/
















































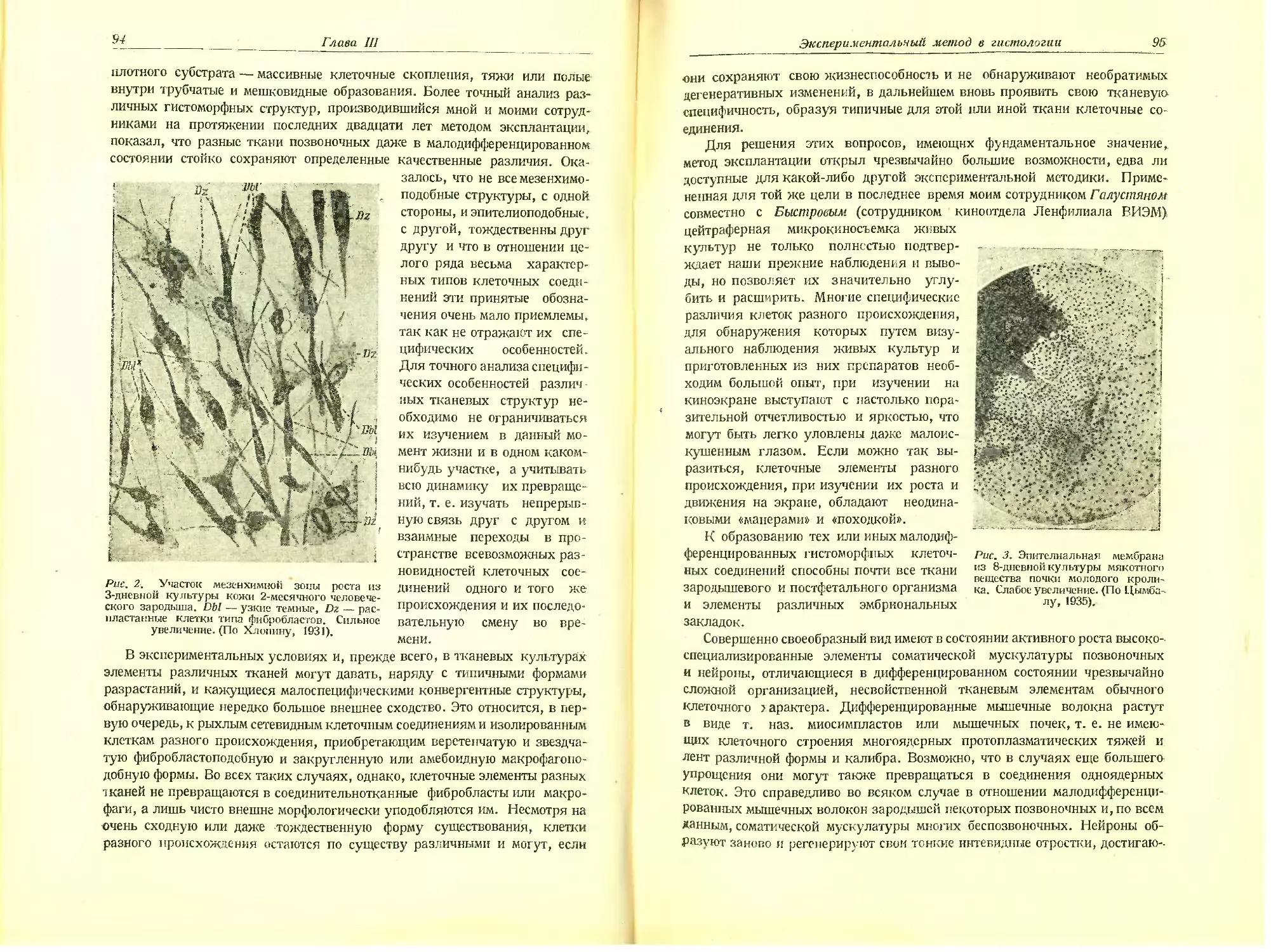




















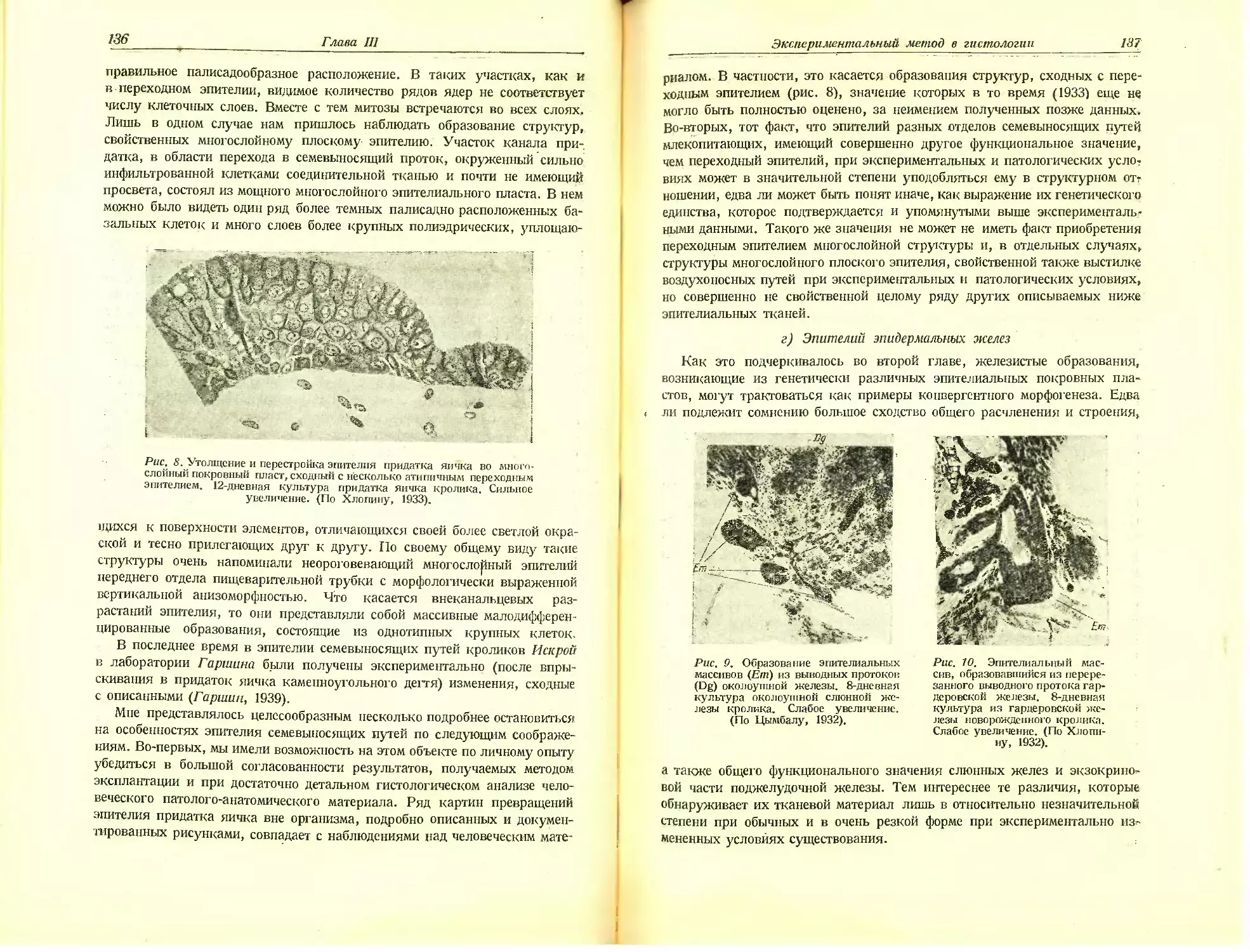
















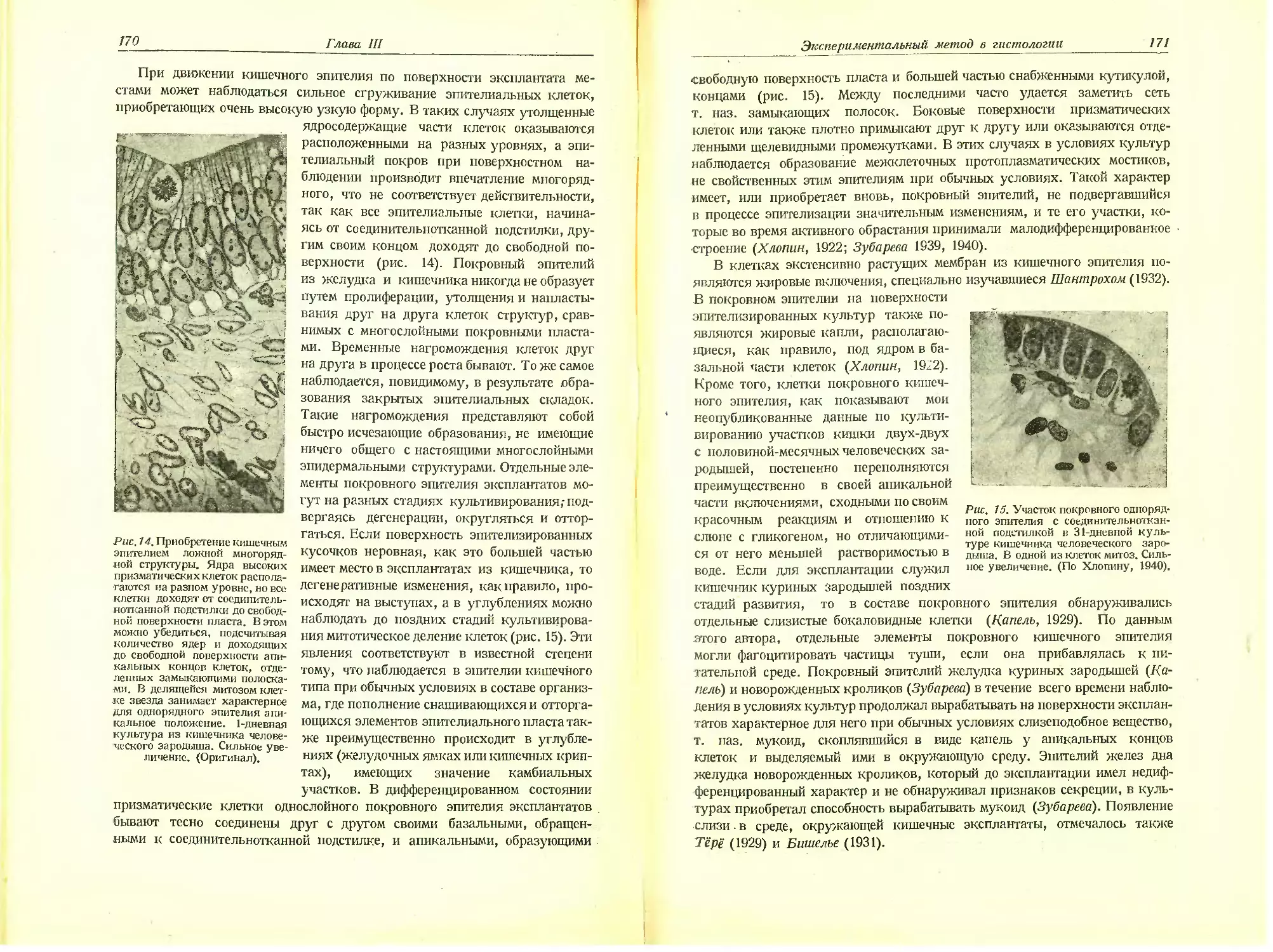




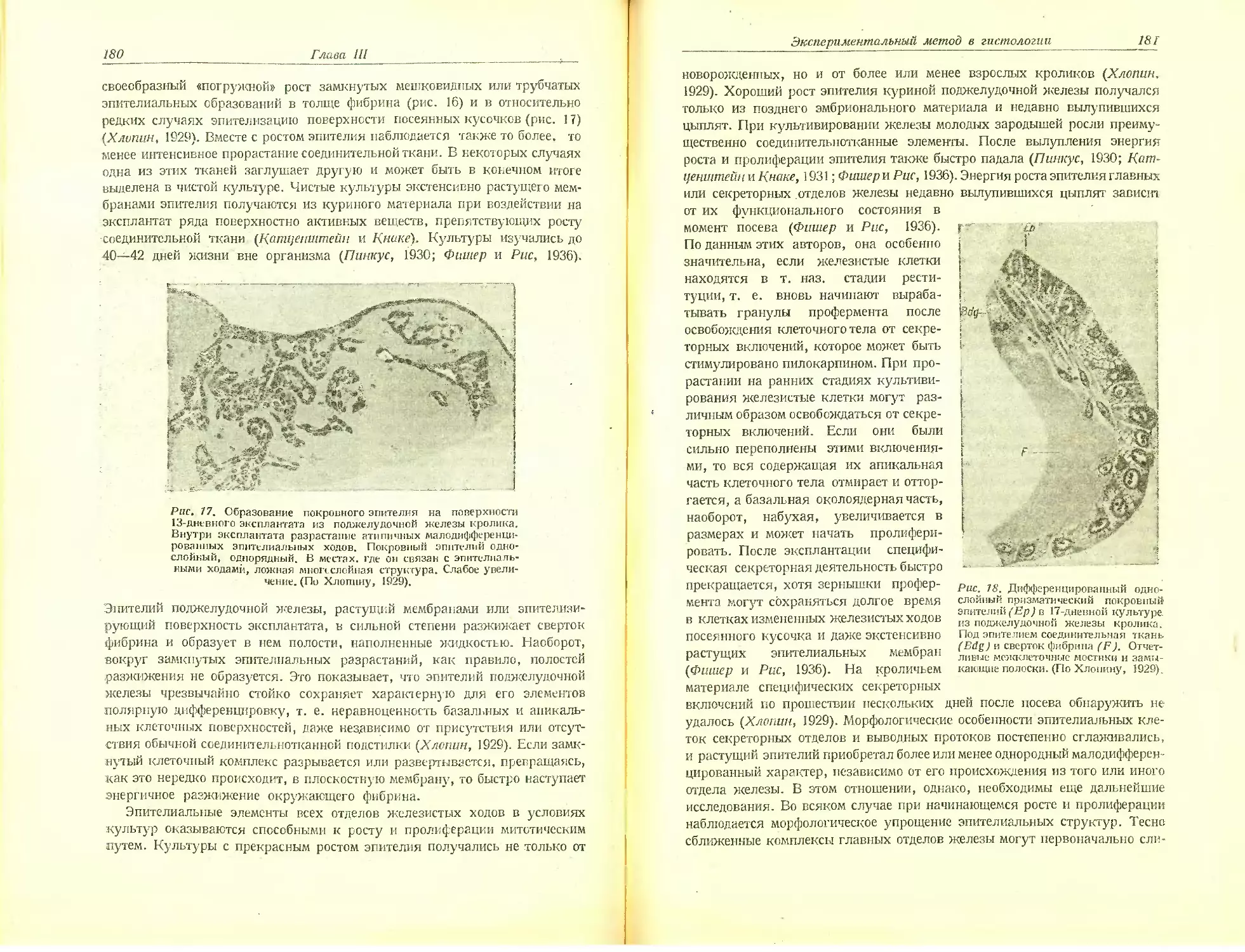














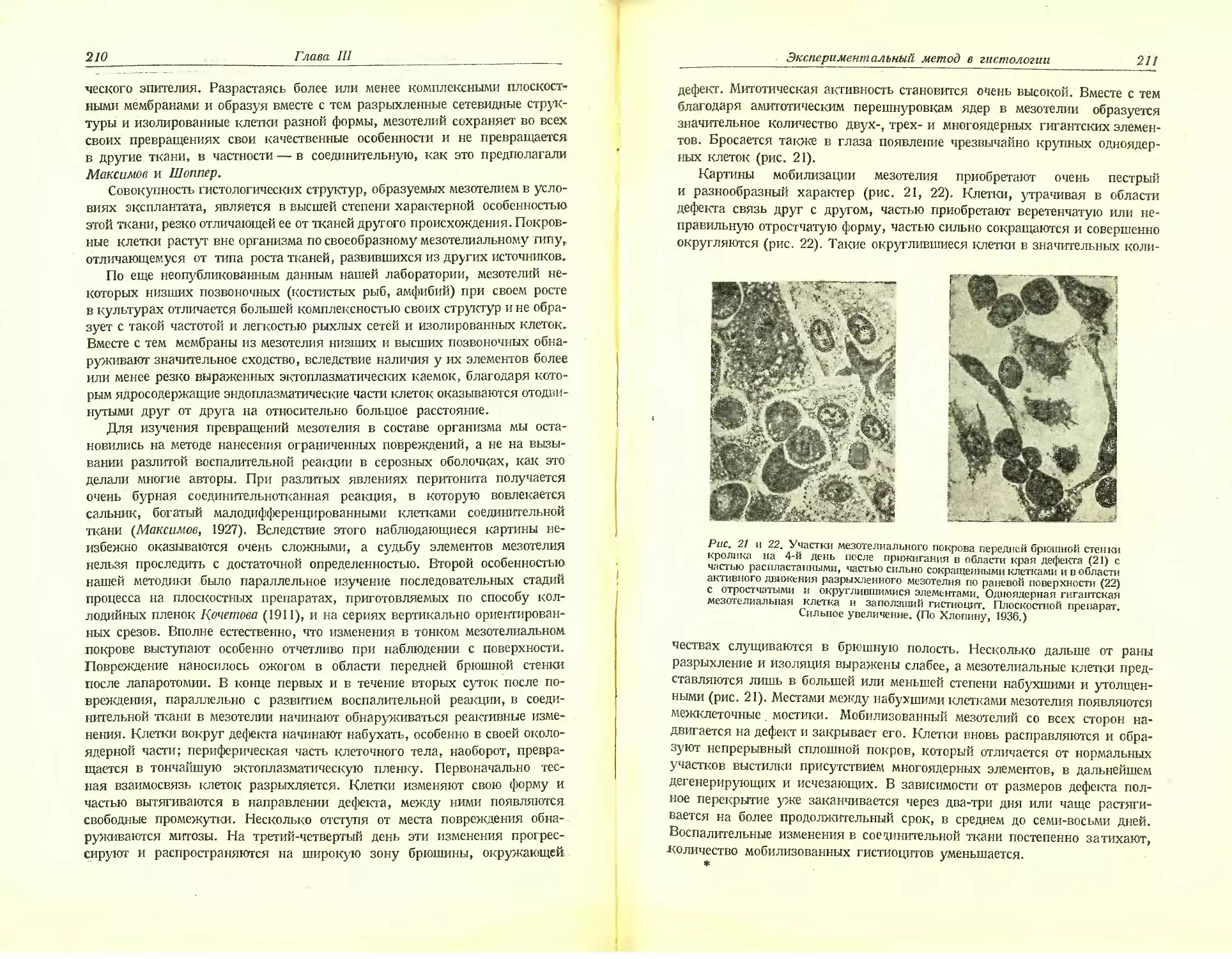



















































































































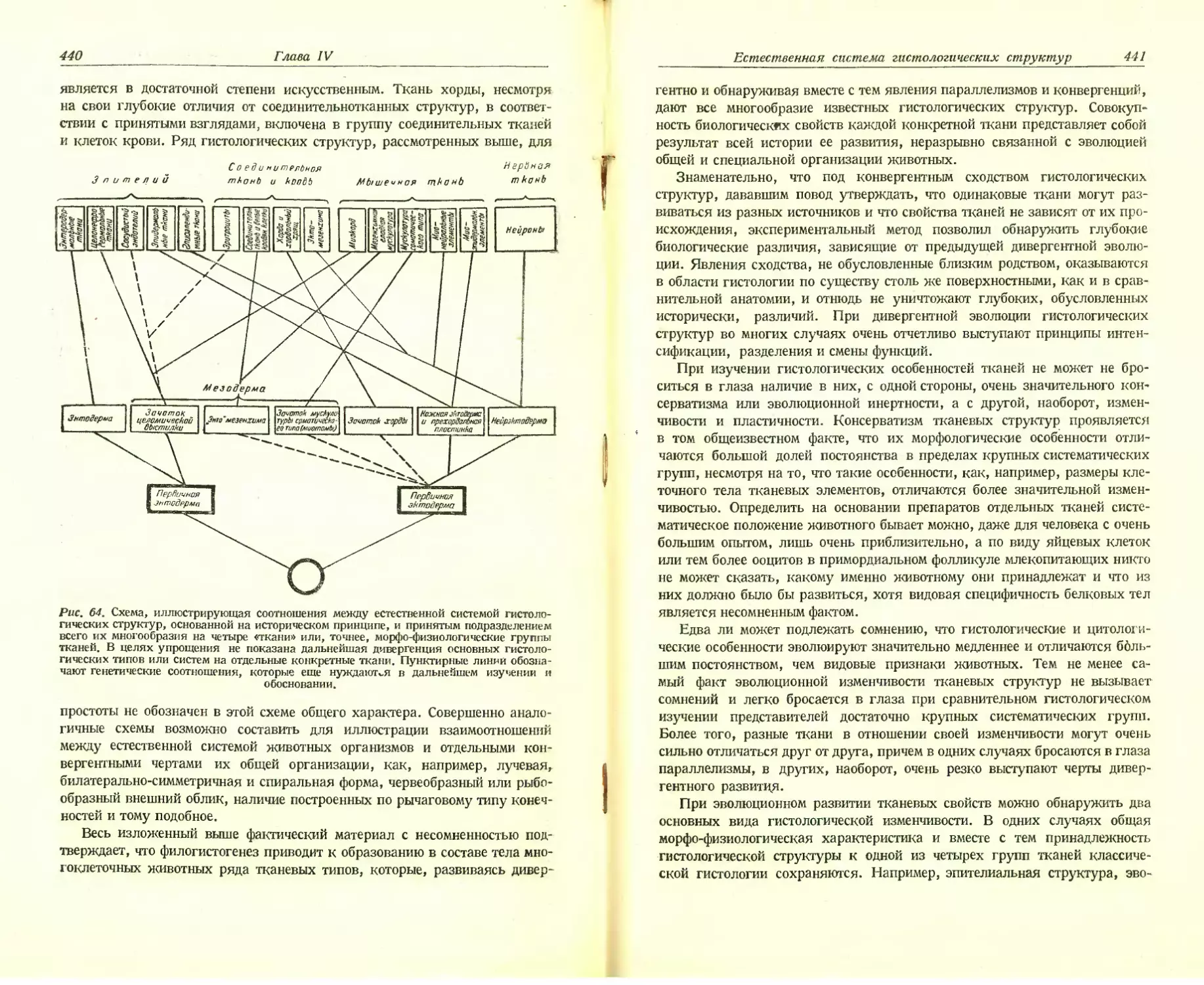


























Автор: Хлопин Н.Г.
Теги: медицина практическая медицина гистология академия наук ссср экспериментальные основы гистологии
Год: 1946



Текст
X л о п и н
ОБЩЕБИОЛОГИЧЕСКИЕ
И ЭКСПЕРИМЕНТАЛЬНЫЕ
ОСНОВЫ
ГИСТ ологи и
академия наук СОЮЗА ССР
и. г. хлоп ин
ОБЩЕБИОЛОГИЧЕСКИЕ
И
ЭКСПЕРИМЕНТАЛЬНЫЕ
ОСНОВЫ ГИСТОЛОГИИ
ИЗДАТЕЛЬСТВО АКАДЕМИИ НАУК СССР
19 4 6
ПРЕДИСЛОВИЕ
Настоящая книга посвящена изложению и обоснованию новой есте-
ственной системы гистологии, построенной на историческом или филогене-
тическом, а не на обычном и общепринятом морфо-физиологическом прин-
ципе. Она является результатом работы автора и научных коллективов, кото-
рыми он руководил на протяжении ряда лет: лабораторий кафедры гисто-
логии и эмбриологии Еоенно-медицинской академии Красной Армии имени
С. М. Кирова, цитологического отделения Онкологического института и
отделения экспериментальной гистологии и тканевых культур ЛФВИЭМ.
Значительная доля участия в этой общей работе принадлежит также иного-
родним товарищам, тесно связанным идейно и тематически с руководимыми
автором лабораториями.
Помимо уже опубликованного в отдельных работах эксперименталь-
ного материала в настоящей книге впервые находят свое отражение много-
численные новые данные автора и его сотрудников. Фактической централь-
ной части книги предпослан очерк развития основных гистологических
представлений и ряд соображений общего характера, позволяющих уни-
чтожить тот разрыв, который существует до последнего времени между
специальными гистологическими проблемами, с одной стороны, и законо-
мерностями эволюционной морфологии, включая эмбриологию и механику
развития, с другой. Вместе с тем уничтожается в значительной степени
искусственное разграничение и даже противопоставление общей гистоло-
гии и микроскопической анатомии.
Лежащий в основе фактической части книги экспериментально-гисто-
логический анализ тканей и использование данных патологической гисто-
' логии позволяют установить тесную органическую связь между основными
закономерностями так называемой нормальной гистологии и специаль-
ными медицинскими проблемами, которые тем самым приобретают новую
прочную общебиологическую, т. е. последовательно материалистическую
эволюционную, основу.
Личный опыт автора и коллективная работа его сотрудников позволили
охватить под общим углом зрения все основные группы тканей классиче-
ской гистологии. Однако не все ткани еще могут получить в настоящее
время одинаково полное и обстоятельное освещение. Поэтому в отношении
некоторых из них задачей автора является не столько решение, сколько
постановка вопросов с единой новой точки зрения.
4 Предисловие ___
Вводная историческая часть книги и часть второй главы писались во
фронтовой обстановке осажденного Ленинграда, а все прочие главы —
в Самарканде. Этими особыми условиями в значительной степени объяс-
няются пробелы в литературе, а также односторонний и неполный подбор
иллюстраций. Автору удалось использовать только тот материал, которым
он располагал во время подготовки к печати рукописи, законченной в
ноябре 1942 г. В настоящее время в текст хотелось бы внести допол-
нения и некоторые изменения. От этого, однако, пришлось отказаться,
чтобы не усложнить печатания и не вызвать его дальнейшей задержки.
Считаю своим долгом принести всем сотрудникам искреннюю благо-
дарность за счастливое время совместной работы, плодом которой, по
существу, является, настоящая книга.
Ленинград Н. Хлопан .
Март, 1946- г. - . • • :
I. ОЧЕРК РАЗВИТИЯ ГИСТОЛОГИЧЕСКОМ СИСТЕМЫ
1. ВВОДНЫЕ ЗАМЕЧАНИЯ
Вопросы систематики и классификации в биологических науках имеют
очень большое значение, так как в них отражаются основные установки
и уровень развития каждой данной дисциплины. Это демонстрируется
с большой отчетливостью историей возникновения и развития зоологи-
ческой и ботанической систем, которые первоначально основывались на
отдельных, более или менее бросающихся в глаза, внешних признаках
животных и растений, а затем с большей или меньшей полнотой и правиль-
ностью стали отражать их генетические отношения и историю развития.
Вместе с общим прогрессом научной мысли, на смену искусственным клас-
сификациям пришла естественная система живых существ, которая в свою
очередь развивалась, изменялась и совершенствовалась в соответствии
с обоснованием и* развитием эволюционного учения, накоплением и изуче -
нием нового фактического материала и развитием новых методов иссле-
дования.
Вместе с тем не подлежит сомнению, что для тех или иных специаль-
ных целей, помимо естественной системы, в биологии могут существовать
и существуют классификации, отражающие не общий путь эволюционного
процесса, а некоторые особенности общей организации или строения и
функции отдельных органов животных, к какой бы естественной группе
эти последние ни относились.
Однако такие классификации вовсе не могут подменять собой есте-
ственной системы, основанной на генетическом принципе. Если с такой точки
зрения подойти к существующим в гистологии взглядам на ткани и их под-
разделению на основные группы, то бросается в глаза, что общепринятая
до последнего времени классификация тканей не может быть названа их
естественной системой, так как генетический момент в ней или совершенно
не отражается *или отражается очень непоследовательно. Это может быть
показано с особенной отчетливостью, если проанализировать некоторые
моменты исторического развития наших представлений об основных тка-
невых группах. Как это ни удивительно, история гистологии очень недо-
статочно освещена в литературе. В курсах гистологии и в специальных
монографиях (Кацнельсон, 1939; Бермель, 1940) много внимания уделяется
истории учения о клетке, которое, несмотря на все свое огромное значение
Для гистологии, отнюдь не может отождествляться с ее историей.
6
Глава I
Настоящий краткий очерк имеет своей целью восполнить в некоторой
степени имеющийся в этом отношении пробел и показать вместе с тем исто-
рические корни современных гистологических представлений.
2. ПРЕДИСТОРИЯ ГИСТОЛОГИЧЕСКИХ ПРЕДСТАВЛЕНИЙ
Как известно, в первом приближении представления о тканях, как
о материалах, из которых построены разные части тела животных, воз-
никли еще в классической Элладе. Аристотель обозначал, как «однород-
ные части» (та 6u.oiop.epTj popta), материал различных по форме костей,
мышц, «жил» и других органов, противопоставляя этот материал органам
или «неоднородным частям» ("“ avopoiop.epTt popta). Он различал влажные
(точнее жидкие) и мягкие части (кровь, соки — iXSpec, жир, семя, мозг,
желчь, молоко и «мясо») и твердые или плотные (кости, жилы — veopa,
вены, внутренности и т. п.), отмечая в то же время их некоторые функ-
циональные особенности. Гален (II век н. э.) обозначал, как similaria или
partes simiiares, материал костей, хрящей, связок, оболочек, мышц и т. д.,
считая вслед за Гиппократом, что человеческое тело образовано такими
составными частями: 1) упомянутыми выше однородными или «сходными»;
2) органами (instrumenta), как, например, мозг, легкие, сердце, внутрен-
ности, и 3) правильным сочетанием однородных и органных частей. Эти
представления о строении тела перешли к арабским последователям антич-
ной философии. Например, Абу Сина (Авиценна, 980—1037, по происхо-
ждению перс, родом из-под Бухары) принимает девять однородных частей:
кости, хрящи, нервы, сухожилия, связки, артерии, вены, оболочки, мясо
(мышцы и внутренности).
Одна из первых монографий старых европейских анатомов, посвящен-
ных «однородным частям» тела, принадлежит Габриэлю Фаллопию (1575,
посмертное издание его ученика Койтера'). «Мы будем рассуждать, — пишет
Фаллопий в первой главе, — об однородных и простых частях, от знания
которых зависит понимание сложных частей и всего тела в целом».
Однородные части получили, по Фаллопию, свое обозначение оттого,
что они делимы на части, свойства которых остаются такими же, какими
были у целого, и не зависят от того, откуда именно эти части взяты. Слож-
ные же части тела не делимы; например, руку нельзя разделить на несколько
рук. Фаллопий подчеркивает разницу между материалом или веществом
(substantia) и оформленным веществом (substantia figurata). Он считает,
что аристотелевские «однородные части» по степени своей плотности могут
быть сведены к четырем классам: 1) жидкие или влажные части (кровь,
желчь, молоко, соки, семя); 2) нежидкие, но мягкие части (жир, костный
мозг, мясо и внутренности); 3) твердые и более прочные части (нервы, вены, f
артерии, кожа, оболочки, волокна, пузырь); 4) самые твердые части (кости,
хрящи, волосы, ногти, а у животных и птиц, кроме того, рога и перья).
Сам Фаллопий подходит к материалу человеческого тела с двух различ-
ных точек зрения. Пользуясь современной терминологией, их можно обо-
значить как генетическую и морфологическую.
Очерк развития гистологической системы 7
Исходя из господствовавших еще в то время представлений, корни
которых восходят к античным мыслителям, что человеческий зародыш раз-
вивается в результате соединения спермы или семени и менструальной
крови, он подразделяет все однородные части по их предполагаемому про-
исхождению на «семенные» (partes seminales — вены, артерии, нервы, кожа
и другие оболочки, кости, связки и т. п.) и на «кровяные» (partes sangui-
neas— мясо, паренхима,* сало и пр.). По форме или внешним призна-
кам (forma) Фаллопий различает «более теплые и более холодные» части
или «влажные» (жидкие) и сухие, поясняя, что, чем тверже части, тем они
суше, чем мягче, тем они влажнее («quanto ad tactum duriores sunt, tanto
sicciores; quanto molliores, tanto humidiores»). По степени сухости или плот-
ности «однородные части» подразделяются на 10 групп и располагаются
в ряд, в начале которого находятся жир и сало, а в конце хрящ, кость и
вещество зубов. Весь фактический материал он излагает в 12 главах: 1) кость,
2) хрящ, 3) нервы, 4) связки, 5) сухожилия, б) оболочки (мембраны), 7) ар-
терии, 8) вены, 9) сало (жир), 10) костный мозг (как разновидность жира),
11) паренхима (вещество внутренностей и мышц), 12) кожа (cutis) и над-
кожица, или кутикула. Относительно кожи подчеркивается, что она состоит
из двух частей — «семенной > (собственно кожа) и самой наружной, очень
тонкой, называемой по-гречески эпидермис, а по-латински кутикула или
«выцвет кожи» (cutis efflorescentia), которая не является «семенным» об-
разованием, а чем-то вроде выделения. Все эти «однородные части» харак-
теризуются по внешнему виду, плотности и значению в организме. Напри-
мер, сухожилие определяется так: «сухожилие является у животных с кро-
вью частью, очень распространенной и однородной, твердой, не содержа-
щей крови, и прочной, разделяемой на прямые волокна, чувствительной,
связанной с головкой мышц, чтобы кости могли в произвольном движе-
нии сгибаться и разгибаться и через них бы мышцы связывались с костями».
Наблюдения, сделанные с помощью микроскопа в течение XVII —
XVIII вв., не играли сколько-нибудь заметной роли в развитии гистоло-
гических представлений, хотя с их помощью и был собран довольно боль-
шой, но разрозненный фактический материал, касающийся тонкого строе-
ния различных частей тела животных и человека.
Вместе с тем анатомы, пользуясь своими обычными методами препа-
ровки или рассечения, часто в сочетании с мацерацией, постепенно соби-
рают новый фактический материал об однородных частях тела.
В XVIII в. уже оформились макроскопические представления об осо-
бой слизистой, клетчатой ткани или клетчатке,** т. е. современной рыхлой
соединительной- ткани. Клетчатка, как особый «элемент» организма, уже
* Термин «паренхима», которым обозначалось вещество мышц и внутренностей,
принадлежит эллинскому врачу и ученому Эразистрату (IIIв. до н. э.), предполагав-
шему, что мышцы и внутренности образуются из излившейся крови (itcpeiZopa— бук-
вально обозначает «излившееся около и внутри»),
** Термин «клетчатка» или «клетчатая ткань» не имеет ничего общего с современными
представлениями о клетке. Он возник оттого, что, вдувая воздух в рыхлую соединитель-
ную ткань, в ней можно вызвать образование многочисленных, долгое время не спадаю-
щихся, наполненных воздухом ячеек или клеток, отграниченных друг от друга тонкими
перепонками межклеточного вещества. Отсюда возник и современный французский тер-
мин для обозначения рыхлой соединительной ткани — tissu lamellaire.
8
Глава 1
вошла в восьмитомную анатомию и физиологию Галлера (1778). Кроме того-
ей, как и другим сходным частям, например кутикуле или эпидермису,
т. е. роговому слою кожи, были посвящены до начала XIX в. специальные
исследования анатомического характера (Борде, 1767). Галлер в первом томе
своего фундаментального труда различает следующие элементы челове-
ческого тела: 1) волокна, объединяющие всевозможные волокнистые струк-
туры с их разновидностью — «пластинками»; 2) клеточную ткань, состоя-
щую из волокон и пластинок; 3) «оболочки», создающиеся или состоящие
из клеточной ткани; 4) жир и водянистую жидкость тела.
В самом начале XVIII в. голландский ученый Ф. Рейш (1701, 1703)
обнаружил, что подобно тому, как кожные сосочки покрыты эпидермисом,
на сосочках переходной части губ и слизистой оболочки различных уча-
стков полости рта имеется тонкая поверхностная пленочка, обозначенная
им термином epithelis или epithelia (в женском роде).
Относительно значения слова «эпителий > имеются разногласия. До послед-
него времени его производили большей частью от предлога — на и
существительного — сосок, считая, что оно обозначает в буквальном
переводе нечто, находящееся на поверхности сосков, так как сам Рейш
назвал словом эпителиа тонкую пленку, обнаруженную им на поверхности
сосочков губ и разных участков ротовой полости.
Более правильным представляется, однако, объяснение этого слова,
предложенное недавно Гейденгайном (1936), согласно которому корень
«thel» следует производить не от бтрл] — сосок, а от глагола — цве-
сти, расти, который обычно у классических авторов имеет форму Ош-
Глагол встречается у Нонна из Паноплиса в Египте (V век н. э.).
Тогда термин «эпителий» приобретает значительно больший смысл и обо-
значает нечто, растущее по поверхности и представляющее собой поверх-
ностный «выцвет», что хорошо гармонирует со всем контекстом самого Рейша
и его биографа и современника Шрейбера (1732), который следующими
словами передает наблюдения Рейша: «uti jam a cutaneis papillis epider-
mis producitur; ita et papillae prolabiorum, genae internae, atque gingiva-
rum efflorescunt, expanduntur in epithelias; velamenta propria; cuticulas;
non epidermides» (стр. 38).
(«Подобно тому, как из кожных сосочков образуется эпидермис, также
и сосочки переходной части губ, внутренней поверхности щек и десен рас-
цветают, распространяются в эпителий, собственные покровы, кутикулы —
не эпидермисы».)
Несколько позже отдельные исследователи отмечали существование
каких-то тонких поверхностных пленок в разных органах и обращали
внимание на черты сходства между эпидермисом и роговыми придатками
кожи (Лудене, 1739). Либеркюн в своей монографии о строе-
нии и работе кишечных ворсинок (1745) отмечает, что ему приходилось
наблюдать тончайшую мембрану на внутренней поверхности кишечника г>
желудка, пищевода и в полости рта, о которой он пишет следующее: «mem
branam hanc epidermidi absimilem non esse inde patet» («Поэтому, очевидно»
эта мембрана не является несходной с эпидермисом»).
Очерк развития гистологической системы
9
Микроскопические и анатомические исследования позволили разгра-
ничить постепенно, хотя часто недостаточно последовательно и отчетливо,
два круга представлений, которые не различались старыми мыслителями
и анатомами. А именно, представление о пока еще гипотетических элемен-
тарных или простых частях (partes simplices, acuv!)e-cr, TtpwToyova), кото-
рые завершились формулировкой клеточного учения Шванном и представле-
ния об «однородных или сходных» частях, получивших в дальнейшем обо-
значение тканей.
3. МАКРОСКОПИЧЕСКАЯ ГИСТОЛОГИЯ НАЧАЛА XIX В.
Развитие современных гистологических представлений начинается,
собственно говоря, с начала XIX в.
В развитии гистологии выдающееся значение принадлежит, как изве-
стно, анатому и физиологу Биша, опубликовавшему в 1801 г. четырехтом-
ный труд под названием «Общая анатомия в приложении к физиологии и
медицине». Капитальный и оригинальный труд Биша основывается в зна-
чительной степени на собственных исследованиях автора, явившегося
вместе с тем последователем и продолжателем своих предшественников,
в частности, известного врача-психиатра Пинеля. Этот последний в своей
«Философской нозографии» (1798) обратил внимание на то, что патологи-
ческие изменения некоторых частей или тканей тела в составе различней-
ших органов имеют очень много общего, например, картина заболеваний
внутренней оболочки желудка, мочевого пузыря, слизистой оболочки носа
п пр.
Задачу своей книги Биша определяет, как «изучение и изложение по-
рознь со всеми их аттрибутами каждой из простых систем, которые, в раз-
личных сочетаниях, образуют наши органы». «Общая анатомия» Биша
по своему содержанию — не медицинская анатомия в современном смысле
слова, а целая энциклопедия, посвященная обширному комплексу дисци-
плин, которые в то время целиком или частично входили в состав анатомии,
а в наше время представляют собой ряд отдельных наук, собственно ана-
томию, гистологию, эмбриологию, физиологию и патологию, в широком
смысле этого слова.
В своих исследованиях Биша руководствуется мыслью, что тело живот-
ных и человека, подобно сложным химическим телам неживой природы,
представляет собой сочетание или соединение простых составных частей.
«Химия имеет свои простые тела, — пишет Биша, — которые образуют
путем различных сочетаний, к которым они способны, сложные тела...
Также и анатомия имеет свои простые ткани, которые, сочетаясь по четыре,
шесть, восемь и т. п., образуют органы». Для обозначения этих простых
частей тела он пользовался терминами «система» и «ткань» иногда как сино-
нимами, а в некоторых случаях вкладывая в них несколько неодинако-
вое содержание, так как в ряде своих простых систем он еще различал вхо-
дящие в их сослав ткани. В отличие от прежних анатомов, он пользовался
для характеристики отдельных простых систем не только их внешним ви-
дом (окраской, консистенцией, различимой простым глазом или с помощью
10
Гпава I
слабой лупы, структурой), но и отношением различных тканей к реактивам
(преимущественно к кислотам и щелочам), к нагреванию, гниению, а также
перевариванию и некоторым другим воздействиям. Микроскопом или силь-
ными увеличительными стеклами он совершенно не пользовался. Всего
Бита различал 21 простую систему, которые частично представляют со-
бой ткани в современном понимании, а иногда более сложные образования,
органы или части органов и даже целые системы органов, а подчас и совер-
шенно гипотетические образования, которые постулировались на основа-
нии существовавших в то время физиологических представлений. Ткани
или системы Биша были следующие: 1) клеточная (клетчатка); 2) нервная
анимальной жизни; 3) нервная органической (вегетативной) жизни; 4) арте-
риальная; 5) венозная; 6) система выделяющих сосудов (systeme des exha-
lants — гипотетических тончайших трубок, являющихся продолжением
кровеносных капилляров); 7) система поглощающих сосудов (systcme des
absorbants — соответствует лимфатической системе); 8) костная; 9) костно-
мозговая; 10) хрящевая; 11) волокнистая (фиброзная); 12) волокнисто-
хрящевая (fibro-cartilagineux); 13) мышечная животной жизни; 14) мы-
шечная органической жизни; 15) слизистая (т. е. система слизистых обо-
лочек); 16) серозная; 17) синовиальная; 18) железистая; 19) кожная (дер-
моидная); 20) эпидермоидная; 21) волосяная.
Все эти системы могут быть разбиты на две группы. Первые семь систем
имеют чрезвычайно широкое распространение в организме (systemes gen£-
ralement distribues) и находятся в составе не только всех главных аппа-
ратов (систем органов), но и других простых систем. Прочие простые сис-
темы не имеют такого общего распространения и даже часто оказываются
почти изолированными.
В составе всех органов тела Биша различает в основном сходную «пита-
тельную паренхиму», состоящую из клетчатки, различных сосудов и нервов.
В этой основе (canevas), через сосуды, в конечном итоге через выделяющие
сосуды, откладывается неодинаковая в разных частях тела «питательная
материя» (matiere nutritive), которой и обусловливаются отличия одних
органов от других. В этом отношении, по мнению Биша, ясность должна
внести химия. Каждая из простых систем Биша отличается рядом призна-
ков: 1) внешней формой, которая, однако, не является существенной осо-
бенностью; 2) организацией или тонкой структурой; 3) тканевыми свой-
ствами — механической растяжимостью, сократимостью, способностью
твердеть и приобретать под влиянием тех или иных воздействий «роговой»
вид (racornissement) и т. п.; 4) жизненными свойствами, т. е. физиологи-
ческими особенностями, как органическая сократимость, органическая
и животная чувствительность.
Описывая отдельные системы с учетом этих признаков, Биша также
отмечает, в состав каких органов они входят. При разборе тонкого строе-
ния систем в их составе приходится различать их собственные части (par-
ties propres), или ткани, и общие части (parties communes), принадлежащие
уже другим системам. Например, клетчатка, являясь общей частью многих
систем, со своей стороны, содержит в своем составе различные сосуды ит. п.
Очерк развития гистологической системы
11
Система слизистых оболочек состоит из собственной ткани, представлен-
ной ее хорионом (основа или tunica propria), ворсинками или сосочками
и общими частями — кровеносными, «выделяющими» и поглощающими
сосудами и железами, ткань которых обозначается также, как паренхима.
К системе серозных оболочек, бывших еще до опубликования общей ана-
томии предметом специального исследования Биша (1800), относится также
паутинная оболочка мозга, что очень напоминает многие современные
ошибочные представления. После чрезвычайно детального морфо-физио-
логического описания каждой системы со всех указанных точек зрения
Биша переходит к наблюдающимся в ней явлениям восстановления, ее
развитию в зародышевом состоянии, возрастным и специально старческим
изменениям. Наконец, он очень подробно останавливается на всевозмож-
ных патологических изменениях, в частности, опухолях. Таким образом,
дается чрезвычайно полная по тому времени и разносторонняя характери-
стика каждой из 21 «ткани». Разумеется, очень многое из высказываний
Биша теперь, через 145 лет, представляется наивным и совершенно непра-
вильным, но в свое время труд Биша, вскоре переведенный на немецкий
язык, имел несомненно выдающееся значение важнейшего этапа во всем
дальнейшем развитии наших представлений о тканях. Вместе с тем сле-
дует подчеркнуть удивительную точность наблюдений и описаний факти-
ческого материала. В этом отношении достаточно, например, указать
на макроскопическое описание щипка из рыхлой соединительной
ткани.
Вальтер (1806), резюмируя представление Биша о генетических отно-
шениях тканей, подчеркивает, что между тканями существуют лишь коли-
чественные различия по степени их сцепления (Cohaesionsgrad), а поэтому
они могут переходить друг в друга. Все ткани образуются из бесструктур-
ной изначальной ткани — клетчатки или слизистой ткани. В конце концов
все ткани могут вновь переходить в клетчатку. Клетчатка может превра-
щаться в двух противоположных направлениях, давая начало следующим
двум генетическим рядам тканей. Один ряд, начинаясь клетчаткой, через
серозные оболочки, слизистые оболочки и кожу с железами, заканчивается
надкожицей с ногтями и волосами; другой ряд представлен, начинаясь
из того же источника, мышечной, фиброзной, хрящевой и костной тканями.
Позднейшие французские анатомы в основном принимали систему
Биша, внося в нее те или иные изменения и большей частью объединяя
некоторые из «тканей», например сосуды и нервы.
Клоке (1815), вместо 21 системы Биша, принимает 13.
Довольно своеобразную, хотя и мало последовательную систему об-
щей и частной анатомии, не имевшую, правда, заметного влияния на об-
щий ход развития гистологических представлений, дает современник Биша,
Малакарне (1803). Он различает следующие системы тела, которые, в свою
очередь, распадаются на более мелкие подразделения: одну общую систему
кожи; четыре «генеральных» системы (клеточную, сосудистую, мышечную
и нервную); семь «универсальных» систем (оболочек, желез, связок, ко-
стей, внутренностей или паренхимную, костного мозга и хрящей); семь
12
Глава I
«частичных» систем (головы, шеи, рук, груди, живота, половых частей и
нижних конечностей).
Известный физиолог и анатом Рудольфи (1814, 1815) был, невидимому^
первым, кто вполне отчетливо подчеркнул сходство целого ряда анатоми-
ческих образований, а именно эпидермиса, ногтей, волос, рогов и когтей,
и высказал мысль, что т. наз. мальпигиева сеть кожи представляет собой
глубокую, более молодую часть эпидермиса.
Большого интереса заслуживает, с точки зрения истории гистологии,
первый том «Руководства по анатомии человека» Меккеля (1815). Он по-
священ общей анатомии и знаменует собой следующий после Биша этап
развития нашей науки.
Фактический материал, излагаемый Меккелем, как и у предшествен-
ников, основывается на макроскопическом изучении, хотя вместе с тем
он пользуется данными микроскопических исследований. Последние имеют,
однако, за редким исключением, гипотетический характер или основы-
ваются, как и у многих других современников и предшественников, на
всевозможных артефактах, имеющих вид микроскопических «шариков»
и отнюдь не соответствующих современному понятию о клетке.*
В живой природе Меккель различает внешнюю форму, или конфигура-
цию, и внутреннюю форму, или тканевую структуру — текстуру. Текстура
не может быть вполне резко отграничена от собственно структуры, т. е.
состава тела от отдельных более крупных частей. Сложные части тела по
своей структуре могут быть сведены к более простым составным частям,
среди которых, смотря по степени их простоты, следует различать бли-
жайшие и более отдаленные.
«Отдаленных» составных частей (entfernte Formbestandtheile) суще-
ствует два вида: «шарики» и свернувшаяся или способная к свертыванию
и поэтому формируемая (gestaltbare) субстанция; шарики значительно
отличаются друг от друга неодинаковой формой и размерами. Даже
т. наз. кровяные шарики имеют не шарообразную, а сплющенную
форму линзы. Эти элементарные шарики отсутствуют в моче, слизистой
ткани, фиброзных частях, хрящах и кости. Зато в большом количестве
встречаются в крови, мышцах, нервном веществе.
Первоначально зародыш не имеет шариков, а весь целиком состоит из
гомогенной, способной к свертыванию жидкости, которая лишь в дальней-
шем «раздваивается» на шарики и жидкую часть. Из шариков и коагули-
рующего вещества, могущего образовать пластинки и волокна, состоят
различные «ближайшие» составные части тела, которые в отношении своей
внутренней структуры обозначаются также как ткани (textus или tela).
Термин partes similares объясняется Меккелем иначе, чем многими из более
старых авторов. Это обозначение, по Меккелю, зависит оттого, что эти
части встречаются в составе организма много раз, в то время как слож-
ные partes dissimilares имеются в количестве, одной или, самое большее,
двух.
* Подробнее см.: Кацнельсон «100 лет учения о клетке» (1939) и Вермель «Основные
этапы в развитии учения о клетке» (1940).
Очерк развития гистологической системы
1.3
Вместо 21 простой системы или ткани Биша, Меккель различает 12,
которые, в свою очередь, могут быть сведены к 11 и, даже возможно, к 10:
1) слизистая; 2) сосудистая; 3) нервная; 4) костная; 5) хрящевая; 6) волок-
нистая; 7) волокнисто-хрящевая; 8) мышечная; 9) серозная и 10) кожная
система. В последнюю можно включить, по мнению Меккеля, системы
собственно кожи, слизистых оболочек, а также, невидимому, желез.
Из всех этих систем три первые являются общими, а семь остальных
особыми (besondere). Слизистая, или клеточная, ткань, в соответствии с пред-
ставлениями, высказывавшимися в менее четкой форме Биша, является,
.по Меккелю, вместе с тем формой существования основной формируемой
субстанции организма; путем ее видоизменения и. усложнения образуются
все прочие ткани тела. В ее составе могут иметься две жидкости: «кровя-
ная вода» (серозная жидкость) и жир. Описывая известные уже в то вре-
мя три главных оболочки сосудистой стенки, Меккель обращает внимание на
то, что внутренняя из них имеет сходство с серозными оболочками и, воз-
можно, представляет собой как бы переход от слизистой ткани к серозной
системе.
Каждая из систем сначала описывается в нормальном (im regelrech-
fen), а затем очень подробно в различных патологических состояниях
(im regelwidrigen Zustande), причем, в соответствии с современным уров-
нем знания, дается также их функциональная характеристика.
До этого времени учение о простых системах или тканях обозначалось,
по почину Биша, как общая анатомия.
Только в 1819 г. появляется в печати вступительная лекция /<. Майера
к курсу анатомии под заглавием «О гистологии и новом подразделении
тканей человеческого тела», в которой для обозначения нашей науки вво-
дится впервые укрепившееся за ней в дальнейшем название, а термин
«ткань» окончательно начинает приобретать права гражданства и посте-
пенно вытесняет применявшийся наряду с ним термин «система». *
Гистологию, или учение о тканях, которое, по мнению Майера, является
аналитической анатомией, он противопоставляет морфологии,** или син-
тетической анатомии.
Он уже подразделяет гистологию, подобно анатомии, на общую и спе-
циальную части, которые, в общем, соответствуют современной общей
гистологии и микроскопической анатомии. Гистология и морфология, или
анатомия, в современном смысле слова, характеризуются им, как две ветви
знания: «одна занимается внутренним, другая внешним строением орга-
нов, как в общем, так и в частном отношениях. Первая стремится описы-
вать текстуру, ткань, textus, «ото? органов, вообще внутренние отноше-
ния животных и органических (т. е. вегетативных) элементов; вторая опи-
сывает форму, положение, forma, pop<fT], вообще внешнее или простран-
* По-гречески «ткань» обозначается словом wriov или iaxot, соответствующим ла-
тинскому textus; Хбуо« — слово, речь, описание, история, книга и т. д. В дальнейшем
некоторые авторы пользовались также термином «гпетиология», который, однако, нс
;получил распространения. ч
** Термин «морфология» ввел Вольфганг Гете (1807) для обозначения науки о живой
форме в ее становлении и изменениях.
14
Глава I
ственное отношение органов. Они являются, таким образом, по существу
различными и распадаются на общую и специальные части. В общей первой
части или в учении о ткани органов, которую автор хочет обозначить сло-
вами «учение о тканях (Gewebelehre), гистология, т. е. в общей гистоло-
гии говорится о животно-органической ткани вообще и об ее подразделе-
нии на различные виды; в частной (besonderen) гистологии проходятся
отдельные ткани и перечисляются их местоположение в различных орга-
нах и их частные особенности') (стр. 6).
Майер различает всего 8 «родов» тканей, большая часть которых, в свою
очередь, распадается на несколько видов. Его подразделение основывается
на макроскопически различимых особенностях тканей с учетом их функци-
онального значения, а также химических особенностей. Система Майера
имеет следующий вид:
I. Пластинчатая (листковая)ткань (Blattergewebe) или белковая система:
1) ткань хрусталика, 2) ткань роговицы, 3) система надкожицы или
эпидермиса (Oberhautsystem), 4) система волос, 5) система ногтей, б) си-
стема зубов (под эпидермисом он понимает не только эпидермис кожи, но
в значительной степени и гипотетическую выстилку многих полых внут-
ренних органов).
II. Клеточно-волокнистая система.
А. Первая ступень: 1) система клетчатки, 2) система жировой оболочки
(Fetthautsystem), 3) система костного мозга, 4) серозная система, 5) сино-
виальная система.
Б. Вторая ступень: б) система оболочек сосудов, 7) система собственно
кожи, 8) система собственно слизистых оболочек (Schleimlederhautsystem),
9) ткань матки.
III. Волокнистая система.
А. Клеточно-волокнистая ткань: 1) особые оболочки желез, селезенки,
почек, 2) белковая оболочка яичка, 3) ткань пещеристых тел, 4) ткань
склеры.
Б. Собственно фиброзные ткани: 5) ткань твердой мозговой оболочки,
6) надкостница, 7) надхрящница, 8) фиброзные суставные капсулы, 9) связки,
10) апоневрозы, 11) сухожилия, 12) неврилемма.
IV. Хрящевая ткань.
1) Хрящевая ткань органической жизни (волокнистый хрящ), 2) хря-
щевая ткань животной жизни (суставные хрящи).
V. Костная ткань.
VI. Железистая ткань.
1) Ткань лимфатических желез, 2) ткань желез капиллярных сосудов,
3) ткань желез кровеносных сосудов, 4) ткань желез секреторных сосудов.,
VII. Мышечная ткань.
1) Мышечная ткань органической жизни, 2) мышечная ткань животной
жизни.
VIII. Нервная ткань.
1) Нервная ткань органической жизни, 2) нервная ткань животной
жизни.
Очерк развития гистологической системы 15
В системе Майера обращает на себя внимание выделение в особую группу
ряда образований, значительная часть которых в дальнейшем получит
общее обозначение эпителиальной ткани. Вместе с тем он относит к этой
группе такое сложное образование, как роговица глаза, включая и ее со-
единительнотканную основу, на том основании, что она расслаивается
при препаровке на пластинки.
Он довольно подробно характеризует химические и макроскопически-
морфологические особенности листковой ткани и ее значение в составе
целого организма, считая, что она находится на низшей ступени организа-
ции, и отмечает ее сильно выраженную способность к восстановлению,
т. е. регенерации.
Семь групп тканей, в свою очередь, могут быть сведены, по Майеру,
к трем «элементарным органам», напоминающим представления XVIII в.:
1) к «клеткам» (в смысле термина «клеточная ткань»), сосудам или сплете-
ниям сосудов, которые он отождествляет с железами; 2) к раздражимым
волокнам или «клеточным» (т. е. соединительнотканным) и мышечным во-
локнам; 3) к чувствительным волокнам, нервам. Особое значение имеет
«листковая ткань», которая накладывается, как покровный слой, там,
где органы тела животных вступают в соприкосновение с внешними влия-
ниями (стр. 39—40), и которая по своему значению является защитной.
Сосудистая стенка у Майера, как и у предшественников, тоже фигури-
рует, как ткань. Она целиком, так же как и ткань матки (собственно миоме-
трий), находится в группе соединительнотканных образований. Хряще-
вая ткань почему-то подразделяется, по аналогии с мышечной и нервной,
на вегетативную и анимальную хрящевые ткани. Непонятно, почему в во-
локнистой системе в качестве подгруппы называется клеточно-волокни-
стая ткань, а наряду с этим имеется особая клеточно-волокнистая система.
Во многих отношениях сходной, но в то же время более гистологиче-
ской по содержанию, является система тканей, принятая в первом томе
учебника физиологии Рудольфа (1821), который, различая в составе живот-
ного организма плотные и жидкие составные части, сводит все многообразие
первых к восьми «простым» частям: 1) клеточная ткань (также клеточное
или слизистое вещество), 2) роговая ткань, 3) хрящевая ткань, 4) костная
ткань, 5) сухожильное волокно, 6) сосудистое волокно, 7) мышечное волокно,
8) нервное волокно. В отличие от других современников Рудольфи считает',
что пищеварительная трубка, а может быть, воздухоносные, мочевые и
половые пути и даже сосуды имеют внутреннюю кожицу — эпителий,
который вместе с эпидермисом и его роговыми придатками выделяется
в особую «роговую ткань». Наличие эпителиальной выстилки внутренних
полых органов Рудольфи скорее постулирует, чем доказывает непосред-
ственными наблюдениями, так как микроскопом он, повидимому, не поль-
зовался. Это предположение подкрепляется и косвенными доказатель-
ствами по сравнению; например, наличием «рогового вещества» на внут-
ренней поверхности желудка некоторых птиц и жвачных животных. Рого-
вая ткань, по мнению Рудольфи, стоит на очень низкой ступени организа-
ции, разлагаема на чешуйки или волокна и не содержит сосудов и нервов.
16 Глава 1
Из сухожильных волокон состоят, кроме сухожилий и связок, такие обра-
зования, как надхрящница, надкостница, твердая мозговая оболочка,
апоневрозы. Сосудистые волокна, которые, с его точки зрения, заметно
отличаются от мышечных, обнаруживаются отчетливее всего как «арте-
. риальные волокна» в составе среднего слоя стенки артерий. Рудольфи
считает, что нет достаточных анатомических и физиологических основа-
ний для отчетливого подразделения мышечных и нервных волокон на орга-
нические (т. е. вегетативные) и анимальные (т. е. соматические). В своем
кратком и сжатом описании тканей Рудольфи пользуется, хотя часто и
очень неправильным, в современном смысле, фактическим материалом из
области макроскопической гистологии беспозвоночных. Например, он
указывает, что «окостеневающие» хрящи (окостенение на почве хряща
в то время всеми понималось как непосредственное превращение хряще-
вой ткани в костную) встречаются у ракообразных (невидимому, хити-
, новые образования), насекомых, моллюсков и некоторых «лучистых» жи-
вотных, а неокостеневающие хрящи — у пиявок. К костной ткани он отно-
сит раковину (спинной щит) сепий, отмечает, что у многих беспозвоночных
встречаются почти исключительно мышечные слои под кожей и что нервы
имеются у всех ракообразных, насекомых и моллюсков, некоторых лучи-
стых и у червя Strongylus gigas. У наиболее просто организованных живот-
ных, не обнаруживающих чувствительности, не удается обнаружить и
нервного вещества.
Все сложные части тела или органы состоят из упомянутых простых
частей и могут быть на них разделены. Все сложные части подразделяются
на четыре группы: сосуды (включая выводные протоки желез), оболочки
(серозные, слизистые, волокнистые, собственно кожа и эпидермис), железы
и внутренности; последние две группы образований являются наиболее
сложными. Из органов образуются системы. Гистологический состав орга-
нов Рудольфи представляет себе еще очень примитивно; например железы
состоят, по его мнению, из слизистого вещества и сосудов.
Совершенно особое и выдающееся место в истории нашей науки при-
надлежит оставшейся незаконченной и незаслуженно забытой монографии
Гейзингера «Система гистологии» (I часть — «Гистография», 1822), которая,
если не по фактическому материалу и по принятой в ней системе тканей,
то по ряду высказываемых автором общих положений и по данному в ней
определению гистологии, как науки, далеко опередила представления
не только всех современников, но даже очень многих гистологов всего XIX
и XX вв. В отношении системы тканей Гейзингер не дал ничего оригиналь-
ного. *
Он принимает следующие 11 тканей (wto?, tela, tissu, Gewebe), каждая
из которых обнимает собой целый ряд разновидностей, обозначенных как
«образования» (Gebilde, textus):
* В сноске на стр. 143 сам Гейзингер подчеркивает, что в подразделениях тканей
не существует какого-нибудь определенного принципа, и что в описательной части он
-сам не может и не хочет дать системы, построенной согласно определенным и последо-
вательно проведенным принципам.
Очерк развития гистологической системы
17
1) клеточная ткань, для которой предлагается термин «образователь-
ная» ткань; этим окончательно подчеркивается общепринятое в то время
представление о клетчатке, как о первичной ткани, из которой в конечном
итоге состоят или возникают все прочие ткани тела; 2) роговая, 3) хряще-
вая, 4) костная, 5) волокнистая ткани, включая все волокнистые соедини-
тельнотканные и мышечные структуры, 6) кожная ткань (собственно кожа
и основа слизистых оболочек), 7) нервная, 8) серозная ткань (ткань сероз-
ных и синовиальных оболочек), 9) сосудистая ткань сложного состава;
за ее первичную составную часть признается внутренняя, относящаяся
к серозной ткани, оболочка, поверх которой лежат один-два слоя, построен-
ные из «образовательной» и особой «волокнистой» тканей; 10) паренхима-
тозная ткань (имеет также сложный состав; из нее состоят лимфатические
узлы, щитовидная и зобная железы, надпочечники, селезенка, яичники);
11) железистая ткань (еще более сложная, имеет в своем составе особые
«сосуды», называемые выводными протоками; из нее состоят различные
железы; здесь же описываются легкие и семенники).
В противоположность Рудольфи, Гейзингер считает, что нет никаких
оснований признавать существование эпителия, как тонкой пленки, пред-
ставляющейся под микроскопом бесструктурной (sic!), на поверхности всех
слизистых оболочек. Он думает, что все разновидности роговой ткани не
обладают обменом веществ и представляют собой выделения организма.
Как видно из приведенных высказываний Гейзингера относительно эпи-
телия и из многих других мест книги, микроскопирование в руках этого
автора отнюдь не имело решающего значения и не давало чего-либо нового
для анализа тонкой структуры тканей. По существу Гейзингер еще относит-
ся к поколению гистологов, для которых основное значение продолжает
сохранять макроскопическая или анатомическая характеристика тканей.
Тем поучительнее, что он смог дать, несмотря на это, такое определение
гистологии как науки, под которым можно почти целиком подписаться
в настоящее время: «гистология есть учение о текстуре
(т. е. тонком строении) так называемых основных систем
животного организма и о причинах и закономер-
ностях их нормального и ненормального разви-
тия» (стр. 20). Пожалуй,единственным недостатком этого определения,
с современной точки зрения, является отсутствие в нем функционального
момента. Оно сказывается также в формальном объединении в единую
волокнистую ткань соединительнотканных и мышечных структур. Гисто-
логия, тем самым, распадается, по Гейзингеру, на три части: описательную,
или гистографию, гистогению, или часть, посвященную развитию тканей
(включая явления регенерации и патологического образования тканей),
и, наконец, гистономию, или изучение «причин и законов развития и суще-
ствования тканей».
Гейзингер совершенно правильно дает определение науки, ориенти-
руясь -на предмет исследования и те закономерности, которым он подчи-
нен, а не на метод, как это делалось после него и часто делается еще в на-
стоящее время, когда ставится знак равенства между гистологией и микро-
2 Хлопни—349
18 Глава I
скопической анатомией и говорится, что это наука о микроскопическом
строении человеческого организма. Гейзингер впервые подчеркивает зна-
чение сравнительного метода в гистологии, говоря, что «мы должны просле-
живать и тщательно сравнивать друг с другом ткани во всех классах и
в возможно большом количестве семейств животных» (стр. 17). При этом
он отмечает, что есть два возможных пути сравнительного изучения тка-
ней — от человека вниз, к низшим животным, или наоборот. Первый путь
удобнее, так как истинные свойства и значение тканей при их первом появ-
лении неясны и вызывают сомнения. Вместе с тем, мне кажется, не будет
ошибочным считать книгу Гейзингера первым опытом в области эволю-
ционной гистологии.
В опубликованной части собственных исследований автора, посвящен-
ной в основном образовательной и роговой тканям, а также жидкостям
тела, пигменту и .минеральным частям, этот принцип и проводится. Ткани
описываются последовательно у человека (разумеется, с нашей точки зре-
ния, крайне несовершенно, неполно и наивно) и млекопитающих, птиц,
различных низших позвоночных, головоногих и некоторых других моллю-
сков, аннелид, различных членистоногих, «лучистых» (голотурии, звезды,
актинии, медузы, полипы, инфузории и коловратки). Во многих случаях,
правда, автор ограничивается одной или несколькими общими фразами.
Все сказанное показывает, что гистология, как самостоятельная наука
с собственными задачами, вполне оформилась до обоснования клеточного
учения и, в сущности, независимо от микроскопического метода исследо-
вания, который в дальнейшем приобрел и для нее основное значение.
Гистогеиетические представления Гейзингера по существу не отличаются
от прежних умозрительных построений и натурфилософских концепций
и в достаточной степени формальны и фантастичны. С другой стороны, они
предваряют в известном отношении гистогеиетические концепции Шванна,
основанные уже на клеточном учении. Гистогенетически ткани могут быть
сведены, по Гейзингеру, к трем основным типам образований: 1) бесструк-
турному веществу, соответствующему образовательной или клеточной
ткани; 2) возникающим в этом веществе шарикам, существующим порознь
или сливающимся в волокна; 3) пузырькам и получающимся путем их
слияния сосудистым образованиям. «В образовательной ткани возникают
капли лимфы (шары), они одеваются «поляризующейся» оболочкой и пре-
вращаются таким образом в пузыри; пузыри взаимно притягиваются и откры-
ваются друг в друга, вот вы и получаете лимфатический сосуд с его клапа-
нами, как следами бывших прежде раздельных пузырей» (стр. 116). По
этому примеру не трудно представить себе полностью уровень гистогене-
тических представлений Гейзингера, а вместе с тем и некоторых его бли-
жайших, а иногда и очень отдаленных последователей.
В элементах общей анатомии Беклара, вышедших вторым, посмертным
изданием в 1828 г., имеется следующее подразделение общих систем орга-
низма: 1) клеточная и жировая ткань (включая костный мозг); 2) серозные
оболочки (включая синовиальные); 3) покровные оболочки (membranes
tegumentairej), т. е. слизистые оболочки и собственно кожа с придатками;
Очерк развития гистологической системы
19
4) сосудистая система (включая лимфатические узлы, селезенку и некото-
рые эндокриновые органы, а также пещеристую или эректильную ткань,
выделяющуюся некоторыми французскими анатомами в особую группу);
5) железы; б) ткань связок (включая сухожилия, фиброзные и волокнистые
хрящи); 7) хрящи; 8) костная система; 9) мышечная система; 10) нервная
система. В особом, одиннадцатом разделе, приводятся различные патоло-
гические образования в тканях и паразиты. Все многообразие тканей (или
органов, во выражению Беклара) может быть сведено на основании их
анатомических, химических, физиологических и патологических свойств
к трем основным группам. Первые восемь перечисленных выше образова-
ний представляют собой лишь видоизменения и усложнения клетчатки.
Второй класс (ordre) органов образован в основном мышечными волокнами,
а третий и последний — нервами и центральными нервными массами.
Как подчеркивает сам Беклар, последняя классификация, основы которой
реально существуют в природе, в некоторых отношениях совпадает со
взглядами, принятыми еще в XVIII в. Галлером. Роговое или эпидерми-
ческое вещество является продуктом выделения и не может рассматриваться,
как анатомическая составная часть. Таким образом, у Беклара довольно
отчетливо высказана мысль об единстве всей обширной группы тканей,
получившей значительно позже обозначение «соединительного вещества».
Эластическая ткань, являющаяся основой сосудистой системы, принадле-
жит, по его мнению, к тому же классу образований, но вместе с тем прибли-
жается к мышечной ткани. Совершенно ошибочно он относит в одну группу
с соединительнотканными образованиями и железы, которые, по его мнению,
образованы системами покровных оболочек и сосудов.
4. ГИСТОЛОГИЯ В ПЕРИОД ОБОСНОВАНИЯ И РАЗВИТИЯ КЛЕТОЧНОГО
УЧЕНИЯ
Тридцатые годы являются переломным этапом в развитии гистологии.
В связи с рядом основных открытий из области микроскопической структуры
тканей животных, сделанных в течение этого десятилетия и подготовив-
ших в конце его формулировку клеточного учения, гистология и в отно-
шении методики перестает быть частью анатомии в самом широком смысле
этого слова.
Выдающееся значение принадлежит в этом отношении классическим
исследованиям блестящей плеяды ученых, принадлежащих к школам
Пуркинье и Мюллера.
Следует отметить, что не только Мюллер и Пуркинье, но и многие из
их учеников являются не анатомами, а физиологами и вместе с тем микро-
скопистами-морфологами. Нарождающаяся микроскопическая гистология
и после подразделения старой всеобъемлющей анатомии на собственно
анатомию и физиологию сохраняет связь со своими морфо-физиологиче-
скими корнями. Упомяну лишь про некоторые из основных открытий этого
времени.
Самому Мюллеру (1830) принадлежит первое обширное монографиче-
ское исследование по микроскопическому строению желез, произведенное
*
20
Глава I
над многочисленными представителями позвоночных и беспозвоночных.
В этом исследовании Мюллеру не удается обнаружить их клеточного строе-
ния, но микроскопическая анатомия желез описывается им с поразительной
точностью и обстоятельностью.
Мюллер и почти одновременно с ним ученик Пуркинье, Дейч, изучают
микроскопическую структуру хорды и хряща (1834) и открывают их кле-
точное строение. Пуркинье совместно с Валентином открывают широкое
распространение в животном мире мерцательного движения (1835), отме-
чая в отдельных случаях, что мерцательные волоски связаны с особыми
зернами, имеющими в центре по маленькому тельцу (имеются в виду покров-
ные клетки сосудистых сплетений мозга; Пуркинье, 1836). При этом Пур-
кинье говорит, что он склонен считать теперь тонкую, состоящую из зерен,
мембрану на поверхности сосудистых сплетений не нервным, а «эпидер-
мидальным» образованием.
Ученик Пуркинье, Рашков (1835), изучает в своей диссертации гисто-
генез зубов. Ремак, представитель школы Мюллера (1836), публикует
свои известные исследования по периферическим нервам.
Пуркинье (1837) впервые описывает нервные клетки позвоночных («ган-
глиозные шары» или «зерна») и железы дна желудка, в составе которых
обнаруживает клетки (зерна). Генле (1837, 1838) дает описание клеточного
строения различных эпителиев и обнаруживает плоский эпителий (эндо-
телий) в сосудах. Предложенные Генле терминология и классификация
эпителиев на долгое время вошли в гистологию.
Наконец, в 1839 г., появляется в полном виде сделавшее эпоху иссле-
дование Шванна, являющегося, подобно Генле и Ремаку, учеником Мюл-
лера. В книге Шванна, помимо формулировки клеточного учения, описы-
вается ряд очень важных новых фактов (например шванновские клетки,
клетки эмбриональной соединительной ткани) и впервые дается класси-
фикация тканей, целиком основанная на клеточном учении. Шванн разли-
чает всего 5 классов тканей: 1) изолированные, самостоятельные клетки
в жидкостях (например кровяные, «слизистые» и гнойные тельца); 2) само-
стоятельные клетки, соединенные в непрерывные ткани — «роговая» ткань,
в смысле Рудольфи, т. е. эпителий с роговыми придатками, «черный пиг-
мент», т. е. т. наз. пигментный эпителий сетчатки, и хрусталик;
3) клетки, у которых слиты друг с другом или с межклеточным веществом
стенки (хрящ, кость, вещество зубов); 4) волокнистые клетки (клетчатка,
ткань сухожилий, эластическая ткань); 5) клетки, у которых слиты друг
с другом стенки и полости (мышцы, нервы, капиллярные сосуды).
В системе Шванна совершенно обойден молчанием ряд структур орга-
низма, например железы. Очень подробно изученная самим Шванном ткань
хорды, равно как и «серозный» листок бластодермы куриного яйца не отно-
сятся им ни к одной из перечисленных групп, хотя известные черты сход-
ства их с эпителием и отмечаются.
Основным отрицательным моментом в системе Шванна является ее резко
выраженный формально морфологический характер, в результате чего
столь различные образования, как мышцы, нервы и капилляры, оказы-
Очерк развития гистологической системы
21
ваются объединенными в одну группу, а уже известный после работы Генле
«плоский эпителий» сосудов и стенки капилляров представляются совер-
шенно различными тканями. Способ развития мышц, нервов и капилляров
из клеток в трактовке Шванна совершенно сходен с гистогене-
тическими представлениями Гейзингера, который еще оперировал не
с реальными клетками, а гипотетическими шариками и пузырьками (см.
выше).
Гистологическая система Шванна не повлияла заметно на две появив-
шиеся вскоре гистологические сводки — классическую «Общую анатомию»
Генле (1841) и «Ткани тела человека и животных» Валентина (1842), в кото-
рых впервые гистологический материал излагается целиком на основе
результатов микроскопических исследований и уже под большим или мень-
шим влиянием клеточного учения.
«Общая анатомия» Генле представляет собой целую энциклопедию, посвя-
щенную физиологической химии, собственно общей анатомии или гистологии,
цитологии, физиологии и даже психологии. В ней подробно излагается
известный в то время фактический материал по «элементарным клеткам»
и анализируются различные, допускавшиеся тогда способы их образования
и превращения в волокнистые структуры. Аморфное клеточное вещество
трактуется, в соответствии с принятой Шванном теорией цитогенеза, как
цитобластема. Вместе с тем Генле не дает определенной системы тканей, поря-
док изложения которых в общем соответствует принятому во время
макроскопического периода гистологии. Различные ткани излагаются по гла-
вам без достаточно четкой рубрикации. Главы, посвященные гистологической
части материала, следующие: 1) надкожица (эпителий); 2) ногти; 3) зернистый
пигмент; 4) волосы; 5) ткань роговицы; 6) хрусталика; 7) стекловидного тела
и соответствующих оболочек; 7) соединительная ткань; 8) жировая ткань;
9) эластическая ткань; 10) питательные соки и соконосные сосуды (хилус,
лимфа, кровь; системы кровеносных, хилоносных и лимфатических сосу-
дов); 11) мышечная ткань; 12) нервная ткань; 13) хрящевая ткань,
14) костная ткань; 15) зубы; 16) слуховые камешки; 17) железы; 18) оболочки.
Введенный, невидимому, впервые И. Мюллером термин «соединительная
ткань» получает здесь впервые права гражданства вместо старых обозна-
чений: клеточная, слизистая или образовательная ткань.
В сводке Валентина впервые дано принятое до настоящего времени,
особенно для целей преподавания, подразделение гистологии на 3 отдела:
учение об общих свойствах тканей (соответствующее современной цитоло-
гической части курса), общая гистология, или описание свойств отдельных
тканей, и частная гистология, посвященная тканевому составу отдельных
органов и аппаратов. Первые две части Валентин объединяет в общую
анатомию. Валентин не проводит клеточного учения сколько-нибудь после-
довательно. В качестве элементарных частей животного организма он упо-
минает «различные кристаллы, бесструктурные вещества, зернышки, ядра,
клетки, волокна и прозрачные мембраны». Все эти образования могут пере-
ходить друг в друга. Общегистологический материал излагается им под
следующими рубриками: 1) элементарные части, имеющие форму неорга-
22
Глава I
нических тел (кристаллы, отолиты, отложения извести); 2) жир (в полостях
клетчатки); 3) пигменты (внутри клеток); 4) роговая ткань (с подразделе-
нием на эпителий, излагаемый в соответствии с классификацией Генле,
и наиболее компактные роговые образования — волосы, ногти); 5) окуты-
вающая ткань (Umhiillungsgewebe)— под эпителием, в мышцах, нервах;
б) эластическая ткань (выйная связка); 7) ткань волокнистых цилиндров
(Fadenzylindergewebe) с подразделением на клетчатку или соединительную,
ткань, ткань сухожилий и связок; 8) сосудистая ткань сосудов и сердца,
находящаяся между внутренней «эпителиальной» выстилкой и наружным
слоем окутывающей ткани и состоящая из эластических, мышечных и клет-
чаточных волокон; 9) нервная ткань (ее главные элементы — нервные
волокна и ганглиозные шары); 10) мышечная ткань (с подразделением на
сложную или поперечнополосатую, простую или органическую и на мы-
шечные волокна сосудов, несколько отличные от простой мышечной
ткани); 11) хрящевая и костная ткань (с указанием на то, что она встречает-
ся также у десятиногих раков и головоногих моллюсков, — очевидно, име-
ются в виду хитин и известковая раковина ); 12) ткань зубов; 13) ткань
желез.
Почти во всех изложенных выше системах различные ткани, составляю-
щие в настоящее время единую группу мезенхимных производных или
тканей межклеточных веществ, не объединялись вместе, а рассматривались
порознь. Это относится не только к различным разновидностям волокнистой
соединительной ткани, с одной стороны, но и к хрящевой и костной тканям,
с другой. Даже рыхлая и плотная волокнистые соединительные ткани раз*
личных частей тела большей частью не объединялись в единую систему.
Предугаданное, но фактически не обоснованное, еще в XVIII в. Галлером
и вновь подчеркнутое Бекларом объединение соединительнотканных обра-
зований не оказало в свое время заметного влияния на развитие гистоло-
гической системы.
Рейхерт в своей монографии, озаглавленной «Замечания о сравнитель-
ном естествознании вообще и сравнительные наблюдения над соединитель-
ной тканью и родственными образованиями» (1845), делает впервые попытку
обосновать генетическое единство большей части т. наз. группы тканей
«соединительного вещества» (Bindesubstanz).
В своих исследованиях Рейхерт (стр. 147) исходит из установленного
им принципа «непрерывности» (Kontinuitat), согласно которому между
однородными, т. е. генетически родственными, тканями можно обнаружить
непрерывный переход, в то время как между неоднородными, чуждыми
друг другу тканями имеется лишь «смежность» или соприкосновение (Коп-
tiguitat). На основании этого он устанавливает генетическое родство
между различными разновидностями рыхлой и плотной коллагеновыми
тканями, хрящевыми и костной тканями, хотя в окончательных выводах,
в конце работы, костная ткань обходится молчанием. Родственными соеди-
нительной ткани образованиями он считает также ткань спинной струны,
капсулу мальпигиевых клубочков почки, а также твердый кожный скелет
и сухожилия речного рака. Он подчеркивает, что капилляры имеют соб-
Очерк развития гистологической системы
23
сгвенную стенку, не однородную с соединительной тканью. Генетическую
близость между соединительной, с одной стороны, и эластической тканью,
а также элементами сосудистой стенки, с другой, он отрицает. В составе
взрослой соединительной ткани ему, в противоположность наблюдениям
Шванна над эмбриональной, удается обнаружить лишь «рудименты» кле-
ток. Вместе с тем он говорит, что «остатки» клеток в соединительной ткани,
хрящевые клетки и костные тельца (костных клеток он не видел) представ-
ляют собой аналогичные образования. Рейхерт предлагает в конце своей
книги подразделение тканей соединительного вещества на следующие четыре
группы, исходя из принципов, очень сходных с теми, которые были поло-
жены Шванном в основу своей системы: 1) ткани, клетки которых еще не
слились вполне с межклеточным веществом (современный клеточный хрящ
и хордоидная ткань); 2) ткани, у которых клеточные оболочки и незначи-
тельная часть содержимого сливаются с межклеточным веществом, а кле-
точные ядра и содержимое ясно различимы (обыкновенный и сетчатый
хрящи, остатки хорды у костистых рыб); 3) ткани, клетки которых в большей
своей части слились с межклеточным веществом (например волокнистый
хрящ); 4) ткани, клетки которых целиком или с сохранением незначитель-
ных рудиментов слились с межклеточным веществом (соединительная ткань
и фиброзные ткани).
Относительно функционального значения всей этой группы тканей Рей-
херт говорит, что они являются механическим связующим средством и
опорой, а также промежуточным звеном, которое обеспечивает взаимо-
влияние и единство близлежащих и удаленных частей организма. Он под-
черкивает, что сосуды и нервы всегда проходят в соединительной ткани.
Какое именно значение имеет соединительное вещество в питании и секре-
ции, хотя бы в смысле их влияния при диффузии кровяной плазмы, на этот
счет, говорит Рейхерт, не имеется определенных данных. Из этого можно
сделать вывод, что Рейхерт в общем правильно, хотя еще и недостаточно
четко, оценивал не только опорное, но и трофическое значение соединитель-
ной ткани. В отношении фактического материала у Рейхерта очень много
ошибок и недочетов; так, например, он считает, в противоположность утке
имевшимся в то время правильным наблюдениям, что межклеточное веще-
ство всех соединительнотканных образований бесструктурно, а то, что
в его составе описывается как волокна, будто бы на самом деле является
лишь складками или морщинами, образуемыми этим гомогенным по суще-
ству межклеточным веществом.
Большой интерес представляют некоторые общие рассуждения^ Рей-
херта в первой части монографии, которые заставляют считать его одним
из основателей сравнительной и эволюционной гистологии. Он подчерки-
вает, что развитие является особенностью органического мира, качественно
отличающей его от мира неорганического, и что все живые существа должны
рассматриваться как «генетические состояния» в цепи общего развития
или дифференцировки. Сравнительная гистология является, по Рейхерту,
частью сравнительного естествознания, которое может производить срав-
нения в смысле генетической связи отдельных органических тел. Особен-
44
Глава 1
ности тканей зависят от генетического фундамента и могут быть оценены
лишь на этой основе. «Сравнительная гистология исходит, так же как срав-
нительное естествознание вообще, из положения, что гистологические
составные части должны рассматриваться, как обусловленные звенья их
цепи развития» (стр. 38). При этом не может подлежать сомнению, что
гистологическая дифференцировка должна быть связана теснейшим обра-
зом с общим и специальным планом развития организмов. Из этих мыслей
Рейхерта видно, какое значение он придавал генетическому принципу
в системе тканей. Именно этот принцип и был им положен в основу объеди-
нения различных соединительнотканных структур.
В настоящее время трудно оценить то значение, которое имело вышед-
шее незадолго до книги Рейхерта краткое сообщение Ремака (1843) об
эмбриональном развитии желез у куриного зародыша. В этом исследова-
нии, которое вошло затем в классическую монографию Ремака «Исследо-
вания над развитием позвоночных» (1855), впервые устанавливается, что
железы развиваются из желточного листка, т. е. эпителиальной выстилки
кишечной трубки зародыша. Столь очевидное теперь развитие желез еще
в общей анатомии Генле (1841) трактовалось совершенно ошибочно. Счита-
лось, что железы возникают независимо от эпителиальной выстилки из окру-
жающей «слизистой» или «образовательной» ткани и вторично вступают
в связь с эпителием, через который и открываются своими выводными про-
токами. Только исследования Ремака дали основы для объединения в еди-
ную группу гистологической системы эпителия и ткани желез, к которым
первоначально, правда ошибочно, относились и такие образования, как,
например, пейеровы бляшки.
Сороковыми годами прошлого столетия заканчивается период развития
гистологии и гистологической системы, предшествующий современному..
5. МОРФО-ФИЗИОЛОГИЧЕСКАЯ СИСТЕМА ГИСТОЛОГИИ ЛЕЙДИГА
И КЁЛЛИКЕРА
В начале пятидесятых годов окончательно устанавливается единствен-
ный, при существующих в настоящее время на нашей планете условиях,
способ новообразования клеток путем их деления. Это положение, подго-
товленное эмбриологическими исследованиями Кёлликера (1844), Бишофа
(1845) и других и впервые установленное Ремаком (1852), окончательно-
формулирует Вирхов (1855) в известном изречении «omnis cellula a cellula»,
которому несколько позже (1857) была дана укрепившаяся окончательно
форма «omnis cellula е cellula».
Следует отметить еще две из многих заслуг Вирхова перед гистологией.
Он окончательно закрепил и правильно обосновал объединение в одну группу
соединительной, костной и хрящевой тканей (1851), состоящих сходным
образом из «клеток и межклеточного вещества, из которых первые пред-
ставляются круглыми, овальными, чечевицеобразными, хвостатыми, раз-
ветвленными и анастомозирующими, последнее же может быть гиалино-
вым, зернистым, полосатым и волокнистым; первые сохраняются при кипя-
чении, последнее становится сначала гомогенным, затем растворяется»
Очерк развития гистологической системы 25
(стр. 156). В 1856 г. Вирхов вводит в науку новый термин «нейроглия»,,
для обозначения открытого им еще в 1846 г. и подробнее описанного в 1854 г.
своеобразного соединительного или склеивающего вещества централь-
ной нервной системы, частью которого является эпендимная выстилка ее
полостей.
В 1853 г. Лейдиг в заключении монографии «Анатомические и гистоло-
гические исследования над рыбами и рептилиями» * дает впервые подразде-
ление всего многообразия гистологических структур на четыре основные
«ткани», или, как нам представляется правильнее говорить в настоящее
время, группы тканей: 1) ткани соединительного вещества (различные виды
соединительной ткани, хрящевая и костная ткани); 2) «оставшиеся самостоя-
тельными клетки» (эпидермис и эпителий с железами, жировые клетки,
хрусталик, клетки крови и лимфы); 3) мышечные волокна; 4) нервные во-
локна (а также нервные клетки или ганглиозные шары и «молекулярное
вещество» нервных центров). Последние две ткани обозначаются, как ани-
мальные.
Эту систему принимает в основном Кёлликер во втором издании своего
руководства (1855), в котором он выделяет, кроме того, как особую пятую
ткань, «ткань истинных желез кровеносных сосудов» (эндокриновых и лим-
фоидных органов). В 3-м издании руководства (1859) эта ткань исчезла,
как самостоятельная группа.
В отличие от Лейдига, «оставшиеся самостоятельными клетки» обозна-
чаются им как клеточная ткань (Zellengewebe), которая подразделяется на
надкожицу (Oberhaut), или эпителий, и на железы; кровь и лимфа вообще
не рассматриваются в общей гистологии, а излагаются в главе, посвящен-
ной сосудистой системе.
В своем «Учебнике гистологии человека и животных» Лейдиг (1857).
говорит о том, на основании каких принципов он принял подразделение
всего многообразия тканей на четыре основные группы: «Я беру поэтому
в качестве руководящей нити физиологические отношения элементарных
частей» (стр. 21).
По существу, однако, система тканей Лейдига и Кёлликера является
не физиологической, а морфо-физиологической, т. е. осуществляет объеди-
нение и разделение структур на основе свойственной всему живому нераз-
рывной связи формы и функции. Функционально сходные образования,
относимые в одну и ту же группу тканей, отличаются вместе с тем и опре-
деленными чертами морфологического сходства. Этим и объясняется тот
факт, что эта система, несмотря на свою почти девяностолетнюю давность,
существует по настоящее время. Наоборот, неоднократно предлагавшиеся
позже односторонние физиологические или морфологические классифика-
* В этой монографии в группу «рептилий» включаются еще и амфибии, которым
в ней, наравне с осетром, уделяется особенно много внимания. Позже (1885) сам Лейдиг
ссылается на свою монографию под названием «Anatomisch-hsstologische Untersuchungen
fiber Fiscbe und Amphibien». Работы Лейдига под таким названием мне найти не удалось.
Невидимому, автор имеет в виду книгу, озаглавленную в свое время «Anatomisch-histolo-
gische Untersuchungen fiber Fische und Reptilien».
26
Глава I
ции, несмотря на их оригинальность и даже, в известном смысле, последо-
вательность, не прививались, так как во всех попытках такого рода допу-
скался сильный разрыв между формой или структурой и функциональным
значением тканей.
Очень знаменательно, что большой и во многих отношениях правильно
понятый фактический материал из области эмбриологии, в частности откры-
тие зародышевых листков и их участия в образовании закладок основных
органов тела, основатели современной гистологии почти или даже совер-
шенно не использовали и не могли органически связать с гистологически-
ми проблемами. Правда, сам Лейдиг (1853) оговаривается, что пред-
лагаемая им система тканей имеет в виду только готовый орга-
низм. «Кто ставит себе задачу, — пишет он, — обработать гистоло-
гию какого-нибудь позвоночного в возможно законченной форме, тот дол-
жен был бы начать просто с яйцеклетки и показать, каким образом клет-
ки — строительный материал организма — образуются, размножаются и
превращаются в ткани и органы после того, как они (т. е. клетки. —Н. А.)
сложились в листоподобные слои зародыша» (стр. 112).
В руководстве Лейдига эмбриологии отводится лишь 18 строчек, а в пер-
вых изданиях руководства Кёлликера вообще ничего не говорится о гене-
тической связи между известными в то время эмбриональными образова-
ниями и тканями взрослого организма. Благодаря Лейдигу, которому
мы обязаны многочисленными и очень точными исследованиями, гистоло-
гия приобретает резко выраженный сравнительный характер и обогащается
очень ценным фактическим материалом, относящимся к низшим позвоноч-
ным и различным типам беспозвоночных. Основываясь, в первую очередь,
на функциональных, но вместе с тем и на морфологических признаках,
Лейдиг относит различные кутикулярные образования, в частности хити-
новые покровы и внутренний скелет членистоногих, к тканям соединитель-
ного вещества. При этом он подчеркивает, особенно в своей, вышедшей
значительно позже, книге «Клетка и ткани» (1885), резко выраженные черты
сходства между хитином и костью.
С пятидесятых годов прошлого столетия и до самого последнего времени
подход к тканям, как чисто морфо-физиологическим категориям, стано-
вится в гистологии доминирующим и общепринятым, а четыре установлен-
ные Лейдигом, а за ним и Кёлликером группы тканевых структур незыб-
лемо входят в гистологический обиход.
Более того, благодаря авторитету Кёлликера, а также Лейдиги, обще-
принятым среди гистологов становится положение, что морфологические
свойства дифференцированных тканей никак не связаны с их развитием
из того или иного источника и что каждый из зародышевых листков может
давать почти все или даже все ткани (Кёлликер, 1876, 1884; Лейдиг, 1885):
«Каждый зародышевый листок, —говорит Кёлликер (1884, стр. 211), — спосо-
бен, если принять во внимание весь ряд животных, у известных существ
дать начало трем и возможно всем тканям, и поэтому зародышевые листки
не являются гистологическими примитивными органами». Эта же мысль
приводится и в последнем издании руководства Кёлликера (1889).
Очерк развития гистологической системы
27
б ГЕНЕТИЧЕСКИЙ ПРИНЦИП В ГИСТОЛОГИЧЕСКОЙ СИСТЕМЕ И УЧЕНИЕ
О СПЕЦИФИЧНОСТИ ТКАНЕЙ
Вместе с тем большие успехи в области эмбриологии, обоснование Дар-
вином эволюционного учения, создание Геккелем на его основе генеалоги-
ческого дерева животных и формулировка биогенетического закона не могли
не повлечь за собой попыток внести в учение о тканях генетический момент
и создать естественную генетическую систему тканей. Попытки такого рода
принадлежали, однако, не гистологам, а в первую очередь эмбриологам
п анатомам Гису, Рауберу и Валъдейеру, зоологу-эволюционисту Геккелю
и патологу Бару. Как это ни удивительно, они не оказали в целом замет-
ного влияния на дальнейшее развитие гистологии, даже были почти совер-
шенно забыты или искажены.
В своей монографии об оболочках животного организма Гис (1865,
1903) предлагает строго различать истинные эпителии и т. наз.
ложные эпителии или эндотелии, на основании их различного происхожде-
ния, с которым совпадают и существенные физиологические особенности.
Истинные эпителии, с точки зрения Г пса, являются производными наруж-
ного п внутреннего пограничных зародышевых листков зародыша, а эндо-
телии (серозных, мозговых оболочек, сосудов и синовиальных оболочек)
возникают при образовании полостей или щелей в соединительной ткани
из мезодермы и поэтому являются тканями, близкими к ней, и не имеют
пограничного значения. Дальнейшие исследования привели Гиса (1866,
1868, 1882) к формулировке учения об архибласте и парабласте, как о двух,
качественно различных, источниках развития тканей позвоночных. Изу-
чая развитие меробластических богатых желтком яиц птиц, Гис пришел
к выводу, что эндотелий, кровь и соединительнотканные образования воз-
никают не из бластомеров, получающихся в результате дробления на ани-
мальном полюсе яйца (т. наз. архибласта), как все прочие ткани
организма, а из каких-то особых клеток «парабласта», проникающих из
белого желтка в бластодерму. Клетки парабласта, по первоначальным пред-
ставлениям Гиса, возникают не из протоплазмы и ядра яйцеклетки после
оплодотворения, а из фолликулярных элементов материнского организма,
мигрировавших в желток.
Представления Гиса встретили жестокую и обоснованную критику.
Однако, несмотря на все свои недостатки и ошибки, теория Гиса
явилась предшественницей учения о мезенхиме, как об едином источ-
нике развития всей группы тканей межклеточного вещества.
В 1877 г. Геккель распространяет гистологическое понятие «эпителий»на
|родышевые листки и выдвигает свое учение об эпителии, как о первич-
ной ткани (см. ниже), а в 1882 г. братья Гертвиги в своей теории целома
противопоставляют мезенхиму всем «эпителиальным» зародышевым листкам
и, в частности, энтероцельной мезодерме. Это резкое противопоставление
друг другу двух основных типов тканевых структур — рыхлой мезенхим-
ной, имеющей опорное значение, и комплексной эпителиальной, характе-
ризующейся своим пограничным значением,—которое с тех пор прочно
28 Глава /
укореняется не только в эмбриологии, но и в гистологии, является даль-
нейшим развитием геккелевской трактовки эпителия.
Мышечная и нервная ткани, с точки зрения Гертвигов, могут в разных
случаях иметь либо «эпителиальное», либо «мезенхимное» происхождение.
Причем мускулатура эпителиального происхождения и морфологически
отличается, в соответствии с сравнительно-гистологическими данными
Вейсмана (1862), от мезенхимной. Для первой характерно строение из т.
паз. «примитивных пучков», вторая же состоит из мышечных кле-
ток.
Сам Гис (1882) и Вальдейер (1883) в дальнейшем ставят знак равенства
между понятием парабласта, как зачатка соединительнотканных образо-
ваний, и представлениями о мезенхиме, сформулированными О. и Р. Гергп-
вигами. Вальдейер подчеркивает, что парабласт, подобно архибласту,
возникает из дробящейся яйцеклетки, а не из гипотетических клеток белого
желтка. Вместе с тем Вальдейер не соглашается с популярными в то время
и разделяемыми Гертвигами представлениями, что из соединительноткан-
ного зачатка могут образоваться также нервы и мышцы, и выступает в пользу
учения о специфичности тканей, полностью обоснованного лишь в настоя-
щее время.
Почти одновременно Раубер (1883) предлагает обозначить зачаток сое-
динительнотканных образований термином «десмобласт» или «гемодесмо-
бласт» и впервые дает развернутый критический анализ основ гистологи-
ческих систем. Мысли и положения, высказанные Раубером, представляют
исключительный интерес и заслуживают подробного изложения. К заро-
дышевым листкам, как источникам развития тканей взрослого организма,
говорит Раубер, можно подходить с двух различных точек зрения: или они
представляют собой лишь имеющие определенное топографическое распо-
ложение клеточные участки тела зародыша, или на них следует смотреть
как на результат более или менее подвинувшейся вперед дифференцировки,
т. е. разделения вещества зародыша на неоднородные части, разделения не
только механического, но также химического и функционального. Сам
Раубер считает более правильной последнюю точку зрения и подчеркивает,
что дальнейшее расчленение тела зародыша, приводящее в конце концов
к образованию тканей и органов, не является чем-то принципиально новым,
а представляет собой продолжение первых стадий развития. Если исходить
из этого, то каждая ткань не может образоваться (вопреки первой из упо-
мянутых выше и господствующей точки зрения) из любого зародышевого
листка, а ткани, возникающие из разных эмбриональных источников, должны
отличаться друг от друга.
Исходя из этих совершенно правильных, хотя и не обоснованных в то
время фактическим материалом, предпосылок, Раубер приходит к выводу,
что, например, эпителии, будучи полифилетическими, в разных частях
организма должны представлять собой по существу очень различные обра-
зования, так как название «эпителий» обозначает лишь внешнюю форму
и определенное взаиморасположение клеток, но не их природу и качествен-
ные особенности. Хотя и не приходится сомневаться, что клетки одинако -
Очерк развития гистологической системы
29
вой формы могут возникать из разных зародышевых листков, однако необ-
ходимо еще изучить, могут ли и одинаковые по существу клетки образо-
ваться из разных источников. «Я остаюсь при своем утверждении, — говорит
он, — что происхождение из какого-нибудь иного зародышевого листка обо-
значает в общем и иную сущность данного объекта», безотносительно к тому,
одинаковая или разная у них форма, так как качество клетки в высокой
степени не зависит от ее формы. Одинаковые же по существу клетки должны
быть монофилетичными, т. е. иметь одно и то же происхождение.
Монофилетической группой, развивающейся из т. наз. «десмаль-
ного» листка, соответствующего мезенхиме братьев Гертвигов, являются
все ткани соединительного вещества, для генетического же обозначения
эпителиев необходимо к существительному «эпителий» прибавлять еще
в качестве прилагательного наименование соответствующего зародышевого
листка или даже еще более точные определения. Целомический эпителий
Раубер предлагает отграничить от собственно «десмального», т. е. соедини-
тельнотканного эпителия сосудов, эпителия остеобластов, одонтобластов
и т. п. Раубер вполне справедливо подчеркивает, что при попытках клас-
сификации необходимо, вообще говоря, руководствоваться определенным
принципом — генетическим, или функциональным, или еще каким-нибудь,
например химическим или формальным. В гистологии же, продолжает
он, еще нет отчетливых принципов подразделения тканей, и многие класси-
фикации представляют собой странную смесь разнообразных принципов.
Это положение остается вполне справедливым в отношении принципов
классификации тканей, проводимым и в современных руководствах и учеб-
никах.
В классификации, принятой Колликером, особое внимание обращается
на внешнюю форму, в генетической же системе форме нельзя приписывать
решающего значения. Генетическая классификация тканей может быть
проведена, по Рауберу, по четырем основным зародышевым листкам:
1) ткани наружного зародышевого листка (нервная и ганглиозная ткань,
нейроглия, эпендима, эпидермальные ткани кожи и их производные, ткани
гипофиза, хрусталика, эмали, эпителий лабиринта, амниона, серозной
оболочки); 2) ткани внутреннего зародышевого листка (эпителий пищева-
рительного и дыхательного аппаратов, мочевого пузыря, барабанной по-
лости и желез пищеварительной трубки, включая бранхиогенные органы,
возможно, также спинную струну); 3) ткани среднего зародышевого листка
(гладкие и поперечнополосатые мышцы, зачатковый и целомический эпите-
лий, эпителий яичка, почек и мочеточника); 4) ткани десмального листка
(десмальные эпителии, быть может, надпочечник, все разновидности соеди-
нительной ткани, хрящ, кость и дентин).
Классификация тканей по функции может быть проведена, по Рауберу,
следующим образом:
1) герминальные ткани яичника и семенника; 2) персональные ткани
с дальнейшим функциональным же подразделением на нервную ткань
с ее центрами, нейроэпителий, дермальный, трофический, респираторный
и секреторный эпителии, ткани крови, лимфы и кроветворных желез, ткани
30
Глава I
поперечнополосатых и гладких мышц, опорные ткани (нейроглия, соеди-
нительная ткань, хрящ, кость).
Классификации по форме Раубер придает следующий вид:
1. Клеточная ткань: а)эпителий, б) ретикулярная ткань, в) «агминп-
рованная» (скученная) ткань серого вещества мозга, гладкие и часть попе-
речнополосатых мышц.
2. Ткани с межклеточным веществом или диаплазматические ткани
(студенистая и фиброзная ткани, хрящ, кость, кровь, лимфа).
3. Плазмодиальные ткани (большая часть поперечнополосатой муску-
латуры, быть может — нейроглия).
Одним из существенных недостатков в положениях Раубера, а за ним
и многих других исследователей, является строгое разделение функцио-
нального и морфологического подхода к тканям, являющимся биологи-
ческими системами и требующим поэтому соблюдения принципа единства
формы и функции. Кроме того, Раубер считает десмальный листок, т. е.
мезенхиму, в соответствии с представлениями Гертвиеос, а также мезо-
дерму за генетически единые зачатки.
В отношении принципов классификации тканей Геккель (1885), разви-
вая свои положения, высказанные прежде (1874, 1877), в основном согла-
шается с Раубером. В качестве одного из возможных примеров подразде-
ления тканей на основе описательного, гистографического принципа он
приводит следующую классификацию: 1) эпителии или «слоевые» ткани
(Scliichtgewebe), состоящие из клеток без межклеточного вещества; 2) «фиб-
розы» или волокнистые ткани (без межклеточного вещества, но имею-
щие волокнистое строение); 3) «коннективы» или соединительные ткани,
содержащие клетки и межклеточное вещество.
Геккель сейчас же оговаривается, что такой описательный под-
ход отличается искусственностью и приводит к противоестественному разде-
лению близко родственных образований. Затем Геккель дает физиологиче-
ское подразделение тканей, которое, как он правильно отмечает, является
руководящим для большей части гистологов, но искусственно и не может
быть проведено последовательно.
В качестве примера приводится следующая физиологическая классифи-
кация: 1) герминальная ткань; 2) покровная ткань (эпителий и эндотелий);
3) опорная ткань (хорда, хрящ, кость, ткань зубов, спикулы, хитин и т. п.);
4) выполняющая ткань (различные виды соединительной ткани); 5) желе-
зистая ткань; б) кровяная ткань, включая ткань кроветворных органов;
7) мышечная ткань; 8) нервная ткань.
Единственной естественной системой тканей, с точки зрения Геккеля,
является филогенетическая система, которая, в силу биогенетического за-
кона, совпадает с системой, построенной на онтогенетическом принципе.
«Историческое развитие тканей из зародышевых листков родоначальной
формы многоклеточных животных (гастреи); —говорит он,— определяет их
классификацию. Эта филогенетическая система является, по моему личному
убеждению, здесь для тканей, так же как и для различных форм орга-
нов и видов животных, единственной естественной системой. По основ-
Очерк развития гистологической системы
31
ному биогенетическому закону она должна совпадать в основном с онтоге-
нетической системой... Каким образом происходило шаг за шагом фило-
генетическое развитие тканей, этому нас учит еще в настоящее время срав-
нительная гистогения низших многоклеточных. . . Если сделать попытку
провести на этой основе, обращая внимание в равной мере на сравнительную
анатомию и онтогению, филогенетическую классификацию тканей, то полу-
чатся такие категории, как и при онтогенетической системе». Развивая
высказанные еще в 1877 г. мысли, Геккель предлагает, исходя из своей
теории гастреи, в общем виде следующую генетическую, но вместе с тем
очень формальную систему тканей.
Первичной тканью является эпителий, который сначала представлен
единой бластодермой, а затем подразделяется на дермальный и гастральный
эпителий экзобласта и энтобласта. Из этой первичной ткани возникают
затем вторичные («апотелиальные», по Геккелю) ткани, которые могут быть,
в свою очередь, подразделены на три группы — птихобластических (полу-
чающихся в результате складкообразования), схизобластических (образую-
щихся путем отщеплений, как, например, мышечные пластинки) и мезен-
химных тканей. Последние образуются путем эмиграции отдельных клеток
и образования межклеточного вещества. Геккель не приписывает общего
значения мезобласту (мезодерме), так как этот последний обнару?живает
в разных группах животных очень большие различия. Геккель впервые
подчеркивает два типа общей организации животных — тип со слабо раз-
членистоногие) и тип с сильно развитой группой тканей соединительного
вещества, несущей основную опорную функцию (например позвоночные).
У позвоночных же можно различать в гистологическом отношении два
пограничных эпителиальных листка (Metlioria), наружный и внутренний,
и расположенный между ними «массовый листок», или «мезодерму», в состав
которой входят все дериваты обоих первичных листков (нервная трубка,
хорда, целомические мешки и мезенхима). Для позвоночных Геккель в сущ-
ности принимает обычную гистологическую систему (эпителиальная, нерв-
ная, мышечная, соединительная и кровяная ткани) с дальнейшим под-
разделением этих групп по генетическому принципу. Эпителиальная ткань
поэтому распадается на 4 группы: 1) экзэпителий (хротальный или экзо-
бластический эпителий); 2) эндэпителий (гастральный или эндобластиче-
ский эпителий); 3) мезэпителий (целомический или мезобластический эпи-
телий); 4) десмэпителий или десмальныйэпителий. Допуская, что у позво-
ночных в составе эт одермы имеются экзобластические стомодеум и прокто-
деум, Геккель относил к экзэпителию также выстилку полости рта и заднего
отдела пищеварительной трубки. Позднейшие эмбриологические данные,
казалось, опровергли наличие у позвоночных развитых стомодеума и прок-
тодеума, и поэтому гистологи стали рассматривать всю выстилку пищевари-
тельной трубки и дыхательного аппарата (кроме преддверия полости рта)
как производные единого эктодермального зачатка.
Нервную ткань Геккель подразделяет на нервные клетки и нервные
волокна и считает, что в ее образовании, кроме экзобласта, могут также
32
Глава I
принимать участие мезенхима и мезобласт. Всей или большей части мышеч-
ной ткани приписывается единое мезобластическое происхождение. Нако-
нец, соединительная ткань (Connectiv — Gewebe), кроме хорды, обра-
зующейся из эпителия, и кровь (Haemalgewebe) производятся из мезен-
химы, причем считается возможным, что мезенхимные клетки могут обра-
зовываться из различных элементов. Различные разновидности соедини-
тельной ткани могут переходить друг в друга и, кроме того, обнаруживают
непосредственную связь с эпителиями, например, с причисляемыми к этим
последним остеобластами и выстилкой синовиальных оболочек. Онтогене-
тический процесс гистологической дифференцировки, по Геккелю, обуслов-
лен филогенетически и передан наследственно от предков современных
форм.
Гистологическая дифференцировка представляет собой рекапитуляцию
имевшего место в филогенезе разделения труда между различными группами
клеток, и поэтому рассматривается как подтверждение биогенетического
закона. В своих статьях Геккель очень резко полемизирует с «уродли-
выми» (monstrose) концепциями Гиса и с Кёлликером и считает этого послед-
него антидарвинистом.
Генетические системы тканей Раубера и Геккеля являются чисто логи-
ческими построениями. Гистологические и эмбриологические представления
того времени объединяются лишь внешне, но не по существу, и не обосно-
вываются новым фактическим материалом. Наиболее удачным, с нашей
точки зрения, является геккелевское подразделение группы эпителиальных
тканей позвоночных, которое во многих отношениях совпадает с нашими
новыми представлениями. Но как раз этой части его системы не было суждено
сыграть в гистологии заметной роли. Зато прочно вошли в науку и внесли
в нее много путаницы формальные представления Геккеля о том, что заро-
дышевые листки представляют собой эпителий и что эпителий вместе с тем
является единой древнейшей и изначальной тканью. Бластодерма, зароды-
шевые листки и зачатки до последнего времени фигурируют в гистологии,
как примеры единой по существу «эпителиальной» ткани.
Такое же в значительной степени отрицательное, с моей точки зрения,
значение имело предложенное Гертвигами противопоставление друг другу,
как тканей, эпителия и мезенхимы. Чисто формальным или, в лучшем слу-
чае, морфо-физиологическим понятиям был придан генетический смысл,
какого они по существу не имели и не могли иметь. По их почину обычным
сделалось, например, противопоставление тканей эпителиального и мезен-
химного происхождения. Тем самым, в противовес забытому и четко сфор-
мулированному теоретическому положению Раубера, что внешняя форма
тканей не определяет их сущности, было принято ставить знак равенства
между самыми разнообразными структурами, имеющими сходный внеш-
ний облик. Всякая рыхлая сетевидная структура стала трактоваться как
единая’ по существу мезенхимная ткань и, наоборот, всякое комплексное
клеточное соединение — как единая эпителиальная ткань. В функциональ-
ном отношении эпителий был- охарактеризован как пограничная ткань
(Гис, 1865; Гертвиги, 1882), а мезенхима — как ткань опорного и выпол-
Очерк развития гистологической системы
33
няющего. (Гертвиги) или, наряду с этим, также и трофического (Пренан,
Бузн и Майар, 1902) значения. Трофическое значение соединительноткан-
ных образований предполагал еще Рейхерт (1845) и подчеркивал в свое
время Гцс(1865). Тем самым, несмотря на то, что сам Геккель был выдаю-
щимся последователем и пропагандистом эволюционного учения Дарвина
и стремился внести эволюционные представления в область тканей, гисто-
логические категории приобрели и сохранили до последнего времени,
в значительной степени именно благодаря Геккелю, ярко выраженный ста-
тический и по существу формальный характер.
Взгляды Геккеля на эпителий, как на филогенетически и онтогенети-
чески первичную единую ткань, и подчеркнутое братьями Гертвигами проти-
воставление тканей «эпителиального и мезенхимного» происхождения были
приняты и детализированы в некоторых отношениях, наряду с другими
авторами, Раблем (1889) и Маурером (1915).
Мысли, развитые Баром (1886), имеют очень большое значение для
гистологической системы с двух точек зрения: с одной стороны, он форму-
лирует четкое представление о генеалогическом дереве тканей, с другой
стороны, впервые в развернутом виде ставит и теоретически решает вопрос
с специфичности тканей. К мысли о строгой специфичности тканей Бар,
будучи по специальности патолого-анатомом, пришел на основе изучения
опухолевого материала.
Допущение широких возможностей межтканевых превращений зароди-
лось, как это отмечалось выше, еще в течение макроскопического периода
гистологии, когда, согласно общепринятой в то время точке зрения, «кле-
точная», слизистая или образовательная ткань (т. е. рыхлая соединитель-
ная ткань) считалась тем первоисточником, из которого могли возникать
все прочие элементарные системы организма. Эта точка зрения в тех или
иных модификациях сохранилась и после перестройки гистологии на основе
клеточного учения.
Для Шванна и его первых последователей источником новообразования
любых клеток сделалась цитобластема. Позже особенной популярностью
«трансформистские» представления пользовались среди патологов. Сам
Вирхов, выставивший положение «omnis cellula е cellula», а за ним и мно-
гие его последователи продолжали приписывать рыхлой соединительной
ткани способность к превращению едва ли не во все прочие ткани тела.
Именно таким образом многие издавна трактовали возникновение рако-
вых опухолей.
Другие, по почину Конгейма, пришли к выводу, что все ткани могут
образоваться вновь из вездесущих белых кровяных клеток, имеющих такое
выдающееся значение в воспалительном процессе. Наконец, не было недо-
статка в высказываниях, что специализированные клетки самых разнооб-
разных тканей млекопитающих и человека могут во взрослом организме
вновь приобретать эмбриональные свойства и превращаться затем в любые
другие ткани. Появление лейкоцитов в эпителиальных пластах, а также
на месте разрушающихся мышц или нервных элементов обычно трактова-
лось, «не мудрствуя лукаво», как их местное новообразование за счет цле-
3 Хлопии—349
34
Глава I
ток других тканей. Эти примитивные, но вполне понятные при общем уровне:
знаний взгляды очень напоминают старые объяснения появления червей
в гниющем мясе или мышей в грязном белье, а позже, уже в XIX в., микро-
организмов в бродящем пивном сусле путем т. наз. самопроиз-
вольного зарождения. Можно лишь удивляться тому, что аналогичные,
безнадежно устаревшие, взгляды в настоящее время не только продол-
жают поддерживаться некоторыми авторами, но и выдаются ими нередко,
за последнее слово «передовой» науки (Корицкий, 1926, 1936; Немилое,
1931; Рихтер, 1939; Астрахан, 1929; Вейцман, 1938; Рывкинд, 1936,
и др.). Против такого упрощенного подхода к тканям уже давно стали
раздаваться голоса отдельных гистологов и патологов. Уже Тирш
в своей монографии об эпителиальном раке (1865), а затем Валъ-
дейер (1867, 1872) доказывали, что эпителиальные структуры раковых
опухолей возникают не из соединительной ткани, а из клеток эпителия.
Вальдейер, картинно резюмируя прежние представления патолого-анато-
мов, говорит, что не существует ни одной ткани тела, ни одной опухоли,
которые бы не производились из соединительнотканных клеток или лей-
коцитов — этих, можно сказать, «универсальных клеток». («Nicht ein
Gewebe des Korpers, nicht ein Neoplasma, vvelche frisch, frei und froh aus
Bindegewebszellen und Leukozyten— diesen Allerweltzellen konnte manfast
sagen — ware abgeleitet worden». 1886, стр. 3). Бар, выставляя положение
«omnis cellula e cellula ejusdem naturae», говорит, что в области патологи-
ческой анатомии «Differenciations, adaptations, metaplasie resument toutes
les descriptionsettiennent la place de toutes les lois de 1’histogenese normale»...
«Evolutions metatypiques les plus variees et parfois les plus inattandues
sont a leur toure les ressources habituelies de 1’histologie patholo-
gique». *
Для Бара весь процесс эмбрионального гистогенеза представляет собой
постепенное, но в конце концов очень полное разъединение различных
клеточных видов, которые у взрослого уже утрачивают способность к пре-
вращениям друг в друга. Клеточная пролиферация при эмбриогенезе бывает,,
согласно Бару, двоякой: или это простое размножение, приводящее к уве-
личению количества равноценных клеток, или особый процесс раздвоения
или расщепления (dedoublement), сравнимый с разложением сложного тела
на более простые. Яйцо является наиболее сложной из существующих
клеток, оно содержит в себе первоначальные элементы всех будущих тка-
ней тела, оно способно образовать все ткани в результате сложных и много-
численных процессов раздвоения. Только будущие половые клетки размно-
жаются без раздвоения.
Весь процесс эмбрионального развития можно представить себе, по
Бару, в виде «гистогенетического дерева», начинающегося единым стволом
и дающего начало множеству различных веток и веточек.
* «Дифференцировки, адаптации, метаплазия являются итогом всех описаний и
заменяют собой все законы нормального гистогенеза»... «Самые разнообразные и подчас
самые неожиданные метатипические превращения являются, в свою очередь, обычными
ресурсами патологической гистологии».
Очерк развития гистологической системы Зо
Концы веточек этого гистогенетического дерева соответствуют всем-
окончательным и строго специфичным разновидностям клеток. Давая пред-
ставление о генетических отношениях всех клеточных форм тела, гисто-
генетическое дерево показывает объективную реальность существования
клеточных семейств и видов, объединяющих в естественные «атависти-
ческие» группы все многообразие структурных элементов организ-
ма. Попытки конкретизировать свои, во многих отношениях правильные
теоретические установки Бар не сделал, да, пожалуй, и не мог сделать
в то время. Поэтому его гистогенетическое дерево осталось совершенно
абстрактным построением, никак не связанным с реальными клетками и
тканями.
В одной из своих последних работ Гис (1901) вводит понятие органооб-
разующих зародышевых участков, в основном совпадающее с позднейшими
презумптивными закладками (Фогт, 1929), и высказывает взгляды на род-
ственные отношения тканей, напоминающие гистогенетическое генеалоги-
ческое дерево Бара.
Представления Бара о раздвоении или расщеплении клеточных свойств
при эмбриональном развитии очень сходны с известным принципом «не-
равнонаследственного» клеточного деления, выдвинутым Вейсманном и->
Ру, против которого позже с резкой критикой выступил О. Гертвиг в
своем популярном и увлекательно написанном руководстве «Клетка и
ткани» (1893, 1898), озаглавленном в последующих изданиях «Общая
биология».
Гертвиг (1906, стр. 428) подчеркивает, что наша гистологическая си-
стема искусственна, хотя она научно оправдана и явно необходима. Вместе <
с тем он очень резко полемизирует, исходя из теоретических посылок,
с учением о специфичности клеток и тканей. Отнюдь не сомневаясь в том,
что в процессах регенерации каждая ткань образуется из себе подобной
и что, например, превращения нервной клетки в мышечное волокно или
соединительнотканной клетки в эпителиальную еще никто не видел, он,
тем не менее, считает неправильным утверждение, что клетки дифферен-.
цированных тканей в силу своей организации вообще не способны ни к ка-
ким другим функциям, кроме тех, которые они выполняют в данный мо-
мент, и не могут превратиться ни во что иное, кроме того, чем они уже
являются. При этом Гертвиг ссылается на лишь недавно открытый факт
регенерации линзы из края глазного бокала. О специфичности клеток,
по мнению Гертвига, возможно говорить только в одном смысле — в смысле
их видовой специфичности.
Дальнейшее развитие экспериментальной эмбриологии и эксперимен-
тальной гистологии показало неправильность этих теоретических устано-
вок О. Гертвига, заставлявших его не соглашаться с учением о специфич-
ности клеток и считать, что при эмбриональном развитии все клетки остаются
ио существу равноценными, а их видимые морфологические и функциональ-
ные особенности определяются лишь положением в организме и взаимо-
отношениями друг с другом, т. е., по сути дела, внешними по отношению
к ним факторами.
36
Глава I
7. ОДНОСТОРОННИЕ ФИЗИОЛОГИЧЕСКИЕ И ФОРМАЛЬНО-МОРФОЛОГИЧЕСКИЕ
ПОСТРОЕНИЯ НОВЕЙШЕГО ВРЕМЕНИ
Как уже отмечалось выше, попытки создания генетической системы
тканевых структур не встретили сочувствия и не могли быть проведены
последовательно, а интереснейшая работа Раубера совершенно выпала из
поля зрения гистологов. Лично мне эта работа и работа Геккеля (1885)
сделались известны лишь недавно, при внимательном ознакомлении с раз-
личными первоисточниками. Прочно вошли в науку представления
Геккеля об эпителии, как изначальной в фило- и онтогенезе ткани, и про-
тивоставление эпителия мезенхиме, обоснованное Гертвигами.
В течение дальнейшего развития гистологии интерес исследователей
к общим вопросам упал и, невидимому, в значительной степени заслонился
блестящим развитием и достижениями техники гистологических исследова-
ний, собиранием и обработкой разрастающегося фактического материала.
Наряду с работами описательного характера появляются и эксперимен-
тальные исследования, проводимые систематически, преимущественно на
соединительной ткани и крови. Пионером в этой области был
А. Максимов, работы которого сделались классическими и легли в основу
современных представлений о мезенхимных производных.
Внимание большей части гистологов концентрируется преимущественно
на тканях и органах позвоночных и, в первую очередь, млекопитающих
и человека. Беспозвоночные интересуют гистологов в этот период относи-
тельно меньше.
, В XX в. в гистологической системе господствует в основном морфо-
физиолргический, а не генетический подход к тканям, причем значительная
часть авторов особенно подчеркивает функциональный момент (К. Шней-
дер, 1902, 1908; Пренан, Буэн и Майар, 1904; Далъгрен, и Гейнер, 1908;
Петерсен, 1922; отчасти Заварзин, 1932—1939, и многие другие).
v Пренан, Буэн и Майар, проводя, в отличие от многих других, очень,
последовательно физиологический принцип, следующим образом форму-
лируют свое отношение к тканям: <у клеток спрашивают, чтобы сделать
из них ткань, не о том, чем они сделаются впоследствии или откуда они
взялись, а лишь о том, чем они являются и что они делают в данный момент»
(стр. 300). Понятие ткань, в их представлении, не включает в себя непре-
менно генетического единства составляющих ее элементов.
В руководствах физиологического направления иногда делаются попытки
.дать классификации тканей и их элементов, сильно отличающиеся от обще-
принятой системы Лейдига и Кёлликера. В основу таких, чисто физиоло-
гических классификаций обычно кладется не тканевая структура, а.функ-
щиональные особенности отдельных тканевых элементов, дифференцирован-
ных определенным образом. Так, например, Шнейдер (1902, 1908) разли-
чает следующие главные типы клеток: покровные (тектоциты), питающие
(нутроциты), железистые (аденоциты), чувствительные (эстоциты), нерв-
ные (нейроциты), почечные (нефррциты), мышечные (миоциты), соединитель-
нотканные (иноциты) и половые (пропагоциты). Глиальные клетки сбли-,
жаются с покровными, а стрекательные — с железистыми элементами. В
Очерк развития гистологической системы 37
соответствии с этим в животном мире различаются следующие ткани: по-
кровная, питающая, нервная, железистая, соединительная, мышечная и
половая и часто также почечная. Покровные, питательные, железистые,
стрекательные, чувствительные, почечные и многие половые клетки обра-
зуют эпителиальные соединения, а соединительнотканные и мышечные —
выполняющую ткань, компактная разновидность которой обозначается,
как плерома.
В руководстве Пренана, Буэна и Майара приняты следующая номен-
клатура и порядок изложения общегистологической части или «специаль-
ной цитологии»: 1) <чувствительная клетка» (нервные элементы и рецеп-
торные аппараты вплоть до кортиева органа); 2) «мышечная клетка»; 3) пи-
тающие (трофические) клетки (cellules nutritives) — железистые и эпите-
лиальные клетки, кровь и различные фагоцитирующие элементы с крове-
творными органами, жировые клетки, клетки Лейдига, плазматические,
тучные клетки и пр.; 4) «опорные клетки» — глия и мезенхимные опорные
структуры, а также эктодермальные опорные клетки кишечно-полостных,
хорда и некоторые другие образования. Затем описываются клеточное деле-
ние и половые клетки. Понятие ткани, при таком подходе к делу, в значи-
тельной степени исчезает.
Одностороннему физиологическому подходу к тканям нельзя отказать
в большой последовательности. Вместе с тем подчеркивание какого-нибудь
одного функционального признака, как руководящего момента класси-
фикации, приводит к настолько большому разрыву между морфологией
и физиологией тканей, что совершенно различные образования оказываются
объединенными в одну группу (например покровные и железистые эпителии,
кровь; кроветворные органы, жировые клетки и прочее), а сходные ткани
и даже разные клеточные элементы одной и той же ткани попадают в раз-
ные рубрики (например кровь, жировые, тучные и плазматические клетки
оказываются среди «трофических клеток», а фибробласты и мезенхимные
структуры механического характера рассматриваются, как «опорные клет-
ки»). Поэтому неудивительно, что попытки построить систему тканей на одно-
стороннем функциональном моменте не имели успеха и не привились, в то
время как классическая морфо-физиологическая система, несмотря на. все
свои недостатки, существует до настоящего времени.
В ряде курсов гистологии, например Петерсена (1922) и Заварзина
(1932—1939), также подчеркивается физиологическая основа классифика-
ции тканей, однако по существу подход этих авторов к тканям является не
чисто физиологическим, а морфо-физиологическим, т. е. в общем соответ-
ствует принятым в гистологии со времени Лейдига нормам. Например,
Заварзин вводит, вместо употребительного обозначения соединительноткан-
ных образований, функциональные понятия трофических- и механических
тканей или, согласно последнему варианту, тканей внутренней среды, но по
существу не изменяет их морфологического содержания. К трофическим
тканям он не относит, как это делается в настоящих физиологических клас-
сификациях, ни кишечный эпителий, ни ткань печени, ни ряд других струк-
тур, трофическое значение которых бросается в глаза. В их состав вклю-
Глава I
-чаются, как это большей частью принято, лишь кровь и некоторые соеди-
нительнотканные образования. Группа механических тканей также объеди-
няет собой в основном соединительнотканные образования. Поэтому замена
-обычного морфо-физиологического термина «соединительная ткань» или
-«ткани межклеточного вещества» другим, более физиологическим, едва ли
.целесообразна, так как физиологическое обозначение никак не отражается
на существе дела. Пожалуй, еще менее удачным следует считать понятие
тканей внутренней среды в применении к одним лишь мезенхимным произ-
водным, хотя оно представляется очень целесообразным в своем настоящем,
более широком смысле. К тканям внутренней среды относятся с полным
-правом не только кровь и все соединительнотканные образования, но боль-
шая часть мышц, ткани нервной системы, в значительной степени ткань
печени высших позвоночных, ткани эндокриновых органов и ряд других
структур, которые, кстати говоря, отнюдь не в меньшей степени регулируют
состав внутренней среды, чем мезенхимные производные.
В некоторых случаях физиологический подход к тканям делается
настолько односторонним, что принимает совершенно уродливые формы.
Например, Каррель высказывает в своей небольшой статье, озаглавленной
«Новая цитология» (1927), точку зрения, что всю классическую описатель-
ную цитологию и гистологию следует признать совершенно устаревшими
и подлежащими полной замене новой цитологией, основанной на изучении
функциональных особенностей различных чистых клеточных штаммов и их
•взаимоотношений с внешней средой, так как морфология клеток и тка-
ней, будучи второстепенной, изменчивой и малопостоянной, целиком зави-
,сит- от внешних условий. Всякое морфологическое исследование клеток
и тканей может быть полностью заменено изучением численных показа-
телей, характеризующих энергию их роста в тканевых культурах.
К столь же уродливым результатам приводит и противоположная край-
ность — выдвигание на первый план формального морфологического мо-
мента при игнорировании не только генетического, но даже и физиологи-
ческого. Примером такого подхода к тканям является довольно известная
статья Петера «Ткани в преподавании» (1929). Совершенно нежизненную
формальную классификацию тканевых структур предлагал сравнительно
недавно Студничка (1911, 1929). С его точки зрения, всякая ткань пред-
.ставляет собой «несущий определенную функцию комплекс элементарных
составных частей и масс». В имеющихся в тканях свободных промежут-
ках находятся жидкости. Под элементарными составными частями Студ-
ничка понимает, во-первых, биосистемы (клетки, гигантские клетки, син-
цитии) и, во-вторых, биоструктуры (фибрилли и волокна), а под «массами»
(и сетями) — плазмодии, внеклеточные протоплазмы, строительные вещества
(Bausubstanzen — основное вещество и кутикулы), пластинки, палочки.
Если сопоставить эти современные.нам высказывания с разобранными
выше представлениями XVIII и начала XIX вв., то нельзя не подметить неко-
торых, достойных удивления черт сходства, обусловленных столь популяр-
ными в наше время «ревизиями» клеточного учения и достаточно формаль-
ным подходом к тканевым структурам.
Очерк развития гистологической системы
г89
Студничка (1929) дает два образца морфологической классификации
/тканей по их «содержанию» или составу из клеток, синцитиев, симплазм
и основных веществ или по форме и расположению частей. Классификация
по «содержанию» имеет следующий вид: 1) клеточные ткани, состоящие:
а) из клеток, расположенных рядом (клеточные паренхимы), и б) из сете-
видно связанных клеток (клеточные сети, сетчатые ткани); 2) синцитиаль-
ные ткани, состоящие из мышечных элементов или волокон; 3) симпласти-
ческие ткани, например, образующиеся из сетевидной клеточной ткани;
4) ткани основного вещества, подразделяющиеся на: а) содержащие клетки
(с небольшим количеством основного вещества, с большим его количеством,
первично-клеточные, как, например, хрящ, и вторичноклеточные, как,
например, влагалище хорды у селахий), б) бесклеточные (первично бескле-
точные, как, например, «мезострома», и вторично бесклеточные, например
•остеоидные).
По форме и расположению частей Студничка различает следующие
основные разновидности тканевых структур: 1) синенхима (компактная
плазмодиальная ткань); 2) паренхима (клеточная ткань типа эпителия);
3) прозенхима (клеточная ткань из веретенчатых или имеющих вид волокон
клеток, как, например, гладкие мышцы); 4) неменхима (ткань, состоящая
исключительно или преимущественно из волокон, как, например, попе-
речнополосатые мышцы и периферические нервы); 5) ретенхима (сетевид-
ная ткань, как, например сетевидные эпителии, мезенхима, ретикулярная
ткань); 6) филленхима (ткань, состоящая из листков, например рыхлая
соединительная ткань); 7) плектенхима (сплетение клеточных отростков
с клеточными телами, например нервная ткань серого вещества); 8) синек-
хима (компактные бесклеточные основные вещества, кутикулярные обра-
зования). Следует оговориться, что сам Студничка не заменяет своим под-
разделением тканей на эти морфологические разновидности общепринятой
системы тканей Лейдига и Кёлликера.
Если у последователей односторонней физиологической классификации
имеются налицо крайний «целлюларизм» и подмена тканей дифференциро-
ванными различным образом клетками, то все без исключения, как более
старые, так и новые формальные морфологические подразделения, по сути
дела, оперируют не с биологическими, а с чисто геометрическими пред-
ставлениями, или даже со странной смесью геометрии и биологии, имею-
щими весьма отдаленное отношение к живым тканевым системам орга-
низма.
8. ЭКЛЕКТИЧЕСКИЕ МОМЕНТЫ В СОВРЕМЕННЫХ ТРАКТОВКАХ
ГИСТОЛОГИЧЕСКОЙ СИСТЕМЫ ЛЕЙДИГА И КЁЛЛИКЕРА
Хотя система тканей Лейдига и Кёлликера и принята сейчас в основном
подавляющим большинством гистологов, многие авторы предпринимают
в ней отдельные изменения или исправления, в большей или меньшей сте-
пени искажающие ее первоначальный последовательный функциональный
или, точнее, морфо-физиологический характер и придающие ей эклекти-
ческое содержание. Прежде всего это касается определения основных тка-
40
Глава 1
невых групп, в основу которого нередко кладутся совершенно различные
принципы. Так, для т. наз. эпителиальной ткани в целом обычно
считается наиболее подходящим формальное морфологическое определе-
ние, учитывающее одновременно и положение в составе организма (Геген-
баур, 1892; Ганземан, 1896; Маршанд, 1899; Максимов, 1914; Шаффер,
1927, и др.). Единство группы тканей «соединительного вещества» обосно-
вывается их происхождением из мезенхимы, а их определение приобретает
тем самым уже не морфо-физиологический, а генетический характер (Мак-
симов, 1914, 1927, и многие другие). Наконец, для «мышечной» и «нервной»
.тканей, как правило, сохраняется их первоначальное функциональное
.определение. Такой эклектический подход к определению основных тка-
невых групп логически явно недопустим, так как всякая классификация,
на каком бы принципе она ни основывалась, должна быть прежде всего
последовательной и выдержанной.
Другие изменения в гистологической системе касаются большей частью
отнесения отдельных тканевых структур не к той группе, к какой они при-
надлежат по своей общей морфо-физиологической характеристике. .
Такое перемещение производится обычно на основе генетического при-
знака, не учитываемого в подразделении на основные четыре группы. Это,
например, касается нейроглии, эпендимы, различных эндотелиев, а часто
и мезотелия, гладкой мускулатуры позвоночных и некоторых других
тканей.
Нейроглия, которая по своей общей морфо-физиологической характе-
ристике может трактоваться с известной точки зрения как разновидность
соединительной ткани (сетевидное строение, волокнистый опорный аппарат,
трофическое и механическое значение), обычно рассматривается отдельно
от собственно соединительной ткани, с генетически родственными ей нерв-
ными элементами. Вместе с тем в понятие «нервная ткань», которое перво-
начально обозначало совокупность нервных клеток с их отростками и
концевыми аппаратами, в настоящее время включаются функционально
совершенно чуждые ей элементы, принимавшиеся в свое время за особого
рода «ткань соединительного вещества». Эпендима рассматривается с из-
вестными оговорками то как эпителий, то как часть нейроглии, при этом,
если собственно эпендиму и включают в состав нейроглии, то покровные
клетки сосудистых сплетений мозга и пигментный слой сетчатки, которые
по существу являются разновидностью эпендимы, всегда обозначают как
эпителий. Эндотелий сосудистой системы или десмальный эпителий (Шаф-
фер, 1921, 1927) в настоящее время или совершенно исключается из состава
эпителиев и рассматривается как производное мезенхимы, вместе с соеди-
нительной тканью, или, по крайней мере, резко противопоставляется всем
прочим эпителиям. Таким же образом до последнего времени многие были
склонны трактовать и мезотелий серозных оболочек (Максимов, 1925, 1927;
Шоппер, 1932), руководствуясь, как и в случае эндотелия, его предпола-
гаемой генетической близостью к соединительной ткани и трактуя одног
временно соответствующим образом его превращения при эксперименталь-
ных условиях. То же самое справедливо и в отношении ряда других эпите-
Очерк развития гистологической системы
41
лиоподобных структур, как, например, покровных клеток мозговых обо-
лочек, десцеметова «эндотелия» и т. п.
Если принимать для эпителиев подчеркнутый еще Гисом (1865) и Герт-
вигами (1882), а за ними и многими другими авторами общий функциональ-
ный признак пограничное™, то эндотелий может рассматриваться как
разновидность эпителиев с неменьшим правом, чем, например, мезотелий
млекопитающих и ряд других эпителиоподобных структур внутренней
среды, организма. С другой стороны, этот же функциональный критерий
помогает исправить формально морфологическую трактовку остеобластов
и одонтобластов, как эпителиев, которая была принята прежде.
Очень непоследовательным является также исключение гладкой вну-
тренностной мускулатуры позвоночных из группы мышечных тканей, на
основе ее генетического родства с соединительной тканью, если оставаться
в отношении классификации тканей на общепринятых морфо-физиологи-
ческих позициях. То же самое было бы справедливо и в отношении т.
наз. миоэпителиальных клеток и мускулатуры радужины. Все. эти
довольно обычные в наше время попытки тех или иных частичных изме-
нений классической системы гистологии имеют своими причинами, с одной
стороны, фактическую, так сказать, стихийную невозможность исключить
из гистологии генетический принцип, который властно стучится в дверь,
как бы его ни изгоняли, а с другой стороны, исторически сложившееся
в нашей науке, недостаточно отчетливое разграничение морфо-физиологи-
ческих и генетических категорий или, наконец, просто игнорирование
логических предпосылок всякой научной классификации.
Вместе с тем нельзя закрывать глаза на то, что последовательное про-
ведение морфо-физиологической классификации тканей, несомненно во
многих отношениях правильно отражающей объективно существующие
в живой природе закономерности, очень часто наталкивается на непреодо-
лимые трудности и даже имеет формальный характер, именно в силу того,
что особенности ее объектов исторически обусловлены и изменчивы.
Поэтому они по самой своей сущности не всегда могут быть уложены
в те или иные рамки, определяющиеся не их развитием, а иными, хотя бы
и очень существенными, признаками. С такими трудностями неизбежно
сталкивается всякая искусственная гистологическая система, основываю-
щаяся не на генетическом, а на любом другом принципе. Существующая же
гистологическая система является в этом смысле искусственной, а не есте-
ственной системой. К таким трудностям и неясностям относится, например,
положение в системе, если не руководиться генетическим моментом, са-
теллитов, шванновских клеток и вообще нейроглии, эпителия многих
эндокриновых органов и, в первую очередь, эпителиальной сети зобной
железы, сетевидной пульпы эмали, ткани спинной струны, а также упомя-
нутых выше покровных клеток мозговых оболочек и многих других ткане-
вых структур.. Даже принятое в настоящее время отнесение крови и лимфы
к группе., соединительнотканных образований, как это справедливо отме-
чалось и отмечается многими (Шаффер, \Q21-, Шмидт, 1930, и.др,), натал-
кивается на серьезные возражения, поскольку и та и другая, строго го-
"42
Глава I
воря, вообще не имеют тканевой структуры, а представляют собой жид-
кость со свободно плавающими в ней форменными элементами. Сближение
крови и лимфы с соединительной тканью оправдывается, в первую очередь,
генетическими соображениями.
Последовательное проведение морфо-физиологической классификации
в ряде случаев неизбежно приводит к грубому формализму и явным несо-
образностям, как это имеет место, например, в случае отнесения эпите-
лиальной сети зобной железы или пульпы эмали к мезенхимным образо-
ваниям.
Еще значительнее становятся трудности, если обратиться к беспозво-
ночным. В какой степени возможно трактовать клетки нефридиев как
эпителиальную ткань? В какую группу относить кутикулярные, панцыр-
ные и скелетные эктодермальные структуры, которые Лейдигом и многими
другими, именно на основе их морфо-физиологических особенностей, отно-
сились к группе тканей соединительного вещества? Чем являются, наконец,
т. наз. паренхима низших червей или эктодерма и энтодерма кишечно-
полостных? Все эти и многие другие аналогичные вопросы неизбежно встают
перед гистологом. Такие же или даже еще более значительные трудности
возникают, если обратиться к экспериментально или патологически изме-
ненным тканевым структурам, в частности, к опухолевым тканям, которые
не могут игнорироваться, поскольку строгое разграничение т. наз. нор-
мальной и патологической гистологии в настоящее время невозможно и
является, по сути дела, совершенно искусственным.
Наконец, нельзя не отметить, что в отношении своего объема и точности
морфо-физиологической характеристики четыре группы тканей вовсе не
являются равноценными. Эпителиальные и мезенхимные структуры харак-
теризуются в целом очень общими и даже подчас малоопределенными при-
знаками, а. мышечные и нервная ткани, наоборот, в высшей степени кон-
кретными специальными особенностями. Однако общая эпителиальная или
мезенхимная характеристика отнюдь не могут противопоставляться функции
сократимости или раздражимости. Элементы пограничных тканей могут
обладать вместе с тем мышечными или нервными особенностями. «Эпите-
лиальность» или «мезенхимность», как общие свойства, совсем не исклю-
чают сократимости или нервной функции. Недаром эпителий и ткани соеди-
нительного вещества, с одной стороны, мышечные и нервная ткани, с дру-
гой, часто противопоставляются друг другу, как ткани общего и специального
значения. Вместе с тем различные конкретные представители эпителиаль-
ных структур, имеющие ту или иную специальную функциональную харак-
теристику, уже не являются тканями общего характера, а могут быть про-
тивопоставлены, как равноценные рубрики, мышцам и нейронам. В качестве
таких, более специальных, функциональных подразделений эпителиальных
и мезенхимных структур, основывающихся на той или иной однородной
специальной дифференцировке их отдельных элементов, можно было бы
указать на следующие: железистые и эндокриновые, связочные и сухо-
жильные, скелетные, защитные, экскреторные, всасывающие, респира-
торные и многие другие, соответствующие различным специальным функ-
Oiepx развития гистологической системы
43
циям организмов. Такими же специальными рубриками являются мышеч-
ные и нервные образования.
Чем проще общая организация животных, тем более универсальный и
мультифункциональный характер имеют их ткани. Чем выше и сложнее
морфологическая организация, тем более специальный характер приобре-
тают отдельные ткани и тем большим становится вместе с тем их разно-
образие.
9. ПРЕДПОСЫЛКИ К ГЕНЕТИЧЕСКОЙ СИСТЕМЕ ТКАНЕВЫХ СТРУКТУР
Естественная генетическая система тканевых структур может иметь
реальный смысл только в том случае, если происхождение тканей из того
или иного источника отражается каким-то определенным образом в их
свойствах. Если же тождественные во всех отношениях гистологические
структуры могут, как правило, возникать из любого источника, то ни о ка-
кой генетической системе тканей, собственно говоря, не может быть и речи,
и их подразделение по происхождению теряет всякий смысл. В пользу
последней точки зрения, казалось, говорил весь фактический материал
описательной гистологии, а потому и все попытки построения генетической
системы тканей не имели и не могли иметь успеха. Действительно, какой
смысл разделять по происхождению эктодермальный эпителий всей по-
лости рта (Геккель, 1886) или, по крайней мере, преддверия полости рта
(Маурер, 1915) и энтодермальный многослойный эпителий всей прочей
передней части пищеварительной трубки, или, наоборот, объединять в еди-
ную энтодермальную группу совершенно различные эпителии пищевода
и кишечника? Это имеет тем меньше смысла, что, по господствовавшим до
последнего времени представлениям, различные эпителии могут переходить
друг в друга, т. е. не обнаруживают каких-либо качественных различий.
Аналогичные соображения справедливы и в отношении многих других
тканевых структур. Кроме того, они показывают, что генетическая система
тканей не может быть построена на основе одних лишь эмбриологических
представлений.
Если бы в природе не было реальных основ для генетической системы
тканей, то приходилось бы допустить, что закономерности органической
эволюции не приложимы в области гистологии, что ткани каким-то образом
оказались вне эволюционного процесса. Последнее допущение, при совре-
менном уровне знаний, представляется совершенно невероятным.
II. ПРОБЛЕМА ЭВОЛЮЦИОННОЙ ГИСТОЛОГИИ J
1. ЭВОЛЮЦИОННАЯ ТРАКТОВКА ГИСТОЛОГИЧЕСКОЙ СИСТЕМЫ „ цч
ЛЕЙДИГА И КЁЛЛИКЕРА !3.
В изложении истории гистологической системы было отмечено, ч^о-
эволюционные представления стали проникать в нашу науку в начале
прошлого столетия. Попытки эволюционного подхода к тканям можйо
найти еще у Гейзингера (1822) и, несколько позже, у Рейхерта (1845).
К семидесятым и восьмидесятым годам относятся исследования ГеккеЛя
и братьев Гертвигов, эволюционные представления которых до настоящего
времени оказывают очень большое влияние на гистологическую мысль.
Затем, однако, многие основные общегистологические вопросы, а вместе
с тем и проблема эволюционной гистологии, как это уже отмечал осы, ото-
шли на задний план. Лишь в последнее время интерес к таким вопросам
снова возрос, причем у нас в Советском Союзе возникли новые эволюцион-
ные направления в гистологии. Одно из них, возглавляемое Заварзиным
и известное как теория параллелизма, посвящено обоснованию, разработке
и объяснению общих эволюционных закономерностей, которым подчинены
четыре морфо-физиологические группы тканей, установленные Лейдигом it
Келлихером. Эта книга посвящается другому направлению, многие основ-
ные положения которого неоднократно затрагивались в отдельных экспе-
риментальных работах, принадлежащих моим сотрудникам и мне, разве-
вались в статьях общего характера и уже были отражены в моей монографии
(1940). :Г"
Не может подлежать сомнению, что тканевой материал, из которого
построены все части тела многоклеточных животных, не оставался неиз-
менным, а эволюировал вместе с развитием органического мира. Ткани
и клетки представляют собой изменяющиеся в эволюционном процессе
интегрированные частные системы организма, находящиеся в сложных
коррелятивных взаимосвязях и, в своем развитии, подчиненные целому
организму. Совокупность их морфологических и функциональных свойств
обусловлена филогенетически и наследственно закрепляется на каждом
этапе развития. Вместе с тем явления изменчивости и наследственности,
лежащие в основе органической эволюции, должны быть применимы и
в области гистологии, а естественный отбор, творческая и элиминирующая
роль которого в развитии органических форм была обоснована Дарвиным,
должен лежать также в основе тканевой эволюции.
Проблема эволюционной гистологии
45
Однако приложение эволюционного учения к области гистологии встре-
чает очень большие трудности. Эти трудности были подчеркнуты Эбнером
(1911) в небольшой статье, многие мысли которой сыграли очень боль-
шую роль в дальнейшем развитии нашей науки и легли в основу теории
гистологических параллелизмов, развиваемой Заварзиным и его сотруд-
никами.
Ввиду этого здесь будет уместно несколько подробнее остановиться на
упомянутой статье Эбнера «Развитие тканей и филогенез», представляющей
собой доклад на 25-м съезде анатомов. Эбнер обращает внимание на пора-
зительный и общеизвестный факт, что некоторые высокодифференцирован-
ице тканевые структуры появляются у представителей различнейших типов
животных, независимо от их систематического положения.
В качестве примеров приводятся поперечнополосатые мышцы позво-
ночных и членистоногих, структура которых совпадает до многих тончай-
ших деталей; хитиновые образования и структура костей, хрящ в ряду
позвоночных и у отдельных представителей членистоногих (Limulus) и
моллюсков (Haliotis и др.;; пузырчатые опорные ткани и студенистая сое-
динительная ткань различных позвоночных и беспозвоночных.
Все подобные явления сходства тканевых структур Эбнер впервые
трактует как явления конвергенции или совпадения структуры, обусло-
вленное функциональным сходством, и совершенно независящее от общ-
ности происхождения («Es handelt sich augenscheinlich um eine Art Kon-
vergenzerscheinungen, um eine ubereinstimmende Struktur wegen iiberein-
stimmender Funktion, ganz unabliangig von gemeinsamer Abstammung aus
dcmselben Urorganismus»).
Совершенно ясно, говорит он, что позвоночные и членистоногие не имеют
общего предка, который бы обладал поперечнополосатой мускулатурой.
Несомненно, что структурная конвергенция наблюдается, несмотря на
химические различия белковых тел, отличающихся видовой специфич-
ностью. Морфологически неразличимые структуры, например, свойствен-
ные мышечным волокнам, несомненно отличаются друг от друга химически
у различных видов животных.
Подробнее всего Эбнер останавливается на явлениях конвергенции
опорных тканей. На основании разбираемых фактов он приходит к выводу,
что зародышевые листки не имеют тканевой специфичности и что «прежние
неясные представления о значении зародышевых листков должны быть,
наконец, оставлены и уступить место взгляду, что гистогенетическая про-
спективная потенция эмбриональной клетки ни в какой мере не опреде-
ляется окончательно ее положением в составе какого-нибудь зародышевого
листка». Вместе с тем и специфичность тканей принимается им лишь с ого-
ворками и исключениями. Совершенно ясно, говорит он, что способы пред-
ставления, справедливые в отношении филогенеза родственных организмов,
не могут быть перенесены безоговорочно на гистогенез. Ткани предста-
вляются гораздо более изменчивыми (leichtflussiger), чем тип организации,
и в своей структуре приспособляются, даже в составе одного и того же
организма, к той задаче, которую они должны выполнять; те же самые
46
Глава 11
структурные типы могут образоваться конвергентно из различной зачат-
ковой плазмы.
Эбнер подчеркивает вместе с тем бесцельность попыток установить
филогенез тканей, независимо от типа организации, так как они неизбежна
должны привести к неразрешимым противоречиям и смешению простран-
ственных переходов одной ткани в другую с истинными процессами раз-
вития. Большое внимание должно быть обращено на взаимное влияние
тканей в течение развития. Существенное филогенетическое значение могут
иметь лишь те ткани, которые определяют развитие того или иного органа
и обусловливают, в порядке зависимой дифференцировки, развитие других
тканей. Большой заслугой Эбнера является то, что он первый обратил вни-
мание на значение конвергенций в тканевой эволюции и на трудности,
встречающиеся при филогенетическом подходе к гистологическим струк-
турам.
Блестящие исследования Заварзина и его сотрудника Орлова зрительных
центров (1912), брюшного мозга (1925) насекомых и внутренностной нерв-
ной системы членистоногих, подытоженные в монографии (1941), показали,
при сопоставлении с литературными данными (касающимися позвоночных
и некоторых других типов животных), что и в области нейрональных взаимо-
отношений в разных отделах нервной системы очень отдаленных друг от
друга типов животных также имеются поразительные явления сходства.
Под таким же углом зрения был затем проведен большой цикл работ За-
варзина и его сотрудников по крови и соединительной ткани (Заварзин^
1934, 1939). На основе всего этого Заварзин (1925, 1934, 1941) высказал
положение, что явления параллелизмов или конвергенций представляют
собой основную закономерность в области эволюции тканей, что тем самым
эволюция гистологических структур принципиально отличается по своим
закономерностям от эволюции целых организмов и что дарвиновская теория
развития должна быть преломлена особым образом в применении к гисто-
логическим проблемам. Резкое противопоставление друг другу организ-
менной и гистологической эволюции, однако, как это будет показано ниже,
неправильно и недостаточно обоснованно. Теория гистологических парал-.
лелизмов, в той эволюционной трактовке, которая ей была дана позже ее
автором (Заварзин, 1934, 1941), делает попытку дать эволюционное объясне-
ние гистологической системе Лейдига и Кёлликера, которая целиком осно-
вывается на морфологическом сходстве тканевых структур, имеющих
сходное функциональное значение в животном организме. Эволюционное
объяснение существования этих универсальных четырех групп тканей
нуждается, однако, в одном существенном дополнении, на которое мной
было недавно (1940) обращено внимание. Высказывания Эбнера и теория
параллелизма ничего не говорят о том, какой общей причиной обусловли-
ваются все бросающиеся в глаза явления сходства в области гистологи-'
ческих структур, независящего во многих случаях от систематического
положения данного вида животных. Мне кажется, что существование сход-
ных во многих отношениях четырех групп тканей у всех многоклеточных
животных, за исключением губок, не имеющих нервной системы и диф;фе--'
Проблема эволюционной гистологии
47
ротированных мышечных элементов, является чрезвычайно важным и
убедительным доказательством единства происхождения животного мира,
подобно тому, как клеточное учение, в основе которого, строго говоря,
также лежит общее всему живому явление параллелизма, говорит нам об
единстве происхождения всех не только животных, но и растительных
организмов нашей планеты. Другого общего научного объяснения ткане-
вых параллелизмов и клеточного учения, с моей точки зрения, не суще-
ствует. Тем самым из области гистологии исчезает основное затруднение —
кажущаяся неприложимость к тканям одного из основных положений
эволюционного учения, согласно которому сходство в строении различных
организмов объясняется, прежде всего, их родством, а различия — дивер-
гентным развитием. Поразительные черты сходства, наблюдаемые в ана-
логичных (имеющих одинаковое функциональное значение) тканевых и
клеточных структурах животных организмов, уже давно сделались обще-
признанным фактом и приобрели большое значение в гистологии. Наоборот,
явления дивергентного развития клеточных и тканевых структур обра-
щали на себя гораздо меньше внимания.
2. ПАРАЛЛЕЛИЗМЫ И КОНВЕРГЕНЦИИ В ОБЛАСТИ ГИСТОЛОГИИ
И СРАВНИТЕЛЬНОЙ АНАТОМИИ
Так как значение явлений сходства для гистологии делается очевидным
уже на основании существования в животном царстве четырех общеприня-
тых групп тканей, то наша первая задача —выяснить, следует ли их трак-
товать как параллелизмы или конвергенции и какое место принадлежит
этим явлениям в области гистологии и сравнительной анатомии.
Понятия параллелизма и конвергенции в применении к тканям нужда-
ются прежде всего в некотором уточнении. Эбнер, а в последнее время
отчасти Шмалъгаузен (1939) трактуют явления гистологического сходства
как конвергенции, поскольку дело идет об организмах, не связанных друг
с другом близким родством. Заварзин, впервые применивший в области
гистологии понятие параллелизма, пользуется им как синонимом понятия
конвергенции.
Мне кажется, что в области эволюционной гистологии следует разли-
чать, подобно тому, как это принято в сравнительной анатомии, паралле-
лизмы и конвергенции, помня, однако, что между ними не всегда можно
провести резкой границы. Разграничение этих понятий в области тканей,
поскольку дело касается очень общих для животных организмов особен-
ностей, не может основываться на большей или меньшей генетической
близости сравниваемых видов животных. Даже у очень далеко отстоящих
друг от друга организмов сходство некоторых гистологических структур
может быть обусловлено как общностью их происхождения в индивидуаль-
ном и филогенетическом развитии, так и в дальнейшем параллельной эво-
люцией. С моей точки зрения, это полностью относится к поразительным
явлениям параллелизма (именно параллелизма, а не конвергенции) в об-
ласти нервной системы членистоногих и позвоночных, установленным За-
варзиным и его сотрудником Орловым. Во-первых, нервная система этих.
48
Гла?а П ~
а.также многих других животных генетически связана с эктодермальным:
кожным покровом примитивных общих предков и наружным зародышевым
листком в онтогенезе, т. е. с гистогенетически единым субстратом^ Во-
вторых, у общих предков этих типов, вероятно, уже успели появиться
какие-то примитивные нервные элементы, находившиеся в определенных
взаимоотношениях друг с другом и с другими элементами их тела. Из этих
примитивных нервных элементов, как из филогенетически общего источ-
ника, у членистоногих и позвоночных развилась «нервная ткань» с. ее
сложными нейрональными взаимоотношениями. Иными словами, следует
считать, что в состав анатомически негомологичных друг другу трубчатого
спинного мозга и брюшной нервной цепочки входит гистологически гомо-
логичный нейрональный материал. В-третьих, следует подчеркнуть, что
насекомые и позвоночные были выбраны Заварзиным, как объекты сравни-
тельного изучения, именно потому, что в отношении общей расчлененности
тела, прекрасно развитых конечностей, построенных по принципу рычагов
и снабженных дифференцированной мускулатурой, способа передвижения
и ряда других особенностей эти животные несомненно обнаруживают при-
знаки конвергенции, которые не могут не иметь значения для параллельной
гистологической эволюции нервной системы.
Весьма возможно, что многие черты сходства, обнаруживаемые муску-
латурой соматического типа членистоногих и позвоночных, а также цело-
мической, в частности, сердечной мускулатурой некоторых животных,
имеют в своей основе не конвергентное, а параллельное развитие, хотя
доказательства их гистогенетической гомологии в связи с существенными
различиями в онтогенезе и не являются в настоящее время достаточно обо-
снованными. Параллелизмом является также сходное у самых разнообраз-
ных животных строение эктодермального пищеварительного и всасывающего
(трофического) эпителия. Последний у представителей различных типов
животных с полостным пищеварением образует мерцательные или кути-
кулярные структуры, а на более низких ступенях эволюции состоит из
фагоцитирующих элементов, способных к внутриклеточному пищеварению.
При переходе к полостному пищеварению из генетически единого прими-
тивного эктодермального субстрата параллельно возникают и новые сход-
ные продукты дифференцировки.
Как параллелизмы, повидимому, можно трактовать, по крайней мере,
часть волокнистых и пузырчатых опорных структур соединительной ткани
у позвоночных и многих беспозвоночных, если считать энтомезенхиму
разных типов животных за гомологичный зачаток.
Наряду с параллелизмами в области гистологических структур мы
встречаемся также с явлениями конвергенции. Они обусловлены единством
элементарных свойств живого субстрата, имеющего в конечном итоге одно
и то же происхождение, сходным функциональным значением и сходными
условиями существования в составе организма соответствующего ткане-
вого и клеточного материала и, наконец, общими физическими или меха-
ническими закономерностями. Типичными примерами гистологических
конвергенций является уже давно замеченное сходство в тонком строении
Проблема эволюционной гистологии
49
хитина и кости, имеющих в составе организма аналогичное значение ске-
летных образований.
Эктодермальный хитин членистоногих и мезенхимальное вещество
кости позвоночных в отношении своего происхождения вовсе не являются
гомологичными друг другу образованиями. По химическому составу они
также весьма различны, хотя их мицеллы, повидимому, имеют сходную
нитевидную конфигурацию, которая в данном случае, вероятно, в значи-
тельной степени определяет микроскопическую структуру этих образований.
Другим примером гистологических конвергенций являются многие
опорные структуры, построенные по тургорному принципу и обнаружи-
вающие известные черты сходства с тканями растений. Так наз. пузырчатая
опорная ткань, распространение которой в животном мире было подчерк-
нуто в новейшее время Эбнером (1911) и детально разобрано в сводке Шаф-
фера (1930), представлена тканью хорды, различными мезенхимальными
хордоидными структурами, включая жировую ткань позвоночных и экто-
дермальными пузырчатыми элементами, образующими опорный аппарат
щупалец некоторых кишечнополостных.
Таким образом, среди пузырчатых опорных тканей следует различать,
с одной стороны, параллельные, а с другой, конвергентные структуры.
Среди сократимых структур мы также встречаемся не только с паралле-
лизмами, но и с конвергентными образованиями. К ним относятся, напри-
мер, гладкая мезенхимальная мускулатура позвоночных и гладкая муску-
латура соматического типа многих беспозвоночных. То же самое справед-
ливо и в отношении некоторых т. наз. эпителиальных структур, обнару-
живающих очень большое функциональное и морфологическое сходство,
но развивающихся у представителей разных типов животных из разных
источников, как, например, стенка эктодермальных нефридиев и целомо-
дуктов, несущих аналогичную выделительную функцию. Одно из порази-
тельных явлений цитологической конвергенции представляют, в частности,
соленоциты нефридиев многих червей и выделительных органов ланцет-
ника, которые часто также обозначаются как нефридии, но, по сути дела,
являются целомодуктами, поскольку они развиваются не из эктодермы,
а из выстилки полости тела.
В настоящее время в очень многих случаях невозможно с достаточной
определенностью трактовать те или иные случаи гистологического сход-
ства, наблюдаемые у отдельных форм животных, как явления конверген-
ции, а не параллелизма или, наоборот; так, в отношении большей части
т. наз. эпителиальных тканей, например, нельзя сказать с полной опре-
деленностью, являются ли они результатом конвергентного или парал-
лельного развития, так как их источники теряются среди вымерших пред-
ков современных многоклеточных животных. Во всяком случае, типичный
общий и очень стойкий признак многих эпителиальных структур — тесное
комплексное взаиморасположение элементов — можно рассматривать и как
результат очень древней конвергенции и как следствие параллельного раз-
вития какой-то единой примитивной эпителиоподобной структуры. К ана-
лизу этих вопросов нам придется вернуться несколько позже. Зато мы
4 Хлопип—349
50
Глава II
встречаемся с многочисленными явлениями несомненной гистологической
конвергенции в составе одного и того же животного организма. Эти кон-
вергентные гистологические дифференцировки, известные уже давно, и
были причиной того, чтобы считать свойства дифференцированных ткане-
вых структур независящими от их развития из того или иного источника.
Собственно говоря, единственными тканевыми элементами, которые, неви-
димому, у всех или почти у всех многоклеточных животных имеют единое
эктодермальное происхождение, являются нейроны, совокупность которых
в классической гистологии обозначается, как нервная ткань. У высоко-
организованных животных, в частности у позвоночных, можно говорить
о развитии всех нейрональных элементов из разных частей единого нейро-
эктодермального или нейрального зачатка, хотя и с известными оговор-
ками. Правда, в отношении источника развития нервных элементов у от-
дельных типов животных, например, у кишечнополостных и даже у игло-
кожих, имеются неясности, так как части нервной системы этих животных
до настоящего времени приписывается неэктодермальное происхождение.
Три прочие группы тканей едва ли не у всех высокоорганизованных жи-
вотных являются, вообще говоря, продуктами конвергентной дифферен-
цировки, развивающимися полифилетически из различных источников.
Эпителиальные структуры, как известно, могут возникать из любой эмбри-
ональной закладки. Сократимые мышечные элементы также могут обра-
зоваться из очень различных источников. Например, у позвоночных они
развиваются из миотомов (мускулатура соматического типа), из миоэпи-
кардиальной пластинки спланхнотомов (миокард), из энтомезенхимы (боль-
шая часть гладкой внутренностной мускулатуры и, по некоторым данным,
часть мускулатуры соматического типа), из эпидермального зачатка или
кожной эктодермы (миоэпителиальные клетки), из нейроэктодермы или
нейрального зачатка (мышцы радужины, построенные у рыб, амфибий и
млекопитающих по гладкому, а у рептилий и птиц по поперечнополоса-
тому типу).
Наконец, т. наз. ткани соединительного вещества, имеющие механиче-
ское и трофическое значение, могут иметь своим источником у позвоноч-
ных не только т. наз. мезенхиму (соединительнотканная группа), но и
нейральный (глиальная группа) и хордальный зачатки. Имеющие известное
сходство с ретикулярной тканью структуры, кроме того, образуются из
зачатка головной кишки (сеть зобной железы, пульпа эмали).
Наконец, само понятие мезенхимы в генетическом отношении не является
однородным. Помимо энтомезенхимы, образующейся из весьма различных
участков мезодермы (дерматом, склеротом, париетальный и висцеральный
листки спланхнотомов), в настоящее время приходится все больше и больше
считаться с эктомезенхимным зачатком («эктомезодерма» или «мезекто-
дерма»), принимающим, по многим данным {Равен, 1931; Стон, 1926;
Гарвей и Бурр, 1926, и др.), участие в образовании некоторых частей ске-
лета и развивающимся из ганглиозной пластинки.
Пигментные клетки соединительной ткани не являются, как это уста-
новлено в настоящее время, мезенхимными или, точнее, энтомезенхимными
Проблема эволюционной гистологии
51
производными, а образуются из клеточного материала нервных валиков
зародыша.
Иными словами, т. наз. пигментная соединительная ткань представляет
собой по своему клеточному составу полифилетическую, а не единую в ге-
нетическом отношении структуру.
Анализируя структурные особенности тканей и в значительной степени
опираясь на исследования Эбнера и на высказанные им взгляды (1911),
Патцелып (1925) приходит к выводу, что можно различать три основных
направления гистологической дифференцировки, которые нередко, кроме
того, сочетаются друг с другом различным образом. Первое характери-
зуется сохранением тесной взаимной связи цитоплазмы клеток посредством
мостиков или разветвленных отростков при более или менее сильном раз-
витии межклеточных промежутков. Такое направление дифференцировки
мы встречаем в типичном виде, например, в эпителиях и тканях нервной
системы. Плазмодиальный характер мышечной ткани представляет собой,
в известной степени, наиболее далеко идущую степень взаимосвязи клеток.
Как второе направление дифференцировки Патцельт рассматривает обра-
зование замыкающих клеточных оболочек, столь характерное, например,
для хордоидных структур. Третий путь гистологической дифференцировки
характеризуется развитием основного или межклеточного вещества того
или иного состава. Наличием в тканях разного происхождения этих трех
основных направлений дифференцировки и объясняются, по Патцелыпу,
столь распространенные в гистологии явления конвергенции, на которые
в свое время обратил внимание Эбнер (см. выше). Как явление, противо-
положное тканеобразованию, рассматривается четвертый путь развития,
приводящий к изоляции отдельных клеток, как это, например, наблюдается
в случае элементов крови и лимфы, а также половых клеток и при оттор-
жении эпителиальных элементов.
Специфичность зародышевых листков, говорит Патцелып, ограничи-
вается проявлением известных характерных различий, при тех или иных
функциональных приспособлениях, приводящих к образованию аналогич-
ных структур.
Являясь в значительной степени последователем Эбнера, Патцелып,
в отличие от него, допускает возможность существования специфических
особенностей у производных различных эмбриональных зачатков на фоне
общих явлений конвергенции.
Таким образом, в основе существующего подразделения тканевых
структур на четыре морфо-физиологические группы лежат широко рас-
пространенные в гистологии явления параллелизмов и конвергенций в
эволюционном развитии и явления конвергентной дифференцировки
в онтогенезе.
Конвергенции и параллелизмы, однако, вовсе не являются специфи-
ческой особенностью тканевых структур. Они имеют не меньшее значение
в эволюции внешней формы, симметрии и расчленения тела и анатомиче-
ского устройства органов, т. е. общей морфо-физиологической организации
Живых существ. Частью параллелизмами, частью конвергенциями очень
52
Глава II
общего характера является развитие в самых различных ветвях генеалоги-
ческого дерева животных основных систем органов, которые, при всем
своем внешнем разнообразии, обладают некоторыми общими морфо-физио-
логическими особенностями, позволяющими обозначать их одинаковыми
для всего животного мира анатомическими терминами. Параллелизмом
очень общего значения являются образование билатеральной симметрии
и дифференцировка тела на головной и хвостовой конец в ряду первично-
ротых и вторичноротых.
Общие, бросающиеся в глаза, морфологические или, вернее, морфо-
физиологические особенности организмов первоначально были положены
в основу их систематики и дали возможность построить ряд искусственных
систем, на смену которым затем пришла единая естественная система,
которая строится и развивается на эволюционном генетическом принципе.
Эта единая естественная система, особенно в своей части, касающейся бес-
позвоночных, имеет еще очень много спорных и неясных моментов, которые
должны быть разрешены дальнейшими исследованиями. На сходстве внеш-
ней формы тела основывалось объединение то в более обширные, то в мел-
кие систематические группы очень отдаленных друг от друга организмов.
Достаточно вспомнить существовавшие некогда типы Radiata и Zoophyfa
и тип моллюсков, включавший первоначально и плеченогих, сборную группу
мшанок и оболочников, единую группу червей, объединявшую в системе
Линнея всех беспозвоночных, за исключением насекомых, а еще не так
давно немертин, плоских, круглых и кольчатых червей со сборной группой
гефиреев, щетииочелюстных и кишечнодышащих.
Лишь сравнительно недавно класс мшанок оказался состоящим из двух
совершенно различных групп организмов — Endoprocta (Kamptozoa), отно-
симых в настоящее время к низшим червям, и Ectoprocta, объединяемых
вместе с брахиоподами и Phoronidea в группу Tentaculata, положение кото-
рой в системе Гроббена, с ее основным подразделением на первично- и
вторичноротых, до настоящего времени остается неясным.
Не может подлежать сомнению, что внешняя форма и симметрия тела
теснейшим образом связаны с образом жизни животных, со всей их биоло-
гией и что, с известной точки зрения, не преследующей целей установления
генетического родства организмов, они могут быть положены в основу их
подразделения на морфо-физиологические группы, подобно тому, как в ги-
стологии, например, можно говорить об эпителиальных или пограничных
и о мышечных тканях. Известными примерами конвергенций из этой об-
ласти являются кишечнополостные и иглокожные, построенные по радиаль-
ному типу с примыкающими к ним в отношении внешней формы тела Pho-
ronis, Cephalodiscus и рядом других форм; фораминиферы, брюхоногие и
некоторые головоногие моллюски, построенные по спиральному типу и обла-
дающие нередко поразительно сходными по внешней форме раковинами;
всевозможные червеобразные и змеевидные, ползающие и роющиеся жи-
вотные, рыбообразные плавающие организмы; «ногатые» животные — чле-
нистоногие и наземные позвоночные, передвигающиеся с помощью конечно-
стей, построенных по типу рычагов.
Проблема эволюционной гистологии
53
Конвергентное сходство внешней формы у отдаленных друг от друга
животных бывает поверхностным, не затрагивает в более или менее значи-
тельной степени их внутренней организации, а у организмов, генетически
более близких, естественным образом, может касаться многих особенно-
стей их строения. Метамерия может возникать параллельно и неодинако-
выми способами в различных группах животных. Не может подлежать
сомнению, что во многих отношениях поразительное сходство между пухо-
едами и вшами, между некоторыми акулами (Lanina), ихтиозаврами и дель-
финами, между птицами и птерозаврами зависит от того, что первые яв-
ляются насекомыми, а вторые и третьи позвоночными и обладают вместе
с тем многими чертами сходства, зависящими не только от явлений кон-
вергенции, но и от их генетического родства. В основе большого сходства
между рядом сумчатых и высших млекопитающих также лежат, с одной
стороны, явления конвергенции, связанные с приспособлением к одина-
ковому образу жизни, а с другой, — их принадлежность к одному и тому
же классу позвоночных и многие унаследованные от общих предков сходные
черты организации.
Чрезвычайно многочисленны и демонстративны примеры конвергенций
и параллелизмов в строении отдельных органов различных животных.
У членистоногих и позвоночных совершенно независимо развиваются ко-
нечности рычагового типа с их бегающими, роющими, прыгающими и хва-
тательными вариантами, в отношении к частям которых даже применяются
те же анатомические термины: бедро, голень, лапка. В отношении внешней
формы передней конечности особенно яркие примеры конвергентного сход-
ства дают роющие варианты, например у медведки, с одной стороны, и
крота, с другой. Хотя крылья насекомых и птиц ни в какой мере не гомо-
логичны друг другу, в них, тем не менее, имеются известные морфологиче-
ские черты сходства, поскольку те и другие являются летательными органами
одного и того же «орнитоптерного» типа, если пользоваться терминологией,
применяющейся в отношении летательных аппаратов, строящихся чело-
веком.
Очень интересно также конвергентное сходство общей формы выдели-
тельных органов нефридального типа у низших червей и сложноразвет-
вленных древовидных «нефридиев», или, точнее, целомодуктов хитонов
и некоторых брюхоногих моллюсков, которое недавно подчеркнул В. До-
гель (1937, 1938).
Хорошие примеры анатомического параллелизма представляют со-
бой мерцательные воронки «урнулы» голотурий — синаптид, пиявок и
сипункулид, являющиеся производными целомической выстилки, цело-
модукты позвоночных, аннелид и моллюсков, сообщающиеся с полостью
тела мерцательными воронками и несущие как выделительную, так и по-
ловую функции, а также трахеи насекомых и паукообразных. Наконец,
пожалуй, к самым поразительным примерам конвергенций относятся «не-
фридии» некоторых червей, например, Phyllodoce, и выделительные органы
(целомодукты) ланцетника, а также камерные глаза моллюсков и позво-
ночных, основанные на том же принципе, что и фотографический аппарат,
54
Глава II
являющийся, так сказать, конвергентным продуктом человеческого твор-
чества. Весьма эффектны также примеры конвергенции мантийных глаз
пластинчатожаберных моллюсков и сложных глаз членистоногих, отме-
ченные В. Догелем. В отношении конвергентного сходства фоторецепторных
органов предметного зрения одним из важнейших моментов являются,
очевидно, законы оптики, которым должны подчиняться любые аппараты,
дающие на каком-либо экране адэкватное изображение окружающего мира.
Наоборот, наземные локомоторные аппараты, построенные человеком, осно-
вываются на принципе свободно вращающихся колес, который, невиди-
мому, не может быть осуществлен в живой природе.
Следует напомнить также примеры конвергентных образований из
области эмбриологии. Например, телолецитальные меробластические яйца
головоногих моллюсков и позвоночных в результате дробления дают сход-
ную в общих чертах дискобластулу. У высших позвоночных и насекомых
развиваются, как конвергентные ценогенезы, зародышевые оболочки (амнио-
тическая и серозная), клеточный материал которых у тех и у других пред-
ставляет собой внезародышевую эктодерму, т. е. в гистологическом отно-
шении может считаться гомологичным.
Не останавливаясь подробно на обычных и достаточно известных при-
мерах конвергенции и параллелизма из области сравнительной анатомии,
которые не имеют прямого отношения к интересующим нас гистологическим
вопросам и частично освещались в нашей литературе (В. Догель, 1937,
1938; Шмальгаузен, 1939), я позволю себе еще коснуться некоторых при-
меров . конвергентного морфогенеза или органообразования в составе од-
ного и того же организма, поскольку явления конвергентного гистогенеза
очень широко распространены в области тканевых структур. Кроме того,
мне кажется, этому вопросу до настоящего времени не уделялось достаточ-
ного внимания.
Старые анатомы на основании очень поверхностных внешних призна-
ков установили понятие желез, которые затем получили физиологиче-
скую характеристику органов, вырабатывающих те или иные продукты.
Еще недавно, а в медицинской практике и до настоящего времени, к желези-
стым органам, кроме собственно желез, почек и гонад, относили также
лимфатические узлы и селезенку, которые, подобно красному костному
мозгу., ничего общего с железами не имеют.
Когда кроветворная роль лимфатических узлов сделалась известной,
их стали обозначать, как glandulae cytogeneae, т. е. «клеткеобразующие
железы». В этом отношении они сопоставлялись с гонадами.
В настоящее время части кроветворного аппарата анатомы и гистологи
.уже не относят к железистым органам. Последний термин применяется
теперь исключительно к эпителиальным образованиям, возникающим
в связи с разрешением общей функциональной задачи увеличения поверх-
ности обмена эпителиальных пластов с внешней или внутренней средой
организма и, в частности, выделяющих те или иные продукты поверхнОг
стей организма. Эта общая задача увеличения поверхности, разрешаемая
путем разрастания элементов эпителиальных пластов в определенном, ба-
Проблема эволюционной гистологии 55
зальном в отношении данного покрова направлении, приводит к образо-
ванию ряда структур — трубчатых ходов, тяжей той или иной формы или
отдельных клеточных комплексов, имеющих, однако, по существу очень
неодинаковое частное функциональное значение, различное происхождение
и способ развития. - >
Легкие в отношении их морфологии и отчасти функционального значе-
ния также принято сравнивать с железами, сопоставляя ацинусы с секре-
торными отделами, а воздухоносные пути с выводными протоками.
Мне представляется, что в состав группы т. наз. железистых органов
входит ряд образований, возникших в результате конвергентного морфо-
генеза из генетически разных источников и нередко совершенно различными
путями. Биологическое значение этих органов очень неодинаково. В отно-
шении генеза и биологического значения прежде всего необходимо строго
разграничить собственно железы, гонады и выделительные органы. Пер-
вичное и основное значение гонад, заключающееся в приготовлении поло-
вых клеток, не имеет ничего общего с процессами секреции в железах,
будь то даже голокриновые железы. Эндокриновая функция появляется
у гонад позвоночных лишь вторично и, насколько известно в настоящее
время, совершенно не характеризует их у сколько-нибудь значительной
части животных. Образование каких-нибудь специфических продуктов само
по себе вовсе не делает клетки или их комплексы железами в полном смысле
этого слова. Нервные клетки некоторых беспозвоночных, дающие хромаф-
финовую реакцию и, повидимому, вырабатывающие адреналин, все-таки
являются нейронами с эндокриновой функцией, а не железами. Очень
вероятным является также признание железистой функции у нервных эле-
ментов позвоночных, концевые аппараты которых, согласно популярным
в настоящее время взглядам, могут выделять вещества, сходные с адрена-
лином, и ацетил-холин.
Мне кажется, что отнесение семенника высших позвоночных, как откры-
той сложной трубчатой железы, в одну рубрику с бруннеровскими, бульбо-
уретральными и рядом других желез, в высшей степени формально и учи-
тывает лишь поверхностное конвергентное сходство органов, а вовсе не их
биологическую специфику.
То же самое полностью справедливо и в отношении яичника, который
принято относить, вместе со щитовидной железой, в одну рубрику закрытых
фолликулярных желез. По сути дела, извитые семенные канальцы семен-
ников и фолликулы, являясь вполне сравнимыми друг с другом по своему
происхождению и значению весьма древними образованиями, не могут быть
поставлены на одну доску с секреторными отделами каких-либо желез.
Гонады представляют собой совершенно особый тип органов, имеющих
свою длительную историю развития, которые и следует рассматривать как
таковые, а не как одну из разновидностей желез.
Большие возражения вызывает также безоговорочное обозначение по-
чек, как сложных трубчатых желез. Выделительные органы типа не-,
фридиев и целомодуктов также являются очень древними и совер-
шенно своеобразными образованиями, которые по существу мало сравнимы
56
Глава II
с железами, хотя и имеют в известных отношениях сходное с ними строе-
ние. В частности, вся история онто- и филогенетического развития выдели-
тельных органов позвоночных с поразительной наглядностью демонстри-
рует их несравнимость по существу с любыми трубчатыми железами. И в
функциональном отношении процесс экскреции, обеспечиваемый особым
фильтрующим гломерулярным аппаратом и избирательной резорбтивной,
а частично и выделительной функцией мочевых канальцев, существенно
отличается от секреции или экскреции желез. Почки позвоночных и чело-
века биологически правильно рассматривать как особые выделительные
органы типа целомодуктов, а не просто как трубчатые железы, принци-
пиально не отличающиеся от других железистых органов, имеющих с ними
известное конвергентное сходство.
Сравнение с железами легких высших позвоночных также мало оправ-
дано, даже если стоять на не очень популярной в настоящее время «эпите-
лиальной» точке зрения на выстилку их дышащих частей. Как показывает
сравнительная анатомия, легкие возникают из мешковидных воздухонос-
ных придатков переднего отдела пищеварительной трубки, не как система
древовидно ветвящихся, сходных с железистыми, ходов, а путем септиро-
вания и усложнения рельефа их первоначально гладкой стенки. Дышащие
отделы, повидимому, с частью внутрилегочных воздухоносных путей соз-
даются до известной степени независимо от ветвящегося бронхиального
дерева из сложной системы ячеек стенки легочных мешков. В онтогенезе,
в котором, наряду с формообразовательными процессами, происходит уси-
ленный рост зачатков легких и воздухоносных путей, различия между
их развитием и развитием обычных желез оказываются в значительной
степени сглаженными. Если легкие млекопитающих в готовом состоянии
обнаруживают конвергентное сходство со сложными альвеолярными или
альвеолярно-трубчатыми железами, а легкие птиц — с сетевидной
трубчатой железой, то эта аналогия в достаточной степени поверхностна
и ни в какой степени не отражает биологической специфики органа ды-
хания.
Среди собственно железистых и среди эндокриновых органов мы также
встречаемся с явлениями конвергентного морфогенеза, поскольку очень
сходные по форме и даже способу развития образования могут возникать
независимо друг от друга из различных зачатков. В качестве примеров-
достаточно указать на трубчатые железы или крипты матки, развивающиеся
из мезодермальной выстилки мюллеровских каналов, и на эктодермальные
кишечные (либеркюновы) крипты; на слюнные железы, в первую очередь
белковые, и на экзокриновую часть поджелудочной железы; наконец, на
эндокриновые органы типа эпителиальных телец, интерренальных органов
и островков Лангерганса и на печень высших позвоночных, характеризую-
щиеся некоторыми общими структурными особенностями. В последнем
случае трудно разграничить, в какой степени мы имеем дело с конвергент-
ным морфогенезом или гистогенезом.
Таким образом, весьма многочисленные и распространенные в орга-
низме позвоночных и, в частности, млекопитающих органы то весьма раз-
Проблема эволюционной гистологии
57
личного, то сходного биологического значения, имеющие «железистый тип»
строения и развивающиеся из разных зачатков, являются примерами ана-
томической или морфогенетической конвергенции.
Изложенные факты, число которых может быть, при желании, значи-
тельно увеличено, показывают, что конвергенции и параллелизмы имеют
большое значение не только в области гистологии, но и сравнительной
анатомии, и являются общебиологической закономерностью, сосущест-
вующей с фундаментальным законом биологии — законом дивергентного
развития.
3. ДИВЕРГЕНТНОЕ РАЗВИТИЕ И ДИВЕРГЕНТНАЯ ДИФФЕРЕНЦИРОВКА
В ОБЛАСТИ ГИСТОЛОГИЧЕСКИХ СТРУКТУР
Так как дивергентное развитие организмов, результатом которого-
является все огромное разнообразие современных и ископаемых живых
существ нашей планеты, достаточно хорошо известно, то здесь нет необ-
ходимости останавливаться на отдельных иллюстрирующих его конкрет-
ных данных. Поэтому мы ограничимся кратким рассмотрением некоторых
гистологических фактов, показывающих, что дивергентное развитие наблю-
дается и в области тканевых структур.
Демонстративными примерами гистологической дивергенции являются
весьма разнообразные направления дифференцировки кожной эктодермы
у представителей различных типов животных. Морфо-физиологические осо-
бенности наружного покрова кожи связаны теснейшим образом как адап-
тивные приспособления с условиями среды обитания, поскольку эпидер-
мис (в широком смысле слова) представляет собой истинно эктосоматиче-
скую ткань, которая непосредственно граничит с внешней средой и нахо-
дится под непосредственным воздействием всевозможных внешних факто-
ров. С другой стороны, эти же особенности несомненно находятся в тесней-
ших коррелятивных взаимоотношениях с рядом других черт организации
животных. В некоторых случаях строение эпидермиса является одним из.
характерных признаков больших систематических групп естественной си-
стемы организмов. Если вспомнить морфо-физиологические особенности
наружного покрова кожи у представителей некоторых конечных звеньев
эволюционных рядов животных, то резко выраженные черты дивергенции
сразу же бросаются в глаза. У позвоночных эпидермис имеет строение много-
слойного эпителия, приобретающего в конце концов у наземных форм,
мощный защитный роговой слой. По своей функциональной характеристике
он является не только разграничительной, но также и изолирующей и за-
щитной тканью, не имеющей, однако, как правило, механической опорной
функции. У членистоногих эпидермис представлен одним слоем клеток
т. наз. гиподермы, образующих на своей свободной поверхности хитиновый
покров, который достигает большой толщины и прочности и несет не только
разграничительную и защитную, но также скелетную и механическую-
функции. Несколько иначе механическая функция эпидермиса осуще-
ствляется у моллюсков с развитой раковиной. Эти различия в морфо-
физиологических особенностях эпидермиса коррелятивно связаны у позво-
38
Глава II
ночных и членистоногих с рядом других глубоких различий в их органи-
зации, подчеркнутых в свое время Геккелем (1885), и прежде всего, с раз-
витием у позвоночных внутреннего скелета, построенного по соединительно-
тканному типу и замещающего в процессе эволюции первичный опорный
аппарат спинной струны.
У туникат мы встречаемся со сменой хорды очень своеоб-
разным наружным скелетом. Подобная дивергенция в развитии эктодер-
мального покрова встречается в самых разнообразных систематических
группах, начиная с кишечнополостных, среди которых мы встречаем формы
с наружным эктодермальным скелетом (многие гидроиды и шестилучевые
полипы) и формы с внутренним скелетом в мезоглее.
Если не считать спинной струны, происхождение которой предста-
вляется по ряду соображений спорным, то энтодерма несет опорную функцию
лишь в виде исключения, образуя у некоторых кишечнополостных хордо-
идный опорный аппарат щупалец. Нужно думать, что основная пищева-
рительная и всасывающая функция эктодермальных производных почти
или в большей части случаев совершенно несовместима с опорным или
скелетным значением. Таким образом, широкое осуществление в животном
мире скелетной функции может быть свойственно лишь наружному покрову
или производным мезодермы в широком смысле, понимая под этим топо-
графическим собирательным термином ткани, которые не граничат непо-
средственно с внешней средой и, в первую очередь, те из них, которые по
своей морфологической характеристике относятся к т. наз. тканям соеди-
нительного вещества. Эти обусловленные основными чертами организации
животных возможности осуществления опорной функции, очевидно, являют-
ся причиной того, что возникновение наружного или внутреннего скелета,
наблюдаемое в различных систематических группах, имеет, с другой
стороны, характер конвергенции или параллелизма. При отсутствии у жи-
вотных специальных скелетных образований опорная функция может в
значительной степени осуществляться наполненным жидкостью целомом
(эхиуриды, аннелиды, кишечнодышащие и другие формы).
Третьим из путей дивергентной эволюции наружного покрова кожи,
связанным с высокой адаптивной специализацией или даже деградацией
(или дегенерацией, по Северцову) всей организации животных, являются
в ряде случаев его изменения при переходе к эндопаразитизму, как это,
например, имеет место у -трематод, цестод, скребней, а также аскарид и
Protomyzostomum из аннелид. У эндопаразитов с редуцированным пище-
варительным аппаратом наружный покров кожи несет, очевидно, несвой-
ственную ему всасывающую функцию, т. е. физиологически замещает собой
отсутствующий у взрослых форм .многих таких животных эктодермальный
эпителий. Так наз. «погруженный эпителий» трематод и цестод,
встречающийся также у бескишечных турбеллярий, представляет собой
структуру, которая по существу лишь с натяжкой укладывается в при-!
вычное понятие эпителий. Синцитиальные покровы скребней и аскарид,
отличающиеся, невидимому, постоянным числом ядер, также являются
очень своеобразными и высокоспециализированными структурами.
Проблема эволюционной гистологии
59
У соединительнотканных энтомезенхимных образований почему-то при-
нято подчеркивать только черты сходства, тогда как они поражают у раз-
ных представителей мелких и крупных систематических групп боль-
шим разнообразием, которое, пожалуй, может смело конкурировать
с коррелятивно связанными с ними во многих случаях эпидермальными
структурами. Достаточно вспомнить опорные образования, построенные по
чипу спикул, с их чрезвычайно простой геометрической или разнообраз-
ной сложной формой, очень разнообразные в отношении ряда особенностей,
волокнистые, содержащие клетки или бесклеточные структуры, опорные
образования тургорного пузырчатого типа, хрящеподобные ткани отдель-
ных животных и, наконец, костную ткань с ее разнообразными модифика-
циями, появляющуюся только в ряду позвоночных.
Результатом дивергентной эволюции следует, повидимому, считать
также большое разнообразие внешней формы сократимых элементов муску-
латуры соматического типа, которое наблюдается у различных предста-
вителей многих систематических групп. Если принять за исходную форму
т. наз. миоэпителиальные или точнее миоэктодермальные клетки кишечно-
полостных, то можно, как в пределах этих типов, так и в ряде других си-
стематических групп, видеть возникновение мышечных элементов веретен-
чатой и сложноотростчатой формы с диффузной или полярно дифференци-
рованной локализацией миофибрилл и, наконец, совершенно своеобразных
элементов, у которых саркоплазматическое клеточное тело соединяется
с сократимой частью посредством отдельных протоплазматических отрост-
ков. На то, что мышечные элементы вместе с тем обнаруживают замеча-
тельные явления параллелизмов и конвергенций, уже указывалось выше.
Если в отношении нейрональных взаимоотношений у представителей
разных типов животных удается подметить поразительные явления парал-
лелизмов, то в отношении формы тела нейронов и общегистологических осо-
бенностей строения нервной системы вместе с тем обнаруживаются явле-
ния дивергенции, которые никак не следует игнорировать, поскольку они
накладывают глубокий отпечаток на гистологическую структуру нервной
системы.
Достаточно напомнить общеизвестную преобладающую мультиполяр-
ную форму центральных нейронов позвоночных и преобладающую. унипо-
лярную форму соответствующих элементов у многих беспозвоночных,
в частности, насекомых, зависящую, по Ленгоссекуи Заварзину (1925),
от их различного положения в отношении синаптических участков ней-
ропиля.
В центральной нервной системе членистоногих и позвоночных характер
и локализация глиальных элементов оказываются также весьма различ-
ными. У первых тела нейроглиальных элементов имеют большей частью
периферическое в отношении нейропиля расположение, в то время как
у вторых они располагаются центрально; или разбросаны во. всей толще
ткани мозга. Едва ли возможно отрицать, что эти различия в строении
нервной; системы зависят от многих дивергентных черт общей организации
соответствующих животных. .... .. . : " ...т. . .
60
Глава П
У элементов стенки целома можно наблюдать в одних случаях развитие-
в направлении фагоцитирующих резервных трофических клеток (например
хлорагогенные клетки аннелид и, быть может, жирового тела насекомых),
которые могут полностью утрачивать морфо-физиологическую характе-
ристику эпителиальных тканей; в других случаях целомический эпителий
отличается в основном свойствами разграничительной или покровной
ткани.
Чрезвычайно широкое распространение в животном мире имеют явле-
ния дивергентной дифференцировки гистологических структур в преде-
лах отдельных крупных систематических групп и повторяющий их во
многих отношениях дивергентный гистогенез генетически единых ткане-
вых типов в индивидуальном развитии отдельных особей. Сюда относится
появление неодинаковых дифференцировок в разных участках генетически
единых эпителиальных пластов, а также изменения покровных эпителиаль-
ных структур при образовании из них многоклеточных желез и эндокри-
новых органов, с примерами которых мы постоянно встречаемся у различ-
ных позвоночных. Выстилка железистых ходов кожных желез, как правило,,
приобретает существенные отличия от эпидермального покровного пласта.
В составе эпидермальной стенки железистых ходов во многих случаях
возникают путем дивергентной дифференцировки сократимые элементы,,
приобретающие совершенно другую морфо-физиологическую характери-
стику» чем их материнская ткань (т. наз. миоэпителиальные и корзинчатые
клетки многих кожных желез). Отчетливо выраженные гистологические
особенности приобретают респираторные отделы эпидермиса жаберного*
аппарата позвоночных. У моллюсков резкая дивергентная дифференцировка,
возникает между образующим раковину наружным эпидермисом кожи и
выстилкой мантийной полости. Принципам разделения и смены функций
в явлениях дивергентного развития тканей несомненно принадлежит перво-
степенное значение.
Достаточно резко выраженные отличия от кожного эпителия приобре-
тает у низших позвоночных эпителий роговицы глаза, который в дальней-
шем уже подвергается лишь относительно незначительным изменениям.
Клетки трахейных капилляров дыхательного аппарата насекомых,
через которые осуществляется газообмен, обнаруживают совершенно осо-
бую специализацию в отношении внешней формы тела, взаимоотношений
с другими тканевыми элементами и функциональных особенностей, резко
отличающую их от хитинизированной гиподермы кожного покрова и возду-
хоносных трахейных трубок.
Аналогичные явления дивергентной дифференцировки обнаруживает
у позвоночных выстилка переднего, в частности жаберного, отдела пище-
варительной трубки. Здесь также специализируются в особых направле-
ниях покров жаберных листков, выстилка плавательного пузыря и, ве-
роятно, дышащих частей легких. Первоначально сходная (амфибии) вы-
стилка глотки и пищевода, с одной стороны, и воздухоносных путей, с дру-
гой, имеющая характер многорядного мерцательного эпителия, приобре-
тает у рептилий, птиц и млекопитающих существенные различия вследствие
Проблема эволюционной, гистологии
61
того, что в переднем отделе пищеварительной трубки развивается много-
слойный ороговевающий или неороговевающий эпителий.
Как известно, при эмбриональном развитии в пищеводе человека наблю-
дается смена многорядного мерцательного эпителия многослойным плоским
(Шридде, 1907), которая не может не трактоваться как пример гистологи-
ческой рекапитуляции пройденного эволюционного пути.
Развитие из выстилки переднего отдела пищеварительной трубки желез
и эндокриновых органов также сопровождается явлениями дивергентного
гистогенеза. Своеобразным путем специализации эпидермиса кожи (напри-
мер ископаемые кистоперые рыбы; Быстрое, 1939) и эпителия
переднего отдела пищеварительной трубки многих позвоночных является
приобретение способности к образованию зубной эмали. Однослойный
эктодермальный эпителий желудочно-кишечного отдела пищеварительного
аппарата у позвоночных, а также многих беспозвоночных, в составе раз-
личных органов обнаруживает ряд особенностей, которые нельзя не трак-
товать как результат дивергентной дифференцировки генетически единого
исходного материала. Один из поучительных примеров этого дает печень
позвоночных, зачаток которой возникает у ланцетника, как слепой вырост
кишечника. У низших позвоночных печень приобретает характер сложной
трубчатой железы, а у высших — совершенно своеобразного органа с резко
•выраженной сетевидной «эндокриновой структурой». При этом элементы
печеночной паренхимы приобретают ряд биологических особенностей, ко-
торыми они резко отличаются от покровного эпителия кишечника и желчных
путей (см. ниже).
С точки зрения классической эмбриологии факт существования у выс-
ших позвоночных в составе пищеварительной трубки двух резко отличаю-
щихся друг от друга типов эпителия — однослойного призматического и
многослойного плоского, из которых последний может практически не от-
личаться от эпидермиса кожи, следовало бы трактовать как замечательный
пример, с одной стороны, дивергентной дифференцировки эктодермального
зачатка, а с другой, конвергентного развития головной части энтодермы
и кожной эктодермы.
На основании ряда экспериментальных данных и теоретических сообра-
жений мне представляется, однако, более правильным давать этому совер-
шенно иную эволюционную трактовку, которая уже излагалась (Хлопин,
1934) и будет еще раз разобрана ниже в фактической части этой
книги.
Выстилка целома у первичноротых и вторичноротых также дает хоро-
шие примеры дивергентной дифференцировки. С усложнением организации
часть стенки вторичной полости тела дифференцируется в сократимые
элементы миокарда, генетическая связь которых, например, наглядно
демонстрируется на сравнительно-анатомическом и эмбриологическом ма-
териале у моллюсков. В ряду позвоночных мы не располагаем формами,
на которых возможно было бы проследить эти отношения. Но эмбриональ-
ное развитие миокарда у позвоночных и взаимоотношения сердца и пери-
карда у кишечнодышащих и оболочников безусловно говорят в пользу
62
Глава II
трактовки сердечной мышцы как специализировавшегося участка цело-
мической выстилки вторичноротых. Стенка целома у позвоночных, кроме
того, дает ряд других дивергентно развивающихся образований: паренхиму
интерренальных органов, мюллеровы каналы с их производными, фолли-
кулярные клетки гонад. Сам целомический эпителий у высших позвоноч-
ных в конечном итоге превращается в очень своеобразную мезотелиальную
выстилку полости тела.
Соединительнотканные образования, производные энтомезенхимы, если
их трактовать в соответствии с существующими взглядами как генетически
единую группу тканей, обнаруживают, в первую очередь у позвоночных,
дивергентное развитие в направлении различных разновидностей волокни-
стой соединительной ткани, хрящевых и костных тканей и большого разно-
образия кровяных клеток. У представителей ряда классов позвоночных
многие из этих структур, невидимому, возникали параллельно. Кроме
того, энтомезенхима позвоночных специализируется в направлении гладко-
мышечных сократимых структур.
Один из наиболее поразительных примеров дивергентной дифференци-
ровки на морфологически и функционально неравноценные гистологические
структуры дает эпендимная нейроглия хордовых, часть которой сохраняет
свой примитивный эпителиоподобный характер, другие же части превра-
щаются в сходную по господствующим взглядам с соединительной тканью
астроцитарную глию, олигодендроглию и, быть может, ортегаглию, чув-
ствительные элементы некоторых органов чувств, гладкую или поперечно-
полосатую мускулатуру радужины, а также некоторые эндотелиоподобные
образования, которые будут подробнее разобраны ниже. Иными словами,
эта, в генетическом отношении, несомненно единая ткань, эволюируя,
дивергентно образует ряд структур, которые, в отношении своей морфо-
физиологической характеристики, входят в состав всех четырех основных
групп тканей классической гистологии (эпителия, соединительной, мышеч-
ной и нервной тканей).
Уже на основании приведенных выше примеров можно высказать поло-
жение, что эволюция тканей происходит под знаком тех же основных зако-
номерностей, которым подчинены эволюционные изменения целых орга-
низмов. Поэтому мы имеем все основания подходить к гистологическим
явлениям и фактам из области сравнительной анатомии и эмбриологии
с одних и тех же основных общебиологических эволюционных позиций.
Как и при эволюции целых организмов и отдельных органов, мы встречаемся
в филогистогенезе со сложными сочетаниями явлений дивергентного, кон-
вергентного и параллельного развития. Эволюционная дивергенция тканей
и органов находится в непосредственных взаимоотношениях с условиями
существования целого организма, а частью опосредствована его внутрен-
ними коррелятивными связями. Конвергенции и параллелизмы в гистоло-
гии и анатомии зависят от общих элементарных свойств живого субстрата,
в первую очередь, вероятно, от некоторых общих основных особенностей
его белковых тел, связанных с генетическим единством всей живой при-
роды, сходного функционального значения, и от общих механических и
Проблема эволюционной гистологии 63
физико-химических условий существования на нашей планете, а также от
особенностей внутренней среды организмов.
Если бросить общий взгляд на огромное, по существу, разнообразие
анатомических форм и тканевых структур в животном мире, то едва ли
справедливо говорить об ограниченных возможностях живой природы
в отношении формообразования и гистологической дифференцировки. Ско-
рее можно удивляться ее почти неограниченным возможностям.
Конвергентные структуры не являются своего рода testimonium pauper-
tatis, как это по сути дела нередко предполагается, а образуются
в результате творческой и элиминирующей роли естественного отбора,
благодаря которой чрезвычайно широкие формообразовательные возмож-
ности живой природы как бы направляются по определенным руслам адап-
тивной эволюции.
Все то, что развивается вне этих русел, оказывается нежизнеспособным
в борьбе за существование и устраняется. Во всей своей широте этот вопрос
выходит за рамки этой книги и должен быть предметом специального иссле-
дования.
В заключение следует подчеркнуть, что роль естественного отбора
в применении к гистологическим структурам опосредствована через целый
организм. Отбираются не те или иные гистологические структуры, а целые
организмы или, вернее, как это справедливо отмечает Шмальгаузен, целые
онтогенезы. Непосредственное перенесение принципа естественного отбора
на частные системы организма, каковыми являются гистологические струк-
туры, и допущение борьбы за существование между частями тела, клетками
и тканями, как это в свое время делал Ру, глубоко ошибочны.
4. ПРОБЛЕМА ВОЗНИКНОВЕНИЯ И РАННИХ ЭТАПОВ ЭВОЛЮЦИИ ТКАНЕЙ
МНОГОКЛЕТОЧНЫХ ЖИВОТНЫХ
Вопрос о возникновении и историческом пути развития тканей на ран-
них этапах эволюции — неотъемлемая часть эволюционной гистологии и
проблемы естественной системы гистологических структур. Он неразрывно
связан с проблемой появления многоклеточности животных организмов
и должен быть включен в цикл подлежащих нашему рассмотрению вопросов
общего характера. Подобно ряду других основных биологических проблем,
вопрос о возникновении тканей может решаться лишь в той или иной ве-
роятной гипотетической форме. В клеточной организации проявляется
противоставление живого существа, как интегрированного целого, особи
или индивидуальности, всему внешнему миру и, в том числе, другим живым
существам. В соответствии с воззрениями Чайлъда (1915, 1921, 1924), инте-
грация связана с появлением в живых существах первичных градиентов,
представляющих собой донервные связи организма.
Существенным моментом в организации первичных Metazoa является
состав их тела не из двух, трех или немногих клеток, а из достаточно много-
численных элементов. С многоклеточностыо (именно много-, а не малокле-
точностыо) связаны теснейшим образом возникновение и эволюция ткане-
вых систем организмов и даже сама возможность тканевой организации
64
Глава II
животных, если не экстраполировать понятия ткани на различный прото-
плазматический материал тех или иных частей тела и органелл Protozoa.
Переход к многоклеточной организации, как известно, трактуется
двояким образом: или путем расчленения единой протоплазматической массы
на клетки или через колониальные одноклеточные формы. Между первой
и второй концепциями резкой границы, строго говоря, провести невоз-
можно, поскольку колония одноклеточных организмов возникает путем
повторного деления из одной особи и уже с момента своего возникновения
должна обладать некоторой степенью интеграции. Существенно различными
в этих двух точках зрения являются, с одной стороны, представление о по-
степенном расчленении одноклеточного тела на все большее и большее
количество клеток, с другой, допущение быстрого или даже одновремен-
ного подразделения первоначальной единой многоядерной массы сразу
на более или менее значительное количество одноядерных элементов.
Взаиморасположение клеток в составе тела первичных многоклеточных
животных можно себе представить различно: или в виде плотного моруло-
образного клеточного комочка («морея», Геккель), полого внутри комплекса
шаровидной или, быть может, сплющенной, а также вытянутой в каком-
нибудь направлении формы («бластея», Геккель), или в виде плоской кле-
точной пластинки («плакула», Бючли) тех или иных очертаний. Теорети-
чески мыслим, хотя и мало вероятен для примитивных Metazoa, линейный
ряд клеток или цепочка клеток. Последняя «стрептоидная» форма скорее
соответствует нашим представлениям о примитивных многоклеточных ра-
стениях типа нитчатых водорослей. Что касается трех упомянутых воз-
можных типов первичных Metazoa, то они едва ли являются резко отгра-
ниченными друг от друга, и, кроме того, вероятно, их внешняя форма должна
была в значительной степени зависеть от условий существования. Дей-
ствительно, между плотным клеточным комочком и шаровидной бластеей
с небольшой центральной полостью почти нет принципиальной разницы.
И тот и другой клеточный комплекс должен был иметь более или менее
шаровидную или лишь незначительно уклоняющуюся от этой последней
форму, если он свободно плавал в воде. Наоборот, в случае жизни на дне
водоема, его вероятнее себе представить, в соответствии с условиями суще-
ствования, сплющенным в вертикальном направлении, т. е. приближаю-
щимся по форме к клеточной пластинке.
Согласно концепции Геккеля (1874, 1877), однослойный полый внутри
клеточный шар бластеи должен был дать путем впячивания двухслойную
«гастрею» с дифференцировкой на эктодерму и энтодерму. Мечников (1874,
1876, 1892) представлял себе, как известно, дальнейшую эволюцию бластеи
иным образом и считал инвагинационную гаструлу вторичным образова-
нием. С его точки зрения, из бластеи должен был возникнуть, первоначально
путем мультиполярной иммиграции клеток внутрь центральной полости,
организм типа паренхимулообразной личиночной стадии современных
кишечнополостных — паренхимелла или фагоцителла. Образование па-
ренхимеллы возможно себе представить и без промежуточной стадии бла-
стеи из морулообразного клеточного комплекса путем процесса, сравни-
Проблема эволюционной гистологии 65
--_—_ ---
мого с морулярной де ламинацией в том виде, как она наблюдается у неко-
торых Gymnoblastida и Calyptoblastida из современных кишечнополостных.
Такие гипотетические организмы типа паренхимеллы должны были
представлять собой комочек тесно расположенных клеток, периферический
слой которых все больше и больше приобретал морфо-физиологическую
характеристику эпителия и постепенно все отчетливее отграничивался от
внутренней клеточной паренхимы. Наружный слой клеток, кинобласт или
эктодерма, в соответствии со взглядами Мечникова, характеризовался
в первую очередь двигательной функцией, а внутренняя клеточная масса,
фагоцитобласт или энтодерма, пищеварительной. Различная дифферен-
цировка элементов кинобласта и фагоцитобласта была первоначально слабо
выражена и лабильна. Фагоцитируя, клетки кинобласта могли мигриро-
вать в состав фагоцитобласта и утрачивать свою морфологическую харак-
теристику эпителия. Наоборот, закончив внутриклеточное пищеварение,
элементы фагоцитобласта могли входить в состав кинобласта.
Специализация элементов кинобласта (эктодермы) и фагоцитобласта
(энтодермы) с течением времени должна была возрастать, а вместе с тем
их способность к превращению друг в друга, вероятно, все более и более
ограничиваться.
Дальнейшее изменение организмов типа паренхимулы могло итти раз-
ными путями, из которых один привел к губкам, а другой — ко всем про-
чим вышестоящим формам.
Способ образования фагоцитобласта путем мультиполярной иммигра-
ции изменился в течение эволюции в униполярную иммиграцию, а затем
заменился впячиванием или инвагинацией. Пищеварительная паренхима
приобрела эпителиоподобный характер и превратилась в клеточный пласт,
ограничивающий внутреннюю пищеварительную полость. Через образо-
вавшийся первичный рот или бластопор клетки пищеварительной парен-
химы или уже успевшая возникнуть пищеварительная полость вступили
в непосредственную связь с внешней средой. Таким образом, должна была
возникнуть, если исходить из теории паренхимеллы, двуслойная органи-
зация примитивных многоклеточных животных.
В концепции Мечникова малопонятным и в гистогенетическом отноше-
нии малооправданным моментом является образование энтодермы (фагоци-
тобласта) в виде внутренней клеточной массы, отграниченной от внешней
среды пластом кинобласта. Неясно, почему фагоцитирующие элементы
этого последнего должны мигрировать внутрь тела для выполнения пище-
варительной функции. При эволюционном развитии тканей и органов,
поскольку возможно судить на основании сравнительного материала, мы
нигде не видим с достаточной убедительностью превращения элементов
внутренней среды организма в пограничные с внешней средой образования.
Трактуемое в таком смысле эволюционное превращение внутренней парен-
химы бескишечных турбелярий в гастроваскулярную систему прямо- и
ветвисто-кишечных форм не свободно от очень серьезных возражений. То
же самое справедливо и в отношении соответствующей трактовки выстилки
дышащих частей легких птиц и млекопитающих. На критике этих взглядов
5 Хлопин—349
66
Глава И
мы остановимся ниже. Наоборот, весьма обычным и частым является обрат-
ный процесс — процесс эволюционного превращения покровных и разгра-
ничительных образований в ткани внутренней среды организма. С этим мы
встречаемся и при развитии тканей нервной системы и, возможно, также
соматической мускулатуры, при развитии эндокриновых органов у позво-
ночных, при эволюционном изменении ткани печени и в ряде других случаев.
Кроме того, все классические представления об образовании многокле-
точных животных не учитывали их взаимоотношений с внешней средой
и значения этих последних для процессов дифференцировки.
Более оправданным, с гистологической точки зрения, способом обра-
зования примитивных форм, подобных гастрее, представляется следующий
гипотетический путь эволюции многоклеточных животных, связанный
с условиями существования и воздействиями внешней среды. Можно думать,
что в связи с существованием на дне водоемов предки многоклеточных
животных должны были приобрести более или менее резко сплющенную
в вертикальном направлении форму или форму плосковыпуклой чечевицы
с уплощенной нижней поверхностью. Вместе с тем в теле таких организмов
должна была возникнуть ориентированная вертикально система градиен-
тов, на основе которой должны были образоваться два клеточных пласта
с неодинаковыми свойствами. Один из пластов все более и более специали-
зировался в направлении пищеварительной функции, приобретая свой-
ства, характеризующие в дальнейшем энтодерму, а другой пласт, соответ-
ствующий области высокого градиента, приобретал особенности наружного
эктодермального покрова тела с присущей этому последнему более высокой
чувствительностью. Свертывание двуслойной пластинки пищеваритель-
ным клеточным пластом внутрь или, что почти одно и то же, инвагинация
дифференцирующейся энтодермы привели в конце концов к образованию
двуслойных организмов типа геккелевской гастреи, являвшихся предками
современных кишечнополостных.
Как и в онтогенезе современных форм, клеточные элементы первичных
многоклеточных должны были постепенно приобрести морфо-физиологиче-
скую характеристик)’ гетерополярных эпителиальных клеток, тесно сомк-
нутых, в первую очередь, своими апикальными концами. Базальные концы
клеток, возможно, образовали цитоплазматические отростки, соединяв-
шиеся друг с другом. За счет этих базальных частей элементов экто- и
энтодермы, очевидно, возникли в дальнейшем образования типа опорной
пластинки и мезоглеи, а также нервные и мышечные продукты дифферен-
цировки цитоплазмы. Неодинаковые свойства апикальной и базальной
частей энто- и эктодермы развиваются, очевидно, в связи с первичной си-
стемой градиентов, ориентированных нормально к поверхности раздела
между организмом и внешней средой.
Естественно возникает вопрос: какая сторона уплощенных придонных
организмов превратилась в энтодерму? Если это была их нижняя, обращен-
ная к плотному субстрату, поверхность, что очень вероятно, то место,
соответствующее будущему бластопору, должно было быть первоначально
обращено вниз, а не вверх, как у ныне живущих сидячих форм кишечно-
Проблема эволюционной гистологии
67
полостных. Это несоответствие можно себе объяснить тем, что двуслойная
плакула или гастрея перешла затем к плавающему образу жизни. Такие
плавающие организмы при переходе к сидячим формам кишечнополостных
стали уже прикрепляться не оральным, а аборальным концом. Это не могло
не быть связано с инверсией системы градиентов в отношении оси тела
животных. Возможно также, что отверстие на оральном конце полипов
образовалось заново, а первичное, обращенное вниз, отверстие или заросло
совершенно или сохранилось в виде незначительного остатка, обнаружи-
вающегося в подошве некоторых современных полипов.
Помимо своих основных специальных функций в составе разграничи-
тельного и защитного, а у свободно плавающих форм игравшего роль мер-
цательного органа движения эктодермального покрова, и в составе пище-
варительной энтодермы, клетки примитивных многоклеточных организ-
мов, очевидно, обладали также и прочими, характеризующими живую про-
топлазму, свойствами, в основе которых лежит обмен веществ, — раз-
дражимостью, активной изменчивостью формы и, в частности, сократи-
мостью, а наряду с этим, известной степенью устойчивости формы н струк-
туры, способностью к размножению, сохранению в отношении внешней
среды известной степени гетерогенности солевого состава и тому подобное.
Полая внутри шарообразная бластула и морула, как стадии онтогенеза
современных форм, должны были возникнуть вторично в связи с переходом
к развитию в пелагических условиях. Эктодерма, соответствующая области
высокого градиента, должна была обладать более высоким уровнем чув-
ствительности и физиологической активности.
В зависимости от изменений в дальнейшем условий существования и
взаимоотношений примитивных многоклеточных организмов с внешней
средой наряду с аборальной областью высокого градиента или вместо нее,
в результате физиологической инверсии, должна была возникнуть вторич-
ная перистомальная область высокой физиологической активности.
Различная степень сложности организации современных кишечнопо-
лостных и пути превращения планулы во взрослый организм могут быть
использованы для объяснения способа возникновения составных частей
будущего мезодермального зачатка (который, как таковой, еще отсутствует
у этого типа животных) и отчасти элементов нервной системы.
У кишечнополостных несомненно прослеживаются известные этапы
эволюционного развития мышечных элементов, представленных у наиболее
низкоорганизованных форм т. наз. миоэпителиальными элементами, нахо-
дящимися в составе эктодермы и энтодермы.
Возникает вопрос: которые из этих сократимых элементов являются
родоначальниками древнейшей сократимой ткани вышестоящих форм —
мускулатуры соматического типа, обеспечивающей движение организмов
во внешней среде, или же филогенетические корни телесной мускулатуры
следует искать и в энтодерме и в эктодерме? Наконец, мыслима и такая
возможность, что в течение эволюции происходила субституция одной
мускулатуры другой, причем в разных филогенетических стволах сомати-
ческие мышечные элементы могли возникнуть из разных источников.
'68
Глава II
В развитии мускулатуры соматического типа необходимо прежде всего
учитывать ее теснейшую связь с элементами эктодермальной нервной си-
стемы. Эта связь, вероятно, устанавливается с самого момента их возник-
новения, проходит красной нитью в течение всей дальнейшей эволюции
и делает очень вероятным образование мышечных элементов во всем живот--
ном мире из эктодермы общих отдаленных предков. Такая взаимосвязь
элементов соматической мускулатуры и нервной системы даже давала
повод думать, что и те и другие возникли путем расщепления или разде-
ления на две неодинаковые части первоначально единых миоэпителиаль-
ных или, точнее, нейромиоэпителиальных клеток (Клейненберг, 1872; За-
варзин, 1925, 1941), находившихся в составе эктодермы. Согласно этой
точке зрения, первоначально единая мионейроэктодермальная клетка
расщепляется на остающуюся в составе эпителиального пласта чувстви-
тельную или нейроэпителиальную клетку и на погружающееся затем во
внутреннюю среду организма мышечное волокно. Таким образом возни-
кает простейший рецепторно-эффекторный аппарат, который мыслим теоре-
тически, но не обнаруживается у современных животных, в противополож-
ность прежним данным О. Гертвига (1878, 1880). Эта гипотеза расщепле-
ния, к которой нам еще придется вернуться в главе, посвященной нервной
системе, не учитывает системности первичной эктодермы и поэтому недооце-
нивает другого и, с моей точки зрения, гораздо более вероятного способа
развития мышечных и нервных элементов путем дивергентной дифферен-
цировки, который будет рассмотрен ниже.
В теснейшей связи с развитием мускулатуры должно было также нахо-
диться образование механических или опорных структур организма, без
которых невозможна эффективная работа мышечных элементов.
Тело простейших современных кишечнополостных уже содержит в своем
составе покровные, железистые, трофические, опорные, мышечные и нерв-
ные образования, соответствующие в функциональном отношении элемен-
там всех четырех основных групп тканей классической гистологии. Вместе
с тем их тело состоит по существу всего из двух первичных тканевых си-
стем — эктодермы и энтодермы, образующих непрерывные клеточные
пласты. Один из этих пластов, эктодерма, представляет собой наружный
покров тела и содержит в своем составе дифференцированные в различных
направлениях клетки — миоэпителиальные, нейроэпителиальные, нервные,
стрекательные, и, по крайней мере у некоторых форм, как, например, у
пресноводной гидры, т. наз. индифферентные i-клетки, способные
к размножению и к дифференцировке во всевозможных направлениях
и являющиеся вместе с тем родоначальниками полового зачатка. Другой
клеточный пласт, энтодерма, образует стенку кишечной полости и состоит
из способных к фагоцитозу или всасыванию трофических эпителиальных-
клеток, миоэпителиальных и т. наз. эктодермальных нервных
элементов. Учитывая тот факт, что все элементы энтодермы могут образо-
ваться из мигрирующих в нее индифферентных клеток, нервное сплетение,
находящееся в ее составе, нельзя считать принципиально отличающимся по
своему происхождению от свойственной всем вышестоящим формам экто-
Проблема эволюционной гистологии
69
дермальной нервной системы. Для полного обоснования этой точки зрения
необходимо детальное изучение гистогенеза нервных элементов кишечно-
полостных. Промежуточную опорную пластинку и студенистую мезоглею
низших кишечнополостных нельзя рассматривать как самостоятельные
тканевые образования, существующие наравне с эктодермой и энтодермой.
Они являются производными одной из двух или обеих первичных тканей
кишечнополостных и должны рассматриваться как продукт особой диффе-
ренцировки базальных поверхностей пограничных клеточных пластов.
В состав основного вещества мезоглеи низших кишечнополостных могут
вторично выселяться в небольшом количестве свободные клеточные эле-
менты, не образующие тканевой структуры.
Основное вещество мезоглеи по способу своего образования является
своеобразным внеклеточным продуктом, сравнимым в известной степени
со стекловидным телом глазного яблока позвоночных и возникающим по
типу базальной мембраны, невидимому, за счет гетерополярно дифферен-
цированных клеток эктодермы. Поэтому ее основная масса не может безо-
говорочно сопоставляться с межклеточным веществом мезенхимных соеди-
нительнотканных образований, с которым она сравнима лишь в известной
степени по своей морфо-физиологической характеристике (положение внутри
организма, опорное значение), но не по способу своего развития. Об истин-
ных соединительных образованиях, мне кажется, можно говорить только
в тех случаях, когда они являются продуктом обособившихся у зародыша
аполярных клеток внутренней среды организма, которые, путем размно-
жения и выделения межклеточного вещества, образуют генетический еди-
ный тканевый комплекс.
Таким образом, эктодерму и энтодерму низших кишечнополостных сле-
дует рассматривать, с точки зрения их морфо-физиологической характери-
стики, не как эпителиальные ткани, а как своеобразные мультифункцио-
иальные тканевые системы, каждая из которых объединяет в себе особен-
ности трех или всех четырех общепринятых основных групп тканей более
высокоорганизованных животных.
Эволюционную дифференцировку примитивной эктодермы гастреепо-
добных гипотетических предков можно себе представить следующим обра-
зом. Клетки эктодермы первоначально в более или менее равной степени
приобрели некоторые морфологические особенности нейроэпителиальных
н миоэпителиальных чувствительных клеток, базальная часть тела которых,,
возможно, имела отросток или отростки, обладавшие известной степенью
сократимости и вместе с тем способностью передавать получаемые апи-
кальным концом клеточного тела раздражения отросткам других таких же
элементов. Часть таких примитивных «мионейроэпителиальных» элементов
в дальнейшем стала развиваться в направлении рецепторных нейроэпите-
лиальных клеток и утратила сократимость, а другая их часть, наоборот,
специализировалась в направлении сократимых миоэпителиальных эле-
ментов. Из рецепторных нейроэпителиальных клеток затем дифференциро-
вались первичные нервные элементы, с которыми мы уже встречаемся во
всех группах современных животных, за исключением губок, эволюционное
70
Глава II
развитие которых шло иными путями и не привело к образованию высоко-
организованных потомков. Возможно, что на основе примитивных связей
между мионейральными отростками первичных мионейроэпителиальных
клеток позже развились более специализированные контакты, а в дальней-
шем настоящие синапсы между чувствительными нейроэпителиальными
клетками и нейронами, с одной стороны, и между нервными и миоэпите-
лиальными, а затем и настоящими мышечными элементами, с другой. Ве-
роятно, эта дивергентная дифференцировка клеток первичной эктодермы
происходила в каких-то определенных количественных соотношениях, за-
висящих от интегрированности частей тела многоклеточных организмов.
У высших представителей кишечнополостных мы видим следующие
изменения в развитии и строении эктодермальных мышечных элементов
У сцифоидных из эктодермы более или менее диффузно образуются настоя-
щие мышечные волокна, обладающие у некоторых форм поперечной исчер-
ченностыо. У гребневиков впервые появляется особый зачаток соматиче-
ской мускулатуры, обособляющийся в виде группы микромер уже в тече-
ние дробления. Затем мышечные микромеры концентрируются у анималь-
ного полюса зародыша и дифференцируются в волокна соматической
мускулатуры.
Вместе с тем у некоторых коралловых полипов происходит в постфе-
тальном периоде дальнейшее усложнение организации, благодаря выкли-
ниванию из эктодермы клеток, которые вырабатывают в мезоглее особые
«опорные образования, т. наз. склериты. У других высших полипов опорные
структуры образуются лишь апикальными поверхностями клеток экто-
дермы, а сравнимые с мезенхимными к четки, вырабатывающие склериты,
отсутствуют.
У сцифомедуз также наблюдается, хотя и в слабой степени, образование
из эктодермы свободных клеток, мигрирующих в мезоглею. Основное ве-
щество этой последней очень обильно, но, должно быть, возникает неза-
висимо от выклинивающихся из состава эктодермы клеток.
У гребневиков, невидимому, уже имеются настоящие мезенхимные
соединительнотканные образования. Клетки, трактуемые как мезенхим-
ные, правда, обособляются на сравнительно поздних стадиях развития из
состава эктодермы и образуют студенистое вещество между эктодермой и
энтодермой.
Таким образом, внутренние опорные образования, являющиеся по су-
ществу первоначально частями эктодермального и, возможно также, экто-
дермального пластов, постепенно обособляются в виде самостоятельных
мезенхимных соединительнотканных структур, генетически связанных
с эктодермой. Элементы этой последней, развиваясь дивергентно в других
направлениях, дают начало элементам соматической мускулатуры и нерв-
ной системы, которые также погружаются во внутреннюю среду организма.
У всех вышестоящих организмов преемственная связь ряда тканей
с примитивными тканевыми системами кишечнополостных не внушает ни-
каких сомнений. Кишечный эпителий со всеми своими производными гене-
тически связан р энтодермой, а всевозможные разновидности эпидермаль-
Проблема эволюционной гистологии
71
tux тканей и элементы нервной системы являются бесспорно в онтогенезе
л филогенезе производными эктодермы. Энтодерма и эктодерма сравнимы
во всем животном мире. Иначе обстоит дело с т. наз. мезодермой,
которая представляет по существу целый комплекс весьма разнообразных
тканевых зачатков.
В способе развития мезодермы в течение эмбрионального развития име-
ются такие большие различия, что ее безоговорочное сопоставление, как
гомологичного у всех животных образования, не представляется возможным
Как известно, различия в способе образования мезодермы в общем совпа-
дают с разделением всех более высокоорганизованных животных на два
главных филогенетических ствола, предложенного Гроббеном (1905). У пред-
ставителей одного из этих стволов, обозначаемых как первичноротые, ро-
говое отверстие генетически связано с бластопором или образуется на том
же конце тела, где имелся последний. У т. наз. вторичноротых бластопору
соответствует анальное отверстие, а рот образуется совершенно заново на
противоположном конце тела. У первичноротых мезодермальный комплекс
обособляется очень рано, большей частью в виде телобластического, т. е.
щедставленного лишь двумя парными крупными клетками, зачатка или,
в некоторых случаях, мелкоклеточного зачатка.
У вторичноротых он образуется, как правило, на более поздних стадиях
развития энтероцельным способом, в виде карманообразных выступов
стенки первичной кишки.
Филогенетически первичноротых вероятнее всего выводить из организ-
мов, сходных с современными гребневиками (Ланг, 1884; Волыперек, 1904),
причем низшие и высшие черви или рассматриваются как две самостоятель-
ные ветви животных, эволюция которых, начавшись из общего корня,
пошла в разных направлениях, или они выводятся из сходных с турбел-
чяриями предков. •
Предками вторичноротых, согласно Шимкевичу (1907, 1923), следует
считать гипотетический организм, тетранейрулу, сходный с личиночной
формой высших полипов, называемой сцифистомой. Согласно этому взгляду,
два главных филогенетических ствола животных расходятся очень рано.
Их общие корни следует искать среди очень низкоорганизованных форм,
которые были предками современных кишечнополостных, а не среди орга-
низмов, сходных с гребневиками, как это предполагается другими авторами.
Если обратить внимание на разные способы образования в ряду хордовых
мезодермального зачатка, который несомненно является у них гомологич-
ным образованием, то становится ясным, что подчас очень большим раз-
личиям в эмбриональных процессах, особенно на ранних стадиях развития,
не приходится придавать принципиального значения в смысле рекапиту-
ляций. Формально энтероцельное образование мезодермальных карманов
у ланцетника, перистомальное развитие мезодермального пласта у амфибий
ч рыб и, наконец, появление мезодермы из утолщенного участка (первичной
полоски) верхнего слоя бластодиста у высших позвоночных отнюдь не мо-
гут трактоваться как принципиально различные способы возникновения
мезодермы у различных представителей хордовых. Во всех этих случаях
72
Глава 11
мы имеем дело лишь с первоначально различным топографическим разме-
щением того же самого гГрезумптивного зачатка мезодермы, который в конце
концов приобретает одинаковое окончательное расположение. При Детер-
минированном развитии со спиральным дроблением у первичноротых и
установкой на трохофорообразную личинку (Иванов, 1937) мы, вероятнее
всего, имеем дело лишь с формально иным способом обособления ряда за-
чатков, входящих в состав мезодермы и по существу гомологичных по
крайней мере основным компонентам мезодермального зачатка вторично-
ротых. То же самое справедливо, повидимому, и в отношении первичноротых
с другими типами дробления. Во всяком случае, едва ли может подлежать
сомнению, что различия в способе образования мезодермы не могут быть
использованы в качестве сколько-нибудь убедительного аргумента против
ее гомологии у представителей отдаленных систематических групп живот-
ного мира.
Между современными формами кишечнополостных, с одной стороны,
и вышестоящими вторичноротыми, с другой, имеется очень большой раз-
рыв. То же самое наблюдается и в отношении отдельных крупных система-
тических групп вторичноротых, отличающихся рядом существенных осо-
бенностей организации и, в ряде случаев, резко выраженными признаками
высокой специализации и даже вторичного упрощения. Характерной чертой
строения вторичноротых является хорошо развитая мезодерма, образующая
целомическую стенку и развивающаяся у их низших представителей ти-
пичным энтероцельным путем.
Вместе с тем анализ предварительной структуры яйца и распределения
зачатков в бластуле асцидий, ланцетника и низших позвоночных показы-
вает, что стенка первичной кишки представляет собой, по сути дела, ком-
плекс нескольких автономных зачатков, которые уже полностью опреде-
лились значительно раньше. Иными словами, образование в онтогенезе
из стенки первичной кишки выстилки вторичной кишечной трубки, хорды
и мезодермы не является бесспорным указанием на происхождение хорды
и мезодермы из кишечника в филогенезе. Весьма возможно, что и у прочих
вторичноротых по существу имеются такие же взаимоотношения между
стенкой вторичной кишки и мезодермой.
Мезодерма вторичноротых представляет собой комплекс зачатков вы-
стилки целома, соматической мускулатуры и мезенхимных соединительно-
тканных образований. Последние, повидимому, почти отсутствуют у Но-
malopterygia (Chaetognatha), а также у кишечнодышащих, слабо выражены
у туникат и ланцетника и достигают высокого развития у иглокожий и
позвоночных.
Мезенхима вторичноротых, по способу своего образования, включая
ларвальную мезенхиму иглокожих, является целенхимой в смысле Зален-
ского и Давыдова, так как ее развитие может быть сведено к бйлатериально
симметричному зачатку целомической мезодермы или энтомезенхимой, свя-
занной в онтогенезе со стенкой первичной кишки.
Так наз. эктбмезенхима (мсзэктодерма), или, точнее, нейроме-
зенхима позвоночных, представляет собой своеобразное производное ней-
Проблема эволюционной гистологии
75
рального зачатка, возникшее йа поздних этапах эволюции и не имеющее,
как показывает экспериментальный гистологический анализ (см. ниже),
ничего общего с энтомезенхимой.
Мезодермальный комплекс вторичноротых представляет собой, вероятно,,
сложное образование, в составе которого вторично объединились возник-
шие первоначально, частью независимо друг от друга, зачатки соматиче-
ской мускулатуры, мезенхимы и выстилки целомической полости, с которой
генетически связан выделительный аппарат типа целомодуктов. Генети-
ческая связь мускулатуры соматического типа с примитивной эктодермой,
отчетливо обнаруживающаяся у кишечнополостных, в онтогенезе всех
вышестоящих форм, в связи с формированием мезодермального зачатка,
полностью утратилась. Если основываться лишь на данных онтогенеза, то
получается впечатление, что соматическая мускулатура вторичноротых
представляет собой ткань не гомологичную эктодермальной мускулатуре
кишечнополостных, а генетически связанную с целомом, который, в свою
очередь, сравним с периферическими частями гастроваскулярной системы
последних. Тогда следовало бы предположить, что соматическая мускула-
гура вторичноротых не развилась преемственно из эктодермальной муску-
латуры кишечнополостных, а филогенетически субституировала ее, возник-
нув из энтодермы с ее миоэпителиальными клетками.
Однако невозможность безоговорочно сводить энтероцельный способ
возникновения мезодермы к рекапитуляции эволюционного развития всех
ее компонентов, как это справедливо подчеркивается особенно в последнее
время (Иванов, 1937), заставляет с известной осторожностью относиться.
К этим эмбриологическим фактам. Существенные различия в способе обра-
зования мезодермы у позвоночных, на которые уже указывалось выше,
демонстрируют то же самое.
Если бы мускулатура соматического типа вторичноротых возникла из
энтодермы кишечнополостных, энтодермальная нервная система которых,
если пока оставить в стороне иглокожих, исчезла у всех вышестоящих форм,
то ее тесные взаимоотношения с эктодермальной нервной системой должны
были бы возникнуть лишь вторично. При этом следовало бы допустить, что
миоэнтодермальные клетки кишечнополостных путем дивергентной диф-
ференцировки дали мышечные элементы, которые, вторично вступив в связг
с эктодермальной нервной системой, приобрели значение общей локомотор-
ной мускулатуры тела.
Это допущение представляется малоправдоподобным. Гораздо вероят-
нее предполагать, в соответствии со всем сказанным выше, преемственную
генетическую связь мускулатуры соматического типа всех вторичноротых
и, в частности, позвоночных и человека с эктодермальной мускулатурой
сходных с современными кишечнополостными предков. Необходимым след-
ствием этого предположения является то, что мускулатура соматического
типа и выстилка вторичной полости теЛа представляют собой разнородные*
образования, которые лишь вторично объединились в онтогенезе современ-
ных форм. Целом вторйчноротых вероятнее всего выводить с точки зрения
энтероцельной теории братьев Гертвигов и гоноцельной теории в ее новой
74
Глава II
трактовке, предложенной Шимкевичем, из эндодермальных каналов гастро-
васкулярной системы кишечнополостных.
Вопрос о первичном возникновении т. наз. энтомезенхимы у вторично-
ротых может быть решен так же предположительно, как и разобранный
вопрос об эволюционных корнях мускулатуры соматического типа. Если
считать мигрирующие в мезоглею Anthozoa эктодермальные клетки, обра
зующие склериты, и эктодермальные же мезенхимные элементы гребневи-
ков за прародителей мезенхимной соединительной ткани стоящих выше
форм, то энтомезенхиму вторичноротых следует себе представлять, как
результат дивергентной дифференцировки элементов первичной эктодермы
предков. Вместе с тем едва ли возможно исключить участие первичной энто-
дермы в образовании некоторых мезенхимных образований, которое пред-
ставлялось столь вероятным Мечникову, выводившему фагоцитирующие
элементы внутренней среды организмов из фагоцитобласта (энтодермы)
предков. В последнем случае связь в онтогенезе многих вторичноротых
энтомезенхимы с целомической мезодермой следовало бы трактовать не
как вторичное явление, а как рекапитуляцию отношений, имевших место
у предков. Наконец, мыслимо также, что в качестве «мезенхимных»
(в смысле братьев Гepmet/гов) клеток, мигрировавших во внутреннюю среду
организма могли фигурировать как эктодермальные, так и энтодермальные
клетки, или что к древнейшему мезенхимному зачатку общих предков в даль-
нейшем присоединялись элементы различного происхождения, т. е. что
морфо-физиологическое понятие мезенхимы у разных представителей живот-
ного мира может иметь, с точки зрения генетической системы тканей, разный
смысл и в каждом отдельном случае нуждается еще в специальном изучении.
Как это будет рассмотрено в главе, посвященной соединительнотканным
образованиям, их, за некоторым исключением, представляется возможным
трактовать вместе с частью (но не со всеми) форменных элементов крови
как генетически единый тканевой комплекс, предположительно разви-
вающийся в обоих главных стволах животного мира из одного и того же
источника. Этим источником могут быть как раз эктодермальные мезенхим-
ные клетки кишечнополостных. С этой точки зрения, древнейший мезенхим-
ный комплекс возник из того же первоисточника, что и мускулатура сома-
тического типа, и лишь вторично объединился в онтогенезе современных
форм с целомической частью мезодермы.
На основании эмбрионального развития первичноротых еще труднее
делать выводы, касающиеся филогенетического развития тканевых зачат-
ков, входящих в состав мезодермы, чем в случае вторичноротых. Широко
распространенный среди первичноротых спиральный тип дробления не
может считаться примитивным способом развития. Детерминированное
спиральное дробление, при котором несомненно более примитивные гра-
диентные отношения между частями развивающегося организма сменились
физиологической мозаичностью, как это убедительно показал Стрелин
•(1940, 1942) на моллюсках, несомненно успело претерпеть глубокие эволю-
ционные изменения, существенно исказившие первоначальные взаимоотно-
шения между тканевыми зачатками.
Проблема эволюционной гистологии
75
С точки зрения видоизмененной Шимкевичем гоноцельной или, точнее,
энтерогоноцельной теории целома, выстилку вторичной полости тела пер-
вичноротых возможно рассматривать как ткань, гомологичную целомиче-
ской части мезодермы вторичноротых. Согласно этому, очень вероятному
предположению, и та и другая являются производными эктодермальной
выстилки радиальных каналов гастроваскулярной (системы кишечно-
полостных, несмотря на то, что онтогенез не дает в этом отношении никаких
определенных указаний.
Последнее в случае детерминированного развития не представляется
удивительным и не может служить аргументом против энтерогоноцельной
или энтероцельной теории происхождения целома у этих форм. В этом от-
ношении весьма поучительно сопоставление с развитием оболочников,
которое в ряду хордовых приобрело, несомненно в результате вторичных
изменений, несвойственную им очень высокую степень детерминирован-
ности, связанную с уменьшением общего количества клеток в теле заро-
дыша. У ряда туникат полностью утратился энтероцельный способ образо-
вания мезодермы, а между эктодермальными бластомерами и хордо-ней-
ральным зачатком несомненно вторично установилась очень тесная связь.
Всех первичноротых, как это отмечалось выше, по многим данным,
выводят из родственных гребневикам ископаемых форм. Однако эта точка
зрения разделяется в настоящее время не всеми.
Многое в организации бескишечных турбеллярий, которые прежде рас-
сматривались как вторично упрощенные формы, обнаруживает несомненные
примитивные черты. На этом основании возможно предположение, что
родоначальниками всех низших червей, а может быть, также и вышестоя-
щих форм, следует считать не многоветвистых, а бескишечных планарий
{Графф, 1882, 1889, 1905, 1913; Беклемишев, 1937). Принятие этой точки
зрения заставило бы не только значительно изменить наши взгляды на
филогению первичноротых, но сделало бы вместе с тем необходимым пере-
смотр вопроса о гипотетических предках многоклеточных животных.
В организации ряда бескишечных турбеллярий существенна особенно
подчеркиваемая Беклемишевым (1937) неясная отграниченность от мезо-
дермальной паренхимы не только пищеварительного синцития, ио и экто-
дермы. Это можно трактовать с двух противоположных точек зрения:
с одной стороны, этот факт может указывать на чрезвычайную примитив-
ность гистологической организации бескишечных планарий. Тогда эти
формы возможно представить себе, как дальнейший этап эволюции самых
примитивных морулообразных или паренхимулообразных гипотетических
предков, индифферентная клеточная масса которых еще только разделяется
на наружный пограничный эпителиальный слой и внутреннюю паренхиму,
соответствующую фагоцитобласту Мечникова и дифференцирующуюся,
в свою очередь, на энтодермальные пищеварительные, мышечные и мезен-
химные элементы. С этой точки зрения, подкрепляемой также примитив-
ными чертами организации нервной системы и отсутствием нефридиального
аппарата, всех прочих планарий, и в частности Polyclada, следовало бы
рассматривать как формы, возникшие путем прогрессивной эволюции из
76
Глава II
бескишечных предков, а не от организмов^ сходных с гребневиками, обла-
давших развитой гастроваскулярной системой и лишь намечающимися
элементами будущего мезодермального зачатка.
С другой точки зрения, которая мне представляется более вероятной и
опирается на детерминированный тип эмбрионального развития Acoela
и Polyclada (не говоря уже о прочих тубелляриях) и на наличие у них
настоящего мезодермального комплекса, отсутствующего у кишечнополост-
ных, в организации Acoela следует различать и примитивные черты и при-
знаки вторичной идиоадаптации и даже деградации. К последним принад-
лежит отсутствие кишечника, или точнее, гастроваскулярной системы,,
превращенной в пищеварительную паренхиму и видоизмененный у ряда
форм характер кожного эпителия, напоминающего в некоторых отношениях
своеобразный эпидермис трематод. Исходя из этого, представляется более
вероятным производить Acoela не от примитивных Parenchymella или
Phagozytella Мечникова, а от кишечнополостных.
Мне кажется, что паренхимуле в развитии многих современных кишечно-
полостных едва ли возможно придавать филогенетическое значение стадии,
рекапитулирующей организацию предков многоклеточных животных. Спо-
соб превращения паренхимулы в планулу путем образования щелевидной
полости в несообщающейся с внешней средой эктодермальной клеточной
массе также не может рассматриваться как сохраненный в онтогенезе
древнейший филогенетический момент.
Гаструляция' путем униполярной иммиграции, наблюдающаяся у не-
которых Hydrozoa, является, по всей вероятности, вторично измененным
способом образования энтодермы. То же самое справедливо и в отношении
различных видов деляминации, а также мультиполярпой иммиграции, ко-
торые, возможно, следует трактовать, в противоположность взглядам Меч-
никова, как результат, с одной стороны, вероятного сдвига момента обосо-
бления энтодермы и эктодермы на более ранние стадии онтогенеза, а с дру-
гой, накопления в яйцах питательного желтка, но не как примитивные
формы их развития.
С аналогичными, имеющими несомненно значение вторичных изменений,
способами образования энтодермы мы часто встречаемся у высокоорганизо-
ванных животных, в частности, в ряду позвоночных. Филогенетическое
развитие энтодермы вне непосредственной связи с окружающей средой,
как это уже подчеркивалось, малопонятно и не объяснимо на основе не-
сомненно имеющего место взаимодействия внутренних и внешних факторов
в процессе организменной эволюции.
Относительно филогенетических корней мускулатуры соматического
типа и мезенхимных соединительнотканных образований первичноротых
можно сделать те же предположения, какие были высказаны в отношении
этих тканевых систем в ряду вторичноротых, хотя в данном случае дело
затрудняется наличием у низших и высших червей мышечных и мезенхим-
ных элементов энтомезодермального и эктомезодермального происхождения.
У червей с вторичной полостью тела зачаток ее выстилки находится только
в составе энтомезодермы. Учитывая особенности спирального дробления.
Проблема эволюционной гистологии
77
отмеченные выше, этой двойственности происхождения в онтогенезе мышеч-
кого и мезенхимного зачатков едва ли можно придавать значение аргу-
мента в пользу их эволюционного развития из энтодермы и эктодермы пред-
ков.
Возможно, что эктомезодерма представляет собой остаток диффузной
эктодермальной мускулатуры и мезенхимы, свойственных многим кишечно-
полостным. В составе энтомезодермы, наоборот, находятся локализирован-
ные зачатки тех же тканей, обособляющиеся на более ранних стадиях раз-
вития и потому утрачивающие видимую связь с эктодермой.
Эволюционное образование мезодермального зачатка, кажущегося еди-
ным, в обоих главных стволах филогенетического дерева животных можно
себе представить следующим образом. Первоначально диффузное развитие
мускулатуры тела и мезенхимы, из эктодермы кишечнополостных в связи
с образованием вторичной перистомальной области высокого градиента
постепенно сконцетрировалось в последней. В результате этого и проис-
шедшего одновременно сдвига момента обособления клеток мышечного и
мезенхимного зачатков на более ранние стадии онтогенеза, в перистомаль-
ной области возникла часть будущей мезодермы в виде особой группы клеток.
В связи с переходом к активно подвижному существованию, формированием
передне-задней оси и головного конца тела, вся организация животных
и вместе с ней мезодермальный зачаток приобрели билатеральную симмет-
рию. В ту же перистомальную область высокого градиента сдвинулось
обособление из состава энтодермы радиальных, а затем билатерально
симметричных каналов гастроваскулярной системы, т. е. родоначальников
вторичной полости тела. Эктодермальные и эктодермальные части мезо-
дермы в конце концов образовали единый зачаток, существующий у всех
целомических животных.
На возможное филогенетическое значение перистомального развития
мезодермы и хордального зачатка в онтогенезе амфибий в свое время ука-
зывал Фогт (1929), который высказал предположение, что у гастреепо-
добных предков позвоночных, помимо дифференцировки на эктодерму
и энтодерму, существовала еще перистомальная область с особыми свой-
ствами. Она и сделалась источником образования мезодермального, а
также хордального зачатков.
Схизоцельный способ образования вторичной полости тела, характер-
ный для первичноротых, следует рассматривать как вторичный. В ряду
хордовых мы наблюдаем с достаточной отчетливостью постепенную замену
более примитивного энтероцельного образования мезодермы (характерного
для ланцетника) схизоцельным, свойственным всем позвоночным, кроме
миноги. У нее по крайней мере в области переднего конца тела энтероцелия
еще сохранилась.
В противоположность зачаткам мускулатуры тела и мезенхимных соеди-
нительнотканных образований, утрачивающим свою связь с вероятным
первоначальным источником своего образования, зачаток нервной системы
сохраняет на всех этапах эволюции теснейшие взаимоотношения с экто-
дермой. Причина этого заключается в том, что возникновение и эволюцион-
Глава II
78
ное развитие нервной системы находятся в двойной зависимости от падаю-
щих извне раздражений и от двигательной функции.
В эмбриональном развитии нервной системы многих или же всех совре-
менных животных нельзя не видеть следов этой двойной зависимости. Она
проявляется, например, в том, что у многих беспозвоночных зачатки голов-
ного и туловищного мозга образуются отдельно друг от друга из разных
участков эктодермы, а также в наличии у позвоночных и беспозвоночных,
помимо централизованного зачатка нервной системы, плакод или сравни-
мых с ними образований.
Таким образом, возможно себе представить, что элементы будущих
тканевых систем должны были первоначально появиться путем дивергент-
ной дифференцировки клеток двух клеточных пластов, эктодермы и энто-
дермы, из которых состояло тело примитивных предков современных много-
клеточных животных.
В течение эволюции первоначальная диффузная дифференцировка клеток
сменилась более ранним обособлением в онтогенезе зачатков отдельных
тканей, древнейшими из которых являются эпидермальный покров тела,
эктодермальная выстилка пищеварительной трубки, мускулатура тела,
нервные элементы, мезенхимные соединительнотканные образования. Не-
сколько позже, вероятно, образовалась путем обособления от эктодермаль-
ной гастроваскулярной системы выстилка вторичной полости тела. Эпидер-
мальный покров тела, эктодермальная выстилка пищеварительной трубки
и стенка целома, представляющие собой различные типы эпителиальных
тканей, на дальнейших этапах эволюции сделались источниками развития
целого ряда других специальных тканей, которые, в отношении своей общей
морфо-физиологической характеристики, частью могут быть отнесены в одну
из четырех тканевых групп классической гистологии, частью представляют
собой структуры, которые не укладываются в общепринятую систему.
Мезенхимные соединительнотканные образования таки е дают начало очеш
большому многообразию дифференцированных в различных направлениях
тканей.
На фоне общей усложняющейся организменной интеграции и парал-
лельно с морфо-физиологической дифференцировкой должна была прогрес-
сивно развиваться частная внутритканевая и межтканевая интеграция,
характеризующая отдельные гистологические структуры, как части целого
организма и тканевые компоненты органов.
Более или менее параллельно с прогрессирующей тканевой дифферен-
цировкой и специализацией должны были развиваться все более резкие
качественные различия соматических клеток от половых и гистологическая
детерминация, т. е. ограничение способности превращения друг в друга
тканевых элементов, имеющих разную историю развития. Распространенные
у ряда низкоорганизованных форм i-клетки вместе с тем утрачивают
значение камбиального резерва для восстановления дифференцированных
соматических элементов и исчезают или превращаются в клетки полового
зачатка, обособляющиеся на ранних стадиях индивидуального развития.
Дифференцированные соматические клетки, которые у некоторых относи-
Проблема эволюционной гистологии
79
1ельно примитивных форм (по некоторым данным), утрачивая свою диф-
ференцировку, могли превращаться в половые элементы, теряют эту спо-
собность. Направление вторичной дифференцировки тканевых элементов,
подвергшихся видимому морфологическому упрощению (т. наз. дедиф-
ференцировке) при бесполом размножении и регенеративных процессах,
все больше ограничивается.
У аннелид еще описывается возникновение элементов нервной системы,
а также части соматической мускулатуры, из кожного эпидермиса и раз-
личных мезодермальных производных (целомической выстилки, соматиче-
ской мускулатуры, выделительного аппарата типа целомодуктов) из особых
индифферентных необластов, которые рассматриваются как мезодермальные
элементы, сохранившие эмбриональные свойства. При регенерации перед-
него конца тела клетки типа необластов заново возникают (по ряду данных)
из дифференцированных мышечных элементов (Иванов, 1903, 1937; Вейц-
маи, 1937; Жинкин 1934). Приписывание более широких потенций необластам
аннелид, как это делают некоторые авторы (Гаммерлинг, 1924; Пробст-,
Шолыпе, 1927), недостаточно обосновано (Иванов, 1937). У наиболее высоко-
организованных животных, например позвоночных, гистологическая детер-
минация достигает наибольшей степени развития.
Их тканевые элементы обнаруживают в лучшем случае способность
к превращениям и дифференцировке лишь в определенных узких границах
и не способны к превращению в ткани, имеющие иное происхождение.
Гистологическая детерминация тканей позвоночных, а также предста-
вителей некоторых других высокоорганизованных типов животных будет
подробнее разобрана ниже, в связи с изложением экспериментального
материала. У некоторых специализированных форм, характеризующихся
постоянством клеточного состава (например коловратки, нематоды), ткане-
вые элементы, невидимому, вообще утрачивают способность к пролиферации
и каким бы то ни было гистологическим превращениям. Хотя в отношении
тканей многих беспозвоночных до настоящего времени имеется еще много
неясностей, общее направление эволюции тканевых систем, связанное с их
прогрессирующей специализацией и гистологической детерминацией, едва
ли подлежит сомнению. Высказывания противоположного характера зиж-
•чятся на более чем слабых и малоубедительных основаниях и не выдержи-
вают (по крайней мере в отношении позвоночных) критической эксперимен-
тальной проверки. Разумеется, следует оговориться (как это, например,
делается в отношении регенеративных возможностей различных животных),
что полного параллелизма между относительной высотой организации и
положением в системе, с одной стороны, и гистологической детерминацией
тканей, с другой, в ряде случаев, невидимому, не наблюдается. Это, однако,
вовсе не противоречит общему положению относительно прогрессирующего-
развития специфичности тканей и их гистологической детерминации.
III. ЭКСПЕРИМЕНТАЛЬНЫЙ МЕТОД В НОРМАЛЬНОЙ
И ПАТОЛОГИЧЕСКОЙ ГИСТОЛОГИИ
1. ПРИНЦИПЫ И ЗНАЧЕНИЕ ЭКСПЕРИМЕНТАЛЬНО-ГИСТОЛОГИЧЕСКОГО
АНАЛИЗА ТКАНЕЙ
Для изучения генетических отношений и эволюционных изменений
организмов биология располагает, как известно, несколькими основными
методами — сравнительно-анатомическим, эмбриологическим и палеонто-
логическим, составляющими т. наз. геккелевскую триаду и позволяющими
с большей или меньшей долей вероятия или даже достоверности делать
выводы филогенетического характера. Однако взятый в отдельности каждый
ai3 этих методов далеко не всегда может привести к правдоподобным выводам
(Северцов, 1931, 1939; Иванов, 1937). В новейшее время делаются также
попытки использовать в том же направлении данные сравнительной био-
химии и механики развития, которые дают во многих отношениях очень
ценные и интересные результаты. Очевидно, те же самые методы могут быть
использованы, с соответствующими оговорками, и для изучения генетических
отношений тканей. Дело в том, что в составе отдельных органов тела одной
из тканей, как правило, принадлежит основное, ведущее, значение, в то
время как прочие их компоненты имеют более или менее вспомогательный
характер. Так, например, в составе кожи, пищеварительной трубки, выде-
лительного аппарата эпителиальные компоненты, безусчовно, определяют
функциональное значение соответствующих органов. В двигательном ап-
парате тела аналогичное значение имеют мышечные волокна, в нервной
системе — нейроны. Что касается соединительной ткани, гладкой му-
скулатуры и сосудов, то они являются общими вспомогательными частями
различных органов тела. Вспомогательное значение имеют, в свою очередь,
во многих случаях, в составе различных органов и элементы нервной си-
стемы. Очевидно, эволюционные изменения тех или иных органов не могут
не быть связаны теснейшим образом с филогистогенезом тканевого материала,
определяющего их функциональное значение. Иными словами, изучение
эволюции тканей нельзя искусственно отрывать от изучения эволюционных
изменений органов тела и естественной генетической системы животных
«организмов.
Филогенетические изменения тканей, или филогистогенез, и филогенети-
ческое развитие организмов и их органов представляют собой не два раз-
личных и в известных отношениях независимых друг от друга процесса,
Экспериментальный метод в гистологии 81
а две стороны единого по существу процесса эволюции, которые неразрывно
связаны друг с другом.
В настоящее время можно считать, что генетические отношения отдель-
ных классов в подтипе позвоночных наиболее точно разработаны. Даже
подразделение позвоночных на большее, чем шесть, количество классов
уже не сможет изменить расположения отдельных групп в естественной
системе (Зенкевич, 1937). Позвоночные вместе с тем наиболее изучены
в гистологическом отношении. Поэтому прежде и подробнее всего мы остано-
вимся на естественной системе их тканей и распространим затем полученные
результаты (поскольку это возможно в настоящее время) на те или иные
генетические ряды беспозвоночных.
Большая часть авторов трактует принципиально одинаковым образом
соотношения групп аннелид-членистоногих, а также соотношения ки-
шечнодышащих, иглокожих и оболочников с позвоночными. К сожалению,
в гистологическом отношении многие из этих групп еще очень недостаточно
изучены современными методами, несмотря на большой накопленный в на-
стоящее время фактический материал.
Как это подчеркивалось в предыдущей главе, эмбриологический метод
имеет для эволюционной морфологии выдающееся значение, когда депо
касается стадии органогенеза и гистологической дифференцировки, особенно
при сопоставлении процессов индивидуального развития с фактами из
области сравнительной анатомии. Что касается самых ранних стадий
онтогенеза, дробления и образования зародышевых листков, то их филоге-
нетическая трактовка особенно трудна и во многих отношениях гипотетична
и спорна. Последнее совершенно понятно, так как древнейшие моменты
филогенеза должны были претерпеть в индивидуальном развитии наиболь-
шие изменения и искажения. Исходя преимущественно из сравнительно-
анатомических данных и используя теорию физиологических градиентов
Чайлъда, мы попытались, тем не менее, дать одно из вероятных объяснений
филогенетического значения этих ранних стадий зародышевого развития.
Ограниченное применение в области гистологии может иметь, по понят-
ным причинам, палеонтологический метод, поскольку с его помощью можно
судить лишь о структуре эмали, межклеточного вещества кости и дентина.
Изменчивость гистологических структур при экспериментальных и
патологических условиях, развитие в составе организма при опухолевых
процессах совершенно уродливых клеток и тканей давно известны. Спо-
собность тканей к разнообразным превращениям не могла не дать повода
думать, что гистологические структуры отличаются большой лабильностью
и изменчивостью, несвойственной, вообще говоря, анатомическим частям
тела и типу организации животных.
Если считать, что все биологические свойства тканей, как живого струк-
турного материала организмов, филогенетически обусловлены, то совер-
шенно естественно распространить это положение не только на морфологи-
ческие и функциональные особенности гистологических структур при
обычных условиях существования в организме, но и на всю совокупность
«х превращений при экспериментальных или патологических условиях.
® X л он и п—. З49
82
Глава III
Иными словами, следует ожидать, что совокупность структурных и функ-
циональных особенностей тканей, которая обнаруживается при разнообраз-
ных условиях существования, должна подчиняться определенным, завися-
щим от всей истории их развития, гистологическим закономерностям.
Каждую же наблюдаемую при обычных условиях гистологическую структуру
организма следует трактовать как одну из форм существования данной
ткани в цепи возможных для нее превращений, совокупность которых и
дает ее наиболее полную и разностороннюю характеристику. Таким обра-
зом, задачей экспериментального и патолого-гистологического методов (как
методов эволюционной морфологии) является систематическое изучение
изменений и становления каждой тканевой структуры организмов и срав-
нение данных, получаемых при этом на разных объектах. Для выводов
филогенетического порядка необходимо критическое сопоставление резуль-
татов экспериментально-гистологического анализа с данными, получен-
ными с помощью других методов эволюционной морфологии. Как это ни
удивительно, систематического и критического экспериментального анализа
тканей под таким углом зрения до последнего времени не было.
Будет уместно остановиться несколько подробнее на том, что предста-
вляет собой экспериментальный метод в гистологии и какие общие процессы
происходят в тканях и их элементах в результате экспериментального
вмешательства.
Очевидно, что свойства каждой тканевой структуры зависят от двоякого
рода факторов: с одной стороны, от совокупности собственных, внутренних
свойств данной ткани, развивающихся в эволюционном процессе и наслед-
ственно закрепляющихся на каждом этапе эволюции, а с другой, от сово-
купности внешних по отношению к данной ткани, но могущих иметь также
значение внутренних факторов целого организма, условий существования..
Экспериментально-гистологический метод заключается в изменении ус-
ловий существования тканей и их элементов, в изменении внутритканевых
и межтканевых связей, которые приводят к явлениям роста, а также,,
в большинстве случаев, к пролиферативным процессам. Одним из резуль-
татов этого бывают более или менее резко выраженные реактивные изме-
нения тканевой и клеточной структуры или дифференцировки. Задача
гистолога, как морфолога-экспериментатора, изучить и дать гистологиче-
ский анализ всей совокупности морфологических изменений, наблюдаемых
в тканях при разных условиях, и вместе с тем углубленно изучить собствен-
ные свойства тканей, как живого материала животного и человеческого-
организма, в первую очередь их морфологические, структурные особенности
во всей динамике превращений.
Другим необходимым следствием тех же самых экспериментальных воз-
действий являются различные изменения тканевого и клеточного обмена
веществ, регистрируемые теми или иными физиологическими или биохими-
ческими методами. Наконец, морфологические и функциональные изме-
нения в отдельных тканях, вызванные теми или иными причинами, могуг
иметь различное общебиологическое и специальное медицинское значение,
в зависимости от того, какая роль им принадлежит в составе организма,
Экспериментальный метод в гистологии
83
где и как именно они происходят в пространстве и во времени и чем в конце
концов завершаются. Все эти вопросы также интересуют в известной сте-
пени гистолога-морфолога, но их детальный анализ уже принадлежит пред-
ставителям других специальностей, в частности, патологии или динамики
развития.
Один и тот же экспериментальный метод может быть ориентирован на
вопросы, интересующие разные специальности; например, любой экспери-
ментально-гистологический метод является одновременно и методом экспе-
риментальной патологии. Вместе с тем экспериментальная гистология
(и методически и по существ}^ подлежащих ее изучению вопросов) тесно свя-
зана с другими общебиологическими и специальными медицинскими дис-
циплинами.
Наибольшее значение в экспериментальной гистологии принадлежит
изучению реактивных изменений тканей в ответ на повреждение, в частности
на нанесение местного дефекта. К таким методам «повреждения» или мето-
дам «дефектов» относится в первую очередь метод регенерации, тесно
связанные с ним методы воспалительных разрастаний и метод эксплантации
или тканевых культур. Все эти методы могут, кроме того, сочетаться с ме-
тодом трансплантации, при котором возможно в большей или меньшей
степени установление новых связей между изучаемой тканью или тканевым
‘комплексом и целым организмом.
Те же самые или аналогичные методы повреждения (разделения на
части, нанесения дефекта, трансплантации и изоляции) используются и в от-
ношении эмбрионального материала для разрешения разных эмбриологи-
ческих вопросов. Они могут быть вместе с тем применены к проблемам
гистогенеза или процессам развития тех или иных специфических дифферен-
цированных тканевых структур из эмбриональных зачатков, частей яйце-
клетки, бластомеров разных стадий дробления или презумптивных участков
бластулы. Таким образом, в зависимости от изучаемого вопроса, эксперимен-
тально-эмбриологическая методика может иметь гистологическую направ-
ленность.
До последнего времени неправильно противопоставляют эксперименталь-
ные физиологические науки описательным морфологическим. Так, напри-
мер, на этом основании, многие считают экспериментальную эмбриологию
или механику развития, в противовес описательной эмбриологии, физиоло-
гической, а не морфологической дисциплиной. Действительно, эксперимен-
тальные методы были прежде всего использованы физиологами для решения
ряда основных физиологических проблем подобно тому, как сравнитель-
ный метод уже давно приобрел фундаментальное значение в области морфо-
логии. На этом основано противопоставление экспериментальных физиоло-
гических дисциплин описательным морфологическим. Но это глубоко оши-
бочный пережиток прежних этапов их развития. В настоящее время экспе-
риментальным методам принадлежит в области морфологии не менее важное
значение, чем в физиологии. Так же неправильно было бы отнести разви-
вающуюся в настоящее время сравнительную физиологию к морфологическим
дисциплинам только на том основании, что она широко использует сравни-
84 Глава III
тельный метод, бывший долгое время прерогативой морфологических
дисциплин. В настоящее время экспериментальная физиология и экспери-
ментальная морфология, включающая в себе, как часть, экспериментальную
гистологию, каждая занимаясь своим кругом проблем, представляют собой
две мощных и равноправных ветви единого по существу комплекса наук
о живой природе.
В применении к позвоночным и, в частности, к млекопитающим патологи
использовали значительную часть экспериментальных гистологических
методов первоначально для решения тех или иных вопросов патологии.
Однако с их помощью попутно выявлялись факты, имеющие большое зна-
чение для гистологии, а затем они уже применялись специально в области
гистологических проблем. Достаточно вспомнить исследования Рибберта,
Подвысоцкого, Циглера с их сотрудниками, которые вызывали различные
гисто-патологические процессы в тканях с помощью нанесения тем или иным
способом повреждений.
Очень многие авторы изучали процессы репаративной или т. наз. пато-
логической регенерации в самых разнообразных тканях: эпителиях, соеди-
нительной ткани, мышцах и нервной системе. В изучении нервной системы,
наряду с регенеративными процессами, очень большое значение приобрели
предшествующие им явления дегенерации, которые широко использовались
для анализа взаимоотношений нервных элементов друг с другом и элемен-
тами других тканей. Современные представления о тонком строении нервной
системы сложились в значительной степени в результате изучения происхо-
дивших в ней дегенеративных и регенеративных процессов специальными
нейрогистологическими методами. Особенно много в этом отношении дали
классические исследования испанской школы Рамон и Кахаля (1928). В но-
вейшее время метод перерезок, широко использованный физиологами (Лен-
гли, 1921, и др.), дал много ценных фактов для анализа нейрональных взаимоот-
ношений в автономной нервной системе (Лаврентьев него сотрудники, 1939).
Как известно, циглеровский метод вызывания асептического воспаления
путем введения стерильных целлоидиновых камер или трубок применил
.Максимов (который по справедливости может считаться пионером в области
экспериментальной гистологии) для гистологического анализа соединитель-
ной ткани и крови млекопитающих и некоторых низших позвоночных.
Сравнительно недавно та же по существу методика была распространена
на других позвоночных и на некоторых представителей беспозвоночных
(Лазаренко, Заварзин, Данини, Крафт, Алфеева).
Обширным циклом работ Гаршин и его сотрудники доказали, что в очаге
воспаления различнейшие эпителиальные ткани млекопитающих претер-
певают ряд в высшей степени интересных структурных изменений и обнару-
живают резко выраженные пролиферативные явления. Хотя основное
внимание в этих работах уделяется общепатологическим вопросам, в част-
ности, отношению этих пролиферативных процессов к проблеме рака, их
фактический материал имеет выдающееся гистологическое значение, осо-
бенно при его сопоставлении с данными, полученными другими методами,
в частности, методом эксплантации.
Экспериментальный метод в гистологии 85
В своей недавно появившейся монографии Гаршин (1939) полностью
принимает нашу систему эпителиальных тканей, построенную первона-
чально на основе работ, сделанных методом эксплантации мной и моими
сотрудниками.
Очень интересные факты, относящиеся к пролиферативным процессам
в кожном эпителии разных беспозвоночных, а также амфибий, установили
Лазаренко, Заварзин, Данини и Алфеева при опытах с асептическим воспа-
лением по методу Циглера и Максимова. Исходя из работ по воспалительным
разрастаниям эпителиев и из своего опыта в области метода тканевых куль-
тур, Лазаренко в последнее время разработал оригинальную методику им-
плантации измельченных органов вместе с кусочками целлоидина в подкож-
ную клетчатку и получил очень ценный и интересный новый фактический
материал, касающийся пролиферативных процессов в эпителиальных тка-
нях, находящихся в искусственно созданном очаге воспаления.
Цикл работ Лазаренко и его сотрудников в отношении общего плана
в значительной своей степени совпадает с нашими исследованиями и полно-
стью подтверждает своим фактическим материалом наши основные выводы.
Интересные в гистологическом отношении данные были также получены
путем трансплантации частиц органов. Из различных методов трансплан-
тации заслуживают быть отмеченными метод пересадки в переднюю камеру
глаза, широко применявшийся в лаборатории ныне покойного Колосова,
и метод Мерфи-Рауса (1914) трансплантации на хорион-аллантоис, давший
ценные результаты в работах Студиукого по гистогенезу костной ткани
(1937).
Остроумная методика, разработанная Сендисоном в лаборатории Кларка
(1923), сделала возможным очень детальное прижизненное изучение воспа-
лительной пролиферации соединительной ткани и сосудов {Кларк и Кларк)
между прозрачными листками целлофана, вставленными в отверстие, про-
деланное в ухе кролика.
Прижизненное наблюдение нормальных процессов роста капилляров и
реактивных изменений в соединительной ткани на некоторые эксперимен-
тальные воздействия производилось в широком масштабе на хвосте голова-
стиков (Кларк и Кларк, 1912—1925). Эта методика дает возможность,
пользуясь сильными увеличениями микроскопа, детально прослеживать
судьбу и изменения отдельных клеток в составе организма.
Пользуясь такой же методикой, Спейделю удалось в последнее время
(1934, 1938) получить ряд интересных данных по гистогенезу и регенерации
периферических нервов и по реактивным изменениям в соматической попе-
речно-полосатой мускулатуре в ответ на повреждение.
Из различных «методов дефекта» выдающееся значение приобрел для
экспериментально-гистологического анализа всевозможных тканей позво-
ночных, в первую очередь млекопитающих и человека, метод эксплантации
или тканевых культур. Его открыл в 1907 г. Гаррисон, усовершенствовали
Каррель с Берроусом и другими сотрудниками и впервые использовали
в нашей стране Авроров и Тимофеевский (1912) для изучения клеток пато-
логически измененной крови и для решения ряда специальных гематоло-
86
Глава III
гических вопросов, В дальнейшем, в лабораториях Максимова и Тимофеев-
ского метод эксплантации был широко применен к гистологическому анализу
соединительной ткани и крови и дал в этом отношении исключительно важ-
ные и ценные результаты.
Затем метод эксплантации был планомерно применен в руководимых
мной лабораториях для гистологического анализа всех основных типов
тканей — соединительной ткани, всевозможных эпителиев, мышечных эле-
ментов разного происхождения и, наконец, вспомогательных и нейрональ-
ных компонентов нервной системы.
Очень важные результаты для патологии и отчасти гистологии дает
в последнее время метод эксплантации в отношении опухолевых тканей
человека.
Факты, установленные мной и моими сотрудниками, в первую очередь
методом эксплантации, легли в основу тех новых представлений в области
гистологии, которым посвящена настоящая книга. Они дали возможность
использовать и обобщить под определенным углом зрения и правильно гисто-
логически трактовать многочисленные, но большей частью разрозненные
данные, полученные в разное время как методом тканевых культур, так
« другими экспериментальными методами, а также данные патологических
работ, сделанных на человеческом секционном и биотическом мате-
риале.
В последнее время, параллельно с методом эксплантации, мы стали так
же планомерно применять и метод регенерации. Параллельное изучение
тканей различными экспериментальными методами одним и тем же научным
коллективом имеет, само собой разумеется, большие преимуще-
ства.
Помимо изменений в тканях, получаемых путем нанесения местного
повреждения или дефекта, очень интересны в гистологическом отношении
тканевые реакции на некоторые общие воздействия на целый организм,
например гормональные или получающиеся в результате авитаминозной
диэты, а также под влиянием введения в организм тех или иных синтетиче-
ских химических продуктов. Помимо изучения гормональных реакций,
получаемых экспериментальным путем, очень интересны также физиологи-
ческие гормональные реакции тканей, наблюдаемые, например, в течение
нормального полового цикла и при беременности.
Экспериментальный анализ тканей имеет выдающееся значение не только
для решения основных проблем гистологии, для установления закономер-
ностей, которым подчинены тканевые структуры в их онтогенетическом и
филогенетическом развитии, но и для ряда сопредельных дисциплин, в пер-
вую очередь для патологической гистологии, изучающей изменения тканей
при различных патологических процессах. Так наз. патологические
изменения в тканях представляют собой, по сути дела, реакции тканевых
структур на экспериментальные воздействия, производимые самой природой
на человеческий или животный организм. В основе этих патологических
изменений лежит та же общая и специфическая реактивность тканей,
которая проявляется и при собственно экспериментальных условиях.
Экспериментальный метод в гистологии
87
Однако обычный секционный или биотический патолого-гистологиче-
ский материал отличается от экспериментального меньшей точностью,
так как патологические процессы в отношении своей этиологии далеко
не всегда достаточно ясны и могут отличаться значительной степенью слож-
ности, а вызванные ими изменения в тканях очень часто не могут быть
детально изучены по стадиям.
Правильное понимание каждой формы существования ткани в цепи
претерпеваемых ею превращений возможно лишь при условии точного учета
последовательности во времени и взаимосвязи в пространстве отдельных
гистологических структур. В этом отношении только экспериментальная
методика и, пожалуй, в первую очередь метод тканевых культур могут
дать достаточно ясные результаты. Правильная же трактовка патолого-
гистологического материала нередко наталкивается на очень большие
трудности. Экспериментальная гистология, устанавливающая общие и
специальные закономерности изменений тканевых структур при различных
условиях существования, дает вместе с тем прочные основы для правильной
трактовки ряда основных гисто-патологических процессов, как, например,
процесса заживления ран, явлений метаплазии, вопросов гистогенеза опу-
холей и т. п.
В настоящее время ряд фактов из области патологии получает новую
прочно обоснованную трактовку именно на основе экспериментально-
гистологического анализа тканей. В свою очередь, биотический и секцион-
ный патолого-гистологический материал имеет существенное значение для
экспериментальной гистологии, но, разумеется, только в тех случаях,
когда он достаточно полноценен в отношении методики обработки и изуче-
ния препаратов и трактовки наблюдаемых картин.
Таким образом, современная гистология (в широком смысле этого слова),
гистология, обогащенная экспериментальной методикой, изучает не только
структуру и функциональное значение тканей при обычных условиях суще-
ствования в составе организма и процессов гистогенеза в течение индиви-
дуального развития, но и всю совокупность превращений гистологических
структур при различных измененных условиях существования, как в со-
ставе, так и вне целого организма, в тканевых культурах, и раскрывает
на этой основе те общие эволюционные закономерности, которым подчинен
тканевой материал органов во всех своих структурных превраще-
ниях.
После этих кратких методических замечаний следует остановиться на
общей характеристике тех процессов, которые происходят в тканях в ре-
зультате повреждения или нанесения дефекта. Непосредственным резуль-
татом местного повреждения достаточной интенсивности являются гибель
большей или меньшей массы живого вещества и образование очага некроза.
При механическом повреждении режущими инструментами сразу обра-
зуется большего или меньшего размера дефект с незначительной зоной
некротизированных тканей вокруг. Очаг некроза является инородным телом
Для окружающих живых тканей и целого организма. Изливающаяся кровь
или тканевая жидкость также образуют в области дефекта некротические
88
Глава III
массы. Так или иначе вследствие любого повреждения достаточной интен-
сивности целость живых тканей нарушается или в них образуется дефект
с некротическими массами, находящимися в его области в большем или
меньшем количестве.
На границе между дефектом и оставшимися неальтерированными замет-
ным образом тканями образуется выраженная в той или иной степени зона
повреждения, ширина и распространение которой зависят от характера
и силы повреждающего агента, времени его воздействия и биологических
особенностей соответствующих тканевых элементов. В этой зоне часть тка-
невых элементов может быть повреждена необратимым образом и постепенно
подвергается некробиозу и гибели, вследствие чего область отмерших тка-
ней или дефекта увеличивается. Другая часть тканевых элементов зоны
повреждения может обнаружить изменения обратимого характера, выражаю-
щиеся, например, в т. наз. паранекротическом комплексе Насонова и Але-
ксандрова (1941), которые в дальнейшем сглаживаются и исчезают. При
очень слабых степенях повреждения, не приводящих к гибели тканевых
элементов и образованию дефекта, дело может ограничиться одними лишь
обратимыми изменениями живой протоплазмы, представляющими большой
интерес с точки зрения цитофизиологии, но не влекущими за собой тех
явлений роста и пролиферации тканевых элементов, изучение которых на-
ходится в центре внимания гистолога-экспериментатора. Метод тканевых
культур также сводится к нанесению дефекта. Разница между опытами
с целым организмом и тканевыми культурами заключается в том, что в пер-
вом случае изучаются тканевые процессы в окружающих дефект тканях,
а во втором случае — в изолированной из организма частице очень малень-
ких размеров, для которой областью дефекта является, по сути дела, место
всех окружающих ее при нормальных условиях тканей и весь целый орга-
низм, из которого она была выделена. При опытах с целым организмом
явления роста и пролиферации тканевых элементов имеют значение воспа-
лительной реакции и процессов регенерации, а в эксплантатах мы имеем
дело с особыми явлениями роста и пролиферации вне организма. Они не
подчинены коррелятивным связям этого последнего и не имеют, строго,
говоря, по своему биологическому значению, ни воспалительного, ни реге-
неративного характера.
В жизнеспособных тканевых элементах по соседству с местом поврежде-
ния или дефектом начинаются реактивные процессы, приводящие к явле-
ниям роста и изменениям исходных нормальных гистологических структур.
Эти изменения могут постепенно захватить значительную область и, в усло-
виях целого организма позвоночных, сопровождаются характерной сосу-
дистой реакцией.
Совокупность местных реактивных процессов в соединительнотканных
образованиях и сосудистом аппарате в ответ на нанесение повреждения или
дефекта представляет собой, по Фишеру—Вазельсу, то, что издавна принято
обозначать в патологии воспалительной реакцией. Однако, как показывают
исследования Гаршина и его сотрудников (1939), в воспалительную реак-
цию, в очень многих случаях, вовлекаются также и эпителиальные ткани»
Экспериментальный метод в гистологии
89
разрастания которых в очаге воспаления могут иметь резко выраженное
защитное, очищающее организм от той или иной вредности, элиминативное
значение.
Многочисленные опыты Гаршина и его сотрудников доказали, что даже
неповрежденные эпителии реагируют на образовавшийся по соседству
воспалительный очаг пролиферативными изменениями, которые имеют,
в зависимости от условий, неодинаковый характер и несомненно воспали-
тельное, а не регенеративное значение.
Согласно представлениям Гаршина, основывающимся не на формальных
признаках воспалительной реакции, а на ее биологическом или патологи-
ческом значении, воспаление есть процесс, который начинается с местного
повреждения организма и протекает, как тканевая реакция пролиферативного
характера. Она обеспечивает регенерацию, завершающую всю цепь взаимо-
связанных явлений, и постепенно переходит в последнюю.
Основное значение воспалительной реакции заключается, так сказать,
в расчистке поля для последующих регенеративных процессов.
Явления тканевого роста заключаются прежде всего в том, что область
дефекта постепенно заполняется клетками или, по крайней мере, их отрост-
ками (осевые цилиндры нейронов), а в некоторых случаях и многоядерными
симпластами, например т. наз. мышечными почками, смещающимися из
соседних неповрежденных или малоповрежденных участков. Это может
зависеть, в свою очередь, от разных причин: от изменения формы и актив-
ного движения тканевых элементов, от увеличения массы, т. е. гипертрофии
или роста протоплазматического тела и, наконец (в большей части случаев),
от увеличения количества клеток в результате размножения или пролифе-
рации митотическим путем.
Имеющиеся во многих тканях при обычных условиях явления роста
и пролиферации, обеспечивающие стационарное поддержание их нормальной
структуры и компенсирующие физиологическое снашивание и гибель отдель-
ных клеток, после нанесения дефекта или повреждения стимулируются и
изменяют в более или менее резкой степени свой характер.
В случае тканевых культур в результате тех же причин, вокруг эксплан-
тированного кусочка либо образуется т. наз. зона экстенсивного роста по
поверхности или в толще окружающего плотного субстрата, либо тканевой
рост происходит непосредственно по наружной поверхности самого эксплан-
тата, не распространяясь за его пределы.
Находящиеся в области дефекта инородные тела рассасываются про-
растающими их тканевыми элементами или окружаются новообразованными
тканями и, в конечном итоге, в ряде случаев могут удаляться из состава
организма.
Рост и пролиферация тканей в условиях целого организма, как правило,
или завершаются заживлением раны (т. е. тканевой регенерацией) или,
продолжаясь после этого, могут привести к более или менее полному вос-
становлению или регенерации поврежденного или удаленного органа.
Специальные проблемы из области регенерации органов уже выходят из
круга подлежащих нашему рассмотрению вопросов. В условиях культур
90
Глава 111
рост и пролиферация тканей имеют разный исход, в зависимости от условий
опытов и свойств тканевого материала. В ряде случаев они могут продол-
жаться в течение месяцев и даже лет (т. е. практически безгранично), не
ослабляя своего темпа. В других случаях они со временем затухают, и
тканевые культуры приходят в своеобразное «покоящееся» состояние,
сохраняя в течение более или менее длительного времени полную жизне-
способность и определенный уровень общего обмена веществ. Во многих
случаях и тот и другой исход процессов роста и пролиферации вне орга-
низма не является окончательным и целиком зависит от воли эксперимен-
татора.
При опытах с трансплантацией или с имплантацией, по Лазаренко, исход
пролиферативных воспалительных процессов может привести в некоторых
случаях к прочному приживлению пересаженного тканевого материала
на новом месте и к установлению новых коррелятивных связей между ним,
окружающими тканями и целым организмом. Весьма нередко, однако,
в пересаженном материале, после известного периода пролиферативной
активности, наступают дистрофические изменения, приводящие к его
постепенной гибели и рассасыванию окружающими тканями. В ряде слу-
чаев при обычной методике пересадок происходит по истечении известного
промежутка времени быстрая гибель трансплантата при бурных реактивных
процессах в окружающих тканях.
Выведенные из нормального состояния относительного покоя растущие
и пролиферирующие ткани изменяются в отношении формы, взаимораспо-
ложения, способа взаимосвязи своих элементов и в отношении особенностей
цитоплазмы и ядерного аппарата последних. Одновременно с морфологиче-
скими изменениями в тканях происходят характерные сдвиги функцио-
нальных показателей, анализ которых выходит за рамки интересующих нас
здесь гисто-морфологических вопросов. Совокупность морфологических
изменений растущих и пролиферирующих тканей может быть в общих чер-
тах охарактеризована как изменение дифференцировки в сторону ее пони-
жения или «дедифференцировка», т. е. как более или менее резко выражен-
ное видимое упрощение или утрата структур, связанных со специфическим
функциональным значением ткани в составе организма. В зависимости
от внешних условий, это видимое упрощение тканевых и протоплазматиче-
ских структур может быть или едва заметно или, наоборот, очень резко.
Во многих случаях устанавливается прямая зависимость между интенсив-
ностью процессов роста и пролиферации, с одной стороны, и степенью сни-
жения дифференцировки, с другой. В условиях энергичного роста и проли-
ферации вне организма видимое морфологическое упрощение тканей во
многих случаях бывает выражено сильнее, чем в составе целого организма.
Повидимому, в этом отношении, помимо интенсивности пролиферативных
процессов, имеют значение и трудно поддающиеся точной характеристике
межтканевые и организменные коррелятивные связи.
При затухании вызванных экспериментально явлений роста и проли-
ферации как в составе, так и вне организма, в тканях, подвергшихся
морфологическому упрощению, уровень дифференцировки, как правило,
Экспериментальный метод в гистологии
91
вновь повышается, т. е. происходит их вторичная дифференцировка, которая
постепенно приводит к исходным гистологическим картинам или к новой
-тканевой структуре, отличающейся в большей или меньшей степени от
первоначальной.
Уровень вторичной дифференцировки и степень ее соответствия тем или
иным нормальным тканевым структурам организма зависят не только от
специфических особенностей данного гистологического материала, но в из-
вестной степени также от внешних условий и, в частности, от межтканевых
корреляций, а также оттогб, в какой степени дифференцирующаяся ткань
может выполнять в данных условиях свои специфические функции. По-
этому неудивительно, что в условиях целого организма явления вторичной
дифференцировки во многих случаях выражаются полнее и отчетливее,
чем в тканевых культурах. При опытах с эмбриональным тканевым мате-
риалом явления дедифференцировки выражаются в меньшей степени или
отсутствуют вовсе. Продолжающаяся митотическая активность клеток
эмбриональных тканей, которая сопровождается более или менее выражен-
ными изменениями их взаиморасположения и способа связи, их активным
движением, может завершаться нормальными или искаженными процессами
гистологической и цитологической дифференцировки. В эмбриональных
закладках органов при экспериментальных условиях наблюдаются не
только явления клеточного и тканевого роста и дифференцировки, но не-
редко более или менее сложные регулятивные и формообразовательные
процессы органогенеза.
В тех случаях, когда одна вполне дифференцированная ткань в резуль-
тате пролиферативных изменений приобретает особенности другой ткани,
в морфологическом и функциональном отношениях вполне дифференциро-
ванной, в патологии принято обозначать конечный результат такого из-
менения дифференцировки, как метаплазия (Вейгерт) или непрямая мета-
плазия. Допускавшуюся старыми патологами т. наз. прямую метаплазию
(Вирхов), т. е. непосредственный переход одной дифференцированной ткани
в другую без явлений роста и пролиферации, в настоящее время, повиди-
мому, следует считать понятием, имеющим лишь историческое значение,
но не соответствующим истинному течению гистологических процессов.
Те немногие факты из области патологии, которые еще до настоящего времени,
в значительной степени в силу традиции, трактуются как примеры прямой
метаплазии, при более тщательном экспериментальном анализе окажутся
относящимися к другой категории явлений.
Гистологические структуры, которые проходят ткани при их возвраще-
нии к исходному состоянию после максимальной степени упрощения и ко-
торые в отношении некоторых особенностей, хотя и не полностью, соответ-
ствуют другим тканевым дифференцировкам, Гаршин (1938) предложил
обозначать термином этранзиторные изменения > или > ателеплазия > (недодиф-
ференцировка), чтобы отличать их от явлений истинной стойкой метаплазии,
имеющих другое патологическое значение. Как показывают многие данные
из области патологии, истинная метаплазия нередко предшествует образо-
ванию злокачественных опухолей.
92
Глава 111
Относительно границ возможных тканевых превращений и, в частности,
процессов метаплазии при патологических условиях до настоящего вре-
мени имеются существенные разногласия, во многих случаях принципиаль-
ного характера, разрешение которых зависит от результатов эксперимен-
тально-гистологического анализа тканей.
Морфологическое упрощение мобилизующихся тканей, переходящих
в состояние усиленного роста и пролиферации, может зависеть от причин
двоякого рода: в одних случаях дедифферевцировка происходит вследствие
ускоренной гибели клеток, высокий уровень специальной дифференцировки
которых связан с необратимой утратой ими жизнеспособности, и усиленного
размножения малодифференцированных, т. наз. камбиальных * элементов,
сохраняющих при этом свой относительно индифферентный характер.
В других случаях сами дифференцированные элементы тканей начинают
энергично делиться митотическим путем, утрачивают при этом в большей
или меньшей степени цитологические признаки своей специализации и
приобретают вид малодифференцированных, сходных с камбиальными,
клеток.
Элементы некоторых высокодифференцированных тканей во многих слу-
чаях реагируют на повреждение не размножением, а лишь ростом своего
протоплазматического тела, которое также может обнаруживать в отно-
шении некоторых своих морфологических особенностей известные признаки
снижения дифференцировки. Высокоспециализированные элементы нервной
системы позвоночных, хотя и являются очень долговечными клетками тела,
невидимому, полностью утрачивают способность к делению при каких бы
то ни было условиях существования.
В условиях энергичного роста и пролиферации клетки соединительной
ткани позвоночных образуют лишь в относительно незначительном коли-
честве наиболее примитивную форму волокнистых межклеточных веществ —
аргирофильные волокна, а в некоторых условиях и вовсе их не образуют.
По своему внешнему облику и способу взаимосвязи тканевые элементы,
находящиеся в малодифференцированном состоянии в условиях роста и
пролиферации, могут быть в общих чертах охарактеризованы следующим
образом.
Наиболее простой, т. наз. цитоморфной или цитотипической, формой
существования тканевых элементов являются изолированные, не соеди-
ненные друг с другом и не образующие никаких закономерных тканевых
структур клетки различной изменчивой формы, зависящей от внешних
условий. Если изолированные клетки свободно взвешены в жидкости (на-
пример в воспалительных эксудатах или в жидкой фазе питательной среды
тканевых культур), то они, как правило, приобретают шарообразный вид,
хотя могут-образовать выступы той или иной формы. Изолированные клетки,
находящиеся на плотном субстрате (в тканях организма, свертке фибрина
* Термин «камбий» был из области ботаники впервые перенесен в гистологию около
середины прошлого столетия Бильротом для обозначения остеогенного слоя надкостницы.
Его применение для обозначения малодиффгренцированных, способных к размножению
клеток различных тканей принадлежит Фишеру — Вазелъсу (1927),
Эк периментальный метод в гистологии
93
и тому подобное), имеют характер амебоидных элементов типа лимфоцитов,
моноцитов или макрофагов соединительной ткани, передвигающихся с по-
мощью псевдоподий, или образуют менее изменчивые, большей частью
заостренные отростки и приобретают веретенчатую, угловатую, лопатовид-
ную, звездчатую и тому подобную форму, напоминающую во многих слу-
чаях т. наз. фибробласты соединительной ткани. Изолированные клетки
этого типа наблюдаются, в основном, в условиях тканевых культур. Форма
отростчатых неамебоидных элементов может зависеть в значительной сте-
пени от плотности окружающей среды. Располагаясь на поверхности плот-
ного субстрата, они, как правило, сильно
уплощаются и приобретают вид очень тон-
ких протоплазматических пластинок. В не-
которых случаях клеточные отростки мо-
гут приобретать нитевидный характер, до-
стигать очень значительной длины и подчас
ветвиться более или менее сложно. В боль-
шинстве случаев существование тканевых эле-
ментов в виде изолированных клеток — пре-
ходящее и малохарактерное явление. Изо-
лированные клетки большей частью, изменяя
свой внешний облик, вновь входят в состав
тех или иных тканевых структур или с тече-
нием времени погибают. Сходные по своему
внешнему облику изолированные клетки мо-
гут образоваться при экспериментальных
условиях из различных тканей животного
организма.
В подавляющем большинстве случаев рас-
тущие и пролиферирующие тканевые элементы оказываются связанными
тем или иным образом в клеточные соединения то более или менее посто-
янной, то, наоборот, изменчивой тканевой структуры. Такую форму су-
ществования клеток можно обозначить как тканевую, гистоморфную или
гистотипическую.
Следует подчеркнуть, что отдельные, а подчас и многочисленные эле-
менты гистоморфных клеточных соединений, утрачивая взаимную связь,
могут переходить в описанное выше цитоморфное состояние. Первоначально,
в соответствии с общепринятой в гистологии номенклатурой, различались
два основных типа клеточных соединений: мезенхимный или соединительно-
тканный (рис. 1 и 2) и эпителиальный (рис. 1, 3 и 4). К первому типу отно-
сились все клеточные соединения с преобладающим сетевидным, более или
менее рыхлым, взаиморасположением элементов отростчатой или веретен-
чатой «фибробластоподобной» формы; ко второму — гистоморфные струк-
туры с преобладающим комплексным тесным расположением клеток угло-
ватой формы, образующих сплошные стелящиеся по поверхности соедини-
тельной ткани или какого-нибудь другого плотного субстрата тонкие .мем-
браны и более толстые пласты, а в условиях «погружного роста» в толще
Рис. /. Комплексная эпители-
альная мембрана (внизу) и рых-
лые соединительнотканные кле-
точные соединения (наверху).
4-дневная культура придатка
яичка молодого кролика. Уве-
личение лупы. (По Хлопину,
1933).
Глава III
плотного субстрата — массивные клеточные скопления, тяжи или полые
внутри трубчатые и мешковидные образования. Более точный анализ раз-
личных гистоморфных структур, производившийся мной и моими сотруд-
никами на протяжении последних двадцати лет методом эксплантации,
показал, что разные ткани позвоночных даже в малодифференцированном
состоянии стойко сохраняют определенные качественные различия. Ока-
Рис. 2-. Участок мезенхимной зоны роста из
З-лневной культуры кожи 2-месячного человече-
ского зародыша. Dbl — узкие темные, Ln — рас-
пластанные клетки типа фибробластов. Сильное
увеличение. (По Хлопину, 1931).
залось, что не все мезенхимо-
подобные структуры, с одной
стороны, и эпителиоподобные.
с другой, тождественны друг
другу и что в отношении це-
лого ряда весьма характер-
ных типов клеточных соеди-
нений эти принятые обозна-
чения очень мало приемлемы,
так как не отражают их спе-
цифических особенностей.
Для точного анализа специфи-
ческих особенностей различ-
ных тканевых структур не-
обходимо не ограничиваться
их изучением в данный мо-
мент жизни и в одном каком-
нибудь участке, а учитывать
всю динамику их превраще-
ний, т. е. изучать непрерыв-
ную связь друг с другом и
взаимные переходы в про-
странстве всевозможных раз-
новидностей клеточных сое-
динений одного и того же
происхождения и их последо-
вательную смену во вре-
мени.
В экспериментальных условиях и, прежде всего, в тканевых культурах
элементы различных тканей могут давать, наряду с типичными формами
разрастаний, и кажущиеся малоспецифическими конвергентные структуры,
обнаруживающие нередко большое внешнее сходство. Это относится, в пер-
вую очередь, к рыхлым сетевидным клеточным соединениями изолированным
клеткам разного происхождения, приобретающим веретенчатую и звездча-
тую фибробластоподобную и закругленную или амебоидную макрофагоно-
добную формы. Во всех таких случаях, однако, клеточные элементы разных
шаней не превращаются в соединительнотканные фибробласты или макро-
фаги, а лишь чисто внешне морфологически уподобляются им. Несмотря на
очень сходную или даже тождественную форму существования, клетки
разного происхождения остаются по существу различными и могут, если
Экспериментальный метод в гистологии
95
они сохраняют свою жизнеспособность и не обнаруживают необратимых
дегенеративных изменений, в дальнейшем вновь проявить свою тканевую
специфичность, образуя типичные для этой или иной ткани клеточные со-
Рис. 3. Эпителиальная мембрана
из 8-дневной культуры мякотного
вещества почки молодого кроли-
ка. Слабое увеличение. (По Цымба-
лу, 1935).
единения.
Для решения этих вопросов, имеющих фундаментальное значение,
метод эксплантации открыл чрезвычайно большие возможности, едва ли
доступные для какой-либо другой экспериментальной методики. Приме-
ненная для той же цели в последнее время моим сотрудником Галустяном
совместно с Быстровым (сотрудником киноотдела Ленфилиала РИЭМ)
цейтраферная микрокиносъемка живых
культур не только полностью подтвер-
ждает наши прежние наблюдения и выво-
ды, но позволяет их значительно углу-
бить и расширить. Многие специфические
различия клеток разного происхождения,
для обнаружения которых путем визу-
ального наблюдения живых культур и
приготовленных из них препаратов необ-
ходим большой опыт, при изучении на
киноэкране выступают с настолько пора-
зительной отчетливостью и яркостью, что
могут быть легко уловлены даже малоис-
кушенным глазом. Если можно так вы-
разиться, клеточные элементы разного
происхождения, при изучении их роста и
движения на экране, обладают неодина-
ковыми «манерами» и «походкой».
К образованию тех или иных малодиф-
ференцированпых гистоморфных клеточ-
ных соединений способны почти все ткани
зародышевого и постфетального организма
и элементы различных эмбриональных
закладок.
Совершенно своеобразный вид имеют в состоянии активного роста высоко-
специализированные элементы соматической мускулатуры позвоночных
и нейроны, отличающиеся в дифференцированном состоянии чрезвычайно
сложной организацией, несвойственной тканевым элементам обычного
клеточного характера. Дифференцированные мышечные волокна растут
в виде т. наз. миосимпластов или мышечных почек, т. е. не имею-
щих клеточного строения многоядерных протоплазматических тяжей и
лент различной формы и калибра. Возможно, что в случаях еще большего
упрощения они могут также превращаться в соединения одноядерных
клеток. Это справедливо во всяком случае в отношении малодифференци-
рованных мышечных волокон зародышей некоторых позвоночных и, по всем
данным, соматической мускулатуры многих беспозвоночных. Нейроны об-
разуют заново и регенерируют свои тонкие нитевидные отростки, достигаю--
96
Глава III
щие при этом весьма значительной длины, могущие ветвиться и образующие
подчас сложнейшие переплеты.
Способность элементов каждой ткани к образованию при разных усло-
виях существования определенной совокупности преемственно связанных
друг с другом гистоморфных структур различного внешнего вида и уровня
дифференцировки может быть обозначена, как их гистобластические, т. е.
тканеобразующие потенции (Хлопин, 1929). Специфичность тканей, выра-
жающаяся в различной совокупности гистобластических потенций, в их
способности образовать строго определенные в каждом отдельном случае
Рис. 4. Участок эпителиальной мембраны с
митозами, из которых один трехполюсный.
S-дневная культура гардеровской железы ново-
рожденного кролика. Сильное увеличение.
(По Хлопииу, 1934).
дифференцированные структуры
и неспособности к превращению
в ткани иного происхождения,
была обозначена мной термином
«гистологическая» или «ткане-
вая детерминация» (1934). Оче-
видно, что совокупность гисто-
бластических потенций и уро-
вень гистологической детерми-
нации являются тканевыми свой-
ствами (и тем самым свойства-
ми целого организма), которые,
развиваясь, изменяются в фило-
генезе и наследственно закреп-
ляются на каждом этапе эволю-
ции. Нормальная структура ка-
ждой ткани в составе сформи-
рованного взрослого организма
является лишь одной из форм
ее существования при опреде-
ленной совокупности внешних
условий. Прочие формы суще-
ствования ткани, зависящие от всей совокупности ее гистобластических
потенций и не обнаруживающиеся при обычных условиях, выявляются
под влиянием тех или иных экспериментальных и патологических воз-
действий.
Таким образом, свойства каждой конкретной ткани должны, с одной
стороны, зависеть от ее функционального значения и от коррелятивных
связей с другими тканями, подчиненных, в свою очередь, общим инте-
грирующим факторам организма. С другой стороны, ее свойства являются
результатом всей предыдущей истории развития. Снимая экспериментально
в большей или меньшей степени первую группу факторов, мы тем самым
можем дать проявиться этому историческому фону, поскольку он сохра-
нился на данном этапе эволюции. В какой степени это возможно в каждом
отдельном случае, будет видно при изложении экспериментальных данных.
Следует ожидать, что ткани, развившиеся конвергентно из разных источ-
ников и приобрезшие сходную морфо-физиологическую дифференцировку,
Экспериментальный метод в гистологии 97
будут отличаться друг от друга в отношении ряда гистобластических потен-
ций, а сходные при обычных условиях и одновременно родственные друг
другу по происхождению гистологические структуры и в отношении дру-
гих гистобластических потенций будут обнаруживать черты сходства.
Если, наоборот, несколько различных по своим морфо-физиологическим
особенностям тканей возникли из одного и того же источника путем дивер-
гентной дифференцировки, то можно предполагать, что они обнаружат
известные черты сходства в отношении своих гистобластических потенций.
Как показывает достаточно большой, имеющийся в настоящее время
в нашем распоряжении фактический материал, все эти предположения
полностью оправдываются в подавляющем большинстве случаев. Лишь
относительно редко позднейшие эволюционные изменения накладывают
на свойства тканей настолько глубокий отпечаток, что он в значительной
степени препятствует выявлению генетического родства тех или иных гис-
тологических структур.
Очень большое значение имеет, само собой разумеется, применение
экспериментально-гистологического метода к тканям различных предста-
вителей животного мира, т. е. его использование в сравнительном аспекте.
Не отрицая большой ценности результатов сравнительного изучения тканей
животных, принадлежащих к отдаленным систематическим группам, подчерк-
нем, что особенное значение, с точки зрения эволюции тканей, принадлежит
(как это отмечалось уже Сееерцовым) гистологическому анализу форм,
связанных друг с другом более или менее близким и прямым родством.
В первую очередь это относится к тканям, пути филогистогенеза которых
неясны и спорны, а поэтому нуждаются в особенно тщательном эксперимен-
тальном анализе.
Гистологические выводы, не подкрепленные сравнительным изучением
гомологичных друг другу тканей родственных форм, могут в ряде случаев
оказаться ошибочными или, в лучшем случае, мало обоснованными. В этом
отношении следует еще раз напомнить высказанное Эбнером (1911) поло-
жение, что попытки установить филогенез тканей, независимо от типа орга-
низации животных, должны неизбежно привести к неразрешимым противо-
речиям и смешению конвергенции, функциональных субституций и про-
странственных переходов одной ткани в другую с истинными процессами
развития.
Последнее, что нуждается в предварительном обсуждении, это вопрос
о том, в какой степени все сказанное применимо в отношении опухолевых
тканей, отличающихся в составе организма автономией роста и могущих
возникать из дифференцированных нормальных тканей или их зачатков на
разных стадиях онтогенеза. По своему гистологическому строению опухо-
левые ткани, как известно, или соответствуют в общих чертах нормальным
дифференцированным тканям организма, отличаясь от них во многих
случаях известной атипичностыо и нередко уродливостью своих клеточных
элементов, или, наоборот, характеризуются неполноценностью своей гисто-
логической дифференцировки и, в определенных случаях, практически ее
полным отсутствием. Существовавшее издавна и сохраняющееся до настоя-
7 Хлопин—349
98
Глава III
щего времени подразделение злокачественных опухолей на саркомы и кар-
циномы, или раки, целиком основывается на формальных признаках.
Если бластоматозные элементы образуют эпителиоподобныс клеточные
соединения, то опухоль относится к карциномам; если эпителиальных
бластоматозных структур в составе опухоли не имеется, то она является
саркомой. Последняя, однако, теперь уже не является синонимом опухоли
соединительнотканного характера, как это иногда неправильно понимается.
Во многих случаях’, особенно если бластоматозная ткань отличается низким
уровнем дифференцировки и не обнаруживает сколько-нибудь типичных
гистологических структур, установление гистогенеза опухоли, на основании
изучения секционного или биоптического материала наталкивается на
непреодолимые трудности. Дело осложняется еще тем, что источником
опухоли могут быть не только нормальные тканевые компоненты данного
органа, но и дистонические клеточные комплексы, оказавшиеся в данном
месте в результате тех или иных неправильностей в ходе онтогенеза. Гисто-
логические особенности опухолей должны быть, очевидно, результатом
взаимодействия свойств исходного . нормального тканевого материала и
приобретенного ими нового опухолевого качества, выражающегося в авто-
номности роста и размножения и в т. наз. опухолевой катаплазии (Бенеке,
Фишер-Вазелъс, 1927), т. е. в определенной степени неполноценности гисто-
логической дифференцировки бластоматозных элементов. Очевидно, у нас
нет ровно никаких оснований ожидать, что элементы опухолевых тканей
приобретают новые гистобластические потенции, т. е. способность к обра-
зованию таких дифференцированных структур, которые не были свой-
ственны материнской нормальной ткани или эмбриональной закладке при
обычных или экспериментально измененных условиях существования.
С нашей точки зрения, хорошо согласующейся с приводимыми ниже фак-
тами, каждая опухолевая структура в отношении своих гистологических
особенностей должна быть сравнима с той или иной формой существования
соответствующей нормальной ткани, т. е. или с ее нормальной структурой
или с теми структурами, которые ей свойственны при неопухолевых проли-
феративных процессах (Хлопин, 1933, 1938).
Наконец, у нас есть все основания считать, что сама опухолевая ткань
при экспериментальных условиях будет обнаруживать такие гистобласти-
ческие потенции, которые помогут выяснить ее истинную гистологическую
природу, если последняя не может быть установлена на обычном патолого-
гистологическом материале. С другой стороны, через изучение опухолевых
тканей как определенных гистологических структур, измененных особого
рода пролиферативными процессами, мы сможем получить факты, важные
для понимания тех или иных свойств соответствующих нормальных тканей
организма. Установление гистогенеза опухолей имеет особенное значение
для их рациональной классификации, поскольку морфо-физиологический
принцип здесь приложим в еще меньшей степени, чем к нормальным неопу-
холевым компонентам организма, так как о функциональном значении
бластоматозной ткани вообще нельзя говорить в том же смысле, как это
говорится в отношении нормальных гистологических структур. Примене-
Экспериментальный метод в гистологии 99
ние же в качестве руководящего принципа классификации только одного
структурного момента неизбежно приводит к грубому формализму.
При изложении фактического материала мы примем обычное подразделе-
ние тканевых структур, в основе которого лежит морфо-физиологическая ха-
рактеристика составляющих их элементов. Изолированные клетки крови,
лимфы, гемолимфы и полостной жидкости мы рассмотрим, как это принято
в настоящее время, в составе группы соединительнотканных образований.
Не образуя сами по себе определенных тканевых структур, свободные
клетки, находящиеся в жидкостях тела или блуждающие в тех или иных
тканях представляют собой части кроветворных тканевых систем, с кото-
рыми они неразрывно связаны в течение всей жизни.
Отдельные конкретные ткани, входящие в состав четырех групп, мы рас-
смотрим, поскольку это возможно в настоящее время, с точки зрения срав-
нительной анатомии, эмбриологии и экспериментальной гистологии, чтобы
восстановить таким образом вероятную историю их филогенетического раз-
вития. Исходя из этого, мы сначала подразделим каждую из четырех гисто-
логических групп на большее или меньшее количество тканевых типов,
объединенных общностью происхождения, а в конце концов попытаемся
дать единую естественную систему тканей на основе генетического, а не
общепринятого морфо-физиологического принципа, распространив его
вместе с тем на опухолевые структуры.
При подразделении тканей на четыре морфо-физиологические группы,
мы встречаемся с рядом трудностей, так как ряд гистологических
структур, в отношении некоторых, а подчас и многих своих особенностей,
не укладывается и не может укладываться в обычную гистологическую
систему, что является вполне понятным, если принять во внимание эволю-
ционные изменения тканевых свойств. Отнесение многих гистологических
структур в ту или иную категорию оказывается поэтому в достаточной
степени произвольным, так как руководящим моментом для этого в таких
случаях часто являются в первую очередь дидактические соображения.
Кроме того, некоторые тканевые структуры вообще принято рассматри-
вать не в общей гистологии, а только в микроскопической анатомии, в ко-
торой общегистологические принципы классификаций тканей проводятся
далеко не всегда.
Если принять для группы эпителиальных структур в качестве общих
основных морфо-физиологических признаков пограничное значение, их
состав из тесно сомкнутых полярно дифференцированных элементов и отсут-
ствие разделяющего их межклеточного вещества, то под эту характеристику
подойдут всевозможные покровные эпителиальные пласты, железистые
ходы и фолликулы, нефроны и собирательные трубки выделительных орга-
нов, отчасти фолликулярные и сертолиевы клетки гонад, эпендима, покровные
клетки сосудистых сплетений мозга и пигментный слой сетчатки, мезотелий,
эндотелий сосудистой системы, покровные клетки мозговых оболочек и
нервных стволиков.
Пожалуй, с таким же правом, как некоторые из перечисленных выше
разнообразных тканей, к эпителиальным структурам следовало бы отнести
100
Глава III
также сателлиты периферических нейронов и даже шванновские
клетки.
Сателлиты по своему общему характеру могут обнаруживать известные
черты сходства с фолликулярным эпителием яичника, а шванновские клетки,
отграничивающие аксоны от соединительной ткани, в известных отношениях
сравнимы с эндотелием.
Разумеется, эти сопоставления представляются в достаточной степени
натянутыми, так как по существу эти структуры не укладываются, как и
многие другие, в рамки четырех принятых групп тканей. Некоторое сходство
вспомогательных компонентов нервной системы с эндотелием подчеркнул
в свое время Лейдиг (1885). Вместе с тем следует отметить, что эндотелий
в этой позднейшей монографии Лейдига уже трактуется, в соответствии
с взглядами Гиса, как уплощенные соединительнотканные клетки и поэтому
не включается в состав эпителиальной ткани. С другой стороны, тканевой
материал таких органов, как аденогипофиз, зобная и околощитовидные же-
лезы, островки Лангерганса, интерренальные органы, желтое тело и в неко-
торых отношениях даже печень высших позвоночных, при всей их несо-
мненной связи с эпителиальными образованиями, никак не может быть вклю-
чен, на основе своих морфо-физиологических особенностей, в группу эпи-
телиальных тканей. То же самое справедливо и в отношении сетевидной
пульпы эмали зубного зачатка. Эти гистологические структуры, быть может
за исключением печеночных балок, не имеют пограничного значения, а по-
лярность у их элементов выражена неясно или совершенно не обнаружи-
вается. Остающийся же отрицательный признак, отсутствие межклеточного
вещества > едва ли является достаточным для их включения в состав эпи-
телиев. Последнее возможно лишь на основе несомненной генетической
связи эндокриновых и сетевидных структур с эпителиальными пластами.
Для их правильной трактовки неизбежно приходится отказаться от после-
довательного проведения морфо-физиологического принципа. Трактовка
нейроглии центральной нервной системы, как особой разновидности тканей
межклеточного вещества, имеющих опорное и механическое значение,
также не является бесспорной, поскольку глиальным структурам свой-
ственна также барьерная функция, а их волокна скорее сравни-
мы с тонофибриллями, но не с волокнистым межклеточным веществом
соединительной ткани. Учитывая, кроме того, несомненную генетическую
связь астроцитарной глии с эпителиоподобным эпендимным аппаратом
низших хордовых, нейроглию можно рассматривать как своеобразную
разграничительную эпителиальную структуру, отнюдь с неменьшим пра-
вом, чем, например, эпителиальную сеть зобной железы. Рассмотрение
нейроглии в составе той или иной тканевой группы классической гистологии
является достаточно произвольным и может оправдываться теми или иными
дополнительными соображениями.
Если желательно привести в первую очередь сопоставление трофиче-
ских и механических структур организма, то рассмотрение нейроглии
в составе соединительнотканной группы представляется вполне целесо-
образным. Если же поставить себе иные цели, например, изучение изме-
Экспериментальный метод в гистологии
101
нения свойств эпителиальных структур в связи с изменением их положения
в составе организма и функционального значения, то имеются все основа-
ния для изучения нейроглии в составе группы эпителиальных тканей,
с которыми она также обнаруживает некоторые черты сходства. Для на-
ших целей будет правильнее рассмотреть нейроглию, вместе с эпендимными
образованиями, в составе эпителиальной группы тканей. Это оправдается
также, как увидим ниже, результатами экспериментально-гистологиче-
ского анализа, при котором некоторые эпителиоподобные черты нейро-
глии выступают особенно отчетливо.
При рассмотрении соединительнотканных структур мы лишь упомянем
нейроглию постольку, поскольку это необходимо для некоторых сопоста-
влений.
Такое же двойственное положение в морфо-физиологической системе
гистологии, иллюстрирующее ее искусственный характер, свойственно
и ряду других гистологических структур, например — миоэпителиальным
н корзинчатым клеткам, которые, с одной стороны, являются компонен-
тами эпителиальных железистых ходов, а, с другой, своеобразными мы-
шечными клетками, ткани спинной струны, у которой есть особенности
механических структур соединительнотканного характера и вместе с тем
несомненные черты сходства с эпителиями, различным нейроэпителиаль-
ным элементам, хроматофорам и некоторым другим образованиям.
В составе эпителиальной группы мы рассмотрим следующие пять типов
тканей, отличающихся друг от друга у позвоночных своим происхождением
из пяти различных источников: 1) энтеродермальные ткани или эпителии
кишечного типа, развивающиеся из энтодермы; 2) эпидермальные ткани
или эпителии кожного типа, развивающиеся в онтогенезе из эктодермы,
или же, по многим данным, связанные с ней филогенетически; 3) эпендимо-
глиальные ткани, возникающие из нервного зачатка; 4) целонефродер-
мальные ткани, генетически связанные с целомической выстилкой; 5) эндо-
телии сосудистой системы, которые, по господствующим взглядам, отно-
сятся к соединительной ткани и образуются из того же мезенхимного за-
чатка, что и последняя. Эндотелиоподобная выстилка синовиальных поло-
стей, повидимому, представлена уплощенными клетками соединительной
ткани, неразрывно связанными с ней и лишь местами имеющими сомкнутое
эпителиоподобное взаимное расположение.
От принятой мной и моими сотрудниками генетической системы эпи-
телиальных структур (Хлопан, 1934) это подразделение отличается объеди-
нением целодермальных и нефродермальных тканей в единую тканевую
систему и добавлением эпендимо-глиального и эндотелиального типов.
Эти два типа мы до последнего времени не рассматривали в составе эпите-
лиев. Однако при последовательном морфо-физиологическом подходе
к гистологическим структурам исключение из состава эпителиальных
тканей эпендимы с родственными ей образованиями и сосудистого эндо-
телия не оправдано и должно быть исправлено. В генетически сборной
группе эпителиальных структур исторически обусловленные различия
между эпидермальными и энтеродермальными тканями выражены по сути
102
Глава III
дела не менее резко, чем между ними, с одной стороны, и эндотелием или
эпендимой, с другой. Если представители каждого из четырех принятых
нами прежде типов эпителиев качественно различны и неспособны к пре-
вращениям друг в друга, то это делает возможным добавление к эпите-
лиальным структурам эндотелия и эпендимных элементов, как двух
самостоятельных и генетически единых групп. Наоборот, принятое до
недавнего времени резкое противопоставление друг другу всех эпителиев,
взятых вместе, как единой ткани, эндотелию или «десмальному эпителию»
неправильно и неоправданно.
Объединение в одну группу целодермальных и нефродермальных тка-
ней, несмотря на значительные морфологические и функциональные раз-
личия между ними у млекопитающих, оправдывается их генетическим
единством и сравнительным экспериментальным анализом целомической
выстилки в ряду позвоночных, подробно разобранным ниже.
Аналогично мы подразделяем группу мышечных тканей на пять типов,
в соответствии с их развитием из того или иного источника (Хлопан, 1936):
1) мускулатура соматического или анимального типа, развивающаяся у по-
звоночных из миотомов, которая представляет собой одну из древнейших тка-
невых систем организма; 2) целомическая или целодермальная мускулатура,
представленная у позвоночных тканью миокарда и развивающаяся из целоми-
ческой выстилки; 3) миодесмальная ткань или гладкая мускулатура ме-
зенхимного происхождения, невидимому, имеющаяся только у позвоноч-
ных; 4) мионейральная ткань, представленная сократимыми элементами
радужины глаза; 5) миоэпидермальные клетки некоторых желез.
Так наз. нервная ткань, т. е. совокупность нейронов живот-
ного организма, представляет собой у всех или почти у всех животных
единый в генетическом отношении очень древний тканевой тип. Так наз.
нейроэпителиальные элементы, которые возможно рассматривать как
нервные или эпителиальные клетки, мы включим в состав эпендимо-гли-
ального типа эпителиальной группы тканей, используя затем полученные
данные для их сравнения с нейронами.
В составе группы тканей соединительного вещества мы рассмотрим, по-
скольку это возможно, мезенхимные или, точнее, «энтомезенхимные» про-
изводные, а у позвоночных и некоторые образования эктомезенхимного
характера. Относительно этих последних, с гистологической точки зрения,
имеется очень много неясностей и спорных моментов.
2. ЭПИТЕЛИАЛЬНЫЕ СТРУКТУРЫ И РОДСТВЕННЫЕ ИМ ОБРАЗОВАНИЯ
С понятием эпителий или эпителиальная ткань в классической гисто-
логии соединяется представление о непрерывных комплексных клеточных
пластах, которые покрывают тело животных снаружи, выстилают пище-
варительную трубку и ряд внутренних полостей организма, сообщающихся
или не сообщающихся с внешней средой, а также о тканевом материале
желез, развивающихся из эпителиальных пластов, и канальцев выдели-
тельных органов.
Экспериментальный метод в гистологии 103
В главе общей гистологии, посвященной эпителию, рассматриваются
только эти образования, к которым приложима общая морфологическая
характеристика данной тканевой группы. Генетический принцип не играет
руководящей роли ни в общей характеристике этой тканевой группы, ни
в ее обычном подразделении на разновидности, хотя и принято отмечать,
что эпителий может развиваться из всех трех зародышевых листков. Ряд
эпителиальных выстилок, которые, согласно принятым в гистологии взгля-
дам, развиваются из мезенхимы и еще со времени Гиса (1865) обозначаются
как эндотелий или десмальный эпителий {Раубер, 1883; Шаффер, 1920,
1927), в настоящее время большей частью рассматриваются как разновид-
ность клеток соединительной ткани и противопоставляются всем другим
эпителиальным структурам. С точки зрения морфо-физиологического под-
хода к тканям, это непоследовательно. Таким же образом многими авто-
рами трактуется и мезотелий серозных оболочек.
На неопределенность понятий эпителий и эпителиальная клетка уже
давно и неоднократно указывалось {Раубер, 1883; Ганземан, 1896; Мар-
шанд, 1899; Фишер-Вазельс, 1927, и др.). Тем не менее принятая до послед-
него времени трактовка эпителиальных структур и их подразделение на
разновидности остаются по существу такими же, какими они были более
ста лет тому назад,, когда Генле (1838) дал впервые описание клеточного
строения самых разнообразных покровных образований. Сводка Меркеля
(1908) не внесла по существу ничего нового в укоренившиеся формальные
представления об эпителиях. Из введенных им новых обозначений было
принято разделение эпителиев на изоморфные и анизоморфные, а номен-
клатура, основанная на общей толщине пласта (лептодермальный, метрио-
дермальный и батидермальный эпителии), не привилась. Новейшая моно-
графическая статья Шаффера (1927) исходит из тех же формальных уста-
новок. Десмальный эпителий резко противопоставляется всем прочим эпи-
телиальным структурам.
Чтобы подчеркнуть преимущественно морфологическое значение поня-
тия эпителий, Маршанд (1899) предложил термин «эпитема» (Epithem,
от греческого emBujua—лежащее на чем-нибудь), который применим да-
леко не ко всем образованиям, обозначаемым как эпителиальные, но от-
личается той же неопределенностью и расплывчатостью.
Попытка Геккеля (1885) подойти к тканям с последовательных эволю-
ционных позиций не только не уменьшила неопределенности понятия
эпителий, но даже усугубила ее. Морфо-физиологическому или даже в пер-
вую очередь морфологическому понятию эпителий был придан несвой-
ственный ему генетический смысл единой изначальной ткани, из которой
в филогенезе возникают все прочие гистологические структуры. Тем са-
мым в состав эпителия были включены все эмбриональные зачатки, имею-
щие комплексную эпителиальную внешность и такие мультифункциональ-
ные образования с разнообразной морфо-физиологической характеристи-
кой, как энтодерма и эктодерма кишечнополостных, которые вовсе не равно-
ценны дифференцированным эпителиальным тканям высокоорганизован-
ных животных. Одним из результатов такой расширенной трактовки группы
104
Глава III
эпителиальных структур и отсутствия дифференцированного подхода к ним
была попытка теоретического обоснования их предполагаемых, но не об-
основанных, необычайно широких возможностей к превращениям и явле-
ниям метаплазии в другие ткани. При этом нередко совершенно игнориро-
вались качественные изменения свойств эпителиальных структур в тече-
ние филогенетического и онтогенетического развития. Тем самым эволю-
ционные идеи Геккеля приводили и приводят еще в настоящее время к взгля-
дам, которые не имеют ничего общего с эволюционным учением, хотя и
именуются «трансформистскими», поскольку они полностью или почти
полностью отрицают специфичность тканей.
Хотя большая часть гистологов и гистопатологов в настоящее время
не идет столь далеко в допущении межтканевых метаплазий, возможность
взаимных переходов и превращений в пределах группы эпителиальных
тканей многими допускается в очень широкой степени. В частности, почти
ни у кого не вызывает сомнений способность едва ли не любой эпителиаль-
ной ткани млекопитающих и человека превращаться в многослойный
эпителий кожного типа, хотя такая возможность, с точки зрения эволю-
ционной гистологии, в большей части случаев столь же невероятна, как
и допущение широких межтканевых метаплазий.
Резко выраженный формализм и расплывчатость принятой трактовки
эпителия заставили меня, опираясь на результаты экспериментально-
гистологического анализа, искать других путей подхода сначала к этим,
а затем и к другим тканевым структурам. Разбираемый ниже эксперимен-
тальный материал показал с полной убедительностью, что т. наз. эпителий
не является единой тканью или единой системой тканей пограничного
значения, а очень обширной и не однородной по своему составу тканевой
группой сборного характера. Принятое подразделение этой тканевой груп-
пы на основании формы клеток или какого-нибудь одного частного функ-
ционального признака, как это иногда делается, не вносит необходимой
ясности и определенности и отличается большой искусственностью.
Наиболее естественной и отвечающей действительному положению вещей
оказалась филогенетическая трактовка эпителиальных, а также и других
тканевых структур и их последовательное подразделение на основе всей
истории развития. Генетический принцип в качестве руководящего мо-
мента не только позволил произвести подразделение эпителиальных струк-
тур на отдельные типы, но дал также возможность найти место в есте-
ственной гистологической системе для многих тканей, которые на основе
приобретенных ими в эволюционном процессе морфо-физиологических
признаков, по сути дела, без больших натяжек, не могут быть вклю-
чены ни в одну из четырех групп тканей классической гистоло-
гии.
Первым результатом работ нашего научного коллектива в этом напра-
влении явилась генетическая система эпителиальных тканей, намеченная
мной в 1932 г., более полно сформулированная в 1934 г. и подробно разби-
раемая в настоящей книге.
Экспериментальный метод в гистологии
105
А. Эпидермальный тканевой тип
а) Краткие эмбриологические и сравнительные данные
Эпидермальные ткани и их сравнение с энтеродермальными требуют
особенно тщательного анализа, так как важнейшие гистологические осо-
бенности многих из них совершенно не вяжутся с господствующими эм-
бриологическими представлениями, в соответствии с которыми гистоло-
гические данные следовало бы трактовать как результат конвергентной
дифференцировки, а не признак генетической близости.
Эпителиальная часть общего кожного покрова у всех существующих
организмов несомненно развилась из одного источника — эктодермы об-
щих предков, независимо оттого, как себе представлять эти гипотетические
родоначальные формы. Кроме того, у большей части кишечнополостных
и у всех планарий более или менее значительная глоточная часть пище-
варительной трубки имеет также эктодермальное происхождение. У всех
прочих первичноротых имеются хорошо развитые эктодермальные стомо-
деум и проктодеум, которые и в отношении гистологических особенностей,
как правило, обнаруживают сходство с кожным эпителием. В частности,
у членистоногих они, как известно, хитинизированы и у некоторых форм
Гнапример высшие ракообразные) могут составлять большую по своей
длине часть пищеварительной трубки. У первичноротых, кроме того, гене-
тически связаны с эктодермой органы дыхания (жабры, трахеи, легкие), а
”акже выделительные органы типа эктодермальных нефридиев и соответ-
ствующих им частей нефромиксий {Гудрич, 1897—1900).
У большей части вторичноротых дело обстоит не так. За исключением
щетинкочелюстных и иглокожих, у всех прочих форм если эктодермаль-
ные участки пищеварительной трубки и обнаруживаются в эмбриогенезе,
то лишь как очень короткие отрезки, которые по своим гистологическим
особенностям, вдобавок, могут совершенно не отличаться от т. наз. экто-
дермальных частей, примыкающих к ним. Особенно резко это бросается
в глаза у позвоночных, у которых эпителии кишечника и кожи достаточно
или даже чрезвычайно резко отличаются друг от друга в отношении гисто-
логических особенностей. Для примера достаточно указать на млекопи-
тающих, характеризующихся многослойным плоским и большей частью
ороговевающим кожным эпителием и однослойным кишечным эпителием
резко отличного от первого строения. Это не могло уже давно не навести
на мысль, что граница между однослойным и многослойным или много-
рядным эпителием в пищеварительной трубке должна совпадать с грани-
цей ее эктодермальной и энтодермальной частей. Параллельно с накоп-
лением эмбриологических фактов предполагаемые границы между экто-
и энтодермой стали сдвигаться в пользу второй и перестали совпадать
с гистологическими особенностями эпителиальных покровов. К области
энтодермы отошли весь пищевод, глотка и сначала относительно неболь-
шая, а затем большая часть выстилки полости рта, за исключением пред-
дверия, соответствующего ротовой впадине зародыша. В исторической
вводной части книги отмечалось, что два очень крупных современника,
106
Глава- HI
Геккель и Раубер, еще сильно расходились в определении границ между
энто- и эктодермой в полости рта. Чтобы уничтожить бросающуюся в глаза
неувязку между эмбриологией и гистологией, Грейль уже давно высказал
весьма правдоподобное предположение, что замещение^первоначально энто-
дермальной части выстилки переднего отдела пищеварительной трубки кож-
ной эктодермой происходит вторично в результате ее врастания через рото-
вое отверстие и жаберные щели и постепенного вытеснения энтодермы в от-
делы пищеварительной трубки, расположенные более каудально. Эта
гипотеза эволюционного смещения границ между экто- и энтодермой или
меторизиса, отражающегося и в онтогенезе, была очень остроумно и ши-
роко использована и развита Шимкевичем (1908, 1913, 1923). Однако не-
которые открытия, сделанные в последнее время методами эксперимен-
тальной эмбриологии, в частности, известные опыты Голътфретера (1933)
с экзогаструляцией у амфибий (см. ниже) показали, что выстилка большей
передней части пищеварительной трубки образуется из зачатка головной
кишки, который инвагинирует при гаструляции и тем самым должен быть
признан за энтодерму. Таково положение вопроса о выстилке переднего
отдела пищеварительной трубки в настоящее время с эмбриологической
точки зрения, совершенно не считающейся с гистологическими фактами.
Гистологами эта точка зрения до последнего времени полностью разде-
лялась. Принципиально сходным образом обстоит дело и с выстилкой
клоаки, где границы между эктодермой и энтодермой, с одной стороны,
и между эпителиями разного строения, с другой, также не совпадают.
Если безоговорочно поддерживать точку зрения, что морфологические
особенности дифференцированных тканей не зависят от способа их раз-
вития, что едва ли не любая ткань может развиваться из любого эмбрио-
нального зачатка и что все эпителии, несмотря на свои морфологические
и гистологические различия являются, по сути дела, единой тканью, в пре-
делах которой возможны любые изменения структуры и явления метапла-
зии, то несовпадению между эмбриологическими и гистологическими фак-
тами нельзя было бы придавать принципиального значения. Тогда един-
ственным более или менее достоверным критерием происхождения ткани
остается ее развитие в онтогенезе. Но если только что упомянутые обще-
принятые положения классической гистологии перестают соответствовать
экспериментально доказуемым гистологическим фактам и, как было пока-
зано выше, представляют собой по сути дела отказ от филогенетического
понимания свойств гистологических структур, то они естественно нуж-
даются в критическом пересмотре.
Разбираемый ниже экспериментальный материал, полученный перво-
начально мной, а затем всем нашим научным коллективом, не мог не за-
ставить меня (Хлопин, 1929, 1932, 1933, 1934) критически просмотреть
известные в то время другие данные и на этой основе притти к выводу, что
гистологическим фактам может и должно придаваться в очень многих слу-
чаях решающее значение для понимания эволюционного развития тканей
и чем самым для правильной трактовки ряда важнейших эмбриологиче-
ских фактов и связанных с ними общих вопросов. С тех пор положение еще
Экспериментальный метод в гистологии 107
изменилось в пользу защищаемой нами точки зрения, благодаря накоп-
лению нового фактического материала, полученного не только в руково-
димых мной, но и в других лабораториях.
Исходя из излагаемых ниже экспериментальных данных -И гистологи-
ческих особенностей соответствующих нормальных эпителиев, мы отно-
сим у позвоночных к единому типу эпидермальных тканей, кроме соб-
ственно кожного эпителия с его производными, также выстилку переднего
отдела пищеварительной трубки, многорядную у некоторых низших по-
звоночных или многослойную у высших, эпителии всех развивающихся
из нее желез и эндокриновых органов, дыхательного аппарата и среднего
уха. Здесь уместно подчеркнуть, что под многорядными эпителиями мы
понимаем только такие структуры, у которых, как и в многослойном эпи-
телии, не все клетки пласта участвуют в образовании его свободной апи-
кальной поверхности, а наряду с длинными клетками имеются также корот-
кие базальные или промежуточные элементы. Кроме того, я умышленно
избегаю обычного обозначения «однослойный многорядный эпителий»,
которое предполагает известное противопоставление многослойного эпителия
многорядному и сближение последнего с однослойным эпителием кишеч-
ного типа, что, с моей точки зрения, по существу неправильно. «Обратно-
многорядный эпителий» органов слухового аппарата будет рассмотрен
среди вспомогательных компонентов нервной системы, как совершенно
своеобразная структура иного, чем обычный многорядный эпителий, зна-
чения.
Таким же образом дело обстоит с областью клоаки и ее производных,
с семевыносящими и мочеотводящими путями, в выстилке которых экспе-
риментальный анализ также позволяет обнаружить существенные черты
сходства с кожным эпителием, но способ эмбрионального развития говорит
против их генетического единства. У всех позвоночных, имеющих во взрос-
лом состоянии клоаку, в ней можно, по имеющимся данным, различать
два отдела. В одном из них эпителиальная выстилка однослойная, сход-
ная с кишечным эпителием, в другой — двуслойная или многослойная,
приближающаяся по своим особенностям в большей или меньшей степени
к эпидермису, хотя и не тождественная с этим последним. У млекопитаю-
щих же из части клоаки, возможно соответствующей ее второму отделу,
при эмбриональном развитии образуется большая часть эпителия моче-
полового синуса и мочевого пузыря.
Аллантоис, образовавшийся на более ранних стадиях развития из соот-
ветствующего клоаке заднего участка вторичной кишечной трубки, остается
в связи с мочевым пузырем и, может быть, участвует своим протоком в об-
разовании его стенки. Все производные этого заднего участка пищевари-
тельной трубки относятся эмбриологами к энтодерме, но по гистологи-
ческим особенностям переходного эпителия (которые разбираются ниже)
обнаруживают несомненное сходство с эпидермисом кожи и на этом осно-
вании относятся нами к тому же эпидермальному тканевому типу.
Вольфовы каналы, из которых в дальнейшем развиваются мочеотво-
дящие и семевыносящие пути, первоначально образуются, как протоки
108
Глава III
предпочки, из передних нефротомов и, разрастаясь в каудальном напра-
влении, в конце концов открываются в клоаку. Несмотря на ряд замеча-
тельных особенностей, установленных в развитии вольфовых каналов,
они в большей части эмбриологических справочников и руководств трак-
туются как мезодермальные производные. Для млекопитающих было
установлено уже давно (Бонне, 1887; Мартин, 1888; Кейбель, 1888, 1897)
и в последнее время подтверждено в отношении человека, что слепой конец
вольфова канала вступает в связь с эктодермой, за счет присоединяющихся
элементов которой и образуется его каудальная часть. Эта эктодермаль-
ная часть вольфова канала и дорастает затем до клоаки. Таким образом,
у зародышей млекопитающих в составе открывающегося в клоаку, а за-
тем в мочеполовой синус вольфова канала следует различать краниальную
мезодермальную и каудальную эктодермальную части. Вопрос о том, в ка-
кой мере каждая из этих частей при дальнейшем росте тела зародыша уве-
личивается в длину и тем самым принимает участие в образовании семе-
выносящих путей, остается до настоящего времени открытым. Гистологи-
ческие данные говорят о том, что эктодермальной части вольфова канала
в этом отношении принадлежит главное значение. Мезодермальная же
часть его в дальнейшем редуцируется. Во всяком случае, эмбриологиче-
ские данные, касающиеся млекопитающих, определенно говорят за участие
эктодермы в образовании вольфова канала. Мочеточник и лоханка, обра-
зующиеся из самого заднего отдела вольфова канала, несомненно
являются производными его эктодермальной части. Трудно сказать точно,
в какой степени элементы эктодермальной, открывающейся в клоаку части
вольфова канала участвуют в образовании ее выстилки и замещают быв-
ший там раньше эпителий.
По существующим взглядам эпителий вольфова канала занимает об-
ласть так наз. треугольника. Так или иначе, вместе с каудальным от-
делом вольфова канала в состав выстилки клоаки млекопитающих прони-
кают элементы эктодермы.
Еще больше неясностей относительно вольфова канала птиц, рептилий
и низших позвоночных, тем более, что экспериментально-гистологического
материала еще совсем нет. Признававшееся прежде (Рюккерт, 1892; Ван-
Вей, 1888 и др.) участие эктодермы в развитии вольфова канала некоторых
низших позвоночных дальнейшими исследованиями не подтвердилось.
Лишь в самое последнее время (Коннор, 1940) появились новые эмбриоло-
гические данные, согласно которым у амфибий клоака образует парные
краниальные выросты, могущие достигать значительной длины, которые
соединяются с вольфовыми каналами. Возможно, но пока не доказано,
что эти выросты клоаки в дальнейшем играют существенную роль в обра-
зовании эпителия вольфова канала. Вместе с тем эпителий, образующийся
из вольфова канала, имеет, по существующим данным, двурядное строение,
по крайней мере у большей части низших позвоночных. Нигде за преде-
лами класса млекопитающих переходного эпителия в мочеотводящих путях
не встречается. Эта разновидность эпителия является, повидимому, одной
из гистологических особенностей млекопитающих.
Экспериментальный метод в гистологии
109
Гистологическая специфика выстилки производных вольфова канала
у рыб, амфибий, рептилий и птиц еще недостаточно известна, чтобы можно
было делать какие-либо выводы филогенетического порядка.
Напомним также, что филогенетически вольфов канал, с точки зрения
Боеери (1898), может быть выведен из перибранхиальной полости ланцет-
ника, т. е. из несомненно эктодермального образования.
Наконец, следует сказать еще несколько слов относительно многослой-
ного эпителия влагалища многих млекопитающих, который должен быть
отнесен к эпидермальной группе тканей. В эмбриологии принято считать,
что выстилка влагалища, так же, как однослойные эпителии матки и яйце-
водов, развивается из мюллеровых каналов. Эти последние, по крайней
мере у большей части позвоночных, возникают из желобообразного углуб-
ления целомической выстилки. По некоторым новым данным (Кофф, 1932;
Вилас, 1932), эпителий влагалища человека образуется не из мюллеровых
каналов, а из выстилки мочеполового синуса, подрастающей навстречу
им и постепенно вытесняющей мюллеровский эпителий. Уже на этой ран-
ней стадии развития достаточно отчетливо заметна граница между эпите-
лиями мюллеровых каналов и мочеполового синуса.
Очень резкая во взрослом состоянии граница между многослойным
влагалищным и однослойным маточным эпителиями у разных млекопитаю-
щих может располагаться на очень разных уровнях, то опускаясь очень
низко (например у лошади большая часть влагалища выстлана одно-
слойным маточным эпителием), то поднимаясь в область матки. Более того,
у одного и того же вида животных, при патологических условиях и даже
в норме, совершенно резкая граница между многослойным эпителием вла-
галища и однослойным маточным может обнаруживать очень заметные
варианты своего положения. На человеческом материале такие сдвиги
наблюдаются нередко (Р. Майер, 1910).
Все это говорит за то, что развитие многослойного эпителия влагалища
из мюллеровских каналов, в лучшем случае, не может считаться достаточно
обоснованным.
Несомненно, что эволюция морфологических и функциональных осо-
бенностей эктодермального наружного покрова наиболее примитивных
форм и развившейся из него, после обособления большей части других
тканевых зачатков (нейрального и, быть может, мышечного), эпидермаль-
ной ткани протекала в теснейших непосредственных взаимоотношениях
с внешней средой. В зависимости от того, в каком направлении шла эволю-
ция организмов, наружный кожный покров приобрел поразительное раз-
нообразие структур. В своем наиболее примитивном виде он, вероятно,
имел характер однослойного мерцательного эпителия (который мы встре-
чаем, например, у турбеллярий, немертин, местами у аннелид и пр.),
имевшего значение и ткани разграничительной, и ткани движения, и ткани
раздражения. Ряд примитивных черт он мог, хотя бы местами, сохранить
у относительно высокоспециализированных в других отношениях
беспозвоночных, например у кишечнодышащих, брахиопод, некоторых
моллюсков и пр. Мерцательный покров очень характерен также для мно-
по
Глава III
гих зародышевых и личиночных форм, что, вероятно, также имеет изве-
стное филогенетическое значение. В таком примитивном виде механиче-
ская защитная функция у наружного покрова в целом была, вероятно,,
выражена слабо или практически вовсе отсутствовала. Тем не менее от-
дельные элементы его могли дифференцироваться в своеобразные орудия
защиты и нападения, которые, например, встречаются у кишечнополост-
ных в виде разнообразных стрекательных клеток. Очень любопытно, что
даже после обособления основной массы нейрального зачатка из первич-
ной эктодермы, у беспозвоночных в составе эпидермиса или в непосред-
ственной близости к нему во многих случаях остаются периферические
чувствительные невроны. Тесная генетическая связь с эпидермальной
тканью характерна и для части мускулатуры (турбеллярий, аннелиды).
Вместе с развитием выраженной механической защитной функции кож-
ный покров приобретает на своей свободной поверхности кутикулярные
образования разной толщины. Для эпидермальной кутикулы при этом
характерна ее непрерывность: она представляет собой продукт превра-
щения слившихся в синцитиальную пластинку апикальных концов клеток,
которые вследствие этого не имеют замыкающих полосок. Кутикуляризи-
рованный кожный покров встречается у многих первичноротых, например,
аннелид, а также у вторичноротых, например, иглокожих. Защитная его
роль может усиливаться развитием хитиновых придатков, могущих обра-
зовать густой войлок на поверхности тела, который стал типичен для ряда
полихет. Во многих случаях заметна несомненная связь между образом
жизни животных и толщиной его кожной кутикулы. Например, у олиго-
хет, живущих в воде, кутикула очень тонкая, а у роющихся в плотном
грунте — сравнительно мощная. Многие, живущие в грубом песке поли-
хеты (например Ophelia), обладают особенно толстой кутикулой. С дру-
гой стороны, из этого правила имеются и видимые исключения, значение
которых остается пока неясным. Достаточно указать на ланцетника, рою-
щегося в очень грубом песке, который, казалось бы, должен сильно травма-
тизировать животное, не обладающее защитным эпидермисом. Правда,
и у ланцетника некоторые авторы описывают подобие поверхностной кути-
кулы-, однако она, несомненно, очень нежна и едва ли может играть роль
механического защитного образования. Детальное сравнительное изу-
чение эпидермиса в связи с образом жизни животного является безусловно
одной из очень интересных задач эволюционной гистологии.
Эволюция наружного покрова у некоторых беспозвоночных пошла
по очень своеобразному пути развития не только защитной, но вместе с тем
и опорной; скелетной функции, с параллельной выработкой весьма твер-
дых и часто очень сложных структур. Такого рода эктодермальный покров
встречается среди беспозвоночных у членистоногих, многих кишечнопо-
лостных, брахиоиод и большей части моллюсков. У гидроидных апикаль-
ные концы эктодермальных элементов образуют похожий на кутикулу
«псевдохитиновый» покров, т. наз. перисарк или тэку, которая,
по мнению Сент-Илера (1930), представляет собой род структурирован-
ного секрета. У коралловых полипов скелет образуется или путем выде-
Экспериментальный метод в гистологии 111
ления эктодермальными элементами сферокристаллов углекислого каль-
ция (как это предполагают Кох, Кремпф и др.) или (по менее популярным
в настоящее время взглядам Гейдера и Оджильви) путем обизвествления
протоплазмы самих клеток. У моллюсков, членистоногих и, повидимому,
брахиопод, скелет представляет собой развившиеся в определенных на-
правлениях эпидермальные кутикулярные образования, состоящие из
органической (хитиновой или «псевдохитиновой») основы и, во многих
случаях, неорганической пропитки из углекислого кальция. Эти скелет-
ные образования всегда имеют закономерную структуру, часто очень слож-
ную и специфическую для отдельных групп. Интересны превращения кож-
ного покрова, связанные с образованием внутреннего скелета у коралло-
вых полипов и особенно у членистоногих. В этих случаях элементы экто-
дермального происхождения врастают в толщу организма, утрачивают
непосредственную связь с внешней средой, а вместе с тем и разграничи-
тельную функцию, и приобретают исключительно механическое опорное
значение скелетных образований или сухожилий. Последнее имеет место
у членистоногих. Развитие эпидермального скелета у артропод, в частно-
сти у насекомых, коррелятивно связано с целым рядом других особенностей
их строения и потому накладывает чрезвычайно глубокий и характерный
отпечаток на всю организацию этих животных, представляющих собой,
во многих отношениях, одних из наиболее высокоорганизованных пред-
ставителей всего животного мира.
Кроме всего этого, непроницаемому хитиновому панцырю у наземных
форм принадлежит еще одна чрезвычайно важная функция — предохра-
нение животного от испарения и высыхания, что является необходимым
условием существования в сухой атмосфере. Вполне независимыми от влаж-
ной среды остались лишь многие брюхоногие моллюски, обладающие хо-
рошо развитой раковиной, и представители панцырных членистоногих,
богатство которых видами превосходит (насекомые) во много раз количе-
ство видов во всех других типах. Строение панцырной хитиновой эпидер-
мальной ткани достигает большой степени сложности и может обнаружи-
вать подчас значительное сходство со структурами пластинчатой кости,
вплоть до наличия концентрических образований, напоминающих гавер-
совы системы. У панцырного эпидермиса, кроме того, имеется налицо резко
выраженная вертикально-анизоморфная структура, представленная одним
слоем способных к пролиферации базальных камбиальных элементов,
безъядерные апикальные концы которых сливаются в синцитиальную
покровную пластинку, подвергающуюся хитинизации и физиологической
дегенерации.
Старый хитин при очередной линьке периодически сбрасывается и
заменяется новым. Очень своеобразна толстая кожистая кутикула оболоч-
ников, состоящая из туницина и содержащая, повидимому, заползающие
в нее мезенхимные элементы. Совершенно своеобразное эволюционное
изменение претерпевают в ряде случаев эпидермальные ткани при пере-
ходе животных к паразитическому образу жизни, превращаясь часто,
вместе с редукцией пищеварительного аппарата, во всасывающую ткань
112
Глава. Ill
внешнего обмена веществ и сохраняя, вместе с тем, во многих случаях
защитный характер (кишечные паразиты). При этом эпидермис вовсе не
приобретает черт сходства с энтеродермальными тканями, а образует очень
своеобразные кутикуляризированные структуры, т. наз. погруженный
эпителий сосальщиков и цестод, «гиподерму» круглых червей и скребней.
В системе эпидермальных тканей кожного покрова беспозвоночных
особое функциональное значение и ряд морфологических особенностей
имеют элементы, приобревшие разнообразный железистый характер. Же-
лезистые эпидермальные клетки или входят в состав покровного пласта
или превращаются в более или менее объемистые одноклеточные, мало-
клеточные и многоклеточные железы, которые погружаются во внутрен-
нюю среду организма и только своим концом, выделяющим секрет, оста-
ются в связи с эпидермальным покровом. Не касаясь ряда железистых
образований, приобретающих очень высокую физиологическую специа-
лизацию (например привлекающие и душистые железы, имеющие половое
значение), упомянем про широко распространенные слизистые и белковые
элементы. Слизистые клетки и железы имеют, вероятно, в большинстве
случаев, защитное или механическое значение, а в некоторых случаях
играют роль при кожном дыхании. Защитный или механический характер
имеют в большинстве случаев, в первую очередь, различные трубкообра-
зующие (например многие полихеты) или прядильные (некоторые анне-
лиды, моллюски, насекомые и пр.) железы, представляющие собой, вероят-
но, продукты эволюционной специализации слизистых и в функциональ-
ном отношении как бы могущие заменять кутикулярные образования.
Так или иначе железистые образования, там, где они имеются налицо, явля-
ются в морфологическом и функциональном отношениях органической
составной частью эпидермальных тканей.
В совершенно ином и своеобразном направлении шла эволюция эпи-
дермального кожного покрова у щетинкочелюстных и особенно у позво-
ночных. Из всех беспозвоночных только представители щетинкочелюстных
приобрели на некоторых участках тела настоящий многослойный эпидер-
мис, физиологическое значение которого неясно. Принимая во внимание
значительную плотность многослойных (на шее до 5—б слоев клеток) уча-
стков эпидермиса и их морфологическое сходство с т. наз. хор-
доидной тканью у Spadella (Гертвиг, 1880), можно высказать предполо-
жение, что они имеют механическое опорное значение для хорошо развитой
и способной к быстрым сокращениям поперечно-полосатой мускулатуры
этих своеобразных животных, которых можно сближать по некоторым
особенностям с хордовыми.
Эпидермис кожи позвоночных имеет в первую очередь защитное зна-
чение, в то время как механическая опорная роль у них переходит, в про-
тивоположность членистоногим, к соединительнотканным образованиям,
достигающим в количественном и качественном отношениях у этих пред-
ставителей животного мира высшей ступени своего развития. У водных
форм, круглоротых и рыб эпидермис, достигающий значительной толщины
и состоящий из многих слоев клеток, имеет характер слизистой оболочки.
Экспериментальный метод в гистологии 113
Преимущественное защитное значение принадлежит, вероятно, находя-
щимся в его составе слизистым и белковым железистым клеткам. Вместе
с тем покровные элементы могут подвергаться более или менее сильно
выраженной кутикуляризации. Своей высшей дифференцировки эпидер-
мис достигает вместе с переходом к наземному существованию и одновре-
менным развитием рогового слоя. Еще в слабой степени ороговение выра-
жено большей частью у амфибий, эпидермис которых приобретает харак-
тер многослойного плоского эпителия. Слабая степень ороговения их эпи-
дермиса не предохраняет организм значительной части этих животных
от высыхания вследствие испарения тканевых соков. Поэтому такие назем-
ные амфибии могут жить только в достаточно влажной среде. Своего наи-
большего совершенства эпидермис достигает у амниот в связи с приобре-
тением ими полной независимости от степени влажности окружающей
атмосферы и с развитием механической защитной роли. В ороговевающем
эпидермисе вертикальная анизоморфность, осуществляющаяся совершенно
иным образом, чем у членистоногих, бывает выражена наиболее резко,
в связи с отчетливым разграничением на базальные, камбиальные слои
клеток и покровные элементы, подвергающиеся ороговению, физиологи-
ческой дегенерации и слущиванию. Любопытно, что участки эпидермиса,
остающиеся у высших позвоночных во влажной среде, например, конъюнк-
тива глаза, могут сохранять гораздо более примитивный характер и напо-
минать, до известной степени, кожный покров водных форм, сохраняя
даже слизистые бокаловидные клетки, не свойственные другим участкам
их эпидермиса. У млекопитающих, перешедших к водному образу жизни
и успевших претерпеть соответствующие изменения всей организации
(например у китообразных), эпидермис, по крайней мере на большей части
кожи, может в значительной степени вторично утратить ороговение, делаясь
похожим на эпидермальную ткань переднего отдела пищеварительного
аппарата у многих других млекопитающих.
Очень характерными производными эпидермиса кожи у высших позво-
ночных являются разнообразные роговые придатки (волосы, перья, когти,
копыта, рога, ногти).
Вместе с исчезновением отдельных железистых клеток у наземных позво-
ночных, начиная с амфибий, во многих случаях особенного развития
достигают многоклеточные железы, образующиеся путем врастания в со-
единительную ткань отдельных участков эпидермиса. При этом, в связи
с приобретением нового функционального значения и ослаблением связи
с (внешним миром, такие участки эпидермальной ткани вторично изменяют
свой характер: многослойность и явления ороговения исчезают (напри-
мер кожные железы амфибий, клубочковые железы млекопитающих)
или ороговение сменяется голокриновой секрецией жироподобных веществ
(сальные железы). Тем не менее, даже в таком измененном виде эпидермаль-
ная ткань желез сохраняет известные качественные особенности: у клу-
бочковых желез несецернирующие отделы трубки сохраняют двуслой-
ность, секреторные делаются двурядными, причем их базальные элементы
приобретают характер сократимых миоэпителиальных клеток, обнаружи-
8 Хлопни—349
114
Глава III
вающих явления функциональной и морфологической конвергенции с глад-
кими мышцами мезенхимного происхождения. Эпидермальная ткань молоч-
ной, гардеровской и, повидимому, слезных желез в выводных протоках
и главных (секреторных) отделах утрачивает многослойность и делается
большей частью двурядной. Стенка выводных протоков состоит из длинных
призматических и коротких базальных клеток, а главных отделов —из
железистых и корзинчатых, повидимому, имеющих характер миоэпители-
альных.
Повидимому, эпидермальные элементы кожных желез наземных позво-
ночных вырабатывали защитный слизистый или белковый секрет (амфи-
бии) и лишь вторично приобрели различные специальные функции -—
выделительную, теплорегуляторную, связанную с обеспечением потом-
ства и пр. Полное или почти полное отсутствие железистых эпидермаль-
ных участков у рептилий, птиц и некоторых млекопитающих является,
повидимому, вторичным.
Эпидермальная ткань передней и задней кишки у первичноротых со-
храняет, как правило, более или менее резко выраженные качественные
особенности кожного покрова. Особенно отчетливо это выражено, напри-
мер, у членистоногих, многие представители которых обладают очень
длинными эпидермальными участками пищеварительного аппарата с ясно
выраженной, хотя и сравнительно тонкой хитиновой кутикулой и весьма
разнообразными железистыми образованиями.
У позвоночных наиболее примитивное строение имеет, повидимому
(по крайней мере местами), выстилка переднего отдела пищеварительного
аппарата, сохраняющая у бесхвостых амфибий и круглоротых многоряд-
ный, призматический, мерцательный характер. В зародышевом состоянии да-
же у человека сходное строение имеет выстилка пищевода, в котором Шридде
подробно проследил любопытные процессы превращения структур эпи-
дермальной ткани. Вместе с тем именно в области передней кишки могут’
появиться филогенетически очень рано роговые образования (например
в виде роговых зубов круглоротых) в несомненной связи с их специфи-
ческим функциональным значением для добывания пищи. У птиц и млеко-
питающих эпидермальная ткань передней кишки во взрослом состоянии
уже приобретает многослойное строение и плоские покровные клетки.
Очень тесные взаимоотношения между характером пищи и защитной нагруз-
кой, с одной стороны, и степенью ороговения, с другой, можно наблю-
дать у млекопитающих. В некоторых случаях эпидермальная выстилка
передней кишки местами приобретает, вследствие разрыхления, своеоб-
разное сетчатое строение, например, по данным Шаффера (1927), у химеры
и Spinax niger. Такое разрыхление может быть связано с развитием осо-
бых лимфоэпителиальных органов — миндалин.
Так же как и в эпидермисе, в выстилке передней кишки у позвоночных
можно проследить исчезновение в процессе эволюции слизистых и зерни-
стых железистых клеток, вероятно белкового характера, и развитие много-
клеточных слюнных желез, которые у многих млекопитающих делаются
особенно многочисленными и достигают большой степени сложности. Be-
Экспериментальный метод в гистологии
ПЪ
роятно, первично железистые элементы эпидермальной выстилки перед-
ней кишки имели так же, как и в кожном покрове, защитное и механиче-
ское значение, появление же пищеварительных ферментов (амилазы) в слюне
имело место лишь вторично и большого физиологического значения
не приобрело.
Интересно, что эпидермальная ткань слизистых, белковых и смешан-
ных слюнных желез млекопитающих в гистологическом отношении имеет
очень большое сходство с кожными железами. Железистые ходы слюнных
желез, за исключением конечных отделов главных выводных протоков
некоторых больших желез, сохранивших многослойное строение, также
приобрели двурядный характер (Циммерман, 1927); базальные же клетки
концевых, а у крупных желез и некоторых других отделов, превратились
в миоэпителиальные (корзинчатые) элементы.
Выстилка переднего отдела пищеварительной трубки образует у мно-
гих позвоночных зубную эмаль. В зубных зачатках эпидермальная пульпа
эмали претерпевает своеобразное сетевидное разрыхление, утрачивая
структурные особенности эпителиальных тканей.
В составе дыхательного аппарата эпидермальный покров частью обна-
руживает известные черты гистологического сходства с другими эпидер-
мальными тканями тех же животных, частью оказывается в высокой сте-
пени специализированным и приобретает совершенно особый характер.
Достаточно будет остановиться на трахеях насекомых и особенно на
легочном аппарате позвоночных, поскольку относительно эпителия этих
дыхательных органов ниже будут приведены экспериментальные данные.
Стенка воздухоносной части трахей, подобно кожному покрову, состоит
из хитинизированной гиподермы, причем хитин образует характерные,
закрученные спиралью утолщения. Респираторные части трахей пред-
ставлены совершенно своеобразными и, очевидно, высокоспециализиро-
ванными трахейными клетками сложной отростчатой формы с внутриклеточ-
ными воздухоносными капиллярами, стенка которых лишена хитина.
Воздухоносные пути позвоночных выстланы на большей части своего про-
тяжения дву-или многорядным мерцательным эпителием с короткими,
не доходящими до свободной поверхности, базальными или промежуточ-
ными клетками. Последние участки воздухоносных путей, примыкающие
к дышащим частям, имеют однослойную, однорядную выстилку. Респи-
раторные отделы у низших позвоночных, согласно большей части данных,
выстланы одним слоем плоских клеток, через которые происходит газо-
обмен. Относительно респираторной выстилки высших позвоночных до
настоящего времени имеются существенные разногласия.
Со времени Цёлликера считалось установленным, что альвеолярный
эпителий выстилает всю дышащую поверхность легких частью в виде со-
держащих ядра полиэдрических или кубических элементов, частью в виде
тонких безъядерных пластинок. Оппель (1905), Озава (1911) и другие счи-
тают, что безъядерные пластинки являются частями ядерных эпителиаль-
ных элементов, а не отдельными от них образованиями. По Лонгу (1925),
выстилка альвеол состоит только из безъядерных пластинок, в то время
&
116 Глава III
как содержащие ядра клетки представляют собой сходные с гистиоцитами
мезенхимальные элементы. Некоторые авторы, особенно в последнее вре-
мя, вовсе отрицают существование безъядерных пластинок, а содержа-
щие ядро элементы считают или эпителиальными (Зееман, 1931; Клара,
1936) или мезенхимальными (Поликар, 1926; Киоди, 1928, и др.). Согласно
этим взглядам, эпителиальная выстилка альвеол бывает или несплошпой
или вовсе отсутствует, причем эпителий, несомненно существовавший
в зародышевом состоянии, будто бы частью или совсем исчезает около
момента рождения.
Наиболее популярны в настоящее время те взгляды, которые допу-
скают существование несплошной эпителиальной выстилки (Баргманн,
1936; Клара, 1936), представленной отдельными или расположенными
группами, содержащими ядра «эпицитами» (Клара, 1936), с обнаженными
от эпителия участками между ними. Респираторные отделы трубчатых
легких птиц, согласно новейшим данным, вовсе лишены эпителия и образо-
ваны соединительнотканными клетками (Баргманн, 1936). Выстилка пла-
вательного пузыря костистых рыб, развивающаяся, подобно легким, из
переднего отдела пищеварительной трубки, образована однорядным плос-
ким или кубическим эпителием. У ганоидных (Мухина, 1941) выстилка
плавательного пузыря представлена высокопризматическим многорядным
мерцательным эпителием с бокаловидными клетками.
В функциональном отношении т. наз. предпузырная часть плаватель-
ного пузыря костистых рыб имеет известное сходство с легкими, поскольку
через ее выстилку происходит обратное поступление части тазов в кровь,
хотя механизм этого процесса, возможно, отличается от легочного газо-
обмена. Область т. наз. газовой железы является, наоборот, специфиче-
ской частью пузыря, как гидростатического аппарата, не имеющей своего
физиологического аналога в составе легких. Если стоять на весьма веро-
ятной точке зрения, что и легкие и плавательные пузыри филогенетически
являются производными задних редуцировавшихся жаберных карманов
(Макушок, 1911, 1912, 1913; Васнецов, 1928), то гистологическое сопоста-
вление выстилки этих органов должно представлять большой
интерес.
Среди производных эпидермальной ткани переднего отдела пищевари-
тельного аппарата мы, кроме того, встречаемся еще с рядом образований,
которые в эволюционном процессе совершенно утратили связь с внешним
миром и целиком погрузились во внутреннюю среду организма. Имеются
в виду эпидермальные элементы эндокриновых органов — гипофиза, зоб-
ной, щитовидной и околощитовидных желез, которые по характеру своих
гистологических особенностей и функциональному значению более или
менее резко отличаются от типичных представителей этой тканевой группы.
Вместе с утратой разграничительного значения и превращения в ткани
внутренней среды организма эпидермальные элементы эндокриновых орга-
нов образуют структуры, мало соответствующие обычному определению
эпителиальных тканей. Это или тяжи или неправильные скопления клеток,
частью не обнаруживающие полярности (паращитовидные железы и адено-
Экспериментальный метод в гистологии
117
гипофиз), но могущие, в случае накопления между ними секрета, обра-
зовать полые внутри пузырьки, которые особенно характерны для щито-
видной железы. В зобной железе, имеющей характер т. наз. лимфо-эпи-
телиальных органов, только тельца Гассаля сохраняют эпидермальный
характер; прочие же эпидермальные элементы ее образуют сетевидные
структуры, имеющие внешнее сходство с ретикулярной тканью.
Сравнительно-гистологический обзор мочевых и семевыносящих путей
пока делать преждевременно, за отсутствием экспериментальных данных,
посвященных низшим позвоночным и беспозвоночным, а также недо-
статочной ясности в отношении гомологии этих образований и имеющихся,
повидимому, в этой области явлений меторизиса.
Вместе с тем следует подчеркнуть, что при обычных условиях эпите-
лию семевыносящих путей даже у млекопитающих далеко не всегда свой-
ствена двурядная структура.
У неполовозрелых молодых кроликов, например, каналы придатка
яичка выстланы настоящим однослойным однорядным эпителием (Хлопан,
1933). То же самое может иметь место и в других случаях в отдельных уча-
стках или даже на большей части протяжения семевыносящих путей. Иными
словами, эпидермальная природа этих образований, в противоположность
большей части эпителиев, относимых нами к эпидермальному тканевому
типу, при обычных условиях существования проявляется не всегда.
б) Многослойные эпидермальные пласты кожи и
пищеварительной трубки позвоночных, волосяные влагалища
Рост и гистологические превращения в культурах эпидермиса кожи
лабораторных млекопитающих и человека изучались в первую очередь,
на эмбриональном материале (Хлопин, 1922, 1932). Имеются также данные
относительно эпидермиса неэмбриональной кожи млекопитающих и чело-
века (Блажо, 1932; Бернштейн, 1930; Пинкус, 1932) и кожи куриных за-
родышей (Стренджуэйс и Фелл, 1926; Мишурский, 1937). На рост эмбрио-
нального и неэмбрионального эпидермиса кожи амфибий и рыб имеются
указания в работах Уленгута (1916), Гольмса (1914), Дедерера (1921) и
других.
Эпидермис может эпителизировать поверхность эксплантатов и давать
картины экстенсивного плоскостного роста по плотному субстрату. Отме-
чался также погружной рост эпидермиса в свертке фибрина в виде массив-
ных тяжей. Сверток фибрина энергично разжижается растущим эпидер-
мисом с образованием наполненных жидкостью полостей.
Наряду с эпидермисом могут расти и соединительнотканные элементы
кожи, то преобладая, то отступая на задний план. При эпителизации поверх-
ности эксплантата и энергичном росте мембранами соединительная ткань
эксплантатов во многих случаях совершенно не прорастает. Наоборот,
если лоскутки эмбриональной кожи свертываются эпидермисом внутрь,
то на их наружной поверхности наблюдается только разрастание соеди-
нительнотканных элементов.
118
Глава III
Экстенсивный плоскостный рост эпидермиса человека (Пинкус, 1932)
двух-двух с половиной-месячных человеческих зародышей (Хлопан, 1932)
происходит не только тонкими однослойными плоскостными мембранами,
но и многослойными более или менее массивными пластами, состоящими
из уплощенных тесно сомкнутых клеток многоугольной или несколько
вытянутой формы. Более массивный характер плоскостные разрастания
эпидермиса имеют обычно в центральных участках зоны роста, вблизи
посеянного кусочка, где количество клеточных слоев нередко достигает 3—4.
В дистальном от этого последнего направлении они приобретают характер
однослойных мембран, края которых большей частью разрыхляются,
образуя атипичные сетевидные структуры и изолированные клетки разной
формы. Повидимому, утолщению в многослойные пласты могут вторично
Рис. 5. Дифференцировка многослойного плоского покровного эпидер-
мального пласта на поверхности 5-дневного эксплантата из кожи
2-месячного человеческого зародыша. Сильное увеличение. (По Хло-
пину, 1932).
подвергаться и более отдаленные от посеянного кусочка участки мембран.
Рост мембран и пластов происходит за счет активного движения и мито-
тического размножения камбиальных клеток эксплантата. Покровные
клетки эмбрионального двуслойного эпидермиса, хотя и не способны к раз-
множению, могут, повидимому, образовать, продвигаясь в зону роста,
быстро отмирающие и распадающиеся мембраны. При замедленном или
приостановившемся росте в эпидермальных пластах наблюдается диффе-
ренцировка характерных для этой ткани волокнистых структур в прото-
плазме, т. наз. тонофибриллей. По данным Пинкуса, тонофибрилли
образуются без связи с хондриозомами, имеющими форму нитей или зер-
нышек. Первоначально двуслойный эмбриональный эпидермис, быстро
эпителизировав всю наружную поверхность эксплантата, постепенно утол-
щается и дифференцируется, превращаясь по истечении пяти-десяти дней
в многослойный плоский эпителий. Утолщение происходит за счет кам-
биального базального слоя путем митотического деления и напластывания
клеток друг на друга. Между клетками образуются правильные межкле-
точные мостики, придающие им характер элементов шиповатого слоя (рис. 5).
Многослойный покровный эпителий эксплантата состоит из неравноценных
друг другу клеточных слоев. Элементы, расположенные близ соединительно-
тканной подстилки, имеют свойства камбиальных и делятся митозами.
Экспериментальный метод в гистологии
119
В выше лежащих слоях клетки утрачивают способность к размножению
и, с приближением к свободной поверхности, начинают обнаруживать
дегенеративные изменения. Несколько позже поверхностные слои клеток
эпидермального покрова эксплантата, утрачивая ядра, подвергаются ати-
пичному ороговению и постепенно слущиваются (рис. 6). Вместе с тем
эпителизированный кусочек с течением времени постепенно уменьшается
в размерах (как бы худеет) и в конце концов отмирает, так как его орого-
вевающий покров, очевидно, делает невозможным обмен веществ между
тканями эксплантата и внешней средой. О явлениях ороговения в куль-
турах из эмбрионального куриного материала упоминали вкратце Стрен-
джуэйс и Фелл (1926). Подробнее они были изучены Майсурским (1937) на
культурах из кожи шестидневных куриных зародышей. По данным этого
автора, особенно удобными для изучения
ороговения были частички кожи, свернув-
шиеся эпидермисом внутрь и находящиеся
вследствие этого в относительно благо-
приятных условиях обмена веществ, так
как питание эпидермиса тогда происхо-
дит со стороны базальной поверхности,
граничащей с соединительной тканью, т.е.
в основном так же, как в составе целого
организма, а не со стороны свободной по-
верхности, как в случае эпителизирован-
ных эксплантатов. Один или несколько
круглой формы эпидермальных компле-
ксов, окруженных со всех сторон эмбрио-
нальной соединительной тканью, подвер-
гаются дифференцировке с образованием
всех слоев, характерных для эпидермиса
взрослых кур, но расположенных кон-
центрически: снаружи с соединительной
тканью граничит призматический базаль-
ный слой, внутри от него располагаются
постепенно уплощающиеся шиповатые,
зернистые и, наконец, ороговевшие клетки.
Рис. 6. Участок дифференцировав-
шегося вне организма покров-
ного эпидермального пласта с оро-
говевающими и слущивающимися
поверхностными клетками. 15-днев-
ная культура из кожи 2-ме-
сячного человеческого зародыша.
Сильное увеличение. (По Xлопи-
ну, 1932).
Последние, слущиваясь, могут образовать массивное скопление в центре
такого эпидермального комплекса или т. наз. жемчужины.
От эпидермальных мембран и пластов, помимо упомянутых выше изо-
лированных клеток, могут также отделяться клеточные комплексы круг-
лой формы, состоящие из двух - трех и более клеток. Клетки таких комп-
лексов бывают тесно сомкнуты и концентрически охватывают друг друга.
Имеются указания на рост в культурах эпителия роговицы амфибий
и млекопитающих (Отель, 1912; Елеонская, 1915; Матсу мото, 1918;
Кирби, 1927; Полев, 1927, и др.). Отель на этом объекте специально изучал
явления эпителизации, а Полеву удалось получить из него чистые культуры.
Гистологического анализа этого материала не дается.
120
Глава III
Здесь же будет уместно упомянуть про рост и превращения в условиях
тканевых культур эпителия волосяных мешков. По еще неопубликован-
ным данным моей сотрудницы Ушаковой, эпителий волосяных мешков
крупных осязательных (синусных) волос новорожденных кроликов обна-
руживает вне организма явления экстенсивного роста мембранами, пла-
стами или массивами, а также эпителизации поверхности эксплантата.
Эпителий волосяных мешков обладает, подобно покровному эпителию
кожи, резко выраженной способностью разжижать фибрин. Отчасти в связи
с этим в экстенсивно растущих мембранах легко наступают разрыхление
и даже полная изоляция элементов и их переход в цитоморфное состояние.
Сетевидные эпителиальные соединения и даже изолированные клетки, как
правило, обнаруживают заметные отличия от растущих в тех же культурах
элементов соединительной ткани. В большей части многослойных эпите-
лизирующих пластов обычно наблюдаются явления не вполне типичного
ороговения или паракератоза, сопровождающиеся характерными морфо-
логическими изменениями клеток и даже утратой ими ядер. Сходные с оро-
говением изменения нередко наблюдаются и в отдельных элементах мало-
:дифференцированных в гистологическом отношении мембран. Аналогич-
ные явления ороговения обнаруживает, как правило, и остающийся внутри
эксплантата эпителий наружного корневого влагалища. Это очень быстро
и легко наступающее ороговение является основным затруднением при
попытках получить более длительные (свыше трех-четырех недель) куль-
туры данного эпителия.
, Репаративная регенерация эпидермиса кожи млекопитающих, чело-
века и, в меньшей степени, низших позвоночных была издавна предметом
многочисленных исследований {Клебс, 1847; Фрайссе, 1855; Барфурт, 1891;
Рибберт, 1881; Маршанд, 1901; Барделебен, 1901, и др.).
До последнего времени при описании гистологических процессов при
заживлении кожных ран в значительной степени руководились данными
Лёба (1898, 1902). В таком виде, например, этот процесс, имеющий фунда-
ментальное значение в человеческой патологии, описывался в четырех первых
изданиях известного учебника гистологии Заварзина (1933—1938). Со-
гласно данным Лёба, после нанесения дефекта происходит первичное пере-
крытие раны клетками шиповатого и зернистого слоев эпидермиса, кото-
рые, изменяя свою форму и расположение, но не размножаясь, очень бы-
стро образуют на поверхности струпа тонкий провизорный защитный
покров. После этого клетки базального слоя, уплощаясь, продвигаются
со всех сторон под струп, образуя на поверхности грануляционной ткани
непрерывный однослойный пласт. Последний, постепенно утолщаясь,
благодаря интенсивному делению клеток, становится многослойным и,
наконец, превращается в полностью дифференцированный новый участок
ороговевающего многослойного плоского эпителия кожи.
Провизорная эпителиальная пленка на поверхности струпа быстро де-
генерирует и исчезает.
Трактовка всего процесса, данная Лёбом, совершенно не гармонировала
с нашими новыми представлениями об эпителиальных тканях и некото-
Экспериментальный метод в гистологии
121
рыми данными, полученными мной методом эксплантации, а также не
была свободна от ряда серьезных принципиальных возражений.
Выполненное под моим руководством гистологическое исследование
заживления ран кожи и слизистой оболочки полости рта кроликов и собак
{Шевченко, 1941) не только дало впервые исчерпывающую и полную кар-
тину превращений в эпителиальном покрове, но и заставило полностью
отказаться от трактовки процесса, данной Лёбом. Очень ценной и по суще-
ству новой в методическом отношении особенностью работы Шевченко
является параллельное изучение регенеративных процессов в многослой-
ном эпителии полости рта на сериях срезов и на тотальных плоскостных
препаратах. Затем исследования Шевченко были распространены на эпи-
телий роговицы глаза и преджелудка крыс (1941). В кожном эпидермисе
и в многослойном эпителии полости рта и преджелудка процессы идут
принципиально сходным образом.
После нанесения повреждения ожогом на раневой поверхности обра-
зуется плотный струп из некротизированных тканей. Никакого первич-
ного перекрытия раны элементами шиповатого и зернистого слоя, в про-
тивоположность описанию Лёба, не происходит. Этот автор, очевидно,
был введен в заблуждение нередко встречающимися на поверхности струпа,
а также в его толще, подсыхающими эритроцитами, которые, интенсивно
закрашиваясь в препаратах, могут действительно симулировать пикно-
тизированные ядра уплощенных клеток. Такие дегенерировавшие эле-
менты в толще струпа Лёб трактовал как активно вросшие в него клетки
эпителия, которые будто бы участвуют в его «организации». Жизнеспособ-
ные клетки окружающего дефект пласта начинают активно продвигаться
со всех сторон под струп, эпителизируя поверхность образуюхцейся одно-
временно грануляционной ткани. Клетки базального, а затем и шипова-
того слоя, уплощаясь, образуют в периферической области раны молодой
эпителиальный регенерат, имеющий на вертикальном разрезе форму за-
остренного клина или несколько утолщенного на конце языка. Форма,
и толщина регенерата зависят в значительной степени от толщины грани-
чащего с дефектом эпидермального пласта, которая даже в разных уча-
стках одного и того же препарата бывает неодинакова из-за характерных
для многослойного эпителия гребешков, вдающихся в соединительную
ткань. Несколько отступя от края раны, уже в течение первых часов после
повреждения резко возрастает митотическая активность.
Митозы встречаются не только в базальном слое, но и в одном-двух,
реже большем количестве слоев шиповатых элементов. Эта большей частью
дву- трехслойная зона многослойного плоского эпителия млекопитаю-
щих, кроме того, характеризуется резкой базофилией своих элементов
и представляет собой, по Шевченко, собственно камбиальную часть эпи-
телиального покрова. Лежащие выше шиповатые клетки, обнаруживаю-
щие возрастающие признаки дифференцировки (которая, в частности,
выражается в появлении постепенно расширяющегося периферического’
оксифильного ободка цитоплазмы), хотя и входят в состав т. наз..
росткового слоя, но уже по сути дела не являются камбиальными или
122
Глава III
«ростковыми» элементами. В ороговевающем эпидермисе в обычных усло-
виях при обычной методике срезов митозы находились настолько редко,
что многие авторы допускали, без достаточных оснований, широкое уча-
стие амитотического деления клеток в регенеративных явлениях. В настоя-
щее время на плоскостных препаратах эпидермиса кожи удалось устано-
вить высокую митотическую активность камбиальных элементов, которая
даже обнаруживает очень интересный и закономерный суточный ритм
(Карлтон, 1934). У белых мышей в коже живота максимум митотической
активности эпидермиса, составляющий 15 %, падает на вечерние и ночные
часы. Аналогичные данные были затем получены и для кож»'
человека.
Продолжающийся процесс эпителизации дефекта сопровождался ин-
тенсивной митотической активностью, область которой приближается
к краям дефекта, а затем вскоре распространяется и на вновь образованный
эпителиальный пласт. В последнем, параллельно с ростом по поверхности,
происходят рост и обычная дифференцировка элементов в вертикальном
направлении, вплоть до образования рогового слоя. В результате этого,
увеличивающийся в размерах регенерат, на большей части своего протя-
жения, оказывается многослойным и позволяет различать в своем составе
все слои, характеризующие данную эпидермальную ткань. Только его
свободный край имеет во многих случаях, и то не всегда, клиновидную
заостренную форму и заканчивается одной-двумя уплощенными клетками
базального слоя. Протекающий таким образом процесс эпителизации за-
канчивается полным перекрытием дефекта, а струп, постепенно умень-
шаясь в размерах, совершенно исчезает. Одновременно с эпителизацией
в соединительной ткани друг друга сменяют все фазы воспалительного
процесса вплоть до восстановления более или менее нормального сосочко-
вого слоя. Постепенно рельеф базальной поверхности эпителиального
регенерата становится неровным, вследствие его неравномерного утолще-
ния и образования гребешков, вдающихся в соединительную ткань. Базаль-
ная мембрана, первоначально отсутствовавшая под новообразованным
эпителием, восстанавливается. Митотическая активность уменьшается.
Существенно важным в гистологическом отношении моментом в иссле-
дованиях Шевченко является доказательство того, что в процессе репара-
тивной регенерации многослойный эпителий полностью сохраняет свою
эпидермальную специфику и что вместе с тем репаративный гистогенез
отличается по своему течению от гистогенеза многослойного эпителия в он-
тогенезе.
Изученный Шевченко (1941) процесс репаративной регенерации эпи-
дермиса аксолотля протекает по несколько иному способу, зависящему
от меньшего уровня его дифференцировки и слабее выраженной вертикаль-
ной анизоморфности. Камбиальная зона распространяется на всю толщу
эпидермиса аксолотля, кроме двух или даже одного поверхностного слоя
уплощенных клеток. Перекрытие дефекта у аксолотля происходит в основ-
ном за счет изменения формы и расположения предсуществовавших по
краям раны элементов.
Экспериментальный метод в гистологии 123
Репаративное митотическое размножение клеток начинается значи-
тельно позже и в конце концов приводит к восстановлению первоначаль-
ных отношений.
При чрезвычайно быстро протекающем процессе эпителизации раны
эпителиальный регенерат первоначально состоит на большей части своего
протяжения из очень немногих, чаще всего двух, слоев весьма сильно упло-
щенных клеток и поэтому представляется гораздо тоньше, чем нормаль-
ный эпидермис. Характерные для кожи аксолотля лейдиговские клетки
также чрезвычайно сильно уплощаются, участвуя в закрытии дефекта.
На основании результатов, полученных Шевченко, делаются понятными
данные тех авторов, которые не видели митозов при репаративной регене-
рации эпидермиса аксолотля, особенно после линейных разрезов (Вейнер,
1930).
Репаративная регенерация эпителия роговицы глаза изучалась уже
давно, преимущественно окулистами (Зальцер, 1910, 1911; Юселиус, 1910;
Ганке, 1915, и др.). Процессы, происходящие при этом, однако, описыва-
лись и трактовались очень по-разному, вплоть до приписывания эпителию
роговицы способности превращаться в элементы соединительной ткани.
Эпителий роговицы представляет собой особый участок эпидермаль-
ного покрова, который, очевидно, очень рано подвергся у предков совре-
менных животных специализации в особом направлении в связи с разви-
тием зрительного аппарата. Поэтому многослойный плоский неорогове-
вающий эпителий роговицы обнаруживает черты большого сходства у всех
позвоночных и появляется в своем типичном виде у таких животных, у ко-
торых в других местах организма многослойного плоского эпителия еще
нё развивается. Вместе с тем в отношении ряда деталей строения эпите-
лий роговицы может заметно отличаться не только у представителей раз-
ных классов позвоночных, но даже в пределах одного и того же класса.
Эти различия касаются как неодинакового количества клеточных слоев,
так и уровня его дифференцировки, выражающегося в различных соот-
ношениях толщины и количества слоев между способными к размножению
камбиальными и утрачивающими эту способность дифференцированными
покровными элементами (Шевченко, 1940).
Исследования, произведенные в последнее время Шевченко (1941), кроме
того, вносят полную ясность и, в частности, позволяют исправить некото-
рые неточности, допущенные недавно Щелкуновым (1938) в этом вопросе.
Шевченко подверг детальному изучению гистологические процессы при
репаративной регенерации после ожога эпителия роговицы крыс, кроли-
ков и лягушек, начиная с первых минут после нанесения дефекта и кончая
полным восстановлением, на сериях вертикальных срезов и на плоско-
стных тотальных препаратах.
Закрытие дефекта эпителием происходит у млекопитающих чрезвы-
чайно быстро в течение первых двух-трех, реже более дней. У летних лягу-
шек оно, при прочих равных условиях, затягивается до пяти-семи дней,
а у зимних — на гораздо более значительный промежуток времени. При
закрытии дефектов роговицы эпителиальный пласт, даже у млекопитаю-
124
Глава 1П
щих, первоначально продвигается по некротизированной соединительной
ткани и местами даже врастает в нее. Молодая соединительная ткань раз-
вивается, как правило, позже, рассасывая и замещая омертвевшие остатки.
При эпителизации происходит сильное уплощение эпителиальных элемен-
тов, сопровождающееся заметным уменьшением толщины образующегося
регенерата, по сравнению с нормальным эпителием.
Количество клеточных слоев тоже уменьшается, доходя в некоторых
случаях до двух. Процесс эпителизации происходит за счет расположен-
ных ближе к соединительнотканной подстилке камбиальных элементов.
Дифференцированные покровные слои, наоборот, вокруг дефекта слущи-
ваются, обнаруживая дегенеративные изменения, часть которых неправиль-
но трактовалась Щелкуноеым как особого рода пролиферация, сопрово-
ждающаяся распадом ядра на мелкие глыбки хроматина. Эпителизации
дефекта одним слоем клеток на самом деле не происходит. Отмечавшиеся
Щелкуноеым картины зависят от примененного им метода повреждения
скарификацией, результаты которой в отдельных случаях с трудом под-
даются учету, и от неоправданного в данном случае метода коллодийных
пленок (Кочетов, 1911), который не позволяет изучать на плоскостных
препаратах всей толщи эпителиального пласта. В зависимости от рельефа
подстилки эпителиальный регенерат может образовать на своем свобод-
ном крае или в отдельных, удаленных от края, участках местные утолще-
ния, как бы натеки. Ничего похожего на превращение эпителиальных
элементов в клетки соединительной ткани, как и во всех других случаях,
не происходит. Эпителизации сопровождается выраженной в разных слу-
чаях в неодинаковой степени митотической пролиферацией камбиальных
клеток эпителия, которая продолжается после закрытия дефекта, в конце
концов приводит к утолщению регенерата и его превращению в нормаль-
ный эпителий роговицы. В соответствии с меньшим уровнем дифференци-
ровки эпителиального покрова роговицы млекопитающих, по сравнению
с эпидермисом кожи и его идиоадаптацией в составе организма, связанной
с особыми условиями существования и функциональным значением, он
обнаруживает в процессе регенерации на фоне общих эпидермальных
особенностей и черты известного своеобразия.
Экспериментальные исследования Гаршина (1927, 1928, 1939) и его
сотрудников над воспалительными разрастаниями эпителия кожи и воло-
сяных мешков в результате введения в соединительную ткань наружных
покровов уха кролика различных агентов (шарлахового масла, кизельгура,
каменноугольного дегтя, холестерина) дают очень богатый материал, иллю-
стрирующий с большой полнотой гистобластические потенции эпидермиса
в этих особых условиях пролиферации в составе целого организма.
Данные Гаршина во многих отношениях подтверждают гистологиче-
ские закономерности, установленные нами первоначально методом эксплан-
тации и дают убедительные дополнительные обоснования предложенной
мной новой естественной системы эпителиальных структур.
Эти опыты не только значительно расширяют и углубляют данные
Б. Фишера (1906), Ручинского (1910), Ларина (1912) и ряда других авто-
Экспериментальный метод в гистологии 125
ров по т. наз. атипичным разрастаниям эпителия кожи под влиянием инъ-
екции шарлахового масла и некоторых других веществ, но позволяют дать
вполне обоснованную правильную трактовку биологического и вместе
с тем патологического значения всего процесса в целом (Гаршин, 1939).
Гаршину удалось доказать с исчерпывающей полнотой воспалительную
природу этих разрастаний и тесную связь пролиферативных процессов
в эпителии с состоянием подстилающей или окружающей его соединитель-
ной ткани. Источником воспалительных разрастаний эпителия может быть,
в зависимости от условий опытов, или наружный эпидермис или эпителий
волосяных мешков или, наконец, и тот и другой одновременно. В своих
первых опытах Гаршин (1925) получал воспалительные разрастания эпи-
телия, после введения взвеси кизельгура в толщу соединительной ткани,
путем срезания эпителия и слоя соединительной ткани, отделяющего первый
от образовавшейся гранулемы. В результате этого окружающий эпидермис
приходил в непосредственное соприкосновение с очагом воспаления и
эпителизировал его, образуя в то же самое время резко выраженные вос-
палительные разрастания вокруг него. В дальнейшем при соответствую-
щих изменениях условий опытов дополнительной эксцизии не производилось.
Если грануляционная ткань остается отделенной от эпителия прослой-
кой старой, хотя и воспалительно измененной ткани с развитым межкле-
точным веществом, то эпителий реагирует на воспалительный процесс
только усиленной пролиферацией и утолщением на месте без образования
погруженных в толщу соединительной ткани разрастаний, с сохранением
основных особенностей кожного эпидермиса. По терминологии Гаршина
такого рода воспалительная пролиферация эпителия обозначается как
рост in situ. Если покровный эпителий или эпителий волосяных мешков
непосредственно граничит с грануляционной тканью, с находящимися
в ней очагами некроза, то он образует разрастания «погружного типа».
Воспалительные разрастания «погружного типа», соответствующие во
многих отношениях разрастаниям в условиях тканевых культур, имеют
характер тяжей или сплошных массивов, образующихся из малодиффе-
ренцированных камбиальных элементов, энергично делящихся митоти-
ческим путем. Из отдельных тяжей вторично могут также образоваться
массивы. Дорастая до окруженных некротическими массами инородных
тел (капель дегтя, холестериновых масс и т. п.), эпителиальные массивы
обычно приобретают колоколообразную форму и своими истончающимися
краями начинают распространяться по поверхности молодой грануляци-
онной ткани, окружающей очаги некроза. Таким образом, а также за счет
сливающихся друг с другом эпителиальных тяжей, возникают многослой-
ные эпителиальные пласты, постепенно обрастающие со всех сторон не-
кротические массы и инородные тела.
В конце концов в толще воспаленной соединительной ткани образуются
кисты большего или меньшего размера с многослойной эпителиальной
стенкой. Первоначально малодифференцированные разрастания, состоя-
щие из однородных камбиальных клеток, начиная с определенной стадии
процесса, обнаруживают явления вторичной дифференцировки. На границе
126
Глава III
с соединительной тканью образуются базальный призматический слой
многослойного эпителия, а ближе к центру массивов или к внутренней
полости кист типичные шиповатые клетки разного уровня дифференци-
ровки, а затем, наконец, и роговой слой. В результате этого малодиффе-
ренцированные массивы превращаются в жемчужины с роговыми массами
внутри, а стенка кист приобретает строение настоящего ороговевающего
эпидермиса. Тем самым достигается отграничение от живых тканей орга-
низма инородных тел и некротических скоплений. Если в эпителиальных
массах, соединяющих стенку кисты с наружным эпителием, в результате
ороговения образуется центральный просвет, то содержимое кисты может
опорожниться наружу. Мелкие инородные тела, например иглы кизель-
зура и небольшие скопления детрита, обросшие базальными слоями эпи-
дермиса, также постепенно элиминируются из организма в результате
нормальных процессов роста и смены клеток многослойного эпителиаль-
ного пласта. Из ороговевающей эпителиальной выстилки крупных кист
могут вновь возникнуть волосяные зачатки, из которых в просвет кисты
вырастают волосы.
Приостановка разрастаний и процессы дифференцировки эпителия
происходят в зависимости от стадий созревания грануляционной ткани.
Слабо выраженные воспалительные разрастания, приводящие местами
к образованию небольших тяжей и языков, вдающихся в соединительно-
тканную подстилку, могут сопровождать в отдельных случаях и обыч-
ную репаративную регенерацию многослойного эпителия (Шевченко, 1941).
Воспалительные разрастания эпидермиса низших позвоночных до на-
стоящего времени почти совершенно не изучены, хотя их получение и обе-
щает много нового и интересного. В работе Бельша (1910) имеются указа-
ния, что у личинок саламандры, после введения под эпидермис шарлахо-
вого масла, удавалось получить эпителиальные разрастания. Явления эпи-
телизации инородного тела (целлоидиновых трубок) в отдельных случаях
получала Алфеева (1927) при изучении асептического воспаления у лягушки.
На основании изучения человеческого секционного и биотического
материала и имеющихся литературных данных, Гаршин пришел к выводу,
что целый ряд пролиферативных процессов в эпидермисе человека должен
трактоваться как воспалительные разрастания.
Штейн (1936) в лаборатории Гаршина показал, что при туберкулезе кожи
(волчанке) можно наблюдать воспалительные разрастания эпителия, при-
водящие в элиминации туберкулезных бугорков. Когда материал из обла-
сти человеческой патологии достаточно тщательно изучен в гистологиче-
ском отношении, получаемые картины полностью совпадают с материалами
эксперимента на животных. Более подробные данные можно найти в моно-
графии Гаршина (1939), на которую я и позволю себе сослаться.
Существенно важным для нас выводом из всех приведенных выше на-
блюдений является то, что эпидермис кожи млекопитающих и человека
при пролиферативных процессах, вызванных самыми разнообразными
агенталш, проделывает принципиально одинаковый цикл характерных
для данной ткани гистологических превращений, т. е. проявляет неспе-
Экспериментальный метод в гистологии
127
цифическую в отношении внешнего агента, но совершенно специфическую
в отношении своих тканевых свойств реактивность. Наблюдающиеся в отдель-
ных случаях различия имеют в основном количественный характер. В част-
ности, интенсивность воспалительных разрастаний эпидермиса зависит
от длительности воспалительного процесса и оттого, насколько быстро или
медленно созревает грануляционная ткань.
Многослойная выстилка влагалища, имеющая типичный эпидермаль-
ный характер, как известно, обнаруживает ряд изменений в течение поло-
вого цикла. У многих млекопитающих эти изменения бывают выражены
в очень резкой степени, сопровождаются вследствие усиленной пролифе-
рации резким утолщением эпителия в периоде течки и образованием рого-
вого слоя, который затем слущивается.
Такие же явления вызываются экспериментально путем введения эстро-
генных веществ. Оказывается, и другие многослойные эпидермальные пла-
сты обнаруживают в той или иной степени аналогичные изменения под
влиянием гормональных воздействий. В частности, это относится к эпите-
лию конъюнктивы глаза, по состоянию которой можно определять совер-
шенно так же, как это делается на основании изменений эпителия влага-
лища, наступление периода течки (Волосков, 1938). Указания на то, что
эпителий влагалища будто бы утолщается за счет присоединения к нему
элементов из соединительнотканной подстилки, которые в дальнейшем
вновь возвращаются в ее состав (Рихтер, 1939), совершенно необоснованны
и не выдерживают никакой критики. То же самое относится и к некоторым
более старым данным (Сабатье и Рувиль 1898; Л. Лёб, 1899; Кромейер, 1902;
Реттере, 1903, и некоторые другие), согласно которым эпидермальные
покровные пласты или пополняются за счет присоединяющихся к ним
клеток соединительной ткани или, наоборот, обладают способностью пре-
вращаться в нее.
Такого рода представления не стоят на уровне современной науки.
в) Покровный эпителий воздухоносных, мочеотводящих и
семевыносящих путей млекопитающих и человека
Данные, касающиеся превращений в эксплантатах этих эпителиев, я
изложу суммарно, на основании своих собственных работ и работ моих
сотрудников, где имеется достаточно полный гистологический анализ на-
блюдавшихся картин.
В условиях тканевых культур выстилка бронхов кроликов (Стрелин,
1930), имеющая строение двурядного призматического мерцательного
эпителия, переходный эпителий мочевого пузыря (Хлопан. 1924; Птохое,
1939), мочеточника (Лежава, 1935) и почечной лоханки (Цымбал, 1935),
а также выстилка протоков придатка яичка (Хлопан, 1934) кроликов
обнаруживают в своих превращениях значительное сходство. Все эти
ткани могут расти или экстенсивно, образуя плоскостные мембраны
и пласты, или эпителизировать поверхность эксплантата, нередко обра-
стая его со всех сторон. Сверток фибрина при этом подвергается сильному
128
Глава III
разжижению с образованием полостей, наполненных жидкостью. Наи-
более интересные в гистологическом отношении изменения наблюдаются
при эпителизации поверхности эксплантатов, сопровождающейся образо-
ванием более или менее дифференцированного покровного эпителия. Перво-
начально эпителизирующий покров может иметь строение более или менее
тонкого, большей частью дву- или однослойного пласта, состоящего из
уплощенных или низкопризматических клеток. В случае переходного
эпителия при этом удается проследить, что рост происходит за счет проли-
ферации и активного движения камбиальных клеток, расположенных
ближе к соединительной ткани, в то время как лежащие выше и, в первую
очередь, покровные элементы, округляясь, отторгаются и погибают {Лежава,
1935; Птохов, 1939). Затем покровный пласт, утолщаясь и дифференци-
руясь, делается дву- или многослойным. Клетки или тесно сомкнуты друг
Рис. 7. Преврашение переход-
ного эпителия мочеточника во
многослойный плоский. 4-днев-
ная культура из мочеточника
6-дневного кролика. Сильное
увеличение. (По Лежава, 1934).
с другом, или между ними появляются тон-
кие щелевидные промежутки с отчетливо
выраженными межклеточными мостиками,
не свойственными этим тканям при обычных
условиях. На свободной поверхности покров-
ных клеток многослойных дифференцирован-
ных пластов можно бывает заметить уплотнен-
ный наподобие кутикулы слой цитоплазмы, а
между их прилегающими друг к другу кра-
ями замыкающие полоски. На некоторых
стадиях культивирования переходный эпите-
лий на поверхности эксплантата приобретал
строение типичного многослойного плоского
неороговевающего эпителия (рис. 7) {Лежа-
ва}. В других случаях он мог вновь обра-
зовать на своей свободной поверхности ха-
рактерные для него при обычных условиях
дву-трехядерные покровные клетки {Лежава, Птохов). Нередко меж-
ду клетками измененного переходного эпителия образовались дости-
гающие подчас значительных размеров полости, наполненные жидкостью.
Обрастающий поверхность эксплантата бронхиальный эпителий, не-
смотря на изменение формы клеток, мог (по крайней мере, местами) со-
хранять в течение более или менее длительного времени оживленно мерцаю-
щие реснички. В дальнейшем они исчезали.
С течением времени нацело эпителизированные эксплантаты постепенно
уменьшались в размерах, а их внутренние части подвергались дегенера-
ции. В конце концов погибал и распадался покровный эпителий, который
до конца сохранял характер многослойного пласта или предварительно
истончался. В разных случаях эти дегенеративные изменения протекали
•с неодинаковой быстротой. Заканчивались смертью они большей частью
через 2—3 недели. Дву- или многослойный эпителиальный покров в куль-
турах из придатка яичка нередко приобретал большое сходство с переход-
-ным эпителием (рис. 8) и отличался тем, что митотическое деление его кле-
Экспериментальный метод в гистологии
129
ток происходило преимущественно в покровном слое, а не в базальном, как
это бывает в измененном переходном эпителии, поверхностные клетки
Которого и в условиях культур, в большинстве случаев, утрачивают спо-
собность к пролиферации. Клетки всех этих тканей могут также, утра-
чивая взаимную связь, существовать некоторое ограниченное время в изо-
лированном виде. В культурах бронхиального эпителия отмечалось обра-
зование изолированных эпителиальных комплексов, состоящих из концен-
трически наслоенных друг на друга клеток (Стрелин, 1930).
В опытах с пролиферативными процессами в составе целого организма
эти эпителии также обнаруживают много общего. Наряду с этим, в усло-
виях целого организма могут отчетливее проявляться более частные спе-
цифические особенности отдельных эпителиев. Поэтому эти эксперимен-
тальные данные будет удобнее изложить порознь по отдельным конкретным
тканям.
Процессы репаративной регенерации эпителия воздухоносных путей
изучались на выстилке трахеи кролика после вырезания небольших
участков слизистой оболочки (Гаршин, 1935) и в моей лаборатории, после
ожога раскаленной иглой (Гольштейн, 1941). Ввиду того, что работа Гар-
шина ориентирована в первую очередь на вопросы патологии, а Гольштейна
на гистологические, положим в основу изложения данные последнего автора.
Очень скоро после повреждения в окружающем дефект эпителии обнару-
живаются в заметном количестве митозы, которые при обычных условиях
практически не встречаются, так как процессы физиологического снаши-
вания и физиологической регенерации, очевидно, протекают очень мед-
ленно. Эти первые митозы локализированы не апикально, как в одноряд-
ных эпителиях, а близ соединительнотканной подстилки в коротких базаль-
ных клетках, что доказывает с полной отчетливостью камбиальное зна-
чение этих элементов (в котором до последнего времени принято было со-
мневаться) и вместе с тем вертикально анизоморфный характер многоряд-
ного эпителия. Мерцательные и бокаловидные клетки вблизи дефекта,
наоборот, слущиваются и исчезают. В более удаленных от места ожога
участках замечается появление необычно большого количества бокаловид-
ных клегок, повидимому, путем превращения в них значительной части
мерцательных элементов, подвергающихся ослизнению и утрачивающих
реснички вместе с базальными тельцами. Пролиферация камбиальных
клеток при одновременном исчезновении мерцательных и бокаловидных
приводит к перестройке эпителиального пласта вокруг дефекта. В та-
ких местах многорядный эпителий превращается в дву- и многослойный
малодифференцированный пласт, в который, без резкой границы, пере-
ходят более удаленные и вместе с тем малоизмененные части эпителиаль-
ной выстилки. Малодифференцированный пласт вокруг дефекта, состоящий
из малоизмененных бывших камбиальных элементов, отличается отсут-
ствием вертикальной анизомфорности и наличием митозов во всех слоях.
Образующаяся в области дефекта обнаженная грануляционная ткань эпи-
телизируется надвигающимся на нее со всех сторон многослойным эпите-
лиальным пластом, дающим одновременно во многих случаях то более»
В Хлопни—349
130
Глава 111
то менее резко выраженные воспалительные разрастания в толщу своей
подстилки. Эпителизирующий пласт в области дефекта приобретает строе-
ние многослойного плоского неороговеваюгцего эпителия. После полного
перекрытия во всей или почти всей области бывшего дефекта может наблю-
даться эпителиальный покров, состоящий из постепенно уплощающихся
к свободной поверхности клеток. В дальнейшем многослойный эпителий
постепенно вновь перестраивается в многорядный мерцательный, пови-
димому, благодаря слущиванию покровных клеточных слоев и дифферен-
цировке в соответствующем направлении пролиферирующих базальных
элементов. В осложненных нагноением случаях эпителиальный покров
мог резко утолщаться, приобретая известное сходство с атипичным пере-
ходным эпителием, лишенным, однако, характерных для него многоядер-
ных покровных клеток.
Так наз. «атипические» разрастания эпителия бронхов получил
впервые Фишер-Вазельс (1922) после интравенозных инъекций кролику
препарата гранугеноль (Granugenol). Их воспалительную природу уста-
новили Гаршин и Пигалев (1927) на основании опытов с введением шпри-
цем в ткань легкого каменноугольного дегтя и «формалинового масла»
(смесь подсолнечного масла с формалином) (Гаршин и Шабад, 1935; Гар-
шин, 1939). Аналогичные результаты были также получены Шабадом (1937)
при интратрахеальном введении каменноугольного дегтя, повторившим
в широком масштабе опыты Кимура (1923). Наиболее резко выраженные
разрастания получались при непосредственном введении каменноуголь-
ного дегтя, а также формалинового масла в ткань легкого. Эти же иссле-
дователи дали гистологическое описание наблюдавшихся картин. Цикл
превращения бронхиального эпителия в очаге воспаления во многих отно-
шениях напоминает описанные выше изменения эпителия трахеи при репа-
ративной регенерации. Расположенная по соседству с воспалительным
очагом многорядная призматическая выстилка бронхов, благодаря размно-
жению своих камбиальных клеток и исчезновению мерцательных и бокало-
видных элементов, постепенно перестраивается в малодифференцирован-
ный, многослойный эпителий.
Такой измененный эпителий бронхов образует, врастая в грануляци-
онную ткань, сплошные клеточные тяжи различной толщины и массивы»
соответствующие структурам, образуемым эпидермисом. Из этих «погруж-
ных» воспалительных разрастаний эпителия (которые, развиваясь значи-
тельно слабее, как побочное явление, наблюдаются и в процессе репара-
тивной регенерации выстилки трахеи) образуются сплошные многослой-
ные эпителиальные пласты, обрастающие со всех сторон очаги некроза»
окружающие капли деггя или масла и отграничивающие их от жизне-
способных тканей. В результате этого образуется киста, стенки которой
состоят из многослойного эпителия с плоскими или местами изопризмати-
ческими покровными клетками. Через канализирующиеся эпителиальные
тяжи киста может приобрести сообщение с бронхами, через которые в конце
концов содержимое выводится наружу.
Рост эпителия пластом, приводящий к образованию кисты, Гаршин
Экспериментальный метод в гистологии
131
рассматривает как регенеративный. Если при таком регенеративном росте
эпителиальный пласт оказывается расположенным непосредственно на
молодой грануляционной ткани, то он вновь образует в толще этой послед-
ней воспалительные разрастания «погружного» типа. Этого не происхо-
дит, когда эпителиальный пласт растет по волокнистой соединительной
ткани.
Параллельно с затуханием воспалительного процесса в соединитель-
ной ткани разрастания эпителия прекращаются. Атипичные эпителиаль-
ные тяжи могут превратиться в трубки, образованные вновь дифферен-
цирующимся многорядным эпителием. Такую вторичную перестройку
претерпевает и выстилка кист, приобретающая в конце концов строение
типичного эпителия воздухоносных путей.
Многие картины, описываемые на основании человеческого секцион-
ного материала при различных патологических процессах в легких (Аска-
нази, 1919; Шмидтманн, 1920: Бракк, 1926, и др.), как отмечает Гаршин
(1939), являются не чем иным, как различными стадиями воспалительных
разрастаний бронхиального эпителия.
При самых разнообразных патологических условиях он дает превра-
щения, которые полностью укладываются в рамки описанных выше при
экспериментальных условиях гистологических структур. То же самое
справедливо и в отношении других частей воздухоносных путей. Напри-
мер, при туберкулезе гортани наблюдается резко выраженная воспалитель-
ная пролиферация эпителия, проникающего даже в хрящ, в результате
которой могут происходить обрастание и, повидимому, элиминация бу-
горков из организма (Гаршин, 1935).
Во всех описанных случаях превращение многорядного эпителия воздухо-
носных путей во многослойный не является метаплазией в полном смысле
этого слова, а относится к категории явлений, обозначаемых Гаршиным
(1938), как транзиторные изменения или ателеплазия. Создающийся при
различных пролиферативных процессах, на месте многорядного, много-
слойный плоский эпителий не обнаруживает признаков ороговения и пред-
ставляется, например, по сравнению с эпидермисом, относительно мало-
дифференцированным и менее сложным. Этот многослойный эпителий пред-
ставляет собой, подобно сплошным тяжам и массивам, одну из форм суще-
ствования бронхиального эпителия, подчеркивающую его большое сход-
ство с кожным эпителием (основанное, с нашей точки зрения, на генети-
ческой близости), но не полное тождество с ним. Вместе с тем ряд данных
из области патологии показывает, что в воздухоносных путях могут встре-
чаться участки настоящего многослойного ороговевающего эпителия.
Хотя гистогенез таких участков и не прослеживался, можно предположить,
с большой долей вероятия, что они возникли путем метаплазии из много-
рядного эпителия, хотя полностью исключить их происхождение
из дистонического эмбрионального зачатка все-таки нельзя. По-
явление многослойного ороговевающего эпителия в воздухоносных
путях при экспериментальном авитаминозе А, хотя не прослеживалось
достаточно детально по стадиям, возможно, также происходит в результате
132
Глава 111
метаплазии (Уолбах и Гау, 1925). В этом случае, однако, нельзя исклю-
чить с полной достоверностью замещения эпителия воздухоносных путей
многослойной плоской выстилкой глотки, врастающей на его место. Учи-
тывая экспериментально доказанное сходство между многорядным эпите-
лием и эпидермисом, следует считать во всех этих случаях истинную мета-
плазию весьма вероятной.
Относительно репаративной регенерации выстилки эпителия мочевого
пузыря мы располагаем в настоящее время очень подробными в гистологи-
ческом отношении данными моего сотрудника Птохова (1940), который
подверг тщательному анализу превращения переходного эпителия после
нанесения дефекта прижиганием и вырезанием.
Данные немногих авторов (Бельуов, 1884; Де-Рувилъ, 1896, 1897; Кор-
пиль и Карно, 1899; Лазио, 1909), изучавших регенеративные процессы
в слизистой оболочке мочевого пузыря, во многом устарели, а в гистоло-
гическом отношении были очень неполны и суммарны.
При том и другом способах повреждения переходный эпителий обна-
руживает, по сути дела, одинаковые гистобластические потенции, хотя
течение процесса регенерации и отличается известными особенностями
количественного порядка.
Весь ход процесса прослеживался параллельно на сериальных срезах
и на тотальных плоскостных препаратах растянутой стенки пузыря взрос-
лых кроликов. Предварительное изучение Птоховым (1940) нормальной
структуры переходного эпителия мочевого пузыря в растянутом и сокра-
щенном состоянии, а также на мацерированных препаратах показало,
в соответствии с данными Лежава (1934, 1935), полученными с выстилкой
мочеточника, что переходный эпителий имеет многорядно-многослойное
строение и обнаруживает определенные черты сходства с многослойным
эпителием кожного типа. В максимально растянутом состоянии он имеет
у новорожденных кроликов три, а у взрослых до пяти слоев более или ме-
нее уплощенных клеток, образующих однослойные базальную и покров-
ную зоны и одно-трехслойную промежуточную. Разделение переходного
эпителия на эти три характерные для него зоны было предложено Лежава
(1934, 1935), на основе изучения выстилки мочеточника. При сокращенной
стенке клетки не только изменяют свою форму и становятся выше, но и
выклиниваются частично своими ядросодержащими участками в лежащие
выше слои, приобретая более или менее длинные отростки, направленные
базально. Эти отростки частью доходят до соединительнотканной подстилки,
частью заканчиваются свободно между телами лежащих ниже клеток эпи-
телия. Поэтому количество видимых рядов клеток и ядер в сокращенном
пузыре превышает истинное количество клеточных слоев, которое у раз-
ных млекопитающих и даже, быть может, в разных местах у одного и того
же животного может варьировать. Покровные клетки, имеющие нередко
несколько ядер и гигантские размеры и элементы т. наз. промежуточной
зоны, находящиеся под ними, своими отростками не доходят до соединитель-
ной ткани, с которой связаны только клетки базальной зоны, имеющие
в первую очередь камбиальное значение. Переходный эпителий обнару-
Экспериментальный метод в гистологии
183
живает, подобно кожному, вертикальную анизоморфность, хотя и в менее
резком виде. Эти наблюдения имеют принципиальное значение, так как
доказывают несостоятельность точки зрения, рассматривающей пе-
реходный эпителий как разновидность однослойного (Данини, 1923,
1924).
При репаративной регенерации переходный эпителий обнаруживает
цикл весьма характерных превращений, которые в еще большей степени
подчеркиваю!' его близость многослойным эпителиям кожного типа. Во-
круг образовавшегося дефекта клетки покровной и промежуточной зон
обнаруживают, в результате повреждения, выраженные в резкой степени
дегенеративные и некробиотические изменения. Отмирающие клетки оттор-
гаются и могут обнажать на большем или меньшем протяжении элементы
базальной зоны, в которой одновременно резко стимулируется митоти-
ческая активность. Количество митозов, регистрируемое на плоскостных
препаратах, бывает очень велико — несколько десятков в одном поле зре-
ния при иммерсионном увеличении. Вследствие этого переходный эпите-
лий, перестраиваясь и приобретая строение малодифференцированного
многослойного, начинает эпителизировать рану, продвигаясь на поверх-
ность молодых грануляций под сохраняющийся некоторое время струп.
В области дефекта эпителизирующий пласт заметно истончается вследствие
сильного распластывания клеток и уменьшения количества их слоев, кото-
рое, однако, как правило, бывает не менее двух. Лишь изредка у свобод-
ного края продвигающегося пласта на вертикальных срезах можно заме-
тить одну-две резко уплощенных клетки, расположенные одним слоем.
В других случаях свободный край представляется, наоборот, несколько
утолщенным. По истечении 5—7 дней вся область дефекта оказывается
закрытой эпителием, который частью сохраняет строение многослойного
с поверхностными уплощенными клетками, частью уже начинает пере-
страиваться, приобретая постепенно вновь характер нормальной выстилки
мочевого пузыря.
Процесс заканчивается восстановлением типичного переходного эпи-
телия на месте бывшего дефекта. Одновременно с перекрытием дефекта,
эпителий в очень многих случаях образует своеобразные утолщенные уча-
стки, вдающиеся в соединительнотканную подстилку. Они, разрастаясь,
принимают вид эпителиальных гнезд круглой формы, могущих полностью
утрачивать связь с покровным пластом и приобретать характер т. наз.
брунновских гнезд, свойственных слизистой оболочке мочевого пузыря
человека, при патологических условиях (например cystitis cystica).
Многослойные структуры, образованные эпителием мочевого пузыря,
при репаративной регенерации, по виду и окрашиваемости в препаратах
своих клеточных элементов сохраняет определенные цитотипические осо-
бенности, свойственные переходному эпителию.
Известное сходство с брунновскими гнездами могут приобретать в усло-
виях тканевых культур эпителиальные комплексы на месте облитериро-
ванного просвета мочеточника, описанные Лежава. Ничего похожего на
превращение элементов соединительной ткани в эпителий мочевого пузыря,
Г34
Глава III
предполагавшееся в свое время де-Рувилем (1896, 1897) при репаративной
регенерации, не происходит.
Относительно воспалительных разрастаний переходного эпителия
имеются лишь указания Захарьевской (1939), которая вводила в почку
кролика каменноугольный деготь. Наряду с пролиферативными процессами
в почечном эпителии, ей удалось получить также разрастания эпителия
лоханки, которые несомненно обнаруживают большое сходство с выше-
описанными изменениями мочевого пузыря и очень резко отличаются от
картин, даваемых эпителием мочевых канальцев. В опытах Захарьевской
из эпителия лоханки возникали пласты, имеющие на большей части сво-
его протяжения дву- или многослойное строение. Эти пласты обрастали
скопления дегтя, окруженные некротическими массами, и образовали
в конце концов сплошную выстилку кистовидных полостей.
Исходя из всех экспериментальных данных, касающихся переходного
эпителия мочевого пузыря, мочеточника и лоханки, можно с очень большой
долей вероятия считать, что описываемые в человеческой патологии при
т. наз. лейкоплакиях явления метаплазии переходного эпителия в орого-
вевающий многослойный плоский, действительно являются таковыми
и что переходный эпителий под влиянием некоторых длительных патоло-
гических процессов может превращаться в настоящий дифференцирован-
ный кожный эпителий со всеми его гистологическими особенностями. То
же самое справедливо и в отношении образования многослойного орого-
вевающего эпителия в мочевых путях при экспериментальном авитами-
нозе А.
Если, кроме того, принять во внимание, что многослойный ороговеваю-
щий эпителий описывался во всех участках мочевого пузыря, мочеточника
и лоханки, то трактовка этих находок как результат истинной метапла-
зии местного переходного эпителия приобретает почти полную достовер-
ность. Это может быть доказано с полной убедительностью только путем
точного прослеживания всего развития процесса по стадиям при непре-
менном учете его последовательности в разных участках мочевых путей.
Такие исследования должны быть одной из задач будущих работ.
Относительно появления в семевыносящих путях человека многослой-
ного эпителия в патолого-анатомической литературе имеются довольно
многочисленные, хотя в каждом отдельном случае основывающиеся на
единичных наблюдениях, указания (Нобль, 1903; Селлеи, 1904; Вольф, 1920;
Коншегг, 1926. и др.). По Вольфу и Коншеггу, при хронических эпидидими-
тах метоплазия может даже доходить до образования ороговевающего
эпителия. Эти данные, однако, оставались неподтвержденными и не вошли
в подробную сводку Оберндорфера (1931). Следует отметить, что все эти
данные (в частности, Вольфа) и описания Коншегга в гистологическом
отношении очень поверхностны и недостаточно убедительны. Во многих
случаях (в частности, в работе Коншегга) иллюстрации совершенно отсут-
ствуют. Экспериментальная работа Кирле и Шоппера (1915), изучавших
регенеративные процессы в придатке яичка после повреждения и также
описавших образование многослойного неороговевающего эпителия, в ги-
Экспериментальный метод в гистологии 135
отологическом отношении также оставляют желать многого. Поэтому мои
данные, полученные методом эксплантации (Xлопин, 1933), подтвержден-
ные в самое последнее время, стояли более или менее особняком. Исходя
из того, что превращения эпителия семевыносящих путей при воспалитель-
ных процессах, как в гистологическом, так и в патологическом отноше-
нии, представляют очень большой интерес, Гаршин совместно со мной
предпринял соответствующее исследование на большом человеческом опе-
рационном материале, обнимающем 49 отдельных случаев. В 31 случае
изученных эпидидимитов нам удалось обнаружить, наряду с дистрофиче-
скими, и резко выраженные гиперпластические изменения в эпителии семе-
выносящих канальцев головки придатка, в канале придатка или в началь-
ном отрезке семевыносящего протока. Изменения во всех изученных от-
делах семевыносящих путей имели, по существу, одинаковый характер
и в большей части случаев относились к типу воспалительной пролифера-
ции эпителия in situ, т. е. в составе стенки канальцев. Внеканальцевые
разрастания эпителия в окружающую соединительную ткань были выра-
жены слабо и встречались значительно реже. В эпителии, расположенном
в области воспалительно измененной соединительной ткани, с более или
менее резко выраженной инфильтрацией лимфоцитами, амебоидными ги-
стиоцитами и специальными лейкоцитами, различия, свойственные разным
отделам семевыносящих путей, постепенно стираются. В семевыносящих
канальцах головки исчезают мерцательные клетки и характерная неравно-
мерность высоты отдельных элементов, а обусловленные ею углубления
в эпителии сглаживаются. Размножаясь митотически и наслаиваясь друг
на друга, эпителиальные клетки принимают частью двуслойное или дву-
слойно-многорядное, частью многослойное расположение. У высоких
элементов двурядного (при обычных условиях) эпителия протока придатка
исчезают сецерненты; появляются двуслойные, многорядные и много-
слойные участки. То же самое отмечается и в хвостовом отделе придатка.
В результате этого в разных отделах семевыносящих путей могут появиться
тождественные по своему строению атипичные участки. В резко утолщен-
ных участках эпителиального покрова наблюдается, кроме того, разная
степень его инфильтрации блуждающими элементами и местами образо-
вание характерных кистевидных полостей неодинакового размера.
Утолщение эпителия приводит к резкому сужению просвета и даже
полной облитерации. Двуслойные более тонкие участки эпителия состоят
из уплощенных или неправильно полиэдрических элементов. При утол-
щении эпителия количество рядов ядер возрастает до трех-четырех, а ча-
сто — до семи и более рядов, причем в нем местами намечается появление
неодинаковых по своему характеру зон. По деталям своего строения такие
утолщенные участки приобретают довольно разнообразный характер и чрез-
вычайно большое сходство с известными участками переходного эпителия.
Покровные клетки бывают полиэдрическими, более или менее изо-
призматическими или уплощенными. Клетки нижележащих слоев имеют
полиэдрическую или местами сильно вытянутую в вертикальном напра-
влении форму. В последнем случае ядра базального слоя имеют довольно
136
Глава Ill
правильное палисадообразное расположение. В таких участках, как и
в переходном эпителии, видимое количество рядов ядер не соответствует
числу клеточных слоев. Вместе с тем митозы встречаются во всех слоях.
Лишь в одном случае нам пришлось наблюдать образование структур,
свойственных многослойному плоскому эпителию. Участок канала при-
датка, в области перехода в семевыносящий проток, окруженный сильно
инфильтрованной клетками соединительной тканью и почти не имеющий
просвета, состоял из мощного многослойного эпителиального пласта. В нем
можно было видеть один ряд более темных палисадно расположенных ба-
зальных клеток и много слоев более крупных полиэдрических, уплощаю-
Рис. 8. Утолщение и перестройка эпителия придатка яичка во много-
слойный покровный пласт, сходный с несколько атипичным переходным
эпителием. 12-дневная культура придатка яичка кролика. Сильное
увеличение. (По Хлопину, 1933).
щихся к поверхности элементов, отличающихся своей более светлой окра-
ской и тесно прилегающих друг к другу. По своему общему виду такие
структуры очень напоминали неороговевающий многослойный эпителий
переднего отдела пищеварительной трубки с морфологически выраженной
вертикальной анизоморфностью. Что касается внеканальцевых раз-
растаний эпителия, то они представляли собой массивные малодифферен-
цированные образования, состоящие из однотипных крупных клеток.
В последнее время в эпителии семевыносящих путей кроликов Искрой
в лаборатории Гаршина были получены экспериментально (после впры-
скивания в придаток яичка каменноугольного дегтя) изменения, сходные
с описанными (Гаршин, 1939).
Мне представлялось целесообразным несколько подробнее остановиться
на особенностях эпителия семевыносящих путей по следующим соображе-
ниям. Во-первых, мы имели возможность на этом объекте по личному опыту
убедиться в большой согласованности результатов, получаемых методом
эксплантации и при достаточно детальном гистологическом анализе чело-
веческого патолого-анатомического материала. Ряд картин превращений
эпителия придатка яичка вне организма, подробно описанных и докумен-
тированных рисунками, совпадает с наблюдениями над человеческим мате-
Экспериментальный метод в гистологии
137
риалом. В частности, это касается образования структур, сходных с пере-
ходным эпителием (рис. 8), значение которых в то время (1933) еще не
могло быть полностью оценено, за неимением полученных позже данных.
Во-вторых, тот факт, что эпителий разных отделов семевыносящих путей
млекопитающих, имеющий совершенно другое функциональное значение,
чем переходный эпителий, при экспериментальных и патологических уело?
виях может в значительной степени уподобляться ему в структурном от*
ношении, едва ли может быть понят иначе, как выражение их генетического
единства, которое подтверждается и упомянутыми выше эксперименталь-
ными данными. Такого же значения не может не иметь факт приобретения
переходным эпителием многослойной структуры и, в отдельных случаях,
структуры многослойного плоского эпителия, свойственной также выстилке
воздухоносных путей при экспериментальных и патологических условиях,
но совершенно не свойственной целому ряду других описываемых ниже
эпителиальных тканей.
г) Эпителий эпидермальных желез
Как это подчеркивалось во второй главе, железистые образования,
возникающие из генетически различных эпителиальных покровных пла-
стов, могут трактоваться как примеры конвергентного морфогенеза. Едва
ли подлежит сомнению большое сходство общего расчленения и строения.
Рис. 9. Образование эпителиальных
массивов (Ет) из выводных протоков
(Dg) околоушной железы. 8-дневная
культура околоушной слюнной же-
лезы кролика. Слабое увеличение.
(По Цымбалу, 1932).
Рис. 10. Эпителиальный мас-
сив, образовавшийся из перере-
занного выводного протока гар-
деровской железы. 8-дневная
культура из гардеровской же-
лезы новорожденного кролика.
Слабое увеличение. (По Хлопи-
ну, 1932).
а также общего функционального значения слюнных желез и экзокрино-
вой части поджелудочной железы. Тем интереснее те различия, которые
обнаруживает их тканевой материал лишь в относительно незначительной
степени при обычных и в очень резкой форме при экспериментально из-
мененных условиях существования.
J38
Глава III
Железы, производные покровных эпидермальных пластов, предста-
вляют собой у млекопитающих разветвленные или, реже, неразветвленные
трубчатые ходы, стенка которых отличается двурядной, а иногда двуслой-
ной эпителиальной выстилкой. Лишь главные выводные протоки некото-
рых из них имеют на известном протяжении многослойную стенку, не отли-
чающуюся принципиально по своему строению от покровного пласта
Расположенные у основания двурядной выстилки базальные клетки во
многих случаях имеют своеобразную вытянутую параллельно соедини-
тельнотканной подстилке или разветвленную форму и отличаются, пови-
димому, подобно гладким мезенхимным элементам, способностью к сокра-
щению. Методом тканевых культур изучались, главным образом на кро-
личьем материале, молочная железа (Максимов, 1925; Хлопан, 1932),
конъюнктивальная, т. наз. гардеровская (Хлопин, 1932), под-
челюстная (Хлопан, 1923; Назу, 1923) и околоушная (Цымбал, 1932) же-
лезы. Превращения эксплантированных частиц этих желез имеют очень
много общего. Их эпителий может расти экстенсивно, наряду с соедини-
тельной тканью, или эпителизировать поверхность эксплантата. При экс-
тенсивном росте по поверхности плотного субстрата образуются тонкие
плоскостные однослойные мембраны и более толстые пласты, а при погруж-
ном росте в толще плотного субстрата — сплошные или полые внутри
тяжи и толстые клеточные массивы (рис. 9, 10). У зародышей и новорожден-
ных животных эпителиальные элементы всех отделов железистых ходов
слюнных и гардеровской желез оказываются способными к росту и про-
лиферации. У выводных протоков эта способность сохраняется у молодых
й взрослых животных; это же оказывается справедливым и в отношении
концевых или главных отделов слюнных желез. У гардеровской железы
главные отделы ко времени прозревания новорожденных животных пере-
полняются характерными липоидными включениями и с возрастом быстро
утрачивают способность к росту' и пролиферации вне организма. Энергия
роста эпителия молочной железы очень сильно зависит от ее функциональ-
ного состояния и бывает наибольшей у беременных или только что родив-
шихся животных (Франческой и Цамбелли, 1933). Из молочной железы бере-
менных мышей Фишеру (1930) удалось выделить чистые культуры эпителия.
Прорастающие экстенсивно железистые клетки, повидимому, перестают вы-
рабатывать свой специфический секрет и утрачивают включения зимогена.
Если апикальные концы клеток, обращенные в просвет концевого отдела же-
лезы, бывают сильно переполнены секреторными включениями, то они отми-
рают и отторгаются, а базальные части клеток, содержащие ядро, могут со-
хранять свою жизнеспособность, расти и делиться митотическим путем. Мио-
эпителиальные корзинчатые клетки постепенно перестают отличаться от
других эпителиальных элементов и наравне с ними участвуют в явлениях
роста и пролиферации. При менее благоприятных условиях существования
железистые ходы нередко нацело отмирают на большем или меньшем про-
тяжении. Внутри эксплантата живые железистые ходы могут превращаться
в системы кистовидно расширенных полостей, эпителиальная выстилка
которых нередко становится двуслойной. Постепенно элементы разных
Экспериментальный метод в гистологии
139
отделов железистых ходов, упрощаясь и уподобляясь друг другу, прини-
мают одинаковый недифференцированный характер. Плоскостные одно-
слойные мембраны, состоящие из уплощенных элементов, могут путем утол-
щения и наслоения клеток друг на друга превращаться в более массивные
пласты. Элементы мембран из функционирующей молочной железы бывают
очень богаты жировыми включениями, которые, по мнению Франческой
и Цамбелли, являются продуктами специфической секреции.
Образующиеся при погружном экстенсивном росте измененных желе-
зистых ходов клеточные массивы, состоящие из сомкнутых многогранных
Рис. 11. Превращение двурядного эпителия (а) млечного хода мо-
лочной железы беременного кролика при эпителизации поверхности
эксплантата в двуслойный пласт с уплощенными клетками (б) и в
тонкую однослойную мембрану (в) в 2-дневной культуре. Утолщение
и вторичная дифференцировка дву- (г) и многослойного (д) покров-
ного эпителия. Тот же исходный материал после 13 дней культи-
вирования. Сильное увеличение. (По Хлопину, 1932).
элементов, во многих случаях подвергаются известной гистологической
дифференцировке. Между их клетками, отделяющимися тонкими щелями,
образуются межклеточные мостики. Местами клетки, концентрически
наслаиваясь друг на друга, образуют структуры, сходные с жемчужинами.
В культурах из молочной железы центральные части таких концентри-
ческих образований даже претерпевают изменения, сходные с ороговением.
От начинающих прорастать измененных железистых ходов посеянного
кусочка и образованных ими при экстенсивном росте мембран, пластов
и клеточных массивов нередко отделяются свободные клетки или их группы,
которые, округляясь, свободно плавают в жидкости или растут среди эле-
ментов соединительной ткани. Среди изолированных эпителиальных кле-
ток в культурах из гардеровской железы обнаруживаются элементы двух
сортов: способные к амебоидному движению или могущие образовать за-
остренные отростки и сохраняющие шаровидную форму, поверхностный
140
Глава III
слой протоплазмы которых представляется уплотненным наподобие кути-
кулы. Иногда клеточные комплексы, разрыхляясь, образуют сетевидные
структуры, по внешнему виду могущие напоминать соединения мезенхим?
ных клеток. .
Первоначально тонкие, часто однослойные, эпителизирующие поверхг
ность эксплантата, пласты постепенно утолщаются и превращаются в дву?
и многослойный покровный эпителий с призматическими или уплощенными
покровными клетками, могущими обнаруживать явления дегенерации
и отторгаться (рис. 11). Базальные клетки дву- и многослойных покров-
ных пластов, образовавшихся в эксплантатах, наоборот, сохраняют свою
жизнеспособность и могут делиться митозом.’
Полученные в последнее время Михайловым и еще не опубликованные
данные, касающиеся сложной клубочковой кожной железы кролика, по-
строенной по типу потовых, показывают, что эпителиальная выстилка ее
железистых ходов способна к пролиферации и экстенсивному росту мем-
бранами, пластами и массивами, а также к эпителизации путем активного
движения поверхности эксплантата. От клеточных комплексов зоны роста,
как и во многих других случаях, изолируются отдельные клетки или группы
тесно соединенных по две-три и более клеток. Растущим эпителием сверток
фибрина сильно разжижается с образованием полостей, наполненных
жидкостью. Среди элементов зоны роста, наряду с вполне жизнеспособ-
ными клетками, делящимися митотическим путем и окрашивающимися
на фиксированных препаратах основными красками, бросаются в глаза
элементы круглой формы с уплотненным поверхностным слоем прото-
плазмы, красящейся кислыми красками. Эти клетки, являющиеся дегене-
ративными, вместе с тем обнаруживают специфические морфологические
изменения, сходные с теми, которые наблюдаются при атипичном орогове-
нии, например при опухолевом росте в целом организме. Сходные резуль-
таты были получены в последнее время Пинкусом (1938) в отношении пото-
вых желез человека. В самое последнее время Михайлову удалось получить
в эксплантатах из сложной клубочковой железы кроликов образование
многослойного покровного эпителия с явлениями ороговения.
Опыты экспериментального изучения пролиферативных процессов эпи-
телия эпидермальных желез в составе организма касаются преимуще-
ственно слюнных желез. Этому вопросу посвящены работы Подвысоцкого
(1886), Рибберта (1897), Марцокки (1904), Левенштейна (1910), Макья-
руло и Бюнглера (1929) и новейшие исследования Искры (1937). Во всех
случаях наблюдались сходные изменения в эпителии, хотя не все авторы
получали то же самое разнообразие гистологических картин.
Подвысоцкий и Рибберт описывали после повреждения регенеративные
процессы преимущественно за-счет эпителия выводных протоков, который,
пролиферируя, образовывал клеточные скопления, превращавшиеся в даль-
нейшем в железистые «ацинусы» или трубки. Марцокки целиком выделял
подчелюстную железу кроликов из окружающих тканей и, после пере-
вязки выводного протока, зашивал ее на старом месте. Одновременно с ги-
белью железистой паренхимы он наблюдал пролиферацию корзинчатых
Экспериментальный метод в гистологии 141
клеток, образовавших первоначально сплошные тяжи, а затем трубчатые
ходы и даже белковые концевые отделы, которые, правда, в дальнейшем
исчезали.
В работах Левенштейна с введением шарлахового масла, Макьяруло
и Бюнглера с инъекциями каменноугольного дегтя не было достаточно
тщательного анализа по стадиям всего процесса в целом. Наиболее полные
данные принадлежат Искре, тщательно изучившему в лаборатории Гар-
шина воспалительные разрастания эпителия подчелюстной железы кро-
ликов после инъекций каменноугольного дегтя. На них мы и остановимся
подробнее.
В развивающемся в результате введения дегтя воспалительном очаге
эпителий разных частей железы (междольковых протоков, слюнных тру-
бок, вставочных и главных отделов) обнаруживает очень резко выражен-
ные пролиферативные изменения. Специальная дифференцировка клетками
утрачивается; они приобретают, как и в условиях эксплантата, сходный
малодифференцированный вид и, разрастаясь, образуют сплошные тяжи
и массивы, состоящие из более или менее однородных полиэдрических
или вытянутых в длину элементов, тесно прилегающих друг к другу. До-
растая до границы между грануляционной тканью и некротическими мас-
сами с каплями дегтя, эпителиальные пролифераты образуют многослой-
ные пласты, которые в конце концов превращаются в стенку кистовидных
полостей.
Эпителиальная выстилка кист, дифференцируясь, приобретает строение
многослойного плоского эпителия, сходного с нормальным покровом ро-
товой полости. Затихание пролиферативных процессов в эпителии и его
вторичная дифференцировка связаны с созреванием грануляционной ткани
и развитием в ней межклеточного вещества. При воспалительных разрас-
таниях эпителия слюнных желез обнаруживаются камбиальное значение
и резко выраженная способность к пролиферации корзинчатых миоэпи-
телиальных клеток.
Очень сходные результаты получаются также в опытах с импланта-
цией частиц слюнной железы с кусочками целлоидина (Лазаренко, 1934).
Согласно этому автору, в концевых отделах и слюнных трубках секретор-
ные и палочковидно исчерченные элементы отмирают, а малодифферен-
цированные эпителиальные разрастания тяжами и массивами образуются
за счет набухающих и усиленно размножающихся митозами корзинчатых
клеток.
Кроме того, Лазаренко наблюдал на поздних стадиях, когда пролифера-
тивные процессы уже закончились, образование в составе многослойной
выстилки кист элементов, напоминающих железистые клетки. Применяв-
шаяся автором методика окраски препаратов не позволила, к сожалению,
изучить детальнее природу сходных со слизью секреторных включений.
Воспалительные разрастания эпителия желез, сходные с описанными
выше, несомненно могут быть обнаружены на человеческом патологическом
материале (Гаршин, 1939). В частности, Захарьевской (1935) удалось по-
дробно изучить процесс эпителизации бугорков при туберкулезе молочной
142
Глава III
железы. Эпителий этой последней также образовал малодифференцирован-
ные тяжи и массивы, за счет которых развивалась сплошная выстилка
вокруг бугорков, построенная по типу многослойного плоского эпителия
кожи.
Как известно, резко выраженные пролиферативные процессы в эпите-
лии молочной железы и образование им свойственных эпидермису много-
слойных структур могут быть вызваны гормональными воздействиями.
Наконец, очень поучительно сопоставление реактивных изменений
в сальных железах, полученных методом эксплантации и при их повре-
ждении в целом организме. Интересным превращениям, сходным с эпидер-
мисом, подвергаются эксплантаты из сложных больших сальных желез
крыс, открывающихся в наружный слуховой проход и впервые подробно
изученных Цымбалом (1933). Сальные железы можно себе представить,
как свернувшиеся и погрузившиеся в толщу соединительной ткани уча-
стки кожного эпидермиса с измененной функцией, сохраняющие в прин-
Рис. 12. Образование ороговевающего многослой-
ного пласта из эпителия концевого отдела саль-
ной железы кролика в 9-дневной культуре. Силь-
ное увеличение. (По Цымбалу, 1933).
ципе основные черты его строе-
ния. В их секреторных от-
делах имеются граничащие с
соединительной тканью кам-
биальные клетки и несколь-
ко слоев элементов, подвер-
гающихся в направлении к
центру, вместо ороговения,
своеобразному жировому пе-
рерождению и постепенной
дегенерации. После эксплан-
тации частиц сальной железы
ее камбиальные клетки
могут, размножаясь, давать
экстенсивный рост мембранами или пластами и эпителизировать
поверхность посеянного кусочка, а также пролиферировать в со-
ставе находящихся внутри его железистых отделов. Ожиревшие же
«сальные» клетки подвергаются полной дегенерации и распаду. Пролифе-
рируя внутри и на поверхности эксплантата, камбиальные элементы саль-
ной железы наслаиваются друг на друга и подвергаются дифференцировке,
но уже не в железистые, а в ороговевающие элементы эпидермиса, обра-
зуя слои зернистых и безъядерных роговых клеток (рис. 12). В тех участ-
ках эпителизирующего пласта, где дифференцировка не заходит так далеко,
образуется по крайней мере дву-трехслойный покров с уплощенными
поверхностными клетками, отличающимися от базальных по своей окраске.
В экстенсивно растущих плоскостных мембранах, отличающихся по взаим-
ному расположению клеток весьма низкой гистологической дифферен-
цировкой, отдельные элементы или их небольшие группы могут приобре-
тать характер клеток, находящихся на ранних стадиях ороговения. Если
таким изменениям подвергаются несколько расположенных рядом клеток,
то они образуют концентрические структуры. Мембраны и эпителизирующие
Экспериментальный метод в гистологии
148
пласты эксплантатов из сальной железы сильно разжижают фибрин и обра-
зуют в нем полости, наполненные жидкостью.
Пролиферативные процессы в сальных железах в составе организма
изучал в свое время Подвысоцкий (1886, 1888), производивший надрезы
в веке кролика. Периферические (камбиальные) клетки поврежденных
меймобовых желез очень быстро реагировали интенсивной митотической
активностью, в результате которой альвеолы, вместо дегенерирующих
сальных клеток, переполнялись малодифференцированными кубическими
элементами эпителия и превращались в сплошные эпителиальные массивы.
В окружающую соединительную ткань врастали сплошные эпителиальные
ночки, которые развивались особенно сильно в направлении организующе-
гося свертка фибрина. Из глубоких слоев эпидермиса также возникали
врастающие в соединительную ткань эпителиальные массивы. За счет
этих эпителиальных разрастаний разного происхождения в конце концов
происходило закрытие дефекта в покровном эпителии.
Результаты, полученные Мизуми (1909) при повреждении больших аналь-
ных сальных желез кролика замораживанием, в некоторых отношениях
очень напоминают описанные выше данные Цымбала. После замораживания
все сальные клетки отмирают и отторгаются, а камбиальные клетки, про-
лиферируя и изменяя направление дифференцировки, образуют на месте
сальных клеток концентрические слои уплощенных элементов. Эти изме-
нения имеют обратимый характер и подходят под категорию явлений, обо-
значаемых Гаршиным как транзиторпые изменения.
д) Эпителий эпидермальных эндокриновых органов
Несмотря на большое количество экспериментальных исследований над
эндокриновыми органами, количество данных, демонстрирующих гисто-
бластические потенции их эпителиев, невелико. Остановимся лишь на тех
немногих работах, в которых имеется фактический материал по интересую-
щим нас ближе вопросам.
Опыты с эксплантацией гипофиза млекопитающих производились Ка-
лахари (1936), Андерсоном и Геймекером (1935), Энгелем и Вербером (1937),
Гайаром (1938) и в нашей лаборатории Цымбалом (неопубликованные дан-
ные). Тщательно обработанный гистологический материал последнего ав-
тора по аденогипофизу новорожденных кроликов я положу в основу даль-
нейшего изложения.
В эксплантатах из передней доли гипофиза имеет место образование
плоскостных мембран, пластов, клеточных массивов и тяжей, а также хо-
рошо выражены явления эпителизации. Разжижение свертка фибрина с обра-
зованием в нем полостей происходит энергично. Разрастающиеся по поверх-
ности плотного субстрата мембраны состоят большей частью из тесно сомк-
нутых клеток и могут, утолщаясь, образовать многослойные пласты. При
образовании полостей разжижения зона роста, как в других случаях, дефор-
мируется, превращаясь в массивный клеточный вал по краю неразжиженного
фибрина. Эпителизации могут подвергаться как поверхность самого экс-
144
Глава III
плантата, так и подсаженные частицы других органов, например мышц
желудка (Цымбал). При эпителизации образуются многослойные покров-
ные пласты, состоящие из полиэдрических элементов. Между клетками
пластов, нередко разделенных узкими щелевидными промежутками, могут
образоваться межклеточные мостики.
Растущие и пролиферирующие митотическим путем эпителиальные эле-
менты гипофиза имеют индифферентный характер. Казахара (1936) описывал
в условиях замедленного роста появление в клетках мембран и пластов
оксифильных зернышек, трактуя это как цитологическую дифференцировку
специфических для этого органа оксифильных клеток. Он же отмечал обра-
зование вне организма, при особых условиях культивирования, элементов,
напоминающих не вполне типичные главные и базофильные клетки. Ука-
зания на дифференцировку специфических элементов гипофиза в условиях
жизни вне организма еще нуждаются в проверке и подтверждении. Обра-
зования гормонов элементами гипофиза, растущими вне организма, уста-
новить не удалось (Кеттинг и Льюис, 1938).
Гистологических данных, касающихся изменений аденогипофиза в усло-
виях целого организма, сравнимых с описанными, мне неизвестно.
Удачные эксплантаты из щитовидной железы куриных зародышей
получили Каррель и Берроуз еще в 1911 г. Длительные чистые культуры
эпителия этого органа на том же объекте изучили Ибелинг (1924) и Демут
(1932). В гистологическом отношении все эти работы недостаточно полны.
На кроличьем материале сделаны работы Шампи (1915), данные которого
в настоящее время устарели, и Галустяна (1940), давшего тщательный гисто-
логический анализ роста и превращений эпителия щитовидной железы вне
организма.
Клетки фолликулов щитовидной железы, энергично делясь митотическим
путем, дают очень разнообразные картины экстенсивного роста. Явления
эпителизации в разных случаях выражены неодинаково и, повидимому,
зависят от возрастного фактора. В то время как у зародышей млекопитаю-
щих и новорожденных эпителизация нередко приводит к полному обраста-
нию эксплантатов эпителиальным покровом, у взрослых животных явления
эпителизации выражены слабо и в большей части случаев отсутствуют со-
вершенно {Галустян). Разжижение фибрина также не постоянно. При
экстенсивном росте мембранами и пластами, а также в случаях эпителиза-
ции, в свертке фибрина может быстро наступать образование полостей
разжижения. В других случаях экстенсивный рост эпителия тяжами в толще
фибрина и среди проросшей соединительной ткани может почти или совсем
не сопровождаться образованием полостей и деформацией зоны роста.
Способность к погружному росту в свертке фибрина без его разжижения
и между' соединительнотканными элементами выражена у эпителия щито-
видной железы в значительно более резкой степени, чем у рассмотренных
выше представителей эпидермального тканевого типа. Образующиеся при
этих условиях подчас очень длинные клеточные тяжи могут ветвиться и
образовать более или менее сложный переплет в зоне роста. Фолликулы
посеянного кусочка частью запустевают, превращаясь в массивные эпи-
Экспериментальный метод в гистологии
145
телиальные комплексы, частью кистовидно вздуваются. В некоторых из
них коллоид, уплотняясь, может сохраняться долгое время.
Новообразование в чисто эпителиальной зоне роста фолликулов с кол-
лоидом, которое описывал Ибелинг, не подтвердилось в опытах Демута
и Галустяна. Повидимому, Ибелинг имел дело не с новообразованием,
а с сохранением предсуществовавших фолликулов, которые при энергичном
росте могли увлекаться разрастающимся эпителием в зону роста. Это явле-
ние на ранних стадиях культивирования с достоверностью проследил Галу-
стян. При эпителизации, специально изучавшейся Галустяном, в эксплан-
татах из эмбрионального свиного материала, на поверхности эксплантата
мог образоваться многорядный и многослойный покровный эпителий с упло-
щающимися на свободной поверхности клетками. В случае эпителизации
эксплантатов из незародышевого материала покровные пласты сохраняли
свое однослойное строение, хотя и, утолщаясь местами, приобретали харак-
тер призматического эпителия. Образование многослойного покровного
эпителия наблюдать не удавалось. От эпителиальных комплексов могли
изолироваться свободные одно- и многоядерные клетки разнообразной
формы.
Превращение эпителиальных элементов щитовидной железы в недиф-
ференцированные соединительнотканные клетки, описанное в свое время
Шампи (1915) и не подтвержденное никем из других исследователей, пови-
димому, объясняется, в первую очередь, его неправильной трактовкой обще-
известного в настоящее время факта изоляции эпителиальных клеток,
приобретающих при этом нередко веретенчатую или отростчатую форму.
Эти обычные в условиях тканевых культур явления уже отмечались не-
однократно в нашем предыдущем изложении. Вместе с тем описанное
им массовое превращение эпителиальных элементов щитовидной железы
в соединительнотканные элементы на ранних стадиях культивирования,
не наблюдавшееся никем из других исследователей, заставляет считать,
что примененная Шампи гистологическая методика была настолько далека
от совершенства, что его препараты не позволили ему отличить эпителия
от соединительной ткани. То же самое справедливо и в отношении данных
Шампи, касающихся эпителия почки, которые будут нами рассмотрены
ниже.
Опыты с регенеративными процессами в щитовидной железе кроликов
и собак после вырезания небольших участков органа были поставлены уже
давно Риббертом (1889), данные которого и в настоящее время сохраняют
значение основных. Правда, сейчас трудно сказать, за неимением соответ-
ствующего новейшего материала, в какой мере пролиферативные процессы,
наблюдавшиеся Риббертом, являются регенеративными и в какой мере
воспалительными.
После повреждения Рибберт отмечал появление митозов в клетках фол-
ликулов. Набухая и наслаиваясь друг на друга, пролиферирующие эпи-
телиальные клетки образовали дву- и многослойные структуры. В неко-
торых случаях фолликулы превращались в сплошные клеточные массивы,
без полости внутри. Кроме того, наблюдалось разрастание эпителиальных
10 X лопин—349
146
Глава III
тяжей и комплексов в грануляционной ткани, образующихся пролиферирую-
щими элементами поврежденных разрезом фолликулов. На более поздних
стадиях процесса Рибберт отмечал появление новых фолликулов. По его
данным, периферические и центральные фолликулы органа реагируют на
травму неодинаковым образом; регенеративные процессы обнаруживаются
преимущественно центральными фолликулами.
Согласно кратким данным Лазаренко (1934), после имплантации частиц
щитовидной железы с кусочками целлоидина, даже по истечении года,
можно наблюдать совершенно жизнеспособные фолликулы с коллоидом,
образовавшиеся заново из разраставшихся в очаге воспаления малодиффе-
ренцированных эпителиальных тяжей и комплексов. Более подробные гисто-
логические данные пока, к сожалению, отсутствуют.
Значительный интерес представляют данные Йошида (1932), изучав-
шего изменения в щитовидной железе морских свинок под влиянием под-
кожного введения раствора о-амидоазотолуола в масле. Продолжитель-
ность опытов достигала 5 месяцев. Автор первоначально отмечал у подопыт-
ных животных резко выраженные дегенеративные процессы в эпителии щито-
видной железы, сменявшиеся затем пролиферативными процессами. В эпи-
телии сохранившихся фолликулов появлялись митозы, в результате кото-
рых происходило резкое утолщение стенки фолликулов, приобретавшей
многослойное строение. В ряде случаев пласты, образовавшиеся из клеток
фолликулов, обнаруживали, по данным Йошида, далеко идущие явления
дифференцировки в типичный многослойный плоский эпителий с появлением
зернышек кератогиалина и резко выраженным ороговением. Ряд фоллику-
лов в конце концов превращался в роговые жемчужины.
Зобную железу млекопитающих изучали методом тканевых культур
Часовников (1927, 1929), Попов (1927) и Галустян (1939). Относительно из-
менений зобной железы амфибий в эксплантате имеются данные Паппен-
геймера (1913) и Вассена (1915). Наиболее полное исследование монографи-
ческого характера превращений тканевых элементов зобной железы, сде-
ланное параллельно методами эксплантации, регенерации, после вырезания
кусочков, ожога, воспалительных разрастаний (после введения шарлахо-
вого масла или каменноугольного дегтя) и имплантации с кусочками цел-
лоидина, принадлежит Галустяну, данные которого мы и положили в основу
дальнейшего изложения. В диссертации Галустяна (1940) имеется, кроме
того, исчерпывающий обзор всей соответствующей литературы. Экспери-
ментально-гистологический анализ зобной железы представляется особенно
интересным, потому что эпителиальные компоненты этого органа, эндо-
криновая функция которого еще не может считаться доказанной, кроме
телец Гассаля, при обычных условиях совершенно утрачивают свое ком-
плексное эпителиообразное взаимное расположение и образуют, подобно
соединительной ткани, сетевидную строму, петли которой заполнены много-
численными лимфоидными клетками (Гаммар, 1926, 1929; Максимов, 1909,
и др.). Поэтому зобная железа по своему строению отнюдь не является
железой, а представляет собой своеобразный лимфоэпителиальный орган
(Экке, 1899). После эксплантации тельца Гассаля погибают, а рыхлая
Экспериментальный метод в гистологии
147
сетевидная эпителиальная строма, состоящая из отростчатых клеток, при-
обретает свойственное тканям эпидермального типа комплексное строение
и обнаруживает очень энергичный экстенсивный рост мембранами, мас-
сивными Пластами и тяжами различной формы. Эпителизации поверхности
эксплантата никогда не наблюдается. Разжижение фибрина с образованием
полостей наступает лишь в виде исключения и в большей части культур
не имеет места, несмотря на энергичный рост эпителия. Лимфоидные эле-
менты органа частью дегенерируют на месте, частью выползают в окружаю-
щий фибрин. В первые дни после посева разрастающийся эпителий может
быть еще в большей или меньшей степени инфильтрирован ими. С течением
времени лимфоциты совершенно исчезают. Наряду с эпителием могут про-
растать обычным образом и элементы соединительной ткани. Во многих
случаях удается получить эксплантаты с чисто эпителиальной зоной роста.
Если зобная железа до эксплантации была подвергнута рентгенизации
{Часовников, 1929; Галустян, 1939), то лимфоидные элементы исчезают
особенно быстро. По наблюдениям Галустяна, быстрее и энергичнее всего
разрастаются и пролиферируют митотическим путем краевые клетки кор-
кового вещества долек зобной железы, граничащие с соединительноткан-
ными прослойками и имеющие, повидимому, значение камбиальных эле-
ментов. Эпителиальная сеть по направлению к центру долек, особенно в их
мякотном веществе, постепенно утрачивает способность к росту и размноже-
нию. Согласно этим данным, Галустян предполагает, что зобную железу
можно представить себе, как эпидермальный массив, проросший соедини-
тельной тканью, с кровеносными сосудами, и инфильтрованный лимфоци-
тами, периферические элементы которого имеют камбиальный характер,
а расположенные ближе к центру находятся на разных стадиях дифферен-
цировки, старения и дегенерации. Последняя проявляется в специфическом
для эпидермальных тканей высших позвоночных виде и сопровождается
образованием гассалевских телец, т. е. образований, напоминающих жем-
чужины и состоящих из концентрически наслаивающихся друг на друга
клеток, которые подвергаются изменениям, сходным в известных отноше-
ниях с ороговением.
Разнообразные опыты над зобной железой, поставленные Галустяном
в условиях целого организма, полностью подтвердили его приведенный
выше вывод, касающийся общего строения органа. Кроме того, они дали
очень ценный новый гистологический материал. При всех методах повре-
ждения паренхима зобной железы освобождается на большем или меньшем
протяжении от лимфоцитов, значительная часть которых погибает и разру-
шается на месте, а часть эмигрирует в соединительную ткань и кровеносные
сосуды. Массовая гибель лимфоцитов наблюдается и после рентгенизации.
Вдали от места повреждения, где ткань зобной железы не подвергается
резким изменениям, в первые дни наблюдается резкое увеличение коли-
чества и размеров гассалевских телец. Вследствие того, что лимфоциты
в первую очередь начинают исчезать на периферии долек, камбиальная зона
эпителия так же, как в условиях эксплантата, начинает выступать осо-
бенно отчетливо. При дальнейшем исчезновении лимфоцитов поврежден-
Глава III
148
ные дольки зобной железы постепенно превращаются в эпителиальные
массивы, состоящие из полиэдрических клеток.
Эпителиальные элементы мякотного вещества оказываются наиболее
чувствительными и при прочих равных условиях погибают первыми. Чем
дальше к периферии дольки, тем жизнеспособнее эпителиальные элементы.
К активной пролиферации и росту в очаге воспаления способны эпителиаль-
ные элементы коркового вещества и, в первую очередь, периферические
камбиальные элементы, в которых обнаруживается очень большое число
митозов. Интенсивность воспалительных разрастаний сплошными клеточ-
ными тяжами разного калибра, как и в других случаях, зависит от течения
(длительности, интенсивности и характера) воспалительной реакции в соеди-
нительной ткани. Эти воспалительные разрастания эпителия зобной железы,
состоящие из вытянутых или полиэдрических малодифференцированных
элементов и нередко сетевидно анастомозирующие друг с другом, в точности
соответствуют по своей морфологии аналогичным разрастаниям других
описанных выше эпидермальных тканей. Эти эпителиальные тяжи, дора-
стая до очагов некроза, образуют эпителизирующие их пласты. Особенно
интересные результаты были получены после введения дегтя, когда в ряде
случаев удавалось наблюдать образование замкнутых кист, выстланных
многослойными эпителиальными пластами. Последние обнаруживали очень
далеко идущие явления дифференцировки, превращаясь в типичный много-
слойный плоский эпителий с палисадообразно расположенными клетками
базального слоя и уплощенными покровными элементами. В ряде случаев
явления дифференцировки завершались совершенно типичным ороговением,
с образованием кератогиалина и мощного рогового слоя, обращенного
в просвет кисты. Ороговевшие клетки, отторгаясь, скоплялись в окаймлен-
ной эпителием полости, располагаясь между некротическими массами и
каплями дегтя. Между клетками шиповатого слоя покровного эпителия
можно было наблюдать межклеточные мостики.
При имплантации по методу Лазаренко первые изменения сохраниввшх
жизнеспособность частиц зобной железы соответствовали описанным выше.
Наблюдались и исчезновение лимфоцитов и образование разрастающихся
в очаге воспаления малодифференцированных эпителиальных тяжей без
последующей эпителизации очагов некроза с кусочками целлоидина. В даль-
нейшем, однако, разросшиеся эпителиальные тяжи, изменяя свою струк-
туру и инфильтрируясь лимфоцитами, могли вновь превращаться, при
затихании воспалительной реакции, в отдельные небольшие участки долек
зобной железы с вновь образованными тельцами Гассаля.
Сопоставление полученных с помощью неодинаковых эксперименталь-
ных методов на одном и том же объекте результатов, из которых мы отме-
тили лишь некоторые, наиболее существенные, для разбираемых здесь во-
просов весьма поучительно в различных отношениях.
Исчезновение лимфоцитов и образование гистоморфных клеточных эпи-
телиальных массивов и тяжей происходят практически тождественным
образом под влиянием всех изучавшихся методов воздействия, как вне,
так и в составе организма. Плоскостные тонкие мембраны являются, как
Экспериментальный метод в гистологии
149
и во многих других случаях, формой существования тех же малодифферент
цированных разрастаний, свойственных в первую очередь особым условиям
жизни вне организма и лишь в виде исключения наблюдающихся при дру-
гих пролиферативных процессах.
Явления эпителизации поверхности эксплантатов и очагов некроза
с инородными телами при воспалительных разрастаниях оказываются по
своему биологическому значению различными процессами. Более того,
явления эпителизации инородных гел в организме обнаруживаются в очень
неодинаковой степени при разных условиях: при дегтярном воспалении
они выступают резче, а в условиях имплантации могут совершенно не
обнаруживаться.
В дальнейшем изложении мы познакомимся с рядом других примеров
этого. Эпителий зобной железы, не эпителизирующий при обычных усло-
виях культивирования поверхности эксплантата, образует прекрасные
эпителизирующие пласты и покровный эпителий при дегтярном воспалении.
Вместе с тем различная способность к эпителизации в условиях культур
несомненно является одним из показателей биологических свойств ткани.
В частности, в этом отношении разные изученные эпидермальные ткани
с большим постоянством заметно отличаются друг от друга. Большая часть
желез хорошо эпителизирует поверхность эксплантата. Из изученных эндо-
криновых органов способность к эпителизации и образованию покровного
эпителия, хорошо выраженная у аденогипофиза, оказывается значительно
более слабой у эпителия щитовидной железы и совершенно отсутствует
у зобной. В данном ряду способность к эпителизации совпадает более или
менее с фибринолитическими свойствами, чего в других случаях, о которых
речь будет ниже, не наблюдается. Наконец, на этом объекте, как и на мно-
гих других, подчеркивается тот факт, что явления дифференцировки в
условиях организма часто удается получить с большей легкостью, чем
в эксплантатах. Направление дифференцировки в условиях дегтярного
воспаления оказывается совершенно иным, чем в имплантатах. В эпителии
сальных желез и отчасти некоторых других производных кожного эпи-
дермиса ороговение или явления паракератоза получаются, наоборот,
с большой легкостью в условиях культур.
Нельзя не поставить в связь с своеобразным положением во внутренней
среде организма ослабление и утрату некоторыми эпидермальными произ-
водными, в частности эпителием упомянутых эндокриновых органов, спо-
собности эпителизировать (в условиях тканевых культур) поверхность
эксплантата и обоазовать дифференцированный покровный эпитепий, на-
ряду с резко выраженной у эпителия щитовидной железы способностьк
к погружному росту в толщу плотного субстрата. В этом случае вероятнее,
всего говорить об особом направлении филогенетического развигия неко-
торых эпидермальных тканей, которые, утратив в эволюционном процессе
барьерное и покровное значение, приобрели свойства особых тканей внут-
ренней среды организма.
Общеизвестные сравнительно-анатомические данные, касающиеся щито-
видной железы и гипофиза, хорошо гармонируют с этими предположениями.
150
Глава III
Щитовидная железа у всех позвоночных имеет во взрослом состоянии строе-
ние и функциональное значение эндокринового органа. У низших хордовых
и у пескоройки ее гомологом является гипобранхиальная борозда, т. е.
определенный участок эпителиальной выстилки переднего, с нашей точки
зрения, эпидермального отдела пищеварительной трубки, который, оче-
видно, затем утрачивает связь с внешней средой, но, как часть эпидермаль-
ного зачатка, эволюирует и приобретает у высших позвоночных некоторые
общие, на данном этапе эволюции, свойства большей части эпидермальных
образований. Аденогипофиз еще у низших позвоночных, круглоротых
(по крайней мере частью) имеет характер эпителиальной трубки (т. наз.
питуитарный отросток), сообщающейся с внешней средой. Только начиная
с рыб, ткань аденогипофиза полностью превращается в особую ткань внут-
ренней среды организма. Что касается зобной железы, то сравнитель-
ная анатомия не дает точных указаний на пути ее филогенетического раз-
вития. Этот орган нам известен только как своеобразное лимфоэпителиаль-
ное образование, встречающееся во всем ряду позвоночных. Попытки искать
его эволюционные корни в антенефридальных канальцах, соответствующих
передним выделительным канальцам ланцетника (Семон), не представляются
достаточно обоснованными. Явно эпидермальный характер эпителия зоб-
ной железы категорически говорит против этого предположения. Наоборот,
филогенетическая связь околощитовидных желез с эпителиальным покровом
жаберного аппарата рыб представляется весьма вероятной.
е) Дышащие части легких и выстилка плавательного пузыря
Относительно выстилки дышащих частей легкого разными описательными
и экспериментальными методами получены очень разноречивые данные,
не позволяющие еще высказать с полной определенностью суждение по
этому вопросу.
Результаты опытов с эксплантацией частиц легкого, кроме того, еще
в значительной степени неполны в гистологическом отношении.
Согласно одним из них, содержащие ядро клетки альвеолярной стенки,
описывавшиеся в классической гистологии, представляют собой соедини-
тельнотканные гистиоциты, которые в эксплантатах превращаются в на-
стоящих макрофагов (Ланг, 1925). Так как большая часть современных
авторов вместе с тем отрицает существование безъядерных плоских клеток
респираторного эпителия, то эти данные, казалось бы, говорят в пользу
точки зрения, защищаемой в настоящее время Поликаром, согласно кото-
рой респираторные отделы легких вообще будто бы лишены эпителиальной
выстилки. Многие, действительно, отмечают в легочных эксплантатах массо-
вое появление макрофагов, которым приписывается так же, как альвеоляр-
ным фагоцитам, то «эпителиальное», то соединительнотканное происхожде-
ние. Другие авторы описывали рост альвеолярного эпителия в культурах
из эмбрионального (Дальтон, 1925) и взрослого (Бинэ и Шампи, 1926)
легкого млекопитающих. Робб-Смит (1936), работавший с. эксплантатами
из легких взрослых кроликов, допускает существование альвеолярного
Экспериментальный метод в гистологии
151
эпителия, но в то же самое время приписывает альвеолярным фагоцитам
мезенхимное происхождение.
Новейшие данные {Сигурдсон, 1940) показывают, что при соблюдении
известных методических предосторожностей (наполнение легкого кровяной
плазмой перед его измельчением для посева) в культурах из легкого удается
получить разрастания своеобразных эпителиоподобных мембран, отличаю-
щихся от клеточных мембран, образуемых бронхиальным эпителием. Такие
мембраны, происходящие по предположению автора из респираторной вы-
зтилки, удается культивировать в чистом виде достаточно длительное
время.
Полученные в нашей лаборатории, еще не оп/бликованные данные
{Галустян) показывают, что характер зоны роста в эксплантатах из
эмбриональных и неэмбриональных легких оказывается очень различным.
В то время как в первом случае наблюдается рост вполне эпителиообразных
мембран, во втором образуются мембраны, элементы которых с первого
взгляда действительно напоминают распластанных по плотному субстрату
макрофагов, но отличаются от этих последних тем, что они своими резко
источенными прозрачными краевыми участками большей частью тесно
сомкнуты друг с другом. Мембраны, состоящие из такого рода элементов,
обнаруживают во многих отношениях внешнее сходство с растущим вне
организма мезотелием серозных оболочек, который будет разобран не-
сколько позже. Явлений эпителизации поверхности эксплантата альвео-
лярная выстилка не дает. При сопоставлении этих фактов с результатами,
полученными Гарлътоном (1925), Каффъе (1931) и Верном (1932) на эмбрио-
нальном кроличьем, кошачьем, человеческом и курином материале, следует
предположить, что выстилка будущих респираторных отделов эмбриональ-
ного легкого ведет себя в условиях культур более или менее сходным с брон-
хиальным эпителием образом, а в функционирующих легких изменяет свои
биологические особенности и становится своеобразной «респираторной»
тканью, отличающейся по ряду свойств от выстилки средних и крупных
бронхов и, в то же самое время, от производных мезенхимы. Это, вероятно,
связано с тем, что выстилка респираторных отделов утрачивает разграни-
чительные свойства, характерные для эпидермальных покровных пластов.
В настоящее время еще не представляется возможным изложить в развер-
нутом виде эту точку зрения и вместе с тем дать описание строения респи-
раторных отделов легкого у разных позвоночных. Во всяком случае, с нашей
точки зрения, представляется маловероятным, чтобы респираторные отделы
легких были или совершенно лишены особой выстилки, как это предпола-
гается у птиц (Баргман, 1936), или чтобы эта последняя была непрерыв-
ной у ряда млекопитающих {Баргман), а состояла из отдельных или рас-
положенных по несколько вместе «эпицитов» {Клара, 1936) с обнаженными
мезенхимными промежутками между ними.
Из разноречивых, но по существу недостаточно убедительных данных,
полученных при экспериментальных воздействиях на легочную ткань
в составе целого организма, упомяну лишь наблюдения Гаршина и Шабада
=(1935), согласно которым в альвеолах, обращенных к расположенному по
152
Глава III
соседству очагу воспаления, удается наблюдать появление непрерывной
изопризматической однослойной эпителиальной выстилки. То же самое
описывалось неоднократно при пневмониях и трактовалось как результат
ателектаза, т. е. непроходимостью для воздуха участка воздухоносных путей,
расположенного по соседству с воспалительным очагом. Согласно предпо-
ложению Гаршина, здесь дело заключается не в ателектазе, а в реактивном
изменении выстилки альвеол под влиянием воспаления.
Изучение методом эксплантации выстилки плавательного пузыря кости-
стых рыб (Хлопан, 1937) показало, что она разрастается вне организма
комплексными тонкими эпителиальными мембранами. По своей способности
к образованию дифференцированных покровных пластов она отличается
от большей части изученных эпидермальных тканей. Образуемые ею в усло-
виях эксплантата структуры всегда отличаются однослойным и однорядным
и, лишь в виде исключения, многорядным характером. С другой стороны,
при опытах Мухиной с трансплантацией участков стенки плавательного
пузыря (неопубликованные данные) удавалось получить заметное утолще-
ние его эпителия и приобретение им многорядной, а местами даже много-
слойной структуры. При сопоставлении всех этих, правда еще очень немного-
численных, данных с упомянутыми выше наблюдениями Мухиной (1941),
касающимися особенностей структуры эпителия плавательного пузыря
у ганоидных и разных костистых рыб при нормальных условиях, можно'
притти к выводу, что эта часть эпидермальных тканей постепенно претер-
певает, в связи со своей специализацией в особом направлении, известное
эволюционное упрощение. Возможно предположить, что выстилка респи-
раторных отделов легких низших и высших позвоночных еще в большей
степени изменяет свои свойства и в конце концов утрачивает (становясь
особой высокоспециализированной тканью внешнего газообмена) морфоло-
гические признаки, характеризующие все прочие эпидермальные пласты
позвоночных. В этом направлении необходимы еще дальнейшие сравни-
тельные экспериментальные исследования над представителями разных
классов позвоночных.
ж) Ткань хорды
Как это уже отмечалось выше, ткань хорды, по своим морфо-физиоло-
гическим особенностям, строго говоря, не может быть отнесена безогово-
рочно ни к одной из четырех тканевых групп классической гистологии.
В соответствии со своим функциональным или, точнее, морфо-физиологиче-
ским подходом к тканям Лейдиг (1855), а за ним и большая часть поздней-
ших гистологов рассматривали и рассматривают ткань хорды как разновид-
ность тканей соединительного вещества. Имеющая опорное значение и
в настоящее время относимая на этом основании и на основании известных
морфологических особенностей, вместе с мезенхимными пузырчатыми хордо-
идными и хондроидными тканями, в особую группу «опорных» и «выполняю-
щих» тканей (Шаффер, 1930), ткань хорды вместе с тем имеет определенные
«эпидермальные» структурные особенности (Студничка, 1931, 1932). У всех
позвоночных хорду можно представить себе как свернутый в сплошную
Экспериментальный метод в гистологии 153
трубку, без просвета внутри, эпидермальный пласт с расположенными на
периферии камбиальными элементами (эпителий хорды) и соответствующими
дифференцированным покровным элементам, превратившимся в высоко-
специализированные пузырчатые клетки, внутри. Часть оболочки хорды,
образованная, по Шафферу (1930), «хордальным» эпителием, должна тракто-
ваться как особого рода мощная базальная мембрана.
Целый ряд морфологических особенностей дифференцированных эле-
ментов хорды, детально изученный Студничкои, подчеркивает ее сходство
с покровными эпидермальными пластами.
Отмечавшиеся у рыб «хордоидные» участки эпидермиса (Кашкаров, 1913),
«хрящеподобные» участки эпителия глотки (Патуельт, 1925), пузырчатые
покровные слои эпидермиса губ и некоторых других частей тела говорят
о том же. К сожалению, экспериментально-гистологический материал, ка-
сающийся хорды, почти отсутствует, а на основании тех немногих данных,,
которые имеются, пока нельзя строить определенных выводов. В условиях
культур (Леви, 1919) ткань спинной струны куриных зародышей образует
плоскостные мембраны, содержащие крупные вакуоли. От хордальных
мембран, имеющих, по этим данным, синцитиальное строение, могут изо-
лироваться многоядерные цитоплазматические скопления и отдельные
клетки. Во всяком случае, даже этих скудных данных достаточно, чтобы
убедиться в эпителиообразном, а отнюдь не сходном с соединительной
тканью, характере материала хорды.
При опытах с регенерацией-хвоста амфибий и рептилий (Барфурт, 1891)
из клеток хорды, повидимому, возникает палочковидное опорное образо-
вание хрящеподобного характера. Частичное превращение хорды в хрящевую
ткань неоднократно описывалось и при обычных условиях (Шаффер, 1930),
причем источником образования хряща являются не дифференцированные
пузырчатые клетки спинной струны, а «хордобласты» или т. наз. эпителий
хорды, имеющий камбиальное значение. Детальный гистологический анализ
превращений ткани хорды и возникающих из нее образований одна из
задач будущего, имеющих, правда, ограниченный практический интерес,
например, с точки зрения изучения редкой опухолевой группы хордом, но
несомненно большое теоретическое и общебиологическое значение. Относи-
тельно филогенетических корней хорды современная эмбриология не дает
определенных указаний. Хорда возникает, как самостоятельный зачаток,
топографически особенно тесно связанный с презумптивной нервной пла-
стинкой и областью первичного рта; но его генетическая связь с тем или
иным зародышевым зачатком позвоночных эмбриологически не устанавли-
вается. У кишечнодышащих хордоподобное образование генетически свя-
зано с выстилкой передней кишки. В эмбриональном развитии амфибий
Фогт (1929) особенно подчеркивает тесную связь дорзальной стенки пер-
вичной кишки, т. е. пластинки хорды, с одной стороны с краем бластопора,
а с другой —с прехордальной пластинкой головной кишки. А область,
прехордальной пластинки, как это будет разобрано ниже, при своей даль-
нейшей гистологической дифференцировке дает начало эпидермальным^
тканям.
154
Глава III
Все это вместе взятое заставляет считать вероятным, что ткань спинной
струны так или иначе связана генетически с наружным эктодермальным
покровом отдаленных предков хордовых. Решение этого вопроса принадле-
жит будущему.
з) Эмбриональные зачатки эпидермиса и выстилки головной кишки
Изолированный презумптивный эпидермис зародышей амфибий в усло-
виях существования без подстилки обнаруживает ограниченную жизне-
способность. В течение некоторого времени он сохраняет комплексное рас-
положение клеток, но не дает никакой гистологической дифференцировки.
Его клетки оказываются лишь способными к цитоморфной дифференцировке
мерцательного аппарата, свойственного ранним этапам развития разных
эмбриональных зачатков. Клеточный, а не тканевой характер этой диффе-
ренцировки обнаруживается в совершенно беспорядочном, не направленном
в определенную сторону, мерцании ресничек отдельных клеток. Эта осо-
бенность ярко бросается в глаза в опытах Гольтфретера (1933) с экзо-
гаструляцией, в которых у зародышей аксолотля, помещенных в гипото-
нический рингеровский раствор (0.35%), наблюдалась не инвагинация,
а эвагинация всей вегетативной и краевой зоны бластулы, а презумптивные
зачатки кожной и нейральной эктодермы оставались в виде полого внутри
пузырька. Этот эктодермальный пузырек спадался в неправильную мор-
щинистую массу, оставаясь связанным с экзоэмбрионом, постепенно сужаю-
щимся стебельком; последний в дальнейшем легко разрывался или распа-
дался. При неполной экзогаструляции эктодерма, оставшаяся на поверх-
ности экзоэмбриоиа в составе полого мешка с большим или меньшим коли-
чеством элементов других зачатков внутри, обнаруживала ту или иную,
в зависимости от условий и индуцирующих воздействий, дифференцировку.
Направления гистологической дифференцировки эктодермы до мозаич-
ной стадии развития зародыша, совпадающей с ранней нейрулой, в зависи-
мости от внешних условий и индуцирующих воздействий, как показывают,
например, опыты с трансплантацией {Мангольд, 1924) или интерплантацией,
очень разнообразны или, вернее, практически безграничны. С момента пер-
вого появления нервных валиков наступает ее детерминация.
Весь приведенный выше экспериментально-гистологический материал
доказывает, что гистологическая детерминация становится окончательно
закрепленным признаком всех развивающихся из кожной эктодермы тканей,
которые в дальнейшем характеризуются строгой специфичностью приобре-
тенных ими свойств и неспособностью превращаться в ткани другого проис-
хождения, образующиеся из других зачатков. С наступлением мозаичной
стадии каждый эмбриональный зачаток, и в частности кожная эктодерма,
становится источником развития строго определенного комплекса тканей, в
большей части случаев характеризующихся некоторыми общими гистоти-
пическими особенностями, на фоне которых проявляется их еще более узкая
специфичность, в каждом отдельном случае имеющая известные качествен-
ные отличия. В отдельных, правда редких, случаях высокая специализа-
Экспериментальный метод в гистологии
155
ция ткани в определенном направлении может, повидимому, совершенно
скрыть или уничтожить общий гистотипический фон.
Изолированная эктодерма, помещенная на плотный субстрат в условиях
тканевых культур в плазме или даже физиологическом растворе (Гольт-
фретер, 1938; Винников, неопубликованные данные), обнаруживает спо-
собность к росту плоскостными комплексными мембранами, не проходя
предварительно стадий тканевой дифференцировки. Даже в состоянии
роста малодифференцированными мембранами кожная эктодерма или эпи-
дермальный зачаток отличается от других зачатков, поставленных в те же
условия существования.
Уже на очень ранней стадии развития, т. наз. прехордальная пластинка
или презумптивный зачаток переднего отдела пищеварительной трубки,
расположенный в области намечающейся дорзальной губы бластопора,
отличается по своим свойствам от презумптивной кишечной эктодермы.
Это с полной убедительностью показывают опыты Куше (1929) с интерплан-
-тацией в глазницу и Гольтфретера (1938) с изоляцией участков бластулы и
ранней гаструлы. При экзогаструляции из зачатка головной кишки диф-
ференцируется характерный в гистологическом отношении эпителий перед-
него отдела пищеварительной трубки, резко отличающийся от дифферен-
цирующегося одновременно энтодермального эпителия желудка и кишек.
В условиях роста малодифференцированными мембранами выстилка
переднего отдела пищеварительной трубки очень сильно отличается от
собственно энтодермы, но вместе с тем и не обнаруживает тождества с кож-
ной эктодермой. Резкие отличия зачатка переднего отдела пищеваритель-
ной трубки от расположенных более каудально собственно энтодермальных
отделов нельзя не сопоставить с резкой гистологической границей, имею-
щейся между дифференцированным эпителием полости рта и пищевода,
с одной стороны, желудка и кишек, с другой. Поэтому в настоящее время
есть все основания различать в составе стенки кишечной трубки зародыша
два резко отличающихся друг от др}та зачатка — зачаток выстилки голов-
ной или передней кишки и собственно энтодерму; их до настоящего времени
принято обозначать по топографическому признаку, несмотря на их раз-
личия, общим собирательным термином < энтодерма'}, который держится в силу
традиций и затемняет истинное положение вещей.
Как обстоит дело с образованием зачатка головной кишки у высших
позвоночных, пока сказать с достоверностью трудно. Его клеточный мате-
риал может находиться в составе обособившегося в течение сильно изме-
ненной первой фазы гаструляции нижнего листка бластодиска. Однако не
исключена возможность, что материал зачатка передней кишки смещается
вторично из верхнего слоя бластодиска в нижний в составе головного от-
ростка. Опыты Уэддингтона (1930, 1932, 1933), мне кажется, не дают опре-
деленного ответа на этот вопрос.
На возможном объяснении особых свойств зачатка головной кишки и
сходства дифференцирующихся из него тканей с производными кожной
эктодермы будет целесообразнее остановиться после изложения фактиче-
ского материала, касающегося собственно эктодермальных производных.
156
Глава III
и) Эпидермальные ткани беспозвоночных
Экспериментальные данные, касающиеся гистологических особенностей:
эпидермальных тканей беспозвоночных, значительно менее полны, несмотря
на то, что опыты с регенерацией ставились и ставятся над различными
представителями в очень широком масштабе. Однако большая часть иссле-
дователей, не будучи гистологами по специальности, не уделяет достаточно
внимания гистологическим вопросам, а главное — не дает тщательного
гистологического анализа наблюдаемых процессов по стадиям, с исполь-
зованием полноценной гистологической методики обработки и изучения
препаратов. Кроме того, многие изучавшиеся объекты представляют исклю-
чительно большие трудности в отношении правильной трактовки тканевых
превращений. Метод эксплантации в отношении беспозвоночных еще недо-
статочно разработан. Лишь в единичных случаях удавалось получитьполно-
ценные явления роста и пролиферации тканей вне организма.
Я остановлюсь лишь на относительно немногих данных, имеющих зна-
чение для интересующих нас вопросов. При регенерации орального конца
тела гидры элементы эктодермы дают путем роста начало только эктодерме-
же, не обнаруживая при этом размножения митотическим путем.
Многочисленные митозы обнаруживаются лишь в мелких недифферен-
цированных 1-клетках, вдали от дефекта, располагающихся в составе
эктодермы и дающих начало не только разным дифференцированным эле-
ментам эктодермы, но и энтодермы (Стрелин, 1928).
Попытка Токина (1934) доказать, что при регенерации аборального
полюса гидры может происходить превращение дифференцированных кле-
ток эктодермы и энтодермы друг в друга, не является достаточно обоснован-
ной, так как в этой области имеются, хотя и в незначительном при нормаль-
ных условиях количестве, тотипотентные i-клетки, из которых, кроме всего-
прочего, развиваются и половые продукты.
В отношении планарий, являющихся одним из излюбленных объектов
для опытов с регенерацией, до настоящего времени имеется много неясно-
стей. Часть авторов описывала регенерацию кожного эпителия за счет или
при участии выползающих в него клеток паренхимы (Стивенс, 1901, 1907;
Кертис, 1902; Бартш, 1923, и др.). Многие авторы не могли обнаружить
митозов при регенерации эпидермиса и предполагали, что деление его клеток
происходит амитотическим путем (Бардин, 1902) или путем распада ядра
на хромидии, из которых в дальнейшем восстанавливаются новые ядра
(Бартш, 1923). Штейнман (1924—1926) даже допускал новообразование
эпидермиса из защитного слизистого покрова на поверхности раны, образую-
щегося из рабдитов, что уже ни в какой степени не является правдоподоб-
ным и вообще не стоит на уровне современной науки, а напоминает популяр-
ные в свое время взгляды старых анатомо-гистологов. Другая группа ав-
торов— Бардин (1902) и, в последнее время, Шаров (1937) — считает, что
регенерирующий эпидермис у триклад происходит из эпидермиса же.
В клетках последнего Шарову удалось наблюдать митозы. Последние отме-
чались и Кёллером (1894) у Stenostomum. Картины, которые некоторыми авто-
Экспериментальный метод в гистологии
157
рами объяснялись, как миграция клеток паренхимы в эпидермис, Шароегргк-
тует, в соответствии с наблюдениями Беклемишева над паразитическими
Acoela (1915), как выклинивание элементов из состава эпидермального
покрова и их миграцию внутрь. Весьма возможно, эти мигрирующие внутрь
клетки кожной эпидермы принимают участие в регенерации нервных и,
по крайней мере, части мышечных элементов, обнаруживающих у планарий
тесные генетические отношения к кожному покрову.
Превращения в процессе регенерации эктодермы в энтодерму, или на-
оборот, у планарий никто не описывал, если не считать неправдоподобного
предположения Ленерта (1891), что новая кожная эктодерма наземных
планарий при регенерации происходит из распадающихся элементов,
мигрирующих главным образом из кишечного эпителия. Эти краткие
данные показывают, во всяком случае, что широко распространенное
мнение об отсутствии гистологической детерминации тканей даже у таких
примитивных форм, как планарии, далеко не достаточно обосновано.
У другой, значительно выше организованной, группы беспозвоночных —
аннелид—данные в отношении регенерации кожного эпителия отличаются
большей определенностью, несмотря на имеющиеся разногласия. Очень
точные в этом отношении наблюдения Иванова (1903, 1908, 1916), а также
работы /Кинкипа (1934), Вейцмана (1927, 1937) показывают, что новообра-
зованный эпидермис возникает только из старого эпидермиса и не из ка-
кого другого источника. Клетки регенерирующего эпидермиса при этом
обнаруживают ряд характерных изменений, набухая, увеличиваются в раз-
мерах, приобретают малодифференцированный эмбриональный характер
п делятся митотическим путем. Предположение Геммерлинга (1924) и Штоль-
те (1927) о регенерации эктодермальных производных за счет мезодермаль-
ных необластов не подтвердилось Вейдманом (1937). Повидимому, эти авторы
были введены в заблуждение поверхностным сходством малодифференци-
рованных клеток регенерирующего эпидермиса и необластов. С другой сто-
роны, согласно большей части данных, элементы нервной системы и кольце-
вой мускулатуры аннелид, имеющих и в онтогенезе эктодермальное проис-
хождение, регенерируют из кожного эпителия. В образовании же генети-
чески чуждых тканей кожный эпителий не принимает участия. Таким
образом, кожный эпителий аннелид является вполне детерминированной
тканью в отношении энтодермальных и мезодермальных производных,
но сохраняет способность образовать элементы нервной системы и кольцевой
мускулатуры, свойственную эмбриональной эктодерме.
Из работ, относящихся к высшим представителям беспозвоночных,
особенного внимания заслуживают исследования Лазаренко (1924, 1925)
над личинкой жука-носорога, Данини над речным раком (1928) и Заварзина
(1927) над беззубкой. Все эти авторы установили, что кожный эпителий
изученных животных при асептическом воспалении, вызванном целлоидино-
выми трубками, во многих случаях начинает пролиферировать и в конце
концов полностью обрастает инородное тело, которое у членистоногих при
очередной линьке удаляется из организма. В свете рассмотренных выше
исследований Гаршина все эти разрастания кожного эпителия беспозвоноч-
158
Глава HI
ных являются воспалительными. В течение всего процесса элементы кож-
ного эпителия, хотя и претерпевают ряд морфологических изменений,
сохраняют свою полную специфичность, не превращаются в элементы соеди-
нительной ткани и не возникают из них, т. е. обнаруживают высокую сте-
пень гистологической детерминации.
Процесс эпителизации инородного тела у беспозвоночных, открытый
первоначально Лазаренко и изученный им особенно детально, протекает
следующим образом. У личинки жука-носорога и у речного рака клетки
гиподермы (у насекомых главная роль принадлежит гиподерме трахей),
располагающиеся в очаге воспаления, набухая, прорастают в новообразую-
щуюся соединительную ткань. Из клеток гиподермы образуются эпители-
зирующие пласты, разрастающиеся между очагом некроза, окружающим
инородное тело, и молодой соединительной тканью.
Как правило, эпителизирующие пласты имеют клиновидно заостренный
край, состоящий из плоских, на вертикальном разрезе, веретенчатых кле-
ток, не образующих хитина. С удалением от свободного нарастающего края,
клетки постепенно приобретают призматическую форму и на своей апикаль-
ной поверхности, обращенной к инородному телу, начинают выделять не-
прерывный слой молодого хитина, который постепенно утолщается. Как
правило, несколько отступя от свободного края пласта, в клетках гипо-
дермы наблюдаются многочисленные митозы. Процесс заканчивается созда-
нием непрерывной эпителиальной выстилки вокруг очага некроза с хорошо
развитым хитиновым панцырем, часть которого при очередной линьке пре-
вращается в участок молодого кожного покрова, а все содержимое эпи-
дермальной кисты, вместе со старым хитином, удаляется из организма.
В опытах с беззубкой эпителизации инородного тела, введенного в ман-
тию, происходила относительно редко, вследствие чего изучение процесса,
по последовательным стадиям встречало значительные трудности. Тем не
менее Заварзину удалось со значительной полнотой, хотя местами и предпо-
ложительно, восстановить весь процесс в целом и подробно описать гисто-
логические изменения в эпидермальном покрове мантии как при регенера-
ции поверхностного дефекта, так и при воспалительных разрастаниях,
приводящих к эпителизации инородного тела.
При эпителизации поверхности раны призматический мерцательный
эпителий, обращенный в полость мантии, сильно уплощается и, большей
частью утрачивая реснички, постепенно закрывает образующуюся в области
дефекта молодую соединительную ткань. Сильная митотическая активность
вспыхивает в регенерирующем эпителии, несколько отступя от края де-
фекта. Постепенно молодой и первоначально малодифференцированный
эпителиальный покров становится выше, его элементы вновь приобретают
призматическую форму и образуют мерцательный аппарат. При воспали-
тельных разрастаниях эпидермальные клетки выклиниваются из состава
эпителиального покрова в подстилающую его воспалительно измененную
соединительную ткань. По данным Заварзина, это происходит в местах,
где в результате воспалительного процесса исчезла и временно не восста-
навливается базальная мембрана. При воспалительных разрастаниях
Экспериментальный метод в гистологии
159
клетки эпидермиса мантии приобретают неправильную звездчатую форму
и, соединяясь друг с другом отростками, образуют в толще воспалительно
измененной соединительной ткани инфильтрирующие ее сетсвидные струк-
туры, большей частью совершенно не имеющие эпителиального характера.
По морфологическим особенностям своих элементов (резкая базофилия
клеточного тела, крупные светлые ядра) эпидермальные разрастания,
однако, ясно отличаются от клеток соединительной ткани. Значительно
труднее бывает отличить такие эпителиальные клетки от элементов мышеч-
ного происхождения. Однако по более точным позднейшим данным Завар-
зина (1927) клетки эпидермиса и измененные мышечные элементы все-таки
удается различать, причем новообразования мышц из эпидермиса не проис-
ходит. В таких случаях, когда атипичные сетевидные соединения клеток
эпидермиса дорастают до окруженного дегенеративными элементами ино-
родного тела, они вновь приобретают вид эпителия. Тесно соединяясь друг
с другом, элементы эпидермиса образуют на поверхности соединительной
ткани, окружающей инородное тело, непрерывный, первоначально мало-
дпфферснцированный пласт, который в конце концов полностью обрастает
некротические массы и в дальнейшем дифференцируется в типичный покров-
ный эпителий. В конце концов целлоидиновая трубка могла прорезаться
наружу и удаляться из организма.
Наблюдения Заварзина, устанавливающие у моллюсков высокую спе-
цифичность клеток эпидермиса, мне кажется, дают возможность правиль-
ной трактовки более старых данных Дрю (1911), который описывает пре-
вращение соединительнотканных клеток другого пластинчатожаберного
моллюска, Pecten, в эпителий. Эти данные до настоящего времени фигури-
руют в литературе (Грей, 1931; Гексли и де-Бер, 1936) в качестве примера
«метаплазии» соединительной ткани в мерцательный эпителий. Дрю транс-
плантировал в запирательную мышцу Pecten кусочки яичника, которые деге-
нерировали и окружались, в результате воспалительной реакции, молодой
соединительной тканью, элементы которой будто бы через три-четыре недели
приобретали сходство с эпителием, а значительно позже, по истечении 98
дней, превращались в типичный призматический мерцательный эпителий.
Рисунки Дрю не оставляют сомнения в том, что описанные им на стадии
трех-четырех недель в качестве сходных с эпителием клетки представляют
собой круглые кровяные фагоциты, которые всегда располагаются на по-
верхности некротических масс и были описаны при асептическом воспалении
у насекомых (Лазаренко, 1923, 1925), ракообразных (Данини, 1925) и мол-
люска беззубки (Заварзин, 1925, 1927). Генетическая связь мерцательного
эпителия, изображенного Дрю, на поздних стадиях опыта с этими круглыми
элементами ничем не обоснована, кроме их весьма отдаленного внешнего
сходства и положения на поверхности некротических масс.
Очевидно, что Дрю, имевший также дело с воспалительными разраста-
ниями эпителия и не проследивший достаточно тщательно всего процесса,
дал совершенно неправильную трактовку своим наблюдениям, призвав
на помощь метаплазию, которой в качестве deus ex machina до последнего
времени охотно объясняются многие, не имеющие с ней ничего общего,
160 Глава III
гистологические и гистопатологические явления. Я умышленно несколько
подробнее остановился на данных Дрю, чтобы на этом примере лишний
раз подчеркнуть необоснованность многих утверждений такого рода.
Описывавшееся у некоторых высших беспозвоночных, в частности
у ракообразных, образование волокон соматической мускулатуры из эпи-
дермальных элементов, недостаточно убедительно. Повидимому, такого
рода утверждения основываются на поверхностном морфологическом сход-
стве пролиферирующих эпидермальных элементов и настоящих миобластов.
Согласно новейшим данным Жинкина (1938), в регенерирующей конечности
рачка Asellus из гиподермы образуется только кожный покров регенерата,
а вся мускулатура его возникает из миобластов, источником которых яв-
ляются лишь старые мышечные волокна.
К сожалению, полноценные гистологические данные по регенерации и
воспалительным разрастаниям эпидермиса беспозвоночных имеют до на-
стоящего времени не систематический, а лишь разрозненный характер.
С одной стороны, они относятся к систематически очень удаленным друг
от друга формам, а с другой, не могут быть сопоставлены с изменениями
других эпителиев, в первую очередь эктодермального эпителия кишечника
тех же форм. Поэтому невозможно проследить эволюционные изменения
свойств эпидермиса в отдельных филогенетических рядах животного мира.
Пожалуй, одним из существенных выводов этого рода, который все-таки
можно сделать, является прогрессирующая гистологическая детерминация
эпидермиса первичноротых в отношении мускулатуры и нервной системы.
Эпидермис, имевший, повидимому, первоначально значение эктодермального
покрова, способного образовать при экспериментальных условиях эле-
менты нервной системы и части мускулатуры, на позднейших этапах эво-
люции утрачивает эту способность и становится полностью детерминиро-
ванной эпидермальной тканью, сравнимой и в этом отношении с эпидермаль-
ными покровами позвоночных. Эпидермис членистоногих, моллюсков и
позвоночных несомненно обнаруживает очень большие различия, завися-
щие, очевидно, от дивергентной эволюции их типовой и тканевой органи-
зации.
Очень важным, с нашей точки зрения, фактом является устанавливае-
мая, начиная с самых примитивных форм, строгая специфичность кожного
и кишечного эпителиев, противостоящих друг другу, несмотря на свою
общую эпителиальную морфо-физиологическую характеристику, как две
строго специфические и неспособные к взаимным превращениям
ткани.
Соотношения между регенерацией кожного эпителия у оболочников и его
развитием при бесполом размножении с процессами эмбрионального гисто-
генеза настолько неясны, что не могут получить в настоящее время опре-
деленной трактовки и настоятельно требуют дальнейших исследо-
ваний.
По той же самой причине пока остается совершенно неясной природа
энергично разрастающихся вне организма элементов столонов асцидии
Clavellina lepadiformis (И. Фишер, 1937).
Экспериментальный метод в гистологии
161
к) Опухоли эпидермального происхождения
Если даже оставить в стороне опухоли, имеющие своим вероятным источ-
ником уродливый эмбриональный клеточный материал, и остановиться
на тех из них, которые возникают, согласно Фишер-Вазельсу (1927), на почве
регенеративных (или, быть может точнее, дегенеративно-пролиферативных
процессов) из дифференцированных эпидермальных тканей, то следует
отметить их очень большое гистологическое разнообразие у людей и живот-
ных. Кожные раки были, кроме того, первыми злокачественными опухолями
млекопитающих, полученными экспериментально, путем длительного сма-
зывания каменноугольным дегтем.
Опухоли, образующиеся первично из эпидермальных компонентов того
или иного органа, сохраняют (хотя и в атипичном, более или менее изу-
родованном виде) гистологические особенности исходного материала и по
своей структуре в значительной степени соответствуют тем картинам,
которые дают эпидермальные ткани при неопухолевых пролиферативных
процессах. В связи с этим уровень дифференцировки опухолевой ткани
может быть в отдельных случаях весьма различен.
При крайней степени опухолевой катаплазии бластоматозные эпидер-
? мальные элементы, как и в случае тканей другого происхождения, полностью
утрачивают способность к какой-либо определенной тканевой взаимосвязи
и разрастаются отдельными или образующими неправильные группы клет-
ками, часто имеющими закругленную форму и неодинаковые, в разных
случаях, размеры. Гистологическая природа этих, т. наз. мелко-
клеточных или крупноклеточных цитобластом (Фишер-Вазельс, 1927), мо-
гущих встречаться в самых разнообразных органах тела, на основании
изучения одной микроскопической картины, не может быть установлена
с достоверностью. Находящиеся в цитоморфном состоянии клетки цито-
бластом соответствуют неоднократно описывавшимся выше изолированным
эпидермальным элементам, наблюдающимся, например, в условиях тка-
невых культур, отличаясь от них своей способностью к длительному
существованию и размножению в таком изолированном состоянии.
Многие раки кожи и переднего отдела пищеварительной трубки (на-
пример языка или пищевода) имеют вид настоящих эпидермальных мас-
сивов с явлениями ороговения и типичными роговыми жемчужинами, ко-
торые не оставляют сомнения в их тканевой природе. Такие же или не об-
наруживающие явлений ороговения, т. наз. плоскоклеточные раки
могут несомненно возникать первично из многих других эпителиев эпидер-
мального типа, не имеющих при нормальных условиях многослойной пло-
ской структуры, но могущих претерпевать сходные превращения при
пролиферации в экспериментальных условиях. Достаточно напомнить о пло-
скоклеточных раках воздухоносных путей, молочной, слюнных, щитовидной
и предстательной желез, мочевого пузыря, влагалища и вагинальной части
матки, которые, в соответствии с приведенными выше данными, мы имеем
все основания считать за новообразования, возникающие путем бласто-
матозного превращения из эпителиев этих органов. Весьма вероятное
11 X лопин-—349
162
Глава III
возникновение плоскоклеточных раков из всевозможных эпителиев эпидер-
мального типа, подтверждая нашу точку зрения об их генетической бли-
зости, в свою очередь, получает объяснение на основе экспериментально-
гистологических данных.
Малодифференцированным и более атипичным видом своих клеточных
элементов, но в гистологическом отношении также несомненным эпидер-
мальным характером отличаются и солидные мягкие раки-мозговики, очень
бедные стромой, свойственные, например, молочной железе и мочевому
пузырю.
Другим, хотя и менее распространенным типом эпидермальных опухолей
являются т. наз. базальноклеточные (Кромпехер, 1903) раки кожи,
воздухоносных путей и некоторых эпидермальных желез, которые по
своему гистологическому характеру напоминают малодифференцированные
воспалительные разрастания эпителиев этих органов.
Изучение локализации этих опухолей в области лица показало, что
места их возникновения поразительно совпадают с линиями срастания лоб-
ного, верхнечелюстных и нижнечелюстных отростков зародыша, что де-
лает очень вероятным их образование из отщепившихся при этих эмбрио-
нальных процессах эпидермальных зачатков (Глазунов, 1933).
Опухолевидные разрастания слюнных желез, известные под названием
смешанных опухолей, несмотря на свойственную им пестроту картин,
как показали исследования Цымбала (1933, 1939), в значительной степени
повторяют картины превращений эпителия этих желез при эксперименталь-
ных условиях. Такой же характер имеют некоторые опухоли молочной
железы. Обозначение их, как смешанные опухоли, поэтому утрачивает
в настоящее время свой смысл, тем более, что описывавшиеся в их составе
хрящеподобные образования, как показывает более тщательное гистоло-
гическое исследование (Цымбал, 1933), имеют очень мало общего с настоящей
хрящевой тканью, а представляют собой своеобразные дегенеративные
участки. То же самое оказалось справедливым и в отношении атипичных
сетевидных опухолевых структур.
В аденомах и т. наз. аденокарциномах эпидермальных желез некоторые
общие гистологические особенности эпидермальных тканей, повидимому,
также сохраняются, хотя в этом отнопюнии необходимо более тщательное
гистологическое исследование и, в частности, детальное сопоставление их
структур со структурами соответствующих эктодермальных опухолей.
Атипичным эпителиальным ходам эпидермальных аденокарцином во мно-
гих случаях свойственна двурядность, характерная для эпидермальных
желез, или многорядность. Изученные в последнее время т. наз. миоте-
лиальные (Гамперль, 1940) опухоли молочной железы оказались, в отно-
шении ряда особенностей своих элементов, построенными из клеток, сход-
ных с миоэпителиальными корзинчатыми, откуда и получили свое новое и
едва ли удачное обозначение.
Ряд опухолей эпидермального происхождения, как это уже отмечалось
выше, в большой степени соответствует нормальным, специфическим для
того или иного органа при обычных условиях, структурам. Для примера
Экспериментальный метод в гистологии
163
достаточно указать на коллоидный рак щитовидной железы, который даже
в матастазах содержит, хотя и атипичные, фолликулы с коллоидом, лимфо-
эпителиальные опухоли (Шминке, 1921) глоточного кольца, папилломы
мочевого пузыря, имеющие в большей или меньшей степени искаженный,
но в то же самое время признаваемый за таковой, переходный эпителий,
т. наз. адамантиномы, имеющие опять-таки искаженные в той или иной
Рис. 13. Эпидермальные опухолевые массивы,
инфильтрирующие соединительнотканную зо-
ну роста. 8-дневная культура мышиного рака
молочной железы. Сильное увеличение. (По
Хлопину, 1932).
степени структуры зубных зачатков.
На противоположном полюсе стоят упомянутые выше опухоли типа
цитобластом, тканевая природа которых устанавливается, с известной долей
вероятия, на основании первичной локализации и с большей точностью
в случаях прогрессирующей
атипизации новообразований,
имевших первоначально до-
статочно определенный тка-
невой характер.
В высокой степени инте-
ресным и многообещающим с
точки зрения гистологии и
патологии является экспери-
ментальное изучение опухо-
левых тканей методом экс-
плантации, дающим возмож-
ность подвергнуть разносто-
роннему экспериментальному
анализу опухолевые ткани
животных и человека.
Из опухолевых тканей,
которые по своему происхож-
дению могут быть отнесены
к эпидермальному типу, ме-
тодом тканевых культур изу-
чены главным образом неко-
торые раки мышей и крыс, в
первую очередь, рак грудной
железы мышей (Дрю, 1923; Фишер с сотрудниками, 1928, 1933; Хлопин,
1932. и др.). Опухолевые элементы, разрастаясь, образуют плоскостные
мембраны, массивные пласты и тяжи, погруженные в толщу плотной среды
(рис. 13). Все эти образования состоят, как правило, из тесно сомкнутых
клеток разной формы, которые могут то в большем, то в меньшем количестве
изолироваться, и по своему общему характеру сходны со структурами,
свойственными соответствующим нормальным тканям. Способность к по-
гружному росту, прорастанию и уничтожению подсаженных частиц нор-
мальных тканей выражена в сильной степени (рис. 13). Чистые культуры
опухолевых раковых клеток, как это уже упоминалось, могут жить и расти
вне организма в течение многих лет, сохраняя свои специфические особен-
ности. Опухолевые комплексы отличаются своим малодифференцированным
164
Глава III
характером, межклеточные мостики в опухолевых комплексах, в отличие
от многих нормальных тканей этого типа, не образуются. В культурах
опухоли крыс Флекснер-Джоблинга, которая, по имеющимся данным, об-
разовалась из кожного покрова, комплексный характер разрастаний выра-
жен слабее, и опухолевые клетки легче изолируются друг от друга (Эрд-
ман, 1923; Улезко-Строгоноеа, 1923, и др.). Имеются указания на рост
элементов эпидермальных раков человека вне организма (Гефер, 1934;
Тимофеееский, 1938). Как показывают новейшие исключительно интерес-
ные данные Беневоленской, различные человеческие опухоли эпидермаль-
ного происхождения (рак кожи и щитовидной железы, опухоли молочной
железы), даже при длительном культивировании, полностью сохраняют
свою тканевую специфичность и обнаруживают превращения, вполне
укладывающиеся в рамки закономерностей, установленных нами для соот-
ветствующих нормальных тканей. Периферические участки мембран из
ракового эпителия кожи могут обнаруживать резко выраженные явления
ороговения.
Путем изучения эксплантатов из т. наз. смешанных опухолей
слюнных желез человека удалось установить, что они не являются смешан-
ными, а представляют собой чрезвычайно разнообразные по структуре раз-
растания эпителия слюнных желез (Цымбал, 1932), дегенеративные участки
которых могут обнаруживать поверхностное сходство с мезенхимными
производными. Опухолевые участки, состоящие из массивных клеточных
скоплений или трубчатых ходов, оказываются в условиях эксплантата спо-
собными к некоторым изменениям, имеющим характер дифференцировки
и специфичным для многих эпидермальных тканей, например к образованию
комплексов, состоящих из концентрически наслоенных друг на друга
клеток, связанных межклеточными мостиками.
Продолжая свои опыты по эксплантации опухолей слюнных желез,
Цымбал в руководимом им в настоящее время цитологическом отделении
Онкологического института получил интересные дополнительные данные.
При культивировании во флаконах ему удалось добиться обширных раз-
растаний мембран и пластов опухолевого эпителия, который обнаруживал
высокую митотическую активность и способность к образованию жемчужин
вне организма. *
Б. Энтеродермальный тканевой тип
К энтеродермальному, или эктодермальному, в узком смысле слова,
тканевому типу мы относим ткани, развивающиеся у позвоночных из за-
чатка вторичной кишечной трубки зародыша. Из состава собственно энто-
дермы мы, однако, исключаем, по ряду соображений, которые будут разо-
браны ниже, зачаток выстилки передней части пищеварительной трубки,
включительно до пищевода, а у ряда форм — до некоторых отделов желудка
* За сообщение новых интересных данных, полученных в цитологическом отделении
Онкологического института, и за любезное разрешение включить их в настоящую моно-
графию пользуюсь случаем выразить профессору В. Е. Цымбалу свою искреннюю благо-
дарность.
Экспериментальный метод в гистологии
165
и зачаток ее небольшого заднего отдела, соответствующего части клоаки.
У первичноротых к этому тканевому типу относится эктодермальная вы-
стилка среднего отдела пищеварительной трубки (mesodaeurn) с ее производ-
ными. Указать точные границы собственно энтодермальных производных
у вторичноротых беспозвоночных, за неимением соответствующих гисто-
логических и гистогенетических данных, пока не представляется возможным.
Филогенетически энтеродермальные ткани всех первичноротых и вто-
ричноротых образовались из примитивной энтодермы гипотетических га-
стрееподобных предков, сходных в некоторых отношениях с современными
кишечнополостными. Эта примитивная мультифункциональная энтодерма,
по своей морфо-физиологической характеристике, как это отмечалось выше,
соединяла в себе свойства различных общепринятых тканевых групп и,
в частности, имела также значение примитивного кишечного эпителия.
Из эпителиальной выстилки кишечной трубки животных с дифферен-
цированными тканями, имеющей в первую очередь значение пищевари-
тельной и всасывающей, но вместе с тем пограничной и защитной ткани,
через которую осуществляется интенсивный внешний обмен веществ орга-
низма, в филогенезе и онтогенезе образуются специализирующиеся в раз-
личных направлениях участки: у позвоночных, например, выстилка же-
лудка, желчных путей и желчного пузыря, утрачивающая всасывающий, но
сохраняющая в первую очередь покровный и защитный характер, много-
клеточные железы различной степени сложности, несущие пищеваритель-
ную функцию и, наконец, органы эндокринового типа, эпителиальные ком-
поненты которых, частью или полностью утрачивая значение пограничных
образований, превращаются в своеобразные ткани внутренней среды орга-
низма, характеризующиеся трофическими или гормональными, а также
своеобразными защитными свойствами.
В сравнительно редких случаях некоторые части энтеродермальных
покровов специализируются как ткани внешнего газообмена.
Генетически единый и вполне определенный энтеродермальный тканевой
тип, дифференцируясь дивергентно и изменяя свои функциональные осо-
бенности, дает в филогенезе и индивидуальном развитии весьма различные
в отношении своей морфо-физиологической характеристики конкретные
ткани, часть которых лишь с большими оговорками и натяжкой может
обозначаться термином «эпителий» и, в соответствии с укоренившейся гисто-
логической традицией, как это отмечалось выше, рассматривается не в об-
щем учении о тканях, а в микроскопической анатомии.
Покровные энтеродермальные пласты и образующиеся из них желези-
стые ходы характеризуются рядом признаков, подчеркивающих генетиче-
ское единство этих образований у всех животных, которые в связи с пара-
зитизмом не утратили вторично пищеварительного аппарата. Только у бески-
шечных турбеллярий энтеродсрмальная часть пищеварительной трубки
лишена просвета и имеет характер пищеварительного синцития или сим-
пласта неэпителиального характера. В соответствии с тем, что было ска-
зано выше, эти своеобразные черты организации Acoela вероятнее тракто-
вать как вторичную специализацию. За этим исключением энтеродермаль-
166
Глава III
ная выстилка пищеварительной трубки и соответствующих железистых
органов в дифференцированном состоянии всегда имеет строение однослой-
ного призматического эпителия, элементы которого образуют на своем
апикальном конце или амебоидные псевдоподии (у животных с внутрикле-
точным пищеварением), или мерцательные реснички, или, в большей части
случаев, своеобразную каемку или кутикулу. Клетки железистого харак-
тера, как правило, бывают лишены упомянутых образований. Несмотря на
наличие кутикулы, между апикальными концами клеток кишечного эпите-
лия обнаруживается отчетливая сеть замыкающих полосок, подчеркиваю-
щих клеточный характер кутикулярного ободка.
Все клетки эпителиального пласта тянутся от его базальной до апикаль-
ной поверхности, т. е. имеют однорядное расположение. Ядра в большей
части случаев располагаются на одном уровне; реже, особенно если клетки
имеют очень высокую и вместе с тем узкую форму, их ядросодержащие
части бывают расположены на разной высоте. В таких случаях иногда гово-
рят о многорядной структуре эпителия, что является совершенно непра-
вильным и вносит терминологическую, а подчас, принципиальную путаницу
в понятия.
Многорядная структура, свойственная, например, эпителию воздухо-
носных путей млекопитающих, в котором ядра также располагаются не на
одном уровне, имеет совершенно другую природу и значение. В данном
случае можно в лучшем случае говорить о ложной многорядности.
В отдельных случаях энтеродермальные покровы могут приобретать
резко выраженное защитное значение, как это, например, имеет место
в мышечном желудке многих птиц. Защитная функция осуществляется
в данном случае совершенно иным способом, чем в многослойных эпителиях
кожного типа, образующих на своей поверхности слои ороговевших клеток.
В мышечном желудке птиц мы видим типичный однослойный однорядный
эпителий, образующий железистые углубления трубчатой формы. Клетки
такого эпителия выделяют на своей апикальной поверхности секрет, кото-
рый, уплотняясь, превращается в твердый и прочный защитный покров.
При нормальных условиях в пищеварительных и защитных энтеродер-
мальных выстилках постоянно или периодически происходит смена эпи-
телия, вследствие замещения снашивающихся и отторгающихся клеток
размножающимися и дифференцирующимися камбиальными элементами,
морфологические особенности и распределение которых еще далеко не во
всех случаях достаточно ясны.
У позвоночных, в первую очередь у млекопитающих, у которых эти
отношения изучены полнее, камбиальные участки имеют вполне опреде-
ленную локализацию в эпителиальных углублениях — либеркюновых крип-
тах кишечника, желудочных ямках или криптах, где постоянно встречаются
митозы (Биццоцеро, 1885—1893; Биццоцеро и Вассале, 1887; Сачердотти,
1896). В составе выступающих в просвет участков эпителиальной выстилки,
митозы, если и встречаются, то как большая редкость; зато здесь нередко
наблюдаются дегенеративные изменения в клетках и их слущивание в про-
свет. Элементы камбиальных участков не имеют отчетливо выраженной
Экспериментальный метод в гистологии
167
кутикулы, а их железистые элементы отличаются меньшим количеством
секреторных включений. Митотическая активность камбиальных участков
кишечного эпителия очень высока. Процент делящихся митозом клеток
достигает у белой крысы, по данным Хрущева (1942), 9.
Таким образом, кишечный и желудочный эпителий и при физиологиче-
ских условиях находится в состоянии движения, продвигаясь из углубле-
ний на выступающие в просвет части слизистой оболочки. У рыб (Биццо-
церо, 1902/03) и у жабы (Кингсбери, 1899) описывалось аналогичное рас-
пределение камбиальных участков. У миноги и у лягушки, в кишечнике
которых ворсинки и крипты отсутствуют, митозы встречаются преиму-
щественно в глубине складок (Биццоцеро, 1893). У некоторых низших
позвоночных (тритона, саламандры) в слизистую оболочку кишечника
местами вдаются, лишенные просвета, эпителиальные почки, имеющие зна-
чение камбиальных участков (Биццоцеро, 1892; Никола, 1894). Сходный
вид малодифференцированных эпителиальных скоплений или почек имеют
также камбиальные участки, описывавшиеся у некоторых насекомых.
Такое распределение биологически неравноценных камбиальных и диффе-
ренцированных участков, в результате которого при физиологической реге-
нерации происходит скольжение элементов энтеродермального пласта, па-
раллельно своей подстилке, было мной в свое время обозначено, как гори-
зонтальная анизоморфность.
Явления роста эпителия кишечного типа при экспериментальных усло-
виях сопровождаются то более, то менее интенсивным митотическим кле-
точным делением.
При пролиферативных процессах энтеродермальным элементам во всех
•изученных случаях свойственна высокая степень комплексности, т. е. их
разрастания, как в составе, так и вне организма, сохраняют тесную «эпи-
телиальную» взаимосвязь клеток.
Сетевидное разрыхление энтеродермальных разрастаний и изоляция
•отдельных клеток округлой, амебоидной или отростчатой формы могут
•быть довольно редко констатированы с достаточной отчетливостью только
в условиях тканевых культур, да и то эпизодически, как проходящее и
малохарактерное явление. Полная изоляция клеток завершается в очень
многих случаях необратимыми дегенеративными изменениями, но иногда,
повидимому, приводит к восстановлению типичной эпителиоподобной
взаимосвязи элементов.
а) Эпителий пищеварительной трубки
Данные, касающиеся роста и превращения вне организма эпителиальной
выстилки кишечника и желудка в течение первых дней культивирования,
а в некоторых случаях до одного-двух месяцев, получены главным образом
на эмбриональном материале — куриных зародышах (Фишер, 1922, 1930;
Льюисы, 1925; Капель, 1929; Тёрё, 1929, 1931; Бишелье, 1931, и др.) и заро-
дышах кролика и человека (Хлопин, 1922, 1934). Относительно эпителия
желудка и кишек молодых и взрослых кроликов имеются лишь работы
Елисеева (1929), Хлопкова (1931) и Зубаревой (1939, 1940). Из эмбрионального.
168
Глава III
материала эксплантирутотся цилиндрические, не вскрытые вдоль отрезки
кишечника и вскрытые и развернутые участки всей толщи стенки этих
органов. При эксплантации неэмбрионального материала для посева служат
частицы слизистой оболочки, которая может быть легко отделена от мы-
шечных слоев и отмыта значительным количеством стерильного физиологи-
ческого раствора от инфицированного микроорганизмами содержимого.
Для стерилизации исходного материала пользовались также освещением
ультрафиолетовыми лучами или слабым раствором риванола.
Эпителий или растет экстенсивно в виде клеточных мембран, стелющихся
по поверхности плотного субстрата, или эпителизирует поверхность эксплан-
тата, нередко обрастая ее со всех сторон. Желудочный и кишечный эпи-
телии очень сильно разжижают сверток фибрина, в котором образуются
полости, наполненные жидкостью. Наряду с экстенсивным ростом эпите-
лия в некоторых случаях может наблюдаться прорастание мезенхимальных
составных частей посеянных кусочков. Мезенхимальная зона роста, состоя-
щая из сетевидно связанных фибробластообразных элементов, является
в течение некоторого времени хоропюй подстилкой для растущих эпите-
лиальных пластов. Во многих случаях получить экстенсивный рост эпи-
телия бывает нелегко, вследствие сильного разжижения им фибрина. По
наблюдениям Хлопана, на человеческом эмбриональном материале получе-
ние экстенсивного роста эпителия облегчается при эксплантации частиц
кишечника на поверхность соединительнотканной культуры. Любопытно
отметить, что эти эпителии через более или менее короткое время губят
проросшую соединительную ткань. Повидимому, это связано с энергичным
разжижением свертка, а частью, быть может, также с выделением каких-то
вредно действующих на клетки продуктов жизнедеятельности. Чистые
культуры эпителия, полученные путем отрезания и пересева на поверхность
свертка участков мембран, могут лишь короткое время сохраняться живыми
и расти. Длительных чистых культур эпителия желудка и кишек еще полу-
чить не удалось.
При экстенсивном росте эпителия по фибрину образование полостей
разжижения вызывает разрывы мембран и их ргруживание в клеточные
тяжи или массивы, которые часто образуют сплошной вал по краю полости
разжижения, могущий оставаться связанным с центральным кусочком
эпителиальными перемычками. В дальнейшем такие эпителиальные ком-
плексы распадаются и погибают. В некоторых случаях центральный ку-
сочек оказывается прижатым к стенке объемистой круглой полости разжи-
жения и образует более или менее значительное утолщение в составе
кольцевого клеточного вала. Такие культуры по своей форме напоминают
перстень. При полной эпителизации и отсутствии экстенсивного роста
эксплантат большей частью оказывается свободно плавающим в полости
разжижения. Нацело эпителизированные эксплантаты из эмбрионального
кишечника могут значительно увеличиваться в объеме, становясь про-
зрачными. Это зависит от сильного набухания или отека находящейся в их
составе эмбриональной соединительной ткани. Их поверхность образует
разной формы выступы и углубления. Покрытые эпителием части таких
Экспериментальный метод в гистологии
169
«кишечных организмов» могут долгое время (один-два месяца) сохраняться
живыми (Фишер, 1922; Хлопин, 1934). Оказавшиеся внутри гладкие мышцы
кишечной стенки обнаруживают при этом самопроизвольные перистальти-
ческие сокращения, благодаря которым весь эксплантат может изменять
свою форму и переворачиваться в окружающей его жидкости. Очевидно,
эпителиальный покров такого, как бы вывернутого наизнанку, участка
кишки вполне удовлетворительно обслуживает обмен веществ эксплантата
и отличается резко выраженной способностью к всасыванию. Частицы
слизистой оболочки неэмбрионального кишечника скручиваются эпителием
наружу и в большей части случаев эпителизируются (Хлопков, 1931). Вор-
синки оказываются торчащими наружу. Частью они дегенерируют и рас-
падаются. Эпителий крипт может разрастаться внутри эксплантата, при-
обретая совершенно атипический характер.
На поверхности эпителизированных эксплантатов из слизистой оболочки
желудка, вследствие неравномерного роста отдельных участков соедини-
тельнотканной подстилки, могут на ранних стадиях образоваться выступы
закругленной формы. С течением времени их поверхность постепенно вырав-
нивается, вследствие исчезновения желудочных ямок и желез, а содержимое,
в противоположность кишечным эксплантатам, подвергается дегенерации
по истечении двух-трех недель.
Это показывает, что желудочный эпителий по своим биологическим
свойствам, в частности, своей всасывающей способности, сильно отли-
чается от кишечного.
В состоянии активного роста, пролиферации и продвижения по поверх-
ности плотного субстрата, высокие призматические эпителиальные эле-
менты, сохраняя непрерывную связь друг с другом, уплощаются и могут
принимать довольно разнообразные очертания. Гистологическое строение
эпителиального пласта подвергается при этом значительному упрощению и
становится индифферентным. Наиболее сильному распластыванию подвер-
гаются элементы экстенсивно растущих по слюде или стеклу мембран,
от краев которых, часто имеющих довольно ровный контур, могут отде-
ляться отдельные эпителиальные клетки различной, часто неправильной,
отростчатой, «фибробластообразной» формы. Если рост и движение эпите-
лия происходят медленно или останавливаются совершенно, что, например,
имеет место по окончании эпителизации поверхности эксплантата или об-
растания стенки полости разжижения, образовавшейся в фибрине, то его
клетки становятся выше и могут вновь приобрести кубическую или высоко-
призматическую форму. Эпителиальные мембраны и пласты отличаются,
как правило, однослойным расположением своих элементов. По данным
Робинова (1936), связь между свободными поверхностями клеток мембран
из кишечного эпителия является наиболее тесной. Именно в этой плоскости
обнаруживаются (как при прижизненном наблюдении, так и на фиксиро-
ванных препаратах) четкие границы между тесно сомкнутыми друг с другом
элементами. В других плоскостях связь между соседними клетками может
быть менее тесной и осуществляется с помощью тонких цитоплазматических
мостиков.
170
Глава III
При движении кишечного эпителия по поверхности эксплантата ме-
стами может наблюдаться сильное сгруживание эпителиальных клеток,
приобретающих очень высокую узкую форму. В таких случаях утолщенные
Рис. 14. Приобретение кишечным
эпителием ложной многоряд-
ной структуры. Ядра высоких
призматических клеток распола-
гаются на разном уровне, но все
клетки доходят от соединитель-
нотканной подстилки до свобод-
ной поверхности пласта. В этом
можно убедиться, подсчитывая
количество ядер и доходящих
до свободной поверхности апи-
кальных концов клеток, отде-
ленных замыкающими полоска-
ми. В делящейся митозом клет-
ке звезда занимает характерное
для однорядного эпителия апи-
кальное положение. 1-дневная
культура из кишечника челове-
ческого зародыша. Сильное уве-
личение. (Оригинал).
ядросодержащие части клеток оказываются
расположенными на разных уровнях, а эпи-
телиальный покров при поверхностном на-
блюдении производит впечатление многоряд-
ного, что не соответствует действительности,
так как все эпителиальные клетки, начина-
ясь от соединительнотканной подстилки, дру-
гим своим концом доходят до свободной по-
верхности (рис. 14). Покровный эпителий
из желудка и кишечника никогда не образует
путем пролиферации, утолщения и напласты-
вания друг на друга клеток структур, срав-
нимых с многослойными покровными пласта-
ми. Временные нагромождения клеток друг
на друга в процессе роста бывают. То же самое
наблюдается, повидимому, в результате обра-
зования закрытых эпителиальных складок.
Такие нагромождения представляют собой
быстро исчезающие образования, не имеющие
ничего общего с настоящими многослойными
эпидермальными структурами. Отдельные эле-
менты покровного эпителия эксплантатов мо-
гут на разных стадиях культивирования .-под-
вергаясь дегенерации, округляться и оттор-
гаться. Если поверхность эпителизированных
кусочков неровная, как это большей частью
имеет место в эксплантатах из кишечника, то
дегенеративные изменения, как правило, про-
исходят на выступах, а в углублениях можно
наблюдать до поздних стадий культивирова-
ния митотическое деление клеток (рис. 15). Эти
явления соответствуют в известной степени
тому, что наблюдается в эпителии кишечного
типа при обычных условиях в составе организ-
ма, где пополнение снашивающихся и отторга-
ющихся элементов эпителиального пласта так-
же преимущественно происходит в углубле-
ниях (желудочных ямках или кишечных крип-
тах), имеющих значение камбиальных
участков. В дифференцированном состоянии
призматические клетки однослойного покровного эпителия эксплантатов
бывают тесно соединены друг с другом своими базальными, обращен-
ными к соединительнотканной подстилке, и апикальными, образующими
Экспериментальный метод в гистологии
171
свободную поверхность пласта и большей частью снабженными кутикулой,
концами (рис. 15). Между последними часто удается заметить сеть
т. наз. замыкающих полосок. Боковые поверхности призматических
клеток или также плотно примыкают друг к другу или оказываются отде-
ленными щелевидными промежутками. В этих случаях в условиях культур
наблюдается образование межклеточных протоплазматических мостиков,
не свойственных этим эпителиям при обычных условиях. Такой характер
имеет, или приобретает вновь, покровный эпителий, не подвергавшийся
в процессе эпителизации значительным изменениям, и те его участки, ко-
торые во время активного обрастания принимали малодифференцированное
строение (Хлопан, 1922; Зубарева 1939, 1940).
В клетках экстенсивно растущих мембран из кишечного эпителия по-
являются жировые включения, специально изучавшиеся Шантрохом (1932).
В покровном эпителии на поверхности
эпителизированных культур также по-
являются жировые капли, располагаю-
щиеся, как правило, под ядром в ба-
зальной части клеток (Хлопин, 19^2).
Кроме того, клетки покровного кишеч-
ного эпителия, как показывают мои
неопубликованные данные по культи-
вированию участков кишки двух-двух
с половиной-месячных человеческих за-
родышей, постепенно переполняются
преимущественно в своей апикальной
Рис. 15. Участок покровного одноряд-
ного эпителия с соединительноткан-
ной подстилкой в 31-дневной куль-
туре кишечника человеческого заро-
дыша. В одной из клеток митоз. Силь-
ное увеличение. (По Хлопину, 1940).
части включениями, сходными по своим
красочным реакциям и отношению к
слюне с гликогеном, но отличающими-
ся от него меньшей растворимостью в
воде. Если для эксплантации служил
кишечник куриных зародышей поздних
стадий развития, то в составе покровного эпителия обнаруживались
отдельные слизистые бокаловидные клетки (Капель, 1929). По данным
этого автора, отдельные элементы покровного кишечного эпителия
могли фагоцитировать частицы туши, если она прибавлялась к пи-
тательной среде. Покровный эпителий желудка куриных зародышей (Ка-
пель) и новорожденных кроликов (Зубарева) в течение всего времени наблю-
дения в условиях культур продолжал вырабатывать на поверхности эксплан-
татов характерное для него при обычных условиях слизеподобное вещество,
т. наз. мукоид, скоплявшийся в виде капель у апикальных концов
клеток и выделяемый ими в окружающую среду. Эпителий желез дна
желудка новорожденных кроликов, который до эксплантации имел недиф-
ференцированный характер и не обнаруживал признаков секреции, в куль-
турах приобретал способность вырабатывать мукоид (Зубарева). Появление
слизи. в среде, окружающей кишечные эксплантаты, отмечалось также
Тёрё (1929) и Бише лье (1931).
172
Глава III
Метод регенерации дает принципиально аналогичные гистологические
результаты, хотя в течение процесса естественно имеется ряд особенностей.
Шевченко (1940) очень детально изучил регенеративные процессы в об-
ласти границы преджелудка и желудка крыс после нанесения ожогом де-
фекта в слизистой оболочке. После нанесения дефекта и в однослойном
желудочном эпителии и в многослойном ороговевающем эпителии пред-
желудка начинаются энергичные пролиферативные процессы, приводящие
в конце концов к заживлению, с образованием нового места стыка между
различными эпителиальными выстилками.
Остановимся только на данных, касающихся эпителия кардиальной
части желудка. После нанесения дефекта в соединительной ткани возникает
воспалительная реакция, а эпителий, мобилизуясь, начинает продвигаться
на область дефекта, постепенно эпителизируя его поверхность. Постепенное
перекрытие дефекта зависит от двух причин: от перераспределения имев-
шегося налицо клеточного материала и от усиленной митотической актив-
ности. Жизнеспособные эпителиальные клетки по краю дефекта, сохраняя
тесную связь друг с другом, постепенно уплощаются, распластываются по
поверхности соединительной ткани, тем самым увеличивают свою крою-
щую поверхность и начинают продвигаться по обнаженной подстилке.
Количество мукоидных включений в них уменьшается. Реактивные изме-
нения в эпителии, начавшись в граничащих с дефектом участках, постепенно
захватывают довольно широкую зону слизистой оболочки и распростра-
няются на желудочные ямки и железы, построенные по кардиальному типу.
Рельеф, граничащей с местом повреждения области слизистой оболочки,
постепенно выравнивается, желудочные ямки, а затем и железы, как бы
развертываясь, укорачиваются и, наконец, в известной зоне, полностью
исчезают. Составлявшие их элементы, если можно так выразиться, расте-
каются по поверхности, приобретая характер малодифференцированного
покровного желудочного эпителия с небольшим количеством мукоидных
включений.
Митотическая активность заметно увеличивается, причем митозы в эпи-
телии первоначально наблюдаются только в окружающей дефект части
слизистой оболочки, а не в новообразованном эпителиальном регенерате.
В последнем они появляются несколько позже, не встречаясь, однако, как
правило, у самого нарастающего края.
После того как более или менее значительная часть дефекта окажется
эпителизированной, в выравнившемся эпителиальном покрове вновь на-
чинают образовываться углубления, типа желудочных ямок, и процесс
гистологического упрощения сменяется вторичной дифференцировкой. По-
следняя, начавшись в более отдаленных от края эпителизирующегося пласта
участках, постепенно приближается к центру бывшего дефекта, захватывая,
после полного перекрытия его, весь новообразованный эпителиальный
покров. В течение и по окончании всего процесса регенерации, точно про-
слеженного по стадиям, однослойный желудочный и многослойный пред-
желудочный эпителии остаются совершенно резко отграниченными друг
от друга. Эпителий желудка ни на одной стадии не образует структур;
Экспериментальный метод в гистологии 173
свойственных даже малодифференцированному, а тем более ороговеваю-
щсму многослойному эпителию.
Данные Гриффина и Вассале (1888) и, в особенности новейшие наблю-
дения Фергюсона (1928), полученные при изучении репаративной регене-
рации слизистой оболочки фундальной области желудка собак, в общем
хорошо согласуются с изложенными выше, хотя эти авторы не очень под-
робно останавливаются на многих гистологических деталях процесса. Из
работы Фергюсона с полной очевидностью явствует, что эпителий желудка
собак в течение всего процесса регенерации сохраняет свойственную ему
однослойность и никогда не приобретает строения многослойного эпителия.
Данные, полученные на человеческом секционном материале (Краевский
и Маят, 1938), говорят то же самое.
К совершенно иным выводам пришел в своих опытах Фюттерер (1905),
вводивший в желудок кроликов едкие вещества (например пирогаллол)
и изучивший конечный результат после нанесенного повреждения. В двух
случаях Фюттерер обнаружил в желудке у подопытных животных участки
многослойного эпителия, вплоть до пилорической области. На этом осно-
вании он приходит к выводу, что в результате репаративной регенерации
желудочный эпителий метаплазировался в многослойный плоский.
Нахождение какого-нибудь эпителия, отличающегося по своей структуре
от свойственного данному органу в норме, до последнего времени в патоло-
гии было принято объяснять метаплазией, несмотря на то, что в большей
части случаев весь ход процесса оставался неизученным. Это объясняется тем,
что, в соответствии с принятыми в классической гистологии взглядами,
эпителий трактуется как единая ткань, которая может быть представлена
то одной, то другой конкретной структурой. Как это ни удивительно, до
наших работ как-то не возникало мысли, что далеко не все различные по
своему строению эпителии могут превращаться друг в друга и, в частности,
в многослойный плоский. В соответствии со всем сказанным выше ясно,
что вывод Фюттерера о метаплазии однослойного эпителия желудка в мно-
гослойный плоский не является сколько-нибудь обоснованным. С нашей
точки зрения, основывающейся на тщательно изученном эксперименталь-
ном материале, который, кроме всего прочего, демонстрирует, что эпителии
обладают огромной способностью к плоскостному росту, нам представляется
более правильным трактовать данные Фюттерера и многие аналогичные
находки не как метаплазию, а как дистопию, т. е. разрастание за свои
обычные границы и замещение одного эпителия другим. Это справедливо,
в частности, в отношении наблюдений Моратти (1930), который, применяя
авитаминозную диэту (без витамина А), описал «метаплазию» эпителия
желудка в многослойный плоский эпителий. Авитаминоз А вызывает
в ряде эпителиев дистрофические изменения и в той или иной степени про-
лиферативную реакцию. Не подлежит сомнению, что в опытах этого автора,
результат которых не был прослежен по стадиям, также имело место
дистоническое разрастание многослойного эпителия пищевода.
Результаты, полученные в нашей лаборатории Колоссом (1941) при изу-
чении по стадиям процесса репаративной регенерации эпителиальной
174
Глава III
выстилки желудка травяной лягушки, показывают, что и у представите-
лей низших позвоночных этот эпителий характеризуется вполне сходным
циклом превращений. После нанесения ожога происходит постепенное
перекрытие дефекта тем же способом, что и у млекопитающих. Вместе с тем
весь процесс в целом протекает медленнее. Благодаря очень значительным
вертикальным размерам клеток покровного эпителия их уплощение вы-
ражено еще резче, чем у млекопитающих. В отличие от последних, эпители-
зация сначала происходит по поверхности некротизированных масс, которые
лишь медленно рассасываются и замещаются в результате гораздо более
вялой, чем у млекопитающих, воспалительной реакции, молодой соедини*
тельной тканью. Выравнивание рельефа слизистой оболочки вокруг де-
фекта, развертывание и растекание по поверхности эпителия желудочных
ямок и желез происходят сходным с описанным выше способом. Повышение
митотической активности выражено очень резко. В отличие от кролика,
дефект, дно которого располагается ниже окружающих частей слизистой
оболочки, первоначально оказывается перекрытым ровным слоем низко-
призматического или изопризматического малодифференцированного эпи-
телия с незначительным количеством мукоидных включений. Только в сред-
ней части области дефекта нередко происходит сгруживание клеток сомк-
нувшегося эпителизирующего пласта. В результате этого здесь образуется
заметное утолщение, вдающееся в просвет и состоящее из неправильно
нагроможденных элементов с более или менее резко выраженными дегене-
ративными изменениями. В дальнейшем этот участок отмирает, его элементы
отваливаются и замещаются окружающими, расположенными одним пра-
вильным слоем, клетками. Только после полного перекрытия дефекта
в эпителии начинается вторичная дифференцировка, параллельно с расса-
сыванием расположенных под эпителием некротических масс и их замеще-
нием молодой соединительной тканью, приводящим к постепенному под-
нятию дна области ожога. Клетки становятся выше, мукоидная секреция
усиливается, и начинается образование углублений — будущих желудоч-
ных ямок, из слепого конца которых постепенно разрастаются желудочные
железы. В конце концов регенерат превращается в участок нормально диф-
ференцированной слизистой оболочки, отличающейся от окружающих
частей менее правильным складчатым рельефом. В железах дифференци-
руются характерные для желудка лягушки слизистые клетки в области
шейки и типичные железистые элементы с мелкими зернами зимогена,
вырабатывающие желудочный сок. Процесс завершается новообразованием,
по всей видимости, функционально полноценного участка слизистой оболочки.
Атипические разрастания эпителия желудка были впервые получены
Гага (1913) путем впрыскивания шарлахового масла, частью с разными
примесями, под слизистую оболочку желудка кроликов, собак и крыс.
Разрастания имели вид трубочек, враставших в подслизистый слой, а от-
туда обратно в слизистую оболочку, давая картины, имеющие внешнее
сходство с аденокарциномой.
Опыты Гаршина (1935), подтвержденные Чурбановым (1937), наглядно
демонстрируют изменения эпителия тела желудка кролика в очаге воспа-
Экспериментальный метод в гистологии
175
ления, вызванного введением в стенку желудка каменноугольного дегтя
или формалинового масла. Вокруг капель инородного тела, располагаю-
щихся обычно в подслизистом слое, начинается интенсивная воспалитель-
ная реакция. Эпителий расположенных по соседству желез реагирует на
воспалительный очаг изменением дифференцировки и пролиферативными
явлениями. Участки очень резко повышенной митотической активности
возникают в области впадения желез в желудочные ямки и в их заканчиваю-
щихся слепо главных отделах. В последних главные клетки утрачивают
свою нормальную дифференцировку и превращаются в элементы мукоид-
ного характера, сходные с теми, которые располагаются в вышележащей
части железы. Обкладочные клетки исчезают не прослеженным детальнее
образом. Возможно, они подвергаются дегенерации. В результате этих
изменений железы превращаются в состоящие из однородных мукоидных
клеток малодифференцированные трубки, слепые концы которых заметно
раздуваются и начинают прорастать через гладкомышечную подстилку
в подслизистый слой. Более удаленные от очага воспаления железы также
превращаются в мукоидные трубчатые ходы, но не вырастают за пределы
слизистой оболочки. Слепые концы дистонических желез, дорастая до
окруженных некротическими массами капель дегтя или масла, «разверты-
ваются», т. е. превращаются в эпителиальные пласты, имеющие свободный
эпителизирующий клиновидно заостренный край и состоящие из одного
слоя уплощенных в большей или меньшей степени элементов, почти или
совершенно не содержащих мукоидных включений. Образовавшиеся из
железистых трубочек эпителиальные пласты, как правило, продвигаются
по воспаленной соединительной ткани, обрастая постепенно инородные
тела и некротические массы. В конце концов края пластов сходятся и об-
разуют сплошную стенку кистовидных полостей, сообщающихся через
бывшие железистые трубки с просветом желудка. В конце концов содержи-
мое кист может опорожниться в просвет желудка через расширившиеся
железистые каналы, а сами кисты превращаются в дивертикулы слизистой
оболочки желудка. По наблюдениям Гаршина окончательная судьба из-
мененного эпителия может быть двоякой. Дистопический эпителий при
затихании воспалительного процесса в окружающей соединительной ткани,
сохраняя мукоидный характер, приобретает вид покровного эпителия же-
лудка. Железы, превратившиеся в мукоидные трубки, но не выросшие за
пределы слизистой оболочки, параллельно с возвращением окружающей
соединительной ткани в исходное состояние, вновь дифференцируются
с образованием в их составе типичных для фундальных желез клеточных
элементов.
При гомопластической имплантации частиц стенки желудка зародышей
кролика вместе с кусочками стерильного целлоидина в подкожную клет-
чатку (Лазаренко, 1939) в создавшемся очаге воспаления эпителий путем
роста и пролиферации постепенно образует большей частью замкнутые
кисты. Первоначально его элементы, уплощаясь, образуют тонкие малодиф-
ференцированные пласты, которые с течением времени снова утолщаются и
приобретают характер изопризматического и высокопризматического по-
176
Глава III
кровного эпителия желудка. Все изменения в эпителии, по данным Лаза-
ренко и в соответствии со взглядами Гаршина, коррелятивно связаны со
стадиями воспалительного процесса в соединительной ткани. Со временем
образования рубцовой ткани совпадают конец пролиферативных процес-
сов в эпителии и его вторичная дифференцировка. Дальнейшая организация
рубца приводит затем к наступлению дистрофических изменений в эпите-
лии, несмотря на васкуляризацию имплантата разросшимися сосудами.
Временно сохраняющие жизнеспособность клетки эпителия вновь упло-
щаются, компенсируя тем самым в известной степени прогрессирующую
дегенерацию и отторжение другой части его клеток. В конце концов имплан-
тат погибает и рассасывается. В условиях своих опытов Лазаренко не наблю-
дал обрастания желудочным эпителием кусочков целлоидина, окруженных
зоной некроза. На всех стадиях процесса имплантированный желудочный
эпителий сохранял однослойное строение и общие гистоморфные свойства
энтеродермальных тканей.
Все изложенные опыты, проделанные очень различными методами на
одном и том же или сходном гистологическом материале, при точном ана-
лизе разыгрывающихся процессов по стадиям, показывают, что специфи-
ческие особенности исходного тканевого материала являются тем основным
моментом, от которого зависит совокупность наблюдаемых гистологических
картин. Различные условия эксперимента представляют собой по существу
факторы, позволяющие реализоваться гистобластическим потенциям, в боль-
шей или меньшей степени свойственным данному тканевому материалу.
Патолого-гистологический материал, относящийся к язвенной болезни
желудка (Любарш, 1906; Чума, 1925; Меркель, 1926; Гаузер, 1926; Конеч-
ный, 1928; Зигмунд, 1929, и др.) и к различным язвенным болезням кишеч-
ника, как, например, дизентерия, тиф, луэс и другие, в отношении описы-
вавшихся для эпителия картин, хорошо согласуется с экспериментальными
данными.
При заживлении дизентерийных язв кишечника происходит типичный
процесс эпителизации (Бейцке, 1918). Эпителий может, врастая в толщу
кишечной стенки, образовать на месте бывших лимфоидных фолликулов
замкнутые кисты, характеризующие т. наз. фолликулярную ди-
зентерию (Лёлейн, 1923). Вследствие усиленного образования бокаловидных
клеток и интенсивного слизеотделения, в содержимом таких кист может
скопляться много слизи. При туберкулезном аппендиците Поппер (1929)
отмечал разрастание либеркюновых крипт. Причем одна из них, достигнув
очага распада, развернулась в пласт, стелющийся на границе некротиче-
ского участка.
Новейшие исследования Галъдман (1940) по экспериментальной пара-
тифозной инфекции показывают, что заживление язв слизистой оболочки
кишечника происходит в результате воспалительных и регенеративных
разрастаний эпителия, обнаруживающего такую же, как и при других
условиях, гистологическую специфику.
Из патолого-анатомических секционных данных остановимся на работе
Захарьевской (1935), посвященной изучению т. наз. псевдомиксомы
Экспериментальный метод в гистологии
177
брюшной полости человека, при которой кишечный эпителий в условиях
целого организма дает чрезвычайно большое, быть может, максимальное
разнообразие гистологических картин. Псевдомиксома представляет собой
довольно редкое заболевание, возникающее в результате прободения черве-
образного отростка и разрастания его эпителия в брюшной полости. Выде-
ляемая кишечным эпителием слизь вызывает воспалительную реакцию
в серозных оболочках, а эта последняя, в свою очередь, стимулирует даль-
нейшие дистопические разрастания эпителия. Получается «порочный круг»
взаимно стимулирующих друг друга процессов, в результате которых брюш-
ная полость переполняется огромными массами слизи и воспалительными
спайками соединительной ткани, по которым разрастается тяжами и пла-
стами энергично пролиферирующий эпителий. Внешне болезнь производит
впечатление неукоснительно развивающегося опухолевого процесса, с ко-
торым она, по существу, не имеет ничего общего.
В зависимости от свойств субстрата, подстилающего эпителий в данном
месте, уровень его дифференцировки оказывается весьма различным. Раз-
растающиеся по скоплениям слизи, нередко инфильтрированным блуждаю-
щими элементами, участки эпителия имеют характер мембран, состоящих
из совершенно плоских малодифференцированных однородных элементов,
между которыми не удается обнаружить замыкающих полосок. В других
местах можно бывает наблюдать утолщение эпителиального покрова и
приобретение его элементами пзопризматической формы. Между их апикаль-
ными концами начинает обнаруживаться сеть замыкающих полосок. На-
конец, на образующихся прослойках соединительной ткани, в которых диф-
ференцируется волокнистое межклеточное вещество, элементы эпителиаль-
ных пластов принимают высокопризматическую форму и дифференцируются
частью в клетки, сходные с кутикулярными, частью в типичные бокаловид-
ные клетки, энергично продуцирующие слизь. Уровень митотической ак-
тивности тем выше, чем менее дифференцирован эпителий. Таким образом,
при псевдомиксоме можно наблюдать весь диапазон дифференцировок ки-
шечного эпителия, начиная от его наименее дифференцированного состоя-
ния, свойственного мембранам тканевых культур, и кончая наиболее спе-
циализированными формами существования. В течение достаточно длитель-
ной болезни кишечный эпителий, поддерживаемый в состоянии пролифе-
рации, обнаруживает совокупность свойственных ему гистобластических
потенций, но никаких намеков на превращение в эпителиальную ткань
иного, например, кожного типа.
Здесь будет уместно остановиться на т. наз. верхних кардиаль-
ных железах или, точнее, островках слизистой оболочки желудка, наблю-
дающихся в 70 % случаев в верхней части человеческого пищевода [Шридде,
1909; Бернер-Питцелып, 1922; Шиффер. 1927). Появление этих образова-
ний в виде резко отграниченных от эпителия пищевода островков можно
констатировать на молодом эмбриональном материале. В дифференцирован-
ном состоянии они также бывают очень резко отграничены от прилегающих
участков многослойного эпителия. Картин, которые можно было бы истол-
ковать как превращение одного вида эпителия в другой, никогда видеть
12 Хлоп пн— 349
178
Глава III
не удается. Все это говорит за то, что в передней части пищеварительной
трубки на ранних стадиях онтогенеза может происходить взаимоперемещение
качественно различного эмбрионального материала; причем часть элемен-
тов энтодермального зачатка эпителия желудка оказывается расположен-
ной среди зачатка головной кишки. Следует подчеркнуть, что первоначаль-
ная длина пищевода у зародыша относительно гораздо меньше и что удале-
ние сместившихся зачатков от кардиальной области желудка происходит
лишь вторично вследствие неравномерного роста.
К сожалению, гистологических данных, касающихся репаративной ре-
генерации или воспалительных разрастаний кишечного эпителия у пред-
ставителей других классов позвоночных, не имеется. Очень скудны и не-
полны в гистологическом отношении данные, относящиеся к различным
беспозвоночным, несмотря на большое количество работ, посвященных
вопросам регенерации. Эктодерма и энтодерма пресноводной гидры, при
опытах с выворачиванием этих животных наизнанку, оказываются неспо-
собными функционально и морфологически замещать друг друга, т. е.
несомненно обнаруживают определенную степень детерминации. Вместе
с тем образование всех дифференцированных клеток гидры, повидимому,
может происходить из одного и того же источника — мелких индифферентных
i-клеток, находящихся в составе эктодермы. При регенерации головного
конца энтодерма и эктодерма претерпевают неодинаковые изменения и вос-
станавливают только подобные себе элементы, не превращаясь друг в друга
(Стрелин, 1928).
По новейшим данным (Бострём, 1936), клетки губок, искусственно дис-
социированные протиранием через мельничный газ, оказываются гораздо
более специализированными, чем это предполагалось прежде. Так, напри-
мер, воротничковые элементы жгутиковых камер (вопрос о том, в какой
мере их можно сравнивать с энтодермой других животных, мы оставляем
открытым) не изменяют своей дифференцировки и не превращаются в дру-
гие клетки, а при воссоздании животного вновь образуют выстилку жгу-
тиковых камер.
Относительно гистогенетических процессов при регенерации трехветви-
стых планарий до настоящего времени остается очень много неясностей и
противоречий. Признававшаяся многими авторами способность элементов
паренхимы превращаться при регенерации в элементы кишечного эпителия
отнюдь не может считаться доказанной. Весьма вероятно, что гистологи-
ческая детерминация по крайней мере некоторых тканей, даже у этих
очень примитивных форм, выражена вполне отчетливо. В частности, ки-
шечник у них регенерирует, повидимому, не метапластическим путем, а из
эндодермальных элементов старого кишечника, делящихся митотическим
путем (Шарое, 1937).
У одной из немертин (Linens ruber) Нуссбаум и Окснер (1910) описали в
свое время образование нового кишечника в отрезках тела, совершенно ли-
шенных элементов энтодермы, из мезодермальной клеточной массы.
По Давыдову (1915, 1928), это совпадаете одной особенностью эмбрио-
нального развития этих животных, при дроблении яиц которых не про-
Экспериментальный метод в гистологии
179
исходит полного разделения эктодермального и мезодермального мате-
риала.
У аннелид эндодермальная стенка кишечника образуется при регенера-
ции из эктодермального же материала, клетки которого, несмотря на неко-
торые морфологические изменения, даже при пролиферации, не утрачивают
свойственных им ресничек (Иванов, 1908). Относительно гистологических
процессов при репаративной ре-
генерации в кишечном эпителии
членистоногих и моллюсков в
известной мне литературе указа-
ний не имеется. При метамор-
фозе генетическая связь дефи-
нитивного кишечного эпителия
с эктодермальным ларвальным
представляется, по имеющимся
данным, несомненной (Поярков,
1900; Генсон, 1932, и др.). Со-
отношение в образовании ки-
шечного эпителия у оболочни-
ков при эмбриональном разви-
тии, с одной стороны, при бес-
полом размножении и регене-
рации, с другой, представляет-
ся неясным и требует новых
фактических данных.
б) Поджелудочная железа
В основу анализа гистобла
стических потенций эпителия
экзокриновой части поджелудоч-
ной железы методом эксплан-
тации положен фактический ма-
териал моей работы (Хлопан.
1929), сделанной на кроличьем
материале, дополненный в от-
дельных местах данными других
авторов, относящимися к экс-
Рис. 16. Образование эпителием вставочного
отдела (St и St') поджелудочной железы зам-
кнутого мешковного выроста с однослойной
стенкой. (Hi — дегенерировавшие главные от-
делы. АК — тангенциально срезанный мало-
дифференцированный эпителиальный комплекс.
3-дневная культура из поджелудочной железы
кролика. Сильное увеличение. (По Хлопипу,
1929).
плантации куриного материала (Капель, 1929; Пинкус, 1930; Катцснштейп
и Кнаке, 1931; Аморозо', 1932; И. Фишер и Рис, 1936, и некоторые другие).
Относительно особенностей превращений вне организма островков
Лангерганса, т. е. эндокриновой части этого органа, сколько-нибудь опре-
деленного фактического материала не имеется. Большая часть авторов
останавливается на плоскостном экстенсивном росте эпителия мембранами.
Однако помимо плоскостного разрастания по поверхности плотного суб-
страта в эксплантатах из поджелудочной железы можно наблюдать также
180
Глава III
своеобразный «погружной» рост замкнутых мешковидных или трубчатых
эпителиальных образований в толще фибрина (рис. 16) и в относительно
редких случаях эпителизацию поверхности посеянных кусочков (рис. 17)
(Хлопан, 1929). Вместе с ростом эпителия наблюдается также то более, то
менее интенсивное прорастание соединительной ткани. В некоторых случаях
одна из этих тканей заглушает другую и может быть в конечном итоге
выделена в чистой культуре. Чистые культуры экстенсивно растущего мем-
бранами эпителия получаются из куриного материала при воздействии на
эксплантат ряда поверхностно активных веществ, препятствующих росту
соединительной ткани (Капщенштейн и 1\наке). Культуры изучались до
40—42 дней жизни вне организма (Пинкус, 1930; Фишер и Рис, 1936).
Рис. 17. Образование покровного эпителия на поверхности
13-дневного эксплантата из поджелудочной железы кролика.
Внутри эксплантата разрастание атипичных малодифференци-
рованных эпителиальных ходов. Покровный эпителий одно-
слойный, однорядный. В местах, где он связан с эпителиаль-
ными ходами, ложная многсслойная структура. Слабое увели-
чение. (По Хлоппну, 1929).
Эпителий поджелудочной железы, растущий мембранами или эпителизи-
рующий поверхность эксплантата, в сильной степени разжижает сверток
фибрина и образует в нем полости, наполненные жидкостью. Наоборот,
вокруг замкнутых эпителиальных разрастаний, как правило, полостей
разжижения не образуется. Это показывает, что эпителий поджелудочной
железы чрезвычайно стойко сохраняет характерную для его элементов
полярную дифференцировку, т. е. неравноценность базальных и апикаль-
ных клеточных поверхностей, даже независимо от присутствия или отсут-
ствия обычной соединительнотканной подстилки (Xлопин, 1929). Если замк-
нутый клеточный комплекс разрывается или развертывается, превращаясь,
как это нередко происходит, в плоскостную мембрану, то быстро наступает
энергичное разжижение окружающего фибрина.
Эпителиальные элементы всех отделов железистых ходов в условиях
культур оказываются способными к росту и пролиферации митотическим
путем. Культуры с прекрасным ростом эпителия получались не только от
Экспериментальный метод в гистологии
181
Рис. 78. Дифференцированный одно-
слойный призматический покровный
эпителий (Ер) в 17-дневной культуре
из поджелудочной железы кролика.
Под эпителием соединительная ткань
(Edg) и сверток фибрина (F). Отчет-
ливые межклеточные мостики и замы-
кающие полоски. (По Хлопину, 1929).
новорожденных, но и от более или менее взрослых кроликов (Xлопин,
1929). Хороший рост эпителия куриной поджелудочной железы получался
только из позднего эмбрионального материала и недавно вылупившихся
цыплят. При культивировании железы молодых зародышей росли преиму-
щественно соединительнотканные элементы. После вылупления энергия
роста и пролиферации эпителия также быстро падала (Пинкус, 1930; Кат-
ценштейн и Кнаке, 1931; Фишер и Рис, 1936). Энергия роста эпителия главных
или секреторных отделов железы недавно вылупившихся цыплят зависит
от их функционального состояния в
момент посева (Фишер и Рис, 1936).
По данным этих авторов, она особенно
значительна, если железистые клетки
находятся в т. наз. стадии рести-
туции, т. е. вновь начинают выраба-
тывать гранулы профермента после
освобождения клеточного тела от секре-
торных включений, которое может быть
стимулировано пилокарпином. При про-
растании на ранних стадиях культиви-
рования железистые клетки могут раз-
личным образом освобождаться от секре-
торных включений. Если они были
сильно переполнены этими включения-
ми, то вся содержащая их апикальная
часть клеточного тела отмирает и оттор-
гается, а базальная околоядерная часть,
наоборот, набухая, увеличивается в
размерах и может начать пролифери-
ровать. После эксплантации специфи-
ческая секреторная деятельность быстро
прекращается, хотя зернышки профер-
мента могут сохраняться долгое время
в клетках измененных железистых ходов
посеянного кусочка и даже экстенсивно
растущих эпителиальных мембран
(Фишер и Рис, 1936). На кроличьем
материале специфических секреторных
включений по прошествии нескольких дней после посева обнаружить не
удалось (Хлопин, 1929). Морфологические особенности эпителиальных кле-
ток секреторных отделов и выводных протоков постепенно сглаживались,
и растущий эпителий приобретал более или менее однородный малодифферен-
цированный характер, независимо от его происхождения из того или иного
отдела железы. В этом отношении, однако, необходимы еще дальнейшие
исследования. Во всяком случае при начинающемся росте и пролиферации
наблюдается морфологическое упрощение эпителиальных структур. Тесно
сближенные комплексы главных отделов железы могут первоначально сли-
18'2
Глава III
ваться в массивные клеточные скопления, вследствие исчезновения тонких
прослоек соединительной ткани. Растущий и пролиферирующий эпителий,
как правило, характеризуется однослойным расположением своих элементов.
Такую же структуру приобретают при своем дальнейшем изменении кле-
точные скопления, возникшие путем слияния. И экстенсивно растущие
мембраны, и стенки полых внутри замкнутых мешковидных или трубчатых
выростов, и пласты, эпителизирующие поверхность эксплантата или ра-
стущую по стенке полости разжижения соединительную ткань, характери-
зуются постоянным однослойным расположением более или менее сильно
уплощенных клеток, плотно сомкнутых своими краями. Удается просле-
дить, как замкнувшиеся перерезанные при посеве материала концы тонких
вставочных отделов железы, постепенно раздуваясь, все больше и больше
выдвигаются в зону роста (рис. 16). Раздуваться могут и разные не прора-
стающие экстенсивно отделы железистых ходов внутри посеянных кусочков.
Сильнее всего уплощение клеток бывает выражено в экстенсивно растущих
мембранах, в которых они могут местами черепицеобразно налегать друг
на друга. Если рост эпителизирующих пластов останавливается, в них удается
наблюдать явления дифференцировки, приводящие к образованию одно-
слойного покровного эпителия. Клетки становятся выше и приобретают
призматическую форму, а между их свободными концами образуется сеть
замыкающих полосок, отсутствующих в условиях энергичного роста и
•пролиферации (рис. 18). Между боковыми поверхностями клеток такого
покровного эпителия возможно бывает заметить появление несвойственных
этой ткани при обычных условиях межклеточных мостиков, а на свободной
поверхности протоплазма, уплотняясь, приобретает характер кутикулы.
Апикальные концы клеток образуют большей частью ровную поверхность,
как и в покровном эпителии в культурах из желудка и кишечника. При
экстенсивном росте, как и в случае других тканей, могут изолироваться
отдельные эпителиальные клетки закругленной, амебоидной, пластинчатой
или неправильно отростчатой формы. Изоляция и округление клеток проис-
ходят также с поверхности эксплантата, если вокруг него образовалась
полость разжижения. Иногда встречаются гигантские эпителиальные много-
ядерные изолированные элементы. В изолированных клетках могут иногда
встречаться митозы.
Фактический материал, касающийся превращений эпителия поджелудоч-
ной железы млекопитающих, при экспериментальных условиях в составе
целого организма в гистологическом отношении изучен недостаточно полно.
Пролиферативные процессы в этом органе удается вызвать как непосред-
ственным повреждением, так и перевязкой выводных протоков. При послед-
ней методике в эпителии различных экзокриновых желез наблюдаются,
с одной стороны, дистрофические изменения, связанные с застоем образуе-
мого ими секрета, а с другой — продолжающиеся ограниченное время
явления пролиферации, приводящие к образованию малодифференцирован-
ных железистых почек и ходов.
В конечном итоге перевязка протока поджелудочной железы приводит
к полному исчезновению экзокриновой части органа при сохранении остров-
Экспериментальный метод в гистологии
183
нового аппарата. В своих опытах над регенеративными процессами в под-
желудочной железе Кирле (1907) описывал новообразование путем проли-
ферации экзокриновой части, а также островков Лангерганса. В противо-
положность этим данным, Мартинотти (1888), Чиполлина (1889) и Б. Фи-
шер (1915) отмечали лишь рост малодифференцированных трубок с одно-
слойной стенкой, но не образование типичных для данного органа и функ-
ционально полноценных долек. Образований структур, сходных с много-
слойным эпителием, эти авторы не отмечали. К сожалению, достаточно
подробных гистологических данных, полученных методами воспалитель-
ных разрастаний в отношении поджелудочной железы, до настоящего
времени нет.
В патолого-анатомической литературе имеются единичные указания на
отдельные находки участков многослойного эпителия в выводных протоках
поджелудочной железы (Гаккель и Нордманн, 1926; Оверлеи, 1921). Безого-
ворочная трактовка таких находок как примеров метаплазии эпителия
поджелудочной железы совершенно не обоснована. Тот факт, что при экс-
периментальных условиях в этом органе не удается получить образования
даже малодифференцированного многослойного эпителия кожного типа,
не может не заставить искать другого объяснения этим наблюдениям.
Возможно, что в поджелудочную железу, как и во многие другие области
организма, могут попадать те или иные дистонические зачатки, дифферен-
цирующиеся на необычном месте с образованием чуждых данному органу
структур, подобно тому, как это часто происходит при образовании упо-
мянутых дистонических кардиальных желез.
Обширный материал из области экспериментальной эмбриологии пока-
зывает, что после трансплантации частиц эмбриональных зачатков, начи-
ная с мозаичной стадии развития (у амфибий — нейрула), пересаженный
материал дифференцируется, независимо от окружающей обстановки, в пол-
ном соответствии со своим происхождением из того или иного источника.
Кроме того, следует подчеркнуть, что далеко не все структуры, описы-
ваемые как многослойный эпителий, являются им на самом деле или срав-
нимы со структурами, свойственными эпителиям кожного типа. Случайные
неправильные нагромождения клеток однослойного эпителия, отмечав-
шиеся выше, начальные стадии врастания однослойного пласта в соедини-
тельнотканную подстилку при недостаточно тщательном анализе гисто-
логических картин могут дать повод к описанию многослойных покровных
структур там, где их на самом деле не имеется. Наблюдение последнего
времени над образованием эндокриновых островковых элементов в составе
выводных протоков поджелудочной железы при нормальных условиях также
может быть причиной ошибочного описания скоплений таких островковых
элементов, как участков многослойного эпителия кожного типа, с которым
они по существу не имеют ничего общего. Наконец, не всякая структура,
имеющая многослойную внешность, может отождествляться даже с мало-
дифференцированным многослойным эпителием кожного типа. В этом отно-
шении достаточно вспомнить т. наз. многослойный эпителий, образуемый
фолликулярными клетками яичника на определенных стадиях развития
184
Глава III
яйцевого фолликула. Едва ли кому придет в голову считать многослойный
фолликулярный эпителий за нечто сходное по существу с кожным эпите-
лием. Все эти соображения и точно анализированный экспериментальный
материал, касающийся различных энтеродермальных тканей, заставляют
меня считать, что случаев образования этим эпите-
лием структур, сравнимых с многослойными э п и-
телиальными структурами кожного типа, никто
и никогда, с малейшей долей достоверности, не
констатировал.
Экспериментальных данных по поджелудочной железе низших позво-
ночных, которые могли бы быть использованы в гистологическом отноше-
нии, в известной мне литературе не имеется. Кроме того, как видно из
сказанного, для выявления вероятных особенностей гистобластических
потенций элементов островкового аппарата еще необходимы специальные
исследования. Как показывают экспериментальные данные, касающиеся
зачатков поджелудочной железы зародышей лягушки, способность к диф-
ференцировке в островки свойственна не всем трем зачаткам этого органа.
ё) Желчные пути и печень
Экспериментально-гистологический материал,, касающийся печени и
желчных путей, также еще нуждается в существенных дополнениях. Между
тем печень является в ряду позвоночных органом, имеющим ряд интерес-
нейших гистологических особенностей.
У ланцетника образование, гомологичное печени позвоночных, имеет'
вид слепого эпителиального выступа пищеварительной трубки. Желези-
стая паренхима печени существенно отличается по своей структуре у пред-
ставителей разных классов позвоночных. У миксины из круглоротых она
имеет строение трубчатой железы с длинными, ветвящимися и слепо закан-
чивающимися секреторными отделами, которые, повидимому, не анасто-
мозируют друг с другом. У миног во взрослом состоянии печень утрачивает
выводные протоки и значение пищеварительной железы. У рыб и амфибий
железистые трубки печени приобретают, вследствие развития анастомозов,
более или менее резко выраженный сетевидный характер. Каждая желези-
стая трубка образована в поперечном сечении несколькими (4—5—6) клет-
ками, причем просвет секреторных отделов, более широкий у рыб, у амфи-
бий приобретает вид тонкого желчного капилляра. Наконец, у высших по-
звоночных и в первую очередь у млекопитающих печеночная паренхима
превращается в сеть балок, состоящих обычно из двух рядов клеток, же-
лобообразные выемки которых образуют в совокупности тончайшие желчт
ные капилляры, пронизывающие всю систему печеночных балок и стоящие
в связи с мелкими разветвлениями желчных протоков. Вместе с тем каж-
дая печеночная клетка вступает с кровеносной системой в особенно тес-
ные взаимоотношения, характеризующие элементы эндокрнновых органов.
При сравнительном изучении печени в ряду позвоночных не могут не
броситься в глаза эволюционные изменения ее гистологической структуры.
Экспериментальный метод в гистологии 185
заключающиеся в ее постепенном превращении из органа обычного желе-
зистого типа в орган с преобладающим эидокриновым строением, напоми-
нающим, например, по своему общему характеру т. наз. кору надпочеч-
ников. Вероятно, эти морфологические изменения связаны с возрастанием
функционального значения печени, как органа внутреннего обмена веществ.
Слизистая оболочка желчного пузыря имеет у низших позвоночных глад-
кую поверхность, а у высших, в частности птиц и млекопитающих, приобре-
тает более или менее сложный складчатый рельеф. У разных животных
в эпителии желчного пузыря в неодинаковой степени выражены явления
мукоидной секреции. На свободной поверхности клеток имеется то более,
то менее отчетливый кутикулярный ободок. У вершины складок клетки
эпителия, как правило, бывают выше, чем у их основания.
По еще неполностью опубликованным данным нашей лаборатории
(Григорьев, 1941), выстилка желчного пузыря у представителей различных
классов позвоночных (рыб, амфибий, рептилий, птиц и млекопитающих)
всегда имеет характер однослойного и однорядного эпителия, в который
могут проникать то в большем, то в меньшем количестве лимфоциты. От-
дельные попытки рассматривать эпителий желчного пузыря как много-
рядный или, точнее, двурядный (Рёзигер, 1930), лишены всякого реального
основания. Описывавшиеся в его составе мелкие клетки являются не мало-
щифференцированными камбиальными элементами эпителия, а заползшими
лимфоцитами, которые в составе эпителиального пласта могут распола-
гаться на разных уровнях.
Эпителии желчных путей, в частности пузыря, и печени обнаруживают
заметные различия в отношении некоторых особенностей. Поэтому их есте-
ственнее рассматривать порознь и начать с желчного пузыря, выстилка
которого во многих отношениях напоминает эпителий пищеварительной
трубки. До последнего времени гистологические данные, касающиеся роста
и превращения эпителия желчного пузыря в условиях тканевых культур,
были очень скудны и исчерпывались кратким сообщением Итикава и Ши-
момура (1925), касающимся желчного пузыря лягушки, и работой Эрдманн
(1931), сделанной на курином эмбриональном материале. Эти авторы отме-
тили экстенсивный рост эпителиальных мембран, который в опытах Эрд-
манн продолжался почти до двух месяцев жизни в организме. По ее данным,
в эпителиальных клетках в течение всего времени культивирования удается
констатировать включения слизи. Григорьев (1941) подверг детальному
сравнительному изучению методом эксплантации эпителий желчного пузыря
костистых рыб, тритона, аксолотля, лягушки, ящерицы, куриных эмбрио-
нов, цыплят, зародышей домашней свиньи, зародышей и молодых кроликов
сроком до одного-полутора месяцев жизни организма. Эпителий желч-
ного пузыря растет экстенсивно, образуя мембраны, или эпителизирует
поверхность эксплантата. Кроме роста эпителия в части культур наблю-
далось также прорастание мезенхимных производных и мезотелия. Рост
эпителия сопровождался разжижением свертка фибрина. Экстенсивного
роста мембранами автору не удалось получить лишь в эксплантатах из
желчного пузыря рыб.
186 1 лава HI,
У всех изученных животных эпителиальные мембраны состоят из одного
слоя полигональных элементов, большей частью уплощенных, реже изо-
призматической формы. При экстенсивном росте по соединительной ткани
эпителиальные элементы приобретают особенно высокую призматическую
форму. У птиц и рептилий на всем протяжении мембран гематоксилином
отчетливо прокрашивались клеточные границы или замыкающие полоски
у апикальной поверхности клеток, что подчеркивает полярную дифферен-
цировку мембран. У млекопитающих и амфибий окраску клеточных границ
удается получить лишь местами — около центрального кусочка, и, особенно,
у клеток, расположенных на соединительнотканной зоне роста. Эпителиаль-
ные мембраны разных животных в отношении своего общего гистотипиче-
ского характера обнаруживают большое сходство. Вместе с тем по ряду
особенностей своих элементов мембраны, как правило, обнаруживают яс-
ную видовую специфичность. Одинаковые по существу функциональное
значение и условия существования эпителия желчного пузыря у разных
позвоночных накладывают свой общий отпечаток на гистоморфную спе-
цифику ткани, на фоне которой уже выступают частные видовые признаки,
характеризующие разные ткани, в том числе эпителий желчного пузыря
отдельных форм.
Эпителиальные пласты на поверхности эксплантатов имеют во всех
случаях однослойное однорядное строение.
Процесс эпителизации происходит у птиц и млекопитающих скорее,
чем у холоднокровных. При эпителизации эпителиальные элементы упло-
щаются на большем или меньшем протяжении от свободного края пласта,
надвигающегося на обнаженную соединительную ткань.
По окончании процесса эпителизации покровный эпителий становится
высокопризматическим и приобретает сходное или тождественное с исход-
ным строение. В апикальной части клеток до самых последних изученных
сроков культивирования образуется мукоидный секрет. Явления роста
эпителия сопровождаются то более, то менее интенсивным митотическим
делением клеток. Внутри эпителизированных кусочков клетки постепенно
дегенерируют, и количество их со временем заметно уменьшается.
Опыты с репаративной регенерацией эпителия желчного пузыря после
сквозного ожога иглой через всю толщу стенки (Григорьев, неопубликован-
ные данные) дали весьма сходные картины. После ожога, параллельно
с развивающейся воспалительной реакцией в соединительной ткани, эпи-
телий желчного пузыря, уплощаясь и упрощаясь, постепенно эпителизиро-
вал дефект. В некоторых случаях эпителиальный пласт продвигался через
незаросшее сквозное отверстие в стенке пузыря на его наружную, обращен-
ную в брюшную полость, поверхность. В удаленных от места ожога участ-
ках слизистой оболочки эпителиальные клетки на ранних стадиях временно
приобретали очень различную высоту. Вследствие этого первоначально
гладкий (у амфибий) рельеф слизистой оболочки становился неровным. По
окончании процесса эпителизации постепенно наступала вторичная диф-
ференцировка. В течение всего процесса эпителий постоянно сохранял одно-
слойное однорядное строение.
Экспериментальный метод в гистологии 187
При изучении распределения митозов после повреждения удалось под-
метить любопытные различия между низшими (амфибии) и высшими (птицы,
млекопитающие) позвоночными. У тритона обнаруживается высокая мито-
тическая активность в области шейки пузыря, у места выхода пузырного
протока. Более того, появление большого количества митозов удается
проследить в печеночном протоке и его разветвлениях внутри печени. Лишь
на более поздних стадиях можно было видеть единичные митозы вблизи
места дефекта, который обычно наносился в средней части пузыря или в об-
ласти его дна. У высших позвоночных с неровным складчатым рельефом
слизистой оболочки митозы могли встретиться в разных участках пузыря,
но с преимущественной локализацией не у вершины складок, а между ними.
В самом регенерате митозы во всех случаях появлялись значительно позже.
Эти наблюдения наводят на мысль, что распределение элементов с различно
выраженными камбиальными свойствами или различной реактивностью
в составе эпителиального пласта претерпевает в филогенезе определенные
и закономерные изменения, связанные, вероятно, в данном случае с изме-
нением рельефа слизистой оболочки, и ставят на очередь дальнейшее изу-
чение этих отношений в энтеродермальных пластах других органов, а также
и в других эпителиях. В качестве параллели к этим данным следует напо-
мнить о наблюдениях Стрелина (1935) над своеобразной локализацией
митозов, возникающих в базальном слое многослойного эпителия роговицы
лягушки после нанесения дефекта. В последнее время Хрущов (1942) уста-
новил в эпителии тонкого кишечника белых крыс наличие двух областей
очень высокой митотической активности в условиях нормального всасы-
вания пищи.
Относительно вызванных экспериментально воспалительных разраста-
ний эпителия желчного пузыря имеются краткие указания в монографии
Гаршина (1939).
Изучение эпителия желчного пузыря при патологических условиях
хорошо согласуется с приведенными экспериментальными данными. При
хронической желчнокаменной болезни и разных воспалительных процессах
в стенке желчного пузыря эпителий обнаруживает описанные выше свойства
энтеродермальных пластов. В его составе описывалось, как метаплазия,
появление типичных бокаловидных клеток, свойственных кишечнику. Отно-
сительно очень редких указаний на обнаружение в желчных путях «мета-
пластического» многослойного эпителия справедливо все сказанное про
аналогичные находки в поджелудочной железе.
Как показывают экспериментальные данные, печеночные элементы выс-
ших позвоночных в отношении ряда особенностей очень заметно отличаются
от эпителия желчных путей.
Большая часть работ, посвященная культивированию печеночных эле-
ментов, выполнена на курином эмбриональном материале, и лишь незначи-
тельное количество их сделано на печени млекопитающих (Смирнова, 1916;
Линч, 1921; Митсуда, 1924; Нордман, 1929; Должанский, 1930; Бишелье,
1931; Шантрох, 1932, и др.). В гистологическом отношении, однако, все
данные неполны. Ряд авторов констатирует экстенсивный рост плоскостных
188
Глава III
мембран из печеночных клеток и отмечает их своеобразный характер.
Должанскому удалось получить чистые культуры экстенсивно растущих
мембранами печеночных клеток и поддержать их существование в течение
нескольких месяцев. Мембраны состоят из однослойно расположенных поли-
гональных уплощенных элементов, сохраняющих независимо от срока
культивирования свой характер, и имеют ровные края. Растущие печеноч-
ные клетки энергично делятся митотическим путем, богаты жировыми
включениями (Шантрох, 1922) и могут также накоплять гликоген (Нордман,
1929). При замедленном росте и пролиферации они ожиревают особенно
сильно. Если при очередном пересеве к чистым печеночным культурам
подсадить эмбриональную соединительную ткань, то может наблюдаться
взаимное прорастание печеночных и соединительнотканных элементов. При
погружном росте в соединительной ткани печеночные элементы образуют
сплошные отпрыски, тяжи и полые внутри трубчатые ходы с однослойной
стенкой, состоящей из клеток кубической или призматической формы
(Долзкапский и Руле, 1934). Вскоре после эксплантации нормальная струк-
тура эксплактированных частиц эмбриональной печени изменяется. Кровя-
ные синусы спадаются, а их эндотелиальные элементы очень быстро высе-
ляются из периферических участков эксплантата в окружающий сверток
фибрина. Вследствие этого печеночные клетки приобретают тесное взаимное
расположение (Леви, 1934). Затем они, прорастая и постепенно уплощаясь,
образуют упомянутые выше мембраны зоны роста. Наиболее благоприятная
для роста и пролиферации печеночных элементов концентрация эмбриональ-
ного экстракта составляет 20 %, а остаточная энергия их роста меньше, чем
у фибробластоподобных клеток (Должанский, 1929).
На основании неопубликованных данных нашей лаборатории (Зуба-
рева, Цымбал) можно сделать вывод, что печеночные элементы новорожден-
ных и молодых кроликов в культурах энергично пролиферируют и растут
характерными тонкими однослойными мембранами, состоящими большей
частью из довольно правильных полигональных сильно уплощенных кле-
ток, тесно связанных своими краями. Разжижение свертка фибрина и обра-
зование в нем полостей выражены слабо и даже могут вообще отсутствовать.
Печеночные клетки никогда не эпителизируют поверхности эксплантата
и не образуют покровных пластов, отграничивающих его от окружающей
среды. Эта особенность, не отмечавшаяся другими авторами, отличает их
в сильной степени от других тканей энтеродермального типа. Мелкие желч-
ные протоки могут, раздуваясь, превращаться в кистовидные полости,
наполненные жидкостью. Наряду с мембранами из печеночных клеток и
эпителия мелких желчных протоков из соединительнотканных элементов
печени образуется обычная сетевидная зона роста с амебоидными макро-
фагами.
Опыты над регенерацией печени млекопитающих наглядно демонстри-
руют очень высокую способность ее элементов к пролиферативным процессам
(Подвысоцкий, 1885; Пенфик, 1889; Мейстер, 1891; Рибберт, 1904; Мас-
сенти, 1924; Фишбек, 1929; Герксгеймер и Тольдте, 1930, и др.). Через
четыре или пять недель после хирургического удаления двух третей массы-
Экспериментальный метод в гистологии
189
печени у крыс общие размеры органа приблизительно возвращаются к норме.
Чрезвычайно быстрая регенерация печени отмечалась также у собак, при-
чем новообразованная ткань приобретала вполне нормальный вид. В отно-
шении источников образования регенерата остается спорным и неясным,
в какой степени в регенерации печеночной паренхимы принимают участие
мелкие желчные протоки. В первую очередь это относится к т. наз.
септальным желчным ходам, высокую регенеративную способность кото-
рых, как своеобразных, сравнимых со вставочными отделами других желез,
частей желчных протоков, едва ли можно отрицать. Несомненно, что сами
печеночные клетки также обладают высокой пролиферативной активностью.
После нанесения дефекта элементы печеночных балок, расположенных
по соседству, претерпевают заметное морфологическое упрощение и могут
в значительной степени уподобляться реактивно измененным клеткам
желчных путей. Это очень сильно затрудняет определение участия желчных
протоков в регенерации печеночных долек. Измененные печеночные балки
разрастаются тяжами и трубками с однослойной стенкой, которые сетевидно
соединяются друг с другом и дают по крайней мере большую часть обра-
зующейся вновь печеночной ткани. Более крупные печеночные протоки,
остающиеся отличимыми от печеночной паренхимы, образуют ветвящиеся
.трубки с однослойной эпителиальной стенкой. Разрастающаяся печеночная
паренхима постепенно вновь дифференцируется в типичные печеночные
балки и приобретает дольчатое строение. В эпителии растущих желчных
протоков и в растущих и пролиферирующих печеночных клетках отмечаются
многочисленные митозы. Ряд авторов указывал на преимущественную
локализацию митозов в периферических частях долек. При эксперименталь-
ном отравлении фосфором в печени также наблюдаются резко выраженные
регенеративные процессы {Клара, 1931).
К сожалению, мы еще не располагаем полностью данными относительно
воспалительных разрастаний ткани печени, получаемых по методу Гар-
шина. Согласно предварительным данным {Гаршин, 1939), печеночные
клетки и эпителий желчных протоков, в ответ на введение каменноугольного
дегтя, энергично растут тяжами и трубками и пролиферируют в очаге
воспаления. Во многих случаях, обрастая очаги некроза с каплями дегтя,
измененные печеночные клетки образуют эпителиальные кисты, стенка
которых построена из однослойного однорядного призматического эпителия.
Параллельно с затуханием воспалительной пролиферации соединительной
ткани, рост эпителия прекращается.
В опытах с воспалительными разрастаниями печеночных имплантатов
по методу Лазаренко {Ржевуцкая, 1937) также наблюдается энергичный
рост и пролиферация эпителия печени и желчных протоков. Печеночные
клетки растут преимущественно трубками и однорядной стенкой. В отли-
чие от опытов с дегтем, в имплантатах не наблюдается обрастания или эпи-
телизации кусочков целлоидина и некротических масс. После периода
более или менее бурной пролиферации в воспалительном очаге постепенно
развивается рубцовая ткань, а в разросшихся эпителиальных трубках
наступают дистрофические изменения, сопровождающиеся накоплением
190 Глава III
темных гранул в цитоплазме. В конце концов эпителиальные пролифераты
погибают. При многих патологических процессах в печени, как известно,,
наблюдаются разрастания в виде т. наз. «ложных желчных ходов», генез
которых до настоящего времени является спорным (Рёссле, 1930; Геркс-
геймер, 1930).
Как видно из изложенного, печеночные элементы млекопитающих,
а также птиц при различной экспериментальной методике обнаруживают
цикл гистологических превращений, сходный по существу с другими энте-
родермальными тканями. Лишь в условиях тканевых культур печеночные
клетки в одном отношении резко отличаются от покровных энтеродер-
мальных пластов; отсутствием эпителизации поверхности эксплантата и
слабо выраженными фибринолитическими свойствами. При других мето-
дах (как это уже отмечалось для некоторых разобранных выше объектов)
эти биологические особенности их не наблюдаются.
Вторая особенность печеночных клеток, также обнаруживаемая мето-
дом эксплантации, заключается в их способности сохранять во многих слу-
чаях свои специфические цитологические признаки в составе очень мало-
дифференцированных в гистологическом отношении структур, каковыми яв-
ляются экстенсивно растущие мембраны. Возможно, что эти биологические
особенности связаны в известной мере с утратой печеночными элементами
высших позвоночных морфо-физиологических признаков эпителиальных
тканей и приобретением ими преобладающего значения особой ткани вну-
тренней среды организма.
Очень большой интерес, с нашей точки зрения, представляет сравни-
тельное изучение печеночной паренхимы в ряду позвоночных. К сожалению,
в этом отношении еще практически ничего не сделано.
г) Свойства собственно энтодермы как эмбрионального зачатка
При изучении способности к дифференцировке энтодермального мате-
риала амфибий, начиная со стадии ранней гаструлы, методом интерплан-
тации в глазницу (Дюркен, 1926), при эксплантации в соляных растворах
(Голътфретер, 1938, 1939) и в плазме (Винников, неопубликованные дан-
ные) энтеродсрмальные бластомеры обнаруживают наименьший объем
проспективных потенций, а затем оказываются способными диф-
ференцироваться лишь в свойственные энтодерме производные — в
желудочный или кишечный эпителий, печеночные клетки или части
поджелудочной железы. В условиях экстенсивного роста вне ор-
ганизма энтодерма, не пройдя предварительно стадий органогенеза,
непосредственно дает начало малодифференцированным гистоморф-
ным разрастаниям однослойными клеточными мембранами, причем их
элементы отличаются богатством желточных включений. Такие мембраны
по способу взаиморасположения клеток напоминают разрастания, полу-
чаемые из дифференцированных эндодермальных тканей. В опытах И. Фи-
шер (1937) наблюдалось образование эксплантированной энтодермой струк-
тур, свойственных секреторным концевым отделам поджелудочной железы.
191
Экспериментальный метод в гистологии
которые при дальнейшем культивировании подвергались дедифференци-
ровке и давали начало экстенсивно растущим эпителиальным мембранам.
Эти данные показывают, что энтодерма представляет собой, начиная
с ранних стадий развития, определенным образом детерминированный кле-
точный материал, который обнаруживает определенную совокупность
гпстобластических потенций, характеризующих разнообразные ткани, раз-
вивающиеся из него в дальнейшем при нормальном онтогенезе.
Опыты Тома (1938) с эксплантацией желточной энтодермы куриных
зародышей, с одной стороны, показали высокую степень специфичности ее
элементов, а с другой — весьма большую изменчивость образуемых ею
гистоморфных структур. Экстенсивно разрастающаяся желточная энто-
дерма образовала комплексные клеточные мембраны, которые в дальней-
шем, разрыхляясь, превращались в сетевидные соединения отростчатых
клеток, напоминающих по внешней форме фибробласты, и изолированные
макрофагоподобные амебоидные элементы.
В этих весьма разнообразных формах существования клетки желточ-
ной энтодермы обнаруживали общую цитотипическую особенность — спо-
собность к образованию и отложению в своей цитоплазме характерных
желточных включений. По данным Тома, желточная энтодерма в состоя-
нии роста мембранами и сетевидными клеточными соединениями образует
волокна межклеточного вещества. Нужно думать, в этом своеобразии по-
ведения при экспериментальных условиях проявляется то, что желточная
энтодерма представляет собой особую часть энтодермального зачатка. Она
претерпевает раннюю специализацию и дифференцировку (т. наз. прежде-
временная дифференцировка Иванова) в особую ткань трофического зна-
чения, имеющую при нормальных условиях морфо-физиологическую ха-
рактеристику эпителия и вместе с тем утрачивает все прочие гистобласти-
ческие потенции, свойственные этому эмбриональному зачатку, т. е. про-
делывает своеобразную эволюционную адаптацию, или, если можно так
выразиться, гистологическую деградацию.
д) Эктодермальные опухолевые ткани
При опухолевой пролиферации энтеродермальных тканей также наблю-
даются гистологические структуры различного уровня дифференцировки.
Наиболее частыми и характерными из злокачественных опухолей этого
типа являются различные модификации т. наз. цилиндроклеточ-
ных раков, бластоматозные элементы которых образуют свойственные эпи-
телиям кишечного типа однослойные однорядные пласты или ходы, напо-
минающие в большей или меньшей степени железы. Такого рода первичные
опухоли, несомненно возникающие из эктодермального зачатка на разных
уровнях его развития или чаще из соответствующих дифференцированных
эпителиев и могущие метастазировать в разные части тела, встречаются
во всех органах пищеварительной системы (Боррман, 1926; Фишер-Ва~
зелье, 1927; Юинг, 1928; Оберндорфер, 1929, и др.). В своем наиболее диф-
ференцированном виде они обнаруживают, как правило, в более или менее
192
Глаза III
типичном или, наоборот, уродливом виде морфологические особенности желу-
дочного или кишечного эпителиев. В их составе нередко встречаются сецер-
нирующие мукоидные или слизистые клетки. Ог^холи типа т. ваз.
гепатобластом состоят из тяжей и скоплений клеток, обнаруживающих,
несмотря на свою уродливость, несомненную дифференцировку печеночных
элементов. Недавно опухоли такого типа удалось получить эксперимен-
тально у мышей под влиянием ортоамидоазотолуола (Морозе некая, 1938).
Другие энтеродермальные опухоли напоминают малодифференцированные
эпителиальные пласты или трубки, образуемые энтеродермальными тка-
нями при пролиферации в экспериментальных условиях. Нередко в составе
одной и той же опухоли можно, бывает, наблюдать участки разного строе-
ния, переходящие друг в друга без резкой границы.
Наименее дифференцированные энтеродермальные опухоли, например
некоторые бластомы желудка т. наз. крупно- и мелкоклеточного типа,
могут быть, наконец, образованы совершенно атипичными недифференци-
рованными клетками, подчас напоминающими малые лимфоциты и не обра-
зующими никакой гистоморфной структуры. В составе таких опухолей,
обозначаемых общим термином щитобластомы» (Фишер-Вазельс, 1927), энте-
родермальные ткани находятся в состоянии наибольшей катаплазии. Их эле-
менты не образуют никаких взаимосвязанных тканевых соединений и пред-
ставляют собой по сути дела изолированные клетки. Элементы таких опу-
холей сравнимы с нормальными элементами энтеродермальных тканей,
находящимися в цитоморфном состоянии. В отличие от последних соответ-
ствующие опухолевые клетки приобретают способность к длительному суще-
ствованию и пролиферации в изолированном цитоморфном состоянии. Оче-
видно, в этом случае высокая степень опухолевой катаплазии препятствует
выявлению каких бы то ни было гистоморфных особенностей ткани. Истин-
ную природу таких опухолей, как и в других случаях, на основании изу-
чения микроскопической картины определить невозможно. Их гистогенез
устанавливается с достаточной долей вероятности в тех случаях, когда
в составе одной и той же опухоли удается проследить постепенную атипи-
зацию характерных энтеродермальных гистоморфных структур, завер-
шающуюся их превращением в несвязанные друг с другом клетки. Изло-
женный выше экспериментальный материал представляется убедительным
обоснованием этой трактовки цитобластом, предложенной Фтиер-Вазель-
сом (1927). Наоборот, совершенно неубедительной и необоснованной пред-
ставляется другая, более старая точка зрения, согласно которой цито-
бластомы трактуются как саркомы соединительнотканного характера,
могущие возникать путем метаплазии из любого тканевого материала.
Говорить в данном случае о метаплазии и тем более о метаплазии в соеди-
нительную ткань совершенно ошибочно уже потому, что опухолевые клетки
цитобластом вообще не образуют никакой дифференцированной или мало-
дифференцированной тканевой структуры.
Сравнительно очень редкими или даже единичными находками являются
первичные опухоли органов пищеварительной системы, содержащих в норме
энтеродермальные ткани, с дифференцированными многослойными плоско-
Экспериментальный, метод в гистологии 193
клеточными или описываемыми, как таковые, эпителиальными структурами.
Если источником возникновения плоскоклеточных раков желудка, по
крайней мере в подавляющем большинстве случаев, является эпителий
пищевода, то редкие находки плоскоклеточных или смешанных раков пе-
чени (Рёзигер, 1930), которых всего отмечено в литературе около 30 слу-
чаев, или совершенно единичные описанные плоскоклеточные раки под-
желудочной железы несомненно образуются первично из тканевых эле-
ментов, бывших до начала опухолевого процесса в составе этих органов.
Исходя из всего того, что мы знаем о гистологической детерминации экто-
дермального зачатка и образующихся из него тканей, мы не имеем никаких
оснований считать их за вероятный источник опухоли. Поскольку дело
идет о настоящих новообразованиях кожного типа, мы имеем все основа-
ния считать, что данные опухолевые зачатки возникли из дистонического
эмбрионального материала, обладающего другими, чем у энтодермы, гисто-
бластическими потенциями. Кроме того, не подлежит сомнению, что ряд
структур, принимаемых за многослойный эпителий или за образования,
сравнимые с жемчужинами, как в случае т. наз. аденоканкроидов пище-
варительной трубки (Герксгеймер, 1907; Боррман, 1926), на самом деле не
имеют признаков эпителиев кожного типа.
До настоящего времени, к сожалению, отсутствует точный гистологи-
4 ческий анализ некоторых малодифференцированных раков, развивающихся,
например, из эпителия желудка, в которых опухолевые клетки образуют
сплошные скопления без полости внутри, напоминающие некоторые мас-
сивные малодифференцированные разрастания кожного эпителия. Эти,
так сказать, вторично солидные карциномы, могущие представлять собой
измененные типичные «цилиндроклеточные» структуры, по способу своего
образования и ряду особенностей, несомненно отличаются от эпителиальных
массивов эпидермального происхождения. Тщательное гистологическое
изучение таких структур, исходя из сказанного выше, должно быть по-
ставлено на очередь.
В отношении экспериментального изучения энтеродермальных раков
методом эксплантации пока сделано еще слишком мало. Кроме отдельных
указаний на рост клеток эпителиальными мембранами (Вейтцмап, 1938),
никаких более обстоятельных данных гистологического характера не
имеется.
Резюмируя все сказанное выше, можно сделать вывод, что весь в гисто-
логическом отношении достаточно точный и изученный в его динамике
фактический материал, которым мы располагаем, доказывает высокую сте-
пень специфичности энтеродермальных тканей в первую очередь у позво-
ночных, а также у большой части беспозвоночных. Энтеродермальные
ткани не способны не только к превращению в соединительнотканные обра-
зования (в чем в настоящее время уже едва ли кто-нибудь сомневается
всерьез), но и в гистологические структуры, свойственные дифференциро-
ванным в той или иной степени эпителиям другого происхождения. Вся
совокупность превращений энтеродермальных тканей в эксперименталь-
ных условиях демонстрирует с полной убедительностью их строгую гисто-
13 X л опин——349
194
Глава III
логическую детерминацию. Они никогда и ни при каких условиях не воз-
никают из тканей другого происхождения и в таковые не превращаются.
е) Филогенетические корни зачатка головной кишки позвоночных
После разбора фактического материала по эпидермальным и энтеро-
дермальным тканям необходимо подвести краткий итог обоснованию на-
шей трактовки происхождения эпителиальной выстилки переднего отде-
ла пищеварительной трубки со всеми ее производными.
Как это неоднократно отмечалось, выстилку переднего отдела пище-
варительной трубки можно трактовать с двух противоположных точек
зрения, рассматривая ее или как результат конвергентной «эпидермо-
идальною дифференцировки части единого эктодермального зачатка или
как сместившийся в филогенезе участок наружного кожного покрова,
который вторично присоединился к эктодермальной части кишечной труб-
ки. Общепринята первая точка зрения. Она основывается в настоящее
время лишь на том, что в онтогенезе ланцетника и низших позвоночных за-
чаток головной кишки инвагинирует или подворачивается вместе с энтодер-
мой и хордой при образовании стенки первичной кишечной трубки, .а не об-
разуется на более поздних стадиях развития путем врастания внутрь клеток
эпидермального зачатка. Этим только и исчерпывается вся аргументация
в пользу взгляда на зачаток головной кишки, как на часть единой энто-
дермы. Все же прочие, разобранные выше факты, определенно говорят
против этой точки зрения. Значение этого эмбриологического факта, однако,
вовсе не следует переоценивать, хотя бы на основании того, что прехор-
дальная пластинка, являющаяся зачатком головной кишки, связана с хор-
дой в такой же степени, как с энтодермой, а хорда представляет собой
самостоятельный зачаток, который в настоящее время уже не приходится
рассматривать как специализированную часть кишечной энтодермы. Это
уже само по себе значительно понижает убедительность эмбриологической
аргументации эволюционного происхождения эпителиальной выстилки пе-
реднего отдела пищеварительной трубки из энтодермы предков. В пользу
филогенетического развития зачатка головной кишки из кожной эктодермы
предков, точки зрения, защищаемой нами, говорят следующие факты.
Во-первых, строгая гистологическая детерминация эпителиев кишеч-
ного и кожного типов и их неспособность превращаться друг в друга
в экспериментальных условиях, которая обнаруживается уже у многих
низкоорганизованных форм и демонстрируется с полной убедительностью
у позвоночных и, в частности, у млекопитающих.
Большие различия в гистобластических потенциях эпидермальных и
энтеродермальных тканей, которые были установлены лишь в последнее
время и позволили противопоставить друг другу эти гистологические
структуры, как две качественно различные тканевые системы, являются,
на основании всего сказанного выше, принципиально новым и, по сути
дела, решающим аргументом в пользу нашей точки зрения. Этим аргумен-
том не обладали прежние исследователи, которые, подобно Геккелю и Рау-
беру, выступали за генетический подход к тканям.
Экспериментальный метод в гистологии
195
Во-вторых, глубокие черты гистологического сходства, доходящего в
ряде случаев до тождества, между эпидермисом кожи и эпителием перед-
него отдела пищеварительной трубки. Этот аргумент делается особенно
убедительным в результате экспериментального обоснования различной
гистологической детерминации эпидермальных и энтеродермальных тканей.
В-третьих, ранняя детерминация зачатка головной кишки у амфибий,
приобретающего уже на ранних стадиях развития качественные отличия
от энтодермы.
В-четвертых, совершенно резкая граница между эпителиями кожного
и кишечного типов, обнаруживающаяся повсеместно, где имеется стык
(именно стык, а не переход друг в друга) между этими тканями.
В-пятых, общеизвестные факты, что эпидермис кожи и эпителий перед-
него отдела пищеварительной трубки участвуют в ряду позвоночных в обра-
зовании зубных зачатков.
В-шестых, осуществление кожным и кишечным эпителиями защитной
функции совершенно различными способами: путем развития многослой-
ное™ с ороговением покровных слоев у высших позвоночных и путем обра-
зования защитного секрета при сохранении однослойное™, как это. на-
пример, имеет место в мышечном желудке птиц. Все это, взятое вместе,
наглядно демонстрирует, что эпителии кожного и кишечного типов пред-
ставляют собой две ткани, имеющие глубокие гистотипические различия,
причиной которых могут быть только неодинаковые пути их исторического
развития и возникновение из разных источников.
Труднее всего дать объяснение, почему в онтогенезе хордовых утрати-
лась видимая генетическая связь между эпидермальным зачатком и за-
чатком головной кишки. Наиболее вероятным представляется поставить
в связь это несомненно вторичное изменение в ходе онтогенеза с развитием
вторичного рта и хордального зачатка. Последний возможно рассматривать
как специализированный участок эктодермы, образовавшийся у пред-
ков современных хордовых на границе между собственно эктодермой и
зачатком эктодермальной же головной кишки, который, в связи с образо-
ванием нового ротового отверстия на аборальном конце, вероятно, занял
положение в области дорзальной губы бластопора и начал инвагинировать
в течение гаструляции вместе с расположенной по соседству прехордаль-
ной пластинкой.
В. Целонефродермалыгый тканевой тип
а) Краткие эмбриологические и сравнительные данные
Генетическая близость эпителиев целома и выделительного аппарата
типа целомодуктов с достаточной отчетливостью вытекает из эмбриоло-
гических и сравнительно-анатомических данных.
В ряду позвоночных от общей полости тела уже у круглоротых обо-
собляется мешочек перикарда, а у млекопитающих завершается разделе-
ние прочей части целома на брюшную и плевральные полости.
У позвоночных и ланцетника выделительный аппарат представлен орга-
нами типа целомодуктов, которые в своем развитии связаны теснейшим
196
Глава III
образом с выстилкой полости тела. У личиночных форм ряда низших позво-
ночных (круглоротые, ганоидные и костистые рыбы, амфибии) между по-
лостью тела и выделительным аппаратом имеются тесные морфологические
и функциональные взаимоотношения, большей частью утрачивающиеся во
взрослом состоянии, в связи с редукцией предпочки. При функционирую-
щей предпочке фильтрация растворимых продуктов обмена происходит
через сосудистый клубок в полость тела, из которого клубочковый фильтрат
через нефростомы поступает в мочевые канальцы, где подвергается изме-
нениям и в конце концов выводится наружу. У костистых рыб функцио-
нально связанный с предпочкой участок целома отделяется от него, пре-
вращаясь в особую камеру предпочки, в стенке которой содержится филь-
трующий аппарат. У селяхий, у которых предпочка рудиментарна, ка-
нальцы вольфова тела сохраняют нефростомы, хотя их функциональное
значение и представляется неясным. Нефростомы первичной почки бес-
хвостых амфибий, хотя и сохраняются частично, но утрачивают связь
с выделительным аппаратом и у взрослых форм превращаются в соединение
полости тела с сосудистой системой, что является в высшей степени инте-
ресным, хотя по существу и мало понятным фактом. У высших позвоноч-
ных нефротомы и развивающийся из них выделительный аппарат полностью
утрачивают связь с целомом, а воронки первичной и вторичной почек даже
не развиваются. Собирательные трубки вторичной почки первоначально
образуются у зародыша как ветвящаяся система выростов лоханки, всту-
пающих в связь с развивающимися из нефрогенной ткани нефронами (Петер,
1909; Феликс, 1905, 1911). Эти первичные собирательные трубки затем ча-
стично редуцируются и превращаются в выстилку чашечек. По новейшим
данным Фукса (1932, 1933), эта редукция первичных собирательных трубок
идет у человека значительно дальше, чем это предполагалось прежде. Не
исключена возможность, что в окончательных собирательных трубках вся
или почти вся выстилка замещается в конце концов эпителием нефронов.
В пользу этого предположения говорят приводимые ниже эксперименталь-
ные данные. У некоторых домашних животных удается обнаружить в со-
сочковых ходах на разном уровне довольно ясную границу между одно-
слойным и двурядным эпителиями, из которых первый, вероятно, при-
надлежит самой почке, а второй, постепенно переходящий в типичный
эпителий мочеотводящих путей, — лоханке.
Другим образованием, тесно связанным с полостью тела, являются
мюллеровы каналы, появляющиеся впервые у рыб и превращающиеся
в женские половые пути. У круглоротых мюллеровы каналы не образуются.
Созревшие женские и мужские половые продукты у круглоротых путем
разрыва стенки гонад попадают в полость тела, которая у половозрелых
форм функционирует, как половые пути, и сообщается с внешней средой
через особые отверстия, абдоминальные поры. Мочеполовой связи, харак-
терной для мужского полового аппарата всех вышестоящих позвоночных,
у них не образуется. Отношения, наблюдаемые у круглоротых, несколько
напоминают способ выведения половых продуктов у ланцетника, у которого
половые пути совершенно отсутствуют, а половые продукты выводятся
Экспериментальный метод в гистологии
197
путем разрыва стенки тела в перибранхиальную полость и через нее
наружу. У рыб выведение женских половых продуктов наружу происхо-
дит довольно различным, в деталях, образом, частью через мюллеровы
каналы, частью через особый участок полости тела, обособляющийся от
всей прочей части ее и превращающийся в особую камеру яичника и боль-
шую или меньшую часть яйцеводов. У живородящих селяхий образуется
плодовместилище, в котором происходит развитие зародышей. У самок ам-
фибий зрелые половые продукты через разрыв стенки яичника поступают
в полость тела, а затем наружу через яйцеводы, образующиеся из пол-
ностью развитых мюллеровых каналов.
У высших позвоночных между воронками яйцеводов (мюллеровых ка-
налов) и яичниками устанавливается настолько близкая связь, что полость
тела фактически выключается из функции выведения половых продуктов,
несмотря на существование открытого сообщения между целомом и жен-
скими половыми путями. Половые клетки непосредственно попадают на
мерцательную бахрому яйцеводов, практически минуя полость тела.
В онтогенезе мюллеровы каналы большей частью образуются, как
желобовидное углубление стенки полости тела, которое постепенно отшну-
ровывается от нее. Их воронки первоначально представляют собой нефро-
стомы предпочки. В течение дальнейшей эволюции первичные воронки
мюллеровых каналов редуцируются и заменяются образующимися вновь
воронковидными сообщениями с полостью тела. В деталях развитие мюл-
лерова канала и его связь с воронкой предпочки у представителей разных
классов варьируют довольно сильно. У селяхий, в отличие от всех других
позвоночных, мюллеров канал, по крайней мере частично, возникает путем
расщепления вольфова канала.
Сопоставляя различные варианты развития мюллеровых каналов, Шим-
кееич пришел к выводу, что в данном случае имеет место меторизис за счет
прогрессирующего удлинения целомической части зачатка. У млекопитаю-
щих из мюллеровых каналов образуется выстилка яйцеводов и матки.
Что касается эпителия влагалища, то, как уже отмечалось выше,
на этот счет имеются разногласия. Гистологические факты заставляют
нас считать, что многослойный эпителий влагалища не является произ-
водным мюллеровых каналов, а в соответствии с эмбриологическими дан-
ными Кофф (1933) и Билас (1934) развивается из выстилки мочеполового
синуса и по всем своим свойствам относится к эпидермальному ткане-
вому типу.
Уже на основании краткого разбора сравнительно-эмбриологического
и анатомического материала и неодинакового в разных случаях функцио-
нального значения полости тела можно высказать предположение, что,
несмотря на генетическую близость эпителиев целома, почек, яйцеводов
и матки, они должны были филогенетически приобрести, в результате ди-
вергентной дифференцировки, очень существенные морфо-физиологические
различия. Установленные нами экспериментально у млекопитающих гисто-
логические особенности этих эпителиев заставили нас первоначально под-
разделить эти эпителии на два резко отличающиеся Друг от друга типа
198 Глава III
целодермальный и нефродермальный, с которыми мы, кроме того, сближали
производные мюллеровых каналов. Законченный в настоящее вре.мя экспе-
риментальный анализ выстилки полости тела на сравнительном материале
и приведенные выше эмбриологические факты заставляют считать более
правильным объединение всех этих эпителиев в единый, но дивергентно
развившийся целонефродермальный тканевой тип.
Помимо упомянутых выше тканей, к тому же тканевому типу должны
быть отнесены, на основе их близких генетических отношений, элементы
паренхимы интерренальных органов (у млекопитающих — коркового веще-
ства надпочечников) и фолликулярные клетки (у млекопитающих — фол-
ликулярные и сертолиевы клетки) гонад, т. е. яичников и семенников.
Все эти тканевые элементы, согласно большей части данных, разви-
ваются из эмбрионального целомического эпителия, хотя, в отдельных слу-
чаях, имеются указания на участие в их развитии, по крайней мере у не-
которых форм, эпителия канальцев вольфова тела. Вероятно, эти разно-
гласия зависят от топографической близости этих зачатков и неодинаковой
ясности в разных случаях эмбриологических картин. Предположение Фи-
шеля (1930), что фолликулярные и сертолиевы клетки млекопитающих раз-
виваются не из целомического эпителия, а из подстилающей его мезен-
химы, стоит особняком и не разделяется другими исследователями. Фак-
тический материал Фишеля и даже приводимые им иллюстрации, мне ка-
жется, отнюдь не убеждают в правильности предложенной им трактовки.
Целомическая выстилка или покровные клетки серозных оболочек
млекопитающих, открытая и обозначенная как эпителий Генле (1841),
получила затем обозначение «эндотелий» (Гис, 1865) или «мезотелий» (Май-
нот, 1901). На большей части своего протяжения она состоит из одного
слоя плоских клеток, а в отдельных местах из приближающихся по форме
к кубическим. Мезотелиальный покров яичника (т. наз. «зачатковый» или,
правильнее, покровный эпителий яичника) также представлен почти изо-
призматическими элементами. На поверхности сердца и легких высота
клеток мезотелиального покрова изменяется в зависимости от степени
растяжения органа. При обычных условиях клетки большей частью бывают
тесно соединены друг с другом краями. Описывавшиеся Колосовым (1902)
межклеточные мостики являются, повидимому, артефактом, но могут воз-
никать при экспериментальных условиях (Хлопан, 1937). Отмечавшиеся
в некоторых случаях на свободной поверхности клеток мезотелии млеко-
питающих, сходные со щеточной каемкой образования (Максимов, 1925.
1927), представляют собой, повидимому, выпадающие при фиксации бел-
ковые свертки (Хлопан, 1937).
Целомическая выстилка брюшной полости у представителей отдельных
классов позвоночных обнаруживает уже давно известные морфологические
различия, которые в пристеночной брюшине в последнее время подверга-
лись специальному сравнительному изучению моей сотрудницей Насоно-
вой (1938, 1940 и неопубликованные данные). У миноги пристеночная цело-
мическая выстилка брюшной полости у обоих полов представлена, почти
на всем своем протяжении, однослойным изопризматическим мерцатель-
Экспериментальный метод в гистологии
199
ным эпителием. У амфибий имеется резко выраженный половой диморфизм.
У самок в составе целомической выстилки есть элементы двух сортов: мер-
цательные и лишенные ресничек. В пристеночной брюшине и те и другие
имеют уплощенную форму, причем несколько более толстые мерцательные
клетки располагаются (при наблюдении с поверхности) группами значи-
тельных размеров, образующими островки или тяжи между элементами,
лишенными ресничек. У самцов мезотелий представлен лишь плоскими,
в пристеночной брюшине, немерцательными клетками. Согласно старым,
очень тщательным исследованиям Никольского (1880) и у самцов удается
обнаружить очень немногочисленные мерцательные клетки. У рептилий
и птиц мезотелий лишен ресничек и на большей части своего протяжения
состоит из плоских клеток. В отдельных участках клетки выше и по форме
приближаются к кубическим, а в виде исключения, например, у некоторых
рептилий, даже становятся высокопризматическими.
Почечные канальцы состоят из одного слоя клеток, имеющих в разных
местах неодинаковую высоту и обнаруживающих у разных позвоночных
в отдельных участках большое сходство цитологических особенностей.
Имеющиеся у низших позвоночных мерцательные отделы канальцев у выс-
ших отсутствуют. Стенка бауменовой капсулы отличается очень сильно
t уплощенными клетками. Недавно даже высказывалось предположение,
что висцеральный листок ее эпителия у млекопитающих совершенно отсут-
ствует и замещен отдельными отростчатыми клетками типа гистиоцитов,
которые образуют на поверхности эндотелия капилляров клубочка не
сплошной слой (Меллендорф, 1927),. По новейшим данным Клара (1936),
эти гистиоцитоподобные клетки («эпициты») представляют собой изменен-
ный, утративший непрерывность эпителий бауменовой капсулы, а не эле-
менты соединительной ткани. Следует подчеркнуть, что состав висцераль-
ного листка бауменовой капсулы из клеток, рыхло связанных друг с дру-
гом отростками, не доказан достаточно убедительно, так как очень тонкие,
тесно сомкнутые друг с другом части сильно уплощенных клеток могут
легко ускользнуть от наблюдения. Это тем более вероятно, что сам Клара,
упомянутая работа которого выполнена очень тщательно, подчеркивает,
что протоплазма эндотелия сосудистого клубочка настолько тонка, что
на большей части своего протяжения не различима и' обнаруживается в пре-
паратах только в утолщенной околоядерной области. Это, однако, не за-
ставляет сомневаться в наличии непрерывной эндотелиальной выстилки
капилляров. В функциональном отношении эпителий выделительного аппа-
рата имеет, как известно, не только экскреторное, но и осморегуляторное
значение.
Эпителий производных целомических мюллеровых каналов (в число
которых не включается многослойная выстилка влагалища млекопитаю-
щих) характеризуется своей однослойностыо и состоит из клеток, имеющих
в разных отделах неодинаковую высоту. У представителей различных
классов позвоночных высота эпителия и его цитологические особенности,
в соответствии с неодинаковым функциональным значением, обнаруживают
большое разнообразие. Вообще говоря, в составе мюллеровского эпителия
200
Глава III
встречаются железистые и мерцательные клетки, причем секреторные
участки могут отличаться высокой и очень разнообразной специализацией,
которая еще очень нуждается в специальных исследованиях сравнитель-
ного характера. Во многих случаях железистые элементы имеют слизи-
стый или слизеподобный характер. У млекопитающих в яйцеводах опи-
сывались в составе эпителия, правда в небольшом количестве, короткие
базальные клетки {Шаффер, 1908). Однако вопрос о двурядности этого
эпителия настоятельно нуждается в пересмотре, так как по своим гистоло-
гическим свойствам выстилка женских половых путей принципиально
отличается от описанных выше многорядных покровов эпидермального
типа. Обнаруженные Галустяном (1935) и некоторыми другими авторами
в составе эпителиальной выстилки яйцеводов своеобразные, расположен-
ные базально мерцательные клетки с ресничками, обращенными внутрь
объемистой вакуоли, отнюдь не равноценны базальным клеткам много-
рядного эпителия эпидермального типа, имеющим камбиальное значение.
По некоторым данным «мерцательная вакуоля» этих клеток имеет сооб-
щение с просветом яйцевода посредством тончайшего капиллярного хода.
Описывавшиеся у селяхий многослойные участки в половых путях самок,
повидимому, не гомологичны типичному мюллеровскому эпителию дру-
гих позвоночных, имеют другой способ развития и, в соответствии с этим,
вероятно, не должны относиться к целонефродермальному тканевому типу.
Замечательной особенностью эпителия женских половых путей млеко-
питающих являются его изменения в течение полового цикла, зависящие
от гормональных влияний и вызываемые также экспериментально. Эти из-
менения изучены еще далеко недостаточно детально в гистологическом отно-
шении. Новый фактический материал моей сотрудницы Жемковой (1941 и
неопубликованные данные) наглядно демонстрирует всю глубину пере-
строек, которые свойственны некоторым участкам этого эпителия при физио-
логических условиях в составе целого организма. Ее исследования ка-
саются пока изменения участков выстилки матки кролика, находящихся
между плацентарными вздутиями и не вступающих в связь с зародышевыми
оболочками. В плацентарных участках картины еще значительно сложнее.
Жемкова детально изучила день за днем изменения в эпителии этих участ-
ков матки, начиная с первого дня беременности и кончая десятым днем
постпуэрперального периода, когда восстанавливаются обычные для не-
беременной матки отношения. По ее данным, эпителий небеременной матки
повсеместно однослойный и однорядный. Его элементы в состоянии физио-
логического покоя имеют невысокую призматическую форму и лишены
ресничек. В течение беременности происходит утолщение всей стенки матки,
в частности, слизистой оболочки, сопровождающееся усложнением ее
рельефа. Пять невысоких первоначально продольных складок ее и слабо
выраженные крипты очень сильно разрастаются и увеличиваются в раз-
мерах. Складки слизистой оболочки приобретают сложно разветвленную
(на поперечных через матку разрезах) древовидную форму. Описываемые
ниже изменения в эпителии начинаются на поверхности складок и посте-
пенно распространяются на крипты, спускаясь вглубь от их устья, но
Экспериментальный метод в гистологии
201
никогда не достигая слепых концов. В области последних эпителий все
время сохраняет свой малодифференцированный камбиальный характер.
В первые дни беременности в эпителии повсеместно наблюдаются много-
численные митозы. Клетки его становятся крупнее и выше. В его составе
дифференцируются мерцательные и содержащие секреторные включения
железистые элементы. Затем волна митотической активности спадает; не-
многочисленные митотические фигуры обнаруживаются лишь в глубине
крипт. Период усиленного размножения клеток эпителия сменяется их
усиленным ростом и амитотическим делением ядер. Количество много-
ядерных гигантских клеток нарастает. Высота клеток или равна или не-
значительно превышает их плоскостные размеры, а многочисленные ядра
располагаются кучкой, занимая почти все клеточное тело. Секреторные
явления и мерцательные реснички постепенно исчезают. Затем клетки
чрезвычайно сильно вытягиваются в длину и приобретают во второй поло-
вине беременности, длящейся у кролика один месяц, высокую столбчатую
форму. Относительное количество цитоплазмы несколько увеличивается,
а ядра приобретают расположение цепочкой, направленной вертикально,
вдоль длинного размера клетки. На скошенных разрезах такой высокий
столбчатый эпителий может симулировать многослойную структуру. Это
зависит оттого, что полигональные в поперечном разрезе клетки, местами
отделенные друг от друга узкими щелевидными промежутками, с замет-
ными в них цитоплазматическими перемычками, кажутся расположенными
в несколько слоев. К концу беременности часть многоядерных клеток утра-
чивает связь с соединительнотканной подстилкой и, изменяя форму, пре-
вращается в неправильные дегенеративные плазмодиальные массы, нагро-
мождающиеся друг на друга и постепенно слущивающиеся в просвет. Ядра
гигантских клеток при этом сморщиваются и утрачивают видимую струк-
туру. В результате усиленной дегенерации и слущивания эпителиальная
выстилка матки после родов резко истончается до одного слоя уплощен-
ных в большей или меньшей степени многоядерных элементов, на смену
которым из глубины крипт начинают выдвигаться новые эпителиальные
клетки. В глубине крипт в это время резко усиливается митотическая
активность. В первые дни постпуэрперального периода молодой эпителий,
продвигающийся на поверхность (вероятно, под влиянием еще циркули-
рующих в организме в большом количестве гормонов), претерпевает сход-
ные с описанными выше, хотя и менее резко выраженные, изменения и
быстро слущивается. Замещающие его новые потоки эпителия, попол-
няющегося митотической активностью камбиальных элементов в глубине
крипт, постепенно приобретают к седьмому-десятому дню внешний вид
и свойства выстилки небеременной матки. Складки слизистой оболочки
и крипты возвращаются к состоянию физиологического покоя.
Эти гистологические процессы, впервые изученные достаточно детально
на примере межплацентарных участков беременной матки кролика, пока-
зывают, какую высокую специализацию приобретает мюллеровский эпи-
телий женских половых путей. Его реактивность в отношении гормональ-
ных влияний представляет собой, повидимому, одну из новейших, приобре-
202
Глава III
тенных в эволюционном процессе, биологических особенностей и подчер-
кивает развитие в составе организма новых интегрирующих факторов, уста-
новление новых коррелятивных связей. Из описанного выше ясно, что весь
цикл изученных гистологических изменений в морфологическом отношении
отличается глубоким своеобразием, а наблюдавшиеся структуры не имеют
по существу ничего общего с многослойными и многорядными эпителиаль-
ными пластами кожного типа. Весьма вероятно, что часть описывавшихся
при патологических и экспериментальных условиях в женских половых
путях структур имеет сходную с описанными выше природу, а не предста-
вляет собой примеров появления тем или иным, путем гетеротопных участ-
ков многослойного эпителия, в качестве которых они трактовались на
основе слишком поверхностного микроскопического изучения. С этой точки
зрения, данные Жемковой безусловно заслуживают очень большого вни-
мания. Задачей дальнейших работ являются углубление и расширение
исследований в этом направлении, имеющем, в равной степени и теорети-
ческий и практический интерес.
За отсутствием экспериментально-гистологических данных, посвя-
щенных производным мюллеровских каналов низших позвоночных,
пока преждевременно давать их более углубленный сравнительный
обзор.
Целомическая выстилка беспозвоночных отличается, как это уже упо-
миналось выше, большим разнообразием. Ее элементы, в отдельных слу-
чаях, очень сильно отличаются в отношении общей формы и высоты, при-
сутствия или отсутствия мерцательного аппарата и ряда других цитоло-
гических и гистологических особенностей. С ней бывают теснейшим обра-
д|ом связаны генетически и функционально выделительные органы типа
целомодуктов или целомические части нефромиксий, а также гомологич-
ные им отделы половых путей, где они имеются. Функциональное значение
целома беспозвоночных также очень разносторонне и, в отдельных случаях,
весьма неодинаково. Как правило, он бывает тесно связан с выделитель-
ным и половым аппаратами, играет вместе с тем. роль тургорного механи-
ческого аппарата организма, приобретающего (например у роющих
форм, как сипункулиды, кишечнодышащие) очень большое функциональное
значение. У ряда форм целом образует более или менее сложную систему
каналов и несет функцию газообмена и сокообращения, субституируя
кровеносную систему, лишенную у беспозвоночных, как правило, эндо-
телиальной выстилки. Такая целомическая кровеносная система дости-
гает высокого уровня развития (например у части пиявок, у сипункулид
и иглокожих). Целомическая система сокообращения характеризуется
у всех беспозвоночных, у которых она имеется, непрерывной эпителиальной
или, если угодно, эндотелиальной выстилкой.
Вторичная редукция целома приводит у членистоногих к образованию
обширной системы щелей и лакун псевдоцеля, лишенных целомической
выстилки, по которым циркулирует гемолимфа. Маленький участок целома
у части ракообразных сохраняется в составе выделительного аппарата,
гомологичного целомодуктам. В других случаях, например у моллюсков,
Экспериментальный метод в гистологии 203
от целома остается лишь перикардиальный синус, включенный в общую,
повидимому, «мезенхимную систему сокообращения.
Ввиду того, что приводимый ниже экспериментальный гистологический
материал относится почти исключительно к позвоночным, не будем оста-
навливаться подробнее на весьма интересных гистологических особенно-
стях целомической выстилки беспозвоночных и на эволюционных изме-
нениях ее свойств, которые вкратце упоминались в одной из предыдущих
глав.
Стенка выделительных органов беспозвоночных может в гистологиче-
ской! отношении очень резко отличаться от мочевых канальцев позвоноч-
ных, обнаруживая в то же самое время подчас удивительное сходство в отно-
шении специальных цитоплазматических структур. Достаточно упомя-
нуть про такие специальные структуры, как палочковидная исчерченность
и сходные со щеточной каемкой образования. Существенные гистологиче-
ские различия касаются в первую очередь количества клеток, образующих
стенку канальцев, которая у позвоночных и некоторых беспозвоночных
(например в зеленой железе десятиногих раков) многоклеточна, но во
многих случаях имеет очень малоклеточную структуру и (в поперечном
сечении) часто бывает образована одной единственной, свернутой в трубку,
клеткой, имеющей как бы внутриклеточный центральный просвет. Одним
из направлений эволюции гистологической структуры канальцев выдели-
тельных органов является, повидимому, увеличение числа составляющих
их клеток и приобретение стенкой канальца эпителиального, в обычном
смысле этого слова, строения. Клетки, образующие трубчатые сегменты
выделительных органов, лишь с большой натяжкой могут быть обозначены,
как эпителий, подобно элементам эктодермальных нефридиев и многих
малоклеточных кожных желез беспозвоночных.
Весьма своеобразными и у всех целомических животных генетически
близко родственными целомической мезодерме элементами являются упо-
минавшиеся выше вспомогательные элементы гонад или фолликулярные,
в широком смысле этого слова, клетки (у половозрелых млекопитающих,
собственно, фолликулярные и сертолиевы клетки). Эти клетки трофиче-
ского характера обеспечивают обмен веществ развивающихся гоноцитов
и, повидимому, создают для них особую внутреннюю среду, отличающуюся
от общей внутренней среды организма, а вместе с тем характеризуются
в известной степени барьерной или пограничной функцией, свойственной
т. наз. эпителиальным тканям. Вместе с тем они являются одной из много-
численных тканей внутренней среды организма, не граничащих непосред-
ственно с какой-либо пеизофизиологической средой.
В отношении морфологических особенностей они у разных животных
и у разных полов одного и того же вида животных отличаются большим
разнообразием. Сертолиевым клеткам у млекопитающих многие припи-
сывают синцитиальную взаимосвязь, хотя отдельные ядерноплазматические
территории т. наз. сертолиева синцития имеют отчетливый клеточный
характер. При исчезновении из состава стенки семенных канальцев поло-
вых элементов (например после облучения рентгеновыми лучами) они
204 Глава 111
приобретают вид типичного однослойного призматического эпителия. При
обычных условиях их форма и взаимоотношения прослеживаются с тру-
дом, так как образованная ими протоплазматическая строма стенки семен-
ного канальца густо заполнена геноцидами на разных стадиях развития.
У неполовозрелых животных будущие семенные канальцы представлены
сплошными тяжами относительно мелких фолликулярных и более крупных
половых клеток, образовавшихся как разрастания целомической выстилки
мочеполовых складок. Сходный в основном общий характер имеют яйце-
носные тяжи или мешки развивающегося яичника, первая (а у человека,
повидимому, частично вторая) волна разрастаний которых из целомиче-
ского эпителия дает начало рудиментарным образованиям мякотного веще-
ства, а третья превращается затем в примордиальные фолликулы. Фолли-
кулярные клетки создают первоначально очень тонкий эндотелиоподобный
покров на поверхности ооцита, затем приобретают в растущем фолликуле
кубическую форму и, энергично пролиферируя, превращаются в толстый
клеточный массив, в котором создаются полости, наполненные жидкостью.
В конце концов накапливающаяся в фолликуле жидкость превращает
фолликулярные клетки в многослойную выстилку граафова пузырька с рас-
положенным в ее утолщенно.м участке ооцитом. По своему внешнему виду
эту многослойную выстилку обозначают как многослойный эпителий.
Как уже отмечалось выше, многослойный фолликулярный эпителий ни
по способу образования, ни по деталям структуры, ни по биологическому
значению не имеет ничего общего даже с малодифференцированными много-
слойными эпителиальными пластами кожного типа, а представляет собой
образование, обнаруживающее весьма поверхностное конвергентное сход-
ство с ними. Этот, вообще говоря, достаточно хорошо известный факт пока-
зывает лишний раз, что не всякое эпителиоподобное многослойное распо-
ложение клеток характерно для эпителиев кожного типа и что каждая
многослойная структура нуждается в тщательном гистологическом ана-
лизе, чтобы по совокупности своих свойств быть отнесенной в ту или иную,
не формальную, а оправданную всей совокупностью своих биологических
особенностей рубрику. К сожалению, эти, достаточно элементарные, рас-
суждения далеко не всегда учитываются при трактовке патологически
измененных тканевых структур, что приводит лишний раз к ошибочному
описанию явлений метаплазии там, где их на самом деле нет.
После овуляции из фолликулярных клеток образуются прорастающие
соединительной тканью и сосудами комплексы лутеиновых клеток желтого
тела, которые представляют собой конечный этап развития этих целоми-
ческих производных.
Последними, относимыми к эпителиальным тканям, производными цело-
мической выстилки позвоночных является, наконец, паренхима интер-
ренальных органов, то образующих самостоятельные анатомические еди-
ницы (костистые рыбы), то срастающихся с нейрогенными адренальными
зачатками в анатомически единые органы, надпочечники. В составе послед-
них интерренальная и адренальная части или интимно прорастают друг
друга клеточными тяжами (например у амфибий) или образуют хорошо
Экспериментальный метод в гистологии
205
отграниченные друг от друга корковое и мякотное вещество органов (млеко-
питающие). Интерренальная паренхима, прорастая сосудами, обнаружи-
вает конвергентную структуру внутренней среды организма с эндокрино-
выми органами иного происхождения и функционального значения.
В какой степени сравнимы половые пути низших первичноротых с цело-
мическими производными выше организованных форм, сказать в настоящее
время трудно. Не исключена возможность, что здесь имела место гистоло-
гическая субституция одной ткани другой. Вместе с тем очень вероятно,
что мезодермальные половые пути турбеллярий представляют собой обо-
собившиеся в первую очередь половые отделы гастроваскулярной системы
предков, из которых развилась или к которым, затем присоединилась цело-
мическая часть .мезодермы. Входить в обсуждение этого вопроса пока прежде-
временно.
Кроме всех упомянутых выше производных, которые с большими или
меньшими оговорками могут быть включены в группу эпителиальных
тканей, стенка целома является несомненно источником развития (как
в филогенезе, так и в онтогенезе) сердечной мускулатуры, которая по своей
морфо-физиологической характеристике никак не может быть включена
в группу эпителиев, а представляет собой своеобразную разновидность
мышечных тканей. В составе их она и будет рассмотрена ниже.
б) Целомическая выстилка
' Покровным клеткам серозных оболочек млекопитающих посвящено
большое количество экспериментальных работ. Это является вполне по-
нятным, так как воспалительные процессы в серозных оболочках имеют
огромное значение в человеческой патологии, а их правильное понимание
зависит в первую очередь от изучения специфических особенностей мезо-
телиального покрова. Относительно целомической выстилки представи-
телей других классов позвоночных, за исключением последних работ нашей
лаборатории, экспериментальных данных почти нет. Первое, по времени,
экспериментальное исследование мезотелия или эпителия (как его тогда
обычно называли) серозных оболочек принадлежит Риндфлейшу (1862),
который вызывал слабый ограниченный перитонит у кроликов и морских
свинок и изучал с помощью весьма несовершенной методики того времени
(без окраски в растворе сахара или после обработки уксусной кислотой)
пленки с поверхности брюшины. Риндфлейш пришел к выводу, что при
перитоните правильная эпителиальная мозаика покровных клеток исче-
зает, вследствие их округления и превращения в элементы, сходные с лим-
фоцитами, являющимися типичными соединительнотканными образова-
ниями, и что, на этом основании, выстилка серозных оболочек предста-
вляет собой, по сути дела, уплощенные соединительнотканные клетки, а не
эпителий, сравнимый с эпителием кожи или слизистых оболочек. Сходная
точка зрения была несколько позже высказана Гисом (1865), на основе
анатомо-физиологических и эмбриологических данных. Подробнее взгляды
Гиса были изложены в исторической вводной части этой книги. Цёлликер,
первоначально принявший эту точку зрения, позже (в 5-м издании своего
206
Глава III
руководства), исходя из более новых эмбриологических и сравнительно-
анатомических представлений, склонился к взгляду, что выстилка сероз-
ных оболочек по своему происхождению должна рассматриваться как эпи-
телий, в противоположность эндотелию сосудистой системы. Точку зре-
ния Кёлликера в дальнейшем приняли Колосов (1893), Вальдейер (1901),
Брунн (1902), а в позднейшее время Менкеберг (1924) и некоторые другие
авторы. Очень большое число исследователей, наоборот, в соответствии со
взглядами Риндфлейша и Гиса рассматривают мезотелий как разновид-
ность соединительной ткани и считают, что его элементы при эксперимен-
тальных и патологических условиях превращаются в настоящие фибро-
бласты и макрофаги (Доминичи, 1902, 1920/21; Шотт, 1909; Вейденрейх,
1911; Кийоно, 1914; Герцог, 1915; Мартане), 1921; Меллендорфы В. и М.,
1926, и др.). Вместе с тем следует подчеркнуть, что некоторые наблюдения
не вяжутся с такой трактовкой. Так, например, Цивидис (1913) при пери-
кардите обнаружил образование мезотелием перикарда трубчатых ходов
в подстилающей соединительной ткани, выстланных кубическими клет-
ками, а Цунода (1909) удавалось вызывать их появление экспериментально
путем введения под грудину инородного тела, которое давлением механи-
чески раздражало перикард. Наконец, Максимов (1925, 1927) и Шоппер
(1932) (главным образом, на основе опытов с эксплантацией) представляют
себе мезотелий млекопитающих, как особую ткань, сохранившую во мно-
гих отношениях свойства эмбриональных спланхнотомов, способную к обра-
зованию эпителиоподобных структур, но при экспериментальных и пато-
логических условиях в конечном итоге превращающуюся в соединительно-
тканные фибробласты. Его способность к превращению в макрофаги послед-
ние авторы отрицают. Вместе с тем Максимов строго отличает мезотелий
серозных оболочек от эндотелия сосудистой системы. Так или иначе, точка
зрения Максимова и Шоппера принципиально мало отличается от взгля-
дов предыдущих авторов. Эти представления, казавшиеся эксперимен-
тально хорошо обоснованными, пользовались до последнего времени наи-
большей популярностью. Предпринятые мной параллельные исследования
на кроличьем материале мезотелия методами эксплантации и регенерации
привели меня к совершенно иной и, по существу, новой трактовке этой
ткани (Хлопин, 1934, 1935, 1936, 1937). Полученный при изучении мезо-
телия фактический материал сыграл очень большую роль в развитии тех
представлений, которые лежат в основе этой книги, так как именно на его
примере с особенной отчетливостью выяснилась невозможность решения
ряда основных проблем гистологии, исходя из общепринятых гистологи-
ческих установок.
Мезотелий разных участков серозных оболочек (брюшной стенки, же-
лудка, сальника, брыжжейки, перикарда и диафрагмы) ведет себя в усло-
виях тканевых культур принципиально сходным, образом. Разрастаясь
и делясь митотически, он образует характерную плоскостную зону экстен-
сивного роста. Мезотелиальные мембраны часто распространяются в двух,
расположенных друг над другом, плоскостях — по свободной поверхности
свертка фибрина и в глубине его у поверхности слюды или покровного
Экспериментальный метод в гистологии
207
стекла. На ранних стадиях культивирования мезотелий может, в некоторых
случаях, первоначально прорастать клеточными тяжами, которые затем,
как правило, превращаются в широкие и плоские мембраны. Разжижение
свертка фибрина с образованием полостей бывает выражено в слабой сте-
пени и часто отсутствует совсем. Лишь в некоторых случаях около централь-
ного кусочка на более ранних стадиях культивирования появляются не-
сколько небольших или одна-две более крупных полости разжижения,
которые могут в большей или меньшей степени деформировать зову роста,
образующую аркады довольно характерного вида. Мезотелиальные мем-
браны могут расти некоторое время по обнаженной соединительной ткани
Рис. 19. Мезотелиальная мембрана в 5-дневной культуре из перикарда кролика
с разрыхленными клеточными соединениями и изолированными клетками на пе-
риферии. Слабое увеличение. (По Хлопину, 1934.)
посеянного кусочка, однако покрытия мезотелием всей наружной поверх-
ности эксплантата и ее отграничения от окружающей среды не наблюдается.
Тонкий мезотелиальный покров исходного материала очень чувствителен
к разного рода внешним воздействиям. Поэтому для получения хороших
культур нужно, по возможности, избегать механического или иного его
повреждения. Наравне с мезотелием с различной интенсивностью может
расти соединительная ткань. Ее рост бывает выражен особенно сильно
в эксплантатах из сальника и брыжжейки, где он может в некоторых слу-
чаях преобладать и даже заглушать рост покровных клеток. В других
случаях (особенно часто в эксплантатах из перикарда, брюшной стенки,
а также сухожильного центра диафрагмы) растущий мезотелий почти или
даже совершенно заглушает соединительную ткань и может быть, особенно
после пересевов, получен в чистых культурах. Мезотелиальные клетки
208
Глава III
энергично делятся митотическим путем. В зависимости от условий суще-
ствования, их форма и взаимное расположение в плоскостных разраста-
ниях бывают неодинаковы. Характерной особенностью мезотелиальных
плоскостных мембран является резко выраженная способность к разрых-
лению и к образованию сетевидных структур и изолированных клеток раз-
ной формы (рис. 19, 20). Мезотелиальные клетки, находящиеся на поверх-
ности центрального кусочка или растущие экстенсивно, могут не только
утрачивать взаимную связь, но и связь со своим, плотным, субстратом.
Округляясь и осыпаясь, они могут свободно плавать в жидкой части пита-
тельной среды. Изолированные и округлившиеся элементы мезотелия при-
обретают макрофагообразный или сходный с лимфоцитами вид. Такое
« я
Рис. 20, а и б. Экстенсивный плоскостной рост мезотелия сероз-
ных оболочек с характерным разрыхлением, выраженным в раз-
личной степени, в 9-дневной культуре из перикарда 28-дневного
кролика (20а) и в 12-дневной культуре из перикарда 6-дневного
кролика (206). Слабое увеличение. (По Хлопину, 1936.)
осыпание имеет нередко массовый характер, в других случаях оно может
быть выражено в слабой степени. Совершенно изолированные или лишь
рыхло соединенные друг с другом отростками мезотелиальные клетки,
хотя в них могут встречаться даже митозы, отличаются в таком состоянии
ограниченной жизнеспособностью. Разрыхление мезотелиальных мем.бран
может быть вызвано различными внешними воздействиями, например трав-
мой при очередном пересеве и охлаждением. В этих случаях удается наблю-
дать, как первоначально сомкнутые своими краями распластанные клетки
мембран сокращаются и отходят друг от друга. Их протоплазма собирается
в околоядерной области, которая становится толще. Такие сократившиеся
клетки могут оставаться во взаимной связи с помощью узких протоплазма-
тических отростков и образовать плоскостную сеть или совершенно изо-
Экспериментальный метод в гистологии
20?
или даже частично распавшиеся на отдельные клетки мембраны, по исте-
чении некоторого промежутка времени, благодаря вновь наступаюше*'
распластыванию клеток приобретают свой первоначальный сомкнутый вид.
В других случаях разрыхление бывает связано с наступлением дегенера-
тивных изменений и приводит к гибели всей культуры или ее части. Эта
нестойкость сомкнутых мезотелиальных клеточных соединений и детально
Изученное лишь в последнее время явление их разрыхления, повидимому,
давали повод считать, что мезотелий в условиях тканевых культур превра-
щается в конечном итоге в соединительную ткань (Максимов, Шоппер),
чего на самом деле не происходит. Как уже отмечалось неоднократно, раз-
рыхление комплексных клеточных соединений может наблюдаться в слу-
чае различных тканей. У мезотелия, которому, вообще говоря, свойственны
более или менее комплексные разрастания, это бросается в глаза в особенно
сильной степени. Если округлившиеся клетки мезотелия имеют макрофаго-
образный или лимфоцитоподобный вид, то изолированные или сетевидно
связанные отростками элементы, как это бывает и в других случаях, могут
приобретать облик, практически не отличимый от соединительнотканных
фибробластов, в то же время отличаясь от них своей тканевой природой,
т. е. по происхождению и дальнейшей судьбе. Более комплексные варианты
плоскостных мезотелиальных структур зоны роста также отличаются зна-
штельным многообразием и, несмотря на свой малодифференцированный
характер, по своей совокупности, ясно отличаются от комплексных струк-
тур, образуемых эпителиальными тканями, часть которых была уже разо-
брана выше. Единственным известным исключением в этом отношении,
повидимому, является выстилка дышащих частей легкого. Характерной
особенностью мезотелиальных мембран, стелющихся по свободной поверх-
ности свертка фибрина, помимо разрыхления периферические участков,
является образование их элементами тончайших краевых эктоплазматиче-
ских каемок, отличающихся от утолщенной околоядерной эндоплазмати-
ческой части клеток. Благодаря этому в мезотелиальных мембранах часто
бросается в глаза относительно редкое расположение ядер. Края соседних
клеток, всегда имеющих однослойное расположение, или бывают тесно
сомкнуты друг с другом или между ними имеются промежутки разной
ширины, в которых можно бывает заметить отдельные межклеточные про-
топлазматические связи. Клетки могут быть или многоугольной или вы-
тянутой в длину формы. Как те, так и другие нередко образуют в составе
мембраны группы или гнезда (рис. 19). При росте мезотелия в толще свертка
мембраны нередко имеют более компактное строение и образуют по своему
периферическому краю выросты, состоящие из сомкнутых клеток. В таких
случаях мезотелиальные элементы нередко приобретают неправильную •
форму и, накладываясь друг на друга отростками, дают картины более
или менее густого переплета. На поверхности центрального кусочка или
по краям полостей разжижения, если они образовались, в местах, где
активного роста в данный момент не происходит, мезотелиальные клетки
могут, тесно смыкаясь своими краями, временно приобретать довольно
высокую призматическую форму, образуя структуру т. наз. куби-
14 X л опин—349
210
Глава III
ческого эпителия. Разрастаясь более или менее комплексными плоскост-
ными мембранами и образуя вместе с тем разрыхленные сетевидные струк-
туры и изолированные клетки разной формы, мезотелий сохраняет во всех
своих превращениях свои качественные особенности и не превращается
в другие ткани, в частности — в соединительную, как это предполагали
Максимов и Шоппер.
Совокупность гистологических структур, образуемых мезотелием в усло-
виях эксплантата, является в высшей степени характерной особенностью
этой ткани, резко отличающей ее от тканей другого происхождения. Покров-
ные клетки растут вне организма по своеобразному мезотелиальному типу,
отличающемуся от типа роста тканей, развившихся из других источников.
По еще неопубликованным данным нашей лаборатории, мезотелий не-
которых низших позвоночных (костистых рыб, амфибий) при своем росте
в культурах отличается большей комплексностью своих структур и не обра-
зует с такой частотой и легкостью рыхлых сетей и изолированных клеток.
Вместе с тем мембраны из мезотелия низших и высших позвоночных обна-
руживают значительное сходство, вследствие наличия у их элементов более
или менее резко выраженных эктоплазматических каемок, благодаря кото-
рым ядросодержащие эндоплазматические части клеток оказываются отодви-
нутыми друг от друга на относительно большое расстояние.
Для изучения превращений мезотелия в составе организма мы оста-
новились на методе нанесения ограниченных повреждений, а не на вызы-
вании разлитой воспалительной реакции в серозных оболочках, как это
делали многие авторы. При разлитых явлениях перитонита получается
очень бурная соединительнотканная реакция, в которую вовлекается
сальник, богатый малодифференцированными клетками соединительной
ткани (Максимов, 1927). Вследствие этого наблюдающиеся картины не-
избежно оказываются очень сложными, а судьбу элементов мезотелия
нельзя проследить с достаточной определенностью. Второй особенностью
нашей методики было параллельное изучение последовательных стадий
процесса на плоскостных препаратах, приготовляемых по способу кол-
лодийных пленок Кочетова (1911), и на сериях вертикально ориентирован-
ных срезов. Вполне естественно, что изменения в тонком мезотелиальном
покрове выступают особенно отчетливо при наблюдении с поверхности.
Повреждение наносилось ожогом в области передней брюшной стенки
после лапаротомии. В конце первых и в течение вторых суток после по-
вреждения, параллельно с развитием воспалительной реакции, в соеди-
нительной ткани в мезотелии начинают обнаруживаться реактивные изме-
нения. Клетки вокруг дефекта начинают набухать, особенно в своей около-
ядерной части; периферическая часть клеточного тела, наоборот, превра-
щается в тончайшую эктоплазматическую пленку. Первоначально тес-
ная взаимосвязь клеток разрыхляется. Клетки изменяют свою форму и
частью вытягиваются в направлении дефекта, между ними появляются
свободные промежутки. Несколько отступя от места повреждения обна-
руживаются митозы. На третий-чствертый день эти изменения прогрес-
сируют и распространяются на широкую зону брюшины, окружающей
Экспериментальный метод в гистологии 211
дефект. Митотическая активность становится очень высокой. Вместе с тем
благодаря амитотическим перешнуровкам ядер в мезотелии образуется
значительное количество двух-, трех- и многоядерных гигантских элемен-
тов. Бросается также в глаза появление чрезвычайно крупных одноядер-
ных клеток (рис. 21).
Картины мобилизации мезотелия приобретают очень пестрый
и разнообразный характер (рис. 21, 22). Клетки, утрачивая в области
дефекта связь друг с другом, частью приобретают веретенчатую или не-
правильную отростчатую форму, частью сильно сокращаются и совершенно
округляются (рис. 22). Такие округлившиеся клетки в значительных коли-
Рис. 21 и 22. Участки мезотелиального покрова передней брюшной стенки
кролика на 4-й день после прижигания в области края дефекта (21) с
частью распластанными, частью сильно сокращенными клетками и в области
активного движения разрыхленного мезотелия по раневой поверхности (22)
с отростчатыми и округлившимися элементами. Одноядерная гигантская
мезотелиальная клетка и заползший гистиоцит. Плоскостной препарат.
Сильное увеличение. (По Хлопину, 1936.)
чествах слущиваются в брюшную полость. Несколько дальше от раны
разрыхление и изоляция выражены слабее, а мезотелиальные клетки пред-
ставляются лишь в большей или меньшей степени набухшими и утолщен-
ными (рис. 21). Местами между набухшими клетками мезотелия появляются
межклеточные. мостики. Мобилизованный мезотелий со всех сторон на-
двигается на дефект и закрывает его. Клетки вновь расправляются и обра-
зуют непрерывный сплошной покров, который отличается от нормальных
участков выстилки присутствием многоядерных элементов, в дальнейшем
дегенерирующих и исчезающих. В зависимости от размеров дефекта пол-
ное перекрытие уже заканчивается через два-три дня или чаще растяги-
вается на более продолжительный срок, в среднем до семи-восьми дней.
Воспалительные изменения в соединительной ткани постепенно затихают,
-количество мобилизованных гистиоцитов уменьшается.
212
Глава HI
Полученные методом регенерации результаты очень хорошо согла-
суются с изложенными выше наблюдениями над ростом и превращениями
мезотелия в условиях тканевых культур. При репаративной регенерации
восстановление мезотелиального покрова происходит только за счет актив-
ного движения и пролиферации мезотелиальных же элементов, а отнюдь не
.в результате уплощения и смыкания друг с другом соединительнотканных
клеток, как это предполагали Льюис (1923), Меллендорфы (1926) и другие.
За счет мезотелия не происходит образования соединительнотканных спаек
между серозными оболочками, как это думал Верещинский (1925). Мезо-
телий не способствует, а, наоборот, закрывая соединительную ткань, пре-
пятствует образованию спаек. Мезотелий при экспериментальных и пато-
логических условиях не возникает ни из какой другой ткани и не превра-
щается в элементы других тканей, в частности—соединительной. Вместе
с тем по своим свойствам он очень резко отличается от большей части
эпителиев. Выстилка серозных оболочек очень чувствительна к различным
повреждающим агентам; ее комплексность, тесная взаимосвязь между ее
клетками легко нарушаются, что, вообще говоря, не свойственно эпите-
лиальным тканям. Все эти своеобразные свойства мезотелия объясняются,
с моей точки зрения, тем, что он представляет собой у млекопитающих не
малодифференцированную, обладающую эмбриональными свойствами (как
это предполагал Максимов), а, наоборот, высокоспециализированную ткань.
Обладая, вероятно, у предков млекопитающих типичной морфо-физиологи-
ческой характеристикой эпителиальных тканей, покровные клетки сероз-
ных оболочек, в связи со своим положением в изофизиологической внутрен-
ней среде организма, утратили многие особенности пограничных эпите-
лиев и сделались своеобразной тканью внутренней среды организма, со-
хранившей покровное значение. Ее разграничительная функция вместе с
тем должна была если не исчезнуть, то в значительной степени ослабиться.
Мезотелий, если можно так выразиться, по совокупности своих свойств,
не является ни соединительной тканью, ни эпителием в обычном смысле
этого слова, чего, с точки зрения классической гистологии, никак и нельзя
было предполагать. Такое новое эволюционное объяснение свойств мезо-
телия лучше всего согласуется с известными фактами и вместе с тем делает
совершенно понятными все отмеченные выше разногласия относительно его
тканевой природы.
Появившаяся вскоре после моего первого сообщения о результатах
опытов с репаративной регенерацией (1935) работа Щелкунова (1936),
выполненная под руководством проф. Заварзина, полностью подтверждает
и в некоторых отношениях дополняет мой фактический материал. Вместе
с тем этот автор, оставаясь целиком на старых гистологических позициях,
приходит к выводу, что мезотелий представляет собой настоящий эпите-
лий, со всеми свойствами этой ткани, что не вытекает, строго говоря, из
его собственных данных и совершенно не учитывает результатов других
экспериментальных исследований.
Для полного обоснования моей, разобранной выше, точки зрения на
мезотелий млекопитающих необходимо было провести достаточно подроб-
Экспериментальный метод в гистологии
213
ные исследования на сравнительном материале в ряду позвоночных. Это
выполнила в моей лаборатории Насонова (1937, 1940, 1942), которая, помимо
отмеченных выше наблюдений над нормальным строением целомической
выстилки, тщательно изучила ее изменения в процессе репаративной реге-
нерации у миноги, амфибий, рептилий и птиц после нанесения дефекта
сквозным ожогом через всю толщу брюшной стенки тела. Весь ход процесса
изучался на плоскостных препаратах, а также на вертикальных срезах.
Результаты опытов Насоновой заслуживают очень большого внимания, так
как они демонстрируют с большой наглядностью эволюционные изменения
гистологических свойств целомического эпителия в связи с изменением
функционального значения полости тела, которое было отмечено выше.
У всех изученных животных заживление дефекта происходит, как и
у млекопитающих, путем активного движения и митотического размно-
жения клеток мезотелия, но наблюдающиеся при этом гистологические
картины существенно отличаются друг от друга у представителей разных
классов позвоночных. Особенно интересные в этом отношении результаты
дал мезотелий миноги. Изопризматический мерцательный целомический
эпителий миноги по краям раны, мобилизуясь, постепенно уплощается
и начинает закрывать дефект сплошным клеточным пластом. Плоскостные,
а также абсолютные размеры клеток и ядер заметно, а местами очень сильно
увеличиваются. Количество появляющихся митозов очень велико. Надви-
гающийся со всех сторон клеточный пласт имеет ровный неразрыхленный
свободный край. Изоляции клеток совершенно не происходит. В разгаре
процесса, в конце первой или в течение второй недели после начала опыта
в эпителиальном пласте можно различать, при наблюдении с поверхности,
три нерезко отграниченные друг от друга концентрические зоны. Свобод-
ный нарастающий край состоит из тесно сомкнутых веретенчатых клеток
удлиненной формы, ориентированных не радиально, а концентрически.
Наиболее широкая средняя зона состоит из очень сильно распластанных и
резко увеличенных в размерах клеток с крупными ядрами и ядрышками.
Периферическая зона, переходящая постепенно в неизмененные окружающие
участки целомической выстилки, характеризуется менее уплощенными
элементами, вытянутыми в радиальном направлении, т. е. в сторону
дефекта. В этой области, как правило, наблюдаются узкие межклеточные
промежутки с заметными в них цитоплазматическими мостиками. Неко-
торое расхождение клеток, обнаруживающихся местами в участках ме-
зотелия, отмечается только на уровне их средней части. Базальные
и особенно апикальные концы остаются тесно сомкнутыми. Весь процесс
протекает значительно медленнее, чем у млекопитающих, и заканчи-
вается лишь в конце первого месяца. Появления многоядерных клеток
не отмечалось.
У самок травяной лягушки между мерцательными и немерцательными
клетками в процессе репаративной регенерации обнаруживаются неко-
торые различия. В составе «мезотелизирующего» пласта мерцательные
клетки располагаются (при рассмотрении с поверхности) ориентирован-
ными преимущественно в радиальном направлении тяжами различной
2J4
Глава III
ширины и в значительной степени сохраняют комплексное расположение,
а если и расходятся, то лишь в очень слабой степени. Вся окаймляющая
дефект зона пласта оказывается состоящей из немерцательных клеток.
Очень вероятно, что в ее образовании, помимо предсуществующих немер-
цательных клеток, принимают также участие и бывшие мерцательные эле-
менты, утратившие свои реснички. Немерцательные клетки в более уда-
ленных от места ожога участках, расположенные между группами реснич-
ных элементов, сохранившими комплексное расположение, набухая, за-
метно отходят друг от друга, оставаясь связанными цитоплазматическими
мостиками различной длины. В области надвигающегося на раневую по-
верхность края мезотелиального пласта разрыхление бывает выражено
еще сильнее. Покровные клетки с несколько набухшей околоядерной и
очень тонкой периферической частью тела принимают неправильную вере-
тенчатую или отростчатую форму, оставаясь лишь местами соединенными
своими длинными отростками и образуя рыхлые сетевидные соединения,
которые продвигаются по обнаженной соединительной ткани, обнаружи-
вающей резкие воспалительные изменения.
Резко бросается в глаза гигантизм отдельных клеточных ядер. Появ-
ляются немногочисленные дву- или трехядерные клетки. Митотическая
активность, постепенно нарастая, достигает своего максимума к концу
первой или началу второй недели. При митозах гигантских ядер количе-
ство хромозом резко увеличивается. Полное закрытие дефекта происходит
в конце третьей недели или позже. Непрерывность мезотелиального по-
крова и комплексная взаимосвязь его элементов постепенно восстанавли-
ваются.
У рептилий (ящерицы) надвигающийся со всех сторон на раневую по-
верхность мезотелий обнаруживает по краю пласта, а также местами и
в несколько удаленных от него участках, то более, то менее резко выражен-
ное разрыхление, местами полную изоляцию отдельных элементов.
Полиморфизм клеток выражен очень сильно, количество дву-, трех- и
многоядерных элементов довольно велико. Помимо разрыхления мезо-
телиального пласта и образования неправильной формы отростчатых эле-
ментов, замечаются картины изоляции клеток особого характера. Набу-
хание отдельных элементов мезотелия в области дефекта приобретает ха-
рактер своеобразного сокращения клеточного тела, приводит к полному
или почти полному округлению их большей околоядерной части, которая
иногда остается связанной с сохраняющейся эктоплазматической частью
посредством тонкой перемычки. Повидимому, часть таких округлившихся
клеток, совершенно отделяясь от соединительнотканной подстилки, слу-
щивается в брюшную полость. После полного перекрытия дефекта в мезо-
телии повсеместно восстанавливаются обычные отношения.
Реактивные изменения в мезотелиальной выстилке голубя, так же как
у млекопитающих, обнаруживают гораздо более разлитой характер и,
протекая интенсивнее, захватывают более значительную площадь, чем
у холоднокровных животных. Набухание околоядерной части клеток и,
наоборот, очень сильное истончение периферической эктоплазматической
Экспериментальный Метод в гистологии
215
части клеточного тела, выражено очень резко. Сетевидное разрыхление
первоначально сплошного покрова и полная изоляция клеток разнообраз-
ной формы прослеживаются хорошо. Даже в местах целомической вы-
стилки, сохранивших значительную степень комплексности, клетки при-
обретают очень неправильную отростчатую форму. Клеточные отростки,
достигающие большой длины, нередко перекрывают соседние элементы.
Местами, в несколько удаленных от места повреждения участках, в резуль-
тате набухания образуются местные утолщения мезотелиального покрова
в виде выступающих в брюшную полость гребешков, состоящих из ряда
веретенчатых (при рассматривании с поверхности) клеток. Часть клеток
этих выступов, сильно сокращаясь, округляется и, повидимому, слущи-
вается. Как правило, встречаются гигантские одно- и многоядерные клетки.
Заживление раны происходит быстро и редко растягивается на две недели
и более. В течение всего процесса регенерации в клетках различных живот-
ных обнаруживаются также определенные видовые цитологические осо-
бенности, на которых мы здесь не будем останавливаться.
Резюмированный в общих чертах материал, касающийся целомической
выстилки позвоночных, полностью обосновывает данную выше трактовку
мезотелия млекопитающих и заставляет считать его тканевую природу
прочно установленной.
Относительно целомической выстилки беспозвоночных имеются лишь
отрывочные данные, касающиеся немногих животных. Наибольшее коли-
чество данных относится к кольчатым червям и, в первую очередь, к оли-
гохетам, репаративные процессы у которых изучали многие авторы (Ива-
нов, 1903, 1907, 1908, 1916; Нусбаум, 1905, 1908; Цреккер, 1923; Вейцман,
1927, 1928, 1937; Жанкин, 1934, и др.). Согласно имеющемуся фактическому
материалу, регенерация целомической выстилкиновообразующихся сегмен-
тов происходит из особых крупных клеток, называемых необластами.
Они или предсуществуют, как таковые, в области диссипиментов, являясь
малодифференцированными потомками мезодермальных бластомеров, или
образуются заново из элементов продольной соматической мускулатуры,
в составе которой имеются волокна неодинакового уровня дифференци-
ровки (Вейцман, 1937). Кроме целомической выстилки, из тех же необла-
стов регенерируют выделительная система и продольная мускулатура,
т. е. весь комплекс мезодермальных производных. Мне кажется, однако,
не исключена возможность, что, помимо образования из мышечных элементов
миобластов, которое проследил в последнее время Заварзин (1938), из старой
целомической выстилки также возникают сходные с ними по внешнему виду
клетки типа необластов, из которых в дальнейшем регенерирует стенка
полости тела. Это предположение потому представляется вероятным, что
регенерирующие элементы, имеющие разное происхождение (например
клетки эпидермиса и те же необласты), обнаруживают настолько большое
морфологическое сходство, что ряд авторов их совсем не различал. В резуль-
тате этого возникли представления, что у этих животных из мезодермальных
необластов могут при регенерации возникать и типичные эктодермальные
ткани (Геммерлинг, 1924; Штольте, 1927). Позднейшие, указанные выше
216
Глава III
более точные исследования не подтвердили этих данных. Мне кажется, что
в отношении регенерации мезодермальных производных аннелид последнее
слово еще не сказано.
в) Гонады и корковое вещество надпочечника
Поразительное сходство с мезотелием серозных оболочек при культи-
вировании вне организма обнаруживают в условиях экстенсивного роста
сертолиевы или, точнее, фолликулярные клетки неполовозрелого семен-
ника кролика (Михайлов, 1937). Данные этого автора вносят ясность в ряд
спорных вопросов и уточняют противоречивые результаты, полученные
в свое время Шампи и Морита (1928), Эзаки (1928) и Вермелем (1931, 1933)
Рис. 23, Участок экстенсивно растущей разрых-
ляющейся мембраны из сертолиевых клеток с
темными эндоплазматическими и светлыми экто-
плазматическими зонами цитоплазмы. Одна клет-
ка сократилась и округляется. 16-дневная куль-
тура из семенника 10-дневного кролика. Сильное
увеличение. (По Михайлову, 1937.)]
на семенниках различных по-
звоночных. Фолликулярные
клетки семенника кролика,
отличаясь способностью к
энергичному росту и проли-
ферации, нередко заглушают
рост соединительной ткани.
Прорастая первоначально не
только мембранами, но и тя-
жами, они в дальнейшем име-
ют исключительно плоскост-
ной рост, общий характер ко-
торого уже не меняется при
повторных пересевах. Мем-
браны из фолликулярных кле-
ток обладают значительным
многообразием структур и
обнаруживают полное морфо-
логическое тождество с мезотелиальными мембранами. Они, подобно послед-
ним, могут сетевидно разрыхляться (рис. 23) и приобретать поверхностное
сходство с соединительнотканными культурами. Фолликулярные клетки,
подобно мезотелию, нередко в массовых количествах округляются, изоли-
руются и осыпаются, приобретая макрофагообразный вид. Своеобразие
свойств и вместе с тем высокую специфичность превращений фолликуляр-
ных клеток семенника не подметили другие авторы, которые или выска-
зывались в пользу возможности превращения их в соединительную ткань
(Эзаки) или, наоборот, лишь отмечали их рост эпителиальными мембранами
(Шампи и Морита, Вермелъ).
Очень интересны также изменения семенных канальцев внутри централь-
ного кусочка эксплантата, прослеженные Михайловым. Эти изменения,
в части случаев, обнаруживают сходство со структурами некоторых опухо-
лей яичка, а в других совершенно точно воспроизводят те картины, которые
свойственны семенным канальцам при атрофических процессах в целом
организме, наблюдаемых, например, в старости (Призель, 1931).
Тщательное изучение методом эксплантации элементов яичника кро-
Экспериментальный метод в гистологии 217
ликов разного возраста (Колесникова, 1940) позволило установить практи-
чески полное тождество структур, образуемых при экстенсивном росте
его фолликулярными и покровными клетками, с описанными выше карти-
нами, характерными для мезотелия и фолликулярных клеток семенника.
Вместе с тем эти данные объяснили картины, которые на том же объекте
неправильно трактовались в смысле превращения фолликулярных клеток
вне организма в соединительнотканные элементы, и уточнили данные авто-
ров (Шампи и Морита, 1928; Улезко-Строганова, 1935; Хрущев и Диоми-
дова, 1937; Жив, 1937), которым не удалось установить специфических
особенностей мезотелиального типа роста фолликулярных клеток и кото-
рые лишь отмечали их разрастание «эпителиальными» мембранами.
Здесь же будет уместно упомянуть относительно судьбы незрелых поло-
вых клеток семенника и яичника в условиях тканевых культур. Некоторые
авторы широко допускают активную эмиграцию в зону роста и даже про-
лиферацию митотическим путем сперматогоний амфибий и птиц (Вермелъ,
1931, 1933) и оогоний млекопитающих (Хрущев и Диомидова, 1937), хотя
и не прослеживают их судьбы в составе центрального кусочка. По другим
данным (Эзаки, 1928; Михайлов, 1937; Колесникова, 1940), сперматогонии
и оогонии млекопитающих не мигрируют активно в зону роста и никаких
изменений прогрессивного характера не обнаруживают.
В первые один - два дня после эксплантации в половых клетках цен-
трального кусочка наблюдается значительное количество митозов, которые
затем исчезают совершенно и больше не возобновляются. Большая часть
половых клеток дегенерирует очень рано, а остальные в небольшом коли-
честве могут переживать более или менее длительное время, не обнаруживая
никаких прогрессивных изменений и не двигаясь активно. Если единичные
половые клетки на ранних стадиях культивирования и увлекаются разра-
стающимися фолликулярными элементами пассивно в зону роста, то они
быстро погибают. Изолированные круглые клетки, описывавшиеся неко-
торыми авторами в зоне роста как половые, согласно исследованиям Михай-
лова и Колесниковой, представляют собой округлившиеся фолликулярные
элементы. Хотя данные, полученные на млекопитающих, и нельзя без-
оговорочно переносить на птиц и амфибий, следует считать предполагаемую
активную эмиграцию половых клеток у последних в лучшем случае недо-
казанной. У млекопитающих же она совершенно не имеет места.
В недавно появившейся работе Мендельсона (1937) дается совершенно
неправильная трактовка картин роста в эксплантатах из семенника. Автор
считает, что эпителиоподобные мембраны представляют собой «герминатив-
ный эпителий», а встречающиеся в их составе отдельные очень крупные
элементы—сертолиевы клетки.
,,С нашей точки зрения, совершенно неправильно само укоренившееся
в гистологии понятие «герминативного эпителия», так как, в соответствии
с теорией зародышевого пути, в составе семенных канальцев имеются двоя-
кие, совершенно различные элементы: клетки Сертоли и расположенные
: между ними половые клетки на разных стадиях развития. Насколько можно
судить по микрофотографиям, приложенным к работе, очевидно, автор
218
Глава III
имел дело с разрастаниями сертолиевых клеток в виде мезотелиальных
мембран, отдельные элементы которых нередко отличаются гигантизмом.
В гонадах зародышей и неполовозрелых млекопитающих (Мартинович,
1938), экснлантированных целиком, наблюдаются некоторые явления
дифференцировки половых клеток, причем нормальные ооциты сохраня-
лись до ста трех дней жизни вне организма.
Относительно роста коркового вещества надпочечника имеются очень
скудные данные (Сиповский, 1929; Кнелль, 1937). Первому из этих авторов
удалось наблюдать лишь рост эпителиообразных мембран без митотической
пролиферации элементов. Второму удалось получить настоящие проли-
ферирующие и растущие мембранами эксплантаты из элементов клубоч-
ковой зоны надпочечника кроликов, быков и коров. К сожалению, послед-
няя работа в смысле обработки фактического материала оставляет желать
многого и не позволяет сделать никаких выводов о гистологических особен-
ностях элементов, растущих вне организма.
Большое количество работ, произведенных различными эксперименталь-
ными методами над гонадами и, в первую очередь, над семенником (подроб-
ная литература приведена в статьях Оберндорфера (1931) и Призеля (1931),
дает очень немного в отношении гистобластических потенций фоллику-
лярных (сертолиевых) клеток. Тщательные исследования Максимова (1898)
над изменениями в семеннике млекопитающих и амфибий после поврежде-
ния не обнаружили в нем заметных регенеративных процессов. Очень много-
численные опыты с трансплантацией семенников в гистологическом отно-
шении не дали интересных результатов (Ситенко, 1939; там же см. подроб-
ную литературу по этому вопросу). В связи с исчезновением половых клеток
из семенных канальцев, наблюдающимся при самых разнообразных экспе-
риментальных воздействиях и патологических процессах, сертолиевы клетки
образуют сетевидные структуры, фагоцитируя отмирающие гоноциты,
в соответствии с тем, что Михайлов описывал в условиях тканевых культур.
После полного или почти полного исчезновения половых клеток серто-
лиевы элементы могут образовать однослойную эпителиальную выстилку
различной толщины, состоящую из призматических или уплощенных клеток.
Данные, согласно которым после удаления у мышей яичников они будто
бы создаются вновь, внушают серьезные сомнения. Здесь уместно остано-
виться в общих чертах на тератоидных разрастаниях семенников петухов,
полученных впервые Михаловским (1926, 1928) и детально изучаемых
в последнее время Фалиным (1939) и его сотрудниками. В результате введе-
ния в ткань семенника солей цинка в очень большом проценте случаев
-наблюдается развитие тератом, состоящих из всевозможных типичным
образом дифференцированных тканей и беспорядочно расположенных
частей органов: кист, выстланных кишечным или кожным эпителием,
островков хрящевой и костной тканей, соматической поперечнополосатой
Мускулатуры, нервной системы и пр. Гистогенез этих очень интересных
тератоидных образований пока еще не прослежен. Вероятнее всего пред-
положить, что в этих случаях происходит атипичный «эфебогенез» за счет
столовых клеток семенника, продукты деления которых развиваются и
Экспериментальный, метод в гистологии
219
дифференцируются при отсутствии организменной интеграции (под ее
знаком протекает нормальное эмбриональное развитие), но при частичном
наличии межтканевых корреляций. Эти опыты заслуживают очень большого
внимания с точки зрения экспериментальной эмбриологии и гистологии.
г) Почечный эпителий
Более или менее подробные данные относительно роста и превращений
.вне организма эпителия канальцев вторичной или окончательной почки
млекопитающих, после первых опытов Карреля и Берроуза (1910), имеются
в работах Шампи (1914), Рингофа (1922), Хлопина (1922), Дрю (1923),
Поликара (1925), Капеля (1929), Н о рдмана (1929), Смирновой и Уразова
(1932), Гросфельда (1932), Маркеса (1933), Эфрусси и Литвака (1934), Роби-
нова (1935) и др. Эти работы сделаны частью на материале, взятом от заро-
дышей, частью на эксплантатах от новорожденных и взрослых животных.
Наиболее полный гистологический анализ почечных эксплантатов сделан
Цымбалом (1935). Опыты с эксплантацией первичной почки амфибий произ-
водили Нишибе (1928), Дрю (1923), Хлопин (1928) и Сунцова (1930).
Различные отделы нефрона и собирательные трубки в эксплантате
способны к росту и пролиферации митотическим путем. Они растут экстен-
сивно в виде плоскостных однослойных клеточных мембран или погружен-
ных в толщу плотного субстрата сплошных, нередко ветвящихся тяжей,
могущих инфильтрировать подсаженные к почечным эксплантатам другие
тканевые частицы (Гросфелъд)-, кроме того, в некоторых случаях, как
и в случае других тканей, комплексные клеточные соединения могут под-
вергаться разрыхлению и образовать изолированные элементы разной
формы. Эти явления, однако, у элементов почечных канальцев выражены
в незначительной степени и, повидимому, имеют преимущественно деге-
неративный характер. Перерезанные при эксплантации концы почечных
канальцев могут зарастать (Чемберс, 1934; Цымбал, 1935) и, кистовидно
расширяясь, выдвигаться в зону роста в виде замкнутых пузырей, которые
превращаются при дальнейшем росте в плоскостные мембраны (Цымбал).
Участие в эпителизации и в экстенсивном росте было констатировано для
разных участков почечных канальцев, за исключением бауменовой капсулы
мальпигиевых телец. Прорастающие элементы почечных канальцев утра-
чивают свою специфическую дифференцировку и приобретают однородный
характер. У мобилизующихся элементов главных отделов можно видеть
отторжение и отмирание более или менее значительной надъядерной части
клеточного тела (Шампи, Цымбал). По наблюдениям Цымбала, экстенсивно
растущие мембраны из собирательных трубок мякотного вещества и нефро-
нов коркового имеют более или менее ровные края, но обнаруживают зна-
чительные различия в отношении формы клеток и ядер, а также взаимо-
отношения элементов. В мембранах из коркового вещества клетки очень
энергично делятся митозом, отличаются полиморфизмом, имеют, как пра-
вило, неправильную форму и подчас длинные отростки, вклинивающиеся
между соседними элементами. Их ядра овальны или снабжены вдавле-
ниямн. В мембранах из мякотного вещества почки, наоборот, преобладают
220
Глава III
правильная полигональная форма клеток и круглые ядра. Размеры тех
и других в направлении от центра к периферии увеличиваются. Митозы
встречаются относительно редко. Эпителизирующие поверхность эксплан-
тата покровные пласты могут в местах активного роста состоять из более
или менее одинаковых уплощенных клеток. По окончании процесса эпите-
лизации, часто приводящего к закрытию всей наружной поверхности
посеянного кусочка сплошным эпителиальным пластом, последний приоб-
ретает весьма характерное, неодинаковое в различных участках, строение
и обнаруживает определенные черты вторичной дифференцировки (рис. 24).
Покровные пласты из почечного эпителия млекопитающих имеют однослой-
Рис. 24. Участки однослойного покровного эпите-
лия неодинаковой высоты, образовавшегося после
полного перекрытия поверхности эксплантата эпи-
телием мочевых канальцев. 25-дневная культура
из коркового вещества почки новорожденного кро-
лика. Сильное увеличение. (По Цымбалу, 1935.)
ный и однорядный характер,
не образуют истинных много-
рядных и многослойных
структур. Лишь местами, в си-
лу сморщивания и уменьше-
ния объема эпителизирован-
ного кусочка и отторжения
его покровных элементов на
поверхности эксплантата, мо-
гут создаваться кратковре-
менные нагромождения кле-
ток, которые никогда не бы-
вают стойки и быстро распа-
даются, оставляя после себя
однослойный покров. Упомя-
нутые в работе Сунцовой мно-
гослойные участки покровно-
го эпителия в эксплантатах
из первичной почки лягушки
также несомненно представ-
ляли собой преходящие кле-
точные нагромождения, а не
истинные и более или менее
стойкие многослойные струк-
туры. Для покровного эпителия эксплантированного кусочка, отли-
чающегося хорошо выраженной полярностью, весьма характерны неоди-
наковая высота элементов и чередование участков различной протяжен-
ности, покрытых плоскими или низко-призматическими клетками с высоко-
призматическими (рис. 24), причем этот полиморфизм выражен в большей
степени у эпителия нефронов в эксплантатах из коркового вещества, чем.
у эпителия собирательных трубок в культурах из мякотного вещества.
Свободные апикальные поверхности клеток имеют, как правило, сильно
выпуклую куполообразную форму и замыкающие полоски. Между их
базальными концами, в большинстве случаев, образуются узкие щелевид-
ные промежутки, в которых заметны немногочисленные, но подчас довольно
длинные межклеточные цитоплазматические связи. Внутренние части
Экспериментальный метод в гистологии
221
нацело эпителизированных эксплантатов постепенно подвергаются деге-
нерации, которая обычно протекает не так быстро, как в случае эпители-
зации эпидермальными покровными пластами. Очевидно, почечные покров-
ные пласты не в достаточной степени несут необходимую для поддержания
жизни эпителизированных эксплантатов всасывающую функцию. Наоборот,
по данным Чемберса, относящимся к первичной почке, в просвет почечных
канальцев с совершенно замкнувшимися перерезанными концами с легкостью
проникают извне вещества, например ряд относительно безвредных красок.
Таким образом, через этот эпителий может свободно происходить передви-
жение растворенных веществ в направлении от базальной к апикальной
поверхности, обращенной при обычных условиях в просвет канальца. Сопо-
ставление последних фактов подчеркивает хорошо выраженную полярность
этой ткани.
Элементы бауменовой капсулы могут становиться в центральных кусоч-
ках выше и приобретать низко-призматическую или кубическую форму
(Хлопин, 1922; Нордман, 1929). Метанефрогенная ткань зародышевой
вторичной почки, повидимому, может, дифференцируясь в составе централь-
ного кусочка, образовать зачатки нефронов (Рингоф, Хлопин). По данным
Рингофа, из нее даже могут образоваться новые мальпигиевы тельца,
включая эндотелий их капилляров. Эти наблюдения в настоящее время
нуждаются в проверке. Предположение Хлопина (1922), что метанефроген-
ная ткань относительно старых зародышей способна к превращению в эле-
менты мезенхимы, не представляется вероятным в соответствии с настоящим
уровнем знаний и подлежит пересмотру. Метанефрогенные элементы почки
новорожденных кроликов, имеющие характер недифференцированных зачат-
ков нефронов, прорастая, ведут себя, повидимому, так же, как подверг-
шийся упрощению эпителий канальцев коркового вещества (Цымбал).
При совместной эксплантации мякотного вещества почки и лоханки
удается получить явления эпителизации частично за счет эпителия соби-
рательных трубок, частично за счет переходного эпителия лоханки, причем
почечный эпителий дает однослойные участки, а переходный — много-
слойные. Те и другие, смыкаясь друг с другом, остаются совершенно четко
отграниченными Щымбал).
В опытах с регенерацией ткани почек после нанесения дефекта многие
авторы описывали рост эпителиальных трубок и тяжей, который в соот-
ветствии с новыми данными, пожалуй, правильнее трактовать как воспали-
тельные разрастания. Эти разрастания почечного эпителия имели малодиф-
ференцированный характер и никогда не превращались в почечные канальцы
(Подвысоцкий, 1888; Мартинотти, 1892; Пейперс, 1895; Пизенти, 1899;
Любарш, 1925). Аналогичные разрастания описывались также по краям
почечных инфарктов (Торель, 1896; Любарш, 1925).
При отравлении сулемой в почечных канальцах происходят поврежде-
ние и гибель эпителия. При частичной гибели нефрона сохранившийся
эпителий, пролиферируя, может восстанавливать значительные участки
поврежденных канальцев (Гейнеке, 1909). При образовании в почке очагов
некроза наблюдается их обрастание эпителием (Пейперс, 1895).
222
Глава III
Во всех этих случаях растущие эпителиальные клетки имеют различную
форму, но всегда располагаются одним слоем.
Новейшие опыты Захарьевской (1939) с введением каменноугольного
дегтя позволили выделить несколько типов воспалительных разрастаний
почечного эпителия.
При частичном повреждении стенки канальцев может наблюдаться
т. наз. внутриканальцевый рост, т. е. регенерация поврежденного участка
in situ, приводящая во многих случаях к его полному восстановлению. При
внеканальцевых разрастаниях в очаге воспаления эпителий канальцев
создает лишь малодифференцированные трубки и тяжи, которые никогда
не превращаются в полноценные почечные канальцы. Эпителиальные
трубки и тяжи, дорастая до границы некротических участков, давали
начало пластам, которые эпителизировали их, образуя стенку кистовидных
полостей. Почечный эпителий всегда сохранял однослойное расположение,
но его клетки обнаруживали неодинаковую высоту.
Никакой дифференцировки внеканальцевых разрастаний эпителия,
с образованием структур, типичных для нормальных почечных канальцев,
не наблюдалось. Ничего похожего на превращение почечного эпителия
в соединительнотканные элементы никогда не происходит.
Согласно кратким данным Лазаренко (1934), при имплантации частиц поч-
ки с кусочками целлоидина в подкожную клетчатку, эпителий почечных ка-
нальцев, делясь митотически, разрастается в очаге воспаления малодиффе-
ренцированными трубками с однослойной стенкой. Высота клеток в разных
участках эпителиальных трубок бывает очень неодинакова. Конечная судьба
таких разрастаний — дегенерация и замещение рубцом. Основной особен-
ностью пролиферативных процессов в почке является то, что регенерация с
новообразованием в конечном итоге дифференцированного почечного эпите-
лия бывает возможна только при частичных повреждениях нефронов и при
внутриканальцевом росте эпителия при обязательном сохранении в целости
мальпигиевых телец, которые не регенерируют сами и даже, согласно боль-
шей части данных, не обнаруживают митотического деления клеток бауме-
новой капсулы. В противном случае получаются лишь малодифференциро-
ванные и неполноценные разрастания. Каждый нефрон реагирует, как це-
лая система, которая или восстанавливается, как таковая, или дает лишь
неполноценные образования, которые в конечном итоге обречены на гибель.
При всех пролиферативных процессах в почечном эпителии никогда
не обнаруживаются образования многослойных структур кожного типа.
В этом отношении он резко отличается от эпителия лоханки (Цымбал, 1935;
Захарьевская, 1939). При самых разнообразных условиях почечный эпите-
лий дает образования (малодифференцированные трубки, покровный эпи-
телий в культурах и в составе организма, нормальные нефроны), которые
отличаются неодинаковой высотой клеток, имеющих часто куполовидно
выпуклую апикальную поверхность. В этом нельзя не видеть одной из
характерных гистоморфных особенностей этой ткани.
Эпителий нефронов, обнаруживая в условиях культур несомненное
сходство с выстилкой собирательных трубок, вместе с тем обнаруживает
Экспериментальный метод в гистологии
223
(например в условиях экстенсивного роста мембранами) известные отли-
чия от нее, отмеченные Цымбалом (1935). Эти два участка почечных канальцев
не вполне идентичны в отношении своих гистологических особенностей.
Еще более резко, однако, выражены различия между эпителиями собира-
тельных трубок и лоханки. Последний при обычных условиях имеет строе-
ние переходного эпителия, а в области сосочков пирамид приобретает
двухрядное строение, сменяясь затем в сосочковых ходах однослойным
эпителием. Высокие призматические клетки сосочковых ходов обнаружи-
вают в отношении своих цитологических особенностей известное сходство
с высокими элементами двухрядного лоханочного эпителия. Во взаимо-
отношениях эпителия собирательных трубок и эпителия лоханки еще имеется,
ряд неясностей. Например, трудно сказать в точности, где, в каждом отдель-
ном случае, располагается граница между ними. Возможно, что в отдель-
ных сосочковых ходах она располагается не вполне одинаково, то совпадая
с устьем трубки, то смещаясь выше, внутрь последней, то, быть может,
находясь уже на поверхности сосочка. Эта область подлежит еще специ-
альному изучению в отношении своих особенностей.
Способность лоханочного эпителия в экспериментальных условиях обра-
зовать многослойные структуры и неспособность к образованию таких
структур у почечного эпителия, включая и эпителий собирательных трубок,
могут быть объяснены двояко. Может быть, эпителий собирательных трубок
настолько специализировался, что, несмотря на свой эпидермальный харак-
тер, совершенно утратил ряд основных гистобластических потенций тканей
этого типа. В пользу такой возможности говорят отмеченные выше эволю-
ционные изменения выстилки дышащих частей легких. С другой стороны,
однако, вторичная почка является филогенетически более новым образо-
ванием, чем легкие. Эпидермальная выстилка каудального отдела вольфова
канала млекопитающих в других своих производных, семевыносящих путях,
сохранила свойственные эпидермальным тканям гистобластические потен-
ции. Между тем, способность к образованию многослойных структур кож-
ного типа эпителием семевыносящих путей при экспериментальных усло-
виях никак не оправдывается его функциональным значением в норме,
которое должно было также или даже в еще большей степени способствовать
эволюционному ослаблению или исчезновению эпидермальных гистобласти-
ческих потенций.
Поэтому мне представляется более вероятным иное объяснение приве-
денных выше результатов экспериментального анализа эпителиев почки
и мочеотводящих путей, заключайщееся в том, что граница между выросшими
из эмбриональной лоханки собирательными трубками и нефронами в сфор-
мированной почке располагается не там, где она первоначально находилась
у зародыша. На этот счет имеются и вполне определенные, упомянутые
выше, эмбриологические данные, согласно которым первичные собиратель-
ные трубки в большей или меньшей степени редуцируются, причем их
эпителий затем входит в состав выстилки лоханки (Нетер, 1909; Феликс,
1905, 1911). По болееновым данным Фукса(1932, 1933), редукция первичных,
собирательных трубок идет значительно дальше, чем это предполагалось
'224
Глава III
Прежними авторами. Все это, вместе взятое, делает вероятным предполо-
жение, что при окончательном формировании почки практически весь
эпителий первичных собирательных трубок, или его большая часть, вытес-
няется обратно в лоханку и замещается нарастающим навстречу нефроген-
ным почечным эпителием.
Таким образом, первичные собирательные трубки эмбриональной почки и
система окончательных собирательных трубок сформированного органа не
тождественны друг другу. Первичные собирательные трубки являются лишь
провизорными образованиями, которым, возможно, принадлежит морфоге-
нетическое значение при формировании дефинитивных собирательных тру-
бок. Нефрогенный эпителий пользуется ими как моделью при своем росте в
направлении к лоханке, до которой он и распространяется в конце концов.
Другими словами, нефроны и собирательные трубки перестают быть генети-
чески различными образованиями, а возникают, повидимому, из того же
источника метонефрогенной ткани или, в конечном итоге, нефротомов. Не-
фрон представляет собой лишь неразветвленный начальный участок системы
почечных канальцев, а собирательные трубки — их древовидно разветвлен-
ные, или, вернее, соединяющиеся друг с другом под острыми углами отделы.
д) Эпителий производных мюллеровых каналов (яйцеводов и матки)
Эпителию яйцеводов и матки посвящены единичные работы, сделанные
методом эксплантации.
Подробный гистологический анализ превращений эпителия яйцеводов
был до настоящего времени на кроличьем материале дан только Галустяном
(1935). Ему же принадлежит и работа по эпителию матки (1932). Данные,
касающиеся маточного эпителия, содержащиеся в работах Гейма (1927),
Каффье (1931), Гирша и Джонса (1933), Азуми (1937) и некоторых других
авторов, настолько неполны, что на их основании невозможно делать какие-
либо гистологические выводы. Эпителий яйцевода растет экстенсивно,
образуя плоскостные мембраны, и эпителизирует, нередко нацело, поверх-
ность эксплантатов. Его элементы энергично делятся митотическим путем.
Энергично разжижая фибрин, он образует в нем полости, наполненные
жидкостью. Вэ многих случаях эпителий совершенно заглушает рост со-
единительной ткани и образует чистые культуры. Периферические участки
эпителиальных мембран могут подвергаться сильному разрыхлению, пре-
вращаясь в сетевидные клеточные соединения и изолированные клетки
неправильной формы с заостренными отростками. При эпителизации поверх-
ности эксплантата расположенные одним слоем эпителиальные элементы
могут подвергаться более или менее сильному уплощению. Дифференци-
рованный покровный эпителий нацело эпителизированных эксплантатов
отличается неодинаковой высотой своих элементов и позволяет большей
частью различать чередующиеся друг с другом участки, состоящие из упло-
щенных, кубических и высокопризматических элементов с куполообраз-
ными выпуклыми апикальными поверхностями и замыкающими полосками
(рис. 25). Между расположенными базально частями боковых поверхностей
клеток покровного эпителия могут образоваться межклеточные промежутки,
Экспериментальный метод в гистологии
225
в которых бывают заметны цитоплазматические связи. В условиях эксплан-
тата эпителиальная выстилка яйцевода подвергается упрощению благодаря
постепенному исчезновению железистых клеток. Более или менее значи-
тельная часть мерцательных элементов может длительное время сохранять
свои реснички как в составе покровного эпителия, так и в энергично раз-
растающихся мембранах, и продолжает оживленно мерцать даже в куль-
турах месячного возраста. Другая
часть мерцательных клеток утра-
чивает свой ресничный аппарат,
а также базальные тельца. Неко-
торые клетки временно сохраня-
ют одну-две или несколько бес-
порядочно мерцающих ресничек.
В особенности на ранних стадиях
культивирования наблюдаются мас-
совое отторжение и дегенерация
отдельных клеток эпителиального
покрова посеянного кусочка. Они
до полного отторжения имеют
очень характерный вид и напоми-
нают по своей форме гвозди со
шляпкой, воткнутые между сосед-
ними элементами и выдающиеся над
их апикальными поверхностями.
Отторгнутые мерцательные эле-
менты совершенно округляются,
причем расположенные на их по-
верхности реснички еще могут
некоторое время продолжать мер-
цать. Любопытно, что единичные
мерцающие реснички могут сохра-
няться даже в совершенно разрых-
ленных участках мембран у клеток,
приобревших неправильную от-
Рис.~'25. Участки однослойного покровного
эпителия различной высоты нацело эпите-
лизированных эксплантатов из яйцеводов
кролика. А—15-дневная, Б — 18-дневная,
В — 23-дневная культуры. Сильное увели-
чение. (По Галустяну, 1936.)
ростчатую форму и напоминающих
по своему внешнему облику т. наз. фибробласты соединительной ткани.
Маточный эпителий кроликов (Галустяи, 1932) ведет себя в эксплантатах
сходным образом, отличаясь лишь отсутствием ресничек. После эпители-
зации поверхности посеянного кусочка в нем также можно наблюдать
однослойные участки неодинаковой высоты. Отмечавшаяся в его составе
многорядность, насколько можно судить в настоящее время, является
ложной и, повидимому, зависит лишь от расположения ядер на разных
уровнях, а не от присутствия в нем коротких базальных клеток, имеющих
камбиальное значение. Последние, еще неопубликованные данные нашей
лаборатории (Жемкова) говорят в пользу однослойного и однорядного
строения маточного эпителия.
15 X лопин—349
226
Глава III
Опыты с эксплантацией эндометриозов {Михайлов, 1936), представлч-
ющих собой, по своему строению, участки слизистой оболочки матки или
яйцеводов и встречающихся как дистопические образования в разных
местах женского организма (преимущественно в серозной оболочке брюш-
ной полости или области пупка), показали, что их эпителий ведет себя
в культурах как эпителий матки и яйцеводов, давая экстенсивный рост
мембранами и эпителизируя поверхность эксплантата. На поверхности
эпителизированных частиц образуются своеобразные тонкостенные пузыри
с эпителиальной стенкой, вследствие сильного отека в отдельных участках
соединительнотканной подстилки. Последнее отмечалось также Жемковой
в эксплантатах из нормального женского эндометрия. Покровный эпителий
в эксплантатах из эндометриозов (в которых удалось до одиннадцати-
двенадцати дней культивирования констатировать секрецию слизи) создает,
повсеместно сохраняя однослойное и однорядное строение, участки очень
неодинаковой высоты и отличается сильно выступающими свободными
апикальными поверхностями своих элементов. В эксплантатах эндометрио-
зов удавалось проследить до двух недель эксплантации мерцательные
движения в тех участках эпителия, которые имели реснички.
Последняя, еще неопубликованная работа Жемковой, посвященная
изучению методом тканевых культур особенностей эпителия матки кролика
на разных стадиях беременности и в послеродовом периоде, существенно
дополняет приведенный выше фактический материал. Для эксплантации
брались участки слизистой оболочки матки, расположенные между плацен-
тами, изменениям эпителия которых в течение беременности была посвящена
предыдущая работа того же автора, рассмотренная выше.
При экстенсивном росте наиболее характерной особенностью эпителия
матки является образование мембран, состоящих из многоядерных гигант-
ских клеток. На более поздних стадиях культивирования в составе мембран
наблюдаются как многоядерные, так и одноядерные элементы. Многоядер-
ные клетки мембран образуются за счет разрастания покровного эпителия
матки, измененного при беременности и состоящего из мнигоядерных эле-
ментов {Жемкова, 1941).
Каждая клетка часто содержит от пяти до двадцати и даже более ядер.
Периферические части мембран, где выражено разрыхление, образуют
сетевидные плоскостные клеточные соединения. Позже, начиная с седьмого
дня эксплантации, среди многоядерных клеток мембран появляются одно-
ядерные элементы, делящиеся митотическим путем и растущие из эпителия
крипт, сохраняющего при беременности свой обычный характер. Эпителий
крипт обычно разрастается в виде веерообразно расширяющихся потоков.
На дальнейших стадиях культивирования мембраны или сохраняют смешан-
ный характер или многоядерные элементы совершенно вымирают и исчезают.
При эпителизации поверхности эксплантатов проявляются те же харак-
терные различия между эпителиями поверхности и крипт. Клетки поверх-
ностного эпителия, наползая на мышцы и соединительную ткань, посте-
пенно уплощаются, сохраняя свою многоядерность. Эпителий крипт,
выдвигаясь на поверхность и эпителизируя кусочек, содержит в своем
Экспериментальный метод в гистологии
227
составе одноядерные клетки различной высоты, в которых часто встре-
чаются митозы, отсутствующие в гигантских элементах. На всех сроках
культивирования слизистой оболочки матки, взятой на третий день после-
родового периода, когда покровный эпителий утрачивает многоядерные
клетки, наблюдался рост эпителиальных мембран, состоящих только из
одноядерных клеток.
Последние данные подчеркивают в еще большей степени, чем это было
возможно сделать раньше, глубокое своеобразие маточного эпителия и его
специализацию в особом, не свойственном другим тканям, направлении.
Все эти экспериментальные исследования вместе с тем заставляют подверг-
нуть коренному пересмотру вопрос о т. наз. явлениях метаплазии, описы-
вавшихся в женском половом аппарате при экспериментальных и пато-
логических условиях.
Имеется очень большое количество фактов, указывающих на то, что
в матке и даже яйцеводах млекопитающих и человека могут появляться
участки многослойного эпителия кожного типа, могущего даже обнару-
живать типичное ороговение. Тейчлендеру (1924) удалось это вызвать путем
внутриматочного введения дегтя. То же самое получается при авитаминозе А,
а в последнее время удалось получить Кленицкому (1938, 1940) под влиянием
различных воздействий. В человеческой патологии находки многослойного
эпителия в матке не представляют большой редкости, а в ряде случаев,
при т. наз. ихтиозе, вся матка оказывается выстланной многослой-
ным ороговевающим эпителием. В свете наших экспериментальных данных,
показывающих глубокое своеобразие эпителия матки и яйцеводов и его
неспособность к превращению ни в клетки соединительной ткани, ни в много-
слойный эпителий кожного типа, все эти наблюдения должны трактоваться
не как метаплазия местного однослойного мюллеровского эпителия, а как
его замещение врастающим из влагалища многослойным эпителием (Хлопин,
1934; Галустян, 1935). Следует подчеркнуть, что во всех решительно слу-
чаях такое замещение не могло быть исключено, а все экспериментальные
данные демонстрируют с большой наглядностью высокую способность
эпителиев к плоскостному росту.
Для решения спора, возникающего по этому вопросу между раковой
лабораторией проф. Шабада и моим научным коллективом, Казанская
выполнила под руководством проф. Шабада следующую работу, доложен-
ную в мае 1941 г. на объединенном заседании общества патологов и общества
анатомов, гистологов и эмбриологов в Ленинграде. У части подопытных
мышей производились перерезка и перевязка маточных рогов для того,
чтобы их полностью изолировать от влагалищного эпителия. Затем опери-
рованным и нормальным мышам длительное время вводили раствор полиа-
нола. Под влиянием этого вещества, в физиологическом отношении сходного
с фолликулином, у мышей в маточных рогах на значительном протяжении
их появляется многослойный плоский эпителий кожного типа. При иссле-
довании маточных рогов, изолированных оперативно от влагалища, в их
эпителии были обнаружены лишь дистрофические изменения, но никакого
намека на появление многослойного эпителия кожного типа. Результаты
228
Глава Ш
опытов полностью совпали со сделанным мной предсказанием и показали
с большой убедительностью правильность нашей точки зрения, а не при-
нятой патологами трактовки этого явления.
Наконец, не исключена возможность, что некоторая часть описанных
случаев находок дистопического многослойного эпителия в женских поло-
вых путях зависит от слишком поверхностного изучения препаратов и что
трактовавшиеся как многослойный эпителий структуры на самом деле не
имеют ничего общего с многослойными эпителиями кожного типа. Поводом
к такого рода ошибочным описаниям могут быть совершенно своеобразные
изменения маточного эпителия, аналогичные тем, которые были детально
изучены Жемковой при беременности. В этом отношении остается еще
широкое поле для систематических гистологических и гисто-патологических
исследований.
В свете установленных нами у млекопитающих гистологических разли-
чий между мезотелием серозных оболочек, с одной стороны, и эпителиями
матки и яйцеводов, с другой, вопрос о природе т. наз. эндометриозов или
эндометриом (Лаухе, 1923; Манкин, 1933, и др.) получает прочно обоснованное
решение. Эндометриозы должны рассматриваться как гетеротопические
участки слизистой оболочки матки или яйцеводов или имплантировавшиеся
вторично (как это предполагают Семпсон, а за ним многие другие авторы),
а может быть, возникшие в отдельных случаях из эмбрионального дистопи-
ческого зачатка мюллеровского эпителия. Во всяком случае нет никаких
оснований считать их результатом метаплазии участка серозных оболочек.
Многочисленные опыты с трансплантацией частиц эндометрия в брюшную
полость, в переднюю камеру глаза и другие места организма, произведен-
ные на животных, преимущественно кроликах (Ш шиллинг, 1909; Якобсон,
1922; Нейман, 1931, 1933; Оллен и Прист, 1932; Верхацкий, 1940, и др.),
показали, что гомотрансплантаты хорошо приживают и сохраняются дли-
тельное время, превращаясь большей частью в замкнутые кисты, выстлан-
ные изнутри маточным эпителием, который обнаруживает свои характер-
ные особенности и реагирует типичным образом на гормональные влияния
в течение астрального цикла. При пересадке в переднюю камеру глаза (Ней-
ман, 1933) наблюдалосьразрастание маточного эпителия по передней поверх-
ности радужной оболочки. Аутотрансплантацией эндометрия в стенку
влагалища или цервикальный канал успешно пользуются также в клинике
для восстановления у женщин месячных (Шмид, 1932, 1934; Фукс, 1935,
1937; Верхацкий, 1940, и др.). Приживший трансплантат сохраняет типичное
для эндометрия строение и обнаруживает характерные для менструального
цикла изменения. Маточный эпителий обнаруживает при этом только
структуры, свойственные производным мюллеровских каналов. Эти опыты,
гистологическая обработка которых, правда, не отличается обстоятельно-
стью, полностью подтверждают высокую специфичность маточного эпителия.
ё) Эмбриональные зачатки целонефродермалъных эпителиев
Презумптивные области бластулы и ранней гаструлы амфибий, соот-
ветствующие будущим зачаткам целонефродермальных эпителиев и распо-
Экспериментальный метод в гистологии
229
лагающиеся в краевой зоне рядом с материалом миотомов, подобно другим
презумптивным областям, не обнаруживают гистологической детерминации
и, например, в опытах с трансплантацией могут изменять свое проспектив-
ное значение, развиваясь в совершенно другие ткани (Мангольд, 1923)
Вместе с тем в опытах с изоляцией (Голыпфретер, 1938) они уже обнару-
живают известные отличия от других презумптивных участков. Со стадии
нейрулы зачатки выстилки целома и выделительных органов, относительно
которых имеются конкретные данные, уже оказываются гистологически детер-
минированными и неспособными развиваться в эктодермальные и энтодер-
мальные производные, а также, повидимому, в элементы соматической
мускулатуры, хотя в экспериментальной эмбриологии и принято считать,
что мезодермальный материал почек конечностей у амфибий будто бы обра-
зуется из соматоплевры, а не из миотомов.
Еще совершенно недифференцированные зачатки вольфова тела амфибий
способны при трансплантации или в изолятах давать типичные или атипич-
ные почечные канальцы (Эрдманн, 1931; Голыпфретер, 1931). То же самое
справедливо и в отношении соответствующих зачатков зародышей в усло-
виях трансплантации на хорион-аллантоис (Данчакоеа, 1934).
Последние работы Голыпфретера (1938) и Винникова (неопубликованные
данные) показывают, что зачатки выделительного аппарата и стенки полости
тела дают в условиях тканевых культур экстенсивные разрастания мембра-
нами, которые по своим особенностям отличаются от мембран, образую-
щихся из других источников. Мембраны из спланхнотомов несомненно
сходны с разрастаниями мезотелия, а мембраны из области нефротомов
с разрастаниями почечного эпителия, разумеется, с поправкой на видовую
специфичность, о которой, кстати говоря, Голыпфретер забывает. Поэтому
он противопоставляет наблюдавшуюся им дифференцировку мерцательных
клеток из нефротомного материала амфибий данным нашей лаборатории,
полученным с почечным эпителием млекопитающих, которому, как известно,
мерцательные элементы совершенно не свойственны. То же самое справед-
ливо и в отношении некоторых рассуждений Голыпфретера, касающихся
выстилки целома.
Эти данные доказывают лишний раз (см. выше) с достаточной убедитель-
ностью, что гистотипические особенности дифференцированных тканей
вполне соответствуют гистотипическим особенностям соответствующих эмбри-
ональных зачатков, которые могут давать сходные с дифференцированными
тканями гистоморфные разрастания, свойственные тканевым культурам,
не пройдя предварительно нормальных этапов морфогенеза и гистологи-
ческой дифференцировки. Этим подчеркивается значение эмбриональных
зачатков, как зачатков определенного гистологического материала, ткане-
вые свойства которого могут проявляться без определенной связи с про-
цессами морфогенеза. Если в филогенезе морфогенез и гистогенез неразрывно
связаны друг с другом, то в индивидуальном развитии осуществление мате-
риалом данного зачатка наследственно закрепленных гистогенетическйх
процессов уже оказывается не связанным с осуществлением им же своих
морфогенетических свойств.
230
Глава III
Приобретенная на ранних этапах индивидуального развития гистоло-
гическая детерминация остается характерным свойством тканей, разви-
вающихся из данного источника, в течение всей жизни организма. На при-
мерах других, разобранных ниже, зачатков и развившихся из них тканей
мы получим дальнейшее подтверждение этих положений.
ж) Опухолевые ткани целонефродермального типа
Первичные опухоли серозных оболочек, обозначенные в свое время
Риббертом термином «целомический рак» (Coelomkrebs), а в настоящее
время называемые часто опухолями покровных клеток (Deckzellentumoren)
или мезотелиомами, имеют очень своеобразную структуру, которая сильно
отличает их от опухолей другого происхождения (Ф. Герцог, 1914; Кузнецов-
ский, 1929; Напп, 1906; Пурман, 1930; Крумбейн, 1924; Фишер-Вазельс,
1929; Затке и Зальцер, 1932; Гроссек, 1932, и др.). Обычно они представляют
собой новообразования, разрастающиеся в пределах самих серозных обо-
лочек и вызывающие их резкое утолщение. По своей микроскопической
структуре они большей частью имеют характер типичных, хотя и очень
своеобразных, эпителиом. Опухолевые клетки располагаются сплошными
гнездами или тяжами, резко отграниченными от стромы, и отличаются
очень большим полиморфизмом. Гигантские одно- и многоядерные элементы
представляют собой обычные находки. Опираясь на исследования Максимова
(1927) и Шоппера (1932), Фишер-Вазельс (1932) выступил с очень резкой
критикой работы Гроссек (1932), в которой, в соответствии с принятой в пато-
логии трактовкой, этот автор считал их источником мезотелий.
Фишер-Вазельс исходил из совершенно правильного положения, что
свойства нормальных тканей должны проявляться в какой-то степени и
в соответствующих опухолевых разрастаниях. Ввиду того, что работами
Максимова и Шоппера казалась установленной близость мезотелия соеди-
нительнотканным образованиям, Фишер-Вазельс стал категорически отри-
цать возможность возникновения первичных опухолей серозных оболочек
типа целомического рака из мезотелия и высказал мысль, что они могут
иметь своим источником только особые эмбриональные зачатки эпителиаль-
ного характера. Наши данные показывают, наоборот, что свойства це-
ломических раков очень хорошо объясняются на основе гистобласти-
ческих потенций мезотелия, выявляющихся при экспериментальных
условиях, и что предположение Фишер-Вазельс не имеет под собой
реальной почвы.
Нет совершенно никаких оснований отрицать возможность развития
первичных опухолей серозных оболочек из дифференцированного мезо-
телия на основе пролиферативных процессов, подобно тому, как это имеет
место для большей части других новообразований. Экспериментальных
данных, касающихся опухолей этого типа, мне неизвестно.
Значительная часть достаточно разнообразных новообразований почки
(Фар и Любарш, 1925) имеет, повидимому, своим источником те или иные
эмбриональные зачатки, оказывающиеся в составе этого органа. Это отно-
сится в первую очередь к т. наз. аденосаркомам почки, встречающимся
Экспериментальный метод в гистологии
231
в детском возрасте, которые нельзя не трактовать как результат бласто-
матозного превращения метанефрогенной ткани. В составе таких опухолей
на фоне саркомоподобных разрастаний веретенчатых клеток имеются нерезко
отграниченные от них полости и трубчатые ходы различной формы с эпите-
лиоподобной стенкой, очень напоминающие атипичные ранние стадии форми-
рования нефронов из метанефрогенной бластомы. Эти картины не могут не
навести на мысль, что пролиферирующая бластоматозно измененная мета-
нефрогенная ткань в составе опухоли сохраняет в известной степени свой-
ственную ей способность к дифференцировке в зачатки почечных канальцев.
Вместе с тем не исключена возможность, что малодифференцированные
зачатки последних могут, пролиферируя, превращаться в веретеноклеточ-
ные метанефрогенные участки. Вопрос о генезе т. наз. опухолей Гравица,
для которых, вместо старого обозначения гипернефромы, предложен термин
«гипернефроидные опухоли», является до настоящего времени спорным. Их
или представляют себе как вторично измененные трубчатые аденомы моче-
вых канальцев, т. е. генетически связывают с почечным эпителием, или
рассматривают как бластоматозно измененную ткань коркового вещества
надпочечников. По Любаршу (1925), следует различать гипернефроидные
опухоли трех разных типов. Учитывая во многих случаях поразительное
морфологическое сходство гипернефроидных опухолей с тканью коркового
вещества надпочечников, по крайней мере некоторые из них нельзя не рас-
сматривать как настоящие гипернефромы, т. е. новообразования, раз-
вивающиеся из интерренального зачатка, но не из почечного эпителия,
дающего разрастания типа трубчатых аденом или папиллярных (трабеку-
лярных) кистом и раковые опухоли, характеризующиеся, как правило,
однослойными эпителиальными структурами.
В литературе имеются экспериментальные данные, касающиеся гипер-
нефромы морской свинки (Роскин, 1931) и гипернефромы человека (Тимо-
феевский, 1939). Последнему автору удалось получить довольно длительные
культуры этой опухоли. К сожалению, ткань нормального надпочечника
изучена методом эксплантации в гистологическом отношении слишком
недостаточно, чтобы можно было сделать какие-нибудь сопоставления,
а соответствующий опухолевый материал нуждается еще в более детальном
гистологическом анализе.
Гистогенез крупноклеточной опухоли яичка т. наз. семиномы
представляется в достаточной степени спорным (Оберндорфер, 1931). По
своему общему характеру она производит впечатление очень сильно ката-
плазированной и малодифференцированной эпителиомы, состоящей из
крупных клеток полиэдрической формы, располагающихся большей частью
гнездами. Данные, полученные Михайловым (1938) методом эксплантации,
показывают, что ее элементы, по совокупности своих превращений в усло-
виях тканевых культур, обнаруживают большое сходство с фолликулярными
клетками неполовозрелого семенника. Это дает возможность в чрезвычайно
запутанном вопросе о гистогенезе этих опухолей высказаться в пользу
их вероятного происхождения из сертолиевых клеток или соответствующих
им элементов эмбрионального организма.
232
Глава Hi
По данным Григорьева (1939), элементы т. наз. опухоли Брауна и Пирс
кроликов, по характеру роста вне организма, также в значительной степени
напоминают фолликулярные клетки семенников. Это противоречит предл
положению Брауна и Пирс, считающих, что опухоль происходит из эпи-
дермиса мошонки, и результатам опытов М. Меллендорф (1938) (полученным
методом эксплантации), согласно которым эта опухоль растет по соеди-
нительнотканному типу.
Результаты, полученные Михайловым (1941) при культивировании
вне организма редкого опухолеподобного разрастания яичника т. наз»
фолликуломы, показали, что ее элементы разрастаются мембранами, напо-
минающими растущие вне организма фолликулярные клетки.
Очень ценный фактический материал дает тщательное в гистоло-
гическом отношении исследование Глазунова (1937) над т. наз. цилиоэпи-
телиальными кистами яичника. Полученные Глазуновым данные, при их
сопоставлении с разобранными выше работами нашей лаборатории (Хлопин,
1936, 1937; Галустян, 1933, 1935; Колесникова, 1940) по мезотелию серозных
оболочек, эпителию матки и яйцеводов и по целомическим компонентам
яичника, показывают с полной очевидностью, что эти опухоли имеют своим
источником мюллеровский эпителий, а не тканевые элементы нормального
яичника. Все разнообразие эпителиальных структур этих кист обнаруживает
несомненное и глубокое сходство с разными структурами, свойственными
эпителиям матки и яйцеводов при обычных и экспериментальных условиях
и не имеют ничего общего со структурами, свойственными мезотелию сероз-
ных оболочек и фолликулярным клеткам яичников.
Последние, неопубликованные данные Михайлова (которые я сообщаю
с любезного разрешения автора) с несомненностью говорят то же самое
и в отношении т. наз. псевдомуцинозных кист. Он изучил их на большом
и тщательно обработанном операционном материале, причем в ряде случаев»
помимо описания присущих им структур, провел успешные опыты с эксплан-
тацией. Эти опыты позволили установить, что в условиях экстенсивного
роста мембранами их эпителий с особенной отчетливостью проявляет
гистобластические потенции производных мюллеровых каналов.
Опухолевая ткань папиллярных кистокарцином яичника при культи-
вировании вне организма обнаруживает большое морфологическое сходство
с эпителием трубы и, вероятно, происходит из него (Беневоленская,.
1939).
Что касается плоскоклеточных опухолей, возникающих в женском
половом аппарате, то они (в соответствии со всем разобранным выше экспе-
риментальным материалом) не являются производными целонефродермаль-
ного типа и были в общих чертах отмечены как представители эпидер-
мального опухолевого типа.
Наибольшие трудности в отношении трактовки представляют встречаемые
в матке опухоли, в составе которых обнаруживаются однослойные и кажу-
щиеся многослойными участки. Без специального и очень детального гисто-
логического анализа (который пока не осуществлен) невозможно высказать,
каких-либо предположений на их счет.
Экспериментальный метод в гистологии
239
Г. Эпендимо-глиальный тканевой тип или вспомогательные ткани
нервной системы
а) Общие замечания. Краткие эмбриологические и сравнительные данные
'Как это уже неоднократно отмечалось выше, положение различных
вспомогательных тканей нервной системы в обычной морфо-физиологической
системе гистологических структур является в достаточной степени неопре-
деленным, противоречивым и даже надуманным. Их рассматривание в составе
эпителиальной группы тканей (которое мы приняли в настоящей книге)
также не свободно от серьезных возражений и в отношении собственно
нейроглии не соответствует общепринятой точке зрения. Наконец, недо-
статочно ясными представляются границы эпендимо-глиального тканевого
типа и то, какие именно конкретные структуры будет целесообразно к нему
отнести. Как это будет подробнее разобрано ниже, мы отнесем пока к эпен-
димо-глиальному тканевому типу все ткани, развивающиеся у позвоночных
из нервной пластинки и плакод и входящие в состав центральной и перифе-
рической нервной системы с органами чувств, элементы которых не имеют
нейрональной дифференцировки, а несут пограничную, опорную, трофи-
ческую, отчасти защитную и, в некоторых случаях, рецепторную функции.
При последовательном проведении генетической системы тканей к еди-
ному нервному или нейральному тканевому типу следовало бы отнести,
как его особые подразделения, нейроны и мышечные элементы нейрального
происхождения наравне с эпендимными и, в широком смысле слова, глиаль-
ными клетками. Поскольку мы, однако, условились придерживаться в
основном морфо-физиологических норм, правда, с некоторыми отступлени-
ями от принятой классификации, сократимые элементы глаза и нейроны
будут рассмотрены в другом месте.
Для того чтобы правильно ориентироваться в современном состоянии
вопроса о вспомогательных тканях нервной системы, необходимо дать
краткий исторический очерк. Честь открытия ненейрональных компонентой
нервной системы принадлежит Пуркинье (1836), который обнаружил у плода
овцы мерцательное движение в стенке внутренних полостей мозга. Он
отметил, что реснички связаны со слоем «зерен» (т. е. клеток эпендимы),,
выстилающих эти полости. Кроме того, он описал на поверхности сосуди-
стых сплетений мозга мембрану, лишенную мерцательного аппарата и
состоящую из правильно расположенных <зерен» (т. е. клеток), каждое
из которых имеет в центре маленькое тельце (т. е. ядро). «Первоначально,—
говорит Пуркинье, — эта мембрана мне представлялась по своему виду, как
нервное образование; теперь же я имею много оснований считать ее эпидер-
моидальной». Двумя годами позже Генле (1838) включает внутреннюю
выстилку полостей мозга в состав эпителиальной ткани, отмечая ее строение
из цилиндрических телец с несколько заостренным концом, содержащих ядро.
Вирхов (1846) описал в человеческом мозгу под эпендимой особую, сходную,
с «соединительным веществом» (т, е. соединительной тканью) мембрану,
с неясной волокнистостью, которая дает начало патологическим выступам.
Они, с его точки зрения, могут рассматриваться как утолщения эпендимы.
334
Глава III
обнаруживающие известное сходство с пахионовыми грануляциями мозго-
вых оболочек. Вместе с тем Вирхов (1846, 1853) полемизирует с Генле,
который в своей «Общей анатомии» предполагал (1841), что эпендима непо-
средственно покрывает нервные волокна. В работе 1854 г. и, особенно,
в примечании к работе 1846 г., перепечатанной в собрании сочинений
(1856), Вирхов резюмирует свои представления следующим образом: эпен-
дима состоит не только из эпителия, но также из покрытого эпителием
слоя соединительной ткани; она не образует отдельного покрова в узком
смысле слова, но лишь выступающий на поверхности слой межуточной
или соединительной ткани мозгового вещества. Эта ткань соединительного
вещества образует в головном и спинном мозгу и в высших органах чувств
подобие клея (Kitt) или связующего вещества (нейроглия), в который погру-
жены нервные элементы. В этом соединительном веществе заметны клетки
« ядра. На поверхности кровеносных сосудов оно сгущается. Таким обра-
зом, основоположник учения о нейроглии Вирхов совершенно правильно
считал, что эпендима и нейроглия представляют собой единую ткань, что
упускается из виду многими позднейшими авторами, которые рассматри-
вают эпендиму как эпителий, а нейроглию как ткань, сходную с соеди-
нительной. Фроманн (1864, 1877) и, особенно, Дейтерс (1865) подробнее
описали отростчатую форму клеток нейроглии (астроцитов), получивших
первоначально название клеток Дейтерса, а позже — клеток Гольджи,
изучавшего их с помощью своей хромосеребряной методики (1885/6, 1894)
и принявшего для их обозначения термин «астроциты» или «звездчатые
клетки». Чрезвычайно точное и во многих отношениях непревзойденное
описание астроцитарных элементов нейроглии млекопитающих сделал
Гирке (1885, 1886), который пользовался параллельно методикой мацерации
и целлоидиновых срезов. Данные этого автора незаслуженно забываются
или недооцениваются. По Гирке (1885), глия представляет собой ткань,
сходную с ретикулярной и состоящую из клеток с отростками и бес-
структурного основного вещества. Соединяясь друг с другом отростками,
достигающими огромной длины и ветвящимися сложным образом, клетки
нейроглии образуют опорный аппарат нервной системы. На мацерирован-
ных препаратах мозга овцы этому автору удалось видеть клетки размером
в 15—-25 длина отростков которых превышала половину, три четверти
н даже 1 мм.
Пользуясь методикой переваривания, по Эвальду — Кюне (1877), Гирке,
кроме того, установил, что протоплазма глиальных клеток подвергается
после рождения особому резкому изменению, сходному с ороговением,
превращаясь в кератин (нейрокератин Эвальда и Кюне). С возрастом про-
цессы ороговения резко усиливаются. Элементы нейроглии отличаются
друг от друга количеством и длиной отростков и степенью ороговения. При
ороговении ядро, уменьшаясь в размерах, сморщивается и затем исчезает,
а от клеточного тела в конце концов остаются роговые нити. У частично
ороговевших клеток отростки представляются пучком нитей с общей прото-
плазматической оболочкой. С отростков процесс ороговения распростра-
няется на клеточное тело.
Экспериментальный метод в гистологии
235
Если сопоставить друг с другом эктодермальный эпидермис, эпителии
органов чувств и нейроглию, то и последнюю следует рассматривать, по
мнению Гирке, как эпителиальную ткань. Это он подкрепляет наблюдениями
над развитием глии в спинном мозгу у зародышей кролика. Глиальные
и нервные клетки развиваются из длинных эпителиальных элементов, из
которых впоследствии остается один слой вокруг центрального канала.
Периферические отростки эпендимных клеток сформированного спинного
мозга также могут ороговевать. В противоположность Фроманну и Гирке,
считавшим, что глиальные клетки анастомозируют, Дейтерс, Болль и
некоторые другие авторы предполагали, что отростки клеток глии пере-
плетаются друг с другом.
Ранвье (1882, 1883), а за ним Вейгерт (1890, 1895) стали различать
в составе глии клетки и волокна. Эту, по существу неправильную, точку
зрения особенно подробно обосновал Вейгерт, очень детально изучивший
нейроглию на человеческом материале с помощью открытой им специаль-
ной методики, позволяющей элективно окрашивать в срезах волокнистые
структуры глии, но не неизмененную протоплазму глиальных элементов.
Поэтому у Вейгерта получилось неправильное представление, что глиаль-
ные клетки лишены отростков и располагаются среди густого войлока воло-
кон. С этим связаны и общие представления этого автора о тканевой при-
роде нейроглии, согласно которым она построена по типу соединительной
ткани. На этот счет Вейгерт (1895) высказывается следующим образом:
«Так как нейроглия построена по типу соединительного вещества, то при-
нятие ее эпителиальной природы очень парадоксально» (стр. 129), и дальше:
«В настоящее время нельзя обойти факта, что нейроглия в морфологическом
и биологическом отношениях имеет свойства соединительного вещества»
(стр. 130). Вместе с тем он правильно подчеркивает, что последнее слово
в этом отношении принадлежит эмбриологам. До Вейгерта аналогичный
взгляд на нейроглию высказывался Лейдигом (1885) и Кёлликером (1889).
Кёлликср в последнем издании своего руководства (1889) подчеркивает
большое сходство нейроглии с простой клеточной соединительной тканью,
отмечая, что одни глиальные элементы иг рают роль опорных образований,
а другие, имеющие уплощенную форму и небольшое количество протоплазмы,
образуют частично выстилки полостей («ложный эпителий»), частично
оболочки ганглиозных клеток и нервных волокон. Высказанная несколько
позже Мюллером (1900) точка зрения, что нейроглия представляет собой
как бы переход от эпителия к соединительной ткани и что ее возможно
правильно понять, отказавшись от «догмы тканевой специфичности», глу-
боко ошибочна. Это совершенно наглядно демонстрируется эксперимен-
тальным анализом этой ткани.
В современный период изучение нейроглии вступило после разработки
новых методов импрегнации золотом и серебром Рамон и Кахалем и его
сотрудниками Ачукарро (1911), Рио Ортега (1916, 1917, 1921) и другими.
Вместе с тем общегистологические вопросы о тканевой природе нейроглии
отошли на задний план и заслонились детализацией цитоморфологических
особенностей ее элементов, что в значительной степени соответствует и
286
Глава III
направлению развития учения о крови и соединительной ткани. Очень
специальная, трудоемкая и часто капризная методика изучения нейроглии,
а также нейронов, привела к тому, что глава гистологии, посвященная
нервной системе, выделилась в полусамостоятельную дисциплину, нейро-
гистологию, и оказалась в значительной степени оторванной от других
отделов нашей науки, а вместе с тем и от ее основных и общих проблем?
Затушеванное исследованиями Вейгерта представление о клеточном строе-
нии нейроглии вновь получило права гражданства в свете новейших работ.
Пользуясь своими новыми методами, Кахаль отчетливо продемонстрировал
три разновидности клеток глии — протоплазматическую и волокнистую
макроглию со сложной системой отростков и т. наз. «третий элемент» (е!
tercer elemento), протоплазматическое тело которого не красилось, и кото-
рый поэтому представлялся лишенным отростков. Волокнистая и прото-
плазматическая глия не всегда четко отграничиваются друг от друга,
так как, кроме того, еще имеются формы промежуточного характера.
К макроглии принято относить также, как ее особые разновидности, берг-
мановские и мюллеровские волокна, перистые клетки и некоторые дру-
гие элементы. Первые два рода клеток соответствовали астроцитам
Гольджи или элементам с довольно крупными светлыми ядрами и не-
большим ободком цитоплазмы, обнаруживающимся методикой Ниссля, а
третий элемент отличался мелкими и темными ядрами без видимого, при
окраске по Нисслю, ободка цитоплазмы. На этот третий элемент еще задолго
до того обратил внимание Робертсон (1900), обозначивший его термином
«мезоглия». Дальнейшие исследования и усовершенствования методов
импрегнации серебром позволили выделить среди клеток, обозначенных
как третий элемент, или мезоглия, еще две разновидности: олигодендроглию
и микроглию. Микроглию особенно подробно изучил Рио Ортега (1921,,
1932), поэтому ее иначе называют «ортегаглия». Макроглия обозначается
также в настоящее время удачным термином «астроглия» (Стронг, 1925).
Нежная сеточка, описанная Гольдом (1909) под названием Gliareticulum,
большей частью позднейших авторов отрицается.
Первоначальное предположение, разделявшееся и Гисом, что нейро-
глия развивается из мигрирующей в нервную трубку зародыша мезенхимы,
было вскоре оставлено. Детальные эмбриологические исследования, среди
которых выдающееся место принадлежит работам самого Гиса (1890), пока-
зали, что эпителиальная выстилка эмбриональной нервной трубки млеко-
питающих, дифференцируясь в двух направлениях, дает начало не только
пейробластам, но и спонгиобластам, превращающимся затем в нейроглию.
Вместе с тем ряд авторов предполагал, что в составе нейроглии имеются
и элементы, развивающиеся из стенки нервной трубки, и мезенхимные
клетки. Эта точка зрения была, в частности, высказана Робертсоном (1900)
в отношении упомянутой выше мезоглии и детально развита в последнее
время Ортега (1919—1932), а за ним и рядом исследователей, резко проти-
вопоставляющих микроглию, которая, по их данным, имеет мезенхимное
происхождение, собственно нейроглии, т. е. макроглии и олигодендроглии
(Пенфилъд, 1932).
Экспериментальный метод в гистологии
237
Многие весьма авторитетные исследователи (Якоб, 1927; Метц и Шпатц,
1924; Шаффер, 1926, и др.), однако,не разделяют этой точки зрения, осно-
вывающейся преимущественно на изменениях микроглии при патологических
условиях и, в частности, на ее способности к фагоцитозу. Они считают,
что микроглия тоже развивается из эмбриональной нервной трубки.
Современное учение о нейроглии млекопитающих и человека вылилось в
настоящее время в сложные генеалогические схемы, демонстрирующие, каким
образом при эмбриональном развитии получается все многообразие ее кле-
точных форм, каждая из которых отличается высокой степенью специфичности
и даже неспособностью к превращениям в другие глиальные же элементы.
Нейроглия центральной нервной системы низших позвоночных изучена
до настоящего времени значительно менее полно. Тем не менее, ее сопоставле-
ние у ланцетника и разных позвоночных в высшей степени поучительно,
так как дает понятие об эволюционном изменении свойств этой ткани.
У ланцетника вся нейроглия представлена только эпендимными клетками
(Ленгоссек, 1895; Э. Мюллер, 1900), апикальные концы которых, снабжен-
ные ресничками, образуют выстилку полости трубчатого мозга, сообщающе-
гося в личиночном состоянии на своем переднем конце с внешней средой
посредством нейропора. Базальные концы клеток эпендимной глии образуют
отростки, частью доходящие до наружной поверхности трубки (т. наз.
пучковые волокна) и образующие ее пограничную перепонку, частью
оплетающие тела нейронов и нервные волокна (т. наз. волокна сплетений).
У круглоротых (Э. Мюллер, 1900) помимо эпендимной глии с ее базаль-
ными нитевидными отростками появляются ближе к наружной поверх-
ности спинного мозга клетки типа астроцитов, утратившие связь с выстил-
кой центрального канала, но соединяющиеся частью своих отростков,
образующих ножки, с мягкой мозговой оболочкой. Эпендима приобретает
вместе с тем двурядное строение: часть клеточных тел ее образует выстилку
центрального канала, часть имеет грушевидную форму и отодвинутое от
просвета центрального канала тело, с которым оно связывается централь-
ным клеточным отростком, вклинивающимся между эпендимными клет-
ками первого ряда и расширяющимся в пластинку.
Может быть, эпендима низших позвоночных имеет значение чувстви-
тельных клеток, образующих особый орган чувства (Третьяков, 1913;
Агдур, 1932). Третьяков (1913) описал в ее составе настоящие нервные
клетки грушевидной формы, вероятно, соответствующие эпендимным клет-
кам второго ряда, частично упомянутым выше. У рыб появляются в зна-
чительном количестве астроцитарные элементы, утратившие связь и
•с наружной поверхностью мозга, а эпендимный слой становится много-
рядным. Первое появление в центральной нервной системе клеток типа
«третьего элемента» (возможно, соответствующих олигодендроглии) отметил
у ящерицы Ачукарро (1915). Относительно микроглии никаких определен-
ных данных сравнительного характера не имеется. За пределами класса
млекопитающих она с достоверностью описана не была.
На основе сравнительного изучения нервной системы ланцетника и
разных позвоночных и процессов гистогенеза в ней Шапер (1894, 1897)
288
Глава Ш
развил следующие взгляды на ее эволюционное развитие в ряду хордовых.
У ланцетника «эпителиальная» стенка эмбриональной нервной трубки
непосредственно дифференцируется на эпендиму и нейробласты, дающие
затем нейроны. У круглоротых эпендимная глия претерпевает дальнейшее
развитие: нейробласты развиваются не непосредственно из первичного
«эпителия» нервной трубки, а из особых круглых зачатковых клеток,
образующихся предварительно из него. Начиная с рыб, первоначальная
эпендимная глия замещается новой генерацией глиальных элементов,
развивающихся не непосредственно из «эпителия» нервной трубки, а из
индифферентных зачатковых клеток, одна часть которых дифференцируется
в нейробласты, а другая — в спонгиобласты.
У млекопитающих роль «зачатковых» клеток Шапера выражена осо-
бенно резко. Смещаясь активно от просвета нервной трубки, где они перво-
начально располагаются, зачатковые клетки образуют мощный плащевой
слой (описанный еще Гисом), элементы которого и дают основную массу
нейроглии и нейронов. В отличие от Гиса, считавшего зачатковые клетки
только предшественниками нейробластов, Шапер признает за ними совер-
шенно индифферентный характер и думает, что они являются бипотентными
родоначальниками и нейрональных и вспомогательных элементов нервной си-
стемы. Согласно Шаперу, индифферентные клетки сохраняются в мозгу дли-
тельное время, быть может, даже пожизненно, однако, начиная с какого-то,
точнее, неопределенного, момента, они утрачивают способность превращаться
в нейробласты, сохраняя свое камбиальное значение в отношении нейроглии
Точку зрения Шапера в настоящее время разделяет большинство нейро-
гистологов и особенно нейропатологов, причем вместо термина «зачат-
ковые» или «индифферентные» клетки теперь большей частью пользуются
новым обозначением, предложенным для них Бейли и Кашингом (1936),
медуллобласты.
Развитые Шапером взгляды не свободны, однако, от очень серьезных
возражений. Прежде всего, мне кажется, нет достаточных оснований гово-
рить о субституции первичного эпендимного опорного аппарата вторичным
астроцитарным. Кроме того, точка зрения на зачатковые клетки, как на
бипотентные родоначальники нейробластов и спонгиобластов, основы-
вается в значительной степени на допущении, что спонгиобласты, подобно
нейробластам, утрачивают способность к размножению митотическим путем.
В отношении спонгиобластов и даже дифференцированной нейроглии это
предположение (как показывают наши экспериментальные данные, разби-
раемые ниже) не соответствует действительности.
Локализация митозов у внутреннего просвета нервной трубки, отме-
ченная, повидимому, впервые Альтманом (1881), особенно подчеркнутая
Раубером (1892) и детально изученная Гисом (1890), является очень харак-
терной особенностью ранних стадий развития нервной системы. Круглой
формы клетки, находящиеся на разных стадиях митотического деления и
располагающиеся у самого центрального канала, Гис обозначил как зачат-
ковые клетки и считал их особой клеточной формой, которая дает в дальней-
шем нейробласты и качественно отличается от элементов, сохраняющих
Экспериментальный метод в гистологии
239
эпителиоподобный вид. Эти же клетки, обозначенные затем Шапером,
как индифферентные, отличаются, по данным этого автора, от эпителио-
подобных элементов, а затем спонгиобластов теми же внешними признаками.
Вместе с тем остается совершенно недоказанным, что клетки, делящиеся
митозами, никогда не переходят, изменяя свой внешний вид, в интеркине-
тическое состояние, и, наоборот, элементы, имеющие вид призматических
клеток эпителия или спонгиобластов, округляясь и приобретая характер
зачатковых клеток, не могут переходить в митотическое состояние. Более
того, такое предположение представляется даже малоправдоподобным.
В ткани любого органа, развивающейся из одного источника и содержащей
малодифференцированные камбиальные элементы, они всегда находятся
частью в интеркинетическом, частью в митотическом состоянии и лишь
постепенно дифференцируются, ослабляя или даже утрачивая свою мито-
тическую активность. Только в извитых семенных канальцах половозре-
лого яичка имеются неделящиеся митозом сертолиевы и делящиеся поло-
вые клетки на разных стадиях развития. Те и другие представляют собой,
однако (в соответствии с наиболее обоснованными данными), два совершенно
различных и не переходящих друг в друга клеточных типа. Что касается
собственно половых клеток, то часть сперматогониев также можно всегда
видеть в состоянии интеркинеза.
Поэтому гораздо более вероятным является предположение, что в нерв-
ной трубке, на первых стадиях ее развития, имеется лишь одна мало диф-
ференцированная клеточная форма, которая в состоянии интеркинеза имеет
вид «медуллярного эпителия», а в митотическом состоянии приобретает
особенности «зачатковых» или «индифферентных» клеток. Затем часть
клеток, находящихся в состоянии интеркинеза, переставая делиться,
дифференцируется в нейробласты, которые появляются, по данным Гелъда
(1909), уже на очень ранних стадиях развития и отличаются по своим цито-
логическим особенностям при специальных методах окраски. Нейробласты
являются уже необратимо дифференцирующимися элементами. Прочие
клетки «медуллярного эпителия» частью дают начало также необратимо
дифференцирующимся спонгиобластам, которые, однако, не утрачивают спо-
собности к делению и, переходя в митотическое состояние, уподобляются
делящимся «индифферентным» клеткам. Вопрос о том, являются ли наи-
менее дифференцированные элементы нервной трубки на поздних стадиях
ее развития только предшественниками нейробластов или также и следующих
поколений спонгиобластов, остается нерешенным.
Некоторые данные, касающиеся эпендимы низших позвоночных, также
говорят против точки зрения Гиса и Шапера на зачатковые или индифферент-
ные клетки, как на особую клеточную форму, которая, будто бы, только
одна и сохраняет способность к пролиферации и дифференцировке в те
или иные элементы нервной системы. По данным Студнички (1900), эпен-
дима взрослых костистых рыб способна к пролиферации и образованию
глиальных и нервных элементов.
То же самое справедливо, по наблюдениям Агдура (1920), в отношении
эпендимы амфибий и даже новорожденных млекопитающих. Симоэс (1925>
240
Глава III
также отмечает, что эпендима амфибий способна к пролиферации, а Вейс-
фейлер (1924) наблюдал резко выраженные регенеративные процессы
в головном мозгу аксолотлей и тритонов после оперативного удаления
значительных масс ткани мозга. Все это говорит за то, что зачатковые клетки
представляют собой малодифференцированные элементы в состоянии митоза,
а не особые камбиальные элементы нервной трубки. В состоянии интер-
кинеза те же камбиальные клетки могут иметь вид эпендимы.
Все приведенное выше говорит, во всяком случае, против точки зрения
Шапера, что астроцитарная нейроглия рыб и, особенно, высших позвоноч-
ных является новой тканью, заместившей собой эпендимную глию примитив-
ных форм. Несомненно, астроцитарная глия не субституировала, а развилась
преемственно из эпендимной глии предков.
Принятое в настоящее время подразделение нейроглии на ряд различных
и как бы отделенных друг от друга китайской стеной клеточных форм произ-
водит впечатление безжизненной и формальной схемы, по моему глубокому
убеждению, не соответствующей действительному положению вещей.
Против ее правильности говорит ряд хорошо известных фактов даже из
области чисто описательной нейрогистологии. Как уже отмечалось выше,
между протоплазматической и волокнистой глией имеются переходные формы.
Это, возможно, указывает на то, что они просто находятся на разном уровне
дифференцировки. Отмеченный уже давно Гирке (1885) своеобразный процесс
кератинизации глиальных элементов нельзя не сопоставить с этими поздней-
шими наблюдениями.
Элементы, обозначаемые как олигодендроглия, при ближайшем рас-
смотрении обнаруживают настолько большое разнообразие, что их отне-
сение к одной и той же категории клеточных форм, четко отграниченной от
других глиальных элементов, представляется малооправдапным и, пожалуй,
доказывается лишь тем, что клетки, относимые к олигодендроглии, более
или менее элективно импрегнируются, если придерживаться определен-
ного рецепта обработки препаратов, непригодного для демонстрации макро-
глии или микроглии. В сером веществе центральной нервной системы эле-
менты олигодендроглии оправдывают свое наименование, так как действи-
тельно обладают небольшим клеточным телом и немногими слабо ветвя-
щимися отростками. Так наз. интерфасцикулярная олигодендроглия
белого вещества, наоборот, демонстрирует все переходы к чрезвычайно
сложно разветвленным крупным элементам, форме тела которых мог бы,
так сказать, позавидовать любой астроцит. Элементы интерфасцикулярной
глии в настоящее время принято сближать со шванновскими клетками.
Наконец, многие авторы отмечали, что в ряде случаев удается обнаружить
любые переходные формы между типичной олигодендроглией и макроглией.
Все это говорит, безусловно, против принятого резкого разграничения
олигодендроглии и макроглии даже при обычных или нормальных условиях
существования. Так же затруднительно, по данным других авторов, при
ближайшем рассмотрении отграничить олигоглию от микроглии. Прежде
всего это относится к элементам типа сателлитов серого вещества, отно-
симых частью к олиго-, частью к микроглии. Кажущиеся столь характер-
Экспериментальный метод в гистологии
241
ними морфологические особенности этих элементов, в том виде, как они
изображаются для педагогических целей, демонстрируются далеко не
всегда и не везде. Кроме того, небезызвестно, что особенно красивые микро-
глиоциты получаются не на препаратах нормального человеческого мозга,
а при некоторых патологических процессах. При других патологических
процессах, наоборот, изменения в микроглии и олигоглии, по некоторым
данным, обнаруживают настолько большое сходство, что авторы, стоящие
на точке зрения мезенхимного происхождения микроглии, склонны при-
писывать его и олигодендроглиоцитам (Белецкий, 1931, 1932, 1938), отгра-
ничивая их тем самым очень резко от макроглии. Если сопоставить эги
немногие разноречивые данные, то, мне кажется, уже на этом основании
с несомненностью выступает несоответствие созданной стройной и вместе
с тем неподвижной схемы и конкретной действительности со всеми выте-
кающими последствиями.
Так или иначе, в головном и спинном мозгу позвоночных к эпендимо-
глиальному тканевому типу относятся, кроме собственно эпендимы и раз-
личных клеточных форм, описываемых в составе нейроглии и развивающихся
из нервной пластинки зародыша, также покровные клетки сосудистых
сплетений, имеющие характер однослойного эпителия. Они представляют
собой, строго говоря, эпендиму стенки тех участков нервной трубки, кото-
рые не развились в вещество мозга и, сохранив очень примитивные в гисто-
логическом отношении черты, вместе с тем специализировались в особом
направлении и приобрели железистую функцию.
Высокоспециализированным участком центральной нервной системы
является сетчатая оболочка органа зрения с т. наз. зрительным нервом
или, вернее, зрительным пучком. Сетчатая оболочка глаза и общее строение
глазного яблока обнаруживают у позвоночных очень высокий уровень
организации и поразительное сходство у представителей разных классов.
Заметные в ряду позвоночных эволюционные изменения гистологических
структур этого органа касаются лишь отдельных деталей. Пути филоге-
нетического становления органа зрения почти совершенно неизвестны,
так как между т. наз. глазками Гесса, разбросанными в нервной трубке
ланцетника, и глазом простейших позвоночных, круглоротых, имеется
огромный разрыв, который лишь в малой степени заполняется, если при-
влечь для сравнения неинвертированные париэтальные глаза, возможно,
представляющие собой сохранившийся более примитивный тип строения
фоторецепторных органов. Очевидно, прослеживающееся у позвоночных
эволюционное усложнение зрительной функции происходило в основном за
счет усложнения других, связанных с ней, частей нервной системы и лишь
в относительно незначительной степени за счет самого фоторецепторного
аппарата. Филогенетические ряды усложняющихся органов зрения, кото-
рые можно изучить на примерах отдельных типов беспозвоночных (хотя бы
моллюсков), не дают, разумеется, представления о том, как это в действи-
тельности происходило у предков современных позвоночных. Фоторе-
цепторная часть сетчатки построена по типу коркового центра и содержит,
кроме нейронов, все известные формы нейроглии. Радиальные мюллеров-
16 Хлсншн—-449
242
Глава III
ские волокна обычно трактуются как особая разновидность макроглии,
хотя они, пожалуй, в большей степени соответствуют специализированным
эпендимным элементам, участвуя в образовании как наружной, так и вну-
тренней пограничной перепонки. Зрительные клетки, обычно обозначавшие-
ся как нейроэпителиальные, по своему положению в сетчатке тоже пред-
ставляют собой эпендиму, специализированную, однако, в другом, фото-
рецепторном, направлении. Некоторые авторы, основываясь на функцио-
нальных особенностях клеток палочек и колбочек, трактуют их как ней-
роны, несмотря на веские возражения против этого, которые, например,
приведены в последнем издании руководства Кёлликера. С этой точки зрения,
принятой также Заварзиным (1913, 1925, 1941), фоторецепторные клетки
представляют собой первый нейрон зрительного пути. Слой клеток палочек
и колбочек играет в развитии сетчатки роль камбия, в котором, как это
подробно изучил в последнее время Винников (1939, 1941, 1943), митозы
встречаются у кролика еще после рождения. Плащевому ядерному
слою других корковых центров в развивающейся сетчатке соответствует
ганглиозный слой, внутренний ядерный слой возникает непосредственно
из малодифференцированных клеток эпендимы (Зеефелъдер, 1910; Р. Кахаль,
1909— 1911, и др.). Иными словами, корковому центру в сетчатке соответ-
ствует только ганглиозный слой, но не вся сетчатка в целом, как это обычно
говорится. У края зрительной части сетчатки мюллеровские волокна посте-
пенно переходят в эпителиоподобный внутренний листок ее слепой части,
не пигментированный в цилиарной и пигментированный у неальбиносов
в радужинной областях. У зрачкового края радужины внутренний листок
глазного бокала переходит в наружный пигментный (у неальбиносов)
слой. Последний состоит из своеобразных, миоэпителиальных клеток задней
поверхности радужины, т. наз. пигментового слоя цилиарной области
и отличающегося от него рядом особенностей пигментного слоя зрительной
части сетчатки или тапетум. Все эти элементы сетчатки являются, по сути
дела, дифференцированной различным образом эпендимой.
Зрительные пучки представляют собой участки белого вещества централь-
ной нервной системы и состоят у млекопитающих из пучков аксонов ган -
глиозных клеток сетчатки и нейроглии, в которой, по имеющимся данным
(Маркезани, 1926; Кон и Мекмиллен, 1932), имеются приблизительно в рав-
ном количестве волокнистая астроглия и олигодендроглия. Олигодендро-
глия представлена сложноразветвленными клетками, которые обнаруживают
несомненное сходство с астроцитами и при перерождении зрительных пучков,
повидимому, превращаются в них (Кон и Мекмиллен). Микроглия пред-
ставлена небольшим количеством клеток, которые выявляются с трудом.
Гистогенез нейроглии зрительных пучков происходит своеобразно, путем
непосредственного превращения в нее пролиферирующих элементов эмбрио-
нальной «эпителиальной» стенки глазного стебелька. Если стоять на точке
зрения Шапера (1897), то следовало бы считать, что нейроглия зрительных
пучков млекопитающих соответствует примитивной эпендимной глии лан-
цетника и круглоротых, но не субституирующей ее астроцитарной глии
других отделов центральной нервной системы высших позвоночных, что
Экспериментальный метод в гистологии
248
является маловероятным и говорит лишний раз против этой кон-
цепции.
Работа Студничка (1897), посвященная сравнительному изучению зри-
тельного нерва различных позвоночных, дает интересный материал по фило-
гистогенезу его глиального аппарата. У миноги имеются наиболее прими-
тивные отношения. Хотя просвет глазного стебелька у нее исчезает довольно
рано, эпендимные клетки образуют сплошной тяж, расположенный централь-
но. Нервные волокна располагаются в виде хорошо отграниченного пери-
ферического слоя между базальными отростками эпендимных клеток,
образующих наружную глиальную мембрану. Аксиальный эпендимный
тяж находится в непосредственной связи с эпендимной выстилкой сетчатки.
Соединительная ткань внутрь зрительного нерва не проникает. У части
хвостатых амфибий также имеется центральный клеточный тяж, у других
форм он во взрослом состоянии разделяется на несколько параллельных
тяжей, а у бесхвостых глиальные элементы приобретают диффузное распо-
ложение. У двудышащих и кистеперых рыб центральный клеточный тяж
сохраняется лишь в проксимальной части зрительного нерва. Затем нерв
подразделяется соединительной тканью на пучки с центрально расположен-
ными глиальными элементами. У ганоидов, селяхий и костистых рыб имеет-
ся диффузная глия при более или менее сложном подразделении нерва
на пучки. У рептилий и птиц глиальные элементы приобретают харак-
терное для млекопитающих расположение продольными рядами.
Париетальные глаза у ряда низших позвоночных, а из рептилий у гат-
терии, как известно, имеют строение и в какой-то степени значение фото-
рецепторных органов (Студничка, 1905). Причем у одних форм сохраняется
в большей степени пинеальный, у других — парапинеальный глаз. Под-
вергаясь рудиментации, пинеальный глаз превращается в т. наз. шишко-
видную железу, которая у птиц имеет действительно железистую структуру
с трубчатыми ходами, выстланными эпителиоподобными клетками, сравни-
мыми с эпендимой. У млекопитающих главная масса эпифиза построена
из клеток, сходных с астроцитами (Ортега, 1932), среди которых имеются
нерезко отграниченные сплошные тяжи или трубчатые ходы, образованные
сходными с эпендимой клетками с большим или меньшим количеством
пигментных включений (И. Хлопана, 1941, и неопубликованные данные).
На примере эпифиза, таким образом, хорошо прослеживается очень своеоб-
разное эволюционное изменение определенной тканевой структуры.
Купффер (1893— 1900,1903) уже давно открыл, что у позвоночных в об-
разовании органов чувств и периферической нервной системы, помимо
нервной пластинки, принимают’ участие особые утолщения эктодермы,
т. наз. плакоды. Плакоды образуются в непосредственном соседстве
с нервной пластинкой или на некотором расстоянии от нее и могут рас-
сматриваться как ее части, в большей или меньшей степени обособившиеся.
Часть плакод развивается в дальнейшем по типу центральной, другая
часть — периферической нервной системы. Участие плакод в развитии
черепномозговых ганглиев подтверждается многочисленными новейшими
исследованиями (Ван-Кампенгаут, 1939; Грот, 1938, 1939, и др.).
*
244
Глава III
Орган обоняния образуется из обонятельной плакоды, соответствующей
самому переднему участку нервной пластинки (Кольмор, 1927; Грот, 1938).
В отличие от всех других соответствующих образований большая часть
обонятельной плакоды остается в составе эктодермального покрова тела,
образуя обонятельные части слизистой оболочки носа, а у низших позво-
ночных — выстилку обонятельных ямок или мешков. Плакода дифферен-
цируется в эпителиальный многорядный пласт с находящимися в его составе
обонятельными клетками, образующими своими базальными отростками
волокна обонятельного нерва. В ряде случаев описывалось образование
скоплений чувствительных нейронов т. наз. обонятельного ганглия путем
выклинивания из состава плакоды отдельных элементов. Обычно опорные
клетки выстилки обонятельной слизистой оболочки рассматриваются просто
как эпителий. Однако, учитывая их способ развития и тесные взаимоот-
ношения с рецепторными обонятельными клетками, их. можно с тем же
правом трактовать как участок эпендимы, оставшийся на поверхности тела.
К сожалению, до настоящего времени мы не располагаем еще эксперимен-
тальными данными, которые помогли бы установить, в какой степени эти
элементы обнаруживают, в отношении своих гистобластических потенций,
сходство с эпидермальным или эпендимо-глиальным тканевым типом.
Не исключена возможность, что т. наз. эпителиальные клетки обонятель-
ной части слизистой оболочки носа займут, в силу своей особой специали-
зации и вместе с тем очень примитивных эпендимных черт, некоторое про-
межуточное положение. Являются ли обонятельные клетки настоящими
нейронами или элементами, сравнимыми со зрительными, пока с достовер-
ностью сказать нельзя.
Слуховая плакода при своем возникновении не может быть отграничена
от нервной пластинки и является первоначально, по существу, ее расширен-
ной боковой частью. Уже позже, при замыкании нервной трубки, слуховая
ямка, а затем слуховой пузырек превращаются в отдельные, первоначально
не связанные с зачатком центральной нервной системы, образования {Поли,
1897; Ретиг и Бругш, 1902; Грот, 1939, и др.). Ряд авторов подчеркивал
или хотя бы изображал на рисунках поразительное сходство строения стенки
слухового пузырька и нервной трубки. И в том и в другой замечаются
многочисленные митозы в округлившихся клетках, расположенных у про-
света, и призматические эпителиальные клетки, достигающие значительной
длины, с ядрами, расположенными ближе к периферии. Иными словами,
слуховые пузырьки имеют сначала совершенно типичное строение зачатка
центральной нервной системы. В течение дальнейшей дифференцировки
выстилка лабиринта становится на большей части своего протяжения тон-
кой и однорядной, сохраняя утолщенные места в рецепторных участках.
Этим утолщенным местам свойственна очень характерная для них «обратная
многорядность» совсем иного типа, чем обычная многорядность эпидер-
мальных тканей. «Обратная многорядность» эпителиоподобного пласта
заключается в том, что он состоит из длинных клеток опорного значения
и коротких чувствительных клеток, находящихся между их апикальными
концами и не достигающих подстилающей ткани. С такого рода структурами
Экспериментальный метод в гистологии
245
мы не встречаемся при обычных условиях в других местах организма.
По новым данным (Грот, 1939), основная масса зачатка статоакустического
ганглия образуется за счет элементов, выселяющихся из слухового пузырька,
а не из ганглиозной пластинки, как это описывалось прежде. Сходная
с соединительной тканью часть перепончатого лабиринта может быть сопо-
ставлена с мягкой пиоарахноидальной мозговой оболочкой глазного яблока,
включая ее суправаскулярный слой. Перилимфатические пространства
ограничены слоем плоских эндотелиоподобных клеток, подобно субарахно-
идальному, а также, повидимому, субдуральному пространствам. Наконец,
в определенных участках стенки перепончатого лабиринта регулярно встре-
чаются типичные хроматофоры {Гассе, 1873; Ретциус, 1884; Александер,
1901), сходные с пигментными клетками сосудистой оболочки глаза и неко-
торых участков собственно мозговых оболочек. Последний, очень интересный
сам по себе, факт в настоящее время малоизвестен, хотя и приводится в руко-
водстве Кёлликера. Мне, лично, также пришлось убедиться на кроличьем
материале в правильности этих указаний. Кроме того, следует напомнить,
что у асцидий статический пузырек располагается в центральной нервной
системе. У личинки асцидии Botryllus schlosseri в центральной нервной
системе описан своеобразный орган двойственного значения, играю-
щий одновременно роль фоторецептора и статорецептора {Грейв и
Вудбридж, 1934).
Наконец, есть указания на то, что часть слухового нерва по своему
строению отличается от периферических нервов, обнаруживая известное
сходство со зрительными пучками. Все эти довольно разнообразные
факты говорят за то, что мы вправе филогенетически рассматривать орган
слуха и равновесия позвоночных как обособившийся участок центральной
нервной системы, а его эпителиоподобную выстилку как специализиро-
вавшуюся в особом направлении эпендиму. Вместе с тем это является
дальнейшим аргументом в пользу того, чтобы рассматривать все плакоды,
наравне с самой нервной пластинкой и ее валиками, как разные части,
по существу, единого нервного зачатка.
Наибольшие трудности в этом отношении представляет хрусталиковая
плакода, развивающаяся в совершенно своеобразную часть органа зрения
позвоночных. В отношении хрусталика следует также подчеркнуть, что его
зачаток на ранних стадиях развития имеет, подобно слуховому пузырьку,
большое сходство с зачатком нервной системы в отношении общей формы
и расположения клеток и первоначальной локализации митозов. Циннова
связка хрусталика, по данным Агабабова и Арнштейна (1904), имеет неко-
торые тинкториальные особенности глиальных волокон. Ткань хрусталика
я также склонен включить в состав эпендимо-глиального типа, хотя ее
гистобластические потенции изучены еще недостаточно, а некоторые факты
из области экспериментальной эмбриологии в настоящее время с трудом
согласуются с этим предположением.
Вопрос о тканевой природе вспомогательных элементов периферической
нервной системы и, в первую очередь, шванновских клеток до настоящего
времени еще вызывает споры, несмотря на то, что их нейроглиальный харак-
246
Гпава III
тер, по существу, едва ли может серьезно оспариваться. Как это уже отме-
чалось выше, классики гистологии Лейдиг и Кёлликер были склонны рас-
сматривать шванновские клетки и сателлиты периферических нейронов
как элементы соединительной ткани, имеющие характер «ложного эпителия»,
или эндотелия. Первым, кто ввел в науку понятие периферической нейро-
глии, был Фритиоф Нансен (1888). Несмотря на многочисленные очень
разнообразные и убедительные доводы в пользу признания сателлитов
и шванновских клеток за особого рода специфические элементы перифери-
ческой нейроглии, качественно отличные от клеток соединительной ткани,
более старая противоположная точка зрения на них, как на элементы соеди-
нительной ткани, все еще имеет приверженцев. Эта точка зрения опирается,
в первую очередь, на то, что шванновские клетки способны при эксперимен-
тальных и патологических условиях, округляясь, превращаться в т. наз.
зернистые или «сетчатые» клетки, участвующие в разрушении миэлина.
Кроме того, некоторые авторы считают очень вероятным, что шванновские
клетки образуют, подобно элементам соединительной ткани, аргирофиль-
ные волокна межклеточного вещества (Лейдлоу, 1930; Нажотт, 1932).
Следует отметить, что, как это ни удивительно, до последнего времени
сохранились также приверженцы третьей точки зрения на шванновские
клетки. Они считают, что шванновские клетки представляют собой пери-
ферические нейробласты, образующие при эмбриональном развитии и при
регенерации отдельные участки осевого цилиндра. Согласно этому, в настоя-
щее время совершенно неправдоподобному и опровергнутому прямыми
наблюдениями взгляду, аксон периферического нерва представляет собой
не разросшийся в длину отросток нейрона (Ремак, 1862; Гис, 1883; Кёл-
ликер, 1900; Рамони Кахаль, 1909—1911, 1928), а цепочку вторично соеди-
нившихся друг с другом сегментов, образованных отдельными шваннов-
скими клетками (Шванн, 1899; Бальфур, 1878; Апати, 1897; О. Шульце,
1905; Шпильмейер, 1922). Оставим в стороне вопросы о тонком строении
периферических нервов и взаимоотношениях шванновских клеток, аксона
и миэлина, которые еще представляются во многих отношениях спорными
и неразрешенными окончательно, поскольку они не имеют прямого отно-
шения к нашей теме.
Классические исследования Гаррисона (1906, 1907/8) показали с полной
очевидностью, что образование аксонов, как отростков нейрона, не связано
со шванновскими клетками и что последние образуются не из окружающей
первоначально аксон мезенхимы, а из ганглиозной пластинки. Последние
прижизненные наблюдения Спейделя (1934) над ростом и регенерацией
нервных волокон в хвосте головастика подтвердили установленные Гар-
рисоном факты и показали воочию, что шванновские клетки, активно дви-
гаясь, лишь вторично накладываются поверх выросшего аксона.
Вместе с тем следует подчеркнуть, что установление генетической связи
шванновских клеток с нервным зачатком само по себе еще отнюдь не гово-
рит за их стойкие и специфические отличия от элементов соединительной
ткани, если придерживаться неоднократно упоминавшегося тезиса клас-
сической гистологии, согласно которому способ развития ткани не опре-
Экспериментальный метод в гистологии 247
деляет ее гистологических особенностей. Решение этого вопроса целиком
зависит от результатов экспериментально-гистологического анализа.
В отношении гистогенеза различных чувствительных периферических
ганглиев и нервов имеются различия в том отношении, что они или разви-
ваются из ганглиозной пластинки, или (в головной области) при большем
или меньшем участии плакод (например тройничный, лицевой, языкогло-
точный и блуждающий нервы; Ван-Кампеигаут, 1929), или даже целиком
за счет них (например слуховой нерв).
С вопросом о тканевой природе шванновских клеток теснейшим образом
связан также вопрос о вспомогательных элементах концевых аппаратов:
двигательных бляшек, которые, по данным Лаврова (1939), принадлежат
целиком нервному волокну, а не являются также частью саркоплазмы
т. наз. клеток внутренней колбы различных чувствительных окончаний.
Нейроглиальная природа всех этих образований представляется в настоящее
время более чем вероятной. На это уже давно указывал, между прочим,
Гельд (1909), говоривший (стр. 244): «Еще остается изучить, путем каких
именно изменений из конечных периферических глиальных клеток обра-
зуются определенные и особые клетки т. наз. периферических конце-
вых нервных аппаратов, как, например, материал бляшки, различных
осязательных телец и так далее». Вместе с тем сам собой возникает и даль-
нейший вопрос: о природе клеток, находящихся в составе эпидермальных
покровных пластов и образующих, например, вкусовые почки, различные
чувствительные бугорки низших позвоночных, меркелевские осязательные
клетки, элементы эймерова органа и тому подобное.
Являются ли эти различные образования измененными клетками эпидер-
мального характера или же они скорее сравнимы с вспомогательными
элементами нервной системы эпендимо-глиального типа, развивающимися
как производные нервные пластинки или плакод? Хотя некоторые факты из
области экспериментальной эмбриологии и говорят в пользу их развития
из клеток эпителиального покрова (Голыпфретер, 1933), ответ на этот
вопрос еще. однако, не является достаточно определенным и бесспорным.
Последнее, на чем мы должны остановиться, является ткань мозговых
и перинейральных оболочек и некоторых других образований, сходных
с ними в известных отношениях.
Мягкие мозговые оболочки еще со времени старых анатомогистологов,
как это отмечалось в нашем историческом введении, было принято сравни-
вать с серозными оболочками. Со времени классических исследований Кея
и Ретциуса (1873, 1875), их ткань рассматривалась как одна из разновид-
ностей рыхлой соединительной ткани, а плоские клетки, выстилающие
субарахноидальное и субдуральное пространства, как уплощенные соеди-
нительнотканные клетки типа фибробластов, которые чаще всего обознача-
лись термином ондотелий» или даже «мезотелий» (У ид, 1917),подобно покров-
ным клеткам серозных оболочек. Хотя Максимов (1927) и подчеркивает
неодинаковое развитие покровных клеток мозговых оболочек, сосудистого
эндотелия и мезотелия, он тем не менее допускает, что по своим биологи-
ческим особенностям покровные клетки мозговых и серозных оболочек,
248
Глава 111
возможно, соответствуют друг другу. Если вспомнить разобранную выше
точку зрения Максимова на мезотелий, то можно утверждать, что близость
покровных клеток мозговых оболочек и соединительной ткани представ-
лялась этому автору несомненной. Эта, представлявшаяся почти само-
очевидной, точка зрения до настоящего времени отражается в большей
части руководств и справочников по гистологии.
Однако изучение эмбрионального развития мозговых оболочек показало
в последнее время, что дело обстоит совсем не так, как это думал Уид (1917),
описавший возникновение субдурального и субарахноидального простран-
ства путем образования щелей и расслоения эмбриональной мезенхимы.
При изучении нормального гистогенеза мозговых оболочек Оверлеи (1922)
высказал предположение, что в их образовании принимают участие клетки,
выселяющиеся в окружающую мезенхиму из нервного зачатка и группи-
рующиеся в пластинку, которая затем расщепляется и дает начало покровным
клеткам мозговых оболочек. Эти выселяющиеся из эмбриональной нервной
системы элементы он обозначил как менингобласты, считая, что от них
зависят своеобразные специфические особенности опухолей мозговых
оболочек, получивших название менингобластомы. Эти же клетки, с точки
зрения Оберлена и Нордмана (1927), образуют т. наз. пахионовы (вернее,
паккионовы) грануляции.
Методами экспериментальной эмбриологии предположение Оберлена,
принятое Массоном (1923), было подтверждено Гарвеем и Бурром (1925),
а также Стоном (1926, 1929, 1932) и Равеном (1931, 1933). Согласно этим
данным, из ганглиозной пластинки наблюдается обильное выселение клеток
эктомезенхимы, дающих в дальнейшем не только обычные соединительно-
тканные образования, но в головной области и некоторые хрящевые
скелетные части.
С точки зрения классической гистологии, не признающей зависимости
свойств дифференцированной ткани от способа ее развития, эти особен-
ности гистогенеза различных «эктомезенхимных» образований головной
области не имеют принципиального значения. Поэтому отнесение покровных
клеток мозговых оболочек к эпендимо-глиальному тканевому типу может
быть обосновано только в случае обнаружения в них свойств, отличающих
их от соединительной ткани и сближающих с другими производными ней-
рального зачатка, что может быть достигнуто только экспериментально-
гистологическим анализом. Основываться же только на своеобразии опу-
холей, возникающих из мозговых оболочек, не достаточно убедительно
для решения этого вопроса, так как их гистогенез до настоящего времени
никем не прослежен. Работы нашей лаборатории, излагаемые ниже (Кан~
негисер, 1938, 1940), позволяют в настоящее время с несомненностью при-
числить загадочные покровные клетки мозговых оболочек к эпендимо-
глиальному тканевому типу, в составе которого они и рассматриваются
в этой книге.
От решения этого вопроса зависит также наш подход к целому ряду
других тканей: т. наз. десцеметову эндотелию роговицы глаза, который
вполне сравним с клетками, выстилающими субдуральное пространство,
Экспериментальный метод в гистологии 249
«эндотелию», покрывающему периферические нервные стволики, и, наконец,
даже к своеобразной «соединительной ткани» капсул различных чувстви-
тельных нервных окончаний (телец Фатер—Почини, Гольджи—Мадзони,
Мейснера и пр.). Хотя мы еще не располагаем в отношении всех этих
образований необходимым и достаточно убедительным экспериментальным
материалом, я позволю себе отнести их (пока, разумеется, предположи-
тельно) к тому же эпендимо-глиальному тканевому типу.
К скелетным образованиям, построенным по типу хрящевой ткани,
но возникающим, согласно ряду экспериментально эмбриологических дан-
ных из «эктомезенхимы» или, точнее, нервного зачатка, мы еще вернемся,
поскольку позволяет фактический материал, в главе о соединительной
ткани, так как по своей морфо-физиологической характеристике они отно-
сятся к последней.
Из этого краткого очерка ясно, что тканям, относимым нами к эпендимо-
глиальному типу, в составе организма позвоночных принадлежит огромное
значение. Эти ткани, по существу, столь же вездесущи и многообразны по
своим структурам, как и производные т. наз. «энтомезенхимы», дающей
различнейшие дифференцировки в составе нашего тела. Рассмотрение их
значительной части в составе морфо-физиологической группы эпителиев
оправдывается лишь в известной степени не только некоторыми особен-
ностями их структуры (даже нейроглию вместе с эпендимой возможно
рассматривать как разрыхленный эпителиальный массив), но и функциональ-
ными особенностями. Едва ли может подлежать сомнению, что «эпителиям»
эпендимо-глиального тканевого типа свойственно в большей или меньшей
степени особое разграничительное значение в составе организма, выра-
жающееся хотя бы в существовании т. наз. гемоэнцефалического или, если
угодно, десмоэнцефалического барьера.
По существу, однако, многие ткани эпендимо-глиального типа, как это
уже отмечалось, настолько своеобразны в отношении своей морфо-физиоло-
гической характеристики, что их отнесение к той или иной из четырех групп
тканей, предусмотренных классической гистологией, представляется в до-
статочной степени неестественным и натянутым.
К сожалению, наши знания о вспомогательных элементах нервной си-
стемы беспозвоночных еще слишком недостаточны для того, чтобы составить
себе достаточно удовлетворительное и мало-мальски полное представление
на их счет. В литературе имеются лишь совершенно разрозненные, очень
неполные и часто неопределенные данные. У кишечнополостных и низших
первичноротых (например турбеллярий) сравнимые с нейроглией вспомо-
гательные элементы нервной системы, возможно, отсутствуют. По крайней
мере, они еще никем, насколько мне известно, не описывались. У аннелид,
в ряду которых прослеживается погружение центральной нервной системы
из состава эпидермального покрова во внутреннюю среду организма, Рет-
циус (1891) и Кахаль (1904) описывали нейроглиальные элементы. Такие же
указания имеются и для пиявок (Апати, 1897; Якубский-, 1908; Санчец,
1909; Менклъ, 1908, 1909), в нервной системе которых даже различали глиаль-
ные клетки и волокна. Можно думать, что у тех аннелид, у которых нерв-
250
Глава III
ная цепочка еще находится в составе эпидермиса, прилегающие к ней
непосредственно эпидермальные клетки в некоторых отношениях
сравнимы с эпендимой. В составе погруженной под эпидермис нервной
цепочки, повидимому, уже имеются вышедшие вместе с ней из состава
эпидермиса нейроглиальные элементы, клеточное тело которых распола-
гается на периферии, а разветвленные отростки находятся в составе вещества
мозга. Такого рода нейроглия сравнима в некоторых отношениях с эпен-
димной глией позвоночных. Сходные отношения имеются, вероятно, в ту-
ловищном мозгу насекомых. В зрительных центрах последних {Рамон
и Кахаль, 1904; Заварзин, 1914; Р. Кахаль и Санчец, 1915) описывались
опорные элементы нейроглиального характера, внешнее сходство которых
с мюллеровскими волокнами сетчатки позвоночных подчеркнул Заварзин
(1925, 1941), а также расположенные поверхностно, сравнимые в некоторых
отношениях с эпендимой, элементы.
То же самое относится к зрительным центрам головоногих моллюсков
(Рамон и Кахаль, 1917), в которых отмечалось также присутствие отрост-
чатых элементов типа астроцитов. Смидт (1900, 1901) описывал у вино-
градной улитки особые сопровождающие и образующие оболочки клетки
в составе нервов, ганглиев и комиссур. Первоначально (1900) он сомневался
в тканевой природе описанных им элементов, считая, что они имеют, воз-
можно, соединительнотканный характер. В своей второй работе (1901)
он уже с определенностью говорит о них как о нейроглии.
Несколько более полные данные получил позже Якубский (1913, 1915)
в отношении пластинчатожаберных, брюхоногих и головоногих моллюсков.
Тело отдельных нейронов, достигающих у моллюсков очень крупных раз-
меров, нейроглиальные элементы окружают наподобие сателлитов, про-
никая даже в их протоплазму.
Этих и некоторых других данных, разбросанных в литературе, явно
недостаточно, чтобы составить себе представление о нейроглии беспозво-
ночных, хотя бы отдаленно соответствующее нашим сведениям на этот счет
у позвоночных.
Даже простое описательное и сравнительное изучение вспомогательных
элементов нервной системы у представителей различных типов и классов
животного мира представляет собой одну из увлекательных, но вместе
с тем очень трудных, задач будущего.
б) Нейроглия и эпендима спинного и головного мозга
Опыты с эксплантацией производились с различными тканями, относи-
мыми нами в настоящее время к генетически единому эпендимо-глиальному
типу. Однако до работы нашей лаборатории никто не сопоставлял получае-
мых в разных случаях картин и не отдавал себе отчета в том, что, несмотря
на очень различную исходную тканевую структуру, в условиях культур
эти ткани обнаруживали черты сходства и по целому ряду вполне стойких
признаков отличаются от тканей другого происхождения. Разбираемые
ниже данные показывают, что в настоящее время имеются все основания
Экспериментальный метод в гистологии
251
говорить об особом едином нейроглиальном (Хлопин, 1938) или эпендимо-
глиальном типе роста в его различных модификациях.
Большая часть работ, выполненных с помощью метода эксплантации,
начиная с классических исследований Гаррисона, произведена на эмбрио-
нальном материале, в составе которого, помимо совершенно индифферент-
ных нейробластов и более или менее дифференцированных нейронов, имеются
спонгиобласты, т. е. будущая нейроглия, и, особенно на более ранних ста-
диях развития, малодифференцированные камбиальные элементы нервной
трубки. Поэтому явления, наблюдаемые в таких эксплантатах, отличаются
большой сложностью. При культивировании частиц эмбриональной централь-
ной нервной системы куриных зародышей, представляющих собой излю-
бленный материал для опытов с тканевыми культурами, описывались более
или менее интенсивное прорастание нервных волокон, т. е. отростков ней-
робластов и эмбриональных нейронов, и выселение последних в зону роста,
рост и эмиграция фибробластообразных клеток и образование плоскостных
комплексных мембран (Оливо, 1927, 1928; Капель, 1929; Лазаренко, 1932;
Михалик, 1935, и др.). Культуры из нервной системы отличаются сильными
фибринолитическими свойствами и образуют полости разжижения. Вслед-
ствие этого периферические части могут отрываться от поверхности эксплан-
татов и оставаться связанными с неразжиженным ободком фибрина. Про-
растающие клетки и нервные волокна при этом нередко сильно растяги-
ваются между центральными кусочками и отставшим от его поверхности
фибрином или связанными с ним частями эксплантата.
Амебоидные клетки, появляющиеся в больших количествах в эксплан-
татах из эмбриональной центральной нервной системы и мозговой коры
новорожденных животных, по почину Костеро (1930), стали обозначаться
как микроглия, которая, по популярным взглядам Ортега, развивается
не из элементов самой нервной трубки, а из проникающих в нее, на извест-
ных этапах эмбрионального развития, мезенхимных клеток. Их внешнее
значительное сходство с настоящими мезенхимными макрофагами считалось
убедительным доказательством этой точки зрения (Ортега, 1932; Уэллс
и Кармикаэл, 1930; Михалик, 1935; Леви, 1934).
Получающиеся в культурах из эмбриональной центральной нервной
системы клеточные мембраны первоначально обозначались просто как
«эпителий» или «мозговой эпителий» (Оливо, 1927; Капель, 1927, 1929;
Фишер, 1930, и др.), причем никакого вопроса об их возможной специфич-
ности и отличии от мембран в культурах различных органов, содержащих
те или иные эпителиальные ткани, вообще не возникало. Такая точка зре-
ния соответствовала принятому до последнего времени в гистологии фор-
мальному взгляду, что все гистологические структуры, имеющие вид сплош-
ных клеточных пластов или трубок, представляют собой единую эпите-
лиальную ткань. С точки зрения Оливо, «эпителиальные мембраны» обра-
зуются за счет наименее дифференцированных камбиальных элементов
эпендимного слоя эмбриональной нервной системы. В противоположность
этому Капель считает элементы этих мембран нейробластами, утратившими
путем дедифференцировки свои специфические особенности, а Костеро
252
Глава III
принимает их за распластанные по поверхности плотного субстрата макро-
фаги. Верн (1930), изучая клеточные мембраны из эмбрионального куриного
мозга, обратил внимание на тот факт, что в протоплазме их клеток, при
более или менее длительном культивировании, удается в некоторых случаях
обнаружить волокна, напоминающие глиальные. Поэтому он, а за ним и
некоторые другие стали обозначать рост «мозгового эпителия», как рост
нейроглии. Указания на появление глиальных клеток в зоне роста без
сколько-нибудь подробного анализа их происхождения и особенностей
имеются и в некоторых других работах, посвященных культивированию
разных участков центральной нервной системы куриных зародышей.
В одной из своих новейших работ Бауэр (1938), изучая зону роста из
эмбриональной нервной системы куриных зародышей, описал образование
структур, напоминающих нитевидные отростки нейронов. Эти наблюдения
он объяснил тем, что «эпителиальная» зона роста в культурах эмбриональ-
ной нервной системы представляет собой не нейроглию, а недифференциро-
ванную ткань, элементы которой способны, в условиях культур, дифферен-
цироваться в нейрональном направлении.
Как это видно из приведенного краткого обзора, только макрофаго-
подобные клетки в эксплантатах из нервной системы сходным образом
трактуют все или почти все авторы, что же касается т. наз. «эпителиаль-
ных мембран», то относительно их тканевой природы высказываются со-
вершенно противоречивые и исключающие друг друга предположения,
причем наблюдаемые при культивировании картины не используются для
анализа тканевых свойств нейроглии.
Мы пошли в наших опытах по линии культивирования достаточно диф-
ференцированной нервной системы млекопитающих (кроликов), когда в ее
составе уже имеется настоящая нейроглия, но процесс миэлинизации еще
почти или совершенно не начинался.
При существующей методике опытов миэлин является в большинстве
случаев очень большой помехой тканевому росту вне организма. Большая
часть данных относится к новорожденным и молодым кроликам (возрастом
от одного до десяти дней). Кроме того, использовался и материал от кро-
личьих зародышей незадолго до рождения, который давал такие же кар-
тины роста. Для того чтобы, по возможности, исключить рост отростков
нейронов и их возможную, по имевшимся данным, эмиграцию, я прежде
всего остановился на изучении нейроглии белого вещества полушарий
мозга (Хлопан, 1941, и дальнейшие неопубликованные данные). В этих
опытах материалом для посева служили довольно тонкие пластинки, вы-
резанные из латеральной стенки боковых желудочков. Для дополнительного
изучения эпендимы, кроме того, эксплантировались частицы из дна чет-
вертого желудочка (Хлопан, 1941).
Культуры изучались до полуторамесячного срока жизни вне организма.
Разжижение фибрина и образование полостей, наполненных жидкостью,
были выражены в очень сильной степени. Нередко в течение первого вре-
мени после эксплантации обнаруживались лишь слабые признаки экстен-
сивного роста отдельными клеточными тяжами, которые при прогрессн-
Экспериментальный метод в гистологии
253
и, лишь в отдельных случаях, на первых стадиях
Рис. 26. Участок белого вещества из стенки бокового же-
лудочка мозга 9-дневного кролика с кровеносным капилля-
ром. Темные и светлые ядра глиальных элементов. Фикса-
ция— Ценкер-фсрмол, окраска — железный гематокси-
лин Гойденгайна. Сильное увеличение. (Оригинал.)
рующем разжижении среды быстро обрывались. Как правило, наблюдалось
также довольно интенсивное осыпание изолированных клеток, которые
или свободно плавали, приобретая шаровидную форму, в жидкости, ок-
ружающей центральный кусочек, или распластывались по поверхности
слюды. Эксплантаты, сжимаясь, заметно уменьшались в объеме и постепенно
округлялись в шарик таким образом, что их покрытая эпендимой поверх-
ность почти всегда оказывалась внутри. Периферическая, травмированная
при подготовке к посеву каемка центрального кусочка обычно дегенериро-
вала и при пересеве отмывалась от жизнеспособной части эксплантата вместе
с отдельными, сохранившимися в ее составе клетками. Обычно, по истече-
нии пяти-восьми дней
кул ьтив ирова н ия,
центральный кусочек
начинал образовы-
вать стелющуюся по
слюде, то очень тон-
кую и нередко разры-
хленную, то, особен-
но на дальнейших
стадиях, более или
менее массивную и
комплексную зону
роста. В некоторых
случаях по истечении
пятнадцати- двадцати
дней весь централь-
ный кусочек мозга,
энергично разраста-
ясь, как бы раство-
рялся в окружающей
его зоне роста. В
уменьшающемся в
размерах округлив-
шемся центральном кусочке происходил ряд интересных изменений. Уже
в течение первых и вторых суток вокруг глиальных ядер появлялись почти
незаметные в исходном материале, при обычных методах окраски, цитоплаз-
матические ободки, то неправильно закругленной или амебоидной формы,
то снабженные заостренными отростками (рис. 26, 27). Различия в размере
и окрашиваемости ядер постепенно сглаживались. С течением времени все
ядра приобретали вид более крупных и светлых ядер, свойственных эле-
ментам макроглии. В цитоплазме глиальных элементов в большинстве
случаев появлялась грубая зернистость. Делящиеся митозом клетки, как
это большей частью имеет место, округлялись. Весь кусочек в целом ста-
новился менее прозрачным. В части эксплантатов глиальные элементы все
больше и больше округлялись и, очень сильно переполняясь включениями,
полностью утрачивали взаимную связь. Такие изменения приводили вскоре
254
Глава 111
Рис. 27. Участок краевой зоны центрального кусоч-
ка эксплантата из стенки бокового мозгового желу-
дочка 1-дневного кролика на 2-й день культивирова-
ния. Выявление цитоплазматических тел глиальных
элементов отростчатой и закругленной амебоидной
формы с темно красящейся зернистостью. 2 митоза.
Такая же обработка. Сильное увеличение.(Оригинал.)
к полной дезинтеграции и гибели посеянных кусочков. В большей части
случаев при сохранении жизнеспособности культур изменения развивались
в другом направлении. На третий-пятый день находящиеся между глиаль-
ными элементами части эксплантата подвергались некоторому разжижению
и частичной резорбции, причем местами образовывались светлые полости.
Вместе с тем все отчетливее выявлялись сетевидно связанные друг с другом
отростками тела глиальных элементов, которые даже при обычных методах
окраски отчетливо выявляли свою типичную для астроцитов форму (рис.
28). Зерна, появившиеся первоначально в цитоплазме, вероятно, в резуль-
тате рассасывания и, быть
может, даже фагоцитоза
отмирающих и дегенери-
рующих частей экспланта-
та, исчезали. Вместо них
в цитоплазме астроцитар-
ных элементов начинала
обнаруживаться то более,
то менее отчетливая волок-
нистость, довольно элек-
тивно окрашивающаяся же-
лезным гематоксилином. В
конце первой или в тече-
ние второй недели, в ре-
зультате сближения кле-
точных тел глиальных эле-
ментов и полного исчезно•:
вения находившихся меж-
ду ними частей эксплан-
тата, весь кусочек приоб-
ретал строение компактно-
го клеточного массива с од-
нотипными, большей частью
светлыми ядрами, в протоплазме клеточных элементов которого в боль-
шинстве случаев образовался густой переплет грубых волокон, интенсивно
чернящихся железным гематоксилином (рис. 29). При соответствующих
специальных методах окраски в толще такого глиального массива могли
долгое время обнаруживаться незначительные остатки волокнистых соеди-
нительнотканных прослоек, отчетливо отличающиеся от элементов нейро-
глии. Такая структура глиального массива сохранялась до предельных
прослеженных сроков культивирования. Форма отдельных клеток в составе
таких глиальных массивов, вследствие неясности клеточных границ, не
могла быть с достоверностью установлена. Однако его образование из
описанной выше пространственной сети астроцитарных элементов и наблю-
давшиеся на более поздних стадиях картины прорастания заставляют ду-
мать, что отдельные элементы в составе массивов сохраняли такую же раз-
ветвленную форму, образуя сплошной переплет клеточных тел и отростков.
Экспериментальный метод в гистологии
255
Пробегающие и взаимно перекрещивающиеся во всех направлениях про-
странства глиальные волокна подтверждают эту точку зрения.
Плоскостные очень многообразные структуры зоны роста в своей сово-
Рис. 28. Участок центрального кусочка эксплантата из стенки
бокового мозгового желудочка 9-дневного кролика на 5-й день
культивирования. Сетевидно связанные друг с другом глиаль-
ные элементы типа астроцитов с волокнистостью в цитоплазме.
Митоз в округлившейся глиальной клетке. Такая же обработка.
Сильное увеличение. (Оригинал.)
купности резко отличались от описанных выше эпидермальных, энтеро-
дермальных и целонефродермальных мембран и пластов. Характеристика
структур ‘Зоны роста
нейроглии как эпите-
лиальных мембран ни
в какой степени не
отражает их специ-
фических особенно-
стей и даже просто
не соответствует дей-
ствительности. Сопо-
ставление общего ви-
да полученных мною
разрастаний с изобра-
жениями экстенсив-
Рис. 29. Участок центрального кусочка эксплантата из
стенки бокового мозгового желудочка 1-дневного кролика
на 40-й день культивирования. Весь центральный кусочек
превратился в сплошной глиальный массив с густым пере-
плетом толстых волокон. Такая же обработка. Сильное
увеличение. (Оригинал.)
ной зоны роста в экс-
плантатах из эмбрио-
нальной нервной си-
стемы куриных заро-
дышей в работах
некоторых других авторов (Оливо, 1927, 1928; Капель, 1929; Михалик, 1935;
Бауэр, 1938, и др.) обнаруживает большое сходство между теми и дру-
гими. Не может подлежать сомнению, что и другие авторы в своих опытах
256
Глава III
имели дело с типичными картинами нейроглиального роста, которые они,
однако, не подвергли достаточно тщательному гистологическому анализу
и поэтому не сумели правильно оценить, считая в большей части случаев,
что они имеют дело с таким же ростом эпителия, с каким приходится встре-
чаться, например, при эксплантации эпидермальных и энтеродермальных
тканей. Если различные разрастающиеся вне организма эпидермальные
ткани по совокупности своих превращений могут быть ясно отличены от
энтеродермальных, то
особенности нейрогли-
альных культур броса-
ются в глаза отнюдь не
меньше, а во многих
случаях даже еще резче.
Особенно мало даже по-
верхностного сходства
между культурами ней-
роглии и различных опи-
санных выше энтеро-
дермальных и целоне-
фродермальных тканей.
Плоскостные разраста-
ния нейроглии, как уже
отмечалось выше, имеют
весьма разнообразный и,
в своем многообразии,
очень типичный вид.
Это — или рыхлые сете-
видные структуры, или
комплексные тонкие
мембраны, или довольно
толстые массивы, или,
наконец, клеточные тя-
жи различной ширины
и длины (рис. 30, 31).
Рис. 30. Экстенсивные разрастания нейроглии стенки
бокового желудочка мозга 9-дневного кролика. Глиаль-
ный массив, переходящий в мембрану и клеточные
тяжи. 23-дпёвная культура. Формол, гематоксилин.
(Оригинал.)
Последние реже образуются непосредственно из центрального кусочка.
Большей частью в тяжи переходят периферические участки пластов или
мембран, что придает им во многих случаях очень своеобразный «лучистый»
или «рогатый» вид. В некоторых случаях значительная часть зоны роста
может состоять из клеточных массивов неправильных или звездчатых
очертаний, соединенных друг с другом тяжами или тонкими цитоплазма-
тическими перемычками. Лишь в относительно редких случаях форма
нейроглиальных клеток в зоне роста становится более или менее пра-
вильной, полигональной. Только состоящие из клеток такой формы
отдельные комплексные участки мембран заслуживают названия
эпителиальных. Значительно чаще нейроглиальные разрастания состоят
из элементов удлиненной, веретенчатой или неправильно звездчатой
Экспериментальный метод в гистологии
275
формы. Поэтому в толстых массивах зоны роста форма отдельных эле-
ментов прослеживается лишь с большим трудом и то далеко не всегда.
В разрыхленных тонких плоскостных разрастаниях, особенно на более
ранних стадиях культивирования, когда они встречаются чаще, все разно-
образие клеточных форм бросается в глаза с особой отчетливостью. В не-
рпе. 31. Глиальные комплексы и тяжи зоны роста, соединенные друг
с другом цитоплазматическими перемычками. Отдельные глиальные
клетки сложной формы с нитевидными отростками. 22-дневная куль-
тура из стенки бокового желудочка мозга 1-дневного кролика. Слабое
увеличение. Формол, гематоксилин. (Оригинал.)
которых случаях, как и элементы других тканей, малодифференцированные
нейроглиальные клетки могут приобретать универсальную фибробласто-
подобную форм)' и утрачивать в значительной степени свои морфологические
особенности. Состоящие из таких элементов рыхлые клеточные соединения
могут, как и в случае других тканей, приобретать поверхностное сходство
с соединительнотканными разрастаниями, отличаясь, однако, от них своим
возникновением из совершенно не свойственных соединительной ткани
других структур и своими дальнейшими превращениями.
Особенно в эксплантатах с незначительной энергией роста глиальные
элементы частью приобретают чрезвычайно своеобразную разветвленную
форму, образуя тонкие нитевидные отростки, придающие им астроцитарный
или даже сходный с нервными клетками вид (рис. 32, 33). Соединяясь друг
с другом или глиальными клетками иной формы своими отростками, такие
астроцитарные или «нейроноидные» элементы образуют в высокой степени
специфичные структуры, никогда не встречающиеся в тканях иного про-
исхождения. Не подлежит сомнению, что ряд авторов, описывавших при
культивировании эмбриональной нервной системы подчас очень интен-
сивную миграцию в зону роста нейронов (Леви, 1934: Михалик, 1935, и др.)
или дифференцировку в нейроны малодифференцированных клеток зоны
17 Хлопин—349
Глава III
258
роста {Бауэр, 1938), был введен в заблуждение такими «нейроноидными», но.
по сути дела, астроцитарными, структурами и описывал в качестве нейронов
нейроглиальные клетки.
Отсутствие в нашем исходном материале нейронов почти во всех, за
единичными исключениями, случаях и детальное изучение возникновения
таких «нейроноидных» клеток доказывают с полной несомненностью их
нейроглиальную природу. Не следует также забывать того, хорошо извест-
ного факта, что астроциты и при обычных условиях существования в орга-
Рис. 32, Разрыхленная глиальная мембрана зоны роста. Распластанные,
округляющиеся н астроцитарные элементы с нитевидными отростками.
11-дневная культура из стенки бокового желудочка мозга 5-дневного
кролика. Сильное увеличение. Формол, гематоксилин. (Оригинал.)
низме могут в значительной степени симулировать своей внешней формой
нейроны. То более, то менее тонкие прямые отростки различной длины
образуют также периферические нейроглиальные элементы клеточных мас-
сивов зоны роста, придающие им зачастую лучистый неровный контур
(рис. 34). В составе клеточных массивов, как это отмечалось, нейроглиаль-
ные клетки нередко приобретают вытянутую форму тела и ядра, образуя
в их составе ориентированные в одном направлении потоки. Перекрещи-
ваясь друг с другом в разных направлениях, такие потоки удлиненных клеток
придают местами зоне роста очень своеобразный характер. Клетки в со-
ставе массивов зоны роста могут обнаруживать то с большей, то с меньшей
отчетливостью нейроглиальную волокнистость цитоплазмы.
Упомянутые выше клеточные тяжи состоят из одного или нескольких
рядов нейроглиальных клеток, более или менее вытянутых в одном напра-
влении. На разных стадиях культивирования (как правило, в большей сте-
пени на более ранних) из состава центрального кусочка и из разных клеточ-
Экспериментальный метод в гистологии
25&
ных соединений зоны роста, изолируются отдельные клетки макрофаго-
образной формы, подобно тому, как это описывалось для разных других
тканей. При этом клеточное ядро обычно становится меньше и темнее.
Такие изолированные клетки, утрачивая связь с плотным субстратом и
свободно плавая в жидкости, совершенно округляются, а на плотном суб-
страте двигаются амебоидно, образуя псевдоподии. Через известное время
такие элементы, вновь изменяя свою форму, могут превратиться в отрост-
чатые или веретенчатые нейроглиальные клетки и войти в состав тех или
Рис. 33. Образование в зоне роста сложно разветвленных астро-
цитарных элементов. Один митоз. 12-дневная культура из стенки
бокового желудочка мозга 5-дневногс кролика. Сильное увеличе-
ние. Формол, гематоксилин. (Оригинал.)
иных клеточных соединений зоны роста. Обратимость таких изменений
клеток нейроглии, которая на основании визуальных наблюдений живых
культур или изучения фиксированных препаратов может лишь предпола-
гаться, доказывается воочию методом цейтраферной микрокиносъемки
(Галустян и Быстров, неопубликованные данные).
В некоторых случаях наблюдалось образование многоядерных глиаль-
ных плазмодиев.
Энергия роста нейроглиальных культур бывает не всегда одинакова.
Часто она бывает очень высока. В эксплантатах из мозга соединительная
ткань не прорастает вовсе. Митозы, которые на более ранних стадиях
культивирования встречаются и в центральном кусочке и в зоне роста,
позже обнаруживаются только в экстенсивно растущих нейроглиальных
элементах.
Изложенный фактический материал следует еще дополнить некоторыми,
правда еще недостаточными, наблюдениями над эпендимой мозговых же-
лудочков.
260
Глава III
При упомянутом выше скручивании и округлении эксплантатов из
стенки боковых желудочков внутри центрального кусочка образовывались
полости, окаймленные эпендимой и имеющие вид т. наз. «розеток». Базаль-
ные части эпендимных клеток без резкой границы терялись в окружающей
массе ткани мозга. В дальнейшем розетки переставали быть различимыми;
эпендимные клетки теряли реснички и базальные тельца, утрачивали свою
первоначально правильную форму и ориентировку, делаясь неотличимыми
от окружающих элементов нейроглиального массива, как бы растворяясь
в нем. Полость внутри розетки при этом исчезала.
Рис. 34. Край глиального пласта зоны роста с тонкими прямыми цито-
плазматическими отростками клеток. 19-дневная культура стенки
бокового желудочка 5-дневного кролика. Сильное увеличение. Фор-
мол, гематоксилин. (Оригинал.)
Эксплантированные частицы из дна ромбовидной ямки имели прибли-
зительно призматическую форму. Одна сторона таких эксплантатов была
покрыта слоем эпендимы, обнаруживающим оживленные мерцательные
движения ресничек. В таких эксплантатах эпендимный слой оставался
на поверхности и выявлял еще более резко выраженные фибринолитические
свойства, чем собственно нейроглия. Вследствие этого, явления экстенсив-
ного роста нейроглиальных элементов наблюдались лишь в редких слу-
чаях и притом только из непокрытых эпендимой участков поверхности
эксплантатов. Экстенсивный рост нейроглии принципиально ничем не от-
личался от описанного выше роста в культурах из полушарий мозга. Раз-
меры эксплантатов с течением времени умешшались, частью вследствие
распада периферических травмированных участков нейроглии, частью в
результате сжатия кусочков и их постепенного превращения в компактные
глиальные массивы. Форма эксплантатов становилась более или менее
шарообразной, а эпендимный слой мог с течением времени занять значитель-
ную часть наружной поверхности. Прижизненное наблюдение оставляло в
некоторых случаях впечатление, что эксплантаты как бы эпителизируются
разрастающимся по их свободной поверхности эпендимным покровом. Однако
в данном случае этот кажущийся процесс эпителизации зависит на самом
Экспериментальный метод в гистологии
261
деле не от активного движения эпендимного покрова, а от общего сжатия и
уменьшения объема эксплантата. К концу второй или в течение третьей не-
дели после эксплантации, а иногда и раньше, глиальный массив централь-
ного кусочка мог настолько уменьшиться в своих размерах, что большая
часть эксплантата оказывалась состоящей из эпендимы, которая в ряде
случаев продолжала обнаруживать мерцательное движение ресничек.
В цитоплазме клеток глиального массива, совершенно так же, как в эксплан-
татах из стенки боковых желудочков мозга, дифференцируется переплет гли-
альных волокон, достигающих значительной толщины. Клетки эпендимного
покрова сливаются без рез-
кой границы своими базаль-
ными концами с подстилаю-
щим его глиальным масси-
вом, составляя с ним одно
целое (рис. 35). Во многих слу-
чаях эпендима утолщалась,
вследствие удлинения верти-
кальных размеров ее клеток,
а ядра принимали многоряд-
ное расположение. Это свя-
зано с сильным уменьшением
объема глиального массива.
В эксплантатах из спин-
ного мозга новорожденных
и молодых кроликов наблю-
дались очень сходные изме-
нения в центральных кусоч-
ках и картины экстенсивного
роста (Хлопин, неопублико-
ванные данные). Центральные
Рис. 35. Покрытая мерцательной эпендимой часть
центрального кусочка 13-дневного эксплантата из
дна 4-го мозгового желудочка 4-дневного кролика.
Центральный кусочек превратился в сплошной
глиальный массив с густым переплетом волокон.
Ценкер-формол, железный гематоксилин Гейден-
гайна. Сильное увеличение. (Оригинал.)
кусочки тоже превращались в сплошные глиальные массивы с типичными
волокнами в цитоплазме клеток. В зоне роста наблюдались мембраны, пласты
и тяжи, сходного с описанными выше вида, которые имели отнюдь не эпи-
телиальный и не соединительнотканный, а нейроглиальный характер.
Также, каки в других случаях, нейроглиальные элементы округлялись
и изолировались, приобретая макрофагоподобный вид. Рыхлые плоскост-
ные соединения астроцитарных или «нейроноидных элементов» были выра-
жены чрезвычайно отчетливо как на более ранних, так и относительно поздних
стадиях культивирования (рис. 36). В отдельных астроцитарных элементах
звездчатой формы, образующих сложный переплет тонких ветвящихся
отростков, появлялась амитотическая перешнуровка и фрагментация ядер,
вследствие чего получались очень своеобразные дву- и многоядерные клетки.
В эксплантатах из спинного мозга, содержащих выстилку центрального
канала, удалось видеть очень демонстративные картины прорастания эпен-
димы. Эпендимные клетки, первоначально обнаруживающие мерцательные
движения ресничек, продвигались в зону роста, образуя пласты и тяжи,
262
Глава III
которые в течение некоторого времени удавалось отличать от окружающих
нейроглиальных разрастаний. С удалением от центрального кусочка про*-
росшие эпендимные клетки, изменяя форму и утрачивая реснички, пере-
ставали быть отличимыми от разрастающейся нейроглии и, повидимому,
сливались с ней в единую глиальную зону роста. В условиях моих опытов,
ориентированных на изучение нейроглии, прогрессивных изменений в
нейронах серого вещества спинного мозга, заметной регенерации их от-
ростков, а тем более активной миграции нервных клеток в зону роста не
наблюдалось.
Имеющийся в распоряжении лаборатории, пока еще ориентировочный
материал по эксплантации коры больших полушарий мозга (И. Хлопина)
Рис. 36. Соединения астроцитарных («нейроноидных») клеток раз-
рыхленной плоскостной зоны роста. Образование 2- и 4-ядерных
клеток в результате амнтотических перешпуровок ядра. 21-дневная
культура из спинного мозга 2-дневного кролика. Формол, гемато-
ксилин. Сильное увеличение. (Оригинал.)
демонстрирует на этом объекте сходные в общих чертах изменения нейро-
глии и образование характерной нейроглиальной зоны экстенсивного роста.
Большого внимания заслуживают наблюдения над ростом и превраще-
ниями вне организма покровных клеток (эпендимы) сосудистых сплетений
мозга (Михайлов, 1938, и неопубликованные данные). Оказывается, что
эти элементы, имеющие при обычных условиях существования в организме
вид изопризматического железистого эпителия, в эксплантатах обнаружи-
вают чрезвычайно большое сходство с нейроглией центральной нервной
системы. Если они образуют тонкие однослойные мембраны путем мас-
сового активного движения по слюде без сколько-нибудь энергичной про-;
лиферации, их элементы сильно уплощаются, сохраняют более или менее
правильные многоугольные очертания и свой эпителиообразный характер,
В других случаях, особенно когда рост покровных клеток сопровождается
энергичной пролиферацией митотическим путем, они образуют типичные
нейроглиальные, а отнюдь не эпителиальные структуры, более или менее
Экспериментальный метод в гистологии
263
толстые пласты, массивы и тяжи, состоящие большей частью из клеток
удлиненной формы и нередко обнаруживающие характерные перекресты
ориентированных в разных направлениях клеточных потоков. В отличие
от собственно нейроглии, описанной выше, покровные клетки не давали
астроцитарных или «нейроноидных» элементов. При разрыхлении комплекс-
ных нейроглиальных структур зоны роста местами образовывались и мало-
характерные сетевидные клеточные соединения (если их рассматривать
вне связи с другими образованиями), а также макрофагоподобные элементы.
Клеточные тяжи в этих эксплантатах имели разный калибр и достигали
значительной длины. Массивные скопления клеток в зоне роста приобретали
к некоторых случаях концентрическое строение, хотя и не очень отчетливо
выраженное, вследствие уплощения наслоенных друг на друга элементов.
Явлений эпителизации поверхности кусочка в эксплантатах из сосудистых
сплетений не наблюдалось.
Сравнение друг с другом культур собственно нейроглии и покровных
клеток сосудистых сплетений демонстрирует, несмотря на некоторые
различия, очень большое сходство гистобластических потенций тканей,
неодинаковых по своей морфо-физиологической характеристике, но по
происхождению близко родственных друг другу, т. е. как раз то, что
совершенно не учитывала и не могла учитывать классическая гисто-
логия.
В отношении изучения гистобластических потенций нейроглии про-
цессы дегенерации и регенерации в нервной системе дают немного. Несмотря
на олень большую литературу, посвященную травме мозга и эксперимен-
тальному изучению процессов репаративной регенерации в центральной
нервной системе (Шпильмейер, 1922; Якоб, 1927; Р. Кахаль, 1928; Ортега
и Пенфильд, 1928; Минкин, 1939, и др.), * многие, даже основные, вопросы
из этой области еще не решены, а если и решаются, то большей частью очень
разноречиво. Это связано с исключительными техническими трудностями
и большой сложностью реактивных процессов, возникающих в глии и
в соединительнотканных образованиях. Некоторые из реактивных изме-
нений в глии и мезенхимных производных обнаруживают, несомненно, более
или менее далеко идущее конвергентное сходство, а некоторые клеточные
формы глиального и соединительнотканного происхождения даже совер-
шенно не могут быть отличены друг от друга. Достаточно напомнить о
т. наз. зернистых или пенистых клетках, богатых жировыми включениями и
описанных впервые Глюге (1841), которые имеют очень малоспецифи-
ческий макрофагоподобный вид и могут возникать при самых разнообразных
патологических процессах в мозгу, а также в периферической нервной систе-
ме. Относительно их происхождения имеются все мыслимые точки зрения.
Кахаль (1928) и другие считают, что они образуются из клеток соедини-
тельной ткани в крови, Якоб (1927), Русси, Лермит и Оберлен (1930) —
из всех элементов нейроглии, а также из соединительной ткани, Ортега
(1932), Ортега и Пенфильд (1928), Лайнел (1929) — только из микроглии,
’Приведенные работы содержат большое количество литературных данных.'
264
Глава III
Белецкий (1939) — из микроглии и олигодендроглии, Фаррар (1908) — из
эндотелия сосудов.
Образование макрофагоподобных клеток из нейроглии в условиях
тканевых культур несомненно говорит против точки зрения Ортега и его
последователей, что зернистые клетки возникают только из микроглии.
После травмы мозга имеется более или менее резко выраженное крово-
излияние, окруженное зоной некробиоза. На границе отмирающей и оста-
ющейся жизнеспособной ткани образуется особенно резко выраженное
скопление из упомянутых зернистых или пенистых клеток. Внутрь очага
некроза врастает соединительная ткань, а на границе жизнеспособной ткани
мозга и организующимися массами детрита с отмирающими элементами
постепенно создается более или менее резко выраженная пограничная
зона волокнистой астроглии или собственно глиальный рубец, в котором
позже развиваются регрессивные изменения. Глиальные и соединительно-
тканные части формирующегося рубца могут в большей или меньшей степени
прорастать друг в друга. Возможно, что в составе целого орга-
низма глиальные рубцы по своему генезу и строению представляют
обой образования, сходные с описанными выше глиальными массивами
центральных кусочков тканевых культур. Относительная интенсивность
соединительнотканной и глиальной реакции зависит от характера и раз-
меров повреждения. После процессов дегенерации, вызванных перерез-
ками, рубец развивается почти исключительно из нейроглии. По данным
Якоба (1922),при появлении глиального рубца очень хорошо прослеживается
дифференцировка волокнистой макроглии из протоплазматической,лпро
текающая так же, как при эмбриональном развитии. Глиальные волокна
развиваются в периферическом слое протоплазмы. Предполагается, что
они могут каким-то образом частично отделяться от цитоплазмы и распо-
лагаться межклеточно. Последнее, исходя из наблюдений над тканевыми
культурами, кажется маловероятным.
Если лишенные цитоплазмы волокна и встречаются, то их образование
вернее всего представлять себе, как конечный этап превращений всей массы
цитоплазмы клеточного отростка по типу отмеченной Гирке «кератинизации»
глиальных элеме нтов.
При гибели отдельных нейронов к нейронофагии способны клетки-
сателлиты, т. е. микроглия и олигодендроглия (Якоб, 1927; Пенфилъд,
1932). Они же образуют на месте погибшего нейрона «розетки». Чрезвы-
чайно странным и не вяжущимся с высокой пролиферативной способностью
глиальных элементов является указание ряда авторов на отсутствие в них
митозов при репаративной регенерации. Это совершенно противоречит
всему, что мы знаем относительно пролиферативных процессов в других
тканях и о значении митоза, как, по существу, единственного полноценного
способа клеточного деления. Все новейшие работы по реактивным изме-
нениям глии, сделанные специальными методами, не дают общей картины
тканевого процесса, а выхватывают из него отдельные клеточные формы,
которые удается импрегнировать с помощью той или иной методики. При-
этом делается допущение, далеко не являющееся самоочевидным, что отдели-
Экспериментальный метод в гистологии
265
ные клеточные формы, несмотря на происходящие в них изменения, сохра-
няют способность элективно окрашиваться такими же методами, что и
в норме. Особенно мало удовлетворяет большая часть результатов, отно-
сящихся к т. наз. олигодендроглии, роль которой в пролиферативных про-
цессах остается, пожалуй, наименее ясной. Последнее, возможно, зависит в
значительной степени оттого, что методика обнаружения олигодендроглии яв-
ляется, пожалуй, наиболее капризной и дает наименее постоянные результаты.
Все результаты опытов с эксплантацией частиц мозга и ряда других
тканей показывают, что образование макрофагоподобных, способных к фаго-
цитозу элементов, является одной из чрезвычайно примитивных и универ-
сальных реакций всевозможных клеток, не имеющей тканевой специфичности
и зависящей, кроме того, от функционального состояния клеточных эле-
ментов. Способность к фагоцитозу представляет собой элементарное свой-
ство живой протоплазмы, которое лишь в редких случаях может исчезать
в связи с высокой специализацией клеток. На способности или неспособ-
ности к фагоцитозу, а также, повидимому, к накоплению в цитоплазме
коллоидных веществ, нельзя делать безоговорочно выводов гистогенети-
ческого характера. Поэтому всякая аргументация в пользу мезенхимного
происхождения тех или иных элементов глии, основанная на способности
макрофагоподобных клеток к фагоцитозу и нефроцитозу, по существу
неправильна и совершенно неубедительна.
е) Зрительные пути, сетчатая оболочка глаза и эпифиз
Сравнительное экспериментальное изучение зрительных путей и сет-
чатой оболочки глаза чрезвычайно интересно, так как дифференцированные
очень различным образом части органа зрения развиваются из единого
рано детерминирующегося зачатка, т. наз. оптико-окулярного аппарата
(Лепла, 1919; Адельман, 1929). Этот зачаток, входя в состав самого
переднего отдела нервной пластинки, уже до замыкания ее в трубку, по
ряду свойств отличается от других частей нейрального зачатка (Шпеман,
1912; Вурдеман, 1929; Менчот, \929;Годлей, 1928, и др.) и характеризуется
хорошо выраженной способностью к самодифференцировке и регулятив-
ным процессам.* Этим подчеркивается особенно большая степень генети-
ческой близости разных отделов зрительного аппарата, который, возникнув
очень давно у предков современных позвоночных, эволюировал, как
единая система. Рассмотрение эпифиза совместно с глазом оправдывается
тем, что он является, как это отмечалось выше, рудиментом париетального
глаза или вероятно точнее оптикоокулярного аппарата, развившегося из
другого участка головного мозга. Поэтому уже заранее возможно было бы
ожидать, что нейроглия зрительных путей, а также-эпифиза, должна отли-
чаться в какой-то степени по своим биологическим свойствам от нейроглии
других отделов центральной нервной системы. В составе зрительного нерва,
как это уже упоминалось, описывается волокнистая астроглия и олигоденд-
* Сводка по экспериментальной эмбриологии, а также гистологии глазного зачатка
имеется в статье Ванникова (Успехи современной биологии, 10, 1939).
766 Глава 111
роглия, причем вторая отличается свойственными белому веществу мозга
крупными размерами и сложно разветвленным клеточным телом (Марке-
зани, 1926). По моим наблюдениям, при обычных гистологических методах
изучения и при обработке по Населю, характерных различий в величине
и окраске ядер элементов нейроглии зрительных пучков обнаружить не
удается. Почти все глиальные ядра отличаются довольно крупными разме-
рами и светлой окраской.
Мы располагаем в настоящее время очень полным экспериментально-
гистологическим материалом, полученным методом эксплантации по эпен-
Рис. 37. Участок зоны роста из 12-дневной культуры зрительного нерва 10-днев-
ного кролика. Массивная и комплексная нейроглиальная зона экстенсивно расту-
щей нейроглии переходит на периферии в клеточные тяжи и изолированные клет-
ки. Большая часть клеток имеет удлиненную форму и образует ориентированные
в разных направлениях потоки. Слабое увеличение. (По Хлопину, 1940.)
димо-глиальным тканям этих отделов нервной системы. Паши данные по
зрительному нерву, хиазме и тракту (Хлопан, 1938, 1939, 1940, и неопубли-
кованные данные) и по эпифизу новорожденных и молодых кроликов (И. Хло-
пана, 1941, и подробные неопубликованные данные) первые и единственные
в литературе. Пигментному слою сетчатки посвящено довольно значительное
число работ (Елеонская, 1915; А. Фишер, 1922; Капель, 1929; Леви, 1934;
И. Фишер, 1938, и др.), сделанных почти исключительно на эмбриональном
курином материале. По рецепторному листку сетчатки имеется всего не-
сколько работ на эмбриональном курином материале (Мейер, 1936; Вейс
и Доррис, 1935) и на сетчатке кролика и черепахи (Шампи, 1914). Однако
ни в одной из этих работ не были не только детально описаны, но даже отме-
Экспериментальный метод в гистологии
267
чены характерные гистотипические свойства изучаемых тканей. По стран-
ному недоразумению, картины экстенсивного роста пигментного слоя
сетчатки со времени работы А. Фишера (1622), Ибелинга и Фишера (1922)
приводились в качестве примера «эпителиальных культур», причем даже
не возникало вопроса, что т. наз. пигментный эпителий является чрезвычайно
отличной по своим гистобластическим потенциям от эпителиев ряда дру! их
органов тканью. Фактический материал по рецепторному листку сетчатки
еще менее полон в гистологическом отношении. Большой цикл работ, выпол-
ненных моим сотрудником Винниковым (1936, 1938, 1939, 1940, 1941, 1943),
не только дает впервые подробный гистологический анализ пигментного
Рис. 38. Участок глиальной зоны роста с круглыми макрофагоподобнымп клет-
ками, которая частично разрыхляется, образуя клеточные тяжи различной шири-
ны. 7-дневпая культура из зрительного нерва 6-дневного кролика. Слабое увели-
чение. (По Хлопину 1940.)
и рецепторного слоев сетчатки, но и открывает с первого взгляда совершенно
неожиданные гистоморфные особенности ряда элементов глазного яблока.
Нейроглия зрительных путей энергично разжижает фибрин с образо-
ванием в нем полостей и, как правило, заглушает рост соединительной ткани,
элементы которой вскоре исчезают без остатка из зоны роста. Поэтому
получение чистых культур нейроглии не представляет труда. Ее экстенсив-
ный рост отличается многообразием форм, которые неизменно повторяются
на различных стадиях культивирования. Она растет комплексными тонкими
мембранами, более толстыми пластами, своеобразными клеточными мас-
сивами и подчас сложно разветвленными тяжами значительной ширины
и толщины и вместе с тем может образовать разрыхленные сетевидные
структуры и изолированные клетки (рис. 37, 38, 39). Энергия пролиферации
растущих глиальных элементов очень велика, а количество наблюдающихся
митотических фигур иногда прямо поражает. В своей совокупности и пре-
268
Глава III
Рис. 39. Экстенсивный рост ней-
роглии клеточными массивами, пе-
реходящими на периферии в заост-
ренные клеточные тяжи. 11-днев-
ная культура из зрительного нер-
ва 6-дневного кролика. Увеличе-
ние лупы. (По Хлопину, 1940.)
вращениях все эти структуры настолько характерны, что могут быть без
труда отличены от картин роста тканей другого происхождения. Глиальные
мембраны, массивы и тяжи могут состоять из элементов разной формы, тс
более или менее полигональных или полиэдрических, то чаще вытянутых
в длину. В последнем случае потоки одинаково ориентированных удлинен-
ных клеток могут взаимно перекрещиваться в разных направлениях, обра-
зуя подчас своеобразные вихревые структуры. Участки мембран, состоящие
из уплощенных полигональных клеток, если их рассматривать вне связи
с другими структурами, обнаруживают несомненное поверхностное сходство
с рядом растущих вне организма эпителиальных,в частности, эпидермальных
тканей.
При наступающем разжижении свертка
фибрина и образовании в нем полостей,
нейроглиальная зона роста может разры-
ваться и деформироваться, нередко стяги-
ваясь по краям неразжиженного ободка
фибрина в сплошной клеточный вал или
отдельные скопления, которые в дальней-
шем могут сделаться новыми центрами
роста. Массивы, мембраны и тяжи в сво-
ей совокупности нередко образуют в зоне
роста лучистые или звездчатые структуры
(рис. 40, 41). Глиальные элементы, остаю-
щиеся в составе первоначально посеянного
кусочка, с течением времени преобразу-
ются в протоплазматические массивы, без
заметных клеточных границ, в которых вза-
имно перекрещивающиеся волокнистые структуры дифференцируются более
отчетливо, чем это имело место в исходном материале. Эти волокнистые струк-
туры могут достигать значительной толщины и имеют такой же характер, что
и в эксплантатах из головного и спинного мозга. В нейроглиальных мас-
сивах зоны роста также можно наблюдать появление волокнистых струк-
тур в протоплазме. Кроме того, в глиальной зоне роста, при более или менее
длительном культивировании, появляются аргирофильные сети волокни-
стого межклеточного вещества, обнаруживаемого при импрегнации сереб-
ром. Изолирующиеся от нейроглиальных комплексов и округляющиеся
клетки могут иметь макрофагообразный вид. На более ранних стадиях
культивирования, большей частью до двух-трех недель, изоляция и осы-
пание клеток принимают нередко массовый характер. Позже количество
таких элементов в культурах, как правило, уменьшается и может даже
сводиться к нулю. Эти макрофагообразные клетки, образование которых
является, повидимому, обратимым процессом, несомненно происходят
из растущих вне организма элементов нейроглии, а не из мезенхимных
производных, которые, как это уже упоминалось, быстро заглушаются
и совершенно исчезают из зоны роста. Эти данные показывают лишний раз,
что нет оснований считать макрофагообразные свободные клетки, описан-
Экспериментальный метод в гистологии
269
ные рядом авторов в культурах из эмбриального и неэмбриального мозга,
элементами мезенхимного происхождения и использовать их как доказа-
тельство взглядов Ортега, касающихся мезенхимного происхождения
микроглии.
Изучение изменений центрального кусочка на ранних стадиях культи-
вирования и возникновения из его элементов зоны роста не оставляет ника-
ких сомнений в том, что практически все глиальные элементы исходного
материала проделывают тождественный цикл превращений и дают по суще-
ству совершенно равнозначные, несмотря на разнообразие их формы,
клетки зоны роста. Иными словами, описываемые в составе зрительного
нерва в приблизительно одинаковом количестве клетки астроглии и олиго-
дендроглии по своим гистобластическим потенциям не отличаются сколько-
нибудь заметно друг от друга и
представляют собой по существу
весьма сходные структурные мо-
дификации одного и того же
клеточного типа нейроглии зри-
тельных путей, а не совершен-
но различные по своим свой-
ствам клеточные формы, как это
предполагается с точки зрения
современной нейрогистологии.
Некоторые данные по измене-
ниям зрительного нерва при его
дегенерации говорят за то, что
элементы олигодендроглии в ко-
Рис. 40. Массивная глиальная зона роста с
клеточными тяжами .и звездчатыми комплекса-
ми, 16-дневная культура из зрительного нер-
ва 6-дневного кролика. Увеличение луцы.
(По Хлопину, 1940.)
нечном итоге превращаются в
волокнистую астроглию (Кон: и
Мекмиллен, 1932).
При сопоставлении наших
данных с последними указани-
ями нельзя не сделать вывода, что олигодендроглия и астроглия, вероятнее
всего, лишь несколько отличаются уровнем своей дифференцировки, причем
астроглия является несколько более дифференцированной, чем олигоден-
дроглия. Эти различия едва ли выражены по существу более резко, чем, на-
пример. различия между базальным и одним-двумя выше лежащими слоями
шиповатых клеток многослойного плоского эпителия кожи. Эти очень простые
и вероятные сопоставления с другими тканями почему-то совершенно упуска-
ются из виду в нейрогистологии. Вместе с тем следует подчеркнуть, что
элементы нейроглии зрительных путей, по существу отождествляемые
с соответствующими им элементами других отделов центральной нерьной
системы, несмотря на несомненное сходство своих превращений в условиях
тканевых культур и картины экстенсивного роста, не оставляющие сомнений
в их принадлежности к единому нейроглиальному типу, тем не менее отли-
чаются рядом особенностей, подобно тому как, например, эпителии единого
эпидермального или энтеродермального типа в составе отдельных органов
270
Глава HI
не полностью идентичны друг другу в отношении биологических
свойств.
.Несмотря на большое общее сходство картин установленного нами
(Хлопин, 1938) нейроглиального типа роста, у глии зрительных путей
можно отметить следующие частные особенности. Прежде всего ее элементы
отличаются преобладающей удлиненной формой тела и ядра. Обладая (по-
добно глии головного и спинного мозга), повидимому, значительной вяз-
костью цитоплазмы, ее элементы могут образовать длинные отростки в двух
взаимно противоположных направлениях, приобретая подчас почти ните-
видную форму, но не образуя, как правило, в зоне роста звездчатых астро-
цитарных или нейроноидных клеток описанного выше типа. Способ взаимо-
связи и комплексность элементов нейроглии зрительных путей также отли-
чаются вполне определенными особенностями. Особенно тесная комплексная
Рис. 4! Лучистые глиальные комплексы.
12-дневная культура из зрительного нерва
13-дневпого кролика. Слабое увеличение.
(По Хлопину, 1940.)
взаимная связь клеток удлинен-
ной формы наблюдается между
концами их клеточных тел, при
значительно менее тесной связи их
обращенных друг к другу длинных
сторон. От этой особенности зави-
сят образование подчас очень длин-
ных клеточных тяжей разного ка-
либра и сетевидных структур, на
которые обычно разрыхляется боль-
шая периферическая часть зоны
роста, а также особенно резко вы-
раженные у этого объекта взаимно-
перекрещивающиеся клеточные по-
токи в составе мембран и массивов,
образующие характерные вихре-
вые структуры и лучистые звездчатые клеточные комплексы. Вме-
сте с тем в отдельных участках уплощенные или полиэдрические
клетки нейроглии зрительных путей, как это отмечалось выше, дают и
довольно типичные для эпидермальных тканей малодифференцировачные
структуры. Митотическая активность этих элементов необычайно высока.
В отношении типа комплексности и способности к образованию клеточных
тяжей особенное сходство с нейроглией зрительных путей обнаруживают
из разобранных выше объектов покровные клетки сосудистых сплетений
мозга.
По исследованиям Винникова (1936, 1940), разные участки сетчатой
оболочки, в частности, ретинальные листки края глазного бокала и т. наз.
«пигментный эпителий» (тапетум) ее зрительной части, обнаруживают в усло-
виях культур некоторые особенности на фоне обгцих картин нейроглиаль-
ного типа роста. Ретинальные листки радужины, цилиарного тела и тапетум
дают только экстенсивный рост, но, в противоположность некоторым друт им
данным (Капель, 1929), никогда не эпителизируют поверхности эксплантата,
обнаруживая в этом отношении сходство с другими структурами эпендими-
Экспериментальный метод в гистологии 27Г
глиального тканевого типа, имеющими морфо-физиологическую характе-
ристику эпителия (эпендима, покровные клетки сосудистых сплетений).
При экстенсивном росте ретинальные листки края глазного бокала
молодых и взрослых кроликов образуют тонкие комплексные мембраны
и рыхлые сетевидные структуры, массивные клеточные пласты, тяжи разно-
образной формы и, наконец, изолированные, частью многоядерные, очень
полиморфные клетки. Клетки пигментированных участков ретинальных
листков неальбинотических животных, в соответствии с данными других
авторов, полученными на эмбриональном курином материале, продолжают
вырабатывать пигмент (рис. 42). В зависимости от условий опытов, интен-
Рис. 42. Участок мембраны в 13-
дпевной культуре из пигментного
слоя слепой части сетчатой оболоч-
ки кролика. В клетках многочис-
ленные пигментные включения.
Сильное увеличение. (По Винни-
кову, 1936.)
Рис. 43. Приобретение внутренним
ретинальным листком «обратно-
многорядной» структуры. 4-днев-
ная культура цилиарного тела 8-
дневного кролика. Сильное увели-
чение. (По Винникову, 1936.)
сивность пигментообразования бывает очень различна. То более, то менее
значительные участки клеточных мембран обнаруживают свойственную
многим эпителиальным тканям правильную мозаику полигональных эле-
ментов, которая, однако, сменяется в пространстве и во времени плоскост-
ными структурами, образованными клетками резко удлиненной фирмы
и рыхлыми сетевидными клеточными соединения.ми. В составе мембран,
имеющих клетки вытянупй формы, последние нередко образуют ориенти-
рованные различным образом потоки. Массивные клеточные пласты состоят
из нескольких слоев клеток, растущих в различных направлениях, и также
обнаруживают в своем составе ориентированные неодинаковым образом
клеточные потоки, могущие перекрещиваться под разными углами. Отли-
чающиеся первоначально друг от друга по некоторым деталям, элементы
ретинальных листков радужины и цилиарного тела еще в составе централь-
ного кусочка, как бы нивелируясь, уподобляются друг другу и дают
совершенно тождественные картины экстенсивного роста. Начинающие
пролиферировать еще в составе центрального кусочка, клетки ретинальных
листов образую'; местами утолщенные покровные участки, которые отли-
272
Глава III
чаются по расположению клеток и локализации митозов более или менее
характерной для нейральных зачатков «обратной многорядностью» (рис. 43).
Чрезвычайно характерные для зоны роста ретинальных листков клеточные
тяжи, отмеченные в свое время Фишером (1922), не обратившим, однако,
внимания на их гистотипическое значение, отличаются большим разно-
образием внешней формы, калибра и строения. То это бывают довольно
короткие выросты центрального кусочка или периферических частей мем-
бран, состоящие из одного-двух рядов клеток полиэдрической формы,
то они достигают очень значительной длины, могут ветвиться и состоят
из более или менее сильно вытянутых в длину элементов, располагающихся
неодинаковым числом рядов. Благодаря сочетанию в зоне роста мембран,
клеточных массивов и тяжей в эксплантатах из ретинальных листков
также наблюдается образование звездчатых своеобразных структур. В цито-
плазме элементов зоны роста местами удается обнаружить дифференцировку
волокнистых структур, которые нельзя не сопоставить с описанными выше
глиальными. Хотя экспериментальный анализ сократимых производных
ретинальных листков будет дан в главе, посвященной мышечным тканям,
здесь все же следует отметить, что т. наз. миоэпителиальные клетки расши-
ряющей зрачок мышцы по своим превращениям в зоне роста ничем не отли-
чаются от внутреннего ретинального листка радужины.
Наружный пигментный слой зрительной части сетчатки зародышей
свиньи и кролика, а также новорожденных и взрослых кроликов, как и
следовало ожидать, обнаруживает, по данным Винникова (1940), большое
сходство превращений в условиях тканевых культур с ретинальными лист-
ками радужины и цилиарного тела, но вместе с тем и некоторые особен-
ности.
На временно непрорастающих частях поверхности эксплантата тапетум
может также заметно утолщаться и приобретать, повидимому, «обратно
многорядное» и даже, возможно, многослойное расположение элементов.
Сильная вакуолизация клеток, обнаруживающаяся местами в таких утол-
щенных участках, а также, особенно при неблагоприятных условиях куль-
тивирования, в зоне роста, напоминает патологические изменения сет-
чатки при retinitis pigmentosa, которые были получены экспериментально
Шрейбером и Венглером (1910) путем введения шарлахового масла. Обра-
зуемая клетками тапетум зона роста в гистологическом отношении полностью
соответствует картинам, полученным при эксплантации радужинной и
цилиарной частей сетчатки. И в данном случае образуются комплексные
мембраны, массивные пласты, клеточные тяжи, разрыхленные сетевидные
структуры и изолированные клетки, со всем многообразием формы и взаимо-
расположения их элементов, описанным выше. Особенности тапетум отно-
сятся в первую очередь к области цитотипических или цитоморфных структур.
Большая часть клеток тапетум, отличающихся, при обычных условиях,
поразительной геометрической правильностью формы, не свойственной
элементам других частей ретинальных листков, характеризуется своей
двуядерностью, которая может более или менее длительное время сохра-
няться ими в зоне роста. Эта двуядерность возникает у них в онтогенезе
Экспериментальный метод в гистологии
273
не амитотическим путем, а путем митоза, не сопровождающегося разделе-
нием клеточного тела. Плазмотомия, однако, совершается рано или поздно
всеми жизнеспособными клетками в условиях тканевых культур. Только
после этого образовавшиеся одноядерные элементы начинают делиться мито-
тическим путем. Другой особенностью дифференцированных клеток тапетум
является характерная игольчатая форма пигментных включений, отли-
чающихся от круглых гранул пигмента элементов слепой части сетчатки.
Эта особенность первое время сохраняется у растущих вне организма
клеток тапетум. Однако, когда пигментообразование вне организма ста-
новится заметным, наряду со старыми игольчатыми пигментными вклю-
чениями появляются во все увеличивающемся относительном количестве
новые включения округлой формы, свойственные эмбриональному тапетум
и ретинальным листкам слепой части сетчатой оболочки.
Исследования Винникова, так же как и работы других авторов, имевших
дело с ретинальными листками, не подтвердили данных Фишера (1922),
который считал, что ему удалось наблюдать в центральном кусочке эксплан-
татов из пигментного слоя радужины явления ороговения. Нужно думать,
Фишер был введен в заблуждение картинами дегенерации, которые, судя
по приведенному им рисунку, имели достаточно отдаленное сходство с
настоящим ороговением или даже с паракератозом.
Особенно интересные данные удалось получить Винникову (1938, 1939,
194 Г, и неопубликованные данные) при экспериментальном анализе рецептор-
ного листка сетчатки, первоначально касавшемся млекопитающих (заро-
дыши кролика и свиньи, новорожденные и молодые кролики), а затем распро-
страненном и на представителей других классов позвоночных (костистых
рыб, амфибий и птиц). У всех изученных позвоночных, помимо разрастаний
глиального типа, наблюдался также ряд изменений в нейронах, на которых
мы остановимся позже. Разрастающиеся вне организма элементы рецептор-
ной части сетчатки обнаруживают большое сходство с другими изученными
частями органа зрения. В образующейся зоне роста наблюдается все много-
образие описанных выше структур: комплексные мембраны и клеточные
массивы, тяжи, разрыхленные сетевидные соединения и изолированные
клетки. Удлиненной формы элементы, как и в других случаях, образуют
характерные, взаимно пересекающиеся клеточные потоки. Участки мембран,
состоящие из полигональных элементов, имеют более или менее выраженный
«эпителиальный» характер. В эксплантатах из сетчатки зародышей местами
можно было наблюдать образование своеобразных «нейроноидных» клеток
с тонкими нитевидными отростками. Я склонен трактовать эти элементы
(в соответствии со всеми приведенными выше данными) как клетки астро-
цитарного характера, хотя их природа нуждается еще в дальнейшем изу-
чении. Гистогенез зоны роста в эксплантатах из сетчатки был прослежен
Винниковым очень детально и в настоящее время может считаться точно
установленным. У зародышей и новорожденных кроликов гистогенез сет-
чатки еще не закончен, а в наружном ядерном слое нередко удается видеть
митозы, которые у выросших животных исчезают. После эксплантации
тонкие частицы сетчатки, как правило, скручиваются таким образом, что
18 Хлопин—349
274
Глава III
ганглиозный слой оказывается внутри, а фоторецепторные элементы
снаружи. Часть последних гибнет; другая, более значительная, претерпе-
вает ряд сложных превращений. Прежде всего у всех фоторецепторных
элементов отторгаются, подвергаясь пластинчатому распаду, членики
палочек и колбочек. На их месте, в течение первых дней после эксплантации,
удается в некоторых случаях наблюдать образование очень тонких, слегка
извивающихся цитоплазматических отростков. Содержащие ядро части
фоторецепторных клеток набухают, а первоначальная своеобразная ядер-
ная структура начинает изменяться. Ядра становятся светлее, отчетливо
выступают ядрышки. У новорожденных кроликов митозы в клетках пало-
чек и колбочек активизируются, а у выросших кроликов с течением времени
появляются вновь. Измененные фоторецепторные клетки в местах, где
наружная пограничная перепонка разрушается, начинают потоками про-
растать экстенсивно, образуя основную массу зоны роста и энергично
делясь митотическим путем. Во внутреннем ядерном слое происходи;
прогрессирующая гибель нервных элементов. Мюллеровские волокна
выявляются все отчетливее, их внешняя форма упрощается и становится
веретенчатой. Из таких измененных мюллеровских волокон, утративших
связь с пограничными перепонками, также образуются потоки делящихся
митозами клеток, которые продвигаются вслед за бывшими фоторецептор-
ными элементами в зону роста. Подвергнувшиеся упрощению и проросшие
мюллеровские волокна и клетки палочек и колбочек ничем не отличаются
друг от друга и образуют единый малодифференцированный клеточный
материал всех нейроглиальных структур зоны роста. Прогрессивных изме-
нений в нейроглиальных элементах внутренних слоев сетчатки пока не
обнаружено. Совершенно сходные изменения в эксплантированных фото-
рецепторных клетках удалось установить у всех упомянутых выше пред-
ставителей других классов позвоночных. Во всех случаях наблюдалось
образование зоны роста нейроглиального типа за счет измененных клеток-
палочек и колбочек. У холоднокровных митотическая активность была
выражена слабо, и экстенсивный рост сопровождался лишь в незначитель-
ной степени пролиферацией. В ряде случаев, так же как в эксплантатах из
мозга, наблюдалось образование клетками глиальных мембран сетчатки
более или менее длинных прямых и даже нитевидных цитоплазматических
отростков.
Изложенные вкратце данные показывают, что, несмотря на высокую
специализацию в двух совершенно различных направлениях, клетки пало-
чек и колбочек и мюллеровские волокна сохраняют в основном характер
эпендимных клеток, способных к пролиферации митотическим путем.
В отношении гистобластических потенций они представляют собой элементы
эпендимо-глиального тканевого типа. Фоторецепторные элементы, пови-
димому, правильнее всего трактовать не как нейроны, обнаруживающие
в экспериментальных условиях другие биологические свойства, а как особую
свето- и цветочувствительную модификацию эпендимы, приобретающей
лишь очень поверхностное сходство с нервными клетками. Возможно»
в свойствах клеток палочек и колбочек сохранились в какой-то степени
Экспериментальный метод в гистологии
275
черты очень примитивных по существу эктодермальных рецепторов, из
которых филогенетически развились высокоспециализированные нервные
клетки со всей полнотой своих совершенно своеобразных биологических
особенностей. Вместе с тем, как это отмечалось, эпендима является в эволю-
ционном процессе источником развития астроцитарной нейроглии. Поэтому
эпендимные и, в частности, фоторецепторные клетки можно также рассматри-
вать как элементы, имеющие характер общих родоначальников вспомога-
тельных и нейрональных элементов нервной системы.
Упоминавшиеся выше наблюдения Шрейбера и Венглера (1910) показы-
вают, что и в условиях целого организма в рецепторной части сетчатки
возможны пролиферативные процессы. Их указание на появление в наруж-
ном ядерном слое митозов прекрасно гармонирует с данными, получен-
ными методом эксплантации. Что касается митозов, встреченных ими в
области ганглиозного слоя, то нет никаких оснований думать, что они при-
надлежали ганглиозным нервным клеткам. Вероятнее всего, авторы имели
дело со смещенными, в результате пролиферативных процессов, бывшими
клетками палочек и колбочек, а, возможно, также и измененными мюл-
леровскими волокнами. Эти интересные опыты в настоящее время настоя-
тельно нуждаются в повторении и дополнительном гистологическом анализе.
Здесь же будет уместно еще раз кратко остановиться на своеобразной
ткани хрусталика. Ряд авторов успешно культивировал вне организма
т. наз. хрусталиковый эпителий (Елеонская, 1915; Керби, 1927; Адачи.
1930). Все авторы указывали на то, что он разрастается «эпителиаль-
ными» мембранами. К сожалению, на основании этих работ, нельзя со-
ставить себе достаточно точного представления о специфических особен-
ностях зоны роста и степени ее вероятного сходства с описанными выше
глиальными структурами. Если принять во внимание, что разрастания
ретинальных листков и эмбриональной нервной системы было также при-
нято описывать как эпителиальные мембраны, то не будет ничего удиви-
тельного, если и эпителий хрусталика окажется нейроглией или очень
сходной с ней тканью. В пользу этого также говорит давно известный факт
регенерации хрусталика из ретинальных листков края глазного бокала
(Колюччи, 1891; Вольф, 1904). Это поразительное с первого взгляда явление,
которое наблюдается также в экспериментально измененном онтогенезе,
считалось одним из веских аргументов против специфичности тканей и неза-
висимости их свойств от способа развития (Гертвиг, 1906). На самом деле
регенерация хрусталика и картины его нормального онтогенеза, мне кажется,
говорят как раз обратное. Более примитивные формы органа зрения позво-
ночных, представленные париетальными глазами, показывают, что способ-
ность к хрусталиковой дифференцировке была приобретена в процессе
эволюции стенкой глазного пузыря, т. е. определенным участком нервного
зачатка. В соответствии с теорией Шимкевича (1923) превращения
камерного неревертированного глаза в ревертированный, линзообразо-
вателъная область глазного пузыря оказалась у края глазного бокала.
Вероятнее всего, вольфовская регенерация хрусталика отражает собой эти
пройденные этапы эволюции. Наблюдающийся в настоящее время у позво-
276
Глава III
ночных способ развитая хрусталика должен был возникнуть позже в связи
с установлением новых коррелятивных зависимостей между глазным бока-
лом и одной их плакод, направление дифференцировки которой при этом
изменилось и пошло по пути образования ткани хрусталика. Установленные
в онтогенезе амфибий различия во взаимодействии зачатка глаза и линзо-
образующих областей эктодермы, вероятно, отражают в какой-то степени
эволюцию свойств этого формообразовательного аппарата (Филатов, 1939).
Во всяком случае, если считать плакоды за участки единого по существу
нервного зачатка, то образование линзы в нормальном онтогенезе и при
экспериментальных условиях происходит из гистогенетически однородного
материала, который в обоих случаях является нервным зачатком. Любо-
пытно, что, поданным Тёрё (1931), имплантированные на место хрусталика
культуры пигментированных ретинальных листков, под воздействием
окружающих частей глаза приобретают внешнюю форму и типичную внутрен-
нюю структуру линзы. При имплантации культур из роговицы такого резуль-
тата не получалось. Это подчеркивает лишний раз генетическую общность
хрусталика и нервного зачатка. Каким образом можно трактовать резуль-
таты опытов с пересадкой у зародышей амфибий других тканевых зачатков,
например миотомов, на место удаленного хрусталика (Попов, 1937), не
вполне ясно. Пересаженный материал, по этим данным, приобретал внешнюю
форму хрусталика. Но ни на основании описаний, ни из рисунков нельзя
представить, в какой степени линзоподобное образование по своей струк-
туре соответствовало настоящей ткани хрусталика, а не являлось лишь ее
имитацией. Учитывая достаточно хорошо обоснованный факт независимости
в онтогенезе морфологических и гистогенетических процессов (Гексли и
де-Беер, 1936), вполне возможно, например, допустить, что в материале
имплантированного миотома, принявшем форму хрусталика, дифферен-
цировались не настоящие хрусталиковые, а более или менее атипичные
мышечные волокна, приобревшие в поле глазного бокала определенную,
свойственную настоящим хрусталиковым волокнам, ориентировку. Анало-
гичные предположения можно высказать и в отношении результатов имплан-
тации другого, чуждого материала. Такая трактовка указанных опытов
не исключается фактическим материалом работ. Было бы очень интересно
пересмотреть этот очень ценный сам по себе экспериментальный материал
под гистологическим углом зрения.
Ткань эпифиза новорожденных кроликов (И. Хлопина, 1941, и неопубли-
кованные данные) дает в условиях тканевых культур чрезвычайно типичные
картины нейроглиального роста в той его модификации, которая стоит
всего ближе к разрастаниям зрительных путей, и, отчасти, ретинальных
листков и покровных клеток сосудистых сплетений мозга. В центральных
кусочках эксплантатов вокруг жизнеспособных, увеличивающихся в раз-
мерах, ядер паренхимы эпифиза постепенно выявляются клеточные тела
полиэдрической, уплощенной или удлиненной формы, плохо различимые,
при обычных методах обработки, в исходном материале. В результате
этих изменений отдельные участки эксплантата приобретают характер
клеточных массивов, которые и дают начало создающейся зоне роста.
Экспериментальный метод в' гистологии
277
Рис. 44. Звездчатые глиальные клеточные комплексы.
17-дневная культура эпифиза новорожденного кро-
лика. Слабое увеличение (По И. Хлопиной,
оригинал.)
Трубчатые ходы и тяжи, образованные в исходном материале эпендимо-
подобными клетками, вскоре исчезают. Их элементы утрачивают свою пра-
вильную форму и ориентировку и перестают быть отличимыми от окружаю-
щих измененных элементов паренхимы эпифиза. Соединительнотканные про-
слойки никакого видимого участия в образовании зоны роста не принимают.
В зоне роста наблюдается все многообразие комплексных эпителиоподобных
и неэпителиоподобных структур, разрыхленных сетевидных клеточных
соединений, тяжей разного калибра и длины и изолированных элементов,
свойственных и другим, изученным в настоящее время, участкам централь-
ной нервной системы. Клеточные комплексы зоны роста с отходящими
от них тяжами отличались характерной звездчатой или лучистой формой.
В некоторых случаях весь центральный кусочек, энергично разрастаясь,
как бы растворялся в ок-
ружающей его зоне роста
и распадался на отдель-
ные или связанные друг
с другом клеточными пе-
ремычками звездчатые ком-
плексы (рис. 44). Различ-
ная ориентировка клеточ-
ных потоков, состоящих
из удлиненных элементов,
приводила к образованию
характерных перекрестов..
Как в центральном кусоч-
ке, так и в зоне роста,
в цитоплазме элементов
эпифиза появлялась харак-
терная фибриллярность,
но переплета толстых гли-
альных волокон, описан-
ных в культурах головно-
го и спинного мозга и зрительных путей, в клеточных массивах центрально! о
кусочка не наблюдалось. Часть элементов эпифиза отличалась в зоне роста
наличием пигментных включений, образование которых, возможно, про-
должалось и в эксплантатах.
Совокупность отмеченных особенностей паренхимы эпифиза нельзя
не поставить в связь с его филогенетический! развитием из париэтального
органа зрения.
г) Оболочки центральной нервной системы
До последнего времени работ по мозговым оболочкам, сделанных методом
эксплантации, не было. Выполненное в моей лаборатории исследование
Каннегисер (1938, 1940) над мягкими оболочками головного мозга кроликов
разного возраста восполняет пробел. Эта работа предпринималась в то
278
Глава HI
время, когда у пас еще не возникало сомнений в обычном соединительно-
тканном характере этих образований. Ее первоначальной целью было
показать неравноценность покровных клеток мозговых оболочек и мезо-
телия, с которым их было принято сближать, несмотря на бросающиеся
в глаза различия в онтогенезе и морфологическом значении. Результаты
первых опытов показали с несомненностью, что в условиях эксплантата
покровные клетки мозговых и серозных оболочек обнаруживают чрезвы-
чайно резко выраженные различия. Но вместо того, чтобы дать ожидав-
шиеся обычные картины роста соединительной ткани, мозговые оболочки
обнаружили какие-то, в то время еще незнакомые и очень своебразные.
гистобластические потенции.
Только после того, как был предпринят систематический эксперимен-
тально-гистологический анализ разных тканей, объединенных затем в эпен-
димо-глиальный тканевой тип, стало возможным правильно понимать
и трактовать полученные результаты.
Прежде всего бросилось в глаза при эксплантации мозговых оболочек
резко выраженное разжижение свертка фибрина и образование зоны роста,
которая состояла из комплексных клеточных соединений, рыхлых сетевидных
структур, клеточных тяжей и многочисленных свободных макрофагоподоб-
ных изолированных клеток. В составе комплексных разрастаний, состоящих
из вытянутых в длину элементов, можно было различить отдельные клеточ-
ные потоки, образующие перекресты и вихревые структуры. Комплексные
участки, с отходящими от них и частично соединяющимися друг с другом
тяжами, образовали характерные звезды и лучи.
Источником возникновения всех этих структур были покровные клетки.
Такой же характер зона роста сохраняла после ряда пересевов до предель-
ных изученных сроков культивирования.
Если соединительнотканная основа мозговых оболочек и прорастала
местами, что наблюдалось лишь в редких случаях, то в дальнейшем рост
соединительной ткани совершенно заглушался покровными клетками.
Только в отдельных эксплантатах, в которые попадали участки довольно
крупных сосудов с обильной окружающей их соединительной тканью при
полном или почти полном отсутствии покровных клеток, удавалось наблю-
дать развитие зоны роста обычного для мезенхимных производных характера.
В большей части случаев эксплантаты из мозговых оболочек обнаруживали
не только большое сходство, но даже полное тождество с картинами роста
нейроглии зрительных путей, эпителия сосудистых сплетений мозга и неко-
торых других тканей эпендимо-глиального типа, дающих часто совершенно
идентичные разрастания. От нейроглии центральной нервной системы
культуры из мозговых оболочек отличались не в большей степени, чем
нейроглия зрительных путей.
Таким образом, в отношении мягких мозговых оболочек, может считаться
установленным, что элементы т. наз. экто.мезенхимы, образующиеся
из нейрального зачатка, дают начало особой дифференцированной ткани,
которая в отношении своих гистобластических потенций чрезвычайно резко
отличается от производных обычной мезенхимы или «энтомезенхимы» и,
Экспериментальный метод в гистологии
279
в соответствии со своим происхождением, обнаруживает поразительное
сходство с другими производными нервного зачатка.
В появившейся почти одновременно работе Бленда (1939) автору не
удалось обнаружить столь резко бросающихся в глаза особенностей роста
мозговых оболочек. Не отдавая себе отчета в специфических особенностях
наблюдавшихся картин и, повидимому, исходя из наличия в зоне роста
сетевидных клеточных соединений и быть может клеточных тяжей, этот
автор не сумел отличить характерных для нейроглии структур от разраста-
ний мезенхимных производных и пришел к выводу, что и мозговые оболочки
и изучавшиеся им параллельно их опухоли растут по соединительноткан-
ному типу. В качестве материала для сопоставления он даже воспользовался
моими данными по эксплантации человеческой мезенхимы, которая не
обнаруживает сколько-нибудь заметного сходства с покровными клетками
мозговых оболочек.
Недостаточно углубленный и точный гистологический анализ, а также,
вероятно, и недостаточный личный опыт нередко приводят к подобного рода
курьезам.
Работа Сальмана, предпринятая у меня в лаборатории, но, к сожалению,
оставшаяся незаконченной, позволяет сделать предварительный вывод,
что и в составе твердой мозговой оболочки имеются элементы, повидимому,
покровные клетки субдурального пространства, которые в отношении
своих гистобластических потенций не обнаруживают заметных отличий
от соответствующих элементов мягких мозговых оболочек. В этом отношении,
однако, необходимо ждать окончательных результатов.
Экспериментальные исследования над мозговыми оболочками в составе
организма, в частности, опыты с прижизненной окраской {Вулард, 1924;
Куби и Шульц, 1925), могли констатировать наличие в их составе клеток
с гранулами краски. При воспалительных процессах наблюдается обра-
зование крупных, энергично воспринимающих краску, макрофагов {Маклин
и Маклин, 1920) и, быть .может, правильнее, макрофагоподобных клеток.
Часть авторов считает, что «макрофаги» образуются за счет плоских покров-
ных клеток {Эссик, 1920; Вулард, 1924).
Принимая во внимание наши данные, полученные методом эксплантации,
участие покровных клеток в образовании макрофагоподобных элементов
представляется вполне вероятным. Это однако, разумеется, отнюдь не
значит, что эти макрофагоподобные клетки тождественны обычным мезен-
химным макрофагам.
При воспалительных процессах в мозговых оболочках у человека,
например при туберкулезном менингите, в субарахноидальном пространстве
может скопиться огромное количество свободных клеток. Однако никаких
указаний на специфические особенности покровных клеток этим путем полу-
чить не удалось.
Единственными, пока известными процессами в составе целого орга-
низма, структурные особенности которых можно объяснить своеобразными
тканевыми свойствами покровных клеток, являются, как это отмечалось
выше, образования т. наз. паккионовых грануляций и некоторых опухолей.
280
Глава III
Паккионовы грануляции сами по себе не представляют образований, имею-
щих какое-нибудь особенное биологическое или патологическое значение.
Но в отношении своей тонкой структуры они заслуживают, как показали
Оверлеи (1922), Оверлеи и Нордман (1927), внимания, отличаясь по свое;
образному концентрическому расположению клеток от образований соеди*
нительнотканного характера. К опухолям мозговых оболочек мы еще вер-
немся в конце этой главы.
Установление нейроглиальной природы покровных клеток мозговых
оболочек заставляет пересмотреть общепринятые взгляды на т. наз. дес-
цеметов эндотелий роговицы (Хлогшн, 1940), который нельзя не сопоставить
с покровными клетками твердой мозговой оболочки, поскольку фиброзная
оболочка глазного яблока, а следовательно, и роговица, являющаяся ее
передней частью, соответствует: этой мозговой оболочке. Как известно,
десцеметов эндотелий принято рассматривать, как соединительнотканное
образование. Однако его вполне эпителиоподобный характер и правильная
форма клеток представляются не вполне обычными для мезенхим-
ных элементов, а сама десцеметова оболочка по своим свойствам отличается
от обычных межклеточных веществ мезенхимных производных. Данные
Мацуи (1929), полученные методом эксплантации, хотя и отличаются
недостаточной полнотой, все-таки позволяют сделать вывод, что в условиях-
тканевых культур десцеметов эндотелий очень заметно отличается по своим
свойствам от клеток соединительной ткани. В условиях репаративной реге-
нерации в десцеметовом «эндотелии» отмечалось появление митозов, причем
его клетки постепенно закрывали дефект на задней поверхности роговицы
(Петер, 1889). Все это, вместе взятое, заставляет думать, что десцеметов
эндотелий, подобно покровным клеткам мозговых, оболочек, представляет
собой структуру нейроглиального характера и нейрогенного происхождения,
резко отличающуюся по своим свойствам от мезенхимных («энтомезенхим-
ных») производных.
Нейроглиальные элементы мозговых оболочек и некоторых других
аналогичных им образований, которые отмечались выше («перинейральный
эндотелий» периферических нервов и, возможно, клетки оболочек инкапсу-
лированных нервных окончаний), можно обозначить собирательным тер-
мином «экзонейральная» или «перинейральная» глия. Нужно думать,
этим элементам свойственно в какой-то степени барьерное значение, харак-
теризующее нейроглию, и участие в образовании или, по крайней .мере,
поддержании вокруг разных частей нервной системы среды особого харак-
тера, отличающейся от общей соединительнотканной внутренней среды
организма.
Напомним кстати, что состав жидкости внутренних полостей мозга.,
субарахноидального и субдурального пространства, а также передней
камеры глаза обнаруживает большое сходство и очень существенно отли-
чается от крови, лимфы и тканевой жидкости соединительной ткани. Наконец,
между внутренними полостями мозга и субарахноидальным пространством
через отверстие Лушка и Мажанди, повидимому, имеется сообщение, что
косвенно также говорит в пользу гистологического сходства между эпен-
Экспериментальный метод в гистологии
281
димой, а следовательно, и глией, и покровными клетками мозговых обо-
лочек.
о) Перепончатый лабиринт
Ткани перепончатого лабиринта представляют очень большие трудности
для экспериментального анализа. Посвященных ему исследований, сделан-
ных методом эксплантации, кроме работы Фелл (1928), имеющей эксперш
ментально-эмбриологический характер и показавшей, что слуховой пузырек
куриных зародышей способен к само дифференцировке, не имеется. В течение
ряда последних лет мной ставились опыты с эксплантацией частиц перепош
чатого лабиринта новорожденных кроликов.
Большой, накопившийся за это время, материал, однако, еще не был
вполне обработан, а в настоящее время неизвестно, сможет ли быть про-
Рис. 45. Край клеточной мембраны, образовавшейся из кортиева
органа, в 4-дневной культуре перепончатой улитки 1-дневного
кролика. Сильное увеличение. (Оригинал.)
должено его изучение. Поэтому я позволю себе в общих чертах изложить
полученные данные, имеющие предварительный характер.
Извлеченный из височной кости новорожденных кроликов перепончатый
лабиринт измельчался и сеялся по методу висячей капли. Благодаря про-
зрачности его тканей удается установить, что в отдельные культуры попа-
дали или участки улитки с хорошо различимым кортиевым органом или
различные части вестибулярного аппарата или эндолимфатический проток.
При интравитальном наблюдении и на препаратах можно бывало обнаружить
экстенсивный рост перилимфатической части лабиринта и его внутренней
эпителиальной или эпендимной выстилки. Явлений эпителизации никогда
не наблюдалось. В некоторых культурах имели место разрастания и эпите-
лия и перелимфатической части стенки, в очень редких случаях был полу-
чен рост чистого эпителия и довольно часто — пролиферация одной лишь
перилимфатической части, причем эпителий сохранялся внутри не вскрытого
широко отрезка лабиринта и не появлялся на его наружной поверхности..
282
Глава III
И перилимфатический покров лабиринта и его внутренняя выстилка энер-
гично разжижают при своем росте сверток фибрина, в котором большей
частью уже вскоре наблюдается образование полостей, связанное с дефор-
мацией зоны роста.
Разрастания внутренней выстилки разных участков лабиринта имеют
вид очень однообразных плоскостных эпителиоподобных мембран, состоящих
из одного слоя тонких распластанных клеток, тесно сомкнутых своими
краями (рис. 45). Лишь периферические части такой плоскостной зоны
роста могут обнаруживать разрыхление и изоляцию отдельных элементов.
Ретракция и округление клеток выстилки лабиринта наблюдались лишь
изредка. Даже в местах, где периферического разрыхления не наблюдается,
краевые клетки часто образуют, наряду с более толстыми цитоплазмати-
ческими выростами, довольно сложную систему тонких нитевидных отрост-
ков, нередко ветвящихся и соединяющихся друг с другом, но не дости-
гающих, как правило, большой длины. В составе мембран форма и размеры
клеток могут заметно варьировать. В значительной части случаев клетки
имеют многогранные очертания, а местами, например при прорастании
элементов кортиева органа, оказываются первоначально сильно вытяну-
тыми в длину. Несмотря на это, в большей части случаев бросается в глаза
преобладающая, очень правильная, совершенно круглая или лишь слегка
овальная форма ядер, на светлом и почти гомогенном фоне содержимого
которых выступают относительно мелкие ядрышки. В элементах мембран
постоянно наблюдались немногочисленные митозы. Изредка в клетках
мембран встречались в небольшом количестве мелкие пигментные зерна.
Заметно отличаясь от многих описанных выше эпендимо-глиальных разра-
станий, мембраны из внутренней выстилки лабиринта в то же самое время
совершенно не обнаруживали сходства с находящимися в аналогичных
экспериментальных условиях эпидермальными тканями.
Разрастания перилимфатической части лабиринта отличались значи-
тельно большим разнообразием и во многих случаях обнаруживали большое
сходство с покровными клетками мозговых оболочек и некоторыми модифи-
кациями нейроглии. Характерными для этих разрастаний структурами
являются мембраны, клеточные пласты и тяжи, состоящие частью из поли-
гональных, частью из вытянутых в длину клеток, и рыхлые сетевидные
клеточные соединения. Изоляция округлившихся и макрофагоподобных
клеток нередко была выражена в сильной степени. Кроме того, в зоне роста
наблюдалась миграция отростчатых хро.матофор с темными включениями
пигмента, которые в очень сильной степени изменяли свою форму. Митоти-
ческая активность клеток перилимфатической части лабиринта была во
многих случаях очень высока. Хотя в настоящее время еще преждевременно
анализировать многие и часто существенные детали, даже то общее впечатле-
ние, которое получается на основании очень кратких и неполных приведен-
ных выше данных, позволяет, мне кажется, сделать некоторые выводы.
Сходство роста вне организма перилимфатической части лабиринта с неко-
торыми модификациями нейроглии безусловно подчеркивает и углубляет
ее отмеченную выше гистологическую гомологию с мягкими мозговыми
Экспериментальный метод в гистологии
283
оболочками и подтверждает точку зрения на перепончатый лабиринт, как
на специализированный в особом направлении участок центральной нервной
системы. Вместе с тем большие различия в гистобластических потенциях
между выстилкой лабиринта, с одной стороны, и нейроглией с эпендимой —
с другой, показывают, что эволюция этой ткани и других вспомогательных
компонентов центральной нервной системы еще у отдаленных предков
позвоночных пошла дивергентно, и ее результаты выразились в приобретении
ими очень стойких гистологических различий. Тот факт, что полость лаби-
ринта еще у некоторых низших позвоночных, например селяхий, сообщается
с внешней средой, а полная изоляция от внешней среды собственно централь-
ной нервной системы произошла еще у предков позвоночных, делает отчасти
понятной резко подчеркнутую у выстилки лабиринта .морфо-физиологиче-
скую характеристику эпителиальных тканей с высокой степенью комплекс-
ности и способностью к росту в экспериментальных условиях эпителио-
подобными мембранами. Кроме того, вероятно, анатомо-физиологические
особенности органа, как стато- и фонорецептора, также благоприятствовали
сохранению у выстилки лабиринта свойств, характеризующих разграни-
чительные и покровные ткани.
е) Нейроглия периферической нервной системы
Совершенно противоположное направление филогенетического развития
характеризует, как показывают экспериментальные данные, нейроглию
периферической нервной системы. Если при опытах с эксплантацией частиц
центральной нервной системы все авторы подчеркивали «эпителиальный»
или, точнее, в значительной мере комплексный характер наблюдавшихся
разрастаний, то в относительно немногочисленных работах, посвященных
культивированию вне организма зародышевых спинальных или симпати-
ческих ганглиев (Мосса, 1928, 1929; Эзаки, 1929; Леви и Делоренци, 1934;
Леви и Млйер, 1940, и некоторые другие), кроме роста «нервных волокон»,
обычно отмечалось появление в зоне роста клеток, обозначавшихся как фибро-
бласты, хотя и высказывалось иногда предположение, что по крайней мере
часть этих «фибробластов» является по существу леммобластами, т. е.
соответствует эмбриональным шванновским клеткам. Никаких попыток
изучить специфические особенности этих фибробластов или даже отметить
их отличия от клеток соединительной ткани не делалось. Правда, в старой
работе Ингебригтсена (1916), которая обычно почему-то не цитируется,
посвященной культивированию вне организма частиц периферических
нервов, отмечается прорастание сплетающихся друг с другом клеточных
нитей, которые, по мнению автора, образуются шванновскими клетками.
Удачные культуры Ингебригтсену удавалось получить только из нервов
в состоянии уоллеровского перерождения, начиная с пятого дня после
перерезки. Нормальные нервы, по его данным, не обнаруживали явлений
роста. Мои наблюдения над культурами частиц блуждающего нерва кро-
ликов (1939, 1940) и данные Сутулова (1941) по эксплантации спинальных
ганглиев того же животного позволяют в настоящее время составить себе
284
Глава 111
достаточно полное представление о гистобластических потенциях шван-
новских и мантийных клеток. *
Предметом моей работы были шванновские клетки блуждающего нерва
зародышей, незадолго до рождения, новорожденных и молодых кроликов.
В части опытов предварительно (за семь-десять дней) производилась
двойная перевязка нерва, причем для эксплантации использовались части
ствола между лигатурами, когда явления уоллеровского перерождения
уже были резко выражены. Эксплантируемые участки нервов по возможности
освобождались от соединительнотканных оболочек. Культуры изучались
до двух месяцев срока жизни. Внутри посеянных частиц происходят обыч-
ные явления перерождения. Осевые цилиндры дегенерируют и исчезают
по истечении нескольких дней. Если нервные волокна уже были заметно
миэлинизированы, то наблюдаются типичные картины перерождения миэли-
новой оболочки с образованием четковидных вздутий и комков миэлина,
подвергающихся рассасыванию, которое, в отличие от картин дегенерации
в- составе организма, происходит без заметного участия блуждающих мезен-
химальных клеток. Отдельные переполненные жировыми каплями шван-
новские элементы, превращаясь в крупные сетчатые или пенистые клетки.,
мигрируют из центрального кусочка в образующуюся зону роста. Большая
же часть шванновских клеток начинает прорастать после их освобождения
от миэлина (около четвертого, пятого дня после посева), которое в нервах
молодых кроликов происходит очень быстро. Если нервные волокна еше
не были заметным образом миэлинизированы, то прорастание шванновских
клеток начинается значительно раньше, часто в течение второго дня. Наряду
со шванновскими клетками активируются и элементы соединительной
ткани, которые, однако, лишь в редких случаях накладывают заметный
отпечаток на характер зоны роста. В большей части изученных культур
структуры зоны роста имели достаточно своеобразный характер, зависевший
от преобладания в ней шванновских клеток. В ряде случаев, после повтор-
ных пересевов, удавалось получить их чистые культуры без сколько-нибудь
заметной примеси растущих элементов соединительной ткани. Рост шван-
новских клеток сопровождался сильным разжижением фибрина и образова-
нием в нем полостей. По относительно слабо выраженной комплексности,
зона роста в целом или ее отдельные участки, при недостаточно внимательном
изучении, действительно нередко могли произвести впечатление соединитель-
нотканных разрастаний, образованных клетками типа фибробластов. Однако
это первое впечатление совершенно не соответствует действительному поло-
жению вещей, хотя и объясняет тот факт, почему ряд авторов говорил про
рост фибробластов в эксплантатах из частей периферической нервной
системы. Подобно тому, как комплексные разрастания различных тканей
по своим особенностям очень сильно отличаются друг от друга и даже по
сути дела далеко не всегда имеют тот вид, который принято связывать
с представлением об эпителиальном характере, и более рыхло связанные
* Мои данные, касающиеся шванновских клеток, были полностью подтверждены
в недавно опубликованной работе Меррей и Стаут (Anat. Rec., 84, 1Q42), выполненной
на человеческом материале.
Экспериментальный метод в гистологии
285
друг с другом клетки далеко не всегда образуют тождественные гисто-
логические структуры, которые, учитывая их различия, было бы целесо-
образно обозначать, как соединительнотканные. Разумеется, здесь речь
идет не об отдельных, вырванных из их взаимосвязи, участках разрастаний
Рис. 46. Чистая 50-дневная культура шванновских клеток блужда-
кщего нерва 8-дневного кролика. Слабое увеличение, (Оригинал.)
в один какой-нибудь момент их жизни, зафиксированный в препарате,
действительно не могущих быть идентифицированными даже очень опытным
исследователем, а о совокупности структур, изучаемых в их взаимосвязи
в пространстве и во времени. Эти, чрезвычайно простые по существу, но
Рис. 47. Рыхлый участок зоны роста, образованной шванновскими
клетками в 13-дневной культуре блуждал щего нерва 1-дневного
кролика. Сильное увеличение. (Оригинал.)
имеющие фундаментальное значение для правильного понимания всех без
исключения явлений из области гистологии соображения, пожалуй, с осо-
бенной отчетливостью иллюстрируются на примере вспомогательных тканей
нервной системы. Поэтому мне представляется правильным обратить на
них в этом месте лишний раз внимание. Шванновские клетки образуют
286
Глава III
в условиях экстенсивного роста густые массивные клеточные соединения
и плоскостные мембраны, отличающиеся различной степенью комплекс-
ности, рыхло расположенные или образующие пучки, большей частью,
узкие клеточные тяжи, сетевидные структуры и изолированные клетки
разной формы. Наиболее характерными для шванновских клеток формами
существования являются узкие нитевидные клеточные тяжи или цепочки,
нередко достигающие очень значительной длины, а также изолированные
нитевидные клетки (рис. 46, 47). В составе массивов и мембран клетки часто
имеют удлиненную форму (рис. 48), хотя отдельные участки комплексных
плоскостных разрастаний могут иметь эпителиоподобный вид мозаики
довольно правильных полигональных элементов. Менее комплексные
плоскостные структуры часто состоят из клеток с многочисленными корот-
кими отростками, отхо-
дящими в разные сторо-
ны от их тела и соеди-
няющимися друг с дру-
гом. При еще более зна-
чительной степени раз-
рыхления образуются
упомянутые выше сете-
видные соединения фиб-
робластоподобных кле-
ток. В очень многих
случаях на долю более
комплексных структур
приходится лишь отно-
сительно небольшая цен-
тральная область зоны
роста, а ее широкая пе-
Рис. 48. Комплексный центральный участок зоны роста,
образованный шванновскими клетками в 14-дневной
культуре блуждающего нерва 0-дневного кролика. В
клетках удлиненной формы заметна фибриллярность.
Сильное увеличение. (Оригинал.)
риферическая часть представлена клеточными тяжами, расходящимися ра-
диально и могущими местами сетевидно соединяться друг с другом.
В шванновских клетках удается наблюдать появление фабриллярной
структуры в цитоплазме,которая, однако, не превращается в грубую волокни-
стость, свойственную описанным выше глиальным массивам в культурах
из головного и спинного мозга (рис. 48). Кроме того, шванновские клетки
обнаружили, подобно нейроглии зрительных путей, способность к обра-
зованию аргирофильных сетей межклеточного вещества. Эти и некоторые
другие аналогичные данные показывают, как это мной уже отмечалось
(Хлопин, 1939, 1940), что способность к образованию межклеточных веществ,
но крайней мере их примитивных форм, является не специфическим призна-
ком какой-нибудь одной тканевойгруппы , а, быть может, общим элементарным
свойством Живой протоплазмы, которое в некоторых случаях достигает
высокого развития, а в других, наоборот, может утрачиваться. Тесная и,
повидимому, относительно прочная взаимная связь удлиненных клеток
своими концами сочетается с незначительной степенью комплексности их
соединения сторонами. Эта особенность, отмечавшаяся выше при рассмотре-
Экспериментальный метод в гистологии
287
нии нейроглии зрительных путей, у шванновских клеток бывает выражена
в еще большей степени.
Если в отношении комплексности структур расположить эпендимо-
глиальные ткани разных отделов нервной системы в ряд, то один конец
его займут выстилка лабиринта, нейроглия головного и спинного мозга,
затем пойдут ретинальные листки, покровные клетки сосудистых сплетений,
ткань эпифиза, нейроглия зрительных путей, экзонейральная глия мозговых
оболочек и, наконец, шванновские клетки. Совершенно несомненно, что
нейроглия разных отделов центральной нервной системы, отличаясь друг
от друга в отношении гистобластических потенций, еще более резко отли-
чается от шванновских клеток, которые, вообще говоря, принято сопостав-
лять, а иногда едва ли не отождествлять с элементами олигодендроглии.
Несомненно, шванновские клетки представляют собой высокоспециализиро-
ванную часть вспомогательных элементов нервной системы, которые в фило-
генезе приобрели ряд стойких отличий, возможно связанных с тем, что они
в большей степени, чем центральная глия, утратили пограничную функцию
и приобрели свойства особой ткани внутренней среды организма с более
слабо выраженным барьерным значением.
В опытах с эксплантацией частиц спинальных ганглиев новорожденных
кроликов (Сутулое, 1941) удалось получить энергичные разрастания не
только отростков чувствительных нейронов, но и вспомогательных элемен-
тов, т. е. сателлитов и шванновских клеток. Сверток фибрина подвергался
очень сильному разжижению с образованием полостей. Наблюдавшиеся
картины роста очень напоминали рост шванновских клеток в культурах
из блуждающего нерва, хотя типичные для последних узкоклеточньд*
тяжи встречались реже и в меньшем количестве. Наиболее типичные для
шванновских клеток разрастания получались, как следовало ожидать,
из концов корешков, из которых создавались характерные узкие, заострен-
ные, веерообразно расходящиеся лучи. Образующаяся из частиц самих
ганглиев зона роста имела преимущественно плоскостной характер клеточ.г
ных мембран и рыхлых сетевидных клеточных соединений, от которых
изолировались то в большем, то в меньшем количестве отростчатые фиброг
бластоподобные клетки. Наблюдалось также образование круглых или
амебоидных макрофагообразных элементов. В составе мембран в относи-
тельно редких случаях наблюдалась эпителиоподобная мозаика полигональ-
ных элементов. Чаще клетки имели удлиненную форму и располагались
ориентированными в разных направлениях потоками, образуя подчас
и вихревые структуры. В более толстых массивных участках зоны роста,
расположенные двумя-тремя слоями клеточные потоки взаимно перекре-
щивались в разных направлениях. На фоне плоскостных мембран и сете-
видных клеточных соединений удавалось наблюдать также характерные
для шванновских клеток длинные нитевидные тяжи. Повидимому, сател-
литы спинальных ганглиев в отношении своих гистобластических потенций
несколько отличаются все-таки от собственно шванновских клеток, хотя
разрастания тех и других обнаруживают много общих черт и представляют
собой две нерезко отграниченные модификации общего для элементов пери-
288
Глава III
ферической нейроглии типа роста, который в свою очередь обнаруживает
и ряд общих для всей нейроглии черт.
В составе целого организма при экспериментальных и патологических
условиях гистобластические потенции шванновских клеток выявляются
с меньшей полнотой и отчетливостью, чем при их пролиферации в тканевых
культурах. При перерождении нервов после перерезки или ранений у млеко-
питающих и человека воспалительная реакция в соединительной ткани
осложняет и затемняет картины изменений в шванновских клетках (Р. Ка-
халь, 1928; Дойников, 1911; Шпильмейер, 1922, и др.). Относительно
происхождения характерных сетчатых или пенистых клеток, заполненных
жировыми включениями, имеются по существу такие же разногласия,
как и относительно их возникновения в центральной нервной системе.
Частично они несомненно представляют собой мезенхимальные макрофаги,
но, как показывают опыты с эксплантацией, в них безусловно могут пре-
вращаться и сами шванновские клетки. Образующиеся из шванновских
клеток после перерождения бюнгнеровские тяжи представляют собой,
в известной степени, структуры, сходные с растущими вне организма клеточг
ными цепочками. В образовании тех и других проявляется, во всяком
случае, одна и та же гистотипическая особенность шванновских клеток —
способность к образованию длинных и узких клеточных тяжей. Еще
отчетливее, пожалуй, проявляется это же цитотипическое свойство при
образовании т. наз. шванном, полученных Нажоттом (1932) при транс-
плантации частиц периферических нервов. Разрастаясь и пролиферируя,
шванновские клетки образовали в этих опытах клубковидно-закрученные
клеточные тяжи. Для изучения шванновских клеток и явлений дегенерации
нервов низшие позвоночные до настоящего времени почти совершенно не
использовались. По данным моего сотрудника Лавренко, отчасти воспол-
няющим этот пробел, дегенерация седалищного нерва лягушки (1941) и
аксолотля (неопубликованные данные) происходит после перерезки в гораздо
более простой, чем у теплокровных позвоночных, обстановке, так как воспа-
лительная реакция в соединительной ткани практически совершенно отсут-
ствует и рассасывание миэлина происходит только благодаря жизнедеятель-
ности самих шванновских клеток. После окончания процесса перерождения
шванновские клетки амфибий превращаются в протоплазматические тяжи
с отчетливо заметной продольной волокнистостью.
ж) Эмбриональные зачатки нервной системы и их гистобластические
потенции
Область презумптивного нервного зачатка амфибий, первоначально
способная к дифференцировке в весьма разнообразные ткани, после обра-
зования подстилающей его дорзальной стенки первичной кишечной трубки
оказывается стойко детерминированной и обнаруживает способность к диф-
ференцировке только в направлении нейрональных и вспомогательных
компонентов нервной системы. Совершенно особыми гистобластичсскими
потенциями обладают нервные валики и образующаяся из них ганглиозная
Экспериментальный метод в гистологии
289
пластинка, дающая начало т. наз. «мезэктодерме» или «эктомезенхиме»
(Стон, 1926, 1929, 1932; Равен, 1931—1936, и др.). Из элементов эктоме-
зенхимы или, точнее, нейромезенхимы возникают, по существующим дан-
ным, совершенно, казалось бы, несвойственные нервному зачатку диф-
ференцировки, и, в частности, образуется значительная часть висцераль-
ного хрящевого скелета головы.
Как уже отмечалось выше, часть нейромезенхимы принимает участие
в образовании мозговых оболочек, нейрогенные элементы которых, в соот-
ветствии со своим происхождением, обнаруживают гистобластические
потенции тканей глиоэпендимного типа, а отнюдь не обычных «энтомезен-
химных» соединительнотканных клеток. Мне представляется более чем
вероятным, что экспериментально-гистологический анализ хрящей нейро-
генного происхождения, которые и при обычных условиях отличаются
известными структурными особенностями от «энто.мезенхимной» хрящевой
ткани, позволит обнаружить их совершенно особые биологические свойства,
связанные с их развитием из нервного зачатка. Правда, мне кажется, что
и самый факт развития хрящевой ткани из нервного зачатка еще нуждается
в дополнительном изучении и уточнении. В разных участках нервного
зачатка возникает резко выраженная анатомическая и отчасти гистоло-
гическая мозаичность. Многочисленными опытами с трансплантацией
разных участков нервной пластинки и плакод была демонстрирована их
способность к формообразованию различных отделов нервной системы,
в составе которых наблюдались и более или менее далеко идущие явления
специфически гистологической дифференцировки.
Согласно данным Гирсберга (1924) и Детлаф (1936), детерминация
нервной пластинки у бесхвостых амфибий сводится к перемещению и кон-
центрации в ее области, под влиянием организатора, элементов глубокого
слоя эктодермы, который и представляет собой зачаток элементов нервной
системы. Клетки наружного слоя эктодермы, вне области нервной пластинки,
не участвуют в ее образовании, а в составе этой последней, по позднейшим
данным Детлаф (1940), повидимому, дифференцируются в эпендиму нервной
трубки. Эти очень интересные наблюдения настоятельно нуждаются в даль-
нейшей разработке, а также в сопоставлении с тем, что известно о детерми-
нации и развитии нервной пластинки у хвостатых амфибий и других позво-
ночных. Только после этого сделается возможным уточнить паши предста-
вления об общих закономерностях дифференцировки нервного зачатка
позвоночных и, в частности, о взаимоотношениях в эмбриогенезе между
спонгиобластами, нейробластами и т. наз. недифференцированными клет-
ками нервной трубки.
Новейшими опытами с удалением, эксплантацией и пересадками участков
нервных валиков (Дюшен, 1938, 1939; И. Фишер, 1938; Доррис, 1940, и др.)
не только подтвердились наблюдения Гаррисона (1906) над развитием
шванновских клеток из ганглиозной пластинки, но быт о также доказано
образование из того же источника хро.матофор, которые до последнего
времени большей частью рассматривались, вопреки старым данным Вей-
денрейха (1912), как клетки соединительной ткани, развивающиеся из общего
19 Хлопни*-349
290
Глава 111
с прочими ее элементами источника — мезенхимы или «энтомезенхимы»
Эти данные ставят хроматофоры в ряд производных нервного зачатка и
заставляют их считать особыми высокоспециализированными элементами/
гистоморфные и цитоморфные особенности которых еще нуждаются в спе-
циальных исследованиях. Структура хроматофор и сложных образуемых
ими хроматофорных органов низших позвоночных (Балловиц, 1931) заста-
вляет думать, что они являются совершенно особыми образованиями,
которые, подобно нейронам и мышечным волокнам, приобрели чрезвычайно
сложную организацию и превратились в очень своеобразные живые системы,
соединяющие в себе признаки клетки и целого органа и подчиненные в своей
функции специальным гуморальным и нервным влияниям. Со временем
учение о хроматофорах, вероятно, составит особую часть главы эксперимен-
тальной гистологии, посвященной тканям нервного происхождения.
Хроматофоры представляют собой лишний пример структур организма,
которые не укладываются в обычную морфо-физиологическую систему
гистологии, так как их рассмотрение в главе о соединительной ткани оправ-
дывается в настоящее время только топографическим признаком.
Опыты с эксплантацией разных частей нервного зачатка амфибий,
произведенные в течение ряда последних лет моим сотрудником Винниковым
(неопубликованные данные), показывают, что их клеточный материал,
способный первоначально к очень своеобразным явлениям цитоморфной
дифференцировки, со временем дает вне организма малодифференцированные
тканевые разрастания, которые по своим гистологическим особенностям
обнаруживают замечательное сходство со структурами глиального типа,
разобранными выше на примере млекопитающих. Это относится к различ-
ным участкам нервной пластинки, в частности, к области глазных зачатков
и нервным валикам. Таким образом, клеточный материал нервного зачатка
может, не проделывая свойственных ему формообразовательных процес-
сов, обнаруживать общие гистоморфные особенности развивающихся из него'
в дальнейшем вспомогательных тканей нервной системы. Это вновь подчер-
кивает значение эмбриональных зачатков, как специфических родоначаль-
ников определенного генетического единого тканевого типа. Осуществляе-
мые частью этого клеточного материала на более ранних стадиях культи-
вирования цитоморфные специальные дифференцировки также представляют
очень большой интерес. В эксплантатах из разных участков нервной пла-
стинки наблюдается дифференцировка своеобразных нейроноидных эле-
ментов, которые в данном случае, повидимому, возможно трактовать как
нейрональные, а не астроцитарные образования. В дальнейшем они деге-
нерируют и исчезают. В культурах из нервных валиков с большой отчет-'
ливостью прослеживается в образующейся плоскостной зоне роста диф-
ференцировка различных хроматофор—типичных звездчатых .меланофор,
переполненных зернами темного пигмента, липофор с характерными жел-
тыми включениями, растворяющимися при обработке спиртом и ксилолом
фиксированных препаратов и тому подобное. Поразительные явления
цитоморфной дифференцировки, на фоне мало дифференцированных гисто-
морфных разрастаний, удалось получить Винникову при эксплантации
Экспериментальный метод в гистологии
291
частиц глазного зачатка. Отдельные клетки зоны роста обнаружили спо-
собность к превращению в фоторецепторные клетки с типичными слоистыми
члениками палочек. Эти наблюдения показывают с большой убедительностью
не только большую степень независимости друг от друга в онтогенезе про-
цессов морфогенеза и гистогенеза, но такую же независимость специфической
цитоморфной дифференцировки от общего хода гистогенеза и морфогенеза.
В изолированных частицах глазных зачатков, развивающихся, как единая
морфогенетическая система, далеко идущие процессы цитоморфной диф-
ференцировки бывают сопряжены с общим ходом гистогенеза разных частей
образующегося глазного бокала.
Мне кажется, достаточно немногих приведенных в этом параграфе
фактов, чтобы показать, что развиваемые нами на основе экспериментально-
гистологического анализа дифференцированных тканей представления об
их естественной генетической системе и лежащее в основе последней
учение о гистологической детерминации полностью подтверждаются и вместе
с тем расширяются путем экспериментального изучения клеточного мате-
риала эмбриональных зачатков. Этим в значительной степени устраняются
кажущиеся противоречия между современной экспериментальной эмбрио-
логией и нашим экспериментальным подходом к проблеме филогистогенеза
или эволюционного развития тканей. Следует пожелать, чтобы эта взаимная
связь между экспериментальной эмбриологией и гистологией продолжала
укрепляться и сделалась бы прочным общим фундаментом этих родственных
друг другу по существу дисциплин.
з) Ткани нейрогенных опухолей
Учение о нейрогенных опухолях стало развиваться после того, как
Вирхов (1864) впервые обратил внимание на особый характер глиом и выде-
лил их в самостоятельную группу новообразований. С тех пор глава пато-
логии об опухолях нервной системы усиленно разрабатывалась и в настоящее
время превратилась в значительной степени самостоятельный отдел онко-
логии или учения об опухолях, разросшийся в целую новую область знания,
которая имеет выдающееся общебиологическое и специальное медицинское
значение.
Современное состояние вопроса о нейрогенных опухолях теснейшим
образом связано с цитоморфологической разработкой и детализацией
представлений о клеточных формах вспомогательных компонентов нервной
системы, их предполагаемыми генетическими отношениями друг к другу
и процессами эмбрионального гистогенеза тканей нервной системы, которые,
как это уже отмечалось выше, принято сводить к достаточно сложным
генеалогическим схемам (Бейли и Кашинг, 1930; Бейли, 1932; Гаккель,
1939, и др.). В учении об опухолях нервной системы, тесно связанном
с ее нормальной гистологией и эмбриологией, почти нет места необоснован-
ным спекуляциям на тему о межтканевых метаплазмах, которые все еще
пользуются известной популярностью в некоторых других отделах пато-
логии. Вместе с тем в существующих классификациях нейрогенных опу-
холей в сильной степени ощущаются большая схематизация и стремление
292
Глава III
вывести все их многообразие только из процессов гистогенеза, без учета
тех гистологических превращений, к которым способна дифференцированная
нейроглия при пролиферативных процессах. Изложенный выше фактический
материал показывает, что различные вспомогательные ткани нервной системы
способны к пролиферативным процессам, а вместе с тем возникает очевид-
ная необходимость все больше считаться в области нервной системы с опу-
холями не только дисонтогенетического характера, но и с возникающими
на почве пролиферативных процессов в дифференцированных вспомога-
тельных тканях. Сопоставление опухолевых структур с разными стадиями
развития нервной системы несомненно очень плодотворно, так как этим
путем действительно удается объяснить гистологические особенности ряда
новообразований. Следует даже пожелать, чтобы этот метод был шире
использован для анализа других бластомотозных тканей. Но желание
увидеть во всех опухолевых структурах нервной системы только аналогов
разных стадий ее эмбрионального гистогенеза безусловно представляется
односторонним, тем более, что в последнее время удалось вызвать экспе-
риментально образование глиом под влиянием концерогенных веществ.
Такие новообразования, как медуллобластома, со всем многообразием
наблюдающихся в ней дифференцировок, ретинобластомы или симпати-
кобластомы, возникающие в очень молодом возрасте и даже в эмбриональной
Жизни, и ряд других новообразований действительно нельзя, пожалуй,
объяснить иначе, как опухолевым превращением сохранившихся дисонто-
генетических клеток эмбриональных закладок. В отношении же различных
глиобластом, астроцитом и даже, быть может, опухолей, которые принято
обозначать, как олигодендроглиомы, менингобластом и различных большей
частью доброкачественных патологических пролифераций периферической
нервной системы нельзя не допускать возможности развития из соответ-
ствующих дифференцированных тканей.
Наряду с большим количеством опухолей, нейрогенная природа кото-
рых может быть установлена на основании свойственных им специфических
гистологических структур, несомненно имеются, как и в случае других
тканей, новообразования, нейроглиальное происхождение которых в высшей
степени вероятно, но структура которых отличается очень малодифферен-
цированным и неопределенным характером и не позволяет поставить точного
гистогенетического диагноза.
Так или иначе, подавляющее большинство опухолей нервной системы
несомненно полностью подтверждает высокую специфичность вспомогатель-
ных тканей нервной системы и неразрывную зависимость их гистологи-
ческих особенностей от развития из определенного детерминированного
эмбрионального зачатка, т. е. то положение, которое является краеуголь-
ным камнем нашей генетической системы тканей.
Одной из очень эффектных иллюстраций зависимости структуры опу-
холей от ее происхождения являются гистологические особенности упоми-
навшихся уже неоднократно опухолей мозговых оболочек, получивших
название менингобластомы (Оверлеи, 1922). Их структура настолько отли-
чается от соединительнотканных образований и обнаруживает настолько
Экспериментальный метод в гистологии
293
большое сходство с тонким строением некоторых опухолей центральной
и периферической нервной системы, что пришлось допустить их нейроген-
ное происхождение, а вместе с тем отказаться от общепринятой точки зрения
на развитие мозговых оболочек из мезенхимы.
В настоящее время упомянутые выше экспериментально-эмбриоло-
гические работы и работы нашей лаборатории, воочию демонстрирующие
нейроглиальную природу покровных клеток мозговых оболочек, подводят
прочные основы под их предполагаемое нейрогенное происхождение. Более
того, гистологические особенности, характерные для растущих вне орга-
низма покровных клеток мозговых оболочек и ряда других тканей эпендимо-
глиального типа, в частности, сосуществование эпителиоподобных клеточных
соединений и своеобразных вихревых структур, обнаруживают несомненное
сходство с аналогичными морфологическими особенностями менингобластом
и многих других нейрогенных опухолей. Это доказывает, что и в случае
последних опухолевые структуры могут быть поняты путем их сопостг вл зния
с гистологическими структурами, свойственными генетически однородной
нормальной ткани в состоянии неопухолевой пролиферации в эксперимен-
тальных условиях, а не только путем подыскивания сходных образований
в нормальном гистогенетическо.м ряду в соответствии с принятыми в совре-
менной нейропатологии установками.
Изучение гистобластических потенций шванновских клеток методом
эксплантации отчетливо демонстрирует несостоятельность точки зрения
Маллори (1920), принятой Пенфильдом (1932), согласно которой опухоле-
подобные разрастания периферических нервов, известные под названием
нейрином, рассматриваются, как производные соединительнотканных обо-
лочек и, в соответствии с этим, обозначаются термином «перинейральные
фибробластомы» и хорошо гармонируют с их вероятным развитием из
элементов периферической нейроглии.
Нейрогенные опухоли уже были предметом ряда работ, сделанных мето-
дом эксплантации (Рессель и Бленд, 1933; Джай и ДжаД 1936; Пинкус,
1936, 1937; Смирнов, по Тимофеевско.му, 1940; Тимофеевский 1939, 1940,
и др.). Однако гистологическая обработка во многих отношениях ценного
материала еще настолько неполна, что не позволяет составить себе пред-
ставлений о специфических тканевых особенностях разрастаний бласто-
матозных клеток вне организма и о степени вероятного сходства гисто-
бластических потенций нейрогенных опухолевых и нормальных тканей.
В большей части случаев авторы лишь констатируют факт роста опухолевых
клеток и ограничиваются очень поверхностным описанием наблюдавшихся
при этом картин. Для примера остановлюсь на одной из наиболее подробных
работ.
Из трех культивировавшихся Пинкусом (1937) человеческих опухолей
две имели нейрогенное происхождение и были диагностированы как эпен-
димома и полиморфная глиобластома. Клетки первой из них, росшей вне
организма почти до трех месяцев, отличались в зоне роста веретенчатой
или многогранной формой, а иногда приобретали, по данным автора, вееро-
образный вид и образовали длинные цитоплазматические отростки. Отно-
294
Глава III
сительно гистотипических особенностей разрастаний . опухолевых клеток
ничего не говорится. Вторая опухоль, культивировавшаяся в течение срока
до девяти месяцев, дала два разных типа роста: мелкоклеточный и крупно-
клеточный. Ряд культур, росших первоначально по крупноклеточному
типу, в дальнейшем превратился в мелкоклеточные разрастания. Этот
факт автор объясняет наличием в исходном опухолевом материале двух
стойких и различных клеточных форм, из которых одна в конце концов
вытесняет и заглушает другую. При мелкоклеточном типе в зоне роста
наблюдается очень сложный сетевидный переплет клеток. При культиви-
ровании в плазме морской свинки сверток фибрина не разжижается, и опу-
холевые клетки приобретают «эпителиальное» расположение.
При крупноклеточном типе роста автор отмечает первоначальный «эпи-
телиальный» характер клеток, который затем изменяется. Опухолевые
элементы приобретают самостоятельность, начинают обнаруживать чрез-
вычайный полиморфизм, приобретая весьма разнообразную удлиненную
форму: то широкую, лентовидную, то очень узкую, нитевидную, до 500 р.
в длину.
В других культурах, наоборот, преобладали клетки с тремя и более
отростками. В протоплазме клеток обнаруживалась волокнистость, тем
более отчетливая, чем слабее становился рост культур. Пинкус делает,
по существ}-, очень правильную, но неудачную по результату, по-
пытку сопоставить наблюдавшиеся им разрастания опухолевых клеток
с культурами из эмбриональной нервной системы куриных зародышей.
Последние им, очевидно, не изучались специально, а потому выбранные для
сравнения, повидимому, более или менее случайные культуры, насколько
можно судить по микрофотограмме, вопреки утверждению автора, обна-
руживают более чем приблизительное сходство. Наоборот, описания автора
и, особенно, приводимые микрофотограммы, убеждают в том, что многие
из наблюдавшихся им картин действительно обнаруживают очень большое
.сходство с изученными мной разрастаниями нейроглии центральной нервной
системы вне организма и. что опухолевая нейроглия двух изученных Пин-
кусом новообразований в отношении своих гистобластических потенций
несомненно стоит очень близко к нормальной. Микрофотограммы, приво-
димые в работе Бленда и Ресселя (1933), вполне согласуются с этим, пока
предварительным, утверждением.
Большого внимания заслуживают краткие данные Тимофеевского (1939,
1940), относящиеся к очень малодифференцированной злокачественной
опухоли периферического нерва, диагносцированной как злокачественная
нейринома, которая длительное время давала нехарактерную зону роста,
а на втором году культивирования обнаружила способность к образованию
длинных клеточных тяжей, которые обнаруживают поразительное сходство
с растущими вне организма нормальными шванновскими клетками (Хлопин,
1939, 1940).
Эти наблюдения имеют очень большое принципиальное значение. С одной
стороны, они позволяют установить экспериментально истинную тканевую
природу опухоли, исходная структура которой не обладала, в силу т. наз.
Экспериментальный метод в гистологии
295
катаплазии, никакими сравнимыми с нормальными тканями гистологи-
ческими особенностями, причем это удалось сделать лишь после очень
длительного культивирования. С другой стороны, они в очень убедитель-
ной форме подтверждают высказанную выше точку зрения, согласно кото-
рой при опухолевом превращении нормальные ткани не приобретают
новых гистобластических потенций, а лишь утрачивают в большей или
меньшей степени способность к их проявлению, разрастаясь малодиф-
ференцированными и потому во многих случаях малохарактерными клеточ-
ными соединениями или даже изолированными клетками. Эта утрата спо-
собности к образованию гистотипических тканевых структур, по крайней
мере в некоторых случаях, оказывается обратимой.
Как это мной уже отмечалось (Хлопин, 1940), изучение в нашей лабора-
тории методом эксплантации ряда тканей эпендимо-глиального типа
позволяет дать иную трактовку данным Хлопина и Цымбала (1934), каса-
ющимся роста вне организма элементов меланотической опухоли человека.
В настоящее время делается вероятным, в соответствии со взглядами Мас-
сона (1923, 1926) и Фута (1932), происхождение по крайней мере части
меланотических опухолей из нервного зачатка, а не из кожной эктодермы.
Такую же трактовку допускают, повидимому, и новейшие данные Вар-
шавской (1939), отметившей, что элементы меланотических опухолей чело-
века не образуют типичных для эпителиальных тканей мембран. То же
самое, повидимому, справедливо и в отношении меланотической опухоли
мышей, удачные культуры которой были получены Гулъдбергом (1935).
Значительный опыт, приобретенный в отношении различных нормаль-
ных эпендимо-глиальных тканей, поставил перед нами на очередь детальный
экспериментально-гистологический анализ тканей нейрогенных опухолей
под тем же углом зрения. Мои опыты с начала 1941 г. позволили накопить
довольно значительный новый экспериментальный материал, который уже
частью достаточно, разработан и поэтому может быть в настоящее время
резюмирован для сопоставления с данными, касающимися нормальной
нейроглии, а также с упомянутыми выше опытами по эксплантации нейро-
генных опухолей человека. Опыты ставились частью по методу висячей
капли, частью во флаконах, в которых человеческие опухоли, по данным
Джай и Джай (1936) и Тимофеевского (1939, 1940), обнаруживают очень
энергичный рост. В своей работе я пошел не по пути попыток длительного
культивирования разнообразного материала (при котором получаемые
результаты, очень эффектные в техническом отношении, неизбежно соче-
таются с недостаточно углубленным гистологическим изучением и с невоз-
можностью детально проследить по стадиям структурные изменения исход-
ного материала), а по пути возможно тщательного гистологического анализа
каждой отдельной опухоли, поскольку вероятность получения двух или
нескольких совершенно идентичных глиом очень мала, и каждый опухо-
левый случай представляет собой, по крайней мере на первых порах, само-
стоятельную и достаточно трудную проблему. Поэтому я большей частью
распределял культивируемый материал таким образом, чтобы продлить
жизнь опухолевых частиц вне организма до полутора, реже до двух или
296
Глава III
трех месяцев жизни вне организма с тем, чтобы иметь достаточно много-
численные тотальные препараты культур и серии срезов по всем стадиям.
Первая из подлежащих нашему рассмотрению опухолей локализиро
валась в левой фронтальной доле больших полушарий, имела клинически
медленное и доброкачественное течение и характер типичной глиобластомы
невысокого уровня дифференцировки (рис. 49). В исходном материале, при
обычных методах обработки, клеточные тела прослеживались только местами
в околоядерной области, а клеточные отростки совершенно терялись в окру-
жающей зернистой,комковатой или волокнистой массе, в которой находились
отдельные интенсивно красящиеся, большей частью извитые волокна,
представляющие собой измененные клеточные отростки, отмечавшиеся
еще Гирке (1885) в нормальной глии.
Рис. 49. Участок малодифференцированной глиобластомы
человека с многоядерными клетками. Ценкер-формол, эозин-
азур. Сильное увеличение.
Ядра имели очень неодинаковые размеры и окрашиваемость. Вокруг
более мелких темных ядер ободка цитоплазмы почти или даже совершенно
не удавалось рассмотреть. В таких же элементах, имевших отчетливое
округлившееся клеточное тело, встречались отдельные митозы. Вокруг части
крупных, более светлых ядер различалось то более, то менее отчетливо
контурированное округленное или неправильной формы клеточное тело.
Обращало на себя внимание значительное количество дву-, трех- и много-
ядерных гигантских клеток с тесно сгруженными, то очень крупными
то более мелкими ядрами. Цитоплазма некоторых элементов подвергалась
своеобразному гиалинозу, превращаясь в крупные ясно контурированные
гомогенные шары с оттесненными на периферию ядрами или ядром. После
эксплантации структура опухоли начинала выявляться все отчетливее,
совершенно также, как это отмечалось выше в отношении нормальной глий.
Часть клеток с мелкими ядрами очень быстро превращалась в округлые
или амебоидные макрофагоподобные элементы, переполнявшиеся жиро-
выми капельками, постепенно мигрировала в окружающий сверток фибрина.
Экспериментальный метод в гистологии
297
приобретая чрезвычайно разнообразную внешнюю форму. В цитоплазме
части таких клеток образовывались светлые пузыри, мешковидно выпячи-
вающиеся на поверхности их тела и достигавшие очень значительных раз-
меров. У другой части мелкоядерных и у крупноядерных элементов начинали
все отчетливее выступать клеточное тело и неразличимые прежде цито-
плазматические отростки. Находившаяся в исходном материале между
отдельными ядерными территориями глыбчатая или комковатая масса
исчезала. К концу первой или в течение второй недели культивирования
в центральном кусочке уже повсеместно совершенно отчетливо выявлялась
сетевидная паренхима опухоли, состоящая из отростчатых астроцитоподоб-
ных элементов с переплетающимися и соединенными друг с другом разветвлен-
ными отростками (рис. 50). За счет этих элементов возникала по истечении
Рис. 50. Та же опухоль на 7-й день культивирования. Выявле-
ние в центральном кусочке сетевидно связанных друг с другом
опухолевых элементов. Ценкер-формол, зозиназур. Сильное увели-
чение. (Оригинал.)
первых трех-четырех, а иногда и большего количества дней зона роста,,
причем во флаконах рост начинался раньше и происходил гораздо энергич-
нее, чем в условиях висячей капли. В некоторых элементах центрального
кусочка продолжались дистрофические и дегенеративные изменения, частью
отмеченные еще в исходном материале и заканчивающиеся смертью. Отростки
дегенерирующих элементов превращались в волокна плотной консистенции,
имеющие разную толщину, и в некоторых случаях извилистый штопоро-
образный ход. После первых пересевов с вырезанием можно было убедиться
в значительной прочности таких измененных клеточных отростков, растя-
гивавшихся и мявшихся под ножом и образовавших вокруг пересеянного
центрального кусочка неправильный переплет нитей. Это своеобразное
изменение цитоплазмы могло распространяться и на околоядерную часть.
Тогда опухолевые клетки превращались в более или менее гомогенные,
утратившие ядра, уродливые образования с округлой или неправильной
центральной частью и отходящими от нее в разном количестве прямыми
или извитыми волокнами и нитями. Это своеобразное перерождение опухо-
левых клеток напоминает старые наблюдения Гирке (1885) над «кератини-
зацией» астроцитарной глии. В части клеток центрального кусочка наблю-
дались картины перешнуровки и фрагментации ядер. Наконец, значитель-
ная часть центрального кусочка могла сохранять до предельных сроков
298
Глава III
культивирования жизнеспособный вид и структуру густой пространствен-
ной сети звездчатых эле-
Рис. 51. Та же опухоль. 22-дневная культура. Крае-
вой участок зоны роста. Формол, гематоксилин
ГСлабое увеличение. (Оригинал.)
!
ментов разной величины,
в цитоплазме которых то
с большей, то с меньшей от-
четливостью обнаружива-
лась фибриллярность. В
ряде случаев при энергич-
ном экстенсивном росте
центральный кусочек це-
ликом или большей своей
частью как бы растворялся,
в окружающей зоне роста.
Его превращения в сплош-
ной грубоволокнистый гли-
альный массив, подобно
тому как это имело место
в культурах из централь-
ной нервной системы, ни-
когда не наблюдалось.
'Разжижение свертка фиб-
рина и образование в нем полостей были выражены в сильной степени.
При этом центральный кусочек мог сильно деформироваться, растягиваться
и даже разрываться на части.
При экстенсивном росте
можно было наблюдать явле-
ния двоякого рода. В одних
случаях более мелкие астро-
цитарные элементы централь-
ного кусочка, втягивая от-
ростки, приобретали вид ма-
лодифференцированных по-
лиэдрических или веретенча-
тых одноядерных элементов,
которые, энергично делясь ми-
тотическим путем, образовали
более или менее комплексную
плоскостную зону роста, в
общем сходную с разраста-
ниями нормальной нейроглии
центральной нервной системы
j(pHC. 51, 52). Макрофагопо-
добные клетки встречались
до предельных прослеженных
Рис. 52. Та же опухоль. 42-дневная культура.
Участок зоны роста. Мембрана из опухолевых гли-
альных клеток с двумя митозами. На фоне мем--
браны макрофагоподобные клетки, образовавшие,
заостренные отростки с пенистой от жировых
включений цитоплазмой. Формол, гематоксилин
Сильное увеличение. (Оригинал.)
сроков. В такого рода культурах митотическая активность была вы.-,
сокой на всех стадиях культивирования. Периферическая часть зоны рот.
Экспериментальной метод в гистологии
299
ста, разрыхляясь, переходила в отдельные клеточные потоки и изоли-
рованные клетки веретенчатой или отростчатой «фибробластоподобной»
формы (рис. 51).
Рис. 53. Та же опухоль. 20-дневная культура. Сетевидный участок
зоны роста, состоящий из астроцитоподобных или нейроноидиых глиаль-
ных опухолевых клеток и отдельных макрсфагоподобных элементов.
Формол, гематоксилин. Сильное увеличение, (Оригинал.)
В отдельных случаях, наряду с такого же рода разрастаниями или без
них, астроцитарные элементы центрального кусочка претерпевали изме-
Рис, 54. Та же опухоль. 15-дневная культура. Нейроноидные
опухолевые глиальные клетки с причудливой системой ните-
видных отростков. Формол, гематоксилин. Слабое увеличение.
(Оригинал.)
нения другого рода, почти или совершенно не обнаруживали митотической
активности, хотя и разрастались достаточно энергично, сильно увеличи-
300
Глава III
ваясь в размерах и приобретая чрезвычайно сложную разветвленную
форму (рис. 53, 54). Такого рода картины наблюдались большей частью
только в течение первых 20—25 дней после эксплантации. Повидимому, при
росте этого второго типа на первый план выступала атипичная дифферен-
цировка опухолевых элементов, которая происходила в том же направлении,
что и в культурах нормальной глии, при образовании вне центрального
кусочка астроцитарных или нейроноподобных элементов. Клетки астро-
цитарной сети центрального кусочка сначала вытягивали в окружающий
фибрин длинные отростки, принимая все более рыхлое взаимное располо-
жение, а затем частично утрачивали взаимную связь и продвигались, мед-
ленно изменяя свою форму, во все стороны, окружая эксплантат чрезвы-
чайно своеобразной сетевидной зоной роста (рис. 53). Часть клеток при
этом сохраняла относительно небольшие размеры, другие вырастали в ги-
Рис. 55. Та >ке опухоль. 15-дневная культура. Чудовищная
гигантская опухолевая клетка. Фэрмол, гематоксилин. Сред-
нее увеличение. (Оригинал.)
гантские одно- и многоядерные протоплазматические тела. И более мелкие
и крупные клетки, образуя в разном количестве длинные нитевидные отро-
стки или же распластываясь, приобретали плоскую веретенчатую или
лентовидную форму. Некоторые элементы частью своего клеточного тела
распластывались в протоплазматическую пластинку различных очертаний,
а с другой стороны вытягивались в один или несколько тонких длинных
отростков. Образованная такого рода элементами зона роста имела вид
цитоплазматической сети с вкрапленными в ее состав отростчатыми клет-
ками разных размеров. В большем или меньшем количестве встречались
и описанные выше макрофагоподобные клетки. На тотальных препаратах
нередко удавалось проследить во всех деталях до тончайших разветвле-
ний форму отдельных клеток, почти или совершенно утративших связь
с другими себе подобными элементами. Многие клетки имели совершенно
нейроноподобный вид, одно- или многоядерное клеточное тело и два, трй
или несколько нитевидных, местами слегка варикозных отростков. общая
Экспериментальный метод в гистологии
301
длина которых могла измеряться одним и более миллиметрами (рис. 54).
Некоторые астроцитарные или нейроноидные клетки гигантских размеров
имели совершенно причудливый, чудовищный облик, быть может, сравнимый
с «Monstergliazellen», описываемыми при некоторых заболеваниях нервной
системы (рис. 55). Образование многоядерных плазмодиев прослеживалось
С полной отчетливостью благодаря многочисленным картинам амитоти-
ческих перешнуровок, причем нередко несколько ядер в одной и той же
клетке оставалось попарно соединенными тонкими нитевидными перемыч-
ками. Только в одном случае мне удалось наблюдать огромную, но несом-
ненно абортивную, митотическую фигуру, с очень многочисленными спу-
танными и частью неясными хромозомами в гигантской нейроноидной
клетке. В цитоплазме клеток могла обнаруживаться нежная мелкая зер-
Рис. 56. 9-дневная культура волокнистой астроцитомы человека. Экстен-
сивный рост нитевидных цитоплазматических отростков опухолевых
глиальных клеток, симулирующих нейриты. Формол, гематоксилин.
Слабое увеличение. (Оригинал.)
нистость и то более, то менее отчетливая фибриллярность. Мне предста-
вляется, что все эти астроцитарные и нейроноидные элементы представляют
собой образования, вполне сравнимые с астроцитарными и нейроноидными
клетками в культурах нормальной нейроглии. Только в опухолевой глии
они оказываются чрезвычайно утрированными и поэтому приобретают не-
редко чудовищный облик. Гистотипические и цитотипические особенности
нормальной ткани проявляются в данном случае на соответствующем опу-
холевом материале в еще более резкой и отчетливой, хотя и несколько урод-
ливой, форме. Если правильная оценка опухолевого материала возможна
только па основе его сравнения с соответствующей нормальной тканью, то,
наоборот, через изучение опухолевой ткани некоторые свойства нормаль-
ной выявляются и подчеркиваются с особенной ясностью.
Мне кажется, что эти наблюдения позволяют также дать правильную
трактовку мелкоклеточного и крупноклеточного типов роста, отмеченных
в своих опытах Пинкусом.
302
Глава III
Другая из анализированных мной методом эксплантации опухолей
развилась в области мозжечка и относилась к типу дифференцированных
волокнистых, относительно бедных клетками астроцитом. Астроцитарные
опухолевые клетки, со сложной системой переплетающихся и сетевидно
соединяющихся отростков, были уже отчетливо заметны в исходном мате-
риале, в противоположность нормальной глии и предыдущей опухоли *
в которых они, при обычных методах обработки, начинали выявляться лишь
по истечении нескольких дней культивирования. В цитоплазме отчетливо
выступали глиальные волокна различной толщины. Часть клеточных отрост-
ков подвергалась полному превращению в вещество волокон. Как и в описан-
ном выше случае, часть опухолевых элементов превращалась в мигрирующие
из центрального кусочка макрофагоподобные клетки. И при культивиро-
вании этой опухоли наблюдались разрастания двух разобранных выше типов.
Разжижение свертка фибрина и образование в нем полостей были также
выражены в сильной степени. Хороший рост этой опухоли удалось получить
только при культивировании во флаконах, причем на первых порах он
в поразительной степени симулировал прорастание нервных волокон.
Через пять-шесть дней на всей или значительной части поверхности цен-
трального кусочка обнаруживались тонкие нитевидные клеточные отростки,
далеко вытягивающиеся в питательную среду и нередко перекрещивающие,
преимущественно в радиальном направлении, образующиеся полости
разжижения. Цитоплазматические нити обычно вырастали более или менее
густыми, веерообразно расходящимися пучками. По своему ходу они вет-
вились и, соединяясь друг с другом, образовали сетевидный переплет (рис. 56).
Вслед за своими, уже успевшими достигнуть значительной длины, отрост-
ками в зону роста начинали смещаться сначала единичные, а затем все
в большем количестве ядерные части опухолевых астроцитарных клеток.
В результате этого первоначальный переплет «нейроидных» волокон пре-
вращался в сложное сочетание клеточных тел и нитевидных отростков,
причем, по сравнению с предыдущей опухолью, в сетевидных, астроцитар-
ных или нейроноидных структурах зоны роста количество тонких цито-
плазматических нитей преобладало над ядросодержащими участками
Часть астроцитарных элементов утрачивала связь с другими клетками и,
особенно в бедных клетками рыхлых местах, позволяла точно проследить
свою чрезвычайно сложную нейроноподобную, биполярную или мульти-
полярную форму со всей системой ветвящихся и причудливо изгибающихся
отростков. Часть астроцитарных или нейроноидных элементов становилась
дву-, трех- или реже многоядерной в результате амитотических перешну-
ровок.
В большем или меньшем количестве клетки такого рода встречались и
на поздних стадиях культивирования, обычно в сочетании с другими
картинами роста.
Другая часть астроцитарных клеток центрального кусочка, наоборот,
втягивая отростки, сближалась друг с другом и, делясь митотическим
путем, образовала густоклеточные более или менее комплексные разраста-
ния, которые лишь на периферии, разрыхляясь, переходили в сетевидные
Экспериментальный метод в гистологии
803
клеточные соединения, клеточные тяжи и изолирующиеся отростчатые и
веретенчатые клетки.
В составе такого рода плоскостных разрастаний клетки также имели
большей частью неправильную форму с отростками и лишь относительно
редко становились полигональными или веретенчатыми. Поэтому в ком-
плексных густых участках клеточные границы и их точная форма почти
или совершенно не прослеживались. Местами, особенно вблизи централь-
ного кусочка, зона роста приобретала довольно массивный характер
У части клеток в составе комплексных разрастаний кое-где также прослежи-
вались довольно длинные нитевидные отростки, которые могли протяги-
ваться в соседние, бедные клетками или лишенные их, участки питательной
среды. При разрыхлении зоны роста более или менее сложная астроци-
тарная форма клеток часто вновь выступала достаточно отчетливо, хотя
местами в таких разрыхленных местах клетки, наоборот, имели или при-
обретали простые малохарактерные «фибробластообразные» очертания.
Клетки-чудовища, столь характерные для предыдущей опухоли, в данном
случае почти не встречались.
Третья из опухолей центральной нервной системы, на которой я позволю
себе еще кратко остановиться, была диагносцирована как олигодендроглиома
с очагами обизвествления. По микроскопической картине она представляла
собой очень богатую клетками, имеющими большей частью удлиненную,
реже полиэдрическую форму, массу, центральные части которой были
переполнены густо расположенными сферическими известковыми конкре-
циями разных размеров. Вокруг известковых сфероидов клетки обычно-
приобретали эпителиоподобный вид. Эта опухоль отличалась очень энер-
гичным ростом, не только во флаконах, но и в висячей капле, при значи-
тельной митотической активности. Сверток фибрина подвергался разжиже-
нию с образованием полостей. Опухолевые клетки изменялись в трех глав-
ных направлениях: превращались в макрофагоподобные или очень мелкие
лимфоцитоподобные элементы и прорастали клеточными потоками, тяжами
или плоскостными мембранами. Центральная часть зоны роста часто при-
обретала массивный характер. Клетки имели большей частью удлиненные,
реже полигональные очертания и могли образовать длинные, почти ните-
видные с варикозными вздутиями отростки. Клеточные ядра имели большей
частью округлую или слегка овальную форму и одно-два крупных ядрышка.
По общему виду зона роста имела преимущественно характер малодиф-
ференцированных глиальных разрастаний, часть элементов которых могла'
приобретать астроцитоподобную, но далеко не столь резко выраженную,
как в предыдущих случаях, форму. Часто встречались двуядерные, реже
трех- и многоядерные клетки относительно небольших размеров. Опухо-
левая ткань, повидимому, отличалась высокой степенью катаплазии, причем
ее гистобластические потенции в течение полуторамесячного культивиро-
вания, которое было прервано подготовкой к эвакуации, не смогли быть
выявлены в достаточно полном объеме.
Наконец, следует еще упомянуть про один случай нейриномы слухо-
вого нерва или опухоли мосто-мозжечкового утла, которая отличалась
304
Глава III
исключительно сильным фибринолизом и в течение почти трехмесячного
срока культивирования во флаконах с постоянными пассажами давала
совершенно типичный для шванновских клеток рост узкоклеточными тяжами.
Для сопоставления были, кроме того, приготовлены из слухового нерва
новорожденных кроликов культуры, которые росли по тому же типу, что
и данная опухоль, а также изученный прежде блуждающий нерв.
Резюмированные в общих чертах результаты культивирования нейро-
генных опухолей при их сопоставлении с приведенным раньше материалом
по нормальным тканям эпендимо-глиального типа с большой отчетливостью
обосновывают точку зрения, согласно которой опухолевые клетки не при-
обретают каких-либо новых гистобластических потенций, по сравнению
с исходными нормальными тканями, а обнаруживают то с большей, то
с меньшей полнотой цикл превращений, свойственный соответствующим
нормальным тканям при неопухолевых пролиферативных процессах. Вместе
с тем проявление опухолевыми клетками общих гистотипических для дан-
ного тканевого типа особенностей может отличаться более или менее значи-
тельным специфическим «опухолевым» своеобразием, анализ которого
выходит за рамки этой книги, посвященной в первую очередь вопросам
общей гистологии, и, кроме того, требует более детального разбора как
собственного, так и литературного опухолевого материала.
Мне кажется, что нейроноидные формы глиальных клеток, открытые
нами при культивировании частиц нормальной центральной нервной системы
и обнаруживающиеся, в особенно утрированном виде, в опухолевом материа-
ле, а также сложно разветвленная форма астроцитов, детально разобранная
Гирке (1885) на мацерированных препаратах мозга, имеют важное прин-
ципиальное значение. В этом нельзя не видеть общих морфологических
особенностей, обусловленных близким родством клеток нервной системы,
имеющих совершенно различное функциональное значение, но развивающих-
ся в фило- и онтогец^зе из одного и того же источника и в своем развитии
теснейшим образом связанных друг с другом.
Д. Эндотелий сосудистой системы
Из приведенного выше материала видно, что сходные по внешнему
виду структуры, которые принято обозначать, по почину Гиса (1865),
термином «эндотелий», по своему происхождению и гистобластическим
потенциям очень сильно отличаются друг от друга и являются продуктами
конвергентной дифференцировки ряда биологически неравноценных тка-
невых типов. Мезотелий высших позвоночных, который долгое время также
рассматривался, а иногда еще и теперь рассматривается, как эндотелий,
оказался высокоспециализированной формой целодермальных тканей. Эндо-
телий, мезотелий или покровные клетки мозговых оболочек, а вместе с ними
и так называемый десцеметов эндотелий находят свое естественное место
среди тканей эпендимо-глиального типа.
Эндотелий сосудистой системы позвоночных, трактуемый как произ-
водное мезенхимы или. точнее, «энтомезенхимы» и входящий, по Шафферу
Экспериментальный метод в гистологии
305
(1921), в группу «десмальных эпителиев» по своему происхождению, биоло-
гическому значению и положению в организме должен быть, естественным
образом, выделен в особую группу тканей и даже, быть может, в самостоя-
тельный тканевой тип. Являясь по своей морфо-физиологической характе-
ристике пограничной тканью эпителиального характера, эндотелий рас-
сматривается обычно в настоящее время как одна из клеточных форм сое-
динительной ткани вместе с последней.
Эта прочно установившаяся традиция, которой следовал до настоящего
времени и автор этой книги, однако, не свободна от серьезных возра-
жений.
При сравнительном обзоре животного царства не может не броситься
в глаза, что сосудистая система, снабженная непрерывной эндотелиальной
стенкой, отграничивающей сосудистое русло от соединительной ткани,
является, по существу, одним из типовых признаков позвоночных. За
пределами типа хордовых система сокообращения, которую принято срав-
нивать с сосудистой системой позвоночных, несмотря на подчас очень
высокий уровень своего развития, не имеет, как правило, непрерывной
эпителиоподобной выстилки. Повидимому, единственным исключением из
этого правила являются немертины, в развитии и строении которых обна-
руживается ряд своеобразных черт, заставляющих предполагать, что они
являются вторично сильно измененными и, вероятно, подвергшимися упро-
щению организмами.
Единственным отделом сосудистой системы, который, например у мол-
люсков, кишечнодышащих и некоторых других форм, имеет эпителиальную
выстилку, является включенный в ее состав участок целомической полости,
представленный околосердечной сумкой. Описывавшиеся неоднократно
у некоторых беспозвоночных отдельные или расположенные группами
клетки на внутренней поверхности сосудов и представляющие собой вре-
менно осевшие клетки гемолимфы не могут сопоставляться с настоящим
сосудистым эндотелием. Вместе с тем у части беспозвоночных, наряду или
вместо сосудистого аппарата безэндотелиального типа, развивается система
замкнутых трубок, снабженных непрерывной эпителиальной выстилкой,
которая функционально субституирует ее, является продуктом особой
специализации целомической полости и поэтому может быть обозначена
как целомическая сосудистая система, или целомическая система сокообра-
щения.
Постепенное развитие такой целомической системы сокообращения очень
хорошо прослеживается у пиявок. У сипункулид она имеет высокий уровень
дифференцировки и несет, как это уже отмечалось, также функцию газооб-
мена. У вторичноротых беспозвоночных очень высокого уровня развития
целомические или генетически связанные с целомом системы сокообращения
достигают у иглокожих, у которых, помимо амбулакральной системы,
имеется еще собственно кровеносная система, представленная осевым ор-
ганом, осевыми синусами и окологлоточным кольцом, и т. наз. перигемаль-
ная система, образующаяся за счет клеток, выселяющихся из состава спланх-
нонлевры заднего целомического пузыря.
20 X лопин—349
306
Глава III
На основе этих сопоставлений не может не возникнуть совершенно есте-
ственного вопроса, с какой же из систем сокообращения, имеющихся у бес-
позвоночных, правильнее сравнивать и гомологизировать сосудистую
систему позвоночных, характернейшей составной частью которой является
непрерывная эпителиальная, в широком смысле слова, или, точнее, эндо-
телиальная выстилка. С точки зрения сравнительной анатомии и сравни-
тельной гистологии мне представляется вполне допустимой трактовка эндо-
телия сосудистой системы позвоночных в соответствии со взглядами,
подробно развитыми в монографии Гаусмана (1935), как высоко спе-
циализированного участка стенки целома, а не с общепринятой точки зре-
ния, как вторично уплощенных и принявших эпителиоподобный облик кле-
ток соединительной ткани.
Отсутствие целомической полости у немертин не может служить бес-
спорным аргументом против сопоставления их сосудистой системы, с по-
лостью тела, так как в их эмбриональном развитии, как известно, обра-
зуются настоящие парные мезодермальные полоски, свойственные целоми-
ческим животным, которые лишь вторично распадаются на рыхло распо-
ложенные и поэтому обозначаемые как «мезенхиматозные» клетки. Очень
возможно, что целомическая полость немертин подверглась вторичной ре-
дукции, а ее оставшаяся часть превратилась в кровеносную систему, если
последняя действительно снабжена настоящим эндотелием, что все-таки не
может считаться совершенно бесспорным.
Общепринятый взгляд на эндотелий сосудистой системы позвоночных,
как на уплощенные клетки соединительной ткани, обосновывается, с одной
стороны, его эмбриональным развитием, с другой, его предполагаемыми
превращениями при экспериментальных и патологических условиях. Кроме
того, следует учитывать, что в составе эндотелия имеются функционально
очень неодинаковые участки. Это заставляет, например, Максимова
(1927), строго различать собственно эндотелий и так называемые «береговые
клетки» (Uferzellen), относящиеся по своим физиологическим особенностям
к ретикулоэндотелиальной системе (купфферовские клетки синусоидов
печени, выстилка капилляров костного мозга и ряда эндокриновых орга-
нов и пр.). Поэтому не исключена также возможность, что в составе
эндотелия позвоночных могут оказаться и в генетическом отношении нерав-
ноценные части, хотя это и не представляется вероятным.
Взгляды на эмбриональное развитие эндотелия позвоночных и на его
генетические отношения с соединительной тканью, подвергались большим
изменениям. В противовес точке зрения Г аса (1866, 1868),* которая
в своей первоначальной форме оказалась несостоятельной, но в измененном
Вольйеггерол«(1883)и бр. Гертвигами (1882) виде господствует и в настоящее
время, был, как известно, выдвинут, по почину того же Гиса (1900), взгляд
(Майнот, 1911; Себин, 1920; Финлей, 1922), согласно которому сосудистая
система развивается из совершенно самостоятельного зачатка т. наз.
ангиобласта, элементы которого не зависимы и от мезодермы и от
См. вводную историческую часть.
Экспериментальный метод в гистологии 307
мезенхимы . Наконец, некоторая часть авторов допускает также возможность
участия энтодермы в образовании первых кровяных островков зародыша
(Рюккерт и Мольер, 1906). У высших позвоночных, согласно наиболее
популярным и тщательным исследованиям (Максимов, 1907, 1909, 1927;
Данчакова, 1908, 1916; Рюккерт и Мольер, 1906,— в отношении сосудов
зародыша), эндотелий разных отделов сосудистой системы, включая и сердца,
развивается, как вне зародыша (в стенке желточного мешка), так и в разных
местах зародышевого организма, из местных клеток мезенхимы. Образующие-
ся в разных местах сосудистые зачатки затем соединяются в единое русло.
Сходным образом происходит образование первых сосудов и в стенке желточ-
ного мешка селахий (Максимов, 1923). Следует, однако, подчеркнуть, что
млекопитающие, птицы, рептилии и селяхии отличаются от прочих позвоноч-
ных очень сильным развитием и ранним возникновением мезенхимы, в со-
ставе которой возможно имеются первоначально неотличимые друг от друга,
но неравноценные и даже различным образом детерминированные элементы.
Не исключена возможность, что одной из детерминированных разновидно-
стей клеток мезенхимы, качественно отличных от клеток будущей соединитель-
ной ткани, является зачаток эндотелия сосудистой системы. То, что при
образовании кровяных островков в стенке желточного мешка называется
мезенхимой, представляет собой, по существу, разрыхленную внезародыше-
вую мезодерму боковых пластинок, которая затем подразделяется и диффе-
ренцируется на кровяные островки, окружающую их мезенхиму и внезаро-
дышевую часть целомической выстилки. Данные из области эксперименталь-
ной эмбриологии безусловно говорят в пользу ранней детерминации мате-
риала кровяных островков (Шиплей, 1916; Меррей, 1932). Особенности
развития мезенхимы высших позвоночных и селяхий являются безусловно
вторично приобретенными и, может быть, искажают или затемняют, в боль-
шей или меньшей степени, первоначальные отношения. У круглоротых,
ганоидных и костистых рыб, а также у амфибий мезенхима образуется зна-
чительно позже и в гораздо более скудном количестве.
У круглоротых и амфибий источником развития первых кровяных элемен-
тов и части сосудистого зачатка вентральной части тела является лежащий
по срединной линии непарный тяж, вилообразно разделяющийся впереди.
У амфибий материал этого непарного тяжа обособляется очень рано в виде
разрозненных клеток по краям обрастания еще во время гаструляции.
Так как развитие эндотелия первых сосудов и эритроцитов неразрывно
связано друг с другом, то данные, касающиеся ранней детерминации оча-
гов эритропоэза, представляют определенный интерес и с точки зрения
образования сосудистой системы.
Помещая изолированные части вентральных отделов мезобласта в ме-
шочки из кожной эктодермы, Слонимский (1930, 1931) показал, что
эритробластическая кроветворная детерминация наступает в этих отделах
спланхнотомов очень рано.
После полного удаления у зародыша лягушки, на стадии ранней
хвостовой ночки этой кроветворной области, развития эритроцитов
в организме вовсе не происходит (Фредеричи, 1926; Госс, 1928). Эндо-
808
Глава HI
телий сердца у амфибий формируется из клеток, выселившихся из
края обрастания мезодермы и скопляющихся между вентральными краями
спланхнотомов и вентральной стенкой кишки. У костистых рыб будущая
кроветворная область, зачатки спинных аорт и кардинальных вен обособ-
ляются в виде т. наз. промежуточных масс или клеточных скоплений,
образующих затем продольный тяж, располагающийся между хордой
и дорзальной стенкой кишечной трубки. Из этого тяжа, так же как у других
позвоночных, в первых очагах кроветворения образуются эндотелиальная
стенка кровеносного сосуда и первичные кровяные клетки (Стоккард,
1915). Из тех же промежуточных масс образуются и клеточные скопления,
из которых возникает эндокард или, во всяком случае, эндотелий сердца.
Все эти эмбриологические данные говорят за то, что у низших позвоночных
красные кровяные клетки и эндотелий некоторых крупных центральных
отделов кровеносной системы имеют совершенно самостоятельный и очень
рано обособляющийся зачаток. Весьма возможно, хотя против этого и го-
ворит общепринятая точка зрения, что с тем же зачатком генетически свя-
зан и эндотелий других отделов сосудистой системы, которому приписы-
вается возникновение на месте из тех или других областей мезенхимного
зачатка, в частности, из склеротомов. Дело в том, что при имеющихся пока
возможностях изучения раннего развития сосудистой системы ни та, ни
другая точка зрения, строго говоря, не могут быть доказаны с достаточной
убедительностью. Гистогенез сосудистой системы ланцетника до настоя-
щего времени изучен еще недостаточно; однако имеются вполне опреде-
ленные указания на развитие эндотелия сосудов непосредственно из
целомической выстилки (Франц, 1927).
Имеющиеся в настоящее время эмбриологические данные по развитию
сосудистой системы и первых кроветворных очагов не позволяют совер-
шенно безоговорочно сближать сосудистый эндотелий с соединительной
тканью и считать т. наз. энтомезенхиму единым и равноценным в своих
разных частях эмбриональным зачатком. К этому нам придется еще вер-
нуться в следующей главе, посвященной соединительной ткани и мезен-
химе.
Рассмотрим, какими гистобластическими потенциями обладает эндоте-
лий на разных стадиях своего развития и в какой мере имеются основания
сближать его с соединительной тканью в течение следующих этапов жизни
организма.
После образования замкнутого круга эмбрионального кровообращения
новообразование дальнейших сосудов происходит в соответствии с обще-
принятыми взглядами исключительно путем образования эндотелиальных
отпрысков. Новообразование кровеносных и лимфатических сосудов исклю-
чительно из эндотелиальных сосудистых почек во многих случаях доказано
с исчерпывающей убедительностью.
Особенно демонстративны, пожалуй, в этом отношении результаты де-
тального прижизненного изучения роста сосудов в хвосте головастика
и прозрачной камере, вставленной в ухо кролика (Кларк и Кларк, 1920,
1932, 1939), а также старые данные Максимова по асептическому воспа-
Экспериментальный метод в гистологии
309
лению (1902). Наоборот, попытки трактовать образование эндотелия но-
вых сосудов иным образом никак нельзя назвать сколько-нибудь убеди-
тельными. В частности, это относится к предполагаемому новообразованию
эндотелия из прилегающих к нему снаружи малодифференцированных
клеток соединительной ткани (Жолли, 1906; Беннингоф, 1926; Щелкунов,
1938, и др.) и к старым представлениям Ранвье (1874) о «вазоформативных»
клетках. Столь же неубедительной и малоправдоподобной является предпо-
лагаемая Щелкуноеым (1936, 1937) регенерация эндотелия артерий
млекопитающих из лежащего под ним подэндотелиального слоя, большое
значение которого для роста и регенерации среднего слоя сосудистой стенки,
согласно данным того же автора, следует считать хорошо обоснованным.
Новейшие данные Щелкуноеа (1939) по сосудистой системе миноги
также говорят против участия подэндотелиального слоя в регенерации
эндотелия и за восстановление его из себе подобных .клеток.
В течение некоторого времени после развития первых кровеносных
сосудов у зародыша, по данным Максимова (1907, 1909), у кролика,
по данным Данчаковой (1909), у птиц может происходить из эндо-
телия новообразование первичных кровяных клеток, эндотелиальных фа-
гоцитов, а также отщепление базофильных блуждающих клеток в окружаю-
щую мезенхиму.
У некоторых низших позвоночных удается наблюдать даже во взрос-
лом состоянии округление и переход в кровь элементов эндотелия, например
в артериальном конусе сердца. Изолирующиеся клетки эндотелия или имеют
малодифференцированный вид или уже приобретают признаки эритробластов.
Из всего обширного экспериментального материала, касающегося разоб-
ранных выше тканей, ясно, что превращение в свободные малодифферен-
цированные макрофагоподобные клетки не может служить само по себе
в настоящее время аргументом в пользу превращения одной ткани в эле-
менты какой-нибудь другой.
Дифференцированному эндотелию взрослых млекопитающих многими
авторами до последнего времени приписываются очень широкие способ-
ности к превращениям при экспериментальных и патологических условиях.
Согласно этим взглядам, эндотелий трактуется как запас совершенно
индифферентных эмбриональных клеток во взрослом организме, который
может быть использован для восстановления элементов самых разнообраз-
ных тканей. Однако приводимые в пользу этого доказательства отнюдь
нельзя считать достаточно убедительными. Кроме того, ряд данных этого
рода не был подтвержден дальнейшими исследованиями. Подобно тому,
как некоторые упомянутые выше авторы допускали новообразование эндо-
телия из прилегающих к нему адвентициальных клеток, другие (Мар-
шанд, 1921, 1924; Г. Герцог, 1915) считают, наоборот, что адвентициаль-
ные клетки, а следовательно, и макрофаги воспалительного очага и даже
лимфоциты, могут возникать из эндотелия. При экспериментальном
туберкулезе (Фут, 1919—1925) эндотелию приписывают широкое участие
в образовании бугорков. То же самое говорится и относительно возникно-
вения зернистых лейкоцитов при воспалении (Г. Герцог, 1921, 1922,
310
Глава III
1923). По некоторым, совершенно неправдоподобным, данным, массовое
образование зернистых лейкоцитов из эндотелия может происходить со-
вершенно молниеносно, буквально в течение немногих минут (Эллер.
1923, 1925; Теппих, 1925, 1926). Эти и некоторые другие, аналогичные
им, взгляды в свое время критиковал Максимов (1927).
Вообще следует подчеркнуть, что при местном появлении каких-либо
свободных кровяных клеток никогда не может быть исключен их занос
через кровяное русло, который несомненно происходит в самых широких
размерах.
Максимов, наиболее умеренный сторонник близких генетических отно-
шений эндотелия с производными мезенхимы, допускает лишь его способ-
ность к превращению в фибробласты соединительной ткани при воспалении
в составе целого организма (1902).
В отношении эндотелия синусоидов печени, селезенки, костного мозга,
надпочечников и некоторых других органов, который, в противоположность
эндотелию большей части сосудистых областей тела, Максимов обо-
значает как «береговые клетки» (1927), им допускаются более широкие воз-
можности превращений. То же самое относится к выстилке синусов лимфа-
тических узлов и находящимся в их просвете сетевидно связанным друг
с другом клеткам.
С несомненностью доказывается во всяком случае резко выраженная
способность этих элементов к фагоцитозу и накоплению прижизненно кра-
сящих веществ (Аничков, 1930) с последующим освобождением из
состава сосудистой стенки и переходом в кровь в виде переполненных вклю-
чениями макрофагов или, быть может вернее, макрофагоподобных клеток.
Кроме того, этим элементам, подобно эндотелию всех прочих частей сосу-
дистой системы, приписывается также способность к превращению в фибро-
бласты соединительной ткани. Из всего изложенного выше мы могли, однако,
убедиться, что, подобно превращению в малодифференцированные базо-
фильные клетки, как раз внешнее уподобление элементам типа макрофагов
и фибробластов еще отнюдь не обозначает действительно превращения в сое-
динительнотканные макрофаги и соединительнотканные фибробласты. Даже
самый факт уподобления эндотелия фибробластам в условиях целого ор-
ганизма едва ли доказуем с достаточной убедительностью. Все утверждения
такого рода представляют собой, по существу, одну из возможных трактовок
наблюдаемых картин, которая в первую очередь зависит от априорного
допущения такой возможности.
К сожалению, данные, полученные в настоящее время методом эксплан-
тации относительно гистобластических потенций эндотелия, далеко не отли-
чаются полнотой, а во многих случаях и достаточной убедительностью.
В богатых капиллярами эксплантатах из некоторых органов куриного
зародыша (У. Льюис, 1931; Тёрё, 1937), сальника, подкожной ткани и
мозговых оболочек кроликов или крыс (Максимов, 1916, 1924, 1927; Зиль-
берберг, 1929; Герцог и Шоппер, 1931; Скраба, 1935, и др.) описывается
прорастание сосудистых почек одновременно с ростом элементов соеди-
нительной ткани. Льюис наблюдал даже образование обширных сетей капил-
Экспериментальный метод в гистологии
311
лярных отпрысков. Растущие капилляры имеют вид полых внутри трубок
разного калибра, образованных тесно сомкнутыми клетками эндотелия,
с замкнутым и большей частью заостренным концом. Образуя боковые
отпрыски, эндотелиальные трубки могут в зоне роста соединяться друг
с другом. В их просвете находится жидкость, в которой бывают взвешены
в разном количестве кровяные клетки, перетекающие пассивно из сосудов
центрального кусочка. Отпрыски капилляров могут быть заметны в зоне
роста, по данным всех авторов, лишь в течение немногих дней после эксплан-
тации; в дальнейшем они исчезают, частью погибая, частью распадаясь,
по их мнению, на отдельные фибробластообразные элементы, не отличи-
мые от находящихся по соседству соединительнотканных клеток. В экс-
плантатах из соматической мускулатуры новорожденных кроликов мне
приходилось видеть появление в разных количествах капиллярных отпрыс-
ков, часть которых могла сохраняться до 10—15 дней в культурах, не
подвергшихся вырезанию.
Действительно ли эндотелий капилляров в конечном итоге превращается
в элементы, во всех отношениях тождественные обычным фибробластообраз-
ным клеткам соединительной ткани, как это предполагается большей частью
авторов, остается, строго говоря, недоказанным, так как на многих
других примерах в настоящее время известно, что фибробластообразный
облик могут приобретать клетки разного происхождения, не утрачивая
в то же самое время своих неодинаковых специфических гистобластических
потенций. По опубликованным недавно наблюдениям Тёрё (1937), эндо-
телий, превратившийся в фибробластообразные элементы, после имплан-
тации культур в переднюю камеру глаза, будто бы не только образует
систему замкнутых полостей, но даже кроветворные очаги, сходные с кро-
вяными островками желточного мешка зародыша. Эти, несомненно, весьма
интересные данные, однако, нуждаются еще в проверке и подтверждении.
Сосудистыми эндотелиальными отпрысками, по данным Беневоленской
(1930), растет также эндотелий эмбриональной человеческой печени. По
данным других авторов (У. Льюис, №22-, Леви, 1923, 1925; Гарофолини, 1927;
Должанский, 1930; Эфруаи, 1930; Бишслье, 1931), эндотелий печеночных
синусоидов куриного зародыша, энергично пролиферируя, образует зону
роста из веретенчатых или уплощенных клеток, которые или образуют более
или менее рыхлые сетевидные соединения или местами довольно тесно
смыкаются друг с другом краями. Насколько возможно судить по имею-
щимся описаниям, элементы такой зоны роста по своему виду и по способу
соединения друг с другом отличаются от обычных фибробластообразных
клеток соединительной ткани. Эндотелиальная природа клеток зоны роста
в эксплантатах из эмбриональной печени достаточно убедительна, если
исходный материал принадлежал молодым зародышам, так как на более
поздних стадиях развития в печени, помимо эндотелия, появляется заметное
количество околососудистой мезенхимы. В эксплантатах из более крупных
сосудов (Максимов, 1927) и эндокарда эмбрионального сердца (Ниши-
йе, 1929) также описывался рост эндотелия, который, по данным послед-
него автора, даже удается выделить в чистых культурах. Максимов, в
312
Глава III
соответствии со своими взглядами на природу эндотелия, описывает его
превращение в фибробласты соединительной ткани. Нишибе также от-
мечает морфологическое тождество изучавшихся им «эндокардиальных»
клеток с фибробластообразными мезенхимными элементами. Последние
данные, однако, в настоящее время не могут считаться достаточно убеди-
тельными, а эндотелиальное происхождение прорастающих в этих куль-
турах элементов доказанным. Эндотелиальная трубка сердечного зачатка
молодых куриных зародышей образует в культурах своеобразные комплекс-
ные мешковидные разрастания, а также изолированные фибробластообраз-
ные и макрофагоподобные клетки (Гарро, 1932). Едва ли можно сомне-
ваться, что описанные Нишибе (1929) эпителиообразные мембраны
в культурах из почки жабы, которым он почему-то приписывает эндоте-
лиальную природу, судя по хорошим микрофотограммам и тексту работы,,
образованы растущим почечным эпителием. Эта работа является удачным
примером того, насколько произвольно и необоснованно некоторые авторы
принимают гистологические обозначения в отношении растущих вне орга-
низма клеток и тканей.
Предпринятое моей сотрудницей Колесниковой эксперименталь-
ное изучение эндотелия было прервано вспыхнувшей Отечественной
войной и отъездом автора на фронт. Поэтому его результаты еще не могут
быть изложены в полном объеме. Однако собранный ею и частично обра-
ботанный большой материал позволяет уже сделать некоторые выводы,
заслуживающие внимания. Опыты были проведены методом эксплантации
на клапанах сердца и разных частях сосудистой системы новорожденных
и молодых кроликов и показали очень значительные технические трудности
изучения сосудистого эндотелия, вследствие его очень легкой повреждае-
мости. Особенно удобным для изучения материалом оказались клапаны..
При эксплантации частиц клапанов, а также крупных сосудов прежде
всего обращали на себя внимание сильное разжижение свертка фибрина
и быстрое образование в нем полостей, что совершенно несвойственно экс-
плантатам из обычных соединительнотканных образований. Если эндотелий
покрывал собой, как это обычно бывает, некоторую часть материнского
кусочка, то разжижение точно соответствовало первоначально покрытой
эндотелием части эксплантата. Сверток фибрина, прилегающий к обна-
женной соединительной ткани или не разжижался вовсе или вовлекался
в этот процесс очень постепенно, вследствие постепенного расширения пер-
воначально образовавшейся полости Обнаженная соединительная ткань
начинала прорастать обычными фибробластообразными клетками, образуя
в то же самое время и свободные блуждающие элементы типа макрофагов.
Тонкий покровный эндотелиальный пласт местами набухал, причем око-
лоядерные части клеток, утолщаясь, образовали, наподобие булыжников
мостовой,. вздутые участки. В эндотелии активировалась митотическая
пролиферация. Пласт эндотелия мог, несколько продвигаясь по обнажен-
ной и не прорастающей еще соединительной ткани, покрывать собой большие
или меньшие части поверхности эксплантата. Наблюдать полное закрытие
всей наружной поверхности центрального кусочка и его отграничение от
Экспериментальный метод в гистологии
313
окружающей среды непрерывным, активно нарастающим эндотелиальным
покровом пока не удавалось с достоверностью. Вместе с тем эндотелиальный
пласт начинал обнаруживать экстенсивный плоскостной рост очень тонкими
и первоначально совершенно комплексными мембранами, состоящими из
тесно сомкнутых плоских клеток различных очертаний. Увеличиваясь
в размерах, эндотелиальные мембраны начинали разрыхляться, постепенно
распадаясь на сетевидно связанные друг с другом отростками клетки. Это
совпадало обычно с далеко заходящим разжижением фибрина и затем могло
уже переходить в ясно выраженные изменения дегенеративного характера.
Для поддержания жизнеспособности культуры приходилось незамедли-
тельно прибегать к пассажу, после которого наблюдавшиеся картины вновь
повторялись. Совершенно такие же картины плоскостного роста эндоте-
лиальных мембран удавалось получить и в эксплантатах из крупных со-
судов.
К сожалению, предельные сроки культивирования пока не превышали
в большей части случаев двух-трех недель, вследствие больших технических
трудностей, связанных со специфическими особенностями данного материала.
Тем не менее, резюмированные выше наблюдения показывают, что эндо-
телий сердца и крупных сосудов, по крайней мере в течение двух-трех не-
дель жизни вне организма, не обнаруживает никакого сходства с обычными
элементами соединительной ткани, а проявляет себя как очень своеобраз-
ный в отношении своих биологических и гистологических свойств детерми-
нированный тканевой тип. Вместе с тем полученные результаты открывают
пути дальнейшего изучения эндотелия сосудистой системы в новом направ-
лении. Из всего изложенного пока еще преждевременно делать какие-либо
определенные выводы. Глава, посвященная эндотелию, в настоящее время
еще далеко не может считаться законченной, а представляет собой скорее
постановку вопроса для дальнейших исследований. Одно мне представляется
совершенно ясным: принятая трактовка эндотелия как элементов близко
родственных фибробластам соединительной ткани, а в некоторых частях
сосудистой системы и ретикулярным клеткам кроветворных органов, не
является точно и незыблемо доказанной. Ряд фактов, начиная от высокой
специфичности, проявляемой эндотелием при образовании сосудистых-
почек, и кончая обнаружением его своеобразных свойств в условиях тка-
невых культур, делает возможным его трактовку как особого высокоспе-
циализированного тканевого типа, обладающего морфо-физиологической
характеристикой т. наз. эпителиальных тканей и качественно отличного
на данном этапе эволюции от соединительной ткани. Дальнейший вопрос
заключается в том, представляет ли собой эндотелий специализировавшуюся
в процессе филогистогенеза часть соединительной ткани, которая вторично
приобрела морфо-физиологические особенности пограничных образований,
или эндотелий развился независимо от собственно мезенхимы и лишь вто-
рично вступил с ней в теснейшие взаимоотношения. Приведенные выше
данные сравнительно-анатомического, а также эмбриологического харак-
тера делают для меня более вероятным предположение, что сосудистая
система позвоночных сравнима с той целомической системой сокообразо-
H14 Глава Ш
вания, которая встречается лишь у относительно небольшого числа беспозво-
ночных, но в эволюции позвоночных достигает чрезвычайно высокого уровня
развития, причем ее генетическая связь с целомом оказывается в большей
или меньшей степени затушеванной, а биологические особенности эндо-
телия обнаруживают, по сравнению с выстилкой полости тела, далеко иду-
щую дивергенцию и специализацию, делающие невозможным их отождест-
вление на данном этапе эволюции. В некоторых случаях между полостью
тела и сосудистой системой у позвоночных может, однако, устанавливаться
вторичная связь через нефростомы выделительных органов, которая, на-
пример, обнаруживается у бесхвостых амфибий.
Пересмотр вопроса об эндотелии равным образом влечет за собой пере-
смотр ряда основных проблем, связанных с кровью, кроветворением и сое-
динительной тканью, так как тесная связь эритроцитов с эндотелием со-
судистой стенки позвоночных и с выстилкой целомической системы сокооб-
ращения беспозвоночных проходит красной нитью и не может не бросаться
в глаза каждому непредубежденному исследователю. Тем самым становится
на очередь вопрос экспериментального изучения, под этим новым углом
зрения, генетических отношений форменных элементов крови, лимфы и
гемолимфы друг с другом и с другими тканевыми типами животного орга-
низма, а также и всего эволюционного развития крови и соединительной
ткани у представителей разных филогенетических стволов животного
царства.
3. ВОПРОС О МЕЗЕНХИМЕ, СОЕДИНИТЕЛЬНЫХ ИЛИ МЕЖУТОЧНЫХ
ТКАНЯХ И СВОБОДНЫХ КЛЕТКАХ СОКОВ ТЕЛА
а) Вводные замечания
Вопрос о мезенхиме, в широком смысле этого слова, и обо всем комплексе
генетически связанных с ней тканей еще не может быть изложен в развер-
нутом виде с точки зрения последовательно проводимых в этой книге общих
установок. То, что здесь может быть дано, представляет собой, в лучшем
случае, попытку критического освещения некоторых существующих в на-
стоящее время взглядов (в которых был в значительной степени воспитан
сам автор этих строк) и логического распространения опыта, приобретен-
ного при изучении ряда тканей, на частный случай т. наз. тканей соедини-
тельного вещества с включаемыми обычно в их состав свободными клетками
соков тела.
Современное учение о соединительной ткани и крови представляет собой
чрезвычайно стройное и цельное здание, возведенное руками бесчисленного
количества исследователей и принявшее свой законченный вид в значитель-
ной степени благодаря экспериментальным работам Максимова и его
статьям обобщающего характера. Составленную им известную сводку по
соединительной ткани и тканям кроветворных органов можно считать
с полным основанием непревзойденной мастерской картиной состояния этой
главы гистологии к 1927 г. Дальнейшему развитию сформулированного
Максимовым учения посвящены у нас в Советском Союзе исследования
Заварзина и его сотрудников, несомненной и большой заслугой которых
Экспериментальный метод в гистологии
315
является распространение экспериментального изучения крови и соеди-
нительной ткани, в первую очередь методикой Циглера—.Максимова, на
отдельных представителей различных типов беспозвоночных, которым
до этого времени уделялось недостаточно внимания, хотя еще наш вели-
кий соотечественник Мечников доказал плодотворность такого рода срав-
нительных исследований. Процессы, разыгрывающиеся в соединительной
ткани в условиях асептического воспаления, Заварзин трактует (1934)
с точки зрения явлений конвергенции или параллелизмов, между которыми
не делает различий. Во взглядах, развиваемых Заварзиным, принимаются
кажущиеся очевидными представления о мезенхиме, как о едином, по
существу, во всем животном мире эмбриональном зачатке, дающем начало
большому многообразию генетически однородных и полностью сравнимым
друг с другом в разных случаях тканей. Исходя из лейдиговского
физиологического или, с моей точки зрения, вернее морфо-физиологиче-
ского подхода к гистологическим структурам, Заварзин дает всему ком-
плексу мезенхимных производных не совсем удачные обозначения опорно-
трофических (1933) или трофических и механических (1934) тканей и,
несколько позже, тканей внутренней среды. Если считать целесообразным
замену общепринятого термина «соединительная ткань», не изменяя по
существу его содержания, каким-нибудь другим, позволяющим ее про-
тивопоставлять эпителиальным тканям, то, пожалуй, целесообразнее, в
соответствии со сказанным выше, вспомнить о старом обозначении «меж-
уточная ткань», «межуточные» или «выполняющие ткани», представляющем
собой по существу свободный перевод греческого слова мезенхима.
С точки зрения сравнения друг с другом с чисто морфо-физиологических
позиций межуточных тканей различных животных вопрос об их генети-
ческом единстве и их гистологической гомологии не представляется
существенным; не имеет принципиального значения и систематическое
положение животных, ткани которых сраниваются. Но для пересмотра
учения о межуточных тканях с точки зрения эволюционной гистологии
и естественной генетической системы гистологических структур эти же
вопросы приобретают исключительно важное принципиальное значение.
Без учета их совершенно невозможно, например, разобраться в том, где
мы имеем дело с действительным эволюционным изменением данной струк-
туры, а где происходит замещение или субституция ее другой тканью,
имеющей другую историю развития, а, следовательно, неизбежно и иные
свойства, иное биологическое значение.
Само собой разумеется, преемственная эволюция какой-нибудь ткани
представляет собой нечто глубоко отличное от ее субституции другой тканью,
тем более, что весь изложенный выше фактический материал доказывает,
в какой степени свойства ткани зависят от всей предыдущей истории ее
развития, отраженной в большей или меньшей степени, а в ряде случаев
с очень большими искажениями, в онтогенезе.
Кроме того, изложенный выше фактический материал заставляет с боль-
шой осторожностью подходить к таким критериям определения родства,
а вместе с тем и других, связанных с последним биологических свойств
316
Глава III
тканевых элементов, как их внешний облик, который у совершенно не-
сходных по существу клеток может обнаруживать в некоторых случаях
далеко идущее сходство. Способность к фагоцитозу также является одним
из элементарных свойств живой протоплазмы и безусловно не имеет тка-
невой специфичности. Даже такая особенность, казалось бы, свойственная
элементам соединительной ткани, являющимся производным единого опре-
деленного зачатка как образование межклеточных веществ, и та (на осно-
вании изложенного выше материала) не может, взятая сама по себе, служить
абсолютным критерием генетического родства клеток и их сходства в
других отношениях. Способность к образованию межклеточных веществ
в той или иной форме и степени также является, повидимому, одним из
элементарных свойств живой протоплазмы, быть может, одной из форм ее
взаимодействия с окружающей внешней средой, наравне с чувствитель-
ностью, сократимостью и рядом других ее свойств, совокупность которых
входит в само понятие жизни. Эти общие элементарные свойства живой
протоплазмы могут, эволюируя, изменяться в широких пределах, раз-
виваясь и достигая высокого совершенства и приобретая вместе с тем новые
качественные особенности или наоборот, подвергаясь далеко идущей ре-
дукции и даже частично исчезая, в той степени, в какой это совместимо
с понятием жизни. Точно так же в прогрессивной эволюции животного мира,
от простейших до человека, и в деградирующих в морфо-физиологическом
отношении отдельных филогенетических ветвях, приводящих, например,
к образованию таких форм, как саккулина среди ракообразных, каждая
из всего многообразия форм, несмотря на все их количественные и качествен-
ные отличия Друг от друга, удовлетворяет некоторым общим признакам,
свойственным всем представителям мира животных.
Все это следует иметь в виду, приступая даже к самому краткому раз-
бору мезенхимы и межуточных тканей с точки зрения естественной генети-
ческой системы гистологических структур.
Произведенный выше анализ ряда тканей показывает, что у позвоночных
в составе того комплекса эмбриональных клеток, который принято обозна-
чать со времени классической монографии бр. Гертвигое (1882) термином
«мезенхима», имеется ряд зачатков, качественно отличающихся друг от
друга и имеющих вместе с тем неодинаковую историю развития. Прежде
всего это относится к разграничению собственно мезенхимы или «энто-
мезенхимы» и так называемой эктомезенхимы, или, точнее, нейромезенхимы.
Часть нейромезенхимпых производных, которая была подвергнута экспе-
риментально-гистологическому анализу, а именно, покровные клетки моз-
говых оболочек обнаруживали совершенно отчетливое сходство с другими
представителями генетически единого эпендимо-глиального тканевого типа
и были включены в его состав. К нейромезенхиме следует также отнести те
клетки, которые, выселяясь из нервных валиков и распространяясь у
позвоночных по всему организму, образуют хроматофоры (Дюшен,
1838, 1939; И. Фишер, 1938, и др.). Последние по своему происхождению
также являются, повидимому, очень высокоспециализированными пред-
ставителями нейрального тина, которые не имеют, по существу, ничего
Экспериментальный метод в гистологии 317
-общего с соединительной тканью и резко отличаются от ее элементов. Как
известно, Вейденрейх (1912) выделил четыре главных области рас-
пространения хроматофор у низших позвоночных: кожную, или дермальную,
перинейральную, т. е. в составе оболочек мозга, органов чувств и перифе-
рических нервов, периваскулярную по ходу сосудистой системы и целоми-
ческую в составе серозных оболочек. Иными словами, хроматофоры фак-
тически повсеместно вступают в теснейшие топографические отношения
с соединительной тканью, хотя и не могут рассматриваться с точки зрения
генетической системы тканей как ее составная часть. С точки же зрения
обычной характеристики тканей хроматофоры, в соответствии с принятыми
в гистологии взглядами, рассматриваются до настоящего времени как одна
из клеточных форм соединительной ткани. У высших позвоночных эта
своеобразная «хроматофорная ткань», как известно, подвергается сильной
редукции, сохраняясь, в конце концов, лишь в отдельных участках орга-
низма, например в сосудистой оболочке глаза, местами в перепончатом
лабиринте, мозговых оболочках и т. д.
У животных альбиносов хроматофоры лишены пигмента, и поэтому
отличать их от клеток собственно сединительной ткани является нелегкой
задачей. Бесцветные хроматофоры представляют собой один из очень лю-
бопытных примеров гистологической мутации, связанной с утратой самого
характерного внешнего признака данных тканевых элементов, которые
тем не менее сохраняют свои прочие, связанные с развитием из нейраль-
ного зачатка, биологические особенности и качественные отличия от клеток
соединительной ткани. Как это уже отмечалось выше, изучение гистоти-
пических и цитотипических свойств хроматофор является одной из интерес-
нейших задач. Весьма интересно также параллельное изучение хроматофор
у беспозвоночных, которые могут также обнаруживать (например у голо-
воногих моллюсков) большую сложность внешней формы и внутренней
структуры. Вопрос о том, из какого именно источника развиваются хро-
матофоры разных беспозвоночных и являются ли они параллельно развив-
шимися из генетически одного и того же источника и родственными или
лишь конвергентными образованиями, также еще нуждается в специальных
исследованиях.
Что именно развивается из нейромезенхимных элементов туловищной
части ганглиозной пластинки (Равен, 1936) помимо хроматофор и, веро-
ятно, также покровных клеток оболочек спинного мозга, в настоящее время
с достоверностью сказать трудно.
Помимо упомянутых выше производных нейромезенхимы, у амфибий
установлено также развитие из того же источника значительной части
хрящевого висцерального скелета (Стон, 1926, 1929; Равен, 1933,
и др.). По крайней мере одностороннее удаление ганглиозной пластинки го-
ловной области влечет за собой недоразвитие соответствующих частей
скелета, а пересадка ее частей на стадии ранней нейрулы в другие места
вызывает на новом месте образование не только участков нервной системы,
но и хряща. Опыты с пересадкой участков соответствующей презумптивной
области на стадии гаструлы не дают развития хряща. Опыты с прижизнен-
318
Глава HI
ной маркировкой ганглиозной пластинки (Стон, 1932) не дали положи-
тельных результатов в смысле наличия в образующемся хряще краски.
По многим данным, участки хряща, которым приписывается развитие
из ганглиозной пластинки, заметно отличаются по своей структуре от обыч-
ного энтомезенхимного хряща. Несмотря на всю убедительность и высокий
интерес данных, касающихся эктомезенхимы, постановка опытов с развитием
хряща все-таки не может полностью исключить возможность того, что
в данном случае имеет место не развитие хряща из материала ганглиозной
пластинки, а лишь индукция его развития в окружающей ткани. Более
убедительными в этом отношении являются данные, полученные методом
изоляции и культивирования отдельных частей нервного зачатка (Вин-
ников, неопубликованный материал).
Если принять полностью все данные, касающиеся эктомезенхимы, то
развитие соединительной ткани и гиалинового хряща из нервного зачатка
следует себе представить как замечательные примеры гистологической
конвергентной дифференцировки, но отнюдь не более удивительные, чем
образование из ретинальных листков глазного бокала гладких или поперечно-
полосатых мышечных волокон радужины, которые будут рассмотрены
ниже в главе о сократимых тканях.
Детальное сравнение тонкой структуры хрящей разного происхожде-
ния и экспериментальный анализ гисто-биологических потенций их эле-
ментов еще принадлежат будущему. Здесь будет уместно указать, что А. Фи-
шер (1923) уже давно поставил удачные опыты с эксплантацией эмбрио-
нального хряща склеры куриных зародышей. В течение довольно длитель-
ного срока культивирования (сколо трех месяцев) хрящевые клетки после
полного исчезновения межклеточного вещества давали очень своеобразные
разрастания, которые, насколько возможно судить по описаниям и рисункам
автора, не обнаруживали никакого сходства с культурами производных
обычной энтомезенхимы. Из какого именно источника развивается этот
своеобразный хрящ, с полной достоверностью сказать трудно. Но
место его образования в составе твердой мозговой оболочки, како-
вой является склера глазного яблока, и своеобразные биологические
свойства его элементов, устанавливаемые методом эксплантации, не могут
не навести на мысль, что изученный Фишером хрящ, возможно, является
одним из хрящей нейрального происхождения, и вызвать желание пере-
исследовать под новым гистологическим и гистогенетическим углом зрения
этот вопрос. Так или иначе, в настоящее время следует считаться с возмож-
ностью наличия у позвоночных трех генетически различных типов хряще-
подобных тканей: обычный энтомезенхимный хрящ в его различных
структурных модификациях, нейральный хрящ и упоминавшийся выше
хордальный хрящ.
Все ткани, развивающиеся из нейромезенхимного зачатка, с точки зре-
ния естественной системы гистологических структур, должны быть исклю-
чены из состава главы о соединительной ткани и крови. Однако генетиче-
ское единство и так называемого энтомезенхимного или собственно мезен-
химного зачатка, в соответствии с разобранными выше фактическими дан-
Экспериментальный метод в гистологии 319
ними и соображениями об эндотелии сосудистой системы, ставит серьез-
ный вопрос об исключении из состава генетически единого комплекса тканей
соединительного вещества не только эндотелия, но и элементов всего гемогло-
бинового кроветворного ряда, который находится (по крайней мере, на
ранних этапах эмбрионального развития) в теснейшем родстве с сосудистой
стенкой и зачаток которого оказывается детерминированным уже на очень,
ранних стадиях онтогенеза. Этот вопрос требует здесь еще дополнительного
обсуждения.
По вполне понятным причинам вопросы кроветворения разрабатывались
в первую очередь на млекопитающих и человеке. Развитие же кровяных
клеток у низших позвоночных изучено до настоящего времени значительно
менее полно.
Основной особенностью гематопоэза у млекопитающих является его
разделение во взрослом состоянии на лимфоидную и миэлоидную части.
Эта особенность давала уже давно повод к жестоким спорам между защит-
никами унитарных и полифилетических, в частности, дуалистических,
теорий гематопоэза. Согласно новейшей, более простой и представляющейся
хорошо обоснованной экспериментально унитарной теории, ярким защит-
ником и, по существу, творцом которой был Максимов, все кровяные
клетки имеют своего общего родоначальника в недифференцированных и по
своим потенциям развития тождественных мезенхимных клетках, которые,
сохраняясь и во взрослом организме, представлены в нем элементами рети-
кулярной стромы кроветворных органов и вездесущими свободными базо-
фильными гемоцитобластами, имеющими морфологический облик лимфо-
цитов, в первую очередь их более крупных модификаций.
Основываясь на своих собственных (1925) работах, имеющих в настоя-
щее время почти двадцатилетнюю давность, я, наряду со многими другими,
также придерживался этой точки зрения, обосновывая и подчеркивая
большое значение в организме малодифференцированных мезенхимных
клеток. Учение о малодифференцированных мезенхимных или камбиальных
клетках в составе соединительнотканных образований позвоночных было
одновременно обосновано и широко развито Максимовым (1925, 1927),
а затем принято и использовано Заварзиным и его сотрудниками (Ясвойн,
1928; Заварзин, 1933, 1935).
Исходя из разделения у млекопитающих кроветворения на лимфоид-
ную и миэлоидную ветви, защитники дуалистической теории, очень темпе-
раментным представителем которой в последнее время являлся прежде
всего Негелц (1930), считают, что миэлоидный и лимфоидный ряды имеют
по самостоятельному родоначальнику—миэлобласту и лимфобласту, каждый
из которых уже детерминирован необратимым образом. Эти две клетки
очень сходны друг с другом, хотя между ними и пытаются находить очень
тонкие, но будто бы постоянные, морфологические различия. Следует,
однако, подчеркнуть, что и новейшие дуалисты (Неге ли) выводят эти
обе формы из еще менее дифференцированных оседлых мезенхимных клеток,
имеющихся в разных местах организма. Поэтому различия между более
новыми вариантами унитарной и дуалистической теорий кроветворения
320
Глава III
сделались, благодаря взаимным уступкам, гораздо менее резкими, чем
они были раньше, когда кроветворные ряды начинались только от свобод-
ных базофильных клеток типа лимфоцитов, а не предполагали еще наличия
единого совершенно недифференцированного мезенхимного резерва. Край-
ние дуалисты (например Шридде, 1923) доводили кроветворное генеа-
логическое дерево вплоть до двух качественно различных оседлых клеточ-
ных форм: эндотелия кровеносных сосудов для клеток миэлоидного ряда
и эндотелия лимфатических сосудов для клеток лимфоидного ряда. Лишь
немногие новейшие авторы (Кеннингем, Себин и Доан, 1925) проводили
резкое разделение кроветворных рядов иным образом и различали гемо-
глобиновый ряд, начинающийся клетками эндотелия, и лимфогранулоцитный
ряд, идущий через общую лимфоидную промежуточную форму от ретикуляр-
ных клеток кроветворных органов, исходя из своей точки зрения, что
эритропоэз, в противоположность лимфогранулопоэзу, всегда развивается
внутрисосудисто.
Я не буду останавливаться на историческом развитии этих взглядов
со времен Эрлиха и на многочисленных их толках и вариантах, которые,
с соответствующей аргументацией, излагаются в курсах гематологии или
в специальных сводках.
При сопоставлении данных, касающихся кроветворения у представи-
телей разных классов позвоночных, уже в настоящее время получается
довольно стройная картина эволюционного развития кроветворного аппа-
рата в этом филогенетическом ряду, которая, мне кажется, убедительно
подкрепляет высказанную выше точку зрения.
Основной особенностью кроветворения у рыб и хвостатых амфибий
является строгое территориальное разделение очагов эритропоэза и лимфо-
гранулопоэза. Эритроциты и тромбоциты образуются у этих форм в селе-
зенке (Биццоцеро нТорре, 1884;Г. Мюллер, 1889;Лопес, 1890;Физали, 1902;
Джордан и Спейдел, 1924, и др.), заканчивая часто свой цикл развития в
циркулирующей крови, а клетки лимфогранулоцитного ряда б. ч. — вне селе-
зенки. У разных животных основные очаги лимфогранулопоэза имеют очень
неодинаковую локализацию: например у ганоидных в соединительной ткани
почки, под серозной оболочкой сердцд, в полостях хрящевого черепа, в стенке
кишечника (Р. Гертвиг, 1879; Држевина, 1905; Дауни, 1909, и др.), у кос-
тистых рыб большей частью в почке, а также в поджелудочной железе
(Биццоцеро и Торре, 1884; Држевина, 1905, и др.), у селахий, главным
образом в слизистой оболочке пищевода и гонадах (Држевина, 1905; Мак-
симов, 1923; Джордан и Спейдел, 1924), у хвостатых амфибий в краевой
зоне печени (Држевина, 1905; Джордан и Спейдел, 1924, и др.). Кроме
этих главных очагов, лимфогранулопоэз может происходить и более или
менее диффузно в разных органах. Частью в норме, частью в эксперимен-
тальных условиях интраваскулярное образование эритробластов или, по
крайней мере, малодифференцированных базофильных клеток, которые, по-
видимому, являются предшественниками элементов гемоглобинового ряда,
неоднократно отмечалось и в других местах сосудистой системы. Мне самому
приходилось это наблюдать с большой отчетливостью в артериальном конусе
Экспериментальный метод в гистологии 321
щуки и аксолотля. К сожалению, до настоящего времени, кроветворени
у круглоротых остается недостаточно полно изученным. Известно, что у ми-
ноги имеется своеобразный кроветворный орган, расположенный вдоль
тела дорзально от спинного мозга. Кроме того, у круглоротых описывалось
кроветворение в стенке кишечника, в частности, в его спиральном клапане
(Джордан и Спейдел, 1930). Возможно, хотя и неизвестно в точности, что
последняя кроветворная область филогенетически предшествует крове-
творению в селезенке, которая у этих животных еще отсутствует.
У бесхвостых амфибий начинается отчетливое образование миэлойдной
ткани кроветворного костного мозга и перемещение в него эритропоэза и час-
тично гранулопоэза, причем красные кровяные клетки и, повидимому, тром-
боциты развиваются исключительно внутрисосудисто, а белые — в окружаю-
щей сосуды ретикулярной ткани (Маркис, 1892; Нейман 1896; Джордан,
1919, и др.). Правда, и селезенка бесхвостых амфибий, повидимому, также
функционирует еще как эритропоэтический орган (Джордан и Спейдел,
1923). Краевая зона печени может в разной степени служить очагом лимфо-
гранулопоэза. У рептилий и птиц селезенка при обычных условиях утрачи-
вает значение эритропоэтического органа, а костный мозг становится основ-
ным очагом кроветворения, причем лимфогранулопоэз, кроме того, совер-
шается и диффузно в соединительной ткани разных участков тела. В
костном мозгу образование эритроцитов и лейкоцитов совершенно отчетливо
разделено территориально. Эритроциты развиваются только внутрисосудисто
в просвете венозных синусов (Биццоцеро и 7 орре, 1884; еан-дер-Штрихт,
1892; Данчакова, 1909, и др.), а лейкоциты — в окружающей сосуды рети-
кулярной ткани. По данным Доан (1923) и Доан, Каннингем и Себин'(1925),
после голодания удается проследить развитие эритробластов из эндотелия,
причем это происходит в особых тонких капиллярах, соединяющих друг
с другом венозные синусы. По этим кроветворным капиллярам постоянного
кровообращения не происходит. У млекопитающих в красном костном
мозгу разделения эритропоэза и гранулопоэза, по многим данным, не про-
слеживается и большей частью признается, что тот и другой происходят
внесосудисто в ретикулярной ткани.
Учитывая очень большие трудности различения в костном мозгу млеко-
питающих внутрисосудистых и внесосудистых участков и очень убедитель-
ные в этом отношении данные, касающиеся рептилий, птиц, а также бес-
хвостых амфибий, мне представляется, что внутрисосудистое развитие
красных кровяных клеток у млеког итающих не может считаться достаточно
обоснованным. Доан, Каннингем и Себин (1925) обнаруживали и у кроли-
ков, хотя и не с такой отчетливостью, как у голубей, существование тонких
капилляров, соединяющих друг с другом венозные синусы и являющихся
с их точки зрения местами интраваскулярного эритропоэза. Кроме того,
не подлежит сомнению, что эритробласты в костном мозгу, как пра-
вило, располагаются очажками. Поэтому территориальное разделение
эоитропоэза и гранулопоэза в костном мозгу млекопитающих, несмотря
на то, что оно выражено значительно менее отчетливо, чем у других позво-
ночных, все-таки представляется в достаточной степени вероятным.
21 Хлопин—-349
322
Глава III
С этой точки зрения было бы очень интересно дополнительно изучить
отношения в печени зародышей млекопитающих, в которой обычно при-
знается внесосудистое образование эритроцитов, хотя имеются и указания
противоположного характера. Выделение лимфоидного кроветворения в
самостоятельные органы и участки тела у позвоночных, несомненно, про-
исходит вторично и своего полного развития достигает только у млекопитаю-
щих с их высоко развитыми лимфоидными очагами. Многочисленные наблю-
дения над т. наз. экстрамедуллярным миэлопоэзом, подробный разбор
которых выходит за рамки поставленной задачи, показывают, что скорее
и чаще всего вне костного мозга наступает развитие специальных и эози-
нофильных лейкоцитов, затем уже могут появляться мегакариоциты и еще
позже, и лишь в редких случаях, эритробласты. Экстрамедуллярный миэло-
поэз наблюдается как при весьма разнообразных патологических состоя-
ниях, так и под влиянием местных или общих экспериментальных воздей-
ствий на целый организм. Самым эффектным методом местного получения
экстрамедуллярного кроветворения является, пожалуй, перевязка почеч-
ных сосудов, после которой в соединительной ткани лоханки образуется
губчатая кость с настоящим кроветворным костным мозгом (Сачердотти
и Фраттен, 1902; Максимов, 1907, и др.). Общими экспериментальными,
методами получения экстрамедуллярного миэлопоэза у млекопитающих
являются повторные кровопускания, введение в организм кровяных ядов,
как то: фенилгидразин или сапонин, холестерин, бактерии и т.п. Прежде
всего, как правило, экстрамедуллярный миэлопоэз получается в селезенке,
затем в печени, лимфатических узлах, надпочечниках и других органах,
из которых и при этих условиях обращают на себя внимание ворота
почки. У амфибий и, вероятно, у других низших позвоночных с селезеноч-
ным эритропоэзом удаление селезенки вызывает резко выраженное
образование эритробластов из эндотелия артериального конуса сердца.
Экстрамедуллярные миэлоидные элементы появляются первоначально, как
правило, внутри сосудов, а затем уже и вне их. Это значит, что большое
значение в образовании экстрамедуллярных очагов кроветворения принад-
лежит заносу клеток через кровяное русло. Местное внесосудистое разви-
тие могло быть установлено только для миэлоцитов, но не для эритробластов.
Некоторые данные, полученные методом эксплантации, подчеркивают
близкие генетические отношения между лимфоидными элементами, с одной
стороны, и гранулоцитами, с другой, но отнюдь не клетками гемоглобино-
вого ряда. Так, например, Максимов (1923) описывал появление в экс-
плантатах из лимфатических узлов специальных миэлоцитов, а Блуму (1937)
удалось в последнее время получить массовое образование специальных
миэлоцитов и лейкоцитов в культурах лимфоидных элементов, взятых из
грудного протока кроликов, правда, под влиянием ряда особых факторов.
Еще раньше Авророву и Тимофеевскому (1912) удалось наблюдать миэ-
лопоэз вне организма в культурах лейкемической крови человека. Согласно
более подробным позднейшим данным Тимофеевского и Беневоленской (1925,
1927), крупные базофильные малодифференцированные клетки крови боль-
ных миэлоидной лейкемией могут ’в эксплантатах превращаться в нейтро-
Экспериментальный метод в гистологии
323
фильные, эозинофильные, базофильные лейкоциты и даже в эритробласты.
Последнее, с нашей, проводимой здесь, точки зрения, может быть истол-
ковано таким образом, что в крови этих больных имеются не только малодиф-
ференцированные родоначальники гранулоцитов, но и особые клетки,
являющиеся предшественниками элементов гемоглобинового ряда и, возмож-
но, образовавшиеся, в свою очередь, из эндотелия. На основании всего
сказанного, следует ожидать, что ткань селезенки низших позвоночных,
а у высших позвоночных — соответствующая ей т. наз. красная пульпа,
должна обладать рядом особых филогенетически обусловленных свойств,
связанных с ее первоначальной эритропоэтической функцией. С этим особым
значением селезенки нельзя также не поставить в связь того факта, что, по
многим данным, в ее развитии большое значение принадлежит целомической
выстилке зародыша, элементы которой, пролиферируя, образуют более или
менее значительную часть зачатка этого органа.
В связи с этим мои прежние данные (1925, 1928), касающиеся сравни-
тельного изучения методом эксплантации элементов соединительной ткани
разных позвоночных, должны быть в настоящее время пересмотрены и
уточнены в той своей части, в которой они касаются ткани селезенки.
То же самое касается и довольно многочисленных работ, сделанных
в разное время методом эксплантации и посвященных специально селезенке
птиц, млекопитающих и человека, которые были недавно кратко резюми-
рованы в моей сводке по культуре тканей (Хлопин, 1940).
Итак, в настоящее время имеются серьезные положительные данные,
заставляющие считать, что в составе т. наз. энтомезенхимы позвоночных
имеются по существу не один, а, по крайней мере, два зачатка. Один из
них представлен эндотелием сосудистой системы и генетически связанными
с ним клетками гемоглобинового ряда, а у всех позвоночных, кроме млеко-
питающих, возможно, также веретенчатыми клетками крови или тромбо-
цитами. Эта часть т. наз. производных мезенхимы, на основании того, что
говорилось в предыдущей главе, повидимому, связана филогенетически
с целомической выстилкой и является по существу особой специализиро-
ванной разновидностью тканей целодермального типа, которая у позво-
ночных настолько резко обособилась и отличается по своим свойствам и
от мезотелия и от других целодермальных тканей, что заслуживает выде-
ления в особый «ангиоэритробластический» тканевой тип. Вопрос о том,
относятся ли к нему же мегакариоциты и кровяные пластинки млекопитаю-
щих, пока остается открытым. Так или иначе, с этой точки зрения прихо-
дится считать эритроциты за свободных высокоспециализированных по-
томков целомического эпителия, а вместе с тем мы встречаемся еще с одним
примером эволюционной дивергентной дифференцировки в пределах не-
когда единого тканевого типа. Вторая часть «энтомезенхимы» позвоночных,
к краткому разбору которой мы теперь должны перейти, представлена теми
ее элементами, которые дают лимфоциты и гранулоциты крови и все много-
образие межуточных тканей.
Здесь уместно еще напомнить, что у ряда низших позвоночных в цирку-
лирующей крови описывались «пигментные лейкоциты». Весьма возможно,
*
324
Глава III
они представляют собой округлившиеся и свободно плавающие в крови
меланофоры, т. е. элементы нейромезенхимного характера. Это подчерки-
вает лишний раз, что форменные элементы крови могут быть, по своему
происхождению, а, следовательно, и биологическим свойствам, клетками,
относящимися к разнообразным тканевым типам.
Другим «соком» тела, содержащим в своем составе клетки разного про-
исхождения, является целомическая полостная жидкость, или т. наз. эксу-
дат серозных полостей млекопитающих и человека, в котором, кроме лей-
коцитов, лимфоцитов, округлившихся малодифференцированных мезен-
химных клеток и гистиоцитов сальника, имеются также мезотелиальные
целомические элементы (Максимов, 1925, 1927).
б) Кровь и волокнистая соединительная ткань позвоночных
Против генетического единства в ряду позвоночных второй части т. наз.
энтомезенхимы, а вместе с тем и ее производных, никаких серьезных дан-
ных пока не имеется. Наоборот, весь обширный экспериментальный и пато-
логический материал, который известен в настоящее время, говорит в пользу
близких генетических отношений всей этой весьма разнообразной группы
тканей, хотя в ее пределах и наблюдается очень высокая специализация
в различных направлениях.
Уже в классических исследованиях Максимова над кровью и сое-
динительной тканью млекопитающих (1902) методом асептического воспа-
ления приводятся убедительные данные в пользу широкого участия в обра-
зовании фагоцитирующих клеток воспалительного очага не только спе-
циальных гранулоцитов, но и лимфоидных клеток, эмигрирующих из кро-
вяного русла. Из гематогенных лимфоидных клеток, наряду с местными
блуждающими клетками в покое или гистиоцитами, возникают одинаковые
(Энергично фагоцитирующие и амебоидно подвижные элементы, которые,
в силу своего происхождения из этих разных источников, были обозначены
Максимовым как полибласты. Эти элементы соответствуют мечниковским
макрофагам и часто обозначаются теперь этим первоначальным термином.
, Блуждающие клетки в покое, клазматоциты или рагиокриновые клетки,
получившие впоследствии обозначение гистиоцитов, считались в то время
одной из двух основных клеточных форм соединительной ткани. На фоне
этой преимущественно эксудативной фазы воспалительной реакции на
повреждение и инородное тело постепенно развиваются пролиферативные
процессы, заключающиеся в первую очередь в размножении и росте второй,
известной в то время, формы местных клеток соединительной ткани типа
т. наз. фибробластов и сосудистых эндотелиальных почек.
Из энергично пролиферирующих оседлых клеток соединительной ткани,
макрофагов и сосудистых почек образуется молодая грануляционная ткань.
Лейкоциты, при обычном асептическом воспалении, не носящем гнойного
характера, погибая, резко уменьшаются в числе и вскоре почти совершенно
исчезают. Пролиферативная фаза постепенно сменяется фазой дифферен-
цировки, в течение которой грануляционная ткань превращается в плотный
Экспериментальный метод в гистологии 325
соединительнотканный рубец с большим количеством межклеточного ве-
щества, окружающий со всех сторон введенное инородное тело, в качестве
которого, по почину Циглера, большей частью служат кусочки цел-
лоидина той Или другой формы. В течение этой поздней фазы воспаления
Максимов отмечал, что часть успокоившихся макрофагов или гистио-
цитов делается неотличимой от оседлых клеток типа фибробластов, а тем
самым сделал вероятным предположение, что лимфоциты крови могут в ко-
нечном итоге превращаться в оседлые клетки соединительной ткани и уча-
ствовать и в образовании межклеточного вещества.
Со времени этих исследований Максимова прошло в настоящее
время 40 лет. Тем не менее, они сохраняют все свое фундаментальное зна-
чение. Вместе с тем последующие исследования заставили внести в концеп-
цию соединительной ткани того времени, страдавшую известным схема-
тизмом, некоторые изменения, сделанные частично уже самим Макси-
мовым (1927). Эти изменения касаются^ прежде всего, понятия оседлых
клеток соединительной ткани, фибробластов, или, как их теперь часто на-
зывают, фиброцитов, которые первоначально трактовались как одинаковые
по существу высокодифференцированные клеточные элементы. Из числа
этих т.наз. фибробластов или фиброцитов пришлось, прежде всего, выделить
внешне сходные с ними недифференцированные или малодифференцирован-
ные мезенхимные клетки. Наличие этих малодифференцированных фибро-
бластообразных клеток в соединительной ткани, на которое мной было обра-
щено внимание еще в 1925 году, тогда же заставило меня высказать мысль,
что тем самым делается очень вероятным предположение о наличии в сое-
динительной ткани разных позвоночных непрерывного клеточного ряда,
разные звенья которого отличаются друг от друга уровнем своей дифферен-
цировки. Этот клеточный ряд начинается от малодифференцированных
мезенхимных клеток, сходных по своим свойствам с элементами ретикуляр-
ной стромы кроветворных органов, и заканчивается зрелыми фибробластами,'
вероятно, утрачивающими постепенно способность к пролиферации и в
конце концов погибающими. Для клеток этого дифференцирующегося
клеточного ряда мной были предложены обозначения: ретикулярная клет-
ка — для малодифференцированных мезенхимных клеток, десмобласты —
для молодых, не достигших окончательной зрелости фибробластов, и
десмоциты — для конечных стадий их дифференцировки.
Эта трактовка оседлых клеточных форм соединительной ткани была
затем проверена мной и полностью подтверждена при изучении эмбрио-
нальной соединительной ткани человеческих зародышей и плодов разного
возраста методом эксплантации (1931), после чего я уже не возвращался
к этим вопросам. Эти представления в значительной степени легли в основу
концепции соединительной ткани, излагаемой в курсе гистологии Завар-
зина (1933—1939), и послужили темой специального сообщения (1935),
В отличие от моей терминологии Заварзин принял обозначения —
камбиальная клетка, фибробласт, фиброцит.
Тогда же я высказал предположение, что в соединительной ткани различ-
ных животных относительное количество клеток разного уровня диффе-
326
Глава III
ренцировки, вероятно, будет неодинаковым. Это предположение получило
свое фактическое подтверждение в очень тщательных исследованиях Яс-
война (1928), которому, кроме того, удалось на оригинальных и в тех-
ническом отношении безукоризненных препаратах — щипках из соеди-
нительной ткани—демонстрировать впервые морфологические различия
фибробластообразных клеток разного уровня дифференцировки. Еще до
этого Максимов (1927) показал, что соединительная ткань разных участков
тела одного и того же животного может отличаться неодинаковым уровнем
своей дифференцировки и, в первую очередь, относительным количеством
«мезенхимных» клеток. Им же был тогда особенно подчеркнут малодиффе-
ренцированный характер соединительной ткани сальника.
Все эти изменения в понимании фибробластообразных клеток соедини-
тельной ткани соответствующим образом отразились и на трактовке части
процессов, наблюдаемых при воспалении в составе целого организма и при
пролиферации соединительной ткани в условиях тканевых культур. В тех
же опытах эти представления получили свое дальнейшее развитие. Ниже
мы еще вернемся к свойствам пролиферирующих соединительнотканных
клеток типа фибробластов и к их взаимоотношениям с макрофагообраз-
ными элементами. Здесь необходимо еще отметить, что не очень давно была
предложена совершенно другая трактовка основных клеточных форм сое-
динительной ткани млекопитающих (В. и М. Меллепдорф, 1926). Со-
гласно этой концепции, развивающейся и в последующих работах тех
же исследователей и их сотрудников, все оседлые клетки рыхлой соедини-
тельной ткани, обозначаемые этими авторами как фиброциты, образуют,
соединяясь своими отростками, связную сеть и представляют собой по су-
ществу недифференцированные или малодифференцированные мезенхимные
элементы, которые под влиянием различных внешних воздействий могут
превращаться не только в гистиоциты, но и в зернистые лейкоциты крови.
Идя еще дальше, те же авторы считают и мезотелий серозных оболочек
такими же эмбриональными мезенхимными клетками, что, в соответствии
со сказанным выше, в главе о целодермальных тканях, ни в какой мере не
соответствует действительному положению вещей.
Эта точка зрения на соединительную ткань, быть может, с первого взгляда
подкупает известной простотой, но она несомненно не выдерживает серьез-
ной и резкой критики, данной в свое время в последней, изданной посмертно,
статье Максимова (1929), в обстоятельных и точных исследованиях Ясвой-
на (1930) и в ряде работ других авторов. Очевидно, супруги Меллен-
дорф были введены в заблуждение рядом картин, которым они дали со-
вершенно неправильную трактовку. Втягивание отростков «фиброцитами»
они безоговорочно принимали за их превращение в макрофагов, дегенера-
тивные макрофаги с фрагментирующими ядрами они считали обра-
зующимися лейкоцитами и допустили ряд других столь же серьезных
ошибок.
На более подробном разборе этой, имеющей в настоящее время, пожа-
луй, лишь исторический интерес, концепции я не буду останавливаться.
Следует лишь отметить, что ошибочные выводы Меллендорфа отчасти объ-
Экспериментальный метод в гистологии
327
ясняются также и материалом: соединительная ткань белых мышей отно-
сительно очень богата малодифференцированными клетками.
Для установления близких генетических отношений свободных клеток
типа лимфоцитов, моноцитов и макрофагов решающие результаты, которые
можно с полным правом охарактеризовать как expermentum crucis, по-
лучены методом эксплантации. В этих опытах в качестве материала для
посева применяли или просто капли крови и полостных эксудатов или чаще
пользовались отцентрифугированными свободными клетками, в частности,
лейкоцитарной пленкой крови.
Превращения в условиях тканевых культур нормальных форменных
элементов крови различных позвоночных и человека (Каррель и Ибелинг,
1922; У. Льюис, 1926; Каффье, 1927, 1928; Максимовой, 1929; Хрущов,
1935, и др.), лимфы млекопитающих (Блум, 1928, 1937) и элементов лейке-
мической крови человека (Авроров и Тимофеевский, 1913; Тимофеевский
и Беневоленская, 1929) в настоящее время изучены очень подробно. Эрит-
роциты не обладают способностью к активному движению и каким-либо
изменениям прогрессивного характера, будучи высокоспециализированными
элементами. Они лишь переживают ограниченное время в условиях экс-
плантата, а затем погибают и исчезают без остатка. Они или фагоцитиру-
ются макрофагами и перевариваются в их протоплазме, оставляя после себя
зернышки желтоватого или бурого цвета, или растворяются внеклеточно. -
Гранулоциты также, несомненно, не обладают способностью к делению и к
каким-либо превращениям. В течение некоторого времени они могут в
культурах активно передвигаться, амебоидно ползая и изменяя форму своего
тела. Постепенно их амебоидная активность затихает, они начинают окру-
гляться, а затем дегенерируют и распадаются.
Указания противоположного характера, имеющиеся в работе Каффье
(1928), повидимому, основаны на каком-то недоразумении. Способность
к превращениям прогрессивного характера в условиях культур незернистых
кровяных клеток была впервые установлена Авроровым и Тимофеевским,
а затем подтверждена многими авторами для нормальной крови и детально
изучена на разных объектах Максимовым, Тимофеевским и Беневоленской.
Сопоставляя данные различных авторов, следует отметить, что судьба
клеток крови типа лимфоцитов и моноцитов в очень сильной степени за-
висит от условий существования и состава питательной среды. При куль-
тивировании в обычных условиях в свертке фибрина моноциты и лимфоциты
частью погибают, частью, увеличиваясь в размерах, приобретают характер
макрофагов, которые могут превращаться в гигантские многоядерные клетки.
Проделывая эти превращения, малые лимфоциты крови и лимфы сначала
делаются неотличимыми от моноцитов (Максимов, Блум). Затем
макрофаги подвергаются раньше или позже дальнейшим превращениям.
Как уже отмечалось выше, макрофаги, полученные из куриной крови,
могут, при известных условиях, довольно длительное время культивиро-
ваться в чистом виде (Каррель и Ибелинг, 1921). Макрофаги из
крови и лимфы млекопитающих, наоборот, по истечении короткого проме-
жутка времени превращаются в связную тканевую систему (Блум, Мак-
828
Глава III
симов, Тимофеевский и Беневоленская). Повидимому, эти разли-
чия зависят в значительной степени от неодинаковых условий
культивирования в опытах разных авторов, хотя не исключена возможность
видовых различий между кровяными макрофагами. Подобно тому, как
не все лимфоциты даже в одной и той же культуре, сохраняющей свою
жизнеспособность, одновременно превращаются в макрофагов, последние
не одновременно подвергаются дальнейшим изменениям, которым благо-
приятствует культивирование без систематических вырезаний. Вместо
амебоидных псевдоподий они начинают образовывать заостренные цито-
плазматические отростки, а ядро становится менее компактным и на фикси-
рованных препаратах окрашивается светлее. Клетки с заостренными от-
ростками утрачивают свойственную макрофагам волнистую перепонку,
приобретают морфологические особенности так называемых фибробластов
тканевых культур и, соединяясь друг с другом, образуют сетевидные струк-
туры. Образовавшиеся фибробластообразные элементы сначала под влия-
нием даже относительно слабых раздражений, например, сильного осве-
щения, сотрясения и т. п., могут, втягивая отростки, вновь приобретать
характер макрофагов (Максимов, 1928). Постепенно, однако, эти
изменения делаются все более стойкими и в конце концов уже оказываются
необратимыми. Немногочисленные у макрофагов фигуры митотического
деления начинают встречаться чаще. Часть макрофагов сохраняется между
образовавшимися фибробластообразными клетками в неизмененном виде,
но перестает активно ползать, приобретая характер гистиоцитов или блуж-
дающих клеток в покое рыхлой соединительной ткани. Соединительно-
тканные клетки фпбробластообразного вида, дифференцированные необра-
тимым образом и приобревшие определенные морфологические особенности,
обозначают в настоящее время как фиброциты.
Одной из характерных особенностей этих элементов являются тонкие,
почти прямые нити, т. наз. тонофибрилли, пробегающие и в их цитоплазме
и могущие быть обнаруженными на фиксированных и окрашенных пре-
паратах. С точки зрения некоторых авторов (Леви и Оливо, 1928;
Леви, 1934), эти структуры представляют собой не нити, а вызванные
растяжением клеточного тела гребневидные выступы и борозды поверхно-
стного уплотненного слоя цитоплазмы.
Эти структуры были впервые описаны в рубцовой ткани Максимо-
вым (1902), а затем обозначены Маллори (1904) как волокна фибро-
глии. Дальнейшее превращение лейкоцитов и моноцитов удалось получить
и подробно изучить Максимову (1929) в культурах из клеток нормальной
крови. Оказалось, что клетки новообразованной ткани постепенно обра-
зуют густую сеть аргирофильных волокон межклеточного вещества, кото-
рые затем, утолщаясь и соединяясь друг с другом, приобретают характер
настоящих коллагеновых пучков соединительной ткани. В конце концов
из лимфоцитов и моноцитов крови вне организма образуется настоящая
соединительная ткань, состоящая из клеток типа фибробластов, гистиоци-
тов и большого количества коллагенового волокнистого межклеточного
вещества. Эти факты, безусловно, имеют фундаментальное значение для
Экспериментальный метод в гистологии
329
гистолетии и патологии, так как решают в положительном смысле вопрос
об образовании соединительнотканных структур свободными кровяными
клетками.
Гюпер и Рессель (1932) описали своеобразные скопления клеток
в зоне роста культур из белых кровяных клеток, которые они обозначают
как капилляроподобные образования. Эти структуры, не имеющие ничего
общего с эндотелиальными сосудистыми трубками, повидимому, представ-
ляют собой скопления кровяных клеток в полостях, образовавшихся в
свертке фибрина, наполненных жидкостью и имеющих иногда трубчатую
форму.
Данные, полученные при культивировании свободных клеток серозных
эксудатов, до настоящего времени во многих отношениях недостаточно
ясны и нуждаются, еще в проверке. В составе исходного материала, по наи-
более обстоятельным данным Максимова (1927), как это отмечалось
выше, следует считаться с присутствием следующих элементов: лимфо-
цитов разного размера, округлившихся недифференцированных мезен-
химных клеток и гистиоцитов сальника, слущившихся и округлившихся
клеток мезотелия, а также больших количеств гранулоцитов. Изолирован-
ные мезотелиальные и недифференцированные мезенхимные клетки эксу-
датов, по данным Максимова, имеют лимфоцитообразный облик и по
внешнему виду не могут быть безошибочно определены. Это делает понят-
ными трудности, связанные с изучением превращений вне организма тех
или иных клеток эксудата. Гистиоциты эксудата в условиях культур обра-
зуют фагоцитирующие макрофаги и гигантские многоядерные клетки,
а лимфоцитоподобные элементы постепенно превращаются в отростчатые
клетки типа фибробластов (Верещинский, 1924; Максимов, 1916, 1927).
Превращение лимфоцитов эксудата в фагоцитирующие макрофаги, которые,
повидимому, являются, как и для лимфоцитов крови и лимфы, промежуточ-
ной стадией образования фибробластообразных клеток, описывалось
Крейбергом (1929), Нордмейер (1932), и др.
В соответствии со своими взглядами на природу мезотелия Макси-
мов и Верещинский считали, что превращение в фибробласты проделывают
слущившиеся и сходные по своему внешнему виду с лимфоцитами мезоте-
лиальные клетки.
Эту точку зрения в настоящее время следует считать неправильной
(Хлопин, 1934, 1937). Вероятнее всего, слущившиеся мезотелиальные
клетки погибают, а лимфоциты и округлившиеся мезенхимные клетки
эксудата действительно превращаются в соединительнотканные элементы
типа фибробластов. Едва ли возможно отрицать, что и гистиоциты эксудата
обладают сходными гистобластическими потенциями. При культивиро-
вании элементов брюшного эксудата по своему методу в непрерывном токе
питательной среды де-Гаан (1926, 1929), де-Гаан и Феринга (1934)
также наблюдали образование сетевидной ткани из свободных недиф-
ференцированных клеток, элементы которой, обозначаемые ими как фибро-
бласты, однако, довольно длительное время сохраняли способность вновь
превращаться в лимфоциты и даже дифференцироваться в гранулоциты
33)
Глава 1П
и красные кровяные клетки, что совершенно несвойственно, согласно наи-
более обоснованным взглядам, настоящим фибробластам соединительной
ткани.
Если считать наблюдения де-Гаана хотя бы частично достаточно
точными в гистологическом отношении, то полученные им сетевидно свя-
занные отростками фибробластообразные клетки следует признать эле-
ментами, сравнимыми с недифференцированными мезенхимными клетками
соединительной ткани или, быть может, элементами ретикулярной стромы
кроветворных органов, а отнюдь не дифференцированными фибробластами
или фиброцитами обычной соединительной ткани. Кроме того, де-Гаан
наблюдал образование многоядерных гигантских клеток. Предполагаемое
им образование лимфоцитов из гигантских клеток представляется неве-
роятным, а вопрос об образовании эритроцитов требует коренного пере-
смотра. В последнее время Вейцман (1939) изучал методом экспланта-
ции клеточные элементы плевральных и перитонеальных эксудатов чело-
века.
Следует вкратце упомянуть о наблюдениях, сделанных многими авто-
рами над превращениями клеток в капле, приготовленной из полостной
жидкости, или гемолимфы ряда беспозвоночных, например червей, рако-
образных и насекомых (Гудрич, 1920; Лазаренко, 1925; Легорн,
1925; Форе-Фремье, 1927, и др.). Согласно этим наблюдениям,
амебоциты могут, быстро изменяя свою форму и образуя заострен-
ные отростки, распластываться по плотному субстрату. Это превращение
неактивных амебоидных клеток в распластанные неамебоидные «хоанолей-
коциты» в условиях кратковременного опыта оказывается обратимым и,
повидимому, сравнимо, как это указывает Форе-Фремье, с превра-
щением макрофагов позвоночных в фибробласты или, точнее, в недиффе-
ренцированные фибробластообразные элементы.
Приведенные наблюдения показывают, что, по крайней мере, часть
незернистых клеток крови лимфы и брюшного эксудата обладает способ-
ностью к превращению в макрофаги и оседлые отростчатые клетки типа
фибробластов, причем последние, в ряде случаев, могут образовать настоя-
щую волокнистую соединительную ткань. С другой стороны, точно такие
же малодифференцированные клетки обнаруживают способность к диф-
ференцировке в направлении зернистых кровяных клеток. В тех случаях,
когда это происходит более или менее одновременно в одной и той же куль-
туре, при практически тождественных условиях существования, этот факт
является по существу столь же поразительным, как, например, дивергент-
ная дифференцировка малодифференцированных родоначальников зерни-
стых кровяных клеток, располагающихся по соседству друг с другом при
обычных условиях в ретикулярной строме кроветворных органов и их
превращение в определенных количественных соотношениях в несколько
разновидностей гранулоцитов. Как осуществляется эта чрезвычайно точ-
ная согласованность друг с другом нормальных процессов дивергентной
дифференцировки, пока остается совершенно неясным. Кроме необходимого
допущения, что в этом и во многих других аналогичных случаях проявляются
Экспериментальный метод в гистологии
331
какие-то организменные и тканевые коррелятивные связи, возможно, гумо-
рального характера, никаких более конкретных объяснений такого рода
фактам из области эмбриональных и постнатальных гистогенезов пока
дать нельзя. Их причинный анализ является одной из увлекательных и
очень сложных задач будущего. То, что, казалось бы, однородный исход-
ный материал образует вне организма разнородные продукты превращения,
сосуществующие друг с другом, во всяком случае говорит за то, что эксплан-
тированные клетки представляют собой не колонии самостоятельных эле-
ментов, а взаимосвязанные тканевые системы. Ряд изложенных выше фак-
тов, касающихся других тканей, доказывает то же самое.
В случае малодифференцированных клеток крови, имеющих внешний
облик лимфоидных элементов или моноцитов, конечно, не может быть пол-
ностью исключена возможность, что они, несмотря на полное или почти пол-
ное морфологическое тождество, все-таки не вполне равноценны, а потому и
обнаруживают различные превращения. Однако, если это можно считать,
на основании сказанного выше, более чем вероятным в отношении мало-
дифференцированных родоначальников клеток гемоглобинового ряда, с
одной стороны, гранулоцитов и соединительной ткани, с другой, которые,
по многим данным, обладают различной историей развития, то допущение
в отношении мезенхимных, в узком смысле слова, лимфоидных клеток
каких-нибудь стойких качественных различий едва ли представляется
правдоподобным.
Вероятнее всего представить себе, что клетки, имеющие малодифферен-
цированный характер, но одну и ту же историю развития, и образующиеся
в составе однородных тканей, какими, например, является ретикулярная
строма костного мозга, могут лишь обладать нестойкими различиями чисто
функционального порядка или, самое большее, обнаруживать известные
явления «старения», о которых так часто говорят гематологи (например
Паппенгейм, 1919) и которые могут быть связаны со сроком их цир-
куляции в крови. В какой-то момент жизни комплекса мало дифференциро-
ванных клеток в части их происходит необратимая детерминация. С мо-
мента этой детерминации первоначально однородные клетки уже становятся
качественно различными и начинают дифференцироваться в строго опре-
деленных направлениях. Это явление сравнимо с тем, что имеет место на
ранних стадиях эмбрионального развития. В какой именно момент жизни
и вследствие каких причин происходит детерминация малодифференцирован-
ных клеток, пока сказать с точностью нельзя. Максимов (1923) в свое
время высказал предположение, что кровяные элементы испытывают в те-
чение митоза какой-то толчок, под влиянием которого обе дочерние клетки
начинают дифференцироваться в каком-нибудь одном определенном на-
правлении. Эта мысль основывалась на том, что однородные клетки, нахо-
дящиеся на ранних стадиях дифференцировки в кроветворных органах,
как правило, встречаются попарно. Если малодифференцированные клетки
находятся в особых условиях существования в кровяном русле и при этом
более или менее длительное время не делятся митозом, то, очень может
быть, они постепенно претерпевают какие-то изменения, которые нельзя
332
Глава III
назвать дифференцировкой в полном смысле этого слова, но которые в то
же самое время понижают их стойкость в отношении внешних воздействий,
их способность к прогрессивным изменениям в других условиях существо-
вания и митотическому делению и завершаются гибелью и распадом. Этот
процесс старения, повидимому, в некоторых случаях, бывает связан с ви-
димыми морфологическими изменениями (например окрашиваемости ци-
топлазмы, формы ядра и т. п,), но в ряде случаев, повидимому, не может
быть уловлен путем простого микроскопического изучения.
До известной степени аналогичный, но по существу не вполне тождествен-
ный процесс старения наблюдается и в дифференцированных клетках крови
и, вероятно, многих тканевых систем организма. Такое старение, приво-
дящее к смерти и большей частью к распаду, является в данных случаях
неизбежным роковым следствием предшествующей дифференцировки. Его
причины находятся, прежде всего, в самих клетках, а не во внешних усло-
виях их существования. Такие изменения свойственны, например, грануло-
цитам с их возрастающей дольчатостью ядра, эритроцитам, отличающимся
друг от друга осмотической стойкостью, ороговевающим клеткам кожного
эпителия и, вероятно, клеткам многих других тканевых систем организма.
Довольно многочисленные данные, касающиеся соединительной ткани
и крови представителей разных классов позвоночных, с моей точки зрения,
еще не позволяют сделать достаточно обоснованных выводов эволюцион-
ного порядка, тем более, если учесть все сказанное выше о нейромезенхиме,
сосудистом эндотелии и клетках гемоглобинового ряда. Интересные сопо-
ставления, сделанные в этом направлении Заварзиным (1934), имеют,
пока еще в значительной степени предварительный характер.
Как это отмечалось Максимовым (1927), а также мной (1925), в соедини-,
тельной ткани ряда низших позвоночных вероятно наличие значительного
количества мало дифференцированных фибробластоподобных элементов.
В пользу этого говорят описательные и экспериментальные, сделанные мето-
дами воспаления и эксплантации, наблюдения самого Максимова (1906),
а также мои (1925) над аксолотлем, Эбергардта (1907/8) над черепахой,
мои (1925) и Крафт (1925) над карасем.
Однако это не является общим правилом, так как, например, у лягушки,
наоборот, по крайней мере преобладающая часть «фибробластов» произ-
водит впечатление дифференцированных клеток (Алфеева, 1927). Данные
Солуха (1908), полученные на курах, говорят в пользу наличия в их сое-
динительной ткани относительно большого количества малодифференци-
рованных элементов. Клетки типа гистиоцитов, частью покоящиеся, частью
в состоянии амебоидного движения, описывались у различных предста-
вителей рыб (Максимов, 1923; Хлопин, 1925; Крафт, 1925; Заварзин, 1938),
амфибий (Максимов, 1906; Беннингоф, 1923; Хлопин, 1925; Алфеева, 1927),
рептилий (Эбергардт, 1907/8; Хлопин, 1925) и птиц (Солуха, 1908; Мясоедов,
1926), причем многими отмечалась нередко неясная отграниченность этой
клеточной формы от элементов типа фибробластов. Кроме того, в составе
соединительной ткани низших позвоночных неоднократно описывались,
частью очень своеобразные, частью сходные с таковыми у млекопитающих
Экспериментальный метод в гистологии
333
другие специальные виды клеток, на которых мы не будем здесь остана-
вливаться.
Разнообразие лейкоцитов крови у низших позвоночных колеблется
в очень широких пределах, причем во многих случаях количественно пре-
обладают малодифференцированные лимфоидные формы, а некоторые гра-
нулоциты могут вовсе отсутствовать. Очень интересно указание Заварзина
(1937) на то, что у некоторых костистых рыб, например у бычка, грануло-
цитов вовсе не встречается, а имеются лишь лимфоидные клетки. При
воспалении у бычка Заварзин (1937) особенно отмечал участие незернистых
кровяных клеток в образовании грануляционной и рубцовой ткани. По
его данным, значительная часть фибробластов в очаге воспаления возни-
кает непосредственно из этих кровяных элементов. Эти очень интересные
наблюдения подтверждают близкие генетические отношения, установлен-
ные у млекопитающих между незернистыми кровяными клетками и фибро-
бластами соединительной ткани, и вместе с тем подчеркивают весьма зна-
менательный факт неодинакового течения воспалительного процесса у раз-
ных представителей позвоночных, который заслуживает большого внимания
и требует еще специального изучения как с точки зрения гистологии, так
и патологии.
Различия между соединительнотканными образованиями в ряду позво-
ночных несомненно не исчерпываются особенностями отдельных клеточных
форм, а должны касаться и ряда общих гистоморфных особенностей, кото-
рые до настоящего времени известны еще слишком недостаточно. Очень
существенные различия в характере роста и способе взаимосвязи оседлых
соединительнотканных клеток разных позвоночных в условиях тканевых
культур, отмечавшиеся мной (1925, 1928, 1937), нуждаются в дополни-
тельных специальных исследованиях.
Что касается некоторых неясных и спорных моментов, относящихся
к оседлым клеточным формам десмобластического или фибробластического
ряда и их взаимоотношений с гистоцитами и макрофагами на разных ста-
диях дифференцировки, то опыты с эксплантацией эмбриональной и взрос-
лой соединительной ткани позволяют уточнить и иллюстрируют некоторые
из высказанных выше положений. Вместе с тем они ставят и ряд новых,
подлежащих разрешению, вопросов.
Экстенсивно растущие культуры соединительнотканного типа могут
быть получены из самых разнообразных органов зародышей, новорожден-
ных и взрослых животных, а также из кровяных клеток. После известного
числа пересевов с вырезанием и делением на части, из мезенхимных тканей
получаются стационарные культуры, состоящие из т. наз. фибробластов
или, точнее, фибробластообразных элементов, которые, как правило, не
образуют сомкнутых комплексных структур.
Их гистологический анализ удобнее всего дать на относительно простых
примерах эмбриональной и неэмбриональной соединительной ткани млеко-
питающих и человека, которые в гистологическом отношении изучены
подробнее всего {Максимов, 1916; М. Меллендорф, 1929, 1931; Хлопин,
1931). На ранних стадиях культивирования в таких эксплантатах, кроме
884
Глава 111
элементов типа фибробластов, встречаются в более или менее значитель-
ном количестве макрофагообразные амебоидные клетки, а также неко-
торые другие разновидности клеточных форм (единичные лимфоциты,
зернистые лейкоциты и пр.), на которых мы здесь не будем останавли-
ваться.
Макрофагообразные элементы, как это уже неоднократно отмечалось,
образуют заостренные отростки и, утрачивая амебоидную подвижность,
могут превращаться в клетки типа фибробластов. В некоторых случаях,
даже на ранних стадиях, количество макрофагов может быть очень незна-
чительным, или они даже практически отсутствуют. Фибробластообразные
элементы, соединяясь друг с другом отростками, образуют сетевидные
структуры, более густые в центральных участках зоны роста, разрыхляю-
щиеся и распадающиеся на отдельные клетки в направлении к ее пери-
ферии, и нередко имеют преимущественно радиальное расположение и
веретенчатую форму. Стелясь по поверхности плотного субстрата, они могут
временно более или менее тесно смыкаться друг с другом, образуя плоскост-
ные комплексы, несколько похожие на эпителиообразные мембраны. Форма
и размеры фибробластообразных элементов довольно изменчивы. Они или
в более или менее сильной степени распластываются по плотному субстрату,
или отличаются узким и относительно темно окрашивающимся клеточным
телом, на поверхности которого могут появляться закругленной формы
выступы. Часть их энергично делится митотическим путем, в других бывают
заметны дегенеративные изменения. По некоторым данным, в соединительно-
тканной зоне роста, при ее тщательном изучении, во многих случаях можно
бывает обнаружить, даже после повторных пересевов, некоторое количе-
ство амебоидных клеток типа макрофагов, связанных непрерывной цепью
переходных форм с фибробластообразными элементами (Хлопан). Иногда
наблюдается массовое превращение фибробластообразных элементов в макро-
фаги, что может быть вызвано искусственно при культивировании в при-
сутствии незначительных количеств трипановой сини (Меллендорф). Пре-
вращение фибробластообразных клеток в макрофаги может, однако, симу-
лироваться их простым округлением и втягиванием отростков, что нередко
предшествует дегенеративным изменениям. Танненберг (1929, 1931) даже
считает, что описанное Меллендорфом образование макрофагов, как пра-
вило, представляет собой такие дегенеративные изменения. В настоящее
время, однако, нельзя категорически отрицать возможность такого
превращения.
Такой же характер имеет, в общем, соединительнотканная зона роста
в культурах из органов куриного зародыша. Но они в гистологическом
отношении, как это ни удивительно, относительно мало изучены. Что
касается наиболее изученных культур из эмбрионального сердца, то их
тканевая природа до настоящего времени, как это будет подробнее разобрано
ниже, остается неясной. В них также описывалось спонтанное и вызывае-
мое искусственно прибавлением поверхностно-активных веществ превра-
щение фибробластообразных клеток в макрофаги (Фишер Д., 1926, 1930;
Тома Д., 1938).
Экспериментальный метод в гистологии
335
В зоне роста эксплантатов соединительнотканного типа при таких
условиях культивирования волокнистое межклеточное вещество или не
успевает образоваться, или лишь наблюдается начало образования арги-
рофильных волокон.
Первоначально было принято обозначать отростчатые соединительно-
тканные клетки таких растущих экстенсивно культур как фибробласты,
приравнивая их к соответствующим дифференцированным элементам сое-
динительной ткани (Максимов, 1916). Такое отождествление, однако, едва ли
правильно, так как зона роста, образующаяся после очередного пересева
с вырезанием, элементы которой энергично делятся митозом, имеет очень
малодифференцированный характер и сильно отличается от дифференци-
рованной соединительной ткани. Кроме того, самый факт энергичного
митотического деления несомненно говорит против сколько-нибудь высо-
кой дифференцировки элементов таких культур. Поэтому естественнее
всего предположить, что энергично растущие и пролиферирующие эксплан-
таты состоят, главным образом, не из клеток, сравнимых с фиброцитами,
а из недифференцированных или малодифференцированных мезенхимных
элементов, лишь некоторая часть которых при таких условиях может созре-
вать и стареть. Тот факт, что в некоторой части клеток энергично растущих
культур наблюдаются различные явления дегенерации, как это уже отме-
чалось выше, вполне соответствует такой точке зрения. Весьма вероятно
также, что упомянутые выше морфологические различия между фибробласто-
подобными элементами, наблюдающиеся часто в одной и той же культуре,
являются в известной степени видимым проявлением их неодинаковой сте-
пени зрелости. Эта своеобразная, энергично пролиферирующая ткань
мезенхимальных эксплантатов, которая может быть обозначена как
эксплантационная бластема (Хлопан, 1931), таким образом, не может счи-
таться колонией совершенно однородных элементов, а представляет собой
своеобразную тканевую систему.
Нужно думать, что условия существования в культурах благоприят-
ствуют фибробластообразной, а не макрофагоподобной форме существования
тканевых элементов; поэтому мезенхимные эксплантаты большей частью
и производят впечатление однородных. Однако макрофагообразные пре-
вращения из клеток, как это уже отмечалось, могут иметь место при извест-
ных условиях. Фибробластообразные клетки соединительнотканных эксплан-
татов образуются, повидимому, за счет недифференцированных мезенхимных
элементов и, возможно, также менее дифференцированных клеток фибро-
бластического ряда исходного материала. Весьма вероятно, что дифферен-
цированные клетки этого ряда после эксплантации погибают. Частичная
дегенерация оседлых соединительнотканных клеток центрального кусочка
действительно имеет место в течение первого времени после посева. Что
касается макрофагов, то они несомненно возникают путем мобилизации
гистиоцитов. Возможно также, что в них превращается и некоторая часть
недифференцированных мезенхимных клеток.
Жировые клетки подкожной клетчатки кролика (Максимов, 1916),
сальника, брыжжейки и жирового костного мозга морских свинок (Буркгардт,
336
Глава III
1934) могут, изменяя свою форму, передвигаться в зону роста. Их жир при
этом разбивается на более мелкие капли и постепенно исчезает. В жировых
клетках с уменьшенным количеством жира наблюдаются митозы. В конце
концов, они могут приобретать характер фибробластообразных элементов
{Максимов) или гистиоцитов (Буркгардт). Тимофеевскому (1939) удалось
получить длительные культуры из подкожной жировой ткани человека,
клетки которых отличались большим количеством жировых включений
и образовали в зоне роста своеобразные, похожие на дольки, скопления.
Насколько возможно судить в настоящее время, культуры из соедини-
тельной ткани самых разнообразных органов имеют сходный внешний вид.
При культивировании без вырезания мезенхимная эксплантационная
бластема, делаясь гуще, дифференцируется с образованием большого коли-
чества волокнистого межклеточного вещества. Часть ее фибробластообраз-
ных элементов при этом сохраняет свой индифферентный характер, более
узкое и темно окрашивающееся клеточное тело, а остальные, распласты-
ваясь в большей или меньшей степени и утрачивая ясные контуры, приобре-
тают вид настоящих фибробластов, или фиброцитов соединительной ткани,
причем в их протоплазме образуются упоминавшиеся выше тонофибрилли,
или фиброглия. Амебоидные макрофаги зоны роста превращаются в типич-
ные покоящиеся гистиоциты. Некоторые, находящиеся вне зоны роста,
в свертке фибрина, гистиоциты подчас образуют очень сложно разветвлен-
ные отростки. Они представляют собой элементы дегенеративного харак-
тера и в дальнейшем погибают. Количество аргирофильных волокон в зоне
роста делается очень значительным, они утолщаются и, соединяясь друг
с другом, приобретают характер коллагеновых пучков. Образование волок-
нистого межклеточного вещества наблюдается в соединительнотканных
эксплантатах из разных органов куриных зародышей и млекопитающих
при соответствующих условиях культивирования. Дифференцировка типич-
ных коллагеновых пучков была подробно описана Максимовым в соедини-
тельнотканных культурах из зобной железы. Бофилъ-Делофё (1932), изучая
образование аргирофильных волокон в мезенхимальных культурах из
разных органов куриного зародыша, не отметил обратной зависимости
между энергией роста и количеством образующегося волокнистого межкле-
точного вещества. По его данным, наоборот, чем энергичнее растут культуры,
тем больше в них появляется аргирофильных волокон.
Если соединительнотканные культуры из подкожной клетчатки куль-
тивируются таким образом, что при каждом пассаже первоначальный кусо-
чек с прилегающей к нему частью зоны роста вырезается и пересеивается
без деления, то их центральная часть становится все плотнее. Ее клетки
частью дегенерируют, а количество межклеточного вещества делается все
больше. Образуются волокнистые колагеновые пучки, неправильные мас-
сивные густые переплеты коллагеновых фибриллей и своеобразные, частью
аморфные, частью волокнистые, располагающиеся концентрическими слоями,
структуры, могущие благодаря обизвествлению приобретать большую
степень твердости (В. Меллендорф, 1932; Дюггели, 1937). Частью концен-
трическая, частью радиальная ориентировка межклеточных структур со-
Экспериментальный метод в гистологии
337
ответствует обычному взаимно перпендикулярному направлению силовых
линий. Сходные обизвествляющиеся коллагеновые структуры были в послед-
нее время получень! Румянцевым и Березкиной (1937) в эксплантатах из
подкожной соединительной ткани кур.
По данным этих авторов, при прибавлении к питательной среде экстракта
из развивающихся костей куриного зародыша межклеточное вещество, не
изменяя своего общего характера, развивается в больших количествах.
Едва ли можно согласиться с указаниями Меллендорфа, а также Румян-
цева и Березкиной, что полученные ими обизвествленные структуры имеют
сходство с костной тканью. Их, несомненно, правильнее всего сравнивать
не с костью, а с наблюдающимися нередко при патологических процессах
в целом организме очагами обизвествленной соединительной ткани. При
культивировании при тех же условиях частиц стенки эмбриональной аорты
в центральном кусочке образуется плотный узелок, состоящий из богатой
волокнистым межклеточным веществом ткани хондроидного характера, но
ясно отличающейся от типичного хряща {Румянцев и Березкина).
Относительно образования эластических волокон в культурах до на-
стоящего времени имеются довольно скудные данные. Впервые их образо-
вание вне организма наблюдалось Блумом (1929) в соединительнотканной
зоне роста при совместном культивировании частиц аорты и сердца заро-
дышей морских свинок. Затем дифференцировка эластических волокон была
описана Порта (1930) и Одиетт (1933) при эксплантации частиц аорты,
кожи и сердца куриных зародышей.
Что касается способа образования и природы волокнистых межклеточ-
ных структур в условиях эксплантации, то на этот счет были высказаны
весьма противоречивые взгляды, которые существуют и существовали
раньше, относительно развития межклеточных веществ в организме. Чего-
либо принципиально нового в отношении этого вопроса метод тканевых
культур не дал. Подавляющее большинство данных, полученных методом
эксплантации, говорит не в пользу внутриклеточного способа образования
аргирофильных волокон. Они появляются или на поверхности или даже
на известном расстоянии от клеточного тела {Бейтсел, 1915; Максимов,
1929; Хлопин, 1931; Момильяно-Леви, 1932; Меллендорф, 1932; Румянцев
и Сунцова, 1935; Румянцев и Березкина, 1937, и др.). Их образование
удается обнаружить не только на соответствующим образом обработанных
препаратах, ио и при прижизненном наблюдении в темном поле зрения
{Момильяно-Леви, 1932). Последним методом, по данным этого автора, их
даже легче обнаружить, чем серебрением, с помощью которого тончайшие
волокна могут и не выявляться. При изучении соединительнотканных куль-
тур из куриного зародыша появление первых аргирофильных структур
удается обнаружить уже к концу первых суток после эксплантации
{Момильяно-Леви). Большинство авторов отрицает прямое превращение
в них фибрина {Бейтсел, Меллендорф) или их кристаллизацию из сыво-
ротки или эмбрионального экстракта, пропитывающих сверток {Максимов,
1929) вдали от клеток, под влиянием выделяемых ими веществ типа фер-
ментов.
22 Хлопин— 349
3S8
Глава III
Попытка Должанского и Руле (1933) доказать образование соединительно-
тканных волокон в свертке фибрина, лишенном клеток, является мало-
убедительной, так как в фибрине и во многих других веществах могут
появляться волокнистые структуры, подчас даже импрегнирующиеся
серебром, которые не имеют ничего общего с межклеточным веществом
соединительной ткани. Независимость образуемых клетками волокон и
волокнистых структур свертка фибрина убедительно показана в работе
Сунцовой (1935). Наиболее вероятным является предположение, что меж-
клеточное вещество соединительной ткани в культурах образуется самими
клетками (Момилъяно-Леви, 1932; Бофиль-Делофё, 1932; Хлопан, 1931.
и др.), первоначально, быть может, в виде коллоидного золя с палочковид-
ными мицеллами, в котором затем путем кристаллизации образуются во-
локнистые структуры. Спор о том, возникает ли оно первоначально как
продукт превращения протоплазмы или, точнее, эктоплазмы клеток или
путем процесса, сходного с секрецией, мне представляется принципиально
маловажным и, при существующих методах исследования, неразрешимым с
достаточной определенностью. Малообоснованным представляется отождест-
вление аргирофильных волокон с коллагеновыми структурами (Момиль-
яно-Леви, 1932; Леви, 1934) и совершенно недопустимым отождествление полу-
чаемых в культурах из разных тканей волокнистых структур друг с другом
на основании того, что они могут быть выявлены при импрегнации серебром.
На опытах с эксплантацией тканей кроветворных органов я не буду
специально останавливаться. Соответствующие данные можно найти в моей
сводке (1940).
Несмотря на то, что культуры фибробластоподобных клеток, выделен-
ные из соединительной ткани разных органов одного и того же животного,
по имеющимся данным, отличаются совершенно сходным внешним видом,
они оказались в ряде случаев нетождественными друг другу в отношении
некоторых физиологических показателей. В частности, энергия роста фибро-
бластоподобных клеток из межмышечной соединительной ткани 17-днев-
ного куриного зародыша превосходит в три раза темп роста таких же клеток
из щитовидной железы того же зародыша и в десять раз выше темпа роста
«фибробластов» из сердца (Паркер, 1932). Правда, следует подчеркнуть,
что, когда речь идет о т. наз. «сердечных фибробластах», то, как
это будет разобрано ниже, остается неясным, какие именно клетки имеются
в виду. Кроме того, оказывается, что культуры фибробластообразных кле-
ток, выделенных из соединительной ткани одного и того же органа на раз-
ных стадиях его развития, также отличаются, хотя и в меньшей степени,
друг от друга энергией роста (Паркер, 1932). Эти различия сохраняются
совершенно стойко при разных условиях культивирования.
Подобно многим известным данным из области нормальной гистологии
соединительной ткани в составе разных органов, эти факты говорят за то,
что волокнистая соединительная ткань в разных местах организма, дифферен-
цируясь дивергентно, приобретает определенные специфические особен-
ности и не является вполне равноценной. Подкожная и межмышечная рых-
лая соединительная ткань, соединительная ткань слизистых оболочек,
Экспериментальный метод в гистологии
339
выстланных разными эпителиями (например в составе переднего отдела
пищеварительной трубки, желудка и кишечника, матки и яйцеводов), сероз-
ных оболочек, внутреннего слоя сосудистой стенки, интерстициальная
ткань разных желез, легких, почек и гонад отличаются друг от друга
подчас очень резкими биологическими и, в частности, морфологическими
особенностями, на которые, правда, далеко не всегда обращается доста-
точно внимания, но которые, безусловно, заслуживают глубокого спе-
циального изучения. Детальное изучение отдельных местных вариантов
соединительной ткани, как, например, показывают обстоятельные иссле-
дования Щелкунова (1935, 1936, 1937) внутреннего слоя сосудистой стенки,
дает очень много интересных и ценных результатов.
Разный уровень дифференцировки и неодинаковое относительное коли-
чество малодифференцированных элементов являются, повидимому, да-
леко не единственными причинами этих местных различий соединительно-
тканных образований. Многое говорит за то, что свойства соединительной
ткани коррелятивно связаны со спецификой главных ведущих тканевых
компонентов разных органов, в частности, например, с глубокими биологи-
ческими различиями, характеризующими эпителии разного происхожде-
ния. О том же свидетельствуют приведенные выше данные, касающиеся
образования межклеточного вещества в соединительнотканных экспланта-
тах из разных органов. Наконец, свойства соединительной ткани,
кроме того, в заметной степени зависят еще от возрастного
фактора.
Глубокое своеобразие характеризует также различные плотные разно-
видности волокнистой соединительной ткани, как, например, сетчатый
слой кожи, ткань сухожилий, разных связок, фасций, апоневрозов и тому
подобное. Обстоятельные исследования Брауна (1939) над сетчатым слоем
кожи показали чрезвычайно закономерную структуру, тесно связанную
с местными механическими факторами этой т. наз. «неоформленной» плот-
ной соединительной ткани.
Не может подлежать сомнению, что соединительная ткань каждого
органа эволюирует вместе и во взаимосвязи с его другими тканевыми ком-
понентами, а потому все местные свойства этих «межуточных» образо-
ваний должны быть филогенетически обусловлены и отличаться теми или
иными достаточно стойкими, наследственно закрепляющимися различиями.
Сравнительное изучение в определенных филогенетических рядах местных
особенностей соединительнотканных образований еще является одной из
интереснейших задач будущего. Следует только подчеркнуть, что оно
должно итти не оторванно, а в тесной связи с изучением филогистогенеза
тех тканевых структур, для которых данная соединительная ткань является
стромой или подстилкой.
Такое систематическое комплексное изучение разных тканей в их
развитии должно сделаться в дальнейшем одной из основ особого
направления в нашей науке, для которого экспериментальный анализ
отдельных тканей является необходимой предварительной сту-
пенью .
*
340
Глава III
в) Скелетные энтомезенхимные ткани позвоночных
Вопрос о генетических отношениях между элементами крови, лимфы
и волокнистой соединительной ткани, с одной стороны, хрящом и костью,
с другой, не может считаться достаточно выясненным.
Наряду со многими данными, наглядно демонстрирующими очень
раннюю детерминацию скелетогенных участков мезенхимы, ряд фактов
из области описательной, экспериментальной и патологической гистологии
говорит против наличия резкой границы между теми или иными разновид-
ностями волокнистой соединительной, хрящевой и костной тканями.
Приводимые ниже данные, полученные методом эксплантации, в своем
подавляющем большинстве подчеркивают специфические качественные осо-
бенности скелетных образований и скелетогенных тканей.
Из скелетогенных участков надкостницы и надхрящницы куриного
зародыша удается получить экстенсивно растущие эксплантаты, которые
при периодических пересевах с вырезанием и делением могут длительное
время культивироваться в чистом виде (Фишер и Паркер, 1929; Паркер,
1934, и др.). По данным этих авторов, такие эксплантаты состоят из фибро-
бластообразных элементов, которые по своему виду и способу соединения
друг с другом тождественны клеткам нескелетогенных мезенхимных
эксплантатов и культур из эмбриональной сердечной мышцы, но отли-
чаются от них по ряду функциональных особенностей.
Однако, сопоставляя друг с другом результаты, полученные Должан-
ским (1929) и Фишером (1932), можно сделать вывод, что культуры из остео-
бластов надкостницы лобной кости, которые Фишер ошибочно называет
хондробластами, по совокупности своих морфологических особенностей
отличаются от обычных соединительнотканных эксплантатов. На ранних
стадиях культивирования они дают очень разнообразные картины, довольно
сильно разжижают фибрин и могут образовать то эпителиообразные мем-
браны, то сетевидные клеточные соединения. На более поздних стадиях
зона роста, как правило, сохраняет более рыхлый вид. По данным Должан-
ского, даже после пятимесячного культивирования (65 пассажей) клетки
зоны роста сохраняют ряд особенностей, свойственных остеобластам. Они
большей частью содержат большую околоядерную вакуолю и богаты вклю-
чениями, часть которых дает микрохимическую реакцию на кальций. По
данным Должанского, в эксплантатах из лобной кости в образовании зоны
роста участвуют не только элементы надкостницы, но и сами костные клетки.
Студицкий (1932), повторивший опыты предыдущего автора, однако, не
обнаружил у растущих вне организма остеобластов сохранения каких-либо
специфических цитологических особенностей. По данным Фишера, хондро-
бласты клиновидной кости образуют культуры, практически не отличимые
от эксплантатов из остеобластов.
Повидимому, гистологические особенности зоны роста, образуемой
скелетогенными элементами, могут варьировать в довольно широких пре-
делах. Наряду со структурами, сходными или даже тождественными с тако-
выми обычных соединительнотканных эксплантатов, они могут давать и не-
Экспериментальный метод в гистологии
34 Г
свойственные последним картины. Остеобласты и хондробласты образуют,
подобно другим мезенхимным элементам, в зоне роста волокна аргирофиль-
ного межклеточного вещества (Бофиль-Делофё, 1932).
Покровные клетки синовиальных оболочек, или «синовиобласты» (Фо-
бель, 1933), по характеру своего роста вне организма и морфологическим
особенностям, больше всего напоминают хондробласты и остеобласты. Они
могут образовать в культурах мукоидное вещество и, повидимому, являются
элементами, родственными производным скелетогенной мезенхимы (Фо-
бель, 1933).
Отчетливее всего особенности скелетогенных элементов проявляются
при замедленном росте, когда на первый план выступают явления диффе-
ренцировки. Как уже указывалось выше, образование хряща и его рост
могут иметь место при эксплантации целых закладок и отсутствии экстен-
сивного роста. Впервые получила вне организма дифференцировку хряща
в экстенсивно растущих эксплантатах из предхрящевой бластемы конеч-
ностей трехдневных куриных зародышей Фелл (1928), в лаборатории кото-
рой затем был выполнен ряд в высшей степени интересных и обстоятельных
работ, посвященных хондрогенезу и остеогенезу вне организма. Несколько
позже ей удалось получить образование хрящевых узелков в эксплантатах
из надкостницы (1932) и эндоста (1933) диафиза трубчатой кости куриных
зародышей. В последнем случае после образования зоны роста костная
частица удалялась, а в зоне остеогенной ткани части культур сначала
наблюдалось образование ткани неопределенного характера, состоящей
из клеток, похожих на остеобласты, и студенистого межклеточного веще-
ства, а затем — дифференцировка последней частью в хрящ, частью в остео-
идную ткань, участки которых без резкой границы переходили друг в друга.
Образовавшийся хрящ отличался невысоким уровнем дифференцировки и
незначительным количеством межклеточного вещества.
Дифференцировку хряща несколько позже получили Нивен (1933),.
Руле (1935), ррушинский (1936), Румянцев и Березкина (1937) в
эксплантатах из скелетогенной мезенхимы разных участков тела ку-
риных зародышей. Последние авторы сначала в течение 10 пассажей
с вырезанием и делением получили чистые культуры фибробластообразных
элементов, которые в дальнейшем столько же времени культивировались
в присутствии экстракта из развивающихся костей без деления на части.
Только при этих условиях в центральном кусочке некоторых культур обра-
зовались плотные узелки, состоящие из типичного гиалинового хряща,
окруженного хондрогенной зоной. В эксплантатах из нескелетогенной
мезенхимы настоящая хрящевая ткань никогда не образовывалась.
Менее убедительны данные А. Фишера и Паркера (1929) и А. Фишера
(1932), которые указывали, что им удалось получить дифференцировку
хрящевого, а затем костного межклеточного вещества в семи- и девяти-
месячных культурах фибробластообразных элементов, выделенных из
надкостницы лобной кости куриного зародыша. Эти авторы ошибочно
обозначали как надхрящницу периост этой накладной кости; их описания
и микрофотограммы заставляют сомневаться в том, имели ли они действи-
342
Глава III
тельно дело с образованием настоящей хрящевой и костной ткани. Дж. Бер-
роуз (1933) также не получил, при вторичной дифференцировке куль-
тур из зачатка нижней челюстной кости куриных зародышей, образования
кости, а лишь коллагеновое волокнистое межклеточное вещество. Недоста-
точно убедительными представляются также опыты Фишера (1931) по сов-
местной эксплантации частиц суставного эмбрионального хряща и чистых
культур фибробластообразных элементов из сердца или надхрящницы
куриных зародышей. По его данным, хрящевая частичка, сама по себе
неспособная к росту, вызывает в обросших ее фибробластообразных эле-
ментах образование хряща и играет роль организатора. Судя по микро-
фотограмме автора, новообразованная ткань имела довольно мало общего
с настоящим хрящем.
Чрезвычайно быстрое образование гиалинового хряща описал Н. Насо-
нов (1934) в культурах из соединительной ткани аксолотля, взятой из об-
ласти плечевого пояса, при условии прибавления к питательной среде
сгущенного экстракта из хрящей того же животного или кролика. Автор
считает возможным, что в исходном материале присутствовали хондро-
генные элементы. Аналогичные опыты, поставленные с лягушачьим мате-
риалом, дали отрицательный результат. В этом направлении очень жела-
тельны дальнейшие исследования.
В первых опытах по эксплантации надкостницы и надхрящницы диф-
ференцировки костной ткани получить не удавалось (Верещинский, 1924;
Поликар и Бушарла, 1925). Несомненное образование перекладин губча-
той кости получила Фелл (1928) в эксплантатах из надкостницы и эндоста
куриных зародышей при условии, если их центральный кусочек не трав-
мировался при пересевах. Выше уже отмечалось, что органотипический
рост хрящевых закладок описывали Стренджуэйс и Фелл (1925), Фелл
(1928) и Демут (1928) при эксплантации предхрящевой бластемы и
целых хрящевых скелетных частей. В этих растущих целиком хрящевых
закладках Фелл наблюдала и подробно изучила вне организма ход пери-
хондрального и эндохондрального окостенения, которое в культурах,
в части случаев, протекает в общих чертах так же, как и в условиях орга-
низма.
При эксплантации зачатков трубчатых костей на самых ранних ста-
диях развития в значительной части случаев окостенения не получается,
особенно если культивирование производится в среде с очень малым содер-
жанием эмбрионального экстракта, задерживающей рост хрящевой за-
кладки (Фелл и Ландауэр, 1935). На более поздних стадиях развития ске-
летных зачатков окостенение наблюдается значительно чаще, а влияние
на него неполноценной среды ослабевает и исчезает. При тех же условиях
окостенение зачатков накладных костей происходит почти во всех случаях.
По данным Гайара (1935), образование кости наблюдается в культурах
остеобластов лобной кости куриных зародышей, если к питательной среде
прибавлять последовательно экстракты из 5-,7-, 15- и 17-дневных зароды-
шей. При культивировании в присутствии обычного эмбрионального
экстракта из семи-, восьмидневных зародышей образования костной ткани
Экспериментальный метод в гистологии
343
не получается {Гайар, 1935; Мишурский, 1938). Нивен (1931) наблюдал
вне организма срастание обломков развивающихся костей куриных заро-
дышей и мышей путем новообразования остеобластами костной ткани.
Весьма интересно, что меккелев хрящ, не подвергающийся при обычных
условиях окостенению и, в соответствии с данными, касающимися амфи-
бий, имеющий по крайней мере в значительной своей части нейромезенхим-
ное происхождение, в эксплантатах никогда не окостеневает. Из окружаю-
щей его скелетогенной ткани нижней челюсти почти без исключения удается
получить окостенение.
Ориентировка образующихся в культурах костных перекладин, по
данным Глюксмана (1938), зависит в значительной степени от механи-
ческих факторов. При появлении окостенения в эксплантированных
закладках удается констатировать образование фосфатазы {Фелл и Робин-
зон, 1929, 1930), т. е. фермента, расщепляющего гексозо-фосфорную кислоту,
за счет которой, повидимому, образуется фосфорнокислый кальций кост-
ного вещества.
Ряд полученных методом эксплантации данных показывает также, что
малодифференцированные скелетогенные элементы по своим физиологи-
ческим показателям очень заметно отличаются от фибробластообразных
клеток из межмышечной соединительной ткани и из сердца куриного за-
родыша. Тканевая природа последних элементов, как это уже отмечалось, не-
достаточно ясна. Энергия роста фибробластообразных элементов из сердца
куриного зародыша, как это показали еще Наррелъ и Ибелинг (1921), резко
увеличивается с повышением концентрации эмбрионального экстракта
в среде до 10—15% его содержания; дальнейшее увеличение его концен-
трации также увеличивает энергию роста, но в меньшей степени. При 40%
содержания экстракта энергия роста достигает своего максимума. Фибро-
бласты из соматической мускулатуры повышают энергию роста при уве-
личении концентрации экстракта до 25%. Выше этого предела легко на-
ступают разжижение свертка и ухудшение роста. Рост хондробластов из
зачатка клиновидной кости и остеобластов из надкостницы лобной кости
активируется при увеличении концентрации эмбрионального экстракта до
15 %. При 25 % его содержания рост этих элементов замедляется, а при 40%
культивирование их делается невозможным, вследствие разжижения свертка
фибрина в течение 24 часов. Различной оказалась и остаточная энергия
роста этих элементов. У клеток, обозначенных как хондробласты, и у остео-
бластов она значительно выше, чем у фибробластообразных элементов
из сердца и соматической мускулатуры. Отличается также в большей или
меньшей степени и форма кривых роста всех этих элементов при различ-
ных условиях.
Эти физиологические данные хорошо совпадают с устанавливаемой мор-
фологически спецификой скелетогенных элементов.
Очень эффектные и обстоятельные экспериментальные исследования
Студицкого (1937) над скелетогенной мезенхимой и развитием хрящевой
и костной тканей, сделанные методом пересадки на хорион-аллантоис
куриных зародышей, также демонстрируют высокую степень специфич-
844
Глава III
ности скелетогенных элементов и очень близкие генетические отношения
между хрящом и костью.
В детализации этих отношений, однако, автор, с моей точки зрения,
идет значительно дальше, чем это допускает фактический материал, а его
предположение о превращении хрящевых клеток, достигших весьма высо-
кого уровня дифференцировки, в костные, принимавшееся и рядом других
исследователей, представляется малообоснованным и маловероятным.
Несмотря на убедительность данных, доказывающих детерминирован-
ность скелетогенных элементов и приобретение ими стойких отличий от
клеток обычной соединительной ткани, ряд авторов и в настоящее время
склонен полностью или частично отрицать их специфичность (Вейденрейх,
1923, 1930; Лериш и Поликар, 1926, 1934, и некоторые другие). Вейден-
рейх считает за высокоспециализированную костную ткань только пластин-
чатую, или, по его терминологии, скорлуповую кость, а образование грубо-
волокнистой кости сближает с неспецифическими процессами обизвествле-
ния соединительной ткани.
Особенно далеко идут в этом отношении Лериш и Поликар, считающие,
что остеобласты вообще являются не костеобразующими элементами, а мало-
специфическими реактивными формами клеток соединительной ткани, воз-
никающими в связи с образованием в данном участке организма какой-то
независящей от местных клеток «остеогенной среды». За гуморальными
факторами костеобразования, значение которых вполне вероятно, эти авторы
перестают видеть клетки с их жизненными функциями и, игнорируя боль-
шой и хорошо обоснованный гистологический материал, резко противоре-
чащий их собственным представлениям, создают свою глубоко ошибочную
концепцию, одной из основ которой являются упомянутые выше неудач-
ные и неубедительные опыты Поликара и Бушарла (1925).
Вместе с тем не подлежит сомнению, что при экспериментальных и
патологических условиях в составе организма приходится считаться с гете-
ротопным появлением дифференцированных скелетных тканей, которые,
без бросающихся в глаза натяжек, никак не удается связать преемственно
с предшествующими скелетогенными зачатками. Если в некоторых слу-
чаях не исключена возможность описания малоспецифических очагов
обизвествления как настоящей костной ткани, и смешения с хрящевой
тканью структур, не имеющих с ней по существу ничего общего, очень
много случаев гетеротопного окостенения и образования хряща все-таки
представляется достаточно несомненным.
Прежде всего это относится к упомянутому выше образованию кости
в области почечных ворот после перевязки почечных сосудов (Сачердотти
и Фратен, 1902; Максимов, 1907; Пожарисский, 1905, и др.). Кроме того,
гетеротопное образование кости получали, хотя и далеко нс так регулярно,
как в этом случае, при пересадках мертвой кости (Нажотт, 1918,
1922; Кристоф, 1923). Поллетини (1922) описывал появление хрящевой
ткани вокруг пересаженных кусочков фиксированной кости, а Данини
(1930, 1936):—вокруг хрящевых трансплантатов. В последнее время
Ю. Петров (1940) отметил образование совершенно типичной губчатой
Экспериментальный метод в гистологии
345
кости и островков ткани, сходных с хрящевой, в мышце бедра кролика
через 8 месяцев после инъекции в нее каменноугольного дегтя. В той же
мышце в результате введения дегтя образовалась злокачественная опухоль
типа полиморфно-клеточной саркомы. Препараты автора, работавшего под
моим руководством, не оставляли никаких сомнений в том, что он дей-
ствительно имел дело с гетеротопным костеобразованием. Слабой сто-
роной всех этих экспериментальных данных, за исключением остеогенеза в
почке, являются их до известной степени случайный характер и невозмож-
ность до настоящего времени вызывать эти процессы достаточно регулярно.
При патологических условиях гетеротопное образование костной ткани
отмечалось неоднократно в стенке артерий (Менкеберг, 1902), в рубцах
(Грубер, 1917; Мебиус, 1924), старых гематомах (Лериш и Поликар, 1926),
в мозгу и мягкой мозговой оболочке (Багров, 1915), в стенке желудка, мыш-
цах и ряде других органов. При своеобразном заболевании, окостеневаю-
щем миозите, наблюдается развитие костной ткани в различных мышцах.
В более редких случаях наблюдалось появление гетеротопного хряща
(Марбург, 1902; Менкеберг, 1902, и др.).
При сопоставлении всех этих данных с тем, что говорилось выше о сое-
динительной ткани, нельзя не притти к выводу, что, с одной стороны, скеле-
тогенная детерминация мезенхимы в течение онтогенеза возникает очень
рано и придает ей особые, стойкие, специфические особенности, отличаю-
щие ее от нескелетогенных элементов. Но, с другой стороны, более чем
вероятно, что превращение элементов обычной соединительной ткани
в скелетогенные, под влиянием некоторых экспериментальных воздей-
ствий и патологических процессов, также может иметь место. Очевидно,
такому превращению подвер! аются малодифферепцированные камбиаль-
ные, а не более или менее специализированные клетки соединительной
ткани. Вместе с тем это скелетогенное превращение, которое, возможно,
является уже необратимым, в большей части случаев (кроме остеогенеза
в почке) совершается с известным трудом, так сказать, с насилием. До на-
стоящего времени не существует метода, который бы позволил вызывать
этот процесс без отказа в любом месте организма в той или иной разновид-
ности волокнистой соединительной ткани.
Невольно приходит в голову сопоставление с тем, что говорилось выше
относительно весьма вероятных случаев истинной метаплазии различных
эпидермальных тканей (например переходного эпителия или многорядной
выстилки дыхательных путей) в настоящий многослойный ороговевающий
эпителий. Для таких превращений в пределах генетически единого тка-
невого типа, разные представители которого отличаются различной дивер-
гентной дифференцировкой, очевидно, как правило, требуется наличие
специальных или, во всяком случае, достаточно сильных или действующих
длительное время внешних факторов, которые, возможно, вызывают про-
лиферативные процессы, а затем, уже на почве их, истинную метаплазию.
Таким образом, имеющийся в настоящее время экспериментальный и
патологический материал, подчеркивая генетическое единство у позво-
ночных части клеток крови, разных волокнистых разновидностей соедини-
346
Глава Ш
тельной ткани, хрящевых и костных тканей, производных «энтомезенхимы»,
говорит в то же самое время за их далеко зашедшую дивергентную дифферен-
цировку, обусловленную всей предыдущей историей развития. При этом
довольно вероятно, что вследствие неодинакового филогенетического воз-
раста разных скелетогенных участков, их элементы при более точном экспе-
риментальном анализе обнаружат известные различия в своих биологи-
ческих свойствах. Очень интересен также вопрос о специфических свой-
ствах дентинообразующей мезенхимы и о генетических отношениях ден-
тина и кости. На этих и на многих других проблемах, связанных со спе-
циализированными в разных направлениях производных «энтомезенхимы»
позвоночных, пока еще преждевременно останавливаться Имеющиеся
в настоящее время указания на развитие дентина из нейромезенхимы,
в частности, данные Гольтфретера (1935), полученные при ксеноплас-
тической пересадке недетерминированного материала гаструлы. не пред-
ставляются достаточно убедительными.
Мною также умышленно оставляется в стороне вопрос о мезенхимных
хондроидных тканях, систематический экспериментальный анализ которых
еще является делом будущего.
г ) Соединительнотканные образования и полостная жидкость беспозвоночных
Сравнение и эволюционная трактовка соединительнотканных образо-
ваний в ряду позвоночных и у разных представителей беспозвоночных
в настоящее время представляются еще более затруднительными и спор-
ными. Очень интересные попытки в этом направлении принадлежат За-
варзину (1934), подошедшему к этому вопросу с точки зрения морфо-физио-
логических явлений конвергенции или параллелизмов. Такого рода сопо-
ставление, не основывающееся на действительном родстве тканей и не
преследующее цели создания естественной генетической системы гистоло-
гических структур, не учитывает их эмбрионального развития, не нуждается -
особенно в прослеживании эволюционных изменений свойств одной и той
же ткани в определенных филогенетических рядах, а также систематиче-
ском экспериментальном изучении специфических особенностей различ-
ных тканей у одного и того же животного. Для наших целей все эти моменты
как раз являются очень важными. А в этом отношении имеющийся в на-
стоящее время материал, к сожалению, еще очень недостаточен.
При сравнении соединительнотканных образований у позвоночных и
беспозвоночных, как это уже отмечалось выше, обычно подчеркиваются
имеющиеся между ними черты сходства, позволяющие издавна объединить
их в морфо-физиологическую группу тканей соединительного вещества или
межуточных тканей. Именно так поступил в своем замечательном обзоре
Эбнер (1911), впервые объяснивший сходство опорных тканей у разных
животных явлениями конвергенции. Эти же по существу моменты подчерк-
нул в своей обстоятельной работе М. Пренан (1922). То же самое проходит
красной нитью в экспериментальных исследованиях Заварзина (1926) и его
статьях обобщающего характера (1925, 1934). Сопоставления по внешнему
сходству делаются также между свободными клетками крови, лимфы,
Экспериментальный метод в гистологии
847
гемолимфы, вообще полостных жидкостей, в которых описываются лим-
фоидные базофильные клетки и разные гранулоциты, зернышки которых
частью обнаруживают сходство в отношении своей окрашиваемости (на-
пример у очень многих животных имеются «эозинофильные лейкоциты»),
частью, правда, отличаются теми или иными специфическими особенно-
стями {Кено, 1891, 1897; Колльман, 1908; М. Пренан, 1922; Заварзин,
1934, и многие другие авторы).
Пожалуй, одним из наиболее поразительных фактов в этом отношении
являются клетки типа эритроцитов, большей частью содержащие ядро и
лишь в единичных случаях безъядерные, которые появляются у отдельных
представителей самых разнообразных типов и классов животного мира,
например у немертин, аннелид, сипункулид, моллюсков, иглокожих.
Этим элементам беспозвоночных особенно много внимания уделяли в послед-
нее время Заварзин-{\934) и Гаусман (1935), причем последний особенно
подчеркнул генетическую связь этих элементов с целомической выстилкой.
Согласно сказанному выше, именно эти элементы с точки зрения естествен-
ной системы гистологических структур должны быть, по всей вероятности,
исключены из числа собственно мезенхимных производных и перенесены
в группу целомических тканей.
Если же подойти к крови и соединительной ткани не с традиционной
точки зрения, устанавливающей совпадения в структуре функционально
сходных частей организма, то в глаза бросается, пожалуй, прежде всего
их большое разнообразие, тесно связанное с рядом других особенностей
организации' животных и, в частности, с развитием опорного значения
эпидермальных образований, на что уже давно обратил внимание Геккель
(1885). Именно в соединительной ткани многих беспозвоночных мы встре-
чаемся с замечательными примерами дивергентного развития, если стоять
на точке зрения генетического родства и их параллельного, а не конвер-
гентного развития в отдельных типах. Последнее, к сожалению, является
еще во многих случаях очень спорным. Прежде всего это относится к ме-
зоглее кишечнополостных, которая, согласно с моей изложенной выше
точкой зрения, в морфо-физиологическом отношении скорее сравнима
с измененной и чрезвычайно сильно развитой во многих случаях базальной
мембраной, сходной в некоторых отношениях со стекловидным телом глаза
позвоночных, но не с настоящими мезенхимными структурами. Даже если
исключить» эти спорные образования, свойственные некоторым очень при-
митивным формам, то и без них разнообразие структур соединительноткан-
ного характера оказывается очень большим. Наиболее резко бросающи-
мися в глаза разными типами их являются, пожалуй, спикульные скелеты,
свойственные, например, в наиболее схематичном виде личиночным фор-
мам иглокожих, бесклеточные волокнистые «пограничные» образования
{Ливанов, 1913) аннелид и сходная с ними бесклеточная соединительная ткань
насекомых {Лазаренко, 1925) и многих других форм и, наконец, волокни-
стые, но содержащие клетки, соединительнотканные структуры, обнару-
живающие некоторое, правда, по существу отдаленное сходство с волок-
нистой соединительной тканью позвоночных, которые, например, известны
348
Глава III
у некоторых моллюсков, голотурий и других животных. Изредка встре-
чающиеся у отдельных беспозвоночных хрящеподобные образования,
являющиеся, возможно, хорошими примерами дивергентной дифферен-
цировки межуточной ткани соответствующих форм, повидимому, сильно
отличаются по своим свойствам друг от друга и от хряща позвоночных.
Так наз. хрящ радулы многих брюхоногих моллюсков скорее на-
поминает сравнительно мало специфическую пузырчатую ткань, но в неко-
торых случаях (у моллюска Haliotis) приобретает в значительной степени
хрящеподобный вид (Новиков, 1912). Очень интересные «хряши»
головоногих моллюсков частью отличаются очень сложной формой тела
своих паукообразных клеток с многочисленными тонкими отростками и,
повидимому, рядом особенностей межклеточного вещества. Другие хря-
щевые части скелета головоногих безусловно обнаруживают значительное
морфологическое сходство с обычным хрящом позвоночных. Ткань, обозна-
чаемая у Limulus как хрящевая, имеет в разных местах довольно разно-
образное строение и лишь частью обнаруживает известное сходство с хря-
щом низших позвоночных.
Сопоставление всех этих тканей с мезенхимным хрящом позвоночных,
с точки зрения генетической системы гистологических структур, далеко
не всегда возможно по существу, так как мезенхимное происхождение
многих из них остается до настоящего времени в лучшем случае спорным.
Например, «хрящ» головоногих моллюсков по многим данным (Мечников,
1867; Боб редкий, 1877; Усов, 1881; Цлаач, 1894) развивается из экто-
дермы, хотя Фаусек (1893) и Бабор (1901) приписывают ему «мезодермальное
происхождение». Что касается «хрящеподобного эндостернита» Limulus,
то его мезенхимное происхождение, с моей точки зрения, не представляется
достаточно бесспорным, если принять во внимание эктодермальное происхо-
ждение эндостернита у сольпуг.
Детальное изучение развития и гистологических особенностей всех этих
образований, с использованием экспериментальной методики, предста-
вляется очень заманчивым. Чего-либо сравнимого с костной тканью позво-
ночных ни у одного из известных представителей беспозвоночных животных
не встречается. При сопоставлении разных структур с соединительноткан-
ной морфо-физиологической характеристикой, в первую очередь, пожалуй,
бросается в глаза их поразительное разнообразие в животном мире, а отнюдь
не подчеркиваемые обычно черты сходства. *
Детальных экспериментальных гистологических данных, касающихся
соединительнотканных образований беспозвоночных, к сожалению, слиш-
ком мало, чтобы на их основании можно было делать выводы филогисто-
гепетического порядка. Кроме того, они относятся к отдельным предста-
вителям очень отдаленных друг от друга в системе животных групп, что
еще более усложняет дело. Тем не менее, исключительного интереса и вни-
мания заслуживают исследования Лазаренко (1925), Цедровского (1925),
Заварзина (1926) и Данини (1928) над асептическим воспалением у насеко-
мых, пластинчатожаберных моллюсков и речного рака, сделанные по мето-
дике Циглера—Максимова.
Экспериментальный метод в гистологии
349
Как известно, у всех этих животных и у позвоночных в некоторых
этапах воспалительной реакции были подчеркнуты такие черты сходства:
скопление на поверхности инородного тела круглых клеток типа макро-
фагов и лейкоцитов, которые в дальнейшем дегенерируют, и появление
вокруг этого дегенеративного слоя большого числа сетевидно связанных
друг с другом отростками веретенчатых или звездчатых клеток, для обозна-
чения которых был принят предложенный мной для незрелых оседлых
клеточных форм соединительной ткани позвоночных термин «десмобласты».
Эта сетевидпая клеточная ткань, окружающая в виде капсулы инородное
тело, была названа десмобластическим синцитием. В петлях этой сети
могут также встречаться свободные круглые клетки. Затем десмобласты
образуют то более густую, то более рыхлую сеть волокон межклеточного
вещества вокруг инородного тела и в некоторых случаях исчезают без
остатка (насекомые), в других (рак) остаются среди волокнистого межкле-
точного вещества в виде т. наз. лейдиговских клеток 1-го рода веретенчатой
формы. У моллюсков в мантии также описываются соединительная ткань
без оседлых клеток и полное исчезновение десмобластов в конце процесса,
но это представляется далеко не столь ясным, как у насекомых. Факты
говорят сами за себя: у несвязанных друг с другом близким родством орга-
низмов отмеченные картины воспалительного процесса несомненно обна-
руживают большое сходство: у всех этих форм имеются клетки, образую-
щие межклеточное волокнистое вещество, сравнимые с десмобластами или
фибробластами позвоночных.
Кроме того, эти данные позволяют также сделать известные сопоста-
вления между клетками крови различных животных. На этом сопоставле-
ние явлений обычно и заканчивается, хотя в работах имеется огромное
количество исключительно ценного и интересного фактического материала.
Я позволю себе провести несколько подробнее сопоставление гистологиче-
ских процессов, наблюдаемых при воспалительной реакции у насекомых
и у позвоночных, не проделывая того же самого для всех других форм.
На насекомых я остановлюсь потому, что их соединительная ткань и гемо-
лимфа были изучены Лазаренко особенно детально, а картина всего вос-
палительного процесса дана с исчерпывающей ясностью. Благодаря лю-
безности автора и с его помощью я имел возможность очень подробно озна-
комиться в свое время с соответствующими препаратами. Поэтому факти-
ческий материал этого исследования мне представляется особенно рельефно.
Наконец, насекомые и позвоночные представляют собой два, во многих
отношениях очень несходных, но в то же самое время очень совершенных,
типа организации животных, которые в первую очередь используются
для демонстрации явлений параллелизма.
Соединительная ткань насекомых и позвоночных обнаруживает и в отно-
шении своей структуры и всего биологического значения при нормальных
условиях огромные различия, которые уже подчеркивались неоднократно.
Бесклеточные сети ветвящихся волокон образуют в составе всех органов
очень тонкие чехлы и прокладки. По сравнению с мощно развитым наруж-
ным хитиновым панцырем, внутренними частями хитинового скелета,
350
Глава III
хитиновыми же сухожилиями соматической мускулатуры, опорное и меха-
ническое значение у этой количественно слабо развитой и очень однородной
в разных местах организма соединительной ткани, вполне оправдывающей
здесь свое название, очень невелико. Она представляет собой упаковочный,
межуточный материал, который в функциональном отношении сравним,
пожалуй, только с рыхлой соединительной тканью позвоночных. Если же
взять для сопоставления всю систему разнообразнейших соединительно-
тканных образований позвоночных в целом и систему волокнистых пере-
понок насекомых, то разница между этими структурами делается столь
же большой, как и между общими чертами организации позвоночных и
насекомых. Следует еще добавить, что по своему характеру волокнистые
структуры насекомых не имеют ничего общего с коллагеновыми пучками
и, повидимому, также преколлагеновыми волокнами. Сходство с эластиче-
скими волокнами ограничивается лишь фактом ветвления волокон насе-
комых, причем в местах ветвления обычно удается заметить трехугольные
площадочки, их видимой гомогенностью и окрашиваемостью резорцин-
фуксином. Для отождествления сравниваемых структур этого явно не-
достаточно.
Вторым компонентом сравниваемой с мезенхимными производными
позвоночных системы личинок жука-носорога является их гемолимфа,
свободно циркулирующая во всех щелях псевдоцеля, форменные элементы
которой развиваются в небольших кроветворных клеточных скоплениях,
сходных во многих отношениях с описанными Павловским (1917) скопле-
ниями «лимфоидной ткани» у скорпионов. В этих своеобразных кроветвор-
ных органах клетки гемолимфы, повидимому, образуются из чрезвычайно
мелких круглых клеток с небольшим ободком цитоплазмы, в которых
удается видеть картины митотического деления. Во всех прочих
клетках, поступающих в полостную жидкость, митозов не наблю-
дается.
Разнообразие клеточных форм гемолимфы исключительно велико, при-
чем далеко не все свободно взвешенные клетки имеют круглую форму. Эти
клетки представлены довольно разнообразными круглыми незернистыми
элементами неодинаковых размеров, элементами веретенчатой формы с базо-
фильной незернистой цитоплазмой и овальным ядром и элементами вытя-
нутой формы с закругленными концами, несколько вздутой средней, содер-
жащей ядро, частью и оксифильной зернистостью в цитоплазме. По внеш-
нему виду эти клетки, в своей совокупности, пожалуй, настолько отли-
чаются от картины крови разных позвоночных, поскольку это мыслимо для
изолированных, свободно плавающих в жидкости элементов. Наконец,
учитывая тот факт, что членистоногие представляют собой по сути дела
целомических животных с вторично исчезнувшей или частично измененной
до неузнаваемости выстилкой полости тела, которой несомненно обладали
их предки, более чем вероятно, что среди многообразия клеток гемолимфы
имеются по существу представители, по крайней мере, двух филогенети-
чески различных тканевых типов более древнего, собственно мезенхимного,
и более молодого, имевшего иную историю развития, целомического. Не-
Экспериментальный метод в гистологии
351
которые факты, установленные Лазаренко, как мы увидим ниже, подтвер-
ждают эту точку зрения.
Теперь обратимся к несколько более обстоятельному анализу процес-
сов, происходящих в псевдоцеле личинки жука-носорога, после введения
целлоидиновой трубки, прослеженных с непревзойденной полнотой в иссле-
дованиях Лазаренко. Прежде всего, на поверхности стерильного инород-
ного тела начинают оседать как круглые незернистые, так и зернистые
оксифильные клетки гемолимфы. Многие из этих элементов, быстро деге-
нерируя, разрушаются, другие сохраняются более длительное время, обра-
зуя более или менее непрерывный слой, сходный, например, со скопле-
ниями свободных макрофагов и лейкоцитов на поверхности стерильного
инородного тела, введенного в соединительную ткань позвоночных.
Так как у насекомых никаких оседлых клеток соединительной ткани
не имеется, то вся местная реакция, столь характерная для позвоночных,
естественно, совсем отсутствует. Вместо этого в области инородного тела
все в больших и больших массах скопляются свободные клетки гемо-
лимфы. Причем поверх макрофагоподобных клеток начинают оседать в пре-
обладающем количестве упомянутые выше веретенчатые элементы. Ни
одного митоза во всех этих клетках в течение всего процесса не встречается.
Увеличение их количества в очаге воспаления происходит, повидимому,
исключительно за счет приноса новых элементов гемолимфой. Оседающие
поверх «дегенеративного слоя» веретенчатые элементы, удлиняясь и обра-
зуя заостренные отростки, соединяются друг с другом в сетевидную «десмо-
бластическую» ткань, окружающую со всех сторон инородное тело и содер-
жащую в своих петлях отдельные клетки гемолимфы с другими морфоло-
гическими признаками.
Приток новых клеток постепенно прекращается, а десмобластическая
ткань переходит к следующей фазе своего развития. Десмобласты начи-
нают, как бы «худеть», причем на месте их бывших отростков, а затем и кле-
точного тела появляются волокна межклеточного вещества, сетевидно свя-
занные друг с другом. Создается впечатление, что за счет постепенно исче-
зающей цитоплазмы, путем ее прямого превращения, образуются волокни-
стые соединительнотканные структуры. Околоядерные части цитоплазмы
десмобластов некоторое время бывают еще различимы в петлях сети меж-
клеточного вещества. Затем остатки клеточных тел и ядра как бы подсы-
хают, оставляя после себя подобие теней, которые в конце концов исчезают
без остатка. В результате этого, вокруг отмирающего полностью дегене-
ративного слоя свободных клеток на поверхности инородного тела обра-
зуется довольно скудная и негустая бесклеточная волокнистая сеть, харак-
терная для соединительной ткани насекомых. Во всем течении этого про-
цесса и свойствах клеток гемолимфы насекомых нельзя не видеть очень
существенных отличий от того, что известно про воспалительную реакцию
и кровяные клетки позвоночных.
Эти различия, с моей точки зрения, заслуживают отнюдь не меньшего
внимания, чем отмеченные выше и особенно подчеркиваемые Заварзиным
черты сходства. Эти, уже достаточно ясно бросающиеся в глаза, различия
усугубляются еще больше, если обратить внимание на свойства клеток,
циркулирующих в гемолимфе.
Бросается в глаза при изучении всего хода процесса чрезвычайно высо-
кая специализация всех без исключения клеточных элементов гемолимфы
насекомых. Эти клетки не делятся митозом и, повидимому, пред-
ставляют собой совокупность нескольких строго детерминированных и
неспособных уже к превращению друг в друга клеточных рядов. Циркули-
рующие в гемолимфе насекомых и скопляющиеся в очаге воспаления клетки
не обнаруживают способности к характерному для крови и соединительной
ткани позвоночных, хорошо известному и особенно подчеркнутому и раз-
витому в курсе гистологии Заварзина развитию, неразрывно связанному
с размножением митотическим путем. Поэтому есть все основания считать
их уже неделящимися митотическим путем высокоспециализированными
формами, которые способны лишь в строго определенных пределах к некото-
рым морфологическим изменениям.
Даже если в гемолимфе происходит превращение части круглых не-
зернистых клеток в веретенчатых предшественников «десмобластов», то,
нужно думать, что эта часть круглых клеток качественно отличается от
другой ее части, изменяющейся в каком-нибудь другом направлении. Если
сопоставлять весь ряд клеток гемолимфы, способный к превращению в дес-
мобластическую сеть с клеточными формами соединительной ткани и крови
позвоночных, то они сравнимы вовсе не со всем десмобластическим или
фибробластическим рядом, включая его возможные лимфоидные звенья,
а только с его конечным звеном — фиброцитом или десмоцитом, который,
в отличие от позвоночных, свободно циркулирует в гемолимфе насекомых
и при соответствующих условиях имеет лишь одну окончательную судьбу:
никогда не делясь митозом, образовать волокнистое межклеточное вещество
и погибнуть. Все предыдущие стадии образования клеток гемолимфы, т. е.
их развитие, связанное с митотическим делением незрелых форм, к сожа-
лению, известны хуже всего. Как отмечалось выше, у данного вида известны
лишь небольшие клеточные скопления, сравниваемые с кроветворными
органами, где удавалось видеть митозы в очень мелких, повидимому, мало-
дифференцированных клетках, которые если и поступают в гемолимфу,
то в совершенно ничтожном количестве. Пожалуй, нет даже достаточных
оснований утверждать, что все клетки гемолимфы образуются только в этих,
известных, в настоящее время, органах.
Как известно, образование свободных клеток гемолимфы взрослых форм
некоторых насекомых описывается в связи с их эктодермальными имаги-
нальными дисками. Без прослеживания эмбрионального гистогенеза гемо-
лимфы и соединительнотканных волокнистых структур, разумеется, очень
трудно делать какие-нибудь определенные и достаточно обоснованные
выводы.
Из всего сказанного ясно, что клеточные формы гемолимфы насекомых
в их взаимоотношениях с образующимся в экспериментальных условиях
межклеточным веществом, при наличии некоторых черт сходства в отдель-
ных этапах процесса, пожалуй, столь же сильно отличаются от взаимо-
Экспериментальный метод в гистологии
353
отношений между кровью и соединительной тканью у позвоночных, как
и общий тип организации этих животных. Различия в биологическом зна-
чении соединительной ткани насекомых и позвоночных особенно подчерки-
ваются, если проследить воспалительную реакцию в ее обычном течении
до конца и учесть сопутствующие ей разрастания кожного эпителия. Обра-
зующаяся у насекомых тонкая и, повидимому, весьма несовершенная кап-
сула из волокон соединительнотканного характера не является по суще-
ству исходом процесса. Настоящим и регулярно наблюдающимся исходом
воспалительной реакции на введение инородного тела у насекомых является,
по данным Лазаренко (1925), образование вокруг него хитиновой эктодер-
мальной капсулы, образующейся в результате разрастания кожного эпите-
лия в очаге воспаления. При очередной линьке инородное тело выводится
из организма и выбрасывается вместе со старым хитином. Эта элиминатив-
ная (Гаршин, 1939) функция кожного эпителия включена у насекомых
в воспалительную реакцию как ее постоянное завершающее звено.
У позвоночных, как это подробно разбиралось выше, элиминативная
функция кожного эпителия также выражена вполне отчетливо, но она
отнюдь не проявляется как постоянно включенное в течение воспалитель-
ного процесса звено. Для осуществления воспалительных разрастаний
кожного эпителия необходим еще ряд дополнительных условий. Обычно
исходом воспалительной реакции позвоночных на введение стерильных
кусочков целлоидина является образование хорошо развитой, изолирую-
щей капсулы из плотной рубцовой ткани, имеющей довольно мало общего
с рыхлой сеточкой волокон, образующейся у насекомых. У последних
хитинизированная гиподерма, мощное развитие которой доминирует во
всей организации, в ходе воспалительного процесса функционально как бы
заменяет, полностью сохраняя свою эпидермальную специфику, роль слабо
развитых межуточных структур.
Следует отметить еще одно очень интересное и важное наблюдение
Лазаренко (1925) над регенерацией поврежденного при введении целлоиди-
новой трубки жирового тела, которое, вероятно, представляет собой спе-
циализированное производное целомической выстилки предков насекомых.
По данным этого автора, молодые дольки жирового тела также образуются
путем трансформации части клеток гемолимфы, которые, повидимому, и
в этом случае не делятся митозом. Мне представляется, что эти наблюдения
говорят в пользу существования в гемолимфе особых свободных целомиче-
ских элементов, не имеющих морфологически выраженной дифференци-
ровки, но качественно отличных от клеток, образующих волокнистые
структуры соединительной ткани. Не исключена возможность того, что
в своеобразной клеточной мозаике гемолимфы будут обнаружены также и
другие специализированные еще в каком-нибудь ином направлении сво-
бодные элементы. Таким образом, мне представляется, что, как и у многих
других форм животных, в полостной жидкости насекомых весьма вероятно
наличие клеток, с которыми связано образование волокнистых межуточных
структур и целомических элементов, имеющих иное биологическое зна-
чение.
23 V лопин—349
354
Глава III
Учитывая все очень существенные различия, которые обнаруживаются
в течение воспалительного процесса, а также в свойствах свободных полост-
ных клеток и соединительнотканных структур насекомых и позвоночных,
мне кажется, нет достаточных оснований переносить способ образования
межклеточного вещества, наблюдаемый у насекомых, на способ его раз-
вития у позвоночных. Различия в этом отношении, пожалуй, предста-
вляются более вероятными, чем черты сходства. Во всяком случае едва ли
можно считать достаточно обоснованной попытку Заварзина (1934) объяс-
нить способ развития межклеточного вещества у позвоночных путем сопо-
ставления с картинами, наблюдаемыми у насекомых и, в гораздо менее
отчетливом виде, у других беспозвоночных.
Последнее, на чем мне хочется здесь остановиться, это на возможном
объяснении некоторых черт сходства очень общего характера между сое-
динительной тканью и кровью позвоночных и насекомых, позволяющих
объединить их в одну и ту же морфо-физиологическую группу тканей и,
в частности, подчеркнутых Заварзиным совпадений в отдельных картинах
воспалительного процесса у этих и у некоторых других животных и тех
глубоких различий, которые, согласно всему сказанному выше, имеются
у представителей этих типов организмов как во всем ходе воспалительного
новообразования соединительной ткани, так и в свойствах клеток их по-
лостной жидкости или крови и соединительнотканных структурах.
Если считать все отмеченные черты сходства достаточно глубокими и
имеющими принципиальное значение, то в них можно видеть доказатель-
ство гистологической гомологичности мезенхимных образований у насеко-
мых и позвоночных, т. е. того, что и те и другие развились преемственно
из древнего примитивного мезенхимного зачатка отдаленных общих пред-
ков, представленного малодифференцированными способными к фагоцитозу
клетками, блуждающими в промежуточном между эктодермой и энтодермой
слое их тела и обладавшими, вероятно, уже тогда, способностью скопляться
вокруг попадающих внутрь этих примитивных форм инородных частиц.
В пользу этого говорят в первую очередь классические наблюдения Меч-
никова над фагоцитозом у многих низших беспозвоночных. Очень глубокие
отмеченные выше и совершенно незаслуженно игнорируемые различия
во всей совокупности биологических свойств свободных клеток полостной
жидкости и крови и соединительнотканных структур позвоночных и насе-
комых являются результатом дивергентного развития всего типа их орга-
низации. Дело здесь не в принципе параллелизма, который Заварзин счи-
тает основным принципом морфологии, а в сочетании связанных с един-
ством происхождения черт сходства с типовыми, приобретенными в про-
цессе дивергентной эволюции, гистологическими различиями, неразрывно
связанными с особенностями общей типовой организацией живот-
ных.
Всего сказанного выше, мне кажется, достаточно для того, чтобы пока-
зать возможность и полную оправданность подхода с наших общих пози-
ций и к соединительнотканным, в широком смысле слова, образованиям
как в ряду позвоночных, так и беспозвоночных. Более детальный разбор
Экспериментальный метод в гистологии
355
этих структур с точки зрения естественной гистологической системы
является преждевременным.
д) Опухолевые соединительнотканные структуры
Опухоли соединительнотканного характера и энтомезенхимного проис-
хождения представляют собой очень обширную и разнообразную группу.
Как и в других случаях, часть опухолевых и опухолеподобных структур
отличается более или менее высоким уровнем дифференцировки и достаточно
характерной для той или иной разновидности межуточных тканей структу-
рой. Плотные фибромы и фибросаркомы с развитым волокнистым межкле-
точным веществом, хондромы и хондросаркомы, остеосаркомы по своей
микроскопической структуре не оставляют сомнений в своей тканевой
природе и в своей генетической связи с обычной энтомезенхимой или ее
специализированной скелетогенной разновидностью.
Иначе обстоит дело со многими новообразованиями, богатыми клетками
и бедными волокнистыми структурами или даже почти или совершенно
лишенными их, большая часть которых носит название сарком с прилага-
тельным, относящимся часто к внешней форме или размерам опухолевых
клеток. Тканевая природа новообразований типа веретеноклеточных,
круглоклеточных, мелкоклеточных или гигантоклеточных сарком в очень
многих случаях представляется совершенно неясной, а их гистогенетиче-
ское обозначение достаточно произвольным и малооправданным.
Правда, наш опыт в области малодифференцированных неопухолевых
разрастаний показывает, что более детальный и точный микроскопический
анализ, чем он зачастую имеет место при обычной патологогистологической
диагностике, позволит обнаружить в ряде опухолевых тканей такие особен-
ности тканевой структуры, которые сделают достаточно вероятным их
отнесение к той или иной гистогенетической группе, к тому или иному
тканевому типу естественной гистологической системы. В этом отношении
следует приветствовать каждый опыт детального изучения этих структур
с применением хороших общих и специальных гистологических методов.
Работа Глазунова (1931), посвященная изучению фибриллообразования
в саркомах, принадлежит к числу таких исследований, хотя, как это уже
подчеркивалось, самый факт образования межклеточного вещества, и тем
более его очень примитивной разновидности, какой являются аргиро-
фильные волокна, взятый сам по себе, еще не может считаться безусловным
гистогенетическим критерием. Зато обнаружение хотя бы в отдельных
вытянутых в длину опухолевых клетках малодифференцированного внеш-
него вида продольной волокнистости, местами обнаруживающей попереч-
ную исчерченность, уже делает более чем вероятным миогенную природу
опухолей и ее принадлежность к мышечным опухолям соматического типа.
Несколько подробнее мы остановимся на такого рода новообразованиях
в главе о мышечных тканях. Едва ли, однако, подлежит сомнению, что
даже после возможно тщательного изучения микроскопической структуры
опухолей тканевая природа достаточного количества их все-таки останется
совсем неясной. В соответствии с нашим подходом к нормальным тканям.
356
Глава IIJ
такого рода новообразования следует трактовать как результат конвер-
гентного бластоматозного патогистогенеза, связанного с низким уровнем
гистоморфной и цитоморфной дифференцировки, которая, в свою очередь,
зависит, в соответствии со взглядами Фишер-Вазелъса (1927), от резко выра-
женной опухолевой катаплазии.
Среди этих опухолей мы имеем все основания предполагать наличие
малоспецифических структур, генетически связанных с очень многими
или, быть может, даже всеми тканевыми типами естественной гистологиче-
ской системы. Здесь окажутся и нейрогенные и эпидермальные, энтеродер-
мальные и цело дермальные, и соматомиогенные и собственно мезенхимные,
или «энтомезенхимные» новообразования типа цитобластом, или
очень малодифференцированные гистобластомы, т. е. опухоли, состоящие
не из несвязанных друг с другом клеток, как это предполагает понятие
цитобластомы, но из клеточных соединений малодифференцировапного и
малоспецифического характера.
Истинную тканевую природу этих опухолей, как это было видно на
некоторых, правда, еще немногочисленных примерах, удается устано-
вить с помощью экспериментально-гистологического анализа методом
эксплантации. Однако уже заранее следует ожидать, что сильно катапла-
зированные соединительнотканные опухоли представят в этом отношении
очень большие трудности, так как именно в их случае можно, пожалуй,
менее всего ожидать появления в экспериментальных условиях достаточно
специфических тканевых структур. Впрочем, решение этого вопроса еще
принадлежит будущему.
Очень сходными во многих отношениях с опухолевыми процессами
являются заболевания кроветворного аппарата и крови типа лейкемий, кото-
рые в гистогенетическом отношении, на основании всего того, что мы
знаем о крови и соединительной ткани, следует сближать с опухолями энто-
мезенхимного происхождения. Лейкемии являются в гистологическом отно-
шении чем-то очень сходным с цитобластомами, поскольку лейкемические
клетки не образуют связных тканевых структур.
У этих «цитобластом», в соответствии со свойствами нормальных кле-
ток крови, полному отсутствию тканевой дифференцировки может сопут-
ствовать высокая цитологическая или цитоморфная специализация эле-
ментов, дифференцирующихся, например, в те или иные формы миэлоцитов
и лейкоцитов. В таких случаях с далеко идущей дифференцировкой опухоле-
подобных или настоящих опухолевых клеток, связанной с утратой ими
способности к размножению митотическим путем, болезнь прогрессирует
за счет пролиферации менее дифференцированных клеток, представляю-
щих собой род «опухолевого камбия». Так наз. миэлобластпые лей-
кемии представляют собой тип заболевания с более катаплазированными
клетками, не обнаруживающими специальной цитоморфной дифференци-
ровки. Опыты Авророва и Тимофеевского (1913) и Тимофеевского и Бенево-
ленской (1927) показывают, что такие катаплазированные клетки способны,
дифференцируясь в условиях тканевых культур, выявлять свои специфи-
ческие, но в данном случае не гистобластические, а цитобластическве потен-
Экспериментальный метод в гистологии
357
ции и превращаться в разные формы гранулоцитов. Те же самые опыты,
как это уже отмечалось выше, с моей точки зрения, делают вероятным суще-
ствование не только смешанных эритролейкемических заболеваний, но,
быть может, и чистых катаплазированных белых эритремических форм,
связанных с опухолеподобным поражением эритробластического аппарата.
В соответствии со всем сказанным выше, патологические клетки эритреми-
ческих заболеваний должны, по всей видимости, рассматриваться, с точки
зрения генетической системы гистологии, вместе с сосудистым, эндотелием
и его патологическими производными.
4. МЫШЕЧНЫЕ ИЛИ СОКРАТИМЫЕ ТКАНИ
а) Общие гистогеиетические и сравнительные соображения
Сократимость в широком смысле этого слова представляет собой одно
из общих элементарных свойств живой протоплазмы, которое обнаружи-
вается, как это неоднократно подчеркивалось в предыдущем изложении,
даже у очень малодифференцированных тканевых элементов,хотя бы в форме
их способности к довольно быстрому втягиванию отростков и округлению.
У собственно мышечных элементов эта элементарная сократимость, раз-
виваясь исторически, совершенствуясь и приобретая те или иные каче-
ственные особенности, становится основной ведущей функцией, с которой
неразрывно связаны и некоторые общие морфологические свойства. Поэтому
неудивительно, что мышечные ткани, развиваясь у самых разнообразных
представителей животного царства, частью из одного и того же, частью
из разных, имеющих за собой неодинаковые пути исторического развития,
источников, во многих случаях обнаруживают большое морфологическое
сходство, которое и позволило их объединить, независимо от происхожде-
ния и систематического положения животных, в единую тканевую группу.
Как это уже отмечалось выше, внешнее сходство мышечных элементов
рассматривалось Эбнером (1911) как один из примеров гистологических
конвергенций, обозначенных затем Заварзиным (1925, 1932, 1941) как
параллелизмы.
При изучении всего многообразия сократимых элементов не могло не
возникнуть попыток вывести одни формы мышц из других путем построения
филогистогенетических рядов, дающих представление об эволюционном
развитии мышечных тканей. Попытка такого рода была сделана еще бр.
Гертвигами (1882), Геккелем (1885), а затем развита Маурером (1906, 1915).
различавшим эпителиальные, эпителогенные и мезенхимные мышцы. Пер-
вые две группы генетически связаны с эпителиальными, третья — с мезен-
химными структурами. Несмотря на ряд положительных сторон и большой
сравнительный материал, собранный и изученный Маурером, его система,
основанная на геккелевском и гервиговском формальном понимании эпи-
телиальных и мезенхимных структур и на смешении морфо-физиологиче-
ских и генетических принципов классификации, не могла внести ясности
во всю проблему сократимых тканей. В ряде случаев несомненно род-
ственные друг другу мышечные элементы оказываются в разных рубриках.
358
Глава III
и наоборот. Весьма ценным и правильным, в общих чертах, является во
взглядах Маурера трактовка поперечной исчерченности, как приобретае-
мого в филогенезе морфологического усложнения, связанного лишь с извест-
ными функциональными признаками, а не как отличительного свойства
определенного типа мышечных тканей. Эта точка зрения была принята
Заварзиным (1932/33) и мной (1933). В блестящей книге Гейденгайна (1911),
богатой ценнейшим описательным фактическим материалом, главное вни-
мание уделяется тонкой структуре мышечных элементов, которая, наряду
с некоторыми функциональными особенностями, и кладется в основу клас-
сификации мышечных тканей с их основным подразделением на гладкие
и поперечнополосатые мышцы. В таком виде глава гистологии, посвящен-
ная мышечным тканям, и фигурирует до настоящего времени почти во всех
учебниках и сводках. Новейшая сводка, принадлежащая Гегквисту (1931),
не содержит ничего принципиально нового и поражает крайним формализ-
мом изложения. Проводимая в этой книге точка зрения на мышечные ткани
была предложена мной в 1936 г., использована в моей монографии (1940)
и частично принята Заварзиным (1941), который вместе с тем вернулся
к принятому старыми авторами, начиная с Биша, подразделению мышц
на анимальные, или соматические, и вегетативные, или внутренностные.
С точки зрения естественной гистологической системы, рассмотрение
группы мышечных тканей, объединяемой на основании очень специального
чисто морфо-физиологического признака, в самостоятельной главе не
является последовательным. В предыдущем изложении нам удалось более
или менее совместить порядок рассмотрения тканей по общепризнанным
морфо-физиологическим группам и в соответствии с нашей генетической
естественной системой. В отношении очень узкоспециализированной руб-
рики сократимых структур этого не удается сделать. Большую часть сокра-
тимых элементов можно или рассматривать, как они по существу этого и
заслуживают, в соответствии с их генетическими отношениями как частные
разновидности разобранных выше тканевых типов, выделив для само-
стоятельного изложения только их наиболее древнюю часть — мускула-
туру соматического типа, или же подвергнуть анализу вместе, как особую
морфо-физиологическую группу тканей, подразделив ее на соответствую-
щие части в связи с их историей развития. Мне представляется, в данном
случае, более целесообразным второй порядок изложения, при котором
с особенной отчетливостью подчеркивается значение экспериментально-
гистологического анализа и на этой основе делается возможным освещение
некоторых вопросов общего характера.
Мышечные элементы представляют собой, особенно у позвоночных, те
гистологические структуры, которые очень часто использовались как
убедительное доказательство против возможности построения и в пользу
безсмысленности даже tнаиболее примитивной, чисто эмбриологической,
генетической системы тканей и как обоснование взгляда, что свойства диф-
ференцированных тканей никак не связаны с их развитием из того или иного
источника. Действительно, у позвоночных мы наблюдаем, как это уже
отмечалось, конвергентную дифференцировку очень типичных веретенча-
i
т
Экспериментальный метод в гистологии
359
тых гладких мышечных волокон или гладких сократимых элементов раз-
ветвленной формы из многослойных эпителиев кожи и переднего отдела
пищеварительной трубки, из «эпителия» края глазного бокала, т. е. нерв-
ного зачатка, и из энтомезенхимы. Образование мускулатуры поперечно-
полосатого типа происходит в этом же филогенетическом ряду из миотомов,
спланхнотомов, по некоторым данным также из мезенхимы, а у птиц и
рептилий!, кроме того, из нервного зачатка. При таком положении вещей,
казалось, представляется совершенно невозможным стоять на точке зре-
ния гистологической специфичности эмбриональных закладок и зависи-
мости свойств дифференцированных тканей от способа их развития. Однако,
как это уже было отмечено мной (1936), экспериментально-гистологический
анализ позволяет обнаружить в мышечных тканях разного происхождения
совершенно отчетливые различия в биологических свойствах и тем самым
использовать именно сократимые элементы для доказательства правиль-
ности нашей противоположной точки зрения и полной обоснованности
генетического подхода к тканям. С точки зрения естественной системы
гистологии мы подразделяем все разнообразие сократимых элементов позво-
ночных на пять уже перечисленных выше типов, которые нам предстоит
рассмотреть здесь подробнее.
Прежде всего мы остановимся на мускулатуре соматического типа, пред-
ставляющей собой древнейшую гистологическую систему с сократимыми
свойствами, с уровнем развития, расчлененностью и общей высотой орга-
низации которой теснейшим образом связаны все морфо-физиологические
особенности животных и в значительной степени их тип организации.
Значение мускулатуры соматического типа в эволюции животных не может
быть рассмотрено здесь во всей широте, так как оно далеко выходит за
рамки наших гистологических задач. Опыт изучения мускулатуры с точки
зрения общей биологии и экологии животных организмов, сделанный
в последнее время Зенкевичем (1944), представляется совершенно ориги-
нальным, исключительно интересным и открывающим широкие перспективы
для дальнейших разносторонних исследований.
Характерной особенностью мускулатуры соматического типа позвоноч-
ных является ее состав из многоядерных элементов с очень сложной вну-
тренней организацией и характерной очень отчетливой поперечной исчер-
ченностью. Для обозначения своеобразных мышечных элементов соматиче-
ского типа весьма целесообразным является предложенный Шнейдером
(1902) термин «мионы», которым мы и будем пользоваться впредь. Несмотря
на высокую специализацию и черты сходства, связанные с единым основным
функциональным значением, элементы соматической мускулатуры позво-
ночных обнаруживают тем не менее очень значительные морфологические
различия, связанные с рядом особенностей гистогенеза, которые до на-
стоящего времени еще недостаточно детально изучены (Маурер, 1906,
1915). Достаточно напомнить своеобразные мышечные коробочки кругло-
ротых, относительно очень массивные волокна многих рыб и заметно отли-
чающиеся от них мышечные элементы высших позвоночных. Даже у одного
и того же животного мышечные элементы в составе разных мышц и в разных
360
Глава III
частях тела обнаруживают заметное разнообразие формы, размеров и
деталей внутреннего строения. Это показывает, что при общей очень боль-
шой морфо-физиологической специализации у мышечных элементов могут
быть обнаружены, во-первых, заметные эволюционные изменения свойств
даже в пределах типа позвоночных и, во-вторых, явления дивергентной
морфологической дифференцировки, связанные с их местными топогра-
фическими и функциональными особенностями.
Не так давно Глюксман (1934) высказал предположение, что в типе
хордовых можно обнаружить эволюционную смену трех типов соматиче-
ской мускулатуры, неразрывно связанную с эволюцией опорных структур
и их взаимоотношений с мышечными волокнами. Первый наиболее прими-
тивный тип встречается у ланцетника, второй — у рыб и третий — у амфи-
бий и высших позвоночных. Гистогенез мышечных волокон при эмбрио-
нальном развитии в некоторых отношениях отражает их филогенез. Хотя
многие данные Глюксмапа, касающиеся гистогенеза мышечных волокон
и взаимоотношений с элементами соединительной ткани, не свободны от
очень серьезных возражений и, с моей точки зрения, должны трактоваться
иным образом, чем это делается автором, такая постановка вопроса безу-
словно заслуживает внимания.
Не следует забывать также, что, помимо своего общего значения сокра-
тимой ткани, соматическая мускулатура несомненно несет еще и другие
важные функции, являясь, например, у ряда низших позвоночных важ-
нейшей трофической тканью, за счет которой может происходить огромная
трата энергии во время нерестового хода лососевых рыб и могут поддержи-
ваться жизненные функции при разных условиях голодания. Как важней-
шему, наряду с печенью, гликогеновому депо организма, мускулатуре
высших позвоночных также принадлежит немаловажное значение одной из
трофических тканей внутренней среды организма. Это доказывает, между
прочим, лишний раз большую условность попыток разделения тканей по
функциональным признакам.
Кроме того, не подлежит сомнению, что самим же мышечным элемен-
там вместе с межуточной соединительной тканью в ряде случаев принад-
лежит и значение опорного аппарата организма, сосуществующего у позво-
ночных со специальными скелетными образованиями, а у некоторых бес-
позвоночных имеющего основное механическое значение. Последняя осо-
бенность также не должна забываться при общей функциональной харак-
теристике мускулатуры.
Часть соматической мускулатуры позвоночных вторично приобретает
значение не локомоторного, а висцеромоторного аппарата. Это относится,
в первую очередь, к т. наз. висцеральной мускулатуре жаберных
дуг низших позвоночных, которую, с гистологической точки зрения, пра-
вильнее рассматривать как особую специализированную© часть мускула-
туры соматического типа, а отнюдь не гладкой висцеральной мезенхимной
мускулатуры, как это обычно делается в сравнительной анатомии. К муску-
латуре соматического типа, повидимому, следует также отнести все по-
строенные по этому типу части мускулатуры пищеварительной трубки,
Экспериментальный метод в гистологии
361.
даже в тех случаях, когда они встречаются, как это имеет место, например
у некоторых рыб, в составе тонкого кишечника.
Хотя в этом отношении еще не имеется определенных эмбриологических
данных, все эти висцеромоторные поперечнополосатые мышцы можно
предположительно рассматривать как вторично смещенные части муску-
латуры соматического типа. Некоторые данные, касающиеся лимфатических
сердец лягушки, делают вероятным отнесение их поперечно исчерченных
мышечных волокон также к мускулатуре соматического типа. Именно по
этой причине я и принял термин «мускулатура соматического типа» (1940),
а не «соматическая мускулатура», как это обычно делается, так как чисто
территориальный или физиологический признак недостаточен для опреде-
ления специфических особенностей ткани и не учитывает весьма вероятных
эволюционных изменений ее локализации и специального функциональ-
ного значения. В ряду позвоночных мы пока не имеем никаких определен-
ных положительных данных, которые бы заставляли считать вероятным,
в данном случае, приобретение гладкой мезенхимной мускулатурой неко-
торых морфологических признаков соматической поперечнополосатой, хотя
теоретически такая возможность не может быть исключена с полной досто-
верностью.
Специфическим тканевым зачатком мускулатуры соматического типа
позвоночных являются миотомы или мышечные пластинки спинных сег-
ментов. презумптивная локализация которых может быть определена с боль-
шой точностью уже на ранних стадиях развития зародышей амфибий.
Экспериментальная эмбриология показывает, что, начиная с «мозаичной
стадии» развития, соответствующей нейруле амфибий, эти эмбриональные
зачатки оказываются вполне детерминированными в гистологическом отно-
шении и при разных внешних условиях обнаруживают способность к диф-
ференцировке в мышечные волокна. То же самое демонстрируется и в отно-
шении недифференцированных или малодифференцированных миобластов
птиц и млекопитающих методом эксплантации (Леви, 1934; Григорьев,
1934; Буркгардт, 1936, и др.). Предположение Буркгардта, что в его
опытах с эксплантацией почек конечностей зародышей крыс источником
миобластов является эндотелий сосудов, не может считаться сколько-
нибудь вероятным и обоснованным.
У некоторых рыб (ганоидные, селяхии) удается проследить с полной
отчетливостью образование мускулатуры плавников за счет особых отпрыс-
ков миотомов. Все литературные указания на развитие мускулатуры сома-
тического типа (например в плавниках морских коньков, в зачатках конеч-
ностей высших позвоночных, в языке, пищеводе и некоторых других ме-
стах организма) из мезенхимы (В. Шмидт, 1927, 1930; Кацнельсон, 1936,
и др.) основаны на неправильной предпосылке, что любые рыхло располо-
женные «мезенхимообразные» клетки могут на этом основании считаться
за элементы вполне определенного генетически единого мезенхимного за-
чатка так, как это понимали в свое время братья Гертвиги (1882). В составе
рыхлых малодифференцированных клеточных соединений зародыша, кото-
рые даже на относительно поздних стадиях его развития неправильно
862
Глава III
трактуются как совершенно равноценные в гистогенетическом отношении
части особого мезенхимного зачатка, как это уже отмечалось выше, сле-
дует различать элементы разного происхождения и с качественно различ-
ными гистобластическими потенциями. Утверждение, что даже такие фило-
генетически несомненно связанные с миотомами части мускулатуры, как
мышцы конечностей, развиваются из «мезенхимы», целиком зависит, с моей
точки зрения, от формального и недифференцированного, хотя и очень
обычного, понимания этого термина. В соответствии со всем сказанным выше,
т. наз. развитие мышечных волокон соматического типа из мезен-
химы следует понимать таким образом, что в составе данных миобластиче-
ских участков организма имеются вполне детерминированные клетки, вы-
шедшие из состава миотомов или генетически связанные с гомологичными
им частями мезодермы, которые уже качественно отличаются от элементов
мезенхимы, дающих начало межуточным тканям и родственным им обра-
зованиям. Эти элементы должны по существу также строго отличаться от
собственно энтомезенхимы, как разобранная выше эктомезенхима и, по-
видимому, «ангиоэритробластический» зачаток.
Особого упоминания заслуживают еще эмбриологические указания на
развитие мускулатуры хвоста амфибий из нервной трубки и мускулатуры
конечностей из соматоплевры боковых пластинок. С точки зрения эволю-
ционной морфологии и, как ее части, эволюционной гистологии, такого
рода указания, очевидно, никак нельзя понимать в буквальном смысле,
так как в противном случае пришлось бы отказаться от принятия более
чем вероятного, преемственного развития в ряду позвоночник конечностей
или хвостовой части тела и входящих в их состав отдельных органов и
тканей и допустить столь широкую возможность субституций, которая
сравнительно-анатомически никак не оправдывается и может лишь при-
вести к полной путанице во всех хорошо обоснованных представлениях и,
в конце концов, даже к отказу от самого понятия гомологии. В отношении
мускулатуры хвоста (Бийтелъ и Вурдеман, 1928; Бийтелъ, 1931; Гексли и
де-Бер, 1936; Иванов, 1937) это кажущееся ее развитие из нервной трубки
принято трактовать совершенно правильно таким образом, что вентраль-
ная часть того образования, которое в хвостовой области представляется
с первого взгляда нервной трубкой, состоит на самом деле из вполне обо-
собленного материала, соответствующего хвостовым спинным сегментам и
располагавшегося на стадии бластулы в непосредственном соседстве
с материалом задних туловищных сомитов. В отношении мускулатуры
конечностей, путь развития которой, кстати говоря, еще нуждается в более
детальном изучении, подобного рода необходимых пояснений обычно не
делается. Если мускулатура конечностей в своем онтогенезе действительно
связана с соматоплеврой, то соответствующие части ее, мне кажется, сле-
дует трактовать как вторично сместившийся материал миотомов, т. е.
общего для позвоночных мышечного зачатка.
Я позволил себе остановиться на всех этих вопросах, так как с точки
зрения естественной системы гистологических структур они имеют прин-
ципиальное значение.
Экспериментальный метод в гистологии
363
Как это отмечалось в главе, посвященной соображениям о возникно-
вении древнейших тканевых систем организма, вопрос о генетических
взаимоотношениях мускулатуры соматического типа у вторичноротых и
у первичноротых не может быть решен в настоящее время в достаточно
определенной форме, так как никаких ясных указаний на это ни сравни-
тельная эмбриология, ни сравнительная анатомия не дают, и теоретически
не исключена возможность субституций одних образований другими. Тем
не менее существование у современных кишечнополостных (которые, по-
видимому, являются близкими родственниками общих вымерших предков
и первичноротых и вторичноротых животных) развитой в различной
степени мускулатуры делает вполне вероятным ее преемственное развитие
из одного источника в разных ветвях филогенетического древа. Раннее
обособление в онтогенезе гребневиков мышечного зачатка в вице последней
серии микромер как бы на границе между эктодермой и энтодермой может
трактоваться как несколько измененный способ их перистомального раз-
вития по краю бластопора, а тем самым может быть использовано как один
из аргументов в пользу филогенетического возникновения этого зачатка
в той же области с последующим вытеснением диффузно развивающихся
в эктодерме и энтодерме сократимых элементов, свойственных другим
кишечнополостным. Эктодермальные элементы тонкой кольцевой муску-
латуры турбелларий и аннелид с этой точки зрения можно трактовать
как филогенетический остаток диффузной соматической мускулатуры,
который в дальнейшем совершенно исчезает. К сожалению, убедительность
такого рода совершенно умозрительных сопоставлений очень недостаточна
и субъективна, хотя к ним и неизбежно приходится прибегать для создания
многих гипотетических представлений общего характера. В качестве рабо-
чей гипотезы, против которой, мне кажется, нельзя привести достаточно
серьезных аргументов, вполне возможно принять преемственное развитие
из одного и того же источника части мезодермального зачатка, дающей
в онтогенезе начало всей или, по крайней мере, основной части мускулатуры
соматического типа во всех филогенетических стволах животного мира;
тем более, что преемственное развитие других очень древних гистоло-
гических систем — энтодермальной, эпидермальной и тесно связан-
ной с последней, нервной системой, прослеживается достаточно хорошо
у всех первичноротых и вторичноротых животных, за совершенно единич-
ными исключениями, не могущими иметь принципиального значения.
Разнообразие размеров, внешней формы и внутренней структуры мышечных
элементов соматического типа у беспозвоночных необычайно велико, пожа-
луй, настолько, насколько это вообще мыслимо для клеток или возникаю-
щих из них своеобразных неклеточных протоплазматических структур,
какими являются многие мышечные волокна.
Клетки «миоэпителиального» характера, удлиненной или сложно раз-
ветвленной формы, элементы с одним или многими ядрами, гигантские
одноядерные мышечные тяжи с односторонним расположением миофибрил-
лей, причудливые протоплазматические образования с отростками, соеди-
няющимися с совершенно обособленными сократимыми частями, ветре-
364
Глава III
чаются во всем своем многообразии у различных представителей животного
мира и являются благодарным материалом для демонстрации явлений дивер-
гентного развития, а также параллелизмов. Для общего объяснения послед-
них, с моей точки зрения, вполне достаточно общих законов эволюционной
биологии, согласно которым глубокие черты сходства имеют своей причиной
генетическое родство, различия зависят от дивергентной эволюции мелких
и крупных систематических групп, а более поверхностные черты сходства
могут быть и результатом конвергентного развития образований, имевших
очень неодинаковые пути эволюции. Детализация филогенетических изме-
нений свойств элементов соматической мускулатуры не входит в задачи
этой книги. Одних данных описательной гистологии для оценки биологиче-
ских особенностей, в частности, черт сходства и различий тканевых эле-
ментов, как это подчеркивалось в общей части, предпосланной фактиче-
скому материалу, в настоящее время уже недостаточно. Углубленный ана-
лиз этих вопросов требует систематического использования эксперимен-
тальных методов на представителях отдельных филогенетических рядов.
Подобно тому, как мышечная дифференцировка, в широком смысле
слова, сама по себе не является отличительной особенностью того или иного
тканевого типа естественной гистологической системы, а может возникать
на фоне весьма различных приобретенных в процессе эволюции свойств
тканевых элементов, гладкое или поперечно исчерченное строение муску-
латуры имеет, как это мной уже неоднократно подчеркивалось, с нашей
точки зрения, еще меньшее принципиальное значение, как типовой гисто-
логический признак, хотя при последовательной морфо-физиологической,
а не генетической, трактовке гистологических явлений ему принадлежит
и должно принадлежать очень почетное место. Поперечнополосатая диф-
ференцировка, как это справедливо отмечено Сееерцовым (1939), является
одним из примеров гистологических ароморфозов и связана, очевидно,
с повышением интенсивности функции сокращения, прежде всего, с ее
резким ускорением. Приобретение поперечнополосатой структуры всей
или некоторой частью мышечных элементов, имеющих в онтогенезе одно
и то же или разное происхождение, наблюдается у многоклеточных живот-
ных, начиная с кишечнополостных.
Поперечно исчерченные мионемы наблюдаются даже, как известно,
у некоторых форм протистов. Поперечнополосатые мышцы имеются у ряда
кишечнополостных. В мускулатуре соматического типа первичноротых
поперечная исчерченность появляется местами у крылоногих моллюсков,
в части мышечных волокон запирающей мышцы у Pecten, причем прочие
и значительные части мускулатуры сохраняют более примитивный глад-
кий вид. Особенно высокого уровня дифференцировки достигает поперечно-
полосатая мускулатура у высших представителей членистоногих, в част-
ности, у насекомых, у которых сложность поперечнополосатой структуры
совпадает с необычайной быстротой сокращений, далеко превосходящей
у летательных мышц самые быстрые сокращения у позвоночных. У форм,
стоящих близко к предкам насекомых, каковыми являются первично-
трахейные, описывается гладкая мускулатура соматического типа. Таким
Экспериментальный метод в гистологии
365
образом, в отдельных филогенетических рядах с большой отчетливостью
прослеживается эволюция телесной мускулатуры от гладкого к поперечно-
полосатому морфо-физиологическому типу. Дальнейшее детальное изуче-
ние эволюции тонкой стрзжтуры мышц в таком разрезе является безусловно
одной из очень интересных задач эволюционной гистологии. У вторично-
ротых, помимо позвоночных и ланцетника, поперечнополосатая мускула-
тура хорошо развита у щетинкочелюстных, в аппарате аристотелева фонаря
морских ежей и у некоторых туникат, по крайней мере, в личиночной
жизни. Все это говорит за то, что не только элементарная сократимость
является универсальным свойством живой протоплазмы, но даже специаль-
ная поперечнополосатая дифференцировка, возникая у очень примитивных
форм, может затем встречаться на самых различных этапах эволюции,
вероятно, в теснейшей связи не только с общим типом организации, но
и с образом жизни и экологическими особенностями животных.
Вместе с тем следует подчеркнуть, что в своих деталях поперечная
исчерченность мышечных волокон может обнаруживать очень большие
различия, на которые, однако, большей частью, не обращается достаточ-
ного внимания. До настоящего времени, при описании тонкого строения
поперечнополосатых мышечных волокон, принято рассматривать одно-
временно мускулатуру высших позвоночных и насекомых, совершенно
забывая, что у этих весьма отдаленных форм несомненно имеются и суще-
ственные различия в одноименных тканях. Многие детали строения попе-
речнополосатых мышц соматического типа насекомых без всяких к тому
оснований безоговорочно переносятся на позвоночных, что, конечно, недо-
пустимо с точки зрения эволюционной гистологии. Дивергентное развитие
гистологических структур при этом полностью игнорируется, а явления
параллелизмов или конвергенций, в силу прочно укоренившихся в гисто-
логии традиций, крайне односторонне выдвигаются на первый план. Не
может подлежать сомнению, что детальному анализу гистологических и
цитологических особенностей и различий строения и функциональных
свойств одноименных мышечных элементов близких и удаленных друг
от друга животных должно принадлежать, с точки зрения эволюционной
гистологий, выдающееся значение. То же самое справедливо и в отноше-
нии конвергентных мышечных структур разного происхождения в составе
организма одного и того же животного.
Вторым типом сократимых тканей является, с точки зрения естествен-
ной системы, мускулатура целомического типа (Хлопни, 1936), предста-
вленная в ряду позвоночных миокардом, который, с нашей точки зрения,
является сократимой модификацией целодермальных тканей и должен рас-
сматриваться как их особым образом специализированная и развившаяся
дивергентно разновидность. Миокард является несомненно филогенети-
чески значительно более молодой, чем мускулатура соматического типа,
сократимой тканью, и вместе с тем имеющей за собой иную историю разви-
тия, которая, с нашей точки зрения, не может не отражаться в ее гистоло-
гических свойствах. Несмотря на свою поперечную полосатость, развив-
шуюся, повидимому, конвергентно с телесной мускулатурой, он очень
366
Глава III
резко отличается от нее сетевидным строением и многими другими струк-
турными особенностями, связанными, очевидно, и с функциональным свое-
образием. В препаратах, как известно, получить «хорошую» поперечную
исчерченность в волокнах миокарда значительно труднее, чем в соматиче-
ской мускулатуре, что не может не говорить об известных различиях их
живого субстрата. Любопытно отметить, что первое, поражающее своей
точностью и ясностью изображение сетевидного строения миокарда нахо-
дится в известной книге «Тайны природы» Левенгука (1695), который в то
время не мог оценить значения своего открытия. В миокарде, пожалуй,,
еще резче, чем в мускулатуре соматического типа, проявляется неразрыв-
ная связь сократимости с функцией проведения импульсов к сокращению.
Сократимость и проводимость соответствующих импульсов представляют
собой две неразрывно связанные друг с другом функциональные стороны
мышечной дифференцировки. В составе миокарда даже специализируются,
повидимому, только у высших позвоночных, особые мышечные центры
возбуждения с системой проводящих волокон, которые до известной сте-
пени функционально субституируют нервный аппарат сердца низших
позвоночных. Если взять проводимость за основную функциональную
характеристику тканевых свойств, то, при последовательном и односто-
роннем физиологическом подходе к тканям, проводящую систему сердца
высших позвоночных можно было бы рассматривать, вместе с нейронами,
как разновидности проводящих тканей.
С точки зрения Гейденгайна (1911), пользующейся в настоящее время
наибольшим распространением, сердечная мускулатура представляет собой
сплошной синцитий без всякого подразделения на клеточные территории.
Согласно более старой точке зрения, проводившейся позже в работах Цим-
мермана (1910) и его учениц Пальчевской и Вернер, миокард имеет клеточное
строение, хотя количество ядер в отдельных его территориях у разных
животных может быть неодинаково. У человека оно равно одному-двум,
а у свиньи доходит до 32. Клеточные границы с этой точки зрения пред-
ставлены т. наз. спаивающими или замыкающими полосками, которые,
кстати говоря, появляются лишь на поздних стадиях развития сердца
млекопитающих, причем с возрастом их количество возрастает. Защитники
противоположного взгляда на сердечную мышцу считают вставочные
участки или блестящие полоски, функциональное значение которых доста-
точно спорно, за утолщенные инофрагмы, через которые миофибрилли
проходят, не прерываясь.
Миокард позвоночных развивается из определенного участка целоми-
ческой выстилки зародыша т. наз. миоэпикардиальной пластинки висцераль-
ного листка сплахнотомов, дающей начало и миокарду и мезотелию эпикарда,
что особенно подчеркивает генетическую близость этих тканей, представ-
ляющихся столь различными. Методами экспериментальной эмбриологии
(Штёр, 1924, 1925) удается получить самодифференцировку миокарда
из соответствующих изолированных участков спланхнотомов зародышей
амфибий при культивировании в эпидермальных мешочках. При частичном
удалении зачатка миокарда соседние участки спланхнотомов, изменяя
Экспериментальный метод в гистологии
367
обычное направление своей дифференцировки, компенсируют в известной
степени недостаток миоэпикардиального материала (Экман, 1921, 1929).
Последнее так же хорошо подчеркивает генетическую близость мезотелия
и миокарда. Несмотря на отдельные исследования, посвященные миокарду
разных позвоночных, достаточно ясной картины филогенетических изме-
нений его гистологических особенностей пока не удается себе составить.
Ряд интересных данных находится в исследованиях Марсо (1904).
У многих целомических беспозвоночных также имеется миокард или
соответствующие ему мышечные участки сосудистой системы, причем во
многих случаях их генетическая близость с выстилкой полости тела, в част-
ности, с перикардиальным мешком выступает с поразительной нагляд-
ностью. Достаточно вспомнить эмбриональное развитие этих образований
и их взаимоотношения у взрослых форм кишечнодышащих из вторично-
ротых и моллюсков из первичноротых. Поэтому мы вправе говорить,,
как это мной уже отмечалось (1936), о наличии особой целомической или це-
лодермальной мускулатуры у беспозвоночных. У животных с «псевдоцелем»,
т. е. с вторично полностью исчезнувшей выстилкой целомической полости
(членистоногие), можно пока лишь говорить о связи сердечной мускулатуры
в эмбриональном развитии с общим мезодермальным комплексом зачатков,
а не с какой-нибудь определенной его частью.
Имеющиеся отдельные работы, посвященные специально некоторым
участкам мускулатуры беспозвоночных, позволяют установить у некоторых
форм, например у насекомых-типулид, поразительные черты сходства
с миокардом в отношении сетевидного строения и даже наличия вставочных
участков (Мазярский, 1926). Не менее важным и интересным является
вместе с тем анализ дивергентных черт строения сердечной мускулатуры
у представителей разных систематических групп. Дальнейшие сравнительно-
гистологические исследования в этом направлении обещают очень много
интересного. Чрезвычайно трудным и с полной определенностью не разре-
шенным в настоящее время вопросом является трактовка филогистоге-
нетических отношений между миокардом позвоночных и других вторично-
ротых и сердечной или соответствующей ей сосудистой мускулатурой первич-
норотых. Если в филогенетическом ряду вторичноротых еще можно говорить
с достаточной определенностью о генетической близости миокарда и о его
параллельном развитии из гистологически единого субстрата, целомической
выстилки общих предков, то генетическая близость сердечной или соот-
ветствующей ей сосудистой мускулатуры в ряду вторичноротых и первично-
ротых является в лучшем случае очень косвенной, и ее признание упирается
прежде всего в неясность генетических отношений целомической выстилки
этих форм. В соответствии с тем, что было сказано в главе, посвященной
вопросу о развитии тканевой организации животных, гистологическая гомо-
логия между целомической выстилкой вторичноротых и первичноротых
не может быть доказана на основании имеющихся эмбриологических срав-
нительно-анатомических фактов, а является лишь гипотезой, против кото-
рой, с моей точки зрения, не имеется убедительных данных.
Принятие или непринятие этой гипотезы в настоящее время не осно-
368
Глава III
вывается на каких-либо прямых и доказательных аргументах. Мне она
представляется достаточно вероятной или, по крайней мере, возможной.
Но даже если ее и принять, то генетические отношения сердечных муску-
латур оказываются весьма косвенными, так как, несомненно, общие предки
первичноротых и вторичноротых не только не обладали сердечной мус-
кулатурой, но даже еще и целомической полостью. Правда, и в данном случае
остается все-таки возможным искать общего филогистогенетического предка
целомической мускулатуры, уже имевшего, по крайней мере отчасти, морфо-
физиологическую характеристику сократимых тканей в миоэнтодермальных
клетках некоторых кишечнополостных, что представляется весьма пробле-
матичным, хотя и не абсолютно невозможным. Следует только подчеркнуть
тот факт, что отмеченные в литературе черты сходства между сердечной
мускулатурой позвоночных и отдельных первичноротых обычно совпадают
с определенным анатомическим конвергентным сходством соответствующих
органов, с которым в данном случае несомненно связано теснейшим образом
и известное совпадение гистологической структуры.
Для внесения хотя бы некоторой ясности во все эти вопросы необходимо
ждать результатов дальнейших, в достаточной степени кропотливых иссле-
дований, в которых, с моей точки зрения, экспериментально-гистологи-
ческому методу должно принадлежать почетное место.
Как это мной уже отмечалось (1936), весьма возможно, что у некоторых
форм беспозвоночных, в частности у насекомых, у которых вся имеющаяся
в их теле мускулатура построена по поперечнополосатому типу, к цело-
мической мускулатуре, кроме мышцы сердца, относится также и внутрен-
ностная мускулатура кишечника. Это предположение получило частичное
подтверждение в работе Кочетова (1940), установившего ряд гистологи-
ческих отличий между сетевидной мускулатурой средней кишки насекомых,
с одной стороны, и их соматической мускулатурой и мышцами хитини-
зированных эктодермальных, переднего и заднего, участков пищеваритель-
ной трубки, с другой.
В свете этих исследований мое первоначальное предположение должно
быть исправлено в том отношении, что к целомической мускулатуре у насе-
комых, возможно, относятся лишь мышцы среднего эктодермального участка
пищеварительной трубки, мускулатура же передней и задней кишки пред-
ставляет собой сместившиеся части мускулатуры соматического типа.
Третий тип мышечных тканей позвоночных представлен мезенхимной
или,точнее, «энтомезенхимной» мускулатурой, генетически теснейшим обра-
зом связанной с соединительной тканью. Близость гладких мышечных
элементов с соединительнотканными клетками, бросавшаяся в глаза старым
авторам [например еще Генле (1841) обозначал гладкую мускулатуру как
сократимую соединительную ткань], подчеркивалась очень многими иссле-
дователями, особенно на протяжении последних 20 лет, причем некоторые
из них, как, например, Штиве (1926), идут настолько далеко, что допускают
возможность развития гладкомышечных клеток в беременной матке не
только из малодифференцированных элементов соединительной ткани,
ио даже из лимфоидных элементов крови, эмигрирующих из сосудов. Близ-
Экспериментальный метод в гистологии 369
кие генетические отношения между малодифференцированным подэндо-
телиальным слоем стенки артерий и мышечными элементами среднего слоя
убедительно доказываются работами Шелку нова (1935, 1936). Чтобы подчерк-
нуть близкие генетические отношения энтомезенхимной гладкой муску-
латуры с соединительной тканью, ее клетки были мной в свое время обозна-
чены как миодесмальные элементы (1936).
В отношении своей структуры при нормальных условиях гладкая
мезенхимная мускулатура очень резко отличается как от соматической,
так и от сердечной. Ее характерной особенностью является отчетливое
клеточное строение, безотносительно от того, существуют или не существуют
описываемые отдельными авторами анастомозы между отдельными мышеч-
,ными волокнами. Со времени первых точных описаний разнообразия форм
гладкомышечных «клеток-волокон», данных Колликером (1849) и за-
тем подтверждавшихся многими позднейшими авторами, в этом едва ли
могут быть сомнения. Гейденгайн (1911) целиком ее поддерживал, хотя
и считал вопрос о существовании анастомозов открытым. Проводимая
Гегквистом- (\93\) точка зрения на гладкомышечные пучки как на сплош-
ную синцитиальную массу с отдельными ядерными эндоплазматическими
участками, мезоплазмой, представленной миофибриллярными частями,
и общей «экзоплазмой», содержащей в себе волокнистые структуры прокол-
лагенового, коллагенового и эластического характера, которые предста-
вляют собой по сути дела межклеточное вещество, является лишь необычной
и нс отвечающей действительному положению вещей трактовкой обычных
. препаратов, не подкрепленной какими-либо убедительными положитель-
. ними доводами. Очень частая веретенчатая форма гладких мышечных клеток
. может варьировать в отдельных случаях в широких пределах от довольно
правильной уплощенной полигональной до сложно разветвленной с отрост-
ками. Клетки последнего рода описываются часто (Беннингоф, 1926) распо-
ложенными изолированно в соединительной ткани. Нужно, однако, отме-
тить, что мышечная природа таких изолированных отростчатых элементов
не всегда бывает достаточно убедительна, так как их отличия от клеток
соединительной ткани далеко не всегда выступают отчетливо, а их трактовка
представляется в значительной степени субъективной. Весьма интересен
тот факт, что в гладкомышечных пучках и пластах между мышечными клет-
ками обнаруживаются тонкие прослойки межклеточного вещества, которым,
как это только что отмечалось, Гегквиап (1931) приписывает, без достаточ-
ных оснований, расположение в экзоплазме, опираясь на эктоплазмати-
ческую теорию образования межклеточного вещества. Наряду с развитием,
.наличие межклеточного вещества соединительнотканного характера между
гладкомышечными клетками, которые, очевидно, и являются его образо-
вателями в процессе своей дифференцировки, также подчеркивает близость
.гладкой мускулатуры и межуточных тканей. Вместестемследует подчеркнуть,
.что в отношении преобладающего характера межклеточного вещества,
степени его развития и некоторых других особенностей между гладкой
мускулатурой разных органов, повидимому, имеются различия, трактовка
которых в настоящее время представляется затруднительной и требует
24 ХлоииЕ*—349
370
Глава III
еще дальнейших исследований. Каких-либо определенных различий, за
исключением общих размеров, между гладкомышечными элементами пред-
ставителей разных классов позвоночных, которые позволяли бы говорить
о постепенных эволюционных изменениях гладкой мускулатуры, насколько
мне известно, не подчеркивалось. За пределами подтипа позвоночных, эле-
ментов, сравнимых с мезенхимной мускулатурой в отношении генеза и
взаимоотношений с соединительной тканью, неизвестно. Все гладкомы-
шечные волокна беспозвоночных, относимые, на основе отсутствия
в них поперечнополосатой структуры, в единую гладкомышечную ткань
с мезенхимной мускулатурой позвоночных, представляют собой, пови-
димому, по сути дела, как это уже отмечалось выше, модификации
мускулатуры соматического типа. Их сближение с гладкой мускула-
турой позвоночных основывается лишь на одном единственном, несуще-
ственном с точки зрения тканевой систематики, формальном признаке. Пови-
димому, сильное развитие в ряду позвоночных гладкой мезенхимной муску-
латуры следует поставить в связь с необычайно высоким уровнем качествен-
ного и количественного развития у них соединительнотканных образований.
Наиболее вероятной точкой зрения является в настоящее время признание
гладкой мезенхимной мускулатуры за развившуюся дивергентно высоко-
специализированную сократимую разновидность соединительнотканных
элементов, которая, вследствие своей мышечной дифференцировки, вместе
с тем обнаруживает конвергентное сходство с некоторыми другими мышеч-
ными структурами иного происхождения. Дальнейшее развитие учения
о гладкомышечных элементах позвоночных, очевидно, связано теснейшим об-
разом с нашими представлениями о межуточныхтканяхис трактовкой соеди-
нительнотканных структур вообще, а также с углубленным эксперименталь-
ным анализом самой гладкой мускулатуры разных органов тела. В этом отно-
шении мы, возможно, еще не гарантированы от некоторых неожиданностей..
Четвертым типом сократимых структур позвоночных является муску-
латура радужины, развитие которой из ретинальных листков глаза, т. е.
по сути дела из зачатка центральной нервной системы, после первых опи-
саний Нусбаума (1899) и ряда исследований позднейших авторов
(Шили, 1902; Форсмарк, 1904; Лепла, 1912, и др.) является прочно уста-
новленным. С точки зрения своего происхождения мышечные элементы
радужины заслуживают названия «мионейральных» (Хлопин, 1936; Вин-
ников, 1938). У млекопитающих мышцы радужины, открытые Валентином
(1835) и довольно подробно описанные Брюкке (1846), построены по глад-
кому типу, а потому их принято считать разновидностями гладкой муску-
латуры. Вместе с тем т. наз. мышца, сужающая зрачок, настолько
отличается по своему характеру от расширяющей мышцы, что их прихо-
дится относить с другой, также формально-морфологической, точки зрения
к двум очень различным типам мышц. Мышца, сужающая зрачок, обнару-
живает настолько поразительное внешнее сходство с гладкой мезенхимной
мускулатурой, что ее, несмотря на глубокие различия в происхождении,
рассматривают до последнего времени вместе с мезенхимной гладкой муску-
латурой (Гегквист, 1931). По наблюдениям Винникова (1938) в толще мышцы.
Экспериментальный метод в гистологии 371
сужающей зрачок, удается обнаружить лишь серебрящиеся волокна.
Коллагеновых и эластических волокнистых структур, свойственных обыч-
ной мезенхимной гладкой мускулатуре, в ней не встречается, равно как
и сосудов. У неальбинотических кроликов в околоядерной части сокра-
тимых элементов встречаются в большем.или меньшем количестве такие
же пигментные зерна, как и в ретинальных листках слепой части сетчатой
оболочки, что подчеркивает генетическую близость этих образований
Мышца, расширяющая зрачок, по своей морфологии относится к т. наз.
миоэпителиальному типу. Ее ядросодержащие части образуют в своей
совокупности непрерывный, большей частью пигментированный, кле-
точный слой, представляющий собой наружный листок т. наз. эпите-
лия радужины, высота которого изменяется в зависимости от степени
ее сокращения или расслабления. Базальные части этих «эпителиальных»
элементов превращены в длинные отростки, богатые миофибрилл ями,
тянущиеся непосредственно под «эпителием» параллельно поверхности
радужины в радиальном направлении и образующие в своей совокупности
т. наз. волокнистую мембрану Бруха, которой Гегквист также приписывает
сплошное синцитиальное строение. Небольшая часть клеток мышцы, рас-
ширяющей зрачок, может целиком погружаться под слой эпителия и при-
обретать веретенчатую форму, свойственную сужающей зрачок мышце.
У разных позвоночных с нередуцированными глазами мышцы радужины
достигают неодинакового развития. У круглоротых в связи с отсутствием
радужины эта мускулатура еще совершенно отсутствует. У разных рыб
и амфибий она развита в неодинаковой степени и имеет сходный с млеко-
питающими гладкий характер. У рептилий и птиц она приобретает
поперечнополосатое строение, причем мышца, сужающая зрачок, оказы-
вается построенной из типичных многоядерных поперечнополосатых волокон.
Иными словами, мышечные элементы радужины, имеющие в ряду позво-
ночных тождественное происхождение, обнаруживают настолько разно-
образную дивергентную дифференцировку, что с точки зрения своих морфо-
физиологических особенностей принадлежат к совершенно разным типам
сократимых структур- С другой стороны, они являются дивергентно диф-
ференцированными и специализированными разновидностями тканей, раз-
вившихся из нервного зачатка.
Мышечные элементы цилиарного тела, которые в отношении своей
структуры обнаруживают совпадение с мускулатурой радужины и у птиц,
также отличаются поперечнополосатым характером, обычно принимают
за мускулатуру мезенхимного или, точнее, энтомезенхимного происхождения.
Однако место их развития — цилиарное тело является по существу особым
участком мягкой мозговой оболочки — делает очень вероятным, в соответ-
ствии со сказанным выше, образование мускулатуры цилиарного тела из
нейромезенхимы, т. е. ее отнесение к эпендимо-глиальному тканевому типу.
В этом отношении необходимы еще специальные исследования.
К пятому и последнему типу сократимых структур, с точки зрения
естественной системы тканей, относятся у многих позвоночных т. наз.
миоэпителиальные или, точнее, миоэпидермальные клетки некоторых
372
Глава III
желез, часть которых была описана еще Кёлликером (1849) и включена
им, на основании сходства в строении, в группу гладких мышц вместе
с мезенхимной гладкой мускулатурой.
В некоторых эпидермальных железах (например потовые железы млеко-1
питающих, кожные железы бесхвостых амфибий) сократимые элементы,
располагающиеся в виде базального ряда в составе эпителиальной выстилки,
действительно обнаруживают большое сходство с веретенчатыми мезен-
химными гладкомышечными клетками. В других случаях им свойственна
более или менее сложно разветвленная форма т. наз. корзинчатых клеток
(молочная и слюнные железы). Естественно возникает вопрос, имеются ли у
беспозвоночных сократимые элементы, сравнимые в каких-нибудь отноше-
ниях с мионейральными и миоэпидермальными клетками позвоночных.
У кишечнополостных с мультнфункциональной эктодермой, которая
объединяет в себе, как это отмечалось выше, свойства всех тканевых групп
классической гистологии, связана значительная часть сократимых элемен-
тов тела. У планарий и у аннелид эктодермальное происхождение имеет
кольцевая мускулазура, причем у аннелид и, возможно, также у планарий,
кожный эпителий является при регенерации источником образования не
только эпителия кожного покрова и кольцевой мускулатуры, но и элементов
нервной системы, т. е. является тканью, объединяющей в себе гистогенети-
ческие особенности и кожной эктодермы и нейрэктодермы позвоночных,
которые у последних детерминируются в разных направлениях уже на
ранних стадиях индивидуального развития. Таким образом, между лионейг
ральными и миоэктодермальными клетками позвоночных, с одной стороны,
и эктодермальной мускулатурой некоторых беспозвоночных устанавливают-
ся столь же отдаленные генетические отношения, как и между соответ-
ствующими животными. Достаточно отдаленной является связь этих гисто-
логических структур с мускулатурой соматического типа, если ее выводить
филогенетически, как это предлагалось еще Маурером, из эктодермальной
мускулатуры общих предков.
Особые свойства мускулатуры соматического типа, обусловленные всей
историей ее развития, подчеркиваются также наличием у нее своеобразных
двигательных нервных окончаний (двигательные бляшки позвоночных
и соответствующие им дуайеровы бугорки насекомых), которые, повидимому,
несвойственны сократимым структурам других типов. В частности, в мио-
карде млекопитающих эффекторные концевые аппараты {Лавров, 1941) по
своей структуре не имеют ничего общего с двигательными бляшками. Эво-
люция нервт омышечных синапсов мускулатуры соматического типа в от-
дельных филогенетических рядах животных еще нуждается в дополнитель-
ном изучении и заслуживает очень большого внимания с точки зрения вза-
имозависимости филогистогенеза нервной и соматической мышечной систем.
Все перечисленные типы сократимых тканей позвоночных являются
конвергентно развившимися образованиями, имевшими очень неодинаковые
пути исторического развития, под общей, хотя и разной в отдельных случаях,
мышечной внешностью которых, в соответствии с нашей точкой зрения,
следует ожидать совершенно отчетливых биологических различий. Для
Экспериментальный метод в гистологии 378
обнаружения их мы прибегли, как и во многих других случаях, к экспери-
ментально-! истологическому анализу сократимых структур.
б) Мускулатура соматического типа
Как это уже отмечалось выше, клеточный материал миотомов и недиф-
ференцированные элементы зачатков мускулатуры соматического типа
более поздних стадий развития у разных позвоночных обнаруживают спо-
собность к самодифференцировке в более или менее типичные для этого
тканевого типа поперечно исчерченные мышечные волокна. Это демон-
стрируют самые разнообразные, достаточно хорошо известные опыты и,
в частности, исследования, произведенные методом эксплантации или
изоляции. При эксплантации частиц дифференцирующейся мускулатуры
соматического типа, взятых от зародышей разного возраста, помимо раз-
растаний малодифференцированных клеток, наблюдается рост своеобразных
симпластических многоядерных тяжей {Льюисы, 1915, 1917; Леей, 1919,
1925, 1934; Фридгейм, 1931; Де-Реньи и Гог, 1931; Хлопин, 1936; Юдиц-
кая, 1938, и др.) По многим данным, эти симпластические тяжи могут в даль-
нейшем распадаться на одноядерные малода} ференцированные клетки
типа миобластов {Льюисы, 1915, 1917; Леви, 1934; Юдицкая, 1938, и др.).
В отношении дифференцированной соматической мускулатуры новоро-
жденных и молодых кроликов происходящие in vitro гистологические про-
цессы впервые прослежены с достаточной полнотой, причем установлен
ряд совершенно новых фактов {Хлопин, 1936, 1940).
По моим данным, миобластическая зона роста в культурах из зародышей
млекопитающих несомненно отличается от обычных мезенхимных и соеди-
нительнотканных эксплантатов своим более резко выраженным плоскостным
характером и несколько большей комплексностью. С увеличением возраста
исходного эмбрионального материала характер экстенсивного роста изме-
няется, и наряду с одноядерными миобластами начинает наблюдаться
прорастание своеобразных многоядерных протоплазматических тяжей или
лент, которые образуются в центральном кусочке из более дифференци-
рованных мышечных волокон и могут быть обозначены как миосимпласты.
На поздних стадиях эмбриональною развития и в случае неэмбриопального
исходного материала мышечные волокна млекопитающих прорастают
исключительно миосимпласгами и, повидимому, утрачивают способность
превращаться в размножающиеся митотическим путем миобласты при
культивировании без периодических вырезаний. Рост миосимпластов,
как правило, наступает позже прорастания элементов межмышечной соеди-
нительной ткани, образующих в окружающем свертке фибрина обычные
сетевидные клеточные соединения. Поэтому миосимпласты обычно врастают
в уже успевшую образоваться соединительнотканную зону роста. С воз-
растом способность к росту вне организма у мышечных волокон заметно
ослабляется, и процент культур с растущими миосимпластами делается
все меньше. В различных мышцах тела, повидимому, в связи с неодинако-
вым темпом их дифференцировки, ослабление способности к росту вне ор-
374
Глава Ill
ганизма выражается в разной степени. Из мускулатуры взрослых животных
до настоящего времени никому не удавалось получить в условиях тканевых
Рис. 57. Ценкеровское перерождение мышечных волокон соматического
типа. 2-дневная культура из мускулатуры брюшной стенки новоро-
жденного кролика. Сильное увеличиние. (По Хлопину, 1940.)
культур прорастания миосимпластов. Чем старше исходный материал,
тем более резко выражены в эксплантированных частицах дегенеративные
изменения и тем большее количество имеющихся налицо мышечных волокон
они захватывают. В эксплантатах из мускулатуры более старых животных
Рис. 58. Два мышечных волокна с увеличенным количеством крае-
вой саркоплазмы, обособляющейся от почти гомогенной дегенериру-
ющей миофибриллярной части и превращающейся в недифференциро-
ванные многоядерные миосимпласты. 3-дневная культура из муску-
латуры передней брюшной стенки 6-дневного кролика. Сильное увеличе-
ние. (По Хлопину, 1940.)
полной дегенерации и распаду подвергаются все мышечные волокна, а жизне-
способной остается лишь часть элементов соединительнотканных прослоек.
Разжижение фибрина с образованием полостей в эксплантатах из муску-
латуры, как правило, совершенно не наблюдается, или оно происходит
лишь в некоторых случаях после длительного культивирования без выре-
зания и прибавления свежей плазмы.
Превращения эмброниальных мышечных волокон, приводящие к обра-
зованию делящихся митотическим путем недифференцированных миобластов,
до настоящего времени недостаточно изучены. Что касается изменения
дифференцированных мышечных волокон и образования из них прорастаю-
Экспериментальный метод в гистологии
375
щих миосимпластов, то эти процессы удалось проследить со значительной
полнотой в эксплантатах из брюшных мышц новорожденных и молодых
кроликов. Уже в течение первых суток после приготовления культур мышеч-
ные волокна подвергаются заметным изменениям, вызванным, повидимому,
их реакцией на механическое повреждение, которое имело место при посеве.
Эти изменения заключаются в исчезновении в большей части волокон попе-
речной исчерченноеги и в появлении в саркоплазме между пучками миофиб-
риллей темно окрашивающихся зернышек. В части волокон затем посте-
пенно наступают дегенеративные изменения, имеющие характер т. наз.
ценкеровского перерождения и заключающиеся в распаде вещества во-
локна на более плотные темно окрашивающиеся и менее плотные светлые
Рис. 59. Миосимпласт зоны роста, в котором, отступя от концевого
булавовидного утолщения, начинают дифференцироваться гладкие
миофибрилли. 7-дневная культура из мускулатуры передней брюш-
ной стенки 5-дневного кролика. Сильное увеличение.
(По Хлопину, 1940.)
участки неправильной формы (рис. 57). В условиях эксплантата эти явле-
ния приводят, как правило, к полной дегенерации. В другой части волокон
дальнейшие изменения протекают иначе и заключаются в постепенном
обособлении друг от друга сохраняющих жизнеспособность элементов
периферической зоны, состоящей из недифференцированной саркоплазмы
с ядрами и центральной дифференцированной массы миофибриллей, кото-
рые постепенно дегенерируют и разрушаются (рис. 58). В незначительном
количестве при этом могут образоваться одноядерные саркоплазматические
участки округлой формы, переполненные измененной миофибриллярной
субстанцией и сходные с т. наз. сарколитами. Наконец, в известной
части волокон новорожденных животных, повидимому, сохранивших
к моменту эксплантации относительно низкий уровень дифференцировки
и осевое расположение ядер, миофибрилли могут исчезать постепенно,
как бы растворяться в окружающей саркоплазме, а не обособляться от нее
в виде дегенеративных масс. В течение второго-четвертого дня, реже не-
сколько позже, в неподвергающихся полному перерождению волокнах
центрального кусочка нередко встречаются митотические фигуры, не сопро-
вождающиеся разделением цитоплазмы. В дальнейшем, митозов в мышечных
волокнах и образующихся из них миосимпластах не наблюдается, и нередко
очень значительное увеличение количества ядер происходит исключительно
путем их простого деления или фрагментации.
Саркоплазматические части мышечных волокон, постепенно увели-
чиваясь в объеме, превращаются в многоядерные миосимпласты, которые
большей частью через четыре-пять дней начинают вырастать за пределы
376
Глава III
центрального кусочка (рис. 59) в успевшую к этому времени образоваться
соединительнотканную зону роста. Количество проросших в зону роста
миосимпластов в течение последующих дней увеличивается.
При пересевах без вырезания их длина продолжает увеличиваться
в течение двух недель и может достигать нескольких миллиметров. Иногда
они растут поодиночке, иногда образуют целые пучки. Большей частью
они отличаются почти прямолинейной или слегка дугообразной формой.
Различные миосимпласты часто имеют весьма неодинаковый калибр. То
они представляют собой широкие цитоплазматические ленты, то узкие тяжи
рис. 60. Дифференцировка миосимпластов, окруженных элемента-
ми соединительной ткани,у края центрального кусочка 8-дневной
культуры из мускулатуры передней брюшной стенки 6-дневного
кролика. Тонкие миосимпласты с почти гомогенной и гладко-фиб-
риллярной цитоплазмой и толстый миосимпласт с миофибриллями,
в которых местами появляется отчетливая поперечная исчерчен-
ность. Сильное увеличение. (По Хлопину, 1940.)
или даже почти нитевидные образования с веретенчатыми вздутиями в обла-
сти ядер. Общий калибр одного и того же миосимпласта и толщина его
отдельных участков могут изменяться как в течение роста, так и вследствие
его сократимости. Нередко их свободный конец бывает булавовидно вздутым
(рис. 59), в других случаях они, наоборот, имеют тонкие заостренные концы.
В редких случаях от миосимпластов, образовавшихся из дифференцирован-
ных неэмбриональных мышц, могут отделяться одно- и двуядерные участки
веретевовидной формы, которые имеют ограниченную жизнеспособность
и вскоре дегенерируют и распадаются. Более узкие миосимпласты, сливаясь
друг с другом, могут образовать широкие ленты или неправильные
саркоплазматические натеки, которые в дальнейшем вновь развет-
вляются.
Образовавшиеся миосимпласты имеют первоначально индифферентный
саркоплазматический характер. Лишь на их поверхности можно бывает
обнаружить тончайшую каемку измененной протоплазмы, представляющую
Экспериментальный метод в гистологии
377
собой подобие оболочки, быть может, сравнимой с той частью сарколеммы,
которая, по многим данным, образуется не соединительнотканными эле-
ментами, а саркоплазмой самих мышечных волокон. Ядра в миосимпластах
местами располагаются поодиночке, но чаще тесно сближенными парами,
рядами и целыми неправильными скоплениями, образующимися путем
амитотических делений или фрагментации (рис. 59). Если свободный конец
миосимпласта бывает колбовидно вздут, то в нем обычно содержится значи-
тельное количество ядер. С течением времени при культивировании с пере-
севами без вырезаний в индифферентных миосимпластах могут дифферен-
цироваться характерные для мышечных волокон структуры — миофибрилли
или даже поперечная исчерченность. Явления дифференцировки миосим-
пластов, по моим наблюдениям, протекают следующим образом: по истс-
чении пяти-семи дней после эксплантации в их протоплазме появляются
Рис. 61. Участок длинного миосимпласта зоны роста с дифферен-
цировкой отчетливых мисфибриллей и поперечной исчерченностью.
19-дневная культура из мускулатуры передней брюшной стенки
6-дневного кролика. Сильное увеличение. (По Хлопину, 1940.)
гладкие продольные фибрилли, становящиеся все более отчетливыми и
достигающие довольно значительной толщины. Как правило, дифферен-
цировка начинается в проксимальной, обращенной к центральному кусочку
части миосимпласта, в то время как его свободный дистальный конец
сохраняет свой более или менее недифференцированный вид (рис. 59).
В начале второй недели в миосимпластах части эксплантатов местами
появляется сначала едва заметная, затем все более отчетливая поперечная
исчерченность отдельных миофибриллей (рис. 60). Двояко преломляющие
и темно окрашивающиеся диски Q правильно чередуются с дисками /, но
могут не совпадать в соседних миофибриллях. По середине дисков / нередко
дифференцируются отчетливые полоски Z, то локализующиеся в единичных
миофибриллях, то перегораживающие поперек большую или меньшую
часть миосимпластов. В результате этого миосимпласты дифференцируются
вне организма в несколько атипичные поперечнополосатые мышечные
волокна (рис. 61). В некоторых эксплантатах столь далеко идущей диф-
ференцировки видеть не удается. В других она захватывает единичные
миосимпласты зоны роста и центрального кусочка или реже значитель-
ную их часть. Свободные дистальные концы миосимпластов, как правило,
сохраняют свой индифферентный, характер. Такие эксплантаты удалось
378
Глава III
наблюдать до трех-четырех недель жизни вне организма. Любопытно отме-
тить, что в ряде мышечных культур от новорожденных кроликов по про-
шествии нескольких дней после эксплантации можно наблюдать появление
самопроизвольных подергиваний и правильных ритмических пульсаций,
достигавших подчас большой частоты (до 180 ударов в 1 минуту), которые
продолжались, иногда с перерывами, в течение трех-четырех недель
жизни вне организма, т. е. в течение срока, когда двигательные нервные
волокна и окончания уже давно должны были исчезнуть.
В миосимпластах и дифференцирующихся из них мышечных волокнах
можно обнаружить довольно короткие, частью нитевидные, частью имеющие
вид зернышек хондриозомы. Их прямого превращения в миофибрилли
[в противоположность данным Дюсберга (1910) и некоторых других авто-
ров, изучавших эмбриональный гистогенез мышечных волокон] обнару-
жить не удается. При образовании миофибриллей в культурах хондриозомы
остаются отчетливо отличимыми от них. Их количество уменьшается, но
они сохраняются в саркоплазме между миофибриллями. Невероятными
представляются также указания Леей (1934) на обратное превращение
миофибриллей в хондриозомы при образовании индифферентных миобла-
стов из эмбриональных мышечных волокон.
Относительно того, какова связь между видимой дифференцировкой
соматических мышечных элементов и их сократимостью, имеются специаль-
ные исследования. В кратковременных эксплантатах из эмбриональной
скелетной мускулатуры еще Льюис (1920) отмечала сокращение мышечных
элементов, лишенных миофибриллей и поперечной исчерченности. Изучая
эксплантированные спинные сегменты куриных зародышей разного воз-
раста, Оливо (1925, 1932) показал, что их элементы начинают реагировать
слабыми сокращениями на переменный ток на стадии 27 — 28 сомитов (60—
65 часов насиживания). В этом возрасте они имеют вид веретенчатых клеток,
не имеющих еще миофибриллей и поперечной исчерченности. При их изуче-
нии в поляризованном свете удается обнаружить слабое и диффузное двой-
ное лучепреломление. Ясная, обнаруживаемая лишь в поляризационном
микроскопе поперечная исчерченность появляется на стадии 32—34 сомитов.
Реньи и Гог (1931), экспериментируя с растущими вне организма в течение
первых' дней после эксплантации индифферентными миосимпластами,
лишенными миофибриллей, обнаружили их сократимость при прикосно-
вении иглой микроманипулятора. Сокращение происходило очень резко
и быстро, но не сопровождалось обычного типа расслаблением. Лишь по
прошествии трех-шести часов сократившийся миосимпласт расправлялся,
причем его удлинение происходило путем роста, а не мышечного рас-
слабления. Фридгейм (1931) наблюдал ритмические сокращения миосимпла-
стов в течение двух-семи дней после эксплантации и вызывал их тетанус
при раздражении переменным током. По его данным, не подтверждаемым
другими исследованиями, сократимость будто бы связана с видимой в поля-
ризованном свете поперечной исчерченностью.
В настоящее время не приходится сомневаться в том, что миофибрилли
представляют собой реально существующие прижизненные структуры,
Экспериментальный метод в гистологии
379
а не образуются, как таковые, в результате фиксации. Что касается попе-
речной исчерченности на ранних этапах дифференцировки, то она не обна-
руживается на окрашенных гистологических препаратах и бывает заметна
лишь в поляризационном микроскопе. При дифференцировке, зашедшей
дальше, она делается вполне отчетливой при наблюдении в обычном неполя-
ризованном свете и видна как на живых объектах, так и на фиксированных
и окрашенных препаратах.
Опыты с тканевыми культурами и изучение миотомов другими экспери-
ментальными методами механики развития доказывают, что миобласты,
миосимпласты и мышечные волокна представляют собой, начиная с ранних
стадий эмбрионального развития, в гистологическом отношении детерми-
нированные элементы, неспособные к превращению в другие ткани. Диф-
ференцированные мышечные волокна млекопитающих представляют собой
своеобразные гистологические элементы, многоядерные симпластические
системы или мионы, состоящие из высокоспециализированной миофибрил-
лярной и более индифферентной саркоплазматической частей. При час-
тичном повреждении мионов их саркоплазматическая, содержащая ядра,
часть отличается резко выраженной способностью к росту и увеличению
массы своего живого вещества, а их дифференцированная миофибрилляр-
ная часть разрушается и погибает. В результате этого мион превращает-
ся в миосимпласт, в котором может вторично дифференцироваться
поперечно исчерченная миофибриллярная часть.
В отличие от данных других авторов, отмечавших сократимость миосим-
пластов в течение немногих первых дней культивирования, когда еще нельзя
исключить частичного сохранения двигательных нервных окончаний,
мои наблюдения доказывают с несомненностью полный автоматизм и даже
способность к самопроизвольным ритмическим сокращениям элементов
мускулатуры соматического типа.
Высокопоучительным представляется сопоставление изложенных выше
данных с многочисленными наблюдениями над репаративной регенерацией
и изменениями при разных патологических процессах мышечных волокон
соматического типа в составе целого организма. При этом, однако, необ-
ходимо учитывать следующие моменты, которые не всегда принимаются
во внимание: возрастные изменения в онтогенезе, особенности, связанные
с систематическим положением животных, характер экспериментального
воздействия или патологического процесса и совокупность внешних условий,
в которых происходят те или иные изменения мышечных волокон.
Одним из основных выводов, получаемых в результате изучения сфор-
мированной соматической мускулатуры вне организма и при репаративной
регенерации, является установление того факта, что рост ее элементов
происходит исключительно за счет предшествующих дифференцированных
мышечных волокон. Никаких недифференцированных запасных кам-
биальных элементов в ее составе, в противоположность эпителиям многих
органов или мезенхимным производным, обнаружить не удается. Это поло-
жение следует считать справедливым не только в отношении соматической
мускулатуры млекопитающих, но и других позвоночных, а, возможно,
380
Глава III
также в отношении достаточно высокоорганизованных беспозвоночных,
как, например, артроподы и моллюски. Противоположные данные Заутера
(1934), касающиеся мускулатуры костистых рыб, отнюдь нельзя признать
сколько-нибудь убедительными. Само собой разумеется, что никаких убеди-
тельных и выдерживающих критику данных в настоящее время нельзя
также привести в пользу новообразования мышечных волокон при регене-
рации из соединительнотканных элементов или лейкоцитов крови, как
это некогда предполагалось Ценкером (1864), Вальдейером (1865), Маслов-
ским (1868), Эрбкамом (1880) и рядом других авторов. Совершенно фанта-
стичны, не обоснованы и не стоят на уровне современной науки пред-
положения о том, что из поврежденных мышечных волокон будто бы могут
возникать соединительная ткань, хрящ, кость, гной (Крёзинг, 1892), или
эритроциты (Вейдман, 1938). В условиях эксплантата наблюдается развитие
тех типичных картин перерождения, которым после классического иссле-
дования Ценкера (1864) было присвоено имя этого автора или которые,
по предложению Неймана (1868), были обозначены, как «глыбчатый распад»
(schollige Zerkliiftung) или «распад на восковидные глыбки». В условиях
эксплантата волокна или участки волокон, подвергшихся ценкеровскому
перерождению, в дальнейшем погибают нацело. Как обстоит дело в целом'
организме, представляется до настоящего времени не вполне ясным. Мно-
гие признают, что в перерожденных участках волокон, наряду с распав-
шимся на глыбки содержимым, могут сохраняться жизнеспособные, содер-
жащие ядра участки саркоплазмы, которые, по некоторым данным, в даль-
нейшем участвуют в регенеративных процессах, превращаясь в недиф-
ференцированные «мышечные клетки», или саркобласты (Асканази, 1891:
Фолькман, 1893; Тома, 1909, и др.). По другим данным, содержащие ядра
участки в подвергшихся глыбчатому распаду волокнах в дальнейшем поги-
бают, не принимая участия в регенеративных явлениях (Шминке, 1909:
Миллар, 1934, и др.). В культурах дальнейшие явления роста мышечных
элементов происходят за счет участков волокон, не подвергшихся глыб-
чатому распаду. В соответствии с этим мне представляется более вероятным,
что источником регенеративных процессов в целом организме также являются
участки волокон, не подвергшиеся глубоким дегенеративным изменениям,
после отторжения от них распавшихся на глыбки и отмирающих частей.
Если сопоставить друг с другом данные, касающиеся ценкеровского пере-
рождения в целом организме (Фолькман, 1893; Шмидт, 1910; Шминке,
1909; Ямин, 1925; Раевская, 1939, и др.), и приведенные выше наблюдения
в эксплантате, то нельзя не признать, что эти изменения являются специ-
фической для мионов соматической мускулатуры реакцией на всевозможные
повреждающие воздействия. Эта реакция свойственна соматическим мышеч-
ным волокнам не только млеко питающих и человека, но и других, изученных
в этом отношении позвоночных (Р. Тома, 1906—1910; Шминке, 1908—1909;
Заварзин, 1938; Крюкова, 1938, и др.). Любопытно отметить, что в миокарде
млекопитающих и мезенхимной мускулатуре позвоночных картин настоящего
ценкеровского перерождения при патологических процессах не наблю-
дается. По данным Цымбала (1937), при дегенерации волокон миокар-
Экспериментальный метод в гистологии
881
да млекопитающих в культурах также не встречается глыбчатого рас-
пада.
В гладких и поперечнополосатых мышечных элементах соматического
топа различных беспозвоночных дегенеративных изменений подобных
ценкеровскому перерождению никто не наблюдал. Описанные в новейших
обстоятельных работах дегенеративные процессы в поврежденных мышеч-
ных волокнах олигохет (Заварзин, 1938), моллюсков (Заварзин, 1925;
Крюкова, 1936) и ракообразных (Жинкин, 1938) резко отличаются от картин,
характерных для соматической мускулатуры позвоночных.
Дальнейшие изменения подвергшихся ценкеровскому перерождению
мышечных волокон в культурах протекают в противоположность тому,
что наблюдается в целом организме, без образования так называемых
Muskelzellenschlauche, относительно происхождения которых было столько
разногласий. Образование Muskelzellenschlauche наблюдается не только
у млекопитающих и человека, но и у других позвоночных. Вслед за Валь-
дейером (1865), который, как известно, является автором этого термина,
многие исследователи считали, что клетки, появляющиеся внутри сарко-
леммных трубок перерожденных мышечных волокон, образуются путем
размножения сохранившихся изолированных содержащих ядра сарко»
плазматических участков. Мечников (1892) также относил миофагов, разру-
шающих при метаморфозе дегенерирующие мышечные волокна хвоста
головастиков бесхвостых амфибий, к элементам миогеиного происхождения.
В противоположность этому Нейман (1868) почти одновременно с Вальдейе-
ром высказал предположение, что клетки внутри сарколемных трубок
представляют собой иммигрировавшие из перимизия элементы. Эта точка
зрения в настоящее время и принимается большей частью исследователей
с той разницей, что в образовании MuskelzellenschlSuche признается уча-
стие не только мобилизованных клеток межмышечной соединительной
ткани, но и лейкоцитов, выползших из кровяного русла (Рахманинов, 1881;
Шминке, 1909; Таратынов, 1914; Миллар, 1934; Крюкова, 1938, и др.).
Таратынов (1914) обстоятельно изучил клеточные элементы Muskelzellen-
schlauche, пользуясь совершенной современной методикой, в частности,
методом прижизненной окраски, и доказал, что сначала в сарколеммных
трубках скопляются специальные лейкоциты, а затем полибласты или
макрофаги. В условиях тканевых культур не может иметь места истинная
воспалительная реакция, одним из результатов которой должно считаться
образование Muskelzellenschlauche или, как их правильнее обозначил
Шминке (1909), Wanderzellenschlauche; поэтому отсутствие этих структур
в наших опытах является вполне естественным и понятным.
В отличие от того, что обычно имеет место в целом организме при меха-
ническом повреждении или ожоге, в эксплантированных частицах мышц
далеко идущие изменения постепенно захватывают все волокна целиком,
которые, в конце концов, перестают существовать, как таковые. Таким
образом, в условиях тканевых культур изменения в мышечных волокнах
должны были бы более всего соответствовать тем, которые происходят
щ целом организме при отсутствии нарушения целости сарколеммы и при
382
Глава III
более или менее элективном повреждении вещества мышечных волокон
как это, например, бывает при их перерождении в случае брюшного тифа
и других токсикозах (Штемплер, 1914), а также в результате заморажива-
ния. Регенеративные процессы после такого рода повреждений особенно
подробно изучил Фолькман (1893) на млекопитающих и сравнил их с тем,
что происходит при значительном нарушении целости мышц.
В дифференцированных мышечных волокнах или их участках, не под-
вергшихся дисковидному или глыбчатому распаду, постепенно исчезает
только миофибриллярная часть, а саркоплазма с ядрами, гипертрофируясь,
превращается в недифференцированные миосимпласты, которые, как мышеч-
ные почки при регенерации, вырастают за пределы посеянного кусочка
в образующуюся вокруг него соединительнотканную зону роста. В условиях
моих опытов (пассажи без вырезания), как это уже неоднократно отме-
чалось, только многоядерные миосимпласты представляют собой жизне-
способные и могущие затем вновь дифференцироваться элементы. Мне
кажется, это подтверждает, что в крайне запутанном вопросе о регенерации
мышечных волокон соматического типа, по крайней мере, у млекопитающих
и человека, следует признать более правильными взгляды тех авторов,
которые, подобно Нейману (1868), придерживаются точки зрения новообра-
зования мышечных волокон не за счет одноядерных клеток-миобластов,
а за счет многоядерных мышечных почек. Нужно отметить, что в пользу
этого, возникшего уже очень давно, но затем многими оспаривавшегося
взгляда, склоняются почти все более новые авторы, работающие над реге-
нерацией мышц у млекопитающих и человека (Шминке, 1909; Бундшу, 1923;
Локателли, 1933; Миллар, 1934, и др.), в противоположность данным
Асканази (1891), Фолъкмана (1893), Даусона (1909); еан-Гелена (1936) и
многих других. Следует отметить, что новейшие данные ван-Гелена очень
суммарны и, повидимому, не основываются на достаточно тщательном анализе
препаратов. Едва ли можно согласиться с Фолькманом, что характер травмы
может в такой сильной степени влиять на способ регенерации, чтобы у одного
и того же вида животных в одних случаях образовались мышечные почки,
а в других миобласты. Если при регенерации мышц и происходит образо-
вание одноядерных миогенных элементов, констатирование которых на
срезах, кстати говоря, никогда не может считаться вполне убедительным,
то такие клетки в сущности являются не миобластами или саркобластами,
а нежизнеспособными изолировавшимися частями или обломками миосим-
пластов. Наблюдения над эксплантатами, в которых той или иной форме
перерождения мышечного волокна подвергаются нацело, пожалуй, говорят
против предположения Шминке (1909), что регенерация миобластами может
выступать на первый план у высших позвоночных в случаях очень сильных
повреждений, разрушающих нацело мышечные волокна. Вместе с тем
мои данные, как это уже отмечалось, не могут вполне исключить возмож-
ности образования миобластов из дифференцированных мышечных волокон
млекопитающих во всех решительно условиях.
Если для высших позвоночных, в соответствии с данными, полученными
методом эксплантации, в настоящее время следует считать характерной
Экспериментальный метод в гистологии
383.
регенерацию соматических мышечных волокон почками или миосимпла-
стами, то в отношении низших позвоночных имеется еще много неясностей,
безусловно требующих дальнейшей разработки вопроса. Из низших позво-
ночных, по данным Шминке (1908), только у хвостатых амфибий имеет
место регенерация миобластами, в то время как у костистых рыб и бес-
хвостых амфибий она происходит мышечными почками. Образование мышеч-
ных почек у мальков костистых рыб, правда, наряду с миобластами, наблю-
дали Нусбаум и Сидориак (1900) и Казанцев (1935) Заутер (1935) у кости-
стых рыб даже пытался обнаружить недифференцированные и плурипо-
тентные регенеративные клетки, за счет которых, по его мнению, могли
заново образоваться мышечные волокна. Данные этой работы нельзя счи-
тать сколько-нибудь обоснованными. Наконец, в последнее время Завар-
зин (1938), изучая регенерацию туловищной мускулатуры у бычка, пришел
к выводу, что, в условиях его опытов, она происходит исключительно
миобластическим путем, причем образовавшиеся из разрушенных мышеч-
ных волокон миобласты энергично делятся митотическим путем. Образо-
вание одноядерных миобластов у хвостатых амфибий отмечается, кроме
Шминке, и другими авторами, значительная часть которых, правда, имела
дело с эмбриональным или личиночным материалом. Барфурт (1891),
а в новейшее время особенно Навилль (1922) показали, что ход регенерации
мышечных волокон у амфибий в большой степени зависит от возрастной
стадии. Навилль нашел, что у молодых головастиков имеет место образование
миобластов, а у более старых — мышечных почек. Это хорошо гармонирует
с приведенными выше данными относительно роста вне организма эмбрио-
нальной и постфетальной мускулатуры кролика, но с трудом сопоставляется
с результатами, полученными на мальках лососевых рыб. Нусбаумом и
Сидориаком и Казанцевым и на взрослых бычках Заварзиным.
У взрослой лягушки Крюкова в последнее время наблюдала при реге-
нерации образование типичных мышечных почек. В описанных Крюковой,
кроме того, изолированных миогенных клетках не приходилось наблюдать
митозов. Поэтому их трактовка как миобластов и участие в образовании
новых мышечных волокон мне представляются недостаточно обоснованными.
Таким образом, способность к росту многоядерными симпластами, даже
если признать в отдельных случаях образование миобластов, является
характерной особенностью дифференцированных мышечных волокон всех
изученных позвоночных. Наоборот, соматическая мускулатура различных
беспозвоночных, по всем имеющимся в настоящее время данным, регене-
рирует исключительно путем образования одноядерных миобластов {Шульц,
1898; Иванов, 1903; Крюкова, 1936; Заварзин, 1938; Жанкин, 1938, и др.).
В отношении деталей структуры мышечных волокон соматического
типа, изучение их вне организма, мне кажется, дает возможность вы-
сказаться с достаточной определенностью о природе инофрагм Z, кото-
рые, с точки зрения Гегквиста (1931), представляют собой внутренние
сарколеммные прослойки, имеющие вид решеток и дающие некоторые красоч-
ные реакции коллагена, сквозь которые продернуты миофибрилли. Изу-
чение хода дифференцировки миосимпластов в культурах показывает,
382
Глава III
более или менее элективном повреждении вещества мышечных волокон,,
как это, например, бывает при их перерождении в случае брюшного тифа
и других токсикозах (Штемплер, 1914), а также в результате заморажива-
ния. Регенеративные процессы после такого рода повреждений особенно
подробно изучил Фолъкман (1893) на млекопитающих и сравнил их с тем,
что происходит при значительном нарушении целости мышц.
В дифференцированных мышечных волокнах или их участках, не под-
вергшихся дисковидному или глыбчатому распаду, постепенно исчезает
только миофибриллярная часть, а саркоплазма с ядрами, гипертрофируясь,
превращается в недифференцированные миосимпласты, которые, как мышеч-
ные почки при регенерации, вырастают за пределы посеянного кусочка
в образующуюся вокруг него соединительнотканную зону роста. В условиях
моих опытов (пассажи без вырезания), как это уже неоднократно отме-
чалось, только многоядерные миосимпласты представляют' собой жизне-
способные и могущие затем вновь дифференцироваться элементы. Мне
кажется, это подтверждает, что в крайне запутанном вопросе о регенерации
мышечных волокон соматического типа, по крайней мере, у млекопитающих
и человека, следует признать более правильными взгляды тех авторов,
которые, подобно Нейману (1868). придерживаются точки зрения новообра-
зования мышечных волокон не за счет одноядерных клеток-миобластов,
а за счет многоядерных мышечных почек. Нужно отметить, что в пользу
этого, возникшего уже очень давно, но затем многими оспаривавшегося
взгляда, склоняются почти все более новые авторы, работающие над реге-
нерацией мышц у млекопитающих и человека (Шминке, 1909; Бундшу, 1923;
Локателли, 1933; Миллар, 1934, и др.), в противоположность данным
Асканази (1891), Фолькмана (1893), Даусона (1909); ван-Гелена (1936) и
многих других. Следует отметить, что новейшие данные ван-Гелена очень
суммарны и, повидимому, не основываются на достаточно тщательном анализе
препаратов. Едва ли можно согласиться с Фолъкманом, что характер травмы
может в такой сильной степени влиять на способ регенерации, чтобы у одного
и того же вида животных в одних случаях образовались мышечные почки,
а в других миобласты. Если при регенерации мышц и происходит образо-
вание одноядерных миогенных элементов, констатирование которых на
срезах, кстати говоря, никогда не может считаться вполне убедительным,
то такие клетки в сущности являются не миобластами или саркобластами,
а нежизнеспособными изолировавшимися частями или обломками миосим-
пластов. Наблюдения над эксплантатами, в которых той или иной форме
перерождения мышечного волокна подвергаются нацело, пожалуй, говорят
против предположения Шминке (1909), что регенерация миобластами может
выступать на первый план у высших позвоночных в случаях очень сильных
повреждений, разрушающих нацело мышечные волокна. Вместе с тем
мои данные, как это уже отмечалось, не могут вполне исключить возмож-
ности образования миобластов из дифференцированных мышечных волокон
млекопитающих во всех решительно условиях.
Если для высших позвоночных, в соответствии с данными, полученными
методом эксплантации, в настоящее время следует считать характерной
Экспериментальный метод в гистологии
383
регенерацию соматических мышечных волокон почками или миосимпла-
стами, то в отношении низших позвоночных имеется еще много неясностей,
безусловно требующих дальнейшей разработки вопроса. Из низших позво-
ночных, по данным Шминке (1908), только у хвостатых амфибий имеет
место регенерация миобластами, в то время как у костистых рыб и бес-
хвостых амфибий она происходит мышечными почками. Образование мышеч-
ных почек у мальков костистых рыб, правда, наряду с миобластами, наблю-
дали Нусбаум и Сидориак (1900) и Казанцев (1935). Заутер (1935) у кости-
стых рыб даже пытался обнаружить недифференцированные и плурипо-
тентные регенеративные клетки, за счет которых, по его мнению, могли
заново образоваться мышечные волокна. Данные этой работы нельзя счи-
тать сколько-нибудь обоснованными. Наконец, в последнее время Завар-
зин (1938), изучая регенерацию туловищной мускулатуры у бычка, пришел
к выводу, что, в условиях его опытов, она происходит исключительно
миобластическим путем, причем образовавшиеся из разрушенных мышеч-
ных волокон миобласты энергично делятся митотическим путем. Образо-
вание одноядерных миобластов у хвостатых амфибий отмечается, кроме
Шминке, и другими авторами, значительная часть которых, правда, имела
дело с эмбриональным или личиночным материалом. Барфурт (1891),
а в новейшее время особенно Навилль (1922) показали, что ход регенерации
мышечных волокон у амфибий в большой степени зависит от возрастной
стадии. Навилль нашел, что у молодых головастиков имеет место образование
миобластов, а у более старых — мышечных почек. Это хорошо гармонирует
с приведенными выше данными относительно роста вне организма эмбрио-
нальной и постфеталыюй мускулатуры кролика, ио с трудом сопоставляется
с результатами, полученными на мальках лососевых рыб. Нусбаумом и
Сидориаком и Казанцевым и на взрослых бычках Заварзиным.
У взрослой лягушки Крюкова в последнее время наблюдала при реге-
нерации образование типичных мышечных почек. В описанных Крюковой,
кроме того, изолированных миогенных клетках не приходилось наблюдать
митозов. Поэтому их трактовка как миобластов и участие в образовании
новых мышечных волокон мне представляются недостаточно обоснованными.
Таким образом, способность к росту многоядерными симпластами, даже
если признать в отдельных случаях образование миобластов, является
характерной особенностью дифференцированных мышечных волокон всех
изученных позвоночных. Наоборот, соматическая мускулатура различных
беспозвоночных, по всем имеющимся в настоящее время данным, регене-
рирует исключительно путем образования одноядерных миобластов {Шульц,
1898; Иванов, 1903; Крюкова, 1936; Заварзин, 1938; Жанкин, 1938, и др.).
В отношении деталей структуры мышечных волокон соматического
типа, изучение их вне организма, мне кажется, дает возможность вы-
сказаться с достаточной определенностью о природе инофрагм Z, кото-
рые, с точки зрения Гегквиста (1931), представляют собой внутренние
сарколеммные прослойки, имеющие вид решеток и дающие некоторые красоч-
ные реакции коллагена, сквозь которые продернуты миофибрилли. Изу-
чение хода дифференцировки миосимпластов в культурах показывает,
384
Глава III
что диски Z могут в виде ясных тончайших полосок появляться сначала
по середине дисков I в отдельных миофибриллях или их столбиках. Если
.волокно достигает более высокого уровня дифференцировки, то они разго-
раживают более или менее значительную часть поперечника волокна или
даже все волокно целиком. Это подтверждает то, что инофрагмы пред-
ставляют собой не решето, а продукт дифференцировки в поперечном напра-
влении всего живого вещества волокна, т. е. сплошные, а не продырявлен-
ные пластинки, в состав которых входят как интерфибриллярные, так и фиб-
риллярные участки. Поэтому следует признать правильными те представле-
ния о природе инофрагм, которые были подробно развиты в свое время Гей-
Зенгайном (1911), а не упомянутую выше более новую точку зренияГегквиста.
Особого обсуждения заслуживает вопрос о делении ядер мышечных
волокон, при эмбриональном развитии которых сначала наблюдаются
митозы, а затем амитотические деления. Еще в настоящее время, несмотря
на огромный и убедительный фактический материал, доказывающий значение
набора хромозом в клетках и говорящий в пользу хромозомного строения
ядра в интеркинезе, многие признают, что амитотическое деление клеток
„может быть полноценным и заменять в известных случаях митоз. То. что
.происходит в мышечных волокнах, казалось бы, как раз подтверждает,
чго ядра, делившиеся амитотически, могут в дальнейшем вновь давать
нормальные митотические картины. С точки зрения современной кариоло-
«тии это не может не представляться парадоксальным. То, что при регене-
рации соматических мышечных волокон даже у млекопитающих могут
наблюдаться митозы, отмечали отдельные авторы (Левен, 1887; Науверк,
1890; Даусон, 1909; Бундшу, 1923, и др.). Большая часть авторов, рабо-
тавших с млекопитающими, однако, отрицает наличие митозов в мышечных
ядрах. Митозы в миогенных элементах при регенерации у низших позво-
ночных и у беспозвоночных описывали многие исследователи, в последнее
время, в частности, Казанцев (1935), Заварзин (1938), Жинкин (1938), и др.
В моих опытах, на ранних стадиях культивирования, довольно много-
численные несомненные митозы наблюдались в мышечных волокнах ново-
рожденных и молодых кроликов (до месячного возраста), т. е. в таком
возрасте, когда в мышечных волокнах предварительно уже должны были
иметь место, согласно господствующим взглядам, амитозы. Мне кажется,
:что эти факты можно примирить с хромозомной теорией строения ядер
,и даже, более того, использовать для правильного понимания природы
соматических мышечных волокон, без того чтобы допускать существование
особого «генеративного амитоза» (Беннингоф, 1922), могущего быть равно-
ценным митозу. .
С другой стороны, несомненно, что амитозы в мышечных волокнах еще
нуждаются в специальных исследованиях, тем более, что Навиллем (1922)
был отмечен их особый характер в мышечных волокнах хвоста головастика.
Разумеется, пока все процессы деления мышечных ядер не будут просле-
жены при жизни в их естественной последовательности, всякие толкования
будут неизбежно спорными и гипотетическими. Новейшие данные Спейделя
(1938) по прижизненному изучению дегенеративных и регенеративных
Экспериментальный метод в гистологии
385
процессов в мышечных волокнах личинок амфибий открывают и в этом
направлении большие возможности.
Весьма возможно, что в мышечных волокнах не все ядра проделывают
амитотические деления, а только более или менее значительная часть их.
Таким образом, в дифференцированных мионах, наряду с ядрами с иска-
женным в результате амитоза хромозомным набором, совокупность которых
в составе мышечного волокна обеспечивает его жизненные функции при
обычных условиях, если можно так выразиться, его «вегетативную» жизнь,
могут сохраняться отдельные, так сказать, «генеративные», или «камбиаль-
ные» ядра с полноценным набором хромозом. Они могут при измененных
условиях делиться вполне нормальными митозами. Только за счет таких
ядер мыслимо образование истинных пролиферирующих митотическим
путем миобластов. Отделившиеся же от массы мышечного волокна клетки
с неполноценными ядрами будут представлять собой лишь нежизнеспособ-
ные обломки мионов, обреченные в дальнейшем на гибель, по крайней
мере, если их изоляция не была лишь кратковременным и преходящим
явлением. Наконец, вполне мыслимо, что сохранение полноценных «гене-
ративных» ядер имеет место не в каждом мышечном волокне, а только
в некоторых из них. Тогда в составе дифференцированной соматической
мускулатуры не все мышечные волокна будут равноценны друг другу в
биологическом отношении. Было бы интересно, например, сравнить под
этим углом зрения друг с другом богатые и бедные миофибриллями волокна.
Весьма возможно, что при специальном изучении в мышечных волокнах
будет возможно обнаружить какие-нибудь закономерные морфологические
различия в ядрах. Возможно также, что эти гипотетические «генеративные»
ядра обнаружат, по крайней мере, в некоторых случаях, определенную
локализацию в волокнах, например, по концам их, отличающимся, по
многим данным, своей менее дифференцированной структурой. Мне уда-
лось найти одну литературную ссылку на малоизвестную работу Левенталя
(1899), который в нормальных мышцах различает 4 типа ядер и указывает
на их возможное неодинаковое функциональное значение. Специальное
изучение этого вопроса в сравнительном аспекте современными методами
представляет безусловно большой интерес. Различиями мышечных волокон
разных животных в отношении количества и распределения «генеративных»
ядер проще всего могут быть объяснены противоречия в вопросе об обра-
зовании саркобластов и мышечных почек при регенерации у разных пред-
ставителей позвоночных. Одним из подтверждений этой точки зрения являются,
в частности, последние данные Заварзина (1938), согласно которым у бычка
при повреждении мышечных волокон только часть их ядер обнаруживает
способность к делению митотическим путем, дру1ая же подвергается деге-
нерации. Таким образом, результаты, получаемые при наблюдении над
мышечными волокнами, могут быть полностью согласованы с той трактовкой
значения митоза и амитотических перешнуровок ядер, которая высказы-
валась мной неоднократно и была, в частности, развита в отношении элемен-
тов соединительной ткани (Хлопин, 1931). Вместе с тем следует признать,
что в мионах млекопитающих способность к митотическому делению гене-
25 Хлопин—349
386
Глава III
ративных ядер находится в подавленном состоянии. В условиях культи-
вирования без вырезания она проявляется лишь в течение первых дней
после эксплантации. Затем митозы исчезают безвозвратно, и при экстенсив-
ном росте миосимпластов в дальнейшем наблюдаются лишь амитотические
деления ядер. Это, однако, вовсе не значит, что митотическая активность
генеративных мышечных ядер не может быть при других условиях или под
влиянием других агентов стимулирована в значительно большей степени.
С этой точки зрения мионы соматической мускулатуры представляют
собой своеобразные симпластические тканевые системы, по крайней мере
часть которых и в дифференцированном состоянии, наряду с высокоспе-
циализированными морфологическими и функциональными особенностями,
обладает особым типом камбиальности, зависящим от наличия в их составе
генеративных ядернопротоплазматических участков. Однако у млеко-
питающих, в условиях репаративной регенерации и в наших опытах, на
первый план выступает лишь способность мышечных волокон к росту путем
увеличения массы саркоплазмы и ядер. Стойкое сохранение мышечными
волокнами своего симпластического характера зависит, вероятно, оттого,
что их саркоплазма и преобладающие количественно ядра с нарушенным
в результате амитозов хромозомным набором могут существовать длитель-
ное время только в виде единой целостной системы, а ее изолировавшиеся
ядернопротоплазматические участки, не обладающие полным хромозомным
набором, будут обладать лишь ограниченной жизнеспособностью.
На основании сопоставления выявляющихся при экспериментальных
условиях особенностей соматических мышечных волокон беспозвоночных
и различных позвоночных можно сделать вывод, что филогистогенез мышеч-
ных волокон позвоночных шел в сторону приобретения и все большего
закрепления свойств своеобразной целостной симпластической системы.
У млекопитающих свойства соматического мышечного волокна, как единой
системы, выражены, пожалуй, наиболее резко. Вместе с тем представляется
возможным трактовать возрастные изменения свойств мышечных волокон
позвоночных на разных этапах эмбрионального развития как пример
рекапитуляции эволюционных изменений гистологических особенностей.
Сопоставление изменений мышечных волокон при репаративной реге-
нерации у беспозвоночных и позвоночных, в первую очередь млекопитающих,
как и в случаях многих других тканей показывает и черты сходства и раз-
личия. Первые возможно трактовать как результат исторического развития
мускулатуры соматического типа из мышечных элементов отдаленных
общих предков, которое происходило параллельно и преемственно в отдель-
ных ветвях филогенетического дерева. Отмечающиеся различия, которые,
с моей точки зрения, нуждаются в более углубленном и детальном изуче-
нии, чем это имело место до настоящего времени, очевидно, следует отнести
за счет дивергентного развития мускулатуры соматического типа в разных
филогенетических рядах.
Во всяком случае, мне представляется односторонней, недостаточно
обоснованной и даже неправильной точка зрения, согласно которой прин-
ципиальное значение при регенеративных процессах в мышцах придается
Экспериментальный метод в гистологии
387
лишь отдельным сходным моментам, а все различия, часть которых
подчеркивалась выше, игнорируются, как нечто совершенно несу-
щественное.
Последним, что нуждается еще в кратком рассмотрении, являются
опухоли соматической мускулатуры млекопитающих и человека. Интересен
тот факт, что в результате внутримышечного введения канцерогенных
веществ удавалось вызвать экспериментально опухоли типа высокодиф-
ференцировапных рабдомиобластом, что косвенно подтверждает развитую
выше точку зрения на существование «генеративных» ядер в составе мышеч-
ных волокон соматического типа и доказывает их высокую способность
к разрастаниям после бластоматозного превращения. К сожалению, эти
наблюдения в гистологическом отношении очень неполны и поэтому не
позволяют детализировать некоторые напрашивающиеся естественным
образом выводы.
В человеческой патологии опухоли, развивающиеся из мускулатуры
соматического типа, играют существенную роль (Абрикосов, 1925). По
своему микроскопическому строению они очень разнообразны и только
более дифференцированные из них имеют настолько характерную попе-
речнополосатую структуру и некоторые другие особенности мышечных
волокон, что их гистогенез представляется достаточно несомненным. Даже
в одной и той же опухоли во многих случаях могут встречаться элементы
очень неодинакового уровня дифференцировки, одно- и многоядерные
клетки то с отчетливой, то с едва заметной продольной гладкой фибрил*
лярностыо, с полным отсутствием поперечной исчерченности или лишь
местами с заметной поперечнополосатой структурой (Абрикосов, 1925;
Раков, 1937).
В настоящее время все опухоли такого рода, независимо от присут-
ствия или отсутствия поперечной исчерченности, с полным основанием
рассматриваются как рабдомиобластомы. Однако, наряду с опухолями,
имеющими те или иные, хотя бы и слабо выраженные, признаки миоген-
ного происхождения, из соматических мышц развиваются и настолько мало-
дифференцированные новообразования, что их гистогенез не может быть
установлен на основании простого микроскопического изучения. Такие
опухоли имеют обычно характер веретеноклеточных или, пожалуй, чаще
полиморфноклеточных сарком. Такого же рода опухоль удалось получить
недавно моему сотруднику Ю- Петрову (1940) после введения дегтя в бедрен-
ную мышцу кролика. О гистогенезе такого рода новообразований возможно
высказывать лишь более или менее вероятные предположения. Если сопо-
ставить различные варианты опухолей телесной мускулатуры с тем, что
известно и было только что описано в отношении гистологических картин,
даваемых в условиях эксплантата эмбриональными и дифференцированными
мышечными волокнами млекопитающих, то разнообразие структур миоген-
ных опухолей делается совершенно понятным. Каждому варианту опухо-
левой структуры можно подобрать вариант соответствующего уровня
дифференцировки из нормальной ткани, поставленной в условия роста
и пролиферации.
388
Глава III
Сопоставление результатов, полученных в последнее время Тимофеев-
ским (1938, 1939) при эксплантации некоторых полиморфноклеточных сар-
ком человека с превращениями вне организма элементов соматической
мускулатуры {Хлопин, 1936, 1940), позволяет высказаться в пользу их
возникновения из детерминированного мышечного зачатка и включить их
с полной достоверностью в группу миогенных опухолей соматической муску-
латуры. Очень большого интереса заслуживают также последние данные
Тимофеевского (1940), согласно которым элементы одной малодифферен-
цированной веретеноклеточной саркомы человека на втором году жизни
вне организма дифференцировались в поперечно исчерченные миосимпласты
и тем самым выявили свою миогенную природу. Совершенно ясно, насколько
велико значение этих наблюдений для подтверждения проводимого в этой
книге учения о гистологической детерминации нормальных и опухолевых
тканей.
в) Мускулатура целомического типа
Хотя сердцем куриного зародыша, после основных работ Карреля и его
сотрудников, широко пользуются в качестве исходного материала для
опытов с тканевыми культурами и, в частности, для получения т. наз.
чистых штаммов «сердечных фибробластов», которые могут месяцами и
годами расти вне организма, относительно тканевой природы элементов
сердечных эксплантатов до настоящего времени имеются весьма существен-
ные разногласия. Очень многие авторы, работавшие с этим исходным мате-
риалом, в соответствии с точкой зрения Карреля, считают, что мускулатура
сердца даже у относительно молодых зародышей в условиях культур поги-
бает, а зона экстенсивного роста образуется исключительно за счет мезен-
химных интерстициальных элементов. В такой общей форме эту точку
зрения, однако, следует считать неправильной, так как ряд исследователей
(Леви, 1925, 1934; Льюисы, 1917; Оливо, 1925, 1929; Гог, 1937, и др.) с
несомненностью доказал превращение волокон сердечной мышцы куриных
зародышей в недифференцированные, энергично пролиферирующие и раз-
растающиеся клетки, т. наз. миобласты. Исходя из этого, Леви (1934) счи-
тает, что вечные фибробластообразные элементы, растущие в длительных
чистых культурах из сердца куриного зародыша, представляют собой не
клетки мезенхимного происхождения, а сердечные миобласты, сходные
с ними лишь по своему внешнему облику. Весьма возможно, что в ряде
случаев эксплантаты из сердца, даже при их длительном росте вне орга-
низма, несмотря на свой кажущийся однородный характер, представляют
собой не чистые культуры, а смесь недифференцированных элементов мио-
карда и клеток мезенхимного происхождения (У. Льюис, 19^3; Нишибе, 1920).
Миокард зародышей млекопитающих и новорожденных животных, кото-
рому посвящено лишь относительно незначительное количество работ
(Конгдон, 1915; Лейк, 1916; Дрю, 1922; Смирнова, 1928; Госс, 1932; Цымбал,
1937), также может энергично расти и пролиферировать вне организма.
Несомненно, кроме того, что элементы миокарда в отдельных эксплан-
татах и на разных этапах своего развития ведут себя неодинаково и даже
Экспериментальный метод в гистологии
389
в тех случаях, когда они оказываются жизнеспособными, наряду с ними
могут прорастать интерстициальные мезенхимные или соединительнотканные
клетки. Поэтому не исключена возможность, что соединительнотканные
элементы могут в течение времени постепенно заглушать рост миобластов
и оказаться, в конце концов, в чистой культуре.
По данным Льюисов (1917), прорастание миокарда в эксплантатах из
сердца куриных зародышей наблюдается лишь в части культур, в то время
как в другой части растут одни мезенхимные элементы. Рост миобластов
наблюдался Льюисами в 11 % случаев из сердца четырехдневного зародыша,
в 8 % шести-семидневного и лишь в 2 % из одиннадцатидневного зародыша
при культивировании в предложенной ими жидкой питательной среде.
Румянцев, описывавший первоначально (1922) образование недифферен-
цированных миобластов из волокон эмбрионального миокарда куриных
зародышей, позже (1927) отмечает, что в кратковременных культурах из
четырех-шестидневных зародышей начинающие прорастать волокна мио-
карда, не утрачивая полностью своей дифференцировки, вскоре дегене-
рируют и заглушаются энергично пролиферирующей мезенхимой. В куль-
турах из сердца двухдневного зародыша, по данным этого автора, мезен-
хима разрасталась настолько энергично, что миокард не обнаруживал
признаков роста и очень быстро погибал, что не соответствует результатам,
которые получил Оливо (1929) на том же материале. Он установил, что
энергия роста миокарда очень велика у зародышей и, постепенно ослабевая
после вылупления, падает до нуля у 25-дневных цыплят. Волокна сердечной
мышцы у них нацело дегенерируют, а жизнеспособной оказывается лишь
соединительная ткань. Наоборот, в эксплантатах из сердца ранних заро-
дышей, по крайней мере в течение первого времени, могут даже наблюдаться
явления прогрессивной дифференцировки.
Прорастание волокон сердечной мышцы зародышей кролика и человека
и новорожденных кроликов, утрату ими своей специфической дифферен-
цировки и превращение в энергично пролиферирующие большей частью
одноядерные клетки с несомненностью установил в последнее время Цымбал,
который вместе с тем отметил, что вскоре после рождения энергия роста
миокарда быстро падает. По его данным, в культурах из сердца десяти-
дневных кроликов элементы миокарда дегенерируют нацело, и наблюдается
лишь рост соединительнотканных элементов.
Изменения элементов миокарда в эксплантатах на разных этапах его
развития по наиболее тщательным и обоснованным данным представляются
следующим образом. В эксплантированных частицах сердца куриного
зародыша 24—30 часов после начала инкубации (6—16 сомитов), миоэпикар-
диальный плащ которого состоит еще из одного слоя ясно различимых
клеток кубической формы, с едва заметными гладкими фибриллями,
в течение первых дней можно наблюдать образование волокон миокарда,
состоящих из сетевидно соединенных друг с другом миобластов. В их про-
топлазме дифференцируются поперечно исчерченные миофибрилли. В таких
эксплантатах вместе с тем наблюдается самопроизвольная пульсация,
которая в исходном материале может или совершенно отсутствовать или
390
Глава III
происходит лишь в ограниченном участке сердечной трубки. Благодаря
митотическому делению ядер масса миокарда постепенно увеличивается
во всех трех измерениях. Таким образом, в эксплантатах из раннего эмбрио-
нального материала наблюдается, собственно говоря, не столько обычный
для тканевых культур экстенсивный рост, сколько увеличение всей массы
и тканевая дифференцировка эксплантированных частиц эмбриональной
закладки. По истечении 10—15 дней уже дифференцировавшиеся поперечно
исчерченные миобласты начинают подвергаться упрощению, вновь утра-
чивают миофибрилли и постепенно превращаются в индифферентные клетки,
образующие обычную зону экстенсивного роста.
Аналогичные явления дифференцировки несколько раньше наблюдал
Экман (1921, 1924) в изолированных закладках сердца зародыша амфибий,
продолжавших развиваться в течение некоторого времени внутри мешочков
из кожной эктодермы.
Рост миокарда более старых зародышей вне организма, в зависимости
от условий, может иметь неодинаковый характер. При культивировании
в обычной питательной среде, содержащей плазму и экстракт, волокна
миокарда куриных зародышей (Оливо), эмбрионов человека и кролика и
новорожденных кроликов (Цымбал) с дифференцированными поперечно-
полосатыми миофибриллями частью дегенерируют и распадаются, частью
постепенно упрощаются, утрачивая поперечную исчерченность и миофиб-
рилли, и превращаются в конце концов в недифференцированные, большей
частью одноядерные клетки, энергично делящиеся митотическим путем.
Эксплантированные частицы целиком или отдельными участками могут
довольно длительное время (до одного и даже до трех месяцев) продолжать
пульсировать. Миобласты, получающиеся в культурах из эмбрионального
куриного миокарда, могут терять взаимную синцитиальную связь до полной
утраты поперечной исчерченности и в таком состоянии продолжать
некоторое время пульсировать.
При культивирований в жидкой среде (Гог, 1937) без периодических
вырезаний частицы миокарда растут очень медленно, распластываясь по
поверхности стекла, причем большая часть его волокон сохраняет Длитель-
ное время (до полугода) свою дифференцировку. Индифферентной зоны
экстенсивного роста при этом, судя по результатам, полученным автором,
строго говоря, почти не образуется, а происходит, хотя и в атипичном
виде, увеличение всей массы эксплантированного кусочка миокарда. Таким
образом, при известных условиях волокна миокарда могут долгое время
сохранять вне организма свою специфическую дифференцировку. Стилуэл
(1938) отмечал дифференцировку миокарда в эксплантатах из сердца восьми-
дневных куриных зародышей. Вторичной дифференцировки подвергшихся
упрощению и пролиферирующих миобластов до настоящего времени никому
с достоверностью получить не удалось. Поэтому Оливо считает, что обра-
зующие зону роста недифференцированные сердечные миобласты предста-
вляют собой клетки, отличающиеся известной неполноценностью от настоя-
щих эмбриональных миобластов, способных к образованию дифференци-
рованных структур миокарда, а процесс упрощения растущей вне организма
Экспериментальный метод в гистологии 391
сердечной мышцы тем самым не представляет собой возврата к эмбриональ-
ному состоянию. Указания на вторичную дифференцировку сердечных
миобластов, имеющиеся в работе Госса (1932), недостаточно убедительны.
Весьма возможно, что этот автор имел дело не с вторичной дифференци-
ровкой, а с сохранением дифференцированных участков миокарда в составе
бывшего центрального кусочка эксплантата.
Чрезвычайно трудно, как это ни удивительно, дать правильную гисто-
логическую характеристику зоны роста, образуемой недифференцирован-
ными сердечными миобластами куриного зародыша. Принято считать,
что, после нескольких пересевов, зоны роста сердечных эксплантатов при-
обретают типичный мезенхимоподобный вид, не отличимый от соединительно-
тканных культур, и состоят из сетевидно связанных отростками фибробла-
стообразных элементов, которые большей частью прямо обозначаются
как фибробласты или как сердечные фибробласты. В ряде случаев, если
принять во внимание упомянутые данные Льюисов, весьма вероятно, что
имеющиеся в составе сердца элементы соединительной ткани или совершенно
заглушают рост миобластов или растут в преобладающем количестве.
Тогда, естественным образом, зона роста будет действительно иметь вполне
мезенхимный или соединительнотканный вид. Вместе с тем имеется ряд
указаний на то, что структуры зоны роста сердечных эксплантатов могут
давать варианты, не свойственные обычным соединительнотканным куль-
турам. Люъис отмечал прорастание сердечных миобластов компактными
тяжами. Он же указывал (1923), что сетевидные структуры, образуемые
сердечными миобластами, могут приобретать вид мезотелиальных мембран.
По его же данным, это, однако, несвойственно мезенхиме других, изучен-
ных им, органов. Из своих очень точных и прекрасно иллюстрированных
наблюдений он даже сделал несомненно неправильный вывод, что «мезен-
хима» может превращаться в мезотелий и наоборот, причем под сердечной
мезенхимой он, очевидно, понимал, на основании совершенно поверхно-
стного формального сходства, сетевидные структуры, образуемые сердечными
миобластами. Значительно позже Гросфельд (1931) отмечал, что зона роста
сердечных эксплантатов имеет, в зависимости от внешних условий, то соеди-
нительнотканный вид, то характер настоящих «эпителиальных» мембран.
Последнее, по его данным, подвергнутым критике Грюнбергом (1932),
наблюдается, как правило, если «сердечные фибробласты» в условиях жид-
кой питательной среды стелются по поверхности стекла. Наблюдавшиеся
изменения в структуре зоны роста Гросфельд использовал для доказатель-
ства того, что эпителий может превращаться в мезенхиму и наоборот,
понимая под «эпителием» не какую-нибудь определенную ткань, а любое
плоскостное клеточное соединение, состоящее из тесно сомкнутых уплощен-
ных клеток, а под мезенхимой, очевидно, любые сетевидные структуры.
€ такой формальной трактовкой гистологических структур, как это неодно-
кратно отмечалось, в настоящее время уже невозможно согласиться, хотя
она и теперь еще не является редкостью. Оливо также указывает на то,
что сердечные миобласты могут образовать эпителиообразные струк~
туры.
392
Глава Ill
В своей большой работе, посвященной образованию т. наз. макро-
фагов из «фибробластов», Тома (1937) приводит микрофотографию
культуры из сердца куриного зародыша, которая в точности соответствует
описанным мною (1936, 1937) мезотелиальным мембранам с характерными
для них ретракцией и округлением отдельных мезотелиальных элементов.
Эти данные он неправильно трактует как пример превращения фибробластов
в макрофаги.
Все эти факты, касающиеся эксплантатов из сердца куриного зародыша,
дше кажется, получают правильную трактовку в работе Цымбала (’937),
посвященной миокарду человека и кролика. Следует отметить, что клеточ-
ные элементы млекопитающих являются, благодаря своим крупным раз-
мерам и лучшей окрашиваемости в препаратах, гораздо более удобным
объектом для решения вопросов гистологического характера.
Анализируя различные варианты зоны роста в сердечных эксплантатах,
нельзя не притти к выводу, что миокард растет не по соединительнотканному,
а по мезотелиальному типу. Подобно мезотелию, он растет комплексными
мембранами, реже тяжами и более рыхлыми сетевидными клеточными
соединениями. Более того, мембраны из сердечных миобластов по ряду
морфологических особенностей оказываются сходными, подчас до полного
тождества, с мембранами, образуемыми в условиях культур покровными
клетками серозных оболочек. При сопоставлении картин, изученных Цым-
балом, мне кажется, что это сходство бывает тем полнее, чем моложе был
исходный материал. Принимая во внимание необыкновенно интенсивный
рост и пролиферацию куриной мезенхимы, которая (по большей части
литературных данных) заглушает рост многих других тканей, что является
особенностью, несвойственной в такой степени соединительной ткани млеко-
питающих, эти результаты нельзя безоговорочно переносить на эксплантаты
из сердца куриных зародышей. Все сказанное заставляет считать, что
эксплантаты из сердца куриных зародышей в отношении особенностей
строения зоны роста нуждаются в каждом отдельном случае в более тщатель-
ном гистологическом анализе, чем это имело место до настоящего времени,
Возможно, что они или, по крайней мере, многие из них до сих пор лишь
по недоразумению фигурируют в качестве образцов мезенхимального типа
роста, подобно тому, как растущие по нейроглиальному типу ретинальные
листки неправильно принято считать образцом эпителиальных культур.
Изучение физиологических особенностей т. наз. чистых клеточных штам-
мов из сердца куриного зародыша тоже показывает с большой убедитель-
ностью их неравноценность с чистыми культурами фибробластообразных
элементов, полученных из соматической мускулатуры, скелетогенных
участков мезенхимы или ретикулярной соединительной ткани.
Миофибрилли сердечной мышцы несомненно представляют собой реально
существующие прижизненные структуры (Леви, 1934), а не образуются,
как это предполагали Льюисы, в результате фиксации. Точка зрения,
согласно которой поперечная исчерченность зависит от более или менее
правильно расположенных хондриозом (Льюисы, Госс), также не представ-
ляется вероятной. Утверждение Леви, что миофибрилли сердечной мышцы
Экспериментальный метод в гистологии
393
при своем исчезновении в культурах превращаются в хондриозомы, не-
достаточно обоснованы. Нужно думать, что в миокарде, как и в сомати-
ческой мускулатуре, между хондриозомами и миофибриллями непосред-
ственной, устанавливаемой микроскопическим наблюдением, морфологи-
ческой преемственности не имеется. В культурах из сердечной мускулатуры,
особенно в условиях замедленного роста, уже давно было отмечено ново-
образование межклеточного вещества в виде сетей так называемых арги-
рофильных волокон. Отмечалось оно и в «чистых штаммах», растущих
в лаборатории Карреля в течение многих лет (Оливо). По исследованиям
Цымбала (1937), аргирофильные волокна несомненно могут образоваться
сердечными миобластами млекопитающих, растущими не по мезенхималь-
ному, а по мезотелиальному типу. Эти факты, а также упомянутое выше
образование волокнистого межклеточного вещества нейроглией и клетками
ретинальных листков показывают, что способность к образованиям арги-
рофильных волокон не является отличительной особенностью лишь мезен-
химных производных, а элементарным свойством тканей, имеющим довольно
широкое распространение. По еще неопубликованным работам нашей
лаборатории, аргирофильные волокна могут также образоваться в условиях
эксплантата мезотелием серозных оболочек (Хлопин) и сертолиевскими
клетками семенника (Михайлов). Недавно появившаяся в печати интересная
работа Румянцева и Березкиной (1937) показывает, что по своей способ-
ности к образованию межклеточного вещества клетки, растущие в эксплан-
татах из сердечной мышцы куриных зародышей, резко отличаются от
элементов мезенхимы эмбриональной аорты и скелетогенных участков,
а также соединительной ткани кожи. Мезенхимные и соединительнотканные
клетки, как это подробнее отмечалось выше, при известных условиях обла-
дают способностью образовать вне организма не только аргирофильные
волокна, но и другие виды межклеточных веществ. По данным же Румянцева
и Березкиной, в эксплантатах из сердечной мускулатуры не удается вызвать
образования никаких других межклеточных структур; количество ново-
образованных аргирофильных волокон в сердечных эксплантатах, как
правило, бывает относительно невелико. Это отметил и Оливо при изучении
длительных клеточных штаммов Карреля, выделенных из частиц эмбриональ-
ного сердца.
Эксплантированные частицы сердца, в зависимости от условий культи-
вирования и возраста исходного материала, могут продолжать более или
менее длительное время пульсировать вне организма. Нередко самостоя-
тельная пульсация временно приостанавливается, чтобы затем возобно-
виться. При эксплантации недифференцированной закладки раннего эмбрио-
нального сердца, которое еще не сокращалось, параллельно с ее дифферен-
цировкой в культурах, появляются самостоятельные сокращения (Оливо).
Сердечные миобласты, утратившие при экстенсивном росте и пролиферации
свою дифференцировку, совершенно утрачивают способность к сокращению.
Вместе с тем, однако, сократимость не связана обязательно, как в случае
соматической мускулатуры, с наличием обнаруживаемых гистологическими
методами поперечно исчерченных миофибриллей. Способност ью к сарко-
394
Глава III
плазматическому сокращению обладают, начиная с определенного этапа
развития, миокард ранней сердечной закладки и его частицы, утратившие
поперечно исчерченные миофибрилли, но сохранившие еще свой сетевидный
синцитиальный характер (Оливо, Цымбал и др.).
Прибавлением к питательной среде хлористого кальция удается вызвать
вне организма фибриллярные подергивания в элементах миокарда, сохра-
нивших свои миофибрилли, но не пульсирующих самостоятельно; в эле-
ментах же миокарда, не содержащих миофибриллей, хлористым кальцием
не удается вызвать подергиваний (Оливо).
При культивировании частиц эмбриональной сердечной мышцы в таких
условиях, когда экстенсивный рост выражен слабо и утрата дифферен-
цировки длительное время не наступает, самостоятельное сокращение
наблюдалось до семи месяцев жизни вне организма (Оливо).
По данным Берроуза (1913), эксплантированные частицы желудочков
сердца куриных зародышей возрастом от шестидесяти часов до десяти
дней пульсируют ритмически с частотой 50—150 ударов в минуту, а частицы
предсердия зародышей любого возраста и вылупившихся цыплят -— с часто-
той 150—220 ударов. Эксплантаты из желудочков более старых куриных
зародышей не пульсируют. Нордман и Рютер (1931) на эмбриональном
курином и крысином материале также показали, что предсердие пульси-
рует чаще, чем желудочки. Между частицами, взятыми из разных участков
желудочков, в отношении ритма тоже наблюдаются различия. Отдельные
участки одного и того же кусочка могут сокращаться с неодинаковой часто-
той. Фишер (1925, 1930) отмечает, что в некоторых случаях при совместном
культивировании двух частиц сердца, пульсирующих с различной частотой,
после их срастания может наступать синхронность сокращений. В других
случаях, наоборот, срастание двух ритмически пульсирующих эксплантатов
приводило к появлению аритмии. При срастании двух частиц эмбриональ-
ного сердца, из которых одна принадлежала куриному, а другая утиному
зародышу, синхронности сокращений никогда не наступало.
Анализ гистологических превращений миокарда представляет интерес
в различных отношениях. Прежде всего он показывает, что излюбленный
объект для всякого рода физиологических исследований (культуры из сердца
куриного зародыша) является далеко не безупречным в смысле ясности
своей тканевой природы. Неизвестно также, с одинаковыми ли по проис-
хождению клеточными штаммами имели дело разные исследователи, и в
какой степени так называемые чистые культуры фибробластов из сердца
куриного зародыша являются в действительности однородными. Весьма
возможно, что данные различных авторов, не подвергавших свои эксплан-
таты достаточно тщательному гистологическому изучению, относятся частью
действительно к чистым культурам элементов мезенхимного происхождения,
частью к недифференцированным сердечным миобластам, частью к куль-
турам смешанного характера.
В высшей степени интересным является тот факт, что соматическая и
сердечная мускулатура, обе построенные по поперечнополосатому типу,
ведут себя у высших позвоночных в условиях культур различным образом.
Экспериментальный метод в гистологии 395
Волокна первой, с известных этапов эмбрионального развития, прорастают
своеобразными миосимпластами; миокард же, пока он сохраняет способ-
ность к экстенсивному росту вне организма, превращается в энергично
пролиферирующие недифференцированные сердечные миобласты, но мио-
симпластов не образует. По своему общему характеру клеточные соеди-
нения сердечных миобластов обнаруживают во многих случаях несомненное
сходство с растущим вне организма мезотелием серозных оболочек. Послед-
ний факт приобретает особое значение потому, что и мезотелий и сердечная
мышца являются производными одной и той же эмбриональной закладки —
спланхнотомов или выстилки полости тела зародыша. В то время как
мышечные волокна соматического типа приобретают в конце концов у высших
позвоночных характер совершенно своеобразных тканевых элементов,
в миокарде, несмотря на его сетевидное синцитиальное строение, сохра-
няется расчленение на ядерно-протоплазматические территории, которые
могут в условиях эксплантата превратиться в делящиеся митотическим
путем малодифференцированные одноядерные клетки. Таким образом, метод
эксплантации позволяет обнаружить большую разницу между этими сокра-
тимыми тканями, построенными по поперечнополосатому типу, и заставляет,
в соответствии с рядом фактов из области сравнительной морфологии,
считать их представителями двух тканевых типов, которые имеют различ-
ную историю развития, но приобрели черты значительного морфологи-
ческого и функционального сходства, сохранив вместе с тем глубокие,
выявляющиеся при экспериментальных условиях, биологические различия.
Для сопоставления с данными, полученными методом эксплантации,
почти не имеется фактов из области превращений волокон дифференцирован-
ного миокарда при экспериментальных условиях в составе целого организма.
Н. Аничков (1912), давший непревзойденное в отношении точности опи-
сание воспалительных изменений в сердечной мышце, вызванных экспери-
ментально, не отмечает заметной пролиферативной реакции волокон мио-
карда в течение воспаления, при котором в них преобладают дистрофические
и дегенеративные процессы. Тем не менее этот автор отметил появление
в очаге воспаления своеобразных довольно крупных клеток, которые воз-
можно образуются из измененных волокон сердечной мышцы. Эти элементы
могут, не обнаруживая прогрессивных изменений, более или менее длитель-
ное время сохраняться в области образовавшегося рубца. Опубликованные
недавно наблюдения Тёрё (1939) над стимулирующим действием экстракта
из сердечной мышцы зародышей на элементы миокарда взрослых животных,
в котором, по его данным, удавалось наблюдать митозы, заставляют наде-
яться, что в дальнейшем удастся изучить гистологические картины проли-
феративных изменений волокон взрослой сердечной мышцы и в условиях
целого организма, а также получить на разных объектах их разрастания
в эксплантатах.
В миокарде в очень редких случаях описывалось образование опухоле-
подобных узелков, т. наз. кордобластом, повидимому, обнаруживающих
заметные структурные отличия от опухолей мускулатуры соматического
типа. Настоящих злокачественных новообразований сердечной мышцы
396
Глава III
неизвестно. Сравнительный экспериментально-гистологический анализ мио-
карда в ряду позвоночных и у разных представителей беспозвоночных
является еще делом будущего.
В отношении сходства, обнаруживаемого разрастаниями в культурах
волокон сердечной мышцы, мезотелия и фолликулярных клеток гонад
млекопитающих, необходимо добавить еще следующее замечание. В настоя-
щее время этот факт можно объяснить тем, что все эти гистологические
структуры развились дивергентно из одного и того же источника, примитив-
ной целомической выстилки общих предков позвоночных, и вместе с тем.
приобрели значение различных тканей внутренней среды организма.
г) Мезенхимная мускулатура позвоночных
Гладким мышцам посвящено относительно небольшое количество работу,
сделанных методом эксплантации, результаты которых довольно противо-
речивы. Согласно имеющимся данным, гладкомышечные клетки некоторых
органов куриного зародыша (амниотической оболочки — М. Льюис, 1920;
М. и У. Льюисы, 1917; мышечного желудка — Эрдман, 1928), ново-
рожденных и взрослых кроликов (мочевого пузыря и артерий — Шампи,
1913; желудка — Хлопин, 1936; Цымбал, 1938) могут, делясь митотическим
путем, прорастать экстенсивно и образовать зону роста соединительноткан-
ного типа. В то время как Шампи считал, что мышечные клетки при этом
приобретают полное морфологическое тождество с обычными фибробласто-
образными элементами соединительной ткани, другие авторы отмечают
сохранение ими известных отличий от соединительнотканных клеток.
При более детальном изучении общий характер зоны роста и морфологи-
ческие особенности экстенсивно растущих гладких мышечных клеток
в ряде случаев могут обнаруживать некоторые особенности (Льюисы, Эрдман,
Леви и Буччанте, 1928; Цымбал). Благодаря довольно правильной, нераз-
ветвленной или малоразветвленной, вытянутой в длину форме и преиму-
щественно радиальному направлению растущих гладкомышечных элемен-
тов, зона роста, имеющая в общем мезенхимоподобный сетевидный характер,
отличается своим, как бы игольчатым видом (Эрдман, Цымбал). Вместе
с тем проросшие гладкомышечные клетки, особенно на периферии зоны
роста, могут, уплощаясь, приобретать, как и элементы другого происхо-
ждения, морфологически не отличимый от соединительнотканных фибро-
бластообразных элементов вид, что, однако, как и в других аналогичных
случаях, не может быть истолковано в смысле полной утраты ими своих
специфических биологических особенностей. Если рассматривать глад-
комышечную зону роста во всех ее вариантах, а не ограничиваться одними
лишь местными особенностями того или иного участка, то она несомненно
по совокупности своих гистологических структур несколько отличается
от обычных соединительнотканных культур, и, что особенно интересно,
совершенно не похожа на картины экстенсивного роста мышечных тканей
другого происхождения, в частности, — гладких сократимых элемен-
тов радужины, развивающихся из нейрального зачатка и т. наз.
Экспериментальный метод в гистологии
397
миоэпителиальных клеток некоторых желез. Вместе с тем пока предста-
вляется не вполне ясным, в какой степени гладкомышечные клетки разных
органов по характеру образуемой ими зоны роста сходны друг с другом.
В частности, это относится к гладкой мускулатуре амниотической оболочки.
В этом отношении необходимы дальнейшие систематические исследования.
Делясь митотическим путем и прорастая, гладкомышечные клетки могут,
как это отмечалось еще Шампи, упрощаться и утрачивать миофибрилли.
Однако миофибрилли или сходная с ними фибриллярная дифференцировка
протоплазмы отмечаются и в гладкомышечных элементах зоны роста (Льюисы,
Леви и Буччанте, Цымбал), протоплазма которых отличается также своим
сильным лучепреломлением. Фибрилли, в противоположность данным
Льюиса, видны не только после фиксации, но и в живых клетках, причем
они располагаются по периферии клеточного тела и по своим свойствам
резко отличаются от нитевидных хондриоконт (Леви, 1934). Гладкомышечные
элементы желудка отличаются довольно сильными фибринолитическими
свойствами. В таких эксплантатах нередко образуются полости разжижения.
При культивировании без периодических вырезаний в гладкомышечной
зоне роста образуется густая сеть аргирофильных волокон межклеточного
вещества (Цымбал).
Гладкомышечные элементы центрального кусочка и зоны роста могут
порознь или целыми группами обнаруживать медленные ритмические
сокращения, которые специально изучали М. Льюис (1920), а в новейшее
время Нордман, Бакстер и Весле (1934).
В культурах из бронхов куриного зародыша сокращения наблюдались
до двух недель жизни вне организма, останавливаясь при охлаждении
культур до 36° (Льюис). При 37° наблюдалось одно сокращение в минуту,
при 38° — одно в 30 секунд, а при 40v — одно в 15 секунд. По данным
Нордман, Бакстер и Весле (1934), сокращения в эксплантированных части-
цах желудка и кишечника куриных зародышей, взятых из разных отделов,
отличались неодинаковой частотой и зависели от количества нервных
элементов ауэрбахова сплетения в данном участке кишечной стенки. Как
уже упоминалось выше, медленные перистальтические сокращения наблю-
дались до месяца и более в нацело эпителизированных культурах из кишеч-
ника куриного и человеческого зародышей (Фишер, 1923; Хлопин, 1934).
Большого внимания заслуживают последние наблюдения Цымбала (1941)
над превращениями вне организма гладкой мускулатуры желудка человека.
Полученный оперативным путем материал эксплантировался во фла сонах
и изучался в течение трехмесячного срока жизни вне организма. Волокна
взрослой мезенхимной мускулатуры обнаружили резко выраженную спо-
собность к пролиферации митотическим путем и образовали зону роста
описанного выше типа, напоминающую разрастания соединительной ткани,
но вместе с тем отличающуюся от них вполне определенным своеобразием.
Автор отмечает также ряд деталей, которых ему не удалось обнаружить
раньше при эксплантации мускулатуры желудка кроликов.
При сопоставлении с изложенными данными, касающимися изучения
мускулатуры соматического типа и сердца, бросаются в глаза чрезвычайно
398
Глава III
резкие различия гистобластических потенций всех этих сократимых тканей,
которые, очевидно, нельзя не поставить в связь с их развитием из разных
источников, неодинаковым филогенетическим возрастом и с разными прой-
денными путями исторического развития.
Из фактического материала, касающегося пролиферативных процессов
и дифференцировки гладких мышечных волокон в составе организма,
я остановлюсь лишь на некоторых наблюдениях. .
Очень отчетливую и резко выраженную пролиферацию гладкой муску-
латуры удалось наблюдать при экспериментальном вызывании внутри-
брюшных спаек у кроликов (Верещинский, 1925). Гладкомышечные клетки,
обнаруживающие миофибриллярную дифференцировку, разрастались целы-
ми потоками и участвовали в образовании спаек между органами брюшной
полости.
Как отмечалось выше, Штиве (1926) описывал новообразование
в беременной матке гладкой мускулатуры из малодифференцированных сое-
динительнотканных клеток и даже лимфоидных элементов крови. Эти
наблюдения не подтвердились в лаборатории Фишер-Вазельса (1932), вслед-
ствие чего возникла острая полемика. Повидимому, более или менее значи-
тельная часть этих данных нуждается еще в пересмотре. Наблюдения
Щелкунова (1935, 1936) над изменениями в стенках артерий при перевязках
и образовании окольного кровообращения у кроликов убедительна дока-
зывают участие малодифференцированных соединительнотканных клеток
подэнтотелиального слоя в новообразовании гладкомышечных элементов
и эластических волокон среднего слоя сосудистой стенки. Способность
к пролиферации гладкомышечных клеток мочеточника отметил недавно
Лежаеа (1940) при перевязке этого органа.
Здесь уместно также упомянуть о наблюдениях Кери (1921), хотя они
и не подтвердились позднейшими исследованиями, сделанными в лабора-
тории Фишер-Вазельса. Кери подвергал мочевой пузырь щенков ритми-
ческому растяжению и сокращению, раздувая его с помощью особого при-
бора физиологическим раствором. После более или менее длительных опытов
он изучил изменения в мышечном слое и обнаружил в его элементах появ-
ление поперечной исчерченности.
Безотносительно к тому, как относиться к точности этих наблюдений,
следует сделать следующую оговорку. Даже если бы наблюдения Кери
в дальнейшем подтвердились, их нельзя было бы трактовать, как это иногда
делалось, как пример метаплазии, т. е. превращение гладкомышечных эле-
ментов пузыря в совершенно другую дифференцированную ткань. Разу-
меется, здесь нет никаких оснований говорить о переходе мезенхимной
мускулатуры в мускулатуру соматического типа или в волокна сердечной
мышцы. В этом случае можно было бы лишь утверждать (если думать,
что наблюдения Кери достаточно точны), что гладкая мускулатура, имеющая
в мочевом пузыре, по всем данным, энтомезенхимное происхождение, может
приобретать при особых функциональных условиях поперечнополосатое
строение, что было бы достаточно интересно и без дальнейших необосно-
ванных выводов. 1
Экспериментальный метод в гистологии 399
Наблюдения над гистологическими особенностями гладкомышечных опу-
холеподобных разрастаний, в частности, очень точные и технически совер-
шенные патолого-гистологические исследования Глазунова (1934) над фибро-
миомами человеческой матки, также подчеркивают генетическую близость
гладкой мускулатуры внутренних органов и соединительной ткани. Гла-
зунову удалось установить, что изученные им опухоли матки образуют
непрерывный ряд, на одном конце которого располагаются миомы, а на
противоположном — фибромы. Между теми и другими имеются всевоз-
можные переходы с преобладанием или гладкомышечных или фиброматоз-
ных особенностей. В составе этих опухолей элементы, имеющие морфоло-
гические признаки гладкомышечных клеток, обнаруживают способность
к образованию не только малодифференцированных аргирофильных, но
и настоящих коллагеновых волокнистых структур.
Все эти данные хорошо согласуются с изложенной выше точкой зрения,
что гладкая мезенхимная мускулатура и ее опухолевые производные у позво-
ночных представляют собой развившуюся дивергентно специализированную
разновидность волокнистой соединительной ткани и что на основании
совокупности своих биологических свойств она чрезвычайно резко отли-
чается от мускулатуры соматического типа и от миокарда.
д) Мионейральные элементы радужины
Для обоснования естественной системы мышечных тканей очень боль-
шое значение имеют исследования моего сотрудника Винникова (1936,
1938) над сократимыми элементами радужины новорожденных и взрослых
кроликов, произведенные методом тка-
невых культур.
При эксплантации частиц радужины,
содержащих ретинальные листки и т.
наз. сократимую мембрану Бруха-
Генле, состоящую из миофибрилляр-
ных частей элементов мышцы, расширя-
ющей зрачок, Винникову (1936) удалось
детально проследить превращения «мио-
эпителиальных клеток» последней.
Клеточная структура наружного рети-
нального листка и мембраны Бруха-
Генле к третьему дню культивирования
Рис. 62. Изменения наружного рети-
нального листка радужины кролика
на 3-й день культивирования. Так на-
зываемая мембрана Бруха-Генле под-
разделяется на клеточные отростки.
Сильное увеличение. (По Винникову,
1936.)
выявляется совершенно отчетливо (рис. 62). Сократимые миофибриллярные
части клеток мышцы, расширяющей зрачок, утрачивают свое параллельное
Друг другу радиальное направление и превращаются в отдельные отростки,
связь которых с ядросодержащей частью клеток прослеживается совер-
шенно отчетливо. В результате этих изменений получаются картины,
сходные с известными стадиями развития этой мышцы, описанными Гер-
цогом (1902). Затем остатки мембраны Бруха совершенно исчезают, и наруж-
ный ретинальный листок приобретает такое же строение, что и внутренний.
400
Глава III
Элементы того и другого образуют единую характерную зону роста описан-
ного выше нейроглиального типа.
Изменения мышцы, сужающей зрачок, которая, по Гегквисту (1931),
имеет типичный «мезенхимный» характер, прослеженные Винниковым
в специальной работе, протекают следующим образом. Миофибриллярная
дифференцировка ее элементов постепенно исчезает в течение четырех-
пяти первых дней после эксплантации. Если материал был взят от неаль-
бинотических животных, то параллельно с исчезновением миофибриллей
в сократимых клетках резко нарастает количество пигментных включений,
свойственных ретинальным клеткам и образующемуся из них у зародышей
«колбовидному органу», т. е. зачатку мышцы, сужающей зрачок. Получается
впечатление, что эмбриональные процессы в экспериментальных условиях
как бы начинают итти вспять, однако их дальнейшее течение уже приоб-
ретает совершенно особое, свойственное условиям тканевых культур тече-
ние. Во все отчетливее и отчетливее отграничивающихся друг от друга
клетках мышцы появляются митозы. Аргирофильные волокна, свойствен-
ные нормальной дифференцированной мышце, исчезают, не обнаруживая
изменений, свойственных в тех же условиях аргирофильным волокнам
мезенхимного происхождения, которые в ряде случаев приобретают харак-
тер коллагеновых структур. Затем измененные мышечные клетки, утратив-
шие свою специальную дифференцировку, энергично разрастаются экстен-
сивно, образуя все многообразие структур, подробно описанных выше
и свойственных ретинальным листкам сетчатой оболочки, а также ряду
других производных нервного зачатка. Зона роста имеет на разных стадиях
культивирования совершенно типичный нейроглиальный характер в той
его модификации, которая прежде всего свойственна различным другим
частям сетчатой оболочки и нейроглии зрительных путей. Если материал
относился к альбиносам, то пигментообразования в эксплантатах не проис-
ходит, и зона роста, лишенная пигмента, совершенно утрачивает свою
единственную цитотипическую особенность, позволяющую безошибочно
отличать ее от эпендимо-глиальных разрастаний, которым несвойственно
пигментообразование.
Картины, характеризующие мышцу, сужающую зрачок, совершенно
безошибочно отличаются с первого взгляда от описывавшихся выше раз-
растаний мезенхимных гладких мышечных волокон, миокарда и муску-
латуры соматического типа.
Не подлежит сомнению, что экспериментально-гистологический анализ
позволяет с поразительной отчетливостью вскрыть истинную тканевую
природу элементов, обнаруживающих при обычных условиях высокую
степень специальной конвергентной дифференцировки мышечного характера.
Имеющиеся, правда, очень скудные и неполные, литературные данные
(Мейер, 1936), касающиеся поперечнополосатой мышцы, сужающей зрачок
куриных зародышей, не позволяют сделать определенного вывода об осо-
бенностях ее превращений вне организма. Неопубликованные наблюдения
Винникова показывают, что эти элементы, утрачивая поперечную исчер-
ченность и миофибриллярную дифференцировку, частью сразу прорастают
Экспериментальный метод в гистологии
401
отдельными одноядерными клетками, частью образуют первоначально
малодифференцированные многоядерные протоплазматические тяжи, обна-
руживающие сходство с миосимпластами соматической мускулатуры.
Однако в данном случае эти образования оказываются лишь промежуточным
этапом к постепенному превращению в одноядерные клетки, которые дают
и на курином материале зону роста, тождественную с разрастанием рети-
нальных листков и совершенно явственно отличающуюся от картин, харак-
теризующих все известные ткани другого происхождения. Таким образом,
данные, полученные в последнее время на птичьем материале, полностью
подтверждают все сказанное выше. Вместе с тем, мне кажется, не исклю-
чена возможность, что дальнейшие исследования с более детальным учетом
возрастного фактора позволят обнаружить у многоядерных поперечно-
полосатых мионейральных волокон птиц какие-нибудь специфические
особенности, связанные с их очень высокой специализацией. Во всяком
случае тщательное сравнение мионейральной мускулатуры радужины
птиц с соматическими мышечными элементами тех же животных,-в част-
ности,вотношениидеталейгистогенеза, еще является одной из задач будущего.
е) Миоэпиде рмалъные элементы
Относительно миоэпидермальных элементов молочной и слюнных желез
здесь остается повторить с некоторыми дополнениями наблюдения, которых
нам неизбежно пришлось коснуться при изложении результатов экспери-
ментально-гистологического анализа эпидермальных тканей. Таких наблю-
дений не очень много, однако, они, как мне кажется, с полной отчетливостью
устанавливают тканевую природу этих элементов и подтверждают воочию
их положение в естественной системе гистологии. В условиях тканевых
культур миоэпидермальные корзинчатые клетки секреторных отделов
молочной {Максимов, 1925) и околоушной (Цымбал, 1932) желез, а также,
повидимому, вставочных отделов и слюнных трубок последней, уже вскоре
после эксплантации набухают и перестают отличаться от изменяющихся
прочих эпителиальных клеток. Утратив свою первоначальную дифферен-
цировку, бывшие миоэпителиальные, железистые и прочие элементы желе-
зистых ходов, ни в чем не отличаясь друг от друга и пролиферируя, обра-
зуют все многообразие свойственных эпидермальным тканям структур
зоны роста и эпителизирующих поверхность эксплантата пластов. Специаль-
ная дифференцировка корзинчатых клеток оказывается особенно непостоян-
ной и быстро исчезающей, причем обнаруживается их одновременное камби-
альное значение в составе железистых ходов.
Еще отчетливее это прослежено в настоящее время при воспалительных
разрастаниях слюнных желез после инъекции в них каменноугольного
дегтя (Искра, 1938) и в имплантатах с кусочками целлоидина (Лазаренко,
1934). И в тех и в других опытах корзинчатые клетки главных, исчерчен-
ных, т. е. слюнных трубок и, повидимому, также вставочных отделов набу-
хали, превращались в клетки обычного эпителиального вида и, пролифе-
рируя митотическим путем, в своих превращениях ничем не отличались
26 Хлопин—349
402
Глава III
от базальных элементов междольковых протоков и других, сохраняющих
свою жизнеспособность клеток железистых ходов.
Из всех пролиферирующих эпителиальных элементов образовались
описанные в соответствующей главе характерные малодифференцированныс
эпидермальные разрастания, из которых во многих случаях возникала
многослойная стенка кистовидных полостей, содержащих в просвете инород-
ные тела (капли дегтя или кусочки целлоидина) с продуктами тканевого
распада. Лазаренко отмечал, в частности, что ему приходилось наблюдать
полную дегенерацию исчерченных эпителиальных элементов слюнных
трубок и одновременно интенсивные пролиферативные процессы в изме-
ненных корзинчатых клетках. Все эти данные с полной убедительностью
демонстрируют эпидермальную природу корзинчатых клеток, на свойства
которых мышечноподобная дифференцировка не накладывает сколько-
нибудь глубокого отпечатка и не изменяет заметным образом их биологи-
ческих свойств. Миоэпителиальные клетки эпидермального характера
столь же резко отличаются от миоэпителиальных клеток радужины, как
и от всех прочих мышечных элементов неэпидермального типа. Очень
желательно было бы изучить под тем же углом зрения более типичные
в отношении мышечной дифференцировки миоэпителиальные клетки неко-
торых кожных желез.
При опухолевом превращении элементов грудной железы, как пока-
зывают последние наблюдения Гампе рля (1940), мышечная дифференци-
ровка корзинчатых клеток может в некоторых случаях очень стойко сохра-
няться, причем общий характер этих разрастаний все-таки несомненно обна-
руживает их эпидермальную природу. По данным этого автора, именно из
несколько измененных корзинчатых или, по его терминологии, «миоте-
лиальных» клеток построены особые новообразования этого органа, состоя-
щие из относительно мелких клеток, образующих массивные ветвящиеся
тяжи. Эти данные можно поставить в параллель с моими, разобранными
выше, наблюдениями над опухолевой нейроглией, которая также обнару-
живает в некоторых случаях в более резко выраженной степени, чем нор-
мальная глия, специальную нейроноидную дифференцировку утрирован-
ного и уродливого характера.
Другое направление дифференцировки эпидермальных тканей прояв-
ляется также в резко утрированном виде в некоторых раках с необычайно
сильным ороговением. В ряде энтеродермальных раков, наоборот, может
проявляться исключительно резкое слизеобразование. Во всех этих фактах
нельзя не видеть некоторых общих, свойственных опухолевым тканям,
закономерностей, которые еще требуют специального изучения.
5. НЕРВНАЯ ТКАНЬ, ИЛИ НЕЙРОНАЛЬНЫЕ ЭЛЕМЕНТЫ НЕРВНОЙ СИСТЕМЫ
а) Общие замечания
Подобно функции сокращения, раздражимость и проведение раздра-
жений также являются одним из элементарных физиологических показателей
живой протоплазмы. В нейронах эта функция, достигающая очень большой
Экспериментальный метод в гистологии
403
степени совершенства и приобретающая ряд новых качественных особен-
ностей, связанных с высокой морфологической специализацией, становится
основным ведущим физиологическим признаком. Как и в прочих высоко-
специализированных в том или другом направлении тканевых элементах,
односторонняя дифференцировка накладывает глубокий отпечаток на все
морфо-физиологические особенности нейронов, к которым полностью прило-
жимо. как к одному из частных случаев, общее высказанное мной положение
(1934), что специфические особенности тканевых элементов представляют
собой развивающиеся в эволюционном процессе и наследственно закреп-
ляющиеся на каждом этапе эволюции признаки многоклеточных организмов.
Как показывают сравнительная анатомия и гистология, образование в составе
тела животных особых нервных элементов восходит к глубокой древности.
Нервная система кишечнополостных уже является настолько высоко-
организованной, что ее возникновение следует отнести на еще более ранние
этапы эволюции, от которых не сохранилось в настоящее время никаких
следов. У губок никаких нервных элементов не имеется, а можно лишь
говорить о какой-то очень примитивной форме мускулатуры. Кроме того,
эти формы стоят, повидимому, в стороне от главного филогенетического
ствола.
Вместе с тем делается очень вероятным, что вымершие предки всех
вышестоящих многоклеточных организмов уже обладали примитивными
нервными и нейробластическими элементами, которые сделались общим
источником развившихся у высокоорганизованных форм весьма разно-
образных нейронов, морфо-физиологические черты сходства которых имеют
своей причиной единство происхождения и появляющиеся в общей органи-
зации животных конвергентные черты сходства, а различия — дивер-
гентную эволюцию стволов филогенетического дерева и дивергентную
дифференцировку в теле отдельных животных, связанную с общим услож-
нением их организации. Большое своеобразие морфо-физиологических
особенностей нейронов хорошо объясняется глубокой филогенетической
древностью нервной системы. В противоположность мышечным тканям,
возникающим во многих случаях конвергентно и параллельно из разных
источников на очень различных этапах эволюции и поэтому относящихся
к совершенно различным тканевым типам естественной гистологической
системы, нейроны всех организмов в филогенезе и онтогенезе связаны
с эктодермой, причем из ее состава все резче и резче выделяются особые
нервные зачатки, а прочие части превращаются в зачатки эпидермальных
тканей. Можно думать, что филогенетический возраст разных нейронов,
генетически связанных с нейробластической эктодермой, а затем с нейробла-
стическими элементами нервных зачатков, неодинаков и что на последующих
этапах эволюции к более древним элементам нервной системы присоеди-
няются и более новые. Более молодые генерации нейронов, возникая из
того же гистогенетического источника, что и более древние, параллельно
в отдельных крупных стволах филогенетического дерева, в своем даль-
нейшем развитии неразрывно связаны с более древними элементами нервной
системы, в состав которой они и включаются как взаимосвязанные части„
4.04
Глава III
При этом происходит постепенное усложнение и уточнение ее нейрональных
взаимоотношений и общей структуры в коррелятивной связи с высотой
организации мускулатуры соматического типа, которая может обнаружи-
вать замечательные явления анатомического параллелизма, например
при дифференцировке из кожно-мускульного мешка высокодифференци-
рованной мускулатуры членистоногих и позвоночных, связанной с конеч-
ностями, построенными по типу расчлененных рычагов. У кишечнополо-
стных, как это отмечалось выше, имеются указания на наличие особой экто-
дермальной нервной системы, гистогенез которой, однако, не может счи-
таться достаточно точно прослеженным. Возможно, что нервные элементы,
находящиеся в составе энтодермы, возникают из мигрирующих в нее индиф-
ферентных клеток, генетически связанных с эктодермой. Принимая во
внимание тот факт, что из таких индифферентных клеток могут возникать
все без исключения клетки тела, эктодермальная часть нервной системы
кишечнополостных не может считаться образованием, существенно отли-
чающимся по своему развитию от ее эктодермальной части. Менее ясно
обстоит дело с частями нервной системы, которым у сравнительно высоко-
организованных форм приписывают неэктодермальное происхождение.
В первую очередь это относится к иглокожим, у которых, наряду
с т. наз. эктонейральной нервной системой, имеется еще ее гипо нейраль-
ная и энтонейральная части. Эктонейральная нервная система несомненно
генетически связана с эктодермой, а менее полно изученной гипонейральной
и энтонейральной некоторые авторы приписывают мезодермальное проис-
хождение. Однако это не может считаться достаточно прочно установленным
и нуждается еще в дополнительных исследованиях.
В последнее время Массон (1932) выдвинул гипотезу, что у позвоночных
в кишечной энтодерме, так же как и в эктодерме, образуются плакоды,
производными которых будто бы являются развивающиеся в составе кишеч-
ного эпителия т. наз. энтерохромаффиновые или базальнозернистые клетки.
Связь этих элементов с нервной системой Массон усматривает, между про-
чим, в том, что в червеобразном отростке наблюдаются своеобразные раз-
растания энтерохромаффиповых элементов, связанные с гиперплазией
мейснерова сплетения. Эту гипотезу нельзя считать сколько-нибудь серьезно
обоснованной.
Нервная система была предметом очень многочисленных и тщательных
исследований описательного характера с точки зрения нейрональных взаимо-
отношений ее элементов, внешняя форма которых известна во многих
случаях с исключительной точностью. На основании богатого собствен-
ного фактического материала и его сопоставления с литературными дан-
ными, Заварзин (1925) пришел к ряду очень интересных теоретических
выводов, которые в несколько измененном и во многих отношениях допол-
ненном виде нашли свое отражение в монографии этого автора (1941),
где сравнительная описательная морфология нервной системы изложена
с исключительным мастерством. Широко используемый в отношении позво-
ночных и в первую очередь млекопитающих метод дегенерации и регене-
рации дал в руках Лаврентьева и его сотрудников богатейший и ценный
Экспериментальный метод в гистологии
405
материал, позволяющий проследить нейрональные соотношения в ряде
случаев, в частности, в автономной нервной системе, когда простое наблю-
дение не может дать ясного ответа на поставленные вопросы (Лаврентьев,
1938). Применяемый с этой целью экспериментальный метод имеет совер-
шенно иную направленность, чем экспериментально-гистологический анализ
тканей в нашем смысле, позволяющий изучать общегистологические зако-
номерности и в первую очередь гистобластические потенции тканей, свой-
ства их камбиальных элементов и тому подобное.
Так как экспериментальный анализ общих гистотипических особен-
ностей нейронов и нейробластических элементов еще находится в довольно
зачаточном состоянии и известные в этом отношении данные касаются,
за немногими исключениями, позвоночных, то в настоящее время невоз-
можно делать подробного разбора нервной системы в разных филогенети-
ческих рядах с этой точки зрения. Достаточно напомнить немногие факты,
которые уже упоминались в главе, посвященной глиоэпендимному тканевому
типу.
У многих аннелид среди первичноротых эпидермис кожного покрова,
как показывают опыты с регенерацией, имеет еще значение по дивалентной
эктодермы, способной даже у взрослых форм дифференцироваться не только
в эпидермальные же клетки, но и в элементы нервной системы и мышц
кольцевого слоя кожно-мускульного мешка. Иными словами, эпителий
кожного покрова сохраняет у них нейробластические потенции. Очень
вероятно, хотя и не может считаться доказанным, что аналогичные соот-
ношения имеются и у ряда турбеллярий, отличающихся организацией,
значительно более примитивной по сравнению с аннелидами.
У низших позвоночных, как это уже отмечалось выше, эпендима даже
у взрослых форм сохраняет общее камбиальное значение для тканей нерв-
ной системы, поскольку имеющиеся данные, полученные методом реге-
нерации, говорят за то, что ее клетки, пролиферируя, могут дифференциро-
ваться в нейрональные и глиальные элементы. У высших позвоночных
эпендима имеет нейробластическое значение, повидимому, только до
определенных этапов онтогенеза, утрачивая его в течение послезародышевой
жизни.
Не останавливаясь на специально нейрологических вопросах, отно-
сящихся, строго говоря, к микроскопической анатомии нервной системы, в
отношении которых я позволю себе сослаться на упомянутые выше моно-
графию Заварзина (1941) и сводку Лаврентьева (1938), я считаю своим дол-
гом разобрать здесь несколько подробнее некоторые вопросы, касающиеся
общего эволюционного усложнения нервной системы и ее нейрональных
соотношений, обнаруживающих замечательные черты сходства у очень
отдаленных друг от друга организмов (Р. Кахаль-, Радлъ, 1912;
Заварзин, 1913, 1925, 1940), обозначенные Заварзиным, как явления парал-
лелизма. С моей точки зрения, эти параллелизмы зависят, как это уже
неоднократно отмечалось, от генетического единства нейронов во всем
животном мире и явлений конвергенции в некоторых сторонах типовой
организации животных, относящихся даже к очень отдаленным друг от
406
Глава III
друга систематическим группам, но вовсе не оттого, что в основе эволюции
нейрональных соотношений лежат какие-то особые закономерности, не-
свойственные филогенетическому развитию целых организмов и их органов.
Для полноты картины необходимо еще раз вернуться к гипотезам,
пытающимся объяснить возникновение элементов нервной системы и их
связь с мускулатурой.
К сожалению, первые этапы эволюции нервной системы, как это неодно-
кратно отмечалось, совершенно неизвестны. По Паркеру (1919), опирающе
муся на появление в наружном покрове губок примитивных сократимых
элементов («независимых эффекторов»), предки всех нейронов появляются
как нейроэпителиальные клетки, связанные своими базальными отростка-
ми с погружающимися внутрь и дифференцирующимися мышечными волок-
нами. Это предположение несколько дополняет гипотезу Гертвига (1878,
1880), согласно которой источниками возникновения мышечных волокон
были клетки, сходные с миоэпителиальными элементами кишечно-полостных.
Возможно, что миоэпителиальные или мышечные элементы действи-
тельно возникли прежде специализированных нейроэпителиальных, однако
весьма вероятно, что они были не только «независимыми эффекторами»
в смысле Паркера, но и элементами, связанными в функциональном отно-
шении с клетками эктодермы, не приобревшими еще специальной нейро-
эпителиальной дифференцировки, но несомненно обладавшими общей
примитивной раздражимостью живой протоплазмы. Как это указывалось
в главе, посвященной вопросам возникновения древнейших тканевых систем,
клеточному слою гипотетических предков многоклеточных животных,
соответствующему эктодерме, можно приписать состав из первоначально
одинаковых малодифференцированных мультифункциональных элементов,
которые в отношении своей морфо-физиологической характеристики имеют
смешанное значение мионейроэпител!ильных клеток. Их апикальные концы,
вероятно, были эпителиоподобно сомкнуты, а базальные имели соединяющие
их друг с другом отростки, обладавшие примитивной сократимостью и про-
водимостью. Путем дивергентной дифференцировки этих клеток затем
возникли, с одной стороны. миоэпителиальные и мышечные, а с другой —
нейроэпителиальные и нервные элементы. Из тех же клеток, обладавших,
вероятно, способностью к примитивной форме образования межклеточного
вещества своими базальными частями, несколько позже возник
и мезенхимный зачаток или, по крайней мере, его часть. Таким образом,
делается понятной первичная связь между мышечными и нервными элемен-
тами без предположения о расщеплении гипотетической бивалентной нерв-
но мышечной клетки на нервную и мышечную части. В приложении к возник-
новению мышечных и нервных элементов эта гипотеза расщепления исходит
из отдельных клеток первичной эктодермы, что, с моей точки зрения, непра-
вильно, а не из эктодермы в целом, как взаимосвязанной примитивной
клеточной или тканевой системы, каковой она была на самом деле. Мне ка-
жется, что наша трактовка возникновения элементов нервной и мышечной
систем стоит ближе к истинному положению вещей, чем гипотеза расщеп-
ления.
Экспериментальный метод в гистологии 407
Трудно сказать, являются т. наз. i - клетки современных гидроид-
ных форм примитивной или возникшей вторично чертой организации.
Если их связывать с обособлением полового зачатка, то они, вероятно,
являются вторичными образованиями, представляющими собой камбиаль-
ный резерв для всех решительно клеток тела и, в частности, для различных
элементов с мышечной или нервной дифференцировкой.
Следующим этапом усложнения нервной системы является появление
тех соотношений, которые свойственны некоторым из изученных в этом
отношении современных кишечнополостных — гидре (Хаджи, 1909), акти-
ниям (Гросселъи, 1909), сцифомедузе корнероту (Бозлер, 1927) и гребне-
вику Вегоё (Гейдер, 1927). Основная особенность современных кишечно-
полостных заключается в наличии в их диффузной нервной системе чувстви-
тельных нейроэпителиальных и собственно нервных клеток, образующих
сплетение, и связанных, в свою очередь, с мышечными элементами. Нервные
клетки сплетений, поскольку они соединены с мышцами, являются прими-
тивными эффекторными, а поскольку они связываются друг с другом,
первичными ассоциативными нейронами. Иными словами, в таких случаях
можно думать о сосуществовании более простой двучленной и более слож-
ной трех- и л ногочленной передачи возбуждения на мышцы, хотя вместе с тем
не совсем исключена возможность того, что часть чувствительных нейро-
эпителиальных клеток сохранила, кроме того, еще наиболее примитивную
прямую связь с сократимыми элементами. По А. Каппе рсу (1920,1921,1932),
клетки сплетений кишечнополостных являются не истинными поляризован-
ными нейронами, а «первичными ганглиозными клетками» с двусторонней
проводимостью, между которыми он допускает синцитиальную связь,
что в свете данных Бозлера (1927) и в соответствии с точкой зрения Заварзина
(1941) едва ли имеет место.
Из клеток сплетений у вышестоящих форм формируется центральная
нервная система, а из чувствительных нейроэпителиальных клеток обра-
зуются чувствительные периферические нейроны. Последнее, в соответ-
ствии со взглядами Заварзина (1941), является более вероятным, чем гипо-
теза центрального возникновения чувствительных нейронов.
У высокоорганизованных форм животных все нейроны могут быть раз-
виты, на основании своих взаимоотношений с другими тканями тела и друг
с другом, на три основных типа: рецепторные, промежуточные, или ассо-
циативные, и эффекторные нейроны. Первые из них непосредственно свя-
заны с чувствительной периферией при помощи концевых аппаратов, сво-
бодных или снабженных особыми клетками ненейронального характера.
С другой стороны, эти же элементы соединены синаптически с другими ней-
ронами. Нейроны второго типа имеют только интернейрональные синапсы,
а эффекторные нейроны, с одной стороны, связаны синаптически с другими
нейрональными элементами, а с другой — образуют концевые аппараты
эффекторного характера, например двигательные окончания на мышцах
или секреторные окончания на железистых клетках. Из трех звеньев,
представленных последовательно соединенными чувствительным, проме-
жуточным и эффекторным нейроном, состоит дуга простого рефлекса спи-
408
Глава HI
нального типа, возникшая, вероятно, из более примитивной двухчленной
и являющаяся, в свою очередь, в ряде случаев источником образования
более сложных, построенных более чем из одного промежуточного звена,
многочленных дуг, характеризующих нервные системы с высокоразви-
тым головным мозгом и его корковыми центрами. Помимо усложнения дуги
рефлекса в длину, происходят его эволюционные изменения «в ширину».
Они заключаются в том, что во многих случаях наблюдается весьма обыч-
ное лавинообразное нарастание рефлекса, при котором импульсы, идущие
с одного чувствительного нейрона, переключаются не на один, а на несколько
включенных параллельно промежуточных нейронов, каждый из которых
может быть в свою очередь связан с несколькими же эффекторными. В дру-
гих случаях, как это, например, бывает в проводящих системах высших
органов чувств, на пути от рецептора к корковому центру может иметь
место сужение русла рефлекса, при котором несколько чувствительных
нейронов или предшествующих им вспомогательных клеток связываются
не с несколькими, а только с одним нейроном и т. д. Наконец, усложне-
ние рефлексов достигается еще с развитием ассоциативных нейронов мест-
ного значения, координирующих работу ряда однородных центральных
нейронов (например клетки Гольджи 2-го типа в их различных моди-
фикациях).
Общегистологический вопрос, подлежащий нашему обсуждению, заклю-
чается в том, каким образом происходят усложнение и, во многих слу-
чаях, уточнение рефлекторной дуги. Подробнее всего это разбирают Завар-
зин (1925, 1941) и Ганстрем (1928) на примерах образования трехчленной
рефлекторной дуги в туловищном мозгу и развития многочленных связей
рецепторных клеток органов зрения с корковыми центрами. Для эволю-
ционного объяснения этих явлений Заварзин прибегает к помощи
гипотезы расщепления, распространяя ее на нейрональные взаимо-
отношения и рассматривая ее как необходимую составную часть теории
параллелизмов. Согласно Заварзину, увеличение количества звеньев
в рефлекторных дугах происходит, вследствие «морфологического» ( 925),
или «эволюционного», согласно новейшей терминологии автора (1941),
поперечного расщепления гистологических структур. Оно мыслится таким
образом, что то или иное звено дуги рефлекса с усложнением структуры
заменяется двумя включенными последовательно элементами. Этот про-
цесс может в случае зрительных центров происходить повторно. Следует
подчеркнуть, что гипотеза расщепления не столько объясняет, сколько
описывает процесс усложнения нейрональных связей, хотя она вместе
с тем противопоставляется другой возможной гипотезе дивергентной диф-
ференцировки. Факты, на которые опирается Заварзин, заключаются в сле-
дующем. В ряду аннелиды — членистоногие у некоторых форм в туло-
вищном мозгу описаны нейроны, имеющие обычно крупные размеры й
по своему значению представляющие собой одновременно двигательные
и ассоциативные нейроны, которые, например у пиявк Aulastomum,
с одной стороны, имеют один или два эфферентных отростка, связанных
с мускулатурой и соответствующих нейритам, а с другой—сложную
Экспериментальный метод в гистологии
409
систему отростков, не выходящих за пределы центральной нервной си-
стемы, представленных дендритами и нейритом, связывающимся с дру-
гими двигательными или двигательно-ассоциативными элементами. Эти
нейроны, связанные посредством афферентных волокон с периферическими
чувствительными клетками, имеют значение двигательных и вместе с тем
промежуточных ассоциативных элементов. У насекомых в туловищном мозгу
Заварзин описывает вместо таких сложных двигательно-ассоциативных
нейронов более простые, одни из которых являются чисто двигательными,
другие — чисто ассоциативными. Этот факт и обозначается как эволю-
ционное расщепление структуры, которое, с одной стороны, противопо-
ставляется принципу дивергентного развития, а с другой—«не предре-
шает никак вопроса о механизме этого явления» (Заварзин, 1941, стр. 113).
Таким же «расщеплением», но только не поперечным, а, так сказать, про-
дольным объясняется и замена одного более сложного ассоциативного
нейрона двумя более простыми, также ассоциативными. Другой пример,
приводимый Заварзиным для подтверждения своей точки зрения, мне
кажется, не подтверждает ее, а скорее ограничивает применимость
гипотезы расщепления и показывает, что эволюционное изменение нейро-
нальных взаимоотношений при общем усложнении организации может
происходить и совершенно иным образом. У краба, стоящего в филогене-
тическом ряду гораздо выше аннелид и отнюдь не являющегося родствен-
ником прямых предков насекомых, в туловищном мозгу имеются, по дан-
ным Бете (1903), поливалентные двигательно-ассоциативные нейроны еще
большей степени сложности, чем у пиявки. Это показывает с несомненностью,
что, при другом направлении прогрессивных изменений в нервной системе,
усложнение нейрональных соотношений может итти по совершенно иному
пути, соответствующему дивергентным чертам в развитии и организации
крабов и насекомых. Следует оговориться, что для объяснения возникно-
вения чувствительных нейронов позвоночных Заварзину (1925, 1941) при-
шлось прибегнуть самому к гипотезе «эволюционного слияния», диамет-
рально противоположного «эволюционному расщеплению».
В ряду позвоночных настоящих поливалентных двигательно-ассоциа-
тивных нейронов с несколькими нейритами не имеется даже у пескоройки
(Третъяков, 1911). Едва ли имеются достаточные основания постулировать
их обязательное присутствие у вымерших предков. Возможно, что в этом
филогенетическом ряду, в соответствии с его специфическими особенностями,
усложнение структуры шло и по иному пути. Было бы глубокой ошибкой
игнорировать бесспорно имеющие место различия, обусловленные дивергент-
ной эволюцией общей организации соответствующих форм'.
Другой и наиболее наглядной иллюстрацией гипотезы расщепления
являются, согласно Заварзину (1941), зрительные центры, среди кото-
рых в разных филогенетических рядах можно найти эволюционное увели-
чение количества звеньев от одного до четырех, в зависимости от высоты
организации и точности работы всего оптического аппарата, который
только на высших этапах эволюции делает возможным предметное
зрение.
410
Глава III
Первоначально {Заварзин, 1925) предпочтительность гипотезы расщеп-
ления перед дивергентной дифференцировкой обосновывалась, между про-
чим, тем, что только при допущении расщепления усложнение структуры
может происходить без уменьшения количества членов данного звена, а
при дивергентной дифференцировке должно неизбежно происходить их
уменьшение, что не соответствует наблюдающимся фактам. Позже {Завар-
зин, 1941) расщепление аргументируется прежде всего трудностью пред-
ставить себе дивергентную эволюцию ассоциативно-двигательных нейро-
нов в направлении ассоциативных элементов путем исчезновения двига-
тельных нейритов. Убедительным аргументом в пользу этой гипотезы
Заварзин считает также поразительное совпадение в распределении синап-
тических областей поливалентных нейронов у одних форм и ряда моно-
валентных нейронов у других.
Мне кажется, что гипотеза расщепления в тех случаях, когда она вообще
представляется приложимой, возникла на основе следующих предпосылок:
во-первых, в результате сопоставления нейрональных взаимоотношений
только у взрослых форм при полном игнорировании гистогенетических
данных даже в тех случаях, когда они известны с той или иной степенью
полноты; во-вторых, в результате допущения, что рефлекторные дуги пред-
ставляют собой какие-то самостоятельные частные системы, начинающиеся
рецепторным звеном и заканчивающиеся эффекторным, в рамках которых
только и происходят все эволюционные изменения; в-третьих, вследствие
неправильной трактовки чувствительных эпендимных клеток спинного
мозга низших позвоночных и фоторецепторных клеток органа зрения как
высокоспециализированных нейронов и, в-четвертых, благодаря игнорирова-
нию факта существования в нервной системе даже во взрослом состоянии (низ-
шие позвоночные) или, по крайней мере, в течение определенных этапов онто-
генеза малодифференцированных камбиальных клеток, способных к про-
лиферации и дифференцировке в нейроны. У таких форм, как аннелиды,
как это отмечалось выше, значение нейробластических камбиальных кле-
ток имеют эпидермальные элементы кожного покрова. Выводы на осно-
вании сопоставления картин, свойственных взрослым формам, без анализа
развития гистологических структур, едва ли могут считаться достаточно
обоснованными, подобно тому как в эволюционной морфологии нельзя
делать выводов только на основании сравнения строения взрослых форм.
Как правильно подчеркивает Шмалъгаузен, естественному отбору подвер-
гаются не только взрослые формы, а целые онтогенезы. Допущение, что
рефлекторные дуги представляют собой как бы самостоятельные частные
системы, может в лучшем случае, да и то далеко не всегда, с известными
оговорками, считаться возможным в отношении взрослого организма, но
никак не развивающейся нервной системы, в которой продолжают диффе-
ренцироваться новые нейроны из камбиальных клеток. Фоторецепторные
элементы у всех изученных позвоночных, согласно экспериментальным дан-
ным нашей лаборатории (Винников, 1939, 1941, 1943), несмотря на свой
весьма специализированный внешний вид, сохраняют способность к про-
лиферации и чрезвычайно резко отличаются в отношении своих биологи-
Экспериментальный метод в гистологии
411
ческих свойств от нейронов. Это хорошо гармонирует со старыми представ-
лениями, согласно которым они являются не нейронами, а нейроэпителиаль-
ными клетками или, точнее, измененной чувствительной эпендимой. Если
учесть все сказанное, то гипотеза расщепления делается излишней и подле-
жит замене другими представлениями. Если использовать факты из области
эмбрионального гистогенеза и экспериментальной гистологии нервной сис-
темы, то эволюционным изменениям нейрональных соотношений может
быть дано следующее объяснение, мне кажется, достаточно обоснованное
и хорошо гармонирующее как с общими закономерностями эволюции,
так и с развиваемыми в этой книге общими идеями.
Если мы обратимся к эмбриональному гистогенезу сетчатки глаза позво-
ночных, на примере млекопитающих, протекающему очень схематично
(Фюрст, 1904; Зеефелъдер, 1910; Винников, 1943), то увидим, что камбиаль-
ный эпендимный слой внутреннего листка глазного бокала, усиленно про-
лиферируя, дает начало растущим в длину клеткам, ядра которых распо-
ложены на разных уровнях, типичным для развивающейся центральной
нервной системы образом. Первоначально все клетки своими ориентиро-
ванными вертикально отростками проходят через всю толщу внутреннего
ретинального листка, соединяя друг с другом обе его поверхности. В таком
утолщенном зачатке будущей сетчатки митозы по преимуществу локализи-
руются в клетках, ядра которых граничат с наружным ретинальным лист-
ком. Прежде всего утрачивают двухстороннюю связь нейробласты буду-
щих ганглиозных клеток, из округлившегося клеточного тела которых,
вместо бывших эмбриональных цитоплазматических отростков, начинают
вырастать нейриты. Они образуют в своей совокупности слой волокон
зрительного нерва и устремляются к стебельку глазного бокала. Затем
уже происходит дифференцировка элементов внутреннего ядерного слоя,
который вместе с тем начинает обособляться от наружного слоя ядер,
в составе которого камбиальные элементы продолжают энергично делиться
митотическим путем, пополняя запас дифференцирующихся элементов. Все
это можно объяснить только тем, что при дивергентной дифференцировке
камбиальных элементов, соответствующих примитивным фоторецепторным
клеткам, прежде всего образуются элементы ганглиозною слоя, а затем
уже двухчленная клеточная цепочка усложняется образованием третьего
звена, биполярных клеток, а также нейронов местного значения, т. е. гори-
зонтальных и амакриновых клеток.
Более чем вероятно, что в эмбриональном гистогенезе рекапитули-
руется, хотя бы в измененном виде, эволюционное усложнение сетчатки,
происходившее путем вставления нового звена между фоторецепторными
и ганглиозными элементами. Последние представляют собой древнейшие
нейроны сетчатки. Следующими по филогенетическому возрасту нервными
элементами являются биполяры и клегки местного значения. Что касается
фоторецепторных клеток, то они, несмотря на высокую специализацию,
сохраняют в то же самое время значение эпендимного камбия, подобно тому
как базальные корзинчатые клетки эпидермальных желез, несмотря на
свою специализацию в направлении сократимых элементов, в то же самое
412 Глава III
время имеют и камбиальное значение. С этой точки зрения, опирающейся
на исследование Винникова, подробно разобранное выше, первыми по счету
истинными нейронами зрительного пути являются биполяры, вторыми —
ганглиозные клетки, а третьими, находящимися за пределами клетчатки,—
нейроны первичной зрительной коры или подкорковых центров промежу-
точного мозга.
Весьма вероятно, что в филогенетическом ряду позвоночных усложне-
ние разных рефлекторных дуг происходит именно путем таких вставок
новых, образующихся из камбиального эпендимного слоя, нейронов, а не
благодаря расщеплению уже успевших образоваться нервных элементов.
У последних могут лишь происходить в той или иной степени изменения
в синапсах. Увеличение включенных параллельно звеньев рефлекторных
дуг может происходить принципиально сходным образом. Иными словами,
усложнение рефлекторного аппарата может совершаться не за счет уже
успевших образоваться звеньев, в частности, древнейших из них, чувстви-
тельного и двигательного или двигательно-ассоциативного, а путем разви-
тия из малодифференцированных клеток нервной системы новых ассоциа-
тивных звеньев или, другими словами, путем дивергентной дифференци-
ровки, связанной с разделением функций. Если стоять на точке зрения пол-
ного' филогенетического параллелизма в различных эволюционных рядах, то
совершенно такой же механизм изменения нейрональных соотношений мыс-
лим в других стволах и ветвях естественной системы животных организмов.
Однако, ввиду того, что наряду с параллелизмами несомненно наблю-
даются и моменты дивергентного развития, полной уверенности в этом нет.
Для решения этого вопроса необходимо прежде всего изучение гистогенеза
нервной системы в разных филогенетических рядах, а также сравнительный
экспериментальный анализ ее элементов. Таким же образом объяснимо
и усложнение нейроглиального аппарата, развивающегося из тех же мало-
дифференцированных эпендимных элементов. Гипотезе эволюционного рас-
щепления элементов нервной системы, мне кажется, можно противопоста-
вить с полным основанием гипотезу эволюционного или филогенетического
камбия, которая может объяснить усложнение нервной системы путем
новообразования нейрональных звеньев, а не путем подразделения на части
уже предсуществующих. Эта же гипотеза хорошо объясняет факт порази-
тельного разнообразия нервных элементов в разных частях нервной системы,
которое не может не быть следствием их дивергентной дифференцировки.
Аналогичным образом можно себе представить замещение мелких чувстви-
тельных нейронов крупными, дифференцирующимися заново из других
частей нейробластического камбия, представленных в гистогенезе позвоноч-
ных нервными валиками с образующейся из них ганглиозной пластинкой,
а также плакодами, и имеющего вид «эволюционного слияния» известного
количества мелких нейронов в один крупный.
Необходимым следствием нашей гипотезы является высокая вероят-
ность того, что образовавшиеся в разное время путем дивергентной диф-
ференцировки нейроны в отношении своих биологических свойств, обна-
руживаемых в экспериментальных условиях, будут проявлять те или иные
Экспериментальный метод в гистологии 413
различия. Ряд особенно бросающихся в глаза биологических особенностей
характеризует, в частности, чувствительные нейроны, большая часть кото-
рых в филогенезе и онтогенезе обнаруживают, начиная с амфибий, заме-
чательное и закономерное изменение первоначальной биполярной формы
в ложную униполярную. Эти же клетки существенно отличаются от прочих
нейронов организма позвоночных тем, что оба их отростка, соответствую-
щие дендриту и нейриту, имеют характер настоящих осевых цилиндров.
Соответствующий дендриту периферический отросток чувствительных нейро-
нов позвоночных дает эффект аксон-рефлекса. Все это подтверждает, что
эти элементы отличаются рядом замечательных особенностей, которые,
вероятно, могут быть раскрыты с большей полнотой с помощью экспери-
ментально-гистологического анализа.
б) Некоторые общегистологические особенности нейронов позвоночных
Несмотря на значительное количество экспериментальных работ, посвя-
щенных нервной системе позвоночных, то, что может быть обозначено
как общегистологические особенности нейронов, в противоположность их
местным особенностям, связям и ходу проводящих путей, т. е. фактам,
относящимся в первую очередь к области микроскопической анатомии нерв-
ной системы, изучено до настоящего времени еще очень неполно. С другой
стороны, многочисленные экспериментальные данные, а также факты из
области патологии дают обильный материал для суждения о цитологических
особенностях нервных элементов при разных условиях их существования.
Из работ последнего времени в этом отношении заслуживают особого вни-
мания исследования, сделанные в области учения о паранекрозе {Насонов
и Александров, 1940) и в области систематического и комплексного изуче-
ния изменений нейронов (а также нейроглии), в разных областях нервной
системы при различных патологических процессах, проводимого Дойни-
ковъш и его сотрудниками {Дойников, 1940). Системное изучение гистоло-
гических процессов в нервной системе имеет безусловно выдающееся
теоретическое и практическое значение и уже в настоящее время дало
исключительно интересные и ценные результаты. Между микроскопической
анатомией нервной системы, с одной стороны, и изучением тонкой струк-
туры отдельных нейронов и ее изменений под влиянием тех или иных внеш-
них агентов ~ с другой, лежит, с моей точки зрения, большая и важная
общегистологическая область, задачей которой является исследование спе-
цифических свойств нервной ткани как материала нервной системы, т. е.
совокупности нейронов с их отростками в их более элементарных ткане-
вых взаимоотношениях. К элементарным тканевым свойствам нейронов,
лежащим в основе специальной морфо-физиологической дифференцировки
нервной системы в целом, относятся способность или неспособность нейро-
нов на разных стадиях развития к пролиферативным процессам и прежде
всего к делению митотическим путем, наличие или отсутствие нейробласти-
ческого камбия и особенности внешней формы клеток и их отростков во
всей динамике ее экспериментально вызываемых изменений, взаимоотно-
шения отростков и их разветвлений, принадлежащих одной и той же клетке
1
414
Глава Ill
или целой совокупности нейронов, гистотипические структуры, образуемые
телами и отростками нервных элементов, взаимоотношения нейронов и
вспомогательных элементов нервной системы, т. е. вся совокупность вопро-
сов, связанных с гистобластическими потенциями нейронального клеточ-
ного материала и с его общими, изученными разнообразными специальными
методами, функциональными особенностями, в отдельности и во взаимо-
связях с нейроглией в широком смысле этого слова. Такого рода кругу
вопросов, позволяющих подходить к нейрональному материалу с таких же
общегистологических позиций, как и к элементам других тканевых типов,
необходимо уделить значительно больше внимания, чем это было до настоя-
щего времени. Само собой разумеется, изучение элементов нервной системы
с такой точки зрения должно производиться, как и в случае других ткане-
вых типов, с филогенетических, эволюционных позиций, возможно широко
используя экспериментально-гистологический анализ и, в частности, метод
эксплантации. Кроме тех, относительно немногочисленных, эксперимен-
тальных данных, которые были приведены в главе, посвященной эпенди-
мо-глиальному тканевому типу, остановимся на некоторых результатах,
полученных в отношении нейробластов и нейронов позвоночных путем
их изучения методом тканевых культур, и на тех поправках, которые необ-
ходимо внести в имеющиеся литературные данные, в результате исследова-
ний нашей лаборатории. Последние успели охватить со значительной пол-
нотой эпендимо-глиальную часть нервной системы, но в отношении нейро-
нов еще находятся в стадии первоначального накопления фактов и первых
выводов теоретического характера.
Характерной особенностью нейробластов и нервных клеток централь-
ной и периферической нервной системы в условиях культур является их
способность к образованию тонких длинных нитевидных цитоплазмати-
ческих отростков, могущих достигать очень значительной длины.
Растущие вне организма отростки нервных клеток обычно принято
не вполне точно обозначать как нервные волокна. От настоящих нервных
волокон целого организма они отличаются отсутствием оболочек и соот-
ветствуют, собственно говоря, лишь проводящей нервные импульсы осево-
цилиндрической части нервных волокон. Эмбриональные шванновские
клетки или леммобласты могут в некоторых случаях (Эзаки, 19 9) сопро-
вождать их рост, тесно прилегать к ним, но не образуют типичной шваннов-
ской оболочки и миэлина. При эксплантации раннего эмбрионального
материала зародышей амфибий, как это впервые установил Гаррисон,
а затем подтвердил ряд авторов на курином эмбриональном материале,
нервные волокна возникают заново в культурах благодаря наступающей
дифференцировке нейробластов, первоначально лишенных отростков в по-
сеянном кусочке. Последние при этом могут сильно увеличиваться в раз-
мерах и, образуя отростки, приобретать характер дифференцированных
нервных элементов (Оливо, 1927; Мосса, 1928, и др.), большей частью не-
типичной мультиполярной формы (Оливо, Лазаренко, 1931). В экспланта-
тах из спинальных и черепных ганглиев зародышей в составе центральных
кусочков, однако, наблюдалась дифференцировка биполярных или типич-
Экспериментальный метод в гистологии
415
них псевдоуниполярных нейронов, с Т-образно ветвящимся клеточным отрост-
ком (Эзаки, 1929; Леей, 1934; Леей и Делоренци, 1934). По некоторым данным,
сами нейробласты, по крайней мере на ранних стадиях культивирования,
могут возникать из недифференцированных камбиальных элементов эмбрио-
нального мозга (Серебряков, 1934). Кроме того, уже дифференцированные
нервные клетки зародышей и молодых животных, отростки которых при
эксплантации были отрезаны на большем или меньшем расстоянии от кле-
точного тела, могут их регенерировать. При эксплантации эмбрионального
материала, содержащего уже более или менее дифференцированные ней-
роны, и частиц неэмбриональной нервной системы, наблюдающийся вне
организма, нередко очень обильный, рост нервных волокон зависит в пер-
вую очередь от явлений их регенерации. При повторных пересевах с выре-
занием или при перерезке части нервных волокон зоны роста регенерация
может повторяться в течение известного промежутка времени (Ингебриг-
тсен, 1913; Леей, 1934).
При пересевах без вырезания увеличение длины нервных волокон про-
должается в течение восьми-двенадцати, а иногда и более дней, причем
сверток фибрина может в конце концов на большем или меньшем протя-
жении, а в некоторых случаях даже целиком (Эзаки, 1929; Лазаренко,
1931), оказаться густо пронизанным ими. Дорастая до периферического
края свертка, конец растущего волокна может загибаться и продолжать
расти или параллельно к краю или в обратном направлении (Лазаренко).
Отдельные волокна могут достигать 17 мм в длину (Эзаки). Кроме того,
что нервные клетки, остающиеся внутри посеянного кусочка, образуют
нервные отростки, многие авторы описывали также эмиграцию в зону
роста самих нервных клеток, которая, однако, никогда не бывает выражена
в сильной степени.
Задача длительного сохранения нейронов живыми вне организма до
сих пор полностью не разрешена. С течением времени они постепенно
вымирают и исчезают. Большей частью это происходит уже на протяжении
первых двух-трех недель. Лишь иногда удается продлить срок их жизни
до месяца или несколько более. Это отмечали: в своих классических опытах,
сделанных на эмбриональном лягушачьем материале, Гаррисон и на мате-
риале, взятом от млекопитающих, Мартинович (1933). В последнее время
Мейер и Яблонскому (1937) удалось сохранить живыми во флаконах Кар-
реля нервные клетки в центральных кусочках спинальных ганглиев кури-
ного зародыша в течение 135 дней.
В эксплантированных участках нервной системы, содержащих лишь
проводники и лишенных нервных клеток, осевые цилиндры быстро дегене-
рируют и исчезают без остатка в течение нескольких дней.
Быстрота начала прорастания нервных волокон зависит от исходного
материала. В культурах из куриных зародышей прорастание начинается
нередко уже по истечении двух-трех часов после эксплантации (Леви,
Мартинович, 1933). По исследованиям Гаррисона и позднейшим данным
Мосса (1928), образование новых отростков у нейробластов, имеющих пер-
воначально более или менее закругленную форму, происходит путем дви-
416
Глава III
Рис. 63. Отростки нейронов в зоне роста 5-дневпой
культуры из головного мозга куриного зародыша.
Метиленовая синь. Сильное увеличение. (По Лаза-
ренко, 1931.)
жения цитоплазмы клетки; в ней образуется местное утолщение, которое
превращается сначала в цитоплазматическую пластинку с зазубренным
краем {Мосса), а затем постепенно вытягивается в тонкий отросток. Сво-
бодный конец новообразующегося или регенерирующего волокна все время
изменяет свою форму и большей частью представляется колбовидно взду-
тым, а в некоторых случаях кистевидно разволокненным на тончайшие
цитоплазматические ниточки. Концевое вздутие, то более, то менее резко
выраженное, образует тончайшие ниточки изменчивой формы и длины
{Леей, 1917), одна из которых, постепенно утолщаясь, становится нарастаю-
щим концом волокна и мо-
жет образовать новое кол-
бовидное вздутие взамен
исчезающего старого. Тон-
чайшие, большей частью
быстро исчезающие, ответ-
вления образуются и по хо-
ду всего растущего волок-
на {Льюисы, 1912, Леей,
1917). По исследованиям
этих же авторов, растущие
концы волокон и в более
редких случаях утолщения
по их ходу могут образо-
вать постоянные разветвле-
ния, а по данным Бауэра
(1932, 1936), — даже син-
цитиальные сети. По дру-
гим данным {Лазаренко,
1931), в культурах могут в
некоторых случаях сохра-
няться морфологические
отличия дендритов от ней-
ритов. В то время как пер-
вые разветвляются, вторые,
как правило, не ветвятся,
хотя большей частью и достигают большой длины. В некоторых
случаях в культурах из центральной нервной системы и спинальных ган-
глиев молодых (четырех-пяти дневных) куриных зародышей отростки нерв-
ных клеток распластываются по плотному субстрату и, сливаясь друг
с другом, приобретают форму широких и тонких лент {Леви, Мосса, 1929).
Такие образования, однако, существуют лишь короткое время и быстро
изменяются.
Растущие нервные волокна представляются большей частью или прямо-
линейными или изгибающимися нитями равномерной толщины, за исклю-
чением концевых вздутий и наблюдающихся в редких случаях отдельных
утолщений по их ходу. Однако под влиянием самых разнообразных воздей-
Экспериментальный метод в гистологий 417
ствий на них могут очень легко образоваться многочисленные, так
называемые варикозные утолщения, которые или исчезают вновь
или, утолщаясь, придают волокнам четкообразный вид. В последнем случае
они являются признаками наступающих дегенеративных изменений, закан-
чивающихся распадом. Варикозности, которые получаются на препаратах,
окрашенных суправитально метиленовой синью, вероятно, являются резуль-
татом предсмертного изменения волокон. Образование резко выраженных
варикозных утолщений может быть вызвано освещением ультрафиолето-
выми лучами (Лазаренко и Бененсон, 1932).
Взаимное расположение нервных волокон в зоне роста может быть
очень разнообразным. Они располагаются поодиночке или образуют пучки
и сложные войлокообразные сплетения (рис. 63) (Лазаренко). Направление
их роста, то более или менее радиальное в отношении посеянного кусочка,
то взаимно перекрещивающееся, зависит, по данным Вейса (1934), в зна-
чительной степени от механических свойств и ультрамикроскопической
структуры среды. При культивировании частиц эмбриональной нервной
системы в свертке фибрина, образованном в стеклянной рамке той или иной
формы и находящемся в состоянии растяжения, рост волокон, так же как
и рост фибробластообразных элементов, происходит преимущественно па-
раллельно силовым линиям, которые, вероятно, вызывают соответствую-
щую ориентировку мицелл свертка (Вейс).
Средняя скорость роста нервных волокон довольно значительна. В раз-
ные моменты наблюдения и в зависимости от внешних условий (степени
плотности среды, температуры и пр.) она может колебаться в широких
пределах. В эксплантатах из эмбриональной нервной системы амфибий
она составляет в среднем около 46 микронов в час (Гаррисон, 1911), а
в случае куриного эмбрионального материала при 38° лишь около 23.5
микрона (Леви). В отдельные моменты она может колебаться, при прочих
равных условиях, от 7.7 до 29 микронов в час (Мосса). Рост может
происходить в температурных границах от 26 до 46°, достигая при 39°
наибольшей скорости, в среднем 33 микрона в час.
После перерезки проросших волокон, центральный отрезок, связанный
с телом клетки, как это уже упоминалось, может регенерировать, а пери-
ферический некоторое время (до 12 часов, по Леви, 1920) еще продолжает
увеличиваться в длину. Затем его рост останавливается, а через 20 часов
(Ингебригтсен, 1913) на нем появляются варикозные вздутия. На про-
тяжении двух суток в периферическом отрезке происходят дегенеративные
изменения, приводящие к его распаду.
Есть указания, будто центральный и периферический участки нервного
волокна, осторожно перерезанного с помощью иглы микроманипулятора,
могут в некоторых случаях, по истечении короткого промежутка времени,
вновь срастаться и восстанавливать свою непрерывность без последующих
явлений дегенерации (Леви, 1925).
Как уже указывалось выше, помимо дифференцировки нейробластов
в посеянном кусочке и экстенсивного роста нервных отростков в эксплан-
татах из нервной системы, многие авторы отмечали продвижение в зону
27 X лопин—-349
418
Глава III
роста и самих нейробластов или более или менее дифференцированных
нейронов. Их форма может быть очень различной и изменяется в широких
границах. Это наблюдается в наиболее сильной степени при их распласты-
вании по поверхности стекла или слюды. В зоне роста нервные клетки не
образуют каких-либо комплексных тканевых структур, а ведут себя неза-
висимо друг от друга, располагаясь совершенно изолированно или соеди-
няясь друг с другом с помощью длинных и тонких отростков. Предполо-
жение Капелл (1929), что нервные клетки, подобно другим тканевым эле-
ментам, могут, упрощаясь и начиная энергично пролиферировать, образо-
вать эпителиоподобные мембраны, получающиеся в конечном итоге в экс-
плантатах из нервной системы, мало доказательно и мало вероятно (Оливо,
1927; Леви, 1934, и др.). В конечном итоге нервные элементы зоны роста
утрачивают нейрофибрилли и погибают. Однако, по некоторым данным,
в них может наблюдаться на ранних стадиях культивирования, хотя и ати-
пичная, но все же дифференцировка (Мосса, 1928). По исследованиям Гри-
горьева (1931—1934), дифференцировка эмигрировавших нейробластов про-
исходит только при том условии, если они находятся в зоне роста, обра-
зованной элементами некоторых других тканей, разрастающихся из под-
саженного к частице нервной системы кусочка другого органа. В некоторых
случаях, по мнению этого автора, может наблюдаться не только диффе-
ренцировка нейробластов, но и образование их отростками окончаний на
поверхности элементов других тканей, растущих в той же культуре, напри-
мер эмбриональной сердечной мышцы или соматических мышечных воло-
кон. Такие картины этот автор склонен считать настоящими нервными окон-
чаниями, сравнимыми с теми, которые свойственны целому организму. С
этим, однако, как справедливо отмечает Леви (1934), едва ли можно
согласиться.
До последнего времени считалось, что даже зародышевые нервные клетки
высших позвоночных неспособны к делению митозом. Только в немногих
опубликованных недавно работах имеются указания, что эмигрировавшие
в зону роста нейроны из коры полушарий новорожденных котят (Марти-
нович, 1933) и продолговатого мозга человеческих и куриных зародышей
(Леви и Мейер, 1933) могут делиться митозом, сохраняя при этом свои
характерные отростки. Деление нейронов — явление очень нерегулярное
(Леви, 1934). В некоторых культурах оно будто бы происходит очень интен-
сивно, но в большей их части отсутствуют совершенно. Дальнейшая судьба
нейронов, которые делятся митозом, не прослежена.
Мартинович (1933), кроме того, описывал картины изменений
мигрировавших в зону роста нервных клеток, которые он трактовал как
явления дедифференцировки и их превращения в элементы, сходные
с фибробластами.
Все данные, касающиеся миграции нейронов и снабженных отростками .
нейробластов в зону роста, их дальнейших изменений и пролиферации
митотическим путем, в свете работ нашей лаборатории должны быть под-
вергнуты очень серьезной критике. При изучении работ соответствующих
авторов и в первую очередь сводки Леви (1934), в которой этим вопросам
Экспериментальный метод в гистологии
419
уделено очень много внимания, прежде всего бросаются в глаза почти пол-
ная голословность и неубедительность большей части утверждений, что
элементы, описываемые в зоне роста как нервные клетки, в действитель-
ности являются таковыми. Изучение соответствующих рисунков заставляет
считать эти утверждения еще менее обоснованными.
В большей части случаев единственным критерием нейрональной при-
роды изучавшихся клеток является наличие у них двух или нескольких
отростков, часто не достигающих значительной длины. По своей форме,
за очень редкими исключениями, клетки, описываемые как нервные, даже не
имеют мало-мальски типичной нейроноидной формы и часто бывают сете-
видно связаны друг с другом отростками. Значительно реже трактуемые
как нейроны клетки действительно отличаются более или менее сложной
нейроноидной формой. Детального изучения изменений в центральном
кусочке эксплантатов, которые бы могли пролить свет на происхождение
наблюдаемых в зоне роста элементов, как правило, не дается. Изучение
специальными нейрологическими методами центральных кусочков, произ-
водившееся в отдельных случаях, не дает картин, которые бы позволяли
утверждать, что типичные нейроны и нейробласты, обнаруживаемые в его
составе, изменяя форму, действительно активно мигрируют в зону роста
и превращаются в, описываемые как нервные, клетки этой последней.
Все это заставляет подходить с большой осторожностью к данным Леви
и ряда других авторов. С другой стороны, почти все без исключения изо-
лированные клетки и все своеобразные сетевидные клеточные соединения
зоны роста, которым приписывается нейрональная природа с большой сте-
пенью точности, соответствуют подробно изученным нами и описанным выше
картинам разрастаний нейроглии центральной или периферической нерв-
ной системы, нейроглиальная природа которых не внушает никаких сомне-
ний. Она доказывается, между прочим, тем, что даже наиболее нейроноид-
ные формы клеток весьма обычны в культурах из белого вещества полу-
шарий мозга, не содержащих, как правило, нейронов, или встречаются
на таких стадиях культивирования, когда все имевшиеся в центральном ку-
сочке нейроны (спинной мозг) уже успевали исчезнуть без остатка, и связь
зоны роста с его измененным нейроглиальным остовом могла быть установле-
на с полной несомненностью (Хлопин, 1941, и неопубликованные данные).
Нейроглиальная, а не нейрональная природа клеток, описанных, в част-
ности, Мартиновичем (1933) в эксплантатах из коры полушарий ново-
рожденных млекопитающих как измененные нейроны, с моей точки зрения,
также не подлежит никакому сомнению. В некоторых из описывавшихся
{Леви, 1934) и документированных рисунками сложных сетевидных струк-
турах зоны роста эксплантатов из эмбриональной нервной системы, пови-
димому, имелось также сочетание выросших из центральных кусочков аксо-
нов и тесно связанных с ними нейроглиальных элементов разнообразной
формы. Клетки, описываемые Леви (1934), Леви и Долоренци (1934), Леви
и Мейер (1940) в зоне роста культур из спинальных ганглиев куриных
зародышей, по крайней мере не относящихся к наиболее ранним стадиям
эмбрионального развития, как нейробласты и нейроны, представляют
*
420 Глава III
собой измененные сателлиты и шванновские клетки, тем более, что они
и в отношении своей величины очень резко отличаются от чувствительных
нейронов, которые, начиная дифференцироваться, очень резко увеличи-
ваются в размерах. В частности, очень мелкие «биполярные нейроны»
Леви являются с почти полной достоверностью не чем иным, как шваннов-
скими клетками. Разобранные выше наблюдения Сутулова (1941) над раз-
растаниями вспомогательных элементов спинальных ганглиев новорожден-
ных кроликов в условиях тканевых культур демонстрируют с полной
несомненностью, что, при совершенном отсутствии миграции нейронов,
уже успевших в исходном материале достигнуть высокого уровня диффе-
ренцировки, сателлиты и шванновские клетки образуют в зоне роста и такие
структуры, которые полностью соответствуют тем, которые Леви и его сотруд-
ники описывали как нейрональные.
Одним из основных источников ошибок авторов, предметом изучения
которых были нейроны, является незнакомство с гистобластическими
потенциями нейроглии, исследование которых является одной из необхо-
димых предпосылок правильного понимания гистологических процессов,
происходящих в эксплантатах из нервной системы. Кроме того, причиной
этих же ошибок была предвзятая и не проверенная точка зрения, что клетки
с отростками, даже в весьма отдаленной степени напоминающие нейроны,
поскольку они наблюдаются в эксплантатах из нервной системы, должны
"быть непременно нервными элементами.
Допущение нейрональной природы отдельных элементов, отмечавшихся
в условиях тканевых культур вне центрального кусочка, имеет до настоя-
щего времени большую или меньшую долю вероятности в относительно
редких случаях. Прежде всего это относится к опытам с эксплантацией
материала эмбриональных закладок ранних стадий онтогенеза, когда нейро-
нальная дифференцировка отдельных, первоначально очень малодиффе-
ренцйрованных клеток, мигрирующих в зону роста, не может быть исклю-
чена. Кроме того, на ранних стадиях культивирования эмбрионального
материала может наблюдаться в некоторых случаях сильное распластыва-
ние всего центрального кусочка, или, по крайней мере, его периферических
участков, по плотному субстрату и вместе с тем смещение имевшихся
в его составе нейробластов, могущее симулировать их активную ми-
грацию, но обусловленное, на самом деле, в первую очередь разрастанием
или даже пассивным растягиванием глиальной или, точнее, спонгио-
бластической стромы. Все. правда, очень немногочисленные литератур-
ные указания на митотическое деление эмбриональных нервных клеток
в условиях тканевых культур, которые стоят совершенно особняком
и не' соответствуют тому, что может считаться достаточно прочно обосно-
ванным в настоящее время, объясняются тем, что делящиеся митотически
клетки были без достаточных оснований приняты за нервные, а на самом
делё представляли собой нейроглиальные элементы. Рисунки, иллюстри-
рующие эти наблюдения, полностью подтверждают настоящую точку зре-
ния. В несомненных нейронах или нейробластах и в условиях тканевых
культур митозов никогда не наблюдается. - . - -
Экспериментальный метод в гистологии 421
Следует отметить, что и сам Леви, с уверенностью описавший совместно
с Г. Мейер митозы в нервных клетках, в одной из следующих работ под-
верг свои собственные данные вполне обоснованному сомнению.
Таким образом, метод эксплантации в настоящее время может полностью
подтвердить положение, что не только нейроны, но и нейробласты, обла-
дающие типичной для них дифференцировкой, представляют собой высо-
коспециализированные клетки, утрачивающие, по всей видимости безвоз-
вратно, способность к пролиферации. Этим они существенно отличаются от
близко-родственных элементов эпендимо-глиального тканевого типа, спо-
собных к размножению митотическим путем. В составе разнообразных
тканей, относящихся к генетически единому типу производных нейральных
зачатков, нейроны в отношении высокого уровня специализации можно,
пожалуй, сравнить с гранулоцитами, которые из «энтомезенхимных» про-
изводных, дифференцируясь, также полностью утрачивают способность
к делению, но в то же самое время отличаются от нейронов кратковремен-
ностью жизни.
Вполне достоверной гистотипической особенностью нейронов и нейро-
бластов являются регенерация и новообразование ими в условиях ткане-
вых культур, сходных с аксонами, цитоплазматических отростков, могущих
образовать в зоне роста очень своеобразные структуры, которые, насколько
возможно судить по имеющимся данным, у разных нервных элементов
отличаются известными особенностями. Достаточно детальных системати-
ческих исследований, ставящих своей целью изучение гистобластических
потенций нейронов разных отделов нервной системы, в смысле особенно-
стей образуемых ими отростков, до настоящего времени очень не-
много.
Упоминавшиеся выше исследования Лазаренко (1931) над ростом аксо-
нов нервных элементов эмбрионального головного мозга куриных зароды-
шей, произведенные в моей лаборатории с применением методики электив-
ного окрашивания метиленовой синью, показали, что нервные отростки
в этом случае никогда не ветвятся в зоне роста, хотя и разрастаются очень
энергично и могут достигать очень большой длины. Они отличаются от на-
чала до конца тонким нитевидным калибром, если не считать нередко наблю-
дающегося концевого колбовидного вздутия и варикозностей, зависящих
на метиленовых препаратах от реактивных изменений нервных элементов
на повреждающие воздействия. Лазаренко очень тщательно изучил и доку-
ментировал рисунками разнообразные картины роста аксонов, которые,
в соответствии со сказанным выше, или росли порознь или чаще целыми,
напоминавшими подчас нервные стволики, пучками, которые могли обра-
зовать сплошные сплетения, но никогда не анастомозировали и не образовали
истинных сетей (рис. 63). Каким именно нейрональным элементам голов-
ного мозга куриных зародышей принадлежали эти аксоны, не могло быть
установлено.
Описанные Леей (1934) сетевидные структуры в эксплантатах из разных
отделов центральной нервной системы куриных зародышей, по крайней
мере в значительной степени, должны быть отнесены за счет не распознан-
422
Глава III
ного им роста нейроглиальных элементов. Поэтому эти данные в настоящее
время недостаточно точны.
Очень детальные исследования провел Винников (1943) над нейронами
сетчатой оболочки глаза разных позвоночных. Они с особой убедитель-
ностью подчеркивают глубокие биологические различия между фоторецеп-
торными клетками и истинными нервными элементами. В сетчатке ново-
рожденных и молодых кроликов биполярные клетки и нейроны местного
значения не обнаружили никаких прогрессивных изменений и довольно
быстро дегенерировали на месте, не образуя нервных отростков и, тем
более, не обнаруживая активной миграции. Наоборот, у части ганглиоз-
ных клеток сетчатки, в условиях тех же опытов, можно было установить
явления регенерации нейритов, которые в составе центрального кусочка
росли параллельно друг другу в направлении, соответствовавшем бывшему
месту выхода зрительного пучка из глазного яблока. Вырастая за пределы
центрального кусочка, аксоны ганглиозных клеток утрачивали свою пра-
вильную ориентировку и могли ветвиться, не образуя, однако, сетевидных
анастомозов друг с другом. Они отличались тонким калибром и не дости-
гали, как правило, значительной длины. В части ганглиозных клеток уже
вскоре после эксплантации наступали дегенеративные изменения, которые
с течением времени захватывали все большую и большую часть их.
Рост аксонов ганглиозных клеток сетчатки Винникову удалось получить
также в эксплантатах из сетчатой оболочки глаза костистых рыб, хвоста-
тых и бесхвостых амфибий и птиц, причем у представителей разных клас-
сов позвоночных он отличался очень резко выраженными особенностями.
В эксплантатах из сетчатки щуки вырастали большей частью аксоны необы-
чайной толщины с хорошо заметной при жизни и даже на препаратах,
обработанных неспециальными методами, фибриллярностью и постепенно
заостряющимся свободным концом.
Постепенно разрастаясь, такие аксоны обычно образовывали отходящие
под острыми углами более тонкие ветви и могли достигать очень значи-
тельной длины, однако, без сетевидных анастомозов друг с другом. Ней-
риты ганглиозных клеток сетчатки амфибий также достигали очень зна-
чительной длины и ветвились довольно сложным образом, причем по харак-
теру ветвлений отчетливо разнились от одноименных аксонов щуки и могли
быть безошибочно отличены от этих последних по своей общей древовидной
или несколько напоминающей причудливый канделябр внешней форме.
Некоторые, менее бросающиеся в глаза, различия обнаруживали также
аксоны лягушки и аксолотля.
Наконец, в эксплантатах из куриной сетчатки нервные отростки отли-
чались тонкой нитевидной формой, не достигали значительной длины и
если и ветвились, то в незначительной степени. Сетевидных анастомозов
между аксонами и миграции нейронов в зону роста во всех случаях не
наблюдалось.
Очень большими особенностями отличается рост вне организма аксо-
нов чувствительных клеток спинальных ганглиев. С особенной отчетли-
востью это показывают исследования Сутулова (1941) над чувствительными
Экспериментальный метод в гистологии
423
нейронами кроличьих зародышей незадолго до рождения и новорожденных
кроликов. Повторяя на млекопитающих опыты, поставленные в разное время
на курином эмбриональном материале, они позволяют исключить отмечен-
ные выше ошибки ряда авторов, в наблюдениях которых во многих слу-
чаях рост периферической нейроглии не мог быть дифференцирован с доста-
точной ясностью от характерных для нейронов картин. Миграции чув-
ствительных нейронов в зону роста и их пролиферации не наблюдалось ни
в одном случае, но рост аксонов мог быть изучен почти до месячного срока
культивирования, причем он нередко возобновлялся после повторных пере-
севов с вырезанием. Спинальные ганглии подобно сетчатке представляют
собой исключительно благодарный объект для изучения гистологических
особенностей одного определенного типа нейронов, которые обнаруживают
при этом совершенно отчетливые отличия как .от нейронов сетчатки, так
и от изученных раньше в опытах Лазаренко нейронов головного мозга кури-
ных зародышей. Рост отростков чувствительных нейронов отличался во
многих случаях на разных стадиях культивирования большой интенсив-
ностью, разнообразием и сложностью наблюдавшихся картин. Аксоны росли
в некоторых случаях изолированно (причем их ветвления и взаимоотноше-
ния друг с другом отдельных веточек могли быть прослежены до мельчай-
ших деталей) или образовали первоначально тянущиеся параллельно друг
другу, веерообразно расходящиеся или перекрещивающиеся под разными
углами цитоплазматические стволы очень значительной толщины. Изоли-
рованные стволы аксонов на неодинаковом расстоянии от центрального
кусочка начинали древовидно ветвиться, причем их более тонкие, а иногда
и более толстые веточки, соединяясь друг с другом, образовали сложные
сетевидные структуры с петлями разных размеров. Часть веточек закан-
чивалась свободно тонким или, в некоторых случаях, несколько утол-
щенным концом или соединялась с отдельными нейроглиальными клет-
ками, другая часть которых не обнаруживала видимой связи с аксонами.
Попытку Леви дифференцировать растущие элементы нейроглии (лем-
мобласты) от соединительнотканных клеток, на основании наличия или
отсутствия связи с аксонами, нельзя считать удачной. В опытах Су ту лова
несомненные нейроглиальные элементы, образуя обширную и очень харак-
терную зону роста, во многих случаях не обнаруживали никакой связи с
аксонами чувствительных нейронов, а рост соединительной ткани, как пра-
вило, вовсе отсутствовал, так как последняя, как и в других случаях,
не выдерживала конкуренции с разрастающейся нейроглией. Если аксоны
разрастались пучками или образовали перекресты, то их веточки, вступая
в связь друг с другом, могли образовать чрезвычайно сложные сети, в кото-
рых совершенно не удавалось установить части, относящиеся к одному или
другому нервному отростку.
Прочность сетевидной связи разветвлений отростков доказывалась тем,
что при образовании полостей разжижения отдельные участки сетей и
аксоны с их ветвями могли заметно растягиваться, не разрываясь сразу.
Таким образом, чувствительные нейроны спинальных ганглиев в экспери-
ментальных условиях несомненно образуют друг с другом, подобно элемен-
424
Глава III
там других тканей, цитоплазматические связи совершенно иного характера,
чем синапсы. Такие же тесные цитоплазматические связи они могут обра-
зовать и с элементами нейроглии. Несомненно, что в течение роста устано-
вившиеся сетевидные связи между отростками нейронов, а также между
ними и другими элементами зоны поста могут напутаться и возникать за-
ново.
Относительно того, каковы взаимоотношения между соединяющимися
отростками нейронов, можно высказать те же предположения, что и отно-
сительно других клеточных элементов, соединяющихся друг с другом с по-
мощью цитоплазматических связей. Здесь может иметь место более или менее
тесное соприкосновение, или слипание, или же непрерывная цитоплазмати-
ческая связь, которую в ряде случаев подчеркивал Леви. Мне кажется,
что этот вопрос не имеет такого принципиального значения, какое ему
придают некоторые исследователи, так как несомненно, что, пожалуй, за
исключением симпластических элементов мускулатуры соматического типа,
элементы зоны роста тканевых культур, подобно тканевым элементам целого
организма, представляют собой достаточно четко отграниченные друг от
друга, большей частью одноядерные протоплазматические территории или
клетки, обладающие в каждой данной системе известной, правда, лишь
относительной, самостоятельностью. Межклеточные границы ь ряде слу-
чаев с полной убедительностью констатируются непосредственным наблю-
дением на гистологических препаратах, а также на живых объектах. Правда,
в случае взаимной связи самых разнообразных элементов с помощью тон-
ких цитоплазматических отростков установить морфологически различи-
мую границу часто бывает невозможно. Связи, устанавливающиеся между
нейронами в условиях тканевых культур, как раз относятся к такого рода
соединениям. Целесообразно напомнить отдельные наблюдения над клет-
ками ненейрональной природы, которые не могут не иметь значения для
понимания сетевидных структур, образуемых разветвлениями аксонов.
Внимательное изучение живых культур показывает, что изоляция так или
иначе соединенных друг с другом клеток может наступать довольно быстро
и без следа какого-либо разрыва (7И. Льюис, 1922). Отростки фибробласто-
образных элементов или клеток мезотелия могут втягиваться и утрачивать
взаимную связь. С образованием новых отростков связь может снова вос-
станавливаться. Если наблюдать за движением мелких оформленных частиц
в живой клеточной цитоплазме, то в подавляющем большинстве случаев
нельзя установить их перетекания из одной клеточной территории в другую
(де-Гари, 1924; Леви, 1925, и др.). Лишь в единичных случаях Леви указы-
вает на их передвижение через клеточные отрос гки из одной клетки в сосед-
нюю. После этого, по его данным, бывшие столь тесно соединенными эле-
менты могут расходиться.
Опыты с повреждением отдельных клеток зоны роста иглой микр )ма-
нипулятора (Чемберс, 1924; Чемберс и Фелл, 193’.) показывают, что даже
в тех случаях, когда межклеточные границы в живом виде незаметны,
клетки, подвергнутые уколу, обособляются от соседних элементов. На
последние, однако, вызванные уколом изменения (коагуляция ядерного
Экспериментальный метод в гистологии 425
содержимого) не распространяются. Это вполне совпадает с данными
(Чемберс и Реньи, 1925; Чемберс и Ледфорд, 1931), полученными на эпите-
лиях расщипанных частиц целого ряда нормальных органов. Лишь в диф-
ференцированном многослойном эпителии целого организма оказалось
возможным доказать прижизненно наличие очень устойчивых протоплазма-
тических мостиков, соединяющих отдельные клетки друг с другом, и распро-
странение на соседние элементы повреждений, вызванных уколом в какой-
нибудь одной клетке.
Под влиянием гипотонической среды элементы комплексных и рыхлых
клеточных соединений могут, утрачивая взаимную связь, приобретать
округленную форму, причем эти изменения оказываются, в известных'
границах, обратимыми (Гросфельд, 1936).
Нужно думать, что в подавляющем большинстве случаев между отдель-
ными клетками имеет место лишь более или менее тесное соприкосновение
или слипание, а не протоплазматическая непрерывность. Такое соединение
осуществляется или между отдельными клеточными отростками или между
плотно прилегающими друг к другу краями или поверхностями соседних
клеток (например в эпителиообразных комплексах); при наступлении раз-
рыхления в таких комплексах могут наблюдаться и межклеточные связи
первого типа. Исчезновение поверхности раздела между протоплазмам.!
двух клеточных элементов, если и происходит, то, повидимому, лишь как
редкий эпизод, не влекущий за собой утраты известной индивидуальности
данных ядерно-протоплазматических территорий. Это — лишь частный
случай наиболее тесной взаимной связи двух элементов, сохраняющих
свою известную индивидуальность. Полное слияние двух или более клеток
с образованием единого гигантского дву - или многоядерного протоплазма-
тического тела или плазмодия может, повидимому, происходить в некото-
рых случаях у свободных амебоидных элементов типа макрофагов.
Едва ли способ взаимосвязи нейронов в условиях тканевых культур
принципиально отличается от того, что известно в отношении других кле-
точных элементов. Бо всяком случае нет никаких причин использовать
данные тканевых культур в качестве аргумента против вполне обоснован-
ной в настоящее время нейронной теории строения нервной системы и
в пользу несоответствующей реальным фактам нейрофибриллярной гипо-
тезы непрерывности или более новой гипотезы «нейренцития» Гелъда (1909),
или, наконец, допускаемой в новейшее время Герриком, по примеру неко-
торых очень старых взглядов, синцитиальной связи между дендритам!!
нервных клеток и защищаемого Штёром мл. (1928—1934) сетевидного строе-
ния автономной нервной системы.
Наблюдения Леви и Делоренци (1934), Леви и Мейер (19^1), касаю-
щиеся клеток ганглиев автономной нервной системы и характера образуе-
мых ими вне организма разрастаний, не могли установить каких-нибудь
ясных отличий от картин, описанных теми же авторами в эксплантатах
из спинальных ганглиев. Имея в виду другой нейрональный
состав ганглиев автономной нервной системы и их значение в отличие от
спинальных ганглиев как нервных центров, трудно принять эти данные
426
Глава III
безоговорочно. Наблюдения над ростом вне организма элементов эксплан-
тированных ганглиев автономной нервной системы отличаются значительно
меньшей полнотой, причем и в этом случае некоторые структуры зоны роста,
описанные в качестве нейрональных, повидимому. не являются таковыми,
а должны быть отнесены за счет нейроглии.
Проанализированные выше экспериментальные данные позволяют утвер-
ждать, что чувствительные нейроны млекопитающих отличаются рядом
гистотипических особенностей от некоторых других достаточно детально
изученных нервных клеток. Это хорошо гармонирует с некоторыми опытами,
произведенными в лаборатории Лаврентьева (1938), согласно которым при
образовании искусственных синапсов клетки спинальных ганглиев ведут
себя иным образом, чем клетки ганглиев автономной нервной системы.
В цитоплазме различных нервных элементов в условиях тканевых куль-
тур на обработанных специальными нейрологическими методами препа-
ратах обнаруживаются то более, то менее типичные нейрофибриллярные
структуры (Леви, 1917, 1934; Эзаки, 1929; Григорьев, 1932, и др.).
В некоторых случаях отчетливые нейрофибриллярные картины отмечались
и в живых нейронах (Вейс и Хси Ванг, 1936; Леви и Мейер, 1937). В отрост-
ках чувствительных нейронов кроликов в условиях тканевых культур
прижизненная видимость нейрофибриллей может быть совершенно ясной.
В некоторых случаях их видимость улучшается, если культуры подверга-
лись охлаждению или воздействию некоторых агентов, вызывающих пара-
некротические изменения.
В заключение представляется целесообразным привести некоторые
наблюдения физиологического характера, полученные с эксплантирован-
ными нейронами методами микрургии, так как, помимо собственного инте-
реса, они уточняют и дополняют некоторые из разобранных выше данных
морфологического характера, касающихся, в частности, образований свя-
зей между отростками нейронов и другими клеточными элементами, расту-
щими вне организма.
Под влиянием электрического тока, непосредственно подводимого к нерв-
ным клеткам или их отросткам неполяризующимися микроэлектродами,
в них наблюдается ряд довольно характерных изменений, обнаруживаю-
щих закономерную зависимость от силы тока, продолжительности раздра-
жения и состояния данной нервной клетки в момент опыта (Петерфи и
Уильямс, 1933). В жизнеспособных нервных клетках, цитоплазма которых
отличается очень большой вязкостью и находится в состоянии плотного
геля (Леви, 1934), при пропускании постоянного тока силой от нескольких
тысячных до сотых миллиампера наблюдается появление зернышек, окра-
шивающихся прижизненно метиленовой синью. В их отростках появляются
четковидные вздутия. Особенно заметным бывает набухание колбовидного
конца нервных волокон. После размыкания тока происходит усиленный
рост нервных отростков, в направлении бывшего анода. Поврежденные или
стареющие нервные элементы реагируют в культурах на электрический ток
иначе, причем в них может ускоряться появление дегенеративных изме-
нений.
Экспериментальный метод в гистологии
427
Нервные отростки, вступающие в связь с элементами других тканей
и снабженные концевыми пуговками или петельками, всегда бывают ясно
отграничены от протоплазмы последних (Григорьев, 1932; Петерфи и Уиль-
ямс, 1934). Вместе с тем связь между нервными отростками и телом ткане-
вого элемента другой природы может быть очень интимной. По упомянутым
исследованиям Петерфи и Уильямса, при раздражении микроэлектродами
нервного отростка, оканчивающегося на глиальной или фибробластооб-
разной клетке, в участке цитоплазмы последних, соединенном с нервным во-
локном, наблюдается заметное набухание. Таким же образом реагирует их
цитоплазма при непосредственном раздражении электрическим током. Еще
этчетливее и вместе с тем иначе реагируют элементы сердечной и сомати-
ческой мускулатуры на раздражения током подросшего к ним нервного
волокна. В них так же, как и при непосредственном раздражении, можно
обнаружить местную волну сокращения. Эти опыты доказывают, что нерв-
ные элементы, растущие в условиях тканевых культур, имеют характерную
раздражимость и могут передавать импульсы тем элементам, с которыми
они вступают в связь.
В заключение следует еще остановиться на том, почему даже такие
опытные исследователи, как например Леви, в ряде случаев несомненно
принимали и описывали нейроглию как нервные элементы. В главе, посвя-
щенной эпендимо-глиальному тканевому типу, неоднократно подчеркива-
лось поразительное сходство в условиях тканевых культур между нейро-
глиальными, астроцитарными элементами и нейронами, которое, в част-
ности, в случае опухолевой глии доходило до полной симуляции роста нерв-
ных отростков. После первых наблюдений за ростом эксплантатов из цен-
тральной нервной системы у меня также не могло не возникнуть мысли,
что и в моих опытах, наряду с нейроглиальными разрастаниями, появля-
лись эмигрировавшие из центрального кусочка или, быть может, даже заново
дифференцирующиеся нейроны. Настолько велико было сходство между
биполярными и мультиполярными клетками зоны роста и- нейронами. Но
тщательное изучение материала показало с несомненностью, что в цент-
ральных кусочках эксплантатов из стенки боковых желудочков, за очень
редкими исключениями, вообще не встречалось нейронов. В таких же
совершенно единичных случаях, когда они были констатированы в течение
первых дней культивирования, в зоне роста удавалось видеть появление
очень немногочисленных, одного, двух или нескольких цитоплазматических
отростков нейрональной природы, которые, отличаясь значительной тол-
щиной, никогда не достигали большой длины. Появление же описанных
выше нейроноидных элементов было самой обычной картиной. Не было
никаких оснований говорить о миграции предсуществующих нейронов в зону
роста. Оставалась еще одна возможность, которую следовало учитывать:
присутствие в составе центральных кусочков эпендимы, элементы которой
могли бы сохранить у очень молодых животных нейробластические потен-
ции. Тщательное изучение центральных кусочков на срезах не обнаружило
картин, которые, хотя бы с натяжкой, могли трактоваться в смысле диф-
ференцировки нейробластов из малодифференцированных камбиальных кле-
428
Глава III
ток. Наконец, во многих случаях рост нейроноидных клеток наблюдался'
и в эксплантатах из стенки боковых желудочков и в культурах спинного
мозга, когда весь целиком центральный кусочек уже успевал приобрести
строение однородного глиального массива, в составе которого никаких
других элементов на непрерывных сериях срезов не обнаруживалось.
Все это заставило меня отказаться от заманчивой мысли, что нейронопо-
добные клетки являются настоящими нейронами или нейробластами, а не
симулирующими их астроцитарными элементами нейроглии. То же самое
говорили и культуры из опухолевой глии, в которых особенно полная симу-
ляция роста нервных отростков была получена в случае высокодифферен-
цированной волокнистой астроцитомы, а не какой-нибудь малодифферен-
цированной опухоли, в составе которой можно было предполагать наличие
каких-нибудь патологических индифферентных клеток.
Все эти данные и очень большой накопившийся материал, исключавший
какие бы то ни было случайности, позволили дать с полной определенностью
правильную нейроглиальную трактовку постоянно наблюдавшихся нейроно-
подобных элементов и вместе с тем заставили подойти критически к соот-
ветствующим наблюдениям ряда авторов. Как это уже вкратце отмечалось
выше, симуляция нейроглией нервных клеток представляет, с моей точки
зрения, очень большой интерес, хотя по сути дела отнюдь не является уди-
вительной. Ведь общая форма элементов астроцитарной глии, в особен-
ности в тех случаях, когда их отростки удается проследить на большом
протяжении, как это, например, делал Гирке (1885) на мацерированных
препаратах, отличается и в норме такой сложностью, которая их делает
во многих отношениях сходными с нейронами, несмотря на глубокие биоло-
гические различия. Факты, установленные методом эксплантации, и наблю-
дения над опухолевым материалом показывают вместе с тем, что сложно
разветвленная форма нейроглиальных элементов при обычных условиях
существования в составе нервной системы не представляет собой пассивного
приспособления формы вспомогательных глиальных элементов к условиям
их существования между нейронами с их сложными системами отростков,
а отражает гистобластические потенции этой ткани, приобретенные ею
в филогенезе и обнаруживающие несомненно очень большое сходство с гисто-
бластическими потенциями нейронов, столь резко отличающихся по своим
физиологическим свойствам. Мне кажется, что основной причиной этого
сходства не может не быть, помимо того, что глия и нейроны представляют
собой взаимосвязанные части единой системы, их генетическое единство,
т. е. их развитие в филогенезе и онтогенезе из одного и того же субстрата —
эктодермы, обладавшей, вероятно, и у предков позвоночных нейробласти-
ческой и глиобластической функцией, а затем специализированного нерв-
ного зачатка, дающего начало главным и вспомогательным элементам нерв-
ной системы. Нейроглия в ее разнообразных модификациях и нейроны
представляют собой, по сути дела, части генетически единого нейрального
тканевого типа, которые, хотя и развились дивергентно, тем не менее.,
в силу своих близких родственных отношений, обнаруживают несомненное
сходство некоторых элементарных гистобластических потенций. В есте-
Экспериментальный метод в гистологии 429
ственной системе гистологических структур эпендимо-глиальные ткани
и нейроны должны рассматриваться как специализированные в разных
направлениях части единого нервного или нейрального тканевого типа,
генетически связанного с эктодермой или эпидермальным покровом отда-
ленных предков, но приобревшего у позвоночных и, вероятно, у ряда дру-
гих высокоорганизованных животных выраженные черты единой общетка-
невой и вместе с тем исключительно разнообразной специальной морфо-
физиологической дифференцировки.
Изложенные в настоящей главе общие соображения и фактический мате-
риал, разумеется, весьма далеки от желаемой полноты и ни в какой степени
не могут претендовать на разрешение многих основных вопросов, связан-
ных с тканевой спецификой нейронов. Задачей нашего изложения было
не столько решение, сколько постановка ряда вопросов с точки зрения
развиваемых и обосновываемых в настоящей книге общих установок. Вместе
с тем того, что было сказано, мне кажется, достаточно для доказательства
положения, что эволюционные изменения нейронов могут быть поняты под
углом зрения общебиологических эволюционных закономерностей.
Для объяснения эволюционных изменений в тканях нервной системы
нет необходимости постулировать какие-то специальные гистологические
закономерности, принципиально отличающиеся от общих закономерностей
эволюционной морфологии. Если в отношении нейрональных взаимоотно-
шений нервная система является очень благодарным материалом для демон-
страции тканевых параллелизмов, существующих наряду с явлениями
тканевой дивергенции, то и эволюция анатомических особенностей нервной
системы в двух основных филогенетических стволах животного мира —
у первичноротых и вторичноротых — также обнаруживает, помимо явле-
ний дивергенции, и резко бросающиеся в глаза параллелизмы очень общего
характера, развивающиеся совершенно независимо друг от друга и свя-
занные с приобретением известных черт сходства в общей организации.
Такими анатомическими параллелизмами в эволюции нервной системы
являются ее все большая и большая концентрация и кефализация, зави-
сящие от параллельно развивающейся билатеральной симметрии свободно
подвижных организмов, дифференцировки у них головной и хвостовой
частей тела и развития дистантных рецепторов на переднем конце. Форми-
рование головного и туловищного мозга и вторичное анатомическое обо-
собление внутренностной или автономной нервной системы происходит
независимо в обоих крупных стволах филогенетического дерева. То же
самое справедливо и в отношении дальнейшей дифференцировки централь-
ной нервной системы, связанной с происходящей параллельно у первично-
ротых и вторичноротых дифференцировкой мускулатуры, которая, разви-
ваясь у членистоногих и позвоночных из кожно-мышечного мешка, дает
сложную систему мышц, антагонистов и синергистов. Последнее связано,
в свою очередь, с образованием у этих, систематически весьма отдаленных
групп конечностей, построенных по типу расчлененных рычагов. Оче-
видно, параллельное развитие анатомических особенностей нервной системы,
связанных с параллельным общим повышением организации животных,
430
Глава 111
находится под контролем творческого и элиминирующего значения есте-
ственного отбора. Параллельная эволюция ряда особенностей нервной си-
стемы, относящихся к области цитологии и микроскопической анатомии,
не может не быть связана неразрывным образом с этими общими анатоми-
ческими параллелизмами и тем бесспорным фактом, что ее гистологический
материал, нейроны, преемственно развивается из одного и того же источ-
ника. Тем самым, через отбор целых организмов или, точнее, целых онто-
генезов, и одновременно с ним совершается отбор и соответствующих гисто-
логических структур или гистогенезов. Нет ничего удивительного, что именно
в нейрональных соотношениях общие анатомические параллелизмы сказы-
ваются наиболее резким и отчетливым образом и заставляют подчас забывать
о существующих одновременно гистологических дивергенциях, связанных
в свою очередь с дивергентными чертами типовой организации. Экспери-
ментальное изучение гистотипических особенностей и гистогенеза ткане-
вого материала нервной системы у представителей разных филогенетических
рядов несомненно не только откроет ряд совершенно новых фактов, с кото-
рыми нельзя будет не считаться в будущем, но и позволит значительно
углубить и уточнить все эти положения общего характера.
IV. ЕСТЕСТВЕННАЯ СИСТЕМА ГИСТОЛОГИЧЕСКИХ
СТРУКТУР
Если сопоставить результаты экспериментально-гистологического ана-
лиза тканей в его филогенетической трактовке с данными сравнительной
анатомии и эмбриологии, а также механики развития, то в составе орга-
низма многоклеточных животных можно установить ряд типов или систем
гистологических структур, развитие которых, при наличии параллелизмов
и явлений конвергенции, отмечавшихся неоднократно и объясняющихся,
в конечном итоге, единством происхождения всего многообразия живот-
ных организмов, происходит по основному закону эволюционной морфо-
логии, схематически изображаемому в виде филогенетического дерева с ди-
вергентно расходящимися ветвями. В ряду позвоночных генетические
отношения большей части тканей вырисовываются уже в настоящее время
достаточно отчетливо. В отношении тканей других филогенетических ря-
дов, в частности тех, которые относятся к другому стволу филогенети-
ческого дерева, к первичноротым, еще остается много неясностей.
Тем не менее й для ряда тканевых типов беспозвоночных, на основании
имеющихся данных, также допустимо сделать некоторые филогенетиче-
ские предположения.
Естественная генетическая система тканей вовсе не заключается в меха-
ническом перенесении принятых эмбриологических данных в область гисто-
логии. Попытки такого рода, принадлежавшие еще Геккелю (1885), а затем
неоднократно повторявшиеся, оказались неудачными, так как не позво-
лили обнаружить в тканях каких-либо качественных особенностей, свя-
занных с их происхождением из разных источников. Кроме того, в настоя-
щее время рекапитуляциям в эмбриональном развитии уже невозможно
придавать, особенно на ранних стадиях онтогенеза, того значения, которое
им приписывалось прежде.
Теория филэмбриогенеза, разработанная школой покойного акаде-
мика А. Н. Северцева, не только демонстрирует на многих конкретных при-
мерах взаимообусловленность онтогенеза и филогенеза, но и устанавли-
вает ряд общих закономерностей этой взаимосвязи.
Необходимым следствием филогенетической системы тканей является,
наоборот, дальнейшее изменение трактовки ряда моментов индивидуаль-
ного развития. Это относится не только к мезодерме, представляющей
собой собирательное понятие и объединяющей ряд неоднородных зачатков,
но отчасти и к стенке вторичной кишечной трубки зародыша позвоночных,
482
Глава IV
в составе которой следует различать, помимо большой собственно эктодер-
мальной части, особые зачатки головной кишки и, повидимому, также
клоаки, представляющие собой сместившиеся в филогенезе части кожной
эктодермы.
Наиболее рано обособившимися друг от друга главными стволами фило-
тистогенетического дерева являются, с нашей точки зрения, первичные
и мультифункциональные энтодерма и эктодерма, дающие в дальнейшем
начало всему многообразию гистологических структур. Часть производ-
ных этих двух основных тканевых систем превращается у всех типов жи-
вотных в ткани энтеродермального и эпидермального типов, имеющие
морфо-физиологическую характеристику кишечного и кожного эпителиев.
Эпителии кишечного типа, являющиеся основной тканью, определяю-
щей функциональное значение пищеварительного аппарата, обнаружи-
вают в соответствии со своей физиологической специализацией в составе
пищеварительной трубки, параллельно развивающейся в различных
филогенетических ветвях, очень демонстративные гистологические па-
раллелизмы. При дивергентной эволюции из однослойных эпителиев
кишечного тина развиваются специализированные в разных направлениях
участки выстилки пищеварительной трубки, железистые и, наконец, эндо-
криновые структуры, утрачивающие в большей или меньшей степени функ-
циональную и морфологическую характеристику эпителиев.
Тип, или система, энтеродермальных тканей представлен в онтогенезе
позвоночных производными большей средней части вторичной кишечной
трубки или энтодермы в узком смысле слова, а у первичноротых — про-
изводными всего эктодермального зачатка. Начиная с ранних этапов эво-
люции, ткани энтеродермального типа приобретают высокую степень гис-
тологической детерминации. Они не образуются из каких-либо других
тканей и не обнаруживают способности к превращениям в ткани других
типов. У позвоночных, в частности млекопитающих и человека, явления
метаплазии энтеродермальных тканей возможны только в пределах опре-
деленных, свойственных им гистологических структур.
Возникновения дифференцированных гистологических структур, свой-
ственных другим тканевым типам, в частности эпителиям другого про-
исхождения, из энтеродермальных тканей не наблюдается. Этим ставятся
определенные границы возможных, но отнюдь не обязательных для эпи-
телиев кишечного типа явлений метаплазии. В состоянии роста и проли-
ферации в экспериментальных и патологических условиях ткани энтеро-
дермального типа у позвоночных и, вероятно, всех достаточно высокоорга-
низованных животных, во всей совокупности своих превращений полностью
сохраняют свои качественные отличия от тканей других типов. Процессы
регенерации энтеродермальных тканей, а также воспалительных разраста-
ний отличаются определенной, свойственной только этому тканевому типу
и разобранной выше спецификой, обнаруживающей определенные вари-
анты у их дифференцированных различным образом представителей и обу-
словленной всей историей их развития. Опухоли энтеродермального типа,
во всем многообразий своих структур, охватывают собой вполне опреде-
Естественная система гистологических структур
433
ленную естественную группу бластоматозных тканей единого происхо-
ждения, гистобластические потенции которых определяются их развитием
из тканей энтеродермального типа и степенью опухолевой катаплазии.
Эпителии кожного типа представляют собой филогенетически очень
удаленный от кишечных эпителиев тканевый тип, несмотря на их принад-
лежность к одной и той же морфо-физиологической эпителиальной группе
тканей классической гистологии.
В разных филогенетических стволах животного мира они приобретают
эволюируя дивергентно, очень неодинаковую морфо-физиологическую
специализацию. В пределах одного и того же филогенетического ряда жи-
вотных они обнаруживают резко выраженные явления дивергентной диф-
ференцировки, образуя покровные пласты очень разнообразной структуры,
железистые и эндокриновые образования, мышечные элементы, а также
поверхности внешнего газообмена. При дивергентной дифференцировке
и высокой функциональной специализации эпидермальные ткани могут
в большей или меньшей степени утрачивать морфо-физиологическую харак-
теристику эпителиев. У позвоночных эпителиальные покровы кожного типа
характеризуются в большей части случаев особыми многослойными, много-
рядными и двурядными структурами и, только в случае особой специали-
зации, однослойными-и однорядными. В состоянии роста и пролиферации
при экспериментальных и патологических условиях почти все эпидермаль-
ные образования млекопитающих отличаются способностью к превраще-
нию в многослойные покровные пласты или погружные клеточные массивы,
характеризующиеся рядом специфических «эпидермальных» особенностей,
и могущие во многих случаях обнаруживать у млекопитающих явления
ороговения. Единственным известным в настоящее время исключением
из этого правила является высокоспециализированная и, повидимому,
претерпевшая эволюционное гистологическое упрощение респираторная
выстилка легких, существование которой, в противоположность некото-
рым популярным в настоящее время взглядам, представляется более чем
вероятным.
Ткани эпидермального типа развиваются в онтогенезе из кожной экто-
дермы и эктодермальных же зачатков выстилки переднего и заднего отде-
лов пищеварительной трубки.
У позвоночных связь зачатка всего переднего эпидермального отдела
пищеварительной трубки с кожной эктодермой в онтогенезе утратилась
и не прослеживается, но подчеркивается его очень ранней эпидермальной
детерминацией и рядом специфических качественных отличий от собственно
эктодермального зачатка.
То же самое справедливо и в отношении заднего клоачного отдела пище-
варительной трубки. У высших позвоночных к эпидермальному тканевому
типу относятся производные внезародышевой эктодермы, а у млекопитаю-
щих —также заднего эктодермального отдела вольфова канала. В отношении
производных вольфова канала низших позвоночных до настоящего времени
имеются неясности. Только эпидермальным образованиям млекопитающих,
sa исключением респираторной выстилки легких, свойственны явления
28 Хлопин—349
434
Глава IV
метаплазии в многослойные и даже ороговевающие покровные пласты,
построенные по типу кожного эпителия. При различных пролиферативных
процессах эпидермальные ткани по динамике своих превращений каче-
ственно отличаются от гистологических структур, образуемых тканями
других типов, не переходя в них и не возникая из них заново. Процессы
заживления ран происходят в эпидермальных покровных пластах особым,
специфическим для них образом. Опухоли эпидермального типа во всем
большом многообразии своих структур охватывают собой единую есте-
ственную группу бластоматозных тканей.
Нейральный или нервный тканевой тип возникает очень рано путем
дивергентной дифференцировки эктодермы и первоначально обнаружи-
вает очень близкие генетические отношения к эпидермальным тканям,
поскольку элементы нервной системы могут образоваться (например еще
у аннелид из первичноротых) из эпидермиса кожи при регенеративных
процессах. Лишь на относительно поздних этапах эволюции дивергентная
дифференцировка и гистологическая детерминация нейроэктодермального
зачатка закрепляются окончательно на ранних этапах онтогенеза и при-
водят к его полному обособлению от кожной эктодермы или эпидермаль-
ного зачатка. Только после этого делается возможным говорить о появлении
в филогенезе особого нейрального тканевого типа, развившегося из нейро-
бластического примитивного эпидермиса. У предков современных позво-
ночных, до образования нервной трубки, нервная система, вероятно, рас-
полагалась в составе дорзального участка эпидермиса, элементы которого,
повидимому, обладали нейробластическими потенциями и имели вместе
с тем значение кожного органа чувств или плакоды. Сходные отношения
сохранились в значительной степени в эпинейральной нервной системе
современных иглокожих. Нервнаятрубка личинки ланцетника еще сохраняет
через нейропор непосредственное сообщение с внешней средой. У современ-
ных позвоночных источником развития тканей нейрального типа являются
нервная пластинка и плакоды, полное обособление которых от эпидермаль-
ного зачатка происходит, повидимому, несколько позже. Нейральный
тканевой тип позвоночных отличается исключительным разнообразием
своих морфо-физиологических модификаций и включает в себя все вспо-
могательные компоненты нервной системы, т. е. ткани эпендимо-глиального
типа с т. наз. нейроэпителиальными и мионейральными сократимыми эле-
ментами, хроматофорами, производными «эктомезенхимы» и свои наиболее
специализированные части — нейроны. Разнообразные ткани нейраль-
ного типа, представляющие собой продукты дивергентной дифференцировки
единого нейрэктодермального зачатка, частью входят в состав всех четы-
рех тканевых групп классической гистологии, частью представлены совер-
шенно особыми структурами, не укладывающимися безоговорочно в рамки
обычной морфо-физиологической системы тканей. При пролиферативных
процессах вспомогательные компоненты нервной системы отличаются
очень большим своеобразием и высокой тканевой специфичностью. Ней-
роглия центральной нервной системы млекопитающих способна к образо-
ванию клеточных массивов особого характера, которые, возможно, указы-
Естественная система гистологических структур
485
вают на некогда близкие генетические отношения между нейральным и
эпидермальным тканевыми типами. Ее структура при обычных условиях
может трактоваться как очень сильно измененный, свернувшийся в трубку
и погрузившийся во внутреннюю среду организма эпидермальный пласт,
приобретающий в ряду позвоночных, параллельно с эпидермисом кожи,
многослойность особого характера. В известных чертах морфологического
сходства между нейронами и астроцитами нельзя не видеть одного из при-
знаков их генетического единства. Группа нейрогенных опухолей пред-
ставляет собой естественный, но очень полиморфный тип бластоматозных
тканей.
Ткань спинной струны, возможно, представляет собой специализиро-
ванную в особом направлении часть примитивного эпидермиса предков
современных хордовых, приобревшую значение особого тканевого типа
со своеобразными гистобластическими потенциями.
Хордомы представляют собой очень своеобразный тип дисонтогенети-
ческих опухолей.
Мускулатура соматического типа является у позвоночных и, повиди-
мому, во всех других филогенетических рядах особым очень древним ткане-
вым типом, который возможно со значительной долей вероятия филогене-
тически выводить из сократимых эктодермальных элементов отдаленных
общих предков. Первоначально мышечные элементы дифференцировались
диффузно, а затем стали возникать из локализированного в области бласто-
пора или первичного рта тканевого зачатка, постепенно вытеснившего
диффузную эктодермальную мускулатуру и сделавшегося древнейшей
частью мезодермального комплекса. Такой путь филогистогенеза муску-
латуры соматического типа, однако, не может считаться бесспорным. Раз-
виваясь из одного и того же источника и сохраняя в своей эволюции морфо-
физиологическую характеристику мышечной ткани, мускулатура сома-
тического типа обнаруживает, наряду с явлениями дивергентного раз-
вития, отчетливые явления параллелизмов у представителей разных
филогенетических рядов. В онтогенезе позвоночных элементы мускула-
туры соматического типа генетически связаны с определенными участками
мезодермы, миотомами, материал которых может вторично смещаться.
В условиях роста вне организма и при регенеративных процессах в со-
ставе целого организма дифференцированные элементы мускулатуры сома-
тического типа характеризуются совершенно специфическим циклом пре-
вращений, завершающихся вторичной дифференцировкой сложных сокра-
тимых структур.
Миогенные опухоли соматической мускулатуры или рабдомиобластомы
могут отличаться очень неодинаковым уровнем дифференцировки, но пред-
ставляют собой генетически единый тип опухолевых тканей.
Целомическая выстилка является в главных филогенетических стволах
многоклеточных животных источником происхождения очень многочис-
ленных целонефродермальных тканей, которые, развиваясь дивергентно,
приобретают на высших этапах эволюции значение ряда самостоятельных
высокоспециализированных тканевых типов.
436 Глава IV
Имеются все основания рассматривать целомическую выстилку у вто-
ричноротых, в соответствии с ее энтороцельным способом развития в онто-
генезе низко организованных форм, как результат дивергентной эволюции
первичного эктодермального тканевого зачатка.
Принимая во внимание вторично резко измененный способ развития
большей части первичноротых, проделывающих характерное спиральное
детерминированное дробление с установкой на трохофорообразную личинку,
нельзя отрицать и в этом филогенетическом ряду возможности энтероцель-
ного возникновения целома, хотя вполне убедительных положительных
данных в этом отношении не имеется.
С этой точки зрения вторичную полость тела в двух главных филогене-
тических стволах можно рассматривать как параллельно возникшие из
одного и того же источника образования, материал стенки которых в ги-
стологическом отношении гомологичен.
Часть производных целомической выстилки развивается, сохраняя
в основном морфо-физиологическую характеристику эпителиальных тка-
ней, другие ее производные в процессе эволюции утрачивают в большей
или меньшей степени эпителиальную характеристику и превращаются
в особые, весьма разнообразным способом дифференцированные ткани
внутренней среды организма. Миокард и сравнимые с ним сократимые
образования беспозвоночных как особый целомический тип сократимых
тканей представляет собой мышечную морфо-физиологическую модифи-
кацию выстилки вторичной полости тела.
У высших позвоночных сама целомическая выстилка с образующимися
из нее мюллеровыми каналами и генетически связанные с ней зачатки вы-
делительного аппарата типа целомодуктов дают начало очень разно-
образным в отношении своих функциональных особенностей тканям цело-
дермального и нефродермального типов: мезотелию серозных оболочек,
фолликулярным клеткам гонад и элементам интерренальных органов,
утратившим в значительной степени морфо-физиологическую характери-
стику эпителиальных тканей, своеобразным эпителиям матки, яйцеводов
и почек. Целодермальные и нефродермальные ткани обнаруживают очень
высокую специализацию в разных направлениях и по своим гистобласти-
ческим потенциям отличаются как друг от друга, так и от тканей всех дру-
гих типов, в которые они никогда не превращаются и из которых не воз-
никают заново. У низших позвоночных различия между цело дермальными
и нефродермальными тканями оказываются выраженными в гораздо менее
значительной степени, и поэтому можно говорить об едином целонефро-
дермальном тканевом типе, дивергентно развивающемся в разных напра-
влениях. Опухолевые производные целонефродермального тканевого типа
представляют собой большую естественную группу, которая распадается
на ряд бластомотозных производных упомянутых выше тканей.
В настоящее время имеются основания рассматривать эндотелий сосу-
дистой системы позвоночных и кровяные клетки гемоглобинового ряда
как представителей особого тканевого типа, развившегося из целомиче-
ской выстилки предков и приобревшего черты очень высокой специали-
Естественная система гистологических структу р
437
зации. С этой точки зрения, сосудистая система позвоночных сравнима
не с лишенной эндотелия сосудистой системой беспозвоночных, а со снаб-
женными непрерывной целомической выстилкой каналами, которые суб-
ституируют ее функционально, достигая у отдельных представителей высо-
кого уровня развития. Генетическая близость эндотелия и соединительной
ткани, признаваемая в настоящее время, не полностью обоснована и допу-
скает, в свете экспериментально-гистологического анализа, совершенно
иную трактовку. Трактовка группы опухолей, обозначаемой как эндо-
телиомы, которая с точки зрения Фишер-Вазельса (1927) в настоящее время
не имеет больше прав гражданства, должна быть, с изложенной точки зре-
ния на эндотелий, вновь пересмотрена. Весь вопрос об эндотелии сосуди-
стой системы нуждается еще для окончательного решения в дополнитель-
ных исследованиях.
Очень пестрая группа мезенхимных или межуточных тканей, объеди-
нявшая до последнего времени, помимо всего многообразия соединитель-
нотканных образований, свободные клетки полостной жидкости, крови
и лимфы, эндотелий сосудистой системы, гладкие мышцы позвоночных
и различные производные т. наз. эктомезенхимы, должна быть с точки зре-
ния естественной системы гистологических структур подвергнута серьёз-
ному пересмотру. Само понятие мезенхимных производных, основываю-
щееся на гертвиговском морфо-физиологическом понимании мезенхимы,
как совокупности свободных или рыхло связанных друг с другом клеток
внутренней среды организма, могущих в онтогенезе и филогенезе возни-
кать из разных источников, не может быть положено в основу генетиче-
ского подхода к тканям. Мезенхима так же, как и эпителий, является по
своему существу морфо-физиологической категорией. Подобно тому, как
нами было показано, что в составе группы эпителиев имеются ткани раз-
ного происхождения, обладающие качественно неодинаковыми биологи-
ческими особенностями и неспособные к превращениям друг в друга, рых-
лые мезенхимные клеточные соединения также не являются равноценными
друг другу. Некоторые из них уже в настоящее время с большой долей
вероятия или даже с достоверностью могут быть включены в состав разных
тканевых типов генетической системы.
Самому по себе факту разрыхления тканевого зачатка или образования
свободных, не связанных друг с другом клеток в онтогенезе не должно при-
даваться решающее значение без дополнительного исследования гисто-
бластических потенций соответствующих эмбриональных клеток и образую-
щихся из них элементов дифференцированных тканей. Хотя в отношении
вопроса о мезенхимных тканях, с точки зрения естественной гистологи-
ческой системы, еще остается много неясностей и филогенетические суб-
ституции в этой области вовсе не могут быть исключены, его предположи-
тельное решение намечается в настоящее время в следующих общих чер-
тах. Часть мезенхимных производных, вероятно, имеет очень глубокие фило-
генетические корни, которые следует искать в мезенхимных клетках жи-
вотных, родственных современным кишечнополостным и сделавшихся пред-
ками всех вышестоящих ископаемых и современных форм. Возможно, что
438
Vitaea IV
источником образования этой древнейшей мезенхимы являлась первичная
мультифункциональная эктодерма, а ее дальнейшее развитие шло в тесной
коррелятивной связи с развитием эпидермальных тканей и мускулатуры
соматического типа. Подобно тому, как образование мускулатуры сома-
тического типа затем сконцентрировалось в области краев первичного рта,
можно допустить дальнейшее развитие соединительнотканных структур
из зачатка, локализировавшегося в этой же области и превратившегося
в «энтомезенхиму» вышестоящих организмов. С этой точки зрения, обнару-
живающаяся связь между целомической мезодермой, с одной стороны, му-
скулатурой соматического типа и «энтомезенхимой», с другой, представляется
вторичной и чисто топографической, не отражающей их действительных
отношений друг к другу, а сам термин «энтомезенхима» — не отражающим
филогенетического пути развития этого тканевого зачатка. Ларвальная
эктомезенхима первичноротых может трактоваться как остаток диффуз-
ного эктодермального мезенхимного зачатка предков, который в даль-
нейшем исчезает без остатка. Древний «энтомезенхимный» зачаток дает
начало исключительно разнообразным дивергентно развивающимся в раз-
ных филогенетических ветвях соединительнотканным структурам и части
свободных клеток полостной жидкости или крови. У позвоночных ткани
этого типа достигают исключительно высокого развития и образуют об-
ширную систему собственно соединительнотканных структур более общего
характера, высокоспециализированных тканей механического значения,
особую внутренностную мезенхимную мускулатуру и различные белые
клетки крови и лимфы.
К этой древнейшей и, повидимому, генетически единой части т. наз. мезен-
химного зачатка у позвоночных дополнительно присоединяются другие
зачатки, имевшие иные пути исторического развития и по своему происхо-
ждению относящиеся к другим тканевым типам естественной гистологиче-
ской системы. Таковыми являются, во-первых, т. наз. эктомезенхима (или
нейромезенхима) и, во-вторых, повидимому, зачаток сосудистого эндо-
телия и клеток гемоглобинового ряда.
Эктомезенхима позвоночных является частью нервного зачатка и по
своим гистобластическим потенциям должна быть отнесена к тканям глио-
эпендимного типа. Это может считаться прочно установленным в отноше-
нии покровных клеток мозговых оболочек и нуждается еще в эксперимен-
тальных доказательствах для некоторых других эктомезенхимных произ-
водных. К нейрогенным производным относятся, по сути дела, также хро-
матофоры позвоночных. Относительно вероятного положения сосудистого
эндотелия и клеток гемоглобинового ряда в естественной системе гисто-
логических структур уже говорилось выше. Наконец, те «мезенхимные»
клетки, которые, по многим данным, дают у позвоночных начало некото-
рой части мускулатуры соматического типа, являются по сути дела не мезен-
химными, а особыми миобластическими элементами, отделившимися от
миотомов, т. е. специфического мышечного зачатка. Таким образом, чрез-
вычайная сложность и многообразие т. паз. группы мезенхимных тканей
позвоночных зависят от двух причин: от очень сильного развития и разно-
Естественная система гистологических структур
439
образной дивергентной дифференцировки той части «энтомезенхимного»
зачатка, которая, в соответствии с современным уровнем знаний, может
трактоваться как генетически единый тканевой тип, или единая тканевая
система, и от присоединения к этой части мезенхимного зачатка генети-
чески чуждых ему образований, относящихся к другим тканевым типам,
но вступающих вторично в тесные топографические и функциональные
взаимоотношения и могущих вместе с тем обнаруживать явления конвер-
гентной морфо-физиологической дифференцировки.
" Все сказанное в отношении нормальных «мезенхимных» тканей отно-
сится в совершенно равной степени к их опухолевым производным, часть
которых, повидимому, образует естественную генетически единую группу
(например многие фибробластомы, лейомиобластомы, хондробластомы,
•остеобластомы в их различных модификациях и, вероятно, также лейкозы
и лейкемические заболевания), а другие должны быть отнесены к соответ-
ствующим другим опухолевым типам (например менингобластомы, мела-
номы и нейриномы к опухолям или опухолеподобным заболеваниям нерв-
ной системы). Как это было отмечено в соответствующей главе, в отноше-
нии мезенхимных производных ряд вопросов может считаться в настоящее
время поставленным, но отнюдь не разрешенным до конца. Только даль-
нейшие исследования приблизят нас к правильному пониманию той исклю-
чительно сложной и важной группы тканей, которую в настоящее время
принято обозначать как группу мезенхимных производных, в отношении
которой мы, по моему глубокому убеждению, еще можем ожидать ряд не
учитываемых в настоящее время неожиданностей.
Самый факт новой постановки вопроса о мезенхимных тканях, с точки
зрения естественной системы гистологических структур, является доста-
точно знаментальным и несомненно открывает новые перспективы в обла-
сти их гистологии и патологии. В отношении этой группы тканей следует
особенно помнить ее вспомогательное, обслуживающее все прочие ткане-
вые типы значение и вместе с тем невозможность правильного эволюцион-
ного решения всей проблемы в целом без учета сложных коррелятивных
взаимосвязей между «мезенхимными» производными и другими тканями
тела, которым принадлежит в составе тех или иных систем органов роль
основных, ведущих компонентов, определяющих их функциональное зна-
чение и вместе с тем воздействующих особенно активно на морфо-физио-
логические особенности вспомогательных частей и, очевидно, на гистогене-
тические процессы в них в течение филогенеза и онтогенеза.
Взаимоотношения между естественной системой гистологических струк-
тур и принятыми четырьмя морфо-физиологическими группами тканей
можно себе представить в виде прилагаемой схемы, составленной приме-
нительно к позвоночным, но с некоторыми частными изменениями, пови-
димому, справедливой и в отношении других типов многоклеточных живот-
ных (рис. 64). В этой схеме нейроглия, являющаяся по существу совокупно-
стью гистологических структур sui generis, не укладывающихся в систему
Лейдига и Кёлликера, включена не в группу соединительнотканных обра-
зований, как это часто делалось и делается, а в группу эпителиев, что также
440
Глава IV
является в достаточной степени искусственным. Ткань хорды, несмотря
на свои глубокие отличия от соединительнотканных структур, в соответ-
ствии с принятыми взглядами, включена в группу соединительных тканей
и клеток крови. Ряд гистологических структур, рассмотренных выше, для
Рис. 64. Схема, иллюстрирующая соотношения между естественной системой гистоло-
гических структур, основанной на историческом принципе, и принятым подразделением
всего их многообразия на четыре «ткани» или, точнее, морфо-физиологические группы
тканей. В целях упрощения не показана дальнейшая дивергенция основных гистоло-
гических типов или систем на отдельные конкретные ткани. Пунктирные линяй обозна-
чают генетические соотношения, которые еще нуждаются в дальнейшем изучении и
обосновании.
простоты не обозначен в этой схеме общего характера. Совершенно анало-
гичные схемы возможно составить для иллюстрации взаимоотношений
между естественной системой животных организмов и отдельными кон-
вергентными чертами их общей организации, как, например, лучевая,
билатерально-симметричная и спиральная форма, червеобразный или рыбо-
образный внешний облик, наличие построенных по рычаговому типу конеч-
ностей и тому подобное.
Весь изложенный выше фактический материал с несомненностью под-
тверждает, что филогистогенез приводит к образованию в составе тела мно-
гоклеточных животных ряда тканевых типов, которые, развиваясь дивер-
Естественная система гистологических структур
441
гентно и обнаруживая вместе с тем явления параллелизмов и конвергенций,
дают все многообразие известных гистологических структур. Совокуп-
ность биологических свойств каждой конкретной ткани представляет собой
результат всей истории ее развития, неразрывно связанной с эволюцией
общей и специальной организации животных.
Знаменательно, что под конвергентным сходством гистологических
структур, дававшим повод утверждать, что одинаковые ткани могут раз-
виваться из разных источников и что свойства тканей не зависят от их про-
исхождения, экспериментальный метод позволил обнаружить глубокие
биологические различия, зависящие от предыдущей дивергентной эволю-
ции. Явления сходства, не обусловленные близким родством, оказываются
в области гистологии по существу столь же поверхностными, как и в срав-
нительной анатомии, и отнюдь не уничтожают глубоких, обусловленных
исторически, различий. При дивергентной эволюции гистологических
структур во многих случаях очень отчетливо выступают принципы интен-
сификации, разделения и смены функций.
При изучении гистологических особенностей тканей не может не бро-
сится в глаза наличие в них, с одной стороны, очень значительного кон-
серватизма или эволюционной инертности, а с другой, наоборот, измен-
чивости и пластичности. Консерватизм тканевых структур проявляется
в том общеизвестном факте, что их морфологические особенности отли-
чаются большой долей постоянства в пределах крупных систематических
групп, несмотря на то, что такие особенности, как, например, размеры кле-
точного тела тканевых элементов, отличаются более значительной измен-
чивостью. Определить на основании препаратов отдельных тканей систе-
матическое положение животного бывает можно, даже для человека с очень
большим опытом, лишь очень приблизительно, а по виду яйцевых клеток
или тем более ооцитов в примордиальном фолликуле млекопитающих никто
не может сказать, какому именно животному они принадлежат и что из
них должно было бы развиться, хотя видовая специфичность белковых тел
является несомненным фактом.
Едва ли может подлежать сомнению, что гистологические и цитологи-
ческие особенности эволюируют значительно медленнее и отличаются боль-
шим постоянством, чем видовые признаки животных. Тем не менее са-
мый факт эволюционной изменчивости тканевых структур не вызывает
сомнений и легко бросается в глаза при сравнительном гистологическом
изучении представителей достаточно крупных систематических групп.
Более того, разные ткани в отношении своей изменчивости могут очень
сильно отличаться друг от друга, причем в одних случаях бросаются в глаза
параллелизмы, в других, наоборот, очень резко выступают черты дивер-
гентного развития.
При эволюционном развитии тканевых свойств можно обнаружить два
основных вида гистологической изменчивости. В одних случаях общая
морфо-физиологическая характеристика и вместе с тем принадлежность
гистологической струкгуры к одной из четырех групп тканей классиче-
ской гистологии сохраняются. Например, эпителиальная структура, эво-
441
Слава IV
люируя и изменяя свои морфо-физиологические особенности, сохраняет
некоторые общие для данного типа эпителиальных тканей признаки, или
та или иная соединительнотканная структура, несмотря »а свои изменения,
вса-таки остается структурой соединительнотканного характера. В других
случаях, наоборот, морфо-физиологическая характеристика гистологической
структуры, изменяясь в каком-нибудь направлении, утрачивает признаки
исходной тканевой группы и приобретает более общие или специальные
особенности тканей, принадлежащих к совершенно другой морфо-физиологи-
ческой группе гистологических структур. В таких случаях условность,
натянутость и даже несостоятельность обычных принципов классифика-
ции тканей, несмотря на всю их привычность и все их дидактические удоб-
ства, часто чрезвычайно резко бросаются в глаза.
Если принять в качестве общей функциональной характеристики группы
эпителиальных тканей подчеркнутый еще Гисом (1865) и бр. Гертвигами^ 1882)
и принятый затем многими авторами признак пограничности, то многие
эпителиальные структуры, эволюируя, по сути дела перестают быть
таковыми. Достаточно напомнить развитие эндокриновых структур из
покровных эпителиальных пластов, утрату пограничных свойств выстил-
кой серозных оболочек в ряду позвоночных или обеспечивающих внешний
газообмен эпидермальных структур, образование астроцитарной и свое-
образной периферической нейроглии и, вероятно, нервных элементов из
эпителиобразной эпендимы и, в конечном итоге, из примитивных эпидер-
мальных тканей, дифференцировку гладких и поперечнополосатых мышеч-
ных волокон и, по некоторым данным, хрящевой ткани из того же источ-
ника и много других, указанных или разобранных выше примеров. Из
межуточных, мезенхимных или соединительнотканных структур, поражаю-
щих своим многообразием, в филогенезе и онтогенезе также вторично
возникают образования, очень резко отличающиеся по своему строению
или функциональному значению от характерных структур этой тканевой
группы, — комплексные хордоидные тургорные образования, группы кле-
ток, напоминающих железистые элементы эндокриновых органов, типич-
ные мышечные структуры позвоночных и некоторые другие ткани. Мезен-
химные гладкомышечные клетки местами, например в артериовенозных
анастомозах, в свою очередь приобретают эпителиоподобный вид. Эле-
менты высокоспециализированных мышечных и нервной тканей, приобрев-
шие в филогенезе одностороннюю дифференцировку, вообще говоря, уже
сохраняют свою общую морфо-физиологическую характеристику, хотя
и из этого правила бывают исключения. Некоторые части мускулатуры
соматического типа у отдельных представителей позвоночных могут вто-
рично специализироваться в особую ткань электрических органов, утра-
чивающую сократимое значение, а нервные элементы, повидимому, пре-
вращаются в своеобразные железистые клетки адренальных органов. При
развитии ткани электрических органов имеют место смена одной очень
специальной и высокодифференцированной тканевой функции другой,
а вместе с тем и структурные изменения мышечных волокон, превращаю-
щихся в электропластинки. • . . , . .
Естественная система гистологических структур
443
Таким образом, филогенетическое развитие отдельных гистологических
структур может происходить или с сохранением их первоначальной при-
надлежности к той или иной морфо-физиологической группе тканей или
же с резко выраженным изменением ее. В последнем случае исходная гисто-
логическая структура, относившаяся по своей морфо-физиологической
характеристике к одной тканевой группе, превращается в эволюционном
процессе в гистологическую структуру, которая, с точки зрения гистоло-
гической системы, основанной на морфологических и функциональных
признаках, имеет другую групповую принадлежность.
Такие эволюционные изменения тканей являются демонстративными
примерами приложимости принципов интенсификации и смены функций
в области гистологии.
Наконец, эволюционные изменения гистологических свойств приводят
к образованию таких структур, которые вообще не укладываются в рамки
обычных общегистологических категорий и, как это отмечалось выше,
обычно рассматриваются не в общей гистологии, а в микроскопической
анатомии. Отнесение таких очень важных многочисленных и интересных
тканевых структур к той или иной тканевой группе классической гисто-
логии часто производится или на основе одних лишь дидактических сооб-
ражений или в результате достаточно произвольного и одностороннего
подчеркивания какой-нибудь одной морфо-физиологической особенности,
позволяющей со значительной натяжкой включить их в рамки принятой
искусственной системы. Нередко, чтобы спасти положение, приходится
отказываться от последовательного проведения морфо-физиологического
принципа классификации и в достал очной степени нелогично переходить
к использованию характеристики тканей по происхождению, которое со-
вершенно игнорируется при установлении основных групп гистологических
слруктур. В результате этого получается двойственность и неопреде-
ленность существующей гистологической системы Лейдига и Кёлликера,
которая, несмотря на наличие частичных исправлений или новые трак-
товки, не может удовлетворить в настоящее время исследователя и властно
требует пересмотра основных гистологических представлений.
История развития гистологических систем, весь фактический материал
и ряд общих соображений, изложенные в этой книге, указывают один из
кутей, по которому может и должна итти в настоящее время гистологи-
ческая мысль, берущая все ценное, что имеется в старых классических
представлениях нашей науки, широко использующая экспериментальные
методы и строящая на основе новых, неизвестных прежним исследователям,
фактов естественную систему тканей животных организмов.
В основе естественной системы гистологии лежит доказанное всем из-
ложенным фактическим материалом положение, что совокупность биоло-
гических свойств каждой ткани, при обязательном учете всей динамики
свойственных ей при экспериментальных, а также патологических усло-
виях превращений, зависит от истории ее развития. Несмотря на большую
кажущуюся изменчивость тканевых структур при экспериментальных
условиях, совокупность приобретенных в филогенезе биологических свойств
444
Глава IV
каждой отдельной ткани является на каждом этапе эволюции стойко за-
крепленной и полностью определяет возможные для нее превращения.
Поэтому естественная система гистологических структур, помимо своего
общебиологического значения, характеризуется прежде всего теснейшей
связью с основными медицинскими проблемами.
То, что сейчас выполнено нашим научным коллективом, позволяет
в одних случаях детальнее, в других — менее полно наметить и обосновать
естественную систему тканей, построенную на историческом принципе,
полная разработка которой потребует еще длительной работы не одного,
а многих научных коллективов.
Плодотворность и правильность избранного нами пути, мне кажется,
достаточно наглядно демонстрируются не только достигнутыми резуль-
татами, но и неоднократно отмечавшимися по ходу изложения перспекти-
вами дальнейших исследований.
ЛИТЕРАТУРА
А. Общий указатель работ
Абрикосов А. 1925. О миомах, развивающихся из поперечнополосатой произвольной
мускулатуры. Тр. 2-го Всеросс. съезда патол.
Абрикосов А. 1939. Основы общей патологической анатомии.
А в р о ров и Тимофеевский. 1913. Опыты культивирования тканей вне орга-
низма. Тр. Общ. естествоисп. и врачей при Томском унив. за 1912 г.
— 1914. Опыты культивирования тканей вне организма. Томск.
Ага ба б о в и А рн ште йн (Aga ba bof f u. Arnstein). 1904. Untersuchungen
liber die Natur der Zonula ciliaris. Arch. mikr. Anat., 63.
Агдур (Agduhr E,). 1920. Цит. по Агдур, 1932.
( — 1932. Chorioid plexus and ependyma. Cytol. a. cell. path, of the nervous system, ed.
by Penfield, 2.
Адачи (Adachi A.). 1931. Uber die Zilchtung von Linsencpithelien in vitro. Arch,
exp. Zellforsch., 10.
Аде л ь ма н (Ade 1 ma n n H.). 1929, 1930. Experimental studies on the Development
of the eye. I—III. J. exp. Zool., 54, 57.
Азум и (Azumi J.). 1920. Mitt. med. Akad. Kiyoto, 20.
Акаматсу (Aka matsu N,). 1922. Uber Gewebskulturen von Lebergewebe. Virch.
Arch., 240.
Александер (Alexander G.). 1901. Das Labyrinthpigment des Menschen und
der hoheren Saugetiere nebst Bemerkungen iiber den feineren Bau des perilymphatischen
Gewebes. Arch. mikr. Anat., 58.
Алфеева (Alfejew S.). 1927. Beitrage zur vergleichenden Histologie des Blutes
und des Bindegewebes. Z. mikr.-anat. Forsch., 9.
Альтман (Altmann). 1881. Uber embryonales Wachstum. Leipzig.
Аморозо (Amoroso E.). 1932. Die Ziichtung von Epithel des embryonalen Hiihner-
pankreas. Arch. exp. Zellforsch., 12.
* Андерсон и Гей мекер (A ndersonE. a. Haymaker W_). 1935. Proc. Sot.
exp. Biol. a. Med., 33.
А н и ч к о в H. 1912. О воспалительных изменениях миокарда. Дисс. СПб.
— 1930. Ретикулоэндотелиальная система.
А п а т и (А р a t h у). 1897. Das leitende Element des Nervensystems. Mitt. Zool. Stat.
Neapel., 12.
Асканази (Askanazy M.). 1891. Zur Regeneration der quergestreiftenMuskel-
fasern. Virch. Arch., 125.
— 1919. Uber die Veranderungen der grossen Luftwege, besonders ihre Epithelmetaplasie
bei der Influenza. Corr.-Bl. schweiz. Arzte.
Астрахан. 1929. Материалы к изучению закономерностей роста в процессе регенера-
ции. Инет, для леч. опухолей I МГУ.
А чу к а р р о (Ac h ис а г го N.). 1911. Nuevo metodo para el estudio de la neuroglia
у del tejido conjuntivo. Boll. Soc. Espan. Biol. Madrid, 1.
Ea6op(Babor J.). 1901. Zur Histogenese der Bindesubstanzen bei Weichtieren. II.
446
Литература
Die Entwicklung des knorpeligen Schadels bei Eledone moschata. Verh. d. 5. internal,
zool. Kongr., Berlin.
Багров H. 1915. О гетеропластическом образовании костей в мозгу и мягкой его
оболочке. Харьк. мед. ж.. 20.
Ба л л о ви ц (Ba 11 о wit z Е.). 1031. Die Pigmentzellen, Chromatophoren und ihre
Vereinigungen (chromatische Organe) in der Haut der Fische, Amphibien und Repti-
lien usw. Handb. d. vergl. Anat. der Wirbeltiere, 1.
Бальфур (Balfour). 1878. A Monograph on the Development of Elasmobranch
Fishes. J. Anat. a. Physiol.
Б a p (B a r d L.). 1886. La specifite cellulaire et I’histogenese chez I’embryon. Arch, de
physiol., 7, ser. 3.
Б a p г м а н (B a r g m a n n). 1936. Die Lungenalveole, Handb. d. mikr. Anat. des Men-
schen, Bd. 5, T. 3.
Барделебен(Вагбе1еЬеп W.). 1901. Die Heilung der Epidermis. Virch. Arch.,
163.
Ба p Д и и (Ba r d e e n), 1902. Embryonic and regenerative development in Planarians.
Biol. Bull., 3.
Бартш (Bartsch O.). 1923. Die Histogenese der Planarienregeneratc. Arch. mikr.
Anat. u. Entwmech., 99.
Барфурт (В a r f u r t h D.). 1891. Zur Regeneration der Gewebe. Arch. mikr.
Anat., 37.
— 1910. Regeneration und Transplantation in der Medizin. Jena.
Бауэр (Bauer K.). 1932. Beobachtungen liber das Wachstum von Nervengewebe iit
vitro. Z. mikr.-anat. Forsch. 28.
— 1936. Ober die kontinuirlichen Gitterbildungen in Nervengewebskulturen. Z. mikr.-anat,
Forsch., 39.
— 1936. Ober Epithelformationen in den Nerve nkulturen und ihre histogenetische Poten-
zem. Roux’Arch., 138.
Бейли (Bailey P.). 1932. Cellular types in primary tumors of the brain. Cytol. a. cell,
path, of the nervous syst., V. 3.
Бейли и Каш инг (Bailey Р. u. Cushing H_). 1930. Gewebs-Verschiedenheit
der Hirngliome und ihre Bedeutung fir die Prognose. Jena.
Бейтсел (Baitsei G.). 1915. The origine and structure of a fibrous tissue which
appeares in living cultures of adult frog tissues. J. exp. med... 21.
Бейцке (Beitzke). 1918. Ober Heilungsprozesse bei der Ruhr. Ziegl. Beitr,, 64.
Бек л a p(Bec la rd F.). 1828. El£mentj d’anatomie g6n6rale. Nouvelle 6d., Bruxelles.
Беклемишев В. 1915. О паразитных турбелляриях Мурманского моря. I. Acoela.
Тр. СПб. общ. естествоисп., 43.
— 1916. О паразитных турбелляриях Мурманского моря. II. Rhabdocoela. Тр. СПб
общ. естествоисп. 45.
— 1937. Класс ресничных червей (Turbellaria). Рук. по зоол., т. I.
Белецкий В. (В elezky W.). 1931. Virch. Arch., 282.
— 1932. Virch. Arch., 284.
— 1938. Virch. Arch., 290.
— 1939. Мезоглия в норме и патологии. Дисс. М.
Бельцов (В eltzow А.). 1894. Zur Regeneration des Epithets der Harnblase. Virch.
Arch., 97.
Беневоленская (Benewolenskaja S.). 1930. Hematopoiesis in culture»
of the embryonic liver of Man. Arch. exp. Zellforsch., 9.
— 1939. Опыты культивирования раковых опухолей человека. Арх. пат. анат. и пат.
физио л., 5.
Бeннинггoф(Benninghoff А.). 1922. Zur Kenntnis und Bedeutung der Ami-
tose und amitoseahnlicher Bilder. Sitz. ber. Ges. Naturwiss. Marburg, № 7.
— 1926. Uber die Formenreihe der glatten Muskulatur und die Bedeutung der Rougetschen
Zellen an den Capiilaren. Z. Zellforsch., 4.
Литература
447
— 1930. BlutgefSsse und Herz. Handb. mikr. Anat. d. Menschen, Bd. 6, T. I.
Бернер-Патцельт(Вогпег-Ра1геИ D.). 1922, Die Entwicklung der Magen-
schlcimhautinseln im cberen Anteil des Oesophagus von ihremersten Auftreten be im
Fetus bis zur Geburt. Anat. Anz., 55.
Берроуз Дж. (В u г г о w s H. J.). 1933. The intercellular product of a pure culture
of osteogenic cells in vitro. Arch. exp. Zellforsch., 14.
Берроуз M. (Burrow s M.). 1913. The cultures as a physiological method. Trans.
Congr. Am. Physiol, a. Surg., 9.
Бете (В e t h e). 1897. Das Zentralnervesystcm von Carcinas moenas. Eine anatomisch-
physiologische Untersuchung. I. Teil. Arch. mikr. Anat., 50.
— 1903. AUgcmeine Anatomie und Physiologic des Nervensystems. Leipzig.
Бийтель (Bijtel J.). 1931. Beitrage zur Schwanzentwicklung der Amphibien.
Roux’Arch., 125.
Бийтель и В у p д e м а н (В i j t e 1 J. u. Woerdemann M.). 1928. Prcc. Kon.
Akad. Weten. Amsterdam, 31.
БинэиШампи(В1ге1 L. etChampyCh.). 1926. Sur les cultures de poumon in
vitro. C. r. Soc. biol., 94.
Биццоцеро (Bizzozero E_). 1888. Uber die Regeneration der Elemente der
Schlauchformigen Drvsen des Epithels des Magen-Darmkanals. Anat. Anz., 3.
— 1892. Uber die schlauchformigen Drusen des Magen-Darmkanals und die Beziehungen
ihres Epithels zu dem Oberflachenepithel der Schleimhaut. Arch. mikr. Anat., 40.
— 1902/3. Sulla rigenerazione dell’epitelio intestinale nei pesci. Atti accad. sc. di
Torino, 38.
Биццоцеро и Вассале (В izzozerou. Vassal e). 1887. Uber die Erzeu-
gung und physiologische Regeneration der Driisenzellen bei den Saugetieren. Virch.
Arch., 110.
Биццоцеро и Торре (Bizzozero u. Torre). 1884. Uber die Entstehung der
roten Blutkorperchen in verschiedenen Wirbeltierklassen. Virch. Arch., 95.
Б и ш a (B i c h a t M. X.). 1800. Traite de membranesen general et de diverses membrans
en particulier. Paris.
Биша. 1801. Anatomie generale, appliquee a la physiologic et a la medecine, v. 1—4.
Paris.
Бише л be (Bisceglie V.). 1931. Studi sui tessuti espiantati. I. Ricerche sulla
morphologia e biologia delle cellule epatiche ed endoteliali in colture di fegato embrio-
nale. Arch. exp. Zellforsch., 10.
— 1931. Id. II. L’attivitA peristaltica degli espianti di intestino embrionale. Arch. exp.
Zellforsch., 12.
— 1931. Id. III. Le modalita di accrescimento e caratteri strutturali delle colture di inte-
stino embrionale. di polio. Arch. exp. Zellforsch., 12.
Бишоф (Bischoff Th.). 1842. Entwicklungsgeschichte der Saugethiere und des Men-
schen. Leipzig.
— 1842. Entwicklungsgeschichte des Kaninchen-Eies. Braunschweig.
— 1845. Entwicklungsgeschichte des Hunde-Eies. Braunschweig.
Бл аи<о (B la s zo A.). 1932. On tissue cultures of the skin of adult rabbit. Arch, exp.
Zellforsch., 12.
5 ленд (Bland J.). 1939. The growth of human meningiomata in culture compared
with that of certain human tissues Arch. exp. Zellforsch., 22.
Блум (Bloom W.). 1928. Mammalian lymph in tissue culture. From lymphocyte to
fibroblast. Arch. exp. Zellforsch., 5.
— 1937. Transformation of lymphocytes into granulocytes in vitro. Anat. Rec., 69.
Бобрецкий 1877. Исследования над развитием головоногих моллюсков. Тр.
Моск. общ. естествоисп., 24.
Бовери (Boveri Th.). 1898. Die Nierenkanalchen des Amphioxus. Ein Beitrag zur
Phylogenie des Urogenitalsystems der Wirbeltiere. Zool. Jahrb., Abt.
Anat., 5.
448
Литература
Б о з л е р (В о z J е г). 1926. Sinnes- und nervenphysiologische UntersuchungenanScypho-
medusen. Z. vergl. Phys., 4.
— 1927. Untersuchungen uber das Nervensystem der Coelenteraten. T. 1. Z. Zellforsch,, 5;
T. II. Z. vergl. Phys., 6.
Бонне (Bonnet R.). 1887. Ober die ektodermale Entstehung des Wolffschen Ganges
bei den Wirbtltieren. Sitz. ber. Ges Morph, u. Physiol. Munchen, 3.
Б о p д ё (В о r d е u). 1767. Recherches sui’ le tissu muqueux ou Porgane cellulaire. Paris.
Бо p p м а н (В о r r ma n n R.). 1926. Geschwulste des Magens und Duodenums. Handb.
spez. path. Anat. u. Hist., Bd. 4, T. 2.
Бофиль-Делофё (Bofill-Delofeu). 1932. DieargyrophilenFaserstrukturen
in mesenchymalen Gewebekulturen von verschiedener Herkunft und verschiedener
Wachstumsgeschwindigkeit. Z. Zellforsch., 14.
Б p а к к (Brack). 1926. Uber echte und falsche Metaplasie des Respirationsepithels.
Virch. Arch., 259.
Брандт (Bra ndt M.). 1926. Uber Regenerationserscheinungen in der Lunge und ihre
Beziehungcn zum primaren Lungenkrehs. Virch. Arch., 262.
Бру hh (v. Brunn M.). 1901. Uber die Entzundung ser6ser Haute usw. Ziegl.
Beitr, 30.
Бундшу (Bundschuh E.). 1923. Regeneration quergestreifter Muskulatur. Ziegl.
Beitr., 71.
Буркгардт(Burkhardt L.). 1934. Beobachtungen an explatiertem Fettgewebe.
Arch. exp. Zellforsch., 16.
— 1936. Differenz-ierung von Skelettmuskulatur im Explantat. Arch. exp. Zellforsch., 18.
Быстров A. (Bystro w). 1939. Zahnstruktur der Crossopterygier. Acta Zool., 20.
Валентин (Valentin G.). 1835. Handbuch der Entwicklungsgeschichte des Men-
schen. Berlin.
— 1842. Gewebe des menschlichen und thierischen Korpers. Handworterbuch d. Physiol,
von. R. Wagner, Bd. I.
Вальдейер (W aideye r). 1865. Uber die Veranderungen der quer gestreiften
Muskeln bei Entziindung und brim Typhusprozess esw. Virch. Arch., 34.
— 1867, 1872. Die Entwicklung der Carcinome. Virch. Arch., 41, 55.
Вальдейер. 1883. Archiblast und Parablast. Arch. mikr. Anat., 22.
— 1901. Kittsubstanz und Grundsubstanz, Epithel und Endothel. Arch. mikr. Anat., 57.
Вальдман A. 1939. Экспериментальные исследования о заживлении паратифозных
поражений лимфатического аппарата кишечника. Арх. биол. наук, 53.
— 1939. Экспериментальные данные о морфологии и патогенезе паратифозных инфек-
ций. Арх. биол. наук, 55.
Вальтер (W а 1 t h е г). 1806. Darstellung des Bichat’schen Systhemes ais erste Fortset-
zung der historischen Umrisse von Frankreichs wissenschaftlicher Kultur. Jahrb. d.
Med. v. Marcus u. Schelling.
Варшавская Б. 1939. Опыты культивирования человеческих меланобластом. Арх
пат. анаг, и пат. физиол., 5.
Вассен (Was sen А.). 1915. Beobachtungen an Thymuskulturen in vitro. Anat.
Hefte, 52.
Ван-Вей (v. Wijhe J.). 1888. Uber die Entwicklung des Exkretionssystems und anderer
Organe bei den Selachiern. Anat. Anz., 3.
— 1898, Uber die Beteiligung des Ektoderms an der Bildung des Vornierenganges bei Seia-
chiern. Verh. anat. Ges.
Вейгерт (W e i g e r t C.). 1890. Berner kungen iiber das Neurogliageriist des menschli-
chen Centralnervensystems. Anat. Anz., Jg. 5.
— 1895. Beitrage zur Kenntnis der normalen menschlichen Neuroglia. Frankf. a. M.
Вейден рейх (W eidenreich F.). 1911. Die Leukozyten und die verwandten
Zellformen. Erg. Anat. u. Entw. gesch., 19.
— 1912. Die Lokalisation des Pigments und ihre Bedeutung in Ontogenie und Phylogenie
der Wirbeltiere. Z. Morph, u. Anthrop. Sonderhcft.
Литература
449
— 1923. Uber den Begriff «Knochen» und die Beziehungen des Knochengewebes zu Bindege-
webe und Knorpel. Anat. Anz., 57, Erg. Heft.
— 1930. Das Knochengewebe. Handb mikr. Anat. d. Menschen, Bd. 2, T. 2.
Вейс (W e i s s P.). 1934. In vitro experiments on the factors determining the course
of outgrowing nerve fibers. J. exp. Zool., 68.
— 1934. Secretory activity of the inner layer of the embryonic mid brain in the chick
as revealed by tissue culture. Anat. Rec., 53.
Вейс и Доррис (W eiss P. aDorris F.). 1935. Mechanical factors determining
cell pattern in retina and cartilage cultivated in vitro. Anat, Rec., 64, Suppl.
Вейс и X с и-В а н г (W e i s s P. a H s i-W a n g). 1936. Neurofibrills in living ganglion
cells cultivated in vitro. Anat. Rec., 67.
Вейсман (W e i s m a n n A.). 1862, Ueber die zwei Typen contractilen Gswebes und
ihre Vcrtheilung in den Gruppen des Thierreichs, sowie fiber die histologische Bedeu-
tung ihrer Formelemente. Z. ration. Med., III. R., 15.
Вейсфейлер (W e i s s f e i 1 e r). 1924. Regenerations des lobes olfactives et de
hemisphferes сёгёЬгаих Chez les batraciens urodeles. C. r. soc. biol, 91. (Thfese. Genfcve).
Be йцман (W e i z m a n n G.). 1938. Ueber die Ziichtung einiger menschlicher malig-
ner Tumoren in vitro. Z. Krebsforsch., 48.
1939. Epithel und Karzinom der erwachsenen Menschen in vitro. Arch. exp.
Zellforsch., 22.
— 1939. Die Zi'.chtung von Kulturen aus menschlichen Pleura- und Peritoncalergiissen.
Arch. exp. Zellforsch., 22.
В e й ц м а н B. (W e i z m a n W.). 1927. Zur Kenntnis der Regenerationserscheinungen
im Mesoderm der Oligochaeten. Roux’Arch., 110.
— 1928. Einige Bemerkungen zur Regeneration der Gonaden bei Lumbriculus. Zool.
Anz., 78.
1937. Происхождение регенерационной ткани в связи со степенью развития организма.
Сб. к 80-летию акад. Н. Насонова. Изд. АН СССР.
— 1938. Индивидуальное развитие организмов и патологические процессы. Усп. совр.
биол., 9.
Вере щинскп й A. (W jereszinski А.). 1924. Vergleichende Untersuchungen
uber Explatation und Transplantation von Knochen, Periost. und Endost. Virch.
Arch., 251.
— 1924. Ueber die freien Zellen der serosen Exsudate, ihren Ursprung, ihre genetische
Wechselbeziehungen und ihre prospektive Potenzen. Haematologica. Pavia. 1 S.
— 1925. Beitrag zur Morphologie und Histogenese der intraperitonealen Verwachsungen
Leipzig.
Верме льЕ (W e r m e 1 E_). 1931. Beobachtungen an Zellen der Samendriise des Fro-
sches. Z. Zellforsch., 13.
— 1933. Kultivieren von Testkulumgeweben eines Hahnes in vitro. Arch. exp.
Zellforsch,, 14.
— 1940. Основные этапы в развитии учения о клетке.
Верн (Verne J.). 1930. La nevroglie dans les cultures du tissu nerveux. C. r. Ass. Anat.,
25-e reun.
— 1932. Cultures d’epithelium pulmonaire de poulet. C. r. Ass. Anat., 27-e reun.
Верхацкий H. 1940. Аутопластическая пересадка эндометрия. Дагест. гос.изд.
Вилас (Vilas Е_). 1934. Zur Entwicklung der menschichen Scheide. Anat. Anz., 79.
Вирхов (Virchow R.). 1846. Ueber das granulierte Aussehen der Wandungen der
Gehirnventrikel. Allg. Z. f. Psych, u. psychgericht. Medizin, 3.
— 1851. Die Identitat von Knochen-, Knorpel- und Bindegewebskorperchen, sowie liber
Schleimgewebe. Wurzburger Verh., 2.
> — 1853. Zur Streitfrage uber die Bindesubstanzen. Virch. Arch., 5.
— 1854. Ueber eine im Gehirn und Rilckenmark des Menschen aufgefundene Substanz mit
der chemischen Reaktion der Cellulose. Virch. Arch., 6.
— 1855. Cellular Pathologic. Virch. Arch., 8.
29 Хлопни—349
450
Литература
— 1856. Gesammelte Abhandlungen zur wissenschaftlichen Medizin. Frankf. a. M.
S. 889—891.
— 1864. Krankhafte Geschwiilste.
Волосков 1938. Дисс. M.
Вольтерек (Woltereck). 1904. Wurmkopf, Wurmrumpf und Trochophora.
Zool. Anz., 28.
Вольф (W о 1 f J.). 1920. Beitrag zur pathologischen Histologie der gonorrhoischen
Epididymitis. Virch. Arch., 228.
Вулард (W collard M.). 1924. Vital staining of the Leptonieninges. J. Anat., 58.
By pдема н (W oerdemann M_). 1929. Experimentelle Untersuchungen i.ber
Lage und Bau der augenbildenden Bezirke in der Medullarplatte beim Axolotli. Roux’
Arch., 116.
де-Г а а н (de Haan J.). 1926. Die Umwandlung von Wanderzellen in Fibroplasten
bei der Gewebezi chtung in vitro. Arch. exp. Zellforsch., 3.
— 1929. Weitere Untersuchungen i ber die Zi chtung von Wanderzellen mittels Durchstro-
mting. Nahrungsbediirfnisse, Phagozytose und Vitalfarbung. Arch. exp. Zellforsch. 7.
— 1929. Das Auftreten verschiedener Zelltypen in Blut und Bindegewebe. Arch. exp.
Zellforsch., 7.
де-Г а а н и Фери нга (de Haan a. F e r i n g a). 1934. On the possibility to force
the growth of perfused cultures in the direction of an adenoid (or haemopcetic) system.
Arch. exp. Zellforsch., 15.
Гага (Haga) 1913. Experimentelle Untersuchungen fiber die Erzeugung atypischer
Epithel- und Schleimhauiwucherungen. Z. Krebsfcrsch., 12.
Га й a p (Gaillard P). 1935. Developmental changes in composition of body fluids
in relation to growth and differentiation of tissue cultures. Protopl., 23.
— 1938. Die Glandula hypophysis das Kaninchens (pars anterior) in vitro und in vivo.
Arch. exp. Zellforsch., 22.
Гаккель В. 1939. Морфология опухолей нервной системы.
Галлер A. (Haller А.). 1778. De partium corporis humani praecipuarum fabrica
et functionibus opus quinquaginta annorum, T. I. Bernae et Lausannae.
Га мма p (H a m ma r.). 1926. Die Menschenthymus. I. Das normale Organ. Z. mikr.-
anat. Forsch., Erganztingsband z. 6. Bd.
— 1929. Das Organ unteranormalen KCrperverhaitnissen. II. Z. mikr-anat. Forsch., Ergiin-
zungsband z. 16. Bd.
Гам мерли hf (H a m m e r 1 i n g). 1924. Die ungeschlechtliche Fortpflanzung und
Regeneration bei Aelcsoma. Zool. Jahrb., Physicl. 41.
Ганзе м ан (Hanseman D.). 1893. Studien uber die Spezifitat, den Altruismus und
die Anaplasie der Zellen mit besonderer Beri cksichtigung der Gesctiw. Iste. Berlin.
— 1896. Ueber die Benennung der bosartigen Geschwi Iste. Centralzeitung № 81.
Г а н к e (H a n k e). 1915. Studien t ber die Regeneration der Hornhaut und die wahre
Natur des Keratoblasten. Graefe’s Arch., 89.
Га ист ре м(На nstr 8 m B.). 1928. Vergleichende Anatomie des Nervensystems der
wirbellosen Tiere. Berlin.
Г a p в e й и Б у p p (H a i v e у a. Burr). 1926. Development of Meninges. Arch, of
Neur. a. Psych., 15.
— 1933. Development of Meninges. Ibid., 29.
ле-Г a p и (de G a r i s Ch ). 1924. Notes on some interrelation of fibroblasts in tissue
culture. Bull. Johns Hopk. Hosp., 35.
Г a p о ф о л и н и (G a г о f о I i n i). 1927. Ricerche su colture di tessuti in vitro. II.
Cellule epatiche eendotelio incolture di fegato embrionale. Boll. soc. ital. Biol, sper., 2.
Гарри сон (Harrison R.). 1904. Neue Versuche und Beobachtungen liber die Ent-
wicklung der peripheren Nerven der Wirbeltiere. Sitz. Ber. d. Niederrheinischen Gesi,
Bonn.
— 1906. Further experiments on the developing of peripheral nerves. Am. J. Anat., 5.
— 1907. Observations on living developing nerve fibres. Proc. Soc. exp. Biol, and Med., 6.
Литература 451
— 1910. The development of peripheral nerve fibres in alteredsurroundings. Roux’Arch., 30.
— 1911. The outgrowth of the nerve fibres as a mode of protoplasmic movement. J. exp.
Zool., 9.
— 1913. The life of tissues outside the organisme from the embryological standpoint
Transact. Congr. Am. Phys. a. Surg., 9.
— 1924. Neuroblast versus sheath cell in the development of peripheral nerves. J. comp.
Neurol., 37.
Гарро (Garrauit H.). 1932. Etude in vitro de i’endothdliumcardiaque embryonnai-
re. C. r. Ass. Anat., 27-e rcun.
Гаршин B. (Garschin W.). 1927. Экспериментальные исследования атипических
разрастаний эпителия кожи. Атипические разрастания эпителия при асептическом
воспалении, вызванном инфузорной землей. Арх. биол. наук, 27; Z. Krebsforsch., 24.
— 1928. Экспериментальные исследования атипических разрастаний эпителия кожи.
Атипические разрастания эпителия при воспалении, вызванном холестерином и
животным углем. Арх. биол. наук, 28; Z. Krebsforsch., 27.
— 1928. Ueber die Bedeutung der atypischen Epithelwucherungen. Z. Krebsforsch., 27.
— 1935. Гетеротопические разрастания эпителия желез дна желудка. Арх. биол. наук;
Frankf. Z. Path., 49.
— 1935. О процессах дифференцировки эпителия дыхательных путей при регенерации
и воспалительной пролиферации. Арх. биол. паук; Frankf. Z. Path., 49.
— 1938. Sur 1’importance pathologique des modifications de la differenciation des voies
respiratoires dans 1’inflammation et sur le rapport de ces modifications avec la meta-
plasie et le cancer. Acta de I’unicn internat. centre le cancer, 3.
— 1939. Воспалительные разрастания эпителия, их биологическое значение и отношение
к проблеме рака. Медгиз.
Гаршин В. и Пига ле в И. (Garschin u. Pigale w). 1931. Эксперименталь-
ные исследования атипических разрастаний эпителия. Атипические разрастания
эпителия бронхов, при воспалении, вызванном введением в ткань легких каменно-
угольного дегтя. Арх. биол. наук, 31; Z. Krebsfors, 33.
Гаршин В. иШабадЛ. (Garschin u. Schabad). 1935. Об атипических раз-
растаниях эпителия бронхов при введении в ткань легких формалина. Арх. пат.
анат. и пат. физиол., 2; Z. Krebsforsch., 43.
Гаузер (Hauser G.). 1926. Peptische Schadigungen des Magens und Darms. Hadb.
spez. path. Anat. u. Hist., Bd. 4, T. I.
Гаусман (Hausmann M.). 1935. Entstehung und Funktion von Gefassystem und
Blut auf cellular-physiologischer Grundlage. Basel.
Гегенбауер (Gegenbauer C.). 1890, Lehrbuch der Anatomie des Menschen. 4.
Aufl. Bd. 1. Leipzig.
Гегквист (Haggquist G.). 1931. Oewebe und Systeme der Muskulatur. Handb.
mikr. Anat. d. Menschen, B. 2, T. 3.
Гейден гай н M. (Н е i d е n h a i n M.). 1911. Plasma und Zelle. 2. Aufl. Jena.
— 1936. Herkunft und Bedeutung der Worte Epithelium und Endothelium. Z. mikr.-anat.
Forsch., 39.
— 1936. Ein Nachwort zu der Frage der Ableitung der Bezeichnungen Epithelium und
Endothelium. Z. mikr. - anat. Forsch., 40.
Ге ii д e p (H e i de r). 1927. Vom Nervensystem der Ctenophoren. Z. Morph, it.
6koi., 9.
Г e й 3 и нт e p (H e u s i n g e r C.). 1822. System der Histologie. I Theil. Histographie.
Eisenach, bei J. Barecke.
Гейм (Heim K.). 1927. Beitrag zur Frage liber die Verschleppungsmoglichkeit und
Wachstumsfahigkeit menschlicher Uterusschleimhaut. Zbl. Gynak., 29.
Гейнеке (Heinecke A). 1909. Die Veranderungen der menschlichen Niere nach
Sublimatvergiftung mit besonderer Beriicksichtigung der Regeneration des Epithets.
Ziegl. Beitr., 45.
Геккель Э. (Haeckel E.). 1874. Die Gastraea-Theorie, die phylogenetische Classi-
*
452
Литература
fication des Thierreichs und die Homologie der Keimblatter. Jena. Z. Naturwiss., 8,
N. F. 1.
— 1877. Histolcgische Bedeutung der Gastraea-Theorie. Nachtrage zur Gastraea-Theorie.
Jena. Z. Naturwiss., 11, N. F. 4.
Геккель Э. 1885. Ursprung und Entwickelung der thierischen Gewebe. Ein histoge-
netischer Beitrag zur Gastraea—Theorie. Jena. Z. Naturwiss., 18, N. F. 11.
Гексли и де-Б ер (Н u х 1 е у J. a. de Beer G.). 1936. Основы экспериментальной
эмбриологии. Биомедгиз.
в а н-Г е л е н (v. G е h 1 е n Н.). 1936. Ueber die Regeneration der quergestreiften Muskula-
tur. Roux’Arch., 135.
Ге л ь д(Н e 1 d H.). 1903. Ueber den Bau der Neuroglia und uber die Wand der Lymphge-
fasse der Haut und Schleimhaut. Abh, Kon. Sachs. Ges. d. Wiss., 28.
— 1909. Die Entwicklung des Ncrvensystems bei den Wirbeltieren. Leipzig.
Генле (Henle J.). 1837. Symbolae ad anatomiam villorum ir.testinalium imprimis
eorum epithelii et vasorum lacteorum. Berolini, apud Aug. Hirschwald.
— 1838. Ueber die Ausbreitung des Epithelium im menschlichen Korper. Muller’s Arch,
f. Anat., Physiol, u. wiss. Med.
___1841. Ailgemcine Anatomie. Lchre von den Mischungs- und Formbestandtheilen des
menschlichen Korpers. Veil. L. Voss.
Генс он (Henson). 1932. Development of alimentary chanel of Pieris brassicae.
Quart. J. micr. Sc., 75.
Герксгеймер (Herxheimer G.). 1907. Ueber heterologe Kankroide. Ziegl.
Beitr., 41.
— 1930. Lebcrgewachse. Handb. spez. path. Anat. u. Hist. Bd. 5, T. I.
Герксгеймер и Тольдте (Herxheimer G. u. Tholldte M.) 1930.
Regeneration und Hypertrophie (Hyperplasie) der Leber. Ibid.
Геррик (Herrick C.). 1924. Neurological foundation of animal behaviour.
Гертвиг О. (Hertwig О.). 1878. Das Nervensystem und die Sinnesorgane der Me-
dusen. Leipzig.
— 1880. Die Actinien anatomisch und histologisch mit besonderer Beriicksichtigung des
Nervensystems untersucht, Jena. Z. Naturwiss., 13, 14.
— 1882. Die Entwicklung des mittleren Keimblattes der Wirbelthiere. Jena. Z. Natur-
wiss., 15, N. F. 8.
— 1883, 1898. Die Zelle und die Gewebe. I und 2 Teil.
— 1906. Allgemeine Biologic.
Гертвиг О. и Г е р т в и г Р. (Н е г t w i g О. u. H e r t w i g R.). 1882. Die Coelom-
theorie. Versuch einer Erklarung des mittleren Keimblattes. Jena. Z. Naturwisss.,
15, N. F. 8.
Гертвиг P. (Hertwig R.), 1873, Die lymphatischen Drusen an der Oberflache des
Stohrherzens. Arch. mikr. Anat., 9.
Г e p ц о г Г. (Herzog H.). 1902. Die Entwicklung der Binnenmuskulatur des Auges.
Arch. mikr. Anat., 40.
— (Herzog G.). 1915. Experimentelle Untersuchungen uber die Einheilung vonFremd-
korpern. Ziegl. Beitr., 61.
— 1921. Zur Frage der Granulocytenbildung bei der Entziindung. Zbi. allg. Path. u. path.
Anat., 31.
— 1922. Zellformen bei Meningoencephalitis. Zbl. allg. Path. u. path. Anat., 33.
—- 1923. Ueber die Bedeutung der Gefasswandzellen in der Pathologic. Klin. Wochen-
schr., 2.
Герцог и Шоппер (H erzog G. u. Schopper W.). 1931. Ueber das Verhalten
der Blutgefasse in der Kultur. Arch. exp. Zellforsch., 11.
Герцог Ф. (Herzog F.). 1914. Maligne Deckzellengeschwiilste des Peritoneums.
Zeigl. Beitr. 58.
Гете В. (G о e t h e W.). 1807. Morphologic. (Цит. no полн. собр. соч.) 32. (Cottasche
Buchhandlung, Gebr. Kroner),
Литература
453
Г е ф е р (Hofer К.). 1934. Beobachtungen an Gewebskulturen menschlicher Haut-
karzinome, kontrolliert durch kinematographische Zeitraumaufnahmen. Arch. exp.
Zellforsch., 16.
Гиккель и Нордман(Н!ске1О. uNordmann J.). 1926. Le role du sy-
stfeme excreteur du pancreas dans la genfese des ilots de Langerhans. Trav. Inst, d’anat,
path, de Strasbourg. Ann. d’anat. path., nomr., tned.-chir., 3.
Гирке (Girke H). 1885, 1886. Die Stiitzsubstanz des Centralnervensystems. I and II.
Arch. mikr. Anat., 25, 26.
Гирсберг(О1е rsber g). 1924. Neurulation bei Rana undTriton. Roux’Aich., 103.
— 1926. Neue Untersuchungen zur Neurulation bei Rana und Triton. Roux’Arch., 107.
Гирш и Джонс (Hirsch E. a. Johns. H.) 1933. Am. J. Obst. a. Gyn., 25.
Г и с В. (H i s W.). 1865. Die Haute und Hohlen des Korpers. Basel. (Wiederabdruck;
Arch. f. Anat. u. Physiol., anat. Abt., 1903).
— 1866. Ueber die erste Anlage des Wirbelthierleibes. Arch. mikr. Anat., 2.
— 1868. Untersuchungen uber die erste Anlage des Wirbelthierleibes. Die erste Entwi-
cklung des Hiihnchens im Ei. Leipzig.
— 1882. Die Lehre von Bindesubstanzkeim (Parablast). Ri ckblick nebst kritischer Be-
sprechung einiger neuererentwicklungsgeschichtlicher Arbeiten. Arch. f. Anat. u. Phy-
siol., anat. Abt.
— 1890. Die Neuroblasten und deren Entstehung im embryonalen Mark. Abh. d. math.-
phys. CI. d. K. Sachsischen Ges. d. Wiss., 15.
— 1900. Lecithoblast und Angioblast der Wirbeltiere. Abh. d. math.-phys. KI. d. K.
Sachs. Ges. d. Viss., 26.
— 1901. Das Prinzip der organbildenden Keimbezirke und die Verwandschaft der Gewebe.
Arch. Anat. u. Physiol., anat. Abt.
Глазунов M. (Glasunow M.). 1934. Ueber die feine Struktur der Uterusmyome.
Arch. Gynak., 157.
— 1931. Ueber die fibrillogenetischen Eigenschaften der Sarkome. Z. Krebsforsch., 34.
— 1937. Гистология и гистогенез так называемых цилиоэпителиальных (серозных)
кистом яичников. Арх. пат. анат. и пат. физиол., 3.
— 1937. Histologie und Histogenese der sog. cilioepithelialen (serosen) Cystome der Ova-
rien. Arch. Gynak., 164.
Гл юг e (G 1 u ge). 1841. Experimente iiber Encephalitis. Abh. z. Phys. u. Path., 2.
Глюксман (G 1 ii c k s m a n). 1934. Z. anat. u. Entw. gesch., 103.
— 1938. Studies on bone mechanics in vitro. I. Anat. Rec., 72.
Г о г (H о g u e, M.)_ 1937. Studies on heart musclein tissue cultures. Anat. Rec., 67.
Гог и д e - P e н ь и (H о g u e M. a. de R e n у i). 1939. Giant muscle cells in tissue
cultures. Arch. exp. Zellforsch., 23.
Г о д л e ii (H о a d 1 e у L.). 1928. Developmental potencies of parts of the early blasto-
derm of the chick. J. exp. Zool., 43.
Гольджи К. (Golgi C.) 1885/86. Sulla fina anatomia degli organi centrali del si-
stema nervoso. Milano. Hcepli.
— 1894. Untersuchungen uber den feineren Bau des Zentralen und peripheren Nerven-
systems. Jena.
Гольме (Hol ms S.). 1914. Observations on isolated living pigment cells from larvae
of amphibians. Univ. publ. zool., 11.
— 1914. Developmental changes of pieces of frog embryos cultivated in lymph. Biol,
bull., 3.
Гол ьтф рете p(Ho 1 tire ter J.). 1929, 1931. Ueber die Aufzucht isolierter Teile
des Amphibienkeims. I and II. Roux’Arch., 117, 124.
— 1933. Die totale Exogastrulation, eine Selbstablosung des Ektoderms vom Entomeso-
derm. Roux’Arch., 129.
— 1935. Morphologische B'tinflussung von Urodelenektoderm bei xenoplastiseher
Transplantation. R ux’Arch., 133.
— 1938. Differenzierungspotenzen isolierter Teile der Urodelengastrula. Roux’Arch., 138.
454
Литература
— 1938, Diffeienzierungspotenzen isolierten Teile der Anurengastrula. Rottx’Arch., 138,
Госс (Goss C.). 1928. Experimental removal of the blood island of Amblystoma pun-
ctatum embryos. J. exp, Zool., 52.
— 1932. The formations of cross-striation in cultures of embryonic heart muscle. Arch,
exp. Zellforsch., 12.
Г рафф (Graff L. v.). 1882, 1889. Monographic der Turbellarien. 1. Rhabdocoelida.
11. Triclada terricola (Landplanarien). Leipzig,
— 1905, 1913. Turbellaria. Bronns Klassen und Ordnungen desTierreichs. Lief. 23(Acoela);
Lief. 35 (Rhabdocoelida).
Г p e й (G г a у ¥.). 1931. Experimental Cytology. Cambridge.
Грей в и В у д бри дж (Gra ve C, aWood bridge Н.). 1934. Botryllus schlo-
sseri: the behaviour and morphology of free swimming larva. J. Morph., 39.
Г p e й л ь (G r e i 1). 1906. Ueber Homologie der Anamnieikiemen. Anat. Anz., 28.
Григорьев Л. (Grigorjeff L.). 1931—1932. Differenzierung des Nervengewebes
ausserhalb des Organismus. Arch. exp. Zellforsch., 11, 13.
— 1934. Взаимоотношения нервной ткани и скелетных мышц в условиях совмест-
ного роста вне организма. Тр. I Гист, конф., М.
Гриффини и Вассале (GriffiniL. uVassale G.). 1888. Ueber Regene-
ration der Magenschleimhaut. Ziegl. Beitr., 3.
Гроб бе н(Ог о bbe n C.), 1905, Lehrbuch der Zoologie begriindet von C. Claus.
7 Aufl.
Грос сек (Grosse k, R.). 1932. Ueber die Deckzellen der serosen Haute und ihre
primaren malignen Geschwi Iste. Z. Krebsfcrsch., 35.
Гроссельи (G г о s s e 1 j i). 1909. Untersuchungenuber das Nervensystem der Acti-
nien. Arb, zool. Inst. Wien, 17.
Гросфельд (Grossfeld H.). 1931. Experimentelle Erzeugung von mesenchy-
malem Epithel aus dem Herzexplantat des Hi hnerembryo. Arch. exp. Zellforsch., 11.
— 1932. In vitro Versuche zum Problem des infiltrativen Tumorwachstums. Z. Krebs-
forsch., 37.
— 1936. Osmotischer Druck, Elektrolyte und Gewebezellen. Protopl., 26.
Грот (Groth W.). 1938. Der Ursprung der Riechzellenneuroblasten und ihre erste
Entwicklung bis zur Ausbildung der Riechnervenanlage beim Kaninchen. Z. mikr.-
anat. Forsch., 43.
— 1939. Der Ursprt ng der Labyrinthplacode und des Ganglion statoacusticum im Ver-
gleich zur Genese des Riechorgans beim Kaninchen. Zugleich ein Beitrag zur Homolo-
gisierung dtr Genese von Gehohr-, Riech- und Sehorgan beim Saugetier. Z. mikr.-anat.
Forsch., 45.
Г p у 6 e p (G r u b e r G.) 1917. Ueber heteroplastische Knochehbildung in der Musku-
latur und ihrer Nachbarschaft. Beitr. Klin. Chir., 106.
Г p юн бе pr (G rii n be rg R,)_ 1932. Ueber Experimente mit der Zweimedienkultur,
Arch. exp. Zellforsch., 13.
Гудрич (Goodrich). 1897—1900. On the nephridia of the Polychaeta. I—II.
Quart. J. micr. Sc. (n. s.), 40, 41, 43.
— 1920. The pseudopodia of leucocytes of invertebrates. Biol. Bull., 46.
Г у л ь д б e p г (G u 1 d b e r g). 1935. Versuche zur Zilchtung von melanotischen Gesch-
wulsten. Arch, exp, Zellforsch., 17.
Гюпер и Peccenb(HueperW. a. Russel M.). 1932. Capillary-like formations
in tissue cultures of leucocytes. Arch exp. Zellforsch., 12.
' Д а в ы д о в К. (D a v у d о f f). 1915. Реституция у немертин. Тр. особ. зоол. лаб., 1.
— 1928. ТгаИё d’embryologie comparee des invertebres. Paris.
Дальгрен и Кепнер (Dahlgren а. Керпег). 1908. Principles of animal
histology. New-York.
Данини (Danini), 1922. К вопросу о строении переходного эпителия. Изв. биол,
• инет., Пермск. унив., 1.
— 1924. Zur Frage uber den Bau des Uebergangsepithels. Z. Anat,, 74.
Литература
455
— 1925. Ueber die entziindliche Bindegewebsneubildung beim Flusskrebs. Z. mikr.-anat-
Forsch., 3.
1928. Untersuchungen iiber die iegenerativen Eigenschaften des Hautepithels. Z. mikr.-
anat. Forsch., 12.
— 1930. Гистологические наблюдения над трансплантированным хрящем. Изв. Псрмск.
биол. инет., 7.
— 1936. Об индукции развития хрящевой ткани. Учен. зап. Пермск. унив., 2.
Дан чакова В. (Dantschakoff). 1908. Untersuchungen iiber die Entwicklung
von Blut und Bindegewebe bei den Vcgeln. 1. Anat. Hefte, 37.
— 1909. Untersuchungen i. ber die Entwicklung von Blut und Bindegewebe bei Vogeln.
Das lockere Bindegewebe des Ht hnchens im fetalen Leben. Arch. mikr. Anat., 73.
1909. Ueber die Entwicklung des Kncchenmarks bei den Vogeln und iiber dessen VerSn-
dcrungen bei Blutentzkhungen und Ernahrungsstorungen. Arch. mikr. Anat., 74.
— 1916. Ueber die Entwicklung des Blutes in den Blutbildungsorganen (Area vasculosa,
Dottersackanlage, Knochenmark, Thymus, Milz und lockeres Bindegewebe) bei Tro-
pidonotus natrix. Arch. mikr. Anat., 87.
— 1924. Roux’Arch., 124.
Дауни (Downey H.). 1910. Phagocytosis of erythrocythe in the lymphorenal tissue
of Polycdon spatula. Fol. haemat., Arch., 9.
Даусон (Dawson J.). 1909. Changes in cross-striped muscle in the healing of incised
wounds. J. Path. a. Bact., 13.
Дегорн (Dehor ne A.). 1925. Leucocytes et Linome. C. r. Sec. Biol., 93.
Дедерер(Ое6егег P.). 1921. The behavior of cells in tissue cultures of Fundulus
hutercclitus with special reference to the ectoderm. Biol. Bull., 41.
Дейтерс (Deiters O.). 1865. Untersuchungen iiber Gehirn und Riickenmark des
Menschen und der Saugethiere. Braunschweig.
Делоренци (De 1 о re n г i E.). 1937. L’origine delle cellule di Kupfer del fegato
studiata col metodo delle colture in vitro. Monit. zool. ital., 48.
Демут (De mu t h F.)_ 1928. Knorpelwachstum in vitro. Mon.schr. Kinderh., 38.
— 1932. Ueber die Zcchtung von Schildrilsenzellen in vitro. Arch. exp. Zellforsch., 13.
Детлаф Г. 1936. Исследование источников материала закладки нервной системы
у Anura в связи с вопросом о действии организатора. Зоолог, ж., 15.
Джай и Д ж а й (G е у G. a. G е у М.). 1936. The maintenance of human normal ceils
and tumor cells in continuous culture. Am. J. Cancer, 27.
Джордан (Jordan H.). 1919. The histology of the blood and the red bone-marrow
of the leopard-frog, Rana pipiens. Am. J. Anat., 25.
Джордан и CnefiflenfJordanH. a. Speidel C.). 1923. Studies on lympho-
cytes. 1. Effect of splenectomy, experimental haemorrhage and a haemolytic toxin
in the frog. Am. J. Anat., 32.
— 1924. Studies on lymphocytes. II. The origin, function, and fate of the lymphocytes
in fisches. J. Morph., 38.
— 1924. Studies on lymphocytes. III. Granulopoiesis in the salamander, with special
reference to the monophyletic theory of blood origin. Am. J. Anat., 33.
— 1924. The fundamental erythropoietic stimulus. Proc. Soc. exp. biol. a. med., 21.
— 1930. Blood formation in cyclostomes. Am. J. Anat., 46.
Доан (Doan C.). 1923. On the intravascular development of erythrocytes in the bone
marrow of the adult pigeon. Proc. Soc. exp. biol. a. med., 20.
Доан, Каннингем иСеб ин (Doan C., Cunningham R. a, Sabin F.).
1925. Experimental studies on the origin and maturation of avian and mammalian
red blood cells. Contr. to Embr., 16.
Догель A. (D ogle! A.). 1881. U?ber das Verhalten nervoser Elemente in dcr Retina
der Ganoiden, Reptitien, Vogel und Saugetiere. Anat. Anz., 3.
Д о г e л ь B. 1937. Некоторые перспективы развития советской сравнительной анатомии
в связи с ее достижениями за последнее время. Природа, № И.
— 1938, 1940. Сравнительная анатомия беспозвоночных, ч. I и II,
456
Литература
Дойников Б. (Doinikow В.). 1911. Beitrage zur Histologic und Histopathologie
der peripheren Nerven. Nissl- Alzheimers hist. u. histopath. Arb., 4.
— 1940. Введение к сборнику «Патогистолсгия нервней системы». Медгиз.
Должанский (Doi janski L.). 1929.Culturespures de tissu hepatique in vitro.
C. r. Soc. Biol., 101.
— 1929. Dauerzrchtung von Knochen und Periostgewebe. Z. Zellforsch., 8.
— 1930. Sur le rapport entre la proliferation et I’activite pigmentogene dans les cultures
de I’Gpithelium de I’iris. C. r. Soc. Biol. 105.
— 1930. Observations sur Involution des elements endotheliaux dans les cultures de foie
in vitro. C. r. Soc. Biol., 105.
Должанский и Ру л e (De 1 j a ns k i L u. Roulet F.). 1933. Studien fiber
die Enstehung der Bindegewebsfibrillen. Virch. Arch., 291.
— 1934. Ueber die gestaltende Wcchselwirkung zwischen dem Epithel und dem Mesen-
ch\m usw. Virch. Arch., 292.
Дом и ничи (Dominici H.). 1902. Polynucleaires et macrophages. Arch. med.
exp. et d’anat. path., 14.
— 1920/21. Etudes sur le tissu conjonctif et les organes hdmatopedtiques des mammiferbs.
Arch. anat. micr., 17.
Доррис (Dorris F.). 1938. Differentiation of the chick eye in vitro. J. exp. Med , 78.
— 1940. Proc. See. exp. biol. a med., 44.
Држе BHHa (Drze wina A.). 1905. Contribution a 1’dtude du tissu lymphoide des
Ichthyopsides. Arch. zool. exp. et gen., str. 4, vol, 3.
Дрю A. (Dre w A.). 1922. A comparative study of normal and malignant tissues grown
in artificial culture. Brit. J. exp. Path., 3.
Д p io A. 1923. Growth and differentiation in tissue cultures. Brit. J. exp. Path., 4.
Д p ю Г. (D r e w G.). 1911. J. exp. Zool., 10.
Д ю г г e л и (D ii g g e 1 i O.). 1937. Ueber den gestaltenden Einfluss von Zugspannungen
auf Bindegewebskulturen. Z. Zellforsch, 26.
Д ю p к e н (D ii r k e n). 1926. Roux’Arch., 107.
flroc6epr(Duesberg J.). 1910. Les chondriosomes des cellules embryonnaires du
poulet et leur role dans la genese des mycfibrilles avec quelques observations sur le
developpement des fibres musculaires strides. Arch. f. Zellforsch., 4.
Дюшен (Du Shane Gr.). 1938. Neural fold derivatives in the Amphibian. Pigment
cells, spinal ganglia and Rohon-Eeard cells J. exp. zool., 78.
— 1939. J. exp. zool., 82.
Елеонская. 1915. О росте оболочек глаза вне организма. Вест, сфт., 32.
Елисеев В. (Elisse jew W_). 1929. Ueber in vitro Ku’turen der Magenschleimhaut
des Kaninchens im postembryonalen Leben. Arch. exp. Zellforsch., 9.
— 1938. ЭкспериА’ентально-гистологическое изучение клеточных форм рыхлой соеди-
нительной ткани некоторых млекопитающих. I и II. Арх. анат., гист, и эмбр., 18.
Ж и в Б. 1937. Кариологическпе исследования овогенеза у человека. Бюлл. эксп. биол.
и мед., 4.
Ж и н к и н Л. (Z h i n k i n). 1934. Ueber die Bedeutung der Mesodermanlagen bei der
Regeneration von Rhynchelmis limosella. Zoo!. Anz., 105.
— 1938. Регенерация мускулатуры у паукообразных. Арх. анат., гист. и эмбр,. 19.
Заварзин A. (Z a w а г z i n). 1913. Гистологические исследования чувствительной
нервной системы и оптических ганглиев насекомых. СПб.
— 1924. Zur Morphologie der Nervenzentren. Das Bauchmark der Insekten. Z. wiss.
Zool., 122.
— 1924. О морфологическом понимании гистологических структур. Новые идеи в биол.
— 1925. Der Parellelismus der Strukturen als ein Grundprinzip der Morphologie. Z. wiss.
Zoo!., 124.
— 1925. О сравнительном методе в гистологии. Изв. биол. инет. Пермск. уннв., 4.
— 1926. Ueber die entziindliche Neubildung von Bindegewebe bei der Teichmuschel (Ano
donta anatina L.)_ Z. mikr.-anat. Forsch., 6.
Литература
457
— 1927. Ueber die reaktiven Veranderungen des Epithels bei der Einfiihrung eines Fremd-
korpers in den Mantel der Anodonta. Z. mikr.-anat. Forsch., 11.
— 1928, О морфологических закономерностях в гистологических структурах. Природа.
— 1932/33—1939. Курс гистологии и микроскопической анатомии. I—V изд.
— 1934. Об эволюционной динамике тканей. Арх. биол. наук, 36.
— 1935. О номенклатуре клеточных форм рыхлой соединительной ткани. Юб. сб. 25-
лет. проф. Н, Аничкова.
— 1937. О реактивном новообразовании соединительной ткани у рыб. Арх. анат., гист.
и эмбр., 15.
— 1938. О регенерации мышечных элементов у Allobophora calliginosa. Арх. анат.,
гист. и эмбр., 19.
— 1938. О регенерации соматических мышц у рыб (Cottus cottus). Арх. апат., гист.
и эмбр., 19.
— 1941. Очерки по эволюционной гистологии нервной системы. Медгиз.
Заленский В. (Salenski W.). 1904. Merphogenetische Studien an Wiirmern.
Mem. de i’Acad. d. Sc. de S-t P6tersbourg, V-me serie, 16.
Зальцер (Salzer F.). 1910. Ueber Implantation konservierter Pferdehornhaut in
normale Cornea des Kaninchens Sitz. ber. Ges. Morph, u. Physiol., 25.
— 1911. Ueber die Regeneration der Kaninchenhornhaut Arch. f. Augenheilk., 69.
3 а т к e и Зальцер (Satke О. u. Salzer G.). 1932. Ueber ein primares Kar-
zinom des Herzbeutels. Virch. Arch., 284.
3 а у т e p (Sa uter V.). 1935. Regeneration und Transplantation bei erwachsenenTiercn.
Roux’Arch., 132.
Захарьевская M. (Zacharjewskaja M.). 1935. Ueber das Epithelwachs-
tum bei einem Pseudomyxom. Z. Krebsforsch., 42.
— 1935. Воспалительные разрастания эпителия при туберкулезе молочной железы.
Юб. сб. 25-лет. проф. Н. Аничкова.
— 1938. О разрастании эпителия почек при очаговом их повреждении. Арх. биол.
наук, 51.
Зееман Г. (See ma n n G.)_ 1931. Histobiologie der Lungenalveole.
Зеефельдер (Seefelder). 1910. Beitrage zur Histogenese und Histologic dvr
Netzhaut, des Pigmentepithels und des Sehnerven. Graefe’s Arch., 73.
Зенкевич JI. 1937. История системы беспозвоночных. Руков. по зоол., 1.
— 1944. Очерки по эволюции двигательного аппарата животных. Ж. общ. биол., 5.
3 и г мунд (Siegmund Н.). 1929. Einfache Entztindungen des Darmrohres. Handb.
spez. path. Anat. u. Hist., Bd. 4, T. 3.
3HHb6ep6epr(Silberberg M.). 1929. Endothei in den Gewebskulturen. Arch,
exp. Zellforsch., 8.
Ибелинг (Ebeling A.). 1924. Cultures pure d’epithelium thyroidien. C. r. Soc.
Biol., 90.
— 1924. Action de I’epithglium thyroidien en cultures pures sur !a croissance des fibro-
blastes. C. r. Soc. Biol., 90.
Ибелинг и Фи шер (Ebeling A. a. Fischer A.). 1922. Mixed cultures of
pure strains of fibroblasts and epithelial cells. J. exp. Med., 36.
Иванов П. (Ivanoff P.). 1903. Die Regeneration von Rumpf- und Kopfsegmenten
bei Lumbriculus variegatus. Z. wiss. Zool.
— 1907. Die Regeneration der Segmente bei den Polychaeten. Z. Morph, u.
Okol., 10.
— 1908. Die Regeneration des vorderen und hinteren Korperendes bei Spirographis spa-
llanzanii. Z. wiss. Zool., 91.
— 1912. Регенеративные процессы у многощетинковых червей.
— 1937. Общая и сравнительная эмбриология. Биомедгиз.
Инге бригтсен (Ingebrigtsen R.). 1913. Studies on the degeneration and
regeneration of the axis cylinders in vitro. J. exp. Med., 17.
— 1916, Confributionto the biologj of peripheral nerves in transplantation. J. exp. Med. ,23.
458
Литерату pa
И о ш и д a (J о s h i d a). 1932. Experiinenteller Beitrag zur Frage der Epithelmetaplasle.
Virch. Arch., 283.
Искра Ф. 19'8. О воспалительных разрастаниях эпигелия слюнных желез. Арх.
биол. наук, 51.
И ш и к а в а и Ш и м о м у р а (I s h i k a w a u. S h i m о m и r a). 1925. Arch,
exp. Zellforsch., 2.
Казанская H. 1941, Экспериментально вызванное полианолом изменение половых
органов мышей-самок, Докл. наобъед. засед. общ. пат. и общ, анат., гист. и эмбр.
Л.
Казанцев В. 1934. Гистологические исследования процессов регенерации ампути-
рованных конечностей аксолотля главным образом для выяснения вопроса о про-
исхождении клеток, дающих начало различным тканям регенерата. Тр. ЛЭЗМ, 3.
— Гистологические исследования процессов регенерации при ампутации хвостового
стебля и грудных плавников у мальков лосося. Тр. ЛЭЗМ.
— 1935. К вопросу о регенерации мускульной ткани у млекопитающих, ДАН.
Ka3axapa(Kasaharn Sh.). 1936. On the cultivation in vitrc of the hypophysis.
Arch. exp. Zellforsch., 18.
ван-К ампенгаут (v, Campe nhout E.)_ 1939. Le r6!e des placodes epiblasti-
ques craniennes chez 1’embryon de canard. C. r. See. Biol., 130.
Каннингем, Себин и Доан (Canningham R.,Sabin F. a. Doan С.)
1925. The development of leucocytes, lymphocytes and monocytes from a specifi
stem cell in adult tissue. Contr. to Embr., 16.
К а п e л ь (К a p e 1 0.1. 1926. Sur la culture in vitro de tissu hepatique et pancreatique.
C. r. Soc. Biol., 95.
— 1927. Ueber Reinkulturen von Epithel in vitro. Arch. exp. Zellforsch., 4.
— 1929. Einige Untersuchungen liber das verhalten des Epithels in vitro. Arch, exp. Zell-
forsch., 8.
Кап перс (Kappers A.). 1920/21. Vergleichende Anatomie des Ncrvensystems der
Wirbeltiere und des Menschen. Haarlem.
— 1932. Principles of development of the nervous system (neurobiotaxis). Cyt. a. cell,
path, of the nerv. syst., v. 1.
Карлтон A. (Carleton A.). 1934. J. Anat., 68.
Карлтон Г. (Carleton H.). 1925. Growth, phagocytosis and other phenomena in
tissue cultures of foetal and adult lung. Phil, trans. Roy. Soc., London, 213.
Каррель (Carrel A.). 1927. La cytologie nouvelle. C. r. Soc. Biol., 96.
Kapp ель и Берроуз (Carrel A. a. Burrows M.). 1910. La culture de tis-
sue adultesen dehors de 1’organisme. C. r. Soc. Biol., 69.
Каррель и Берроуз. 1911. Cultivation in vitro of the thyroid gland. J. exp.
Med., 13.
Каррель и Ибелинг (Carrel A. a. Ebeling A.). 1921. The multiplication
of fibroblasts in vitro. J. exp. Med., 34.
— 1922. Pure cultures of large mononuclear leucocytes. J. exp. Med., 36.
— 1924. Au sujet d’une famille de fibroblastes se multipliant in vitro depuis douze ans.
C. r. Soc. Biol., 90.
— 1924. The fundamental properties of the fibroblasts and the macrophages. 1. The fibro-
blasts. II. The macrophages. J. exp. Med., 44.
Катценштейн и Кваке (Ka tze ns te 1 n W. u. Knake E.). 1931. Die
Anrcgung des Epithel wachstums bei gleichzeitiger Storung des Bindegewebswachstums
durch oberflachlich aktive Stoffe in Gewebekultur. Z. Krebsforsch., 33.
Каффье(Са11 ier P.). 1927. Ueber die Umwandlungsfahigkeit der weissen Elemente
des nornialen menschlichen Blutes bei in vitro Kultivierung. Arch. exp. Zellforsch. 4.
— 1928. Die prospektiven Potenzen des normalen Menschehblutes. Arch, exp, Zellforsch. 4.
— 1931. Das Mesenchymgewebe der menschlichen embryonalen Lunge, besondersauf Wuchs-
form und Chromosomenbestand untersucht. Arch. exp. Zellforsch., 11.
Кахаль Рамон и (Cajal R. у) 1894. Die Retina der Wirbeltiere. Wiesbaden.
Литература
459
-* 1904. Neuroglia und Neurifobrillen bti Lumbricus. Trab. Lab. inv. biol. univ. Mad-
rid, 3.
— 1909. Nota sobre la estructura de la retina de la Mcsca (Musca domestica). Trab. Mad-
rid., 7.
— 1909—1911. Histologie du systfeme nerveux, Paris.
— 1919. Contribution al conocimiento de la retina у centres opticos de los cefalopodes.
Trab. Madrid.
— 1923. Quelques methodes simples pour la coloration de la neuroglie. Schweiz. Arch.
Neuv. u. Psych., 13.
— 1928. Degeneration and regeneration of the nervous system. Oxford Univ. Press.
Кахаль P. и СанчецД. (Cajal R.ySanchez D.). 1909. El sistemo nervioso
de los Hirudineos. Trab. Madrid, 7, 10.
— 1915. Contribucion al conocimiento del sistema nervioso de los insectos. Trab. Mad-
rid, 13.
— 1917. Dates para el conocimiento histogenetico de los centres opticos de los insectos.
Trab. Madrid, 14.
Кацнельсон 3. (К aznelson Z.). 1936. Entwicklung der myotomalen Musku-
latur bei Urodelen. Z. mikr.-anat. Forsch., 40.
— 1936. Гистогенез поперечнополосатой мышечной ткани. Усп. совр. биол., 5.
— 1939. Сто лет учения о клетке. Изд. АН СССР.
Каш к аров (Kaschkaroff В.). 1913. Ueber die Epidermis bei Trachypterus
taenia. Anat. Anz., 44.
КедровскийБ. (Kedrowsky B.). 1925. Reaktive Veranderungen in den Gewe-
ben der Teichmuschel (Anodonta sp.) bei Einfiihrung von sterilem Сelloidin, Virch.
Arch., 257.
Ke й и Ретциус (Key A. u. Retzius G.). 1873, 1875. Studien iiber die Anatomie
des Nervensystems und des Bindegewebes. Bd. 1 u. 2. Stockholm.
К e й б e л ь (K e i b e 1 F.). 1888. Die Entwicklungvorgange am hinteren Ende des Meer-
schweinchenembryos. Arch. Anat. u. Entw. gesch.
— 1897. Normentafel zur Entwicklungsgeschichte des Schweines. Jena.
Келлер (Keller). 1894. Die ungeschlechtliche Fortpflanzung der Siisswassertur-
bellarien. Z. Natuiwiss., 28.
Кёлликер A. (KOlliker A.). 1844. Entwicklungsgeschichte der Cephalopoden.
Zurich.
— 1849. Beitrage zur Kenntnis der glatten Muskeln. Z. wiss. Zool., 1.
— 1855. 1859, 1889—1902. Handbuch der Gewebelehre des Menschen. 2, 3 und
6 Aufl.
— 1876. Entwicklungsgeschichte des Menschen und der hcheren Thiere. 2 Aufl.
— 1884. Die embryonalen Keimblatter und die Gewebe. Z. wiss. Zool., 40.
FC e н о (С u ё n о t L.). 1891. Etudes sur le sang et les glandes lymphatiques dans la serie
animale. Arch. zool. exp. et g€n., S6r. 2, v. 9.
— 1897. Les globules sanguins et les organes lymphoides des Invertebres. Arch. anat.
micr., 1.
Керби (Kirby D.). 1927. The cultivation of lens epithelium in vitro. J. exp. Med., 45.
К e p и (С а г e у E.). 1921. Am. J. Physiol., 58.
Кертис (Curtis W.). 1902. The life history, the normal fission and the reproductive
organs of Planaria maeulata. Proc. Bost. Soc. Nat. Hist., 30.
Кеттинг и Льюис (C Utting W. a. Lewis M.). 1938. Failure of the anterior
lobe pituitary to produce hormones in tissue cultures. Arch. exp. Zellforsch., 21.
К и й о н о (К i у о п о К.). 1914. Die vitale Karminspeicherung.
Кингсбери (Kingsbury В.). 1899. The regeneration of the intestinal epithe-
lium of the Toad (Bufo lentiginosus americanus) during transformation. Transact.
Am. micr. Soc., 20.
К и о д и (C h i о d i V.). 1928. Sulla natura delle cellule libere del polmone e del rive-
stimento dell’alveolo polmonare. Arch, d’anat., hist, et embr., 8.
460
Литература
Кирле (Kyrle J.). 1907. Ueber die Regenerationsvorgange im tierischen Pancreas.
Arch. mikr. Anat., 72.
Кирле и Шоппер (Kyrle J. и Schopper К.). 1915. Ueber Regenerations-
vorgange im tierischen Nebenhoden. Virch. Arch., 220.
К л а а ч (Klaatsch H.). 1894. Ueber die Herkunft der Skleroblasten. Ein Beitrag
zur Lehre von der Osteogenese. Morph. Jahrb., 21.
Клара (Clara M.)„ 1927. Die arterio-venosen Anastomosen der Vogel und Sauge-
tiere. Erg. Anat., 27.
— 1931. Ueber den Bau der Leber beim Kaninchen und die Regenerationserscheinun-
gen an diesem Gewebe bei experimenteller Phosphorvergiftung. Z. mikr.-anat.
Forsch., 26.
— 1936. Vergleichende Histobiologie des Nierenglomerulus und der Lungenalveole. Z.
mikr.-anat. Forsch., 40.
Кларк Э. (Clark E.). 1912. Further observations on living growing lymphatics: their
relation to mesenchyme cells. J. Anat., 13.
Кларки Кларк (Clark E. a. Clark E.). 1920. Reaction of cells in the tail
of amphibian larvae to injected croton oil (aseptic inflammation). Am. J. Anat., 27.
— 1932. Observation on the new growth of lymphatic vessels as seen in transparent
chambers introduced into the rabbitsear. Am. J. Anat., 51.
— 1939. Microscopic observations on the grewth of blood Capillaries in the living
mammal. Am. J. Anat., 64.
— 1940. Microscopical observation on the extra-endothelial cells of living mammalian
blood-vessels. Am. J. Anat., 66.
Клебс (Klebs E.). 1874. Die Regeneration des Plattenepithels. Arch. exp. Path., 3.
Клейпенберг (К 1 e i n e n b e r g). 1872. Hydra. F.ine anatomisch-entwicklungs-
geschichtliche Untersuchung. Leipzig.
— 1886. Die Entstehung des Annelids aus der Larve von Lepadorhynchus. Z. wiss.
Zool., 44.
К. л e н и ц к и й Я. 1938. Об изменениях, полученных при введении бластомогенпого
агента в половые пути мышей-самок. Арх. биол. наук, 51.
— 1940. Действие 3 : 4 : 8 : 9 дибензпирена на половые пути мышей-самок. Арх. биол.
наук, 58.
К н ё л л ь (К п б 1 1 Е.). 1937. Ueber die Ziichtung von Nebennierenepithelzellen. Arch,
exp. Zellforsch., 20.
Колль м ан (K oilman n M.). 1908. Recherches sui les leucocytes et le tissu lym-
phoide des invertebres. Ann. Sc. nat., zool., 8.
Колосов A. (Kolossow A.). 1893. Ueber die Struktur des Pleuroperitoneal- und
Gefassepithels (Endothels). Arch. mikr. Anat., 42.
К о л ь м е р (Ко 1 iner W.). 1927. Geruchsorgan. Gehororgan. Handb. mikr. Anat.
des Menschen, Bd. 3, T. I.
Кон и Макмил л ен(С one W. a. Macmillan J.). 1932. The optic nerve and
papilla. Cyt. a. cell. path, of the nerv. syst., T. 2.
Конгдон (Congdon E.). 1915. . The identification of tissues in artificial culture
Anat. Rec., 9.
Конечный (Konjetzny G.). 1928. Die Entziindung des Magens. Handb. spez.
path. Anat. und Hist., Bd. 4, T. 2.
Коннор (O’Conner R.) 1940. An experimental study of the development of the
amphibian cloaca. J. Anat., 74.
Кон ше г г (Konschegg Th.). 1926. Zur Epithelmetaplasie. Virch. Arch., 259.
К о p и п к и й Г. 1926. Опыт трансформационной клеточной патологии: трансформа-
ционная теория опухолей. Изд. 2-го МГУ.
— 1936. О так называемых «первичных адамантиномах» костей. Арх. пат. анат. и пат.
физио л., 2.
Корн и ль и Карно (Cornil V. et Carnot Р.). 1899. R6generation cicatri-
cielle des cayites tnuqueuses et de leur revetement 6pithelial. Arch. med. exp.
Литература
461
К о с т е р о (Costero J.). 1930. Studien an Mikrogliazelien (s. g. Hortega-Zellen)
in Gewebskulturen von Gehirn. Arb. Staatsinst. exp. Ther. Frank!., 23.
— 1930. Estudio del comportamento de la microglia cultivada in vitro. Dat. concernien-
tes a su histogenesis. Mem. R. Soc. esp. Hist, nat., 14.
К о ф ф (К о f f). 1933. Contr. to Embr. 140.
Кочетов. 1911. Исследования над пигментным эпителием сетчатки в связи с вопросом
о клеточном делении. Тр. СПб. имп. общ. естествоисп., 39.
Кочетов Н. 1940. Исследования по сравнительной гистологии мышечной ткани.
Дисс. 1 ЛМИ.
Краевски й Н. и М а я т В. 1938. К учению о регенерации слизистой оболочки
желудка. Арх. пат. анат. и пат. физиол., 4.
Краске (Kraske). 1878. Experimentelle Untersuchungen uber Regeneration dec
quergestreiften Muskulatur. Fiabil.-Schr. Halle.
Крафт И. 1925. О воспалительном новообразовании соединительной ткани у кости-
стых рыб. Изв. биол. инет. Пермск. унив., 4.
Крёзинг (К rosing R.). 1892. Ueber die Ruckbildung und Entwicklung der quer-
gestreiften Muskulatur. Virch. Arch., 128.
Kpeft6epr(Kreybcrg L.). 1929. The prospective potencies of the human lym-
phocyte. Arch. exp. Zellforsch., 8.
К p e к к e p (К r e c k e r). 1923. J. exp. Zool., 37.
Кристоф (Cristophe). 1923. Recherches sur les greffes d’os fixe a I’alcohol et
sur le mecanisme de I’osteogenese. Arch, franco-beige de chir., 22.
К p о м а й e p (К г о m а у c r). 1902. Neue biologische Beziehungen zwischen Epithel
und Bindegewebe. Desmoplasie. Arch. Dermat. u. Syph., 62.
Кромпехер (Kro mpe c her), 1903. Der Basalzellenkrebs. Jena.
К p у м б e й н (Kr u mb e i n), 1924. Primfires Pleurakarzinom. Virch. Arch. 249.
Круши HCKHH(Kruszynski J.). 1936. Recherches sur le cartilage. Trav. inst.
d’hist, et d’embr. de 1’univ. Wilno, 10.
Крюкова 3. 1936. Регенерация запирательной мышцы у беззубки (Anodonta anatina).
Арх. биол. наук, 37.
— 1938. Регенерация m. sartcrius у лягушки. Арх. анат., гист. и эмбр. 19.
Куби п Шульц(КиЬ1е9. a. Schultz G.). 1925. Vital and supravital studies
of the cells in the cerebrospinal fluid and of the meninges in cat. Bull. Johns Hopk.
hosp., 37.
Кузнецовский H. 1929. О гистологических особенностях злокачественных опу-
холей покровных клеток (мезотелия) серозных полостей. Вести, хир., 48/49.
Купффер (v. Kupffer С.). 1893—1960. Studien zur vergleichenden Entwicklungs-
geschichte des Kcpfes der Kranioten. 1—4.
— 1903. Die Morphogenie des Zentralnervensystems. Handb. d. vergl. u. exper. Entw.
gesch.
Куше (Kusche). 1929. Interplantation umschriebener Zellbezirke aus der Blastula
und der Gastrula der Amphibien. Roux’Arch., 120.
•Лаврентьев Б. 1938. Антагонистическая иннервация автономной нервной системы
и методы ее исследования. Статья в сборнике «Морфология автономной нервной
системы». Изд. ВИЭМ.
Лазаренко Ф. (Lazarenko Th). 1925. Die morphologische Bedeutung der
Blut- und Bindegewebsekmente der Insekten. Z. mikr-anat. Forsch., 3.
— 1928. Experimentelle Untersuchungen uber das Hypodeimisepithel der Insekten. Z.
mikr.-anat. Forsch., 12.
— 1934. Опыт применения нового метода к экспериментальному изучению тканей и его
предварительные результаты. Арх. биол. наук, 37.
— 1935. Метод культивирования тканей в организме и первые результаты его приме-
нения. Тр. ВИЭМ.
— 1939. Опыты культивирования тканей и органов в организме. II. Слизистая оболочка
желудка. Арх. анат., гист. и эмбр., 21.
462
Литература
Jla3Ho(Lasio G.). 1909. Ueber die Regeneration der Schleimhaut der Harnblase usw.
Virch. Arch., 178.
Ла й вел (L i n e 1 1). 1929. The histology of neuroglial changes following cerebral
trauma. Arch. Neur. und Psych., 22.
Ланг A. (Lang A.). 1884. Die Polycladen (Seeplanarien) des Golfes von Neapel und
der angrenzenden Meeresabschnitte. Eine Monographic. Fauna und Flora des Golfes
von Neapel, XI. Moncgraphie.
Ланг Ф. (Lang F.). 1925. Ueber Gewebekulturen der Lunge. Arch. exp. Zellforsch., 2.
Ланг П. (Lang. P.). 1912. Ueber Regeneration bei Planarien. Arch. mikr. Anat., 79.
Л а у x e (La uc he A.). 1923. Die extragenitalen heterotopen Epithelwucherungen vom
Bau der Uterusschleimhaut (Fibroadencmatosis seroepithelialis). Virch. Arch., 243.
Лёб Л. (L о e b L.l. 1898. Ueber Regeneration des Epithels. Roux’Arch., 6.
— 1899. An experimental study of the transformation of epithelium to connective tissue,
Medecine, 5.
— 1902. Ueber das Wachstum des Epithels. Roux’Arch., 13.
Левен (L e v e n). 1887. Experimentelle Untersuchungen liber die Regeneration
der quergestreiften Muskeln. Inaug.-Diss. Halle.
Левенгук (Leeuwenhuek A.). 1695. Arcana naturae. Delphis Batav.
Левенталь (L 6 w e n t h a 1). Цит. no Lubarsch-Ostertag, Ergebnisse, Jahrg.
6, 1899.
Левен штейн (LOwenstein). 1910. Ueber atypische Epithelwucherungen und
Tumoren der Speicheldriisen, besonders der Parotis. Frankf. Z. Path., 4.
Леви (Levi G.). 1917. Connessioni e struttura degli elementi nervosi sviluppati fuori
dell’organismo. Mem. Accad. Lincei, 12, IV.
— 1919. Nuovi studi su cellule coltivati in vitro. Attivita biologiche, intima struttwa,
caratteri mcrfologici specifici. Arch. ital. Anat., 16.
— 1920. Sulla persistenza dei caratteri specifici nolle cellule coltivate in vitro. Mon. Zool.
ital., 31.
— 1923. Struttura e propriety degli endoteli vascolari. Ricerche su coltura in vitro. Giorn.
Biol, e Med. sper., 1.
— 1925. Conservazione e perdita dell’indipendenza delle cellule dei tessuti. Arch. exp.
Zellforsch., 1.
— 1925. Ricerche sperimentali sopra elementi nervosi sviluppati in vitro. Arch. exp. Zell-
forsch., 2.
— 1934. Explantation, besonders die Struktur und die biologischen Eigenschaften der in
vitro gezi chteten Zellen und Gewebe. Erg. Anat. u. Entw. gesch., 31.
Леви и Буччанте (LeviG. eBucciante). 1928. Sulla natura delle coio-
razioni vitali studiata sulle cellule coltivate in vitro. Arch. Zellforsch., 7.
Леви и Делоренци (Levi G. e Delorenzi E.). 1934. Trasformazioni
degli elementi dei gangli spinali e. simpatici coltivati in vitro. Arch. ital. di anat. e
di embr., 33.
Леви и M e й e p (L e v i G. u. M e у e r H.). 1933. Die Struktur der lebenden Neurone.
Anat. Anz., 83.
— 1937. Nuove ricerche sulle propriefa morfologiche e biologiche del protoplasma del
neurone. Riv. Pat. sper., 8.
— 1941. Nouvelles recherches sur le tissu nerveux cultive in vitro. Morphologie, crois-
sance et relations reciproques des neurones. Arch, de biol., L. 2.
Леви и Оливо (LeviG. eOlivo О.). 1928. Le proprieta strutturali delle cellule
e dei tessuti coltivati in vitro. Arch. exp. Zellforsch., 6.
Лежава A. 1940. О митотическом делении диференцированных гладких мышечных
клеток мезенхимной системы. Сообщ. груз. фил. АН СССР, 1.
Лейдиг Ф. (Leydig F.). 1853. Anatomisch-histologische Untersuchungen uber
Fische und Reptilien. Berlin.
— 1855. Vom Bau des thierischen Korpers.
— 1857. Lehrbuch der Histologie des Menschen und der Thiere. Frankf. a. M.
Литература
468
— 1885. Zelle und Gewebe. Neue Beitrage zur Histologie des Thierkorpers. Berlin.
Ле й д л о у (L a i d 1 о w G.). 1930. Silver staining of the endoneurial fibers of the cere-
brospinal nerves. Am. J. Path., 6.
Лейк (Lake N.). 1916. Observations upon the growth of tissues in vitro relating to
origin of the heart beat, J. Physiol., 50.
Л e л e й н (L о h 1 e i n). 1923. Ueber die s. g. follikulare Ruhr. Jena.
Ленгли (Langley J.). 1921. The autonomic nervous system. Cambridge.
Ленгоссек (LenhossekM. v.). 1893. Der feinere Bau des Nervensystems im
Lichte neuester Forschungen. Berlin.
Ленерт (Lehnert G.). 1891. Beobachtungen an Landplanarien. Arch.
Naturgesch., 57.
Л e п л a (L e p 1 a t G.). 1912. Recherches sur le developpement de la membrane vascu-
laire de Poeil des oiseaux. Arch, de biol. 27.
— 1919. Action du milieu sur le developpement des larves d’amphibiens. Localisation
et differenciation des premieres 6bauches oculaires chez les vertebres. Cyclopie et
anophthalmie. Arch, de Biol., 30.
Лериш и Пол и кар (L eriche R. etPolicard A.). 1926. Les problfemcs
de la physiologie normale et pathologique de Pos. Paris.
— 1934. Position actuelle du probleme de I’osteogenese. La presse med., 99.
Ливанов H. 1913. Пограничные образования Polychaeta etc. Тр. общ. естеств.,
Казань.
Линч (Lynch). 1921. Am. J. Anat., 29.
Локателли (Locate I li P.). 1933. Sulla rigenerazione del tessuto muscolare
striato. Pathologica, 25.
Льюис M. (Lewis M.). 1915. Rhytmical contraction of the skeletal muscle tissue
observed in tissue cultures. Am.J.Physiol.
— 1919. The development of cross-striation in the heart muscle of the chick embryo. Bull.
Johns Hopk. hosp., 30.
— 1920. Muscular contraction in tissue cultures. Contr. to Embr,, 9.
— 1929. The development of the cross-striated myofibrils in the heart muscle of the chick
embryo. Anat. Rec., 16.
Льюис M. иЛью и сУ. (Lewis M. a. Lewis W.). 1912. The cultivation of sym-
pathetic nerves from the intestine of chick embryos in saline solutions. Anat. Rec., 6.
— 1917. The contraction of smooth muscle cells in tissue cultures. Am. J. Physio!., 44.
___ 1917. Behaviour of cross-striated muscle in tissue cultures. Am. J. Anat., 28.
Льюи с У. (Le wi s W.)_ 1922. Endothelium in tissue cultures. Am. J. Anat., 30.
— 1923. The transformation of mesenchyme into mesothelium in tissue cultures. Anat.
Rec., 25.
— 1926. The transformation of mononuclear blood cells into macrophages, epithelioid
cells and giant-cells. Harvey Leet, of the Harvey See, N. Y. C.
— 1931. The outgrowth of endothelium and capillaries in tissue cultures. Bull. Johns
Hopk. hosp., 49.
Любар ш (Lubarsch O.). 1906. Ueber heterotope Epithelwucherung und Krebs.
Verb, dtsch. path. Ges., X. Tag.
— 1925. Die hypertrophischen, hyperplastischen und regenerativen Vorgange. (Niere).
Handb. spez. path. Anat. u. Hist., Bd. 6, T. I.
Мазярский C. (Maziarski S.). 1926. Sur le tissu musculaire des Insectes. Les
cellules et les fibres musculaires dans les gaines ovariques et dans la tunique perito-
neale chez les Tipulides. Bull. Acad, polon.
Майер K. (Maye r C_). 1819. Ueber Histologie und eine neue Eintheilung der Gewebe
des menschlichen Korpers. Bonn bei Ad. Marcus.
Майнот (Minot Ch.). 1901. The embryological basis of pathology. Science, N. S. 13.
__ 1911. Die Entstehung des Angioblastes und die Entwicklung des Blutes. Handb. d.
Entw. gesch. d. Menschen von Keibel u. Mall, Bd. 2.
M а к л -i н и M а к л и н (M a c k 1 i n C. a, Macklin M.). 1920. A study of brain
464 ________________________Литература
repair in the rat by the use of trypan blue, with special reference to the vital staining
of the macrophages. Arch. Neur. a. Psych., 3.
Максимов (Maxi mo w A.). 1898. Die histologischen Vorgange bei der Heilung
von Hodenverletzungen und Regenera tionsfahigkeit des Hodengewebes. Ziegl. Beitr.,26.
— 1902. Experimentelle Untersuchungen i.ber entzfindliche Neubildung von Bindegewebe.
Ziegl. Beitr, Suppl. 5.
— 1906. Ueber entzlndliche Bindegewebsneubildung beim Axolotl. Ziegl. Beitr., 39.
— 1907. Ueber die Entwicklung der Blut- und Bindegewebszellen beim Saugetierembryo.
Fol. hacmat., 4.
— 1907. Experimentelle Untersuchungen zur postfetalen Histogenese des myeloiden Ge-
webes. Ziegl. Beitr., 4.
— 1909. Untersuchungen uber Blut und Bindegewebe. 1, Die friihesten Entwicklungssta-
dien der Blut- und Bindegewebszellenbeim Saugetierembryousw. Arch. mikr. Anat., 73.
— 1909. 11. Ueber die Histogenese der Thymus bei den Saugetieren. Arch, mikr, Anat., 74.
— 1914. Основы гистологии. СПб.
— 1916. О культивировании in vitro соединительной ткани взрослых млекопитающих.
Русск. арх. анат., Гист, и эмбр., 1.
— 1923. Untersuchungen tber Blut und Bindegewebe, VIII. Die cytologischen Eigen-
schaften der Fibroblasten, Reticulumzellen und Lymphocyten des lymphoiden Gewebes
ausserhalb des Organismus, ihre genetischen Wechselbeziehungen und prospektiven
Entwicklungspotenzen. Arch. mikr. Anat., 97.
— 1923. Untersuchungen uber Blut und Bindegewebe. IX. Ueber die experimentelle Er-
zeugung von myeloiden Zellen in Kulturen des lymphoiden Gewebes. Arch. mikr.
Anat., 97.
— 1923. Untersuchungen liber Blut und Bindegewebe. X. Ueber die Blutbildung bei den
Selachiern im erwachsenen und embryonalen Zustande. Arch. mikr. Anat., 97.
— 1924. Relation of blood cells to connective tissue and endothelium. Physiol. Rev., 4.
— 1925.. Ueber Krebsahniiche Verwandlung der Milchdriise in Gewebskulturen. Virch.
Arch., 256.
— 1927. Ueber das Mesothel (Deckzellcn der serosen Haute) und die Zellen der serosen
Exsudate. Untersuchungen an entziindetem Gewebe und an Gewebskulturen. Arch,
exp. Zellforsch., 4.
— 1927. Bindegewebe und blutbildende Gewebe, Handb. d. mikr. Anat. d. Menschen,
Bd. 2.
— 1928. Ueber die Entstehung von argyrophilen und kollagenen Fasern in Kulturen von
Bindegewebe und von Blutleucocyten. Zbl. allg. Path., 47. i
— 1928. Cultures of blood leucocytes. From, lymphocyte and monocyte to connective
tissue. Arch. exp. Zellforsch., 5.
— 1929. Ueber die Entwicklung argyrophiler und kollagener Fasern in Kulturen von
erwachsenem Saugetiergewebe. Z. mikr.-anat. Forsch., 17.
Маку ш о к (Ma k use ho k). 1911, 1912, 1913. Anat. Anz., 39, 42, 44.
Макьяруло и Бюнглер (Macchiarulo u. Bungler). 1929. Durch
Teer erzeugte eigenartige Veranderungen der Parotisdriise. Frankf. Z. Path., 37.
Малакарне(Ма!асагпе V.). 1803. I sistemi e la reciproca influenza loro inda-
gati. Padua.
Маллори (Mallory F.). 1904. A hitherto undescribed fibrillar substance produced
by connective tissue cells. J. Med. Res., 10.
Маллори (Mallory A.). 1920. The type cell of the socalled dural endothelioma. J.
Metab. Res., 41. g
Мангольд (Mangold O.). 1923. Transplantationsversuche zur Frage der Spezi-
fitat und Bildung der Keimblatter bei Triton. Arch. mikr. Anat. u. Entw. mech., 100.
M а н к и н 3. (M a n k i n Z.). 1933. Beitrage zur Lehre von den endometrioiden Hete-
rotopien. Arch. Klin. Chir., 175.
Марбург (Marburg). 1902. Endarteriitis cartilaginosa der grossen Hirngefasse.
Zbl. Path., 13.
Литература
465
Маркезанп (Marchesa ni О.). 1026. Die Morphologie der Glia im Nervus opti-
cus und in der Retina, dargesteilt nachden neuesten Untersuchungsmethoden und Un-
tersuchungsergebnissen. I. Mitt. Graefe’s Arch., 117.
Маркес (Markets S.). 1933. Versuche der Zilchtung von Nierengewebe in vitro.
Arch. exp. Zellforsch., 14.
Маркис (Marquis C.). 1892. Das Knochenmark der Amphibien in den verschiedenen
Jahreszeiten. Inaug.-Diss., Dorpat.
Марсо (Marceau F.). 1904. Recherches sur la structure et le d6veloppement compa-
res des fibres cardiaques dans la serie des vertebres. Ann. Sc. nat., 8, ser. zool., 19.
— 1905. Recherches sur la structure du coeur chez les mollusques suivies d’une etude Spe-
ciale des coeurs branchiauxet de lews appendices glandulaires chez les Cephalopodes.
Arch. anat. micr., 7.
Мартин (Martin E.). 1888. Ueber die Anlage der Urniere bei Kaninchen. Arch,
mikr. Anat., 32.
Мартинович (Martinovic P.). 1933. Survival in vitro of explants of the cerebral
cortex of the cat cultivated in the cerebro-spinal fluid of the young animal. Arch. exp.
Zellforsch., 12.
— 1938. Development in vitro of the mammalian gonad. Arch. exp. Zellforsch., 22.
Мартинотти (Martinotti). 1888. Sulla estirpazione del pancreas. Giorn. R. accad.
med. di Torino, N 4.
— 1890. Ueber Hyperplasie und Regeneration der driisigen Elemente in Beziehung auf
ihre Funktionsfahigkeit. Zbl. Path., 1.
Марцокки (Marzocchi V.). 1904. Sui processi rigenerativi nelle ghiandole sotto
mascellari del coniglio. Arch. sc. med., 28.
— 1904. Sui processi rigenerativi delle ghiandole salivari sierose. Sperimentale, 57.
Маршанд (Marc ha nd F.)_ 1899. Ueber die Beziehungen der pathologischen Ana-
tomie zur Entwicklungsgeschichte. Verh. dtsch. path., Ges., 2 Tag., Munchen.
— 1901. Der Prozess der Wundheilung. Stuttgart.
— 1921. Die Veranderungen der peritonealen Deckzellen nach Einfuhrung kleiner Fremd-
korper. Ziegl. Beitr., 69.
— 1924. Die drtlichen reaktiven Vorgange (Lehre von der Entziindung). Handb. d. allg.
Path., Bd. 4, Abt. 1.
Масловский (Ma s 1 о w s k y). 1868. Ueber die Neubildung und Heilung des quer-
gestreiften Muskelgewebes nach traumatischen Verletzungen. Wien. med. Wochenschr.
MacceHTn(Massenti G.). 1924. Sulla rigenerazione del fegato. Arch. Sc. biol,, 6.
Массон (Masson P_). 1923. Diagnostics de laboratoire. 11. Tumeurs.
— 1932. Neural proliferations in the vermiform appendix. Cyt. a. cell. path, of the
nerv. syst., v. 3.
Матсу мото (Matsumoto S.). 1918. Contribution to the study of epithelial mo-
vement. The cornea epithelium of the frog in tissue culture. J. exp. Zool., 86.
Маурер (Maurer). 1906. Die Entwicklung des Muskelsystems. Handb. d. vergl. u.
exp. Entw. gesch. d. Wirbeltiere, Bd. 3, T. I.
— <1915. Grundziige der vergleichenden Gewebelehre.
Ma ц у и (Ma t s u i J.). 1929. Ueber die in vitro Kulturen des Endothels der Membrana
Descemetii. Arch. exp. Zellforsch., 8.
M e б и у с (M e b i u s). 1924. Die formale Genese der Knochenbildung in Bauchnarben.
Virch. Arch., 248.
Мейер Г. (Meyer H.). 1936. Kulturen von Ciliar- und Irismuskeln bei Hiihnerembryo-
nen. Z. mikr.-anat. Forsch., 39.
— 1936. Ziichtung der Retina des Huhnes in vitro. Z. mikr.-anat. Forsch., 39.
Мейер Г. и Яблонский (Meyer Н. and Jablonski W.). 1937. Cultiva-
tion of nerve cells in vitro over a long period. J. Anat., 72.
M e й e p P. (Meyer R.). 1910. Ueber Erosio portionis uteri. Verh. dtsch. path. Ges.,
14 Tag.
— 1911. Ueber embryonale Gewebsanomalien und ihre pathoiogische Bedeutung im allge-
30 Хлопин—349
466
Литература
meinen und solche des mannlichen Genitalapparats im besonderen. Erg. Path., 15.
Мейзегеймер (Meisenheimer). 1898. Entwicklungsgeschichte von Limax
maximus. Z. wiss. Zool., 63.
Me й с т e p (Me i s te r). 1891. Ueber die Regeneration der Leberdriise nach Entfernung
ganzer Lappen und fiber die Beteiligung der Leber an der Harnstoffbildung. Zbl.
Path., 2.
Меккель (Meckel J.). 1815. Handbuch der menschlichen Anatomie. Bd. I. Allge-
meine Anatomie. Halle und Berlin.
Меллендорф B. (M Ollendorff W. v.). 1927. Einige Beobachtungen iiber den
Aufbau des Nierenglomerulus. Z. Zellforsch., 6.
— 1930. Der Exkretionsapparat. Handb. d. mikr. Anat. d. Menschen, Bd. 7, T. I.
— 1932. Das Mutterstiick von Bindegewebskulturen. Ein Beitrag zur Frage, wie konstruk-
tive Fasersysteme und Hartsubstanzen entstehen. Z. Zellforsch., 15.
Меллендорф В. и M. (М о I 1 е n d о г f f W. u. M.). 1926. Das Fibrozytennetz im
lockeren Bindegewebe, seine Veranderungsfahigkeit und Anteilnahme am Stoffwechsel.
Z. Zellforsch., 3.
Меллендорф M. (M Ollendorff M. v.). 1931. Beobachtungen bei der Dauer-
ziichtung von Bindegewebe erwachsener Kaninchen. Z. Zellforsch., 12.
— 1931. Die Entstehung von Histiozyten in Kulturen erwachsenen Bindegewebes. Arch,
exp. Zellforsch., II.
— 1938. Der Brown-Pearce Tumor in der Gewebekultur. Arch. exp. Zellforsch., 21.
Меллендорф M. и В. (M Ollendorff M. u. W.). 1929. Bindegewebestudien.
VIII. Ueber die Potenzen der Fibrozyten des erwachsenen Bindegewebes in vitro.
Z. Zellforsch., 9.
Мендельсон (Me n de 1 sohn W.). 1937. The cultivation of adult rabbit testicle
in roller tubes. Anat. Rec., 69.
Менкеберг (M 6 n k e berg). 1902. Ueber Knochenbildung in der Arterienwand.
Virch. Arch., 167.
— 1924. Die Erkrankungen des Herzbeutels. Handb. spez. path. Anat. u. Hist.,
Bd. 2.
Мен к л ь (Men cl E.). 1908. Ueber die Histologie und Histogenese der sogenannten
Punktsubstanz Leydigs in dem Bauchstrange der Hirudineen. Z. wiss. Zool., 89.
— 1909. Zur Kenntnis der Neuroglia bei Nephelis. Zool. Anz., 34.
Менчот (Me n c ho t E.). 1929. Abgrenzung des Augenmatherials und anderer Teil-
bezirkc der Medullarplatte, die Teilbewegung wahrend der Aufspaltung. Roux’Arch.,
116.
Меркель Г. (Merkel H.). 1926. Die Magenveratzungen. Handb. d. spez. path. Anat.
u. Hist., Bd. 4, T. I.
Меркель Ф. (Merkel F.). 1908. Epithelium. Erg. Anat. u. Entw. gesch., 18.
M e p p e й (M u г г а у P.). 1932. The development in vitro of the blood of the early chick
embryo. Proc. Roy. Soc. Lond., B, III.
Меррей и Стаут (Murray M. a. Stout A.). 1942. Characteristics of human
Schwann cells in vitro. Anat. Rec., 84. л
Метц и ILInaTn(Metz u, Spatz). 1924. Die Hortegaschen Zellen(das s. g. «dritte
Element») und fiber ihre funktionelle Bedeutung. Z. ges. Neur. u. Psych., 89.
Мечников И. (Metschnikoff E.). 1874. Studien fiber die Entwicklung der
Medusen und Siphonophoren. Z. wiss. Zool., 24.
— 1886. Embryologische Studien an Medusen. Wien.
— 1892. La phagocytose musculaire. Ann. Inst. Pasteur.
— 1892. Legons sur la pathologie comparee de rimflammation. Paris.
Ми зум и (M i s u m i). 1909. Ueber Riickbildung an Talgdriiscn, Viich. Arch., 197.
Минкин C. 1939. Структурные изменения головного мозга после ранения. Дисс
Харьков.
Миллар (Millar W.). 1934. Regeneration of skeletal muscles in young rabbits. J.
Path. a. Bact., 38.
Литература
467
Мите у да (Mitsuda). 1924. Untersuchungen iiber Transplantation und Explanta-
tion von Lebergewebe usw. Virch. Arch., 248.
Михайлов В. 1941. Опыт экспериментально-гистологического анализа опухолей
яичника. Докл. на сессии Гос. Онкол. инет.
Михал и к (v. Mihalik Р.). 1935. Ueber die Nervengewebekulturen mit beson-
derer Berixksichtigung der Neuronenlehre und der Mikrogliafrage. Arch. exp.
Zellforsch., 17.
МихаловскийИ. (Michalowsky). 1926. Zbl, allg. Path. u. path. Anat., 38.
— 1928. Virch. Arch., 267.
Мишу p с к и й (Miszur s ki B.). 1937. Researches on the keratinisation of the epi-
thelium in tissue cultures. Arch. exp. Zellforsch., 20.
— 1938. Recherches sur 1’influence des extraits des embryons de diff6rent age sur la
croissance et la differenciation du cartilage et de 1’os en culture. Arch. exp.
Zellforsch., 22.
— 1939. Further conti ibution to the influence of extracts from embryos of different age
on the growth of cartilage and ossification in vitro. Arch. exp. Zellforsch., 23.
Мом ильян о-Л e в и (M о m i g 1 i a n o-L e v i Q.). 1932. Formazione e maturazione
delle fibre collagene nelle colture dei tessuti significato della distribuzione reticolare
e fascicolare della stroma collagene nei vari organi. Z. Zellforsch., 16.
Монтальчини и Сачердоте (Monta lei ni e Sacer dote). 1937. .For-
mazione di fibrille reticolare e collagene in colture di epiteli e di elementi miocardici.
Monit. Zool., 47.
Морозенская JI. 1938. Экспериментальные опухоли печени, вызванные ортоамид-
азотолуолом. Арх. биол. наук, 51.
Мосса (М ossa S.). 1928, 1929. Ulteriori studi sulla rigenerazione dei neuriti e sulla
modificazione dei neuroblasti dei gangli spinali di embrione di polio coltivate in vitro.
Arch. exp. Zellforsch., 7.
Мюллер Г. (M ii J 1 e r H.). 1889. Zur Frage der Blutbildung. Sitz. ber. d. K. Akad.
d. Wiss. Wien, mathem.-naturwiss. KI. Ill, Bd. 98.
Мюллер И. (M ii 1 1 e r J.). 1830. De glandularum secernentium structura penitiori.
Lipsiae.
M ю л л e p E. (M u 1 1 e r E_). 1900. Studien iiber Neuroglia. Arch. mikr. Anat., 55.
Мясоедов C. (Mjassoj edoff S.). 1926. Die Zellformen des Bindegewebs in des
Blutes und die Blutbildung beim erwachsenen Huhn. Fol. haemat., Arch., 32.
H а в и л л ь (Naville А.). 1922. Recherches sur I’histogenese de la rdgdndration
chez les Batraciens Anoures. Arch, de biol., 34.
H а жотт (Nageotte J.). 1918. Formation de pifeces squelettiques surnumeraires
provoqu£es par la presence de greffons morts dans 1’oreille du lapin adulte. C. r. Soc.
biol., 81.
— 1922. reorganisation de la matiferc vivante dans ses rapports avec la vie. Paris.
— 1932. Sheaths of peripheral nerves. Degeneration and Regeneration. Cyt. a, cell. path,
of the nerv. syst.
H а з у (N a su S.). 1923. Beitrage zur Frage der Ueberlebensfahigkeit der Gewebe. Eine
Untersuchung iiber die Veranderungen an Zellen, die von der normalen Zirkulation
abgeschnitten sind. Virch, Arch., 243.
На ясен (Nansen F.). 1888. Die Nervenelemente, ihre Struktur und Verbindung im
Zentralnervensystem. Anat. Anz., 3.
Hann (N a p p). 1906. Drei Faile von primarem Karzinom des Bauchfeils. Z.
Krebsforsch. 4.
Насонов Д. и Александров В. 1940. Реакция живого вещества на внешние
воздействия. Изд. АН СССР.
Н а С о н о в Н. 1934. Образование хряща in vitro у аксолотля. ДАН СССР, 3.
Нау верк (Na u we г k G.). 1890. Ueber Muskelregeneration nach Verletzungen.
Jena.
He re ли (Nageli). 1930. Blutkrankheiten und Blutdiagnostik.
468
Литература
Н е и ман (Ne u ma n n E.)_ 1868. Ueber die von Zenker beschriebenen Veranderungen
der willkiirlichen Muskeln beim Typhusprozess. Arch. f. Heilkttnde, 9.
— 1868. Ueber die Heilungsprozesse nach Muskelverletzungen. Arch. mikr. Anat., 4.
— 1896. Hamatologische Studien. Virch. Arch., 143.
— (Neumann). 1931, 1933. Arch. Gyn., 151, 152.
H e м и л о в A. 1931. Что такое ткань? Природа.
Ни вен (Niven J.). 1931. The repaire in vitro of embryonic bones after experimental
fractures. Arch. exp. Zellforsch., 17.
— 1933, The development in vivo and in vitro of avian patella. Roux’Arch., 128.
Никола (Nicolas A.). 1894. Les «bourgeons germinatifs» dans 1’intestin de la larve
de salamandre. Bibl. anat., 2.
Никольский. 1880. О мерцательных элементах на серозных оболочках лягушки.
Дисс. СПб.
Ни с с л b(N i ssl F.). 1894. Ueber eine neue Untersuchungsmethode der Zentralorgane.
Zbl. Nervenheilk. u. Psych., 18.
Нишибе (N i s h i b e M.). 1928/1929). On the cultivation of kidney tissue from the
adult toad. Arch. exp. Zellforsch., 7.
— 1928, 1929. Growth of endocardial cells from the chick embryo heart in vitro. Arch. exp.
Zellforsch., 7.
H о б л ь (N о b 1). 1903. Zur Histologie der gonorrhoischen Deferentitis und Epididymitis.
Arch. f. Derm., 67.
Новиков M. (Now ikoff M.). 1912. Исследование о хрящевой и костной тканях
Дисс.
— 1912. Studien iiber das Knorpelgcwebe der Wlrbellosen. Z. Zool., 103.
НордманМ. (Nordmann M.)_ 1929. The behaviour of adult mammalian kidney
in tissue cultures. Arch. exp. Zellforsch., 9.
— 1929. Wachstum und Stoffwechsel der Leberzellen in der Gewebskultur. Arch. exp.
Zellforsch., 8.
Норд м ан, Бакстер и Весле (Nordmann, Baxter u. Wesle). 1934.
Kontraktionen der Magen- und Darmmuskulatur des Hiihnerembryos in der Gewebs-
kultur. Arch. exp. Zellforsch., 15.
Нордман И P ют e p (N ordman n u. Riither). 1931. Ueber die Schlagtatigkeit
des explantierten Herzmuskels von Huhn und Ratte und ihre Beziehungen zum Reiz-
leitungssystem. Arch. exp. Zellforsch., II.
Норд ме й ep (Nordmeyer N.). 1932. Ueber Beziehungen von Exsudatzellen zu
Fibrozyten verschiedener Tierarten. Aich. exp. Zellforsch., 13.
Нусбаум (Nussbaum M.). 1899. Entwicklungsgeschichte des Menschenauges.
Graefe-Saemisch Hanbd. d. Augenheilkunde.
Нусбаум (Nussbaum J.). 1905, 1908. Z. wiss. Zool., 79, 89.
Нусбаум иОкснер (Nussbaum u. Oxner). 1910. Roux’Arch. 30.
Нусбаум и Сидор и ак (Nussbaum u. Sidoriak). 1900. Regeneration
nach kiinstlicher Verletzung bei Forellenembryonen. Roux’Arch., 10.
О б e p л e н (O'b e r 1 i n g Ch.). 1921. Bull, de 1’assoc. franc, pour l’6tude du
cancer.
— 1922. Les tumeurs des meninges. Bull, assoc, fr. pour l’6tude du cancer, Juin.
Оберлени Нордман (О berli ng Ch. et Nordmann J.). 1927. Tumeurs
du nerf optique. Ann. d’ocul., 164.
Оберндорф ep (Oberndorfer S.) 1929. Die Geschwiilste des Darmes. Handb.
spez. path. Anat. u. Hist., Bd. 4, T. 3.
— 1931. Die inneren mannlichen Geschlechtsorgane. Handb. spez. path. Anat. u. Hist.,
Bd. 6, T. 3.
О г а в a (Ogawa C_). 1920. Contribution to the histology of the respiratory spaces in
the vertabrate lungs. Am. J. Anat., 27.
О 3 а в a (O s a w a). 1911. Nissin Igaku. Bd. 2.
О л л e и и П p и с т (A 1 1 e n a. P r i e s t). 1932. Surg. gyn. a. obst., 55.
Литература 469
Оливо (О live О.). 1S25. Sui caratteri morfologici di unceppo di elementi del miocardo
embrionale di polio, coltivati in vitro per 6 mesi. Monit. Zool. ital., 36.
— 1927. Differenziazione e dedifferenziazione del tessuto nervoso embrionale di polio
coltivato per piu settimane in vitro. Arch. exp. Zellforsch., 5.
— 1929. Sulle modificazioni strutturaii e funzionali de) miocardo di polio coltivato in
vitro. Arch. exp. Zellforsch., 8.
— 1930. Differenziazione morfologica e funzionale de tessuto miccardico coltivato in vitro
e conservazione prolongata delle sue propriety specifiche. Monit. Zool. ital., 40.
— 1930. Differenziazione in vitro di fibre collagene. Boll. See. itai. Biol, sper., 5.
— 1932. Birefrangenza e miofibrille nei mioblasti di polio. Boll. Soc. ital. Biol,
sper., 7.
ОппельА. (Oppel A.). 1905. Lehrbuch der vergleichenden mikroskopischen Anato-
mie, Bd. 6.
— 1912. Kausal-morphologische Zellstudien. IV. Die Explantation von Saugetiergewebe,
ein der Regulation von Seiten des Organismus nicht unterworfenes Gestaltungsgesche-
hen. Roux’Arch., 34.
— 1912. Die aktive Epithel be we gung, ein Faktor beim Gestaltungs-und Ernaltungsge-
schehen. Roux’Arch., 35.
— 1912. Die aktive Epithelbewegung. Anat. Anz. 41.
Орлов Ю. (Or loff J.). 1924. Ueber den histologischen Bau der Ganglien des Mund-
magennervensystems der Insekten. Z. mikr.-anat. Forsch., 2.
— 1927. Innervation des Darmes des Flusskrebses. Z. mikr.-anat. Forsch., 4.
— 1929. Ueber den histologischen Bau der Ganglien des Mundmagennervensystems der
Krustazeen. Z. Zellforsch., 8.
— 1930. Ueber die Innervation des Anodontamantels. Z. mikr.-anat. Forsch., 20.
Ортега дел ь-Р ио (H ortega del Rio). 1916. Nuevas reglas para la colora-
cion constante de las formacionvs connectivas, por el metodo de Achucarro. Trab. Lab.
inv. biol. univ. Madiid.
— 1919. Coloiacion rapida de tejidos normales у pathologicos con carbonate de plata ammo-
niacal. Trab. Madrid, 17.
— 1921. El tercer elemento de los centros nerviosos: histogenesis у evolucion normal;
exodo у distribucion regional de la microglia. Mem. R. Soc. Esp. Hist. Nat., 11.
— 1921. El tercer elemento de los centros nerviosos. Poder fagocitario у movilidad de la
microglia. Bol. de la Soc. esp. de biol. 9.
— 1932. Microglia. Cyt. a. cell. path, of the nervous syst., v. 2.
ОртегаиПенф и л ьд (H ortega del Rio a. Penfield W.). 1927. Cere-
bral cicatrix. The reaction of neuroglia and microglia to brain wounds. Bull. John.
Hopk. hosp., 41.
Oct (Ost). 1906. Zur Kenntnis der Regeneration der Extremitaten bei den Arthropoden.
Roux’Arch., 22.
Пав до вскийЕ. 1917. Материалы к сравнительной анатомии и к истории развития
скорпионов. Дисс. Петроград.
— 1924. К строению лимфоидного аппарата и соединительной ткани с ее производными
у скорпионов. Арх. анат., гист. и эмбр., 3.
Паппенгейм (Pappenheim А.). 1919. Morphologische Haematologie,
Паппе и геймер (Pappenheimer). 1913. Further studies of the histology
of the thymus. Am. J. Anat., 14.
П a p и н B. 1912. К вопросу сб экспериментальных атипических разрастаниях эпите-
лия. Дисс. Казань.
П а р к е р (Р а г к е г). 1919. The elementary nervous system. Philadelphia and
London.
— 1932, 1933. The races that constitute the group of common fibroblasts. I and II. J. exp.
Med., 55 a. 58. (также Science, 76).
— 1934. Studies on Organcgenesis. I. The ability of iselatid blcod cells to form organised
vessels in vitro. J. exp. Med., 64.
470
Литература
Пат це л ьт(Р a t z е 1 t V.). 1S25. Zellen, Gewebe. Fasern und die Spezifitat der Keim-
blatter. Z. mikr.-anat. Forsch., 3.
Пейперс (Peipers). 1895. Beitrage zur kompensatorischen Hypertrophie und zur
Regeneration. Roux’Arch., I.
Пенфильд (Penfield Wj. 1932. Neuroglia, normal and pathological. Cyt. a.
cell. path, of the nervous syst.
— 1932. Tumors of the sheathes of the nervous system. Cyt. a. cell. path, of the nerv. syst.,
v. 3.
Петер A. (Peter A.). 1889. Ueber die Regeneration des Endothels der Cornea. Arch,
mikr. Anat., 33.
— 1909. Untersuchungen iiber Bau und Entwicklung der Niere. Jena.
Петер К. (Peter К.). 1909. Die Gewebe im Unterricht. Anat. Anz., 68.
— 1931. Ueber anatomische und physiologische Einheiten des Korpers. Anat. Anz., 71.
— 1933. Ueber die Begriffsbildung von Organ und Gewebe und iiber das Lehren von Ana-
tomie und Histologie. Anat. Anz., 76.
Петерсен (Petersen). 1922, 1924. Histologie und mikroskopische Anatomie. I
und II. Munchen und Wiesbaden.
Петерфи и Уи ль HMC(PeterfiT. u. Williams S.). 1933, 1934. Elektrische
Reizversuche an geziichteten Gewebezellen. I, II, III. Arch. exp. Zellforsch., 14
and 16.
П и з e н т и (P i s e n t i). 1899. Sur la cicatrisation des blessures du rein et sur la rdge-
пё rat ion partielle de cet organe. Arch. ital. du biol., 4.
Пи не ль (Pinel Ph.). 1798. Nosographie philosophique ou la methode d’analyse
appliqufie й la medccine. 6-me 6d. Paris, 1818.
П и H к у с Г. (P i n k u s H.). 1931. Untersuchungen iiber des Verhalten des Hiihnerpan-
kreas in der Gewebeziichtung. Arch. exp. Zellforsch. 10.
— 1932. Ueber Gewebekulturen menschlicher Epidermis. Arch. Derm. u. Syph., 165.
— 1936, 1937. The isolation of pure strains of cells from human turners. I. II. Growth
characteristics of a sarcoma and two brain tumors in tissue culture. Conclusions. Am.
J. Cancer, 20, 26 and 29.
— 1938. Notes on structure and biological properties of human epidermis and sweat glands
cells in tissue cultures and in the organism. Arch. exp. Zellforsch., 22.
По д в ы с о цк и й В. (Podwyssotzky W.). 1886, 1887. Experimentele Unter-
suchungen iiber die Regeneration der Driisengewebe. 1. Die Regeneration des Leber-
gewebes. II. Die Regeneration des Nierenepithels. III. Die Regeneration an den Mei-
bomschen Drusen. IV. Die Regeneration an den Speicheldriisen. Ziegl. Beitr.
1 und 2.
П о ж a p и с с к и й (P о s c h a r i s s k у). 1905. Ueber heteroplastische Knochenbil-
dung. Ziegl. Beitr., 38.
По лев. 1927. Explantationsversuche mit Augengeweben. Ber. 46. Zusammenkunft
dtsch. ophthal. Ges., Heidelberg.
Поли (P о I i C.). 1897. Zur Entwicklung der Gehorblase bei den Wirbeltieren. Arch,
mikr. Anat., 48.
Поликар (Policard A.). 1925. Recherches sur les cultures de tissu renal des mam-
mifferes. Bull. hist, appl., 2.
— 1925. Recherches sur la culture in vitro de tissu renal. Signification des cordons epithfi-
liaux neoformes. C. r. Soc. biol., 92.
— 1926. Sur la nature du revStement des alveoles pulmonaires des mammiferes. Bull,
d’hist., 3.
— 1926. A propos du revetemant des alveoles pulmonaires. C. r. Ass. des Anat., 21-e
reunion, Liege.
— 1928. Precis d’histologie physiologique. Coll. Testut, Paris.
Поликар иБу шарла (Policard A. etBoucharlat M.). 1925. Caractfe-
res des cultures de tissus des mammiftres dans du plasma d’animaux сапсёгеих. C. r.
Soc. biol., 92.
Литература
471
Поллетини (Р о I 1 е t i n i). 1922. Su neoformazione cartilaginee e ossee determi-
nate da innesto di frammenti di cartilagine e d’ossa fissati. Arch. ital. chir., 6.
По нфи к (P о nf i c k E.). 1889. Ueber das Mass der Entbehrlichkeit und der Wieder-
stellungsfahigkeit des Leberorgans. Zbl. med. Wiss. № 35.
Попов В. 1937. О морфо генном влиянии глазной чаши на разные эмбриональные ткани
и на зачатки некоторых органов. Арх. анат., гист. и эмбр., 16.
Попов Н. (Р о р о f f N.). 1927. The histogenesis of the thymus as shown by tissue
cultures. Arch. exp. Zellforsch., 4.
Поппе p (Popper). 1929. Zur Kenntnis der Tuberkulose des Wurmfortsatzes. Wien,
med. Wochenschr., № 15.
Поярков (Poyarkoff). 1900. Recherches histologiqu;s sur la mitamarphase
d’un Coleoptfcre (Callerucella). Arch. anat. micr., 12.
ПренанМ. (P Tenant M.). 1922. Recherches sur le parenchyme des Plathelmi nthes.
Arch, morph, gen. et exp.
П p e h а н А., Буэн П. и МайарЛ. (PrenantA. BouinP. et Maillard L.).
1904. Traitd d’histologie. T. 1. Cytologie generale et speciale. Paris.
П p и з e л ь (P r i e s e 1). 1931. Die Missbildungen der mannlichen Geschlechtsorgane.
Handb. spez. path. Anat. u. Hist., Bd. 6, T. 3.
Пробст (Probst). 1931. Roux’Arch., 124.
Пуркинье (Purkinje J.). 1836. Ueber Flimmerbewegungen im Gehirn. Miill.
Arch.
— 1838. Bericht iiber die Versammlung deutscher Naturforschcr und Aerzte in Prag im
September 1837 von Graf K. Sternberg und Prof. J. v. Kromholz. Prag.
Пу p м а н (P urman n Th.). 1930. Ueber einen Fall von peritonea lei Deckzellenge-
« schwulst bei einem Kalbe. Ziegl. Beitr., 84.
П фул ь (Pf uh 1 W.). 1932. Die Leber. Handb. d. mikr. Anat. des Menschen, Bd. 5.
T. 2.
Pa б л ь (R a bl C.). 1889. Ueber die Prinzipien der Histologie. Verh. anat. Ges., Berlin.
3 Vers.
Равен (Raven C.)_ 1931—1936. Zur Entwircklung der Ganglienleiste. I—V. Roux-
Arch., 125, 129, 130, 132, 134.
P а д л ь (R a d 1). 1912. Neue Lehre vom Zentralnervensystem. Leipzig.
Раевская. 1939. Цит. по Насонову и Александрову, 1940.
Раков A. (Rakov). 1937. Malignant rhabdomyoblastomas of the skeletal muscula-
ture. Am. J. Cancer, 30.
Ранвье (Ranvier L.). 1874. Du dGveloppement et de i’accroissement des vaisseaux
sanguins. Arch, de Phys., I.
— 1882. De la nevroglie. C. r. Acad. Sc., 94.
Раубер A. (Rauber A.). 1882. Ueber das Dickenwachstum des Gehirns. Sitz. ber.
naturforsch. Ges. Leipzig;
— 1883. Die Entwicklung der Gewebe des Saugethierkorpers und die histologischen Systeme
Sitz. ber. naturforsch. Ges. Leipzig, 10 Jahrgang ^Leipzig, 1884).
Раус н M e p ф и (R о u s P. a. Murphy J.). 1914. On immunity to transplantable chic-
ken tumors. J. exp. Med., 20.
Рахманинов И. 1881. К вопросу о регенерации поперечнополосатой мышечной
ткани. Дисс, М.
Pe3Hrep(Roesiger W.). 1930. Ziegl. Beitr., 85.
Рейхерт (Reichert С.). 1845. Bemerkungen zur vergleichenden Naturforschung
im Allgemeinen und vergleichende Beobachtungen iiber das Bindegewebe und die
verwandten Gebilde. Dorpat.
P e й ш Ф. (R uy s c h Fr.). 1701, 1703. Thesaurus anatomicus I.
— 1744. Opera omnia. Amstelodami.
P e м а к P. (R e m a k R.). 1836. Vorlaufige Mitteilung mikroskopischer Beobachtungen
iiber den inneren Bau der Cerebrospinalnerven und iiber die Entstehung ihrer For-
melemente. Miill. Arch.
472
Литература
— 1843. Ueber die Entwicklung des Hiinchens im Ei. Miill. Arch.
— 1852. Ueber extrazellulare Entstehung thierischer Zelien und iiber Vermebrung dersel-
ben durch Theilung. Mull. Arch.
— 1855. Untersuchungen iiber die Entwicklung der Wirbelthiere. Berlin.
д e-P e н ь и и Гог (de R е п у i G. a. Hogue M.). 1931. Studies made by means of
microdissection on skeletal muscle grown in tissue culture. Arch. exp. Zellf., II.
— 1938. Tissue cultures of heart muscle. Anat. Rec.; 70.
РёссельиБленд (Russel a. Bland). 1933. A study of gliomas by the method
of tissue culture. J. Path. a. Bact., 36.
P ё с с л e (Rossie). 1930. Entzilndung der Leber. Handb. spez. path. Anat. u. Hist.,
Bd. 5, T. 1.
Рётиги Б p у г in (R c t h i g P. u. В r u g s c h Th.). 1902. Die Entwicklung des
Labyrinthes beim Huhn. Arch. mikr. Anat., 59.
Реттере (Retterer E.). 1903. Recherches expel imentales sur 1’hyperplasie epithe-
liale et sur la transformation de I’fepith^lium en tissu conjonctif. C. r. hept. Acad.
Sc., 136.
— 1916. Les relations genetiques entre derme et cpiderme. C. r. See. biol., 72.
Ретциус (Retzius). 1893. Studien iiber Ependym und Glia. Biol. Untersuch.,
N. F. 5.
P ж e в у ц к а я О. 1938. О микроскопическом строении печени крупного рогатого скота
и регенерации эпителиальных элементов печени в экспериментальных условиях.
Диес. Зоовет. инет., Оренбург.
Рибберт (Ribbert Н.). 1889. Zur Regeneration des Epithels der Cornea. Berl.
Klin. Wochenschr., 26, № 38.
— 1889. Ueber die Regeneration des Schilddriisengewebee. Virch. Arch., 117.
— 1891. Ueber die Regeneration der Mamilla nebst Bemerkungen iiber ihre Entwicklung.
Arch. mikr. Anat., 37.
— 1895. Beitrage zur Regeneration der Speicheldriisen. Roux’Arch., 1.
— 1897 (1898). Die Veranderungen transplantierter Gewebe. Roux’Arch., 6.
— 1904. Zur Regeneration der Leber und Niere. Roux’Arch. 18.
P и д (R e e d M.). 1904. The regeneration of the first leg of a crayfish. Roux’Arch., 18.
Рингоф (Rienhoff W.). 1922. Development and growth of the metanephros or
permanent kidney of chick embryos. Bull. Johns Hopk. hosp., 33.
P и н д ф л e й in (R i n d f 1 e i s c h E.). 1862. Beitrage zur Lehre von den Entztindungen
seroser Membranen. Virch. Arch., 23.
Рихтер И. 1939. Об особенностях смены клеток вагинального эпителия грызунов.
Бюлл. эксп. биол. и мед., 8.
Робб-С мит (Rob b-S m i t h A.). 1936. Gewebekultur aus der erwachsenen Kanin-
chenlunge. Ziegl. Beitr., 97.
Робертсон (Robertson). 1900. A text-book of Pathology in relation to mental
Diseases by Ford-Robertson, Edinborough.
Робинов (Ro bi now K.). 1935. Ueber das Verhalten der Marksubstanz der Niere
erwachsener Kaninchen und Ratten in der Gewebekultur. Z. Zellforsch., 22.
— 1936. On the structure of epithelial membranes in tissue cultures. Protopl., 27.
— 1938. Experimental investigation into the structure of simple epithelium sheets in
tissue cultures. Arch. exp. Zellforsch., 22.
Роек и h(Ro ski n G.). 1931, Veigleichend-cytologische Beobachtungen an den Hyper-
nephromen von Meerschweinchen und an dessen Explantaten in vitro. Arch. exp. Zell-
forsch., 11.
д e-P у в и л ь (de R о u v i I J е E.). 1896. De la regenetation de I’epithGlium vesical.
C. r. hdbd. Acad. Sc., 123.
— 1896. Du tissu conjonctif commc rGgenerateur des epithGlium. Thfcse. Paris.
Рудольфи (Rudolphi K.). 1814—1815. Ueber Hornbildung. Abh. d. phys. Cl.
Kngl. preuss. Acad. d. Wiss., I. Berlin, 1818.
— 1821. Grundriss der Physiologie. Bd. I. Berlin.
Литература
478
Руле (Roulet R.). 1935. Studien iiber Knorpel- und Knochenbildung in Gewebekul-
turen. Arch. exp. Zellforsch., 17.
Румянцев A. (Rum jantzew A.). 1922. Культивирование ткани сердца куриного
зародыша. Русск. зоол. ж., 3.
— 1927. Beobachtungen fiber die Entwicklung des Herzmuskelgewebes der Embryonen
von Hiihnern in vitro. Arch. exp. Zellforsch., 11.
Румянцев А. и Березкина. 1937. Наблюдения над развитием основного ве-
щества хряша в опытах in vitro. Арх. анат., гист. и эмбр., 17.
Румянцев и Сунцова. 1935. Наблюдения над развитием и дифференцировкой
волокнистых структур в культурах тканей. Тр. Инет. эксп. морфоген. 3; Arch. exp.
Zellforsch., 17.
Русси, Лер ми тиКорни ль (Roussy G., Lhermitte J. etCornil L.l.
1924. Essai de classification des tumeurs cerebrates. Ann. anat. path., '1.
P у ч и и с к и й Б. 1910. К вопросу об экспериментальном атипическом разрастании
эпителиальной ткани. Русск. врач, № 23.
Р ы в к и н д А. 1936. К вопросу об участии эпителия в образовании костной ткани.
Арх. пат. анат. и пат. физиол., 2.
Рюккерт (Ruckert J.). 1892. Die Entwicklung der Exkretionsorgane.
Erg. Anat.
Рюккерт и Monnbep(RilckertJ. und M о 1 1 i e r S.). 1906. Die erste Entste-
hung der Gefasse und des Blutes bei Wirbeltieren. Handb. d. vergl. u. exp. Entw.
gesch. d. Wirbeltiere, Bd. I, T. 1, Hiilfte 2.
Сабатье и д e-P у в и л л ь (S a b a t i e r A. et de R о u v i 1 1 e E.). 1898. Sur la gc-
nese des epitheliums. C. r. hebd. Acad. Sc., 127.
Санчец (Sanchez D.). 1918, 1919. Sobre el desarollo de los elementos nerviosps
en la retina del Pieris brassicae. Trab. Lab. in biol. univ. Madrid, 16 у 17.
— 1925. L’histogenese dans les centres nerveux des insectes pendant la metamorphose.
Trav. lab. rech. biol. Madrid, 23.
Сачердотти (Sacerdotti C.). 1886. Sulla rigenerazione dell’epitelio muciparo
del tubo gastro-intestinale degli Anfibi. Atti. R. accad. Sc. Torino, 31; Arch. mikr.
Anat., 48.
Сачердотти и Фраттен (Sacerdotti C. u. Frattin G.). 1902. Ueber
die heteroplastische Knochenbildung. Experimentelle Untersuchungen. Virch.
Arch., 168.
Себин (Sabin F.). 1920. Studies on the origin of blood-vessels and of red blood cor-
puscles as seen in the living blastoderm of chicks during the second day of incubation.
Contr. to Embr., 9.
Северцов A. (Sewertzoff A.). 1931. Die morphologischen Gesetzmassigkeiten
der Evolution. Jena.
— 1934. Главные направления эволюционного процесса. Бномедгиз.
— 1939. Морфологические закономерности эволюции.
С е л л е и (S е 1 1 е i). 1904. Beitrage zur Histologie der Epididymitis gonorrhoica. Der-
mat. Z., 11.
Семон (Senion R.). 1891. Studien iiber den Bauplan des Urogenitalsystems der Wir-
beltiere. Jena. Z. Naturwiss., 19.
Сендисон (Sandison J.). 1924. A new method for the microscopic study of li-
ving growing tissues by the introduction of a transparent chamber in the rabbit’s ear.
Anat. Rec., 28.
Сен т-И лер К. (Sain t-H i 1 a i r e). 1930. Morphogenetische Untersuchungen der
nichtzellularen Gebilde der Tiere. Periderm der Hydroiden. Zool. Jahrb., 47.
Серебряков. 1934. Рост и дифференцировка нервной ткани вне организма in vitro.
Тр. I Гист, конф., М.
CHrypflCOH(Sigurdsson). 1940. ZCchtung von Lungengewebe nach einem neuen
Verfahren. Arch. exp. Zellf., 24.
Симоэнс Pa п о 30 (S i mo e ns Rapo so D.). 1925. La regeneration de la moelle
474
Литература
epinifcre et des ganglions rhachidiens chez les amphibiensadultes. Trav. Lab, rech.
biol. univ. Madrid, 23.
Ситенко В. 1939. Гомопластические пересадки яичка. Дисс. ВМА. Л.
Слонимский (Slonimski Р.). 1930. Sur le pouvoir d’auto-differentiation stride
de I’ebauche sanguine presomptive des amphibiens. C. r. Soc. biol., 104.
— 1930. Sur la o.ualite d’origine des elements rouges et blancs du sang chez les amphibiens.
C. r. Soc. biol., 104.
С ми дт (S m i d t H.). 1900. Ueber die Darsteliung der Begleit- und Gliazellen im Ner-
vensystem von Helix mit der Golgimethode. Arch. mikr. Anat., 55.
— 1901. Weitere Untersuchungen uber die Glia von Helix. Anat. Anz., 19.
Смирнова B. (Smirnow a). 1910. Sur la culture des tissue en dehors de 1’organisme.
C. r. Soc. biol., 79.
— 1928—1929. Zur Kenntnis der Entdifferenzierung des Muskelgewebes in vitro. Arch,
exp. Zellforsch., 7.
Смирнова и Уразов. 1932. Превращения эпителия почки in vitro. Арх. анат.,
гист. и эмбр., 2.
С о л у х а Н. 1908. О клеточных формах соединительной ткани птиц в нормальном
состоянии и при воспалении. Дисс. СПб.
Спей дел (Speidel С.). 1932. Studies on living nerves. 1. J. exp. Zool., 61.
— Studies on living nerves. II. Am. J. Anat., 52.
— 1934. Growth, irritation and repaire of nerves. Arch. exp. Zellforsch., 15.
— 1935. Studies on living nerves. IV. Biol. Bull., 58.
— 1938. Studies on living muscles. I. Am. J. Anat., 62.
Стивенс (Stevens N.). 1907. A histological study of regeneration in Planaria simp-
licissima, Pl. morgani and Pl. maculata. Roux’Arch., 24.
— 1930. Notes on regeneration in Planaria lugubris. Roux’Arch., 131.
Стилуэлл (Stilwell E.). 1938. Cytological study of chick heart muscle in tissue
culture. Arch. exp. Zellforsch., 21.
CTOKKapfl(Stockard C.). 1915. The origin of blood and vascular endothelium in
embryos without a circulation of the blood and in the normal embryo. Am. J.
Anat., 18.
Стон (Stone L.). 1926. J. exp. Zool., 95.
— 1929. Roux’Arch., 118.
— 1932. Anat. Rec., 51.
Стрелин Г. (Strelin G.). 1928. Rontgenologische Untersuchungen an Hydren. II.
Roux’Arch., 115.
— 1928. Zur Frage iiber den morphologischen Bau und die Herkunft einiger Zellelemente
von Pelmatohydra oligactis. Zool. Anz., 79.
— 1935. О физиологическом градиенте. I. Градиент реактивности роговицы лягушек
и причины его возникновения. Арх. биол. наук, 37.
Стренджуэйси Фелл (StrangewaysT. a. Fell Н.). 1925. Experimen-
tal studies on the differentiation of the embryonic tissues growing in vivo and in vitro.
I. The development of the undifferentiated limb bud when subcutaneously grafted
into the postembryonic chick and when cultivated in vitro. Proc. Roy. Soc.
London, 99.
— 1926. The development of the isolated early embryonic eye in the fowl when cultivated
in vitro. Proc. Roy. Soc. London, 100.
СтудицкийА. (Studizky A.). 1932. Ueber das Wachstum des Knochengewebes
und Periosts in vitro und auf der Allantois. Arch. exp. Zellforsch., 13.
— 1937. Потенциал скелетогенной клетки. Юб. сб. 80-лет. акад. Н. Насонова. Изд. АН
СССР.
Студн и чка (Studnicka F.). 1897. Untersuchungen iiber den Bau des Sehnerven
der Wirbeltiere. Jena. Z. Naturwiss., 31.
— 1900. Untersuchungen iiber den Bau des Ependyms der nervesen Zentralorgane. Anat.
Hefte, 15.
Литература
475
— 1905. Die Parietalorgane. Oppel’s Lehrbuch der vergleichenden mikroskopischen Anato-
mic der Wirbeitiere, Bd. 5.
— 1929. Die Organisation der lebendigem Masse. Handb. mikr. Anat. d. Menschen,
Bd. I, T. 1.
— 1931. Das Gewebe der Chorda dorsalis von Esox lucius L. Z. Zellforsch., 13.
Сунцова B. (S unzoff). 1930. Analyse der Gewebe beim Wachstum der Froschniere
in vitro. Arch. exp. Zellforsch., 10.
— 1935. Die Beziehung zwischen den Fibrinfasern und den argyrophilen Fasern in vitro.
Arch. exp. Zellforsch., 17.
Танненберг (Tannenberg J.). 1929. Ueber die Entwicklungspotenzen der
Fibroblasten in der Gewebskultur. 1—111. Arch. exp. Zellforsch., 9.
— 1931. Ueber die Umwandlungsfiihigkeit der Fibroblasten in Makrophagen. Arch. exp.
Zellforsch., 11.
T a p а т ы н о в H. 1914. О рассасывании мышц при повреждении их и о происхождении
миофагов. Дисс. Казань.
Те й чл е н де р (Те u t sc h I a nde г О.). 1924. Ueber experimentelle Erzeugung der
Cholesteatome und Kankroid des Uterus. Dtsch. med. Wochenschr., № 50.
T e о д о p Л. 1938. Аутотрансплантация яичника в переднюю камеру глаза. Ростиздат.
Теппих (Toppich G.). 1925. Die zellularen Abwehrvorgange in der Lunge bei Erst-
und Wiederinfektion mit Tuberkelbazillen, Krankheitsforsch., 2.
— 1926. Der Abbau der Tuberkelbazillen in der Lunge durch Zellvorgange und ihr Wie-
derauftreten in veranderter Form. Krankheitsforsch., 3.
Тёрё (Toro E.). 1929. Das organoide Wachstum der Darmkulturen. Arch. exp. Zell-
forsch., 9.
— 1931. Ueber die linsenfaserbildende Fahigkeit der Iriszellen bei Hiihnern und Ratten
auf Grund von Implantationsversuchen von Cornea- und Iriskulturen ins linsenlose
Auge. Verh. anat. Ges., 40 Tag.
— 1934. Ueber die Einpflanzung von Gewebekulturen. Arch. exp. Zellforsch., 15.
- — 1937. Ueber die Potenz der Endothelzellen in der Gefassbildung. Arch. exp.
Zellforsch., 19.
— 1939. Neue Untersuchungen zur Wirkung des embryonalen Herzextrakts. Arch. exp.
Zellforsch., 22.
T и м о ф e e в с к и й A. (T i m о f e j e w s k у A.). 1928. Ueber Leukozytenkulturen
des Menschenblutes in vitro. Arch. exp. Zellforsch., 6.
— 1938. К вопросу о способности клеток злокачественных опухолей к дифференцировке.
Врач, дело, № 8.
— 1939. Continuous cultures of tumors of man. Acta med. URSS, 2.
— 1940. Эксплантация опухолей. Общая и частная онкология. 1. Медгиз.
Т и мофеевск и й А. и Беневоленская С. (Timofejewskyu. Ben е-
woienskaja). 1925. Explantationsversuche von weissen Blutkorperchen mit
Tuberkelbazillen. Arch. exp. Zellforsch., 2.
— 1927. Zur Frage iiber die Reaktion pathologischer Leukozytenformen des Menschen-
blutes in vitro auf Tuberkelbazillen. Virch. Arch., 264.
— 1929. Neue Beobachtungen an lymphoiden Zellen der myeloiden und lymphatischen
Leukamie im Explantationsversuche. Arch. exp. Zellforsch., 8.
Тимофеевский, Беневоленская и Варшавская. 1936. О культи-
вировании злокачественных опухолей человека. Вопр. онкол. 10.
Тирш (Thiersch С.). 1865. Epithelialkrebs namentlich der Haut. Leipzig.
T о к и н. 1934. Онтогения клетки. Тр. I Гист, копфер., М.
Т о ма А. (Т h о ma s А.). 1937. Recherches sur les transformations des cellules en dehors
de 1’organisme. Ann. Sc. Nat., ser. Zool.
— 1938. Des epitheliums en culture et dans 1’organisme. Arch. exp. Zell-
forsch., 22.
Тома P. (Thoma R.). 1906. Untersuchungen iiber die wachsartige Umwandlung der
Muskelfasern. I. Mitt. Virch. Arch., 186.
476
Литература
z* <*> л I I I I G в G G GOG
To ма P, 1909. Die spateren Scliicksale der maximal kontrahierten Wiilste und die Muskel-
regeneration nach Verletzungen. II. Mitt., Virch. Arch., 195.
— 1910. Die Zerreissungen der Muskulatur, die wachsartige Umwandlung ilberle-
bender Muskelfasern, die bei Massenligaturen entstehenden wachsartigen Umwandlun-
gen derselben und ihre Beziehungen zu den bei Abdominaltyphus beobachteten Veran-
derungen. III. Mitt. Virch. Arch., 200.
T о p e л ь (T h о r e 1 Ch.). 1896. Ueber typische und Pseudoregeneration bei Nierenin-
farkten. Virch. Arch., 146.
Третьяков Д. 1915. Органы чувств речной миноги. СПб.
— 1911. Мозг пескоройки. СПб.
У и д (W е е d L.). 1917. The development of the cerebrospinal spaces in pig and in man,
Contr. to Embr., 5.
— 1920. The cells of the arachnoid. Bull. Johns Hopk. hosp., 31.
Улезко-СтрогановаК. (Ulesko-Stroganowa). 1932. Zur Histogenese
der Flexner-Joblingschen Neubildungen. Arch. exp. Zellforsch., 13.
— 1935. Рост яичников in vitro Тр. Ак.-гин. инет.
Уленгут (Uhlenhuth). 1916. Die Zellvermehrung in den Hautkulturen von
Rana pipiens. Roux’Arch., 42.
о л 6 a x и Г ay (W a 1 1 b a c h a. Howe). 1925. J. exp. Med., 42.
cob. 1881. Untersuchungen iiber die Entwicklung der Cephalopoden. Arch. d. Biol., 2.
эддингтон (W a d d i n g t о n C.). 1931. Experiments on early chick and duck
embryos cultivated in vitro. Arch. exp. Zellforsch., 11.
1932. Experiments on the development of chick and duck embryos, cultivated in vitro.
Phil. Trans. Roy. Soc. London, 22.
1933. Induction by the primitive streak and its derivatives in the chick. J. exp.
Biol., 10.
1933. Induction by the entoderm in birds. Roux’Arch., 128.
1935. The development of isolated parts of the chick blastoderm. J. exp. Zool.. 71.
элс и К армикаэл (Wells A. a. Carmichael E.) 1930. Microglia:
An experimental study by means of tissue culture end vital stainig. Brain, 53.
алии Л. 1939. Усп. совр. биол., № 12.
а л л о п и й (Fallopius Gabr.). 1575. De partibns similiaribus humani corporis
ex diversis exemplaribus a V. Coiter summa cum diligentia collectae etc. Norimbrgaee.
ар и Любарш (Fahr Th. u. Lubars ch O.). 1925. Die Nierengewilchse. Handb.
d. spez. path. Anat. u. Hist., Bd. 6, T. I.
a p p a p (F a r r a r). 1908. On the phenomena of repair in the cerebral cortex. Hist,
ti. histopath. Arb., 2.
а у с e к. 1893. О так назыв. «белом хряще» и т. д. Тр. Акад. Наук, СПб., 41.
е д о т о в Д. 1935. Очерк эволюции животного мира. Тр. Палеозоя, инет. АН СССР, 4.
е л и к с (F е 1 i х W.). 1905. Die Entwicklung der Harnorgane. Handb. vergl, u, exp.
Entw. gesch., Bd. 3,
— 1911. Die Entwicklung der Harn- und Geschlechtsorgane. Haudb. d. Entw. gesch, d.
Menschen von Keibel und Mall. Bd. 2.
Ф e л л (F e 1 1 H.). 1928. The development in vitro of the isolated tocyste ot the embryo-
nic fowl. Arch. exp. Zellforsch., 7.
— 1928. Experiments on the differentiation in vitro of cartilage and bone. Arch. exp.
Zellforsch., 7.
— 1931. Osteogenesis in vitro. Arch. exp. Zellforsch., 11.
— 1932. The osteogenic capacity in vitro of periosteum ar.d tudosteum isolated from the
limb skeleton of fowl embryos and young chicken. J, ct-Auat., 66.
— 1933. Chondrogenesis in cultures of endosteum. Prcc. Roy. Soc. B. 112.
Фелл иЛандауэр(Ре!1Н. a. Landauer W.) 1935, Experiments ou skeleltal
growth and development in vitro in relation to the problem ot aviau Phoeomelia. Proc.
Roy. Soc., 118.
Фелл и P о б и n с о н (F e 1 1 H. a. R о b i n s о n R.). 1929. Th? growth, development
Литература
477
and phosphates activity of embryonic avian femora and limb-buds cultivated in vitro.
Biochem. J., 23.
— 1930. The development and phosphates activity in vivo and in vitro of mandibular
skeletal tissue of the embryonic fowl. Biochem., 24.
Фергюсон (Ferguson A.). 1928. A cytological study of the regeneration of gastric
glands following the experimental removal of large areas of mucosa. Am. J. Anat., 42.
Ф и з а л и (P h i s a 1 i x). 1902. Role de la rate dans la formation des hematies chez les
vertebres inferieurs. C. r. Soc. biol., 54.
Ф и л а т о в Д. 1939. Сравнительно-морфологическое направление в механике развития,
его объект, цели и пути. Изд. АН СССР.
Финлей (Finley В.). 1922. The development of the subcutaneous vascular plexus
in the head of the human embryo. Contr. to Emb., 14.
Фишбек (F ischback F.). 1929. A morphologic study of regeneration of liver after
partial removal. Arch, of Path., 7.
ФишельА. (Fischel A.). 1930. Ueber die Entwicklung der Keimdriisen des Men-
schen. Z. Anat., 92.
Фишер A. (Fischer A.). 1922. A three months old strain of epithelium. J. exp.
Med., 36.
— 1922. Cultures of organized tissues. J. exp. Med., 36.
— 1922. A pure strain of cartilage cells in vitro. J. exp. Med., 36.
— 1924. The differentiation and keratinization of epithelium in vitro. J. exp. med., 39.
— 1925. Beitrage zur Biologie der Gewebezellen. Eine vergleichende biologische Studie
der normalen und malignen Gewebezellen in vitro. Roux’Arch., 104.
— 1926. Umwandlung von Fibroblasten zu Makrophagen in vitro. Arch. exp.
Zellforsch., 3.
— 1926. DauerzLchtung reiner Stamme von Carzinomzellen in vitro. Z. Krebsforsch., 25.
— 1928. Zur Biologie der in vitro geziichteten Krebszellen. Z. Krebsforsch., 28.
— 1930. Gewebezi.chtung. Munchen. Miiller und Steinicke.
— 1931. Wachstum von hyalinem Knorpel in vitro. Roux’Arch., 125.
Фи ше jr А. и Паркер P. (Fischer u. Parker). 1929. Dauerziichtung in vitro
ohne Wachstumsbeschleunigung. Arch. exp. Zellforsch., 8.
— 1929. Proliferation und Differenzierung. Arch. exp. Zellforsch., 8.
Фишер Б. (Fischer B.)_ 1906. Die experimentelle Erzeugungatypischer Epithel-
wucherungen und die Entstehung bosartiger Geschwiilste. Munch, med. Woch. schr.
№ 42; Verh. dtsch. path. Ges., X. Tag., Stuttgart.
— 1915. Frankf. Z. Path., 17.
— 1922. Ueber experimentelle Erzeugung grosser Flimmerepithelblasen der Lunge.
Frankf. Z. Path., 27.
4>Hmep-Ba3eAbc(Fischer-Wasels B_). 1927. Metaplasie und Gewebsmiss-
bildung. Allgemeime Geschwul'.tlehre. Handb. norm. u. path. Physiol., Bd. 14/2.
— 1929. Primares Karzinom des Herzbeutels. Verh. dtsch. path. Ges., 24. Tag.
— 1932. Ueber die primaren malignenGeschwiilste der Serosadeckzellen. Z. Krebsforsch., 37.
Фишер И. (Fischer Use). 1937. Ueber die Differenzierung der Pancreaszelle in
vitro. Z. Zellforsch., 27.
— 1937. Ueber das verhalten des stolonialen Gewebes der Ascidia Clavellina lepadiformis
in vitro. Roux’Arch., 137.
— 1937. Die Pigmentbildung des Irisepithels in vitro. Arch. exp. Zellforsch., 21.
— 1938. Ueber die Differenzierung von Melanophoren und Lipophoren in Ektodermexplan-
taten von Amblystoma. Arch. exp. Zellforsch., 22.
Ф и ш e p И. и P и с Э. (F i s c h e r J. u. R i e s E.). 1936. Das Verhalten der Pancreas-
zelle des Hiihnchens in d^r Gewebekultur. Arch. exp. Zellforsch., 18.
Ф о б e л ь (V a u b e 1 E.). 1933. The form and function of synovial cells in tissue cultu-
res. I and II. J. exp. Med., 58.
Фогт (Vogt W.). 1929. Gestaltungsanalyse am Amphibienkeime mit ortlicher Vital-
farbung. Roux’Arch., 120.
478
Литература
Фолькман (Volkmann R.). 1893. Ueber die Regeneration des quergestreiften
Muskelgewebes beim Menschen und SSugetier. Ziegl. Beitr., 12.
Форе-Фремье(Раи16-Ргеппе1 E.). 1927. Lesamibocytesdes invert£brescU’etat
quiescent et й I’etat actif. Arch. anat. micr., 23.
Фо рема рк (Fo rsmar k E). 1904. Zur Kenntnis der Irismuskulatur des Menschen,
ihr Bau und ihre Entwicklung. Akad. Abh. Stockholm.
Фра йссе (F ra isse P.). 1885. Die Regeneration von Geweben und Organen bei den
Wirbelthieren, besonders Amphibien und Reptilien. Kassel und Berlin.
Франческой и Цамбе л л и (F rancescon А. е Z a m b е 1 1 i G.). 1933.
Ricerche sulla ghiandola mammaria di adu.to coltivata in vitro. Boll. soc. ital. Biol,
sper., 8.
Франц (Franz V.). 1927. Morphologie der Acranier. Erg. Anat. u. Entw. gesch., 27.
Фредеричи (Fredericl E_). 1926. Arch, de biol., 36.
Ф p и д г e й м (F r i e d h e i m E.). 1931. Morphologische und funktionelle Untersuchungen
an isolierten in vitro gezuchteten Skeletmuskeifasern. Arch. exp. Zellforsch., 11.
Фроман и (F г о m a n n). 1864, 1877. Untersuchungen iiber die normale und patho-
logische Anatomie des Riickenmarks. I und II.
Фукс (Fuchs F.). 1932. Die Entwicklung der Fornices calicisder menschlichcn Niere.
Z. Anat. u. Entw. gesch., 97.
— 1933. Zur Entwicklung der Sammelrohren der menschlichen Niere. Z. Anat. u. Entw.
gesch., 101.
— 1935, 1937. Zbi. Gyn., № 16 u. № 18.
Фу т (F о о t N.). 1919. Studies on endothelial reactions. I. J. med. Res. 40.
— 1920—1923. Studies on endothelial reactions. II—VII. J. exp. Med., 32, 33, 34, 36, 37.
— 1925. The endothelial phagocyte. Anat. Rec., 30.
Ф ю p с т (F ii r s t). 1904. Zur Kenntnis der Histogenese und des Wachstums der Retina.
Lunds Univ. Arskr., 40.
Фютте p e p (F ii t te re r G.). 1905, Ueber Epithelmetaplasie. Erg. Path., II, 9.
X а д ж и (H a d z i). 1909. Ueber das Nervensystem von Hydra. Jena. Z. Naturwiss., 31.
Хлопков A. (Chlopkow A.). 1931. Intestinal epithelium of the adult rabbit in
cultures in vitro. Arch. exp. Zellforsch., 10.
Хрущев Г. 1935. Цитологические исследования на культурах нормальной крови
человека. Тр. Мед.-биол. инет. им. Горького, 3.
— 1942. Физиологическая регенерация эпителия тонкой кишки и факторы ее регули-
рующие. Докл. на ассоц. биол. г. Самарканда.
Хрущев и Берлин (Chruschtschoff a. Berlin). 1935. Cytological inve-
stigations on cultures of normal human blood. J. Genet., 31.
X p у що в и Ди о ми д ов а. 1937. Эпителий яичников взрослых млекопитающих вне
организма. Биол. ж., 5.
Ценкер (Zenker). 1864. Ueber die Veranderungen der willkiirlichen Muskeln im
Typhus abdominalis. Leipzig.
Цивиди c (Tsi vi di s A.). 1913. Ueber driisenahnliche Epithelbildungen bei Pericar-
ditis. Virch. Arch., 211.
Циммерман(ПальчевскаяиВернер) (Zimmerman K.). 1910. Ueber
den Bau der Herzmuskulatur. Arch. mikr. Anat., 75.
— 1927. Die Speicheldriisen der Mundhehle und die Bauchspeicheldriise. Handb. mikr.
Anat. d. Menschen., Bd. 5, T. I.
Цунода (Tsunoda T.) 1909. Histologische und experimentelle Untersuchungen zur
Pathogenese der Sehnenflecke des Herzens. Frankf. Z. Path., 3.
Ц ы м б а л B. 1941. Некоторые задачи экспериментально-гистологического исследования
опухолей в свете современных данных по эксплантации. Докл. на сессии Гос. онкол.
инет. Л.
Чайльд (Child, С.). 1915. Individuality in Organisms. Chicago.
— 1921. The origin and development of the nervous system from a physiological point
of view. Chicago.
Литература
479
— 1924. Physiological foundation of behaviour. New York.
Часовников (Tschassownikow N.). 1926. Ueber in vitro Kulturen des
Thymus. Arch. exp. Zellforsch., 3.
Часовников. 1929. Ueber in vitro Zichtung von rontgenisiertem Thymus. Arch,
exp. Zellforsch., 8.
Чемберс (Chambers). 1924. The physical structure of protoplasma as determinated
by micro-dissection and injection. Cowdry Gen. Cytol.
— 1934. Some features of cell permeability in relation to kidney function. Arch. exp.
Zellforsch., 15.
Чемберс и Ледфорд (Chambers a. Ludford). 1931. Microdissection stu-
dies on malignant and non-malignant tissue cells. Arch. exp. Zellforsch., 12.
Чемберс и Реньи (Chambers a. Renyi). 1925. The structure of cells in
tissues as revealed by microdissection. I. The physical relationship of the cells in epi-
thelia. Am. J. Anat., 35.
Чемберс иФелл (Chambers a. Fell). 1931. Micro-operations on cells in tissue
cultures. Proc. Roy. See. London, 109,
Ч и n о л л и н a (C i p о 1 1 i n a). 1889. Experimentaluntersuchungen iiber die partielle
Regeneration des Pancreas. Rif. med., 2.
Ч у м a (C h u m a). 1923. Zur normalen und pathologischen Histologie der Magenschleim-
haut. Virch. Arch., 247.
Чурбанов H. 1937. Гетеротопические воспалительные разрастания эпителия же-
лудка. Дисс. ВИЭМ. Л.
ШабадЛ. (Schabad L.). 1935. Об альвеолярном эпителии. Сб. 25-лет. проф. Н.
Аничкова.
— 1937. Experimentelle atypische Epithelwucherungen nach intratrachealer Einfiihrung des
Steinkohlenteers in die Lungen. Z. Krebsforsch., 38.
Шамбо (Cham ba u gh G.). 1928. Cytology of the internal ear. Spec. cyt. ed. by Cowdry,
v. 2.
Шампи (ChampyC h_). 1912. Sur les ph6nomenes cytologiques qui s’observent dans
les tissus cultives en dehors de 1’organismc. I. Tissus epitheliaux et glandulaires. C. r.
Soc. biol., 72.
— 1913. La dediffGrentiation des tissus cultives en dehors de 1’organisme. Bibl. anat., 33.
— 1914. Quelques resultats de la methode de culture des tissus. I. G6n6ra!ites. II. Le muscle
lisse. Arch. Zool. exp. et gen., 53.
— 1914. Quelques resultats de la methode de culture des tissus. III. Le rein. Arch. zool.
exp. et gen., 54.
— 1914. Id. IV. La retine. Arch. zool. exp. et gen., 55.
— 1915. Id. V. La glande thyroide. Arch. zool. exp. et gen., 55.
— 1920. Id. VI. Le testicule. Arch. zool. exp. et gen., 60.
Шампи и Морита(СЬашру Ch. et Morita J.). 1928. Recherches sur les
cultures des tissus. Observations sur les cultures de testicule et d’ovaire chez les mammi-
feres, les oiseaux et les batraciens. Arch. exp. Zellforsch., 5.
Шантрох (Santroch Z.). 1932. Untersuchungen iiber die Fettsubstanzen in den
Gewebekulturen. Arch. exp. Zellforsch., 13.
Шапер (Schaper A.). 1894. Die morphologische und histologische Entwicblung
des Kleinhirns der Teleostier. Morph. Jahrb., 21.
— 1897. Fie frilhesten Differenzierungsvorgiinge im Zentralnervensystem. Ri их’Arch., 5.
Шапери Коген (Schaper A. u. Cohen C.). 1905. Beitrage zur Analyse des
tierischen Wachstums. Roux’Arch., 19.
Шаров И. 1937. К вопросу о гистогенезе при регенерации у планарий. Юб. сб. 80-лет.
акад. Н. Насонова.
Шаффер И. (Schaffer J.). 1908. Ueber Bau und Funktion des Eileiterepithels
beim Menschen. Mschr. Geburtsh., 28.
— 1921. Vorschlage zur Verbesserung der histologischen Nomenklatur nebst Bemerkungen
iiber die Begriffe «Endothel» und «Vorknorpel». Erg. Anat. u. Entw. gesch., 23.
480
Литература
— 1927. Das Epithelgewebe. Handb. mikr. Anat. des Menschen, Bd. 2, T. I.
— 1930. Die Stiitzgewebe. Handb. mikr. Anat. des Menschen, Bd. 2, T. 2.
Шаффер К. (Schaffer К.). 1926. Z. ges. Anat. u. Entw. gesch., 81.
Шванн (Schwann Th.). 1839. Mikroskopische Untersuchungen iiber die Uebereinstim-
mung in der Struktur und dem Wachstum der Thiere und Pflanzen. (Русск. перевод
под ред. 3. Кацнельсона, 1939, изд. АН СССР).
Ш и л n(S z i 1 у А.). 1902. Beitragzur Kenntnis der Anatomie und Entwicklungsgeschichte
der hinteren Irisschichten. Arch. Ophth., 53.
Шимкевич В. 1922. Курс сравнительной анатомии позвоночных животных.
3-е изд.
— 1923. Биологические основы зоологии. 4-е изд.
Шип л ей (Shipley Р.). 1916. The development of erythrocytes from hemoglobine
free cells and the differentiation of heart muscle fibres in tissue cultures. Anat.
Rec., 10.
Шмальгаузен И. 1938. Организм как целое в индивидуальном и историческом
развитии. Изд. АН СССР.
— 1939. Пути и закономерности эволюционного процесса. Изд. АН СССР.
Шмид (Schmid). 1932. Med. Klin.. № 42.
— 1934. Arch. Gyn., 155.
Шмидт В. (Schmidt W.)_ 1930. Ueber die Definition von «Organ» und «Gewebe».
Z. Zellforsch., 10.
Ш м и д т В. K. (S c h m i d t V.). 1927. Die Histogenese der quergestreiften Muskulatur
und des Muskelsehnenuberganges. Z. mikr.-anat. Forsch., 8.
— 1930. Die Entwicklung der Flossen von Hippocampus und die Histogenese in denselben.
Z. mikr.-anat. Forsch., 20.
Ш м и д т M. (Schmidt M.). 1910. Ueber Starkstromverletzungen. Verh. dtsch. anat.
Ges.
Ill м и д т м а н (S c h m i d t m a n n). 1920. Einige bemerkenswerte Beobachtungen
zur Pathologie der Grippe. Virch. Arch., 228.
Ш м и н к e (S c h m i n k e A.). 1908. Die Regeneration der quergestreiften Muskelfasern
bei den Wirbeltieren. I. Ichthyopsiden. Verh. physiol.-med. Ges. Wurzburg,
N. F. 39.
— 1908. Regeneration der quergestreiften Muskelfasern bei den Sauropsiden. Ziegl.
Beitr., 43.
— 1909. Die Regeneration der quergestreiften Muskelfasern bei den Saugetieren. Ziegl.
Beitr., 45.
— (Schmincke A.). 1921. Ueber lymphoepitheliale Geschwiilste. Ziegl. Beitr., 68.
Шнейдер К. (Schneider C.). 1902. Lehrbuch der vergleichenden Histologie.
Jena.
— 1908. Prakticum der vergleichenden Histologie. Jena.
Шоппер (S chopper W.), 1932. Explantationsstudien an Blutgefassen und serosen
Hauten. Untersuchungen am normalen und pathologisch veranderten Netz, an weichen
Hirnhauten und an grossen Gefassen junger Meerschweinchcn. Ziegl. Beitr., 88.
Ш о т т (S c h о 11 E.). 1909. Morphologische und experimentelle Untersuchungen iiber
Bedeutung und Herkunft der Zellen der serosen Hohlen usw. Arch. mikr. Anat., 74.
Шпеман (Spemann H.). 1912. Zur Entwicklung des Wirbeltierauges. Zool.
Jahrb., 23.
— 1936. Experimentelle Beitrage zu einer Theorie der Entwicklung. Berlin.
Шпильмейер (Spielmeyer W.). 1922. Histopathologie des Nerve nsystems.
I Band. Allgemeiner Teil. Berlin.
Шрейбер (Schreiber J.). 1732. Historia vitae et meritorum Frederici Ruysch.
Amstelaedami.
Шре йбери Венгле p(Schre ibe rt, u, We ngler F.). 1910. Ueber Wirkun-
gen des Scharlachols auf das Auge, speziell auf die Netzhaut. Mitosenbildung der Gan-
glienzellen. Arch. Ophth., 74.
Литература
481
Шридде (Schridde Н.). 1907. Die Entwicklung des menschlichen Speiserohrene-
pithels und ihre Bedeutung fiir die Metaplasielehre. Wiesbaden.
— 1909. Die ortsfremden Epithelgewebe des Menschen. Jena.
Штейн A. (Stein A.). 1936. Ueber entziindliche Epithelwucherungen bei einigen
Fofmen der Hauttuberkulose. Arch. Derm. u. Syph., 174.
Штейнман(51е1пшапп P.). 1924—1925. Das Verhalten der Zellen und Gewebe
im regenerierenden Tricladenkorper. Verh. d. naturforsch. Ges. Basel, 36.
— 1926. Prospektive Analyse von Restitutionsvorgangen. Roux’Arch., 108.
Штёрмл. (Stohr Ph. jun.). 1928. Das peripherische Nervensystem. Handb. d. mikr.
Anat. des Menschen, Bd. 4, T. I.
— 1930, 1932, 1934. Mikroskopische Studien zur Innervation des Magendarmkanals. I,
II, III. Z. Zellforsch., 12, 16, 21.
i 1тиве (Stieve H.). 1926. Die regelmassigen.Veranderungen der Muskulatur und
des Bindegewebes in der menschlichen Gebarmutter etc. Z. mikr.-anat.
Forsch., 6.
Ш г и л л и н г (Stilling). 1909. Ziegl. Beitr., 47.
Шт ольте (Stolte). 1927, Z. wiss. zool., 129.
в а н-д e p-111 т p и x т (van der S t r i c h t O). 1892. Nouvelles recherches sur la genfese
des globules rouges et des globules blancs du sang. Arch, de bioi., 12.
Шульц E. 1898. О регенерации ног у пауков. Тр. СПб. общ. естествоисп., 29.
Шульце О. (Schultze О). 1905. Beitrage zur Histogenese des Nervensystems. I.
Ueber die multizellulare Entstehung der peripheren sensiblen Nervenfaser und das
Vorhandensein eines allgemeinen Endnetzes sensibler Neuroblasten bei Amphibien-
larven. Arch.mikr. Anat., 66.
Щелкунов C. (SCelkunow S.). 1935. Интима мелких артерий и вен. Арх. биол.
наук, 37.
— 1936. L’intima des petites artferes et des veines et le mesenchyme vasculaire. Arch,
anat.-micr., 32.
— 1936. Beitrage zur Frage des Baues der Blutgefasse unter normalen und experimentellen
Bedingungen. Z. Anat. u. Entw. gesch., 106.
— 1936. Строение брюшины и ее производных при нормальных и экспериментальных
условиях. Арх. анат., гист. и эмбр., 15.
— 1937. Прогрессивное и регрессивное развитие капилляров. Арх. анат., гист. и
эмбр., 17.
— 1937. Сосудистая мезенхима и ее роль в постэмбриональном морфогенезе сосудистой
системы. Дисс. ВИЭМ, Л.
— 1938. Регенерация роговицы. Арх. анат., гист. и эмбр., 18.
— 1939. Строение кровеносных сосудов круглоротых. Докл. в общ. анат., гист. и эмбр.
Л.
__ 1940. Строение синовиальной оболочки. Тр. ВМАКА им. Кирова, 24.
Эбергардт (Eberhardt J.). 1907/8. О клеточных формах крови и соединитель-
ной ткани черепахи в нормальном состоянии и при воспалении. Дисс. СПб.
Эбнер (v. Ebner V.). 1911. Gewebeentwicklung und Phylogenese. Anat. Anz., Erg.
hefte zum Bd. 38 (Verh. anat. Ges., 25. Versam., Leipzig.).
Эвальд и Июне (Ewald A. u. Kiihne W.). 1877. Ueber einen neuen Bestandt-
heil des Nervengewebes. Verh. d. nat.-med. Vereins zu Heidelberg, N. F. I.
9rreniiHr(Eggeling H.). 1931 Handb. vergl. Anat. d. Wirbeltiere, Bd. I.
Эзаки (Esaki S.). 1928. Ueber Kulturen des Hodengewebes der Saugetiere und iiber
die Natur des interstitiellen Hodengewebes und der Zwischenzellen. Z. mikr.-anat.
Forsch., 15.
— 1929. New studies of nervous tissues cultures in vitro. C. r. Ass. Anat., 24. reunion.
Э к к e (E c k e). 1899. Structure et modifications fonctionelles du thymus de la grenouille.
Bull. Acad. r. med. Belgique, 4-e serie, 13.
Экман (Ekman G.). 1921. Experimentelle Beitrage zur Entwicklung des Bombi-
nator-Herzens. Oetversikt at Finskevetens, Soc. 63.
31 Хлопин—349
482
Литература
— 1929. Experimentelle Untersuchungen iiber die friiheste Herzentwicklung bei Rana
fusca. Roux’Arch., 110.
Эллер (Oeller H.). 1923. Die funktionelle Bedeutung der Gefasswandzellen bei aku-
ten Infektionen. Med. Klin., 19.
— 1925. Experimentelle Studien zur pathologischen Physiologic des Mesenchyms und
seiner Stoffwechselleistungen bei Infektionen. Krankheitsforsch., I.
Эмме л (E m me 1 V.). 1910. A study of the differentiation of tissues in the regenerating
crustacean limb. Am. J. Anat., 10.
Энгель и Вербер (E ngel P. u. Werber E.). 1937. Ueber das Wachstum der
Mausehypophyse in der Gewebekultur. Arch. exp. Zellforsch., 20.
Э p б кам (Er b ka m). 1880. Beitrage zur Kenntnis der Degeneration und Regeneration
der quergestreiften Muskulatur nach Quetschungen. Virch. Arch., 79. ]
Эрдман В. (Erdmann W.). 1931. Ueber das Selbstdifferenzierungsvermogen von
Amphibienkeimteilen bekannter prospektiver Bedeutung im Explantat. Roux’
Arch., 124.
Эрдман P. (Erdmann R.). 1928. Ueber die Wachstumsformen verschiedener in
vitro gezichteter Gewebe. Ein Beitrag zum Differenzierungsproblem der Zellen. Verh.
dtsch. Zool. Ges., 32.
— 1931. Epithelstudien, I. DicZiichtungdes Gallenblasenepithels. Arch.exp.Zellforsch., 11.
Эсси к (Essik C.). 1920. Formation of macrophages by the cells lining the subara-
chnoid cavity in response to the stimulus of particulate matter. Contr. to
Embr. 9.
9 Ф p У с с и (E p h r u s s i B.). 1930. Sur la culture de I’endothelium de foie embryonnaire.
C. r. Soc. biol., 103.
Эф py с си и Литвак (Ephrussi В. et Litvac A.). 1934. Quelques resultats
de la statistique des mitoses dans les cultures d’epithelium renal du lapin. Arch. exp.
Zellforsch., 16.
Ю д и цк а я С. 1938. Рост и дифференцировка эмбриональной мускулатуры конечно-
стей цыпленка. ДАН СССР, 21.
Ю и н г (Е w i n g J.). 1928. Neoplastic diseases. 3-е ed.
Юсе л ну с (J u s е 1 i u s). 1910. Experimentelle Untersuchungen iiber die Regenera-
tion des Epithels der Cornea unter normalen Verhaltnissen. Graefe’s Arch., 75.
Яблонский и Me йе p (Ja b lo nsk i W. a. Meyer H.). 1938. The occurence
of wandering cells in cultures of nervous tissue etc. J. Anat., 73.
Якоб (Jakob A.). 1927. Normale und pathologische Anatomie und Histologie des
Grosshirns. I Bd. Normale Anatomie und Histologie und allgemeine Histopathologic
des Grosshirns. Handb. d. Psychiatric, Bd. I, T. I.
Якобсон (J a ko bso n). 1923. Am. J. Obst. a. Gyn., 6.
Яку беки ii (Ja k u bs ki A.). 1908. Untersuchungen iiber das Stiltzgewebe des Ner-
vensystems im vorderen und hinteren Korperende der Hirudineen nebst Bemerkungen
iiber deren Neuromerie. Bull. Inst. Acad. Sci. Cracovie, 9.
— 1908. Untersuchungen iiber das Stiltzgewebe des Nervensystems bei den Hirudineen.
Bull. Inst. Acad. Sci. Cracovie, 10.
— 1913. Studien iiber das Gliagewebe der Mollusken. T. I. Lamellibranchiata und Gastro-
poda. Z. wiss. Zool., 104.
— 1915, Studien iiber das Gliagewebe der Mollusken. T. II, Cephalopoda. Z. wiss.
Zool., 113.
Я ми н (Ja m i n F.). 1925. Degeneration und Regeneration. Handb. d. norm, und path.
Physiol., Bd. 8, T. I.
Яс войн Г. (Jasswoin G.). 1928. Vergleichende Studien iiber einige Zellformen
des lockeren Bindegewebes der Saugetiere. Z. mikr.-anat., Forsch., 15.
— 1930. Experimenteil-morphologische Studien iiber einige Zellformen des lockeren Bin-
degewebes der Saugetiere. Z. mikr.-anat. Forsch., 19.
?— 1934. О возникновении основного вещества кости у млекопитающих. Арх. биол.
наук, 37.
Литература
483
— 1934. Sur 1’origine de la substance fondamentale de 1’os chez les mammiferes. Arch,
anat. micr., 21.
— 1935. On the formation of the ground substance of loose connective tissue. Quart. J.
micr. Sc., 78.
Б. Указатель работ H. Г. Хлопина и руководимого им научного коллектива!
Винников Я. (W i nniko w J.). 1936. Experimentell-histologische Untersuchungen
iiber die retinalen Anteile der Regenbogenhaut und der Ziliarfortsatze, Arch.- exp.
Zellforsch., 19.
— 1936. Экспериментально-гистологические исследования над ретинальной частью
радужки и цилиарных отростков. Арх. анат., гист. и эмбр., 15. (То же, дисс. ВИЭМ,
Л., 1938).
— 1938. Рост и превращения in vitro мионейральвых элементов (сфинктер и дилататср
зрачка). ДАН, 18.
— 1938. Экспериментально-гистологические исследования над сократимыми элемен-
тами радужины. Биол. ж., 7.
— 1939. Реет и превращения в тканевых культурах наружного пигментного слоя зри-
тельной части сетчатки (тапетум). ДАН, 20.
— 1939. Рост и превращения в тканевых культурах перцепторного листка сетчатки.
ДАН, 24.
— 1939. Экспериментальная эмбриология глазного бокала и его производных. Усп.
совр. биол., 10.
i — 1940. Экспериментально-гистологические исследования над зрительной частью сет-
чатки. I. Наружный пигментный слой. Арх. анат., гист. и эмбр., 23.
— 1941. Рост и превращения вне организма элементов сетчатки на разных стадиях ее
развития в ряду позвоночных. Докл. на засед. общ. анат., гист. и эмбр. Л.
— 1943. Экспериментальные исследования развития и строения сетчатки позвоночных.
Дисс. ВИЭМ.
ГалустянШ. (Галстян, Galustjan Sch.). 1933. Ueber die Verwandlungen
des Epithels der Gebarmutter im Explantat, Arch. exp. Zellforsch., 13.
— 1935. Experimentell-histologische Untersuchungen fiber das Eileiterepithel. Arch. exp.
Zellforsch., 17.
— 1935. О превращениях эпителия яйцеводов вне организма в связи с его гистологиче-
скими особенностями при обычных условиях. Арх. биол. наук, 38.
— 1939. Опыт культивирования in vitro эпителия щитовидной железы. ДАН СССР, 23.
— 1939. Общий характер роста и превращения вне организма элементов рентгенизи-
рованной зобной железы. ДАН СССР, 23.
— 1940. Экспериментально-гистологические исследования над эпителием щитовидной
железы. Арх. анат., гист. и эмбр., 23.
— 1940. Морфология зобной железы в свете экспериментального анализа. Дисс.
ВИЭМ. Л.
Гольштейн Н. 1939. Превращения многорядного эпителия трахеи при репаративной
регенерации. ДАН СССР, 23.
Григорьев Н. 1940. Экспериментальные исследования над опухолью Брауна
Пирс. Бюлл. экспер. биол. и мед., 9.
— 1941. Рост и превращения вне организма эпителия желчного пузыря. ДАН СССР, 31.
— 1942. Сравнительные наблюдения над явлениями роста и пролиферации эпителия
желчного пузыря в ряду позвоночных. Докл. на ассоц. биол. гор. Самарканда.
Же мкова 3. 1941. Циклические изменения маточного эпителия кроликов при бере-
менности. Докл. на объед. засед. общ. анат., гист. и эмбр. и общ. патол. Л.
— 1941. Рост и превращения эпителия беременной матки кролика вне организма. Докл.
там же.
Зубарева А. 1939, 1940. Рост и превращения эпителия желудка вне организма!
ДАН СССР, 23; Арх. анат., гист. и эмбр., 23.
*
484
Литература
Каннегисер Н. 1938, 1940. Рост и превращения элементов мягких мозговых обо-
лочек головного мозга вне организма. ДАН, 18; Арх. анат., гист. и эмбр., 23.
Колесникова Н. 1939, 1940. Рост и превращения фолликулярных и покровных
клеток яичника вне организма. ДАН СССР, 23; Арх. анат., гист. и эмбр., 23.
Колосс Е. 1941. О превращениях эпителия желудка лягушки при репаративной
регенерации. ДАН СССР, 31.
ЛавренкоВ. 1941. Об изменениях периферических нервных волокон лягушки после
перерезки. ДАН СССР, 30.
Лазаренко Ф. (Lazarenko Th.). 1931. Ein Beitrag zur Morphologie des Wach-
stums von embryonalem Nervengewebe in vitro. Arch. exp. Zellforsch., 11.
Лазаренко Ф. и Бененсон M. (Lazarenko u. Benenson). 1932. Zur
Frage iiber den Einfluss der ultravioletten Strahlen auf das Wachstum der Nerven-
fasern in vitro. Arch. exp. Zellforsch., 13.
Лежава A. (Lezava A.). 1934. Experimentell-histologische Untersuchungeniiber
das Uebergangsepithel. Z. Anat. u. Entw. gesch., 103.
— 1935. Возрастные изменения переходного эпителия и его превращения при экспери-
ментальных условиях. Арх. биол. наук, 37.
Михайлов В. (Michailow W.). 1937. Experimentell-histologische Untersuchun-
gen iiber die Elemente der Hodenkanalchen. Z. Zellforsch., 26.
— 1938. Экспериментально-гистологические исследования над элементами семенных
канальцев. Дисс. ВИЭМ, Л.
— 1938. Рост и превращения in vitro покровных клеток сосудистых сплетений мозга.
ДАН СССР, 18.
— 1938. Опыт экспериментального анализа элементов семиномы. Юб. сборн. Гос. Ле-
нингр. онкол. инет.
— 1938. Экспериментально-гистологические исследования над эпителием эндометриомы.
Труды I Укр. съезда онкол., Киев.
Насонова С. 1937. Мезэтел гй лягушки в экспериментальных условиях. Арх. анат.,
гист. и эмбр., 16.
— 1940. О репаративной регенераци i мезотелия миноги. Арх. анат., гист. и эмбр., 23.
Петр о в Ю. 1940. «Дегтярная саркома» у кролика. Арх. биол. наук, 57.
П т о х о в М. 1940. К вопросу о строении переходного эпителия мочевого пузыря кро-
лика. Арх. анат., гнет, и эмбр., 23.
— 1940. Экспериментально-гистологические исследования над переходным эпителием
мочевого пузыря кролика. Арх. анат., гист. и эмбр., 23.
— 1940. О некоторых морфологических особенностях эпителиальных опухолей моче-
вого пузыря. Арх. биол. наук, 57.
— 1940. Превращения переходного эпителия мочевого пузыря при репаративной реге-
нерации. Дисс. ВИЭМ. Л.
С и п о в с к и й П. (Ssipowsky Р.). Ueber in vitro Kulturen der Kaninchenneben-
nieren. Arch. exp. Zellforsch., 8.
Стрелин Г. (Strelin G.). 1930. Ueber in vitro Kulturen der Bronchen des Kanin-
chens mit besonderer Beriicksichtigung des Epithels. Arch. exp. Zellforsch., 9.
— 1940. Градиенты и мозаичность в развитии моллюсков. Арх. анат., гист. и
эмбр., 25.
— 1943. О природе чайльдовских градиентов и их значение в динамике развития орга-
низмов. Дисс.
Сутул о в Л. 1941. Особенности роста вне организма отростков чувствительных ней-
ронов. ДАН СССР, 30.
___ 1941. Рост и превращения вне организма нейроглиальных элементов спинальных
ганглиев. ДАН СССР, 30.
Хлопин Н. (Chlopin N.). 1922. Ueber in vitro Kulturen der embryonalen Gewebe
der Saugetiere. Arch. mikr. Anat., 96.
— 1923. Исследозания над тканевыми культурами из органов млекопитающих. Дисс.
ВМА.
Литература
485
— 1923. Исследования над тканевыми культурами из органов аксолотля. Тр. I Съезда
зоол., анат. и гист.
— 1923. Ueber in vitro Kulturen von Geweben der Saugetiere mit besonderer Berilcksi-
chtigung des Epithels. I. Kulturen der Submaxillaris. Virch. Arch., 243.
— 1924. Ueber in vitro Kulturen von Geweben der Saugetiere mit besonderer Beriickei-
chtigung des Epithels. II. Kulturen der Harnblasenschleimhaut. Virch. Arch., 252.
— 1924. Einige Betrachtungen iiber das Bindegewebe und das Blut. Virch. Arch., 252.
— 1924. Кровь, как особый вид соединительной ткани. Тр. 1-го всерссс. съезда патол.,
Л., Сентябрь 1923 г.
— 1925. Studien iiber Gewebskulturen im artfremden Blutplasma. I. Allgemeines. II.
Bindegewebe der Wirbeltiere. Z. mikr. - anat. Forsch., 2.
Хлопин H. иХлопйнаА. 1925. Studien iiber Gewebskulturen im artfremden Blut-
plasma. III. Die Histogenese der Zellformen in den explantaten der blutbildenden
Organe des Axolotls. IV. Ein Beitrag zur Vitalfarbung explantierter ZJielemente.
Arch. exp. Zellforsch., 1.
X л о п и н H. 1925. О культивировании in vitro соединительнотканных элементов рыб.
Изв. Биол. инет. Пермск. у нив., 4.
— 1925. Морфологическое значение клеточных элементов соединительной ткани и
крови в свете современных экспериментальных исследований. Изв. Биол.
инет. Пермск. увив., 4.
— 1927. Experimentelle Untersuchungen i ber die sekretorischen Prozessc im Zytoplasma.
Arch. exp. Zellforsch., 4.
— 1928. Gewebskultur niederer Vertebraten. Arch. exp. Zellforsch., 6.
— 1928. Experimentelle Untersuchungen uber die Vitalfarbung tierischer Zellen. Arch,
exp. Zellforsch., 6.
— 1929. Ueber die Verwandlungen des Epithels der Bauchspeicheldriise in Gewebskul-
turen. Arch. exp. Zellforsch. 9.
— 1930. Ein Beitrag zur Morphologie und zum Mechanismus der Eisenspeicherung.
Z. Zellforsch., 11.
— 1930. Значение метода эксплантации для морфологии. Природа, № 9.
— 1931. Ueber in vitro-Kulturen des menschlichen Mesenchyms. Arch. exp. Zellforsch., 11.
— 1931. Studien iiber Gewebskulturen im artfremden Blutplasma. V. Das Verhalten und
die Verwandlungen des menschlichen Mesenchyms im Explantat. Arch, exp, Zell-
forsch., 12.
— 1932. Experimentell-histologische Untersuchungen uber das Epithelgewebe der Har-
derschen Druse. Z. mikr-anat. Forsch., 30.
— 1932. Ueber einige Wachstums und Differenzierungserscheinungen an der embryonalen
menschlichen Epidermis im Explantat. Roux’Arch., 126.
— 1932. Ueber die Verwandlungen des Mausekarzinomepithels im Explantat. Z. Krebs-
forsch., 37.
— 1932. Ueber das Wachstum und die Organisationsfahigkeit einiger Epithelgewebe aus-
serhalb des Organismus. Z. Krebsforsch., 37.
— 1933. Специфичность и развитие эпителиальных тканей в Свете экспериментально-
гистологических исследований. Арх. биол. наук, 34.
— 1933. О превращениях вне организма производных вольфова и мюллерова канала
и о специфичности эпителиальных тканей мезодермального происхождения.
Арх. биол. наук, 34.
— 1933. Ueber einige Probleme der modernen Histclcgie. Z. mikr.-anat. Fcrsch., 34.
— 1933. Experimentell-histologische Untersuchungen uber das Epithelgewebe des Neben-
hodens. Z. Zellforsch., 20.
— 1933. La importancia de las investigaciones histoldgicas, histogeneticas у embrioWgicas
experimentales en cancerologia. Accion med., 5.
— 1934. Эволюция эпителиальных тканей и их взаимоотношения, с внутренней и внеш-
ней средой организма. Арх. биол. наук, 36.
— 1934. Ueber die Verwandlungsfahigkeit verschiedener Epithelgewebe ausserhalb des
486
Литера тура
Organismus und ihre Bedeutung fur das Problem der histologischen Determination.
Arch. exp. Zellforsch., 15.
— 1934. Динамика эпителиальных тканей и ее значение для понимания роли эпителиев
в воспалительных процессах. Докл. на Всесоюзн. конфер. патол., М.
— 1935. О некоторых особенностях и значении пограничных участков между покров-
ными эпителиальными пластами разного строения в пищеварительном аппарате.
Юб. сб. 25-лет. проф. Н. Аничкова.
— 1935. Система и пути эволюции эпителиальных тканей в свете экспериментального
анализа. Усп. совр. биол., 4.
— 1935. Превращения мезотелия серозных оболочек при экспериментальных условиях
и вопрос об его тканевой специфичности. Бклл. ВИЭМ, № 3.
— 1936. Гистологические элементы мышечных тканей. Арх. анат., гист. и эмбр., 15.
— 1936. Ueber das Verhalten und Wachstum seroser Haute im Explantat. Arch. exp.
Zellforsch., 19.
— 1936. Ueber Regenerationsprezesse im Mesothel und die Bedeutung der Serosadeckzel-
len. Ziegl. Beitr., 98.
— 1937. О покровных клетках серозных оболочек и их положении в системе гистоло-
гических структур. Арх. анат., гист. и эмбр., 16.
— 1937, Рост и превращения вне организма выстилки плавательного пузыря костистых
рыб. Арх. анат., гист. и эмбр., 17.
— 1937. Experimentell-histologishe Untersuchungen iiber das Coelomepithel. Acta
zool., 18.
— 1938. Опыт экспериментального анализа вспомогательных компонентов нервной
системы. ДАН СССР, 18.
* — 1938. Гистологическая детерминация, гистогенез и проблема естественной системы
опухолевых тканей. Тр. I Съезда онкологов УССР.
— 1938. Вопросы гистогенеза опухолевых тканей в свете учения о гистологической
детерминации. Юб. сб. Онкол. инет.
— 1939. Рост и превращения вне организма тканевых элементов зрительных путей.
Арх. анат., гист. и эмбр., 21.
— 1939. О культивировании in vitro частиц периферических нервов. ДАН СССР, 23.
— 1940. Материалы к построению естественной системы гистологических структур.
Арх. анат., гист. и эмбр., 23.
— 1940. Экспериментально-гистологические исследования над мускулатурой сомати-
ческого типа. Арх. анат., гист. и эмбр., 23.
— 1940. Рост и превращения вне организма элементов периферических нервов. Арх.
агат., гист. и эмбр., 23.
— 1940. Культура тканей. Медгиз. Л.
— 1940. Das allgemeine Verhalten und Wachstum der postfoetalen Neuroglia im Explan-
tat. Acta neerl. morph, norm, et path., 3.
— 1941. Опыты с эксплантацией белого вещества полушарий головного мозга. ДАН
СССР, 31.
— 1943. Морфологические классификации и генетическая система тканевых структур.
Усп. совр. биол., 16.
— 1943. О некоторых закономерностях эволюции тканей. Журн. общ. биол., 4.
ХлопинН. иГар ш и н В. 1937. Об изменениях эпителия семевыносящих путей
при эпидидимитах. Арх. биол. наук, 47.
Хлопни Н., Лазаренко Ф. и Цымбал В. 1929. Опыт культивирования in
vitro некоторых сарком в гетерологической плазме. Вести, хирургии, 53.
Хлоп и нН. и Цымбал В. (Chiopin u. Zymbal) 1934. О превращениях вне
организма элементов меланотических опухолей. Бюлл. ВИЭМ, № 3.
— 1934. Ueber das Verhalten und die Verwandlungen melanotischer Geschwulstelemente
im Explantat. Acta cancr., 1.
X л о п и н а И. 1911. Рост и превращения элементов эпифиза вне организма. ДАН
СССР. 31
Литература 487
ЦымбалВ. (Zymbal W.) 1932. Ueber die Vcrwandlungen des Epithels des Ohrspci-
cheldriise im Explantat. Arch. exp. Zellforsch., 12.
— 1933. Histologische und experimentelle Untersuchungen am Epithelgewebe der Talg-
driisen (Gehorgangdruse der Ratte). Z. Zellforsch,, 18.
— 1933. Histologische und experimentelle Untersuchungen iiber die Geschwiilste derSpei-
cheldriisen. Zeigl. Beitr., 91.
— 1935. Рост и превращения вне организма почечного эпителия млекопитающих. Арх.
биол. наук, 38.
— 1935. Experimentell-histologische Untersuchungen iiber das Epithelgewebe des Meta-
nephros. Arch. exp. Zellforsch., 18.
— 1937. Превращения in vitro сердечной мышцы. Арх. анат., гист. и эмбр., 17.
— 1938. Опухоли слюнных желез. Дисс. ВИЭМ. Л.
— 1938. О культивировании вне организма гладкой мезенхимной мускулатуры. Докл.
на конф. ВИЭМ по мышечным тканям. Л.
— Экспериментально-гистологические исследования над эпителием гипофиза (рукопись).
Шевченко Н. 1940. Динамика структур многослойных эпидермальных тканей
в процессе репаративной регенерации. Дисс. ВИЭМ. Л.
— 1941. Репаративная регенерация эпидермальной ткани слизистой оболочки ротовой
полости. Арх. анат., гист. и эмбр., 26.
— Заживление ран кожного эпителия. Зоол. ж. (в печати).
— Репаративная регенерация эпителия роговицы млекопитающих. Офтальмол. вести,
(в печати).
— О репаративной регенерации эпидермиса аксолотля. ДАН СССР (в печати).
— Репаративная регенерация многослойного плоского эпителия преджелудка на
месте стыка его с однослойным призматическим (Рукопись).
ОГЛАВЛЕНИЕ
Стр.
Предисловие ............................................................. 3
I. Очерк развития гистологической системы
1. Вводные замечания.................................................... 5
2. Предистория гистологических представлений ........................... 6
3. Макроскопическая гистология начала XIX в............................. 9
4. Гистология в период обоснования и развития клеточного учения ....... 19
5. Морфо-физиологическая система гистологии Лейдига и Кёлликера........ 24
6. Генетический принцип в гистологической системе и учение о специфичности
тканей................................................................27
7. Односторонние физиологические и формально-морфологические построения
новейшего времени...................................................... 36
8. Эклектические моменты в современных трактовках гистологической системы
Лейдига и Кёлликера ................................................... 39
9. Предпосылки к генетической системе тканевых структур................ 43
II. Проблема эволюционной гистологии
1. Эволюционная трактовка гистологической системы Лейдига и Кёлликера .... 44
2. Параллелизмы и конвергенции в области гистологии и сравнительной анатомии 47
3. Дивергентное развитие и дивергентная дифференцировка в области гистологи-
ческих структур.......................................................57
4. Проблема возникновения и ранних этапов эволюции тканей многоклеточных
животных .............................................................. 63
III. Экспериментальный метод в нормальной и патологической гистологии
1. Принципы и значение экспериментально-гистологического анализа тканей . . 81
2. Эпителиальные структуры и родственные им образования............... 102
А. Эпидермальный тканевой тип ....................... 105
а) Краткие эмбриологические и сравнительные данные ...............—
б) Многослойные эпидермальные пласты кожи и пищеварительной трубки
позвоночных, волосяные влагалища............................. 117
в) Покровный эпителий воздухоносных, мочеотводящих и семевынося-
щих путей млекопитающих и человека......................... 127
г) Эпителий эпидермальных желез.............................. 137
д) Эпителий эпидермальных эндокриновых органов............... 143
е) Дышащие части легких и выстилка плавательного пузыря...... 150
ж) Ткань хорды .............................................. 152
з) Эмбриональные зачатки эпидермиса и выстилки головной кишки . . . 154
и) Эпидермальные ткани беспозвоночных......................... 156
к) Опухоли эпидермального происхождения....................... 161
490
Оглавление
Б. Энтеродермальный тканевой тип I .................... . 164
а) Эпителий пищеварительной трубки............................. 167
б) Поджелудочная железа........................................ 179
в) Желчные пути и печень....................................... 184
г) Свойства собственно энтодермы как эмбрионального зачатка.... 190
д) Эктодермальные опухолевые ткани............................. 191
е) Филогенетические корни зачатка головной кишки позвоночных .... 194
В. Целонефрздермальный тканевой тип............................... 195
а) Краткие эмбриологические и сравнительные данные ............ •—
б) Целомическая выстилка ...................................... 205
в) Гонады и корковое вещество надпочечника..................... 216
г) Почечный эпителий .......................................... 219
д) Эпителий производных мюллеровых каналов (яйцеводов и матки) . . . 224
е) Эмбриональные зачатки целонефродермальных эпителиев........ 228
ж) Опухолевые ткани целонефродермального типа................. 230
Г. Эпендимо-глиальный тканевой тип или вспомогательные ткани нервной
системы......................................................... 23з
а) Общие замечания. Краткие эмбриологические и сравнительные данные —
б) Нейроглия и эпендима спинного и головного мозга............. 250
в) Зрительные пути, сетчатая оболочка глаза и эпифиз... 265
г) Оболочки центральной нервной системы........................ 277
д) Перепончатый лабиринт....................................... 281
е) Нейроглия периферической нервной системы ...... 283
ж) Эмбриональные зачатки нервной системы и их гистобластические по-
тенции ......................................................... 288
з) Ткани нейрогенных опухолей.................................. 291
Д. Эндотелий сосудистой системы................................... 304
3. Вопрос о мезенхиме, соединительных или межуточных тканях и свободных
клетках соков тела...................................................... 314
а) Вводные замечания.......................................... —
б) Кровь и волокнистая соединительная ткань позвоночных....... 324
в) Скелетные энтомезенхимные ткани позвоночных................. 340
г) Соединительнотканные образования и полостная жидкость беспозво-
ночных ........................................................ 34
д) Опухолевые соединительнотканные структуры................... 355
4. Мышечные или сократимые ткани....................................... 357
а) Общие гисгогенетические и сравнительные соображения......... •—
б) Мускулатура соматического типа.............................. 373
в) Мускулатура целомического типа.............................. 388
г) Мезенхимная мускулатура позвоночных ........................ 396
д) Мионейральные элементы радужины............................. 399
е) Миоэпидермальные элементы................................... 401
5. Нервная ткань или нейрональные элементы нервной системы............. 402
а) Общие замечания............................................. —
б) Некоторые общегистологические особенности нейронов позвоночных . 413
Оглавление 491
IV. Естественная система гистологических структур........... 431
Литература.................................................................. 445
А. Общий указатель работ......................................................—
Б. Указатель работ Н. Г. Хлспина и руководимого им научного коллек-
тива ...................................................................483
Н. Г. ХЛОПИН. ОБЩЕБИОЛОГИЧЕСКИЕ и
ЭКСПЕРИМЕНТАЛЬНЫЕ ОСНОВЫ ГИСТО-
ЛОГИИ
*
Печатается по постановлению
Редакционно-издательского со-
вета Академии Наук СССР
Редактор проф. Г. К. Хрущев
*
Подписано к печати 13/VI 1946 г. РИСО АН СССР
№ 2091. Печ. л. 31’/,. Уч.-изд. л. 40,5. Тираж
3000 экз. Зак. № 349
2-я типография «Печатный Двор» им.
А. М. Горького треста «Полиграфкнига» ОГИЗа при
Совете Министров РСФСР, Ленинград, Гатчин-
ская, 26.