
Возбудители бабезиоза — простейшие, представители рода Babesia, человек
заражается бэбезидозом редко, в основном В. divergens Этот вид паразитирует
обычно у крупного рогатого скота, овец и иногда передается человеку клещами.
Заболевание развивается только у лиц со сниженным иммунитетом, в частности
после спленэктомии. По характеру течения инфекции бабезиоз напоминает тропи-
ческую малярию. Паразиты развиваются в эритроцитах и размножаются простым
делением. При поздней диагностике часты летальные исходы.
Лабораторная диагностика основывается на исследовании мазков и толстых
капель крови, окрашенных по Романовскому-Гимзе.
Идентификация бабезии. Бабезии расположены внутри эритроцитов в виде
одиночных кольцевидных образований, иногда по 2 или 4 особи в одном эритро-
ците, и напоминают кольцевидные трофозоиты Р. falciparum. Однако они мельче,
не столь правильной кольцевидной формы. В цитоплазме паразитов отсутствует
пигмент. Шизонты и гаметоциты не встречаются.
778 ЛАБОРА ГОРНАЯ ДИАГНОСТИКА ПАРАЗИТАРНЫХ БОЛЕЗНЕЙ
ПАРАЗИТЫ В КОСТНОМ МОЗГЕ
Объекты исследования — возбудители висцеральных лейшманиозов — Leishma-
nia infantum и Leishmania donovani.
Лейшмании
Лейшмании — группа жгутиковых простейших, вызывающих различные виды
лейшманиозов. В крови обнаруживают только висцеротропные лейшмании —
Leishmania donovani подвида infantum Leishmania donovani — возбудители висце-
ральных лейшманиозов. Наиболее часто завозимый в Россию паразит этой груп-
пы, Leishmania donovani подвида infantum, встречается в странах Средиземноморья
(Греции, Тунисе, Турции, Франции, Испании, Португалии, Италии и др.), в
ряде стран - бывших республик СССР (Таджикистане, Туркмении, Узбекистане,
Казахстане, Грузии, Азербайджане), а также на южном берегу Крыма, в предгорьях
Кавказа. Средиземноморско-среднеазиатский (детский) висцеральный лейшмани-
оз встречается очень редко. Это тяжелая протозойная инфекция, характеризуемая
хроническим течением, поражением лимфоидно-макрофагальной системы, спле-
номегалией, анемией и вторичной иммуносупрессией.
Биология возбудителей
Переносчиками возбудителей лейшманиоза являются москиты. Лейшмании в
организме человека (и некоторых животных — шакалов, лисиц) захватываются
моноцитами, где развиваются в амастиготы, вызывая генерализованный ретику-
логистиоцитоз. Амастигота — безжгутиковая стадия развития лейшманий оваль-
ной или круглой формы, размером 3-5х1-3 мкм, с круглым ядром размером 0,7-
0,8 мкм. В цитоплазме можно видеть кинетопласт палочковидной формы.
Лейшмании являются внутриклеточными паразитами и размножаются простым
делением. На месте укуса москита формируется воспалительный бугорок, из кото-
рого размножившиеся лейшмании разносятся по органам и тканям, локализуясь в
селезенке, костном мозге, крови, лимфатических узлах.
Лабораторная диагностика основывается на обнаружении амастигот лейш-
маний в аспирате костного мозга. Исследование крови не проводят из-за низкой
чувствительности этого метода. В некоторых случаях лейшмании могул быть
обнаружены в пунктате увеличенных лимфатических узлов.
Сбор проб костного мозга проводят путем аспирации с помощью шприца из
грудины, гребня подвздошной кости или из эпифиза большеберцовой кости (у
детей до 3 лет). Аспират немедленно переносят на чистые обезжиренные предмет-
ные стекла и готовят тонкие мазки, не разрушая клетки. Мазки быстро высушива-
ют на воздухе или с помощью фена, красят по Романовскому-Гимзе аналогично
тонким мазкам крови.
Идентификация лейшмании. Препараты исследуют в иммерсионной систе-
ме, лейшмании в них расположены обычно вне клеток, но нередко их обнару-
живают в цитоплазме мононуклеаров и нейтрофильных лейкоцитов; количество
лейшманий в одной клетке может достигать нескольких десятков. Они имеют
овальную или круглую форму размером 3-5x1-3 мкм. Цитоплазма окрашивается
в серовато-голубой, ядро — в красный или красно-фиолетовый, кинетопласт — в
темно-фиолетовый цвет.
ПАРАЗИТЫ В ЛИКВОРЕ
В спинномозговой жидкости (СМЖ) можно обнаружить трипаносомы, ток-
соплазмы, сапрофитные амебы — Naegleria fowleri. Acanthamoeba spp., Balamuthia
ЛАБОРАТОРНАЯ ДИАГНОСТИКА ПАРАЗИТАРНЫХ БОЛЕЗНЕЙ 779
mandrillaris. Иногда в спинномозговой жидкости обнаруживают личинки трихи-
нелл, Trichinella spp., личинки кишечной угрицы — Strongyloidesstercoralis, элементы
эхиноккоковой кисты (Echinococcus spp.у Однако исследование СМЖ не является
основным методом выявления указанных возбудителей.
При паразитарных болезнях в спинномозговой жидкости формируется ком-
плекс неспецифических признаков. При макроскопическом исследовании СМЖ
обращают внимание на ее цвет и прозрачность: нормальная жидкость бесцветна,
прозрачна, содержит 0,12-0,2 г/л белка и 1-2 клетки/мкл. Увеличение количества
клеток (цитоз) и изменение их состава могут свидетельствовать о заболеваниях, в
том числе паразитарной этиологии. Так, обнаружение в СМЖ лимфоидного плео-
цитоза с наличием гистиоцитов, макрофагов или эозинофильных гранулоцитов
может свидетельствовать о цистицеркозе головного мозга.
Эозинофильный плеоцитоз может наблюдаться при цистицеркозе, ангиострон-
гилезе, гнатостомозе, парагонимозе, аскаридозе, трихинеллезе, эхинококкозе,
фасциолезе. шистосомозе, токсокарозе.
Акантамебы, баламутим и неглерии
Это свободноживущие простейшие, амебы, способные, при проникновении
в мозг (чаще всего при попадании с водой в носовые полости) вызвать нейтро-
фильный и гранулематозный энцефалит и острый менингоэнцефалит, харак-
теризуемый высокой летальностью. Возбудители живут в почве, воде, воздуш-
ных фильтрах, экскрементах, разлагающихся тканях животных, промышленных
охлаждающих устройствах, воде бассейнов. Неглерии в виде цист проникают в
головной мозг через эпителий носовой полости, клетки которого способны к фаго-
цитозу. Неглерии размножаются, вызывая кровоизлияния и некрозы, развивается
первичный амебный менингоэнцефалит. Болезнь поражает детей и молодых здо-
ровых людей.
Акантамебы и баламутии проникают в головной мозг через носоглотку и вызы-
вают гранулематозный амебный энцефалит в основном у ослабленных людей,
получивших иммуносупрессивную терапию, в том числе у реципиентов органов и
тканей и ВИЧ-инфицированных. Акантамебы могут проникать через поврежден-
ную роговицу с последующим развитием кератита, а также через кожу, на которой
появляются язвенно-некротические изменения. В этих случаях их обнаруживают
при исследовании соскобов с конъюнктивы и жидкости из передней камеры глаза.
Встречаются диссеминированные формы акантамебиаза с поражением разных
органов, включая легкие.
Лабораторная диагностика основывается на обнаружении трофозоитов диа-
метром 10-35 мкм, с округлыми псевдоподиями и крупным ядром и цист диаме-
тром 7-13 мкм в нативных или окрашенных препаратах СМЖ. Применяют также
культуральные методы, иммунодиагностику, гистологические исследования.
Идентификация. Трофозоиты сапрофитных амеб в ликворе весьма схожи с
макрофагами. Трофозоиты неглерий концентрируются вокруг кровеносных сосу-
дов в ЦНС, напоминают слизней с однонаправленным движением. При помеще-
нии в воду они приобретают жгутиковую форму. При акантамебиазе и баламутна -
зе, помимо трофозоитов, в СМЖ выявляются цисты.
Трипаносомы в СМЖ появляются на поздней стадии болезни, когда возникают
симптомы поражения ЦНС. Исследуют нативный мазок из осадка СМЖ после
ее центрифугирования. Препарат просматривают под большим увеличением,
начиная с краев, поскольку трипаносомы часто концентрируются по самому
краю препарата. Можно также исследовать фиксированный и окрашенный по
Романовскому-Гимзе мазок.
780 ЛАБОРАТОРНАЯ ДИАГНОМИКА ПАРАЗИТАРНЫХ БОЛЕЗНЕЙ
Токсоплазмы в СМЖ могут быть обнаружены у больных с острым токсоплаз-
мозом. Исследуют препарат из осадка СМЖ после центрифугирования. При окра-
ске по Романовскому-Гимзе цитоплазма токсоплазмы приобретает синий цвет, а
ядро — темно-красный. Токсоплазма сохраняет полулунную форму, один конец ее
заострен, другой закруглен, размеры 3x7 мкм.
Трихинеллы на стадии личинки иногда проникают в СМЖ, в частности у боль-
ных с тяжелым течением трихинеллеза. Осадок СМЖ на наличие личинок трихи-
нелл исследуют при малом увеличении микроскопа.
Эхинококк в виде эхинококкового пузыря может развиваться в любых органах
и тканях, в ЦНС — редко.
ПАРАЗИТЫ В ЛИМФАТИЧЕСКИХ УЗЛАХ
Объектами исследования в увеличенных лимфатических узлах при трипаносо-
мозе яе ляются Trypanosoma spp.
В некоторых случаях в аспирате лимфатических узлов при висцеральном лейш-
маниозе можно обнаружить амастиготы лейшманий, однако основным методом
диагностики висцерального лейшманиоза является исследование аспиратов кост-
ного мозга.
Трипаносомы
Возбудителями трипаносомоза являются жгутиковые простейшие. В Африке
встречаются два вида трипаносомозов, их возбудители — Trypanosoma brucei
gambiense (гамбийский трипаносомоз) и Г. b. rhodesiense (родезийский трипа-
носомоз), которые вызывают соответственно антропонозную и зоонозную
формы африканского трипаносомоза, или сонной болезни. Переносчиком воз-
будителя африканского трипаносомоза является муха цеце. При африканском
трипаносомозе возбудит ели поражают лимфатические узлы, клетки ретикуло-
эндотелиальной системы, в дальнейшем происходит диссеми нация трипаносом
с поражением ЦНС. В России трипаносомоз выявляется крайне редко, в виде
завозных случаев.
Лабораторная диагностика африканского трипаносомоза основывается
на исследовании аспиратов лимфатических узлов (гамбийский трипаносомоз).
Возможно выявление возбудителей африканского (родезийского) трипаносомоза
в крови, СМЖ, пунктате костного мозга.
Сбор проб проводят путем биопсии лимфатического узла. Пунктат лимфатиче-
ского узла переносят из иглы на середину предметного стекла в каплю физраство-
ра, накрывают покровным стеклом и исследуют при увеличении х400. В препарате
должны быть эритроциты и лейкоциты. Препарат просматривают, начиная с
периферически к участков, поскольку трипаносомы стремятся проникнуть к краю
покровного стекла.
Тонкий мазок из биоптата фиксируют и окрашивают по Романовскому-Гимзе.
Трипаносомы обнаруживают в виде изогнутых образований, цитоплазма окра-
шивается в синий цвет, ядро и кинетопласт, расположенный вблизи от одного
из концов трипаносомы, — в красный или фиолетовый. Рядом с кинетопластом
начинается жгут, прикрепленный к клеточной стенке на всем протяжении, кроме
передней части, которая заканчивается свободно.
В нативном препарате трипаносомы обраща ют на себя внимание энергичными
движениями, их размер п ревышает размер клеток в биоптате.
Идентификация трипаносом до вида ш возможна. В окрашенных препаратах
среди эритроцитов видны характерные по форме и по размеру кинетопласты и
ЛАБ0оАТ0РНАЯ ДИАГНОСТИКА ПАРАЗИТАРНЫХ БОЛЕЗНЕЙ 781
жгутиковые образования. Размеры колеблются в пределах 14-33x1,5-3,5 мкм.
Основные элементы строения:
< тело удлиненное;
> ядро расположено в центре:
< > на одном из полюсов находится кинетопласт;
< вдоль клетки расположена ундулируюшая мембрана, заканчивающаяся жгу-
тиком.
Чувствительность метода составляет 70-75%.
ИССЛЕДОВАНИЯ ПАРАЗИТОВ В КАЛЕ
Паразиты пищеварительного тракта
Фекалии — сложный по составу экскрет. В них могут быть обнаружены пропа-
гативные стадии множества паразитов и других симбионтов, обитающих в пище-
варительном тракте.
Фауну пищеварительного тракта обычно объединяют в две группы: простейшие
кишечника и кишечные гельминты. В группу простейших кишечника включают
не только патогенные виды, но и непатогенные, морфологически иногда трудно-
отличимые от патогенных. С фека днями выделяются яйца гельминтов, взрослые
особи которых обитают в кишечнике, а также вне кишечника, например пара-
гонима, обитающего в респираторном тракте, кишечной и японской шистосом,
локализующихся в мезентериальных венах. В фекалиях могул быть обнаружены
вегетативные стадии и цисты более 20 видов простейших, яйца и личинки многих
видов гельминтов, основные из которых представлены в табл. 26-2.
Стеаторею с повышенным содержанием нейтрофильного жира, жирных кис-
лот или мыла наблюдают чаще всего при инфекциях, протекающих с синдромом
мальабсорбции, — лямблиозе, стронгилоидозе, криптоспоридиозе, циклоспорозе,
изоспорозе. Часто обнаруживаю^ непереваренные мышечные волокна, кристаллы
Шарко-Лейдена. Примесь крови и слизи в фекалиях выявляется при амебиазе и
балантидиазе. При амебной дизентерии эритроциты, как правило, склеены в кучки
разной величины, лишь небольшая их часть встречается в свободном состоянии.
При амебиазе в фекалиях среди небольшого количества нейтрофилов обнаружи-
вают большое количество эозинофилов.
Методы исследования
Сбор, доставка и консервирование проб
На результаты исследования влияет срок поступления и хранения материала
в лаборатории. При подозрении на амебную дизентерию фекалии должны быть
исследованы немедленно (не позже получаса) после дефекации. В пробе сохраня-
ются и обнаруживаются вегетативные стадии и других амеб и жгутиковых. Пробы
оформленных фекалий можно до исследования хранить в течение 3-4 ч в холо-
дильнике или поместить в консервирующую жидкость.
Собирают пробы фекалий в сухие водонепроницаемые емкости из стекла или
полимеров. В них не должно быть следов химических веществ, губительно дей-
ствующих на вегетативные стадии простейших и личинки гельминтов. Пробы
фекалий не должны содержать мочу. Взятие проб фекалий нежелательно прово-
дить у пациентов, принимающих противопаразитарные лекарственные препараты,
подвергаемых незадолго до обследования рентгеноскопии с применением бария
или после масляных клизм. Доставка проб фекалий в лабораторию должна быть по
Таблица 2Б-2. Клинико-лабораторные характеристики паразитов пищеварительного тракта
Возбудитель/инвазия Патогенная /инфитивная стадия и ее размеры Локализация Симптомы, признаки, показатели го Лабораторная диагностика (метод, диа- гностическая стадия)
Anisakidae spp. Личинка 1,5-3,0 см Разные участки ЖКТ Боль на месте проникновения личинки, рвота, лихорадка, эозинофилия Гистологическое исследование извлечен- ной личинки, иммуноферментный анализ ° (ИФА) 5
Ancylostoma duodenaiei Анкилостомоз Взрослая особь 1-2 см Тонкий кишечник Астения, микроцитарная гипохромная анемия Копроовоскопия 5
Necator атепсапиз/некалороз Взрослая особь 1-2 см Тонкий кишечник Астения, микроцитарная гипохромная анемия Копроовоскопия 3
Ascads lumbricoides/аскарироз Взрослая особь 20—40 см Тонкий кишечник Боли в животе, метеоризм, интоксикация непроходимость кишечника Копроовоскопия
Hymenolepis лала/гименоле- ПИДОЗ Взрослая особь 4 см Тонкий кишечник Боль в животе, диарея, астения Копроовоскопия =)
Hymenolepis diminutadvwteHO- лепидоз крысиный Взрослая особь до 60 см Тонкий кишечник Боль в животе, диспепсия G»J Копроовоскопия 5
Dicrocoeiium lanceatum/рмро- целиоз* (Примечание: перенесено в трематоды) Взрослая особь 1 см Желчные протоки Боль в животе, диспепсия Копроовоскопия g СП
Dipylidiwn caninumJflymnw- диоз* Взрослая особь до 70 см Тонкий кишечник Энтеоит, боли в животе Копроовоскопия
Diphyllobothriidae д. spp. Дифиллоботриозы Взрослая особь до 12 м Тонкий кишечник Боль в животе, метеоризм, макроцитарная анемия Копроовоскопия. идентификация члеников
Metagonimus yokogawai/меуа- гонимоз* Взрослая особь 1 мм Тонкий кишечник Энтерит, диспепсия Копроовоскопия
franophyetus sohiciiobatowil нанофиетоз Взрослая особь 1 мм Тонкий кишечник Энтерит, диспепсия Копроовоскопия
Opisthorcbis felineus/o п ис го р- хоз Clonorchis з/лалз/а/клонорхоз Взрослая особь 8-13 мм Взрослая особь 10-20 мм Желчные и панкреатические протоки Желчные и панкреатические протоки Лихорадка, эозинофилия, боль в правом подреберье, дискинезия желчных путей Лихорадка, эозинофилия, боль в правом подреберье, дискинезия желчных путей Копроовоскопия, исследование дуоде- нального содержимого, ИФА Копроовоскопия, исследование дуоде- нального содержимого, ИФА
Продолжение табл. 26-2
Возбудмтель/инвазия Патогенная /инфитивная стадия и ее размеры Локализация Симптомы, признаки, показатели Лабораторная диагностика (метод, диа- гностическая стадия)
Strongyloides stercoralis/строн- гилоидоз Взрослая особь 2 мм. Личинка до 250 мкм ЖКТ, другие органы и ткани при диссеминиро- ванной форме Линейная крапивница, кашель, диарея, мальабсорбция Копроларвоскопия. Специальные методы (Бермана и его модификации). Обнаружение личинок
Taeniarhynclws saginatus! тениаринхоз Взрослая особь до 12 м Тонкий кишечник Метеоризм, боль в животе, отхождение члеников Копроовоскопия. Яйца в фекалиях и перианальном соскобе или отпечатке, идентификация члеников
Taenia solium/aemoz Взрослая особь до 7 м Тонкий кишечник Метеоризм, боль в животе, отхождение члеников. Цистицеркоз Копроовоскопия. Яйца в фекалиях и перианальном соскобе или отпечатке, идентификация члеников
Trichostrongylidae spp Атp ихон- стронгилидоз* Взрослая особь до 6 мм Тонкий кишечник Энтерит, боли в животе Копроовоскопия :=]
Tnchocephaius (richiurus. /три- хоцефалез Взрослая особь до 5 см Толстая кишка Боли в животе, нарушения функции ЖКТ, астения Копроовоскопия о
Fasciola /?₽ра(/са/фасциолез Взрослая особь 2-3 см Желчные протоки Лихорадка, крапивница, эозинофилия, гепатомегалия, желтуха Копроовоскопия, исследование дуоде- 5 нального содержимого, ИФА 5
Schistosoma mansoni, S. japonictim, S. mekongi, S. inter calatum/i№KT0C0»t3 кишечный** Взрослая особь 0,2-2,5 см Вены, разные органы Зуд, эритема, эозинофилия, кровь в фекалиях, гепатоспленомегалия, портальная гипертензия Копроовоскопия, ИФА S —1 ^4
Enterobius vermicularisi энтеробиоз Взрослая особь до 1,3 см Тонкий и толстый кишеч- ник Лерианальный зуд, боль в животе, астения Обнаружение яиц в перианальном соско- бе или отпечатке
Протозоозы
Entamoeba histolytica/амебмаз 15-50 мкм Толстый кишечник, реже другие органы и ткани Колит, диарея, примесь слизи и крови в фекалиях, тенезмы, лихорадка Трофозоиты и цисты в фекапиях, ИФА %
Balantidium со/г/балантидиаз Трофозоит 30-150 мкм Циста 40-65 мкм Толстый кишечник Диарея, примесь слизи и крови в фекалиях, боли в животе Трофозоиты в фекалиях О
Blastocystis hominis/бпалто- ЦИСТОЗ Трофозоит 5-30 мкм Кишечник Диарея, боли в животе Трофозоиты в фекалиях
Dienthamoeba fragifis/pyidma- мебиаз Тросрозоит 5-15 мкм Толстый кишечник Диарея, боли в животе Трофозоиты в фекалиях
GO
Окончание табл. 26-2
Возбудктель/инвазия Патогенная /инфитивная стадия и ее размеры Локализация Симптомы, признаки, показатели Лабораторная диагностика (метод, диа- гностическая стадия)
Cryptosporidium parvum/крт- тоспоридиоз Трофозоит 10-15 мкм Тонкий кишечник Диарея, анорексия, потеря массы тела Ооцисты в фекалиях
Cyclospora caetanensis/uyiKno- спороз Трофозоит 8—10 мкм Тонкий кишечник Диарея, анорексия, потеря массы тела Ооцисты в фекалиях
Isospora де////изоспороз Трофозоит 20-30x10- 20 мкм Тонкий кишечник Диарея, анорексия, потеря массы тела Ооцисты в фекалиях
Lamblia intsstinalis (Giardia /атМаулямблиоз Трофозоит 10-20 мкм Двенадцатиперстная кишка Диарея, диспепсия боли в животе Трофозоиты в дуоденальном содержи- мом, трофизоиты и цисты в фекалиях
' Встречаются редко.
“ Возможны только завозные случаи из тропических стран.
ЛАБОРАТОРНАЯ ДИАГНОСТИКА ПАРАЗИТАРНЫХ
ЛАБОРАТОРНАЯ ДИАГНОСТИКА ПАРАЗИТАРНЫХ БОЛЕЗНЕЙ 785
возможности немедленной после взятия, то есть в теплом виде. В случае задержки
доставки проб в лабораторию или невозможности исследования в короткие сроки
пробы должны быть помешены в химические консерванты или храниться при тем-
пературе 0-4°С не более 1 сут. В качестве консервантов можно использовать 4%
раствор формалина, 3-10% раствор уксусной кислоты, 1,0-1,5% раствор детерген-
тов (моющие порошки без биоактивных добавок), а также жидкость Барбагалло
(3 мл формалина 40% + 97 мл физраствора или 1 л дистиллированной воды +
30 мл 40% формалина + 8,5 г поваренной соли). Для сохранения простейших
кишечника применяют консервант Турдыева (80 мл 0,2% раствора NaNO2 + 2 мл
глицерина + 10 мл концентрированного формалина + 8 мл концентрированного
раствора Люголя). Соотношение проб фекалий и консерванта составляет 1:3. Для
сохранения взрослых особей гельминтов и их фрагментов используют жидкость
Барбагалло, 10% формалин, 70% спирт, глицерин.
Длительное сохранение вегетативных стадии и цист простейших, яиц гельмин-
тов в консервантах позволяет в затруднительных случаях переслать законсерви-
рованный материал на консультацию в специализированную лабораторию. Такой
материал, содержащий точно идентифицированные простейшие или гельминты,
можно использовать в учебных целях, а также для контроля знаний врачей клини-
ческой лабораторной диагностики.
Вероятность наличия и обнаружения паразитарных объектов в пробах фекалий
в значительной степени определяется консистенцией стула. Вегетативные стадии
простейших обнаруживают обычно в жидких фекалиях и редко в оформленных.
Наоборот, цисты простейших в неоформленном стуле присутствуют редко, за
исключением тех случаев, когда жидкие фекалии получены после очистительной
клизмы; при этом в пробе могут быть обнаружены как вегетативные стадии, так и
цисты. Обычно цисты обнаруживают только в оформленном стуле.
Макроскопические методы исследования
Макроскопические методы исследования направлены на поиск взрослых особей
гельминтов (например, остриц, аскарид) или их фрагментов (сколексов, члеников
и частей стробилы цестод). Применяют два метода: отмучивания и отстаивания.
Метод отмучивания
Простым и эффективным является просмотр разжиженных водой фекалий
в чашках Петри. Фекалии размешивают с водой до образования равномерной
суспензии, после чего исследуют небольшими порциями на темном фоне в кюветах
или чашках Петри. Пинцетом или препаровальными иглами извлекают все подо-
зрительные частицы белого цвета, помещают на предметное стекло в каплю 50%
раствора глицерина или воды, накрывают другим предметным стеклом и исследу-
ют под лупой, а в случае необходимости — под микроскопом.
Ленточные гельминты определяют по строению членика. Узкий длинный чле-
ник характерен д ля бычьего и свиного цепня, короткий, плоский — для лентеца
широкого. Вид пенточного гельминта устанавливают по строению матки:
❖ если она имее г форму розетки — это лентец широкий;
< $• если она имеет 20-30 боковых ответвлений — это цепень бычий;
> если 8-12 — цепень свиной.
Установить факт полного отхождения крупных цестод после дегельминтизации
можно, исследуя при малом увеличении узкий головной конец:
❖ наличие четырех присосок с крючьями — свиной;
без крючьев — бычий;
< > с двумя щелями (ботриями) — лентец широкий.
Метод отстаивания
Фекалии помещают в цилиндр и заливают 3-4 объемами воды, перемешивают
до образования гомогенной суспензии и дают отстояться 10-15 мин надосадоч-
ный слой жидкости сливают. Процедуру повторяют трижды и исследуют осадок.
786 ЛАБОРАТОРНАЯ ДИАГНОСТИКА ПАРАЗИТАРНЫХ БОЛЕЗНЕЙ
Метод флотации
Используют для выявления яиц гельминтов. Метод основан на том, что япца
гельминтов всплывают на поверхность растворов с высоким удельным весом
(нитрата аммония, нитрата натрия, сульфата натрия, хлорида натрия). Метод
эффективен только для выявления яиц нематод и цестод, неэффективен для выяв-
ления яиц трематод.
Метод осаждения
Дчя одновременного поиска возбудителей гельминтозов и протозоозов целе-
сообразно применять наиболее простой, универсальный и эффективный метод —
формалин-эфирное осаждение. В качестве основных реактивов используют 10%
формалин, этиловый эфир, препараты докрашивают раствором Люголя. Метод
осаждения эффективно выявляет инвазии разной степени интенсивности. Его
применяют для выявления яиц и личинок гельминтов кишечника и печени, цист
и ооцист простейших. Трофозоиты простейших в используемых реактивах быстро
погибают и деформируются, поэтому не выявляются методом осаждения.
Микроскопические методы
Микроскопические методы исследования направлены в основном на обнару-
жение пропагативных стадий возбудителей (яиц, личинок, цист, ооцист). В связи
с низкой интенсивностью инвазии целесообразно применять для диагностики
методы обогащения. Отрицательные результаты исследований методом нативного
мазка не считаются достоверными.
Нативный (свежий неокрашенный) мазок. Он представляет собой взвесь
фекалий в капле воды, физраствора либо 50% раствора глицерина (последний
обеспечивает просветление материала, облегчая обнаружение яиц гельминтов, и
предохраняет препарат от высыхания). Достоинство метода — возможность обна-
ружения, помимо яиц гельминтов, вегетативных стадий простейших (иногда и их
идентификацию по характеру движения).
В нативном мазке простейшие кишечника и цисты бесцветны, отличаются от
фона лишь светопреломлением. Однако очень существенно, что вететативные
стадии проявляют в препарате, приготовленном на физрастворе, характерную для
данного вида подвижность, чем способствуют их обнаружению. В нативном мазке
могут быть распознаны гематофаги Е. histolytica, цисты амеб и лямблий, яйца и
дичинки гельминтов.
При низкой интенсивности инвазии чувствительность метода недостаточная,
поэтому исследование материала с применением методов обогащения следует про-
водить в обязательном порядке.
Нативный мазок, обработанный раствором Люголя, предназначен для
обнаружения и идентификации цист простейших.
На предметном стекле готовят два нативных мазка: один на физрастворе, дру-
гой с раствором Люголя. Если мазок один, его исследуют вначале в неокрашенном
виде для выявления подвижных вегетативных стадий простейших, а затем добав-
ляют каплю раствора Люголя. При двух мазках один исследуют на наличие веге-
тативных стадий простейших и яиц гельминтов, другой обрабатывают раствором
Люголя. В последнем случае каплю раствора наносят у края покровного стекла,
йод при этом диффундирует в мазок, окрашивая его с разной интенсивностью по
мере проникновения в разные участки мазка.
Достоинство мазка, обработанного раствором Люголя, — возможность обна-
ружения в цистах простейших важных морфологических деталей, невидимых в
нативном (неокрашенном) мазке.
Метод толстого мазка фекалий по Като и Миури
Принцип метода — исследование на яйца гельминтов большего объема фекалий
за счет просветления мазка смесью Като. На предметное стекло наносят комочек
фекалий величиной с горошину, покрывают замоченной в смеси Като целлофано-
ЛАБОРАТОРНАЯ ДИАГНОСТИКА ПАРАЗИТАРНЫХ БОЛЕЗНЕЙ 787
вой пластинкой, придавливают резиновой пробкой для получения равномерной
толщины мазка. Препарат оставляют для просветления на 40-50 мин при ком-
натной температуре. Просветленный мазок микроскопируют через целлофановую
пленку. В толстом мазке по Като обнаруживают яйца аскарид, власоглава, анкило-
стоматид, лентецов (D. latum и др.), трематод (О. fdbieus, F. hepatica, D. lanceatum
и др.). Эффективность метода для выявления яиц гельминтов достигает 84%.
Методы обогащения
Исследование проб фекалий в мазках любого типа не всегда достаточно эффек-
тивно из-за низкой численности цист или яиц в исследуемом материале, непосто-
янства их выделения из кишечника и других причин. Отрицательный результат
анализа фекалий на простейшие и гельминты методом нативного мазка не может
считаться достоверным. Это обстоятельство вынуждает проводить обследование
пациента повторно и применять методы обогащения. Методы основаны на кон-
центрации яиц и личинок гельминтов или цист простейших в пробе благодаря
отделению собственно фекальных частиц и накоплению паразитарных объектов
в препаратах. Процедуре обогащения могут подвергаться как свежие фекалии, так
и материал из консервирующих жидкостей. Стандартными методами являются
следующие:
❖ толстый мазок фекалий на предметном стекле под гигроскопическим цел-
лофаном, пропитанным раствором глицерина и фенола, — метод Като и
Миури, эффективность которого вы ше, чем исследование нативного мазка,
за счет большего объема исследуемых фекалий, однако при этом все-таки
не выявляются случаи гельминтозов с низкой интенсивностью, простейшие
кишечника;
< > формалин-эфирное осаждение;
❖ уксусно-эфирный метод осаждения;
❖ химико-седиментационный метод (редко применяется из-за трудоемкости).
СПЕЦИАЛЬНЫЕ МЕТОДЫ
Обнаружение яиц остриц на перианальных складках
Принцип метода основан на взятии яиц остриц с перианальной области, отло-
женных там самкой в период ее гибели. 11омимо яиц остриц, этим методом могут
быть выявлены онкосферы бычьего цепня, редко личинки кишечной угрицы.
Перианальный отпечаток на клейкую ленту (по Грэхему)
Взятие материала проводят путем прикосновения кусочком ленты к перианаль-
ным складкам, яйца гельминтов прилипают к ней. Ленту приклеивают к пред-
метному стеклу и просматривают под микроскопом. Перед исследованием под
микроскопом приподнимают ленту и наносят каплю иммерсионного масла под ее
середину — это улучшит прозрачность ленты.
Перианальный отпечаток на липкую поверхность глазных лопаток
(метод Б.Е. Рабиновича)
Набор для исследования состоит из стеклянных глазных лопаток со стержнем,
уложенных в пенал; специальною клея из клеола, касторового масла, эфира и
спирта. Взятие материала проводят стеклянной глазной лопаточкой, покрытой
клеем, которую исследуют с обеих сторон при малом увеличении после установки
в кассете-держателе.
Метод в 2 раза чувствительнее, менее трудоемок и значительно дешевле, чем
метод липкой ленты. Преимуществом метода является удобство для персонала и
пациентов.
Перианальный соскоб по Кеворковой
По Кеворковой используют шпатель с ватным тампоном, по Тор1ушину — шпа-
тель с ватным тампоном, смоченным в глицерине. Эти методы менее эффективны
788 ЛАБОРАТОРНАЯ ДИАГНОСТИКА ПАРАЗИТАРНЫХ БОЛЕЗНЕЙ
по сравнению с методами исследования по Грэхему и Рабиновичу, в последние
годы их применяют редко
Обнаружение личинок Strongyloides stercoralis — кишечной ут рицы
Выделяемые с фекалиями личинки обнаруживают, используя термотропизм
личинок — выход их из остывающих фекалий в среду (воду) с более высокой тем-
пературой. Применяют метод исследования фекалий по Берману. Для проведения
этого исследования лечащий врач в направлении в лабораторию должен указать,
какой именно гельминтоз подозревается у пациента. Эффективность однократно-
го исследования этим методом составляет 20- 30% и возрастает при повторных (не
менее трех) исследованиях.
Простейшие
Стандартным методом вы явления цист (ооцист) простейших в пробах фекалий
является формалин-эфирное осаждение. При этом трофозоиты погибают и не
выявляются. Временное хранение проб в стандартных консервантах не снижает
эффективности выявления пист. Трофозоиты могут быть обнаружены в нативном
мазке, в основном в пробах, исследование которых проводят немедленно после
взятия. В отношении цист лямблий чувствительность однократного обследова-
ния методом формалин-эфирного осаждения составляет около 65%, троекратное
исследование повышает чувствительность метода до Я0-90%. Для выявления
простейших кишечника требуется проведение, по крайней мере, 3 исследований с
интервалом несколько дней.
Некоторые виды простейших выявляются с помощью специальных окрасок,
например по Цилю-Нильсену, сафранином (криптоспоридии, циклоспоры).
Обитатели кишечника относятся к одно i из 4 групп: амебам, жгутиковым, рес-
нитчатым и кокцидиям.
Амеба дизентерийная
Entamoeba histolytica является возбудителем важнейшей кишечной инфекции
амебной дизентерии (амебиаза). Локализуется в толстом кишечнике, выделяет-
ся в форме трофозоита (с жидкими фекалиями) и/или цисты (в оформленных
фекалиях). Поскольку большинство видов амеб, обитающих в кишечнике, непа-
тогенны, следует быть очень осмотрительными при лабораторной диагностике.
Трофозоиты дизентерийных амеб в нативном препарате имеют зеленоватый цвет,
благодаря преломлению света они поблескивают, их диаметр может быть раз-
личным — от 15 до 60 мкм. Важнейшим диагностическим признаком амебиаза
(амебной дизентерии) является обнаружение трофозоитов с поглощенными ими
эритроцитами — гематофагов, а также характерное однонаправленное движение
трофозоитов в нативном препарате. Во всех остальных случаях обнаружение
Е. AiWyfca-подобных трофозоитов, не содержащих эритроцитов, не является
основанием для диагноза амебиаза как болезни. В равной степени это относится
и к случаям обнаружения только цист Е. histolytica, что характерно для цистоноси-
телей или реконвалесцентов. В связи с этим особую важность приобретает диф-
ференциация трофозоитов и цист Е. histolytica от других видов амеб, обитающих
в кишечнике. Трофозоиты амеб различаются по наличию или отсутствию пери-
ферического хроматина в ядре. Трофозоиты Е. histolytica имеют периферический
ядерный хроматин, маленькую кариосому в ядре, мелкозернистую цитоплазму, в
которой отсутствуют бактерии, могут находиться (или отсутствовать) эритроци -
ты. Трофозоиты непатогенных амеб Entamoeba coli и Entamoeba hartmannl, так же
как и дизентерийная амеба, имеют периферический ядерный хроматин, однако
у Е. coli цитоплазма имеет грубую зернистость, в ней обнаруживают бактерии и
дрожжи, которые никогда не обнаруживают в цитоплазме дизентерийной амебы,
В цитоплазме трофозоита Е harbnanni могут находиться поглощенные эритроци-
ЛАБОРАТОРНАЯ ДИАГНОСТИКА ПАРАЗИТАРНЫХ БОЛЕЗНЕЙ 789
ты, однако размер трофозоитов составляет 8-15 мкм. тогда как у дизентерийной
амебы — 15-60 мкм. У непатогенных амеб Endolimax папа, lodamoeba butschiii
и Dientamoeba fragilis периферического хроматина в ядре трофозоитов нет, они
имеют меньший диаметр (до 15 мкм) и большую кариосому в ядре. Более 50% тро-
фозоитов D. fragilis имеют 2 ядра. Дифференциацию и идентификацию трофозои-
тов Е. histolytica осуществляют по влажным (нативным) препаратам, ориентируясь
на характер их движения (однонаправленное), наличие в цитоплазме трофозоитов
эритроцитов. Для более точной идентификации трофозоитов и цист проводят
экспресс-окраску препаратов буферным метиленовым синим, а также окраску
постоянных препаратов трихромовым красителем или железным гематоксилином
по Гайденгайну.
Лямблия
(Синоним Lamblia intestinalis — Giardia lambbd) — единственная среди остальных
5 видов, обитающих в кишечнике жгутиковых, вызывающая у людей симпто-
мы заболевания ЖКТ, в основном диарею, диспепсию, боли в животе, особенно
у первично зараженных детей. Это распространенный кишечный протозооз,
которым в неь оторых регионах заражено до 30% детей, обратившихся за меди-
цинской помощью с симптомами заболевания желудочно-кишечного тракта.
У трофозоитов лямблий имеется 4 пары жгутиков, дифференцировать их необхо-
димо от Pentatruhomonas (Trichomonas) hominis. Цисты лямблии имеют овальную
форму с 2-4 ядрами и несколькими аксонемами, расположенными продольно.
Размеры цист — 8-12x7-10 мкм. Трофозоиты лямблий выявляются в нативном
мазке и отличаются характерным движением (подобно падаюшему листу, коле-
блясь то вперед, то назад), наличием жгутиков. Размеры трофозоита составляют
10-20 мкм. Для изучения структуры цист нативный препарат обрабатывают
раствором Люголя, Для детальной дифференциации мазки в некоторых случаях
окрашивают трихромом.
Балангидии
Balantidium coli — единственный патогенный представитель из группы рес-
ничных. Люди заболевают балантидиозом редко, заражаясь главным образом от
свиней. Возбудитель обнаруживают в фекалиях в форме трофозоитов или цист.
Трофозоиты крупные, 50-200x40-70 мкм, очень подвижные, хорошо заметны
в препаратах с физраствором при малом увеличении. Они покрыты короткими
волосками (ресничками), имеют макро- и микронуклеус. В окружающей среде
неустойчивы, исследовать фекалии необходимо как можно быстрее. Диаметр
цист — 45-75 мкм.
Криптоспоридии
Cryptosporidium spp. — важнейший представитель кокцидий кишечника.
Cryptosporidium parvum вызывает диарею у детей младше 5-летнего возраста, одна-
ко его значение многократно возросло как возбудителя СПИД-ассоциируемой
инфекции криптоспоридиоза. Криптоспоридий развивается в ворсинках эпителия
кишечника, располагаясь в особой вакуоле.
Метод исследования — обнаружение окрашенных ооцист в фиксированном
мазке фекалий. Мазок фекалий высушивают на воздухе, фиксируют в смеси
Никифорова 10-15 мин или над пламенем горелки и красят по Цилю-Нильсену.
Ооцисты криптоспоридий имеют круглую форму, диаметр 5-6 мкм и окрашива-
ются в разные оттенки ярко-красного цвета. Сопутствующая микрофлора окраши-
вается в зеленый цвет. Характерный признак ооцист криптоспоридий — наличие
внутри черных гранул.
В красный цвет также могут окрашиваться капли жироподобных веществ, гра-
нулы детрита. Даже при случайном сходстве по размерам эти образования можно
отличить от ооцист по от сутствию у них отчетливой оболочки и структурирован-
ного содержимого. В сомнительных случаях применяют модификацию окраски по
790 ЛАБОРАТОРНАЯ ДИАГНОСТИКА ПАРАЗИТАРНЫХ БОЛЕЗНЕЙ
Цилю-Нильсену — уменьшают концентрацию малахитового зеленого, что облег-
чает видовую идентификацию ооцист Cryptosporidium spp., Cyclospora caetan&isis,
Isospora belli.
В последние годы применяют окраску 1% водным раствором сафранина или
1% метиленовым синим. Эти способы более надежны, просты, требуют меньше
времени по сравнению с методом окраски по Цилю-Нильсену.
Изоспора
Изоспора — возбудитель изоспороза, протозойной инфекции, значение которой
в патологии человека стало возрастать в связи с эпидемией ВИЧ/СПИДа.
Ооцисты Isospora belli могут быть обнаружены в нативных препаратах или в
препаратах, приготовленных из осадка фекалий, полученных методом формалин-
эфирного осаждения. При обработке мазка раствором йода ядра ооцист хорошо
прокрашиваются. Ооцисты I. belli имеют овальную форму, размер 32x16 мкм, в
центре находится неразделенная масса цитоплазмы.
Циклоспора
Циклоспора — возбудитель циклоспороза, актуальность этой протозойной
инфекции возрастает в связи с эпидемией ВИЧ/СПИДа. В нативных препаратах
в ооцистах Cyclospora caetanensis при малом увеличении хорошо видны многочис-
ленные сферические тельца, а при большом увеличении — 2 спороцисты с 2 споро-
зоитами в каждой. В окрашенных по Цилю-Нильсену препаратах ооцисты могут
быть бесцветными или окрашенными с разной интенсивностью. Это свойство, а
также их больший размер (8-10 мкм) служат отличительными признаками при
сравнении с ооцистами криптоспоридий.
Гельминты
Б фекалиях могут быть обнаружены яйца и/или личинки многих видов гель-
минтов, относящихся к трем классам: нематодам, цестодам и трематодам. Для
обнаружения чаще всего используют универсальный метод формалин-эфирного
осаждения, который позволяет одновременно выявить цисты и ооцисты простей-
ших.
Нематоды
Нематоды, обитающие в кишечнике, относятся в большинстве своем к группе
геогельминтов, их пропагативные стадии (яйца, личинки) приобретают инва-
зивность лишь после пребывания в почве Исключением является острица, яйца
которой становятся инвазивными через 3-4 ч после откладки их самкой на
перианальной области. Личинки кишечной угрицы инвазивны уже в кишечнике,
вследствие этого возможно развитие диссеминированных форм стронгилиидоза с
проникновением личинок в разные органы и ткани.
Лабораторная диагностика нематод основана на обнаружении яиц и/или личи-
нок в препаратах из фекалий или в перианальном соскобе (отпечатке) при энте-
робиозе.
Идентификация яиц аскарид, власоглава, анкилостоматид, трихостронгилид,
остриц и личинок кишечной угрицы проводят, ориентируясь на опыт, атласы и
справочники. Некоторые затруднения может представить идентификация опло-
дотворенных и неоплодотворенных яиц Ascaris lumbricoides. Первые имеют оваль-
ную или круглую форму, покрыты толстой наружной фестончатой многослойной
оболочкой, окрашенной пигментами фекалий в темно-желтый или коричневый
цвет: вторые — удлиненные по форме, покрыты грубой оболочкой с неровными
фестонами, более прозрачны, чем оплодотворенные (видны крупные желточные
клетки). Наличие в препарате только неоплодотворенных яиц свидетельствует о
низкой интенсивности инвазии (отсутствии самцов). Следует иметь в виду, что
ЛАБОРАТОРНАЯ ДИАГНОСТИКА ПАРАЗИТАРНЫХ БОЛЕЗНЕЙ 791
яйца Ancylosloma duodenale и Necator amerioanus морфологически неразличимы,
поэтому при их выявлении следует писать: «Обнаружены яйца анкилостоматид
или Ancylostomatidea g. spp.>>.
Нематодозы зоонозной природы — токсокароз, анизакидоз, трихинеллез, диро-
филяриоз — диагностируют иммунологическими и гистологическими методами.
Цестоды
Цестоды — биогельминты, в их жизненном цикле участвуют животные. Человек
для большинства цестод является окончательным хозяином (паразитируют взрос-
лые особи), для эхинококков — промежуточным (паразитируют в личиночной
стадии). Взрослые стадии цестод продуцируют яйца (диагностические стадии), о
наличии личиночных стадий эхинококков судят по результатам иммунологиче-
ского исследования.
Выявляемость яиц Т. saginatus и Н. папа существенно повышается при комбина-
ции методов обогащения по Калантаряну и толстого мазка по Като. D. latum выде-
ляет яиц так много, что они надежно выявляются в нативном мазке.
Идентификацию яиц цестод (цепней — Hymenolepis папа. Taenia solium,
Taeniarhynchus saginatus и лентеца широкого — biphyllobothrium latum) проводят,
ориентируясь на морфологические признаки. Следует иметь в виду, что онкосферы
Т. solium и Т. saginatus морфологически неразличимы, при их обнаружении следует
писать: «Обнаружены онкосферы тениид или Taeniidae spp.». При инвазии кар-
ликовым цепнем возможно размножение гельминтов в кишечнике человека без
выхода в окружающую среду, особенно у лиц со сниженным иммунитетом.
Трематоды
Трематоды — биогельминты, человек для них является единственным оконча-
тельным хозяином (шистосомы) или одним из окончательных наряду с животны-
ми (описторх, фасциола и др.). Все виды трематод, паразитирующие в организме
человека, продуцируют яйца, выделяемые с фекалиями (исключение составляет
Schistosoma haematobium, яйца которой выделяются с мочой).
Вследствие слабой интенсивности трематодных инвазий рекомендуют всегда
применять метод формалин-эфирного обогащения или метод Като. Следует также
иметь в виду важную особенность диагностики трематодозов: в острой стадии
инвазии, которая может продолжаться в течение нескольких недель или месяцев,
яйца паразитами не продуцируются и копроовоскопия в это время бесполезна.
Наличие возбудителя в острой стадии может быть установлено только косвенно,
иммунологическими методами.
Морфологические признаки яиц трематод приведены в табл. 26-2. в которой
также представлены отличительные признаки морфологически сходных яиц опи-
сторха, метагонима и клонорха. Яйца Fasciola hepatic» и bicrocelium lanceatum могут
быть «транзитными» (находились в съеденной печени крупного рогатого скота
или овец), для исключения ложного диагноза исследование фекалий необходимо
повторить через несколько дней, исключив из рациона пациента сырую или тер-
мически слабо обработанную печень.
ПАРАЗИТЫ В ДУОДЕНАЛЬНОМ СОДЕРЖИМОМ
В дуоденальном содержимом можно обнаружить яйца гельминтов, паразити-
рующих в печени, — O.fehncus, D. lancearum, С. sinensis, F. hepatica, личинки S. sterc-
oral^, ооцисты — Isospora belli, Cryptosporidium spp., трофозоиты лямблий.
Полученное при дуоденальном зондировании содержимое необходимо подвер-
гать немедленному микроскопическому исследованию, так как через 30 мин состав
его меняется. Для исследования дуоденальное содержимое выливают в чашки
Петри и микроскопируют при малом увеличении. Затем каждую порцию дуоде-
нального содержимого соединяют с равным количеством эфира, смесь взбалтыва-
792 ЛАБОРАТОРНАЯ ДИАГНОСТИКА ПАРАЗИТАРНЫХ БОЛЕЗНЕЙ
ют и центрифугируют. Надосадочную жидкость выливают, а осадок помещают на
предметное стекло и просматривают под микроскопом. В нативных мазках можно
увидеть увеличенное количество эпителиальных клеток желчных протоков, лей-
коцитов. а также обнаружить эозинофилы (свидетельство острого или хрониче-
ского холангита или холецистита паразитарной этиологии), трофозоиты лямблий,
яйца и личинки гельминтов. Помимо нативных препаратов, можно исследовать
окрашенные мазки для выявления ооцист изоспоры и криптоспоридий. Чаще
всего применяют модифицированную окраску по Цилю-Нильсену.
ПАРАЗИТЫ В МОЧЕ
В норме моча соломенно-желтого цвета, прозрачная. Для диагностики многих
паразитарных поражений мочевыделительной системы имеет значение изменение
цвета и прозрачности мочи.
В моче могут быть обнаружены яйца шистосом (Schistosoma haematobium) и
диоктофим (Dioctophymerenale), изредка — трофозоиты Trichomonas vaginalis, цисты
Entamoeba histolytica, микрофилярии Wuchcreria bancrofti и Onchocerca volvulus.
Следует иметь в виду, что некоторые виды простейших, обнаруживаемые в про-
бах мочи, могут быть следствием загрязнения ее фекалиями или влагалищными
выделениями. В емкостях из-под напитков с содержанием углекислоты, сахаров
часто развиваются свободноживущие личинки мочекислой угрицы. которые под-
вижны и способны захватывать лейкоциты и эритроциты в моче. Эти объекты,
не имеющие отношения к паразитарным болезням, привлекают внимание специ-
алистов лабораторий, чаще всего их ошибочно принимают за личинки кишечной
угрицы. Для того чтобы исключить их как причину поражений мочевыделитель-
ной системы, следует сделать повторный анализ пробы мочи, взятой стерильно.
Неспецифические признаки паразитарных ботезней в осадке мочи наблюда-
ются при следующих паразитозах: малярии, мочеполовом шистосомозе и лимфа-
тических филяриидозах, диоктофимозе. При осложненной тропической малярии
(остром гемолизе и гемоглобинурийной лихорадке) могут быть обнаружены
повышенное содержание белка, неизмененные или выщелоченные эритроциты,
зернистые и гиалиновые цилиндры, выявляются также альбуминурия, уробилину-
рия, билирубинурия. В моче обнаруживают глыбки и желтые крошковатые массы
аморфного гемоглобина, оксигемоглобин, метгемоглобин.
При мочеполовом шистосомозе часто наблюдается эозинофилурия, при кото-
рой в осадке мочи можно обнаружить до 15-95% эозинофилов, причем их коли-
чество в моче всегда выше, чем в крови.
Лимфатические филяриидозы часто сопровождаются хилурией, моча стано-
вится «жирной», мутной, опалесцирующей, напоминающей молоко. В сосуде
собранная моча разделяется на три слоя: верхний — молочный хилус, средний —
обесцвеченная моча со сгустками и детритом, нижний - клеточный осадок. Для
диоктофимоза характерны гематурия, олигурия, анурия, пиурия, резистентные к
общепринятым методам терапии.
Для исследования на мочеполовой шистосомоз мочу собирают в период наи-
большей активности обследуемого. В качестве пробы для исследования исполь-
зуют последнюю порцию мо ш объемом до 10 мл. Количество яиц S. haematobium
в пробе мочи увеличивается после нагрузки на мышцы брюшного пресса, что
действует на стенку мочевого пузыря, стимулирует выход яиц S. haematobium в
просвет мочевого пузыря и повышает вероятность их обнаружения. Исследуют
только свежую порцию мочи. В тех случаях, когда это невозможно, в мочу реко-
мендуют добавлять консерванты, способствующие длительному сохранению яиц
5. haematobium (несколько суток). Наиболее простым и доступным консервантом
является формалин, концентрация которого после добавления в мочу должна быть
ЛАБОРАТОРНАЯ ДИАГНОСТИКА ПАРАЗИТАРНЫХ БОЛЕЗНЕЙ 793
в пределах 5%. Сбор проб для обнаружения возбудителя диоктофимоза проводят
обычным способом.
Для обнаружения в моче яиц или личинок гельминтов применяют два метода:
ot аждение и фильтрацию.
• Метод осаждения менее чувствительный, но более дешевый и простой в
выполнении. Пробы мочи центрифугируют и из осадка готовят препараты
на покровных стеклах. Препараты исследуют в нативном или окрашенном
виде.
• Метод фильтрации благодаря его высокой чувствительности используют для
количественной оценки инвазии. При фильтрации мочи через фильтр не про-
пускаются яйца и личинки.
Мочеполовой шистосомоз
Заражение происходит при проникновении через кожу и слизистые оболочки
находящихся в воде пресноводных водоемов личинок — церкарий. В организме
человека шистосомы паразитируют в венах мочевого пузыря, с мочой выделяются
яйца гельминтов, имеющие удлиненную овальную форму, длину до 150 мкм, на
одном из полюсов расположен большой шип.
Диоктофимоз
Редкая и тяжелая паразитарная болезнь. Заражение происходит при случайном
проглатывании промежуточных хозяев — червей-олигохет. В организме человека,
обычно в правой почке, развивается крупная особь гельминта (длиной до 1 м,
толщиной до 1 см), приводя к необратимым изменениям. В моче выявляются яйца
эллипсоидной формы, коричневого цвета, с толстой оболочкой, покрытой вдавле-
ниями, на полюсах имеются крышечки, размеры яиц — 60-80x40-44 мкм.
ПАРАЗИТЫ МОКРОТЫ
Мокрота — отделяемое дыхательных путей: легких, бронхов, трахеи, гортани,
она выделяется при кашле или отхаркивании. К мокроте, как правило, приме-
шивается слизь из горла или слюна, поэтому мокроту очень важно правильно
собирать.
В мокроте обнаруживают цисты пневмоцист (Pneumocystis carinii), яйца пара-
гонима (Paragonimus westermani), личинки Strongyloides stercoralis (при диссеми-
нированной форме стронги яоидоза). В некоторых случаях при образовании
эхинококковой кисты в легком можно обнаружить оболочку и содержимое кисты
(сколексы и крючья эхинококкового пузыря). Редкими находками при микро-
скопическом исследовании мокроты и лаважной жидкости являются трофозоиты
амеб (Acanthamoeba spp.. Entamoeba histolytica, Entamoeba gingivalis), трофозоиты
токсоплазмы (T. gondii), личинки аскарид (A. lumbricoides) при миграции в раннем
периоде инвазии, яйца Thonunx aerophylus (напоминают яйца власоглава).
При пневмоцистозе мокрота вязкая, с большим количеством слизи, серовато-
стекловидная. При парагонимозе и в миграционной фазе аскаридоза мокрота
становится жидкой в связи с наличием серозной жидкости. В случаях длительного
течения парагонимоза мокрота приобретает темно-коричневый цвет и сливкоо-
бразную консистенцию. При развитии амебного абсцесса легкого в мокроте может
быть гной. В некоторых случаях (при аскаридозе, токсокарозе, стронгилоидозе)
в мокроте обнаруживают эозинофилы, кристаллы Шарко-Лейдена, спирали
Куршмана.
794 ЛАБОРАТОРНАЯ ДИАГНОСТИКА ПАРАЗИТАРНЫХ БОЛЕЗНЕЙ
Свежевыделенную мокроту собирают в чистую сухую посуду чаще всего в
чашку Петри. Мокроту лучше собирать утром натощак, желательно еще до приема
пищи. Перед сбором мокроты необходимо тщательно прополоскать рот. В слу-
чае затруднения получения мокроты используют паровые или ультразвуковые
ингаляции. Они особенно показаны при исследовании мокроты на наличие пнев-
моцист. Пациента в этих случаях необходимо предупреждать о нежелательности
использования зубных паст, так как входящие в состав последних консерванты
препятствуют хорошему окрашиванию препаратов мокроты. В целях обнаружения
пневмоцист мокроту нужно просматривать немедленно, в крайнем случае через
1-4 ч, поместив ее при этом в холодильник при 4 °C, так как в мокроте быстро раз-
вивается бактериальная флора и разрушаются клеточные элементы. Материал для
исследования на наличие пневмоцист желательно получать методом бронхоальве-
олярного лаважа. Во время бронхоскопии после установления бронхоскопа в устье
одного из субсегментарных бронхов осуществляют инстилляцию в дистальные
отделы бронхиального дерева теплого физиологического раствора с последующей
его аспирацией. Аспират может быть разделен на 3 порции: бронхиальный, брон-
хоальвеолярный и альвеолярный. Полученный аспират (лаважную жидкость)
также рекомендуют исследовать немедленно. Описано два способа временного
сохранения материала:
<> лаважную жидкость сливают в стоящую во льду посуду с 50 мл раствора
Хенкса (коммерческого; pH = 7,2+7,4);
о лаважную жидкость охлаждают до 4 °C. после чего в течение некоторого вре-
мени ее можно использовать для дополнительных исследований.
Мокроту разливают в чашки Петри тонким слоем, рассматривают на светлом и
черном фоне. Отбирают частицы из «ржавых» образований, обрывки ткани и тому
подобное на предметное стекло, расправляют осторожно препаровальной иглой,
накрывают покровным стеклом и просматривают под микроскопом. При большом
содержании гноя и слизи мокроту смешивают с равным объемом 2% соды или
0,5% раствором КОН или NaOH. перемешивают, встряхиваю! в течение 5 мин,
помещают в термостат или на водяную баню на 3 мин, центрифугируют 2-3 мин
со скоростью 1000-1500 об./мин; из осадка приготавливают препарат и исследу-
ют под микроскопом. Проводят макроскопические (в целях изучения физических
свойств мокроты) и микроскопические паразитологические исследования.
Для микроскопического исследования необходимо просматривать мокроту
методом нативного мазка. Для выявления пневмоцист мокроту подвергают специ-
фическим методам окраски.
Обычно применяют скрининговые окраски, позволяющие увидеть содержимое
8-ядерной цисты (так называемые внутрицисз ные тельца), однако не выявляющие
клеточную стенку пневмоцист. К ним относят окраску по Романовскому-Гимзе,
гематоксилин-эозином, по Райту.
Для окончательной верификации пневмоцист применяют сложные, многоэтап-
ные, зачастую требующие дорогостоящих реактивов окраски: окраска толуидино-
вым синим, методом серебрения по Гомори-Грокотту, крезиловым фиолетовым. С
их помощью четко визуализируется клеточная стенка вист Pneumocystis carinii.
ПАРАЗИТЫ ОТДЕЛЯЕМОГО МОЧЕПОЛОВЫХ ПУТЕЙ
Тги homonas vaginalis, трихомонада — единственное простейшее, обитающее в
мочеполовых путях.
Trichomonas vaginalis у части инфицированных им индивидов вызывает острый
или хронический трихомоноз. Т. vaginalis — жгутиковое простейшее, характерной
овальной или грушевидной формы Размеры трофозоита — 4-13x2-9 мкм, имеет
3-5 жгутиков, по краю тела расположена ундулирующая мембрана, доходящая
ЛАБОРАТОРНАЯ ДИАГНОСТИКА ПАРАЗИТАРНЫХ БОЛЕЗНЕЙ 795
примерно до половины тела. Ядро одно, расположено в передней части тела.
Размножается паразит простым делением, цист не образует.
Исследованию на наличие этого простейшего подлежат отделяемое из уретры,
цервикального канала, влагалища, простатический секрет, содержимое парауре-
тральных ходов, бартолиниевых желез.
Лабораторная диагностика основана на микроскопии нативных влажных пре-
паратов из экссудатов половых органов или осадка мочи на наличие подвижных
трофозоитов, а также окрашенных препаратов. В свежем материале хорошо видны
трофозоиты, передвигающиеся «нервными», дергающимися или прыгающими,
движениями. Чувствительность этого метода достигает 75-80%, специфичность
высокая — до 100%. На эффективность выявления трихомонад влияют техниче-
ски правильное взятие материала и своевременный его просмотр.
Выделения, собранные из уретры, влагалища, шейки матки, переносят в каплю
теплого физиологического раствора или помещают в пробирку, содержащую 3 мл
такого раствора. Центрифугируют пробирку в течение 2 мин. Удаляют пипеткой
надосадочную жидкость, из осадка готовят нативные или окрашенные мазки.
Просмотр нативных препаратов чаще всего проводят методом раздавленной
капли-суспензии и реже методом висячей капли.
В дополнение к препаратам, приготовленным из свежего материала, готовят
фиксированные и окрашенные постоянные препараты. Мазки фиксируют в смеси
Никифорова в течение 3-5 мин и окрашивают различными методами. В препара-
тах, окрашенных железным гематоксилином, по Романовскому-Гимзе, акридином
оранжевым или трихромовым красителем, трофозоиты легко распознаются благо-
даря наличию жгутиков и короткой ундулирующей мембраны.
Микроскопия окрашенных препаратов является простым и дешевым методом,
не требующим немедленного проведения исследования, специального оборудова-
ния, однако его специфичность и чувствительность низкие. Полезны повторные
исследования.
В нативных препаратах трихомонады следует дифференцировать от свободно -
живущих жгутиковых, попадающих в препарат из воды или контаминированной
посуды. Эти сапрофитные жгутиковые имеют лишь 2 жгутика и двигаются прямо-
линейно. В окрашенных препаратах у влагалищных трихомонад отчетливо видны
оболочка, ядро, цитоплазма, блефаропласт, жгутики.
Культуральный метод является наиболее чувствительным, специфичным, одна-
ко и наиболее дорогим методом. Влагалищные трихомонады дают хороший рост
на искусственных питательных средах, содержащих антибиотики для подавления
роста сопутствующей микрофлоры. Материал обязательно следует забирать сте-
рильным зондом или бактериальной петлей. Микроскопическое исследование
культур и идентификацию Т. vaginalis следует проводить на 3-5-й день. Поскольку
длительность развития трихомонад в культуре зависит от величины посевной дозы,
отрицательный результат выставляют после исследования на 7-9-й и 11-17-й день
посева. Из придонного осадка готовят нативный препарат для микроскопии.
ПАРАЗИТЫ В БИОПТАТАХ ТКАНЕЙ
Биоптаты берут из здоровых или патологически измененных тканей в целях
обнаружения в них паразитических простейших и яиц или личинок гельминтов.
СРЕЗ КОЖИ НА НАЛИЧИЕ МИКРОФИЛЯРИЙ
Onchocerca volvulus и другие получают с помощью иглы и скальпеля или брит-
венного лезвия.
Принцип метода — обнаружение микрофилярий, вышедших из биоптата, поме-
шенного в физраствор.
796 ЛАБОРАТОРНАЯ ДИАГНОСТИКА ПАРАЗИТАРНЫХ БОЛЕЗНЕЙ
Исследуемый участок кожи обрабатывают спиртом или эфиром, после их
испарения вводят стерильную тонкую иглу (трого поверхностно-горизонтально и
приподнимают кожу над иглой. Срезают кусочек кожи, приподнятый с помощью
иглы, острым глазным скальпелем, удерживая его горизонтально и двигая против
иглы. При правильно выполненной процедуре получают кусочек размером около
2 мм, без следов крови. Затем помещают биоптат в каплю физиологического рас-
твора на предметное стекло. Исследуют препарат под микроскопом (малое увели-
чение) на наличие живых подвижных микрофилярий сразу после приготовления
препарата.
Микрофилярии начинают выходить из биоптата в раствор немедленно, через
30 мин его покидают около 70% и через час практически все, находившиеся в
биоптате. Если биоптат сразу после его получения взвесить на торсионных весах,
через час можно определить интенсивность инвазии (количество микрофилярий
в 1 мг кожи). При очень большом числе микрофилярий для облегчения подсчета
препарат обезвоживают, добавляя каплю формалина.
БИОПТАТ ИЗ КОЖНОГО ИНФИЛЬТРАТА, ЛЕЙШМАНИОМЫ НА НАЛИЧИЕ АМАСТИГОТ
ДЕРМАТОТРОПНЫХ ЛЕЙШМАНИЙ
Возбудителями являются Leishmania tropica (антропонозная форма кожного
лейшманиоза) и Leishmania major (зоонозная фирма кожного лейшманиоза).
Проводят микроскопию клеток краевого инфильтрата вокруг язвы на наличие
лейшманий в макрофагах или вне их. Биоптаг необходимо получить с минималь-
ным количеством крови и максимальным количеством макрофагов Для этого уча-
сток инфильтративного валика анемизируют, сжимая его двумя пальцами левой
руки или пинцетом, делают надрез острым скальпелем и соскабливают кусочек
ткани. Биоптат можно получить, откусив кусочек ткани пинцетом, или ввести
в инфильтрат стерильную зубоврачебную иглу с насечкой и вращать ее в ткани
инфильтрата, а затем извлечь вместе с захваченным материалом. Биоптат вместе с
сукровичной жидкостью переносят на предметное стекло, размазывают получен-
ный мазок, фиксируют, красят по Романовскому-Гимзе и исследуют с иммерсион-
ной системой, как при исследовании мазка костного мозга.
БИОПТАТ МЫШЦ НА НАЛИЧИЕ ТРИХИНЕЛЛ (ВСПОМОГАТЕЛЬНЫЙ МЕТОД)
Обнаружение инкапсулированных или живых личинок трихинелл в мышцах
приводят компрессионным методом или методом переваривания.
Порядок приготовления и исследования препаратов. Для исследования
компрессионным методом берут небольшой биоптат дельтовидной, двуглавой или
икроножной мышцы, делают не менее 24 срезов вдоль мышечных волокон, сдав-
ливают срез между двумя стеклянными пластинами и исследуют под микроскопом
при малом увеличении. При интенсивной иньазии в препарате обнаруживают
личинки трихинелл; при слабой интенсивности инвазии требуется исследование
большого количества срезов (70-100). Размеры личинок — 0,2-0,6 мм.
БИОПСИЯ ТКАНЕЙ НА ЦИСТИЦЕРКИ
При гениозе у пациента может развиться цистииеркоз — паразитирование в
органах и тканях (глазу, конъюнктиве, мозгу, подкожной клетчатке, мышцах)
личиночной стадии свиного цепня — Таета solium. Биоптат ткани (например,
подкожного узла) раздвигают препаровальными иглами в целях выделения бело-
ватого пузырька 0,5-2,0 см в диаметре — цистицерка. Его раздавливают между
двумя предметными стеклами и исследуют под микроскопом. Наличие в пузырьке
сколекса с 4 присосками и венчиком крючьев подтверждает диагноз цистицеркоза.
Если цистицерк пропитан известью, для обнаружения головки ею необходимо
предварительно декальцинировать в 4% растворе азотной кислоты в течение 1 ч.
ЛАБОРАТОРНАЯ ДИАГНОСТИКА ПАРАЗИТАРНЫХ БОЛЕЗНЕЙ 797
БИОПТАТЫ ИЗ ЕРХНЕГО ОТДЕЛА ЖЕЛУДОЧНО-КИШЕЧНОГ О ТРАКТА
В них можно обнаружить личинки Anisakidae spp., которые проникают в сли-
зистые и более глубокие ткани полости рта, пищевода, желудка (иногда и в более
дистальные отделы ЖКТ). Размеры личинок — 0,3x1-3 см. Вокруг личинки быстро
формируется гранулема, в некоторых случаях инвазия осложняется флегмоной.
ИССЛЕДОВАНИЕ СОДЕРЖИМОГО КИСТЫ
Материал получают во время оперативных вмешательств и исследуют при подо-
зрении на эхинококкоз. Диагностическими признаками эхинококкоза являются
слоистая структура стенки кисты, обнаружение в жидкости кисты сколексов и
крючьев эхинококка.
ИССЛЕДОВАНИЕ СОДЕРЖИМОГО АБСЦЕССА
Осложнением кишечного амебиаза является развитие внекишечных поражений
и в частности амебного абсцесса (чаще печени, легких). В содержимом амебного
абсцесса возбудители амебиаза не выявляются. Содержимое представляет собой
клеточный детрит сливкообразной консистенции, темно-коричневого или серого
цвета.
Осложнением фасциолеза является развитие фасциолезного абсцесса. В содер-
жимом абсцесса можно обнаружить яйца Е hepatlea, кристаллы Шарко-Лейдена.
ИССЛЕДОВАНИЕ ГЕЛЬМИНТОВ, ИЗВЛЕЧЕННЫХ ИЗ-ПОД КОЖИ И СЛИЗИСТЫХ
ОБОЛОЧЕК, ВКЛЮЧАЯ ОРГАН ЗРЕНИЯ
Возбудитель — нематода Dirofilaria repens, основными хозяевами которой явля-
ются собаки, переносчиком — комары. В России регистрируются местные случаи
заражения дирофиляриозом. Под кожей и слизистыми оболочками (часто в орга-
не зрения) развиваются единичные незрелые особи. Характерными признаками
гельминтов являются белый цвет, жесткая структура, напоминающая леску или
скрипичную струну, длина до 30 см, толщина 1-2 мм. При микроскопическом
исследовании выявляют продольную исчерченность тела гельминта и кутикуляр-
ные шипы.
МЕТОДЫ ИММУНОДИАГНОСТИКИ ПАРАЗИТАРНЫХ
ЗАБОЛЕВАНИЙ
Методы иммунологической диагностики относятся к непрямым методам и
основаны на выявлении антигенов возбудителей или антител, вырабатываемых
против антигенов возбудителей. Эти методы в клинической паразитологии при-
обретают все большее значение. Необходимость применения иммунологических
методов возникает при невозможности получения материала для микроскопии
неинвазивными способами на протяжении всего течения инфекции/инвазии (ток-
соплазмоза, внекишечного амебиаза, трихинеллеза, токсокароза, эхинококкоза,
цистицеркоза, ларвального парагонимоза) или в острой фазе, когда диагностиче-
ские стадии возбудителей еще не появились (описторхоза, легочного парагони-
моза, фасциолеза). Иногда иммунологические методы применяют в дополнение
к паразитологическим, если чувствительность последних в хронической фазе
недостаточна из-за слабой интенсивности инфекции/инвазии (трипаносомозы,
криптоспоридиоз, стронгилоидоз, шистосомозы).
При анализе и интерпретации результатов серологической диагностики следует
считаться с явлением иммунологической инкубации, то есть появлением антител
или формированием кожной реакции на антигены паразита лишь через опреде-
ленный латентный период, в течение которого антитела применяемым методом
еще не определяются, а кожная проба еще не стала положительной. Следует пом-
798 ЛАБОРАТОРНАЯ ДИАГНОСТИКА ПАРАЗИТАРНЫХ БОЛЕЗНЕЙ
нить также о возможности перекрестных серологических реакций при некоторых
паразитарных заболеваниях (токсокарозе, описторхозе и др.).
ТОКСОПЛАЗМОЗ
В целях лабораторной диагностики этой широко распространенной протозой-
ной инфекции применяют очень широкий набор иммунологических методов:
кожную пробу с токсоплазмином. РСК, РИГА, НРИФ, ИФА, а также оригинальную
реакцию с красителем (реакцию Сэбина-Фельдмана). В последние годы ведущее
место в иммунодиагностике токсоплазмоза занимают НРИФ в полуколичествен-
ном варианте (титр IgG) и ИФА в количественном варианте (IgG и IgM, в МЕ/мл).
Интерпретация результатов представлена в табл. 26-3.
Таблица 26-3. Интерпретация результатов иммунологического обследования пациента на токсо-
плазменную инфекцию (по Лысенко А.Я., 1995)
Метод исследования Результат Интерпретация
Серологическое исследование на антитоксоплазменные IgG Низкие титры (низкая кон- центрация, в МЕ/мкл) Токсоплазмоносительство или субклиниче- ская хроническая инфекция
Высокие титры (высокие показатели, в МЕ/мкл) Острая инфекция, клинические проявления обусловлены токсоплазмозам
Серологическое исследование на антитоксоплазменные IgM Положительный у ново- рожденного Врожденный токсоплазмоз
Положительный у детей и взрослых Свежая токсоплазменная инфекция или рецидив
АМЕБИАЗ ВНЕКИШЕЧНЫЙ
Применяют тест-системы НРИФ и ИФА.
ЛЯМБЛИОЗ
Применяют тест-систему ИФА для определения антител против лямблий. Имеет
вспомогательное значение при массовых противоэпидемических мероприятиях. В
последние годы применяют иммунохроматографический метод для определения
антигена лямблий в фекалиях. Чувствительность этого метода высокая, однако
снижается при исследовании проб фекалий, хранившихся до исследования в кон-
серванте.
ТОКСОКАРОЗ
Применяют тест-систему ИФА для определения антител против личинок токсо-
кар. Является основным методом диагностики токсокароза.
ТРИХИНЕЛЛЕЗ
Применяют тест-систему ИФА для определения антител против личинок трихи-
нелл. Является основным методом диагностики трихинеллеза,
0ПИСТ0РХ03
Применяют иммуноферментные тест-системы для определения антител против
возбудителей описторхоза. Является методом диагностики острой фазы инва-
зии, продолжающейся до 1,5 мес. В дальнейшем диагноз можно поставить путем
копроовоскопического исследования.
ЭХИНОКОККОЗЫ
Применяют тест-системы ИФА для выявления антител к Echinococcus
multilocularis, а также ИЭФ и РСК с лиофилизированным антигеном Е. granulosus.
Является основным методом диагностики, необходимо сопоставление его резуль-
татов с инструментальными методами исследования новообразования, а также
паразитологического исследования содержимого эхинококковой кисты, полу-
ЛАБОРАТОРНАЯ ДИАГНОСТИКА ПАРАЗИТАРНЫХ БОЛЕЗНЕЙ 799
ченной при оперативном вмешательстве. Пунктировать кисту в диагностических
целях нельзя ввиду риска анафилактического шока и диссеминирования возбуди-
теля.
ТРИХОМОНОЗ
Применяют НРИФ. Отпечатки материала из различных очагов поражения
наносят на предметное стекло со специальными лунками одноразовыми зондами с
хорошо сорбирующим ватным тампоном. Трихомонады видны в виде полиморф-
ных образований с характерным ярко-зеленым свечением. Бактериальная флора
окрашивается в оранжевый цвет, лейкоциты, эпителиальные клетки, сперматозои-
ты — в красно-бурый. Результат считается отрицательным при отсутствии клеточ-
ных элементов со специфическим свечением.
Чувствительность и специфичность иммунологического метода сравнимы с
таковыми при кульгуральном методе. Он сравнительно дешев, прост в выполне-
нии, позволяет получить ответ в течение 40-60 мин.
ЛЕЙШМАНИОЗ ВИСЦЕРАЛЬНЫЙ
Применяют ИФА и непрямую реакцию флюоресцирующих антител — НРИФ.
ЭКСПРЕСС-ДИАГНОСТИКА МАЛЯРИИ
Позволяют поставить диагноз без микроскопа. Применение экспресс-методов
допускается в особых случаях по специальному решению органов здравоохране-
ния, ответственных за противомалярийные программы.
Экспресс-методы основаны на выявлении биохимических субстанций паразита
в крови пациента. Разработаны и применяются на практике следующие тест-
системы, содержащие антитела:
к богатому гистидином белку HRP2, специфичному для трофозоитов
Р. falciparum (выявляют бесполую паразитемию при тропической малярии);
> лактатдег идрогеназе pLDH, специфичной для Р falciparum (выявляют тропи-
ческую малярию, не делая различий между бесполыми паразитами и гамето-
цитами);
лактатдегидрогеназе, специфичной для Р. vivax (выявляют трехдневную
малярию);
• о панспецифичной лактатдегидрогеназе или альдолазе (выявляют малярию
без указания вида).
Чувствительность и специфичность этих тестов достаточно высоки (более
90%), если численность паразитов превышает 100 в 1 мкл, однако ниже, чем чув-
ствительность стандартного исследования толстой капли крови. Преимущества
быстрых методов перед микроскопией заключаются в том, что они просты, зани-
мают меньше времени, в меньшей степени зависят от субъективных факторов, не
требуют специального оборудования. Недостатками экспресс-методов являются:
< невозможность выявить гаметоциты Р. falciparum (методами, основанными
на выявлении HRP2) или отличить выявленные гаметоциты от бесполых
форм (при использовании pLDH);
• > невозможность оценить уровень паразитемии;
• о невозможность использования их для оценки эффективности лечения,
поскольку тесты остаются положительными (в течение до 2 нед при HRP2)
после прекращения шизогонии.
ПРОЧИЕ ПАРАЗИТАРНЫЕ БОЛЕЗНИ
Методы иммунологической диагностики паразитарных болезней постоянно рас-
ширяются и совершенствуются. Активно развиваются молекулярно-биологические
методы с выявлением нуклеиновых кислот возбудителей паразитарных болезней.
Однако еще не накоплен достаточный материал для широких рекомендаций этих
800 ЛАБОРАТОРНАЯ ДИАГНОСТИКА ПАРАЗИТАРНЫХ БОЛЕЗНЕЙ
методов к использованию в клинико-диагностических лабораториях при диагно-
стике паразитарных заболеваний.
СПИСОК РЕКОМЕНДУЕМОЙ ЛИТЕРАТУРЫ
Авдюхина Т.Н., Константинова Т.Н., Горбунова Ю.П, Лабораторная диагностика гель-
минтозов. Ч. I. Нематодозы: Учебное пособие. — М.: РМАПО, 2007. — 51 с.
Авдюхина Т.Н., Константинова Т.Н.. Горбунова Ю.П Лабораторная диагностика гель-
минтозов. Ч. II. Цестодозы и трематодозы: Учебное пособие. — М.: РМАПО, 2007. — 48 с.
Беляев А.Е., Званцов А.Б., Авдюхина Т.И. Практическое руководство по эпидемиологи-
ческому надзору за малярией для стран Европейского региона, столкнувшихся с возвратом
малярии. — Копенгаген. ВОЗ, 2006. — 119 с.
Лысенко А.Я., Владимова М.Г., Кондрашин А.В., Майори Дж. Клиническая паразитоло-
гия. — Женева: ВОЗ. 2002. — С. 15-78.
Лысенко А.Я., Горбунова Ю.П., Константинова Т.Н., Авдюхина Т.И. Лабораторная диа-
гностика кишечных протозоозов: Учебное пособие. — М.: РМАПО, 2007. — 42 с.
Основные методы лабораторной диагностики паразитарных болезней. — Женева: ВОЗ,
1994. - 131 с.
Bench Aids for the diagnosis of filarial infections. — Geneva: WHO, 1997.
Bench Aids for the diagnosis of intestinal parasites. — Geneva: WHO, 1994.
ПРЕДМЕТНЫЙ УКАЗАТЕЛЬ
А
Аденовирусы 641
Акантамебы 779
Амеба дизентерийная 788
Амебиаз 798
Анализ
- иммуноферментный 288
- иммунофлюореснентный 287
- иммунохромотографический 290
Анемия иммунная гемолитическая 188
Антигены эритроцитов 177
Антитела
- антинуклеарные 81
- антифосфолипидные 96
- антицитруллиновые 102
- Доната-Ландштейнера 189
- к RA-33 106
- к ацетилхолиновому рецептору 140
- к базальной мембране клубочка 111
- к гладким мышцам 117
- к глиадину 122
- к гликопротеину-1 99
- к глутамат-декарбоксилазе 130
- к кардиолипину 97
- к микросомам печени-почек 118
- к митохондриям 119
- к островковым клеткам 128
- к пероксидазе 124
- к скелетным мышцам и миокарду 142
- к тиреоглобулину 126
- к фактору Кастла 120
- к эндомизию 121
Апробация крови доноров 180
Аспергиллез 756. 769
Астровирусы 644
Аутоантитела
- при пемфигоиде 144
- при пузырчатке 143
Б
Бабезии 777
Бактерии
- гемофильные 375
- грамположительные 406
- неферментирующие 380
Баламутии 779
Балантидиоз 789
Бластомикоз 769
Болезнь
- Аддисона 67,125.131
- Аддисонова 38
- Ауески 572
- Баттона 130
- Бехчета 107, 108
- Бюргера 108
- гемолитическая плода
и новорожденного 185
- Грейвса 67, 92,127
- Дюринга 145
- Иценко-Кушинга 18
- Кавасаки 107
- Крона 67
- Лайма 659
- Такаясу 107
- Чагаса 143
Бордетеллы 323
В
Вальденстрема
макроглобулинемия 113
Вассермана реакция 96
Вирус
- бешенсз ва 645
- гепатита
— А 673
— В 674
— С 676
— D 678
— Е 679
- гриппа 649
— H1N1 652
- иммунодефицита 653
- клещевого энцефалита 656
- кори 659
- краснухи 663
- парагриппа 665
- респираторно-синцитиальный 699
- эпидемического паротита 667
- Эпштейна-Барр 25, 55,139, 542
Вирусология 542
- методы исследования вирусов 5б1
— вирусоскопия 571
— гемагглютинация 575
— гемадсорбции 575
— иммуноферментный анализ 607
802 ПРЕДМЕТНЫЙ УКАЗАТЕЛЬ
— иммунофлуоресценция 582
— клеточные культуры 562
— молекулярно-биологические 613
---амплификационные 619
---гибридизационные 615
— обеспечение качества
исследований 633
— преаналитический этап 627
— прямая микроскопия 571
— развивающиеся куриные
эмбрионы 561
— реакция
---гемагглютинации 590,599
--- нейтрализации 591
---связывания комплемента 601
---торможения гемагглютинации 596
— серодиагност ика 589
— цитология 571
— электронная микроскопия 578
- обшие сведения о вирусах 548
- орга низация работы
лабораторий 545
- таксономия вирусов 550
Возбудители микозов 728
Волчанка системная красная 23, 67
Г
Гемолиз 24
Герпесвирусы 680
Гиалогифомикоз 770
1истоплазмоз 770
Гонококк 519
Гранулематоз Вегенера 67,106
Д
Дерматит Дюринга 115
Дерматомиозит 67, 76
Дерматофитии 759
Диагностика
- антифосфолипидного синдрома 150
- аутоиммунных васкулитов 157
- аутоиммунных заболеваний 66
- бактериологическая 230
— TORCH-комплекс 233
— исследование
--- желчи 257
---крови 239
---материала 269, 272, 273,281
--- мокроты 247
---отделяемого из мочеполовых
органов 265
---поомывных вод 281
---рвотных масс 281
---спинномозговой жидкости 242
--- фекалий 277
— лабораторная 230
---мокроты 252
— методы исследования 283
— раневые инфекции 244
— этиологическая 234
- заболеваний кожи 172
- полиэндокринопагий 170
- ревматических заболеваний 146
- ревматоидного артрита 154
- сахарного диабета 169
- системной красной волчанки 149
Ди октофимоз 793
3
Заболевания
- аутоиммунные 66
— антитела 70
— лабораторные показатели 74
Зигомикозы 759
Зоб Хашимото 67, 94,127
И
Иерсинии 365
Изоспора 790
Иммунитет
- алг оритм лабораторного
исследования 61
- антиген-неспецифические
факторы 48
- антиген-специфические факторы 51
- врожденный 17
— гуморальные факторы 21
— нарушения 36
гуморального иммунитета 39
дефекты Т-лимфоцитов 36
комбинированные 40
- и воспаление 48
- приобретенный 25
— Fc-рецепторы 31
— иммуноглобулины 31
— система
В-лимфоцитов 30
Т-лимфоцитов 25
Иммуногематология 176
- лабораторные исследования 189
- терапия концентратом
тромбоцитов 190
Иммунодефицит
- вторичный 47
- первичный 35
Иммунология лабораторная 17
Индикация
- антигенов возбудителей 287
- молекулярно-генетическая
бактерий 291
- специфических антител 295
Интерлейкины 221
Интерферон 226
К
Калицивирусы 691
Кампилобактеры 388
Кандидоз 757
Клетки
- врожденного иммунитета 17
— NK-клетки 20
— моноциты и макрофаги 20
— полиморфноядерные лейкоциты 17
— эозинофилы 19
- Лангерганса 20
Клостридии 451
Коксиеллы 335
Кокцидиоидомикоз 769
Комплексы
- иммунные
— циркулирующие 112
--криоглобулины 113
- мембрано-атакующий 22
Коринебактерии 468
Коронавирусы 694
Криптококкоз 758, 770
Криптоспоридии 789
Кровепаразиты 773
Культуры клеток
- диплоидные 565
- первично-трипсинизированные 564
- перевиваемые 569
Л
Лаваж бронхоальвеолярный 731
Легионеллы 329
Лейшмании 778
Лептоспиры 460
Лимфома Ходжкина 19
Листерии 445
Лихорадка геморрагическая 669
Лямблиоз 789, 798
ПРЕДМЕТНЫЙ УКАЗАТЕЛЬ 803
М
Малярия 774
Менингококк 438
Микобактерии 484
Микозы
- кистей и стоп 767
- кожи 767
- легких 768
- слизистых оболочек 767
Микология 726
- культуральное исследование 754
- лабораторная диагностика 729
- лампа Вуда 735
- макроскопические исследования 735
Микрофилярии 777
Микрочипы биологические 625
Мукормикоз 770
Н
Неглерии 779
Нематоды 790
О
Описторхоз 798
Определение чувствительности к
антимикробным препаратам 300
П
Папилломавирусы 708
Паразиты
- в биоптатах тканей 795
- в дуоденальном содержи мом 791
- в мокроте 793
- в моче 792
- отделяемого мочеполовых
путей 794
Паракокцидиоидомикоз 770
Парвовирус 706
Пенициллиоз 770
Полиовирусы 714
Проба Кумбса 184
Р
Реакции посттрансфузионные 184
Реакция
- агглютинации 290
Риновирусы 700
Ротавирусы 702
804 ПРЕДМЕТНЫЙ УКАЗАТЕЛЬ
С - Кауфмана 346, 362
Синдром - аллергический 36 - антифосфолипидный 150 - Бехчета 116 - Вискотта-Олдрича 43 - Гийена-Барре 67,138 - голых лимфоцит ов 44 - Гуда 43 - ГХдпасчера 67,107.111 - Ди Джорджи 37 - Дресслера 142 - инфекционный 36 - Карпентера 170 - Костманна 19 - Ламберта-Итона 137 - Леттерера-Зиве 45 - Луи Барр 45 - Миллера-Фишера 138 - Незелофа 45 - Оменна 45 - пролиферативный 36 - Рейно 87, 93 - Рейтера 55 - системного воспалительного ответа 216 - Фелти 18 - Чедиака-Хигаси 37 - Черджа-Стросс 106 - Шарпа 87, 148 - Шегрена 75 - Шмидта 170 - Ядассона-Левандовского 38 Система -АВО 177 — определение группы крови 179 - резус 177 — определение резус- принадлежности 179 Склеродермия 67 Склероз - рассеянный 133 - системный 75 Споротрихоз 770 Среда - ВСУЕ-а 330 -НТМ 305 -MRSA 307 - Борде-Жангу 327 - Игла 563 - Китта-Тароцци 454 - Клиглера 347 - Кэри-Блеер 278 - Левина 344 - Мак-Конки 344 - Мюллера 345. 362 - Олькеницкого 371 - Плоскирева 345 - Раппопорта 346 - Ресселя 347 - Симмонса 349 - СКС 278 - Стюарта 343, 389 - Хеттингера 371 - Хью-Лейфсона 346 - Цейсслера 455 - Эймса 343, 389 - Эндо 344 Стафилококки 406 Стрептококки 417 - пневмококковый 435 Т Тельца - Бабеша-Негри 572 - Боллингера 572 - Донована 268 Терапия трансфузионная 179 Тест Ваалера-Розе 100 Токсокароз 798 Трематоды 791 Трепонемы 496 Трипаносомы 780 Трихинеллез 798 Трихомоноз 799 Тромбопоэтин 228 Ф Фактор - антинуклеарный 74,75 - Виллебранда 116 - Касла 70 - Кастла 120 - колониестимулирующий 227 - ревматоидный 74, 100 Феогифомикозы 770 Физиология грибов 727 Франсиселлы 530 Фузариоз 770 Фунгемия 768
ПРЕДМЕТНЫЙ УКАЗАТЕЛЬ 805
X
Хеликобактеры 394
Хламидии 312
Ц
Цсстоды 791
Циклоспора 790
Цитокины 193
- диагностическое значение 220
- методы оценки 198
— биологический 198
— иммуногистохимический 212
— иммунофррментный 201
— иммунохимический 200
— определение продукции
клетками 207
- при аллергии 219
- при аутоиммунной патологии 217
- при сепсисе 216
- роль в патогенезе заболеваний 213
Ш
Шенлейна-Геноха пурпура 107
Шистосомоз 793
Э
Энтеробактерии 342
Энтеровирусы 720
Эритропоэтин 227
Эхинококкоз 798
Abe
Bartonella (Rochalimaea) spp. 242
Borrelia spp. 242
Brucella spp. 242
Cardiobacterium hominis 242
Legionella spp. 242
Leptosp.ra spp. 242
Mycobacterium spp. 242
Streptobacillus moniliformis 242